WO2009093703A1 - L-アミノ酸の製造法 - Google Patents
L-アミノ酸の製造法 Download PDFInfo
- Publication number
- WO2009093703A1 WO2009093703A1 PCT/JP2009/051104 JP2009051104W WO2009093703A1 WO 2009093703 A1 WO2009093703 A1 WO 2009093703A1 JP 2009051104 W JP2009051104 W JP 2009051104W WO 2009093703 A1 WO2009093703 A1 WO 2009093703A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- gene
- amino acid
- culture
- microalgae
- medium
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/08—Lysine; Diaminopimelic acid; Threonine; Valine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/14—Glutamic acid; Glutamine
Definitions
- the present invention relates to a method for producing an L-amino acid using a microorganism.
- L-amino acids are used in various fields such as seasonings, food additives, feed additives, chemical products, and pharmaceuticals.
- L-amino acids such as L-threonine and L-lysine are industrially produced by fermentation using L-amino acid-producing bacteria such as Escherichia bacteria having the ability to produce L-amino acids.
- L-amino acid-producing bacteria strains isolated from the natural world, artificial mutants of the strains, recombinants in which L-amino acid biosynthetic enzymes are enhanced by gene recombination, and the like are used.
- Examples of the method for producing L-threonine include the methods described in Patent Documents 1 to 4.
- examples of the method for producing L-lysine include the methods described in Patent Documents 5 to 8.
- saccharides that is, glucose, fructose, sucrose, molasses, starch hydrolysate, and the like are used as carbon sources.
- a carbon source in the L-amino acid fermentation method is a saccharified starch derived from higher plants such as corn and cassava. Since these have low moisture content and high starch content, it is easy to obtain starch industrially.
- starch contained in microalgae has a content comparable to corn and cassava per dry weight, but the dry alga body weight per algal culture is less than 1%.
- Patent Documents 9 to 10 or Non-Patent Document 1 describe performing ethanol fermentation using microalgal starch, but the results of ethanol fermentation are not shown. In addition, no examples of saccharification of microalgal starch used for amino acid production have been shown so far.
- Escherichia coli which is a typical amino acid-producing bacterium, can grow using glycerol as a sole carbon source (Non-patent Document 2), and a long chain fatty acid having 12 or more carbon chains as a sole carbon source. It is known that it can grow as (Non-patent Document 3). Therefore, Escherichia coli can assimilate both long-chain fatty acids and glycerol, which are hydrolysates of fats and oils, but does not have lipase activity and cannot assimilate fats and oils directly. It is described in Patent Document 4. Furthermore, it is generally known that the solubility of long-chain fatty acids is extremely low.
- Non-Patent Document 5 the solubility is 0.1 g / L or more for lauric acid, but 0.0003 g / L or less for oleic acid.
- palmitic acid is 0.00000003 g / L or less is described. Therefore, it is difficult to assimilate glycerol and fatty acids with high water solubility at the same time.
- L-amino acids by direct fermentation using a hydrolyzate of fats and oils, which is a mixture of long-chain fatty acids and glycerol, as a carbon source. Has not been reported so far.
- Soybean seeds and oil palm fruits which are oil plants generally used as edible fats and oils, contain about 20% fat.
- microalgae are known to produce fats and oils, and the yield of fats and oils per area greatly exceeds oil plants.
- the steps of alga body separation, dehydration, cell disruption, and purification are complicated and difficult. Therefore, there has been no report on the production of L-amino acids by direct fermentation using algae-derived fats and oils as a carbon source.
- the present invention provides a more efficient L-amino acid production method, and in particular, L-amino acid fermentation production using microorganisms that have been conventionally carried out mainly using sugars derived from higher plants as a carbon source.
- a carbon source derived from microalgae for the method, a more inexpensive method for producing L-amino acids is provided.
- the present inventors have obtained a saccharified product obtained by hydrolyzing a starch obtained from microalgae with a bacterium having L-amino acid-producing ability, Cultivation in a medium using a carbon source without completely purifying a hydrolyzate obtained by hydrolyzing an extract containing oil or fat or a fraction obtained by hydrolyzing alga bodies of microalgae
- L-amino acids can be produced efficiently. Based on this finding, the present invention has been completed.
- a bacterium having L-amino acid-producing ability is cultured in a medium containing a processed product of microalgae, L-amino acid is produced and accumulated in the culture, and L-amino acid is collected from the culture.
- the treated product is, for example, (1) a crushed product of the microalgae culture, (2) an extract or fraction of the crushed product containing a mixture of organic matter derived from the microalgae, or (3 ) A hydrolyzate of the crushed product, the extract or the fraction.
- the treated product is preferably a crushed alga body of a microalga that produces starch, or a saccharified product obtained by hydrolyzing an extract containing starch or a fraction of the extract.
- the saccharified product is preferably a reaction product obtained by an enzymatic reaction using amylase from a crushed product of microalga or a fraction containing starch. Said amylase is preferably glucoamylase.
- the treated product is preferably a hydrolyzate obtained by hydrolyzing an algal body crushed product of microalgae that produces oil or fat, an extract containing oil or fat, or a fraction of the extract.
- the hydrolyzate is preferably a reaction product obtained by enzymatic reaction using lipase from a crushed algal body of a microalga or a fraction containing oils and fats.
- the hydrolyzate may be subjected to an emulsification treatment.
- the method for obtaining the crushed material is one or more methods selected from the group consisting of high temperature treatment, organic solvent treatment, boiling treatment, and strong alkali treatment.
- An example of the high temperature treatment is treatment at a temperature of 150 ° C. or higher.
- the microalgae is preferably an algae belonging to the class of green algae, treboxya algae or diatoms, and more preferably an algae belonging to the class of Chlorophyceae.
- the bacterium is preferably a bacterium belonging to the family Enterobacteriaceae or a coryneform bacterium, and more preferably a bacterium belonging to the genus Escherichia.
- the L-amino acid is, for example, one or more L-amino acids selected from the group consisting of L-lysine, L-threonine, and L-glutamic acid.
- the bacterium is dihydrodipicolinate reductase, diaminopimelate decarboxylase, diaminopimelate dehydrogenase, phosphoenolpyruvate carboxylase, aspartate aminotransferase, diaminopimelate epimerase, aspartate semi
- the activity of one or more enzymes selected from the group consisting of aldehyde dehydrogenase, tetrahydrodipicolinate succinylase, and succinyldiaminopimelate deacylase is enhanced and / or the activity of lysine decarboxylase is weakened It is preferable that
- the bacterium is selected from the group consisting of aspartate semialdehyde dehydrogenase, aspartokinase I encoded by the thr operon, homoserine kinase, aspartate aminotransferase, and threonine synthase.
- the activity of one or more selected enzymes is preferably enhanced.
- the bacterium is one or more selected from the group consisting of glutamate dehydrogenase, citrate synthase, phosphoenolpyruvate carboxylase, and methyl citrate synthase. It is preferred that the activity of the enzyme is enhanced and / or the activity of ⁇ -ketoglutarate dehydrogenase is weakened.
- the medium preferably contains the treated product as a carbon source.
- the present invention also provides a method for producing an L-amino acid comprising the following steps.
- the treated product is, for example, (1) a crushed product of the microalgae culture, (2) an extract or fraction of the crushed product containing a mixture of organic matter derived from the microalgae, or (3 ) A processed product which is a hydrolyzate of the crushed product, the extract or the fraction.
- the crushing method is preferably one or more methods selected from the group consisting of high temperature treatment, organic solvent treatment, boiling treatment, and strong alkali treatment.
- the treatment product preparation step preferably includes a step of saccharification by crushing and / or extracting and fractionating microalgae producing starch and hydrolyzing the treatment product.
- the saccharification step preferably includes performing an enzyme reaction using amylase. Said amylase is preferably glucoamylase.
- the treated product preparation step preferably includes a step of crushing and / or extracting / fractionating fine algae producing fats and oils and hydrolyzing the treated oil. It is preferable that the hydrolysis step includes performing an enzymatic reaction using lipase. The hydrolyzate may be emulsified.
- the microalgae is preferably an algae belonging to a green plant gate or an irregular planta, and more preferably, the microalgae is an algae belonging to a green alga, Trepoxya algae, or a diatom. Particularly preferably, the microalgae are algae belonging to the class Chlorophyceae.
- the bacterium is preferably a bacterium belonging to the family Enterobacteriaceae or a coryneform bacterium, and more preferably Escherichia coli.
- L-amino acids can be produced more efficiently.
- L-amino acids can be produced at low cost by using an inexpensive carbon source derived from microalgae.
- Microalgae used in the present invention and its culture method Any microalgae can be used in the present invention, but it is a microalgae that accumulates starch and / or fats and oils in the algae. It is preferable.
- Algae refers to all organisms that perform oxygen-generating photosynthesis, excluding moss plants, fern plants, and seed plants that inhabit the ground. Algae includes prokaryotes, cyanobacteria, eukaryotes, Glaucophyta, red plant algae (Rhodophyta), green plant gate (Chlorophyta), cryptophyte Gates (Cryptophyta), Haptophyta (Haptophyta), Hetero sparklephyta, Dinophyta, Dinophyta, Euglenophyta, Euglenaphyta Included are various unicellular and multicellular organisms that are classified as Chlorarachniophyta. Microalgae refers to algae with a microscopic structure excluding seaweeds that are multicellular organisms from these algae (Biodiversity Series (3) Diversity and strains of algae: edited by Mitsuo Senbara 1999)).
- Plants including algae often use starch as storage polysaccharide (Ball, S. G. and Morell, M. K. 2003. Annual Review of Plant Biology, 54: 207-233).
- algae that accumulate starch and typical algae include the Plasinophyceae, Chlorophyceae, Trebouxiophyceae, and Aosa algae that belong to the green plant kingdom. (Ulvophyceae) and axle algae (Charophyceae).
- algae belonging to the Chlorophyceae and Trebouxiophyceae are well studied, and the algae belonging to the Chlorophyceae is Chlamydomonas, and the algae belonging to the Trevorxia algae is Chlorella. The genus is mentioned.
- Chlamydomonas genus Chlamydomonas reinhardtii (Ball, SG 1998. The Molecular Biology of Chloroplasts and Mitochondria in Chlamydomonas, pp. 549-567. Rochaix J.-D. M., and Merchant S. (Eds), Kluwer Academic Publishers), and Chlorella kessleri (formerly Chlorella ⁇ vulgaris) (Izumo, A. et al. 2007. Plant Science 172: 1138-1147) ).
- Chlamydomonas reinhardtii CC125 strain is exemplified as Chlamydomonas reinhardi
- Chlorellalorkessleri 11h strain is exemplified as Chlorella kessarelli.
- These strains can be found, for example, in the University of Texas Algae Culture Collection (The University Texas at Austin, The Culture Collection of Algae (UTEX), 1 University Station A6700, Austin, TX 78712-0183, USA), respectively. It is stored with a UTEX 263 accession number and can be obtained from UTEX.
- Chlorella Quesarelli 11h strain was stored in the IAM Culture Collection at the University of Tokyo Institute for Molecular Cell Biology with the storage number C-531, and then transferred to the National Institute for Environmental Studies, Microbial System Storage Facility (NIES) .
- the shares are also stored in the American Type Culture Collection (ATCC, PO Box 1549, Manassas, VA 20108, 1, United States of America) with the acceptance number of ATCC11468 and may be sold by ATCC. it can.
- Neochloris oleo abundance include Neochloris oleoabundans UTEX 1185, Nanochloris SP includes Nannochloris sp.
- UTEX LB 1999 and Thalassiosira pseudonana UTEX LB FD2. These strains can be obtained from the University of Texas Algae Culture Collection (The University of Texas, Austin, The Culture Collection of Algae (UTEX), University, Station A6700, Austin, TX 78712-0183, USA).
- Neochloris Oreo abundance and Nanochloris SP are modified NORO medium (Yamaberi, K. et al. 1998. J. Mar. Biotechnol. 6: 44-48; Takagi, M. et al. 2000. Appl. Microbiol. Biotechnol) .54: 112-117) and Bold's Basal Medium (Tornabene, T. G. et al. 1983. Enzyme and Microb. Technol. 5: 435-440; Archibald, P. A. and Bold, H. C. 1970. Phytomorphology 20: 383-389).
- F / 2 medium (Lie, C.-P. and Lin, L.-P. 2001. Bot. Bull. Acad. Sin. 42: 207-214) Etc. can be used suitably.
- a photobioreactor can also be used for culturing microalgae (WO2003 / 094598 pamphlet).
- the initial pH is preferably around 7-9 neutral, and pH adjustment is often not performed during culturing, but it may be done as needed.
- the culture temperature is preferably 25-35 ° C., and particularly around 28 ° C. is a commonly used temperature, but the culture temperature may be any temperature suitable for the algae used.
- air is blown into the culture medium, and an aeration rate of 0.1-2 vvm (volume per volume per minute) per one minute of the culture solution volume is often used as the aeration rate. Further, CO 2 is blown in order to accelerate the growth, but it is preferable to blow about 0.5-5% with respect to the aeration amount.
- the optimal intensity of light irradiation varies depending on the type of microalgae, but about 1,000-10,000 lux is often used.
- a white fluorescent lamp is generally used indoors, but is not limited thereto. It is also possible to incubate outdoors with sunlight. If necessary, the culture solution may be stirred or circulated with an appropriate strength.
- Algae are known to accumulate fats and oils in the algae when the nitrogen source is depleted (Thompson GA Jr. 1996. Biochim. Biophys. Acta 1302: 17-45), which limits the concentration of the nitrogen source.
- the medium can also be used for the main culture.
- the culture of microalgae includes a culture solution containing algal bodies and algal bodies recovered from the culture solution.
- the method for recovering the algal cells from the culture solution is possible by general centrifugation, filtration, or sedimentation by gravity using a flocculant (Grima, E. M. et al. 2003). Biotechnol. Advances 20: 491-515).
- the microalgae treatment product is a treatment product containing a mixture of organic substances derived from cells of crushed microalgae, and produces L-amino acids. Which promotes the production and accumulation of L-amino acids by bacteria having ability, and specifically includes (1) a crushed culture of the microalgae, and (2) a mixture of organic substances derived from the microalgae And an extract or fraction of the crushed product, or (3) a hydrolyzate of the crushed product, the extract or the fraction.
- “Promoting the production and accumulation of L-amino acids” means that a mixture of organic substances derived from cells of crushed microalgae contained in the treated product is used for bacterial growth and production of L-amino acids. -It means that it contributes substantially as a source of carbon constituting the amino acid, and if it is a processed product that can make such a contribution, it will be treated as a "processed product that promotes production and accumulation of L-amino acids" of the present invention. included.
- the treated product promotes the production and accumulation of L-amino acid is confirmed by culturing the bacterium under the same conditions except for the presence or absence of the treated product, and comparing the production and accumulation amount of L-amino acid in the culture it can.
- L-amino acid accumulation may be any as long as it is improved compared to L-amino acid accumulation in the culture to which the treated product is not added, but it is preferably 10% or more, preferably 20% or more, more preferably compared to the culture without addition. It is desirable that L-amino acid accumulation is improved by 30% or more.
- the growth rate of microorganisms and the increase in the amount of microorganisms in the medium are also included in the “promoting the production and accumulation of L-amino acids” of the present invention. It is desirable that the rate and the amount of bacterial cells are increased by 10% or more, preferably 20% or more, and more preferably 30% or more compared to the culture without addition.
- the L of the present invention can be used as long as it can substantially contribute as a source of carbon constituting the bacterial cell component and L-amino acid in the growth of bacteria and the production of L-amino acid. -Included in processed products that promote production accumulation of amino acids. Therefore, when the amount of L-amino acid produced and accumulated is increased as compared with the condition where no treated product is added, it is also included in the treated product of the present invention, but from the same amount of purified material as the contained carbon source. It is preferable that the L-amino acid production accumulation amount is improved as compared with the case where a carbon source is added.
- L-amino acid production accumulation is improved when the treatment process for purifying the carbon source is shortened as compared with the case where the carbon source comprising the purified substance is used.
- the shortening time of the treatment process is preferably shortened by 10% or more, preferably 20% or more, more preferably 30% or more.
- the target processed product can be obtained by crushing, extraction, fractionation, hydrolysis, and any combination thereof using the promotion of L-amino acid production and accumulation as an index. it can.
- any method may be used for disrupting the culture as long as the algal bodies are sufficiently disrupted.
- high-temperature treatment for example, a temperature of 100 ° C. or higher (preferably 150 ° C. or higher, further Preferably 175 to 215 ° C.), organic solvent treatment (eg, treatment with a mixed solvent of methanol: chloroform), boiling treatment, strong alkali treatment, ultrasonic treatment, French press, etc., and any combination thereof.
- the high temperature treatment includes a high temperature and high pressure reaction under conditions called hydrothermal reaction.
- the algal bodies can be crushed by a physical method.
- the crushed algae-derived organic solution can be used as a crude extract as it is or subjected to a hydrolysis reaction, but it can be removed by filtration or centrifugation to remove insoluble matter such as cell walls or freeze-drying. It can also be concentrated. Furthermore, you may use the solution containing the starch which performed a certain fraction. As for the starch fraction from the algal crushed material, the protein fraction can be separated and recovered based on the difference in specific gravity, for example, by the sedimentation rate from the suspension. Moreover, fats and oils can also be fractionated from alga body crushed material.
- 80% methanol or 80% acetone is added to the crushed alga body or the concentrated crushed material, and insoluble oils and fats are extracted with a solvent such as hexane or chloroform. Oils and fats can be extracted as minutes.
- the mixture of organic substances derived from the microalgae of the present invention preferably contains what can be used as a carbon source.
- the carbon source added separately to the medium for amino acid fermentation can be reduced or eliminated.
- the carbon source that can be used include starch and / or a hydrolyzate of fats and oils.
- the mixture of organic substances derived from the microalgae of the present invention contains starch produced by the microalgae
- this saccharified product can be added to the medium as a carbon source.
- the saccharified starch can be obtained from, for example, an organic solution derived from microalgae or a fraction containing starch by a chemical method such as acid hydrolysis or an enzymatic reaction using amylase.
- Starch is composed of amylose in which glucose is linearly linked by ⁇ -1,4-glucoside bonds and amylopectin having both ⁇ -1,4-glucoside bonds and ⁇ -1,6-glucoside bonds in the branches. It is a high molecular polysaccharide.
- Amylase cocoon is a general term for enzymes that hydrolyze glucoside bonds such as starch.
- ⁇ -amylase ⁇ -amylase EC 3.2.1.1
- ⁇ -amylase ⁇ -amylase EC 3.2.1.2
- glucoamylase glucoamylase EC 3.2.1.3
- the ⁇ -Amylase is an endo-type enzyme that randomly cleaves ⁇ -1,4-glucoside bonds such as starch and glycogen.
- ⁇ -amylase is an exo-type enzyme that sequentially degrades ⁇ -1,4-glucoside bonds in maltose units from the non-reducing end of starch.
- Glucoamylase also called amyloglucosidase
- amyloglucosidase is an exo-type enzyme that sequentially degrades ⁇ -1,4-glucoside bonds in units of glucose from the non-reducing end of starch, and also degrades ⁇ -1,6-linkages contained in amylopectin. To do. Since glucoamylase produces glucose directly from starch, it is widely used in the production of glucose and is also a preferred enzyme in the present invention.
- a saccharified product can be obtained from an algal body by an enzymatic reaction.
- a combination of boiling, ultrasonic treatment, alkali treatment, etc. as pretreatment (Izumo, A. et al. 2007. Plan Science 172: 1138) -1147).
- the conditions for the enzyme reaction can be appropriately set according to the properties of the enzyme used.
- amyloglucosidase Sigma-Aldrich A-9228
- an enzyme concentration of 2 to 20 U / mL, a temperature of 40 to 60 ° C., and a pH of 4 to 6 are preferable.
- the pH adjustment when an organic acid that can be assimilated by bacteria used in the production of L-amino acid is used as a buffer, the organic acid can be used as a carbon source together with a saccharified product of starch.
- the enzyme reaction product can be added to the medium as it is.
- the saccharified product of starch produced by microalgae hydrolyzed starch to produce oligosaccharides or monosaccharides such as maltose or glucose that can be assimilated by bacteria.
- the saccharified product of starch produced by microalgae may be substantially saccharified, but may be partially saccharified.
- 50% by weight or more, more preferably 70% by weight or more, particularly preferably 90% by weight or more of starch is converted to glucose.
- the saccharified product of starch produced by microalgae may contain a carbohydrate other than starch produced by microalgae or a saccharified product thereof.
- this hydrolyzate can be added to the medium as a carbon source.
- a crude extract obtained by crushing algal bodies of microalgae by heat treatment, etc. hydrolyze a mixed solution of organic substances extracted with a solvent such as ethanol, a mixture of methanol and chloroform, or acetone. You can also These solutions can be used as they are, but can also be concentrated by a treatment such as lyophilization or evaporation.
- This solution contains components that can be used as an organic nitrogen source such as amino acids and components that are effective for the growth of bacteria having amino acid-producing ability such as metals, and can also be used as a medium component that is not a carbon source.
- the fats and oils produced by the microalgae may be in any form as long as hydrolysis of fats and oils, preferably hydrolysis of fats and oils by enzymes is possible. , Extracts of crushed cell bodies containing oil and fat, fractions containing oil and fat obtained from the extract, and the like. Moreover, it is preferable that the said extract or fraction contains the organic substance effective for amino acid fermentation other than fats and oils.
- Oils and fats are esters of fatty acids and glycerol, also called triglycerides.
- the fatty acid species produced by hydrolysis are preferably those that can be assimilated as a carbon source by the bacteria used in the method of the present invention, and those having a high content are more preferable. preferable.
- Examples of long-chain fatty acid species that can be assimilated by bacteria having L-amino acid-producing ability include lauric acid, myristic acid, palmitic acid, stearic acid, and oleic acid.
- organisms include lipids that liberate fatty acids by hydrolysis in addition to fats and oils, and fatty acids generated by hydrolysis of lipids can also be used as a carbon source.
- lipids include simple lipids such as wax and ceramide, and examples of complex lipids include phospholipids and glycolipids.
- the oil / fat hydrolyzate is a hydrolyzate obtained by hydrolyzing the fine algal fat / oil by a chemical method or an enzymatic method.
- a chemical hydrolysis method a continuous high-temperature hydrolysis method in which oil and fat and water are brought into countercurrent contact under high temperature (250-260 ° C.) and high pressure (5-6 MPa) is generally performed. It is also known that fats and oils are hydrolyzed in the presence of a strong acid or an acid catalyst (US Pat. No. 4,218,386).
- the reaction is carried out industrially at a low temperature (around 30 ° C.) using an enzyme (Jaeger, K. E. et al. 1994. FEMS Microbiol. Rev. 15: 29-63).
- the enzyme lipase which catalyzes the hydrolysis reaction of fats and oils can be used.
- a hydrolysis rate of about 70-80% can be obtained.
- Industrially, high temperature (250-260 ° C) and high pressure (5-6 MPa) conditions are used.
- the enzymatic method can perform hydrolysis under milder conditions. It is easy for those skilled in the art to perform the enzyme reaction at a temperature suitable for the lipase reaction while stirring water and fats and oils.
- Lipase is an industrially important enzyme and has various industrial uses (Hasan, F. et al. 2006. Enzyme and Microbiol. Technol. 39: 235-251). One type or two or more types of enzymes may be used.
- Lipase is an enzyme that hydrolyzes fats and oils into fatty acids and glycerol, and is also called triacylglycerol lipase or triacylglyceride lipase.
- Lipase has been found in various organisms, but any species of lipase can be used as long as it catalyzes the above reaction.
- various attempts have been made to produce biodiesel fuel, which is a fatty acid ester, from fats and alcohols using lipase enzymes (Fukuda, H., Kondo, A., and Noda, H. 2001. J. Biosci. Bioeng. 92, 405-416).
- lipases derived from microorganisms many lipases derived from the genera Bacillus, Burkholderia, Pseudomonas, and Staphylococcus are known (Jaeger, K. E., and Eggert, T. 2002. Curr. Opin. Biotechnol). . 13: 390-397).
- the base sequence of the gene encoding LipA (GenBank Accession No. M74010) derived from Bacillus subtilis is shown in SEQ ID NO: 1, and the amino acid sequence is shown in SEQ ID NO: 2.
- the base sequence of the gene encoding LipA (GenBank Accession No. X70354) derived from Burkholderia glumae is shown in SEQ ID NO: 3, and the amino acid sequence is shown in SEQ ID NO: 4.
- the base sequence of the gene encoding LipA (GenBank Accession No. D50587) derived from Pseudomonas aeruginosa is shown in SEQ ID NO: 5, and the amino acid sequence is shown in SEQ ID NO: 6.
- the base sequence of the lipase derived from Staphylococcus aureus (GenBank Accession No. M12715) is shown in SEQ ID NO: 7, and the amino acid sequence is shown in SEQ ID NO: 8.
- the lipase derived from the yeast Candida ⁇ antarctica (GenBank Accession No.Z30645) is one of the commonly used lipases (Breivik, H., Haraldsson, G. G. and Kristinsson, B. 1997. J. Am. Oil Chem . Soc. 74: 1425-1429).
- the base sequence of the gene encoding the lipase is shown in SEQ ID NO: 9, and the amino acid sequence is shown in SEQ ID NO: 10.
- yeast Candida rugosa (Candida cylindracea) is known to have five or more lipases encoded by different genes (Alberghina, L. and Lotti, M. 1997. Methods Enzymol. 284: 246- 260).
- LIP1 and LIP2 are known as major lipases.
- the nucleotide sequence of lip1 (GenBank Accession No. X64703) encoding LIP1 is shown in SEQ ID NO: 11, and the amino acid sequence is shown in SEQ ID NO: 12.
- the base sequence of the gene of lip2 (GenBank Accession No. X64703) encoding LIP2 is shown in SEQ ID NO: 13, and the amino acid sequence is shown in SEQ ID NO: 14.
- Candida genus yeast such as Candida cylindracea
- the CTG codon encoding leucine encodes serine in the universal code (Kawaguchi, Y. et al. 1989. Nature 341: 164-166; Ohama, T. et al. 1993. Nucleic Acids Res. 21: 4039-4045).
- SEQ ID NOs: 11 to 14 the amino acid corresponding to CTG is described as Leu for convenience, but is actually Ser.
- the above lipase can be prepared from the cells or cultures of the above microorganisms, but it can be expressed in other host microorganisms using genetic engineering techniques using genes encoding each lipase. It may be prepared by.
- a yeast-derived gene whose CTG codon encodes serine, such as Candida rugosa (Candida cylindracea)
- CTG must be changed to another universal codon encoding serine (Schmidt -Dannert, C. 1999. Bioorg. Med. Chem. 7: 2123-2130).
- the lipase is characterized by having a GXSXG motif called lipase box around the Ser at the active center, and three residues of Ser, Asp, and His called catalytic traid that are commonly found in lipases, esterases, and serine proteases.
- the preservation of the group is mentioned.
- lipase ⁇ box corresponds to positions 106 to 110
- catalytic traid represents Ser at position 108, Asp at position 164, and His at position 187. Two residues correspond.
- Oil and fat hydrolyzate is a mixture of fatty acid and glycerol, and it is known that the weight ratio of glycerol to fatty acid contained in general oil and fat hydrolyzate is about 10%.
- the hydrolyzate may be the reaction product itself after the hydrolysis reaction, and is a product obtained by fractionating or purifying the reaction product as long as it contains a carbon source that can be assimilated by bacteria such as fatty acids derived from lipids and glycerol. There may be.
- the weight ratio of glycerol to fatty acid is preferably 2 to 50: 100, and more preferably 5 to 20: 100.
- the oil / fat hydrolyzate is generally separated into a lower layer containing glycerol (aqueous phase) and an upper layer containing fatty acid (oil phase) at a temperature around room temperature.
- a fraction mainly containing glycerol can be obtained.
- the upper layer is collected, a fraction mainly containing fatty acids can be obtained.
- any of these may be used as the carbon source, but it is preferable to use both glycerol and fatty acids.
- the emulsification treatment examples include emulsification accelerator addition, stirring, homogenization, ultrasonic treatment and the like. It is considered that the emulsification treatment makes it easier for bacteria to assimilate glycerol and fatty acids, and L-amino acid fermentation becomes more effective.
- the emulsification treatment may be any treatment as long as the bacteria having L-amino acid-producing ability make the mixture of fatty acid and glycerol easy to assimilate.
- an emulsification accelerator or a surfactant may be added as an emulsification method.
- examples of the emulsification promoter include phospholipids and sterols.
- nonionic surfactant polyoxyethylene sorbitan fatty acid ester such as poly (oxyethylene) sorbitan monooleate (Tween ⁇ ⁇ 80), alkyl glucoside such as n-octyl ⁇ -D-glucoside, Examples thereof include sucrose fatty acid esters such as sucrose stearate and polyglycerin fatty acid esters such as polyglycerin stearate.
- the zwitterionic surfactant include N, N-dimethyl-N-dodecylglycine betaine which is an alkylbetaine.
- Triton X-100 Triton X-100
- polyoxyethylene (20) cetyl ether Brij-58
- nonylphenol ethoxylate Tegitol NP-40
- This operation may be any operation that promotes emulsification and homogenization of a mixture of fatty acid and glycerol.
- stirring treatment, homogenizer treatment, homomixer treatment, ultrasonic treatment, high pressure treatment, high temperature treatment and the like can be mentioned, and stirring treatment, homogenizer treatment, ultrasonic treatment and combinations thereof are more preferable.
- the treatment with the above emulsification accelerator with the stirring treatment, the homogenizer treatment, and / or the ultrasonic treatment, and these treatments are desirably performed under alkaline conditions where fatty acids are more stable.
- the alkaline condition is preferably pH 9 or higher, more preferably pH 10 or higher.
- Glycerol concentration can be measured with kits such as F-kit glycerol (Roche Diagnostics) and various biosensors.
- concentration of fatty acid or oil is determined by gas chromatography (Hashimoto, K. et al. 1996. Biosci. Biotechnol. Biochem. 70: 22-30) or HPLC (Lin, J. T. et al. 1998. J. Chromatogr A. 808: 43-49).
- Bacteria used in the present invention bacteria having L-amino acid-producing ability are used. Bacteria are not particularly limited as long as they can efficiently produce L-amino acids from organic substances produced by microalgae, in particular, saccharified starches or hydrolyzed oils and fats, such as Escherichia, Pantoea, Examples include, but are not limited to, bacteria belonging to the family Enterobacteriaceae such as Enterobacter, and so-called coryneform bacteria belonging to the genus Brevibacterium, Corynebacterium, and Microbacterium.
- the L-amino acid-producing bacterium in the present invention may be modified so as to enhance the assimilation ability of the hydrolyzate of fats and oils. For example, deletion of a gene encoding a transcription factor FadR that has a DNA binding ability to regulate fatty acid metabolism found in the intestinal bacteria group (DiRusso, C. C. et al. 1992. J. Biol. Chem. 267: 8685-8691; DiRusso, C. C. et al. 1993. ol Mol. Microbiol. 7: 311-322).
- the Escherichia coli fadR gene is located at base numbers 1,234,161 to 1,234,880 on the genome sequence of Escherichia coli MG1655 registered under Genbank Accession No. U00096, and GenBank accession No. It is a gene encoding a protein registered in AAC74271.
- the fadR gene sequence is shown in SEQ ID NO: 15.
- a gene involved in glycerol metabolism may be modified.
- glpR gene As genes involved in glycerol metabolism, the expression of glpR gene (EP1715056) is weakened to increase the utilization of glycerol, or glpA, glpB, glpC, glpD, glpE, glpF, glpG, glpK, glpQ, Expression of glycerol metabolic genes (EP1715055A) such as glpT, glpX, tpiA, gldA, dhaK, dhaL, dhaM, dhaR, fsa and talC genes may be enhanced.
- glycerol dehydrogenase gene gldA
- dhaKLM PEP-dependent dihydroxyacetone kinase gene
- dak ATP-dependent dihydroxyacetone kinase gene
- glycerol kinase In glycerol kinase (glpK), it is preferable to use a desensitized glpK gene in which feedback inhibition by fructose-1,6-phosphate is released. (WO2008 / 081959, WO2008 / 107277)
- the Enterobacteriaceae family includes bacteria belonging to genera such as Escherichia, Enterobacter, Erbinia, Klebsiella, Pantoea, Photohubadus, Providencia, Salmonella, Serratia, Shigella, Morganella, and Yersinia.
- the bacteria belonging to the genus Escherichia that can be used in the present invention are not particularly limited.
- Neidhardt et al. Neidhardt, F. C. Ed. 1996. Escherichia coli and Salmonella: Cellular and Molecular Biology / Second Edition pp 2477-2483.
- Table 1 1. American Society for Microbiology Press, Washington, DC).
- Specific examples include Escherichia coli W3110 (ATCC 273325) and Escherichia coli MG1655 (ATCC 47076) derived from the wild type K-12 strain of the prototype.
- strains can be sold, for example, from the American Type Culture Collection (address P.O. Box 1549 Manassas, VA 20108, United States of America). That is, the registration number corresponding to each strain is given, and it can receive distribution using this registration number. The registration number corresponding to each strain is described in the catalog of American Type Culture Collection. The same applies to strains with the following ATCC numbers.
- the bacterium belonging to the genus Pantoea means that the bacterium is classified into the genus Pantoea according to the classification known to microbiologists. Certain types of Enterobacter agglomerans were recently reclassified as Pantoea agglomerans, Pantoea ananatis, Pantoea stewarti and others (Int. J. Syst. Bacteriol. 1993) . 43: 162-173).
- the bacteria belonging to the genus Pantoea include bacteria that have been reclassified to the genus Pantoea in this way.
- Pantoea citrea Pantoea citrea
- Pantoea Ananatis AJ13355 (FERM BP-6614) (European Patent Application Publication No. 0952221)
- Pantoea Ananatis AJ13356 (FERM BP-6615) (European Patent Application Publication No. 0952221)
- Enterobacter bacteria examples include Enterobacter agglomerans, Enterobacter aerogenes, and the like. Specifically, strains exemplified in European Patent Application Publication No. 952221 can be used. A representative strain of the genus Enterobacter is Enterobacter agglomerans ATCC12287.
- Examples of the genus Erwinia include Erbinia amylobola and Erwinia carotobola, and examples of the Klebsiella bacterium include Klebsiella planticola. Specifically, the following strains are mentioned.
- the “coryneform bacterium” has been conventionally classified into the genus Brevibacterium, but includes bacteria that are currently classified into the genus Corynebacterium (Liebl, W. et al. 1991. Int. J. Syst. Bacteriol., 41: 255-260), and Brevibacterium spp. Closely related to the genus Corynebacterium. Examples of such coryneform bacteria include the following.
- strains can be exemplified.
- Corynebacterium acetoacidophilum ATCC13870 Corynebacterium acetoglutamicum ATCC15806 Corynebacterium alkanolyticum ATCC21511 Corynebacterium carnae ATCC15991 Corynebacterium glutamicum ATCC13020, ATCC13032, ATCC13060
- Corynebacterium herculis ATCC13868 Brevibacterium divaricatam ATCC14020 Brevibacterium flavum ATCC13826, ATCC14067 Brevibacterium immariophilum ATCC14068 Brevibacterium lactofermentum ATCC13869 (Corynebacterium glutamicum ATCC13869) Brevibacterium rose ATCC13825 Brevibacterium saccharolyticum AT
- a bacterium having an amino acid-producing ability refers to a bacterium having an ability to produce an L-amino acid and secrete it into the medium when cultured in the medium.
- it refers to a bacterium capable of accumulating the target L-amino acid in the medium in an amount of preferably 0.5 g / L or more, more preferably 1.0 g / L or more.
- L-amino acids include L-alanine, L-arginine, L-asparagine, L-aspartic acid, L-cysteine, L-glutamic acid, L-glutamine, glycine, L-histidine, L-isoleucine, L-leucine, L- Includes lysine, L-methionine, L-phenylalanine, L-proline, L-serine, L-threonine, L-tryptophan, L-tyrosine and L-valine. In particular, L-threonine, L-lysine and L-glutamic acid are preferable.
- an auxotrophic mutant, an L-amino acid analog resistant strain or a metabolically controlled mutant, or a recombinant strain with enhanced expression of an L-amino acid biosynthetic enzyme can be applied to the breeding of amino acid-producing bacteria such as coryneform bacteria or Escherichia bacteria (amino acid fermentation, Academic Publishing Center, Inc., May 30, 1986, first edition) Issue, see pages 77-100).
- amino acid-producing bacteria such as coryneform bacteria or Escherichia bacteria (amino acid fermentation, Academic Publishing Center, Inc., May 30, 1986, first edition) Issue, see pages 77-100).
- the auxotrophy, analog resistance, metabolic control mutation and other properties imparted may be singly or may be two or more.
- L-amino acid biosynthesis enzymes whose expression is enhanced may be used alone or in combination of two or more.
- imparting properties such as auxotrophy, analog resistance, and metabolic regulation mutation may be combined with enhancement of biosynthetic enzymes.
- an auxotrophic mutant an analog resistant strain, or a metabolically controlled mutant having L-amino acid production ability
- the parent strain or the wild strain is subjected to normal mutation treatment, that is, irradiation with X-rays or ultraviolet rays, or N-methyl.
- the L-amino acid-producing ability can be imparted or enhanced by enhancing the enzyme activity by gene recombination.
- the enhancement of enzyme activity include a method of modifying a bacterium so that expression of a gene encoding an enzyme involved in L-amino acid biosynthesis is enhanced.
- an amplified plasmid in which a DNA fragment containing the gene is introduced into an appropriate plasmid for example, a plasmid vector containing at least a gene responsible for the replication replication function of the plasmid in a microorganism
- these genes can be achieved by making multiple copies on the chromosome by joining, transferring, etc., or by introducing mutations into the promoter regions of these genes (see International Publication No. 95/34672). .
- the promoter for expressing these genes may be any promoter that functions in coryneform bacteria, and the promoter of the gene itself used. Or may be modified.
- the expression level of the gene can also be controlled by appropriately selecting a promoter that functions strongly in coryneform bacteria, or by bringing the -35 and -10 regions of the promoter closer to the consensus sequence.
- the method for enhancing the expression of the enzyme gene as described above is described in International Publication No. 00/18935, European Patent Application Publication No. 1010755, and the like.
- L-threonine-producing bacteria Preferred as microorganisms having L-threonine-producing ability include bacteria in which one or more activities of L-threonine biosynthetic enzymes are enhanced.
- L-threonine biosynthesis enzymes include aspartokinase III (lysC), aspartate semialdehyde dehydrogenase (asd), aspartokinase I (thrA) encoded by the thr operon, homoserine kinase (thrB), threonine synthase ( thrC), aspartate aminotransferase (aspartate transaminase) (aspC).
- the parentheses are abbreviations for the genes (the same applies to the following description). Of these enzymes, aspartate semialdehyde dehydrogenase, aspartokinase I, homoserine kinase, aspartate aminotransferase, and threonine synthase are particularly preferred.
- the L-threonine biosynthetic gene may be introduced into a bacterium belonging to the genus Escherichia in which threonine degradation is suppressed. Examples of the Escherichia bacterium in which threonine degradation is suppressed include, for example, the TDH6 strain lacking threonine dehydrogenase activity (Japanese Patent Laid-Open No. 2001-346578).
- the enzyme activity of the L-threonine biosynthetic enzyme is suppressed by the final product, L-threonine. Therefore, in order to construct an L-threonine-producing bacterium, it is desirable to modify the L-threonine biosynthetic gene so that it is not subject to feedback inhibition by L-threonine.
- the thrA, thrB, and thrC genes constitute the threonine operon, but the threonine operon forms an attenuator structure, and the expression of the threonine operon inhibits isoleucine and threonine in the culture medium. The expression is suppressed by attenuation.
- This modification can be achieved by removing the leader sequence or attenuator of the attenuation region (Lynn, S. P. et al. 1987. J. Mol. Biol. 194: 59-69; 02/26993 pamphlet; see the International Publication No. 2005/049808 pamphlet).
- a threonine operon as governed by a presser and promoter may be constructed. (See European Patent No. 0593792)
- a strain resistant to ⁇ -amino- ⁇ -hydroxyvaleric acid (AHV) may be selected. Is possible.
- the threonine operon modified so as not to be subjected to feedback inhibition by L-threonine has an increased copy number in the host or is linked to a strong promoter to improve the expression level. Is preferred.
- the increase in copy number can be achieved by transferring the threonine operon onto the genome by transposon, Mu-phage, etc., in addition to amplification by plasmid.
- L-threonine biosynthetic enzyme it is also preferable to enhance the glycolytic system, TCA cycle, genes related to the respiratory chain, genes controlling gene expression, and sugar uptake genes.
- these genes effective for L-threonine production include transhydronase (pntAB) gene (European Patent 733712), phosphoenolpyruvate carboxylase gene (pepC) (International Publication No. 95/06114 pamphlet), Phosphoenolpyruvate synthase gene (pps) (European Patent No. 877090), pyruvate carboxylase gene of Coryneform bacterium or Bacillus genus bacteria (International Publication No. 99/18228, European Patent Publication No. 1092776) Is mentioned.
- pntAB transhydronase
- pepC phosphoenolpyruvate carboxylase gene
- pps Phosphoenolpyruvate synthase gene
- pps European Patent No. 8
- genes that confer resistance include rhtA gene (Livshits, V. A. et al. 2003. Res. Microbiol. 154: 123-135), rhtB gene (European Patent Application Publication No. 0994190), rhtC gene ( European Patent Application Publication No. 1013765), yfiK, yeaS gene (European Patent Application Publication No. 1016710).
- rhtA gene Livshits, V. A. et al. 2003. Res. Microbiol. 154: 123-135)
- rhtB gene European Patent Application Publication No. 0994190
- rhtC gene European Patent Application Publication No. 1013765
- yfiK European Patent Application Publication No. 101710
- European Patent Application Publication No. 1016710 European Patent Application Publication No. 1016710
- L-threonine-producing bacteria or parent strains for inducing them examples include E. coli TDH-6 / pVIC40 (VKPM B-3996) (US Patent No. 5,175,107, US Patent No. 5,705,371), E. coli 472T23. / pYN7 (ATCC 98081) (U.S. Pat.No. 5,631,157), E.coli NRRL-21593 (U.S. Pat.No. 5,939,307), E.coli FERM BP-3756 (U.S. Pat.No. 5,474,918), E.coli FERM BP-3519 And FERM BP-3520 (U.S. Patent No. 5,376,538), E.
- E. coli MG442 (Gusyatiner et al., 1978. Genetika (in Russian), 14: 947-956), E. coli VL643 and VL2055 (European Patent Application Publication No. Strains belonging to the genus Escherichia, such as, but not limited to, 1149911).
- the TDH-6 strain lacks the thrC gene, is sucrose-utilizing, and the ilvA gene has a leaky mutation. This strain also has a mutation in the rhtA gene that confers resistance to high concentrations of threonine or homoserine.
- the B-3996 strain carries the plasmid pVIC40 in which the thrA * BC operon containing the mutated thrA gene is inserted into the RSF1010-derived vector. This mutant thrA gene encodes aspartokinase homoserine dehydrogenase I which is substantially desensitized to feedback inhibition by threonine.
- E. coli VKPM B-5318 (EP 0593792B) can also be used as an L-threonine producing bacterium or a parent strain for inducing it.
- the B-5318 strain is isoleucine non-required, and the control region of the threonine operon in the plasmid pVIC40 is replaced by a temperature sensitive lambda phage C1 repressor and a PR promoter.
- VKPM B-5318 was assigned to Russian National Collection of Industrial Microorganisms (VKPM) (1 Dorozhny proezd., 1 Moscow 117545, Russia) on May 3, 1990 under the accession number VKPM B-5318. Has been deposited internationally.
- the thrA gene encoding Escherichia coli aspartokinase homoserine dehydrogenase I is located at base numbers 337 to 2,799 on the genome sequence of Escherichia coli MG1655 strain registered in Genbank Accession No. ⁇ U00096, and GenBank accession No. AAC73113 It is a gene encoding a protein registered in.
- the thrB gene encoding the homoserine kinase of Escherichia coli is located at base numbers 2,801-3,733 on the genome sequence of Escherichia coli MG1655 registered under Genbank Accession No. U00096, and is registered under GenBank accession No. AAC73114 It is a gene that encodes the protein.
- the thrC gene encoding the threonine synthase of Escherichia coli is located at base numbers 3,734-5,020 on the genome sequence of Escherichia coli MG1655 registered in Genbank Accession No. U00096, and is registered under GenBank accession No. AAC73115. It is a gene that encodes the protein. These three genes are encoded as a threonine operon consisting of thrLABC downstream of the thrL gene encoding the leader peptide. In order to increase the expression of the threonine operon, it is effective to remove the attenuator region that affects transcription, preferably from the operon (WO 2005/049808, WO2003 / 097839).
- mutant thrA gene encoding aspartokinase homoserine dehydrogenase I resistant to feedback inhibition by threonine, and the thrB and thrC genes are one operon from the well-known plasmid pVIC40 present in the threonine producing strain E. coli VKPM B-3996. Can be obtained as Details of plasmid pVIC40 are described in US Pat. No. 5,705,371.
- the rhtA gene has nucleotide numbers 848,433-849,320 on the genome sequence of Escherichia coli MG1655 strain registered in Genbank Accession No. U00096 acquired as a gene that gives resistance to homoserine and threonine (rht: resistant to threonine / homoserine). It is a gene that codes for a protein located in (complementary strand) and registered in GenBank Accession No. AAC73900. It has also been found that the rhtA23 mutation that improves rthA expression is a G ⁇ A substitution at position -1 relative to the ATG start codon (Livshits, V. A. et al. 2003. Res Microbiol. 154 : 123-135, European Patent Application No. 1013765).
- the asd gene of Escherichia coli is located at base numbers 3,571,798 to 5723,572,901 (complementary strand) on the genome sequence of Escherichia coli MG1655 strain registered in Genbank Accession No. U00096, and registered as GenBank accession No. AAC76458 It is a gene that encodes a protein. It can be obtained by PCR using primers prepared based on the nucleotide sequence of the gene (see White, T. J. et al. 1989. Trends Genet. 5: 185-189). The asd gene of other microorganisms can be obtained similarly.
- the aspC gene of Escherichia coli is located at base numbers 983, 742 to 984,932 (complementary strands) on the genome sequence of Escherichia coli MG1655 strain registered in Genbank Accession No. U00096, and registered with GenBank accession No. AAC74014 It is a gene that encodes a protein that can be obtained by PCR.
- the aspC gene of other microorganisms can be obtained similarly.
- L-Lysine-producing bacteria include L-lysine analog resistant strains and metabolic control mutants.
- L-lysine analogs include oxalysine, lysine hydroxamate, S- (2-aminoethyl) -L-cysteine (hereinafter sometimes abbreviated as “AEC”), ⁇ -methyllysine, ⁇ -chloro.
- AEC S- (2-aminoethyl) -L-cysteine
- caprolactam etc. are mentioned, it is not limited to these.
- Mutants having resistance to these lysine analogs can be obtained by subjecting bacteria belonging to the family Enterobacteriaceae or coryneform bacteria to ordinary artificial mutation treatment.
- L-lysine-producing bacteria include Escherichia coli AJ11442 (FERM BP-1543, NRRL B-12185; see JP-A-56-18596 and US Pat. No. 4,346,170), Escherichia coli VL611. Strains (JP 2000-189180 A) and the like.
- WC196 strain see International Publication No. 96/17930 pamphlet
- L-lysine-producing bacteria can be constructed by increasing the enzyme activity of the L-lysine biosynthesis system. These increases in enzyme activity can be achieved by increasing the copy number of the gene encoding the enzyme in the cell or by modifying the expression regulatory sequence.
- the modification for enhancing the expression of the gene can be performed, for example, by increasing the copy number of the gene in the cell using a gene recombination technique.
- a DNA fragment containing the gapA gene may be ligated with a vector that functions in a host bacterium, preferably a multicopy vector, to produce a recombinant DNA, which is introduced into the bacterium and transformed.
- Increasing the gene copy number can also be achieved by having multiple copies of the above genes on the bacterial genomic DNA.
- homologous recombination is performed using a sequence present in multiple copies on the genomic DNA as a target.
- a sequence present in multiple copies on genomic DNA repetitive DNA and inverted repeats present at the end of a transposable element can be used.
- each gene may be linked in tandem beside the gapA gene present on the genome, or may be redundantly incorporated on an unnecessary gene on the genome.
- the gene expression can be enhanced by the method described in the pamphlet of International Publication No. 00/18935 using expression control sequences such as each promoter of the gene on genomic DNA or plasmid.
- Regulators that replace powerful genes, bring the -35 and -10 regions of each gene closer to consensus sequences, amplify regulators that increase gene expression, or reduce gene expression It can also be achieved by deleting or weakening.
- lac promoter for example, lac promoter, trp promoter, trc promoter, tac promoter, araBA promoter, lambda phage PR promoter, PL promoter, tet promoter, T7 promoter, ⁇ 10 promoter and the like are known as strong promoters. It is also possible to introduce a base substitution or the like into the promoter region or SD region of the gapA gene and modify it to a stronger one. Methods for evaluating promoter strength and examples of strong promoters are described in Goldstein et al. (Prokaryotic promoters in biotechnology. Biotechnol. Annu. Rev. 1995. 1: 105-128) and the like.
- L-lysine biosynthetic enzymes include dihydrodipicolinate synthase gene (dapA), aspartokinase gene (lysC), dihydrodipicolinate reductase gene (dapB), diaminopimelate decarboxylase gene (lysA) , Diaminopimelate dehydrogenase gene (ddh) (international publication No.
- the parent strain also encodes a gene (cyo) ((EP 1170376 A) involved in energy efficiency, a gene encoding nicotinamide nucleotide transhydrogenase (pntAB) (US Pat. No. 5,830,716), and a protein having L-lysine excretion activity.
- cyo a gene involved in energy efficiency
- pntAB nicotinamide nucleotide transhydrogenase
- pntAB nicotinamide nucleotide transhydrogenase
- gdhA glutamate dehydrogenase
- Wild-type dihydrodipicolinate synthase derived from Escherichia coli is known to undergo feedback inhibition by L-lysine, and wild-type aspartokinase derived from Escherichia coli is subject to inhibition and feedback inhibition by L-lysine. It has been known. Therefore, when using the dapA gene and the lysC gene, these genes are preferably mutant genes that are not subject to feedback inhibition by L-lysine.
- DNA encoding a mutant dihydrodipicolinate synthase that is not subject to feedback inhibition by L-lysine examples include DNA encoding a protein having a sequence in which the histidine residue at position 118 is substituted with a tyrosine residue.
- the threonine residue at position 352 is replaced with an isoleucine residue
- the glycine residue at position 323 is replaced with an asparagine residue
- 318 Examples include DNA encoding AKIII having a sequence in which the methionine at the position is replaced with isoleucine (see US Pat. Nos. 5,610,010 and 6,040,160 for these variants). Mutant DNA can be obtained by site-specific mutagenesis such as PCR.
- RSFD80, pCAB1, and pCABD2 are known as plasmids containing mutant dapA encoding mutant mutant dihydrodipicolinate synthase and mutant lysC encoding mutant aspartokinase (USA) Patent No. 6040160).
- Escherichia coli JM109 strain US Pat. No. 6,040,160 transformed with this plasmid was named AJ12396, and this strain was established on 28 October 1993 at the Institute of Biotechnology, Ministry of International Trade and Industry. Deposited to the National Institute of Advanced Industrial Science and Technology (AIST) as Deposit No. FERM P-13936, transferred to an international deposit based on the Budapest Treaty on November 1, 1994, with the deposit number of FERM BP-4859 It is deposited with.
- RSFD80 can be obtained from AJ12396 strain by a known method.
- examples of such enzymes include homoserine dehydrogenase, lysine decarboxylase (cadA, ldcC), malic enzyme, etc., and a strain in which the activity of the enzyme is reduced or absent is disclosed in International Publication No. WO95 / 23864, It is described in WO96 / 17930 pamphlet, WO2005 / 010175 pamphlet and the like.
- both the cadA gene and ldcC gene encoding lysine decarboxylase it is preferable to reduce the expression of both the cadA gene and ldcC gene encoding lysine decarboxylase. Decrease in the expression of both genes can be performed according to the method described in WO2006 / 078039 pamphlet.
- a mutation that reduces or eliminates the activity of the enzyme in the cell is applied to the gene of the enzyme on the genome by a usual mutation treatment method or gene recombination technique. What is necessary is just to introduce.
- Such mutations can be introduced, for example, by deleting a gene encoding an enzyme on the genome by genetic recombination or by modifying an expression regulatory sequence such as a promoter or Shine-Dalgarno (SD) sequence. Achieved.
- a modified gene in which a partial sequence of the target gene is modified so that it does not produce a normally functioning enzyme is prepared, and a bacterium belonging to the family Enterobacteriaceae is transformed with the DNA containing the gene.
- a bacterium belonging to the family Enterobacteriaceae is transformed with the DNA containing the gene.
- the gene replacement using such homologous recombination is a method called “Red-driven integration” (Datsenko, K. A, and Wanner, B. L. 2000. Proc. Natl. Acad. Sci U S A.
- a preferred L-lysine-producing bacterium includes Escherichia coli WC196 ⁇ cadA ⁇ ldcC / pCABD2 (WO2006 / 078039). This strain was constructed by disrupting the cadA and ldcC genes encoding lysine decarboxylase and introducing plasmid pCABD2 (US Pat. No. 6,040,160) containing a lysine biosynthesis gene from WC196 strain. The WC196 strain was obtained from the W3110 strain derived from E. coli K-12, and encodes aspartokinase III in which feedback inhibition by L-lysine was released by replacing threonine at position 352 with isoleucine.
- the WC196 strain was named Escherichia coli AJ13069.
- WC196 ⁇ cadA ⁇ ldcC was named AJ110692, and was deposited internationally on October 7, 2008, at the National Institute of Advanced Industrial Science and Technology Patent Biological Deposit Center (1-6 Chuo, 1-chome, 1-chome, Tsukuba, Ibaraki, 305-8566, Japan) And the accession number FERM BP-11027 is assigned.
- pCABD2 is a mutant dapA gene encoding dihydrodipicolinate synthase (DDPS) derived from Escherichia coli having a mutation that is desensitized to feedback inhibition by L-lysine, and a mutation that is desensitized to feedback inhibition by L-lysine.
- a mutant lysC gene encoding aspartokinase III derived from Escherichia coli, dapB gene encoding dihydrodipicolinate reductase derived from Escherichia coli, and ddh encoding a diaminopimelate dehydrogenase derived from Brevibacterium lactofermentum Contains genes (International Publication Nos. WO95 / 16042 and WO01 / 53459).
- Coryneform bacteria having the ability to produce L-lysine include AEC-resistant mutant strains (Brevibacterium lactofermentum AJ11082 (NRRL B-11470), etc .: Japanese Patent Publication Nos. 56-1914 and 56-1915 No. 57-14157, No. 57-14158, No. 57-30474, No. 58-10075, No. 59-4993, No. 61-35840, No. 62-24074, JP-B 62-36673, JP-B 5-11958, JP-B 7-112437, JP-B 7-112438); amino acids such as L-homoserine for its growth (See Japanese Patent Publication No. 48-28078, Japanese Patent Publication No.
- L-cysteine producing bacteria examples include E. coli JM15 (US Pat. No. 6,218,168) transformed with a different cysE allele encoding a serine acetyltransferase resistant to feedback inhibition. , Russian Patent Application No. 2003121601), E. coli W3110 (US Pat.No. 5,972,663) having an overexpressed gene encoding a protein suitable for excretion of a substance toxic to cells, cysteine desulfohydrase activity E. coli strains such as reduced E. coli strains (JP-A-11-155571) and E. coli W3110 (international publication No. 0127307) with increased activity of transcription regulators of the positive cysteine regulon encoded by the cysB gene. Examples include, but are not limited to, the strains to which they belong.
- L-leucine-producing bacteria examples include leucine-resistant E. coil strains (eg, 57 strains (VKPM B-7386, US Pat. No. 6,124,121)) or ⁇ E. coli strains resistant to leucine analogs such as 2-thienylalanine, 3-hydroxyleucine, 4-azaleucine, and 5,5,5-trifluoroleucine (Japanese Patent Publication No. 62-34397 and JP-A-8-70879), Although strains belonging to the genus Escherichia such as E. coli strains obtained by the genetic engineering method described in International Publication No. 96/06926, E. coli H-9068 (Japanese Patent Laid-Open No. 8-70879) can be mentioned, It is not limited to these.
- the bacterium used in the present invention may be improved by increasing the expression of one or more genes involved in L-leucine biosynthesis.
- a gene of leuABCD operon represented by a mutant leuA gene (US Pat. No. 6,403,342) encoding isopropyl malate synthase which is preferably desensitized to feedback inhibition by L-leucine can be mentioned.
- the bacterium used in the present invention may be improved by increasing the expression of one or more genes encoding proteins that excrete L-amino acids from bacterial cells. Examples of such genes include b2682 gene and b2683 gene (ygaZH gene) (European Patent Application Publication No. 1239041).
- Coryneform bacteria producing L-isoleucine include coryneform bacteria (JP 2001-169788) in which a brnE gene encoding a branched-chain amino acid excretion protein is amplified, and L-isoleucine production by protoplast fusion with L-lysine producing bacteria.
- Coryneform bacterium imparted with ability JP-A 62-74293
- coryneform bacterium with enhanced homoserine dehydrogenase JP-A 62-91193
- threonine hydroxamate resistant strain JP-A 62-195293
- ⁇ - Examples include ketomarone resistant strains (Japanese Patent Laid-Open No. 61-15695) and methyllysine resistant strains (Japanese Patent Laid-Open No. 61-15696).
- L-histidine producing bacteria examples include E. coli 24 strain (VKPM B-5945, Russian Patent No. 2003677), E. coli 80 strain (VKPM B- 7270, Russian patent 2119536), E. coli NRRL B-12116-B12121 (US Pat.No. 4,388,405), E. coli H-9342 (FERM BP-6675) and H-9343 (FERM BP-6676) (US) Patent No. 6,344,347), E. coli H-9341 (FERM BP-6674) (European Patent Application Publication No. 1085087), E. coli AI80 / pFM201 (US Pat.No. 6,258,554), and other strains belonging to the genus Escherichia However, it is not limited to these.
- L-histidine-producing bacteria or parent strains for inducing them include strains in which expression of one or more genes encoding L-histidine biosynthetic enzymes are increased.
- genes include ATP phosphoribosyltransferase gene (hisG), phosphoribosyl AMP cyclohydrolase gene (hisI), phosphoribosyl-ATP pyrophosphohydrolase gene (hisI), phosphoribosylformimino-5- Examples include aminoimidazole carboxamide ribotide isomerase gene (hisA), amide transferase gene (hisH), histidinol phosphate aminotransferase gene (hisC), histidinol phosphatase gene (hisB), and histidinol dehydrogenase gene (hisD). It is done.
- L-histidine biosynthetic enzymes encoded by hisG and hisBHAFI are known to be inhibited by L-histidine, and therefore L-histidine production ability is feedback-inhibited by the ATP phosphoribosyltransferase gene (hisG). Can be efficiently increased by introducing mutations that confer resistance to (Russian Patent Nos. 2003677 and 2119536).
- strains having L-histidine-producing ability include E. coli FERM-P 5038 and 5048 introduced with a vector carrying a DNA encoding an L-histidine biosynthesis enzyme (Japanese Patent Laid-Open No. 56-005099).
- E. coli strain (European Patent Application Publication No. 1016710) into which an amino acid transport gene was introduced, E. coli 80 imparted resistance to sulfaguanidine, DL-1,2,4-triazole-3-alanine and streptomycin Strains (VKPM B-7270, Russian Patent No. 2119536).
- L-glutamic acid-producing bacteria examples include, but are not limited to, strains belonging to the genus Escherichia such as E. coli VL334thrC + (EP 1172433).
- E. coli VL334 (VKPM B-1641) is an L-isoleucine and L-threonine auxotrophic strain having mutations in the thrC gene and the ilvA gene (US Pat. No. 4,278,765).
- the wild type allele of the thrC gene was introduced by a general transduction method using bacteriophage P1 grown on cells of wild type E. coli K-12 strain (VKPM B-7).
- VKPM B-8961 L-isoleucine-requiring L-glutamic acid producing bacterium VL334thrC +
- L-glutamic acid-producing bacteria or parent strains for deriving the same include, but are not limited to, strains with enhanced activity of one or more L-glutamic acid biosynthetic enzymes.
- examples of such genes include glutamate dehydrogenase (gdhA), glutamine synthetase (glnA), glutamate synthetase (gltAB), isocitrate dehydrogenase (icdA), aconitate hydratase (acnA, acnB), citrate synthase (gltA), Methyl citrate synthase (prpC), phosphoenolpyruvate carbocilase (ppc), pyruvate dehydrogenase (aceEF, lpdA), pyruvate kinase (pykA, pykF), phosphoenolpyruvate synthase (ppsA), enolase ( eno),
- strains modified to increase expression of citrate synthetase gene, phosphoenolpyruvate carboxylase gene, and / or glutamate dehydrogenase gene include European Patent Application Publication No. 1078989, European Patent Application Publication No. 955368. And those disclosed in European Patent Application No. 952221.
- L-glutamic acid-producing bacteria or parent strains for deriving the same are those in which the activity of an enzyme that catalyzes the synthesis of a compound other than L-glutamic acid by diverging from the biosynthetic pathway of L-glutamic acid is reduced or absent Stocks are also mentioned.
- Examples of such enzymes include isocitrate triase (aceA), ⁇ -ketoglutarate dehydrogenase (sucA), phosphotransacetylase (pta), acetate kinase (ack), acetohydroxy acid synthase (ilvG), Examples include acetolactate synthase (ilvI), formate acetyltransferase (pfl), lactate dehydrogenase (ldh), glutamate decarboxylase (gadAB), and the like.
- aceA isocitrate triase
- sucA ⁇ -ketoglutarate dehydrogenase
- pta phosphotransacetylase
- ack acetate kinase
- ilvG acetohydroxy acid synthase
- Examples include acetolactate synthase (ilvI), formate acetyltransferase (pfl), lactate dehydrogenase (ld
- E. coli W3110sucA Km r
- E. coli AJ12624 (FERM BP-3853)
- E. coli AJ12628 (FERM BP-3854)
- E. coli AJ12949 (FERM BP-4881)
- E. coli W3110sucA is a strain obtained by disrupting the ⁇ -ketoglutarate dehydrogenase gene (hereinafter also referred to as “sucA gene”) of E. coli W3110. This strain is completely deficient in ⁇ -ketoglutarate dehydrogenase.
- examples of coryneform bacteria having reduced ⁇ -ketoglutarate dehydrogenase activity include the following strains.
- Brevibacterium lactofermentum L30-2 strain Japanese Unexamined Patent Publication No. 2006-340603
- Brevibacterium lactofermentum strain ⁇ S International pamphlet No.
- L-glutamic acid-producing bacteria include those belonging to the genus Escherichia and having resistance to an aspartic acid antimetabolite. These strains may be deficient in ⁇ -ketoglutarate dehydrogenase, for example, E. coli AJ13199 (FERM BP-5807) (US Patent No. 5,908,768), and FFRM P- with reduced L-glutamate resolution 12379 (US Pat. No. 5,393,671); AJ13138 (FERM BP-5565) (US Pat. No. 6,110,714) and the like.
- Pantoea ananatis AJ13355 strain An example of an L-glutamic acid-producing bacterium of Pantoea ananatis is Pantoea ananatis AJ13355 strain. This strain was isolated from the soil of Iwata City, Shizuoka Prefecture as a strain that can grow on a medium containing L-glutamic acid and a carbon source at a low pH. Pantoea Ananatis AJ13355 was commissioned on February 19, 1998 at the National Institute of Advanced Industrial Science and Technology, the Patent Biological Deposit Center (address: 1st, 1st, 1st, 1-chome, Tsukuba, Ibaraki, Japan, 305-8566).
- ⁇ KGDH ⁇ -ketoglutarate dehydrogenase
- Such strains include AJ13356 (US Pat. No. 6,331,419) in which the ⁇ KGDH-E1 subunit gene (sucA) of AJ13355 strain is deleted, and sucA derived from SC17 strain selected from AJ13355 strain as a low mucus production mutant.
- SC17sucA US Pat. No. 6,596,517) which is a gene-deficient strain.
- AJ13356 was founded on February 19, 1998 at the Institute of Biotechnology, National Institute of Advanced Industrial Science and Technology (currently the National Institute of Advanced Industrial Science and Technology, Patent Biological Deposit Center, 1-chome, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan 305-8566 No. 6) was deposited under the deposit number FERM P-16645, transferred to an international deposit under the Budapest Treaty on January 11, 1999, and given the deposit number FERM BP-6616.
- AJ13355 and AJ13356 are deposited as Enterobacter agglomerans in the above depository organization, but are described as Pantoea ananatis in this specification.
- the SC17sucA strain has been assigned a private number AJ417, and deposited on February 26, 2004 at the above-mentioned National Institute of Advanced Industrial Science and Technology as the accession number FERM BP-08646.
- SC17sucA / RSFCPG + pSTVCB strain As L-glutamic acid-producing bacteria of Pantoea ananatis, SC17sucA / RSFCPG + pSTVCB strain, AJ13601 strain, NP106 strain, and NA1 strain can be mentioned.
- the SC17sucA / RSFCPG + pSTVCB strain is different from the SC17sucA strain in that the plasmid RSFCPG containing the citrate synthase gene (gltA), the phosphoenolpyruvate carboxylase gene (ppsA), and the glutamate dehydrogenase gene (gdhA) derived from Escherichia coli, This is a strain obtained by introducing a plasmid pSTVCB containing a citrate synthase gene (gltA) derived from bacteria lactofermentum.
- the AJ13601 strain was selected from the SC17sucA / RSFCPG + pSTVCB strain as a strain exhibiting resistance to a high concentration of L-glutamic acid at low pH.
- the NP106 strain is a strain obtained by removing the plasmid RSFCPG + pSTVCB from the AJ13601 strain.
- AJ13601 shares were registered with the National Institute of Advanced Industrial Science and Technology, Patent Biological Deposit Center (305-1856, Ibaraki, Japan, 1st-chome, 1st-chome, 1st-chome, 1st-centre, 6th). Deposited as 17516, transferred to an international deposit under the Budapest Treaty on July 6, 2000, and assigned the deposit number FERM BP-7207.
- a method for conferring L-glutamic acid-producing ability to coryneform bacteria a method of amplifying the yggB gene encoding mechanosensitive channel (International Publication WO2006 / 070944), a mutation introducing a mutation in the coding region It is also possible to use a method for introducing a type yggB gene.
- the yggB gene is located at base numbers 1,337,692 to 1,336,091 (complementary strands) on the genome sequence of Corynebacterium glutamicum ATCC 13030 registered in Genbank Accession No. NC_003450, and GenBank Accession No. NP_600492, also called NCgl1221 It is a gene that encodes a registered membrane protein.
- a method for imparting resistance to an organic acid analog or a respiratory inhibitor and a method for imparting sensitivity to a cell wall synthesis inhibitor may be mentioned.
- a method of imparting monofluoroacetic acid resistance Japanese Patent Laid-Open No. 50-113209
- a method of imparting adenine resistance or thymine resistance Japanese Patent Laid-Open No. 57-065198
- a method of weakening urease Japanese Patent Laid-Open No. 52-038088
- a method for imparting resistance to malonic acid Japanese Patent Laid-Open No.
- Brevibacterium flavum AJ3949 (FERM BP-2632: see JP-A-50-113209) Corynebacterium glutamicum AJ11628 (FERM P-5736; see JP 57-065198) Brevibacterium flavum AJ11355 (FERM P-5007; see JP-A-56-1889) Corynebacterium glutamicum AJ11368 (FERM P-5020; see JP 56-1889) Brevibacterium flavum AJ11217 (FERM P-4318; see JP-A-57-2689) Corynebacterium glutamicum AJ11218 (FERM P-4319; see JP-A-57-2689) Brevibacterium flavum AJ11564 (FERM P-5472; see JP 56-140895 A) Brevibacterium flavum AJ11439 (FERM P-5136; see JP-A-56-35981) Corynebacterium glutamicum H7684 (FERM BP-3004
- L-phenylalanine-producing bacteria examples include E. coli AJ12739 (tyrA :: Tn10, tyrR) lacking chorismate mutase-prefenate dehydrogenase and tyrosine repressor ( VKPM B-8197) (WO 03/044191), E. coli HW1089 (ATCC 55371) carrying a mutant pheA34 gene encoding chorismate mutase-prefenate dehydratase with desensitized feedback inhibition (US Pat.No. 5,354,672) Strains belonging to the genus Escherichia such as E.
- E. coli MWEC101-b KR8903681
- E. coli NRRL B-12141 E. coli NRRL B-12141
- NRRL B-12145 E. coli NRRL B-12146
- NRRL B-12147 US Pat.No. 4,407,952
- E. coli K-12 [W3110 (tyrA) / pPHAB]
- E. coli K that retains the gene encoding chorismate mutase-prefenate dehydratase whose feedback inhibition has been released.
- -12 [W3110 (tyrA) / pPHAD] (FERM BP-12659)
- E. coli K-12 [W3110 (tyrA) / pPHATerm] (FERM BP-12662) and E. coli K-12 named AJ 12604 [W3110 (tyrA) / pBR-aroG4, pACMAB] (FERM BP-3579) can also be used (EP 488424 B1).
- L-phenylalanine producing bacteria belonging to the genus Escherichia having an increased activity of the protein encoded by the yedA gene or the yddG gene can also be used (US Patent Application Publication Nos. 2003/0148473 and 2003/0157667, International Publication No. 03/044192). .
- Coryneform bacteria that produce phenylalanine include Corynebacterium glutamica BPS-13 strains with reduced phosphoenolpyruvate carboxylase or pyruvate kinase activity (FERM BP-1777, K77 (FERM BP-2062) and K78 (FERM BP -2063) (European Patent Publication No. 331145, JP-A No. 02-303495), a tyrosine-requiring strain (JP-A No. 05-049489) and the like can be used.
- L-tryptophan-producing bacteria examples include E. coli JP4735 / pMU3028 (DSM10122) and JP6015 / pMU91 lacking tryptophanyl-tRNA synthetase encoded by the mutant trpS gene (DSM10123) (U.S. Pat.No. 5,756,345), E. coli having a serA allele encoding phosphoglycerate dehydrogenase not subject to feedback inhibition by serine and a trpE allele encoding an anthranilate synthase not subject to feedback inhibition by tryptophan.
- SV164 pGH5 (US Pat.No.
- E. coli AGX17 (pGX44) (NRRL B-12263) and AGX6 (pGX50) aroP (NRRL B-12264) lacking tryptophanase (US Pat.No. 4,371,614)
- Escherichia coli such as E. coli AGX17 / pGX50, pACKG4-pps (WO9708333, US Pat.No. 6,319,696) with increased phosphoenolpyruvate production capacity
- Strains include belonging to Rihia genus, but is not limited thereto.
- L-tryptophan-producing bacteria belonging to the genus Escherichia with increased activity of the protein encoded by the yedA gene or the yddG gene can also be used (US Patent Application Publications 2003/0148473 and 2003/0157667).
- L-tryptophan-producing bacteria or parent strains for inducing them examples include anthranilate synthase (trpE), phosphoglycerate dehydrogenase (serA), 3-deoxy-D-arabinohepturonic acid-7-phosphorus Acid synthase (aroG), 3-dehydroquinate synthase (aroB), shikimate dehydrogenase (aroE), shikimate kinase (aroL), 5-enolic acid pyruvylshikimate 3-phosphate synthase (aroA), chorismate synthase (aroC ), Prephenate dehydratase, chorismate mutase and tryptophan synthase (trpAB).
- trpE anthranilate synthase
- serA phosphoglycerate dehydrogenase
- aroG 3-deoxy-D-arabinohepturonic acid-7-phosphorus Acid synthas
- Prefenate dehydratase and chorismate mutase are encoded by the pheA gene as a bifunctional enzyme (chorismate mutase / prephenate dehydrogenase (CM / PDH)).
- phosphoglycerate dehydrogenase 3-deoxy-D-arabinohepturonic acid-7-phosphate synthase, 3-dehydroquinate synthase, shikimate dehydratase, shikimate kinase, 5-enolic acid Pyruvylshikimate 3-phosphate synthase, chorismate synthase, prefenate dehydratase, chorismate mutase-prefenate dehydrogenase are particularly preferred.
- strains having such mutations include E. coli SV164 carrying a desensitized anthranilate synthase and a mutant serA gene encoding phosphoglycerate dehydrogenase with desensitized feedback inhibition Examples include a transformant obtained by introducing the plasmid pGH5 ⁇ (International Publication No. 94/08031) into E. coli SV164.
- L-tryptophan-producing bacteria or parent strains for deriving the same examples include strains into which a tryptophan operon containing a gene encoding an inhibitory anthranilate synthase has been introduced (Japanese Patent Laid-Open Nos. 57-71397 and 1994). 62-244382, US Pat. No. 4,371,614). Furthermore, L-tryptophan-producing ability may be imparted by increasing the expression of a gene encoding tryptophan synthase in the tryptophan operon (trpBA). Tryptophan synthase consists of ⁇ and ⁇ subunits encoded by trpA and trpB genes, respectively. Furthermore, L-tryptophan production ability may be improved by increasing the expression of the isocitrate triase-malate synthase operon (WO 2005/103275).
- Coryneform bacteria include corynebacterium glutamicum AJ12118 (FERM BP-478 patent No. 01688102) which is resistant to sulfaguanidine, coryneform bacteria introduced with tryptophan operon (Japanese Patent Laid-Open No. 63240794), coryneform bacteria Coryneform bacteria (Japanese Patent Laid-Open No. 01994749) into which a gene encoding shikimate kinase derived therefrom has been introduced can be used.
- L-proline-producing bacteria examples include E. coli 702ilvA (VKPM B-8012) that lacks the ilvA gene and can produce L-proline (European Patent Publication) Examples include, but are not limited to, strains belonging to the genus Escherichia such as No. 1,172,433).
- the bacterium used in the present invention may be improved by increasing the expression of one or more genes involved in L-proline biosynthesis.
- An example of a gene preferable for L-proline-producing bacteria includes a proB gene (German Patent No. 3127361) encoding glutamate kinase that is desensitized to feedback inhibition by L-proline.
- the bacterium used in the present invention may be improved by increasing the expression of one or more genes encoding proteins that excrete L-amino acids from bacterial cells. Examples of such genes include b2682 gene and b2683 gene (ygaZH gene) gene (European Patent Publication No. 1,239,041).
- bacteria belonging to the genus Escherichia having L-proline-producing ability include NRRL B-12403 and NRRL B-12404 (British Patent No. 2075056), VKPM B-8012 (Russian Patent Application 2000124295), German Patent No. 3127361 And E. coli strains such as the plasmid variants described in Bloom FR et al (The 15th Miami winter symposium, 1983, p.34).
- L-arginine-producing bacteria examples include E. coli 237 strain (VKPM B-7925) (US Patent Application Publication No. 2002/058315) and mutant N- Derived strains carrying acetylglutamate synthase ( Russian Patent Application No. 2,001,112,869), E. coli 382 strain (VKPM B-7926) (European Patent Publication No.1,170,358), argA gene encoding N-acetylglutamate synthetase introduced Strains belonging to the genus Escherichia such as, but not limited to, arginine producing strains (European Patent Publication No. 1,170,361).
- L-arginine-producing bacteria or parent strains for deriving the same also include strains in which expression of one or more genes encoding L-arginine biosynthetic enzymes are increased.
- genes include N-acetylglutamylphosphate reductase gene (argC), ornithine acetyltransferase gene (argJ), N-acetylglutamate kinase gene (argB), acetylornithine transaminase gene (argD), ornithine carbamoyltransferase gene ( argF), arginosuccinate synthetase gene (argG), arginosuccinate lyase gene (argH), carbamoylphosphate synthetase gene (carAB).
- argC N-acetylglutamylphosphate reductase gene
- argJ ornithine acetyltransferase gene
- argB
- L-valine-producing bacteria examples include, but are not limited to, strains modified to overexpress the ilvGMEDA operon (US Pat. No. 5,998,178). Not. It is preferable to remove the ilvGMEDA operon region required for attenuation so that the operon expression is not attenuated by the produced L-valine. Furthermore, it is preferred that the ilvA gene of the operon is disrupted and the threonine deaminase activity is reduced. Examples of L-valine-producing bacteria or parent strains for deriving the same also include mutants having aminoacyl t-RNA synthetase mutations (US Pat. No. 5,658,766).
- E. coli VL1970 having a mutation in the ileS gene encoding isoleucine tRNA synthetase can be used.
- E. coli VL1970 was registered with Russian National Collection of Industrial Microorganisms (VKPM) (1 Dorozhny proezd., 1 Moscow 117545, Russia) on June 24, 1988 under the accession number VKPM B-4411. It has been deposited.
- VKPM Russian National Collection of Industrial Microorganisms
- a mutant strain International Publication No. 96/06926, that requires lipoic acid for growth and / or lacks H + -ATPase can be used as a parent strain.
- L-valine-producing bacteria of coryneform bacteria include, for example, a strain modified so that expression of a gene encoding an enzyme involved in L-valinate biosynthesis is enhanced.
- an enzyme involved in L-valinate biosynthesis for example, an enzyme encoded by the ilvBNC operon, that is, an acetohydroxyacid synthase encoded by ilvBN and an isomeroreductase encoded by ivlC (International Publication No. 00/50624) Is mentioned.
- the attenuation is released to release the suppression of the expression by the produced L-valine. Is desirable.
- the coryneform bacterium having L-valine-producing ability may be performed by reducing or eliminating the activity of at least one enzyme involved in a substance metabolic pathway that reduces L-valine production. For example, it is conceivable to reduce the activity of threonine dehydratase involved in L-leucine synthesis or the enzyme involved in D-pantosenate synthesis (WO 00/50624).
- Another method for imparting L-valine-producing ability includes a method for imparting resistance to amino acid analogs and the like.
- L-isoleucine and L-methionine auxotrophs, and mutant strains (FERM P-1841, FERM P having resistance to D-ribose, purine ribonucleoside or pyrimidine ribonucleoside and capable of producing L-valine) -29, JP-B 53-025034), mutants resistant to polyketoids (FERM P-1763, FERM P-1764, JP-B 06-065314), and in a medium containing acetic acid as the only carbon source.
- -Mutants (FERM BP-3006, FERM BP-3007, Patent No. 3006929) that are resistant to valine and are sensitive to pyruvate analogs (fluoropyruvate, etc.) in a medium containing glucose as the only carbon source. .
- L-isoleucine-producing bacteria and L-isoleucine-producing bacteria or parent strains for inducing them include mutants having resistance to 6-dimethylaminopurine (Japanese Patent Laid-Open No. 5-304969), thiisoleucine, isoleucine hydroxamate Mutants having resistance to isoleucine analogs such as the above, and mutants having resistance to DL-ethionine and / or arginine hydroxamate (Japanese Patent Laid-Open No. 5-130882), but are not limited thereto.
- a recombinant strain transformed with a gene encoding a protein involved in L-isoleucine biosynthesis such as threonine deaminase and acetohydroxy acid synthase can also be used as a parent strain (JP-A-2-458, FR 0356739, and US Pat. No. 5,998,178).
- Coryneform bacteria producing L-isoleucine include coryneform bacteria (JP 2001-169788) in which a brnE gene encoding a branched-chain amino acid excretion protein is amplified, and L-isoleucine production by protoplast fusion with L-lysine producing bacteria.
- Coryneform bacterium imparted with ability JP-A 62-74293
- coryneform bacterium with enhanced homoserine dehydrogenase JP-A 62-91193
- threonine hydroxamate resistant strain JP-A 62-195293
- ⁇ - Examples include ketomarone resistant strains (Japanese Patent Laid-Open No. 61-15695) and methyllysine resistant strains (Japanese Patent Laid-Open No. 61-15696).
- L-methionine-producing bacteria or parent strains for deriving L-methionine-producing bacteria include, but are not limited to, L-threonine-requiring strains and mutant strains resistant to norleucine. 139471).
- a strain lacking a methionine repressor and a recombinant strain transformed with a gene encoding a protein involved in L-methionine biosynthesis such as homoserine transsuccinylase and cystathionine ⁇ -synthase can also be used as a parent strain.
- Japanese Patent Laid-Open No. 2000-139471 Japanese Patent Laid-Open No. 2000-139471.
- the gene used is not limited to the gene having the above-mentioned genetic information or a gene having a known sequence, and the function of the encoded protein is impaired.
- genes having conservative mutations such as homologues and artificially modified variants of the genes can also be used. That is, it may be a gene encoding a protein having a sequence including substitution, deletion, insertion or addition of one or several amino acids at one or several positions in the amino acid sequence of a known protein.
- “one or several” differs depending on the position of the protein in the three-dimensional structure of the amino acid residue and the type of amino acid residue, but specifically, preferably 1 to 20, more preferably 1 to 10 Means, more preferably 1-5.
- a conservative mutation is a polar amino acid between Phe, Trp, and Tyr when the substitution site is an aromatic amino acid, and between Leu, Ile, and Val when the substitution site is a hydrophobic amino acid. Is an amino acid having a hydroxyl group between Gln and Asn, in the case of a basic amino acid, between Lys, Arg, and His, and in the case of an acidic amino acid, between Asp and Glu. In some cases, it is a mutation that substitutes between Ser and Thr.
- substitutions considered as conservative substitutions include substitution from Ala to Ser or Thr, substitution from Arg to Gln, His or Lys. , Asn to Glu, Gln, Lys, His or Asp, Asp to Asn, Glu or Gln, Cys to Ser or Ala, Gln to Asn, Glu, Lys, His, Asp or Arg Substitution, Glu to Gly, Asn, Gln, Lys or Asp substitution, Gly to Pro substitution, His to Asn, Lys, Gln, Arg or Tyr substitution, Ile to Leu, Met, Val or Phe Substitution, Leu to Ile, Met, Val or Phe, Lys to Asn, Glu, Gln, His or Arg, Met to Ile, Leu, Val or Phe, Phe to Trp, Tyr, Met, Ile or Leu substitution, Ser to Thr or Ala substitution, Thr to Ser or Ala substitution, Trp to Phe or Tyr substitution
- amino acid substitutions, deletions, insertions, additions, or inversions as described above include naturally occurring mutations (mutants or variants) such as those based on individual differences or species differences of the microorganism from which the gene is derived. Also included by Such a gene can be modified, for example, by site-directed mutagenesis so that the amino acid residue at a specific site of the encoded protein contains substitutions, deletions, insertions or additions. Can be obtained by:
- the gene having a conservative mutation as described above has a homology of 80% or more, preferably 90% or more, more preferably 95% or more, particularly preferably 97% or more with respect to the entire encoded amino acid sequence.
- each codon in the gene sequence may be replaced with a codon that is easy to use in the host into which the gene is introduced.
- the gene having a conservative mutation may be one obtained by a method usually used for mutation treatment such as treatment with a mutation agent.
- a gene is a DNA that hybridizes with a probe complementary to a known gene sequence or a probe that can be prepared from the complementary sequence under stringent conditions and encodes a protein having a function equivalent to that of a known gene product. Also good.
- stringent conditions refers to conditions under which so-called specific hybrids are formed and non-specific hybrids are not formed.
- DNAs having high homology for example, 80% or more, preferably 90% or more, more preferably 95% or more, particularly preferably 97% or more, are hybridized to each other.
- Conditions under which DNAs with low homology do not hybridize or conditions for washing of ordinary Southern hybridization, 60 ° C., 1 ⁇ SSC, 0.1% SDS, preferably 0.1 ⁇ SSC, 0.1% SDS, more preferably The conditions include washing once at a salt concentration and temperature corresponding to 68 ° C., 0.1 ⁇ SSC, and 0.1% SDS, more preferably 2 to 3 times.
- a part of the complementary sequence of the gene can also be used.
- Such a probe can be prepared by PCR using an oligonucleotide prepared on the basis of a known gene sequence as a primer and a DNA fragment containing these base sequences as a template.
- hybridization washing conditions include 50 ° C., 2 ⁇ SSC, and 0.1% SDS.
- Preparing a treated product of the microalgae that promotes the production and accumulation of amino acids (b) culturing the bacterium in a medium containing the treated product of the microalgae, and producing and accumulating L-amino acids in the culture; c) L-amino acids are collected from the culture.
- the treated product is preferably contained as a carbon source, and in this case, a treated product containing a saccharified starch or a hydrolyzed oil is particularly preferable.
- the above “as a carbon source” means that it can substantially contribute as a source of carbon constituting the cell components and L-amino acids in the growth of bacteria and the production of L-amino acids. If the growth of bacteria or the production and accumulation of L-amino acids is better when cultured in a medium containing a hydrolyzate than in a medium not containing organic matter produced by microalgae, the hydrolyzate is Evaluated as a carbon source.
- the culture medium may contain only the organic substance produced by microalgae as a carbon source, and may contain another carbon source.
- any of batch culture, fed-batch culture, and continuous culture can be used, and the oil and fat hydrolyzate in the medium is used as the initial medium. It may be contained, may be contained in a feeding medium, and may be contained in both of these.
- Fed-batch culture refers to a culture method in which a medium is fed continuously or intermittently into a culture container and the medium is not removed from the container until the end of the culture.
- Continuous culture refers to a method in which a medium is fed continuously or intermittently into a culture container and the medium (usually equivalent to the medium to be fed) is extracted from the container.
- the initial medium means a medium (medium at the start of the culture) used for batch culture (batch culture) before feeding the fed-batch medium in fed-batch culture or continuous culture. It means a medium supplied to a fermenter when fed-batch culture or continuous culture is performed.
- batch culture means a method in which a new medium is prepared every time, a strain is planted there, and no medium is added until harvest.
- the organic matter produced by the microalgae used may be used at any concentration that is suitable for producing L-amino acids.
- concentration of glucose which is a saccharified starch is preferably about 0.05 w / v% to 50 w / v%, more preferably about 0.1 w / v% to 40 w / v%, particularly preferably 0.2 w / v. About 20% w / v% is contained in the medium.
- the amount of glycerol and fatty acid which are hydrolysates of fats and oils is 0.01 to 10 w / v%, preferably 0.02 to 5 w / v%, more preferably 0.05 to 2 w / v%. It is desirable to make it.
- Organic substances produced by microalgae can be used alone or in combination with other carbon sources such as glucose, fructose, sucrose, waste molasses, and starch hydrolysate.
- the organic matter produced by the microalgae and other carbon sources can be mixed in any ratio, but the proportion of the organic matter produced by the microalgae in the carbon source is 10% by weight or more.
- the amount is preferably 50% by weight or more, more preferably 70% by weight.
- Other preferred carbon sources include glucose, fructose, sucrose, lactose, galactose, molasses, sugars such as sugar hydrolyzate obtained by hydrolysis of starch and biomass, alcohols such as ethanol and glycerol, fumar Organic acids such as acid, citric acid and succinic acid.
- the organic acid when the enzymatic hydrolysis of starch is performed in the presence of an organic acid as a buffer, the organic acid may be added to the medium as a carbon source together with a saccharified product.
- the ratio of starch saccharified product to organic acid in the carbon source may be arbitrary, but is preferably 1: 1 to 1: 3.
- the organic acid may be any as long as the bacteria to be used can assimilate, and examples include acetic acid, citric acid, succinic acid, fumaric acid and the like, and acetic acid is particularly preferable.
- starch saccharified product and oil hydrolyzate produced by microalgae at the start of culture are as described above, but starch saccharified product and oil hydrolyzed by microalgae in culture are hydrolyzed.
- a mixture of organic matter produced by microalgae may be added.
- the algae-derived organic matter may be contained at a constant concentration throughout the entire culture process, or may be added only to the fed-batch medium or the initial medium, and other carbon sources are sufficient. If so, there may be a period in which the hydrolyzate of oils and fats is insufficient for a certain period of time.
- temporary means that the hydrolyzate of fats and oils may be deficient within a time of within 10%, within 20%, or within a maximum of 30% of the entire fermentation time. Thus, even if the concentration of the hydrolyzate of fats and oils may temporarily become zero, if there is a culture period in a medium containing organic matter produced by algae, Included in the phrase “includes organic matter produced as a carbon source”.
- a medium conventionally used in the fermentation production of L-amino acids using microorganisms can be used except that it contains an organic substance produced by algae. That is, in addition to a carbon source, a normal medium containing a nitrogen source, inorganic ions, and other organic components as required can be used.
- a nitrogen source inorganic ammonium salts such as ammonium sulfate, ammonium chloride, ammonium phosphate, ammonium acetate, and urea, or organic nitrogen such as nitrate and soybean hydrolysate, ammonia gas, aqueous ammonia, and the like can be used.
- peptone, yeast extract, meat extract, malt extract, corn steep liquor, soybean hydrolyzate and the like can also be used. Only 1 type of these nitrogen sources may be contained in the culture medium, and it may contain 2 or more types. These nitrogen sources can be used for both the initial medium and the fed-batch medium. In addition, the same nitrogen source may be used for both the initial culture medium and the feed medium, or a different nitrogen source from the initial culture medium may be used.
- the medium of the present invention preferably contains a phosphate source and a sulfur source in addition to a carbon source and a nitrogen source.
- phosphoric acid source phosphoric acid polymers such as potassium dihydrogen phosphate, dipotassium hydrogen phosphate and pyrophosphoric acid can be used.
- the sulfur source may be any one containing sulfur atoms, but sulfates such as sulfates, thiosulfates and sulfites, and sulfur-containing amino acids such as cysteine, cystine and glutathione are desirable. However, ammonium sulfate is desirable.
- the medium may contain a growth promoting factor (a nutrient having a growth promoting effect) in addition to the above components.
- a growth promoting factor a nutrient having a growth promoting effect
- trace metals include iron, manganese, magnesium, calcium and the like
- vitamins include vitamin B1, vitamin B2, vitamin B6, nicotinic acid, nicotinamide, and vitamin B12.
- L-lysine-producing bacteria that can be used in the present invention have many L-lysine biosynthetic pathways as described later, and L-lysine resolution is weakened. It is desirable to add one or more selected from homoserine, L-isoleucine, and L-methionine.
- the initial medium and fed-batch medium may have the same or different medium composition.
- the initial culture medium and the fed-batch medium may have the same or different sulfur concentration.
- the composition of each feeding medium may be the same or different.
- the medium used in the present invention may be either a natural medium or a synthetic medium as long as it contains a carbon source, a nitrogen source, and other components as necessary.
- Organic substances produced by algae contain components used for amino acids in addition to carbon sources.
- the nitrogen source and other components can be reduced as compared with a normal medium as necessary.
- Cultivation is preferably carried out under aerobic conditions for 1 to 7 days, and the culture temperature is preferably 20 ° C. to 45 ° C., preferably 24 ° C. to 45 ° C., particularly preferably 33 to 42 ° C.
- the culture is preferably aeration culture, and the oxygen concentration is preferably adjusted to 5 to 50%, desirably about 10% of the saturation concentration.
- the pH during the culture is preferably 5-9.
- an inorganic or organic acidic or alkaline substance such as calcium carbonate, ammonia gas, aqueous ammonia or the like can be used for pH adjustment.
- the concentration of the accumulated L-amino acid may be any concentration as long as it can be collected and recovered from the medium or cells, but is preferably 1 g / L or more, more preferably 50 g / L or more, and even more preferably 100 g / L or more. is there.
- the pH during the cultivation is controlled to 6.5 to 9.0, and the pH of the medium at the end of the cultivation is controlled to 7.2 to 9.0.
- the fermenter pressure during the fermentation is controlled to be positive, or carbon dioxide gas or a mixed gas containing carbon dioxide gas is supplied to the medium so that at least 2 g of bicarbonate ions and / or carbonate ions in the medium are present.
- L-glutamic acid fermentation it is possible to perform culture while precipitating L-glutamic acid in the medium using a liquid medium adjusted to conditions under which L-glutamic acid is precipitated.
- conditions under which L-glutamic acid precipitates include pH 5.0 to 4.0, preferably pH 4.5 to 4.0, more preferably pH 4.3 to 4.0, and particularly preferably pH 4.0. Can do. (European Patent Application Publication No. 1078989)
- the L-amino acid can be collected from the culture solution by combining an ion exchange resin method, a precipitation method and other known methods.
- L-amino acid accumulates in the microbial cells, for example, the microbial cells are crushed by ultrasonic waves and the microbial cells are removed by centrifugation, and the L-amino acid is removed from the supernatant obtained by ion exchange resin method or the like.
- the recovered L-amino acid may be a free L-amino acid or a salt containing sulfate, hydrochloride, carbonate, ammonium salt, sodium salt, or potassium salt.
- the L-amino acid collected in the present invention may contain microbial cells, medium components, moisture, and microbial metabolic byproducts in addition to the target L-amino acid.
- the purity of the collected L-amino acid is 50% or more, preferably 85% or more, particularly preferably 95% or more (US5,431,933, JP1214636B, US4,956,471, US4,777,051, US4946654, US5,840358, US6 , 238,714, US2005 / 0025878).
- Chlorella kessleri 11h strain obtained from University of Texas Algae Culture Collection (The University of Texas at Austin, The Culture Collection of Algae (UTEX), 1 University Station A6700, Austin, TX 78712-0183, USA) ( UTEX 263), Neochloris oleoabundans UTEX 1185 strain, Nannochloris sp. UTEX LB 1999 strain and Thalassiosira pseudonana UTEX LB FD2 strain were used.
- Example 1 Culture of microalgae Chlorella kessleri Chlorella kessleri in a 500 mL Erlenmeyer flask containing 100 mL of 0.2 x Gamborg B5 medium (Nippon Pharmaceutical Co., Ltd.), light intensity 10,000 lux (TOMY) The culture was performed with a culture apparatus CL-301) for 6 days, and this was used as a preculture solution. Note that white light from a fluorescent lamp was used as the light source. Add 16 mL of pre-culture solution to 1L medium bottle containing 800 mL of 0.2 ⁇ Gambog B5 medium, and CO 2 concentration is 3% at 500 mL / min at a culture temperature of 30 °C and light intensity of 10,000 lux. Culturing was carried out for 12 days while blowing a mixed gas of air and CO 2 so as to be.
- Example 1 (2) Preparation of starch saccharified solution from Chlorella kessleri
- the 1 L culture broth of Example 1 (1) was precipitated by centrifugation, and 80 mL of ethanol was added to the precipitate. After suspending the algal bodies, it was boiled for 30 minutes. The suspension was centrifuged to precipitate algal bodies, washed twice with 80 mL of ethanol, and then dried in a desiccator. 40 mL of 0.2 M KOH was added to the dried algae, boiled for 30 minutes, and further treated with 15,000 W for 10 minutes using an ultrasonic device (Kubota INSONATOR 201MA) to disrupt the cells.
- Kubota INSONATOR 201MA an ultrasonic device
- Example 1 (3) L-lysine production culture using starch saccharified solution derived from Chlorella kessleri as a carbon source Escherichia coli WC196 ⁇ cadA ⁇ ldcC / pCABD2 described in International Publication No. 2006/078039 as an L-lysine producing bacterium was used.
- WC196 ⁇ cadA ⁇ ldcC / pCABD2 was cultured at 37 ° C. for 20 hours in an LB agar medium (Tryptone 10 g / L, Yeast extract 5 g / L, NaCl 10 g / L, agar 15 g / L) containing 20 mg / L of streptomycin sulfate.
- the cells on the agar medium are scraped off, inoculated into a 500 mL Erlenmeyer flask containing 40 mL of L-lysine production medium containing 20 mg / L of streptomycin sulfate, and cultured at 37 ° C for 24 hours. went.
- the saccharified solution of Example 1 (2) prepared from starch produced by Chlorella kessleri (2.75 g / L as the amount of glucose) and 4.8 g / L of acetic acid used as a buffer during the preparation thereof were carbonized.
- the source As a control, the culture was carried out in a medium containing one of glucose and acetic acid at the same concentration and a medium containing both glucose and acetic acid as carbon sources.
- Example 2 (1) Preparation of organic material liquid without starch from Chlorella kessleri Algae bodies for 3.6 L of the culture liquid of Example 1 (1) were precipitated by centrifugation, and 100 mL of the algae was deposited on the precipitate. Ethanol was added to suspend the algal cells and allowed to stand at room temperature for 24 hours. Thereafter, filtration was performed to collect the filtrate, and the alga body residue was extracted twice with 100 mL of ethanol to obtain a total of 300 mL of ethanol extract.
- the ethanol extract was ethanol-removed with a rotary evaporator, dissolved in sterilized water, and adjusted to pH 7.0 with an aqueous potassium hydroxide solution to prepare 45 mL of an organic substance liquid derived from algal cells.
- the organic liquid was sterilized by autoclaving at 115 ° C. for 10 minutes and then used as an alternative to the medium component for L-lysine fermentation. Since this organic liquid does not contain starch, it does not contain a carbon source that can be used for amino acid-producing bacteria.
- Example 2 (2) L-lysine production culture using Chlorella kessleri-free organic liquid without starch as a medium component other than carbon source Escherichia coli WC196 ⁇ cadA ⁇ ldcC / pCABD2, which is an L-lysine-producing bacterium, was added to streptomycin sulfate.
- the cells were cultured at 37 ° C. for 20 hours on an LB agar medium containing 20 mg / L of salt.
- the cells on the agar medium were scraped, inoculated into a thick test tube containing 4 mL of L-lysine production medium containing 20 mg / L of streptomycin sulfate, and cultured at 37 ° C. for 24 hours.
- the B component in the L-lysine production medium was extracted from Chlorella kessleri algae into a medium in which the components of the B section were halved (hereinafter, this medium is referred to as ⁇ L-lysine production medium (half amount B section) ''). Culturing was carried out by adding 75 ⁇ L of the organic substance liquid of Example 2 (1) (corresponding to 6 mL in terms of culture solution). As controls, L-lysine production medium, L-lysine production medium (half amount B section), and L-lysine production medium (half amount B section) medium without glucose, 75 ⁇ L of organic liquid was added, Culture was performed.
- [L-lysine production medium composition] (A ward) Glucose 20 g / L (B ward) (NH 4 ) 2 SO 4 24 g / L KH 2 PO 4 1 g / L Yeast Extract 2 g / L FeSO 4 ⁇ 7H 2 O 10 mg / L MnSO 4 ⁇ 4H 2 O 10 mg / L (C ward) Calcium carbonate 30 g / L
- Zones A and B were sterilized by autoclave at 115 ° C for 10 minutes, and zone C was sterilized by dry heat at 180 ° C for 3 hours. After cooling to room temperature, the three were mixed and a MgSO 4 .7H 2 O solution was added to a final concentration of 1 g / L.
- Example 3 Culture of Chlorella kessleri with jar fermenter Chlorella kessleri was placed at 30 ° C in a 2 L jar fermenter (manufactured by ABLE) containing 500 mL of 0.2 x Gamborg B5 medium (Nippon Pharmaceutical). The mixture was stirred and cultured at a light intensity of 20,000 lux for 7 days, and this was used as a preculture solution.
- this jar fermenter is a light irradiation type jar fermenter in which an annular fluorescent lamp that emits white light surrounds a glass vessel as a light source.
- Example 3 (2) Preparation of a mixed solution of organic matter containing starch and high-temperature treatment from Chlorella kessleri and saccharified solution by amylase 300 mL of the algal culture solution of Example 3 (1) was placed in a reaction vessel in a high-temperature reactor. Then, the mixture was heated to 175 ° C. or 215 ° C. over 60 minutes with stirring, and then held at each temperature for 5 minutes.
- Example 3 (3) L-glutamic acid-producing culture using a saccharified solution obtained by amylase treatment of a mixed solution of organic matter containing starch derived from Chlorella kessleri as a high-temperature treatment.
- Brevibacterium lactofermentum strain L30-2 described in No. 340603 was used.
- the L30-2 strain was seeded on a CM-Dex plate medium and cultured at 31.5 ° C. for 24 hours.
- the cells on the plate medium were scraped off by one platinum loop, inoculated into a thick test tube containing 4 mL of L-glutamic acid production medium, and cultured at a culture temperature of 31.5 ° C. for 13 hours.
- Example 3 (2) As the carbon source in the main culture, the saccharified solution of Example 3 (2) (1.0 g / L as the amount of glucose) prepared from the algal starch degradation product of Chlorella kessleri, and the reagent glucose at the same concentration as the control were used.
- L-glutamic acid production medium and a medium in which the components of B section in L-glutamic acid production medium are halved hereinafter, this medium is referred to as ⁇ L-glutamic acid production medium (half amount B section) ''). Incubation was performed with the components.
- [CM-Dex medium] Glucose 5 g / L Polypeptone 10 g / L Yeast extract 10 g / L KH 2 PO 4 1 g / L MgSO 4 ⁇ 7H 2 O 0.4 g / L FeSO 4 ⁇ 7H 2 O 0.01 g / L MnSO 4 ⁇ 7H 2 O 0.01 g / L Urea 3 g / L Soy hydrolyzate 1.2 g / L Biotin 10 ⁇ g / L Adjust to pH 7.5 using NaOH. Autoclave sterilization conditions at 120 ° C for 20 minutes.
- composition of L-glutamic acid production medium (A ward) Carbon source Algae-derived starch degradation product 1.0 g / L (glucose) (B ward) (NH 4 ) 2 SO 4 15 g / L KH 2 PO 4 1 g / L MgSO 4 ⁇ 7H 2 O 0.4 g / L FeSO 4 ⁇ 7H 2 O 10 mg / L MnSO 4 ⁇ 4H 2 O 10 mg / L VB1 ⁇ HCl 200 ⁇ g / L Biotin 300 ⁇ g / L Soy hydrolyzate 0.48 g / L (C ward) Calcium carbonate 50 g / L
- the pH was adjusted to 7.8 and pH 8.0 using KOH, respectively, and autoclaved at 115 ° C. for 10 minutes.
- the C zone was sterilized by dry heat at 180 ° C. for 3 hours. The three were mixed after each cooled to room temperature.
- L-glutamic acid production medium and L-glutamic acid production medium (half amount B) contain L-glutamic acid derived from soybean hydrolyzate originally 0.45 g / L, 0.24 g / L.
- Table 3 shows values obtained by subtracting the amount of L-glutamic acid from the soybean hydrolyzate in the medium components.
- Example 4 Preparation of degradation product of amylase and lipase from organic mixed solution containing starch by high-temperature treatment from Chlorella kessleri Crushing alga bodies treated at high temperature of 215 ° C prepared in Example 3 (2) After adjusting the pH of the solution to 6.0 with 1N HCl, 400 U of amyloglucosidase alone (Sigma-Aldrich A-9228) or the same amount of amyloglucosidase and 1000 U of lipase (Sigma-Aldrich L1754) Enzyme was added and reacted at 50 ° C. for 24 hours.
- each reaction solution was concentrated to 10 mL, and then the residue in the amyloglucosidase single treatment solution was removed by centrifugation.
- the pH of the supernatant was adjusted to 7.0 using 1N NaOH, and the volume was increased to 15 ml. Thereafter, autoclaving was performed at 120 ° C. for 20 minutes.
- 1.13 mL of 10% Tween80 aqueous solution was added to the treatment solutions of both enzymes, and the mixture was heated to 60 ° C. and then stirred by vortexing.
- Example 4 (2) Construction of Escherichia coli L-lysine-producing bacteria deficient in fadR
- the transcription factor FadR that regulates fatty acid metabolism in Escherichia coli is encoded by the fadR gene (SEQ ID NO: 15) (DiRusso, CC et al. 1992. J. Biol. Chem. 267: 8685-8691).
- the WC196 ⁇ cadA ⁇ ldcC strain described in International Patent Publication WO2006 / 078039 was used as an L-lysine producing strain of Escherichia coli.
- a synthetic oligonucleotide designed with a part of the target gene on the 5 ′ side of the synthetic oligonucleotide and a part of the antibiotic resistance gene on the 3 ′ side is used as a primer.
- a gene disrupted strain can be constructed in one step.
- the antibiotic resistance gene incorporated into the gene disruption strain can be removed (JP 2005-058227, WO 2005/010175).
- Plasmid pMW118-attL-kan-attR (JP 2005-058227, WO2005 / 010175) was used as a PCR template.
- pMW118-attL-kan-attR is a plasmid in which attL and attR genes, which are attachment sites of ⁇ phage, and kan gene, which is an antibiotic resistance gene, are inserted into pMW118 (Takara Bio Inc.). Inserted in order.
- the synthetic oligonucleotides shown in SEQ ID NOS: 16 and 17 having the sequences corresponding to both ends of attL and attR at the 3 ′ end of the primer and the 5 ′ end of the primer corresponding to a part of the fadR gene as the target gene are used as the primer. PCR was performed.
- the amplified PCR product was purified on an agarose gel and introduced into an Escherichia coli WC196 ⁇ cadA ⁇ ldcC strain containing a plasmid pKD46 having a temperature-sensitive replication ability by electroporation.
- Plasmid pKD46 (Datsenko, K. A. and Wanner, B. L. 2000. Proc. Natl. Acad. Sci. USA. 97: 6640-6645) is a ⁇ Red homologous recombination system controlled by the arabinose-inducible ParaB promoter.
- the plasmid pKD46 is necessary for integrating the PCR product into the chromosome of the WC196 ⁇ cadA ⁇ ldcC strain.
- a competent cell for electroporation was prepared as follows. Specifically, Escherichia coli WC196 strain cultured overnight at 30 ° C. in LB medium (trypton 10 g / L, Yeast extract 5 g / L, NaCl 10 g / L) containing 100 mg / L ampicillin (100 mg / L) and L-arabinose (1 mM) were diluted 100-fold with 5 mL of LB medium. The obtained dilution was allowed to grow until the OD600 was about 0.6 while aerated at 30 ° C., then concentrated 100 times, and washed three times with 10% glycerol so that it could be used for electroporation.
- Electroporation was performed using 70 ⁇ L of competent cells and approximately 100 ng of PCR product. After electroporation, add 1 mL of SOC medium (Sambrook, J. and Russell, DW 2001, Molecular Cloning, A Laboratory, Manual, Third Edition, Cold Spring, Harbor Laboratory, Press, and New York) at 37 ° C for 1 hour. Then, the plate was cultured on an LB agar medium containing Km (kanamycin) (40 mg / L) at 37 ° C. to select a Km-resistant recombinant.
- SOC medium Standardbrook, J. and Russell, DW 2001, Molecular Cloning, A Laboratory, Manual, Third Edition, Cold Spring, Harbor Laboratory, Press, and New York
- the deletion of the mutant fadR gene that could be identified by the kanamycin resistance gene was confirmed by PCR.
- the obtained fadR-deficient strain was named WC196 ⁇ cadA ⁇ ldcC ⁇ fadR :: att-kan strain.
- pMW-intxis-ts is a plasmid carrying a gene encoding ⁇ phage integrase (Int) and gene encoding excisionase (Xis) and having temperature-sensitive replication ability.
- the WC196 ⁇ cadA ⁇ ldcC ⁇ fadR att-kan strain competent cell obtained above was prepared according to a conventional method, transformed with the helper plasmid pMW-intxis-ts, and an LB agar medium containing 100 ⁇ g / L ampicillin at 30 ° C. Plated above, ampicillin resistant strains were selected.
- the WC196LC ⁇ fadR strain was transformed with a lysine production plasmid pCABD2 (WO95 / 16042) carrying the dapA, dapB, lysC and ddh genes according to a conventional method to obtain a WC196 ⁇ cadA ⁇ ldcC ⁇ fadR / pCABD2 strain.
- Example 4 (3) L-lysine production culture using amylase and lipase degradation product of organic matter containing starch from Chlorella kessleri as a carbon source Escherichia coli WC196 ⁇ cadA ⁇ ldcC ⁇ fadR / pCABD22 was cultured at 37 ° C. for 20 hours on an LB agar medium containing 20 mg / L of streptomycin sulfate. Cells on the agar medium were scraped off, inoculated into a large test tube containing 4 mL of L-lysine production medium containing 20 mg / L of streptomycin sulfate, and cultured at a culture temperature of 37 ° C. for 24 hours.
- Example 4 (1) prepared from starch produced by Chlorella kessleri (1.33 g / L as the amount of glucose) and the hydrolyzed solution of Example 4 (1) prepared from starch and fats and oils were used.
- Carbon source As a control, culture was performed in a medium containing the same concentration of glucose as a carbon source.
- Zones A and B were sterilized by autoclave at 115 ° C for 10 minutes, and zone C was sterilized by dry heat at 180 ° C for 3 hours. After cooling to room temperature, the three were mixed and a MgSO 4 .7H 2 O solution was added to a final concentration of 1 g / L.
- Example 5 Culture of microalgae Neochloris oleoabundans Neochloris oleoabundans UTEX 1185 strain was cultured at 30 ° C in a 500 mL Erlenmeyer flask containing 100 mL of Modified NORO medium at a light intensity of 10,000 lux (cultivated by TOMY) CL-301) was cultured with shaking for 6 days, and this was used as a preculture. Note that white light from a fluorescent lamp was used as the light source.
- the obtained extract was divided into equal amounts, and each extract was concentrated to obtain a crude extract.
- the crude extract was suspended in 100% mL of 80% methanol, and liquid-liquid distribution was performed three times in a separatory funnel to which the suspension and 100 mL of hexane were added, and a hexane layer containing oil and fat (referred to as an oil and fat fraction)
- An 80% methanol layer (referred to as a water-soluble fraction) containing a large amount of water-soluble organic matter was obtained.
- the crude extract and the oil / fat fraction were mixed with 45 ml of hot water pre-warmed, and then lipase koji (Sigma-Aldrich Co.
- Example 5 (3) L-lysine production culture using fat and oil hydrolyzate derived from Neochloris oleoabundans L-lysine-producing Escherichia coli WC196 ⁇ cadA ⁇ ldcC / pCABD2 glycerol stock was thawed and 100 ⁇ L each.
- Approximately 1/8 amount of the cells on the obtained plate was inoculated into a Sakaguchi flask containing 20 mL of the fermentation medium described below containing 25 mg / L of streptomycin, and 24 hours at 37 ° C in a reciprocating shaker.
- Example 5 Incubate for hours. After each sample derived from the algae of Example 5 (2) was suspended in distilled water, Tween 80 was added to a concentration of 1%, pH was adjusted to 7.0 using 3N KOH, and 120 ° C. What was autoclaved for 20 minutes was used as a carbon source solution. The medium composition (final concentration) used for the culture is shown below. The hydrolyzate of the crude extract derived from algae, the hydrolyzate of the oil and fat fraction derived from algae, or the water-soluble fraction derived from algae was used as the carbon source. For each carbon source, the sum of the measured fatty acid concentration and glycerol concentration is listed.
- a carbon source prepared by mixing reagent fatty acids and reagent glycerol and adjusting the concentration to the same was used.
- the pH was adjusted to 7.0 with KOH and autoclaved at 115 ° C. for 10 minutes.
- the carbon source and MgSO 4 .7H 2 O were separately sterilized and then mixed.
- Calcium carbonate is sterilized by dry heat at 180 ° C for 3 hours and added separately.
- the amount of L-lysine in the culture supernatant was measured with Biotech Analyzer AS310 (Sakura Seiki). In this medium, the degree of growth was determined by counting the number of viable bacteria. Table 5 shows the average value of the results of the culture performed in two flasks.
- the L-lysine concentration when only the target surfactant 0.5% Tween 80 solution was used as the carbon source was L-lysine derived from the medium, and L-lysine production was not observed, but the algae-derived crude extract When the enzyme hydrolyzate was used, good L-lysine production was observed.
- L-lysine production equivalent to or higher than the control carbon source of reagent fatty acid and reagent glycerol was observed, and almost no fatty acid was used as the carbon source. L-lysine production was also observed when using a water-soluble fraction derived from a crude extract not containing it. From this, it is possible to produce L-lysine not only from the hydrolyzate of the oil and fat fraction extracted from the algal bodies of microalgae, but also from water-soluble organic substances, and from the hydrolyzate of the crude algal extract. It has been shown that L-lysine production can be significantly greater than the control carbon source of reagent fatty acids and reagent glycerol.
- Neochloris oleoabundans Culture of Neochloris oleoabundans
- UTEX 1185 strain in a 500 mL Erlenmeyer flask containing 100 mL of Modified NORO medium at 30 ° C. and light intensity of 5,000 lux (cultivation device CL-301 manufactured by TOMY) )
- CL-301 manufactured by TOMY light intensity of 5,000 lux
- the obtained crude extract was suspended in 80 mL of 80% methanol, and liquid-liquid distribution was performed three times in a separatory funnel to which the suspension and 100 mL of hexane were added. Called min).
- the lipid fraction was mixed with 50 ml of hot water pre-warmed, and then 700 U of lipase (Sigma-Aldrich L1754) was added and reacted at 42 ° C. for 18 hours. After completion of the reaction, the solution was concentrated to 30 mL, and it was divided into 15 mL portions, and one of them was made into a fat and oil hydrolyzate.
- alkaline hydrolysis was performed by the following method. First, a solution of the fat and oil decomposition product was prepared so as to have a final molar concentration of 0.1 N NaOH, and the solution was stirred for 90 minutes while being heated to 95 ° C. on a hot plate to perform an alkali hydrolysis treatment. After this lipase treatment, the alkaline hydrolyzed solution was used as a total lipid hydrolyzate.
- Example 6 L-lysine production culture using lipid hydrolyzate derived from Neochloris oleoabundans as a carbon source
- the glycerol stock of Escherichia coli WC196 ⁇ cadA ⁇ ldcC / pCABD2 strain was thawed, and 100 ⁇ L of each was diluted to 25 mg / L.
- the solution was evenly applied to an L plate containing streptomycin and cultured at 37 ° C. for 20 hours.
- [Escherichia bacterium L-lysine production medium] (A ward) Oil hydrolyzate derived from algae 1.22 g / L (fatty acid concentration + glycerol concentration) Hydrolyzate of total lipids derived from algae 2.14 g / L (fatty acid concentration + glycerol concentration) One (B ward) (NH 4 ) 2 SO 4 24 g / L KH 2 PO 4 1.0 g / L FeSO 4 ⁇ 7H 2 O 0.01 g / L MnSO 4 ⁇ 7H 2 O 0.01 g / L Yeast Extract 2.0 g / L (C ward) Calcium carbonate 30 g / L
- Zones A and B were sterilized by autoclave at 115 ° C for 10 minutes, and zone C was sterilized by dry heat at 180 ° C for 3 hours. After cooling to room temperature, the A section, the B section and the C section were mixed, and a MgSO 4 .7H 2 O solution was added to a final concentration of 1 g / L.
- Algae-derived oil and fat decomposition products are treated with lipase alone, so that glycolipids and phospholipids in lipids are hardly decomposed, and are a carbon source in which only fats and oils are decomposed.
- the alga-derived total lipid degradation products are further subjected to alkaline hydrolysis treatment after lipase treatment, not only fats and oils but also glycolipids and phospholipids are degraded. Comparing the sum of the amount of fatty acid and glycerol in these carbon sources, the fat hydrolyzate is 1.22 g / L, whereas the total lipid hydrolyzate decomposes glycolipids and phospholipids in addition to fats and oils.
- L-lysine can be produced not only from oils and fats extracted from algal bodies of microalgae, but also from glycolipids and phospholipids, and L-lysine can also be produced from all lipids of algal bodies. It was shown that.
- Example 7 Culture of microalgae Nannochloris sp. Nannochloris sp. UTEX LB 1999 strain was cultivated at 30 ° C in a 500 mL Erlenmeyer flask containing 100 mL of Modified NORO medium at a light intensity of 10,000 lux (TOMY) The culture was performed with a culture apparatus CL-301) for 6 days, and this was used as a preculture solution. Note that white light from a fluorescent lamp was used as the light source.
- an ultrasonic device Kubota INSONATOR 201MA
- the obtained extract was concentrated and suspended in 100 mL of 80% methanol, and the liquid was partitioned three times in a separatory funnel to which the suspension and 100 mL of hexane were added. An oil / fat fraction) and an 80% methanol layer (water-soluble fraction) containing a large amount of water-soluble organic matter were obtained. Next, the oil and fat fraction was suspended in 40 mL of hot water pre-warmed, 500 U of lipase (Sigma L1754) was added, and the mixture was reacted at 42 ° C. for 20 hours. The fatty acid concentration and glycerol concentration of the reaction solution and water-soluble fraction were measured, and each was used as a carbon source for amino acid fermentation.
- Example 7 (3) L-lysine production culture using fat hydrolyzate derived from Nannochloris sp.
- the glycerol stock of Escherichia coli WC196 ⁇ cadA ⁇ ldcC / pCABD2 strain was thawed, and 100 ⁇ L of each was diluted to 25 mg / L.
- Approximately 1/8 amount of the cells on the obtained plate was inoculated into a Sakaguchi flask containing 20 mL of the following fermentation medium containing 25 mg / L of streptomycin, and the reciprocal shake culture apparatus was used at 16 ° C. for 16 hours.
- the pH was adjusted to 7.0 with KOH and autoclaved at 115 ° C. for 10 minutes.
- the carbon source and MgSO 4 .7H 2 O were separately sterilized and then mixed.
- Calcium carbonate is sterilized by dry heat at 180 ° C for 3 hours and added separately.
- the amount of L-lysine in the culture supernatant was measured with Biotech Analyzer AS310 (Sakura Seiki). In this medium, the degree of growth was determined by counting the number of viable bacteria. Table 7 shows the average value of the results of the culture performed in two flasks.
- the L-lysine concentration when only the target surfactant 0.5% Tween 80 solution was used as the carbon source was L-lysine derived from the medium, and L-lysine production was not observed, but the oil and fat fraction derived from Nannochloris sp. When the hydrolyzate was used, good L-lysine production was observed. In addition, L-lysine production was also observed when water-soluble organic substances containing no fatty acid as a carbon source were used.
- an ultrasonic device Kubota INSONATOR 201MA
- Example 8 (2) L-lysine production culture using crude organic matter derived from Nannochloris sp.
- a carbon source Melt the glycerol stock of Escherichia coli WC196LC / pCABD2 strain, and add 100 ⁇ L each of 25 mg / L streptomycin. It was uniformly applied to the contained L plate and cultured at 37 ° C. for 20 hours. Approximately 1/8 amount of the cells on the obtained plate was inoculated into a Sakaguchi flask containing 20 mL of the following fermentation medium containing 25 mg / L of streptomycin, and the reciprocal shake culture apparatus was used at 16 ° C. for 16 hours. Or it culture
- Example 8 (1) serving as a carbon source Each sample derived from the algae of Example 8 (1) serving as a carbon source was suspended in distilled water, Tween 80 was added to a concentration of 1%, pH was adjusted to 7.0 using 3N KOH, What was autoclaved at 120 ° C. for 20 minutes was used as a carbon source solution.
- the medium composition used for the culture is shown below. The sum of the fatty acid concentration and the glycerol concentration measured for the hydrolyzate of the algae-derived crude extract used as the carbon source is described.
- the pH was adjusted to 7.0 with KOH and autoclaved at 115 ° C. for 10 minutes.
- a carbon source MgSO? ⁇ 7H 2 O, after separately sterilized respectively, and mixed.
- Calcium carbonate is sterilized by dry heat at 180 ° C for 3 hours and added separately.
- Example 9 Cultivation of diatom Thalassiosira pseudonana UTEX LB FD2 strain
- Thalassiosira pseudonana UTEX LB FD2 strain was cultured in a 1 L medium bottle containing 500 mL of F / 2 medium at a culture temperature of 25 ° C and a light intensity of 7,000.
- the mixture was cultured for 3 days while blowing a mixed gas of air and CO 2 so that the CO 2 concentration became 1% at 200 mL / min at lux, and this was used as a preculture solution.
- Aquamarine S manufactured by YASHIMA PURE CHEMICALS
- Example 9 (3) L-lysine production culture using oil hydrolyzate derived from Thalassiosira pseudonana as a carbon source Melt the glycerol stock of Escherichia coli WC196LC / pCABD2 strain, and add 100 ⁇ L each of 25 mg / L The solution was evenly applied to an L plate containing streptomycin and cultured at 37 ° C. for 20 hours. Approximately 1/40 amount of the cells on the obtained plate was inoculated into 4 mL of the fermentation medium described below containing 25 mg / L of streptomycin in a thick test tube, and was cultured at 37 ° C in a reciprocating shake culture apparatus. Cultured for 24 hours.
- Example 9 (2) which is a carbon source
- Tween 80 is added to the hydrolyzed solution so that the concentration becomes 2%, and the pH is adjusted to 7.0 using 3N KOH, and 120 ° C. Then, an autoclave for 20 minutes was used as a carbon source solution.
- the medium composition used for the culture is shown below. Zone A is a carbon source, and the sum of the measured fatty acid concentration and glycerol concentration is described.
- Zones A and B were sterilized by autoclave at 115 ° C for 10 minutes, and zone C was sterilized by dry heat at 180 ° C for 3 hours. After cooling to room temperature, the A section, the B section and the C section were mixed, and a MgSO 4 .7H 2 O solution was added to a final concentration of 1 g / L.
- SEQ ID NO: 1 Base sequence of LipA gene derived from Bacillus subtilis
- SEQ ID NO: 2 Amino acid sequence of LipA derived from Bacillus subtilis
- SEQ ID NO: 3 Base sequence of LipA gene derived from Burkholderia glumae
- SEQ ID NO: 4 Amino acid sequence of LipA derived from Burkholderia glumae
- Pseudomonas aeruginosa-derived LipA gene base sequence number 6 Pseudomonas aeruginosa-derived LipA amino acid sequence number 7: Staphylococcus aureus lipase gene sequence number 8: Staphylococcus aureus lipase amino acid sequence number 9: Candida antarctica-derived lipase SEQ ID NO: 10: Candida antarctica derived lipase amino acid sequence
- SEQ ID NO: 11 Candida rugosa derived lipase gene lip1 SEQ ID NO:
- L-amino acids can be produced efficiently.
- L-amino acids can be produced at a lower cost by using a carbon source derived from microalgae.
Abstract
Description
(a)微細藻類を培地で培養し、該培養物を、破砕、抽出、分画及び加水分解から選ばれる1以上の方法により処理して、L-アミノ酸生産能を有する細菌によるL-アミノ酸の生産蓄積を促進する該微細藻類の処理物を調製し、
(b)該細菌を、該微細藻類の処理物を含む培地に培養し、培養物中にL-アミノ酸を生産蓄積させ、
(c)該培養物からL-アミノ酸を採取することを特徴とするL-アミノ酸の製造法。
<1>本発明で使用する微細藻類とその培養法
本発明における微細藻類(microalgae)は、どのようなものでも用いることが出来るが、スターチ及び/または油脂を藻体内に蓄積する微細藻類であることが好ましい。
本発明において、微細藻類の処理物とは、破砕した微細藻類の細胞に由来する有機物の混合物を含む処理物であって、L-アミノ酸生産能を有する細菌によるL-アミノ酸の生産蓄積を促進するものを指し、具体的には、(1)該微細藻類の培養物の破砕物、(2)該微細藻類に由来する有機物の混合物を含む、該破砕物の抽出物もしくは分画物、又は、(3)該破砕物、該抽出物もしくは該分画物の加水分解物が挙げられる。
従って、処理物を添加していない条件に比べ、L-アミノ酸の生産蓄積量が増大している場合も本発明の処理物に含まれるが、含まれる炭素源と同量の精製された物質からなる炭素源を加えた場合と比べてL-アミノ酸生産蓄積量が向上していることが好ましい。
Burkholderia glumae由来のLipA(GenBank Accession No. X70354)をコードする遺伝子の塩基配列は配列番号3に、アミノ酸配列を配列番号4に示す。
Pseudomonas aeruginosa由来のLipA(GenBank Accession No. D50587)をコードする遺伝子の塩基配列を配列番号5に、アミノ酸配列を配列番号6に示す。
Staphylococcus aureus由来のリパーゼ(GenBank Accession No. M12715)の塩基配列を配列番号7に、アミノ酸配列を配列番号8に示す。
本発明においては、L-アミノ酸生産能を有する細菌を使用する。細菌としては、微細藻類により生産される有機物、特に、スターチの糖化物あるいは油脂の加水分解物からL-アミノ酸を効率よく製造し得るものであれば特に制限されず、例えばエシェリヒア属、パントエア属、エンテロバクター属等の腸内細菌科に属する細菌、及び、ブレビバクテリウム属、コリネバクテリウム属、ミクロバクテリウム属に属するいわゆるコリネ型細菌等が挙げられるが、これらに制限されない。
パントエア・アナナティスAJ13356株(FERM BP-6615)(欧州特許出願公開0952221号明細書)
エルビニア・カロトボーラ ATCC15713株
クレブシエラ・プランティコーラAJ13399株(FERM BP-6600)(欧州特許出願公開955368号明細書)
クレブシエラ・プランティコーラAJ13410株(FERM BP-6617)(欧州特許出願公開955368号明細書)
コリネバクテリウム・アセトグルタミカム
コリネバクテリウム・アルカノリティカム
コリネバクテリウム・カルナエ
コリネバクテリウム・グルタミカム
コリネバクテリウム・リリウム
コリネバクテリウム・メラセコーラ
コリネバクテリウム・サーモアミノゲネス (コリネバクテリウム・エフィシエンス)
コリネバクテリウム・ハーキュリス
ブレビバクテリウム・ディバリカタム
ブレビバクテリウム・フラバム
ブレビバクテリウム・インマリオフィラム
ブレビバクテリウム・ラクトファーメンタム(コリネバクテリウム・グルタミカム)
ブレビバクテリウム・ロゼウム
ブレビバクテリウム・サッカロリティカム
ブレビバクテリウム・チオゲニタリス
コリネバクテリウム・アンモニアゲネス
ブレビバクテリウム・アルバム
ブレビバクテリウム・セリヌム
ミクロバクテリウム・アンモニアフィラム
コリネバクテリウム・アセトアシドフィラム ATCC13870
コリネバクテリウム・アセトグルタミカム ATCC15806
コリネバクテリウム・アルカノリティカム ATCC21511
コリネバクテリウム・カルナエ ATCC15991
コリネバクテリウム・グルタミカム ATCC13020, ATCC13032, ATCC13060
コリネバクテリウム・リリウム ATCC15990
コリネバクテリウム・メラセコーラ ATCC17965
コリネバクテリウム・サーモアミノゲネス AJ12340(FERM BP-1539)
コリネバクテリウム・ハーキュリス ATCC13868
ブレビバクテリウム・ディバリカタム ATCC14020
ブレビバクテリウム・フラバム ATCC13826, ATCC14067
ブレビバクテリウム・インマリオフィラム ATCC14068
ブレビバクテリウム・ラクトファーメンタム ATCC13869(コリネバクテリウム・グルタミカムATCC13869)
ブレビバクテリウム・ロゼウム ATCC13825
ブレビバクテリウム・サッカロリティカム ATCC14066
ブレビバクテリウム・チオゲニタリス ATCC19240
コリネバクテリウム・アンモニアゲネス ATCC6871、ATCC6872
ブレビバクテリウム・アルバム ATCC15111
ブレビバクテリウム・セリヌム ATCC15112
ミクロバクテリウム・アンモニアフィラム ATCC15354
L-スレオニン生産能を有する微生物として好ましいものは、L-スレオニン生合成系酵素の1種又は2種以上の活性が増強された細菌が挙げられる。L-スレオニン生合成系酵素としては、アスパルトキナーゼIII(lysC)、アスパルテートセミアルデヒドデヒドロゲナーゼ(asd)、thrオペロンにコードされるアスパルトキナーゼI(thrA)、ホモセリンキナーゼ(thrB)、スレオニンシンターゼ(thrC)、アスパルテートアミノトランスフェラーゼ(アスパルテートトランスアミナーゼ)(aspC)が挙げられる。カッコ内は、その遺伝子の略記号である(以下の記載においても同様)。これらの酵素の中では、アスパルテートセミアルデヒドデヒドロゲナーゼ、アスパルトキナーゼI、ホモセリンキナーゼ、アスパルテートアミノトランスフェラーゼ、及びスレオニンシンターゼが特に好ましい。L-スレオニン生合成系遺伝子は、スレオニン分解が抑制されたエシェリヒア属細菌に導入してもよい。スレオニン分解が抑制されたエシェリヒア属細菌としては、例えば、スレオニンデヒドロゲナーゼ活性が欠損したTDH6株(特開2001-346578号)等が挙げられる。
以下、L-リジン生産菌及びその構築方法を例として示す。
例えば、L-リジン生産能を有する株としては、L-リジンアナログ耐性株又は代謝制御変異株が挙げられる。L-リジンアナログの例としては、オキサリジン、リジンヒドロキサメート、S-(2-アミノエチル)-L-システイン(以下、「AEC」と略記することがある。)、γ-メチルリジン、α-クロロカプロラクタムなどが挙げられるが、これらに限定されない。これらのリジンアナログに対して耐性を有する変異株は、腸内細菌科に属する細菌やコリネ型細菌を通常の人工変異処理に付すことによって得ることができる。L-リジン生産菌として具体的には、エシェリヒア・コリAJ11442株(FERM BP-1543、NRRL B-12185;特開昭56-18596号公報及び米国特許第4346170号明細書参照)、エシェリヒア・コリ VL611株(特開2000-189180号公報)等が挙げられる。また、エシェリヒア・コリのL-リジン生産菌として、WC196株(国際公開第96/17930号パンフレット参照)を用いることも出来る。
L-システイン生産菌又はそれを誘導するための親株の例としては、フィードバック阻害耐性のセリンアセチルトランスフェラーゼをコードする異なるcysEアレルで形質転換されたE. coli JM15(米国特許第6,218,168号、ロシア特許出願第2003121601号)、細胞に毒性の物質を排出するのに適したタンパク質をコードする過剰発現遺伝子を有するE. coli W3110 (米国特許第5,972,663号)、システインデスルフォヒドラーゼ活性が低下したE. coli株 (特開平11-155571号)、cysB遺伝子によりコードされる正のシステインレギュロンの転写制御因子の活性が上昇したE. coli W3110 (国際公開第0127307号)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。
L-ロイシン生産菌又はそれを誘導するための親株の例としては、ロイシン耐性のE. coil株 (例えば、57株 (VKPM B-7386, 米国特許第6,124,121号))またはβ-2-チエニルアラニン、3-ヒドロキシロイシン、4-アザロイシン、5,5,5-トリフルオロロイシンなどのロイシンアナログ耐性のE. coli株(特公昭62-34397号及び特開平8-70879号)、国際公開第96/06926号に記載された遺伝子工学的方法で得られたE. coli株、E. coli H-9068 (特開平8-70879号)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。
L-ヒスチジン生産菌又はそれを誘導するための親株の例としては、E. coli 24株 (VKPM B-5945、ロシア特許第2003677号)、E. coli 80株 (VKPM B-7270、ロシア特許第2119536号)、E. coli NRRL B-12116 - B12121 (米国特許第4,388,405号)、E. coli H-9342 (FERM BP-6675)及びH-9343 (FERM BP-6676) (米国特許第6,344,347号)、E. coli H-9341 (FERM BP-6674) (欧州特許出願公開第1085087号)、E. coli AI80/pFM201 (米国特許第6,258,554号)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。
L-グルタミン酸生産菌又はそれを誘導するための親株の例としては、E. coli VL334thrC+ (EP 1172433)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。E. coli VL334 (VKPM B-1641)は、thrC遺伝子及びilvA遺伝子に変異を有するL-イソロイシン及びL-スレオニン要求性株である(米国特許第4,278,765号)。thrC遺伝子の野生型アレルは、野生型E. coli K-12株 (VKPM B-7)の細胞で増殖したバクテリオファージP1を用いる一般的形質導入法により導入された。この結果、L-イソロイシン要求性のL-グルタミン酸生産菌VL334thrC+ (VKPM B-8961) が得られた。
E. coli W3110sucA::Kmr
E. coli AJ12624 (FERM BP-3853)
E. coli AJ12628 (FERM BP-3854)
E. coli AJ12949 (FERM BP-4881)
ブレビバクテリウム・ラクトファーメンタムL30-2株(特開2006-340603号明細書)
ブレビバクテリウム・ラクトファーメンタムΔS株(国際公開95/34672号パンフレット)
ブレビバクテリウム・ラクトファーメンタムAJ12821(FERM BP-4172;フランス特許公報9401748号明細書参照)
ブレビバクテリウム・フラバムAJ12822 (FERM BP-4173;フランス特許公報9401748号明細書)
コリネバクテリウム・グルタミカムAJ12823(FERM BP-4174;フランス特許公報9401748号明細書)
コリネバクテリウム・グルタミカムL30-2株(特開2006-340603号)
このような耐性菌の具体例としては、下記のような菌株が挙げられる。
ブレビバクテリウム・フラバムAJ3949 (FERM BP-2632:特開昭50-113209参照)
コリネバクテリウム・グルタミカムAJ11628 (FERM P-5736;特開昭57-065198参照)
ブレビバクテリウム・フラバムAJ11355(FERM P-5007;特開昭56-1889号公報参照)
コリネバクテリウム・グルタミカムAJ11368(FERM P-5020;特開昭56-1889号公報参照)
ブレビバクテリウム・フラバムAJ11217(FERM P-4318;特開昭57-2689号公報参照)
コリネバクテリウム・グルタミカムAJ11218(FERM P-4319;特開昭57-2689号公報参照)
ブレビバクテリウム・フラバムAJ11564(FERM P-5472;特開昭56-140895公報参照)
ブレビバクテリウム・フラバムAJ11439(FERM P-5136;特開昭56-35981号公報参照)
コリネバクテリウム・グルタミカムH7684(FERM BP-3004;特開平04-88994号公報参照)
ブレビバクテリウム・ラクトファーメンタムAJ11426(FERM P-5123;特開平56-048890号公報参照)
コリネバクテリウム・グルタミカムAJ11440(FERM P-5137;特開平56-048890号公報参照)
ブレビバクテリウム・ラクトファーメンタムAJ11796(FERM P-6402;特開平58-158192号公報参照)
L-フェニルアラニン生産菌又はそれを誘導するための親株の例としては、コリスミ酸ムターゼ-プレフェン酸デヒドロゲナーゼ及びチロシンリプレッサーを欠損したE. coli AJ12739 (tyrA::Tn10, tyrR) (VKPM B-8197)(国際公開03/044191号)、フィードバック阻害が解除されたコリスミ酸ムターゼ-プレフェン酸デヒドラターゼをコードする変異型pheA34遺伝子を保持するE. coli HW1089 (ATCC 55371) (米国特許第 5,354,672号)、E. coli MWEC101-b (KR8903681)、E. coli NRRL B-12141, NRRL B-12145, NRRL B-12146及びNRRL B-12147 (米国特許第4,407,952号)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。また、親株として、フィードバック阻害が解除されたコリスミ酸ムターゼ-プレフェン酸デヒドラターゼをコードする遺伝子を保持するE. coli K-12 [W3110 (tyrA)/pPHAB] (FERM BP-3566)、E. coli K-12 [W3110 (tyrA)/pPHAD] (FERM BP-12659)、E. coli K-12 [W3110 (tyrA)/pPHATerm] (FERM BP-12662)及びAJ 12604と命名されたE. coli K-12 [W3110 (tyrA)/pBR-aroG4, pACMAB] (FERM BP-3579)も使用できる(EP 488424 B1)。さらに、yedA遺伝子またはyddG遺伝子にコードされるタンパク質の活性が増大したエシェリヒア属に属するL-フェニルアラニン生産菌も使用できる(米国特許出願公開2003/0148473号及び2003/0157667、国際公開03/044192号)。
L-トリプトファン生産菌又はそれを誘導するための親株の例としては、変異trpS遺伝子によりコードされるトリプトファニル-tRNAシンテターゼが欠損したE. coli JP4735/pMU3028 (DSM10122)及びJP6015/pMU91 (DSM10123) (米国特許第5,756,345号)、セリンによるフィードバック阻害を受けないフォスフォグリセリレートデヒドロゲナーゼをコードするserAアレル及びトリプトファンによるフィードバック阻害を受けないアントラニレートシンターゼをコードするtrpEアレルを有するE. coli SV164 (pGH5) (米国特許第6,180,373号)、トリプトファナーゼが欠損したE. coli AGX17 (pGX44) (NRRL B-12263)及びAGX6(pGX50)aroP (NRRL B-12264) (米国特許第4,371,614号)、フォスフォエノールピルビン酸生産能が増大したE. coli AGX17/pGX50,pACKG4-pps (WO9708333, 米国特許第6,319,696号)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。yedA遺伝子またはyddG遺伝子にコードされるタンパク質の活性が増大したエシェリヒア属に属するL-トリプトファン生産菌も使用できる(米国特許出願公開2003/0148473及び2003/0157667)。
L-プロリン生産菌又はそれを誘導するための親株の例としては、ilvA遺伝子が欠損し、L-プロリンを生産できるE. coli 702ilvA (VKPM B-8012) (欧州特許公開公報1,172,433号)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。
L-アルギニン生産菌又はそれを誘導するための親株の例としては、E. coli 237株 (VKPM B-7925) (米国特許出願公開2002/058315号)、及び、変異N-アセチルグルタメートシンターゼを保持するその誘導株(ロシア特許出願第2,001,112,869号)、E. coli 382株 (VKPM B-7926) (欧州特許公開公報1,170,358号)、N-アセチルグルタメートシンテターゼをコードするargA遺伝子が導入されたアルギニン生産株(欧州特許公開公報1,170,361号)などのエシェリヒア属に属する株が挙げられるが、これらに限定されない。
ノコハク酸リアーゼ遺伝子(argH)、カルバモイルフォスフェートシンテターゼ遺伝子(carAB)が挙げられる。
L-バリン生産菌又はそれを誘導するための親株の例としては、ilvGMEDAオペロンを過剰発現するように改変された株(米国特許第5,998,178号)が挙げられるが、これらに限定されない。アテニュエーションに必要なilvGMEDAオペロンの領域を除去し、生産されるL-バリンによりオペロンの発現が減衰しないようにすることが好ましい。さらに、オペロンのilvA遺伝子が破壊され、スレオニンデアミナーゼ活性が減少することが好ましい。
L-バリン生産菌又はそれを誘導するための親株の例としては、アミノアシルt-RNAシンテターゼの変異を有する変異株(米国特許第5,658,766号)も挙げられる。例えば、イソロイシンtRNAシンテターゼをコードするileS 遺伝子に変異を有するE. coli VL1970が使用できる。E. coli VL1970は、1988年6月24日、ロシアン・ナショナル・コレクション・オブ・インダストリアル・マイクロオルガニズムズ(VKPM) (1 Dorozhny proezd., 1 Moscow 117545, Russia)に、受託番号VKPM B-4411で寄託されている。
さらに、生育にリポ酸を要求する、及び/または、H+-ATPaseを欠失している変異株(国際公開96/06926号)を親株として用いることができる。
例えば、L-イソロイシンおよびL-メチオニン要求性,ならびにD-リボ-ス,プリンリボヌクレオシドまたはピリミジンリボヌクレオシドに耐性を有し,かつL-バリン生産能を有する変異株(FERM P-1841、FERM P-29、特公昭53-025034) や、ポリケトイド類に耐性を有する変異株(FERM P-1763、FERM P-1764、特公平06-065314) 、更には酢酸を唯一の炭素源とする培地でL-バリン耐性を示し、且つグルコースを唯一の炭素源とする培地でピルビン酸アナログ(フルオロピルビン酸等)に感受性を有する変異株(FERM BP-3006、FERM BP-3007、特許3006929号)が挙げられる。
L-イソロイシン生産菌又はそれを誘導するための親株の例としては、6-ジメチルアミノプリンに耐性を有する変異株(特開平5-304969号)、チアイソロイシン、イソロイシンヒドロキサメートなどのイソロイシンアナログに耐性を有する変異株、さらにDL-エチオニン及び/またはアルギニンヒドロキサメートに耐性を有する変異株(特開平5-130882号).が挙げられるが、これらに限定されない。さらに、スレオニンデアミナーゼ、アセトヒドロキシ酸シンターゼなどのL-イソロイシン生合成に関与するタンパク質をコードする遺伝子で形質転換された組換え株もまた親株として使用できる(特開平2-458号, FR 0356739, 及び米国特許第5,998,178号)。
L-メチオニン生産菌又はそれを誘導するための親株の例としては、L-スレオニン要求株、ノルロイシンに耐性を有する変異株が挙げられるが、これらに限定されない(特開2000-139471号)。さらに、メチオニンリプレッサーを欠損した株や、ホモセリントランスサクシニラーゼ、シスタチオニンγ-シンテースなどのL-メチオニン生合成に関与するタンパク質をコードする遺伝子で形質転換された組換え株もまた親株として使用できる(特開2000-139471号)。
また、遺伝子の配列におけるそれぞれのコドンは、遺伝子が導入される宿主で使用しやすいコドンに置換したものでもよい。
本発明のL-アミノ酸の製造法においては、微細藻類の前記処理物を含む培地で、L-アミノ酸生産能を有する細菌を培養して、培養物中にL-アミノ酸を生産蓄積させ、該培養物からL-アミノ酸を採取する。あるいは、(a)微細藻類を培地で培養し、該培養物を、破砕、抽出、分画及び加水分解から選ばれる1以上の方法により処理して、L-アミノ酸生産能を有する細菌によるL-アミノ酸の生産蓄積を促進する該微細藻類の処理物を調製し、(b)該細菌を、該微細藻類の処理物を含む培地に培養し、培養物中にL-アミノ酸を生産蓄積させ、(c)該培養物からL-アミノ酸を採取する。前記処理物は炭素源として含まれることが好ましく、この場合には、特にスターチの糖化物あるいは油脂の加水分解物を含む処理物であることが好ましい。
Chlorella kessleri を、100 mLの0.2×ガンボーグB5培地(日本製薬)を入れた500 mL容三角フラスコにて30℃、光強度10,000 lux(TOMY社製培養装置CL-301)で6日間振とう培養し、これを前培養液とした。尚、光源には、蛍光灯からの白色光を用いた。0.2×ガンボーグB5 培地800 mLを入れた1L容メディウムビンに、前培養液16 mLを添加し、培養温度30℃、光強度10,000 luxにて、500 mL/minで、CO2濃度が3%となるように空気とCO2の混合ガスを吹き込みながら、12日間培養を行った。
KNO3 500 mg/L
MgSO4・7H2O 50 mg/L
NaH2PO4・H2O 30 mg/L
CaCl2・2H2O 30 mg/L
(NH4)2SO4 26.8 mg/L
Na2-EDTA 7.46 mg/L
FeSO4・7H2O 5.56 mg/L
MnSO4・H2O 2 mg/L
H3BO3 0.6 mg/L
ZnSO4・7H2O 0.4 mg/L
KI 0.15 mg/L
Na2MoO2・2H2O 0.05 mg/L
CuSO4・5H2O 0.005 mg/L
CoCl2・6H2O 0.005 mg/L
120℃ 15分 オートクレーブ殺菌
実施例1(1)の培養液1 L分の藻体を遠心分離にて沈殿させ、その沈殿物に80 mLのエタノールを加え、藻体を懸濁させた後に30分煮沸した。その懸濁液を遠心分離し、藻体を沈殿させた後に、80 mLのエタノールで2回洗浄した後、デシケーター内で乾燥させた。その乾燥藻体に40 mLの0.2 M KOHを加え、30分煮沸し、さらに超音波装置(Kubota社INSONATOR 201MA)により15,000W、10分の処理を行い、細胞を破砕した。その破砕液に8 mLの1 M酢酸溶液、250 Uのアミログルコシダーゼ(シグマ-アルドリッチ(Sigma-Aldrich)社A-9228)を加え、55℃にて18時間反応させた。反応終了後、反応物中のグルコースをバイオテックアナライザーAS210(サクラ精機)にて定量した。酵素濃度を2倍にしても、生成するグルコース量はほとんど変化がなかったことから、スターチのうち、大部分がグルコースに変換されていると推定された。藻体破砕物(残渣)を遠心分離によって除去し、その上清をアミノ酸発酵の炭素源として用いた。
L-リジン生産菌として国際公開第2006/078039号パンフレットに記載されているエシェリヒア・コリWC196ΔcadAΔldcC/pCABD2を用いた。WC196ΔcadAΔldcC/pCABD2をストレプトマイシン硫酸塩20mg/Lを含有するLB寒天培地(Tryptone 10 g/L、Yeast extract 5 g/L、NaCl 10g /L、寒天15g/L)にて37℃で20時間培養した。寒天培地上の細胞を掻き取り、ストレプトマイシン硫酸塩20 mg/Lを含有するL-リジン生産培地40 mLを入れた500 mL容三角フラスコに植菌し、培養温度37℃にて、24時間培養を行った。本培養においては、Chlorella kessleriが生産するスターチから調製した実施例1(2)の糖化液(グルコース量として2.75 g/L)とその調製の際に、バッファーとして利用した酢酸4.8 g/Lを炭素源とした。対照として、同濃度のグルコース、又は酢酸の一方を含む培地と、グルコースと酢酸の両方を炭素源として含む培地にて培養を実施した。
(A区)
炭素源
グルコース 2.75 g/L
酢酸 4.8 g/L
(B区)
(NH4)2SO4 24 g/L
KH2PO4 1 g/L
Yeast Extract 2 g/L
FeSO4・7H2O 10 mg/L
MnSO4・4H2O 10 mg/L
(C区)
炭酸カルシウム 15 g/L
A区、B区は115℃にて10分間、オートクレーブ殺菌し、C区は180℃にて3時間、乾熱滅菌した。室温に冷却後3者を混合して、MgSO4・7H2O溶液を終濃度1 g/Lとなるように加えた。

実施例1(1)の培養液3.6 L分の藻体を遠心分離にて沈殿させ、その沈殿物に100 mLのエタノールを加え、藻体を懸濁させた後、室温で24 時間静置した。その後、濾過を行って濾液を回収し、藻体残渣を100 mLのエタノールで2回抽出操作を行い、計300 mLのエタノール抽出液とした。そのエタノール抽出液をロータリーエバポレーターによりエタノール除去した後、滅菌水に溶解し、水酸化カリウム水溶液でpH7.0に合わせ、藻体由来の有機物液45 mLを調製した。その有機物液を、115℃にて10分間オートクレーブ滅菌した後、L-リジン発酵の培地成分の代替として用いた。本有機物液はスターチを含んでいないため、アミノ酸生産菌に利用できる炭素源は、含まれていない。
L-リジン生産菌であるエシェリヒア・コリWC196ΔcadAΔldcC/pCABD2を、ストレプトマイシン硫酸塩20 mg/Lを含有するLB寒天培地にて37℃で20時間培養した。寒天培地上の細胞を掻き取り、ストレプトマイシン硫酸塩20 mg/Lを含有するL-リジン生産培地4 mLを入れた太試験管に植菌し、培養温度37℃にて、24時間培養を行った。本培養においては、L-リジン生産培地中のB区の成分を半減した培地(以下、本培地を「L-リジン生産培地(半量B区)」と呼ぶ)にChlorella kessleri藻体から抽出した実施例2(1)の有機物液75 μL(培養液に換算すると、6 mL分に相当)を加えて、培養を実施した。対照としては、L-リジン生産培地、L-リジン生産培地(半量B区)、及びグルコースが入っていないL-リジン生産培地(半量B区)培地に有機物液75 μLを添加した条件にて、培養を行った。
(A区)
グルコース 20 g/L
(B区)
(NH4)2SO4 24 g/L
KH2PO4 1 g/L
Yeast Extract 2 g/L
FeSO4・7H2O 10 mg/L
MnSO4・4H2O 10 mg/L
(C区)
炭酸カルシウム 30 g/L
室温に冷却後、3者を混合して、MgSO4・7H2O溶液を終濃度1 g/Lとなるように加えた。

Chlorella kessleri を、0.2×ガンボーグB5培地(日本製薬)500 mLを入れた2 L容ジャーファーメンター(ABLE社製)にて30℃、光強度20,000 luxで7日間攪拌培養し、これを前培養液とした。尚、本ジャーファーメンターは、光源として、白色光を発する環形蛍光灯がガラスベッセルの周りを取り囲む光照射型のジャーファーメンターである。0.2×ガンボーグB5 培地1.5 Lを入れた2 L容ミニジャーファーメンターに、前培養液30 mLを添加し、培養温度30℃、光強度20,000 luxにて、500 mL/minでCO2濃度を3%に保ちながら空気-CO2混合ガスを吹き込み、14日間培養を行った。
実施例3(1)の藻類培養液300 mLを高温反応装置内の反応容器に投入し、攪拌しつつ60分間かけて175℃もしくは215℃まで昇温させた後、それぞれの温度で5分間保持させた。
L-グルタミン酸生産菌としては、特開2006-340603号に記載のブレビバクテリウム・ラクトファーメンタムL30-2株を用いた。L30-2株をCM-Dexプレート培地に播種し31.5℃で24時間培養した。そのプレート培地上の細胞を1白金耳分掻き取り、L-グルタミン酸生産培地4 mLを入れた太試験管に植菌し、培養温度31.5℃にて、13時間培養を行った。本培養における炭素源として、Chlorella kessleriの藻類スターチ分解物から調製した実施例3(2)の糖化液(グルコース量として1.0 g/L)、ならびに対照として同濃度の試薬グルコースを用いた。また、L-グルタミン酸生産培地、およびL-グルタミン酸生産培地中のB区の成分を半減した培地(以下、本培地を「L-グルタミン酸生産培地(半量B区)」と呼ぶ)の2種類の培地成分にて、培養を実施した。
グルコース 5 g/L
ポリペプトン 10 g/L
酵母エキス 10 g/L
KH2PO4 1 g/L
MgSO4・7H2O 0.4 g/L
FeSO4・7H2O 0.01 g/L
MnSO4・7H2O 0.01 g/L
尿素 3 g/L
大豆加水分解物 1.2 g/L
ビオチン 10 μg/L
NaOHを用いて、pH7.5に調整。120℃にて20分間オートクレーブ殺菌条件。
(A区)
炭素源
藻類由来のスターチ分解物 1.0 g/L(グルコース)
(B区)
(NH4)2SO4 15 g/L
KH2PO4 1 g/L
MgSO4・7H2O 0.4 g/L
FeSO4・7H2O 10 mg/L
MnSO4・4H2O 10 mg/L
VB1・HCl 200 μg/L
Biotin 300 μg/L
大豆加水分解物 0.48 g/L
(C区)
炭酸カルシウム 50 g/L

実施例3(2)にて調製した215℃の高温処理した藻体破砕液80 mLのpHを1N HClを用いて6.0に調整した後、400 Uのアミログルコシダーゼ単独(シグマ-アルドリッチ社A-9228)もしくは同量のアミログルコシダーゼと1000Uのリパーゼ(シグマ-アルドリッチ社 L1754)両酵素 を加え、50℃にて24時間反応させた。反応終了後の各反応液を10 mLまで濃縮した後、アミログルコシダーゼ単独処理液中の残渣を遠心分離で取り除き、その上清のpHを1N NaOHを用いて7.0に調整し、15mlにメスアップした後、120℃にて20分の条件にて、オートクレーブ処理した。一方、両酵素の処理液は、1.13mLの10%Tween80水溶液を加え、60℃に加温した後、ボルテックスにて攪拌した。次に、その両酵素の処理液中の残渣を遠心分離で取り除き、その上清のpHを1N NaOHを用いて7.0に調整し、15 mLにメスアップした後、120℃、20分の条件にて、オートクレーブ処理した。各炭素源中のグルコースと脂肪酸の量を測定し、培養評価に用いた。
エシェリヒア・コリの脂肪酸代謝を調節する転写因子FadRはfadR遺伝子(配列番号15)によってコードされている(DiRusso, C. C. et al. 1992. J. Biol. Chem. 267: 8685-8691)。本遺伝子破壊の親株は、エシェリヒア・コリのL-リジン生産株として、国際特許公報WO2006/078039に記載のWC196ΔcadAΔldcC株を用いた。
L-リジン生産菌であるエシェリヒア・コリWC196ΔcadAΔldcCΔfadR/pCABD22を、ストレプトマイシン硫酸塩20 mg/Lを含有するLB寒天培地にて37℃で20時間培養した。寒天培地上の細胞を掻き取り、ストレプトマイシン硫酸塩20mg/Lを含有するL-リジン生産培地4 mLを入れた太試験管に植菌し、培養温度37℃にて、24時間培養を行った。本培養においては、Chlorella kessleriが生産するスターチから調製した実施例4(1)の糖化液(グルコース量として1.33 g/L)とスターチと油脂から調製した実施例4(1)の加水分解液を炭素源とした。対照として、同濃度のグルコースを炭素源として含む培地にて培養を実施した。
(A区)
炭素源
藻類由来のスターチ分解物 1.33 g/L(グルコース)
(B区)
(NH4)2SO4 24 g/L
KH2PO4 1 g/L
Yeast Extract 2 g/L
FeSO4・7H2O 10 mg/L
MnSO4・4H2O 10 mg/L
(C区)
炭酸カルシウム 15 g/L

Neochloris oleoabundans UTEX 1185株を、100 mLのModified NORO培地を入れた500 mL容三角フラスコにて30℃、光強度10,000 lux(TOMY社製培養装置CL-301)で6日間振とう培養し、これを前培養液とした。尚、光源には、蛍光灯からの白色光を用いた。Modified NORO培地800 mLを入れたL容メディウムビンに、前培養液32 mLを添加し、培養温度30℃、光強度10,000 luxにて、500 mL/minでCO2濃度が3%となるように空気とCO2の混合ガスを吹き込みながら、14日間培養を行った。
NaCl 29.22 g/L
KNO3 1.0 g/L
MgCl2・6H2O 1.5 g/L
MgSO4・7H2O 0.5 g/L
KCl 0.2 g/L
CaCl2・2H2O 0.2 g/L
K2HPO4 0.045 g/L
Tris(hydroxymethyl)aminomethane 2.45 g/L
Na2-EDTA 1.89 mg/L
ZnSO4・7H2O 0.087 mg/L
H3BO3 0.61 mg/L
CoCl2・6H2O 0.015 mg/L
CuSO4・5H2O 0.06 mg/L
MnCl2 0.23 mg/L
(NH4)6Mo7O24・4H2O 0.38 mg/L
Fe(III)・EDTA 3.64 mg/L
Vitamin B1 0.1 mg/L
Vitamin B12 0.5 mg/L
Biotin 0.5 mg/L
pHを1N HClにて8.0に調整後、120℃にて10分間オートクレーブ殺菌
実施例5(1)の培養液3.6 L分の藻体を遠心分離にて沈殿させ、得られた藻体を用いて、下記の抽出操作を3回繰り返した。まず、藻体を100 mLのメタノール:クロロホルム=2:1に懸濁させた後に、超音波装置(Kubota社INSONATOR 201MA)により15,000W、10 分の処理を行い、細胞を破砕した。その破砕液を一晩、静置抽出した後、濾過により固形物を取り除き、抽出液を回収した。
L-リジン生産菌であるエシェリヒア・コリWC196ΔcadAΔldcC/pCABD2株のグリセロールストックを融解し、各100 μLを、25 mg/Lのストレプトマイシンを含むLプレートに均一に塗布し、37℃にて20時間培養した。得られたプレートのおよそ1/8量の菌体を、25 mg/Lのストレプトマイシンを含む以下に記載の発酵培地20 mLを入れた坂口フラスコに接種し、往復振とう培養装置で37℃において24時間培養した。実施例5(2)の藻類由来の各サンプルを、蒸留水に懸濁させた後、1%濃度になるようにTween80を加え、pHを3N KOHを用いて7.0に調整し、120℃にて20分オートクレーブを行ったものを炭素源溶液として用いた。培養に用いた培地組成(終濃度)を以下に示す。藻類由来の粗抽出物の加水分解物、藻類由来の油脂画分の加水分解物、あるいは、藻類由来の水溶性画分を炭素源として用いた。それぞれの炭素源について、測定された脂肪酸濃度とグリセロール濃度の和が記載されている。また、対照として、試薬脂肪酸と試薬グリセロールを混合し、同濃度になるように調整した炭素源を用いた。微細藻類Neochloris oleoabundansより抽出した油脂の加水分解物の脂肪酸とグリセロールの分析を行い、主要な脂肪酸として含まれていたオレイン酸(純正化学社製一級)、リノール酸(ナカライテスク社製)及びパルミチン酸(ナカライテスク社製特級)を重量組成比4.2 : 3.8 : 1.0で含み、脂肪酸とグリセロールの重量組成比が11.4:1となるように調整した。
炭素源として
試薬脂肪酸+試薬グリセロール 1.09 g/L(脂肪酸濃度+グリセロール濃度)
藻類由来の粗抽出物の加水分解物 1.09 g/L(脂肪酸濃度+グリセロール濃度)
藻類由来の油脂画分の加水分解物 1.19 g/L(脂肪酸濃度+グリセロール濃度)
藻類由来の水溶性画分 0.04 g/L(脂肪酸濃度+グリセロール濃度)
のいずれか
MgSO4・7H2O 1.0 g/L
(NH4)2SO4 24 g/L
KH2PO4 1.0 g/L
FeSO4・7H2O 0.01 g/L
MnSO4・7H2O 0.01 g/L
Yeast Extract 2.0 g/L
炭酸カルシウム 30 g/L

Neochloris oleoabundans UTEX 1185株を、100 mLのModified NORO培地を入れた500mL容三角フラスコにて30℃、光強度5,000 lux(TOMY社製培養装置CL-301)で6日間振とう培養し、これを前培養液とした。尚、光源には、蛍光灯からの白色光を用いた。Modified NORO培地800mLを入れたL容メディウムビンに、前培養液32mLを添加し、培養温度30℃、光強度5,000 luxにて、500 mL/minにてCO2濃度が3%となるように空気とCO2の混合ガスを吹き込みながら、14日間培養を行った。
NaCl 29.22 g/L
KNO3 1.0 g/L
MgCl2・6H2O 1.5 g/L
MgSO4・7H2O 0.5 g/L
KCl 0.2 g/L
CaCl2・2H2O 0.2 g/L
K2HPO4 0.045 g/L
Tris(hydroxymethyl)aminomethane 2.45 g/L
Na2-EDTA 1.89 mg/L
ZnSO4・7H2O 0.087 mg/L
H3BO3 0.61 mg/L
CoCl2・6H2O 0.015 mg/L
CuSO4・5H2O 0.06 mg/L
MnCl2 0.23 mg/L
(NH4)6Mo7O24・4H2O 0.38 mg/L
Fe(III)・EDTA 3.64 mg/L
Vitamin B1 0.1 mg/L
Vitamin B12 0.5 μg/L
Biotin 0.5 μg/L
1N HClにてpH8.0に調整後、120℃にて10分 オートクレーブ殺菌
実施例6(1)の培養液1.8L分の藻体を遠心分離にて沈殿させ、その沈殿物に50 mLのメタノール:クロロホルム=2:1を加え、下記の抽出操作を3回繰り返した。まず、藻体を50 mLのメタノール:クロロホルム=2:1に懸濁させ、一晩、静置抽出した後、濾過により固形物を取り除き、抽出液を回収した。
エシェリヒア・コリWC196ΔcadAΔldcC/pCABD2株のグリセロールストックを融解し、各100 μLを、25 mg/Lのストレプトマイシンを含むLプレートに均一に塗布し、37℃にて20時間培養した。得られたプレートのおよそ1/40量の菌体を、太試験管の、25 mg/Lのストレプトマイシンを含む以下に記載の発酵培地の4 mLに接種し、往復振とう培養装置で37℃において24時間培養した。炭素源となる実施例6(2)の藻類由来の各サンプルは、加水分解後の溶液に、2%濃度になるようにTween80を加え、pHを3N KOHを用いて7.0に調整し、120℃にて20分オートクレーブを行ったものを炭素源溶液として用いた。培養に用いた培地組成を以下に示す。A区は炭素源であり、測定された脂肪酸濃度とグリセロール濃度の和が記載されている。
(A区)
藻類由来の油脂加水分解物 1.22 g/L(脂肪酸濃度+グリセロール濃度)
藻類由来の全脂質類の加水分解物 2.14 g/L(脂肪酸濃度+グリセロール濃度)
のいずれか
(B区)
(NH4)2SO4 24 g/L
KH2PO4 1.0 g/L
FeSO4・7H2O 0.01 g/L
MnSO4・7H2O 0.01 g/L
Yeast Extract 2.0 g/L
(C区)
炭酸カルシウム 30 g/L

Nannochloris sp. UTEX LB 1999株を、100 mLのModified NORO培地を入れた500 mL容三角フラスコにて30℃、光強度10,000 lux(TOMY社製培養装置CL-301)で6日間振とう培養し、これを前培養液とした。尚、光源には、蛍光灯からの白色光を用いた。Modified NORO培地800 mLを入れた1L容メディウムビンに、前培養液32mLを添加し、培養温度30℃、光強度10,000 luxにて、500 mL/minでCO2濃度が3%となるように空気とCO2の混合ガスを吹き込みながら、14日間培養を行った。
実施例7(2)の培養液3.6 L分の藻体を遠心分離にて沈殿させ、得られた藻体を用いて、下記の抽出操作を3回繰り返した。まず、藻体を100 mLのメタノール:クロロホルム=2:1に懸濁させた後に、超音波装置(Kubota社INSONATOR 201MA)により15,000Wにて10分間処理を行い、細胞を破砕した。その破砕液を一晩静置抽出した後、濾過により固形物を取り除き、抽出液を回収した。得られた抽出液を濃縮した後、80%メタノール100 mLに懸濁させ、その懸濁液とヘキサン100 mLを加えた分液ロート内で液々分配を3回行い、油脂を含むヘキサン層(油脂画分)と水溶性有機物を多く含む80%メタノール層(水溶性画分)を得た。次に、油脂画分を予め加温した40 mLの熱水に懸濁させた後に、500Uのリパーゼ (シグマ L1754)を加え、42℃にて20時間反応させた。その反応液と水溶性画分の脂肪酸濃度とグリセロール濃度を測定し、それぞれをアミノ酸発酵の炭素源として用いた。
エシェリヒア・コリWC196ΔcadAΔldcC/pCABD2株のグリセロールストックを融解し、各100 μLを、25 mg/Lのストレプトマイシンを含むLプレートに均一に塗布し、37℃にて20時間培養した。得られたプレートのおよそ1/8量の菌体を、25 mg/Lのストレプトマイシンを含む以下に記載の発酵培地20 mLを入れた坂口フラスコに接種し、往復振とう培養装置で37℃において16もしくは20時間培養した。実施例7(2)の藻類由来の各サンプルを、蒸留水に懸濁させた後、1%濃度になるようにTween80を加え、pHを3N KOHを用いて7.0に調整し、120℃にて20分オートクレーブを行ったものを炭素源溶液として用いた。培養に用いた培地組成を以下に示す。藻類由来の油脂画分加水分解物あるいは藻類由来の水溶性画分を炭素源として用いた。それぞれの炭素源について測定された脂肪酸濃度とグリセロール濃度の和が記載されている。
炭素源として
藻類由来の油脂画分の加水分解物 1.06 g/L(脂肪酸濃度+グリセロール濃度)
藻類由来の水溶性画分 0.00 g/L(脂肪酸濃度+グリセロール濃度)
のいずれか
MgSO4・7H2O 1.0 g/L
(NH4)2SO4 24 g/L
KH2PO4 1.0 g/L
FeSO4・7H2O 0.01 g/L
MnSO4・7H2O 0.01 g/L
Yeast Extract 2.0 g/L
炭酸カルシウム 30 g/L

実施例7(1)と同様にして培養したNannochloris sp. UTEX LB 1999株の培養液1.2L分の藻体を遠心分離にて沈殿させ、得られた藻体を用いて、下記の抽出操作を3回繰り返した。まず、藻体を100mLのメタノール:クロロホルム=2:1に懸濁させた後に、超音波装置(Kubota社INSONATOR 201MA)により15,000Wにて10分間処理を行い、細胞を破砕した。その破砕液を一晩静置抽出した後、濾過により固形物を取り除き、抽出液を回収し、濃縮した。次に、予め加温した40 mLの熱水に懸濁した後、リパーゼ (シグマ-アルドリッチ社 L1754)500Uを加え、42℃にて20時間反応させた。その反応液の脂肪酸濃度とグリセロール濃度を測定し、それぞれをアミノ酸発酵の炭素源として用いた。
エシェリヒア・コリWC196LC/pCABD2株のグリセロールストックを融解し、各100μLを、25 mg/Lのストレプトマイシンを含むLプレートに均一に塗布し、37℃にて20時間培養した。得られたプレートのおよそ1/8量の菌体を、25 mg/Lのストレプトマイシンを含む以下に記載の発酵培地20 mLを入れた坂口フラスコに接種し、往復振とう培養装置で37℃において16もしくは20時間培養した。炭素源となる実施例8(1)の藻類由来の各サンプルは、蒸留水に懸濁させた後、1%濃度になるようにTween80を加え、pHを3N KOHを用いて7.0に調整し、120℃にて20分オートクレーブを行ったものを炭素源溶液として用いた。培養に用いた培地組成を以下に示す。炭素源として用いた藻類由来の粗抽出物の加水分解物について測定された脂肪酸濃度とグリセロール濃度の和が記載されている。
藻類由来の粗抽出物の加水分解物 0.37 g/L(脂肪酸濃度+グリセロール濃度)
MgSO4・7H2O 1.0 g/L
(NH4)2SO4 24 g/L
KH2PO4 1.0 g/L
FeSO4・7H2O 0.01 g/L
MnSO4・7H2O 0.01 g/L
Yeast Extract 2.0 g/L
炭酸カルシウム 30 g/L

Thalassiosira pseudonana UTEX LB FD2株を、500 mLのF/2培地を入れた1L容メディウムビンにて、培養温度25℃、光強度7,000 luxにて、200 mL/minでCO2濃度が1%となるように空気とCO2の混合ガスを吹き込みながら、3日間培養し、これを前培養液とした。F/2培地の海水成分としては、人工海水であるアクアマリンS(八洲薬品 社(YASHIMA PURE CHEMICALS)製)を用いた。F/2培地800 mLを入れた1L容メディウムビンに、前培養液32mLを添加し、培養温度25℃、光強度7,000 luxにて、200 mL/minでCO2濃度が1%となるように空気との混合ガスを吹き込みながら、7日間培養を行った。尚、光源には、蛍光灯からの白色光を用いた。
NaNO3 75 mg/L
NaH2PO4・H2O 5 mg/L
Na2SiO3・9H2O 20 mg/L
FeCl3・6H2O 6.4 mg/L
MnSO4・H2O 0.304 mg/L
ZnSO4・7H2O 0.046 mg/L
Na2MoO4・2H2O 14.6 μg/L
CuCl2・2H2O 13.6 μg/L
Na2EDTA・2H2O 8.8 mg/L
CoCl2・6H2O 23.8 μg/L
Vitamin B12 0.135 mg/L
Biotin 0.025 mg/L
Vitamin B1 1.1 mg/L
アクアマリンS(人工海水) 40 g/L
120℃にて10分 オートクレーブ殺菌
実施例9(1)の培養液1.8L分の藻体を遠心分離にて沈殿させ、その沈殿物に50 mLのメタノール:クロロホルム=2:1を加え、下記の抽出操作を3回繰り返した。まず、藻体を50mLのメタノール:クロロホルム=2:1に懸濁させ、一晩、静置抽出した後、濾過により固形物を取り除き、抽出液を回収した。
得られた抽出液を濃縮した後、40 mLの熱水に懸濁させた後に、リパーゼ (シグマ L1754)500Uを加え、42℃にて18時間反応させた。その反応液の脂肪酸濃度とグリセロール濃度を測定し、それぞれをアミノ酸発酵の炭素源として用いた。
エシェリヒア・コリWC196LC/pCABD2株のグリセロールストックを融解し、各100 μLを、25 mg/Lのストレプトマイシンを含むLプレートに均一に塗布し、37℃にて20時間培養した。得られたプレートのおよそ1/40量の菌体を、太試験管の、25 mg/Lのストレプトマイシンを含む以下に記載の発酵培地の4 mLに接種し、往復振とう培養装置で37℃において24時間培養した。炭素源となる実施例9(2)の藻類由来の各サンプルは、加水分解後の溶液に、2%濃度になるようにTween80を加え、pHを3N KOHを用いて7.0に調整し、120℃、20分オートクレーブを行ったものを炭素源溶液として用いた。培養に用いた培地組成を以下に示す。A区は炭素源であり、測定された脂肪酸濃度とグリセロール濃度の和が記載されている。
[A区]
藻類由来の粗抽出物の加水分解物 1.09 g/L(脂肪酸濃度+グリセロール濃度)
[B区]
(NH4)2SO4 24g/L
KH2PO4 1.0g/L
FeSO4・7H2O 0.01g/L
MnSO4・7H2O 0.01g/L
Yeast Extract 2.0g/L
[C区]
炭酸カルシウム 30g/L
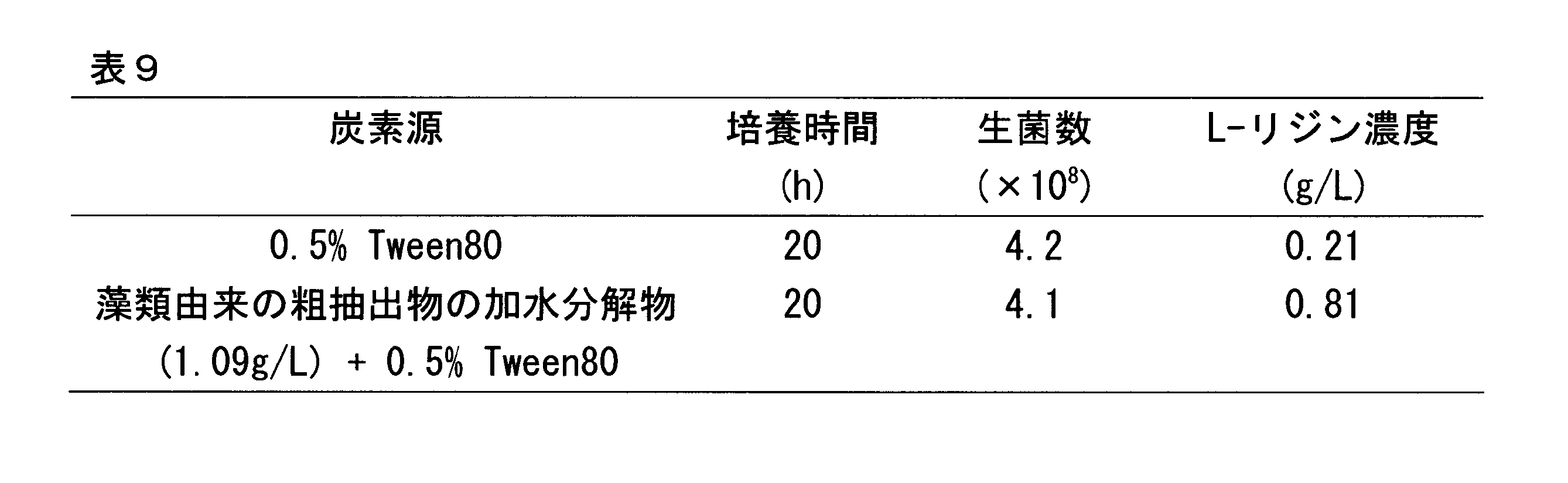
配列番号1:Bacillus subtilis由来 LipA遺伝子の塩基配列
配列番号2:Bacillus subtilis由来 LipAのアミノ酸配列
配列番号3:Burkholderia glumae由来LipA遺伝子の塩基配列
配列番号4:Burkholderia glumae由来LipAのアミノ酸配列
配列番号5:Pseudomonas aeruginosa 由来LipA遺伝子の塩基配列
配列番号6:Pseudomonas aeruginosa 由来LipAのアミノ酸配列
配列番号7:Staphylococcus aureus由来リパーゼ遺伝子の塩基配列
配列番号8:Staphylococcus aureus由来リパーゼのアミノ酸配列
配列番号9:Candida antarctica由来リパーゼ遺伝子の塩基配列
配列番号10:Candida antarctica由来リパーゼのアミノ酸配列
配列番号11:Candida rugosa由来リパーゼ遺伝子lip1の塩基配列
配列番号12:Candida rugosa由来リパーゼLIP1のアミノ酸配列
配列番号13:Candida rugosa由来リパーゼ遺伝子lip2の塩基配列
配列番号14:Candida rugosa由来リパーゼLIP2のアミノ酸配列
配列番号15:Eschrichia coli転写因子遺伝子fadRの塩基配列
配列番号16:fadR増幅用プライマー
配列番号17:fadR増幅用プライマー
Claims (34)
- L-アミノ酸生産能を有する細菌を、微細藻類の処理物を含む培地に培養し、培養物中にL-アミノ酸を生産蓄積させ、該培養物からL-アミノ酸を採取することを特徴とするL-アミノ酸の製造法であって、該処理物が、前記細菌によるL-アミノ酸の生産蓄積を促進するものである、L-アミノ酸の製造法。
- 前記処理物が、(1)該微細藻類の培養物の破砕物、(2)該微細藻類に由来する有機物の混合物を含む、該破砕物の抽出物もしくは分画物、又は、(3)該破砕物、該抽出物もしくは該分画物の加水分解物である請求項1記載の方法。
- 前記処理物が、スターチを産生する微細藻類の藻体破砕物、又はスターチを含むその抽出物もしくは該抽出物の分画物を加水分解して得られる糖化物である、請求項1又は2に記載の方法。
- 前記糖化物が、微細藻類の藻体破砕物又はスターチを含むその分画物から、アミラーゼを用いた酵素反応により得られた反応産物である、請求項3に記載の方法。
- 前記アミラーゼがグルコアミラーゼである請求項4に記載の方法。
- 前記処理物が、油脂を産生する微細藻類の藻体破砕物、又は油脂を含むその抽出物もしくは該抽出物の分画物を加水分解して得られる加水分解物である、請求項1又は2に記載の方法。
- 前記加水分解物が、微細藻類の藻体破砕物又は油脂を含むその分画物から、リパーゼを用いた酵素反応により得られた反応産物である、請求項6記載の方法。
- 前記加水分解物が乳化処理を施されたものであることを特徴とする請求項6~7のいずれか一項に記載の方法。
- 前記微細藻類の破砕法が高温処理であることを特徴とする請求項1~8のいずれか一項に記載の方法。
- 前記微細藻類の破砕法が100℃以上の温度にて処理することを特徴とする請求項1~9のいずれか一項に記載の方法。
- 前記微細藻類が緑色植物門、不等毛植物門に属する藻類である請求項1~10のいずれか一項に記載の方法。
- 前記微細藻類が緑藻綱、トレボキシア藻綱、又は珪藻綱に属する藻類である、請求項11に記載の方法。
- 前記微細藻類が緑藻綱(Chlorophyceae)に属する藻類である、請求項1~12のいずれか一項に記載の方法。
- 前記細菌が腸内細菌科に属する細菌またはコリネ型細菌である請求項1~13のいずれか一項に記載の方法。
- 前記腸内細菌科に属する細菌がエシェリヒア・コリである請求項14に記載の方法。
- 前記L-アミノ酸がL-リジン、L-スレオニン、L-グルタミン酸からなる群より選ばれる1種又は2種以上のL-アミノ酸である、請求項1~15のいずれか一項に記載の方法。
- 前記L-アミノ酸がL-リジンであり、前記細菌がジヒドロジピコリン酸レダクターゼ、ジアミノピメリン酸デカルボキシラーゼ、ジアミノピメリン酸デヒドロゲナーゼ、フォスフォエノールピルベートカルボキシラーゼ、アスパラギン酸アミノトランスフェラーゼ、ジアミノピメリン酸エピメラーゼ、アスパルテートセミアルデヒドデヒドロゲナーゼ、テトラヒドロジピコリン酸スクシニラーゼ、及び、スクシニルジアミノピメリン酸デアシラーゼからなる群より選択される1種または2種以上の酵素の活性が増強されている、及び/または、リジンデカルボキシラーゼの活性が弱化されている、請求項15に記載の方法。
- 前記L-アミノ酸がL-スレオニンであり、前記細菌がアスパラギン酸セミアルデヒドデヒドロゲナーゼ、thrオペロンにコードされるアスパルトキナーゼI、ホモセリンキナーゼ、アスパラギン酸アミノトランスフェラーゼ、及び、スレオニンシンターゼからなる群より選択される1種または2種以上の酵素の活性が増強されている請求項15に記載の方法。
- 前記L-アミノ酸がL-グルタミン酸であり、前記細菌がグルタメートデヒドロゲナーゼ、クエン酸シンターゼ、ホスホエノールピルビン酸カルボキシラーゼ、及び、メチルクエン酸シンターゼからなる群より選択される1種または2種以上の酵素の活性が増強されている、及び/または、α-ケトグルタル酸デヒドロゲナーゼの活性が弱化されている請求項15に記載の方法。
- 前記培地が前記処理物を炭素源として含む請求項1~19のいずれか一項に記載の方法。
- L-アミノ酸の製造方法であって、
(a)微細藻類を培地で培養し、該培養物を、破砕、抽出、分画及び加水分解から選ばれる1以上の方法により処理して、L-アミノ酸生産能を有する細菌によるL-アミノ酸の生産蓄積を促進する該微細藻類の処理物を調製し、
(b)該細菌を、該微細藻類の処理物を含む培地に培養し、培養物中にL-アミノ酸を生産蓄積させ、
(c)該培養物からL-アミノ酸を採取することを特徴とするL-アミノ酸の製造法。 - 前記処理物が、(1)該微細藻類の培養物の破砕物、(2)該微細藻類に由来する有機物の混合物を含む、該破砕物の抽出物もしくは分画物、又は、(3)該破砕物、該抽出物もしくは該分画物の加水分解物である請求項21記載の方法。
- 前記破砕が、高温処理、有機溶媒処理、煮沸処理、強アルカリ処理からなる群から選択される1以上の方法によって行われる請求項21記載の方法。
- 前記処理物調製工程が、スターチを産生する微細藻類を破砕及び/または抽出・分画し、その処理物を加水分解することによって糖化する工程を含む請求項21~23のいずれか一項に記載の方法。
- 前記糖化する工程が、アミラーゼを用いた酵素反応を施すことを特徴とする、請求項24に記載の方法。
- 前記アミラーゼがグルコアミラーゼである請求項25に記載の方法。
- 前記処理物調製工程が、油脂を産生する微細藻類を破砕及び/または抽出・分画し、その油処理物を加水分解する工程を含む、請求項21~23のいずれか一項に記載の方法。
- 前記加水分解工程が、リパーゼを用いた酵素反応を施すことを特徴とする、請求項27記載の方法。
- 前記加水分解物が乳化処理を施されたものであることを特徴とする請求項27または28のいずれか一項に記載の方法。
- 前記微細藻類が緑色植物門、不等毛植物門に属する藻類である請求項21~29のいずれか一項に記載の方法。
- 前記微細藻類が緑藻綱、トレボキシア藻綱、又は珪藻綱に属する藻類である、請求項30に記載の方法。
- 前記微細藻類が緑藻綱(Chlorophyceae)に属する藻類である、請求項31に記載の方法。
- 前記細菌が腸内細菌科に属する細菌またはコリネ型細菌である請求項21~32のいずれか一項に記載の方法。
- 前記腸内細菌科に属する細菌がエシェリヒア・コリである請求項33に記載の方法。
Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN2009801028942A CN102348806A (zh) | 2008-01-23 | 2009-01-23 | L-氨基酸的生产方法 |
| EP09704195A EP2248906A4 (en) | 2008-01-23 | 2009-01-23 | PROCESS FOR THE PREPARATION OF L-AMINO ACID |
| BRPI0906795-7A BRPI0906795A2 (pt) | 2008-01-23 | 2009-01-23 | Método para produzir um l-aminoácido |
| JP2009550578A JP5526785B2 (ja) | 2008-01-23 | 2009-01-23 | L−アミノ酸の製造法 |
| US12/842,355 US8354254B2 (en) | 2008-01-23 | 2010-07-23 | Method for producing an L-amino acid |
| US13/707,950 US8728772B2 (en) | 2008-01-23 | 2012-12-07 | Method for producing an L-amino acid |
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008-012553 | 2008-01-23 | ||
| JP2008012553 | 2008-01-23 | ||
| JP2008-135265 | 2008-05-23 | ||
| JP2008135265 | 2008-05-23 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| US12/842,355 Continuation US8354254B2 (en) | 2008-01-23 | 2010-07-23 | Method for producing an L-amino acid |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2009093703A1 true WO2009093703A1 (ja) | 2009-07-30 |
Family
ID=40901206
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2009/051104 WO2009093703A1 (ja) | 2008-01-23 | 2009-01-23 | L-アミノ酸の製造法 |
Country Status (7)
| Country | Link |
|---|---|
| US (2) | US8354254B2 (ja) |
| EP (1) | EP2248906A4 (ja) |
| JP (1) | JP5526785B2 (ja) |
| KR (1) | KR20100120663A (ja) |
| CN (1) | CN102348806A (ja) |
| BR (1) | BRPI0906795A2 (ja) |
| WO (1) | WO2009093703A1 (ja) |
Cited By (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2011013707A1 (ja) * | 2009-07-29 | 2011-02-03 | 味の素株式会社 | L-アミノ酸の製造法 |
| KR101115357B1 (ko) | 2010-09-03 | 2012-03-06 | 대상 주식회사 | 습식 상태의 미세조류로부터 조오일을 추출하는 방법 |
| WO2012077739A1 (ja) * | 2010-12-10 | 2012-06-14 | 味の素株式会社 | L-アミノ酸の製造法 |
| WO2013008931A1 (ja) * | 2011-07-14 | 2013-01-17 | 味の素株式会社 | 脂肪酸類の製造法 |
| JP2013039085A (ja) * | 2011-08-18 | 2013-02-28 | Ihi Corp | エタノールの製造方法 |
| JP2013520196A (ja) * | 2010-02-25 | 2013-06-06 | コルゲート・パーモリブ・カンパニー | 炭素供給源としてキャッサバ(Cassava)・バガスまたはジャックフルーツ(jackfruit)種子を含む発酵培地中で、コリネバクテリウム・グルタミクム(Corynebacteriumglutamicum)ATCC21831またはコリネバクテリウム・グルタミクムATCC21493を使用してアルギニンを産生するプロセス |
| WO2013176261A1 (ja) * | 2012-05-25 | 2013-11-28 | 味の素株式会社 | 微細藻類を用いた栄養添加剤の製造法 |
| JP2014500031A (ja) * | 2010-12-21 | 2014-01-09 | サンボ インターナショナル エスタブリッシュメント | 光合成微生物中のCoQ10およびCoQH2の含有量を増加させる方法 |
| JP2015198662A (ja) * | 2015-06-01 | 2015-11-12 | コルゲート・パーモリブ・カンパニーColgate−Palmolive Company | 炭素供給源としてキャッサバ(Cassava)・バガスまたはジャックフルーツ(jackfruit)種子を含む発酵培地中で、コリネバクテリウム・グルタミクム(Corynebacteriumglutamicum)ATCC21831またはコリネバクテリウム・グルタミクムATCC21493を使用してアルギニンを産生するプロセス |
| CN103911419B (zh) * | 2014-03-20 | 2017-01-11 | 广东肇庆星湖生物科技股份有限公司 | 一种双菌株联合发酵生产l-缬氨酸的方法 |
Families Citing this family (34)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR20100120663A (ko) | 2008-01-23 | 2010-11-16 | 아지노모토 가부시키가이샤 | L-아미노산의 제조법 |
| JP5488467B2 (ja) | 2008-09-05 | 2014-05-14 | 味の素株式会社 | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| JP2012223092A (ja) | 2009-08-28 | 2012-11-15 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| JP2013074795A (ja) | 2010-02-08 | 2013-04-25 | Ajinomoto Co Inc | 変異型rpsA遺伝子及びL−アミノ酸の製造法 |
| WO2012019168A2 (en) | 2010-08-06 | 2012-02-09 | Moderna Therapeutics, Inc. | Engineered nucleic acids and methods of use thereof |
| US20120237975A1 (en) | 2010-10-01 | 2012-09-20 | Jason Schrum | Engineered nucleic acids and methods of use thereof |
| JP2014500020A (ja) * | 2010-11-26 | 2014-01-09 | ビーエーエスエフ ソシエタス・ヨーロピア | 異性体として純粋な置換シクロヘキサノールの調製 |
| US8765425B2 (en) | 2011-03-23 | 2014-07-01 | Butamax Advanced Biofuels Llc | In situ expression of lipase for enzymatic production of alcohol esters during fermentation |
| US8759044B2 (en) | 2011-03-23 | 2014-06-24 | Butamax Advanced Biofuels Llc | In situ expression of lipase for enzymatic production of alcohol esters during fermentation |
| AU2012236099A1 (en) | 2011-03-31 | 2013-10-03 | Moderna Therapeutics, Inc. | Delivery and formulation of engineered nucleic acids |
| RU2496867C2 (ru) | 2011-04-25 | 2013-10-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО "АГРИ") | Способ получения l-аминокислоты семейства глутамата с использованием коринеформной бактерии |
| JP2014521338A (ja) * | 2011-07-28 | 2014-08-28 | グリコス バイオテクノロジーズ インコーポレイテッド | 油含有供給原料のバイオ発酵のための組成物及び方法 |
| US9464124B2 (en) | 2011-09-12 | 2016-10-11 | Moderna Therapeutics, Inc. | Engineered nucleic acids and methods of use thereof |
| EP3682905B1 (en) | 2011-10-03 | 2021-12-01 | ModernaTX, Inc. | Modified nucleosides, nucleotides, and nucleic acids, and uses thereof |
| RS63244B1 (sr) | 2011-12-16 | 2022-06-30 | Modernatx Inc | Kompozicije modifikovane mrna |
| US10501512B2 (en) | 2012-04-02 | 2019-12-10 | Modernatx, Inc. | Modified polynucleotides |
| US9572897B2 (en) | 2012-04-02 | 2017-02-21 | Modernatx, Inc. | Modified polynucleotides for the production of cytoplasmic and cytoskeletal proteins |
| EP2833892A4 (en) | 2012-04-02 | 2016-07-20 | Moderna Therapeutics Inc | MODIFIED POLYNUCLEOTIDES FOR THE PRODUCTION OF PROTEINS AND PEPTIDES ASSOCIATED WITH ONCOLOGY |
| US9283287B2 (en) | 2012-04-02 | 2016-03-15 | Moderna Therapeutics, Inc. | Modified polynucleotides for the production of nuclear proteins |
| ES2921623T3 (es) | 2012-11-26 | 2022-08-30 | Modernatx Inc | ARN modificado terminalmente |
| US8980864B2 (en) | 2013-03-15 | 2015-03-17 | Moderna Therapeutics, Inc. | Compositions and methods of altering cholesterol levels |
| WO2014154787A2 (fr) | 2013-03-29 | 2014-10-02 | Roquette Freres | Procédé d'enrichissement en protéines de la biomasse de microalgues |
| JP2016527912A (ja) | 2013-08-23 | 2016-09-15 | ロケット フレールRoquette Freres | 酸素アベイラビリティを制御することによる「オフノート」なしの脂質に富む微細藻類バイオマスからの粉末の工業的製造方法 |
| JP2016192903A (ja) * | 2013-09-17 | 2016-11-17 | 味の素株式会社 | 海藻由来バイオマスからのl−アミノ酸の製造方法 |
| AU2014329452B2 (en) | 2013-10-03 | 2019-06-20 | Moderna Therapeutics, Inc. | Polynucleotides encoding low density lipoprotein receptor |
| US20160340640A1 (en) * | 2014-01-20 | 2016-11-24 | Roquette Freres | Method for the protein enrichment of microalgal biomass |
| JP2017131111A (ja) | 2014-06-03 | 2017-08-03 | 味の素株式会社 | L−アミノ酸の製造法 |
| CN104357503A (zh) * | 2014-11-23 | 2015-02-18 | 吉林大学 | 一种提高异亮氨酸产量的方法 |
| CN108699581B (zh) | 2016-02-08 | 2023-02-28 | 科比恩生物技术有限公司 | 微藻生物质的蛋白质富集方法 |
| CN110004192A (zh) * | 2018-10-17 | 2019-07-12 | 许传高 | 一种制取颗粒型苏氨酸的方法 |
| CN109439702A (zh) * | 2018-10-18 | 2019-03-08 | 许传高 | 处理苏氨酸高浓度发酵废水的工艺 |
| CN109628518B (zh) * | 2018-12-23 | 2021-11-02 | 新疆阜丰生物科技有限公司 | 一种生产和提取l-谷氨酰胺的方法 |
| CN116536227A (zh) * | 2022-01-26 | 2023-08-04 | 廊坊梅花生物技术开发有限公司 | 一种生产苏氨酸的修饰的棒状杆菌属微生物及其构建方法与应用 |
| CN116622597A (zh) * | 2022-02-10 | 2023-08-22 | 廊坊梅花生物技术开发有限公司 | 高产苏氨酸工程菌的构建方法 |
Citations (175)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3708395A (en) | 1969-07-23 | 1973-01-02 | Kyowa Hakko Kogyo Kk | Process for producing l-lysine |
| JPS4828078A (ja) | 1971-08-14 | 1973-04-13 | ||
| US3825472A (en) | 1972-04-27 | 1974-07-23 | Ajinomoto Kk | Method of producing l-lysine by fermentation |
| JPS5031093A (ja) | 1973-07-26 | 1975-03-27 | ||
| JPS5053588A (ja) | 1973-09-22 | 1975-05-12 | ||
| JPS50113209A (ja) | 1974-02-13 | 1975-09-05 | ||
| JPS5238088A (en) | 1975-09-19 | 1977-03-24 | Ajinomoto Co Inc | Preparation of l-glutamic acid |
| JPS52102498A (en) | 1976-02-20 | 1977-08-27 | Ajinomoto Co Inc | Preparation of l-lysine |
| JPS531833A (en) | 1976-06-28 | 1978-01-10 | Shin Kobe Electric Machinery | Method of producing battery separator |
| JPS539394A (en) | 1976-07-09 | 1978-01-27 | Kyowa Hakko Kogyo Co Ltd | Preparation of l-lysine by fermentation |
| JPS5325034A (en) | 1976-08-19 | 1978-03-08 | Fujii Denko | Anti precipitation device for horizontal movement |
| JPS5343591A (en) | 1976-10-01 | 1978-04-19 | Hitachi Ltd | Atomizing device for flameless atomic absorption analysis |
| JPS5386089A (en) | 1976-12-29 | 1978-07-29 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS5386090A (en) | 1976-12-29 | 1978-07-29 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS559784A (en) | 1978-07-10 | 1980-01-23 | Ajinomoto Co Inc | Preparation of l-lysine |
| JPS559759A (en) | 1978-07-07 | 1980-01-23 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS559783A (en) | 1978-07-10 | 1980-01-23 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| US4218386A (en) | 1978-06-15 | 1980-08-19 | The Procter & Gamble Company | Hydrolysis of triglycerides |
| JPS561914A (en) | 1979-06-15 | 1981-01-10 | Leitz Ernst Gmbh | Camera mounted on microscope |
| JPS561915A (en) | 1979-06-21 | 1981-01-10 | Nec Corp | Hologram and production of hologram and light beam scanner |
| JPS561889A (en) | 1979-06-20 | 1981-01-10 | Ajinomoto Co Inc | Preparation of l-glutamic acid by fermentation |
| JPS565099A (en) | 1979-06-25 | 1981-01-20 | Ajinomoto Co Inc | Production of l-histidine through fermentation process and microorganism used therefor |
| JPS566499A (en) | 1979-06-26 | 1981-01-23 | Sanyo Electric Co | Hybrid integrated circuit unit |
| JPS568692A (en) | 1979-07-03 | 1981-01-29 | Kyowa Hakko Kogyo Co Ltd | Preparation of l-lysine by fermentation |
| JPS5618596A (en) | 1979-07-23 | 1981-02-21 | Ajinomoto Co Inc | Production of l-lysine through fermentation process |
| JPS5632995A (en) | 1979-08-28 | 1981-04-02 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS5635981A (en) | 1979-08-31 | 1981-04-08 | Ajinomoto Co Inc | Novel variant |
| JPS5639778A (en) | 1979-09-10 | 1981-04-15 | Ajinomoto Co Inc | Novel modified strain |
| JPS5648890A (en) | 1979-08-10 | 1981-05-02 | Ajinomoto Co Inc | Preparation of l-glutamic acid by fermentation |
| US4278765A (en) | 1978-06-30 | 1981-07-14 | Debabov Vladimir G | Method for preparing strains which produce aminoacids |
| JPS56140895A (en) | 1980-04-02 | 1981-11-04 | Ajinomoto Co Inc | Preparation of l-glutamic acid by fermentation |
| GB2075056A (en) | 1980-04-14 | 1981-11-11 | Ajinomoto Kk | L-proline-producing Microorganisms |
| JPS572689A (en) | 1981-03-23 | 1982-01-08 | Ajinomoto Co Inc | Preparation of l-glutamic acid |
| JPS5714158A (en) | 1980-07-01 | 1982-01-25 | Nihon Radiator Co | Refrigeration cycle |
| JPS5714157A (en) | 1980-06-27 | 1982-01-25 | Matsushita Electric Ind Co Ltd | Refrigerant flow rate control for airconditioner |
| JPS5730474A (en) | 1980-07-31 | 1982-02-18 | Victor Co Of Japan Ltd | Slow-motion reproduction system of magnetic reproducing device |
| JPS5765198A (en) | 1980-10-09 | 1982-04-20 | Ajinomoto Co Inc | Fermentative production of l-glutamic acid |
| JPS5771397A (en) | 1980-08-22 | 1982-05-04 | Ajinomoto Co Inc | Preparation of l-tryptophan by fermentation method |
| JPS5810075A (ja) | 1981-07-09 | 1983-01-20 | 安藤 雅威 | 賽を「おう」用した遊具 |
| DE3127361A1 (de) | 1981-07-08 | 1983-02-03 | Schering Ag, 1000 Berlin Und 4619 Bergkamen | Herstellung und anwendung von plasmiden mit genen fuer die biosynthese von l-prolin |
| JPS58158192A (ja) | 1982-03-15 | 1983-09-20 | Ajinomoto Co Inc | 発酵法によるl−グルタミン酸の製造方法 |
| US4407952A (en) | 1979-06-15 | 1983-10-04 | Ajinomoto Company Incorporated | Method for producing L-phenylalanine by fermentation |
| US4411997A (en) | 1980-12-29 | 1983-10-25 | Ajinomoto Company Incorporated | Method for producing L-lysine by fermentation |
| JPS594993A (ja) | 1982-07-01 | 1984-01-11 | Nippon Kokan Kk <Nkk> | 調質型鋼管の溶接方法 |
| JPS6087788A (ja) | 1983-08-29 | 1985-05-17 | Ajinomoto Co Inc | 発酵法によるl―アミノ酸の製造法 |
| JPS6115695A (ja) | 1984-06-29 | 1986-01-23 | Ajinomoto Co Inc | 発酵法によるl−イソロイシンの製造方法 |
| JPS6115696A (ja) | 1984-06-29 | 1986-01-23 | Ajinomoto Co Inc | 発酵法によるl−イソロイシンの製造法 |
| JPS6135840A (ja) | 1984-07-30 | 1986-02-20 | Sofutaade Kogyo Kk | 触媒の交換方法 |
| JPS6224074A (ja) | 1985-07-25 | 1987-02-02 | Toyota Motor Corp | 潤滑装置 |
| JPS6236673A (ja) | 1985-08-09 | 1987-02-17 | Konishiroku Photo Ind Co Ltd | 感光性平版印刷版の処理方法 |
| JPS6274293A (ja) | 1985-09-28 | 1987-04-06 | Kyowa Hakko Kogyo Co Ltd | L−イソロイシンの製造法 |
| JPS6291193A (ja) | 1985-06-05 | 1987-04-25 | Kyowa Hakko Kogyo Co Ltd | L−スレオニンおよびl−イソロイシンの製造法 |
| JPS6234397B2 (ja) | 1979-11-15 | 1987-07-27 | Ajinomoto Kk | |
| JPS62195293A (ja) | 1986-02-22 | 1987-08-28 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−イソロイシンの製造法 |
| JPS62244382A (ja) | 1986-04-16 | 1987-10-24 | Ajinomoto Co Inc | 新規プロモーター及び該プロモーターを用いた遺伝子発現方法 |
| JPS63240794A (ja) | 1987-03-30 | 1988-10-06 | Ajinomoto Co Inc | L−トリプトフアンの製造法 |
| US4777051A (en) | 1986-06-20 | 1988-10-11 | Ajinomoto Co., Inc. | Process for the production of a composition for animal feed |
| JPH01214636A (ja) | 1988-02-23 | 1989-08-29 | Toto Ltd | 和風便器の施工方法 |
| EP0331145A2 (en) | 1988-03-04 | 1989-09-06 | Kyowa Hakko Kogyo Co., Ltd. | Process for producing amino acids |
| KR890003681B1 (ko) | 1987-03-26 | 1989-09-30 | 주식회사 미원 | 미생물에 의한 l-페닐 알라닌의 제조방법 |
| JPH02458A (ja) | 1987-10-12 | 1990-01-05 | Ajinomoto Co Inc | 発酵法によるl―イソロイシンの製造法 |
| JPH02109985A (ja) | 1988-02-22 | 1990-04-23 | Eurolysine | 細菌染色体上ヘの目的遺伝子の組み込み方法及び該方法によって得られた細菌 |
| WO1990004636A1 (en) | 1988-10-25 | 1990-05-03 | Vsesojuzny Nauchno-Issledovatelsky Institut Genetiki I Selektsii Promyshlennykh Mikroorganizmov (Vniigenetika) | Strain of bacteria escherichia coli, producer of l-threonine |
| US4946654A (en) | 1984-04-07 | 1990-08-07 | Bayer Aktiengesellschaft | Process for preparing granulates |
| US4956471A (en) | 1986-04-28 | 1990-09-11 | Ajinomoto Company, Inc. | Process for isolating and purifying amino acids |
| JPH02303495A (ja) | 1989-05-17 | 1990-12-17 | Kyowa Hakko Kogyo Co Ltd | 芳香族アミノ酸の製造法 |
| JPH0488994A (ja) | 1990-07-30 | 1992-03-23 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl―グルタミン酸の製造法 |
| JPH057491A (ja) | 1990-10-15 | 1993-01-19 | Ajinomoto Co Inc | 温度感受性プラスミド |
| JPH0511958A (ja) | 1991-07-01 | 1993-01-22 | N T T Data Tsushin Kk | サーバ・クライアント型ウインドウシステム |
| JPH0549489A (ja) | 1991-08-22 | 1993-03-02 | Ajinomoto Co Inc | 発酵法によるl−フエニルアラニンの製造法 |
| JPH05130882A (ja) | 1991-11-11 | 1993-05-28 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−イソロイシンの製造法 |
| JPH05227977A (ja) | 1991-09-20 | 1993-09-07 | Ajinomoto Co Inc | エシェリヒア・コリによる新規l−スレオニン生産菌の創成とそれによるl−スレオニンの生産方法 |
| JPH05304969A (ja) | 1992-02-25 | 1993-11-19 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるアミノ酸の製造法 |
| RU2003677C1 (ru) | 1992-03-30 | 1993-11-30 | Всесоюзный научно-исследовательский институт генетики и селекции промышленных микроорганизмов | Штамм бактерий ESCHERICHIA COLI - продуцент L-гистидина |
| JPH0665314A (ja) | 1992-01-31 | 1994-03-08 | Spherilene Srl | オレフィンの重合用の成分及び触媒 |
| EP0593792A1 (en) | 1992-10-14 | 1994-04-27 | Ajinomoto Co., Inc. | Novel L-threonine-producing microbacteria and a method for the production of L-threonine |
| JPH06125779A (ja) | 1984-09-27 | 1994-05-10 | Ajinomoto Co Inc | 組換えdnaを有するコリネ型細菌を用いる芳香族アミノ酸の製造法 |
| US5354672A (en) | 1992-11-24 | 1994-10-11 | Ian Fotheringham | Materials and methods for hypersecretion of amino acids |
| JPH06102028B2 (ja) | 1985-10-04 | 1994-12-14 | 協和醗酵工業株式会社 | アミノ酸の製造法 |
| US5376538A (en) | 1991-09-04 | 1994-12-27 | Kyowa Hakko Kogyo Co., Ltd. | Process for producing L-threonine with strains of E coli resistant to phenylalanine and leucine |
| US5378616A (en) | 1991-08-07 | 1995-01-03 | Ajinomoto Co., Inc. | Mutant Escherichia coli capable of enhanced L-glutamic acid production by fermentation |
| WO1995006114A1 (fr) | 1993-08-24 | 1995-03-02 | Ajinomoto Co., Inc. | Allele de phosphenolpyruvate carboxylase, gene de cet allele et procede de production de l'acide amine |
| JPH0787983A (ja) * | 1993-09-27 | 1995-04-04 | Mitsubishi Heavy Ind Ltd | 微細藻からのエタノール製造方法及び装置 |
| JPH07112437A (ja) | 1993-10-19 | 1995-05-02 | Tajiri:Kk | プラスチック廃棄物の減容機 |
| JPH07112438A (ja) | 1993-10-18 | 1995-05-02 | Nissei Plastics Ind Co | 複数種の成形材料供給装置 |
| WO1995016042A1 (fr) | 1993-12-08 | 1995-06-15 | Ajinomoto Co., Inc. | Procede de production de l-lysine par fermentation |
| US5431933A (en) | 1991-09-17 | 1995-07-11 | Degussa Aktiengesellschaft | Animal feed supplement based on a fermentation broth amino acid, a process for its production and its use |
| WO1995023864A1 (fr) | 1994-03-04 | 1995-09-08 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| WO1995034672A1 (fr) | 1994-06-14 | 1995-12-21 | Ajinomoto Co., Inc. | GENE A DESHYDROGENASE α-CETOGLUTARIQUE |
| WO1996006926A1 (fr) | 1994-08-30 | 1996-03-07 | Ajinomoto Co., Inc. | Procede pour produire de la l-valine et de la l-leucine |
| JPH0870879A (ja) | 1994-06-30 | 1996-03-19 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−ロイシンの製造法 |
| WO1996017930A1 (fr) | 1994-12-09 | 1996-06-13 | Ajinomoto Co., Inc. | Nouveau gene de decarboxylase de lysine et procede de production de lysine l |
| US5573945A (en) | 1994-01-10 | 1996-11-12 | Ajinomoto Co., Inc. | Mutant and method for producing L-glutamic acid by fermentation |
| WO1996040934A1 (fr) | 1995-06-07 | 1996-12-19 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| EP0488424B1 (en) | 1990-11-30 | 1997-03-05 | Ajinomoto Co., Inc. | Recombinant DNA sequences encoding feedback inhibition released enzymes, plasmids comprising the recombinant DNA sequences, transformed microorganisms useful in the production of aromatic amino acids, and a process for preparing aromatic amino acids by fermentation |
| WO1997008333A1 (fr) | 1995-08-30 | 1997-03-06 | Ajinomoto Co., Inc. | Procede de production d'acides amines levogyres |
| US5658766A (en) | 1991-05-30 | 1997-08-19 | Ajinomoto Co., Inc. | Strains of Escherichia coli which produce isoleucine or valine and a method for their production |
| US5661012A (en) | 1992-11-10 | 1997-08-26 | Ajinomoto Co., Inc. | Method for the production of L-threonine by fermentation, using mutated DNA encoding aspartokinase III |
| US5705371A (en) | 1990-06-12 | 1998-01-06 | Ajinomoto Co., Inc. | Bacterial strain of escherichia coli BKIIM B-3996 as the producer of L-threonine |
| WO1998004715A1 (en) | 1996-07-30 | 1998-02-05 | Archer-Daniels-Midland Company | Novel strains of escherichia coli, methods of preparing the same and use thereof in fermentation processes for l-threonine production |
| US5756345A (en) | 1995-09-05 | 1998-05-26 | Degussa Aktiengesellschaft | Production of tryptophan by the bacterium Escherichia coli |
| JPH10165180A (ja) | 1996-12-05 | 1998-06-23 | Ajinomoto Co Inc | L−リジンの製造法 |
| RU2119536C1 (ru) | 1997-01-21 | 1998-09-27 | Государственный научно-исследовательский институт генетики и селекции промышленных микроорганизмов | Штамм escherichia coli - продуцент l-гистидина |
| US5830716A (en) | 1993-10-28 | 1998-11-03 | Ajinomoto Co., Inc. | Increased amounts of substances by modifying a microorganism to increase production of NADPH from NADH |
| US5840358A (en) | 1996-05-31 | 1998-11-24 | Degussa Aktiengesellschaft | Process for the preparation of an animal feed supplement based on fermentation broth |
| WO1999018228A2 (de) | 1997-10-04 | 1999-04-15 | Forschungszentrum Jülich GmbH | Verfahren zur mikrobiellen herstellung von aminosäuren der aspartat- und/oder glutamatfamilie und im verfahren einsetzbare mittel |
| US5908768A (en) | 1996-04-23 | 1999-06-01 | Ajinomoto Co., Inc. | Process for producing L-glutamic acid by fermentation with E. coli resistant to aspartic acid antimetabolite |
| JPH11155571A (ja) | 1997-11-25 | 1999-06-15 | Ajinomoto Co Inc | L−システインの製造法 |
| US5972663A (en) | 1997-06-19 | 1999-10-26 | Consortium Fur Elektrochemische Industrie Gmbh | Microorganisms and processes for the fermentative preparation of L-cysteine, L-cystine, N-acetylserine or thiazolidine derivatives |
| EP0952221A2 (en) | 1998-03-18 | 1999-10-27 | Ajinomoto Co., Ltd. | L-Glutamic acid-producing bacterium and method for producing L-glutamic acid |
| EP0955368A2 (en) | 1998-03-18 | 1999-11-10 | Ajinomoto Co., Ltd. | L-glutamic acid-producing bacterium and method for producing l-glutamic acid |
| US5998178A (en) | 1994-05-30 | 1999-12-07 | Ajinomoto Co., Ltd. | L-isoleucine-producing bacterium and method for preparing L-isoleucine through fermentation |
| JP3006929B2 (ja) | 1990-09-18 | 2000-02-07 | 協和醗酵工業株式会社 | 発酵法によるl−バリンの製造法 |
| WO2000018935A1 (fr) | 1998-09-25 | 2000-04-06 | Ajinomoto Co.,Inc. | Procede de construction d'une bacterie produisant des acides amines, et procede de production d'acides amines par une technique de fermentation utilisant ladite bacterie |
| EP0994190A2 (en) | 1998-10-13 | 2000-04-19 | Ajinomoto Co., Inc. | DNA conferring L-homoserine resistance to bacteria, and its use |
| JP2000139471A (ja) | 1998-11-17 | 2000-05-23 | Ajinomoto Co Inc | 発酵法によるl−メチオニンの製造法 |
| JP2000157276A (ja) | 1998-11-24 | 2000-06-13 | Ajinomoto Co Inc | サーマス属細菌のl−リジン生合成系遺伝子 |
| EP1010755A1 (en) | 1998-12-18 | 2000-06-21 | Ajinomoto Co., Inc. | Method for producing L-Glutamic acid by fermentation |
| EP1013765A1 (en) | 1998-12-23 | 2000-06-28 | Ajinomoto Co., Ltd. | Gene and method for producing L-amino acids |
| EP1016710A2 (en) | 1998-12-30 | 2000-07-05 | Ajinomoto Co., Inc. | Method for producing L-amino acids |
| US6110714A (en) | 1995-08-23 | 2000-08-29 | Ajinomoto Co., Inc. | Process for producing L-glutamic acid by fermentation |
| WO2000050624A1 (de) | 1999-02-22 | 2000-08-31 | Forschungszentrum Jülich GmbH | Verfahren zur mikrobiellen herstellung von l-valin |
| JP2000253879A (ja) | 1999-03-09 | 2000-09-19 | Ajinomoto Co Inc | L−リジンの製造法 |
| US6124121A (en) | 1997-10-29 | 2000-09-26 | Ajinomoto Co., Inc. | Method for producing L-leucine |
| WO2000061723A1 (fr) | 1999-04-09 | 2000-10-19 | Ajinomoto Co., Inc. | Bacteries produisant du l-amino acide et procede de production de l-amino acide |
| US6180373B1 (en) | 1992-09-28 | 2001-01-30 | Consortium f{umlaut over (u)}r elektrochemische Industrie GmbH | Microorganisms for the production of tryptophan and process for the preparation thereof |
| EP1078989A2 (en) | 1999-08-20 | 2001-02-28 | Ajinomoto Co., Ltd. | Method for producing L-glutamic acid by fermentation accompanied by precipitation |
| JP2001057896A (ja) | 1999-06-15 | 2001-03-06 | Ajinomoto Co Inc | L−リジンの製造法 |
| EP1085087A2 (en) | 1999-09-20 | 2001-03-21 | Kyowa Hakko Kogyo Co., Ltd. | Method for producing amino acids by fermentation using aminoquinoline resistant bacterial strains |
| US6218168B1 (en) | 1995-10-26 | 2001-04-17 | CONSORTIUM FüR ELEKTROCHEMISCHE INUDSTRIE GMBH | Process for preparing O-acetylserine, L-cysteine and L-cysteine-related products |
| EP1092776A1 (en) | 1999-10-14 | 2001-04-18 | Ajinomoto Co., Inc. | Method for producing L-amino acid by fermentation |
| WO2001027307A1 (de) | 1999-10-14 | 2001-04-19 | Consortium für elektrochemische Industrie GmbH | Verfahren zur fermentativen herstellung von l-cystein oder l-cystein-derivaten |
| US6238714B1 (en) | 1999-05-05 | 2001-05-29 | Degussa-Huls Ag | Feedstuff additive which contains D-pantothenic acid and/or its salts and a process for the preparation thereof |
| JP2001169788A (ja) | 1999-10-27 | 2001-06-26 | Degussa Huels Ag | 分枝アミノ酸の排出をコードするヌクレオチド配列、その単離法および使用 |
| US6258554B1 (en) | 1998-07-03 | 2001-07-10 | Kyowa Hakko Kogyo Co., Ltd. | Method for producing metabolites biologically synthesized via phosphoribosyl pyrophosphate |
| WO2001053459A1 (fr) | 2000-01-21 | 2001-07-26 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| US6303383B1 (en) | 1999-03-16 | 2001-10-16 | Ajinomoto Co., Inc. | Temperature sensitive plasmid for coryneform bacteria |
| EP1149911A2 (en) | 2000-04-26 | 2001-10-31 | Ajinomoto Co., Ltd. | Amino acid producing strains belonging to the genus Escherichia and method for producing amino acid |
| EP1170358A1 (en) | 2000-07-06 | 2002-01-09 | Ajinomoto Co., Ltd. | L-arginine producing Escherichia coli and method of producing L-arginine |
| EP1170376A1 (en) | 2000-07-05 | 2002-01-09 | Ajinomoto Co., Inc. | Method for producing substances utilizing microorganisms |
| EP1170361A2 (en) | 2000-06-28 | 2002-01-09 | Ajinomoto Co., Inc. | New mutant N-Acetylglutamate synthase and method for L-Arginine production |
| EP1172433A1 (en) | 2000-07-06 | 2002-01-16 | Ajinomoto Co., Inc. | Bacterium having ability to produce L-glutamic acid, L-proline or L-arginine and method for producing L-glutamic acid, L-proline or L-arginine |
| US6344347B1 (en) | 1999-09-20 | 2002-02-05 | Kyowa Hakko Kogyo Co., Ltd. | Method for producing L-amino acids by fermentation |
| US20020025564A1 (en) | 2000-08-24 | 2002-02-28 | Ajinomoto Co., Inc. | Method for producing basic amino acid |
| WO2002026993A1 (en) | 2000-09-28 | 2002-04-04 | Archer-Daniels-Midland Company | Escherichia coli strains which over-produce l-threonine and processes for the production of l-threonine by fermentation |
| US20020058315A1 (en) | 2000-09-26 | 2002-05-16 | Ajinomoto Co., Inc. | Bacterium having ability to produce L-glutamic acid, L-proline or L-arginine and method for producing L-glutamic acid, L-proline or L-arginine |
| US6403342B1 (en) | 1999-07-09 | 2002-06-11 | Anjinomoto Co., Inc. | DNA coding for mutant isopropylmalate synthase L-leucine-producing microorganism and method for producing L-leucine |
| US20020110876A1 (en) | 2000-08-11 | 2002-08-15 | Ajinomoto Co., Inc. | Method for producing threonine and isoleucine |
| EP1239041A2 (en) | 2001-02-13 | 2002-09-11 | Ajinomoto Co., Inc. | Method for producing L-amino acid using bacteria belonging to the genus Escherichia |
| RU2000124295A (ru) | 2000-09-26 | 2003-02-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Бактерия, обладающая способностью к продукции l-глутаминовой кислоты, l-пролина или l-аргинина, и способ получения l-глутаминовой кислоты, l-пролина или l-аргинина |
| JP2003135066A (ja) | 1999-03-19 | 2003-05-13 | Ajinomoto Co Inc | L−リジンの製造法 |
| WO2003044191A1 (fr) | 2001-11-23 | 2003-05-30 | Ajinomoto Co.,Inc. | Procede de production de l-aminoacides a l'aide d'escherichia |
| WO2003044192A1 (fr) | 2001-11-23 | 2003-05-30 | Ajinomoto Co.,Inc. | Procede de production de l-aminoacides a l'aide d'escherichia |
| US6596517B2 (en) | 2001-02-20 | 2003-07-22 | Ajinomoto Co., Inc. | Method for producing L-glutamic acid |
| WO2003094598A1 (en) | 2002-05-13 | 2003-11-20 | Greenfuel Technologies Corporation | Photobioreactor and process for biomass production and mitigation of pollutants in flue gases |
| EP1484410A1 (en) | 2003-06-05 | 2004-12-08 | Ajinomoto Co., Ltd. | Fermentation methods and genetically modified bacteria with increased substrate and byproduct uptake. |
| US20050025878A1 (en) | 2003-07-11 | 2005-02-03 | Degussa Ag | Process for the granulation of an animal feedstuff additive |
| WO2005010175A1 (en) | 2003-07-29 | 2005-02-03 | Ajinomoto Co., Inc. | Method for producing l-lysine or l-threonine using escherichia bacteria having attnuated malic enzyme activity |
| RU2003121601A (ru) | 2003-07-16 | 2005-02-27 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО "АГРИ") (RU) | Мутантная серинацетилтрансфераза |
| WO2005049808A1 (en) | 2003-11-21 | 2005-06-02 | Ajinomoto Co., Inc. | Method for producing l-amino acid by fermentation |
| WO2005073390A2 (en) | 2004-01-30 | 2005-08-11 | Ajinomoto Co., Inc. | L-amino acid-producing microorganism and method for producing l-amino acid |
| WO2005103275A1 (ja) | 2004-04-26 | 2005-11-03 | Ajinomoto Co., Ltd. | 発酵法によるl-トリプトファンの製造法 |
| US20060135308A1 (en) | 2003-07-04 | 2006-06-22 | Samuel Abraham | Differential for motor vehicles with device for locking thereof |
| WO2006070944A2 (en) | 2004-12-28 | 2006-07-06 | Ajinomoto Co., Inc. | L-glutamic acid-producing microorganism and a method for producing l-glutamic acid |
| WO2006078039A1 (en) | 2005-01-18 | 2006-07-27 | Ajinomoto Co., Inc. | L-amino acid producing microorganism and a method for producing l-amino acid |
| EP1715056A1 (de) | 2005-04-23 | 2006-10-25 | Degussa AG | Verfahren zur Herstellung von L-Aminosäuren unter Verwendung verbesserter Stämme der Familie Enterobacteriaceae |
| EP1715055A2 (de) | 2005-04-22 | 2006-10-25 | Degussa GmbH | Verfahren zur Herstellung von L-Aminosäuren unter Verwendung verbesserter Stämme der Familie Enterobacteriaceae |
| JP2006340603A (ja) | 2003-06-23 | 2006-12-21 | Ajinomoto Co Inc | L−グルタミン酸の製造法 |
| EP1813677A1 (en) | 2004-10-07 | 2007-08-01 | Ajinomoto Co., Inc. | Process for producing basic substance |
| US20070202582A1 (en) | 2006-02-28 | 2007-08-30 | Bush Ronnie A | Process for the production of ethanol from algae |
| WO2007100009A1 (ja) * | 2006-03-03 | 2007-09-07 | Ajinomoto Co., Inc. | L-アミノ酸の製造法 |
| WO2008002053A1 (en) * | 2006-06-26 | 2008-01-03 | Cj Cheiljedang Corporation | Method for producing amino acids using glycerol |
| WO2008081959A1 (en) | 2006-12-22 | 2008-07-10 | Ajinomoto Co., Inc. | A method for producing an l-amino acid by fermentation using a bacterium having an enhanced ability to utilize glycerol |
| WO2008102861A1 (ja) | 2007-02-22 | 2008-08-28 | Ajinomoto Co., Inc. | L-アミノ酸の製造法 |
| WO2008107277A1 (de) | 2007-03-05 | 2008-09-12 | Evonik Degussa Gmbh | Verfahren zur herstellung von l-aminosäuren unter verwendung von stämmen der familie enterobacteriaceae |
Family Cites Families (30)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS5326391A (en) * | 1976-07-26 | 1978-03-11 | Kanegafuchi Chem Ind Co Ltd | Preparation of l-lysine by fermentation |
| JPS5455786A (en) * | 1977-10-05 | 1979-05-04 | Kanegafuchi Chem Ind Co Ltd | Preparation of l-glutamic acid by fermentation |
| JPS5718990A (en) * | 1980-07-10 | 1982-01-30 | Ajinomoto Co Inc | Production of l-glutamic acid through fermentation |
| JPS5692794A (en) * | 1979-12-27 | 1981-07-27 | Ajinomoto Co Inc | Preparation of l-glutamic acid by fermentation method |
| EP0645456B1 (en) | 1993-09-27 | 2001-04-25 | Mitsubishi Jukogyo Kabushiki Kaisha | Process and system for the production of ethanol from microalgae |
| RU2215783C2 (ru) | 2001-05-15 | 2003-11-10 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото - Генетика" | МУТАНТНАЯ N-АЦЕТИЛГЛУТАМАТ СИНТАЗА (ВАРИАНТЫ), ФРАГМЕНТ ДНК, ШТАММ БАКТЕРИИ Escherichia coli - ПРОДУЦЕНТ АРГИНИНА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА |
| JP2003250544A (ja) | 2002-03-04 | 2003-09-09 | National Institute Of Technology & Evaluation | タンパク質の性質を改変する方法 |
| US6911332B2 (en) | 2002-06-12 | 2005-06-28 | Ajinomoto Co., Inc. | Isolated polynucleotides encoding d-arabino-3-hexulose-6-phosphate synthases from Methylophilus methylotrophus |
| US7060475B2 (en) | 2003-02-28 | 2006-06-13 | Ajinomoto Co., Inc. | Polynucleotides encoding polypeptides involved in intermediates metabolism of central metabolic pathway in methylophilus methylotrophus |
| US7026149B2 (en) | 2003-02-28 | 2006-04-11 | Ajinomoto Co., Inc. | Polynucleotides encoding polypeptides involved in the stress response to environmental changes in Methylophilus methylotrophus |
| US7029893B2 (en) | 2003-02-28 | 2006-04-18 | Ajinomoto Co., Inc. | Polynucleotides encoding polypeptides involved in amino acid biosynthesis in methylophilus methylotrophus |
| US7468262B2 (en) | 2003-05-16 | 2008-12-23 | Ajinomoto Co., Inc. | Polynucleotides encoding useful polypeptides in corynebacterium glutamicum ssp. lactofermentum |
| JP4836786B2 (ja) * | 2003-07-11 | 2011-12-14 | − ツァチェルニュウク、タティアナ、アー. エゴロワ | 生体化合物の安定同位体標識のための組成物及び方法 |
| JP2007117082A (ja) | 2005-09-27 | 2007-05-17 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| DE102005056667A1 (de) * | 2005-11-28 | 2007-05-31 | Basf Ag | Fermentative Herstellung organischer Verbindungen |
| US7135308B1 (en) | 2006-02-28 | 2006-11-14 | Propulsion Logic, Llc | Process for the production of ethanol from algae |
| JP5407858B2 (ja) | 2006-07-19 | 2014-02-05 | 味の素株式会社 | 腸内細菌科の細菌を用いたl−アミノ酸の製造方法 |
| JP2010017081A (ja) | 2006-10-10 | 2010-01-28 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| EP2094858B1 (en) | 2006-12-11 | 2015-02-18 | Ajinomoto Co., Inc. | Method for producing an l-amino acid |
| JP2010130899A (ja) | 2007-03-14 | 2010-06-17 | Ajinomoto Co Inc | L−グルタミン酸系アミノ酸生産微生物及びアミノ酸の製造法 |
| WO2008133131A1 (ja) | 2007-04-16 | 2008-11-06 | Ajinomoto Co., Inc. | 有機酸の製造方法 |
| CN101688176B (zh) | 2007-04-17 | 2013-11-06 | 味之素株式会社 | 具有羧基的酸性物质的生产方法 |
| JP2010263790A (ja) | 2007-09-04 | 2010-11-25 | Ajinomoto Co Inc | アミノ酸生産微生物及びアミノ酸の製造法 |
| JP2010226957A (ja) | 2007-10-17 | 2010-10-14 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| KR20100120663A (ko) | 2008-01-23 | 2010-11-16 | 아지노모토 가부시키가이샤 | L-아미노산의 제조법 |
| JP2011167071A (ja) | 2008-05-22 | 2011-09-01 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| JP5488467B2 (ja) | 2008-09-05 | 2014-05-14 | 味の素株式会社 | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| CN102471790B (zh) | 2009-07-29 | 2014-10-29 | 味之素株式会社 | 产生l-氨基酸的方法 |
| JP2012223092A (ja) | 2009-08-28 | 2012-11-15 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| JP2012239452A (ja) | 2011-05-24 | 2012-12-10 | Ajinomoto Co Inc | 澱粉高蓄積微細藻類及びそれを用いたグルコースの製造法、並びに目的物質の製造法 |
-
2009
- 2009-01-23 KR KR1020107018301A patent/KR20100120663A/ko not_active Application Discontinuation
- 2009-01-23 JP JP2009550578A patent/JP5526785B2/ja not_active Expired - Fee Related
- 2009-01-23 BR BRPI0906795-7A patent/BRPI0906795A2/pt not_active Application Discontinuation
- 2009-01-23 WO PCT/JP2009/051104 patent/WO2009093703A1/ja active Application Filing
- 2009-01-23 CN CN2009801028942A patent/CN102348806A/zh active Pending
- 2009-01-23 EP EP09704195A patent/EP2248906A4/en not_active Withdrawn
-
2010
- 2010-07-23 US US12/842,355 patent/US8354254B2/en active Active
-
2012
- 2012-12-07 US US13/707,950 patent/US8728772B2/en active Active
Patent Citations (198)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3708395A (en) | 1969-07-23 | 1973-01-02 | Kyowa Hakko Kogyo Kk | Process for producing l-lysine |
| JPS4828078A (ja) | 1971-08-14 | 1973-04-13 | ||
| US3825472A (en) | 1972-04-27 | 1974-07-23 | Ajinomoto Kk | Method of producing l-lysine by fermentation |
| JPS5031093A (ja) | 1973-07-26 | 1975-03-27 | ||
| JPS5053588A (ja) | 1973-09-22 | 1975-05-12 | ||
| JPS50113209A (ja) | 1974-02-13 | 1975-09-05 | ||
| JPS5238088A (en) | 1975-09-19 | 1977-03-24 | Ajinomoto Co Inc | Preparation of l-glutamic acid |
| JPS52102498A (en) | 1976-02-20 | 1977-08-27 | Ajinomoto Co Inc | Preparation of l-lysine |
| JPS531833A (en) | 1976-06-28 | 1978-01-10 | Shin Kobe Electric Machinery | Method of producing battery separator |
| JPS539394A (en) | 1976-07-09 | 1978-01-27 | Kyowa Hakko Kogyo Co Ltd | Preparation of l-lysine by fermentation |
| JPS5325034A (en) | 1976-08-19 | 1978-03-08 | Fujii Denko | Anti precipitation device for horizontal movement |
| JPS5343591A (en) | 1976-10-01 | 1978-04-19 | Hitachi Ltd | Atomizing device for flameless atomic absorption analysis |
| JPS5386089A (en) | 1976-12-29 | 1978-07-29 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS5386090A (en) | 1976-12-29 | 1978-07-29 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| US4218386A (en) | 1978-06-15 | 1980-08-19 | The Procter & Gamble Company | Hydrolysis of triglycerides |
| US4278765A (en) | 1978-06-30 | 1981-07-14 | Debabov Vladimir G | Method for preparing strains which produce aminoacids |
| JPS559759A (en) | 1978-07-07 | 1980-01-23 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS559784A (en) | 1978-07-10 | 1980-01-23 | Ajinomoto Co Inc | Preparation of l-lysine |
| JPS559783A (en) | 1978-07-10 | 1980-01-23 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS561914A (en) | 1979-06-15 | 1981-01-10 | Leitz Ernst Gmbh | Camera mounted on microscope |
| US4407952A (en) | 1979-06-15 | 1983-10-04 | Ajinomoto Company Incorporated | Method for producing L-phenylalanine by fermentation |
| JPS561889A (en) | 1979-06-20 | 1981-01-10 | Ajinomoto Co Inc | Preparation of l-glutamic acid by fermentation |
| JPS561915A (en) | 1979-06-21 | 1981-01-10 | Nec Corp | Hologram and production of hologram and light beam scanner |
| JPS565099A (en) | 1979-06-25 | 1981-01-20 | Ajinomoto Co Inc | Production of l-histidine through fermentation process and microorganism used therefor |
| US4388405A (en) | 1979-06-25 | 1983-06-14 | Ajinomoto Company Incorporated | Method for producing L-histidine by fermentation |
| JPS566499A (en) | 1979-06-26 | 1981-01-23 | Sanyo Electric Co | Hybrid integrated circuit unit |
| JPS568692A (en) | 1979-07-03 | 1981-01-29 | Kyowa Hakko Kogyo Co Ltd | Preparation of l-lysine by fermentation |
| JPS5618596A (en) | 1979-07-23 | 1981-02-21 | Ajinomoto Co Inc | Production of l-lysine through fermentation process |
| US4346170A (en) | 1979-07-23 | 1982-08-24 | Ajinomoto Company, Incorporated | Method for producing L-lysine by fermentation |
| JPS5648890A (en) | 1979-08-10 | 1981-05-02 | Ajinomoto Co Inc | Preparation of l-glutamic acid by fermentation |
| JPS5632995A (en) | 1979-08-28 | 1981-04-02 | Ajinomoto Co Inc | Preparation of l-lysine by fermentation |
| JPS5635981A (en) | 1979-08-31 | 1981-04-08 | Ajinomoto Co Inc | Novel variant |
| JPS5639778A (en) | 1979-09-10 | 1981-04-15 | Ajinomoto Co Inc | Novel modified strain |
| JPS6234397B2 (ja) | 1979-11-15 | 1987-07-27 | Ajinomoto Kk | |
| JPS56140895A (en) | 1980-04-02 | 1981-11-04 | Ajinomoto Co Inc | Preparation of l-glutamic acid by fermentation |
| GB2075056A (en) | 1980-04-14 | 1981-11-11 | Ajinomoto Kk | L-proline-producing Microorganisms |
| JPS5714157A (en) | 1980-06-27 | 1982-01-25 | Matsushita Electric Ind Co Ltd | Refrigerant flow rate control for airconditioner |
| JPS5714158A (en) | 1980-07-01 | 1982-01-25 | Nihon Radiator Co | Refrigeration cycle |
| JPS5730474A (en) | 1980-07-31 | 1982-02-18 | Victor Co Of Japan Ltd | Slow-motion reproduction system of magnetic reproducing device |
| JPS5771397A (en) | 1980-08-22 | 1982-05-04 | Ajinomoto Co Inc | Preparation of l-tryptophan by fermentation method |
| US4371614A (en) | 1980-08-22 | 1983-02-01 | Ajinomoto Co., Inc. | E.Coli bacteria carrying recombinant plasmids and their use in the fermentative production of L-tryptophan |
| JPS5765198A (en) | 1980-10-09 | 1982-04-20 | Ajinomoto Co Inc | Fermentative production of l-glutamic acid |
| US4411997A (en) | 1980-12-29 | 1983-10-25 | Ajinomoto Company Incorporated | Method for producing L-lysine by fermentation |
| JPS572689A (en) | 1981-03-23 | 1982-01-08 | Ajinomoto Co Inc | Preparation of l-glutamic acid |
| DE3127361A1 (de) | 1981-07-08 | 1983-02-03 | Schering Ag, 1000 Berlin Und 4619 Bergkamen | Herstellung und anwendung von plasmiden mit genen fuer die biosynthese von l-prolin |
| JPS5810075A (ja) | 1981-07-09 | 1983-01-20 | 安藤 雅威 | 賽を「おう」用した遊具 |
| JPS58158192A (ja) | 1982-03-15 | 1983-09-20 | Ajinomoto Co Inc | 発酵法によるl−グルタミン酸の製造方法 |
| JPS594993A (ja) | 1982-07-01 | 1984-01-11 | Nippon Kokan Kk <Nkk> | 調質型鋼管の溶接方法 |
| JPS6087788A (ja) | 1983-08-29 | 1985-05-17 | Ajinomoto Co Inc | 発酵法によるl―アミノ酸の製造法 |
| US4946654A (en) | 1984-04-07 | 1990-08-07 | Bayer Aktiengesellschaft | Process for preparing granulates |
| JPS6115695A (ja) | 1984-06-29 | 1986-01-23 | Ajinomoto Co Inc | 発酵法によるl−イソロイシンの製造方法 |
| JPS6115696A (ja) | 1984-06-29 | 1986-01-23 | Ajinomoto Co Inc | 発酵法によるl−イソロイシンの製造法 |
| JPS6135840A (ja) | 1984-07-30 | 1986-02-20 | Sofutaade Kogyo Kk | 触媒の交換方法 |
| JPH06125779A (ja) | 1984-09-27 | 1994-05-10 | Ajinomoto Co Inc | 組換えdnaを有するコリネ型細菌を用いる芳香族アミノ酸の製造法 |
| JPS6291193A (ja) | 1985-06-05 | 1987-04-25 | Kyowa Hakko Kogyo Co Ltd | L−スレオニンおよびl−イソロイシンの製造法 |
| JPS6224074A (ja) | 1985-07-25 | 1987-02-02 | Toyota Motor Corp | 潤滑装置 |
| JPS6236673A (ja) | 1985-08-09 | 1987-02-17 | Konishiroku Photo Ind Co Ltd | 感光性平版印刷版の処理方法 |
| JPS6274293A (ja) | 1985-09-28 | 1987-04-06 | Kyowa Hakko Kogyo Co Ltd | L−イソロイシンの製造法 |
| JPH06102028B2 (ja) | 1985-10-04 | 1994-12-14 | 協和醗酵工業株式会社 | アミノ酸の製造法 |
| JPS62195293A (ja) | 1986-02-22 | 1987-08-28 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−イソロイシンの製造法 |
| JPS62244382A (ja) | 1986-04-16 | 1987-10-24 | Ajinomoto Co Inc | 新規プロモーター及び該プロモーターを用いた遺伝子発現方法 |
| US4956471A (en) | 1986-04-28 | 1990-09-11 | Ajinomoto Company, Inc. | Process for isolating and purifying amino acids |
| US4777051A (en) | 1986-06-20 | 1988-10-11 | Ajinomoto Co., Inc. | Process for the production of a composition for animal feed |
| KR890003681B1 (ko) | 1987-03-26 | 1989-09-30 | 주식회사 미원 | 미생물에 의한 l-페닐 알라닌의 제조방법 |
| JPS63240794A (ja) | 1987-03-30 | 1988-10-06 | Ajinomoto Co Inc | L−トリプトフアンの製造法 |
| JPH02458A (ja) | 1987-10-12 | 1990-01-05 | Ajinomoto Co Inc | 発酵法によるl―イソロイシンの製造法 |
| JPH02109985A (ja) | 1988-02-22 | 1990-04-23 | Eurolysine | 細菌染色体上ヘの目的遺伝子の組み込み方法及び該方法によって得られた細菌 |
| JPH01214636A (ja) | 1988-02-23 | 1989-08-29 | Toto Ltd | 和風便器の施工方法 |
| EP0331145A2 (en) | 1988-03-04 | 1989-09-06 | Kyowa Hakko Kogyo Co., Ltd. | Process for producing amino acids |
| WO1990004636A1 (en) | 1988-10-25 | 1990-05-03 | Vsesojuzny Nauchno-Issledovatelsky Institut Genetiki I Selektsii Promyshlennykh Mikroorganizmov (Vniigenetika) | Strain of bacteria escherichia coli, producer of l-threonine |
| US5631157A (en) | 1988-10-25 | 1997-05-20 | Ajinomoto Co., Inc. | Bacterial strain of Escherichia coli VNII genetika 472T23 as the producer of L-threonine |
| US5175107A (en) | 1988-10-25 | 1992-12-29 | Ajinomoto Co., Inc. | Bacterial strain of escherichia coli bkiim b-3996 as the producer of l-threonine |
| JPH02303495A (ja) | 1989-05-17 | 1990-12-17 | Kyowa Hakko Kogyo Co Ltd | 芳香族アミノ酸の製造法 |
| US5705371A (en) | 1990-06-12 | 1998-01-06 | Ajinomoto Co., Inc. | Bacterial strain of escherichia coli BKIIM B-3996 as the producer of L-threonine |
| JPH0488994A (ja) | 1990-07-30 | 1992-03-23 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl―グルタミン酸の製造法 |
| JP3006929B2 (ja) | 1990-09-18 | 2000-02-07 | 協和醗酵工業株式会社 | 発酵法によるl−バリンの製造法 |
| JPH057491A (ja) | 1990-10-15 | 1993-01-19 | Ajinomoto Co Inc | 温度感受性プラスミド |
| EP0488424B1 (en) | 1990-11-30 | 1997-03-05 | Ajinomoto Co., Inc. | Recombinant DNA sequences encoding feedback inhibition released enzymes, plasmids comprising the recombinant DNA sequences, transformed microorganisms useful in the production of aromatic amino acids, and a process for preparing aromatic amino acids by fermentation |
| US5658766A (en) | 1991-05-30 | 1997-08-19 | Ajinomoto Co., Inc. | Strains of Escherichia coli which produce isoleucine or valine and a method for their production |
| JPH0511958A (ja) | 1991-07-01 | 1993-01-22 | N T T Data Tsushin Kk | サーバ・クライアント型ウインドウシステム |
| US5378616A (en) | 1991-08-07 | 1995-01-03 | Ajinomoto Co., Inc. | Mutant Escherichia coli capable of enhanced L-glutamic acid production by fermentation |
| US5393671A (en) | 1991-08-07 | 1995-02-28 | Ajinomoto Co., Inc. | Mutant Escherichia coli capable of enhanced L-glutamic acid production |
| JPH0549489A (ja) | 1991-08-22 | 1993-03-02 | Ajinomoto Co Inc | 発酵法によるl−フエニルアラニンの製造法 |
| US5376538A (en) | 1991-09-04 | 1994-12-27 | Kyowa Hakko Kogyo Co., Ltd. | Process for producing L-threonine with strains of E coli resistant to phenylalanine and leucine |
| US5431933A (en) | 1991-09-17 | 1995-07-11 | Degussa Aktiengesellschaft | Animal feed supplement based on a fermentation broth amino acid, a process for its production and its use |
| JPH05227977A (ja) | 1991-09-20 | 1993-09-07 | Ajinomoto Co Inc | エシェリヒア・コリによる新規l−スレオニン生産菌の創成とそれによるl−スレオニンの生産方法 |
| JPH05130882A (ja) | 1991-11-11 | 1993-05-28 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−イソロイシンの製造法 |
| JPH0665314A (ja) | 1992-01-31 | 1994-03-08 | Spherilene Srl | オレフィンの重合用の成分及び触媒 |
| US5474918A (en) | 1992-02-25 | 1995-12-12 | Kyowa Kakko Kogyo Co., Ltd. | Process for the production of L-threonine and L-isoleucine by fermentation of Escherichia coli |
| JPH05304969A (ja) | 1992-02-25 | 1993-11-19 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるアミノ酸の製造法 |
| RU2003677C1 (ru) | 1992-03-30 | 1993-11-30 | Всесоюзный научно-исследовательский институт генетики и селекции промышленных микроорганизмов | Штамм бактерий ESCHERICHIA COLI - продуцент L-гистидина |
| US6180373B1 (en) | 1992-09-28 | 2001-01-30 | Consortium f{umlaut over (u)}r elektrochemische Industrie GmbH | Microorganisms for the production of tryptophan and process for the preparation thereof |
| EP0593792A1 (en) | 1992-10-14 | 1994-04-27 | Ajinomoto Co., Inc. | Novel L-threonine-producing microbacteria and a method for the production of L-threonine |
| US5661012A (en) | 1992-11-10 | 1997-08-26 | Ajinomoto Co., Inc. | Method for the production of L-threonine by fermentation, using mutated DNA encoding aspartokinase III |
| US5354672A (en) | 1992-11-24 | 1994-10-11 | Ian Fotheringham | Materials and methods for hypersecretion of amino acids |
| WO1995006114A1 (fr) | 1993-08-24 | 1995-03-02 | Ajinomoto Co., Inc. | Allele de phosphenolpyruvate carboxylase, gene de cet allele et procede de production de l'acide amine |
| JPH0787983A (ja) * | 1993-09-27 | 1995-04-04 | Mitsubishi Heavy Ind Ltd | 微細藻からのエタノール製造方法及び装置 |
| JPH07112438A (ja) | 1993-10-18 | 1995-05-02 | Nissei Plastics Ind Co | 複数種の成形材料供給装置 |
| JPH07112437A (ja) | 1993-10-19 | 1995-05-02 | Tajiri:Kk | プラスチック廃棄物の減容機 |
| US5830716A (en) | 1993-10-28 | 1998-11-03 | Ajinomoto Co., Inc. | Increased amounts of substances by modifying a microorganism to increase production of NADPH from NADH |
| EP0733712B1 (en) | 1993-10-28 | 2001-12-12 | Ajinomoto Co., Inc. | Process for producing substance |
| US6040160A (en) | 1993-12-08 | 2000-03-21 | Ajinomoto Co., Inc. | Method of producing L-lysine by fermentation |
| WO1995016042A1 (fr) | 1993-12-08 | 1995-06-15 | Ajinomoto Co., Inc. | Procede de production de l-lysine par fermentation |
| JPH11192088A (ja) | 1993-12-08 | 1999-07-21 | Ajinomoto Co Inc | 発酵法によるl−リジンの製造法 |
| US5573945A (en) | 1994-01-10 | 1996-11-12 | Ajinomoto Co., Inc. | Mutant and method for producing L-glutamic acid by fermentation |
| WO1995023864A1 (fr) | 1994-03-04 | 1995-09-08 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| US5998178A (en) | 1994-05-30 | 1999-12-07 | Ajinomoto Co., Ltd. | L-isoleucine-producing bacterium and method for preparing L-isoleucine through fermentation |
| WO1995034672A1 (fr) | 1994-06-14 | 1995-12-21 | Ajinomoto Co., Inc. | GENE A DESHYDROGENASE α-CETOGLUTARIQUE |
| JPH0870879A (ja) | 1994-06-30 | 1996-03-19 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−ロイシンの製造法 |
| WO1996006926A1 (fr) | 1994-08-30 | 1996-03-07 | Ajinomoto Co., Inc. | Procede pour produire de la l-valine et de la l-leucine |
| WO1996017930A1 (fr) | 1994-12-09 | 1996-06-13 | Ajinomoto Co., Inc. | Nouveau gene de decarboxylase de lysine et procede de production de lysine l |
| US5827698A (en) | 1994-12-09 | 1998-10-27 | Ajinomoto Co., Inc. | Lysine decarboxylase gene and method of producing l-lysine |
| WO1996040934A1 (fr) | 1995-06-07 | 1996-12-19 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| US6110714A (en) | 1995-08-23 | 2000-08-29 | Ajinomoto Co., Inc. | Process for producing L-glutamic acid by fermentation |
| WO1997008333A1 (fr) | 1995-08-30 | 1997-03-06 | Ajinomoto Co., Inc. | Procede de production d'acides amines levogyres |
| EP0877090B1 (en) | 1995-08-30 | 2005-11-23 | Ajinomoto Co., Inc. | Process for producing l-amino acids |
| US6319696B1 (en) | 1995-08-30 | 2001-11-20 | Ajinomoto Co., Inc. | Process for producing L-amino acids |
| US5756345A (en) | 1995-09-05 | 1998-05-26 | Degussa Aktiengesellschaft | Production of tryptophan by the bacterium Escherichia coli |
| US6218168B1 (en) | 1995-10-26 | 2001-04-17 | CONSORTIUM FüR ELEKTROCHEMISCHE INUDSTRIE GMBH | Process for preparing O-acetylserine, L-cysteine and L-cysteine-related products |
| US5908768A (en) | 1996-04-23 | 1999-06-01 | Ajinomoto Co., Inc. | Process for producing L-glutamic acid by fermentation with E. coli resistant to aspartic acid antimetabolite |
| US5840358A (en) | 1996-05-31 | 1998-11-24 | Degussa Aktiengesellschaft | Process for the preparation of an animal feed supplement based on fermentation broth |
| US5939307A (en) | 1996-07-30 | 1999-08-17 | The Archer-Daniels-Midland Company | Strains of Escherichia coli, methods of preparing the same and use thereof in fermentation processes for l-threonine production |
| WO1998004715A1 (en) | 1996-07-30 | 1998-02-05 | Archer-Daniels-Midland Company | Novel strains of escherichia coli, methods of preparing the same and use thereof in fermentation processes for l-threonine production |
| JPH10165180A (ja) | 1996-12-05 | 1998-06-23 | Ajinomoto Co Inc | L−リジンの製造法 |
| RU2119536C1 (ru) | 1997-01-21 | 1998-09-27 | Государственный научно-исследовательский институт генетики и селекции промышленных микроорганизмов | Штамм escherichia coli - продуцент l-гистидина |
| US5972663A (en) | 1997-06-19 | 1999-10-26 | Consortium Fur Elektrochemische Industrie Gmbh | Microorganisms and processes for the fermentative preparation of L-cysteine, L-cystine, N-acetylserine or thiazolidine derivatives |
| WO1999018228A2 (de) | 1997-10-04 | 1999-04-15 | Forschungszentrum Jülich GmbH | Verfahren zur mikrobiellen herstellung von aminosäuren der aspartat- und/oder glutamatfamilie und im verfahren einsetzbare mittel |
| US6124121A (en) | 1997-10-29 | 2000-09-26 | Ajinomoto Co., Inc. | Method for producing L-leucine |
| JPH11155571A (ja) | 1997-11-25 | 1999-06-15 | Ajinomoto Co Inc | L−システインの製造法 |
| EP0952221A2 (en) | 1998-03-18 | 1999-10-27 | Ajinomoto Co., Ltd. | L-Glutamic acid-producing bacterium and method for producing L-glutamic acid |
| EP0955368A2 (en) | 1998-03-18 | 1999-11-10 | Ajinomoto Co., Ltd. | L-glutamic acid-producing bacterium and method for producing l-glutamic acid |
| US6331419B1 (en) | 1998-03-18 | 2001-12-18 | Ajinomoto Co., Inc. | L-glutamic acid-producing bacterium and method for producing L-glutamic acid |
| US6258554B1 (en) | 1998-07-03 | 2001-07-10 | Kyowa Hakko Kogyo Co., Ltd. | Method for producing metabolites biologically synthesized via phosphoribosyl pyrophosphate |
| WO2000018935A1 (fr) | 1998-09-25 | 2000-04-06 | Ajinomoto Co.,Inc. | Procede de construction d'une bacterie produisant des acides amines, et procede de production d'acides amines par une technique de fermentation utilisant ladite bacterie |
| EP0994190A2 (en) | 1998-10-13 | 2000-04-19 | Ajinomoto Co., Inc. | DNA conferring L-homoserine resistance to bacteria, and its use |
| JP2000139471A (ja) | 1998-11-17 | 2000-05-23 | Ajinomoto Co Inc | 発酵法によるl−メチオニンの製造法 |
| JP2000157276A (ja) | 1998-11-24 | 2000-06-13 | Ajinomoto Co Inc | サーマス属細菌のl−リジン生合成系遺伝子 |
| EP1010755A1 (en) | 1998-12-18 | 2000-06-21 | Ajinomoto Co., Inc. | Method for producing L-Glutamic acid by fermentation |
| EP1013765A1 (en) | 1998-12-23 | 2000-06-28 | Ajinomoto Co., Ltd. | Gene and method for producing L-amino acids |
| JP2000189180A (ja) | 1998-12-30 | 2000-07-11 | Ajinomoto Co Inc | L―アミノ酸の製造法 |
| EP1016710A2 (en) | 1998-12-30 | 2000-07-05 | Ajinomoto Co., Inc. | Method for producing L-amino acids |
| WO2000050624A1 (de) | 1999-02-22 | 2000-08-31 | Forschungszentrum Jülich GmbH | Verfahren zur mikrobiellen herstellung von l-valin |
| JP2000253879A (ja) | 1999-03-09 | 2000-09-19 | Ajinomoto Co Inc | L−リジンの製造法 |
| US6303383B1 (en) | 1999-03-16 | 2001-10-16 | Ajinomoto Co., Inc. | Temperature sensitive plasmid for coryneform bacteria |
| JP2003135066A (ja) | 1999-03-19 | 2003-05-13 | Ajinomoto Co Inc | L−リジンの製造法 |
| WO2000061723A1 (fr) | 1999-04-09 | 2000-10-19 | Ajinomoto Co., Inc. | Bacteries produisant du l-amino acide et procede de production de l-amino acide |
| US6238714B1 (en) | 1999-05-05 | 2001-05-29 | Degussa-Huls Ag | Feedstuff additive which contains D-pantothenic acid and/or its salts and a process for the preparation thereof |
| JP2001057896A (ja) | 1999-06-15 | 2001-03-06 | Ajinomoto Co Inc | L−リジンの製造法 |
| US6403342B1 (en) | 1999-07-09 | 2002-06-11 | Anjinomoto Co., Inc. | DNA coding for mutant isopropylmalate synthase L-leucine-producing microorganism and method for producing L-leucine |
| EP1078989A2 (en) | 1999-08-20 | 2001-02-28 | Ajinomoto Co., Ltd. | Method for producing L-glutamic acid by fermentation accompanied by precipitation |
| EP1085087A2 (en) | 1999-09-20 | 2001-03-21 | Kyowa Hakko Kogyo Co., Ltd. | Method for producing amino acids by fermentation using aminoquinoline resistant bacterial strains |
| US6344347B1 (en) | 1999-09-20 | 2002-02-05 | Kyowa Hakko Kogyo Co., Ltd. | Method for producing L-amino acids by fermentation |
| EP1092776A1 (en) | 1999-10-14 | 2001-04-18 | Ajinomoto Co., Inc. | Method for producing L-amino acid by fermentation |
| WO2001027307A1 (de) | 1999-10-14 | 2001-04-19 | Consortium für elektrochemische Industrie GmbH | Verfahren zur fermentativen herstellung von l-cystein oder l-cystein-derivaten |
| JP2001169788A (ja) | 1999-10-27 | 2001-06-26 | Degussa Huels Ag | 分枝アミノ酸の排出をコードするヌクレオチド配列、その単離法および使用 |
| WO2001053459A1 (fr) | 2000-01-21 | 2001-07-26 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| JP2001346578A (ja) | 2000-04-26 | 2001-12-18 | Ajinomoto Co Inc | アミノ酸生産菌及びアミノ酸の製造法 |
| EP1149911A2 (en) | 2000-04-26 | 2001-10-31 | Ajinomoto Co., Ltd. | Amino acid producing strains belonging to the genus Escherichia and method for producing amino acid |
| EP1170361A2 (en) | 2000-06-28 | 2002-01-09 | Ajinomoto Co., Inc. | New mutant N-Acetylglutamate synthase and method for L-Arginine production |
| EP1170376A1 (en) | 2000-07-05 | 2002-01-09 | Ajinomoto Co., Inc. | Method for producing substances utilizing microorganisms |
| EP1172433A1 (en) | 2000-07-06 | 2002-01-16 | Ajinomoto Co., Inc. | Bacterium having ability to produce L-glutamic acid, L-proline or L-arginine and method for producing L-glutamic acid, L-proline or L-arginine |
| EP1170358A1 (en) | 2000-07-06 | 2002-01-09 | Ajinomoto Co., Ltd. | L-arginine producing Escherichia coli and method of producing L-arginine |
| US20020110876A1 (en) | 2000-08-11 | 2002-08-15 | Ajinomoto Co., Inc. | Method for producing threonine and isoleucine |
| JP2002065287A (ja) | 2000-08-24 | 2002-03-05 | Ajinomoto Co Inc | 塩基性アミノ酸の製造方法 |
| US20020025564A1 (en) | 2000-08-24 | 2002-02-28 | Ajinomoto Co., Inc. | Method for producing basic amino acid |
| US20020058315A1 (en) | 2000-09-26 | 2002-05-16 | Ajinomoto Co., Inc. | Bacterium having ability to produce L-glutamic acid, L-proline or L-arginine and method for producing L-glutamic acid, L-proline or L-arginine |
| RU2000124295A (ru) | 2000-09-26 | 2003-02-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Бактерия, обладающая способностью к продукции l-глутаминовой кислоты, l-пролина или l-аргинина, и способ получения l-глутаминовой кислоты, l-пролина или l-аргинина |
| WO2002026993A1 (en) | 2000-09-28 | 2002-04-04 | Archer-Daniels-Midland Company | Escherichia coli strains which over-produce l-threonine and processes for the production of l-threonine by fermentation |
| EP1239041A2 (en) | 2001-02-13 | 2002-09-11 | Ajinomoto Co., Inc. | Method for producing L-amino acid using bacteria belonging to the genus Escherichia |
| US6596517B2 (en) | 2001-02-20 | 2003-07-22 | Ajinomoto Co., Inc. | Method for producing L-glutamic acid |
| RU2001112869A (ru) | 2001-05-15 | 2003-03-20 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Новая мутантная n-ацетилглутамат синтаза и способ получения l-аргинина |
| WO2003044191A1 (fr) | 2001-11-23 | 2003-05-30 | Ajinomoto Co.,Inc. | Procede de production de l-aminoacides a l'aide d'escherichia |
| US20030148473A1 (en) | 2001-11-23 | 2003-08-07 | Ajinomoto Co., Inc. | Method for producing L-amino acid using bacteria belonging to the genus escherichia |
| US20030157667A1 (en) | 2001-11-23 | 2003-08-21 | Ajinomoto Co., Inc. | Method for producing L-amino acids using bacteria belonging to the genus escherichia |
| WO2003044192A1 (fr) | 2001-11-23 | 2003-05-30 | Ajinomoto Co.,Inc. | Procede de production de l-aminoacides a l'aide d'escherichia |
| WO2003094598A1 (en) | 2002-05-13 | 2003-11-20 | Greenfuel Technologies Corporation | Photobioreactor and process for biomass production and mitigation of pollutants in flue gases |
| EP1484410A1 (en) | 2003-06-05 | 2004-12-08 | Ajinomoto Co., Ltd. | Fermentation methods and genetically modified bacteria with increased substrate and byproduct uptake. |
| JP2006340603A (ja) | 2003-06-23 | 2006-12-21 | Ajinomoto Co Inc | L−グルタミン酸の製造法 |
| US20060135308A1 (en) | 2003-07-04 | 2006-06-22 | Samuel Abraham | Differential for motor vehicles with device for locking thereof |
| US20050025878A1 (en) | 2003-07-11 | 2005-02-03 | Degussa Ag | Process for the granulation of an animal feedstuff additive |
| RU2003121601A (ru) | 2003-07-16 | 2005-02-27 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО "АГРИ") (RU) | Мутантная серинацетилтрансфераза |
| WO2005010175A1 (en) | 2003-07-29 | 2005-02-03 | Ajinomoto Co., Inc. | Method for producing l-lysine or l-threonine using escherichia bacteria having attnuated malic enzyme activity |
| JP2005058227A (ja) | 2003-07-29 | 2005-03-10 | Ajinomoto Co Inc | L−リジンまたはl−スレオニンの製造法 |
| WO2005049808A1 (en) | 2003-11-21 | 2005-06-02 | Ajinomoto Co., Inc. | Method for producing l-amino acid by fermentation |
| WO2005073390A2 (en) | 2004-01-30 | 2005-08-11 | Ajinomoto Co., Inc. | L-amino acid-producing microorganism and method for producing l-amino acid |
| WO2005103275A1 (ja) | 2004-04-26 | 2005-11-03 | Ajinomoto Co., Ltd. | 発酵法によるl-トリプトファンの製造法 |
| EP1813677A1 (en) | 2004-10-07 | 2007-08-01 | Ajinomoto Co., Inc. | Process for producing basic substance |
| WO2006070944A2 (en) | 2004-12-28 | 2006-07-06 | Ajinomoto Co., Inc. | L-glutamic acid-producing microorganism and a method for producing l-glutamic acid |
| WO2006078039A1 (en) | 2005-01-18 | 2006-07-27 | Ajinomoto Co., Inc. | L-amino acid producing microorganism and a method for producing l-amino acid |
| EP1715055A2 (de) | 2005-04-22 | 2006-10-25 | Degussa GmbH | Verfahren zur Herstellung von L-Aminosäuren unter Verwendung verbesserter Stämme der Familie Enterobacteriaceae |
| EP1715056A1 (de) | 2005-04-23 | 2006-10-25 | Degussa AG | Verfahren zur Herstellung von L-Aminosäuren unter Verwendung verbesserter Stämme der Familie Enterobacteriaceae |
| US20070202582A1 (en) | 2006-02-28 | 2007-08-30 | Bush Ronnie A | Process for the production of ethanol from algae |
| WO2007101172A2 (en) * | 2006-02-28 | 2007-09-07 | Propulsion Logic Llc | Process for the production of ethanol from algae |
| WO2007100009A1 (ja) * | 2006-03-03 | 2007-09-07 | Ajinomoto Co., Inc. | L-アミノ酸の製造法 |
| WO2008002053A1 (en) * | 2006-06-26 | 2008-01-03 | Cj Cheiljedang Corporation | Method for producing amino acids using glycerol |
| WO2008081959A1 (en) | 2006-12-22 | 2008-07-10 | Ajinomoto Co., Inc. | A method for producing an l-amino acid by fermentation using a bacterium having an enhanced ability to utilize glycerol |
| WO2008102861A1 (ja) | 2007-02-22 | 2008-08-28 | Ajinomoto Co., Inc. | L-アミノ酸の製造法 |
| WO2008107277A1 (de) | 2007-03-05 | 2008-09-12 | Evonik Degussa Gmbh | Verfahren zur herstellung von l-aminosäuren unter verwendung von stämmen der familie enterobacteriaceae |
Non-Patent Citations (49)
| Title |
|---|
| ALBERGHINA, L.; LOTTI, M., METHODS ENZYMOL., vol. 284, 1997, pages 246 - 260 |
| ARCHIBALD, P.A.; BOLD, H.C., PHYTOMORPHOLOGY, vol. 20, 1970, pages 383 - 389 |
| BALL, S.G.; MORELL, M.K., ANNUAL REVIEW OF PLANT BIOLOGY, vol. 54, 2003, pages 207 - 233 |
| BLOOM F.R. ET AL., THE 15TH MIAMI WINTER SYMPOSIUM, 1983, pages 34 |
| BREIVIK, H.; HARALDSSON, G.G.; KRISTINSSON, B., J. AM. OIL CHEM. SOC., vol. 74, 1997, pages 1425 - 1429 |
| CHISTI Y., BIOTECHNOL. ADV., vol. 25, 2007, pages 294 - 306 |
| CHISTI, Y. ET AL.: "Biodiesel From Microalgae.", BIOTECHNOL. ADV., vol. 25, no. 3, 2007, pages 294 - 306, XP026983679 * |
| CHO E.H.; GUMPORT R.I.; GARDNER J.F., J. BACTERIOL., vol. 184, 2002, pages 5200 - 5203 |
| CHO, E.H.; GUMPORT, R.I.; GARDNER, J.F., J. BACTERIOL., vol. 184, 2002, pages 5200 - 5203 |
| DATSENKO, K.A.; WANNER, B.L., PROC. NATL. ACAD. SCI. USA, vol. 97, 2000, pages 6640 - 6645 |
| DATSENKO, K.A.; WANNER, B.L., PROC. NATL. ACAD. SCI. USA., vol. 97, 2000, pages 6640 - 6645 |
| DATSENKO, K.A; WANNER, B.L., PROC. NATL. ACAD. SCI. USA, vol. 97, 2000, pages 6640 - 6645 |
| DIRUSSO, C.C. ET AL., J. BIOL. CHEM., vol. 267, 1992, pages 8685 - 8691 |
| DIRUSSO, C.C. ET AL., MOL. MICROBIOL., vol. 7, 1993, pages 311 - 322 |
| FUKUDA, H.; KONDO, A.; NODA, H., J. BIOSCI. BIOENG., vol. 92, 2001, pages 405 - 416 |
| GOLDSTEIN ET AL.: "Prokaryotic promoters in biotechnology", BIOTECHNOL. ANNU. REV., vol. 1, 1995, pages 105 - 128 |
| GRIMA, E.M. ET AL., BIOTECHNOL. ADVANCES, vol. 20, 2003, pages 491 - 515 |
| GUSYATINER ET AL., GENETIKA, vol. 14, 1978, pages 947 - 956 |
| HASAN, F. ET AL., ENZYME AND MICROBIOL. TECHNOL., vol. 39, 2006, pages 235 - 251 |
| HASHIMOTO, K. ET AL., BIOSCI. BIOTECHNOL. BIOCHEM., vol. 70, 1996, pages 22 - 30 |
| INT. J. SYST. BACTERIOL., vol. 43, 1993, pages 162 - 173 |
| IZUMO A. ET AL., PLANT SCIENCE, vol. 172, 2007, pages 1138 - 1147 |
| JAEGER, K.E. ET AL., FEMS MICROBIOL. REV., vol. 15, 1994, pages 29 - 63 |
| JAEGER, K.E.; EGGERT, T., CURR. OPIN. BIOTECHNOL., vol. 13, 2002, pages 390 - 397 |
| KAWAGUCHI, Y. ET AL., NATURE, vol. 341, 1989, pages 164 - 166 |
| LIE, C.-P.; LIN, L.-P., BOT. BULL. ACAD. SIN., vol. 42, 2001, pages 207 - 214 |
| LIEBL; W. ET AL., INT. J. SYST. BACTERIOL., vol. 41, 1991, pages 255 - 260 |
| LIN, J.T. ET AL., J. CHROMATOGR. A., vol. 808, 1998, pages 43 - 49 |
| LIVSHITS, V.A. ET AL., RES. MICROBIOL., vol. 154, 2003, pages 123 - 135 |
| LYNN, S.P.; BURTON, W.S.; DONOHUE, T.J.; GOULD, R.M.; GUMPORT, R.L; GARDNER, J.F., J. MOI. BIOL., vol. 194, 1987, pages 59 - 69 |
| MATSUMOTO, M. ET AL., APPL. BIOCHEM. BIOTECHNOL., vol. 105-108, 2003, pages 247 - 254 |
| OHAMA, T. ET AL., NUCLEIC ACIDS RES., vol. 21, 1993, pages 4039 - 4045 |
| OYAMA Y. ET AL., PLANTA, vol. 224, 2006, pages 646 - 654 |
| QIU Z.; GOODMAN M.F., J. BIOL. CHEM., vol. 272, 1997, pages 8611 - 8617 |
| ROBERTSON, G.H. ET AL., J. AGRIC. FOOD CHEM., vol. 54, 2006, pages 353 - 365 |
| SCHMIDT-DANNERT, C., BIOORG. MED. CHEM., vol. 7, 1999, pages 2123 - 2130 |
| See also references of EP2248906A4 |
| SPOLAORE, P. ET AL., J. BIOSCI. BIOENG., vol. 101, 2006, pages 87 - 96 |
| TAKAGI, M. ET AL., APPL. MICROBIOL. BIOTECHNOL., vol. 54, 2000, pages 112 - 117 |
| THOMPSON G.A. JR., BIOCHIM. BIOPHYS. ACTA, vol. 1302, 1996, pages 17 - 45 |
| TONON, T. ET AL., PHYTOCHEMISTRY, vol. 61, 2002, pages 15 - 24 |
| TORNABENE, T.G. ET AL., ENZYME AND MICROB. TECHNOL., vol. 5, 1983, pages 435 - 440 |
| VALLE F. ET AL., GENE, vol. 23, 1983, pages 199 - 209 |
| VORUM, H. ET AL., BIOCHIMICA ET BIOPHYSICA ACTA, vol. 1126, 1992, pages 135 - 142 |
| WANG, J.P ET AL., J. AGRIC. FOOD CHEM., vol. 54, 2006, pages 9405 - 9410 |
| WENTE, S.R.; SCHACHMAN, H.K., J. BID. CHEM., vol. 266, 1991, pages 20833 - 20839 |
| WHITE, T.J. ET AL., TRENDS GENET, vol. 5, 1989, pages 185 |
| WINKLER W.C., CURR. OPIN. CHEM. BIOL., vol. 9, 2005, pages 594 - 602 |
| YAMABERI, K. ET AL., J. MAR. BIOTECHNOL., vol. 6, 1998, pages 44 - 48 |
Cited By (16)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP2460883A4 (en) * | 2009-07-29 | 2013-01-16 | Ajinomoto Kk | PROCESS FOR THE PREPARATION OF L-AMINO ACID |
| WO2011013707A1 (ja) * | 2009-07-29 | 2011-02-03 | 味の素株式会社 | L-アミノ酸の製造法 |
| EP2460883A1 (en) * | 2009-07-29 | 2012-06-06 | Ajinomoto Co., Inc. | Method for producing l-amino acid |
| US8771981B2 (en) | 2009-07-29 | 2014-07-08 | Ajinomoto Co., Inc. | Method for producing an L-amino acid |
| US9150892B2 (en) | 2010-02-25 | 2015-10-06 | Colgate-Palmolive Company | Process of producing arginine employing Corynebacterium glutamicum ATCC 21831 or Corynebacterium glutamicum ATCC 21493 in an afermantation medium comprising cassava bagasse or jackfruit seed as a carbon source |
| JP2013520196A (ja) * | 2010-02-25 | 2013-06-06 | コルゲート・パーモリブ・カンパニー | 炭素供給源としてキャッサバ(Cassava)・バガスまたはジャックフルーツ(jackfruit)種子を含む発酵培地中で、コリネバクテリウム・グルタミクム(Corynebacteriumglutamicum)ATCC21831またはコリネバクテリウム・グルタミクムATCC21493を使用してアルギニンを産生するプロセス |
| KR101115357B1 (ko) | 2010-09-03 | 2012-03-06 | 대상 주식회사 | 습식 상태의 미세조류로부터 조오일을 추출하는 방법 |
| WO2012077739A1 (ja) * | 2010-12-10 | 2012-06-14 | 味の素株式会社 | L-アミノ酸の製造法 |
| US8951760B2 (en) | 2010-12-10 | 2015-02-10 | Ajinomoto Co., Inc. | Method for producing an L-amino acid |
| JP2014500031A (ja) * | 2010-12-21 | 2014-01-09 | サンボ インターナショナル エスタブリッシュメント | 光合成微生物中のCoQ10およびCoQH2の含有量を増加させる方法 |
| US9376660B2 (en) | 2010-12-21 | 2016-06-28 | Sanbo International Establishment | Method for increasing the CoQ10 and CoQH2 content in phototrophic microorganisms |
| WO2013008931A1 (ja) * | 2011-07-14 | 2013-01-17 | 味の素株式会社 | 脂肪酸類の製造法 |
| JP2013039085A (ja) * | 2011-08-18 | 2013-02-28 | Ihi Corp | エタノールの製造方法 |
| WO2013176261A1 (ja) * | 2012-05-25 | 2013-11-28 | 味の素株式会社 | 微細藻類を用いた栄養添加剤の製造法 |
| CN103911419B (zh) * | 2014-03-20 | 2017-01-11 | 广东肇庆星湖生物科技股份有限公司 | 一种双菌株联合发酵生产l-缬氨酸的方法 |
| JP2015198662A (ja) * | 2015-06-01 | 2015-11-12 | コルゲート・パーモリブ・カンパニーColgate−Palmolive Company | 炭素供給源としてキャッサバ(Cassava)・バガスまたはジャックフルーツ(jackfruit)種子を含む発酵培地中で、コリネバクテリウム・グルタミクム(Corynebacteriumglutamicum)ATCC21831またはコリネバクテリウム・グルタミクムATCC21493を使用してアルギニンを産生するプロセス |
Also Published As
| Publication number | Publication date |
|---|---|
| US8354254B2 (en) | 2013-01-15 |
| EP2248906A4 (en) | 2012-07-11 |
| US20130084609A1 (en) | 2013-04-04 |
| US8728772B2 (en) | 2014-05-20 |
| KR20100120663A (ko) | 2010-11-16 |
| BRPI0906795A2 (pt) | 2015-08-18 |
| JPWO2009093703A1 (ja) | 2011-05-26 |
| CN102348806A (zh) | 2012-02-08 |
| JP5526785B2 (ja) | 2014-06-18 |
| EP2248906A1 (en) | 2010-11-10 |
| US20110014663A1 (en) | 2011-01-20 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5526785B2 (ja) | L−アミノ酸の製造法 | |
| US8771981B2 (en) | Method for producing an L-amino acid | |
| EP2003209B1 (en) | Method for producing l-amino acid | |
| EP2094858B1 (en) | Method for producing an l-amino acid | |
| WO2009142286A1 (ja) | L-アミノ酸の製造法 | |
| JP2009165355A (ja) | L−アミノ酸を生産する微生物及びl−アミノ酸の製造法 | |
| WO2018030507A1 (ja) | L-アミノ酸の製造法 | |
| JP5303918B2 (ja) | L−アミノ酸の製造法 | |
| JP2016192903A (ja) | 海藻由来バイオマスからのl−アミノ酸の製造方法 | |
| WO2015064648A1 (ja) | 脂肪酸を生成する緑藻類 | |
| US20140127761A1 (en) | Method for Producing Fatty Acid | |
| US8975045B2 (en) | Mutant RpsA gene and method for producing L-amino acid | |
| JP2016208852A (ja) | 脂肪酸を生成する緑藻改変株 | |
| WO2014061805A1 (ja) | L-アミノ酸の製造法 | |
| WO2014061804A1 (ja) | L-アミノ酸の製造法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WWE | Wipo information: entry into national phase |
Ref document number: 200980102894.2 Country of ref document: CN |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 09704195 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2009550578 Country of ref document: JP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2009704195 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: 20107018301 Country of ref document: KR Kind code of ref document: A |
|
| ENP | Entry into the national phase |
Ref document number: PI0906795 Country of ref document: BR Kind code of ref document: A2 Effective date: 20100723 |