RU2562862C2 - Конъюгаты меченных радиоактивным цирконием сконструированных антител с цистеиновыми заменами - Google Patents
Конъюгаты меченных радиоактивным цирконием сконструированных антител с цистеиновыми заменами Download PDFInfo
- Publication number
- RU2562862C2 RU2562862C2 RU2012123007/10A RU2012123007A RU2562862C2 RU 2562862 C2 RU2562862 C2 RU 2562862C2 RU 2012123007/10 A RU2012123007/10 A RU 2012123007/10A RU 2012123007 A RU2012123007 A RU 2012123007A RU 2562862 C2 RU2562862 C2 RU 2562862C2
- Authority
- RU
- Russia
- Prior art keywords
- antibody
- cysteine
- antibodies
- zirconium
- protein
- Prior art date
Links
- 235000018417 cysteine Nutrition 0.000 title claims abstract description 219
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 title claims abstract description 218
- 230000002285 radioactive effect Effects 0.000 title abstract description 23
- -1 cysteine amino acids Chemical class 0.000 claims abstract description 73
- 235000001014 amino acid Nutrition 0.000 claims abstract description 64
- 229910052726 zirconium Inorganic materials 0.000 claims abstract description 62
- QCWXUUIWCKQGHC-UHFFFAOYSA-N Zirconium Chemical compound [Zr] QCWXUUIWCKQGHC-UHFFFAOYSA-N 0.000 claims abstract description 54
- 150000001413 amino acids Chemical class 0.000 claims abstract description 46
- 238000003384 imaging method Methods 0.000 claims abstract description 30
- 239000000126 substance Substances 0.000 claims abstract description 14
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 claims description 159
- 230000027455 binding Effects 0.000 claims description 150
- 238000006467 substitution reaction Methods 0.000 claims description 138
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 claims description 128
- 238000000034 method Methods 0.000 claims description 121
- 108090000623 proteins and genes Proteins 0.000 claims description 120
- 239000000427 antigen Substances 0.000 claims description 101
- 108091007433 antigens Proteins 0.000 claims description 101
- 102000036639 antigens Human genes 0.000 claims description 101
- 102000004169 proteins and genes Human genes 0.000 claims description 94
- 229960000575 trastuzumab Drugs 0.000 claims description 91
- 235000018102 proteins Nutrition 0.000 claims description 90
- 239000003153 chemical reaction reagent Substances 0.000 claims description 76
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 claims description 62
- 229940024606 amino acid Drugs 0.000 claims description 62
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 claims description 61
- 206010028980 Neoplasm Diseases 0.000 claims description 60
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 56
- UBQYURCVBFRUQT-UHFFFAOYSA-N N-benzoyl-Ferrioxamine B Chemical group CC(=O)N(O)CCCCCNC(=O)CCC(=O)N(O)CCCCCNC(=O)CCC(=O)N(O)CCCCCN UBQYURCVBFRUQT-UHFFFAOYSA-N 0.000 claims description 48
- 241000282414 Homo sapiens Species 0.000 claims description 43
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 42
- 102000005962 receptors Human genes 0.000 claims description 38
- 108020003175 receptors Proteins 0.000 claims description 38
- 239000000203 mixture Substances 0.000 claims description 33
- 201000011510 cancer Diseases 0.000 claims description 30
- 238000001514 detection method Methods 0.000 claims description 28
- 229960000958 deferoxamine Drugs 0.000 claims description 27
- 108060003951 Immunoglobulin Proteins 0.000 claims description 23
- 102000018358 immunoglobulin Human genes 0.000 claims description 23
- 238000004519 manufacturing process Methods 0.000 claims description 20
- 238000000746 purification Methods 0.000 claims description 20
- 230000014509 gene expression Effects 0.000 claims description 19
- 239000003446 ligand Substances 0.000 claims description 19
- 241001465754 Metazoa Species 0.000 claims description 18
- 102100033423 GDNF family receptor alpha-1 Human genes 0.000 claims description 16
- 230000015572 biosynthetic process Effects 0.000 claims description 15
- 102100022430 Melanocyte protein PMEL Human genes 0.000 claims description 14
- 125000000539 amino acid group Chemical group 0.000 claims description 14
- 238000001727 in vivo Methods 0.000 claims description 14
- 101000579425 Homo sapiens Proto-oncogene tyrosine-protein kinase receptor Ret Proteins 0.000 claims description 13
- 102100028286 Proto-oncogene tyrosine-protein kinase receptor Ret Human genes 0.000 claims description 13
- 238000002703 mutagenesis Methods 0.000 claims description 13
- 231100000350 mutagenesis Toxicity 0.000 claims description 13
- 101000997961 Homo sapiens GDNF family receptor alpha-1 Proteins 0.000 claims description 12
- 101000628535 Homo sapiens Metalloreductase STEAP2 Proteins 0.000 claims description 12
- 102100026711 Metalloreductase STEAP2 Human genes 0.000 claims description 12
- 101710120037 Toxin CcdB Proteins 0.000 claims description 12
- 230000006870 function Effects 0.000 claims description 12
- 238000011161 development Methods 0.000 claims description 11
- 101000620359 Homo sapiens Melanocyte protein PMEL Proteins 0.000 claims description 10
- 102100032780 Semaphorin-5B Human genes 0.000 claims description 10
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 10
- 241000699670 Mus sp. Species 0.000 claims description 9
- 102100026159 Tomoregulin-1 Human genes 0.000 claims description 9
- 102100027203 B-cell antigen receptor complex-associated protein beta chain Human genes 0.000 claims description 8
- 101001065550 Homo sapiens Lymphocyte antigen 6K Proteins 0.000 claims description 8
- 101000654679 Homo sapiens Semaphorin-5B Proteins 0.000 claims description 8
- 101000713169 Homo sapiens Solute carrier family 52, riboflavin transporter, member 2 Proteins 0.000 claims description 8
- 101000835745 Homo sapiens Teratocarcinoma-derived growth factor 1 Proteins 0.000 claims description 8
- 102100032129 Lymphocyte antigen 6K Human genes 0.000 claims description 8
- 102100036862 Solute carrier family 52, riboflavin transporter, member 2 Human genes 0.000 claims description 8
- 102100026404 Teratocarcinoma-derived growth factor 1 Human genes 0.000 claims description 8
- 102100029690 Tumor necrosis factor receptor superfamily member 13C Human genes 0.000 claims description 8
- 102100039094 Tyrosinase Human genes 0.000 claims description 8
- 210000000988 bone and bone Anatomy 0.000 claims description 8
- 239000000969 carrier Substances 0.000 claims description 8
- 101000834937 Homo sapiens Tomoregulin-1 Proteins 0.000 claims description 7
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 claims description 7
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 claims description 7
- 230000000527 lymphocytic effect Effects 0.000 claims description 7
- 102100031511 Fc receptor-like protein 2 Human genes 0.000 claims description 6
- 101000914491 Homo sapiens B-cell antigen receptor complex-associated protein beta chain Proteins 0.000 claims description 6
- 101001063456 Homo sapiens Leucine-rich repeat-containing G-protein coupled receptor 5 Proteins 0.000 claims description 6
- 101000958332 Homo sapiens Lymphocyte antigen 6 complex locus protein G6d Proteins 0.000 claims description 6
- 101000853730 Homo sapiens RING finger and transmembrane domain-containing protein 2 Proteins 0.000 claims description 6
- 101000834948 Homo sapiens Tomoregulin-2 Proteins 0.000 claims description 6
- 102100031036 Leucine-rich repeat-containing G-protein coupled receptor 5 Human genes 0.000 claims description 6
- 102100038210 Lymphocyte antigen 6 complex locus protein G6d Human genes 0.000 claims description 6
- 102100032131 Lymphocyte antigen 6E Human genes 0.000 claims description 6
- 102100035928 RING finger and transmembrane domain-containing protein 2 Human genes 0.000 claims description 6
- 102100026160 Tomoregulin-2 Human genes 0.000 claims description 6
- 230000004913 activation Effects 0.000 claims description 6
- 230000001086 cytosolic effect Effects 0.000 claims description 6
- 230000004069 differentiation Effects 0.000 claims description 6
- 238000002955 isolation Methods 0.000 claims description 6
- 150000007523 nucleic acids Chemical group 0.000 claims description 6
- 210000002307 prostate Anatomy 0.000 claims description 6
- 102100024220 CD180 antigen Human genes 0.000 claims description 5
- 101000980829 Homo sapiens CD180 antigen Proteins 0.000 claims description 5
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 5
- 239000003102 growth factor Substances 0.000 claims description 5
- 239000011734 sodium Substances 0.000 claims description 5
- 102100025218 B-cell differentiation antigen CD72 Human genes 0.000 claims description 4
- 108010001445 CD79 Antigens Proteins 0.000 claims description 4
- 102000000796 CD79 Antigens Human genes 0.000 claims description 4
- 102100031517 Fc receptor-like protein 1 Human genes 0.000 claims description 4
- 101710120224 Fc receptor-like protein 1 Proteins 0.000 claims description 4
- 101000846911 Homo sapiens Fc receptor-like protein 2 Proteins 0.000 claims description 4
- 101000623901 Homo sapiens Mucin-16 Proteins 0.000 claims description 4
- 101000825475 Homo sapiens Protein shisa-2 homolog Proteins 0.000 claims description 4
- 101000844504 Homo sapiens Transient receptor potential cation channel subfamily M member 4 Proteins 0.000 claims description 4
- 101000606090 Homo sapiens Tyrosinase Proteins 0.000 claims description 4
- 102000004310 Ion Channels Human genes 0.000 claims description 4
- 108090000862 Ion Channels Proteins 0.000 claims description 4
- 102100023123 Mucin-16 Human genes 0.000 claims description 4
- 102100037603 P2X purinoceptor 5 Human genes 0.000 claims description 4
- 206010060862 Prostate cancer Diseases 0.000 claims description 4
- 102100036735 Prostate stem cell antigen Human genes 0.000 claims description 4
- 208000000236 Prostatic Neoplasms Diseases 0.000 claims description 4
- 102100022938 Protein shisa-2 homolog Human genes 0.000 claims description 4
- 102100038437 Sodium-dependent phosphate transport protein 2B Human genes 0.000 claims description 4
- 101710178300 Tumor necrosis factor receptor superfamily member 13C Proteins 0.000 claims description 4
- 108060008724 Tyrosinase Proteins 0.000 claims description 4
- 210000004901 leucine-rich repeat Anatomy 0.000 claims description 4
- 230000005945 translocation Effects 0.000 claims description 4
- 108010046304 B-Cell Activation Factor Receptor Proteins 0.000 claims description 3
- 101710129514 B-cell differentiation antigen CD72 Proteins 0.000 claims description 3
- 102100025473 Carcinoembryonic antigen-related cell adhesion molecule 6 Human genes 0.000 claims description 3
- 101000914326 Homo sapiens Carcinoembryonic antigen-related cell adhesion molecule 6 Proteins 0.000 claims description 3
- 101000628547 Homo sapiens Metalloreductase STEAP1 Proteins 0.000 claims description 3
- 101000716102 Homo sapiens T-cell surface glycoprotein CD4 Proteins 0.000 claims description 3
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 claims description 3
- 102000052922 Large Neutral Amino Acid-Transporter 1 Human genes 0.000 claims description 3
- 102100026712 Metalloreductase STEAP1 Human genes 0.000 claims description 3
- 206010035226 Plasma cell myeloma Diseases 0.000 claims description 3
- 108010029485 Protein Isoforms Proteins 0.000 claims description 3
- 102000001708 Protein Isoforms Human genes 0.000 claims description 3
- 108091006232 SLC7A5 Proteins 0.000 claims description 3
- 102100036011 T-cell surface glycoprotein CD4 Human genes 0.000 claims description 3
- 230000033228 biological regulation Effects 0.000 claims description 3
- 230000001419 dependent effect Effects 0.000 claims description 3
- 201000000050 myeloid neoplasm Diseases 0.000 claims description 3
- 229910052709 silver Inorganic materials 0.000 claims description 3
- 239000004332 silver Substances 0.000 claims description 3
- 229910052708 sodium Inorganic materials 0.000 claims description 3
- 230000001052 transient effect Effects 0.000 claims description 3
- 102000035160 transmembrane proteins Human genes 0.000 claims description 3
- 108091005703 transmembrane proteins Proteins 0.000 claims description 3
- 208000030507 AIDS Diseases 0.000 claims description 2
- 102100020998 Aspartate beta-hydroxylase domain-containing protein 1 Human genes 0.000 claims description 2
- 102100022108 Aspartyl/asparaginyl beta-hydroxylase Human genes 0.000 claims description 2
- 101710140787 Aspartyl/asparaginyl beta-hydroxylase Proteins 0.000 claims description 2
- 230000024704 B cell apoptotic process Effects 0.000 claims description 2
- 108091008875 B cell receptors Proteins 0.000 claims description 2
- 102000007536 B-Cell Activation Factor Receptor Human genes 0.000 claims description 2
- 102100027205 B-cell antigen receptor complex-associated protein alpha chain Human genes 0.000 claims description 2
- 101710166261 B-cell antigen receptor complex-associated protein beta chain Proteins 0.000 claims description 2
- 102100038080 B-cell receptor CD22 Human genes 0.000 claims description 2
- 230000003844 B-cell-activation Effects 0.000 claims description 2
- 208000011691 Burkitt lymphomas Diseases 0.000 claims description 2
- 102100031658 C-X-C chemokine receptor type 5 Human genes 0.000 claims description 2
- 102000002110 C2 domains Human genes 0.000 claims description 2
- 108050009459 C2 domains Proteins 0.000 claims description 2
- 108091005462 Cation channels Proteins 0.000 claims description 2
- 102100037677 Cell surface hyaluronidase Human genes 0.000 claims description 2
- 101710125063 Cell surface hyaluronidase Proteins 0.000 claims description 2
- 108010008955 Chemokine CXCL13 Proteins 0.000 claims description 2
- 102000006574 Chemokine CXCL13 Human genes 0.000 claims description 2
- 101100004180 Chironomus tentans BR3 gene Proteins 0.000 claims description 2
- 102100030886 Complement receptor type 1 Human genes 0.000 claims description 2
- 102100026245 E3 ubiquitin-protein ligase RNF43 Human genes 0.000 claims description 2
- 102000012545 EGF-like domains Human genes 0.000 claims description 2
- 108050002150 EGF-like domains Proteins 0.000 claims description 2
- 102000010180 Endothelin receptor Human genes 0.000 claims description 2
- 108050001739 Endothelin receptor Proteins 0.000 claims description 2
- 102100031507 Fc receptor-like protein 5 Human genes 0.000 claims description 2
- 102000016970 Follistatin Human genes 0.000 claims description 2
- 108010014612 Follistatin Proteins 0.000 claims description 2
- 108091006027 G proteins Proteins 0.000 claims description 2
- 101710105157 GDNF family receptor alpha-1 Proteins 0.000 claims description 2
- 102000030782 GTP binding Human genes 0.000 claims description 2
- 108091000058 GTP-Binding Proteins 0.000 claims description 2
- 102000034615 Glial cell line-derived neurotrophic factor Human genes 0.000 claims description 2
- 108091010837 Glial cell line-derived neurotrophic factor Proteins 0.000 claims description 2
- 208000031886 HIV Infections Diseases 0.000 claims description 2
- 102100031546 HLA class II histocompatibility antigen, DO beta chain Human genes 0.000 claims description 2
- 102000018713 Histocompatibility Antigens Class II Human genes 0.000 claims description 2
- 108010027412 Histocompatibility Antigens Class II Proteins 0.000 claims description 2
- 101000783987 Homo sapiens Aspartate beta-hydroxylase domain-containing protein 1 Proteins 0.000 claims description 2
- 101000914489 Homo sapiens B-cell antigen receptor complex-associated protein alpha chain Proteins 0.000 claims description 2
- 101000884305 Homo sapiens B-cell receptor CD22 Proteins 0.000 claims description 2
- 101000922405 Homo sapiens C-X-C chemokine receptor type 5 Proteins 0.000 claims description 2
- 101000727061 Homo sapiens Complement receptor type 1 Proteins 0.000 claims description 2
- 101000692702 Homo sapiens E3 ubiquitin-protein ligase RNF43 Proteins 0.000 claims description 2
- 101100119857 Homo sapiens FCRL2 gene Proteins 0.000 claims description 2
- 101000846908 Homo sapiens Fc receptor-like protein 5 Proteins 0.000 claims description 2
- 101000866281 Homo sapiens HLA class II histocompatibility antigen, DO beta chain Proteins 0.000 claims description 2
- 101001065568 Homo sapiens Lymphocyte antigen 6E Proteins 0.000 claims description 2
- 101000576802 Homo sapiens Mesothelin Proteins 0.000 claims description 2
- 101001024605 Homo sapiens Next to BRCA1 gene 1 protein Proteins 0.000 claims description 2
- 101000829779 Homo sapiens Probable G-protein coupled receptor 19 Proteins 0.000 claims description 2
- 101000604039 Homo sapiens Sodium-dependent phosphate transport protein 2B Proteins 0.000 claims description 2
- 101100425948 Homo sapiens TNFRSF13C gene Proteins 0.000 claims description 2
- 206010020853 Hypertonic bladder Diseases 0.000 claims description 2
- 108010006444 Leucine-Rich Repeat Proteins Proteins 0.000 claims description 2
- 101710157879 Lymphocyte antigen 6E Proteins 0.000 claims description 2
- 206010025323 Lymphomas Diseases 0.000 claims description 2
- 102000018697 Membrane Proteins Human genes 0.000 claims description 2
- 108010052285 Membrane Proteins Proteins 0.000 claims description 2
- 108090000015 Mesothelin Proteins 0.000 claims description 2
- 102000003735 Mesothelin Human genes 0.000 claims description 2
- 102100025096 Mesothelin Human genes 0.000 claims description 2
- 101100182721 Mus musculus Ly6e gene Proteins 0.000 claims description 2
- 101100042271 Mus musculus Sema3b gene Proteins 0.000 claims description 2
- 101100365741 Mus musculus Shisa2 gene Proteins 0.000 claims description 2
- 102400000058 Neuregulin-1 Human genes 0.000 claims description 2
- 108090000556 Neuregulin-1 Proteins 0.000 claims description 2
- 101710189969 P2X purinoceptor 5 Proteins 0.000 claims description 2
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 claims description 2
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 claims description 2
- 102100033237 Pro-epidermal growth factor Human genes 0.000 claims description 2
- 102100023417 Probable G-protein coupled receptor 19 Human genes 0.000 claims description 2
- 108010080192 Purinergic Receptors Proteins 0.000 claims description 2
- 102000014400 SH2 domains Human genes 0.000 claims description 2
- 108050003452 SH2 domains Proteins 0.000 claims description 2
- 108091058557 SILV Proteins 0.000 claims description 2
- 108091006576 SLC34A2 Proteins 0.000 claims description 2
- 102000009203 Sema domains Human genes 0.000 claims description 2
- 108050000099 Sema domains Proteins 0.000 claims description 2
- 101710199399 Semaphorin-5B Proteins 0.000 claims description 2
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 2
- 102000003618 TRPM4 Human genes 0.000 claims description 2
- 108060008245 Thrombospondin Proteins 0.000 claims description 2
- 102000002938 Thrombospondin Human genes 0.000 claims description 2
- 101710175559 Tomoregulin-1 Proteins 0.000 claims description 2
- 102100031228 Transient receptor potential cation channel subfamily M member 4 Human genes 0.000 claims description 2
- 241000269368 Xenopus laevis Species 0.000 claims description 2
- 101100365738 Xenopus laevis shisa1 gene Proteins 0.000 claims description 2
- 230000005540 biological transmission Effects 0.000 claims description 2
- 230000007123 defense Effects 0.000 claims description 2
- 230000009266 disease activity Effects 0.000 claims description 2
- 230000005284 excitation Effects 0.000 claims description 2
- 208000032839 leukemia Diseases 0.000 claims description 2
- 210000004698 lymphocyte Anatomy 0.000 claims description 2
- 210000003593 megakaryocyte Anatomy 0.000 claims description 2
- 230000005012 migration Effects 0.000 claims description 2
- 238000013508 migration Methods 0.000 claims description 2
- 230000000921 morphogenic effect Effects 0.000 claims description 2
- 230000004766 neurogenesis Effects 0.000 claims description 2
- 230000007310 pathophysiology Effects 0.000 claims description 2
- 239000001488 sodium phosphate Substances 0.000 claims description 2
- 229910000162 sodium phosphate Inorganic materials 0.000 claims description 2
- 208000035782 susceptibility to 1 Hirschsprung disease Diseases 0.000 claims description 2
- 210000000225 synapse Anatomy 0.000 claims description 2
- 201000000596 systemic lupus erythematosus Diseases 0.000 claims description 2
- 208000001608 teratocarcinoma Diseases 0.000 claims description 2
- 108010037277 thymic shared antigen-1 Proteins 0.000 claims description 2
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 claims description 2
- 102100034845 KiSS-1 receptor Human genes 0.000 claims 6
- 108010023729 Complement 3d Receptors Proteins 0.000 claims 4
- 102000011412 Complement 3d Receptors Human genes 0.000 claims 4
- 101001091205 Homo sapiens KiSS-1 receptor Proteins 0.000 claims 4
- 102100032768 Complement receptor type 2 Human genes 0.000 claims 2
- 101001136592 Homo sapiens Prostate stem cell antigen Proteins 0.000 claims 2
- 108010076800 Kisspeptin-1 Receptors Proteins 0.000 claims 2
- 239000002299 complementary DNA Substances 0.000 claims 2
- 108010009392 Cyclin-Dependent Kinase Inhibitor p16 Proteins 0.000 claims 1
- 101000941929 Homo sapiens Complement receptor type 2 Proteins 0.000 claims 1
- 108091007643 Phosphate carriers Proteins 0.000 claims 1
- 102100023087 Protein S100-A4 Human genes 0.000 claims 1
- DAWBXZHBYOYVLB-UHFFFAOYSA-J oxalate;zirconium(4+) Chemical compound [Zr+4].[O-]C(=O)C([O-])=O.[O-]C(=O)C([O-])=O DAWBXZHBYOYVLB-UHFFFAOYSA-J 0.000 claims 1
- 238000013414 tumor xenograft model Methods 0.000 claims 1
- 230000000694 effects Effects 0.000 abstract description 25
- 239000003814 drug Substances 0.000 abstract description 20
- 238000003745 diagnosis Methods 0.000 abstract description 4
- 238000006880 cross-coupling reaction Methods 0.000 abstract 1
- 125000005647 linker group Chemical group 0.000 description 79
- 150000003573 thiols Chemical class 0.000 description 72
- 210000004027 cell Anatomy 0.000 description 66
- 230000009257 reactivity Effects 0.000 description 61
- 108010090804 Streptavidin Proteins 0.000 description 58
- 239000000562 conjugate Substances 0.000 description 54
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 52
- 230000021615 conjugation Effects 0.000 description 48
- 238000004458 analytical method Methods 0.000 description 36
- 229920001184 polypeptide Polymers 0.000 description 34
- 125000003396 thiol group Chemical group [H]S* 0.000 description 31
- 238000006243 chemical reaction Methods 0.000 description 30
- 239000012634 fragment Substances 0.000 description 30
- 238000003556 assay Methods 0.000 description 28
- 125000000415 L-cysteinyl group Chemical group O=C([*])[C@@](N([H])[H])([H])C([H])([H])S[H] 0.000 description 27
- 229960002685 biotin Drugs 0.000 description 27
- 239000011616 biotin Substances 0.000 description 27
- 235000020958 biotin Nutrition 0.000 description 26
- 125000003275 alpha amino acid group Chemical group 0.000 description 25
- 102000009027 Albumins Human genes 0.000 description 24
- 108010088751 Albumins Proteins 0.000 description 24
- 108020004414 DNA Proteins 0.000 description 24
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 24
- 239000000243 solution Substances 0.000 description 24
- 102000001301 EGF receptor Human genes 0.000 description 22
- 238000002600 positron emission tomography Methods 0.000 description 22
- 239000000523 sample Substances 0.000 description 21
- 239000000543 intermediate Substances 0.000 description 20
- 238000002965 ELISA Methods 0.000 description 19
- 230000003287 optical effect Effects 0.000 description 19
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 18
- 230000003993 interaction Effects 0.000 description 18
- 108060006698 EGF receptor Proteins 0.000 description 17
- 150000001875 compounds Chemical class 0.000 description 17
- 229940079593 drug Drugs 0.000 description 17
- 238000010521 absorption reaction Methods 0.000 description 16
- 239000000499 gel Substances 0.000 description 16
- 238000002372 labelling Methods 0.000 description 16
- 239000002904 solvent Substances 0.000 description 16
- CIVGYTYIDWRBQU-UFLZEWODSA-N 5-[(3as,4s,6ar)-2-oxo-1,3,3a,4,6,6a-hexahydrothieno[3,4-d]imidazol-4-yl]pentanoic acid;pyrrole-2,5-dione Chemical compound O=C1NC(=O)C=C1.N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 CIVGYTYIDWRBQU-UFLZEWODSA-N 0.000 description 14
- 230000035772 mutation Effects 0.000 description 14
- 238000012216 screening Methods 0.000 description 14
- 108091034117 Oligonucleotide Proteins 0.000 description 13
- 238000012879 PET imaging Methods 0.000 description 13
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 13
- 210000004369 blood Anatomy 0.000 description 13
- 239000008280 blood Substances 0.000 description 13
- 239000007850 fluorescent dye Substances 0.000 description 13
- 125000000524 functional group Chemical group 0.000 description 13
- 239000002245 particle Substances 0.000 description 13
- 238000002360 preparation method Methods 0.000 description 13
- 239000000047 product Substances 0.000 description 13
- 210000002966 serum Anatomy 0.000 description 13
- 239000000758 substrate Substances 0.000 description 13
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 13
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 12
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 12
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 12
- 239000002202 Polyethylene glycol Substances 0.000 description 12
- 125000004429 atom Chemical group 0.000 description 12
- 238000002347 injection Methods 0.000 description 12
- 239000007924 injection Substances 0.000 description 12
- 238000005259 measurement Methods 0.000 description 12
- 239000012070 reactive reagent Substances 0.000 description 12
- 210000001519 tissue Anatomy 0.000 description 12
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 11
- ZMXDDKWLCZADIW-UHFFFAOYSA-N N,N-Dimethylformamide Chemical compound CN(C)C=O ZMXDDKWLCZADIW-UHFFFAOYSA-N 0.000 description 11
- MUBZPKHOEPUJKR-UHFFFAOYSA-N Oxalic acid Chemical compound OC(=O)C(O)=O MUBZPKHOEPUJKR-UHFFFAOYSA-N 0.000 description 11
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 11
- 230000000269 nucleophilic effect Effects 0.000 description 11
- 239000013612 plasmid Substances 0.000 description 11
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 11
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 10
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 10
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical group NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 10
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Chemical group NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 10
- PEEHTFAAVSWFBL-UHFFFAOYSA-N Maleimide Chemical compound O=C1NC(=O)C=C1 PEEHTFAAVSWFBL-UHFFFAOYSA-N 0.000 description 10
- 241000699666 Mus <mouse, genus> Species 0.000 description 10
- 239000000090 biomarker Substances 0.000 description 10
- 239000000872 buffer Substances 0.000 description 10
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 10
- 230000004048 modification Effects 0.000 description 10
- 238000012986 modification Methods 0.000 description 10
- 238000007254 oxidation reaction Methods 0.000 description 10
- 239000002953 phosphate buffered saline Substances 0.000 description 10
- 229920001223 polyethylene glycol Polymers 0.000 description 10
- 230000002829 reductive effect Effects 0.000 description 10
- 238000011160 research Methods 0.000 description 10
- 230000001225 therapeutic effect Effects 0.000 description 10
- 238000011282 treatment Methods 0.000 description 10
- 239000004472 Lysine Substances 0.000 description 9
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 9
- 241000283973 Oryctolagus cuniculus Species 0.000 description 9
- 230000004071 biological effect Effects 0.000 description 9
- 230000018109 developmental process Effects 0.000 description 9
- 238000001962 electrophoresis Methods 0.000 description 9
- 102000052116 epidermal growth factor receptor activity proteins Human genes 0.000 description 9
- 108700015053 epidermal growth factor receptor activity proteins Proteins 0.000 description 9
- 150000002148 esters Chemical class 0.000 description 9
- 238000002474 experimental method Methods 0.000 description 9
- 239000000463 material Substances 0.000 description 9
- 230000003647 oxidation Effects 0.000 description 9
- 238000000163 radioactive labelling Methods 0.000 description 9
- 150000003839 salts Chemical class 0.000 description 9
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 8
- DJQYYYCQOZMCRC-UHFFFAOYSA-N 2-aminopropane-1,3-dithiol Chemical group SCC(N)CS DJQYYYCQOZMCRC-UHFFFAOYSA-N 0.000 description 8
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 8
- 102000004190 Enzymes Human genes 0.000 description 8
- 108090000790 Enzymes Proteins 0.000 description 8
- 241000588724 Escherichia coli Species 0.000 description 8
- XEKOWRVHYACXOJ-UHFFFAOYSA-N Ethyl acetate Chemical compound CCOC(C)=O XEKOWRVHYACXOJ-UHFFFAOYSA-N 0.000 description 8
- 108010087819 Fc receptors Proteins 0.000 description 8
- 102000009109 Fc receptors Human genes 0.000 description 8
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 8
- 239000002253 acid Substances 0.000 description 8
- 238000007413 biotinylation Methods 0.000 description 8
- 230000006287 biotinylation Effects 0.000 description 8
- 229940098773 bovine serum albumin Drugs 0.000 description 8
- 239000002738 chelating agent Substances 0.000 description 8
- 239000003795 chemical substances by application Substances 0.000 description 8
- 210000000349 chromosome Anatomy 0.000 description 8
- 238000010276 construction Methods 0.000 description 8
- 239000012636 effector Substances 0.000 description 8
- 229940088598 enzyme Drugs 0.000 description 8
- 239000002609 medium Substances 0.000 description 8
- YOHYSYJDKVYCJI-UHFFFAOYSA-N n-[3-[[6-[3-(trifluoromethyl)anilino]pyrimidin-4-yl]amino]phenyl]cyclopropanecarboxamide Chemical compound FC(F)(F)C1=CC=CC(NC=2N=CN=C(NC=3C=C(NC(=O)C4CC4)C=CC=3)C=2)=C1 YOHYSYJDKVYCJI-UHFFFAOYSA-N 0.000 description 8
- 238000012636 positron electron tomography Methods 0.000 description 8
- QCWXUUIWCKQGHC-YPZZEJLDSA-N zirconium-89 Chemical compound [89Zr] QCWXUUIWCKQGHC-YPZZEJLDSA-N 0.000 description 8
- WXTMDXOMEHJXQO-UHFFFAOYSA-N 2,5-dihydroxybenzoic acid Chemical compound OC(=O)C1=CC(O)=CC=C1O WXTMDXOMEHJXQO-UHFFFAOYSA-N 0.000 description 7
- 102000000844 Cell Surface Receptors Human genes 0.000 description 7
- 108010001857 Cell Surface Receptors Proteins 0.000 description 7
- JGFZNNIVVJXRND-UHFFFAOYSA-N N,N-Diisopropylethylamine (DIPEA) Chemical compound CCN(C(C)C)C(C)C JGFZNNIVVJXRND-UHFFFAOYSA-N 0.000 description 7
- 108700020796 Oncogene Proteins 0.000 description 7
- 229920002684 Sepharose Polymers 0.000 description 7
- 239000004480 active ingredient Substances 0.000 description 7
- 125000003277 amino group Chemical group 0.000 description 7
- 238000013459 approach Methods 0.000 description 7
- 230000001588 bifunctional effect Effects 0.000 description 7
- 239000013078 crystal Substances 0.000 description 7
- 238000000338 in vitro Methods 0.000 description 7
- 238000004949 mass spectrometry Methods 0.000 description 7
- 239000003068 molecular probe Substances 0.000 description 7
- 229910052757 nitrogen Inorganic materials 0.000 description 7
- 229920002401 polyacrylamide Polymers 0.000 description 7
- 238000012546 transfer Methods 0.000 description 7
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 7
- 101710132601 Capsid protein Proteins 0.000 description 6
- 101710094648 Coat protein Proteins 0.000 description 6
- MHAJPDPJQMAIIY-UHFFFAOYSA-N Hydrogen peroxide Chemical compound OO MHAJPDPJQMAIIY-UHFFFAOYSA-N 0.000 description 6
- 108010058683 Immobilized Proteins Proteins 0.000 description 6
- 101710125418 Major capsid protein Proteins 0.000 description 6
- 101710141454 Nucleoprotein Proteins 0.000 description 6
- 101710083689 Probable capsid protein Proteins 0.000 description 6
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical compound [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 6
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Chemical compound CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 6
- 238000007792 addition Methods 0.000 description 6
- 239000011324 bead Substances 0.000 description 6
- 238000012512 characterization method Methods 0.000 description 6
- 238000002591 computed tomography Methods 0.000 description 6
- 238000009826 distribution Methods 0.000 description 6
- 238000009472 formulation Methods 0.000 description 6
- 238000002523 gelfiltration Methods 0.000 description 6
- 229940072221 immunoglobulins Drugs 0.000 description 6
- 229910052751 metal Inorganic materials 0.000 description 6
- 239000002184 metal Substances 0.000 description 6
- 210000000822 natural killer cell Anatomy 0.000 description 6
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 6
- 238000002823 phage display Methods 0.000 description 6
- 238000012552 review Methods 0.000 description 6
- 238000002741 site-directed mutagenesis Methods 0.000 description 6
- 239000001632 sodium acetate Substances 0.000 description 6
- 235000017281 sodium acetate Nutrition 0.000 description 6
- 239000011780 sodium chloride Substances 0.000 description 6
- 239000007787 solid Substances 0.000 description 6
- 239000011550 stock solution Substances 0.000 description 6
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 5
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 5
- 102000004506 Blood Proteins Human genes 0.000 description 5
- 108010017384 Blood Proteins Proteins 0.000 description 5
- 206010006187 Breast cancer Diseases 0.000 description 5
- 206010009944 Colon cancer Diseases 0.000 description 5
- YMWUJEATGCHHMB-UHFFFAOYSA-N Dichloromethane Chemical compound ClCCl YMWUJEATGCHHMB-UHFFFAOYSA-N 0.000 description 5
- XTHFKEDIFFGKHM-UHFFFAOYSA-N Dimethoxyethane Chemical compound COCCOC XTHFKEDIFFGKHM-UHFFFAOYSA-N 0.000 description 5
- 238000012286 ELISA Assay Methods 0.000 description 5
- 239000004471 Glycine Substances 0.000 description 5
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 5
- 241001529936 Murinae Species 0.000 description 5
- QPCDCPDFJACHGM-UHFFFAOYSA-N N,N-bis{2-[bis(carboxymethyl)amino]ethyl}glycine Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(=O)O)CCN(CC(O)=O)CC(O)=O QPCDCPDFJACHGM-UHFFFAOYSA-N 0.000 description 5
- 239000000611 antibody drug conjugate Substances 0.000 description 5
- 229940127089 cytotoxic agent Drugs 0.000 description 5
- 238000013461 design Methods 0.000 description 5
- 230000004927 fusion Effects 0.000 description 5
- 229940127121 immunoconjugate Drugs 0.000 description 5
- 238000011534 incubation Methods 0.000 description 5
- 230000004807 localization Effects 0.000 description 5
- 238000009206 nuclear medicine Methods 0.000 description 5
- 238000001556 precipitation Methods 0.000 description 5
- 241000894007 species Species 0.000 description 5
- JJAHTWIKCUJRDK-UHFFFAOYSA-N succinimidyl 4-(N-maleimidomethyl)cyclohexane-1-carboxylate Chemical compound C1CC(CN2C(C=CC2=O)=O)CCC1C(=O)ON1C(=O)CCC1=O JJAHTWIKCUJRDK-UHFFFAOYSA-N 0.000 description 5
- 239000000725 suspension Substances 0.000 description 5
- 238000003786 synthesis reaction Methods 0.000 description 5
- 238000012360 testing method Methods 0.000 description 5
- DTQVDTLACAAQTR-UHFFFAOYSA-N trifluoroacetic acid Substances OC(=O)C(F)(F)F DTQVDTLACAAQTR-UHFFFAOYSA-N 0.000 description 5
- ASOKPJOREAFHNY-UHFFFAOYSA-N 1-Hydroxybenzotriazole Chemical compound C1=CC=C2N(O)N=NC2=C1 ASOKPJOREAFHNY-UHFFFAOYSA-N 0.000 description 4
- VHYFNPMBLIVWCW-UHFFFAOYSA-N 4-Dimethylaminopyridine Chemical compound CN(C)C1=CC=NC=C1 VHYFNPMBLIVWCW-UHFFFAOYSA-N 0.000 description 4
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 4
- 208000026310 Breast neoplasm Diseases 0.000 description 4
- 0 CC(CC(N1CC(CC2)CCC2C(NCCCCCN(*(CCC(NCCCCCN(C(CCC(NCCCCCN(C(C)=O)O)=O)=O)O)=O)=O)O)=O)=O)C1=O Chemical compound CC(CC(N1CC(CC2)CCC2C(NCCCCCN(*(CCC(NCCCCCN(C(CCC(NCCCCCN(C(C)=O)O)=O)=O)O)=O)=O)O)=O)=O)C1=O 0.000 description 4
- QOSSAOTZNIDXMA-UHFFFAOYSA-N Dicylcohexylcarbodiimide Chemical compound C1CCCCC1N=C=NC1CCCCC1 QOSSAOTZNIDXMA-UHFFFAOYSA-N 0.000 description 4
- GKQLYSROISKDLL-UHFFFAOYSA-N EEDQ Chemical compound C1=CC=C2N(C(=O)OCC)C(OCC)C=CC2=C1 GKQLYSROISKDLL-UHFFFAOYSA-N 0.000 description 4
- 102100031968 Ephrin type-B receptor 2 Human genes 0.000 description 4
- 241001524679 Escherichia virus M13 Species 0.000 description 4
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 4
- FXHOOIRPVKKKFG-UHFFFAOYSA-N N,N-Dimethylacetamide Chemical compound CN(C)C(C)=O FXHOOIRPVKKKFG-UHFFFAOYSA-N 0.000 description 4
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 description 4
- 229910019142 PO4 Inorganic materials 0.000 description 4
- 241000700159 Rattus Species 0.000 description 4
- 102100029981 Receptor tyrosine-protein kinase erbB-4 Human genes 0.000 description 4
- 101710100963 Receptor tyrosine-protein kinase erbB-4 Proteins 0.000 description 4
- CDBYLPFSWZWCQE-UHFFFAOYSA-L Sodium Carbonate Chemical compound [Na+].[Na+].[O-]C([O-])=O CDBYLPFSWZWCQE-UHFFFAOYSA-L 0.000 description 4
- 239000007983 Tris buffer Substances 0.000 description 4
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 4
- 230000003213 activating effect Effects 0.000 description 4
- 150000001408 amides Chemical class 0.000 description 4
- 150000001412 amines Chemical class 0.000 description 4
- 230000001188 anti-phage Effects 0.000 description 4
- 238000004422 calculation algorithm Methods 0.000 description 4
- 230000008859 change Effects 0.000 description 4
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 4
- 230000001268 conjugating effect Effects 0.000 description 4
- 238000007796 conventional method Methods 0.000 description 4
- 231100000433 cytotoxic Toxicity 0.000 description 4
- 239000002254 cytotoxic agent Substances 0.000 description 4
- 230000001472 cytotoxic effect Effects 0.000 description 4
- 230000029087 digestion Effects 0.000 description 4
- 239000000539 dimer Substances 0.000 description 4
- 239000002552 dosage form Substances 0.000 description 4
- 238000002330 electrospray ionisation mass spectrometry Methods 0.000 description 4
- 230000029142 excretion Effects 0.000 description 4
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 4
- 230000036541 health Effects 0.000 description 4
- 239000008241 heterogeneous mixture Substances 0.000 description 4
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 4
- 230000002998 immunogenetic effect Effects 0.000 description 4
- 238000011065 in-situ storage Methods 0.000 description 4
- PGLTVOMIXTUURA-UHFFFAOYSA-N iodoacetamide Chemical compound NC(=O)CI PGLTVOMIXTUURA-UHFFFAOYSA-N 0.000 description 4
- 125000005439 maleimidyl group Chemical group C1(C=CC(N1*)=O)=O 0.000 description 4
- 239000003550 marker Substances 0.000 description 4
- 230000001404 mediated effect Effects 0.000 description 4
- 239000003094 microcapsule Substances 0.000 description 4
- HBAYEVATSBINBX-UHFFFAOYSA-N n-[5-[acetyl(hydroxy)amino]pentyl]-n'-hydroxy-n'-[5-[[4-[hydroxy-[5-[(4-isothiocyanatophenyl)carbamothioylamino]pentyl]amino]-4-oxobutanoyl]amino]pentyl]butanediamide Chemical compound CC(=O)N(O)CCCCCNC(=O)CCC(=O)N(O)CCCCCNC(=O)CCC(=O)N(O)CCCCCNC(=S)NC1=CC=C(N=C=S)C=C1 HBAYEVATSBINBX-UHFFFAOYSA-N 0.000 description 4
- 230000036961 partial effect Effects 0.000 description 4
- 239000000546 pharmaceutical excipient Substances 0.000 description 4
- 230000003285 pharmacodynamic effect Effects 0.000 description 4
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 4
- 239000010452 phosphate Substances 0.000 description 4
- 230000008569 process Effects 0.000 description 4
- QELSKZZBTMNZEB-UHFFFAOYSA-N propylparaben Chemical compound CCCOC(=O)C1=CC=C(O)C=C1 QELSKZZBTMNZEB-UHFFFAOYSA-N 0.000 description 4
- 230000009145 protein modification Effects 0.000 description 4
- 238000011002 quantification Methods 0.000 description 4
- 230000005855 radiation Effects 0.000 description 4
- 230000035945 sensitivity Effects 0.000 description 4
- 239000007790 solid phase Substances 0.000 description 4
- 239000000375 suspending agent Substances 0.000 description 4
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 4
- 210000004881 tumor cell Anatomy 0.000 description 4
- 241001515965 unidentified phage Species 0.000 description 4
- 239000013598 vector Substances 0.000 description 4
- BDNKZNFMNDZQMI-UHFFFAOYSA-N 1,3-diisopropylcarbodiimide Chemical compound CC(C)N=C=NC(C)C BDNKZNFMNDZQMI-UHFFFAOYSA-N 0.000 description 3
- VOXZDWNPVJITMN-ZBRFXRBCSA-N 17β-estradiol Chemical compound OC1=CC=C2[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CCC2=C1 VOXZDWNPVJITMN-ZBRFXRBCSA-N 0.000 description 3
- 101100067974 Arabidopsis thaliana POP2 gene Proteins 0.000 description 3
- WVDDGKGOMKODPV-UHFFFAOYSA-N Benzyl alcohol Chemical compound OCC1=CC=CC=C1 WVDDGKGOMKODPV-UHFFFAOYSA-N 0.000 description 3
- 241000282693 Cercopithecidae Species 0.000 description 3
- KRKNYBCHXYNGOX-UHFFFAOYSA-K Citrate Chemical compound [O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O KRKNYBCHXYNGOX-UHFFFAOYSA-K 0.000 description 3
- 108020004705 Codon Proteins 0.000 description 3
- 241000699802 Cricetulus griseus Species 0.000 description 3
- 102100025698 Cytosolic carboxypeptidase 4 Human genes 0.000 description 3
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 3
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- 102100027285 Fanconi anemia group B protein Human genes 0.000 description 3
- 241000724791 Filamentous phage Species 0.000 description 3
- 101000932590 Homo sapiens Cytosolic carboxypeptidase 4 Proteins 0.000 description 3
- 101100118549 Homo sapiens EGFR gene Proteins 0.000 description 3
- 101001064462 Homo sapiens Ephrin type-B receptor 2 Proteins 0.000 description 3
- 108010073807 IgG Receptors Proteins 0.000 description 3
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 3
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 3
- 102100029185 Low affinity immunoglobulin gamma Fc region receptor III-B Human genes 0.000 description 3
- 241000699660 Mus musculus Species 0.000 description 3
- 101001033003 Mus musculus Granzyme F Proteins 0.000 description 3
- 108010021466 Mutant Proteins Proteins 0.000 description 3
- 102000008300 Mutant Proteins Human genes 0.000 description 3
- 108091005804 Peptidases Proteins 0.000 description 3
- 102000035195 Peptidases Human genes 0.000 description 3
- 101710100969 Receptor tyrosine-protein kinase erbB-3 Proteins 0.000 description 3
- 102100029986 Receptor tyrosine-protein kinase erbB-3 Human genes 0.000 description 3
- 101100123851 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) HER1 gene Proteins 0.000 description 3
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 3
- PZBFGYYEXUXCOF-UHFFFAOYSA-N TCEP Chemical compound OC(=O)CCP(CCC(O)=O)CCC(O)=O PZBFGYYEXUXCOF-UHFFFAOYSA-N 0.000 description 3
- WDLRUFUQRNWCPK-UHFFFAOYSA-N Tetraxetan Chemical compound OC(=O)CN1CCN(CC(O)=O)CCN(CC(O)=O)CCN(CC(O)=O)CC1 WDLRUFUQRNWCPK-UHFFFAOYSA-N 0.000 description 3
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 3
- 239000004473 Threonine Substances 0.000 description 3
- 238000002835 absorbance Methods 0.000 description 3
- 239000008186 active pharmaceutical agent Substances 0.000 description 3
- 238000010171 animal model Methods 0.000 description 3
- 229940049595 antibody-drug conjugate Drugs 0.000 description 3
- 230000006907 apoptotic process Effects 0.000 description 3
- 239000007900 aqueous suspension Substances 0.000 description 3
- 125000003118 aryl group Chemical group 0.000 description 3
- 230000001580 bacterial effect Effects 0.000 description 3
- 230000008901 benefit Effects 0.000 description 3
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 3
- 238000004113 cell culture Methods 0.000 description 3
- 230000004663 cell proliferation Effects 0.000 description 3
- 239000013522 chelant Substances 0.000 description 3
- 238000007385 chemical modification Methods 0.000 description 3
- 239000003638 chemical reducing agent Substances 0.000 description 3
- 238000004587 chromatography analysis Methods 0.000 description 3
- 239000003593 chromogenic compound Substances 0.000 description 3
- 229920001577 copolymer Polymers 0.000 description 3
- 239000010949 copper Substances 0.000 description 3
- 238000004132 cross linking Methods 0.000 description 3
- 239000003431 cross linking reagent Substances 0.000 description 3
- 238000012217 deletion Methods 0.000 description 3
- 230000037430 deletion Effects 0.000 description 3
- 238000011033 desalting Methods 0.000 description 3
- 235000014113 dietary fatty acids Nutrition 0.000 description 3
- BGRWYRAHAFMIBJ-UHFFFAOYSA-N diisopropylcarbodiimide Natural products CC(C)NC(=O)NC(C)C BGRWYRAHAFMIBJ-UHFFFAOYSA-N 0.000 description 3
- 239000003085 diluting agent Substances 0.000 description 3
- 150000002019 disulfides Chemical class 0.000 description 3
- 239000000975 dye Substances 0.000 description 3
- 235000019439 ethyl acetate Nutrition 0.000 description 3
- 238000011156 evaluation Methods 0.000 description 3
- 239000000194 fatty acid Substances 0.000 description 3
- 229930195729 fatty acid Natural products 0.000 description 3
- 150000004665 fatty acids Chemical class 0.000 description 3
- 238000000855 fermentation Methods 0.000 description 3
- 230000004151 fermentation Effects 0.000 description 3
- 238000007429 general method Methods 0.000 description 3
- 229960005219 gentisic acid Drugs 0.000 description 3
- 238000004128 high performance liquid chromatography Methods 0.000 description 3
- 238000013537 high throughput screening Methods 0.000 description 3
- 238000003780 insertion Methods 0.000 description 3
- 230000037431 insertion Effects 0.000 description 3
- XMBWDFGMSWQBCA-OIOBTWANSA-N iodane Chemical compound [124IH] XMBWDFGMSWQBCA-OIOBTWANSA-N 0.000 description 3
- 150000002500 ions Chemical class 0.000 description 3
- 229910052742 iron Inorganic materials 0.000 description 3
- 239000007788 liquid Substances 0.000 description 3
- 238000004811 liquid chromatography Methods 0.000 description 3
- 238000004895 liquid chromatography mass spectrometry Methods 0.000 description 3
- 238000011068 loading method Methods 0.000 description 3
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 3
- 210000004962 mammalian cell Anatomy 0.000 description 3
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 3
- 231100000252 nontoxic Toxicity 0.000 description 3
- 230000003000 nontoxic effect Effects 0.000 description 3
- 102000039446 nucleic acids Human genes 0.000 description 3
- 108020004707 nucleic acids Proteins 0.000 description 3
- 238000011580 nude mouse model Methods 0.000 description 3
- 210000001672 ovary Anatomy 0.000 description 3
- 239000007800 oxidant agent Substances 0.000 description 3
- 239000008194 pharmaceutical composition Substances 0.000 description 3
- 150000008300 phosphoramidites Chemical class 0.000 description 3
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 3
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 3
- 238000011363 radioimmunotherapy Methods 0.000 description 3
- 239000000376 reactant Substances 0.000 description 3
- 239000011541 reaction mixture Substances 0.000 description 3
- 230000009467 reduction Effects 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 230000004044 response Effects 0.000 description 3
- PYWVYCXTNDRMGF-UHFFFAOYSA-N rhodamine B Chemical compound [Cl-].C=12C=CC(=[N+](CC)CC)C=C2OC2=CC(N(CC)CC)=CC=C2C=1C1=CC=CC=C1C(O)=O PYWVYCXTNDRMGF-UHFFFAOYSA-N 0.000 description 3
- 102220206292 rs1057521851 Human genes 0.000 description 3
- 239000007974 sodium acetate buffer Substances 0.000 description 3
- 229940074404 sodium succinate Drugs 0.000 description 3
- ZDQYSKICYIVCPN-UHFFFAOYSA-L sodium succinate (anhydrous) Chemical compound [Na+].[Na+].[O-]C(=O)CCC([O-])=O ZDQYSKICYIVCPN-UHFFFAOYSA-L 0.000 description 3
- 239000007858 starting material Substances 0.000 description 3
- 239000008362 succinate buffer Substances 0.000 description 3
- KDYFGRWQOYBRFD-UHFFFAOYSA-L succinate(2-) Chemical compound [O-]C(=O)CCC([O-])=O KDYFGRWQOYBRFD-UHFFFAOYSA-L 0.000 description 3
- UMGDCJDMYOKAJW-UHFFFAOYSA-N thiourea Chemical compound NC(N)=S UMGDCJDMYOKAJW-UHFFFAOYSA-N 0.000 description 3
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 3
- 239000004474 valine Substances 0.000 description 3
- 230000003612 virological effect Effects 0.000 description 3
- 150000003754 zirconium Chemical class 0.000 description 3
- PUPZLCDOIYMWBV-UHFFFAOYSA-N (+/-)-1,3-Butanediol Chemical compound CC(O)CCO PUPZLCDOIYMWBV-UHFFFAOYSA-N 0.000 description 2
- VRDGQQTWSGDXCU-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 2-iodoacetate Chemical compound ICC(=O)ON1C(=O)CCC1=O VRDGQQTWSGDXCU-UHFFFAOYSA-N 0.000 description 2
- GEYOCULIXLDCMW-UHFFFAOYSA-N 1,2-phenylenediamine Chemical compound NC1=CC=CC=C1N GEYOCULIXLDCMW-UHFFFAOYSA-N 0.000 description 2
- XQUPVDVFXZDTLT-UHFFFAOYSA-N 1-[4-[[4-(2,5-dioxopyrrol-1-yl)phenyl]methyl]phenyl]pyrrole-2,5-dione Chemical compound O=C1C=CC(=O)N1C(C=C1)=CC=C1CC1=CC=C(N2C(C=CC2=O)=O)C=C1 XQUPVDVFXZDTLT-UHFFFAOYSA-N 0.000 description 2
- HZAXFHJVJLSVMW-UHFFFAOYSA-N 2-Aminoethan-1-ol Chemical compound NCCO HZAXFHJVJLSVMW-UHFFFAOYSA-N 0.000 description 2
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 2
- FPQQSJJWHUJYPU-UHFFFAOYSA-N 3-(dimethylamino)propyliminomethylidene-ethylazanium;chloride Chemical compound Cl.CCN=C=NCCCN(C)C FPQQSJJWHUJYPU-UHFFFAOYSA-N 0.000 description 2
- PBVAJRFEEOIAGW-UHFFFAOYSA-N 3-[bis(2-carboxyethyl)phosphanyl]propanoic acid;hydrochloride Chemical compound Cl.OC(=O)CCP(CCC(O)=O)CCC(O)=O PBVAJRFEEOIAGW-UHFFFAOYSA-N 0.000 description 2
- SFYDWLYPIXHPML-UHFFFAOYSA-N 3-nitro-1-(2,4,6-trimethylphenyl)sulfonyl-1,2,4-triazole Chemical compound CC1=CC(C)=CC(C)=C1S(=O)(=O)N1N=C([N+]([O-])=O)N=C1 SFYDWLYPIXHPML-UHFFFAOYSA-N 0.000 description 2
- ZCYVEMRRCGMTRW-UHFFFAOYSA-N 7553-56-2 Chemical compound [I] ZCYVEMRRCGMTRW-UHFFFAOYSA-N 0.000 description 2
- HBAQYPYDRFILMT-UHFFFAOYSA-N 8-[3-(1-cyclopropylpyrazol-4-yl)-1H-pyrazolo[4,3-d]pyrimidin-5-yl]-3-methyl-3,8-diazabicyclo[3.2.1]octan-2-one Chemical class C1(CC1)N1N=CC(=C1)C1=NNC2=C1N=C(N=C2)N1C2C(N(CC1CC2)C)=O HBAQYPYDRFILMT-UHFFFAOYSA-N 0.000 description 2
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 2
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 2
- 108090000672 Annexin A5 Proteins 0.000 description 2
- 102000004121 Annexin A5 Human genes 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- 102100032312 Brevican core protein Human genes 0.000 description 2
- CPELXLSAUQHCOX-UHFFFAOYSA-M Bromide Chemical compound [Br-] CPELXLSAUQHCOX-UHFFFAOYSA-M 0.000 description 2
- PJUNKUSWDHNJGX-UHFFFAOYSA-N CC(CC(N1CC(CC2)CCC2C(NCCCCCN(C(CCC(NCCCCCN(C(CCC(NCCCCCN(C(C)=O)O)=O)=O)O)=O)=O)O)=O)=O)C1=O Chemical compound CC(CC(N1CC(CC2)CCC2C(NCCCCCN(C(CCC(NCCCCCN(C(CCC(NCCCCCN(C(C)=O)O)=O)=O)O)=O)=O)O)=O)=O)C1=O PJUNKUSWDHNJGX-UHFFFAOYSA-N 0.000 description 2
- UGEVPPINYSCSQY-UHFFFAOYSA-N CCC(NCCCCCN(C(CCC(NCCCCCN(C(CCC(NCCCCCN(C(C)=O)O)=O)=O)O)=O)=O)O)=O Chemical compound CCC(NCCCCCN(C(CCC(NCCCCCN(C(CCC(NCCCCCN(C(C)=O)O)=O)=O)O)=O)=O)O)=O UGEVPPINYSCSQY-UHFFFAOYSA-N 0.000 description 2
- 241000283707 Capra Species 0.000 description 2
- 102100020743 Dipeptidase 1 Human genes 0.000 description 2
- AOJJSUZBOXZQNB-TZSSRYMLSA-N Doxorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)[C@H](O)[C@H](C)O1 AOJJSUZBOXZQNB-TZSSRYMLSA-N 0.000 description 2
- IAYPIBMASNFSPL-UHFFFAOYSA-N Ethylene oxide Chemical compound C1CO1 IAYPIBMASNFSPL-UHFFFAOYSA-N 0.000 description 2
- 101150064015 FAS gene Proteins 0.000 description 2
- 108010021468 Fc gamma receptor IIA Proteins 0.000 description 2
- 108010021472 Fc gamma receptor IIB Proteins 0.000 description 2
- VTLYFUHAOXGGBS-UHFFFAOYSA-N Fe3+ Chemical class [Fe+3] VTLYFUHAOXGGBS-UHFFFAOYSA-N 0.000 description 2
- GYHNNYVSQQEPJS-YPZZEJLDSA-N Gallium-68 Chemical compound [68Ga] GYHNNYVSQQEPJS-YPZZEJLDSA-N 0.000 description 2
- 108010010803 Gelatin Proteins 0.000 description 2
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 2
- 239000007821 HATU Substances 0.000 description 2
- 239000007995 HEPES buffer Substances 0.000 description 2
- 101000851181 Homo sapiens Epidermal growth factor receptor Proteins 0.000 description 2
- OAKJQQAXSVQMHS-UHFFFAOYSA-N Hydrazine Chemical compound NN OAKJQQAXSVQMHS-UHFFFAOYSA-N 0.000 description 2
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 2
- WTDRDQBEARUVNC-UHFFFAOYSA-N L-Dopa Natural products OC(=O)C(N)CC1=CC=C(O)C(O)=C1 WTDRDQBEARUVNC-UHFFFAOYSA-N 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 2
- 102100029205 Low affinity immunoglobulin gamma Fc region receptor II-b Human genes 0.000 description 2
- AFVFQIVMOAPDHO-UHFFFAOYSA-N Methanesulfonic acid Chemical compound CS(O)(=O)=O AFVFQIVMOAPDHO-UHFFFAOYSA-N 0.000 description 2
- 101100335081 Mus musculus Flt3 gene Proteins 0.000 description 2
- NQTADLQHYWFPDB-UHFFFAOYSA-N N-Hydroxysuccinimide Chemical compound ON1C(=O)CCC1=O NQTADLQHYWFPDB-UHFFFAOYSA-N 0.000 description 2
- RHGKLRLOHDJJDR-UHFFFAOYSA-N Ndelta-carbamoyl-DL-ornithine Natural products OC(=O)C(N)CCCNC(N)=O RHGKLRLOHDJJDR-UHFFFAOYSA-N 0.000 description 2
- 206010029260 Neuroblastoma Diseases 0.000 description 2
- 108091005461 Nucleic proteins Chemical group 0.000 description 2
- 102000004316 Oxidoreductases Human genes 0.000 description 2
- 108090000854 Oxidoreductases Proteins 0.000 description 2
- 108010067902 Peptide Library Proteins 0.000 description 2
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Natural products OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 2
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 2
- 239000004793 Polystyrene Substances 0.000 description 2
- 101710193132 Pre-hexon-linking protein VIII Proteins 0.000 description 2
- 239000004365 Protease Substances 0.000 description 2
- 101001010820 Rattus norvegicus Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 2
- 241000283984 Rodentia Species 0.000 description 2
- 108010071390 Serum Albumin Proteins 0.000 description 2
- 102000007562 Serum Albumin Human genes 0.000 description 2
- 239000000589 Siderophore Substances 0.000 description 2
- 229930006000 Sucrose Natural products 0.000 description 2
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 2
- 102000004142 Trypsin Human genes 0.000 description 2
- 108090000631 Trypsin Proteins 0.000 description 2
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Natural products NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 2
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 2
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 2
- 125000002777 acetyl group Chemical group [H]C([H])([H])C(*)=O 0.000 description 2
- 150000007513 acids Chemical class 0.000 description 2
- 238000001042 affinity chromatography Methods 0.000 description 2
- 239000003570 air Substances 0.000 description 2
- 101150115889 al gene Proteins 0.000 description 2
- 150000001299 aldehydes Chemical class 0.000 description 2
- 125000000217 alkyl group Chemical group 0.000 description 2
- 230000029936 alkylation Effects 0.000 description 2
- 238000005804 alkylation reaction Methods 0.000 description 2
- 230000001093 anti-cancer Effects 0.000 description 2
- 230000002494 anti-cea effect Effects 0.000 description 2
- 230000000259 anti-tumor effect Effects 0.000 description 2
- 230000000890 antigenic effect Effects 0.000 description 2
- 239000007864 aqueous solution Substances 0.000 description 2
- 235000010323 ascorbic acid Nutrition 0.000 description 2
- 239000011668 ascorbic acid Substances 0.000 description 2
- WPYMKLBDIGXBTP-UHFFFAOYSA-N benzoic acid Chemical compound OC(=O)C1=CC=CC=C1 WPYMKLBDIGXBTP-UHFFFAOYSA-N 0.000 description 2
- 230000029918 bioluminescence Effects 0.000 description 2
- 238000005415 bioluminescence Methods 0.000 description 2
- 238000001815 biotherapy Methods 0.000 description 2
- 150000001720 carbohydrates Chemical class 0.000 description 2
- 235000014633 carbohydrates Nutrition 0.000 description 2
- 238000012219 cassette mutagenesis Methods 0.000 description 2
- YCIMNLLNPGFGHC-UHFFFAOYSA-N catechol Chemical compound OC1=CC=CC=C1O YCIMNLLNPGFGHC-UHFFFAOYSA-N 0.000 description 2
- 230000030833 cell death Effects 0.000 description 2
- 230000024245 cell differentiation Effects 0.000 description 2
- 230000006037 cell lysis Effects 0.000 description 2
- 229960005395 cetuximab Drugs 0.000 description 2
- 239000011248 coating agent Substances 0.000 description 2
- 238000000576 coating method Methods 0.000 description 2
- 230000024203 complement activation Effects 0.000 description 2
- 230000000295 complement effect Effects 0.000 description 2
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 2
- 238000004590 computer program Methods 0.000 description 2
- 239000007859 condensation product Substances 0.000 description 2
- 238000011109 contamination Methods 0.000 description 2
- 229910000365 copper sulfate Inorganic materials 0.000 description 2
- ARUVKPQLZAKDPS-UHFFFAOYSA-L copper(II) sulfate Chemical compound [Cu+2].[O-][S+2]([O-])([O-])[O-] ARUVKPQLZAKDPS-UHFFFAOYSA-L 0.000 description 2
- 230000008878 coupling Effects 0.000 description 2
- 238000010168 coupling process Methods 0.000 description 2
- 238000005859 coupling reaction Methods 0.000 description 2
- 238000005520 cutting process Methods 0.000 description 2
- NZNMSOFKMUBTKW-UHFFFAOYSA-N cyclohexanecarboxylic acid Chemical compound OC(=O)C1CCCCC1 NZNMSOFKMUBTKW-UHFFFAOYSA-N 0.000 description 2
- 231100000599 cytotoxic agent Toxicity 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- FAMRKDQNMBBFBR-BQYQJAHWSA-N diethyl azodicarboxylate Substances CCOC(=O)\N=N\C(=O)OCC FAMRKDQNMBBFBR-BQYQJAHWSA-N 0.000 description 2
- HPNMFZURTQLUMO-UHFFFAOYSA-N diethylamine Chemical compound CCNCC HPNMFZURTQLUMO-UHFFFAOYSA-N 0.000 description 2
- 229960005156 digoxin Drugs 0.000 description 2
- MHUWZNTUIIFHAS-CLFAGFIQSA-N dioleoyl phosphatidic acid Chemical compound CCCCCCCC\C=C/CCCCCCCC(=O)OCC(COP(O)(O)=O)OC(=O)CCCCCCC\C=C/CCCCCCCC MHUWZNTUIIFHAS-CLFAGFIQSA-N 0.000 description 2
- 201000010099 disease Diseases 0.000 description 2
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 2
- 125000002228 disulfide group Chemical group 0.000 description 2
- 238000000132 electrospray ionisation Methods 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 239000013604 expression vector Substances 0.000 description 2
- 238000001914 filtration Methods 0.000 description 2
- 108020001507 fusion proteins Proteins 0.000 description 2
- 102000037865 fusion proteins Human genes 0.000 description 2
- 238000001641 gel filtration chromatography Methods 0.000 description 2
- 239000008273 gelatin Substances 0.000 description 2
- 229920000159 gelatin Polymers 0.000 description 2
- 235000019322 gelatine Nutrition 0.000 description 2
- 235000011852 gelatine desserts Nutrition 0.000 description 2
- 239000011521 glass Substances 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 2
- 150000004820 halides Chemical class 0.000 description 2
- 125000005179 haloacetyl group Chemical group 0.000 description 2
- 210000004408 hybridoma Anatomy 0.000 description 2
- XMBWDFGMSWQBCA-UHFFFAOYSA-N hydrogen iodide Chemical compound I XMBWDFGMSWQBCA-UHFFFAOYSA-N 0.000 description 2
- NPZTUJOABDZTLV-UHFFFAOYSA-N hydroxybenzotriazole Substances O=C1C=CC=C2NNN=C12 NPZTUJOABDZTLV-UHFFFAOYSA-N 0.000 description 2
- 229960001001 ibritumomab tiuxetan Drugs 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 239000012535 impurity Substances 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 208000015181 infectious disease Diseases 0.000 description 2
- 230000002458 infectious effect Effects 0.000 description 2
- 238000001802 infusion Methods 0.000 description 2
- 108091008042 inhibitory receptors Proteins 0.000 description 2
- 238000001990 intravenous administration Methods 0.000 description 2
- 239000011630 iodine Substances 0.000 description 2
- 229910052740 iodine Inorganic materials 0.000 description 2
- 150000002540 isothiocyanates Chemical class 0.000 description 2
- 150000002576 ketones Chemical class 0.000 description 2
- 210000000265 leukocyte Anatomy 0.000 description 2
- RGLRXNKKBLIBQS-XNHQSDQCSA-N leuprolide acetate Chemical compound CC(O)=O.CCNC(=O)[C@@H]1CCCN1C(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](CC(C)C)NC(=O)[C@@H](NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H]1NC(=O)CC1)CC1=CC=C(O)C=C1 RGLRXNKKBLIBQS-XNHQSDQCSA-N 0.000 description 2
- 229960004502 levodopa Drugs 0.000 description 2
- 108020001756 ligand binding domains Proteins 0.000 description 2
- 238000001972 liquid chromatography-electrospray ionisation mass spectrometry Methods 0.000 description 2
- 238000002595 magnetic resonance imaging Methods 0.000 description 2
- 230000003211 malignant effect Effects 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- 239000002207 metabolite Substances 0.000 description 2
- 229930182817 methionine Natural products 0.000 description 2
- 230000003020 moisturizing effect Effects 0.000 description 2
- 238000010369 molecular cloning Methods 0.000 description 2
- 238000012544 monitoring process Methods 0.000 description 2
- 229940125645 monoclonal antibody drug Drugs 0.000 description 2
- 210000001616 monocyte Anatomy 0.000 description 2
- 230000004899 motility Effects 0.000 description 2
- OWIUPIRUAQMTTK-UHFFFAOYSA-M n-aminocarbamate Chemical compound NNC([O-])=O OWIUPIRUAQMTTK-UHFFFAOYSA-M 0.000 description 2
- 230000009826 neoplastic cell growth Effects 0.000 description 2
- 210000000440 neutrophil Anatomy 0.000 description 2
- ZQPPMHVWECSIRJ-KTKRTIGZSA-N oleic acid Chemical compound CCCCCCCC\C=C/CCCCCCCC(O)=O ZQPPMHVWECSIRJ-KTKRTIGZSA-N 0.000 description 2
- 210000000056 organ Anatomy 0.000 description 2
- 150000002894 organic compounds Chemical class 0.000 description 2
- 235000006408 oxalic acid Nutrition 0.000 description 2
- 150000002923 oximes Chemical class 0.000 description 2
- 238000007911 parenteral administration Methods 0.000 description 2
- 230000001575 pathological effect Effects 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 239000008188 pellet Substances 0.000 description 2
- 230000035515 penetration Effects 0.000 description 2
- 230000035699 permeability Effects 0.000 description 2
- 229920002223 polystyrene Polymers 0.000 description 2
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 239000003755 preservative agent Substances 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 230000005180 public health Effects 0.000 description 2
- CWZZGQMHJJMMPM-UHFFFAOYSA-N pyrrole-2,5-dione;zirconium Chemical compound [Zr].O=C1NC(=O)C=C1 CWZZGQMHJJMMPM-UHFFFAOYSA-N 0.000 description 2
- 238000003156 radioimmunoprecipitation Methods 0.000 description 2
- 239000012217 radiopharmaceutical Substances 0.000 description 2
- 229940121896 radiopharmaceutical Drugs 0.000 description 2
- 230000002799 radiopharmaceutical effect Effects 0.000 description 2
- 230000035484 reaction time Effects 0.000 description 2
- 230000001105 regulatory effect Effects 0.000 description 2
- 238000010405 reoxidation reaction Methods 0.000 description 2
- 125000006853 reporter group Chemical group 0.000 description 2
- GHMLBKRAJCXXBS-UHFFFAOYSA-N resorcinol Chemical compound OC1=CC=CC(O)=C1 GHMLBKRAJCXXBS-UHFFFAOYSA-N 0.000 description 2
- 230000000717 retained effect Effects 0.000 description 2
- 206010039073 rheumatoid arthritis Diseases 0.000 description 2
- DFEYYRMXOJXZRJ-UHFFFAOYSA-N sevoflurane Chemical compound FCOC(C(F)(F)F)C(F)(F)F DFEYYRMXOJXZRJ-UHFFFAOYSA-N 0.000 description 2
- 229960002078 sevoflurane Drugs 0.000 description 2
- 238000002603 single-photon emission computed tomography Methods 0.000 description 2
- 238000003998 size exclusion chromatography high performance liquid chromatography Methods 0.000 description 2
- 229910000029 sodium carbonate Inorganic materials 0.000 description 2
- 239000012453 solvate Substances 0.000 description 2
- 230000003595 spectral effect Effects 0.000 description 2
- 239000003381 stabilizer Substances 0.000 description 2
- 230000000087 stabilizing effect Effects 0.000 description 2
- 125000001424 substituent group Chemical group 0.000 description 2
- KZNICNPSHKQLFF-UHFFFAOYSA-N succinimide Chemical group O=C1CCC(=O)N1 KZNICNPSHKQLFF-UHFFFAOYSA-N 0.000 description 2
- 239000005720 sucrose Substances 0.000 description 2
- 235000000346 sugar Nutrition 0.000 description 2
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 description 2
- 230000004083 survival effect Effects 0.000 description 2
- 238000013268 sustained release Methods 0.000 description 2
- 239000012730 sustained-release form Substances 0.000 description 2
- 238000001308 synthesis method Methods 0.000 description 2
- 239000003826 tablet Substances 0.000 description 2
- 230000008685 targeting Effects 0.000 description 2
- 231100001274 therapeutic index Toxicity 0.000 description 2
- 238000004809 thin layer chromatography Methods 0.000 description 2
- SRVJKTDHMYAMHA-WUXMJOGZSA-N thioacetazone Chemical compound CC(=O)NC1=CC=C(\C=N\NC(N)=S)C=C1 SRVJKTDHMYAMHA-WUXMJOGZSA-N 0.000 description 2
- 239000003053 toxin Substances 0.000 description 2
- 231100000765 toxin Toxicity 0.000 description 2
- 108700012359 toxins Proteins 0.000 description 2
- VZCYOOQTPOCHFL-UHFFFAOYSA-N trans-butenedioic acid Natural products OC(=O)C=CC(O)=O VZCYOOQTPOCHFL-UHFFFAOYSA-N 0.000 description 2
- 102000027257 transmembrane receptors Human genes 0.000 description 2
- 108091008578 transmembrane receptors Proteins 0.000 description 2
- 239000012588 trypsin Substances 0.000 description 2
- 210000003462 vein Anatomy 0.000 description 2
- 238000001262 western blot Methods 0.000 description 2
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 description 1
- DLNKOYKMWOXYQA-VXNVDRBHSA-N (+)-norephedrine Chemical group C[C@@H](N)[C@@H](O)C1=CC=CC=C1 DLNKOYKMWOXYQA-VXNVDRBHSA-N 0.000 description 1
- JKHVDAUOODACDU-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 3-(2,5-dioxopyrrol-1-yl)propanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCN1C(=O)C=CC1=O JKHVDAUOODACDU-UHFFFAOYSA-N 0.000 description 1
- PVGATNRYUYNBHO-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-(2,5-dioxopyrrol-1-yl)butanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCN1C(=O)C=CC1=O PVGATNRYUYNBHO-UHFFFAOYSA-N 0.000 description 1
- BQWBEDSJTMWJAE-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-[(2-iodoacetyl)amino]benzoate Chemical compound C1=CC(NC(=O)CI)=CC=C1C(=O)ON1C(=O)CCC1=O BQWBEDSJTMWJAE-UHFFFAOYSA-N 0.000 description 1
- PMJWDPGOWBRILU-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-[4-(2,5-dioxopyrrol-1-yl)phenyl]butanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCC(C=C1)=CC=C1N1C(=O)C=CC1=O PMJWDPGOWBRILU-UHFFFAOYSA-N 0.000 description 1
- VLARLSIGSPVYHX-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 6-(2,5-dioxopyrrol-1-yl)hexanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCN1C(=O)C=CC1=O VLARLSIGSPVYHX-UHFFFAOYSA-N 0.000 description 1
- WCMOHMXWOOBVMZ-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 6-[3-(2,5-dioxopyrrol-1-yl)propanoylamino]hexanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCNC(=O)CCN1C(=O)C=CC1=O WCMOHMXWOOBVMZ-UHFFFAOYSA-N 0.000 description 1
- IHVODYOQUSEYJJ-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 6-[[4-[(2,5-dioxopyrrol-1-yl)methyl]cyclohexanecarbonyl]amino]hexanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCNC(=O)C(CC1)CCC1CN1C(=O)C=CC1=O IHVODYOQUSEYJJ-UHFFFAOYSA-N 0.000 description 1
- LNAZSHAWQACDHT-XIYTZBAFSA-N (2r,3r,4s,5r,6s)-4,5-dimethoxy-2-(methoxymethyl)-3-[(2s,3r,4s,5r,6r)-3,4,5-trimethoxy-6-(methoxymethyl)oxan-2-yl]oxy-6-[(2r,3r,4s,5r,6r)-4,5,6-trimethoxy-2-(methoxymethyl)oxan-3-yl]oxyoxane Chemical compound CO[C@@H]1[C@@H](OC)[C@H](OC)[C@@H](COC)O[C@H]1O[C@H]1[C@H](OC)[C@@H](OC)[C@H](O[C@H]2[C@@H]([C@@H](OC)[C@H](OC)O[C@@H]2COC)OC)O[C@@H]1COC LNAZSHAWQACDHT-XIYTZBAFSA-N 0.000 description 1
- XMQUEQJCYRFIQS-YFKPBYRVSA-N (2s)-2-amino-5-ethoxy-5-oxopentanoic acid Chemical compound CCOC(=O)CC[C@H](N)C(O)=O XMQUEQJCYRFIQS-YFKPBYRVSA-N 0.000 description 1
- MZOFCQQQCNRIBI-VMXHOPILSA-N (3s)-4-[[(2s)-1-[[(2s)-1-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-methyl-1-oxopentan-2-yl]amino]-5-(diaminomethylideneamino)-1-oxopentan-2-yl]amino]-3-[[2-[[(2s)-2,6-diaminohexanoyl]amino]acetyl]amino]-4-oxobutanoic acid Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@@H](N)CCCCN MZOFCQQQCNRIBI-VMXHOPILSA-N 0.000 description 1
- LTDQGCFMTVHZKP-UHFFFAOYSA-N (4-bromophenyl)-(4,6-dimethoxy-3-methyl-1-benzofuran-2-yl)methanone Chemical compound O1C2=CC(OC)=CC(OC)=C2C(C)=C1C(=O)C1=CC=C(Br)C=C1 LTDQGCFMTVHZKP-UHFFFAOYSA-N 0.000 description 1
- GHOKWGTUZJEAQD-ZETCQYMHSA-N (D)-(+)-Pantothenic acid Chemical compound OCC(C)(C)[C@@H](O)C(=O)NCCC(O)=O GHOKWGTUZJEAQD-ZETCQYMHSA-N 0.000 description 1
- WRIDQFICGBMAFQ-UHFFFAOYSA-N (E)-8-Octadecenoic acid Natural products CCCCCCCCCC=CCCCCCCC(O)=O WRIDQFICGBMAFQ-UHFFFAOYSA-N 0.000 description 1
- DNIAPMSPPWPWGF-GSVOUGTGSA-N (R)-(-)-Propylene glycol Chemical compound C[C@@H](O)CO DNIAPMSPPWPWGF-GSVOUGTGSA-N 0.000 description 1
- 125000003088 (fluoren-9-ylmethoxy)carbonyl group Chemical group 0.000 description 1
- YYMCVDNIIFNDJK-XFQWXJFMSA-N (z)-1-(3-fluorophenyl)-n-[(z)-(3-fluorophenyl)methylideneamino]methanimine Chemical compound FC1=CC=CC(\C=N/N=C\C=2C=C(F)C=CC=2)=C1 YYMCVDNIIFNDJK-XFQWXJFMSA-N 0.000 description 1
- TZCPCKNHXULUIY-RGULYWFUSA-N 1,2-distearoyl-sn-glycero-3-phosphoserine Chemical compound CCCCCCCCCCCCCCCCCC(=O)OC[C@H](COP(O)(=O)OC[C@H](N)C(O)=O)OC(=O)CCCCCCCCCCCCCCCCC TZCPCKNHXULUIY-RGULYWFUSA-N 0.000 description 1
- LMDZBCPBFSXMTL-UHFFFAOYSA-N 1-Ethyl-3-(3-dimethylaminopropyl)carbodiimide Substances CCN=C=NCCCN(C)C LMDZBCPBFSXMTL-UHFFFAOYSA-N 0.000 description 1
- SGVWDRVQIYUSRA-UHFFFAOYSA-N 1-[2-[2-(2,5-dioxopyrrol-1-yl)ethyldisulfanyl]ethyl]pyrrole-2,5-dione Chemical compound O=C1C=CC(=O)N1CCSSCCN1C(=O)C=CC1=O SGVWDRVQIYUSRA-UHFFFAOYSA-N 0.000 description 1
- DIYPCWKHSODVAP-UHFFFAOYSA-N 1-[3-(2,5-dioxopyrrol-1-yl)benzoyl]oxy-2,5-dioxopyrrolidine-3-sulfonic acid Chemical compound O=C1C(S(=O)(=O)O)CC(=O)N1OC(=O)C1=CC=CC(N2C(C=CC2=O)=O)=C1 DIYPCWKHSODVAP-UHFFFAOYSA-N 0.000 description 1
- CULQNACJHGHAER-UHFFFAOYSA-N 1-[4-[(2-iodoacetyl)amino]benzoyl]oxy-2,5-dioxopyrrolidine-3-sulfonic acid Chemical compound O=C1C(S(=O)(=O)O)CC(=O)N1OC(=O)C1=CC=C(NC(=O)CI)C=C1 CULQNACJHGHAER-UHFFFAOYSA-N 0.000 description 1
- IXPNQXFRVYWDDI-UHFFFAOYSA-N 1-methyl-2,4-dioxo-1,3-diazinane-5-carboximidamide Chemical compound CN1CC(C(N)=N)C(=O)NC1=O IXPNQXFRVYWDDI-UHFFFAOYSA-N 0.000 description 1
- HRBGUGQWTMBDTR-UHFFFAOYSA-N 2,3,4-tri(propan-2-yl)benzenesulfonyl chloride Chemical compound CC(C)C1=CC=C(S(Cl)(=O)=O)C(C(C)C)=C1C(C)C HRBGUGQWTMBDTR-UHFFFAOYSA-N 0.000 description 1
- LDGWQMRUWMSZIU-LQDDAWAPSA-M 2,3-bis[(z)-octadec-9-enoxy]propyl-trimethylazanium;chloride Chemical compound [Cl-].CCCCCCCC\C=C/CCCCCCCCOCC(C[N+](C)(C)C)OCCCCCCCC\C=C/CCCCCCCC LDGWQMRUWMSZIU-LQDDAWAPSA-M 0.000 description 1
- KGLPWQKSKUVKMJ-UHFFFAOYSA-N 2,3-dihydrophthalazine-1,4-dione Chemical compound C1=CC=C2C(=O)NNC(=O)C2=C1 KGLPWQKSKUVKMJ-UHFFFAOYSA-N 0.000 description 1
- LSYXXLMBRSSBGS-UHFFFAOYSA-N 2,4,6-tris(hydroxymethyl)phenol Chemical group OCC1=CC(CO)=C(O)C(CO)=C1 LSYXXLMBRSSBGS-UHFFFAOYSA-N 0.000 description 1
- KUMMBDBTERQYCG-UHFFFAOYSA-N 2,6-bis(hydroxymethyl)-4-methylphenol Chemical group CC1=CC(CO)=C(O)C(CO)=C1 KUMMBDBTERQYCG-UHFFFAOYSA-N 0.000 description 1
- QEVGZEDELICMKH-UHFFFAOYSA-L 2-(carboxylatomethoxy)acetate Chemical compound [O-]C(=O)COCC([O-])=O QEVGZEDELICMKH-UHFFFAOYSA-L 0.000 description 1
- FALRKNHUBBKYCC-UHFFFAOYSA-N 2-(chloromethyl)pyridine-3-carbonitrile Chemical compound ClCC1=NC=CC=C1C#N FALRKNHUBBKYCC-UHFFFAOYSA-N 0.000 description 1
- FZDFGHZZPBUTGP-UHFFFAOYSA-N 2-[[2-[bis(carboxymethyl)amino]-3-(4-isothiocyanatophenyl)propyl]-[2-[bis(carboxymethyl)amino]propyl]amino]acetic acid Chemical compound OC(=O)CN(CC(O)=O)C(C)CN(CC(O)=O)CC(N(CC(O)=O)CC(O)=O)CC1=CC=C(N=C=S)C=C1 FZDFGHZZPBUTGP-UHFFFAOYSA-N 0.000 description 1
- JUIKUQOUMZUFQT-UHFFFAOYSA-N 2-bromoacetamide Chemical compound NC(=O)CBr JUIKUQOUMZUFQT-UHFFFAOYSA-N 0.000 description 1
- LSTRKXWIZZZYAS-UHFFFAOYSA-N 2-bromoacetyl bromide Chemical compound BrCC(Br)=O LSTRKXWIZZZYAS-UHFFFAOYSA-N 0.000 description 1
- NEAQRZUHTPSBBM-UHFFFAOYSA-N 2-hydroxy-3,3-dimethyl-7-nitro-4h-isoquinolin-1-one Chemical compound C1=C([N+]([O-])=O)C=C2C(=O)N(O)C(C)(C)CC2=C1 NEAQRZUHTPSBBM-UHFFFAOYSA-N 0.000 description 1
- 125000003903 2-propenyl group Chemical group [H]C([*])([H])C([H])=C([H])[H] 0.000 description 1
- KGIGUEBEKRSTEW-UHFFFAOYSA-N 2-vinylpyridine Chemical compound C=CC1=CC=CC=N1 KGIGUEBEKRSTEW-UHFFFAOYSA-N 0.000 description 1
- LQJBNNIYVWPHFW-UHFFFAOYSA-N 20:1omega9c fatty acid Natural products CCCCCCCCCCC=CCCCCCCCC(O)=O LQJBNNIYVWPHFW-UHFFFAOYSA-N 0.000 description 1
- KIUMMUBSPKGMOY-UHFFFAOYSA-N 3,3'-Dithiobis(6-nitrobenzoic acid) Chemical compound C1=C([N+]([O-])=O)C(C(=O)O)=CC(SSC=2C=C(C(=CC=2)[N+]([O-])=O)C(O)=O)=C1 KIUMMUBSPKGMOY-UHFFFAOYSA-N 0.000 description 1
- ALKYHXVLJMQRLQ-UHFFFAOYSA-N 3-Hydroxy-2-naphthoate Chemical compound C1=CC=C2C=C(O)C(C(=O)O)=CC2=C1 ALKYHXVLJMQRLQ-UHFFFAOYSA-N 0.000 description 1
- HVCOBJNICQPDBP-UHFFFAOYSA-N 3-[3-[3,5-dihydroxy-6-methyl-4-(3,4,5-trihydroxy-6-methyloxan-2-yl)oxyoxan-2-yl]oxydecanoyloxy]decanoic acid;hydrate Chemical compound O.OC1C(OC(CC(=O)OC(CCCCCCC)CC(O)=O)CCCCCCC)OC(C)C(O)C1OC1C(O)C(O)C(O)C(C)O1 HVCOBJNICQPDBP-UHFFFAOYSA-N 0.000 description 1
- FEWJPZIEWOKRBE-UHFFFAOYSA-M 3-carboxy-2,3-dihydroxypropanoate Chemical compound OC(=O)C(O)C(O)C([O-])=O FEWJPZIEWOKRBE-UHFFFAOYSA-M 0.000 description 1
- NCPQROHLJFARLL-UHFFFAOYSA-N 4-(2,5-dioxopyrrol-1-yl)butanoic acid Chemical compound OC(=O)CCCN1C(=O)C=CC1=O NCPQROHLJFARLL-UHFFFAOYSA-N 0.000 description 1
- OBWSOTREAMFOCQ-UHFFFAOYSA-N 4-(4-amino-3,5-dimethylphenyl)-2,6-dimethylaniline;hydrochloride Chemical compound Cl.CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 OBWSOTREAMFOCQ-UHFFFAOYSA-N 0.000 description 1
- XZKIHKMTEMTJQX-UHFFFAOYSA-N 4-Nitrophenyl Phosphate Chemical compound OP(O)(=O)OC1=CC=C([N+]([O-])=O)C=C1 XZKIHKMTEMTJQX-UHFFFAOYSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- ZMRMMAOBSFSXLN-UHFFFAOYSA-N 4-[4-(2,5-dioxopyrrol-1-yl)phenyl]butanehydrazide Chemical compound C1=CC(CCCC(=O)NN)=CC=C1N1C(=O)C=CC1=O ZMRMMAOBSFSXLN-UHFFFAOYSA-N 0.000 description 1
- 125000004042 4-aminobutyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])C([H])([H])N([H])[H] 0.000 description 1
- 229960000549 4-dimethylaminophenol Drugs 0.000 description 1
- KIUMMUBSPKGMOY-UHFFFAOYSA-L 5-[(3-carboxylato-4-nitrophenyl)disulfanyl]-2-nitrobenzoate Chemical compound C1=C([N+]([O-])=O)C(C(=O)[O-])=CC(SSC=2C=C(C(=CC=2)[N+]([O-])=O)C([O-])=O)=C1 KIUMMUBSPKGMOY-UHFFFAOYSA-L 0.000 description 1
- DNXDWMHPTVQBIP-ZQIUZPCESA-N 5-[(3as,4s,6ar)-2-oxo-1,3,3a,4,6,6a-hexahydrothieno[3,4-d]imidazol-4-yl]-n-[6-[(2-iodoacetyl)amino]hexyl]pentanamide Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)NCCCCCCNC(=O)CI)SC[C@@H]21 DNXDWMHPTVQBIP-ZQIUZPCESA-N 0.000 description 1
- QLPHBNRMJLFRGO-YDHSSHFGSA-N 5-[(3as,4s,6ar)-2-oxo-1,3,3a,4,6,6a-hexahydrothieno[3,4-d]imidazol-4-yl]-n-[6-[3-(pyridin-2-yldisulfanyl)propanoylamino]hexyl]pentanamide Chemical compound C([C@H]1[C@H]2NC(=O)N[C@H]2CS1)CCCC(=O)NCCCCCCNC(=O)CCSSC1=CC=CC=N1 QLPHBNRMJLFRGO-YDHSSHFGSA-N 0.000 description 1
- NJYVEMPWNAYQQN-UHFFFAOYSA-N 5-carboxyfluorescein Chemical compound C12=CC=C(O)C=C2OC2=CC(O)=CC=C2C21OC(=O)C1=CC(C(=O)O)=CC=C21 NJYVEMPWNAYQQN-UHFFFAOYSA-N 0.000 description 1
- BZTDTCNHAFUJOG-UHFFFAOYSA-N 6-carboxyfluorescein Chemical compound C12=CC=C(O)C=C2OC2=CC(O)=CC=C2C11OC(=O)C2=CC=C(C(=O)O)C=C21 BZTDTCNHAFUJOG-UHFFFAOYSA-N 0.000 description 1
- 102100031126 6-phosphogluconolactonase Human genes 0.000 description 1
- 108010029731 6-phosphogluconolactonase Proteins 0.000 description 1
- QSBYPNXLFMSGKH-UHFFFAOYSA-N 9-Heptadecensaeure Natural products CCCCCCCC=CCCCCCCCC(O)=O QSBYPNXLFMSGKH-UHFFFAOYSA-N 0.000 description 1
- 244000215068 Acacia senegal Species 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- QGZKDVFQNNGYKY-UHFFFAOYSA-O Ammonium Chemical compound [NH4+] QGZKDVFQNNGYKY-UHFFFAOYSA-O 0.000 description 1
- 206010002091 Anaesthesia Diseases 0.000 description 1
- 102100021569 Apoptosis regulator Bcl-2 Human genes 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 208000023275 Autoimmune disease Diseases 0.000 description 1
- 108090001008 Avidin Proteins 0.000 description 1
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 1
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 1
- 238000011729 BALB/c nude mouse Methods 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 108090000363 Bacterial Luciferases Proteins 0.000 description 1
- 206010051728 Bone erosion Diseases 0.000 description 1
- 102100027052 Bone morphogenetic protein receptor type-1B Human genes 0.000 description 1
- 244000056139 Brassica cretica Species 0.000 description 1
- 235000003351 Brassica cretica Nutrition 0.000 description 1
- 235000003343 Brassica rupestris Nutrition 0.000 description 1
- 101710140080 Brevican core protein Proteins 0.000 description 1
- FERIUCNNQQJTOY-UHFFFAOYSA-N Butyric acid Natural products CCCC(O)=O FERIUCNNQQJTOY-UHFFFAOYSA-N 0.000 description 1
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 1
- 108050005493 CD3 protein, epsilon/gamma/delta subunit Proteins 0.000 description 1
- 101150013553 CD40 gene Proteins 0.000 description 1
- 108060001253 CD99 Proteins 0.000 description 1
- 102000024905 CD99 Human genes 0.000 description 1
- 108010021064 CTLA-4 Antigen Proteins 0.000 description 1
- 229940045513 CTLA4 antagonist Drugs 0.000 description 1
- 101710205625 Capsid protein p24 Proteins 0.000 description 1
- OKTJSMMVPCPJKN-NJFSPNSNSA-N Carbon-14 Chemical compound [14C] OKTJSMMVPCPJKN-NJFSPNSNSA-N 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 102000003952 Caspase 3 Human genes 0.000 description 1
- 108090000397 Caspase 3 Proteins 0.000 description 1
- 108010076667 Caspases Proteins 0.000 description 1
- 102000011727 Caspases Human genes 0.000 description 1
- 108010067225 Cell Adhesion Molecules Proteins 0.000 description 1
- 102000016289 Cell Adhesion Molecules Human genes 0.000 description 1
- VEXZGXHMUGYJMC-UHFFFAOYSA-M Chloride anion Chemical compound [Cl-] VEXZGXHMUGYJMC-UHFFFAOYSA-M 0.000 description 1
- 208000030808 Clear cell renal carcinoma Diseases 0.000 description 1
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 1
- 229920002785 Croscarmellose sodium Polymers 0.000 description 1
- 206010011732 Cyst Diseases 0.000 description 1
- 102000004127 Cytokines Human genes 0.000 description 1
- 108090000695 Cytokines Proteins 0.000 description 1
- 102100039498 Cytotoxic T-lymphocyte protein 4 Human genes 0.000 description 1
- 102100037579 D-3-phosphoglycerate dehydrogenase Human genes 0.000 description 1
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 1
- IGXWBGJHJZYPQS-SSDOTTSWSA-N D-Luciferin Chemical compound OC(=O)[C@H]1CSC(C=2SC3=CC=C(O)C=C3N=2)=N1 IGXWBGJHJZYPQS-SSDOTTSWSA-N 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 1
- RGHNJXZEOKUKBD-SQOUGZDYSA-M D-gluconate Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C([O-])=O RGHNJXZEOKUKBD-SQOUGZDYSA-M 0.000 description 1
- AEMOLEFTQBMNLQ-AQKNRBDQSA-N D-glucopyranuronic acid Chemical compound OC1O[C@H](C(O)=O)[C@@H](O)[C@H](O)[C@H]1O AEMOLEFTQBMNLQ-AQKNRBDQSA-N 0.000 description 1
- 102000053602 DNA Human genes 0.000 description 1
- 239000003298 DNA probe Substances 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- SBJKKFFYIZUCET-JLAZNSOCSA-N Dehydro-L-ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(=O)C1=O SBJKKFFYIZUCET-JLAZNSOCSA-N 0.000 description 1
- CYCGRDQQIOGCKX-UHFFFAOYSA-N Dehydro-luciferin Natural products OC(=O)C1=CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 CYCGRDQQIOGCKX-UHFFFAOYSA-N 0.000 description 1
- SBJKKFFYIZUCET-UHFFFAOYSA-N Dehydroascorbic acid Natural products OCC(O)C1OC(=O)C(=O)C1=O SBJKKFFYIZUCET-UHFFFAOYSA-N 0.000 description 1
- 239000004375 Dextrin Substances 0.000 description 1
- 229920001353 Dextrin Polymers 0.000 description 1
- FEWJPZIEWOKRBE-JCYAYHJZSA-N Dextrotartaric acid Chemical compound OC(=O)[C@H](O)[C@@H](O)C(O)=O FEWJPZIEWOKRBE-JCYAYHJZSA-N 0.000 description 1
- LTMHDMANZUZIPE-AMTYYWEZSA-N Digoxin Natural products O([C@H]1[C@H](C)O[C@H](O[C@@H]2C[C@@H]3[C@@](C)([C@@H]4[C@H]([C@]5(O)[C@](C)([C@H](O)C4)[C@H](C4=CC(=O)OC4)CC5)CC3)CC2)C[C@@H]1O)[C@H]1O[C@H](C)[C@@H](O[C@H]2O[C@@H](C)[C@H](O)[C@@H](O)C2)[C@@H](O)C1 LTMHDMANZUZIPE-AMTYYWEZSA-N 0.000 description 1
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 1
- 101150029707 ERBB2 gene Proteins 0.000 description 1
- 229910052693 Europium Inorganic materials 0.000 description 1
- 108010007457 Extracellular Signal-Regulated MAP Kinases Proteins 0.000 description 1
- 108050001049 Extracellular proteins Proteins 0.000 description 1
- 108090000331 Firefly luciferases Proteins 0.000 description 1
- BJGNCJDXODQBOB-UHFFFAOYSA-N Fivefly Luciferin Natural products OC(=O)C1CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 BJGNCJDXODQBOB-UHFFFAOYSA-N 0.000 description 1
- VZCYOOQTPOCHFL-OWOJBTEDSA-N Fumaric acid Chemical compound OC(=O)\C=C\C(O)=O VZCYOOQTPOCHFL-OWOJBTEDSA-N 0.000 description 1
- 108010015133 Galactose oxidase Proteins 0.000 description 1
- 108010073178 Glucan 1,4-alpha-Glucosidase Proteins 0.000 description 1
- 102100022624 Glucoamylase Human genes 0.000 description 1
- 239000004366 Glucose oxidase Substances 0.000 description 1
- 108010015776 Glucose oxidase Proteins 0.000 description 1
- 108010018962 Glucosephosphate Dehydrogenase Proteins 0.000 description 1
- ZWZWYGMENQVNFU-UHFFFAOYSA-N Glycerophosphorylserin Natural products OC(=O)C(N)COP(O)(=O)OCC(O)CO ZWZWYGMENQVNFU-UHFFFAOYSA-N 0.000 description 1
- 229930186217 Glycolipid Natural products 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 102000009465 Growth Factor Receptors Human genes 0.000 description 1
- 108010009202 Growth Factor Receptors Proteins 0.000 description 1
- 229920000084 Gum arabic Polymers 0.000 description 1
- 102100031573 Hematopoietic progenitor cell antigen CD34 Human genes 0.000 description 1
- 102100028999 High mobility group protein HMGI-C Human genes 0.000 description 1
- 241000282412 Homo Species 0.000 description 1
- 101000971171 Homo sapiens Apoptosis regulator Bcl-2 Proteins 0.000 description 1
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 1
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 1
- 101000984546 Homo sapiens Bone morphogenetic protein receptor type-1B Proteins 0.000 description 1
- 101000731086 Homo sapiens Brevican core protein Proteins 0.000 description 1
- 101000932213 Homo sapiens Dipeptidase 1 Proteins 0.000 description 1
- 101000777663 Homo sapiens Hematopoietic progenitor cell antigen CD34 Proteins 0.000 description 1
- 101000986379 Homo sapiens High mobility group protein HMGI-C Proteins 0.000 description 1
- 101001044893 Homo sapiens Interleukin-20 receptor subunit alpha Proteins 0.000 description 1
- 101000738771 Homo sapiens Receptor-type tyrosine-protein phosphatase C Proteins 0.000 description 1
- 101000946843 Homo sapiens T-cell surface glycoprotein CD8 alpha chain Proteins 0.000 description 1
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 1
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 1
- 102100022420 Inactive rhomboid protein 1 Human genes 0.000 description 1
- 101710136320 Inactive rhomboid protein 1 Proteins 0.000 description 1
- 108010008212 Integrin alpha4beta1 Proteins 0.000 description 1
- 102100025390 Integrin beta-2 Human genes 0.000 description 1
- 102000008607 Integrin beta3 Human genes 0.000 description 1
- 108010020950 Integrin beta3 Proteins 0.000 description 1
- 108010064593 Intercellular Adhesion Molecule-1 Proteins 0.000 description 1
- 102100037877 Intercellular adhesion molecule 1 Human genes 0.000 description 1
- 238000012695 Interfacial polymerization Methods 0.000 description 1
- 108091029795 Intergenic region Proteins 0.000 description 1
- 102100022706 Interleukin-20 receptor subunit alpha Human genes 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 1
- RHGKLRLOHDJJDR-BYPYZUCNSA-N L-citrulline Chemical group NC(=O)NCCC[C@H]([NH3+])C([O-])=O RHGKLRLOHDJJDR-BYPYZUCNSA-N 0.000 description 1
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- JVTAAEKCZFNVCJ-UHFFFAOYSA-M Lactate Chemical compound CC(O)C([O-])=O JVTAAEKCZFNVCJ-UHFFFAOYSA-M 0.000 description 1
- 102100038609 Lactoperoxidase Human genes 0.000 description 1
- 108010023244 Lactoperoxidase Proteins 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 108010000817 Leuprolide Proteins 0.000 description 1
- 206010024612 Lipoma Diseases 0.000 description 1
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- DDWFXDSYGUXRAY-UHFFFAOYSA-N Luciferin Natural products CCc1c(C)c(CC2NC(=O)C(=C2C=C)C)[nH]c1Cc3[nH]c4C(=C5/NC(CC(=O)O)C(C)C5CC(=O)O)CC(=O)c4c3C DDWFXDSYGUXRAY-UHFFFAOYSA-N 0.000 description 1
- 208000007433 Lymphatic Metastasis Diseases 0.000 description 1
- 108010064548 Lymphocyte Function-Associated Antigen-1 Proteins 0.000 description 1
- 102000008072 Lymphokines Human genes 0.000 description 1
- 108010074338 Lymphokines Proteins 0.000 description 1
- 241001082241 Lythrum hyssopifolia Species 0.000 description 1
- 241000282560 Macaca mulatta Species 0.000 description 1
- 239000004907 Macro-emulsion Substances 0.000 description 1
- 108010026217 Malate Dehydrogenase Proteins 0.000 description 1
- OFOBLEOULBTSOW-UHFFFAOYSA-L Malonate Chemical compound [O-]C(=O)CC([O-])=O OFOBLEOULBTSOW-UHFFFAOYSA-L 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 206010027459 Metastases to lymph nodes Diseases 0.000 description 1
- 102000007474 Multiprotein Complexes Human genes 0.000 description 1
- 108010085220 Multiprotein Complexes Proteins 0.000 description 1
- 102000016943 Muramidase Human genes 0.000 description 1
- 108010014251 Muramidase Proteins 0.000 description 1
- 101000623899 Mus musculus Mucin-13 Proteins 0.000 description 1
- HSHXDCVZWHOWCS-UHFFFAOYSA-N N'-hexadecylthiophene-2-carbohydrazide Chemical compound CCCCCCCCCCCCCCCCNNC(=O)c1cccs1 HSHXDCVZWHOWCS-UHFFFAOYSA-N 0.000 description 1
- APGLTERDKORUHK-LURJTMIESA-N N,N-dimethyl-L-Valine Chemical group CC(C)[C@H](N(C)C)C(O)=O APGLTERDKORUHK-LURJTMIESA-N 0.000 description 1
- 108010062010 N-Acetylmuramoyl-L-alanine Amidase Proteins 0.000 description 1
- WHNWPMSKXPGLAX-UHFFFAOYSA-N N-Vinyl-2-pyrrolidone Chemical compound C=CN1CCCC1=O WHNWPMSKXPGLAX-UHFFFAOYSA-N 0.000 description 1
- BAQMYDQNMFBZNA-UHFFFAOYSA-N N-biotinyl-L-lysine Natural products N1C(=O)NC2C(CCCCC(=O)NCCCCC(N)C(O)=O)SCC21 BAQMYDQNMFBZNA-UHFFFAOYSA-N 0.000 description 1
- HDFGOPSGAURCEO-UHFFFAOYSA-N N-ethylmaleimide Chemical group CCN1C(=O)C=CC1=O HDFGOPSGAURCEO-UHFFFAOYSA-N 0.000 description 1
- 229910002651 NO3 Inorganic materials 0.000 description 1
- NHNBFGGVMKEFGY-UHFFFAOYSA-N Nitrate Chemical compound [O-][N+]([O-])=O NHNBFGGVMKEFGY-UHFFFAOYSA-N 0.000 description 1
- 239000000020 Nitrocellulose Substances 0.000 description 1
- 208000008589 Obesity Diseases 0.000 description 1
- 239000005642 Oleic acid Substances 0.000 description 1
- ZQPPMHVWECSIRJ-UHFFFAOYSA-N Oleic acid Natural products CCCCCCCCC=CCCCCCCCC(O)=O ZQPPMHVWECSIRJ-UHFFFAOYSA-N 0.000 description 1
- 108020005187 Oligonucleotide Probes Proteins 0.000 description 1
- 206010061535 Ovarian neoplasm Diseases 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 241001504519 Papio ursinus Species 0.000 description 1
- 102000057297 Pepsin A Human genes 0.000 description 1
- 108090000284 Pepsin A Proteins 0.000 description 1
- 108010081690 Pertussis Toxin Proteins 0.000 description 1
- 206010057249 Phagocytosis Diseases 0.000 description 1
- BELBBZDIHDAJOR-UHFFFAOYSA-N Phenolsulfonephthalein Chemical compound C1=CC(O)=CC=C1C1(C=2C=CC(O)=CC=2)C2=CC=CC=C2S(=O)(=O)O1 BELBBZDIHDAJOR-UHFFFAOYSA-N 0.000 description 1
- 101710177166 Phosphoprotein Proteins 0.000 description 1
- 241000276498 Pollachius virens Species 0.000 description 1
- 239000004698 Polyethylene Substances 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 101710098940 Pro-epidermal growth factor Proteins 0.000 description 1
- 101710120463 Prostate stem cell antigen Proteins 0.000 description 1
- 102000007066 Prostate-Specific Antigen Human genes 0.000 description 1
- 108010072866 Prostate-Specific Antigen Proteins 0.000 description 1
- 101800004937 Protein C Proteins 0.000 description 1
- 102000017975 Protein C Human genes 0.000 description 1
- 102000004022 Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000412 Protein-Tyrosine Kinases Proteins 0.000 description 1
- 108010067787 Proteoglycans Proteins 0.000 description 1
- 101100501691 Rattus norvegicus Erbb2 gene Proteins 0.000 description 1
- 101100501698 Rattus norvegicus Erbb4 gene Proteins 0.000 description 1
- 102000004278 Receptor Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000873 Receptor Protein-Tyrosine Kinases Proteins 0.000 description 1
- 102100037422 Receptor-type tyrosine-protein phosphatase C Human genes 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 208000006265 Renal cell carcinoma Diseases 0.000 description 1
- 206010062104 Renal mass Diseases 0.000 description 1
- 108050002653 Retinoblastoma protein Proteins 0.000 description 1
- 101800001700 Saposin-D Proteins 0.000 description 1
- 229920005654 Sephadex Polymers 0.000 description 1
- 239000012507 Sephadex™ Substances 0.000 description 1
- 101710149279 Small delta antigen Proteins 0.000 description 1
- 241001137868 Streptomyces pilosus Species 0.000 description 1
- QAOWNCQODCNURD-UHFFFAOYSA-L Sulfate Chemical compound [O-]S([O-])(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-L 0.000 description 1
- 239000012505 Superdex™ Substances 0.000 description 1
- 102100034922 T-cell surface glycoprotein CD8 alpha chain Human genes 0.000 description 1
- 239000012317 TBTU Substances 0.000 description 1
- JVHROZDXPAUZFK-UHFFFAOYSA-N TETA Chemical compound OC(=O)CN1CCCN(CC(O)=O)CCN(CC(O)=O)CCCN(CC(O)=O)CC1 JVHROZDXPAUZFK-UHFFFAOYSA-N 0.000 description 1
- 229920002253 Tannate Polymers 0.000 description 1
- 108010000499 Thromboplastin Proteins 0.000 description 1
- 102100030859 Tissue factor Human genes 0.000 description 1
- 229920001615 Tragacanth Polymers 0.000 description 1
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 102100022563 Tubulin polymerization-promoting protein Human genes 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 1
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 1
- 206010053613 Type IV hypersensitivity reaction Diseases 0.000 description 1
- 108090000848 Ubiquitin Proteins 0.000 description 1
- 102000044159 Ubiquitin Human genes 0.000 description 1
- 108010092464 Urate Oxidase Proteins 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 102100033220 Xanthine oxidase Human genes 0.000 description 1
- 108010093894 Xanthine oxidase Proteins 0.000 description 1
- CLZISMQKJZCZDN-UHFFFAOYSA-N [benzotriazol-1-yloxy(dimethylamino)methylidene]-dimethylazanium Chemical compound C1=CC=C2N(OC(N(C)C)=[N+](C)C)N=NC2=C1 CLZISMQKJZCZDN-UHFFFAOYSA-N 0.000 description 1
- 235000010489 acacia gum Nutrition 0.000 description 1
- 239000000205 acacia gum Substances 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- VJHCJDRQFCCTHL-UHFFFAOYSA-N acetic acid 2,3,4,5,6-pentahydroxyhexanal Chemical compound CC(O)=O.OCC(O)C(O)C(O)C(O)C=O VJHCJDRQFCCTHL-UHFFFAOYSA-N 0.000 description 1
- DPXJVFZANSGRMM-UHFFFAOYSA-N acetic acid;2,3,4,5,6-pentahydroxyhexanal;sodium Chemical compound [Na].CC(O)=O.OCC(O)C(O)C(O)C(O)C=O DPXJVFZANSGRMM-UHFFFAOYSA-N 0.000 description 1
- 230000023445 activated T cell autonomous cell death Effects 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 238000012382 advanced drug delivery Methods 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 238000001261 affinity purification Methods 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- HAXFWIACAGNFHA-UHFFFAOYSA-N aldrithiol Chemical compound C=1C=CC=NC=1SSC1=CC=CC=N1 HAXFWIACAGNFHA-UHFFFAOYSA-N 0.000 description 1
- 125000003545 alkoxy group Chemical group 0.000 description 1
- 125000003282 alkyl amino group Chemical group 0.000 description 1
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 description 1
- 239000012080 ambient air Substances 0.000 description 1
- 230000037005 anaesthesia Effects 0.000 description 1
- 238000000540 analysis of variance Methods 0.000 description 1
- 230000033115 angiogenesis Effects 0.000 description 1
- 230000001772 anti-angiogenic effect Effects 0.000 description 1
- 229940046836 anti-estrogen Drugs 0.000 description 1
- 230000001833 anti-estrogenic effect Effects 0.000 description 1
- 230000003064 anti-oxidating effect Effects 0.000 description 1
- 230000009830 antibody antigen interaction Effects 0.000 description 1
- 238000011230 antibody-based therapy Methods 0.000 description 1
- 239000002246 antineoplastic agent Substances 0.000 description 1
- 239000003963 antioxidant agent Substances 0.000 description 1
- 235000006708 antioxidants Nutrition 0.000 description 1
- 239000003418 antiprogestin Substances 0.000 description 1
- 230000001640 apoptogenic effect Effects 0.000 description 1
- 238000003782 apoptosis assay Methods 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 125000000732 arylene group Chemical group 0.000 description 1
- 229940072107 ascorbate Drugs 0.000 description 1
- 229960005070 ascorbic acid Drugs 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 239000000305 astragalus gummifer gum Substances 0.000 description 1
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 1
- 230000001363 autoimmune Effects 0.000 description 1
- 239000000022 bacteriostatic agent Substances 0.000 description 1
- 229960000686 benzalkonium chloride Drugs 0.000 description 1
- 229940077388 benzenesulfonate Drugs 0.000 description 1
- SRSXLGNVWSONIS-UHFFFAOYSA-M benzenesulfonate Chemical compound [O-]S(=O)(=O)C1=CC=CC=C1 SRSXLGNVWSONIS-UHFFFAOYSA-M 0.000 description 1
- 229960001950 benzethonium chloride Drugs 0.000 description 1
- UREZNYTWGJKWBI-UHFFFAOYSA-M benzethonium chloride Chemical compound [Cl-].C1=CC(C(C)(C)CC(C)(C)C)=CC=C1OCCOCC[N+](C)(C)CC1=CC=CC=C1 UREZNYTWGJKWBI-UHFFFAOYSA-M 0.000 description 1
- 235000019445 benzyl alcohol Nutrition 0.000 description 1
- CADWTSSKOVRVJC-UHFFFAOYSA-N benzyl(dimethyl)azanium;chloride Chemical compound [Cl-].C[NH+](C)CC1=CC=CC=C1 CADWTSSKOVRVJC-UHFFFAOYSA-N 0.000 description 1
- 230000005255 beta decay Effects 0.000 description 1
- 230000005266 beta plus decay Effects 0.000 description 1
- WQZGKKKJIJFFOK-FPRJBGLDSA-N beta-D-galactose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-FPRJBGLDSA-N 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 102000005936 beta-Galactosidase Human genes 0.000 description 1
- 108010005774 beta-Galactosidase Proteins 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 229960000397 bevacizumab Drugs 0.000 description 1
- 230000002146 bilateral effect Effects 0.000 description 1
- 210000003445 biliary tract Anatomy 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 238000012742 biochemical analysis Methods 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- BAQMYDQNMFBZNA-MNXVOIDGSA-N biocytin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)NCCCC[C@H](N)C(O)=O)SC[C@@H]21 BAQMYDQNMFBZNA-MNXVOIDGSA-N 0.000 description 1
- 238000012925 biological evaluation Methods 0.000 description 1
- 230000031018 biological processes and functions Effects 0.000 description 1
- 108091006004 biotinylated proteins Proteins 0.000 description 1
- QKSKPIVNLNLAAV-UHFFFAOYSA-N bis(2-chloroethyl) sulfide Chemical compound ClCCSCCCl QKSKPIVNLNLAAV-UHFFFAOYSA-N 0.000 description 1
- HUTDDBSSHVOYJR-UHFFFAOYSA-H bis[(2-oxo-1,3,2$l^{5},4$l^{2}-dioxaphosphaplumbetan-2-yl)oxy]lead Chemical compound [Pb+2].[Pb+2].[Pb+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O HUTDDBSSHVOYJR-UHFFFAOYSA-H 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 230000036760 body temperature Effects 0.000 description 1
- 239000007853 buffer solution Substances 0.000 description 1
- 244000309464 bull Species 0.000 description 1
- DQXBYHZEEUGOBF-UHFFFAOYSA-N but-3-enoic acid;ethene Chemical compound C=C.OC(=O)CC=C DQXBYHZEEUGOBF-UHFFFAOYSA-N 0.000 description 1
- SNCZNSNPXMPCGN-UHFFFAOYSA-N butanediamide Chemical compound NC(=O)CCC(N)=O SNCZNSNPXMPCGN-UHFFFAOYSA-N 0.000 description 1
- LRHPLDYGYMQRHN-UHFFFAOYSA-N butyl alcohol Substances CCCCO LRHPLDYGYMQRHN-UHFFFAOYSA-N 0.000 description 1
- 125000000484 butyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])C([H])([H])[H] 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 239000002775 capsule Substances 0.000 description 1
- 150000001718 carbodiimides Chemical class 0.000 description 1
- 125000002915 carbonyl group Chemical group [*:2]C([*:1])=O 0.000 description 1
- 239000001768 carboxy methyl cellulose Substances 0.000 description 1
- 150000001732 carboxylic acid derivatives Chemical class 0.000 description 1
- 230000022131 cell cycle Effects 0.000 description 1
- 230000010261 cell growth Effects 0.000 description 1
- 210000000170 cell membrane Anatomy 0.000 description 1
- 230000015861 cell surface binding Effects 0.000 description 1
- 230000019522 cellular metabolic process Effects 0.000 description 1
- 230000007541 cellular toxicity Effects 0.000 description 1
- 125000003636 chemical group Chemical group 0.000 description 1
- 238000001311 chemical methods and process Methods 0.000 description 1
- 238000002512 chemotherapy Methods 0.000 description 1
- 125000002603 chloroethyl group Chemical group [H]C([*])([H])C([H])([H])Cl 0.000 description 1
- 229940001468 citrate Drugs 0.000 description 1
- 229960002173 citrulline Drugs 0.000 description 1
- 235000013477 citrulline Nutrition 0.000 description 1
- 238000004140 cleaning Methods 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000003759 clinical diagnosis Methods 0.000 description 1
- 238000005354 coacervation Methods 0.000 description 1
- 239000000084 colloidal system Substances 0.000 description 1
- 210000001072 colon Anatomy 0.000 description 1
- 239000003086 colorant Substances 0.000 description 1
- 238000012875 competitive assay Methods 0.000 description 1
- 230000004154 complement system Effects 0.000 description 1
- 230000000536 complexating effect Effects 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 239000000356 contaminant Substances 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 238000012937 correction Methods 0.000 description 1
- 239000006184 cosolvent Substances 0.000 description 1
- 229960005168 croscarmellose Drugs 0.000 description 1
- 239000001767 crosslinked sodium carboxy methyl cellulose Substances 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 125000004122 cyclic group Chemical group 0.000 description 1
- HPXRVTGHNJAIIH-UHFFFAOYSA-N cyclohexanol Chemical compound OC1CCCCC1 HPXRVTGHNJAIIH-UHFFFAOYSA-N 0.000 description 1
- 125000000058 cyclopentadienyl group Chemical group C1(=CC=CC1)* 0.000 description 1
- 208000031513 cyst Diseases 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 1
- 238000002784 cytotoxicity assay Methods 0.000 description 1
- 231100000263 cytotoxicity test Toxicity 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 238000000354 decomposition reaction Methods 0.000 description 1
- 235000020960 dehydroascorbic acid Nutrition 0.000 description 1
- 239000011615 dehydroascorbic acid Substances 0.000 description 1
- 238000005831 deiodination reaction Methods 0.000 description 1
- 230000003111 delayed effect Effects 0.000 description 1
- 238000004925 denaturation Methods 0.000 description 1
- 230000036425 denaturation Effects 0.000 description 1
- 239000000412 dendrimer Substances 0.000 description 1
- 229920000736 dendritic polymer Polymers 0.000 description 1
- 229940099217 desferal Drugs 0.000 description 1
- 235000019425 dextrin Nutrition 0.000 description 1
- 238000002050 diffraction method Methods 0.000 description 1
- 238000009792 diffusion process Methods 0.000 description 1
- LTMHDMANZUZIPE-PUGKRICDSA-N digoxin Chemical compound C1[C@H](O)[C@H](O)[C@@H](C)O[C@H]1O[C@@H]1[C@@H](C)O[C@@H](O[C@@H]2[C@H](O[C@@H](O[C@@H]3C[C@@H]4[C@]([C@@H]5[C@H]([C@]6(CC[C@@H]([C@@]6(C)[C@H](O)C5)C=5COC(=O)C=5)O)CC4)(C)CC3)C[C@@H]2O)C)C[C@@H]1O LTMHDMANZUZIPE-PUGKRICDSA-N 0.000 description 1
- LTMHDMANZUZIPE-UHFFFAOYSA-N digoxine Natural products C1C(O)C(O)C(C)OC1OC1C(C)OC(OC2C(OC(OC3CC4C(C5C(C6(CCC(C6(C)C(O)C5)C=5COC(=O)C=5)O)CC4)(C)CC3)CC2O)C)CC1O LTMHDMANZUZIPE-UHFFFAOYSA-N 0.000 description 1
- 150000002016 disaccharides Chemical class 0.000 description 1
- DLNKOYKMWOXYQA-UHFFFAOYSA-N dl-pseudophenylpropanolamine Natural products CC(N)C(O)C1=CC=CC=C1 DLNKOYKMWOXYQA-UHFFFAOYSA-N 0.000 description 1
- 238000004980 dosimetry Methods 0.000 description 1
- BJAJDJDODCWPNS-UHFFFAOYSA-N dotp Chemical compound O=C1N2CCOC2=NC2=C1SC=C2 BJAJDJDODCWPNS-UHFFFAOYSA-N 0.000 description 1
- 230000002222 downregulating effect Effects 0.000 description 1
- 230000003828 downregulation Effects 0.000 description 1
- 229960004679 doxorubicin Drugs 0.000 description 1
- 238000012377 drug delivery Methods 0.000 description 1
- ZSWFCLXCOIISFI-UHFFFAOYSA-N endo-cyclopentadiene Natural products C1C=CC=C1 ZSWFCLXCOIISFI-UHFFFAOYSA-N 0.000 description 1
- 230000006862 enzymatic digestion Effects 0.000 description 1
- 230000002255 enzymatic effect Effects 0.000 description 1
- 229940116977 epidermal growth factor Drugs 0.000 description 1
- 229960005309 estradiol Drugs 0.000 description 1
- 229940011871 estrogen Drugs 0.000 description 1
- 239000000262 estrogen Substances 0.000 description 1
- 239000000328 estrogen antagonist Substances 0.000 description 1
- CCIVGXIOQKPBKL-UHFFFAOYSA-M ethanesulfonate Chemical compound CCS([O-])(=O)=O CCIVGXIOQKPBKL-UHFFFAOYSA-M 0.000 description 1
- 125000001495 ethyl group Chemical group [H]C([H])([H])C([H])([H])* 0.000 description 1
- FAMRKDQNMBBFBR-UHFFFAOYSA-N ethyl n-ethoxycarbonyliminocarbamate Chemical compound CCOC(=O)N=NC(=O)OCC FAMRKDQNMBBFBR-UHFFFAOYSA-N 0.000 description 1
- 239000005038 ethylene vinyl acetate Substances 0.000 description 1
- OGPBJKLSAFTDLK-UHFFFAOYSA-N europium atom Chemical compound [Eu] OGPBJKLSAFTDLK-UHFFFAOYSA-N 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 239000010685 fatty oil Substances 0.000 description 1
- 210000003754 fetus Anatomy 0.000 description 1
- 229950003499 fibrin Drugs 0.000 description 1
- 239000000796 flavoring agent Substances 0.000 description 1
- UZCGKGPEKUCDTF-UHFFFAOYSA-N fluazinam Chemical compound [O-][N+](=O)C1=CC(C(F)(F)F)=C(Cl)C([N+]([O-])=O)=C1NC1=NC=C(C(F)(F)F)C=C1Cl UZCGKGPEKUCDTF-UHFFFAOYSA-N 0.000 description 1
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 1
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 1
- 238000001215 fluorescent labelling Methods 0.000 description 1
- 235000013355 food flavoring agent Nutrition 0.000 description 1
- 235000003599 food sweetener Nutrition 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 229940050411 fumarate Drugs 0.000 description 1
- 210000001035 gastrointestinal tract Anatomy 0.000 description 1
- 238000005227 gel permeation chromatography Methods 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 229940050410 gluconate Drugs 0.000 description 1
- 229940116332 glucose oxidase Drugs 0.000 description 1
- 235000019420 glucose oxidase Nutrition 0.000 description 1
- 229940097042 glucuronate Drugs 0.000 description 1
- 229930195712 glutamate Natural products 0.000 description 1
- 229940049906 glutamate Drugs 0.000 description 1
- 229960002989 glutamic acid Drugs 0.000 description 1
- 150000002334 glycols Chemical class 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- 239000008187 granular material Substances 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 229940093915 gynecological organic acid Drugs 0.000 description 1
- 201000010536 head and neck cancer Diseases 0.000 description 1
- 208000014829 head and neck neoplasm Diseases 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 229940022353 herceptin Drugs 0.000 description 1
- 125000005549 heteroarylene group Chemical group 0.000 description 1
- 125000005842 heteroatom Chemical group 0.000 description 1
- 125000000623 heterocyclic group Chemical group 0.000 description 1
- ALBYIUDWACNRRB-UHFFFAOYSA-N hexanamide Chemical compound CCCCCC(N)=O ALBYIUDWACNRRB-UHFFFAOYSA-N 0.000 description 1
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 102000051957 human ERBB2 Human genes 0.000 description 1
- 238000009396 hybridization Methods 0.000 description 1
- 239000000017 hydrogel Substances 0.000 description 1
- 229910052739 hydrogen Inorganic materials 0.000 description 1
- QAOWNCQODCNURD-UHFFFAOYSA-M hydrogensulfate Chemical compound OS([O-])(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-M 0.000 description 1
- 229920001477 hydrophilic polymer Polymers 0.000 description 1
- 229920001600 hydrophobic polymer Polymers 0.000 description 1
- 229920003063 hydroxymethyl cellulose Polymers 0.000 description 1
- 229940031574 hydroxymethyl cellulose Drugs 0.000 description 1
- 239000001866 hydroxypropyl methyl cellulose Substances 0.000 description 1
- 235000010979 hydroxypropyl methyl cellulose Nutrition 0.000 description 1
- 229920003088 hydroxypropyl methyl cellulose Polymers 0.000 description 1
- 239000012216 imaging agent Substances 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 238000010166 immunofluorescence Methods 0.000 description 1
- 230000002163 immunogen Effects 0.000 description 1
- 229940027941 immunoglobulin g Drugs 0.000 description 1
- 238000011532 immunohistochemical staining Methods 0.000 description 1
- 238000001114 immunoprecipitation Methods 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 238000011503 in vivo imaging Methods 0.000 description 1
- ADFCQWZHKCXPAJ-UHFFFAOYSA-N indofine Natural products C1=CC(O)=CC=C1C1CC2=CC=C(O)C=C2OC1 ADFCQWZHKCXPAJ-UHFFFAOYSA-N 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 238000013383 initial experiment Methods 0.000 description 1
- 229940102223 injectable solution Drugs 0.000 description 1
- 229940102213 injectable suspension Drugs 0.000 description 1
- 238000011081 inoculation Methods 0.000 description 1
- 125000001905 inorganic group Chemical group 0.000 description 1
- 229910052500 inorganic mineral Inorganic materials 0.000 description 1
- 210000000936 intestine Anatomy 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 230000010189 intracellular transport Effects 0.000 description 1
- 238000007913 intrathecal administration Methods 0.000 description 1
- 230000002601 intratumoral effect Effects 0.000 description 1
- PNDPGZBMCMUPRI-UHFFFAOYSA-N iodine Chemical compound II PNDPGZBMCMUPRI-UHFFFAOYSA-N 0.000 description 1
- 238000005342 ion exchange Methods 0.000 description 1
- 239000012948 isocyanate Substances 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 125000000741 isoleucyl group Chemical group [H]N([H])C(C(C([H])([H])[H])C([H])([H])C([H])([H])[H])C(=O)O* 0.000 description 1
- TWBYWOBDOCUKOW-UHFFFAOYSA-M isonicotinate Chemical compound [O-]C(=O)C1=CC=NC=C1 TWBYWOBDOCUKOW-UHFFFAOYSA-M 0.000 description 1
- QXJSBBXBKPUZAA-UHFFFAOYSA-N isooleic acid Natural products CCCCCCCC=CCCCCCCCCC(O)=O QXJSBBXBKPUZAA-UHFFFAOYSA-N 0.000 description 1
- 239000000644 isotonic solution Substances 0.000 description 1
- 230000000155 isotopic effect Effects 0.000 description 1
- 238000001948 isotopic labelling Methods 0.000 description 1
- 230000000366 juvenile effect Effects 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 238000011005 laboratory method Methods 0.000 description 1
- 229940057428 lactoperoxidase Drugs 0.000 description 1
- 210000002429 large intestine Anatomy 0.000 description 1
- 229960004338 leuprorelin Drugs 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 239000013541 low molecular weight contaminant Substances 0.000 description 1
- 239000008176 lyophilized powder Substances 0.000 description 1
- 230000006674 lysosomal degradation Effects 0.000 description 1
- 210000003712 lysosome Anatomy 0.000 description 1
- 230000001868 lysosomic effect Effects 0.000 description 1
- 239000004325 lysozyme Substances 0.000 description 1
- 229960000274 lysozyme Drugs 0.000 description 1
- 235000010335 lysozyme Nutrition 0.000 description 1
- 210000002540 macrophage Anatomy 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- VZCYOOQTPOCHFL-UPHRSURJSA-N maleic acid Chemical compound OC(=O)\C=C/C(O)=O VZCYOOQTPOCHFL-UPHRSURJSA-N 0.000 description 1
- 210000005075 mammary gland Anatomy 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 230000008774 maternal effect Effects 0.000 description 1
- 108010082117 matrigel Proteins 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 230000010534 mechanism of action Effects 0.000 description 1
- 229940126601 medicinal product Drugs 0.000 description 1
- 210000002780 melanosome Anatomy 0.000 description 1
- 238000005374 membrane filtration Methods 0.000 description 1
- 229910021645 metal ion Inorganic materials 0.000 description 1
- 150000002739 metals Chemical class 0.000 description 1
- 229920000609 methyl cellulose Polymers 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 235000010981 methylcellulose Nutrition 0.000 description 1
- 239000001923 methylcellulose Substances 0.000 description 1
- 229960002900 methylcellulose Drugs 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- 239000004530 micro-emulsion Substances 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 108010029942 microperoxidase Proteins 0.000 description 1
- 239000004005 microsphere Substances 0.000 description 1
- 239000011707 mineral Substances 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 238000000302 molecular modelling Methods 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 150000002772 monosaccharides Chemical class 0.000 description 1
- 238000000465 moulding Methods 0.000 description 1
- 238000011512 multiplexed immunoassay Methods 0.000 description 1
- 235000010460 mustard Nutrition 0.000 description 1
- UPSFMJHZUCSEHU-JYGUBCOQSA-N n-[(2s,3r,4r,5s,6r)-2-[(2r,3s,4r,5r,6s)-5-acetamido-4-hydroxy-2-(hydroxymethyl)-6-(4-methyl-2-oxochromen-7-yl)oxyoxan-3-yl]oxy-4,5-dihydroxy-6-(hydroxymethyl)oxan-3-yl]acetamide Chemical compound CC(=O)N[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1O[C@H]1[C@H](O)[C@@H](NC(C)=O)[C@H](OC=2C=C3OC(=O)C=C(C)C3=CC=2)O[C@@H]1CO UPSFMJHZUCSEHU-JYGUBCOQSA-N 0.000 description 1
- 239000002088 nanocapsule Substances 0.000 description 1
- 239000002105 nanoparticle Substances 0.000 description 1
- 239000013642 negative control Substances 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 239000002547 new drug Substances 0.000 description 1
- GUCVJGMIXFAOAE-NJFSPNSNSA-N niobium-95 Chemical compound [95Nb] GUCVJGMIXFAOAE-NJFSPNSNSA-N 0.000 description 1
- 229920001220 nitrocellulos Polymers 0.000 description 1
- 239000002736 nonionic surfactant Substances 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 239000000346 nonvolatile oil Substances 0.000 description 1
- 238000000655 nuclear magnetic resonance spectrum Methods 0.000 description 1
- 239000012434 nucleophilic reagent Substances 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 125000003835 nucleoside group Chemical group 0.000 description 1
- 239000002773 nucleotide Substances 0.000 description 1
- 125000003729 nucleotide group Chemical group 0.000 description 1
- GYCKQBWUSACYIF-UHFFFAOYSA-N o-hydroxybenzoic acid ethyl ester Natural products CCOC(=O)C1=CC=CC=C1O GYCKQBWUSACYIF-UHFFFAOYSA-N 0.000 description 1
- 235000020824 obesity Nutrition 0.000 description 1
- 229940049964 oleate Drugs 0.000 description 1
- 239000002751 oligonucleotide probe Substances 0.000 description 1
- 238000012634 optical imaging Methods 0.000 description 1
- 150000007524 organic acids Chemical class 0.000 description 1
- 235000005985 organic acids Nutrition 0.000 description 1
- 125000000962 organic group Chemical group 0.000 description 1
- 125000002524 organometallic group Chemical group 0.000 description 1
- 230000001590 oxidative effect Effects 0.000 description 1
- 239000001301 oxygen Substances 0.000 description 1
- 229910052760 oxygen Inorganic materials 0.000 description 1
- LXCFILQKKLGQFO-UHFFFAOYSA-N p-hydroxybenzoic acid methyl ester Natural products COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 1
- 229940014662 pantothenate Drugs 0.000 description 1
- 235000019161 pantothenic acid Nutrition 0.000 description 1
- 239000011713 pantothenic acid Substances 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 230000001717 pathogenic effect Effects 0.000 description 1
- 229960003330 pentetic acid Drugs 0.000 description 1
- 229940111202 pepsin Drugs 0.000 description 1
- 125000001151 peptidyl group Chemical group 0.000 description 1
- 230000010412 perfusion Effects 0.000 description 1
- 210000001322 periplasm Anatomy 0.000 description 1
- 102000013415 peroxidase activity proteins Human genes 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 230000008782 phagocytosis Effects 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- 239000012071 phase Substances 0.000 description 1
- 229960003531 phenolsulfonphthalein Drugs 0.000 description 1
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 1
- 230000004962 physiological condition Effects 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- 229920001983 poloxamer Polymers 0.000 description 1
- 229920001200 poly(ethylene-vinyl acetate) Polymers 0.000 description 1
- 229920000747 poly(lactic acid) Polymers 0.000 description 1
- 229920000728 polyester Polymers 0.000 description 1
- 229920002338 polyhydroxyethylmethacrylate Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 239000000244 polyoxyethylene sorbitan monooleate Substances 0.000 description 1
- 235000010482 polyoxyethylene sorbitan monooleate Nutrition 0.000 description 1
- 229920000053 polysorbate 80 Polymers 0.000 description 1
- 229920002451 polyvinyl alcohol Polymers 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 229940069328 povidone Drugs 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 230000002028 premature Effects 0.000 description 1
- 238000004321 preservation Methods 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 230000003623 progesteronic effect Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- VVWRJUBEIPHGQF-MDZDMXLPSA-N propan-2-yl (ne)-n-propan-2-yloxycarbonyliminocarbamate Chemical compound CC(C)OC(=O)\N=N\C(=O)OC(C)C VVWRJUBEIPHGQF-MDZDMXLPSA-N 0.000 description 1
- IVRIRQXJSNCSPQ-UHFFFAOYSA-N propan-2-yl carbonochloridate Chemical compound CC(C)OC(Cl)=O IVRIRQXJSNCSPQ-UHFFFAOYSA-N 0.000 description 1
- 239000004405 propyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010232 propyl p-hydroxybenzoate Nutrition 0.000 description 1
- 229960003415 propylparaben Drugs 0.000 description 1
- 235000019833 protease Nutrition 0.000 description 1
- 235000019419 proteases Nutrition 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 239000013636 protein dimer Substances 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 238000001742 protein purification Methods 0.000 description 1
- 238000000734 protein sequencing Methods 0.000 description 1
- 239000012460 protein solution Substances 0.000 description 1
- 229940024999 proteolytic enzymes for treatment of wounds and ulcers Drugs 0.000 description 1
- 150000003254 radicals Chemical class 0.000 description 1
- 239000000941 radioactive substance Substances 0.000 description 1
- 229940051022 radioimmunoconjugate Drugs 0.000 description 1
- 230000003439 radiotherapeutic effect Effects 0.000 description 1
- 229910052761 rare earth metal Inorganic materials 0.000 description 1
- 150000002910 rare earth metals Chemical class 0.000 description 1
- 238000001525 receptor binding assay Methods 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000026267 regulation of growth Effects 0.000 description 1
- 239000011347 resin Substances 0.000 description 1
- 229920005989 resin Polymers 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 238000004007 reversed phase HPLC Methods 0.000 description 1
- 238000005096 rolling process Methods 0.000 description 1
- CVHZOJJKTDOEJC-UHFFFAOYSA-N saccharin Chemical compound C1=CC=C2C(=O)NS(=O)(=O)C2=C1 CVHZOJJKTDOEJC-UHFFFAOYSA-N 0.000 description 1
- 229940081974 saccharin Drugs 0.000 description 1
- 235000019204 saccharin Nutrition 0.000 description 1
- 239000000901 saccharin and its Na,K and Ca salt Substances 0.000 description 1
- YGSDEFSMJLZEOE-UHFFFAOYSA-M salicylate Chemical compound OC1=CC=CC=C1C([O-])=O YGSDEFSMJLZEOE-UHFFFAOYSA-M 0.000 description 1
- 229960001860 salicylate Drugs 0.000 description 1
- 238000013391 scatchard analysis Methods 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 150000003958 selenols Chemical class 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 238000004904 shortening Methods 0.000 description 1
- 230000008054 signal transmission Effects 0.000 description 1
- 238000001542 size-exclusion chromatography Methods 0.000 description 1
- 210000000813 small intestine Anatomy 0.000 description 1
- 235000010413 sodium alginate Nutrition 0.000 description 1
- 239000000661 sodium alginate Substances 0.000 description 1
- 229940005550 sodium alginate Drugs 0.000 description 1
- 235000019812 sodium carboxymethyl cellulose Nutrition 0.000 description 1
- 229920001027 sodium carboxymethylcellulose Polymers 0.000 description 1
- MKNJJMHQBYVHRS-UHFFFAOYSA-M sodium;1-[11-(2,5-dioxopyrrol-1-yl)undecanoyloxy]-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].O=C1C(S(=O)(=O)[O-])CC(=O)N1OC(=O)CCCCCCCCCCN1C(=O)C=CC1=O MKNJJMHQBYVHRS-UHFFFAOYSA-M 0.000 description 1
- ULARYIUTHAWJMU-UHFFFAOYSA-M sodium;1-[4-(2,5-dioxopyrrol-1-yl)butanoyloxy]-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].O=C1C(S(=O)(=O)[O-])CC(=O)N1OC(=O)CCCN1C(=O)C=CC1=O ULARYIUTHAWJMU-UHFFFAOYSA-M 0.000 description 1
- VUFNRPJNRFOTGK-UHFFFAOYSA-M sodium;1-[4-[(2,5-dioxopyrrol-1-yl)methyl]cyclohexanecarbonyl]oxy-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].O=C1C(S(=O)(=O)[O-])CC(=O)N1OC(=O)C1CCC(CN2C(C=CC2=O)=O)CC1 VUFNRPJNRFOTGK-UHFFFAOYSA-M 0.000 description 1
- 239000000600 sorbitol Substances 0.000 description 1
- 230000009870 specific binding Effects 0.000 description 1
- 238000004611 spectroscopical analysis Methods 0.000 description 1
- 238000010183 spectrum analysis Methods 0.000 description 1
- 238000012453 sprague-dawley rat model Methods 0.000 description 1
- 230000006641 stabilisation Effects 0.000 description 1
- 238000011105 stabilization Methods 0.000 description 1
- 230000003068 static effect Effects 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- SFVFIFLLYFPGHH-UHFFFAOYSA-M stearalkonium chloride Chemical compound [Cl-].CCCCCCCCCCCCCCCCCC[N+](C)(C)CC1=CC=CC=C1 SFVFIFLLYFPGHH-UHFFFAOYSA-M 0.000 description 1
- 238000011146 sterile filtration Methods 0.000 description 1
- 238000000547 structure data Methods 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- KDYFGRWQOYBRFD-UHFFFAOYSA-N succinic acid Chemical compound OC(=O)CCC(O)=O KDYFGRWQOYBRFD-UHFFFAOYSA-N 0.000 description 1
- 229940014800 succinic anhydride Drugs 0.000 description 1
- 229960002317 succinimide Drugs 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 125000000446 sulfanediyl group Chemical group *S* 0.000 description 1
- 125000002653 sulfanylmethyl group Chemical group [H]SC([H])([H])[*] 0.000 description 1
- 239000003774 sulfhydryl reagent Substances 0.000 description 1
- BDHFUVZGWQCTTF-UHFFFAOYSA-M sulfonate Chemical compound [O-]S(=O)=O BDHFUVZGWQCTTF-UHFFFAOYSA-M 0.000 description 1
- YBBRCQOCSYXUOC-UHFFFAOYSA-N sulfuryl dichloride Chemical compound ClS(Cl)(=O)=O YBBRCQOCSYXUOC-UHFFFAOYSA-N 0.000 description 1
- 239000003765 sweetening agent Substances 0.000 description 1
- 230000008409 synovial inflammation Effects 0.000 description 1
- 238000004885 tandem mass spectrometry Methods 0.000 description 1
- 229940095064 tartrate Drugs 0.000 description 1
- 125000005931 tert-butyloxycarbonyl group Chemical group [H]C([H])([H])C(OC(*)=O)(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- ABZLKHKQJHEPAX-UHFFFAOYSA-N tetramethylrhodamine Chemical compound C=12C=CC(N(C)C)=CC2=[O+]C2=CC(N(C)C)=CC=C2C=1C1=CC=CC=C1C([O-])=O ABZLKHKQJHEPAX-UHFFFAOYSA-N 0.000 description 1
- MPLHNVLQVRSVEE-UHFFFAOYSA-N texas red Chemical compound [O-]S(=O)(=O)C1=CC(S(Cl)(=O)=O)=CC=C1C(C1=CC=2CCCN3CCCC(C=23)=C1O1)=C2C1=C(CCC1)C3=[N+]1CCCC3=C2 MPLHNVLQVRSVEE-UHFFFAOYSA-N 0.000 description 1
- 239000002562 thickening agent Substances 0.000 description 1
- 125000000101 thioether group Chemical group 0.000 description 1
- 150000003568 thioethers Chemical class 0.000 description 1
- 230000025366 tissue development Effects 0.000 description 1
- JOXIMZWYDAKGHI-UHFFFAOYSA-N toluene-4-sulfonic acid Chemical compound CC1=CC=C(S(O)(=O)=O)C=C1 JOXIMZWYDAKGHI-UHFFFAOYSA-N 0.000 description 1
- PYHOFAHZHOBVGV-UHFFFAOYSA-N triazane Chemical compound NNN PYHOFAHZHOBVGV-UHFFFAOYSA-N 0.000 description 1
- 230000001960 triggered effect Effects 0.000 description 1
- 230000007306 turnover Effects 0.000 description 1
- 125000001493 tyrosinyl group Chemical group [H]OC1=C([H])C([H])=C(C([H])=C1[H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 238000005199 ultracentrifugation Methods 0.000 description 1
- 238000002604 ultrasonography Methods 0.000 description 1
- 230000003827 upregulation Effects 0.000 description 1
- 125000005500 uronium group Chemical group 0.000 description 1
- 230000004862 vasculogenesis Effects 0.000 description 1
- 210000005048 vimentin Anatomy 0.000 description 1
- 238000012800 visualization Methods 0.000 description 1
- 238000002424 x-ray crystallography Methods 0.000 description 1
- QCWXUUIWCKQGHC-RNFDNDRNSA-N zirconium-95 Chemical compound [95Zr] QCWXUUIWCKQGHC-RNFDNDRNSA-N 0.000 description 1
Images



















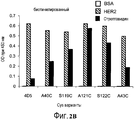
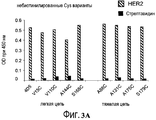
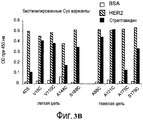



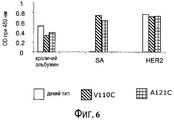


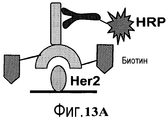
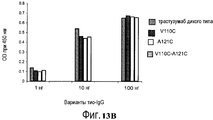
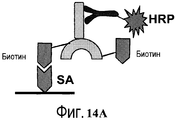
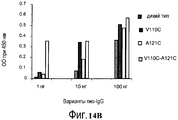











Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K51/00—Preparations containing radioactive substances for use in therapy or testing in vivo
- A61K51/02—Preparations containing radioactive substances for use in therapy or testing in vivo characterised by the carrier, i.e. characterised by the agent or material covalently linked or complexing the radioactive nucleus
- A61K51/04—Organic compounds
- A61K51/08—Peptides, e.g. proteins, carriers being peptides, polyamino acids, proteins
- A61K51/10—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody
- A61K51/1093—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody conjugates with carriers being antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- A61K38/17—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/30—Macromolecular organic or inorganic compounds, e.g. inorganic polyphosphates
- A61K47/42—Proteins; Polypeptides; Degradation products thereof; Derivatives thereof, e.g. albumin, gelatin or zein
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K48/00—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K49/00—Preparations for testing in vivo
- A61K49/06—Nuclear magnetic resonance [NMR] contrast preparations; Magnetic resonance imaging [MRI] contrast preparations
- A61K49/08—Nuclear magnetic resonance [NMR] contrast preparations; Magnetic resonance imaging [MRI] contrast preparations characterised by the carrier
- A61K49/10—Organic compounds
- A61K49/14—Peptides, e.g. proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K49/00—Preparations for testing in vivo
- A61K49/06—Nuclear magnetic resonance [NMR] contrast preparations; Magnetic resonance imaging [MRI] contrast preparations
- A61K49/08—Nuclear magnetic resonance [NMR] contrast preparations; Magnetic resonance imaging [MRI] contrast preparations characterised by the carrier
- A61K49/10—Organic compounds
- A61K49/14—Peptides, e.g. proteins
- A61K49/16—Antibodies; Immunoglobulins; Fragments thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K51/00—Preparations containing radioactive substances for use in therapy or testing in vivo
- A61K51/02—Preparations containing radioactive substances for use in therapy or testing in vivo characterised by the carrier, i.e. characterised by the agent or material covalently linked or complexing the radioactive nucleus
- A61K51/04—Organic compounds
- A61K51/08—Peptides, e.g. proteins, carriers being peptides, polyamino acids, proteins
- A61K51/10—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody
- A61K51/1027—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody against receptors, cell-surface antigens or cell-surface determinants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K51/00—Preparations containing radioactive substances for use in therapy or testing in vivo
- A61K51/02—Preparations containing radioactive substances for use in therapy or testing in vivo characterised by the carrier, i.e. characterised by the agent or material covalently linked or complexing the radioactive nucleus
- A61K51/04—Organic compounds
- A61K51/08—Peptides, e.g. proteins, carriers being peptides, polyamino acids, proteins
- A61K51/10—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody
- A61K51/1045—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody against animal or human tumor cells or tumor cell determinants
- A61K51/1051—Antibodies or immunoglobulins; Fragments thereof, the carrier being an antibody, an immunoglobulin or a fragment thereof, e.g. a camelised human single domain antibody or the Fc fragment of an antibody against animal or human tumor cells or tumor cell determinants the tumor cell being from breast, e.g. the antibody being herceptin
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07C—ACYCLIC OR CARBOCYCLIC COMPOUNDS
- C07C259/00—Compounds containing carboxyl groups, an oxygen atom of a carboxyl group being replaced by a nitrogen atom, this nitrogen atom being further bound to an oxygen atom and not being part of nitro or nitroso groups
- C07C259/04—Compounds containing carboxyl groups, an oxygen atom of a carboxyl group being replaced by a nitrogen atom, this nitrogen atom being further bound to an oxygen atom and not being part of nitro or nitroso groups without replacement of the other oxygen atom of the carboxyl group, e.g. hydroxamic acids
- C07C259/06—Compounds containing carboxyl groups, an oxygen atom of a carboxyl group being replaced by a nitrogen atom, this nitrogen atom being further bound to an oxygen atom and not being part of nitro or nitroso groups without replacement of the other oxygen atom of the carboxyl group, e.g. hydroxamic acids having carbon atoms of hydroxamic groups bound to hydrogen atoms or to acyclic carbon atoms
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07C—ACYCLIC OR CARBOCYCLIC COMPOUNDS
- C07C323/00—Thiols, sulfides, hydropolysulfides or polysulfides substituted by halogen, oxygen or nitrogen atoms, or by sulfur atoms not being part of thio groups
- C07C323/50—Thiols, sulfides, hydropolysulfides or polysulfides substituted by halogen, oxygen or nitrogen atoms, or by sulfur atoms not being part of thio groups containing thio groups and carboxyl groups bound to the same carbon skeleton
- C07C323/51—Thiols, sulfides, hydropolysulfides or polysulfides substituted by halogen, oxygen or nitrogen atoms, or by sulfur atoms not being part of thio groups containing thio groups and carboxyl groups bound to the same carbon skeleton having the sulfur atoms of the thio groups bound to acyclic carbon atoms of the carbon skeleton
- C07C323/60—Thiols, sulfides, hydropolysulfides or polysulfides substituted by halogen, oxygen or nitrogen atoms, or by sulfur atoms not being part of thio groups containing thio groups and carboxyl groups bound to the same carbon skeleton having the sulfur atoms of the thio groups bound to acyclic carbon atoms of the carbon skeleton with the carbon atom of at least one of the carboxyl groups bound to nitrogen atoms
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/32—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against translation products of oncogenes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K19/00—Hybrid peptides, i.e. peptides covalently bound to nucleic acids, or non-covalently bound protein-protein complexes
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; CARE OF BIRDS, FISHES, INSECTS; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2267/00—Animals characterised by purpose
- A01K2267/03—Animal model, e.g. for test or diseases
- A01K2267/0331—Animal model for proliferative diseases
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07C—ACYCLIC OR CARBOCYCLIC COMPOUNDS
- C07C2601/00—Systems containing only non-condensed rings
- C07C2601/12—Systems containing only non-condensed rings with a six-membered ring
- C07C2601/14—The ring being saturated
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/51—Complete heavy chain or Fd fragment, i.e. VH + CH1
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/55—Fab or Fab'
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/62—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising only variable region components
- C07K2317/624—Disulfide-stabilized antibody (dsFv)
Abstract
Изобретение относится к области биотехнологии, конкретно к получению меченых антител, и может быть использовано для диагностики. Антитела конструируют с заменой одной или нескольких аминокислот исходного антитела на не являющиеся перекрестно связанными, высокореакционноспособные цистеиновые аминокислоты. Сконструированные антитела с цистеиновыми заменами (Ab) конъюгируют с одной или несколькими метками с комплексом циркония (Z) через линкер (L) для получения конъюгатов меченных цирконием сконструированных антител с цистеиновыми заменами, обладающих формулой I: Ab-(L-Z)p, где p представляет собой 1-4. Изобретение позволяет повысить эффективность диагностики за счет использования меченных радиоактивным цирконием сконструированных антител с цистеиновыми заменами для получения изображений. 5 н. и 10 з.п. ф-лы, 29 ил., 12 табл., 23 пр.
Description
По настоящей заявке испрашивается приоритет заявки США, имеющей серийный № 12/612912 и поданной 5 ноября 2009 г, являющейся частичным продолжением заявки США, имеющей серийный № 12/399241 и поданной 6 марта 2009 г, являющейся продолжением заявки США, имеющей серийный № 11/233258 и поданной 22 сентября 2005 г., в настоящее время заявка США 7521541, поданная 21 апреля 2009 г., а также по пункту 35 USC § 119(e) испрашивается приоритет Предварительной патентной заявки США, имеющей серийный № 60/612468 и поданной 23 сентября 2004 г., и Предварительной патентной заявки США, имеющей серийный № 60/696353 и поданной 30 июня 2005 г., полное содержание каждой из которых приведено в настоящем документе в качестве ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится, главным образом, к сконструированным антителам с реакционноспособными остатками цистеина, и более конкретно, к антителам с терапевтическими или диагностическими применениями. Сконструированные антитела с цистеиновыми заменами можно конъюгировать с химиотерапевтическими средствами, токсинами, аффинными лигандами, такими как биотин, и метками для детекции, такими как радиоактивные изотопы и флуорофоры. Изобретение также относится к способам использования антител и соединений - конъюгатов антитело-лекарственное средство для диагностики или лечения клеток млекопитающих или связанных с ними патологических состояний in vitro, in situ и in vivo.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Получение изображений на молекулярном уровне является важным инструментом для разработки и оценки новых лекарственных средств. Иммуно-позитронная эмиссионная томография (ИммуноПЭТ) представляет собой быстро развивающийся способ отслеживания и количественной оценки моноклональных антител (mAb) in vivo, поскольку в нем эффективно скомбинирована высокая чувствительность ПЭТ с высокой чувствительностью mAb. ИммуноПЭТ приближается к тому, чтобы стать клиническим способом выбора для неинвазивной диагностики, обеспечивающим «всеобъемлющее иммуногистохимическое окрашивание in vivo» (van Dongen GA, et al. «ImmunoPET: a navigator in monoclonal antibody development and applications» Oncologist 2007;12: 1379-89). Поскольку для ИммуноПЭТ необходимо присоединение позитронно-активного радиоизотопа к намеченной специфической молекуле, необходимо, чтобы биологическое время полужизни молекулы совпадало со временем полужизни радиоактивного изотопа (Verel I, et al. «The promise of immuno-PET in radioimmunotherapy» J Nucl Med 2005; 46 Suppl 1: 164S-71S). Хотя антитела (~150 кДа) обладают временем полужизни в сыворотке в диапазоне от суток до недель, получение изображений, как правило, обеспечивает максимальные соотношения сигнала к фону через 2-6 суток после введения метки на основе антител, что требует использования радиоактивных изотопов, таких как 89Zr и 124I со временем полужизни 3,3 суток и 4,2 суток, соответственно. К сожалению, время полужизни легко доступного 64Cu (12,7 ч) является слишком коротким для получения изображений с хорошим контрастом в эти временные рамки.
Разработка средств на основе Mab для получения изображений при позитронно-эмиссионной томографии (ПЭТ) (Иммуно-ПЭТ) продолжает являться многообещающим инструментом для локализации и количественного определения молекулярных мишеней и может улучшать клиническую диагностику патологических состояний (van Dongen et al (2007) Oncologist 12;1379-89; Williams et al (2001) Cancer Biother Radiopharm 16:25-35; Holliger et al (2005) Nat Biotechnol 23: 1126-36). ПЭТ представляет собой способ получения изображений на молекулярном уровне, все больше используемый для детекции заболеваний. Системы получения изображений ПЭТ создают изображения на основе распределения позитронно-активных изотопов в ткани пациента. Изотопы, как правило, вводят пациенту посредством инъекции молекул-зондов, содержащих позитронно-активный изотоп, такой как F-18, C-11, N-13 или O-15, ковалентно присоединенный к молекуле, поддающейся легкому метаболизму или локализации в организме (например, глюкоза), или химически связывающейся с участками рецепторов внутри организма. В некоторых случаях изотоп вводят пациенту в виде ионного раствора или посредством ингаляции. Небольшие средства для получения изображения иммуно-ПЭТ, такие как Fab-фрагменты антител (50 кДа) или диатела, спаренные димеры ковалентно связанной VH-VL области Mab, 55 кДа (Shively et al (2007) J Nucl Med 48: 170-2), могут являться особенно пригодными, поскольку они обладают коротким временем полужизни, высоким проникновением в ткани, и достигают оптимального соотношения опухоли к фону между двумя и четырьмя часами после инъекции, облегчая использование изотопов с коротким временем полужизни, таких как широкодоступный 18F (109,8 мин).
Иод 124 (124I) присоединяли к антителу 3F9 и использовали для дозиметрической оценки радиммунотерапии нейробластомы (Larson SM, et al «PET scanning of iodine-124-3F9 as an approach to tumor dosimetry during treatment planning for radioimmunotherapy in a child with neuroblastoma» J Nucl Med 1992;33:2020-3). Позже, с возникновением более сложных инструментов ПЭТ и улучшенных способов мечения радиоактивным иодом, 124I использовали в многочисленных исследованиях иммуноПЭТ (Verel I, et al «High-quality 124I-labelled monoclonal antibodies for use as PET scouting agents prior to 131I-radioimmunotherapy» European journal of nuclear medicine и molecular imaging 2004; 31:1645-52; Lee FT et al «Immuno-PET of human colon xenograft-bearing BALB/c nude mice using 124I-CDR-grafted humanized A33 monoclonal antibody» J Nucl Med 2001;42:764-9; Sundaresan G, et al. «124I-labeled engineered anti-CEA minibodies and diabodies allow high-contrast, antigen-specific small-animal PET imaging of xenografts in athymic mice» J Nucl Med 2003; 44:1962-9; Jain M and Batra SK. «Genetically engineered antibody fragments and PET imaging: a new era of radioimmunodiagnosis» J Nucl Med 2003; 44:1970-2; Gonzalez Trotter DE et al. «Quantitation of small-animal (124)I activity distributions using a clinical PET/CT scanner» J Nucl Med 2004; 45:1237-44; Robinson MK, et al. «Quantitative immuno-positron emission tomography imaging of HER2-positive tumor xenografts with an iodine-124 labeled anti-HER2 diabody» Cancer Res 2005; 65:1471-8; Jayson GC et al. «Molecular imaging and biological evaluation of HuMV833 anti-VEGF antibody: implications for trial design of antiangiogenic antibodies» J Natl Cancer Inst 2002; 94:1484-93; Divgi CR, et al. «Preoperative characterisation of clear-cell renal carcinoma using iodine-124-labelled antibody chimeric G250 (124I-cG250) and PET in patients with renal masses: a phase I trial» Lancet Oncol 2007; 8:304-10). Несмотря на относительно простые способы мечения радиоактивным иодом, доступные для присоединения 124I к mAb, важные ограничения замедляют повсеместное доклиническое использование этого радиоактивного изотопа. Следует отметить, что схема распада комплекса включает позитроны с высокой энергией (β+ макс. 1,5 и 2,1 МэВ), что отрицательно влияет на разрешение микроПЭТ для мелких животных. Кроме того, интернализованные меченные иодом белки подвергаются ферментативному деиодированию с быстрым выведением свободного иодида из клеток-мишеней, что приводит к тому, что изображения ПЭТ не отражают действительного поглощения mAb «Perera RM et al. «Internalization, intracellular trafficking, and biodistribution of monoclonal antibody 806: a novel anti-epidermal growth factor receptor antibody» Neoplasia (New York, N.Y 2007;9: 1099-110). Использование 89Zr преодолевает эти недостатки, поскольку позитроны, испускаемые при распаде 89Zr (β макс. 897 кэВ), обеспечивают разрешение микроПЭТ, сравнимое с 18F и 11C (приблизительно 1 мм). Метаболиты интернализованных 89Zr-mAb отслеживают также внутри клеток в лизосомах, обеспечивая лучшую корреляцию действительного поглощения mAb с получением изображений ПЭТ (van Dongen GA, et al. «Immuno-PET: a navigator in monoclonal antibody development and applications» Oncologist 2007;12: 1379-89).
Общепринятые средства присоединения, т.е., связывания посредством ковалентных связей, метки, такой как радиоактивный изотоп, флуоресцентный краситель или группа лекарственного средства, к антителу, как правило, приводит к гетерогенной смеси молекул, где группы метки присоединены к ряду участков антитела. Например, цитотоксические средства как правило, конъюгируют с антителами через многочисленные остатки лизина в антителе, получая гетерогенную смесь конъюгата антитело-лекарственное средство. В зависимости от условий реакции, гетерогенная смесь, как правило, содержит распределение антител с присоединенными от 0 до приблизительно 8 или более групп лекарственного средства. Кроме того, в пределах каждой подгруппы конъюгатов с определенным целочисленным соотношением групп лекарственного средства к антителу, присутствует потенциально гетерогенная смесь, где группа лекарственного средства присоединена к различным участкам антитела. Аналитические и препаративные способы являются неадекватными для разделения и характеризации видов молекул конъюгата антитело-лекарственное средство внутри гетерогенной смеси, полученной в результате реакции конъюгации. Антитела представляют собой большие, сложные и структурно разнообразные биомолекулы, часто со множеством реакционноспособных функциональных групп. Их реакционная способность по отношению к линкерным реагентам и промежуточным соединениям лекарственное средство-линкер зависит от таких факторов, как pH, концентрация, концентрация соли и сорастворители. Более того, способ многоступенчатой конъюгации может являться невоспроизводимым из-за сложностей в контролировании условий реакции и характеризации реагентов и промежуточных соединений.
Тиолы цистеина являются реакционноспособными при нейтральном pH, в отличие от большинства аминов, которые являются протонированными и менее нуклеофильными при pH около 7. Поскольку свободные тиоловые (RSH, сульфгидрильные) группы являются относительно реакционноспособными, белки с остатками цистеина часто существуют в их окисленной форме в виде связанных дисульфидными мостиками олигомеров или имеют дисульфидные группы, образующие внутренние мостики. Внеклеточные белки, как правило, не имеют свободных тиолов (Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London, at page 55). Количество свободного тиола в белке можно оценивать общепринятым анализом Элмана. Иммуноглобулин M является примером пентамера, связанного дисульфидными связями, в то время как иммуноглобулин G является примером белка с внутренними дисульфидными мостиками, связывающими субъединицы вместе. В белках, подобных этим, необходимо восстановление дисульфидных связей с помощью такого реагента, как дитиотреитол (DTT) или селенол (Singh et al (2002) Anal. Biochem. 304: 147-156), для получения реакционноспособного свободного тиола. Этот способ может приводить к потере третичной структуры и антигенсвязывающей специфичности антитела.
Тиоловые группы остатков цистеина антител, как правило, являются более реакционноспособными, т.е. более нуклеофильными, по отношению к электрофильным реагентам для конъюгации, чем аминогруппы или гидроксильные группы антител. Остатки цистеина вводили в белки способами генной инженерии для формирования ковалентных присоединений к лигандам или для формирования новых внутримолекулярных дисульфидных связей (Better et al (1994) J. Biol. Chem. 269(13):9644-9650; Bernhard et al (1994) Bioconjugate Chem. 5: 126-132; Greenwood et al (1994) Therapeutic Immunology 1:247-255; Tu et al (1999) Proc. Natl. Acad. Sci USA 96:4862-4867; Kanno et al (2000) J. of Biotechnology, 76:207-214; Chmura et al (2001) Proc. Nat. Acad. Sci. USA 98(15):8480-8484; US 6248564). Однако, конструирование тиоловых групп цистеина посредством мутации различных аминокислотных остатков белка до цистеиновых аминокислот потенциально является проблематичным, в частности, в случае неспаренных (свободных Cys) остатков или остатков, относительно доступных для реакции или окисления. В концентрированных растворах белка, в периплазме E. coli, культуральных супернатантах, или частично или полностью очищенном белке, неспаренные остатки Cys на поверхности белка могут спариваться и окисляться с формированием межмолекулярных дисульфидных мостиков, и таким образом, димеров или мультимеров белка. Формирование дисульфидных димеров делает новые Cys нереакционноспособными для конъюгации с лекарственным средством, лигандом или другой меткой. Более того, если в белке с помощью окисления образуется внутримолекулярная дисульфидная связь между новым сконструированным Cys и существующим остатком Cys, обе группы Cys недоступны для участия в активном участке и взаимодействий. Более того, белок может становиться неактивным или неспецифическим из-за неправильного сворачивания или потери третичной структуры (Zhang et al (2002) Anal. Biochem. 311:1-9).
Сайт-специфическая конъюгация является предпочтительной по сравнению с модификацией случайных аминогрупп, поскольку она позволяет химическую модификацию в участке, отдаленном от участка связывания, обеспечивая полное сохранение биологической активности и позволяя контроль над возможным количеством добавленных простетических групп. Сконструированные антитела с цистеиновыми заменами разработаны в форме FAB-фрагментов антител (тиоFab) и экспрессированы в форме полноразмерных, моноклональных (тиоMab) антител IgG. См.: US 7521541; Junutula JR et al. «Rapid identification of reactive cysteine residues for site-specific labeling of antibody-Fabs» J Immunol Methods 2008; 332:41-52; Junutula JR et al. «Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index» (2008) Nat Biotechnol. 26:925-32, содержание которых приведено в качестве ссылки. Антитела ТиоFab и ТиоMab конъюгировали посредством линкеров на тиолах нового введенного цистеина с помощью вступающих в реакцию с тиолом линкерных реагентов и реагентов лекарственное средство-линкер для получения конъюгатов с лекарственным средством сконструированного антитела с цистеиновыми заменами (Тио ADC) с противораковыми свойствами, включая анти-MUC16 (US 2008/0311134), анти-CD22 (US 2008/0050310), анти-ROBO4 (US 2008/0247951), анти-TENB2 (US 2009/0117100), анти-CD79B (US 2009/0028856; US 2009/0068202) Тио ADC.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Соединения по изобретению включают сконструированные антитела с цистеиновыми заменами, где одна или несколько аминокислот исходного антитела заменены на свободную аминокислоту цистеин. Сконструированное антитело с цистеиновыми заменами содержит одну или несколько свободных цистеиновых аминокислот, обладающих значением реакционной способности тиола в диапазоне от 0,6 до 1,0. Свободная аминокислота цистеин представляет собой остаток цистеина, введенный при конструировании в исходное антитело и не являющийся частью дисульфидного мостика.
Сконструированные антитела с цистеиновыми заменами могут являться пригодными для диагностики и лечения злокачественных опухолей и включают антитела, специфические для рецепторов поверхности клеток и трансмембранных рецепторов, и для опухолеассоциированных антигенов (TAA). Такие антитела можно использовать в форме голых антител (не конъюгированных с группой лекарственного средства или метки) или в форме конъюгатов антитело-цирконий (AZC).
Варианты осуществления способов получения и скрининга сконструированных антител с цистеиновыми заменами включают варианты, где исходное антитело представляет собой фрагмент антитела, такой как hu4D5Fabv8. Исходное антитело может представлять собой также слитый белок, содержащий альбуминсвязывающую пептидную последовательность (ABP). Исходное антитело может представлять собой также гуманизированное антитело, выбранное из huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 (трастузумаб).
Сконструированные антитела с цистеиновыми заменами по изобретению можно сайт-специфическим и эффективным образом присоединять к вступающему в реакцию с тиолом реагенту. Вступающий в реакцию с тиолом реагент может представлять собой содержащий радиоактивный изотоп реагент, мультифункциональный линкерный реагент, связывающий метку реагент, содержащий флуорофор реагент или промежуточное соединение лекарственное средство-линкер.
Сконструированное антитело с цистеиновыми заменами можно метить поддающейся детекции меткой, иммобилизовывать на твердофазной подложке и/или конъюгировать с группой лекарственного средства.
Другой аспект изобретения относится к меченному цирконием сконструированному антителу с цистеиновыми заменами, содержащему сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающему формулой I:

где p представляет собой 1-4.
Другой аспект изобретения относится к меченному десферриоксамином сконструированному антителу с цистеиновыми заменами, содержащему сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающему формулой II:
где L-Df выбран из:

где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4.
Другой аспект изобретения относится к реагенту для мечения десферриоксамином, выбранному из структур:

где R выбран из:

Другой аспект изобретения относится к способу получения меченного десферриоксамином сконструированного антитела с цистеиновыми заменами, содержащего сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающего формулой II:
где L-Df выбран из:

где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4;
где способ включает реакцию композиции, выбранной из структур:

где R выбран из:

со сконструированным антителом с цистеиновыми заменами, имеющим одну или несколько свободных цистеиновых аминокислот, посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
Другой аспект изобретения относится к способу получения меченного цирконием сконструированного антитела с цистеиновыми заменами, содержащего сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающего формулой I:
где p представляет собой 1-4;
где способ включает образование комплекса содержащего цирконий реагента с меченным десферриоксамином сконструированным антителом с цистеиновыми заменами, содержащим сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающим формулой II:
где L-Df выбран из:

где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4;
посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
Другой аспект изобретения относится к способу получения изображения, включающему:
введение меченного цирконием сконструированного антитела с цистеиновыми заменами животному; и
детекцию in vivo присутствия меченного цирконием сконструированного антитела с цистеиновыми заменами посредством получения изображений,
где меченное цирконием сконструированное антитело с цистеиновыми заменами содержит сконструированное антитело с цистеиновыми заменами (Ab), имеющее одну или несколько свободных цистеиновых аминокислот, конъюгированных с одним или несколькими комплексами циркония (Z) посредством линкера (L) и имеющее формулу I:

где p представляет собой 1-4.
Другой аспект изобретения относится к диагностическим применениям соединений и композиций, описанных в настоящем документе.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фигуре 1A показано трехмерное представление фрагмента антитела hu4D5Fabv7 в координатах кристалла, полученных с помощью рентгеновского излучения. Положения в структуре иллюстративных сконструированных остатков Cys тяжелой и легкой цепей пронумерованы (в соответствии с последовательной системой нумерации).
На фигуре 1B показана схема последовательной нумерации (верхний ряд), начиная с N-конца, по сравнению со схемой нумерации Kabat (нижний ряд) для 4D5v7fabH. Вставки с нумерацией по Kabat обозначены a, b, c.
На фигурах 2A и 2B показаны измерения связывания с детекцией оптической плотности при 450 нм для hu4D5Fabv8 и Cys мутантных (ТиоFab) фаговых вариантов hu4D5Fabv8: (A) небиотинилированного фаг-hu4D5Fabv8 и (B) биотинилированного фаг-hu4D5Fabv8 (B) посредством анализа PHESELECTOR для взаимодействий с BSA (незакрашенный столбец), HER2 (заштрихованный столбец) или стрептавидином (закрашенный столбец).
На фигурах 3A и 3B показаны измерения связывания с детекцией оптической плотности при 450 нм для hu4D5Fabv8 (слева) и Cys мутантных (ТиоFab) вариантов hu4D5Fabv8: (A) небиотинилированного фаг-hu4D5Fabv8 и (B) биотинилированного фаг-hu4D5Fabv8 посредством анализа PHESELECTOR для взаимодействий с: BSA (незакрашенный столбец), HER2 (заштрихованный столбец) и стрептавидином (незакрашенный столбец). Варианты легкой цепи расположены на левой стороне, а варианты тяжелой цепи расположены на правой стороне. Реакционная способность тиола = OD450нм для связывания стрептавидина ÷ OD450нм для связывания HER2 (антитела).
На фигуре 4A показаны значения относительной доступности поверхности остатков в hu4D5Fabv8 дикого типа. Участки легкой цепи расположены на левой стороне, а участки тяжелой цепи расположены на правой стороне.
На фигуре 4B показаны измерения связывания с детекцией оптической плотности при 450 нм для вариантов биотинилированного hu4D5Fabv8 (слева) и Cys мутанта (ТиоFab) hu4D5Fabv8 при взаимодействиях с HER2 (сутки 2), стрептавидином (SA) (сутки 2), HER2 (сутки 4) и SA (сутки 4). Cys варианты фаг-hu4D5Fabv8 выделяли и хранили при 4°C. На сутки 2 или сутки 4 проводили конъюгацию с биотином с последующими анализами PHESELECTOR для мониторирования их взаимодействия с Her2 и стрептавидином, как описано в примере 2, и тестировали стабильность реакционноспособных тиоловых групп на сконструированных вариантах ТиоFab.
На фигуре 5 показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированного с биотин-малеимидом-hu4D5Fabv8 (A121C) и небиотинилированного hu4D5Fabv8 дикого типа при связывании со стрептавидином и HER2. Каждый Fab тестировали при 2 нг и 20 нг.
На фигуре 6 показан анализ ELISA с детекцией оптической плотности при 450 нм для биотинилированного ABP-hu4D5Fabv8 дикого типа (wt), и цистеиновых мутантов ABP-hu4D5Fabv8 V110C и A121C при связывании с кроличьим альбумином, стрептавидином (SA) и HER2.
На фигуре 7 показан анализ ELISA с детекцией оптической плотности при 450 нм для цистеиновых мутантов (вариантов ТиоFab) биотинилированного ABP-hu4D5Fabv8: (слева направо) варианты с одним Cys ABP-V110C, ABP-A121C и варианты с двойным Cys ABP-V110C-A88C и ABP-V110C-A121C при связывании с кроличьим альбумином, HER2 и стрептавидином (SA), и использовании в качестве зондов Fab-HRP или SA-HRP.
На фигуре 8 показано связывание фага с биотинилированным ТиоFab и антитела против фага с HRP с HER2 (вверху) и стрептавидином (внизу).
На фигуре 13A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2, со связыванием меченного HRP вторичного антитела для детекции оптической плотности.
На фигуре 13B показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированных с биотином-малеимидом вариантов тио-трастузумаба и небиотинилированного трастузумаба дикого типа при связывании с иммобилизованным HER2. Слева направо: V110C (один cys), A121C (один cys), V110C/A121C (двойной cys) и трастузумаб. Каждый вариант тио-IgG и трастузумаб тестировали при 1, 10 и 100 нг.
На фигуре 14A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2, со связыванием биотина с анти-IgG-HRP для детекции оптической плотности.
На фигуре 14B показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированных с биотин-малеимидом вариантов тио-трастузумаба и небиотинилированного трастузумаба дикого типа при связывании с иммобилизованным стрептавидином. Слева направо: V110C (один cys), A121C (один cys), V110C/A121C (двойной cys) и трастузумаб. Каждый вариант тио-IgG и трастузумаб тестировали при 1, 10 и 100 нг.
На фигуре 15 показан общий способ подготовки сконструированного антитела с цистеиновыми заменами (ТиоMab), экспрессированного из культуры клеток, для конъюгации.
На фигуре 16 показан анализ электрофорезом в невосстанавливающем (вверху) и восстанавливающем (внизу) денатурирующем полиакриламидном геле вариантов 2H9 ТиоMab Fc (слева направо, дорожки 1-9): A339C; S337C; S324C; A287C; V284C; V282C; V279C; V273C, и 2H9 дикого типа после очистки на иммобилизованном белке A. Дорожка справа представляет собой маркер размера, указывающий на то, что интактные белки составляют приблизительно 150 кДа, фрагменты тяжелой цепи приблизительно 50 кДа, и фрагменты легкой цепи приблизительно 25 кДа.
На фигуре 17A показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (+DTT) (справа) денатурирующем полиакриламидном геле вариантов 2H9 ТиоMab варианты (слева направо, дорожки 1-4): L-V15C; S179C; S375C; S400C, после очистки на иммобилизованном белке A.
На фигуре 17B показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (+DTT) (справа) денатурирующем полиакриламидном геле вариантов 2H9 и 3A5 ТиоMab, после очистки на иммобилизованном белке A.
На фигуре 18 показан анализ вестерн-блоттингом вариантов биотинилированного Тио-IgG. Варианты ТиоMab 2H9 и 3A5 анализировали электрофорезом в восстанавливающем денатурирующем полиакриламидном геле, белки переносили на нитроцеллюлозную мембрану. В качестве зондов для анализа присутствия антитела и конъюгированного биотина служили анти-IgG-HRP (вверху) и стрептавидин-HRP (внизу), соответственно. Дорожка 1: 3A5 H-A121C. Дорожка 2: 3A5 L-V110C. Дорожка 3: 2H9 H-A121C. Дорожка 4: 2H9 L-V110C. Дорожка 5: 2H9 дикого типа.
На фигуре 19 показан анализ ELISA связывания вариантов биотинилированного 2H9 со стрептавидином посредством использования в качестве зонда анти-IgG-HRP и измерения оптической плотности при 450 нм (верхняя столбчатая диаграмма). Нижняя схематическая диаграмма изображает дизайн эксперимента, используемый для анализа ELISA.
На фигуре 20 показаны бифункциональные реагенты для присоединения хелатирующего агента для 89Zr десферриоксамина B (Df, вверху) к белкам с использованием реакционноспособных по отношению к аминогруппам линкеров, TFP-N-SucDf и Df-Bz-NCS (в центре), и реакционноспособных по отношению к тиолу линкеров, Df-Chx-Mal, Df-Bac и Df-lac (внизу).
На фигуре 21 показано получение Df-Chx-Mal, Df-Bac, Df-Iac и конъюгация с тио-трастузумабом через остатки Cys, включенные в тяжелую цепь Fab. Условия реакции: i. DIEA, DMF/H2O (10:1), RT, 0,5-1 ч; ii. DIEA, DMF, 0°C, 4 ч; iii. pH 7,5, RT, 1 ч; iv. pH 9, RT, 5 ч; v. pH 9, RT, 2 ч.
На фигуре 22 показано образование хелатов оксалата циркония-89 с меченным десферриоксамином сконструированным антителом с цистеиновыми заменами, таких как варианты Df-линкер-трастузумаба, содержащие четыре линкера: N-Suc, Bz-SCN, Chx-малеимид (CHx-Mal), или ацетил (Ac).
На фигуре 23 показан масс-спектрометрический анализ восстановленных антител, показывающий отдельные сигналы от легкой и тяжелой цепей. A: тио-трастузумаб, B: Df-Ac-тио-трастузумаб (с использованием Df-Bac), и C: Df-Ac-тио-трастузумаб (с использованием Df-Iac), и D: Df-Chx-Mal-тио-трастузумаб.
На фигуре 24 показана стабильность 89Zr-Chx-Mal-тио-трастузумаба (незакрашенные круги) и 89Zr-Df-Ac-тио-трастузумаба (закрашенные круги) в сыворотке мыши при 37°C (n=3).
На фигуре 25 показаны репрезентативные изображения целого организма (проекция максимальной интенсивности), полученные через 96 часов после болюсной инъекции в хвостовую вену 100 мКи 89Zr-трастузумаба, полученного с использованием четырех различных линкеров (Bz-SCN, N-Suc, Chx-Mal и Ac).
На фигуре 26 показано поглощение in vivo в избранных тканях через 24, 96 и 144 часа после инъекции, как измерено посредством ПЭТ.
ПОДРОБНОЕ ОПИСАНИЕ ИЛЛЮСТРАТИВНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
В настоящее время приведены подробные ссылки на конкретные варианты осуществления изобретения, примеры которых проиллюстрированы в сопровождающих структурах и формулах. В то время как изобретение описано в отношении перечисленных вариантов осуществления, понятно, что они не предназначены для ограничения изобретения этими вариантами осуществления. С другой стороны, изобретение предназначено, чтобы включать все альтернативы, модификации и эквиваленты, которые могут быть включены в объем настоящего изобретения, как определено формулой изобретения.
Специалисту в данной области известно множество способов и материалов, подобных или эквивалентных описанным в настоящем документе, которые можно использовать в практике настоящего изобретения. Настоящее изобретение никаким образом не ограничено описанными способами и материалами.
Если не определено иначе, технические и научные термины, используемые в настоящем документе, обладают таким же значением, какое является общепринятым для специалиста в области, к которой относится это изобретение, и соответствует: Singleton et al (1994) Dictionary of Microbiology and Molecular Biology, 2nd Ed., J. Wiley & Sons, New York, NY; и Janeway, C, Travers, P., Walport, M., Shlomchik (2001) Immunobiology, 5th Ed., Garland Publishing, New York.
ОПРЕДЕЛЕНИЯ
Если не указано иначе, следующие термины и фразы, как применяют в настоящем документе, предназначены, чтобы обладать следующими значениями:
При использовании торговых наименований в настоящем документе, авторы настоящего изобретения подразумевают независимое включение состава продукта под торговым наименованием, аналога патентованного лекарственного средства и активного фармацевтического ингредиента(ингредиентов) продукта под торговым наименованием.
Термин «антитело» в настоящем документе используют в самом широком смысле, и он конкретно охватывает моноклональные антитела, поликлональные антитела, димеры, мультимеры, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антитела, пока они обладают желательной биологической активностью (Miller et al (2003) Jour. of Immunology 170:4854-4861). Антитела могут являться мышиными, человеческими, гуманизированными, химерными или полученными из других видов. Антитело представляет собой белок, образуемый иммунной системой, способный узнавать и связывать специфический антиген. (Janeway, C, Travers, P., Walport, M., Shlomchik (2001) Immuno Biology, 5th Ed., Garland Publishing, New York). Антиген-мишень, как правило, обладает несколькими участками связывания, называемыми также эпитопами, узнаваемыми CDR множества антител. Каждое антитело, которое специфически связывается с отличным эпитопом, обладает отличной структурой. Таким образом, один антиген может иметь более одного соответствующего антитела. Антитело включает полноразмерную молекулу иммуноглобулина или иммунологически активную часть полноразмерной молекулы иммуноглобулина, т.е., молекулы, содержащей антигенсвязывающий участок, содержащий антигенсвязывающий участок, иммуноспецифически связывающий антиген интересующей мишени или его часть, где такие мишени включают в качестве неограничивающих примеров, клетку или клетки злокачественных опухолей, продуцирующие аутоиммунные антитела, ассоциированные с аутоиммунным заболеванием. Иммуноглобулин, описанный в настоящем документе, может относиться к любому типу (например, IgG, IgE, IgM, IgD, и IgA), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу молекулы иммуноглобулина. Иммуноглобулины могут происходить из любых видов. В одном аспекте, однако, иммуноглобулин имеет человеческое, мышиное или кроличье происхождение.
«Фрагменты антител» содержат часть полноразмерного антитела, как правило, его антигенсвязывающую или вариабельную область. Примеры фрагментов антител включают Fab, Fab', F(ab')2 и Fv фрагменты; диатела; линейные антитела; миниантитела (Olafsen et al (2004) Protein Eng. Design & Sel. 17(4):315-323), фрагменты, полученные из экспрессирующей библиотеки Fab, анти-идиотипические (анти-Id) антитела, CDR (определяющую комплементарность область) и эпитоп-связывающие фрагменты любого из вышеуказанного, специфически связывающиеся с антигенами клеток злокачественных опухолей, вирусными антигенами или антигенами микроорганизмов, молекулы одноцепочечных антител; и мультиспецифические антитела, сформированные из фрагментов антител.
Термин «моноклональное антитело», как применяют в настоящем документе, относится к антителу, полученному из популяции по существу однородных антител, т.е., отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, являясь направленными против одного антигенного участка. Более того, в отличие от препаратов поликлональных антител, включающих различные антитела, нацеленные против различных детерминант (эпитопов), каждое моноклональное антитело нацелено против отдельной детерминанты антигена. В дополнение к их специфичности, моноклональные антитела являются преимущественными в том смысле, что их можно синтезировать без контаминации другими антителами. Определение «моноклональные» указывает на характер антитела, как полученного из по существу гомогенной популяции антител, и его не следует истолковывать как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела для использования в соответствии с настоящим изобретением можно получать способом гибридомы, впервые описанным Kohler et al (1975) Nature 256:495, или их можно получать способами рекомбинантной ДНК (см., например: US 4816567; US 5807715). Моноклональные антитела можно выделять также из фаговых библиотек антител с использованием способов, описанных, например, в Clackson et al (1991) Nature, 352:624-628; Marks et al (1991) J. Mol. Biol, 222:581-597.
Моноклональные антитела в настоящем документе конкретно включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как остаток цепи(цепей) является идентичным или гомологичным соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих к другому классу или подклассу антител, так же как фрагменты таких антител, пока они обладают желательной биологической активностью (US 4816567; и Morrison et al (1984) Proc. Natl. Acad. Sci. USA, 81:6851-6855). Интересующие химерные антитела в настоящем документе включают «приматизированные» антитела, содержащие антигенсвязывающие последовательности вариабельного домена, полученные из не относящегося к человеку примата (например, обезьяны Старого Света, обезьяны и т.д.), и человеческие последовательности константной области.
«Интактное антитело» в настоящем документе представляет собой антитело, содержащее домены VL и VH, а также константный домен (CL) легкой цепи и константные домены тяжелой цепи CH1, CH2 и CH3. Константные домены могут представлять собой константные домены с природной последовательностью (например, константные домены с человеческой природной последовательностью) или вариант их аминокислотной последовательности. Интактное антитело может обладать одной или несколькими «эффекторными функциями», которые относятся к тем видам биологической активности, которую можно приписать константной области Fc (природной последовательности Fc-области или варианту аминокислотной последовательности Fc-области) антитела. Примеры эффекторных функций антител включают связывание C1q; комплементзависимую цитотоксичность; связывание Fc рецептора; антителозависимую опосредуемую клетками цитотоксичность (ADCC); фагоцитоз; и понижающую регуляцию рецепторов поверхности клеток, таких как B-клеточный рецептор и BCR.
В зависимости от аминокислотных последовательностей константных доменов их тяжелых цепей, интактные антитела можно отнести к различным «классам». Существует пять главных классов интактных иммуноглобулинов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно далее разделить на «подклассы» (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, соответствующие различным классам иммуноглобулинов, называют α, δ, ε, γ и μ, соответственно. Субъединичные структуры и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны. Формы Ig включают модификации шарнира или бесшарнирные формы (Roux et al (1998) J. Immunol. 161:4083-4090; Lund et al (2000) Eur. J. Biochem. 267:7246-7256; US 2005/0048572; US 2004/0229310).
«Рецептор ErbB» представляет собой рецепторную протеинтирозинкиназу, принадлежащую к семейству рецепторов ErbB, члены которого являются важными медиаторами роста, дифференцировки и выживаемости клеток. Семейство рецепторов ErbB включает четыре отдельных члена, включая рецептор эпидермального фактора роста (EGFR, ErbB1, HER1), HER2 (ErbB2 или p185neu), HER3 (ErbB3) и HER4 (ErbB4 или tyro2). Панель анти-ErbB2 антител охарактеризована с использованием линии клеток опухоли молочной железы человека SKBR3 (Hudziak et al (1989) Mol. Cell. Biol. 9(3):1165-1172. Максимальное ингибирование получено с антителом, называемым 4D5, ингибирующим пролиферацию клеток на 56%. Другие антитела в панели в этом анализе уменьшали пролиферацию клеток в меньшей степени. Кроме того, обнаружено, что антитело 4D5 сенсибилизирует линии клеток опухоли молочной железы со сверхэкспрессией ErbB2 к цитотоксическим эффектам TNF-α (US 5677171). Анти-ErbB2 антитела, обсуждаемые в Hudziak et al., дополнительно охарактеризованы в Fendly et al (1990) Cancer Research 50: 1550-1558; Kotts et al. (1990) In Vitro 26(3):59A; Sarup et al. (1991) Growth Regulation 1:72-82; Shepard et al. J. (1991) Clin. Immunol. 11(3):117-127; Kumar et al. (1991) Mol. Cell. Biol. 11(2):979-986; Lewis et al. (1993) Cancer Immunol. Immunother. 37:255-263; Pietras et al. (1994) Oncogene 9: 1829-1838; Vitetta et al. (1994) Cancer Research 54:5301-5309; Sliwkowski et al. (1994) J. Biol. Chem. 269(20): 14661-14665; Scott et al. (1991) J. Biol. Chem. 266: 14300-5; D'souza et al. Proc. Natl. Acad. Sci. (1994) 91:7202-7206; Lewis et al. (1996) Cancer Research 56:1457-1465; и Schaefer et al. (1997) Oncogene 15:1385-1394.
Рецептор ErbB, как правило, содержит внеклеточный домен, который может связывать лиганд ErbB; липофильный трансмембранный домен; консервативный внутриклеточный домен тирозинкиназы; и карбоксил-концевой сигнальный домен, несущий несколько остатков тирозина, которые могут быть фосфорилированы. Рецептор ErbB может представлять собой рецептор ErbB с «природной последовательностью» или «вариант его аминокислотной последовательности». Предпочтительно, рецептор ErbB представляет собой рецептор ErbB человека с природной последовательностью. Соответственно, «член семейства рецепторов ErbB» представляет собой EGFR (ErbB1), ErbB2, ErbB3, ErbB4 или любой другой рецептор ErbB, который известен в настоящее время или который будет идентифицирован в будущем.
Термины «ErbB1», «рецептор эпидермального фактора роста», «EGFR» и «HER1» используют в настоящем документе взаимозаменяемо, и они относятся к EGFR, как описано, например, в Carpenter et al (1987) Ann. Rev. Biochem., 56:881-914, включая их встречающиеся в природе мутантные формы (например, делеционный мутант EGFR, как в Humphrey et al (1990) Proc. Nat. Acad. Sci. (USA) 87:4207-4211). Термин erbB1 относится к гену, кодирующему белковый продукт EGFR. Антитела против HER1 описаны, например, в Murthy et al (1987) Arch. Biochem. Biophys., 252:549-560 и в WO 95/25167.
Термин «ERRP», «родственный EGF-рецептору белок», «родственный EGFR белок» и «родственный рецептору эпидермального фактора роста белок» используют в настоящем документе взаимозаменяемо, и они относятся к ERRP, как описано, например, в US 6399743 и в Публикации США № 2003/0096373.
Выражения «ErbB2» и «HER2» используют в настоящем документе взаимозаменяемо, и они относятся к белку HER2 человека, описанному, например, в Semba et al (1985) Proc. Nat. Acad. Sci. (USA) 82:6497-6501 и Yamamoto et al (1986) Nature, 319:230-234 (номер доступа в Genebank X03363). Термин «erbB2» относится к гену, кодирующему ErbB2 человека, и «neu» относится к гену, кодирующему p185neu крысы. Предпочтительным ErbB2 является природная последовательность ErbB2 человека.
«ErbB3» и «HER3» относятся к полипептиду рецептора, как описано, например, в Патентах США №№ 5183884 и 5480968, а также в Kraus et al (1989) Proc. Nat. Acad. Sci. (USA) 86:9193-9197. Антитела против ErbB3 известны в данной области и описаны, например, в Патентах США №№ 5183884, 5480968 и в WO 97/35885.
Термины «ErbB4» и «HER4» в настоящем документе относятся к полипептиду рецептора, как описано, например, в Патентной заявке EP № 599274; Plowman et al (1993) Proc. Natl. Acad. Sci. USA 90: 1746-1750; и Plowman et al (1993) Nature 366:473-475, включая их изоформы, например, как описано в WO 99/19488. Антитела против HER4 описаны, например, в WO 02/18444.
Антитела против рецепторов ErbB являются коммерчески доступными из ряда источников, включая, например, Santa Cruz Biotechnology, Inc., California, USA.
Термин «вариант аминокислотной последовательности» относится к полипептидам, обладающим аминокислотными последовательностями, отличающимися до некоторой степени от полипептида с природной последовательностью. Обычно варианты аминокислотной последовательности обладают по меньшей мере приблизительно 70% идентичностью последовательности по меньшей мере с одним связывающим рецептор доменом природного лиганда ErbB или по меньшей мере с одним связывающим лиганд доменом природного рецептора ErbB, и предпочтительно, они по меньшей мере приблизительно на 80%, более предпочтительно, по меньшей мере приблизительно на 90% гомологичны по последовательности таким связывающим рецептор или лиганд доменам. Варианты аминокислотной последовательности имеют замены, делеции и/или вставки в конкретных положениях внутри аминокислотной последовательности природной аминокислотной последовательности. Аминокислоты обозначают общепринятыми наименованиями, в однобуквенном и трехбуквенном кодах.
«Идентичность последовательности» определяют как процент остатков в варианте аминокислотной последовательности, являющихся идентичными после выравнивания последовательностей и введения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей. Способы и компьютерные программы для выравнивания хорошо известны в данной области. Одной такой компьютерной программой является «Align 2», авторство которой принадлежит Genentech, Inc. и подано с пользовательской документацией в Бюро регистрации авторских прав США, Washington, DC 20559, 10 декабря 1991 г.
«Антителозависимая опосредуемая клетками цитотоксичность» и «ADCC» относятся к опосредованной клетками реакции, при которой неспецифические цитотоксические клетки, экспрессирующие Fc-рецепторы (FcR) (например, клетки естественные киллеры (NK), нейтрофилы, и макрофаги) узнают связанное антитело на клетке-мишени и затем вызывают лизис клетки-мишени. Первичные клетки, опосредующие ADCC, NK клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках обобщена в таблице 3 на странице 464 в Ravetch and Kinet, (1991) «Annu. Rev. Immunol.» 9:457-92. Для оценки активности ADCC интересующей молекулы можно проводить анализ ADCC in vitro, такой как описан в US 5500362 и US 5821337. Пригодные эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (PBMC) и естественные клетки-киллеры (NK). Альтернативно или дополнительно, активность ADCC интересующей молекулы можно оценивать in vivo, например, в модели на животных, такой, как описана в Clynes et al (1998) PROC. NAT. ACAD. SCI. (USA) (USA) 95:652-656.
«Эффекторные клетки человека» представляют собой лейкоциты, которые экспрессируют один или несколько рецепторов константной области (FcR) и выполняют эффекторные функции. Предпочтительно, клетки экспрессируют по меньшей мере FcyRIII и выполняют эффекторную функцию(функции) ADCC. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (PBMC), естественные клетки-киллеры (NK), моноциты, цитотоксические T-клетки и нейтрофилы; где PBMC и NK клетки являются предпочтительными. Эффекторные клетки можно выделить из их природного источника, например, из крови или PBMC, как описано в настоящем документе.
Термины «Fc рецептор» или «FcR» используют для описания рецептора, который связывается с константной областью Fc антитела. Предпочтительный FcR представляет собой FcR человека с природной последовательностью. Более того, предпочтительным FcR является FcR, который связывается с антителом IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcyRII, и FcγRIII, включая аллельные варианты и формы альтернативного сплайсинга этих рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), обладающие сходными аминокислотными последовательностями, которые отличаются, в первую очередь, в их цитоплазматических доменах. Активирующий рецептор FcγRIIA содержит иммунорецепторный активирующий мотив, основанный на тирозине (ITAM) в его цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит иммунорецепторный ингибирующий мотив, основанный на тирозине (ITIM), в его цитоплазматическом домене. (См. обзор M. в Daлron, «Annu. Rev. Immunol.» 15:203-234 (1997)). Обзор FcR приведен в Ravetch and Kinet, «Annu. Rev. Immunol»., 9:457-92 (1991); Capel et al (1994) Immunomethods 4:25-34; и de Haas et al (1995) J. Lab. Clin. Med. 126:330-41. Другие FcR, включая те, которые будут идентифицированы в будущем, включены в термин «FcR» в настоящем документе. Термин включает также неонатальный рецептор, FcRn, который является ответственным за перенос материнских IgG к плоду (Guyer et al (1976) J. Immunol, 117:587 и Kim et al (1994) J. Immunol. 24:249).
«Комплементзависимая цитотоксичность» или «CDC» относится к способности молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента запускается связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом) в комплексе с родственными ей антигеном. Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al J. Immunol. Methods, 202: 163 (1996).
«Нативные антитела» обычно представляют собой гетеротетрамерные гликопротеины приблизительно 150000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (H) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как количество дисульфидных связей меняется среди тяжелых цепей различных изотипов иммуноглобулинов. В каждой тяжелой и легкой цепи также равномерно расположены внутрицепьевые дисульфидные мостики. Каждая тяжелая цепь на одном конце имеет вариабельный домен (VH) с последующими несколькими константными доменами. Каждая легкая цепь на одном конце имеет вариабельный домен (VL) и на другом конце - константный домен. Константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, и вариабельный домен легкой цепи выровнен с вариабельным доменом тяжелой цепи. Считают, что конкретные аминокислотные остатки формируют поверхность раздела между вариабельными доменами легкой цепи и тяжелой цепи.
Термин «вариабельный» относится к факту, что конкретные части вариабельных доменов сильно отличаются по последовательности среди антител и используются для связывания и специфичности каждого конкретного антитела для его конкретного антигена. Однако вариабельность не является равномерно распределенной на всем протяжении вариабельных доменов антител. Она сконцентрирована в трех участках, называемых гипервариабельными областями, в вариабельных доменах как на легкой цепи, так и на тяжелой цепи. Наиболее высоко консервативные части вариабельных доменов называют каркасными областями (FR). Каждый из вариабельных доменов природных тяжелых и легких цепей содержит четыре FR, преимущественно принимающие конфигурацию β-листа, соединенные тремя гипервариабельными областями, формирующими петли, соединяющие структуры β-листа, и в некоторых случаях формирующие их часть. Гипервариабельные области на каждой цепи удерживаются вместе в непосредственной близости посредством FR и, вместе с гипервариабельными областями из другой цепи, участвуют в формировании антигенсвязывающего участка антител (см. Kabat et al (1991) Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD). Константные домены не являются непосредственно вовлеченными в связывание антитела с антигеном, но обладают различными эффекторными функциями, такими как участие антитела в антителозависимой клеточной токсичности (ADCC).
Термин «гипервариабельная область» при использовании в настоящем документе относится к аминокислотным остаткам антитела, ответственным за связывание антигена. Гипервариабельная область обычно содержит аминокислотные остатки из «определяющей комплементарность области» или «CDR» (например, остатки 24-34 (LI), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al выше) и/или остатки из «гипервариабельной петли» (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (HI), 53-55 (H2) и 96-101 (H3) в вариабельном домене тяжелой цепи; Chothia and Lesk (1987) J. Mol. Biol, 196:901-917). Остатки «каркасной области» или «FR» представляют собой остатки вариабельного домена, отличные от остатков гипервариабельной области, как определено в настоящем документе.
Расщеплением антител папаином получают два идентичных антигенсвязывающих фрагмента, называемых «Fab» фрагментами, каждый с одним антигенсвязывающим участком, и оставшийся «Fc» фрагмент, наименование которого отражает его способность легко кристаллизоваться. Обработкой пепсином получают F(ab')2 фрагмент, обладающий двумя антигенсвязывающими участками и еще способный к перекрестному связыванию антигена.
«Fv» представляет собой минимальный фрагмент антитела, содержащий полный узнающий антиген и антигенсвязывающий участок. Эта область состоит из димера одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи в тесной, нековалентной связи. В этой конфигурации три гипервариабельные области каждого вариабельного домена взаимодействуют для определения антигенсвязывающего участка на поверхности димера VH-VL. Шесть гипервариабельных областей совместно придают антигенсвязывающую специфичность антителу. Однако, даже отдельный вариабельный домен (или половина Fv, содержащая только три гипервариабельные области, специфичные для антигена) обладает способностью узнавать и связывать антиген, хотя и с более низкой аффинностью, чем целый связывающий участок.
Fab-фрагмент содержит также константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на карбокси-конце CH1 домена тяжелой цепи, включая один или несколько остатков цистеина из шарнирной области антитела. Fab'-SH в настоящем документе является обозначением Fab', в котором остаток(остатки) цистеина константных доменов несут по меньшей мере одну свободную тиоловую группу. F(ab')2 фрагменты антитела первоначально были получены в форме пар Fab'-фрагментов с шарнирными цистеинами между ними. Известны также другие химические соединения фрагментов антител.
«Легкие цепи» антител из любых видов позвоночных можно отнести к одному из двух явно различимых типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотных последовательностей их константных доменов.
«Одноцепочечные Fv» или «scFv» фрагменты антител содержат VH и VL домены антитела, где эти домены присутствуют на одной полипептидной цепи. Предпочтительно, полипептид Fv дополнительно содержит полипептидный линкер между VH и VL доменами, позволяющий scFv формировать желательную структуру для связывания антигена. Обзор scFv см. в Pluckthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994). scFv фрагменты анти-ErbB2 антитела описаны в WO 93/16185; Патентах США №№ 5571894; и 5587458.
Термин «диатела» относится к малым фрагментам антитела с двумя антигенсвязывающими участками, где фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной и той же полипептидной цепи (VH-VL). С использованием линкера, слишком короткого, чтобы позволить спаривание двух доменов одной и той же цепи, домены вынуждают спариваться с комплементарными доменами другой цепи и формировать два антигенсвязывающих участка. Диатела более подробно описаны, например, в EP 404,097; WO 93/11161; и Hollinger et al (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448.
«Гуманизированные» формы не относящихся к человеку антител» (например, грызунов) представляют собой химерные антитела, содержащие минимальную последовательность, полученную из не относящегося к человеку иммуноглобулина. Гуманизация является способом переноса информации для связывания антигена от мыши к неиммуногенному акцепторному антителу человека, и приводит к получению множества терапевтически пригодных лекарственных средств. Способ гуманизации, как правило, начинается с переноса всех шести мышиных определяющих комплементарность областей (CDR) в каркас человеческого антитела (Jones et al, (1986) Nature 321:522-525). Эти CDR-привитые антитела, как правило, не сохраняют исходную аффинность для связывания антигена, и фактически, аффинность часто серьезно нарушена. Помимо CDR, избранные каркасные остатки не относящегося к человеку антитела также необходимо включать для поддержания правильной конформации CDR (Chothia et al (1989) Nature 342:877). Показано, что перенос ключевых мышиных остатков в человеческий акцептор для поддержания структурной конформации привитых CDR сохраняет связывание антигена и аффинность (Riechmann et al (1992) J. Mol. Biol. 224, 487-499; Foote and Winter, (1992) J. Mol. Biol. 224:487-499; Presta et al (1993) J. Immunol. 151, 2623-2632; Werther et al (1996) J. Immunol. Methods 157:4986-4995; и Presta et al (2001) Thromb. Haemost. 85:379-389). По большей части, гуманизированные антитела представляют собой иммуноглобулины человека (антитело-реципиент), в котором остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области не относящихся к человеку видов (антитела-донора), таких как мышь, крыса, кролик или не относящийся к человеку примат, обладающими желаемой специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими не присутствующими у человека остатками. Более того, гуманизированные антитела могут содержать остатки, не обнаруженные в антителе-реципиенте или в антителе-доноре. Эти модификации выполняют для дальнейшего усовершенствования характеристик антитела. Как правило, гуманизированное антитело содержит в основном все из по меньшей мере одного, и как правило, двух вариабельных доменов, в которых все или в основном все гипервариабельные петли соответствуют петлям не относящегося к человеку иммуноглобулина, и все или в основном все FR являются FR с последовательностью человеческого иммуноглобулина. Гуманизированное антитело, необязательно, может содержать также по меньшей мере часть константной области (Fc) иммуноглобулина, как правило, иммуноглобулина человека. Более подробно см. US 6407213; Jones et al (1986) Nature, 321:522-525; Riechmann et al (1988) Nature 332:323-329; и Presta, (1992) Curr. Op. Struct. Biol, 2:593-596.
«Свободная аминокислота цистеин» относится к остатку аминокислоты цистеина, который введен при конструировании в исходное антитело, имеет тиоловую функциональную группу (-SH), и не является спаренным в качестве внутримолекулярного или межмолекулярного дисульфидного мостика.
Термин «значение реакционной способности тиола» представляет собой количественную характеристику реакционной способности свободных цистеиновых аминокислот. Значение реакционной способности тиола представляет собой процент свободной аминокислоты цистеина в сконструированном антителе с цистеиновыми заменами, вступающий в реакцию с реакционноспособным по отношению к тиолу реагентом, и преобразованный до максимального значения 1. Например, свободная аминокислота цистеин в сконструированном антителе с цистеиновыми заменами, которая вступает в реакцию со 100% выходом с реакционноспособным по отношению к тиолу реагентом, таким как биотин-малеимидный реагент, с формированием меченного биотином антитела, имеет значение реакционной способности тиола 1,0. Другая аминокислота цистеин, введенная при конструировании в такое же или другое исходное антитело, которая вступает в реакцию с 80% выходом с реакционноспособным по отношению к тиолу реагентом, имеет значение реакционной способности тиола 0,8. Другая аминокислота цистеин, введенная при конструировании в такое же или другое исходное антитело, которая совсем не вступает в реакцию с реакционноспособным по отношению к тиолу реагентом, имеет значение реакционной способности тиола 0. Определение значения реакционной способности тиола конкретного цистеина можно проводить анализом ELISA, масс-спектроскопией, жидкостной хроматографией, радиоавтографией или другими количественными аналитическими тестами.
«Исходное антитело» представляет собой антитело, содержащее аминокислотную последовательность, в которой один или несколько аминокислотных остатков заменяют на один или несколько остатков цистеина. Исходное антитело может содержать природную последовательность или последовательность дикого типа. Исходное антитело может обладать предсуществующими модификациями аминокислотной последовательности (такими как добавления, делеции и/или замены) по сравнению с другими природными формами, формами дикого типа или модифицированными формами антитела. Исходное антитело может быть нацелено против интересующего антигена-мишени, например, биологически важного полипептида. Предусмотрены также антитела, нацеленные против неполипептидных антигенов (таких как опухолеассоциированные гликолипидные антигены; см. US 5091178).
Иллюстративные исходные антитела включают антитела, обладающие аффинностью и избирательностью для рецепторов поверхности клеток и трансмембранных рецепторов, и опухолеассоциированных антигенов (TAA).
Другие иллюстративные исходные антитела включают антитела, выбранные, и без ограничения, из антитела против рецептора эстрогена, антитела против рецептора прогестерона, анти-p53 антитела, анти-HER-2/neu антитела, анти-EGFR антитела, антитела против катепсина D, анти-Bcl-2 антитела, антитела против E-кадгерина, анти-CA125 антитела, анти-CA15-3 антитела, анти-CA19-9 антитела, анти-c-erbB-2 антитела, антитела против P-гликопротеина, анти-CEA антитела, антитела против белка ретинобластомы, антитела против онкобелка ras, антитела против Льюиса X, анти-Ki-67 антитела, анти-PCNA антитела, анти-CD3 антитела, анти-CD4 антитела, анти-CD5 антитела, анти-CD7 антитела, анти-CD8 антитела, анти-CD9/p24 антитела, анти-CD10 антитела, анти-CD11c антитела, анти-CD13 антитела, анти-CD14 антитела, анти-CD15 антитела, анти-CD19 антитела, анти-CD20 антитела, анти-CD22 антитела, анти-CD23 антитела, анти-CD30 антитела, анти-CD31 антитела, анти-CD33 антитела, анти-CD34 антитела, анти-CD35 антитела, анти-CD38 антитела, анти-CD41 антитела, анти-LCA/CD45 антитела, анти-CD45RO антитела, анти-CD45RA антитела, анти-CD39 антитела, анти-CD100 антитела, анти-CD95/Fas антитела, анти-CD99 антитела, анти-CD106 антитела, антитела против убиквитина, анти-CD71 антитела, анти-c-myc антитела, антитела против цитокератинов, антитела против виментинов, антитела против белков HPV, антитела против легких цепей каппа, антитела против легких цепей лямбда, антитела против меланосом, антитела против специфического антигена простаты, анти-S-100 антитела, антитела против антигена tau, антитела против фибрина, антитела против кератинов и антитела против Tn-антигена.
«Выделенное» антитело представляет собой антитело, которое идентифицировано и отделено и/или выделено из компонента его природного окружения. Загрязняющие компоненты его натурального окружения представляют собой материалы, которые могут мешать исследовательским, диагностическим или терапевтическим применениям антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления антитело является очищенным (1) до более чем 95% по массе антитела, как определено способом Лоури, и наиболее предпочтительно, до более чем 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности при использовании секвенатора с вращающимся стаканом, или (3) до гомогенности в SDS-PAGE в невосстанавливающих или восстанавливающих условиях с использованием окраски Кумасси синим или, предпочтительно, серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, с тех пор, когда перестает присутствовать по меньшей мере один из компонентов природного окружения антитела. Обычно, однако, выделенное антитело получают посредством по меньшей мере одной стадии очистки.
Антитело, «которое связывает» интересующую молекулярную мишень или антиген, например, антиген ErbB2, представляет собой антитело, способное связывать этот антиген с достаточной аффинностью, так что антитело является пригодным для нацеливания на клетку, экспрессирующую антиген. Когда антитело представляет собой антитело, связывающее ErbB2, оно обычно предпочтительно связывает ErbB2 в отличие от других рецепторов ErbB, и может представлять собой антитело, которое не вступает в значительную перекрестную реакцию с другими белками, такими как EGFR, ErbB3 или ErbB4. В таких вариантах осуществления степень связывания антитела с этими не относящимися к ErbB2 белками (например, связывания поверхности клеток с эндогенным рецептором) составляет менее 10%, как определено анализом активированной флуоресценцией сортировки клеток (FACS) или радиоиммунопреципитацией (RIA). Иногда анти-ErbB2 антитело не вступает в значительную перекрестную реакцию с белком neu крысы, например, как описано в Schecter et al. (1984) Nature 312:513 и Drebin et al (1984) Nature 312:545-548.
Молекулярные мишени для антител, включенные в настоящее изобретение, включают CD белки и их лиганды, такие как, в качестве неограничивающих примеров: (i) CD3, CD4, CD8, CD19, CD20, CD22, CD34, CD40, CD79α (CD79a) и CD79β (CD79b); (ii) члены семейства рецепторов ErbB, такие как рецептор EGF, рецептор HER2, HER3 или HER4; (iii) молекулы клеточной адгезии, такие как LFA-1, Macl, p150,95, VLA-4, ICAM-1, VCAM и αν/β3 интегрин, включая их альфа- или бета-субъединицы (например, анти-CD11a, анти-CD18 или анти-CD11b антитела); (iv) факторы роста, такие как VEGF; IgE; антигены группы крови; рецептор flk2/flt3; рецептор ожирения (OB); рецептор mpl; CTLA-4; белок C, BR3, c-met, тканевой фактор, β7 и т.д.; и (v) опухолеассоциированный антигены (TAA) поверхности клеток и трансмембранные опухолеассоциированные антигены (TAA).
Если нет иных указаний, термин «моноклональное антитело 4D5» обозначает антитело, обладающее антигенсвязывающими остатками мышиного антитела 4D5 (ATCC CRL 10463), или полученное из него. Например, моноклональное антитело 4D5 может представлять собой мышиное моноклональные антитело 4D5 или его вариант, такой как гуманизированное 4D5. Иллюстративные гуманизированные антитела 4D5 включают huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 (трастузумаб, ГЕРЦЕПТИН®), как в Патенте США № 5821337.
«Фаговый дисплей» представляет собой способ, посредством которого варианты полипептидов экспонируют в виде белков, слитых с белком оболочки, на поверхности частиц фага, например, нитевидного фага. Применимость фагового дисплея основана на факте, что большие библиотеки рандомизированных вариантов белка можно быстро и эффективно отсортировать по тем последовательностям, которые связывают антиген-мишень с высокой аффинностью. Экспонирование пептидных и белковых библиотек на фаге использовали для скрининга миллионов полипептидов для отбора полипептидов со специфическими свойствами связывания. Способы поливалентного фагового дисплея использовали для экспонирования малых случайных пептидов и малых белков, как правило, посредством слияния с pIII или pVIII нитевидного фага (Wells and Lowman, (1992) Curr. Opin. Struct. Biol, 3:355-362, и процитированные в этом документе ссылки). В моновалентном фаговом дисплее белковую или пептидную библиотеку сливают с белком оболочки фага или его частью, и экспрессируют на низких уровнях в присутствии белка дикого типа. Эффекты авидности снижены по сравнению с поливалентным фагом, так что отбор проходит на основе собственной аффинности лиганда, и используют фагмидные векторы, упрощающие манипуляции с ДНК. Lowman and Wells, Methods: A companion to Methods in Enzymology, 3:205-0216 (1991). Фаговый дисплей включает способы получения антителоподобных молекул (Janeway, C, Travers, P., Walport, M., Shlomchik (2001) Immunobiology, 5th Ed., Garland Publishing, New York, p.627-628; Lee et al ).
«Фагмида» представляет собой плазмидный вектор, обладающий бактериальной точкой начала репликации, например, ColE1, и копией межгенной области бактериофага. Фагмиду можно использовать для любого известного бактериофага, включая нитевидный бактериофаг и лямбдоидный бактериофаг. Плазмида также, как правило, содержит селективный маркер для устойчивости к антибиотику. Фрагменты ДНК, клонированные в эти векторы, можно размножать в форме плазмид. Когда клетки, несущие эти векторы, обеспечивают всеми генами, необходимыми для продукции фаговых частиц, тип репликации плазмиды меняется на репликацию по типу катящегося кольца для получения копий одной цепи плазмидной ДНК и упаковки фаговых частиц. Фагмиды могут формировать инфекционные или неинфекционные фаговые частицы. Этот термин включает фагмиды, содержащие ген белка оболочки фага или его фрагмент, связанный с геном гетерологичного полипептида в форме слитого гена, так что гетерологичный полипептид экспонирован на поверхности фаговых частиц.
«Линкер», «линкерное звено», или «связующее звено» обозначает химическую группу, содержащую ковалентную связь или цепь атомов, которая ковалентно присоединяет антитело к группе лекарственного средства. В различных вариантах осуществления линкер обозначают как L. Линкеры включают двухвалентный радикал, такой как алкилдииловые, ариленовые, гетероариленовые группы, такие как: -(CR2)nO(CR2)n-, повторяющиеся звенья алкилокси (например, полиэтиленокси, PEG, полиметиленокси) и алкиламино (например, полиэтиленамино, Джеффамин™); и сложные эфиры и амиды двухосновной кислоты, включая сукцинат, сукцинамид, дигликолят, малонат и капроамид.
Термин «метка» обозначает любую группу, которую можно ковалентно присоединять к антителу и которая функционирует для: (i) обеспечения поддающегося детекции сигнала; (ii) взаимодействия со второй меткой для модификации поддающегося детекции сигнала, обеспечиваемого первой или второй меткой, например, FRET (резонансный перенос энергии флуоресценции); (iii) стабилизации взаимодействий или увеличения аффинности связывания с антигеном или лигандом; (iv) влияния на подвижность, например, электрофоретическую подвижность, или клеточную проницаемость, посредством заряда, гидрофобности, формы или других физических параметров, или (v) предоставления связывающей группы, для модуляции аффинности лиганда, связывания антитело/антиген или образования комплексных ионов.
Стереохимические определения и обозначения, используемые в настоящем документе, в основном, следуют S. P. Parker, Ed., McGraw-Hill Dictionary of Chemical Terms (1984) McGraw-Hill Book Company, New York; и Eliel, E. and Wilen, S., Stereochemistry of Organic Compounds (1994) John Wiley & Sons, Inc., New York. Многие органические соединения существуют в оптически активных формах, т.е., они обладают способностью вращать плоскость плоскополяризованного света. При описании оптически активного соединения, префиксы D и L или R и S, используют для обозначения абсолютной конфигурации молекулы вокруг ее хирального центра(центров). Префиксы d и l или (+) и (-) используют для обозначения знака вращения плоскополяризованного света соединением, где (-) или l обозначает, что соединение является левовращающим. Соединение с префиксом (+) или d является правовращающим. Для данной химической структуры, эти стереоизомеры являются идентичными, за исключением того, что они являются зеркальными отражениями друг друга. Конкретный стереоизомер можно обозначать также, как энантиомер, и смесь таких изомеров часто называют энантиомерной смесью. Смесь энантиомеров 50:50 обозначают рацемической смесью или рацематом, которые могут возникать в отсутствие стереоизбирательности или стереоспецифичности химической реакции или процесса. Термины «рацемическая смесь» и «рацемат» относятся к эквимолярной смеси двух видов энантиомеров, лишенной оптической активности.
Фраза «фармацевтически приемлемая соль», как применяют в настоящем документе, относится к фармацевтически приемлемым органическим или неорганическим солям AZC. Иллюстративные соли включают соли сульфат, цитрат, ацетат, оксалат, хлорид, бромид, иодид, нитрат, бисульфат, фосфат, кислый фосфат, изоникотинат, лактат, салицилат, кислый цитрат, тартрат, олеат, таннат, пантотенат, битартрат, аскорбат, сукцинат, малеат, гентизинат, фумарат, глюконат, глюкуронат, сахарат, формат, бензоат, глутамат, метансульфонат, этансульфонат, бензолсульфонат, п-толуолсульфонат и памоат (т.е., 1,1'-метилен-бис-(2-гидрокси-3-нафтоат)), но не ограничены ими. Фармацевтически приемлемая соль может предполагать включение другой молекулы, такой как ион ацетата, ион сукцината или другой противоион. Противоион может представлять собой любую органическую или неорганическую группу, стабилизирующую заряд исходного соединения. Более того, фармацевтически приемлемая соль может иметь в структуре более одного заряженного атома. В случаях, когда несколько заряженных атомов составляют часть фармацевтически приемлемой соли, она может иметь несколько противоионов. Таким образом, фармацевтически приемлемая соль может иметь один или несколько заряженных атомов и/или один или несколько противоионов.
«Фармацевтически приемлемый сольват» обозначает ассоциацию одной или нескольких молекул растворителя и AZC. Растворители, формирующие фармацевтически приемлемые сольваты, включают в качестве неограничивающих примеров, воду, изопропанол, этанол, метанол, DMSO, этилацетат, уксусную кислоту и этаноламин.
Следующие аббревиатуры использованы в настоящем документе и имеют следующие определения: BME представляет собой бета-меркаптоэтанол, Boc представляет собой N-(трет-бутоксикарбонил), cit представляет собой цитруллин (2-амино-5-уреидопентановая кислота), dap представляет собой долапроин, DCC представляет собой 1,3-дициклогексилкарбодиимид, DCM представляет собой дихлорметан, DEA представляет собой диэтиламин, DEAD представляет собой диэтилазодикарбоксилат, DEPC представляет собой диэтилфосфорилцианидат, DIAD представляет собой диизопропилазодикарбоксилат, DIEA представляет собой N,N-диизопропилэтиламин, dil представляет собой долаизолейцин, DMA представляет собой диметилацетамид, DMAP представляет собой 4-диметиламинопиридин, DME представляет собой диметиловый эфир этиленгликоля (или 1,2-диметоксиэтан), DMF представляет собой N,N-диметилформамид, DMSO представляет собой диметилсульфоксид, doe представляет собой долафенин, dov представляет собой N,N-диметилвалин, DTNB представляет собой 5,5'-дитиобис(2-нитробензойную кислоту), DTPA представляет собой диэтилентриаминпентауксусную кислоту, DTT представляет собой дитиотреитол, EDCI представляет собой гидрохлорид 1-(3-диметиламинопропил)-3-этилкарбодиимида, EEDQ представляет собой 2-этокси-1-этоксикарбонил-1,2-дигидрохинолин, ES-MS представляет собой масс-спектрометрию с электрораспылением, EtOAc представляет собой этилацетат, Fmoc представляет собой N-(9-флуоренилметоксикарбонил), gly представляет собой глицин, HATU представляет собой гексафторфосфат O-(7-азабензотриазол-1-ил)-N,N,N',N'-тетраметилурония, HOBt представляет собой 1-гидроксибензотриазол, HPLC представляет собой высокоэффективную жидкостную хроматографию, ile представляет собой изолейцин, lys представляет собой лизин, MeCN (CH3CN) представляет собой ацетонитрил, MeOH представляет собой метанол, Mtr представляет собой 4-анизилдифенилметил (или 4-метокситритил), nor представляет собой (1S,2R)-(+)-норэфедрин, PAB представляет собой п-аминобензилкарбамоил, PBS представляет собой фосфатно-солевой буфер (pH 7), PEG представляет собой полиэтиленгликоль, Ph представляет собой фенил, Pnp представляет собой п-нитрофенил, MC представляет собой 6-малеимидокапроил, phe представляет собой L-фенилаланин, PyBrop представляет собой гексафторфосфат бром-трис-пирролидинофосфония, SEC представляет собой эксклюзионную хроматографию, Su представляет собой сукцинимид, TFA представляет собой трифторуксусную кислоту, TLC представляет собой тонкослойную хроматографию, UV представляет собой ультрафиолет, и val представляет собой валин.
СКОНСТРУИРОВАННЫЕ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
Соединения по изобретению включают сконструированные антитела с цистеиновыми заменами, в которых одна или несколько аминокислот антитела дикого типа или исходного антитела заменены на аминокислоту цистеин. Любую форму антитела можно модифицировать таким образом, т.е. подвергать мутации. Например, исходный Fab-фрагмент антитела можно модифицировать для получения сконструированного Fab с цистеиновыми заменами, обозначенного в настоящем документе как «ТиоFab». Подобным образом, исходное моноклональное антитело можно модифицировать для получения «ТиоMab». Следует отметить, что мутация в одном участке приводит к одному сконструированному остатку цистеина в ТиоFab, в то время как мутация в одном участке приводит к двум сконструированным остаткам цистеина в ТиоMab, из-за димерной природы антитела IgG. Мутанты с замененными («сконструированными») остатками цистеина (Cys) оценивали по реакционной способности вновь введенных сконструированных тиоловых групп цистеина. Значение реакционной способности тиола является относительным числовым термином в диапазоне от 0 до 1,0, и его можно измерять для любого сконструированного антитела с цистеиновыми заменами. Значения реакционной способности тиола сконструированных антител с цистеиновыми заменами по изобретению лежат в диапазонах 0,6-1,0; 0,7-1,0; или 0,8-1,0.
Способы дизайна, отбора и получения по изобретению предоставляют сконструированные антитела с цистеиновыми заменами, реакционноспособные по отношению к электрофильной функциональной группе. Эти способы, кроме того, предоставляют соединения конъюгата с антителом, такие как соединения конъюгата антитело-цирконий (AZC) с атомами циркония в обозначенных сконструированных избранных участках. Реакционноспособные остатки цистеина на поверхности антитела позволяют специфическую конъюгацию группы циркония через реакционноспособную по отношению к тиолу группу, такую как малеимид или галоацетил. Реакционная способность нуклеофильной тиоловой функциональной группы остатка Cys по отношению к малеимидной группе приблизительно в 1000 выше по сравнению с любой другой функциональной группой аминокислот в белке, такой как аминогруппа остатков лизина или N-концевая аминогруппа. Специфические для тиола функциональные группы в иодацетиловых и малеимидных реагентах могут вступать в реакцию с аминогруппами, однако, необходимы более высокий pH (>9,0) и более длительные периоды времени реакции (Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London).
Сконструированные антитела с цистеиновыми заменами по изобретению предпочтительно сохраняют антигенсвязывающую способность эквивалентных им антител дикого типа, исходных антител. Таким образом, сконструированные антитела с цистеиновыми заменами способны связываться, предпочтительно, специфически, с антигенами. Такие антигены включают, например, опухолеассоциированные антигены (TAA), рецепторные белки поверхности клеток и другие молекулы поверхности клеток, трансмембранные белки, белки передачи сигнала, регуляторные факторы выживаемости клеток, регуляторные факторы пролиферации клеток, молекулы, ассоциированные с развитием или дифференцировкой тканей (например, как известно или как предполагают, вносящие в них функциональный вклад), лимфокины, цитокины, молекулы, вовлеченные в регуляцию клеточного цикла, молекулы, вовлеченные в васкулогенез, и молекулы, ассоциированные с ангиогенезом (например, как известно или как предполагают, вносящие в него функциональный вклад). Опухолеассоциированный антиген может представлять собой фактор кластера дифференцировки (т.е., CD белок). Антиген, с которым способно связываться сконструированное антитело с цистеиновыми заменами, может являться членом подгруппы одной из вышеупомянутых категорий, где другая подгруппа(подгруппы) указанной категории содержит другие молекулы/антигены, обладающие отличной характеристикой (по отношению к интересующему антигену).
Исходное антитело может представлять собой также гуманизированное антитело, выбранное из huMAb4D5-1, huMAb4D5-2, huMAb4D5-3, huMAb4D5-4, huMAb4D5-5, huMAb4D5-6, huMAb4D5-7 и huMAb4D5-8 (Трастузумаб, ГЕРЦЕПТИН®), как описано в таблице 3 US 5821337, включенной в настоящее описание в качестве ссылки в полном объеме; антитела гуманизированное 520C9 (WO 93/21319) и гуманизированное 2C4, как описано в настоящем описании.
Сконструированные антитела с цистеиновыми заменами по изобретению можно сайт-специфически и эффективно присоединять к реакционноспособному по отношению к тиолу реагенту. Реакционноспособный по отношению к тиолу реагент может представлять собой мультифункциональный линкерный реагент, реагент с меткой связывания, т.е. аффинности, (например, биотин-линкерный реагент), метку для детекции (например, реагент с флуорофором), реагент для иммобилизации на твердой фазе (например, SEPHAROSE™, полистироле или стекле), или промежуточное соединение цирконий-линкер. Одним из примеров реакционноспособного по отношению к тиолу реагента является N-этилмалеимид (NEM). В иллюстративном варианте осуществления реакция ТиоFab с реагентом биотин-линкер предоставляет биотинилированное ТиоFab, посредством которого можно детектировать и измерять присутствие и реакционную способность сконструированного остатка цистеина. Реакция ТиоFab с мультифункциональным линкерным реагентом предоставляет ТиоFab с функционализированным линкером, который далее может вступать в реакцию с реагентом с группой циркония или с другой меткой. Реакция ТиоFab с промежуточным соединением цирконий-линкер предоставляет конъюгат ТиоFab с цирконием.
Иллюстративные способы, описанные в настоящем документе, можно широко использовать для идентификации и продукции антител, и более широко, для других белков, посредством использования стадий разработки и скрининга, описанных в настоящем документе.
Такой способ можно использовать для конъюгации других реакционноспособных по отношению к тиолу средств, в которых реакционноспособная группа представляет собой, например, малеимид, иодацетамид, пиридилдисульфид, или другой реакционноспособный по отношению к тиолу партнер для конъюгации (Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, 1997, Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Hermanson, G. in Bioconjugate Techniques (1996) Academic Press, San Diego, pp. 40-55, 643-671). Партнер может представлять собой цитотоксическое средство (например, токсин, такой как доксорубицин или токсин коклюша), флуорофор, такой как флуоресцентный краситель, подобный флуоресцеину или родамину, хелатирующий агент для получения изображений или металлсодержащее радиотерапевтическое средство, пептидильную или непептидильную метку или метку для детекции, или модифицирующее выведение средство, такое как различные изомеры полиэтиленгликоля, пептид, связывающий третий компонент, или другой углевод или липофильное средство.
Участки, идентифицированные в иллюстративном фрагменте антитела, hu4D5Fabv8, в настоящей работе, находятся в первую очередь в константном домене антитела, который является достаточно консервативным во всех видах антител. Эти участки можно широко использовать для других антител, без дальнейшей необходимости структурной разработки или знаний о структурах конкретных антител и без влияния на свойства связывания антигена, присущие вариабельным доменам антитела.
Сконструированные антитела с цистеиновыми заменами, которые могут являться пригодными для лечения злокачественной опухоли, включают в качестве неограничивающих примеров, антитела против рецепторов поверхности клеток и опухолеассоциированных антигенов (TAA). Такие антитела можно использовать в форме голых антител (не конъюгированных с группой метки) или в форме конъюгатов антитело-цирконий (AZC) формулы I. Опухолеассоциированные антигены известны в данной области и могут быть получены для использования в получении антител с использованием способов и информации, хорошо известных в данной области. В попытках открыть эффективные клеточные мишени для диагностики и терапии злокачественных опухолей, исследователи стремились к идентификации трансмембранных или иным образом ассоциированных с опухолями полипептидов, специфически экспрессирующихся на поверхности одного или нескольких конкретных типа(типов) клеток злокачественных опухолей по сравнению с одной или несколькими нормальными незлокачественными клеткой(клетками). Часто, такие опухолеассоциированные полипептиды более сильно экспрессированы на поверхности клеток злокачественных опухолей по сравнению с поверхностью незлокачественных клеток. Идентификация таких опухолеассоциированных антигенных полипептидов на поверхности клеток привела к возможности специфического нацеливания на клетки злокачественных опухолей для их разрушения способами терапии на основе антител.
TAA включают в качестве неограничивающих примеров TAA (1)-(36), перечисленные ниже. Для удобства, информация относительно этих антигенов, все из которых известны в данной области, перечислена ниже и включает наименования, альтернативные наименования, номера доступа в Genbank и основополагающую ссылку(ссылки), следующие идентификационным обозначениям последовательностей нуклеиновых кислот и белков Национального центра биотехнологической информации (NCBI). Последовательности нуклеиновых кислот и белков, соответствующие TAA (1)-(36), доступны в публичных базах данных, таких как GenBank. Опухолеассоциированные антигены, являющиеся мишенями для антител, включают все варианты и изоформы аминокислотной последовательности, обладающие по меньшей мере приблизительно 70%, 80%, 85%, 90% или 95% идентичностью последовательности по отношению к последовательности, идентифицированной в процитированных ссылках, или обладающие по существу такими же биологическими свойствами или характеристиками, как TAA, обладающие последовательностью, обнаруженной в процитированных ссылках. Например, TAA, обладающие вариантом последовательности, как правило, способны специфически связываться с антителом, специфически связывающимся с TAA с соответствующими перечисленными последовательностями. Последовательности и описание в ссылке, конкретно процитированной в настоящем документе, в явной форме приведены в качестве ссылки.
ОПУХОЛЕАССОЦИИРОВАННЫЕ АНТИГЕНЫ (1)-(36):
(1) BMPR1B (рецептор костного морфогенного белка типа IB, номер доступа в Genbank NM_001203)
ten Dijke,P., et al Science 264 (5155): 101-104 (1994), Oncogene 14 (11): 1377-1382 (1997)); WO2004063362 (пункт формулы изобретения 2); WO2003042661 (пункт формулы изобретения 12); US2003134790-A1 (страницы 38-39); WO2002102235 (пункт формулы изобретения 13; страница 296); WO2003055443 (страницы 91-92); WO200299122 (Пример 2; страницы 528-530); WO2003029421 (пункт формулы изобретения 6); WO2003024392 (пункт формулы изобретения 2; Фиг.112); WO200298358 (пункт формулы изобретения 1; страница 183); WO200254940 (страницы 100-101); WO200259377(страницы 349-350); WO200230268 (пункт формулы изобретения 27; страница 376); WO200148204 (Пример; Фиг.4)
NP_001194 bone morphogenetic protein receptor, type IB /pid=NP_001194.1
Перекрестные ссылки: MIM:603248; NP_001194.1; AY065994
(2) E16 (LAT1, SLC7A5, номер доступа в Genbank NM_003486)
Biochem. Biophys. Res. Commun. 255 (2), 283-288 (1999), Nature 395 (6699):288-291 (1998), Gaugitsch, H.W., et al (1992) J. Biol. Chem. 267 (16): 11267-11273); WO2004048938 (Пример 2); WO2004032842 (Пример IV); WO2003042661 (пункт формулы изобретения 12); WO2003016475 (пункт формулы изобретения 1); WO200278524 (Пример 2); WO200299074 (пункт формулы изобретения 19; страницы 127-129); WO200286443 (пункт формулы изобретения 27; страницы 222, 393); WO2003003906 (пункт формулы изобретения 10; страницы 293); WO200264798 (пункт формулы изобретения 33; страницы 93-95); WO200014228 (пункт формулы изобретения 5; страницы 133-136); US2003224454 (Фиг.3); WO2003025138 (пункт формулы изобретения 12; страница 150);
NP_003477 solute carrier family 7 (cationic amino acid transporter, y+ system), member 5 /pid=NP_003477.3 - Homo sapiens
Перекрестные ссылки: MIM:600182; NP_003477.3; NM_015923; NM_003486_1
(3) STEAP1 (эпителиальный антиген простаты, содержащий шесть трансмембранных доменов, номер доступа в Genbank NM_012449)
Cancer Res. 61 (15), 5857-5860 (2001), Hubert, R.S., et al (1999) Proc. Natl. Acad. Sci. U.S.A. 96 (25): 14523-14528); WO2004065577 (пункт формулы изобретения 6); WO2004027049 (Фиг.1L); EP1394274 (Пример 11); WO2004016225 (пункт формулы изобретения 2); WO2003042661 (пункт формулы изобретения 12); US2003157089 (Пример 5); US2003185830 (Пример 5); US2003064397 (Фиг.2); WO200289747 (Пример 5; страницы 618-619); WO2003022995 (Пример 9; Фиг.13A, Пример 53; страница 173, Пример 2; Фиг.2A);
NP_036581 six transmembrane epithelial antigen of the prostate
Перекрестные ссылки: MIM:604415; NP_036581.1; NM_012449_1
(4) 0772P (CA125, MUC16, номер доступа в Genbank AF361486)
J. Biol. Chem. 276 (29):27371-27375 (2001)); WO2004045553 (пункт формулы изобретения 14); WO200292836 (пункт формулы изобретения 6; Фиг.12); WO200283866 (пункт формулы изобретения 15; страницы 116-121); US2003124140 (Пример 16); Перекрестные ссылки: GI:34501467; AAK74120.3; AF361486_1
(5) MPF (MPF, MSLN, SMR, фактор активации мегакариоцитов, мезотелин, номер доступа в Genbank NM_005823) Yamaguchi, N., et al Biol. Chem. 269 (2), 805-808 (1994), Proc. Natl. Acad. Sci. U.S.A. 96 (20): 11531-11536 (1999), Proc. Natl. Acad. Sci. U.S.A. 93 (1): 136-140 (1996), J. Biol. Chem. 270 (37):21984-21990 (1995)); WO2003101283 (пункт формулы изобретения 14); (WO2002102235 (пункт формулы изобретения 13; страницы 287-288); WO2002101075 (пункт формулы изобретения 4; страницы 308-309); WO200271928 (страницы 320-321); WO9410312 (страницы 52-57); Перекрестные ссылки: MIM:601051; NP_005814.2; NM_005823_1
(6) Napi3b (NAPI-3B, NPTIIb, SLC34A2, семейство переносчиков растворенных веществ 34 (фосфат натрия), член 2, натрий-зависимый переносчик фосфата 3b, типа II, номер доступа в Genbank NM 006424)
J. Biol. Chem. 277 (22): 19665-19672 (2002), Genomics 62 (2):281-284 (1999), Feild, J.A., et al (1999) Biochem. Biophys. Res. Commun. 258 (3):578-582); WO2004022778 (пункт формулы изобретения 2); EP1394274 (Пример 11); WO2002102235 (пункт формулы изобретения 13; страница 326); EP875569 (пункт формулы изобретения 1; страницы 17-19); WO200157188 (пункт формулы изобретения 20; страница 329); WO2004032842 (Пример IV); WO200175177 (пункт формулы изобретения 24; страницы 139-140);
Перекрестные ссылки: MIM:604217; NP_006415.1; NM_006424_1
(7) Sema 5b (FLJ10372, KIAA1445, Mm.42015, SEMA5B, SEMAG, Семафорин 5b Hlog, домен sema, семь повторов тромбоспондина (типа 1 и подобный типу 1), трансмембранный домен (TM) и короткий цитоплазматический домен, (семафорин) 5B, номер доступа в Genbank AB040878)
Nagase T., et al (2000) DNA Res. 7 (2): 143-150); WO2004000997 (пункт формулы изобретения 1); WO2003003984 (пункт формулы изобретения 1); WO200206339 (пункт формулы изобретения 1; страницы 50); WO200188133 (пункт формулы изобретения 1; страницы 41-43, 48-58); WO2003054152 (пункт формулы изобретения 20); WO2003101400 (пункт формулы изобретения 11);
Номера доступа: Q9P283; EMBL; AB040878; BAA95969.1. Genew; HGNC: 10737;
(8) PSCA hlg (2700050C12Rik, C530008O16Rik, кДНК RIKEN 2700050C12, кДНК RIKEN гена 2700050C12, номер доступа в Genbank AY358628); Ross et al (2002) Cancer Res. 62:2546-2553; US2003129192 (пункт формулы изобретения 2); US2004044180 (пункт формулы изобретения 12); US2004044179 (пункт формулы изобретения 11); US2003096961 (пункт формулы изобретения 11); US2003232056 (Пример 5); WO2003105758 (пункт формулы изобретения 12); US2003206918 (Пример 5); EP1347046 (пункт формулы изобретения 1); WO2003025148 (пункт формулы изобретения 20);
Перекрестные ссылки: GI:37182378; AAQ88991.1; AY358628_1
(9) ETBR (рецептор эндотелина типа B, номер доступа в Genbank AY275463);
Nakamuta M., et al Biochem. Biophys. Res. Commun. 177, 34-39, 1991; Ogawa Y., et al Biochem. Biophys. Res. Commun. 178, 248-255, 1991; Arai H., et al Jpn. Circ. J. 56, 1303-1307, 1992; Arai H., et al J. Biol. Chem. 268, 3463-3470, 1993; Sakamoto A., Yanagisawa M., et al Biochem. Biophys. Res. Commun. 178, 656-663, 1991; Elshourbagy N.A., et al J. Biol. Chem. 268, 3873-3879, 1993; Haendler B., et al J. Cardiovasc. Pharmacol. 20, s1-S4, 1992; Tsutsumi M., et al Gene 228, 43-49, 1999; Strausberg R.L., et al Proc. Natl. Acad. Sci. U.S.A. 99, 16899-16903, 2002; Bourgeois C, et al J. Clin. Endocrinol. Metab. 82, 3116-3123, 1997; Okamoto Y., et al Biol. Chem. 272, 21589-21596, 1997; Verheij J.B., et al Am. J. Med. Genet. 108, 223-225, 2002; Hofstra R.M.W., et al Eur. J. Hum. Genet. 5, 180-185, 1997; Puffenberger E.G., et al Cell 79, 1257-1266, 1994; Attie T., et al, Hum. Mol. Genet. 4, 2407-2409, 1995; Auricchio A., et al Hum. Mol. Genet. 5:351-354, 1996; Amiel J., et al Hum. Mol. Genet. 5, 355-357, 1996; Hofstra R.M.W., et al Nat. Genet. 12, 445-447, 1996; Svensson P.J., et al Hum. Genet. 103, 145-148, 1998; Fuchs S., et al Mol. Med. 7, 115-124, 2001; Pingault V., et al (2002) Hum. Genet. 111, 198-206; WO2004045516 (пункт формулы изобретения 1); WO2004048938 (Пример 2); WO2004040000 (пункт формулы изобретения 151); WO2003087768 (пункт формулы изобретения 1); WO2003016475 (пункт формулы изобретения 1); WO2003016475 (пункт формулы изобретения 1); WO200261087 (Фиг.1); WO2003016494 (Фиг.6); WO2003025138 (пункт формулы изобретения 12; страницы 144); WO200198351 (пункт формулы изобретения 1; страницы 124-125); EP522868 (пункт формулы изобретения 8; Фиг.2); WO200177172 (пункт формулы изобретения 1; страницы 297-299); US2003109676; US6518404 (Фиг.3); US5773223 (пункт формулы изобретения 1a; кол. 31-34); WO2004001004;
(10) MSG783 (RNF124, гипотетический белок FLJ20315, номер доступа в Genbank NM_017763);
WO2003104275 (пункт формулы изобретения 1); WO2004046342 (Пример 2); WO2003042661 (пункт формулы изобретения 12); WO2003083074 (пункт формулы изобретения 14; страница 61); WO2003018621 (пункт формулы изобретения 1); WO2003024392 (пункт формулы изобретения 2; Фиг.93); WO200166689 (Пример 6);
Перекрестные ссылки: LocusID:54894; NP_060233.2; NM_017763_1
(11) STEAP2 (HGNC_8639, IPCA-1, PCANAP1, STAMP1, STEAP2, STMP, ассоциированный с раком предстательной железы ген 1, ассоциированный с раком предстательной железы белок 1, эпителиальный антиген простаты 2, содержащий шесть трансмембранных доменов, белок простаты, содержащий шесть трансмембранных доменов, номер доступа в Genbank AF455138)
Lab. Invest. 82 (11): 1573-1582 (2002)); WO2003087306; US2003064397 (пункт формулы изобретения 1; Фиг.1); WO200272596 (пункт формулы изобретения 13; страницы 54-55); WO200172962 (пункт формулы изобретения 1; Фиг.4B); WO2003104270 (пункт формулы изобретения 11); WO2003104270 (пункт формулы изобретения 16); US2004005598 (пункт формулы изобретения 22); WO2003042661 (пункт формулы изобретения 12); US2003060612 (пункт формулы изобретения 12; Фиг.10); WO200226822 (пункт формулы изобретения 23; Фиг.2); WO200216429 (пункт формулы изобретения 12; Фиг.10);
Перекрестные ссылки: GI:22655488; AAN04080.1; AF455138_1
(12) TrpM4 (BR22450, FLJ20041, TRPM4, TRPM4B, катионный канал транзиторного рецепторного потенциала, подсемейство M, член 4, номер доступа в Genbank NM_017636)
Xu, X.Z., et al Proc. Natl. Acad. Sci. U.S.A. 98 (19): 10692-10697 (2001), Cell 109 (3):397-407 (2002), J. Biol. Chem. 278 (33):30813-30820 (2003)); US2003143557 (пункт формулы изобретения 4); WO200040614 (пункт формулы изобретения 14; страницы 100-103); WO200210382 (пункт формулы изобретения 1; Фиг.9A); WO2003042661 (пункт формулы изобретения 12); WO200230268 (пункт формулы изобретения 27; страница 391); US2003219806 (пункт формулы изобретения 4); WO200162794 (пункт формулы изобретения 14; Фиг.1A-D);
Перекрестные ссылки: MIM:606936; NP_060106.2; NM_017636_1
(13) CRIPTO (CR, CR1, CRGF, CRIPTO, TDGF1, полученный из тератокарциномы фактор роста, номер доступа в Genbank NP 003203 или NM_003212)
Ciccodicola, A., et al EMBO J. 8 (7): 1987-1991 (1989), Am. J. Hum. Genet. 49 (3):555-565 (1991)); US2003224411 (пункт формулы изобретения 1); WO2003083041 (Пример 1); WO2003034984 (пункт формулы изобретения 12); WO200288170 (пункт формулы изобретения 2; страницы 52-53); WO2003024392 (пункт формулы изобретения 2; Фиг.58); WO200216413 (пункт формулы изобретения 1; страницы 94-95, 105); WO200222808 (пункт формулы изобретения 2; Фиг.1); US5854399 (Пример 2; кол. 17-18); US5792616 (Фиг.2);
Перекрестные ссылки: MIM:187395; NP_003203.1; NM_003212_1
(14) CD21 (CR2 (рецептор комплемента 2) или C3DR (рецептор C3d/вируса Эпштейна-Барр) или Hs.73792 номер доступа в Genbank M26004)
Fujisaku et al (1989) J. Biol. Chem. 264 (4):2118-2125); Weis J.J., et al J. Exp. Med. 167, 1047-1066, 1988; Moore M., et al Proc. Natl. Acad. Sci. U.S.A. 84, 9194-9198, 1987; Barel M., et al Mol. Immunol. 35, 1025-1031, 1998; Weis J.J., et al Proc. Natl. Acad. Sci. U.S.A. 83, 5639-5643, 1986; Sinha S.K., et al (1993) J. Immunol. 150, 5311-5320; WO2004045520 (Пример 4); US2004005538 (Пример 1); WO2003062401 (пункт формулы изобретения 9); WO2004045520 (Пример 4); WO9102536 (Фиг.9.1-9.9); WO2004020595 (пункт формулы изобретения 1);
Номера доступа: P20023; Q13866; Q14212; EMBL; M26004; AAA35786.1.
(15) CD79b (CD79B, CD79β, IGb (иммуноглобулин-ассоциированная молекула бета), B29, номер доступа в Genbank NM_000626 или 11038674)
Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (7):4126-4131, Blood (2002) 100 (9):3068-3076, Muller et al (1992) Eur. J. Immunol. 22 (6): 1621-1625); WO2004016225 (пункт формулы изобретения 2, Фиг.140); WO2003087768, US2004101874 (пункт формулы изобретения 1, страница 102); WO2003062401 (пункт формулы изобретения 9); WO200278524 (Пример 2); US2002150573 (пункт формулы изобретения 5, страница 15); US5644033; WO2003048202 (пункт формулы изобретения 1, страницы 306 и 309); WO 99/558658, US6534482 (пункт формулы изобретения 13, Фиг.17A/B); WO200055351 (пункт формулы изобретения 11, страницы 1145-1146);
Перекрестные ссылки: MIM:147245; NP_000617.1; NM_000626_1
(16) FcRH2 (IFGP4, IRTA4, SPAP1A (содержащий SH2 домен фосфатазный якорный белок 1a), SPAP1B, SPAP1C, номер доступа в Genbank NM_030764, AY358130)
Genome Res. 13 (10):2265-2270 (2003), Immunogenetics 54 (2):87-95 (2002), Blood 99 (8):2662-2669 (2002), Proc. Natl. Acad. Sci. U.S.A. 98 (17):9772-9777 (2001), Xu, M.J., et al (2001) Biochem. Biophys. Res. Commun. 280 (3):768-775; WO2004016225 (пункт формулы изобретения 2); WO2003077836; WO200138490 (пункт формулы изобретения 5; Фиг.18D-1-18D-2); WO2003097803 (пункт формулы изобретения 12); WO2003089624 (пункт формулы изобретения 25);
Перекрестные ссылки: MIM:606509; NP_110391.2; NM_030764_1
(17) HER2 (ErbB2, номер доступа в Genbank M11730)
Coussens L., et al Science (1985) 230(4730): 1132-1139); Yamamoto T., et al Nature 319, 230-234, 1986; Semba K., et al Proc. Natl. Acad. Sci. U.S.A. 82, 6497-6501, 1985; Swiercz J.M., et al J. Cell Biol. 165, 869-880, 2004; Kuhns J.J., et al J. Biol. Chem. 274, 36422-36427, 1999; Cho H.-S., et al Nature 421, 756-760, 2003; Ehsani A., et al (1993) Genomics 15, 426-429; WO2004048938 (Пример 2); WO2004027049 (Фиг.1I); WO2004009622; WO2003081210; WO2003089904 (пункт формулы изобретения 9); WO2003016475 (пункт формулы изобретения 1); US2003118592; WO2003008537 (пункт формулы изобретения 1); WO2003055439 (пункт формулы изобретения 29; Фиг.1A-B); WO2003025228 (пункт формулы изобретения 37; Фиг.5C); WO200222636 (Пример 13; страницы 95-107); WO200212341 (пункт формулы изобретения 68; Фиг.7); WO200213847 (страницы 71-74); WO200214503 (страницы 114-117); WO200153463 (пункт формулы изобретения 2; страницы 41-46); WO200141787 (страница 15); WO200044899 (пункт формулы изобретения 52; Фиг.7); WO200020579 (пункт формулы изобретения 3; Фиг.2); US5869445 (пункт формулы изобретения 3; Кол. 31-38); WO9630514 (пункт формулы изобретения 2; страницы 56-61); EP1439393 (пункт формулы изобретения 7); WO2004043361 (пункт формулы изобретения 7); WO2004022709; WO200100244 (Пример 3; Фиг.4);
Номера доступа: P04626; EMBL; M11767; AAA35808.1. EMBL; М11761; AAA35808.1.
(18) NCA (CEACAM6, номер доступа в Genbank M18728);
Barnett T., et al Genomics 3, 59-66, 1988; Tawaragi Y., et al Biochem. Biophys. Res. Commun. 150, 89-96, 1988; Strausberg R.L., et al Proc. Natl. Acad. Sci. U.S.A. 99: 16899-16903, 2002; WO2004063709; EP1439393 (пункт формулы изобретения 7); WO2004044178 (Пример 4); WO2004031238; WO2003042661 (пункт формулы изобретения 12); WO200278524 (Пример 2); WO200286443 (пункт формулы изобретения 27; страница 427); WO200260317 (пункт формулы изобретения 2);
Номера доступа: P40199; Q14920; EMBL; M29541; AAA59915.1. EMBL; M18728;
(19) MDP (DPEP1, номер доступа в Genbank BC017023)
Proc. Natl. Acad. Sci. U.S.A. 99 (26): 16899-16903 (2002)); WO2003016475 (пункт формулы изобретения 1); WO200264798 (пункт формулы изобретения 33; страницы 85-87); JP05003790 (Фиг.6-8); W09946284 (Фиг.9);
Перекрестные ссылки: MIM:179780; AAH17023.1; BC017023_1
(20) IL20Rα (IL20Ra, ZCYTOR7, номер доступа в Genbank AF184971); Clark H.F., et al Genome Res. 13, 2265-2270, 2003; Mungall A.J., et al Nature 425, 805-811, 2003; Blumberg H., et al Cell 104, 9-19, 2001; Dumoutier L., et al J. Immunol. 167, 3545-3549, 2001; Parrish-Novak J., et al J. Biol. Chem. 277, 47517-47523, 2002; Pletnev S., et al (2003) Biochemistry 42: 12617-12624; Sheikh F., et al (2004) J. Immunol. 172, 2006-2010; EP1394274 (Пример 11); US2004005320 (Пример 5); WO2003029262 (страницы 74-75); WO2003002717 (пункт формулы изобретения 2; страница 63); WO200222153 (страницы 45-47); US2002042366 (страницы 20-21); WO200146261 (страницы 57-59); WO200146232 (страницы 63-65); W09837193 (пункт формулы изобретения 1; страницы 55-59);
Номера доступа: Q9UHF4; Q6UWA9; Q96SH8; EMBL; AF184971; AAF01320.1.
(21) Бревикан (BCAN, BEHAB, номер доступа в Genbank AF229053)
Gary S.C., et al Gene 256, 139-147, 2000; Clark H.F., et al Genome Res. 13, 2265-2270, 2003; Strausberg R.L., et al Proc. Natl. Acad. Sci. U.S.A. 99, 16899-16903, 2002; US2003186372 (пункт формулы изобретения 11); US2003186373 (пункт формулы изобретения 11); US2003119131 (пункт формулы изобретения 1; Фиг.52); US2003119122 (пункт формулы изобретения 1; Фиг.52); US2003119126 (пункт формулы изобретения 1); US2003119121 (пункт формулы изобретения 1; Фиг.52); US2003119129 (пункт формулы изобретения 1); US2003119130 (пункт формулы изобретения 1); US2003119128 (пункт формулы изобретения 1; Фиг.52); US2003119125 (пункт формулы изобретения 1); WO2003016475 (пункт формулы изобретения 1); WO200202634 (пункт формулы изобретения 1);
(22) EphB2R (DRT, ERK, Hek5, EPHT3, Tyro5, номер доступа в Genbank NM_004442)
Chan,J. и Watt, V.M., Oncogene 6 (6), 1057-1061 (1991) Oncogene 10 (5):897-905 (1995), Annu. Rev. Neurosci. 21:309-345 (1998), Int. Rev. Cytol. 196: 177-244 (2000)); WO2003042661 (пункт формулы изобретения 12); WO200053216 (пункт формулы изобретения 1; страница 41); WO2004065576 (пункт формулы изобретения 1); WO2004020583 (пункт формулы изобретения 9); WO2003004529 (страницы 128-132); WO200053216 (пункт формулы изобретения 1; страница 42);
Перекрестные ссылки: MIM:600997; NP_004433.2; NM_004442_1
(23) ASLG659 (B7h, номер доступа в Genbank AX092328)
US20040101899 (пункт формулы изобретения 2); WO2003104399 (пункт формулы изобретения 11); WO2004000221 (Фиг.3); US2003165504 (пункт формулы изобретения 1); US2003124140 (Пример 2); US2003065143 (Фиг.60); WO2002102235 (пункт формулы изобретения 13; страница 299); US2003091580 (Пример 2); WO200210187 (пункт формулы изобретения 6; Фиг.10); WO200194641 (пункт формулы изобретения 12; Фиг.7b); WO200202624 (пункт формулы изобретения 13; Фиг.1A-1B); US2002034749 (пункт формулы изобретения 54; страницы 45-46); WO200206317 (Пример 2; страницы 320-321, пункт формулы изобретения 34; страницы 321-322); WO200271928 (страницы 468-469); WO200202587 (Пример 1; Фиг.1); WO200140269 (Пример 3; страницы 190-192); WO200036107 (Пример 2; страницы 205-207); WO2004053079 (пункт формулы изобретения 12); WO2003004989 (пункт формулы изобретения 1); WO200271928 (страницы 233-234, 452-453); WO 0116318;
(24) PSCA (Предшественник антигена стволовых клеток простаты, номер доступа в Genbank AJ297436)
Reiter R.E., et al Proc. Natl. Acad. Sci. U.S.A. 95, 1735-1740, 1998; Gu Z., et al Oncogene 19, 1288-1296, 2000; Biochem. Biophys. Res. Commun. (2000) 275(3):783-788; WO2004022709; EP1394274 (Пример 11); US2004018553 (пункт формулы изобретения 17); WO2003008537 (пункт формулы изобретения 1); WO200281646 (пункт формулы изобретения 1; страница 164); WO2003003906 (пункт формулы изобретения 10; страница 288); WO200140309 (Пример 1; Фиг.17); US2001055751 (Пример 1; Фиг.1b); WO200032752 (пункт формулы изобретения 18; Фиг.1); WO9851805 (пункт формулы изобретения 17; страница 97); W09851824 (пункт формулы изобретения 10; страница 94); WO9840403 (пункт формулы изобретения 2; Фиг.1B);
Номер доступа: O43653; EMBL; AF043498; AAC39607.1.
(25) GEDA (номер доступа в Genbank AY260763);
AAP14954 lipoma HMGIC fusion-partner-like protein /pid=AAP14954.1 - Homo sapiens Species: Homo sapiens (human)
WO2003054152 (пункт формулы изобретения 20); WO2003000842 (пункт формулы изобретения 1); WO2003023013 (Пример 3, пункт формулы изобретения 20); US2003194704 (пункт формулы изобретения 45);
Перекрестные ссылки: GI:30102449; AAP14954.1; AY260763_1
(26) BAFF-R (рецептор фактора активации B-клеток, BLyS рецептор 3, BR3, номер доступа в Genbank AF116456); BAFF receptor /pid=NP_443177.1 - Homo sapiens
Thompson, J.S., et al Science 293 (5537), 2108-2111 (2001); WO2004058309; WO2004011611; WO2003045422 (Пример; страницы 32-33); WO2003014294 (пункт формулы изобретения 35; Фиг.6B); WO2003035846 (пункт формулы изобретения 70; страницы 615-616); WO200294852 (Кол. 136-137); WO200238766 (пункт формулы изобретения 3; страница 133); WO200224909 (Пример 3; Фиг.3);
Перекрестные ссылки: MIM:606269; NP_443177.1; NM_052945_1; AF132600
(27) CD22 (изоформа B-клеточного рецептора CD22-B, BL-CAM, Lyb-8, Lyb8, SIGLEC-2, FLJ22814, номер доступа в Genbank AK026467);
Wilson et al (1991) J. Exp. Med. 173: 137-146; WO2003072036 (пункт формулы изобретения 1; Фиг.1);
Перекрестные ссылки: MIM:107266; NP_001762.1; NM_001771_1
(28) CD79a (CD79A, CD79α, иммуноглобулин-ассоциированная молекула альфа, специфический для B-клеток белок, ковалентно взаимодействующий с Ig бета (CD79B) и формирующий комплекс на поверхности с молекулами IgM, передает сигнал, вовлеченный в дифференцировку B-клеток), pI: 4,84, MW: 25028 TM: 2 [P] Хромосома гена: 19q13.2, номер доступа в Genbank NP_001774.10)
WO2003088808, US20030228319; WO2003062401 (пункт формулы изобретения 9); US2002150573 (пункт формулы изобретения 4, страницы 13-14); W09958658 (пункт формулы изобретения 13, Фиг.16); WO9207574 (Фиг.1); US5644033; Ha et al (1992) J. Immunol. 148(5): 1526-1531; Mueller et al (1992) Eur. J. Biochem. 22: 1621-1625; Hashimoto et al (1994) Immunogenetics 40(4):287-295; Preud'homme et al (1992) Clin. Exp. Immunol. 90(1): 141-146; Yu et al (1992) J. Immunol. 148(2) 633-637; Sakaguchi et al (1988) EMBO J. 7(11):3457-3464;
(29) CXCR5 (Рецептор лимфомы Беркитта 1, связанный с G белком рецептор, активируемый хемокином CXCL13, функционирует в миграции лимфоцитов и гуморальной защите, играет роль в инфекции HIV-2 и, возможно, развитии СПИД, лимфомы, миеломы и лейкоза); 372 ак, pI: 8,54 MW: 41959 TM: 7 [P] Хромосома гена: 11q23.3, номер доступа в Genbank NP_001707.1)
WO2004040000; WO2004015426; US2003105292 (Пример 2); US6555339 (Пример 2); WO200261087 (Фиг.1); WO200157188 (пункт формулы изобретения 20, страница 269); WO200172830 (страницы 12-13); WO200022129 (Пример 1, страницы 152-153, Пример 2, страницы 254-256); W09928468 (пункт формулы изобретения 1, страница 38); US5440021 (Пример 2, кол. 49-52); W09428931 (страницы 56-58); W09217497 (пункт формулы изобретения 7, Фиг.5); Dobner et al (1992) Eur. J. Immunol. 22:2795-2799; Barella et al (1995) Biochem. J. 309:773-779;
(30) HLA-DOB (Бета-субъединица молекулы MHC класса II (Ia антиген), связывающая пептиды и представляющая их CD4+ T-лимфоцитам); 273 ак, pI: 6,56 MW: 30820 TM: 1 [P] Хромосома гена: 6p21.3, номер доступа в Genbank NP_002111.1)
Tonnelle et al (1985) EMBO J. 4(11):2839-2847; Jonsson et al (1989) Immunogenetics 29(6):411-413; Beck et al (1992) J. Mol. Biol. 228:433-441; Strausberg et al (2002) Proc. Natl. Acad. Sci USA 99: 16899-16903; Servenius et al (1987) J. Biol. Chem. 262:8759-8766; Beck et al (1996) J. Mol. Biol. 255: 1-13; Naruse et al (2002) Tissue Antigens 59:512-519; W09958658 (пункт формулы изобретения 13, Фиг.15); US6153408 (Кол. 35-38); US5976551 (кол. 168-170); US6011146 (кол. 145-146); Kasahara et al (1989) Immunogenetics 30(1):66-68; Larhammar et al (1985) J. Biol. Chem. 260(26): 14111-14119;
(31) P2X5 (управляемый лигандом пуринергического рецептора P2X ионный канал 5, ионный канал, управляемый внеклеточным ATP, может быть вовлечен в передачу возбуждения по синапсам и нейрогенез, недостаточность может вносить вклад в патофизиологию идиопатической нестабильности детрузора); 422 ак), pI: 7,63, MW: 47206 TM: 1 [P] Хромосома гена: 17p13.3, номер доступа в Genbank NP_002552.2)
Le et al (1997) FEBS Lett. 418(1-2): 195-199; WO2004047749; WO2003072035 (пункт формулы изобретения 10); Touchman et al (2000) Genome Res. 10: 165-173; WO200222660 (пункт формулы изобретения 20); WO2003093444 (пункт формулы изобретения 1); WO2003087768 (пункт формулы изобретения 1); WO2003029277 (страница 82);
(32) CD72 (антиген дифференцировки B-клеток CD72, Lyb-2) БЕЛКОВАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ Полная maeaity...tafrfpd (1..359; 359 ак), pI: 8,66, MW: 40225 TM: 1 [P] Хромосома гена: 9p13.3, номер доступа в Genbank NP_001773.1)
WO2004042346 (пункт формулы изобретения 65); WO2003026493 (страницы 51-52, 57-58); WO200075655 (страницы 105-106); Von Hoegen et al (1990) J. Immunol. 144(12):4870-4877; Strausberg et al (2002) Proc. Natl. Acad. Sci USA 99: 16899-16903;
(33) LY64 (Лимфоцитарный антиген 64 (RP105), мембранный белок типа I семейства богатых лейцином повторов (LRR), регулирует активацию и апоптоз B-клеток, потеря функции ассоциирована с увеличенной активностью заболевания у пациентов с системной красной волчанкой); 661 ак, pI: 6,20, MW: 74147 TM: 1 [P] Хромосома гена: 5q12, номер доступа в Genbank NP_005573.1)
US2002193567; WO9707198 (пункт формулы изобретения 11, стр. 39-42); Miura et al (1996) Genomics 38(3):299-304; Miura et al (1998) Blood 92:2815-2822; WO2003083047; W09744452 (пункт формулы изобретения 8, страницы 57-61); WO200012130 (страницы 24-26);
(34) FcRH1 (подобный Fc-рецептору белок 1, предположительный рецептор для Fc-домена иммуноглобулинов, содержащий Ig-подобный домен типа C2 и домен ITAM, может играть роль в дифференцировке B-лимфоцитов); 429 ак, pI: 5,28, MW: 46925 TM: 1 [P] Хромосома гена: 1q21-1q22, номер доступа в Genbank NP_443170.1)
WO2003077836; WO200138490 (пункт формулы изобретения 6, Фиг.18E-1-18-E-2); Davis et al (2001) Proc. Natl. Acad. Sci USA 98(17):9772-9777; WO2003089624 (пункт формулы изобретения 8); EP1347046 (пункт формулы изобретения 1); WO2003089624 (пункт формулы изобретения 7);
(35) IRTA2 (ассоциированный с транслокацией рецептор 2 суперсемейства иммуноглобулинов, предположительный иммунорецептор с возможной ролью в развитии B-клеток и лимфомагенезе; нарушение регуляции гена посредством транслокации происходит при некоторых злокачественных новообразованиях B-клеток); 977 ак, pI: 6,88 MW: 106468 TM: 1 [P] Хромосома гена: 1q21, номер доступа в Genbank Человек: AF343662, AF343663, AF343664, AF343665, AF369794, AF397453, AK090423, AK090475, AL834187, AY358085; Мышь: AK089756, AY158090, AY506558; NP_112571.1
WO2003024392 (пункт формулы изобретения 2, Фиг.97); Nakayama et al (2000) Biochem. Biophys. Res. Commun. 277(1): 124-127; WO2003077836; WO200138490 (пункт формулы изобретения 3, Фиг.18B-1-18B-2);
(36) TENB2 (TMEFF2, томорегулин, TPEF, HPP1, TR, предположительный трансмембранный протеогликан, родственный семейству факторов роста EGF/херегулина и фоллистатину); 374 ак, Номера доступа в NCBI: AAD55776, AAF91397, AAG49451, NCBI RefSeq: NP_057276; Ген в NCBI: 23671; OMIM: 605734; SwissProt Q9UIK5; номер доступа в Genbank AF179274; AY358907, CAF85723, CQ782436
WO2004074320 (SEQ ID NO 810); JP2004113151 (SEQ ID NO 2, 4, 8); WO2003042661 (SEQ ID NO 580); WO2003009814 (SEQ ID NO 411); EP1295944 (страницы 69-70); WO200230268 (страница 329); WO200190304 (SEQ ID NO 2706); US2004249130; US2004022727; WO2004063355; US2004197325; US2003232350; US2004005563; US2003124579; Horie et al (2000) Genomics 67: 146-152; Uchida et al (1999) Biochem. Biophys. Res. Commun. 266:593-602; Liang et al (2000) Cancer Res. 60:4907-12; Glynne-Jones et al (2001) Int J Cancer. Oct 15;94(2): 178-84;
(37) PMEL17 (гомолог silver; SILV; D12S53E; PMEL17; (SI); (SIL); ME20; gp100) BC001414; BT007202; M32295; M77348; NM_006928; McGlinchey,R.P. et al (2009) Proc. Natl. Acad. Sci. U.S.A. 106 (33), 13731-13736; Kummer,M.P. et al (2009) J. Biol. Chem. 284 (4), 2296-2306;
(38) TMEFF1 (трансмембранный белок с EGF-подобным доменом и двумя фоллистатин-подобными доменами 1; Томорегулин-1; H7365; C9orf2; C9ORF2; U19878; X83961) NM_080655; NM_003692; Harms, P.W. (2003) Genes Dev. 17 (21), 2624-2629; Gery, S. et al (2003) Oncogene 22 (18):2723-2727;
(39) GDNF-Ra1 (рецептор альфа 1 семейства GDNF; GFRA1; GDNFR; GDNFRA; RETL1; TRNR1; RET1L; GDNFR-alpha1; GFR-ALPHA-1; U95847; BC014962; NM_145793)
NM_005264; Kim, M.H. et al (2009) Mol. Cell. Biol. 29 (8), 2264-2277; Treanor, J.J. et al (1996) Nature 382 (6586): 80-83;
(40) Ly6E (комплекс лимфоцитарного антигена 6, локус E; Ly67,RIG-E,SCA-2,TSA-1)
NP_002337.1; NM_002346.2; de Nooij-van Dalen, A.G. et al (2003) Int. J. Cancer 103 (6), 768-774; Zammit, D.J. et al (2002) Mol. Cell. Biol. 22 (3):946-952;
(41) TMEM46 (гомолог shisa 2 (Xenopus laevis); SHISA2) NP_001007539.1;
NM_001007538,1; Furushima,K. et al (2007) Dev. Biol. 306 (2), 480-492; Clark,H.F. et al (2003) Genome Res. 13 (10):2265-2270;
(42) Ly6G6D (комплекс лимфоцитарного антигена 6, локус G6D; Ly6-D, MEGT1) NP_067079.2; NM_021246.2; Mallya, M. et al (2002) Genomics 80 (1): 113-123; Ribas, G. et al (1999) J. Immunol. 163 (1):278-287;
(43) LGR5 (содержащий богатые лейцином повторы связанный с белком G рецептор 5; GPR49, GPR67) NP_003658.1; NM_003667.2; Salanti,G. et al (2009) Am. J. Epidemiol. 170 (5):537-545; Yamamoto,Y. et al (2003) Hepatology 37 (3):528-533;
(44) RET (протоонкоген ret; MEN2A; HSCR1; MEN2B; MTC1; (PTC); CDHF12; Hs.168114; RET51; RET-ELE1) NP_066124.1; NM_020975.4; Tsukamoto, H. et al (2009) Cancer Sci. 100 (10): 1895-1901; Narita,N. et al (2009) Oncogene 28 (34):3058-3068;
(45) LY6K (комплекс лимфоцитарного антигена 6, локус K; LY6K; HSJ001348; FLJ35226) NP_059997.3; NM_017527.3; Ishikawa,N. et al (2007) Cancer Res. 67 (24): 11601-11611; de Nooij-van Dalen,A.G. et al (2003) Int. J. Cancer 103 (6):768-774;
(46) GPR19 (связанный с белком G рецептор 19; Mm.4787) NP_006134.1; NM_006143.2;
Montpetit, A. and Sinnett,D. (1999) Hum. Genet. 105 (1-2): 162-164; O'Dowd, B.F. et al (1996) FEBS Lett. 394 (3):325-329;
(47) GPR54 (рецептор KISS1; KISS1R; GPR54; HOT7T175; AXOR12) NP_115940.2; NM_032551.4; Navenot, J.M. et al (2009) Mol. Pharmacol. 75 (6): 1300-1306; Hata, K. et al (2009) Anticancer Res. 29 (2):617-623;
(48) ASPHD1 (содержащий домен аспартат-бета-гидроксилазы белок 1; LOC253982) NP_859069.2; NM_181718.3; Gerhard, D.S. et al (2004) Genome Res. 14 (10B):2121-2127;
(49) Тирозиназа (TYR; OCAIA; OCA1A; тирозиназа; SHEP3) NP_000363.1; NM_000372.4; Bishop, D.T. et al (2009) Nat. Genet. 41 (8):920-925; Nan, H. et al (2009) Int. J. Cancer 125 (4):909-917;
(50) TMEM118 (белок с «безымянным пальцем», трансмембранный белок 2; RNFT2; FLJ14627)
NP_001103373.1; NM_001109903.1; Clark, H.F. et al (2003) Genome Res. 13 (10):2265-2270; Scherer,S.E. et al (2006) Nature 440 (7082):346-351
(51) GPR172A (связанный с белком G рецептор 172A; GPCR41; FLJ11856; D15Ertd747e)
NP_078807.1; NM_024531.3; Ericsson, T.A. et al (2003) Proc. Natl. Acad. Sci. U.S.A. 100 (11):6759-6764; Takeda, S. et al (2002) FEBS Lett. 520 (1-3):97-101.
Исходное антитело может представлять собой также слитый белок, содержащий последовательность альбуминсвязывающего пептида (ABP) (Dennis et al. (2002) «Albumin Binding As A General Strategy For Improving The Pharmacokinetics Of Proteins» J Biol Chem. 277:35035-35043; WO 01/45746). Антитела по изобретению включают слитые белки с последовательностями ABP, объясняемые в: (i) Dennis et al (2002) J Biol Chem. 277:35035-35043 в таблицах III и IV, страница 35038; (ii) US 20040001827 в SEQ ID NO: 9-22; и (iii) WO 01/45746 на страницах 12-13, в SEQ ID NO: z1-z14, и содержание всех из них приведено в настоящем документе в качестве ссылки.
МУТАГЕНЕЗ
ДНК, кодирующую вариант аминокислотной последовательности исходного полипептида, получают множеством способов, известных в данной области. Эти способы включают, без ограничения, получение посредством сайт-специфического (или опосредованного олигонуклеотидом) мутагенеза, ПЦР-мутагенеза и кассетного мутагенеза ранее полученной ДНК, кодирующей полипептид. Варианты рекомбинантных антител можно также конструировать посредством манипуляций с рестрикционными фрагментами или посредством ПЦР с удлинением перекрывающихся фрагментов с использованием синтетических олигонуклеотидов. Праймеры для мутагенеза кодируют замену(замены) кодонов на кодоны для цистеина. Общепринятые способы мутагенеза можно использовать для получения ДНК, кодирующей такие мутантные сконструированные антитела с цистеиновыми заменами. Общее руководство можно найти в Sambrook et al Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989; и Ausubel et al Current Protocols in Molecular Biology, Greene Publishing and Wiley-Interscience, New York, N.Y., 1993.
Сайт-специфический мутагенез является одним из способов получения вариантов с заменами, т.е. мутантных белков. Этот способ хорошо известен в данной области (см., например, Carter (1985) et al Nucleic Acids Res. 13:4431-4443; Ho et al (1989) Gene (Amst.) 77:51-59; и Kunkel et al (1987) Proc. Natl. Acad. Sci. USA 82:488). Кратко, при проведении сайт-специфического мутагенеза ДНК, исходную ДНК изменяют, сначала проводя гибридизацию олигонуклеотида, кодирующего желательную мутацию, с одной цепью такой исходной ДНК. После гибридизации используют ДНК-полимеразу для синтеза полной второй цепи с использованием прогибридизовавшегося олигонуклеотида в качестве праймера, и с использованием одной цепи исходной ДНК в качестве матрицы. Таким образом олигонуклеотид, кодирующий желательную мутацию, включают в полученную двухцепочечную ДНК. Сайт-специфический мутагенез можно проводить в гене, экспрессирующем белок, подлежащий мутагенезу, в экспрессирующей плазмиде, и полученную плазмиду можно секвенировать для подтверждения вставки желательных мутаций замены на цистеин (Liu et al (1998) J. Biol. Chem. 273:20252-20260). Способы и форматы сайт-специфического мутагенеза включают коммерчески доступные способы и форматы, например, набор для множественного сайт-специфического мутагенеза QuikChange® (Stratagene, La Jolla, CA).
ПЦР-мутагенез также пригоден для получения вариантов аминокислотной последовательности исходного полипептида. См. Higuchi, (1990) in PCR Protocols, pp.177-183, Academic Press; Ito et al (1991) Gene 102:67-70; Bernhard et al (1994) Bioconjugate Chem. 5: 126-132; и Vallette et al (1989) Nuc. Acids Res. 17:723-733. Кратко, когда малые количества ДНК-матрицы используют в качестве исходного материала для ПЦР, праймеры, которые немного отличаются по последовательности от соответствующей области в ДНК-матрице, можно использовать для получения относительно больших количеств специфического фрагмента ДНК, отличающегося от последовательности матрицы только в положениях, в которых праймеры отличаются от матрицы.
Другой способ получения вариантов, кассетный мутагенез, основан на способе, описанном Wells et al (1985) Gene 34:315-323. Исходный материал представляет собой плазмиду (или другой вектор), содержащий ДНК исходного полипептида, подлежащего мутагенезу. Идентифицируют кодон(ы) в исходной ДНК, подлежащие мутагенезу. Должны присутствовать участки для уникальной рестрикционной эндонуклеазы с каждой стороны идентифицированного участка(участков) мутации. Если таких участков рестрикции не существует, их можно получить с использованием вышеописанного способа опосредованного олигонуклеотидами мутагенеза для введения их в подходящие положения ДНК исходного полипептида. Плазмидную ДНК разрезают в этих участках, чтобы линеаризовать ее. Двухцепочечный олигонуклеотид, кодирующий последовательность ДНК между участками рестрикции, но содержащий желательную мутацию(мутации), синтезируют с использованием общепринятых способов, когда две цепи олигонуклеотида синтезируют по отдельности и затем гибридизуют вместе с использованием общепринятых способов. Этот двухцепочечный олигонуклеотид обозначают кассетой. Эту кассету конструируют с наличием 5'- и 3'-концов, совместимых с концами линеаризованной плазмиды, так что ее можно напрямую лигировать в плазмиду. Эта плазмида теперь содержит последовательность ДНК с мутацией. Мутантную ДНК, содержащую закодированные цистеиновые замены, можно подтверждать секвенированием ДНК.
Отдельные мутации получают также направляемым олигонуклеотидами мутагенезом с использованием двухцепочечной плазмидной ДНК в качестве матрицы, посредством основанного на ПЦР мутагенеза (Sambrook and Russel, (2001) Molecular Cloning: A Laboratory Manual, 3rd edition; Zoller et al (1983) Methods Enzymol. 100:468-500; Zoller, M.J. and Smith, M. (1982) Nucl. Acids Res. 10:6487-6500).
По настоящему изобретению, hu4D5Fabv8, экспонированное на фаге M13 (Gerstner et al (2002) «Sequence Plasticity In The Antigen-Binding Site Of A Therapeutic Anti-HER2 Antibody», J Mol Biol. 321:851-62) использовали во всех экспериментах в качестве модельной системы. Цистеиновые мутации вводили в конструкции hu4D5Fabv8-фаг, hu4D5Fabv8 и ABP-hu4D5Fabv8. Выделения hu4D5-ТиоFab-фага проводили с использованием способа преципитации полиэтиленгликолем (PEG), как описано ранее (Lowman, Henry B. (1998) Methods in Molecular Biology (Totowa, New Jersey) 87 (Combinatorial Peptide Library Protocols) 249-264).
Олигонуклеотиды получают способом фосфорамидитного синтеза (US 4415732; US 4458066; Beaucage, S. and Iyer, R. (1992) «Advances in the synthesis of oligonucleotides by the phosphoramidite approach», Tetrahedron 48:2223-2311). Фосфорамидитный способ включает циклическое добавление нуклеотидных мономерных звеньев с реакционноспособной 3'-фосфорамидитной группой к олигонуклеотидной цепи, растущей на твердой подложке, состоящей из стекла с контролируемым размером пор или полистирола с высокой степенью сшивки, и, наиболее часто, в направлении от 3' к 5', где 3'-концевой нуклеозид присоединяют к твердой подложке в начале синтеза (US 5047524; US 5262530). Способ обычно осуществляют на практике с использованием автоматических, коммерчески доступных синтезаторов (Applied Biosystems, Foster City, CA). Олигонуклеотиды можно химически метить с помощью неизотопных групп для детекции, захвата, стабилизации или других целей (Andrus, A. «Chemical methods for 5' non-isotopic labelling of PCR probes and primers» (1995) in PCR 2: A Practical Approach, Oxford University Press, Oxford, pp. 39-54; Hermanson, G. in Bioconjugate Techniques (1996) Academic Press, San Diego, pp. 40-55, 643-671; Keller, G. and Manak, M. in DNA Probes Second Edition (1993), Stockton Press, New York, pp. 121-23).
АНАЛИЗ PHESELECTOR
Анализ PHESELECTOR (фаговый ELISA для отбора реакционноспособных тиолов) позволяет детекцию реакционноспособных групп цистеина в антителах в формате фагового ELISA (US 7521541; Junutula JR et al. «Rapid identification of reactive cysteine residues for site-specific labeling of antibody-Fabs» J Immunol Methods 2008; 332:41-52). Способ покрытия интересующим белком (например, антителом) поверхностей лунок, последующей инкубации с фаговыми частицами и затем с меченным HRP вторичным антителом с детекцией оптической плотности подробно описан в примере 2. Можно проводить скрининг мутантных белков, экспонированных на фагах, быстрым, точным и высоко производительным способом. Библиотеки сконструированных антител с цистеиновыми заменами можно получать и подвергать отбору по связыванию с использованием такого же способа для идентификации подходящих реакционноспособных участков включения свободного Cys из случайных белково-фаговых библиотек антител или других белков. Этот способ включает реакцию мутантных по цистеину белков, экспонированных на фаге, с аффинным реагентом или репортерной группой, которая также является реакционноспособной по отношению к тиолу. Фигура 8 иллюстрирует анализ PHESELECTOR посредством схематического представления, изображающего связывание Fab или ТиоFab с HER2 (вверху) и биотинилированного ТиоFab со стрептавидином (внизу).
ЭКСПРЕССИЯ И ОЧИСТКА БЕЛКА
ДНК, кодирующую сконструированные антитела с цистеиновыми заменами, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Клетки гибридомы служат источником такой ДНК. После выделения ДНК можно помещать в экспрессирующие векторы, которыми затем трансфицируют клетки-хозяева, такие как клетки E. coli, клетки обезьяны COS, клетки яичников китайского хомяка (CHO) или другие клетки-хозяева млекопитающих, такие как клетки миеломы (US 5807715; US 2005/0048572; US 2004/0229310), которые в ином случае не продуцируют белка антитела, для получения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Выходы сконструированных антител с цистеиновыми заменами hu4D5Fabv8 являлись сходными с выходами hu4D5Fabv8 дикого типа. Обзорные статьи по рекомбинантной экспрессии в бактериях ДНК, кодирующей антитело, включают Skerra et al (1993) Curr. Opinion in Immunol. 5:256-262 и Pluckthun (1992) Immunol. Revs. 130: 151-188.
После конструирования и отбора сконструированные антитела с цистеиновыми заменами, например, ТиоFab, с неспаренными остатками Cys с высокой реакционной способностью, можно получать посредством: (i) экспрессии в бактериальной системе, например, E. coli, или в системе культуры клеток млекопитающих (WO 01/00245), например, клеток яичников китайского хомяка (CHO); и (ii) очистки с использованием общепринятых способов очистки белка (Lowman et al (1991) J. Biol. Chem. 266(17): 10982-10988).
ТиоFab экспрессировали при индукции в 34B8, несупрессорном штамме E. coli (Baca et al (1997) Journal Biological Chemistry 272(16): 10678-84). См. Пример 3a. Собранный осадок клеток ресуспендировали в PBS (фосфатно-солевом буфере), общий лизис клеток проводили пропусканием через микрофлюидайзер, и ТиоFab очищали аффинной хроматографией на SEPHAROSE™ с белком G (Amersham). ТиоFab конъюгировали с биотин-PEO-малеимидом, как описано выше, и биотинилированые-ТиоFab дополнительно очищали гель-фильтрационной хроматографией на Superdex-200™ (Amersham), отсекающей свободный биотин-PEO-малеимид и олигомерную фракцию ТиоFab.
АНАЛИЗ МАСС-СПЕКТРОСКОПИИ
Масс-спектрометрический анализ жидкостной хроматографией с ионизацией электрораспылением (LC-ESI-MS) использовали для точного определения молекулярной массы конъюгированных с биотином Fab (Cole, R.B. Electro Spray Ionization Mass Spectrometry: Fundamentals, Instrumentation And Applications. (1997) Wiley, New York). Аминокислотную последовательность биотинилированного пептида hu4D5Fabv8 (A121C) определяли расщеплением трипсином с последующим анализом LC-ESI-тандемной MS (таблица 4, пример 3b).
Fab-фрагмент антитела hu4D5Fabv8 содержит приблизительно 445 аминокислотных остатков, включая 10 остатков Cys (пять на легкой и пять на тяжелой цепи). Определена структура с высоким разрешением вариабельного фрагмента гуманизированного 4D5 (Fv4D5), см.: Eigenbrot et al «X-Ray Structures Of The Antigen-Binding Domains From Three Variants Of Humanized Anti-P185her2 Antibody 4D5 And Comparison With Molecular Modeling» (1993) J Mol Biol. 229:969-995). Все остатки Cys присутствуют в форме дисульфидных связей, таким образом, эти остатки не имеют каких-либо реакционноспособных тиоловых групп для конъюгации с цирконий-малеимидом (если только не обработаны восстанавливающим агентом). Таким образом, вновь сконструированный остаток Cys может оставаться неспаренным, и способным вступать в реакцию, т.е., образовывать конъюгат с электрофильным линкерным реагентом или промежуточным соединением цирконий-линкер, таким как цирконий-малеимид. На фигуре 1A показано трехмерное представление фрагмента антитела hu4D5Fabv8 в координатах кристалла, полученных с помощью рентгеновского излучения. Положения в структуре сконструированных остатков Cys тяжелой и легкой цепей пронумерованы в соответствии с последовательной системой нумерации. Эта последовательная система нумерации коррелирует с системой нумерации Kabat (Kabat et al., (1991) Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD) для варианта 4d5v7fabH трастузумаба в соответствии с фигурой 1B, на которой показана схема последовательной нумерации (верхний ряд), начиная с N-конца, отличающаяся от схемы нумерации по Kabat (нижний ряд) вставками, обозначенными a,b,c. При использовании системы нумерации Kabat, действительная линейная аминокислотная последовательность может содержать меньше аминокислот или может содержать дополнительные аминокислоты, соответствующие укорочению FR или CDR вариабельного домена или вставке в них. Участки сконструированного варианта тяжелой цепи с цистеиновыми заменами идентифицированы по схеме последовательной нумерации и схеме нумерации Kabat в следующей таблице:

Для M13 фагмиды-Cys мутантных Fab (фигуры 3A и 3B) можно проводить быстрый скрининг по сравнению с Fab белками. Связывание фагмиды-ТиоFab с антигеном и со стрептавидином можно тестировать покрытием планшетов для ELISA HER2 и стрептавидином, соответственно, с последующим использованием в качестве зонда анти-Fab-HRP (Пероксидаза хрена), как описано в примере 2 и изображено на фигуре 8. Этот способ позволяет одновременное мониторирование эффекта на связывание антигена и на реакционную способность тиоловой группы сконструированного остатка Cys/конъюгированной молекулы биотина. Способ можно использовать также для скрининга реакционноспособных тиоловых групп любого белка, экспонированного на фаге M13. Конъюгированные или неконъюгированные фагмиды-ТиоFab очищают простой преципитацией PEG.
Антигенсвязывающий фрагмент гуманизированного 4D5 (hu4D5Fab) хорошо экспрессируется в E. Coli и был экспонирован на бактериофаге (Garrard et al (1993) Gene 128: 103-109). Fab-фрагмент антитела hu4D5Fabv8 был экспонирован на фаге M13 в качестве модельной системы в анализе на основе ELISA для тестирования реакционной способности тиола. Фигура 8 представляет собой графическое изображение анализа PHESELECTOR, показывающее связывание биотинилированного ТиоFab фага и антитела против фага с HRP с HER2 (вверху) и стрептавидином (внизу). Пять аминокислотных остатков (L-Ala43, H-Ala40, H-Ser119, H-Ala121 и H-Ser122) первоначально выбраны по информации о кристаллической структуре как удаленные от антигенсвязывающей поверхности (Eigenbrot et al. (1993) J Mol Biol. 229:969-995). Рентгеновская кристаллическая структура из базы данных белков обозначена 1FVC. Остатки Cys сконструированы в этих положениях посредством сайт-специфического мутагенеза. Препараты ТиоFab-фага выделяли и подвергали реакции с реагентом для биотинилирования.
Конъюгированные и неконъюгированные с биотином варианты тестировали по связыванию с HER2 и стрептавидином с использованием анализа PHESELECTOR на основе ELISA (фигура 8, пример 2) с HRP (пероксидаза хрена)-конъюгированным антителом против фага. Взаимодействие небиотинилированного фаг-hu4D5Fabv8 (фигура 2A) и биотинилированного фаг-hu4D5Fabv8 (Фигура 2B) с BSA (незакрашенный прямоугольник), HER2 (серый прямоугольник) или стрептавидином (закрашенный прямоугольник) мониторировали с помощью антитела анти-M13-пероксидаза хрена (HRP) посредством проявления стандартной реакции HRP и измерения оптической плотности при 450 нм. Оптическую плотность, полученную при превращении колориметрического субстрата, измеряли при 450 нм. Реакционная способность ТиоFab по отношению HER2 является показателем связывания антигена. Реакционная способность ТиоFab по отношению к стрептавидину является показателем степени биотинилирования. Реакционная способность ТиоFab по отношению к BSA является отрицательным контролем для неспецифического взаимодействия. Как видно на фигуре 2A, все варианты ТиоFab-фага обладали сходным связыванием с HER2 по сравнению со связыванием hu4D5Fabv8-фага дикого типа. Более того, конъюгация с биотином не влияла на связывание ТиоFab с HER2 (Фигура 2B).
Удивительно и неожиданно, для образцов ТиоFab-фагов показали изменяющиеся уровни активности связывания стрептавидина. Из всех протестированных фаг-ТиоFab, для сконструированного антитела с цистеиновыми заменами A121C показали максимальную реакционную способность тиола. Даже несмотря на то, что hu4D5Fabv8-фаг дикого типа инкубировали с такими же количествами биотин-малеимида, этот фаг обладал небольшим связыванием стрептавидина, что указывает на то, что предсуществующие остатки цистеина (вовлеченные в формирование дисульфидных связей) из hu4D5Fabv8 и белков оболочки фага M13 не мешают сайт-специфической конъюгации биотин-малеимида. Эти результаты показывают, что анализ фагового ELISA можно успешно использовать для скрининга реакционноспособных тиоловых групп на поверхности Fab.
Анализ PHESELECTOR позволяет скрининг реакционноспособных тиоловых групп в антителах. Идентификация варианта A121C этим способом является иллюстративной. Можно проводить эффективный поиск по целой молекуле Fab для идентификации большего числа вариантов ТиоFab с реакционноспособными тиоловыми группами. Параметр относительную доступность поверхности использовали для идентификации и количественной оценки доступности для растворителя аминокислотных остатков в полипептиде. Доступность поверхности можно выражать как площадь поверхности (Е2), которая может вступать в контакт с молекулой растворителя, например, воды. Пространство, занимаемое водой, составляет сферу радиусом приблизительно 1,4 Е. Программное обеспечение является свободно доступным или лицензируемым (Secretary to CCP4, Daresbury Laboratory, Warrington, WA44 AD, United Kingdom, Fax: (+44) 1925 603825, или доступным в интернет: www.ccp4.ac.uk/dist/html/INDEX.html) как пакет программ для кристаллографии CCP4, использующий алгоритмы для расчета доступности поверхности каждой аминокислоты белка с известными координатами, полученными рентгеновской кристаллографией («The CCP4 Suite: Programs for Protein Crystallography» (1994) Acta. Cryst. D50:760-763). Два иллюстративных модуля программного обеспечения, осуществляющие расчеты доступности поверхности «AREAIMOL» и «SURFACE», основаны на алгоритмах B.Lee and F. M. Richards (1971) J. Mol. Biol. 55:379-400.
AREAIMOL определяет поверхность белка, доступную для растворителя, как локус в центре сферы зонда (представляющей молекулу растворителя), по мере того, как она катится по Ван-дер-Ваальсовой поверхности белка. AREAIMOL рассчитывает площадь поверхности, доступной для растворителя, посредством получения точек поверхности на расширенной сфере вокруг каждого атома (на расстоянии от центра атома, равном сумме радиусов атома и зонда), и исключения точек, лежащих внутри одних и тех же сфер, ассоциированных с соседними атомами. AREAIMOL находит площадь атомов, доступных для растворителя, в файле координат PDB и суммирует доступную площадь по остатку, по цепи и по целой молекуле. Доступные площади (или разности площадей) для отдельных атомов можно записать в файл выходных данных псевдо-PDB. AREAIMOL присваивает один радиус каждому элементу и распознает только ограниченное количество различных элементов. Неизвестным типам атомов (т.е., не присутствующим во внутренней базе данных AREAIMOL) по умолчанию присваивается радиус 1,8 Å. Список распознаваемых атомов представляет собой:

AREAIMOL и SURFACE выдают абсолютные значения доступности, т.е. количество квадратных ангстрем (Å). Относительную доступность поверхности рассчитывают на основе стандартного состояния, соответствующего аминокислоте внутри полипептида. Стандартное состояние представляет собой трипептид Gly-X-Gly, где X представляет собой интересующую аминокислоту, и стандартное состояние должно представлять собой «растянутую» конформацию, т.е. подобную конформации в бета-цепях. Растянутая конформация максимизирует доступность X. Рассчитанную доступную площадь делят на доступную площадь в стандартном состоянии трипептида Gly-X-Gly и регистрируют частное, представляющее собой относительную доступность. Процент доступности представляет собой относительную доступность, умноженную на 100.
Другой иллюстративный алгоритм для расчета доступности поверхности основан на модуле SOLV программы xsae (Broger, C, F. Hoffman-LaRoche, Basel), рассчитывающем относительную доступность аминокислотного остатка для водяной сферы на основании рентгеновских координат полипептида.
Относительную доступность поверхности для каждой аминокислоты в hu4D5Fabv7 рассчитывали с использованием информации о кристаллической структуре (Eigenbrot et al. (1993) J Mol Biol. 229:969-995). Значения относительной доступности поверхности для аминокислот легкой цепи и тяжелой цепи hu4D5Fabv7 показаны в нисходящем порядке в таблице 1.




Следующие два критерия использовали для идентификации остатков hu4D5Fabv8, которые можно конструировать с заменой на остатки Cys:
1. Исключены аминокислотные остатки, которые полностью скрыты, т.е. менее чем с 10% относительной доступностью поверхности. В таблице 1 показано, что существует 134 (легкая цепь) и 151 (тяжелая цепь) остатков в hu4D5Fabv8, доступных более чем на 10% (относительная доступность поверхности). Выбрано десять наиболее доступных остатков Ser, Ala и Val из-за их близкого структурного свойства с Cys по сравнению с другими аминокислотами, так что вновь сконструированный Cys вносит только минимальные структурные ограничения в антитело. Можно также проводить скрининг других участков замены на цистеин, и они могут быть пригодны для конъюгации.
2. Остатки сортируют на основании их роли в функциональных и структурных взаимодействиях Fab. Затем отбирали остатки, не вовлеченные во взаимодействия с антигеном и удаленные от существующих дисульфидных связей. Вновь сконструированные остатки Cys должны быть удалены от участка связывания антигена, и не должны ни мешать связыванию антигена, ни ошибочно спариваться с остатками цистеина, вовлеченными в формирование дисульфидных связей.
Следующие остатки hu4D5Fabv8 удовлетворяли вышеуказанным критериям и были выбраны для замены на Cys: L-V15, L-A43, L-V110, L-A144, L-S168, H-A88, H-A121, H-S122, H-A175 и H-S179 (показаны на фигуре 1).
Реакционную способность тиола можно распространять на любое антитело, в котором можно выполнять замену аминокислот на реакционноспособные цистеиновые аминокислоты в пределах легкой цепи, выбранных из: L-10 - L-20; L-38 - L-48; L-105 - L-115; L-139 - L-149; L-163 - L-173; и в пределах тяжелой цепи, выбранных из: H-35 - H-45; H-83 - H-93; H-114 - H-127; и H-170 - H-184, и в пределах Fc-области, выбранных из H-268 - H-291; H-319 - H-344; H-370 - H-380; и H-395 - H-405.
Реакционную способность тиола можно распространять также на конкретные домены антитела, такие как константный домен легкой цепи (CL) и константные домены тяжелой цепи, CH1, CH2 и CH3. Замены на цистеин, приводящие к значению реакционной способности тиолов 0,6 и выше, можно выполнять в константных доменах тяжелых цепей α, δ, ε, γ и μ интактных антител: IgA, IgD, IgE, IgG и IgM, соответственно, включая подклассы IgG: IgG1, IgG2, IgG3, IgG4, IgA и IgA2.
Из данных кристаллической структуры очевидно, что выбранные 10 мутантов Cys находятся далеко от антигенсвязывающего участка, такого как поверхность контакта с HER2 в этом случае. Эти мутанты можно тестировать экспериментально по непрямым эффектам на функциональные взаимодействия. Реакционную способность тиола всех Cys вариантов Fab измеряли и рассчитывали, как описано в примерах 1 и 2, и представлено в таблице 2. Остатки L-V15C, L-V110C, H-A88C и H-A121C имели реакционноспособные и стабильные тиоловые группы (Фигуры 3A и 3B). Мутанты V15C, V110C, A144C, S168C представляют собой Cys варианты легкой цепи. Мутанты A88C, A121C, A175C, S179C представляют собой Cys варианты тяжелой цепи. Удивительно и неожиданно, что участки с высокой относительной доступностью поверхности не обладают наиболее высокой реакционной способностью тиола, как рассчитано посредством анализа PHESELECTOR (таблица 2). Иными словами, относительная доступность поверхности (таблицы 1, 2) не коррелирует с реакционной способностью тиола (таблица 2). Фактически, остатки Cys, сконструированные в участках с умеренной доступностью поверхности 20%-80% (фигура 4A, таблица 1), или частично экспонированные участки, подобные остаткам Ala или Val, обладают лучшей реакционной способностью тиола, т.е. >0,6, (фигура 3B, таблица 2), чем Cys, введенный вместо остатков Ser, таким образом, необходимость использования анализа PHESELECTOR для скрининга реакционноспособных участков тиола только по информации о кристаллической структуре является недостаточной для отбора этих участков (фигура 3B и 4A).
Данные по реакционной способности тиола показаны на фигурах 3A и 3B для аминокислотных остатков Cys мутантов 4D5 ТиоFab: (3A) небиотинилированный (контроль) и (3B) биотинилированные фаг-ТиоFab. Реакционноспособные тиоловые группы на поверхности антитела/Fab идентифицировали анализами PHESELECTOR по взаимодействию небиотинилированного фаг-hu4D5Fabv8 (3A) и биотинилированного фаг-hu4D5Fabv8 (3B) с BSA (незакрашенный прямоугольник), HER2 (серый прямоугольник) или стрептавидином (закрашенный прямоугольник). Анализ проводили, как описано в примере 2. Варианты легкой цепи расположены на левой стороне, и варианты тяжелой цепи расположены на правой стороне. Связывание небиотинилированных Cys мутантов 4D5 ТиоFab является низким, как ожидалось, однако, сильное связывание с HER2 сохраняется. Соотношение связывания со стрептавидином и с HER2 биотинилированных Cys мутантов 4D5 ТиоFab дает значение реакционной способности тиолов в таблице 2. Фоновая оптическая плотность при 450 нм или малые количества неспецифического связывания белка Cys мутантов биотинилированного 4D5 ТиоFab с BSA также видны на фигуре 3B. Значения относительной доступности поверхности избранных аминокислотных остатков, замененных на остаток Cys, показаны на фигуре 4A. Относительная доступность поверхности рассчитана по доступной структуре hu4D5Fabv7 и показана в таблице 1 (Eigenbrot et al. (1993) J Mol Biol. 229:969-995). Конформационные параметры структур hu4D5Fabv7 и hu4D5Fabv8 хорошо согласуются и позволяют определение любой корреляции между относительной доступностью поверхности hu4D5Fabv7 и реакционной способностью тиола цистеиновых мутантов hu4D5Fabv8. Измеренная реакционная способность тиола остатков Cys на фаге с ТиоFab, введенных в частично экспонированные остатки (Ala или Val), показывает лучшую реакционную способность тиола по сравнению с остатками Cys, введенными в остатки Ser (Таблица 2). В таблице 2 можно видеть, что для Cys мутантов ТиоFab корреляция между значениями реакционной способности тио- и относительной доступностью поверхности мала или отсутствует.
Аминокислоты в положениях L-15, L-43, L-110, L-144, L-168, H-40, H-88, H-119, H-121, H-122, H-175 и H-179 антитела можно, как правило, подвергать мутации (заменять) с помощью свободных цистеиновых аминокислот. Диапазоны в пределах приблизительно 5 аминокислотных остатков с каждой стороны от этих положений также можно заменять на свободные цистеиновые аминокислоты, т.е. L-10 - L-20; L-38 - L-48; L-105 - L-115; L-139 - L-149; L-163 - L-173; H-35 - H-45; H-83 - H-93; H-114 - H-127; и H-170 - H-184, так же как в диапазонах Fc-области, выбранных из H-268 - H-291; H-319 - H-344; H-370 - H-380; и H-395 - H-405, для получения сконструированных антител с цистеиновыми заменами по изобретению.

Два Cys варианта для легкой цепи (L-V15C и L-V110C) и два для тяжелой цепи (H-A88C и H-A121C) выбраны для дальнейшего анализа, поскольку для этих вариантов показана наивысшая реакционная способность тиола (таблица 2).
В отличие от очистки фага, получение Fab может требовать 2-3 суток, в зависимости от масштаба продукции. На протяжении этого времени, тиоловые группы могут терять реакционную способность из-за окисления. Для тестирования стабильности тиоловых групп на hu4D5Fabv8-фаге, измеряли реакционную способность тиола фаг-тиоFab (фигура 4B). После выделения ТиоFab-фага, на сутки 1, сутки 2 и сутки 4, все образцы конъюгировали с биотин-PEO-малеимидом и тестировали в анализе фагового ELISA (PHESELECTOR) для тестирования связывания HER2 и стрептавидина. L-V15C, L-V110C, H-A88C и H-A121C сохраняли значительные уровни реакционной способности тиола по сравнению с другими вариантами ТиоFab (фигура 4B).
СПОСОБЫ ПОЛУЧЕНИЯ СКОНСТРУИРОВАННЫХ АНТИТЕЛ С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
Соединения по изобретению включают сконструированные антитела с цистеиновыми заменами, в которых одна или несколько аминокислот исходного антитела заменены на свободную аминокислоту цистеин. Сконструированное антитело с цистеиновыми заменами содержит одну или несколько свободных цистеиновых аминокислот, обладающих значением реакционной способности тиола в диапазоне 0,6-1,0. Свободная аминокислота цистеин представляет собой остаток цистеина, введенный при конструировании в исходное антитело и не являющийся частью дисульфидного мостика.
В одном аспекте сконструированное антитело с цистеиновыми заменами получено способом, включающим:
(a) замену одного или нескольких аминокислотных остатков исходного антитела на цистеин; и
(b) определение реакционной способности тиола сконструированного антитела с цистеиновыми заменами посредством реакции сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу реагентом.
Сконструированное антитело с цистеиновыми заменами может являться более реакционноспособным, чем исходное антитело, по отношению к реакционноспособному по отношению к тиолу реагенту.
Остатки свободной аминокислоты цистеина могут быть локализованы в тяжелой или легкой цепях, или в константном или вариабельном доменах. Фрагменты антитела, например, Fab, также можно конструировать с заменой аминокислот фрагментов антитела на одну или нескольких цистеиновых аминокислот для получения сконструированных фрагментов антитела с цистеиновыми заменами.
Другой аспект изобретения относится к способу получения (изготовления) сконструированного антитела с цистеиновыми заменами, включающему:
(a) введение одной или нескольких цистеиновых аминокислот в исходное антитело для получения сконструированного антитела с цистеиновыми заменами; и
(b) определение реакционной способности тиола сконструированного антитела с цистеиновыми заменами с помощью реакционноспособного по отношению к тиолу реагента;
где сконструированное антитело с цистеиновыми заменами является более реакционноспособным, чем исходное антитело, по отношению к реакционноспособному по отношению к тиолу реагенту.
Стадия (a) способа получения сконструированного антитела с цистеиновыми заменами может включать:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновыми заменами;
(ii) экспрессию сконструированного антитела с цистеиновыми заменами; и
(iii) выделение и очистку сконструированного антитела с цистеиновыми заменами.
Стадия (b) способа получения сконструированного антитела с цистеиновыми заменами может включать экспрессию сконструированного антитела с цистеиновыми заменами на вирусной частице, выбранной из частицы фага или фагмиды.
Стадия (b) способа получения сконструированного антитела с цистеиновыми заменами может включать также:
(i) реакцию сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу аффинным реагентом для получения аффинно меченного сконструированного антитела с цистеиновыми заменами; и
(ii) измерение связывания аффинно меченного сконструированного антитела с цистеиновыми заменами со связывающей средой.
Другой аспект по изобретению относится к способу скрининга сконструированных антител с цистеиновыми заменами с высоко реакционноспособными, неспаренными цистеиновыми аминокислотами по реакционной способности тиола, включающему:
(a) введение одной или нескольких цистеиновых аминокислот в исходное антитело для получения сконструированного антитела с цистеиновыми заменами;
(b) реакцию сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу аффинным реагентом для получения аффинно меченного сконструированного антитела с цистеиновыми заменами; и
(c) измерение связывания аффинно меченного сконструированного антитела с цистеиновыми заменами со связывающей средой; и
(d) определение реакционной способности тиола сконструированного антитела с цистеиновыми заменами с помощью реакционноспособного по отношению к тиолу реагента.
Стадия (a) способа скрининга сконструированных антител с цистеиновыми заменами может включать:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновыми заменами;
(ii) экспрессию сконструированного антитела с цистеиновыми заменами; и
(iii) выделение и очистку сконструированного антитела с цистеиновыми заменами.
Стадия (b) способа скрининга сконструированного антитела с цистеиновыми заменами может включать экспрессию сконструированного антитела с цистеиновыми заменами на вирусной частице, выбранной из частицы фага или фагмиды.
Стадия (b) способа скрининга сконструированного антитела с цистеиновыми заменами может включать также:
(i) реакцию сконструированного антитела с цистеиновыми заменами с реакционноспособным по отношению к тиолу аффинным реагентом для получения аффинно меченного сконструированного антитела с цистеиновыми заменами; и
(ii) измерение связывания аффинно меченного сконструированного антитела с цистеиновыми заменами со связывающей средой.
МЕЧЕНЫЕ СКОНСТРУИРОВАННЫЕ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
Сконструированные антитела с цистеиновыми заменами по изобретению можно конъюгировать с любой группой метки, которую можно ковалентно присоединять к антителу посредством тиоловой группы цистеина (Singh et al (2002) Anal. Biochem. 304: 147-15; Harlow E. и Lane, D. (1999) Using Antibodies: A Laboratory Manual, Cold Springs Harbor Laboratory Press, Cold Spring Harbor, NY; Lundblad R.L. (1991) Chemical Reagents for Protein Modification, 2nd ed. CRC Press, Boca Raton, FL). Присоединенная метка может функционировать для: (i) обеспечения поддающегося детекции сигнала; (ii) взаимодействия со второй меткой для модификации поддающегося детекции сигнала, обеспечиваемого первой или второй меткой, например, для получения FRET (резонансного переноса энергии флуоресценции); (iii) стабилизации взаимодействий или увеличения аффинности связывания с антигеном или лигандом; (iv) влияния на подвижность, например, электрофоретическую подвижность, или клеточную проницаемость, посредством заряда, гидрофобности, формы или других физических параметров, или (v) предоставления связывающей группы, для модуляции аффинности лиганда, связывания антитело/антиген, или образования комплексных ионов.
Меченые сконструированные антитела с цистеиновыми заменами могут являться полезными для диагностических анализов, например, для детекции экспрессии интересующего антигена в специфических клетках, тканях или сыворотке. Для диагностических применений антитело, как правило, метят поддающейся детекции группой.
Доступно множество меток, которые можно в общем сгруппировать по следующим категориям:
(a) Радиоизотопы (радиоактивные изотопы), такие как 3H, 11C, 14C, 18F, 32P, 35S, 64Cu, 68Ga, 86Y, 89Zr, 99Tc, 111In, 123I, 124I, 125I, 131I, 133Xe, 177LU, 211At или 213Bi. Меченные радиоизотопами антитела являются полезными в экспериментах получения изображений, направленных на рецепторы. Антитело можно метить с помощью реагентов с лигандами, которые связывают, образуют хелат или другой комплекс с радиоизотопом металла, где реагент является реакционноспособным по отношению к сконструированному тиолу антитела, с использованием способов, описанных в Current Protocols in Immunology, Volumes 1 и 2, Coligen et al, Ed. Wiley-Interscience, New York, NY, Pubs. (1991). Хелатирующие лиганды, которые могут образовывать комплексы с ионом металла, включают DOTA, DOPA, DOTP, DOTMA, DTPA и TETA (Macrocyclics, Dallas, TX). Радиоактивные изотопы можно нацеливать посредством образования комплексов с сконструированными антителами с цистеиновыми заменами в форме конъюгатов антитело-цирконий по изобретению (Wu et al (2005) Nature Biotechnology 23(9): 1137-1146).
Металл-хелатные комплексы, пригодные в качестве меток для антитела для экспериментов по получению изображений, описаны в: US 5342606; US 5428155; US 5316757; US 5480990; US 5462725; US 5428139; US 5385893; US 5739294; US 5750660; US 5834456; Hnatowich et al (1983) J. Immunol. Methods 65: 147-157; Meares et al (1984) Anal. Biochem. 142:68-78; Mirzadeh et al (1990) Bioconjugate Chem. 1:59-65; Meares et al (1990) J. Cancerl990, Suppl. 10:21-26; Izard et al (1992) Bioconjugate Chem. 3:346-350; Nikula et al (1995) Nucl. Med. Biol. 22:387-90; Camera et al (1993) Nucl. Med. Biol. 20:955-62; Kukis et al (1998) J. Nucl. Med. 39:2105-2110; Verel et al (2003) J. Nucl. Med. 44:1663-1670; Camera et al (1994) J. Nucl. Med. 21:640-646; Ruegg et al (1990) Cancer Res. 50:4221-4226; Verel et al (2003) J. Nucl. Med. 44:1663-1670; Lee et al (2001) Cancer Res. 61:4474-4482; Mitchell, et al (2003) J. Nucl. Med. 44:1105-1112; Kobayashi et al (1999) Bioconjugate Chem. 10:103-111; Miederer et al (2004) J. Nucl. Med. 45:129-137; DeNardo et al (1998) Clinical Cancer Research 4:2483-90; Blend et al (2003) Cancer Biotherapy & Radiopharmaceuticals 18:355-363; Nikula et al (1999) J. Nucl. Med. 40:166-76; Kobayashi et al (1998) J. Nucl. Med. 39:829-36; Mardirossian et al (1993) Nucl. Med. Biol. 20:65-74; Roselli et al (1999) Cancer Biotherapy & Radiopharmaceuticals, 14:209-20.
(b) Флуоресцентные метки, такие как хелаты редкоземельных элементов (хелаты европия), типы флуоресцеина, включая FITC, 5-карбоксифлуоресцеин, 6-карбоксифлуоресцеин; типы родамина, включая TAMRA; данзил; лиссамин; цианины; фикоэритрины; Техасский Красный; и их аналоги. Флуоресцентные метки можно конъюгировать с антителами с использованием способов, описанных, например, в Current Protocols in Immunology, выше. Реагенты с флуоресцентными красителями и флуоресцентной меткой включают реагенты, коммерчески доступные из Invitrogen/Molecular Probes (Eugene, OR) и Pierce Biotechnology, Inc. (Rockford, IL).
(c) Различные фермент-субстратные метки доступны или описаны (US 4275149). Фермент, как правило, катализирует химическое изменение хромогенного субстрата, которое можно измерять с использованием различных способов. Например, фермент может катализировать изменение окраски субстрата, которое можно измерять спектрофотометрически. Альтернативно, фермент может изменять флуоресценцию или хемилюминесценцию субстрата. Способы количественного определения изменения флуоресценции описаны выше. Хемилюминесцентный субстрат становится электронно-возбужденным посредством химической реакции и может затем испускать свет, который можно измерять (например, с использованием хемилюминометра) или передавать энергию акцептору флуоресценции. Примеры ферментых меток включают люциферазы (например, люциферазу светлячков и бактериальную люциферазу; US 4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназу, уриназу, пероксидазу, такую как пероксидаза хрена (HRP), щелочную фосфатазу (AP), β-галактозидазу, глюкоамилазу, лизоцим, оксидазы сахаридов (например, оксидазу глюкозы, оксидазу галактозы и глюкоза-6-фосфатдегидрогеназу), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидазу, микропероксидазу и т.п. Способы конъюгации ферментов с антителами описаны в O'Sullivan et al (1981) «Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay», in Methods in Enzym. (ed J. Langone & H. Van Vunakis), Academic Press, New York, 73:147-166.
Примеры комбинаций фермент-субстрат включают, например:
(i) пероксидазу хрена (HRP) с перекисью водорода в качестве субстрата, где перекись водорода окисляет предшественник красителя (например, ортофенилендиамин (OPD) или 3,3',5,5'-тетраметилбензидингидрохлорид (TMB));
(ii) щелочную фосфатазу (AP) с пара-нитрофенилфосфатом в качестве хромогенного субстрата; и
(iii) β-D-галактозидазу (β-D-Gal) с хромогенным субстратом (например, п-нитрофенил-β-D-галактозидазой) или флуорогенным субстратом 4-метилумбеллиферил-β-D-галактозидазой.
Множество других комбинаций фермент-субстрат доступно специалистам в данной области. Общий обзор см. в US 4275149 и US 4318980.
Метку можно опосредованно конъюгировать с сконструированным антителом с цистеиновыми заменами. Например, антитело можно конъюгировать с биотином и любую метку из трех широких категорий, указанных выше, можно конъюгировать с авидином или стрептавидином, или наоборот. Биотин избирательно связывается со стрептавидином, и, таким образом, метку можно конъюгировать с антителом данным опосредованным образом. Альтернативно для достижения опосредованной конъюгации метки с вариантом полипептида, вариант полипептида конъюгируют с небольшим гаптеном (например, дигоксином) и одну из различных типов меток, указанных выше, конъюгируют с полипептидным вариантом против гаптена (например, антителом против дигоксина). Таким образом можно достигать опосредованной конъюгации метки с вариантом полипептида (Hermanson, G. (1996) in Bioconjugate Techniques Academic Press, San Diego).
Вариант полипептида по настоящему изобретению можно использовать в любом известном способе анализа, таком как ELISA, анализы конкурентного связывания, прямом и непрямом сэндвич-анализах, и анализах иммунопреципитации (Zola, (1987) Monoclonal Antibodies: A Manual of Techniques, pp.147-158, CRC Press, Inc.).
Метка для детекции может являться пригодной для локализации, визуализации и количественной оценки события связывания или узнавания. Меченые антитела по изобретению могут детектировать рецепторы поверхности клеток. Другим применением для меченных поддающейся детекции меткой антител является способ иммунного связывания с помощью бусин, включающий конъюгацию бусин с флуоресцентно меченным антителом и детекцию сигнала флуоресценции при связывании лиганда. В сходных способах детекции связывания используют эффект поверхностного плазмонного резонанса (SPR) для измерения и детекции взаимодействий антитело-антиген.
Такие метки для детекции, как флуоресцентные красители и хемилюминесцентные красители (Briggs et al (1997) «Synthesis of Functionalised Fluorescent Dyes and Their Coupling to Amines and Amino Acids», J. Chem. Soc, Perkin-Trans. 1: 1051-1058), обеспечивают поддающийся детекции сигнал и являются общепринятыми для мечения антитела, предпочтительно, со следующими свойствами: (i) меченое антитело должно производить очень сильный сигнал с низким фоном, так что небольшие количества антител можно с большой чувствительностью детектировать как в бесклеточных, так и в основанных на клетках анализах; и (ii) меченое антитело должно являться фотостабильным, так чтобы флуоресцентный сигнал можно было наблюдать, мониторировать и регистрировать без значительного фотообесцвечивания. Для применений, включающих связывание на поверхности клетки меченого антитела с мембранами или поверхностями клеток, особенно живых клеток, метки предпочтительно (iii) обладают хорошей растворимостью в воде для достижения эффективных концентраций конъюгата и чувствительности детекции и (iv) являются нетоксичными для живых клеток, так чтобы не нарушать нормальных процессов метаболизма клеток или не вызывать преждевременной гибели клеток.
Прямую количественную оценку интенсивности клеточной флуоресценции и подсчет событий флуоресцентного мечения, например, связывания на поверхности клеток конъюгатов пептид-краситель, можно проводить в системе (FMAT® 8100 HTS System, Applied Biosystems, Foster City, Calif.), автоматизирующей нерадиоактивные анализы смешивания и считывания с живыми клетками или бусинами (Miraglia, «Homogeneous cell- and bead-based assays for high throughput screening using fluorometric microvolume assay technology», (1999) J. of Biomolecular Screening 4: 193-204). Применения меченых антител включают также анализы связывания с рецептором на поверхности клеток, анализы иммунного связывания, твердофазные иммунофлуоресцентные анализы (FLISA), анализы расщепления каспазой (Zheng, «Caspase-3 controls both cytoplasmic and nuclear events associated with Fas-mediated apoptosis in vivo», (1998) Proc. Natl. Acad. Sci. USA 95:618-23; US 6372907), анализы апоптоза (Vermes, «A novel assay for apoptosis. Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V» (1995) J. Immunol. Methods 184:39-51) и анализы цитотоксичности. Способ флуориметрического анализа в микрообъемах можно использовать для идентификации повышающей или понижающей регуляции посредством молекулы, нацеленной на поверхность клеток (Swartzman, «A homogeneous and multiplexed immunoassay for high-throughput screening using fluorometric microvolume assay technology», (1999) Anal. Biochem. 271:143-51).
Меченые сконструированные антитела с цистеиновыми заменами по изобретению являются пригодными в качестве биомаркеров и зондов для получения изображений различными методами и способами получения биомедицинских и молекулярных изображений, такими как: (i) МРТ (магнитно-резонансная томография); (ii) МикроКТ (компьютерная томография); (iii) SPECT (однофотонная эмиссионная компьютерная томография); (iv) ПЭТ (позитронная эмиссионная томография) Chen et al (2004) Bioconjugate Chem. 15:41-49; (v) биолюминесценция; (vi) флуоресценция; и (vii) ультразвук. Иммуносцинтиография представляет собой способ получения изображений при котором антитела, меченные радиоактивными веществами, вводят пациенту - животному или человеку, и получают изображение в участках организма, где локализовано антитело (US 6528624). Биомаркеры для получения изображений можно объективно измерять и оценивать в качестве показателя нормальных биологических процессов, патогенных процессов или фармакологических ответов на терапевтическое вмешательство. Биомаркеры могут принадлежать к нескольким типам: маркеры типа 0 представляют собой маркеры естественного течения заболевания и по длительности проявления коррелируют с известными клиническими показателями, например, оценкой ЯМР синовиального воспаления при ревматоидном артрите; маркеры типа I фиксируют эффект вмешательства вне зависимости от механизма действия, даже несмотря на то, что механизм может не быть ассоциирован с клиническим исходом; маркеры типа II функционируют как суррогатные конечные точки, где по изменению биомаркера или по сигналу от биомаркера прогнозируют клиническое преимущество для «подтверждения» намеченного ответа, например, измерение эрозии костной ткани при ревматоидном артрите посредством КТ. Биомаркеры для получения изображений таким образом могут предоставлять фармакодинамическую (PD) терапевтическую информацию относительно: (i) экспрессии белка-мишени, (ii) связывания лекарственного средства с белком-мишенью, т.е., избирательности, и (iii) фармакокинетических данных о выведении и времени полужизни. Преимущества биомаркеров для получения изображений in vivo по сравнению с биомаркерами для лабораторных исследований включают: неинвазивную обработку, количественную оценку целого организма, повторяющиеся введение и оценку, т.е. множественные временные точки, и эффекты, потенциально переносимые от доклинических результатов (для мелких животных) к клиническим результатам (для человека). Для некоторых применений получение изображений биологических объектов заменяет эксперименты на животных или минимизирует количество экспериментов на животных в доклинических исследованиях.
Метки для получения изображений с радиоактивными изотопами включают такие радиоактивные изотопы, как 3H, 11C, 14C, 18F, 32P, 35S, 64Cu, 68Ga, 86Y, 89Zr, 99Tc, 111In, 123I, 124I, 125I, 131I, 133Xe, 177Lu, 211At или 213Bi. Можно получать комплекс иона радиоактивного изотопа металла с хелатирующим линкером, таким как DOTA. Линкерные реагенты, такие как DOTA-малеимид (4-малеимидобутирамидобензил-DOTA), можно получать реакцией аминобензил-DOTA с 4-малеимидомасляной кислотой (Fluka), активированной изопропилхлорформатом (Aldrich), с последующим использованием способа Axworthy et al (2000) Proc. Natl. Acad. Sci. USA 97(4): 1802-1807). DOTA-малеимидные реагенты вступают в реакцию со свободными цистеиновыми аминокислотами сконструированных антител с цистеиновыми заменами и обеспечивают наличие на антителе образующего комплексы с металлами лиганда (Lewis et al (1998) Bioconj. Chem. 9:72-86). Реагенты для мечения с хелатирующим линкером, такие как DOTA-NHS (сложный моно(N-гидроксисукцинимидный) эфир 1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусной кислоты), являются коммерчески доступными (Macrocyclics, Dallas, TX). Нацеленное на рецепторы получение изображений с помощью меченных радиоактивным изотопом антител может обеспечивать маркер активации пути посредством детекции и количественной оценки возрастающего накопления антител в ткани опухоли (Albert et al (1998) Bioorg. Med. Chem. Lett. 8: 1207-1210). Конъюгированные радиоактивные металлы могут оставаться внутри клеток после лизосомальной деградации.
Способы мечения пептидов хорошо известны. См. Haugland, 2003, Molecular Probes Handbook of Fluorescent Probes and Research Chemicals, Molecular Probes, Inc.; Brinkley, 1992, Bioconjugate Chem. 3:2; Garman, (1997) Non-Radioactive Labelling: A Practical Approach, Academic Press, London; Means (1990) Bioconjugate Chem. 1:2; Glazer et al (1975) Chemical Modification of Proteins. Laboratory Techniques in Biochemistry and Molecular Biology (T. S. Work и E. Work, Eds.) American Elsevier Publishing Co., New York; Lundblad, R. L. and Noyes, C. M. (1984) Chemical Reagents for Protein Modification, Vols. I and II, CRC Press, New York; Pfleiderer, G. (1985) «Chemical Modification of Proteins», Modern Methods in Protein Chemistry, H. Tschesche, Ed., Walter DeGryter, Berlin and New York; и Wong (1991) Chemistry of Protein Conjugation and Cross-linking, CRC Press, Boca Raton, Fla.); De Leon-Rodriguez et al (2004) Chem.Eur. J. 10: 1149-1155; Lewis et al (2001) Bioconjugate Chem. 12:320-324; Li et al (2002) Bioconjugate Chem. 13: 110-115; Mier et al (2005) Bioconjugate Chem. 16:240-237.
Пептиды и белки, меченные двумя группам, флуоресцентным репортером и тушителем в достаточной близости, подвергаются резонансному переносу энергии флуоресценции (FRET). Репортерные группы, как правило, представляют собой флуоресцентные красители, возбуждаемые светом определенной длины волны и переносящие энергию на группу акцептора, или тушителя, с соответствующим сдвигом Стокса для излучения с максимальной яркостью. Флуоресцентные красители включают молекулы с большими ароматическими группами, такие как флуоресцеин и родамин, и их производные. Флуоресцентный репортер может подвергаться частичному или полному тушению группой тушителя в интактном пептиде. При расщеплении пептида пептидазой или протеазой можно измерять поддающееся детекции увеличение флуоресценции (Knight, C. (1995) «Fluorimetric Assays of Proteolytic Enzymes», Methods in Enzymology, Academic Press, 248: 18-34).
Меченые антитела по изобретению можно использовать также в качестве средства для аффинной очистки. По этому способу, меченое антитело иммобилизуют на твердой фазе, такой как смола Sephadex или фильтровальная бумага, с использованием способов, хорошо известных в данной области. Иммобилизованное антитело приводят в контакт с образцом, содержащим антиген, подлежащий очистке, и затем подложку промывают подходящим растворителем, удаляющим в основном весь материал образца, за исключением антигена, подлежащего очистке, который является связанным с иммобилизованным вариантом полипептида. Наконец, подложку промывают другим подходящим растворителем, таким как глициновый буфер, pH 5,0, высвобождающим антиген с варианта полипептида.
Реагенты для мечения, как правило, несут реакционноспособную функциональную группу, которая может вступать в реакцию (i) неспосредственно с тиолом цистеина сконструированного антитела с цистеиновыми заменами с образованием меченого антитела, (ii) с линкерным реагентом с образованием промежуточного соединения линкер-метка, или (iii) с имеющим линкер антителом с образованием меченого антитела. Реакционноспособные функциональные группы реагентов для мечения включают: малеимид, галоацетил, сложный эфир иодацетамида и сукцинимидила (например, NHS, N-гидроксисукцинимид), изотиоцианат, сульфонилхлорид, 2,6-дихлортриазинил, сложный эфир пентарфторфенила и фосфорамидит, хотя можно использовать также другие функциональные группы.
Иллюстративной реакционноспособной функциональной группой является N-гидроксисукцинимидиловый сложный эфир (NHS) карбоксильной группы заместителя поддающейся детекции метки, например, биотина или флуоресцентного красителя. NHS сложный эфир метки может являться заранее полученным, выделенным, очищенным и/или охарактеризованным, или он может образовываться in situ и вступать в реакцию с нуклеофильной группой антитела. Как правило, карбоксильную форму метки активируют посредством реакции с некоторой комбинацией карбодиимидного реагента, например, дициклогексилкарбодиимида, диизопропилкарбодиимида или урониевого реагента, например, TSTU (тетрафторборат (O-(N-сукцинимидил)-N,N,N',N'-тетраметилурония), HBTU (гексафторфосфат O-бензотриазол-1-ил)-N,N,N',N'-тетраметилурония) или HATU (гексафторфосфат O-(7-азабензотриазол-1-ил)-N,N,N',N'-тетраметилурония), активатора, такого как 1-гидроксибензотриазол (HOBt), и N-гидроксисукцинимида для получения NHS сложного эфира метки. В некоторых случаях, метку и антитело можно связывать активацией метки in situ и реакцией с антителом с образованием конъюгата метка-антитело в одну стадию. Другие реагенты для активации и связывания включают TBTU (гексафторфосфат 2-(1H-бензотриазо-1-ил)-1-1,3,3-тетраметилурония), TFFH (2-фтор-гексафторфосфат N,N',N'',N'''-тетраметилурония), PyBOP (гексафторфосфат бензотриазол-1-ил-окси-трис-пирролидинофосфония), EEDQ (2-этокси-1-этоксикарбонил-1,2-дигидрохинолин), DCC (дициклогексилкарбодиимид); DIPCDI (диизопропилкарбодиимид), MSNT (1-(мезитилен-2-сульфонил)-3-нитро-1H-1,2,4-триазол и арилсульфонилгалогениды, например, триизопропилбензолсульфонилхлорид.
КОНЪЮГАЦИЯ БИОТИН-МАЛЕИМИДА С ТИОFAB
Вышеописанные свойства ТиоFab определены в присутствии фага, поскольку слияние Fab с белком оболочки фага потенциально может изменять доступность или реакционную способность тиола Cys. Таким образом, конструкции ТиоFab клонировали в экспрессирующий вектор под промотором щелочной фосфатазы (Chang et al (1987) Gene 55: 189-196), и экспрессию ТиоFab индуцировали посредством выращивания клеток E. coli в бесфосфатной среде. ТиоFab очищали на колонке с SEPHAROSE™ с белком G и анализировали в восстанавливающих и невосстанавливающих гелях для SDS-PAGE. Эти анализы позволяют оценку того, сохранили ли ТиоFab свою реакционноспособную тиоловую группу или сделаны неактивными посредством формирования внутримолекулярных или межмолекулярных дисульфидных связей. ТиоFab L-V15C, L-V110C, H-A88C и H-A121C экспрессировали и очищали хроматографией на колонке с SEPHAROSE™ с белком-G (подробно см. в разделе методы). Очищенные белки анализировали в геле для SDS-PAGE в восстанавливающих (с DTT) и невосстанавливающих (без DTT) условиях. Другие восстанавливающие средства, такие как BME (бета-меркаптоэтанол), можно использовать в геле для расщепления межцепочечных дисульфидных групп. Из анализа SDS-PAGE в геле очевидно, что главная (~90%) фракция ТиоFab присутствует в мономерной форме, в то время как hu4D5Fabv8 дикого типа присутствует в основном в мономерной форме (47 кДа).
ТиоFab (A121C) и hu4D5Fabv8 дикого типа инкубировали со 100-кратным избытком биотин-малеимида в течение 3 часов при комнатной температуре, и биотинилированные Fab наносили на колонку для гель-фильтрации Superdex-200™. Эта стадия очистки являлась пригодной для отделения мономерных Fab от олигомерных Fab, а также от избыточного свободного биотин-малеимида (или свободного циркониевого реагента).
На фигуре 5 показано подтверждение свойств вариантов ТиоFab в отсутствие фагового окружения. Белки без слияния с фагом, hu4D5Fabv8 и hu4D5Fabv8-A121C (ТиоFab-A121C), экспрессировали и очищали с использованием бусин с агарозой с белком-G с последующей инкубацией со 100-кратным молярным избытком биотин-малеимида. Сравнивали связывание стрептавидина и HER2 с биотинилированным сконструированным ТиоFab с cys заменами и небиотинилированным Fab дикого типа. Степень конъюгации с биотином (взаимодействие со стрептавидином) и способность связывания с HER2 мониторировали посредством анализов ELISA. Каждый Fab тестировали при 2 нг и 20 нг.
Биотинилированный A121C ТиоFab сохранял связывание с HER2, сравнимое со связыванием hu4D5Fabv8 дикого типа (фигура 5). Fab дикого типа и A121C-ТиоFab очищали гель-хроматографией на колонке. Два образца тестировали по связыванию HER2 и стрептавидина посредством ELISA с использованием анти-Fab-HRP козы в качестве вторичного антитела. Как Fab дикого типа (незакрашенный прямоугольник), так и ТиоFab (прямоугольник с точками) обладают сходным связыванием с HER2, но только ТиоFab сохранял связывание со стрептавидином. Только фоновый уровень взаимодействия со стрептавидином наблюдали для небиотинилированного hu4D5Fabv8 дикого типа (фигура 5). Масс-спектральный (LC-ESI-MS) анализ биотинилированного-ТиоFab (A121C) приводил к главному пику 48294,5 дальтон в отличие от hu4D5Fabv8 дикого типа (47737 дальтон). Разница в 537,5 дальтон между двумя молекулами точно соответствует одному биотин-малеимиду, конъюгированному с ТиоFab. Результаты масс-спектрального секвенирования белка (LC-ESI-тандемного масс-спектрального анализа) дополнительно подтвердили, что конъюгированная молекула биотина присутствует на вновь сконструированном остатке Cys (таблица 4, пример 3).
САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ БИОТИН-МАЛЕИМИДА С АЛЬБУМИНСВЯЗЫВАЮЩИЙ ПЕПТИД (ABP)-ТИОFAB
Связывание с белком плазмы может являться эффективным способом улучшения фармакокинетических свойств короткоживущих молекул. Альбумин является наиболее распространенным белком в плазме. Связывающие сывороточный альбумин пептиды (ABP) могут изменять фармакодинамику слитых с ними активных доменов белков, включая изменение поглощения, проникновения и диффузии в тканях. Эти фармакодинамические параметры можно регулировать специфическим выбором подходящей последовательности связывающего сывороточный альбумин пептида (US 20040001827). Серию альбуминсвязывающих пептидов идентифицировали скринингом фагового дисплея (Dennis et al. (2002) «Albumin Binding As A General Strategy For Improving The Pharmacokinetics Of Proteins» J Biol Chem. 277:35035-35043; WO 01/45746). Соединения по изобретению включают ABP последовательности, объясняемые в: (i) Dennis et al (2002) J Biol Chem. 277:35035-35043 в таблицах III и IV, страница 35038; (ii) US 20040001827 в SEQ ID NO: 9-22; и (iii) WO 01/45746 на страницах 12-13, SEQ ID NO: z1-z14, и содержание всех из них приведено в настоящем документе в качестве ссылки.
Альбуминсвязывающие белки (ABP)-Fab конструировали слиянием альбуминсвязывающего пептида с C-концом тяжелой цепи Fab в стехиометрическом соотношении 1:1 (1 ABP/1 Fab). Показано, что связывание этих ABP-Fab с альбумином увеличивало время их полужизни более чем в 25 раз у кроликов и мышей. Вышеописанные реакционноспособные остатки Cys можно, таким образом, вводить в эти ABP-Fab и использовать для сайт-специфической конъюгации с циркониевыми реагентами с последующими исследованиями на животных in vivo.
Последовательности альбуминсвязывающих пептидов включают в качестве неограничивающих примеров аминокислотные последовательности, перечисленные в SEQ ID NO: 1-5:

Последовательности альбуминсвязывающих пептидов (ABP) связывают альбумин множества видов (мыши, крысы, кролика, быка, макака-резуса, бабуина и человека) с Kd (кролик) = 0,3 мкМ. Альбуминсвязывающий пептид не конкурирует с лигандами, как известно, связывающими альбумин и обладает временем полужизни (T½) у кролика 2,3 ч. Белки ABP-ТиоFab очищали на BSA-SEPHAROSE™ с последующей конъюгацией с биотин-малеимидом и очисткой хроматографией на колонке Superdex-S200, как описано в предыдущих разделах. Очищенные биотинилированные белки являлись гомогенными и лишенными каких-либо олигомерных форм (пример 4).
На фигуре 6 показаны свойства вариантов альбуминсвязывающий пептид (ABP)-ТиоFab. Анализы ELISA проводили для тестирования способности ABP-hu4D5Fabv8-wt, ABP-hu4D5Fabv8-V110C и ABP-hu4D5Fabv8-A121C связываться с кроличьим альбумином, стрептавидином и HER2. Биотинилированные ABP-ТиоFab способны связывать альбумин и HER2 с аффинностью, сходной с аффинностью ABP-hu4D5Fabv8 дикого типа, как подтверждено ELISA (фигура 6) и анализом кинетики BIAcore (таблица 3). Планшет для ELISA покрывали альбумином, HER2 и SA, как описано. Для связывания биотинилированного ABP-ТиоFab с альбумином, HER2 и SA в качестве зонда использовали анти-Fab HRP. Биотинилированные ABP-ТиоFab были способны связываться со стрептавидином по сравнению с небиотинилированным контрольным ABP-hu4D5Fabv8-wt, что указывает на то, что ABP-ТиоFab являлись конъюгированными с биотин-малеимидом, подобно ТиоFab, сайт-специфическим образом, поскольку одни и те же мутанты Cys использовали для обоих вариантов (фигура 6).

Альтернативно, альбуминсвязывающий пептид можно связывать с антителом посредством ковалентного присоединения через линкерную группу.
КОНСТРУИРОВАНИЕ ABP-ТИОFAB С ДВУМЯ СВОБОДНЫМИ ТИОЛОВЫМИ ГРУППАМИ НА FAB
Вышеуказанные результаты показывают, что все четыре варианта (L-V15C, L-V110C, H-A88C и H-A121C) тиоFab (сконструированных Fab антител с цистеиновыми заменами) обладают реакционноспособными тиоловыми группами, которые можно использовать для сайт-специфической конъюгации с реагентом для мечения, линкерным реагентом или промежуточным соединением цирконий-линкер. L-V15C можно экспрессировать и очищать, но с относительно низкими выходами. Однако, выходы экспрессии и очистки вариантов L-V110C, H-A88C и H-A121C являлись сходными с выходами hu4D5Fabv8. Таким образом, эти мутанты можно использовать для дальнейшего анализа и рекомбинации для получения более одной тиоловой группы на Fab. Для этой цели конструировали одну тиоловую группу на легкой и одну на тяжелой цепи для получения двух тиоловых групп на молекулу Fab (L-V110C/H-A88C и L-V110C/H-A121C). Эти два варианта с двойным Cys экспрессировали в экспрессирующей системе E. coli и очищали. Обнаружено, что гомогенность очищенных биотинилированных ABP-ТиоFab является сходной с гомогенностью вариантов с отдельным Cys.
Исследовали эффекты конструирования двух реакционноспособных остатков Cys на Fab (фигура 7). Присутствие второго биотина тестировали посредством исследования связывания биотинилированного ABP-ТиоFab с SA с использованием стрептавидин-HRP (фигура 7). Для анализа HER2/Fab планшет для ELISA покрывали HER2, и в качестве зонда использовали анти-Fab HRP. Для анализа SA/Fab планшет для ELISA покрывали SA, и в качестве зонда использовали анти-Fab HRP. Для анализа SA/SA планшет для ELISA покрывали SA, и в качестве зонда использовали SA-HRP. Фигура 7. Анализы ELISA взаимодействия cys вариантов биотинилированного ABP-hu4D5Fabv8 с HER2, стрептавидином (SA). Для HER2/Fab, SA/Fab и SA/SA показали, что их взаимодействия выявлены посредством анти-Fab-HRP, SA-HRP, соответственно. SA/Fab выявляет присутствие одного биотина на Fab, и более одного биотина на Fab выявляют анализом SA/SA. Связывание HER2 с мутантами с двойным cys является сходным со связыванием с вариантами с одним Cys (фигура 7). Однако, степень биотинилирования мутантов с двойным Cys была выше по сравнению с вариантами с одним Cys благодаря более одной свободной тиоловой группе на молекулу Fab (фигура 7).
КОНСТРУИРОВАНИЕ ТИО-IgG ВАРИАНТОВ ТРАСТУЗУМАБА
Цистеин вводили в конкретные остатки полноразмерного моноклонального антитела, трастузумаба (ГЕРЦЕПТИН®, Genentech Inc.). Мутанты трастузумаба с одним cys H-A88C, H-A121C и L-V110C, и мутанты трастузумаба с двойным cys V110C-A121C и V110C-A121C экспрессировали в клетках CHO (яичников китайского хомяка) посредством нестационарной ферментации в среде, содержащей 1 мМ цистеин. Последовательность мутантной тяжелой цепи A88C (450 ак) представляет собой SEQ ID NO:6. Последовательность мутантной тяжелой цепи A121C (450 ак) представляет собой SEQ ID NO:7. Последовательность мутантной легкой цепи V110C (214 ак) представляет собой SEQ ID NO:8.

В одном из вариантов осуществления сконструированные антитела с цистеиновыми заменами тио-трастузумаб содержат одну или несколько из следующих последовательностей вариабельной области тяжелой цепи со свободной аминокислотой цистеином (SEQ ID NO: 9-16).

В другом варианте осуществления сконструированные антитела с цистеиновыми заменами тио-трастузумаб содержат одну или несколько из следующих последовательностей вариабельных областей легкой цепи со свободной аминокислотой цистеином (SEQ ID NO: 17-27).

Полученные полноразмерные варианты IgG тио-трастузумаба анализировали по реакционной способности тиола и активности связывания HER2. На фигуре 13A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2 и меченным HRP вторичным антителом для детекции оптической плотности. На фигуре 13B показаны измерения связывания с иммобилизованным HER2 с детекцией оптической плотности при 450 нм (слева направо): небиотинилированный трастузумаб дикого типа (Wt), конъюгированные с биотином-малеимидом варианты тио-трастузумаба V110C (один cys), A121C (один cys) и V110C-A121C (двойной cys). Каждый вариант тио-IgG и трастузумаб тестировали при 1, 10 и 100 нг. Измерения показывают, что биотинилированные анти-HER2 ТиоMab сохраняют активность связывания HER2.
На фигуре 14A показано графическое изображение связывания биотинилированного антитела с иммобилизованным HER2 со связыванием биотина с анти-IgG-HRP для детекции оптической плотности. На фигуре 14B показаны измерения связывания с детекцией оптической плотности при 450 нм для конъюгированных с биотином-малеимидом вариантов тио-трастузумаба и небиотинилированного трастузумаба дикого типа при связывании со стрептавидином. Слева направо: V110C (один cys), A121C (один cys), V110C/A121C (двойной cys) и трастузумаб. Каждый вариант тио IgG и исходный трастузумаб тестировали при 1, 10 и 100 нг. Измерения показывают, что ТиоMab против HER2 обладают высокой реакционной способностью тиола.
Цистеин вводили в конкретные остатки полноразмерного анти-EphB2R антитела 2H9. Мутант с одним cys H-A121C 2H9 экспрессировали в клетках CHO (яичников китайского хомяка) посредством нестационарной ферментации в среде, содержащей 1 мМ цистеин. Мутантная последовательность тяжелой цепи A121C 2H9 (450 ак) представляет собой SEQ ID NO:28.

Сконструированные тио-2H9 антитела с цистеиновыми заменами содержат следующие последовательности константной области тяжелой цепи Fc со свободной аминокислотой цистеином (SEQ ID NO: 29-38).

На фигуре 16 показан анализ электрофорезом в невосстанавливающем (вверху) и восстанавливающем (внизу) денатурирующем SDS-PAGE (электрофорезом в полиакриламидном геле) вариантов 2H9 ТиоMab Fc (слева направо, дорожки 1-9): A339C; S337C; S324C; A287C; V284C; V282C; V279C; и V273C, вместе с 2H9 дикого типа, после очистки на иммобилизованном белке A. Дорожка справа представляет собой маркер размера, указывающий на то, что интактные белки составляют приблизительно 150 кДа, фрагменты тяжелой цепи приблизительно 50 кДа и фрагменты легкой цепи приблизительно 25 кДа. На фигуре 17A показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (справа) денатурирующем полиакриламидном геле вариантов 2H9 ТиоMab (слева направо, дорожки 1-4): L-V15C; S179C; S375C; S400C, после очистки на иммобилизованном белке A. На фигуре 17B показан анализ электрофорезом в невосстанавливающем (слева) и восстанавливающем (+DTT) (справа) денатурирующем полиакриламидном геле дополнительных вариантов ТиоMab 2H9 и 3A5 после очистки на иммобилизованном белке A. Варианты ТиоMab 2H9 (в области Fab, так же как в Fc-области) экспрессировали и очищали, как описано. Как показано на фигурах 16, 17A и 17B, все белки являются гомогенными в SDS-PAGE после процедуры восстановления и окисления из примера 11 для получения реакционноспособных ТиоMab для конъюгации (пример 12).
Цистеин вводили в конкретные остатки полноразмерного анти-MUC16 антитела 3A5. Мутант 3A5 с одним cys H-A121C экспрессировали в клетках CHO (яичников китайского хомяка) посредством нестационарной ферментации в среде, содержащей 1 мМ цистеин. Мутантная последовательность тяжелой цепи A121C 3A5 (446 ак) содержит SEQ ID NO:39.

Сконструированные антитела с цистеиновыми заменами тио-3A5 анти-MUC16 содержат следующие последовательности вариабельной области тяжелой цепи со свободной аминокислотой цистеином (SEQ ID NO: 40-44).

Сконструированные тио-3A5 анти-MUC16 антитела с цистеиновыми заменами содержат следующие последовательности вариабельной области легкой цепи со свободной аминокислотой цистеином (SEQ ID NO: 45-49).

РЕАКЦИОННАЯ СПОСОБНОСТЬ ТИОЛА ТИОMAB
Реакционную способность тиола полноразмерных сконструированных антител с цистеиновыми заменами IgG (ТиоMab) измеряли по биотинилированию и связыванию стрептавидина. Анализ вестерн-блоттингом разрабатывали для скрининга ТиоMab, специфически конъюгированного с биотин-малеимидом. В этом анализе антитела анализировали восстанавливающим SDS-PAGE, и присутствие биотина специфически тестировали инкубацией со стрептавидином-HRP. Как видно на фигуре 18, наблюдают взаимодействие стрептавидина-HRP с тяжелой цепью или легкой цепью в зависимости от того, какой сконструированный с заменами cys вариант используют, и не наблюдают взаимодействия для дикого типа, что указывает на специфическую конъюгацию вариантов ТиоMab с биотином на сконструированном остатке Cys. На фигуре 18 показан анализ электрофорезом в денатурирующем геле восстановленных биотинилированных вариантов Тио-IgG после связывания с иммобилизованным анти-IgG-HRP (верхний гель) и стрептавидином-HRP (нижний гель). Дорожка 1: 3A5 H-A121C. Дорожка 2: 3A5 L-V110C. Дорожка 3: 2H9 H-A121C. Дорожка 4: 2H9 L-V110C. Дорожка 5: исходное анти-EphB2R 2H9, дикого типа. Каждый мутант (дорожки 1-4) связывался с анти-IgG с детекцией HRP (вверху), что указывает на сохранение избирательности и аффинности. Связывание с иммобилизованным стрептавидином с детекцией HRP (внизу) подтвердило локализацию биотина на тяжелой и легкой цепях. Локализацией цистеиновой мутации на сконструированных антителах с цистеиновыми заменами в дорожках 1 и 3 является тяжелая цепь. Локализацией цистеиновой мутации на сконструированных антителах с цистеиновыми заменами в дорожках 2 и 4 является легкая цепь. Участок цистеиновой мутации подвергается конъюгации с биотин-малеимидным реагентом.
Анализ ТиоMab сконструированных антител с цистеиновыми заменами на фигуре 18 и варианта 2H9 V15C посредством LC/MS дает количественные показатели реакционной способности тиола (Таблица 5).

Конструирование цистеиновых замен проводили в константном домене, т.е. Fc-области, IgG антител. Множество аминокислотных участков переводили в цистеиновые участки, и экспрессированные мутанты, т.е. сконструированные антитела с цистеиновыми заменами, оценивали по реакционной способности тиола.
Варианты биотинилированного Fc ТиоMab 2H9 оценивали по реакционной способности тиола посредством количественной оценки с помощью HRP посредством связывания с иммобилизованным стрептавидином в анализе ELISA (фигура 19). Анализ ELISA разрабатывали для быстрого скрининга остатков Cys с реакционноспособными тиоловыми группами. Как показано на схематической диаграмме на фигуре 19, взаимодействие стрептавидин-биотин мониторировали с использованием в качестве зонда анти-IgG-HRP с последующим измерением оптической плотности при 450 нм. Эти результаты подтвердили, что 2H9-ТиоFc варианты V282C, A287C, A339C, S375C и S400C обладали реакционной способностью тиола, от средней до наиболее высокой. Степень конъюгации с биотином вариантов 2H9 ТиоMab Fc количественно оценивали анализом LS/MS, как указано в таблице 6. Анализ LS/MS подтвердил, что варианты A282C, S375C и S400C обладали 100% конъюгацией с биотином, и V284C и A339C обладали 50% конъюгацией, что указывает на присутствие реакционноспособной тиоловой группы цистеина. Другие варианты ТиоFc, и исходное 2H9 дикого типа либо обладали очень малым биотинилированием, либо не были биотинилированы.

РЕАКЦИОННАЯ СПОСОБНОСТЬ ТИОЛА ВАРИАНТОВ ЛЕГКОЙ ЦЕПИ ТИО-4D5 FAB
Скринингом множества сконструированных вариантов легкой цепи Fab анти-ErbB2 антитела 4D5 с цистеиновыми заменами получили ряд вариантов с значением реакционной способности тиола 0,6 и выше (таблица 7), как измерено анализом PHESELECTOR на фигуре 8. Значения реакционной способности тиола из таблицы 7 нормализованы по варианту тяжелой цепи 4D5 ТиоFab (HC-A121C), который принимали за 100%, принимая полное биотинилирование варианта HC-A121C, и представлены как значения в процентах.

РЕАГЕНТЫ ДЛЯ МЕЧЕНИЯ ЦИРКОНИЕМ
Иллюстративные бифункциональные реагенты на основе десферриоксамина B (Df) использовали для образования комплексов 89Zr с антителами, включая моноклональные антитела (mAb). Десферриоксамин B (Ν'-{5-[ацетил(гидрокси)амино]пентил}-N-[5-({4-[(5-аминопентил)(гидрокси)амино]-4-оксобутаноил}амино)пентил]-N-гидроксисукцинамид (CAS Reg. No. 70-51-9); и известный также как дефероксамин, десфероксамин B, DFO-B, DFOA, DFB или десферал), представляет собой бактериальный сидерофор, продуцируемый actinobacter Streptomyces pilosus (фигура 20, вверху). Десферриоксамин B имеет применение в медицине в качестве хелатирующего агента, используемого для удаления избытка железа из организма (Miller, Marvin J. «Syntheses and therapeutic potential of hydroxamic acid based siderophores and analogs» (1989) Chemical Reviews 89 (7): 1563-1579). Соль мезилат DFO-B является коммерчески доступной. Первоначальные эксперименты проводили с N-(S-ацетил)тиоацетил-Df (SATA-Df) и mAb с добавленными малеимидными группами, 4-[N-малеимидометил]циклогексан-1-карбоксилатом (mAb-SMCC), присоединенными к ε-аминогруппе в боковой цепи лизина (Meijs WE et al. «Zirconium-labeled monoclonal antibodies and their distribution in tumor-bearing nude mice» (1997) J. Nucl. Med. 38: 112-8; Meijs WE et al. «A facile method for the labeling of proteins with zirconium isotopes» (1996) Nucl Med Biol. 23:439-48). Однако, полученные тиоэфирные конъюгаты (mAb-SMCC-SATA-Df) были нестабильными в сыворотке человека при 37°C (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» (2003) J Nucl Med 44: 1271-81). Другие иллюстративные реакционноспособные по отношению к аминогруппам бифункциональные хелаторы на основе Df, модифицированного янтарным ангидридом (Suc), использовали для превращения аминогруппы Df в карбоновую кислоту и последующей активации в форме сложного эфира 2,3,5,6-тетрафторфенила (TFP). TFP-N-Suc-Df (фигура 20, в центре) присоединяли к ε-аминогруппам лизина mAb, и получали хелаты очищенного mAb-N-Suc-Df с 89Zr. Полученный 89Zr-mAb-N-Suc-Df являлся стабильным в физиологических условиях, и его биораспределение сравнивали с mAb-SMCC-SATA-Df у мышей (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» (2003) J Nucl Med. 44: 1271-81). Однако, получение TFP-N-Suc-Df требует защиты гидроксамидных групп в форме комплекса Fe(III). Железо удаляют обработкой ЭДТА до образования хелатов с 89Zr, но многоступенчатый способ является трудоемким и подвержен опасности неполного удаления железа из десферриоксамина и/или неполного удаления ЭДТА из буфера для конъюгации, что может отрицательно влиять на выход образования 89Zr-хелатов. Таким образом, гетеробифункциональный реакционноспособный по отношению к аминогруппам реагент, п-изотиоцианатбензил-десферриоксамин (Df-Bz-NCS), недавно разработан для включения Df в белки посредством связи с тиомочевиной, фигура 20, в центре (Perk LR et al. «Facile radiolabeling of monoclonal antibodies and other proteins with zirconium-89 or gallium-68 for PET Imaging using p-isothiocyanatobenzyl-desferrioxamine» (2008) Nature Protocols, published online:DOI: 10.1038/nprot.2008.22; Perk LR et al. «p-Isothiocyanatobenzyl-desferrioxamine: a new bifunctional chelate for facile radiolabeling of monoclonal antibodies with zirconium-89 for immuno-PET imaging» (2009) European Journal Of Nuclear Medicine And Molecular Imaging). Для конъюгатов антител, полученных с использованием Df-Bz-NCS, показали стабильность и свойства получения изображений, сравнимые с контрольными конъюгатами, полученными с использованием TFP-N-Suc-Df. Поскольку разработаны надежные способы присоединения 89Zr к антителам через ε-аминогруппы лизина, число опубликованных доклинических и клинических исследований иммуноПЭТ с меченными 89Zr антителами быстро увеличивается (Verel I, et al. «Long-lived positron emitters zirconium-89 and iodine-124 for scouting of therapeutic radioimmunoconjugates with PET» (2003) Cancer Biother Radiopharm. 18:655-61; Nagengast WB et al. «In vivo VEGF imaging with radiolabeled bevacizumab in a human ovarian tumor xenograft» (2007) J Nucl Med. 48:1313-9; Perk LR, et al. «(89)Zr as a PET surrogate radioisotope for scouting biodistribution of the therapeutic radiometals (90)Y and (177)Lu in tumor-bearing nude mice after coupling to the internalizing antibody cetuximab» (2005) J Nucl Med. 46: 1898-906; Perk LR et al. «Quantitative PET imaging of Met-expressing human cancer xenografts with (89)Zr-labelled monoclonal antibody DN30» (2008) European Journal Of Nuclear Medicine And Molecular Imaging 35:1857-67; Perk LR et al. «Preparation and evaluation of (89)Zr-Zevalin for monitoring of (90)Y-Zevalin biodistribution with positron emission tomography» (2006) European Journal Of Nuclear Medicine And Molecular Imaging 33:1337-45; Borjesson PK et al. «Performance of immuno-positron emission tomography with zirconium-89-labeled chimeric monoclonal antibody U36 in the detection of lymph node metastases in head and neck cancer patients» (2006) Clin Cancer Res. 12:2133-40; Aerts HJ et al. «Disparity between in vivo EGFR expression and 89Zr-labeled cetuximab uptake assessed with PET» (2009) J Nucl Med. 50:123-31; Dijkers EC et al. «Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging» (2009) J Nucl Med 50(6):974-981).
Варианты осуществления комплексов циркония включают также связывающие цирконий (образующие с ним хелаты) лиганды, такие как DTPA (CAS Reg. No. 67-43-6), DOPA (1,4,7,10-тетраазациклододекан-1,4,7,10-тетрауксусная кислота) (Liu, Shuang (2008) Advanced Drug Delivery Reviews 60(12):1347-1370), циклопентадиенильные и аллильные группы (Erker, G. (1991) Pure and Applied Chemistry 63(6):797-806; Erker, G. (1990) Jour, of Organometallic Chem. 400(1-2):185-203), содержание каждого из которых приведено в настоящем документе в качестве ссылки.
Комплексы циркония (Z) и других радиоактивных изотопов можно конъюгировать с антителами (Ab), включая моноклональные антитела (mAb), через ε-аминогруппу боковой цепи лизина или через тиоловую группу цистеина. Поскольку приблизительно 40 боковых цепей лизина (Wang L et al «Structural characterization of the maytansinoid-monoclonal antibody immunoconjugate, huN901-DM1, by mass spectrometry» (2005) Protein Sci. 14:2436-46) или 8 остатков цистеина (Hamblett KJ et al. «Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate» (2004) Clin Cancer Res. 10:7063-70) доступны для конъюгации в mAb, оба способа обеспечивают гетерогенность по отношению к соотношениям конъюгатов mAb и участкам конъюгации. Модификация остатка лизина внутри участка связывания может уменьшать биологическую активность конъюгата (Cai W et al. «PET imaging of colorectal cancer in xenograft-bearing mice by use of an 18F-labeled T84.66 anti-carcinoembryonic antigen diabody» (2007) J Nucl Med. 48:304-10; Shively JE. «18F labeling for immuno-PET: where speed and contrast meet» (2007) J Nucl Med. 48: 170-2; Tait JF et al «Improved detection of cell death in vivo with annexin V radiolabeled by site-specific methods» (2006) J Nucl Med. 47: 1546-53; Schellenberger EA et al «Optical imaging of apoptosis as a biomarker of tumor response to chemotherapy» (2003) Neoplasia (New York, N.Y) 5: 187-92), в то время как модификация остатков цистеина в шарнирной области обеспечивает уменьшенное время полужизни в плазме (Hamblett KJ et al. «Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate» (2004) Clin Cancer Res. 10:7063-70). Этих ограничений можно избежать с использованием mAb, сконструированных с содержанием цистеина, расположенного избирательно, с целью сайт-специфической конъюгации, с помощью биохимического анализа PHESELECTOR (US 7521541; Junutula JR et al. «Rapid identification of reactive cysteine residues for site-specific labeling of antibody-Fabs» J Immunol Methods 2008;332:41-52) для быстрой идентификации предпочтительных аминокислот в антителе для мутации с заменой на цистеин. Полученное антитело (ТИОMAB) затем химически избирательно и сайт-специфически конъюгируют с цитотоксическими лекарственными средствами без какой-либо потери аффинности связывания или неблагоприятных эффектов на стабильность каркаса антитела (Junutula JR et al. «Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index» (2008) Nat Biotechnol. 26:925-32).
С точки зрения получения изображений, высокая аффинность к мишени и минимальное неспецифическое поглощение необходимы для оптимального качества изображения. Соответственно, сайт-специфически радиоактивно меченные сконструированные антитела с цистеиновыми заменами (ТИОMAB) могут обеспечивать метки с неизмененной аффинностью связывания и стабильностью каркаса, которые могут минимизировать неспецифическое поглощение метаболитов вне ткани-мишени. Один аспект настоящего изобретения относится к способу сайт-специфического радиоактивного мечения ТИОMAB с использованием новых реакционноспособных по отношению к тиолу бифункциональных реагентов на основе Df малеимидоциклогексил-десферриоксамина (Df-Chx-Mal), бромацетил-десферриоксамина (Df-Bac) и иодацетил-десферриоксамина (Df-Iac) (фигура 20). Иллюстративные варианты осуществления включают варианты осуществления, в которых эти реагенты сайт-специфически конъюгируют с ТИОMAB трастузумабом (тио-трастузумаб), получают хелаты с 89Zr и оценивают in vitro и in vivo.
Одним из метастабильных изомеров циркония является 89Zr со временем полужизни 78,4 часов с типами распада бета (испускание электронов), испускания позитронов (бета плюс) и гамма-излучения.
Радиоактивный изотоп или другие метки можно включать в конъюгат известными способами (Fraker et al (1978) Biochem. Biophys. Res. Commun. 80: 49-57; «Monoclonal Antibodies in Immunoscintigraphy» Chatal, CRC Press 1989). Меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой иллюстративный хелатирующий агент для конъюгации радиоактивного изотопа с антителом (WO 94/11026).
ЛИНКЕРЫ
«Линкер» (L) представляет собой бифункциональную или мультифункциональную группу, которую можно использовать для соединения одной или нескольких групп комплекса с цирконием (Z) и молекулы антитела (Ab) с образованием конъюгатов антитело-цирконий (AZC) формулы I. Конъюгаты антитело-цирконий (AZC) можно удобно получать с использованием линкера, обладающего реакционноспособной функциональной группой для связывания с цирконием и с антителом. Тиол цистеина сконструированного антитела с цистеиновыми заменами (Ab) может образовывать связь с функциональной группой линкерного реагента, группой циркониевой метки или промежуточным соединением цирконий-линкер.
В одном аспекте линкер имеет реакционноспособный участок с электрофильной группой, являющейся реакционноспособной по отношению к нуклеофильному цистеину, присутствующему в антителе. Тиол цистеина антитела является реакционноспособным по отношению к электрофильной группе линкера и образует ковалентную связь с линкером. Пригодные электрофильные группы включают в качестве неограничивающих примеров малеимидные и галоацетамидные группы.
Сконструированные антитела с цистеиновыми заменами могут вступать в реакцию с линкерными реагентами или промежуточными соединениями цирконий-линкер, с электрофильными функциональными группами, такими как малеимид или α-галокарбонил, согласно способу конъюгации на странице 766 в Klussman, et al (2004), Bioconjugate Chemistry 15(4):765-773, и согласно способам из примеров 17-19.
В другом варианте осуществления группы Z являются одинаковыми.
В другом варианте осуществления группы Z являются различными.
Иллюстративные варианты осуществления соединений конъюгатов антитело-цирконий (AZC) формулы I включают:

где X представляет собой:

где Y представляет собой:

R независимо представляет собой H или C1-C6 алкил; и n представляет собой 1-12.
В другом варианте осуществления линкер обладает реакционноспособной функциональной группой, обладающей нуклеофильной группой, реакционноспособной по отношению к электрофильной группе, присутствующей в антителе. Пригодные электрофильные группы антитела включают в качестве неограничивающих примеров, карбонильные группы альдегидов и кетонов. Гетероатом нуклеофильной группы линкера может вступать в реакцию с электрофильной группой антитела и образовывать ковалентную связь с молекулой антитела. Пригодные нуклеофильные группы линкера включают в качестве неограничивающих примеров, гидразид, оксим, амино, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид. Электрофильная группа антитела обеспечивает удобный участок для присоединения к линкеру.
В другом варианте осуществления линкер может быть замещен группами, модулирующими растворимость или реакционную способность. Например, заряженный заместитель, такой как сульфонат (-SO3 -) или аммоний, может увеличивать растворимость реагента в воде и облегчать реакцию присоединения линкерного реагента к антителу или группе циркония, или облегчать реакцию присоединения Ab-L (промежуточного соединения антитело-линкер) к Z или Z-L (промежуточного соединения цирконий-линкер) к Ab, в зависимости от способа синтеза, используемого для получения AZC.
Соединения по изобретению конкретно включают, но без ограничения, AZC, полученные с помощью линкерных реагентов: BMPEO, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), и включая бис-малеимидные реагенты: DTME, BMB, BMDB, BMH, BMOE, BM(PEO)2 и BM(PEO)43, коммерчески доступные от Pierce Biotechnology, Inc., Customer Service Department, P.O. Box 117, Rockford, IL. 61105 U.S.A. Бис-малеимидные реагенты позволяют присоединение тиоловой группы сконструированного антитела с цистеиновыми заменами к содержащей тиол циркониевой группе, метке или линкерному промежуточному соединению, последовательным или одновременным образом. Другие функциональные группы помимо малеимида, реакционноспособные по отношению к тиоловой группе сконструированного антитела с цистеиновыми заменами, циркониевой группе, метке или линкерному промежуточному соединению, включают иодацетамид, бромацетамид, винилпиридин, дисульфид, пиридилдисульфидизоцианат и изотиоцианат.

Пригодные линкерные реагенты можно получать также из других коммерческих источников, таких как Molecular Biosciences Inc. (Boulder, CO), или синтезировать способами, описанными в Toki et al (2002) J. Org. Chem. 67: 1866-1872; Walker, M.A. (1995) J. Org. Chem. 60:5352-5355; Frisch et al (1996) Bioconjugate Chem. 7: 180-186; US 6214345; WO 02/088172; US 2003130189; US2003096743; WO 03/026577; WO 03/043583; и WO 04/032828.
Иллюстративные линкерные реагенты включают:

где n представляет собой целое число в диапазоне 1-10, и T представляет собой -H или -SO3Na;

где n представляет собой целое число в диапазоне 0-3;

В другом варианте осуществления линкер L может представлять собой линкер древовидного типа для ковалентного присоединения более одной группы циркония через разветвленную, мультифункциональную линкерную группу к антителу (Sun et al (2002) Bioorganic & Medicinal Chemistry Letters 12:2213-2215; Sun et al (2003) Bioorganic & Medicinal Chemistry 11:1761-1768). Линкеры древовидного типа могут увеличивать молярное соотношение циркония к антителу, т.е. нагрузку AZC. Таким образом, когда сконструированное антитело с цистеиновыми заменами несет только одну реакционноспособную тиоловую группу цистеина, множество циркониевых групп можно присоединять через линкер древовидного типа.
Следующие иллюстративные варианты осуществления линкерных реагентов древовидного типа позволяют конъюгацию вплоть до девяти содержащих циркониевую группу нуклеофильных реагентов посредством реакции с содержащими хлорэтил и азотистый иприт функциональными группами:


Другие варианты осуществления разветвленных, древовидных линкеров включают линкеры с самоуничтожающимися дендримерными звеньями 2,6-бис(гидроксиметил)-п-крезола и 2,4,6-трис(гидроксиметил)-фенола (WO 2004/01993; Szalai et al (2003) J. Amer. Chem. Soc. 125: 15688-15689; Shamis et al (2004) J. Amer. Chem. Soc. 126: 1726-1731; Amir et al (2003) Angew. Chem. Int. Ed. 42:4494-4499).
МЕЧЕННЫЕ ДЕСФЕРРИОКСАМИНОМ СКОНСТРУИРОВАННЫЕ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ
В одном аспекте изобретение относится к меченному десферриоксамином сконструированному антителу с цистеиновыми заменами, содержащему сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающему формулой II:

где L-Df выбран из:

где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4.
ПОЛУЧЕНИЕ КОНЪЮГАТОВ АНТИТЕЛО-ЦИРКОНИЙ
Конъюгаты антитело-цирконий (AZC) формулы I можно получать несколькими способами с использованием реакций, условий и реагентов органической химии, известных специалистам в данной области, включая: (1) реакцию цистеиновой группы сконструированного антитела с цистеиновыми заменами с линкерным реагентом для образования промежуточного соединения антитело-линкер Ab-L, посредством ковалентной связи, с последующей реакцией с активированной группой циркониевой метки Z; и (2) реакцию нуклеофильной группы циркониевой группы с линкерным реагентом для образования промежуточного соединения циркониевая метка-линкер Z-L, посредством ковалентной связи, с последующей реакцией с цистеиновой группой сконструированного антитела с цистеиновыми заменами. Способы конъюгации (1) и (2) можно использовать со множеством сконструированных антител с цистеиновыми заменами, групп циркониевой метки и линкеров для получения конъюгатов антитело-цирконий формулы I.
Тиоловые группы цистеина антитела являются нуклеофильными и способными вступать в реакцию для формирования ковалентных связей с электрофильными группами линкерных реагентов и промежуточных соединений цирконий-линкер, включая: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; (ii) алкил и бензилгалиды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы; и (iv) дисульфиды, включая пиридилдисульфиды, посредством сульфидного обмена. Нуклеофильные группы на группе циркониевой метки включают в качестве неограничивающих примеров: группы амина, тиола, гидроксила, гидразида, оксима, гидразина, тиосемикарбазона, гидразинкарбоксилата и арилгидразида, способные вступать в реакцию для формирования ковалентных связей с электрофильными группами линкерных групп и линкерных реагентов.
В конкретных условиях сконструированные антитела с цистеиновыми заменами можно сделать реакционноспособными для конъюгации с линкерными реагентами посредством обработки восстанавливающим агентом, таким как DTT (реактив Клеланда, дитиотреитол) или TCEP (трис(2-карбоксиэтил)фосфингидрохлорид; Getz et al (1999) Anal. Biochem. Vol 273:73-80; Soltec Ventures, Beverly, MA). Полноразмерные сконструированные моноклональные антитела с цистеиновыми заменами (ТиоMab), экспрессированные в клетках CHO, восстанавливали с помощью приблизительно 50-кратного избытка TCEP в течение 3 часов при 37°C для восстановления дисульфидных связей, которые могут формироваться между вновь введенными остатками цистеина и цистеином, присутствующим в культуральной среде. Восстановленное ТиоMab разводили и наносили на колонку HiTrap S в 10 мМ ацетате натрия, pH 5, и элюировали с помощью PBS, содержащего 0,3 М хлорид натрия. Дисульфидные связи вновь образовывались между остатками цистеина, присутствующими в исходном Mab, с помощью разведенного (200 нМ) водного сульфата меди (CuSO4) при комнатной температуре, в течение ночи. Можно использовать другие окислители, т.е. окисляющие агенты и окисляющие условия, известные в данной области. Окисление атмосферным воздухом также является эффективным. Эта стадия мягкого, частичного повторного окисления формирует внутрицепочечные дисульфиды эффективно с высокой точностью. Приблизительно 10-кратный избыток промежуточного соединения цирконий-линкер добавляют, перемешивают и оставляют приблизительно на один час при комнатной температуре для осуществления конъюгации и образования конъюгата ТиоMab антитело-цирконий. Смесь после конъюгации подвергают гель-фильтрации, наносят на колонку HiTrap S и проводят элюцию для удаления избытка промежуточного соединения цирконий-линкер и других примесей.
На фигуре 15 показан общий способ подготовки сконструированного антитела с цистеиновыми заменами, экспрессированного из культуры клеток, для конъюгации. Цистеиновые аддукты, предположительно с различными межцепьевыми дисульфидными связями, подвергают восстановительному отщеплению для получения восстановленной формы антитела. Межцепочечные дисульфидные связи между спаренными остатками цистеина вновь формируют в условиях частичного окисления, таких как воздействие атмосферного кислорода. Вновь введенные сконструированные и неспаренные остатки цистеина остаются доступными для реакции с линкерными реагентами или промежуточными соединениями цирконий-линкер для образования конъюгатов антител по изобретению. С ТиоMab, экспрессированных в линиях клеток млекопитающих, получали внешний аддукт Cys, конъюгированный с конструированным Cys посредством формирования -S-S- связи. Таким образом, очищенные ТиоMab необходимо обрабатывать способами восстановления и окисления, как описано в примере 11, для получения реакционноспособных ТиоMab. Эти ТиоMab используют для конъюгации с содержащими малеимид радиоактивными метками, цитотоксическими лекарственными средствами, флуорофорами и другими метками.
ПОЛУЧЕНИЕ И АНАЛИЗ КОНЪЮГАТОВ
89
Zr-Df-ТРАСТУЗУМАБА
Защищенный активный сложный эфир TFP-N-SucDf-Fe получали согласно описанному ранее способу (Verel I et al «89Zr Immuno-PET: Comprehensive Procedures For The Production Of 89Zr-Labeled Monoclonal Antibodies» (2003) J Nucl Med 44: 1271-81) и конъюгировали с трастузумабом с использованием 5-кратного молярного избытка TFP-N-SucDf-Fe для получения N-SucDf-трастузумаба в среднем с 1,6 молекулами десферриоксамина (Таблица 8). Df-Bz-SCN-трастузумаб получали объединением с 8-кратным молярным избытком Df-Bz-SCN при pH 8,5 (Perk LR, et al. «Facile radiolabeling of monoclonal antibodies and other proteins with zirconium-89 or gallium-68 for PET Imaging using p-isothiocyanatobenzyl-desferrioxamine» (2008) Nature Protocols; published online:DOI: 10.1038/nprot.2008.22). Реакция приводит к Df-Bz-SCN-трастузумабу с присоединенными в среднем 2,4 молекулами десферриоксамина (Таблица 1).
Новый реакционноспособный по отношению к тиолу бифункциональный линкер на основе малеимида Df-Chx-Mal получали из эквимолярных количеств мезилата десферриоксамина и SMCC (фигура 21, пример 13). Реакция завершалась через 30 мин при комнатной температуре, и продукт выделяли преципитацией при добавлении воды с 45% выходом и более чем 95% чистотой. Реакцией 8,5-кратного молярного избытка Df-Chx-Mal со свежеприготовленым тио-трастузумабом (фигура 21, пример 17) получали конъюгат Df-Chx-Mal-тио-трастузумаб точно с 2 молекулами десферриоксамина за 1 час (таблица 1, фигура 21). Бромацетилдесферриоксамин (BDf-Bac) получали реакцией с эквимолярными количествами мезилата десферриоксамина и бромацетилбромида при 0°C (пример 14). Продукт получали с 14% выходом после очистки HPLC. Алкилированием свежеполученного тио-трастузумаба (фигура 21, пример 16) с помощью 12-кратного молярного избытка Df-Bac получали конъюгат (Df-Ac-тио-трастузумаб) с 1,8 молекулами Df на антитело через 5 часов (таблица 8, фигура 21, пример 18). Низкая реакционная способность бромида побудила авторов настоящего изобретения исследовать более реакционноспособное производное иодацетила (Df-Iac). Df-Iac получали с 53% выходом реакцией мезилата десферриоксамина с небольшим избытком иодацетата N-гидроксисукцинимидила (фигура 21, пример 15). Продукт получали более чем с 95% чистотой посредством преципитации из реакционной смеси. Последующей реакцией с 11-кратным избытком Df-Iac получали Df-Ac-тио-трастузумаб с 1,8 молекулами Df через 2 часа (таблица 1, фигура 21, пример 19). На основании опыта авторов настоящего изобретения, Df-Chx-Mal является предпочтительным реагентом из трех исследованных соединений. Следует отметить, что реакция Df-Chx-Mal завершалась при среднем pH в пределах 1 часа, в отличие от более высокого pH и более длительного времени реакции, необходимых для алкилирования тиоловых групп с помощью Df-Bac и Df-Iac. Кроме того, более низкая реакционная способность галогенацетамидов может вызывать неполную загрузку обеих доступных остатков цистеина тио-трастузумаба.

89Zr образовывал хелаты в форме оксалата 89-циркония со всеми четырьмя вариантами Df-трастузумаба с использованием ранее описанного экспериментального способа (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» (2003) J Nucl Med. 44: 1271-81). Белки с радиоактивной меткой очищали на обессоливающей колонке, и конечный раствор концентрировали до необходимого объема мембранной фильтрацией. Выход, чистота и конечная сецифическая активность конъюгатов 89Zr обобщены в таблице 9. Как правило, выход хелатов превышал 80%, за исключением Df-N-Suc линкера, полученного с низким выходом, предположительно, из-за более низкого количества Df на молекулу антитела и/или неполного удаления Fe(III), используемого для защиты хелатирующего агента в ходе активации и конъюгации. После очистки вариантов Df-трастузумаба с использованием обессоливающей колонки, чистота продукта составляла более 90% с малым количеством (1-6%) высокомолекулярных агрегатов, детектированных в каждом образце. Df-Chx-Mal-тио-трастузумаб обеспечивал комплекс 89Zr с чистотой 99% (Таблица 9), в отличие от конъюгата Df-Ac, загрязненного приблизительно 8% низкомолекулярных загрязнений и 2% высокомолекулярных агрегатов. Загрязнения выдерживали удаление с использованием колонки NAP-10, но удаление являлось возможным с использованием повторяющейся смены буфера на фильтре Amicon.
| Таблица 9 Выходы, специфическая активность и чистота меченного радиоактивной меткой 89Zr-Df-линкер-трастузумаба |
|
 |
БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ КОНЪЮГАТОВ
89
Zr-Df-ТРАСТУЗУМАБА
Биологическую активность вновь полученных сайт-специфических конъюгатов Df-линкер-тио-трастузумаб определяли с использованием анализа Скэтчарда, анализа связывания с линией клеток рака молочной железы BT474. Полученные значения KD сравнивали с немодифицированным трастузумабом (0,91±0,20 нМ). KD для конгъюгата тио-трастузумаба, содержащего Chx-Mal линкер, составляла 0,93±0,15 нМ, и значения для конъюгатов, содержащих Ac линкер, составляли 1,22±0,22 нМ для конъюгата, полученного с использованием Df-Bac, и 0,87±0,15 нМ для конъюгата, полученного с использованием Df-Iac. Результаты анализов биологической активности указывают на то, что модификация тио-трастузумаба не влияет на аффинность связывания антитела с HER2.
СТАБИЛЬНОСТЬ СЫВОРОТКИ IN VITRO
Ранее опубликованные конъюгаты Df-антитела с линкерами N-Suc и Bz-SCN, содержащие связи амида или тиомочевины, являлись стабильными in vitro в течение периода 6 суток в сыворотке при 37°C (Verel I et al «89Zr immuno-PET: comprehensive procedures for the production of 89Zr-labeled monoclonal antibodies» J Nucl Med 2003;44: 1271-81; Perk LR et al, (2009) European Journal Of Nuclear Medicine And Molecular Imaging 35(10): 1857-1867). Стабильность конъюгатов 89Zr-тио-трастузумаба с Chx-Mal и Ac линкерами определяли в сыворотке мыши при 37°C. Значительной потери связанного с антителом 89Zr не наблюдали в течение периода 5 суток. Оба тио-конъюгата являлись стабильными со средней потерей связанного с антителом 89Zr 1,8% в сутки для 89Zr-Df-Chx-Mal-тио-трастузумаба и 1,4% в сутки для 89Zr-Df-Ac-тио-трастузумаба (Фигура 24). Медленное формирование высокомолекулярных молекул, предположительно, агрегатов, наблюдали для обоих линкеров Df-Chx-Mal и Df-Ac.
ПОЛУЧЕНИЕ ИЗОБРАЖЕНИЙ МИКРОПЭТ IN VIVO
Двадцати животным (5 животных на группу), несущим подкожные ксенотрансплантаты BT474M1 (размер ~200 мм3), инъецировали внутривенно 89Zr-трастузумаб. Количество антитела, инъецированного на животное, составляло 1,4±0,29 мг/кг. Изображения для проекции максимальной интенсивности для репрезентативных животных (через 96 ч p.i.) показаны на фиг.3. Поглощение 89Zr-трастузумаба избранными тканями обобщено на фиг.4. В изображениях через 1 час (не показаны) преобладало сильное поглощение в кровяных депо, за исключением Df-Bac, где быстрое выделение липофильных загрязнений из печени и желчных путей приводило к увеличенному поглощению в кишечнике. Загрязнения полностью выводились в течение первых 24 часов, и увеличенное поглощение в тонком и толстом кишечнике не детектировали через 24 часа или в более позднее время после инъекции метки. Хотя поглощение тканями конъюгата Df-Ac в результате являлось немного меньшим (~8%), на соотношения опухоли к крови (таблица 10) не влияла потеря инъецированной радиоактивности. В изображениях через 96 часов преобладало сильное поглощение в опухолях с небольшими различиями, обнаруженными среди четырех различных вариантов 89Zr-трастузумаба (фигура 25). Поглощение в опухоли было идентичным для каждой метки, достигая максимальных значений через 24 часа после инъекции и максимальных соотношений для опухоли и крови через 144 часа из-за выведения из крови (таблица 10). Для конъюгата на основе тиола 89Zr-Df-Chx-Mal-тио-трастузумаба показали увеличенное поглощение в костной ткани (P<0,05) по сравнению с конъюгатами на основе амина (Df-Bz-SCN и Df-N-Suc) через 96 и 144 часов p.i. Поглощение в костной ткани Df-Ac-тио-трастузумаба не было значимо повышенным (P=0,20) по сравнению с линкерами Df-Bz-SCN и Df-N-Suc, но может становиться значимым при коррекции на 8% потери радиоактивности в течение первых 24 часов. Поглощение в почках для каждой метки являлось низким (фигура 26), как ожидали для меток на основе антитела, но для 89Zr-Df-Chx-Mal-тио-трастузумаба было немного выше по сравнению с другими линкерами через 24, 96 и 144 часа (P<0,05).

Для ксенотрансплантов BT474 (3+ уровень экспрессии HER2) показали более низкое абсолютное поглощение метки (15 %ID/г), чем ранее измерено Dijkers et al для SKOV3 (3+ уровень экспрессии HER2) 33,4±7,7%ID/г (Dijkers EC, et al. «Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging» (2009) J Nucl Med 50(6):974-981). Однако, соотношение опухоли к крови 5,7-7,1 (таблица 10) является сравнимым со значением, полученным для SKOV3 (соотношение опухоли к крови 7,6). Различия в поглощении в опухоли можно отнести к модели опухоли и общей дозы трастузумаба. Использовали материал с более высокой специфической активностью, таким образом, инъецировали значительно меньше антитела (35 мкг, 1,4 мг/кг) по сравнению с исследованием Dijkers et al с SKOV3 (100 мкг, 4 мг/кг). Различия в специфической активности также можно отнести к более низкому поглощению в костях свободного 89Zr в эксперименте в настоящем документе 2-3 %ID/г по сравнению с моделью SKOV3 (5-10 %ID/г). К сожалению, Dijkers et al не приводят объяснений относительно повышенного поглощения в костной ткани. Известно, что цирконий связывается с белками плазмы (Mealey J, Jr. «Turn-over of carrier-free zirconium-89 in man» (1957) Nature 179:673-4 и позже депонируется в костных минералах (Fletcher CR. «The radiological hazards of zirconium-95 and niobium-95» (1969) Health Phys. 16:209-20; Shiraishi Y and Ichikawa R. «Absorption and retention of 144 Ce and 95 Zr-95 Nb in newborn, juvenile and adult rats» (1972) Health Phys. 22:373-8). Поскольку инъецированный материал не содержит свободного 89Zr, поглощение в костной ткани может происходить из-за разрушения 89Zr антитела или из-за 89Zr, неспецифически ассоциированного с антителом, который может затем образовывать транс-хелаты с белками плазмы в отличие от 89Zr, связанного с Df.
Три специфичных для тиола реагента приведены в настоящем документе в качестве примера для хемоселективной конъюгации десферриоксамина (Df) с моноклональными антителами через тиоловую группу цистеина сконструированного антитела с цистеиновыми заменами. Тиол-специфические Df-реагенты получали ацилированием аминогруппы десферриоксамина B с выходами 14% (Df-Bac), 53% (Df-Iac) и 45% (Df-Chx-Mal), и конъюгировали с тио-трастузумабом, получая сайт-специфическую модификацию обоих сконструированных остатков цистеина за 1-5 ч. Активность связывания сайт-специфических конъюгатов тио-трастузумаба с HER2 были идентичны активности немодифицированного трастузумаба. Для Df-модифицированного тио-трастузумаба (Df-Ac-тио-трастузумаб и Df-Chx-Mal-тио-трастузумаб) получали хелаты с 89Zr (фигура 22) с выходами, превышающими 80%, в течение 1 часа, что сравнимо с лизиновыми конъюгатами, полученными с использованием ранее описанных линкеров Df-Bz-SCN и Df-N-Suc. Как для 89Zr-Df-Ac-тио-трастузумаба, так и для 89Zr-Df-Chx-Mal-тио-трастузумаба показали сравнимую стабильность в сыворотке мыши. Для обоих соединений показали также возможность получения изображений ПЭТ на модели рака молочной железы BT474M1, что сравнимо с лизиновыми конъюгатами, достигающими поглощения в опухоли 10-15% ID/г с соотношением опухоли к крови в диапазоне 6,1-7,1. В целом, новые реагенты являются легко доступными, обладая хорошей реакционной способностью по отношению к тиоловым группам белка, и обладают очень хорошими хелатирующими свойствами по отношению к 89Zr. 89Zr-меченные антитела являлись стабильными в сыворотке и обладали отличными свойствами получения изображений ПЭТ. Df-Chx-Mal является пригодным реагентом для конъюгации Df с антителами через боковую цепь цистеина и обладает несколькими преимуществами по сравнению с Df-Bac и Df-Iac. Во-первых, умеренный pH 7,5 необходим для полной конъюгации Df-Chx-Mal в течение 1 часа по сравнению с pH 9 и 2 или 5 часами, необходимыми для Df-Bac и Df-Iac. Кроме того, сайт-специфически меченные 89Zr конъюгаты сконструированного ТИОMAB можно использовать сходным образом с 18F меченными конъюгатами ТИОFAB (Gill HS, et al. «A modular platform for the rapid site-specific radiolabeling of proteins with 18F exemplified by quantitative positron emission tomography of human epidermal growth factor receptor 2» (2009) Jour, of Med. Chem. 52:5816-25) в качестве полезных инструментов для применений получения изображений ПЭТ в биомедицинских исследованиях.
ВВЕДЕНИЕ КОНЪЮГАТОВ АНТИТЕЛО-ЦИРКОНИЙ
Конъюгаты антитело-цирконий (AZC) по изобретению можно вводить любым способом, подходящим для состояния, подлежащего лечению. AZC, как правило, вводят парентерально, т.е. инфузией, подкожно, внутримышечно, внутривенно, внутрикожно, интратекально и эпидурально.
ФАРМАЦЕВТИЧЕСКИЕ СОСТАВЫ
Фармацевтические составы диагностических конъюгатов антитело-цирконий (AZC) по изобретению, как правило, получают для парентерального введения, т.е. болюсной, внутривенной, внутриопухолевой инъекции вместе с фармацевтически приемлемым парентеральным носителем и в пригодной для инъекции единичной лекарственной форме. Конъюгат антитело-цирконий (AZC), обладающий желательной степенью чистоты, необязательно смешивают с фармацевтически приемлемыми разбавителями, носителями, наполнителями или стабилизаторами (Remington's Pharmaceutical Sciences (1980) 16th edition, Osol, A. Ed.), в форме лиофилизированного состава или водного раствора.
Приемлемые разбавители, носители, наполнители и стабилизаторы в используемых дозах и концентрациях не токсичны для реципиентов и включают буферы, такие как фосфатный, цитратный и относящиеся к другим органическим кислотам; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметэтония; хлорид бензалкония, хлорид бензэтония; феноловый, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); полипептиды с низкой молекулярной массой (менее чем приблизительно 10 остатков); белки, сывороточный альбумин, желатин, или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металла (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как ТВИН™, ПЛЮРОНИК™ или полиэтиленгликоль (PEG). Например, лиофилизированные составы анти-ErbB2 антитела описаны в WO 97/04801, содержание которой приведено в настоящем документе в качестве ссылки в полном объеме.
Активные фармацевтические ингредиенты можно заключать также в микрокапсулы, полученные, например, способами коацервации или межповерхностной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметацилатные) микрокапсулы, соответственно, в коллоидные системы для доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получать препараты с замедленным высвобождением. Пригодные примеры препаратов с замедленным высвобождением включают содержащие AZC полупроницаемые матрицы из твердых гидрофобных полимеров, где матрицы находятся в виде изделий с заданной формой, например, пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат), или поливиниловый спирт)), полилактиды (US 3773919), сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, неразлагаемый ацетат этиленвинила, разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOT™ (пригодные для инъекции микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту.
Подлежащие использованию для введения in vivo составы должны быть стерильными, что легко осуществить фильтрацией через стерильные мембраны для фильтрации.
Составы включают составы, пригодные для вышеуказанных способов введения. Составы можно для удобства предоставлять в единичной дозированной форме, и их можно получать любыми способами, хорошо известными в данной области фармацевтики. Способы и составы в основном можно найти в Remington's Pharmaceutical Sciences (Mack Publishing Co., Easton, PA). Такие способы включают стадию объединения активного ингредиента с носителем, представляющим собой один или несколько вспомогательных ингредиентов. Как правило, составы получают посредством однородного и тесного объединения активного ингредиента с жидкими носителями или тонкоизмельченными твердыми носителями, или и с теми, и с другими, и затем, при необходимости, формования продукта.
Водные суспензии по изобретению содержат активные вещества в смеси с наполнителями, пригодными для изготовления водных суспензий. Такие наполнители включают суспендирующее вещество, такое как карбоксиметилцеллюлоза натрия, кроскармеллоза, повидон, метилцеллюлоза, гидроксипропилметилцеллюлоза, альгинат натрия, поливинилпирролидон, трагакантовая камедь и гуммиарабик, и диспергирующие или увлажняющие средства, такие как природный фосфатид (например, лецитин), продукт конденсации алкиленоксида с жирной кислотой (например, полиоксиэтиленстеарат), продукт конденсации этиленоксида с алифатическим спиртом с длинными цепями (например, гептадекаэтиленоксицетанол), продукт конденсации этиленоксида с неполным сложным эфиром, полученным из жирной кислоты и ангидрид гексита (например, моноолеат полиоксиэтиленсорбитана). Водная суспензия может содержать также один или несколько консервантов, таких как этил- или н-пропил-п-гидроксибензоат, одно или несколько окрашивающих средств, одно или несколько придающих вкус средств и один или несколько подсластителей, таких как сахароза или сахарин.
Фармацевтические композиции AZC могут присутствовать в форме стерильного пригодного для инъекции препарата, такого как стерильная пригодная для инъекции водная или масляная суспензия. Эту суспензию можно составлять, как известно в данной области, с использованием тех пригодных диспергирующих или увлажняющих средств и суспендирующих средств, которые упомянуты выше. Стерильный пригодный для инъекции препарат может представлять собой также стерильные пригодные для инъекции раствор или суспензию в нетоксичном пригодном для парентерального введения разбавителе или растворителе, например, раствор в 1,3-бутандиоле, или может представлять собой лиофилизированный порошок. Среди пригодных носителей и растворителей, которые можно использовать, присутствуют вода, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, стерильные жирные масла являются общепринятыми в качестве растворителя или суспендирующего вещества. Для этой цели можно применять любое легкое жирное масло, включая синтетические моно- или диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, можно подобным образом использовать для получения пригодных для инъекции препаратов.
Количество активного ингредиента, которое можно объединять с материалом носителя для получения единичной лекарственной формы, меняется в зависимости от обрабатываемого хозяина и конкретного способа введения. Например, водный раствор, предназначенный для внутривенной инфузии, может содержать приблизительно от 3 до 500 мкг активного ингредиента на миллилитр раствора, чтобы могла происходить инфузия подходящего объема со скоростью приблизительно 30 мл/ч.
Составы, пригодные для парентерального введения, содержат водные и неводные стерильные растворы для инъекции, которые могут содержать анти-окислители, буферы, бактериостатические средства и растворенные вещества, делающие состав изотоническим с кровью предполагаемого реципиента; и водные и неводные стерильные суспензии, которые могут включать суспендирующие средства и загустители.
Хотя пероральное введение белковых лекарственных средств нежелательно из-за гидролиза или денатурации в пищеварительном тракте, составы AZC, пригодные для перорального введения, можно получать в форме единичных лекарственных форм, таких как капсулы, саше или таблетки, каждая из которых содержит предопределенное количество AZC.
Составы можно упаковывать в контейнеры для единичных доз или множественных доз, например, герметично закрытые ампулы и флаконы, и можно хранить в высушенном замораживанием (лиофилизированном) состоянии, требующем только добавления стерильного жидкого носителя, например, воды, для инъекции, непосредственно перед использованием. Приготовленные для немедленного использования растворы и суспензии для инъекций получают из стерильных порошков, гранул и таблеток описанного ранее вида. Предпочтительными единичными дозированными составами являются составы, содержащие ежесуточную дозу активного ингредиента или единичную часть ежесуточной дозы, как перечислено в настоящем документе выше, или ее подходящую часть.
Изобретение, кроме того, относится к ветеринарным композициям, содержащим по меньшей мере один активный ингредиент, как определено выше, вместе с ветеринарным носителем, соответственно. Ветеринарные носители представляют собой материалы, пригодные для цели введения композиции, и могут представлять собой твердые, жидкие или газообразные материалы, которые в ином случае являются инертными или приемлемыми в области ветеринарии и являются совместимыми с активным ингредиентом. Эти ветеринарные композиции можно вводить парентерально, перорально или любым другим желательным способом.
СПОСОБЫ ПОЛУЧЕНИЯ ИЗОБРАЖЕНИЙ МЕЧЕНОГО АНТИТЕЛА
В другом варианте осуществления изобретения сконструированные антитела с цистеиновыми заменами могут являться меченными через тиол цистеина радиоактивными изотопами, флуоресцентными красителями, превращаемыми под действием биолюминесценции субстратными группами, превращаемыми под действием хемилюминесценции субстратными группами, ферментами и другими детектируемыми метками для экспериментов по получению изображений для диагностических, фармакодинамических и терапевтических применений. Как правило, меченое сконструированное антитело с цистеиновыми заменами, т.е. «биомаркер» или «зонд», вводят посредством инъекции, перфузии или перорального приема внутрь в живой организм, например, человеку, грызуну или другому мелкому животному, в перфузируемый орган или образец ткани. Распределение зонда детектируют с течением времени и представляют в виде изображения.
ПРИМЕРЫ
Препараты растворителей и химикатов закупали в Aldrich (Milwaukee, WI), если не указано иначе. Следующие системы для обращеннофазовой HPLC использовали для анализа и очистки продуктов. Система A: Phenomenex BioSep-SEC-S 3000 (300×4,60 мм, 5 мкм) 50 мМ PBS 0,5 мл/мин, оборудованная детектором УФ-поглощения и радиоактивности (PMT); Система B Altima C-18 (100×22,0 мм, 5 мкм) 0,05% TFA+10-50% ацетонитрил, 0-30 мин, 24 мл/мин, оборудованная УФ-детектором. Масс-спектрометрический анализ низкомолекулярных продуктов проводили в системе PE Sciex API 150EX LCMS, оборудованной колонкой Onyx Monolithic C18. LCMS анализ белков проводили на тройном квадрупольном масс-спектрометре TSQ Quantum с расширенным диапазоном массовых чисел (Thermo Electron, Thermo Fisher Scientific Inc., USA). Образцы белка для анализа LCMS восстанавливали обработкой 20 мМ дитиотреитолом (DTT) при 37°C в течение 1 часа для разделения тяжелых и легких цепей. Образцы подвергали хроматографии на микроколонке PRLP-S 1000 A (50×2,1 мм, Polymer Laboratories, Varian Inc., USA), нагретой до 75°C. Использовали линейный градиент от 30-40% B (растворитель A, 0,05% TFA в воде; растворитель B, 0,04% TFA в ацетонитриле), и элюат напрямую ионизировали с использованием источника электрораспыления. Данные собирали в системе данных Xcalibur и деконволюцию проводили с использованием программного обеспечения ProMass (Novatia, Monmouth Junction, NJ). Спектры ЯМР регистрировали на спектрометре Bruker Avance II 400 при 298K, и регистрировали химические сдвиги относительно TMS. Концентрации белка измеряли при 280 нм с использованием Eppendorf BioPhotometer (Westbury, NY). 89Zr получали из Memorial Sloan-Kettering Cancer Center (New York, NY) в форме оксалата 89Zr(IV) в 1M растворе щавелевой кислоты с удельной активностью 470-1195 Ки/ммоль (Holland JP, et al (2009) «Standardized methods for the production of high specific-activity zirconium-89» Nucl Med Biol. 36:729-39). Гетеробифункциональный линкер сукцинимидил-4-[N-малеимидометил]циклогексан-1-карбоксилат (SMCC) закупали в Pierce (Rockford, IL) и иодацетат N-гидроксисукцинимидила получали из Indofine Chemical Company (Hillsborough, NJ). Колонки NAP-10 получали от GE Healthcare, USA и фильтры для ультрацентрифугирования Amicon Ultra-4 (10000 MWCO) от Millipore (Billerica, MA). Df-Bz-SCN закупали в Macrocyclics (Dallas, TX).
Пример 1 Получение биотинилированного ТиоFab-фага
Проводили реакцию ТиоFab-фага (5×1012 фаговых частиц) с 150-кратным избытком биотин-PEO-малеимида ((+)-биотинил-3-малеимидопропионамидил-3,6-диоксаоктандиамин, Oda et al (2001) Nature Biotechnology 19:379-382, Pierce Biotechnology, Inc.) в течение 3 часов при комнатной температуре. Избыток биотин-PEO-малеимида удаляли из биотин-конъюгированного фага повторяющимися преципитациями PEG (3-4 раза). Можно использовать другие коммерчески доступные реагенты для биотинилирования с электрофильными группами, реакционноспособными по отношению к тиоловым группам цистеина, включая биотин-BMCC, PEO-иодацетил-биотин, иодацетил-LC-биотин и биотин-HPDP (Pierce Biotechnology, Inc.), и N α-(3-малеимидилпропионил)биоцитин (MPB, Molecular Probes, Eugene, OR). Другие коммерческие источники биотинилированных, бифункциональных и мультифункциональных линкерных реагентов включают Molecular Probes, Eugene, OR, и Sigma, St. Louis, MO.

Пример 2 Анализ PHESELECTOR
Бычьим сывороточным альбумином (BSA), внеклеточным доменом erbB2 (HER2) и стрептавидином (100 мкл при 2 мкг/мл) по отдельности покрывали 96-луночные планшеты Maxisorp. После блокирования с помощью 0,5% Твин-20 (в PBS), биотинилированный и небиотинилированный hu4D5Fabv8-ТиоFab-фаг (2×1010 фаговых частиц) инкубировали в течение 1 часа при комнатной температуре с последующей инкубацией с меченным пероксидазой хрена (HRP) вторичным антителом (антитело против белка оболочки фага M13, белка pVIII). Фигура 8 иллюстрирует анализ PHESELECTOR посредством схематического представления, изображающего связывание Fab или ТиоFab с HER2 (вверху) и биотинилированного ТиоFab со стрептавидином (внизу).
Проводили стандартную реакцию с HRP и измеряли оптическую плотность при 450 нм. Реакционную способность тиола измеряли, рассчитывая соотношение OD450 для стрептавидина/OD450 для HER2. Значение реакционной способности тиола 1 указывает на полное биотинилирование тиола цистеина. В случае измерений связывания белка Fab, использовали hu4D5Fabv8 (2-20 нг) с последующей инкубацией с меченными HRP поликлональными анти-Fab антителами козы.
Пример 3a Экспрессия и очистка ТиоFab
ТиоFab экспрессировали при индукции в 34B8, несупрессорном штамме E. coli (Baca et al (1997) Journal Biological Chemistry 272(16): 10678-84). Собранный осадок клеток ресуспендировали в PBS (фосфатно-солевом буфере), общий лизис клеток проводили пропусканием через микрофлюидайзер, и ТиоFab очищали аффинной хроматографией на SEPHAROSE™ с белком G (Amersham).
ТиоFab L-V15C, L-V110C, H-A88C и H-A121C экспрессировали и очищали хроматографией на колонке с SEPHAROSE™ с белком G. Олигомерный-Fab присутствовал в фракциях 26-30, и большая часть мономерной формы находилась в фракциях 31-34. Фракции, состоящие из мономерной формы, объединяли и анализировали посредством SDS-PAGE вместе с hu4D5Fabv8 дикого типа, и анализировали в геле для SDS-PAGE в восстанавливающих (с DTT или BME) и невосстанавливающих (без DTT или BME) условиях. Фракции после гель-фильтрации A121C-ТиоFab анализировали невосстанавливающим SDS-PAGE.
ТиоFab конъюгировали с биотин-PEO-малеимидом, как описано выше, и биотинилированные-ТиоFab дополнительно очищали гель-фильтрационной хроматографией на Superdex-200™ (Amersham), отсекающей свободный биотин-PEO-малеимид и олигомерную фракцию ТиоFab. Каждый из hu4D5Fabv8 дикого типа и hu4D5Fabv8 A121C-ТиоFab (в количестве 0,5 мг) по отдельности инкубировали со 100-кратным молярным избытком биотин-PEO-малеимида в течение 3 часов при комнатной температуре и наносили на колонку для гель-фильтрации Superdex-200 для отделения свободного биотина, так же как олигомерных Fab, от мономерной формы.
Пример 3b Анализ ТиоFab
Фрагменты после ферментативного расщепления биотинилированного hu4D5Fabv8 (A121C) ТиоFab и hu4D5Fabv8 дикого типа анализировали масс-спектрометрическим анализом жидкостной хроматографией с ионизацией электрораспылением (LS-ESI-MS). Различие между исходной массой 48294,5 биотинилированного hu4D5Fabv8 (A121C) и исходной массой 47737,0 hu4D5Fabv8 дикого типа составляло 557,5 единиц массы. Этот фрагмент указывает на присутствие одной биотин-PEO-малеимидной группы (C23H36N5O7S2). В таблице 4 показано распределение значений фрагментации, подтверждающее последовательность.
| Таблица 4 LC-ESI-Масс-спектрометрический анализ биотинилированного hu4D5Fabv8 ТиоFab A121C после расщепления трипсином |
||
| Аминокислота | Фрагмент b | Фрагмент y |
| A (Аланин) | 72 | |
| M (Метионин) | 203 | 2505 |
| D (Аспарагиновая кислота) | 318 | 2374 |
| Y (Тирозин) | 481 | 2259 |
| W (Триптофан) | 667 | 2096 |
| G (Глицин) | 724 | 1910 |
| Q (Глутамин) | 852 | 1853 |
| G (Глицин) | 909 | 1725 |
| T (Треонин) | 1010 | 1668 |
| L (Лейцин) | 1123 | 1567 |
| V (Валин) | 1222 | 1454 |
| T (Треонин) | 1323 | 1355 |
| V (Валин) | 1422 | 1254 |
| S (Серин) | 1509 | 1155 |
| S (Серин) | 1596 | 1068 |
| С (Цистеин)+биотин | 2242 | 981 |
| S (Серин) | 2329 | 335 |
| T (Треонин) | 2430 | 248 |
| К (Лизин) | 175 |
До и после гель-фильтрации на Superdex-200 проводили анализы SDS-PAGE в гелях, с восстановлением и без восстановления посредством DTT или BME, биотинилированного ABP-hu4D5Fabv8-A121C, биотинилированного ABP-hu4D5Fabv8-V110C, биотинилированного ABP-hu4D5Fabv8-(V110C-A88C) с двойным Cys и биотинилированного ABP-hu4D5Fabv8-(V110C-A121C) с двойным Cys.
Масс-спектрометрический анализ (MS/MS) hu4D5Fabv8-(V110C)-BMPEO-DM1 (после очистки гель-фильтрацией на Superdex-200): Fab+1 51607,5, Fab 50515,5. Эти данные показывают 91,2% конъюгацию. Анализ MS/MS hu4D5Fabv8-(V110C)-BMPEO-DM1 (восстановленного): LC 23447,2, LC+1 24537,3, HC (Fab) 27072,5. Эти данные показывают, что вся конъюгация DM1 произошла на легкой цепи Fab.
Пример 11 Восстановление/окисление ТиоMab для конъюгации
Полноразмерные сконструированные моноклональные антитела с цистеиновыми заменами (ТиоMab), экспрессированные в клетках CHO, восстанавливали с помощью приблизительно 50-кратного избытка TCEP (трис(2-карбоксиэтил)фосфингидрохлорида; Getz et al (1999) Anal. Biochem. Vol 273:73-80; Soltec Ventures, Beverly, MA) в течение 3 часов при 37°C. Восстановленный ТиоMab (Фигура 15) разводили и наносили на колонку HiTrap S в 10 мМ ацетате натрия, pH 5, и элюировали с помощью PBS, содержащего 0,3 M хлорид натрия. Элюированный восстановленный ТиоMab обрабатывали 200 нМ водным сульфатом меди (CuSO4) при комнатной температуре, в течение ночи. Окисление окружающим воздухом также являлось эффективным.
Пример 13 N-[4-(N-малеимидометил)циклогексан-1-карбоксил]десферриоксамин (Df-Chx-Mal)

4-[N-малеимидометил]циклогексан-1-карбоксилат (SMCC, 40 мг, 0,12 ммоль), мезилат десферриоксамина (78 мг, 0,12 ммоль) и N,N-диизопропилэтиламин (22 мкл, 0,13 ммоль) растворяли в смеси DMF (2,0 мл) и 0,2 мл воды (фигура 21). Полученный мутный раствор перемешивали при комнатной температуре в течение 30 мин. Добавляли воду (8 мл), и преципитированный продукт выделяли посредством фильтрации, промывали водой и высушивали под пониженным давлением с получением 42 мг (45%) N-[4-(N-малеимидометил)циклогексан-1-карбоксил]десферриоксамина (Df-Chx-Mal) в виде белого твердого вещества (фигура 20, внизу).
1H ЯМР (400 МГц, d6-DMSO) δ 0,87-0,90 (м, 2H), 1,20-1,26 (м, 8Н), 1,35-1,41 (м, 6Н), 1,45-1,55 (м, 8H), 1,60-1,70 (м, 4H), 1,97 (с, 3H, ацетил), 2,26-2,29 (м, 4H), 2,56-2,60 (м, 4H), 2,95-3,05 (м, 6H), 3,24 (д, J=7,0 Гц, 2H), 3,44-3,48 (м, 6H), 7,00 (с, 2H, малеимид), 7,62 (т, J=5,4 Гц, 1H, амид), 7,75 (м, 2H, амид), 9,59 (с, 2H, гидроксил), 9,64 (с, 1H, гидроксил). MS ESI (m/z): [M+H]+ рассчитано для C37H62N7O11 780,44; обнаружено 780.6.
Пример 14 N-бромацетилдесферриоксамин (Df-Bac)

Раствор бромацетилбромида (27 мкл, 0,30 ммоль) в DMF (1 мл) добавляли по каплям за 5 минут в охлажденную (0°C) смесь мезилата десферриоксамина (200 мг, 0,30 ммоль) и N,N-диизопропилэтиламина (106 мкл, 0,60 ммоль) в DMF (5 мл), после чего реакционную смесь перемешивали при 0°C в течение 4 часов (фигура 21). Добавляли воду (10 мл), и продукт выделяли с использованием HPLC (Система B, время удержания 7,5 мин) с получением 29 мг (14%) N-бромацетилдесферриоксамина (Df-Bac) в виде белого твердого вещества (фигура 20, внизу).
1H ЯМР (400 МГц, d6-DMSO) δ 1,18-1,26 (м, 6H), 1,35-1,42 (м, 6H), 1,45-1,55 (м, 6H), 1,97 (с, 3Н, ацетил), 2,24-2,30 (м, 4H), 2,54-2,59 (м, 4Н), 2,96-3,07 (м, 6Н), 3,44-3,47 (м, 6Н), 3,82 (c, 2Н, бромацетил), 7,74 (м, 2H, амид), 8,21 (т, 1H, амид), 9,59 (с, 2H, гидроксил), 9,63 (с, 1H, гидроксил). MS ESI (m/z): [M+H]+ рассчитано для C27H50BrN6O9 681,27, 683,27; обнаружено 681,1, 683,0.
Пример 15 N-иодацетилдесферриоксамин (Df-Iac)

Мезилат десферриоксамина (200 мг, 0,30 ммоль) и N,N-диизопропилэтиламин (53 мкл, 0,30 ммоль) смешивали в DMF (4 мл) и воде (0,4 мл). Добавляли иодацетат N-гидроксисукцинимидила (93 мг, 0,33 ммоль) и полученную смесь перемешивали при комнатной температуре в течение 1 часа (фигура 21). Добавляли воду (8 мл) и преципитированный продукт отделяли, промывали водой и высушивали под пониженным давлением с получением 115 мг (53%) N-иодацетилдесферриоксамина (Df-Iac) в виде белого твердого вещества (фигура 20, внизу).
1H ЯМР (400 МГц, d6-DMSO) δ 1,20-1,25 (м, 6H), 1,35-1,42 (м, 6H), 1,47-1,54 (м, 6H), 1,97 (с, 3H, ацетил), 2,25-2,29 (м, 4H), 2,56-2,59 (м, 4Н), 2,98-3,03 (м, 6Н), 3,45 (м, 6Н), 3,61 (с, 2Н, иодацетил), 7,75 (м, 2H, амид), 8,17 (т, 1H, амид), 9,57 (с, 2H, гидроксил), 9,61 (с, 1H, гидроксил). MS ESI (m/z): [M+H]+ рассчитано для C27H50IN6O9 729,26; обнаружено 729,1.
Пример 16 Тио-трастузумаб
Конструирование, экспрессия и очистка ТИОMAB с заменой Cys вместо Ala114 (нумерация Kabat) в тяжелой цепи описаны ранее (Junutula JR, et al «Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index» (2008) Nat Biotechnol 26:925-32). Выделенный тио-трастузумаб подготавливали для конъюгации способом восстановления и повторного окисления для удаления дисульфидных аддуктов, связанных с Cys114. Сначала белок восстанавливали в течение 24 часов обработкой 40-кратным молярным избытком DTT и 2 мМ ЭДТА в 88 мМ Трис-буфере pH 7,5. Для удаления DTT перед повторным окислением, pH раствора тио-трастузумаба доводили до 5 добавлением 10 мМ буфера сукцината натрия. Затем раствор наносили на ионообменную колонку (HiTrap SP FF, GE Healthcare), которую стерилизовали и уравновешивали с помощью 10 мМ буфера сукцината натрия pH 5. Колонку промывали 10 мМ буфером сукцинатом натрия (10 мл), и затем тио-трастузумаб элюировали 3 мл буфера 50 мМ Трис, 150 мМ NaCl с pH 7,5. Повторное окисление тио-трастузумаба осуществляли обработкой 25-кратным молярным избытком дегидроаскорбиновой кислоты (100 мМ в N,N-диметилацетамиде (DMA)) в буфере 75 мМ Трис, 150 мМ NaCl pH 7,5 при 25°C в течение 3,5 часов. После повторного окисления тио-трастузумаб конъюгировали с десферриоксамином без дополнительной очистки. MS ESI (m/z): обнаружено для легкой цепи 23440,0, для тяжелой цепи 50627,3.
Пример 17 Df-Chx-Mal-тио-трастузумаб

2 мМ исходный раствор бифункционального хелатирующего средства получали растворением Df-Chx-Mal (1,5 мг, 2 мкмоль) в 1:1 смеси (1 мл) DMF и DMA посредством нагревания при 44°C в течение 30 мин, затем исходный раствор разделяли на аликвоты и хранили при -80°C (фигура 21). Затем аликвоту исходного раствора (220 мкл, 0,440 мкмоль) добавляли к раствору тио-трастузумаба (7,5 мг, 52 нмоль) в буфере 50 мМ Трис, 150 мМ NaCl pH 7,5 (1,5 мл) и инкубировали при комнатной температуре в течение 1 часа. Затем раствор подвергали замене буфера на фильтре Amicon Ultra-4 на буфер 0,25 M ацетат натрия с получением 1 мл раствора конъюгата Df-Chx-Mal-тио-трастузумаба в концентрации 6 мг/мл. MS ESI (m/z): обнаружено для легкой цепи 23440,2, для тяжелой цепи 51407,3 (фигура 23, D).
Пример 18 Df-Ac-тио-трастузумаб с использованием Df-Bac

12 мМ исходный раствор бифункционального хелатирующего средства получали растворением Df-Bac (8 мг, 12 мкмоль) в 1 мл DMA (фигура 21). Затем исходный раствор разделяли на аликвоты и хранили при -80°C. Повторно окисленный тио-трастузумаб подвергали замене буфера на фильтре Amicon Ultra-4 на буфер 0,05 M борат натрия pH 9. Аликвоту исходного раствора Df-Bac (35 мкл, 0,410 мкмоль) добавляли к раствору тио-трастузумаба (4,9 мг, 34 нмоль) в буфере 0,05 M борате натрия pH 9 (1 мл) и инкубировали при комнатной температуре в течение 5 часов. Реакционную смесь наносили на колонку NAP-10, и Df-Ac-тио-трастузумаб элюировали с помощью 1,5 мл буфера 0,25 M ацетата натрия с получением продукта в концентрации 3,2 мг/мл. MS ESI (m/z): обнаружено для легкой цепи 23440,1, для тяжелой цепи 51228,1 (фигура 23, B).
Пример 19 Df-Ac-тио-трастузумаб с использованием Df-Iac

11 мМ исходный раствор бифункционального хелатирующего средства получали растворением Df-Iac (8 мг, 11 мкмоль) в DMSO (1 мл), затем исходный раствор разделяли на аликвоты и хранили при -80°C (фигура 21). pH раствора тио-трастузумаба (3,2 мл) доводили до 9 добавлением 0,5 мл 0,1 M карбоната натрия. Затем аликвоту исходного раствора (110 мкл, 1,20 мкмоль) добавляли к раствору тио-трастузумаба (16 мг, 110 нмоль) в буфере 50 мМ Трис, 150 мМ NaCl, 0,0125 M карбонат натрия с pH 9 (4 мл) и инкубировали при комнатной температуре в течение 2 часов. Затем раствор подвергали замене буфера на фильтре Amicon Ultra-4 на буфер 0,25 M ацетат натрия буфер с получением 1 мл раствора конъюгата Df-Ac-тио-трастузумаба в концентрации 8 мг/мл. MS ESI (m/z): обнаружено для легкой цепи 23440,1, для тяжелой цепи 51228,3 (фигура 23, C).
Пример 20 Общий способ получения хелатов 89Zr
Раствор оксалата 89Zr(IV) (2-4 мКи, 100 мкл) в 1 M щавелевой кислоте смешивали с 2 M раствором Na2CО3 (45 мкл) и инкубировали при комнатной температуре в течение 3 мин, после чего добавляли буфер 0,5 M HEPES (0,15 мл) (фигура 22). Конъюгат Df-тио-трастузумаба (1 мг, 7 нмоль) разводили 0,25 M ацетатом натрия/0,5% гентизиновой кислотой до конечного объема 0,356 мл и добавляли к раствору 89Zr. Наконец, добавляли вторую часть буфера HEPES (0,350 мл), получая общий объем 1 мл. Смесь инкубировали при комнатной температуре в течение 1 часа. Для удаления свободного 89Zr белок с радиоактивной меткой очищали с использованием обессоливающей колонки NAP-10. Колонку NAP-10 уравновешивали с помощью 20 мл 0,25 M ацетата натрия/0,5% гентизиновой кислоты. Реакционную смесь наносили на колонку NAP-10, и 89Zr-Df-тио-трастузумаб элюировали с помощью 1,5 мл буфера 0,25 M ацетата натрия/0,5% гентизиновой кислоты (1,5 мл). При необходимости 89Zr-Df-тио-трастузумаб концентрировали с использованием фильтра Amicon Ultra-4 до желательного объема. Продукт анализировали посредством SEC HPLC (Система A).
Пример 21 Стабильность в сыворотке in vitro
Раствор конъюгата 89Zr-Df-тио-трастузумаба 0,5-1,5 мКи (1 мг) в буфере 0,25 M ацетате натрия/0,5% гентизиновой кислоте (0,1 мл) добавляли к свежей сыворотке мыши (0,9 мл) и инкубировали при 37°C в течение 0-96 часов. Образцы (20 мкл) раствора сыворотки анализировали с использованием SEC HPLC (Система A), результаты показаны на фигуре 24.
Пример 22 Модели на животных
Бежевых голых мышей XID в возрасте 6-8 недель получали из Harlan Sprague Dawley (Livermore, CA). За трое суток до инокуляции клеток, мышам имплантировали (s.c, левый бок) 0,36 мг частиц 17β-эстрадиола с замедленным высвобождением в течение 60-суток (Innovative Research of America) для поддержания уровня эстрогена в сыворотке. Мышам инокулировали в жировую прослойку молочной железы 5×106 клеток BT474M1 в 50% матригеле без содержания фенола красного. BT474M1 представляет собой субклон линии клеток опухоли молочной железы человека BT474, полученный из California Pacific Medical Center. При содержании животных и манипуляциях с ними следовали протоколам, одобренным Genentech's Institutioned Animal Care and Use Committee, аккредитованным Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC).
Пример 23 Получение изображений микроПЭТ
Мышей подвергали анестезии приблизительно 3% севофлураном до получения эффекта и инъецировали мышам i.v. через хвостовую вену приблизительно 0,1 мКи меченного радиоактивным 89Zr трастузумаба в изотоническом растворе (100-130 мкл) и возвращали их в клетку для восстановления. Получение изображений ПЭТ (фигура 25) проводили на сканере Inveon PET/CT через 1, 24, 96 и 144 часа после инъекции метки следующим образом. Животных после подвергания анестезии севофлураном располагали в положении лежа, головой вперед на площадке сканера и проводили статическое сканирование 15 или 30 мин. Температуру тела измеряли ректальным зондом и поддерживали с помощью теплого воздуха. Реконструкцию повторяющихся изображений целого организма получали с использованием алгоритма максимальной апостериорной вероятности (MAP, гиперпараметр бета (β) 0,05) и корректировали на затухание сигнала с использованием плотности тканей, полученной при КТ. Проекции получали с помощью программного обеспечения ASIPro (Siemens Preclinical Solutions) и использовали для получения количественных уровней активности в каждом интересующем органе с использованием анализа интересующих областей.
Статистический анализ: Графики на фигуре 26 получали с помощью программного обеспечения R версии 2.4.1 (R Foundation for Statistical Computing, Vienna, Austria). Статистическую значимость определяли с использованием двустороннего критерия Стьюдента или ANOVA, и значения P менее 0,05 считали значимыми; данные представлены как среднее ± s.d., если не указано иначе.
Объем настоящего изобретения не является ограниченным конкретными вариантами осуществления, описанными в примерах, которые предназначены в качестве иллюстраций некоторых аспектов изобретения, и любые варианты осуществления, являющиеся функционально эквивалентными, входят в объем этого изобретения. Действительно, различные модификации по изобретению в дополнение к показанным и описанным в настоящем документе очевидны специалистам в данной области и предназначены для включения в объем прилагаемой формулы изобретения.
Содержание всех патентов, патентных заявок и ссылок, процитированных на протяжении описания, конкретно приведено в качестве ссылки.
Claims (15)
1. Меченное цирконием сконструированное антитело с цистеиновыми заменами, содержащее сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающее формулой I:

где p представляет собой 1-4;
где (L-Z) выбран из
 ,
,
где волнистая линия обозначает присоединение к антителу; и где цирконий образует комплекс с десферриоксаминовой группой; и
где сконструированное антитело содержит последовательность в тяжелой цепи, выбранную из SEQ ID NO: 11, 12, 13 и 15:


где цистеин в SEQ ID NO: 11, 12, 13 и 15 представляет собой свободную аминокислоту цистеин, где сконструированное антитело с цистеиновыми заменами в легкой цепи содержит последовательность, выбранную из SEQ ID NO: 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 45 и 46:

где цистеин в SEQ ID NO: 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 45 и 46 представляет собой свободную аминокислоту цистеин.
где p представляет собой 1-4;
где (L-Z) выбран из
где волнистая линия обозначает присоединение к антителу; и где цирконий образует комплекс с десферриоксаминовой группой; и
где сконструированное антитело содержит последовательность в тяжелой цепи, выбранную из SEQ ID NO: 11, 12, 13 и 15:
где цистеин в SEQ ID NO: 11, 12, 13 и 15 представляет собой свободную аминокислоту цистеин, где сконструированное антитело с цистеиновыми заменами в легкой цепи содержит последовательность, выбранную из SEQ ID NO: 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 45 и 46:
где цистеин в SEQ ID NO: 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 45 и 46 представляет собой свободную аминокислоту цистеин.
2. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где p равно 2.
3. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами получают способом, включающим:
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновыми заменами;
(ii) экспрессию сконструированного антитела с цистеиновыми заменами; и
(iii) выделение и очистку сконструированного антитела с цистеиновыми заменами.
(i) мутагенез последовательности нуклеиновой кислоты, кодирующей сконструированное антитело с цистеиновыми заменами;
(ii) экспрессию сконструированного антитела с цистеиновыми заменами; и
(iii) выделение и очистку сконструированного антитела с цистеиновыми заменами.
4. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами выбрано из моноклонального антитела, биспецифического антитела, химерного антитела, человеческого антитела, гуманизированного антитела и Fab-фрагмента.
5. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами представляет собой А121С тио-трастузумаб.
6. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами получают способом, включающим замену одного или нескольких аминокислотных остатков исходного антитела на одну или несколько свободных цистеиновых аминокислот, где исходное антитело избирательно связывается с антигеном и сконструированное антитело с цистеиновыми заменами избирательно связывается с тем же антигеном, что и исходное антитело.
7. Меченное цирконием сконструированное антитело с цистеиновыми заменами по п. 1, где сконструированное антитело с цистеиновыми заменами или исходное антитело связывается с одним или несколькими из рецепторов (1)-(51):
(1) BMPR1B (рецептор костного морфогенного белка типа IB);
(2) Е16 (LAT1, SLC7A5);
(3) STEAP1 (эпителиальный антиген простаты, содержащий шесть трансмембранных доменов);
(4) 0772Р (СА125, MUC16);
(5) MPF (MPF, MSLN, SMR, фактор активации мегакариоцитов, мезотелин);
(6) Napi3b (NAPI-3B, NPTIIb, SLC34A2, семейство переносчиков растворенных веществ 34 (фосфат натрия), член 2, натрий-зависимый переносчик фосфата 3b, типа II);
(7) Sema 5b (FLJ10372, KIAA1445, Mm. 42015, SEMA5B, SEMAG, Семафорин 5b Hlog, домен sema, семь повторов тромбоспондина (типа 1 и подобный типу 1), трансмембранный домен (ТМ) и короткий цитоплазматический домен, (семафорин) 5В);
(8) PSCA hlg (2700050C12Rik, C530008O16Rik, кДНК RIKEN 2700050С12, кДНК RIKEN гена 2700050С12);
(9) ETBR (рецептор эндотелина типа В);
(10) MSG783 (RNF124, гипотетический белок FLJ20315);
(11) STEAP2 (HGNC_8639, IPCA-1, PCANAP1, STAMP1, STEAP2, STMP, ассоциированный с раком предстательной железы ген 1, ассоциированный с раком предстательной железы белок 1, эпителиальный антиген простаты 2, содержащий шесть трансмембранных доменов, белок простаты, содержащий шесть трансмембранных доменов);
(12) TrpM4 (BR22450, FLJ20041, TRPM4, TRPM4B, катионный канал транзиторного рецепторного потенциала, подсемейство М, член 4);
(13) CRIPTO (CR, CR1, CRGF, CRIPTO, TDGF1, полученный из тератокарциномы фактор роста);
(14) CD21 (CR2 (рецептор комплемента 2) или C3DR (рецептор C3d/вируса Эпштейна-Барр) или Hs. 73792);
(15) CD79b (CD79B, CD79β, IGb (иммуноглобулин-ассоциированная молекула бета), В29);
(16) FcRH2 (IFGP4, IRTA4, SPAP1A (содержащий SH2 домен фосфатазный якорный белок 1a), SPAP1B, SPAP1C);
(17) HER2;
(18) NCA;
(19) MDP;
(20) IL20Rα;
(21) Бревикан;
(22) EphB2R;
(23) ASLG659;
(24) PSCA;
(25) GEDA;
(26) BAFF-R (рецептор фактора активации В-клеток, BLyS рецептор 3, BR3;
(27) CD22 (изоформа B-клеточного рецептора CD22-B);
(28) CD79a (CD79A, CD79α, иммуноглобулин-ассоциированная молекула альфа, специфический для В-клеток белок, ковалентно взаимодействующий с Ig бета (CD79B) и формирующий комплекс на поверхности с молекулами IgM, передает сигнал, вовлеченный в дифференцировку В-клеток);
(29) CXCR5 (рецептор лимфомы Беркитта 1, связанный с G белком рецептор, активируемый хемокином CXCL13, функционирует в миграции лимфоцитов и гуморальной защите, играет роль в инфекции HIV-2 и, возможно, развитии СПИД, лимфомы, миеломы и лейкоза);
(30) HLA-DOB (бета-субъединица молекулы МНС класса II (1а антиген), связывающая пептиды и представляющая их CD4+ Т-лимфоцитам);
(31) Р2Х5 (управляемый лигандом пуринергического рецептора Р2Х ионный канал 5, ионный канал, управляемый внеклеточным АТР, может быть вовлечен в передачу возбуждения по синапсам и нейрогенез, недостаточность может вносить вклад в патофизиологию идиопатической нестабильности детрузора);
(32) CD72 (антиген дифференцировки В-клеток CD72, Lyb-2);
(33) LY64 (лимфоцитарный антиген 64 (RP105), мембранный белок типа I семейства богатых лейцином повторов (LRR), регулирует активацию и апоптоз В-клеток, потеря функции ассоциирована с увеличенной активностью заболевания у пациентов с системной красной волчанкой);
(34) FcRH1 (подобный Fc-рецептору белок 1, предположительный рецептор для Fc-домена иммуноглобулинов, содержащий Ig-подобный домен типа С2 и домен ITAM, может играть роль в дифференцировке В-лимфоцитов);
(35) IRTA2 (ассоциированный с транслокацией рецептор 2 суперсемейства иммуноглобулинов, предположительный иммунорецептор с возможной ролью в развитии В-клеток и лимфомагенезе; нарушение регуляции гена посредством транслокации происходит при некоторых злокачественных новообразованиях В-клеток);
(36) TENB2 (предположительный трансмембранный протеогликан, родственный семейству факторов роста EGF/херегулина и фоллистатину);
(37) PMEL17 (гомолог silver; SILV; D12S53E; PMEL17; (SI); (SIL); МЕ20; gp100);
(38) TMEFF1 (трансмембранный белок с EGF-подобным доменом и двумя фоллистатин-подобными доменами 1; Томорегулин-1; Н7365; C9orf2; C90RF2; U19878; Х83961;
(39) GDNF-Ra1 (рецептор альфа 1 семейства GDNF; GFRA1; GDNFR; GDNFRA; RETL1; TRNR1; RET1L; GDNFR-alpha1; GFR-ALPHA-1; U95847; ВС014962);
(40) Ly6E (комплекс лимфоцитарного антигена 6, локус Е; Ly67, RIG-E, SCA-2, TSA-1);
(41) ТМЕМ46 (гомолог shisa 2 (Xenopus laevis); SHISA2);
(42) Ly6G6D (комплекс лимфоцитарного антигена 6, локус G6D; Ly6-D, MEGT1);
(43) LGR5 (содержащий богатые лейцином повторы, связанный с белком G рецептор 5; GPR49, GPR67);
(44) RET (протоонкоген ret; MEN2A; HSCR1; MEN2B; МТС1; (РТС); CDHF12; Hs. 168114; RET51; RET-ELE1);
(45) LY6K (комплекс лимфоцитарного антигена 6, локус K; LY6K; HSJ001348; FLJ35226);
(46) GPR19 (связанный с белком G рецептор 19; Mm. 4787);
(47) GPR54 (рецептор KISS1; KISS1R; GPR54; НОТ7Т175; AXOR12);
(48) ASPHD1 (содержащий домен аспартат-бета-гидроксилазы белок 1; LOC253982);
(49) Тирозиназа (TYR; OCAIA; OCA1A; тирозиназа; SHEP3);
(50) TMEM118 (белок с «безымянным пальцем», трансмембранный белок 2; RNFT2; FLJ14627); и
(51) GPR172A (связанный с белком G рецептор 172А; GPCR41; FLJ11856; D15Ertd747e).
(1) BMPR1B (рецептор костного морфогенного белка типа IB);
(2) Е16 (LAT1, SLC7A5);
(3) STEAP1 (эпителиальный антиген простаты, содержащий шесть трансмембранных доменов);
(4) 0772Р (СА125, MUC16);
(5) MPF (MPF, MSLN, SMR, фактор активации мегакариоцитов, мезотелин);
(6) Napi3b (NAPI-3B, NPTIIb, SLC34A2, семейство переносчиков растворенных веществ 34 (фосфат натрия), член 2, натрий-зависимый переносчик фосфата 3b, типа II);
(7) Sema 5b (FLJ10372, KIAA1445, Mm. 42015, SEMA5B, SEMAG, Семафорин 5b Hlog, домен sema, семь повторов тромбоспондина (типа 1 и подобный типу 1), трансмембранный домен (ТМ) и короткий цитоплазматический домен, (семафорин) 5В);
(8) PSCA hlg (2700050C12Rik, C530008O16Rik, кДНК RIKEN 2700050С12, кДНК RIKEN гена 2700050С12);
(9) ETBR (рецептор эндотелина типа В);
(10) MSG783 (RNF124, гипотетический белок FLJ20315);
(11) STEAP2 (HGNC_8639, IPCA-1, PCANAP1, STAMP1, STEAP2, STMP, ассоциированный с раком предстательной железы ген 1, ассоциированный с раком предстательной железы белок 1, эпителиальный антиген простаты 2, содержащий шесть трансмембранных доменов, белок простаты, содержащий шесть трансмембранных доменов);
(12) TrpM4 (BR22450, FLJ20041, TRPM4, TRPM4B, катионный канал транзиторного рецепторного потенциала, подсемейство М, член 4);
(13) CRIPTO (CR, CR1, CRGF, CRIPTO, TDGF1, полученный из тератокарциномы фактор роста);
(14) CD21 (CR2 (рецептор комплемента 2) или C3DR (рецептор C3d/вируса Эпштейна-Барр) или Hs. 73792);
(15) CD79b (CD79B, CD79β, IGb (иммуноглобулин-ассоциированная молекула бета), В29);
(16) FcRH2 (IFGP4, IRTA4, SPAP1A (содержащий SH2 домен фосфатазный якорный белок 1a), SPAP1B, SPAP1C);
(17) HER2;
(18) NCA;
(19) MDP;
(20) IL20Rα;
(21) Бревикан;
(22) EphB2R;
(23) ASLG659;
(24) PSCA;
(25) GEDA;
(26) BAFF-R (рецептор фактора активации В-клеток, BLyS рецептор 3, BR3;
(27) CD22 (изоформа B-клеточного рецептора CD22-B);
(28) CD79a (CD79A, CD79α, иммуноглобулин-ассоциированная молекула альфа, специфический для В-клеток белок, ковалентно взаимодействующий с Ig бета (CD79B) и формирующий комплекс на поверхности с молекулами IgM, передает сигнал, вовлеченный в дифференцировку В-клеток);
(29) CXCR5 (рецептор лимфомы Беркитта 1, связанный с G белком рецептор, активируемый хемокином CXCL13, функционирует в миграции лимфоцитов и гуморальной защите, играет роль в инфекции HIV-2 и, возможно, развитии СПИД, лимфомы, миеломы и лейкоза);
(30) HLA-DOB (бета-субъединица молекулы МНС класса II (1а антиген), связывающая пептиды и представляющая их CD4+ Т-лимфоцитам);
(31) Р2Х5 (управляемый лигандом пуринергического рецептора Р2Х ионный канал 5, ионный канал, управляемый внеклеточным АТР, может быть вовлечен в передачу возбуждения по синапсам и нейрогенез, недостаточность может вносить вклад в патофизиологию идиопатической нестабильности детрузора);
(32) CD72 (антиген дифференцировки В-клеток CD72, Lyb-2);
(33) LY64 (лимфоцитарный антиген 64 (RP105), мембранный белок типа I семейства богатых лейцином повторов (LRR), регулирует активацию и апоптоз В-клеток, потеря функции ассоциирована с увеличенной активностью заболевания у пациентов с системной красной волчанкой);
(34) FcRH1 (подобный Fc-рецептору белок 1, предположительный рецептор для Fc-домена иммуноглобулинов, содержащий Ig-подобный домен типа С2 и домен ITAM, может играть роль в дифференцировке В-лимфоцитов);
(35) IRTA2 (ассоциированный с транслокацией рецептор 2 суперсемейства иммуноглобулинов, предположительный иммунорецептор с возможной ролью в развитии В-клеток и лимфомагенезе; нарушение регуляции гена посредством транслокации происходит при некоторых злокачественных новообразованиях В-клеток);
(36) TENB2 (предположительный трансмембранный протеогликан, родственный семейству факторов роста EGF/херегулина и фоллистатину);
(37) PMEL17 (гомолог silver; SILV; D12S53E; PMEL17; (SI); (SIL); МЕ20; gp100);
(38) TMEFF1 (трансмембранный белок с EGF-подобным доменом и двумя фоллистатин-подобными доменами 1; Томорегулин-1; Н7365; C9orf2; C90RF2; U19878; Х83961;
(39) GDNF-Ra1 (рецептор альфа 1 семейства GDNF; GFRA1; GDNFR; GDNFRA; RETL1; TRNR1; RET1L; GDNFR-alpha1; GFR-ALPHA-1; U95847; ВС014962);
(40) Ly6E (комплекс лимфоцитарного антигена 6, локус Е; Ly67, RIG-E, SCA-2, TSA-1);
(41) ТМЕМ46 (гомолог shisa 2 (Xenopus laevis); SHISA2);
(42) Ly6G6D (комплекс лимфоцитарного антигена 6, локус G6D; Ly6-D, MEGT1);
(43) LGR5 (содержащий богатые лейцином повторы, связанный с белком G рецептор 5; GPR49, GPR67);
(44) RET (протоонкоген ret; MEN2A; HSCR1; MEN2B; МТС1; (РТС); CDHF12; Hs. 168114; RET51; RET-ELE1);
(45) LY6K (комплекс лимфоцитарного антигена 6, локус K; LY6K; HSJ001348; FLJ35226);
(46) GPR19 (связанный с белком G рецептор 19; Mm. 4787);
(47) GPR54 (рецептор KISS1; KISS1R; GPR54; НОТ7Т175; AXOR12);
(48) ASPHD1 (содержащий домен аспартат-бета-гидроксилазы белок 1; LOC253982);
(49) Тирозиназа (TYR; OCAIA; OCA1A; тирозиназа; SHEP3);
(50) TMEM118 (белок с «безымянным пальцем», трансмембранный белок 2; RNFT2; FLJ14627); и
(51) GPR172A (связанный с белком G рецептор 172А; GPCR41; FLJ11856; D15Ertd747e).
8. Меченное десферриоксамином сконструированное антитело с цистеиновыми заменами, содержащее сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающее формулой II:

где L-Df выбран из:

где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4.
где L-Df выбран из:
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4.
9. Способ получения меченного десферриоксамином сконструированного антитела с цистеиновыми заменами по п. 8,
где способ включает реакцию композиции, выбранной из структур:

где R выбран из:

со сконструированным антителом с цистеиновыми заменами, имеющим одну или несколько свободных цистеиновых аминокислот, посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
где способ включает реакцию композиции, выбранной из структур:
где R выбран из:
со сконструированным антителом с цистеиновыми заменами, имеющим одну или несколько свободных цистеиновых аминокислот, посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
10. Способ получения меченного цирконием сконструированного антитела с цистеиновыми заменами, содержащего сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и комплексом циркония (Z), обладающего формулой I:

где p представляет собой 1-4;
где способ включает образование комплекса циркониевого реагента с меченным десферриоксамином сконструированным антителом с цистеиновыми заменами, содержащим сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающим формулой II:

где L-Df выбран из:

где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4;
посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
где p представляет собой 1-4;
где способ включает образование комплекса циркониевого реагента с меченным десферриоксамином сконструированным антителом с цистеиновыми заменами, содержащим сконструированное антитело с цистеиновыми заменами (Ab), конъюгированное через свободную аминокислоту цистеин с линкером (L) и группой десферриоксамина (Df), обладающим формулой II:
где L-Df выбран из:
где волнистая линия обозначает присоединение к антителу (Ab); и
p представляет собой 1-4;
посредством чего получают меченное десферриоксамином сконструированное антитело с цистеиновыми заменами.
11. Способ по п. 10, где циркониевый реагент представляет собой оксалат 89циркония.
12. Способ получения изображений, включающий:
введение меченного цирконием сконструированного антитела с цистеиновыми заменами по п. 1 животному; и
детекцию in vivo присутствия меченного цирконием сконструированного антитела с цистеиновыми заменами посредством получения изображений.
введение меченного цирконием сконструированного антитела с цистеиновыми заменами по п. 1 животному; и
детекцию in vivo присутствия меченного цирконием сконструированного антитела с цистеиновыми заменами посредством получения изображений.
13. Способ по п. 12, где Z содержит цирконий в комплексе с десферриоксамином В.
14. Способ по п. 12, где меченное цирконием сконструированное антитело с цистеиновыми заменами связывается с антигеном.
15. Способ по п. 12, где животное представляет собой модель ксенотрансплантата опухоли на мышах.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US12/612,912 US20100111856A1 (en) | 2004-09-23 | 2009-11-05 | Zirconium-radiolabeled, cysteine engineered antibody conjugates |
| US12/612,912 | 2009-11-05 | ||
| PCT/US2010/055465 WO2011056983A1 (en) | 2009-11-05 | 2010-11-04 | Zirconium-radiolabeled, cysteine engineered antibody conjugates |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| RU2012123007A RU2012123007A (ru) | 2013-12-10 |
| RU2562862C2 true RU2562862C2 (ru) | 2015-09-10 |
Family
ID=43383591
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2012123007/10A RU2562862C2 (ru) | 2009-11-05 | 2010-11-04 | Конъюгаты меченных радиоактивным цирконием сконструированных антител с цистеиновыми заменами |
Country Status (10)
| Country | Link |
|---|---|
| US (2) | US20100111856A1 (ru) |
| EP (1) | EP2496270B1 (ru) |
| JP (1) | JP5850843B2 (ru) |
| KR (2) | KR20170136652A (ru) |
| CN (1) | CN102596260B (ru) |
| BR (1) | BR112012007774A2 (ru) |
| CA (1) | CA2780216A1 (ru) |
| MX (1) | MX340674B (ru) |
| RU (1) | RU2562862C2 (ru) |
| WO (1) | WO2011056983A1 (ru) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US11779649B2 (en) | 2018-12-21 | 2023-10-10 | Novartis Ag | Antibodies to PMEL17 and conjugates thereof |
Families Citing this family (145)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6136311A (en) | 1996-05-06 | 2000-10-24 | Cornell Research Foundation, Inc. | Treatment and diagnosis of cancer |
| US9701754B1 (en) | 2002-10-23 | 2017-07-11 | City Of Hope | Covalent disulfide-linked diabodies and uses thereof |
| CA2646329C (en) | 2006-03-20 | 2018-07-03 | The Regents Of The University Of California | Engineered anti-prostate stem cell antigen (psca) antibodies for cancer targeting |
| CN101802013B (zh) | 2007-07-16 | 2014-07-02 | 健泰科生物技术公司 | 人源化抗cd79b抗体和免疫偶联物及使用方法 |
| NZ583367A (en) | 2007-07-16 | 2012-10-26 | Genentech Inc | Anti-cd79b antibodies and immunoconjugates and methods of use |
| CA2698343C (en) | 2007-09-04 | 2018-06-12 | The Regents Of The University Of California | High affinity anti-prostate stem cell antigen (psca) antibodies for cancer targeting and detection |
| CA2700410C (en) * | 2007-10-03 | 2020-10-06 | Cornell University | Treatment of proliferative disorders using antibodies to psma |
| MX2010008437A (es) | 2008-01-31 | 2010-11-25 | Genentech Inc | Anticuerpos anti-cd79b e inmunoconjugados y metodos de uso. |
| GB0803076D0 (en) | 2008-02-20 | 2008-03-26 | Univ Ghent | Mucosal Membrane Receptor and uses thereof |
| US20100009390A1 (en) * | 2008-05-09 | 2010-01-14 | The Regents Of The University Of California | Mutant antibodies with high affinity for egfr |
| US20100008978A1 (en) * | 2008-05-09 | 2010-01-14 | The Regents Of The University Of California | Nanoparticles effective for internalization into cells |
| US8999661B2 (en) * | 2008-07-30 | 2015-04-07 | Dana-Farber Cancer Institute, Inc. | Compositions for detecting cell death and methods of use thereof |
| ES2712732T3 (es) | 2009-02-17 | 2019-05-14 | Cornell Res Foundation Inc | Métodos y kits para el diagnóstico de cáncer y la predicción de valor terapéutico |
| ES2547712T3 (es) | 2009-02-27 | 2015-10-08 | Genentech, Inc. | Métodos y composiciones para el marcado de proteínas |
| MX354143B (es) | 2009-12-02 | 2018-02-14 | Imaginab Inc | Minicuerpos j591 y diacuerpos-cys para apuntar con especificidad de objetivo a un antígeno de membrana específico para próstata de humano (psma) y métodos para su uso. |
| US8877897B2 (en) | 2010-02-23 | 2014-11-04 | Genentech, Inc. | Compositions and methods for the diagnosis and treatment of tumor |
| RU2613886C2 (ru) * | 2010-06-03 | 2017-03-21 | Дженентек, Инк. | Антитела и иммуноконъюгаты, визуализируемые при помощи иммуно-позитрон-эмиссионной томографии, и способы их применения |
| SG185428A1 (en) | 2010-06-08 | 2012-12-28 | Genentech Inc | Cysteine engineered antibodies and conjugates |
| TW201302793A (zh) | 2010-09-03 | 2013-01-16 | Glaxo Group Ltd | 新穎之抗原結合蛋白 |
| JP6253987B2 (ja) | 2010-12-20 | 2017-12-27 | ジェネンテック, インコーポレイテッド | 抗メソテリン抗体及びイムノコンジュゲート |
| MX350807B (es) | 2011-11-29 | 2017-09-20 | Genentech Inc | Composiciones y metodos para el analisis de cancer de prostata. |
| AR090549A1 (es) | 2012-03-30 | 2014-11-19 | Genentech Inc | Anticuerpos anti-lgr5 e inmunoconjugados |
| BR112014027166A2 (pt) | 2012-05-01 | 2017-06-27 | Genentech Inc | anticorpo, ácido nucleico, célula hospedeira, método para produzir um anticorpo, imunoconjugado, formulação farmacêutica, método de tratamento, método de inibir proliferação e métodos de detecção. |
| MA37538B1 (fr) | 2012-05-21 | 2017-11-30 | Genentech Inc | Anticorps et immunoconjugués anti-ly6e et procédés d'utilisation |
| KR102006997B1 (ko) * | 2012-07-03 | 2019-08-02 | 한국생명공학연구원 | IgG Fc 위치선택적 결합 펩티드 및 이를 포함하는 하이브리드 분자 |
| CN107973856B (zh) | 2012-07-04 | 2021-11-23 | 弗·哈夫曼-拉罗切有限公司 | 共价连接的抗原-抗体缀合物 |
| BR112014030843A2 (pt) | 2012-07-04 | 2019-10-15 | Hoffmann La Roche | anticorpo anti-teofilina, formulação farmacêutica e uso do anticorpo |
| CA2872192A1 (en) | 2012-07-04 | 2014-01-09 | F. Hoffmann-La Roche Ag | Anti-biotin antibodies and methods of use |
| WO2014011327A1 (en) * | 2012-07-12 | 2014-01-16 | Dana-Farber Cancer Institute, Inc. | Radiolabeled probes for the non-invasive detection and imaging of cell death |
| CA2879670A1 (en) * | 2012-08-02 | 2014-02-06 | Genentech, Inc. | Anti-etbr antibodies and immunoconjugates |
| MX2015001399A (es) * | 2012-08-02 | 2015-09-07 | Genentech Inc | Anticuerpos anti-etbr e inmunoconjugados. |
| CN104797599A (zh) * | 2012-11-05 | 2015-07-22 | 全药工业株式会社 | 抗体或抗体组合物的制备方法 |
| US20140147381A1 (en) * | 2012-11-29 | 2014-05-29 | Gregory David Espenan | 89zr compounds, to include somatostatin, apparatus and products comprising such compounds, methods of making same, and methods of using same for radio imaging and/or treatment |
| EP2934602B1 (en) | 2012-12-19 | 2019-02-27 | Genentech, Inc. | Methods and compositions for radiohalogen protein labeling |
| KR102447350B1 (ko) | 2013-02-08 | 2022-09-23 | 노파르티스 아게 | 면역접합체의 제조를 위한 항체의 변형에 사용되는 특정 부위 |
| WO2014164988A1 (en) * | 2013-03-13 | 2014-10-09 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Tetrahydroxamate chelators of zirconium89 and niobium90 for use in diagnostic applications |
| MX2015010777A (es) | 2013-03-14 | 2016-04-25 | Genentech Inc | Anticuerpos e inmunoconjugados anti-b7-h4. |
| US9562099B2 (en) | 2013-03-14 | 2017-02-07 | Genentech, Inc. | Anti-B7-H4 antibodies and immunoconjugates |
| ES2699599T3 (es) * | 2013-03-15 | 2019-02-11 | Abbvie Biotherapeutics Inc | Variantes de Fc |
| EP3008212A4 (en) * | 2013-06-10 | 2017-05-24 | Millennium Pharmaceuticals, Inc. | Methods of treatment of cancer |
| CN105518027A (zh) | 2013-09-17 | 2016-04-20 | 豪夫迈·罗氏有限公司 | 使用抗lgr5抗体的方法 |
| AR098126A1 (es) | 2013-10-21 | 2016-05-04 | Genentech Inc | Anticuerpos anti-ly6e (locus e del complejo del antígeno 6 linfocítico), inmunoconjugados y métodos para usarlos |
| WO2015089344A1 (en) | 2013-12-13 | 2015-06-18 | Genentech, Inc. | Anti-cd33 antibodies and immunoconjugates |
| WO2015095124A1 (en) | 2013-12-16 | 2015-06-25 | Genentech Inc. | Peptidomimetic compounds and antibody-drug conjugates thereof |
| CR20160271A (es) | 2013-12-16 | 2016-12-02 | Genentech Inc | Compuestos peptidométicos y sus conjugados de anticuerpo-fármaco |
| MX2016007958A (es) | 2013-12-17 | 2016-08-03 | Genentech Inc | Anticuerpos anti-cd3 y metodos de uso. |
| CN105873616B (zh) | 2014-01-03 | 2020-06-05 | 豪夫迈·罗氏有限公司 | 共价连接的多肽毒素-抗体缀合物 |
| CN105873615B (zh) | 2014-01-03 | 2020-12-25 | 豪夫迈·罗氏有限公司 | 共价连接的helicar-抗helicar抗体缀合物及其用途 |
| WO2015101586A1 (en) | 2014-01-03 | 2015-07-09 | F. Hoffmann-La Roche Ag | Bispecific anti-hapten/anti-blood brain barrier receptor antibodies, complexes thereof and their use as blood brain barrier shuttles |
| BR112016015693A2 (pt) | 2014-01-24 | 2017-10-24 | Genentech Inc | método para tratar um câncer de próstata e anticorpo |
| EA201691827A1 (ru) * | 2014-03-12 | 2017-01-30 | Новартис Аг | Конкретные участки для модификации антител с целью получения иммуноконъюгатов |
| RU2016150370A (ru) | 2014-05-22 | 2018-06-26 | Дженентек, Инк. | Антитела и иммуноконъюгаты против GPC3 |
| CA2949982A1 (en) | 2014-06-11 | 2015-12-17 | Genentech, Inc. | Anti-lgr5 antibodies and uses thereof |
| JP7286267B2 (ja) | 2014-08-28 | 2023-06-05 | バイオアトラ インコーポレイテッド | 修飾t細胞に対する条件的活性型キメラ抗原受容体 |
| JP7085837B2 (ja) | 2014-09-12 | 2022-06-17 | ジェネンテック, インコーポレイテッド | 抗her2抗体及び免疫複合体 |
| PE20170670A1 (es) | 2014-09-12 | 2017-06-06 | Genentech Inc | Anticuerpos anti-cll-1 e inmunoconjugados |
| SG11201701128YA (en) | 2014-09-12 | 2017-03-30 | Genentech Inc | Cysteine engineered antibodies and conjugates |
| TW201625689A (zh) | 2014-09-12 | 2016-07-16 | 建南德克公司 | 抗-b7-h4抗體及免疫結合物 |
| LT3262071T (lt) | 2014-09-23 | 2020-06-25 | F. Hoffmann-La Roche Ag | Anti-cd79b imunokonjugatų naudojimo būdai |
| EP3207027B1 (en) * | 2014-10-16 | 2019-06-19 | The University of Melbourne | Novel imaging composition and uses thereof |
| CN107250358B (zh) * | 2014-11-05 | 2021-03-30 | 伊卢米纳剑桥有限公司 | 在样品制备和测序期间使用铁载体螯合剂减少dna损伤 |
| KR20170094216A (ko) | 2014-12-17 | 2017-08-17 | 에프. 호프만-라 로슈 아게 | 소르타제를 사용하는 효소 매개되는 폴리펩티드 접합을 위한 신규한 방법 |
| CN104645364A (zh) * | 2015-01-27 | 2015-05-27 | 南京江原安迪科正电子研究发展有限公司 | 89Zr标记地诺单抗的标记产物及其制备方法、应用 |
| EP3277314A4 (en) | 2015-04-03 | 2018-08-29 | Eureka Therapeutics, Inc. | Constructs targeting afp peptide/mhc complexes and uses thereof |
| WO2016179003A1 (en) | 2015-05-01 | 2016-11-10 | Genentech, Inc. | Masked anti-cd3 antibodies and methods of use |
| EP3307780A1 (en) | 2015-06-15 | 2018-04-18 | Genentech, Inc. | Antibodies and immunoconjugates |
| JP2018526972A (ja) | 2015-06-16 | 2018-09-20 | ジェネンテック, インコーポレイテッド | 抗cd3抗体及び使用方法 |
| EP4137158A1 (en) | 2015-08-07 | 2023-02-22 | Imaginab, Inc. | Antigen binding constructs to target molecules |
| JP6861702B2 (ja) | 2015-09-25 | 2021-04-21 | エフ.ホフマン−ラ ロシュ アーゲーF. Hoffmann−La Roche Aktiengesellschaft | ソルターゼ・コンジュゲート化ループを含む組換え免疫グロブリン重鎖およびそのコンジュゲート |
| EP3353291B1 (en) | 2015-09-25 | 2021-06-09 | F. Hoffmann-La Roche AG | Novel soluble sortase a |
| CN108138204B (zh) | 2015-09-25 | 2021-12-31 | 豪夫迈·罗氏有限公司 | 使用分选酶a生产硫酯的方法 |
| CN108026560A (zh) | 2015-09-25 | 2018-05-11 | 豪夫迈·罗氏有限公司 | 在低共熔溶剂中使用分选酶的转酰胺反应 |
| MA43345A (fr) | 2015-10-02 | 2018-08-08 | Hoffmann La Roche | Conjugués anticorps-médicaments de pyrrolobenzodiazépine et méthodes d'utilisation |
| US10758634B2 (en) | 2016-03-18 | 2020-09-01 | Wake Forest University | Compounds, compositions and associated methods using zirconium-89 in immuno-positron emission tomography |
| WO2017167712A1 (en) | 2016-03-30 | 2017-10-05 | F. Hoffmann-La Roche Ag | Improved sortase |
| KR102001821B1 (ko) | 2016-04-06 | 2019-07-19 | (주)알테오젠 | 변형항체를 포함하는 항체-약물 접합체 |
| EP3442999A2 (en) | 2016-04-15 | 2019-02-20 | Alpine Immune Sciences, Inc. | Icos ligand variant immunomodulatory proteins and uses thereof |
| SG11201808994YA (en) | 2016-04-15 | 2018-11-29 | Bioatla Llc | Anti-axl antibodies, antibody fragments and their immunoconjugates and uses thereof |
| KR20190006495A (ko) | 2016-04-15 | 2019-01-18 | 알파인 이뮨 사이언시즈, 인코포레이티드 | Cd80 변이체 면역조절 단백질 및 그의 용도 |
| ES2930255T3 (es) | 2016-05-13 | 2022-12-09 | Bioatla Inc | Anticuerpos anti-Ror2, fragmentos de anticuerpos, sus inmunoconjugados y usos de los mismos |
| JP7133477B2 (ja) | 2016-06-24 | 2022-09-08 | ジェネンテック, インコーポレイテッド | 抗ポリユビキチン多重特異性抗体 |
| US11834490B2 (en) | 2016-07-28 | 2023-12-05 | Alpine Immune Sciences, Inc. | CD112 variant immunomodulatory proteins and uses thereof |
| US11471488B2 (en) | 2016-07-28 | 2022-10-18 | Alpine Immune Sciences, Inc. | CD155 variant immunomodulatory proteins and uses thereof |
| EP3491013A1 (en) | 2016-07-28 | 2019-06-05 | Alpine Immune Sciences, Inc. | Cd155 variant immunomodulatory proteins and uses thereof |
| US20240018268A1 (en) | 2016-07-29 | 2024-01-18 | Juno Therapeutics, Inc. | Anti-idiotypic antibodies against anti-cd19 antibodies |
| TWI782930B (zh) | 2016-11-16 | 2022-11-11 | 美商再生元醫藥公司 | 抗met抗體,結合met之雙特異性抗原結合分子及其使用方法 |
| US10517966B2 (en) | 2016-11-18 | 2019-12-31 | Astellas Pharma Inc. | Anti-human MUC1 antibody Fab fragment |
| CN110023334B (zh) | 2016-11-21 | 2023-11-14 | 科雅博有限责任公司 | 抗gp73抗体和免疫偶联物 |
| AU2017367734A1 (en) | 2016-12-01 | 2019-06-20 | Regeneron Pharmaceuticals, Inc. | Radiolabeled anti-PD-l1 antibodies for immuno-PET imaging |
| KR20190104528A (ko) | 2016-12-03 | 2019-09-10 | 주노 쎄러퓨티크스 인코퍼레이티드 | Car-t 세포들 투여를 결정하는 방법 |
| US11274157B2 (en) | 2017-01-12 | 2022-03-15 | Eureka Therapeutics, Inc. | Constructs targeting histone H3 peptide/MHC complexes and uses thereof |
| US11266745B2 (en) | 2017-02-08 | 2022-03-08 | Imaginab, Inc. | Extension sequences for diabodies |
| SG11201907208XA (en) | 2017-02-10 | 2019-09-27 | Regeneron Pharma | Radiolabeled anti-lag3 antibodies for immuno-pet imaging |
| WO2018170026A2 (en) | 2017-03-16 | 2018-09-20 | Alpine Immune Sciences, Inc. | Cd80 variant immunomodulatory proteins and uses thereof |
| AU2018235835A1 (en) | 2017-03-16 | 2019-09-05 | Alpine Immune Sciences, Inc. | PD-L2 variant immunomodulatory proteins and uses thereof |
| PT3596116T (pt) | 2017-03-16 | 2023-12-04 | Alpine Immune Sciences Inc | Proteínas imunomoduladoras de pd-l1 variante e suas utilizações |
| CN110475779A (zh) | 2017-04-12 | 2019-11-19 | 豪夫迈·罗氏有限公司 | 标记含醛靶分子的方法 |
| KR20200012860A (ko) | 2017-04-26 | 2020-02-05 | 유레카 쎄라퓨틱스, 인코포레이티드 | 글리피칸 3을 특이적으로 인식하는 구축물 및 그의 용도 |
| CA3064597A1 (en) | 2017-06-02 | 2018-12-06 | Juno Therapeutics, Inc. | Articles of manufacture and methods related to toxicity associated with cell therapy |
| TWI795415B (zh) | 2017-07-07 | 2023-03-11 | 日商安斯泰來製藥股份有限公司 | 新穎的抗人類CEACAM5抗體Fab片段 |
| KR20200027522A (ko) | 2017-07-13 | 2020-03-12 | 에프. 호프만-라 로슈 아게 | Pivka에 대한 신규 결합제 및 어세이 |
| US10730944B2 (en) | 2017-07-24 | 2020-08-04 | Regeneron Pharmaceuticals, Inc. | Anti-CD8 antibodies and uses thereof |
| EP4219540A3 (en) | 2017-10-10 | 2023-12-06 | Alpine Immune Sciences, Inc. | Ctla-4 variant immunomodulatory proteins and uses thereof |
| US20200283500A1 (en) | 2017-10-18 | 2020-09-10 | Alpine Immune Sciences, Inc. | Variant icos ligand immunomodulatory proteins and related compositions and methods |
| EP3697813A1 (en) | 2017-10-20 | 2020-08-26 | H. Hoffnabb-La Roche Ag | Copy protection for antibodies |
| WO2019089858A2 (en) | 2017-11-01 | 2019-05-09 | Juno Therapeutics, Inc. | Methods of assessing or monitoring a response to a cell therapy |
| WO2019089848A1 (en) | 2017-11-01 | 2019-05-09 | Juno Therapeutics, Inc. | Methods associated with tumor burden for assessing response to a cell therapy |
| MA51210A (fr) | 2017-12-01 | 2020-10-07 | Juno Therapeutics Inc | Procédés de dosage et de modulation de cellules génétiquement modifiées |
| WO2019165143A1 (en) * | 2018-02-21 | 2019-08-29 | Cytomx Therapeutics, Inc. | Positron emission tomography imaging of activatable binding polypeptides and related compositions thereof |
| WO2019175131A1 (en) | 2018-03-14 | 2019-09-19 | F. Hoffmann-La Roche Ag | Method for affinity maturation of antibodies |
| CN111971300A (zh) | 2018-03-14 | 2020-11-20 | 豪夫迈·罗氏有限公司 | 新颖的抗-肌钙蛋白t抗体 |
| CN111936522A (zh) | 2018-04-18 | 2020-11-13 | 豪夫迈·罗氏有限公司 | 新型抗胸苷激酶抗体 |
| US20210363219A1 (en) | 2018-06-15 | 2021-11-25 | Alpine Immune Sciences, Inc. | Pd-1 variant immunomodulatory proteins and uses thereof |
| EP3844503A1 (en) | 2018-08-31 | 2021-07-07 | F. Hoffmann-La Roche AG | Thymidine kinase (tk-1) in prognostic indices for dlbcl |
| MX2021004146A (es) | 2018-10-10 | 2021-06-08 | Astellas Pharma Inc | Composicion farmaceutica que contiene complejo de fragmento fab de anticuerpo antihumano con sitio marcado. |
| AU2019356573A1 (en) | 2018-10-11 | 2021-05-27 | Inhibrx, Inc. | PD-1 single domain antibodies and therapeutic compositions thereof |
| EP3863721A1 (en) | 2018-10-11 | 2021-08-18 | Inhibrx, Inc. | 5t4 single domain antibodies and therapeutic compositions thereof |
| WO2020076977A2 (en) | 2018-10-11 | 2020-04-16 | Inhibrx, Inc. | Dll3 single domain antibodies and therapeutic compositions thereof |
| TW202028246A (zh) | 2018-10-11 | 2020-08-01 | 美商英伊布里克斯公司 | B7h3單域抗體及其治療性組合物 |
| AU2019387497A1 (en) | 2018-11-30 | 2021-06-24 | Juno Therapeutics, Inc. | Methods for treatment using adoptive cell therapy |
| CA3120868A1 (en) | 2018-11-30 | 2020-06-04 | Alpine Immune Sciences, Inc. | Cd86 variant immunomodulatory proteins and uses thereof |
| WO2020117257A1 (en) | 2018-12-06 | 2020-06-11 | Genentech, Inc. | Combination therapy of diffuse large b-cell lymphoma comprising an anti-cd79b immunoconjugates, an alkylating agent and an anti-cd20 antibody |
| CN113396230A (zh) | 2019-02-08 | 2021-09-14 | 豪夫迈·罗氏有限公司 | 癌症的诊断和治疗方法 |
| CN115943306A (zh) | 2019-03-29 | 2023-04-07 | 乐天医药生技股份有限公司 | 用于光免疫疗法的方法和相关生物标志物 |
| AU2020275415A1 (en) | 2019-05-14 | 2021-11-25 | Genentech, Inc. | Methods of using anti-CD79B immunoconjugates to treat follicular lymphoma |
| WO2020236679A1 (en) * | 2019-05-17 | 2020-11-26 | Cytomx Therapeutics, Inc. | Methods and compositions for determining the biodistribution of activatable anti-cd166 antibody conjugates |
| US20200399371A1 (en) | 2019-06-21 | 2020-12-24 | Regeneron Pharmaceuticals, Inc. | Use of bispecific antigen-binding molecules that bind muc16 and cd3 in combination with 4-1bb co-stimulation |
| AU2020296181A1 (en) | 2019-06-21 | 2021-12-16 | Regeneron Pharmaceuticals, Inc. | Use of bispecific antigen-binding molecules that bind PSMA and CD3 in combination with 4-1BB co-stimulation |
| MX2022002886A (es) | 2019-09-16 | 2022-04-06 | Regeneron Pharma | Proteinas de union met radiomarcadas para la obtencion de imagenes por inmuno-pet. |
| TW202122114A (zh) | 2019-10-18 | 2021-06-16 | 美商建南德克公司 | 使用抗CD79b 免疫偶聯物治療彌漫性大B 細胞淋巴瘤的方法 |
| JP2023504736A (ja) | 2019-12-06 | 2023-02-06 | ジュノー セラピューティクス インコーポレイテッド | Gprc5d標的結合ドメインに対する抗イディオタイプ抗体ならびに関連する組成物および方法 |
| US20230028050A1 (en) | 2019-12-06 | 2023-01-26 | Juno Therapeutics, Inc. | Anti-idiotypic antibodies to bcma-targeted binding domains and related compositions and methods |
| BR112022014667A2 (pt) | 2020-01-29 | 2022-09-20 | Inhibrx Inc | Anticorpos de domínio único de cd28 e construtos multivalentes e multiespecíficos dos mesmos |
| WO2021217051A1 (en) | 2020-04-24 | 2021-10-28 | Genentech, Inc. | Methods of using anti-cd79b immunoconjugates |
| US20230324408A1 (en) | 2020-08-05 | 2023-10-12 | Juno Therapeutics, Inc. | Anti-idiotypic antibodies to ror1-targeted binding domains and related compositions and methods |
| WO2022036146A1 (en) | 2020-08-12 | 2022-02-17 | Genentech, Inc. | Diagnostic and therapeutic methods for cancer |
| JP2023550278A (ja) | 2020-10-30 | 2023-12-01 | エフ. ホフマン-ラ ロシュ アーゲー | 胆管癌のマーカーとしてのtimp1 |
| WO2022147463A2 (en) | 2020-12-31 | 2022-07-07 | Alamar Biosciences, Inc. | Binder molecules with high affinity and/ or specificity and methods of making and use thereof |
| WO2022241446A1 (en) | 2021-05-12 | 2022-11-17 | Genentech, Inc. | Methods of using anti-cd79b immunoconjugates to treat diffuse large b-cell lymphoma |
| WO2023019092A1 (en) | 2021-08-07 | 2023-02-16 | Genentech, Inc. | Methods of using anti-cd79b immunoconjugates to treat diffuse large b-cell lymphoma |
| WO2023034750A1 (en) | 2021-08-30 | 2023-03-09 | Genentech, Inc. | Anti-polyubiquitin multispecific antibodies |
| WO2023111168A1 (en) | 2021-12-17 | 2023-06-22 | F. Hoffmann-La Roche Ag | A novel antibody for detection of amyloid beta 42 (aβ42) |
| WO2023172883A1 (en) | 2022-03-07 | 2023-09-14 | Alpine Immune Sciences, Inc. | Immunomodulatory proteins of variant cd80 polypeptides, cell therapies thereof and related methods and uses |
| WO2023178357A1 (en) | 2022-03-18 | 2023-09-21 | Evolveimmune Therapeutics, Inc. | Bispecific antibody fusion molecules and methods of use thereof |
| CN117198390B (zh) * | 2023-09-08 | 2024-03-12 | 中国科学院广州生物医药与健康研究院 | 通过设计和改造二硫键交联位点的slc膜蛋白复合物的制备方法 |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2006034488A2 (en) * | 2004-09-23 | 2006-03-30 | Genentech, Inc. | Cysteine engineered antibodies and conjugates |
Family Cites Families (290)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3773919A (en) | 1969-10-23 | 1973-11-20 | Du Pont | Polylactide-drug mixtures |
| US4275149A (en) | 1978-11-24 | 1981-06-23 | Syva Company | Macromolecular environment control in specific receptor assays |
| US4318980A (en) | 1978-04-10 | 1982-03-09 | Miles Laboratories, Inc. | Heterogenous specific binding assay employing a cycling reactant as label |
| US4458066A (en) | 1980-02-29 | 1984-07-03 | University Patents, Inc. | Process for preparing polynucleotides |
| US4415732A (en) | 1981-03-27 | 1983-11-15 | University Patents, Inc. | Phosphoramidite compounds and processes |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| JPS6098584A (ja) | 1983-11-02 | 1985-06-01 | Canon Inc | カメラ―体形vtr |
| US5807715A (en) | 1984-08-27 | 1998-09-15 | The Board Of Trustees Of The Leland Stanford Junior University | Methods and transformed mammalian lymphocyte cells for producing functional antigen-binding protein including chimeric immunoglobulin |
| US5342606A (en) | 1984-10-18 | 1994-08-30 | Board Of Regents, The University Of Texas System | Polyazamacrocyclic compounds for complexation of metal ions |
| US5316757A (en) | 1984-10-18 | 1994-05-31 | Board Of Regents, The University Of Texas System | Synthesis of polyazamacrocycles with more than one type of side-chain chelating groups |
| US4737456A (en) | 1985-05-09 | 1988-04-12 | Syntex (U.S.A.) Inc. | Reducing interference in ligand-receptor binding assays |
| US5091178A (en) | 1986-02-21 | 1992-02-25 | Oncogen | Tumor therapy with biologically active anti-tumor antibodies |
| IL85035A0 (en) | 1987-01-08 | 1988-06-30 | Int Genetic Eng | Polynucleotide molecule,a chimeric antibody with specificity for human b cell surface antigen,a process for the preparation and methods utilizing the same |
| GB8720833D0 (en) * | 1987-09-04 | 1987-10-14 | Celltech Ltd | Recombinant dna product |
| WO1989006692A1 (en) | 1988-01-12 | 1989-07-27 | Genentech, Inc. | Method of treating tumor cells by inhibiting growth factor receptor function |
| US5047524A (en) | 1988-12-21 | 1991-09-10 | Applied Biosystems, Inc. | Automated system for polynucleotide synthesis and purification |
| US5262530A (en) | 1988-12-21 | 1993-11-16 | Applied Biosystems, Inc. | Automated system for polynucleotide synthesis and purification |
| DE3920358A1 (de) | 1989-06-22 | 1991-01-17 | Behringwerke Ag | Bispezifische und oligospezifische, mono- und oligovalente antikoerperkonstrukte, ihre herstellung und verwendung |
| AU6355090A (en) | 1989-08-23 | 1991-04-03 | Scripps Clinic And Research Foundation | Compositions and methods for detection and treatment of epstein-barr virus infection and immune disorders |
| US5183884A (en) | 1989-12-01 | 1993-02-02 | United States Of America | Dna segment encoding a gene for a receptor related to the epidermal growth factor receptor |
| US5256643A (en) | 1990-05-29 | 1993-10-26 | The Government Of The United States | Human cripto protein |
| AU9016591A (en) | 1990-10-25 | 1992-05-26 | Tanox Biosystems, Inc. | Glycoproteins associated with membrane-bound immunoglobulins as antibody targets on B cells |
| US5571894A (en) | 1991-02-05 | 1996-11-05 | Ciba-Geigy Corporation | Recombinant antibodies specific for a growth factor receptor |
| US5543503A (en) | 1991-03-29 | 1996-08-06 | Genentech Inc. | Antibodies to human IL-8 type A receptor |
| US5440021A (en) | 1991-03-29 | 1995-08-08 | Chuntharapai; Anan | Antibodies to human IL-8 type B receptor |
| ATE194385T1 (de) | 1991-03-29 | 2000-07-15 | Genentech Inc | Menschliche pf4a rezeptoren und ihre verwendung |
| LU91067I2 (fr) | 1991-06-14 | 2004-04-02 | Genentech Inc | Trastuzumab et ses variantes et dérivés immuno chimiques y compris les immotoxines |
| JP3050424B2 (ja) | 1991-07-12 | 2000-06-12 | 塩野義製薬株式会社 | ヒトエンドセリンリセプター |
| US5264557A (en) | 1991-08-23 | 1993-11-23 | The United States Of America As Represented By The Department Of Health And Human Services | Polypeptide of a human cripto-related gene, CR-3 |
| US5587458A (en) | 1991-10-07 | 1996-12-24 | Aronex Pharmaceuticals, Inc. | Anti-erbB-2 antibodies, combinations thereof, and therapeutic and diagnostic uses thereof |
| US6011146A (en) | 1991-11-15 | 2000-01-04 | Institut Pasteur | Altered major histocompatibility complex (MHC) determinant and methods of using the determinant |
| US6153408A (en) | 1991-11-15 | 2000-11-28 | Institut Pasteur And Institut National De La Sante Et De La Recherche Medicale | Altered major histocompatibility complex (MHC) determinant and methods of using the determinant |
| CA2122732C (en) | 1991-11-25 | 2008-04-08 | Marc D. Whitlow | Multivalent antigen-binding proteins |
| US5428139A (en) | 1991-12-10 | 1995-06-27 | The Dow Chemical Company | Bicyclopolyazamacrocyclophosphonic acid complexes for use as radiopharmaceuticals |
| US5480990A (en) | 1991-12-10 | 1996-01-02 | The Dow Chemical Company | Bicyclopolyazamacrocyclocarboxylic acid complexes for use as contrast agents |
| US5739294A (en) | 1991-12-10 | 1998-04-14 | The Dow Chemical Company | Bicyclopol yazamacrocyclophosphonic acid complexes for use as contrast agents |
| ATE503496T1 (de) | 1992-02-06 | 2011-04-15 | Novartis Vaccines & Diagnostic | Biosynthetisches bindeprotein für tumormarker |
| AU4025193A (en) | 1992-04-08 | 1993-11-18 | Cetus Oncology Corporation | Humanized C-erbB-2 specific antibodies |
| IL107366A (en) | 1992-10-23 | 2003-03-12 | Chugai Pharmaceutical Co Ltd | Genes coding for megakaryocyte potentiator |
| ES2091684T3 (es) | 1992-11-13 | 1996-11-01 | Idec Pharma Corp | Aplicacion terapeutica de anticuerpos quimericos y radiomarcados contra el antigeno de diferenciacion restringida de los linfocitos b humanos para el tratamiento del linfoma de las celulas b. |
| CA2103323A1 (en) | 1992-11-24 | 1994-05-25 | Gregory D. Plowman | Her4 human receptor tyrosine kinase |
| US5644033A (en) | 1992-12-22 | 1997-07-01 | Health Research, Inc. | Monoclonal antibodies that define a unique antigen of human B cell antigen receptor complex and methods of using same for diagnosis and treatment |
| US5869445A (en) | 1993-03-17 | 1999-02-09 | University Of Washington | Methods for eliciting or enhancing reactivity to HER-2/neu protein |
| US5801005A (en) | 1993-03-17 | 1998-09-01 | University Of Washington | Immune reactivity to HER-2/neu protein for diagnosis of malignancies in which the HER-2/neu oncogene is associated |
| US5385893A (en) | 1993-05-06 | 1995-01-31 | The Dow Chemical Company | Tricyclopolyazamacrocyclophosphonic acids, complexes and derivatives thereof, for use as contrast agents |
| US5462725A (en) | 1993-05-06 | 1995-10-31 | The Dow Chemical Company | 2-pyridylmethylenepolyazamacrocyclophosphonic acids, complexes and derivatives thereof, for use as contrast agents |
| US6214345B1 (en) | 1993-05-14 | 2001-04-10 | Bristol-Myers Squibb Co. | Lysosomal enzyme-cleavable antitumor drug conjugates |
| US5773223A (en) | 1993-09-02 | 1998-06-30 | Chiron Corporation | Endothelin B1, (ETB1) receptor polypeptide and its encoding nucleic acid methods, and uses thereof |
| CZ292061B6 (cs) | 1994-03-17 | 2003-07-16 | Merck Patent Gmbh | Jednořetězcové fragmenty protilátek a protilátky proti receptoru epidermálního růstového faktoru, způsob jejich přípravy a farmaceutický prostředek, který je obsahuje |
| US5750370A (en) | 1995-06-06 | 1998-05-12 | Human Genome Sciences, Inc. | Nucleic acid encoding human endothlein-bombesin receptor and method of producing the receptor |
| US5707829A (en) | 1995-08-11 | 1998-01-13 | Genetics Institute, Inc. | DNA sequences and secreted proteins encoded thereby |
| AU716785B2 (en) | 1995-07-27 | 2000-03-09 | Genentech Inc. | Stabile isotonic lyophilized protein formulation |
| US20020193567A1 (en) | 1995-08-11 | 2002-12-19 | Genetics Institute, Inc. | Secreted proteins and polynucleotides encoding them |
| DK0853487T3 (da) * | 1995-09-18 | 2000-10-23 | Us Health | Neutraliserende monoklonale antistoffer mod respiratorisk syncytial virus |
| US5834456A (en) | 1996-02-23 | 1998-11-10 | The Dow Chemical Company | Polyazamacrocyclofluoromonoalkylphosphonic acids, and their complexes, for use as contrast agents |
| JP3646191B2 (ja) | 1996-03-19 | 2005-05-11 | 大塚製薬株式会社 | ヒト遺伝子 |
| DK0896586T4 (en) | 1996-03-27 | 2015-08-24 | Genentech Inc | ErbB3 antibodies. |
| NZ332598A (en) | 1996-05-17 | 2000-04-28 | Schering Corp | Human BAS-1 protein, and use as an antagonist for modulating physiology or development of a cell |
| US5945511A (en) | 1997-02-20 | 1999-08-31 | Zymogenetics, Inc. | Class II cytokine receptor |
| US20030185830A1 (en) | 1997-02-25 | 2003-10-02 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of prostate cancer |
| US7033827B2 (en) | 1997-02-25 | 2006-04-25 | Corixa Corporation | Prostate-specific polynucleotide compositions |
| US6541212B2 (en) | 1997-03-10 | 2003-04-01 | The Regents Of The University Of California | Methods for detecting prostate stem cell antigen protein |
| ES2221982T3 (es) | 1997-03-10 | 2005-01-16 | The Regents Of The University Of California | Antigeno de celulas madre de la prostata (psca). |
| US6261791B1 (en) | 1997-03-10 | 2001-07-17 | The Regents Of The University Of California | Method for diagnosing cancer using specific PSCA antibodies |
| US6555339B1 (en) | 1997-04-14 | 2003-04-29 | Arena Pharmaceuticals, Inc. | Non-endogenous, constitutively activated human protein-coupled receptors |
| US6319688B1 (en) | 1997-04-28 | 2001-11-20 | Smithkline Beecham Corporation | Polynucleotide encoding human sodium dependent phosphate transporter (IPT-1) |
| WO1998051824A1 (en) | 1997-05-15 | 1998-11-19 | Abbott Laboratories | Reagents and methods useful for detecting disease of the urinary tract |
| US6890749B2 (en) | 1997-05-15 | 2005-05-10 | Abbott Laboratories | Reagents and methods useful for detecting diseases of the prostate |
| US6753165B1 (en) * | 1999-01-14 | 2004-06-22 | Bolder Biotechnology, Inc. | Methods for making proteins containing free cysteine residues |
| US6248564B1 (en) * | 1997-08-29 | 2001-06-19 | Harvard University | Mutant MHC class I molecules |
| WO1999019488A1 (en) | 1997-10-15 | 1999-04-22 | Children's Medical Center Corporation | Novel human egf receptors and use thereof |
| US20030060612A1 (en) | 1997-10-28 | 2003-03-27 | Genentech, Inc. | Compositions and methods for the diagnosis and treatment of tumor |
| US20020034749A1 (en) | 1997-11-18 | 2002-03-21 | Billing-Medel Patricia A. | Reagents and methods useful for detecting diseases of the breast |
| US6110695A (en) | 1997-12-02 | 2000-08-29 | The Regents Of The University Of California | Modulating the interaction of the chemokine, B Lymphocyte Hemoattractant, and its Receptor, BLR1 |
| WO1999046284A2 (en) | 1998-03-13 | 1999-09-16 | The Burnham Institute | Molecules that home to various selected organs or tissues |
| US6528624B1 (en) | 1998-04-02 | 2003-03-04 | Genentech, Inc. | Polypeptide variants |
| US6534482B1 (en) | 1998-05-13 | 2003-03-18 | Epimmune, Inc. | Expression vectors for stimulating an immune response and methods of using the same |
| US20030064397A1 (en) | 1998-05-22 | 2003-04-03 | Incyte Genomics, Inc. | Transmembrane protein differentially expressed in prostate and lung tumors |
| US20020187472A1 (en) | 2001-03-09 | 2002-12-12 | Preeti Lal | Steap-related protein |
| WO2000012130A1 (en) | 1998-08-27 | 2000-03-09 | Smithkline Beecham Corporation | Rp105 agonists and antagonists |
| JP4689781B2 (ja) | 1998-09-03 | 2011-05-25 | 独立行政法人科学技術振興機構 | アミノ酸輸送蛋白及びその遺伝子 |
| AU5963699A (en) | 1998-10-02 | 2000-04-26 | Mcmaster University | Spliced form of (erb)b-2/neu oncogene |
| US6468546B1 (en) | 1998-12-17 | 2002-10-22 | Corixa Corporation | Compositions and methods for therapy and diagnosis of ovarian cancer |
| US20030091580A1 (en) | 2001-06-18 | 2003-05-15 | Mitcham Jennifer L. | Compositions and methods for the therapy and diagnosis of ovarian cancer |
| US6858710B2 (en) | 1998-12-17 | 2005-02-22 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of ovarian cancer |
| US6962980B2 (en) | 1999-09-24 | 2005-11-08 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of ovarian cancer |
| US20020119158A1 (en) | 1998-12-17 | 2002-08-29 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of ovarian cancer |
| US20030190669A1 (en) | 1998-12-30 | 2003-10-09 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| AU776672B2 (en) | 1998-12-30 | 2004-09-16 | Beth Israel Deaconess Medical Center | Characterization of a calcium channel family |
| CZ20012587A3 (cs) | 1999-01-29 | 2002-05-15 | Corixa Corporation | Izolovaný protein, nukleová kyselina, virový vektor, farmaceutický prostředek, izolovaná populace T buněk, způsob posílení imunitní odpovědi, způsob odstranění nádorových buněk, způsob stimulace a/nebo namnoľení T buněk a způsob přípravy fúzního proteinu |
| GB9905124D0 (en) | 1999-03-05 | 1999-04-28 | Smithkline Beecham Biolog | Novel compounds |
| AU3395900A (en) | 1999-03-12 | 2000-10-04 | Human Genome Sciences, Inc. | Human lung cancer associated gene sequences and polypeptides |
| US7304126B2 (en) | 1999-05-11 | 2007-12-04 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| US6399743B1 (en) | 1999-05-14 | 2002-06-04 | Dept. Of Veterans Affairs | Isolation and characterization of a rat epidermal growth factor related protein |
| US7049410B2 (en) | 1999-05-14 | 2006-05-23 | Majumdar Adhip P N | Antibodies to a novel EGF-receptor related protein (ERRP) |
| WO2000075655A1 (fr) | 1999-06-03 | 2000-12-14 | Takeda Chemical Industries, Ltd. | Procede de criblage avec cd100 |
| HU226742B1 (en) | 1999-06-25 | 2009-08-28 | Genentech Inc | Humanized anti-erbb2 antibodies and treatment with anti-erbb2 antibodies |
| AU784157B2 (en) | 1999-06-25 | 2006-02-16 | Genentech Inc. | Methods of treatment using anti-ErbB antibody-maytansinoid conjugates |
| US7589172B2 (en) | 1999-07-20 | 2009-09-15 | Genentech, Inc. | PRO256 polypeptides |
| US7297770B2 (en) | 1999-08-10 | 2007-11-20 | Genentech, Inc. | PRO6496 polypeptides |
| US7294696B2 (en) | 1999-08-17 | 2007-11-13 | Genentech Inc. | PRO7168 polypeptides |
| CA2380355A1 (en) | 1999-09-01 | 2001-03-08 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| US20030129192A1 (en) | 1999-09-10 | 2003-07-10 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of ovarian cancer |
| US20030206918A1 (en) | 1999-09-10 | 2003-11-06 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of ovarian cancer |
| US20030232056A1 (en) | 1999-09-10 | 2003-12-18 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of ovarian cancer |
| ATE400587T1 (de) | 1999-10-29 | 2008-07-15 | Genentech Inc | Gegen das prostata-stammzellantigen (psca) gerichtete antikörper und deren verwendung |
| US6372907B1 (en) | 1999-11-03 | 2002-04-16 | Apptera Corporation | Water-soluble rhodamine dye peptide conjugates |
| ES2591102T3 (es) | 1999-11-29 | 2016-11-24 | The Trustees Of Columbia University In The City Of New York | Aislamiento de cinco genes novedosos que codifican nuevos melanomas de tipo receptor de Fc implicados en la patogénesis del linfoma/melanoma |
| WO2001040269A2 (en) | 1999-11-30 | 2001-06-07 | Corixa Corporation | Compositions and methods for therapy and diagnosis of breast cancer |
| AU2087401A (en) | 1999-12-10 | 2001-06-18 | Epimmune, Inc. | Inducing cellular immune responses to her2/neu using peptide and nucleic acid compositions |
| EP2248828A1 (en) | 1999-12-23 | 2010-11-10 | ZymoGenetics, Inc. | Method for treating inflammation |
| CA2395539C (en) | 1999-12-23 | 2009-08-25 | Zymogenetics, Inc. | Soluble interleukin-20 receptor |
| NZ502058A (en) | 1999-12-23 | 2003-11-28 | Ovita Ltd | Isolated mutated nucleic acid molecule for regulation of ovulation rate |
| US6610286B2 (en) | 1999-12-23 | 2003-08-26 | Zymogenetics, Inc. | Method for treating inflammation using soluble receptors to interleukin-20 |
| ATE337403T1 (de) | 1999-12-24 | 2006-09-15 | Genentech Inc | Verfahren und verbindungen zur verlängerung der halbwertzeiten bei der ausscheidung von biowirksamen verbindungen |
| US20040001827A1 (en) | 2002-06-28 | 2004-01-01 | Dennis Mark S. | Serum albumin binding peptides for tumor targeting |
| US7294695B2 (en) | 2000-01-20 | 2007-11-13 | Genentech, Inc. | PRO10268 polypeptides |
| US20030224379A1 (en) | 2000-01-21 | 2003-12-04 | Tang Y. Tom | Novel nucleic acids and polypeptides |
| AU2001234493A1 (en) | 2000-01-21 | 2001-07-31 | Corixa Corporation | Compounds and methods for prevention and treatment of her-2/neu associated malignancies |
| AU2001243142A1 (en) | 2000-02-03 | 2001-08-14 | Hyseq, Inc. | Novel nucleic acids and polypeptides |
| US20030186372A1 (en) | 2000-02-11 | 2003-10-02 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| US20020142377A1 (en) | 2000-02-22 | 2002-10-03 | Glucksmann Maria Alexandra | 18607, a novel human calcium channel |
| US20030219806A1 (en) | 2000-02-22 | 2003-11-27 | Millennium Pharmaceuticals, Inc. | Novel 18607, 15603, 69318, 12303, 48000, 52920, 5433, 38554, 57301, 58324, 55063, 52991, 59914, 59921 and 33751 molecules and uses therefor |
| US20040005561A1 (en) | 2000-03-01 | 2004-01-08 | Corixa Corporation | Compositions and methods for the detection, diagnosis and therapy of hematological malignancies |
| US20040002068A1 (en) | 2000-03-01 | 2004-01-01 | Corixa Corporation | Compositions and methods for the detection, diagnosis and therapy of hematological malignancies |
| EP1261743A2 (en) | 2000-03-07 | 2002-12-04 | Hyseq, Inc. | Novel nucleic acids and polypeptides |
| US7097840B2 (en) * | 2000-03-16 | 2006-08-29 | Genentech, Inc. | Methods of treatment using anti-ErbB antibody-maytansinoid conjugates |
| WO2001072962A2 (en) | 2000-03-24 | 2001-10-04 | Fahri Saatcioglu | Novel prostate-specific or testis-specific nucleic acid molecules, polypeptides, and diagnostic and therapeutic methods |
| US20030186889A1 (en) | 2000-03-31 | 2003-10-02 | Wolf-Georg Forssmann | Diagnostic and medicament for analysing the cell surface proteome of tumour and inflammatory cells and for treating tumorous and inflammatory diseases, preferably using a specific chemokine receptor analysis and the chemokine receptor-ligand interaction |
| AU2001253140A1 (en) | 2000-04-03 | 2001-10-15 | The Government Of The United States Of America, As Represented By The Secretary Of The Department Of Health And Human Services | Tumor markers in ovarian cancer |
| US6806054B2 (en) | 2000-04-07 | 2004-10-19 | Arena Pharmaceuticals, Inc. | Non-endogenous, constitutively activated known G protein-coupled receptors |
| WO2001088133A2 (en) | 2000-05-18 | 2001-11-22 | Lexicon Genetics Incorporated | Human semaphorin homologs and polynucleotides encoding the same |
| WO2001090304A2 (en) | 2000-05-19 | 2001-11-29 | Human Genome Sciences, Inc. | Nucleic acids, proteins, and antibodies |
| AU2001275437A1 (en) | 2000-06-09 | 2001-12-17 | Idec Pharmaceuticals Corporation | Gene targets and ligands that bind thereto for treatment and diagnosis of ovarian carcinomas |
| JP2004523203A (ja) | 2000-06-16 | 2004-08-05 | インサイト・ゲノミックス・インコーポレイテッド | Gタンパク質結合受容体 |
| AU2001271714A1 (en) | 2000-06-30 | 2002-01-14 | Human Genome Sciences, Inc. | B7-like polynucleotides, polypeptides, and antibodies |
| AU2001273194A1 (en) | 2000-06-30 | 2002-01-14 | Amgen Inc. | B7-Like Molecules and Uses Thereof |
| JP2004528003A (ja) | 2000-06-30 | 2004-09-16 | インサイト・ゲノミックス・インコーポレイテッド | 細胞外マトリクスおよび細胞接着分子 |
| AU2002214531A1 (en) | 2000-07-03 | 2002-01-30 | Curagen Corporation | Proteins and nucleic acids encoding same |
| US20040044179A1 (en) | 2000-07-25 | 2004-03-04 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| AU2001283507A1 (en) | 2000-07-27 | 2002-02-13 | Mayo Foundation For Medical Education And Research | B7-h3 and b7-h4, novel immunoregulatory molecules |
| WO2002010382A2 (en) | 2000-07-28 | 2002-02-07 | Ulrich Wissenbach | Trp8, trp9 and trp10, markers for cancer |
| US7229623B1 (en) | 2000-08-03 | 2007-06-12 | Corixa Corporation | Her-2/neu fusion proteins |
| WO2002013847A2 (en) | 2000-08-14 | 2002-02-21 | Corixa Corporation | Methods for diagnosis and therapy of hematological and virus-associated malignancies |
| BR0113235A (pt) | 2000-08-14 | 2004-06-08 | Corixa Corp | Composições e métodos para a terapia e a diagnose de malignidades associadas com her-2/neu |
| GB0020953D0 (en) | 2000-08-24 | 2000-10-11 | Smithkline Beecham Biolog | Vaccine |
| AU2001270118A1 (en) | 2000-08-24 | 2002-03-04 | Genentech, Inc. | Compositions and methods for the diagnosis and treatment of tumor |
| US20020119148A1 (en) | 2000-09-01 | 2002-08-29 | Gerritsen Mary E. | ErbB4 antagonists |
| AU2001290548A1 (en) | 2000-09-11 | 2002-03-26 | Hyseq, Inc. | Novel nucleic acids and polypeptides |
| US20030119121A1 (en) | 2000-09-15 | 2003-06-26 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| DE60141546D1 (de) | 2000-09-15 | 2010-04-22 | Zymogenetics Inc | Polypeptide enthaltend extrazellulären Domäne von IL-20RA und/oderr IL-20RB |
| US6613567B1 (en) | 2000-09-15 | 2003-09-02 | Isis Pharmaceuticals, Inc. | Antisense inhibition of Her-2 expression |
| AU2001292724B2 (en) | 2000-09-18 | 2005-05-26 | Biogen Idec Ma Inc. | CRIPTO mutant and uses thereof |
| UA83458C2 (ru) | 2000-09-18 | 2008-07-25 | Байоджен Айдек Ма Інк. | Выделенный полипептид baff-r (рецептор фактора активации в-клеток семейства tnf) |
| WO2002030268A2 (en) | 2000-10-13 | 2002-04-18 | Eos Biotechnology, Inc. | Methods of diagnosis of prostate cancer, compositions and methods of screening for modulators of prostate cancer |
| ES2329012T3 (es) | 2000-11-07 | 2009-11-20 | Zymogenetics, Inc. | Receptor del factor de necrosis tumoral humano. |
| US20020150573A1 (en) | 2000-11-10 | 2002-10-17 | The Rockefeller University | Anti-Igalpha-Igbeta antibody for lymphoma therapy |
| WO2002061087A2 (en) | 2000-12-19 | 2002-08-08 | Lifespan Biosciences, Inc. | Antigenic peptides, such as for g protein-coupled receptors (gpcrs), antibodies thereto, and systems for identifying such antigenic peptides |
| US20020159986A1 (en) | 2001-01-12 | 2002-10-31 | John Langenfeld | Bone morphogenetic protein-2 in the treatment and diagnosis of cancer |
| US20030119133A1 (en) | 2001-01-16 | 2003-06-26 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| US20030119119A1 (en) | 2001-01-16 | 2003-06-26 | Genentech, Inc. | Secreted and transmembrane polypeptides and nucleic acids encoding the same |
| US7754208B2 (en) | 2001-01-17 | 2010-07-13 | Trubion Pharmaceuticals, Inc. | Binding domain-immunoglobulin fusion proteins |
| CA2440703A1 (en) | 2001-01-24 | 2002-08-01 | Protein Design Labs, Inc. | Methods of diagnosis of breast cancer, compositions and methods of screening for modulators of breast cancer |
| WO2002060317A2 (en) | 2001-01-30 | 2002-08-08 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of pancreatic cancer |
| WO2002064780A1 (en) | 2001-02-12 | 2002-08-22 | Bionomics Limited | Dna sequences for human tumour suppressor genes |
| US20030087250A1 (en) | 2001-03-14 | 2003-05-08 | Millennium Pharmaceuticals, Inc. | Nucleic acid molecules and proteins for the identification, assessment, prevention, and therapy of ovarian cancer |
| AU2002311787A1 (en) | 2001-03-28 | 2002-10-15 | Zycos Inc. | Translational profiling |
| WO2003008537A2 (en) | 2001-04-06 | 2003-01-30 | Mannkind Corporation | Epitope sequences |
| US6820011B2 (en) | 2001-04-11 | 2004-11-16 | The Regents Of The University Of Colorado | Three-dimensional structure of complement receptor type 2 and uses thereof |
| WO2002083866A2 (en) | 2001-04-17 | 2002-10-24 | The Board Of Trustees Of The University Of Arkansas | Repeat sequences of the ca125 gene and their use for diagnostic and therapeutic interventions |
| CA2444691A1 (en) | 2001-04-18 | 2002-10-31 | Protein Design Labs, Inc. | Methods of diagnosis of lung cancer, compositions and methods of screening for modulators of lung cancer |
| EA007469B1 (ru) | 2001-04-26 | 2006-10-27 | Байоджен Айдек Эмэй Инк. | Антитела, блокирующие cripto, и их применения |
| US6884869B2 (en) | 2001-04-30 | 2005-04-26 | Seattle Genetics, Inc. | Pentapeptide compounds and uses related thereto |
| CA2446788A1 (en) | 2001-05-09 | 2002-11-14 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of prostate cancer |
| WO2002092836A2 (en) | 2001-05-11 | 2002-11-21 | Sloan-Kettering Institute For Cancer Research | Nucleic acid sequence encoding ovarian antigen, ca125, and uses thereof |
| EP1436003B3 (en) | 2001-05-24 | 2012-03-14 | ZymoGenetics, Inc. | Taci-immunoglobulin fusion proteins |
| JP2005518185A (ja) | 2001-06-04 | 2005-06-23 | キュラジェン コーポレイション | 新規タンパク質およびそれをコード化する核酸 |
| AU2002314901A1 (en) | 2001-06-04 | 2002-12-16 | Eos Biotechnology, Inc. | Methods of diagnosis and treatment of androgen-dependent prostate cancer, prostate cancer undergoing androgen-withdrawal, and androgen-independent prostate cancer |
| US20030087266A1 (en) | 2001-06-05 | 2003-05-08 | Lori Friedman | IGs as modifiers of the p53 pathway and methods of use |
| ATE483976T1 (de) | 2001-06-05 | 2010-10-15 | Exelixis Inc | Gfats als modifikatoren des p53-wegs und verwendungsverfahren |
| US7235358B2 (en) | 2001-06-08 | 2007-06-26 | Expression Diagnostics, Inc. | Methods and compositions for diagnosing and monitoring transplant rejection |
| US7125663B2 (en) | 2001-06-13 | 2006-10-24 | Millenium Pharmaceuticals, Inc. | Genes, compositions, kits and methods for identification, assessment, prevention, and therapy of cervical cancer |
| CA2451465A1 (en) | 2001-06-18 | 2002-12-27 | Eos Biotechnology Inc. | Methods of diagnosis of ovarian cancer, compositions and methods of screening for modulators of ovarian cancer |
| US7189507B2 (en) | 2001-06-18 | 2007-03-13 | Pdl Biopharma, Inc. | Methods of diagnosis of ovarian cancer, compositions and methods of screening for modulators of ovarian cancer |
| WO2003004989A2 (en) | 2001-06-21 | 2003-01-16 | Millennium Pharmaceuticals, Inc. | Compositions, kits, and methods for identification, assessment, prevention, and therapy of breast cancer |
| US20030108958A1 (en) | 2001-06-28 | 2003-06-12 | Rene De Waal Malefyt | Biological activity of AK155 |
| US20030120040A1 (en) | 2001-06-29 | 2003-06-26 | Genentech, Inc. | Secreted and Transmembrane polypeptides and nucleic acids encoding the same |
| WO2003004529A2 (en) | 2001-07-02 | 2003-01-16 | Licentia Ltd. | Ephrin-tie receptor materials and methods |
| US20040076955A1 (en) | 2001-07-03 | 2004-04-22 | Eos Biotechnology, Inc. | Methods of diagnosis of bladder cancer, compositions and methods of screening for modulators of bladder cancer |
| WO2003003984A2 (en) | 2001-07-05 | 2003-01-16 | Curagen Corporation | Novel proteins and nucleic acids encoding same |
| WO2003055439A2 (en) | 2001-07-18 | 2003-07-10 | The Regents Of The University Of California | Her2/neu target antigen and use of same to stimulate an immune response |
| AU2002337657A1 (en) | 2001-07-25 | 2003-02-17 | Millennium Pharmaceuticals, Inc. | Novel genes, compositions, kits, and methods for identification, assessment, prevention, and therapy of prostate cancer |
| JP2005510208A (ja) | 2001-08-03 | 2005-04-21 | ジェネンテック・インコーポレーテッド | TACIs及びBR3ポリペプチドとその用途 |
| US20070015145A1 (en) | 2001-08-14 | 2007-01-18 | Clifford Woolf | Nucleic acid and amino acid sequences involved in pain |
| US20030092013A1 (en) | 2001-08-16 | 2003-05-15 | Vitivity, Inc. | Diagnosis and treatment of vascular disease |
| WO2003018621A2 (en) | 2001-08-23 | 2003-03-06 | Oxford Biomedica (Uk) Limited | Genes |
| AU2002357643A1 (en) | 2001-08-29 | 2003-04-14 | Vanderbilt University | The human mob-5 (il-24) receptors and uses thereof |
| US20040235068A1 (en) * | 2001-09-05 | 2004-11-25 | Levinson Arthur D. | Methods for the identification of polypeptide antigens associated with disorders involving aberrant cell proliferation and compositions useful for the treatment of such disorders |
| US20030124579A1 (en) | 2001-09-05 | 2003-07-03 | Eos Biotechnology, Inc. | Methods of diagnosis of ovarian cancer, compositions and methods of screening for modulators of ovarian cancer |
| ES2537074T3 (es) | 2001-09-06 | 2015-06-02 | Agensys, Inc. | Ácido nucleico y proteína correspondiente denominados STEAP-1 útiles en el tratamiento y la detección de cáncer |
| JP2005518782A (ja) | 2001-09-17 | 2005-06-30 | プロテイン デザイン ラブス, インコーポレイテッド | ガンの診断方法、ガンのモジュレータのスクリーニング組成物及び方法 |
| NZ573831A (en) | 2001-09-18 | 2010-07-30 | Genentech Inc | Compositions and methods for the diagnosis and treatment of tumor, particularly breast tumor - TAT193 |
| IL160933A0 (en) | 2001-09-18 | 2004-08-31 | Proteologics Inc | Methods and compositions for treating ?cap associated diseases |
| WO2003025148A2 (en) | 2001-09-19 | 2003-03-27 | Nuvelo, Inc. | Novel nucleic acids and polypeptides |
| US7091186B2 (en) | 2001-09-24 | 2006-08-15 | Seattle Genetics, Inc. | p-Amidobenzylethers in drug delivery agents |
| WO2003026577A2 (en) | 2001-09-24 | 2003-04-03 | Seattle Genetics, Inc. | P-amidobenzylethers in drug delivery agents |
| US20030077644A1 (en) | 2001-09-28 | 2003-04-24 | Bing Yang | Diagnosis and treatment of diseases caused by mutations in CD72 |
| US20050130117A1 (en) | 2001-10-03 | 2005-06-16 | Davis Cong L. | Modulators of lymphocyte activation and migration |
| US20040249144A1 (en) | 2001-10-03 | 2004-12-09 | Zairen Sun | Regulated breast cancer genes |
| WO2003034984A2 (en) | 2001-10-19 | 2003-05-01 | Genentech, Inc. | Compositions and methods for the diagnosis and treatment of inflammatory bowel disorders |
| US20040241703A1 (en) | 2002-08-19 | 2004-12-02 | Genentech, Inc. | Compositions and methods for the diagnosis and treatment of tumor |
| US20050123925A1 (en) | 2002-11-15 | 2005-06-09 | Genentech, Inc. | Compositions and methods for the diagnosis and treatment of tumor |
| US7825089B2 (en) | 2001-10-24 | 2010-11-02 | National Jewish Health | Three-dimensional structures of TALL-1 and its cognate receptors and modified proteins and methods related thereto |
| CN1685055A (zh) | 2001-10-31 | 2005-10-19 | 爱尔康公司 | 骨形态发生蛋白(bmp)、bmp受体和bmp结合蛋白及它们在诊断和治疗青光眼中的用途 |
| WO2003042661A2 (en) | 2001-11-13 | 2003-05-22 | Protein Design Labs, Inc. | Methods of diagnosis of cancer, compositions and methods of screening for modulators of cancer |
| US20030232350A1 (en) | 2001-11-13 | 2003-12-18 | Eos Biotechnology, Inc. | Methods of diagnosis of cancer, compositions and methods of screening for modulators of cancer |
| US20050123536A1 (en) | 2001-11-20 | 2005-06-09 | Che-Leung Law | Treatment of immunological disorders using anti-dc30 antibodies |
| WO2003045422A1 (en) | 2001-11-29 | 2003-06-05 | Genset S.A. | Agonists and antagonists of prolixin for the treatment of metabolic disorders |
| AU2002349784A1 (en) | 2001-12-03 | 2003-06-17 | Asahi Kasei Pharma Corporation | Nf-kappab activating genes |
| AU2002366951A1 (en) | 2001-12-10 | 2003-07-09 | Nuvelo,Inc. | Novel nucleic acids and polypeptides |
| US20030134790A1 (en) | 2002-01-11 | 2003-07-17 | University Of Medicine And Dentistry Of New Jersey | Bone Morphogenetic Protein-2 And Bone Morphogenetic Protein-4 In The Treatment And Diagnosis Of Cancer |
| US7452675B2 (en) | 2002-01-25 | 2008-11-18 | The Queen's Medical Center | Methods of screening for TRPM4b modulators |
| DE60334364D1 (de) | 2002-02-21 | 2010-11-11 | Univ California | Behandlungsverfahren unter verwendung von anti-cd22-antikörpern |
| EP1575480A4 (en) | 2002-02-22 | 2008-08-06 | Genentech Inc | COMPOSITIONS AND METHODS FOR TREATING DISEASES RELATED TO THE IMMUNE SYSTEM |
| WO2003073823A2 (en) | 2002-03-01 | 2003-09-12 | Exelixis, Inc. | PDPK1s AS MODIFIERS OF THE p53 PATHWAY AND METHODS OF USE |
| WO2003104399A2 (en) | 2002-06-07 | 2003-12-18 | Avalon Pharmaceuticals, Inc | Cancer-linked gene as target for chemotherapy |
| WO2003097803A2 (en) | 2002-05-15 | 2003-11-27 | Avalon Pharmaceuticals | Cancer-linked gene as target for chemotherapy |
| EP2258712A3 (en) | 2002-03-15 | 2011-05-04 | Multicell Immunotherapeutics, Inc. | Compositions and Methods to Initiate or Enhance Antibody and Major-histocompatibility Class I or Class II-restricted T Cell Responses by Using Immunomodulatory, Non-coding RNA Motifs |
| CA2486490A1 (en) | 2002-03-19 | 2003-12-31 | Curagen Corporation | Therapeutic polypeptides, nucleic acids encoding same, and methods of use |
| JP2005534286A (ja) | 2002-03-21 | 2005-11-17 | サネシス ファーマシューティカルズ, インコーポレイテッド | キナーゼインヒビターの同定 |
| EP1494693B1 (en) | 2002-03-22 | 2010-12-08 | Biogen Idec MA Inc. | Cripto-specific antibodies |
| US7193069B2 (en) | 2002-03-22 | 2007-03-20 | Research Association For Biotechnology | Full-length cDNA |
| AU2003245239A1 (en) | 2002-03-25 | 2003-11-03 | Uab Research Foundation | FC receptor homolog, reagents, and uses thereof |
| AU2003222103A1 (en) | 2002-03-28 | 2003-10-13 | Idec Pharmaceuticals Corporation | Novel gene targets and ligands that bind thereto for treatment and diagnosis of colon carcinomas |
| US20030194704A1 (en) | 2002-04-03 | 2003-10-16 | Penn Sharron Gaynor | Human genome-derived single exon nucleic acid probes useful for gene expression analysis two |
| BR0308953A (pt) | 2002-04-05 | 2006-03-14 | Agensys Inc | composições, proteìna, polinucleotìdeo, método de geração de uma resposta imune, método de detecção, composição farmacêutica, anticorpo ou seu fragmento, animal transgênico, hibridoma, método de fornecimento de um agente citotóxico ou agente de diagnóstico e método de inibição do crescimento de células cancerosas |
| US20040101874A1 (en) | 2002-04-12 | 2004-05-27 | Mitokor Inc. | Targets for therapeutic intervention identified in the mitochondrial proteome |
| CA2481507A1 (en) | 2002-04-16 | 2003-10-30 | Genentech, Inc. | Compositions and methods for the diagnosis and treatment of tumor |
| WO2003089904A2 (en) | 2002-04-17 | 2003-10-30 | Baylor College Of Medicine | Aib1 as a prognostic marker and predictor of resistance to encocrine therapy |
| AU2003228869A1 (en) | 2002-05-03 | 2003-11-17 | Incyte Corporation | Transporters and ion channels |
| AU2003232453A1 (en) | 2002-05-30 | 2003-12-19 | David K. Bol | Human solute carrier family 7 member 11 (hslc7a11) |
| AU2003239969A1 (en) | 2002-06-04 | 2003-12-19 | Avalon Pharmaceuticals, Inc. | Cancer-linked gene as target for chemotherapy |
| WO2003101283A2 (en) | 2002-06-04 | 2003-12-11 | Incyte Corporation | Diagnostics markers for lung cancer |
| WO2003104270A2 (en) | 2002-06-06 | 2003-12-18 | Ingenium Pharmaceuticals Ag | Dudulin 2 genes, expression products, non-human animal model: uses in human hematological disease |
| SG145559A1 (en) | 2002-06-06 | 2008-09-29 | Oncotherapy Science Inc | Genes and polypeptides relating to human colon cancers |
| AU2003245441A1 (en) | 2002-06-12 | 2003-12-31 | Avalon Pharmaceuticals, Inc. | Cancer-linked gene as target for chemotherapy |
| EP1552002A4 (en) | 2002-06-18 | 2006-02-08 | Archemix Corp | APTAMER TOXIN MOLECULES AND METHOD FOR THEIR USE |
| US20040249130A1 (en) | 2002-06-18 | 2004-12-09 | Martin Stanton | Aptamer-toxin molecules and methods for using same |
| JP4364121B2 (ja) | 2002-06-20 | 2009-11-11 | スナップトラック・インコーポレーテッド | 結合されたgps受信器と通信システムにおける相互干渉の低減 |
| WO2004000221A2 (en) | 2002-06-20 | 2003-12-31 | The Regents Of The University Of California | Compositions and methods for modulating lymphocyte activity |
| EP1534331B1 (en) | 2002-06-21 | 2014-10-29 | Johns Hopkins University School of Medicine | Membrane associated tumor endothelium markers |
| WO2004009622A2 (en) | 2002-07-19 | 2004-01-29 | Cellzome Ag | Protein complexes of cellular networks underlying the development of cancer and other diseases |
| WO2004011611A2 (en) | 2002-07-25 | 2004-02-05 | Genentech, Inc. | Taci antibodies and uses thereof |
| CA2494104A1 (en) | 2002-07-31 | 2004-04-22 | Seattle Genetics, Inc. | Anti-cd20 antibody-drug conjugates for the treatment of cancer and immune disorders |
| AU2003251471A1 (en) | 2002-08-06 | 2004-02-25 | Bayer Healthcare Ag | Diagnostics and therapeutics for diseases associated with human cxc chemokine receptor 5(cxcr5) |
| JP2004121218A (ja) | 2002-08-06 | 2004-04-22 | Jenokkusu Soyaku Kenkyusho:Kk | 気管支喘息または慢性閉塞性肺疾患の検査方法 |
| AU2003278725A1 (en) | 2002-08-27 | 2004-03-19 | Bristol-Myers Squibb Company | Polynucleotide predictor set for identifying protein tyrosine kinase modulators |
| WO2004020595A2 (en) | 2002-08-29 | 2004-03-11 | Five Prime Therapeutics, Inc. | Novel human polypeptides encoded by polynucleotides |
| AU2002951346A0 (en) | 2002-09-05 | 2002-09-26 | Garvan Institute Of Medical Research | Diagnosis of ovarian cancer |
| CA2496888A1 (en) | 2002-09-06 | 2004-03-18 | Mannkind Corporation | Epitope sequences |
| AU2003300776A1 (en) | 2002-09-09 | 2004-05-25 | Omeros Corporation | G protein coupled receptors and uses thereof |
| JP2004113151A (ja) | 2002-09-27 | 2004-04-15 | Sankyo Co Ltd | 癌遺伝子及びその用途 |
| CA2500978A1 (en) | 2002-10-03 | 2004-04-15 | Mcgill University | Antibodies and cyclic peptides which bind cea (carcinoembryonic antigen) and their use as cancer therapeutics |
| AU2003288918A1 (en) | 2002-10-04 | 2004-05-04 | Van Andel Research Institute | Molecular sub-classification of kidney tumors and the discovery of new diagnostic markers |
| WO2004042017A2 (en) * | 2002-10-31 | 2004-05-21 | Genentech, Inc. | Methods and compositions for increasing antibody production |
| JP2006516094A (ja) | 2002-11-08 | 2006-06-22 | ジェネンテック・インコーポレーテッド | ナチュラルキラー細胞関連疾患の治療のための組成物と方法 |
| WO2004044178A2 (en) | 2002-11-13 | 2004-05-27 | Genentech, Inc. | Methods and compositions for diagnosing dysplasia |
| US8007804B2 (en) | 2002-11-15 | 2011-08-30 | Musc Foundation For Research Development | Complement receptor 2 targeted complement modulators |
| AU2003294355A1 (en) | 2002-11-15 | 2004-06-15 | The Board Of Trustees Of The University Of Arkansas | Ca125 gene and its use for diagnostic and therapeutic interventions |
| AU2003297300A1 (en) | 2002-11-20 | 2004-06-15 | Biogen Idec Inc. | Novel gene targets and ligands that bind thereto for treatment and diagnosis of carcinomas |
| AU2003294462C1 (en) | 2002-11-21 | 2011-06-30 | University Of Utah Research Foundation | Purinergic modulation of smell |
| WO2004048938A2 (en) | 2002-11-26 | 2004-06-10 | Protein Design Labs, Inc. | Methods of detecting soft tissue sarcoma, compositions and methods of screening for soft tissue sarcoma modulators |
| WO2004053079A2 (en) | 2002-12-06 | 2004-06-24 | Diadexus, Inc. | Compositions, splice variants and methods relating to ovarian specific genes and proteins |
| US20040157278A1 (en) | 2002-12-13 | 2004-08-12 | Bayer Corporation | Detection methods using TIMP 1 |
| ES2388280T3 (es) | 2002-12-20 | 2012-10-11 | Abbott Biotherapeutics Corp. | Anticuerpos que reaccionan frente a GPR64 y utilización de los mismos |
| WO2004058309A1 (en) | 2002-12-23 | 2004-07-15 | Human Genome Sciences, Inc. | Neutrokine-alpha conjugate, neutrokine-alpha complex, and uses thereof |
| CA2512536A1 (en) | 2003-01-08 | 2004-07-29 | Bristol-Myers Squibb Company | Biomarkers and methods for determining sensitivity to epidermal growth factor receptor modulators |
| US20050181375A1 (en) | 2003-01-10 | 2005-08-18 | Natasha Aziz | Novel methods of diagnosis of metastatic cancer, compositions and methods of screening for modulators of metastatic cancer |
| US20050227301A1 (en) | 2003-01-10 | 2005-10-13 | Polgen | Cell cycle progression proteins |
| US20040171823A1 (en) | 2003-01-14 | 2004-09-02 | Nadler Steven G. | Polynucleotides and polypeptides associated with the NF-kappaB pathway |
| WO2004065576A2 (en) | 2003-01-15 | 2004-08-05 | Millennium Pharmaceuticals, Inc. | Methods and compositions for the treatment of urological disorder using differential expressed polypeptides |
| WO2004065417A2 (en) * | 2003-01-23 | 2004-08-05 | Genentech, Inc. | Methods for producing humanized antibodies and improving yield of antibodies or antigen binding fragments in cell culture |
| US20060228710A1 (en) | 2003-02-14 | 2006-10-12 | Morris David W | Novel therapeutic targets in cancer |
| US20030224411A1 (en) | 2003-03-13 | 2003-12-04 | Stanton Lawrence W. | Genes that are up- or down-regulated during differentiation of human embryonic stem cells |
| CA2652945C (en) * | 2006-05-30 | 2015-06-02 | Genentech, Inc. | Antibodies and immunoconjugates and uses therefor |
| CA2676766A1 (en) * | 2007-02-09 | 2008-08-21 | Genentech, Inc. | Anti-robo4 antibodies and uses therefor |
| AU2008251608B2 (en) * | 2007-05-08 | 2014-03-27 | Genentech, Inc. | Cysteine engineered anti-MUC16 antibodies and antibody drug conjugates |
| CN101802013B (zh) * | 2007-07-16 | 2014-07-02 | 健泰科生物技术公司 | 人源化抗cd79b抗体和免疫偶联物及使用方法 |
| NZ583367A (en) * | 2007-07-16 | 2012-10-26 | Genentech Inc | Anti-cd79b antibodies and immunoconjugates and methods of use |
| CA2698541C (en) * | 2007-10-19 | 2018-01-09 | Genentech, Inc. | Cysteine engineered anti-tenb2 antibodies and antibody drug conjugates |
-
2009
- 2009-11-05 US US12/612,912 patent/US20100111856A1/en not_active Abandoned
-
2010
- 2010-11-04 KR KR1020177034613A patent/KR20170136652A/ko not_active Application Discontinuation
- 2010-11-04 CN CN201080050134.4A patent/CN102596260B/zh active Active
- 2010-11-04 RU RU2012123007/10A patent/RU2562862C2/ru active
- 2010-11-04 CA CA2780216A patent/CA2780216A1/en not_active Abandoned
- 2010-11-04 BR BR112012007774A patent/BR112012007774A2/pt not_active IP Right Cessation
- 2010-11-04 MX MX2012005211A patent/MX340674B/es active IP Right Grant
- 2010-11-04 JP JP2012537224A patent/JP5850843B2/ja active Active
- 2010-11-04 EP EP10776064.7A patent/EP2496270B1/en active Active
- 2010-11-04 KR KR1020127011607A patent/KR20120102625A/ko not_active Application Discontinuation
- 2010-11-04 WO PCT/US2010/055465 patent/WO2011056983A1/en active Application Filing
-
2014
- 2014-06-10 US US14/300,630 patent/US20150017094A1/en not_active Abandoned
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2006034488A2 (en) * | 2004-09-23 | 2006-03-30 | Genentech, Inc. | Cysteine engineered antibodies and conjugates |
Non-Patent Citations (1)
| Title |
|---|
| DIJKERS E.C.F. et al, Development and Characterization of Clinical-Grade 89Zr-Trastuzumab for HER2/neu ImmunoPET Imaging, The Journal of Nuclear Medicine, 2009, v. 50, n. 6, p. 974-981. GOVINDAN S.V. et al, Deferoxamine as a chelator for 67Ga in the preparation of antibody conjugates, 2005, v. 32, p. 513-519. РОЙТ А. и др., Иммунология, Москва, "Мир", 2000, стр. 110-111 * |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US11779649B2 (en) | 2018-12-21 | 2023-10-10 | Novartis Ag | Antibodies to PMEL17 and conjugates thereof |
Also Published As
| Publication number | Publication date |
|---|---|
| EP2496270B1 (en) | 2018-01-31 |
| RU2012123007A (ru) | 2013-12-10 |
| KR20170136652A (ko) | 2017-12-11 |
| JP2013510093A (ja) | 2013-03-21 |
| US20100111856A1 (en) | 2010-05-06 |
| MX340674B (es) | 2016-07-20 |
| CA2780216A1 (en) | 2011-05-12 |
| WO2011056983A1 (en) | 2011-05-12 |
| CN102596260A (zh) | 2012-07-18 |
| MX2012005211A (es) | 2012-06-13 |
| KR20120102625A (ko) | 2012-09-18 |
| JP5850843B2 (ja) | 2016-02-03 |
| CN102596260B (zh) | 2017-04-05 |
| BR112012007774A2 (pt) | 2016-11-22 |
| US20150017094A1 (en) | 2015-01-15 |
| EP2496270A1 (en) | 2012-09-12 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| RU2562862C2 (ru) | Конъюгаты меченных радиоактивным цирконием сконструированных антител с цистеиновыми заменами | |
| US11873330B2 (en) | Cysteine engineered antibodies and conjugates | |
| JP4948413B2 (ja) | システイン操作抗体および結合体 | |
| MX2007003404A (es) | Anticuerpos y conjugados diseñados de cisteina |