ES2909143T3 - Microbios de Chlorella o Prototheca modificados genéticamente y aceite producido a partir de estos - Google Patents
Microbios de Chlorella o Prototheca modificados genéticamente y aceite producido a partir de estos Download PDFInfo
- Publication number
- ES2909143T3 ES2909143T3 ES18212337T ES18212337T ES2909143T3 ES 2909143 T3 ES2909143 T3 ES 2909143T3 ES 18212337 T ES18212337 T ES 18212337T ES 18212337 T ES18212337 T ES 18212337T ES 2909143 T3 ES2909143 T3 ES 2909143T3
- Authority
- ES
- Spain
- Prior art keywords
- oil
- microbe
- prototheca
- gene
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B1/00—Production of fats or fatty oils from raw materials
- C11B1/02—Pretreatment
- C11B1/025—Pretreatment by enzymes or microorganisms, living or dead
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/64—Fats; Fatty oils; Ester-type waxes; Higher fatty acids, i.e. having at least seven carbon atoms in an unbroken chain bound to a carboxyl group; Oxidised oils or fats
- C12P7/6436—Fatty acid esters
- C12P7/6445—Glycerides
- C12P7/6463—Glycerides obtained from glyceride producing microorganisms, e.g. single cell oil
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23D—EDIBLE OILS OR FATS, e.g. MARGARINES, SHORTENINGS, COOKING OILS
- A23D9/00—Other edible oils or fats, e.g. shortenings, cooking oils
- A23D9/007—Other edible oils or fats, e.g. shortenings, cooking oils characterised by ingredients other than fatty acid triglycerides
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23D—EDIBLE OILS OR FATS, e.g. MARGARINES, SHORTENINGS, COOKING OILS
- A23D9/00—Other edible oils or fats, e.g. shortenings, cooking oils
- A23D9/02—Other edible oils or fats, e.g. shortenings, cooking oils characterised by the production or working-up
-
- C—CHEMISTRY; METALLURGY
- C10—PETROLEUM, GAS OR COKE INDUSTRIES; TECHNICAL GASES CONTAINING CARBON MONOXIDE; FUELS; LUBRICANTS; PEAT
- C10M—LUBRICATING COMPOSITIONS; USE OF CHEMICAL SUBSTANCES EITHER ALONE OR AS LUBRICATING INGREDIENTS IN A LUBRICATING COMPOSITION
- C10M169/00—Lubricating compositions characterised by containing as components a mixture of at least two types of ingredient selected from base-materials, thickeners or additives, covered by the preceding groups, each of these compounds being essential
- C10M169/04—Mixtures of base-materials and additives
- C10M169/041—Mixtures of base-materials and additives the additives being macromolecular compounds only
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B1/00—Production of fats or fatty oils from raw materials
- C11B1/06—Production of fats or fatty oils from raw materials by pressing
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B1/00—Production of fats or fatty oils from raw materials
- C11B1/10—Production of fats or fatty oils from raw materials by extracting
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B3/00—Refining fats or fatty oils
- C11B3/001—Refining fats or fatty oils by a combination of two or more of the means hereafter
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B3/00—Refining fats or fatty oils
- C11B3/006—Refining fats or fatty oils by extraction
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B3/00—Refining fats or fatty oils
- C11B3/008—Refining fats or fatty oils by filtration, e.g. including ultra filtration, dialysis
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B3/00—Refining fats or fatty oils
- C11B3/02—Refining fats or fatty oils by chemical reaction
- C11B3/06—Refining fats or fatty oils by chemical reaction with bases
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B3/00—Refining fats or fatty oils
- C11B3/10—Refining fats or fatty oils by adsorption
-
- C—CHEMISTRY; METALLURGY
- C11—ANIMAL OR VEGETABLE OILS, FATS, FATTY SUBSTANCES OR WAXES; FATTY ACIDS THEREFROM; DETERGENTS; CANDLES
- C11B—PRODUCING, e.g. BY PRESSING RAW MATERIALS OR BY EXTRACTION FROM WASTE MATERIALS, REFINING OR PRESERVING FATS, FATTY SUBSTANCES, e.g. LANOLIN, FATTY OILS OR WAXES; ESSENTIAL OILS; PERFUMES
- C11B3/00—Refining fats or fatty oils
- C11B3/12—Refining fats or fatty oils by distillation
- C11B3/14—Refining fats or fatty oils by distillation with the use of indifferent gases or vapours, e.g. steam
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
- C12N9/2405—Glucanases
- C12N9/2434—Glucanases acting on beta-1,4-glucosidic bonds
- C12N9/2442—Chitinase (3.2.1.14)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/64—Fats; Fatty oils; Ester-type waxes; Higher fatty acids, i.e. having at least seven carbon atoms in an unbroken chain bound to a carboxyl group; Oxidised oils or fats
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/64—Fats; Fatty oils; Ester-type waxes; Higher fatty acids, i.e. having at least seven carbon atoms in an unbroken chain bound to a carboxyl group; Oxidised oils or fats
- C12P7/6409—Fatty acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y301/00—Hydrolases acting on ester bonds (3.1)
- C12Y301/02—Thioester hydrolases (3.1.2)
- C12Y301/02014—Oleoyl-[acyl-carrier-protein] hydrolase (3.1.2.14), i.e. ACP-thioesterase
-
- H—ELECTRICITY
- H01—ELECTRIC ELEMENTS
- H01B—CABLES; CONDUCTORS; INSULATORS; SELECTION OF MATERIALS FOR THEIR CONDUCTIVE, INSULATING OR DIELECTRIC PROPERTIES
- H01B3/00—Insulators or insulating bodies characterised by the insulating materials; Selection of materials for their insulating or dielectric properties
- H01B3/18—Insulators or insulating bodies characterised by the insulating materials; Selection of materials for their insulating or dielectric properties mainly consisting of organic substances
- H01B3/20—Insulators or insulating bodies characterised by the insulating materials; Selection of materials for their insulating or dielectric properties mainly consisting of organic substances liquids, e.g. oils
-
- H—ELECTRICITY
- H01—ELECTRIC ELEMENTS
- H01F—MAGNETS; INDUCTANCES; TRANSFORMERS; SELECTION OF MATERIALS FOR THEIR MAGNETIC PROPERTIES
- H01F27/00—Details of transformers or inductances, in general
- H01F27/28—Coils; Windings; Conductive connections
- H01F27/32—Insulating of coils, windings, or parts thereof
- H01F27/321—Insulating of coils, windings, or parts thereof using a fluid for insulating purposes only
-
- H—ELECTRICITY
- H01—ELECTRIC ELEMENTS
- H01F—MAGNETS; INDUCTANCES; TRANSFORMERS; SELECTION OF MATERIALS FOR THEIR MAGNETIC PROPERTIES
- H01F27/00—Details of transformers or inductances, in general
- H01F27/08—Cooling; Ventilating
- H01F27/10—Liquid cooling
- H01F27/105—Cooling by special liquid or by liquid of particular composition
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Wood Science & Technology (AREA)
- Oil, Petroleum & Natural Gas (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Microbiology (AREA)
- Health & Medical Sciences (AREA)
- Zoology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biotechnology (AREA)
- General Chemical & Material Sciences (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Power Engineering (AREA)
- Physics & Mathematics (AREA)
- Polymers & Plastics (AREA)
- Food Science & Technology (AREA)
- Medicinal Chemistry (AREA)
- Cell Biology (AREA)
- Spectroscopy & Molecular Physics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Lubricants (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Fats And Perfumes (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Immobilizing And Processing Of Enzymes And Microorganisms (AREA)
- Enzymes And Modification Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
icrobio de Chlorella o Prototheca modificado genéticamdente para eliminar o regular a la acil-ACP tioesterasa.
Description
DESCRIPCIÓN
Microbios de Chlorella o Prototheca modificados genéticamente y aceite producido a partir de estos
CAMPO DE LA INVENCIÓN
[0001] La presente invención se refiere a la producción de aceites a partir de microorganismos.
ANTECEDENTES
[0002] Combustible fósil es un término general para los depósitos geológicos combustibles enterrados de materiales orgánicos, formados a partir de plantas y animales en descomposición que se han convertido en petróleo crudo, carbón, gas natural o aceites pesados por exposición al calor y a la presión en la corteza terrestre durante cientos de de millones de años. Los combustibles fósiles son un recurso finito y no renovable.
[0003] Muchas industrias, incluyendo las de la fabricación de plásticos y productos químicos, dependen en gran medida de la disponibilidad de hidrocarburos como materia prima para sus procesos de fabricación.
[0004] En la publicación PCT n.02008/151149 se describen métodos y materiales para el cultivo de microalgas para la producción de aceite, la extracción de aceite microbiano y la producción de alimentos, aceite para alimentos, combustibles y otros productos oleoquímicos a partir del aceite producido por microbios oleaginosos.
[0005] Una aplicación oleoquímica importante es la producción de fluidos dieléctricos industriales, que se utilizan para el aislamiento eléctrico y la refrigeración o disipación de calor en transformadores y otros dispositivos eléctricos. Entre estos dispositivos eléctricos se incluyen transformadores de potencia y distribución, disyuntores, condensadores, aparamenta, máquinas de rayos X y cables aislantes.
[0006] El aceite de origen biológico, en particular el aceite de soja con alto contenido de ácido oleico, se ha utilizado como fluido dieléctrico en transformadores herméticos desde la década de 1990 (véase Srivastava (2009) Int'l J Computer Electrical Eng, v. 1(2) págs. 212-216). Los fluidos dieléctricos de origen biológico actuales son triacilgliceroles (TAG) de alto contenido de ácido oleico purificados con aditivos incorporados (véase la patente de EE. UU. n.o 6,274,067 y la patente de EE. UU. n.o 8,043,0607). Por ejemplo, los principales beneficios del fluido dieléctrico de aceite de soja con alto contenido de ácido oleico en comparación con el fluido dieléctrico a base de aceite mineral son (i) un mayor punto de inflamación (2x), (ii) una mayor vida útil del transformador (4-8x) y (iii) un menor coste en remediar vertidos debido a la alta biodegradabilidad del aceite de origen biológico (>3x) y su menor toxicidad (véase Schneider (2006) J Sci Food Agric, v. 86 págs.: 1769-1780). En W02010/063031 describe un método para producir un aceite, que comprende cultivar Prototheca que contiene un gen exógeno y aislar los lípidos de esta. En W02011/008565 se describe un método para producir un producto de ácido graso, que comprende cultivar, por ejemplo, Chlorella o Prototheca que comprende una molécula de ácido nucleico que codifica acil-ACP tioesterasa exógena. En W02010/120939 se describe aceite microbiano de Chlorella o Prototheca que contiene al menos un 60 % de ácido graso C18:1.
[0007] Las principales desventajas de los aceites de origen biológico en comparación con los aceites de origen mineral son la inestabilidad oxidativa de los aceites de origen biológico, el mayor coste de la adquisición de aceites de origen biológico y la transición de los equipos de aceites de origen mineral a aceites de origen biológico (véase Schneider (2006), supra). Aunque los fluidos dieléctricos de origen biológico ocupan una parte importante del mercado de fluidos dieléctricos, los fluidos dieléctricos de aceite mineral dominan el mercado en la actualidad. 0tra desventaja significativa es el coste de producción de estos aceites a base de soja y su desviación de una importante fuente de alimentos hacia aplicaciones no alimentarias.
RESUMEN
[0008] La invención está definida por el objeto de las reivindicaciones. En algunas formas de realización, la presente invención proporciona un microbio de Chlorella o Prototheca modificado genéticamente para eliminar o regular a la baja un gen endógeno de acil-ACP tioesterasa. En una forma de realización preferida, el microbio se manipula para alterar una acil-ACP tioesterasa endógena. En una forma de realización preferida, la acil-ACP tioesterasa endógena se elimina mediante recombinación homóloga. En una forma de realización preferida, el microbio se manipula para que exprese un ARN inhibidor que se dirige a una acil-ACP tioesterasa endógena. En una forma de realización preferida, el microbio comprende además uno o más genes exógenos que son expresados por la célula. En una forma de realización preferida, el gen exógeno codifica una sacarasa, una acil-ACP tioesterasa o una desaturasa. En una forma de realización preferida, los genes exógenos codifican: una sacarasa; una acil-ACP tioesterasa; o una
sacarasa y una acil-ACP tioesterasa. En una forma de realización preferida, el microbio es Prototheca moriformis o Chlorella protothecoides. Más preferiblemente, el microbio es Prototheca moriformis.
[0009] En algunas formas de realización, la invención proporciona un método para producir un aceite microbiano, donde el método comprende: a) cultivar un microbio como se ha definido anteriormente; y b) separar el aceite del microbio. En una forma de realización preferida, el método incluye además el paso de: c) someter el aceite a refinado, blanqueo, desodorización o desgomado para producir aceite microbiano RBD. En una forma de realización preferida, el método comprende además añadir al aceite microbiano un antioxidante, un desactivador de iones metálicos, un inhibidor de la corrosión, un desemulsionante, un aditivo antidesgaste, un inhibidor del punto de fluidez o un compuesto antihidrólisis.
[0010] En determinadas formas de realización, la invención proporciona un aceite producido mediante un método como se ha definido anteriormente, en el que el aceite comprende menos del 7,5 % de C18:2 y en el que: i) el número de copias del gen exógeno es superior a 2; ii) el gen exógeno es el gen de tioesterasa FatB2 de Cuphea wrightii; y iii) el microbio es Prototheca moriformis.
BREVE DESCRIPCIÓN DE LOS DIBUJOS
[0011]
Figura 1. Perfil de enfriamiento típico para el fraccionamiento de aceite RBD (Tf = temperatura de filtración). Figura 2. Perfil de enfriamiento típico para el fraccionamiento de oleína de algas (Tf = temperatura de filtración). Figura 3. Efecto de VPL 10-310 en el punto de fluidez del aceite de algas y aceites fraccionados. "Aceite desodorizado" es aceite RBD; "oleína" es oleína # 1; "súper oleína" es súper oleína #1".
DESCRIPCIÓN DETALLADA
[0012] La presente invención surge, en parte, del descubrimiento de que Prototheca y otros microorganismos oleaginosos presentan, en determinadas formas de realización, propiedades inesperadamente ventajosas para la producción de fluidos dieléctricos, entre otras aplicaciones, tales como lubricantes biodegradables, especialmente aceites de motor y fluidos hidráulicos, que anteriormente estaban hechos principalmente a base de aceites minerales. Los lubricantes a base de aceite microbiano se pueden usar para reemplazar los lubricantes de petróleo en espadas de motosierra, lodos y aceites de perforación, fluidos puros para trabajar metales, lubricantes para la industria alimentaria, aceites para engranajes abiertos, grasas biodegradables, fluidos hidráulicos, aceites marinos y lubricantes para motores fuera de borda, aceites para bombas de agua y subterráneas, lubricantes para raíles, lubricantes para amortiguadores, aceites para tractores, lubricantes para equipos agrícolas, aceites para ascensores, aceites para desmoldeo, lubricantes para motores de dos tiempos y otros lubricantes.
[0013] La presente invención surge también, en parte, del descubrimiento de procesos de modificación de aceites microbianos para reducir su punto de fluidez. La transesterificación de lípidos produce ésteres de ácidos grasos de cadena larga. Se pueden adaptar otros procesos enzimáticos y químicos para producir ácidos grasos, aldehídos, alcoholes, alcanos y alquenos. En algunas aplicaciones, se producen compuestos de hidrocarburos útiles en fluidos dieléctricos.
[0014] Esta descripción detallada se divide en secciones para la comodidad del lector. En la sección I se proporcionan definiciones de los términos utilizados en este documento. En la sección II se proporciona una descripción de las condiciones de cultivo útiles en las formas de realización de los métodos de la invención. En la sección III se proporciona una descripción de los métodos y materiales de ingeniería genética. En la sección IV se proporciona una descripción de la ingeniería genética de microbios para permitir la utilización de sacarosa, con referencia específica a las microalgas, como por ejemplo Prototheca. En la sección V se proporciona una descripción de la ingeniería genética para modificar la biosíntesis de lípidos. En la sección VI se describen métodos para fabricar aceites microbianos de formas de realización de la invención y productos derivados de estos, tales como fluidos dieléctricos. En la sección VII se describen ejemplos que ilustran las diversas formas de realización de la invención.
I. DEFINICIONES
[0015] A menos que se definan de otro modo, todos los términos técnicos y científicos utilizados en este documento tienen el significado comúnmente entendido por una persona experta en la materia a la que pertenece esta invención. Las siguientes referencias proporcionan a la persona experta una definición general de muchos de los términos utilizados en esta invención: Singleton y col., Dictionary of Microbiology and Molecular Biology (2nd ed.
1994); The Cambridge Dictionary of Science and Technology (Walker ed., 1988); The Glossary of Genetics, 5th Ed.,
R. Rieger y col. (eds.), Springer Verlag (1991); y Hale & Marham, The Harper Collins Dictionary of Biology (1991). Tal como se usan en este documento, los siguientes términos tienen los significados que se les atribuyen a menos que se especifique lo contrario.
[0016] "Activo en las microalgas" se refiere a un ácido nucleico que es funcional en las microalgas. Por ejemplo, de un promotor que se ha utilizado para activar un gen de resistencia a los antibióticos para dotar de resistencia a los antibióticos a una microalga transgénica, se dice que es activo en las microalgas.
[0017] La "proteína transportadora de acilo" o "ACP" es una proteína que se une a una cadena de acilo en crecimiento durante la síntesis de ácidos grasos como un tioéster en el tiol distal del resto 4'-fosfopanteteína y comprende un componente del complejo de la ácido graso sintasa.
[0018] "Molécula de acil-CoA" o "acil-CoA" es una molécula que comprende un resto acilo unido covalentemente a la coenzima A a través de un enlace tioéster en el tiol distal del resto 4'-fosfopanteteína de la coenzima A.
[0019] "Antioxidante" es una molécula que es capaz de inhibir la oxidación de otras moléculas. Los antioxidantes se suelen añadir a los productos industriales. Un uso común es como estabilizadores en combustibles y lubricantes para evitar la oxidación, y en gasolinas para evitar la polimerización que conduce a la formación de residuos que ensucian el motor. También se usan mucho para evitar la degradación oxidativa de polímeros como cauchos, plásticos y adhesivos que provoca una pérdida de resistencia y flexibilidad en estos materiales.
[0020] "Compuesto antihidrólisis" es una molécula que inhibe la descomposición de un compuesto químico por reacción con el agua. Las carbodiimidas, por ejemplo, se pueden emplear como compuestos antihidrólisis. Los compuestos antihidrólisis están disponibles en el mercado, por ejemplo, en SpecialChem, entre otros.
[0021] El "aditivo antidesgaste" es un aditivo para un fluido (por ejemplo, un aceite lubricante) que da como resultado una vida útil más prolongada de la máquina debido a una mayor resistencia al desgaste y al gripado de los componentes. Los aditivos antidesgaste evitan el contacto directo de metal con metal entre las piezas de la máquina cuando desaparece la película de aceite. Por lo general, el aditivo reacciona con el metal en la superficie de la pieza y forma una película que puede deslizarse sobre la superficie de fricción. Los aditivos antidesgaste suelen contener compuestos de zinc y fósforo. Los ejemplos de aditivos antidesgaste incluyen ditiofosfato de zinc (ZDP), dialquilditiofosfato de zinc (ZDDP, que también actúa como inhibidor de la corrosión y antioxidante), fosfato de tricresilo (TCP, utilizado para operaciones a alta temperatura), halocarbonos (parafinas cloradas, para operaciones con presión extrema), monooleato de glicerol, ácido esteárico (que se adhiere a las superficies a través de un proceso de adsorción reversible por debajo de 150 °C, útil para condiciones de contacto moderado).
[0022] "Porcentaje de área" se refiere al área de los picos observados utilizando los métodos de detección FAME GC/FID en los que cada ácido graso de la muestra se convierte en un éster metílico de ácido graso (FAME) antes de la detección. Por ejemplo, se observa un pico separado para un ácido graso de 14 átomos de carbono sin instauración (C14:0) en comparación con cualquier otro ácido graso como C14:1. El área del pico para cada clase de FAME es directamente proporcional a su composición porcentual en la mezcla y se calcula en función de la suma de todos los picos presentes en la muestra (es decir, [área bajo el pico específico/área total de todos los picos medidos] X 100). Cuando se hace referencia a los perfiles de lípidos (ácidos grasos) de los aceites y las células descritos en el presente documento, "al menos 4 % C8-C14" significa que al menos el 4 % de los ácidos grasos totales de la célula o de la composición de glicerolípidos extraída tienen una longitud de cadena que incluye 8, 10, 12 o 14 átomos de carbono.
[0023] "Axénico" se refiere a un cultivo de un organismo sin contaminación por otros organismos vivos.
[0024] "Biodiésel" es un éster alquílico de ácido graso producido de forma biológica que es apto para el uso como combustible en un motor diésel.
[0025] "Biomasa" es material producido por crecimiento y/o propagación de células. La biomasa puede contener células y/o contenidos intracelulares, así como material extracelular, incluidos, entre otros, compuestos secretados por una célula.
[0026] "Biorreactor" es un recinto o recinto parcial en el que se cultivan células, opcionalmente en suspensión.
[0027] La “tensión de ruptura" de un fluido dieléctrico es el voltaje al cual el fluido dieléctrico pierde sus propiedades aislantes.
[0028] "Catalizador" es un agente, como una molécula o un complejo macromolecular, capaz de facilitar o propiciar una reacción química de un reactivo a un producto sin convertirse en parte del producto. Un catalizador aumenta la velocidad de una reacción, después de lo cual, el catalizador puede actuar sobre otro reactivo para formar el producto. Un catalizador reduce por lo general la energía de activación total requerida para la reacción, de modo que avanza más rápido o a una temperatura más baja. Por lo tanto, se puede alcanzar más rápidamente un equilibrio de reacción. Entre los ejemplos de catalizadores se incluyen las enzimas, que son catalizadores biológicos; el calor, que es un catalizador no biológico; y metales utilizados en los procesos de refinado de combustible fósil.
[0029] "Material celulósico" es el producto de la digestión de celulosa, incluidas la glucosa y la xilosa y, opcionalmente, compuestos adicionales tales como disacáridos, oligosacáridos, lignina, furfurales y otros compuestos. Los ejemplos no excluyentes de fuentes de material celulósico incluyen bagazo de caña de azúcar, pulpa de remolacha azucarera, rastrojo de maíz, astillas de madera, serrín y pasto varilla.
[0030] "Cocultivo", y sus variantes como "cultivo conjunto" y "cofermentación”, se refieren a la presencia de dos o más tipos de células en el mismo biorreactor. Ambos de los dos o más tipos de células pueden ser microorganismos, tales como microalgas, o pueden ser células de microalgas cultivadas con un tipo de célula diferente. Las condiciones de cultivo pueden ser aquellas que fomenten el crecimiento y/o la propagación de los dos o más tipos de células o aquellas que faciliten el crecimiento y/o la proliferación de una, o un subconjunto, de las dos o más células mientras se mantiene el crecimiento celular para el resto.
[0031] "Cofactor" es cualquier molécula, distinta del sustrato, necesaria para que una enzima lleve a cabo su actividad enzimática.
[0032] El "ADN complementario" o "ADNc" es una copia de ADN del ARNm, generalmente obtenido por transcripción inversa del ARN mensajero (ARNm) o amplificación (por ejemplo, a través de la reacción en cadena de la polimerasa ("PCR")).
[0033] “Inhibidor de la corrosión" es una molécula que, cuando se añade a un fluido, disminuye la velocidad de corrosión de un metal o una aleación en contacto con el fluido.
[0034] "Cultivado", y sus variantes como "fermentado", se refieren al fomento intencional del crecimiento (aumentos del tamaño celular, del contenido celular y/o de la actividad celular) y/o propagación (aumentos del número de células a través de la mitosis) de una o más células mediante el uso de condiciones seleccionadas y/o controladas. La combinación de crecimiento y propagación puede denominarse proliferación. Entre los ejemplos de condiciones seleccionadas y/o controladas se incluyen el uso de un medio definido (con características conocidas como pH, fuerza iónica y fuente de carbono), temperatura especificada, tensión de oxígeno, niveles de dióxido de carbono y crecimiento en un biorreactor. Cultivar no se refiere al crecimiento o propagación de microorganismos en la naturaleza o de otro modo sin intervención humana; por ejemplo, el crecimiento natural de un organismo que finalmente se fosiliza para producir crudo geológico no es un cultivo.
[0035] La "citólisis" es la lisis de células en un ambiente hipotónico. La citólisis es causada por una ósmosis excesiva, o movimiento de agua, hacia el interior de una célula (hiperhidratación). La célula no puede soportar la presión osmótica del agua en su interior, por lo que explota.
[0036] La "harina deslipidada" y la "biomasa microbiana deslipidada" es la biomasa microbiana después de que el aceite (incluidos los lípidos) se haya extraído o aislado de ella, ya sea mediante el uso de extracción mecánica (es decir, ejercida por una prensa extrusora) o con disolventes, o ambas. La harina deslipidada tiene una cantidad reducida de aceite/lípidos en comparación con antes de la extracción o el aislamiento de aceite/lípidos de la biomasa microbiana, pero contiene algo de aceite/lípido residual.
[0037] "Desemulsionante" es una molécula que rompe las emulsiones (generalmente emulsiones líquido-líquido) o evita que se formen. Los desemulsionantes se basan típicamente en las siguientes químicas: resinas de fenolformaldehído catalizadas por ácido, resinas de fenol-formaldehído catalizadas por base, poliaminas, diepóxidos, polioles. Estas moléculas suelen estar etoxiladas (y/o propoxiladas) para proporcionar el grado deseado de solubilidad en agua/aceite. La adición de óxido de etileno aumenta la solubilidad en agua, mientras que el óxido de propileno la disminuye. Las formulaciones desemulsionantes disponibles en el mercado suelen ser una mezcla de dos a cuatro compuestos químicos diferentes, en disolvente(s) portador(es) como xileno, nafta aromática pesada (HAN), isopropanol, metanol, 2-etilhexanol o diésel.
[0038] "Dieléctrico" o un "fluido dieléctrico" es un fluido que no tiene conductividad, o que tiene un nivel muy bajo de conductividad, de una corriente eléctrica en circunstancias normales (o en las circunstancias de uso previstas). Los
fluidos dieléctricos se utilizan para el aislamiento eléctrico, la refrigeración y la lubricación, por ejemplo, en transformadores y otros dispositivos eléctricos. Entre los dispositivos eléctricos que utilizan fluidos dieléctricos se incluyen los transformadores de potencia y distribución, disyuntores, condensadores, interruptores, máquinas de rayos X y cables aislantes.
[0039] La "rigidez dieléctrica" de un material (por ejemplo, un aislante) es el voltaje máximo requerido para producir una ruptura dieléctrica, esto es, un fallo de sus propiedades aislantes, expresado en voltios por unidad de espesor. La rigidez dieléctrica de un material se puede determinar según los métodos estándar, por ejemplo, los métodos de prueba ASTM D1816, D877, D3300, D117, D2413, D6180, D6181 o D1310.
[0040] "Vector de expresión" o "construcción de expresión" o "plásmido" o "construcción de ADN recombinante" se refiere a un ácido nucleico que se ha generado mediante la intervención humana, incluyendo por medios recombinantes o síntesis química directa, con una serie de elementos de ácido nucleico especificados que permiten la transcripción y/o traducción de un ácido nucleico concreto en una célula hospedadora. El vector de expresión puede formar parte de un plásmido, virus o fragmento de ácido nucleico. Típicamente, el vector de expresión incluye un ácido nucleico que se va a transcribir unido operativamente a un promotor.
[0041] "Gen exógeno" es un ácido nucleico que codifica la expresión de un ARN y/o una proteína que se ha introducido ("transformado") en una célula, y también se denomina "transgén". Una célula transformada puede denominarse célula recombinante, en la que pueden introducirse genes exógenos adicionales. El gen exógeno puede ser de una especie diferente (y, por tanto, heterólogo), o de la misma especie (y, por tanto, homólogo), con respecto a la célula que se está transformando. Por lo tanto, un gen exógeno puede incluir un gen homólogo que ocupa una ubicación diferente en el genoma de la célula o que está bajo un control diferente, en relación con la copia endógena del gen. Un gen exógeno puede estar presente en más de una copia en la célula. Un gen exógeno puede mantenerse en una célula como una inserción en el genoma (nuclear o plásmido) o como una molécula episomal.
[0042] “Proporcionado exógenamente’’ se refiere a una molécula proporcionada al medio de cultivo de un cultivo celular.
[0043] El "prensado con extrusión" es un método mecánico para extraer aceite de materias primas como la soja y la colza. Una prensa extrusora es una máquina de tipo tornillo, que presiona el material a través de una cavidad similar a un barril enjaulado. Las materias primas entran por un lado de la prensa y la torta prensada sale por el otro lado mientras el aceite se filtra entre las barras de la jaula y se recoge. La máquina utiliza la fricción y la presión continua de los tornillos para mover y comprimir la materia prima. El aceite se filtra a través de pequeñas aberturas que no permiten el paso de sólidos. A medida que se presiona la materia prima, la fricción generalmente hace que se caliente.
[0044] "Ácido graso" es un ácido carboxílico con una cola (cadena) alifática larga. La porción alifática del ácido graso puede estar completamente saturada (sin enlace(s) doble(s)) o puede estar insaturada en una o más porciones de la molécula. La mayoría de los ácidos grasos naturales tienen una cadena con un número par de átomos de carbono, de 4 a 28. Los ácidos grasos pueden ser componentes de triglicéridos u otros lípidos, por ejemplo, fosfolípidos, esfingolípidos. Los ácidos grasos se pueden caracterizar por "números de lípidos". Los números de lípidos toman la forma C:D, donde C es el número de átomos de carbono del ácido graso y D es el número de enlaces dobles del ácido graso. En consecuencia, "C18:1" se refiere a un ácido graso con 18 carbonos y 1 enlace doble, mientras que "C18:2" se refiere a un ácido graso con 18 carbonos y 2 enlaces dobles.
[0045] La "acil-ACP tioesterasa" es una enzima que cataliza la escisión de un ácido graso de una proteína transportadora de acilo (ACP) durante la síntesis de lípidos.
[0046] La "acil-CoA/aldehído reductasa" es una enzima que cataliza la reducción de una molécula de acil-CoA a un alcohol primario.
[0047] La "acil-CoA reductasa" es una enzima que cataliza la reducción de una molécula de acil-CoA a un aldehído.
[0048] La "aldehído graso descarbonilasa" es una enzima que cataliza la conversión de un aldehído graso en un alcano.
[0049] La "aldehído graso reductasa" es una enzima que cataliza la reducción de un aldehído a un alcohol primario.
[0050] El punto de inflamación de un material es la temperatura a la que continuará ardiendo durante al menos 5 segundos después de la ignición por una llama abierta. El punto de inflamación se puede determinar según métodos estándar, por ejemplo, métodos de prueba ASTM D92 o D1310.
[0051] El "punto de inflamación" es la temperatura más baja a la que un material puede vaporizarse para formar una mezcla inflamable en el aire. En el punto de inflamación, el material puede encenderse, pero es posible que los vapores producidos durante la ignición no se produzcan a una velocidad suficiente para sostener la combustión. El punto de inflamación se puede determinar según métodos estándar, por ejemplo, métodos de prueba ASTM D3278, D3828, D56 o D93.
[0052] "Fuente de carbono fijo" es una molécula o moléculas que contienen carbono, normalmente una molécula orgánica, que está presente a temperatura y presión ambiente en forma sólida o líquida en un medio de cultivo y que puede ser utilizada por un microorganismo que se cultiva en este.
[0053] "Heterótrofo”, en lo que respecta a las condiciones de cultivo, es un cultivo en ausencia sustancial de luz mientras se utiliza o metaboliza una fuente de carbono fijo.
[0054] "Homogeneizado" es biomasa que se ha disgregado físicamente.
[0055] "Fluido hidráulico" es el fluido que sirve como medio de transmisión de potencia en un sistema hidráulico.
[0056] "Hidrocarburo" es una molécula que contiene únicamente átomos de hidrógeno y de carbono en la que los átomos de carbono están unidos covalentemente para formar una cadena lineal, ramificada, cíclica o parcialmente cíclica a la que están unidos los átomos de hidrógeno. La estructura molecular de los compuestos de hidrocarburos varía desde la más simple, en forma de metano (CH4), que es un componente del gas natural, hasta las muy pesadas y muy complejas, como algunas moléculas como los asfaltenos que se encuentran en el petróleo crudo, el petróleo y los betunes. Los hidrocarburos pueden estar en forma gaseosa, líquida o sólida, o cualquier combinación de estas formas, y pueden tener uno o más enlaces dobles o triples entre átomos de carbono adyacentes en cadena principal. Por consiguiente, el término incluye parafina, lípidos, alcanos y alquenos lineales, ramificados, cíclicos o parcialmente cíclicos. Entre los ejemplos se incluyen propano, butano, pentano, hexano, octano y escualeno.
[0057] "Relación hidrógeno:carbono" es la relación de átomos de hidrógeno a átomos de carbono en una molécula sobre una base de átomo a átomo. La relación puede usarse para referirse al número de átomos de carbono e hidrógeno de una molécula de hidrocarburo. Por ejemplo, el hidrocarburo con la relación más alta es el metano CH4 (4:1).
[0058] "Fracción hidrófoba" es la porción, o fracción, de un material que es más soluble en una fase hidrófoba en comparación con una fase acuosa. Una fracción hidrófoba es sustancialmente insoluble en agua y normalmente no polar.
[0059] "Aumentar la producción de lípidos" se refiere a un aumento en la productividad de lípidos de un cultivo microbiano mediante, por ejemplo, el aumento del peso en seco de las células por litro de cultivo, el aumento del porcentaje de células que constituyen lípidos o el aumento de la cantidad total de lípidos por litro de volumen de cultivo por unidad de tiempo.
[0060] "Promotor inducible" es un promotor que media la transcripción de un gen unido operativamente en respuesta a un estímulo particular.
[0061] El "aceite industrial" es un aceite que es útil en la industria. Los aceites industriales comunes incluyen lubricantes para espadas de motosierra, fluidos para trabajar metales, lubricantes de calidad alimentaria, aceites para engranajes, aceites marinos, lubricantes para motores, aceites para tractores, lubricantes para equipos agrícolas, aceites para ascensores, aceites para desmoldeo y similares. El "lubricante para espadas de motosierra" se utiliza para la lubricación externa de la espada y la cadena de las motosierras. Los "fluidos para trabajar metales" son fluidos que se usan para enfriar y/o lubricar el proceso de dar forma a una pieza de metal para convertirla en un objeto útil. Los "lubricantes de calidad alimentaria" son lubricantes aceptables para su uso en equipos, aplicaciones y plantas de procesamiento de carne de res, carne de ave y otros alimentos. Los "aceites para engranajes" son aceites que son útiles para lubricar engranajes, por ejemplo, en transmisiones, cajas de transferencia y diferenciales en automóviles, camiones y otra maquinaria. Los "aceites marinos" son aceites que son útiles para lubricar las partes móviles de los equipos marinos. Los "lubricantes de motor" se utilizan para la lubricación de varios motores de combustión interna. Si bien la función principal es lubricar las piezas móviles, los lubricantes para motores también pueden limpiar, inhibir la corrosión, mejorar la estanqueidad y enfriar el motor al alejar el calor de las piezas móviles.
Los "aceites para tractores" son aceites útiles para lubricar las partes móviles de los tractores. Los "lubricantes para equipos agrícolas" son lubricantes que son útiles para lubricar las partes móviles de los equipos agrícolas. Los "aceites para ascensores" son aceites utilizados como fluido hidráulico en ascensores hidráulicos. Los "aceites de desmoldeo" son aceites útiles en la producción de artículos formados usando un molde. Los aceites de desmoldeo facilitan el desmoldeo del artículo formado y pueden tener características de acondicionamiento superficial que proporcionan un acabado superficial deseado.
[0062] “En unión operativa" es una unión funcional entre dos secuencias de ácido nucleico, como una secuencia de control (típicamente un promotor) y la secuencia enlazada (típicamente una secuencia que codifica una proteína, también denominada secuencia codificante). Un promotor está en unión operativa con un gen exógeno si puede mediar en la transcripción del gen.
[0063] "ln situ" significa "en su lugar" o "en su posición original". El "índice de yodo" (o “número de yodo") es una medida del grado de insaturación de un aceite. Es la masa de yodo que consumen los enlaces insaturados de un aceite. Por ejemplo, un aceite con un índice de yodo de 50 es un aceite en el que 100 gramos de aceite consumirían 50 gramos de yodo. Los valores de yodo se determinan de manera rutinaria en la técnica. Los métodos estándar para determinar los valores de yodo incluyen ASTM D5768-02 (2006) y DIN 53241.
[0064] La “concentración limitante de un nutriente" es una concentración de un compuesto en un cultivo que limita la propagación de un organismo cultivado. Una "concentración no limitante de un nutriente" es una concentración que permite la máxima propagación durante un período de cultivo dado. Por lo tanto, el número de células producidas durante un período de cultivo dado es menor en presencia de una concentración limitante de un nutriente que cuando el nutriente no es limitante. Se dice que un nutriente está "en exceso" en un cultivo cuando el nutriente está presente en una concentración mayor que la que permite la máxima propagación.
[0065] La "lipasa" es una enzima soluble en agua que cataliza la hidrólisis de enlaces éster en sustratos lipídicos insolubles en agua. Las lipasas catalizan la hidrólisis de los lípidos en gliceroles y ácidos grasos.
[0066] Una “enzima que modifica los lípidos" se refiere a una enzima que altera la estructura covalente de un lípido. Entre los ejemplos de enzimas que modifican los lípidos se incluyen una lipasa, una acil-ACP tioesterasa, una desaturasa, incluyendo una estearoil-ACP desaturasa (SAD) y una acilo graso desaturasa (FAD), y una aldehído graso descarbonilasa.
[0067] "Enzima de la ruta de los lípidos" es cualquier enzima que desempeñe una función en el metabolismo de los lípidos, es decir, la síntesis, modificación o degradación de los lípidos, y cualquier proteína que modifique químicamente los lípidos, así como las proteínas transportadoras.
[0068] “Lípido" o "lípidos" son una clase de moléculas que son solubles en disolventes no polares (como el éter y el cloroformo) y son relativa o completamente insolubles en agua. Las moléculas de lípidos tienen estas propiedades porque consisten en gran parte en largas colas hidrocarbonadas que son de naturaleza hidrófoba. Entre los ejemplos de lípidos se incluyen los ácidos grasos (saturados e insaturados); glicéridos o glicerolípidos (tales como monoglicéridos, diglicéridos, triglicéridos o grasas neutras y fosfoglicéridos o glicerofosfolípidos); no glicéridos (esfingolípidos, lípidos de esteroles incluyendo colesterol y hormonas esteroides, lípidos de prenol incluyendo terpenoides, alcoholes grasos, ceras y policétidos); y derivados de lípidos complejos (lípidos unidos a azúcares, o glicolípidos, y lípidos unidos a proteínas). Las "grasas" son un subgrupo de lípidos llamados "triacilglicéridos".
[0069] "Lubricante" es una sustancia capaz de reducir la fricción, el calor y/o el desgaste cuando se introduce como una película entre superficies sólidas.
[0070] “Lisado" es una solución que contiene el contenido de las células lisadas.
[0071] "Lisar" o "lisis" es romper la membrana celular y, opcionalmente, la pared celular de un organismo biológico o una célula lo suficiente para liberar al menos algo de contenido intracelular.
[0072] "Desactivador de iones metálicos", también conocido como "desactivador de metales" o "agente desactivador de metales (MDA)", es un aditivo de combustible y/o aceite que se utiliza para estabilizar fluidos mediante la desactivación (normalmente mediante el secuestro) de iones metálicos. Los iones metálicos se pueden producir por la acción de ácidos naturales en el combustible y ácidos generados en lubricantes por procesos oxidativos con las partes metálicas de los sistemas.
[0073] "Microalga" es un organismo microbiano eucariota que contiene un cloroplasto o plástido y, opcionalmente, que es capaz de hacer la fotosíntesis, o un organismo microbiano procariota capaz de hacer la fotosíntesis. Las microalgas incluyen las fotoautótrofas obligadas, que no pueden metabolizar una fuente de carbono fijo como energía, así como las heterótrofas, que pueden vivir únicamente de una fuente de carbono fijo. Las microalgas incluyen organismos unicelulares que se separan de sus células hermanas poco después de la división celular, como Chlamydomonas, así como microbios como, por ejemplo, Volvox, que es un microbio fotosintético multicelular simple de dos tipos de células distintas. Las microalgas incluyen células como Chlorella, Dunaliella, y Prototheca. Las microalgas también incluyen otros organismos fotosintéticos microbianos en los que se da la adhesión célula-célula, como Agmenellum, Anabaena, y Pyrobotrys. "Microalgas" también se refiere a microorganismos heterótrofos obligados que han perdido la capacidad de realizar la fotosíntesis, como algunas especies de algas dinoflageladas y especies del género Prototheca.
[0074] "Microorganismo" y "microbio" son organismos microscópicos unicelulares.
[0075] "Coexpresado de manera natural”, con referencia a dos proteínas o genes, significa que las proteínas o sus genes se coexpresan de manera natural en un tejido u organismo del que derivan, por ejemplo, porque los genes que codifican las dos proteínas están bajo el control de una secuencia reguladora común o porque se expresan en respuesta a un mismo estímulo.
[0076] "Aceite" se refiere a cualquier aceite de triacilglicéridos, producido por organismos, incluyendo levaduras oleaginosas, plantas y/o animales. "Aceite", a diferencia de "grasa", se refiere, a menos que se indique lo contrario, a lípidos que generalmente, pero no siempre, son líquidos a temperaturas y presiones ambiente ordinarias. Por ejemplo, "aceite" incluye aceites vegetales o de semillas derivados de plantas, incluidos, entre otros, un aceite derivado de aguacate, nueces de Brasil, caléndula, camelina, camelina sativa, canola, anacardo, ricino, manteca de cacao (también conocida como cacao, que es un aceite de triacilglicéridos derivado del grano de cacao que es sólido a temperaturas y presiones ambiente típicas), coco, café, copra, cilantro, maíz, semilla de algodón, cuphea, euforbia, avellana, cáñamo, jatrofa, jojoba, kenaf, linaza, altramuz, macadamia, semillas de mostaza, avena, oliva, adormidera, palma, palmiste, cacahuete, nuez pecán, semilla de calabaza, colza, arroz, cártamo, sésamo, soja, girasol y árbol de tung, así como combinaciones de estos. "Aceite microbiano" se refiere a un aceite derivado de un microbio.
[0077] "Levadura oleaginosa" significa una levadura que puede acumular de manera natural más del 20 % de su peso celular en seco en forma de lípidos y son del subreino de los hongos-Di>arya. La levadura oleaginosa incluye, pero no se limita a, organismos tales como Yarrowia lipolytica, Rhodotorula glutinis, Cryptococcus curvatus, y Lipomyces starkeyi.
[0078] El "choque osmótico" es la ruptura de células en una solución después de una reducción repentina de la presión osmótica. A veces se induce un choque osmótico para liberar componentes celulares de tales células en una solución.
[0079] "Enzima degradante de polisacáridos" es cualquier enzima capaz de catalizar la hidrólisis, o la sacarificación, de cualquier polisacárido. Por ejemplo, las celulasas catalizan la hidrólisis de la celulosa.
[0080] Los "polisacáridos" o "glicanos" son carbohidratos formados por monosacáridos unidos por enlaces glucosídicos. La celulosa es un polisacárido que forma parte de algunas paredes celulares de las plantas. La celulosa se puede despolimerizar mediante enzimas para producir monosacáridos como xilosa y glucosa, así como disacáridos y oligosacáridos más grandes.
[0081] El "punto de fluidez" es la temperatura más baja a la que un líquido se verterá o fluirá en un conjunto específico de condiciones. Los estándares de punto de fluidez ejemplares incluyen ASTM D97-11, D5853-11 y D5949-10, pero se pueden emplear otros conocidos o desarrollados por los expertos en la técnica para realizar determinaciones de punto de fluidez en relación con los métodos descritos en este documento.
[0082] Los "depresores del punto de fluidez”, o "PPD”, son polímeros que controlan la formación de cristales de cera en aceites o lubricantes, lo que da como resultado un punto de fluidez más bajo y un mejor rendimiento del flujo a baja temperatura.
[0083] "Promotor" es una secuencia de control de ácido nucleico que dirige la transcripción de un ácido nucleico. Como se usa en este documento, un promotor incluye secuencias de ácido nucleico necesarias cerca del sitio de inicio de la transcripción, tal como, en el caso de un promotor de tipo polimerasa II, un elemento TATA. Un promotor también incluye opcionalmente elementos potenciadores o represores distales, que pueden ubicarse hasta varios miles de pares de bases desde el sitio de inicio de la transcripción.
[0084] "Recombinante" se refiere a una célula, ácido nucleico, proteína o vector que se ha modificado debido a la introducción de un ácido nucleico exógeno o la alteración de un ácido nucleico nativo. Por lo tanto, por ejemplo, las células recombinantes expresan genes que no se encuentran dentro de la forma nativa (no recombinante) de la célula o que expresan genes nativos de manera diferente a como los expresa una célula no recombinante. Un "ácido nucleico recombinante" es un ácido nucleico formado originalmente in vitro, en general, mediante la manipulación del ácido nucleico, por ejemplo, utilizando polimerasas y endonucleasas, o que de otro modo se encuentra en una forma que no se da de manera natural en la naturaleza. Pueden producirse ácidos nucleicos recombinantes, por ejemplo, para colocar dos o más ácidos nucleicos en una unión operativa. Por lo tanto, un ácido nucleico aislado o un vector de expresión formado mediante la unión in vitro de moléculas de ADN que normalmente no están unidas en la naturaleza se considera recombinante. Una vez que se produce un ácido nucleico recombinante y se introduce en una célula u organismo hospedador, puede replicarse usando la maquinaria celular in vivo de la célula hospedadora; sin embargo, dichos ácidos nucleicos, una vez producidos de forma recombinante, aunque posteriormente se repliquen intracelularmente, todavía se consideran recombinantes. De manera similar, una "proteína recombinante" es una proteína elaborada utilizando técnicas recombinantes, es decir, mediante la expresión de un ácido nucleico recombinante.
[0085] El "aceite RBD" es un aceite que se ha sometido a refinado, blanqueo o desodorización.
[0086] El "diésel renovable" es una mezcla de alcanos (como C10:0, C12:0, C14:0, C16:0 y C18:0) producidos por hidrogenación y desoxigenación de lípidos.
[0087] La "sacarificación" es un proceso de conversión de biomasa, normalmente biomasa celulósica o lignocelulósica, en azúcares monoméricos, como glucosa y xilosa. Material celulósico o biomasa "sacarificado/a” o "despolimerizado/a” se refiere a material celulósico o biomasa que se ha convertido en azúcares monoméricos mediante sacarificación.
[0088] La "sonicación" es un proceso de alteración de materiales biológicos, como una célula, mediante el uso de energía de ondas sonoras.
[0089] "Especie de furfural" es 2-furancarboxaldehído o un derivado que conserva las mismas características estructurales básicas.
[0090] “Rastrojo” son los tallos secos y las hojas de un cultivo que quedan después de la cosecha de un grano.
[0091] El "gen de utilización de sacarosa" es un gen que, cuando se expresa, ayuda a la capacidad de una célula para utilizar sacarosa como fuente de energía. Las proteínas codificadas por un gen de utilización de sacarosa se denominan en la presente "enzimas de utilización de sacarosa" e incluyen transportadores de sacarosa, sacarasas y hexocinasas tales como glucocinasas y fructocinasas.
[0092] "Transformador" es un dispositivo que transfiere energía eléctrica de un circuito a otro a través de conductores acoplados de manera inductiva, típicamente las bobinas del transformador.
[0093] Los términos "preparación para el invierno" del aceite o "preparación del aceite para el invierno" se refieren a un proceso que incluye la eliminación de los componentes de mayor punto de fusión de un aceite y/o la adición de uno o más depresores del punto de fluidez.
II. CULTIVO Y CONDICIONES DE CULTIVO
[0094] En algunas formas de realización, la presente invención generalmente está relacionada con el cultivo de microbios oleaginosos, tales como microalgas recombinantes, incluyendo especies y cepas de Chlorella y Prototheca, para la producción de aceite microbiano (lípidos). Para comodidad del lector, esta sección se subdivide en subsecciones. En la subsección 1 se describen especies y cepas de Prototheca y cómo identificar nuevas especies y cepas de Prototheca y microalgas relacionadas por comparación de ADN genómico, así como otras microalgas, levaduras, hongos y bacterias útiles en los métodos descritos en este documento. En la subsección 2 se describen biorreactores útiles para el cultivo. En la subsección 3 se describen medios para el cultivo. En la subsección 4 se describe la producción de aceite (lípidos) según métodos de cultivo ilustrativos descritos en este documento. En la subsección 5 se describen los tipos de levadura oleaginosa adecuados para usar en los métodos descritos en este documento, las condiciones de cultivo para generar biomasa de levadura y los perfiles de lípidos y la composición química de la biomasa preparada según los métodos ilustrativos descritos en este documento.
1. Especies v cepas de Prototheca v otros microbios oleaginosos
[0095] Prototheca es un microorganismo notable para su uso en la producción de lípidos, porque puede producir altas cantidades de lípidos, en particular lípidos adecuados para la producción de fluidos dieléctricos y otros lubricantes. El lípido producido por Prototheca tiene un mayor grado de saturación que el producido por otras microalgas. Es más, el lípido de Prototheca generalmente está libre de pigmento (niveles bajos o indetectables de clorofila y ciertos carotenoides) y, en cualquier caso, contiene mucho menos pigmento que los lípidos de otras microalgas. Además, las células de Prototheca recombinantes proporcionadas para su uso en los métodos descritos en el presente documento pueden usarse para producir lípidos con un mayor rendimiento y eficacia, y con un coste reducido, con respecto a la producción de lípidos a partir de otros microorganismos. Entre las especies y cepas ilustrativas de Prototheca para usar en los métodos descritos en este documento se incluyen Prototheca wickerhamii, Prototheca stagnora (incluida UTEX 327), Prototheca portoricensis, Prototheca moriformis (incluidas las cepas UTEX 1441, 1435), y Prototheca zopfii. Las especies del género Prototheca son heterótrofos obligados.
[0096] Las especies de Prototheca para su uso en los métodos descritos en el presente documento pueden identificarse mediante la amplificación de algunas regiones concretas del genoma. Por ejemplo, la identificación de una determinada especie o cepa de Prototheca se puede lograr a través de la amplificación y secuenciación del ADN nuclear y/o de los cloroplastos usando cebadores y metodología usando cualquier región del genoma, por ejemplo usando los métodos descritos en Wu y col., Bot. Bull. Acad. Sin. (2001) 42:115-121 Identification of Chlorella spp. isolates using ribosomal DNA sequences. Los expertos en la técnica pueden utilizar métodos establecidos de análisis filogenético, como la amplificación y la secuenciación del espaciador transcrito interno ribosomal (ITS1 e ITS2 rDNA), ARNr 23S, ARNr 18S y otras regiones genómicas conservadas para identificar especies no solo de Prototheca, sino de otros organismos productores de hidrocarburos y lípidos con perfiles de lípidos y capacidad de producción similares. Para ver ejemplos de métodos de identificación y clasificación de algas, véase también, por ejemplo, Genetics, agosto de 2005; 170(4):1601-10 y RNA, abril de 2005; 11(4):361-4.
[0097] Por lo tanto, la comparación de ADN genómico se puede usar para identificar especies adecuadas de microalgas para usar en los métodos descritos en este documento. Las regiones de ADN genómico conservado, tales como, entre otros, el ADN que codifica el ARNr 23S, pueden amplificarse a partir de especies de microalgas y compararse con secuencias de consenso para detectar especies de microalgas que estén relacionadas desde el punto de vista taxonómico con las microalgas preferidas utilizadas en los métodos descritos en el presente documento. A continuación, se muestran ejemplos de tales comparaciones de secuencias de ADN para especies dentro del género Prototheca. La comparación de ADN genómico también puede ser útil para identificar especies de microalgas que se han identificado erróneamente en una colección de cepas. A menudo, una colección de cepas identificará especies de microalgas en función de las características fenotípicas y morfológicas. El uso de estas características puede dar lugar a una categorización errónea de la especie o el género de una microalga. El uso de la comparación de ADN genómico puede ser un método mejor para clasificar las especies de microalgas en función de su relación filogenética.
[0098] Las microalgas ilustrativas para su uso en los métodos descritos en el presente documento suelen tener secuencias de ADN genómico que codifican el ARNr 23S que tienen al menos un 99 %, al menos un 95 %, al menos un 90 % o al menos un 85 % de identidad de nucleótidos con al menos una de las secuencias enumeradas en la SEQ ID N.011-19.
[0099] Para la comparación de secuencias para determinar el porcentaje de identidad de nucleótidos o aminoácidos, normalmente una secuencia actúa como secuencia de referencia, con la que se comparan las secuencias de prueba. Cuando se usa un algoritmo de comparación de secuencias, las secuencias de prueba y de referencia se introducen en un ordenador, se designan las coordenadas de la subsecuencia, si es necesario, y se designan los parámetros del programa del algoritmo de secuencia. A continuación, el algoritmo de comparación de secuencias calcula el porcentaje de identidad de secuencia para la(s) secuencia(s) de prueba en relación con la secuencia de referencia, en función de los parámetros de programa designados.
[0100] La alineación óptima de las secuencias para la comparación se puede realizar, por ejemplo, mediante el algoritmo de homología local de Smith & Waterman, Adv. Appl. Math. 2:482 (1981), por el algoritmo de alineación de homología de Needleman y Wunsch, J. Mol. Biol.48:443 (1970)), por el método de búsqueda de similitud de Pearson y Lipman, Proc. Nat'l. Acad. Sci. USA 85:2444 (1988), mediante implementaciones computarizadas de estos algoritmos (GAP, BESTFIT, FASTA y TFASTA en Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI), o mediante inspección visual (véase en general Ausubel y col., supra).
[0101] Otro ejemplo de un algoritmo adecuado para determinar el porcentaje de identidad de secuencia y la similitud de secuencia es el algoritmo BLAST, que se describe en Altschul y col., J. Mol. Biol. 215:403-410 (1990). El
programa informático para realizar análisis BLAST está disponible para el público a través del Centro Nacional para la Información Biotecnológica de EE. UU. (National Center for Biotechnology Information, en la dirección web www.ncbi.nlm.nih.gov). Este algoritmo implica identificar primero pares de secuencias de puntuación alta (HSP) mediante la identificación de palabras cortas de longitud W en la secuencia de consulta, que coinciden o satisfacen una puntuación de umbral T de valor positivo cuando se alinean con una palabra de la misma longitud en una secuencia de base de datos. T se conoce como el umbral de puntuación de palabras adyacentes (Altschul y col., supra.). Estas coincidencias iniciales de palabras adyacentes actúan como semillas para iniciar búsquedas para encontrar HSP más largos que las contengan. Las coincidencias de palabras se extienden en ambas direcciones a lo largo de cada secuencia hasta donde se puede aumentar la puntuación de alineación acumulada. Las puntuaciones acumulativas se calculan usando, para las secuencias de nucleótidos, los parámetros M (puntuación de recompensa para un par de residuos coincidentes; siempre > 0) y N (puntuación de penalización para residuos coincidentes; siempre < 0). Para las secuencias de aminoácidos, se utiliza una matriz de puntuación para calcular la puntuación acumulativa. La extensión de las coincidencias de palabras en cada dirección se detiene cuando: la puntuación de alineación acumulativo desciende en la cantidad X de su valor máximo alcanzado; la puntuación acumulativa llega a cero o menos debido a la acumulación de uno o más alineamientos de residuos de puntuación negativa; o se llega al final de cualquiera de las secuencias. Para identificar si un ácido nucleico o un polipéptido está dentro del alcance de la invención, son adecuados los parámetros predeterminados de los programas BLAST. El programa BLASTN (para secuencias de nucleótidos) usa por defecto una longitud de palabra (W) de 11, una expectativa (E) de 10, M=5, N=-4 y una comparación de ambas cadenas. Para las secuencias de aminoácidos, el programa BLASTP utiliza por defecto una longitud de palabra (W) de 3, una expectativa (E) de 10 y la matriz de puntuación BLOSUM62. El programa TBLATN (que usa la secuencia de proteínas para la secuencia de nucleótidos) usa por defecto una longitud de palabra (W) de 3, una expectativa (E) de 10 y una matriz de puntuación BLOSUM 62. (véase Henikoff y Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1989).
[0102] Además de calcular el porcentaje de identidad de secuencia, el algoritmo BLAST también realiza un análisis estadístico de la similitud entre dos secuencias (véase, por ejemplo, Karlin & Altschul, Proc. Nat'l. Acad. Sci. USA 90:5873-5787 (1993)). Una medida de similitud proporcionada por el algoritmo BLAST es la probabilidad de suma más pequeña (P(N)), que proporciona una indicación de la probabilidad por la cual se produciría por casualidad una coincidencia entre dos secuencias de nucleótidos o aminoácidos. Por ejemplo, un ácido nucleico se considera similar a una secuencia de referencia si la probabilidad de la suma más pequeña en una comparación del ácido nucleico de prueba con el ácido nucleico de referencia es menor que aproximadamente 0,1, más preferiblemente menor que aproximadamente 0,01 y, de la manera más preferible, menor que aproximadamente 0,001.
[0103] Se puede utilizar una amplia variedad de microbios oleaginosos además de Prototheca en los métodos descritos en el presente documento. Por ejemplo, Chlorella, que incluye pero no se limita a las cepas de la especie protothecoides de Chlorella, es una microalga excelente para usar en los métodos descritos en el presente documento. Las consideraciones que afectan la selección de microorganismos para su uso en los métodos descritos en este documento, además de la producción de lípidos o hidrocarburos adecuados para la producción de aceites, combustibles y sustancias oleoquímicas, pueden incluir uno o más de las siguientes: (1) alto contenido de lípidos como porcentaje de peso celular; (2) facilidad de crecimiento; (3) facilidad para la modificación por ingeniería genética; y (4) facilidad de procesamiento de biomasa. En formas de realización particulares, el microorganismo de tipo salvaje o modificado genéticamente produce células que tienen al menos un 40 %, al menos un 45 %, al menos un 50 %, al menos un 55 %, al menos un 60 %, al menos un 65 % o al menos un 70 % o más de aceite microbiano (es decir, lípidos y ácidos grasos). Los organismos preferidos crecen (y se cultivan) de manera heterótrofa (en azúcares, en ausencia sustancial de luz). Las microalgas por lo general son microbios excelentes para usar en los métodos descritos en este documento. Los ejemplos de microalgas que se pueden usar para poner en práctica los métodos incluyen, entre otros, las siguientes algas enumeradas en la Tabla 1.
T l 1. E m l mi r l l in .


2. Biorreactor
[0104] Los microorganismos se cultivan tanto para fines de manipulación genética como para la producción de aceite microbiano (por ejemplo, hidrocarburos como lípidos, ácidos grasos, aldehidos, alcoholes y alcanos). El primer tipo de cultivo se lleva a cabo a pequeña escala e inicialmente, al menos, en condiciones en las que el microorganismo de partida pueda crecer. El cultivo con fines de producción de hidrocarburos normalmente se lleva a cabo a gran escala en un biorreactor (por ejemplo, en biorreactores de 10000 l, 40000 l, 100000 l o más grandes). Las microalgas, incluyendo especies de Prototheca, así como los otros microbios oleaginosos descritos en este documento, se cultivan típicamente en los métodos descritos en este documento en medios líquidos dentro de un biorreactor. Normalmente, el biorreactor no permite la entrada de cantidades sustanciales de luz o de ninguna cantidad de luz. Toda(s) la(s) etapa(s) de cultivo del microbio oleaginoso, incluidas las microalgas, se puede(n) realizar en ausencia sustancial de luz.
[0105] El biorreactor o fermentador se utiliza para cultivar células de microalgas durante las diversas fases de su ciclo fisiológico. Los biorreactores ofrecen muchas ventajas para su uso en métodos de crecimiento y propagación heterótrofos. Las microalgas y otros microbios oleaginosos descritos en el presente documento normalmente se fermentan en líquido en grandes cantidades, como, por ejemplo, en cultivos en suspensión. Los biorreactores, como los fermentadores de acero, pueden albergar volúmenes de cultivo muy grandes (se pueden utilizar biorreactores de 40 000 litros y de mayor capacidad). Los biorreactores también suelen permitir el control de las condiciones de cultivo, como la temperatura, el pH, la tensión de oxígeno y los niveles de dióxido de carbono. Por ejemplo, los biorreactores suelen ser configurables, por ejemplo, utilizando puertos conectados al sistema de conductos, para permitir que los componentes gaseosos, como oxígeno o nitrógeno, burbujeen a través de un cultivo líquido. Otros parámetros de cultivo, como el pH de los medios de cultivo, la identidad y concentración de elementos traza y otros componentes de los medios, también se pueden manipular con mayor facilidad usando un biorreactor.
[0106] Los biorreactores se pueden configurar para que los medios de cultivo fluyan a través del biorreactor durante todo el período de tiempo durante el cual las microalgas se reproducen y aumentan en número. Por ejemplo, los medios se pueden infundir en el biorreactor después de la inoculación pero antes de que las células alcancen la densidad deseada. En otros casos, un biorreactor se llena con medios de cultivo al comienzo de un cultivo y no se infunde más medio de cultivo después de inocular el cultivo. En otras palabras, la biomasa de microalgas (u otros microbios) se cultiva en un medio acuoso durante un período de tiempo durante el cual las microalgas se reproducen y aumentan en número; sin embargo, no fluyen cantidades de medio de cultivo acuoso a través del biorreactor durante todo el período de tiempo. Por lo tanto, no es necesario que el medio de cultivo acuoso fluya a través del biorreactor después de la inoculación.
[0107] Los biorreactores equipados con dispositivos tales como palas giratorias e impulsores, mecanismos basculantes, barras agitadoras o medios para la infusión de gas presurizado pueden usarse para someter los cultivos de microalgas a mezcla. La mezcla puede ser continua o intermitente. Por ejemplo, no se mantiene un régimen de flujo turbulento de entrada de gas y entrada de medios para la reproducción de microalgas hasta que se haya logrado un aumento deseado del número de dichas microalgas.
[0108] Los puertos del biorreactor se pueden utilizar para introducir o extraer gases, sólidos, semisólidos y líquidos en la cámara del biorreactor que contiene las microalgas. Si bien muchos biorreactores tienen más de un puerto (por ejemplo, uno para la entrada de medios y otro para el muestreo), no es necesario que solo una sustancia entre o salga de un puerto. Por ejemplo, se puede usar un puerto para hacer fluir medios de cultivo al biorreactor y luego usarlo para el muestreo, como entrada de gas, como salida de gas o para otros fines. Preferiblemente, un puerto de muestreo se puede usar repetidamente sin alterar y comprometer la naturaleza axénica del cultivo. Un puerto de muestreo se puede configurar con una válvula u otro dispositivo que permita detener e iniciar el flujo de muestras o proporcionar un medio de muestreo continuo. Los biorreactores suelen tener al menos un puerto que permite la inoculación de un cultivo, y dicho puerto también se puede utilizar para otros fines, como la introducción de medios o gases.
[0109] Los puertos de los biorreactores permiten manipular el contenido de gas del cultivo de microalgas. A modo ilustrativo, parte del volumen de un biorreactor puede ser gas en lugar de líquido, y las entradas de gas del biorreactor permiten bombear gases al biorreactor. Los gases que pueden bombearse beneficiosamente a un biorreactor incluyen aire, mezclas de aire/CO2, gases nobles, como el argón, y otros gases. Los biorreactores suelen
estar equipados para permitir que el usuario controle la velocidad de entrada de un gas en el biorreactor. Como se ha indicado anteriormente, se puede usar el aumento del flujo de gas en un biorreactor para aumentar la mezcla del cultivo.
[0110] El aumento del flujo de gas también afecta a la turbidez del cultivo. La turbulencia se puede lograr colocando un puerto de entrada de gas por debajo del nivel del medio de cultivo acuoso para que el gas que entra al biorreactor burbujee hacia la superficie del cultivo. Uno o más puertos de salida de gas permiten que escape el gas, evitando así la acumulación de presión en el biorreactor. Preferiblemente, un puerto de salida de gas conduce a una válvula "unidireccional" que evita que los microorganismos contaminantes entren en el biorreactor.
3. Medios
[0111] Las microalgas, así como otros medios de cultivo microbiano, normalmente contienen componentes como una fuente de nitrógeno fijo, una fuente de carbono fijo, oligoelementos, opcionalmente una solución amortiguadora para el mantenimiento del pH y fosfato (normalmente proporcionado como una sal de fosfato). Otros componentes pueden incluir sales como el cloruro de sodio, particularmente para las microalgas de agua de mar. Las fuentes de nitrógeno incluyen fuentes de nitrógeno orgánico e inorgánico, incluyendo, por ejemplo, sin limitación, nitrógeno molecular, nitrato, sales de nitrato, amoníaco (puro o en forma de sal, como, (NH4)2SO4 y NH4OH), proteína, harina de soja, licor de maíz y extracto de levadura. Los ejemplos de oligoelementos incluyen zinc, boro, cobalto, cobre, manganeso y molibdeno, por ejemplo, en las formas respectivas de ZnCl.2, H3BO3, CoCl2 ■ 6 H2O, CuCl2 ■ 2 H2O, MnCh ■ 4 H2O y (NH4)6Mo7O24 ■ 4H2O
[0112] Los microorganismos adecuados se pueden encontrar en varios lugares y entornos de todo el mundo. Como consecuencia de su aislamiento de otras especies y su divergencia evolutiva resultante, puede ser difícil predecir el medio de crecimiento particular para el crecimiento óptimo y la generación de constituyentes de lípidos y/o hidrocarburos. En algunos casos, algunas cepas de microorganismos pueden ser incapaces de crecer en un medio de cultivo concreto debido a la presencia de algún componente inhibidor o a la ausencia de algún requisito nutricional esencial requerido por la cepa concreta del microorganismo.
[0113] Los medios de crecimiento sólidos y líquidos generalmente están disponibles en una amplia variedad de fuentes, y se pueden encontrar instrucciones para la preparación de medios particulares que sean adecuados para una amplia variedad de cepas de microorganismos, por ejemplo, en línea en www.utex.org/, un sitio web mantenido por la Universidad de Texas en Austin, 1 University Station a 6700, Austin, Texas, 78712-0183, para su colección de cultivos de algas (UTEX). Por ejemplo, varios medios de agua dulce y agua salada incluyen los descritos en la publicación PCT n.02008/151149, que se incorpora a la presente por referencia.
[0114] En un ejemplo particular, el medio de proteosa es adecuado para cultivos axénicos, y se puede preparar un volumen de 1 litro del medio (pH ~6,8) mediante la adición de 1 g de proteosa peptona a 1 litro de medio de cultivo Bristol. El medio Bristol comprende 2,94 mM NaNO3, 0,17 mM CaCl2 ■ 2 H2O, 0,3 mM MgSO4 ■ 7 H2O, 0,43 mM, 1,29 mM KH2PO4, y 1,43 mM NaCl en una solución acuosa. Para medio de agar al 1,5 %, se pueden agregar 15 g de agar a 1 L de la solución. La solución se cubre y se esteriliza en autoclave, y luego se almacena a temperatura refrigerada antes de su uso. Otro ejemplo es el medio para el aislamiento de Prototheca (PIM), que comprende 10 g/l de ftalato de hidrógeno y potasio (KHP), 0,9 g/l de hidróxido de sodio, 0,1 g/l de sulfato de magnesio, 0,2 g/l de hidrógeno fosfato de potasio, 0,3 g/l de cloruro de amonio, 10 g/l de glucosa 0,001 g/l de clorhidrato de tiamina, 20 g/L de agar, 0,25 g/l de 5-fluorocitosina, a un pH en el rango de 5,0 a 5,2 (véase Pore, 1973, App. Microbiology, 26: 648-649). Se pueden identificar fácilmente otros medios adecuados para usar con los métodos descritos en este documento consultando la URL que se ha identificado antes o consultando otras organizaciones que mantienen cultivos de microorganismos, como SAG, CCAP o CCALA. SAG se refiere a la Colección de Cultivos de Algas de la Universidad de Gottingen (Gottingen, Alemania), CCAP se refiere a la colección de cultivos de algas y protozoos gestionada por la Asociación Escocesa de Ciencias Marinas (Scottish Association for Marine Science, Escocia, Reino Unido), y CCALA se refiere a la colección de cultivos colección del laboratorio de algas del Instituto de Botánica (Institute of Botany, Treboñ, República Checa). Adicionalmente, en la patente de EE. UU. n.o 5,900,370 se describen formulaciones de medios y condiciones adecuadas para la fermentación heterótrofa de especies de Prototheca.
[0115] Para una producción rentable, la selección de una fuente de carbono fijo es importante, ya que el coste de la fuente de carbono fijo debe ser lo suficientemente bajo para que la producción de aceite sea económica. Las fuentes de carbono adecuadas incluyen, por ejemplo, acetato, floridósido, fructosa, galactosa, ácido glucurónico, glucosa, glicerol, lactosa, manosa, N-acetilglucosamina, ramnosa, rafinosa, estaquiosa, sacarosa y/o xilosa. Las materias primas adecuadas útiles según los métodos descritos en este documento incluyen, por ejemplo, licor negro, almidón de maíz, material celulósico despolimerizado, suero de leche, azúcar invertido (glucosa/fructosa), melaza, patata, sorgo, sacarosa, remolacha azucarera, caña de azúcar, jugo de caña espeso, arroz y trigo. Las fuentes de carbono
también se pueden proporcionar como una mezcla, tal como una mezcla de sacarosa y pulpa de remolacha azucarera despolimerizada.
[0116] La fuente o fuentes de carbono se pueden suministrar a una concentración de al menos aproximadamente 50 pM, al menos aproximadamente 100 pM, al menos aproximadamente 500 pM, al menos aproximadamente 5 mM, al menos aproximadamente 50 mM y al menos aproximadamente 500 mM, de una o más fuentes de carbono fijos proporcionadas exógenamente. Se prefieren fuentes de carbono altamente concentradas como materia prima para la fermentación, y la fuente de carbono se puede proporcionar en una materia prima a una concentración cercana a su solubilidad máxima (es decir, a una concentración que excede el 90 % de solubilidad, como una concentración del 95 % o más, es decir, del 99 % de solubilidad).
[0117] Por ejemplo, se utilizan concentraciones de glucosa de al menos 300 g/l, al menos 400 g/l, al menos 500 g/l o al menos 600 g/l o más en la materia prima en un cultivo discontinuo, en el que la fuente de carbono fijo altamente concentrado se alimenta a las células con el tiempo a medida que las células crecen y acumulan aceite microbiano (lípido). Alternativamente, pueden usarse concentraciones de sacarosa de al menos 500 g/L, al menos 600 g/L, al menos 700 g/L, al menos 800 g/L o más en la materia prima en un cultivo discontinuo. Los ejemplos no excluyentes de fuentes de carbono de sacarosa altamente concentradas incluyen jugo de caña espeso, jugo de caña de azúcar, jugo de remolacha azucarera y melaza. Las fuentes de carbono de particular interés para los fines de los métodos descritos en este documento incluyen celulosa (en forma despolimerizada), glicerol, sacarosa y sorgo, cada una de las cuales se analiza con más detalle a continuación.
[0118] Según los métodos descritos en el presente documento, los microorganismos pueden cultivarse usando biomasa celulósica despolimerizada como materia prima. La biomasa celulósica (por ejemplo, rastrojo, como el rastrojo de maíz) es económica y se puede obtener con facilidad; sin embargo, los intentos de utilizar este material como materia prima para la levadura han fracasado. En particular, se ha descubierto que dichas materias primas inhiben el crecimiento de la levadura, y la levadura no puede usar los azúcares de 5 carbonos producidos a partir de materiales celulósicos (por ejemplo, xilosa de hemicelulosa). Por el contrario, las microalgas pueden crecer en material celulósico procesado. Los materiales celulósicos generalmente incluyen alrededor de un 40-60 % de celulosa; aproximadamente un 20-40 % de hemicelulosa; y un 10-30 % de lignina.
[0119] Entre los materiales celulósicos adecuados se incluyen los residuos de cultivos energéticos herbáceos y leñosos, así como cultivos agrícolas, es decir, las partes de la planta, principalmente tallos y hojas, que no se extraen de los campos con el alimento principal o el producto de fibra. Los ejemplos incluyen desechos agrícolas como bagazo de caña de azúcar, cascarillas de arroz, fibra de maíz (incluidos tallos, hojas, cáscaras y zuros), harina de soja, paja de trigo, paja de arroz, pulpa de remolacha azucarera, pulpa de cítricos, cáscaras de cítricos; desechos forestales tales como los de entresacas de maderas duras y blandas, y residuos de maderas duras y blandas procedentes de operaciones madereras; desechos de madera tales como desechos de aserraderos (astillas de madera, serrín) y desechos de plantas de celulosa; desechos urbanos tales como las fracciones de papel de los desechos sólidos municipales, desechos urbanos de madera y desechos verdes urbanos tales como recortes de césped municipal; y residuos de construcción de madera. Los celulósicos adicionales incluyen cultivos celulósicos dedicados, como el pasto varilla, la madera de álamo híbrido y el miscanto, la caña de fibra y el sorgo de fibra. Entre los azúcares de cinco carbonos que se producen a partir de dichos materiales se incluye la xilosa.
[0120] Los materiales celulósicos se procesan para aumentar la eficacia con la que el microbio puede utilizar los azúcares contenidos en dichos materiales. Los métodos descritos en este documento se pueden poner en práctica para aprovechar los nuevos métodos para el tratamiento de materiales celulósicos después de la explosión ácida, de modo que los materiales sean adecuados para su uso en un cultivo heterótrofo de microbios (por ejemplo, microalgas y levaduras oleaginosas). Como se ha mencionado antes, la biomasa lignocelulósica se compone de varias fracciones, incluyendo la celulosa, un polímero cristalino de glucosa con enlaces beta 1,4 (un azúcar de seis carbonos), hemicelulosa, un polímero asociado más libremente compuesto predominantemente de xilosa (un azúcar de cinco carbonos) y, en menor medida, manosa, galactosa, arabinosa, lignina, un polímero aromático complejo compuesto por alcohol sinapílico y sus derivados, y pectinas, que son cadenas lineales de un ácido poligalacturónico con enlaces alfa 1,4. Debido a la estructura polimérica de la celulosa y la hemicelulosa, los azúcares (por ejemplo, glucosa monomérica y xilosa) que contienen no se encuentran en una forma que muchos microbios puedan usar (metabolizar) de manera eficiente. Para tales microbios, el procesamiento adicional de la biomasa celulósica para generar los azúcares monoméricos que forman los polímeros puede ser muy útil para garantizar que los materiales celulósicos se utilicen de manera eficiente como materia prima (fuente de carbono).
[0121] La celulosa o biomasa celulósica se somete a un proceso, denominado "explosión", en el que la biomasa se trata con ácido sulfúrico diluido (u otro) a temperatura y presión elevadas. Este proceso condiciona la biomasa de manera que pueda someterse de manera eficiente a la hidrólisis enzimática de las fracciones celulósica y
hemicelulósica en monómeros de glucosa y xilosa. Los azúcares monoméricos resultantes se denominan azúcares celulósicos. Los azúcares celulósicos pueden ser utilizados posteriormente por microorganismos para producir una variedad de metabolitos (por ejemplo, lípidos). El paso de explosión ácida da como resultado una hidrólisis parcial de la fracción de hemicelulosa en monosacáridos constituyentes. Estos azúcares pueden liberarse completamente de la biomasa con un tratamiento adicional. El tratamiento adicional puede ser un tratamiento hidrotérmico que incluye lavar el material explotado con agua caliente, lo que elimina contaminantes como sales. Este paso no es necesario para las fermentaciones de etanol celulósico debido a las concentraciones de azúcar más diluidas que se utilizan en dichos procesos. Alternativamente, el tratamiento adicional puede ser un tratamiento con ácido adicional; o bien, el tratamiento adicional puede ser la hidrólisis enzimática del material explotado. Estos tratamientos también se pueden utilizar en cualquier combinación. El tipo de tratamiento puede afectar al tipo de azúcares liberados (por ejemplo, azúcares de cinco carbonos frente a azúcares de seis carbonos) y la etapa en la que se liberan en el proceso. Como consecuencia, se pueden crear diferentes corrientes de azúcares, ya sean predominantemente de cinco o seis carbonos. Estas corrientes enriquecidas de cinco o seis carbonos pueden dirigirse a microorganismos específicos con diferentes capacidades de utilización de carbono.
[0122] Los métodos descritos en este documento implican típicamente la fermentación a densidades celulares más altas que las que se logran en la fermentación con etanol. Debido a las mayores densidades de los cultivos para la producción heterótrofa de lípidos, la fuente de carbono fijo (por ejemplo, la(s) corriente(s) de azúcar(es) derivado(s) de la celulosa) está preferiblemente en forma concentrada. El contenido de glucosa del material celulósico despolimerizado es preferentemente de al menos 300 g/litro, al menos 400 g/litro, al menos 500 g/litro o al menos 600 g/litro antes del paso de cultivo, que es opcionalmente un cultivo discontinuo en el que el material se suministra a las células a lo largo del tiempo a medida que las células crecen y acumulan lípidos. Las corrientes de azúcar celulósico no se utilizan en o cerca de este rango de concentración en la producción de etanol celulósico. Por lo tanto, con el fin de generar y mantener muy altas las densidades de células durante la producción de aceite lignocelulósico, la(s) materia(s) prima(s) de carbono debe(n) suministrarse a los cultivos heterótrofos de forma muy concentrada. Sin embargo, cualquier componente de la corriente de alimentación que no sea un sustrato y no sea metabolizado por el microorganismo oleaginoso se acumulará en el biorreactor, lo que puede generar problemas si el componente es tóxico o inhibe la producción del producto final deseado. Mientras que la lignina y los subproductos derivados de la lignina, los subproductos derivados de carbohidratos como los furfurales y los hidroximetilfurfurales, y las sales derivadas de la generación de los materiales celulósicos (tanto en el proceso de explosión como en el posterior proceso de neutralización), e incluso los azúcares pentosas/hexosas no metabolizados pueden presentar problemas en las fermentaciones etanólicas, estos efectos se amplifican significativamente en un proceso en el que su concentración en la materia prima inicial es alta. Para lograr concentraciones de azúcar de materiales celulósicos de 300 g/L, 400 g/L, 500 g/L o más para azúcares de seis carbonos que pueden usarse en aplicaciones de producción a gran escala, la concentración de estos materiales tóxicos puede ser 20 veces superior a las concentraciones típicamente presentes en las fermentaciones etanólicas de biomasa celulósica.
[0123] En el tratamiento del proceso de explosión del material celulósico se utilizan cantidades significativas de ácido sulfúrico, calor y presión, liberando así subproductos de carbohidratos, a saber, furfurales e hidroximetilfurfurales. Los furfurales y los hidroximetilfurfurales se producen durante la hidrólisis de la hemicelulosa a través de la deshidratación de la xilosa en furfural y agua. Estos subproductos (por ejemplo, furfurales e hidroximetilfurfurales) se pueden eliminar del material lignocelulósico sacarificado antes de introducirlo en el biorreactor. El proceso para la eliminación de los subproductos de los carbohidratos puede ser el tratamiento hidrotérmico de los materiales celulósicos sometidos a la explosión. Además, pueden usarse para la producción cepas capaces de tolerar compuestos tales como furfurales o hidroximetilfurfurales. Otra posibilidad es usar microorganismos que no solo sean capaces de tolerar los furfurales en los medios de fermentación, sino que también puedan metabolizar estos subproductos durante la fermentación.
[0124] El proceso de explosión también genera niveles significativos de sales. Por ejemplo, las condiciones típicas de explosión pueden generar conductividades superiores a 5 mS/cm cuando la biomasa celulósica explotada se resuspende en una proporción de 10:1 agua:sólidos (peso en seco). La biomasa explotada diluida se puede someter a sacarificación enzimática y el sobrenadante resultante se puede concentrar hasta 25 veces para su uso en el biorreactor. El nivel de sal (medido por conductividad) en la(s) corriente(s) de azúcar concentrado puede ser inaceptablemente alto (hasta 1,5 M Na+ equivalentes). También se generan sales adicionales tras la neutralización de los materiales explotados para el posterior proceso de sacarificación enzimática. Según los métodos descritos en el presente documento, estas sales se pueden eliminar para que la(s) corriente(s) de azúcar celulósico concentrado resultante(s) se pueda(n) usar en procesos heterótrofos para producir lípidos. El método para eliminar estas sales puede ser la desionización con resinas, como, entre otras, DOWEX Marathon MR3. El paso de desionización con resina puede ocurrir antes de la concentración de azúcar o el ajuste del pH y el tratamiento hidrotérmico de la biomasa antes de la sacarificación, o cualquier combinación de los anteriores; o bien, el paso se lleva a cabo después de uno o más de estos procesos. Alternativamente, el propio proceso de explosión se modifica para evitar la
generación de sales a niveles inaceptablemente altos. Por ejemplo, una alternativa adecuada a la explosión con ácido sulfúrico (u otro ácido) de la biomasa celulósica es la reducción mecánica a pulpa para hacer que la biomasa celulósica sea receptiva a la hidrólisis enzimática (sacarificación). Como alternativa, pueden usarse cepas nativas de microorganismos resistentes a niveles elevados de sales o cepas modificadas genéticamente con resistencia a niveles elevados de sales.
[0125] El proceso de preparación de biomasa celulósica explotada para su uso en la producción heterótrofa de aceite microbiano usando microbios oleaginosos se puede realizar de la siguiente manera. Un primer paso comprende el ajuste del pH de la biomasa celulósica explotada resuspendida al rango de 5,0-5,3 seguido de lavado de la biomasa celulósica tres veces. Este paso de lavado se puede lograr por una variedad de medios que incluyen el uso de resinas de intercambio de iones y desalinización, ósmosis inversa, tratamiento hidrotérmico (como se ha descrito anteriormente) o simplemente resuspensión y centrifugación repetidas en agua desionizada. Este paso de lavado da como resultado una corriente celulósica cuya conductividad está entre 100-300 pS/cm y la eliminación de cantidades significativas de furfurales e hidroximetilfurfurales. Los decantados de este paso de lavado se pueden guardar para concentrar azúcares de cinco carbonos liberados de la fracción de hemicelulosa. Un segundo paso comprende la sacarificación enzimática de la biomasa celulósica lavada. Por ejemplo, se puede utilizar Accellerase (Genencor). Un tercer paso comprende la recuperación de azúcares mediante centrifugación o decantación y lavado de la biomasa sacarificada. La biomasa resultante (sólidos) es un componente rico en lignina y denso en energía que puede usarse como combustible o desecharse. Se recoge la corriente de azúcar recuperada en el proceso de centrifugado/decantación y aclarado. Un cuarto paso comprende la microfiltración para eliminar los sólidos contaminantes con la recuperación del permeado. Un quinto paso comprende un paso de concentración que se puede lograr utilizando un evaporador al vacío. Este paso puede incluir opcionalmente la adición de agentes antiespumantes como P'2000 (Sigma/Fluka), que a veces es necesario debido al contenido de proteínas de la materia prima de azúcar resultante.
[0126] La fuente de carbono puede ser glicerol, incluido el subproducto de glicerol acidulado y no acidulado de la transesterificación de biodiésel. La fuente de carbono puede incluir glicerol y al menos otra fuente de carbono. En algunos casos, todo el glicerol y dicha al menos otra fuente de carbono fijo se proporcionan al microorganismo al comienzo de la fermentación. En algunos casos, el glicerol y dicha al menos otra fuente de carbono fijo se proporcionan al microorganismo de manera simultánea en una proporción predeterminada. En algunos casos, el glicerol y dicha al menos otra fuente de carbono fijo se proporcionan a los microbios a una velocidad predeterminada durante el transcurso de la fermentación.
[0127] Algunas microalgas experimentan una división celular más rápida en presencia de glicerol que en presencia de glucosa (véase la publicación PCT N.02008/151149). En estos casos, los procesos de crecimiento en dos etapas, en los que las células se alimentan primero con glicerol para aumentar rápidamente la densidad celular y luego se alimentan con glucosa para acumular aceite microbiano (lípidos), pueden mejorar la eficiencia con la que se produce el aceite. El uso del subproducto de glicerol del proceso de transesterificación proporciona ventajas económicas significativas cuando se vuelve a poner en un proceso de producción de aceite microbiano. También se proporcionan otros métodos de alimentación, como los que emplean mezclas de glicerol y glucosa como fuente de carbono fijo. Con el uso de tales mezclas para la alimentación también se obtienen beneficios económicos similares. Además, se puede proporcionar a las microalgas azúcares alternativos, como la sacarosa, en varias combinaciones con glicerol.
[0128] La fuente de carbono puede ser azúcar invertido. El azúcar invertido se produce dividiendo la sacarosa en los monosacáridos que lo componen, fructosa y glucosa. La producción de azúcar invertido se puede lograr mediante varios métodos que se conocen en la técnica. Uno de estos métodos es calentar una solución acuosa de sacarosa. A menudo, se emplean catalizadores para acelerar la conversión de sacarosa en azúcar invertido. Estos catalizadores pueden ser biológicos; por ejemplo, se pueden agregar enzimas como invertasas y sacarasas a la sacarosa para acelerar la reacción de hidrólisis para producir azúcar invertido. El ácido es un ejemplo de un catalizador no biológico que, cuando se combina con el calor, puede acelerar la reacción de hidrólisis. Una vez que se produce el azúcar invertido, es menos propenso a la cristalización en comparación con la sacarosa y, por lo tanto, brinda ventajas para el almacenamiento y las fermentaciones discontinuas, donde, en el caso del cultivo heterótrofo de microbios, incluidas las microalgas, existe la necesidad de una fuente concentrada de carbono. La fuente de carbono puede ser azúcar invertido, preferiblemente en forma concentrada (al menos el 90 % de su máxima solubilidad en las condiciones utilizadas, como se ha descrito anteriormente), es decir, al menos 800 g/litro, al menos 900 g/litro, al menos 1000 g/litro o al menos 1100 g/litro. El azúcar invertido, preferiblemente en forma concentrada, se suministra a las células con el tiempo a medida que las células crecen y acumulan lípidos.
[0129] La fuente de carbono puede ser sacarosa, incluyendo una materia prima compleja que contiene sacarosa, como el jugo de caña espeso del procesamiento de la caña de azúcar. Como se ha señalado anteriormente, debido a las densidades más altas de los cultivos para la producción de aceite heterótrofo, la fuente de carbono fijo (por
ejemplo, sacarosa, glucosa, etc.) se encuentra en forma concentrada, es decir, al menos 500 g/litro, al menos 600 g/litro, al menos 700 g/litro o al menos 800 g/litro de la fuente de carbono fijo antes del paso de cultivo, que es opcionalmente un cultivo discontinuo en el que el material se suministra a las células a lo largo del tiempo a medida que las células crecen y acumulan lípidos. En algunos casos, la fuente de carbono es sacarosa en forma de jugo de caña espeso, típicamente en forma concentrada, es decir, al menos 60 % de sólidos o aproximadamente 770 g/litro de azúcar, al menos 70 % de sólidos o aproximadamente 925 g/litro de azúcar, o al menos 80 % de sólidos o aproximadamente 1125 g/litro de azúcar antes de la etapa de cultivo, que es opcionalmente un cultivo discontinuo. El jugo de caña espeso concentrado se suministra a las células con el tiempo a medida que las células crecen y acumulan lípidos.
[0130] El medio de cultivo puede incluir además al menos una enzima de utilización de sacarosa. En algunos casos, el medio de cultivo incluye una sacarasa. La enzima sacarasa puede ser una enzima sacarasa secretable codificada por un gen exógeno de sacarasa expresado por la población de microorganismos. Por lo tanto, en algunos casos, como se describe con más detalle en la sección IV, a continuación, el microbio utilizado en los métodos descritos en este documento se ha diseñado genéticamente para que exprese una enzima de utilización de sacarosa, como un transportador de sacarosa, una sacarasa, una hexoquinasa, una glucocinasa o una fructocinasa.
[0131] Las materias primas complejas que contienen sacarosa incluyen la melaza de desecho del procesamiento de la caña de azúcar; el uso de este producto de desecho de bajo valor del procesamiento de la caña de azúcar puede proporcionar ahorros significativos en los costes de producción de hidrocarburos y otros aceites. Otra materia prima compleja que contiene sacarosa que es útil en los métodos descritos en este documento es el sorgo, incluyendo jarabe de sorgo y sorgo puro. El jarabe de sorgo se produce a partir del jugo de la caña de sorgo dulce. Su perfil de azúcares se compone principalmente de glucosa (dextrosa), fructosa y sacarosa.
4. Producción de aceite
[0132] Para la producción de aceite (lípido) según los métodos descritos en este documento, es preferible cultivar células en la oscuridad, como es el caso, por ejemplo, cuando se utilizan fermentadores extremadamente grandes (de 40000 litros y más) que no permiten que la luz llegue al cultivo. Por ejemplo, Prototheca y otras especies de microalgas pueden cultivarse y propagarse para la producción de aceite en un medio que contenga una fuente de carbono fijo y en ausencia de luz; dicho crecimiento se conoce como crecimiento heterótrofo.
[0133] Como ejemplo, se introduce en el medio un inóculo de células de microalgas productoras de lípidos; hay un período de retraso (fase de retraso) antes de que las células comiencen a propagarse. Después del período de retraso, la tasa de propagación aumenta constantemente y entra en la fase logarítmica o exponencial. A la fase exponencial le sigue, a su vez, una ralentización de la propagación debido a la disminución de nutrientes como el nitrógeno, el aumento de sustancias tóxicas y los mecanismos de percepción de quórum. Después de esta desaceleración, la propagación se detiene y las células entran en una fase estacionaria o en un estado de crecimiento constante, según el entorno particular que se les proporcione. Para obtener una biomasa rica en lípidos, el cultivo generalmente se cosecha mucho después del final de la fase exponencial, que puede terminarse antes de tiempo permitiendo que el nitrógeno u otro nutriente clave (que no sea carbono) se agote, obligando a las células a convertir las fuentes de carbono, presentes en exceso, en lípidos. Los parámetros de las condiciones de cultivo se pueden manipular para optimizar la producción total de aceite, la combinación de ácidos grasos del aceite producido y/o la producción de un ácido graso específico y los lípidos correspondientes.
[0134] Preferiblemente, los microorganismos cultivados usando las condiciones descritas en este documento y otras conocidas en la técnica comprenden al menos aproximadamente el 20 % en peso de lípido, preferiblemente al menos aproximadamente el 40 % en peso, más preferiblemente al menos aproximadamente el 50 % en peso y, de la manera más preferible, al menos aproximadamente 60 % en peso. Las condiciones del proceso se pueden ajustar para aumentar la producción de lípidos adecuados para un uso particular y/o para reducir el coste de producción. Por ejemplo, se puede cultivar una microalga en presencia de una concentración limitante de uno o más nutrientes, como, por ejemplo, nitrógeno, fósforo o azufre, mientras se proporciona un exceso de energía de carbono fijo, como la glucosa. La limitación del nitrógeno tiende a aumentar la producción de lípidos microbianos en comparación con la producción de lípidos microbianos en un cultivo en el que se proporciona nitrógeno en exceso. El aumento en la producción de lípidos puede ser de al menos aproximadamente: 10 %, 50 %, 100 %, 200 % o 500 %. El microbio se puede cultivar en presencia de una cantidad limitante de un nutriente durante una parte del período de cultivo total o durante todo el período. La concentración de nutrientes se puede alternar entre una concentración limitante y una concentración no limitante al menos dos veces durante el período de cultivo total. El contenido de lípidos de las células puede incrementarse continuando el cultivo durante periodos de tiempo prolongados mientras se proporciona un exceso de carbono, pero limitando o no el nitrógeno.
[0135] Alternativamente, la producción de lípidos aumenta mediante el cultivo de un microbio productor de lípidos (por ejemplo, microalgas) en presencia de uno o más cofactores para una enzima de la ruta de los lípidos (por ejemplo, una enzima sintética de ácidos grasos). En general, la concentración de los cofactores es suficiente para aumentar la producción de aceite microbiano (por ejemplo, lípidos y ácidos grasos) en comparación con la producción de aceite microbiano en ausencia de los cofactores. El o los cofactores pueden proporcionarse al cultivo incluyendo en el cultivo un microbio (por ejemplo, microalgas) que contenga un gen exógeno que codifique el o los cofactores. Alternativamente, se pueden proporcionar cofactor(es) a un cultivo incluyendo un microbio (por ejemplo, microalgas) que contenga un gen exógeno que codifique una proteína que participa en la síntesis del cofactor. Los cofactores adecuados pueden incluir una vitamina requerida por una enzima de la ruta de los lípidos, como, por ejemplo: biotina y pantotenato. Los genes que codifican cofactores adecuados para su uso en los métodos descritos en este documento o que participan en la síntesis de dichos cofactores son ampliamente conocidos y pueden introducirse en microbios (por ejemplo, microalgas u otros microbios oleaginosos descritos en este documento), utilizando construcciones y técnicas como las descritas anteriormente.
[0136] Los ejemplos específicos de biorreactores, condiciones de cultivo y métodos de crecimiento y propagación heterótrofos descritos en el presente documento se pueden combinar de cualquier manera adecuada para mejorar las eficiencias del crecimiento microbiano y la producción de lípidos y/o proteínas.
[0137] Se ha generado biomasa de microalgas con un alto porcentaje de acumulación de aceite/lípidos en peso en seco (véase la publicación PCT n.02008/151149). La biomasa de microalgas generada por los métodos de cultivo descritos en este documento y útil según los métodos descritos en este documento comprende al menos un 10 % de aceite de microalgas en peso en seco. La biomasa de microalgas puede comprender al menos un 25 %, al menos un 50 %, al menos un 55 % o al menos un 60 % de aceite de microalgas en peso en seco. La biomasa de microalgas puede contener del 10 al 90 % de aceite de microalgas, del 25 al 75 % de aceite de microalgas, del 40 al 75 % de aceite de microalgas o del 50 al 70 % de aceite de microalgas en peso en seco.
[0138] El aceite de microalgas de la biomasa que se describe en este documento, o extraído de la biomasa para su uso en los métodos y composiciones descritos en este documento, puede comprender glicerolípidos con una o más cadenas laterales de éster de ácido graso distintas. Los glicerolípidos se componen de una molécula de glicerol esterificada a una, dos o tres moléculas de ácidos grasos, que pueden tener longitudes variables y grados de saturación variables. Las características de longitud y saturación de las moléculas de ácidos grasos (y los aceites de microalgas que las contienen) se pueden manipular para modificar las propiedades o proporciones de las moléculas de ácidos grasos en los aceites de microalgas descritos en este documento a través de condiciones de cultivo o mediante ingeniería de rutas de lípidos, como se describe en más detalles e la sección V, a continuación. Por lo tanto, se pueden preparar mezclas específicas de aceite de algas (u otros microorganismos) dentro de una sola especie de algas o mezclando la biomasa o el aceite de algas de dos o más especies de microalgas, o mezclando el aceite de algas descrito en este documento con aceites de otras fuentes como soja, colza, canola, palma, palmiste, coco, maíz, residuos vegetales, árbol de sebo, aceituna, girasol, semilla de algodón, grasa de pollo, sebo bovino, sebo porcino, microalgas, macroalgas, microbios, cuphea, lino, cacahuete, grasa blanca de primera, manteca de cerdo, Camellina sativa, semilla de mostaza, anacardo, avena, lupino, kenaf, caléndula, socorro, café, linaza (lino), avellana, euforbia, semilla de calabaza, cilantro, camelia, sésamo, cártamo, arroz, árbol de tung, cacao, copra, adormidera, ricino, nuez pecán, jojoba, macadamia, nueces de Brasil, aguacate, petróleo o una fracción destilada de cualquiera de los aceites anteriores.
[0139] Como se ha señalado antes, la composición del aceite, es decir, las propiedades y proporciones de los constituyentes de ácidos grasos de los glicerolípidos, también pueden manipularse combinando biomasa o aceite de al menos dos especies distintas de microalgas. Al menos dos de las distintas especies de microalgas pueden tener diferentes perfiles de glicerolípidos. Las distintas especies de microalgas se pueden cultivar juntas o por separado como se describe en este documento, preferiblemente en condiciones heterótrofas, para generar los respectivos aceites. Diferentes especies de microalgas pueden contener diferentes porcentajes de constituyentes de ácidos grasos distintos en los glicerolípidos de la célula.
[0140] Por lo general, las cepas de Prototheca tienen perfiles de lípidos con ácidos grasos C16 y C18 como especies predominantes. Dichos ácidos grasos de cadena más larga, especialmente los ácidos grasos monosaturados C16 y C18 (es decir, C16:1 y C18:1) son generalmente preferidos para la producción de fluidos dieléctricos (véase, por ejemplo, la patente de EE. UU. n.o 6,274,067). Por ejemplo, Prototheca moriformis (UTEX 1435), Prototheca stagnora (UTEX 327), y Prototheca moriformis (UTEX 1441) contienen entre un 12 % y un 30 % de ácidos grasos C16 y entre un 50 % y un 58 % de ácidos grasos C18:1. Chlorella protothecoides (UTEX 250) contiene aproximadamente un 73 % de ácidos grasos C18:1 y otras cepas de Chlorella protothecoides, incluidas, entre otras, UTEX 25, UTEX 249, UTEX 256, UTEX 264, UTEX 411, CCAP 211/17, CCAP 221/8D y SAG 221 10d, pueden contener entre un 7 % y un 18 % de ácidos grasos C 16 y entre un 55 % y un 75 % de ácidos grasos C18:1. El aceite microbiano (lípido) útil en
los productos descritos en el presente documento (como los fluidos dieléctricos) puede tener al menos un 50 % de C18:1, por ejemplo, al menos un 55 %, al menos un 60 %, al menos un 65 %, al menos un 70 %, al menos alrededor de un 75 %, al menos alrededor de un 80 %, al menos alrededor de un 85 % y al menos alrededor de un 90 % de C18:1. En algunas formas de realización, el aceite microbiano (lípido) es menos del 7,5 %, por ejemplo de menos del 5 %, menos del 2,5 % y menos del 1 % de C18:2. El aceite microbiano puede tener cualquier combinación de porcentajes de C18:1 y C18:2 que sumen 100 % o menos. Por ejemplo, el aceite microbiano puede tener al menos un 50 % de C18:1 y menos de un 10 % de C18:2 o al menos un 80 % de C18:1 y menos de un 5 % de C18:2.
[0141] El aceite (lípido) de microalgas (u otro microorganismo) también puede incluir otros constituyentes producidos por las microalgas, o incorporados en el aceite de microalgas del medio de cultivo. Estos otros componentes pueden estar presentes en cantidades variables según las condiciones de cultivo, la especie, el método de extracción utilizado para recuperar el aceite de la biomasa y otros factores que pueden afectar la composición del aceite. Entre los ejemplos no limitativos de tales constituyentes se incluyen los carotenoides, presentes en menos de 0,4 microgramos/ml; el licopeno, presente en menos de 0,001 microgramos/ml; el betacaroteno, presente en menos de 0. 02.microgramos/ml; la clorofila, presente en menos de 0,02 miligramos por kilogramo de aceite; el gamma tocoferol, presente de 0,40 a 0,60 miligramos por 100 gramos de aceite; el campesterol, presente de 3 a 9 miligramos por 100 gramos de aceite; y los tocotrienoles, presentes en menos de 0,5 miligramos por gramo de aceite.
[0142] Los otros constituyentes pueden incluir, sin limitación, fosfolípidos, tocoferoles, tocotrienoles, carotenoides (por ejemplo, alfacaroteno, betacaroteno, licopeno, etc.), xantofilas (por ejemplo, luteína, zeaxantina, alfacriptoxantina y beta-critoxantina), y varios compuestos orgánicos o inorgánicos. En algunos casos, el aceite extraído de especies de Prototheca comprende entre 0,003 y 0,039 microgramos de luteína/gramo de aceite, menos de 0,003 microgramos de licopeno/gramo de aceite; y menos de 0,003 microgramos de betacaroteno/gramo de aceite.
III. MÉTODOS Y MATERIALES DE INGENIERÍA GENÉTICA
[0143] Los métodos descritos en este documento se pueden poner en práctica usando microalgas recombinantes u otros microbios oleaginosos recombinantes. En esta sección se describen métodos y materiales para modificar genéticamente microbios oleaginosos, como microalgas, ejemplificando específicamente las células de Prototheca, para hacer que las células hospedadoras recombinantes sean útiles en los métodos descritos en este documento, incuyendo, entre otros, células hospedadoras de Prototheca moríformis, Prototheca zopfii, Prototheca krugani, y Prototheca stagnora recombinantes. La descripción de estos métodos y materiales se divide en subsecciones para la comodidad del lector. En la subsección 1 se describen los métodos de transformación. En la subsección 2 se describen métodos de ingeniería genética que utilizan recombinación homóloga. En la subsección 3 se describen los vectores de expresión y los componentes.
1. Métodos de ingeniería - transformación
[0144] Las células se pueden transformar mediante cualquier técnica adecuada, incluyendo, por ejemplo, biolística, electroporación (véase Maruyama y col. (2004), Biotechnology Techniques 8:821-826), transformación de perlas de vidrio y transformación de filamentos de carburo de silicio. Otro método que se puede utilizar implica la formación de protoplastos y el uso de CaCl2 y polietilenglicol (PEG) para introducir ADN recombinante en microalgas u otras células microbianas (véase Kim y col. (2002), Mar. Biotechnol. 4:63-73, que describe el uso de este método para la transformación de Chorella ellipsoidea). La cotransformación de microalgas se puede utilizar para introducir dos moléculas de vector distintas en una célula de manera simultánea (véase, por ejemplo, Protist, 2004 Dec; 155 (4): 381-93).
[0145] También pueden utilizarse métodos biolísticos (véase, por ejemplo, Sanford, Trends ln Biotech. (1988) 6:299 302, patente de EE. UU. n.04,945,050); electroporación (Fromm y col., Proc. Nat'l. Acad. Sci. (EE. UU.) (1985) 82:5824 5828), el uso de un rayo láser, la microinyección o cualquier otro método capaz de introducir ADN en una microalga para la transformación de microbios oleaginosos, como una célula de Prototheca.
2. Métodos de ingeniería - recombinación homologa
[0146] La recombinación homóloga está relacionada con la capacidad de secuencias de ADN complementarias para alinear e intercambiar regiones de homología. En el proceso de recombinación homóloga, el ADN transgénico ("donante") que contiene secuencias homólogas a las secuencias genómicas a las que se dirige ("plantilla") se introduce en el organismo y luego se recombina en el genoma en el sitio de las secuencias homólogas genómicas correspondientes. Los pasos mecánicos de este proceso, en la mayoría de los casos, incluyen: (1) el emparejamiento de segmentos de ADN homólogos; (2) la introducción de roturas de doble cadena en la molécula de ADN donante; (3) la invasión de la molécula de ADN plantilla por los extremos libres del ADN del donante seguida de síntesis de
ADN; y (4) la resolución de eventos de reparación de ruptura de doble cadena que dan como resultado productos de recombinación finales.
[0147] La capacidad de llevar a cabo una recombinación homóloga en un organismo hospedador tiene muchas implicaciones prácticas para lo que se puede hacer a nivel genético molecular y es útil en la generación de un microbio oleaginoso que pueda producir aceites personalizados (lípidos). Por su propia naturaleza, la recombinación homóloga es un evento preciso de selección génica; por lo tanto, la mayoría de las líneas transgénicas generadas con la misma secuencias de direccionamiento serán esencialmente idénticas en términos de fenotipo, lo que requerirá la selección de muchos menos eventos de transformación. La recombinación homóloga también se dirige a eventos de inserción de genes en el cromosoma hospedador, lo que da como resultado una excelente estabilidad genética, incluso en ausencia de selección genética. Debido a que diferentes locus cromosómicos pueden afectar a la expresión génica, incluso de promotores/UTR heterólogos, la recombinación homóloga puede ser un método para consultar locus en un entorno genómico desconocido y para evaluar el impacto de un entorno genómico particular en la expresión génica.
[0148] Las aplicaciones de ingeniería genética particularmente útiles que utilizan la recombinación homóloga cooptan elementos reguladores específicos del hospedador, como promotores/UTR, para impulsar la expresión de genes heterólogos de una manera altamente específica. Por ejemplo, puede usarse la ablación o inactivación de genes/familias de genes de desaturasa con un gen heterólogo que codifica un marcador selectivo para aumentar el porcentaje global de ácidos grasos saturados producidos en la célula hospedadora. En el ejemplo 4 se describen las construcciones de direccionamiento de recombinación homóloga y un ejemplo práctico de tales ablaciones del gen de la desaturasa (inactivaciones o knockouts) generadas en Prototheca moriformis. Otro enfoque para disminuir la expresión de un gen endógeno es usar un método inducido por ARN de regulación a la baja o silenciamiento de la expresión génica que incluye, entre otros, un enfoque de ARNi o antisentido, así como un enfoque de ARNbc. Los enfoques antisentido, ARNi, ARNbc y ARN horquillado son ampliamente conocidos en la técnica e incluyen la introducción de una construcción de expresión que, cuando se expresa como ARNm, conduce a la formación de un ARN horquillado o una construcción de expresión que contiene una porción del gen diana que se transcribe en la orientación antisentido. Todos estos enfoques dan como resultado la disminución de la expresión del gen diana. En el ejemplo 4 también se describen construcciones de expresión y un ejemplo práctico de la regulación a la baja de un gen endógeno de desaturasa de ácidos grasos delta 12 (FADc) de Prototheca moriformis mediante un enfoque de ARN horquillado.
[0149] Gracias a que la recombinación homóloga es un evento preciso de direccionamiento génico, se puede usar para modificar cualquier nucleótido dentro de un gen o región de interés con precisión, siempre que se hayan identificado suficientes regiones flanqueantes. Por lo tanto, la recombinación homóloga puede usarse como medio para modificar secuencias reguladoras que afectan a la expresión génica de ARN y/o proteínas. También se puede usar para modificar regiones codificantes de proteínas para modificar actividades enzimáticas tales como especificidad de sustrato, afinidad y Km, efectuando así el cambio deseado en el metabolismo de la célula hospedadora. La recombinación homóloga proporciona un medio poderoso para manipular el genoma del hospedador, lo que da como resultado la selección de genes, la conversión de genes, la deleción de genes, la duplicación de genes y la inversión de genes, y el intercambio de elementos reguladores de la expresión de genes, como promotores, potenciadores y 3'UTR.
[0150] La recombinación homóloga se puede lograr utilizando construcciones de direccionamiento que contienen fragmentos de secuencias endógenas para “dirigirse a” o actuar sobre el gen o la región de interés dentro del genoma de la célula hospedadora endógena. Dichas secuencias de direccionamiento pueden ubicarse en el extremo 5' del gen o región de interés, en el extremo 3' del gen/región de interés, o incluso flanquear el gen/la región de interés. Tales construcciones de direccionamiento pueden transformarse en la célula hospedadora como un ADN plasmídico superenrollado con un esqueleto de vector adicional, un producto de PCR sin un esqueleto de vector o como una molécula linealizada. En algunos casos, puede ser ventajoso exponer primero las secuencias homólogas dentro del ADN transgénico (ADN donante) con una enzima de restricción. Este paso puede aumentar la eficacia de la recombinación y disminuir la aparición de eventos no deseados. Otros métodos para aumentar la eficacia de la recombinación incluyen el uso de la PCR para generar ADN transgénico transformante que contiene extremos lineales homólogos a las secuencias genómicas sobre las que se desea actuar.
[0151] Para fines ilustrativos y sin intención limitativa, las regiones de secuencias de ADN donadoras que son útiles para la recombinación homóloga incluyen la región KE858 de ADN de Prototheca-moriformis. KE858 es un fragmento genómico de 1,3 kb que abarca parte de la región codificante de una proteína que comparte homología con la familia de proteínas del ARN de transferencia (ARNt). Mediante transferencias de Southern se ha demostrado que la secuencia KE858 está presente en una sola copia en el genoma de Prototheca moriformis (UTEX 1435). Esta región
y los ejemplos del uso de esta región para el direccionamiento de la recombinación homologa se han descrito en la Solicitud PCT N.0 PCT/US2009/66142. Otra región útil del ADN donador es la secuencia genómica 6S.
3. Vectores v componentes de vectores
[0152] Los vectores para la transformación de microorganismos se pueden preparar mediante técnicas conocidas y familiares para los expertos en la materia a la vista de la descripción del presente documento. Un vector normalmente contiene uno o más genes, en donde cada gen codifica la expresión de un producto deseado (el producto génico) y está unido operativamente a una o más secuencias de control que regulan la expresión génica o dirigen el producto génico hacia una ubicación particular en la célula recombinante. Para ayudar al lector, esta subsección se divide en subsecciones. En la subsección A se describen secuencias de control que pueden estar contenidas en vectores. En la subsección B se describen genes típicamente contenidos en vectores, así como métodos de optimización de codones y genes preparados con ellos.
A. Secuencias de control
[0153] Las secuencias de control son ácidos nucleicos que regulan la expresión de una secuencia codificante o que dirigen un producto génico a una ubicación particular dentro o fuera de una célula. Las secuencias de control que regulan la expresión incluyen, por ejemplo, promotores que regulan la transcripción de una secuencia codificante y terminadores que terminan la transcripción de una secuencia codificante. Otra secuencia de control es una secuencia no traducida en 3' ubicada en el extremo de una secuencia codificante que codifica una señal de poliadenilación. Las secuencias de control que dirigen productos génicos a ubicaciones particulares incluyen aquellas que codifican péptidos señal, que dirigen la proteína a la que están unidas a una ubicación particular dentro o fuera de la célula.
[0154] Por lo tanto, un diseño de vector ejemplar para la expresión de un gen exógeno en una microalga u otro microbio oleaginoso contiene una secuencia de codificación para un producto génico deseado (por ejemplo, un marcador seleccionable, una enzima de modificación de la ruta de lípidos o una enzima de utilización de sacarosa) en unión operativa con un promotor activo de la microalga u otro microbio oleaginoso. Alternativamente, si el vector no contiene un promotor en unión operativa con la secuencia de codificación de interés, la secuencia de codificación se puede transformar en las células de manera que se una operativamente a un promotor endógeno en el punto de integración del vector. Se ha demostrado que el método de transformación sin promotores funciona en microalgas (véase, por ejemplo, Plant Journal 14: 4, (1998), págs. 441-447) y otros microbios.
[0155] Muchos promotores son activos en las microalgas, incluyendo promotores que son endógenos a las algas que se transforman, así como promotores que no son endógenos a las algas que se transforman (es decir, promotores de otras algas, promotores de plantas superiores y promotores de virus de plantas o virus de algas). Los promotores exógenos y/o endógenos ilustrativos que son activos en las microalgas (así como los genes de resistencia a los antibióticos funcionales en las microalgas) se describen en la publicación PCT N.o 2008/151149 y las referencias citadas en ella.
[0156] El promotor usado para expresar un gen exógeno puede ser el promotor unido de manera natural a ese gen o puede ser un promotor de gen heterólogo. Algunos promotores son activos en más de una especie de microalgas. Otros promotores son específicos para una especie. Los promotores ilustrativos incluyen promotores como la P-tubulina de Chlamydomonas reinhardtii, utilizada en los Ejemplos siguientes, y promotores virales, como los promotores derivados del virus del mosaico de la coliflor (CMV) y el virus de la Chlorella, que han demostrado ser activos en múltiples especies de microalgas (véase, por ejemplo, Plant Cell Rep. 2005 Mar;23(10-11):727-35; J Microbiol. 2005 Aug;43(4):3S1-5; Mar Biotechnol (New York). 2002 Jan;4(1):S3-73). Otro promotor que es apto para el uso en la expresión de genes exógenos en Prototheca es el promotor de glutamato deshidrogenasa/5'UTR de Chlorella sorokiniana. Normalmente, se utilizan al menos 10, 20, 30, 40, 50 o 60 nucleótidos o más de estas secuencias que contienen un promotor. Los promotores ilustrativos útiles para la expresión de genes exógenos en Prototheca se enumeran en el listado de secuencias de esta solicitud, como el promotor del gen HUP1 de Chlorella (SEQ ID N.o 1) y el promotor de nitrato reductasa de Chlorella ellipsoidea (SEQ ID N.02). Los promotores de virus de Chlorella también se pueden usar para expresar genes en Prototheca, como la SEQ ID N.o 1-7 de la patente de EE. UU. n.o 6,395,965. Puede encontrarse promotores adicionales activos en Prototheca, por ejemplo, en Biochem Biophys Res Comun. 1994;Oct 14204 (1): 187-94; Plant Mol Biol. 1994 Oct; 26 (1): 85-93; Virology. 2004 Aug 15; 326 (1): 150-9; y Virology. 2004 Jan 5; 318 (1): 214-23.
[0157] Un promotor generalmente se puede caracterizar como constitutivo o inducible. Los promotores constitutivos son generalmente activos o funcionan para activar la expresión en todo momento (o en ciertos momentos del ciclo de vida celular) al mismo nivel. Los promotores inducibles, por el contrario, están activos (o se vuelven inactivos) o se regulan significativamente al alza o a la baja solo en respuesta a un estímulo. Ambos tipos de promotores tienen un
uso en los métodos descritos en este documento. Entre los promotores inducibles útiles en los métodos descritos en este documento se incluyen aquellos que median la transcripción de un gen unido operativamente en respuesta a un estímulo, como una molécula pequeña proporcionada exógenamente (por ejemplo, glucosa, como en la SEQ ID N.0 1), temperatura (calor o frío), falta de nitrógeno en los medios de cultivo, etc. Los promotores adecuados pueden activar la transcripción de un gen esencialmente silencioso o regular al alza, preferiblemente sustancialmente, la transcripción de un gen unido operativamente que se transcribe a un nivel bajo.
[0158] La inclusión de la secuencia de control de la región de terminación es opcional y, si se emplea, la elección será principalmente por conveniencia, ya que las regiones de terminación son relativamente intercambiables. La región de terminación puede ser nativa de la región de iniciación de la transcripción (el promotor), puede ser nativa de la secuencia de ADN de interés o puede obtenerse de otra fuente. Véase, por ejemplo, Chen y Orozco, Nucleic Acids Res. (1988) 16:8411.
[0159] Los métodos descritos en el presente documento también pueden hacer uso de vectores que contienen secuencias de control y genes recombinantes que proporcionan la expresión compartimentada de un gen de interés. Los orgánulos para el direccionamiento son los cloroplastos, los plastidios, las mitocondrias y el retículo endoplásmico. Además, los métodos descritos en este documento también pueden hacer uso de secuencias de control y genes recombinantes y vectores que los contienen descritos en este documento que permiten la secreción de una proteína fuera de la célula.
[0160] Las proteínas expresadas en el genoma nuclear de Prototheca se pueden dirigir al plastidio utilizando señales de direccionamiento a plastidios. Se conocen secuencias de direccionamiento a plastidios que son endógenas de Chlorella, como los genes del genoma nuclear de Chlorella que codifican proteínas que se dirigen al plastidio; véase, por ejemplo, los números de acceso de GenBank AYS4S197 y AF499S84, y los vectores que contienen dichas secuencias de control se pueden usar en los métodos descritos en el presente documento para dirigir la expresión de una proteína a un plastidio de Prototheca.
[0161] Los ejemplos siguientes describen el uso de secuencias de direccionamiento a plastidios de algas para dirigir proteínas heterólogas al compartimento correcto de la célula hospedadora. Se crearon bibliotecas de ADN utilizando células de Prototheca moríformis y Chlorella protothecoides, y se describen en la Solicitud PCT N.o PCT/US2009/066142.
[0162] La expresión de un polipéptido en Prototheca o en otro microbio oleaginoso puede dirigirse al retículo endoplásmico. La inclusión de una señal de retención o clasificación adecuada en un vector de expresión garantiza que las proteínas se retengan en el retículo endoplásmico (RE) y no sigan hacia el aparato de Golgi. Por ejemplo, el vector IMPACTVECTOR1.3, de Wageningen UR-Plant Research International, incluye la conocida señal de clasificación o retención KDEL. Con este vector, la retención en el RE tiene una ventaja práctica porque se ha observado que mejora los niveles de expresión por quintuplicado. La razón principal de esto parece ser que el RE contiene concentraciones más bajas y/o diferentes proteasas responsables de la degradación postraduccional de las proteínas expresadas que las que están presentes en el citoplasma. Se conocen señales de retención en el RE funcionales en microalgas verdes. Por ejemplo, véase Proc Natl Acad Sci EE. UU. 2005 Apr 26; 102 (17): 6225-30.
[0163] Alternativamente, se puede direccionar un polipéptido para la secreción fuera de la célula en el medio de cultivo. Véase Hawkins y col., Current Microbiology vol. 38 (1999), págs. 335-341 para ver ejemplos de señales de secreción activas en Chlorella que también se pueden utilizar en otras microalgas, como Prototheca.
B. Genes v optimización de codones
[0164] Normalmente, un gen incluye un promotor, una secuencia codificante y secuencias de control de la terminación. Cuando se ensambla mediante tecnología de ADN recombinante, un gen puede denominarse casete de expresión y puede estar flanqueado por sitios de restricción para una inserción conveniente en un vector que se usa para introducir el gen recombinante en una célula hospedadora. El casete de expresión puede estar flanqueado por secuencias de ADN del genoma u otro ácido nucleico diana para facilitar la integración estable del casete de expresión en el genoma mediante recombinación homóloga. Alternativamente, el vector y su casete de expresión pueden permanecer sin integrar, en cuyo caso el vector normalmente incluye un origen de replicación, que es capaz de proporcionar la replicación del ADN del vector heterólogo.
[0165] Un gen común presente en un vector es un gen que codifica una proteína, cuya expresión permite diferenciar la célula recombinante que contiene la proteína de las células que no expresan la proteína. Tal gen, o su producto génico correspondiente, se denomina marcador seleccionable. Cualquiera de una amplia variedad de marcadores seleccionables puede emplearse en una construcción transgénica útil para transformar Prototheca o cualquier otro
microbio oleaginoso útil en los métodos descritos en este documento. Entre los ejemplos de marcadores seleccionables adecuados se incluyen el gen de resistencia G418, el gen de la nitrato reductasa (véase Dawson y col. (1997), Current Microbiology 35:356-362), el gen de la higromicina fosfotransferasa (HPT; véase Kim y col. (2002), Mar. Biotechnol. 4:63-73), el gen de la neomicina fosfotransferasa y el gen ble, que confiere resistencia a la fleomicina (Huang y col. (2007), Appl. Microbiol. Biotechnol. 72:197-205). Los métodos para determinar la sensibilidad de las microalgas y otros microbios oleaginosos a los antibióticos son ampliamente conocidos. Por ejemplo, véase Mol Gen Genet. 1996 Oct 16; 252 (5): 572-9.
[0166] También se pueden emplear otros marcadores seleccionables que no están basados en antibióticos en una construcción transgénica útil para transformar microalgas en general, incluyendo especies de Prototheca. Los genes que confieren la capacidad de utilizar algunas fuentes de carbono que antes no podían ser utilizadas por las microalgas también se pueden utilizar como marcador seleccionable. A modo de ilustración, las cepas de Prototheca moriformis suelen texner dificultades para crecer, si es que lo hacen, con sacarosa. El uso de una construcción que contiene un gen de sacarasa puede conferir la capacidad de los transformantes positivos para crecer en sacarosa como sustrato de carbono.
[0167] En los métodos descritos en el presente documento, el vector de expresión se puede usar para preparar una célula hospedadora recombinante que incluirá al menos dos, y a menudo tres, genes, si uno de los genes es un marcador seleccionable. Por ejemplo, una Prototheca modificada genéticamente puede crearse por transformación con vectores que comprenden, además de un marcador seleccionable, uno o más genes exógenos, como, por ejemplo, un gen de sacarasa o un gen de acil ACP-tioesterasa. Uno o ambos genes pueden expresarse usando un promotor inducible, lo que permite controlar el tiempo relativo de expresión de estos genes para mejorar la producción de lípidos y la conversión a ésteres de ácidos grasos. La expresión de dos o más genes exógenos puede estar bajo el control del mismo promotor inducible o bajo el control de diferentes promotores inducibles (o constitutivos). En esta última situación, se puede inducir la expresión de un primer gen exógeno durante un primer período de tiempo (durante el cual se puede inducir o no la expresión de un segundo gen exógeno), y se puede inducir la expresión de un segundo gen exógeno durante un segundo período de tiempo (durante el cual la expresión de un primer gen exógeno puede o no inducirse).
[0168] Los dos o más genes exógenos (además de cualquier marcador seleccionable) pueden ser: una acil-ACP tioesterasa y una acil-CoA/aldehído reductasa, cuya acción combinada produce un producto alcohólico. Además se proporcionan otras combinaciones de genes exógenos, incluyendo, sin limitación, una acil-ACP tioesterasa y una acil-CoA reductasa para generar aldehídos. El vector puede proporcionar la combinación de una acil-ACP tioesterasa, una acil-CoA reductasa y una aldehído graso descarbonilasa para generar alcanos. En cada uno de estos casos, uno o más de los genes exógenos pueden expresarse utilizando un promotor inducible.
[0169] Otros vectores ilustrativos que expresan dos o más genes exógenos incluyen los que codifican tanto un transportador de sacarosa como una enzima sacarasa y los que codifican tanto un marcador seleccionable como una sacarasa secretada. La Prototheca recombinante u otra microalga o célula microbiana transformada con cualquier tipo de vector produce lípidos a un menor coste de fabricación debido a la capacidad diseñada para usar caña de azúcar (y azúcares derivados de la caña de azúcar) como fuente de carbono. La inserción de los dos genes exógenos descritos anteriormente se puede combinar con la interrupción de la biosíntesis de polisacáridos a través de mutagénesis dirigida y/o aleatoria, que dirige un flujo de carbono aún mayor hacia la producción de lípidos. Individualmente y en combinación, la conversión trófica, la ingeniería para alterar la producción de lípidos y el tratamiento con enzimas exógenas alteran la composición lipídica producida por un microorganismo. La alteración puede ser un cambio en la cantidad de lípidos producidos, la cantidad de una o más especies de lípidos (ácidos grasos) producida en relación con otras especies de lípidos y/o los tipos de especies de lípidos producidos en el microorganismo. Por ejemplo, las microalgas pueden diseñarse para que produzcan una mayor cantidad y/o porcentaje de TAG.
[0170] Para la expresión óptima de una proteína recombinante, es beneficioso emplear secuencias de codificación que produzcan ARNm con codones utilizados preferentemente por la célula hospedadora que se va a transformar. Por lo tanto, la expresión adecuada de los transgenes puede requerir que el uso de codones del transgén coincida con el sesgo de codones específico del organismo en el que se expresa el transgén. Los mecanismos precisos que subyacen a este efecto son muchos, pero incluyen el equilibrio adecuado de los grupos de ARNt aminoacilados disponibles con las proteínas que se sintetizan en la célula, junto con una traducción más eficiente del ARN mensajero (ARNm) transgénico cuando se satisface esta necesidad. Cuando el uso de codones en el transgén no está optimizado, las reservas de ARNt disponibles no son suficientes para permitir una traducción eficiente del ARNm heterólogo, lo que da como resultado el estancamiento y la terminación ribosomal y la posible inestabilidad del ARNm transgénico.
[0171] En este documento se describen ácidos nucleicos con codones optimizados útiles para la expresión exitosa de proteínas recombinantes en Prototheca. El uso de codones en especies de Prototheca se analizó mediante el estudio de secuencias de ADNc aisladas de Prototheca moriformis. Este análisis representa la interrogación sobre 24000 codones, que dio como resultado la Tabla 2 siguiente.
Tabla 2. Uso preferido de codones en cepas de Prototheca.
Ala GCG 345 (0,36) Asn AAT 8 (0,04)
GCA 66 (0,07) AAC 201 (0,96)
GCT 101 (0,11)
GCC 442 (0,46) Pro CCG 161 (0,29)
CCA 49 (0,09)
Cys TGT 12(0,10) CCT 71(0,13)
TGC 105 (0,90) CCC 267 (0,49)
Asp GAT 43 (0,12) Gin CAG 226 (0,82)
GAC 316 (0,88) CAA 48 (0,18)
Glu GAG 377 (0,96) Arg AGG 33 (0,06)
GAA 14 (0,04) AGA 14 (0,02)
CGG 102 (0,18)
Phe TTT 89 (0,29) CGA 49 (0,08)
TTC 216 (0,71) CGT 51(0,09)
CGC 331 (0,57)
Gly GGG 92 (0,12)
GGA 56 (0,07) Ser AGT 16(0,03)
GGT 76 (0,10) AGC 123 (0,22)
GGC 559 (0,71) TCG 152 (0,28)
TCA 31(0,06)
His CAT 42(0,21) TCT 55 (0,10)
CAC 154 (0,79) TCC 173 (0,31)
Ile ATA 4(0,01) Thr ACG 184 (0,38)
ATT 30 (0,08) ACA 24 (0,05)
ATC 338 (0,91) ACT 21 (0,05)
ACC 249 (0,52)
Lys AAG 284 (0,98)
AAA 7(0,02) Val GTG 308 (0,50)
GTA 9 (0,01)
Leu TTG 26(0,04) GTT 35 (0,06)
TTA 3 (0,00) GTC 262 (0,43)
CTG 447 (0,61)
CTA 20(0,03) Trp TGG 107 (1,00)
CTT 45 (0,06)
CTC 190 (0,26) Tyr TAT 10 (0,05)
TAC 180 (0,95)
Met ATG 191 (1,00)
Terminador TGA/T AG/T AA
[0172] El gen del vector recombinante puede haberse sometido a optimización de codones con referencia a una cepa de microalgas que no sea una cepa de Prototheca u otra cepa microbiana. Por ejemplo, los métodos de
recodificación de genes para la expresión en microalgas se describen en la patente de EE.UU. n.07,135,290. Hay información adicional para la optimización de codones que está disponible, por ejemplo, en la base de datos de uso de codones de GenBank.
[0173] Si bien los métodos y materiales descritos en este documento permiten la introducción de cualquier gen exógeno en Prototheca u otras microalgas u otros microbios oleaginosos, los genes relacionados con la utilización de sacarosa y la modificación de la ruta de los lípidos son de particular interés para los microbios que no pueden utilizarla de forma natural o para los microbios que la utilizan de manera ineficaz, como se analiza en las siguientes secciones.
IV. UTILIZACIÓN DE SACAROSA
[0174] La célula de Prototheca recombinante o de otra microalga puede contener uno o más genes de utilización de sacarosa exógenos. Dichos uno o más genes que codifican una o más proteínas pueden seleccionarse del grupo que consiste en una fructoquinasa, una glucoquinasa, una hexoquinasa, una sacarasa, un transportador de sacarosa. Por ejemplo, la expresión de un transportador de sacarosa y una sacarasa permite a Prototheca o cualquier otra microalga u otra célula microbiana transportar sacarosa al interior de la célula desde el medio de cultivo e hidrolizar sacarosa para producir glucosa y fructosa. Opcionalmente, también se puede expresar una fructoquinasa en los casos en que la actividad de hexoquinasa endógena es insuficiente para la fosforilación máxima de fructosa. Los números de registro de Genbank CAD91334, CAB92307 y CAA53390 son ejemplos de transportadores de sacarosa adecuados. Los números de registro de Genbank P26984, P26420 y CAA43322 son ejemplos de fructoquinasas adecuadas.
[0175] En una forma de realización, los métodos descritos en este documento se ponen en práctica con una célula hospedadora de Prototheca que secreta una sacarasa. La secreción de una sacarasa obvia la necesidad de expresión de un transportador que pueda transportar sacarosa al interior de la célula. Esto se debe a que una invertasa secretada cataliza la conversión de una molécula de sacarosa en una molécula de glucosa y una molécula de fructosa, las cuales pueden ser transportadas y utilizadas por microbios útiles en los métodos descritos en este documento. Por ejemplo, la expresión de una sacarasa (como la SEQ ID N.o 3) con una señal de secreción (como la de SEQ ID N.o 4 (de levadura), la SEQ ID N.o 5 (de plantas superiores), la SeQ ID NO :6 (señal de secreción de consenso eucariota), y la SEQ ID N.07 (combinación de secuencia señal de plantas superiores y consenso eucariota) genera actividad invertasa fuera de la célula. La expresión de dicha proteína, tal como lo permite la metodología de ingeniería genética descrita en este documento, permite que las células que ya son capaces de utilizar glucosa extracelular como fuente de energía utilicen sacarosa como fuente de energía extracelular.
[0176] Las especies de Prototheca que expresan una invertasa que se secreta en un medio que contiene sacarosa son una especie de microalgas preferida para la producción de aceite microbiano para su uso como fluido dieléctrico u otro lubricante (para la producción de aceites alimentarios, algunos consumidores pueden preferir el aceite producido con microbios no recombinantes). La expresión y el direccionamiento extracelular de esta proteína completamente activa permite que las células hospedadoras resultantes crezcan en sacarosa, mientras que sus equivalentes no transformadas no pueden. Por lo tanto, en la práctica de los métodos descritos en este documento se pueden utilizar células recombinantes de Prototheca con un gen de invertasa con codones optimizados, incluyendo, entre otros, el gen de invertasa de levadura, integrado en su genoma de manera que el gen de invertasa se exprese según lo evaluado por la actividad de invertasa y la hidrólisis de sacarosa. Los genes de invertasa son útiles como marcadores seleccionables en Prototheca y otras células recombinantes de microalgas, ya que tales células pueden crecer en sacarosa, mientras que sus equivalentes no transformadas no pueden; y los métodos para seleccionar células hospedadoras recombinantes usando una invertasa son un marcador seleccionable poderoso para la genética molecular de algas.
[0177] La expresión exitosa de una sacarasa en Prototheca también demuestra que las proteínas heterólogas (recombinantes) pueden expresarse en una célula de alga y transitar con éxito fuera de la célula y al medio de cultivo en una forma completamente activa y funcional. Por lo tanto, están disponibles métodos y reactivos para expresar una amplia y diversa gama de proteínas heterólogas en microalgas y secretarlas al exterior de la célula hospedadora. Dichas proteínas incluyen, por ejemplo, enzimas industriales tales como, por ejemplo, lipasas, proteasas, celulasas, pectinasas, amilasas, esterasas, oxidorreductasas, transferasas, lactasas, isomerasas e invertasas.
[0178] Entre los ejemplos de sacarasas adecuadas se incluyen las identificadas por los números de acceso de Genbank CAB95010, NP012104 y CAA0S839. Los ejemplos no limitantes de invertasas adecuadas se enumeran a continuación en la Tabla 3. Las secuencias de aminoácidos para cada invertasa enumerada se incluyen en el Listado de secuencias que aparece más adelante. En algunos casos, el gen de utilización de sacarosa exógeno adecuado
para usar en los métodos y vectores descritos en el presente documento codifica una sacarasa que tiene al menos un 40, 50, 60, 75 o 90 % o más de identidad de aminoácidos con una sacarasa seleccionada de la Tabla 3.
Tabla 3. Sacarasa.
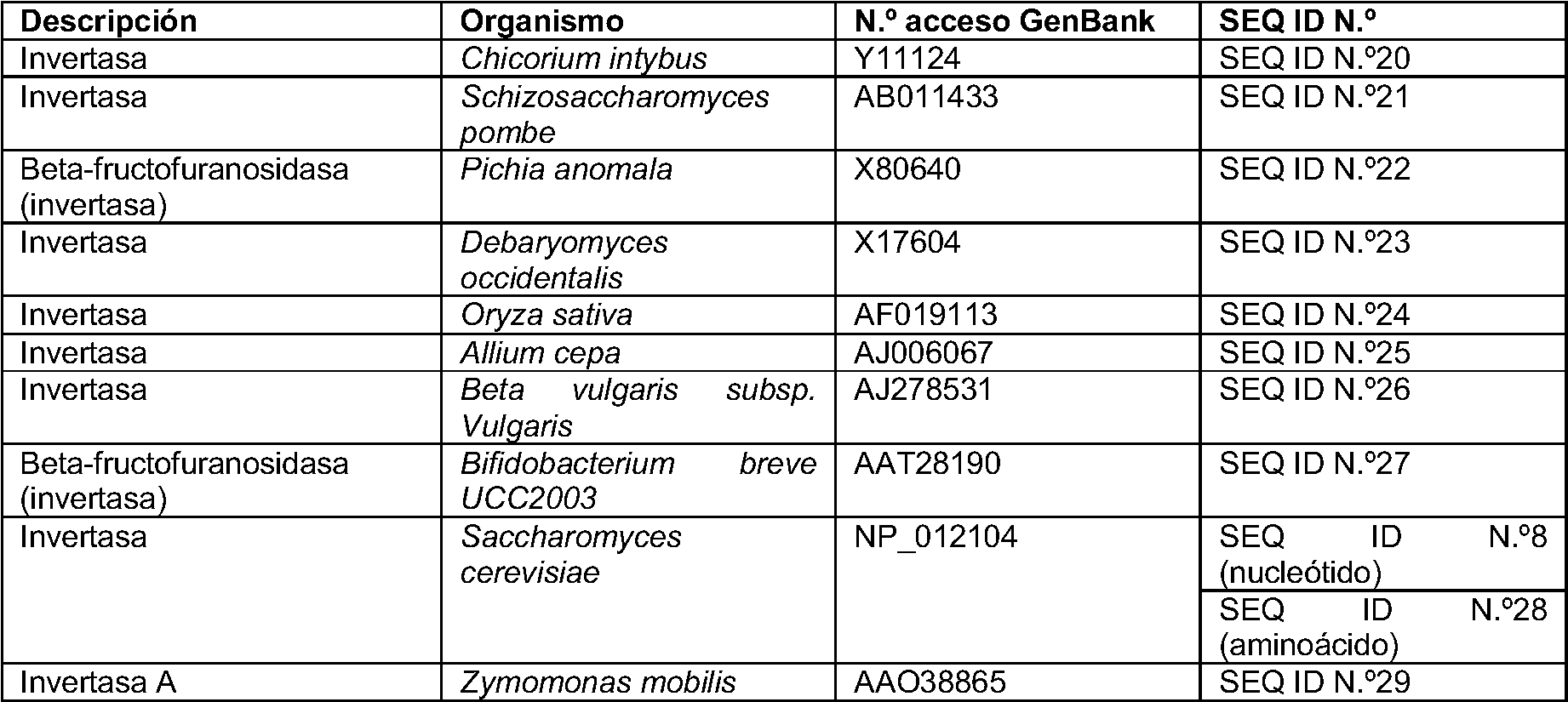
[0179] La secreción de una invertasa al medio de cultivo por Prototheca permite que las células crezcan tanto en la melaza de desecho del procesamiento de la caña de azúcar como en la glucosa pura de grado reactivo; el uso de este producto de desecho de bajo valor del procesamiento de la caña de azúcar puede proporcionar ahorros significativos en los costes de producción de lípidos y otros aceites. Por lo tanto, los métodos descritos en este documento pueden implicar el uso de un cultivo microbiano que contenga una población de Prototheca u otros microorganismos de microalgas, y un medio de cultivo que comprenda (i) sacarosa y (ii) una enzima sacarasa. La sacarosa del cultivo puede provenir de sorgo, remolacha azucarera, caña de azúcar, melaza o material celulósico despolimerizado (que opcionalmente puede contener lignina). Si bien los microbios ejemplificados aquí se modifican de manera que puedan utilizar sacarosa, los métodos y reactivos descritos en este documento se pueden aplicar de manera que las materias primas, como la celulosa, sean utilizables por un microbio hospedador diseñado con la capacidad de secretar celulasas, pectinasas, isomerasas o similares, de modo que los productos de descomposición de las reacciones enzimáticas ya no sean simplemente tolerados, sino que el hospedador los utilice como fuente de carbono.
V. INGENIERÍA DE LA RUTA DE LOS LÍPIDOS
[0180] Además de alterar la capacidad de Prototheca (u otras microalgas u otras células microbianas) para utilizar materias primas como materias primas que contienen sacarosa, las Prototheca recombinantes (u otras microalgas u otras células microbianas) que se han modificado para alterar las propiedades y/o proporciones de los lípidos producidos son útiles en los métodos descritos en el presente documento. La ruta puede modificarse además, o alternativamente, para alterar las propiedades y/o proporciones de diversas moléculas lipídicas producidas a través del procesamiento enzimático de lípidos e intermedios en la ruta de los ácidos grasos. En varios casos, las células recombinantes tienen, en comparación con sus equivalentes no transformadas, una producción de lípidos aumentada u optimizada por unidad de volumen y/o por unidad de tiempo, longitud de cadena de carbono (por ejemplo, para productos químicos industriales, incluidos, entre otros, fluidos dieléctricos, y otras aplicaciones que requieren materia prima lipídica), número reducido de enlaces dobles o triples, opcionalmente a cero, y aumento de la relación hidrógeno:carbono de una especie particular de lípido (ácido graso) o de una población de lípidos distintos.
[0181] En casos particulares, una o más enzimas clave que controlan los puntos de ramificación en el metabolismo para la síntesis de ácidos grasos se han regulado al alza o a la baja para mejorar la producción de lípidos. La regulación al alza se puede lograr, por ejemplo, mediante la transformación de células con construcciones de expresión en las que se expresa un gen que codifica la enzima de interés, por ejemplo, utilizando un promotor fuerte y/o elementos potenciadores que aumentan la transcripción. Dichas construcciones pueden incluir un marcador seleccionable de modo que los transformantes puedan someterse a selección, que también puede usarse para la amplificación de la construcción y un aumento concomitante en el nivel de expresión de la enzima codificada. Los ejemplos de enzimas adecuadas para la regulación al alza según los métodos descritos en el presente incluyen la
piruvato deshidrogenasa, que desempeña un papel en la conversión del piruvato en acetil-CoA (entre los ejemplos, algunos de microalgas, se incluyen los números de acceso de Genbank NP 415392; AAA53047; Q1XDM1; y CAF05587). La regulación al alza de la piruvato deshidrogenasa puede aumentar la producción de acetil-CoA y, por lo tanto, aumentar la síntesis de ácidos grasos. La Acetil-CoA carboxilasa cataliza el paso inicial en la síntesis de ácidos grasos. En consecuencia, esta enzima se puede regular para aumentar la producción de ácidos grasos (entre los ejemplos, algunos de microalgas, se incluyen los números de acceso de Genbank BAA94752; AAA75528; AAA81471; YP 537052; YP53S879; NP 045833 y BAA57908). La producción de ácidos grasos también se puede aumentar mediante la regulación al alza de la proteína transportadora de acilo (ACP), que transporta las cadenas de acilo en crecimiento durante la síntesis de ácidos grasos (entre los ejemplos, algunos de microalgas, se incluyen los números de acceso de Genbank A0T0F8; P51280; NP 849041; YP 874433). La glicerol-3-fosfato aciltransferasa cataliza el paso que limita la tasa de la síntesis de ácidos grasos. La regulación al alza de esta enzima puede aumentar la producción de ácidos grasos (los ejemplos, algunos de microalgas incluyen los números de acceso de Genbank AAA74319; AAA33122; AAA37647; P44857; y AB094442).
[0182] La regulación al alza y/o a la baja de genes se puede aplicar a reguladores globales que controlan la expresión de los genes de las rutas biosintéticas de ácidos grasos. En consecuencia, uno o más reguladores globales de la síntesis de ácidos grasos pueden regularse al alza o a la baja, según corresponda, para inhibir o mejorar, respectivamente, la expresión de una pluralidad de genes sintéticos de ácidos grasos y, en última instancia, para aumentar la producción de lípidos. Entre los ejemplos se incluyen proteínas de unión a elementos reguladores de esteroles (SREBP), como SREBP-1a y SREBP-1c (para ver ejemplos, consúltense los números de acceso de Genbank NP_035S10 y Q9WTN3).
[0183] Los métodos descritos en este documento también se pueden poner en práctica con células de Prototheca recombinantes (u otras microalgas u otros microbios) que se han modificado para que contengan uno o más genes exógenos que codifican enzimas que modifican los lípidos como, por ejemplo, acil-ACP tioesterasas (véase la Tabla 4), acil-CoA/aldehído reductasas (véase la Tabla 6), acil-CoA reductasas, aldehido graso descarbonilasa, aldehido graso reductasas, desaturasas (tales como estearoil-ACP desaturasas y acil desaturasas) y escualeno sintasas (consúltese el número de acceso de GenBank AF205791). En algunos casos, los genes que codifican una acil-ACP tioesterasa y una proteína transportadora de acilo expresada de forma natural se transforman en una célula de Prototheca (u otra microalga u otra célula microbiana), opcionalmente con uno o más genes que codifican otras enzimas que modifican los lípidos. En otros casos, la ACP y la acil-ACP tioesterasa pueden tener una afinidad entre sí que otorga una ventaja cuando las dos se usan juntas en los microbios y los métodos descritos en este documento, independientemente de si se expresan o no de forma conjunta de forma natural en un tejido u organismo particular. Por lo tanto, en algunas formas de realización, la presente invención contempla tanto pares coexpresados de forma natural de estas enzimas como aquellos que comparten una afinidad para interactuar entre sí para facilitar la escisión de una cadena de carbono de longitud específica de la ACP.
[0184] En otros casos, un gen exógeno que codifica una desaturasa se transforma en la célula de Prototheca (u otra microalga u otra célula microbiana) junto con uno o más genes que codifican otras enzimas que modifican los lípidos para proporcionar modificaciones con respecto a la saturación de lípidos. En otro caso, se sobreexpresa un gen endógeno de desaturasa (por ejemplo, mediante la introducción de copias adicionales del gen) en una célula de Prototheca (u otra microalga u otra célula microbiana). La estearoil-ACP desaturasa (véanse, por ejemplo, los números de acceso de GenBank AAF15308; ABM45911 y AAY8S08S), por ejemplo, cataliza la conversión de estearoil-ACP en oleoil-ACP. La regulación al alza de este gen puede aumentar la proporción de ácidos grasos monoinsaturados producidos por una célula, mientras que la regulación a la baja puede reducir la proporción de monoinsaturados. Con fines ilustrativos, las estearoil-ACP desaturasas (SAD) son responsables de la síntesis de ácidos grasos C18:1 a partir de precursores C18:0. Otra familia de desaturasas son las desaturasasde acilos grasos (FAD), incluidas las desaturasas de ácidos grasos delta 12. Estas desaturasas también proporcionan modificaciones con respecto a la saturación de lípidos. Con fines ilustrativos, las desaturasas de ácidos grasos delta 12 son responsables de la síntesis de ácidos grasos C18:2 a partir de precursores C18:1. De manera similar, la expresión de una o más glicerolípido desaturasas puede controlarse para alterar la relación de ácidos grasos insaturados a saturados tales como ácido graso w-6 desaturasa, ácido graso w-3 desaturasa o oleato w-6 desaturasa. En algunos casos, la desaturasa se puede seleccionar con referencia a una longitud de cadena de carbono deseada, de modo que la desaturasa sea capaz de realizar modificaciones específicas de ubicación dentro de un sustrato con una longitud de carbono específica, o sustratos que tengan una longitud de carbono dentro de un rango específico. En otro caso, si el perfil de ácidos grasos deseado es un aumento de los monoinsaturados (como C16:1 y/o C18:1), la sobreexpresión de una SAD o la expresión de una SAD heteróloga puede combinarse con el silenciamiento o la inactivación (por ejemplo, a través de mutación, ARNi, ARN horquillado, inactivación de un gen endógeno de desaturasa, etc.) de una acilo graso desaturasa (FAD). En el ejemplo 4 siguiente se describe la ablación dirigida o la inactivación de estearoil-ACP desaturasas y desaturasas de ácidos grasos delta 12 y también describe el uso de construcciones antisentido de ARN horquillado para disminuir la expresión de un gen de desaturasa endógeno.
[0185] Por lo tanto, en casos particulares, los microbios de la presente invención se manipulan genéticamente para que expresen uno o más genes exógenos seleccionados de una acil-ACP tioesterasa, una acil-CoA/aldehído reductasa, una acil-CoA reductasa, una aldehido graso reductasa, una desaturasa, una aldehido graso descarbonilasa o una proteína portadora de acilo expresada de forma natural. Los métodos de expresión adecuados se describen anteriormente para la expresión de un gen de lipasa, incluidos, entre otros métodos, la expresión inducible y la expresión compartimentada. Una acil-ACP tioesterasa escinde un ácido graso de una proteína transportadora de acilo (ACP) durante la síntesis de lípidos. A través de un procesamiento enzimático adicional, el ácido graso escindido se combina luego con una coenzima para producir una molécula de acil-CoA. Esta acil-CoA es el sustrato para la actividad enzimática de una acil-CoA reductasa para producir un aldehído, así como para una acil-CoA/aldehído reductasa para producir un alcohol. El aldehído producido por la acción de la acil-CoA reductasa identificada anteriormente es el sustrato para una mayor actividad enzimática por una aldehído graso reductasa para producir un alcohol, o una aldehído graso descarbonilasa para producir un alcano o alqueno.
[0186] En algunos casos, los ácidos grasos, los glicerolípidos o los correspondientes alcoholes primarios, aldehídos, alcanos o alquenos, generados por los métodos descritos en este documento, contienen 16 o 18 átomos de carbono. Los ácidos grasos preferidos para la producción de fluidos dieléctricos o los correspondientes alcoholes, aldehídos, alcanos y alquenos contienen 16-18 átomos de carbono. En algunos casos, los ácidos grasos anteriores son saturados (sin enlaces dobles o triples carbono-carbono; monoinsaturados (enlace doble simple); poliinsaturados (dos o más enlaces dobles; y pueden ser lineales (no cíclicos) o ramificados o una mezcla de los dos tipos). Para los fluidos dieléctricos, se prefieren los ácidos grasos monoinsaturados, especialmente el ácido oleico (C18:1).Para aumentar la producción de lípidos que tengan la longitud de cadena y/o el grado de saturación deseados, se puede diseñar la célula de la microalga para que sobreexprese una tioesterasa con la especificidad de longitud de cadena deseada, elimine la producción de tioesterasas con especificidad de longitud de cadena más corta o reduzca la expresión de tales genes, y/o inactive los genes de desaturasa responsables del grado de saturación en los lípidos deseados.
[0187] Varias enzimas descritas anteriormente suelen tener una especificidad preferencial para la hidrólisis de un sustrato que contiene un número específico de átomos de carbono. Por ejemplo, una acil-ACP tioesterasa puede tener preferencia por escindir un ácido graso que tiene 12 átomos de carbono de la ACP. En algunos casos, la ACP y la tioesterasa de longitud específica pueden tener una afinidad entre sí que las hace particularmente útiles como combinación (por ejemplo, los genes exógenos de ACP y tioesterasa pueden coexpresarse de forma natural en un tejido u organismo en particular del cual se derivan). Por lo tanto, en varios casos, la célula de Prototheca recombinante (u otra microalga u otra célula microbiana) de la invención puede contener un gen exógeno que codifica una proteína con especificidad para catalizar una actividad enzimática (por ejemplo, escisión de un ácido graso de una ACP, reducción de una acil-CoA a un aldehído o un alcohol, o conversión de un aldehído en un alcano) con respecto al número de átomos de carbono contenidos en el sustrato. La especificidad enzimática puede ser, en varios casos, para un sustrato que tiene de 8 a 34 átomos de carbono y, preferiblemente, de 16 a 18 átomos de carbono.
[0188] Otras acil-ACP tioesterasas de adecuadas para usar con los microbios y los métodos descritos en este documento incluyen, entre otros, las enumeradas en la Tabla 4.
Tabla 4. Acil-ACP tioesterasas rasas números de acceso de GenBank.


[0189] Los productos químicos a base de bioaceite, como los fluidos dieléctricos, tienen composiciones de ácidos grasos con alto contenido de ácido oleico (C18:1) que se originan a partir de ésteres naturales (es decir, aceites de semillas) como el aceite de girasol y el aceite de canola. En la Tabla 5 se muestran los perfiles de ácidos grasos de los aceites de semillas comerciales comunes. Todos los datos de aceites de semillas comerciales que aparecen a continuación se recopilaron de US Pharmacopeias Food and Chemicals Codes, 7th Ed. 2010-2011.
© © TJ
;gQ.
© Q 3.

[0190] Las acil-CoA/aldehído reductasas adecuadas para su uso con los microbios y los métodos descritos en el presente documento incluyen, entre otros, las enumeradas en la Tabla 6.
________Tabla 6. acil-CoA/aldehído reductasas enumeradas por sus números de acceso de GenBank.________
AAC45217, YP_0478S9, BAB8547S, YP_00108S217, YP_580344, YP_001280274, YP_2S4583, YP_43S109,
YP 9597S9, ZP_0173S9S2, ZP_01900335, ZP_0189209S, ZP_01103974, ZP_01915077, YP_92410S,
YP_130411, ZP 01222731, YP_550815, YP_983712, YP_001019S88, YP_5247S2, YP_85S798,
ZP_01115500, YP_001141848, NP_33S047, NP_21S059, YP_882409, YP_70S15S, YP_00113S150,
YP 9523S5, ZP 01221833, YP_13007S, NP_5S793S, AAR887S2, ABK2858S, NP_197S34, CAD30S94,
NP_0010S39S2, BAD4S254, NP_001030809, EAZ10132, EAZ43S39, EAZ07989, NP_0010S2488, CAB88537,
NP_001052541, CAHSS597, CAE02214, CAHSS590, CAB88538, EAZ39844, AAZ0SS58, CAAS8190,
CAA52019 y BAC84377________________________________________________________________________
[0191] Las acil-ACP tioesterasas son los terminadores de la biosíntesis de ácidos grasos de las plantas superiores (y algunas especies de microalgas) y, en la mayoría de las especies de plantas, esto lo llevan a cabo miembros de la familia de genes FatA, cuyo papel es terminar el alargamiento en la etapa de C16:0 a C18:0. En especies que sintetizan ácidos grasos de cadena más corta (como Cuphea, Elaeis, Myrística, o Umbelullaría), un grupo diferente de acil-ACP tioesterasas codificadas por genes FatB lleva a cabo este paso de terminación.
[0192] Otras enzimas adecuadas para usar en los métodos descritos en este documento incluyen aquellas que tienen al menos un 70 % de identidad de aminoácidos con una de las proteínas enumeradas en las Tablas 6 y 8, y que muestran la actividad enzimática deseada correspondiente (por ejemplo, la escisión de un ácido graso de una proteína portadora de acilo, la reducción de un acil-CoA a un aldehído o un alcohol, o la conversión de un aldehído en un alcano). En casos adicionales, la actividad enzimática está presente en una secuencia que tiene al menos alrededor del 75 %, al menos alrededor del 80 %, al menos alrededor del 85 %, al menos alrededor del 90 %, al menos alrededor del 95 % o al menos alrededor del 99 % de identidad con una de las secuencias descritas anteriormente, todas las cuales se incorporan al presente documento por referencia.
[0193] Al seleccionar la combinación deseada de genes exógenos que expresar (o genes endógenos que inactivar o ambos), se puede adaptar el aceite generado por el microbio, que luego se puede extraer de la biomasa acuosa. Por ejemplo, el microbio puede contener: (i) un gen exógeno que codifica una acil-ACP tioesterasa; (ii) opcionalmente, una proteína transportadora de acilo coexpresada de manera natural o una proteína transportadora de acilo que tiene afinidad por la acil-ACP tioesterasa de ácido graso; (iii) un gen de desaturasa endógeno mutado, en el que la mutación inactiva el gen de desaturasa o la proteína desaturasa, tal como una desactivación de desaturasa; (iv) la sobreexpresión de una proteína transportadora de estearoil acil desaturasa endógena o la expresión de una SAD heteróloga; y (v) cualquier combinación de los anteriores.
[0194] Los genes que codifican tales enzimas, como las acil-ACP tioesterasas, se pueden obtener de células que ya se sabe que poseen una producción significativa de lípidos, como Chlorella protothecoides. Los genes que ya se sabe que tienen una función en la producción de lípidos, por ejemplo, un gen que codifica una enzima que satura los enlaces dobles, pueden transformarse individualmente en células receptoras. Los métodos para identificar genes que pueden alterar (mejorar) la producción de lípidos en microalgas se describen en la publicación PCT N.02008/151149, que se incorpora a la presente por referencia.
[0195] Por lo tanto, en algunos casos, en la práctica de la presente invención se puede utilizar una célula de Prototheca u otra microalga u otra célula microbiana que haya sido manipulada genéticamente para expresar una enzima de la ruta de los lípidos a un nivel alterado en comparación con una célula de tipo salvaje de la misma especie. En algunos casos, la célula produce más lípidos en comparación con la célula de tipo salvaje cuando ambas células se cultivan en las mismas condiciones. En algunos casos, la célula ha sido manipulada genéticamente y/o seleccionada para que exprese una enzima de la ruta de los lípidos a un nivel más alto que la célula de tipo salvaje. En algunos casos, la enzima de la ruta de los lípidos se selecciona del grupo que consiste en piruvato deshidrogenasa, acetil-CoA carboxilasa, proteína transportadora de acilo y glicerol-3 fosfato aciltransferasa. En algunos casos, la célula ha sido manipulada genéticamente y/o seleccionada para que exprese una enzima de la ruta de los lípidos a un nivel más bajo que la célula de tipo salvaje. En un caso en el que la célula expresa la enzima de la ruta de los lípidos a un nivel más bajo, la enzima de la ruta de los lípidos comprende citrato sintasa.
[0196] En algunos casos, la célula ha sido manipulada genéticamente y/o seleccionada para que exprese un regulador global de la síntesis de ácidos grasos a un nivel alterado en comparación con la célula de tipo salvaje, por lo que los niveles de expresión de una pluralidad de genes sintéticos de ácidos grasos están alterados en comparación a la célula de tipo salvaje. En algunos casos, la enzima de la ruta de los lípidos comprende una enzima
que modifica un ácido graso. En algunos casos, la enzima de la ruta de los lípidos se selecciona de una estearoil-ACP desaturasa y una glicerolípido desaturasa. En algunos casos, la célula ha sido manipulada genéticamente y/o seleccionada para que exprese un nivel más bajo de una enzima de la ruta de los lípidos, o para que no exprese una enzima específica de la ruta de los lípidos (es decir, cuando una enzima de la ruta de los lípidos ha sido inaplicada o reemplazada con un gen exógeno).
[0197] En otro caso se utiliza un microbio productor de aceite que contiene uno o más genes exógenos y/o uno o más genes endógenos inactivados, en el que los genes exógenos o endógenos codifican proteína(s) seleccionada(s) del grupo que consiste en una acil-ACP tioesterasa, una acil-CoA reductasa, una aldehído graso reductasa, una acil-CoA/aldehído reductasa, una aldehído graso descarbonilasa, una desaturasa y una proteína portadora de acilo. En otro caso, un gen de desaturasa endógeno se sobreexpresa en un microbio que contiene uno o más de los genes exógenos anteriores. El gen exógeno puede estar en unión operativa con un promotor, que es inducible o reprimible en respuesta a un estímulo. En algunos casos, el estímulo se selecciona del grupo que consiste en una molécula pequeña proporcionada exógenamente, calor, frío y nitrógeno limitado o nulo en los medios de cultivo. En algunos casos, el gen exógeno se expresa o se dirige de otro modo a un compartimento celular. El compartimento celular se puede seleccionar del grupo que consiste en un cloroplasto, un plastidio y una mitocondria. En algunas formas de realización, el microbio es Prototheca moríformis, Prototheca krugani, Prototheca stagnora o Prototheca zopfii.
[0198] El gen exógeno o el gen endógeno inactivado puede codificar una acil-ACP tioesterasa de ácido graso. En algunos casos, la tioesterasa codificada por el gen exógeno o endógeno inactivado cataliza la escisión de un ácido graso de 8 a 18 carbonos de una proteína transportadora de acilo (ACP). En algunos casos, la tioesterasa codificada por el gen exógeno o el gen endógeno inactivado cataliza la escisión de un ácido graso de 10 a 14 carbonos de una ACP. La tioesterasa codificada por el gen exógeno o el gen endógeno inactivado puede catalizar la escisión de un ácido graso de 12 carbonos de una ACP. En algunos casos, la tioesterasa codificada por el gen exógeno cataliza la escisión de un ácido graso de 16-18 carbonos de una ACP.
[0199] El gen exógeno puede codificar una grasa acil-CoA/aldehído reductasa. En algunos casos, la reductasa codificada por el gen exógeno cataliza la reducción de una acil-CoA de 8 a 18 carbonos a un alcohol primario correspondiente. En algunos casos, la reductasa codificada por el gen exógeno o el gen endógeno inactivado cataliza la reducción de una acil-CoA de 10 a 14 carbonos a un alcohol primario correspondiente. La reductasa codificada por el gen exógeno o el gen endógeno inactivado puede catalizar la reducción de una acil-CoA de 12 carbonos a dodecanol.
[0200] En la aplicación de los métodos descritos en este documento se puede utilizar una célula de Prototheca recombinante (u otra célula microalgal o microbiana) que contenga dos genes exógenos (o dos genes endógenos inactivados), en donde un primer gen exógeno o un gen endógeno inactivado codifica una acil-ACP tioesterasa y un segundo gen exógeno o un gen endógeno inactivado codifica una proteína seleccionada del grupo consiste en una acil-CoA reductasa, una acil-CoA/aldehído reductasa y una proteína transportadora de acilo. En algunos casos, los dos genes exógenos están cada uno en unión operativa con un promotor, que es inducible en respuesta a un estímulo. En algunos casos, cada promotor es inducible en respuesta a un estímulo idéntico, como nitrógeno limitado o nulo en los medios de cultivo. La limitación o falta total de nitrógeno en los medios de cultivo estimula la producción de aceite en algunos microorganismos como Prototheca y otras microalgas y otras especies microbianas, y se puede utilizar como desencadenante para inducir la producción de aceite (lípidos) a niveles elevados. Cuando se usa en combinación con los métodos de ingeniería genética descritos en el presente documento, el lípido como porcentaje del peso en seco de las células se puede llevar a niveles altos, como al menos el 30 %, al menos el 40 %, al menos el 50 %, al menos el 60 %, al menos el 70 % y al menos el 75 %.
[0201] Los nuevos aceites (lípidos) y los fluidos dieléctricos derivados de ellos descritos en este documento son distintos de otros aceites naturales que tienen un alto contenido de ácidos grasos C16 y C18, como el aceite de girasol y de canola.
[0202] La tioesterasa codificada por el primer gen exógeno puede catalizar la escisión de un ácido graso de 8 a 18 carbonos de una ACP. Además, en aquellos casos en los que se desean aceites de cadena más larga, la expresión de uno o más genes con una longitud de cadena más corta (es decir, por debajo de C14, como C12, C10 y/o C8) y/o genes de ACP correspondientes, se reduce (a través de la alteración de su expresión) o se elimina (por inactivación, por ejemplo).
[0203] En los diversos casos descritos anteriormente, la Prototheca (u otra microalga u otra célula microbiana) puede contener al menos un gen exógeno o al menos un gen endógeno inactivado (o diseñado para reducir la expresión) que codifica una enzima de la ruta de los lípidos. En algunos casos, la enzima de la ruta de los lípidos se selecciona del grupo que consiste en una estearoil-ACP desaturasa, una ácido graso desaturasa, una glicerolípido desaturasa,
una piruvato deshidrogenasa, una acetil-CoA carboxilasa, una proteína transportadora de acilo y una glicerol-3 fosfato aciltransferasa. En otros casos, la célula de Prototheca u otra célula contiene una enzima que modifica los lípidos seleccionada del grupo que consiste en una acil-ACP tioesterasa, una acil-CoA/aldehído reductasa, una acil-CoA reductasa, una aldehido graso reductasa, una aldehido graso descarbonilasa y/o una proteína transportadora de acilo.
VI. PRODUCCIÓN DE ACEITE MICROBIANO Y PRODUCTOS DERIVADOS DE ESTE
1. Producción de aceite microbiano
[0204] Para la producción de aceite microbiano según los métodos descritos en el presente documento, el aceite crudo (lípidos) sin procesar producido por las células microbianas se recolecta, o se recoge de otro modo, por cualquier medio conveniente. El aceite se puede aislar por extracción de células completas, por ejemplo. En este método, las células primero se rompen y luego los lípidos y ácidos grasos intracelulares y asociados a la membrana celular/pared celular, así como los hidrocarburos extracelulares, pueden separarse de la masa celular, por ejemplo mediante el uso de centrifugación como se ha descrito antes. Los lípidos intracelulares producidos en los microorganismos se extraen, en muchos casos, después o durante el proceso de lisis de las células microbianas.
[0205] Más específicamente, después de completar el cultivo, los microorganismos normalmente se separan del caldo de fermentación. A menudo, la separación se realiza mediante centrifugación para generar una pasta concentrada de biomasa microbiana. La biomasa se puede lavar opcionalmente con una solución de lavado (por ejemplo, agua DI) para eliminar el caldo de fermentación y los residuos. Opcionalmente, la biomasa microbiana lavada también se puede secar (secar en horno, liofilizar, etc.) antes de la ruptura celular. Alternativamente, las células se pueden lisar sin separación de parte o todo el caldo de fermentación cuando se completa la fermentación. Por ejemplo, las células pueden estar en una proporción de menos de 1:1 v:v de células a líquido extracelular cuando se lisan las células.
[0206] Los microorganismos que contienen un lípido pueden lisarse para producir un lisado. Como se detalla en el presente documento, el paso de lisis de un microorganismo (también denominado lisis celular) se puede lograr por cualquier medio conveniente, incluida la lisis inducida por calor, la adición de una base, la adición de un ácido, el uso de enzimas como proteasas y enzimas de degradación de polisacáridos como como amilasas, ultrasonidos, lisis mecánica, choque osmótico, infección con un virus lítico y/o expresión de uno o más genes líticos. La lisis se realiza para liberar moléculas intracelulares que han sido producidas por el microorganismo. Cada uno de estos métodos para lisar un microorganismo se puede usar como un solo método o en combinación de manera simultánea o secuencial. La extensión de la ruptura celular se puede observar mediante análisis microscópico. Utilizando uno o más de los métodos descritos en el presente documento, normalmente se observa más del 70 % de ruptura celular. Preferiblemente, la ruptura celular es superior al 80 %, más preferiblemente superior al 90 % y más preferiblemente alrededor del 100 %.
[0207] El microorganismo se puede lisar después del crecimiento, por ejemplo, para aumentar la exposición del aceite microbiano para la extracción o el procesamiento posterior. Si se utiliza un gen de lipasa exógeno, el momento de la expresión de la lipasa (por ejemplo, a través de un promotor inducible) o la lisis celular puede ajustarse para optimizar la producción de lípidos y/o hidrocarburos. A continuación, se describen varias técnicas de lisis. Estas técnicas se pueden utilizar individualmente o en combinación.
[0208] El paso de lisis de un microorganismo puede comprender el calentamiento de una suspensión celular que contiene el microorganismo. En este caso, el caldo de fermentación que contiene los microorganismos (o una suspensión de microorganismos aislados del caldo de fermentación) se calienta hasta que los microorganismos, es decir, las paredes celulares y las membranas de los microorganismos, se degraden o se rompan. Por lo general, las temperaturas aplicadas son de al menos 50 °C. Para una lisis celular más eficiente, se pueden utilizar temperaturas más altas, como al menos 30 °C, al menos 60 °C, al menos 70 °C, al menos 80 °C, al menos 90 °C, al menos 100 °C, al menos 110 °C, al menos 120 °C, o al menos 130 °C o superiores. La lisis de células por tratamiento térmico se puede realizar poniendo en ebullición el microorganismo. Alternativamente, el tratamiento térmico (sin ebullición) se puede realizar en un autoclave. El lisado tratado con calor se puede enfriar para un tratamiento posterior. La ruptura celular también se puede realizar mediante tratamiento con vapor, es decir, mediante la adición de vapor presurizado. El tratamiento con vapor de microalgas para la ruptura celular se describe, por ejemplo, en la patente de EE. UU. 6,750,048. En algunos casos, el tratamiento con vapor se puede lograr rociando vapor en el fermentador y manteniendo el caldo a la temperatura deseada durante menos de 90 minutos, preferiblemente menos de 60 minutos y más preferiblemente menos de 30 minutos.
[0209] El paso de lisis de un microorganismo puede comprender añadir una base a una suspensión celular que contiene el microorganismo. La base debe ser lo suficientemente fuerte para hidrolizar al menos una parte de los compuestos proteicos de los microorganismos utilizados. Las bases que son útiles para solubilizar proteínas se conocen en la técnica de la química. Las bases ejemplares que son útiles en los métodos descritos en el presente documento incluyen, entre otros, hidróxidos, carbonatos y bicarbonatos de litio, sodio, potasio, calcio y mezclas de los mismos. Una base preferida es KOH. El tratamiento de base de microalgas para la ruptura celular se describe, por ejemplo, en la patente de EE. UU. 6,750,048.
[0210] El paso de lisis de un microorganismo puede comprender agregar un ácido a una suspensión celular que contiene el microorganismo. La lisis ácida se puede efectuar usando un ácido a una concentración de 10-500 mN o preferiblemente 40-160 nM. La lisis ácida se realiza preferiblemente por encima de la temperatura ambiente (por ejemplo, a 40-160°, es decir, una temperatura de 50-130°). Para temperaturas moderadas (por ejemplo, de temperatura ambiente a 100 °C y particularmente de temperatura ambiente a 65°), el tratamiento ácido se puede combinar de manera útil con sonicación u otros métodos de ruptura celular.
[0211] El paso de lisis de un microorganismo puede comprender lisar el microorganismo usando una enzima. Las enzimas preferidas para lisar un microorganismo son las proteasas y las enzimas que degradan los polisacáridos, como la hemicelulasa (por ejemplo, la hemicelulasa de Aspergillus niger, Sigma Aldrich, St. Louis, MO; #H2125), pectinasa (por ejemplo, pectinasa de Rhizopus sp.; Sigma Aldrich, St. Louis, MO; #P2401), Mannaway 4.0 L (Novozymes), la celulasa (por ejemplo, celulosa de Trichoderma viride; Sigma Aldrich, St. Louis, MO; #C9422) y la driselasa (por ejemplo, driselasa de Basidiomicetos sp.; Sigma Aldrich, St. Louis, MO; #D9515).
[0212] La lisis se puede lograr utilizando una enzima como, por ejemplo, una celulasa como una enzima degradadora de polisacáridos, opcionalmente de Chlorella o un virus de Chlorella, y/o una proteasa, tal como proteasa de Streptomyces griseus, quimotripsina, proteinasa K, proteasas enumeradas en Degradation of Polylactide by Commercial Proteases, Oda Y y col., Journal of Polymers and the Environment, vol. 8, 2000 Jan 1, pp. 29-32(4), Alcalase 2.4 FG (Novozymes) y Flavourzyme 100 L (Novozymes). También se puede usar cualquier combinación de una proteasa y una enzima degradadora de polisacáridos, incluida cualquier combinación de las proteasas anteriores y enzimas degradadoras de polisacáridos.
[0213] En otro caso, la lisis se puede realizar utilizando una prensa extrusora. En este proceso, la biomasa se hace pasar a través de un dispositivo de tipo tornillo a alta presión, lisando las células y haciendo que el lípido intracelular se libere y se separe de la proteína y la fibra (y otros componentes) de la célula.
[0214] El paso de lisis de un microorganismo se puede realizar usando ultrasonido, es decir, mediante sonicación. Por lo tanto, las células también pueden lisarse con sonido de alta frecuencia. El sonido se puede producir electrónicamente y transportar a través de una punta metálica a una suspensión celular adecuadamente concentrada. Esta sonicación (o ultrasonicación) rompe la integridad celular en función de la creación de cavidades en la suspensión celular.
[0215] El paso de lisis de un microorganismo se puede realizar mediante lisis mecánica. Las células pueden lisarse mecánicamente y, opcionalmente, homogeneizarse para facilitar la recolección de hidrocarburos (por ejemplo, lípidos). Por ejemplo, se puede usar un disruptor de presión para bombear una suspensión que contiene células a través de una válvula de orificio restringido. Se aplica alta presión (hasta 1500 bar), seguida de una expansión instantánea a través de una boquilla de salida. La ruptura de la célula se logra mediante tres mecanismos diferentes: impacto en la válvula, alto cizallamiento del líquido en el orificio y caída repentina de la presión en la descarga, lo que provoca una explosión de la célula. El método libera moléculas intracelulares. Alternativamente, se puede utilizar un molino de bolas. En un molino de bolas, las células se agitan en suspensión con pequeñas partículas abrasivas, como perlas. Las células se rompen debido a las fuerzas de cizallamiento, la molienda entre las perlas y las colisiones con las perlas. Las perlas rompen las células para liberar el contenido celular. Las células también pueden romperse por fuerzas de cizallamiento, como con el uso de mezclas (por ejemplo, con una licuadora Waring o de alta velocidad), la prensa francesa o incluso la centrifugación en caso de paredes celulares débiles, para romper las células.
[0216] El paso de lisis de un microorganismo se puede realizar aplicando un choque osmótico (es decir, suspendiendo las células del microorganismo en una solución hipotónica).
[0217] El paso de lisis de un microorganismo puede comprender la infección del microorganismo con un virus lítico. Se conoce una amplia variedad de virus para la lisis de microorganismos adecuados para usar en los métodos descritos en el presente documento, y la selección y el uso de un virus lítico particular para un microorganismo particular está dentro del nivel de experiencia en la técnica. Por ejemplo, el virus de paramecium bursaria chlorella
(PBCV-1) es el prototipo de un grupo (familia Phycodnaviridae, género Chlorovirus) de virus de ADN de doble cadena grandes, icosaédricos, formadores de placa, que se replican y lisan algunas algas verdes unicelulares eucariotas similares a la Chlorella. En consecuencia, cualquier microalga susceptible puede lisarse infectando el cultivo con un virus de Chlorella adecuado. Los métodos para infectar especies de Chlorella con un virus de Chlorella se conocen en la técnica. Véase, por ejemplo, Adv. Virus Res 2006;66:293-336; Virology, 1999 Apr 25; 257 (1): 15-23; Virology, 2004 Jan 5; 318 (1): 214-23; Nucleic Acids Symp. Ser. 2000;(44):161-2; J.Virol. 2006 March ;80(5):2437-44; y Annu. Rev. Microbiol. 1999;53:447-94.
[0218] El paso de lisis de un microorganismo puede comprender la autólisis. En este caso, se manipula genéticamente un microorganismo para que produzca una proteína lítica que lisará el microorganismo. Este gen lítico se puede expresar usando un promotor inducible de modo que las células puedan crecer primero hasta una densidad deseable en un fermentador, seguido de la inducción del promotor para expresar el gen lítico para lisar las células. El gen lítico puede codificar una enzima degradadora de polisacáridos. En ciertos otros casos, el gen lítico es un gen de un virus lítico. Así, por ejemplo, un gen lítico de un virus de Chlorella puede expresarse en una célula de alga; véase Virology 260, 308-315 (1999); FEMS Microbiology Letters 180 (1999) 45-53; Virology 263, 376-387 (1999); y Virology 230, 361-368 (1997). La expresión de genes líticos se realiza preferiblemente usando un promotor inducible, como un promotor activo en microalgas que es inducido por un estímulo como la presencia de una molécula pequeña, luz, calor y otros estímulos.
[0219] Están disponibles varios métodos para separar lípidos de lisados celulares producidos por los métodos anteriores. Por ejemplo, los lípidos y los derivados de lípidos como los aldehídos grasos, los alcoholes grasos y los hidrocarburos como los alcanos se pueden extraer con un disolvente hidrófobo como el hexano (véase Frenz y col.
1989, Enzyme microb. Technol., 11:717). Los lípidos y derivados de lípidos también se pueden extraer mediante licuefacción (véase, por ejemplo, Sawayama y col. 1999, Biomass and Bioenergy 17:33-39 y Inou y col. 1993, Biomass and Bioenergy 6(4):269-274); licuefacción de petróleo (véase, por ejemplo, Minowa y col. 1995, Fuel 74(12): 1735-1738); y extracción con CO2 supercrítico (véase, por ejemplo, Mendes y col. 2003, Inorganica Chimica Acta 356:328-334). Miao y Wu describen un protocolo de recuperación de lípidos de microalgas de un cultivo de Chlorella prototheocoides en el que las células se recolectaron por centrifugación, se lavaron con agua destilada y se secaron por liofilización. El polvo celular resultante se pulverizó en un mortero y luego se extrajo con n-hexano. Miao y Wu, Biosource Technology (2006) 97:841-846.
[0220] Así, los lípidos, derivados de lípidos e hidrocarburos generados por los microorganismos descritos en este documento pueden recuperarse mediante extracción con un disolvente orgánico. En algunos casos, el disolvente orgánico preferido es el hexano. Normalmente, el disolvente orgánico se añade directamente al lisado sin separación previa de los componentes del lisado. El lisado generado por uno o más de los métodos descritos anteriormente se puede poner en contacto con un disolvente orgánico durante un período de tiempo suficiente para permitir que los componentes lipídicos y/o hidrocarburos formen una solución con el disolvente orgánico. En algunos casos, la solución se puede refinar aún más para recuperar los componentes de lípidos o hidrocarburos específicos deseados. Los métodos de extracción con hexano son ampliamente conocidos en la técnica.
[0221] Otros métodos para extraer lípidos de microorganismos se describen en la solicitud PCT n.0 US10/031108, que se incorpora a la presente por referencia.
[0222] Los lípidos y derivados de lípidos tales como aldehídos grasos, alcoholes grasos e hidrocarburos tales como alcanos producidos por células como se describe en este documento pueden modificarse mediante el uso de una o más enzimas, incluida una lipasa. Cuando los hidrocarburos están en el entorno extracelular de las células, se pueden añadir una o más enzimas a ese entorno en condiciones en las que la enzima modifica el hidrocarburo o completa su síntesis a partir de un precursor de hidrocarburo. Alternativamente, los hidrocarburos pueden aislarse parcial o completamente del material celular antes de la adición de uno o más catalizadores tales como enzimas. Dichos catalizadores se agregan exógenamente y su actividad ocurre fuera de la célula o in vitro.
2. Procesamiento adicional de aceite microbiano
[0223] De este modo, los lípidos e hidrocarburos producidos por las células in vivo, o modificados enzimáticamente in vitro, como se describe en este documento, pueden seguir procesándose opcionalmente por medios convencionales. El procesamiento puede incluir el "craqueo" para reducir el tamaño y, por lo tanto, aumentar la relación hidrógeno:carbono de las moléculas de hidrocarburo. Los métodos de craqueo catalítico y térmico se utilizan habitualmente en el procesamiento de aceites de hidrocarburos y triglicéridos. Los métodos catalíticos implican el uso de un catalizador, como un catalizador ácido sólido. El catalizador puede ser sílice-alúmina o una zeolita, que da como resultado la rotura heterolítica o asimétrica de un enlace carbono-carbono para dar como resultado un carbocatión y un anión hidruro. Estos productos intermedios reactivos luego se reordenan o se transfieren con otro
hidrocarburo. De este modo, las reacciones pueden regenerar los intermedios para dar como resultado un mecanismo de cadena de autopropagación. Los hidrocarburos también pueden procesarse para reducir, opcionalmente a cero, el número de enlaces dobles o triples carbono-carbono que tienen. Los hidrocarburos también se pueden procesar para quitarles o eliminarles un anillo o una estructura cíclica. Los hidrocarburos también se pueden procesar para aumentar la relación hidrógeno:carbono. Esto puede incluir la adición de hidrógeno ("hidrogenación") y/o el "craqueo" de hidrocarburos en hidrocarburos más pequeños.
[0224] Una vez que se extraen los lípidos, estos pueden someterse, según los métodos descritos en el presente documento, a uno o más pasos de procesamiento. Estos pasos de procesamiento son distintos de los pasos de refinado realizados en el petróleo crudo (por ejemplo, petróleo y otras fuentes) cuando se producen combustibles. Estos pasos de procesamiento son, en algunos aspectos, comparables a los que se realizan en los aceites de semillas durante la producción para el consumo humano. En algunas formas de realización, los lípidos extraídos se desgoman para extraer lecitina y otros fosfolípidos. En otras formas de realización, los lípidos extraídos se refinan usando una base o un metal alcalino. En otras formas de realización más, los lípidos extraídos se pasan a través de una arcilla blanqueadora, normalmente una arcilla ácida. En otras formas de realización, los lípidos extraídos se desodorizan para eliminar o reducir las impurezas volátiles como aldehídos y cetonas. En otros casos más, los lípidos extraídos se acondicionan para el invierno para eliminar o reducir ceras o grasas saturadas. Los pasos de procesamiento anteriores se pueden realizar en cualquiera y todas las combinaciones en los lípidos extraídos, dependiendo de las características del producto deseado. Los lípidos extraídos que se han refinado (por ejemplo, con una base o un metal alcalino), blanqueado (por ejemplo, con una arcilla blanqueadora) y/o desodorizado generalmente se denominan aceite RBD. El aceite RBD producido a partir de lípidos extraídos de microalgas y/o levaduras oleaginosas descritos en este documento es útil en una variedad de aplicaciones industriales, incluida la producción de fluidos dieléctricos.
[0225] En algunos casos, se realiza el desgomado para eliminar contaminantes como los fosfolípidos del aceite. En algunos casos, el desgomado del aceite extraído es parte del refinado, blanqueo y desodorización (o RBD). El proceso de RBD elimina o reduce el olor, color y/o sabor del aceite extraído. En algunos casos, el proceso de refinado suele constar de dos pasos, el desgomado y un paso de neutralización que elimina los ácidos grasos libres (FFA) del aceite mediante extracción cáustica con hidróxido de sodio. El paso de blanqueo puede implicar mezclar el aceite con varias arcillas blanqueadoras para absorber el color, los metales traza y los compuestos de azufre. El paso de desodorización puede ser un proceso de destilación que ocurre a baja presión y alta temperatura. En un proceso de destilación ilustrativo, el aceite se pone al vacío y se calienta con vapor para eliminar cualquier resto de sabor u olor y ácidos grasos libres. La desodorización también se puede lograr mediante el tratamiento con carbón activado.
[0226] Los pasos mencionados anteriormente pueden servir para reducir el punto de fluidez. En varios casos, el punto de fluidez del aceite microbiano (lípido) se puede reducir a aproximadamente -10 grados C, aproximadamente -15 grados C, aproximadamente -20 grados C, aproximadamente -25 grados C, aproximadamente 30 grados C, aproximadamente -35 grados C, o aproximadamente -40 grados C. Además, el punto de fluidez del aceite microbiano puede estar dentro de cualquier rango limitado por cualquiera de estos valores, por ejemplo, alrededor de -10 grados C a -40 grados C o alrededor de -15 grados C a aproximadamente -35 grados C, etc. La reducción en el punto de fluidez puede ocurrir debido a que estos pasos reducen la proporción relativa de la fracción saturada, que consiste principalmente en triglicéridos palmíticos y esteáricos, conocida como fracción de estearina. El fraccionamiento del aceite reduce la concentración de triglicéridos saturados del aceite. El fraccionamiento puede realizarse mediante fraccionamiento en seco, como en el proceso de preparación para el invierno conocido en la industria del aceite vegetal. En este proceso, el aceite microbiano (por ejemplo, de algas) primero se refina, se blanquea y se desodoriza mediante métodos similares a los que se utilizan en la industria del aceite vegetal. Esto da como resultado un aceite con un punto de fluidez en el rango de -5 a -10 grados C, por ejemplo -8 grados C.
[0227] A continuación, la temperatura del aceite RBD puede reducirse de forma controlada hasta que se formen núcleos cristalinos. A continuación, el aceite se puede mantener a esa temperatura de cristalización durante varias horas para facilitar el crecimiento de los cristales. Luego, los cristales se eliminan por filtración para dar como resultado dos fracciones: una fase sólida que contiene parte o la mayor parte de la fracción de estearina y una fase líquida que contiene principalmente la fracción de oleína. Esto da como resultado un aceite con un punto de fluidez en el rango de -8 a -15 grados C, por ejemplo -11 grados C. La fase líquida se puede someter a fraccionamiento nuevamente a una temperatura de cristalización más baja para efectuar una mayor eliminación de estearina. La fracción líquida purificada resultante, equivalente a una súper oleína, como se conoce comúnmente en la industria del aceite vegetal, tiene mejores propiedades térmicas que el aceite microbiano nativo. Por ejemplo, un segundo fraccionamiento puede dar como resultado aceite con un punto de fluidez en el rango de -15 grados a -25 grados C, por ejemplo -20 grados C. El aceite resultante es excepcionalmente útil en una variedad de aplicaciones, incluyendo,
de manera importante, aplicaciones alimentarias, en las que el aceite microbiano se puede utilizar como un sustitutivo más barato y, a menudo, más saludable, en su totalidad o en parte, de los aceites animales y vegetales.
3. Productos derivados de aceites microbianos
[0228] Los aceites microbianos descritos en este documento también se pueden usar para producir productos, como lubricantes, fluidos hidráulicos, aceites industriales o fluidos dieléctricos. Entre los aceites industriales comunes se incluyen lubricantes para espadas de motosierras, fluidos para trabajar metales, lubricantes de calidad alimentaria, aceites para engranajes, aceites marinos, lubricantes para motores, aceites para tractores, lubricantes para equipos agrícolas, aceites para ascensores, aceites para desmoldeo y similares. Los fluidos dieléctricos se utilizan típicamente para enfriar y/o aislar eléctricamente componentes eléctricos (especialmente en equipos de distribución de energía eléctrica de alta tensión), como, por ejemplo, reconectadores automáticos, condensadores, disyuntores, cables de transmisión llenos de fluido de alta tensión, componentes de distribución de energía, equipos de conmutación (por ejemplo, un interruptor de ruptura de carga de alta tensión, como los descritos en USPN 6,797,909), transformadores, componentes de transmisión y reguladores de tensión.
[0229] Los fluidos dieléctricos tradicionales incluyen los lubricantes a base de aceite mineral. Estos incluyen los aceites de base del Grupo 1, II y II+, que son aceites de base de petróleo que se han refinado convencionalmente o se han hidrotratado ligeramente y que tienen un índice de viscosidad (VI) de menos de 120. Estos también incluyen los aceites de base del Grupo III (incluyendo " aceite de motor sintético" en los EE. UU.) que son productos derivados del petróleo convencional altamente refinados. Los aceites de base del Grupo III pueden fabricarse mediante hidroprocesamiento (hidrocraqueo y/o hidroisomerización) de aceites de base del Grupo 1 o Grupo II/II+ y contienen menos saturados, azufre y nitrógeno que los aceites de base del Grupo I, II o II+ y tienen un VI superior a 120. La ASTM (American Society of Testing and Materials) establece especificaciones para fluidos dieléctricos y otras composiciones de hidrocarburos (como combustible diésel (ASTM Dp75), combustible para aviones (ASTM D1655) y biodiésel (ASTM D6751) según cualquier de una serie de factores, como el punto de ebullición, el índice de cetano, el punto de turbidez, el punto de inflamación, la viscosidad, el punto de anilina, el contenido de azufre, el contenido de agua, el contenido de cenizas, la corrosión de tira de cobre y los residuos de carbono.
[0230] Los fluidos dieléctricos de origen biológico se pueden preparar mediante una variedad de procesos. Por ejemplo, un proceso, que comienza con aceite vegetal crudo, incluye los pasos de desgomado, refinación alcalina, blanqueo, desodorización, hidrogenación, preparación para el invierno (para producir aceite vegetal RBD), tratamiento con arcilla para eliminar trazas de compuestos polares y materiales ácidos (véase la patente de EE. UU. n.06,274,067), y combinación con aditivos para producir fluidos dieléctricos de origen biológico.
[0231] Entre las propiedades clave de los fluidos dieléctricos se incluyen la viscosidad, la inflamabilidad, la reactividad, la miscibilidad, la capacidad de aislamiento eléctrico, la biodegradabilidad y el coste de fabricación. Si bien estas y otras propiedades se explican a continuación, el lector puede apreciar mejor algunas de las ventajas de ciertos aceites descritos en este documento al comprender algunas de las ventajas y desventajas de los fluidos dieléctricos de origen biológico tradicionales frente a los fluidos dieléctricos de base de aceite mineral. En cuanto a la viscosidad, los fluidos dieléctricos de origen biológico generalmente tienen una viscosidad y un punto de fluidez más altos y, por lo tanto, propiedades a baja temperatura más pobres en comparación con los fluidos dieléctricos a base de aceite mineral. Sin embargo, la viscosidad de estos últimos puede variar de lote a lote debido a la inconsistencia y complejidad de los compuestos en varias fuentes de aceite mineral. Los fluidos dieléctricos de origen biológico generalmente tienen puntos de inflamación y fuego más altos (al menos dos veces) en comparación con los fluidos dieléctricos de base de aceite mineral. Los fluidos dieléctricos de origen biológico generalmente tienen una estabilidad hidrolítica, térmica y oxidativa inferior, y un índice de acidez más alto (alrededor de dos veces) en comparación con los fluidos dieléctricos a base de aceite mineral. Los fluidos dieléctricos de origen biológico generalmente son más biodegradables y tienen menor toxicidad en comparación con los fluidos dieléctricos de base de aceite mineral y están hechos de un recurso renovable, en lugar de no renovable. Los fluidos dieléctricos de origen biológico generalmente cuestan más de producir y requieren más aditivos en comparación con los fluidos dieléctricos con base de aceite mineral.
[0232] Los nuevos fluidos dieléctricos que se proporcionan en este documento, en algunos casos, tienen todas las ventajas de los fluidos dieléctricos tradicionales de origen biológico con menos, y en algunos casos, ninguna de las desventajas. Estas y otras ventajas de los presentes métodos pueden apreciarse mejor después de tener en cuenta la siguiente descripción de las propiedades generales de los fluidos dieléctricos.
[0233] De manera idónea, la viscosidad de un fluido dieléctrico debería variar lo menos posible con la temperatura. La viscosidad es una medida de la resistencia de un fluido al flujo o corte ("espesor") y se mide en cinemática (kv) y absoluta (dinámica) (cSt o mm2/s @ 40 y 100 °C). (ASTM D2270-04; ASTM D445; ASTM D88). Generalmente, se
desea el lubricante menos viscoso que fuerce adecuadamente la separación de dos superficies móviles. La viscosidad a veces se considera la característica más importante de un fluido hidráulico. Si la viscosidad es demasiado alta, aumentan la fricción, la caída de presión, el consumo de energía y la generación de calor. Si la viscosidad es demasiado baja, es posible que aumenten las fugas internas a temperaturas de funcionamiento más altas. La película de aceite puede ser insuficiente para evitar un desgaste excesivo o un posible gripado de las piezas móviles. Algunas viscosidades ilustrativas (en unidades cSt) de fluido dieléctrico derivado de varias fuentes son: derivado de aceite mineral: 20 a 40 °C y 4 a 100 °C; derivados del aceite de soja: 30 a 40 °C y 7,6 a 100 °C; derivados del aceite de girasol: 40 a 40 °C y 8,7 a 100 °C; y derivados de aceite de colza (canola): 33 a 40 °C. (Siniawski y col.; J. Synthetic Lubrication; 24, 101-110 (2007); Schneider; J. Sci. Food Agric., 86, 1769-1780 (2006)). En casos particulares, los fluidos dieléctricos pueden tener viscosidades similares a las de los fluidos dieléctricos derivados de las fuentes anteriores. En casos ilustrativos, el fluido dieléctrico tiene una viscosidad a 40 °C de menos de aproximadamente 110 cSt, por ejemplo, en el rango de 20-30 cSt y/o una viscosidad a 100 °C en el rango de aproximadamente 2 a aproximadamente 15 cSt, por ejemplo, 4-8 cSt.
[0234] El índice de viscosidad (VI, un número sin unidades) es una medida de la variación de la viscosidad con la variación de la temperatura. Para VI, se compara el kv del aceite a 40 °C con dos aceites de referencia (con VI de 0 y 100), donde todos los aceites tienen el mismo kv a 100 °C (ASTM D2270). El valor VI generalmente debe ser lo más alto posible. Los valores altos de VI indican que la viscosidad del aceite cambia poco con la temperatura. En general: un VI bajo está por debajo de 35; un VI medio es de 35 a 80; un VI alto es de 80 a 110; un VI muy alto es de 110 a 125; un super VI es de 125 a 160; y un VI súper alto es igual o superior a 160. Los VI de fluidos dieléctricos derivados de varios materiales de partida incluyen: derivados de aceite mineral: 103; derivados del aceite de soja: 246; y derivados del aceite de girasol: 206. (Siniawski y col.; J. Synthetic Lubrication; 24, 101-110 (2007)). En casos particulares, los fluidos dieléctricos pueden tener VI similares a los de los fluidos dieléctricos derivados de las fuentes anteriores.
[0235] El punto de fluidez es la temperatura más baja a la que se verterá o fluirá un líquido (°C) (ASTM D97). El punto de fluidez debe ser al menos 10 °C inferior a la temperatura ambiente prevista más baja a la que se utilizará el fluido dieléctrico. Los puntos de fluidez de los fluidos dieléctricos derivados de diversos materiales de partida incluyen: derivados del aceite mineral: -50 °C; derivados del aceite de soja: -9°C; derivados del aceite de girasol: -12°C; y derivados del aceite de colza (canola): -21°C. (Siniawski y col.; J. Synthetic Lubrication; 24, 101-110 (2007)). En casos particulares, los fluidos dieléctricos pueden tener puntos de fluidez similares a los de los fluidos dieléctricos derivados de las fuentes anteriores. En varios casos, el punto de fluidez de un fluido dieléctrico con base de aceite microbiano puede ser de aproximadamente -10 grados C, aproximadamente -15 grados C, aproximadamente -20 grados C, aproximadamente -25 grados C, aproximadamente 30 grados C, aproximadamente -35 grados C, o alrededor de -40 grados C. Además, el punto de fluidez del fluido dieléctrico con base de aceite microbiano puede estar dentro de cualquier rango limitado por cualquiera de estos valores, por ejemplo, aproximadamente -10 grados C a -40 grados C o aproximadamente - 15 grados C a aproximadamente -35 grados C, etc.
[0236] Por ejemplo, y como se ha descrito anteriormente, el aceite RBD producido según los métodos descritos en este documento se puede producir fácilmente con puntos de fluidez de aproximadamente -8 °C o menos. Este punto de fluidez se puede reducir aún más mezclando el aceite RBD con un depresor del punto de fluidez para lograr aceites con puntos de fluidez en el rango de -15 a -20 °C o menos en función de la cantidad de depresor del punto de fluidez que se agregue al aceite. La fracción de oleína de un solo fraccionamiento produce fácilmente aceite con un punto de fluidez de aproximadamente -11 °C, que puede reducirse mezclando la fracción de oleína con un depresor del punto de fluidez para lograr aceites con puntos de fluidez en el rango de -16 a -20 °C o menos en función de la cantidad de depresor del punto de fluidez añadido al aceite. La fracción de oleína de un segundo fraccionamiento ("super oleína") produce fácilmente aceite con un punto de fluidez de aproximadamente -20 °C, que puede reducirse mezclando la fracción de super oleína con un depresor del punto de fluidez para lograr aceites con puntos de fluidez por debajo de - 20 °C, es decir, -26 °C o menos en función de la cantidad de depresor del punto de fluidez que se haya agregado al aceite. Existe en el mercado una amplia variedad de depresores del punto de fluidez, que se pueden adquirir de Chevron, Oronite, Infineum, General Electric, RohmMax Evonik y otros. Los depresores del punto de fluidez ilustrativos para usar con los aceites microbianos (lípidos) descritos en este documento incluyen VISCOPLEX® 10-310 o 1-133 (Rohmax-Evonik Additives GmbH), u otros poli(alquil) acrilatos y poli(metil)acrilatos como INFINEUM® V-351 (Infineum UK limited), PMA-D110 y PMA D.
[0237] La lubricidad (propiedades antidesgaste) de un fluido dieléctrico es importante, ya que se produce un desgaste prematuro cuando la viscosidad del fluido es insuficiente y la película del fluido no evita el contacto con la superficie (ASTM D2882). En algunos casos, los fluidos dieléctricos pueden tener buena lubricidad (equivalente o mejor que la norma ASTM D2882).
[0238] La volatilidad, o la tendencia de un aceite a vaporizarse (atm de vapor frente a °C), también es importante para un fluido dieléctrico. En general, se prefiere una volatilidad más baja. En algunos casos, los fluidos dieléctricos
pueden tener una volatilidad tan baja como la de los fluidos dieléctricos tradicionales con base de aceite mineral y de origen biológico, e incluso más baja.
[0239] La inflamabilidad del fluido dieléctrico es importante. En general, se prefiere una inflamabilidad más baja (véase "Bio-Based Lubricante A Market Opportunity Study Update" United Soybean Board, Nov. 2008, Omni Tech International, Ltd., www.soynewuses.org/downloads/reports/BioBasedLubricantsMarketStudy.pdf). En casos particulares, los fluidos dieléctricos pueden tener una inflamabilidad tan baja como los fluidos dieléctricos basados en aceite mineral y biodieléctricos tradicionales, e incluso más baja.
[0240] El punto de inflamación es la temperatura más baja (°C) a la que un aceite se vaporiza para formar una mezcla inflamable en el aire. En las normas ASTM D3278, d 3828, D56 y D93 se describen las especificaciones de punto de inflamación adecuadas para fluidos dieléctricos. Para evitar la ignición del aceite, el punto de inflamación generalmente debe ser lo más alto posible. Los puntos de inflamación de los fluidos dieléctricos derivados de diversas fuentes incluyen: derivados del aceite mineral: 147 °C; y derivados de TAG (típico): 324 °C. (New Safety Dielectric Coolants for Distribution and Power Transformers, www.cooperpower.com/Library/pdf/00048.pdf). En algunos casos, los fluidos dieléctricos pueden tener puntos de inflamación similares a los de los fluidos dieléctricos derivados de las fuentes anteriores e iguales o superiores a las especificaciones ASTM D1310 y ASTM D92.
[0241] El punto de ignición es la temperatura más baja (°C) a la que un aceite continuará ardiendo durante al menos 5 segundos después de la ignición por una llama abierta. En las normas ASTM D1310 y ASTM D92 se describen las especificaciones del punto de inflamación adecuadas para fluidos dieléctricos. Para evitar la ignición del aceite, el punto de combustión debe ser lo más alto posible. Los puntos de inflamación de los fluidos dieléctricos derivados de varias fuentes incluyen: derivados del aceite mineral: 165 °C; y derivados de TAG (típicos): 360 °C. (New Safety Dielectric Coolants for Distribution and Power Transformers, www.cooperpower.com/Library/pdf/00048.pdf) En algunos casos, los fluidos dieléctricos pueden tener puntos de ignición similares a los de los fluidos dieléctricos derivados de las fuentes anteriores e iguales o superiores a las especificaciones ASTM D1310 y ASTM D92. En algunos casos, ese punto de combustión está por encima de los 300 °C, por ejemplo, de 300 °C a 450 °C.
[0242] La reactividad de un fluido dieléctrico es importante; el fluido dieléctrico no debe reaccionar (o debe tener una baja reactividad) con ácidos/bases, calor y aire.
[0243] La reactividad hidrolítica se refiere a la susceptibilidad de un fluido a la descomposición en presencia de ácidos o bases. En las normas ASTM D2619 y ASTM D943 se describe la reactividad hidrolítica adecuada para fluidos dieléctricos. En los TAG, los grupos funcionales susceptibles son los ésteres y los grupos funcionales sensibles al ácido/base. En casos particulares, los fluidos dieléctricos pueden tener una baja reactividad hidrolítica (equivalente o mejor que en ASTM D2619 y/o ASTM D943).
[0244] La estabilidad térmica se refiere a la susceptibilidad de un fluido dieléctrico a la descomposición térmica. En los fluidos dieléctricos derivados del bioaceite, la inestabilidad térmica generalmente se debe a los hidrógenos p del glicerol, lo que finalmente da como resultado productos de eliminación. En casos particulares, los fluidos dieléctricos pueden tener una alta estabilidad térmica (igual o superior a la de los fluidos dieléctricos tradicionales derivados de bioaceites).
[0245] La susceptibilidad oxidativa se refiere a la susceptibilidad de un fluido dieléctrico a reaccionar con oxígeno para formar productos de oxidación. En ASTM D943 y ASTM D2272 se describe la estabilidad oxidativa adecuada para fluidos dieléctricos. Se desea una baja susceptibilidad a la oxidación; los valores más altos indican más lubricantes oxidativos. En algunos casos, los fluidos dieléctricos pueden tener una susceptibilidad oxidativa baja (por ejemplo, ASTM D943 o ASTM D2272).
[0246] El número de neutralización (índice de acidez/valor ácido) es una medida de la cantidad de ácido en un aceite o fluido dieléctrico. Los ácidos se forman cuando los aceites (o fluidos dieléctricos) se oxidan con el tiempo y el uso. Los ácidos surgen en los lubricantes de origen biológico a partir de la oxidación, la termólisis de ésteres o la hidrólisis de ácido/base. En las normas ASTM D947, ASTM D3487 y ASTM D6871 se describen números de neutralización adecuados para fluidos dieléctricos. Por lo general, el índice de acidez debe ser lo más bajo posible. El índice de acidez del aceite mineral estándar es 0,03 y el del aceite de origen biológico es 0,06. (Ester Transformer Fluids, IEEE/PES Transformer Committee Meeting, October 7, 2003, www.transformerscommittee.org/info/F03/F03-EsterFluids.pdf). En casos particulares, los fluidos dieléctricos pueden tener índices de acidez bajos (por ejemplo, ASTM D947, ASTM D3487 o ASTM D6871).
[0247] La miscibilidad se refiere a la capacidad de un fluido para mezclarse con otros fluidos. De manera idónea, un fluido dieléctrico debe poder mezclarse bien con otros lubricantes, fluidos y aditivos, pero no con agua. La
demulsibilidad se refiere a cómo de bien un fluido hidráulico se resiste a mezclarse con agua. La demulsibilidad es óptima en un fluido dieléctrico. La miscibilidad con los lubricantes y aditivos deseados es óptima en un fluido dieléctrico. En algunos casos, los fluidos dieléctricos pueden tener buena miscibilidad y demulsibilidad.
[0248] Los fluidos dieléctricos deben tener buenas propiedades de aislamiento eléctrico, es decir, deben evitar la disipación de corriente eléctrica. Las pruebas del factor de potencia de aislamiento se realizan en transformadores para medir las pérdidas dieléctricas (medidas en %). Este valor informa sobre el estado del transformador: humedad, sequedad, deterioro del aislante, condición de los devanados, barreras, cambiadores de tomas, aisladores pasantes y aceite. Los valores del factor de potencia asociados con un fluido dieléctrico deben ser lo más bajos posible, típicamente del 0,5 % o menos. Por ejemplo, el factor de potencia del aceite nuevo enviado desde una refinería no debe superar el 0,05 % a 25 °C ni el 0,3 % a 100 °C. (Norma IEEE C57, 106-1991 citada en www.nttworldwide.com/tech2209.htm). Para aceite nuevo en equipos nuevos que funcionan a 69 kV o menos, el factor de potencia no debe ser superior al 0,15 % a 25 °C y no superior al 1,5 % a 100 °C; operando de 69 kV a 288 kV o menos, el factor de potencia no debe ser superior al 0,10 % a 25 °C y no superior al 1,0 % a 100 °C; funcionando a 345 kV o más, el factor de potencia no debe ser superior al 0,05 % a 25 °C y no superior al 0,3 % a 100 °C. El aceite nuevo para interruptores automáticos debe tener un factor de potencia no superior al 0,05 % a 25 °C y no superior al 0,3 % a 100 °C. El aceite utilizado en los interruptores automáticos no debe tener un factor de potencia superior al 1,0 % a 25 °C. algunos casos, los fluidos dieléctricos pueden tener requisitos de factor de potencia favorables.
[0249] La rigidez dieléctrica se refiere a la máxima fuerza de campo eléctrico que el fluido dieléctrico (aislante eléctrico) puede resistir antes de romperse. La rigidez dieléctrica se mide en unidades de MV/m (permisividad relativa) y en la norma ASTM D877 se proporcionan especificaciones adecuadas para fluidos dieléctricos. Para su uso como aislante eléctrico, la rigidez dieléctrica del lubricante debe ser lo más alta posible. En casos particulares, los fluidos dieléctricos pueden tener fuerzas dieléctricas iguales o superiores a las especificadas por la norma ASTM D877.
[0250] El factor de disipación es una medida de la pérdida eléctrica debida al fluido dieléctrico cuando se utiliza como aislante eléctrico y se mide en unidades de % a 25 °C. En la norma ASTM D924 se proporcionan especificaciones adecuadas para los fluidos dieléctricos. Como aislante eléctrico, el valor del factor de disipación debe ser lo más bajo posible. En algunos casos, los fluidos dieléctricos pueden tener factores de disipación iguales o superiores a los especificados por la norma ASTM D924.
[0251] La conductividad eléctrica es una medida de la capacidad de un fluido dieléctrico, cuando se utiliza como aislante eléctrico, para conducir una corriente eléctrica y se mide en unidades de S*m.-1. En la norma ASTM D2624 se proporcionan especificaciones adecuadas para fluidos dieléctricos. Como aislante, el valor de la conductividad eléctrica del fluido dieléctrico debe ser lo más bajo posible. Los fluidos dieléctricos pueden tener una conductividad eléctrica favorable en comparación con los especificados en la norma ASTM D2624.
[0252] Para su uso en transformadores eléctricos y otras aplicaciones, las propiedades térmicas del fluido dieléctrico deben ser tales que el calor se transfiera de manera eficiente. El calor específico se refiere a la capacidad térmica de una sustancia y se mide en unidades de cal/gm/°C. En la norma ASTM D-2766 se proporcionan especificaciones adecuadas para fluidos dieléctricos. Los valores de calor específico más altos permiten una transferencia de calor y un enfriamiento más eficientes. Los valores de calor específico para los fluidos dieléctricos derivados del aceite mineral son generalmente de alrededor de 0,39 y, para los fluidos dieléctricos derivados de TAG, alrededor de 0,45. (Safety Dielectric Coolants for Distribution and Power Transformers, www.cooperpower.com/Library/pdf/00048.pdf). Los fluidos dieléctricos pueden tener valores de calor específico iguales o superiores a 0,39 y/o que cumplan con las especificaciones de la norma ASTM D2624.
[0253] Las propiedades ambientales de un fluido dieléctrico son importantes. Generalmente, se deben emplear fluidos dieléctricos seleccionados para mitigar los efectos ambientales de un vertido u otro accidente. La biodegradabilidad se refiere a la propiedad de un fluido dieléctrico de descomponerse en dióxido de carbono y agua en el medio ambiente y generalmente se mide en unidades de % por 28 días. En las normas OECD 301B y ASTM D-6046 se proporcionan especificaciones de biodegradabilidad adecuadas para fluidos dieléctricos. Los valores de biodegradabilidad fácilmente biodegradables son generalmente ~ 100 %; los valores de biodegradabilidad inherentemente biodegradables son generalmente 20-70 %; y los valores de biodegradabilidad no biodegradables son generalmente insignificantes al 0 %. Los fluidos dieléctricos derivados del aceite mineral generalmente tienen valores de biodegradabilidad en el rango de 15 a 35 %, y los fluidos dieléctricos derivados de bioaceite generalmente tienen valores de biodegradabilidad en el rango de 70 a 100 %. Los fluidos dieléctricos pueden tener valores de biodegradabilidad en el rango de 70-100 % (véase Renewable Lubricants Manual: Biobased Oils, Fluids, & Greases www.renewablelubricants.com/ RenewableLubricantsManual_Biodegradable.html#lntroduction).
[0254] El índice de yodo (o número de yodo) es una medida del grado de insaturación de un aceite. Más específicamente, el índice de yodo es la masa de yodo que consumen los enlaces insaturados de un aceite. Los aceites secantes tienen índices de yodo relativamente altos, de alrededor de 175 o más. Los aceites de soja tienen valores de aproximadamente 130 y los aceites de oliva tienen valores de yodo de aproximadamente 80. Los valores de yodo se determinan de forma rutinaria en la técnica. Los métodos estándar para determinar los valores de yodo incluyen ASTM D5768-02 (2006) y DIN 53241. En varios casos, un aceite microbiano en un producto a base de aceite microbiano, por ejemplo, un fluido dieléctrico, puede tener un índice de yodo de entre aproximadamente 25 y aproximadamente 200, por ejemplo, alrededor de 50, alrededor de 75, alrededor de 100, alrededor de 125, alrededor de 150 o alrededor de 175. Además, el índice de yodo puede estar dentro de cualquier rango limitado por cualquiera de estos valores, por ejemplo, alrededor de 25 a alrededor de 175, alrededor de 50 a alrededor de 200, alrededor de 50 a alrededor de 175, etc.
[0255] La insaturación de ácidos grasos también se puede modificar. El aumento de la instauración disminuye los puntos de congelación/vertido. La monoinsaturación, como la que se observa en los biolubricantes con alto contenido de ácido oleico, actualmente es óptima y representa un equilibrio entre el punto de fluidez y la reactividad oxidativa. Los aceites monoinsaturados reaccionan con el aire, pero mucho más lentamente que los ácidos grasos poliinsaturados o PUFA. Los ejemplos de PUFA incluyen ácido araquidónico (ARA), ácido eicosapentaenoico (EPA) y ácido docosahexaenoico (DHA). Los ácidos grasos diinsaturados y poliinsaturados son muy susceptibles a la oxidación y no son adecuados para aplicaciones eléctricas. Un problema con los fluidos dieléctricos derivados de aceites vegetales es la presencia de ácidos grasos poliinsaturados (por ejemplo, ácido linoleico y ácido linolénico). Una ventaja de los fluidos dieléctricos descritos en este documento es que el aceite microbiano que comprenden (o del que se derivan) contiene menos ácidos grasos di- y poliinsaturados que los fluidos dieléctricos derivados de otros bioaceites y, en algunos casos, nada de esos ácidos.
[0256] El perfil de lípidos del fluido dieléctrico suele ser muy similar al perfil de lípidos del aceite de alimentación. Se prefieren altas cantidades de ácidos grasos monoinsaturados de cadena más larga (C16-C18) para su uso como fluidos dieléctricos. Los ácidos grasos poliinsaturados (como C18:2, C18:3, ARA, EPA y DHA) no son los preferidos debido a la oxidación y la producción de productos de oxidación. Los ácidos grasos saturados tienden a ser sólidos o líquidos con un alto punto de congelación, lo que hace que los ácidos grasos saturados no sean deseables en grandes cantidades en los fluidos dieléctricos. En varios casos, el aceite microbiano (lípido) útil en los fluidos dieléctricos tiene al menos alrededor del 50 % de C18:1, por ejemplo, al menos alrededor del 55 %, al menos alrededor del 60 %, al menos alrededor del 65 %, al menos alrededor del 70 %, como mínimo al menos alrededor del 75 %, al menos alrededor del 80 %, al menos alrededor del 85 % y al menos alrededor del 90 % C18:1. En estos u otros casos, el aceite microbiano (lípido) tiene menos del 10 % de C18:2, por ejemplo, menos del 7,5 %, menos del 5 %, menos del 2,5 % y menos del 1 % de C18: 2. El aceite microbiano puede tener cualquier combinación de porcentajes de C18:1 y C18:2 que suman 100 % o menos. Por ejemplo, el aceite microbiano puede tener al menos un 50 % de C18:1 y menos de un 10 % de C18:2 o al menos un 80 % de C18:1 y menos de un 5 % de C18:2.
[0257] Con fines ilustrativos, se proporcionan aceites de TAG de microbios oleaginosos que contienen menos del 2 % de C18:2 (véase el Ejemplo 4), en comparación con el 20-75 % del aceite de girasol y el 48-65 % del aceite de soja. También se proporcionan aceites de TAG con menos del 0,5 % de C18:3, en comparación con el 5-10 % del aceite de soja.
[0258] Estas y otras propiedades de un fluido dieléctrico se pueden obtener, manipular y/o variar según los métodos descritos en este documento para proporcionar un producto, como un lubricante, un fluido hidráulico, un aceite industrial o un fluido dieléctrico, adecuado para cualquier aplicación. Por ejemplo, se puede manipular genéticamente microbios oleaginosos, como se ha descrito anteriormente, para alterar la longitud de la cadena, la saturación y/o la composición de los diversos ácidos grasos del lípido. En algunos casos, un aceite microbiano útil como se describe en este documento lo produce un microbio manipulado genéticamente que se ha manipulado para que exprese uno o más genes exógenos. Por ejemplo, el microbio modificado genéticamente puede ser Prototheca (por ejemplo, Prototheca moriformis) o Chlorella. Los genes exógenos ilustrativos incluyen aquellos que codifican sacarasa y/o acil-ACP tioesterasa.
[0259] Además, los lípidos extraídos de microalgas o levaduras oleaginosas pueden someterse a diversas modificaciones químicas para lograr una propiedad deseada en un fluido dieléctrico. Las alteraciones típicas incluyen la alteración de la longitud de la cadena de ácidos grasos (FA). Los ácidos grasos de cadena más corta tienen puntos de fluidez reducidos. Las modificaciones químicas también pueden usarse para reducir la insaturación e incluyen alquilación, adición de radicales, acilación, reacciones alder-eno, hidroformilación, hidrogenación selectiva, oligomerización, hidroaminometilación, aciloxilación y epoxidación. Además, o como alternativa, se puede mezclar un
aditivo, como un depresor del punto de fluidez, con el aceite microbiano procesado para lograr una propiedad deseada, por ejemplo, el punto de fluidez. Los aditivos ilustrativos se analizan con mayor detalle a continuación.
[0260] Como se ha mencionado antes, en casos particulares, el aceite microbiano crudo extraído de un microbio oleaginoso típicamente se "enriquece" antes de incorporarlo a un producto descrito en este documento. Por ejemplo, puede haber contaminantes en los lípidos microbianos que pueden cristalizarse y/o precipitarse y caer de la solución como sedimento. La formación de sedimentos es particularmente un problema cuando se usa un fluido dieléctrico a temperaturas más bajas. El sedimento o los precipitados pueden causar problemas tales como disminución del flujo, obstrucción, etc. En la técnica se conocen procesos que se ocupan específicamente de la eliminación de estos contaminantes y sedimentos para producir un producto de mayor calidad. Entre los ejemplos de dichos procesos se incluyen el pretratamiento del aceite para eliminar contaminantes tales como fosfolípidos y ácidos grasos libres (por ejemplo, desgomado, refinado cáustico y filtración con adsorbente de sílice).
[0261] La preparación para el invierno se puede utilizar para enriquecer el aceite microbiano. Existen varios enfoques para preparar un fluido dieléctrico para el invierno. Un enfoque es mezclar el fluido con otros fluidos dieléctricos. Otro enfoque es utilizar aditivos que pueden reducir el punto de congelación. El fraccionamiento en seco también se puede utilizar para reducir la proporción relativa de la fracción saturada (la fracción de estearina). Al enfriar el aceite, se pueden cristalizar los saturados y luego filtrar los cristales. El fraccionamiento separa selectivamente un fluido en componentes o fracciones individuales, lo que permite la eliminación o inclusión de fracciones específicas. Otros métodos de fraccionamiento incluyen el fraccionamiento de urea, el fraccionamiento de solventes y la destilación térmica.
[0262] Luego se puede agregar tierra de diatomeas u otro material filtrante como arcilla blanqueadora al líquido enfriado para formar una suspensión, que luego se puede filtrar a través de una hoja de presión u otro tipo de filtro para eliminar las partículas. Luego, el líquido filtrado puede pasar a través de un filtro de residuos para eliminar los sedimentos restantes y la tierra de diatomeas, con el fin de producir un producto final. Alternativamente, o además, este producto, o el aceite microbiano producido al final de cualquiera de los pasos del proceso anterior, se puede mezclar con un depresor del punto de fluidez para producir un producto, como un fluido dieléctrico.
[0263] Un método para producir un aceite lubricante o un fluido dieléctrico puede ser un método que comprende los pasos de (a) cultivar un microorganismo que contiene lípidos usando métodos descritos en este documento, (b) lisar un microorganismo que contiene lípidos para producir un lisado, (c) aislar la composición lipídica del microorganismo lisado, y (d) enriquecer la composición lipídica aislada, con lo que se produce un aceite lubricante o fluido dieléctrico. Típicamente, el paso (d) incluirá uno o más pasos de refinado, blanqueo y/o desodorización y uno o más pasos de fraccionamiento para reducir la proporción relativa de la fracción saturada eliminando los triglicéridos palmítico y/o esteárico. En otro caso, el aceite lubricante o fluido dieléctrico resultante del paso (d) se mezcla con un depresor del punto de fluidez.
[0264] Opcionalmente, se pueden mezclar otros aditivos para aumentar la estabilidad oxidativa de los lípidos aislados con el aceite microbiano, lubricante o fluido dieléctrico producido por estos métodos. Los ejemplos de tales aditivos incluyen antioxidantes tales como tocoferoles (vitamina E, por ejemplo, alfa-, beta- y/o delta-tocoferol), ácido ascórbico (vitamina C). Los antioxidantes adecuados están disponibles en el mercado. La empresa BASF comercializa una línea de antioxidantes adecuados a base de fenol y a base de aminas con la marca IRGANOX®. IRGANOX L109, IRGANOX L64, IRGANOX L57, otros antioxidantes IRGANOX y otros compuestos a base de aminas y fenol son adecuados como aditivos antioxidantes para los aceites y productos, incluidos los fluidos dieléctricos. Otros ejemplos no excluyentes de antioxidantes incluyen butilhidroxianisol (BHA), butilhidroxitolueno (BHT), monobutilhidroquinona terciaria (TBHQ), butilhidroanisol, tetrahidrobutrofenona, palmitato de ascorbilo y galato de propilo. En algunos casos, un producto a base de aceite microbiano, por ejemplo, un fluido dieléctrico, incluye adicionalmente un antioxidante del 0,1 % al 5 % en peso, y preferiblemente del 0,5 % al 2 %.
[0265] Otros aditivos que se pueden añadir opcionalmente a los lípidos aislados para su uso como productos tales como fluidos dieléctricos son desactivadores de iones metálicos, inhibidores de corrosión, aditivos antidesgaste y/o protectores contra la hidrólisis. Algunos aditivos ampliamente utilizados en fluidos dieléctricos se describen en Schneider, 2006, J Science Food and Agricultura; 86: 1769-1780.). Los desactivadores de iones metálicos tienen dos funciones principales. Suprimen el ataque químico en la superficie del metal y también pasivan la superficie del metal para suprimir cualquier residuo que pueda actuar como catalizador para la formación de radicales (electrones desapareados). Los desactivadores de metales están disponibles en el mercado. Por ejemplo, la empresa BASF ofrece una línea de desactivadores de metales, incluida la línea de desactivadores de metales IRGAMET®. La empresa RTVANDERBILT vende la línea de desactivadores de metales CUVAN®. Otros ejemplos de desactivadores de metales incluyen triazoles derivatizados incluyendo 1-(di-isooctilaminometil)-1,2,4-triazol, 1-(2-metoxiprop-2-il)toliltriazol, 1 -(1-ciclohexiloxipropil)toliltriazol, 1 -(1-ciclohexiloxiheptil)toliltriazol, 1 -(1-ciclohexiloxibutil)toliltriazol, 1 [b¡s(2-et¡lhex¡l)am¡nomet¡l-4-metilbenzotr¡azol, boros derivatizados incluyendo borato de trietilo, borato de tripropilo, borato de triisopropilo, borato de tributilo, borato de tripentilo, borato de trihexilo , borato de triciclohexilo, borato de trioctilo, borato de triisooctilo y otros desactivadores metálicos de hidrazina derivatizados, por ejemplo, 2',3-bis[[3-[3,5-d¡-tert-butil-4-h¡drox¡fen¡l]prop¡on¡l]]propon¡oh¡draz¡na, y similares). En algunos casos, un producto a base de aceite microbiano, por ejemplo, un fluido dieléctrico, incluye adicionalmente uno o más desactivadores de metales en una proporción del 0,1 % al 5 % en peso, y preferiblemente del 0,5 % al 2 %.
[0266] Por lo tanto, los fluidos dieléctricos preparados según los métodos descritos en este documento pueden contener varios aditivos, incluidos, entre otros, uno o más de los siguientes aditivos: (a) un antioxidante, incluidos, entre otros, BHT y otros fenoles; (b) un desactivador de iones metálicos tales como Cu, Zn y similares, incluidos, entre otros, benzotriazol; (c) inhibidores de la corrosión, incluidos, entre otros, sulfonatos de éster y ésteres de ácido succínico; (d) desemulsionantes; (e) aditivos antidesgaste, incluidos, entre otros, ditiofosfato de zinc; (f) aditivos para reducir el punto de fluidez, incluidos, entre otros, copolímeros de anhídrido maleico y estireno, poli(alquil)acrilatos, incluidos, entre otros, polimetacrilatos; y (g) compuestos que protegen contra la hidrólisis, incluyendo pero sin limitarse a carbodiimidas.
[0267] En algunos casos, un método produce un producto que incluye un aceite microbiano que tiene un punto de fluidez entre aproximadamente -10 °C y aproximadamente -40 °C, y en el que la composición de ácidos grasos del aceite microbiano es al menos 50 % de C18:1 y menos del 10 % de C18:2. El método implica cultivar un microbio modificado genéticamente diseñado para expresar uno o más genes exógenos hasta que el microbio tenga al menos un 10 % de aceite en peso en seco. Los microbios modificados genéticamente ilustrativos incluyen Prototheca (por ejemplo, Prototheca moriformis) o Chlorella. Los genes exógenos ilustrativos incluyen aquellos que codifican sacarasa y/o acil-ACP tioesterasa. En algunos casos, el microbio manipulado genéticamente expresa al menos dos genes exógenos, por ejemplo, que codifican la sacarasa y acil-ACP tioesterasa, que codifican dos acil-ACP tioesterasas diferentes, o que codifican sacarasa y dos acil-ACP tioesterasas diferentes. Una vez que el microbio tiene al menos un 10 % de aceite en peso en seco, el aceite se separa del microbio y se somete a refinado, blanqueo, desodorización o desgomado para producir aceite RBD. Opcionalmente, se puede agregar al aceite RBD un antioxidante, un desactivador de iones metálicos, un inhibidor de la corrosión, un desemulsionante, un aditivo antidesgaste, un depresor del punto de fluidez y/o un compuesto antihidrólisis para producir el producto deseado.
[0268] En casos particulares, un método de fraccionamiento produce un aceite microbiano adecuado para su incorporación en productos (por ejemplo, un fluido dieléctrico) que tiene un punto de fluidez entre aproximadamente -10 °C y aproximadamente -40 °C, y en donde la composición de ácidos grasos del aceite microbiano es al menos 50 % de C18:1 y menos de 10 % de C18:2. El método implica someter un aceite microbiano inicial (es decir, "primero") a refinado, blanqueo, desodorización o desgomado para producir aceite RBD, en el que el aceite RBD se caracteriza por un punto de fluidez inicial y una primera temperatura, lo que reduce la temperatura del aceite RBD a una segunda temperatura, y filtrar el aceite RBD a la segunda temperatura para proporcionar un segundo aceite microbiano caracterizado por un segundo punto de fluidez que es más bajo que el punto de fluidez inicial, donde el segundo punto de fluidez está entre aproximadamente -10° C y aproximadamente -40 °C, y en donde la composición de ácidos grasos del segundo aceite microbiano es al menos 50 % de C18:1 y menos de 10 % de C18:2. Una primera temperatura ilustrativa está entre más de 15 °C y aproximadamente 50 °C, y una segunda temperatura ilustrativa está entre aproximadamente -15 °C y aproximadamente 15 °C. Se puede añadir un aditivo antidesgaste, un depresor del punto de fluidez y/o un compuesto antihidrólisis al segundo aceite microbiano para producir un producto deseado. En variaciones de estos casos, el primer aceite microbiano se produce cultivando un microbio modificado genéticamente para que exprese uno o más genes exógenos hasta que el microbio tenga al menos un 10 % de aceite por peso en seco y luego separando el aceite del microbio para producir el primer aceite microbiano. Este método se puede emplear para producir, por ejemplo, un lubricante, un fluido hidráulico, un aceite industrial o un fluido dieléctrico. En algunos casos, cuando el producto es un fluido dieléctrico, el fluido incluye uno o más de un antioxidante, un desactivador de iones metálicos, un inhibidor de la corrosión, un desemulsionante, un aditivo antidesgaste, un depresor del punto de fluidez o un compuesto antihidrólisis.
[0269] En un caso, se produce un fluido dieléctrico mezclando aceites y/o fluidos dieléctricos derivados de microbios oleaginosos con aceites o fluidos dieléctricos existentes. Los aceites y fluidos dieléctricos existentes pueden ser de origen vegetal o animal (o ambos, por ejemplo, petróleo).
[0270] Por lo tanto, se puede usar una variedad de métodos en los que se utilizan lípidos de microbios oleaginosos para producir fluidos dieléctricos y otros productos útiles en una variedad de aplicaciones industriales y de otro tipo. Los ejemplos de procesos para modificar el aceite producido por los métodos descritos en este documento incluyen, pero no se limitan a, hidrólisis del aceite, hidroprocesamiento del aceite y esterificación del aceite. Otras modificaciones químicas de lípidos de microalgas incluyen, sin limitación, epoxidación, oxidación, hidrólisis, sulfatación, sulfonación, etoxilación, propoxilación, amidación y saponificación. La modificación del aceite de
microalgas produce productos oleoquímicos básicos que pueden modificarse aún más para dar productos oleoquímicos derivados seleccionados para una función deseada. De manera similar a la descrita anteriormente con referencia a los procesos de producción de combustible, estas modificaciones químicas también se pueden realizar en los aceites generados a partir de los cultivos microbianos descritos en este documento.
[0271] En algunos casos, un fluido dieléctrico descrito en este documento se emplea en un sistema eléctrico, como un transformador, que incluye un tanque que alberga un conjunto de núcleo/bobina de transformador, en el que el fluido dieléctrico rodea el conjunto de núcleo/bobina. En variaciones de tales casos, el tanque también incluye un material absorbente de oxígeno que está en contacto con los gases del tanque, pero aislado del contacto con el fluido aislante dieléctrico. Los materiales absorbentes de oxígeno adecuados son aquellos que son capaces de reducir la concentración de oxígeno libre en la atmósfera que rodea al fluido dieléctrico dentro del tanque y que a su vez reducen la presencia de oxígeno disuelto en el propio fluido. Dichos compuestos pueden denominarse compuestos captadores de oxígeno. Los compuestos captadores de oxígeno útiles incluyen los comúnmente empleados en la industria del envasado de alimentos. Entre los representantes de compuestos útiles de captación de oxígeno se incluyen los siguientes: sulfito de sodio; sulfato de cobre pentahidratado; una combinación de carbón y polvo de hierro activado; mezclas de hidrosulfito, hidróxido de calcio, bicarbonato de sodio y carbón activado; un polvo de haluro metálico que recubre la superficie de un polvo metálico; y combinaciones de compuestos alcalinos, tales como hidróxido de calcio, con carbonato de sodio o bicarbonato de sodio. También se consideran útiles las mezclas y combinaciones de una o más de las composiciones anteriores. También son útiles como compuestos captadores de oxígeno aquellas composiciones proporcionadas según la patente de EE. UU. n.02,825,651, que se incorpora por referencia, que incluye una composición captadora de oxígeno que comprende una mezcla de una sal de sulfito y un acelerador tal como sulfato de cobre hidratado, cloruro estannoso u óxido de cobalto. Otra clase útil de compuestos captadores de oxígeno incluye aquellas composiciones que comprenden una sal de manganeso, hierro, cobalto o níquel, un compuesto alcalino y un sulfito o compuesto delicuescente, tal como se describe en la patente de EE. UU. n.o 4,384,972, que también se incorpora por referencia. Los compuestos captadores de oxígeno preferidos incluyen (o incluyen como su componente base) al menos un óxido de hierro básico, tal como un óxido de hierro ferroso, o están hechos de mezclas de materiales de óxido de hierro. Las composiciones útiles que contienen óxido de hierro están disponibles en el mercado, por ejemplo, con el nombre comercial "Ageless" de Mitsubishi Gas Chemical Company de Duncan, Carolina del Sur (EE. UU.) y con el nombre comercial "Freshmax" de Multisorb Technologies, Inc. de Buffalo, Nueva York (EE. UU.). Son útiles los agentes absorbentes de oxígeno que comprenden una mezcla de sales ferrosas y un modificador de oxidación y/o un compuesto de sulfito o sulfato metálico.
[0272] La invención, tal como se define en las reivindicaciones, se ejemplifica en los siguientes ejemplos, que se ofrecen con el fin de ilustrar, pero no limitar, la invención reivindicada.
VIII. EJEMPLOS
EJEMPLO 1: Métodos para cultivar P ro to th e c a
[0273] Se cultivaron cepas de Prototheca para lograr un alto porcentaje de aceite por peso celular seco. Las células criopreservadas se descongelaron a temperatura ambiente y se añadieron 500 ul de células a 4,5 ml de medio (4,2 g/L de K2HPO4, 3,1 g/l de NaH2PO4, 0,24 g/L de MgSO4-7H2O, 0,25 g/L de ácido cítrico monohidrato, 0,025 g/L de CaCl2 2 H2O, 2 g/L de extracto de levadura) más glucosa al 2 % y se cultivaron durante 7 días a 28 °C con agitación (200 r.p.m.) en una placa de 6 pocillos. Los pesos celulares en seco se determinaron centrifugando 1 ml de cultivo a 14000 r.p.m. durante 5 minutos en un tubo Eppendorf pesado previamente. Se descartó el sobrenadante del cultivo y se lavó el sedimento celular resultante con 1 ml de agua desionizada. El cultivo se centrifugó de nuevo, se descartó el sobrenadante y los sedimentos celulares se pusieron a -80 °C hasta que se congelaron. A continuación, las muestras se liofilizaron durante 24 horas y se calcularon los pesos celulares en seco. Para la determinación de los lípidos totales en los cultivos, se extrajeron 3 ml de cultivo y se sometieron a análisis utilizando un sistema Ankom (Ankom Inc., Macedon, NY) según el protocolo del fabricante. Las muestras se sometieron a extracción por solvente con un extractor Amkom XT10 según el protocolo del fabricante. Los lípidos totales se determinaron como la diferencia de masa entre las muestras secas hidrolizadas con ácido y las muestras secas extraídas con disolvente. Las medidas de porcentaje del peso celular en seco de aceite se muestran en la Tabla 9.

[0274] Se genotiparon muestras de microalgas de múltiples cepas del género Prototheca. El ADN genómico se aisló de la biomasa de algas como sigue. Se centrifugaron células (aproximadamente 200 mg) de cultivos líquidos durante 5 minutos a 14000 x g. A continuación, las células se resuspendieron en agua destilada estéril, se centrifugaron durante 5 minutos a 14000 x g y se descartó el sobrenadante. Se añadió una sola perla de vidrio de ~2 mm de diámetro a la biomasa y los tubos se pusieron a -80 °C durante al menos 15 minutos. Se retiraron las muestras y se añadieron 150 pl de tampón de trituración (Sarkosyl al 1 %, sacarosa 0,25 M, NaCI 50 mM, EDTA 20 mM, Tris-HCI 100 mM, pH 8,0, 0,5 pg/ul de RNasa A). Los gránulos se resuspendieron mediante agitación vorticial breve, seguido de la adición de 40 ul de NaCl 5M. Las muestras se agitaron brevemente en un agitador vorticial, seguido de la adición de SS pl de CTAB al 5 % (bromuro de cetiltrimetilamonio) y una breve agitación vorticial final. A continuación, las muestras se incubaron a 65 °C durante 10 minutos, después de lo cual se centrifugaron a 14000 x g durante 10 minutos. El sobrenadante se transfirió a un tubo nuevo y se extrajo una vez con 300 pl de fenol:cloroformo:alcohol isoamílico 12:12:1, seguido de centrifugación durante 5 minutos a 14000 x g. La fase acuosa resultante se transfirió a un tubo nuevo que contenía 0,7 vol de isopropanol ( 190 pl), se mezcló por inversión y se incubó a temperatura ambiente durante 30 minutos o durante la noche a 4 °C. El ADN se recuperó mediante centrifugación a 14000 x g durante 10 minutos. A continuación, el sedimento resultante se lavó dos veces con etanol al 70 %, seguido de un lavado final con etanol al 100 %. Los sedimentos se secaron al aire durante 20-30 minutos a temperatura ambiente seguido de resuspensión en 50 pl de TrisCl 10 mM, EDTA 1 mM (pH 8,0).
[0275] Cinco pl de ADN de algas total, preparado como se ha descrito anteriormente, se diluyeron 1:50 en Tris 10 mM, pH 8,0. Las reacciones de PCR, volumen final de 20 pl, se establecieron como sigue. Se añadieron diez pl de master mix 2 x iProof HF (BIO-RAD) a 0,4 pl de cebador SZ02S13 (5'-TGTTGAAGAATGAGCCGGCGAC-3‘ (SEQ ID N.09) a una concentración de solución madre de 10 mM). Esta secuencia de cebadores va desde la posición 567 588 en el número de acceso de Gen Bank L43357 y está altamente conservada en plantas superiores y genomas de plastidios de algas. A esto le siguió la adición de 0,4 pl del cebador SZ02S15 (5'-CAGTGAGCTATTACGCACTC-3' (SEQ ID N.010) a una concentración de reserva de 10 mM). Esta secuencia de cebador es complementaria a la posición 1112-1093 en el número de acceso de Gen Bank L43357 y está altamente conservada en plantas superiores y genomas de plastidios de algas. A continuación, se agregaron 5 pl de ADN total diluido y 3,2 pl de dH2O. Las reacciones de PCR se realizaron de la siguiente manera: 98 °C, 45"; 98 °C, 8"; 53 °C, 12"; 72 °C, 20" durante 35 ciclos seguidos de 72 °C durante 1 min y mantenimiento a 25 °C. Para la purificación de los productos de PCR, se añadieron 20 pl de Tris 10 mM, pH 8,0 a cada reacción, seguido de extracción con 40 pl de fenol:cloroformo:alcohol isoamílico 12:12:1, agitación en agitador vorticial y centrifugación a 14000 xg durante 5 minutos. Las reacciones de PCR se aplicaron a columnas S-400 (GE Healthcare) y se centrifugaron durante 2 minutos a 3000 x g. Los productos de PCR purificados se clonaron posteriormente con TOPO en PCR8/GW/TOPO y se seleccionaron los clones positivos en placas LB/Spec. El ADN plasmídico purificado se secuenció en ambas direcciones usando cebadores directos e inversos M13. En total, se seleccionaron doce cepas de Prototheca para obtener las secuencias de ADN de su ARNr 23S, y las secuencias se enumeran en el Listado de secuencias. A continuación, se incluye un resumen de las cepas y los números de lista de secuencias. Se analizó la divergencia global de las secuencias con respecto a la secuencia UTEX 1435 (SEQ ID N.o 15). Surgieron dos pares (UTEX 329/UTEX 1533 y UTEX 329/UTEX 1440) como los más divergentes. En ambos casos, la alineación por pares dio como resultado una identidad de secuencia por pares del 75,0 %. El porcentaje de identidad de secuencia con UTEX 1435 también se incluye a continuación:
Especie Cepa % identidad de SEQ ID N.0
nucleótidos
Prototheca kruegani UTEX 329 75,2 SEQ ID N.011 Prototheca wickerhamii UTEX 1440 99 SEQ ID N.012 Prototheca stagnora UTEX 1442 75,7 SEQ ID N.013 Prototheca moriformis UTEX 288 75,4 SEQ ID N.014 Prototheca moriformis UTEX 1439; 1441; 1435; 1437 100 SEQ ID N.015 Prototheca wikerhamii UTEX 1533 99,8 SEQ ID N.016 Prototheca moriformis UTEX 1434 75,9 SEQ ID N.017 Prototheca zopfii UTEX 1438 75,7 SEQ ID N.018 Prototheca moriformis UTEX 1436 88,9 SEQ ID N.019
[0276] Se analizó el perfil de lípidos de las muestras de lípidos de un subconjunto de las cepas enumeradas anteriormente mediante HPLC. Los resultados se muestran a continuación en la Tabla 10.
T l 1 . Div r i n li í i n i Pr h

| UTEX 1435 | 1,09 I 25,77 | 0 | 2,75 | 54,01 | 11,90 | 2,44 | 0  |
|
[0277] Aceite extraído de Prototheca moriformis UTEX 1435 (mediante extracción con solvente o utilizando una prensa extrusora) se analizó para determinar los carotenoides, clorofila, tocoferoles, otros esteroles y tocotrienoles. Los resultados se resumen a continuación en la Tabla 11.
Tabla 11. Análisis de carotenoides, clorofila, tocoferol/esteroles y tocotrienol en aceite extraído de Prototheca moriformis UTEX 1435.

[0278] Se refinó y blanqueó aceite extraído de Prototheca moriformis, de cuatro lotes separados, usando métodos estándar de procesamiento de aceite vegetal. Brevemente, el aceite crudo extraído de Prototheca moriformis se clarificó en un decantador horizontal, donde se separaron los sólidos del aceite. El aceite clarificado se transfirió luego a un tanque con ácido cítrico y agua y se dejó reposar por aproximadamente 24 horas. Después de 24 horas, la mezcla en el tanque formó 2 capas separadas. La capa inferior estaba compuesta por agua y gomas que luego se eliminaban por decantación antes de transferir el aceite desgomado a un tanque de blanqueo. Luego se calentó el aceite junto con otra dosis de ácido cítrico. A continuación, se añadió arcilla de blanqueo al tanque de blanqueo y la mezcla se calentó más al vacío para evaporar el agua que estaba presente. Luego, la mezcla se bombeó a través de un filtro de hojas para eliminar la arcilla blanqueadora. Luego, el aceite filtrado se pasó a través de un filtro de pulido final de 5 pm y luego se recogió para su almacenamiento hasta su uso. Luego se analizó el aceite refinado y blanqueado (RB) para detectar los carotenoides, clorofila, esteroles, tocotrienoles y tocoferoles. Los resultados de estos análisis se resumen en la Tabla 12 a continuación. "Nd" denota que no se detectó y la sensibilidad de detección se enumera a continuación:
Sensibilidad de detección
[0279]
Carotenoides (mcg/g) nd = <0,003 mcg/g
Clorofila (mcg/g) nd = <0,03 mcg/g
Esteroles ( %) nd = 0,25 %
Tocoferoles (mcg/g); nd = 3 mcg/g
Tabla 12. Análisis de carotenoides, clorofila, esteróles, tocotrienoles y tocoferoles de aceite de Prototheca moriformis refinado blan ueado.^

[0280] También se analizaron los mismos cuatro lotes de aceite de Prototheca moriformis en busca de oligoelementos y los resultados se resumen a continuación en la Tabla 13.
Tabla 13. Análisis elemental de aceite de Prototheca moriformis refinado blan ueado.


EJEMPLO 2: Métodos generales para la transformación biolística de P ro to th e c a
[0281] Se prepararon microportadores de oro de Seashell de 550 nanómetros según el protocolo del fabricante. El plásmido (20 pg) se mezcló con 50 gl de tampón de unión y SO gl (30 mg) de portadores de oro S550d y se incubó en hielo durante 1 min. Se añadió tampón de precipitación (100 gl) y la mezcla se incubó en hielo durante 1 min más. Después de agitar con agitador vorticial, las partículas recubiertas de ADN se sedimentaron por rotación a 10000 r.p.m. en una microcentrífuga Eppendorf 5415C durante 10 segundos. El sedimento de oro se lavó una vez con 500 gl de etanol frío al 100 %, se sedimentó mediante centrifugación breve en la microcentrífuga y se resuspendió con 50 gl de etanol enfriado con hielo. Después de una breve sonicación (1-2 segundos), se transfirieron inmediatamente 10 gl de partículas recubiertas de ADN a la membrana transportadora.
[0282] Las cepas de Prototheca se cultivaron en medio de proteosa (2 g/l de extracto de levadura, NaN032,94 mM, CaCl22H20 0,17 mM, MgS04-7H20 0,3 mM, K2HP04 0,4 mM, KH2P04 1,28 mM, NaCl 0,43 mM) con glucosa al 2 % en un agitador giratorio hasta alcanzar una densidad celular de 2x106células/ml. Las células se recogieron, se lavaron una vez con agua destilada estéril y se resuspendieron en 50 gl de medio. Se extendieron 1x107 células en el tercio central de una placa de medio de proteosa no selectiva. Las células se bombardearon con el sistema de suministro de partículas biolísticas PDS-1000/He (Bio-Rad). Se usaron discos de ruptura (1350 psi) y las placas se colocaron 6 cm por debajo del conjunto pantalla/macroportador. Se permitió que las células se recuperaran a 25 °C durante 12-24 h. Tras la recuperación, las células se rasparon de las placas con una espátula de goma, se mezclaron con 100 gl de medio y se extendieron sobre placas que contenían la selección de antibiótico adecuada. Después de 7-10 días de incubación a 25 °C, eran visibles en las placas colonias que representaban células transformadas. Las colonias se recogieron y se colocaron en placas de agar selectivas (fuente de antibiótico o de carbono) para una segunda ronda de selección.
EJEMPLO 3: Expresión de genes heteróloaos de acil-ACP tioesterasa en células de microalaas
[0283] Los métodos y resultados de la expresión del gen heterólogo de la tioesterasa en células de microalgas, incluidas especies de Prototheca, se han descrito previamente en la Solicitud PCT N.0 PCT/US2009/66412, que se incorpora a la presente por referencia. En este ejemplo se describen resultados utilizando otros genes/productos genéticos de tioesterasa de especies de plantas superiores.
[0284] Una acil-ACP tioesterasa de Ricinus communis se introdujo en un contexto genético de Prototheca moríformis UTEX 1435 y la secuencia de ADNc con codones optimizados (SEQ ID N.o 87) y las secuencias de aminoácidos (del N.o de acceso GenBank ABS30422.1) (SEQ ID N.o 88) se enumeran en el Listado de secuencias. La construcción de expresión contenía secuencias de direccionamiento de recombinación homóloga 5' (SEQ ID N.0100) y 3' (SEQ ID N.0101) (que flanquean la construcción) a la región genómica 6S para la integración en el genoma nuclear y una región codificante sucH de sacarasa de S. cerevisiae bajo el control del promotor de P-tubulina/5'UTR de C. reinhardtii y la 3' UTR de nitrato reductasa de Chlorella vulgaris. Este casete de expresión sucH de S. cerevisiae se enumera como la SEQ ID N.o 78 y sirvió como marcador de selección. La región de codificación de R. communis estaba bajo el control del promotor Amt03/5'UTR de Prototheca moriformis (SEQ ID N.084) y la 3'UTR de nitrato reductasa de C. vulgaris (SEQ ID N.085). El péptido de tránsito nativo de Ricinus communis también se reemplazó con el péptido de tránsito de estearoil desaturasa de C.protothecoides (SEQ ID N° 86) y la secuencia de ADNc de la tioesterasa con el péptido de tránsito reemplazado se enumera como la SEQ ID N.087. La totalidad del casete de expresión de Ricinus communis se denominó pSZ1375 y se transformó en un conexto genético de Prototheca moriformis. Los clones positivos se cribaron en placas con sacarosa como única fuente de carbono. Se seleccionó un subconjunto de los clones positivos y se cultivó en condiciones de producción de lípidos y se determinaron los perfiles de lípidos (ácidos grasos) usando métodos de transesterificación directa como se ha descrito anteriormente. Los perfiles de ácidos grasos de los clones seleccionados se resumen en la Tabla 14 siguiente.
Tabla 14. Perfiles de ácidos rasos de Prototheca trans énica con ACP-tioesterasa de Ricinus communis

[0285] Los resultados muestran que los transformantes con el transgén de tioesterasa de Ricinus-communis tienen niveles alterados de ácidos grasos C16:0 y, en menor medida, de ácidos grasos C18:0, en comparación con la cepa de tipo salvaje. Además, hubo un aumento concomitante en el nivel de ácidos grasos C18:1 en comparación con el nivel de tipo salvaje.
EJEMPLO 4: Alteración de los niveles de ácidos grasos saturados en las microalgas P r o to th e c a m o r ifo rm is
A. Disminución de la expresión de estearoil ACP desaturasa v desaturasa de ácidos grasos delta 12 mediante un enfoque de inactivación génica
[0286] Como parte de un cribado genómico utilizando un enfoque bioinformático basado en ADNc, transcriptoma Illumina y secuenciación Roche 454 de ADN genómico de Prototheca moriformis (UTEX 1435), se identificaron dos grupos específicos de genes implicados en la desaturación de ácidos grasos: estearoil ACP desaturasas (SAD) y desaturasas de ácidos grasos delta 12 (A12 FAD). Las enzimas estearoil ACP desaturasa forman parte de la ruta de síntesis de lípidos y funcionan para introducir enlaces dobles en las cadenas de acilo graso, por ejemplo, la síntesis de ácidos grasos C18:1 a partir de ácidos grasos C18:0. Las desaturasas de ácidos grasos delta 12 también forman parte de la ruta de síntesis de lípidos y funcionan para introducir enlaces dobles en ácidos grasos ya insaturados, por ejemplo, la síntesis de ácidos grasos C18:2 a partir de ácidos grasos C18:1. El análisis de transferencia Southern utilizando sondas basadas en las dos clases de genes de desaturasa de ácidos grasos identificados durante los procesos bioinformáticos indicó que cada clase de genes de desaturasa probablemente estaba compuesta por varios miembros de la familia. Además, los genes que codifican estearoil ACP desaturasas se dividieron en dos familias distintas. Sobre la base de estos resultados, se diseñaron tres construcciones de interrupción génica para interrumpir múltiples miembros de la familia de genes al dirigirse a regiones codificantes más conservadas dentro de cada familia de enzimas desaturasas.
[0287] Se diseñaron tres construcciones de direccionamiento de recombinación homóloga utilizando: (1) porciones altamente conservadas de la secuencia codificante de los miembros de la familia de la desaturasa de ácidos grasos
delta 12 (d12FAD) y (2) dos construcciones dirigidas a cada una de las dos familias distintas de SAD, cada una con regiones conservadas de las secuencias codificantes de cada familia. Esta estrategia está diseñada para incrustar un gen marcador seleccionable (el casete tal2 de sacarasa de S. cerevisiae que confiere la capacidad de hidrolizar la sacarosa) en estas regiones codificantes altamente conservadas (dirigidas a varios miembros de la familia) en lugar de una estrategia clásica de reemplazo de genes en la que la recombinación homologa se dirigiría a las regiones flanqueantes del gen deseado.
[0288] Todas las construcciones se introdujeron en las células mediante transformación biolística usando los métodos descritos anteriormente y las construcciones se linealizaron antes de inyectarlas en las células. Los transformantes se seleccionaron en placas/medios que contenían sacarosa y los cambios en el perfil de ácidos grasos se analizaron usando el método descrito anteriormente. Las secuencias relevantes de cada una de las tres construcciones de direccionamiento se enumeran a continuación.
Descripción________________________________________________ SEQ ID N.° Secuencia 5' de la construcción de direccionamiento a d12fad SEQ ID N.° 30 Secuencia 3' de la construcción de direccionamiento a d12fad SEQ ID N.° 31 Secuencia de ADNc de la construcción de direccionamiento a d12fad SEQ ID N.° 32 Secuencia 5' de la construcción de direccionamiento a SAD2A SEQ ID N.° 33 Secuencia 3' de la construcción de direccionamiento a SAD2A SEQ ID N.° 34 Secuencia de ADNc de la construcción de direccionamiento a SAD2A SEQ ID N.° 35 Secuencia 5' de la construcción de direccionamiento a SAD2B SEQ ID N.° 36 Secuencia 3' de la construcción de direccionamiento a SAD2B SEQ ID N.° 37 Secuencia de adnc de la construcción de direccionamiento a SAD2B SEQ ID N.° 38
[0289] Se seleccionaron clones positivos representativos de transformaciones con cada una de las construcciones y se determinaron los perfiles de ácidos grasos para estos clones (expresados en % de área), que se resumen en la Tabla 15 a continuación.

[0290] Cada una de las construcciones tuvo un impacto medible en la clase deseada de ácido graso, y en los tres casos los niveles de C18:0 aumentaron notablemente, particularmente con las dos SADm inactivadas. La comparación adicional de múltiples clones de SAD inactivadas indicó que las líneas de SAD2B inactivadas tenían reducciones significativamente mayores en ácidos grasos C18:1 que los niveles de ácidos grasos C18:1 observados con las líneas de SAD2A inactivadas.
[0291] Se generaron inactivaciones adicionales de desaturasa de ácidos grasos A12(FAD) en una Prototheca moriformis (UTEX 1435) utilizando los métodos descritos anteriormente. Para identificar posibles homólogos de A12FAD, se usaron los siguientes cebadores para amplificar una región genómica que codifica una supuesta FAD:
Cebador 15-TCACTTCATGCCGGCGGTCC-3' SEQ ID N.°74
Cebador 25'- GCGCTCCTGCTTGGCTCGAA-3' SEQ ID N°:75
[0292] Las secuencias resultantes de la amplificación genómica del ADN genómico de Prototheca moriformis mediante los cebadores anteriores fueron muy similares, pero esto indicó la existencia de múltiples genes o alelos de A12FAD en Prototheca moriformis.
[0293] En función de este resultado, se diseñaron dos construcciones de interrupción génica para eliminar uno o más genes de A12FAD. La estrategia fue incorporar un casete de sacarasa (suc2 de S. cerevisiae), lo que confiere la
capacidad de hidrolizar la sacarosa como un marcador seleccionable, en regiones codificantes altamente conservadas en lugar de utilizar una estrategia clásica de reemplazo génico. La primera construcción, denominada pSZ1124, contenía secuencias genómicas de direccionamiento 5' y 3' que flanqueaban un promotor de P-tubulina de C. reinhardtii que impulsa la expresión del gen suc2S de S. cerevisiae y una 3'UTR de nitrato reductasa de Chlorella vulgarís (casete suc2 de S. cerevisiae). La segunda construccción, denominada pSZ1125, contenía secuencias genómicas de direccionamiento 5' y 3' que flanqueaban un promotor de P-tubulina de C. reinhardtii que impulsa la expresión del gen suc2S de S. cerevisiae y una 3'UTR de nitrato reductasa de Chlorella vulgaris. Las secuencias relevantes de las construcciones se enumeran en el Listado de secuencias:
Secuencia genómica de direccionamiento 5' pSZ1124 (FAD2B) SEQ ID N.076
Secuencia genómica de direccionamiento 3' pSZ1124 (FAD2B) SEQ ID N.o 77
Casete suc2 de S. cerevisiae SEQ ID N.o 78
Secuencia genómica de direccionamiento 5' pSZ1125 (FAD2C) SEQ ID N.o 79
Secuencia genómica de direccionamiento 3' pSZ1125 (FAD2C) SEQ ID N.o 80
[0294] pSZ1124 y pSZ1125 se introdujeron cada una en un contexto de Prototheca moriformis y se seleccionaron clones positivos en función de la capacidad para hidrolizar sacarosa. En la Tabla 16 se resumen los perfiles de ácidos grasos (en % de área, generados utilizando los métodos descritos anteriormente) obtenidos en dos líneas transgénicas en las que se utilizaron los vectores de direccionamiento pSZ1124 y pSZ1125.
Tabla 1S. Perfiles de ácidos rasos de A12 FAD inactivadas.

[0295] El clon transgénico que contenía la construcción FAD2B (pSZ1124) dio un resultado muy interesante e inesperado en el perfil de lípidos, ya que los niveles de C18:2, que se esperaría que disminuyeran, solo disminuyeron en aproximadamente un % de área. Sin embargo, los niveles de ácidos grasos C18:1 aumentaron significativamente, casi exclusivamente a expensas de los niveles de C16:0, que disminuyeron significativamente. El clon transgénico que contenía la construcción FAD2C (pSZ1125) también produjo un cambio en el perfil de ácidos grasos: los niveles de C18:2 se redujeron significativamente junto con un aumento correspondiente en los niveles de C18:1.
B. Enfoque de ARN horquillado para la regulación a la baja de desaturasa de ácidos grasos delta 12 (FADc) en células de P ro to th e c a
[0296] Se introdujeron vectores que regulaban a la baja la expresión génica de FADC (gen de desaturasa de ácidos grasos delta 12) en un contexto genético de Prototheca moriformis UTEX 1435. El gen de sacarasa suc2 de Saccharomyces cerevisiae se utilizó como marcador seleccionable, confiriendo la capacidad de crecer en sacarosa como única fuente de carbono a los clones positivos, y se usaron dos tipos de construcciones. El primer tipo de construcción utilizó una porción del primer exón de la región codificante de FADc unida en cis a su primer intrón seguida de una unidad repetida del primer exón en orientación inversa. Este tipo de construcción se diseñó para formar una horquilla cuando se expresa como ARNm. Se crearon dos construcciones de este primer tipo, una activada por el promotor de Amt03 de Prototheca moriformis (SEQ ID N.o 84), denominado pSZ1468, y una segunda activada por el promotor de P-tubulina de Clamidomomas reinhardtii (SEQ ID NT89), denominado pSZ1469. El segundo tipo de construcción utilizó el gran exón 2 de FADc en la orientación antisentido activado por el promotor Amt03 de Prototheca moriformis (SEQ ID N.084), denominado pSZ1470, o activado por el promotor de P-tubulina de Clamidomomas reinhardtii (SEQ ID N.o 89), denominado pSZ1471. Las cuatro construcciones tenían un casete de sacarasa suc2 de S. cerevisiae (SEQ ID N.o 78) y secuencias de direccionamiento de recombinación homóloga 5' (SEQ ID N.o 100) y 3' (SEQ ID N.o 101) (que flanquean la construcción) a la región genómica 6S para la integración en el genoma nuclear. Las secuencias de las porciones FADc de cada construcción de ARN horquillado junto con las porciones relevantes de cada construcción se enumeran en el Listado de secuencias como:
Descripción_____________________________________ SEQ ID N.°
Casete de ARN horquillado FADc pSZ1468 SEQ ID N.o 90 Porciones relevantes de la construcción pSZ1468 SEQ ID N.o 91 Casete de ARN horquillado FADc pSZ1469 SEQ ID N.o 92 Porciones relevantes de la construcción pSZ1469 SEQ ID N.o 93 Exón 2 de un casete de ARN horquillado FADc pSZ1470 SEQ ID N.o 94 Porciones relevantes de la construcción pSZ1470 SEQ ID N.o 95 Exón 2 de un casete de ARN horquillado FADc pSZ1471 SEQ ID N.o 96
Porciones relevantes de la construcción pSZ1471 SEQ ID N.097
[0297] Cada una de las cuatro construcciones se transformó en un contexto de Prototheca moríformis y los clones positivos se examinaron utilizando placas con sacarosa como única fuente de carbono. Se seleccionaron clones positivos de cada transformación y se seleccionó un subconjunto para determinar el impacto de los casetes de horquilla y antisentido contenidos en pSZ1468, pSZ1469, pSZ1470 y pSZ1471 en los perfiles de ácidos grasos. Los clones seleccionados de cada transformación se cultivaron en condiciones de producción de lípidos y los perfiles de ácidos grasos se determinaron usando métodos de transesterificación directa como se ha descrito anteriormente. Los perfiles de ácidos grasos representativos de cada una de las transformaciones se resumen a continuación en la Tabla 17. Las células de tipo salvaje 1 y 2 eran células de Prototheca moriformis sin transformar que se analizaron con cada uno de los transformantes como control negativo.
Tabla 17. Perfiles de ácidos grasos de células de Prototheca moriformis que contienen construcciones de ARN

[0298] Los resultados anteriores muestran que las construcciones de horquilla pSZ1468 y pSZ1469 mostraron los fenotipos esperados: una reducción en los niveles de ácidos grasos C18:2 y un aumento en los niveles de ácidos grasos C18:1 en comparación con el tipo salvaje 1 y el tipo salvaje 2, respectivamente. Las construcciones antisentido, pSZ1470 y pSZ1471 no dieron como resultado una disminución en los niveles de ácidos grasos C18:2, sino que mostraron un ligero aumento en comparación con el tipo salvaje 1 y el tipo salvaje 2, respectivamente, y una ligera disminución en los niveles de ácidos grasos C16:0.
C. Expresión de una estearoil-ACP desaturasa exóaena
[0299] La estearoil-ACP desaturasa de Olea europea se introdujo (número de acceso de GenBank AAB67840.1) en un contexto genético de Prototheca moriformis UTEX1435. La construcción de expresión contenía secuencias de direccionamiento de recombinación homóloga en 5' (SEQ ID NT100) y 3' (SEQ ID NT101) (que flanquean la
construcción) a la región genómica 6S para la integración en el genoma nuclear y una región suc2 codificante de sacarasa S. cerevisiae bajo el control del promotor de P-tubulina/5'UTR de . Reinhardtii y nitrato reductasa 3' UTR de Chlorella vulgaris. Este casete de expresión suc2 de S. cerevisiae se enumera como la SEQ ID N.078 y sirvió como marcador de selección. La región de codificación de estearoil-ACP desaturasa de Olea europea estaba bajo el control del promotor Amt03/5'UTR de Prototheca moriformis (SEQ ID N.°84) y 3’UTR de nitrato reductasa de C. vulgaris, y el péptido de tránsito nativo se reemplazó con el péptido de tránsito de estearoil-ACP desaturasa de Chlorella protothecoides (SEQ ID N.o 86). Las secuencias de ADNc con codones optimizados y las secuencias de aminoácidos (con el péptido de tránsito reemplazado) se enumeran en el Listado de secuencias como la SEQ ID N.o 98 y la SEQ ID N.o 99, respectivamente. La totalidad del casete de expresión SAD de O. europaea se denominó pSZ1377 y se transformó en un contexto genético de Prototheca moriformis. Los clones positivos se cribaron en placas con sacarosa como única fuente de carbono. Se seleccionó un subconjunto de los clones positivos y se cultivó en condiciones de producción de lípidos y se determinaron los perfiles de ácidos grasos utilizando métodos de transesterificación directa como se ha descrito anteriormente. Los perfiles de ácidos grasos de los clones seleccionados se resumen en la Tabla 18 a continuación.
Tabla 18. Perfil de ácidos grasos de células de Prototheca moriformis transgénicas con estearoil-ACP desaturasa de Olea euro ea.

[0300] Los resultados anteriores demuestran que la introducción de una desaturasa heteróloga, en este caso una estearoil-ACP desaturasa de Olea europea, puede resultar en niveles más altos de ácidos grasos C18:1 y una disminución concomitante en los niveles de ácidos grasos C18:0 y C16:0.
EJEMPLO COMPARATIVO 5: Cultivo de levadura oleaginosa
[0301] Las cepas de levaduras oleaginosas usadas en este Ejemplo y posteriores se obtuvieron de la Deutsche Sammlung von Mikroorganismen un Zellkulturen GmbH (DSMZ), ubicada en Inhoffenstrabe 7B, 38124 Braunschweig, Alemania, o del Centraalbureau voor Schimmelscultures (CBS) ubicado en P.O. Box 85167, 3508 Utrecht, Países Bajos. Ciento ochenta y cinco cepas de levaduras oleaginosas se analizaron para detectar su tasa de crecimiento y producción de lípidos.
[0302] Todas las cepas se convirtieron en axénicas por siembra en estría en colonias individuales en placas de agar YPD (medio YPD como se describe a continuación con 2 % de agar añadido). Se recogieron colonias individuales de las placas YPD de cada cepa y se cultivaron hasta la fase logarítmica tardía en medio YPD (10 g de extracto de bacto-levadura, 20 g de bacto-peptona y 20 g de glucosa/1 L de volumen final en agua destilada) en un agitador rotatorio a 200 r.p.m. a 30 °C.
[0303] Para la evaluación de la productividad de lípidos, se añadieron 2 ml de medio YPD a un tubo de biorreactor tarado de 50 ml (MidSci, Inc.) y se inoculó a partir de una reserva congelada de cada cepa. Luego, los tubos se colocaron en una incubadora a 30 °C y se cultivaron durante 24 horas, agitando a 200 r.p.m. para generar un cultivo de semillas. Después de 24 horas, se añadieron 8 ml de medio Y1 (base nitrogenada de levadura sin aminoácidos, Difco) que contenía tampón de ftalato 0,1 M, pH 5,0 y se mezcló bien pipeteando con cuidado. El cultivo resultante se dividió en partes iguales en un segundo tubo de biorreactor tarado. Cada uno de los cultivos duplicados resultantes de 5 ml se colocaron luego en una incubadora a 30 °C con agitación a 200 r.p.m. durante 5 días. A continuación, se recogieron las células para determinar la productividad de lípidos y el perfil de lípidos. Se usaron 3 ml del cultivo para la determinación del peso celular en seco y el contenido de lípidos totales (productividad de lípidos) y se usó 1 ml para la determinación del perfil de ácidos grasos. En cualquier caso, los cultivos se colocaron en tubos y se centrifugaron a 3500 r.p.m. durante 10 minutos para sedimentar las células. Después de decantar el sobrenadante, se agregaron 2 ml de agua desionizada a cada tubo y se usaron para lavar el sedimento celular resultante. Los tubos se volvieron a centrifugar a 3500 r.p.m. durante 10 minutos para sedimentar las células lavadas, luego se decantó el sobrenadante y los sedimentos celulares se colocaron en un congelador a -70 °C durante 30 minutos. A continuación, los tubos se transfirieron a un liofilizador durante la noche para que se secaran. Al día siguiente, se registró el peso del tubo cónico más la biomasa seca resultante del cultivo de 3mL y el sedimento celular resultante
se sometió a extracción de lípidos totales utilizando un sistema de hidrólisis ácida de Ankom (según las instrucciones del fabricante) para determinar el contenido de lípidos totales.
[0304] De las 185 cepas seleccionadas, se eligieron 30 en función de la tasa de crecimiento y la productividad de lípidos. La productividad de lípidos (expresada como porcentaje de lípidos del peso celular seco) de estas 30 cepas se resume a continuación en la Tabla 19.
Tabla 19. Productividad de lí idos de ce as de levaduras olea inosas.

[0305] Los sedimentos celulares resultantes de 1 ml de cultivo se sometieron a transesterificación directa y análisis por GC para la determinación del perfil de ácidos grasos. Un resumen de los perfiles de ácidos grasos para 17 de las cepas de levadura anteriores se lista a continuación en la Tabla 20.

[0306] Se realizó un análisis del perfil de ácidos grasos de cepas adicionales de levadura oleaginosa y se descubrió que varias cepas producían un alto porcentaje de ácido graso C16:1, incluyendo Torulaspora delbruekii CBS 2924. Esta cepa de levadura oleaginosa tenía una productividad de lípidos de aproximadamente un 40 % de lípidos como porcentaje de DCW y un perfil de ácidos grasos de: C12:0 (0,36 %); C14:0 (1,36 %); C15:0 (0,16 %); C16:0 (10,82 %); C 16:1 (42,9 %); C17:0 (0,11 %); C18:0 (2,1 %); C18:1 (35,81 %); C18:2 (4,62 %). Se descubrió que esta cepa tenía un porcentaje particularmente alto de C16:1 (ácido palmitoleico) como parte de su perfil de ácidos grasos. Se identificaron cuatro cepas adicionales que producían un alto porcentaje de 16:1: Yarrowia lipolytica CBS 6012 (10,10 %); Yarrowia lipolytica CBS 6331 (14,80 %), Yarrowia lipolytica CBS 10144 (12,90 %) y Yarrowia lipolytica CBS 5589 (14,20 %).
EJEMPLO COMPARATIVO 6: Genotipado de cepas de levaduras oleaginosas
[0307] Se realizó el genotipado de 48 cepas diferentes de levaduras oleaginosas. Se aisló el ADN genómico de cada una de las 48 cepas diferentes de biomasa de levadura oleaginosa como sigue. Se centrifugaron células (aproximadamente 200 mg) de cultivos líquidos durante 5 minutos a 14000 x g. A continuación, las células se resuspendieron en agua destilada estéril, se centrifugaron durante 5 minutos a 14000 x g y se descartó el sobrenadante. Se añadió una sola perla de vidrio de ~2 mm de diámetro a la biomasa y los tubos se pusieron a -80 °C durante al menos 15 minutos. Se retiraron las muestras y se añadieron 150 pl de tampón de trituración (Sarkosyl al 1 %, sacarosa 0,25 M, NaCl 50 mM, EDTA 20 mM, Tris-HCl 100 mM, pH 8,0, 0,5 pg/ul de ARNasa A). Los gránulos se resuspendieron mediante agitación vorticial breve, seguido de la adición de 40 ul de NaCl 5M. Las muestras se agitaron brevemente en un agitador vorticial, seguido de la adición de SS pl de CTAB al 5 % (bromuro de cetiltrimetilamonio) y una breve agitación vorticial final. A continuación, las muestras se incubaron a 65 °C durante 10 minutos, después de lo cual se centrifugaron a 14000 x g durante 10 minutos. El sobrenadante se transfirió a un tubo nuevo y se extrajo una vez con 300 pl de fenol: cloroformo: alcohol isoamílico 12:12:1, seguido de centrifugación durante 5 minutos a 14000 x g. La fase acuosa resultante se transfirió a un tubo nuevo que contenía 0,7 vol de isopropanol ( 190 pl), se mezcló por inversión y se incubó a temperatura ambiente durante 30 minutos o durante la noche a 4 °C. El ADN se recuperó mediante centrifugación a 14000 x g durante 10 minutos. A continuación, el sedimento resultante se lavó dos veces con etanol al 70 %, seguido de un lavado final con etanol al 100 %. Los sedimentos se secaron al aire durante 20-30 minutos a temperatura ambiente seguido de resuspensión en 50 pl de TrisCl 10 mM, EDTA 1 mM (pH 8,0).
[0308] Cinco pl de ADN de algas total, preparado como se ha descrito anteriormente, se diluyeron en proporción 1:50 en Tris 10 mM, pH 8,0. Las reacciones de PCR, volumen final de 20 pl, se establecieron como sigue. Se agregaron diez pl de 2 x master mix iProof HF (BIO-RAD) a 0,4 pl de cebador directo SZ5434 (5' GTCCCTGCCCTTTGTACACAC -3' (SEQ ID N.°39) a una concentración de solución madre de 10 mM) y 0,4 pl de cebador inverso SZ5435 ( 5-TTGATATGCTTAAGTTCAGCGGG -3' (SEQ ID N.040) a una concentración madre de 10 mM). Los cebadores se seleccionaron en función de la conservación de secuencias entre tres regiones principales de 18S y cinco regiones principales de genes de ARNr 26S de hongos. El cebador directo es idéntico a los nucleótidos 1S32-1S52 del n.° de acceso Genbank AY550243 y el cebador inverso es idéntico a los nucleótidos 4S4271-4S4293 del n.° de acceso Genbank NC 001144. A continuación, se agregaron 5 pl de ADN total diluido y 3,2 pl de dH2Ü. Las reacciones de PCR se realizaron de la siguiente manera: 98°C, 45"; 98°C, 8"; 53°C, 12"; 72°C, 20" durante 35 ciclos seguido de 72 °C durante 1 min y mantenimiento a 25 °C. Para la purificación de los productos de PCR, se añadieron 20 pl de Tris 10 mM, pH 8,0 a cada reacción, seguido de extracción con 40 pl de fenol:cloroformo:alcohol isoamílico 12:12:1, agitación en agitador vorticial y centrifugación a 14000 xg durante 5 minutos. Las reacciones de PCR se aplicaron a columnas S-400 (GE Healthcare) y se centrifugaron durante 2 minutos a 3000 x g. Los productos de p Cr purificados resultantes se clonaron y transformaron en E. coli utilizando el kit de vector ZeroBlunt PCR4Blunt-TOPO (Invitrogen) según las instrucciones del fabricante. Las reacciones de secuenciación se llevaron a cabo directamente en colonias resistentes a ampicilina. El ADN plasmídico purificado se secuenció en ambas direcciones usando cebadores directos e inversos M13. Los productos de PCR purificados se clonaron posteriormente con TOPO en PCR8/GW/TOPO y se seleccionaron los clones positivos en placas LB/Spec. El ADN plasmídico purificado se secuenció en ambas direcciones usando cebadores directos e inversos M13.
[0309] Una lista de las 48 cepas de levadura oleaginosa que se genotiparon se encuentra en la Tabla 21 junto con las SEQ ID N.o correspondientes.
Tabla 21. Cepas de levaduras oleaginosas genotipadas.
Nombre de la cepa_______________________ Número de cepa_________ SEQ ID N.°_________ Rhodotorula glutinis DSMZ-DSM 7098 SEQ ID NT41
Lipomyces tetrasporus CBS 5911 SEQ ID NT41 Rhodotorula glutinis var. glutinis CBS 3044 SEQ ID NT42
Lipomyces tetrasporus CBS 8664 SEQ ID N.°42
Lipomyces tetrasporus CBS1808 SEQ ID N.°43
Lipomyces tetrasporus CBS1810 SEQ ID N.°43
Lipomyces starkeyi CBS1809 SEQ ID N.°44
Trichosporon montevideense CBS 8261 SEQ ID N.°44
Yarrowia lipolytica CBS 6331 SEQ ID N.°45
Cryptococcus curvatus CBS 5324 SEQ ID N.°46
Rhodotorula mucilaginosa var. mucilaginosa CBS 316 SEQ ID N.°46
Cryptococcus curvatus CBS 570 SEQ ID N.°46
Cryptococcus curvatus CBS2176 SEQ ID N.°46
Cryptococcus curvatus CBS 2744 SEQ ID N.°46
Cryptococcus curvatus CBS 2754 SEQ ID N.°46
Cryptococcus curvatus CBS 2829 SEQ ID N.°46
Cryptococcus curvatus CBS 5163 SEQ ID N.°46
Cryptococcus curvatus CBS 5358 SEQ ID N.°46
Trichosporon sp. CBS 7617 SEQ ID N.°47 Spororobolomyces alborubescens CBS 482 SEQ ID N.°48
Rhodotorula glutinis var. glutinis CBS 324 SEQ ID N.°49
Rhodotorula glutinis var. glutinis CBS 4476 SEQ ID N.°50
Trichosporon behrend CBS 5581 SEQ ID N.°51
Geotrichum histeridarum CBS 9892 SEQ ID N.°52
Rhodotorula aurantiaca CBS 8411 SEQ ID N.°53
Cryptococcus curvatus CBS 8126 SEQ ID N.°53
Trichosporon domesticum CBS 8111 SEQ ID N.°54
Rhodotorula toruloides CBS 8761 SEQ ID N.°55
Rhodotorula terpendoidalis CBS 8445 SEQ ID N.°56
Yarrowia lipolytica CBS 10144 SEQ ID N.°57
Rhodotorula glutinis var. glutinis CBS 5805 SEQ ID N.°58
Yarrowia lipolytica CBS 10143 SEQ ID N.°59
Lipomyces tetrasporus CBS 5607 SEQ ID N.°60
Yarrowia lipolytica CBS 5589 SEQ ID N.°61
Lipomyces tetrasporus CBS 8724 SEQ ID N.°62
Rhodosporidium sphaerocarpum CBS2371 SEQ ID N.°63
Trichosporon brassicae CBS 6382 SEQ ID N.°64
Cryptococcus curvatus CBS2755 SEQ ID N.°65
Lipomyces tetrasporus CBS 7656 SEQ ID N.°65
Lipomyces starkeyi CBS 7786 SEQ ID N.°66
Yarrowia lipolytica CBS 6012 SEQ ID N.°67
Trichosporon loubieri var. loubieri CBS 8265 SEQ ID N.°68
Geotrichum vulgare CBS10073 SEQ ID N.°69
Rhodosporidium toruloides CBS 14 SEQ ID N.°70
Rhodotorula glutinis var. glutinis CBS 6020 SEQ ID N.°71
Lipomyces orientalis CBS10300 SEQ ID N.°71
Rhodotorula aurantiaca CBS 317 SEQ ID N.°72
Torulaspora delbrueckii CBS 2924 SEQ ID N.°73
EJEMPLO COMPARATIVO 7: Cultivo de R h o d o c o c c u s o o a c u s para lograr un alto contenido de aceite
[0310] Se generó un cultivo de siembra de Rhodococcus opacus PD630 (DSM 44193, Deutsche Sammlung von Mikroorganismen und Zellkuttwen GmbH) utilizando 2 ml de una solución madre crioconservada inoculada en 50 ml de medio MSM con sacarosa al 4 % (véase Schlegel, y col., (1961) Arch Mikrobiol 38, 209-22) en un matraz deflector de 250 ml. El cultivo de siembra se hizo crecer a 30 °C con agitación de 200 r.p.m. hasta que alcanzó una densidad óptica de 1,16 a 600 nm. Se utilizaron 10 ml del cultivo del matraz de siembra para inocular cultivos para la producción de lípidos en dos condiciones de nitrógeno diferentes: NH4O 10 mM y NHiCl 18,7 mM (cada uno por duplicado). Los cultivos se cultivaron a 30 °C con agitación a 200 r.p.m. durante 6 días. Las células cultivadas en la condición de NH4O 10 mM alcanzaron un máximo de 57,2 % (promedio) de lípidos por DCW después de 6 días de cultivo. Las células cultivadas en la condición de NH4Cl 18,7 mM alcanzaron un máximo de 51,8 % (promedio) de lípidos por DCW después de 5 días en cultivo.
[0311] Una muestra de biomasa de Rhodococcus opacus se sometió a transesterificación directa y se analizó mediante GC/FID para determinar un perfil de ácidos grasos. Los resultados fueron: C14:0 (2,33); C15:0 (9.08);
C16:0 (24.56); C16:1 (11.07); C17:0 (10.50); 2 especies de C17 equivalentes de enlace doble (2DBE) (19,90); C18:0 (2.49); C18:1 (17.41); C18:2 (0,05); C19:0 (0,75) y especies 2DBE de C19 (1,87).
EJEMPLO COMPARATIVO 8: Extracción de aceite de microorganismos
A. Extracción de aceite de microalqas usando una prensa extrusora v una ayuda para el prensado
[0312] La biomasa de microalgas que contenía un 38 % de aceite por DCW se secó utilizando un secador de tambor, lo que resultó en un contenido de humedad resultante de 5-5,5 %. La biomasa se introdujo en una prensa francesa L250. Se introdujeron 30,4 kg (67 libras) de biomasa a través de la prensa y no se recuperó aceite. La misma biomasa microbiana seca combinada con un porcentaje variable de pasto varilla como ayuda para el prensado se hizo pasar a través de la prensa. La combinación de biomasa microbiana seca y 20 % p/p de pasto varilla produjo el mejor porcentaje general de recuperación de aceite. A continuación, las tortas prensadas se sometieron a extracción con hexano y el rendimiento final para la condición de 20 % de pasto varilla fue del 61,6 % del aceite total disponible (calculado en peso). La biomasa con más del 50 % del peso de las células secas de aceite no requirió el uso de una ayuda para el prensado como el pasto varilla para extraer el aceite. Otros métodos de extracción de un aceite de microalgas mediante una prensa extrusora se describen en la Solicitud PCT N.0 PCT/US2010/31108, que se incorpora al presente documento por referencia.
B. Extracción de aceite de levaduras oleaginosas utilizando una prensa extrusora
[0313] Se obtuvo una cepa de levadura Rhodotorula glutinis (DSMZ-DSM 70398) de la Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (Colección alemana de cultivos de microorganismos y células, InhoffenstraRe 7B, 38124 Braunschweig, Alemania). Las células crioconservadas se descongelaron y se agregaron a 50 ml de medio YPD (descrito anteriormente) con 1x solución vitamínica DAS (1000x: 9g/L tricina; 0,S7g/L tiamina-HCl; 0,01 g/L d-biotina; 0,008 cianocobalamina; 0,02 pantotenato cálcico; y 0,04 g/L ácido p-aminobenzoico) y se cultivaron a 30 °C con agitación a 200 r.p.m. durante 18-24 horas hasta que la lectura de DO fue superior a 5 DO (AS00). Luego, el cultivo se transfirió a fermentadores de 7 L y se cambió a medio YP1 (8,5 g/L Difco Yeast Nitrogen Base sin aminoácidos y sulfato de amonio, 3 g/l de sulfato de amonio, 4 g/l de extracto de levadura) con una solución de vitamina 1x DAS. Se tomaron muestras de los cultivos dos veces al día y se analizó la DO (A600), el peso celular en seco (DCW) y la concentración de lípidos. Cuando los cultivos alcanzaron más de 50 g/L DCW, los cultivos se cosecharon. En función del peso celular en seco, la biomasa de levadura contenía aproximadamente un 50 % de aceite. Dos muestras de biomasa de levadura se sometieron a transesterificación directa y se analizaron mediante GC/FID para determinar un perfil de ácidos grasos. Los resultados se expresan en porcentaje de área y se muestran en la Tabla 22, a continuación.
Tabla 22. Perfil de ácidos rasos de muestras de biomasa de levadura transesterificada.

[0314] El caldo de levadura cosechado se secó usando tres métodos diferentes para la comparación: (1) secado en bandeja en un horno de aire forzado a 75 °C durante la noche; (2) secado en un secador de tambor sin concentración; y (3) el caldo de levadura se concentró hasta un 22 % de sólidos y luego la suspensión se secó en un secador de tambor. El material de cada una de las tres diferentes condiciones de secado se acondicionó con calor y se hizo pasar por una prensa de tornillo para la extracción de aceite. La temperatura de la prensa fue de 150 °F y la biomasa de levadura seca acondicionada se mantuvo a aproximadamente 190 °F hasta que estuvo lista para introducirla en la prensa.
[0315] El contenido de humedad de la levadura secada en bandeja fue del 1,45 % y después la levadura seca se acondicionó en un horno a 90 °C durante 10 minutos. El contenido de humedad después del acondicionamiento fue del 0,9 %. El material secado en bandeja acondicionado se introdujo luego en una prensa de tornillo Taby de sobremesa (prensa de aceite Taby Pressen Tipo 70 con un motor de 2,2 Hp y un diámetro de tornillo de 70 mm) para la extracción de aceite. Este material no produjo ninguna cantidad significativa de aceite y se observó un fuerte ensanchamiento de los extremos con el prensado.
[0316] El contenido de humedad del caldo de levadura secado en tambor sin concentración fue del 5,4 % y la levadura secada en tambor se acondicionó luego en un horno a 90 °C durante 20 minutos. El contenido de humedad después del acondicionamiento fue del 1,4 %. La levadura secada en tambor acondicionada se introdujo luego en una prensa de tornillo Taby de sobremesa para la extracción de aceite. El material expulsó bien el aceite, con un ensanchamiento mínimo de los extremos.
[0317] El contenido de humedad del caldo de levadura concentrada secada en tambor fue del 2,1 % y la levadura concentrada secada en tambor se acondicionó luego en un horno a 900C durante 20 minutos. El contenido de humedad después del acondicionamiento fue del 1,0 %. La levadura concentrada secada en tambor acondicionada se introdujo luego en una prensa de tornillo Taby de sobremesa para la extracción de aceite. El material expulsó bien el aceite, con un ensanchamiento mínimo de los extremos.
C. Secado y extracción de aceite de bacterias oleaginosas
[0318] La cepa de bacterias oleaginosas Rhodococcus opacus PD630(DSMZ-DSM 44193) se cultivó según los métodos proporcionados en el presente documento para producir biomasa de bacterias oleaginosas con aproximadamente un 32 % de lípidos por DCW.
[0319] El caldo de Rhodococcus opacus cosechado se concentró mediante centrifugación y luego se lavó con agua desionizada y se resuspendió en 1,8 l de agua desionizada. Se añadieron 50 gramos de celulosa purificada (PB20-Pre-co-Floc, EP Minerals, Nevada, EE. UU.) a la biomasa resuspendida y los sólidos totales se ajustaron con agua desionizada al 20 %. Luego, la biomasa de Rhodococcus- se secó en un secador de tambor y el contenido de humedad de Rhodococcus-después del secado en tambor fue de aproximadamente 3 %.
[0320] El material secado en tambor se acondicionó luego con calor en un horno a 130 °C durante 30 minutos con un contenido de humedad resultante de aproximadamente 1,2 %. A continuación, la biomasa acondicionada con calor se pasó por una prensa Taby de sobremesa (prensa de tornillo) para la extracción de aceite. La temperatura de la prensa fue de 209 °F y la biomasa de levadura seca acondicionada se mantuvo a aproximadamente 240 °F hasta que estuvo lista para ser introducida en la prensa. La recuperación del aceite estuvo acompañada de un fuerte ensanchamiento de los extremos del material.
EJEMPLO 9: Procesamiento de aceite extraído: descenso del punto de fluidez
Resumen
[0321] El aceite microbiano preparado según los ejemplos anteriores se puede procesar según los métodos descritos en este documento para mejorar sus propiedades para el uso en alimentos y lubricantes. Además de los microbios descritos en los ejemplos anteriores, las microalgas Chlorella protothecoides son excelentes productoras de aceite microbiano. Para métodos de cultivo de especies y cepas de Chlorella para obtener un alto contenido de aceite y extraer aceite de ellas, véanse las publicaciones PCT N.o 2008/151149, 2010/120939, y 2010/138,620, que se incorporan a la presente por referencia.
[0322] El punto de fluidez se redujo en el aceite obtenido de Chlorella protothecoides mediante la reducción de la proporción relativa de la fracción saturada, que consiste principalmente en triglicéridos palmíticos y esteáricos conocidos en la técnica como la fracción de estearina. Esto se logró fraccionando el aceite para reducir la concentración de triglicéridos saturados del aceite. Esto se hizo mediante cristalización o fraccionamiento en seco, de manera similar al proceso de preparación para el invierno conocido en la industria del aceite vegetal. El aceite de algas se refinó, blanqueó y desodorizó primero mediante los métodos descritos anteriormente (también podrían emplearse métodos similares a los utilizados en la industria del aceite vegetal) para producir "aceite RBD".
[0323] La temperatura del aceite RBD se redujo de manera controlada hasta que se formaron núcleos cristalinos. A continuación, el aceite se mantuvo a esa temperatura de cristalización durante varias horas para facilitar el crecimiento de los cristales. A continuación, los cristales se eliminaron por filtración para dar como resultado dos fracciones: una fase sólida que contenía parte o la mayor parte de la fracción de estearina y una fase líquida que contenía principalmente la fracción de oleína. La fase líquida se sometió a fraccionamiento de nuevo a una temperatura de cristalización más baja para efectuar una eliminación adicional de la estearina. La fracción líquida purificada resultante, equivalente a una súper oleína como se conoce comúnmente en la industria del aceite vegetal, tiene mejores propiedades térmicas que el aceite de algas nativo.
Materiales y métodos
Materiales
[0324] El aceite de algas (refinado, blanqueado y desodorizado) lo produjo Solazyme, Inc (South San Francisco, CA). En la Tabla 23 se resumen las propiedades del aceite utilizado en el estudio.
Tabla 23. Propiedades del aceite de algas utilizado en el estudio
Análisis Valor
Humedad [ %] 0,01
Ácido graso libre [ % como oleico] 0,03
Indice de yodo 83,5
Perfil de ácidos grasos
8:0 0,00
10:0 0,00
12:0 0,03
14:0 1,12
16:0 14,02
18:0 3,24
18:1 67,73
18:2 11,18
18:3 0,62
20:0 0,32
20:1 0,20
[0325] El depresor del punto de fluidez (PPD) a base de copolímero de metacrilato de polialquilo VISCOPLEX® 10 310 que contiene ~50 % (p/p) de vehículo de aceite de colza y VISCOPLEX® 1-133 que contiene un vehículo de aceite mineral refinado los suministró RohmMax Evonik (Horsham, PA).
Métodos
A. Fraccionamiento en seco: cristalización
[0326] Se colocaron alrededor de 2,5 kg de aceite de algas en un recipiente de doble pared de 3 L conectado a un baño de agua circulante de temperatura controlada, que sirvió para calentar y enfriar el producto (Crystallization & Degumming, Charleroi, Bélgica). El reactor estaba equipado con un agitador de velocidad variable. El enfriamiento se controló monitoreando las temperaturas del aceite y del agua que circula entre las paredes dobles del reactor. Se tomó una muestra de una gota de suspensión de cristales del reactor con una varilla y se depositó en un cubreobjetos para monitorear la formación de cristales al final del enfriamiento. La muestra se analizó inmediatamente bajo un microscopio antes de que los cristales tuvieran la oportunidad de derretirse.
[0327] El patrón general de enfriamiento se muestra en la Fig. 1. La velocidad del agitador fue de 30 r.p.m. durante la primera fase y de 15 r.p.m. hasta el final del programa de enfriamiento.
B. Fraccionamiento en seco: Filtración
[0328] Al final de la cristalización, la suspensión de cristales se filtró utilizando un filtro prensa de membrana de 1 L (Choquenet SA, Chauny, Francia). La filtración se realizó en una cámara que se mantuvo a la temperatura final de enfriamiento. El tiempo de filtración fue de 20 min y la presión de alimentación del filtro fue de 4 barg.
[0329] Al final del paso de separación, se pesaron las fracciones de estearina y oleína, se calcularon los rendimientos de las fracciones y se apartó una muestra de cada fracción para su posterior análisis. La súper oleína de algas #1 se produjo procesando la oleína del primer fraccionamiento y repitiendo el proceso de cristalización y filtración descrito anteriormente siguiendo el programa de enfriamiento que se muestra en la Fig. 2. Las súper oleínas de algas #2 y #3 se produjeron fraccionando primero el aceite desodorizado y repitiendo el proceso de cristalización y filtración usando un programa de enfriamiento similar al que se muestra en la Fig. 2.
C. Punto de fluidez _PP!
[0330] Se pesaron depresores del punto de fluidez (0,5 y 1,0 gramos) en matraces. Se añadieron a cada matraz fracciones de aceite de algas, oleína y superoleína (100 gramos). Las mezclas se mezclaron completamente. Cada muestra se analizó según el método estándar D 97 ASTM (American Society for Testing and Materials). La muestra se vertió en un tubo de ensayo y se calentó sin agitación en un baño de agua donde la temperatura se fijó a 48,0 °C. La muestra se calentó hasta alcanzar los 46,0 °C. Después de calentarla, la muestra se enfrió a 25,0 °C (en un baño de agua). Luego, la muestra se colocó en un cilindro de metal en un baño de metanol. La temperatura del baño de metanol se fijó entre -1,0 °C y -2,0 °C hasta que la temperatura de la muestra alcanzó los 10,0 °C. Luego, la temperatura del baño de metanol se redujo a -17,0 °C hasta que la temperatura de la muestra alcanzó -7,0 °C. Cuando la temperatura de la muestra estuvo aproximadamente 11,0 °C por encima del punto de fluidez esperado, se
sacó la muestra del baño de metanol en cada reducción de 3,0 °C, para comprobar la capacidad de fluidez. El punto de fluidez de la muestra se determinó como la temperatura cuando la muestra en el tubo de ensayo dejó de verterse cuando se sacó del baño de metanol. A la temperatura registrada, se añadieron 3,0 °C, para dar el valor real del punto de fluidez de la muestra.
[0331] Las propiedades del aceite producido en cada paso podrían mejorarse aún más según los métodos descritos en este documento mediante la adición de un depresor químico del punto de fluidez que redujera aún más el punto de fluidez. Los depresores del punto de fluidez utilizados para este ejemplo fueron VISCOPLEX® 10-310 y 1-133, ambos producidos por Evonik, pero se pueden obtener resultados similares usando cualquier depresor del punto de fluidez estándar. Los resultados se muestran en la Tabla 24, a continuación, y en la Fig. 3.
T l 24. Ef l fr i n mi n r r n fl i z1 n i l °
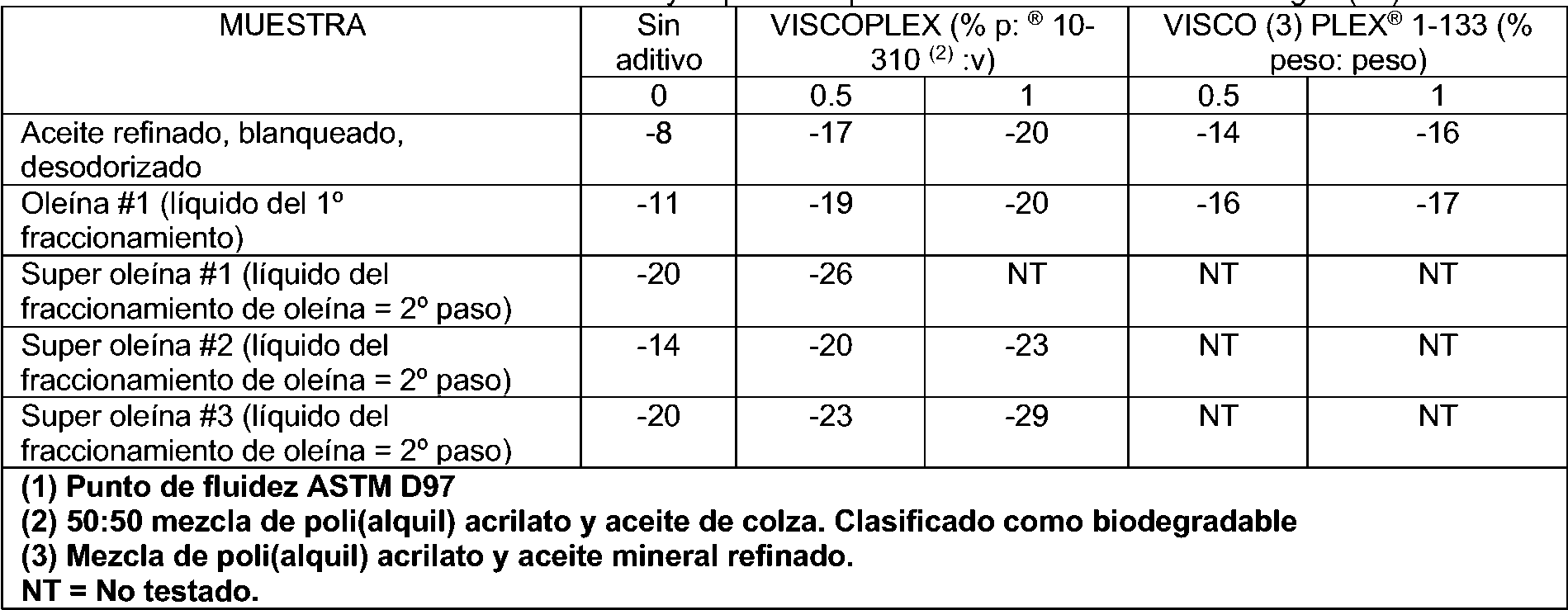
EJEMPLO 10: Puntos de fluidez del aceite producido a partir de microalqas modificadas
[0332] Protheca moriformis (UTEX 1435) se transformó con una de las siguientes construcciones plasmídicas de la Tabla 25 usando los métodos del Ejemplo 2.

[0333] Cada una de las construcciones contenía una región para la integración en el genoma nuclear y una región suc2 codificante de sacarasa de S. cerevisiae bajo el control del promotor de (3-tubulina/5'UTR de C. Reinhardtii y nitrato reductasa 3' UTR de Chlorella vulgaris. Este casete de expresión suc2 de S. cerevisiae se enumera como la SEQ ID N.° 78 y sirvió como marcador de selección. Las secuencias relevantes para las regiones de direccionamiento utilizadas para la integración del genoma nuclear se muestran a continuación.
secuencia 5' para la secuencia genómica de direccionamiento 6S SEQ ID N.° 100
secuencia 3' para la secuencia genómica de direccionamiento 6S SEQ ID N.° 101
secuencia 5' para la integración genómica en el locus FADc SEQ ID N.° 102
secuencia 3' para la integración genómica en el locus FADc SEQ ID N.° 103
secuencia 5' para la integración genómica en el locus SAD2B SEQ ID N.° 36
secuencia 3' para la integración genómica en el locus SAD2B SEQ ID N.037
[0334] Además del marcador seleccionable de sacarosa, tres de las cuatro construcciones también contenían secuencias adicionales diferentes para la expresión de proteínas o ARN. En la Tabla 26 se enumeran enzimas importantes o casetes de ARN horquillado que están codificados por la secuencia de ADN en la construcción indicada. Todas las regiones de codificación de proteínas tuvieron codones optimizados para reflejar el sesgo de codones inherente a los genes nucleares de Prototheca moríformis UTEX 1435 (véase la Tabla 2). Tanto las secuencias de aminoácidos como las secuencias de ADNc para la construcción utilizada se enumeran en el listado de secuencias.
Tabla 26. construcciones plasmídicas para la expresión de tioesterasas o ARN horquillado utilizados para transformar Protheca moriformis UTEX 1435.

[0335] Las regiones codificantes tanto de ACP tioesterasa de Carthamus tinctorius (CtOTE en la construcción 3) como de Tioesterasa FatB2 de Cuphea wrightii (CwTE2 en la construcción 4) estaban bajo el control del promotor Amt03/5'UTR de Prototheca moriformis (SEQ ID N.°84) y 3’UTR de nitrato reductasa de C. vulgaris (SEQ ID NT85). El péptido de tránsito nativo de la ACP tioesterasa de C. tintorius fue reemplazado por el péptido de tránsito de estearoil-ACP desaturasa de Chlorella protothecoides (SEQ ID N.o 86). Las secuencias de ADNc con codones optimizados y las secuencias de aminoácidos (con el péptido de tránsito reemplazado) de la ACP tioesterasa de C. tintorius se incluyen en el Listado de secuencias como la SEQ ID N.o 106 y la SEQ ID N.o 104, respectivamente. Las secuencias de ADNc con codones optimizados y las secuencias de aminoácidos de Tioesterasa FatB2 de Cuphea wrightii se incluyen en el Listado de secuencias como la SEQ ID N.o 107 y la SEQ ID N.o 105, respectivamente. La construcción 1 que contiene el ARN de horquilla FADc se describe en el Ejemplo 4.
[0336] Cada construcción se transformó en un contexto genético de Prototheca moriformis. Los clones positivos se cribaron en placas con sacarosa como única fuente de carbono. Se seleccionó un subconjunto de los clones positivos y se cultivó en condiciones de producción de lípidos. UTEX 1435 de tipo salvaje se cultivó usando glucosa, mientras que todas las demás líneas transgénicas se cultivaron en sacarosa. Para cada construcción, se cultivaron transformantes y se aisló el aceite. Se analizaron los perfiles de ácidos grasos de los aceites aislados y se determinaron los puntos de fluidez como se describe en este documento. Los puntos de fluidez se determinaron utilizando el método de prueba estándar ASTM D97 para la evaluación del punto de fluidez. Los perfiles de ácidos grasos y los puntos de fluidez de los aceites para las cepas transgénicas se muestran en la Tabla 27 a continuación. La Tabla 27 revela los datos para la manipulación exitosa de los puntos de fluidez de los aceites producidos por microalgas modificadas genéticamente. El punto de fluidez del aceite transformado con la construcción 3 se redujo de -10,5 °C a -19,5 °C.
Tabla 27. Perfiles de ácidos grasos y temperaturas de punto de fluidez de células de Prototheca moriformis que contienen diferentes construcciones.

EJEMPLO 11: Microalaas modificadas genéticamente con perfiles de ácidos grasos alterados
[0337] Como se ha descrito anteriormente, la integración de genes heterólogos para inactivar o atenuar enzimas endógenas específicas de la ruta de los lípidos en especies de Prototheca pueden alterar los perfiles de ácidos grasos. Como las acil-ACP tioesterasas endógenas catalizan la escisión de un ácido graso de una proteína transportadora de acilo durante la síntesis de lípidos, son enzimas importantes de la ruta de los lípidos para establecer el perfil de lípidos del organismo hospedador. Se crearon construcciones plasmídicas para evaluar si el perfil de lípidos de una célula hospedadora puede verse afectado como resultado de una inactivación o inactivación de un gen endógeno de acil-ACP tioesterasa, FATA1.
A. Alteración de los perfiles de ácidos grasos por inactivación de un gen de tioesterasa endógeno de Prototheca moriformis
[0338] Un derivado clásicamente mutagenizado de Protheca moriformis UTEX 1435, S1920, se transformó con una de las siguientes construcciones plasmídicas de la Tabla 28 utilizando los métodos del Ejemplo 2. Cada construcción contenía una región para la integración en el genoma nuclear para interrumpir el gen FATA1 endógeno y un región codificante de sacarasa suc2 de S. cerevisiae bajo el control del promotor de P-tubulina/5'UTR de C. Reinhardtii y 3' UTR de nitrato reductasa de Chlorella vulgaris. Este casete de expresión suc2 de S. cerevisiae se enumera como la SEQ ID N.078 y sirvió como marcador de selección. Todas las regiones de codificación de proteínas tuvieron cordones optimizados para reflejar el sesgo de codones inherente a los genes nucleares de Prototheca moriformis UTEX 1435 (véase la Tabla 2). Las secuencias relevantes para las regiones de direccionamiento del gen FATA1 utilizadas para la integración del genoma nuclear se muestran a continuación.
Descripción____________________________________ SEQ ID N.°
secuencia 5' para la integración en el locus FATA1 SEQ ID N.0108 secuencia 3' para la integración en el locus FATA1 SEQ ID N.0109
Ta l 2 . n r i n l mí i iliz r r n f rm r Pr h m rif rmi TEX 14 1 2 .

[0339] Los sitios de restricción relevantes en la construcción FATA1-CrbTub_ylnv_nr-FATA1 se indican en minúsculas en la secuencia que aparece a continuación, en negrita y subrayados, y son 5'-3' BspQ 1, Kpn I, Asc I, Mfe I, Sac I, BspQ I, respectivamente. Los sitios BspQI delimitan los extremos 5' y 3' del ADN transformante. Las secuencias en minúsculas y en negrita representan el ADN genómico de S1920 que permite la integración dirigida en el locus FATA1 a través de la recombinación homóloga. Procediendo en la dirección 5' a 3', el promotor de P-tubulina de . Reinhardtii que impulsa la expresión del gen de sacarasa de levadura (que confiere la capacidad de S1920 para metabolizar sacarosa) se indica mediante recuadros de texto. El iniciador ATG y el terminador TGA para la invertasa se indican en cursiva mayúscula y negrita, mientras que la región codificante se indica en cursiva minúscula. La 3' UTR de nitrato reductasa de Chlorella-vulgaris se indica con texto subrayado en minúsculas seguido de la región genómica S190 FATA1 indicada con texto en negrita y minúsculas.

g »ca u ca u ca u c tc »u a ta »ta tcu a ca ca c tc t»u a c »c tR u tcu tu tu a t»u a c t»ttR C C u cca ca c tt»c tu cc ttu a c c tu tu a atatccctücc<->cttttatcaaacaücctca»tutütttüatcttutütütac»cücttttucüauttuctauct<->cttütüctatttücüaata ccaccccca u ca tcccc ttccc tcu tttca ta tcu c ttu ca tcccaaccu caac tta tc ta cu c tu tcc tu c ta tccc tcau cu ctu c tcc t g c tc c tg c tc a c tg c c c c tc g c a c a g c c ttg g tt tg g g c tc c g c c tg ta ttc tc c tg g ta c tg c a a c c tg ta a a c c a g c a c tg c a a tg c tg a tg c a c g g g a a g ta g tg g g a tg g g a a c a c a a a tg g a g g a tc g ta g a g c t c a c ta g ta tc g a t t tc g a a g a c a g g g tg g t tg g c tg g atggggaaacgctggtcgcgggattcgatcctgctgcttatatcctccctggaagcacacccacgactctgaagaagaaaacg tgcacacacacaacccaaccggccgaatatttgcttccttatcccgggtccaagagagactgcgatgcccccctcaatcagcat cctectccctgccgcttcaatcttccctgcttgcctgcgcccgcggtgcgccgtctgcccgcccagtcagtcactcctgcacaggc cccttgtgcgcagtgctcctgtaccctttaccgctccttccattctgcgaggccccctattgaatgtattcgttgcctgtgtggcca agcgggctgctgggcgcgccgccgtcgggcagtgctcggcgactttggcggaagccgattgttcttctgtaagccacgcgcttg ctgctttgggaagagaagggggggggtactgaatggatgaggaggagaaggaggggtattggtattatctgagttgggtgaa g a g c (S E Q ID N O : 111 )
[0340] para introducir el gen de ACP-tioesterasa 2 (CwFatB2) de Cuphea wrightii (número de acceso: U56104) en S1920 en el locus FATA1, se generó una construcción para expresar la región codificante de la proteína del gen CwFatB2 bajo el control del promotor Amt03/5'UTR de Prototheca moriformis (SEQ ID N.° 84) y 3’UTR de nitrato reductasa de C. vulgaris (SEQ ID N.°85). La construcción que se ha expresado en S1920 se puede escribir como FATA1-CrbTub_ylnv_nr::amt03_CwTE2_nr-FATA1.
[0341] Los sitios de restricción relevantes en la construcción FATA1-CrbTub_ylnv_nr::amt03_CwTE2_nr-FATA1 se indican en minúsculas, en negrita y subrayados en la secuencia a continuación y son 5'-3' BspQ 1, Kpn I, Ase I, Mfe I, BamH I, EcoR I, Spe I, Ase I, Pac I, Sac I, BspQ I, respectivamente. Los sitios BspQI delimitan los extremos 5' y 3' del ADN transformante. Las secuencias en minúsculas y en negrita representan el ADN genómico de S1920 que permite la integración dirigida en el locus FATA1 a través de la recombinación homologa. Procediendo en la dirección 5' a 3', el promotor de (3-tubulina de C. Reinhardtii que impulsa la expresión del gen de sacarasa de levadura (que confiere la capacidad de S1920 para metabolizar sacarosa) se indica mediante recuadros de texto. El iniciador ATG y el terminador TGA para la invertasa se indican en cursiva mayúscula y negrita, mientras que la región codificante se indica en cursiva minúscula. La 3' UTR de nitrato reductasa de Chlorella vulgaris se indica mediante un texto subrayado en minúsculas seguido de un promotor amt03 endógeno de Prototheca moriformis, indicado por un recuadro de texto en cursiva. Los codones iniciador ATG y terminador TGA de ACP-tioesterasa de C. wrightii se indican con mayúsculas y negrita cursiva, mientras que el resto de la región codificante de ACP-tioesterasa se indica con negrita y cursiva. La 3' UTR de nitrato reductasa de C. vulgaris se indica de nuevo con texto subrayado en minúsculas seguido de la región genómica S1920 FATA1 indicada con texto en negrita y minúsculas. Las secuencias de ADNc con codones optimizados y las secuencias de aminoácidos de Tioesterasa FatB2 de Cuphea wrightii se incluyen en el Listado de secuencias como la SEQ ID N.° 107 y la SEQ ID N.° 105, respectivamente.
g c t c t t c g g a g t c a c t g t g c c a c tg a g t t c g a c t g g t a g c tg a a t g g a g tc g c t g c t c c a c t a a a c g a a t t g t c a g c a c c g c c a g c c g g c c g a g g a c c c g a g t c a ta g c g a g g g t a g t a g c g c g c c a t g g c a c c g a c c a g c c t g c t t g c c a g t a c t g g c g t c t c t t c c g c t t c t c t g t g g t c c t c t g c g c g c t c c a g c g c g t g c g c t t t t c c g g t g g a t c a t g c g g t c c g t g g c g c a c c g c a g c g g c c g c t g c c c a t g c a g c g c c g c t g c t t c c g a a c a g tg g c g g t c a g g g c c g c a c c c g c g g ta g c c g t c c g t c c g g a a c c c g c c c a a g a g t t t t g g g a g c a g c t t g a g c c c t g c a a g a t g g c g g a g g a c a a g c g c a t c t t c c t g g a g g a g c a c c g g t g c g t g g a g g t c c g g g g c t g a c c g g c c g t c g c a t t c a a c g t a a t c a a t c g c a t g a t g a t c a g a g g a c a c g a a g tc t t g g t g g c g g tg g c c a g a a a c a c t g t c c a t t g c a a g g g e a ta g g g a t g c g t t c c t t c a c c t c t c a t t t c t c a t t t c t g a a tc c c t c c c t g c t c a c t c t t t c t c c t c c t c c t t c eegtteaegeagcattcggggtaedctttcttgcgctatgacacttccagcaaaaggtagggcgggctgcgagacggcttcccggc gctgcatgcaacaccgatgatgcttcgaccccccgaagctccttcggggctgcatgggcgctccgatgccgctccagggcgagcgc tgtttaaatagccaggcccccgattgcaaagacattatagcgagctaccaaagccatattcaaacacctagatcactaccacttctacac aggccactcgagcttgtgatcgcactccgctaagggggcgcctcttcctcttcgtttcagtcacaacccgcaaacggcgcgcc/1 TG

ccaccaacaccacgttgacgaaggccaacagctacaacgtcgacctgtccaacagcaccggcaccctggagttcgagctggtg tacgccgtcaacaccacccagacgatctccaagtccgtgttcgcggacctctccctctggttcaagggcctggaggaccccgagg aglacctccgcatgggcltcgagglgtccgcgícctccuctlcctggaccgcgggaacagcaagglgaagtícgígaaggagaa cccctacttcaccaaccgcatgagcgtgaacaaccagccctícaagagcgagaacgacctgtcctactacaaggtgtacggctlg ctggaccagaacatcctggagctgtacttcaacgacggcgacgtcgtgtccaccaacacciacttcatgaccaccgggaacgcc ctgggctccgtgaacatgacgacgggggtggacaacctgttctacatcgacaagttccaggígcgcgaggtcaagTGAcsisiU ggcagcagcagctcggatagtatcgacacactctggacgctggtcgtgtgatggactgttgccgccacacttgctgccttgacctgtga atatccctgccgcttttatcaaacagcctcagtgtgtttgatcttgtgtgtacgcgcttttgcgagttgctagctgcttgtgctatttgcgaata ccacccccagcatccccttccctcgtttcatatcgcttgcatcccaaccgcaacttatctacgctgtcctgctatccctcagcgctgctcct gctcctgctcactgcccctcgcacagccttggtttgggctccgcctgtattctcctggtactgcaacctgtaaaccagcactgcaatgctg atgcacgggaagtagtgggatgggaacacaaatggaggatcccgcgtctcgaacagagcgcgcagaggaacgctgaaggtctcg cctctgtcgcacctcagcgcggcatacaccacaataaccacctgacgaatgcgcttggttcttcgtccattagcgaagcgtccggttca cacacgtgccacgttggcgaggtggcaggtgacaatgatcggtggagctgatggtcgaaacgttcacagcctagggatatcgaattc

cccccgccccacccccaagcccggcaagttcggcaactggcccagcagcctgagccagcccttcaagcccaagagcaacccc aacggccgcttccaggtgaaggccaacgtgugcccccacggssssssccccaaggccaacggcagcgccgtgagcctgaag tccggcagcctgaacaccctggaggacccccccagcagcccccccccccgcaccttcctgaaccagctgcccgactggagccg cctgcgcaccgccatcaccaccgtgttcgtggccgccgagaagcagttcacccgcctggaccgcaagugcaagcgccccgaca tgctggtggaetggttcggcagcgagaccatcgtgcaggaeggcctggtgttccgcgagcgcttcagcatccgcagctacgagat cggcgccgaccgcaccgccagcatcgagaccctgatgaaccacctgcaggacaccagcctgaaccactgcaagagcgtggg cctgctgaacgacggcttcggccgcacccccgagatgtgcacccgcgacctgatctgggtgctgaccaagatgcagatcgtggtg aaccgctaccccacctggggcgacaccgtggagatcaacagctggttcagccugagcggcaagatcggcatgggccgcgagt ggctgatcagcgactgcaacaccggcgugatcctggtgcgcgccaccagcgcctgggccatgatgaaccagaagacccgccg cttcagcaagctgccctgcgaggtgcgccaggagatcgccccccacttcgtggacgccccccccgtgatcgaggacaacgacc gcaagctgcacaagttcgacgtgaagaccggcgacagcatctgcaagggcctgacccccggctggaacgacttcgacgtgaac cagcacgtgagcaacgtgaagtacatcggctggattctggagagcatgcccaccgaggtgctggagacccaggagctgtgcag cctgaccctggagtaccgccgcgagtgcggccgcgagagcgtggtggagagcgtgaccagcatgaaccccagcaaggtgggc gaccgcagccagtaccagcacctgctgcgccíggaggacggcgccgacatcatgaagggccgcaccgagtggcgccccaag aacgccggcaccaaccgcgccatcagcaccTGA t t a a t t a a c tc g a a g c a g c a g c a g c tc g g a ta g ta tc g a c a c a c tc ta g a c g c tg g tc g tg tg a tg g a c tg ttg c c g c c a c a c t tg c tg c c ttg a c c tg tg a a ta tc c c tg c c g c t t t ta tc a a a c a g c c tc a g tg tg tt tg
a tc ttg tg tg ta c g c g c t t t tg c g a g t tg c ta g c tg c ttg tg c ta tt tg c g a a ta c c a c c c c c a g c a tc c c c ttc c c tc g t t tc a ta tc g c t tg catcccaaccgcaacttatctacgctgtcctgctatccctcagcgctgctcctgctcctgctcactgcccctcgcacagccttggtttggg c tc c g c c tg ta t tc tc c tg g ta c tg c a a c c tg ta a a c c a g c a c tg c a a tg c tg a tg c a c g g g a a g ta g tg g g a tg g g a a c a c a a a tg g
a a a g c t tg a g c tc ttg tt t tc c a g a a g g a g ttg c tc c t tg a g c c tt tc a t tc tc a g c c tc g a ta a c c tc c a a a g c c g c tc ta a t tg tg g a g g
g g g t t c g a a g a c a g g g t g g t t g g c t g g a t g g g g a a a c g c t g g t c g c g g g a t t c g a t c c t g c t g c t t a t a t c c t c c c t g g a a g c a
c a c c c a c g a c t c t g a a g a a g a a a a c g t g c a c a c a c a c a a c c c a a c c g g c c g a a ta t t t g c t t c c t ta t c c c g g g t c c a a g a g
a g a c t g c g a t g c c c c c c t c a a tc a g c a t c c t c c t c c c tg c c g c t t c a a tc t t c c c t g c t t g c c tg c g c c c g c g g t g c g c c g t c t g c
c c g c c c a g t c a g t c a c t c c t g c a c a g g c c c c t t g t g c g c a g t g c t c c t g ta c c c t t t a c c g c t c c t t c c a t t c t g c g a g g c c c c c t
a t t g a a t g t a t t c g t t g c c t g t g t g g c c a a g c g g g c t g c t g g g c g c g c c g c c g t c g g g c a g t g c t c g g c g a c t t t g g c g g a a g c
c g a t t g t t c t t c t g t a a g c c a c g c g c t lg c t g c t t t g g g a a g a g a a g g g g g g g g g t a c t g a a t g g a t g a g g a g g a g a a g g a g g g g t a t t g g t a t t a t c t g a g t t g g g t g a a g a g c (S E Q ID N O : 112)
[0342] Tras la transformación de FATA1-CrbTub_ylnv_nr-FATA1 en S1920, los transformantes primarios se purificaron clonalmente y se cultivaron en condiciones estándar de producción de lípidos a pH 5,0 similares a las condiciones descritas en el Ejemplo 1. Los perfiles de ácidos grasos se analizaron mediante cromatografía de gases / detección de ionización de llama de éster metílico de ácidos grasos estándar (FAME GC/FID). En la Tabla 29 siguiente se proporcionan los perfiles de ácidos grasos de varios transformantes.
Tabla 29. Perfiles de ácidos grasos de células de Prototheca moriformis que contienen un marcador seleccionable r in rr m ir n l l FATA1 n n

[0343] Estos resultados muestran que la ablación del alelo FATA1 endógeno del hospedador altera el perfil de lípidos de las microalgas modificadas. El impacto de dirigir un marcador seleccionable al alelo FATA1 endógeno es una
clara disminución de la producción de ácidos grasos C16:0 con un aumento en la producción de ácidos grasos C18:1.
[0344] Tras la transformación de FATA1-CrbTub_ylnv_nr::amt03_CwTE2_nr-FATA1 en S1920, los transformantes primarios se purificaron clonalmente y se cultivaron en condiciones estándar de producción de lípidos a pH 7,0 con diferentes fuentes de carbono hasta una concentración total de 40 g/l. La concentración de sacarosa fue de 40 g/l. Cuando solo se usó glucosa como fuente de carbono, se proporcionó glucosa a 40 g/l. Cuando se utilizó glucosa y fructosa como fuente de carbono, se proporcionó glucosa a 20 g/L y fructosa a 20 g/L. Los perfiles de ácidos grasos se evaluaron mediante GC-FID. Los perfiles de ácidos grasos resultantes se enumeran en la Tabla 30.
Tabla 30. Perfiles de ácidos grasos de células de Prototheca moriformis que contienen un marcador seleccionable y una tioesterasa exó ena ara interrum ir un alelo FATA1 endó eno.

[0345] Según el direccionamiento de solo un marcador seleccionable al alelo FATA1 del hospedador, la integración de un marcador seleccionable concomitante con una tioesterasa exógena altera el perfil de lípidos de las microalgas modificadas. Como antes, la orientación de un gen exógeno al alelo FATA1 da como resultado una clara disminución de la producción de ácidos grasos C16:0. La expresión adicional de la tioesterasa CwTE2 en el locus FATA1 también afecta a los ácidos grasos de cadena media y a la producción de ácidos grasos C18:1 en una medida que depende del nivel de actividad tioesterasa exógena presente en los transformantes analizados. Los genes flanqueados por unidades repetidas como la 3' UTR de nitrato reductasa de C. vulgaris en construcciones como FATA1-CrbTub_ylnv_nr::amt03_CwTE2_nr-FATA1 puede amplificarse tras la integración en el genoma hospedador. Existe una buena concordancia entre el número de copias del transgén amplificado en el sitio de integración diana y los niveles de tioesterasa, como lo revelan los impactos en los perfiles de ácidos grasos o la acumulación de proteínas recombinantes, según lo evaluado por transferencia Western.
[0346] Las líneas transgénicas en las que el gen CwTE2 ha sufrido amplificación muestran un marcado aumento en los ácidos grasos de cadena media (C10:0-C14:0) y una disminución simultánea en los ácidos grasos C18:1. Por el contrario, aquellos transformantes en los que CwTE2 ha sufrido poca o ninguna amplificación (probablemente 1-2 copias) concuerdan con una menor expresión de la tioesterasa exógena, lo que resulta en un ligero aumento en los ácidos grasos de cadena media y un impacto mucho mayor en el aumento de los ácidos grasos C18:1.
[0347] En conjunto, estos datos demuestran que la ablación del alelo FATA1 endógeno del hospedador altera el perfil de lípidos de las microalgas modificadas.
B. Alteración de los perfiles de lipidos por eliminación de un gen de tioesterasa endógeno de Prototheca moriformis
[0348] Una construcción, pSZ1773, para regular a la baja la expresión del gen FATA1 de Prototheca moriformis por un ARN de horquilla se introdujo en un contexto genético de Prototheca moriformis UTEX 1435 S1920. El gen de sacarasa suc2 de Saccharomyces cerevisiae se utilizó como marcador seleccionable, confiriendo la capacidad de crecer en la sacarosa como única fuente de carbono. La parte de la construcción que codifica el ARN horquillado utilizó el primer exón de la región codificante de FatAl, seguido del intrón endógeno y una unidad repetida del primer exón en la orientación inversa. Se incluyeron secuencias de direccionamiento de recombinación homóloga 5' y 3' (que flanquean la construcción) a la región genómica 6S, enumeradas como la SEQ ID N.0 100 y 101 respectivamente, para la integración de la construcción de horquilla en el genoma nuclear. Esta construcción se denomina SS::P-Tub:suc2:nr::P-tub:hairpinFatA:nr::SS.
[0349] Los sitios de restricción relevantes en SS::P-Tub:suc2:nr::P-tub:hairpin FatA:nr::SS se indican en minúsculas, en negrita y subrayados en la secuencia siguiente y son 5'-3' BspQ 1, Kpn I, Mfe I, BamH I, EcoR I, Spe I, Xho I, Sac I, BspQ I, respectivamente. Los sitios BspQI delimitan los extremos 5' y 3' del ADN transformante. Las secuencias en minúsculas y en negrita representan el ADN genómico de S1920 que permite la integración dirigida en el locus 6s a través de la recombinación homóloga. Procediendo en la dirección 5' a 3', el promotor de P-tubulina de . Reinhardtii que impulsa la expresión del gen de sacarasa de levadura (que confiere la capacidad de S1920 para metabolizar sacarosa) se indica mediante recuadros de texto. El iniciador ATG y el terminador TGA para la invertasa se indican en cursiva mayúscula y negrita, mientras que la región codificante se indica en cursiva minúscula. La 3' UTR de nitrato reductasa de Chlorella vulgaris se indica mediante un texto subrayado en minúsculas seguida del segundo promotor de P-tubulina de . Reinhardtii que impulsa la expresión de la horquilla FatAl, indicada por texto en cursiva en recuadros. El codón ATG iniciador de FatAl se indica en mayúsculas, negrita y cursiva, mientras que el resto del primer exón de la región codificante de FatAl se indica en mayúsculas. El intrón del gen FatA se indica en mayúsculas subrayadas, y se creó una región conectora que se muestra en mayúsculas subrayadas, cursiva y negrita en el intrón FatAl/primer exón invertido para ayudar en el corte y empalme del ARN en estos vectores. El primer exón invertido de FatAl se indica en mayúsculas. La 3' UTR de nitrato reductasa de . vulgaris se indica de nuevo con texto subrayado en minúsculas seguida de la región genómica S19206S indicada con texto en negrita y minúsculas. Las secuencias de las porciones FATA de esta construcción de ARNi se enumeran como la SEQ ID N.0110.
aacatcagcaacgccggcccctggagccggttcgccaccaacaccacgttgacgaaggccaacagctacaacgtcgacctgtc caacagcaccggcaccctggagttcgagctggtgtacgccgtcaacaccacccagacgatctccaagtccgtgttcgcggacctc tccctclgg/lcaagggcctggüggaccccgüggagtacctccgcatgggc/tcgaggtgtccgcgtcctccttcltcctggaccgc gggaacagcaaggtgaagttcgtgaaggagaacccctacttcaccaaccgcatgagcgtgaacciaccagcccttcaagagcg agaacgcicctgtcctactacaaggtgtacggcttgctggaccagaacatcctggagctgtacttcaacgacggcgacgtcgtgtcc accaacacctacttcatgaccaccgggaacgccctgggctccgtgaacatgacgacgggggtggacaacctgttctacatcgac rt¿/g//c¿flgg/gig6g¿/gg/i(/m., 7Y¿4caattggcagcagcagctcggatagtatcgacacactctggacgctggtcgtgtgat ggactgttgccgccacacttgctgccttgacctgtgaatatccctgccgcttttatcaaacagcctcagtgtgtttgatcttgtgtgtacgcg cttttgcgagttgctagctgcttgtgctatttgcgaataccacccccagcatccccttccctcgtttcatatcgcttgcatcccaaccgcaac ttatctacgctgtcctgctatccctcagcgctgctcctgctcctgctcactgcccctcgcacagccttggtttgggctccgcctgtattctcc tggtactgcaacctgtaaaccagcactgcaatgctgatgcacgggaagtagtgggatgggaacacaaatggaggatcccgcgtctcg aacagagcgcgcagaggaacgctgaaggtctcgcctctgtcgcacctcagcgcggcatacaccacaataaccacctgacgaatgcg

G T G G T C C T C T G C G C G C T C C A G C G C G T G C G C T T T T C C G G T G G A T C A T G C G G T C C G T GGCGCACCGCAGCGGCCGCTGCCCATGCAGCGCCGCTGCTTCCGAACAGTGGCG G T C A G G G C C G C A C C C G C G G T A G C C G T C C G T C C G G A A C C C G C C C A A G A G T T T T G G G A G C A G C T T G A G C C C T G C A A G A T G G C G G A G G A C A A G C G C A T C T T C C T G G A G G A G C A C C G G T G C G T G G A G G T C C G G G G C T G A C C G G C C G T C G C A T T C A A C G T A A T C A A T C G C A T G A T G A T C A G A G G A C A C G A A G T C T T G G T G G C G G T G G C C A G A A A C A C T G T C C A T T G C A A G G G C A T A G G G A T G C G T T C C T T C A C C T C T C A T T T C T C A T T T C T G A A T C C C T C C C T G C T C A C T C T T T C T C C T C C T C C T T C C C G T T C A C G C A G O i7 T C Y y Y fY *T ;C A 4 C Y ; ,4 (> (/T (/ (> (/C C C G T G C T C C T C C A G G A A G A T G C G C T T G T C C T C C G C C A T C T T G C A G G G C T C A A G C T G C T C C C A A A A C T C T T G G G C G G G T T C C G G A C G G A C G G C T A C C G C G G G T G C G G C C C T G A C C G C C A C T G T T C G G A A G C A G C G G C G C T G C A T G G G C A G C G G C C G C T G C G G T G C G C C A C G G A C C G C A T G A T C C A C C G G A A A A G C G C A C G C G C T G G A G C G C G C A G A G G A C C A C A G A G A A G C G G A A G A G A C G C C A G T A C T G G C A A G C A G G C T G G T C G G T G C C A T a tcga tagatc tcttaaggcagcagcagctcggatagtatcgacacactctggacgctggtcgtgtgatggact gttgccgccacacttgctgccttgacctgtgaatatccctgccgcttttatcaaacagcctcagtgtgtttgatcttgtgtgtacgcgcttttg
Claims (13)
1. Microbio de Chlorella o Prototheca modificado genéticamente para eliminar o regular a la baja un gen endógeno de acil-ACP tioesterasa.
2. Microbio de la reivindicación 1, en donde el microbio está diseñado para alterar una acil-ACP tioesterasa endógena.
3. Microbio de la reivindicación 1 o la reivindicación 2, en donde la acil-ACP tioesterasa endógena se elimina por recombinación homóloga.
4. Microbio de la reivindicación 1 o la reivindicación 2, en donde el microbio está diseñado para expresar un ARN inhibidor que actúa sobre una acil-ACP tioesterasa endógena.
5. Microbio de una cualquiera de las reivindicaciones 1 a 4, en donde el microbio comprende además uno o más genes exógenos que son expresados por la célula.
6. Microbio de la reivindicación 5, en donde el gen exógeno codifica una sacarasa, una acil-ACP tioesterasa o una desaturasa.
7. Microbio de la reivindicación 6, en donde el/los gen(es) exógeno(s) codifica(n): una sacarasa; una acil-ACP tioesterasa; o una sacarasa y una acil-ACP tioesterasa.
8. Microbio de cualquiera de las reivindicaciones 1 a 7, en donde el microbio es Prototheca moríformis o Chlorella protothecoides.
9. Microbio de la reivindicación 8, en donde el microbio es Prototheca moriformis.
10. Método para producir un aceite microbiano, donde el método comprende:
a. cultivar un microbio según cualquiera de las reivindicaciones 1 a 9; y
b. separar el aceite del microbio.
11. Método de la reivindicación 10, que además incluye el paso de:
c. someter el aceite a refinado, blanqueo, desodorización o desgomado para producir aceite microbiano RBD.
12. Método de la reivindicación 10 o la reivindicación 11, en donde el método comprende adicionalmente añadir un antioxidante, un desactivador de iones metálicos, un inhibidor de la corrosión, un desemulsionante, un aditivo antidesgaste, un depresor del punto de fluidez o un compuesto antihidrólisis al aceite microbiano.
13. Aceite producido mediante el método de cualquiera de las reivindicaciones 10 a 12, en donde el aceite comprende menos del 7,5 % de C18:2, y en donde:
i) el número de copias del gen exógeno es superior a 2;
ii) el gen exógeno es la tioesterasa FatB2 de Cuphea wrightii; y
iii) el microbio es Prototheca moriformis.
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US40990210P | 2010-11-03 | 2010-11-03 | |
| US201161438966P | 2011-02-02 | 2011-02-02 | |
| US201161522231P | 2011-08-10 | 2011-08-10 | |
| US201161546932P | 2011-10-13 | 2011-10-13 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2909143T3 true ES2909143T3 (es) | 2022-05-05 |
Family
ID=45003066
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES11785851T Active ES2746074T3 (es) | 2010-11-03 | 2011-11-03 | Aceites microbianos con puntos de fluidez reducidos, fluidos dieléctricos producidos a partir de estos y métodos relacionados |
| ES18212337T Active ES2909143T3 (es) | 2010-11-03 | 2011-11-03 | Microbios de Chlorella o Prototheca modificados genéticamente y aceite producido a partir de estos |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES11785851T Active ES2746074T3 (es) | 2010-11-03 | 2011-11-03 | Aceites microbianos con puntos de fluidez reducidos, fluidos dieléctricos producidos a partir de estos y métodos relacionados |
Country Status (13)
| Country | Link |
|---|---|
| US (4) | US9066527B2 (es) |
| EP (2) | EP2635663B1 (es) |
| JP (3) | JP5970463B2 (es) |
| KR (2) | KR102065526B1 (es) |
| CN (2) | CN108823254A (es) |
| AU (2) | AU2011323288B2 (es) |
| BR (1) | BR112013011039A8 (es) |
| CA (2) | CA3024641A1 (es) |
| ES (2) | ES2746074T3 (es) |
| MX (1) | MX354145B (es) |
| SG (2) | SG10201509035WA (es) |
| WO (1) | WO2012061647A2 (es) |
| ZA (1) | ZA201303271B (es) |
Families Citing this family (83)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2008151149A2 (en) | 2007-06-01 | 2008-12-11 | Solazyme, Inc. | Production of oil in microorganisms |
| US20100303989A1 (en) | 2008-10-14 | 2010-12-02 | Solazyme, Inc. | Microalgal Flour |
| ES2583639T3 (es) | 2008-11-28 | 2016-09-21 | Terravia Holdings, Inc. | Producción de aceites específicos en microorganismos heterótrofos |
| US8475660B2 (en) | 2010-04-06 | 2013-07-02 | Heliae Development, Llc | Extraction of polar lipids by a two solvent method |
| US8308951B1 (en) | 2010-04-06 | 2012-11-13 | Heliae Development, Llc | Extraction of proteins by a two solvent method |
| US8115022B2 (en) | 2010-04-06 | 2012-02-14 | Heliae Development, Llc | Methods of producing biofuels, chlorophylls and carotenoids |
| US8273248B1 (en) * | 2010-04-06 | 2012-09-25 | Heliae Development, Llc | Extraction of neutral lipids by a two solvent method |
| US8313648B2 (en) * | 2010-04-06 | 2012-11-20 | Heliae Development, Llc | Methods of and systems for producing biofuels from algal oil |
| EP2556138B1 (en) | 2010-04-06 | 2014-06-11 | Heliae Development LLC | Methods of and systems for producing biofuels |
| JP5996527B2 (ja) | 2010-05-28 | 2016-09-21 | テラヴィア ホールディングス, インコーポレイテッド | 用途に応じた油を含む食品成分 |
| EP2576801B1 (en) | 2010-06-01 | 2019-10-02 | DSM IP Assets B.V. | Extraction of lipid from cells and products therefrom |
| SG10201509035WA (en) | 2010-11-03 | 2015-12-30 | Solazyme Inc | Microbial Oils With Lowered Pour Points, Dielectric Fluids Produced Therefrom, And Related Methods |
| JP6071904B2 (ja) | 2011-02-02 | 2017-02-01 | テラヴィア ホールディングス, インコーポレイテッド | 組み換え油産生微生物から生成される用途に応じた油 |
| IT1403878B1 (it) * | 2011-02-14 | 2013-11-08 | A & A Flii Parodi Srl | Fluido dielettrico vegetale per trasformatori elettrici |
| BR112013028621A2 (pt) | 2011-05-06 | 2016-11-29 | Solazyme Inc | "microalgas oleaginosas recombinantes, método para produzir biomassa de microalga ou um produto produzido da biomassa, e método para produzir uma composição lipídica de microalga". |
| US8236194B1 (en) * | 2011-06-17 | 2012-08-07 | Lubrigreen Biosynthetics, Llc | Refrigerating fluid compositions comprising estolide compounds |
| EP2702125B1 (en) * | 2011-06-17 | 2019-06-19 | Biosynthetic Technologies, LLC | Dielectric fluids comprising estolide compounds and methods of making and using the same |
| EP2702126A1 (en) * | 2011-06-17 | 2014-03-05 | Biosynthetic Technologies, LLC | Estolide compositions exhibiting high oxidative stability |
| WO2013003268A1 (en) * | 2011-06-27 | 2013-01-03 | Dow Global Technologies Llc | Genetically-engineered microbial oil dielectric fluid |
| ES2694823T3 (es) * | 2011-06-30 | 2018-12-27 | Dow Global Technologies Llc | Compuestos de bifenol-éter como marcadores para hidrocarburos líquidos y otros combustibles y aceites |
| WO2013075116A2 (en) | 2011-11-17 | 2013-05-23 | Heliae Development, Llc | Omega 7 rich compositions and methods of isolating omega 7 fatty acids |
| US20140047762A1 (en) * | 2012-07-09 | 2014-02-20 | Real Technologies Llc | Fuel additive and method for use |
| US9719114B2 (en) | 2012-04-18 | 2017-08-01 | Terravia Holdings, Inc. | Tailored oils |
| SG11201406711TA (en) | 2012-04-18 | 2014-11-27 | Solazyme Inc | Tailored oils |
| EP2867355A1 (en) * | 2012-06-29 | 2015-05-06 | BP Biofuels UK Ltd. | Process for separation of renewable materials from microorganisms |
| WO2014041553A1 (en) * | 2012-09-12 | 2014-03-20 | Savita Oil Technologies Limited | Mustard oil based insulating fluid composition and process for preparation thereof |
| WO2014100649A1 (en) | 2012-12-20 | 2014-06-26 | Cargill, Incorporated | Enzymatically-degummed oil and uses thereof |
| JP6491881B2 (ja) * | 2012-12-27 | 2019-03-27 | 花王株式会社 | アシル−acpチオエステラーゼ |
| US10098371B2 (en) | 2013-01-28 | 2018-10-16 | Solazyme Roquette Nutritionals, LLC | Microalgal flour |
| US9567615B2 (en) | 2013-01-29 | 2017-02-14 | Terravia Holdings, Inc. | Variant thioesterases and methods of use |
| US9816079B2 (en) | 2013-01-29 | 2017-11-14 | Terravia Holdings, Inc. | Variant thioesterases and methods of use |
| US9783836B2 (en) | 2013-03-15 | 2017-10-10 | Terravia Holdings, Inc. | Thioesterases and cells for production of tailored oils |
| US9290749B2 (en) | 2013-03-15 | 2016-03-22 | Solazyme, Inc. | Thioesterases and cells for production of tailored oils |
| US9249252B2 (en) | 2013-04-26 | 2016-02-02 | Solazyme, Inc. | Low polyunsaturated fatty acid oils and uses thereof |
| WO2015005139A1 (ja) | 2013-07-12 | 2015-01-15 | 花王株式会社 | アシル-acpチオエステラーゼ |
| FR3009619B1 (fr) | 2013-08-07 | 2017-12-29 | Roquette Freres | Compositions de biomasse de microalgues riches en proteines de qualite sensorielle optimisee |
| WO2015051319A2 (en) | 2013-10-04 | 2015-04-09 | Solazyme, Inc. | Tailored oils |
| US9458407B2 (en) * | 2013-12-10 | 2016-10-04 | T2e Energy Holdings, LLC | Algal oil based bio-lubricants |
| US9499846B2 (en) | 2013-12-10 | 2016-11-22 | Mark Randall | Method for recycling flue gas |
| KR102435269B1 (ko) | 2013-12-20 | 2022-08-22 | 디에스엠 아이피 어셋츠 비.브이. | 미생물 세포로부터의 미생물 오일의 수득 방법 |
| US11124736B2 (en) | 2013-12-20 | 2021-09-21 | Dsm Ip Assets B.V. | Processes for obtaining microbial oil from microbial cells |
| AR098896A1 (es) | 2013-12-20 | 2016-06-22 | Dsm Ip Assets Bv | Proceso para obtener aceite microbiano a partir de células microbianas |
| AR098890A1 (es) | 2013-12-20 | 2016-06-22 | Dsm Ip Assets Bv | Proceso para obtener aceite microbiano a partir de células microbianas |
| NZ721409A (en) | 2013-12-20 | 2022-10-28 | Dsm Ip Assets Bv | Processes for obtaining microbial oil from microbial cells |
| BR112016014519B1 (pt) * | 2013-12-20 | 2022-07-12 | Dsm Ip Assets B.V | Método de extração de lipídeos apropriados para uso na produção de biocombustíveis |
| EP3120368A4 (en) * | 2014-03-17 | 2017-11-22 | Novvi LLC | Dielectric fluid and coolant made with biobased base oil |
| US9394550B2 (en) | 2014-03-28 | 2016-07-19 | Terravia Holdings, Inc. | Lauric ester compositions |
| CN104130952B (zh) * | 2014-04-03 | 2016-06-08 | 河南师范大学 | 一株胶红酵母及其在发酵生产类胡萝卜素和油脂中的应用 |
| AU2015277857B2 (en) | 2014-06-20 | 2021-07-22 | Kao Corporation | Method for producing lipid |
| CN106574255A (zh) | 2014-07-10 | 2017-04-19 | 泰拉瑞亚控股公司 | 酮脂酰acp合酶基因及其用途 |
| EP3172320B1 (en) | 2014-07-24 | 2019-11-20 | Corbion Biotech, Inc. | Variant thioesterases and methods of use |
| BR112017005370A2 (pt) | 2014-09-18 | 2017-12-12 | Terravia Holdings Inc | acil-acp tioesterases e mutantes das mesmas |
| WO2016130863A2 (en) * | 2015-02-11 | 2016-08-18 | T2e Energy Holdings, LLC | Algal oil based bio-lubricants |
| DK3274430T3 (da) | 2015-03-24 | 2022-10-03 | Corbion Biotech Inc | Mikroalgesammensætninger og deres anvendelser |
| BR112017020906A2 (pt) | 2015-03-31 | 2018-07-10 | Terravia Holdings Inc | microalgas adaptadas para condições de cultura heterotróficas |
| CN107960101A (zh) | 2015-04-06 | 2018-04-24 | 柯碧恩生物技术公司 | 具有lpaat消融的产油微藻 |
| ES2948463T3 (es) * | 2015-05-19 | 2023-09-12 | Quaker Chem Corp | Esteres sintéticos obtenidos a partir de ácido oleico de alta estabilidad |
| US10256007B2 (en) * | 2015-08-14 | 2019-04-09 | Dr. Ambedkar Institute of Technology | Process to extract liquid dielectric coolant from the sesame oil |
| US10032694B2 (en) | 2016-03-08 | 2018-07-24 | Toyota Motor Engineering & Manufacturing North America, Inc | Power electronics assemblies having a semiconductor cooling chip and an integrated fluid channel system |
| CN107346866A (zh) * | 2016-05-04 | 2017-11-14 | 熊启兵 | 一种配电柜冷却系统 |
| CN109415654A (zh) * | 2016-07-20 | 2019-03-01 | 玛拉可再生能源公司 | 用于对油进行冬化的两步分馏方法 |
| US10121729B2 (en) * | 2016-07-25 | 2018-11-06 | Toyota Motor Engineering & Manufacturing North America, Inc. | Power electronics assemblies having a semiconductor device with metallized embedded cooling channels |
| US20180142218A1 (en) | 2016-10-05 | 2018-05-24 | Terravia Holdings, Inc. | Novel acyltransferases, variant thioesterases, and uses thereof |
| EP3538628B1 (en) | 2016-11-09 | 2022-05-18 | Novvi LLC | Synthetic oligomer compositions and methods of manufacture |
| US10844276B2 (en) * | 2017-03-03 | 2020-11-24 | Locus Oil Ip Company, Llc | Composition and methods for microbial enhanced digestion of polymers in fracking wells |
| WO2018204859A1 (en) | 2017-05-05 | 2018-11-08 | Purissima, Inc. | Neurotransmitters and methods of making the same |
| US11473028B2 (en) | 2017-07-14 | 2022-10-18 | Novvi Llc | Base oils and methods of making the same |
| EP3652281A4 (en) | 2017-07-14 | 2021-04-07 | Novvi LLC | BASE OILS AND MANUFACTURING PROCESSES FOR THEM |
| JP6647593B2 (ja) * | 2017-07-19 | 2020-02-14 | バイオックス化学工業株式会社 | 緑藻を用いた有用脂質製造法 |
| US11814598B2 (en) | 2018-03-21 | 2023-11-14 | Cargill, Incorporated | Synthetic ester and mineral oil dielectric fluids with increased stability |
| CN108849854B (zh) * | 2018-07-13 | 2021-02-09 | 深圳市润科生物科技有限公司 | 一种外周血单个核细胞冻存方法 |
| EP3844293A4 (en) | 2018-08-30 | 2022-06-08 | Checkerspot, Inc. | HYDROFORMYLATED TRIGLYCERIDES AND THEIR USES |
| FR3085962B1 (fr) * | 2018-09-14 | 2021-06-18 | Fermentalg | Procede d'extracton d'une huile riche en pufa |
| BR112021015352A2 (pt) | 2019-02-11 | 2021-10-05 | Checkerspot, Inc. | Composições de óleo de triglicerídeo |
| EP4077439A4 (en) | 2019-12-18 | 2023-12-13 | Checkerspot, Inc. | USES OF MICROBE-DERIVED MATERIALS IN POLYMER APPLICATIONS |
| WO2021163194A1 (en) * | 2020-02-10 | 2021-08-19 | C16 Biosciences, Inc. | Microbially produced palm oil substitutes |
| EP3882928B1 (en) * | 2020-03-17 | 2023-11-15 | Hitachi Energy Ltd | Mfc/nc in transformer boards used in power transformers |
| WO2022104046A1 (en) * | 2020-11-12 | 2022-05-19 | C16 Biosciences, Inc. | Edible microbial oil |
| EP4208528A4 (en) | 2021-09-17 | 2023-10-11 | Checkerspot, Inc. | HIGH OLEIC OIL COMPOSITIONS AND THEIR USES |
| CN113982691B (zh) * | 2021-11-11 | 2023-12-15 | 安徽理工大学 | 一种煤矿井下有害气体检测预警系统 |
| WO2023091669A1 (en) | 2021-11-19 | 2023-05-25 | Checkerspot, Inc. | Recycled polyurethane formulations |
| WO2023102069A1 (en) | 2021-12-01 | 2023-06-08 | Checkerspot, Inc. | Polyols, polyurethane dispersions, and uses thereof |
| NL2032949B1 (en) * | 2022-09-05 | 2024-03-21 | Nopalm Ingredients B V | Providing an oil composition through fermentation of biomass with a yeast |
Family Cites Families (311)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US2235056A (en) | 1938-03-04 | 1941-03-18 | Ici Ltd | Process for the recovery of glycerol from still residues from fermentation processes |
| US2383602A (en) | 1943-03-04 | 1945-08-28 | Colgate Palmolive Peet Co | Process for treatment of fatty glycerides |
| US2967700A (en) | 1955-03-01 | 1961-01-10 | Morris B Kallison | Whipping and aerating apparatus |
| US2874171A (en) | 1957-02-20 | 1959-02-17 | Upjohn Co | Recovery of ergosterol |
| US2825651A (en) | 1957-07-01 | 1958-03-04 | Carnation Co | In-package oxygen remover |
| US3142135A (en) | 1962-02-13 | 1964-07-28 | Grain Processing Corp | Production of carotenoids by the cultivation of algae |
| US3280502A (en) | 1962-11-07 | 1966-10-25 | Hoffmann La Roche | Process for the preparation of lutein |
| US3320693A (en) | 1964-09-11 | 1967-05-23 | Kk | Method of industral cultivation of unicellular green algae such as chlorella |
| US3475274A (en) | 1967-08-07 | 1969-10-28 | Commercial Solvents Corp | Production of riboflavin |
| US3962466A (en) | 1972-11-10 | 1976-06-08 | Dai-Nippon Sugar Manufacturing Co., Ltd. | Method for treatment of microorganisms |
| JPS5328989B2 (es) | 1974-05-27 | 1978-08-17 | ||
| US4005062A (en) | 1974-08-16 | 1977-01-25 | Standard Oil Company (Indiana) | Process of preparing water-soluble whippable extract from microorganism protein material |
| US3957578A (en) | 1975-01-03 | 1976-05-18 | Hokkaido Sugar Co., Ltd. | Method for manufacture of α-galactosidase by microorganism |
| US4103039A (en) | 1976-08-18 | 1978-07-25 | Fuji Oil Company, Limited | Method for producing improved shea fat |
| FR2375319A1 (fr) | 1976-12-23 | 1978-07-21 | British Petroleum Co | Procede de traitement d'extraits lipidiques |
| US4182777A (en) | 1977-03-01 | 1980-01-08 | Standard Oil Company (Indiana) | Co-dried yeast whey food product and process |
| US4384972A (en) | 1977-06-21 | 1983-05-24 | Toppan Printing Co., Ltd. | Foodstuff freshness keeping agents |
| IL57712A (en) | 1979-07-03 | 1984-02-29 | Yissum Res Dev Co | Cultivation of halophilic algae of the dunaliella species for the production of fuel-like product |
| US4273790A (en) | 1979-11-19 | 1981-06-16 | Standard Brands Incorporated | Low-fat liquid spread and process |
| US4373434A (en) | 1980-11-24 | 1983-02-15 | Simon-Rosedowns Limited | Apparatus for the expansion of oil bearing seeds |
| JPS57150379A (en) | 1981-03-14 | 1982-09-17 | Kikujiro Ishiwatari | Crushing of cell membrane of chlorella |
| US4390561A (en) | 1981-11-04 | 1983-06-28 | The Procter & Gamble Company | Margarine oil product |
| JPS606799A (ja) | 1983-06-07 | 1985-01-14 | 北畠 三之丞 | 汚れ落しに効果を発揮する米糠を原料とする洗剤の製法 |
| JPS6071904A (ja) | 1983-09-28 | 1985-04-23 | Matsushita Electric Ind Co Ltd | 光ディスク評価装置 |
| US4519845A (en) | 1984-02-09 | 1985-05-28 | Uop Inc. | Separation of sucrose from molasses |
| US4945050A (en) | 1984-11-13 | 1990-07-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues and apparatus therefor |
| US4627192B1 (en) * | 1984-11-16 | 1995-10-17 | Sigco Res Inc | Sunflower products and methods for their production |
| US4755467A (en) | 1985-06-03 | 1988-07-05 | Unisearch Limited | Method for the production of sorbitol and gluconate |
| US5001059A (en) | 1985-07-01 | 1991-03-19 | Bio-Technical Resources, Inc. | L-ascorbic acid production in microorganisms |
| US5900370A (en) | 1985-07-01 | 1999-05-04 | Bio-Technical Resources | Process for the production of ascorbic acid with prototheca |
| US4673490A (en) | 1985-08-23 | 1987-06-16 | Fluor Corporation | Process for separating crude oil components |
| JPH0775557B2 (ja) | 1986-04-25 | 1995-08-16 | 富士レビオ株式会社 | 脂質結合性シアル酸の測定法 |
| US5091116A (en) | 1986-11-26 | 1992-02-25 | Kraft General Foods, Inc. | Methods for treatment of edible oils |
| US5360730A (en) | 1987-06-05 | 1994-11-01 | Universal Foods Corporation | Zeaxanthin producing strains of Neospongiococcum Excentricum |
| FR2626584B1 (fr) | 1988-01-28 | 1990-07-13 | Agronomique Inst Nat Rech | Sequence ars efficace chez yarrowia lipolytica et procede pour sa preparation |
| US4992605A (en) | 1988-02-16 | 1991-02-12 | Craig Wayne K | Production of hydrocarbons with a relatively high cetane rating |
| US4901635A (en) | 1988-04-08 | 1990-02-20 | Anderson International Corp. | Apparatus and method for the continuous extrusion and partial deliquefaction of oleaginous materials |
| JPH078217Y2 (ja) | 1988-07-07 | 1995-03-01 | 日立電線株式会社 | 発光ダイオードヘッド |
| US20060094089A1 (en) | 1988-09-07 | 2006-05-04 | Martek Biosciences Corporation | Process for the heterotrophic production of microbial products with high concentrations of omega-3 highly unsaturated fatty acids |
| US5340742A (en) | 1988-09-07 | 1994-08-23 | Omegatech Inc. | Process for growing thraustochytrium and schizochytrium using non-chloride salts to produce a microfloral biomass having omega-3-highly unsaturated fatty acids |
| US5130242A (en) | 1988-09-07 | 1992-07-14 | Phycotech, Inc. | Process for the heterotrophic production of microbial products with high concentrations of omega-3 highly unsaturated fatty acids |
| US6680426B2 (en) | 1991-01-07 | 2004-01-20 | Auburn University | Genetic engineering of plant chloroplasts |
| US5693507A (en) | 1988-09-26 | 1997-12-02 | Auburn University | Genetic engineering of plant chloroplasts |
| WO1990013612A1 (en) | 1989-05-10 | 1990-11-15 | Davy Mckee (London) Limited | Multi-step hydrodesulphurisation process |
| DE4004800C2 (de) | 1990-02-13 | 2000-12-21 | Aventis Cropscience Gmbh | Im Habitus und Ertrag veränderte transgene Pflanzen |
| US5407957A (en) | 1990-02-13 | 1995-04-18 | Martek Corporation | Production of docosahexaenoic acid by dinoflagellates |
| US7053267B2 (en) | 1990-03-16 | 2006-05-30 | Calgene Llc | Plant seed oils |
| US5298421A (en) | 1990-04-26 | 1994-03-29 | Calgene, Inc. | Plant medium-chain-preferring acyl-ACP thioesterases and related methods |
| US5164308A (en) | 1990-05-21 | 1992-11-17 | Martek Corporation | Preparation of labelled triglyceride oils by cultivation of microorganisms |
| US6483008B1 (en) | 1990-08-15 | 2002-11-19 | Calgene Llc | Methods for producing plants with elevated oleic acid content |
| US5945585A (en) | 1990-12-20 | 1999-08-31 | E. I. Du Pont De Nemours And Company | Specific for palmitoyl, stearoyl and oleoyl-alp thioesters nucleic acid fragments encoding acyl-acp thiosesterase enzymes and the use of these fragments in altering plant oil composition |
| EP0563191B2 (en) | 1990-12-20 | 2000-01-19 | E.I. Du Pont De Nemours And Company | Nucleotide sequences of soybean acyl-acp thioesterase genes |
| CU22292A1 (es) | 1991-05-07 | 1995-01-31 | Cigb | Procedimiento para la obtencion a escala industrial de licores de fructosa-glucosa a partir de sacarosa e instalacion para el mismo |
| US5455167A (en) | 1991-05-21 | 1995-10-03 | Calgene Inc. | Medium-chain thioesterases in plants |
| US5270175A (en) | 1991-07-12 | 1993-12-14 | Dna Plant Technology Corporation | Methods and compositions for producing metabolic products for algae |
| JP3143636B2 (ja) | 1991-09-11 | 2001-03-07 | 株式会社サン・クロレラ | 細胞破裂によるクロレラ細胞壁の破砕方法 |
| DE4130986A1 (de) | 1991-09-18 | 1993-03-25 | Bayer Ag | Pinosylvinsynthase-gene |
| US6355861B1 (en) | 1991-10-10 | 2002-03-12 | Rhone-Poulenc Agrochimie | Production of gamma linolenic acid by a Δ6-desaturase |
| PH31293A (en) | 1991-10-10 | 1998-07-06 | Rhone Poulenc Agrochimie | Production of y-linolenic acid by a delta6-desaturage. |
| FR2686619B1 (fr) | 1992-01-28 | 1995-07-13 | Commissariat Energie Atomique | Procede de production selective de lipides poly-insatures a partir d'une culture de micro-algues du type porphyridium et cuve utilisee dans ce procede. |
| US5395455A (en) | 1992-03-10 | 1995-03-07 | Energy, Mines And Resources - Canada | Process for the production of anhydrosugars from lignin and cellulose containing biomass by pyrolysis |
| DE4209779C1 (es) | 1992-03-26 | 1993-07-15 | Oelmuehle Leer Connemann Gmbh & Co., 2950 Leer, De | |
| WO1994000009A1 (en) | 1992-06-19 | 1994-01-06 | Nonomura Arthur M | Methods and compositions for enhancing carbon fixation in plants |
| US6410281B1 (en) | 1992-07-10 | 2002-06-25 | Omegatech, Inc. | Reducing corrosion in a fermentor by providing sodium with a non-chloride sodium salt |
| JPH078217B2 (ja) | 1992-08-11 | 1995-02-01 | 昭 遠藤 | 固形味付酢もずくの製造方法 |
| JP3090810B2 (ja) | 1993-03-09 | 2000-09-25 | 日本碍子株式会社 | パルミトオレイン酸の製造方法 |
| GB2277052A (en) | 1993-04-14 | 1994-10-19 | Du Pont Canada | Polyurethane foam laminates |
| JPH078215A (ja) | 1993-04-30 | 1995-01-13 | Kawasaki Steel Corp | ドコサヘキサエン酸含有海洋性微細藻類食品素材およびその製造方法 |
| JPH078217A (ja) | 1993-06-29 | 1995-01-13 | Kawasaki Steel Corp | ドコサヘキサエン酸含有健康食品およびその製造方法 |
| JP3506740B2 (ja) | 1993-09-09 | 2004-03-15 | 日清オイリオ株式会社 | ドコサヘキサエン酸含有藻類の培養方法 |
| EP0728212A1 (en) | 1993-11-10 | 1996-08-28 | Calgene, Inc. | Plant acyl acp thioesterase sequences |
| US5563058A (en) | 1994-04-06 | 1996-10-08 | Calgene, Inc. | Plant lysophosphatidic acid acyltransferases |
| US5910630A (en) | 1994-04-06 | 1999-06-08 | Davies; Huw Maelor | Plant lysophosphatidic acid acyltransferases |
| ATE368118T1 (de) | 1994-05-18 | 2007-08-15 | Bayer Bioscience Gmbh | Für enzyme, die die fähigkeit besitzen lineare alpha 1,4-glucane in pflanzen, pilzen und mikroorganismen zu synthesieren, kodierende dna sequenzen |
| US5756135A (en) | 1994-09-15 | 1998-05-26 | Robert D. Seeley Trust | Water insoluble yeast solids product and process of making same |
| US5680812A (en) | 1995-01-23 | 1997-10-28 | Linsgeseder; Helmut | Apparatus and method for the extraction of vegetable oils |
| WO1996039476A1 (en) | 1995-06-06 | 1996-12-12 | Agro Management Group, Inc. | Vegetable based biodegradable liquid lubricants |
| US5685218A (en) | 1995-07-14 | 1997-11-11 | The French Oil Mill Machinery Co. | Method for treating oil-bearing material |
| US6255505B1 (en) | 1996-03-28 | 2001-07-03 | Gist-Brocades, B.V. | Microbial polyunsaturated fatty acid containing oil from pasteurised biomass |
| AR006830A1 (es) | 1996-04-26 | 1999-09-29 | Du Pont | Aceite de soja con alta estabilidad oxidativa |
| US5595965A (en) * | 1996-05-08 | 1997-01-21 | The Lubrizol Corporation | Biodegradable vegetable oil grease |
| US5949017A (en) | 1996-06-18 | 1999-09-07 | Abb Power T&D Company Inc. | Electrical transformers containing electrical insulation fluids comprising high oleic acid oil compositions |
| US6312623B1 (en) * | 1996-06-18 | 2001-11-06 | Abb Power T&D Company Inc. | High oleic acid oil compositions and methods of making and electrical insulation fluids and devices comprising the same |
| US7109392B1 (en) | 1996-10-09 | 2006-09-19 | Cargill, Incorporated | Methods for increasing oleic acid content in seeds from transgenic plants containing a mutant delta 12 desaturase |
| US6200788B1 (en) | 1997-01-24 | 2001-03-13 | Kirin Beer Kabushiki Kaisha | β-ketoacyl-ACP synthetase II enzymes and genes coding for same |
| US6395965B1 (en) | 1997-03-21 | 2002-05-28 | Restoragen, Inc. | Plant containing a gene construct comprising a chlorella virus promoter and a lac operator |
| US6243725B1 (en) | 1997-05-21 | 2001-06-05 | Premier International, Ltd. | List building system |
| JP2001512029A (ja) | 1997-08-01 | 2001-08-21 | マーテック・バイオサイエンスィズ・コーポレーション | Dha含有栄養組成物およびそれらの製造方法 |
| WO1999016860A1 (fr) | 1997-09-29 | 1999-04-08 | Nihon Tobacco Inc. | Composition a l'extrait de levure, levure utilisee pour l'obtention de ladite composition et procede de production de ladite composition |
| WO1999037166A1 (en) | 1998-01-21 | 1999-07-29 | University Of Maryland Biotechnology Institute | Methods for the enrichment of live feed with nutrients essential for fish larvae |
| US6139897A (en) | 1998-03-24 | 2000-10-31 | Kao Corporation | Oil or fat composition containing phytosterol |
| US20020012979A1 (en) | 1998-06-08 | 2002-01-31 | Alan Berry | Vitamin c production in microorganisms and plants |
| US6531303B1 (en) | 1998-07-06 | 2003-03-11 | Arkion Life Sciences Llc | Method of producing geranylgeraniol |
| DK1947189T3 (da) | 1998-07-06 | 2011-03-14 | Dcv Inc | Fremgangsmåde til vitaminfremstilling |
| US7511190B2 (en) | 1999-11-17 | 2009-03-31 | Mendel Biotechnology, Inc. | Polynucleotides and polypeptides in plants |
| JP2000136199A (ja) | 1998-10-29 | 2000-05-16 | Asahi Glass Co Ltd | シゾサッカロミセス・ポンベで使用可能なシグナルペプチド、分泌型発現ベクター、およびそれらを用いたタンパク質生産方法 |
| US6762345B1 (en) | 1998-12-03 | 2004-07-13 | E. I. Du Pont De Nemours And Company | Plant stearoyl desaturases |
| JP2000175696A (ja) | 1998-12-14 | 2000-06-27 | Yoshio Tanaka | ドナリエラ藻体の抽出方法 |
| US6166231A (en) | 1998-12-15 | 2000-12-26 | Martek Biosciences Corporation | Two phase extraction of oil from biomass |
| US6534261B1 (en) | 1999-01-12 | 2003-03-18 | Sangamo Biosciences, Inc. | Regulation of endogenous gene expression in cells using zinc finger proteins |
| US7211418B2 (en) | 1999-01-14 | 2007-05-01 | Martek Biosciences Corporation | PUFA polyketide synthase systems and uses thereof |
| WO2000061740A1 (en) | 1999-04-10 | 2000-10-19 | Maxygen, Inc. | Modified lipid production |
| ATE446370T1 (de) | 1999-04-12 | 2009-11-15 | Monsanto Technology Llc | Öl, das brassicastanol enthält |
| ES2265945T3 (es) | 1999-06-04 | 2007-03-01 | Consejo Superior De Investigaciones Cientificas | Plantas, semillas y aceites de girasol con un contenido alto de oleico y alto de estearico. |
| DE19926456A1 (de) | 1999-06-10 | 2000-12-14 | Norddeutsche Pflanzenzucht Han | Verfahren zur Erhöhung des Fettsäuregehalts in Pflanzensamen |
| DE10000978A1 (de) | 2000-01-12 | 2001-07-26 | Gvs Ges Fuer Erwerb Und Verwer | Verfahren zur Erhöhung des Gehalts an Fettsäuren in Pflanzen und Mikroorganismen |
| MX350779B (es) | 2000-01-19 | 2017-09-18 | Dsm Ip Assets B V * | Procedimiento de extraccion sin solvente. |
| WO2010120939A2 (en) * | 2009-04-14 | 2010-10-21 | Solazyme, Inc. | Methods of microbial oil extraction and separation |
| US6338866B1 (en) | 2000-02-15 | 2002-01-15 | Applied Food Biotechnology, Inc. | Pet foods using algal or fungal waste containing fatty acids |
| GB0007651D0 (en) | 2000-03-29 | 2000-05-17 | Ascorbex Ltd | Gene sequence |
| AU2001253699B2 (en) | 2000-04-21 | 2006-04-27 | Martek Biosciences Corporation | Trophic conversion of obligate phototrophic algae through metabolic engineering |
| US20020178467A1 (en) | 2000-05-12 | 2002-11-28 | Katayoon Dehesh | Plastid transit peptide sequences for efficient plastid targeting |
| EP1303616A2 (en) | 2000-07-25 | 2003-04-23 | Calgene LLC | Nucleic acid sequences encoding beta-ketoacyl-acp synthase and uses thereof |
| EP1178118A1 (en) | 2000-08-02 | 2002-02-06 | Dsm N.V. | Isolation of microbial oils |
| JP2002125601A (ja) | 2000-10-25 | 2002-05-08 | Kurorera Kogyo Kk | 動物性プランクトン用餌料とその製造方法及び動物性プランクトンの培養方法 |
| AU2002224445A1 (en) | 2000-10-26 | 2002-05-06 | Joe E. Guyer | Method of generating and recovering gas from subsurface formations of coal, carbonaceous shale and organic-rich shales |
| AU2002220655B2 (en) | 2000-11-21 | 2006-05-25 | Upfield Europe B.V. | Edible spread containing a natural fat phase |
| JP2002161294A (ja) | 2000-11-29 | 2002-06-04 | Kanegafuchi Chem Ind Co Ltd | 可塑性油脂組成物 |
| US7081567B2 (en) | 2000-12-03 | 2006-07-25 | Lexun Xue | Transgenic dunaliella salina as a bioreactor |
| US20020144455A1 (en) | 2001-01-06 | 2002-10-10 | Bertrand Jerome C. | Non sooting candle composition |
| WO2002085293A2 (en) | 2001-04-20 | 2002-10-31 | Cargill, Incorporated | Production of alpha-lipoic acid |
| US6620427B2 (en) * | 2001-04-24 | 2003-09-16 | Abbott Laboratories | Method for improving bone mineralization |
| US6974893B2 (en) | 2001-06-29 | 2005-12-13 | Brookhaven Science Associates, Llc | Isoform of castor oleate hydroxylase |
| WO2003012071A2 (en) | 2001-08-03 | 2003-02-13 | Elitra Pharmaceuticals, Inc. | Nucleic acids of aspergillus fumigatus encoding industrial enzymes and methods of use |
| WO2003037884A2 (en) | 2001-09-19 | 2003-05-08 | Archer-Daniels-Midland Company | Process for separation of tocopherols |
| JP3816774B2 (ja) | 2001-10-01 | 2006-08-30 | 独立行政法人科学技術振興機構 | 炭化水素資化微細藻類およびそれを用いたバイオレメディエーション方法 |
| JP4334352B2 (ja) | 2002-01-23 | 2009-09-30 | ロイヤル ネダルコ ベスローテン フェンノートシャップ | ペントース糖の発酵 |
| US7314974B2 (en) | 2002-02-21 | 2008-01-01 | Monsanto Technology, Llc | Expression of microbial proteins in plants for production of plants with improved properties |
| US20030229237A1 (en) | 2002-04-02 | 2003-12-11 | Haas Michael J. | In situ production of fatty acid alkyl esters |
| ES2391381T3 (es) | 2002-05-03 | 2012-11-23 | Martek Biosciences Corporation | Lípidos de alta calidad y métodos para su producción mediante liberación enzimática a partir de biomasa |
| US20030211594A1 (en) | 2002-05-07 | 2003-11-13 | Rosebrook Donald Ian | Microalgae for remediation of waste and method of culturing the same |
| CA2490771A1 (en) | 2002-06-21 | 2003-12-31 | Monsanto Technology Llc | Thioesterase-related nucleic acid sequences and methods |
| US7232935B2 (en) | 2002-09-06 | 2007-06-19 | Fortum Oyj | Process for producing a hydrocarbon component of biological origin |
| US20040074760A1 (en) | 2002-10-17 | 2004-04-22 | Carnegie Mellon University | Production of biofuels |
| US7135290B2 (en) | 2003-04-12 | 2006-11-14 | Solazyme, Inc. | Methods and compositions for evolving hydrogenase genes |
| BR0316380A (pt) | 2002-11-18 | 2005-10-04 | Monsanto Technology Llc | Produção de proteìna e óleo aumentada em plantas pelo rompimento da trilha de fenilpropanóide |
| WO2004065537A2 (de) | 2003-01-20 | 2004-08-05 | Sungene Gmbh & Co. Kgaa | Expressionskassette mit promotoren der stärkesynthase 3 zur expression von nukleinsäuren in stärkehaltigen geweben von pflanzen |
| JP4966006B2 (ja) | 2003-01-28 | 2012-07-04 | セレクティス | カスタムメイドメガヌクレアーゼおよびその使用 |
| CA2518400A1 (en) | 2003-03-03 | 2004-09-16 | Meiji Seika Kaisha, Ltd. | Transgenic plants modified to accumulate fructooligosaccharides and production thereof |
| US7238482B2 (en) | 2003-05-07 | 2007-07-03 | E. I. Du Pont De Nemours And Company | Production of polyunsaturated fatty acids in oleaginous yeasts |
| US7125672B2 (en) | 2003-05-07 | 2006-10-24 | E. I. Du Pont De Nemours And Company | Codon-optimized genes for the production of polyunsaturated fatty acids in oleaginous yeasts |
| US7032664B2 (en) | 2004-06-02 | 2006-04-25 | Halliburton Energy Services, Inc. | Nanocomposite particulates and methods of using nanocomposite particulates |
| US7259255B2 (en) | 2003-06-25 | 2007-08-21 | E. I. Du Pont De Nemours And Company | Glyceraldehyde-3-phosphate dehydrogenase and phosphoglycerate mutase promoters for gene expression in oleaginous yeast |
| US7267976B2 (en) | 2003-07-02 | 2007-09-11 | E.I. Du Pont De Nemours And Company | Acyltransferases for alteration of polyunsaturated fatty acids and oil content in oleaginous yeasts |
| EP1642959B1 (en) | 2003-07-09 | 2010-05-19 | The Nisshin OilliO Group, Ltd. | Process for producing symmetrical triglyceride |
| CA2536635C (en) | 2003-08-25 | 2015-05-12 | Funzyme Biotechnologies Sa | Novel fungal proteins and nucleic acids encoding same |
| PE20050398A1 (es) | 2003-09-22 | 2005-06-03 | Rosales Jose Antonio Socla | Proceso y purificacion de las xantofilas de marigold |
| US20060048240A1 (en) | 2004-04-01 | 2006-03-02 | Nickolai Alexandrov | Sequence-determined DNA fragments and corresponding polypeptides encoded thereby |
| EP1670880A4 (en) | 2003-10-02 | 2007-04-04 | Univ Mississippi | METHOD FOR PRODUCING BIODIESEL AND OTHER VALUABLE CHEMICALS FROM SLUDGE FROM WASTEWATER TREATMENT PLANTS |
| US7504259B2 (en) | 2003-11-12 | 2009-03-17 | E. I. Du Pont De Nemours And Company | Δ12 desaturases suitable for altering levels of polyunsaturated fatty acids in oleaginous yeast |
| CN100577622C (zh) | 2003-11-20 | 2010-01-06 | 索尔维公司 | 从甘油生产二氯丙醇的方法,甘油最终来自生物柴油生产中动物脂肪的转化 |
| CA2805816A1 (en) | 2003-12-23 | 2005-07-14 | Basf Plant Science Gmbh | Transgenic plants expressing a putative palmitoyl protein thioesterase |
| CN1238469C (zh) | 2004-01-16 | 2006-01-25 | 清华大学 | 有机介质反应体系中脂肪酶转化油脂生产生物柴油新工艺 |
| US7063957B2 (en) | 2004-03-26 | 2006-06-20 | The University Of Hong Kong | Methods for production of astaxanthin from the green microalgae Chlorella in dark-heterotrophic cultures |
| US20080194029A1 (en) | 2004-05-07 | 2008-08-14 | Peter Hegemann | Method for Increasing the Ratio of Homologous to Non-Homologous Recombination |
| US7364883B2 (en) | 2004-05-07 | 2008-04-29 | Yeastern Biotech Co., Ltd. | Process for producing poly-unsaturated fatty acids by oleaginous yeasts |
| EP1766037B1 (en) | 2004-05-12 | 2015-07-01 | Transworld Technologies Limited | Generation of hydrogen from hydrocarbon-bearing materials |
| US7214297B2 (en) | 2004-06-28 | 2007-05-08 | Applied Materials, Inc. | Substrate support element for an electrochemical plating cell |
| US20060075522A1 (en) | 2004-07-31 | 2006-04-06 | Jaclyn Cleveland | Genes and uses for plant improvement |
| US7279617B2 (en) | 2004-09-22 | 2007-10-09 | Ceres, Inc. | Promoter, promoter control elements, and combinations, and uses thereof |
| GB0421937D0 (en) | 2004-10-02 | 2004-11-03 | Univ York | Acyl CoA synthetases |
| WO2006047445A2 (en) | 2004-10-22 | 2006-05-04 | Martek Biosciences Corporation | Process for preparing materials for extraction |
| US7189559B2 (en) | 2004-11-04 | 2007-03-13 | E. I. Du Pont De Nemours And Company | Mortierella alpina lysophosphatidic acid acyltransferase homolog for alteration of polyunsaturated fatty acids and oil content in oleaginous organisms |
| US7273746B2 (en) | 2004-11-04 | 2007-09-25 | E.I. Dupont De Nemours And Company | Diacylglycerol acyltransferases for alteration of polyunsaturated fatty acids and oil content in oleaginous organisms |
| US8685679B2 (en) | 2004-11-04 | 2014-04-01 | E I Du Pont De Nemours And Company | Acyltransferase regulation to increase the percent of polyunsaturated fatty acids in total lipids and oils of oleaginous organisms |
| US7588931B2 (en) | 2004-11-04 | 2009-09-15 | E. I. Du Pont De Nemours And Company | High arachidonic acid producing strains of Yarrowia lipolytica |
| US7879591B2 (en) | 2004-11-04 | 2011-02-01 | E.I. Du Pont De Nemours And Company | High eicosapentaenoic acid producing strains of Yarrowia lipolytica |
| US20060225341A1 (en) | 2004-12-20 | 2006-10-12 | Rodolfo Rohr | Production of biodiesel |
| CA2784248C (en) | 2004-12-30 | 2015-02-10 | Sun Drilling Products Corporation | Thermoset nanocomposite particles, processing for their production, and their use in oil and natural gas drilling applications |
| ES2356086T5 (es) | 2005-01-14 | 2021-03-04 | Neste Oyj | Procedimiento para la producción de hidrocarburos |
| DE102005003624A1 (de) | 2005-01-26 | 2006-07-27 | Nutrinova Nutrition Specialties & Food Ingredients Gmbh | Herstellung und Anwendung eines antioxidativ wirksamen Extraktes aus Crypthecodinium sp. |
| US7692049B2 (en) | 2005-01-31 | 2010-04-06 | Exxonmobil Chemical Patents Inc. | Hydrocarbon compositions useful for producing fuels and methods of producing the same |
| EP1871883A1 (en) | 2005-03-02 | 2008-01-02 | Metanomics GmbH | Process for the production of fine chemicals |
| WO2006102342A2 (en) | 2005-03-18 | 2006-09-28 | Microbia, Inc. | Production of carotenoids in oleaginous yeast and fungi |
| US20060223153A1 (en) | 2005-04-05 | 2006-10-05 | Luca Technologies, Llc | Generation of materials with enhanced hydrogen content from anaerobic microbial consortia |
| US7426960B2 (en) | 2005-05-03 | 2008-09-23 | Luca Technologies, Inc. | Biogenic fuel gas generation in geologic hydrocarbon deposits |
| CN101227900A (zh) | 2005-05-11 | 2008-07-23 | 高级生物营养公司 | 稳定形式的鱼油 |
| US8048652B2 (en) | 2005-05-12 | 2011-11-01 | Martek Biosciences Corporation | Biomass hydrolysate and uses and production thereof |
| FI1741767T4 (fi) | 2005-07-04 | 2023-04-25 | Menetelmä dieselalueen hiilivetyjen valmistamiseksi | |
| MX2008002633A (es) | 2005-08-25 | 2008-09-26 | A2Be Carbon Capture Llc | Metodo, aparato y sistema para produccion de biodiesel a partir de algas. |
| WO2007027669A1 (en) | 2005-08-29 | 2007-03-08 | Cps Biofuels, Inc. | Improved biodiesel fuel, additives, and lubbricants |
| FR2890961B1 (fr) | 2005-09-21 | 2007-11-23 | Inst Francais Du Petrole | Procede perfectionne de fabrication d'esters ethyliques a partir de corps gras d'origine naturelle |
| AU2006294680A1 (en) | 2005-09-27 | 2007-04-05 | Cornell University | Invertase and inhibitors from coffee |
| US8163675B2 (en) | 2005-10-20 | 2012-04-24 | Akzo Nobel N.V. | Emulsifier based on polyamines and fatty acid/maleic anhydride |
| US20070167396A1 (en) | 2006-01-19 | 2007-07-19 | Solazyme, Inc. | Methods and compositions for cholesterol reduction in mammals |
| US20090274736A1 (en) | 2006-01-19 | 2009-11-05 | Solazyme Inc. | Nutraceutical Compositions From Microalgae And Related Methods of Production And Administration |
| WO2007101862A1 (en) * | 2006-03-09 | 2007-09-13 | Centre National De La Recherche Scientifique (Cnrs) | Method of producing sialylated oligosaccharides |
| US20070218183A1 (en) | 2006-03-14 | 2007-09-20 | Bunge Oils, Inc. | Oil composition of conjugated linoleic acid |
| AU2007226511A1 (en) | 2006-03-15 | 2007-09-20 | Dsm Ip Assets B.V. | Plant seed oils containing polyunsaturated fatty acids |
| CN100590186C (zh) | 2006-03-17 | 2010-02-17 | 中国科学院大连化学物理研究所 | 一种生产生物油脂和生物柴油的方法 |
| BRPI0710665A2 (pt) | 2006-04-03 | 2011-08-16 | Advanced Bionutrition Corp | formulações de ração contendo ácido docosa-hexaenóico |
| US20100248322A1 (en) | 2006-04-05 | 2010-09-30 | Luca Technologies, Inc. | Chemical amendments for the stimulation of biogenic gas generation in deposits of carbonaceous material |
| US7309602B2 (en) | 2006-04-13 | 2007-12-18 | Ambrozea, Inc. | Compositions and methods for producing fermentation products and residuals |
| CA2648934A1 (en) | 2006-04-13 | 2007-10-25 | Ambrozea, Inc. | Compositions and methods for producing fermentation products and residuals |
| US20100021968A1 (en) | 2006-05-12 | 2010-01-28 | Arizona Board of Regents, a body corporate of the state of Arizona acting for and on behalf of | Novel chlorella species and uses therefor |
| WO2007141257A1 (en) | 2006-06-06 | 2007-12-13 | Total Raffinage Marketing | Lysophosphatidic acid acyltransferase genes and uses thereof |
| RU2483057C2 (ru) * | 2006-06-28 | 2013-05-27 | Ньюселис Инк. | Смеси жирных кислот и их применение |
| CN101108997B (zh) * | 2006-07-19 | 2010-07-21 | 中国科学院大连化学物理研究所 | 一种微生物油脂的制备方法 |
| US20100228068A1 (en) | 2006-08-16 | 2010-09-09 | Bioecon International Holding N.V. | Production of linear alkanes by hydrotreating mixtures of triglycerides with vacuum gasoil |
| CN101611125A (zh) | 2006-09-14 | 2009-12-23 | 生物燃料箱公司 | 细胞类脂向生物燃料的充分且高效的转换的方法 |
| JP2010503416A (ja) | 2006-09-18 | 2010-02-04 | アリゾナ ボード オブ リージェンツ, ア ボディー コーポレイト オブ ザ ステート オブ アリゾナ アクティング フォー アンド オン ビハーフ オブ アリゾナ ステート ユニバーシティー | 藻類の中鎖長脂肪酸および炭化水素 |
| JP2008081559A (ja) | 2006-09-26 | 2008-04-10 | Nippon Shokubai Co Ltd | バイオディーゼル燃料組成物およびその製造方法 |
| WO2008130372A2 (en) | 2006-09-28 | 2008-10-30 | Microbia, Inc. | Production of sterols in oleaginous yeast and fungi |
| US9101161B2 (en) | 2006-11-02 | 2015-08-11 | The Coca-Cola Company | High-potency sweetener composition with phytoestrogen and compositions sweetened therewith |
| WO2008060571A2 (en) | 2006-11-13 | 2008-05-22 | Aurora Biofuels, Inc. | Methods and compositions for production and purification of biofuel from plants and microalgae |
| ITMI20062193A1 (it) | 2006-11-15 | 2008-05-16 | Eni Spa | Processo per produrre frazioni idrocarburiche da miscele di origine biologica |
| JP4820277B2 (ja) | 2006-12-20 | 2011-11-24 | 花王株式会社 | ケトン体及び/又は2級アルコールの製造法 |
| US9637714B2 (en) | 2006-12-28 | 2017-05-02 | Colorado State University Research Foundation | Diffuse light extended surface area water-supported photobioreactor |
| WO2008081898A1 (ja) | 2006-12-28 | 2008-07-10 | Tohoku Techno Arch Co., Ltd. | 強塩基性陰イオン交換樹脂の再生方法 |
| WO2008083351A2 (en) | 2006-12-29 | 2008-07-10 | Genifuel Corporation | Controlled growth environments for algae cultivation |
| US7977076B2 (en) | 2006-12-29 | 2011-07-12 | Genifuel Corporation | Integrated processes and systems for production of biofuels using algae |
| BRPI0807237A2 (pt) | 2007-02-02 | 2014-04-29 | Kirin Holdings Kk | Dna que codifica xilitol desidrogenase, construção de ácido nucléico, vetores, microorganismos e métodos para produzir xilitol desigrogenase e etanol |
| US8129512B2 (en) | 2007-04-12 | 2012-03-06 | Pioneer Hi-Bred International, Inc. | Methods of identifying and creating rubisco large subunit variants with improved rubisco activity, compositions and methods of use thereof |
| MX2009007356A (es) | 2007-05-02 | 2009-10-08 | Ouro Fino Participacoes E Empr | Proceso para producir biodiesel y/o aceite combustible. |
| WO2008144224A1 (en) | 2007-05-15 | 2008-11-27 | Dow Global Technologies Inc. | High resilience foams |
| KR20100022473A (ko) * | 2007-05-17 | 2010-03-02 | 쿠퍼 인더스트리스, 인코포레이티드 | 식물성 오일 유전성 유체 조성물 |
| US8801975B2 (en) | 2007-05-17 | 2014-08-12 | Cooper Industries, Llc | Vegetable oil dielectric fluid composition |
| WO2008151149A2 (en) | 2007-06-01 | 2008-12-11 | Solazyme, Inc. | Production of oil in microorganisms |
| EP2463370B1 (en) | 2007-06-01 | 2013-07-31 | Sapphire Energy, Inc. | Use of genetically modified organisms to generate biomass degrading enzymes |
| US7914832B2 (en) | 2007-06-01 | 2011-03-29 | Kyoto Eiyo Co., Ltd. | Method for producing chlorella fermented food |
| ES2607795T3 (es) | 2007-06-12 | 2017-04-04 | Cps Biofuels, Inc. | Producción de gasolina a partir de materias primas fermentables |
| AU2008274551B2 (en) * | 2007-07-09 | 2014-05-29 | BASF Agricultural Solutions Seed US LLC | Brassica plant comprising mutant fatty acyl-ACP thioesterase alleles |
| US20090018300A1 (en) | 2007-07-11 | 2009-01-15 | Archer-Daniels-Midland Company | Monomers and polymers from bioderived carbon |
| CN101092353B (zh) | 2007-07-12 | 2010-12-01 | 上海交通大学 | 动植物油脂转化脂肪酸单酯的制备方法 |
| CN101130513A (zh) | 2007-08-24 | 2008-02-27 | 北京科技大学 | 一种从小球藻藻粉中提取纯化叶黄素的方法 |
| KR101650584B1 (ko) | 2007-08-31 | 2016-08-23 | 디에스엠 아이피 어셋츠 비.브이. | 다중불포화된 지방산 함유 고체 지방 조성물 및 그의 용도 및 생산 |
| US20090176272A1 (en) | 2007-09-12 | 2009-07-09 | Kuehnle Agrosystems, Inc. | Expression of nucleic acid sequences for production of biofuels and other products in algae and cyanobacteria |
| NZ584460A (en) | 2007-09-12 | 2012-10-26 | Martek Biosciences Corp | Biological oil produced by a microorganism of the kingdom Stramenopil by heterotropic fermentation |
| JP5711968B2 (ja) | 2007-10-03 | 2015-05-07 | イー・アイ・デュポン・ドウ・ヌムール・アンド・カンパニーE.I.Du Pont De Nemours And Company | 高エイコサペンタエン酸を生成するためのヤロウィア・リポリティカ(YarrowiaLipolytica)の最適化株 |
| US7883822B2 (en) | 2007-10-17 | 2011-02-08 | Texas Instruments Incorporated | Graded lithographic mask |
| WO2009058799A1 (en) | 2007-11-01 | 2009-05-07 | Wake Forest University School Of Medicine | Compositions and methods for prevention and treatment of mammalian diseases |
| FR2924126B1 (fr) | 2007-11-28 | 2011-04-15 | Roquette Freres | Nouveau procede de culture d'une microalgue heterotrophe |
| US20090145392A1 (en) | 2007-11-30 | 2009-06-11 | Clark Richard Hugh | Fuel formulations |
| US8815567B2 (en) | 2007-11-30 | 2014-08-26 | E I Du Pont De Nemours And Company | Coenzyme Q10 production in a recombinant oleaginous yeast |
| MX2010006539A (es) | 2007-12-11 | 2011-02-23 | Synthetic Genomics Inc | Secrecion de acidos grasos por parte de microorganismos fotosinteticos. |
| GB0724720D0 (en) | 2007-12-19 | 2008-01-30 | Ici Plc | Triglyceride macromonomers |
| WO2009105620A1 (en) | 2008-02-20 | 2009-08-27 | Cco Technology, Ltd. | Selective short chain monounsaturated oils |
| CN101230364A (zh) | 2008-02-25 | 2008-07-30 | 清华大学 | 一种利用异养小球藻高密度发酵生产生物柴油的方法 |
| US8048654B2 (en) | 2010-06-09 | 2011-11-01 | Joule Unlimited Technologies, Inc. | Methods and compositions for the recombinant biosynthesis of fatty acids and esters |
| US8598378B2 (en) | 2008-03-14 | 2013-12-03 | University Of Hawaii | Methods and compositions for extraction and transesterification of biomass components |
| US8043496B1 (en) | 2008-03-18 | 2011-10-25 | Peter Allen Schuh | System for extracting oil from algae |
| EP2271742A4 (en) | 2008-03-31 | 2011-05-25 | Kuehnle Agrosystems Inc | GENE EXPRESSION ON A NUCLEAR BASIS FOR BIOFUELS PRODUCTION AND TREATMENT OF COPRODUCTS IN ALGAE |
| US20100170144A1 (en) | 2008-04-09 | 2010-07-08 | Solazyme, Inc. | Hydroprocessing Microalgal Oils |
| RU2542374C2 (ru) | 2008-04-09 | 2015-02-20 | Солазим, Инк. | Способ химической модификации липидов микроводорослей, способ получения мыла и мыло, включающее соли жирных кислот омыленных липидов микроводорослей |
| WO2009140354A1 (en) | 2008-05-13 | 2009-11-19 | Cargill, Incorporated | Polyol made from partially hydrogenated, fully epoxidized natural oils |
| CN101280328B (zh) | 2008-05-27 | 2011-06-29 | 清华大学 | 一种从自养到异养两步培养小球藻生产生物柴油的方法 |
| US8435790B2 (en) | 2008-07-25 | 2013-05-07 | The Regents Of The University Of California | Methods of modulating lipid concentrations in eukaryotic cells |
| US8273694B2 (en) | 2008-07-28 | 2012-09-25 | Jeffrey A Brown | Synthetic compositions obtained from algae |
| AU2009276720A1 (en) | 2008-07-28 | 2010-02-04 | Qteros, Inc. | Methods and compositions for improving the production of products in microorganisms |
| US20100035309A1 (en) | 2008-08-06 | 2010-02-11 | Luca Technologies, Inc. | Analysis and enhancement of metabolic pathways for methanogenesis |
| US8927475B2 (en) | 2008-08-08 | 2015-01-06 | The Dial Corporation | Consumer products comprising algae derived ingredients |
| WO2010019813A2 (en) | 2008-08-13 | 2010-02-18 | Sapphire Energy, Inc. | Production of fatty actds by genetically modified photosynthetic organisms |
| NZ591972A (en) | 2008-10-03 | 2013-04-26 | Agrisoma Biosciences Inc | Production of modified fatty acids in plants |
| EP2975122B1 (en) | 2008-10-07 | 2018-06-06 | REG Life Sciences, LLC | Method and compositions for producing fatty aldehydes |
| US20100303957A1 (en) | 2008-10-14 | 2010-12-02 | Solazyme, Inc. | Edible Oil and Processes for Its Production from Microalgae |
| US20120128851A1 (en) | 2008-10-14 | 2012-05-24 | Solazyme, Inc | Novel microalgal food compositions |
| US20100297296A1 (en) | 2008-10-14 | 2010-11-25 | Solazyme, Inc. | Healthier Baked Goods Containing Microalgae |
| US20100303989A1 (en) | 2008-10-14 | 2010-12-02 | Solazyme, Inc. | Microalgal Flour |
| US20100297325A1 (en) | 2008-10-14 | 2010-11-25 | Solazyme, Inc. | Egg Products Containing Microalgae |
| US20100297295A1 (en) | 2008-10-14 | 2010-11-25 | Solazyme, Inc. | Microalgae-Based Beverages |
| US20130122180A1 (en) | 2008-10-14 | 2013-05-16 | Solazyme, Inc. | Microalgal Food Compositions |
| BRPI0920280B8 (pt) | 2008-10-14 | 2022-11-16 | Corbion Biotech Inc | Composição alimentícia, e, método para fabricar uma composição alimentícia |
| US20100297323A1 (en) | 2008-10-14 | 2010-11-25 | Solazyme, Inc. | Gluten-free Foods Containing Microalgae |
| US20100303961A1 (en) | 2008-10-14 | 2010-12-02 | Solazyme, Inc. | Methods of Inducing Satiety |
| US20100303990A1 (en) | 2008-10-14 | 2010-12-02 | Solazyme, Inc. | High Protein and High Fiber Algal Food Materials |
| US20100297292A1 (en) | 2008-10-14 | 2010-11-25 | Solazyme, Inc. | Reduced Pigmentation Microalgae Strains and Products Therefrom |
| US20100297331A1 (en) | 2008-10-14 | 2010-11-25 | Solazyme, Inc. | Reduced Fat Foods Containing High-Lipid Microalgae with Improved Sensory Properties |
| ES2583639T3 (es) | 2008-11-28 | 2016-09-21 | Terravia Holdings, Inc. | Producción de aceites específicos en microorganismos heterótrofos |
| CN101748134B (zh) * | 2008-11-28 | 2012-07-04 | 中国科学院上海生命科学研究院 | 一种使微生物在高于其极限生长温度下生长的方法 |
| MX2011006081A (es) | 2008-12-11 | 2011-06-24 | Bio Architecture Lab Inc | Biosintesis de productos quimicos de uso generalizado. |
| CN102325864B (zh) | 2008-12-23 | 2016-01-27 | Reg生命科学有限责任公司 | 硫酯酶相关的方法和组合物 |
| US8075797B2 (en) | 2009-01-29 | 2011-12-13 | Honeywell International Inc. | Azeotrope-like compositions of pentafluoropropane, chlorotrifluoropropene, and hydrogen fluoride |
| US20100196575A1 (en) * | 2009-01-30 | 2010-08-05 | Solae, Llc | Melting Vegetable Protein Based Substitute Cheese |
| JP5477011B2 (ja) | 2009-02-03 | 2014-04-23 | セントラル硝子株式会社 | (z)−1−クロロ−3,3,3−トリフルオロプロペンの精製方法 |
| CN101824440A (zh) * | 2009-03-04 | 2010-09-08 | 中国科学院大连化学物理研究所 | 一种微生物油脂的分离方法 |
| CN102365343B (zh) | 2009-03-27 | 2016-08-03 | 纳幕尔杜邦公司 | 介电传热流体 |
| KR101899933B1 (ko) | 2009-04-14 | 2018-09-19 | 테라비아 홀딩스 인코포레이티드 | 신규의 미세 조류 식품 조성물 |
| CA2760326A1 (en) | 2009-05-13 | 2010-11-18 | Basf Plant Science Company Gmbh | Acyltransferases and uses thereof in fatty acid production |
| AU2010254104A1 (en) | 2009-05-26 | 2011-12-15 | Solazyme, Inc. | Fractionation of oil-bearing microbial biomass |
| WO2010150411A1 (ja) | 2009-06-26 | 2010-12-29 | オーエスジー株式会社 | 硬質被膜、および硬質被膜被覆工具 |
| EP2448955B1 (en) * | 2009-06-29 | 2015-09-30 | Synthetic Genomics, Inc. | Acyl-acp thioesterase genes and uses therefor |
| WO2011026008A1 (en) | 2009-08-28 | 2011-03-03 | Phycal Llc | Biofuel from recombinant oleginous algae using sugar carbon sources |
| US20110162259A1 (en) | 2009-09-25 | 2011-07-07 | Ls9, Inc. | Production of fatty acid derivatives |
| EP2327776A1 (en) | 2009-11-30 | 2011-06-01 | Institut National De La Recherche Agronomique | Method for the production of Very Long Chain Fatty Acids (VLCFA) by fermentation with a recombinant Yarrowia sp |
| CA2782423C (en) | 2009-12-18 | 2019-06-18 | Cargill Incorporated | Brassica plants yielding oils with a low total saturated fatty acid content |
| WO2011090730A1 (en) | 2009-12-28 | 2011-07-28 | Martek Biosciences Corporation | Recombinant thraustochytrids that grow on xylose, and compositions, methods of making, and uses thereof |
| WO2011082253A2 (en) * | 2009-12-30 | 2011-07-07 | Board Of Trustees Of Michigan State University | A method to produce acetyldiacylglycerols (ac-tags) by expression ofan acetyltransferase gene isolated from euonymus alatus (burning bush) |
| US8553906B2 (en) | 2010-02-02 | 2013-10-08 | Creative Technology Ltd | Apparatus for enabling karaoke |
| WO2011130576A1 (en) | 2010-04-14 | 2011-10-20 | Solazyme, Inc. | Oleaginous yeast food compositions |
| US20110252696A1 (en) | 2010-04-14 | 2011-10-20 | Solazyme, Inc. | Fuel And Chemical Production From Oleaginous Yeast |
| WO2011130578A2 (en) | 2010-04-14 | 2011-10-20 | Solazyme, Inc. | Lipid-rich microalgal flour food compositions |
| JP5996527B2 (ja) | 2010-05-28 | 2016-09-21 | テラヴィア ホールディングス, インコーポレイテッド | 用途に応じた油を含む食品成分 |
| SG10201509035WA (en) | 2010-11-03 | 2015-12-30 | Solazyme Inc | Microbial Oils With Lowered Pour Points, Dielectric Fluids Produced Therefrom, And Related Methods |
| US8530207B2 (en) | 2010-12-23 | 2013-09-10 | Exxonmobil Research And Engineering Company | Photosynthetic microorganisms comprising exogenous prokaryotic acyl-ACP thioesterases and methods for producing fatty acids |
| JP6071904B2 (ja) * | 2011-02-02 | 2017-02-01 | テラヴィア ホールディングス, インコーポレイテッド | 組み換え油産生微生物から生成される用途に応じた油 |
| BR112013028621A2 (pt) | 2011-05-06 | 2016-11-29 | Solazyme Inc | "microalgas oleaginosas recombinantes, método para produzir biomassa de microalga ou um produto produzido da biomassa, e método para produzir uma composição lipídica de microalga". |
| CN104126006B (zh) | 2011-11-28 | 2017-06-30 | 泰拉瑞亚控股公司 | 包括原藻脂质通路基因的基因工程化的微生物株系 |
| US20130157917A1 (en) | 2011-12-20 | 2013-06-20 | Rhodia Operations | Industrial cleaning compositions and methods for using same |
| AU2012358203A1 (en) | 2011-12-23 | 2014-07-24 | Terravia Holdings, Inc. | Algal thermoplastics, thermosets, paper, adsorbants and absorbants |
| SG11201406711TA (en) | 2012-04-18 | 2014-11-27 | Solazyme Inc | Tailored oils |
| US9719114B2 (en) | 2012-04-18 | 2017-08-01 | Terravia Holdings, Inc. | Tailored oils |
| EA201591667A1 (ru) | 2013-03-08 | 2016-05-31 | Солазайм, Инк. | Смазочные материалы, содержащие микробы, накапливающие масло |
| US9249252B2 (en) | 2013-04-26 | 2016-02-02 | Solazyme, Inc. | Low polyunsaturated fatty acid oils and uses thereof |
| WO2015051319A2 (en) | 2013-10-04 | 2015-04-09 | Solazyme, Inc. | Tailored oils |
| US10280431B2 (en) | 2013-12-18 | 2019-05-07 | Nutech Ventures | Acyltransferases and methods of using |
| US9394550B2 (en) | 2014-03-28 | 2016-07-19 | Terravia Holdings, Inc. | Lauric ester compositions |
| CN106574255A (zh) | 2014-07-10 | 2017-04-19 | 泰拉瑞亚控股公司 | 酮脂酰acp合酶基因及其用途 |
| EP3172320B1 (en) * | 2014-07-24 | 2019-11-20 | Corbion Biotech, Inc. | Variant thioesterases and methods of use |
| CN107960101A (zh) | 2015-04-06 | 2018-04-24 | 柯碧恩生物技术公司 | 具有lpaat消融的产油微藻 |
| US20180216144A1 (en) | 2015-09-28 | 2018-08-02 | Corbion Biotech, Inc. | Triglyceride oils having asymmetric triglyceride molecules |
| US20180142218A1 (en) | 2016-10-05 | 2018-05-24 | Terravia Holdings, Inc. | Novel acyltransferases, variant thioesterases, and uses thereof |
-
2011
- 2011-11-03 SG SG10201509035WA patent/SG10201509035WA/en unknown
- 2011-11-03 JP JP2013537836A patent/JP5970463B2/ja not_active Expired - Fee Related
- 2011-11-03 CN CN201810544646.4A patent/CN108823254A/zh active Pending
- 2011-11-03 US US13/288,815 patent/US9066527B2/en active Active
- 2011-11-03 CA CA3024641A patent/CA3024641A1/en not_active Abandoned
- 2011-11-03 KR KR1020197008866A patent/KR102065526B1/ko active IP Right Grant
- 2011-11-03 EP EP11785851.4A patent/EP2635663B1/en active Active
- 2011-11-03 WO PCT/US2011/059224 patent/WO2012061647A2/en active Application Filing
- 2011-11-03 BR BR112013011039A patent/BR112013011039A8/pt not_active Application Discontinuation
- 2011-11-03 SG SG2013034368A patent/SG190154A1/en unknown
- 2011-11-03 CA CA2816125A patent/CA2816125C/en active Active
- 2011-11-03 MX MX2013004631A patent/MX354145B/es active IP Right Grant
- 2011-11-03 AU AU2011323288A patent/AU2011323288B2/en active Active
- 2011-11-03 EP EP18212337.2A patent/EP3521408B1/en active Active
- 2011-11-03 ES ES11785851T patent/ES2746074T3/es active Active
- 2011-11-03 KR KR1020137013979A patent/KR101964886B1/ko active IP Right Grant
- 2011-11-03 ES ES18212337T patent/ES2909143T3/es active Active
- 2011-11-03 CN CN2011800532582A patent/CN103282473A/zh active Pending
-
2013
- 2013-05-06 ZA ZA2013/03271A patent/ZA201303271B/en unknown
-
2015
- 2015-06-04 US US14/730,671 patent/US9388435B2/en active Active
-
2016
- 2016-01-21 JP JP2016009933A patent/JP6453257B2/ja active Active
- 2016-05-10 AU AU2016202999A patent/AU2016202999B2/en active Active
- 2016-06-10 US US15/179,253 patent/US10167489B2/en active Active
-
2018
- 2018-04-05 JP JP2018073024A patent/JP2018104722A/ja active Pending
- 2018-11-09 US US16/186,316 patent/US10344305B2/en active Active
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2909143T3 (es) | Microbios de Chlorella o Prototheca modificados genéticamente y aceite producido a partir de estos | |
| US10100341B2 (en) | Tailored oils produced from recombinant oleaginous microorganisms | |
| ES2898678T3 (es) | Composiciones alimentarias que comprenden aceites a medida |