RU2238325C2 - Способ получения l-аминокислоты c использованием микроорганизма - Google Patents
Способ получения l-аминокислоты c использованием микроорганизма Download PDFInfo
- Publication number
- RU2238325C2 RU2238325C2 RU2001118542A RU2001118542A RU2238325C2 RU 2238325 C2 RU2238325 C2 RU 2238325C2 RU 2001118542 A RU2001118542 A RU 2001118542A RU 2001118542 A RU2001118542 A RU 2001118542A RU 2238325 C2 RU2238325 C2 RU 2238325C2
- Authority
- RU
- Russia
- Prior art keywords
- microorganism
- respiratory chain
- ndh
- producing
- amino acid
- Prior art date
Links
Images

Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P1/00—Preparation of compounds or compositions, not provided for in groups C12P3/00 - C12P39/00, by using microorganisms or enzymes
- C12P1/04—Preparation of compounds or compositions, not provided for in groups C12P3/00 - C12P39/00, by using microorganisms or enzymes by using bacteria
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/22—Tryptophan; Tyrosine; Phenylalanine; 3,4-Dihydroxyphenylalanine
- C12P13/222—Phenylalanine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0012—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7)
- C12N9/0036—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on NADH or NADPH (1.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0012—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7)
- C12N9/0036—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on NADH or NADPH (1.6)
- C12N9/0038—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on NADH or NADPH (1.6) with a heme protein as acceptor (1.6.2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/08—Lysine; Diaminopimelic acid; Threonine; Valine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/22—Tryptophan; Tyrosine; Phenylalanine; 3,4-Dihydroxyphenylalanine
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Biomedical Technology (AREA)
- Mycology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Organic Low-Molecular-Weight Compounds And Preparation Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
Изобретение относится к биотехнологии и представляет собой способ получения L-аминокислоты с использованием микроорганизма, принадлежащего к роду Escherichia или коринеформной бактерии. Данный способ включает выращивание указанного микроорганизма в среде для продуцирования и накопление в среде и сбор накопленной аминокислоты. При этом микроорганизм создан из родительского штамма, имеющего путь дыхательной цепи с высоким энергетическим выходом и путь дыхательной цепи с низким энергетическим выходом в качестве путей дыхательной цепи, и этот микроорганизм представляет собой мутантный штамм или штамм генетического рекомбинанта, имеющего одну или обе из следующих характеристик: путь дыхательной цепи с высоким энергетическим выходом усилен; путь дыхательной цепи с низким энергетическим выходом является дефицитным. Данный способ позволяет увеличить выход L-аминокислоты. 7 з.п. ф-лы, 5 табл., 2 ил.
Description
Область техники
Настоящее изобретение относится к способу получения вещества с использованием микроорганизма. В настоящем изобретении микроорганизм обычно выбирают из бактерий, принадлежащих к роду Escherichia, или коринеформных бактерий, которые традиционно используют для производства веществ. Получаемое вещество может быть выбрано из веществ, обычно получаемых с использованием микроорганизмов, например L-аминокислот, нуклеиновых кислот, антибиотиков, витаминов, факторов роста, физиологически активных веществ и т.д. Настоящее изобретение раскрывает средства улучшения продуктивности по конечным целевым веществам в способах получения веществ с использованием микроорганизмов.
Описание близкой области техники
Многие организмы накапливают энергию, необходимую для жизнедеятельности, за счет дыхания. При дыхании микроорганизмов различные ферментные комплексы обычно функционируют соответственно видам или среде развития, и эффективность накопления энергии также меняется значительно. Углеводы, белки и алифатические кислоты преобразуются в ацетил-КоА в результате гликолиза, β -окисления и т.д. и подвергаются катаболизму в цикле лимонной кислоты. Энергия, сохраненная в этом случае в форме HAD.H, используется для "выбрасывания" протона из микробных клеток при содействии HAD.H-дегидрогеназы (NDH) и следующей системы электронного переноса, включающей оксидоредуктазы, и посредством чего образуется градиент концентрации протонов между внутренней стороной и внешней стороной цитоплазматической мембраны. Этот градиент концентрации протонов используется в качестве движущей силы синтеза аденозинтрифосфата (АТФ). В это же время существует путь, проявляющий высокую способность к протонному выбору, и путь, проявляющий низкую способность к протонному выбору, наряду с путями электронного переноса, зависящими от комбинации NDH и оксидоредуктаз. Считают, что путь с высокой способностью к протонному выбору проявляет высокий энергетический выход, а путь с низкой способностью к протонному выбору показывает низкий энергетический выход. Следовательно, один вид микроорганизма одновременно содержит множество путей электронного переноса в дыхательной цепи параллельно, и эти пути включают пути с высоким энергетическим выходом и низким энергетическим выходом.
Существует два вида NDH и два вида концевых оксидаз в дыхательной цепи Escherichia coli в аэробных условиях. То есть для NDH известны NDH-1 (кодируемая nuo-опероном) высокого энергетического выхода и NDH-II (кодируемая ndh-опероном) низкого энергетического выхода. Кроме того, что касается концевой оксидазы, то известны цитохромоксидаза bo (кодируемая cyoABCD-опероном), отнесенная к SохМ типу (Castresana, J., Saraste, M., Trends in Biochem. Sci., 20, 443-448, 1995) и проявляющая высокий энергетический выход, и цитохром bd-оксидаза (кодируемая cydAB-опероном), проявляющая низкий энергетический выход. Хотя известно, что экспрессируемые количества этих ферментов дыхательной цепи меняются в ответ на среду развития (Minagawa et al., The Journal of Biological Chemistry, 265:11198-11203, 1990; Tseng et al., Journal of Bacteriology, 178:1094-1098, 1996; Green et al., Molecular Microbiology, 12:433-444, 1994; Bongaerts et al., Molecular Microbiology, 16:521-534, 1995, все еще существует много неизвестных моментов относительно физиологического значения особенностей их экспрессии.
Кроме того, в Corynebacterium glutamicum существует комплекс цитохром bc1 и подтверждено наличие, по меньшей мере, двух типов концевых оксидаз, SохМ-оксидазы и цитохромоксидазы bd (The Second Symposium Concerning Metabolic Engineering, Lecture Abstracts, 1999). Это показывает, что путь электронного переноса от хинонового пула к молекуле кислорода включает два типа пути, путь, использующий комплекс цитохром bc1 и SoxM-оксидазу, и путь, использующий только цитохромоксидазы bd. Считают, что первый представляет собой путь электронного переноса высокого энергетического выхода, в котором число протонного переноса для переноса одного электрона является высоким, а последний представляет собой путь, в котором число протонного переноса для переноса одного электрона является низким.
Что касается концевой оксидазы Е.coli, то если сравнивать урожай биомассы в аэробной культуре мутантного штамма, имеющего только цитохромоксидазу bo, мутантного штамма, имеющего только цитохромоксидазу bd, и исходного штамма, имеющего обе оксидазы, урожай биомассы будет самым низким в мутантном штамме, имеющем только цитохромоксидазу bd, и это зависит от вида и эффективности энергонакопления концевой оксидазы (Annual Meeting of the Society for fermentation and Bioengineering Japan, 1995, Lecture Abstracts, №357).
Кроме того, описан энергетический выход мутантов с дефицитом некоторых ферментов дыхательной цепи (Calhoun et al., Journal of Bacteriology, 175:3020-3925, 1993).
Однако отсутствуют указания относительно изменения энергетического выхода путем амплификации гена дыхательной цепи, обеспечивающего высокую эффективность, такого как ген для NDH-I и SохМ-оксидазы, и даже не известны попытки использовать ее для получения веществ. Более того, для получения веществ не были предприняты попытки использовать делецию ферментов дыхательного пути низкой эффективности, таких как NDH-II и цито-хромоксидаза bd.
Сущность изобретения
В живых организмах для биосинтеза веществ, таких как L-аминокислоты и нуклеиновые кислоты, требуется энергия. Большая часть энергии, используемой в дальнейшем, заключается в восстанавливающей способности HAD.H, HADФ.H и т.д. и энергии, сохраненной в виде АТОФ. Таким образом, заявители настоящего изобретения предположили, что если в способе производства целевых веществ с использованием микроорганизмов запас энергии, используемый при продуцировании целевых веществ, был увеличен, продуктивность по целевым веществам будет улучшена. Исходя из этой концепции, объектом настоящего изобретения является создание микроорганизма, показывающего улучшенный энергетический выход, и разработка способа получения целевого вещества путем его использования.
Заявители настоящего изобретения заключили, что микроорганизм, имеющий повышенный запас энергии, мог бы быть создан путем усиления пути дыхательной цепи, проявляющего высокую эффективность энергонакопления, или путем придания дефицитности пути дыхательной цепи, проявляющего низкую эффективность энергонакопления. Более конкретно, что касается Е.coli, то штаммы, которые, как считают, имеют улучшенный энергетический выход, получены путем амплификации гена, кодирующего цитохромоксидазу bо как фермента дыхательной цепи с высоким энергетическим выходом, или путем делеции гена, кодирующего NDH-II как фермента дыхательной цепи с низким энергетическим выходом. Затем с их использованием осуществили получение L-аминокислоты и установили, что продуктивность по L-аминокислоте улучшается в штаммах, энергетический выход которых был улучшен. Следовательно, реализуется настоящее изобретение.
То есть, настоящее изобретение предлагает следующее.
(1) Способ получения целевого вещества с использованием микроорганизма, включающий выращивание микроорганизма в среде с целью продуцирования и накопления целевого вещества, где микроорганизм создан из родительского штамма микроорганизма, имеющей в качестве путей дыхательной цепи путь дыхательной цепи с высоким энергетическим выходом и путь дыхательной цепи с низким энергетическим выходом, и этот микроорганизм представляет собой мутантный штамм или штаммы генетического рекомбинанта, имеющий или одну, или обе из следующих характеристик:
(А) путь дыхательной цепи с высоким энергетическим выходом усилен,
(Б) путь дыхательной цепи с низким энергетическим выходом является дефицитным.
(2) Способ получения целевого вещества в соответствии с пунктом (1), где путь дыхательной цепи с высоким энергетическим выходом усилен путем увеличения копийности гена, кодирующего фермент, вовлеченный в дыхательную цепь, или модификации регулирующий экспрессию последовательности гена.
(3) Способ получения целевого вещества в соответствии с пунктами (1) или (2), где путь дыхательной цепи с низким энергетическим выходом сделан дефицитным путем разрушения гена, кодирующего фермент, вовлеченный в дыхательную цепь.
(4) Способ получения целевого вещества в соответствии с любым из пунктов (1)-(3), где ферменты дыхательной цепи с высоким энергетическим выходом включают SохМ-оксидазу, bc1-комплекс, NDH-1 или два, или три вида из них.
(5) Способ получения целевого вещества в соответствии с любым из пунктов (1)-(4), где ферменты дыхательной цепи с низким энергетическим выходом включают цитохромоксидазу bd, NDH-II или оба из них.
(6) Способ получения целевого вещества в соответствии с любым из пунктов (1)-(5), где в микроорганизме активность SoxM-оксидазы усилена, а NDH-II сделана дефицитной.
(7) Способ получения целевого вещества в соответствии с любым из пунктов (1)-(6), где SoxM-оксидаза представляет собой цитохромоксидазу bо.
(8) Способ получения целевого вещества в соответствии с любым из пунктов (1)-(7), где микроорганизм представляет собой бактерии, принадлежащие роду Escherichia, или коринеформные бактерии.
(9) Способ получения целевого вещества в соответствии с любым из пунктов (1)-(8), где целевое вещество представляет собой L-аминокислоту или нуклеиновую кислоту.
В соответствии с настоящим изобретением в способе получения целевого вещества с использованием микроорганизма, включающем выращивание микроорганизма в среде с целью получения и накопления целевого вещества в среде и сбор целевого вещества, продуктивность по целевому веществу может быть улучшена на основе принципов, отличных от обычной стратегии.
Краткое описание чертежей
На фиг.1 представлено создание плазмиды pTS-Δ ndh для получения линии с разрушенным геном для NDH-II.
На фиг.2 показано создание плазмиды pMAN997.
Подробное описание изобретения
Далее настоящее изобретение объяснено более подробно.
Вещество, получаемое способом получения настоящего изобретения, особенно не ограничено пока оно представляет собой вещество, которое может быть продуцировано микроорганизмом. Примерами таких веществ являются различные L-аминокислоты, такие как L-треонин, L-лизин, L-глутаминовая кислота, L-лейцин, L-изолейцин, L-валин и L-фенилаланин; нуклеиновые кислоты, такие как гуаниловая кислота и инозиновая кислота; витамины; антибиотики; факторы роста; физиологически активные вещества и т.д.
Микроорганизм, используемый для настоящего изобретения, представляет собой микроорганизм, обладающий способностью продуцировать целевое вещество, которое указано выше, и созданный из родительского штамма микроорганизма, имеющего в качестве путей дыхательной цепи путь дыхательной цепи с высоким энергетическим выходом и путь дыхательной цепи с низким энергетическим выходом и имеющий или одну, или обе из следующих характеристик:
(А) путь дыхательной цепи с высоким энергетическим выходом усилен,
(Б) путь дыхательной цепи с низким энергетическим выходом является дефицитным.
В общем случае микроорганизмы, включая Е.coli и коринеформные бактерии, одновременно содержат множество путей электронного переноса дыхательной цепи параллельно, и эти пути включают пути с высоким числом протонного переноса и пути с низким числом протонного переноса на один электрон. В Е.coli, например, что касается донора электрона HAD.H, то существуют NDHI и NDHII в качестве HAD.H-дегидрогеназ, которые катализируют протонный перенос от HAD.H к хиноновому пулу. Из них NDHI проявляет высокий энергетический выход, а NDHII проявляет низкий энергетический выход. То есть NDHII имеет молекулярное число протонов, которое может быть выделено с одним электроном (число протонного переноса), равное 0, тогда как число протонного переноса для NDHI, как считают, равно 2.
В настоящем изобретении такой путь, показывающий высокое число протонного переноса на один электрон, которое описано выше, например путь дыхательной цепи с высоким энергетическим выходом, усилен, а путь дыхательной цепи с низким энергетическим выходом сделан дефицитным. Путь дыхательной цепи с высоким энергетическим выходом может быть усилен путем усиления активности фермента дыхательной цепи, вовлеченного в путь дыхательной цепи. Путь дыхательной цепи с низким энергетическим выходом может быть сделан дефицитным за счет уменьшения или исключения активности фермента дыхательной цепи, вовлеченного в этот путь дыхательной цепи.
Фермент дыхательной цепи, вовлеченный в путь дыхательной цепи, не имеет особых ограничений пока он представляет собой фермент, составляющий путь дыхательной цепи. Конкретно примерами таких ферментов являются дегидрогеназы, которые катализируют электронный перенос от донора электрона к хиноновому пулу, такому как убихинон, диметилменахинон и менахинон, и оксидазы, которые катализируют электронный перенос от хинонового пула к донору электрона.
Оксидазы, которые катализируют реакцию получения молекулы воды путем электронного переноса от хинонового пула, классифицированы как SoxM-тип (bo-тип) и bd-тип.Число протонного переноса bo-типа равно 2, тогда как число протонного переноса bd-типа равно 1. Следовательно, bo-тип проявляет более высокий энергетический выход.
В настоящем изобретении определения “высокий” и “низкий”, используемые для энергетического выхода, не применяются в абсолютном значении, а приняты для обозначения относительных понятий, описанных выше.
Средства усиления активности фермента дыхательной цепи с высоким энергетическим выходом и средства снижения или исключения активности фермента дыхательного пути с низким энергетическим выходом будут объяснены ниже.
Для того чтобы усилить активность фермента дыхательной цепи с высоким энергетическим выходом, например, рекомбинантная ДНК может быть получена путем лигирования фрагмента гена, кодирующего фермент, с вектором, функционирующим в клетке микроорганизма, предпочтительно с мультикопийным вектором, и введена в микроорганизм для трансформации клетки. Копийность гена, кодирующего фермент в клетке трансформированного штамма, за счет этого повышается, и в результате ферментативная активность увеличивается. Этот способ будет объяснен далее на примере суо-оперона (cyoABCDE), кодирующего цитохромоксидазу bo как гена фермента дыхательной пути с высоким энергетическим выходом.
Последовательность суо-оперона Е.coli уже описана (Chepuri et al., The Journal of Biological Chemistry, 265:11185-11192, 1990), поэтому этот оперон может быть клонирован на основе этой последовательности. Также в качестве суо-оперона можно использовать ген бактерии, принадлежащей к роду Escherichia, или ген, полученный из других организмов, таких как коринеформные бактерии.
В качестве вектора для клонирования гена и введения гена в микроорганизм может быть использована, например, плазмида, способная к спонтанной репликации в клетках Е.coli. Конкретными примерами таких векторов являются pUC19, pUC18, pBR322, PHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pSTV29 и т.д. В случае введения гена в коринеформные бактерии предпочтительно может быть использован шаттл-вектор, способный к спонтанной репликации в коринеформных бактериях и Е.coli. Примеры плазмид, спонтанно реплицирующихся в коринеформных бактериях и Е.coli, перечислены ниже.
рАМ 330 (см. нерассмотренную публикацию японского патента (Kokai) №58-67699)
рНМ 1519 (см. нерассмотренную публикацию японского патента №58-77895)
pAJ 655 (см. нерассмотренную публикацию японского патента №58-192900)
pAJ 611 (см. нерассмотренную публикацию японского патента №58-192900)
pAJ 1844 (см. нерассмотренную публикацию японского патента №58-192900)
pCG 1 (см. нерассмотренную публикацию японского патента №57-134500)
pCG 2 (см. нерассмотренную публикацию японского патента №58-35197)
pCG 4 (см. нерассмотренную публикацию японского патента №57-183799)
pCG11 (см. нерассмотренную публикацию японского патента №57-183799)
рНК4 (см. нерассмотренную публикацию японского патента №5-7491)
Для того чтобы лигировать фрагмент ДНК, содержащий суо-оперон, и вектор с образованием рекомбинантной ДНК, вектор вначале расщепляют рестрикционным ферментом, приемлемым для концов суо-оперона. Лигирование обычно осуществляют с использованием лигазы, такой как ДНК лигаза фага Т4.
Чтобы ввести в микроорганизм рекомбинантную ДНК, полученную, как описано выше, может быть использован любой известный метод трансформации. Например, можно использовать способ обработки реципиентных клеток хлоридом кальция, чтобы повысить проницаемость ДНК, который был описан для Е.coli K-12 (Mandel М., Higa A., J.Mol.Biol., 53, 159, 1970); и способ получения компетентных клеток из клеток, которые находятся в фазе роста, с последующим введением в них ДНК, который описан для Bacillus subtilis (Duncan C.H., Wilson G.A., Young F.E., Gene, 1, 153, 1977). Помимо этих способов также можно использовать способ получения ДНК реципиентных клеток в протопластах или сферопластах, которые могут легко принимать рекомбинантную ДНК с последующим введением рекомбинантной ДНК в клетки; этот способ, как известно, применим для Bacillus subtilis, актиномицетов и дрожжей (Chang S., Choen S.N., Molec. Gen. Genet., 168, 111, 1979; Bibb M.J., Ward J.M., Hopwood O.A., Natute, 274, 398, 1978; Hinnen A., Hicks J.B., Fink G.R., Proc. Natl. Sci., USA, 75, 1929, 1978). Трансформация коринеформных бактерий может быть достигнута с помощью метода электропробы (см. нерассмотренную публикацию японского патента №2-207791).
Амплификация активности цитохромоксидазы bo также может быть достигнута за счет того, что делается возможным существование множества копий суо-оперона на хромосомной ДНК реципиента. Для того чтобы ввести множество копий суо-оперона в хромосомную ДНК микроорганизма, такого как бактерии, принадлежащие роду Escherichia, или коринеформные бактерии, осуществляют гомологическую рекомбинацию с использованием последовательности, множественные копии которой существуют в хромосомной ДНК в качестве мишеней. В качестве последовательностей, множественные копии которых присутствуют в хромосомной ДНК, могут быть использованы повторяющаяся ДНК или инвертированные повторы, присутствующие на конце перемещающегося генетического элемента. Кроме того, как описано в нерассмотренной публикации японского патента №2-109985, также можно вводить суо-оперон в транспозон и получать возможность его переноса, чтобы ввести множество копий суо-оперона в хромосомную ДНК. Любым способом повышается число копий суо-оперона внутри клетки, и в результате повышается активность цитохромоксидазы bo.
Усиление активности цитохромоксидазы bo также может быть достигнуто помимо описанной выше амплификации гена путем замены регулирующей экспрессии на последовательности суо-оперона, такой как промотор, более сильным промотором (см. нерассмотренную публикацию японского патента №1-215280). Например, lac промотор, trp промотор, trc промотор, tac промотор, PR промотор и PL промотор лямбда фага, tet промотор, аmуЕ промотор и т.д. известны как сильные промоторы. Замещение этими промоторами усиливает экспрессию суо-оперона и, следовательно, активность цитохромоксидазы bo увеличивается. Усиление регулирующей экспрессии последовательности может быть объединено с повышением числа копий суо-оперона.
Повышение активности фермента дыхательной цепи с высоким энергетическим выходом также может быть достигнуто путем введения такой мутации, что внутриклеточная активность фермента будет повышена через мутагенез микроорганизма. Примерами таких мутаций являются мутации кодирующей области, повышающие удельную активность фермента, мутации в регулирующих экспрессию последовательностях, увеличивающие экспрессию гена, и т.д. В качестве мутагенной обработки можно упомянуть способы, в которых используется обработка ультрафиолетовым излучением или обработка мутагенным агентом, таким как N-метил-N’-нитро-N-нитрозогуанидин (NTG) и азотистая кислота.
Для того чтобы уменьшить или устранить активность фермента дыхательной цепи с низким энергетическим выходом, в ген вводят мутацию так, что внутриклеточная активность фермента будет уменьшаться или устраняться, или ген на хромосоме микроорганизма разрушают так, что ген не будет нормально функционировать. Далее на примере гена ndh, кодирующего NDH-II как гена фермента дыхательной цепи с низким энергетическим выходом, будет объяснен способ разрушения ndh гена.
Последовательность ndh гена Е.coli уже описана (Young et al., European Journal of Biochemistry, 116:165-170, 1981), и поэтому ген может быть клонирован на основе этой последовательности. Также в качестве ndh гена можно использовать ген бактерии, принадлежащей к роду Escherichia, или ген, полученный из других организмов, таких как кориноформные бактерии.
Ген ndh на хромосоме может быть разрушен путем трансформации микроорганизма с помощью ДНК, содержащей ndh ген, модифицированный путем внутренней делеции так, чтобы он не продуцировал нормально функционирующую NDH-II (ndh ген делеционного типа), и создания возможности рекомбинации между ndh геном делеционного типа и ndh геном на хромосоме. Такое разрушение гена путем гомологической рекомбинации уже было представлено, и существуют способы, использующие линейную ДНК, плазмиду, которая содержит чувствительную к температуре, контролирующую репликацию область и т.д. В настоящем изобретении предпочтителен способ, использующий плазмиду, которая содержит чувствительную к температуре контролирующую репликацию область.
Ген ndh на хромосоме реципиента может быть замещен ndh геном делеционного типа следующим образом. Вначале получают рекомбинантную ДНК путем вставки чувствительной к температуре контролирующей репликацию области, ndh гена делеционного типа и маркерного гена устойчивости к лекарственному средству, полученной рекомбинантной ДНК трансформируется микроорганизм. Затем полученный трансформант культивируют при температуре, при которой чувствительная к температуре, контролирующая репликацию область не функционирует, и далее трансформант выращивают в среде, содержащей лекарственное средство, чтобы получить трансформированный штамм, в которой рекомбинантная ДНК введена в хромосомную ДНК.
В такой линии, где рекомбинантная ДНК введена в хромосомную ДНК, ndh ген делеционного типа рекомбинирован с ndh геном, изначально присутствующем на хромосоме, и два слитых гена хромосомного ndh гена и ndh гена делеционного типа введены в хромосому так, что другие части рекомбинантной ДНК (векторный сегмент, чувствительная к температуре контролирующая репликацию область и устойчивый к лекарству маркер) будут присутствовать между двумя слитыми генами. Следовательно, трансформант экспрессирует NDH-II, так как в этом состоянии нормальный ndh ген является доминантным.
Затем для того чтобы на хромосомной ДНК оставить только ndh ген делеционного типа, одну копию ndh гена из хромосомной ДНК удаляют вместе с векторным сегментом (включая чувствительную к температуре, контролирующую репликацию область и маркер лекарственной устойчивости) путем рекомбинации двух ndh генов. В этом случае нормальный ndh ген остается на хромосомной ДНК, а ndh ген делеционного типа вырезается из хромосомной ДНК или наоборот ndh ген делеционного типа остается на хромосомной ДНК, а нормальный ndh ген вырезается из хромосомной ДНК. В обоих случаях вырезаемая ДНК может быть сохранена в клетке как плазмида, когда клетку культивируют при температуре, при которой может функционировать чувствительная к температуре контролирующая репликацию область. Впоследствии клетку выращивают при температуре, при которой чувствительная к температуре, контролирующая репликацию область не может функционировать, чтобы элиминировать плазмидную ДНК, и может быть получен мутант с ndh геном делеционного типа.
Примерами вектора, имеющего чувствительную к температуре точку инициации репликации для Е.coli, являются плазмида pMAN997, описанная в Международной патентной публикации WO 99/03988, и т.д., а примером вектора, имеющего чувствительную к температуре точку инициации репликации для коринеформных бактерий, является плазмида pHSC4, описанная в нерассмотренной публикации японского патента №5-7491, и т.д. Однако плазмиды не ограничены этими плазмидами, и могут быть также использованы другие векторы.
Конкретными примерами таких микроорганизмов, полученных способом, описанным выше, являются микроорганизмы, у которых SoxM-оксидаза или NDH-I, или обе из них усилены, микроорганизмы, у которых активность цитохромоксидазы bd, или NDH-II, или активности обеих уменьшены, и микроорганизмы, у которых SoxM-оксидаза, или NDH-I, или обе из них усилены, а активность цитохромоксидазы bd, или NDH-II, или активности обеих понижены или устранены. Более конкретно можно назвать, например, Е.coli, где активность SохМ-оксидазы усилена, а NDH-II сделана дефицитной. Примером SохМ-оксидазы является цитохромоксидаза bo.
Микроорганизм, использованный в настоящем изобретении, не имеет особенных ограничений, пока ему могут быть приданы упомянутые выше свойства, и примерами такого микроорганизма являются бактерии, принадлежащие роду Eschirichia, такие как Е.coli, коринеформные бактерии, такие как Brevibacterium lactofermentum (Corynebacterium glutamicum), бактерии Bacillus, такие как Bacillus subtilis, бактерии Serratia, такие как Serratia marcescens, дрожжи, такие как Saccharomyces cerevisae, и т.д.
Если говорить более конкретно, то когда продуктом ферментации является L-треонин, можно назвать Е.coli VKPM В-3996 (RIA 1867) (см. патент США №5175107), Corynebacterium acetoacidophilum AJ12318 (FERM BP-1172) (см. патент США №5188949) и т.д.; в случае L-лизина можно назвать Е. coli AJ11442 (NRRL В-12185, FERM ВР-1543) (см. патент США №4346170), Е.coli W3110 (tyrA) (эта линия получена путем удаления плазмиды pHATerm из Е.coli W3110 (tyrA)/pHATerm (FERM ВР-3653) (см. Международную патентную публикацию WO 95/16042), Brevibacterium lactofermentum AJ12435 (FERM BP-2294) (см. патент США №5304476), Brevibacterium lactofermentum AJ3990 (АТСС31269) (см. патент США №4066501) и др.; в случае L-глутаминовой кислоты можно назвать Е.coli AJ12624 (FERM ВР-3853) (см. нерассмотренную публикацию французского патента №2680178), Brevibacterium lactofermentum AJ12821 (FERM BP-4172) (см. нерассмотренную публикацию японского патента №5-26811, нерассмотренную публикацию французского патента №2701489), Brevibacterium lactofermentum AJ12475 (FERM BP-2922) (см. патент США №5272067), Brevibacterium lactofermentum AJ13029 (FERM BP-5189) (см. Международную патентную публикацию JP(?) 95/01586) и др.; в случае L-лейцина можно назвать Е.coli АJ1478 (FERM P-5274) (см. Публикацию японского патента (Kokoku) №62-34397), Brevibacterium lactofermentum AJ3718 (FERM P-2516) (см. патент США №3970519) и др.; в случае L-изолейцина можно назвать Е.coli KX141 (VKPM B-4781) (см. нерассмотренную европейскую патентную публикацию №519113), Brevibacterium flavum AJ12149 (FERM BP-759) (см. патент США №4656135) и др.; в случае L-валина можно назвать Е.coli VL1970 (VKPM В-4411) (см. нерассмотренную европейскую патентную публикацию №519113), Brevibacterium lactofermentum AJ12341 (FERM BP-1763) (см. патент США №5188948) и др.; в случае L-фенилаланина можно назвать Е.coli AJ12604 (FERM BP-3579) (см. нерассмотренную публикацию японского патента №5-236947, нерассмотренную европейскую патентную публикацию №488424), Brevibacterium lactofermentum AJ12673 (FERM BP-4160) (см. нерассмотренную публикацию патента Франции №2686898) и т.д.
В микроорганизме использованный в настоящем изобретении, в зависимости от целевого вещества активность фермента, вовлеченного в биосинтез целевого вещества, может быть усилена. Кроме того, активность фермента, неблагоприятного для синтеза целевого соединения, может быть уменьшена или исключена.
Целевое вещество может быть получено путем выращивания такого микроорганизма, как микроорганизм, описанный выше, в среде с целью продуцирования и накопления целевого вещества в среде и сбора целевого вещества.
Среда, используемая для получения целевого вещества, может быть обычно используемой хорошо известной средой, выбранной в зависимости от применяемого микроорганизма. То есть среда может представлять собой обычную среду, содержащую источник углерода, источник азота, неорганические ионы, а также другие органические компоненты, если это необходимо. Никакая специальная среда в настоящем изобретении не требуется.
В качестве источника углерода можно использовать сахара, такие как глюкоза, лактоза, галактоза, фруктоза или крахмальный гидролизат; спирты, такие как глицерин или сорбит; органические кислоты, такие как фумаровая кислота, лимонная кислота или янтарная кислота и т.д.
В качестве источника азота возможно использование неорганических аммонийных солей, таких как сульфат аммония, хлорид аммония или фосфат аммония; органического азота, такого как гидролизат соевых бобов; газообразный аммиак; водный аммиак и др.
Желательно, чтобы требуемые вещества, такие как витамин B1, L-гомосерин и L-тирозин или дрожжевой экстракт, содержались в соответствующем количестве в качестве питательных органических веществ, добавленных в незначительных количествах. Помимо вышеназванных веществ в небольших количествах добавляют фосфат калия, сульфат магния, ион железа, ион марганца и т.п., если это необходимо.
Выращивание может быть осуществлено при обычно используемых хорошо известных условиях, выбранных в соответствии с применяемым микроорганизмом. Например, выращивание предпочтительно осуществлять в аэробных условиях в течение 16-120 часов. Температура выращивания предпочтительно регулируется так, чтобы она находилась в интервале от 25 до 45°С, а величина рН во время выращивания предпочтительно устанавливается при 5-8. Неорганические или органические, кислые или щелочные вещества, а также газообразный аммиак и подобные вещества могут быть использованы для доведения величины рН.
Для сбора метаболических продуктов из среды после выращивания в настоящем изобретение не требуется никаких специальных методов. То есть настоящее изобретение может быть осуществлено на практике с использованием сочетания обычных хорошо известных ионнообменных методик, методик треципитации и других методик.
Наилучший способ осуществления изобретения
Далее настоящее изобретение объясняется более подробно с помощью следующих примеров.
Пример 1: Клонирование гена цитохромоксидазы bo
Последовательность суо-оперона (cyoABCDE), кодирующая цитохромоксидазы bо Е.coli, уже описана (Chepure et al., The Journal of Biological Chemistry, 265:11185-11192, 1990), и поэтому оперон был клонирован на основе этой последовательности.
Более конкретно, ген целевого суо-оперона получали из библиотеки фагов Kohara (Kohara et al., Cell, 50:495-508, 1987), содержащей суо-оперон. Фаговую ДНК получали из фагового клона 147[2Н5] Kohara, содержащего оперон, с использованием лямбда-препарата Wizard (Promega). Полученную фаговую ДНК 147[2Н5] расщепляли с помощью PshBI, и полученный фрагмент размером 5.5 т.п.н., содержащий суо-оперон, “затупляли по концам” и вставляли в SmaI-сайт вектора pMW119 (Nippon Gene), чтобы клонировать суо-оперон, содержащий промоторную область. В полученной плазмиде суо-оперон был вставлен в обратном направлении относительно промотора лактозного оперона вектора pMW119. Эту плазмиду обозначали как pMW(CYO)B.
Плазмиду pMW(CYO)B вводили в штамм Е.coli W3110 (полученный из National Institute of Genetics, Mishima, Shizuoka, Япония) с получением штамма W3110/pMW(CYO)В. Активность убихинолоксидазы, присутствующей в клеточных экстрактах штамма W3110 и штамма W3110/pMW(CYO)B, измеряли как активность концевой оксидазы с использованием известного метода (Kita et al., The Journal of Biological Chemistry, 259:3368-3374, 1984). Полученные результаты представлены в таблице 1.

Установлено, что активность концевой оксидазы повышалась в штамме, в который введена плазмида pMW(CYO)B, как это видно из данных таблицы 1. Это усиление активности концевой оксидазы, как считают, вызвано усилением активности цитохромоксидазы bo за счет усиления суо-оперона.
Пример 2: Создание NDH-II-дефицитной линии
Для того чтобы получить NDH-II-дефицитную линию, получали внутренне-расщепленную частичную последовательность гена NDH-II (разрушенный ген NDH-II). Частичную последовательность гена NDH-II клонировали на основе известной последовательности ndh гена Е.coli, кодирующего NDH-II (Young et al. European Journal of Biochemistry, 116:165-170, 1981).
Более конкретно, разрушенный ген NDH-II получают следующим образом (фиг.1). Во-первых, фрагмент ДНК размером приблизительно 2.4 т.п.н., содержащий частичную последовательность гена NDH-II, получали путем амплификации хромосомной ДНК Е.coli с помощью ПЦР с использованием в качестве праймеров ndh-1 (SEQ ID NO: 1) и ndh-2 (SEQ ID NO: 2). Этот фрагмент клонировали в вектор pGEM-T (Promega) с получением плазмиды pGEM-ndh. Плазмиду pGEM-ndh расщепляли рестрикционными ферментами EcoRI и StuI, и полученный фрагмент ДНК размером 0.5 т.п.н. отбирали и лигировали с pTWV229 (Takara Shuzo), расщепленной ферментами EcoRI и StuI, с получением плазмиды pTWV-ndh.
Затем плазмиду pTWV-ndh расщепляли рестрикционным ферментом StuI, и полученный фрагмент размером 0.9 т.п.н. отбирали и вводили в HincII сайт плазмиды pTWV-ndh. Таким образом, получали плазмиду pTWVΔ ndh, содержащую часть множества сайтов рестрикции плазмиды pTWV229 в частичной последовательности ndh-гена. Плазмида pTWVΔ ndh содержит ndh-последовательность, вставленную с последовательностью из 17 пар оснований, берущую начало от pTWV299 при StuI-сайте, в ndh-последовательности. Затем фрагмент размером 1.5 т.п.н., полученный расщеплением pTWVΔ ndh с помощью HindII- и EcoRI, вставляли между HindII и EcoRI-сайтами чувствительной к температуре плазмиды pMAN997 (см. Международную патентную публикацию WO 99/03988) с получением плазмиды pTS-Δ ndh. Гомологическую рекомбинацию проводили между этой плазмидой pTS-Δ ndh и геномом штамма W3110 относительно ndh-гена с помощью обычной методики рекомбинации с использованием чувствительной к температуре плазмиды pTS-Δ ndh (Matuyama et al., Journal of Bacteriology, 162:1196, 1985) с получением штамма W3110(ndh), который не экспрессировал нормальный белок NDH-II, потому что последовательность из 17 пар оснований, происходящая от pTWV299, вставлена в кодирующую область ndh на геноме. Из штамма W3110 (tyrA) в штамм W3110 (ndh) вводили дефицитность по гену tyrA путем P1 трансдукции с использованием устойчивости к тетрациклину в качестве маркера с получением штамма W3110 (ndh, tyrA).
Упомянутая выше плазмида pMAN997 получена путем обмена VspI-HindIII фрагментов pMAN031 (J. Bacteriol., 162, 1196, 1985) и вектора pUC19 (Takara Shuzo) (фиг.2).
Кроме того, хотя штамм W3110 (tyrA) подробно описан в нерассмотренной публикации европейского патента №488424/1992, способ его получения кратко описан ниже.
Штамм Е.coli W3110 получен из National Institute of Genetics (Mishima, Shizuoka). Этот штамм высевали на LB-чашку, содержащую стрептомицин, и отбирали штамм, который образует колонию, получая штамм, устойчивый к стрептомицину. Отобранный устойчивый к стрептомицину штамм и штамм Е.coli K-12 ME8424 смешивали и выращивали в полной среде (L-жидкая питательная среда: 1% бактотриптона, 0,5% дрожжевого экстракта, 0,5% NaCl) при 37°С в течение 15 минут как стационарную культуру, чтобы индуцировать конъюгацию. Штамм Е.coli K-12 ME8424 имеет генетические признаки (HfrPО45, thi, relAl, tyrA:Tn10, ung-1, nadB), и его можно получить в National Institute of Genetics. Затем культуру высевали в полную среду (L-жидкая питательная среда: 1% бактотриптона, 0,5% дрожжевого экстракта, 0,5% NaCl, 1,5% агара), содержащую стрептомицин, тетрациклин и L-тирозин, и отбирали штамм, который образовывал колонию. Этот штамм обозначали как Е.coli W3110 (tyrA).
Нерассмотренная публикация европейского патента №488424/1992 раскрывает много штаммов, полученных путем введения плазмиды в вышеописанный штамм. Например, штамм, полученный путем введения плазмиды pHATerm, обозначен как Е.coli W3110 (tyrA)/pHATerm, сдан на хранение 16 ноября 1991 г. в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (1-3 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan, почтовый код: 305 (в настоящее время независимая административная корпорация, National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary (Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan, почтовый код: 305-5466) в качестве международного банка хранения, предусмотренного Будапештским Договором, и получила номер по каталогу FERM ВР-3653. Штамм Е.coli W3110 (tyrA) может быть получен путем элиминации плазмиды pHATerm из описанного выше штамма обычным способом.
Пример 3: Получение L-лизина
Плазмиду pMW(CYO)B, полученную в примере 1, вводили в штамм W3110 (tyrA) и штамм W3110 (ndh, tyrA), полученные в примере 2, с получением соответственно штаммов W3110 (tyrA)/pMW(CYO)B и W3110(ndh, tyrA)/pMW(CYO)В. Аналогично плазмиду pMW119 вводили в штамм W3110 (tyrA) с получением штамма W3110 (tyrA)/pMWl19. Путем выращивания в колбе оценивали продуктивность по L-лизину штамма W3110 (tyrA)/pMW(CYO)В, штамма W3110 (ndh, tyrA)/pMW(CYO)В и линии W3110 (tyrA)/pMW119 в качестве контроля. Выращивание проводили с использованием среды, имеющей приведенный ниже состав, при 37°С в течение от 24 до 48 часов при встряхивании. Полученные результаты приведены в таблице 2.
Состав среды
Глюкоза 40 г/л
MgSO4• 7H2O 1 г/л
KH2PO4 1 г/л
FeSO4• 7H2O 0,01 г/л
MnSO4• 5H2O 0,01 г/л
Дрожжевой экстракт (Difco) 2 г/л
L-тирозин 0,1 г/л или 0,05 г/л
Величину рН среды доводили до 7,0 с помощью КОН, и среду выдерживали в автоклаве при 115°С в течение 10 минут. Однако глюкозу и МgSO4• 7Н2O стерилизовали отдельно. Кроме того, в соответствии с Фармакопеей Японии перед выращиванием к среде добавляли 30 г/л СаСОз, который подвергнут сухой стерилизации при 180°С, и 100 мкг/л антибиотиков, ампициллина.
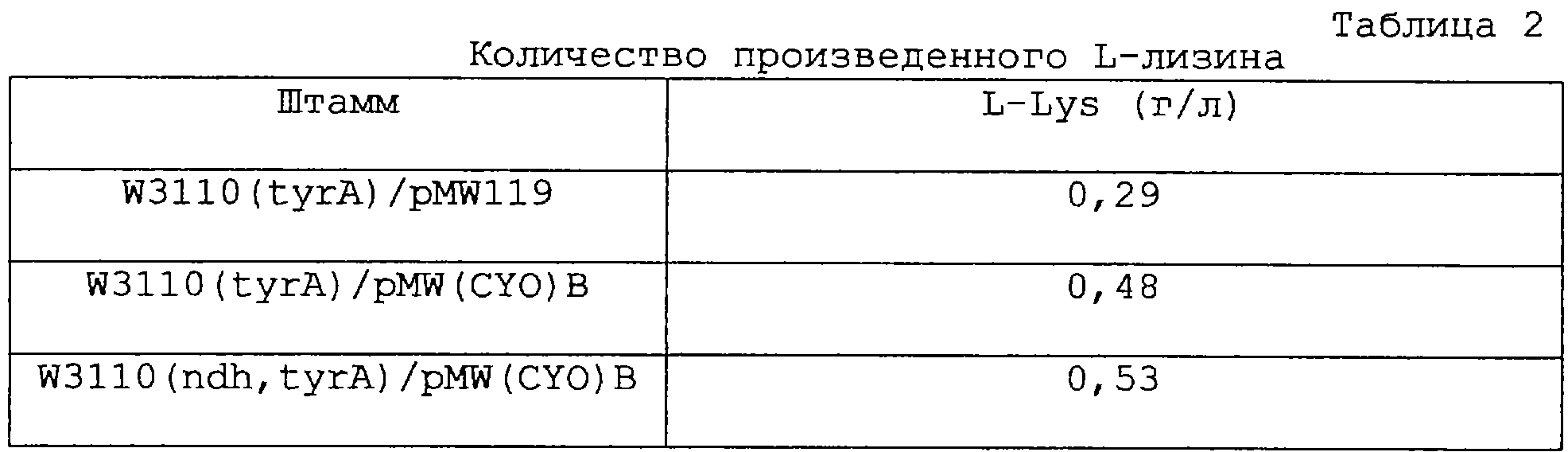
Установлено, что продуктивность по L-лизину улучшается в продуцирующих L-лизин бактериях Е.coli за счет усиления активности цитохромоксидазы bo. Это, как считают, происходит из-за того, что эффективность энергонакопления улучшается за счет усиления пути дыхательной цепи с высоким энергетическим выходом, и эта энергия используется для производства L-лизина.
Также установлено, что продуктивность по L-лизину улучшается в продуцирующих L-лизин бактериях Е.coli, сделанных дефицитными по NDH-II. Это, как считают, происходит из-за того, что эффективность энергонакопления улучшается за счет дефицитности пути дыхательной цепи с низким энергетическим выходом, и энергия используется для производства L-лизина.
Пример 4: Получение L-треонина
Плазмиду pMW(CYO)B, полученную описанным выше способом, вводили в продуцирующую L-треонин бактерию Е.coli VKPM В-3996 (RIA 1867) (см. патент США №5175107, далее называемую штаммом “В-3996”) с получением штамма B-3996/pMW(CYO)В. Штамм В-3996 нес плазмиду pVIC40 (Международная патентная публикация WO 90/04636), полученную путем вставки оперона треонина в векторную плазмиду pAYC32 широкого круга хозяев, содержащую устойчивый к стрептомицину маркер (см. Chistorerdov A.Y., Tsygankov Y.D., Plasmid, 1986, 16, 161-167). Штамм В-3996 депонирован в Институте исследования антибиотиков (СССР) (ВНИИА) под регистрационным номером RIA1867.
В качестве контроля получен штамм B-3996/pMW119 путем введения в штамм В-3996 плазмиды pMW119. Продуктивность по L-треонину этих штаммов B-3996/pMW (CYO)В и B-3996/pMW119 оценивали при выращивании в колбе. Выращивание проводили с использованием среды, имеющей состав, приведенный в таблице 3, при температуре 37°С в течение 38 часов при перемешивании со скоростью 114-116 об/мин. Компонент А, Компонент В и Компонент С, которые приведены в таблице 3, приготовлены и стерилизованы по отдельности, а затем охлаждены и смешаны в соотношении 16/20 объема Компонента А, 4/20 объема Компонента В и 30 г/л Компонента С. Полученные результаты представлены в таблице 4.


Установлено, что продуктивность по L-треонину продуцирующих L-треонин бактерий Е.coli может быть улучшена путем повышения активности цитохром bо-оксидазы.
Пример 5: Получение L-фенилаланина
Плазмиду рАСМАВ выделяли из штамма Е.coli W3110 (tyrA)/pACMAB, pBR-aroG4 в соответствии с обычным способом очистки плазмиды. Плазмида представляла собой плазмиду, полученную путем вставки фрагмента ДНК, содержащего ген для десенсибилизированной хоризматмутаза/префенатдегидротазы (СМ-PDH) в соответствующей системе биосинтеза L-фенилаланина, между BamHI- и HindIII-рестрикционными сайгами векторной плазмиды pACYC184 (Арr) (см. Международную патентную публикацию WO 97/08333). Штамм W3110 (tyrA)/pACMAB, pBR-aroG4 (обозначаемый как AJ12604) сдан на хранении в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (1-3 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan, почтовый код: 305) и получил номер по каталогу FERM Р-11975. Затем он был передан в международный банк хранения в соответствии с Будапештским Договором от 26 сентября 1991 г. и получил номер по каталогу FERM BP-3579.
Плазмиду pACMAB “затупляли по концам” путем расщепления ферментом SalI. В нее вставляли “затупленный по концам” фрагмент ДНК, содержащий суо-оперон размером 5.5 т.п.н., который получен из ДНК фага 147[2Н5] Kohara, упомянутого выше, путем расщепления с помощью PshBI. Полученную плазмиду рАСМАВ-суо вводили в штамм W3110 (tyrA/pBR-aroG4). Полученный трансформант выращивали в среде для получения L-фенилаланина (содержащую 20 г глюкозы, 29,4 г гидрофосфата натрия, 6 г дигидрофосфата калия, 1 г хлорида натрия, 2 г хлорида аммония, 10 г цитрата натрия, 0,4 г глутамата натрия, 3 г семиводного гидрата сульфата магния, 0,23 г хлорида кальция, 2 мг гидрохлорида тиамина и 100 мг L-тирозина в 1 л воды, рН 7,0) при 37°С в течение 40 часов. L-фенилаланин, содержащийся в среде, количественно оценивали с помощью высокоэффективной жидкостной хроматографии. Полученные результаты представлены в таблице 5.

Установлено, что продуктивность по L-фенилаланину продуцирующих L-фенилаланин бактерий Е.coli улучшается за счет усиления активности цитохромоксидазы bo.


Claims (8)
1. Способ получения L-аминокислоты с использованием микроорганизма, включающий стадии выращивания микроорганизма в среде для продуцирования и накопления L-аминокислоты в среде и сбора L-аминокислоты, где микроорганизм создан из родительского штамма микроорганизма, имеющего путь дыхательной цепи с высоким энергетическим выходом и путь дыхательной цепи с низким энергетическим выходом в качестве путей дыхательной цепи, и этот микроорганизм представляет собой мутантный штамм или штамм генетического рекомбинанта, имеющего одну или обе из следующих характеристик: A) Путь дыхательной цепи с высоким энергетическим выходом усилен; B) Путь дыхательной цепи с низким энергетическим выходом является дефицитным, где микроорганизм выбирают из группы, состоящей из бактерии, принадлежащей к роду Escherichia, и коринеформной бактерии.
2. Способ получения L-аминокислоты по п.1, где путь дыхательной цепи с высоким энергетическим выходом усилен путем увеличения копийности гена, кодирующего фермент, вовлеченный в дыхательную цепь, или модификации, регулирующей экспрессию последовательности гена.
3. Способ получения L-аминокислоты по п.1 или 2, где путь дыхательной цепи с низким энергетическим выходом сделан дефицитным путем разрушения гена, кодирующего фермент, вовлеченный в дыхательную цепь.
4. Способ получения L-аминокислоты по любому из пп.1-3, где ферментами дыхательной цепи с высоким энергетическим выходом у коринеформных бактерий являются bс 1-комплекс и SoxM-оксидаза, а у бактерий рода Escherichia - NDH-I и SoxM-оксидаза.
5. Способ получения L-аминокислоты по любому из пп.1-4, где ферментами дыхательной цепи с низким энергетическим выходом у бактерий рода Escherichia являются цитохромоксидаза bd и NDH-II, а у коринеформных бактерий - цитохромоксидаза bd.
6. Способ получения L-аминокислоты по любому из пп.1-5, где в микроорганизме активность SoxM-оксидазы усилена, а NDH-II сделана дефицитной.
7. Способ получения L-аминокислоты по любому из пп.1-6, где фермент пути дыхательной цепи с высоким энергетическим выходом представляет собой цитохромоксидазу bo.
8. Способ получения L-аминокислоты по любому из пп.1-7, где микроорганизм представляет собой бактерию, принадлежащую к роду Escherichia.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2000-204252 | 2000-07-05 | ||
| JP2000204252A JP4380029B2 (ja) | 2000-07-05 | 2000-07-05 | 微生物を利用した物質の製造法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| RU2001118542A RU2001118542A (ru) | 2003-06-20 |
| RU2238325C2 true RU2238325C2 (ru) | 2004-10-20 |
Family
ID=18701548
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2001118542A RU2238325C2 (ru) | 2000-07-05 | 2001-07-04 | Способ получения l-аминокислоты c использованием микроорганизма |
Country Status (16)
| Country | Link |
|---|---|
| US (2) | US8586334B2 (ru) |
| EP (2) | EP1170376B1 (ru) |
| JP (1) | JP4380029B2 (ru) |
| KR (1) | KR100805644B1 (ru) |
| CN (1) | CN1280419C (ru) |
| AT (2) | ATE501265T1 (ru) |
| AU (1) | AU782560B2 (ru) |
| BR (1) | BRPI0102666B1 (ru) |
| DE (2) | DE60144205D1 (ru) |
| DK (2) | DK2067864T3 (ru) |
| ES (2) | ES2331601T3 (ru) |
| MY (1) | MY127434A (ru) |
| PE (1) | PE20020172A1 (ru) |
| PL (1) | PL206590B1 (ru) |
| RU (1) | RU2238325C2 (ru) |
| TW (1) | TWI238192B (ru) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2819270C1 (ru) * | 2019-12-23 | 2024-05-16 | СиДжей ЧеилДжеданг Корпорейшн | Микроорганизм для продуцирования L-аминокислоты, обладающий повышенной активностью цитохрома С, и способ получения L-аминокислоты с его использованием |
Families Citing this family (94)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2175351C2 (ru) | 1998-12-30 | 2001-10-27 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО "АГРИ") | Фрагмент днк из escherichia coli, определяющий повышенную продукцию l-аминокислот (варианты), и способ получения l-аминокислот |
| JP2001046067A (ja) * | 1999-08-04 | 2001-02-20 | Ajinomoto Co Inc | 好熱性バチルス属細菌由来のl−リジン生合成系遺伝子 |
| PL341895A1 (en) * | 1999-08-12 | 2001-02-26 | Ajinomoto Kk | Plasmide autonomously replicable in corynebacter bacteria |
| BRPI0016995B1 (pt) * | 2000-01-21 | 2016-03-08 | Ajinomoto Kk | bactéria escherichia coli geneticamente modificada, e, método para produzir l-lisina |
| WO2003072785A2 (en) * | 2002-02-27 | 2003-09-04 | Dsm Ip Assets B.V. | Fermentation process |
| WO2004033421A2 (en) | 2002-10-04 | 2004-04-22 | Genencor International, Inc. | Improved production of bacterial strains |
| KR101148255B1 (ko) | 2002-10-04 | 2012-08-08 | 다니스코 유에스 인크. | 고수율을 갖는 1,3-프로판디올의 생물학적 제조 방법 |
| BR0304860A (pt) * | 2002-11-11 | 2004-08-31 | Ajinomoto Kk | Método para produzir uma substância alvo pela utilização de uma-bactéria pertencente ao gênero escherichia |
| DE10254074A1 (de) * | 2002-11-19 | 2004-06-17 | Forschungszentrum Jülich GmbH | Verfahren zur mikrobiellen Herstellung von Stoffwechselprodukten |
| MXPA05008857A (es) * | 2003-02-18 | 2006-03-09 | Metabolic Explorer Sa | Procedimiento de preparacion de microorganismos evolucionados que permite la creacion o la modificacion de vias metabolicas. |
| US7335496B2 (en) * | 2003-06-05 | 2008-02-26 | Ajinomoto Co., Inc. | Method for producing target substance |
| PL1651758T3 (pl) * | 2003-07-29 | 2009-04-30 | Ajinomoto Kk | Sposób wytwarzania L-lizyny lub L-treoniny przy użyciu bakterii Escherichia o atenuowanej aktywności enzymu jabłczanowego |
| FR2862068B1 (fr) * | 2003-11-06 | 2007-10-12 | Metabolic Explorer Sa | Souches de microorganismes optimisees pour des voies de biosyntheses consommatrices de nadph |
| JP4380305B2 (ja) * | 2003-11-21 | 2009-12-09 | 味の素株式会社 | 発酵法によるl−アミノ酸の製造法 |
| PL1664318T3 (pl) * | 2004-01-30 | 2010-03-31 | Ajinomoto Kk | Mikroorganizm wytwarzający L-aminokwas i sposób wytwarzania L-aminokwasu |
| JP4780783B2 (ja) * | 2004-02-27 | 2011-09-28 | 協和発酵バイオ株式会社 | アミノ酸の製造法 |
| US7344874B2 (en) * | 2004-03-04 | 2008-03-18 | Ajinomoto Co., Inc. | L-glutamic acid-producing microorganism and a method for producing L-glutamic acid |
| US7300776B2 (en) | 2004-04-26 | 2007-11-27 | Ajinomoto Co., Inc. | L-amino acid-producing bacterium and a method for producing L-amino acid |
| US7482140B2 (en) * | 2004-06-15 | 2009-01-27 | Ajinomoto Co., Inc. | L-tyrosine-producing bacterium and a method for producing L-tyrosine |
| US7205132B2 (en) * | 2004-09-10 | 2007-04-17 | Ajinomoto Co., Inc. | L-glutamic acid-producing microorganism and a method for producing L-glutamic acid |
| US7794989B2 (en) * | 2004-12-28 | 2010-09-14 | Ajinomoto Co., Inc. | L-glutamic acid-producing microorganism and a method for producing L-glutamic acid |
| US7547531B2 (en) | 2005-01-18 | 2009-06-16 | Ajinomoto Co., Inc. | L-amino acid producing microorganism which has been modified to inactive the fimH gene, and a method for producing I-amino acid |
| JP4985407B2 (ja) * | 2005-05-30 | 2012-07-25 | 味の素株式会社 | ポリ−γ−グルタミン酸の製造法及びその製造法に用いられる微生物 |
| US20070004014A1 (en) * | 2005-06-29 | 2007-01-04 | Yuichiro Tsuji | Method for producing l-threonine |
| JP2008283863A (ja) | 2005-08-26 | 2008-11-27 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造方法 |
| JP2007185184A (ja) * | 2005-12-16 | 2007-07-26 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| EP1979486B1 (en) | 2006-01-30 | 2013-04-17 | Ajinomoto Co., Inc. | L-amino acid producing bacterium and method of producing l-amino acid |
| JP2009095237A (ja) * | 2006-02-02 | 2009-05-07 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| JP2009118740A (ja) * | 2006-03-03 | 2009-06-04 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| WO2007119574A2 (en) | 2006-03-23 | 2007-10-25 | Ajinomoto Co., Inc. | A method for producing an l-amino acid using bacterium of the enterobacteriaceae family with attenuated expression of a gene coding for small rna |
| JP2009060791A (ja) | 2006-03-30 | 2009-03-26 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| JP2009165355A (ja) | 2006-04-28 | 2009-07-30 | Ajinomoto Co Inc | L−アミノ酸を生産する微生物及びl−アミノ酸の製造法 |
| KR101109402B1 (ko) | 2006-05-09 | 2012-01-30 | 미쓰이 가가쿠 가부시키가이샤 | 보효소 재생에 의한 히드록시카르복실산류의 생산방법 |
| JP4954985B2 (ja) * | 2006-05-09 | 2012-06-20 | 三井化学株式会社 | 補酵素合成強化によるグリコール酸の生産方法 |
| WO2007136133A1 (en) | 2006-05-23 | 2007-11-29 | Ajinomoto Co., Inc. | A method for producing an l-amino acid using a bacterium of the enterobacteriaceae family |
| JP5407858B2 (ja) * | 2006-07-19 | 2014-02-05 | 味の素株式会社 | 腸内細菌科の細菌を用いたl−アミノ酸の製造方法 |
| US20080293101A1 (en) * | 2006-07-27 | 2008-11-27 | Peters Matthew W | Engineered microorganisms for increasing product yield in biotransformations, related methods and systems |
| US9562224B2 (en) * | 2006-09-07 | 2017-02-07 | William Marsh Rice University | Reduced activity of ubiCA in E. coli |
| JP2010017081A (ja) * | 2006-10-10 | 2010-01-28 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| WO2008072761A2 (en) * | 2006-12-11 | 2008-06-19 | Ajinomoto Co., Inc. | Method for producing an l-amino acid |
| RU2006143864A (ru) | 2006-12-12 | 2008-06-20 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) (RU) | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, В КОТОРОЙ ОСЛАБЛЕНА ЭКСПРЕССИЯ ГЕНОВ cynT, cynS, cynX, ИЛИ cynR, ИЛИ ИХ КОМБИНАЦИИ |
| RU2006145712A (ru) * | 2006-12-22 | 2008-06-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) (RU) | Способ получения l-аминокислот методом ферментации с использованием бактерий, обладающих повышенной способностью к утилизации глицерина |
| CN101627110B (zh) | 2007-01-22 | 2014-08-13 | 味之素株式会社 | 生产l-氨基酸的微生物和l-氨基酸的生产方法 |
| JP2010088301A (ja) * | 2007-02-01 | 2010-04-22 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| JP2010110217A (ja) | 2007-02-22 | 2010-05-20 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| JP2010226956A (ja) * | 2007-07-23 | 2010-10-14 | Ajinomoto Co Inc | L−リジンの製造法 |
| CN101939412B (zh) | 2007-09-04 | 2016-01-20 | 味之素株式会社 | 生产氨基酸的微生物以及氨基酸的生产方法 |
| JP2010263790A (ja) * | 2007-09-04 | 2010-11-25 | Ajinomoto Co Inc | アミノ酸生産微生物及びアミノ酸の製造法 |
| RU2395579C2 (ru) * | 2007-12-21 | 2010-07-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia |
| JP2011067095A (ja) | 2008-01-10 | 2011-04-07 | Ajinomoto Co Inc | 発酵法による目的物質の製造法 |
| EP2248906A4 (en) | 2008-01-23 | 2012-07-11 | Ajinomoto Kk | PROCESS FOR THE PREPARATION OF L-AMINO ACID |
| RU2008105793A (ru) | 2008-02-19 | 2009-08-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) (RU) | Способ конструирования оперонов, содержащих трансляционно сопряженные гены, бактерия, содержащая такой оперон, способ продукции полезного метаболита и способ мониторинга экспрессии гена |
| JP2011167071A (ja) | 2008-05-22 | 2011-09-01 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| EP2336347B1 (en) | 2008-09-08 | 2017-03-15 | Ajinomoto Co., Inc. | An l-amino acid-producing microorganism and a method for producing an l-amino acid |
| KR101123213B1 (ko) * | 2008-09-11 | 2012-03-19 | 부산대학교 산학협력단 | 미생물 전자전달계 및 탄소원 대사경로가 재설계된 재조합 대장균 생촉매 |
| JP2012029565A (ja) | 2008-11-27 | 2012-02-16 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| JP2010142200A (ja) | 2008-12-22 | 2010-07-01 | Ajinomoto Co Inc | L−リジンの製造法 |
| BRPI1007069A2 (pt) | 2009-01-23 | 2015-08-25 | Ajinomoto Kk | Método para produzir um l-aminoácido. |
| JP5521347B2 (ja) | 2009-02-16 | 2014-06-11 | 味の素株式会社 | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| JPWO2011013707A1 (ja) | 2009-07-29 | 2013-01-10 | 味の素株式会社 | L−アミノ酸の製造法 |
| JP2012223092A (ja) | 2009-08-28 | 2012-11-15 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| JP2013013329A (ja) | 2009-11-06 | 2013-01-24 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| RU2460793C2 (ru) | 2010-01-15 | 2012-09-10 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО АГРИ) | Способ получения l-аминокислот с использованием бактерий семейства enterobacteriaceae |
| RU2010101135A (ru) | 2010-01-15 | 2011-07-20 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО АГРИ) (RU) | Бактерия семейства enterobacteriaceae - продуцент l-аспартата или метаболитов, производных l-аспартата, и способ получения l-аспартата или метаболитов, производных l-аспартата |
| JP2013074795A (ja) | 2010-02-08 | 2013-04-25 | Ajinomoto Co Inc | 変異型rpsA遺伝子及びL−アミノ酸の製造法 |
| RU2471868C2 (ru) | 2010-02-18 | 2013-01-10 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО АГРИ) | Мутантная аденилатциклаза, днк, кодирующая ее, бактерия семейства enterobacteriaceae, содержащая указанную днк, и способ получения l-аминокислот |
| RU2471870C2 (ru) | 2010-06-03 | 2013-01-10 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО АГРИ) | СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА И L-ЦИТРУЛЛИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, В КОТОРОЙ ОСЛАБЛЕНА ЭКСПРЕССИЯ ГЕНА pepA |
| RU2010122646A (ru) | 2010-06-03 | 2011-12-10 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО АГРИ) (RU) | Способ получения l-аминокислоты с использованием бактерии семейства enterobacteriaceae, в которой ослаблена экспрессия генов, кодирующих транспортер лизина/аргинина/орнитина |
| RU2482188C2 (ru) | 2010-07-21 | 2013-05-20 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО АГРИ) | СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ РОДА Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ОПЕРОН astCADBE |
| RU2501858C2 (ru) | 2010-07-21 | 2013-12-20 | Закрытое акционерное общество "Научно-исследовательский институт "Аджиномото-Генетика" (ЗАО АГРИ) | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae |
| JP2014036576A (ja) | 2010-12-10 | 2014-02-27 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| RU2011134436A (ru) | 2011-08-18 | 2013-10-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО "АГРИ") | Способ получения l-аминокислоты с использованием бактерии семейства enterobacteriaceae, обладающей повышенной экспрессией генов каскада образования флагелл и клеточной подвижности |
| JP2015013812A (ja) | 2011-11-01 | 2015-01-22 | 味の素株式会社 | 植物ウイルスの感染抑制剤およびそれを用いた植物ウイルス感染抑制方法 |
| RU2013118637A (ru) | 2013-04-23 | 2014-10-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО "АГРИ") | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, В КОТОРОЙ РАЗРЕГУЛИРОВАН ГЕН yjjK |
| PE20150681A1 (es) | 2013-05-13 | 2015-05-15 | Ajinomoto Kk | Metodo para producir l-aminoacidos |
| WO2015014829A1 (en) * | 2013-07-31 | 2015-02-05 | F. Hoffmann-La Roche Ag | Method for the recombinant production of a polypeptide in prokaryotic cells |
| RU2013140115A (ru) | 2013-08-30 | 2015-03-10 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО "АГРИ") | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae, В КОТОРОЙ НАРУШЕНА ЭКСПРЕССИЯ КЛАСТЕРА ГЕНОВ znuACB |
| JP2016192903A (ja) | 2013-09-17 | 2016-11-17 | 味の素株式会社 | 海藻由来バイオマスからのl−アミノ酸の製造方法 |
| JP5958653B2 (ja) | 2013-10-02 | 2016-08-02 | 味の素株式会社 | アンモニア制御装置およびアンモニア制御方法 |
| RU2013144250A (ru) | 2013-10-02 | 2015-04-10 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО "АГРИ") | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae, В КОТОРОЙ ОСЛАБЛЕНА ЭКСПРЕССИЯ ГЕНА, КОДИРУЮЩЕГО ФОСФАТНЫЙ ТРАНСПОРТЕР |
| JP6459962B2 (ja) | 2013-10-21 | 2019-01-30 | 味の素株式会社 | L−アミノ酸の製造法 |
| JP6519476B2 (ja) | 2013-10-23 | 2019-05-29 | 味の素株式会社 | 目的物質の製造法 |
| RU2015120052A (ru) | 2015-05-28 | 2016-12-20 | Аджиномото Ко., Инк. | Способ получения L-аминокислоты с использованием бактерии семейства Enterobacteriaceae, в которой ослаблена экспрессия гена gshA |
| BR112018017227A2 (pt) | 2016-02-25 | 2019-02-05 | Ajinomoto Kk | método para produzir um l-aminoácido |
| JP7066977B2 (ja) | 2017-04-03 | 2022-05-16 | 味の素株式会社 | L-アミノ酸の製造法 |
| WO2020071538A1 (en) | 2018-10-05 | 2020-04-09 | Ajinomoto Co., Inc. | Method for producing target substance by bacterial fermentation |
| WO2020138178A1 (en) | 2018-12-27 | 2020-07-02 | Ajinomoto Co., Inc. | Method for producing basic l-amino acids or salts thereof by fermentation of an enterobacteriaceae bacterium |
| BR112021014194A2 (pt) | 2019-02-22 | 2021-12-28 | Ajinomoto Kk | Método para a produção de um l-aminoácido |
| BR112021017870A2 (pt) | 2019-04-05 | 2021-12-07 | Ajinomoto Kk | Método para produzir um l-aminoácido |
| CN111909917B (zh) * | 2019-05-10 | 2022-10-14 | 中国科学院微生物研究所 | 一种内溶素Lysmeta1及其编码基因与应用 |
| JP2022550084A (ja) | 2019-09-25 | 2022-11-30 | 味の素株式会社 | 細菌の発酵によるl-アミノ酸の製造方法 |
| WO2021133030A1 (ko) * | 2019-12-23 | 2021-07-01 | 씨제이제일제당 (주) | 사이토크롬 c 활성이 강화된 l-아미노산 생산 미생물 및 이를 이용한 l-아미노산 생산방법 |
| CN113106075B (zh) * | 2021-02-25 | 2022-06-14 | 齐鲁工业大学 | 一种细胞色素氧化酶突变体及其应用 |
| JP2024003706A (ja) * | 2022-06-27 | 2024-01-15 | 花王株式会社 | 芳香族化合物の製造方法 |
Family Cites Families (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4104124A (en) * | 1976-08-30 | 1978-08-01 | Louisiana State University Foundation | Process for the production of single cell protein and amino acids |
| DE4027453A1 (de) * | 1990-08-30 | 1992-03-05 | Degussa | Neue plasmide aus corynebacterium glutamicum und davon abgeleitete plasmidvektoren |
| US5830716A (en) * | 1993-10-28 | 1998-11-03 | Ajinomoto Co., Inc. | Increased amounts of substances by modifying a microorganism to increase production of NADPH from NADH |
| FI980551A (fi) * | 1998-03-11 | 1999-09-12 | Valtion Teknillinen | Transformoidut mikro-organismit, joilla on parannettuja ominaisuuksia |
| US6074830A (en) * | 1998-06-09 | 2000-06-13 | E. I. Du Pont De Nemours & Company | 3,4-dihydroxy-2-butanone 4-phosphate synthase |
| JPH11346776A (ja) * | 1998-06-11 | 1999-12-21 | Ajinomoto Co Inc | ブレビバクテリウム・ラクトファーメンタムのシトクロムbd型キノールオキシダーゼ遺伝子 |
| US7220571B2 (en) * | 2000-09-28 | 2007-05-22 | Archer-Daniels-Midland Company | Escherichia coli strains which over-produce L-threonine and processes for the production of L-threonine by fermentation |
| ATE498691T1 (de) * | 2003-10-21 | 2011-03-15 | Cargill Inc | Herstellung von monatin und monatinvorstufen |
-
2000
- 2000-07-05 JP JP2000204252A patent/JP4380029B2/ja not_active Expired - Lifetime
-
2001
- 2001-07-02 EP EP01116050A patent/EP1170376B1/en not_active Expired - Lifetime
- 2001-07-02 ES ES01116050T patent/ES2331601T3/es not_active Expired - Lifetime
- 2001-07-02 ES ES09003058T patent/ES2360174T3/es not_active Expired - Lifetime
- 2001-07-02 DK DK09003058.6T patent/DK2067864T3/da active
- 2001-07-02 DE DE60144205T patent/DE60144205D1/de not_active Expired - Lifetime
- 2001-07-02 AU AU54169/01A patent/AU782560B2/en not_active Expired
- 2001-07-02 AT AT09003058T patent/ATE501265T1/de not_active IP Right Cessation
- 2001-07-02 AT AT01116050T patent/ATE442453T1/de not_active IP Right Cessation
- 2001-07-02 DE DE60139838T patent/DE60139838D1/de not_active Expired - Lifetime
- 2001-07-02 EP EP09003058A patent/EP2067864B1/en not_active Expired - Lifetime
- 2001-07-02 DK DK01116050T patent/DK1170376T3/da active
- 2001-07-03 MY MYPI20013161A patent/MY127434A/en unknown
- 2001-07-04 BR BRPI0102666A patent/BRPI0102666B1/pt active IP Right Grant
- 2001-07-04 PE PE2001000664A patent/PE20020172A1/es not_active Application Discontinuation
- 2001-07-04 PL PL348448A patent/PL206590B1/pl unknown
- 2001-07-04 KR KR1020010039675A patent/KR100805644B1/ko active IP Right Grant
- 2001-07-04 RU RU2001118542A patent/RU2238325C2/ru active
- 2001-07-05 CN CNB01125954XA patent/CN1280419C/zh not_active Expired - Lifetime
- 2001-07-05 TW TW090116463A patent/TWI238192B/zh not_active IP Right Cessation
- 2001-07-05 US US09/897,988 patent/US8586334B2/en active Active
-
2013
- 2013-10-16 US US14/055,302 patent/US20140045219A1/en not_active Abandoned
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2819270C1 (ru) * | 2019-12-23 | 2024-05-16 | СиДжей ЧеилДжеданг Корпорейшн | Микроорганизм для продуцирования L-аминокислоты, обладающий повышенной активностью цитохрома С, и способ получения L-аминокислоты с его использованием |
Also Published As
| Publication number | Publication date |
|---|---|
| US20140045219A1 (en) | 2014-02-13 |
| KR20020005441A (ko) | 2002-01-17 |
| MY127434A (en) | 2006-12-29 |
| EP1170376B1 (en) | 2009-09-09 |
| DE60144205D1 (de) | 2011-04-21 |
| JP4380029B2 (ja) | 2009-12-09 |
| KR100805644B1 (ko) | 2008-02-26 |
| EP1170376A1 (en) | 2002-01-09 |
| DE60139838D1 (de) | 2009-10-22 |
| ES2331601T3 (es) | 2010-01-11 |
| TWI238192B (en) | 2005-08-21 |
| ATE442453T1 (de) | 2009-09-15 |
| ATE501265T1 (de) | 2011-03-15 |
| AU5416901A (en) | 2002-01-10 |
| BR0102666A (pt) | 2002-02-26 |
| US20020160461A1 (en) | 2002-10-31 |
| PL206590B1 (pl) | 2010-08-31 |
| US8586334B2 (en) | 2013-11-19 |
| AU782560B2 (en) | 2005-08-11 |
| DK2067864T3 (da) | 2011-06-27 |
| EP2067864A1 (en) | 2009-06-10 |
| EP2067864B1 (en) | 2011-03-09 |
| CN1335401A (zh) | 2002-02-13 |
| CN1280419C (zh) | 2006-10-18 |
| PE20020172A1 (es) | 2002-03-22 |
| PL348448A1 (en) | 2002-01-14 |
| ES2360174T3 (es) | 2011-06-01 |
| JP2002017363A (ja) | 2002-01-22 |
| DK1170376T3 (da) | 2009-12-14 |
| BRPI0102666B1 (pt) | 2017-05-02 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| RU2238325C2 (ru) | Способ получения l-аминокислоты c использованием микроорганизма | |
| JP4265093B2 (ja) | スレオニン及びイソロイシンの製造法 | |
| JP4380305B2 (ja) | 発酵法によるl−アミノ酸の製造法 | |
| US7833761B2 (en) | Amino acid producing microorganism and a method for producing an amino acid | |
| EP1740694B1 (en) | L-tryptophan-producing bacterium and a method for producing l-tryptophan | |
| JP3861290B2 (ja) | ストレス耐性微生物及び発酵生産物の製造法 | |
| CZ121396A3 (en) | Process for preparing chemical compounds | |
| WO2007037460A9 (en) | An l-amino acid-producing bacterium and a method for producing l-amino acids | |
| US7163810B2 (en) | Method for producing target substance | |
| US20050106688A1 (en) | Method for producing L-amino acid | |
| ES2659513T3 (es) | Microorganismo que produce L-treonina teniendo el gen tyrR inactivado, procedimiento de producción del mismo y procedimiento de producción de L-treonina usando el microorganismo | |
| WO2001005959A1 (fr) | Obtention d'une substance cible par un procede de fermentation | |
| JP2007117076A (ja) | L−アミノ酸生産菌及びl−アミノ酸の製造法 | |
| JP2000050894A (ja) | 発酵生産物の製造法及びストレス耐性微生物 | |
| JP2007117077A (ja) | L−アミノ酸生産菌及びl−アミノ酸の製造法 | |
| BamHI | S kkkk I | |
| WO2001005979A1 (fr) | Methode d'elaboration d'une substance cible par fermentation |