KR20230065381A - 유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 - Google Patents
유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 Download PDFInfo
- Publication number
- KR20230065381A KR20230065381A KR1020237014846A KR20237014846A KR20230065381A KR 20230065381 A KR20230065381 A KR 20230065381A KR 1020237014846 A KR1020237014846 A KR 1020237014846A KR 20237014846 A KR20237014846 A KR 20237014846A KR 20230065381 A KR20230065381 A KR 20230065381A
- Authority
- KR
- South Korea
- Prior art keywords
- tale
- sequence
- domain
- dna
- crispr
- Prior art date
Links
Images







































































































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
- C12N15/902—Stable introduction of foreign DNA into chromosome using homologous recombination
- C12N15/907—Stable introduction of foreign DNA into chromosome using homologous recombination in mammalian cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
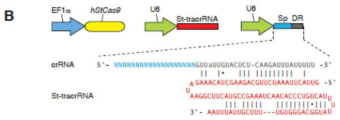
- C12N15/635—Externally inducible repressor mediated regulation of gene expression, e.g. tetR inducible by tetracyline
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
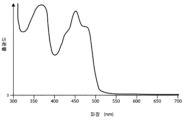
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/102—Mutagenizing nucleic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
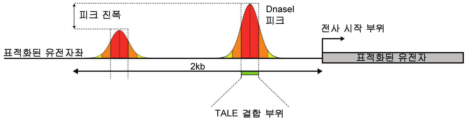
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/62—DNA sequences coding for fusion proteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/01—Fusion polypeptide containing a localisation/targetting motif
- C07K2319/09—Fusion polypeptide containing a localisation/targetting motif containing a nuclear localisation signal
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/16011—Human Immunodeficiency Virus, HIV
- C12N2740/16041—Use of virus, viral particle or viral elements as a vector
- C12N2740/16043—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14111—Dependovirus, e.g. adenoassociated viruses
- C12N2750/14141—Use of virus, viral particle or viral elements as a vector
- C12N2750/14143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Wood Science & Technology (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Medicinal Chemistry (AREA)
- Crystallography & Structural Chemistry (AREA)
- Virology (AREA)
- Mycology (AREA)
- Cell Biology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Enzymes And Modification Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Epidemiology (AREA)
- Pharmacology & Pharmacy (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Abstract
본 발명은 일반적으로 유도 전사 효과기를 사용할 수 있는 유전자 발현의 공간적이고 시간적인 제어에 사용되는 방법 및 조성물에 관한 것이다. 본 발명은 특히 세포에서의 관심 게놈 유전자좌의 발현을 변형하거나 교란하는 유도 방법에 관한 것이며, 이때 상기 게놈 유전자좌는 데옥시리보핵산 (DNA) 결합 폴리펩티드를 포함하는 비-자연발생적 또는 조작된 조성물과 접촉될 수 있다.
Description
관련 명세서 및 참고문헌
이 명세서는 미국 가출원 일련 번호 61/675,778 (2012 년 7 월 25 일 출원), 61/721,283 (2012 년 11 월 1 일 출원), 61/736,465 (2012 년 12 월 12 일 출원), 61/794,458 (2013 년 3 월 15 일 출원) 및 61/835,973 (2013 년 6 월 17 일 출원) (발명의 명칭: 유도성 DNA 결합 단백질 및 게놈 교란 도구 및 이의 적용 (INDUCIBLE DNA BINDING PROTEINS AND GENOME PERTURBATION TOOLS AND APPLICATIONS THEREOF)) 에 우선권을 주장한다.
미국 가출원 번호 61/565,171 (2011 년 11 월 30 일 출원) 및 미국 출원 번호 13/554,922 (2012 년 7 월 30 일 출원) 및 13/604,945 (2012 년 9 월 6 일 출원) (발명의 명칭: 설계자 TAL 효과기에 대한 뉴클레오티드-특이적 인지 서열 (NUCLEOTIDE-SPECIFIC RECOGNITION SEQUENCES FOR DESIGNER TAL EFFECTORS)) 를 또한 참고한다.
미국 가출원 번호 61/736,527 (2012 년 12 월 12 일 출원); 61/748,427 (2013 년 1 월 2 일 출원); 61/757,972 (2013 년 1 월 29 일 출원), 61/768,959 (2013 년 2 월 25 일 출원) 및 61/791,409 (2013 년 3 월 15 일 출원) (발명의 명칭: 서열 조작을 위한 시스템 방법 및 조성물 (SYSTEMS METHODS AND COMPOSITIONS FOR SEQUENCE MANIPULATION)) 를 또한 참고한다.
미국 가출원 번호 61/758,468 (2013 년 1 월 30 일 출원) 및 61/769,046 (2013 년 3 월 15 일 출원) (발명의 명칭: 서열 조작을 위한 시스템, 방법 및 조성물의 조작 및 최적화 (ENGINEERING AND OPTIMIZATION OF SYSTEMS, METHODS AND COMPOSITIONS FOR SEQUENCE MANIPULATION)) 을 또한 참고한다.
미국 가출원 번호 61/835,931; 61/835,936; 61/836,080; 61/836,101; 61/836,123 및 61/836,127 (2013 년 6 월 17 일 출원) 을 또한 참고한다.
미국 가출원 번호 61/842,322 (2013 년 7 월 2 일 출원) (발명의 명칭: 유전자 생성물의 발현 변형을 위한 CRISPR-CAS 시스템 및 방법 (CRISPR-CAS SYSTEMS AND METHODS FOR ALTERING EXPRESSION OF 유전자 산물S)) 및 미국 가출원 번호 61/847,537 (2013 년 7 월 17 일 출원) (발명의 명칭: 서열 조작 및 적용을 위한 시스템, 방법 및 조성물의 전달, 조작 및 최적화 (DELIVERY, ENGINEERING AND OPTIMIZATION OF SYSTEMS, METHODS AND COMPOSITIONS FOR SEQUENCE MANIPULATION AND APPLICATIONS)) 을 또한 참고한다.
전술한 출원 및 이에 인용되거나 그의 실행 동안 인용된 모든 문헌 ("출원 인용 문헌") 및 출원 인용 문헌에서의 인용되거나 참고된 모든 문헌, 및 본원에 인용되거나 참고된 모든 문헌 ("본원에 인용된 문헌"), 및 본원에 인용된 문헌에서 인용되거나 참고된 모든 문헌은 본원에 언급되거나 본원에 참고로 포함되는 임의의 문헌에서 언급된 임의의 제품에 대한 임의의 제조사 지시사항, 설명서, 제품 사양 및 제품 시트와 함께 본원에 참고로 포함되고, 본 발명의 실행에서 이용될 수 있다. 보다 특히, 모든 참고 문헌은 개별적 문헌이 참고로 포함되는 것으로 구체적이고 개별적으로 표시되는 바와 동일한 정도로 참고로 포함된다.
발명의 분야
본 발명은 일반적으로 유도 전사 효과기를 사용할 수 있는 게놈 교란과 같은 유전자 발현의 공간적이고 시간적인 제어에 사용되는 방법 및 조성물에 관한 것이다.
연방 자금 지원 설명
본 발명은 미국 국립 보건원에 의해 수여된 R01NS073124 및 개척자 상 (Pioneer Award) 1 DP IMH 100706 하 정부 지원으로 이루어졌다. 정부는 본 발명에 특정 권리를 갖는다.
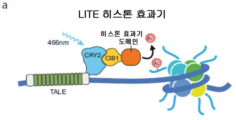
정상 유전자 발현은 조심스럽게 조직된 시간적이고 공간적인 성분, 유기체의 정상 발생, 항상성 유지 및 향상에 필요한 정확도를 갖는 동적 프로세스이다. 결과적으로, 유전자 또는 유전자 집합의 증가, 감소 또는 변형된 기능에 의한 필요한 유전자 발현 패턴의 조절 장애는 광범위한 병리학과 연결된다. 시공간적으로 정확한 방식으로 유전자 발현을 조정할 수 있는 기술은 정상 생물학적 프로세스 및 질환 메커니즘을 책임지는 유전자적 신호의 설명을 가능하게 할 것이다. 이러한 기술적 필요성을 다루기 위해서, 출원인은 유전자 발현, 특히 내재적 유전자 발현의 광-매개 제어를 제공하는 광-유도 전사 효과기 (LITE) 를 조절할 수 있는 유도 분자 도구를 개발하였다.
유도 유전자 발현 시스템은 삽입된 개방 판독 프레임 또는 shRNA 서열의 화학적 유도 활성화가 가능하도록 통상 설계되어, 각각 유전자 과발현 또는 억제를 초래한다. 과발현에 대한 개방 판독 프레임을 사용하는 것의 불리한 점은 스플라이스 변이의 손실 및 유전자 크기 제한을 포함한다. RNA 간섭을 통한 유전자 발현은, 이의 인간 생물학에 있어서의 형질전환적 동력에도 불구하고, 복잡한 오프-타겟 (off-target) 효과에 의해 방해될 수 있다. 에스트로겐, 엑디손 및 FKBP12/FRAP 기반 시스템을 포함하는 특정 유도 시스템은 내재적 유전자의 오프-타겟을 활성화시키는 것으로 알려져있다. 장기간 항생체 처리의 잠재적으로 유해한 효과는 테트라사이클린 전사활성제 (TET) 기반 시스템의 사용을 복잡하게 할 수 있다. 이러한 화학적 유도 시스템의 시간적 정확도는 유도물질 흡수 및 제거의 속도에 따라 좌우된다. 또한, 유도물질이 일반적으로 전신 전달되기 때문에, 이러한 시스템의 공간적 정확도는 외재적 벡터 전달의 정확도에 의해 결부된다.
미국 특허 공개 번호 20030049799 는 사전선택된 자극에 반응하여 검출가능한 산출물이 생기도록 조작된 자극-반응성 스위치에 관한 것이다.
관심 게놈 유전자좌의 효율적이고 정확한 공간적 및 시간적 제어를 가능하게 하는 방법 및 조성물에 대한 명백한 필요성이 존재한다. 이들 방법 및 조성물은 생체내 및 시험관내 모두에서의 게놈 발현의 조절 및 조정을 제공할 수 있을 뿐 아니라 수많은 질환 병리학에 대한 신규한 치료 방법을 제공할 수 잇다.
상기 출원에서의 임의의 문헌의 인용 또는 확인은 그러한 문헌이 본 발명에 대해 선행 기술로서 이용가능하다는 것을 허용하는 것은 아니다.
한 양태에서 본 발명은 하나 이상의 스위치를 포함할 수 있는 비-자연 발생적 또는 조작된 TALE 또는 CRISPR-Cas 시스템을 제공하며, 이때 상기 TALE 또는 CRISPR-Cas 시스템의 활성은 스위치에 대한 하나 이상의 유도물질 에너지원과의 접촉에 의해 제어된다. 본 발명의 구현예에서 상기 TALE 또는 CRISPR-Cas 시스템의 활성 또는 하나 이상의 스위치에 관한 제어가 활성화되고, 증진되고, 종료되거나 억제될 수 있다. 하나 이상의 유도물질 에너지원과의 접촉은 제 1 효과 및 제 2 효과를 초래할 수 있다. 제 1 효과는 핵 이입, 핵 이출, 2 차 성분 (예컨대 효과기 분자) 의 영입, 형태적 변화 (단백질, DNA 또는 RNA 의), 절단, 카고 (cargo) (예컨대 갇혀있는 분자 또는 보조-인자) 의 배출, 결합 또는 해리 중 하나 이상일 수 있다. 제 2 효과는 상기 TALE 또는 CRISPR-Cas 시스템의 활성 또는 하나 이상의 스위치에 관한 제어의 활성화, 증진, 종료 또는 억제 중 하나 이상일 수 있다. 한 구현예에서 제 1 효과 및 제 2 효과는 캐스케이드로 발생할 수 있다.
본 발명의 또 다른 양태에서 TALE 또는 CRISPR-Cas 시스템은 하나 이상의 핵 국부화 신호 (NLS), 핵 이출 신호 (NES), 기능적 도메인, 유연성 링커, 돌연변이, 결실, 변형 또는 절두를 추가로 포함할 수 있다. NLS, NES 또는 기능적 도메인 중 하나 이상은 조건적으로 활성화되거나 불활성화될 수 있다. 또 다른 구현예에서, 돌연변이는 전사 인자 상동 부위에서의 돌연변이, DNA 결합 도메인에서의 돌연변이 (예컨대 염기성 헬릭스 루프 헬릭스의 염기성 잔기 돌연변이화), 내인성 NLS 에서의 돌연변이 또는 내인성 NES 에서의 돌연변이 중 하나 이상일 수 있다. 본 발명은 유도물질 에너지원이 열, 초음파, 전자기 에너지 또는 화학 물질일 수 있다는 것을 이해한다. 본 발명의 바람직한 구현예에서, 유도물질 에너지원은 항생제, 소분자, 호르몬, 호르몬 유도체, 스테로이드 또는 스테로이드 유도체일 수 있다. 보다 바람직한 구현예에서, 유도물질 에너지원은 아브시스산 (ABA), 독시사이클린 (DOX), 큐메이트, 라파마이신, 4-히드록시타목시펜 (4OHT), 에스트로겐 또는 엑디손일 수 있다. 본 발명은 항생제 기반 유도 시스템, 전자기 에너지 기반 유도 시스템, 소분자 기반 유도 시스템, 핵 수용체 기반 유도 시스템 및 호르몬 기반 유도 시스템으로 이루어지는 군에서 선택될 수 있는 하나 이상의 스위치를 제공한다. 보다 바람직한 구현예에서 하나 이상의 스위치는 테트라사이클린 (Tet)/DOX 유도 시스템, 광 유도 시스템, ABA 유도 시스템, 큐메이트 억제제/오퍼레이터 시스템, 4OHT/에스트로겐 유도 시스템, 엑디손-기반 유도 시스템 및 FKBP12/FRAP (FKBP12-라파마이신 복합체) 유도 시스템으로 이루어지는 군에서 선택될 수 있다.
본 발명의 한 양태에서 유도물질 에너지원은 전자기 에너지이다. 전자기 에너지는 450nm-700nm 범위의 파장을 갖는 가시광의 성분일 수 있다. 바람직한 구현예에서 가시광의 성분은 450nm-500nm 범위의 파장을 가질 수 있으며 청색광일 수 있다. 청색광은 0.2 mW/cm2 이상, 또는 보다 바람직하게는 4 mW/cm2 이상의 강도를 가질 수 있다. 또 다른 구현예에서, 가시광의 성분은 620-700nm 범위의 파장을 가질 수 있으며 적색광이다.
본 발명은 하나 이상의 기능적 도메인이 하기로 이루어지는 군에서 선택될 수 있는 시스템을 포함한다: 트랜스포사아제 도메인, 인테그라아제 도메인, 재조합효소 도메인, 리졸바아제 도메인, 인버타아제 도메인, 프로테아제 도메인, DNA 메틸트랜스퍼라아제 도메인, DNA 히드록실메틸라아제 도메인, DNA 데메틸라아제 도메인, 히스톤 아세틸라아제 도메인, 히스톤 데아세틸라아제 도메인, 뉴클레아제 도메인, 억제제 도메인, 활성제 도메인, 핵-국부화 신호 도메인, 전사-조절 단백질 (또는 전사 복합체 영입) 도메인, 세포 흡수 활성 관련 도메인, 핵산 결합 도메인, 항체 제시 도메인, 히스톤 개질 효소, 히스톤 개질 효소의 영입제; 히스톤 개질 효소의 저해제, 히스톤 메틸트랜스퍼라아제, 히스톤 데메틸라아제, 히스톤 키나아제, 히스톤 포스파타아제, 히스톤 리보실라아제, 히스톤 데리보실라아제, 히스톤 유비퀴티나아제, 히스톤 데유비퀴티나아제, 히스톤 비오티나아제 및 히스톤 테일 프로테아제.
본 발명은 또한 유전적 또는 비유전적 관심 유전자좌의 교란을 위한 시스템의 용도를 제공한다. 또한, 약학 화합물의 제조를 위한 시스템의 용도가 제공된다.
추가 양태에서, 본 발명은 하나 이상의 스위치를 포함하는 비-자연 발생적 또는 조작된 TALE 또는 CRISPR-Cas 시스템을 제공하는 것을 포함하는, 상기 TALE 또는 CRISPR-Cas 시스템의 제어 방법을 제공하며, 이때 상기 TALE 또는 CRISPR-Cas 시스템의 활성은 스위치에 대한 하나 이상의 유도물질 에너지원과의 접촉에 의해 제어된다.
본 발명의 한 구현예에서, 본 발명은 상기 TALE 또는 CRISPR-Cas 시스템의 활성 또는 하나 이상의 스위치에 관한 제어가 활성화되고, 증진되고, 완료되거나 억제될 수 있는 방법을 제공한다. 하나 이상의 유도물질 에너지원과의 접촉으로 제 1 효과 및 제 2 효과가 생길 수 있다. 제 1 효과는 핵 이입, 핵 이출, 2 차 성분 (예컨대 효과기 분자) 의 유도, 형태적 변화 (단백질, DNA 또는 RNA 의), 카고 (예컨대 갇힌 분자 또는 보조-인자) 의 절단, 배출, 결합 또는 해리 중 하나 이상일 수 있다. 제 2 효과는 상기 TALE 또는 CRISPR-Cas 시스템의 활성 또는 하나 이상의 스위치에 관한 제어의 활성화, 증진, 완료 또는 억제 중 하나 이상일 수 있다. 한 구현예에서 제 1 효과 및 제 2 효과는 캐스케이드로 발생할 수 있다.
본 발명의 방법의 또 다른 양태에서 TALE 또는 CRISPR-Cas 시스템은 하나 이상의 핵 국부화 신호 (NLS), 핵 이출 신호 (NES), 기능적 도메인, 유연성 링커, 돌연변이, 결실, 변형 또는 절두를 추가로 포함할 수 있다. NLS, NES 또는 기능적 도메인 중 하나 이상은 조건적으로 활성화되거나 불활성화될 수 있다. 또 다른 구현예에서, 돌연변이는 전사 인자 상동 부위에서의 돌연변이, DNA 결합 도메인에서의 돌연변이 (예컨대 염기성 헬릭스 루프 헬릭스의 염기성 잔기의 돌연변이화), 내인성 NLS 에서의 돌연변이 또는 내인성 NES 에서의 돌연변이 중 하나 이상일 수 있다. 본 발명은 유도물질 에너지원이 열, 초음파, 전자기 에너지 또는 화학 물질일 수 있다는 것을 이해한다. 본 발명의 바람직한 구현예에서, 유도물질 에너지원은 항생제, 소분자, 호르몬, 호르몬 유도체, 스테로이드 또는 스테로이드 유도체일 수 있다. 보다 바람직한 구현예에서, 유도물질 에너지원은 아브시스산 (ABA), 독시사이클린 (DOX), 큐메이트, 라파마이신, 4-히드록시타목시펜 (4OHT), 에스트로겐 또는 엑디손일 수 있다. 본 발명은 항생제 기반 유도 시스템, 전자기 에너지 기반 유도 시스템, 소분자 기반 유도 시스템, 핵 수용체 기반 유도 시스템 및 호르몬 기반 유도 시스템으로 이루어지는 군에서 선택될 수 있는 하나 이상의 스위치를 제공한다. 보다 바람직한 구현예에서 하나 이상의 스위치는 테트라사이클린 (Tet)/DOX 유도 시스템, 광 유도 시스템, ABA 유도 시스템, 큐메이트 억제제/오퍼레이터 시스템, 4OHT/에스트로겐 유도 시스템, 엑디손-기반 유도 시스템 및 FKBP12/FRAP (FKBP12-라파마이신 복합체) 유도 시스템으로 이루어지는 군에서 선택될 수 있다.
본 발명의 방법의 한 양태에서 유도물질 에너지원은 전자기 에너지이다. 전자기 에너지는 450nm-700nm 범위의 파장을 갖는 가시광의 성분일 수 있다. 바람직한 구현예에서 가시광의 성분은 450nm-500nm 범위의 파장을 가질 수 있고 청색광일 수 있다. 청색광은 0.2 mW/cm2 이상, 또는 보다 바람직하게는 4 mW/cm2 이상의 강도를 가질 수 있다. 또 다른 구현예에서, 가시광의 성분은 620-700nm 범위의 파장을 가질 수 있고 적색광이다.
본 발명은 하나 이상의 기능적 도메인이 하기로 이루어지는 군에서 선택될 수 있는 방법을 포함한다: 트랜스포사아제 도메인, 인테그라아제 도메인, 재조합효소 도메인, 리졸바아제 도메인, 인버타아제 도메인, 프로테아제 도메인, DNA 메틸트랜스퍼라아제 도메인, DNA 히드록실메틸라아제 도메인, DNA 데메틸라아제 도메인, 히스톤 아세틸라아제 도메인, 히스톤 데아세틸라아제 도메인, 뉴클레아제 도메인, 억제제 도메인, 활성제 도메인, 핵-국부화 신호 도메인, 전사-조절 단백질 (또는 전사 복합체 영입) 도메인, 세포 흡수 활성 관련 도메인, 핵산 결합 도메인, 항체 제시 도메인, 히스톤 개질 효소, 히스톤 개질 효소의 영입제; 히스톤 개질 효소의 저해제, 히스톤 메틸트랜스퍼라아제, 히스톤 데메틸라아제, 히스톤 키나아제, 히스톤 포스파타아제, 히스톤 리보실라아제, 히스톤 데리보실라아제, 히스톤 유비퀴티나아제, 히스톤 데유비퀴티나아제, 히스톤 비오티나아제 및 히스톤 테일 프로테아제.
본 발명의 추가 양태는 TALE 시스템이 하기를 포함하는 DNA 결합 폴리펩티드를 포함하는 본원에 기재한 바와 같은 시스템 또는 방법을 제공한다:
(i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 또는 그 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함).
본 발명의 시스템 및 방법은 하기를 포함하는 DNA 결합 폴리펩티드를 제공하며: (a) N-말단 캡핑 부위 (b) 관심 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 내지 40 개의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인, 및 (c) C-말단 캡핑 부위, 여기서 (a), (b) 및 (c) 가 사전결정된 N-말단에서 C-말단의 배향으로 배열될 수 있고, 이때 게놈 유전자좌가 표적 DNA 서열 5'-T0N1N2....Nz Nz+1-3' (여기서 T0 및 N = A, G, T 또는 C 임) 을 포함하고, 상기 표적 DNA 서열은 DNA 결합 도메인에 결합하고, DNA 결합 도메인은 (X1-11-X12X13-X14-33 또는 34 또는 35)z (여기서 X1-11 은 11 개의 인접한 아미노산 사슬이고, X12X13 은 반복 가변 이중잔기 (RVD) 이고, X14-33 또는 34 또는 35 는 21, 22 또는 23 개의 인접한 아미노산 사슬이고, z 는 적어도 5 내지 40 일 수 있음) 를 포함할 수 있고, 이때 상기 폴리펩티드는 코돈 최적화된 핵산 분자에 의해 인코딩되고 이로부터 번역되어 폴리펩티드가 바람직하게는 관심 유전자좌의 DNA 에 결합할 수 있다.
추가 구현예에서, 본 발명의 시스템 또는 방법은 야생형 N-말단 캡핑 부위의 147 개의 인접한 아미노산을 포함하는 N-말단 캡핑 부위 또는 이의 단편, 또는 야생형 C-말단 캡핑 부위의 68 개의 인접합 아미노산을 포함하는 C-말단 캡핑 부위 또는 이의 단편, 또는 야생형 N-말단 캡핑 부위의 136 개의 인접한 아미노산을 포함하는 N-말단 캡핑 부위 또는 이의 단편 및 야생형 C-말단 캡핑 부위의 183 개의 인접한 아미노산을 포함하는 C-말단 캡핑 부위 또는 이의 단편을 제공한다. 또 다른 구현예에서, 하나 이상의 RVD 는 (a) 구아닌 (G) 의 인지를 위한 HH, KH, NH, NK, NQ, RH, RN, SS, NN, SN, KN; (b) 아데닌 (A) 의 인지를 위한 NI, KI, RI, HI, SI; (c) 티민 (T) 의 인지를 위한 NG, HG, KG, RG; (d) 시토신 (C) 의 인지를 위한 RD, SD, HD, ND, KD, YG; (e) A 또는 G 의 인지를 위한 NV, HN; 및 (f) A 또는 T 또는 G 또는 C 의 인지를 위한 H*, HA, KA, N*, NA, NC, NS, RA, S* (여기서 (*) 는 X13 에서의 아미노산이 부재함을 의미함) 로 이루어지는 군에서 선택될 수 있다.
또 다른 구현예에서 하나 이상의 RVD 는 (a) 구아닌 (G) 의 인지를 위한 HH, KH, NH, NK, NQ, RH, RN, SS; (b) 아데닌 (A) 의 인지를 위한 SI; (c) 티민 (T) 의 인지를 위한 HG, KG, RG; (d) 시토신 (C) 의 인지를 위한 RD, SD; (e) A 또는 G 의 인지를 위한 NV, HN 및 (f) A 또는 T 또는 G 또는 C 의 인지를 위한 H*, HA, KA, N*, NA, NC, NS, RA, S* (여기서 (*) 는 X13 에서의 아미노산이 부재함을 의미함) 로 이루어지는 군에서 선택될 수 있다. 바람직한 구현예에서, G 의 인지를 위한 RVD 는 RN, NH, RH 또는 KH 이거나; A 의 인지를 위한 RVD 는 SI 이거나; T 의 인지를 위한 RVD 는 KG 또는 RG 이고; C 의 인지를 위한 RVD 는 SD 또는 RD 이다. 또 다른 구현예에서, 하기 중 하나 이상이 존재한다: X1-4 에서의 [LTLD] 또는 [LTLA] 또는 [LTQV], 또는 위치 X30-33 또는 X31-34 또는 X32-35 에서의 [EQHG] 또는 [RDHG].
본 발명의 한 양태에서 TALE 시스템은 AAV 또는 렌티바이러스 (lentivirus) 벡터 내로 패키징된다.
본 발명의 추가 양태는 CRISPR 시스템이 하기를 포함하는 벡터 시스템을 포함할 수 있는, 본원에 기재한 바와 같은 시스템 또는 방법을 제공하며: a) 관심 유전자좌를 표적화하는 CRISPR-Cas 시스템 안내 RNA 에 작동가능하게 연결된 제 1 조절 요소, b) Cas 단백질에 작동가능하게 연결된 제 2 조절 유도 요소, 이때 성분 (a) 및 (b) 는 시스템의 동일하거나 상이한 벡터 상에 위치할 수 있고, 안내 RNA 는 관심 유전자좌의 DNA 를 표적화하고, Cas 단백질 및 안내 RNA 는 함께 자연적으로 발생하지 않는다. 본 발명의 바람직한 구현예에서, Cas 단백질은 Cas9 효소이다. 본 발명은 또한 AAV 또는 렌티바이러스인 벡터를 제공한다.
본 발명은 특히, 관심 게놈 유전자좌의 발현 변형의 유도 방법 및 게놈 관심 게놈 유전자좌 (게놈 유전자좌는 데옥시리보핵산 (DNA) 결합 폴리펩티드를 포함하는 비-자연 발생적 또는 조작된 조성물과 접촉할 수 있음) 의 발현을 유도가능하게 변형하는 조성물에 관한 것이다.
이러한 폴리펩티드는 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 또는 그 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인을 포함할 수 있다. 상기 에너지 민감성 단백질 또는 이의 단편은 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너에 결합할 수 있다. 폴리펩티드는 또한, 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 변이체 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인을 포함할 수 있으며, 이때 에너지 민감성 단백질 또는 이의 단편은 에너지원에 의한 유도시 상호작용 파트너에 결합할 수 있다. 상기 방법은 또한 에너지원을 적용하고 게놈 유전자좌의 발현이 변형되는 것을 측정하는 것을 포함할 수 있다. 본 발명의 바람직한 구현예에서 게놈 유전자좌는 세포 내에 있을 수 있다.
본 발명은 또한 관심 게놈 유전자좌 발현 억제의 유도 방법 및 관심 게놈 유전자좌의 발현을 유도가능하게 억제하는 조성물에 관한 것이며, 이때 상기 게놈 유전자좌는 DNA 결합 폴리펩티드를 포함하는 비-자연 발생적 또는 조작된 조성물과 접촉될 수 있다.
폴리펩티드는 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 억제제 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 또는 그 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인을 포함할 수 있다. 에너지 민감성 단백질 또는 이의 단편은 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있다. 폴리펩티드는 또한, 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 변이체 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인을 포함할 수 있으며, 이때 에너지 민감성 단백질 또는 이의 단편은 에너지원에 의한 유도시 상호작용 파트너에 결합할 수 있다. 상기 방법은 또한 에너지원을 적용하고, 게놈 유전자좌의 발현이 억제되는 것을 측정하는 것을 포함할 수 있다. 본 발명의 바람직한 구현예에서 게놈 유전자좌는 세포 내에 있을 수 있다.
본 발명은 또한 관심 게놈 유전자좌의 발현 활성화 유도 방법 및 관심 게놈 유전자좌의 발현을 유도가능하게 활성화하는 조성물에 관한 것이며, 여기서 게놈 유전자좌는 DNA 결합 폴리펩티드를 포함하는 비-자연 발생적 또는 조작된 조성물과 접촉될 수 있다.
폴리펩티드는 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 활성제 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 또는 그 이상의 TALE 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인을 포함할 수 있다. 에너지 민감성 단백질 또는 이의 단편은 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너에 결합할 수 있다. 폴리펩티드는 또한 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 변이체 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인을 포함할 수 있으며, 여기서 에너지 민감성 단백질 또는 이의 단편은 에너지원에 의한 유도시 상호작용 파트너에 결합할 수 있다. 상기 방법은 또한 에너지원을 적용하고, 게놈 유전자좌의 발현이 활성화되는 것을 측정하는 것을 포함할 수 있다. 본 발명의 바람직한 구현예에서 게놈 유전자좌는 세포 내에 있을 수 있다.
본 발명의 또 다른 바람직한 구현예에서, 유도 효과기는 광 유도 전사 효과기 (LITE) 일 수 있다. LITE 시스템의 모듈성은 전사 모듈화에 임의 수의 효과기 도메인이 이용될 수 있게 한다.
본 발명의 또 다른 바람직한 구현예에서, 유도 효과기는 화학 물질일 수 있다.
본 발명은 또한 CRISPR (군집화 규칙적 공간화 단길이 회귀성 반복물 (clustered regularly interspaced short palindromic repeats))/Cas 시스템을 사용하는 유도 멀티플렉스 게놈 조작을 고려한다.
본 발명은 또한 본 발명의 폴리펩티드를 인코딩하는 핵산을 포함한다. 핵산은 프로모터, 유리하게는 인간 시냅신 I 프로모터 (hSyn) 를 포함할 수 있다. 특히 유리한 구현예에서, 핵산은 아데노 관련 바이러스 벡터 (AAV) 내에 패키징될 수 있다.
본 발명은 추가로 또한, 본원에 기재한 방법 및 조성물을 포함하는 치료 방법 또는 치료요법에 관한 것이다.
따라서, 본 발명 내에 임의의 이전에 공지된 생성물, 상기 생성물 제조 공정, 또는 상기 생성물 사용 방법을 포함하지 않아, 출원인이 권한을 남겨 둠으로써 임의의 이전에 공지된 생성물, 공정 및 방법의 권리불요구를 개시하는 것이 본 발명의 목적이다. 본 발명이 본 발명의 범주 내에, USPTO (35 U.S.C. §112, 첫 번째 단락) 또는 EPO (EPC 의 제 83 조) 의 서면 설명 및 실시가능 필요사항을 충족하지 않는 임의의 생성물, 상기 생성물 제조 공정 또는 상기 생성물 사용 방법을 포함하는 것으로 의도되지 않아, 출원인이 권한을 남겨 둠으로써 이전에 기재된 생성물, 상기 생성물 제조 공정 또는 상기 생성물 사용 방법의 권리불요구를 개시하는 것에 또한 주목한다.
이 개시물 및 특히 특허청구범위 및/또는 단락에서, 용어 예컨대 "포함하다", "포함된", "포함하는" 등은 미국 특허법에 부여된 의미를 가질 수 있다는 것에 주목한다; 예를 들어 이는 "함유하다", "함유된", "함유하는" 등을 의미할 수 있고; 용어 예컨대 "~로 본질적으로 이루어지는" 및 "~로 본질적으로 이루어진다" 는 미국 특허법에서 기인한 의미를 갖고, 예를 들어 요소가 명백하게 인용되지 않게 하나, 선행 기술에서 발견되거나 본 발명의 기본적으로 신규한 특징에 영향을 주는 요소는 배제한다.
구현예 및 기타 구현예가 하기 상세한 설명으로부터 개시되거나 명백하고 그에 의해 포함된다.
예로써 주어졌으나 기재한 특정 구현예에만 본 발명을 제한하는 것으로 의도되지 않는 하기의 상세한 설명은 동반하는 도면과 함께 가장 최상으로 이해될 수 있다.
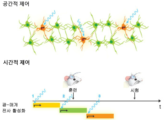
도 1 은 공간적 및 시간적 정확도의 필요성을 나타내는 모식도를 나타낸다.
도 2 는 전사 활성제 유사 효과기 (TALE) 를 나타낸다. TALE 는 그의 서열의 코어에서 34 개 aa 반복물로 이루어진다. 각각의 반복물은 TALE 에 의해 결합되는 표적 DNA 에서의 염기에 상응한다. 반복물은 위치 12 및 13 에서 2 개의 가변적인 아미노산에 의해서만 상이하다. 이러한 대응의 코드가 설명된 바 있으며 (Boch, J et al. , Science, 2009 및 Moscou, M et al. , Science, 2009) 이 도면에서 나타낸다. 출원인은 이러한 코드를 통합하며 게놈 내에 선택 서열을 결합시킬 수 있는 설계자 TALE 의 합성 방법을 개발하였다 (Zhang, F et al. , Nature Biotechnology, 2011).
도 3 은 LITE: TALE/크립토크롬 전사 활성화의 설계를 나타낸다. 각각의 LITE 는 CRY2 에 융합된 TALE 및 VP64 (전사 인자) 에 융합된 크립토크롬 결합 파트너 CIB1 을 포함할 수 있는 2-성분 시스템이다. 불활성 상태에서, TALE 은 이의 융합된 CRY2 도메인을 관심 유전자의 프로모터 부위에 국부화시킨다. 이때, CIB1 은 CRY2 에 결합할 수 없고, CIB1-VP64 가 핵 공간 내에서 미결합된 채로 있다. 488 nm (청색) 광으로 자극시, CRY2 는 형태적 변화를 거쳐, 이의 CIB1 결합 부위가 밝혀진다 (Liu, H et al. , Science, 2008). CIB1 의 신속한 결합은 융합된 VP64 도메인의 영입을 초래하여, 표적 유전자의 전사를 유도한다.
도 4 는 LITE 활성에 대한 크립토크롬 이량체 절두의 효과를 나타낸다. CRY2 및 CIB1 의 활성을 변경시키는 것으로 알려져 있는 절두를 (Kennedy M et al, Nature Methods 2010) 전체 길이 단백질에 대해 비교하였다. Neurog2 의 프로모터에 대해 표적화된 LITE 를 도메인의 각 조합에 대해 Neuro-2a 세포에서 시험하였다. 488 nm 광으로 자극한 후, Neurog2 의 전사 수준을 자극된 샘플 및 미자극된 샘플에 대해 qPCR 을 사용하여 정량하였다.
도 5 는 KLF4 LITE 의 광-강도 의존적 반응을 나타낸다.
도 6 은 Neurog2 LITE 의 활성화 반응속도 및 Neurog2 LITE 의 불활성화 반응속도를 나타낸다.
도 7A 는 출원인의 RVD 스크리닝 시스템을 사용하여 측정한 바와 같은 다양한 RVD 의 염기-선호도를 나타낸다.
도 7B 는 출원인의 RVD 스크리닝 시스템을 사용하여 측정한 바와 같은 추가적인 RVD 의 염기-선호도를 나타낸다.
도 8A-D 는 다음을 나타낸다: (a) 잔토모나스 종 (Xanthomonas sp.) 에서 유래한 TALE 의 자연적 구조. 각각의 DNA-결합 모듈은 34 개의 아미노산으로 이루어지는데, 각각의 반복물의 12 번째 및 13 번째 아미노산 위치에서의 RVD 는 부호 NG = T, HD = C, NI = A 및 NN = G 또는 A 에 따라 표적화되는 DNA 염기를 명시한다. DNA-결합 모듈은 비반복적 N 및 C 말단에 측면 위치하여, 전좌, 핵 국부화 (NLS) 및 전사 활성화 (AD) 도메인을 수반한다. N 말단 내 암호성 신호는 표적 부위의 제 1 염기로서 티민을 명시한다. (b) TALE 툴박스는 커스텀 TALE-TF 및 TALEN 의 신속하고 값싼 구성을 가능하게 한다. 키트는 총 12 개 플라스미드로 이루어진다: 4 개 단량체 플라스미드가 PCR 증폭용 주형으로서 사용되고, 4 개의 TALE-TF 및 4 개의 TALEN 클로닝 백본은 0.5 반복물에 의해 표적화된 4 개의 상이한 염기에 상응한다. CMV, 사이토메갈로바이러스 프로모터; N 말단, Hax3 TALE 로부터의 비반복적 N 말단; C 말단, Hax3 TALE 로부터의 비반복적 C 말단; BsaI, 커스텀 TALE DNA-결합 도메인의 삽입에 사용된 유형 II 제한 부위; ccdB + CmR, ccdB 음성 선택 유전자 및 클로람페니콜 저항성 유전자를 포함하는 음성 선택 카세트; NLS, 핵 국부화 신호; VP64, 단순 포진 바이러스의 VP 16 단백질에서 유래한 합성 전사 활성제; 2A, 2A 자체-절단 링커; EGFP, 증강된 녹색 형광 단백질; 폴리A 신호, 폴리아데닐화 신호; FokI, FokI 엔도뉴클레아제로부터의 촉매적 도메인. (c) TALE 은 커스텀 TALE-TF 를 생성시키고 게놈으로부터의 내인성 유전자의 전사를 조정하는데 사용될 수 있다. TALE DNA-결합 도메인은 합성 VP64 전사 활성제에 융합되는데, 이는 RNA 폴리머라아제 및 전사 개시에 필요한 기타 인자를 영입한다. (d) TALEN 은 비상동 쌍 또는 상동성 유도 쌍을 통해 게놈 편집을 촉진시키기 위해 위치-특이적 이중가닥 파단점을 생성시키는데 사용될 수 있다. 2 개의 TALEN 은 측면 위치 16-bp 스페이서의 결합 부위 쌍을 표적화한다. 좌측 및 우측 TALEN 은 각각 표적 부위의 상부 및 하부 가닥을 인지한다. 각각의 TALE DNA-결합 도메인은 FokI 엔도뉴클레아제의 촉매적 도메인에 융합되고; FokI 이 이량체화될 때, 이는 좌측 및 우측 TALEN-결합 도메인 사이의 부위에서 DNA 를 절단한다.
도 9A-F 는 단량체 서열 (위치 12 및 13 에서의 RVD 제외) 및 특정 서열을 갖는 단량체가 발생하는 빈도를 열거하는 표를 나타낸다.
도 10 은 TALE 활성에 대한 비-RVD 아미노산의 효과 비교를 나타낸다.
도 11 은 VP64, p65 와 VP16 사이의 활성화 수준을 비교하는 활성제 스크린을 나타낸다.
도 12A-D 는 TALE 전사 억제제 구조의 개발을 나타낸다. (a) TALE 억제제 스크리닝에 대한 SOX2 TALE 의 설계. 인간 게놈의 SOX2 유전자좌 내의 14bp 서열을 표적화하는 TALE 을 합성하였다. (b) 스크리닝한 모든 억제제 및 그의 숙주 기원의 목록 (좌측). 8 개의 상이한 후보물 억제제 도메인은 SOX2 TALE 의 C-말단에 융합되었다. (c) 내인성 SOX2 mRNA 의 감소 배수를 모의 트랜스펙션 세포에서의 SOX2 mRNA 수준을, 각각의 후보 TALE 억제제로 트랜스펙션시킨 세포에서의 SOX2 mRNA 수준에 의해 나누어, qRTPCR 을 사용하여 측정한다. (d) 내인성 CACNA1C 의 전사 억제. G-표적화 RVD 로서 NN, NK 및 NH 를 사용하는 TALE 을 구축하여 인간 CACNA1C 유전자좌 내 18 bp 표적 부위를 표적화하였다. 각각의 TALE 을 SID 억제 도메인에 융합시킨다. NLS, 핵 국부화 신호; KRAB, 크루펠 (Kruppel)-관련 박스; SID, mSin 상호작용 도메인. 모든 결과를 HEK 293FT 세포에서 3 개의 독립적 실험으로부터 수집하였다. 오류 막대는 s.e.m. 을 나타내고; n = 3 이다. * p < 0.05, 스튜던트 (Student) t 시험.
도 13A-C 는 SID 및 SID4X 를 사용하는 TALE 전사 억제제 구조의 최적화를 나타낸다. (a) TALE 억제제 구조의 시험을 위한 p11 TALE 의 설계. 마우스 (Mus musculus) 게놈의 p11 (s100a10) 유전자좌 내의 20bp 서열을 표적화하는 TALE 을 합성하였다. (b) 내인성 마우스 p11 mRNA 의 전사 억제. 야생형 TALE 구조의 2 개 상이한 절두부를 갖는 마우스 p11 유전자좌를 표적화하는 TALE 을 x-축에 나타낸 바와 같이 상이한 억제제 도메인에 융합시켰다. 괄호 안의 값은 DNA 결합 반복물에 측면 위치하며 상기 구축물에서 사용한 억제제 도메인이 그 뒤에 있는 TALE DNA 결합 도메인의 N-말단 및 C-말단에서의 아미노산 수를 나타낸다. 내인성 p11 mRNA 수준을 qRT-PCR 을 사용하여 측정하고, GFP-인코딩 구축물로 트랜스펙션시킨 음성 대조군 세포에서의 수준으로 정규화시켰다. (c) 내인성 마우스 p11 의 전사 억제 배수. 내인성 p11 mRNA 의 감소 배수를, 음성 대조군 GFP 구축물로 트랜스펙션시킨 세포에서의 p11 mRNA 수준을 각각의 후보 TALE 억제제로 트랜스펙션시킨 세포에서의 p11 mRNA 수준에 의해 나누어, qRT-PCR 을 사용하여 측정한다. x-축에 따른 구축물의 표지는 이전 패널에서와 동일하다. NLS, 핵 국부화 신호; SID, mSin 상호작용 도메인; SID4X, 단길이 펩티드 링커에 의해 연결된 SID 도메인의 최적화된 4-회 탠덤 (tandem) 반복물. 모든 결과는 Neuro2A 세포에서의 3 개 독립적 실험에서 수집하였다. 오류 막대는 s.e.m. 을 나타내고; n = 3 이다. *** /p< 0.001, 스튜던트 t 시험.
도 14 는 2 가지 상이한 유형의 TALE 구조의 비교를 나타낸다.
도 15A-C 는 화학적 유도 TALE ABA 유도 시스템을 나타낸다. ABI (ABA 비-민감성 1) 및 PYL (PYL 단백질: 피라박틴 저항성 (PYR)/PYRl-유사 (PYL)) 은 식물 호르몬 아브시스산 (ABA) 의 결합시 이량체화하는 하기 열거한 2 개 단백질로부터의 도메인이다. 이러한 식물 호르몬은 출원인이 출원인의 유도 TALE 시스템에서 사용한 소분자 화학 물질이다. 이러한 시스템에서, TALE DNA-결합 폴리펩티드는 ABI 도메인에 융합되는 반면, VP64 활성화 도메인 또는 SID 억제제 도메인 또는 임의의 효과기 도메인은 PYL 도메인에 연결된다. 따라서 ABA 분자의 존재에 의해 유도시, 2 개 상호작용 도메인, ABI 및 PYL 은 이량체화되고 TALE 이 효과기 도메인에 연결되게 하여 표적 유전자 발현을 조절하는데 있어서의 이의 활성을 수행할 수 있게 한다.
도 16A-B 는 화학적 유도 TALE 4OHT 유도 시스템을 나타낸다.
도 17 은 LITE 기능성에 대한 크립토크롬2 이형이량체 배향의 효과를 나타낸다.
도 18 은 마우스 피질 뉴런 배양액에서의 mGlur2 LITE 활성을 나타낸다.
도 19 는 LITE AAV 벡터을 사용하는 1 차 마우스 뉴런의 형질도입을 나타낸다.
도 20 은 생체내 LITE 성분의 발현을 나타낸다.
도 21 은 사용한 특정 NES 펩티드가 LDLASLIL 인 구축물의 향상된 설계를 나타낸다.
도 22 는 40H 타목시펜의 부재 및 존재 하 Sox2 mRNA 수준을 나타낸다.
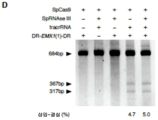
도 23A-E 는 스트렙토코쿠스 피오게네스 (Streptococcus pyogenes) SF370 으로부터의 유형 II CRISPR 유전자좌가 DNA 의 표적화된 DSB 가 촉진되도록 포유동물 세포에서 재구성될 수 있다는 것을 나타낸다. (A) NLS 를 갖는 SpCas9 및 SpRNase III 의 조작은 포유동물 핵 내로의 이입을 가능하게 한다. (B) SpCas9 및 SpRNase III 의 포유동물 발현은 EFla 프로모터에 의해 유도되는 반면, tracrRNA 및 예비-crRNA 어레이 (DR-스페이서-DR) 는 U6 프로모터에 의해 유도된다. PAM 을 갖는 인간 EMX1 유전자좌로부터의 프로토스페이서 (청색 하이라이트) 를 예비-crRNA 어레이에서 스페이서에 대한 주형으로서 사용한다. (C) 표적 유전자좌와 EMX1-표적화 crRNA 사이의 염기 페어링의 모식적 표현. 적색 화살표는 추정상 절단 부위를 나타낸다. (D) SpCas9-매개 삽입-결실 (indel) 에 대한 SURVEYOR 검정. (E) 187 클론 암플리콘으로부터 확인된 돌연변이화된 대립형질의 대표 서열을 나타내는 예시적 크로마토그램. 적색 대시 기호는 결실된 염기를 나타내고; 적색 염기는 삽입 또는 돌연변이를 나타낸다. 스케일 바 = 10 ㎛.
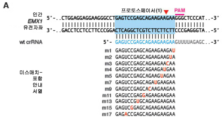
도 24A-C 는 SpCas9 가 포유동물 세포에서의 다수 게놈 유전자좌를 표적화하도록 재프로그래밍될 수 있다는 것을 나타낸다. (A) 청색 선으로 표시한 5 개 프로토스페이서의 위치를 나타내는 인간 EMX1 유전자좌의 모식도, 상응하는 PAM 은 자홍색으로 나타냄. (B) 예비-crRNA 및 tracrRNA 의 직접 반복 (회색) 부위 사이의 하이브리드화를 나타내는 예비-crRNA:tracrRNA 복합체 (상부) 의 모식도. 키메라 RNA 설계의 모식도 (M. Jinek et al., A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816 (Aug 17, 2012)) (하부). tracrRNA 서열을 적색으로 나타내고 20bp 스페이서를 청색으로 나타낸다. (C) 인간 EMX1 유전자좌에서의 5 개 프로토스페이서에서의 Cas9-매개 절단의 효능을 비교하는 SURVEYOR 검정. 각각의 프로토스페이서를 처리된 예비-crRNA:tracrRNA 복합체 (crRNA) 또는 키메라 RNA (chiRNA) 를 사용하여 표적화한다.
도 25A-D 는 SpCas9 특이성의 평가 및 TALEN 과의 효율 비교를 나타낸다. (A) 단일 점 돌연변이를 갖는 EMX1-표적화 키메라를 생성시켜 스페이서-프로토스페이서 미스매치의 효과를 평가하였다. (B) 상이한 돌연변이체 키메라 RNA 의 절단 효율을 비교하는 SURVEYOR 검정. (C) TALEN 표적화 EMX1 의 설계를 나타내는 모식도. (D) TALEN 및 SpCas9 의 효율을 비교하는 SURVEYOR 겔 (N = 3).
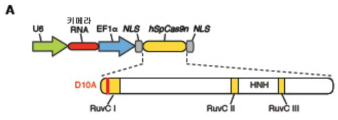
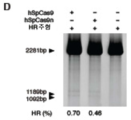
도 26A-G 는 상동 재조합 및 멀티플렉스 게놈 조작을 위한 Cas9 의 적용을 나타낸다. (A) RuvC I 도메인의 돌연변이는 Cas9 를 절단 효소 (SpCas9n) 로 전환시킨다. (B) EMX1-표적화 키메라 RNA 와 SpCas9 와의 동시-발현은 삽입-결실을 유발하는 반면, SpCas9n 은 그렇지 않다 (N = 3). (C) 재조합 계획의 모식적 표시. 수복 주형은 제한 부위가 EMX1 유전자좌에 삽입되도록 설계된다. 변형된 부위를 증폭하는데 사용한 프라이머를 적색 화살표로 나타낸다. (D) 제한 단편 길이 다형성 겔 분석. 화살표는 HindIII 소화에 의해 생성된 단편을 나타낸다. E) 성공적인 재조합을 나타내는 예시적 크로마토그램. (F) SpCas9 는 EMX1 및 PVALB 를 표적화하는 2 개 스페이서를 포함하는 crRNA 어레이를 사용하여 멀티플렉스 게놈 변형을 촉진시킬 수 있다. crRNA 어레이의 설계를 나타내는 모식도 (상부). 스페이서 모두 효율적인 프로토스페이서 절단을 매개한다 (하부). (G) SpCas9 를 사용하여 정확한 게놈 결실을 수행할 수 있다. EMX1 을 표적화하는 2 개 스페이서 (상부) 는 118bp 게놈 결실을 매개하였다 (하부).
도 27 은 유형 II CRISPR-매개 DNA 이중가닥 파단의 모식도를 나타낸다. 스트렙토코쿠스 피오게네스 SF370 으로부터의 유형 II CRISPR 유전자좌는 4 개 유전자, Cas9, Casl, Cas2 및 Csnl 의 클러스터 뿐 아니라 2 개의 비-코딩 RNA 요소, tracrRNA, 및 비-반복 서열의 단길이 스트레치 (스페이서, 각각 30bp) 에 의해 공간이 남겨진 반복 서열의 특징적 어레이 (직접 반복물) 을 포함한다 (15-18, 30, 31). 각각의 스페이서는 통상 외래 유전적 물질 (프로토스페이서) 에서 유래하고, CRISPR-매개 핵산 절단의 특이성을 유도한다. 표적 핵산에서, 각각의 프로토스페이서는 그의 인지가 개별적 CRISPR 시스템에 특이적인 프로토스페이서 인접 모티프 (PAM) 와 연관된다 (22, 23). 유형 II CRISPR 시스템은 순차적 단계로 표적화 DNA 이중가닥 파단 (DSB) 을 실행한다 (M. Jinek et al, Science 337, 816 (Aug 17, 2012); Gasiunas, R. et al. Proc Natl Acad Sci USA 109, E2579 (Sep 25, 2012); J. E. Garneau et al, Nature 468, 67 (Nov 4, 2010); R. Sapranauskas et al, Nucleic Acids Res 39, 9275 (Nov, 2011); A. H. Magadan et al. PLoS One 7, e40913 (2012)). 먼저, 예비-crRNA 어레이 및 tracrRNA 를 CRISPR 유전자좌로부터 전사한다. 두 번째로, tracrRNA 를 예비-crRNA 의 직접 반복물에 하이드리드화하고 이중체로서 Cas9 와 연관시킨다 (예비-crRNA 의, 개별적인, 절두된 스페이서 서열을 포함하는 성숙 crRNA 로의 처리를 매개함). 세 번째로, 성숙 crRNA:tracrRNA 이중체는 Cas9 를 crRNA 의 스페이서 부위와 프로토스페이서 DNA 사이의 이형이중체 형성을 통해 프로토스페이서 및 필수적 PAM 으로 이루어지는 DNA 표적으로 유도한다. 마지막으로, Cas9 는 PAM 의 표적 DNA 업스트림의 절단을 매개하여 프로토스페이서 내에 DSB 를 생성시킨다.
도 28A-C 는 Cas9-매개 유전자 표적화를 위한 상이한 tracrRNA 전사체의 비교를 나타낸다. (A) 시험한 2 개 (단길이 및 장길이) tracrRNA 전사체의 설계 및 서열을 나타내는 모식도. 각각의 전사체는 U6 프로모터에 의해 주도된다. 전사 시작 부위를 +1 로 표기하고 전사 터미네이터는 표시한 바와 같다. 청색 선은 그의 역-상보 서열이 tracrRNA 검출용 노던 블롯 프로브 생성에 사용되는 부위를 나타낸다. (B) EMX1 유전자좌의 hSpCas9-매개 절단의 효율을 비교하는 SURVEYOR 검정. 2 개의 생물학적 레플리카를 각각의 tracrRNA 전사체에 대해 나타낸다. (C) 장길이 또는 단길이 tracrRNA 뿐 아니라 SpCas9 및 DR-EMX1(1)-DR 을 운반하는 U6 발현 구축물로 트랜스펙션된 293FT 세포로부터 추출한 총 RNA 의 노던 블롯 분석. 좌측 및 우측 패널은 각각 SpRNase III 의 부재 또는 존재 하에 트랜스펙션된 293FT 세포로부터의 것이다. U6 은 인간 U6 snRNA 를 표적화하는 프로브로 블롯팅된 로딩 대조군을 나타낸다. 단길이 tracrRNA 발현 구축물의 트랜스펙션은 tracrRNA 의 처리 형태의 과잉 수준을 이끌어낸다 (~75bp) (E. Deltcheva et al., Nature 471, 602 (Mar 31, 2011)). 매두 소량의 장길이 tracrRNA 를 노던 블롯에서 검출한다. 이들 실험의 결과로서, 출원인은 포유동무루 세포에서의 적용을 위해 단길이 tracrRNA 사용을 선택하였다.
도 29 는 이중가닥 파단-유도 미세 삽입 및 결실의 검출을 위한 SURVEYOR 검정을 나타낸다 (D. Y. Guschin et al. Methods Mol Biol 649, 247 (2010)). Cas9-매개 절단 효율을 측정하는데 사용한 SURVEYOR 검정의 모식도. 먼저, 게놈 PCR (gPCR) 을 사용하여 변형 및 미변형 세포의 불균일 집단으로부터의 Cas9 표적 부위를 증폭하고, gPCR 생성물을 천천히 재어닐링하여 이형이중체를 생성한다. 재어닐링한 이형이중체를 SURVEYOR 뉴클레아제에 의해 절단하는 반면, 동종이중체는 미손상인 채로 둔다. Cas9-매개 절단 효율 (% 삽입-결실) 을 절단된 DNA 분획물을 기준으로 계산한다.
도 30A-B 는 포유동물 세포에서의 crRNA 처리의 노던 블롯 분석을 나타낸다. (A) 2 개 직접 반복물 (DR-EMX1(l)-DR) 에 의해 측면 위치한 단일 스페이서에 대한 발현 벡터를 나타내는 모식도. 인간 EMX1 유전자좌 프로토스페이서 1 을 표적화하는 30bp 스페이서 (표 1) 를 청색으로 나타내고 직접 반복물을 회색으로 나타낸다. 주황색 선은 그의 역-상보 서열을 사용하여 EMX1(1) crRNA 검출용 노던 블롯 프로브를 생성시키는 부위를 나타낸다. (B) DR-EMX(1)-DR 을 운반하는 U6 발현 구축물로 트랜스펙션된 293FT 세포로부터 추출한 총 RNA 의 노던 블롯 분석. 좌측 및 우측 패널은 각각 SpRNase III 의 부재 또는 존재 하에 트랜스펙션된 293FT 세포로부터의 것이다. DR-EMX(1)-DR 은 SpCas9 및 단길이 tracrRNA 의 존재 하에서만 성숙 crRNA 로 처리되었으며, SpRNase III 의 존재에 의해서는 좌우되지 않았다. 트랜스펙션된 293FT 총 RNA 로부터 검출된 성숙 crRNA 는 ~33bp 이고 S. 피오게네스 (S. pyogenes) 로부터의 39-42bp 성숙 crRNA 보다 더 짧은데 (E. Deltcheva et al., Nature 471, 602 (Mar 31, 2011)), 이는 인간 293FT 세포에서의 처리된 성숙 crRNA 가 S. 피오게네스에서의 박테리아 성숙 crRNA 와 상이할 것으로 예상된다는 것을 제시한다.
도 31A-B 는 Cas9 를 갖는 키메라 crRNA 또는 예비-crRNA 어레이에 대한 바이시스트로닉 발현 벡터를 나타낸다. (A) 예비-crRNA 어레이에 대한 발현 벡터의 설계를 나타내는 모식도. 스페이서는 어닐링된 올리고뉴클레오티드를 사용하여 2 개 BbsI 부위 사이에 삽입될 수 있다. 올리고뉴클레오티드에 대한 서열 설계를 하기에 나타내며, 적절한 라이게이션 어댑터를 함께 표시하였다. (B) 키메라 crRNA 에 대한 발현 벡터의 모식도. 안내 서열은 어닐링된 올리고뉴클레오티드를 사용하여 2 개 BbsI 부위 사이에 삽입될 수 있다. 벡터는 이미 부분적 직접 반복물 (회색) 및 부분적 tracrRNA (적색) 서열을 포함한다. WPRE, 마못 (Woodchuck) 간염 바이러스 전사후 조절 요소.
도 32A-B 는 인간 PVALB 및 마우스 Th 유전자좌에서의 프로토스페이서의 선택을 나타낸다. 각각 PVALB 및 Th 유전자의 마지막 엑손 내의 3 개 프로토스페이서의 위치 및 인간 PVALB (A) 및 마우스 Th (B) 유전자좌의 모식도. 30bp 프로토스페이서를 흑색 선으로 나타내고 인접한 PAM 서열을 자홍색 막대로 나타낸다. 센스 및 안티-센스 가닥 상의 프로토스페이서를 각각 DNA 서열 위 및 아래에 나타낸다.
도 33A-C 는 인간 게놈에서의 PAM 서열의 발생을 나타낸다. 인간 게놈에서의 인접한 스트렙토코쿠스 피오게네스 SF370 유전자좌 1 PAM (NGG) (A) 와 스트렙토코쿠스 써모필루스 (Streptococcus thermophilus) LMD9 유전자좌 1 PAM (NNAGAAW) (B) 사이 거리의 히스토그램. (C) 염색체에 의한 각각의 PAM 에 대한 거리. Chr, 염색체. 인간 염색체 서열의 플러스 및 마이너스 가닥 모두를 사용하여 추정상 표적을 확인하였다. 일부 프로토스페이서 표적에서 절단 활성을 제한할 수 있는 크로마틴, DNA 메틸화-, RNA 구조 및 기타 인자를 고려하면, 실제 표적화 능력이 이러한 컴퓨터적 분석 결과보다 덜 할 수 있다는 것에 주목하는 것이 중요하다.
도 34A-D 는 스트렙토코쿠스 써모필루스 LMD-9 로부터의 유형 II CRISPR 가 또한 진핵세포에서 기능할 수 있다는 것을 나타낸다. (A) 스트렙토코쿠스 써모필루스 LMD-9 로부터의 CRISPR 유전자좌 2 의 모식도. (B) S. 써모필루스 (S. thermphilus) CRISPR 시스템에 대한 발현 시스템의 설계. 항시성 EF1a 프로모터를 사용하여 인간 코돈-최적화 hStCas9 를 발현시킨다. 정확한 전사 개시를 확실히 하기 위해, U6 프로모터를 사용하여 tracrRNA 및 crRNA 의 성숙한 버전을 발현시킨다. 성숙 crRNA 및 tracrRNA 에 대한 서열을 나타낸다. crRNA 서열에서 소문자 "a" 로 나타낸 단일 염기를 사용하여, RNA Pol III 전사 터미네이터로서 역할하는 폴리U 서열을 제거하였다. (C) 인간 EMX1 유전자좌에서의 프로토스페이서 및 상응하는 PAM 서열 표적을 나타내는 모식도. 2 개의 프로토스페이서 서열을 하이라이트 표시하고 NNAGAAW 모티프를 만족시키는 그의 상응하는 PAM 서열을 자홍색 선으로 표시한다. 프로토스페이서 모두는 안티-센스 가닥을 표적화한다. (D) 표적 유전자좌에서의 StCas9-매개 절단을 나타내는 SURVEYOR 검정. RNA 안내 스페이서 1 및 2 는 각각 14% 및 6.4% 유도되었다. 이들 2 개 프로토스페이서 부위에서의 생물학적 레플리카 전체에 걸친 절단 활성의 통계적 분석을 표 1 에서 발견할 수 있다.
도 35 는 AAV-프로모터-TALE-효과기 구축물의 예를 나타내며, 여기서 hSyn = 인간 시냅신 1 프로모터, N+136 = TALE N-말단, AA +136 절두, C63 = TALE C-말단, AA +63 절두, vp = VP64 효과기 도메인, GFP = 녹색 형광 단백질, WPRE = 마못 간염 바이러스 전사후 조절 요소, bGH = 소 성장 호르몬 폴리A, ITR = AAV 역전 말단 반복물 및 AmpR = 암피실린 저항성 유전자이다.
도 36A-C 는 LITE 시스템의 설계 및 최적화를 나타낸다. (a) TALE DNA-결합 도메인은 CRY2 에 융합되고 전사 효과기 도메인은 CIB1 에 융합된다. 불활성 상태에서, TALE-CRY2 는 표적 유전자의 프로모터 부위에 결합하는 한편, CIB1-효과기는 핵 내에서 결합하지 않은 채로 남아있다. VP64 전사 활성제는 상기에 나타낸다. 청색으로 조명시, TALE-CRY2 및 CIB1-효과기는 빠르게 이량체화하고, CIB1-효과기를 표적 프로모터에 영입한다. 상기 효과기는 표적 유전자의 전사를 조정한다. (b) 그의 광-민감성 결합 파트너의 기능적 절두를 포함하는 LITE 를 갖는 내인성 표적 Neurog2 mRNA 의 광-의존적 상향조절. LITE-트랜스펙션된 Neuro-2a 세포를 7% 의 사용률로 5 mW/cm2 의 강도에서 466 nm 광으로 24 시간 동안 자극하였다 (0.066 Hz 에서의 1 초 펄스). (c) TALE-CRY2PHR 및 CIB1-VP64 LITE 에 의한 광-의존적 Neurog2 상향조절의 시간 경과. LITE-트랜스펙션된 Neuro-2a 세포를 7% 의 사용률로 5 mW/cm2 의 강도에서 466 nm 광으로 자극하였고 (0.066 Hz 에서 1 초 펄스), Neurog2 mRNA 수준은 광 자극 6 시간 후 감소한다. 모든 Neurog2 mRNA 수준을 발현하는 GFP 대조군 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3-4) (*, p < 0.05; 및 ***, p < 0.001).
도 37A-F 는 뉴런에서의 내인성 유전자좌를 표적화하는 시험관내 및 생체내 AAV-매개 TALE 전달을 나타낸다. (a) AAV 내로 패키징된 항시성 TALE 전사 활성제의 일반적 모식도. 효과기 도메인 VP64 를 하이라이트 표시하였다. hSyn: 인간 시냅신 프로모터; 2A: 구제역-유래 2A 펩티드; WPRE: 마못 간염 전사후 반응 요소; bGH pA: 소 성장 호르몬 폴리-A 신호, (b) 1 차 피질 뉴런에서의 (a) 로부터의 AAV-TALE-VP64 구축물로의 형질도입을 나타내는 대표 이미지. 세포를 GFP 및 뉴런 마커 NeuN 에 대해 염색하였다. 스케일 바 = 25 ㎛. (c) 다양한 내인전 유전자좌를 표적화하는 AAV-TALE-VP64 구축물을 1 차 피질 뉴런에서 전사 활성화에 대해 스크리닝하였다 (*, p < 0.05; **, p < 0.01 ; p < 0.001). (d) AAV 에 의한 TALE-VP64 의 마우스 ILC 로의 효율적 전달. 스케일 바 = 100 ㎛. (Cg1 = 대상 피질, PLC = 변연전 피질, ILC = 변연계 아래 피질), (e) ILC 에서의 뉴런의 효율적 형질도입의 고배율 이미지. (f) ILC 에서의 생체내 TALE-VP64 에 의한 Grm2 mRNA 상향조절 (평균 ± s.e.m.; n = 조건 당 3 마리 동물), 300 ㎛ 조직 펀치를 사용하여 측정함.
도 38A-I 은 1 차 뉴런 및 생체내에서의 내인성 전사의 LITE-매개 광유전학적 조절을 나타낸다. (a) 교환된 CRY2PHR 및 CIB1 구조를 갖는 AAV-LITE 활성제 구축물, (b) 1 차 뉴런에서의 AAV-전달 LITE 구축물의 동시-형질도입을 나타내는 대표적 이미지. GFP, HA-tag 및 DAPI 에 대해 세포를 염색하였다. (스케일 바 = 25 ㎛). (c) 0.8% 사용률로 466 nm 광 펄스로 (0.033Hz 에서 250 ms 펄스 또는 0.016Hz 에서 500 ms 펄스; 5 mW/cm2) 자극 24 시간 후 1 차 뉴런에서의 Grm2 발현의 광-유도 활성화. (d) 4 시간 및 24 시간점에서의 광 자극의 존재 및 부재 하 1 차 피질 뉴런에서의 Grm2 mRNA 의 상향 조절. 발현 수준을 GFP 로만 형질도입된 뉴런에 대해 나타낸다. (e) GFP 단독 대조군 형질도입, LITE 를 갖는 미자극 뉴런, 및 LITE 를 갖는 광-자극 뉴런에서의 mGluR2 단백질 수준의 정량. 대표적인 웨스턴 블롯을 로딩 대조군으로서 β-튜불린-III 을 사용하여 나타낸다. (f) LITE 시스템, 광학 섬유 이식물, 및 조직 단리에 사용한 0.35 mm 직경 뇌 펀치를 사용한 형질도입을 나타내는 모식도. (g) 두 LITE 성분 모두로 동시-형질도입된 ILC 의 대표적 이미지. 염색을 HA-tag (적색), GFP (녹색) 및 DAPI (청색) 로 나타낸다. (스케일 바 = 25 ㎛). (h) ILC 로 형질도입된 LITE 를 사용하는 내인성 Grm2 발현의 광-유도 활성화. **, p < 0.05; 각 실험 조건에 대해 4 마α리의 상이한 마우스로부터 데이터를 생성시켰다. (i) LITE 1.0 및 최적화 LITE 2.0 을 사용하는 Neurog2 발현의 광 유도 및 증가 배수. LITE2.0 은 높은 수준의 활성화를 유지하면서 최소의 배경을 제공한다. NLSα-임포틴 및 NLSsv40, 각각-임포틴 및 원숭이 바이러스 40 으로부터의 핵 국부화 신호; GS, Gly-Ser 링커; NLS*, 표시한 잔기가 Ala 로 치환되어 핵 국부화 활성을 방지하는 돌연변이화된 NLS; △318-334; 고등 식물 헬릭스-루프-헬릭스 전사 인자 상동성 부위의 결실.
도 39A-H 는 TALE- 및 LITE-매개 후생적 변형을 나타낸다. (a) LITE 후생적 변형제 (epiLITE) 의 모식도. (b) AAV 벡터 내의 조작된 후생적 전사 억제제 SID4X 의 모식도. 효율적인 AAV 패키징을 확실히 하기 위해 GFP (800bp) 보다는 phiLOV2.1 (330bp) 을 형광 마커로서 사용하였다. (c) 광 자극의 존재 및 부재 하 1 차 피질 뉴런에서의 내인성 Grm2 발현의 epiLITE-매개 억제. GFP 단독으로 형질도입된 뉴런에 대해 하향조절 배수를 나타낸다. (d) 광-자극의 존재 및 부재 하 Grm2 프로모터에서의 H3K9 히스톤 잔기 아세틸화에서의 epiLITE-매개 감소. (e, f) epiTALE-메틸트랜스퍼라아제 (epiTALE-KYP, -TgSET8 및 -NUE) 에 의한 Grm2 mRNA 의 감소 배수, 및 Grm2 프로모터에서의 히스톤 메틸화 마크 H3K9mel, H4K20me3 및 H3K27me3 의 상응하는 풍족. (g, h) epiTALE 히스톤 데아세틸라아제 (epiTALE-HDAC8, -RPD3, -Sir2a 및 -Sin3a) 에 의한 Grm2 mRNA 의 감소 배수, 및 Grm2 프로모터에서의 히스톤 잔기 아세틸화 마크 H4K8Ac 및 H3K9Ac 에 있어서의 상응하는 감소. 모든 패널에서 나타낸 값은 평균 ± s.e.m., n = 3-4 이다.
도 40 은 시험관내 CRY2 의 흡수 스펙트럼의 설명을 나타낸다. 크립토크롬 2 를 350-475 nm 광에 의해 최적으로 활성화시켰다1. 흡수 및 활성화에서의 급격한 감소가 480 nm 초과의 파장에 대해 나타났다. 스펙트럼을 [Banerjee, R. et al. The Signaling State of Arabidopsis Cryptochrome 2 Contains Flavin Semiquinone. Journal of 'Biological Chemistry 282, 14916-14922, doi: 10.1074/jbc.M700616200 (2007)] 로부터 조정하였다.
도 41 은 LITE-매개 유전자 발현에 대한 조명 사용률의 효과를 나타낸다. 가변적인 사용률 (총 시간의 % 로서의 조명) 을 사용하여 KLF4 유전자를 표적화하는 LITE 를 발현하는 293FT 세포를 자극하여, LITE 활성에 대한 사용률의 효과를 조사하였다. KLF4 발현 수준을 GFP 만을 발현하는 세포와 비교하였다. 자극 매개변수는 다음과 같다: 466 nm, 5 mW/cm2, 24 시간 동안. 펄스를 0.067 Hz 에서 하기 지속 기간으로 수행하였다: 1.7% = 0.25 초 펄스, 7% = 1 초 펄스, 27% = 4 초 펄스, 100% = 연속 조명. (평균 ± s.e.m.; n = 3-4).
도 42A-B 는 LITE-매개 유전자 발현 및 세포 생존에 대한 광 강도의 효과를 나타낸다. (a) CRY2PHR :: CIB1 LITE 의 전사 활성은 466 nm 청색광의 강도에 따라 가변적이라는 것이 발견되었다. Neuro 2a 세포를 24 시간 동안 7% 사용률 (0.066Hz 에서 1 초 펄스) 에서 자극하였다. (b) 적색-형광 에티디움 동종이량체-1 에 대해 양성인 세포 대 칼세인-양성 세포의 % 로서 광-유도 독성을 측정하였다. 모든 Neurog2 mRNA 수준을 GFP 만을 발현하는 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3-4).
도 43 은 LITE-매개 유전자 발현에 대한 전사 활성화 도메인의 영향을 나타낸다. 상이한 전사 활성화 도메인 (VP16, VP64 및 p65) 을 사용하는 LITE 에 의한 광의 존재 및 부재 하 Neurog2 상향조절. LITE 로 트랜스펙션한 Neuro-2a 세포를 24 시간 동안 466 nm 광으로 5 mW/cm2 의 강도 및 7% 의 사용률에서 (0.066 Hz 에서 1 초 펄스) 자극하였다. (평균 ± s.e.m.; n = 3-4)
도 44A-C 는 내인성 유전자 전사의 화학적 유도를 나타낸다. (a) 아브시스산 (ABA) 수용체 시스템을 기반으로 하는 화학 물질 유도 2 개 하이브리드 TALE 시스템의 설계를 나타내는 모식도. ABI 및 PYL 은 ABA 첨가시 이량체화하며 ABA 철회시 해리된다. (b) ABA-의존적 Neurog2 상향조절의 시간 경과. 250 μM 의 ABA 를 TALE(Neurog2)-ABI 및 PYL-VP64 를 발현하는 HEK 293FT 세포에 첨가하였다. ABA 첨가 후 나타낸 시점에서 mRNA 증가 배수를 측정하였다. (c) ABA 자극 24 시간 후 Neurog2 mRNA 수준의 감소. 모든 Neurog2 mRNA 수준을 발현하는 GFP 대조군 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3-4).
도 45A-C 는 AAV 상청액 생성을 나타낸다. (a) 렌티바이러스 및 AAV 벡터 (GFP 운반) 를 사용하여 형질유도 효율을 시험하였다. (b) 1 차 배아 피질 뉴런을 동일 수의 AAV 또는 렌티바이러스-트랜스펙션된 293FT 세포로부터 유래한 300 및 250 μL 상청액으로 형질도입하였다. GFP 발현의 대표적 이미지를 7 d.p.i. 에서 수집하였다. 스케일 바 = 50 ㎛. (c) 나타낸 공정을 AAV 상청액의 생성 및 1 차 뉴런의 후속적 형질도입을 위해 개발하였다. 293FT 세포를 관심 유전자를 운반하는 AAV 벡터, AAVl 혈청형 패키징 벡터 (pAAV1), 및 PEL 을 사용하는 헬퍼 플라스미드 (pDF6) 로 트랜스펙션하였다. 48 시간 후, 상청액을 수확하고 0.45 ㎛ PVDF 멤브레인을 통해 여과하였다. 1 차 뉴런을 이후 상청액으로 형질도입하고 남아 있는 분취액을 -80℃ 에서 보관하였다. 5-6 일 후 AAV 구축물 발현의 안정한 수준에 도달하였다. 이러한 공정에 따른 AAV 상청액 생성을 96-웰 포맷으로 96 개 이하의 상이한 바이러스 구축물의 생성에 사용할 수 있다 (도 37C 에서 나타낸 뉴런에서의 TALE 스크린에 이용).
도 46 은 DNaseI-민감성 크로마틴 부위에 의해 유도된 TALE 표적 부위의 선택을 나타낸다. ENCODE (http://genome.ucsc.edu) 로부터의 마우스 피질 조직 데이터를 기반으로 한 높은 DNaseI 민감성을 사용하여 개방 크로마틴 부위를 확인하였다. 전사 시작 부위의 2 kb 업스트림 부위 내의 최고 진폭을 갖는 피크를 표적화를 위해 선택하였다. TALE 결합 표적을 이후 피크의 중심에서 200 bp 부위 내에서 골랐다.
도 47 은 1 차 뉴런 건강에 대한 광 사용률의 영향을 나타낸다. 1 차 피질 뉴런 건강에 대한 광 자극의 효과를 사용률 7%, 0.8%, 및 무광 조건에 대해 비교하였다. 칼세인을 사용하여 뉴런 생존도를 평가하였다. 명시야 이미지를 캡쳐하여 형태 및 세포 무결성을 나타내었다. 466 nm 광의 5 mW/cm2 로 24 시간 동안 표시 사용률로 1 차 피질 뉴런을 자극하였다. 대표적 이미지, 스케일 바 = 50 ㎛. 펄스를 하기의 방식으로 수행하였다: 7% 사용률 = 0.067 Hz 에서 1 초 펄스, 0.8% 사용률 = 0.0167 Hz 에서 0.5 초 펄스.
도 48 은 광유전학적 자극 동안의 마우스 이미지를 나타낸다. 잠에서 깬, 자유로이 거동하는, LITE-주입 마우스를 뇌정위적으로 이식한 캐뉼라 및 광학 섬유로 사진찍었다.
도 49 는 마우스 변연계 아래 피질에서의 AAVl/2 에 의한 LITE 성분의 동시-형질도입 효율을 나타낸다. TALE(Grm2)-CIB1 단독, CRY2PHR-VP64 단독에 의해 형질도입되거나 동시-형질도입된 세포를, 모든 형질도입된 세포의 % 로서 계산하였다.
도 50 은 기준선 전사 조절에 대한 개별적인 LITE 성분의 기여도를 나타낸다. Grm2 mRNA 수준을 개별적인 LITE 성분으로 트랜스펙션된 1 차 뉴런에서 측정하였다. Grm2 TALE 1-CIB1 단독을 발현하는 1 차 뉴런은 전체 LITE 시스템을 발현하는 미자극 세포에서와 같이 Grm2 mRNA 수준에 있어서 유사한 증가를 초래하였다 (평균 ± s.e.m.; n = 3-4).
도 51 은 활성화, 배경 신호 및 감소 배수에 대한 LITE 성분 조작의 효과를 나타낸다. 단백질 변형을 이용하여, LITE 성분이 감소된 배경 전사 활성화를 초래하면서 광에 의한 유도 비율을 향상시킨다는 것을 발견하였다. 단백질 변형을 하기에서 상세히 토의한다. 간략하게, 내인성 핵 이출 신호에서의 돌연변이 및 핵 국부화 신호를 사용하여 CRY2PHR-VP64 성분의 핵 이입을 향상시켰다. 핵 국부화 또는 CIB1 전사 활성화를 감소시키는 것으로 의도된 CIB1 의 여러 변이를 추구하여 TALE-CIB1 성분의 배경 활성에 대한 기여도를 감소시켰다. 시험한 CRY2PHR-VP64 및 TALE-CIB1 의 모든 조합의 결과를 상기에 나타낸다. 막대 그래프의 좌측에 있는 표는 각 조합에 사용한 도메인/돌연변이의 특정 조합을 표시한다. 표 및 막대 그래프의 각 행은 성분 세부사항, 광/무광 활성, 및 특정 CRY2PHR/CIB1 조합에 대한 광에 의한 유도 비율을 포함한다. LITE 1.0 에 비해 감소된 배경 및 증가된 유도 배수를 모두 초래한 조합을 "+" 표시한 표 열에서 녹색으로 하이라이트 표시한다 (t-시험 p<0.05). CRY2PHR-VP64 구축물: 3 개의 새로운 구축물을 CRY2PHR-VP64 핵 이입을 향상시키는 목적으로 설계하였다. 먼저, CRY2PHR 의 예측된 내인성 핵 이출 서열 내의 돌연변이 L70A 및 L74A 를 유도하여 단백질의 핵 이출을 제한하였다 (효과기 열에서 '*' 로 나타냄). 두 번째로, α-임포틴 핵 국부화 서열을 CRY2PHR-VP64 의 N-말단에 융합시켰다 (효과기 열에서 'A' 로 나타냄). 세 번째로, SV40 핵 국부화 서열을 CRY2PHR-VP64 의 C-말단에 융합시켰다 (효과기 열에서 'P' 로 나타냄). TALE-CIB1 링커: LITE 1.0 에서 사용한 TALE 과 CIB1 사이의 SV40 NLS 링커를 TALE-CIB1 단백질의 핵 이출을 증가시키도록 설계된 여러 링커 중 하나로 대체하였다 (CIB1 링커 열에서 사용한 기호를 괄호 안에 나타냄): 유연성 글리신-세린 링커 (G), 아데노바이러스 유형 5 E1B 핵 이출 서열 (W), HIV 핵 이출 서열 (M), MAPK 핵 이출 서열 (K) 및 PTK2 핵 이출 서열 (P). NLS* 내인성 CIB1 핵 국부화 서열 돌연변이: 핵 국부화 신호는 야생형 CIB1 서열 내에 존재한다. 이러한 신호는 NLS* 구축물 내에서 K92A, R93A, K105A 및 K106A 에서 돌연변이화되어 TALE-CIB1 핵 국부화가 약화되었다 (NLS* 열에서 'N' 으로 나타냄). △CIB1 전사 인자 상동성 결실: 가능한 기저 CIB1 전사 활성화를 제거하려는 노력으로, 결실 구축물을 고등 식물에서의 염기성 헬릭스-루프-헬릭스 전사 인자에 대한 높은 상동성 부위가 제거되게 설계하였다. 이들 결실된 부위는 Aaa230-256, Aaa276-307, Aaa308-334 로 이루어졌다 (ACIB1 열에서 '1', '2' 및 '3' 으로 나타냄). 각각의 경우, 결실된 부위를 3 개 잔기 GGS 링크로 대체하였다. CIB1 내로의 NES 삽입: TALE- CIB1 의 광-의존적 핵 이입을 촉진하기 위한 하나의 계획은 CRY2PHR 과의 이량체화 접촉면에서 CIB1 에 NES 를 삽입하여, CRY2PHR 과 결합시 신호가 숨겨질 수 있는 것이다. 이를 위해, NES 를 공지된 CRY2 상호작용 도메인 CIBN 내 상이한 위치 (aa 1-170) 에서 삽입하였다. 위치는 하기와 같다 (NES 열에서 사용한 기호를 괄호 안에 나타냄): aa28 (1), aa52 (2), aa73 (3), aal20 (4), aal40 (5), aal60 (6). *bHLH 염기성 헬릭스-루프-헬릭스 돌연변이: 직접적 CIB1-DNA 상호작용을 감소시키기 위해, CIB1 에서의 염기성 헬릭스-루프-헬릭스 부위의 여러 염기성 잔기를 돌연변이시켰다. 하기 돌연변이는 모든 *bHLH 구축물 내에 존재한다 (도 51 의 *bHLH 컬럼에서 'B' 로 나타냄): R175A, G176A, R187A 및 R189A.
도 52 는 TALE-CIB1 의 광 매개 동시-의존적 핵 이입의 설명을 나타낸다. (a) 광의 부재 하, TALE-CIB1 LITE 성분은 핵 국부화 신호, NLS 의 부재 (또는 약한 핵 이출 신호, NES 의 첨가) 로 인해 세포질에서 존재한다. 한편 NLS 를 함유하는 CRY2PHR-VP64 성분은 저절로 핵에 능동적으로 이입된다. (b) 청색광의 존재 하에, TALE-CIB1 은 CRY2PHR 에 결합한다. CRY2PHR-VP64 에 존재하는 강력한 NLS 는 이제 LITE 성분 모두의 복합체의 핵 이입을 매개하여, 이들이 표적화된 유전자좌에서 전사를 활성화시킬 수 있게 한다.
도 53 은 주목할만한 LITE 1.9 조합을 나타낸다. LITE 2.0 구축물에 추가로, 조작된 LITE 성분 스크린으로부터의 여러 CRY2PHR-VP64 :: TALE-CIB1 조합에 특히 주목하였다. 돌연변이화된 내인성 NLS 를 갖는 α-임포틴 NLS 효과기 구축물 및 △276-307 TALE-CIB1 구축물이 조합된 LITE 1.9.0 은 9 초과의 유도 비율 및 180 초과의 절대 광 활성화를 나타내었다. 미변형된 CRY2PHR-VP64 와 돌연변이화된 NLS, △318-334, AD5 NES TALE-CIB1 구축물이 조합된 LITE 1.9.1 은, 4 의 유도 비율을 1.06 의 배경 활성화로 얻었다. 2 미만의 배경 활성화 및 7 내지 12 범위의 유도 비율을 갖는 다른 LITE 1.9 조합의 선택을 또한 하이라이트 표시하였다.
도 54A-D 는 뉴런에서의 TALE SID4X 억제제 분석 및 적용을 나타낸다. a) 합성적 억제제를 4 개의 SID 도메인 (SID4X) 을 연쇄 연결시켜 구축하였다. 최적의 TALE -억제제 구조를 확인하기 위해, SID 또는 SID4X 를 마우스 p11 유전자를 표적화하도록 설계된 TALE 에 융합시켰다. (b) p11 mRNA 에서의 감소 배수를 qRT-PCR 을 사용하여 검정하였다. (c) AAV 에 패키징된 항시성 TALE 전사 억제제의 일반적 모식도. hSyn: 인간 시냅신 프로모터; 2A: 구제역-유래 2 A 펩티드; WPRE: 마못 간염 전사후 반응 요소; bGH pA: 소 성장 호르몬 폴리-A 신호. phiLOV2.1 (330bp) 을 효율적 AAV 패키징을 확실히 하기 위한 단길이 형광 마커로서 선택하였다. (d) 내인성 마우스 유전자좌 Grm5 및 Grm2 를 표적화하는 2 개 TALE 을 SID4X 에 융합시키고 1 차 뉴런에 바이러스 형질도입하였다. SID4X 를 통한 표적 유전자 하향 조절을 GFP 만을 발현하는 뉴런에서의 수준에 대하여 각각의 TALE 에 대해 나타낸다. (평균 ± s.e.m.; n = 3-4).
도 55A-B 는 다양한 집합의 epiTALE 이 뉴런 및 Neuro2a 세포에서의 전사 억제를 매개한다는 것을 나타낸다. a) 상이한 히스톤 효과기 도메인에 융합된 총 24 개 Grm2 표적화 TALE 을 AAV 를 사용하여 1 차 피질 마우스 뉴런에 형질도입하였다. Grm2 mRNA 수준을, GFP 단독으로 형질도입된 뉴런에 대해 RT-qPCR 을 사용하여 측정하였다. * 는 p < 0.05 인 억제를 나타낸다. b) 총 32 개 epiTALE 을 Neuro2A 세포에 트랜스펙션시켰다. 이들 중 20 개는 표적화된 Neurog2 유전자좌의 유의한 억제를 매개하였다 (* = p<0.05).
도 56A-D 는 Neuro 2A 세포에서의 히스톤 개질과 함께 epiTALE 매개된 전사 억제를 나타낸다. (a) Neuro 2A 세포에서 쥐과 Neurog2 유전자좌를 표적화하는 SIRT3 및 히스톤 탈아세틸화 후생적 효과기 NcoR 에 융합된 TALE 을 Neurog2 전사체 수준에 대한 억제 활성에 대해 검정하였다. (b) ChIP RT-qPCR 은 NcoR 및 SIRT3 epiTALE 에 대한 Neurog2 프로모터에서의 H3K.9 아세틸화에 있어서의 감소를 나타낸다. (c) 공지된 히스톤 메틸트랜스퍼라아제 결합 활성을 갖는 후생적 효과기 PHF19 를 Neurog2 mRNA 수준의 Neurog2 매개 억제를 표적화하는 TALE 에 융합시켰다. (d) ChIP RT-qPCR 은 PHF19 epiTALE 에 대한 Neurog2 프로모터에서의 H3K27me3 수준에 있어서 증가를 나타낸다.
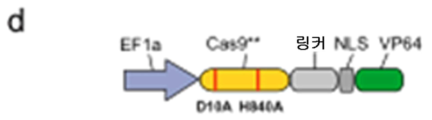
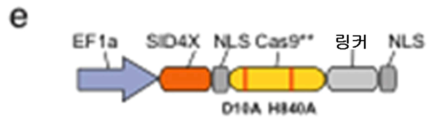
도 57A-G 는 RNA-안내 DNA 결합 단백질 Cas9 가 전사 효과기 도메인을 특정 게놈 유전자좌에 표적화하는데 사용될 수 있다는 것을 나타낸다. (a) 유형 II 스트렙토코쿠스 피오게네스 CRISPR/Cas 시스템으로부터의 RNA-안내 뉴클레아제 Cas9 는 2 개의 알라닌 치환 (D10A 및 H840A) 을 도입함으로써 핵산분해적으로 불활성인 RNA-안내 DNA 결합 단백질 (Cas9**) 로 전환될 수 있다. 모식도는 합성적 안내 RNA (sgRNA) 가 인간 게놈에서의 특정 유전자좌에 대한 Cas9**-효과기 융합을 지시할 수 있다는 것을 나타낸다. sgRNA 는 표적 서열을 명기하는 5' 말단에서 20bp 안내 서열을 포함한다. 표적 게놈 DNA 상에서, 20bp 표적 부위는 그 뒤에 5'-NGG PAM 모티프가 뒤따를 수 있다. (b, c) 각각 인간 KLF4 및 SOX2 유전자좌에서의 sgRNA 표적 부위를 나타내난 모식도. 각각의 표적 부위를 청색 막대로 표시하고 상응하는 PAM 서열을 자홍색 막대로 표시한다. (d, e) Cas9**-VP64 전사 활성제 및 SID4X-Cas9** 전사 억제제 구축물의 모식도. (f, g) 각각 Cas9**-VP64 및 SID4X-Cas9** 매개된 KLF4 의 활성화 및 SOX2 억제. 모든 mRNA 수준을 GFP 모의 트랜스펙션된 대조군 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3).
도 58 은 2 개 TALE 이 각각의 내인성 마우스 유전자좌 Grm5, Grin2a 및 Grm2 를 표적화하는, 설계된 6 개 TALE 을 나타낸다. TALE 을 전사 활성제 도메인 VP64 또는 억제제 도메인 SID4X 에 융합시키고, 1 차 뉴런에 바이러스 형질도입하였다. VP64 를 통한 표적 유전자 상향조절 및 SID4X 를 통한 하향조절 모두를, GFP 만을 발현하는 뉴런에서의 수준에 대하여 각각의 TALE 에 대해 나타낸다.
도 59A-B 는 하기를 나타낸다: (A) SID4X 억제제 도메인을 강조하는 LITE 억제제 구축물. (B) Grm2 T1-LITE 및 Grm2 T2-LITE 를 사용하는 1 차 피질 뉴런에서의 내인성 Grm2 발현의 광-유도 억제. 하향조절 배수를 GFP 단독으로 형질도입된 뉴런에 대해 나타낸다 (평균 ± s.e.m.; 모든 서브패널에 대해 n = 3-4).
도 60A-B 는 CRY2PHR 및 CIB1 성분 교환을 나타낸다. (A) TALE-CIB1::CRY2PHR-VP64 는 TALE-CRY2PHR::CIB1-VP64 보다 더 높은 수준에서 Ngn2 를 활성화시킬 수 있었다. (B) Ngn2 LITE 의 활성화 비율 (광 대 무광) 배수는 두 설계 모두에 대해 유사한 효율을 나타낸다. 자극 매개변수는 도 36B 에서 사용한 것들과 동일하였다.
도 61 은 유도 Cas9 에 대한 Tet Cas9 벡터 설계를 나타낸다.
도 62 는 Cas9 및 EGFP 의 독시사이클린 유도 후 293FT 세포에서의 EGFP 발현 및 벡터를 나타낸다.
도 1 은 공간적 및 시간적 정확도의 필요성을 나타내는 모식도를 나타낸다.
도 2 는 전사 활성제 유사 효과기 (TALE) 를 나타낸다. TALE 는 그의 서열의 코어에서 34 개 aa 반복물로 이루어진다. 각각의 반복물은 TALE 에 의해 결합되는 표적 DNA 에서의 염기에 상응한다. 반복물은 위치 12 및 13 에서 2 개의 가변적인 아미노산에 의해서만 상이하다. 이러한 대응의 코드가 설명된 바 있으며 (Boch, J et al. , Science, 2009 및 Moscou, M et al. , Science, 2009) 이 도면에서 나타낸다. 출원인은 이러한 코드를 통합하며 게놈 내에 선택 서열을 결합시킬 수 있는 설계자 TALE 의 합성 방법을 개발하였다 (Zhang, F et al. , Nature Biotechnology, 2011).
도 3 은 LITE: TALE/크립토크롬 전사 활성화의 설계를 나타낸다. 각각의 LITE 는 CRY2 에 융합된 TALE 및 VP64 (전사 인자) 에 융합된 크립토크롬 결합 파트너 CIB1 을 포함할 수 있는 2-성분 시스템이다. 불활성 상태에서, TALE 은 이의 융합된 CRY2 도메인을 관심 유전자의 프로모터 부위에 국부화시킨다. 이때, CIB1 은 CRY2 에 결합할 수 없고, CIB1-VP64 가 핵 공간 내에서 미결합된 채로 있다. 488 nm (청색) 광으로 자극시, CRY2 는 형태적 변화를 거쳐, 이의 CIB1 결합 부위가 밝혀진다 (Liu, H et al. , Science, 2008). CIB1 의 신속한 결합은 융합된 VP64 도메인의 영입을 초래하여, 표적 유전자의 전사를 유도한다.
도 4 는 LITE 활성에 대한 크립토크롬 이량체 절두의 효과를 나타낸다. CRY2 및 CIB1 의 활성을 변경시키는 것으로 알려져 있는 절두를 (Kennedy M et al, Nature Methods 2010) 전체 길이 단백질에 대해 비교하였다. Neurog2 의 프로모터에 대해 표적화된 LITE 를 도메인의 각 조합에 대해 Neuro-2a 세포에서 시험하였다. 488 nm 광으로 자극한 후, Neurog2 의 전사 수준을 자극된 샘플 및 미자극된 샘플에 대해 qPCR 을 사용하여 정량하였다.
도 5 는 KLF4 LITE 의 광-강도 의존적 반응을 나타낸다.
도 6 은 Neurog2 LITE 의 활성화 반응속도 및 Neurog2 LITE 의 불활성화 반응속도를 나타낸다.
도 7A 는 출원인의 RVD 스크리닝 시스템을 사용하여 측정한 바와 같은 다양한 RVD 의 염기-선호도를 나타낸다.
도 7B 는 출원인의 RVD 스크리닝 시스템을 사용하여 측정한 바와 같은 추가적인 RVD 의 염기-선호도를 나타낸다.
도 8A-D 는 다음을 나타낸다: (a) 잔토모나스 종 (Xanthomonas sp.) 에서 유래한 TALE 의 자연적 구조. 각각의 DNA-결합 모듈은 34 개의 아미노산으로 이루어지는데, 각각의 반복물의 12 번째 및 13 번째 아미노산 위치에서의 RVD 는 부호 NG = T, HD = C, NI = A 및 NN = G 또는 A 에 따라 표적화되는 DNA 염기를 명시한다. DNA-결합 모듈은 비반복적 N 및 C 말단에 측면 위치하여, 전좌, 핵 국부화 (NLS) 및 전사 활성화 (AD) 도메인을 수반한다. N 말단 내 암호성 신호는 표적 부위의 제 1 염기로서 티민을 명시한다. (b) TALE 툴박스는 커스텀 TALE-TF 및 TALEN 의 신속하고 값싼 구성을 가능하게 한다. 키트는 총 12 개 플라스미드로 이루어진다: 4 개 단량체 플라스미드가 PCR 증폭용 주형으로서 사용되고, 4 개의 TALE-TF 및 4 개의 TALEN 클로닝 백본은 0.5 반복물에 의해 표적화된 4 개의 상이한 염기에 상응한다. CMV, 사이토메갈로바이러스 프로모터; N 말단, Hax3 TALE 로부터의 비반복적 N 말단; C 말단, Hax3 TALE 로부터의 비반복적 C 말단; BsaI, 커스텀 TALE DNA-결합 도메인의 삽입에 사용된 유형 II 제한 부위; ccdB + CmR, ccdB 음성 선택 유전자 및 클로람페니콜 저항성 유전자를 포함하는 음성 선택 카세트; NLS, 핵 국부화 신호; VP64, 단순 포진 바이러스의 VP 16 단백질에서 유래한 합성 전사 활성제; 2A, 2A 자체-절단 링커; EGFP, 증강된 녹색 형광 단백질; 폴리A 신호, 폴리아데닐화 신호; FokI, FokI 엔도뉴클레아제로부터의 촉매적 도메인. (c) TALE 은 커스텀 TALE-TF 를 생성시키고 게놈으로부터의 내인성 유전자의 전사를 조정하는데 사용될 수 있다. TALE DNA-결합 도메인은 합성 VP64 전사 활성제에 융합되는데, 이는 RNA 폴리머라아제 및 전사 개시에 필요한 기타 인자를 영입한다. (d) TALEN 은 비상동 쌍 또는 상동성 유도 쌍을 통해 게놈 편집을 촉진시키기 위해 위치-특이적 이중가닥 파단점을 생성시키는데 사용될 수 있다. 2 개의 TALEN 은 측면 위치 16-bp 스페이서의 결합 부위 쌍을 표적화한다. 좌측 및 우측 TALEN 은 각각 표적 부위의 상부 및 하부 가닥을 인지한다. 각각의 TALE DNA-결합 도메인은 FokI 엔도뉴클레아제의 촉매적 도메인에 융합되고; FokI 이 이량체화될 때, 이는 좌측 및 우측 TALEN-결합 도메인 사이의 부위에서 DNA 를 절단한다.
도 9A-F 는 단량체 서열 (위치 12 및 13 에서의 RVD 제외) 및 특정 서열을 갖는 단량체가 발생하는 빈도를 열거하는 표를 나타낸다.
도 10 은 TALE 활성에 대한 비-RVD 아미노산의 효과 비교를 나타낸다.
도 11 은 VP64, p65 와 VP16 사이의 활성화 수준을 비교하는 활성제 스크린을 나타낸다.
도 12A-D 는 TALE 전사 억제제 구조의 개발을 나타낸다. (a) TALE 억제제 스크리닝에 대한 SOX2 TALE 의 설계. 인간 게놈의 SOX2 유전자좌 내의 14bp 서열을 표적화하는 TALE 을 합성하였다. (b) 스크리닝한 모든 억제제 및 그의 숙주 기원의 목록 (좌측). 8 개의 상이한 후보물 억제제 도메인은 SOX2 TALE 의 C-말단에 융합되었다. (c) 내인성 SOX2 mRNA 의 감소 배수를 모의 트랜스펙션 세포에서의 SOX2 mRNA 수준을, 각각의 후보 TALE 억제제로 트랜스펙션시킨 세포에서의 SOX2 mRNA 수준에 의해 나누어, qRTPCR 을 사용하여 측정한다. (d) 내인성 CACNA1C 의 전사 억제. G-표적화 RVD 로서 NN, NK 및 NH 를 사용하는 TALE 을 구축하여 인간 CACNA1C 유전자좌 내 18 bp 표적 부위를 표적화하였다. 각각의 TALE 을 SID 억제 도메인에 융합시킨다. NLS, 핵 국부화 신호; KRAB, 크루펠 (Kruppel)-관련 박스; SID, mSin 상호작용 도메인. 모든 결과를 HEK 293FT 세포에서 3 개의 독립적 실험으로부터 수집하였다. 오류 막대는 s.e.m. 을 나타내고; n = 3 이다. * p < 0.05, 스튜던트 (Student) t 시험.
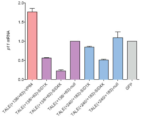
도 13A-C 는 SID 및 SID4X 를 사용하는 TALE 전사 억제제 구조의 최적화를 나타낸다. (a) TALE 억제제 구조의 시험을 위한 p11 TALE 의 설계. 마우스 (Mus musculus) 게놈의 p11 (s100a10) 유전자좌 내의 20bp 서열을 표적화하는 TALE 을 합성하였다. (b) 내인성 마우스 p11 mRNA 의 전사 억제. 야생형 TALE 구조의 2 개 상이한 절두부를 갖는 마우스 p11 유전자좌를 표적화하는 TALE 을 x-축에 나타낸 바와 같이 상이한 억제제 도메인에 융합시켰다. 괄호 안의 값은 DNA 결합 반복물에 측면 위치하며 상기 구축물에서 사용한 억제제 도메인이 그 뒤에 있는 TALE DNA 결합 도메인의 N-말단 및 C-말단에서의 아미노산 수를 나타낸다. 내인성 p11 mRNA 수준을 qRT-PCR 을 사용하여 측정하고, GFP-인코딩 구축물로 트랜스펙션시킨 음성 대조군 세포에서의 수준으로 정규화시켰다. (c) 내인성 마우스 p11 의 전사 억제 배수. 내인성 p11 mRNA 의 감소 배수를, 음성 대조군 GFP 구축물로 트랜스펙션시킨 세포에서의 p11 mRNA 수준을 각각의 후보 TALE 억제제로 트랜스펙션시킨 세포에서의 p11 mRNA 수준에 의해 나누어, qRT-PCR 을 사용하여 측정한다. x-축에 따른 구축물의 표지는 이전 패널에서와 동일하다. NLS, 핵 국부화 신호; SID, mSin 상호작용 도메인; SID4X, 단길이 펩티드 링커에 의해 연결된 SID 도메인의 최적화된 4-회 탠덤 (tandem) 반복물. 모든 결과는 Neuro2A 세포에서의 3 개 독립적 실험에서 수집하였다. 오류 막대는 s.e.m. 을 나타내고; n = 3 이다. *** /p< 0.001, 스튜던트 t 시험.
도 14 는 2 가지 상이한 유형의 TALE 구조의 비교를 나타낸다.
도 15A-C 는 화학적 유도 TALE ABA 유도 시스템을 나타낸다. ABI (ABA 비-민감성 1) 및 PYL (PYL 단백질: 피라박틴 저항성 (PYR)/PYRl-유사 (PYL)) 은 식물 호르몬 아브시스산 (ABA) 의 결합시 이량체화하는 하기 열거한 2 개 단백질로부터의 도메인이다. 이러한 식물 호르몬은 출원인이 출원인의 유도 TALE 시스템에서 사용한 소분자 화학 물질이다. 이러한 시스템에서, TALE DNA-결합 폴리펩티드는 ABI 도메인에 융합되는 반면, VP64 활성화 도메인 또는 SID 억제제 도메인 또는 임의의 효과기 도메인은 PYL 도메인에 연결된다. 따라서 ABA 분자의 존재에 의해 유도시, 2 개 상호작용 도메인, ABI 및 PYL 은 이량체화되고 TALE 이 효과기 도메인에 연결되게 하여 표적 유전자 발현을 조절하는데 있어서의 이의 활성을 수행할 수 있게 한다.
도 16A-B 는 화학적 유도 TALE 4OHT 유도 시스템을 나타낸다.
도 17 은 LITE 기능성에 대한 크립토크롬2 이형이량체 배향의 효과를 나타낸다.
도 18 은 마우스 피질 뉴런 배양액에서의 mGlur2 LITE 활성을 나타낸다.
도 19 는 LITE AAV 벡터을 사용하는 1 차 마우스 뉴런의 형질도입을 나타낸다.
도 20 은 생체내 LITE 성분의 발현을 나타낸다.
도 21 은 사용한 특정 NES 펩티드가 LDLASLIL 인 구축물의 향상된 설계를 나타낸다.
도 22 는 40H 타목시펜의 부재 및 존재 하 Sox2 mRNA 수준을 나타낸다.
도 23A-E 는 스트렙토코쿠스 피오게네스 (Streptococcus pyogenes) SF370 으로부터의 유형 II CRISPR 유전자좌가 DNA 의 표적화된 DSB 가 촉진되도록 포유동물 세포에서 재구성될 수 있다는 것을 나타낸다. (A) NLS 를 갖는 SpCas9 및 SpRNase III 의 조작은 포유동물 핵 내로의 이입을 가능하게 한다. (B) SpCas9 및 SpRNase III 의 포유동물 발현은 EFla 프로모터에 의해 유도되는 반면, tracrRNA 및 예비-crRNA 어레이 (DR-스페이서-DR) 는 U6 프로모터에 의해 유도된다. PAM 을 갖는 인간 EMX1 유전자좌로부터의 프로토스페이서 (청색 하이라이트) 를 예비-crRNA 어레이에서 스페이서에 대한 주형으로서 사용한다. (C) 표적 유전자좌와 EMX1-표적화 crRNA 사이의 염기 페어링의 모식적 표현. 적색 화살표는 추정상 절단 부위를 나타낸다. (D) SpCas9-매개 삽입-결실 (indel) 에 대한 SURVEYOR 검정. (E) 187 클론 암플리콘으로부터 확인된 돌연변이화된 대립형질의 대표 서열을 나타내는 예시적 크로마토그램. 적색 대시 기호는 결실된 염기를 나타내고; 적색 염기는 삽입 또는 돌연변이를 나타낸다. 스케일 바 = 10 ㎛.
도 24A-C 는 SpCas9 가 포유동물 세포에서의 다수 게놈 유전자좌를 표적화하도록 재프로그래밍될 수 있다는 것을 나타낸다. (A) 청색 선으로 표시한 5 개 프로토스페이서의 위치를 나타내는 인간 EMX1 유전자좌의 모식도, 상응하는 PAM 은 자홍색으로 나타냄. (B) 예비-crRNA 및 tracrRNA 의 직접 반복 (회색) 부위 사이의 하이브리드화를 나타내는 예비-crRNA:tracrRNA 복합체 (상부) 의 모식도. 키메라 RNA 설계의 모식도 (M. Jinek et al., A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816 (Aug 17, 2012)) (하부). tracrRNA 서열을 적색으로 나타내고 20bp 스페이서를 청색으로 나타낸다. (C) 인간 EMX1 유전자좌에서의 5 개 프로토스페이서에서의 Cas9-매개 절단의 효능을 비교하는 SURVEYOR 검정. 각각의 프로토스페이서를 처리된 예비-crRNA:tracrRNA 복합체 (crRNA) 또는 키메라 RNA (chiRNA) 를 사용하여 표적화한다.
도 25A-D 는 SpCas9 특이성의 평가 및 TALEN 과의 효율 비교를 나타낸다. (A) 단일 점 돌연변이를 갖는 EMX1-표적화 키메라를 생성시켜 스페이서-프로토스페이서 미스매치의 효과를 평가하였다. (B) 상이한 돌연변이체 키메라 RNA 의 절단 효율을 비교하는 SURVEYOR 검정. (C) TALEN 표적화 EMX1 의 설계를 나타내는 모식도. (D) TALEN 및 SpCas9 의 효율을 비교하는 SURVEYOR 겔 (N = 3).
도 26A-G 는 상동 재조합 및 멀티플렉스 게놈 조작을 위한 Cas9 의 적용을 나타낸다. (A) RuvC I 도메인의 돌연변이는 Cas9 를 절단 효소 (SpCas9n) 로 전환시킨다. (B) EMX1-표적화 키메라 RNA 와 SpCas9 와의 동시-발현은 삽입-결실을 유발하는 반면, SpCas9n 은 그렇지 않다 (N = 3). (C) 재조합 계획의 모식적 표시. 수복 주형은 제한 부위가 EMX1 유전자좌에 삽입되도록 설계된다. 변형된 부위를 증폭하는데 사용한 프라이머를 적색 화살표로 나타낸다. (D) 제한 단편 길이 다형성 겔 분석. 화살표는 HindIII 소화에 의해 생성된 단편을 나타낸다. E) 성공적인 재조합을 나타내는 예시적 크로마토그램. (F) SpCas9 는 EMX1 및 PVALB 를 표적화하는 2 개 스페이서를 포함하는 crRNA 어레이를 사용하여 멀티플렉스 게놈 변형을 촉진시킬 수 있다. crRNA 어레이의 설계를 나타내는 모식도 (상부). 스페이서 모두 효율적인 프로토스페이서 절단을 매개한다 (하부). (G) SpCas9 를 사용하여 정확한 게놈 결실을 수행할 수 있다. EMX1 을 표적화하는 2 개 스페이서 (상부) 는 118bp 게놈 결실을 매개하였다 (하부).
도 27 은 유형 II CRISPR-매개 DNA 이중가닥 파단의 모식도를 나타낸다. 스트렙토코쿠스 피오게네스 SF370 으로부터의 유형 II CRISPR 유전자좌는 4 개 유전자, Cas9, Casl, Cas2 및 Csnl 의 클러스터 뿐 아니라 2 개의 비-코딩 RNA 요소, tracrRNA, 및 비-반복 서열의 단길이 스트레치 (스페이서, 각각 30bp) 에 의해 공간이 남겨진 반복 서열의 특징적 어레이 (직접 반복물) 을 포함한다 (15-18, 30, 31). 각각의 스페이서는 통상 외래 유전적 물질 (프로토스페이서) 에서 유래하고, CRISPR-매개 핵산 절단의 특이성을 유도한다. 표적 핵산에서, 각각의 프로토스페이서는 그의 인지가 개별적 CRISPR 시스템에 특이적인 프로토스페이서 인접 모티프 (PAM) 와 연관된다 (22, 23). 유형 II CRISPR 시스템은 순차적 단계로 표적화 DNA 이중가닥 파단 (DSB) 을 실행한다 (M. Jinek et al, Science 337, 816 (Aug 17, 2012); Gasiunas, R. et al. Proc Natl Acad Sci USA 109, E2579 (Sep 25, 2012); J. E. Garneau et al, Nature 468, 67 (Nov 4, 2010); R. Sapranauskas et al, Nucleic Acids Res 39, 9275 (Nov, 2011); A. H. Magadan et al. PLoS One 7, e40913 (2012)). 먼저, 예비-crRNA 어레이 및 tracrRNA 를 CRISPR 유전자좌로부터 전사한다. 두 번째로, tracrRNA 를 예비-crRNA 의 직접 반복물에 하이드리드화하고 이중체로서 Cas9 와 연관시킨다 (예비-crRNA 의, 개별적인, 절두된 스페이서 서열을 포함하는 성숙 crRNA 로의 처리를 매개함). 세 번째로, 성숙 crRNA:tracrRNA 이중체는 Cas9 를 crRNA 의 스페이서 부위와 프로토스페이서 DNA 사이의 이형이중체 형성을 통해 프로토스페이서 및 필수적 PAM 으로 이루어지는 DNA 표적으로 유도한다. 마지막으로, Cas9 는 PAM 의 표적 DNA 업스트림의 절단을 매개하여 프로토스페이서 내에 DSB 를 생성시킨다.
도 28A-C 는 Cas9-매개 유전자 표적화를 위한 상이한 tracrRNA 전사체의 비교를 나타낸다. (A) 시험한 2 개 (단길이 및 장길이) tracrRNA 전사체의 설계 및 서열을 나타내는 모식도. 각각의 전사체는 U6 프로모터에 의해 주도된다. 전사 시작 부위를 +1 로 표기하고 전사 터미네이터는 표시한 바와 같다. 청색 선은 그의 역-상보 서열이 tracrRNA 검출용 노던 블롯 프로브 생성에 사용되는 부위를 나타낸다. (B) EMX1 유전자좌의 hSpCas9-매개 절단의 효율을 비교하는 SURVEYOR 검정. 2 개의 생물학적 레플리카를 각각의 tracrRNA 전사체에 대해 나타낸다. (C) 장길이 또는 단길이 tracrRNA 뿐 아니라 SpCas9 및 DR-EMX1(1)-DR 을 운반하는 U6 발현 구축물로 트랜스펙션된 293FT 세포로부터 추출한 총 RNA 의 노던 블롯 분석. 좌측 및 우측 패널은 각각 SpRNase III 의 부재 또는 존재 하에 트랜스펙션된 293FT 세포로부터의 것이다. U6 은 인간 U6 snRNA 를 표적화하는 프로브로 블롯팅된 로딩 대조군을 나타낸다. 단길이 tracrRNA 발현 구축물의 트랜스펙션은 tracrRNA 의 처리 형태의 과잉 수준을 이끌어낸다 (~75bp) (E. Deltcheva et al., Nature 471, 602 (Mar 31, 2011)). 매두 소량의 장길이 tracrRNA 를 노던 블롯에서 검출한다. 이들 실험의 결과로서, 출원인은 포유동무루 세포에서의 적용을 위해 단길이 tracrRNA 사용을 선택하였다.
도 29 는 이중가닥 파단-유도 미세 삽입 및 결실의 검출을 위한 SURVEYOR 검정을 나타낸다 (D. Y. Guschin et al. Methods Mol Biol 649, 247 (2010)). Cas9-매개 절단 효율을 측정하는데 사용한 SURVEYOR 검정의 모식도. 먼저, 게놈 PCR (gPCR) 을 사용하여 변형 및 미변형 세포의 불균일 집단으로부터의 Cas9 표적 부위를 증폭하고, gPCR 생성물을 천천히 재어닐링하여 이형이중체를 생성한다. 재어닐링한 이형이중체를 SURVEYOR 뉴클레아제에 의해 절단하는 반면, 동종이중체는 미손상인 채로 둔다. Cas9-매개 절단 효율 (% 삽입-결실) 을 절단된 DNA 분획물을 기준으로 계산한다.
도 30A-B 는 포유동물 세포에서의 crRNA 처리의 노던 블롯 분석을 나타낸다. (A) 2 개 직접 반복물 (DR-EMX1(l)-DR) 에 의해 측면 위치한 단일 스페이서에 대한 발현 벡터를 나타내는 모식도. 인간 EMX1 유전자좌 프로토스페이서 1 을 표적화하는 30bp 스페이서 (표 1) 를 청색으로 나타내고 직접 반복물을 회색으로 나타낸다. 주황색 선은 그의 역-상보 서열을 사용하여 EMX1(1) crRNA 검출용 노던 블롯 프로브를 생성시키는 부위를 나타낸다. (B) DR-EMX(1)-DR 을 운반하는 U6 발현 구축물로 트랜스펙션된 293FT 세포로부터 추출한 총 RNA 의 노던 블롯 분석. 좌측 및 우측 패널은 각각 SpRNase III 의 부재 또는 존재 하에 트랜스펙션된 293FT 세포로부터의 것이다. DR-EMX(1)-DR 은 SpCas9 및 단길이 tracrRNA 의 존재 하에서만 성숙 crRNA 로 처리되었으며, SpRNase III 의 존재에 의해서는 좌우되지 않았다. 트랜스펙션된 293FT 총 RNA 로부터 검출된 성숙 crRNA 는 ~33bp 이고 S. 피오게네스 (S. pyogenes) 로부터의 39-42bp 성숙 crRNA 보다 더 짧은데 (E. Deltcheva et al., Nature 471, 602 (Mar 31, 2011)), 이는 인간 293FT 세포에서의 처리된 성숙 crRNA 가 S. 피오게네스에서의 박테리아 성숙 crRNA 와 상이할 것으로 예상된다는 것을 제시한다.
도 31A-B 는 Cas9 를 갖는 키메라 crRNA 또는 예비-crRNA 어레이에 대한 바이시스트로닉 발현 벡터를 나타낸다. (A) 예비-crRNA 어레이에 대한 발현 벡터의 설계를 나타내는 모식도. 스페이서는 어닐링된 올리고뉴클레오티드를 사용하여 2 개 BbsI 부위 사이에 삽입될 수 있다. 올리고뉴클레오티드에 대한 서열 설계를 하기에 나타내며, 적절한 라이게이션 어댑터를 함께 표시하였다. (B) 키메라 crRNA 에 대한 발현 벡터의 모식도. 안내 서열은 어닐링된 올리고뉴클레오티드를 사용하여 2 개 BbsI 부위 사이에 삽입될 수 있다. 벡터는 이미 부분적 직접 반복물 (회색) 및 부분적 tracrRNA (적색) 서열을 포함한다. WPRE, 마못 (Woodchuck) 간염 바이러스 전사후 조절 요소.
도 32A-B 는 인간 PVALB 및 마우스 Th 유전자좌에서의 프로토스페이서의 선택을 나타낸다. 각각 PVALB 및 Th 유전자의 마지막 엑손 내의 3 개 프로토스페이서의 위치 및 인간 PVALB (A) 및 마우스 Th (B) 유전자좌의 모식도. 30bp 프로토스페이서를 흑색 선으로 나타내고 인접한 PAM 서열을 자홍색 막대로 나타낸다. 센스 및 안티-센스 가닥 상의 프로토스페이서를 각각 DNA 서열 위 및 아래에 나타낸다.
도 33A-C 는 인간 게놈에서의 PAM 서열의 발생을 나타낸다. 인간 게놈에서의 인접한 스트렙토코쿠스 피오게네스 SF370 유전자좌 1 PAM (NGG) (A) 와 스트렙토코쿠스 써모필루스 (Streptococcus thermophilus) LMD9 유전자좌 1 PAM (NNAGAAW) (B) 사이 거리의 히스토그램. (C) 염색체에 의한 각각의 PAM 에 대한 거리. Chr, 염색체. 인간 염색체 서열의 플러스 및 마이너스 가닥 모두를 사용하여 추정상 표적을 확인하였다. 일부 프로토스페이서 표적에서 절단 활성을 제한할 수 있는 크로마틴, DNA 메틸화-, RNA 구조 및 기타 인자를 고려하면, 실제 표적화 능력이 이러한 컴퓨터적 분석 결과보다 덜 할 수 있다는 것에 주목하는 것이 중요하다.
도 34A-D 는 스트렙토코쿠스 써모필루스 LMD-9 로부터의 유형 II CRISPR 가 또한 진핵세포에서 기능할 수 있다는 것을 나타낸다. (A) 스트렙토코쿠스 써모필루스 LMD-9 로부터의 CRISPR 유전자좌 2 의 모식도. (B) S. 써모필루스 (S. thermphilus) CRISPR 시스템에 대한 발현 시스템의 설계. 항시성 EF1a 프로모터를 사용하여 인간 코돈-최적화 hStCas9 를 발현시킨다. 정확한 전사 개시를 확실히 하기 위해, U6 프로모터를 사용하여 tracrRNA 및 crRNA 의 성숙한 버전을 발현시킨다. 성숙 crRNA 및 tracrRNA 에 대한 서열을 나타낸다. crRNA 서열에서 소문자 "a" 로 나타낸 단일 염기를 사용하여, RNA Pol III 전사 터미네이터로서 역할하는 폴리U 서열을 제거하였다. (C) 인간 EMX1 유전자좌에서의 프로토스페이서 및 상응하는 PAM 서열 표적을 나타내는 모식도. 2 개의 프로토스페이서 서열을 하이라이트 표시하고 NNAGAAW 모티프를 만족시키는 그의 상응하는 PAM 서열을 자홍색 선으로 표시한다. 프로토스페이서 모두는 안티-센스 가닥을 표적화한다. (D) 표적 유전자좌에서의 StCas9-매개 절단을 나타내는 SURVEYOR 검정. RNA 안내 스페이서 1 및 2 는 각각 14% 및 6.4% 유도되었다. 이들 2 개 프로토스페이서 부위에서의 생물학적 레플리카 전체에 걸친 절단 활성의 통계적 분석을 표 1 에서 발견할 수 있다.
도 35 는 AAV-프로모터-TALE-효과기 구축물의 예를 나타내며, 여기서 hSyn = 인간 시냅신 1 프로모터, N+136 = TALE N-말단, AA +136 절두, C63 = TALE C-말단, AA +63 절두, vp = VP64 효과기 도메인, GFP = 녹색 형광 단백질, WPRE = 마못 간염 바이러스 전사후 조절 요소, bGH = 소 성장 호르몬 폴리A, ITR = AAV 역전 말단 반복물 및 AmpR = 암피실린 저항성 유전자이다.
도 36A-C 는 LITE 시스템의 설계 및 최적화를 나타낸다. (a) TALE DNA-결합 도메인은 CRY2 에 융합되고 전사 효과기 도메인은 CIB1 에 융합된다. 불활성 상태에서, TALE-CRY2 는 표적 유전자의 프로모터 부위에 결합하는 한편, CIB1-효과기는 핵 내에서 결합하지 않은 채로 남아있다. VP64 전사 활성제는 상기에 나타낸다. 청색으로 조명시, TALE-CRY2 및 CIB1-효과기는 빠르게 이량체화하고, CIB1-효과기를 표적 프로모터에 영입한다. 상기 효과기는 표적 유전자의 전사를 조정한다. (b) 그의 광-민감성 결합 파트너의 기능적 절두를 포함하는 LITE 를 갖는 내인성 표적 Neurog2 mRNA 의 광-의존적 상향조절. LITE-트랜스펙션된 Neuro-2a 세포를 7% 의 사용률로 5 mW/cm2 의 강도에서 466 nm 광으로 24 시간 동안 자극하였다 (0.066 Hz 에서의 1 초 펄스). (c) TALE-CRY2PHR 및 CIB1-VP64 LITE 에 의한 광-의존적 Neurog2 상향조절의 시간 경과. LITE-트랜스펙션된 Neuro-2a 세포를 7% 의 사용률로 5 mW/cm2 의 강도에서 466 nm 광으로 자극하였고 (0.066 Hz 에서 1 초 펄스), Neurog2 mRNA 수준은 광 자극 6 시간 후 감소한다. 모든 Neurog2 mRNA 수준을 발현하는 GFP 대조군 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3-4) (*, p < 0.05; 및 ***, p < 0.001).
도 37A-F 는 뉴런에서의 내인성 유전자좌를 표적화하는 시험관내 및 생체내 AAV-매개 TALE 전달을 나타낸다. (a) AAV 내로 패키징된 항시성 TALE 전사 활성제의 일반적 모식도. 효과기 도메인 VP64 를 하이라이트 표시하였다. hSyn: 인간 시냅신 프로모터; 2A: 구제역-유래 2A 펩티드; WPRE: 마못 간염 전사후 반응 요소; bGH pA: 소 성장 호르몬 폴리-A 신호, (b) 1 차 피질 뉴런에서의 (a) 로부터의 AAV-TALE-VP64 구축물로의 형질도입을 나타내는 대표 이미지. 세포를 GFP 및 뉴런 마커 NeuN 에 대해 염색하였다. 스케일 바 = 25 ㎛. (c) 다양한 내인전 유전자좌를 표적화하는 AAV-TALE-VP64 구축물을 1 차 피질 뉴런에서 전사 활성화에 대해 스크리닝하였다 (*, p < 0.05; **, p < 0.01 ; p < 0.001). (d) AAV 에 의한 TALE-VP64 의 마우스 ILC 로의 효율적 전달. 스케일 바 = 100 ㎛. (Cg1 = 대상 피질, PLC = 변연전 피질, ILC = 변연계 아래 피질), (e) ILC 에서의 뉴런의 효율적 형질도입의 고배율 이미지. (f) ILC 에서의 생체내 TALE-VP64 에 의한 Grm2 mRNA 상향조절 (평균 ± s.e.m.; n = 조건 당 3 마리 동물), 300 ㎛ 조직 펀치를 사용하여 측정함.
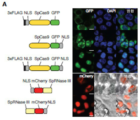
도 38A-I 은 1 차 뉴런 및 생체내에서의 내인성 전사의 LITE-매개 광유전학적 조절을 나타낸다. (a) 교환된 CRY2PHR 및 CIB1 구조를 갖는 AAV-LITE 활성제 구축물, (b) 1 차 뉴런에서의 AAV-전달 LITE 구축물의 동시-형질도입을 나타내는 대표적 이미지. GFP, HA-tag 및 DAPI 에 대해 세포를 염색하였다. (스케일 바 = 25 ㎛). (c) 0.8% 사용률로 466 nm 광 펄스로 (0.033Hz 에서 250 ms 펄스 또는 0.016Hz 에서 500 ms 펄스; 5 mW/cm2) 자극 24 시간 후 1 차 뉴런에서의 Grm2 발현의 광-유도 활성화. (d) 4 시간 및 24 시간점에서의 광 자극의 존재 및 부재 하 1 차 피질 뉴런에서의 Grm2 mRNA 의 상향 조절. 발현 수준을 GFP 로만 형질도입된 뉴런에 대해 나타낸다. (e) GFP 단독 대조군 형질도입, LITE 를 갖는 미자극 뉴런, 및 LITE 를 갖는 광-자극 뉴런에서의 mGluR2 단백질 수준의 정량. 대표적인 웨스턴 블롯을 로딩 대조군으로서 β-튜불린-III 을 사용하여 나타낸다. (f) LITE 시스템, 광학 섬유 이식물, 및 조직 단리에 사용한 0.35 mm 직경 뇌 펀치를 사용한 형질도입을 나타내는 모식도. (g) 두 LITE 성분 모두로 동시-형질도입된 ILC 의 대표적 이미지. 염색을 HA-tag (적색), GFP (녹색) 및 DAPI (청색) 로 나타낸다. (스케일 바 = 25 ㎛). (h) ILC 로 형질도입된 LITE 를 사용하는 내인성 Grm2 발현의 광-유도 활성화. **, p < 0.05; 각 실험 조건에 대해 4 마α리의 상이한 마우스로부터 데이터를 생성시켰다. (i) LITE 1.0 및 최적화 LITE 2.0 을 사용하는 Neurog2 발현의 광 유도 및 증가 배수. LITE2.0 은 높은 수준의 활성화를 유지하면서 최소의 배경을 제공한다. NLSα-임포틴 및 NLSsv40, 각각-임포틴 및 원숭이 바이러스 40 으로부터의 핵 국부화 신호; GS, Gly-Ser 링커; NLS*, 표시한 잔기가 Ala 로 치환되어 핵 국부화 활성을 방지하는 돌연변이화된 NLS; △318-334; 고등 식물 헬릭스-루프-헬릭스 전사 인자 상동성 부위의 결실.
도 39A-H 는 TALE- 및 LITE-매개 후생적 변형을 나타낸다. (a) LITE 후생적 변형제 (epiLITE) 의 모식도. (b) AAV 벡터 내의 조작된 후생적 전사 억제제 SID4X 의 모식도. 효율적인 AAV 패키징을 확실히 하기 위해 GFP (800bp) 보다는 phiLOV2.1 (330bp) 을 형광 마커로서 사용하였다. (c) 광 자극의 존재 및 부재 하 1 차 피질 뉴런에서의 내인성 Grm2 발현의 epiLITE-매개 억제. GFP 단독으로 형질도입된 뉴런에 대해 하향조절 배수를 나타낸다. (d) 광-자극의 존재 및 부재 하 Grm2 프로모터에서의 H3K9 히스톤 잔기 아세틸화에서의 epiLITE-매개 감소. (e, f) epiTALE-메틸트랜스퍼라아제 (epiTALE-KYP, -TgSET8 및 -NUE) 에 의한 Grm2 mRNA 의 감소 배수, 및 Grm2 프로모터에서의 히스톤 메틸화 마크 H3K9mel, H4K20me3 및 H3K27me3 의 상응하는 풍족. (g, h) epiTALE 히스톤 데아세틸라아제 (epiTALE-HDAC8, -RPD3, -Sir2a 및 -Sin3a) 에 의한 Grm2 mRNA 의 감소 배수, 및 Grm2 프로모터에서의 히스톤 잔기 아세틸화 마크 H4K8Ac 및 H3K9Ac 에 있어서의 상응하는 감소. 모든 패널에서 나타낸 값은 평균 ± s.e.m., n = 3-4 이다.
도 40 은 시험관내 CRY2 의 흡수 스펙트럼의 설명을 나타낸다. 크립토크롬 2 를 350-475 nm 광에 의해 최적으로 활성화시켰다1. 흡수 및 활성화에서의 급격한 감소가 480 nm 초과의 파장에 대해 나타났다. 스펙트럼을 [Banerjee, R. et al. The Signaling State of Arabidopsis Cryptochrome 2 Contains Flavin Semiquinone. Journal of 'Biological Chemistry 282, 14916-14922, doi: 10.1074/jbc.M700616200 (2007)] 로부터 조정하였다.
도 41 은 LITE-매개 유전자 발현에 대한 조명 사용률의 효과를 나타낸다. 가변적인 사용률 (총 시간의 % 로서의 조명) 을 사용하여 KLF4 유전자를 표적화하는 LITE 를 발현하는 293FT 세포를 자극하여, LITE 활성에 대한 사용률의 효과를 조사하였다. KLF4 발현 수준을 GFP 만을 발현하는 세포와 비교하였다. 자극 매개변수는 다음과 같다: 466 nm, 5 mW/cm2, 24 시간 동안. 펄스를 0.067 Hz 에서 하기 지속 기간으로 수행하였다: 1.7% = 0.25 초 펄스, 7% = 1 초 펄스, 27% = 4 초 펄스, 100% = 연속 조명. (평균 ± s.e.m.; n = 3-4).
도 42A-B 는 LITE-매개 유전자 발현 및 세포 생존에 대한 광 강도의 효과를 나타낸다. (a) CRY2PHR :: CIB1 LITE 의 전사 활성은 466 nm 청색광의 강도에 따라 가변적이라는 것이 발견되었다. Neuro 2a 세포를 24 시간 동안 7% 사용률 (0.066Hz 에서 1 초 펄스) 에서 자극하였다. (b) 적색-형광 에티디움 동종이량체-1 에 대해 양성인 세포 대 칼세인-양성 세포의 % 로서 광-유도 독성을 측정하였다. 모든 Neurog2 mRNA 수준을 GFP 만을 발현하는 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3-4).
도 43 은 LITE-매개 유전자 발현에 대한 전사 활성화 도메인의 영향을 나타낸다. 상이한 전사 활성화 도메인 (VP16, VP64 및 p65) 을 사용하는 LITE 에 의한 광의 존재 및 부재 하 Neurog2 상향조절. LITE 로 트랜스펙션한 Neuro-2a 세포를 24 시간 동안 466 nm 광으로 5 mW/cm2 의 강도 및 7% 의 사용률에서 (0.066 Hz 에서 1 초 펄스) 자극하였다. (평균 ± s.e.m.; n = 3-4)
도 44A-C 는 내인성 유전자 전사의 화학적 유도를 나타낸다. (a) 아브시스산 (ABA) 수용체 시스템을 기반으로 하는 화학 물질 유도 2 개 하이브리드 TALE 시스템의 설계를 나타내는 모식도. ABI 및 PYL 은 ABA 첨가시 이량체화하며 ABA 철회시 해리된다. (b) ABA-의존적 Neurog2 상향조절의 시간 경과. 250 μM 의 ABA 를 TALE(Neurog2)-ABI 및 PYL-VP64 를 발현하는 HEK 293FT 세포에 첨가하였다. ABA 첨가 후 나타낸 시점에서 mRNA 증가 배수를 측정하였다. (c) ABA 자극 24 시간 후 Neurog2 mRNA 수준의 감소. 모든 Neurog2 mRNA 수준을 발현하는 GFP 대조군 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3-4).
도 45A-C 는 AAV 상청액 생성을 나타낸다. (a) 렌티바이러스 및 AAV 벡터 (GFP 운반) 를 사용하여 형질유도 효율을 시험하였다. (b) 1 차 배아 피질 뉴런을 동일 수의 AAV 또는 렌티바이러스-트랜스펙션된 293FT 세포로부터 유래한 300 및 250 μL 상청액으로 형질도입하였다. GFP 발현의 대표적 이미지를 7 d.p.i. 에서 수집하였다. 스케일 바 = 50 ㎛. (c) 나타낸 공정을 AAV 상청액의 생성 및 1 차 뉴런의 후속적 형질도입을 위해 개발하였다. 293FT 세포를 관심 유전자를 운반하는 AAV 벡터, AAVl 혈청형 패키징 벡터 (pAAV1), 및 PEL 을 사용하는 헬퍼 플라스미드 (pDF6) 로 트랜스펙션하였다. 48 시간 후, 상청액을 수확하고 0.45 ㎛ PVDF 멤브레인을 통해 여과하였다. 1 차 뉴런을 이후 상청액으로 형질도입하고 남아 있는 분취액을 -80℃ 에서 보관하였다. 5-6 일 후 AAV 구축물 발현의 안정한 수준에 도달하였다. 이러한 공정에 따른 AAV 상청액 생성을 96-웰 포맷으로 96 개 이하의 상이한 바이러스 구축물의 생성에 사용할 수 있다 (도 37C 에서 나타낸 뉴런에서의 TALE 스크린에 이용).
도 46 은 DNaseI-민감성 크로마틴 부위에 의해 유도된 TALE 표적 부위의 선택을 나타낸다. ENCODE (http://genome.ucsc.edu) 로부터의 마우스 피질 조직 데이터를 기반으로 한 높은 DNaseI 민감성을 사용하여 개방 크로마틴 부위를 확인하였다. 전사 시작 부위의 2 kb 업스트림 부위 내의 최고 진폭을 갖는 피크를 표적화를 위해 선택하였다. TALE 결합 표적을 이후 피크의 중심에서 200 bp 부위 내에서 골랐다.
도 47 은 1 차 뉴런 건강에 대한 광 사용률의 영향을 나타낸다. 1 차 피질 뉴런 건강에 대한 광 자극의 효과를 사용률 7%, 0.8%, 및 무광 조건에 대해 비교하였다. 칼세인을 사용하여 뉴런 생존도를 평가하였다. 명시야 이미지를 캡쳐하여 형태 및 세포 무결성을 나타내었다. 466 nm 광의 5 mW/cm2 로 24 시간 동안 표시 사용률로 1 차 피질 뉴런을 자극하였다. 대표적 이미지, 스케일 바 = 50 ㎛. 펄스를 하기의 방식으로 수행하였다: 7% 사용률 = 0.067 Hz 에서 1 초 펄스, 0.8% 사용률 = 0.0167 Hz 에서 0.5 초 펄스.
도 48 은 광유전학적 자극 동안의 마우스 이미지를 나타낸다. 잠에서 깬, 자유로이 거동하는, LITE-주입 마우스를 뇌정위적으로 이식한 캐뉼라 및 광학 섬유로 사진찍었다.
도 49 는 마우스 변연계 아래 피질에서의 AAVl/2 에 의한 LITE 성분의 동시-형질도입 효율을 나타낸다. TALE(Grm2)-CIB1 단독, CRY2PHR-VP64 단독에 의해 형질도입되거나 동시-형질도입된 세포를, 모든 형질도입된 세포의 % 로서 계산하였다.
도 50 은 기준선 전사 조절에 대한 개별적인 LITE 성분의 기여도를 나타낸다. Grm2 mRNA 수준을 개별적인 LITE 성분으로 트랜스펙션된 1 차 뉴런에서 측정하였다. Grm2 TALE 1-CIB1 단독을 발현하는 1 차 뉴런은 전체 LITE 시스템을 발현하는 미자극 세포에서와 같이 Grm2 mRNA 수준에 있어서 유사한 증가를 초래하였다 (평균 ± s.e.m.; n = 3-4).
도 51 은 활성화, 배경 신호 및 감소 배수에 대한 LITE 성분 조작의 효과를 나타낸다. 단백질 변형을 이용하여, LITE 성분이 감소된 배경 전사 활성화를 초래하면서 광에 의한 유도 비율을 향상시킨다는 것을 발견하였다. 단백질 변형을 하기에서 상세히 토의한다. 간략하게, 내인성 핵 이출 신호에서의 돌연변이 및 핵 국부화 신호를 사용하여 CRY2PHR-VP64 성분의 핵 이입을 향상시켰다. 핵 국부화 또는 CIB1 전사 활성화를 감소시키는 것으로 의도된 CIB1 의 여러 변이를 추구하여 TALE-CIB1 성분의 배경 활성에 대한 기여도를 감소시켰다. 시험한 CRY2PHR-VP64 및 TALE-CIB1 의 모든 조합의 결과를 상기에 나타낸다. 막대 그래프의 좌측에 있는 표는 각 조합에 사용한 도메인/돌연변이의 특정 조합을 표시한다. 표 및 막대 그래프의 각 행은 성분 세부사항, 광/무광 활성, 및 특정 CRY2PHR/CIB1 조합에 대한 광에 의한 유도 비율을 포함한다. LITE 1.0 에 비해 감소된 배경 및 증가된 유도 배수를 모두 초래한 조합을 "+" 표시한 표 열에서 녹색으로 하이라이트 표시한다 (t-시험 p<0.05). CRY2PHR-VP64 구축물: 3 개의 새로운 구축물을 CRY2PHR-VP64 핵 이입을 향상시키는 목적으로 설계하였다. 먼저, CRY2PHR 의 예측된 내인성 핵 이출 서열 내의 돌연변이 L70A 및 L74A 를 유도하여 단백질의 핵 이출을 제한하였다 (효과기 열에서 '*' 로 나타냄). 두 번째로, α-임포틴 핵 국부화 서열을 CRY2PHR-VP64 의 N-말단에 융합시켰다 (효과기 열에서 'A' 로 나타냄). 세 번째로, SV40 핵 국부화 서열을 CRY2PHR-VP64 의 C-말단에 융합시켰다 (효과기 열에서 'P' 로 나타냄). TALE-CIB1 링커: LITE 1.0 에서 사용한 TALE 과 CIB1 사이의 SV40 NLS 링커를 TALE-CIB1 단백질의 핵 이출을 증가시키도록 설계된 여러 링커 중 하나로 대체하였다 (CIB1 링커 열에서 사용한 기호를 괄호 안에 나타냄): 유연성 글리신-세린 링커 (G), 아데노바이러스 유형 5 E1B 핵 이출 서열 (W), HIV 핵 이출 서열 (M), MAPK 핵 이출 서열 (K) 및 PTK2 핵 이출 서열 (P). NLS* 내인성 CIB1 핵 국부화 서열 돌연변이: 핵 국부화 신호는 야생형 CIB1 서열 내에 존재한다. 이러한 신호는 NLS* 구축물 내에서 K92A, R93A, K105A 및 K106A 에서 돌연변이화되어 TALE-CIB1 핵 국부화가 약화되었다 (NLS* 열에서 'N' 으로 나타냄). △CIB1 전사 인자 상동성 결실: 가능한 기저 CIB1 전사 활성화를 제거하려는 노력으로, 결실 구축물을 고등 식물에서의 염기성 헬릭스-루프-헬릭스 전사 인자에 대한 높은 상동성 부위가 제거되게 설계하였다. 이들 결실된 부위는 Aaa230-256, Aaa276-307, Aaa308-334 로 이루어졌다 (ACIB1 열에서 '1', '2' 및 '3' 으로 나타냄). 각각의 경우, 결실된 부위를 3 개 잔기 GGS 링크로 대체하였다. CIB1 내로의 NES 삽입: TALE- CIB1 의 광-의존적 핵 이입을 촉진하기 위한 하나의 계획은 CRY2PHR 과의 이량체화 접촉면에서 CIB1 에 NES 를 삽입하여, CRY2PHR 과 결합시 신호가 숨겨질 수 있는 것이다. 이를 위해, NES 를 공지된 CRY2 상호작용 도메인 CIBN 내 상이한 위치 (aa 1-170) 에서 삽입하였다. 위치는 하기와 같다 (NES 열에서 사용한 기호를 괄호 안에 나타냄): aa28 (1), aa52 (2), aa73 (3), aal20 (4), aal40 (5), aal60 (6). *bHLH 염기성 헬릭스-루프-헬릭스 돌연변이: 직접적 CIB1-DNA 상호작용을 감소시키기 위해, CIB1 에서의 염기성 헬릭스-루프-헬릭스 부위의 여러 염기성 잔기를 돌연변이시켰다. 하기 돌연변이는 모든 *bHLH 구축물 내에 존재한다 (도 51 의 *bHLH 컬럼에서 'B' 로 나타냄): R175A, G176A, R187A 및 R189A.
도 52 는 TALE-CIB1 의 광 매개 동시-의존적 핵 이입의 설명을 나타낸다. (a) 광의 부재 하, TALE-CIB1 LITE 성분은 핵 국부화 신호, NLS 의 부재 (또는 약한 핵 이출 신호, NES 의 첨가) 로 인해 세포질에서 존재한다. 한편 NLS 를 함유하는 CRY2PHR-VP64 성분은 저절로 핵에 능동적으로 이입된다. (b) 청색광의 존재 하에, TALE-CIB1 은 CRY2PHR 에 결합한다. CRY2PHR-VP64 에 존재하는 강력한 NLS 는 이제 LITE 성분 모두의 복합체의 핵 이입을 매개하여, 이들이 표적화된 유전자좌에서 전사를 활성화시킬 수 있게 한다.
도 53 은 주목할만한 LITE 1.9 조합을 나타낸다. LITE 2.0 구축물에 추가로, 조작된 LITE 성분 스크린으로부터의 여러 CRY2PHR-VP64 :: TALE-CIB1 조합에 특히 주목하였다. 돌연변이화된 내인성 NLS 를 갖는 α-임포틴 NLS 효과기 구축물 및 △276-307 TALE-CIB1 구축물이 조합된 LITE 1.9.0 은 9 초과의 유도 비율 및 180 초과의 절대 광 활성화를 나타내었다. 미변형된 CRY2PHR-VP64 와 돌연변이화된 NLS, △318-334, AD5 NES TALE-CIB1 구축물이 조합된 LITE 1.9.1 은, 4 의 유도 비율을 1.06 의 배경 활성화로 얻었다. 2 미만의 배경 활성화 및 7 내지 12 범위의 유도 비율을 갖는 다른 LITE 1.9 조합의 선택을 또한 하이라이트 표시하였다.
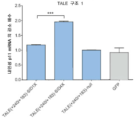
도 54A-D 는 뉴런에서의 TALE SID4X 억제제 분석 및 적용을 나타낸다. a) 합성적 억제제를 4 개의 SID 도메인 (SID4X) 을 연쇄 연결시켜 구축하였다. 최적의 TALE -억제제 구조를 확인하기 위해, SID 또는 SID4X 를 마우스 p11 유전자를 표적화하도록 설계된 TALE 에 융합시켰다. (b) p11 mRNA 에서의 감소 배수를 qRT-PCR 을 사용하여 검정하였다. (c) AAV 에 패키징된 항시성 TALE 전사 억제제의 일반적 모식도. hSyn: 인간 시냅신 프로모터; 2A: 구제역-유래 2 A 펩티드; WPRE: 마못 간염 전사후 반응 요소; bGH pA: 소 성장 호르몬 폴리-A 신호. phiLOV2.1 (330bp) 을 효율적 AAV 패키징을 확실히 하기 위한 단길이 형광 마커로서 선택하였다. (d) 내인성 마우스 유전자좌 Grm5 및 Grm2 를 표적화하는 2 개 TALE 을 SID4X 에 융합시키고 1 차 뉴런에 바이러스 형질도입하였다. SID4X 를 통한 표적 유전자 하향 조절을 GFP 만을 발현하는 뉴런에서의 수준에 대하여 각각의 TALE 에 대해 나타낸다. (평균 ± s.e.m.; n = 3-4).
도 55A-B 는 다양한 집합의 epiTALE 이 뉴런 및 Neuro2a 세포에서의 전사 억제를 매개한다는 것을 나타낸다. a) 상이한 히스톤 효과기 도메인에 융합된 총 24 개 Grm2 표적화 TALE 을 AAV 를 사용하여 1 차 피질 마우스 뉴런에 형질도입하였다. Grm2 mRNA 수준을, GFP 단독으로 형질도입된 뉴런에 대해 RT-qPCR 을 사용하여 측정하였다. * 는 p < 0.05 인 억제를 나타낸다. b) 총 32 개 epiTALE 을 Neuro2A 세포에 트랜스펙션시켰다. 이들 중 20 개는 표적화된 Neurog2 유전자좌의 유의한 억제를 매개하였다 (* = p<0.05).
도 56A-D 는 Neuro 2A 세포에서의 히스톤 개질과 함께 epiTALE 매개된 전사 억제를 나타낸다. (a) Neuro 2A 세포에서 쥐과 Neurog2 유전자좌를 표적화하는 SIRT3 및 히스톤 탈아세틸화 후생적 효과기 NcoR 에 융합된 TALE 을 Neurog2 전사체 수준에 대한 억제 활성에 대해 검정하였다. (b) ChIP RT-qPCR 은 NcoR 및 SIRT3 epiTALE 에 대한 Neurog2 프로모터에서의 H3K.9 아세틸화에 있어서의 감소를 나타낸다. (c) 공지된 히스톤 메틸트랜스퍼라아제 결합 활성을 갖는 후생적 효과기 PHF19 를 Neurog2 mRNA 수준의 Neurog2 매개 억제를 표적화하는 TALE 에 융합시켰다. (d) ChIP RT-qPCR 은 PHF19 epiTALE 에 대한 Neurog2 프로모터에서의 H3K27me3 수준에 있어서 증가를 나타낸다.
도 57A-G 는 RNA-안내 DNA 결합 단백질 Cas9 가 전사 효과기 도메인을 특정 게놈 유전자좌에 표적화하는데 사용될 수 있다는 것을 나타낸다. (a) 유형 II 스트렙토코쿠스 피오게네스 CRISPR/Cas 시스템으로부터의 RNA-안내 뉴클레아제 Cas9 는 2 개의 알라닌 치환 (D10A 및 H840A) 을 도입함으로써 핵산분해적으로 불활성인 RNA-안내 DNA 결합 단백질 (Cas9**) 로 전환될 수 있다. 모식도는 합성적 안내 RNA (sgRNA) 가 인간 게놈에서의 특정 유전자좌에 대한 Cas9**-효과기 융합을 지시할 수 있다는 것을 나타낸다. sgRNA 는 표적 서열을 명기하는 5' 말단에서 20bp 안내 서열을 포함한다. 표적 게놈 DNA 상에서, 20bp 표적 부위는 그 뒤에 5'-NGG PAM 모티프가 뒤따를 수 있다. (b, c) 각각 인간 KLF4 및 SOX2 유전자좌에서의 sgRNA 표적 부위를 나타내난 모식도. 각각의 표적 부위를 청색 막대로 표시하고 상응하는 PAM 서열을 자홍색 막대로 표시한다. (d, e) Cas9**-VP64 전사 활성제 및 SID4X-Cas9** 전사 억제제 구축물의 모식도. (f, g) 각각 Cas9**-VP64 및 SID4X-Cas9** 매개된 KLF4 의 활성화 및 SOX2 억제. 모든 mRNA 수준을 GFP 모의 트랜스펙션된 대조군 세포에 대해 측정하였다 (평균 ± s.e.m.; n = 3).
도 58 은 2 개 TALE 이 각각의 내인성 마우스 유전자좌 Grm5, Grin2a 및 Grm2 를 표적화하는, 설계된 6 개 TALE 을 나타낸다. TALE 을 전사 활성제 도메인 VP64 또는 억제제 도메인 SID4X 에 융합시키고, 1 차 뉴런에 바이러스 형질도입하였다. VP64 를 통한 표적 유전자 상향조절 및 SID4X 를 통한 하향조절 모두를, GFP 만을 발현하는 뉴런에서의 수준에 대하여 각각의 TALE 에 대해 나타낸다.
도 59A-B 는 하기를 나타낸다: (A) SID4X 억제제 도메인을 강조하는 LITE 억제제 구축물. (B) Grm2 T1-LITE 및 Grm2 T2-LITE 를 사용하는 1 차 피질 뉴런에서의 내인성 Grm2 발현의 광-유도 억제. 하향조절 배수를 GFP 단독으로 형질도입된 뉴런에 대해 나타낸다 (평균 ± s.e.m.; 모든 서브패널에 대해 n = 3-4).
도 60A-B 는 CRY2PHR 및 CIB1 성분 교환을 나타낸다. (A) TALE-CIB1::CRY2PHR-VP64 는 TALE-CRY2PHR::CIB1-VP64 보다 더 높은 수준에서 Ngn2 를 활성화시킬 수 있었다. (B) Ngn2 LITE 의 활성화 비율 (광 대 무광) 배수는 두 설계 모두에 대해 유사한 효율을 나타낸다. 자극 매개변수는 도 36B 에서 사용한 것들과 동일하였다.
도 61 은 유도 Cas9 에 대한 Tet Cas9 벡터 설계를 나타낸다.
도 62 는 Cas9 및 EGFP 의 독시사이클린 유도 후 293FT 세포에서의 EGFP 발현 및 벡터를 나타낸다.
발명의 상세한 설명
용어 "핵산" 또는 "핵산 서열" 은 단일- 또는 이중-가닥 형태인 데옥시리보핵산 또는 리보핵산 올리고뉴클레오티드를 의미한다. 상기 용어는 핵산, 즉 자연적 뉴클레오티드의 공지된 유사체를 포함하는 올리고뉴클레오티드를 포함한다. 상기 용어는 또한 합성 백본을 갖는 핵산-유사 구조를 포함한다: 예를 들어 Eckstein, 1991; Baserga et al, 1992; Milligan, 1993; WO 97/03211; WO 96/39154; Mata, 1997; Strauss-Soukup, 1997; 및 Samstag, 1996 을 참고한다.
본원에서 사용하는 바와 같이, "재조합체" 는 합성되거나 시험관내 조작된 폴리뉴클레오티드 (예를 들어, "재조합체 폴리뉴클레오티드"), 세포 또는 기타 생물학적 시스템에서 유전자 산물을 생성시키기 위해 재조합체 폴리뉴클레오티드를 사용하는 방법, 또는 재조합체 폴리뉴클레오티드에 의해 인코딩된 폴리펩티드 ("재조합체 단백질") 를 의미한다. "재조합체 수단" 은 상이한 근원으로부터의 다양한 코딩 부위 또는 도메인 또는 프로모터 서열을 갖는 핵산을 이의 발현을 위해, 예를 들어 본 발명의 벡터 내 폴리펩티드 코딩 서열의 유도 또는 항시성 발현을 위해 발현 카세트 또는 벡터에 라이게이션하는 것을 포함한다.
핵산에 대해 사용하는 경우 용어 "이종" 은 상기 핵산이 자연계에서 보통 발견되지 않는 세포 또는 바이러스 내에 존재하는 것을 나타내거나; 자연계에서 보통 발견되는 바처럼 서로 동일한 관계로 발견되지 않거나, 재조합적으로 조작되어 그의 발현 수준, 또는 세포 또는 구조에서의 다른 핵산 또는 다른 분자에 대한 물리적 관계가 자연계에서 보통 발견되지 않는, 둘 이상의 하위서열을 포함한다. 이러한 맥락에서 사용된 유사한 용어는 "외인성" 이다. 예를 들어, 이종 핵산은 통상 재조합적으로 생성되어, 자연계에서 발견되지 않는 방식으로 배열된 관련없는 유전자로부터의 둘 이상의 서열을 가지며; 그 예로, 본 발명의 아데노바이러스-기반 벡터 내로 삽입된 프로모터 서열에 작동가능하게 연결된 인간 유전자를 들 수 있다. 예를 들어, 관심 이종 핵산은 면역원성 유전자 생성물을 인코딩할 수 있으며, 이때 아데노바이러스를 담체 또는 약물-백신 조성물로서 치료적 또는 예방적으로 투여한다. 이종 서열은 다양한 조합의 프로모터 및 서열을 포함할 수 있으며, 이의 예를 본원에서 상세히 기재한다.
"치료적 리간드" 는 치료 효과를 갖는 표적 세포의 수용체에 결합할 수 있는 물질일 수 있다.
"치료 효과" 는 임의 종류의 의학적 치료의 결과일 수 있으며, 이의 결과는 당업자에 의해 바람직하고 이로운 것인지 판단된다. "치료 효과" 는 의학적 치료에 대한 반응으로서 발생하는 거동 또는 생리적 변화일 수 있다. 그 결과는 예측된, 미예측된, 또는 심지어 의도되지 않은 의학적 치료의 결과일 수 있다. "치료 효과" 는 예를 들어, 병원체에 의한 감염을 겪는 대상에서의 증상 감소를 포함할 수 있다.
"표적 세포" 는 그의 활성에 있어서의 변형이 원하는 결과 또는 반응을 유도할 수 있는 세포일 수 있다. 본원에서 사용하는 바와 같이, 세포는 시험관내 세포일 수 있다. 상기 세포는 전체 유기체로 발생할 수 없을 수 있는 단리된 세포일 수 있다.
"리간드" 는 생물학적 목적용으로 역할하도록 생분자에 결합하고 생분자와 복합체를 형성하는 임의의 물질일 수 있다. 본원에서 사용하는 바와 같이, "리간드" 는 또한 "항원" 또는 "면역원" 을 의미할 수 있다. 본원에서 사용하는 바와 같이 "항원" 및 "면역원" 은 상호교환가능하게 사용된다.
유전자 또는 핵산의 "발현" 은 세포적 유전자 발현 뿐 아니라 클로닝 시스템 및 임의의 다른 맥락에서의 핵산(들) 의 전사 및 번역을 포함한다.
본원에서 사용하는 바와 같이, "벡터" 는 한 환경에서 다른 환경으로 개체를 이동시킬 수 있거나 이를 촉진시키는 도구이다. 예를 들어, 재조합 DNA 기술에서 사용한 일부 벡터는 개체, 예컨대 DNA 조각 (예컨대 이종 DNA 조각, 예컨대 이종 cDNA 조각) 이 표적 세포 내로 이동될 수 있게 한다. 본 발명은 바이러스 벡터, 박테리아 벡터, 원생동물 벡터, DNA 벡터 또는 이의 재조합체를 포함할 수 있는 재조합체 벡터를 포괄한다.
벡터에서의 발현을 위한 외인성 DNA (예를 들어, 관심 에피토프 및/또는 항원 및/또는 치료제를 인코딩함) 및 이러한 외인성 DNA 를 제공하는 문헌 뿐 아니라 핵산 분자의 발현 증강을 위한 전사 및/또는 번역 인자의 발현에 대해서, 그리고 "관심 에피토프", "치료적", "면역 반응", "면역학적 반응", "보호성 면역 반응", "면역학적 조성물", "면역원성 조성물" 및 "백신 조성물" 과 같은 용어에 관해서, 그 중에서도, 미국 특허 번호 5,990,091 (1999 년 11 월 23 일 발행), 및 WO 98/00166 및 WO 99/60164, 및 이에 인용된 문헌 및 특허 및 이들 PCT 출원의 공소제기에 있어서의 기록 문헌을 참조하고; 이들 모두는 본원에 참고로 포함된다. 따라서, 미국 특허 번호 5,990,091 및 WO 98/00166 및 WO 99/60164 및 이에 인용된 문헌 및 특허 및 이들 PCT 출원의 공소제기에 있어서의 기록 문헌, 및 이에 인용된 기타 문헌 또는 본원에 참고로 포함된 문헌은 본 발명의 실행에서 참조될 수 있고; 이에 인용된 모든 외인성 핵산 분자, 프로모터 및 벡터는 본 발명의 실행에서 사용될 수 있다. 이와 관련하여, 미국 특허 번호 6,706,693; 6,716,823; 6,348,450; 미국 특허 출원 일련 번호 10/424,409; 10/052,323; 10/116,963; 10/346,021; 및 WO 99/08713 (PCT/US98/16739 로부터, 1999 년 2 월 25 일 공개) 이 또한 언급된다.
본 발명의 양태는 전달 시스템을 통해 관심 유전자좌 또는 세포 또는 유기체에 전달되는 본 발명의 TALE 및 CRISPR-Cas 시스템을 포괄한다. 한 전달 수단은 벡터를 통한 것이며, 여기서 벡터는 바이러스 벡터, 예컨대 렌티- 또는 바큘로- 또는 바람직하게는 아데노-바이러스/아데노-관련 바이러스 벡터이나, 다른 전달 수단이 공지되어 있으며 (예컨대 효모 시스템, 미소낭포, 유전자 총/금 나노입자에 대한 부착 벡터) 제공된다. 일부 구현예에서, 하나 이상의 바이러스 또는 플라스미드 벡터는 나노입자, 엑소좀, 미소낭포 또는 유전자 총을 통해 전달될 수 있다.
본원에서 사용하는 바와 같이, 용어 "약물 조성물" 및 "약물", "백신 조성물", "백신", "백신 조성물", "치료적 조성물" 및 "치료적-면역학적 조성물" 은 항원 또는 병원체에 대항한 보호를 유도하는 임의의 조성물을 포함한다. 일부 구현예에서, 보호는 병원체에 의한 감염의 억제 또는 예방으로 인한 것일 수 있다. 다른 구현예에서, 보호는 관심 항원(들) 에 대항한 면역 반응에 의해 유도될 수 있거나, 항원에 대항하여 효과적으로 보호하는 것일 수 있고; 예를 들어, 대상에 대한 투여 또는 주사 후, 표적화된 항원 또는 면역원에 대항하여 보호성 면역 반응을 이끌어내거나 본 발명의 발명적인 아데노바이러스 벡터로부터 발현된 항원 또는 면역원에 대항하여 효과적인 보호를 제공한다. 용어 "약학 조성물" 은 대상에게 전달되는 임의의 조성물을 의미한다. 일부 구현예에서, 상기 조성물은 병원체에 의한 감염을 억제하거나 방지하도록 전달될 수 있다.
"치료적 유효량" 은 대상에게 투여시, 관심 유전자 생성물에 대해 치료적 반응 또는 면역 반응을 생성시키는, 관심 유전자를 인코딩하는 재조합 벡터의 양 또는 농도이다.
본원에서 사용하는 바와 같은 용어 "바이러스 벡터" 는 비제한적으로, 레트로바이러스, 아데노바이러스, 아데노-관련 바이러스, 알파바이러스 및 단순 포진 바이러스를 포함한다.
본 발명은 에너지 형태를 사용하는 내인성 또는 외인성 유전자 발현의 시공간적 제어를 포괄한다. 에너지 형태는 비제한적으로, 전자기 조사, 음파 에너지, 화학적 에너지 및 열 에너지를 포함할 수 있다. 본 발명의 바람직한 구현예에서, 에너지 형태는 전자기 조사, 바람직하게는 광 에너지이다. DNA 결합 아연 핑거 단백질에 연결된 전사 활성제와 같은 내인성 유전자의 발현 제어를 위한 이전의 접근방식은 시간적 또는 공간적 제어를 위한 메커니즘을 제공하지 않았다. 본원에 기재된 시스템의 광활성화를 위한 능력은 세포의 국부화 집단 내에서 정확한 시간에 시작하도록 유전자 발현 조절의 유도를 가능하게 한다.
상기 출원에서 상세히 설명한 바와 같은 제어의 양태는 적어도 하나 이상의 스위치(들) 에 관한 것이다. 본원에서 사용하는 바와 같은 용어 "스위치" 는 생물학적 기능의 활성화, 억제, 증강 또는 완료와 같은 상기 기능의 모든 양태를 포함하여, 변화에 영향을 주는 공동작용 방식으로 작용하는 성분의 집합 또는 시스템을 의미한다. 한 양태에서 용어 스위치는 유전자 조절 단백질의 기본 성분 및 이들 단백질이 인지하는 특이적 DNA 서열을 포함하는 유전적 스위치를 포함한다. 한 양태에서, 스위치는 유전자 조절에 사용되는 유도 및 억제 시스템에 관한 것이다. 일반적으로, 유전자 발현을 가능하게 하는 일부 분자 (유도물질로 지칭함) 가 존재하지 않으면 유도 시스템은 작동이 불가할 수 있다. 상기 분자는 "발현을 유도" 하는 것으로 일컬어진다. 이러한 일이 발생하는 방식은 제어 메커니즘 뿐 아니라 세포 유형에 있어서의 차이에 따라 좌우된다. 억제 시스템은 유전자 발현을 억제하는 일부 분자 (보조억제제로 지칭함) 가 존재하는 경우를 제외하고는 작동 가능하다. 상기 분자는 "발현을 억제" 하는 것으로 일컬어진다. 이러한 일이 발생하는 방식은 제어 메커니즘 뿐 아니라 세포 유형에 있어서의 차이에 따라 좌우된다. 본원에서 사용하는 바와 같은 용어 "유도" 는 수반된 분자 메커니즘에 관계없이 스위치의 모든 양태를 포함할 수 있다. 따라서 본 발명에 의해 이해되는 바와 같은 스위치는 비제한적으로 항생제 기반 유도 시스템, 전자기 에너지 기반 유도 시스템, 소분자 기반 유도 시스템, 핵 수용체 기반 유도 시스템 및 호르몬 기반 유도 시스템을 포함할 수 있다. 바람직한 구현예에서 스위치는 테트라사이클린 (Tet)/DOX 유도 시스템, 광 유도 시스템, 아브시스산 (ABA) 유도 시스템, 큐메이트 억제제/오퍼레이터 시스템, 4OHT/에스트로겐 유도 시스템, 엑디손-기반 유도 시스템 또는 FKBP12/FRAP (FKBP12-라파마이신 복합체) 유도 시스템일 수 있다.
본 발명의 한 양태에서 하나 이상의 스위치는 TALE 또는 CRISPR-Cas 시스템과 연관될 수 있으며, 여기서 TALE 또는 CRISPR-Cas 시스템의 활성은 스위치에 대한 하나 이상의 유도물질 에너지원과의 접촉에 의해 제어된다. 본 발명의 양태에 대해 본원에서 사용되는 바와 같은 용어 "접촉" 은 스위치와 유도물질 에너지원 사이의 임의의 관련적 관계를 의미하며, 이는 성분과의 물리적 상호작용 (함께 결합하는 분자 또는 단백질에서와 같이) 또는 에너지원에 의해 방사된 에너지의 경로에 있거나 상기 에너지에 의해 부딪치는 물리적 상호작용일 수 있다 (빛, 열 또는 소리의 흡수 또는 반사의 경우에서와 같이). 본 발명의 일부 양태에서 스위치와 유도물질 에너지원과의 접촉은 유도물질 에너지원의 적용에 의해 이루어진다. 본 발명은 또한 수동적 피드백 시스템을 통한 접촉을 포괄한다. 이는 TALE 또는 CRISPR-Cas 시스템 활성이, 이미 존재하며 따라서 적용할 필요가 없는 유도물질 에너지원과의 접촉에 의해 제어되는 임의의 수동적 조절 메커니즘을 비제한적으로 포함한다. 예를 들어 상기 에너지원은 세포 또는 세포 환경에 이미 존재하는 분자 또는 단백질일 수 있다. 수동적 접촉을 초래하는 상호작용은 비제한적으로 수용체/리간드 결합, 수용체/화학적 리간드 결합, 수용체/단백질 결합, 항체/단백질 결합, 단백질 이량체화, 단백질 이형이량체화, 단백질 다량체화, 핵 수용체/리간드 결합, 전사-후 변형 예컨대 인산화, 탈인산화, 유비퀴틴화 또는 탈유비퀴틴화를 포함할 수 있다.
2 개의 주요 분자 도구가 광반응성 전사 활성제-유사 (TAL) 효과기 시스템의 설계에 있어서 영향력을 행사하였다. 먼저, 조작된 TAL 효과기의 DNA 결합 특이성을 이용하여 게놈 내 특정 부위에 대해 복합체를 국부화시킨다. 두 번째로, 광-유도 단백질 이량체화를 사용하여 TAL 효과기에 의해 명시된 부위에 대해 활성 또는 억제 도메인을 유도시켜, 다운스트림 유전자를 조절한다.
유도 효과기는 시간적 또는 공간적으로 특이적인 유전자 발현 제어가 필요한 시험관내 또는 생체내 적용에 고려된다. 시험관내 예: 발생 신호의 시기를 밝히기 위한 발생 유전자의 시간적으로 정확한 유도/억제, 세포-유형 패턴화 조직의 생성을 위한 세포 운명 재프로그래밍 인자의 공간적으로 제어된 유도. 시험관내 예: 특정 뇌 부위 내의 유전자 발현의 조합된 시간적 및 공간적 제어.
본 발명의 바람직한 구현예에서, 유도 효과기는 광 유도 전사 효과기 (LITE) 이다. LITE 시스템의 모듈성으로 인해, 임의 수의 효과기 도메인이 전사 조절에 이용될 수 있다. 특히 유리한 구현예에서, 전사 활성제 유사 효과기 (TALE) 및 활성화 도메인 VP64 가 본 발명에서 사용된다.
LITE 는 시간적 및 공간적으로 정확한 방식으로 개별적 내인성 유전자의 발현을 조정하거나 변형시키도록 설계된다. 각각의 LITE 는 커스텀화된 DNA-결합 전사 활성제 유사 효과기 (TALE) 단백질, 아라바돕시스 탈리아나 (Arabadopsis thaliana) 로부터의 광-반응성 크립토크롬 이형이량체, 및 전사 활성화/억제 도메인으로 이루어지는 2 성분 시스템을 포함할 수 있다. TALE 은 관심 유전자의 프로모터 서열에 결합하도록 설계된다. TALE 단백질은 크립토크롬 이형이량체의 절반 (크립토크롬-2 또는 CIB1) 에 융합되면서, 남아있는 크립토크롬 파트너가 전사 효과기 도메인에 융합된다. 효과기 도메인은 활성제 예컨대 VP16, VP64 또는 p65, 또는 억제제 예컨대 KRAB, EnR 또는 SID 일 수 있다. LITE 의 미자극 상태에서, TALE-크립토크롬2 단백질은 관심 유전자의 프로모터에 대해 국부화되나, CIB1-효과기 단백질에 결합하지는 않는다. 청색 스펙트럼 광으로 LITE 자극시, 크립토크롬-2 는 활성화되고, 형태적 변화를 거치며, 이의 결합 도메인이 밝혀진다. 다음으로 CIB1 은 크립토크롬-2 에 결합하여 관심 유전자의 프로모터 부위에 대해 효과기 도메인을 국부화시키고 유전자 과발현 또는 침묵화 (silencing) 를 개시시킨다.
활성제 및 억제제 도메인은 종류, 강도, 메커니즘, 지속기간, 크기, 또는 임의 수의 기타 매개변수를 기반으로 선택될 수 있다. 바람직한 효과기 도메인은 비제한적으로, 트랜스포사아제 도메인, 인테그라아제 도메인, 재조합효소 도메인, 리졸바아제 도메인, 인버타아제 도메인, 프로테아제 도메인, DNA 메틸트랜스퍼라아제 도메인, DNA 데메틸라아제 도메인, 히스톤 아세틸라아제 도메인, 히스톤 데아세틸라아제 도메인, 뉴클레아제 도메인, 억제제 도메인, 활성제 도메인, 핵-국부화 신호 도메인, 전사-단백질 영입 도메인, 세포 흡수 활성 관련 도메인, 핵산 결합 도메인 또는 항체 제시 도메인을 포함한다.
LITE 또는 임의의 기타 유도 효과기에서의 유전자 표적화는 커스텀화된 TALE DNA 결합 단백질의 특이성을 통해 이루어질 수 있다. 관심 유전자의 프로모터 부위에서 표적 서열이 선택되며 상기 서열에 대해 커스텀화된 TALE 이 설계된다. TALE 의 중심 부분은 34 개 아미노산 길이의 탠덤 반복물로 이루어진다. 이들 반복물의 서열은 거의 동일하나, 각각의 반복물의 12 번째 및 13 번째 아미노산 (반복 가변성 2 잔기로 지칭함) 은 가변적이어서, 각각의 반복물의 뉴클레오티드-결합 특이성을 결정한다. 따라서, TALE 단량체 반복물의 적절한 배치를 갖는 구축물을 합성하여, 표적 프로모터 서열에 특이적인 DNA 결합 단백질을 제작한다.
본 발명의 유리한 구현예에서, 본원에 제공된 방법은 단리된, 비-자연 발생적인 재조합 또는 조작된 DNA 결합 단백질을 사용한다 (이는 향상된 효율 및 확장된 특이성을 갖는 핵산 서열의 표적화를 가능하게 하는 그의 조직 구조의 일부로서 TALE 단량체 또는 TALE 단량체 또는 반-단량체를 포함함).
자연 발생적 TALE 또는 "야생형 TALE" 은 수많은 종류의 프로테오박테리아에 의해 분비된 핵산 결합 단백질이다. TALE 폴리펩티드는 대부분 33, 34 또는 35 개 아미노산 길이이며 서로 주로 아미노산 위치 12 및 13 에 있어서 상이한 고도로 보존된 단량체 폴리펩티드의 탠덤 반복물로 구성된 핵산 결합 도메인을 포함한다. 유리한 구현예에서 핵산은 DNA 이다. 본원에서 사용하는 바와 같이, 용어 "폴리펩티드 단량체", "TALE 단량체" 또는 "단량체" 는 TALE 핵산 결합 도메인 내의 고도로 보존된 반복적 폴리펩티드 서열을 의미하는데 사용되며, 용어 "반복 가변성 2-잔기" 또는 "RVD" 는 폴리펩티드 단량체의 위치 12 및 13 에서의 고도로 가변적인 아미논산을 의미하는데 사용된다. DNA 결합 도메인 내에 포함되는 TALE 단량체의 일반적 표시는 X1-11-(X12X13)-X14-33 또는 34 또는 35 이며, 여기서 첨자는 아미노산 위치를 나타내고, X 는 임의의 아미노산을 나타낸다. X12X13 은 RVD 를 나타낸다. 일부 폴리펩티드 단량체에서, 위치 13 에서의 가변적 아미노산은 결실되거나 부재하고, 이러한 단량체에서, RVD 는 단일 아미노산으로 이루어진다. 이러한 경우 RVD 는 대안적으로 X* 로 표시될 수 있으며, 여기서 X 는 X12 를 나타내고, (*) 는 X13 이 부재함을 나타낸다. DNA 결합 도메인은 TALE 단량체의 여러 반복물을 포함하며 이는 (X1-11-(X12X13)-X14-33 또는 34 또는 35)z 로서 표시되고, 이때 유리한 구현예에서, z 는 적어도 5 내지 40 이다. 추가로 유리한 구현예에서, z 는 적어도 10 내지 26 이다.
TALE 단량체는 그의 RVD 에서의 아미노산 동일성에 의해 결정되는 뉴클레오티드 결합 친화성을 갖는다. 예를 들어, NI 의 RVD 를 갖는 폴리펩티드 단량체는 바람직하게는 아데닌 (A) 에 결합하고, NG 의 RVD 를 갖는 단량체는 바람직하게는 티민 (T) 에 결합하고, HD 의 RVD 를 갖는 단량체는 바람직하게는 시토신 (C) 에 결합하고, NN 의 RVD 를 갖는 단량체는 바람직하게는 아데닌 (A) 및 구아닌 (G) 둘 모두에 결합한다. 본 발명의 또 다른 구현예에서, IG 의 RVD 를 갖는 단량체는 바람직하게는 T 에 결합한다. 따라서, TALE 의 핵산 결합 도메인에서의 폴리펩티드 단량체 반복물의 수 및 순서는 이의 핵산 표적 특이성을 결정한다. 본 발명의 추가 구현예에서, NS 의 RVD 를 갖는 단량체는 모든 4 개 염기 쌍을 인지하며 A, T, G 또는 C 에 결합할 수 있다. TALE 의 구조 및 기능은 예를 들어, Moscou et al, Science 326: 1501 (2009); Boch et al, Science 326:1509-1512 (2009); 및 Zhang et al, Nature Biotechnology 29: 149-153 (2011) 에 추가로 기재되어 있으며, 이들 각각은 그 전체가 본원에 참고로 포함된다.
본 발명의 방법에서 사용한 폴리펩티드는 단리된, 비-자연 발생적인 재조합 또는 조작된 핵산 결합 단백질이며 이는 특정 핵산 서열을 표적화하도록 설계되는 폴리펩티드 단량체 반복물을 함유하는 핵산 또는 DNA 결합 부위를 갖는다.
본원에서 기재한 바와 같이, HN 또는 NH 의 RVD 를 갖는 폴리펩티드 단량체는 바람직하게는 구아닌에 결합하며, 이로써 표적 핵산 서열을 함유하는 구아닌에 대한 높은 결합 특이성을 갖는 TALE 폴리펩티드가 생성되게 한다. 본 발명의 바람직한 구현예에서, RVD RN, NN, NK, SN, NH, KN, HN, NQ, HH, RG, KH, RH 및 SS 를 갖는 폴리펩티드 단량체는 바람직하게는 구아닌에 결합한다. 본 발명의 훨씬 더 유리한 구현예에서, RVD RN, NK, NQ, HH, KH, RH, SS 및 SN 을 갖는 폴리펩티드 단량체는 바람직하게는 구아닌에 결합하며 이로써 표적 핵산 서열을 함유하는 구아닌에 대한 높은 결합 특이성을 갖는 TALE 폴리펩티드가 생성되게 한다. 본 발명의 보다 더 유리한 구현예에서, RVD HH, KH, NH, NK, NQ, RH, RN 및 SS 를 갖는 폴리펩티드 단량체는 바람직하게는 구아닌에 결합하며 이로써 표적 핵산 서열을 함유하는 구아닌에 대한 높은 결합 특이성을 갖는 TALE 폴리펩티드가 생성되게 한다. 추가로 유리한 구현예에서, 구아닌에 대한 높은 결합 특이성을 갖는 RVD 는 RN, NH, RH 및 KH 이다. 또한, NV 의 RVD 를 갖는 폴리펩티드 단량체는 바람직하게는 아데닌 및 구아닌에 결합한다. 본 발명의 보다 바람직한 구현예에서, H*, HA, KA, N*, NA, NC, NS, RA 및 S* 의 RVD 를 갖는 단ㄹ야체는 비슷한 친화성으로 아데닌, 구아닌, 시토신 및 티민에 결합한다.
본 발명의 보다 더 유리한 구현예에서 아데닌에 대한 특이성을 갖는 RVD 는 NI, RI, KI, HI 및 SI 이다. 본 발명의 보다 바람직한 구현예에서, 아데닌에 대한 특이성을 갖는 RVD 는 HN, SI 및 RI 이고, 가장 바람직하게는 아데닌 특이성에 대한 RVD 는 SI 이다. 본 발명의 보다 더 바람직한 구현예에서 티민에 대한 특이성을 갖는 RVD 는 NG, HG, RG 및 KG 이다. 본 발명의 추가로 유리한 구현예에서, 티민에 대한 특이성을 갖는 RVD 는 KG, HG 및 RG 이고, 가장 바람직하게는 티민 특이성에 대한 RVD 는 KG 또는 RG 이다. 본 발명의 보다 더 바람직한 구현예에서 시토신에 대한 특이성을 갖는 RVD 는 HD, ND, KD, RD, HH, YG 및 SD 이다. 본 발명의 추가로 유리한 구현예에서, 시토신에 대한 특이성을 갖는 RVD 는 SD 및 RD 이다. 본 발명의 가장 바람직한 구현예에 포함되는 대표적 RVD 및 이들이 표적화하는 뉴클레오티드에 대해서는 도 7B 를 참고한다. 추가로 유리한 구현예에서 변이체 TALE 단량체는 도 7A 에서 나타낸 바와 같이 뉴클레오티드에 대한 특이성을 나타내는 임의의 RVD 를 포함할 수 있다. 모든 이러한 TALE 단량체는 관련되나 동일하지는 않은 표적 핵산 서열의 레퍼토리에 결합할 수 있는 퇴행적 TALE 폴리펩티드가 생성될 수 있게 한다. 본 발명의 추가 구현예에서, RVD NT 는 G 및 A 에 결합할 수 있다. 본 발명의 추가 구현예에서, RVD NP 는 A, T 및 C 에 결합할 수 있다. 본 발명의 보다 유리한 구현예에서, 하나 이상의 선택된 RVD 는 NI, HD, NG, NN, KN, RN, NH, NQ, SS, SN, NK, KH, RH, HH, KI, HI, RI, SI, KG, HG, RG, SD, ND, KD, RD, YG, HN, NV, NS, HA, S*, N*, KA, H*, RA, NA 또는 NC 일 수 있다.
핵산 또는 DNA 결합 도메인의 하나 이상의 폴리펩티드 단량체의 사전결정된 N-말단에서 C-말단 순서는 본 발명의 폴리펩티드가 결합할 상응하는 사전결정된 표적 핵산 서열을 결정한다. 본원에서 사용하는 바와 같이 단량체 및 적어도 하나 이상의 반-단량체는 관심 유전자 또는 게놈 유전자좌를 "표적화하도록 특이적으로 순서화된다". 식물 게놈에서, 자연적 TALE-결합 부위는 항상 티민 (T) 으로 시작하는데, 이는 TALE 폴리펩티드의 비-반복성 N-말단 내의 암호성 신호에 의해 명시될 수 있고; 일부 경우 상기 부위는 반복물 0 으로서 지칭될 수 있다. 동물 게놈에서, TALE 결합 부위는 반드시 티민 (T) 으로 시작할 필요는 없으며 본 발명의 폴리펩티드는 T, A, G 또는 C 로 시작하는 DNA 서열을 표적화할 수 있다. TALE 단량체의 탠덤 반복물은 항상, 반복성 전체 길이 TALE 단량체의 처음 20 개 아미노산과만 동일성을 공유할 수 있는 서열의 스트레치 또는 반-길이 반복물로 종료되며, 상기 반-반복물은 반-단량체로서 지칭될 수 있다 (도 8). 그러므로, 이는 표적화되는 핵산 또는 DNA 의 길이가 전체 단량체 수 + 2 와 동일하다는 것에 따른다.
예를 들어 핵산 결합 도메인은, 사전결정된 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 개 뉴클레오티드 길이 핵산 서열에 결합시키기 위해 N-말단에서 C-말단 방향으로 배열된 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 개 이상의 폴리펩티드 단량체를 함유하도록 조작될 수 있다. 본 발명의 보다 유리한 구현예에서, 핵산 결합 도메인은 각각 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27 및 28 개 뉴클레오티드 길이의 핵산 서열을 표적화하기 위해 특이적으로 순서화되거나 배열된 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 개 이상의 전체 길이 폴리펩티드 단량체를 함유하도록 조작될 수 있다. 특정 구현예에서 폴리펩티드 단량체는 인접하다. 일부 구현예에서, 특히 TALE 폴리펩티드의 C-말단에서 존재하는 경우, 반-단량체가 하나 이상의 단량체 대신에 사용될 수 있다.
폴리펩티드 단량체는 일반적으로 33, 34 또는 35 개 아미노산 길이이다. RVD 를 제외하고, 폴리펩티드 단량체의 아미노산 서열은 고도로 보존되거나 본원에 기재된 바와 같이, RVD 를 제외하고 폴리펩티드 단량체에서의 아미노산은 TALE 활성에 영향을 주는 패턴을 나타내며, 이의 확인이 본 발명의 바람직한 구현예에서 사용될 수 있다. 단량체 서열 중의 아미노산의 대표적 조합 (RVD 제외) 은 TALE 활성에 대한 효과를 갖는 것으로 출원인에 의해 나타내어진다 (도 10). 본 발명의 보다 바람직한 구현예에서, DNA 결합 도메인이 (X1-11-X12X13-X14-33 또는 34 또는 35)z 를 포함하는 경우 (여기서 X1-11 은 11 개의 인접한 아미노산의 사슬이고, X12X13 은 반복 가변성 2 잔기 (RVD) 이고, X14-33 또는 34 또는 35 는 21, 22 또는 23 개의 인접한 아미노산의 사슬이고, z 는 적어도 5 내지 26 임), 아미노산의 바람직한 조합은 X1-4 에서의 [LTLD] 또는 [LTLA] 또는 [LTQV], 또는 위치 X30-33 또는 X31-34 또는 X32-35 에서의 [EQHG] 또는 [RDHG] 이다. 또한, 단량체에서의 다른 관심 아미노산 조합은, 단량체가 34 개 아미노산 길이인 경우, X1-4 에서의 [LTPD] 및 X16-20 에서의 [NQALE] 및 X32-34 에서의 [DHG] 이다. 단량체가 33 또는 35 개 아미노산 길이인 경우, 상응하는 이동이 인접한 아미노산 [NQALE] 및 [DHG] 의 위치에서 발생하고; 바람직하게는, 본 발명의 구현예는 X15-19 또는 X17-21 에서 [NQALE] 및 X31-33 또는 X33-35 에서 [DHG] 를 가질 수 있다.
본 발명의 추가 구현예에서 단량체에서의 관심 아미노산 조합은, 단량체가 34 개 아미노산 길이인 경우, X1-4 에서의 [LTPD] 및 X16-20 에서의 [KRALE] 및 X32-34 에서의 [AHG] 또는 X1-4 에서의 [LTPE] 및 X16-20 에서의 [KRALE] 및 X32-34 에서의 [DHG] 이다. 단량체가 33 또는 35 개 아미노산 길이인 경우, 상응하는 이동이 인접한 아미노산 [KRALE], [AHG] 및 [DHG] 의 위치에서 발생한다. 바람직한 구현예에서, 인접한 아미노산의 위치는 (X1-4 에서의 [LTPD] 및 X15-19 에서의 [KRALE] 및 X31-33 에서의 [AHG]) 또는 (X1-4 에서의 [LTPE] 및 X15-19 에서의 [KRALE] 및 X31-33 에서의 [DHG]) 또는 (X1-4 에서의 [LTPD] 및 X17-21 에서의 [KRALE] 및 X33-35 에서의 [AHG]) 또는 (X1-4 에서의 [LTPE] 및 X17-21 에서의 [KRALE] 및 X33-35 에서의 [DHG]) 일 수 있다. 본 발명의 추가 구현예에서, 인접한 아미노산 [NGKQALE] 는 위치 X14-20 또는 X13-19 또는 X15-21 에서 존재한다. 이들 대표적 위치는 본 발명의 다양한 구현예를 제안하며 본원에서 기재한 모든 TALE 단량체에서의 관심 아미노산의 조합 또는 추가적인 관심 아미노산을 확인하기 위한 안내를 제공한다 (도 9A-F 및 10).
하기 제공된 것은 폴리펩티드 단량체의 보존된 부분의 예시적 아미노산 서열이다. 각 서열에서의 RVD 의 위치를 XX 또는 X* 로 나타낸다 (여기서 (*) 는 RVD 가 단일 아미노산이고 잔기 13 (X13) 이 부재하다는 것을 나타냄).

서열 (X1-11-X14-34 또는 X1-11-X14-35) (여기서 X 는 임의의 아미노산이고 첨자는 아미노산 위치임) 로 나타낼 수 있는 RVD 를 제외하고 TALE 단량체의 추가적인 목록을 도 9A-F 에 제공한다.
[Zhang et al, Nature Biotechnology 29:149-153 (2011)] 에서 기재한 바와 같이, TALE 폴리펩티드 결합 효율은, 자연 발생적 TALE 의 DNA 결합 부위의 직접적 N-말단 또는 C-말단인 "캡핑 부위" 로부터의 아미노산 서열을 조작된 TALE DNA 결합 부위의 위치 N-말단 또는 C-말단에서의 조작된 TALE 에 포함시킴으로써 증가될 수 있다. 따라서 특정 구현예에서, 본원에서 기재된 TALE 폴리펩티드는 N-말단 캡핑 부위 및/또는 C-말단 캡핑 부위를 추가로 포함한다.
N-말단 캡핑 부위의 예시적 아미노산 서열은 하기와 같다:

C-말단 캡핑 부위의 예시적 아미노산 서열은 하기와 같다:

본원에서 사용하는 바오 같이 사전결정된 N-말단 캡핑 부위, 반복 TALE 단량체를 포함하는 DNA 결합 도메인 및 C-말단 캡핑 부위의 "N-말단" 에서 "C-말단" 배향은 본 발명의 d-TALE 또는 폴리펩티드에서의 상이한 도메인의 구성을 위한 구조적 기초를 제공한다.
전체 N-말단 및/또는 C-말단 캡핑 부위는 DNA 결합 부위의 결합 활성을 증강시키는데 필요하지 않다. 따라서 특정 구현예에서, N-말단 및/또는 C-말단 캡핑 부위의 단편은 본원에 기재된 TALE 폴리펩티드에 포함된다.
특정 구현예에서, 본원에 기재된 TALE 폴리펩티드는 N-말단 캡핑 부위의 적어도 10, 20, 30, 40, 50, 54, 60, 70, 80, 87, 90, 94, 100, 102, 110, 117, 120, 130, 140, 147, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240, 250, 260 또는 270 개 아미노산을 포함하는 N-말단 캡핑 부위 단편을 함유한다. 특정 구현예에서, N-말단 캡핑 부위 단편 아미노산은 N-말단 캡핑 부위의 C-말단 (DNA-결합 부위 근부 말단) 의 것이다. [Zhang et al., Nature Biotechnology 29:149-153 (2011)] 에서 기재한 바와 같이, C-말단 240 개 아미노산을 포함하는 N-말단 캡핑 부위 단편은 전체 길이 캡핑 부위와 동일하게 결합 활성을 증강시키는 한편, C-말단 147 개 아미노산을 포함하는 단편은 전체 길이 캡핑 부위 효능의 80% 초과를 유지하고, C-말단 117 개 아미노산을 포함하는 단편은 전체 길이 캡핑 부위 활성의 50% 초과를 유지한다.
일부 구현예에서, 본원에 기재된 TALE 폴리펩티드는 C-말단 캡핑 부위의 적어도 6, 10, 20, 30, 37, 40, 50, 60, 68, 70, 80, 90, 100, 110, 120, 127, 130, 140, 150, 155, 160, 170, 180 개 아미노산을 포함하는 C-말단 캡핑 부위 단편을 함유한다. 특정 구현예에서, C-말단 캡핑 부위 단편 아미노산은 C-말단 캡핑 부위의 N-말단 (DNA-결합 부위 근부 말단) 의 것이다. [Zhang et al, Nature Biotechnology 29: 149-153 (2011)] 에서 기재한 바와 같이, C-말단 68 개 아미노산을 포함하는 C-말단 캡핑 부위 단편은 전체 길이 캡핑 부위와 동일하게 결합 활성을 증강시키는 한편, C-말단 20 개 아미노산을 포함하는 단편은 전체 길이 캡핑 부위 효능의 50% 초과를 유지한다.
특정 구현예에서, 본원에 기재된 TALE 폴리펩티드의 캡핑 부위는 본원에 제공된 캡핑 부위 서열에 대해 동일한 서열을 가질 필요는 없다. 따라서 일부 구현예에서, 본원에 기재된 TALE 폴리펩티드의 캡핑 부위는 본원에 제공된 캡핑 부위 아미노산 서열에 대해 적어도 50%, 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 동일하거나 동일성을 공유하는 서열을 갖는다. 서열 동일성은 서열 상동성과 관련된다. 상동성 비교는 육안으로, 또는 통상 용이하게 이용가능한 서열 비교 프로그램을 보조로 사용하여 수행될 수 있다. 이들 시판 컴퓨터 프로그램은 둘 이상의 서열 사이의 상동성 퍼센트 (%) 를 계산할 수 있으며 또한 둘 이상의 아미노산 또는 핵산 서열에 의해 공유된 서열 동일성을 계산할 수 있다. 일부 바람직한 구현예에서, 본원에 기재된 TALE 폴리펩티드의 캡핑 부위는 본원에 제공된 캡핑 부위 아미노산 서열에 대해 95% 이상 동일하거나 동일성을 공유하는 서열을 갖는다.
서열 상동성은, BLAST 또는 FASTA 를 비제한적으로 포함하는 당업계에 공지된 임의의 다수 컴퓨터 프로그램에 의해 생성될 수 있다. GCG Wisconsin Bestfit 패키지와 같은 정렬을 실행하기에 적합한 컴퓨터 프로그램을 또한 사용할 수 있다. 소프트웨어가 최적 정렬을 생성시키고 나면, 상동성 %, 바람직하게는 서열 동일성 % 를 계산할 수 있다. 소프트웨어는 통상 서열 비교의 일부로서 이를 수행하며 수치적 결과를 산출해낸다.
본원에 기재된 유리한 구현예에서, 본 발명의 TALE 폴리펩티드는 하나 이상의 효과기 도메인에 연결된 핵산 결합 도메인을 포함한다. 용어 "효과기 도메인" 또는 "조절 또는 기능적 도메인" 은 핵산 결합 도메인에 의해 인지된 핵산 서열에 결합하는 것 외의 활성을 갖는 폴리펩티드 서열을 의미한다. 핵산 결합 도메인을 하나 이상의 효과기 도메인과 조합시킴으로써, 본 발명의 폴리펩티드를 사용하여, 핵산 결합 도메인이 특이적으로 결합하는 특정 표적 DNA 서열에 대해 효과기 도메인에 의해 매개된 하나 이상의 기능 또는 활성을 표적화할 수 있다. 용어 "효과기 도메인" 및 "기능적 도메인" 은 본 출원에서 상호교환가능하게 사용된다.
본원에서 기재된 TALE 폴리펩티드의 일부 구현예에서, 효과기 도메인에 의해 매개된 활성은 생물학적 활성이다. 예를 들어, 일부 구현예에서 효과기 도메인은 전사 저해제 (즉, 억제제 도메인), 예컨대 mSin 상호작용 도메인 (SID), SID4X 도메인 또는 크루펠 (Kruppel)-연관 상자 (KRAB) 또는 KRAB 도메인의 단편이다. 일부 구현예에서 효과기 도메인은 전사의 증강제 (즉, 활성화 도메인), 예컨대 VP16, VP64 또는 p65 활성화 도메인이다. 이들 상이한 활성화 도메인이 Sox2 mRNA 수준에 대해 갖는 효과의 그래프적 비교를 도 11 에 제공한다.
본원에서 사용하는 바와 같이, VP16 은 헤르페스바이러스 단백질이다. 이는 바이러스 전초기 유전자 발현을 특이적으로 활성화시키는 매우 강력한 전사 활성제이다. VP16 활성화 도메인은 산성 잔기가 풍부하며 고전적 산성 활성화 도메인 (AAD) 으로서 간주된다. 본원에서 사용하는 바와 같이, VP64 활성화 도메인은 VP16 의 최소 활성화 도메인의 사량체 반복물이다. 본원에서 사용하는 바와 같이, p65 는 NF-카파 B 전사 인자 복합체를 구성하는 2 개 단백질 중 하나이다. 다른 단백질은 p50 이다. p65 활성화 도메인은 p65 서브유닛의 일부이며 이는 p50 의 부재 하에서도 강력한 전사 활성제이다. 특정 구현예에서, 효과기 도메인은 포유동물 단백질 또는 이의 생물학적 활성 단편이다. 이러한 효과기 도메인은 "포유동물 효과기 도메인" 으로서 지칭한다.
일부 구현예에서 핵산 결합은 예를 들어, 트랜스포사아제 도메인, 인테그라아제 도메인, 재조합효소 도메인, 리졸바아제 도메인, 인버타아제 도메인, 프로테아제 도메인, DNA 메틸트랜스퍼라아제 도메인, DNA 히드록실메틸라아제 도메인, DNA 데메틸라아제 도메인, 히스톤 아세틸라아제 도메인, 히스톤 데아세틸라아제 도메인, 뉴클레아제 도메인, 억제제 도메인, 활성제 도메인, 핵-국부화 신호 도메인, 전사-조절 단백질 (또는 전사 복합체 영입) 도메인, 세포 흡수 활성 관련 도메인, 핵산 결합 도메인, 항체 제시 도메인, 히스톤 개질 효소, 히스톤 개질 효소의 영입제; 히스톤 개질 효소의 저해제, 히스톤 메틸트랜스퍼라아제, 히스톤 데메틸라아제, 히스톤 키나아제, 히스톤 포스파타아제, 히스톤 리보실라아제, 히스톤 데리보실라아제, 히스톤 유비퀴티나아제, 히스톤 데유비퀴티나아제, 히스톤 비오티나아제 및 히스톤 테일 프로테아제를 비제한적으로 포함하는 효과기 도메인 또는 기능적 도메인과 연결된다.
일부 구현예에서, 효과기 도메인은 트랜스포사아제 활성, 인테그라아제 활성, 재조합효소 활성, 리졸바아제 활성, 인버타아제 활성, 프로테아제 활성, DNA 메틸트랜스퍼라아제 활성, DNA 데메틸라아제 활성, 히스톤 아세틸라아제 활성, 히스톤 데아세틸라아제 활성, 뉴클레아제 활성, 핵-국부화 신호전달 활성, 전사 억제제 활성, 전사 활성제 활성, 전사 인자 영입 활성 또는 세포 흡수 신호전달 활성을 비제한적으로 포함하는 활성을 나타내는 단백질 도메인이다. 본 발명의 다른 바람직한 구현예는 본원에 기재된 활성의 임의 조합을 포함할 수 있다.
[Zhang et al, Nature Biotechnology 29: 149-153 (2011)] 에서 기재된 바와 같이, 핵산 결합 도메인 및 효과기 도메인을 갖는 TALE 폴리펩티드를 사용하여, 핵산 결합 도메인에 의해 인지된 사전결정된 핵산 서열을 갖는 게놈 위치에 대해 효과가 도메인의 활성을 표적화할 수 있다. 본원에 기재된 본 발명의 일부 구현예에서, TALE 폴리펩티드는 유전자 조절 활성, 예컨대 조절, 코딩 및/또는 유전자간 부위에 대한 전사 또는 번역 변형제 활성, 예컨대 인핸서 및/또는 억제제 활성을 표적화하기 위해 설계되고 사용되며, 이는 코딩 부위의 전사 업스트림 및 다운스트림에 영향을 줄 수 있고 유전자 발현을 증강시키거나 억제하는데 사용될 수 있다. 예를 들어, TALE 폴리펩티드는 전사 인자로부터의 DNA-결합 도메인을 갖는 효과기 도메인, 전사 인자 (활성제, 억제제, 보조-활성제, 보조-억제제) 로부터의 효과기 도메인, 사일런서 (silencer), 핵 호르몬 수용체 및/또는 크로마틴 연관 단백질 및 그의 변형제 (예를 들어 메틸라아제, 키나아제, 포스파타아제, 아세틸라아제 및 데아세틸라아제) 를 포함할 수 있다. 바람직한 구현예에서, TALE 폴리펩티드는 뉴클레아제 도메인을 포함할 수 있다. 보다 바람직한 구현예에서 뉴클레아제 도메인은 비-특이적 FokI 엔도뉴클레아제 촉매 도메인이다.
추가 구현예에서, 유전자 발현 조절을 위해 유용한 도메인은 또한 암유전자의 유전자 산물로부터 수득될 수 있다. 본 발명의 추가로 유리한 구현예에서, 인테그라아제 또는 트랜스포사아제 활성을 갖는 효과기 도메인은 외인성 핵산 서열의 특정 핵산 서열 부위 내로의 통합을 촉진시키고, 특정 내인성 핵산 서열을 제거 (녹-아웃 (knock-out)) 하고/하거나 예컨대 DNA 메틸트랜스퍼라아제, DNA 데메틸라아제, 히스톤 아세틸라아제 및 히스톤 데아세틸라아제 활성을 촉진시킴으로써 후생적 신호 및 결과적인 유전자 조절을 변형시키는데 사용될 수 있다. 다른 구현예에서, 뉴클레아제 활성을 갖는 효과기 도메인은 본 발명의 폴리펩티드가 특이적으로 결합하는 표적 서열을 잘라내거나 소화시켜 게놈 구조를 변형시키는데 사용될 수 있으며, 이들 부위에 외인성 유전자를 도입시킬 수 있다. 추가 구현예에서, 인버타아제 활성을 갖는 효과기 도메인은 DNA 단편의 배향을 바꾸어 게놈 구조를 변형시키는데 사용될 수 있다.
특히 유리한 구현예에서, 본 발명의 방법에서 사용된 폴리펩티드는 전사 활성을 표적화하는데 사용될 수 있다. 본원에서 사용하는 바와 같이, 용어 "전사 인자" 는 전사 제어를 위해 관심 유전자 또는 게놈 유전자좌와 연관된 특정 DNA 서열에 결합하는 단백질 또는 폴리펩티드를 의미한다. 전사 인자는 RNA 중합효소의 관심 유전자로의 영입을 촉진 (활성제로서) 하거나 차단 (억제제로서) 할 수 있다. 전사 인자는 그의 기능을 단독으로 또는 더 큰 단백질 복합체의 일부로서 수행할 수 있다. 전사 인자에 의해 사용되는 유전자 조절의 메커니즘은 비제한적으로, a) RNA 중합효소 결합의 안정화 또는 탈안정화, b) 히스톤 단백질의 아세틸화 또는 탈아세틸화 및 c) 보조-활성제 또는 보조-억제제 단백질의 영입을 포함한다. 또한, 전사 인자는 기본 전사, 전사의 증강, 발생, 세포간 신호전달에 대한 반응, 환경적 신호에 대한 반응, 세포-주기 제어 및 발병과정을 비제한적으로 포함하는 생물학적 활성에 있어서 그 역할을 수행한다. 전사 인자에 대한 정보와 관련하여, [Latchman and DS (1997) Int. J. Biochem. Cell Biol. 29 (12): 1305-12; Lee TI, Young RA (2000) Annu. Rev. Genet. 34: 77-137 및 Mitchell PJ, Tjian R (1989) Science 245 (4916): 371-8] 가 언급되고, 이는 본원에 그 전체가 참고로 포함된다.
LITE 의 광 반응성은 크립토크롬-2 및 CIB1 의 활성화 및 결합을 통해 이루어진다. 상기 언급한 바와 같이, 청색광 자극은 크립토크롬-2 에 있어서의 활성화 형태적 변화를 유도하여, 이의 결합 파트너 CIB1 의 영입을 초래한다. 이러한 결합은 빠르고 가역적이며, 펄스화된 자극 후 <15 초 내에 포화가 이루어지고 자극 종료 후 <15 분에 기준선으로 복귀된다. 이러한 신속한 결합 속도는 유도제의 흡수 및 청소율보다는 전사/번역 및 전사체/단백질 분해의 속도에 의해서만 일시적으로 결합한 LITE 시스템을 초래한다. 크립토크롬-2 활성화는 또한 매우 민감하여, 낮은 광 강도 자극의 사용을 가능하게 하며 광독성의 위험성을 완화시킨다. 추가로, 미손상 포유동물 뇌와 같은 맥락에서, 가변적인 광 강도는 LITE 자극 부위의 크기를 제어하는데 사용될 수 있어, 벡터 단독 전달이 제공할 수 있는 것보다 더 큰 정확성을 부여한다.
LITE 시스템의 모듈성은 임의 수의 효과기 도메인이 전사 조정에 사용될 수 있게 한다. 따라서, 활성제 및 억제제 도메인은 종류, 강도, 메커니즘, 지속기간, 크기, 또는 임의 수의 기타 매개변수를 기반으로 선택될 수 있다.
출원인은 그 다음으로 LITE 시스템의 2 가지 원형적 시현을 제시한다. 제 1 예는 마우스 유전자 NEUROG2 의 전사를 활성화시키도록 설계된 LITE 이다. 마우스 NEUROG2 의 업스트림 프로모터 부위에 위치한 서열 TGAATGATGATAATACGA 를 표적으로서 선택하고, 상기 서열에 매치되도록 TALE 을 설계하고 합성하였다. TALE 서열을 핵 국부화 신호 (아미노산: SPKKKRKVEAS) 를 통해 크립토크롬-2 에 대한 서열에 연결시켜, 사이토졸로부터의 단백질의 핵 공간으로의 이동을 촉진시킨다. 동일한 핵 국부화 신호를 사용하여, 전사 활성제 도메인 VP64 에 연결된 CIB1 도메인을 포함하는 제 2 벡터를 합성하였다. 이러한 제 2 벡터, 또한 GFP 서열을 2A 전사 스킵 신호에 의해 CIB1-VP64 융합 서열로부터 분리하였다. 각 구축물의 발현을 편재성, 항시성 프로모터 (CMV 또는 EF1-α) 에 의해 유도하였다. Neuro 2A 세포주로부터의 마우스 신경아세포종 세포를 2 개 벡터로 동시-트랜스펙션시켰다. 벡터 발현이 가능하도록 인큐베이션한 후, 샘플을 488 nm LED 의 어레이로부터의 주기적 펄스의 청색광에 의해 자극하였다. 미자극된 동시-트랜스펙션 샘플 및 형광 리포터 YFP 로만 트랜스펙션된 샘플을 대조군으로서 사용하였다. 각 실험 완료시, mRNA 를 qPCR 을 통해 분석한 샘플로부터 정제하였다.
크립토크롬-2 및 CIB1 의 절두 형태를 클로닝하고 전체 길이 형태의 크립토크롬-2 및 CIB1 과 조합으로 시험하여, 각각의 이형이량체 쌍의 효과를 측정하였다. 크립토크롬-2 단백질의 보존된 광반응성 부위 및 전체 길이 형태 CIB1 로 이루어지는 CRY2PHR 도메인의 조합은 Neurog2 mRNA 수준의 최고 상향조절을 초래하였다 (YFP 샘플의 ~22 배 초과 및 미자극 동시-트랜스펙션 샘플의 ~7 배 초과). 전체 길이 크립토크롬-2 (CRY2) 와 전체 길이 CIB1 의 조합은 낮은 절대 활성화 수준 (YFP 의 ~4.6 배 초과) 뿐 아니라 낮은 기준선 활성화 (미자극 동시-트랜스펙션 샘플에 대한 YFP 의 ~1.6 배 초과) 를 초래하였다. 이들 크립토크롬 단백질 페어링은 필요한 유도의 절대 수준 및 LITE 시스템의 기준선 "누수도" 를 최소화하기 위한 필요성에 따라 특정 용도를 위해 선택될 수 있다.
가역성 및 활성화의 속도는 LITE 시스템에 대한 중요한 설계 매개변수이다. LITE 시스템의 속도를 분석하기 위해, Neurog2 TALE-CRY2PHR 및 CIB1-VP64 형태 시스템으로 이루어지는 구축물을 시험하여 이의 활성화 및 불활성화 속도를 측정하였다. 추출하기 전 짧게는 0.5 시간 내지 길게는 24 시간 동안 샘플을 자극하였다. Neurog2 발현의 상향조절은 최단 시점, 0.5 시간에서 관찰되었다 (YFP 샘플에 대해 ~5 배). Neurog2 발현은 자극의 12 시간에서 최대치였다 (YFP 샘플에 대해 ~19 배). 불활성화 속도를 6 시간 동안 동시-트랜스펙션 샘플을 자극하여 분석하여, 이때 자극을 중단하고, 샘플을 0 내지 12 시간 동안 배양물 중 유지시켜 mRNA 가 분해되게 하였다. Neurog2 mRNA 수준은 자극 완료 후 0.5 시간에서 최대치였고 (YFP 샘플에 대해 ~16 배), 그 후 상기 수준은 ~3 시간 반감기로 저하된 후 12 시간까지 기준선 수준 근처로 복구되었다.
제 2 의 원형적 예는 인간 유전자 KLF4 의 전사를 활성화시키도록 설계된 LITE 이다. 인간 KLF4 의 업스트림 프로모터 부위에 위치한 서열 TTCTTACTTATAAC 를 표적으로서 선택하고, 상기 서열에 매치되도록 TALE 을 설계하고 합성하였다. TALE 서열을 핵 국부화 신호 (아미노산: SPKKKRKVEAS) 를 통해 CRY2PHR 에 대한 서열에 연결시켰다. 상기 기재된 동일한 CIB1-VP64 활성제 단백질을 또한 LITE 시스템의 이러한 시현에 사용하였다. HEK293FT 세포주로부터의 인간 배아 신장 세포를 2 개 벡터로 동시-트랜스펙션하였다. 벡터 발현이 가능하도록 인큐베이션한 후, 488 nm LED 의 어레이로부터의 주기적 펄스 청색광에 의해 샘플을 자극하였다. 미자극 동시-트랜스펙션한 샘플 및 형광 리포터 YFP 로만 트랜스펙션한 샘플을 대조군으로서 사용하였다. 각 실험의 종료시, mRNA 를 qPCR 을 통해 분석한 샘플로부터 정제하였다.
LITE 시스템의 광-강도 반응을, 샘플을 증가한 광 동력 (0-9 mW/cm2) 으로 자극하여 시험하였다. KLF4 mRNA 수준의 상향조절이 0.2 mW/cm2 의 낮은 자극에 대해 관찰되었다. KLF4 상향조절은 5 mW/cm2 에서 포화되었다 (YFP 샘플에 대해 2.3 배). 세포 생존도 시험을 또한 9 mW/cm2 이하의 동력에 대해 수행하였고, >98% 세포 생존도를 나타내었다. 유사하게, 자극 사용률 가변화에 대한 KLF4 LITE 반응을 시험하였다 (1.6-100%). 상이한 사용률 간에 KLF4 활성화에 있어서의 차이가 관찰되지 않았는데, 이는 15 초 마다 0.25 초의 낮은 자극 패러다임이 최대 활성화를 초래한다는 것을 나타낸다.
본 발명은 에너지원 예컨대 전자기 조사, 음파 에너지 또는 열 에너지를 고려한다. 유리하게는, 전자기 조사는 가시광의 성분이다. 바람직한 구현예에서, 광은 약 450 내지 약 495 nm 파장의 청색광이다. 특히 바람직한 구현예에서, 파장은 약 488 nm 이다. 또 다른 바람직한 구현예에서, 광 자극은 펄스를 통한 것이다. 광 동력은 약 0-9 mW/cm2 범위일 수 있다. 바람직한 구현예에서, 15 초 마다 0.25 초의 낮은 자극 패러다임은 최대 활성화를 초래한다.
본 발명은 특히, 세포에서의 관신 게놈 유전자좌의 발현을 변형시키거나 게놈 또는 비유전적 유전자좌를 교란하는 유도 방법에 관한 것이며, 이때 상기 게놈 또는 비유전적 유전자좌는 데옥시리보핵산 (DNA) 결합 폴리펩티드를 포함하는 비-자연 발생적 또는 조작된 조성물과 접촉될 수 있다.
본 발명의 세포는 원핵세포 또는 진핵세포, 유리하게는 동물 세포, 보다 유리하게는 포유동물 세포일 수 있다.
상기 폴리펩티드는 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체 (화학적 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도됨) 를 포함하는 DNA 결합 도메인을 포함할 수 있다. 화학적 또는 에너지 민감성 단백질 또는 이의 단편은 화학적 원천의 결합에 의해 유도시 형태적 변화를 거쳐 이를 상호작용 파트너에 결합시킬 수 있다. 폴리펩티드는 또한 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 변이체 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인을 포함할 수 있으며, 이때 화학적 또는 에너지 민감성 단백질 또는 이의 단편은 화학적 원천에 의한 유도시 상호작용 파트너에 결합할 수 있다. 상기 방법은 또한 화학적 원천을 적용하고 게놈 유전자좌의 발현이 변형되는 것을 측정하는 것을 포함할 수 있다.
여러 상이한 설계의 이러한 화학적 유도 시스템이 존재한다: 1. 아브시스산 (ABA) 에 의해 유도가능한 ABI-PYL 기반 시스템 (예를 들어, http://stke.sciencemag.org/cgi/content/abstract/sigtrans;4/164/rs2 참고), 2. 라파마이신 (또는 라파마이신 기반의 관련 화학 물질) 에 의해 유도가능한 FKBP-FRB 기반 시스템 (예를 들어, http://www.nature.com/nmeth/joumal/v2/n6/full/nmeth763.html 참고), 3. 지베렐린 (GA) 에 의해 유도가능한 GID1-GAI 기반 시스템 (예를 들어, http://www.nature.com/nchembio/joumal/v8/n5/full/nchembio.922.htm 참고).
본 발명에 의해 고려되는 또 다른 시스템은 하위-세포 국부화에 있어서의 변화를 기반으로 한 화학적 유도 시스템이다. 출원인은 또한, 폴리펩티드가 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체 (화학적 또는 에너지 민감성 단백질에 대한 추가의 링커인 적어도 하나 이상의 효과기 도메인에 연결된 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도됨) 를 포함하는 DNA 결합 도메인을 포함하는 시스템을 개발하였다. 상기 단백질은 화학적 또는 에너지 민감성 단백질에 대한 에너지 이동 또는 화학 물질 결합시 전체 폴리펩티드의 하위-세포 국부화 (즉, 세포질로부터의 전체 폴리펩티드의 세포 핵 내로의 이동) 에 있어서의 변화를 초래할 것이다. 하나의 하위-세포 부분 또는 세포소기관 (그의 활성이 효과기 도메인에 대한 기질의 결핍으로 인해 격리됨) 으로부터의 전체 폴리펩티드의 또 다른 곳 (기질이 존재함) 으로의 이동은 전체 폴리펩티드가 이의 바람직한 기질 (즉 포유동물 핵 내의 게놈 DNA) 과 접촉될 수 있게 하며 표적 유전자 발현의 활성화 또는 억제를 초래한다.
이러한 유형의 시스템은 또한 효과기 도메인이 뉴클레아제인 경우 세포 내 관심 게놈 유전자좌의 절단을 유도하는데 사용될 수 있다.
이러한 화학적 유도 시스템에 대한 설계는 4-히드록시타목시펜 (4OHT) 에 의해 유도가능한 에스트로겐 수용체 (ER) 기반 시스템이다 (예를 들어, http://www.pnas.Org/content/104/3/1027.abstract 참고). ERT2 로 지칭되는 에스트로겐 수용체의 돌연변이화된 리간드-결합 도메인은 4-히드록시타목시펜 결합시 세포 핵 내로 전좌된다. 2 개의 탠덤 ERT2 도메인을 유연성 펩티드 링커와 함께 연결시킨 후, 포유동물 게놈 내의 특정 서열을 표적화하는 TALE 단백질에 융합시키고 하나 이상의 효과기 도메인에 연결시켰다. 상기 폴리펩티드는 효과기 도메인에 연결된 TALE 단백질이 불활성이 되게 하는 4OHT 의 부재 하에 세포의 세포질 내에 존재할 것이다. 4OHT 의 존재 하에, 탠덤 ERT2 도메인에 대한 4OHT 의 결합은 전체 펩티드가 세포 핵 내로 이동하도록 유도하여, 효과기 도메인에 연결된 TALE 단백질이 활성이 되게 한다.
4-히드록시타목시펜 (4OHT) 에 의해 유도가능한 에스트로겐 수용체 (ER) 기반 시스템의 또 다른 구현예에서, 본 발명은 핵 이출 신호 (NES) 를 포함할 수 있다. 유리하게는, NES 는 LDLASLIL 의 서열을 가질 수 있다. 본 발명의 추가 구현예에서 임의의 핵 수용체, 갑상선 호르몬 수용체, 레티노산 수용체, 에스트로겐 수용체, 에스트로겐-관련 수용체, 글루코코르티코이드 수용체, 프로게스테론 수용체, 안드로겐 수용체의 임의의 자연 발생적 또는 조작된 유도체는 ER 기반 유도 시스템과 유사한 유도 시스템에서 사용될 수 있다.
또 다른 유도 시스템은 에너지, 열 또는 전파에 의해 유도가능한 일시적 수용체 포텐셜 (TRP) 이온 채널을 사용하는 설계를 기반으로 한다 (예를 들어, http://www.sciencemag.org/content/336/6081/604 참고). 이들 TRP 패밀리 단백질은 광 및 열을 포함하는 상이한 자극에 반응한다. 상기 단백질이 광 또는 열에 의해 활성화되는 경우, 이온 채널이 개방되며 칼슘과 같은 이온이 혈장 멤브레인 내로 유입되게 된다. 이러한 이온 유입물은 TALE 단백질 및 하나 이상의 효과기 도메인을 포함하는 폴리펩티드에 연결된 세포내 이온 상호작용 파트너에 결합할 것이며, 결합은 폴리펩티드의 하위세포 국부화의 변화를 유도하여, 전체 폴리펩티드가 세포 핵에 유입되게 한다. 핵 내로 들어오고나면, 효과기 도메인에 연결된 TALE 단백질은 활성이 되며 세포 내 표적 유전자 발현을 조정한다.
이러한 유형의 시스템은 또한, 효과기 도메인이 뉴클레아제인 경우 세포 내 관심 게놈 유전자좌의 절단을 유도하는데 사용될 수 있다. 레이저 또는 기타 형태의 에너지원으로 광이 생성될 수 있다. 전파 형태로 전달된 에너지원으로부터의 에너지 흡수 후 열을 방출하는 나노-입자로부터 또는 에너지원으로부터의 온도 상승 결과로 열이 생성될 수 있다.
광 활성화가 유리한 구현예일 수 있으나, 때때로 이는 특히 광이 피부 또는 기타 기관을 관통할 수 없는 생체내 적용에 대해 불리하다. 이러한 경우, 에너지 활성화의 다른 방법, 특히, 유사한 효과를 갖는 전계 에너지 및/또는 초음파가 고려된다. 필요한 경우, 또 다른 에너지원에 의한 최대 효과를 위해 LITE 시스템의 단백질 페어링이 변경되고/되거나 변형될 수 있다.
전계 에너지는 생체내 조건 하에 약 1 볼트/cm 내지 약 10 k볼트/cm 의 하나 이상의 전기 펄스를 사용하여, 바람직하게는 실질적으로 당업계에서 기재된 바와 같이 투여된다. 펄스 대신 또는 이에 추가로, 전계는 연속 방식으로 전달될 수 있다. 전기 펄스는 1 μs 내지 500 밀리초, 바람직하게는 1 μs 내지 100 밀리초 동안 적용될 수 있다. 전계는 약 5 분 동안 펄스 방식으로 또는 연속적으로 적용될 수 있다.
본원에서 사용하는 바와 같이, '전계 에너지' 는 세포가 노출되는 전기 에너지이다. 바람직하게는 전계는 생체내 조건 하에 약 1 볼트/cm 내지 약 10 k볼트/cm 또는 그 이상의 강도를 갖는다 (WO97/49450 참고).
본원에서 사용하는 바와 같이, 용어 "전계" 는 가변적 커패시턴스 및 전압에서의 하나 이상의 폴스를 포함하며 지수형 및/또는 구형파 및/또는 변조파 및/또는 변조 구형파 형태를 포함한다. 전계 및 전기에 대한 지시대상은 세포 환경에서의 전기 전위 차이의 존재에 대한 지시대상을 포함하는 것으로 의도되어야 한다. 이러한 환경은 당업계에서 알려져 있는 바와 같이 정전기, 교류 (AC), 직류 (DC) 등에 의해 설정될 수 있다. 전계는 균일하고, 비-균일하거나, 시간 의존적 방식으로 강도 및/또는 방향에 있어서 가변적일 수 있다.
전계의 단일 또는 다수 적용 뿐 아니라 초음파의 단일 도는 다수 적용이 또한 임의 순서 및 임의 조합으로 가능하다. 초음파 및/또는 전계는 단일 또는 다수 연속 적용으로서 또는 펄스로서 (박동성 전달) 전달될 수 있다.
전기천공법은 외래 물질을 살아있는 세포 내로 도입하기 위해 시험관내 및 생체내 절차 모두에서 사용되어왔다. 시험관내 적용으로, 살아 있는 세포의 샘플을 먼저 관심 작용제와 혼합하고 평행 플레이트와 같은 전극 사이에 둔다. 이후, 전극이 전계를 세포/이식물 혼합물에 적용한다. 시험관내 전기천공법을 수행하는 시스템의 예는 Electro Cell Manipulator ECM600 제품 및 Electro Square Porator T820 (둘 모두 BTX Division of Genetronics, Inc 사제 (미국 특허 번호 No 5,869,326 참고)) 을 포함한다.
공지된 전기천공법 기술 (시험관내 및 생체내 모두) 은 짧은 고전압 펄스를 처리 부위 근처에 위치한 전극에 적용함으로써 작용한다. 전극 사이에서 생성된 전계는 세포 멤브레인은 일시적으로 다공성이 되게 하여, 관심 작용제가 세포에 유입되게 한다. 공지된 전기천공법 적용에서, 이러한 전계는 1000 V/cm 에 따라, 약 100 .mu.s 지속기간의 단일 구형파 펄스를 포함한다. 이러한 펄스는 예를 들어 Electro Square Porator T820 의 공지된 적용으로 생성될 수 있다.
바람직하게는, 전계는 시험관내 조건 하에 약 1 V/cm 내지 약 10 kV/cm 의 강도를 갖는다. 따라서, 전계는 1 V/cm, 2 V/cm, 3 V/cm, 4 V/cm, 5 V/cm, 6 V/cm, 7 V/cm, 8 V/cm, 9 V/cm, 10 V/cm, 20 V/cm, 50 V/cm, 100 V/cm, 200 V/cm, 300 V/cm, 400 V/cm, 500 V/cm, 600 V/cm, 700 V/cm, 800 V/cm, 900 V/cm, 1 kV/cm, 2 kV/cm, 5 kV/cm, 10 kV/cm, 20 kV/cm, 50 kV/cm 또는 그 이상의 강도를 가질 수 있다. 보다 바람직하게는 시험관내 조건 하에 약 0.5 kV/cm 내지 약 4.0 kV/cm 이다. 바람직하게는 전계는 생체내 조건 하에 약 1 V/cm 내지 약 10 kV/cm 의 r아도를 갖는다. 그러나, 전계 강도는 표적 부위에 전달된 펄스의 수가 증가될 때 감소될 수 있다. 따라서, 낮은 전계 강도에서의 전계의 박동성 전달이 예상된다.
바람직하게는 전계의 적용은 다수 펄스 예컨대 동일 강도 및 커패시턴스의 이중 펄스 또는 가변적 강도 및/또는 커패시턴스의 순차적 펄스의 형태로의 적용이다. 본원에서 사용하는 바와 같이, 용어 "펄스" 는 가변적 커패시턴스 및 전압에서의 하나 이상의 전기 펄스를 포함하며 지수형 및/또는 구형파 및/또는 변조파/구형파 형태를 포함한다.
바람직하게는 전기 펄스는 지수형파 형태, 구형파 형태, 변조파 형태 및 변조 구형파 형태에서 선택되는 파형으로서 전달된다.
바람직한 구현예는 낮은 전압에서 직류를 이용한다. 따라서, 출원인은 1V/cm 내지 20V/cm 의 전계 강도에서, 100 밀리초 이상, 바람직하게는 15 분 이상 동안 세포, 조직 또는 조직 덩어리에 적용되는 전계의 사용을 개시하고 있다.
초음파는 유리하게는 약 0.05 W/cm2 내지 약 100 W/cm2. 의 동력 수준에서 투여된다. 진단용 또는 치료용 초음파가 사용될 수 있거나, 이의 조합물이 사용될 수 있다.
본원에서 사용하는 바와 같이, 용어 "초음파" 는 인간 청력의 범위를 넘는 높은 주파수의 기계적 진동으로 이루어지는 에너지 형태를 의미한다. 초음파 스펙트럼의 낮은 주파수 한계는 일반적으로 약 20 kHz 에서 선택될 수 있다. 초음파의 대부분의 진단용 적용은 1 및 15 MHz' 범위의 주파수를 이용한다 (Ultrasonics in Clinical Diagnosis, P. N. T. Wells, ed., 2nd. Edition, Publ. Churchill Livingstone [Edinburgh, London & NY, 1977] 로부터).
초음파는 진단적 및 치료적 적용 모두에서 사용되어왔다. 진단 도구로서 사용되는 경우 ("진단용 초음파"), 750 mW/cm2 이하의 에너지 밀도가 사용되어왔으나, 초음파는 통상 약 100 mW/cm2 이하의 에너지 밀도 범위 (FDA 권고사항) 로 사용된다. 물리치료에서, 초음파는 통상 약 3 내지 4 W/cm2 까지의 범위 (WHO 권고사항) 로 에너지원으로서 사용된다. 다른 치료적 적용에서, 높은 강도의 초음파, 예를 들어 100 W/cm 에서 1 kW/cm2 이하 (또는 더 높음) 에서의 HIFU 를 단기간 동안 이용할 수 있다. 본 명세서에서 사용하는 바와 같은 용어 "초음파" 는 진단적, 치료적 및 초점성 초음파를 포함하는 것으로 의도된다.
초점성 초음파 (FUS) 는 열 에너지가 침습성 프로브 없이 전달되게 한다 (Morocz et al 1998 Journal of Magnetic Resonance Imaging Vol.8, No. 1, pp.136-142 참고). 또 다른 형태의 초점성 초음파는 고강도 초점성 초음파 (HIFU) 이며 이는 [Moussatov et al in Ultrasonics (1998) Vol.36, No.8, pp.893-900] 및 [TranHuuHue et al in Acustica (1997) Vol.83, No.6, pp.1103-1106] 에 의해 검토된다.
바람직하게는, 진단용 초음파 및 치료용 초음파의 조합이 이용된다. 이러한 조합은 제한되는 것으로 의도되지 않으나, 당업자는 임의의 다양한 조합의 초음파가 사용될 수 있다는 것을 이해할 것이다. 추가적으로, 에너지 밀도, 초음파 주파수 및 노출 기간은 가변적일 수 있다.
바람직하게는 초음파 에너지원에 대한 노출은 약 0.05 내지 약 100 Wcm-2 의 동력 밀도에서 이루어진다. 보다 더 바람직하게는, 초음파 에너지원에 대한 노출은 약 1 내지 약 15 Wcm-2 의 동력 밀도에서 이루어진다.
바람직하게는 초음파 에너지원에 대한 노출은 약 0.015 내지 약 10.0 MHz 의 주파수에서 이루어진다. 보다 바람직하게는 초음파 에너지원에 대한 노출은 약 0.02 내지 약 5.0 MHz 또는 약 6.0 MHz 의 주파수에서 이루어진다. 가장 바람직하게는, 초음파는 3 MHz 의 주파수에서 적용된다.
바람직하게는 노출은 약 10 밀리초 내지 약 60 분의 기간 동안이다. 바람직하게는 노출은 약 1 초 내지 약 5 분의 기간 동안이다. 보다 바람직하게는, 초음파는 약 2 분 동안 적용된다. 그러나 와해될 특정 표적 세포에 따라서, 노출은 장기간, 예를 들어 15 분 동안일 수 있다.
유리하게는, 표적 세포는 약 0.05 Wcm-2 내지 약 10 Wcm-2 의 음향 동력 밀도에서 약 0.015 내지 약 10 MHz 범위의 주파수로 초음파 에너지원에 노출된다 (WO 98/52609 참고). 그러나 예를 들어, 100 Wcm-2 초과의 음향 동력 에너지에서, 감소된 기간, 예를 들어 밀리초 범위 또는 그 이하 동안 1000 Wcm-2 의 음향 동력 에너지에서 초음파 에너지원에 대한 노출과 같은 대안물이 또한 가능하다.
바람직하게는 초음파의 적용은 다수 펄스의 형태이며; 따라서 연속파 및 펄스형파 (초음파의 박동성 전달) 모두 임의의 조합으로 이용될 수 있다. 예를 들어, 연속파 초음파가 적용된 후 펄스형파 초음파가 적용될 수 있으며, 그 반대도 가능하다. 이는 임의 횟수로, 임의 순서 및 조합으로 반복될 수 있다. 펄스형파 초음파는 연속파 초음파의 배경에 반하여 적용될 수 있으며, 임의 수의 펄스가 임의 수의 군에서 사용될 수 있다.
바람직하게는, 초음파는 펄스형파 초음파를 포함할 수 있다. 매우 바람직한 구현예에서, 초음파는 0.7 Wcm-2 또는 1.25 Wcm-2 의 동력 밀도에서 연속파로서 적용된다.
초음파의 사용은 광과 같이, 표적에 정확히 초점화될 수 있으므로 유리하다. 또한, 초음파는 광과 달리 조직 내로 보다 깊숙히 초점화될 수 있으므로 유리하다. 따라서, 이는 전체-조직 침투 (예컨대 간엽에 제한되지는 않음) 또는 전체 기관 (예컨대 전체 간 또는 전체 근육, 예컨대 심장에 제한되지는 않음) 치료요법에 보다 양호하게 적합하다. 또 다른 중요한 이점은 초음파가 광범위한 진단적 및 치료적 적용에서 사용되는 비-침습성 자극이라는 것이다. 예를 들어, 초음파는 의료 영상화 기술, 및 추가적으로, 정형외과 치료요법에 있어서 널리 공지된다. 추가로, 척추동물 대상에 대한 초음파 적용에 적합한 기구가 광범위하게 이용가능하며 그의 용도가 당업계에 널리 공지되어있다.
LITE 의 신속한 전사 반응 및 내인성 표적화는 전사 역학의 연구에 이상적인 시스템에 기여한다. 예를 들어, LITE 는 표적 유전자의 유도된 발현시 mRNA 스플라이스 변이체 생성의 역학 연구에 사용될 수 있다. 전사 주기의 다른 한편, mRNA 분해 연구는 종종, 과잉 유전자에서의 발현 수준 변화를 일으키는 강력한 세포외 자극에 대한 반응으로 수행된다. LITE 는 내인성 표적의 전사를 가역적으로 유도하는데 이용될 수 있으며, 그 후 자극이 중단될 수 있고 고유 표적의 분해 속도가 추적될 수 있다.
LITE 의 시간적 정확도는 실험적 개입과 맞물려 시간 유전적 조절에 대한 동력을 제공할 수 있다. 예를 들어, 장기간 강화작용 (LTP) 에서 예상된 관여를 갖는 표적은 기관형 또는 해리성 뉴런 배양물에 있어서, 그러나 LTP 를 유도하는 자극 동안에만 조정되어, 세포의 정상 발생의 간섭이 방지될 수 있다. 유사하게, 질환 표현형을 나타내는 세포 모델에서, 특정 치료요법의 유효성에 관여되는 것으로 예상되는 표적은 처리 동안에만 조정될 수 있다. 반대로, 유전적 표적은 병리적 자극 동안에만 조정될 수 있다. 외부 실험 자극에 대한 유전적 신호의 시기가 관련되는 임의 수의 실험은 LITE 조정의 효용으로부터 잠재적으로 유익할 수 있다.
생체내 맥락은 유전자 발현 제어를 위한 LITE 의 용도에 대해 동등하게 많은 기회를 제공한다. 상기 언급한 바와 같이, 광유도성은 이전에 성취불가능한 공간적 정확도에 대한 가능성을 제공한다. 광전극 기술의 발전을 이용하여, 자극 광 섬유 리드 (lead) 는 정확한 뇌 부위 내에 위치할 수 있다. 자극 부위 크기는 광 강도에 의해 조정될 수 있다. 이는 미국 가출원 번호 61/671,615 의 바이러스 벡터 또는 분자 슬레드 (sled) 를 통한 LITE 의 전달과 함께 수행될 수 있거나, 트랜스제닉 LITE 동물이 이용가능해진 경우, 정확한 뇌 부위에서의 유전자 발현을 여전히 조정가능하게 하면서 바이러스의 사용을 없앨 수 있다. LITE 는 투명한 유기체, 예컨대 고정 제브라피시에서 사용되어 매우 정확한 레이저 유도 국부 유전자 발현 변화를 가능하게 할 수 있다.
본 발명은 또한 CRISPPv/Cas 시스템을 사용하는 멀티플렉스 게놈 조작을 고려한다. 평상적인 유전적 변이체 및 요소의 기능적 해명은 정확한 게놈 편집 기술을 필요로 한다. 유형 II 원핵세포 CRISPR (군집화된 규칙적 공간화 단길이 군집화 규칙적 공간화 단길이 회귀성 반복물) 적응 면역계가 RNA-안내 부위-특이적 DNA 절ㄹ단을 촉진시키는 것으로 나타난 바 있다. 출원인은 2 개 상이한 유형 II CRISPR 시스템을 조작하였고 Cas9 뉴클레아제가 단길이 RNA 에 의해 유도되어 인간 및 마우스 세포에서 내인성 게놈 유전자좌에서의 정확한 절단을 유도할 수 있다는 것을 입증한다. Cas9 는 또한 최소 돌연변이유발 활성을 갖는 상동성-유도 복구가 촉진되도록 절단 효소로 전환될 수 있다. 마지막으로, 다수의 안내 서열은 단일 CRISPR 어레이 내로 인코딩되어 포유동물 게놈 내 여러 부위의 동시 편집이 가능하게 하는데, 이는 CRISPR 기술의 용이한 프로그램가능성 및 광범위한 적용가능성을 입증한다.
일반적으로, "CRISPR 시스템" 은 전사체 및 CRISPR-연관 ("Cas") 유전자의 활성을 유도하거나 이의 발현에 관여된 다른 요소, 예컨대 Cas 유전자 인코딩 서열, tracr (트랜스-활성화 CRISPR) 서열 (예를 들어 tracrRNA 또는 활성 부분 tracrRNA), tracr-mate 서열 ("직접 반복물" 및 내인성 CRISPR 시스템의 맥락에 있어서 tracrRNA-처리 부분 직접 반복물 포함), 안내 서열 (또한 내인성 CRISPR 시스템의 맥락에 있어서 "스페이서" 로서 지칭함), 또는 CRISPR 유전자좌로부터의 기타 서열 및 전사체를 집합적으로 의미한다. 일부 구현예에서, CRISPR 시스템의 하나 이상의 요소는 유형 I, 유형 II 또는 유형 III CRISPR 시스템에서 유래한다. 일부 구현예에서, CRISPR 시스템의 하나 이상의 요소는 내인성 CRISPR 시스템을 포함하는 특정 유기체, 예컨대 스트렙토코쿠스 피오게네스에서 유래한다. 일반적으로, CRISPR 시스템은 표적 서열 부위에서의 CRISPR 복합체 형성을 촉진하는 요소를 특징으로 한다 (또한 내인성 CRISPR 시스템의 맥락에서 프로토스페이서로서 지칭함). CRISPR 복합체 형성의 맥락에서, "표적 서열" 은 안내 서열이 상보성을 갖도록 설계된 서열을 의미하며, 이때 표적 서열과 안내 서열 사이의 하이브리드화가 CRISPR 복합체의 형성을 촉진한다. 표적 서열은 임의의 폴리뉴클레오티드, 예컨대 DNA 또는 RNA 폴리뉴클레오티드를 포함할 수 있다. 일부 구현예에서, 표적 서열은 세포의 핵 또는 세포질 내에 위치한다.
통상, 내인성 CRISPR 시스템의 맥락에서, CRISPR 복합체 (하나 이상의 Cas 단백질과 복합화되며 표적 서열에 하이브리드화된 안내 서열을 포함함) 의 형성은 표적 서열 내 또는 표적 서열 근처 (예를 들어 표적 서열로부터 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 또는 그 이상의 염기쌍) 하나 또는 둘 모두의 가닥의 절단을 초래한다. 이론에 속박되지 않고, 모든 또는 일부의 tracr 서열은 또한 예컨대 안내 서열에 작동가능하게 연결되는 tracr mate 서열의 전부 또는 일부에 대한 하이브리드화에 의해 CRISPR 복합체의 일부를 형성할 수 있다. 일부 구현예에서, CRISPR 시스템의 하나 이상의 요소의 발현을 유도하는 하나 이상의 벡터가 숙주 세포 내로 도입되어, CRISPR 시스템의 요소의 발현이 하나 이상의 표적 부위에서 CRISPR 복합체 형성을 유도한다. 예를 들어, Cas 효소, tracr-mate 서열에 연결된 안내 서열, 및 tracr 서열은 각각 개별 벡터 상 개별 조절 요소에 작동가능하게 연결될 수 있다. 대안적으로, 동일하거나 상이한 조절 요소로부터 발현된 둘 이상의 요소는 단일 벡터 내에서 조합될 수 있으며, CRISPR 시스템의 임의 성분을 제공하는 하나 이상의 추가적 벡터는 제 1 벡터에 포함되지 않는다. 단일 벡터 내에서 조합되는 CRISPR 시스템 요소는 임의의 적합한 배향으로 배열될 수 있으며, 예를 들어 하나의 요소가 제 2 의 요소 (의 "업스트림") 에 대해 5' 또는 제 2 의 요소 (의 "다운스트림") 에 대해 3' 로 배열될 수 있다. 하나의 요소의 코딩 서열은 제 2 의 요소의 코딩 서열의 동일 또는 반대 가닥 상에 위치할 수 있고, 동일 또는 반대 방향으로 배향될 수 있다. 일부 구현예에서, 단일 프로모터는 안내 서열, tracr mate 서열 (임의로는 안내 서열에 작동가능하게 연결됨) 및 하나 이상의 인트론 서열 (예를 들어 각각 상이한 인트론, 하나 이상의 인트론 중 둘 이상 또는 모두 단일 인트론) 내에 포매된 tracr 서열 중 하나 이상 및 CRISPR 효소를 인코딩하는 전사체의 발현을 유도한다. 일부 구현예에서, CRISPR 효소, 안내 서열, tracr mate 서열 및 tracr 서열은 동일 프로모터에 작동가능하게 연결되며 이로부터 발현된다.
일부 구현예에서, 벡터는 하나 이상의 삽입 부위, 예컨대 제한 엔도뉴클레아제 인지 서열 (또한 "클로닝 부위" 로서 지칭함) 을 포함한다. 일부 구현예에서, 하나 이상의 삽입 부위 (예를 들어 약 또는 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 이상의 삽입 부위) 는 하나 이상의 벡터의 하나 이상의 서열 요소의 업스트림 및/또는 다운스트림에 위치한다. 일부 구현예에서, 벡터는 tracr mate 서열의 업스트림, 및 임의로는 tracr mate 서열에 작동가능하게 연결된 조절 요소의 다운스트림 삽입 부위를 포함하여, 안내 서열의 삽입 부위 내로의 삽입 후 발현시 안내 서열이 진핵세포에서의 표적 서열에 대한 CRISPR 복합체의 서열-특이적 결합을 유도한다. 일부 구현예에서, 벡터는 둘 이상의 삽입 부위를 포함하며, 각각의 삽입 부위는 2 개의 tracr mate 서열 사이에 위치하여 각 부위에서의 안내 서열 삽입을 가능하게 한다. 이러한 배열에서, 둘 이상의 안내 서열은 단일 안내 서열의 둘 이상의 카피, 둘 이상의 상이한 안내 서열, 또는 이들의 조합을 포함할 수 있다. 다수의 상이한 안내 서열을 사용하는 경우, 단일 발현 구축물을 사용하여 CRISPR 활성을 세포 내 다수의 상이한, 상응하는 표적 서열에 대해 표적화할 수 있다. 예를 들어, 단일 벡터는 약 또는 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 이상의 안내 서열을 포함할 수 있다. 일부 구현예에서, 약 또는 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 이상의 이러한 안내-서열-함유 벡터가 제공될 수 있으며, 임의로는 세포에 전달된다.
일부 구현예에서, 벡터는 Cas 단백질과 같은 CRISPR 효소를 인코딩하는 효소-코딩 서열에 작동가능하게 연결된 조절 요소를 포함한다. Cas 단백질의 비제한적 예는 Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9 (또한 Csn1 및 Csx12 으로 공지됨), Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, 이의 동족체, 또는 이의 변형된 형태를 포함한다. 일부 구현예에서, 미변형된 CRISPR 효소는 DNA 절단 활성을 갖는다 (예컨대 Cas9). 일부 구현예에서, CRISPR 효소는 표적 서열의 위치에서, 예컨대 표적 서열 내 및/또는 표적 서열의 보체 내 하나 또는 둘 모두의 가닥의 절단을 유도한다. 일부 구현예에서, CRISPR 효소는 표적 서열의 첫 번째 또는 마지막 뉴클레오티드로부터 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 50, 100, 200, 500 이상의 염기쌍 내 하나 또는 둘 모두의 가닥의 절단을 유도한다. 일부 구현예에서, 벡터는 상응하는 야생형 효소에 대하여 돌연변이화되어 상기 돌연변이화된 CRISPR 효소가 표적 서열을 함유하는 표적 폴리뉴클레오티드의 하나 또는 둘 모두의 가닥을 절단하는 능력이 소실되는, CRISPR 효소를 인코딩한다. 예를 들어, S. 피오게네스로부터의 Cas9 의 RuvC I 효소 도메인에서의 아스파르테이트-알라닌 치환 (D10A) 은 두 가닥 모두를 절단하는 뉴클레아제로부터의 Cas9 를 닉카아제 (nickase) (단일 가닥을 절단함) 로 전환시킨다. Cas9 가 닉카아제가 되도록 하는 돌연변이의 다른 예는 비제한적으로, H840A, N854A 및 N863A 를 포함한다. 추가예로서, Cas9 의 둘 이상의 촉매 도메인 (RuvC I, RuvC II 및 RuvC III) 은, 모든 DNA 절단 활성이 실질적으로 결핍되는 돌연변이화된 Cas9 를 생성시키도록 돌연변이화될 수 있다. 일부 구현예에서, D10A 돌연변이는 H840A, N854A 또는 N863A 돌연변이 중 하나 이상과 조합되어 모든 DNA 절단 활성이 실질적으로 결핍되는 Cas9 효소를 생성시킨다. 일부 구현예에서, CRISPR 효소는 돌연변이화된 효소의 DNA 절단 활성이 이의 비-돌연변이화 형태에 대하여 약 25%, 10%, 5%, 1%, 0.1%, 0.01% 이하인 경우 모든 DNA 절단 활성이 실질적으로 결핍되는 것으로 고려된다.
일부 구현예에서, CRISPR 효소를 인코딩하는 효소 코딩 서열은 특정 세포, 예컨대 진핵세포에서의 발현을 위해 최적화된 코돈이다. 진핵세포는 특정 유기체, 예컨대 인간, 마우스, 랫트, 토끼, 개 또는 비-인간 영장류를 비제한적으로 포함하는 포유동물의 것들 또는 이로부터 유래한 것들일 수 있다. 일반적으로 코돈 최적화는, 선천적 서열의 하나 이상의 코돈 (예를 들어 약 또는 약 1, 2, 3, 4, 5, 10, 15, 20, 25, 50 이상의 코돈) 을 선천적 아미노산 서열을 유지하면서 숙주 세포의 유전자에서 보다 자주 또는 가장 자주 사용되는 코돈으로 대체함으로써, 관심 숙주 세포에서의 증강된 발현을 위해 핵산 서열을 변형시키는 프로세스를 의미한다. 다양한 종류는 특정 아미노산의 특정 코돈에 대한 특정한 편향을 나타낸다. 코돈 편향 (유기체 사이의 코돈 사용빈도에 있어서의 차이) 은 종종 메신저 RNA (mRNA) 의 번역 효율과 상호관련되는데, 이는 다른 것들 중에서도, 번역되는 코돈의 특성 및 특정 트랜스퍼 RNA (tRNA) 분자의 이용가능성에 의존적인 것으로 여겨진다. 세포 내에서의 선택된 tRNA 의 우세는 일반적으로 펩티드 합성에서 가장 자주 사용된 코돈의 반영이다. 따라서, 유전자는 코돈 최적화를 기반으로 주어진 유기체에서의 최적 유전자 발현에 맞추어질 수 있다. 코돈 사용빈도 표는 예를 들어 www.kazusa.orjp/codon/ 에서 이용가능한 "코돈 사용빈도 데이터베이스" (2002 년 7 월 9 일 방문) 에서 용이하게 이용가능하며, 이들 표는 수많은 방식으로 적합화될 수 있다. [Nakamura, Y., et al. " Codon usage tabulated from the international DNA sequence databases: status for the year 2000"Nucl. Acids Res. 28:292 (2000)] 를 참고한다. 특정 숙주 세포에서의 발현을 위한 특정 서열을 최적화하는 코돈에 대한 컴퓨터 알고리즘이 또한 이용가능하며, 예컨대 Gene Forge (Aptagen; Jacobus, PA) 가 또한 이용가능하다. 일부 구현예에서, CRISPR 효소를 인코딩하는 서열에서의 하나 이상의 코돈 (예를 들어 1, 2, 3, 4, 5, 10, 15, 20, 25, 50 이상, 또는 모든 코돈) 은 특정 아미노산에 대해 가장 자주 사용된 코돈에 상응한다.
일부 구현예에서, 벡터는 하나 이상의 핵 국부화 서열 (NLS), 예컨대 약 또는 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 이상의 NLS 를 포함하는 CRISPR 효소를 인코딩한다. 일부 구현예에서, CRISPR 효소는 아미노-말단에서 또는 근처에서 약 또는 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 이상의 NLS 를, 카르복시-말단에서 또는 근처에서 약 또는 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 이상의 NLS 를, 또는 이들의 조합을 포함한다 (예를 들어 아미노-말단에서 하나 이상의 NLS 및 카르복시-말단에서 하나 이상의 NLS). 하나 초과의 NLS 가 존재하는 경우, 각각은 서로에게 독립적으로 선택될 수 있어, 단일 NLS 가 하나 초과의 카피로 존재할 수 있고/있거나 하나 초과의 다른 NLS 와 조합으로 하나 이상의 카피로 존재할 수 있다. 일부 구현예에서, NLS 의 가장 가까운 아미노산이 N- 또는 C-말단으로부터의 폴리펩티드 사슬을 따라 약 1, 2, 3, 4, 5, 10, 15, 20, 25, 30, 40, 50 개 이상의 아미노산 내에 있는 경우, NLS 는 N- 또는 C-말단 근처에 있는 것으로 고려된다. NLS 의 비제한적 예는 하기에서 유래한 NLS 서열을 포함한다: 아미노산 서열 PKKKRKV 를 갖는 SV40 바이러스 거대 T-항원의 NLS; 뉴클레오플라스민으로부터의 NLS (예를 들어 서열 KRPAATKKAGQAKKKK 를 갖는 뉴클레오플라스민 이연 NLS); 아미노산 서열 PAAKRVKLD 또는 RQRRNELKRSP 를 갖는 c-myc NLS; 서열 NQSSNFGPMKGGNFGGRSSGPYGGGGQYFAKPRNQGGY 를 갖는 hRNPA1 M9 NLS; 임포틴-알파로부터의 IBB 도메인의 서열 RMRIZFKNKGKDTAELRRRRVEVSVELRKAKKDEQILKRRNV; 자궁근종 T 단백질의 서열 VSRKRPRP 및 PPKKARED; 인간 p53 의 서열 POPKKKPL; 마우스 c-abl IV 의 서열 SALIKKKKKMAP; 인플루엔자 바이러스 NS1 의 서열 DRLRR 및 PKQKKRK; 간염 바이러스 델타 항원의 서열 RKLKKKIKKL; 마우스 Mx1 단백질의 서열 REKKKFLKRR; 인간 폴리(ADP-리보오스) 중합효소의 서열 KRKGDEVDGVDEVAKKKSKK; 및 스테로이드 호르몬 수용체 (인간) 글루코코르티코이드의 서열 RKCLQAGMNLEARKTKK.
일반적으로, 하나 이상의 NLS 는 진핵세포의 핵에서 검출가능한 양으로 CRISPR 효소를 축적하는 것을 유도하기에 충분한 강도의 것이다. 일반적으로, 핵 국부화 활성의 강도는 CRISPR 효소에서의 NLS 의 수, 특히 사용한 NLS(들), 또는 이들 인자의 조합에서 유래할 수 있다. 핵 내 축적의 검출은 임의의 적합한 기술에 의해 수행될 수 있다. 예를 들어, 검출가능 마커는 CRISPR 효소에 융합될 수 있어, 핵의 위치를 검출하기 위한 수단과의 조합으로 (예를 들어 핵에 대한 특이적 염색 예컨대 DAPI) 세포 내 위치가 가시화될 수 있다. 세포 핵은 또한 세포로부터 단리될 수 있으며, 이의 내용물은 단백질 검출을 위한 임의의 적합한 프로세스, 예컨대 면역조직화학, 웨스턴 블롯 또는 효소 활성 검정에 의해 분석될 수 있다. 핵 내 축적은 또한 예컨대 CRISPR 효소 또는 복합체에 노출되지 않은 대조군, 또는 하나 이상의 NLS 가 결핍되는 CRISPR 효소에 노출된 대조군과 비교하여 CRISPR 복합체 형성 효과에 대한 검정 (예를 들어 DNA 절단 또는 표적 서열에서의 돌연변이에 대한 검정, 또는 CRISPR 복합체 형성 및/또는 CRISPR 효소 활성에 의해 영향받은 변경된 유전자 발현 활성에 대한 검정) 에 의해 간접적으로 검출될 수 있다.
본 발명의 또 다른 구현예에서, 본 발명은 유도 Cas9 를 포함할 수 있는 유도 CRISPR 에 관한 것이다.
CRISPR 시스템은 하기: I. CRISPR/Cas 시스템 키메라 RNA (chiRNA) 폴리뉴클레오티드 서열에 작동가능하게 연결된 제 1 조절 요소 (이때, 상기 폴리뉴클레오티드 서열은 (a) 진핵세포에서의 표적 서열에 하이브리드화할 수 있는 안내 서열, (b) tracr mate 서열 및 (c) tracr 서열을 포함할 수 있음), 및 II. 하나 이상의 핵 국부화 서열을 포함할 수 있는 CRISPR 효소를 인코딩하는 효소-코딩 서열에 작동가능하게 연결된 제 2 조절 요소를 포함할 수 있는 하나 이상의 벡터를 포함할 수 있는 벡터 시스템 내에서 인코딩될 수 있으며, 여기서 (a), (b) 및 (c) 는 5' 에서 3' 배향으로 배열되고, 성분 I 및 II 는 시스템의 동일하거나 상이한 벡터 상에 위치하고, 전사되는 경우, tracr mate 서열은 tracr 서열에 하이브리드화되고 안내 서열은 표적 서열에 대한 CRISPR 복합체의 서열-특이적 결합을 유도하고, CRISPR 복합체는 (1) 표적 서열에 대해 하이브리드화되는 안내 서열, 및 (2) tracr 서열에 대해 하이브리드화되는 tracr mate 서열로 복합화되는 CRISPR 효소를 포함할 수 있고, CRISPR 효소를 인코딩하는 효소 코딩 서열은 추가로 이종 기능적 도메인을 인코딩한다.
유리한 구현예에서, 유도 Cas9 를 렌티바이러스에서 제조할 수 있다. 예를 들어, 도 61 은 Tet Cas9 벡터 설계를 나타내고 도 62 는 293FT 세포에서의 벡터 및 EGFP 발현을 나타낸다. 특히, 유도 테트라사이클린 시스템은 유도 CRISPR 에 대해 고려된다. 벡터는 [Markusic et al, Nucleic Acids Research, 2005, Vol. 33, No. 6 e63] 에서 기재된 바와 같이 설계될 수 있다. 테트라사이클린-의존적 전사 조절 시스템은 테트라사이클린 억제제 단백질 (TetR) 및 특이적 DNA-결합 부위, 테트라사이클린 오퍼레이터 서열 (TetO) 로 이루어지는 대장균 Tn10 테트라사이클린 저항성 오퍼레이터를 기반으로 한다. 테트라사이클린의 부재 하, TetR 은 이량체화하며 TetO 에 결합한다. 테트라사이클린 또는 독시사이클린 (테트라사이클린 유도체) 은 TetR 에 결합하고 형태적 변화를 유도할 수 있어, TetO 로부터의 이의 해리를 초래한다. 유리한 구현예에서, 벡터는 CRISPR 복합체의 조절된 발현을 위해 자가조절된 rtTA 발현을 갖는 단일 Tet-On 렌티바이러스 벡터일 수 있다. 테트라사이클린 또는 독시사이클린은 유도 CRISPR 복합체 활성화를 위해 고려될 수 있다.
또 다른 구현예에서, 큐메이트 유전자-스위치 시스템은 유도 CRISPR 에 대해 고려된다. 유사한 시스템은 [Mullick et al., BMC Biotechnology 2006, 6:43 doi: 10.1186/1472-6750-6-43] 에서 기재된 바와 같다. 유도 큐메이트 시스템은 3 가지 상이한 전략을 사용하여 포유동물 세포에서의 유전자 발현을 조절하기 위한 박테리아 오페론 (cmt 및 cym) 의 조절 메커니즘을 포함한다. 억제제 구성에서, 조절은 강력한 항시성 프로모터의 다운스트림에 위치한 오퍼레이터 부위 (CuO) 에 대한 억제제 (CymR) 의 결합에 의해 매개된다. 큐메이트, 소분자의 첨가로, 억제가 완화된다. 전사촉진제 구성에서, CymR 과 VP16 의 활성화 도메인과의 융합에 의해 형성된 키메라 전사촉진제 (cTA) 단백질은 CMV 최소 프로모터의 업스트림에 위치한 여러 카피의 CuO 에 결합하는 경우 전사를 활성화시킬 수 있다. 큐메이트 첨가는 DNA 결합을 없애며, 따라서 cTA 에 의한 전사촉진을 없앤다. 본 발명은 또한 역 큐메이트 활성제 (rcTA) 를 고려하는데, 이는 큐메이트의 부재 하에서보다는 존재 하에 전사를 활성화시킨다. CymR 은 강력한 프로모터, 예컨대 CMV 로부터의 발현을 가역적으로 차단하는 억제제로서 사용될 수 있다. 큐메이트 억제제/오퍼레이터 시스템의 특정 양태는 미국 특허 번호 7745592 에서 추가로 기재된다.
다른 유도 시스템은 예컨대 비제한적으로, 중금속 [Mayo KE et al, Cell 1982, 29:99-108; Searle PF et al, Mol Cell Biol 1985, 5: 1480-1489 and Brinster RL et al, Nature (London) 1982, 296:39-42], 스테로이드 호르몬 [Hynes NE et al, Proc Natl Acad Sci USA 1981, 78:2038-2042; Klock G et al, Nature (London) 1987, 329:734-736 and Lee F et al, Nature (London) 1981, 294:228-232.], 열 충격 [Nouer L: Heat Shock Response. Boca Raton, FL: CRC; 1991] 및 개발된 기타 시약 [Mullick A, Massie B: Transcription, translation and the control of gene expression. In Encyclopedia of Cell Technology Edited by: Speir RE. Wiley; 2000: 1140-1164 and Fussenegger M, . Biotechnol Prog 2001, 17: 1-51] 에 의한 조절을 고려한다. 그러나, 이러한 유도 포유동물 프로모터로는 "오프 (off)" 상태의 "누수성" 및 유도물질 (열 충격, 중금속, 글루코코르티코이드 등) 의 다면발현성 효과와 같은 한계가 존재한다. 포유동물 세포에서 세포적 프로세스로의 간섭을 감소시키기 위한 시도로, 곤충 호르몬 (엑디손) 의 사용이 제시된 바 있다 [No D et al., Proc Natl Acad Sci USA 1996, 93:3346-3351]. 또 다른 명쾌한 시스템은 유도물질로서 라파마이신을 사용하나 [Rivera VM et al, Nat Med 1996, 2: 1028-1032], 면역억제제로서의 라파마이신의 역할은 이의 생체내 용도에 대한 주요 한계이며, 따라서 유전자 발현 제어를 위해 생물학적으로 불활성인 화합물을 발견할 필요가 있었다 [Saez E et al., Proc Natl Acad Sci USA 2000, 97: 14512-14517].
본 발명은 또한 본 발명의 폴리펩티드를 인코딩하는 핵산을 포함한다. 핵산은 프로모터, 유리하게는 인간 시냅신 I 프로모터 (hSyn) 를 포함할 수 있다. 특히 유리한 구현예에서, 핵산은 아데노 연관 바이러스 벡터 (AAV) 내로 패키징될 수 있다.
또한 본 발명에 의해 고려되는 것은 하나 초과의 아데노바이러스 혈청형으로부터의 바이러스 준입자 (subviral particle) 를 포함할 수 있는 재조합 아데노바이러스 및 재조합 벡터이다. 예를 들어, 아데노바이러스 벡터가 특정 조직 또는 세포 유형에 대해 변경된 향성 (tropism) 을 나타낼 수 있다는 것이 공지되어 있으며 (Havenga, M.J.E. et al., 2002), 따라서, 상이한 아데노바이러스 캡시드, 즉 섬유 또는 각종 아데노바이러스 혈청형으로부터의 펜톤 단백질의 혼합 및 매칭이 유리할 수 있다. 섬유 및 펜톤을 포함하는 아데노바이러스 캡시드의 변형은 미변형된 아데노바이러스와 상이한 향성을 갖는 아데노바이러스 벡터를 초래할 수 있다. 표적 세포를 감염시키는 능력에 있어서 최적화되고 변형되는 아데노바이러스 벡터는 치료적 및 예방적 용량이 유의하게 감소될 수 있게 하여, 감소된 국부적 및 전파 독성을 초래한다.
바이러스 벡터 유전자 전달 시스템은 유전자 전달 및 유전자 치료요법 적용에서 흔히 사용된다. 상이한 바이러스 벡터 시스템은 그만의 고유한 이점 및 불리한 점을 갖는다. 본 발명의 병원체-유래 리간드를 발현시키는데 사용될 수 있는 바이러스 벡터는 비제한적으로, 하기에 보다 상세히 기재된 아데노바이러스 벡터, 아데노-연관 바이러스 벡터, 알파바이러스 벡터, 단순포진 바이러스 벡터 및 레트로바이러스 벡터를 포함한다.
아데노바이러스의 추가적인 일반적 특징은 아데노바이러스의 생물학이 상세히 분석된다는 것이며; 상기 아데노바이러스는 중증 인간 병리학과 연관되지 않고; 아데노바이러스는 이의 DNA 를 숙주 세포 내로 도입하는데 있어서 매우 효과적이고; 아데노바이러스는 광범위한 세포를 감염시킬 수 있으며 넓은 숙주 범위를 갖고; 아데노바이러스는 상대적으로 쉽게 다량으로 생성될 수 있고; 아데노바이러스는 바이러스 게놈의 초기 부위 1 ("E1") 에서의 결실에 의해 결손이 되고/되거나 비-복제형이 될 수 있다.
아데노바이러스는 비-외피형 DNA 바이러스이다. 아데노바이러스의 게놈은 각 가닥의 5'-말단에 공유 결합된 55-kDa 말단 단백질을 갖는 대략 36,000 개 염기쌍 ("bp") 의 선형 이중가닥 DNA 분자이다. 아데노바이러스 DNA 는 약 100 bp 의 동일한 반전 말단 반복물 ("ITR") 을 함유하는데, 정확한 길이는 혈청형에 따라 좌우된다. 복제의 바이러스 기원은 게놈 말단에서 정확히 ITR 내에 위치한다. DNA 합성은 2 단계로 발생한다. 먼저, 복제는 가닥 교체에 의해 진행되고, 딸 듀플렉스 분자 및 부모 대체된 가닥을 생성시킨다. 대체된 가닥은 단일 가닥이며 "팬핸들 (panhandle)" 중간체를 형성할 수 있는데, 이는 복제가 개시되게 하고 딸 듀플렉스 분자를 생성시킨다. 대안적으로, 복제는 게놈의 양쪽 말단으로부터 진행될 수 있으며 동시에, 팬핸들 구조 형성에 대한 필요사항이 제거된다.
생산적 감염 주기 동안, 바이러스 유전자는 2 개 상으로 발현된다: 바이러스 DNA 복제까지의 기간인 초기상, 및 바이러스 DNA 복제의 개시와 동시에 일어나는 후기상. 초기상 동안, 부위 E1, E2, E3 및 E4 에 의해 인코딩되는 초기 유전자 산물만이 발현되며, 이는 바이러스 구조 단백질의 합성을 위해 세포를 준비시키는 수많은 기능을 수행한다 (Berk, A.J., 1986). 후기상 동안, 후기 바이러스 유전자 산물이 초기 유전자 산물에 추가로 발현되며 숙주 세포 DNA 및 단백질 합성이 정지된다. 결과적으로, 세포는 바이러스 DNA 및 바이러스 구조 단백질의 생성에 역점을 두게 된다 (Tooze, J., 1981).
아데노바이러스의 E1 부위는 표적 세포의 감염 후 발현된 아데노바이러스의 첫 번째 부위이다. 상기 부위는 2 개 전사 단위, E1A 및 E1B 유전자로 이루어지며, 이들 둘 모두는 1 차 (배아) 설치류 배양물의 종양성 형질전환에 필요하다. E1A 유전자 산물의 주요 기능은 휴지기 세포가 세포 주기에 들어가게끔 유도하고 세포 DNA 합성을 재개시키며, E1B 유전자 및 바이러스 게놈의 기타 초기 부위 (E2, E3 및 E4) 를 전사적으로 활성화시키는 것이다. E1A 유전자 단독을 갖는 1 차 세포의 트랜스펙션은 제한되지 않은 증식 (불멸화) 을 유도할 수 있으나, 완전한 형질전환을 초래하지는 않는다. 그러나 대부분의 경우 E1A 의 발현은 프로그래밍된 세포 사멸 (세포자멸사) 의 유도를 초래하며, 가끔만 불멸화가 수득된다 (Jochemsen et al., 1987). E1B 유전자의 동시발현은 세포자멸사의 유도를 방지하고 완전한 형태적 형질전환이 발생하도록 하기 위해 필요하다. 구축된 불멸 세포주에서, E1A 의 높은 수준의 발현은 E1B 의 부재 하 완전한 형질전환을 일으킬 수 있다 (Roberts, B.E. et al, 1985).
E1B 인코딩 단백질은 바이러스 복제가 가능하도록 세포 기능을 적용하는데 있어서 E1A 를 보조한다. 본질적으로 핵 내에서 국부화되는 복합체를 형성하는 E1B 55 kD 및 E4 33 kD 단백질은 숙주 단백질의 합성을 저해하고 바이러스 유전자 발현을 촉진하는데 있어서 그 기능을 수행한다. 이들의 주요 영향은 감염의 후기상의 발병에 부수적으로, 바이러스 mRNA 를 핵으로부터 세포질에 선택적으로 이동시키는 것을 확립하는 것이다. E1B 21 kD 단백질은 생산적 감염 주기의 올바른 시간적 제어를 위해 중요하며, 이로써 바이러스 생명 주기가 완료되기 전에 숙주 세포의 때이른 사멸을 방지한다. E1B 21 kD 유전자 산물을 발현할 수 없는 돌연변이체 바이러스는 숙주 세포 염색체 DNA (deg-표현형) 의 과도한 분해에 의해, 그리고 증강된 세포변성 효과 (cyt-표현형; Telling et al, 1994) 에 있어서 동반되는 단축된 감염 주기를 나타낸다. deg 및 cyt 표현형은 추가로 E1A 유전자가 돌연변이화되는 경우 억제되는데, 이는 이러한 표현형이 E1A 와 상관관계에 있다는 것을 나타낸다 (White, E. et al., 1988). 추가로, E1B 21 kDa 단백질은 E1A 가 다른 바이러스 유전자를 스위치 온 (switch on) 하는 속도를 감속시킨다. E1B 21 kD 가 이러한 E1A 의존적 기능을 해소하는 메커니즘은 아직 알려져 있지 않다.
예를 들어 레트로바이러스와 반대로, 아데노바이러스는 숙주 세포의 게놈에 효율적으로 통합되지 않고, 미분화 세포를 감염시킬 수 있으며, 재조합 유전자를 생체내 효율적으로 이동시킬 수 있다 (Brody et al., 1994). 이들 특징은 아데노바이러스로 하여금 예를 들어 관심 항원 또는 면역원의, 이를 필요로 하는 세포, 조직 또는 대상 내로의 생체내 유전자 이동에 대한 효과적인 후보물로 만든다.
다수 결실을 함유하는 아데노바이러스 벡터는 벡터의 운반 수용력을 증가시키고 복제 적격 아데노바이러스 (RCA) 를 생성시키는 재조합 가능성을 감소시킨다. 아데노바이러스가 다수의 결실을 함유하는 경우, 각각의 결실이, 단독으로 존재하는 경우, 복제 결함 및/또는 미-복제 아데노바이러스를 초래할 필요는 없다. 결실 중 하나가 아데노바이러스가 복제 결함이 되거나 미-복제가 되게 하는 한, 추가적인 결실이 다른 목적, 예를 들어 이종 뉴클레오티드 서열에 대한 아데노바이러스 게놈의 운반 수용력을 증가시키기 위해 포함될 수 있다. 바람직하게는, 결실 중 하나 초과는 기능적 단백질의 발현을 방지하고 아데노바이러스가 복제 결함 및/또는 미-복제 및/또는 약화되게 한다. 보다 바람직하게는, 모든 결실은 아데노바이러스가 복제-결함 및/또는 미-복제 및/또는 약화되게 할 수 있는 결실이다. 그러나, 본 발명은 또한 복제 적격 및/또는 야생형인 아데노바이러스 및 아데노바이러스 벡터를 포함하는데, 즉 대상에서의 감염 및 복제를 위해 필요한 모든 아데노바이러스 유전자를 포함한다.
아데노바이러스 재조합체를 이용하는 본 발명의 구현예는 E1-결함 또는 결실, 또는 E3-결함 또는 결실, 또는 E4-결함 또는 결실, 또는 E1 및 E3, 또는 E1 및 E4, 또는 E3 및 E4, 또는 E1, E3 및 E4 결실을 포함하는 아데노바이러스 벡터, 또는 모든 바이러스 유전자가 결실되는 "헬퍼 의존적 (gutless)" 아데노바이러스 벡터를 포함할 수 있다. 아데노바이러스 벡터는 E1, E3 또는 E4 유전자에서의 돌연변이, 또는 이들 또는 모든 아데노바이러스 유전자에서의 결실을 포함할 수 있다. E1 돌연변이는, E1-결함 아데노바이러스 돌연변이체가 복제-결함 및/또는 비-허용 세포에서 미-복제성인 것으로 일컬어지며 적어도 매우 약화되기 때문에, 벡터의 안전역 (safety margin) 을 증가시킨다. E3 돌연변이는 메커니즘을 방해함으로써 항원의 면역원성을 증강시켜, 아데노바이러스가 MHC 클래스 I 분자를 하향 조절한다. E4 돌연변이는 후기 유전자 발현을 억제함으로써 아데노바이러스의 면역원성을 감소시켜, 그에 따라 동일한 벡터를 이용하는 반복된 백신 재접종을 가능하게 할 수 있다. 본 발명은 E1, 또는 E3, 또는 E4, 또는 E1 및 E3, 또는 E1 및 E4 에 있어서 결실되거나 돌연변이화되는 임의의 혈청형 또는 혈청군의 아데노바이러스 벡터를 포괄한다. 이들 아데노바이러스 유전자의 결실 또는 돌연변이는 이들 단백질 활성을 손상 또는 실질적으로 완전히 손실시킨다.
"헬퍼 의존적" 아데노바이러스 벡터는 아데노바이러스 벡터 패밀리에서의 또 다른 유형의 벡터이다. 이의 복제에는 헬퍼 바이러스 및 E1a 및 Cre 모두를 발현하는 특이적 인간 293 세포주, 자연적 환경에서 존재하지 않는 조건이 필요하며; 벡터는 모든 바이러스 유전자가 없고, 따라서 백신 운반체로서의 상기 벡터는 비-면역원성이고 백신 재접종을 위해 수회 접종될 수 있다. "헬퍼 의존적" 아데노바이러스 벡터는 또한 관심 항원 또는 면역원(들) 을 수용하기 위해 36 kb 공간을 포함하며, 따라서 다수의 항원 또는 면역원을 세포 내로 동시 전달할 수 있다.
아데노-연관 바이러스 (AAV) 는 인간 집단에 대해 내인성인 단일 가닥 DNA 파르보바이러스이다. 다양한 종류로부터의 세포에서 생산적 감염이 가능함에도 불구하고, AAV 는 데펜도바이러스 (dependovirus) 여서, 이의 고유 복제를 위해 아데노바이러스 또는 포진 바이러스로부터 헬퍼 기능을 필요로 한다. 이들 헬퍼 바이러스로부터의 헬퍼 기능 부재 하에서, AAV 는 세포를 감염시키고, 핵에서 탈피되고, 이의 게놈이 숙주 염색체 내로 통합되나, 새로운 바이러스 입자를 복제하거나 생성시키지는 않을 것이다.
AAV 의 게놈은 박테리아 플라스미드로 클로닝되며 잘 분석된다. 바이러스 게놈은 각각 145 개 염기의 2 개 말단 반복물을 포함하는 4682 개 염기로 이루어진다. 이들 말단 반복물은 바이러스에 대한 DNA 복제의 기원으로서 역할한다. 일부 조사자들이 또한 이들이 인핸서 기능을 갖는다는 것을 제안한 바 있다. 게놈의 나머지는 2 개의 기능적 도메인으로 나뉜다. 게놈의 좌측 부분은 바이러스 DNA 복제 및 생명유지 유전자 발현을 조절하는 rep 기능을 코딩한다. 생명유지 게놈의 우측 면은 구조적 캡시드 단백질 VP1, VP2 및 VP3 을 인코딩하는 cap 유전자를 함유한다. rep 및 cap 유전자 모두에 의해 인코딩되는 단백질은 생산적 AAV 복제 동안 trans 에서 기능한다.
AAV 는 형질도입 벡터로서 사용하기 위한 이상적인 후보물로 고려되며, 이러한 방식으로 사용되고 있다. 이러한 AAV 형질도입 벡터는 trans 에서 주어진 아데노바이러스 또는 포진 바이러스 헬퍼 기능의 존재 하에 복제하기에 충분한 cis-작용 기능을 포함한다. 재조합 AAV (rAAV) 는 많은 실험실에서 구축되고 있으며 다양한 혈통의 세포 내로 외인성 유전자를 운반하는데 사용되고 있다. 이들 벡터에서, AAV cap 및/또는 rep 유전자는 바이러스 게놈으로부터 결실되고 선택 DNA 조각으로 대체된다. 현재 벡터는 4300 개 이하 염기의 삽입된 DNA 를 수용할 수 있다.
rAAV 를 생성시키기 위해서, 원하는 바이러스 구축물을 함유하는 플라스미드를 아데노바이러스-감염된 세포 내로 트랜스펙션한다. 추가로, 제 2 헬퍼 플라스미드를 이들 세포 내로 동시트랜스펙션하여 재조합 바이러스 구축물의 복제 및 패키징에 의무적인 AAV rep 및 cap 유전자를 제공한다. 이러한 조건 하에, AAV 의 rep 및 cap 단백질은 trans 에서 rAAV 구축물의 복제 및 패키징을 자극하는 작용을 한다. 트랜스펙션 3 일 후, rAAV 를 아데노바이러스와 함께 세포로부터 수확한다. 오염 아데노바이러스를 이후 열 처리에 의해 불활성화시킨다.
단순 포진 바이러스 1 (HSV-1) 은 80 개 초과의 유전자를 인코딩하는 153 kb 의 게놈을 갖는 외피형, 이중가닥 DNA 바이러스이다. 이의 광범위한 숙주 범위는 세포 멤브레인에서 발견된 세포외 헤파린 술페이트 분자에 대한 바이러스 외피 당단백질의 결합으로 인한 것이다 (WuDunn & Spear, 1989). 바이러스의 부화는 외피 당단백질 gD 및 섬유모세포 성장 인자 수용체를 필요로 한다 (Kaner, 1990). HSV 는 세포를 용해적으로 감염시킬 수 있거나 잠복성을 구축할 수 있다. HSV 벡터를 사용하여 광범위한 세포 유형을 감염시킨다 (Lowenstein, 1994; Huard, 1995; Miyanohara, 1992; Liu, 1996; Goya, 1998).
재조합 HSV 벡터 및 암플리콘 벡터로 지칭되는 2 개 유형의 HSV 벡터가 존재한다. 재조합 HSV 벡터는 상동 재조합 이벤트를 통해 전사 단위를 HSV 게놈 내로 직접 삽입하여 생성된다. 암플리콘 벡터는 선택 전사 단위, 복제 기원 및 패키징 신호를 갖는 플라스미드를 기반으로 한다.
HSV 벡터는 외래 유전자의 삽입을 위한 큰 수용력, 뉴런에서의 잠복성 구축력, 넓은 숙주 범위 및 18 개월까지 CNS 에 이식유전자 발현을 부여하는 능력의 명백한 이점을 갖는다 (Carpenter & Stevens, 1996).
레트로바이러스는 유전자 이동 프로토콜에서 광범위하게 사용되는 외피형 단일가닥 RNA 바이러스이다. 레트로바이러스는 gag, pro, pol 및 env 로 지칭되는 4 개 유전자 부위로 이루어진 약 7-10 kb 의 이배체 게놈을 갖는다. 이들 유전자 부위는 구조적 캡시드 단백질, 바이러스 프로테아제, 인테그라아제 및 바이러스 역전사효소 및 외피 당단백질을 각각 인코딩한다. 게놈은 또한 패키징 신호 및 cis-작용 서열 (장길이-말단 반복물 (LTR) 로 지칭) 을 각각의 말단에 갖는데, 이는 전사 제어 및 통합에 있어서 역할을 갖는다.
가장 흔하게 사용되는 레트로바이러스 벡터는 몰로니 (Moloney) 쥐과 백혈병 바이러스 (Mo-MLV) 를 기반으로 하며 외피 당단백질의 수용체 결합 표면 도메인에 따라 가변적 세포 향성을 갖는다.
재조합 레트로바이러스 벡터는 모든 레트로바이러스 유전자로부터 결실되는데, 이는 마커 또는 치료적 유전자, 또는 둘 모두로 대체된다. 재조합 레트로바이러스를 증식시키기 위해, 바이러스 유전자, gag, pol 및 env 를 trans 에서 제공하는 것이 필요하다.
렌티바이러스는 유사분열 및 유사분열 후 세포 모두를 감염시키고 그의 유전자를 발현시키는 능력을 갖는 복잡한 레트로바이러스이다. 가장 흔하게 공지된 렌티바이러스는 인간 면역결핍 바이러스 (HIV) 이며, 이는 다른 바이러스의 외피 당단백질을 사용하여 광범위한 세포 유형을 표적화한다.
원형 신드비스 바이러스 (Sindbis virus) (SIN), 셈리키 삼림 바이러스 (Semliki Forest virus) (SFV) 및 베네수엘라 말 뇌염 바이러스 (Venezuelan equine encephalitis virus) (VEE) 를 포함하는 알파바이러스는 20 면체 캡시드 내에 + 가닥 RNA 게놈을 함유하는 외피형 바이러스의 군을 구성한다.
본 발명의 바이러스 벡터는 시험관내 및 생체내 모두에서 세포에 항원 또는 면역원 발현 핵산을 전달하는데 유용하다. 특히, 본 발명의 벡터는 핵산을 세포, 보다 바람직하게는 포유동물 세포에 전달하거나 이동시키는데 유리하게 이용될 수 있다. 관심 핵산은 펩티드 및 단백질, 바람직하게는 치료적 (예를 들어, 의료용 또는 수의학용) 또는 면역원성 (예를 들어, 백신용) 펩티드 또는 단백질을 인코딩하는 핵산을 포함한다.
바람직하게는, 관심 항원 또는 면역원을 인코딩하는 코돈은 "최적화된" 코돈이며, 즉, 상기 코돈은 예를 들어, 인플루엔자 바이러스에 의해 종종 사용되는 코돈 대신, 대상의 종류에서 고도로 발현된 유전자에서 종종 나타나는 것들이다. 이러한 코돈 사용빈도는 동물 세포에서 항원 또는 면역원의 효율적 발현을 제공한다. 다른 구현예에서, 예를 들어 관심 항원 또는 면역원이 박테리아, 효모 또는 또 다른 발현 시스템에서 발현되는 경우, 코돈 사용빈도 패턴은 항원 또는 면역원이 발현되는 유기체에서의 고도로 발현된 유전자에 대한 코돈 편향을 나타내도록 변경된다. 코돈 사용빈도 패턴은 많은 종류의 고도로 발현된 유전자에 대한 문헌에서 공지되어있다 (예를 들어, Nakamura et al, 1996; Wang et al, 1998; McEwan et al. 1998).
추가적인 대안물로서, 바이러스 벡터는 배양물에서 세포를 감염시켜 원하는 유전자 산물을 발현하는데, 예를 들어 관심 단백질 또는 펩티드를 생성시키는데 사용될 수 있다. 바람직하게는, 단백질 또는 펩티드는 배지로 분비되고 당업계에 공지된 일상적 기술을 사용하여 그로부터 정제될 수 있다. 단백질의 세포외 분비를 유도하는 신호 펩티드 서열은 당업계에 공지되어 잇으며 이를 인코딩하는 뉴클레오티드 서열은 당업계에 공지된 일상적 기술에 의해 관심 펩티드 또는 단백질을 인코딩하는 뉴클레오티드 서열에 작동가능하게 연결될 수 있다. 대안적으로, 세포는 용해될 수 있으며 발현된 재조합 단백질은 세포 용해물로부터 정제될 수 있다. 바람직하게는 세포는 동물 세포이고, 보다 바람직하게는 포유동물 세포이다. 또한, 특정 관심 바이러스 벡터에 의한 형질도입에 적격인 세포가 바람직하다. 이러한 세포는 PER.C6 세포, 911 세포 및 HEK293 세포를 포함한다.
숙주 세포 배양을 위한 배양 배지는 조직 배양에 흔히 사용되는 배지, 그 중에서도 예컨대 M199-얼 (earle) 베이스, 이글 MEM (E-MEM), 둘베코 MEM (DMEM), SC-UCM102, UP-SFM (GIBCO BRL), EX-CELL302 (Nichirei), EX-CELL293-S (Nichirei), TFBM-01 (Nichirei), ASF 104 를 포함한다. 특정 세포 유형에 적합한 배양 배지는 미국 미생물 보존 센터 (American Type Culture Collection) (ATCC) 또는 유럽 세포 배양물 보존 센터 (European Collection of Cell Cultures) (ECACC) 에서 발견될 수 있다. 배양 배지에는 아미노산 예컨대 L-글루타민, 염, 항진균 또는 항박테리아제 예컨대 Fungizone®, 페니실린-스트렙토마이신, 동물 혈청 등이 보충될 수 있다. 세포 배양 배지는 임의로는 혈청-불포함일 수 있다.
본 발명은 또한 LITE 또는 보노 발명에서 유용한 하나 이상의 작용제를 발현 또는 과발현할 수 있는 세포주 또는 트랜스제닉 동물에 관한 것이다. 바람직하게는 세포주 또는 동물은 하나 이상의 LITE 를 발현하거나 과발현한다.
트랜스제닉 동물은 통상 척추동물, 보다 바람직하게는 설치류, 예컨대 랫트 또는 마우스일 뿐 아니라 기타 포유동물 예컨대 인간, 염소, 돼지 또는 소 등을 포함한다.
이러한 트랜스제닉 동물은 새로운 유용한 화합물에 대한 스크리닝 검정에서 및 질환의 동물 모델로서 유용하다. 상기 정의한 바와 같이 하나 이상의 폴리펩티드를 특이적으로 발현시켜, 질환 발전에 대한 이러한 폴리펩티드의 효과를 연구할 수 있다. 추가로, 유전자 치료요법 및 각종 약물을 포함하는 치료요법을 트랜스제닉 동물에서 시험할 수 있다. 트랜스제닉 동물의 생성 방법은 당업계에 공지되어있다. 예를 들어, 유전자를 배아 내로 도입하기 위한 여러 가능한 경로가 존재한다. 이는 (i) 배아 줄기 세포의 직접 트랜스펙션 또는 레트로바이러스 감염, 이후 발생의 배반포 단계에서의 이들 세포의 배아 내로의 도입; (ii) 초기 배아의 레트로바이러스 감염; 및 (iii) DNA 의 접합자 또는 초기 배아 세포 내로의 직접 미세주입을 포함한다. 유전자 및/또는 전이유전자는 또한 당업계에 공지된 유전적 조절 요소 및/또는 구조적 요소를 포함할 수 있다. 전이유전자 도입을 위한 표적 세포의 유형은 배아 줄기 세포 (ES) 이다. ES 세포는 시험관내 배양된 사전-이식 배아로부터 수득될 수 있으며 배아에 융합된다 (Evans et al, 1981, Nature 292: 154-156; Bradley et al, 1984, Nature 309:255-258; Gossler et al, 1986, Proc. Natl. Acad. Sci. USA 83:9065-9069; and Robertson et al, 1986 Nature 322:445-448). 이식유전자는 DNA 트랜스펙션, 미세주입, 또는 레트로바이러스-매개 형질도입과 같은 다양한 표준 기술에 의해 ES 세포 내로 효율적으로 도입될 수 있다. 생성된 형질전환 ES 세포는 이후 비-인간 동물로부터의 배반포와 조합될 수 있다. 도입된 ES 세포는 이후 배아를 군집화시키며 생성된 키메라 동물의 생식세포 계열에 기여한다 (Jaenisch, 1988, Science 240: 1468-1474).
LITE 는 또한 가치있는 생체내 시간적 정확도를 제공할 수 있다. LITE 는, 예를 들어 C. 엘레간스 (C. elegans) 성장의 특정 단계 동안에만 특정 세포자멸사 유전자를 억제하여, 발생의 특정 단계 동안 유전자 발현을 변형시키는데 사용될 수 있다. LITE 는 유전적 신호를 특정 실험 창 (window) 에 대해 시기 조정하는데 사용될 수 있다. 예를 들어, 학습에 연루된 유전자는 미손상 설치류 또는 영장류 뇌의 정확도 부위에서의 학습 자극 동안에만 과발현되거나 억제될 수 있다. 또한, LITE 는 질환 발전의 특정 단계 동안에만 유전자 발현 변화를 유도하는데 사용될 수 있다. 예를 들어, 암유전자는 종양이 특정 크기 또는 전이 단계에 도달하고 단 한번 과발현될 수 있다. 반대로, 알츠하이머병 발전에서 의심된 단백질은 특정 뇌 부위 내에서 및 동물 생 (life) 에서 규정된 시점에서만 녹다운될 수 있다. 이들 예가 LITE 시스템의 잠재적 적용을 철저히 열거하지는 않으나, 이들은 LITE 가 강력한 기술일 수 있는 일부 분야를 강조한다.
본 발명의 치료적 및 진단적 조성물은 장애를 치료하거나 진단하기에 충분한 양으로 개인에게 투여된다. 유효량은 개인의 상태, 체중, 성별 및 연령과 같은 다양한 인자에 따라 가변적일 수 있다. 기타 인자는 투여 방식을 포함한다.
약학 조성물은 다양한 경로 예컨대 피하, 국소, 경구 및 근육내에 의해 개인에게 제공될 수 있다.
본원에 개시된 방법에 따라 확인된 화합물은 적절한 투약량에서 단독으로 사용될 수 있다. 대안적으로, 기타 작용제의 동시-투여 또는 순차적 투여가 바람직할 수 있다.
본 발명은 또한 본 발명의 신규한 치료 방법에서 사용하기 위해 적합한 국소, 경구, 전신 및 비경구 약학 제형을 제공하는 목적을 갖는다. 활성 성분으로서 본 발명에 따라 확인된 화합물을 함유하는 조성물은 투여용의 종래 비히클 중 광범위한 치료 투약 형태로 투여될 수 있다. 예를 들어, 화합물은 정제, 캡슐 (각각 시간적 방출형 및 서방형 제형 포함), 알약, 분말, 과립, 엘릭시르, 팅크, 용액, 현탁액, 시럽 및 에멀젼과 같은 경구 투약 형태로, 또는 주사에 의해 투여될 수 있다. 마찬가지로, 이는 또한 정맥내 (볼루스 및 주입물 모두), 복강내, 피하, 국소 (흡장의 존재 또는 부재 하), 또는 근육내 형태로 투여될 수 있다 (모든 사용 형태는 약학 업계의 당업자에게 널리 공지됨).
유리하게는, 본 발명의 화합물은 단일 1 일 용량으로 투여될 수 있거나, 총 1 일 투약량이 1 일 2, 3 또는 4 회의 분할 용량으로 투여될 수 있다. 추가로, 본 발명의 화합물은 적합한 비강내 비히클의 국소 사용을 통해, 또는 경피 경로를 통해 비강내 형태로 투여될 수 있으며, 경피 피부 패치의 이들 형태를 사용하는 것은 당업자에게 널리 공지되어있다. 경피 전달 시스템의 형태로 투여되기 위해서는, 투약량의 투여는 물론 투약 계획 전체에 걸쳐 간헐식 보다는 연속식일 것이다.
하나 초과의 활성제로의 병용 치료를 위해서, 활성제가 별개의 투약량 제형 중에 존재하는 경우, 상기 활성제는 동시에 투여될 수 있거나, 각각 별도 시차 시간으로 투여될 수 있다.
본 발명의 화합물을 이용하는 투약 계획은 환자의 유형, 인종, 연령, 체중, 성별 및 의학적 상태; 치료하는 병상의 중증도; 투여 경로; 한 환자의 신장, 간 및 심혈관 기능; 및 이용한 이의 특정 화합물을 포함하는 다양한 인자에 따라 선택된다. 통상의 지식을 갖는 의사는 병상의 진행을 방지하고, 대응하거나 막기 위해 필요한 약물의 유효량을 용이하게 결정하고 처방할 수 있다. 독성을 갖지 않는 효능을 산출해내는 범위 내의 약물 농도를 얻는데 있어서 최적의 정확도는 표적 부위에 대한 약물의 이용가능성 속도를 기반으로 한 계획을 필요로 한다. 이는 약물의 분포도, 평형 및 제거에 대한 고려를 포함한다.
본 발명 및 이의 이점이 상세히 기재되어 있으나, 첨부된 특허청구범위에서 정의한 바와 같이 본 발명의 취지 및 범주에서 벗어나지 않고 다양한 변화, 대용 및 변형이 본원에서 이루어질 수 있다는 것이 이해되어야 한다.
하기 실시예에서 본 발명을 추가로 설명하며, 이는 설명의 목적으로만 제공되며 어떠한 식으로도 본 발명을 제한하는 것으로 의도되지 않는다.
실시예
실시예 1
내인성 포유동물 게놈으로부터의 유전자 발현을 직접적으로 조정하는 능력은 정상 유전자 기능 및 질환 메커니즘을 해명하기 위해 중요하다. 세포 집단 내에서 유전자 발현의 공간적 및 시간적 제어를 추가로 개선시키는 진전은 유전자 조정의 유용성을 확장시키는 가능성을 갖는다. 출원인은 이전에 잔타모나스 오리제 (Xanthamonas oryze) 로부터 전사 활성제-유사 효과기 (TALE) 를 개발하여, 부위-특이적 DNA 결합 단백질의 신속한 설계 및 구축을 가능하게 하였다. 출원인은 내인성 포유동물 게놈에서의 광-조절 유전자 발현을 가능하게 하는 분자 도구 세트를 개발하였다. 상기 시스템은 아라비도프시스 탈리아나 (Arabidopsis thaliana) 로부터의 광-민감성 이량체화 단백질 도메인에 연결된 조작된 인공 전사 인자로 이루어진다. 상기 시스템은 450nm -500nm 범위의 광에 반응하며 포유동물 세포에서 6.2 mW/cm2 강도로의 광 자극 후 전분화능 인자의 발현에 있어서 유의한 증가를 유도할 수 있다. 출원인은 광범위한 유전자의 표적화를 위한 도구를 개발하고 있다. 출원인은 유전자 발현의 광-매개 제어를 위한 도구상자가 현존하는 광유전학적 방법을 보완할 수 있으며 장차 뇌에서의 특정 유전자의 시기-, 세포 유형- 및 농도 의존적 역할을 해명하는 것을 도울 수 있다고 믿는다.
내인성 포유동물 게놈으로부터의 유전자 발현을 직접적으로 조정하는 능력은 정상 유전자 기능 및 질환 메커니즘을 해명하기 위해 중요하다. 출원인은 내인성 포유동물 게놈에서의 광-조절 유전자 발현을 가능하게 하는 분자 도구 세트 개발을 제시한다. 이러한 시스템은 아라비도프시스 탈리아나로부터의 광-민감성 이량체화 단백질 도메인 크립토크롬 2 (CRY2) 및 CIB1 에 연결된 전사 활성제 유사 효과기 (TALE) 및 활성화 도메인 VP64 로 이루어진다. 출원인은 KLF4 및 Neurog2 의 프로모터 부위를 표적화하도록 설계된 이들 LITE 구축물로 트랜스펙션한 HEK293FT 및 Neuro-2a 세포의 청색광 자극이 표적 발현에 있어서 유의한 증가를 초래한다는 것을 나타내는데, 이는 TALE-기재 광학 유전자 발현 조정 기술의 기능성을 입증한다.
도 1 은 공간적 및 시간적 정확도에 대한 필요성을 설명하는 모식도를 나타낸다.
도 2 는 전사 활성제 유사 효과기 (TALE) 를 나타낸다. TALE 는 그의 서열의 코어에서 34 개 aa 반복물로 이루어진다. 각각의 반복물은 TALE 에 의해 결합되는 표적 DNA 에서의 염기에 상응한다. 반복물은 위치 12 및 13 에서 2 개의 가변적인 아미노산에 의해서만 상이하다. 이러한 대응의 코드가 설명된 바 있으며 (Boch, J et al. , Science, 2009 and Moscou, M et al. , Science, 2009) 이 도면에서 나타낸다. 출원인은 이러한 코드를 통합하며 게놈 내에 선택 서열을 결합시킬 수 있는 설계자 TALE 의 합성 방법을 개발하였다 (Zhang, F et al. , Nature Biotechnology, 2011).
도 3 은 LITE: TALE/크립토크롬 전사 활성화의 설계를 나타낸다. 각각의 LITE 는 CRY2 에 융합된 TALE 및 VP64 (전사 인자) 에 융합된 크립토크롬 결합 파트너 CIB1 을 포함할 수 있는 2-성분 시스템이다. 불활성 상태에서, TALE 은 이의 융합된 CRY2 도메인을 관심 유전자의 프로모터 부위에 국부화시킨다. 이때, CIB1 은 CRY2 에 결합할 수 없고, CIB1-VP64 가 핵 공간 내에서 미결합된 채로 있다. 488 nm (청색) 광으로 자극시, CRY2 는 형태적 변화를 거쳐, 이의 CIB1 결합 부위가 밝혀진다 (Liu, H et al. , Science, 2008). CIB1 의 신속한 결합은 융합된 VP64 도메인의 영입을 초래하여, 표적 유전자의 전사를 유도한다.
도 4 는 LITE 활성에 대한 크립토크롬 이량체 절두의 효과를 나타낸다. CRY2 및 CIB1 의 활성을 변경시키는 것으로 알려져 있는 절두를 전체 길이 단백질에 대해 비교하였다. Neurog2 의 프로모터에 대해 표적화된 LITE 를 도메인의 각 조합에 대해 Neuro-2a 세포에서 시험하였다. 488 nm 광으로 자극한 후, Neurog2 의 전사 수준을 자극된 샘플 및 미자극된 샘플에 대해 qPCR 을 사용하여 정량하였다.
도 5 는 KLF4 LITE 의 광-강도 의존적 반응을 나타낸다.
도 6 은 Neurog2 LITE 의 활성화 반응속도 및 Neurog2 LITE 의 불활성화 반응속도를 나타낸다.
실시예 2
정상 유전자 발현은 주의깊게 조직된 시간적 및 공간적 성분을 갖는 동적 프로세스이며, 이의 정확도는 정상 발생, 항상성 유지 및 유기체의 진보를 위해 필요하다. 그 다음, 유전자 또는 유전자 세트의 증가, 감소 또는 변경된 기능에 의한 필요한 유전자 발현 패턴의 조절장애는 광범위한 집합의 병리학과 연결된다. 시공간적 정확도 방식으로 유전자 발현을 조정할 수 있는 기술은 정상 생물학적 프로세스 및 질환 메커니즘을 책임지는 유전적 신호의 해명을 가능하게 할 것이다. 이러한 기술적 필요성을 다루기 위해, 출원인은 내인성 유전자 발현의 광-매개 제어를 제공하는 광-유도 전사 효과기 (LITE) 를 개발하였다.
유도 유전자 발현 시스템은 통상 삽입된 개방 판독 프레임 또는 shRNA 서열의 화학적 유도 활성화가 가능하도록 설계되어, 각각 유전자 과발현 또는 억제를 초래한다. 과발현을 위한 개방 판독 프레임 사용의 불리한 점은 스플라이스 변이의 손실 및 유전자 크기의 제한을 포함한다. RNA 간섭을 통한 유전자 억제는, 인간 생물학에 있어서의 이의 형질전환적 영향력에도 불구하고, 복합한 오프-타겟 효과에 의해 저해될 수 있다. 에스트로겐, 엑디손 및 FKBP12/FRAP 기반 시스템을 포함하는 특정 유도 시스템은 오프-타겟 내인성 유전자를 활성화시키는 것으로 공지되어있다. 장기간 항생제 치료의 잠재적 유해 효과는 테트라사이클린 전사촉진제 (TET) 기반 시스템의 사용을 복잡하게 만들 수 있다. 이러한 화학적 유도 시스템의 생체내, 시간적 정확도는 유도제 흡수 및 제거의 속도에 따라 좌우된다. 또한, 유도제가 일반적으로 전신 전달되므로, 이러한 시스템의 공간적 정확도는 외인성 벡터 전달의 정확도에 의해 제약된다.
이러한 제한에 대응하여, LITE 는 시간적 및 공간적으로 정확한 방식으로 개별적 내인성 유전자의 발현을 조정하도록 설계된다. 각각의 LITE 는 커스텀화된 DNA-결합 전사 활성제 유사 효과기 (TALE) 단백질, 아라바돕시스 탈리아나로부터의 광-반응성 크립토크롬 이형이량체 및 전사 활성화/억제 도메인으로 이루어지는 2 성분 시스템이다. TALE 은 관심 유전자의 프로모터 서열에 결합하도록 설계된다. TALE 단백질은 크립토크롬 이형이량체의 절반 (크립토크롬-2 또는 CIB1) 에 융합되면서, 남아있는 크립토크롬 파트너가 전사 효과기 도메인에 융합된다. 효과기 도메인은 활성제 예컨대 VP16, VP64 또는 p65, 또는 억제제 예컨대 KRAB, EnR 또는 SID 일 수 있다. LITE 의 미자극 상태에서, TALE-크립토크롬2 단백질은 관심 유전자의 프로모터에 대해 국부화되나, CIB1-효과기 단백질에 결합하지는 않는다. 청색 스펙트럼 광으로 LITE 자극시, 크립토크롬-2 는 활성화되고, 형태적 변화를 거치며, 이의 결합 도메인이 밝혀진다. 다음으로 CIB1 은 크립토크롬-2 에 결합하여 관심 유전자의 프로모터 부위에 대해 효과기 도메인을 국부화시키고 유전자 과발현 또는 침묵화를 개시시킨다.
LITE 에서의 유전자 표적화는 커스텀화된 TALE DNA 결합 단백질의 특이성을 통해 이루어질 수 있다. 관심 유전자의 프로모터 부위에서 표적 서열이 선택되며 상기 서열에 대해 커스텀화된 TALE 이 설계된다. TALE 의 중심 부분은 34 개 아미노산 길이의 탠덤 반복물로 이루어진다. 이들 반복물의 서열은 거의 동일하나, 각각의 반복물의 12 번째 및 13 번째 아미노산 (반복 가변성 2 잔기로 지칭함) 은 가변적이어서, 각각의 반복물의 뉴클레오티드-결합 특이성을 결정한다. 따라서, TALE 단량체 반복물의 적절한 배치를 갖는 구축물을 합성하여, 표적 프로모터 서열에 특이적인 DNA 결합 단백질을 제작한다.
LITE 의 광 반응성은 크립토크롬-2 및 CIB1 의 활성화 및 결합을 통해 이루어진다. 상기 언급한 바와 같이, 청색광 자극은 크립토크롬-2 에 있어서의 활성화 형태적 변화를 유도하여, 이의 결합 파트너 CIB1 의 영입을 초래한다. 이러한 결합은 빠르고 가역적이며, 펄스화된 자극 후 <15 초 내에 포화가 이루어지고 자극 종료 후 <15 분에 기준선으로 복귀된다. 이러한 신속한 결합 속도는 유도제의 흡수 및 청소율보다는 전사/번역 및 전사체/단백질 분해의 속도에 의해서만 일시적으로 결합한 LITE 시스템을 초래한다. 크립토크롬-2 활성화는 또한 매우 민감하여, 낮은 광 강도 자극의 사용을 가능하게 하며 광독성의 위험성을 완화시킨다. 추가로, 미손상 포유동물 뇌와 같은 맥락에서, 가변적인 광 강도는 LITE 자극 부위의 크기를 제어하는데 사용될 수 있어, 벡터 단독 전달이 제공할 수 있는 것보다 더 큰 정확성을 부여한다.
LITE 시스템의 모듈성은 임의 수의 효과기 도메인이 전사 조정에 사용될 수 있게 한다. 따라서, 활성제 및 억제제 도메인은 종류, 강도, 메커니즘, 지속기간, 크기, 또는 임의 수의 기타 매개변수를 기반으로 선택될 수 있다.
출원인은 그 다음으로 LITE 시스템의 2 가지 원형적 시현을 제시한다. 제 1 예는 마우스 유전자 NEUROG2 의 전사를 활성화시키도록 설계된 LITE 이다. 마우스 NEUROG2 의 업스트림 프로모터 부위에 위치한 서열 TGAATGATGATAATACGA 를 표적으로서 선택하고, 상기 서열에 매치되도록 TALE 을 설계하고 합성하였다. TALE 서열을 핵 국부화 신호 (아미노산: SPKKKRKVEAS) 를 통해 크립토크롬-2 에 대한 서열에 연결시켜, 사이토졸로부터의 단백질의 핵 공간으로의 이동을 촉진시킨다. 동일한 핵 국부화 신호를 사용하여, 전사 활성제 도메인 VP64 에 연결된 CIB1 도메인을 포함하는 제 2 벡터를 합성하였다. 이러한 제 2 벡터, 또한 GFP 서열을 2A 전사 스킵 신호에 의해 CIB1-VP64 융합 서열로부터 분리하였다. 각 구축물의 발현을 편재성, 항시성 프로모터 (CMV 또는 EF1-α) 에 의해 유도하였다. Neuro 2A 세포주로부터의 마우스 신경아세포종 세포를 2 개 벡터로 동시-트랜스펙션시켰다. 벡터 발현이 가능하도록 인큐베이션한 후, 샘플을 488 nm LED 의 어레이로부터의 주기적 펄스의 청색광에 의해 자극하였다. 미자극된 동시-트랜스펙션 샘플 및 형광 리포터 YFP 로만 트랜스펙션된 샘플을 대조군으로서 사용하였다. 각 실험 완료시, mRNA 를 qPCR 을 통해 분석한 샘플로부터 정제하였다.
크립토크롬-2 및 CIB1 의 절두 형태를 클로닝하고 전체 길이 형태의 크립토크롬-2 및 CIB1 과 조합으로 시험하여, 각각의 이형이량체 쌍의 효과를 측정하였다. 크립토크롬-2 단백질의 보존된 광반응성 부위 및 전체 길이 형태 CIB1 로 이루어지는 CRY2PHR 도메인의 조합은 Neurog2 mRNA 수준의 최고 상향조절을 초래하였다 (YFP 샘플의 ~22 배 초과 및 미자극 동시-트랜스펙션 샘플의 ~7 배 초과). 전체 길이 크립토크롬-2 (CRY2) 와 전체 길이 CIB1 의 조합은 낮은 절대 활성화 수준 (YFP 의 ~4.6 배 초과) 뿐 아니라 낮은 기준선 활성화 (미자극 동시-트랜스펙션 샘플에 대한 YFP 의 ~1.6 배 초과) 를 초래하였다. 이들 크립토크롬 단백질 페어링은 필요한 유도의 절대 수준 및 LITE 시스템의 기준선 "누수도" 를 최소화하기 위한 필요성에 따라 특정 용도를 위해 선택될 수 있다.
가역성 및 활성화의 속도는 LITE 시스템에 대한 중요한 설계 매개변수이다. LITE 시스템의 속도를 분석하기 위해, Neurog2 TALE-CRY2PHR 및 CIB1-VP64 형태 시스템으로 이루어지는 구축물을 시험하여 이의 활성화 및 불활성화 속도를 측정하였다. 추출하기 전 짧게는 0.5 시간 내지 길게는 24 시간 동안 샘플을 자극하였다. Neurog2 발현의 상향조절은 최단 시점, 0.5 시간에서 관찰되었다 (YFP 샘플에 대해 ~5 배). Neurog2 발현은 자극의 12 시간에서 최대치였다 (YFP 샘플에 대해 ~19 배). 불활성화 속도를 6 시간 동안 동시-트랜스펙션 샘플을 자극하여 분석하여, 이때 자극을 중단하고, 샘플을 0 내지 12 시간 동안 배양물 중 유지시켜 mRNA 가 분해되게 하였다. Neurog2 mRNA 수준은 자극 완료 후 0.5 시간에서 최대치였고 (YFP 샘플에 대해 ~16 배), 그 후 상기 수준은 ~3 시간 반감기로 저하된 후 12 시간까지 기준선 수준 근처로 복구되었다.
제 2 의 원형적 예는 인간 유전자 KLF4 의 전사를 활성화시키도록 설계된 LITE 이다. 인간 KLF4 의 업스트림 프로모터 부위에 위치한 서열 TTCTTACTTATAAC 를 표적으로서 선택하고, 상기 서열에 매치되도록 TALE 을 설계하고 합성하였다. TALE 서열을 핵 국부화 신호 (아미노산: SPKKKRKVEAS) 를 통해 CRY2PHR 에 대한 서열에 연결시켰다. 상기 기재된 동일한 CIB1-VP64 활성제 단백질을 또한 LITE 시스템의 이러한 시현에 사용하였다. HEK293FT 세포주로부터의 인간 배아 신장 세포를 2 개 벡터로 동시-트랜스펙션하였다. 벡터 발현이 가능하도록 인큐베이션한 후, 488 nm LED 의 어레이로부터의 주기적 펄스 청색광에 의해 샘플을 자극하였다. 미자극 동시-트랜스펙션한 샘플 및 형광 리포터 YFP 로만 트랜스펙션한 샘플을 대조군으로서 사용하였다. 각 실험의 종료시, mRNA 를 qPCR 을 통해 분석한 샘플로부터 정제하였다.
LITE 시스템의 광-강도 반응을, 샘플을 증가한 광 동력 (0-9 mW/cm2) 으로 자극하여 시험하였다. KLF4 mRNA 수준의 상향조절이 0.2 mW/cm2 의 낮은 자극에 대해 관찰되었다. KLF4 상향조절은 5 mW/cm2 에서 포화되었다 (YFP 샘플에 대해 2.3 배). 세포 생존도 시험을 또한 9 mW/cm2 이하의 동력에 대해 수행하였고, >98% 세포 생존도를 나타내었다. 유사하게, 자극 사용률 가변화에 대한 KLF4 LITE 반응을 시험하였다 (1.6-100%). 상이한 사용률 간에 KLF4 활성화에 있어서의 차이가 관찰되지 않았는데, 이는 15 초 마다 0.25 초의 낮은 자극 패러다임이 최대 활성화를 초래한다는 것을 나타낸다.
LITE 가 유전자 발현 제어를 위한 유리한 선택을 나타내는 잠재적 적용이 존재한다. LITE 가 특히 유리한 수많은 시험관내 적용이 존재한다. 이러한 모든 경우, LITE 는 올바른 스플라이스 변이체 발현을 위한 가능성을 갖는 내인성 유전자 발현 유도의 이점을 갖는다.
LITE 활성화가 광유도성이기 때문에, 차폐 또는 래스터화 레이저 스캐닝을 통해 생성된 공간적으로 규정된 광 패턴을 사용하여 국한된 세포 하위집합에서의 발현 수준을 변형시킬 수 있다. 예를 들어, 공간적으로 제약된 세포 집합에서만 세포간 신호전달 분자를 과발현하거나 침묵화시켜, 자극 부위로부터 그의 거리에 관련하여 가까운 세포의 반응은 세포 비-자율적 프로세스의 공간적 특징을 해명하는 것을 도울 수 있다. 추가적으로, 세포 재프로그래밍 생물학에 있어서의 최근 진전은 전사 인자 집합의 과발현이 하나의 세포 유형, 예컨대 섬유모세포를 또 다른 세포 유형, 예컨대 뉴런 또는 심장근육세포로 형질전환하는데 이용될 수 있다는 것을 나타내고 있다. 추가로, 조직 내 세포 유형의 올바른 공간적 분포는 적절한 기관형 기능에 있어서 중요하다. LITE 를 사용하는 재프로그래밍 인자의 과발현은 조직 조작 적용을 위해 공간적으로 정확한 방식으로 다수의 세포 혈통을 재프로그래밍하는데 이용될 수 있다.
LITE 의 신속한 전사 반응 및 내인성 표적화는 전사 역학의 연구에 이상적인 시스템에 기여한다. 예를 들어, LITE 는 표적 유전자의 유도된 발현시 mRNA 스플라이스 변이체 생성의 역학 연구에 사용될 수 있다. 전사 주기의 다른 한편, mRNA 분해 연구는 종종, 과잉 유전자에서의 발현 수준 변화를 일으키는 강력한 세포외 자극에 대한 반응으로 수행된다. LITE 는 내인성 표적의 전사를 가역적으로 유도하는데 이용될 수 있으며, 그 후 자극이 중단될 수 있고 고유 표적의 분해 속도가 추적될 수 있다.
LITE 의 시간적 정확도는 실험적 개입과 맞물려 시간 유전적 조절에 대한 동력을 제공할 수 있다. 예를 들어, 장기간 강화작용 (LTP) 에서 예상된 관여를 갖는 표적은 기관형 또는 해리성 뉴런 배양물에 있어서, 그러나 LTP 를 유도하는 자극 동안에만 조정되어, 세포의 정상 발생의 간섭이 방지될 수 있다. 유사하게, 질환 표현형을 나타내는 세포 모델에서, 특정 치료요법의 유효성에 관여되는 것으로 예상되는 표적은 처리 동안에만 조정될 수 있다. 반대로, 유전적 표적은 병리적 자극 동안에만 조정될 수 있다. 외부 실험 자극에 대한 유전적 신호의 시기가 관련되는 임의 수의 실험은 LITE 조정의 효용으로부터 잠재적으로 유익할 수 있다.
생체내 맥락은 유전자 발현 제어를 위한 LITE 의 용도에 대해 동등하게 많은 기회를 제공한다. 상기 언급한 바와 같이, 광유도성은 이전에 성취불가능한 공간적 정확도에 대한 가능성을 제공한다. 광전극 기술의 발전을 이용하여, 자극 광 섬유 리드는 정확한 뇌 부위 내에 위치할 수 있다. 자극 부위 크기는 광 강도에 의해 조정될 수 있다. 이는 바이러스 벡터를 통한 LITE 의 전달과 함께 수행될 수 있거나, 트랜스제닉 LITE 동물이 이용가능해진 경우, 정확한 뇌 부위에서의 유전자 발현을 여전히 조정가능하게 하면서 바이러스의 사용을 없앨 수 있다. LITE 는 투명한 유기체, 예컨대 고정 제브라피시에서 사용되어 매우 정확한 레이저 유도 국부 유전자 발현 변화를 가능하게 할 수 있다.
LITE 는 또한 가치있는 생체내 시간적 정확도를 제공할 수 있다. LITE 는, 예를 들어 C. 엘레간스 성장의 특정 단계 동안에만 특정 세포자멸사 유전자를 억제하여, 발생의 특정 단계 동안 유전자 발현을 변형시키는데 사용될 수 있다. LITE 는 유전적 신호를 특정 실험 창에 대해 시기 조정하는데 사용될 수 있다. 예를 들어, 학습에 연루된 유전자는 미손상 설치류 또는 영장류 뇌의 정확도 부위에서의 학습 자극 동안에만 과발현되거나 억제될 수 있다. 또한, LITE 는 질환 발전의 특정 단계 동안에만 유전자 발현 변화를 유도하는데 사용될 수 있다. 예를 들어, 암유전자는 종양이 특정 크기 또는 전이 단계에 도달하고 단 한번 과발현될 수 있다. 반대로, 알츠하이머병 발전에서 의심된 단백질은 특정 뇌 부위 내에서 및 동물 생에서 규정된 시점에서만 녹다운될 수 있다. 이들 예가 LITE 시스템의 잠재적 적용을 철저히 열거하지는 않으나, 이들은 LITE 가 강력한 기술일 수 있는 일부 분야를 강조한다.
실시예 3
포유동물 TALE 전사 억제제의 개발
출원인은 조사자가 내인성 유전자의 전사를 억제할 수 있도록 포유동물 TALE 억제제 구조를 개발하였다. TALE 억제제는 유전자 뿐 아니라 미-코딩 전사체 예컨대 마이크로RNA 를 억제하는 가능성을 가져, 이들이 특정 유전적 요소의 평상적 역할을 시험하기 위한 매우 바람직한 도구가 되게 한다. 포유동물 세포에서 TALE 과 함께 사용하기 위한 적합한 억제 도메인을 확인하기 위해, 인간 SOX2 유전자의 프로모터를 표적화하는 TALE 을 사용하여 억제 도메인 후보물 무리의 전사 억제 활성을 평가하였다 (도 12a). 진핵세포 숙주 종류 범위에 걸친 억제 도메인을 선택하여, 카에노르합디티스 엘레간스 (Caenorhabditis elegans) 로부터의 PIE-1 억제 도메인 (PIE-1) (Batchelder, C. et al. Transcriptional repression by the Caenorhabditis elegans germ-line protein PIE-1. Genes Dev. 13, 202-212 (1999)), 드로소필라 멜라노가스테르 (Drosophila melanogaster) 로부터의 Ubx 유전자 내 QA 도메인 (Ubx-QA) (Tour, E., Hittinger, C.T. & McGinnis, W. Evolutionarily conserved domains required for activation and repression functions of the Drosophila Hox protein Ultrabithorax. Development 132, 5271-5281 (2005)), 아라비도프시스 탈리아나로부터의 IAA28 억제 도메인 (IAA28-RD)(4), mSin 상호작용 도메인 (SID) (Ayer, D.E., Laherty, CD., Lawrence, Q.A., Armstrong, A.P. & Eisenman, R.N. Mad proteins contain a dominant transcription repression domain. Mol. Cell. Biol. 16, 5772-5781 (1996)), Tbx3 억제 도메인 (Tbx3-RD), 및 호모 사피엔스 (Homo Sapiens) 로부터의 크루펠-연관 박스 (KRAB) (Margolin, J.F. et al. Kruppel-associated boxes are potent transcriptional repression domains. Proc. Natl. Acad. Sci. U S A 91, 4509-4513 (1994)) 억제 도메인을 포함하는 강력한 합성 억제제 발견 기회를 증가시켰다. KRAB 의 상이한 절두가 가변적 수준의 전사 억제를 나타내는 것으로 공지되어있기 때문에 (Margolin, J.F. et al. Kruppel-associated boxes are potent transcriptional repression domains. Proc. Natl. Acad. Sci. U S A 91, 4509-4513 (1994)), KRAB 의 3 개 상이한 절두를 시험하였다 (도 12c). 이러한 TALE 억제제 후보물을 HEK 293FT 세포에서 발현시켰고, 이는 2 개의 광범위하게 사용되는 포유동물 전사 억제 도메인, SID (Ayer, D.E., Laherty, CD., Lawrence, Q.A., Armstrong, A.P. & Eisenman, R.N. Mad proteins contain a dominant transcription repression domain. Mol. Cell. Biol. 16, 5772-5781 (1996)) 및 KRAB (Margolin, J.F. et al. Kruppel-associated boxes are potent transcriptional repression domains. Proc. Natl. Acad. Sci. U S A 91, 4509-4513 (1994)) 도메인을 운반하는 TALE 이 내인성 SOX2 발현을 억제할 수 있는 한편, 다른 도메인은 전사 활성에 대해 적은 효과를 갖는다는 것이 발견되었다 (도 12c). TALE 결합으로 인한 SOX2 전사의 잠재적 교란을 제어하기 위해서, 임의 효과기 도메인을 갖지 않는 SOX2-표적화 TALE DNA 결합 도메인 단독은 SOX2 의 전사 활성에 대해 효과를 갖지 않았다 (GFP 의 발현 또는 모의물과 유사) (도 12c, Null 조건). SID 도메인이 KRAB 도메인보다 내인성 SOX2 유전자좌를 26% 더 전사 억제시킬 수 있었기 때문에 (도 12c), 후속 연구를 위해 SID 도메인을 사용하는 것으로 결정되었다.
내인성 전사의 하향 조절을 위한 SID 억제제 도메인의 유효성을 추가로 시험하기 위해서, SID 를 이전의 실험으로부터의 CACNA1C-표적 TALE 과 조합하였다 (도 12d). qRT-PCR 을 사용하여, CACNA1C-표적화 TALE 상의 VP64 도메인의 SID 로의 대체가 CACNA1C 전사를 억제할 수 있었다는 것이 발견되었다. NH-함유 TALE 억제제는 NN-함유 TALE 과 유사한 수준의 전사 억제를 달성할 수 있었던 한편 (~4 배 억제), NK 를 사용하는 TALE 억제제는 상당히 덜 활성이었다 (~2 배 억제) (도 12d). 이러한 데이터는 SID 가 실제로 적합한 억제 도메인이라는 것을 입증하면서, 또한 NK 보다 더 적합한 G-표적화 RVD 로서 NH 를 추가로 지지한다.
TALE 은 내인성 게놈 상의 특정 서열을 인지하도록 용이하게 커스텀화될 수 있다. 본원에서, TALE 도구상자의 두 가지 중요한 한계를 다루기 위해 일련의 스크리닝을 수행하였다. 함께, 강건한 TALE 억제제 구조 뿐 아니라 미절충된 활성 강도를 갖는 보다 엄중한 G-특이적 RVD 의 확인은 포유동물 전사 및 게놈 기능을 조사하기 위한 TALE 의 유용성을 추가로 확장시킨다.
TALE 과 함께 사용할 강건한 신규 억제제 도메인으로서 SID (mSin 상호작용 도메인) 를 확인한 후, 포유동물 세포에서 TALE 과 함께 사용하기 위한 SID 도메인을 기반으로 하는 보다 활성인 억제제 도메인 구조를 또한 설계하고 확인하였다. 상기 도메인은 SID4X 로 지칭되며, 이는 짧은 펩티드 링커에 의해 연결된 4 개 SID 도메인의 탠덤 반복물이다. 상이한 TALE 억제제 구조를 시험하기 위해, 마우스 (Mus musculus) p11 (s1OOa1O) 유전자의 프로모터를 표적화하는 TALE 을 사용하여, 일련의 후보 TALE 억제제 구조의 전사 억제 활성을 평가하였다 (도 13a). TALE 의 상이한 절두가 가변적 수준의 전사 활성화 활동을 나타내는 것으로 공지되어 있기 때문에, SID 또는 SID4X 도메인에 융합된 TALE 의 2 개 상이한 절두를 시험하였는데, 한 형태는 DNA 결합 탠덤 반복물에 측면 위치하는 N- 및 C-말단에서 136 및 183 개 아미노산을 갖고, 또 다른 형태는 N- 및 C-말단에서 240 및 183 개 아미노산을 보유한다 (도 13b, c). 후보 TALE 억제제를 마우스 Neuro2A 세포에서 발현시켰고, SID 및 SID4X 도메인 모두를 운반하는 TALE 이 내인성 p11 발현을 4.8 배까지 억제할 수 있는 한편, GFP-인코딩 음성 대조군 구축물은 표적 유전자의 전사에 효과를 갖지 않는다는 것이 발견되었다 (도 13b, c). TALE 결합으로 인한 p11 전사의 잠재적 교란을 제어하기 위해, 임의의 효과기 도메인을 갖지 않는 p11-표적화 TALE DNA 결합 도메인 (시험한 구축물과 동일한 N- 및 C-말단 절두를 가짐) 은 내인성 p11 의 전사 활성에 대해 효과를 갖지 않았다 (도 13b, c, null 구축물).
SID4X 도메인을 포함하는 구축물이 TALE DNA 결합 도메인의 절두에 따라 SID 도메인보다 내인성 p11 유전자좌를 167% 및 66% 더 전사 억제할 수 있었기 때문에 (도 13c), SID4X 도메인에 융합된, 각각 N- 및 C-말단에서 136 및 183 개 아미노산을 포함하는 절두된 TALE DNA 결합 도메인이 표적 유전자 발현의 하향 조절을 가능하게 하는 강력한 TALE 억제제 구조이며 SID 도메인을 이용하는 이전 설계보다 더 활성이라는 결론이 내려졌다.
mSin 상호작용 도메인 (SID) 및 SID4X 도메인은 포유동물 발현에 대해 코돈 최적화되었으며 측면위치 NheI 및 XbaI 제한 부위 (Genscript) 로 합성되었다. TALE DNA 결합 도메인의 절두 변이체를 PCR 증폭하고 NheI 및 XbaI 제한 부위를 사용하여 SID 또는 SID4X 도메인에 융합시킨다. TALE 결합으로 생긴 전사에 대한 임의의 효과를 제어하기 위해서, PCR 클로닝을 사용하여 TALE DNA 결합 도메인 단독만을 운반하는 발현 벡터를 구축하였다. 모든 구축물의 코딩 부위를 Sanger 서열분석을 사용하여 완전히 확인하였다. 2 개 상이한 유형의 TALE 구조의 비교를 도 14 에 나타낸다.
실시예 4
포유동물 TALE 전사 활성제 및 뉴클레아제의 개발
커스텀화된 TALE 은 전사 조정 및 게놈 편집을 포함하는 광범위한 게놈 조작 적용에 사용될 수 있다. 본원에서, 출원인은 계층적 라이게이션 절차를 사용하는 커스텀 TALE 전사 인자 (TALE-TF) 및 뉴클레아제 (TALEN) 의 신속한 구축을 위한 도구상자를 기재하고있다. 상기 도구상자는 1 주일 내 커스텀 TALE-TF 및 TALEN 의 알맞고 신속한 구축을 촉진시키며, 동시에 다수의 표적에 대한 TALE 을 구축하기 위해 용이하게 규모 확대될 수 있다. 출원인은 또한 각각 정량 역전사 PCR 및 Surveyor 뉴클레아제를 사용하여 커스텀 TALE-TF 및 TALEN 의 포유동물 세포에서의 활성을 시험하기 위한 세부사항을 제공한다. TALE 도구상자는 광범위한 생물학적 적용을 가능하게 할 것이다.
TALE 은 숙주 식물에서 유전자 전사를 조정하여 박테리아 군집화를 촉진시키기 위해 잔토모나스 종에 의해 사용된 자연적 박테리아 효과기 단백질이다 (Boch, J. & Bonas, U. Xanthomonas AvrBs3 family-type III effectors: discovery and function. Annu. Rev. Phytopathol. 48, 419-436 (2010) and Bogdanove, A. J., Schornack, S. & Lahaye, T. TAL effectors: finding plant genes for disease and defense. Curr. Opin. Plant Biol. 13, 394-401 (2010)). 단백질의 중심 부위는 34-aa 서열의 탠덤 반복물을 함유하며 (단량체로 지칭) 이는 DNA 인지 및 결합에 필요하다 (Romer, P. et al. Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science 318, 645-648 (2007); Kay, S., Hahn, S., Marois, E., Hause, G. & Bonas, U. A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science 318, 648-651 (2007); Kay, S., Hahn, S., Marois, E., Wieduwild, R. & Bonas, U. Detailed analysis of the DNA recognition motifs of the Xanthomonas type III effectors AvrBs3 and AvrBs3Deltarepl6. Plant J. 59, 859-871 (2009) and Romer, P. et al. Recognition of AvrBs3-like proteins is mediated by specific binding to promoters of matching pepper Bs3 alleles. Plant Physiol. 150, 1697-1712 (2009).) (도 8). 자연 발생적 TALE 은 1.5 내지 33.5 개 범위의 가변적 수의 단량체를 갖는 것으로 발견되었다 (Boch, J. & Bonas, U. Xanthomonas AvrBs3 family-type III effectors: discovery and function. Annu. Rev. Phytopathol. 48, 419-436 (2010)). 각각의 단량체의 서열이 고도로 보존됨에도 불구하고, 이들은 반복 가변성 2 잔기로 지칭되는 2 개 위치에 있어서 주로 상이하다 (RVD, 12 번째 및 13 번째 위치). 최근 보고서는 이들 2 개 잔기의 동일성이 각각의 TALE 반복물의 뉴클레오티드-결합 특이성을 결정하며 간단한 암호가 각각의 RVD 의 표적 염기를 명시한다는 것을 발견한 바 있다 (NI = A, HD = C, NG = T, NN = G 또는 A) (Boch, J. et al. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509-1512 (2009) and Moscou, M.J. & Bogdanove, A.J. A simple cipher governs DNA recognition by TAL effectors. Science 326, 1501 (2009)). 따라서, 각각의 단량체는 하나의 뉴클레오티드를 표적화하고, TALE 에서의 단량체 선형 서열은 5' 에서 3' 배향으로 표적 DNA 서열을 명시한다. 식물 게놈 내의 자연적 TALE-결합 부위는 항상 티민으로 시작하는데 (Boch, J. et al. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509-1512 (2009) and Moscou, M.J. & Bogdanove, A.J. A simple cipher governs DNA recognition by TAL effectors. Science 326, 1501 (2009)), 이는 아마도 TALE 의 비반복적 N 말단 내 암호성 신호에 의해 명시된다. 탠덤 반복물 DNA-결합 도메인은 항상 반-길이 반복물로 종료된다 (0.5 반복물, 도 8). 따라서, 표적화되는 DNA 서열의 길이는 전체 반복 단량체의 수 + 2 와 동일하다.
식물에서, 병원체는 종종 숙주-특이적이다. 예를 들어, 푸사리움 옥시스포룸 (Fusarium oxysporum) f. sp. 리코페르시시 (lycopersici) 는 토마토 시듬병을 일으키나 오직 토마토만을 공격하고, F. 옥시스포룸 f. 디안티이 (dianthii) 푸시니아 그라미니스 (Puccinia graminis) f. sp. 트리티시 (tritici) 는 오직 밀만을 공격한다. 식물은 대부분의 병원체에 저항하기 위한 현존하며 유도된 방어 기작을 갖는다. 식물 세대에 걸친 돌연변이 및 재조합 이벤트는, 특히 병원체가 식물보다 더 많은 빈도로 복제하는 바, 민감성이 생기게 하는 유전적 변이성을 일으킨다. 식물에서는 비-숙주 저항성이 존재할 수 있는데, 예를 들어 숙주 및 병원체는 양립불가하다. 또한 수평 저항성, 예를 들어, 통상 많은 유전자에 의해 제어되는 모든 종류의 병원체에 대한 부분적 저항성, 및 수직 저항성, 예를 들어, 통상 소수 유전자에 의해 제어되는 일부 종류의 병원체에 대한 전체 저항성 (다른 종류에 대해서는 아님) 이 존재할 수 있다. 유전자 대 유전자 (Gene-for-Gene) 수준에 있어서, 식물 및 병원체는 함께 진화하며, 한 나머지에서의 유전적 변화는 다른 것에서 변화한다. 따라서, 자연적 가변성을 사용하여, 육종자는 수율, 품질, 균일성, 강인성, 저항성에 대해 가장 유용한 유전자를 조합한다. 저항성 유전자의 근원은 선천적 또는 외래 품종, 조상 전래 (Heirloom) 품종, 야생 식물 동류 및 유도된 돌연변이 (예를 들어 돌연변이유발제로 식물 물질 처리) 를 포함한다. 본 발명을 사용하여, 식물 육종자에게 돌연변이를 유도하는 새로운 도구가 제공된다. 따라서, 당업자는 저항성 유전자 근원의 게놈을 분석할 수 있으며, 원하는 특징 또는 특성을 갖는 품종에서 본 발명을 이용하여 이전의 돌연변이유발제보다 더 많은 정확도로 저항성 유전자의 발생을 유도하여, 이에 따라 식물 육종 프로그램을 가속화하고 향상시킨다.
출원인은, 잘못된 라이게이션이 최소화되도록 하는 라이게이션 어댑터의 비유사성 최대화 및 별개의 소화 및 라이게이션 단계의 단일 골든 게이트 (Golden Gate) (Engler, C, Kandzia, R. & Marillonnet, S. A one pot, one step, precision cloning method with high throughput capability. PLoS ONE 3, e3647 (2008); Engler, C, Gruetzner, R., Kandzia, R. & Marillonnet, S. Golden gate shuffling: a one-pot DNA shuffling method based on type lis restriction enzymes. PLoS ONE 4, e5553 (2009) and Weber, E., Engler, C, Gruetzner, R., Werner, S. & Marillonnet, S. A modular cloning system for standardized assembly of multigene constructs. PLoS ONE 6, el6765 (2011)) 반응으로의 조합을 비롯한 소수의 최적화를 갖는 TALE 어셈블리 시스템을 추가로 향상시킨다. 간단하게, 각각의 뉴클레오티드-특이적 단량체 서열은 TALE 탠덤 반복물 내의 단량체 위치를 고유하게 명시하는 라이게이션 어댑터로 증폭된다. 이러한 단량체 라이브러리가 생성되고나면, 이는 많은 TALE 의 어셈블리를 위해 편리하게 재사용될 수 있다. 필요한 각각의 TALE 에 대해서, 적절한 단량체가 먼저 육량체에 라이게이션되고, 그 다음 PCR 을 통해 증폭된다. 이후, 적절한 TALE 클로닝 백본을 갖는 두 번째 골든 게이트 소화-라이게이션 (도 8) 으로, 완전히 어셈블리된, 서열-특이적 TALE 이 산출된다. 백본은 TALE N 및 C 말단에 의해 측면 위치한 ccdB 음성 선택 카세트를 함유하는데, 이는 TALE 이 성공적으로 구축되는 경우 탠덤 반복물 DNA-결합 도메인에 의해 대체된다. ccdB 는 빈 백본으로 형질전환된 세포에 비하여 선택하여, 이로써 탠덤 반복물이 삽입된 클론이 산출된다 (Cermak, T. et al. Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res. 39, e82 (2011)).
단량체 DNA-결합 도메인의 어셈블리는 적절한 TALE-TF 또는 TALEN 클로닝 백본 내로 삽입되어 커스텀화 TALE-TF 및 TALEN 이 구축될 수 있다. TALE-TF 는 TALE C 말단 내 자연적 활성화 도메인을 합성 전사 활성화 도메인 VP64 로 대체함으로써 구축된다 (Zhang, F. et al. Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription. Nat. Biotechnol. 29, 149-153 (2011); 도 8). 전사 시작 부위의 결합 부위 업스트림을 표적화함으로써, TALE-TF 는 부위-특이적 방식으로 전사 복합체를 영입하고 유전자 전사를 개시한다. TALEN 은 TALE DNA-결합 도메인의 C-말단 절두 (+63 aa) (Miller, J.C. et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 29, 143-148 (2011)) 를 비특이적 FokI 엔도뉴클레아제 촉매 도메인과 융합하여 구축된다 (도 14). +63-aa C-말단 절두는 또한 전사 조정에 충분한 최소 C 말단으로서 기능하는 것으로 나타난다 (Zhang, F. et al. Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription. Nat. Biotechnol. 29, 149-153 (2011)). TALEN 은 ~17 개 염기에 의해 분리되는 2 개 표적 서열에 대한 결합을 통해 이량체를 형성한다. 결합 부위쌍 사이에서, FokI 촉매 도메인은 이량체화되며 이중 가닥 파단물 (DSB; 도 8) 을 도입함으로써 분자 가위로서 기능한다. 보통, DSB 는 비상동 말단-연결 (Huertas, P. DNA resection in eukaryotes: deciding how to fix the break. Nat. Struct. Mol. Biol. 17, 11-16 (2010)) 경로 (NHEJ) 에 의해 보수되어, 소 결실 및 기능적 유전자 녹아웃을 초래한다. 대안적으로, TALEN-매개 DSB 는 상동 재조합을 자극할 수 있어, 외인성 공여체 DNA 주형의 부위-특이적 삽입이 가능하게 한다 (Miller, J.C. et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 29, 143-148 (2011) and Hockemeyer, D. et al. Genetic engineering of human pluripotent cells using TALE nucleases. Nat. Biotechnol. 29, 731-734 (2011)).
VP64 활성화 도메인으로 구축되는 TALE-TF 와 함께, 본 발명의 다른 구현예는 VP16 및 p65 활성화 도메인으로 구축되는 TALE 폴리펩티드에 관한 것이다. 이러한 상이한 활성화 도메인의 Sox2 mRNA 수준에 대한 효과의 그래프적 비교를 도 11 에서 제공한다.
실시예 5
도 17 은 LITE 기능성에 대한 크립토크롬2 이형이량체 배향의 효과를 나타낸다. 뉴로제닌 (Neurogenin) 2 (Neurog2) LITE 의 2 개 형태를 합성하여 크립토크롬 2 포토리아제 상동성 부위 (CRY2PHR)/칼슘 및 인테그린-결합 단백질 1 (CIB1) 이량체 배향의 효과를 조사하였다. 한 형태에서, CIB1 도메인을 TALE (Neurog2) 도메인의 C-말단에 융합시킨 한편, CRY2PHR 도메인을 VP64 도메인의 N-말단에 융합시켰다. 반대 형태에서, CRY2PHR 도메인을 TALE (Neurog2) 도메인의 C-말단에 융합시킨 한편, CIB1 도메인을 VP64 도메인의 N-말단에 융합시켰다. 각각의 플라스미드 집합을 Neuro2a 세포에서 트랜스펙션시키고 자극한 후 (466 nm, 5 mW/cm2, 15 초 당 1 초 펄스, 12 시간) qPCR 분석을 위해 수확하였다. 자극 LITE 및 미자극 LITE Neurog2 발현 수준을 자극 GFP 대조군 샘플로부터의 Neurog2 수준으로 정규화시켰다. TALE-CRY2PHR/CIB1-VP64 LITE 는 상승된 기저 활성 및 높은 광 유도 Neurog2 발현을 나타내었는데, 이는 더 높은 절대 활성화가 필요한 상황에 대한 이의 적합성을 제시하였다. TALE-CIB1/CRY2PHR-VP64 LITE 의 상대적 광 유도 활성이 그의 대응물보다 낮았으나, 낮은 기저 활성은 최소 기준선 활성화를 필요로 하는 적용에서의 이의 유용성을 제시하였다. 또한, TALE-CIB1 구축물은 TALE-CRY2PHR 구축물에 비해 크기가 더 작았고, 바이러스 패키징과 같은 적용에 대해 잠재적 이점을 갖는다.
도 18 은 마우스 피질 뉴런 배양물에서의 대사자극성 글루타메이트 수용체 2 (mGlur2) LITE 활성을 나타낸다. mGluR2 표적화 LITE 를 플라스미드 pAAV-인간 시냅신 I 프로모터 (hSyn)-HA-TALE(mGluR2)-CIB1 및 pAAV-hSyn-CRY2PHR-VP64-2A-GFP 를 통해 구축하였다. 이러한 융합 구축물을 이후 아데노 연관 바이러스 벡터 (AAV) 내로 패키징하였다. 추가적으로, hSyn-TALE-VP64-2A-GFP 및 GFP 만을 운반하는 AAV 가 생성되었다. 배아 마우스 (E16) 피질 배양물을 폴리-L-리신 코팅된 24 웰 플레이트 상에 플레이팅하였다. 5 일 후 시험관내 뉴런 배양물을 TALE(mGluR2)-CIB1 및 CRY2PHR-VP64 AAV 스톡 (stock) 의 혼합물로 동시 형질도입하였다. 대조군 샘플을 TALE(mGluR2)-VP64 AAV 또는 GFP AAV 로 형질도입하였다. AAV 형질도입 6 일 후, 실험 샘플을 2 가지 광 펄스 패러다임 중 어느 하나를 사용하여 자극하였다: 분 당 0.5 초 및 30 초 당 0.25 초. 뉴런을 24 시간 동안 자극하고 qPCR 분석을 위해 수확하였다. 모든 mGluR2 발현 수준을 각각의 자극 GFP 대조군에 대해 정규화하였다. 데이터는 LITE 시스템이 시험관내 1 차 뉴런 배양물 중 표적 유전자의 광-의존적 활성화를 유도하는데 사용될 수 있다는 것을 제시하였다.
도 19 는 LITE AAV 벡터로의 1 차 마우스 뉴런의 형질도입을 나타낸다. 1 차 마우스 피질 뉴런 배양물을 LITE 시스템의 2 개 성분인 hSyn-CRY2PHR-VP64-2A-GFP 및 hSyn-HA-TALE-CIB1 을 인코딩하는 AAV 벡터로 5 일에 시험관내 동시-형질도입하였다. 좌측 패널: 형질도입 6 일 후, 뉴런 배양물은 hSyn-CRY2PHR-VP64-2A-GFP 벡터로부터 GFP 의 높은 발현을 나타내었다. 우측 패널: 동시-형질도입된 뉴런 배양물을 고정시키고 hSyn-HA-TALE-CIB1 에서의 TALE 도메인의 N-말단 상 HA 에피토프에 특이적인 항체로 염색하였다. 적색 신호는 HA 발현을 나타내었는데, 특히 핵 신호가 강력하였다 (청색 채널에서의 DAPI 에 의해 염색된 DNA). 이러한 영상은 함께, 각 LITE 성분의 발현이 1 차 마우스 뉴런 배양물에서 이루어질 수 있다는 것을 제시하였다 (스케일 바=50 um).
도 20 은 생체내 LITE 성분의 발현을 나타낸다. hSyn-CRY2PHR-VP64 를 운반하는 혈청형 1/2 의 AAV 벡터를 HEK293FT 세포의 트랜스펙션을 통해 생성시키고, 헤파린 컬럼 결합을 통해 정제하였다. 미손상 마우스 뇌 내로의 주입을 위해 벡터를 농축하였다. 1 uL 의 정제된 AAV 스톡을, 정위 수술 및 주사에 의해 8 주령 수컷 C57BL/6 마우스의 해마 및 변연계 아래 피질에 주사하였다. 생체내 형질도입 7 일 후, 마우스를 안락사시키고 뇌 조직을 파라포름알데히드 관류에 의해 고정하였다. 뇌 박편을 바이브라톰 상에서 제조하고, 영상화를 위해 장착하였다. 해마 및 변연계 아래 피질에서의 강력하고 광범위한 GFP 신호는 LITE 성분 CRY2PHR-VP64 의 효율적 형질도입 및 높은 발현을 제시하였다.
실시예 6
NES 요소 사용에 의한 향상된 설계
에스트로겐 수용체 T2 (ERT2) 는 누수 쟁점을 갖는다. ERT2 도메인은 4-히드록시테스토스테론 (4OHT) 부재 하에서도 핵에 유입될 수 있어, TAL 에 의한 표적 유전자 활성화의 배경 수준을 초래한다. NES (핵 이출 신호) 는 살아있는 세포의 세포질에 대해 단백질을 표적화하는 펩티드 신호이다. NES 를 현존 구축물에 첨가함으로써, 출원인은 4OHT 의 부재 하 ERT2-TAL 단백질이 핵 내로의 유입을 방지하여, ERT2 도메인의 "누수" 로 인한 배경 활성화 수준을 저하시키는 것을 목적으로 한다.
도 21 은 사용한 특정 NES 펩티드 서열이 LDLASLIL 인 구축물의 향상된 설계를 나타낸다.
도 22 는 40H 타목시펜의 부재 및 존재 하에서의 Sox2 mRNA 수준을 나타낸다. Y-축은 qRT-PCR 에 의해 측정된 바와 같은 Sox2 mRNA 수준이다. X-축은 상부에서 기재한 상이한 구축물 설계의 패널이다. + 및 - 표시는 0.5uM 4OHT 의 존재 또는 부재를 나타낸다.
실시예 7
CRISPR/Cas 시스템을 사용하는 멀티플렉스 게놈 조작
일상적 유전자 변이체 및 요소의 기능적 해명은 정확한 게놈 편집 기술을 필요로 한다. 유형 II 원핵생물 CRISPR (군집화 규칙적 공간화 단길이 회귀성 반복물) 조정 면역계는 RNA-안내 부위-특이적 DNA 절단을 촉진시키는 것으로 나타난 바 있다. 출원인은 2 개 상이한 유형 II CRISPR 시스템을 조작하였고, Cas9 뉴클레아제가 단길이 RNA 에 의해 유도되어 인간 및 마우스 세포에서 내인성 게놈 유전자좌에서의 정확한 절단을 유도할 수 있다는 것을 입증한다. Cas9 는 또한 절단 효소로 전환되어 최소의 돌연변이유발 활성으로 상동성-유도 수복을 촉진시킬 수 있다. 마지막으로, 다수의 안내 서열은 단일 CRISPR 어레이에 인코딩되어 포유동물 게놈 내의 여러 부위의 동시 편집을 가능하게 하는데, 이는 CRISPR 기술의 용이한 프로그램성 및 광범위한 적용성을 입증한다.
원핵생물 CRISPR 적응 면역계는 재구성되고 조작되어 포유동물 세포에서의 멀티플렉스 게놈 편집을 매개할 수 있다.
정확하고 효율적인 게놈 표적화 기술이 개별적 유전 요소의 선택적 교란을 가능하게 함으로써 일상적 유전적 변이의 조직적 역조작을 가능하게 하기 위해서 필요하다. 게놈-편집 기술 예컨대 설계자 아연 핑거 (ZF) (M. H. Porteus, D. Baltimore, Chimeric nucleases stimulate gene targeting in human cells. Science 300, 763 (May 2, 2003); J. C. Miller et al., An improved zinc-finger nuclease architecture for highly specific genome editing. Nat Biotechnol 25, 778 (Jul, 2007); J. D. Sander et al, Selection-free zinc-finger-nuclease engineering by context-dependent assembly (CoDA). Nat Methods 8, 67 (Jan, 2011) and A. J. Wood et al, Targeted genome editing across species using ZFNs and TALENs. Science 333, 307 (Jul 15, 2011)), 전사 활성제-유사 효과기 (TALE) (A. J. Wood et al, Targeted genome editing across species using ZFNs and TALENs. Science 333, 307 (Jul 15, 2011); M. Christian et al., Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 186, 757 (Oct, 2010); F. Zhang et al, Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription. Nat Biotechnol 29, 149 (Feb, 2011); J. C. Miller et al., A TALE nuclease architecture for efficient genome editing. Nat Biotechnol 29, 143 (Feb, 2011); D. Reyon et al, FLASH assembly of TALENs for high-throughput genome editing. Nat Biotechnol 30, 460 (May, 2012); J. Boch et al., Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509 (Dec 11, 2009) and M. J. Moscou, A. J. Bogdanove, A simple cipher governs DNA recognition by TAL effectors. Science 326, 1501 (Dec 11, 2009)), 및 호밍 (homing) 메가뉴클레아제 (B. L. Stoddard, Homing endonuclease structure and function. Quarterly reviews of biophysics 38, 49 (Feb, 2005)) 가 표적화 게놈 변형이 가능하도록 시작함에도 불구하고, 확장가능하고, 알맞고, 조작하기 용이한 새로운 기술에 대한 필요성이 존재한다. 여기서, 출원인은 유형 II 원핵생물 CRISPR 적응 면역계 (H. Deveau, J. E. Garneau, S. Moineau, CRISPR/Cas system and its role in phage-bacteria interactions. Annual review of microbiology 64, 475 (2010); P. Horvath, R. Barrangou, CRISPR/Cas, the immune system of bacteria and archaea. Science 327, 167 (Jan 8, 2010); K. S. Makarova et al., Evolution and classification of the CRISPR-Cas systems. Nat Rev Microbiol 9, 467 (Jun, 2011) and D. Bhaya, M. Davison, R. Barrangou, CRISPR-Cas systems in bacteria and archaea: versatile small RNAs for adaptive defense and regulation. Annu Rev Genet 45, 273 (2011)) 로부터의 RNA-유도 Cas9 뉴클레아제 기반의 정확도 게놈 조작 도구의 새로운 클래스의 개발을 보고하고있다 (M. Jinek et al, A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816 (Aug 17, 2012); G. Gasiunas, R. Barrangou, P. Horvath, V. Siksnys, Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc Natl Acad Sci USA 109, E2579 (Sep 25, 2012) and J. E. Garneau et al, The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 468, 67 (Nov 4, 2010)).
스트렙토코쿠스 피오게네스 SF370 유형 II CRISPR 유전자좌는 Cas9 뉴클레아제 뿐 아니라 2 개 비-코딩 RNA: tracrRNA 및 예비-crRNA 어레이 (동일한 직접적 반복물 (DR) 에 의해 공간이 남겨진 뉴클레아제 안내 서열 (스페이서) 을 함유함) 를 포함하는 4 개 유전자로 이루어진다 (도 27) (E. Deltcheva et al, CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 471, 602 (Mar 31, 2011)). 출원인은 핵심 성분의 이종 발현을 통해 포유동물 염색체에 표적화 이중 가닥 파단물 (DSB) 을 도입하기 위해 이러한 원핵생물 RNA-프로그램가능 뉴클레아제 시스템을 이용하는 것을 탐색하였다. tracrRNA, 예비-crRNA, 숙주 인자 RNase III 및 Cas9 뉴클레아제의 발현이 시험관내 DNA 절단 (M. Jinek et al., A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816 (Aug 17, 2012) and G. Gasiunas, R. Barrangou, P. Horvath, V. Siksnys, Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc Natl Acad Sci US A 109, E2579 (Sep 25, 2012)) 및 원핵세포 (R. Sapranauskas et al, The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic Acids Res 39, 9275 (Nov, 2011) and A. H. Magadan, M. E. Dupuis, M. Villion, S. Moineau, Cleavage of phage DNA by the Streptococcus thermophilus CRISPR3-Cas system. PLoS One 7, e40913 (2012)) 에서 필요하며 그에 충분하다는 것이 이전에 나타난 바 있다. 출원인은 S. 피오게네스 Cas9 (SpCas9) 및 RNase III (SpRNase III) 를 코돈 최적화하고 핵 국부화 신호 (NLS) 를 부착하여 포유동물 세포에서의 핵 구획화를 확실히 하였다. 인간 293FT 세포에서의 이러한 구축물의 발현이 SpCas9 를 핵에 대해 표적화하는데 필요하다는 것이 밝혀졌다 (도 23A). CRISPR 의 미-코딩 RNA 성분을 재구성하기 위해, 출원인은 RNA 중합효소 III U6 프로모터 하 (도 23B) 에 89-뉴클레오티드 (nt) tracrRNA (도 28) 를 발현시켰다. 유사하게, 출원인은 DR 에 측면 위치한 단일 안내 스페이서를 포함하는 예비-crRNA 어레이의 발현을 유도하기 위해 U6 프로모터를 사용하였다 (도 23B). 출원인은 NGG 에 선행하는 인간 EMX1 유전자좌에서 30-염기쌍 (bp) 부위를 표적화하기 위해 필요한 프로토스페이서 인접 모티프 (PAM) 인 초기 스페이서 (프로토스페이서) 를 설계하였다 (도 23C 및 도 27) (H. Deveau et ah, Phage response to CRISPR-encoded resistance in Streptococcus thermophilus. J Bacteriol 190, 1390 (Feb, 2008) and F. J. Mojica, C. Diez-Villasenor, J. Garcia-Martinez, C. Almendros, Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology 155, 733 (Mar, 2009)).
CRISPR 시스템 (SpCas9, SpRNase III, tracrRNA 및 예비-crRNA) 의 이종 발현이 포유동물 염색체의 표적화 절단을 달성할 수 있는지를 시험하기 위해, 출원인은 CRISPR 성분의 상이한 조합으로 293FT 세포를 트랜스펙션시켰다. 포유동물 DNA 에서의 DSB 가 삽입결실-형성 비-상동 말단 연결 (NHEJ) 경로에 의해 부분적으로 수복되기 때문에, 출원인은 내인성 표적 절단이 검출되도록 SURVEYOR 검정 (도 29) 을 사용하였다 (도 23D 및 도 28B). 모든 4 개의 필요한 CRISPR 성분의 동시-트랜스펙션은 프로토스페이서의 효율적 절단을 초래하였으며 (도 23D 및 도 28B), 이는 Sanger 서열분석에 의해 이후에 확인되었다 (도 23E). 흥미롭게도, SpRNase III 은 프로토스페이서의 절단에 필요하지 않았으며 (도 23D), 89-nt tracrRNA 는 이의 부재 하에 진행된다 (도 28C). 유사하게, 예비-crRNA 의 성숙은 RNase III (도 23D 및 도 30) 을 필요로 하지 않으며, 이는 예비-crRNA 성숙을 보조하는 내인성 포유동물 RNase가 존재할 수 있다는 것을 제시한다 (M. Jinek, J. A. Doudna, A three-dimensional view of the molecular machinery of RNA interference. Nature 457, 405 (Jan 22, 2009); C. D. Malone, G. J. Hannon, Small RNAs as guardians of the genome. Cell 136, 656 (Feb 20, 2009) and G. Meister, T. Tuschl, Mechanisms of gene silencing by double-stranded RNA. Nature 431, 343 (Sep 16, 2004)). 임의의 남아 있는 RNA 또는 Cas9 성분을 제거하는 것은 CRISPR 시스템의 게놈 절단 활성을 없앴다 (도 23D). 이러한 결과는 포유동물 세포에서 효율적 CRISPR-매개 게놈 변형을 위한 최소의 3-성분 시스템을 규정한다.
다음으로, 출원인은 EMX1 유전자좌 내의 추가적인 프로토스페이서를 표적화함으로써 진핵세포에서의 CRISPR-매개 절단의 일반화가능성을 탐색하였다 (도 24A). 동시-전달을 향상시키기 위해, 예비-crRNA 및 SpCas9 모두를 유도하도록 발현 벡터를 설계하였다 (도 31). 동시에, 출원인은 최근 시험관내 인증된 키메라 crRNA-tracrRNA 하이브리드 (도 24B, 상부) 설계를 조정하였는데 (M. Jinek et ah, A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816 (Aug 17, 2012)), 이때 자연적 crRNA:tracrRNA 듀플렉스를 모방하기 위해 성숙 crRNA 가 합성 줄기-루프를 통해 부분적 tracrRNA 에 융합된다 (도 24B, 하부). 출원인은 SpCas9 가 예비-crRNA (DR-스페이서-DR) 및 tracrRNA 와 동시-발현되는 경우 모든 프로토스페이서 표적의 절단을 관찰하였다. 그러나, 모든 키메라 RNA 설계가 그의 게놈 표적의 절단을 촉진시킬 수 있는 것은 아니다 (도 24C, 표 1). 출원인은 이후 인간 PVALB 및 마우스 Th 유전자좌를 표적화하는 예비-crRNA 및 키메라 RNA 를 설계함으로써 인간 및 마우스 세포 모두에서 추가적인 게놈 유전자좌의 표적화를 시험하였다 (도 32). 출원인은 crRNA:tracrRNA 설계를 사용하여 모든 3 개의 마우스 Th 및 1 개의 PVALB 표적에서 효율적 변형을 달성하였으며, 따라서 다수 유기체에 걸쳐 상이한 유전자좌를 변형시키는데 있어서 CRISPR 시스템의 광범위한 적용가능성을 입증한다 (표 1). 동일한 프로토스페이서 표적에 대해서, 키메라 RNA 의 절단 효율은 crRNA:tracrRNA 듀플렉스의 효율보다 더 낮거나 검출가능하지 않았다. 이는 RNA 의 발현 및 안정성에 있어서의 차이, 내인성 RNAi 장비에 의한 분해, 또는 비효율적 Cas9 로딩 또는 표적 인지를 초래하는 2 차 구조로 인한 것일 수 있다.
효율적 게놈 편집은 높은 정확도 및 효율 모두를 갖는 뉴클레아제 표적 특이적 게놈 유전자좌를 필요로 한다. CRISPR-매개 절단의 특이성을 조사하기 위해서, 출원인은 스페이서 및 이의 포유동물 프로토스페이서 표적 사이의 단일-뉴클레오티드 미스매치를 분석하였다 (도 25A). 출원인은 PAM 의 5' 12-bp 이하의 단일-염기 미스매치가 SpCas9 에 의한 게놈 절단을 완전히 없앤 반면, 더 먼 업스트림 돌연변이를 갖는 스페이서가 프로토스페이서 표적에 대한 활성을 보유하였다는 것을 관찰하였다 (도 25B). 이는 Cas9 특이성의 이전의 박테리아 및 시험관내 연구와 일치한다 (M. Jinek et ah, A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816 (Aug 17, 2012) and R. Sapranauskas et al, The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic Acids Res 39, 9275 (Nov, 2011)). 추가로, CRISPR 은 동일한 EMX1 프로토스페이서를 표적화하는 TALE 뉴클레아제 (TALEN) 쌍만큼 효율적으로 게놈 절단을 매개할 수 있다 (도 25, C 및 D).
게놈의 표적화된 변형은 이상적으로 실수-유발 NHEJ 메커니즘에서 발생하는 돌연변이를 방지한다. 야생형 SpCas9 는 부위-특이적 DSB 를 매개할 수 있는데, 이는 NHEJ 또는 상동성-유도 수복 (HDR) 을 통해 수복될 수 있다. 출원인은 SpCas9 의 RuvC I 도메인에서의 아스파르테이트-알라닌 치환 (D10A) 을 조작하여 뉴클레아제를 DNA 닉카아제로 전환시켰는데 (SpCas9n, 도 26A) (M. Jinek et al., A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816 (Aug 17, 2012); G. Gasiunas, R. Barrangou, P. Horvath, V. Siksnys, Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc Natl Acad Sci U S A 109, E2579 (Sep 25, 2012) and R. Sapranauskas et al, The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic Acids Res 39, 9275 (Nov, 2011)), 이는 니킹된 게놈 DNA 가 균일하게 또는 고-적합도 HDR 을 통해 통상 수복되기 때문이다. SURVEYOR (도 26B) 및 327 개 암플리콘의 서열분석은 SpCas9n 에 의해 유도된 임의의 삽입-결실을 검출하지 못했다. 그러나, 니킹된 DNA 가 드문 경우 DSB 중간체를 통해 진행될 수 있으며 NHEJ 이벤트를 초래한다는 것은 주목할만 하다 (M. T. Certo et al., Tracking genome engineering outcome at individual DNA breakpoints. Nat Methods 8, 671 (Aug, 2011)). 출원인은 프로토스페이서 근처의 제한 부위 쌍을 도입하기 위해 상동성 수복 주형을 갖는 동일한 EMX1 유전자좌에서의 Cas9-매개 HDR 을 시험하였다 (도 26C). SpCas9 및 SpCas9n 은 유사한 수준에서의 EMX1 유전자좌 내로의 수복 주형의 통합을 촉진하였는데 (도 26D), 출원인은 Sanger 서열분석을 통해 추가로 확인하였다 (도 26E). 이러한 결과는 표적화된 게놈 삽입을 촉진시키기 위한 CRISPR 의 유용성을 입증한다. 야생형 SpCas9 의 14-bp (시드 (seed) 서열로부터 12-bp 및 PAM 으로부터 2-bp) 표적 특이성 (도 25B) 을 고려하여, 닉카아제의 사용은 오프-타겟 돌연변이를 감소시킬 수 있다.
마지막으로, 어레이된 스페이서를 갖는 CRISPR 유전자좌의 자연적 구조 (도 27) 는 멀티플렉스화 게놈 조작의 가능성을 제시한다. EMX1- 및 PVALB-표적화 스페이서 쌍을 인코딩하는 단일 CRISPR 어레이를 사용하여, 출원인은 유전자좌 모두에서 효율적 절단을 검출하였다 (도 26F). 출원인은 119-bp 간격의 EMX1 내 2 개 표적에 대한 스페이서를 사용하여 동시발생 DSB 를 통해 더 큰 게놈 부위의 표적화된 결실을 추가로 시험하였고, 1.6% 결실 효능을 관찰하였으며 (182 개 암플리콘 중 3 개; 도 26G), 그에 따라 CRISPR 시스템이 단일 게놈 내에 멀티플렉스화 편집을 매개할 수 있다는 것을 입증하였다.
서열-특이적 DNA 절단을 프로그래밍하도록 RNA 를 사용하는 능력은 새로운 클래스의 게놈 조작 도구를 규정한다. 여기서, 출원인은 효율적 게놈 편집이 촉진되도록 S. 피오게네스 CRISPR 시스템이 포유동물 세포에서 이종으로 재구성될 수 있다는 것을 나타내며; 동반 연구는 여러 인간 세포주에서 고효율 CRISPR-매개 게놈 표적화를 독립적으로 확인하였다 (Mali et al.). 그러나, CRISPR 시스템의 여러 양태는 이의 효율 및 다재다능성이 증가되도록 추가로 향상될 수 있다. NGG PAM 에 대한 필요조건은 크로마틴 및 DNA 메틸화 상태로 인한 crRNA 2 차 구조 또는 게놈 접근성에 의해 제기된 잠재적 제약을 해명하지 않고 인간 게놈에서 평균 8-bp 마다로 S. 피오게네스 CRISPR 표적 공간을 제한한다 (도 33). 이러한 제한 중 일부는 미생물적 다양성에 걸친 (K. S. Makarova et al., Evolution and classification of the CRISPR-Cas systems. Nat Rev Microbiol 9, 467 (Jun, 2011)) 이의 상이한 PAM 필요조건 및 Cas9 효소 패밀리를 이용함으로써 극복될 수 있다 (H. Deveau et al., Phage response to CRISPR-encoded resistance in Streptococcus thermophilus. J Bacteriol 190, 1390 (Feb, 2008) and F. J. Mojica, C. Diez-Villasenor, J. Garcia-Martinez, C. Almendros, Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology 155, 733 (Mar, 2009)). 실제로, 다른 CRISPR 유전자좌는 포유동물 세포 내로 이식될 가능성이 있으며; 예를 들어, 스트렙토코쿠스 써모필루스 LMD-9 CRISPR1 은 또한 포유동물 게놈 절단을 매개할 수 있다 (도 34). 마지막으로, 포유동물 세포에서의 멀티플렉스 게놈 편집을 실행하는 능력은 기초 과학, 생물공학 및 의학에 걸쳐 강력한 적용을 가능하게 한다 (P. A. Carr, G. M. Church, Genome engineering. Nat Biotechnol 27, 1151 (Dec, 2009)).
실시예 8
CRISPR/Cas 시스템을 사용하는 멀티플렉스 게놈 조작: 보충 물질
세포 배양 및 트랜스펙션. 인간 배아 신장 (HEK) 세포주 293FT (Life Technologies) 를 10% 소 태아 혈청 (HyClone), 2mM GlutaMAX (Life Technologies), 100U/mL 페니실린 및 100㎍/mL 스트렙토마이신이 보충된 둘베코 개질 이글 배지 (DMEM) 에서 37℃ 에서 5% C02 인큐베이션하면서 유지시켰다. 마우스 neuro2A (N2A) 세포주 (ATCC) 를 37℃ 에서 5% C02 와 함께, 5% 소 태아 혈청 (HyClone), 2mM GlutaMAX (Life Technologies), 100U/mL 페니실린 및 100㎍/mL 스트렙토마이신이 보충된 DMEM 으로 유지시켰다.
웰 당 200,000 개 세포의 밀도로 트랜스펙션 1 일 전 293FT 또는 N2A 세포를 24-웰 플레이트 (Corning) 에 시딩하였다. 세포를 제조사의 권고 프로토콜에 따라 리포펙타민 2000 (Life Technologies) 을 사용하여 트랜스펙션하였다. 24-웰 플레이트의 각 웰에 대해 총 800ng 플라스미드를 사용하였다.
게놈 변형에 대한 Surveyor 검정 및 서열분석. 293FT 또는 N2A 세포를 상기 기재한 바와 같이 플라스미드 DNA 로 트랜스펙션하였다. 게놈 DNA 추출 전 트랜스펙션후 72 시간 동안 37℃ 에서 세포를 인큐베이션하였다. 게놈 DNA 를 제조사의 프로토콜에 따라 QuickExtract DNA 추출 키트 (Epicentre) 를 사용하여 추출하였다. 간략하게, 세포를 QuickExtract 용액에 재현탁하고 65℃ 에서 15 분 동안 및 98℃ 에서 10 분 동안 인큐베이션하였다.
각 유전자에 대한 CRISPR 표적 부위를 둘러싸고 있는 게놈 부위를 PCR 증폭하고, 생성물을 제조사의 프로토콜에 따라 QiaQuick Spin Column (Qiagen) 을 사용하여 정제하였다. 총 400ng 의 정제된 PCR 생성물을 2㎕ 10X Taq 중합효소 PCR 완충액 (Enzymatics) 및 초순수와 총 부피 20㎕ 로 혼합하고, 재-어닐링 프로세스를 거치게 하여 이형이중체가 형성되게 하였다: 95℃ 에서 10 분, 95℃ 에서 85℃ (-2℃/초로 램핑 (ramping)), 85℃ 에서 25℃ (-0.25℃/초), 및 25℃ 에서 1 분 고정. 재어닐링 후, 생성물을 제조사의 권고 프로토콜에 따라 SURVEYOR 뉴클레아제 및 SURVEYOR 인핸서 S (Transgenomics) 로 처리하고, 4-20 Novex TBE 폴리-아크릴아미드 겔 (Life Technologies) 에서 분석하였다. 겔을 SYBR Gold DNA 염색제 (Life Technologies) 로 30 분 동안 염색하고 Gel Doc 겔 영상화 시스템 (Biorad) 으로 영상화하였다. 정량은 상대적 밴드 강도를 기반으로 하였다.
상동 재조합의 검출을 위한 제한 단편 길이 다형성 검정. HEK 293FT 및 N2A 세포를 플라스미드 DNA 로 트랜스펙션하고, 37℃ 에서 72 시간 동안 인큐베이션한 후 상기 기재한 바와 같이 게놈 DNA 를 추출하였다. 표적 게놈 부위를 상동 재조합 (HR) 주형의 상동성 암 (arm) 외부의 프라이머를 사용하여 PCR 증폭하였다. PCR 생성물을 1% 아가로오스 겔 상에서 분리하고 MinElute GelExtraction 키트 (Qiagen) 를 사용하여 추출하였다. 정제된 생성물을 HindIII (Fermentas) 로 소화시키고 6% Novex TBE 폴리-아크릴아미드 겔 (Life Technologies) 에서 분석하였다.
RNA 추출 및 정제. HEK 293FT 세포를 이전에 언급한 바와 같이 유지시키고 트랜스펙션하였다. 세포를 트립신화에 의해 수확한 후 인산 완충 식염수 (PBS) 로 세척하였다. 총 세포 RNA 를 제조사의 프로토콜에 따라 TRI 시약 (Sigma) 으로 추출하였다. 추출한 총 RNA 를 Naonodrop (Thermo Scientific) 을 사용하여 정량하고 동일 농도로 정규화하였다.
포유동물 세포에서의 crRNA 및 tracrRNA 발현의 노던 블롯 분석. RNA 를 동일 부피의 2X 로딩 완충액 (Ambion) 과 혼합하고, 5 분 동안 95℃ 로 가열하고, 1 분 동안 얼음 상에서 냉각시킨 후, 8% 변성 폴리아크릴아미드 겔 (SequaGel, National Diagnostics) 상에, 30 분 이상 동안 겔을 사전-런닝시킨 후 로딩하였다. 샘플을 40 W 한도에서 1.5 시간 동안 전기영동하였다. 그 후, RNA 를 실온에서 1.5 시간 동안 반-건조 이동 장치 (Bio-rad) 내에서 300 mA 에서 Hybond N+ 멤브레인 (GE Healthcare) 에 옮겼다. RNA 를 Stratagene UV Crosslinker the Stratalinker (Stratagene) 상의 자동가교 버튼을 사용하여 멤브레인에 가교시켰다. 멤브레인을 ULTRAhyb-Oligo Hybridization 완충액 (Ambion) 중에서 30 분 동안, 42℃ 에서 회전시키면서 사전-하이브리드화한 후, 프로브를 첨가하고 밤새 하이브리드화하였다. 프로브를 IDT 에서 주문하고 T4 폴리뉴클레오티드 키나아제 (New England Biolabs) 와 함께 [감마-32P] ATP (Perkin Elmer) 로 표지하였다. 멤브레인은 사전가온된 (42℃) 2xSSC, 0.5% SDS 로 1 분 동안 1 회 세척한 후 42℃ 에서 2 회 30 분 세척하였다. 멤브레인은 1 시간 동안 또는 밤새 실온에서 인광체 스크린에 노출시킨 후 phosphorimager (Typhoon) 로 스캔하였다.
표 1. 포유동물 게놈 표적의 프로토스페이서 서열 및 변형 효율. 프로토스페이서 표적을 스트렙토코쿠스 피오게네스 유형 II CRISPR 및 스트렙토코쿠스 써모필루스 CRISPR1 유전자좌 (인간 및 마우스 게놈에서 3 개의 상이한 유전자에 대하여 그의 필요한 PAM 을 가짐) 를 기반으로 하여 설계하였다. 세포를 Cas9 및 precrRNA/tracrRNA 또는 키메라 RNA 로 트랜스펙션하였다. 세포를 트랜스펙션 72 시간 후 분석하였다. 삽입-결실 % 를 나타낸 세포주로부터의 SURVEYOR 검정 결과를 기반으로 계산하고, 모든 스페이서 표적에 대해 N = 3 이고, 오류는 S.E.M. 이고, N.D. 는 SURVEYOR 검정을 사용하여 검출불가능이고; N.T. 는 이 연구에서 시험하지 않은 것이다.

표 2. SURVEYOR 검정, RFLP 검정, 게놈 서열분석 및 노던 블롯에 사용한 프라이머 및 프로브에 대한 서열.

보충적 서열
> U6-단길이 tracrRNA (스트렙토코쿠스 피오게네스 SF370)

> U6-장길이 tracrRNA (스트렙토코쿠스 피오게네스 SF370)

> U6-DR-BbsI 백본-DR (스트렙토코쿠스 피오게네스 SF370)

> U6-키메라 RNA-BbsI 백본 (스트렙토코쿠스 피오게네스 SF370)

> 3xFLAG-NLS-SpCas9-NLS



> SpRNase3-mCherry-NLS


> 3xFLAG-NLS-SpCas9n-NLS (D10A 닉카아제 돌연변이에 밑줄 그음)



> hEMX1-HRTemplate-HindIII-NheI


> NLS-StCsn1-NLS




> U6-St_tracrRNA(7-97)

> EMX1_TALEN_좌측


>EMX1_TALEN_우측


실시예 9
AAV 구축물의 클로닝 (구축)
AAV-프로모터-TALE-효과기 백본의 구축. AAV-프로모터-TALE-효과기의 구축을 위해 백본을 표준 서브클로닝 방법에 의해 클로닝하였다. 특히, 벡터는 항생제 저항성 유전자, 예컨대 암피실린 저항성 및 프로모터-TALE-효과기 삽입물에 측면 위치한 2 개 AAV 역전 말단 반복물 (itr's) (서열, 하기 참조) 을 함유하였다. 프로모터 (hSyn), 효과기 도메인 (이 실시예에서 VP64, SID4X 또는 CIB1)/2 개 유형 IIS 제한 부위 (이 경우 BsaI) 를 갖는 스페이서를 함유하는 TALE 유전자의 N- 및 C-말단 부분을 상기 벡터 내로 서브클로닝하였다. 서브클로닝을 수행하기 위해, 각각의 DNA 성분을 중합효소-연쇄 반응을 사용하여 증폭한 후 특정 제한 효소로 소화시켜 매치되는 DNA 점성 말단을 생성시켰다. 벡터를 DNA 제한 효소로 유사하게 소화시켰다. 모든 DNA 단편을 이후 매치되는 말단에서 어닐링시키고 리가아제 효소를 사용하여 함께 융합하였다.
개별적 TALE 의 AAV-프로모터-TALE-효과기 백본 내로의 어셈플리.
상이한 TALE 단량체 서열을 상기 기재한 AAV-프로모터-TALE-효과기 백본 내로 통합하기 위한 계획은 유형 IIS 제한 효소로의 개별 단량체의 제한 및 그의 고유한 오버행 (overhang) 의 라이게이션으로 12 내지 16 개 단량체의 어셈블리를 형성시켜 최종 TALE 을 형성시키고 이를 N- 및 C-말단 사이의 스페이서에 존재하는 유형 IIS 부위를 사용하여 AAV-프로모터-TALE-효과기 백본 내로 이를 라이게이션하는 것을 기반으로 하였다 (골든 게이트 어셈블리로 지칭함). TALE 단량체 어셈블리의 이러한 방법은 이전에 기재된 바 있다 (NE Sanjana, L Cong, Y Zhou, M M Cunniff, G Feng & F Zhang A transcription activator-like effector toolbox for genome engineering Nature Protocols 7, 171-192 (2012) doi: 10.1038/nprot.2011.431).
상기 요약한 일반적인 클로닝 계획을 사용하여, 상이한 프로모터, 효과기 도메인 및 TALE 단량체를 함유하는 AAV 벡터를 용이하게 구축할 수 있다.
뉴클레오티드 서열:
좌측 AAV ITR
우측 AAV ITR
hSyn 프로모터

TALE N-말단 (+136 AA 절두)

TALE C-말단 (+63 AA 절두)

암피실린 저항성 유전자

실시예 10
내인성 포유동물 전사의 광학적 제어
내인성 포유동물 게놈의 전사를 직접적으로 조정하는 능력은 정상 유전자 기능 및 질환 메커니즘을 해명하기 위해 중요하다. 여기서, 출원인은 광-유도 전사 효과기 (LITE), 커스텀화가능한 TALE DNA-결합 도메인을 아라비도프시스 탈리아나로부터의 광-민감성 크립토크롬 2 단백질 및 이의 상호작용 파트너 CIB1 과 통합시키는 2-하이브리드 시스템의 개발을 기재하고 있다. LITE 는 곧 활성화될 수 있어, 내인성 포유동물 유전자 발현 뿐 아니라 표적화 후생적 크로마틴 변형의 가역성 양방향 조절을 매개한다. 출원인은 이러한 시스템을 1 차 마우스 뉴런 뿐 아니라 각성 상태의, 거동 중인 마우스의 뇌 내에 생체내 적용하였다. LITE 시스템은 내인성 세포 프로세스의 광유전학적 제어의 신규한 방식을 확립하며, 유전적 및 후생적 조절의 일상적 역할의 직접적인 시험을 가능하게 한다.
유전자 발현의 동적 성질은 생존계에서의 세포 프로그래밍, 항상성유지 및 환경적 적응을 가능하게 한다. 세포 및 유기체 기능에 대한 유전자의 기여도를 분석하는 것은 따라서 유전자 발현의 공간적 및 시간적 제어 조정을 가능하게 하는 접근 방식을 필요로 한다. 미생물 및 식물-유래 광-민감성 단백질은 광유전학적 활성제로서 조작되어 높은 시공간적 분해능을 제공하는 광의 사용이 많은 세포 기능을 제어할 수 있게 한다 (Deisseroth, K. Optogenetics. Nature methods 8, 26-29, doi: 10.1038/nmeth.f.324 (2011); Zhang, F. et al. The microbial opsin family of optogenetic tools. Cell 147, 1446-1457, doi: 10.1016/j .cell.2011.12.004 (2011); Levskaya, A., Weiner, O. D., Lim, W. A. & Voigt, C. A. Spatiotemporal control of cell signalling using a light-switchable protein interaction. Nature 461, 997-1001, doi: 10.1038/nature08446 (2009); Yazawa, M., Sadaghiani, A. M., Hsueh, B. & Dolmetsch, R. E. Induction of protein-protein interactions in live cells using light. Nature biotechnology 27, 941-945, doi: 10.1038/nbt. l569 (2009); Strickland, D. et al. TULIPs: tunable, light-controlled interacting protein tags for cell biology. Nature methods 9, 379-384, doi: 10.1038/nmeth.l904 (2012); Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi: 10.1038/nmeth. l524 (2010); Shimizu-Sato, S., Huq, E., Tepperman, J. M. & Quail, P. H. A light-switchable gene promoter system. Nature biotechnology 20, 1041-1044, doi: 10.1038/nbt734 (2002); Ye, H., Daoud-El Baba, M., Peng, R. W. & Fussenegger, M. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice. Science 332, 1565-1568, doi:10.1126/science.1203535 (2011); Polstein, L. R. & Gersbach, C. A. Light-inducible spatiotemporal control of gene activation by customizable zinc finger transcription factors. Journal of the American Chemical Society 134, 16480-16483, doi: 10.1021/ja3065667 (2012); Bugaj, L. J., Choksi, A. T., Mesuda, C. K., Kane, R. S. & Schaffer, D. V. Optogenetic protein clustering and signaling activation in mammalian cells. Nature methods (2013) and Zhang, F. et al. Multimodal fast optical interrogation of neural circuitry. Nature 446, 633-639, doi: 10.1038/nature05744 (2007)). 그러나, 광을 사용하여 내인성 전사 조절을 직접적으로 조정하기 위한 다목적이고 강건한 기술은 달성하기 어려운 채로 남아 있다.
여기서, 출원인은 광-유도 전사 효과기 (LITE), 포유동물 세포에서의 내인성 유전적 및 후생적 프로세스의 시공간적으로 정확한 제어를 가능하게 하는 모듈식 광유전학적 시스템의 개발을 보고하고 있다. LITE 는 잔토모나스 종으로부터의 전사 활성제-유사 효과기 (TALE) 의 프로그래밍가능한 DNA-결합 도메인 (Boch, J. et al. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509-1512, doi: 10.1126/science.l 178811 (2009) and Moscou, M. J. & Bogdanove, A. J. A simple cipher governs DNA recognition by TAL effectors. Science 326, 1501, doi: 10.1126/science.1178817 (2009)) 을 아라비도프시스 탈리아나로부터의 광-유도 이형이량체 단백질 크립토크롬 2 (CRY2) 및 CIB1 (Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi: 10.1038/nmeth.l524 (2010) and Liu, H. et al. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 322, 1535-1539, doi: 10.1126/science. 1163927 (2008)) 과 조합한다. 이들은 이종 유전자 요소의 도입을 필요로 하지 않고, 외인성 화학적 보조 인자에 의존적이지 않으며, 빠르고 가역성인 이량체화 반응속도를 나타낸다 (Levskaya, A., Weiner, O. D., Lim, W. A. & Voigt, C. A. Spatiotemporal control of cell signalling using a light-switchable protein interaction. Nature 461, 997-1001, doi: 10.1038/nature08446 (2009); Yazawa, M., Sadaghiani, A. M., Hsueh, B. & Dolmetsch, R. E. Induction of protein-protein interactions in live cells using light. Nature biotechnology 27, 941-945, doi: 10.1038/nbt.l569 (2009), Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi: 10.1038/nmeth. l524 (2010); Shimizu-Sato, S., Huq, E., Tepperman, J. M. & Quail, P. H. A light-switchable gene promoter system. Nature biotechnology 20, 1041-1044, doi: 10.1038/nbt734 (2002) and Liu, H. et al. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 322, 1535-1539, doi: 10.1126/science. 1163927 (2008)). 다른 광유전학적 도구와 같이, LITE 는 바이러스 벡터 내로 패키징될 수 있으며 특정 세포 집단을 탐색하도록 유전적으로 표적화될 수 있다. 출원인은 1 차 뉴런 뿐 아니라 마우스 뇌 (생체내) 에서의 이러한 시스템의 적용을 입증한다.
LITE 시스템은 2 개의 독립적 성분을 함유한다 (도 36A): 제 1 성분은 게놈 앵커 (anchor) 이며 광-민감성 CRY2 단백질에 융합된 커스텀화 TALE DNA-결합 도메인 (TALE-CRY2) 으로 이루어진다. 제 2 성분은 원하는 전사 효과기 도메인에 융합된 CIB1 으로 이루어진다 (CIB1-효과기). 효율적 핵 표적화를 확실히 하기 위해, 출원인은 핵 국부화 신호 (NLS) 를 양 모듈 모두에 부착시켰다. 광의 부재 하 (불활성 상태), TALE-CRY2 는 표적 유전자의 프로모터 부위에 결합하는 한편 CIB1-효과기는 핵 구획 내에 자유로이 남아있다. 청색광으로의 조명 (피크 ~450 nm) 은 CRY2 에서 형태적 변화를 촉발시키고 이후 CIB1-효과기 (도 36A 에서 나타낸 VP64) 를 표적 유전자좌에 영입하여 전사 조정을 매개한다. 이러한 모듈식 설계는 각각의 LITE 성분이 독립적을 조작될 수 있게 한다. 예를 들어, 동일한 게놈 앵커는 활성화 또는 억제 효과기와 조합되어 (Beerli, R. R., Segal, D. J., Dreier, B. & Barbas, C. F., 3rd. Toward controlling gene expression at will: specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proceedings of the National Academy of Sciences of the United States of America 95, 14628-14633 (1998) and Cong, L., Zhou, R., Kuo, Y.-c, Cunniff, M. & Zhang, F. Comprehensive interrogation of natural TALE DNA-binding modules and transcriptional repressor domains. Nat Commun 3, 968) 동일한 내인성 게놈 유전자좌에 걸쳐 양성적 및 음성적 전사 제어를 가할 수 있다.
가장 효율적인 LITE 구조를 확인하기 위해서, 출원인은 TALE 및 전사 활성제 VP64 (Beerli, R. R., Segal, D. J., Dreier, B. & Barbas, C. F., 3rd. Toward controlling gene expression at will: specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proceedings of the National Academy of Sciences of the United States of America 95, 14628-14633 (1998); Zhang, F. et al. Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription. Nat Biotechnol 29, 149-153, doi: 10.1038/nbt. l775 (2011); Miller, J. C. et al. A TALE nuclease architecture for efficient genome editing. Nature biotechnology 29, 143-148, doi: 10.1038/nbt. l755 (2011) and Hsu, P. D. & Zhang, F. Dissecting neural function using targeted genome engineering technologies. ACS chemical neuroscience 3, 603-610, doi: 10.1021/cn300089k (2012).) 를 각각 CRY2 및 CIB1 의 상이한 절두 (Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi: 10.1038/nmeth.l524 (2010)) 에 융합시키고, 신경 계통-명시 전사 인자 뉴로제닌 2 (Neurog2) 의 청색광 조명 유도된 전사 변화를 측정하여 각 설계의 효능을 평가하였다 (도 36B). 출원인은 전체 길이 CRY2 뿐 아니라 포토리아제 상동성 부위 단독으로 이루어지는 절두 (CRY2PHR, 아미노산 1-498) 를 평가하였다 (Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi: 10.1038/nmeth. l524 (2010)). CIB1 에 대해서, 출원인은 전체 길이 단백질 뿐 아니라 N-말단 도메인-단독 단편을 시험하였다 (CIBN, 아미노산 1-170) (Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi:10.1038/nmeth.l524 (2010)). 4 개 초기 LITE 페어링 중 3 개는 Neuro 2a 세포에서 유의한 광-유도 Neurog2 mRNA 상향조절을 생성시켰다 (p < 0.001, 도 36B). 이들 중에서, TALE-CRY2PHR::CIB1-VP64 는 GFP-단독 대조군 또는 미자극 LITE 샘플로 정규화되는 경우 가장 높은 완전 광-매개 mRNA 증가를 산출하였으며 (도 36B), 따라서 후속 실험에 적용되었다.
효율적 LITE 구조를 확립하여, 출원인은 파장 (도 40), 사용률 (도 41) 및 광 강도 (도 42 및 실시예 11) 를 포함하는 광 자극 매개변수를 체계적으로 최적화하였다 (Banerjee, R. et al. The signaling state of Arabidopsis cryptochrome 2 contains flavin semiquinone. The Journal of biological chemistry 282, 14916-14922, doi: 10.1074/jbc.M700616200 (2007)). 출원인은 또한 활성화 도메인 VP16 및 p65 를 VP64 에 추가로 비교하여 LITE CIB1-효과기 성분의 모듈성을 시험하였다. 모든 3 개 도메인은 유의한 광-의존적 Neurog2 mRNA 상향조절을 생성시켰다 (p < 0.001, 도 43). 출원인은 광-자극의 부재 하 그의 낮은 기저 활성으로 인해, 후속 실험을 위해 VP64 를 선택하였다.
발현 속도가 조절 요소, mRNA 처리 및 전사체 안정성을 포함하는 많은 인자에 의존하는 바, 내인성 유전자 발현의 조작은 다양한 도전과제를 제시한다 (Moore, M. J. & Proudfoot, N. J. Pre-mRNA processing reaches back to transcription and ahead to translation. Cell 136, 688-700, doi: 10.1016/j.cell.2009.02.001 (2009) and Proudfoot, N. J., Furger, A. & Dye, M. J. Integrating mRNA processing with transcription. Cell 108, 501-512 (2002)). CRY2 와 CIB1 사이의 상호작용이 초단위 이하의 기간으로 발생함에도 불구하고 (Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi: 10.1038/nmeth.l524 (2010)), LITE-매개 활성화는 전사의 내재 반응속도에 의해 제한될 가능성이 있다. 출원인은 30 분 내지 24 시간의 광 자극 시간 경과 동안 mRNA 수준을 측정함으로써 LITE-매개 Neurog2 발현의 온-키네틱스 (on-kinetics) 를 조사하였다 (도 36C). Neurog2 mRNA 의 상대적 수준은 광 자극 개시 30 분 후 만큼 상당히 일찍 증가하였으며 GFP-트랜스펙션된 음성 대조군에 비해 대략 20 배 상향조절되어 12 시간에서 포화될 때까지 꾸준히 상승하였다. 유사하게, 출원인은 6 시간 동안 세포를 자극하고 조명 중단 후 여러 시점에서의 Neurog2 전사체의 수준을 측정함으로써 시스템의 오프-키네틱스 (off-kinetics) 를 평가하였다 (도 36D). Neurog2 mRNA 수준은 자극후 30 분까지 짧게 증가하였으며, 효과는 잔류 CRY2PHR-CIB1 이량체화 또는 이전에 영입된 RNA 중합효소로 인한 것일 수 있다. 그 후, Neurog2 발현은 ~3 시간의 반감기로 감소하였는데, 이는 전사체가 광 자극의 부재 하 자연적 수준으로 복귀하는 것을 입증한다. 반대로, 식물 호르몬 아브시스산 수용체를 기반으로 한 소분자 유도 TALE 시스템 (Liang, F.-S., Ho, W. Q. & Crabtree, G. R. Engineering the ABA Plant Stress Pathway for Regulation of Induced Proximity. Sci. Signal. 4, rs2-, doi: 10.1126/scisignal.2001449 (2011)) 은 약물 확산, 대사 또는 청소 (clearance) 에 의해 잠재적으로 제한되는 더 느린 온- 및 오프-키네틱스를 나타내었다 (도 44).
출원인은 다음으로 바이러스 형질도입을 통한 뉴런 적용을 위한 LITE 의 유용성을 탐색하였다. 출원인은 TALE 유전자의 전달을 위한 아데노-연관 바이러스 (AAV)-기반 벡터 및 AAV 생성을 위한 단순화된 프로세스를 개발하였다 (도 37A 및 B, 도 45 및 실시예 11). AAV 의 ssDNA-기반 게놈은 재조합에 덜 취약하여, 렌티바이러스 벡터에 비하여 이점을 제공한다 (Holkers, M. et al. Differential integrity of TALE nuclease genes following adenoviral and lentiviral vector gene transfer into human cells. Nucleic acids research 41, e63, doi: 10.1093/nar/gksl446 (2013)).
1 차 마우스 피질 뉴런에서의 전사를 조정하기 위한 AAV-매개 TALE 전달을 특징화하기 위해, 출원인은 신경전달 또는 뉴런 분화에 관여된 유전자, 이온 채널 서브유닛, 및 신경학적 질환에 연루된 유전자를 포함하는 총 28 개 쥐과 유전자좌를 표적화하는 TALE-VP64 전사 활성제 패널을 구축하였다. 각 표적 유전자의 프로모터에서의 DNase I-민감성 부위는 TALE 결합 서열 선택에 대한 안내를 제공하였다 (도 46). 출원인은 TALE 활성이 출원인의 AAV-TALE 생성 프로세스를 사용하여 효율적으로 스크리닝될 수 있다는 것을 확인하였으며 (도 45), 이러한 방식으로 선택되고 AAV 벡터를 사용하여 1 차 뉴런에 전달된 TALE 이 다양한 어레이의 유전자 표적을 가변적 정도로 활성화시켰다는 것을 발견하였다 (도 37C). 또한, AAV-TALE 의 입체공간적 전달은 마우스 전전두엽 피질에서의 강건한 생체내 발현을 매개하였다 (도 37D, E). 마우스 변연계 아래 피질 (ILC) 에서의 TALE(Grm2)-VP64 의 발현은 GFP-주입 대조군에 비해 Grm2 mRNA 수준에 있어서 2.5 배 증가를 유도하였다 (도 37F).
TALE 활성제를 배양된 1 차 뉴런에 전달하여, 출원인은 다음으로 LITE 성분의 전달을 위한 벡터로서 AAV 를 사용하는 것을 탐색하였다. 이를 위해, 출원인은 LITE 전이유전자가 포함된 각각의 재조합 AAV 의 총 바이러스 게놈 크기가 4.8 kb 의 패키징 한계를 초과하지 않았다는 것을 확실히 할 필요가 있었다 (Wu, Z., Yang, H. & Colosi, P. Effect of Genome Size on AAV Vector Packaging. Mol Ther 18, 80-86 (2009)). 출원인은 TALE N- 및 C-말단을 단축시키고 (N-말단에서 136 aa 를, C-말단에서 63 aa 를 유지), CRY2PHR (1.5kb) 및 CIB1 (1kb) 도메인을 교환하였다 (TALE-CIB1 및 CRY2PHR-VP64; 도 38A). 이러한 LITE 를 2 개 AAV 벡터의 조합에 의해 동시-형질도입을 통해 1 차 피질 뉴런에 전달시켰다 (도 38B; 개별 성분에 대해 83-92% 의 전달 효율, >80% 동시-형질도입 효율). 출원인은 0.8% 의 감소된 사용률로 2 광 펄스 주파수에서 Grm2-표적화 LITE 를 시험하여 뉴런 건강성을 확인하였다 (도 47). 두 자극 조건 모두 Grm2 mRNA 수준에 있어서 ~7 배 광 의존적 증가를 달성하였다 (도 38C). 추가의 연구는 실질적 표적 유전자 발현 증가가 신속히 이루어질 수 있다는 것을 확인하였다 (4 시간 내 4 배 상향조절; 도 38D). 또한, 출원인은 자극 후 mGluR2 단백질의 유의한 상향조절을 관찰하였는데, 이는 mRNA 수준에서 LITE 에 의해 영향받은 변화가 단백질 수준으로 옮겨지는 것을 입증한다 (GFP 수준에 대해 p < 0.01, 무광 조건에 대해 p < 0.05; 도 38E).
LITE 시스템을 생체내 적용하기 위해서, 출원인은 CRY2PHR-VP64 LITE 성분 및 Grm2-표적화 TALE-CIB1 을 운반하는 고농도 AAV 벡터 (1012 DNAseI 저항성 입자/mL) 의 1:1 혼합물을 야생형 C57BL/6 마우스의 ILC 에 입체공간적으로 전달하였다. 생체내 LITE-발현 뉴런의 광학적 자극을 제공하기 위해서, 출원인은 주입 부위에서 광섬유 캐뉼라를 이식하였다 (도 38F 및 도 48) (Zhang, F. et al. Optogenetic interrogation of neural circuits: technology for probing mammalian brain structures. Nat Protoc 5, 439-456, doi: 10.1038/nprot.2009.226 (2010)). 주입 부위에서의 뉴런은 TALE(Grm2)-CIB1 및 CRY2PHR-VP64 모두를 발현하는 >80% 의 형질도입된 세포를 갖는 바이러스 모두에 의해 효율적으로 동시-형질도입되었다 (도 38G 및 도 49). 수술 후 8 일에, 출원인은 고체-상태 473 nm 레이저를 이식된 섬유 캐뉼라에 연결시켜 거동 중인 마우스의 ILC 를 자극하였다. 12 시간 자극 기간 후 (5 mW, 0.8% 사용률, 0.0167 Hz 에서 0.5 초 광 펄스 사용), Grm2 mRNA 에서의 변화에 대해 광 섬유 캐뉼라 이식 부위로부터의 뇌 조직을 분석하였다 (도 38H). 출원인은 미자극 ILC (p≤0.01) 에 비해 광 자극 후 Grm2 mRNA 에 있어서 유의한 증가를 관찰하였다. 종합하여, 이러한 결과로, LITE 가 배양된 뉴런에서 및 생체내에서 내인성 유전자 발현의 광학적 제어를 가능하게 한다는 것이 확인된다.
생체내 LITE 활성제의 무광 조건에서 관찰된 기저 상향조절의 지속성으로 인해, 출원인은 배경의 근원을 확인하고 약화시키며 광-매개 유전자 유도의 효율 (유전자 발현의 광/무광 비) 을 향상시키는 것을 목표로 하는 또 다른 라운드의 최적화를 수행하였다. LITE 표적화 성분 TALE-CIB1 만을 발현하는 뉴런은 LITE 성분 모두 (모두 GFP 대조군에 대해 p < 0.001) 를 발현하는 미자극 뉴런에서 발견된 바와 유사한 Grm2 mRNA 증가를 생성시킨 한편, 효과기 성분 CRY2PHR-VP64 단독은 전사에 유의하게 영향을 미치지 않았는데 (p > 0.05, 도 50), 이는 LITE 에 의한 배경 전사 활성화가 오직 DNA 표적화 성분으로부터만 발생할 수 있다는 것을 시사한다.
따라서, 출원인은 TALE-CIB1 에 의한 기저 표적 상향-조절을 감소시키기 위해 포괄적 스크리닝을 실행하였다 (도 51). 최적화는 다음의 2 가지 계획에 초점을 맞추었다: 첫 번째로, CIB1 은 식물 전사 인자이며 포유동물 세포에서도 고유 조절 효과를 가질 수 있다 (Liu, H. et al. Photoexcited CRY2 Interacts with CIB1 to Regulate Transcription and Floral Initiation III Arabidopsis. Science 322, 1535-1539, doi: 10.1126/science.1163927 (2008)). 출원인은 고등 식물의 염기성 헬릭스-루프 헬릭스 전사 인자 중 보존된 3 개 CIB1 부위를 결실시켜 이러한 효과를 없애는 것을 모색하였다 (도 51). 두 번째로, 출원인은 TALE-CIB1 이 광의 부재 하 표적 유전자좌에 결합하는 것을 방지하는 것을 목표로 하였다. 이를 달성하기 위해, 출원인은 LS-함유 CRY2PHR-VP64 로의 광-유도 이량체화때까지 세포질에서 국부화되도록 TALE-CIB1 을 조작하였다 (도 52). 2 가지 계획 모두를 시험하기 위해서, 출원인은 73 개의 별개 LITE 구조를 평가하고 12 개의 효과기-표적화 도메인 쌍을 확인하였는데 (도 51 및 도 53 에서 "+" 열로 나타냄) 이는 향상된 광-유도 효율 및 감소된 전체 기초선 모두를 가졌다 (본래 LITE1.0 과 비교한 무광 조건에서의 mRNA 증가 배수; p<0.05). LITE2.0 로 지정된, 2 가지 계획 모두를 통합하는 하나의 구조는 최고의 광 유도를 입증하였으며 (광/무광 = 20.4) 본래 구조와 비교하여 6 배 초과로 감소된 배경 활성화가 초래되었다 (도 38I). 또 다른 - LITE1.9.1 - 은 4 배 광 유도를 유지하면서 최소의 배경 mRNA 증가 (1.06) 를 생성시켰다 (도 53).
출원인은 TALE 및 LITE 조정에 의해 접근가능한 프로세스의 범위를 더 확장시키는 것을 모색하였다. 내인성 전사 억제는 종종 히스톤 메틸 메틸트랜스퍼라아제 (HMT) 및 데아세틸라아제 (HDAC) 와 같은 크로마틴 변형 효소에 의해 매개된다. 출원인은 이전에 mSin3 상호작용 도메인 (SID), mSin3-HDAC 복합체의 부분이 293FT 세포에서 표적 유전자를 하향조절하기 위하여 TALE 과 융합될 수 있다는 것을 나타낸 바 있다 (Beerli, R. R., Segal, D. J., Dreier, B. & Barbas, C. F., 3rd. Toward controlling gene expression at will: specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proceedings of the National Academy of Sciences of the United States of America 95, 14628-14633 (1998) and Cong, L., Zhou, R., Kuo, Y.-c., Cunniff, M. & Zhang, F. Comprehensive interrogation of natural TALE DNA-binding modules and transcriptional repressor domains. Nat Commun 3, 968, doi:http://www.nature.com/ncomms/journal/v3/n7/suppinfo/ncomms1962_S1.html (2012)). 이러한 TALE 억제제를 추가로 향상시키기 위해, 출원인은 VP64 의 쿼드러플 (quadruple) VP16 탠덤 반복물 구조와 유사한 SID 의 4 개 반복물 (Beerli, R. R., Segal, D. J., Dreier, B. & Barbas, C. F., 3rd. Toward controlling gene expression at will: specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proceedings of the National Academy of Sciences of the United States of America 95, 14628-14633 (1998)) 이 유전자 전사를 억제하는 이의 효력을 증가시킬 수 있다고 추론하였다. 실제로, TALE-SID4X 구축물은 293FT 세포에서 TALE-SID 만큼 2 배 효율적이었으며 (도 54A 및 54B) 또한 뉴런에서의 효율적 유전자 억제를 매개하였다 (도 54C 및 54D).
출원인은 내인성 유전자좌에 대한 히스톤 효과기의 TALE-매개 표적화가 특이적 후생적 변형을 유도할 수 있어, 후생 뿐 아니라 전사 역학의 탐색을 가능하게 한다는 가설을 세웠다 (도 39A). 출원인은 CRY2PHR-SID4X 구축물을 생성시켰으며, 표적화된 Grm2 프로모터에서의 H3K9 아세틸화에 있어서 ~2 배 감소에 부수적으로 (도 39D), 뉴런에서의 Grm2 의 광-매개 전사 억제를 입증하였다 (도 39B 및 도 39C). 유전자좌 특이적 히스톤 개질에 대한 히스톤 잔기 표적의 다양성을 확장시키려는 노력으로, 출원인은 문헌으로부터 일련의 억제성 히스톤 효과기 도메인을 유도하였다 (표 6). 광범위한 계통발생적 스펙트럼에 걸쳐 끌어내어, 도메인은 HDAC, 히스톤 메틸트랜스퍼라아제 (HMT) 및 히스톤 아세틸트랜스퍼라아제 (HAT) 저해제 뿐 아니라 HDAC 및 HMT 영입 단백질을 포함하였다. 효율적 AAV 패키징이 촉진되도록 작은 크기의 기능적 절두 및 단백질이 바람직하다. 생성된 후생적-변형 TALE-히스톤 효과기 융합 구축물 (epiTALE) 을 1 차 뉴런 및 Neuro 2a 세포에서 각각 Grm2 및 Neurog2 전사를 억제하는 그의 능력에 대해 시험하였다 (도 39E, 도 39F 및 도 55). 1 차 뉴런에서, 24 개 epiTALE 중 23 개가 p < 0.05 의 통계적 기준을 사용하여 grm2 의 전사를 성공적으로 억제하였다. 유사하게, Neuro 2a 세포에서의 epiTALE 발현은 시험한 32 개 히스톤 효과기 도메인 중 20 개에 대해 Neurog2 발현 감소를 초래하였다 (p < 0.05). 유망한 epiTALE 의 하위집합을 1 차 뉴 런 및 Neuro 2a 세포에서 발현시키고, 표적화된 내인성 프로모터에서의 상대적 히스톤 잔기 마크 (mark) 수준을 ChIP-RT-qPCR 에 의해 정량하였다 (도 39G, 도 39H 및 도 56). 1 차 뉴런 또는 Neuro 2a 세포에서, H3K9me1, H4K20me3, H3K27me3, H3K9ac 및 H4K8ac 의 수준은 각각, KYP (A. 탈리아나), TgSET8 (T, 곤디), NUE 및 PHF19 (C. 트라초마티스 (trachomatis) 및 H. 사피엔스), Sin3a, Sirt3 및 NcoR (모두 H. 사피엔스) 및 hdac8, RPD3 및 Sir2a (X. 라에비스 (laevis), S. 세레비지에, P. 팔시파룸 (falciparum)) 에서 유래한 epiTALE 에 의해 변형되었다. 이러한 도메인은 준비된 후생적 효과기의 근원을 제공하여 LITE 에 의한 전사 및 후생적 제어 범위를 확장시킨다.
뇌와 같은 이종 조직에서의 시공간적으로 정확한 생체내 유전자 조절을 달성하는 능력은 조사자로 하여금 발생, 학습, 기억 및 질환 진행과 같이 다양한 프로세스엣서의 동적 유전자 조절의 역할에 대해 의문을 갖게끔 한다. LITE 는 시간적으로 정확하고, 공간적으로 표적화된, 세포주, 1 차 뉴런 및 마우스 뇌에서의 생체내 내인성 유전자 발현의 바이모달 (bimodal) 제어를 가능하게 하는데 사용될 수 있다. LITE 의 TALE DNA 결합 성분은 광범위한 게놈 유전자좌를 표적화하도록 커스텀화될 수 있으며, 다른 DNA 결합 도메인 예컨대 RNA-안내 Cas9 효소 (Cong, L. et al. Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819823 (2013)) 를 TALE 대신 사용하여 다재다능한 유전자좌-특이적 표적화를 가능하게 할 수 있다 (도 57). 신규한 방식의 LITE 조정은 또한 효과기 모듈을 후생적 변형 효소와 같은 새로운 관능기로 대체함으로써 이루어질 수 있다 (de Groote, M. L., Verschure, P. J. & Rots, M. G. Epigenetic Editing: targeted rewriting of epigenetic marks to modulate expression of selected target genes. Nucleic acids research 40, 10596-10613, doi:10.1093/nar/gks863 (2012)). 따라서, LITE 시스템은 현존하는 광유전학적 도구상자에 대한 새로운 집합의 능력이 생길 수 있게 하며, 광을 사용하는 내인성 유전자 조절을 변형시키기 위한 매우 일반화가능하고 다재다능한 플랫폼을 구축한다.
방법 요약. LITE 구축물을 GenJet 를 사용하여 Neuro 2A 세포에 트랜스펙션하였다. TALE 또는 LITE 구축물을 운반하는 AAV 벡터를 사용하여 마우스 1 차 배아 피질 뉴런 뿐 아니라 마우스 뇌 (생체내) 를 형질도입하였다. RNA 를 추출하고 역전사하고 TaqMan-기반 RT-qPCR 을 사용하여 mRNA 수준을 측정하였다. 광 방사 다이오드 또는 고체 상태 레이저를 각각 조직 배양물 및 생체내 광 전달에 사용하였다.
LITE 의 설계 및 구축. 모든 LITE 구축물 서열은 실시예 11 에서 발견될 수 있다.
Neuro 2a 배양 및 실험. Neuro 2a 세포 (Sigma-Aldrich) 를 5% HyClone 열-불활성화 FBS (Thermo Scientific), 1% 페니실린/스트렙토마이신 (Life Technologies) 이 보충된, 1:1 비의 OptiMEM (Life Technologies) 대 GlutaMax 및 나트륨 피루베이트를 갖는 고-글루코오스 DMEM (Life Technologies) 을 함유하는 배지에서 성장시키고, 2 일마다 1:5 에서 계대하였다. 120,000 개 세포를 24-웰 플레이트의 각 웰에 트랜스펙션 18-20 시간 전에 플레이팅하였다. 트랜스펙션 1 시간 전에, 배지를 5% HyClone 열-불활성화 FBS 및 1% 페니실린/스트렙토마이신이 보충된 DMEM 으로 바꾸었다. 세포를, 제조사의 지시사항에 따라 1.5 ㎕ 의 GenJet (SignaGen Laboratories) 트랜스펙션 시약을 사용하여 웰 당 1.0 ㎍ 의 총 구축물 DNA (등몰비) 로 트랜스펙션하였다. 배지를 트랜스펙션 후 24 시간 및 44 시간에 교환하고, 광 자극을 48 시간에 시작하였다. 자극 매개변수는 하기와 같았다: 5 mW/cm2, 466 nm, 7% 사용률 (1 초 광 펄스 0.067 Hz), 도면 범례에서 다르게 나타내지 않는 한 24 시간 동안. RNA 를 제조사의 지시사항에 따라 RNeasy 키트 (Qiagen) 를 사용하여 추출하고, 샘플 당 1 ㎍ 의 RNA 를 qScript (Quanta Biosystems) 를 사용하여 역전사하였다. 상대적 mRNA 수준을, 표적화 유전자 뿐 아니라 내인성 대조군으로서 GAPDH 에 대해 특이적인 TaqMan 프로브 (Life Technologies, Taqman 프로브 ID 에 대해서는 표 3 참조) 를 사용하여 정량 실시간 PCR (qRT-PCR) 에 의해 측정하였다. △△Ct 분석을 사용하여, GFP 단독으로 형질도입되고 광 자극 처리된 음성 대조군에 대한 변화 배수를 수득하였다. LIVE/DEAD 검정 키트 (Life Technologies) 를 지시사항에 따라 사용하여 독성 실험을 수행하였다.
AAV 벡터 생성. 293FT 세포 (Life Technologies) 를 항생제 불포함 D10 배지 (GlutaMax 및 나트륨 피루베이트를 갖는 DMEM 고 글루코오스, 10% 열-불활성화된 Hyclone FBS 및 1% 1M HEPES) 에서 성장시키고, 매일 1:2-2.5 에서 계대하였다. 계대 총 수를 10 이하로 유지시키고, 세포가 절대 85% 밀집도 (confluence) 를 넘지 않도록 성장시켰다. 트랜스펙션 전 날에, 21.5 mL 의 D10 배지 중 1x106 세포를 15 cm 디쉬에 플레이팅하고 18-22 시간 동안 또는 ~80% 밀집도가 될 때까지 인큐베이션항였다. 트랜스펙션 시약으로서 사용하기 위해, 1 mg/mL 의 PEl "Max" (Polysciences) 를 물에 용해하고 용액의 pH 를 7.1 로 조정하였다. AAV 생성을 위해서, 10.4 ㎍ 의 pDF6 헬퍼 플라스미드, 8.7 ㎍ 의 pAAV1 혈청형 패키징 벡터, 및 5.2 ㎍ 의 pAAV 벡터 (관심 유전자 운반) 를 434 ㎕ 의 혈청-불포함 DMEM 에 첨가하고, 130 ㎕ 의 PEl "Max" 용액을 DMEM-희석 DNA 혼합물에 첨가하였다. DNA/DMEM/PEI 칵테일을 볼텍싱하고 실온에서 15 분 동안 인큐베이션하였다. 인큐베이션한 후, 트랜스펙션 혼합물을 22 mL 의 완전 배지에 첨가하고, 짧게 볼텍싱하고, 293FT 세포의 15 cm 디쉬에 대해 배지를 교체하는데 사용하였다. 상청액 생성을 위해, 트랜스펙션 산청액을 48 시간에 수확하고, 0.45 ㎛ PVDF 필터 (Millipore) 를 통해 여과하고, 분취액으로 분배하고, 저장을 위해 -80℃ 에서 동결시켰다.
1 차 피질 뉴런 배양. 해리된 피질 뉴런을 E16 (Charles River Labs) 상에서 C57BL/6N 마우스 배아로부터 제조하였다. 피질 조직을 빙냉 HBSS - (50 mL 10x HBSS, 435 mL dH20, 0.3 M HEPES pH 7.3, 및 1% 페니실린/스트렙토마이신) 중에서 해부하였다. 피질 조직을 20 mL 의 빙냉 HBSS 로 3 회 세척한 후 37℃ 에서 20 분 동안, 8 mL 의 HBSS 중에서 240 ㎕ 의 2.5% 트립신 (Life Technologies) 과 함께 소화시켰다. 이후 피질을 1 mL FBS 를 함유하는 20 mL 의 가온된 HBSS 로 3 회 세척하였다. 피질을 2 ml 의 HBSS 중에서 분말화하고, 폴리-D-리신 코팅된 24-웰 플레이트 (BD Biosciences) 에서 150,000 개 세포/웰로 플레이팅하였다. 뉴런을 1X B27 (Life Technologies), GlutaMax (Life Technologies) 및 1% 페니실린/스트렙토마이신이 보충된 Neurobasal 배지 (Life Technologies) 중에서 유지시켰다.
1 차 뉴런 형질도입 및 광 자극 실험. 1 차 피질 뉴런을 DIV 5 상에서 250 ㎕ 의 AAVI 상청액으로 형질도입하였다. 배지 및 상청액을 그 다음날 보통의 완전 neurobasal 로 대체하였다. Neurobasal 을 AAV 형질도입 6 일 후 1X B27, GlutaMax (Life Technologies) 및 1% 페니실린/스트렙토마이신을 함유하는 최소 필수 배지 (Life Technologies) 로 교환하여, 광 자극 동안 Neurobasal 에 함유된 HEPES 및 리보플라빈으로부터의 광독성 생성물의 형성을 방지하였다.
광 자극을 5 mW/cm2 의 강도, 0.8% 의 사용률 (0.033Hz 에서의 250 ms 펄스 또는 0.016Hz 에서의 500 ms 펄스), 24 시간 동안 466 nm 청색광 (도면 범례에서 다르게 나타내지 않는 한) 으로, AAV 형질도입 6 일 후 시작하였다 (DIV 11). Cells-to-Ct 키트를 제조사 지시사항에 따라 사용하여 (Life Technologies) RNA 추출 및 역전사를 수행하였다. Neuro 2a 세포에 대해 상기 기재한 바와 같은 TaqMan 프로브를 사용하여 정량 실시간 PCR (qRT-PCR) 에 의해 상대적 mRNA 수준을 측정하였다.
1 차 뉴런의 면역조직화학. 1 차 뉴런의 면역조직화학을 위해, 세포를 수확 후 폴리-D-리신/라미닌 코팅된 커버슬립 (BD Biosciences) 상에 플레이팅하였다. AAV1-형질도입을 상기 기재한 바와 같이 수행하였다. 뉴런을 4% 파라포름알데히드 (Sigma Aldrich) 로 15 분 동안 실온에서 형질도입 후 7 일 고정시켰다. 블로킹 및 침투화를 DPBS (Life Technologies) 중 0.5% Triton-X100 (Sigma-Aldrich) 및 10% 정상 염소 혈청 (Life Technologies) 으로 1 시간 동안 실온에서 수행하였다. 뉴런을 1 차 항체와 함께 밤새 4℃ 에서 인큐베이션하고, DPBS 로 3 회 세척하고 2 차 항체와 함께 90 분 동안 실온에서 인큐베이션하였다. 항체 제공처 및 사용 농도에 대해서는 표 4 를 참조한다. 커버슬립을 마지막으로, DAPI 와 함께 Prolong Gold Antifade 시약 (Life Technologies) 사용하여 고정하고 X-Cite 120Q 광원 (Lumen Dynamics) 을 갖는 Axio Scope A.1 (Zeiss) 상에서 영상화하였다. AxioCam MRm 카메라 및 AxioVision 4.8.2 를 사용하여 영상을 획득하였다.
웨스턴 블롯. 총 단백질 용해물의 제조를 위해, 빙냉 용해 완충액 (RIPA, 세포 신호전달; 0.1% SDS, Sigma-Aldrich; 및 complete ultra protease inhibitor mix, Roche Applied Science) 중에서 1 차 피질 뉴런을 광 자극 (상기 참조) 후 수확하였다. 세포 용해물을 Bioruptor 초음파발생장치 (sonicator) (Diagenode) 에서 'M' 설정으로 5 분 동안 초음파처리하고, 21,000 x g 에서 10 분 동안 4℃ 에서 원심분리하였다. RC DC 단백질 검정 (Bio-Rad) 을 사용하여 단백질 농도를 측정하였다. 레인 당 30-40 ㎍ 의 총 단백질을, Precision Plus Protein Dual Color Standard (Bio-Rad) 와 함께 4-15% Tris-HCl 겔 (Bio-Rad) 상에서 비환원 조건 하에 분리하였다. 폴리비닐리덴 디플루오라이드 멤브레인 (Millipore) 으로의 습식 전기이동 후, Tris 완충 식염수 (TBS, Bio-Rad) 중 5% BLOT-QuickBlocker (Millipore) 중에서 45 분 동안 멤브레인을 블로킹하고, 웨스턴 블롯을 항-mGluR2 (Abcam, 1:1.000) 및 항-α-튜불린 (Sigma-Aldrich 1:20,000) 으로 밤새 4℃ 에서 프로브 처리한 후, 세척하고 항-마우스-IgG HRP 항체 인큐베이션하였다 (Sigma-Aldrich, 1:5,000 - 1:10,000). 추가의 항체 세부사항에 대해서는 표 4 를 참조한다. ECL 웨스턴 블롯 기질 (SuperSignal West Femto 키트, Thermo Scientific) 을 통해 검출을 수행하였다. AlphaImager (Innotech) 시스템으로 블롯을 영상화하고, ImageJ 소프트웨어 1.46r 을 사용하여 정량하였다.
농축되고 정제된 AAV1/2 벡터의 생성. 생체내 입체공간적 주입을 위한 농축되고 정제된 AAV 의 생성을, AAV1 상청액 생성에 대해 상기 요약한 동일한 초기 단계를 사용하여 수행하였다. 그러나 트랜스펙션에 대해서는, 동일 비의 AAV1 및 AAV2 혈청형 플라스미드를 AAV1 단독 대신 사용하였다. 구축물 ekdd 5 개 플레이트를 트랜스펙션하고 세포를 트랜스펙션 후 48 시간에 세포-스크래퍼로 수확하였다. HiTrap 헤파린 친화성 컬럼 (GE Healthcare) 을 사용하여 AAV1/2 입자의 정제를 수행하였다 (McClure, c., Cole, K. L., Wulff, P., Klugmann, M. & Murray, A. J. Production and titering of recombinant adeno-associated viral vectors. J Vis Exp, e3348, doi:l0.3791/3348 (2011)). 출원인은 500 ㎕ 농축 컬럼 (100 kDa 절삭값, Millipore) 을 사용하여 구축물 당 100 ㎕ 의 최종 부피에 이르기까지 제 2 농축 단계를 추가하여, 더 높은 바이러스 역가를 수득하였다. WPRE 에 대한 커스텀 Taqman 프로브를 사용하여 (Life Technologies) qRT-PCR 에 의해 AAV 적정을 수행하였다. qRT-PCR 전에, 농축된 AAV 를 DNaseI (New England Biolabs) 으로 처리하여 DNaseI-저항성 입자만에 대한 측정값을 획득하였다. DNaseI 열-불활성화 후, 바이러스 외피를 프로테이나아제 K 소화 (New England Biolabs) 에 의해 분해시켰다. 바이러스 역가를, 공지된 WPRE 카피수를 갖는 표준 곡선을 기반으로 계산하였다.
AAV1/2 의 입체공간적 주입 및 광학적 이식. 모든 동물 절차는 동물 관리에 대한 MIT 위원회에 의해 승인받았다. 성체 (10-14 주령) 수컷 C57BL/6N 마우스를 케타민/자일라진 (100 mg/kg 케타민 및 10 mg/kg 자일라진) 을 복강내 (i.p.) 주입하여 마취하였고, 선제성 무통증이 주어졌다 (Buprenex, 1 mg/kg, i.p.). 승인된 절차에 따라 개두술을 수행하고 1 ㎕ 의 AAV1/2 을 0.35/1.94/-2.94 (브레그마 (bregma) 에 대해 mm 으로 측면, 전면 및 하면 좌표) 에서 ILC 에 주입하였다. 동일한 수술적 절차 동안, 섬유를 갖는 광학적 캐뉼라 (Doric Lenses) 를, 브레그마에 대하여 0.35/1.94/-2.64 에 위치한 광학 섬유의 말단에 단독으로 ILC 에 이식하였다. 상기 캐뉼라를 Metabond 치아용 시멘트 (Parkell Inc) 및 Jet 의치 보수물 (Lang dental) 을 사용하여 두개골에 부착시켜, 이를 둘러싼 안정한 원뿔형을 구축하였다. 절개부를 봉합하고 적절한 수술-후 진통제를 수술 후 3 일 동안 투여하였다.
ILC 뇌 섹션에 대한 면역조직화학. 마우스에 케타민/자일라진 마취제의 치사량을 주입하고 PBS 및 4% 파라포름알데히드 (PFA) 로 경심 관류하였다. 뇌를 4% PFA 중에서 4℃ 에서 밤새 추가적으로 고정시킨 후, 동결보호를 위해 밤새 실온에서 30% 수크로오스에 이동시켰다. 뇌를 이후 Tissue-Tek 최적 절단 온도 (Optimal Cutting Temperature) (OCT) 화합물 (Sakura Finetek) 에 옮기고 -80℃ 에서 동결시켰다. 18 ㎛ 섹션을 cryostat (Leica Biosystems) 상에서 절단하고 Superfrost Plus 유리 슬라이드 (Thermo Fischer) 에 탑재시켰다. 섹션을 15 분 동안 4% PFA 로 후-고정하고, 1 차 뉴런에 대해 상기 기재한 바와 같이 면역조직화학을 수행하였다.
ILC 에서의 광 자극 및 mRNA 수준 분석. 수술 후 8 일에, 각성 상태이며 자유로이 움직이는 마우스를 섬유 패치 케이블 및 회전식 연결부를 통해 광학 이식물에 연결된 473 nm 레이저원 (OEM Laser Systems) 을 사용하여 자극하였다. 자극 매개변수는 1 차 뉴런에 사용한 바와 동일하였다: 5 mW (총 출력), 0.8% 사용률 (0.016 Hz 에서 500 ms 광 펄스), 총 12 시간 동안. 형질도입된 구축물 및 광 자극을 포함하는 실험 조건을 표 5 에 열거한다.
광 자극 종료 후, 마우스를 CO2 를 사용하여 안락사시키고, 전액골 피질 (PFC) 을 얼음 상에서 신속히 해부하고 RNA later (Qiagen) 중에서 4℃ 에서 밤새 인큐베이션하였다. 200 ㎛ 섹션을 바이브라톰 (vibratome) (Leica Biosystems) 상에서 4℃ 에서 RNA later 중에서 절단하였다. 이후 섹션을 드라이 아이스 상 유리 커버슬라이드에서 동결시키고, 바이러스 형질도입된 ILC 를 형광 입체현미경 (Leica M165 FC) 하에 확인하였다. 광학 섬유 트랙 종료부에 직접적으로 배 쪽에 위치한 ILC 의 0.35 mm 직경 펀치를 추출하였다 (Harris uni-core, Ted Pella). 뇌 펀치 샘플을 이후 50 ㎕ Cells-to-Ct RNA 용해 완충액 중에서 RNase-불포함 펠렛-막자 그라인더 (Kimble Chase) 를 사용하여 균질화하고, RNA 추출, 역전사 및 qRT-PCR 을 1 차 뉴런 샘플에 대해 기재한 바와 같이 수행하였다.
크로마틴 면역침전. 뉴런 또는 Neuro2a 세포를 상기 기재한 바와 같이 배양하고 형질도입하거나 트랜스펙션하였다. ChIP 샘플을, 세포 수 및 세포 유형에 대해 소수 조정하여, 이전에 기재된 바와 같이 제조하였다 (Blecher-Gonen, R. et al. High-throughput chromatin immunoprecipitation for genome-wide mapping of in vivo protein-DNA interactions and epigenomic states. Nature protocols 8, 539-554 (2013)). 세포를 24-웰 포맷으로 수확하고, 96-웰 포맷으로 세척하고, 용해를 위해 미세원심분리 튜브에 옮겼다. 샘플을 이후, 30 초 온/오프 주기를 사용하여 21 분 동안 Biorupter 초음파처리 장치 (Diagenode) 로 수조 초음파처리에 의해 직접적으로 용해하였다. qPCR 을 사용하여 표적화된 유전자좌에서의 히스톤 마크의 축적을 평가하였다.
통계적 분석. 모든 실험을 최소 2 개의 독립적인 생물학적 복제품로 수행하였다. 2 개 조건 비교시 스튜던트 양방향 t-검정, 여러 샘플을 서로 비교시 터키 사후검정 분석과 함께 ANOVA, 및 여러 샘플을 음성 대조군에 대해 비교시 던넷 사후검정 분석과 함께 ANOVA 를 사용하여, Prism (GraphPad) 으로 통계적 분석을 수행하였다.
실시예 11
실시예 10 에 대한 보충적 정보: 내인성 포유동물 전사의 광학적 제어
광자극 하드웨어 - 시험관내. 커스텀 빌트 LED 광자극 장치를 사용하여 시험관내 광 자극 실험을 수행하였다. 모든 전자 부품을 커스텀 프린트 회로 보드 (ExpressPCB) 상에 탑재하였다. 피크가 466 nm 인 청색 LED (모델 #: YSL-R542B5C-All, China Young Sun LED Technology; 'LED - Super Bright Blue' COM-00529 로서 SparkFun Electronics 에 의해 배포됨) 를 Corning 24-웰 플레이트의 웰과 함께 정렬된 3 개 군에서 어레이화하였다. LED 전류 흐름을 25 mA DynaOhm 드라이버 (LEDdymanics #4006-025) 에 의해 조절하였다. LED 어레이의 열을 Arduino UNO 마이크로컨트롤러 보드를 통해 TTL 컨트롤 (Fairchild Semiconductor PN2222BU-ND) 에 의해 어드레스하였다. Thorlabs PM100D 파워 미터 및 S120VC 광다이오드 검출기를 이용하여 광 출력량을 어레이 위 80 mm 의 거리로부터 측정하였다. 환기 공간을 제공하고 광 필드 균일성을 최대화하기 위해, 80 mm 길이 환기 스페이서를 LED 어레이와 24-웰 샘플 플레이트 사이에 두었다. 팬 (Evercool EC5015M12CA) 을 스페이서 유닛의 한 웰에 따라 탑재하면서, 반대쪽 웰을 기류 증가가 가능하도록 간격을 두어 제작하였다.
ImageJ 소프트웨어를 사용하는 LIVE/DEAD® 검정의 정량. LIVE/DEAD (Life Technologies) 염색 세포의 영상을 형광 현미경에 의해 캡쳐하고 하기와 같이 처리하였다: 배경을 제한다 (Process → Subtract Background). 형광 부위를 기반으로 한 역치를 세포 상태의 정확한 확인이 가능하도록 설정한다 (Image → Adjust → Threshold). 분할 분석을 수행하여 개별 세포의 자동화된 계수가 가능하게 한다 (Process → Binary → Watershed). 마지막으로, 잔해 신호를 필터링하고 세포를 계수한다 (Analyze → Analyze Particles). 사멸 세포의 % 로서 독성을 측정하였다.
화학적 유도 TALE. Neuro2A 세포를 5% HyClone 열-불활성화 FBS (Thermo Scientific), 1% 페니실린/스트렙토마이신 (Life Technologies) 및 25mM HEPES (Sigma Aldrich) 가 보충된, 1:1 비의 OptiMEM (Life Technologies) 대 GlutaMax 및 나트륨 피루베이트를 갖는 고-글루코오스 DMEM (Life Technologies) 을 함유하는 배지에서 성장시켰다. 150,000 개 세포를 24-웰 플레이트의 각 웰에 트랜스펙션 18-20 시간 전에 플레이팅하였다. 세포를, 웰 당 1 ㎍ 의 총 구축물 DNA (등몰비) 및 2 ㎕ 의 리포펙타민 2000 (Life Technologies) 으로, 제조사의 권고 프로토콜에 따라 트랜스펙션하였다. 배지를 트랜스펙션 후 12 시간에 교환하였다. 반응속도 시험을 위해서, 화학적 유도를 트랜스펙션 후 24 시간에 시작하였고, 이때 아브시스산 (ABA, Sigma Aldrich) 을 250 μM 의 최종 농도로 새 배지에 첨가하였다. RNeasy 키트 (Qiagen) 를 제조사의 지시사항에 따라 사용하여 RNA 를 추출하고, 샘플 당 1 ㎍ 의 RNA 를 qScript (Quanta Biosystems) 를 사용하여 역전사하였다. 상대적 mRNA 수준을, 표적화 유전자 뿐 아니라 내인성 대조군으로서 GAPDH 에 대해 특이적인 TaqMan 프로브 (Life Technologies, Taqman 프로브 ID 에 대해서는 보충적 표 2 참조) 를 사용하여 정량 실시간 PCR (qRT-PCR) 에 의해 측정하였다. △△Ct 분석을 사용하여, GFP 로 모의 트랜스펙션한 세포인 음성 대조군에 대한 변화 배수를 수득하였다.
Cas9 전사 효과기. HEK 293FT 세포를, 24 웰 디쉬에 시딩하고 24 시간 후 리포펙타민 2000 (Life Technologies) 을 사용하여 돌연변이체 Cas9 융합 단백질 및 합성 안내 RNA (sgRNA) 와 함께 동시-트랜스펙션하였다. 트랜스펙션 후 72 시간에, 총 RNA 를 정제하였다 (RNeasy Plus, Qiagen). 1ug 의 RNA 를 cDNA 로 역전사하였다 (qScript, Quanta BioSciences). 정량 실시간 PCR 을 제조사의 지시사항 (Life Technologies) 에 따라 수행하고, hKlf4 (Hs00358836_m1), hSox2 (Hs01053049_s1) 및 내인성 대조군 GAPDH (Hs02758991_g1) 에 대해 TaqMan 검정을 사용하여 3 반복물로 수행하였다.
hSpCas9 활성제 플라스미드를 hEF1a 프로모터의 발현 하에 렌티바이러스 벡터 내로 클로닝하였다 (pLenti-EF1a-Cas9-NLS-VP64). hSpCas9 억제제 플라스미드를 동일 벡터 내로 클로닝하였다 (pLenti-EF1a-SID4x-NLS-Cas9-NLS). KLF4 유전자좌에 대해 표적화된 안내 서열 (20bp) 은 GCGCGCTCCACACAACTCAC, GCAAAAATAGACAATCAGCA, GAAGGATCTCGGCCAATTTG 이다. SOX2 유전자좌에 대해 표적화된 안내 RNA 에 대한 스페이서 서열은 GCTGCCGGGTTTTGCATGAA, CCGGGCCCGCAGCAAACTTC,GGGGCTGTCAGGGAATAAAT 이다.
광유전학적 작동기: 미생물 및 식물-유래 광 민감성 단백질을 광유전학적 작동기로서 조작하여, 멤브레인 전위 (520/Deisseroth, K. Optogenetics. Nature methods 8, 26-29, doi:10.1038/nmeth.f.324 (2011); Zhang, F. et al. The microbial opsin family of optogenetic tools. Cell 147, 1446-1457, doi:10.1016/j.cel1.2011.12.004 (2011) and Yizhar, 0., Fenno, L. E., Davidson, T. J., Mogri, M. & Deisseroth, K. Optogenetics in neural systems. Neuron 71, 9-34, doi: 10.1016/j.neuron.2011.06.004 (2011)), 세포내 생화학적 신호전달 (Airan, R. D., Thompson, K. R., Fenno, L. E., Bernstein, H. & Deisseroth, K. Temporally precise in vivo control of intracellular signalling. Nature 458, 1025-1029, doi:10.1038/nature07926 (2009)), 단백질 상호작용 (Levskaya, A., Weiner, O. D., Lim, W. A. & Voigt, C. A. Spatiotemporal control of cell signalling using a light-switchable protein interaction. Nature 461, 997-1001, doi:10.1038/nature08446 (2009); Yazawa, M., Sadaghiani, A. M., Hsueh, B. & Dolmetsch, R. E. Induction of protein-protein interactions in live cells using light. Nat Biotechnol 27, 941-945, doi:10.1038/nbt.1569 (2009); Strickland, D. et al. TULIPs: tunable, light-controlled interacting protein tags for cell biology. Nature methods 9, 379-384, doi:10.1038/nmeth.1904 (2012) and Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi: 10. 1038/nmeth. 1524 (2010)) 및 이종 유전자 발현 (Yazawa, M., Sadaghiani, A. M., Hsueh, B. & Dolmetsch, R. E. Induction of protein-protein interactions in live cells using light. Nat Biotechnol 27,941-945, doi:10.1038/nbt.1569 (2009); Kennedy, M. J. et al. Rapid blue-light-mediated induction of protein interactions in living cells. Nature methods 7, 973-975, doi:10.1038/nmeth.1524 (2010); Shimizu-Sato, S., Huq, E., Tepperman, J. M. & Quail, P. H. A light-switchable gene promoter system. Nat Biotechnol 20, 1041-1044, doi:10.1038/nbt734 (2002); Ye, H., Daoud-El Baba, M., Peng, R. W. & Fussenegger, M. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice. Science 332, 1565-1568, doi:10.ll26/science.1203535 (2011); Wang, X., Chen, X. & Yang, Y. Spatiotemporal control of gene expression by a light-switchable transgene system. Nature methods 9, 266-269, doi:10.1038/nmeth.1892 (2012) and Polstein, L. R. & Gersbach, C. A. Light-inducible spatiotemporal control of gene activation by customizable zinc finger transcription factors. J Am Chem Soc 134, 16480-16483, doi:10.1021/ja3065667 (2012)) 을 포함하는 세포 기능의 광학적 제어를 가능하게 하였다.
주변 광 노출: 모든 세포를 자극 동안을 제외하고 모든 시기에 낮은 광 수준 (<0.01 mW/cm2) 에서 배양하였다. 실내에서의 주변 광 (0.1-0.2 mW/cm2) 으로서 선택한 이러한 예방책은 LITE 시스템을 상당히 활성화시키는 것으로 발견되었다 (도 36D). 생체내 실험 동안 광으로부터 동물을 보호하기 위해서는 특별한 예방책이 선택되지 않았다 - 이식된 광학 섬유 내 이상적 증식을 가정하더라도, 주변 광으로 인한 섬유 말단에서의 광 전달의 추정치는 <0.01 mW 였다 (200 ㎛ 섬유 코어 직경 및 0.22 개구수를 기준으로 함).
Neuro2A 세포에서의 광 자극 매개변수의 최적화: 근UV 유도 세포독성을 최소화하기 위해서, 출원인은 TALE-CRY2 가 활성화되도록 466 nm 청색 LED 를 선택하였고, 파장은 450 nm 의 CRY2 흡수 최대치로부터 약간 적색-이동되었으나 여전히 80% 초과의 활성을 유지하였다 (Banerjee, R. et al. The signaling state of Arabidopsis cryptochrome 2 contains flavin semiquinone. J Biot Chem 282, 14916-14922, doi:l0.1074/jbc.M700616200 (2007)) (도 42). 광 노출을 최소화하기 위해서, 출원인은 강하지 않은 자극 프로토콜을 선택하였다 (0.067 Hz 에서 1 초 광 펄스, ~7% 사용률). 이는 광 사용률이 광범위한 사용률 매개변수 (1.7% 내지 100% 사용률, 도 41) 에 대한 LITE-매개 전사 활성화에 대해 유의한 효과를 갖지 않았다는 출원인의 발견을 기반으로 하였다. 0 내지 10 mW/cm2 의 광 강도 범위를 갖는 조명으로, Ngn2 mRNA 수준이 5 mW/cm2 이하의 강도의 함수로서 증가하였다는 것이 밝혀졌다. 그러나, Ngn2 mRNA 수준의 증가는 10 mW/cm2 에서 감소하였는데 (도 36C), 이는 고강도 광이 LITE 기능 또는 세포 생리학에 대해 유해한 효과를 가질 수 있다는 것을 제시한다. 이러한 관찰을 더 양호하게 특징화하기 위해, 출원인은 살아있는 세포에 대해 칼세인 대조염색하는 에티디움 동종이량체-1 세포독성 검정을 수행하였고, 10 mW/cm2 의 높은 자극 강도에서 상당히 더 높은 % 의 에티디움-양성 세포를 발견하였다. 반대로, 5 mW/cm2 자극으로부터의 에티디움-양성 세포 계수는 비자극 대조군과 구별가능하지 않았다. 따라서 5 mW/cm2 는 낮은 세포독성을 유지하면서 강건한 LITE 활성화를 획득하기에 최적인 것으로 나타났다.
1 차 뉴런에서의 광-유도 독성의 감소: 뉴런에서 LITE 의 초기 적용으로, 배양된 뉴런이 Neuro 2a 세포보다 청색광에 훨씬 더 민감하다는 것이 밝혀졌다. 출원인이 Neuro 2a 세포에 대해 이전에 최적화한 자극 매개변수 (466 nm, 5 mW/cm2 강도, 7% 사용률, 0.067 Hz 에서 1 초 광 펄스, 총 24 시간 동안) 는 1 차 뉴런에서 >50% 독성을 초래하였다. 따라서 출원인은, 출원인이 광범위한 사용률이 LITE-매개 전사 활성화에 대해 약간의 효과를 갖는다는 것을 이전에 관찰했던 바 (도 41), 더 낮은 사용률로 생존을 시험하였다. 동일한 광 강도 (5 mW/cm2) 에서의 0.8% 의 감소된 사용률 (0.0167 Hz 에서 0.5 초 광 펄스) 은 미자극 배양물과 구별불가능한 높은 생존률을 유지하기에 충분하였다 (도 47).
생체내 실험에서의 광 전파 및 독성: 이전의 연구는 뇌 조직에서의 상이한 파장의 광의 전파 효율을 조사하였다. 473 nm 광 (이 연구에서 사용한 파장) 에 대해, 0.35 mm 의 조직을 통과한 후 >90% 감쇠가 존재하였다 (Witten, Hana B. et at. Recombinase-Driver Rat Lines: Tools, Techniques, and Optogenetic Application to Dopamine-Mediated Reinforcement. Neuron 72, 721-733, doi:http://dx.doi.org/10.1016/j.neuron.2011.1O.028 (2011)). 추정된 5 mW/cm2 광력 밀도를, 조직의 0.35 mm 조직 깊이 (이 연구에서 사용한 뇌 펀치의 직경) 및 5 mW 의 총 전력 출력을 기준으로 추정하였다. 생체내 사용한 광 자극 사용률은 1 차 뉴런에 사용한 바 (도 47) 와 동일하였다 (0.8%, 0.0167 Hz 에서 0.5 초).
CRY2 흡수 스펙트럼: CRY2 의 흡수 스펙트럼 조명을 도 42 에 나타내었다. 상기 스펙트럼은 480 nm 초과의 흡수에서 첨예한 감소를 나타내었다 (Banerjee, R. et al. The Signaling State of Arabidopsis Cryptochrome 2 Contains Flavin Semiquinone. Journal of Biological Chemistry 282, 14916-14922, doi:10.1074/jbc.M700616200 (2007)). 500 nm 초과의 파장은 사실상 흡수되지 않았는데, 이는 황색 또는 적색광 민감성 단백질로의 추후 멀티모달 광학적 제어에 유용할 수 있다.
AAV1 상청액 프로세스의 발전: 종래의 AAV 입자 생성은 번거로운 생성 및정제 프로세스를 필요로 하며, 많은 구축물을 동시에 시험하는 것이 비현실적이 되게 하였다 (Grieger, J. C., Choi, V. W. & Samulski, R. J. Production and characterization of adeno-associated viral vectors. Nat Protoc 1, 1412-1428, doi: 10.1038/nprot.2006.207 (2006)). 이러한 연구에서, 간단하지만 상당히 효율적인 AAV 생성 프로세스는 트랜스펙션된 293FT 세포로부터의 여과된 상청액을 사용하였다 (도 43). 최근 보고는 293FT 세포에서 생성된 AAV 입자가 세포질에서 뿐만 아니라 배양 배지에서도 상당량으로 발견될 수 있었다는 것을 나타낸다 (Lock M, A. M., Vandenberghe LH, Samanta A, Toelen J, Debyser Z, Wilson JM. Rapid, Simple, and Versatile Manufacturing of Recombinant Adeno-Associated Viral Vectors at Scale. Human Gene Therapy 21, 1259-1271, doi:10.1089/hum.2010.055 (2010)). 숙주 세포의 사이토졸과 상청액 사이의 바이러스 입자 비는 AAV 혈청형에 따라 가변적이며, 폴리에틸렌이민 (PEI) 을 바이러스 패키징 플라스미드를 트랜스펙션하는데 사용한 경우 분비가 증진되었다 (Lock M, A. M., Vandenberghe LH, Samanta A, Toelen J, Debyser Z, Wilson JM. Rapid, Simple, and Versatile Manufacturing of Recombinant Adeno-Associated Viral Vectors at Scale. Human Gene Therapy 21, 1259-1271, doi:10.1089/hum.2010.055 (2010)). 현 연구에서, TALE 을 운반하는 AAV 벡터 (도 38A) 로 트랜스펙션되고 AAV1 혈청형을 사용하여 패키징된 2x105 개 293FT 세포가 1 mL 당 5.6 ± 0.24 x 1010 DNAseI 저항성 게놈 카피 (gc) 의 농도에서 250 ㎕ 의 AAV1 을 생성시킬 수 있었다는 것이 발견되었다. 250 ㎕ 의 여과된 상청액은 80-90% 의 효율로 150,000 개 1 차 피질 뉴런을 형질도입시킬 수 있었다 (도 38B 및 도 43). 이러한 프로세스는 또한 96-웰 포맷에 성공적으로 적합화되어, 동시에 96 개 이하의 상이한 구축물로부터 125 ul AAV1 상청액의 생성을 가능하게 하였다. 35 ul 의 상청액을 이후 96-웰 포맷으로 배양된 1 차 뉴런의 1 개 웰을 형질도입하는데 사용할 수 있어, 단일 웰로부터의 생물학적 3 반복물에서의 형질도입을 가능하게 하였다.
표 3. 모든 Taqman 프로브에 대한 제품 정보 (Life Technologies)

표 4. 이 연구에서 사용한 항체에 대한 클론, 제품 번호 및 농도

표 5. CHIP-qPCR 에 사용한 qPCR 프라이머

표 6. TALE 에 의해 표적화된 게놈 서열

표 7. 마우스 변연계 아래 피질 (ILC) 에서의 Grm2 의 생체내 LITE-매개 활성화에 대한 바이러스 형질도입 및 광 자극 매개변수. 동측성 LITE-발현 반구에서의 Grm2 mRNA 수준을, 도 39J 에서 나타낸 모든 3 가지 실험 조건에 대해 대측성 mCherry-발현 대조군 반구와 비교한다.

표 8. HDAC 영입 효과기 도메인

Nan, X. et al. Transcriptional repression by the methyl-CpG-binding protein MeCP2 involves a histone deacetylase complex. Nature 393, 386-389 (1998).
Boeke, J., Ammerpohl, 0., Kegel, S., Moehren, U. & Renkawitz, R. The minimal repression domain of MBD2b overlaps with the methyl-CpG-binding domain and binds directly to Sin3A. Journal ofBiological Chemistry 275, 34963-34967 (2000).
Laherty, C. D. et al. Histone deacetylases associated with the mSin3 corepressor mediate mad transcriptional repression. Cell 89, 349-356 (1997).
Zhang, J., Kalkum, M., Chait, B. T. & Roeder, R. G. The N-CoR-HDAC3 nuclear receptor corepressor complex inhibits the JNK pathway through the integral subunit GPS2. Molecular cell 9, 611-623 (2002).
Lauberth, S. M. & Rauchman, M. A conserved 12-amino acid motif in Salll recruits the nucleosome remodeling and deacetylase corepressor complex. Journal of Biological Chemistry 281, 23922-23931 (2006).
Gu, H. & Roizman, B. Herpes simplex virus-infected cell protein 0 blocks the silencing of viral DNA by dissociating histone deacetylases from the CoREST, AiREST complex.
Ouyang, J., Shi, Y, Valin, A., Xuan, Y & Gill, G. Direct binding of CoREST1 to SUMO-2/3 contributes to gene-specific repression by the LSD1/CoREST1/HDAC complex. Molecular cell 34, 145-154 (2009)
표 9. HDAC 효과기 도메인

Vannier, D., Balderes, D. & Shore, D. Evidence that the transcriptional regulators SIN3 and RPD3, and a novel gene (SDS3) with similar functions, are involved in transcriptional silencing in S. cerevisiae. Genetics 144, 1343-1353 (1996).
Gregoretti, I., Lee, Y.-M. & Goodson, H. V. Molecular evolution of the histone deacetylase family: functional implications of phylogenetic analysis. Journal ofmolecular biology 338,17-31 (2004).
Gao, L., Cueto, M. A., Asselbergs, F. & Atadja, P. Cloning and functional characterization of HDAC11, a novel member of the human histone deacetylase family. Journal of Biological Chemistry 277,25748-25755 (2002).
Wu, K., Tian, L., Malik, K., Brown, D. & Miki, B. Functional analysis of HD2 histone deacetylase homologues in Arabidopsis thaliana. The Plant Journal 22, 19-27 (2000).
Scher, M. B., Vaquero, A. & Reinberg, D. SirT3 is a nuclear NAD+-dependent histone deacetylase that translocates to the mitochondria upon cellular stress. Genes & development 21,920-928 (2007).
Hnisz, D., Schwarzmuller, T. & Kuchler, K. Transcriptional loops meet chromatin: a dual, Aelayer network controls white,Aiopaque switching in Candida albicans. Molecular microbiology 74, 1-15 (2009).
Landry, J. et al. The silencing protein SIR2 and its homologs are NAD-dependent protein deacetylases. Proceedings ofthe National Academy ofSciences 97, 5807-5811 (2000).
Wilson, J. M., Le, V. Q., Zimmerman, c., Marmorstein, R. & Pillus, L. Nuclear export modulates the cytoplasmic Sir2 homologue Hst2. EMBO reports 7,1247-1251 (2006).
Gertz, M. & Steegborn, C. Function and regulation of the mitochondrial Sirtuin isoform Sirt5 in Mammalia. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 1804, 1658-1665 (2010).
Zhu, A. Y. et al. Plasmodium falciparum Sir2A preferentially hydrolyzes medium and long chain fatty acyl lysine. ACS chemical biology 7, 155-159 (2011).
Tennen, R. I., Berber, E. & Chua, K. F. Functional dissection of SIRT6: identification of domains that regulate histone deacetylase activity and chromatin localization. Mechanisms ofageing and development 131, 185-192 (2010).
표 10. 히스톤 메틸트랜스퍼라아제 (HMT) 효과기 도메인

Pennini, M. E., Perrinet, S. p., Dautry-Varsat, A. & Subtil, A. Histone methylation by NUE, a novel nuclear effector of the intracellular pathogen Chlamydia trachomatis. PLoS pathogens 6, el000995 (2010).
Mujtaba, S. et al. Epigenetic transcriptional repression of cellular genes by a viral SET protein. Nature cell biology 10, 1114-1122 (2008).
Tachibana, M., Matsumura, Y, Fukuda, M., Kimura, H. & Shinkai, Y G9a/GLP complexes independently mediate H3K9 and DNA methylation to silence transcription. The EMBO journal 27, 2681-2690 (2008).
Snowden, A. W., Gregory, P. D., Case, C. C. & Pabo, C. O. Gene-specific targeting of H3K9 methylation is sufficient for initiating repression in vivo. Current biology 12,2159-2166 (2002).
Rathert, P., Zhang, X., Freund, C., Cheng, X. & Jeltsch, A. Analysis of the substrate specificity of the Dim-5 histone lysine 메틸transferase using peptide arrays. Chemistry & biology 15,5-11 (2008).
Jackson, J. P. et al. Dimethylation of histone H3 lysine 9 is a critical mark for DNA methylation and gene silencing in Arabidopsis thaliana. Chromosoma 112,308-315 (2004).
Thorstensen, T. et al. The Arabidopsis SUVR4 protein is a nucleolar histone methyltransferase with preference for monomethylated H3K9. Nucleic acids research 34, 5461-5470 (2006).
Vielle, A. et al. H4K20me1 Contributes to Downregulation of X-Linked Genes for C. elegans Dosage Compensation. PLoS Genetics 8, e1002933 (2012).
Couture, J.-F., Collazo, E., Brunzelle, J. S. & Trievel, R. C. Structural and functional analysis of SET8, a histone H4 Lys-20 methyltransferase. Genes & development 19, 1455-1465 (2005).
Sautel, C. l. F. et al. SET8-mediated methylations of histone H4 lysine 20 mark silent heterochromatic domains in apicomplexan genomes. Molecular and cellular biology 27, 5711-5724 (2007).
표 11. 히스톤 메틸트랜스퍼라아제 (HMT) 영입 효과기 도메인

Hathaway, N. A. et al. Dynamics and memory of heterochromatin in living cells. Cell (2012).
Ballare, C. et al. Phf19 links methylated Lys36 of histone H3 to regulation of Polycomb activity. Nature structural & molecular biology 19, 1257-1265 (2012).
Jin, Q. et al. The protein phosphatase-1 (PP1) regulator, nuclear inhibitor of PP1 (NIPP1), interacts with the polycomb group protein, embryonic ectoderm development (EED), and functions as a transcriptional repressor. Journal of Biological Chemistry 278, 30677-30685 (2003).
표 12. 히스톤 아세틸트랜스퍼라아제 저해제 효과기 도메인

Cervoni, N., Detich, N., Seo, S.-B., Chakravarti, D. & Szyf, M. The oncoprotein Set/TAF-1beta, an inhibitor of histone acetyltransferase, inhibits active demethylation of DNA, integrating DNA methylation and transcriptional silencing. Journal of Biological Chemistry 277,25026-25031 (2002)
보충적 서열
> TALE(Ngn2)-NLS-CRY2


>TALE(Ngn2)-NLS-CRY2PHR


> CIB1-NLS-VP64_2A_GFP

> CIBN-NLS-VP64_2A_GFP

> CIB1-NLS-VP16_2A_GFP

> CIB1-NLS-p65_2A_GFP


> HA-TALE(12mer)-NLS-VP64_2A_GFP

> HA-TALE(12mer)-NLS-SID4X_2A_phiLOV2.1


> HA-TALE(12mer)-NLS-CIB1

> CRY2PHR-NLS-VP64_2A_GFP

> CRY2PHR-NLS-SID4X_2A_phiLOV2.1

> TALE(KLF4)-NLS-CRY2PHR

> HA-NLS-TALE(p11, N136)-SID


> HA-NLS-TALE(p11, N136)-SID4X

>HA-TALE(ng2, C63)-GS-cib1-mutNLS

> HA-TALE(ng2, C63)-wNES-cib1-mutNLS


> HA-TALE(ng2, C63)-mNES-cib1-mutNLS


> HA-TALE(ng2, C63)-ptk2NES-cib1-mutNLS

> HA-TALE(ng2, C63)-mapkkNES-cib1-mutNLS

> HA-TALE(ng2, C63)-GS-cib1△3-mutNLS


> HA-TALE(ng2, C63)-wNLS-cib1△3-mutNLS

> HA-TALE(ng2, C63)-mNLS-cib1△3-mutNLS

> HA-TALE(ng2, C63)-GS-cib1-mutNLS-mutbHLH


> HA-TALE(ng2, C63)-wNES-cib1-mutNLS-mutbHLH

> HA-TALE(ng2, C63)-GS-cib1△1-mutNLS

> HA-TALE(ng2, C63)-wNLS-cib1△1-mutNLS


> HA-TALE(ng2, C63)-GS-cib1△2-mutNLS


> HA-TALE(ng2, C63)-wNLS-cib1△2-mutNLS

> HA-TALE(ng2, C63)-NLS-cibl-mutNLS-mutbHLH)


> HA-TALE(ng2, C63)-NLS-cib1△1-mutNLS


> HA-TALE(ng2, C63)-NLS-cib1△2-mutNLS

> HA-TALE(ng2, C63)-GS-iNES1-cib1-mutNLS

> HA-TALE(ng2, C63)-GS-iNES2-cib1-mutNLS


> HA-TALE(ng2, C63)-GS-iNES3-cib1-mutNLS

> HA-TALE(ng2, C63)-GS-iNES4-cib1-mutNLS

> HA-TALE(ng2, C63)-GS-iNES5-cib1-mutNLS


> HA-TALE(ng2, C63)-GS-iNES6-cib1-mutNLS


> HA-TALE(ng2, C63)-NLS-cib1△1

> HA-TALE(ng2, C63)-NLS-cib1△2


> 알파-임포틴-NLS-CRY2PHR-NLS-VP64_2A_GFP

> mutNES-CRY2PHR-NLS-VP64_2A_GFP

> CRY2PHR-NLS-VP64-NLS_2A_GFP


> Neurog2-TALE(N240,C63)-PYL

>ABI-NLS-VP64

> hSpCas9(D10A, H840A)-링커-NLS-VP64

>SID4X-NLS-FLAG-링커-hSpCas9(D10A, H840A)-NLS


후생적 효과기 도메인 서열
>hs_NCoR
>pf_Sir2A


>nc_DIM5

>sc_HST2

>hs_SIRT3

>hs_NIPP1

>ct_NUE

>hs_MBD2b

>ca_HST2

>hs_PHF19

>hs_HDAC11


>ml_MesoLo4

>pbcv1_vSET

>at_KYP

>tg_TgSET8

>hs_SIRT6


>ce_Set1

>mm_G9a

>hs_SIRT5

>xl_HDAC8

>mm_HP1a

>at_HDT1

>mm_SAll
>hs_SETD8

>sc_RPD3

>ec_CobB

>hs_SUV39H1

>hs_RCOR1

>hs_sin3a

>at_SUVR4

>rn_MeCP2_NLS


>mm_SET-TAF1B

>ce_Set4

광자극 하드웨어 제어 스트립트
하기의 Arduino 스트립트를 광 자극된 24-웰 플레이트의 각 4-웰 열의 개별 제어가 가능하도록 사용하였다.





실시예 12
내인성 포유동물 전사의 광학적 제어
1 차 마우스 피질 뉴 런에서의 전자를 조정하기 위한 AAV-매개 TALE 전달의 효능을 시험하기 위해서, 출원인은 각각 mGluR5, NMDA 서브유닛 2A 및 mGluR2 를 인코딩하는 3 개 마우스 신경전달물질 수용체: Grm5, Grin2a 및 Grm2 의 유전자좌를 표적화하는 6 개 TALE-DNA 결합 도메인을 구축하였다 (도 58). 표적 부위 접근 가능성을 증가시키기 위해, 출원인은 UCSC 게놈 브라우저로부터 마우스 피질 DNase I 민감성 데이터를 사용하여 추정상 개방형 크로마틴 부위를 확인하였다. 각각의 표적 유전자의 프로모터 내 DNase I 민감성 부위는 TALE 결합 서열의 선택을 위한 안내를 제공하였다 (도 46). 각각의 TALE 에 대해서, 출원인은 mSin3 상호작용 도메인 (SID) 의 쿼드러플 탠덤 반복물 또는 전사 활성제로서 VP64 를 이용하였다 (Beerli, R.R., Segal, D.J., Dreier, B. & Barbas, C.F., 3rd Toward controlling gene expression at will: specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proc Natl Acad Sci USA 95, 14628-14633 (1998) and Ayer, D.E., Laherty, C.D., Lawrence, Q.A., Armstrong, A.P. & Eisenman, R.N. Mad proteins contain a dominant transcription repression domain. Molecular and Cellular Biology 16, 5772-5781 (1996)) as a repressor. Applicants have previously shown that a single SID fused to TALE downregulated a target gene effectively in 293FT cells (Cong, L., Zhou, R., Kuo, Y.-c., Cunniff, M. & Zhang, F. Comprehensive interrogation of natural TALE DNA-binding modules and transcriptional repressor domains. Nat Commun 3, 968 (2012)). 이러한 TALE 억제제를 추가로 향상시키기 위해, 출원인은 VP64 의 성공적 쿼드러플 VP16 반복물 구조와 유사한 SID 의 4 개 반복물 (Beerli, R.R., Segal, D.J., Dreier, B. & Barbas, C.F., 3rd Toward controlling gene expression at will: specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proc Natl Acad Sci USA 95, 14628-14633 (1998)) 이 이의 억제 활성을 증가시킬 수 있다고 추론하였다. 이는 실제로 TALE-SID4X 구축물이 293FT 세포에서 TALE-SID 에 대해 억제를 ~2 배 증진시킨 바와 같은 경우이다 (도 54).
출원인은 6 개 TALE-VP64 구축물 중 4 개 (T1, T2, T5 및 T6) 가 AAV-형질도입된 1 차 뉴런에서의 그의 표적 유전자 Grm5 및 Grm2 를, 각각 2- 및 8-배까지 효율적으로 활성화시켰다는 것을 발견하였다 (도 58). 유사하게, 6 개 TALE-SID4X 억제제 중 4 개 (T9, T10, T1l, T12) 가 그의 내인성 표적 Grin2a 및 Grm2 의 발현을 각각 2- 및 8-배까지 감소시켰다 (도 58). 함께, 이러한 결과는 구성적 TALE 이 뉴런에서의 내인성 표적 유전자 발현을 양상적 또는 음성적으로 조정할 수 있다는 것을 나타낸다. 특히, 주어진 TALE 에 의한 효율적인 활성화 또는 억제는 반대편 방향에서의 전사 조정에 있어서 그의 효율을 예측하지 않았다. 따라서, 다수의 TALE 은 특정 유전자좌에 대한 가장 효율적 TALE 을 확인하기 위해 스크리닝될 필요가 있을 수 있다.
LITE 의 뉴런 적용을 위해, 출원인은 출원인의 6 개 구성적 TALE 활성제 비교를 기반으로 (도 58), 1 차 뉴런에서 가장 강력한 수준의 표적 상향조절을 나타낸 Grm2 TALE (T6) 을 선택하였다. 출원인은 0.8% 의 동일한 사용률로 2 광 펄스 주파수를 사용하여 이의 기능을 조사하였다. 자극 조건 모두 Grm2 mRNA 수준에 있어서 ~7 배 광-의존적 증가를 획득하였다 (도 38C). 추가 연구로, 유의한 표적 유전자 발현 증가가 빠르게 이루어질 수 있다는 것이 확인되었다 (4 시간 내에 4-배 상향조절; 도 38D). 또한, 출원인은 자극 후 mGluR2 단백질의 유의한 상향조절을 관찰하였는데, 이는 mRNA 수준에서 LITE 에 의해 영향받은 변화가 단백질 도메인으로 옮겨지는 것을 입증한다 (도 38E). 종합하여, 이러한 결과로, LITE 가 뉴런에서의 내인성 유전자 발현의 시간적으로 정확한 광학적 제어를 가능하게 한다는 것이 확인된다.
출원인의 이전에 실행한 LITE 활성제에 대한 치하로서, 출원인은 다음으로 TALE-SID4X 구축물을 기반으로 LITE 억제제를 조작하였다. 구성적 Grm2 TALE (T11 및 T12, 도 59A) 은 최고 수준의 전사 억제를 매개하였으며, LITE 억제제로서 선택되었다 (도 59A, B). 광-유도 억제제 모두가 각각 T11 및 T12 에 대해 1.95 배 및 1.75 배 감소로 Grm2 발현의 유의한 하향조절을 매개하였는데, 이는 뉴런에서의 광학적으로 제어된 억제의 실행가능성을 입증한다 (도 38G).
AAV 를 사용하여 뉴런 내에 LITE 를 전달하기 위해, 출원인은 LITE 전이유전자가 포함된 총 바이러스 게놈 크기가 4.8 kb 를 초과하지 않았다는 것을 확실히 해야만 했다 (Wu, Z., Yang, H. & Colosi, P. Effect of Genome Size on AAV Vector Packaging. Mol Ther 18, 80-86 (2009) and Dong JY, F.P., Frizzell RA Quantitative analysis of the packaging capacity of recombinant adeno-associated virus. Human Gene Therapy 7, 2101-2112 (1996)). 이를 위해, 출원인은 TALE N- 및 C-말단을 단축시키고 (N-말단에서 136 aa 를, C-말단에서 63 aa 를 유지), CRY2PHR 및 CIB1 도메인을 교환하였다 (TALE-CIB1 및 CRY2PHR-VP64; 도 38A). 이러한 스위치는 LITE 의 각 성분이 AAV 벡터 내로 들어맞게 하였으며 광-매개 전사 조정의 효능을 감소시키지 않았다 (도 60). 이러한 LITE 는 2 개 AAV 벡터의 조합에 의한 동시-형질도입을 통해 1 차 피질 뉴런 내로 효율적으로 전달될 수 있다 (도 38B; >80% 동시-형질도입 효율을 갖는 ㄴ개별 성분에 대해 83-92% 의 전달 효율).
실시예 13
유도성 렌티바이러스 Cas9
렌티바이러스 제조. pCasES10 (렌티바이러스 이동 플라스미드 백본을 함유함) 을 클로닝한 후, 10% 소 태아 혈청을 포함하고 항생제를 포함하지 않는 DMEM 중에서 트랜스펙션하기 전날에 낮은 계대 (p=5) 에서의 HEK293FT 를 T-75 플라스크에 50% 밀집도로 시딩하였다. 20 시간 후, 배지를 OptiMEM (혈청 불포함) 배지로 교환하고 4 시간 후 트랜스펙션을 수행하였다. 세포를 10 ug 의 렌티바이러스 이동 플라스미드 (pCasES10) 및 다음의 패키징 플라스미드로 트랜스펙션하였다: 5 ug 의 pMD2.G (VSV-g 위형) 및 7.5 ug 의 psPAX2 (gag/pol/rev/tat). 양이온 지질 전달제 (50uL 리포펙타민 2000 및 100ul Plus 시약) 와 함께 4 mL OptiMEM 중에서 트랜스펙션을 수행하였다. 6 시간 후, 배지를 10% 소 태아 혈청을 갖는 항생제-불포함 DMEM 으로 교환하였다.
렌티바이러스 정제. 바이러스 상청액을 48 시간 후 수확하였다. 상청액을 먼저 잔해가 없도록 하고 0.45um 저 단백질 결합 (PVDF) 필터를 통해 여과하였다. 이를 2 시간 동안 24,000 rpm 에서 초원심분리로 회전시켰다. 바이러스 펠렛을 50ul 의 DMEM 중에서 밤새 4℃ 에서 재현탁하였다. 이를 분취하고 즉시 -80℃ 에서 동결시켰다.
FACS 를 사용하는 클론 단리. HEK293FT 및 HUES64 인간 배아 줄기 세포의 클론 단리를 위해, 세포를 현탁액 중 1ul 또는 5ul 의 정제된 바이러스로 감염시켰다. 감염 후 24 시간에, 1uM 독시사이클린을 세포 배양 배지에 첨가하였다. 24 시간 또는 48 시간 이상 후, 세포를 BD FACSAria IIu 기기 상에서 형광-보조 세포 분류 (FACS) 처리하여, 독시사이클린 처리 후 EGFP (및 따라서 Cas9) 를 강건하게 발현한 단일 세포를 단리하였다. 추가 사용을 위해 세포를 벌크로 또는 개별 웰에 플레이팅하여 통합된 유도 Cas9 를 갖는 클론 집단이 선택되게 하였다. 분류 효율은 항상 >95% 였고, EGFP 형광을 확인하기 위해 플레이팅한 직후 세포가 가시화되었다.
도 61 은 Tet Cas9 벡터 설계를 나타낸다.
도 62 는 293FT 세포에서의 벡터 및 EGFP 발현을 나타낸다.
pCasES020 유도 Cas9 의 서열:







1. 하기를 포함하는, 세포에서의 관심 게놈 유전자좌의 발현을 변형하는 유도 방법:
(a) (i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 데옥시리보핵산 (DNA) 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함)
을 포함하는 DNA 결합 폴리펩티드를 포함하는 비-자연발생적 또는 조작된 조성물과 게놈 유전자좌를 접촉시키고;
(b) 에너지원을 적용시키고;
(c) 게놈 유전자좌의 발현이 변형된 것을 측정함.
2. 단락 1 에 있어서, 적어도 하나 이상의 효과기 도메인이 트랜스포사아제 도메인, 인테그라아제 도메인, 재조합효소 도메인, 리졸바아제 도메인, 인버타아제 도메인, 프로테아제 도메인, DNA 메틸트랜스퍼라아제 도메인, DNA 데메틸라아제 도메인, 히스톤 아세틸라아제 도메인, 히스톤 데아세틸라아제 도메인, 뉴클레아제 도메인, 억제제 도메인, 활성제 도메인, 핵-국부화 신호 도메인, 전사-단백질 영입 도메인, 세포 흡수 활성 관련 도메인, 핵산 결합 도메인 또는 항체 제시 도메인으로 이루어지는 군에서 선택되는 방법.
3. 단락 2 에 있어서, 적어도 하나 이상의 효과기 도메인이 뉴클레아제 도메인 또는 재조합효소 도메인인 방법.
4. 단락 3 에 있어서, 뉴클레아제 도메인이 비-특이적 FokI 엔도뉴클레아제 촉매 도메인인 방법.
5. 단락 1-4 중 어느 하나에 있어서, 에너지 민감성 단백질이 크립토크롬-2 (CRY2) 인 방법.
6. 단락 1-5 중 어느 하나에 있어서, 상호작용 파트너가 크립토크롬-상호작용 염기성 헬릭스-루프-헬릭스 (CIB1) 인 방법.
7. 단락 1-6 중 어느 하나에 있어서, 에너지원이 전자기 조사, 음파 에너지 또는 열 에너지로 이루어지는 군에서 선택되는 방법.
8. 단락 7 에 있어서, 전자기 조사가 가시광의 성분인 방법.
9. 단락 8 에 있어서, 가시광 성분이 450nm-500nm 범위의 파장을 갖는 방법.
10. 단락 8 에 있어서, 가시광 성분이 청색광인 방법.
11. 단락 1 에 있어서, 에너지원을 적용하는 것이 6.2 mW/cm2 이상 강도로의 청색광 자극을 포함하는 방법.
12. 단락 1-11 중 어느 하나에 있어서, DNA 결합 도메인이 (X1-11-X12X13-X14-33 또는 34 또는 35)z 를 포함하는 방법으로서, 여기서 X1-11 이 11 개의 인접한 아미노산 사슬이고, X12X13 이 반복 가변 이중잔기 (RVD) 이고, X14-33 또는 34 또는 35 가 21, 22 또는 23 개의 인접한 아미노산 사슬이고, z 가 적어도 5 내지 40 이고, 하나 이상의 RVD 가 NI, HD, NG, NN, KN, RN, NH, NQ, SS, SN, NK, KH, RH, HH, HI, KI, RI, SI, KG, HG, RG, SD, ND, KD, RD, YG, HN, NV, NS, HA, S*, N*, KA, H*, RA, NA 및 NC 로 이루어지는 군에서 선택되고, 이때 (*) 가 X13 에서의 아미노산이 부재함을 의미하는 방법.
13. 단락 12 에 있어서, z 가 적어도 10 내지 26 인 방법.
14. 단락 12 에 있어서:
X1-ll 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 1-11 로 나타내는 12 개의 인접한 아미노산의 서열이거나,
X14-34 또는 X14-35 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 12-32 또는 12-33 으로 나타내는 21 또는 22 개의 인접한 아미노산의 서열인 방법.
15. 단락 12 에 있어서, 하나 이상의 RVD 가 (a) 구아닌 (G) 의 인지를 위한 HH, KH, NH, NK, NQ, RH, RN, SS; (b) 아데닌 (A) 의 인지를 위한 SI; (c) 티민 (T) 의 인지를 위한 HG, KG, RG; (d) 시토신 (C) 의 인지를 위한 RD, SD; (e) A 또는 G 의 인지를 위한 NV; 및 (f) A 또는 T 또는 G 또는 C 의 인지를 위한 H*, HA, KA, N*, NA, NC, NS, RA, S* (여기서 (*) 는 X13 에서의 아미노산이 부재함을 의미함) 로 이루어지는 군에서 선택되는 방법.
16. 단락 15 에 있어서:
G 의 인지를 위한 RVD 가 RN, NH, RH 또는 KH 이거나;
A 의 인지를 위한 RVD 가 SI 이거나;
T 의 인지를 위한 RVD 가 KG 또는 RG 이고;
C 의 인지를 위한 RVD 가 SD 또는 RD 인 방법.
17. 단락 12 에 있어서, 하기 중 하나 이상이 존재하는 방법:
X1-4 에서의 [LTLD] 또는 [LTLA] 또는 [LTQV], 또는
위치 X30-33 또는 X31-34 또는 X32-35 에서의 [EQHG] 또는 [RDHG].
18. 단락 1-17 중 어느 하나에 있어서:
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 147 개의 인접한 아미노산을 포함하거나,
C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 68 개의 인접한 아미노산을 포함하거나,
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 136 개의 인접한 아미노산을 포함하고 C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 183 개의 인접한 아미노산을 포함하는 방법.
19. 단락 1-18 중 어느 하나에 있어서, 관심 게놈 유전자좌가 분화 인자, 전사 인자, 신경전달물질 이동체, 신경전달물질 신타아제, 시냅스 단백질, 가소성 단백질, 시냅스전 활성 구역 단백질, 시냅스후 밀도 단백질, 신경전달물질 수용체, 후생적 변형제, 신경 운명 선별화 인자, 액손 안내 분자, 이온 채널, CpG 결합 단백질, 유비퀴틴화 단백질, 호르몬, 호메오박스 단백질, 성장 인자, 발암유전자 또는 원발암유전자를 인코딩하는 유전자와 관련되는 방법.
20. 하기를 포함하는, 세포에서의 관심 게놈 유전자좌의 발현을 억제하는 유도 방법:
(a) (i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함)
을 포함하는 DNA 결합 폴리펩티드를 포함하는 비-자연발생적 또는 조작된 조성물과 게놈 유전자좌를 접촉시키고;
(b) 에너지원을 적용시키고;
(c) 게놈 유전자좌의 발현이 억제된 것을 측정함.
21. 단락 20 에 있어서, 폴리펩티드가 하나 이상의 SID 억제제 도메인을 포함하는 방법.
22. 단락 21 에 있어서, 폴리펩티드가 4 개 이상의 SID 억제제 도메인을 포함하는 방법.
23. 단락 21 에 있어서, 폴리펩티드가 SID4X 억제제 도메인을 포함하는 방법.
24. 단락 20 에 있어서, 폴리펩티드가 KRAB 억제제 도메인을 포함하는 방법.
25. 단락 20-24 중 어느 하나에 있어서, 에너지 민감성 단백질이 크립토크롬-2 (CRY2) 인 방법.
26. 단략 20-25 중 어느 하나에 있어서, 상호작용 파트너가 크립토크롬-상호작용 염기성 헬릭스-루프-헬릭스 (CIB1) 인 방법.
27. 단락 20-26 중 어느 하나에 있어서, 에너지원이 전자기 조사, 음파 에너지 또는 열 에너지로 이루어지는 군에서 선택되는 방법.
28. 단락 20 에 있어서, 전자기 조사가 가시광의 성분인 방법.
29. 단락 28 에 있어서, 가시광 성분이 450nm-500nm 범위의 파장을 갖는 방법.
30. 단락 28 에 있어서, 가시광 성분이 청색광인 방법.
31. 단락 20 에 있어서, 에너지원을 적용하는 것이 6.2 mW/cm2 이상 강도로의 청색광 자극을 포함하는 방법.
32. 단락 20-31 중 어느 하나에 있어서, DNA 결합 도메인이 (X1-11-X12X13-X14-33 또는 34 또는 35)z 를 포함하는 방법으로서, 여기서 X1-11 이 11 개의 인접한 아미노산 사슬이고, X12X13 이 반복 가변 이중잔기 (RVD) 이고, X14-33 또는 34 또는 35 가 21, 22 또는 23 개의 인접한 아미노산 사슬이고, z 가 적어도 5 내지 40 이고, 하나 이상의 RVD 가 NI, HD, NG, NN, KN, RN, NH, NQ, SS, SN, NK, KH, RH, HH, HI, KI, RI, SI, KG, HG, RG, SD, ND, KD, RD, YG, HN, NV, NS, HA, S*, N*, KA, H*, RA, NA 및 NC 로 이루어지는 군에서 선택되고, 이때 (*) 가 X13 에서의 아미노산이 부재함을 의미하는 방법.
33. 단락 32 에 있어서, z 가 적어도 10 내지 26 인 방법.
34. 단락 32 에 있어서:
X1-ll 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 1-11 로 나타내는 11 개의 인접한 아미노산의 서열이거나,
X14-34 또는 X14-35 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 12-32 또는 12-33 으로 나타내는 21 또는 22 개의 인접한 아미노산의 서열인 방법.
35. 단락 20-34 중 어느 하나에 있어서:
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 147 개의 인접한 아미노산을 포함하거나,
C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 68 개의 인접한 아미노산을 포함하거나,
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 136 개의 인접한 아미노산을 포함하고 C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 183 개의 인접한 아미노산을 포함하는 방법.
36. 단락 20-35 중 어느 하나에 있어서, 관심 게놈 유전자좌가 분화 인자 또는 이온 채널 성분을 인코딩하는 유전자와 관련되는 방법.
37. 단락 36 에 있어서, 분화 인자가 SRY-박스-2 (SOX2) 이며 유전자 SOX2 에 의해 인코딩되는 방법.
38. 단락 36 에 있어서, 분화 인자가 p11 이며 유전자 p11 에 의해 인코딩되는 방법.
39. 단락 36 에 있어서, 이온 채널 성분이 CACNA1C 이며 유전자 CACNA1C 에 의해 인코딩되는 방법.
40. 하기를 포함하는, 세포에서의 관심 게놈 유전자좌의 발현을 활성화하는 유도 방법:
(a) (i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 이상의 TALE 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함)
을 포함하는 DNA 결합 폴리펩티드를 포함하는 비-자연발생적 또는 조작된 조성물과 게놈 유전자좌를 접촉시키고;
(b) 에너지원을 적용시키고;
(c) 게놈 유전자좌의 발현이 활성화된 것을 측정함.
41. 단락 40 에 있어서, 폴리펩티드가 하나 이상의 VP16 또는 VP64 활성제 도메인을 포함하는 방법.
42. 단락 40 에 있어서, 폴리펩티드가 하나 이상의 p65 활성제 도메인을 포함하는 방법.
43. 단락 40-42 중 어느 하나에 있어서, 에너지 민감성 단백질이 CRY2 인 방법.
44. 단락 40-43 중 어느 하나에 있어서, 상호작용 파트너가 CIB1 인 방법.
45. 단락 40 에 있어서, 에너지원이 전자기 조사, 음파 에너지 또는 열 에너지로 이루어지는 군에서 선택되는 방법.
46. 단락 45 에 있어서, 전자기 조사가 가시광의 성분인 방법.
47. 단락 46 에 있어서, 가시광 성분이 450nm-500nm 범위의 파장을 갖는 방법.
48. 단락 46 에 있어서, 가시광 성분이 청색광인 방법.
49. 단락 40 에 있어서, 에너지원을 적용하는 것이 6.2 mW/cm2 이상 강도로의 청색광 자극을 포함하는 방법.
50. 단락 40-49 중 어느 하나에 있어서, DNA 결합 도메인이 (X1-11-X12X13-X14-33 또는 34 또는 35)z 를 포함하는 방법으로서, 여기서 X1-11 이 11 개의 인접한 아미노산 사슬이고, X12X13 이 반복 가변 이중잔기 (RVD) 이고, X14-33 또는 34 또는 35 가 21, 22 또는 23 개의 인접한 아미노산 사슬이고, z 가 적어도 5 내지 40 이고, 하나 이상의 RVD 가 NI, HD, NG, NN, KN, RN, NH, NQ, SS, SN, NK, KH, RH, HH, HI, KI, RI, SI, KG, HG, RG, SD, ND, KD, RD, YG, HN, NV, NS, HA, S*, N*, KA, H*, RA, NA 및 NC 로 이루어지는 군에서 선택되고, 이때 (*) 가 X13 에서의 아미노산이 부재함을 의미하는 방법.
51. 단락 50 에 있어서, z 가 적어도 10 내지 26 인 방법.
52. 단락 50 에 있어서:
X1-ll 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 1-11 로 나타내는 11 개의 인접한 아미노산의 서열이거나,
X14-34 또는 X14-35 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 12-32 또는 12-33 으로 나타내는 21 또는 22 개의 인접한 아미노산의 서열인 방법.
53. 단락 40-52 중 어느 하나에 있어서:
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 147 개의 인접한 아미노산을 포함하거나,
C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 68 개의 인접한 아미노산을 포함하거나,
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 136 개의 인접한 아미노산을 포함하고 C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 183 개의 인접한 아미노산을 포함하는 방법.
54. 단락 40-53 중 어느 하나에 있어서, 관심 게놈 유전자좌가 분화 인자, 후생적 변형제 또는 이온 채널 성분을 인코딩하는 유전자와 관련되는 방법.
55. 단락 54 에 있어서, 분화 인자가 뉴로제닌-2 이며 유전자 NEUROG2 에 의해 인코딩되는 방법.
56. 단락 54 에 있어서, 분화 인자가 크루펠 유사 인자 4 이며 유전자 KLF-4 에 의해 인코딩되는 방법.
57. 단락 54 에 있어서, 후생적 변형제가 Tet 메틸시토신 디옥시게나아제 1 이며 유전자 tet-1 에 의해 인코딩되는 방법.
58. 단락 54 에 있어서, 이온 채널 성분이 CACNA1C 이며 유전자 CACNA1C 에 의해 인코딩되는 방법.
59. 세포에서의 게놈 유전자좌의 발현을 유도가능하게 변형하기 위한 비-자연발생적 또는 조작된 조성물로서, 상기 조성물이 하기를 포함하는 DNA 결합 폴리펩티드를 포함하고:
(i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함);
상기 폴리펩티드가 코돈 최적화된 핵산 분자에 의해 인코딩되고 이로부터 번역되어, 폴리펩티드가 바람직하게는 게놈 유전자좌의 DNA 에 결합하고,
상기 폴리펩티드가 에너지원의 적용시 게놈 유전자좌의 발현을 변형시키는 조성물.
60. 단락 59 에 있어서, 적어도 하나 이상의 효과기 도메인이 트랜스포사아제 도메인, 인테그라아제 도메인, 재조합효소 도메인, 리졸바아제 도메인, 인버타아제 도메인, 프로테아제 도메인, DNA 메틸트랜스퍼라아제 도메인, DNA 데메틸라아제 도메인, 히스톤 아세틸라아제 도메인, 히스톤 데아세틸라아제 도메인, 뉴클레아제 도메인, 억제제 도메인, 활성제 도메인, 핵-국부화 신호 도메인, 전사-단백질 영입 도메인, 세포 흡수 활성 관련 도메인, 핵산 결합 도메인 또는 항체 제시 도메인으로 이루어지는 군에서 선택되는 조성물.
61. 단락 59 에 있어서, 적어도 하나 이상의 효과기 도메인이 뉴클레아제 도메인 또는 재조합효소 도메인인 조성물.
62. 단락 61 에 있어서, 뉴클레아제 도메인이 비-특이적 FokI 엔도뉴클레아제 촉매 도메인인 조성물.
63. 단락 59-62 중 어느 하나에 있어서, 에너지 민감성 단백질이 크립토크롬-2 (CRY2) 인 조성물.
64. 단락 59-63 중 어느 하나에 있어서, 상호작용 파트너가 크립토크롬-상호작용 염기성 헬릭스-루프-헬릭스 (CIB1) 인 조성물.
65. 단락 59-64 중 어느 하나에 있어서, 에너지원이 전자기 조사, 음파 에너지 또는 열 에너지로 이루어지는 군에서 선택되는 조성물.
66. 단락 65 에 있어서, 전자기 조사가 가시광의 성분인 조성물.
67. 단락 66 에 있어서, 가시광 성분이 450nm-500nm 범위의 파장을 갖는 조성물.
68. 단락 66 에 있어서, 가시광 성분이 청색광인 조성물.
69. 단락 59 에 있어서, 에너지원을 적용하는 것이 6.2 mW/cm2 이상 강도로의 청색광 자극을 포함하는 조성물.
70. 단락 59-69 중 어느 하나에 있어서, DNA 결합 도메인이 (X1-11-X12X13-X14-33 또는 34 또는 35)z 를 포함하는 조성물로서, 여기서 X1-11 이 11 개의 인접한 아미노산 사슬이고, X12X13 이 반복 가변 이중잔기 (RVD) 이고, X14-33 또는 34 또는 35 가 21, 22 또는 23 개의 인접한 아미노산 사슬이고, z 가 적어도 5 내지 40 이고, 하나 이상의 RVD 가 NI, HD, NG, NN, KN, RN, NH, NQ, SS, SN, NK, KH, RH, HH, HI, KI, RI, SI, KG, HG, RG, SD, ND, KD, RD, YG, HN, NV, NS, HA, S*, N*, KA, H*, RA, NA 및 NC 로 이루어지는 군에서 선택되고, 이때 (*) 가 X13 에서의 아미노산이 부재함을 의미하는 조성물.
71. 단락 70 에 있어서, z 가 적어도 10 내지 26 인 조성물.
72. 단락 70 에 있어서:
X1-ll 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 1-11 로 나타내는 12 개의 인접한 아미노산의 서열이거나,
X14-34 또는 X14-35 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 12-32 또는 12-33 으로 나타내는 21 또는 22 개의 인접한 아미노산의 서열인 조성물.
73. 단락 70 에 있어서, 하나 이상의 RVD 가 (a) 구아닌 (G) 의 인지를 위한 HH, KH, NH, NK, NQ, RH, RN, SS; (b) 아데닌 (A) 의 인지를 위한 SI; (c) 티민 (T) 의 인지를 위한 HG, KG, RG; (d) 시토신 (C) 의 인지를 위한 RD, SD; (e) A 또는 G 의 인지를 위한 NV; 및 (f) A 또는 T 또는 G 또는 C 의 인지를 위한 H*, HA, KA, N*, NA, NC, NS, RA, S* (여기서 (*) 는 X13 에서의 아미노산이 부재함을 의미함) 로 이루어지는 군에서 선택되는 조성물.
74. 단락 73 에 있어서:
G 의 인지를 위한 RVD 가 RN, NH, RH 또는 KH 이거나;
A 의 인지를 위한 RVD 가 SI 이거나;
T 의 인지를 위한 RVD 가 KG 또는 RG 이고;
C 의 인지를 위한 RVD 가 SD 또는 RD 인 조성물.
75. 단락 70 에 있어서, 하기 중 하나 이상이 존재하는 조성물:
X1-4 에서의 [LTLD] 또는 [LTLA] 또는 [LTQV], 또는
위치 X30-33 또는 X31-34 또는 X32-35 에서의 [EQHG] 또는 [RDHG].
76. 단락 59-75 중 어느 하나에 있어서:
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 147 개의 인접한 아미노산을 포함하거나,
C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 68 개의 인접한 아미노산을 포함하거나,
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 136 개의 인접한 아미노산을 포함하고 C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 183 개의 인접한 아미노산을 포함하는 조성물.
77. 단락 59-76 중 어느 하나에 있어서, 관심 게놈 유전자좌가 분화 인자, 전사 인자, 신경전달물질 이동체, 신경전달물질 신타아제, 시냅스 단백질, 가소성 단백질, 시냅스전 활성 구역 단백질, 시냅스후 밀도 단백질, 신경전달물질 수용체, 후생적 변형제, 신경 운명 선별화 인자, 액손 안내 분자, 이온 채널, CpG 결합 단백질, 유비퀴틴화 단백질, 호르몬, 호메오박스 단백질, 성장 인자, 발암유전자 또는 원발암유전자를 인코딩하는 유전자와 관련되는 조성물.
78. 세포에서의 게놈 유전자좌의 발현을 유도가능하게 억제하기 위한 비-자연발생적 또는 조작된 조성물로서, 상기 조성물이 하기를 포함하는 DNA 결합 폴리펩티드를 포함하고:
(i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함);
상기 폴리펩티드가 코돈 최적화된 핵산 분자에 의해 인코딩되고 이로부터 발현되어, 폴리펩티드가 바람직하게는 게놈 유전자좌의 DNA 에 결합하고,
상기 폴리펩티드가 에너지원의 적용시 게놈 유전자좌의 발현을 억제하는 조성물.
79. 단락 78 에 있어서, 폴리펩티드가 1 개 이상의 SID 억제제 도메인을 포함하는 조성물.
80. 단락 79 에 있어서, 폴리펩티드가 4 개 이상의 SID 억제제 도메인을 포함하는 조성물.
81. 단락 78 에 있어서, 폴리펩티드가 SID4X 억제제 도메인을 포함하는 조성물.
82. 단락 78 에 있어서, 폴리펩티드가 KRAB 억제제 도메인을 포함하는 조성물.
83. 단락 78-82 중 어느 하나에 있어서, 에너지 민감성 단백질이 크립토크롬-2 (CRY2) 인 조성물.
84. 단락 78-83 중 어느 하나에 있어서, 상호작용 파트너가 크립토크롬-상호작용 염기성 헬릭스-루프-헬릭스 (CIB1) 인 조성물.
85. 단락 78-84 중 어느 하나에 있어서, 에너지원이 전자기 조사, 음파 에너지 또는 열 에너지로 이루어지는 군에서 선택되는 조성물.
86. 단락 78 에 있어서, 전자기 조사가 가시광의 성분인 조성물.
87. 단락 86 에 있어서, 가시광 성분이 450nm-500nm 범위의 파장을 갖는 조성물.
88. 단락 86 에 있어서, 가시광 성분이 청색광인 조성물.
89. 단락 78 에 있어서, 에너지원을 적용하는 것이 6.2 mW/cm2 이상 강도로의 청색광 자극을 포함하는 조성물.
90. 단락 78-89 중 어느 하나에 있어서, DNA 결합 도메인이 (X1-11-X12X13-X14-33 또는 34 또는 35)z 를 포함하는 방법으로서, 여기서 X1-11 이 11 개의 인접한 아미노산 사슬이고, X12X13 이 반복 가변 이중잔기 (RVD) 이고, X14-33 또는 34 또는 35 가 21, 22 또는 23 개의 인접한 아미노산 사슬이고, z 가 적어도 5 내지 40 이고, 하나 이상의 RVD 가 NI, HD, NG, NN, KN, RN, NH, NQ, SS, SN, NK, KH, RH, HH, HI, KI, RI, SI, KG, HG, RG, SD, ND, KD, RD, YG, HN, NV, NS, HA, S*, N*, KA, H*, RA, NA 및 NC 로 이루어지는 군에서 선택되고, 이때 (*) 가 X13 에서의 아미노산이 부재함을 의미하는 조성물.
91. 단락 90 에 있어서, z 가 적어도 10 내지 26 인 조성물.
92. 단락 90 에 있어서:
X1-ll 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 1-11 로 나타내는 11 개의 인접한 아미노산의 서열이거나,
X14-34 또는 X14-35 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 12-32 또는 12-33 으로 나타내는 21 또는 22 개의 인접한 아미노산의 서열인 조성물.
93. 단락 78-92 중 어느 하나에 있어서:
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 147 개의 인접한 아미노산을 포함하거나,
C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 68 개의 인접한 아미노산을 포함하거나,
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 136 개의 인접한 아미노산을 포함하고 C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 183 개의 인접한 아미노산을 포함하는 조성물.
94. 단락 78-93 중 어느 하나에 있어서, 관심 게놈 유전자좌가 분화 인자 또는 이온 채널 성분을 인코딩하는 유전자와 관련되는 조성물.
95. 단락 94 에 있어서, 분화 인자가 SRY-박스-2 (SOX2) 이며 유전자 SOX2 에 의해 인코딩되는 조성물.
96. 단락 94 에 있어서, 분화 인자가 p11 이며 유전자 p11 에 의해 인코딩되는 조성물.
97. 단락 95 에 있어서, 이온 채널 성분이 CACNA1C 이며 유전자 CACNA1C 에 의해 인코딩되는 조성물.
98. 세포에서의 게놈 유전자좌의 발현을 유도가능하게 활성화하기 위한 비-자연발생적 또는 조작된 조성물로서, 상기 조성물이 하기를 포함하는 DNA 결합 폴리펩티드를 포함하고:
(i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함);
상기 폴리펩티드가 코돈 최적화된 핵산 분자에 의해 인코딩되고 이로부터 발현되어, 폴리펩티드가 바람직하게는 게놈 유전자좌의 DNA 에 결합하고,
상기 폴리펩티드가 에너지원의 적용시 게놈 유전자좌의 발현을 활성화하는 조성물.
99. 단락 98 에 있어서, 폴리펩티드가 하나 이상의 VP16 또는 VP64 활성제 도메인을 포함하는 조성물.
100. 단락 98 에 있어서, 폴리펩티드가 하나 이상의 p65 활성제 도메인을 포함하는 조성물.
101. 단락 98-100 중 어느 하나에 있어서, 에너지 민감성 단백질이 CRY2 인 조성물.
102. 단락 98-101 중 어느 하나에 있어서, 상호작용 파트너가 CIB1 인 조성물.
103. 단락 98 에 있어서, 에너지원이 전자기 조사, 음파 에너지 또는 열 에너지로 이루어지는 군에서 선택되는 조성물.
104. 단락 103 에 있어서, 전자기 조사가 가시광의 성분인 조성물.
105. 단락 104 에 있어서, 가시광 성분이 450nm-500nm 범위의 파장을 갖는 조성물.
106. 단락 104 에 있어서, 가시광 성분이 청색광인 조성물.
107. 단락 98 에 있어서, 에너지원을 적용하는 것이 6.2 mW/cm2 이상 강도로의 청색광 자극을 포함하는 조성물.
108. 단락 98-107 중 어느 하나에 있어서, DNA 결합 도메인이 (X1-11-X12X13-X14-33 또는 34 또는 35)z 를 포함하는 방법으로서, 여기서 X1-11 이 11 개의 인접한 아미노산 사슬이고, X12X13 이 반복 가변 이중잔기 (RVD) 이고, X14-33 또는 34 또는 35 가 21, 22 또는 23 개의 인접한 아미노산 사슬이고, z 가 적어도 5 내지 40 이고, 하나 이상의 RVD 가 NI, HD, NG, NN, KN, RN, NH, NQ, SS, SN, NK, KH, RH, HH, HI, KI, RI, SI, KG, HG, RG, SD, ND, KD, RD, YG, HN, NV, NS, HA, S*, N*, KA, H*, RA, NA 및 NC 로 이루어지는 군에서 선택되고, 이때 (*) 가 X13 에서의 아미노산이 부재함을 의미하는 조성물.
109. 단락 108 에 있어서, z 가 적어도 10 내지 26 인 조성물.
110. 단락 108 에 있어서:
X1-ll 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 1-11 로 나타내는 11 개의 인접한 아미노산의 서열이거나,
X14-34 또는 X14-35 중 하나 이상이 도 9 의 서열 (X1-11-X14-34 또는 X1-11-X14-35) 에서의 아미노산 12-32 또는 12-33 으로 나타내는 21 또는 22 개의 인접한 아미노산의 서열인 조성물.
111. 단락 98-110 중 어느 하나에 있어서:
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 147 개의 인접한 아미노산을 포함하거나,
C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 68 개의 인접한 아미노산을 포함하거나,
N-말단 캡핑 부위 또는 이의 단편이 야생형 N-말단 캡핑 부위의 136 개의 인접한 아미노산을 포함하고 C-말단 캡핑 부위 또는 이의 단편이 야생형 C-말단 캡핑 부위의 183 개의 인접한 아미노산을 포함하는 조성물.
112. 단락 98-111 중 어느 하나에 있어서, 관심 게놈 유전자좌가 분화 인자, 후생적 변형제, 이온 채널 성분 또는 수용체를 인코딩하는 유전자와 관련되는 조성물.
113. 단락 112 에 있어서, 분화 인자가 뉴로제닌-2 이며 유전자 NEUROG2 에 의해 인코딩되는 조성물.
114. 단락 112 에 있어서, 분화 인자가 크루펠 유사 인자 4 이며 유전자 KLF-4 에 의해 인코딩되는 조성물.
115. 단락 112 에 있어서, 후생적 변형제가 Tet 메틸시토신 디옥시게나아제 1 이며 유전자 tet-1 에 의해 인코딩되는 조성물.
116. 단락 112 에 있어서, 이온 채널 성분이 CACNA1C 이며 유전자 CACNA1C 에 의해 인코딩되는 조성물.
117. 단락 112 에 있어서, 수용체가 대사자극성 글루타메이트 수용체이며 유전자 mGlur2 에 의해 인코딩되는 조성물.
118. 단락 98-117 중 어느 하나에 있어서, 발현이 화학적으로 유도가능한 조성물.
119. 단락 118 에 있어서, 화학적으로 유도가능한 발현 시스템이 4-히드록시타목시펜 (4OHT) 에 의해 유도가능한 에스트로겐 수용체 (ER) 기반 시스템인 조성물.
120. 단락 119 에 있어서, 핵 이출 신호 (NES) 를 추가로 포함하는 조성물.
121. 단락 120 에 있어서, NES 가 LDLASLIL 의 서열을 갖는 조성물.
122. 단락 98-121 중 어느 하나에 따른 조성물을 인코딩하는 핵산.
123. 단락 122 에 있어서, 프로모터를 포함하는 핵산.
124. 단락 123 에 있어서, 프로모터가 인간 시냅신 I 프로모터 (hSyn) 인 핵산.
125. 단락 122-124 중 어느 하나에 있어서, 아데노 연관 바이러스 벡터 (AAV) 내로 패키징되는 핵산.
126. 하기를 포함하는, 관심 게놈 유전자좌의 발현을 변형하는 유도 방법:
(a) (i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함)
을 포함하는 DNA 결합 폴리펩티드를 포함하는 비-자연발생적 또는 조작된 조성물과 게놈 유전자좌를 접촉시키고;
(b) 에너지원을 적용시키고;
(c) 게놈 유전자좌의 발현이 변형된 것을 측정함.
127. 게놈 유전자좌의 발현을 유도가능하게 변형하기 위한 비-자연발생적 또는 조작된 조성물로서, 상기 조성물이 하기를 포함하는 DNA 결합 폴리펩티드를 포함하고:
(i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함);
상기 폴리펩티드가 바람직하게는 게놈 유전자좌의 DNA 에 결합하고,
상기 폴리펩티드가 에너지원의 적용시 게놈 유전자좌의 발현을 변형하는 조성물.
128. 하기를 포함하는, 세포에서의 관심 게놈 유전자좌의 발현을 교란하기 위한 유도 방법:
(a) 데옥시리보핵산 (DNA) 결합 폴리펩티드를 포함하는 비-자연발생적 또는 조작된 조성물과 게놈 유전자좌를 접촉시키고;
(b) 유도물질 근원을 적용하고;
(c) 게놈 유전자좌의 발현 교란이 발생하는 것을 측정함.
129. 단락 128 에 있어서, 발현을 교란하는 것이 발현을 변형하고 (상향 또는 하향), 발현 결과를 변형하거나 (예컨대 뉴클레아제를 사용) 발현 이동을 제거하고, 예를 들어 의존적 선택사항에 대하여 발현을 변형하는 것인 방법.
130. 단락 128 또는 129 에 있어서, 유도물질 근원이 에너지원 (예컨대 파동 또는 열) 또는 소분자인 방법.
131. 단락 126-130 중 어느 하나에 있어서, DNA 결합 폴리펩티드가 하기를 포함하는 방법:
(i) 에너지 민감성 단백질 또는 이의 단편에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 5 개 이상의 전사 활성제-유사 효과기 (TALE) 단량체 및 적어도 하나 이상의 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 형태적 변화를 거쳐, 상호작용 파트너와 결합할 수 있음), 및/또는
(ii) 상호작용 파트너에 연결된 적어도 하나 이상의 효과기 도메인 또는 관심 게놈 유전자좌를 표적화하도록 특이적으로 유도된 적어도 하나 이상의 TALE 단량체 또는 반-단량체를 포함하는 DNA 결합 도메인 (이때 상기 에너지 민감성 단백질 또는 이의 단편은 유도물질 에너지원에 의한 유도시 상호작용 파트너에 결합함).
132. 하기를 포함하는, 세포에서의 관심 게놈 유전자좌의 발현을 교란하기 위한 유도 방법:
(a) I. CRISPR/Cas 시스템 키메라 RNA (chiRNA) 폴리뉴클레오티드 서열에 작동가능하게 연결된 제 1 조절 요소 (이때, 상기 폴리뉴클레오티드 서열은 (a) 진핵세포에서의 표적 서열에 하이브리드화할 수 있는 안내 서열, (b) tracr mate 서열 및 (c) tracr 서열을 포함함), 및
II. 적어도 하나 이상의 핵 국부화 서열을 포함하는 CRISPR 효소를 인코딩하는 효소-코딩 서열에 작동가능하게 연결된 제 2 조절 유도 요소를 포함하는 하나 이상의 벡터를 포함하는 벡터 시스템을 게놈 유전자좌와 접촉시키고
{여기서 (a), (b) 및 (c) 는 5' 에서 3' 배향으로 배열되고,
성분 I 및 II 는 시스템의 동일하거나 상이한 벡터 상에 위치하고,
전사되는 경우, tracr mate 서열은 tracr 서열에 하이브리드화되고 안내 서열은 표적 서열에 대한 CRISPR 복합체의 서열-특이적 결합을 유도하고,
CRISPR 복합체는 (1) 표적 서열에 대해 하이브리드화되는 안내 서열, 및 (2) tracr 서열에 대해 하이브리드화되는 tracr mate 서열로 복합화되는 CRISPR 효소를 포함하고,
CRISPR 효소를 인코딩하는 효소 코딩 서열은 추가로 이종 기능적 도메인을 인코딩함};
(b) 유도물질 근원을 적용하고;
(c) 게놈 유전자좌의 발현 교란이 발생하는 것을 측정함.
133. 단락 132 에 있어서, 발현을 교란하는 것이 발현을 변형하고 (상향 또는 하향), 발현 결과를 변형하거나 (예컨대 뉴클레아제를 사용) 발현 이동을 제거하고, 예를 들어 의존적 선택사항에 대하여 발현을 변형하는 것인 방법.
134. 단락 132 또는 133 에 있어서, 유도물질 근원이 화학 물질인 방법.
135. 단락 132 내지 134 중 어느 하나에 있어서, 벡터가 렌티바이러스인 방법.
136. 단락 132 내지 135 중 어느 하나에 있어서, 제 2 조절 유도 요소가 테트라사이클린-의존적 조절 시스템을 포함하는 방법.
137. 단락 132 내지 135 중 어느 하나에 있어서, 제 2 조절 유도 요소가 큐메이트 유전자 스위치 시스템을 포함하는 방법.
138. 세포가 원핵세포 또는 진핵세포인, 단락 1-137 중 어느 하나에 따른 조성물, 핵산 또는 방법.
139. 단락 138 에 있어서, 진핵세포가 동물 세포인 조성물, 핵산 또는 방법.
140. 단락 139 에 있어서, 동물 세포가 포유동물 세포인 조성물, 핵산 또는 방법.
* * *
본 발명의 바람직한 구현예를 이에 따라 상세히 기재하였으며, 본 발명의 취지 또는 범주를 벗어나지 않고 그의 많은 명백한 변형이 가능하므로, 상기 단락에 의해 정의된 본 발명이 상기 상세한 설명에서 나타낸 특정 세부사항에 제한되지 않는다는 것이 이해될 것이다.
SEQUENCE LISTING
<110> THE BROAD INSTITUTE, INC.
MASSACHUSETTS INSTITUTE OF TECHNOLOGY
<120> INDUCIBLE DNA BINDING PROTEINS AND GENOME PERTURBATION TOOLS AND
APPLICATIONS THEREOF
<130> 44790.99.2005
<140> PCT/US2013/051418
<141> 2013-07-21
<150> 61/835,973
<151> 2013-06-17
<150> 61/794,458
<151> 2013-03-15
<150> 61/736,465
<151> 2012-12-12
<150> 61/721,283
<151> 2012-11-01
<150> 61/675,778
<151> 2012-07-25
<160> 513
<170> PatentIn version 3.5
<210> 1
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 1
Leu Thr Leu Asp
1
<210> 2
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 2
Leu Thr Leu Ala
1
<210> 3
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 3
Leu Thr Gln Val
1
<210> 4
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 4
Glu Gln His Gly
1
<210> 5
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 5
Arg Asp His Gly
1
<210> 6
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 6
Leu Asp Leu Ala Ser Leu Ile Leu
1 5
<210> 7
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 7
Leu Thr Pro Asp
1
<210> 8
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 8
Asn Gln Ala Leu Glu
1 5
<210> 9
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 9
Lys Arg Ala Leu Glu
1 5
<210> 10
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 10
Leu Thr Pro Glu
1
<210> 11
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 11
Asn Gly Lys Gln Ala Leu Glu
1 5
<210> 12
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 12
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 13
<211> 33
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(12)
<223> Any amino acid
<400> 13
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Xaa Gly Gly Lys Gln
1 5 10 15
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His
20 25 30
Gly
<210> 14
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 14
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Ala Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 15
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 15
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 16
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 16
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 17
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 17
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Ala Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 18
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 18
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 19
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 19
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Val Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 20
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 20
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly
<210> 21
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 21
Leu Thr Pro Tyr Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Ser Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 22
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 22
Leu Thr Arg Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp
20 25 30
His Gly
<210> 23
<211> 35
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 23
Leu Ser Thr Ala Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Gly Ile Gly Glu Gln Leu Leu Lys Leu Arg Thr Ala
20 25 30
Pro Tyr Gly
35
<210> 24
<211> 35
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (12)..(13)
<223> Any amino acid
<400> 24
Leu Ser Thr Ala Gln Val Val Ala Val Ala Ser Xaa Xaa Gly Gly Lys
1 5 10 15
Pro Ala Leu Glu Ala Val Arg Ala Gln Leu Leu Ala Leu Arg Ala Ala
20 25 30
Pro Tyr Gly
35
<210> 25
<211> 288
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 25
Met Asp Pro Ile Arg Ser Arg Thr Pro Ser Pro Ala Arg Glu Leu Leu
1 5 10 15
Ser Gly Pro Gln Pro Asp Gly Val Gln Pro Thr Ala Asp Arg Gly Val
20 25 30
Ser Pro Pro Ala Gly Gly Pro Leu Asp Gly Leu Pro Ala Arg Arg Thr
35 40 45
Met Ser Arg Thr Arg Leu Pro Ser Pro Pro Ala Pro Ser Pro Ala Phe
50 55 60
Ser Ala Asp Ser Phe Ser Asp Leu Leu Arg Gln Phe Asp Pro Ser Leu
65 70 75 80
Phe Asn Thr Ser Leu Phe Asp Ser Leu Pro Pro Phe Gly Ala His His
85 90 95
Thr Glu Ala Ala Thr Gly Glu Trp Asp Glu Val Gln Ser Gly Leu Arg
100 105 110
Ala Ala Asp Ala Pro Pro Pro Thr Met Arg Val Ala Val Thr Ala Ala
115 120 125
Arg Pro Pro Arg Ala Lys Pro Ala Pro Arg Arg Arg Ala Ala Gln Pro
130 135 140
Ser Asp Ala Ser Pro Ala Ala Gln Val Asp Leu Arg Thr Leu Gly Tyr
145 150 155 160
Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val
165 170 175
Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His
180 185 190
Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val
195 200 205
Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala
210 215 220
Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala
225 230 235 240
Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp
245 250 255
Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val
260 265 270
Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn
275 280 285
<210> 26
<211> 183
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 26
Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu Ser Arg Pro Asp Pro
1 5 10 15
Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val Ala Leu Ala Cys Leu
20 25 30
Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys Gly Leu Pro His Ala
35 40 45
Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile Pro Glu Arg Thr Ser
50 55 60
His Arg Val Ala Asp His Ala Gln Val Val Arg Val Leu Gly Phe Phe
65 70 75 80
Gln Cys His Ser His Pro Ala Gln Ala Phe Asp Asp Ala Met Thr Gln
85 90 95
Phe Gly Met Ser Arg His Gly Leu Leu Gln Leu Phe Arg Arg Val Gly
100 105 110
Val Thr Glu Leu Glu Ala Arg Ser Gly Thr Leu Pro Pro Ala Ser Gln
115 120 125
Arg Trp Asp Arg Ile Leu Gln Ala Ser Gly Met Lys Arg Ala Lys Pro
130 135 140
Ser Pro Thr Ser Thr Gln Thr Pro Asp Gln Ala Ser Leu His Ala Phe
145 150 155 160
Ala Asp Ser Leu Glu Arg Asp Leu Asp Ala Pro Ser Pro Met His Glu
165 170 175
Gly Asp Gln Thr Arg Ala Ser
180
<210> 27
<211> 18
<212> DNA
<213> Mus sp.
<400> 27
tgaatgatga taatacga 18
<210> 28
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 28
Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser
1 5 10
<210> 29
<211> 14
<212> DNA
<213> Homo sapiens
<400> 29
ttcttactta taac 14
<210> 30
<211> 7
<212> PRT
<213> Simian virus 40
<400> 30
Pro Lys Lys Lys Arg Lys Val
1 5
<210> 31
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown: Nucleoplasmin bipartite
NLS peptide"
<400> 31
Lys Arg Pro Ala Ala Thr Lys Lys Ala Gly Gln Ala Lys Lys Lys Lys
1 5 10 15
<210> 32
<211> 9
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown: C-myc NLS peptide"
<400> 32
Pro Ala Ala Lys Arg Val Lys Leu Asp
1 5
<210> 33
<211> 11
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown: C-myc NLS peptide"
<400> 33
Arg Gln Arg Arg Asn Glu Leu Lys Arg Ser Pro
1 5 10
<210> 34
<211> 38
<212> PRT
<213> Homo sapiens
<400> 34
Asn Gln Ser Ser Asn Phe Gly Pro Met Lys Gly Gly Asn Phe Gly Gly
1 5 10 15
Arg Ser Ser Gly Pro Tyr Gly Gly Gly Gly Gln Tyr Phe Ala Lys Pro
20 25 30
Arg Asn Gln Gly Gly Tyr
35
<210> 35
<211> 42
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown: IBB domain from importin-alpha
NLS polypeptide"
<400> 35
Arg Met Arg Ile Glx Phe Lys Asn Lys Gly Lys Asp Thr Ala Glu Leu
1 5 10 15
Arg Arg Arg Arg Val Glu Val Ser Val Glu Leu Arg Lys Ala Lys Lys
20 25 30
Asp Glu Gln Ile Leu Lys Arg Arg Asn Val
35 40
<210> 36
<211> 8
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown: Myoma T NLS peptide"
<400> 36
Val Ser Arg Lys Arg Pro Arg Pro
1 5
<210> 37
<211> 8
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown: Myoma T NLS peptide"
<400> 37
Pro Pro Lys Lys Ala Arg Glu Asp
1 5
<210> 38
<211> 7
<212> PRT
<213> Homo sapiens
<400> 38
Pro Gln Pro Lys Lys Lys Pro
1 5
<210> 39
<211> 12
<212> PRT
<213> Mus sp.
<400> 39
Ser Ala Leu Ile Lys Lys Lys Lys Lys Met Ala Pro
1 5 10
<210> 40
<211> 5
<212> PRT
<213> Influenza virus
<400> 40
Asp Arg Leu Arg Arg
1 5
<210> 41
<211> 7
<212> PRT
<213> Influenza virus
<400> 41
Pro Lys Gln Lys Lys Arg Lys
1 5
<210> 42
<211> 10
<212> PRT
<213> Hepatitis virus
<400> 42
Arg Lys Leu Lys Lys Lys Ile Lys Lys Leu
1 5 10
<210> 43
<211> 10
<212> PRT
<213> Mus sp.
<400> 43
Arg Glu Lys Lys Lys Phe Leu Lys Arg Arg
1 5 10
<210> 44
<211> 20
<212> PRT
<213> Homo sapiens
<400> 44
Lys Arg Lys Gly Asp Glu Val Asp Gly Val Asp Glu Val Ala Lys Lys
1 5 10 15
Lys Ser Lys Lys
20
<210> 45
<211> 17
<212> PRT
<213> Homo sapiens
<400> 45
Arg Lys Cys Leu Gln Ala Gly Met Asn Leu Glu Ala Arg Lys Thr Lys
1 5 10 15
Lys
<210> 46
<211> 33
<212> DNA
<213> Homo sapiens
<400> 46
ggaagggcct gagtccgagc agaagaagaa ggg 33
<210> 47
<211> 33
<212> DNA
<213> Homo sapiens
<400> 47
cattggaggt gacatcgatg tcctccccat tgg 33
<210> 48
<211> 33
<212> DNA
<213> Homo sapiens
<400> 48
ggacatcgat gtcacctcca atgactaggg tgg 33
<210> 49
<211> 33
<212> DNA
<213> Homo sapiens
<400> 49
catcgatgtc ctccccattg gcctgcttcg tgg 33
<210> 50
<211> 33
<212> DNA
<213> Homo sapiens
<400> 50
ttcgtggcaa tgcgccaccg gttgatgtga tgg 33
<210> 51
<211> 33
<212> DNA
<213> Homo sapiens
<400> 51
tcgtggcaat gcgccaccgg ttgatgtgat ggg 33
<210> 52
<211> 33
<212> DNA
<213> Homo sapiens
<400> 52
tccagcttct gccgtttgta ctttgtcctc cgg 33
<210> 53
<211> 33
<212> DNA
<213> Homo sapiens
<400> 53
ggagggaggg gcacagatga gaaactcagg agg 33
<210> 54
<211> 33
<212> DNA
<213> Homo sapiens
<400> 54
aggggccgag attgggtgtt cagggcagag agg 33
<210> 55
<211> 33
<212> DNA
<213> Homo sapiens
<400> 55
atgcaggagg gtggcgagag gggccgagat tgg 33
<210> 56
<211> 33
<212> DNA
<213> Homo sapiens
<400> 56
ggtggcgaga ggggccgaga ttgggtgttc agg 33
<210> 57
<211> 33
<212> DNA
<213> Mus musculus
<400> 57
caagcactga gtgccattag ctaaatgcat agg 33
<210> 58
<211> 33
<212> DNA
<213> Mus musculus
<400> 58
aatgcatagg gtaccaccca caggtgccag ggg 33
<210> 59
<211> 33
<212> DNA
<213> Mus musculus
<400> 59
acacacatgg gaaagcctct gggccaggaa agg 33
<210> 60
<211> 37
<212> DNA
<213> Homo sapiens
<400> 60
ggaggaggta gtatacagaa acacagagaa gtagaat 37
<210> 61
<211> 37
<212> DNA
<213> Homo sapiens
<400> 61
agaatgtaga ggagtcacag aaactcagca ctagaaa 37
<210> 62
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 62
aaaaccaccc ttctctctgg c 21
<210> 63
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 63
ggagattgga gacacggaga g 21
<210> 64
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 64
ctggaaagcc aatgcctgac 20
<210> 65
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 65
ggcagcaaac tccttgtcct 20
<210> 66
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 66
gtgctttgca gaggcctacc 20
<210> 67
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 67
cctggagcgc atgcagtagt 20
<210> 68
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 68
accttctgtg tttccaccat tc 22
<210> 69
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 69
ttggggagtg cacagacttc 20
<210> 70
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 70
ggctccctgg gttcaaagta 20
<210> 71
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 71
agaggggtct ggatgtcgta a 21
<210> 72
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
probe"
<400> 72
tagctctaaa acttcttctt ctgctcggac 30
<210> 73
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
probe"
<400> 73
ctagccttat tttaacttgc tatgctgttt 30
<210> 74
<211> 335
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 74
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
cgaaacaccg gaaccattca aaacagcata gcaagttaaa ataaggctag tccgttatca 300
acttgaaaaa gtggcaccga gtcggtgctt ttttt 335
<210> 75
<211> 423
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 75
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
cgaaacaccg gtagtattaa gtattgtttt atggctgata aatttctttg aatttctcct 300
tgattatttg ttataaaagt tataaaataa tcttgttgga accattcaaa acagcatagc 360
aagttaaaat aaggctagtc cgttatcaac ttgaaaaagt ggcaccgagt cggtgctttt 420
ttt 423
<210> 76
<211> 339
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 76
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
cgaaacaccg ggttttagag ctatgctgtt ttgaatggtc ccaaaacggg tcttcgagaa 300
gacgttttag agctatgctg ttttgaatgg tcccaaaac 339
<210> 77
<211> 309
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 77
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
cgaaacaccg ggtcttcgag aagacctgtt ttagagctag aaatagcaag ttaaaataag 300
gctagtccg 309
<210> 78
<211> 4269
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 78
atggactata aggaccacga cggagactac aaggatcatg atattgatta caaagacgat 60
gacgataaga tggccccaaa gaagaagcgg aaggtcggta tccacggagt cccagcagcc 120
gacaagaagt acagcatcgg cctggacatc ggcaccaact ctgtgggctg ggccgtgatc 180
accgacgagt acaaggtgcc cagcaagaaa ttcaaggtgc tgggcaacac cgaccggcac 240
agcatcaaga agaacctgat cggagccctg ctgttcgaca gcggcgaaac agccgaggcc 300
acccggctga agagaaccgc cagaagaaga tacaccagac ggaagaaccg gatctgctat 360
ctgcaagaga tcttcagcaa cgagatggcc aaggtggacg acagcttctt ccacagactg 420
gaagagtcct tcctggtgga agaggataag aagcacgagc ggcaccccat cttcggcaac 480
atcgtggacg aggtggccta ccacgagaag taccccacca tctaccacct gagaaagaaa 540
ctggtggaca gcaccgacaa ggccgacctg cggctgatct atctggccct ggcccacatg 600
atcaagttcc ggggccactt cctgatcgag ggcgacctga accccgacaa cagcgacgtg 660
gacaagctgt tcatccagct ggtgcagacc tacaaccagc tgttcgagga aaaccccatc 720
aacgccagcg gcgtggacgc caaggccatc ctgtctgcca gactgagcaa gagcagacgg 780
ctggaaaatc tgatcgccca gctgcccggc gagaagaaga atggcctgtt cggcaacctg 840
attgccctga gcctgggcct gacccccaac ttcaagagca acttcgacct ggccgaggat 900
gccaaactgc agctgagcaa ggacacctac gacgacgacc tggacaacct gctggcccag 960
atcggcgacc agtacgccga cctgtttctg gccgccaaga acctgtccga cgccatcctg 1020
ctgagcgaca tcctgagagt gaacaccgag atcaccaagg cccccctgag cgcctctatg 1080
atcaagagat acgacgagca ccaccaggac ctgaccctgc tgaaagctct cgtgcggcag 1140
cagctgcctg agaagtacaa agagattttc ttcgaccaga gcaagaacgg ctacgccggc 1200
tacattgacg gcggagccag ccaggaagag ttctacaagt tcatcaagcc catcctggaa 1260
aagatggacg gcaccgagga actgctcgtg aagctgaaca gagaggacct gctgcggaag 1320
cagcggacct tcgacaacgg cagcatcccc caccagatcc acctgggaga gctgcacgcc 1380
attctgcggc ggcaggaaga tttttaccca ttcctgaagg acaaccggga aaagatcgag 1440
aagatcctga ccttccgcat cccctactac gtgggccctc tggccagggg aaacagcaga 1500
ttcgcctgga tgaccagaaa gagcgaggaa accatcaccc cctggaactt cgaggaagtg 1560
gtggacaagg gcgcttccgc ccagagcttc atcgagcgga tgaccaactt cgataagaac 1620
ctgcccaacg agaaggtgct gcccaagcac agcctgctgt acgagtactt caccgtgtat 1680
aacgagctga ccaaagtgaa atacgtgacc gagggaatga gaaagcccgc cttcctgagc 1740
ggcgagcaga aaaaggccat cgtggacctg ctgttcaaga ccaaccggaa agtgaccgtg 1800
aagcagctga aagaggacta cttcaagaaa atcgagtgct tcgactccgt ggaaatctcc 1860
ggcgtggaag atcggttcaa cgcctccctg ggcacatacc acgatctgct gaaaattatc 1920
aaggacaagg acttcctgga caatgaggaa aacgaggaca ttctggaaga tatcgtgctg 1980
accctgacac tgtttgagga cagagagatg atcgaggaac ggctgaaaac ctatgcccac 2040
ctgttcgacg acaaagtgat gaagcagctg aagcggcgga gatacaccgg ctggggcagg 2100
ctgagccgga agctgatcaa cggcatccgg gacaagcagt ccggcaagac aatcctggat 2160
ttcctgaagt ccgacggctt cgccaacaga aacttcatgc agctgatcca cgacgacagc 2220
ctgaccttta aagaggacat ccagaaagcc caggtgtccg gccagggcga tagcctgcac 2280
gagcacattg ccaatctggc cggcagcccc gccattaaga agggcatcct gcagacagtg 2340
aaggtggtgg acgagctcgt gaaagtgatg ggccggcaca agcccgagaa catcgtgatc 2400
gaaatggcca gagagaacca gaccacccag aagggacaga agaacagccg cgagagaatg 2460
aagcggatcg aagagggcat caaagagctg ggcagccaga tcctgaaaga acaccccgtg 2520
gaaaacaccc agctgcagaa cgagaagctg tacctgtact acctgcagaa tgggcgggat 2580
atgtacgtgg accaggaact ggacatcaac cggctgtccg actacgatgt ggaccatatc 2640
gtgcctcaga gctttctgaa ggacgactcc atcgacaaca aggtgctgac cagaagcgac 2700
aagaaccggg gcaagagcga caacgtgccc tccgaagagg tcgtgaagaa gatgaagaac 2760
tactggcggc agctgctgaa cgccaagctg attacccaga gaaagttcga caatctgacc 2820
aaggccgaga gaggcggcct gagcgaactg gataaggccg gcttcatcaa gagacagctg 2880
gtggaaaccc ggcagatcac aaagcacgtg gcacagatcc tggactcccg gatgaacact 2940
aagtacgacg agaatgacaa gctgatccgg gaagtgaaag tgatcaccct gaagtccaag 3000
ctggtgtccg atttccggaa ggatttccag ttttacaaag tgcgcgagat caacaactac 3060
caccacgccc acgacgccta cctgaacgcc gtcgtgggaa ccgccctgat caaaaagtac 3120
cctaagctgg aaagcgagtt cgtgtacggc gactacaagg tgtacgacgt gcggaagatg 3180
atcgccaaga gcgagcagga aatcggcaag gctaccgcca agtacttctt ctacagcaac 3240
atcatgaact ttttcaagac cgagattacc ctggccaacg gcgagatccg gaagcggcct 3300
ctgatcgaga caaacggcga aaccggggag atcgtgtggg ataagggccg ggattttgcc 3360
accgtgcgga aagtgctgag catgccccaa gtgaatatcg tgaaaaagac cgaggtgcag 3420
acaggcggct tcagcaaaga gtctatcctg cccaagagga acagcgataa gctgatcgcc 3480
agaaagaagg actgggaccc taagaagtac ggcggcttcg acagccccac cgtggcctat 3540
tctgtgctgg tggtggccaa agtggaaaag ggcaagtcca agaaactgaa gagtgtgaaa 3600
gagctgctgg ggatcaccat catggaaaga agcagcttcg agaagaatcc catcgacttt 3660
ctggaagcca agggctacaa agaagtgaaa aaggacctga tcatcaagct gcctaagtac 3720
tccctgttcg agctggaaaa cggccggaag agaatgctgg cctctgccgg cgaactgcag 3780
aagggaaacg aactggccct gccctccaaa tatgtgaact tcctgtacct ggccagccac 3840
tatgagaagc tgaagggctc ccccgaggat aatgagcaga aacagctgtt tgtggaacag 3900
cacaagcact acctggacga gatcatcgag cagatcagcg agttctccaa gagagtgatc 3960
ctggccgacg ctaatctgga caaagtgctg tccgcctaca acaagcaccg ggataagccc 4020
atcagagagc aggccgagaa tatcatccac ctgtttaccc tgaccaatct gggagcccct 4080
gccgccttca agtactttga caccaccatc gaccggaaga ggtacaccag caccaaagag 4140
gtgctggacg ccaccctgat ccaccagagc atcaccggcc tgtacgagac acggatcgac 4200
ctgtctcagc tgggaggcga caagcgtcct gctgctacta agaaagctgg tcaagctaag 4260
aaaaagaaa 4269
<210> 79
<211> 1449
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 79
atgaagcagc tggaggagtt actttctacc tctttcgaca tccagtttaa tgacctgacc 60
ctgctggaaa ccgccttcac tcacacctcc tacgcgaatg agcaccgcct actgaatgtg 120
agccacaacg agcgcctgga gtttctgggg gatgctgtct tacagctgat catctctgaa 180
tatctgtttg ccaaataccc taagaaaacc gaaggggaca tgtcaaagct gcgctccatg 240
atagtcaggg aagagagcct ggcgggcttt agtcgttttt gctcattcga cgcttatatc 300
aagctgggaa aaggcgaaga gaagtccggc ggcaggaggc gcgatacaat tctgggcgat 360
ctctttgaag cgtttctggg cgcacttcta ctggacaaag ggatcgacgc agtccgccgc 420
tttctgaaac aagtgatgat ccctcaggtc gaaaagggaa acttcgagag agtgaaggac 480
tataaaacat gtttgcagga atttctccag accaagggag atgtagcaat agattatcag 540
gtaataagtg agaaaggacc agctcacgcc aaacaattcg aagttagcat cgttgttaat 600
ggcgcagtgt tgtcgaaggg cttgggtaaa tcaaaaaaac tggccgagca ggacgctgct 660
aaaaacgccc tcgctcagct cagcgaggta ggatccgtga gcaagggcga ggaggataac 720
atggccatca tcaaggagtt catgcgcttc aaggtgcaca tggagggctc cgtgaacggc 780
cacgagttcg agatcgaggg cgagggcgag ggccgcccct acgagggcac ccagaccgcc 840
aagctgaagg tgaccaaggg tggccccctg cccttcgcct gggacatcct gtcccctcag 900
ttcatgtacg gctccaaggc ctacgtgaag caccccgccg acatccccga ctacttgaag 960
ctgtccttcc ccgagggctt caagtgggag cgcgtgatga acttcgagga cggcggcgtg 1020
gtgaccgtga cccaggactc ctccctgcag gacggcgagt tcatctacaa ggtgaagctg 1080
cgcggcacca acttcccctc cgacggcccc gtaatgcaga agaagaccat gggctgggag 1140
gcctcctccg agcggatgta ccccgaggac ggcgccctga agggcgagat caagcagagg 1200
ctgaagctga aggacggcgg ccactacgac gctgaggtca agaccaccta caaggccaag 1260
aagcccgtgc agctgcccgg cgcctacaac gtcaacatca agttggacat cacctcccac 1320
aacgaggact acaccatcgt ggaacagtac gaacgcgccg agggccgcca ctccaccggc 1380
ggcatggacg agctgtacaa gaagcgtcct gctgctacta agaaagctgg tcaagctaag 1440
aaaaagaaa 1449
<210> 80
<211> 4269
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 80
atggactata aggaccacga cggagactac aaggatcatg atattgatta caaagacgat 60
gacgataaga tggccccaaa gaagaagcgg aaggtcggta tccacggagt cccagcagcc 120
gacaagaagt acagcatcgg cctggccatc ggcaccaact ctgtgggctg ggccgtgatc 180
accgacgagt acaaggtgcc cagcaagaaa ttcaaggtgc tgggcaacac cgaccggcac 240
agcatcaaga agaacctgat cggagccctg ctgttcgaca gcggcgaaac agccgaggcc 300
acccggctga agagaaccgc cagaagaaga tacaccagac ggaagaaccg gatctgctat 360
ctgcaagaga tcttcagcaa cgagatggcc aaggtggacg acagcttctt ccacagactg 420
gaagagtcct tcctggtgga agaggataag aagcacgagc ggcaccccat cttcggcaac 480
atcgtggacg aggtggccta ccacgagaag taccccacca tctaccacct gagaaagaaa 540
ctggtggaca gcaccgacaa ggccgacctg cggctgatct atctggccct ggcccacatg 600
atcaagttcc ggggccactt cctgatcgag ggcgacctga accccgacaa cagcgacgtg 660
gacaagctgt tcatccagct ggtgcagacc tacaaccagc tgttcgagga aaaccccatc 720
aacgccagcg gcgtggacgc caaggccatc ctgtctgcca gactgagcaa gagcagacgg 780
ctggaaaatc tgatcgccca gctgcccggc gagaagaaga atggcctgtt cggcaacctg 840
attgccctga gcctgggcct gacccccaac ttcaagagca acttcgacct ggccgaggat 900
gccaaactgc agctgagcaa ggacacctac gacgacgacc tggacaacct gctggcccag 960
atcggcgacc agtacgccga cctgtttctg gccgccaaga acctgtccga cgccatcctg 1020
ctgagcgaca tcctgagagt gaacaccgag atcaccaagg cccccctgag cgcctctatg 1080
atcaagagat acgacgagca ccaccaggac ctgaccctgc tgaaagctct cgtgcggcag 1140
cagctgcctg agaagtacaa agagattttc ttcgaccaga gcaagaacgg ctacgccggc 1200
tacattgacg gcggagccag ccaggaagag ttctacaagt tcatcaagcc catcctggaa 1260
aagatggacg gcaccgagga actgctcgtg aagctgaaca gagaggacct gctgcggaag 1320
cagcggacct tcgacaacgg cagcatcccc caccagatcc acctgggaga gctgcacgcc 1380
attctgcggc ggcaggaaga tttttaccca ttcctgaagg acaaccggga aaagatcgag 1440
aagatcctga ccttccgcat cccctactac gtgggccctc tggccagggg aaacagcaga 1500
ttcgcctgga tgaccagaaa gagcgaggaa accatcaccc cctggaactt cgaggaagtg 1560
gtggacaagg gcgcttccgc ccagagcttc atcgagcgga tgaccaactt cgataagaac 1620
ctgcccaacg agaaggtgct gcccaagcac agcctgctgt acgagtactt caccgtgtat 1680
aacgagctga ccaaagtgaa atacgtgacc gagggaatga gaaagcccgc cttcctgagc 1740
ggcgagcaga aaaaggccat cgtggacctg ctgttcaaga ccaaccggaa agtgaccgtg 1800
aagcagctga aagaggacta cttcaagaaa atcgagtgct tcgactccgt ggaaatctcc 1860
ggcgtggaag atcggttcaa cgcctccctg ggcacatacc acgatctgct gaaaattatc 1920
aaggacaagg acttcctgga caatgaggaa aacgaggaca ttctggaaga tatcgtgctg 1980
accctgacac tgtttgagga cagagagatg atcgaggaac ggctgaaaac ctatgcccac 2040
ctgttcgacg acaaagtgat gaagcagctg aagcggcgga gatacaccgg ctggggcagg 2100
ctgagccgga agctgatcaa cggcatccgg gacaagcagt ccggcaagac aatcctggat 2160
ttcctgaagt ccgacggctt cgccaacaga aacttcatgc agctgatcca cgacgacagc 2220
ctgaccttta aagaggacat ccagaaagcc caggtgtccg gccagggcga tagcctgcac 2280
gagcacattg ccaatctggc cggcagcccc gccattaaga agggcatcct gcagacagtg 2340
aaggtggtgg acgagctcgt gaaagtgatg ggccggcaca agcccgagaa catcgtgatc 2400
gaaatggcca gagagaacca gaccacccag aagggacaga agaacagccg cgagagaatg 2460
aagcggatcg aagagggcat caaagagctg ggcagccaga tcctgaaaga acaccccgtg 2520
gaaaacaccc agctgcagaa cgagaagctg tacctgtact acctgcagaa tgggcgggat 2580
atgtacgtgg accaggaact ggacatcaac cggctgtccg actacgatgt ggaccatatc 2640
gtgcctcaga gctttctgaa ggacgactcc atcgacaaca aggtgctgac cagaagcgac 2700
aagaaccggg gcaagagcga caacgtgccc tccgaagagg tcgtgaagaa gatgaagaac 2760
tactggcggc agctgctgaa cgccaagctg attacccaga gaaagttcga caatctgacc 2820
aaggccgaga gaggcggcct gagcgaactg gataaggccg gcttcatcaa gagacagctg 2880
gtggaaaccc ggcagatcac aaagcacgtg gcacagatcc tggactcccg gatgaacact 2940
aagtacgacg agaatgacaa gctgatccgg gaagtgaaag tgatcaccct gaagtccaag 3000
ctggtgtccg atttccggaa ggatttccag ttttacaaag tgcgcgagat caacaactac 3060
caccacgccc acgacgccta cctgaacgcc gtcgtgggaa ccgccctgat caaaaagtac 3120
cctaagctgg aaagcgagtt cgtgtacggc gactacaagg tgtacgacgt gcggaagatg 3180
atcgccaaga gcgagcagga aatcggcaag gctaccgcca agtacttctt ctacagcaac 3240
atcatgaact ttttcaagac cgagattacc ctggccaacg gcgagatccg gaagcggcct 3300
ctgatcgaga caaacggcga aaccggggag atcgtgtggg ataagggccg ggattttgcc 3360
accgtgcgga aagtgctgag catgccccaa gtgaatatcg tgaaaaagac cgaggtgcag 3420
acaggcggct tcagcaaaga gtctatcctg cccaagagga acagcgataa gctgatcgcc 3480
agaaagaagg actgggaccc taagaagtac ggcggcttcg acagccccac cgtggcctat 3540
tctgtgctgg tggtggccaa agtggaaaag ggcaagtcca agaaactgaa gagtgtgaaa 3600
gagctgctgg ggatcaccat catggaaaga agcagcttcg agaagaatcc catcgacttt 3660
ctggaagcca agggctacaa agaagtgaaa aaggacctga tcatcaagct gcctaagtac 3720
tccctgttcg agctggaaaa cggccggaag agaatgctgg cctctgccgg cgaactgcag 3780
aagggaaacg aactggccct gccctccaaa tatgtgaact tcctgtacct ggccagccac 3840
tatgagaagc tgaagggctc ccccgaggat aatgagcaga aacagctgtt tgtggaacag 3900
cacaagcact acctggacga gatcatcgag cagatcagcg agttctccaa gagagtgatc 3960
ctggccgacg ctaatctgga caaagtgctg tccgcctaca acaagcaccg ggataagccc 4020
atcagagagc aggccgagaa tatcatccac ctgtttaccc tgaccaatct gggagcccct 4080
gccgccttca agtactttga caccaccatc gaccggaaga ggtacaccag caccaaagag 4140
gtgctggacg ccaccctgat ccaccagagc atcaccggcc tgtacgagac acggatcgac 4200
ctgtctcagc tgggaggcga caagcgtcct gctgctacta agaaagctgg tcaagctaag 4260
aaaaagaaa 4269
<210> 81
<211> 2012
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 81
gaatgctgcc ctcagacccg cttcctccct gtccttgtct gtccaaggag aatgaggtct 60
cactggtgga tttcggacta ccctgaggag ctggcacctg agggacaagg ccccccacct 120
gcccagctcc agcctctgat gaggggtggg agagagctac atgaggttgc taagaaagcc 180
tcccctgaag gagaccacac agtgtgtgag gttggagtct ctagcagcgg gttctgtgcc 240
cccagggata gtctggctgt ccaggcactg ctcttgatat aaacaccacc tcctagttat 300
gaaaccatgc ccattctgcc tctctgtatg gaaaagagca tggggctggc ccgtggggtg 360
gtgtccactt taggccctgt gggagatcat gggaacccac gcagtgggtc ataggctctc 420
tcatttacta ctcacatcca ctctgtgaag aagcgattat gatctctcct ctagaaactc 480
gtagagtccc atgtctgccg gcttccagag cctgcactcc tccaccttgg cttggctttg 540
ctggggctag aggagctagg atgcacagca gctctgtgac cctttgtttg agaggaacag 600
gaaaaccacc cttctctctg gcccactgtg tcctcttcct gccctgccat ccccttctgt 660
gaatgttaga cccatgggag cagctggtca gaggggaccc cggcctgggg cccctaaccc 720
tatgtagcct cagtcttccc atcaggctct cagctcagcc tgagtgttga ggccccagtg 780
gctgctctgg gggcctcctg agtttctcat ctgtgcccct ccctccctgg cccaggtgaa 840
ggtgtggttc cagaaccgga ggacaaagta caaacggcag aagctggagg aggaagggcc 900
tgagtccgag cagaagaaga agggctccca tcacatcaac cggtggcgca ttgccacgaa 960
gcaggccaat ggggaggaca tcgatgtcac ctccaatgac aagcttgcta gcggtgggca 1020
accacaaacc cacgagggca gagtgctgct tgctgctggc caggcccctg cgtgggccca 1080
agctggactc tggccactcc ctggccaggc tttggggagg cctggagtca tggccccaca 1140
gggcttgaag cccggggccg ccattgacag agggacaagc aatgggctgg ctgaggcctg 1200
ggaccacttg gccttctcct cggagagcct gcctgcctgg gcgggcccgc ccgccaccgc 1260
agcctcccag ctgctctccg tgtctccaat ctcccttttg ttttgatgca tttctgtttt 1320
aatttatttt ccaggcacca ctgtagttta gtgatcccca gtgtccccct tccctatggg 1380
aataataaaa gtctctctct taatgacacg ggcatccagc tccagcccca gagcctgggg 1440
tggtagattc cggctctgag ggccagtggg ggctggtaga gcaaacgcgt tcagggcctg 1500
ggagcctggg gtggggtact ggtggagggg gtcaagggta attcattaac tcctctcttt 1560
tgttggggga ccctggtctc tacctccagc tccacagcag gagaaacagg ctagacatag 1620
ggaagggcca tcctgtatct tgagggagga caggcccagg tctttcttaa cgtattgaga 1680
ggtgggaatc aggcccaggt agttcaatgg gagagggaga gtgcttccct ctgcctagag 1740
actctggtgg cttctccagt tgaggagaaa ccagaggaaa ggggaggatt ggggtctggg 1800
ggagggaaca ccattcacaa aggctgacgg ttccagtccg aagtcgtggg cccaccagga 1860
tgctcacctg tccttggaga accgctgggc aggttgagac tgcagagaca gggcttaagg 1920
ctgagcctgc aaccagtccc cagtgactca gggcctcctc agcccaagaa agagcaacgt 1980
gccagggccc gctgagctct tgtgttcacc tg 2012
<210> 82
<211> 3462
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 82
atgaaaaggc cggcggccac gaaaaaggcc ggccaggcaa aaaagaaaaa gtccgacctg 60
gtacttggac tggatattgg tatcggttcg gtgggagtcg gaatcctcaa caaggtcacg 120
ggggagatca ttcacaagaa ctcgcggatc ttccccgcag ctcaggctga gaacaacttg 180
gtgcggagaa cgaataggca gggcaggcga ctggcgagga ggaagaaaca caggagagtc 240
cgattgaacc ggctgttcga ggagtccggt ttgatcaccg actttacgaa aatctcgatt 300
aaccttaatc cctatcagct tcgggtgaaa ggcctgacag acgaactttc gaatgaggaa 360
cttttcatcg cgctgaaaaa catggtcaag cacagaggga tttcctacct cgatgacgcc 420
tcggatgacg gaaattcctc agtaggagat tatgcacaga tcgtgaaaga gaactcaaag 480
caactggaaa caaagacacc ggggcagatc caacttgaaa gataccagac atacggacag 540
ctcagaggag attttacggt ggagaaggac ggtaaaaagc acagactcat taacgtattt 600
cccacgtcgg cgtacagatc cgaagcgctc cgcatccttc agactcaaca ggagttcaac 660
ccgcaaatta ctgatgagtt catcaaccgc tatttggaaa tcttgaccgg aaagcgcaag 720
tattatcatg ggccgggtaa tgagaaatcc agaacagatt acggccgata cagaacttcg 780
ggggaaacct tggataacat ctttggtatt ttgattggaa agtgcacctt ttacccggac 840
gagtttcgag cggccaaggc gtcatacaca gcacaagagt ttaatctctt gaatgatttg 900
aacaacttga cggtccccac ggagacaaag aagctctcca aagagcaaaa gaaccaaatc 960
atcaactacg tcaagaacga gaaggctatg gggccagcga agctgttcaa gtatatcgct 1020
aaacttctca gctgtgatgt ggcggacatc aaagggtacc gaatcgacaa gtcgggaaaa 1080
gcggaaattc acacgtttga agcatatcga aagatgaaaa cgttggaaac actggacatt 1140
gagcagatgg accgggaaac gctcgacaaa ctggcatacg tgctcacgtt gaatactgaa 1200
cgagagggaa tccaagaggc ccttgaacat gagttcgccg atggatcgtt cagccagaag 1260
caggtcgacg aacttgtgca attccgcaag gcgaatagct ccatcttcgg gaagggatgg 1320
cacaactttt cggtcaaact catgatggag ttgatcccag aactttatga gacttcggag 1380
gagcaaatga cgatcttgac gcgcttgggg aaacagaaaa cgacaagctc atcgaacaaa 1440
actaagtaca ttgatgagaa attgctgacg gaagaaatct ataatccggt agtagcgaaa 1500
tcggtaagac aagcgatcaa aatcgtgaac gcggcgatca aggaatatgg tgactttgat 1560
aacatcgtaa ttgaaatggc tagagagacg aacgaagatg acgagaaaaa ggcaatccag 1620
aagatccaga aggccaacaa ggatgaaaaa gatgcagcga tgcttaaagc ggccaaccaa 1680
tacaatggaa aggcggagct gccccattca gtgtttcacg gtcataaaca gttggcgacc 1740
aagatccgac tctggcatca gcagggtgag cggtgtctct acaccggaaa gactatctcc 1800
atccatgact tgattaacaa ttcgaaccag tttgaagtgg atcatattct gcccctgtca 1860
atcacctttg acgactcgct tgcgaacaag gtgctcgtgt acgcaacggc aaatcaggag 1920
aaaggccagc ggactccgta tcaggcgctc gactcaatgg acgatgcgtg gtcattccgg 1980
gagctgaagg cgttcgtacg cgagagcaag acactgagca acaaaaagaa agagtatctg 2040
ctgacagagg aggacatctc gaaattcgat gtcaggaaga agttcatcga gcggaatctt 2100
gtcgacactc gctacgcttc cagagtagta ctgaacgcgc tccaggaaca ctttagagcg 2160
cacaaaattg acacgaaggt gtcagtggtg agagggcagt tcacatccca actccgccga 2220
cattggggca tcgaaaagac gcgggacaca tatcaccatc atgcggtgga cgcgctgatt 2280
attgccgctt cgtcccagtt gaatctctgg aaaaagcaga agaacacgct ggtgtcgtat 2340
tcggaggatc agcttttgga catcgaaacc ggggagctga tttccgacga tgaatacaaa 2400
gaatcggtgt ttaaggcacc atatcagcat ttcgtggaca cgctgaagag caaagagttt 2460
gaggacagca tcctcttttc gtaccaagtg gactcgaagt ttaatcgcaa gatttcagac 2520
gccacaatct acgcgacgag gcaggcgaag gtgggcaaag ataaagcaga tgaaacctac 2580
gtccttggta aaatcaagga catctacact caggacgggt acgatgcgtt catgaaaatc 2640
tacaagaagg ataagtcgaa gtttctcatg taccgccacg atccacagac tttcgaaaaa 2700
gtcattgagc ctattttgga gaactaccct aacaagcaaa tcaacgagaa agggaaagaa 2760
gtcccgtgca acccctttct gaagtacaag gaagagcacg gttatatccg caaatactcg 2820
aagaaaggaa atgggcctga gattaagtcg cttaagtatt acgactcaaa gttgggtaac 2880
cacatcgaca ttaccccgaa agactccaac aacaaagtcg tgttgcagtc cgtctcgccc 2940
tggcgagcag atgtgtattt taataagacg accggcaaat atgagatcct tggactcaaa 3000
tacgcagacc ttcaattcga aaaggggacg ggcacttata agatttcaca agagaagtac 3060
aacgacatca agaaaaagga aggggtcgat tcagattcgg agttcaaatt caccctctac 3120
aaaaacgacc tcctgcttgt gaaggacaca gaaacgaagg agcagcagct ctttcggttc 3180
ctctcacgca cgatgcccaa acaaaaacat tacgtcgaac ttaaacctta cgataagcaa 3240
aagtttgaag ggggagaggc actgatcaaa gtattgggta acgtagccaa tagcggacag 3300
tgtaagaaag ggctgggaaa gtccaatatc tcgatctata aagtacgaac agatgtattg 3360
ggaaaccagc atatcatcaa aaatgagggg gataaaccca aactcgattt caagcgtcct 3420
gctgctacta agaaagctgg tcaagctaag aaaaagaaat aa 3462
<210> 83
<211> 340
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 83
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
cgaaacaccg ttacttaaat cttgcagaag ctacaaagat aaggcttcat gccgaaatca 300
acaccctgtc attttatggc agggtgtttt cgttatttaa 340
<210> 84
<211> 2601
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 84
atggactata aggaccacga cggagactac aaggatcatg atattgatta caaagacgat 60
gacgataaga tggccccaaa gaagaagcgg aaggtcggta tccacggagt cccagcagcc 120
gtagatttga gaactttggg atattcacag cagcagcagg aaaagatcaa gcccaaagtg 180
aggtcgacag tcgcgcagca tcacgaagcg ctggtgggtc atgggtttac acatgcccac 240
atcgtagcct tgtcgcagca ccctgcagcc cttggcacgg tcgccgtcaa gtaccaggac 300
atgattgcgg cgttgccgga agccacacat gaggcgatcg tcggtgtggg gaaacagtgg 360
agcggagccc gagcgcttga ggccctgttg acggtcgcgg gagagctgag agggcctccc 420
cttcagctgg acacgggcca gttgctgaag atcgcgaagc ggggaggagt cacggcggtc 480
gaggcggtgc acgcgtggcg caatgcgctc acgggagcac ccctcaacct gaccccagag 540
caggtcgtgg caattgcgag caaccacggg ggaaagcagg cactcgaaac cgtccagagg 600
ttgctgcctg tgctgtgcca agcgcacgga cttacgccag agcaggtcgt ggcaattgcg 660
agcaaccacg ggggaaagca ggcactcgaa accgtccaga ggttgctgcc tgtgctgtgc 720
caagcgcacg gactaacccc agagcaggtc gtggcaattg cgagcaacat cgggggaaag 780
caggcactcg aaaccgtcca gaggttgctg cctgtgctgt gccaagcgca cgggttgacc 840
ccagagcagg tcgtggcaat tgcgagcaac cacgggggaa agcaggcact cgaaaccgtc 900
cagaggttgc tgcctgtgct gtgccaagcg cacggcctga ccccagagca ggtcgtggca 960
attgcgagca accacggggg aaagcaggca ctcgaaaccg tccagaggtt gctgcctgtg 1020
ctgtgccaag cgcacggact gacaccagag caggtcgtgg caattgcgag caacatcggg 1080
ggaaagcagg cactcgaaac cgtccagagg ttgctgcctg tgctgtgcca agcgcacgga 1140
cttacacccg aacaagtcgt ggcaattgcg agcaaccacg ggggaaagca ggcactcgaa 1200
accgtccaga ggttgctgcc tgtgctgtgc caagcgcacg gacttacgcc agagcaggtc 1260
gtggcaattg cgagcaacca cgggggaaag caggcactcg aaaccgtcca gaggttgctg 1320
cctgtgctgt gccaagcgca cggactaacc ccagagcagg tcgtggcaat tgcgagcaac 1380
atcgggggaa agcaggcact cgaaaccgtc cagaggttgc tgcctgtgct gtgccaagcg 1440
cacgggttga ccccagagca ggtcgtggca attgcgagca acatcggggg aaagcaggca 1500
ctcgaaaccg tccagaggtt gctgcctgtg ctgtgccaag cgcacggcct gaccccagag 1560
caggtcgtgg caattgcgag caaccacggg ggaaagcagg cactcgaaac cgtccagagg 1620
ttgctgcctg tgctgtgcca agcgcacgga ctgacaccag agcaggtcgt ggcaattgcg 1680
agcaaccacg ggggaaagca ggcactcgaa accgtccaga ggttgctgcc tgtgctgtgc 1740
caagcgcacg gactcacgcc tgagcaggta gtggctattg catccaacaa cgggggcaga 1800
cccgcactgg agtcaatcgt ggcccagctt tcgaggccgg accccgcgct ggccgcactc 1860
actaatgatc atcttgtagc gctggcctgc ctcggcggac gacccgcctt ggatgcggtg 1920
aagaaggggc tcccgcacgc gcctgcattg attaagcgga ccaacagaag gattcccgag 1980
aggacatcac atcgagtggc aggttcccaa ctcgtgaaga gtgaacttga ggagaaaaag 2040
tcggagctgc ggcacaaatt gaaatacgta ccgcatgaat acatcgaact tatcgaaatt 2100
gctaggaact cgactcaaga cagaatcctt gagatgaagg taatggagtt ctttatgaag 2160
gtttatggat accgagggaa gcatctcggt ggatcacgaa aacccgacgg agcaatctat 2220
acggtgggga gcccgattga ttacggagtg atcgtcgaca cgaaagccta cagcggtggg 2280
tacaatcttc ccatcgggca ggcagatgag atgcaacgtt atgtcgaaga aaatcagacc 2340
aggaacaaac acatcaatcc aaatgagtgg tggaaagtgt atccttcatc agtgaccgag 2400
tttaagtttt tgtttgtctc tgggcatttc aaaggcaact ataaggccca gctcacacgg 2460
ttgaatcaca ttacgaactg caatggtgcg gttttgtccg tagaggaact gctcattggt 2520
ggagaaatga tcaaagcggg aactctgaca ctggaagaag tcagacgcaa gtttaacaat 2580
ggcgagatca atttccgctc a 2601
<210> 85
<211> 2600
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 85
atggactata aggaccacga cggagactac aaggatcatg atattgatta caaagacgat 60
gacgataaga tggccccaaa gaagaagcgg aaggtcggta tccacggagt cccagcagcc 120
gtagatttga gaactttggg atattcacag cagcagcagg aaaagatcaa gcccaaagtg 180
aggtcgacag tcgcgcagca tcacgaagcg ctggtgggtc atgggtttac acatgcccac 240
atcgtagcct tgtcgcagca ccctgcagcc cttggcacgg tcgccgtcaa gtaccaggac 300
atgattgcgg cgttgccgga agccacacat gaggcgatcg tcggtgtggg gaaacagtgg 360
agcggagccc gagcgcttga ggccctgttg acggtcgcgg gagagctgag agggcctccc 420
cttcagctgg acacgggcca gttgctgaag atcgcgaagc ggggaggagt cacggcggtc 480
gaggcggtgc acgcgtggcg caatgcgctc acgggagcac ccctcaacct gaccccagag 540
caggtcgtgg caattgcgag caaccacggg ggaaagcagg cactcgaaac cgtccagagg 600
ttgctgcctg tgctgtgcca agcgcacgga cttacgccag agcaggtcgt ggcaattgcg 660
agcaaccacg ggggaaagca ggcactcgaa accgtccaga ggttgctgcc tgtgctgtgc 720
caagcgcacg gactaacccc agagcaggtc gtggcaattg cgagcaacca cgggggaaag 780
caggcactcg aaaccgtcca gaggttgctg cctgtgctgt gccaagcgca cgggttgacc 840
ccagagcagg tcgtggcaat tgcgagcaac atcgggggaa agcaggcact cgaaaccgtc 900
cagaggttgc tgcctgtgct gtgccaagcg cacggcctga ccccagagca ggtcgtggca 960
attgcgagca accacggggg aaagcaggca ctcgaaaccg tccagaggtt gctgcctgtg 1020
ctgtgccaag cgcacggact gacaccagag caggtcgtgg caattgcgag ccatgacggg 1080
ggaaagcagg cactcgaaac cgtccagagg ttgctgcctg tgctgtgcca agcgcacgga 1140
cttacacccg aacaagtcgt ggcaattgcg agccatgacg ggggaaagca ggcactcgaa 1200
accgtccaga ggttgctgcc tgtgctgtgc caagcgcacg gacttacgcc agagcaggtc 1260
gtggcaattg cgagccatga cgggggaaag caggcactcg aaaccgtcca gaggttgctg 1320
cctgtgctgt gccaagcgca cggactaacc ccagagcagg tcgtggcaat tgcgagcaac 1380
ggagggggaa agcaggcact cgaaaccgtc cagaggttgc tgcctgtgct gtgccaagcg 1440
cacgggttga ccccagagca ggtcgtggca attgcgagca acggaggggg aaagcaggca 1500
ctcgaaaccg tccagaggtt gctgcctgtg ctgtgccaag cgcacggcct gaccccagag 1560
caggtcgtgg caattgcgag ccatgacggg ggaaagcagg cactcgaaac cgtccagagg 1620
ttgctgcctg tgctgtgcca agcgcacgga ctgacaccag agcaggtcgt ggcaattgcg 1680
agcaacggag ggggaaagca ggcactcgaa accgtccaga ggttgctgcc tgtgctgtgc 1740
caagcgcacg gactcacgcc tgagcaggta gtggctattg catccaacgg agggggcaga 1800
cccgcactgg agtcaatcgt ggcccagctt tcgaggccgg accccgcgct ggccgcactc 1860
actaatgatc atcttgtagc gctggcctgc ctcggcggac gacccgcctt ggatgcggtg 1920
aagaaggggc tcccgcacgc gcctgcattg attaagcgga ccaacagaag gattcccgag 1980
aggaatcaca tcgagtggca ggttcccaac tcgtgaagag tgaacttgag gagaaaaagt 2040
cggagctgcg gcacaaattg aaatacgtac cgcatgaata catcgaactt atcgaaattg 2100
ctaggaactc gactcaagac agaatccttg agatgaaggt aatggagttc tttatgaagg 2160
tttatggata ccgagggaag catctcggtg gatcacgaaa acccgacgga gcaatctata 2220
cggtggggag cccgattgat tacggagtga tcgtcgacac gaaagcctac agcggtgggt 2280
acaatcttcc catcgggcag gcagatgaga tgcaacgtta tgtcgaagaa aatcagacca 2340
ggaacaaaca catcaatcca aatgagtggt ggaaagtgta tccttcatca gtgaccgagt 2400
ttaagttttt gtttgtctct gggcatttca aaggcaacta taaggcccag ctcacacggt 2460
tgaatcacat tacgaactgc aatggtgcgg ttttgtccgt agaggaactg ctcattggtg 2520
gagaaatgat caaagcggga actctgacac tggaagaagt cagacgcaag tttaacaatg 2580
gcgagatcaa tttccgctca 2600
<210> 86
<211> 141
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 86
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc t 141
<210> 87
<211> 141
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 87
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 120
gagcgcgcag ctgcctgcag g 141
<210> 88
<211> 485
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 88
gtgtctagac tgcagagggc cctgcgtatg agtgcaagtg ggttttagga ccaggatgag 60
gcggggtggg ggtgcctacc tgacgaccga ccccgaccca ctggacaagc acccaacccc 120
cattccccaa attgcgcatc ccctatcaga gagggggagg ggaaacagga tgcggcgagg 180
cgcgtgcgca ctgccagctt cagcaccgcg gacagtgcct tcgcccccgc ctggcggcgc 240
gcgccaccgc cgcctcagca ctgaaggcgc gctgacgtca ctcgccggtc ccccgcaaac 300
tccccttccc ggccaccttg gtcgcgtccg cgccgccgcc ggcccagccg gaccgcacca 360
cgcgaggcgc gagatagggg ggcacgggcg cgaccatctg cgctgcggcg ccggcgactc 420
agcgctgcct cagtctgcgg tgggcagcgg aggagtcgtg tcgtgcctga gagcgcagtc 480
gagaa 485
<210> 89
<211> 408
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 89
gtagatttga gaactttggg atattcacag cagcagcagg aaaagatcaa gcccaaagtg 60
aggtcgacag tcgcgcagca tcacgaagcg ctggtgggtc atgggtttac acatgcccac 120
atcgtagcct tgtcgcagca ccctgcagcc cttggcacgg tcgccgtcaa gtaccaggac 180
atgattgcgg cgttgccgga agccacacat gaggcgatcg tcggtgtggg gaaacagtgg 240
agcggagccc gagcgcttga ggccctgttg acggtcgcgg gagagctgag agggcctccc 300
cttcagctgg acacgggcca gttgctgaag atcgcgaagc ggggaggagt cacggcggtc 360
gaggcggtgc acgcgtggcg caatgcgctc acgggagcac ccctcaac 408
<210> 90
<211> 164
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 90
cggaccccgc gctggccgca ctcactaatg atcatcttgt agcgctggcc tgcctcggcg 60
gacgacccgc cttggatgcg gtgaagaagg ggctcccgca cgcgcctgca ttgattaagc 120
ggaccaacag aaggattccc gagaggacat cacatcgagt ggca 164
<210> 91
<211> 858
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 91
atgagtattc aacatttccg tgtcgccctt attccctttt ttgcggcatt ttgccttcct 60
gtttttgctc acccagaaac gctggtgaaa gtaaaagatg ctgaagatca gttgggtgca 120
cgagtgggtt acatcgaact ggatctcaac agcggtaaga tccttgagag ttttcgcccc 180
gaagaacgtt ttccaatgat gagcactttt aaagttctgc tatgtggcgc ggtattatcc 240
cgtattgacg ccgggcaaga gcaactcggt cgccgcatac actattctca gaatgacttg 300
gttgagtact caccagtcac agaaaagcat cttacggatg gcatgacagt aagagaatta 360
tgcagtgctg ccataaccat gagtgataac actgcggcca acttacttct gacaacgatc 420
ggaggaccga aggagctaac cgcttttttg cacaacatgg gggatcatgt aactcgcctt 480
gatcgttggg aaccggagct gaatgaagcc ataccaaacg acgagcgtga caccacgatg 540
cctgtagcaa tggcaacaac gttgcgcaaa ctattaactg gcgaactact tactctagct 600
tcccggcaac aattaataga ctggatggag gcggataaag ttgcaggacc acttctgcgc 660
tcggcccttc cggctggctg gtttattgct gataaatctg gagccggtga gcgtgggtct 720
cgcggtatca ttgcagcact ggggccagat ggtaagccct cccgtatcgt agttatctac 780
acgacgggga gtcaggcaac tatggatgaa cgaaatagac agatcgctga gataggtgcc 840
tcactgatta agcattgg 858
<210> 92
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 92
gcgcgctcca cacaactcac 20
<210> 93
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 93
gcaaaaatag acaatcagca 20
<210> 94
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 94
gaaggatctc ggccaatttg 20
<210> 95
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 95
gctgccgggt tttgcatgaa 20
<210> 96
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 96
ccgggcccgc agcaaacttc 20
<210> 97
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 97
ggggctgtca gggaataaat 20
<210> 98
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 98
ctgtgctgaa ggatctgggg 20
<210> 99
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 99
atgctgcagg cataggacaa 20
<210> 100
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 100
gagggggaga gggactaaag a 21
<210> 101
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
primer"
<400> 101
gctctccctc cccagctta 19
<210> 102
<211> 14
<212> DNA
<213> Mus sp.
<400> 102
tatctgaact ctcc 14
<210> 103
<211> 14
<212> DNA
<213> Mus sp.
<400> 103
tgtctgtctt gcat 14
<210> 104
<211> 14
<212> DNA
<213> Mus sp.
<400> 104
tggctgttgc cagg 14
<210> 105
<211> 14
<212> DNA
<213> Mus sp.
<400> 105
tacctggagc tagc 14
<210> 106
<211> 14
<212> DNA
<213> Mus sp.
<400> 106
tacacaggat gtcc 14
<210> 107
<211> 14
<212> DNA
<213> Mus sp.
<400> 107
ttggccctgt gcag 14
<210> 108
<211> 14
<212> DNA
<213> Mus sp.
<400> 108
tagcgcagcg atcg 14
<210> 109
<211> 14
<212> DNA
<213> Mus sp.
<400> 109
tattgccaag agag 14
<210> 110
<211> 14
<212> DNA
<213> Mus sp.
<400> 110
tgactggaac atac 14
<210> 111
<211> 14
<212> DNA
<213> Mus sp.
<400> 111
tgatggactt gtat 14
<210> 112
<211> 14
<212> DNA
<213> Mus sp.
<400> 112
tggaccttct ccct 14
<210> 113
<211> 14
<212> DNA
<213> Mus sp.
<400> 113
taggtctcct ggag 14
<210> 114
<211> 14
<212> DNA
<213> Mus sp.
<400> 114
tggctcagga actt 14
<210> 115
<211> 14
<212> DNA
<213> Mus sp.
<400> 115
ttctctaagc tgcc 14
<210> 116
<211> 14
<212> DNA
<213> Mus sp.
<400> 116
tgagccctgg agga 14
<210> 117
<211> 14
<212> DNA
<213> Mus sp.
<400> 117
tgcctaagat ggag 14
<210> 118
<211> 14
<212> DNA
<213> Mus sp.
<400> 118
tgtagtgagt gttc 14
<210> 119
<211> 14
<212> DNA
<213> Mus sp.
<400> 119
tgtctaggtg atgt 14
<210> 120
<211> 14
<212> DNA
<213> Mus sp.
<400> 120
tctctgctag aagg 14
<210> 121
<211> 14
<212> DNA
<213> Mus sp.
<400> 121
tctaggtcaa gtgt 14
<210> 122
<211> 14
<212> DNA
<213> Mus sp.
<400> 122
tcctctgctc cgct 14
<210> 123
<211> 14
<212> DNA
<213> Mus sp.
<400> 123
tctaggagtg tagc 14
<210> 124
<211> 14
<212> DNA
<213> Mus sp.
<400> 124
tgcctggctg ctgg 14
<210> 125
<211> 14
<212> DNA
<213> Mus sp.
<400> 125
tatctgaact ctcc 14
<210> 126
<211> 14
<212> DNA
<213> Mus sp.
<400> 126
tcagagctgt cctc 14
<210> 127
<211> 14
<212> DNA
<213> Mus sp.
<400> 127
tgcaagagta ggag 14
<210> 128
<211> 14
<212> DNA
<213> Mus sp.
<400> 128
tagtgactga ttcc 14
<210> 129
<211> 14
<212> DNA
<213> Mus sp.
<400> 129
ttggaggagc acca 14
<210> 130
<211> 16
<212> DNA
<213> Mus sp.
<400> 130
tgaatgatga taatac 16
<210> 131
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 131
Gly Gly Ser Gly
1
<210> 132
<211> 1603
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 132
Met Ser Arg Thr Arg Leu Pro Ser Pro Pro Ala Pro Ser Pro Ala Phe
1 5 10 15
Ser Ala Asp Ser Phe Ser Asp Leu Leu Arg Gln Phe Asp Pro Ser Leu
20 25 30
Phe Asn Thr Ser Leu Phe Asp Ser Leu Pro Pro Phe Gly Ala His His
35 40 45
Thr Glu Ala Ala Thr Gly Glu Trp Asp Glu Val Gln Ser Gly Leu Arg
50 55 60
Ala Ala Asp Ala Pro Pro Pro Thr Met Arg Val Ala Val Thr Ala Ala
65 70 75 80
Arg Pro Pro Arg Ala Lys Pro Ala Pro Arg Arg Arg Ala Ala Gln Pro
85 90 95
Ser Asp Ala Ser Pro Ala Ala Gln Val Asp Leu Arg Thr Leu Gly Tyr
100 105 110
Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val
115 120 125
Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His
130 135 140
Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val
145 150 155 160
Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala
165 170 175
Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala
180 185 190
Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp
195 200 205
Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val
210 215 220
Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn
225 230 235 240
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
245 250 255
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
260 265 270
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
275 280 285
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
290 295 300
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
305 310 315 320
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
325 330 335
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
340 345 350
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
355 360 365
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
370 375 380
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
385 390 395 400
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
405 410 415
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
420 425 430
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
435 440 445
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
450 455 460
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
465 470 475 480
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
485 490 495
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
500 505 510
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
515 520 525
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
530 535 540
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
545 550 555 560
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
565 570 575
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
580 585 590
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
595 600 605
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
610 615 620
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
625 630 635 640
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
645 650 655
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
660 665 670
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
675 680 685
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
690 695 700
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
705 710 715 720
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
725 730 735
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
740 745 750
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
755 760 765
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
770 775 780
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Arg
785 790 795 800
Pro Ala Leu Glu Ser Ile Val Ala Gln Leu Ser Arg Pro Asp Pro Ala
805 810 815
Leu Ala Ala Leu Thr Asn Asp His Leu Val Ala Leu Ala Cys Leu Gly
820 825 830
Gly Arg Pro Ala Leu Asp Ala Val Lys Lys Gly Leu Pro His Ala Pro
835 840 845
Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile Pro Glu Arg Thr Ser His
850 855 860
Arg Val Ala Asp His Ala Gln Val Val Arg Val Leu Gly Phe Phe Gln
865 870 875 880
Cys His Ser His Pro Ala Gln Ala Phe Asp Asp Ala Met Thr Gln Phe
885 890 895
Gly Met Ser Arg His Gly Leu Leu Gln Leu Phe Arg Arg Val Gly Val
900 905 910
Thr Glu Leu Glu Ala Arg Ser Gly Thr Leu Pro Pro Ala Ser Gln Arg
915 920 925
Trp Asp Arg Ile Leu Gln Ala Ser Gly Met Lys Arg Ala Lys Pro Ser
930 935 940
Pro Thr Ser Thr Gln Thr Pro Asp Gln Ala Ser Leu His Ala Phe Ala
945 950 955 960
Asp Ser Leu Glu Arg Asp Leu Asp Ala Pro Ser Pro Met His Glu Gly
965 970 975
Asp Gln Thr Arg Ala Ser Ala Ser Pro Lys Lys Lys Arg Lys Val Glu
980 985 990
Ala Ser Lys Met Asp Lys Lys Thr Ile Val Trp Phe Arg Arg Asp Leu
995 1000 1005
Arg Ile Glu Asp Asn Pro Ala Leu Ala Ala Ala Ala His Glu Gly
1010 1015 1020
Ser Val Phe Pro Val Phe Ile Trp Cys Pro Glu Glu Glu Gly Gln
1025 1030 1035
Phe Tyr Pro Gly Arg Ala Ser Arg Trp Trp Met Lys Gln Ser Leu
1040 1045 1050
Ala His Leu Ser Gln Ser Leu Lys Ala Leu Gly Ser Asp Leu Thr
1055 1060 1065
Leu Ile Lys Thr His Asn Thr Ile Ser Ala Ile Leu Asp Cys Ile
1070 1075 1080
Arg Val Thr Gly Ala Thr Lys Val Val Phe Asn His Leu Tyr Asp
1085 1090 1095
Pro Val Ser Leu Val Arg Asp His Thr Val Lys Glu Lys Leu Val
1100 1105 1110
Glu Arg Gly Ile Ser Val Gln Ser Tyr Asn Gly Asp Leu Leu Tyr
1115 1120 1125
Glu Pro Trp Glu Ile Tyr Cys Glu Lys Gly Lys Pro Phe Thr Ser
1130 1135 1140
Phe Asn Ser Tyr Trp Lys Lys Cys Leu Asp Met Ser Ile Glu Ser
1145 1150 1155
Val Met Leu Pro Pro Pro Trp Arg Leu Met Pro Ile Thr Ala Ala
1160 1165 1170
Ala Glu Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu Gly Leu Glu
1175 1180 1185
Asn Glu Ala Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg Ala Trp
1190 1195 1200
Ser Pro Gly Trp Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe Ile
1205 1210 1215
Glu Lys Gln Leu Ile Asp Tyr Ala Lys Asn Ser Lys Lys Val Val
1220 1225 1230
Gly Asn Ser Thr Ser Leu Leu Ser Pro Tyr Leu His Phe Gly Glu
1235 1240 1245
Ile Ser Val Arg His Val Phe Gln Cys Ala Arg Met Lys Gln Ile
1250 1255 1260
Ile Trp Ala Arg Asp Lys Asn Ser Glu Gly Glu Glu Ser Ala Asp
1265 1270 1275
Leu Phe Leu Arg Gly Ile Gly Leu Arg Glu Tyr Ser Arg Tyr Ile
1280 1285 1290
Cys Phe Asn Phe Pro Phe Thr His Glu Gln Ser Leu Leu Ser His
1295 1300 1305
Leu Arg Phe Phe Pro Trp Asp Ala Asp Val Asp Lys Phe Lys Ala
1310 1315 1320
Trp Arg Gln Gly Arg Thr Gly Tyr Pro Leu Val Asp Ala Gly Met
1325 1330 1335
Arg Glu Leu Trp Ala Thr Gly Trp Met His Asn Arg Ile Arg Val
1340 1345 1350
Ile Val Ser Ser Phe Ala Val Lys Phe Leu Leu Leu Pro Trp Lys
1355 1360 1365
Trp Gly Met Lys Tyr Phe Trp Asp Thr Leu Leu Asp Ala Asp Leu
1370 1375 1380
Glu Cys Asp Ile Leu Gly Trp Gln Tyr Ile Ser Gly Ser Ile Pro
1385 1390 1395
Asp Gly His Glu Leu Asp Arg Leu Asp Asn Pro Ala Leu Gln Gly
1400 1405 1410
Ala Lys Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln Trp Leu Pro
1415 1420 1425
Glu Leu Ala Arg Leu Pro Thr Glu Trp Ile His His Pro Trp Asp
1430 1435 1440
Ala Pro Leu Thr Val Leu Lys Ala Ser Gly Val Glu Leu Gly Thr
1445 1450 1455
Asn Tyr Ala Lys Pro Ile Val Asp Ile Asp Thr Ala Arg Glu Leu
1460 1465 1470
Leu Ala Lys Ala Ile Ser Arg Thr Arg Glu Ala Gln Ile Met Ile
1475 1480 1485
Gly Ala Ala Pro Asp Glu Ile Val Ala Asp Ser Phe Glu Ala Leu
1490 1495 1500
Gly Ala Asn Thr Ile Lys Glu Pro Gly Leu Cys Pro Ser Val Ser
1505 1510 1515
Ser Asn Asp Gln Gln Val Pro Ser Ala Val Arg Tyr Asn Gly Ser
1520 1525 1530
Lys Arg Val Lys Pro Glu Glu Glu Glu Glu Arg Asp Met Lys Lys
1535 1540 1545
Ser Arg Gly Phe Asp Glu Arg Glu Leu Phe Ser Thr Ala Glu Ser
1550 1555 1560
Ser Ser Ser Ser Ser Val Phe Phe Val Ser Gln Ser Cys Ser Leu
1565 1570 1575
Ala Ser Glu Gly Lys Asn Leu Glu Gly Ile Gln Asp Ser Ser Asp
1580 1585 1590
Gln Ile Thr Thr Ser Leu Gly Lys Asn Gly
1595 1600
<210> 133
<211> 1492
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 133
Met Ser Arg Thr Arg Leu Pro Ser Pro Pro Ala Pro Ser Pro Ala Phe
1 5 10 15
Ser Ala Asp Ser Phe Ser Asp Leu Leu Arg Gln Phe Asp Pro Ser Leu
20 25 30
Phe Asn Thr Ser Leu Phe Asp Ser Leu Pro Pro Phe Gly Ala His His
35 40 45
Thr Glu Ala Ala Thr Gly Glu Trp Asp Glu Val Gln Ser Gly Leu Arg
50 55 60
Ala Ala Asp Ala Pro Pro Pro Thr Met Arg Val Ala Val Thr Ala Ala
65 70 75 80
Arg Pro Pro Arg Ala Lys Pro Ala Pro Arg Arg Arg Ala Ala Gln Pro
85 90 95
Ser Asp Ala Ser Pro Ala Ala Gln Val Asp Leu Arg Thr Leu Gly Tyr
100 105 110
Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val
115 120 125
Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His
130 135 140
Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val
145 150 155 160
Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala
165 170 175
Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala
180 185 190
Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp
195 200 205
Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val
210 215 220
Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn
225 230 235 240
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys
245 250 255
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
260 265 270
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
275 280 285
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
290 295 300
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
305 310 315 320
Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
325 330 335
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
340 345 350
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
355 360 365
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
370 375 380
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
385 390 395 400
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
405 410 415
Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val
420 425 430
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
435 440 445
Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu
450 455 460
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
465 470 475 480
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
485 490 495
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
500 505 510
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
515 520 525
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
530 535 540
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
545 550 555 560
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
565 570 575
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
580 585 590
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
595 600 605
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
610 615 620
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
625 630 635 640
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
645 650 655
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
660 665 670
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
675 680 685
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
690 695 700
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
705 710 715 720
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
725 730 735
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
740 745 750
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
755 760 765
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
770 775 780
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Arg
785 790 795 800
Pro Ala Leu Glu Ser Ile Val Ala Gln Leu Ser Arg Pro Asp Pro Ala
805 810 815
Leu Ala Ala Leu Thr Asn Asp His Leu Val Ala Leu Ala Cys Leu Gly
820 825 830
Gly Arg Pro Ala Leu Asp Ala Val Lys Lys Gly Leu Pro His Ala Pro
835 840 845
Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile Pro Glu Arg Thr Ser His
850 855 860
Arg Val Ala Asp His Ala Gln Val Val Arg Val Leu Gly Phe Phe Gln
865 870 875 880
Cys His Ser His Pro Ala Gln Ala Phe Asp Asp Ala Met Thr Gln Phe
885 890 895
Gly Met Ser Arg His Gly Leu Leu Gln Leu Phe Arg Arg Val Gly Val
900 905 910
Thr Glu Leu Glu Ala Arg Ser Gly Thr Leu Pro Pro Ala Ser Gln Arg
915 920 925
Trp Asp Arg Ile Leu Gln Ala Ser Gly Met Lys Arg Ala Lys Pro Ser
930 935 940
Pro Thr Ser Thr Gln Thr Pro Asp Gln Ala Ser Leu His Ala Phe Ala
945 950 955 960
Asp Ser Leu Glu Arg Asp Leu Asp Ala Pro Ser Pro Met His Glu Gly
965 970 975
Asp Gln Thr Arg Ala Ser Ala Ser Pro Lys Lys Lys Arg Lys Val Glu
980 985 990
Ala Ser Lys Met Asp Lys Lys Thr Ile Val Trp Phe Arg Arg Asp Leu
995 1000 1005
Arg Ile Glu Asp Asn Pro Ala Leu Ala Ala Ala Ala His Glu Gly
1010 1015 1020
Ser Val Phe Pro Val Phe Ile Trp Cys Pro Glu Glu Glu Gly Gln
1025 1030 1035
Phe Tyr Pro Gly Arg Ala Ser Arg Trp Trp Met Lys Gln Ser Leu
1040 1045 1050
Ala His Leu Ser Gln Ser Leu Lys Ala Leu Gly Ser Asp Leu Thr
1055 1060 1065
Leu Ile Lys Thr His Asn Thr Ile Ser Ala Ile Leu Asp Cys Ile
1070 1075 1080
Arg Val Thr Gly Ala Thr Lys Val Val Phe Asn His Leu Tyr Asp
1085 1090 1095
Pro Val Ser Leu Val Arg Asp His Thr Val Lys Glu Lys Leu Val
1100 1105 1110
Glu Arg Gly Ile Ser Val Gln Ser Tyr Asn Gly Asp Leu Leu Tyr
1115 1120 1125
Glu Pro Trp Glu Ile Tyr Cys Glu Lys Gly Lys Pro Phe Thr Ser
1130 1135 1140
Phe Asn Ser Tyr Trp Lys Lys Cys Leu Asp Met Ser Ile Glu Ser
1145 1150 1155
Val Met Leu Pro Pro Pro Trp Arg Leu Met Pro Ile Thr Ala Ala
1160 1165 1170
Ala Glu Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu Gly Leu Glu
1175 1180 1185
Asn Glu Ala Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg Ala Trp
1190 1195 1200
Ser Pro Gly Trp Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe Ile
1205 1210 1215
Glu Lys Gln Leu Ile Asp Tyr Ala Lys Asn Ser Lys Lys Val Val
1220 1225 1230
Gly Asn Ser Thr Ser Leu Leu Ser Pro Tyr Leu His Phe Gly Glu
1235 1240 1245
Ile Ser Val Arg His Val Phe Gln Cys Ala Arg Met Lys Gln Ile
1250 1255 1260
Ile Trp Ala Arg Asp Lys Asn Ser Glu Gly Glu Glu Ser Ala Asp
1265 1270 1275
Leu Phe Leu Arg Gly Ile Gly Leu Arg Glu Tyr Ser Arg Tyr Ile
1280 1285 1290
Cys Phe Asn Phe Pro Phe Thr His Glu Gln Ser Leu Leu Ser His
1295 1300 1305
Leu Arg Phe Phe Pro Trp Asp Ala Asp Val Asp Lys Phe Lys Ala
1310 1315 1320
Trp Arg Gln Gly Arg Thr Gly Tyr Pro Leu Val Asp Ala Gly Met
1325 1330 1335
Arg Glu Leu Trp Ala Thr Gly Trp Met His Asn Arg Ile Arg Val
1340 1345 1350
Ile Val Ser Ser Phe Ala Val Lys Phe Leu Leu Leu Pro Trp Lys
1355 1360 1365
Trp Gly Met Lys Tyr Phe Trp Asp Thr Leu Leu Asp Ala Asp Leu
1370 1375 1380
Glu Cys Asp Ile Leu Gly Trp Gln Tyr Ile Ser Gly Ser Ile Pro
1385 1390 1395
Asp Gly His Glu Leu Asp Arg Leu Asp Asn Pro Ala Leu Gln Gly
1400 1405 1410
Ala Lys Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln Trp Leu Pro
1415 1420 1425
Glu Leu Ala Arg Leu Pro Thr Glu Trp Ile His His Pro Trp Asp
1430 1435 1440
Ala Pro Leu Thr Val Leu Lys Ala Ser Gly Val Glu Leu Gly Thr
1445 1450 1455
Asn Tyr Ala Lys Pro Ile Val Asp Ile Asp Thr Ala Arg Glu Leu
1460 1465 1470
Leu Ala Lys Ala Ile Ser Arg Thr Arg Glu Ala Gln Ile Met Ile
1475 1480 1485
Gly Ala Ala Pro
1490
<210> 134
<211> 665
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 134
Met Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn Phe Pro Asp Met
1 5 10 15
Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr Leu Asn Pro Thr
20 25 30
Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser Ser Met Ile Thr
35 40 45
Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly Leu Asn Leu Pro
50 55 60
Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser Arg Leu Ser Ile
65 70 75 80
Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Lys Arg Lys Phe
85 90 95
Asp Thr Glu Thr Lys Asp Cys Asn Glu Lys Lys Lys Lys Met Thr Met
100 105 110
Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile
115 120 125
Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His
130 135 140
Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
145 150 155 160
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg Arg
165 170 175
Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg Arg Glu
180 185 190
Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val Pro Gly Cys
195 200 205
Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu Ile Ile Asn Tyr
210 215 220
Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu Ser Met Lys Leu Ala
225 230 235 240
Ile Val Asn Pro Arg Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys
245 250 255
Glu Val Ala Ser Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val
260 265 270
Leu Ser Gly Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu
275 280 285
Met Val Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn
290 295 300
Thr Ser Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser
305 310 315 320
Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val Ala
325 330 335
Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Gly Ser Gly Arg Ala
340 345 350
Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu
355 360 365
Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe
370 375 380
Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp
385 390 395 400
Met Leu Ile Asn Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu
405 410 415
Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu
420 425 430
Glu Leu Phe Thr Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp
435 440 445
Val Asn Gly His Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala
450 455 460
Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu
465 470 475 480
Pro Val Pro Trp Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln
485 490 495
Cys Phe Ser Arg Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys
500 505 510
Ser Ala Met Pro Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys
515 520 525
Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp
530 535 540
Thr Leu Val Asn Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp
545 550 555 560
Gly Asn Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn
565 570 575
Val Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe
580 585 590
Lys Ile Arg His Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His
595 600 605
Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp
610 615 620
Asn His Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu
625 630 635 640
Lys Arg Asp His Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile
645 650 655
Thr Leu Gly Met Asp Glu Leu Tyr Lys
660 665
<210> 135
<211> 500
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 135
Met Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn Phe Pro Asp Met
1 5 10 15
Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr Leu Asn Pro Thr
20 25 30
Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser Ser Met Ile Thr
35 40 45
Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly Leu Asn Leu Pro
50 55 60
Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser Arg Leu Ser Ile
65 70 75 80
Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Lys Arg Lys Phe
85 90 95
Asp Thr Glu Thr Lys Asp Cys Asn Glu Lys Lys Lys Lys Met Thr Met
100 105 110
Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile
115 120 125
Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His
130 135 140
Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
145 150 155 160
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile Ala Ser Pro Lys Lys Lys
165 170 175
Arg Lys Val Glu Ala Ser Gly Ser Gly Arg Ala Asp Ala Leu Asp Asp
180 185 190
Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu
195 200 205
Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu
210 215 220
Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Ile Asn Ser
225 230 235 240
Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val
245 250 255
Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu Glu Leu Phe Thr Gly
260 265 270
Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn Gly His Lys
275 280 285
Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu
290 295 300
Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro Val Pro Trp Pro
305 310 315 320
Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg Tyr
325 330 335
Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala Met Pro Glu
340 345 350
Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn Tyr
355 360 365
Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn Arg
370 375 380
Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu Gly
385 390 395 400
His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Met Ala
405 410 415
Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe Lys Ile Arg His Asn
420 425 430
Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr Gln Gln Asn Thr
435 440 445
Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu Ser
450 455 460
Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His Met
465 470 475 480
Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met Asp
485 490 495
Glu Leu Tyr Lys
500
<210> 136
<211> 693
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 136
Met Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn Phe Pro Asp Met
1 5 10 15
Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr Leu Asn Pro Thr
20 25 30
Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser Ser Met Ile Thr
35 40 45
Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly Leu Asn Leu Pro
50 55 60
Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser Arg Leu Ser Ile
65 70 75 80
Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Lys Arg Lys Phe
85 90 95
Asp Thr Glu Thr Lys Asp Cys Asn Glu Lys Lys Lys Lys Met Thr Met
100 105 110
Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile
115 120 125
Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His
130 135 140
Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
145 150 155 160
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg Arg
165 170 175
Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg Arg Glu
180 185 190
Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val Pro Gly Cys
195 200 205
Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu Ile Ile Asn Tyr
210 215 220
Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu Ser Met Lys Leu Ala
225 230 235 240
Ile Val Asn Pro Arg Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys
245 250 255
Glu Val Ala Ser Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val
260 265 270
Leu Ser Gly Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu
275 280 285
Met Val Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn
290 295 300
Thr Ser Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser
305 310 315 320
Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val Ala
325 330 335
Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Ala Pro Pro Thr Asp
340 345 350
Val Ser Leu Gly Asp Glu Leu His Leu Asp Gly Glu Asp Val Ala Met
355 360 365
Ala His Ala Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Asp
370 375 380
Gly Asp Ser Pro Gly Pro Gly Phe Thr Pro His Asp Ser Ala Pro Tyr
385 390 395 400
Gly Ala Leu Asp Met Ala Asp Phe Glu Phe Glu Gln Met Phe Thr Asp
405 410 415
Ala Leu Gly Ile Asp Glu Tyr Gly Gly Glu Phe Pro Gly Ile Arg Arg
420 425 430
Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp
435 440 445
Val Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu Glu Leu Phe Thr
450 455 460
Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn Gly His
465 470 475 480
Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys
485 490 495
Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro Val Pro Trp
500 505 510
Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg
515 520 525
Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala Met Pro
530 535 540
Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn
545 550 555 560
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn
565 570 575
Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu
580 585 590
Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Met
595 600 605
Ala Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe Lys Ile Arg His
610 615 620
Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr Gln Gln Asn
625 630 635 640
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
645 650 655
Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
660 665 670
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
675 680 685
Asp Glu Leu Tyr Lys
690
<210> 137
<211> 792
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 137
Met Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn Phe Pro Asp Met
1 5 10 15
Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr Leu Asn Pro Thr
20 25 30
Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser Ser Met Ile Thr
35 40 45
Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly Leu Asn Leu Pro
50 55 60
Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser Arg Leu Ser Ile
65 70 75 80
Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Lys Arg Lys Phe
85 90 95
Asp Thr Glu Thr Lys Asp Cys Asn Glu Lys Lys Lys Lys Met Thr Met
100 105 110
Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile
115 120 125
Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His
130 135 140
Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
145 150 155 160
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg Arg
165 170 175
Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg Arg Glu
180 185 190
Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val Pro Gly Cys
195 200 205
Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu Ile Ile Asn Tyr
210 215 220
Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu Ser Met Lys Leu Ala
225 230 235 240
Ile Val Asn Pro Arg Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys
245 250 255
Glu Val Ala Ser Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val
260 265 270
Leu Ser Gly Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu
275 280 285
Met Val Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn
290 295 300
Thr Ser Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser
305 310 315 320
Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val Ala
325 330 335
Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Pro Ser Gly Gln Ile
340 345 350
Ser Asn Gln Ala Leu Ala Leu Ala Pro Ser Ser Ala Pro Val Leu Ala
355 360 365
Gln Thr Met Val Pro Ser Ser Ala Met Val Pro Leu Ala Gln Pro Pro
370 375 380
Ala Pro Ala Pro Val Leu Thr Pro Gly Pro Pro Gln Ser Leu Ser Ala
385 390 395 400
Pro Val Pro Lys Ser Thr Gln Ala Gly Glu Gly Thr Leu Ser Glu Ala
405 410 415
Leu Leu His Leu Gln Phe Asp Ala Asp Glu Asp Leu Gly Ala Leu Leu
420 425 430
Gly Asn Ser Thr Asp Pro Gly Val Phe Thr Asp Leu Ala Ser Val Asp
435 440 445
Asn Ser Glu Phe Gln Gln Leu Leu Asn Gln Gly Val Ser Met Ser His
450 455 460
Ser Thr Ala Glu Pro Met Leu Met Glu Tyr Pro Glu Ala Ile Thr Arg
465 470 475 480
Leu Val Thr Gly Ser Gln Arg Pro Pro Asp Pro Ala Pro Thr Pro Leu
485 490 495
Gly Thr Ser Gly Leu Pro Asn Gly Leu Ser Gly Asp Glu Asp Phe Ser
500 505 510
Ser Ile Ala Asp Met Asp Phe Ser Ala Leu Leu Ser Gln Ile Ser Ser
515 520 525
Ser Gly Gln Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr
530 535 540
Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu Glu
545 550 555 560
Leu Phe Thr Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val
565 570 575
Asn Gly His Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr
580 585 590
Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro
595 600 605
Val Pro Trp Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys
610 615 620
Phe Ser Arg Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser
625 630 635 640
Ala Met Pro Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp
645 650 655
Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr
660 665 670
Leu Val Asn Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly
675 680 685
Asn Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val
690 695 700
Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe Lys
705 710 715 720
Ile Arg His Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr
725 730 735
Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn
740 745 750
His Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys
755 760 765
Arg Asp His Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr
770 775 780
Leu Gly Met Asp Glu Leu Tyr Lys
785 790
<210> 138
<211> 967
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (158)..(159)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (192)..(193)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (226)..(227)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (260)..(261)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (294)..(295)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (328)..(329)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (362)..(363)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (396)..(397)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (430)..(431)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (464)..(465)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (498)..(499)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (532)..(533)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (566)..(567)
<223> Any amino acid
<400> 138
Met Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Val Asp Leu Arg Thr Leu
1 5 10 15
Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser
20 25 30
Thr Val Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His
35 40 45
Ala His Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val
50 55 60
Ala Val Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His
65 70 75 80
Glu Ala Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu
85 90 95
Glu Ala Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln
100 105 110
Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr
115 120 125
Ala Val Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro
130 135 140
Leu Asn Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly
145 150 155 160
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
165 170 175
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa
180 185 190
Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
195 200 205
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
210 215 220
Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
225 230 235 240
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa
450 455 460
Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Xaa Xaa Gly Gly Arg Pro Ala Leu Glu Ser Ile
565 570 575
Val Ala Gln Leu Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn
580 585 590
Asp His Leu Val Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp
595 600 605
Ala Val Lys Lys Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr
610 615 620
Asn Arg Arg Ile Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Pro
625 630 635 640
Lys Lys Lys Arg Lys Val Glu Ala Ser Gly Ser Gly Arg Ala Asp Ala
645 650 655
Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp
660 665 670
Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu
675 680 685
Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu
690 695 700
Ile Asn Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys
705 710 715 720
Gly Asp Val Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu Glu Leu
725 730 735
Phe Thr Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn
740 745 750
Gly His Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr Tyr
755 760 765
Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro Val
770 775 780
Pro Trp Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe
785 790 795 800
Ser Arg Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala
805 810 815
Met Pro Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp
820 825 830
Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu
835 840 845
Val Asn Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn
850 855 860
Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr
865 870 875 880
Ile Met Ala Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe Lys Ile
885 890 895
Arg His Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr Gln
900 905 910
Gln Asn Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His
915 920 925
Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg
930 935 940
Asp His Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu
945 950 955 960
Gly Met Asp Glu Leu Tyr Lys
965
<210> 139
<211> 922
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (158)..(159)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (192)..(193)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (226)..(227)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (260)..(261)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (294)..(295)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (328)..(329)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (362)..(363)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (396)..(397)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (430)..(431)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (464)..(465)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (498)..(499)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (532)..(533)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (566)..(567)
<223> Any amino acid
<400> 139
Met Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Val Asp Leu Arg Thr Leu
1 5 10 15
Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser
20 25 30
Thr Val Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His
35 40 45
Ala His Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val
50 55 60
Ala Val Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His
65 70 75 80
Glu Ala Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu
85 90 95
Glu Ala Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln
100 105 110
Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr
115 120 125
Ala Val Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro
130 135 140
Leu Asn Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly
145 150 155 160
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
165 170 175
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa
180 185 190
Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
195 200 205
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
210 215 220
Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
225 230 235 240
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa
450 455 460
Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Xaa Xaa Gly Gly Arg Pro Ala Leu Glu Ser Ile
565 570 575
Val Ala Gln Leu Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn
580 585 590
Asp His Leu Val Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp
595 600 605
Ala Val Lys Lys Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr
610 615 620
Asn Arg Arg Ile Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Pro
625 630 635 640
Lys Lys Lys Arg Lys Val Glu Ala Ser Pro Lys Lys Lys Arg Lys Val
645 650 655
Glu Ala Ser Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala
660 665 670
Asp Tyr Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser
675 680 685
Met Leu Pro Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala
690 695 700
Asp Tyr Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser
705 710 715 720
Met Leu Pro Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala
725 730 735
Asp Tyr Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser
740 745 750
Met Leu Pro Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala
755 760 765
Asp Tyr Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser
770 775 780
Met Leu Pro Ser Arg Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu
785 790 795 800
Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Ile Glu Lys Ser
805 810 815
Phe Val Ile Thr Asp Pro Arg Leu Pro Asp Tyr Pro Ile Ile Phe Ala
820 825 830
Ser Asp Gly Phe Leu Glu Leu Thr Glu Tyr Ser Arg Glu Glu Ile Met
835 840 845
Gly Arg Asn Ala Arg Phe Leu Gln Gly Pro Glu Thr Asp Gln Ala Thr
850 855 860
Val Gln Lys Ile Arg Asp Ala Ile Arg Asp Gln Arg Glu Thr Thr Val
865 870 875 880
Gln Leu Ile Asn Tyr Thr Lys Ser Gly Lys Lys Phe Trp Asn Leu Leu
885 890 895
His Leu Gln Pro Val Arg Asp Arg Lys Gly Gly Leu Gln Tyr Phe Ile
900 905 910
Gly Val Gln Leu Val Gly Ser Asp His Val
915 920
<210> 140
<211> 983
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<220>
<221> MOD_RES
<222> (158)..(159)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (192)..(193)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (226)..(227)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (260)..(261)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (294)..(295)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (328)..(329)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (362)..(363)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (396)..(397)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (430)..(431)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (464)..(465)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (498)..(499)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (532)..(533)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (566)..(567)
<223> Any amino acid
<400> 140
Met Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Val Asp Leu Arg Thr Leu
1 5 10 15
Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser
20 25 30
Thr Val Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His
35 40 45
Ala His Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val
50 55 60
Ala Val Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His
65 70 75 80
Glu Ala Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu
85 90 95
Glu Ala Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln
100 105 110
Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr
115 120 125
Ala Val Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro
130 135 140
Leu Asn Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly
145 150 155 160
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
165 170 175
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa
180 185 190
Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
195 200 205
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
210 215 220
Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
225 230 235 240
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa Xaa Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Xaa
450 455 460
Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Xaa Xaa Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Xaa Xaa Gly Gly Arg Pro Ala Leu Glu Ser Ile
565 570 575
Val Ala Gln Leu Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn
580 585 590
Asp His Leu Val Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp
595 600 605
Ala Val Lys Lys Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr
610 615 620
Asn Arg Arg Ile Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Pro
625 630 635 640
Lys Lys Lys Arg Lys Val Glu Ala Ser Asn Gly Ala Ile Gly Gly Asp
645 650 655
Leu Leu Leu Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala
660 665 670
His Leu Lys Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe
675 680 685
Phe Ala Asp Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu
690 695 700
Ser Thr Ala Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val
705 710 715 720
Glu Gly Asp Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr
725 730 735
Gly Asn Phe Lys Lys Arg Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn
740 745 750
Glu Lys Lys Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu
755 760 765
Gly Glu Glu Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr
770 775 780
Lys Ser Ile Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn
785 790 795 800
Phe Ser Asn Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp
805 810 815
Tyr Ile His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser
820 825 830
Ile Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
835 840 845
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly
850 855 860
Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile
865 870 875 880
Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe
885 890 895
Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met Thr
900 905 910
Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His Glu Met
915 920 925
Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu His
930 935 940
Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu Ser Cys
945 950 955 960
Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His Val Gln Asn
965 970 975
Leu Tyr Gly Asn Leu Gly Val
980
<210> 141
<211> 830
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 141
Met Lys Met Asp Lys Lys Thr Ile Val Trp Phe Arg Arg Asp Leu Arg
1 5 10 15
Ile Glu Asp Asn Pro Ala Leu Ala Ala Ala Ala His Glu Gly Ser Val
20 25 30
Phe Pro Val Phe Ile Trp Cys Pro Glu Glu Glu Gly Gln Phe Tyr Pro
35 40 45
Gly Arg Ala Ser Arg Trp Trp Met Lys Gln Ser Leu Ala His Leu Ser
50 55 60
Gln Ser Leu Lys Ala Leu Gly Ser Asp Leu Thr Leu Ile Lys Thr His
65 70 75 80
Asn Thr Ile Ser Ala Ile Leu Asp Cys Ile Arg Val Thr Gly Ala Thr
85 90 95
Lys Val Val Phe Asn His Leu Tyr Asp Pro Val Ser Leu Val Arg Asp
100 105 110
His Thr Val Lys Glu Lys Leu Val Glu Arg Gly Ile Ser Val Gln Ser
115 120 125
Tyr Asn Gly Asp Leu Leu Tyr Glu Pro Trp Glu Ile Tyr Cys Glu Lys
130 135 140
Gly Lys Pro Phe Thr Ser Phe Asn Ser Tyr Trp Lys Lys Cys Leu Asp
145 150 155 160
Met Ser Ile Glu Ser Val Met Leu Pro Pro Pro Trp Arg Leu Met Pro
165 170 175
Ile Thr Ala Ala Ala Glu Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu
180 185 190
Gly Leu Glu Asn Glu Ala Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg
195 200 205
Ala Trp Ser Pro Gly Trp Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe
210 215 220
Ile Glu Lys Gln Leu Ile Asp Tyr Ala Lys Asn Ser Lys Lys Val Val
225 230 235 240
Gly Asn Ser Thr Ser Leu Leu Ser Pro Tyr Leu His Phe Gly Glu Ile
245 250 255
Ser Val Arg His Val Phe Gln Cys Ala Arg Met Lys Gln Ile Ile Trp
260 265 270
Ala Arg Asp Lys Asn Ser Glu Gly Glu Glu Ser Ala Asp Leu Phe Leu
275 280 285
Arg Gly Ile Gly Leu Arg Glu Tyr Ser Arg Tyr Ile Cys Phe Asn Phe
290 295 300
Pro Phe Thr His Glu Gln Ser Leu Leu Ser His Leu Arg Phe Phe Pro
305 310 315 320
Trp Asp Ala Asp Val Asp Lys Phe Lys Ala Trp Arg Gln Gly Arg Thr
325 330 335
Gly Tyr Pro Leu Val Asp Ala Gly Met Arg Glu Leu Trp Ala Thr Gly
340 345 350
Trp Met His Asn Arg Ile Arg Val Ile Val Ser Ser Phe Ala Val Lys
355 360 365
Phe Leu Leu Leu Pro Trp Lys Trp Gly Met Lys Tyr Phe Trp Asp Thr
370 375 380
Leu Leu Asp Ala Asp Leu Glu Cys Asp Ile Leu Gly Trp Gln Tyr Ile
385 390 395 400
Ser Gly Ser Ile Pro Asp Gly His Glu Leu Asp Arg Leu Asp Asn Pro
405 410 415
Ala Leu Gln Gly Ala Lys Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln
420 425 430
Trp Leu Pro Glu Leu Ala Arg Leu Pro Thr Glu Trp Ile His His Pro
435 440 445
Trp Asp Ala Pro Leu Thr Val Leu Lys Ala Ser Gly Val Glu Leu Gly
450 455 460
Thr Asn Tyr Ala Lys Pro Ile Val Asp Ile Asp Thr Ala Arg Glu Leu
465 470 475 480
Leu Ala Lys Ala Ile Ser Arg Thr Arg Glu Ala Gln Ile Met Ile Gly
485 490 495
Ala Ala Pro Ala Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Gly
500 505 510
Ser Gly Arg Ala Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly
515 520 525
Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala
530 535 540
Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp
545 550 555 560
Phe Asp Leu Asp Met Leu Ile Asn Ser Arg Gly Ser Gly Glu Gly Arg
565 570 575
Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Val
580 585 590
Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu Val Glu
595 600 605
Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly Glu Gly
610 615 620
Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr
625 630 635 640
Thr Gly Lys Leu Pro Val Pro Trp Pro Thr Leu Val Thr Thr Leu Thr
645 650 655
Tyr Gly Val Gln Cys Phe Ser Arg Tyr Pro Asp His Met Lys Gln His
660 665 670
Asp Phe Phe Lys Ser Ala Met Pro Glu Gly Tyr Val Gln Glu Arg Thr
675 680 685
Ile Phe Phe Lys Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys
690 695 700
Phe Glu Gly Asp Thr Leu Val Asn Arg Ile Glu Leu Lys Gly Ile Asp
705 710 715 720
Phe Lys Glu Asp Gly Asn Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr
725 730 735
Asn Ser His Asn Val Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly Ile
740 745 750
Lys Val Asn Phe Lys Ile Arg His Asn Ile Glu Asp Gly Ser Val Gln
755 760 765
Leu Ala Asp His Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro Val
770 775 780
Leu Leu Pro Asp Asn His Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys
785 790 795 800
Asp Pro Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe Val Thr
805 810 815
Ala Ala Gly Ile Thr Leu Gly Met Asp Glu Leu Tyr Lys Val
820 825 830
<210> 142
<211> 774
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 142
Met Lys Met Asp Lys Lys Thr Ile Val Trp Phe Arg Arg Asp Leu Arg
1 5 10 15
Ile Glu Asp Asn Pro Ala Leu Ala Ala Ala Ala His Glu Gly Ser Val
20 25 30
Phe Pro Val Phe Ile Trp Cys Pro Glu Glu Glu Gly Gln Phe Tyr Pro
35 40 45
Gly Arg Ala Ser Arg Trp Trp Met Lys Gln Ser Leu Ala His Leu Ser
50 55 60
Gln Ser Leu Lys Ala Leu Gly Ser Asp Leu Thr Leu Ile Lys Thr His
65 70 75 80
Asn Thr Ile Ser Ala Ile Leu Asp Cys Ile Arg Val Thr Gly Ala Thr
85 90 95
Lys Val Val Phe Asn His Leu Tyr Asp Pro Val Ser Leu Val Arg Asp
100 105 110
His Thr Val Lys Glu Lys Leu Val Glu Arg Gly Ile Ser Val Gln Ser
115 120 125
Tyr Asn Gly Asp Leu Leu Tyr Glu Pro Trp Glu Ile Tyr Cys Glu Lys
130 135 140
Gly Lys Pro Phe Thr Ser Phe Asn Ser Tyr Trp Lys Lys Cys Leu Asp
145 150 155 160
Met Ser Ile Glu Ser Val Met Leu Pro Pro Pro Trp Arg Leu Met Pro
165 170 175
Ile Thr Ala Ala Ala Glu Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu
180 185 190
Gly Leu Glu Asn Glu Ala Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg
195 200 205
Ala Trp Ser Pro Gly Trp Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe
210 215 220
Ile Glu Lys Gln Leu Ile Asp Tyr Ala Lys Asn Ser Lys Lys Val Val
225 230 235 240
Gly Asn Ser Thr Ser Leu Leu Ser Pro Tyr Leu His Phe Gly Glu Ile
245 250 255
Ser Val Arg His Val Phe Gln Cys Ala Arg Met Lys Gln Ile Ile Trp
260 265 270
Ala Arg Asp Lys Asn Ser Glu Gly Glu Glu Ser Ala Asp Leu Phe Leu
275 280 285
Arg Gly Ile Gly Leu Arg Glu Tyr Ser Arg Tyr Ile Cys Phe Asn Phe
290 295 300
Pro Phe Thr His Glu Gln Ser Leu Leu Ser His Leu Arg Phe Phe Pro
305 310 315 320
Trp Asp Ala Asp Val Asp Lys Phe Lys Ala Trp Arg Gln Gly Arg Thr
325 330 335
Gly Tyr Pro Leu Val Asp Ala Gly Met Arg Glu Leu Trp Ala Thr Gly
340 345 350
Trp Met His Asn Arg Ile Arg Val Ile Val Ser Ser Phe Ala Val Lys
355 360 365
Phe Leu Leu Leu Pro Trp Lys Trp Gly Met Lys Tyr Phe Trp Asp Thr
370 375 380
Leu Leu Asp Ala Asp Leu Glu Cys Asp Ile Leu Gly Trp Gln Tyr Ile
385 390 395 400
Ser Gly Ser Ile Pro Asp Gly His Glu Leu Asp Arg Leu Asp Asn Pro
405 410 415
Ala Leu Gln Gly Ala Lys Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln
420 425 430
Trp Leu Pro Glu Leu Ala Arg Leu Pro Thr Glu Trp Ile His His Pro
435 440 445
Trp Asp Ala Pro Leu Thr Val Leu Lys Ala Ser Gly Val Glu Leu Gly
450 455 460
Thr Asn Tyr Ala Lys Pro Ile Val Asp Ile Asp Thr Ala Arg Glu Leu
465 470 475 480
Leu Ala Lys Ala Ile Ser Arg Thr Arg Glu Ala Gln Ile Met Ile Gly
485 490 495
Ala Ala Pro Ala Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Gly
500 505 510
Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu
515 520 525
Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu Pro Gly
530 535 540
Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu
545 550 555 560
Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu Pro Gly
565 570 575
Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu
580 585 590
Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu Pro Gly
595 600 605
Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu
610 615 620
Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu Pro Ser
625 630 635 640
Arg Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly
645 650 655
Asp Val Glu Glu Asn Pro Gly Pro Ile Glu Lys Ser Phe Val Ile Thr
660 665 670
Asp Pro Arg Leu Pro Asp Tyr Pro Ile Ile Phe Ala Ser Asp Gly Phe
675 680 685
Leu Glu Leu Thr Glu Tyr Ser Arg Glu Glu Ile Met Gly Arg Asn Ala
690 695 700
Arg Phe Leu Gln Gly Pro Glu Thr Asp Gln Ala Thr Val Gln Lys Ile
705 710 715 720
Arg Asp Ala Ile Arg Asp Gln Arg Glu Thr Thr Val Gln Leu Ile Asn
725 730 735
Tyr Thr Lys Ser Gly Lys Lys Phe Trp Asn Leu Leu His Leu Gln Pro
740 745 750
Val Arg Asp Arg Lys Gly Gly Leu Gln Tyr Phe Ile Gly Val Gln Leu
755 760 765
Val Gly Ser Asp His Val
770
<210> 143
<211> 1356
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 143
Met Ser Arg Thr Arg Leu Pro Ser Pro Pro Ala Pro Ser Pro Ala Phe
1 5 10 15
Ser Ala Asp Ser Phe Ser Asp Leu Leu Arg Gln Phe Asp Pro Ser Leu
20 25 30
Phe Asn Thr Ser Leu Phe Asp Ser Leu Pro Pro Phe Gly Ala His His
35 40 45
Thr Glu Ala Ala Thr Gly Glu Trp Asp Glu Val Gln Ser Gly Leu Arg
50 55 60
Ala Ala Asp Ala Pro Pro Pro Thr Met Arg Val Ala Val Thr Ala Ala
65 70 75 80
Arg Pro Pro Arg Ala Lys Pro Ala Pro Arg Arg Arg Ala Ala Gln Pro
85 90 95
Ser Asp Ala Ser Pro Ala Ala Gln Val Asp Leu Arg Thr Leu Gly Tyr
100 105 110
Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val
115 120 125
Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His
130 135 140
Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val
145 150 155 160
Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala
165 170 175
Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala
180 185 190
Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp
195 200 205
Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val
210 215 220
Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn
225 230 235 240
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
245 250 255
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
260 265 270
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly
275 280 285
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
290 295 300
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
305 310 315 320
Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
325 330 335
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
340 345 350
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
355 360 365
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
370 375 380
Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
385 390 395 400
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
405 410 415
Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val
420 425 430
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
435 440 445
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
450 455 460
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
465 470 475 480
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala
485 490 495
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
500 505 510
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
515 520 525
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
530 535 540
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
545 550 555 560
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
565 570 575
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
580 585 590
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
595 600 605
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
610 615 620
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
625 630 635 640
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
645 650 655
Ile Ala Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala
660 665 670
Gln Leu Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His
675 680 685
Leu Val Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val
690 695 700
Lys Lys Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg
705 710 715 720
Arg Ile Pro Glu Arg Thr Ser His Arg Val Ala Asp His Ala Gln Val
725 730 735
Val Arg Val Leu Gly Phe Phe Gln Cys His Ser His Pro Ala Gln Ala
740 745 750
Phe Asp Asp Ala Met Thr Gln Phe Gly Met Ser Arg His Gly Leu Leu
755 760 765
Gln Leu Phe Arg Arg Val Gly Val Thr Glu Leu Glu Ala Arg Ser Gly
770 775 780
Thr Leu Pro Pro Ala Ser Gln Arg Trp Asp Arg Ile Leu Gln Ala Ser
785 790 795 800
Gly Met Lys Arg Ala Lys Pro Ser Pro Thr Ser Thr Gln Thr Pro Asp
805 810 815
Gln Ala Ser Leu His Ala Phe Ala Asp Ser Leu Glu Arg Asp Leu Asp
820 825 830
Ala Pro Ser Pro Met His Glu Gly Asp Gln Thr Arg Ala Ser Ala Ser
835 840 845
Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Lys Met Asp Lys Lys Thr
850 855 860
Ile Val Trp Phe Arg Arg Asp Leu Arg Ile Glu Asp Asn Pro Ala Leu
865 870 875 880
Ala Ala Ala Ala His Glu Gly Ser Val Phe Pro Val Phe Ile Trp Cys
885 890 895
Pro Glu Glu Glu Gly Gln Phe Tyr Pro Gly Arg Ala Ser Arg Trp Trp
900 905 910
Met Lys Gln Ser Leu Ala His Leu Ser Gln Ser Leu Lys Ala Leu Gly
915 920 925
Ser Asp Leu Thr Leu Ile Lys Thr His Asn Thr Ile Ser Ala Ile Leu
930 935 940
Asp Cys Ile Arg Val Thr Gly Ala Thr Lys Val Val Phe Asn His Leu
945 950 955 960
Tyr Asp Pro Val Ser Leu Val Arg Asp His Thr Val Lys Glu Lys Leu
965 970 975
Val Glu Arg Gly Ile Ser Val Gln Ser Tyr Asn Gly Asp Leu Leu Tyr
980 985 990
Glu Pro Trp Glu Ile Tyr Cys Glu Lys Gly Lys Pro Phe Thr Ser Phe
995 1000 1005
Asn Ser Tyr Trp Lys Lys Cys Leu Asp Met Ser Ile Glu Ser Val
1010 1015 1020
Met Leu Pro Pro Pro Trp Arg Leu Met Pro Ile Thr Ala Ala Ala
1025 1030 1035
Glu Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu Gly Leu Glu Asn
1040 1045 1050
Glu Ala Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg Ala Trp Ser
1055 1060 1065
Pro Gly Trp Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe Ile Glu
1070 1075 1080
Lys Gln Leu Ile Asp Tyr Ala Lys Asn Ser Lys Lys Val Val Gly
1085 1090 1095
Asn Ser Thr Ser Leu Leu Ser Pro Tyr Leu His Phe Gly Glu Ile
1100 1105 1110
Ser Val Arg His Val Phe Gln Cys Ala Arg Met Lys Gln Ile Ile
1115 1120 1125
Trp Ala Arg Asp Lys Asn Ser Glu Gly Glu Glu Ser Ala Asp Leu
1130 1135 1140
Phe Leu Arg Gly Ile Gly Leu Arg Glu Tyr Ser Arg Tyr Ile Cys
1145 1150 1155
Phe Asn Phe Pro Phe Thr His Glu Gln Ser Leu Leu Ser His Leu
1160 1165 1170
Arg Phe Phe Pro Trp Asp Ala Asp Val Asp Lys Phe Lys Ala Trp
1175 1180 1185
Arg Gln Gly Arg Thr Gly Tyr Pro Leu Val Asp Ala Gly Met Arg
1190 1195 1200
Glu Leu Trp Ala Thr Gly Trp Met His Asn Arg Ile Arg Val Ile
1205 1210 1215
Val Ser Ser Phe Ala Val Lys Phe Leu Leu Leu Pro Trp Lys Trp
1220 1225 1230
Gly Met Lys Tyr Phe Trp Asp Thr Leu Leu Asp Ala Asp Leu Glu
1235 1240 1245
Cys Asp Ile Leu Gly Trp Gln Tyr Ile Ser Gly Ser Ile Pro Asp
1250 1255 1260
Gly His Glu Leu Asp Arg Leu Asp Asn Pro Ala Leu Gln Gly Ala
1265 1270 1275
Lys Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln Trp Leu Pro Glu
1280 1285 1290
Leu Ala Arg Leu Pro Thr Glu Trp Ile His His Pro Trp Asp Ala
1295 1300 1305
Pro Leu Thr Val Leu Lys Ala Ser Gly Val Glu Leu Gly Thr Asn
1310 1315 1320
Tyr Ala Lys Pro Ile Val Asp Ile Asp Thr Ala Arg Glu Leu Leu
1325 1330 1335
Ala Lys Ala Ile Ser Arg Thr Arg Glu Ala Gln Ile Met Ile Gly
1340 1345 1350
Ala Ala Pro
1355
<210> 144
<211> 1001
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 144
Met Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Pro Lys Lys Lys Arg
1 5 10 15
Lys Val Glu Ala Ser Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln
20 25 30
Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val Ala Gln His
35 40 45
His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His Ile Val Ala
50 55 60
Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln
65 70 75 80
Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly
85 90 95
Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr
100 105 110
Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln
115 120 125
Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val
130 135 140
His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro
145 150 155 160
Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu
165 170 175
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
180 185 190
Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln
195 200 205
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
210 215 220
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly
225 230 235 240
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
245 250 255
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly
260 265 270
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
275 280 285
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
290 295 300
Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
305 310 315 320
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
325 330 335
Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
340 345 350
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val
355 360 365
Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
370 375 380
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
385 390 395 400
Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr
405 410 415
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
420 425 430
Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu
435 440 445
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
450 455 460
Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln
465 470 475 480
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
485 490 495
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly
500 505 510
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
515 520 525
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile
530 535 540
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
545 550 555 560
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
565 570 575
Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
580 585 590
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
595 600 605
Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
610 615 620
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val
625 630 635 640
Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
645 650 655
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
660 665 670
Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr
675 680 685
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
690 695 700
Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu
705 710 715 720
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
725 730 735
Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln
740 745 750
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
755 760 765
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly
770 775 780
Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu Ser Arg Pro Asp Pro
785 790 795 800
Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val Ala Leu Ala Cys Leu
805 810 815
Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys Gly Leu Pro His Ala
820 825 830
Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile Pro Glu Arg Thr Ser
835 840 845
His Arg Val Ala Asp His Ala Gln Val Val Arg Val Leu Gly Phe Phe
850 855 860
Gln Cys His Ser His Pro Ala Gln Ala Phe Asp Asp Ala Met Thr Gln
865 870 875 880
Phe Gly Met Ser Arg His Gly Leu Leu Gln Leu Phe Arg Arg Val Gly
885 890 895
Val Thr Glu Leu Glu Ala Arg Ser Gly Thr Leu Pro Pro Ala Ser Gln
900 905 910
Arg Trp Asp Arg Ile Leu Gln Ala Ser Gly Met Lys Arg Ala Lys Pro
915 920 925
Ser Pro Thr Ser Thr Gln Thr Pro Asp Gln Ala Ser Leu His Ala Phe
930 935 940
Ala Asp Ser Leu Glu Arg Asp Leu Asp Ala Pro Ser Pro Met His Glu
945 950 955 960
Gly Asp Gln Thr Arg Ala Ser Ala Ser Gly Ser Gly Met Asn Ile Gln
965 970 975
Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu Arg Arg Glu Arg Glu Ala
980 985 990
Glu His Gly Tyr Ala Ser Met Leu Pro
995 1000
<210> 145
<211> 1099
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 145
Met Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Pro Lys Lys Lys Arg
1 5 10 15
Lys Val Glu Ala Ser Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln
20 25 30
Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val Ala Gln His
35 40 45
His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His Ile Val Ala
50 55 60
Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln
65 70 75 80
Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly
85 90 95
Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr
100 105 110
Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln
115 120 125
Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val
130 135 140
His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro
145 150 155 160
Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu
165 170 175
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
180 185 190
Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln
195 200 205
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
210 215 220
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly
225 230 235 240
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
245 250 255
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly
260 265 270
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
275 280 285
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
290 295 300
Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
305 310 315 320
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
325 330 335
Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
340 345 350
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val
355 360 365
Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
370 375 380
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
385 390 395 400
Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr
405 410 415
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
420 425 430
Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu
435 440 445
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
450 455 460
Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln
465 470 475 480
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
485 490 495
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly
500 505 510
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
515 520 525
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile
530 535 540
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
545 550 555 560
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
565 570 575
Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
580 585 590
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
595 600 605
Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
610 615 620
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val
625 630 635 640
Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
645 650 655
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
660 665 670
Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr
675 680 685
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
690 695 700
Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln Ala Leu
705 710 715 720
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
725 730 735
Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Lys Gln
740 745 750
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
755 760 765
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly
770 775 780
Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu Ser Arg Pro Asp Pro
785 790 795 800
Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val Ala Leu Ala Cys Leu
805 810 815
Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys Gly Leu Pro His Ala
820 825 830
Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile Pro Glu Arg Thr Ser
835 840 845
His Arg Val Ala Asp His Ala Gln Val Val Arg Val Leu Gly Phe Phe
850 855 860
Gln Cys His Ser His Pro Ala Gln Ala Phe Asp Asp Ala Met Thr Gln
865 870 875 880
Phe Gly Met Ser Arg His Gly Leu Leu Gln Leu Phe Arg Arg Val Gly
885 890 895
Val Thr Glu Leu Glu Ala Arg Ser Gly Thr Leu Pro Pro Ala Ser Gln
900 905 910
Arg Trp Asp Arg Ile Leu Gln Ala Ser Gly Met Lys Arg Ala Lys Pro
915 920 925
Ser Pro Thr Ser Thr Gln Thr Pro Asp Gln Ala Ser Leu His Ala Phe
930 935 940
Ala Asp Ser Leu Glu Arg Asp Leu Asp Ala Pro Ser Pro Met His Glu
945 950 955 960
Gly Asp Gln Thr Arg Ala Ser Ala Ser Gly Ser Gly Met Asn Ile Gln
965 970 975
Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu Arg Arg Glu Arg Glu Ala
980 985 990
Glu His Gly Tyr Ala Ser Met Leu Pro Gly Ser Gly Met Asn Ile Gln
995 1000 1005
Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu Arg Arg Glu Arg Glu
1010 1015 1020
Ala Glu His Gly Tyr Ala Ser Met Leu Pro Gly Ser Gly Met Asn
1025 1030 1035
Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu Arg Arg Glu
1040 1045 1050
Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu Pro Gly Ser Gly
1055 1060 1065
Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr Leu Glu Arg
1070 1075 1080
Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu Pro Ser
1085 1090 1095
Arg
<210> 146
<211> 1187
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 146
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp
995 1000 1005
Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His
1010 1015 1020
Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala
1025 1030 1035
Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu
1040 1045 1050
Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly
1055 1060 1065
Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln
1070 1075 1080
Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro
1085 1090 1095
Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr
1100 1105 1110
Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr
1115 1120 1125
Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val Asn
1130 1135 1140
Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr Ser
1145 1150 1155
Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met
1160 1165 1170
Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1175 1180 1185
<210> 147
<211> 1188
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 147
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Tyr Pro Glu Arg
835 840 845
Leu Arg Arg Ile Leu Thr Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val
1130 1135 1140
Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr
1145 1150 1155
Ser Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser
1160 1165 1170
Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1175 1180 1185
<210> 148
<211> 1188
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 148
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Gln Leu Pro Pro
835 840 845
Leu Glu Arg Leu Thr Leu Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val
1130 1135 1140
Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr
1145 1150 1155
Ser Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser
1160 1165 1170
Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1175 1180 1185
<210> 149
<211> 1185
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 149
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Asp Leu Ala Ser
835 840 845
Leu Ile Leu Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn Phe Pro
850 855 860
Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr Leu Asn
865 870 875 880
Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser Ser Met
885 890 895
Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly Leu Asn
900 905 910
Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser Arg Leu
915 920 925
Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Ala Ala
930 935 940
Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys Lys Met
945 950 955 960
Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser
965 970 975
Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met
980 985 990
Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser
995 1000 1005
Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg
1010 1015 1020
Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg
1025 1030 1035
Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp
1040 1045 1050
Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu
1055 1060 1065
Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile Glu
1070 1075 1080
Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe
1085 1090 1095
Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met
1100 1105 1110
Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His
1115 1120 1125
Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly
1130 1135 1140
Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp
1145 1150 1155
Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp
1160 1165 1170
Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1175 1180 1185
<210> 150
<211> 1187
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 150
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Gln Lys Lys Leu
835 840 845
Glu Glu Leu Glu Leu Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp
995 1000 1005
Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His
1010 1015 1020
Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala
1025 1030 1035
Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu
1040 1045 1050
Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly
1055 1060 1065
Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln
1070 1075 1080
Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro
1085 1090 1095
Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr
1100 1105 1110
Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr
1115 1120 1125
Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val Asn
1130 1135 1140
Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr Ser
1145 1150 1155
Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met
1160 1165 1170
Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1175 1180 1185
<210> 151
<211> 1159
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 151
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp
995 1000 1005
Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His
1010 1015 1020
Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala
1025 1030 1035
Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu
1040 1045 1050
Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly
1055 1060 1065
Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln
1070 1075 1080
Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro
1085 1090 1095
Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr
1100 1105 1110
Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr
1115 1120 1125
Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val Asn
1130 1135 1140
Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr Ser
1145 1150 1155
Ser
<210> 152
<211> 1160
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 152
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Tyr Pro Glu Arg
835 840 845
Leu Arg Arg Ile Leu Thr Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val
1130 1135 1140
Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr
1145 1150 1155
Ser Ser
1160
<210> 153
<211> 1160
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 153
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Ser Pro Lys Lys Lys
835 840 845
Arg Lys Val Glu Ala Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val
1130 1135 1140
Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr
1145 1150 1155
Ser Ser
1160
<210> 154
<211> 1187
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 154
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp
995 1000 1005
Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His
1010 1015 1020
Val Arg Ala Arg Ala Ala Gln Ala Thr Asp Ser His Ser Ile Ala
1025 1030 1035
Glu Ala Val Ala Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu
1040 1045 1050
Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly
1055 1060 1065
Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln
1070 1075 1080
Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro
1085 1090 1095
Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr
1100 1105 1110
Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr
1115 1120 1125
Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val Asn
1130 1135 1140
Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr Ser
1145 1150 1155
Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met
1160 1165 1170
Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1175 1180 1185
<210> 155
<211> 1179
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 155
Ser Arg Thr Arg Leu Pro Ser Pro Pro Ala Pro Ser Pro Ala Phe Ser
1 5 10 15
Ala Asp Ser Phe Ser Asp Leu Leu Arg Gln Phe Asp Pro Ser Leu Phe
20 25 30
Asn Thr Ser Leu Phe Asp Ser Leu Pro Pro Phe Gly Ala His His Thr
35 40 45
Glu Ala Ala Thr Gly Glu Trp Asp Glu Val Gln Ser Gly Leu Arg Ala
50 55 60
Ala Asp Ala Pro Pro Pro Thr Met Arg Val Ala Val Thr Ala Ala Arg
65 70 75 80
Pro Pro Arg Ala Lys Pro Ala Pro Arg Arg Arg Ala Ala Gln Pro Ser
85 90 95
Asp Ala Ser Pro Ala Ala Gln Val Asp Leu Arg Thr Leu Gly Tyr Ser
100 105 110
Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val Ala
115 120 125
Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His Ile
130 135 140
Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val Lys
145 150 155 160
Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala Ile
165 170 175
Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala Leu
180 185 190
Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp Thr
195 200 205
Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val Glu
210 215 220
Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn Leu
225 230 235 240
Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln
245 250 255
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
260 265 270
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly
275 280 285
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
290 295 300
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile
305 310 315 320
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
325 330 335
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
340 345 350
Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
355 360 365
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
370 375 380
Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
385 390 395 400
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val
405 410 415
Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
420 425 430
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
435 440 445
Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr
450 455 460
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
465 470 475 480
Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu
485 490 495
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
500 505 510
Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln
515 520 525
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
530 535 540
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly
545 550 555 560
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
565 570 575
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly
580 585 590
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
595 600 605
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
610 615 620
Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
625 630 635 640
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
645 650 655
Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
660 665 670
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val
675 680 685
Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
690 695 700
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
705 710 715 720
Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr
725 730 735
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
740 745 750
Glu Gln Val Val Ala Ile Ala Ser His Asp Gly Gly Arg Pro Ala Leu
755 760 765
Glu Ser Ile Val Ala Gln Leu Ser Arg Pro Asp Pro Ala Leu Ala Ala
770 775 780
Leu Thr Asn Asp His Leu Val Ala Leu Ala Cys Leu Gly Gly Arg Pro
785 790 795 800
Ala Leu Asp Ala Val Lys Lys Gly Leu Pro His Ala Pro Ala Leu Ile
805 810 815
Lys Arg Thr Asn Arg Arg Ile Pro Glu Arg Thr Ser His Arg Val Ala
820 825 830
Ala Ser Leu Tyr Pro Glu Arg Leu Arg Arg Ile Leu Thr Asn Gly Ala
835 840 845
Ile Gly Gly Asp Leu Leu Leu Asn Phe Pro Asp Met Ser Val Leu Glu
850 855 860
Arg Gln Arg Ala His Leu Lys Tyr Leu Asn Pro Thr Phe Asp Ser Pro
865 870 875 880
Leu Ala Gly Phe Phe Ala Asp Ser Ser Met Ile Thr Gly Gly Glu Met
885 890 895
Asp Ser Tyr Leu Ser Thr Ala Gly Leu Asn Leu Pro Met Met Tyr Gly
900 905 910
Glu Thr Thr Val Glu Gly Asp Ser Arg Leu Ser Ile Ser Pro Glu Thr
915 920 925
Thr Leu Gly Thr Gly Asn Phe Lys Ala Ala Lys Phe Asp Thr Glu Thr
930 935 940
Lys Asp Cys Asn Glu Ala Ala Lys Lys Met Thr Met Asn Arg Asp Asp
945 950 955 960
Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile Thr Glu Gln Asn
965 970 975
Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His Lys Ala Lys Lys
980 985 990
Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val Thr Lys Glu Leu
995 1000 1005
Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg Ala Ala Gln Ala
1010 1015 1020
Thr Asp Ser His Ser Ile Ala Glu Ala Val Ala Arg Glu Lys Ile
1025 1030 1035
Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val Pro Gly Cys Asp
1040 1045 1050
Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu Ile Ile Asn Tyr
1055 1060 1065
Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu Ser Met Lys Leu
1070 1075 1080
Ala Ile Val Asn Pro Arg Pro Asp Phe Asp Met Asp Asp Ile Phe
1085 1090 1095
Ala Lys Glu Val Ala Ser Thr Pro Met Thr Val Val Pro Ser Pro
1100 1105 1110
Glu Met Val Leu Ser Gly Tyr Ser His Glu Met Val His Ser Gly
1115 1120 1125
Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu His Val Asn Pro
1130 1135 1140
Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu Ser Cys Phe Asn
1145 1150 1155
Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His Val Gln Asn Leu
1160 1165 1170
Tyr Gly Asn Leu Gly Val
1175
<210> 156
<211> 1163
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 156
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp
995 1000 1005
Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His
1010 1015 1020
Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala
1025 1030 1035
Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu
1040 1045 1050
Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly
1055 1060 1065
Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gly
1070 1075 1080
Gly Ser Val Ala Ser Thr Pro Met Thr Val Val Pro Ser Pro Glu
1085 1090 1095
Met Val Leu Ser Gly Tyr Ser His Glu Met Val His Ser Gly Tyr
1100 1105 1110
Ser Ser Glu Met Val Asn Ser Gly Tyr Leu His Val Asn Pro Met
1115 1120 1125
Gln Gln Val Asn Thr Ser Ser Asp Pro Leu Ser Cys Phe Asn Asn
1130 1135 1140
Gly Glu Ala Pro Ser Met Trp Asp Ser His Val Gln Asn Leu Tyr
1145 1150 1155
Gly Asn Leu Gly Val
1160
<210> 157
<211> 1277
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 157
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Tyr Pro Glu Arg
835 840 845
Leu Arg Arg Ile Leu Thr Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gly Gly Ser Gly Glu Glu Glu Lys Ser Lys Ile Thr Glu Gln Asn
1085 1090 1095
Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His Lys Ala Lys
1100 1105 1110
Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val Thr Lys
1115 1120 1125
Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg Arg Gly
1130 1135 1140
Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg Arg Glu
1145 1150 1155
Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val Pro Gly
1160 1165 1170
Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu Ile Ile
1175 1180 1185
Asn Tyr Val Gln Ser Leu Gln Arg Gly Gly Ser Val Ala Ser Thr
1190 1195 1200
Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr
1205 1210 1215
Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val Asn
1220 1225 1230
Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr Ser
1235 1240 1245
Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met
1250 1255 1260
Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1265 1270 1275
<210> 158
<211> 1158
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 158
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp
995 1000 1005
Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His
1010 1015 1020
Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala
1025 1030 1035
Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu
1040 1045 1050
Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly
1055 1060 1065
Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln
1070 1075 1080
Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro
1085 1090 1095
Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr
1100 1105 1110
Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr
1115 1120 1125
Gly Gly Ser Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser
1130 1135 1140
Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1145 1150 1155
<210> 159
<211> 1159
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 159
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Tyr Pro Glu Arg
835 840 845
Leu Arg Arg Ile Leu Thr Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Gly Gly Ser Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro
1130 1135 1140
Ser Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly
1145 1150 1155
Val
<210> 160
<211> 1188
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 160
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Ser Pro Lys Lys Lys
835 840 845
Arg Lys Val Glu Ala Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Ala Ala Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Ala Val Ala Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Ser His Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val
1130 1135 1140
Asn Ser Gly Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr
1145 1150 1155
Ser Ser Asp Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser
1160 1165 1170
Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1175 1180 1185
<210> 161
<211> 1164
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 161
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Ser Pro Lys Lys Lys
835 840 845
Arg Lys Val Glu Ala Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gly Gly Ser Val Ala Ser Thr Pro Met Thr Val Val Pro Ser Pro
1085 1090 1095
Glu Met Val Leu Ser Gly Tyr Ser His Glu Met Val His Ser Gly
1100 1105 1110
Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu His Val Asn Pro
1115 1120 1125
Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu Ser Cys Phe Asn
1130 1135 1140
Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His Val Gln Asn Leu
1145 1150 1155
Tyr Gly Asn Leu Gly Val
1160
<210> 162
<211> 1159
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 162
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Leu Tyr Pro Glu Arg
835 840 845
Leu Arg Arg Ile Leu Thr Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Gly Gly Ser Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro
1130 1135 1140
Ser Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly
1145 1150 1155
Val
<210> 163
<211> 1198
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 163
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Leu Tyr Pro Glu Arg Leu Arg Arg Ile Leu Thr Asn Pro Thr Phe
885 890 895
Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser Ser Met Ile Thr Gly
900 905 910
Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly Leu Asn Leu Pro Met
915 920 925
Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser Arg Leu Ser Ile Ser
930 935 940
Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Ala Ala Lys Phe Asp
945 950 955 960
Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys Lys Met Thr Met Asn
965 970 975
Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile Thr
980 985 990
Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His Lys
995 1000 1005
Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
1010 1015 1020
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg
1025 1030 1035
Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg
1040 1045 1050
Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val
1055 1060 1065
Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu
1070 1075 1080
Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu
1085 1090 1095
Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe Asp Met
1100 1105 1110
Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met Thr Val
1115 1120 1125
Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His Glu Met
1130 1135 1140
Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu
1145 1150 1155
His Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu
1160 1165 1170
Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His
1175 1180 1185
Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1190 1195
<210> 164
<211> 1198
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 164
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Leu Tyr Pro Glu Arg Leu Arg
900 905 910
Arg Ile Leu Thr Ser Tyr Leu Ser Thr Ala Gly Leu Asn Leu Pro Met
915 920 925
Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser Arg Leu Ser Ile Ser
930 935 940
Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Ala Ala Lys Phe Asp
945 950 955 960
Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys Lys Met Thr Met Asn
965 970 975
Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile Thr
980 985 990
Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His Lys
995 1000 1005
Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
1010 1015 1020
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg
1025 1030 1035
Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg
1040 1045 1050
Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val
1055 1060 1065
Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu
1070 1075 1080
Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu
1085 1090 1095
Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe Asp Met
1100 1105 1110
Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met Thr Val
1115 1120 1125
Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His Glu Met
1130 1135 1140
Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu
1145 1150 1155
His Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu
1160 1165 1170
Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His
1175 1180 1185
Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1190 1195
<210> 165
<211> 1198
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 165
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Leu Tyr
915 920 925
Pro Glu Arg Leu Arg Arg Ile Leu Thr Asp Ser Arg Leu Ser Ile Ser
930 935 940
Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys Ala Ala Lys Phe Asp
945 950 955 960
Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys Lys Met Thr Met Asn
965 970 975
Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu Lys Ser Lys Ile Thr
980 985 990
Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys His Lys
995 1000 1005
Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
1010 1015 1020
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg
1025 1030 1035
Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg
1040 1045 1050
Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val
1055 1060 1065
Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu
1070 1075 1080
Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu
1085 1090 1095
Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe Asp Met
1100 1105 1110
Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met Thr Val
1115 1120 1125
Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His Glu Met
1130 1135 1140
Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu
1145 1150 1155
His Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu
1160 1165 1170
Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His
1175 1180 1185
Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1190 1195
<210> 166
<211> 1200
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 166
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Ala
945 950 955 960
Ala Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Leu
965 970 975
Tyr Pro Glu Arg Leu Arg Arg Ile Leu Thr Glu Glu Glu Lys Ser Lys
980 985 990
Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys Lys Met Lys
995 1000 1005
His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser
1010 1015 1020
Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg
1025 1030 1035
Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg
1040 1045 1050
Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp
1055 1060 1065
Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu
1070 1075 1080
Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile Glu
1085 1090 1095
Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe
1100 1105 1110
Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met
1115 1120 1125
Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His
1130 1135 1140
Glu Met Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly
1145 1150 1155
Tyr Leu His Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp
1160 1165 1170
Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp
1175 1180 1185
Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1190 1195 1200
<210> 167
<211> 1198
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 167
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Leu Tyr Pro Glu Arg Leu Arg Arg Ile Leu Thr Met Lys His Lys
995 1000 1005
Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp Ser Ser Lys Val
1010 1015 1020
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg
1025 1030 1035
Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg
1040 1045 1050
Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val
1055 1060 1065
Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu
1070 1075 1080
Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu
1085 1090 1095
Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe Asp Met
1100 1105 1110
Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met Thr Val
1115 1120 1125
Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His Glu Met
1130 1135 1140
Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu
1145 1150 1155
His Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu
1160 1165 1170
Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His
1175 1180 1185
Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1190 1195
<210> 168
<211> 1198
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 168
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Gly Gly Gly Gly Ser
835 840 845
Gly Gly Gly Gly Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu Asn
850 855 860
Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys Tyr
865 870 875 880
Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp Ser
885 890 895
Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala Gly
900 905 910
Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp Ser
915 920 925
Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe Lys
930 935 940
Ala Ala Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Ala Ala Lys
945 950 955 960
Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu Glu
965 970 975
Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile Lys
980 985 990
Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn Asp
995 1000 1005
Ser Ser Lys Val Thr Leu Tyr Pro Glu Arg Leu Arg Arg Ile Leu
1010 1015 1020
Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile His Val Arg Ala Arg
1025 1030 1035
Arg Gly Gln Ala Thr Asp Ser His Ser Ile Ala Glu Arg Val Arg
1040 1045 1050
Arg Glu Lys Ile Ser Glu Arg Met Lys Phe Leu Gln Asp Leu Val
1055 1060 1065
Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala Gly Met Leu Asp Glu
1070 1075 1080
Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg Gln Ile Glu Phe Leu
1085 1090 1095
Ser Met Lys Leu Ala Ile Val Asn Pro Arg Pro Asp Phe Asp Met
1100 1105 1110
Asp Asp Ile Phe Ala Lys Glu Val Ala Ser Thr Pro Met Thr Val
1115 1120 1125
Val Pro Ser Pro Glu Met Val Leu Ser Gly Tyr Ser His Glu Met
1130 1135 1140
Val His Ser Gly Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu
1145 1150 1155
His Val Asn Pro Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu
1160 1165 1170
Ser Cys Phe Asn Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His
1175 1180 1185
Val Gln Asn Leu Tyr Gly Asn Leu Gly Val
1190 1195
<210> 169
<211> 1165
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 169
Met Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro
1 5 10 15
Ser Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp
20 25 30
Leu Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp
35 40 45
Ser Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu
50 55 60
Trp Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro
65 70 75 80
Thr Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro
85 90 95
Ala Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala
100 105 110
Gln Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys
115 120 125
Ile Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu
130 135 140
Val Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His
145 150 155 160
Pro Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala
165 170 175
Ala Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln
180 185 190
Trp Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu
195 200 205
Leu Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile
210 215 220
Ala Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg
225 230 235 240
Asn Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val
245 250 255
Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
260 265 270
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
275 280 285
Val Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr
290 295 300
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
305 310 315 320
Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu
325 330 335
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
340 345 350
Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln
355 360 365
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
370 375 380
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly
385 390 395 400
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
405 410 415
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn
420 425 430
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
435 440 445
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
450 455 460
Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
465 470 475 480
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
485 490 495
Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu
500 505 510
Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val
515 520 525
Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln
530 535 540
Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln
545 550 555 560
Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr
565 570 575
Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro
580 585 590
Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu
595 600 605
Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu
610 615 620
Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln
625 630 635 640
Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His
645 650 655
Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly
660 665 670
Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln
675 680 685
Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly
690 695 700
Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu
705 710 715 720
Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser
725 730 735
Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro
740 745 750
Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile
755 760 765
Ala Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln
770 775 780
Leu Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu
785 790 795 800
Val Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys
805 810 815
Lys Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg
820 825 830
Ile Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Ser Pro Lys Lys
835 840 845
Lys Arg Lys Val Glu Ala Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu
850 855 860
Leu Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu
865 870 875 880
Lys Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala
885 890 895
Asp Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr
900 905 910
Ala Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly
915 920 925
Asp Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn
930 935 940
Phe Lys Lys Arg Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Lys
945 950 955 960
Lys Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu
965 970 975
Glu Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser
980 985 990
Ile Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser
995 1000 1005
Asn Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr
1010 1015 1020
Ile His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser
1025 1030 1035
Ile Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys
1040 1045 1050
Phe Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys
1055 1060 1065
Ala Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln
1070 1075 1080
Arg Gly Gly Ser Val Ala Ser Thr Pro Met Thr Val Val Pro Ser
1085 1090 1095
Pro Glu Met Val Leu Ser Gly Tyr Ser His Glu Met Val His Ser
1100 1105 1110
Gly Tyr Ser Ser Glu Met Val Asn Ser Gly Tyr Leu His Val Asn
1115 1120 1125
Pro Met Gln Gln Val Asn Thr Ser Ser Asp Pro Leu Ser Cys Phe
1130 1135 1140
Asn Asn Gly Glu Ala Pro Ser Met Trp Asp Ser His Val Gln Asn
1145 1150 1155
Leu Tyr Gly Asn Leu Gly Val
1160 1165
<210> 170
<211> 1159
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 170
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Ser Arg Thr Arg Leu Pro Ser
1 5 10 15
Pro Pro Ala Pro Ser Pro Ala Phe Ser Ala Asp Ser Phe Ser Asp Leu
20 25 30
Leu Arg Gln Phe Asp Pro Ser Leu Phe Asn Thr Ser Leu Phe Asp Ser
35 40 45
Leu Pro Pro Phe Gly Ala His His Thr Glu Ala Ala Thr Gly Glu Trp
50 55 60
Asp Glu Val Gln Ser Gly Leu Arg Ala Ala Asp Ala Pro Pro Pro Thr
65 70 75 80
Met Arg Val Ala Val Thr Ala Ala Arg Pro Pro Arg Ala Lys Pro Ala
85 90 95
Pro Arg Arg Arg Ala Ala Gln Pro Ser Asp Ala Ser Pro Ala Ala Gln
100 105 110
Val Asp Leu Arg Thr Leu Gly Tyr Ser Gln Gln Gln Gln Glu Lys Ile
115 120 125
Lys Pro Lys Val Arg Ser Thr Val Ala Gln His His Glu Ala Leu Val
130 135 140
Gly His Gly Phe Thr His Ala His Ile Val Ala Leu Ser Gln His Pro
145 150 155 160
Ala Ala Leu Gly Thr Val Ala Val Lys Tyr Gln Asp Met Ile Ala Ala
165 170 175
Leu Pro Glu Ala Thr His Glu Ala Ile Val Gly Val Gly Lys Gln Trp
180 185 190
Ser Gly Ala Arg Ala Leu Glu Ala Leu Leu Thr Val Ala Gly Glu Leu
195 200 205
Arg Gly Pro Pro Leu Gln Leu Asp Thr Gly Gln Leu Leu Lys Ile Ala
210 215 220
Lys Arg Gly Gly Val Thr Ala Val Glu Ala Val His Ala Trp Arg Asn
225 230 235 240
Ala Leu Thr Gly Ala Pro Leu Asn Leu Thr Pro Glu Gln Val Val Ala
245 250 255
Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
260 265 270
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
275 280 285
Val Ala Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val
290 295 300
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
305 310 315 320
Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu
325 330 335
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
340 345 350
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
355 360 365
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
370 375 380
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
385 390 395 400
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
405 410 415
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Asn Gly
420 425 430
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
435 440 445
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
450 455 460
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
465 470 475 480
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
485 490 495
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
500 505 510
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
515 520 525
Ile Ala Ser Asn Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
530 535 540
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
545 550 555 560
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
565 570 575
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
580 585 590
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
595 600 605
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
610 615 620
Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala
625 630 635 640
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
645 650 655
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
660 665 670
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
675 680 685
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
690 695 700
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
705 710 715 720
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
725 730 735
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
740 745 750
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
755 760 765
Ser His Asp Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala Gln Leu
770 775 780
Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His Leu Val
785 790 795 800
Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val Lys Lys
805 810 815
Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg Arg Ile
820 825 830
Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Ser Pro Lys Lys Lys
835 840 845
Arg Lys Val Glu Ala Ser Asn Gly Ala Ile Gly Gly Asp Leu Leu Leu
850 855 860
Asn Phe Pro Asp Met Ser Val Leu Glu Arg Gln Arg Ala His Leu Lys
865 870 875 880
Tyr Leu Asn Pro Thr Phe Asp Ser Pro Leu Ala Gly Phe Phe Ala Asp
885 890 895
Ser Ser Met Ile Thr Gly Gly Glu Met Asp Ser Tyr Leu Ser Thr Ala
900 905 910
Gly Leu Asn Leu Pro Met Met Tyr Gly Glu Thr Thr Val Glu Gly Asp
915 920 925
Ser Arg Leu Ser Ile Ser Pro Glu Thr Thr Leu Gly Thr Gly Asn Phe
930 935 940
Lys Lys Arg Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Lys Lys
945 950 955 960
Lys Lys Met Thr Met Asn Arg Asp Asp Leu Val Glu Glu Gly Glu Glu
965 970 975
Glu Lys Ser Lys Ile Thr Glu Gln Asn Asn Gly Ser Thr Lys Ser Ile
980 985 990
Lys Lys Met Lys His Lys Ala Lys Lys Glu Glu Asn Asn Phe Ser Asn
995 1000 1005
Asp Ser Ser Lys Val Thr Lys Glu Leu Glu Lys Thr Asp Tyr Ile
1010 1015 1020
His Val Arg Ala Arg Arg Gly Gln Ala Thr Asp Ser His Ser Ile
1025 1030 1035
Ala Glu Arg Val Arg Arg Glu Lys Ile Ser Glu Arg Met Lys Phe
1040 1045 1050
Leu Gln Asp Leu Val Pro Gly Cys Asp Lys Ile Thr Gly Lys Ala
1055 1060 1065
Gly Met Leu Asp Glu Ile Ile Asn Tyr Val Gln Ser Leu Gln Arg
1070 1075 1080
Gln Ile Glu Phe Leu Ser Met Lys Leu Ala Ile Val Asn Pro Arg
1085 1090 1095
Pro Asp Phe Asp Met Asp Asp Ile Phe Ala Lys Glu Val Ala Ser
1100 1105 1110
Thr Pro Met Thr Val Val Pro Ser Pro Glu Met Val Leu Ser Gly
1115 1120 1125
Tyr Gly Gly Ser Pro Leu Ser Cys Phe Asn Asn Gly Glu Ala Pro
1130 1135 1140
Ser Met Trp Asp Ser His Val Gln Asn Leu Tyr Gly Asn Leu Gly
1145 1150 1155
Val
<210> 171
<211> 846
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 171
Met Lys Arg Pro Ala Ala Thr Lys Lys Ala Gly Gln Ala Lys Lys Lys
1 5 10 15
Lys Lys Met Asp Lys Lys Thr Ile Val Trp Phe Arg Arg Asp Leu Arg
20 25 30
Ile Glu Asp Asn Pro Ala Leu Ala Ala Ala Ala His Glu Gly Ser Val
35 40 45
Phe Pro Val Phe Ile Trp Cys Pro Glu Glu Glu Gly Gln Phe Tyr Pro
50 55 60
Gly Arg Ala Ser Arg Trp Trp Met Lys Gln Ser Leu Ala His Leu Ser
65 70 75 80
Gln Ser Leu Lys Ala Leu Gly Ser Asp Leu Thr Leu Ile Lys Thr His
85 90 95
Asn Thr Ile Ser Ala Ile Leu Asp Cys Ile Arg Val Thr Gly Ala Thr
100 105 110
Lys Val Val Phe Asn His Leu Tyr Asp Pro Val Ser Leu Val Arg Asp
115 120 125
His Thr Val Lys Glu Lys Leu Val Glu Arg Gly Ile Ser Val Gln Ser
130 135 140
Tyr Asn Gly Asp Leu Leu Tyr Glu Pro Trp Glu Ile Tyr Cys Glu Lys
145 150 155 160
Gly Lys Pro Phe Thr Ser Phe Asn Ser Tyr Trp Lys Lys Cys Leu Asp
165 170 175
Met Ser Ile Glu Ser Val Met Leu Pro Pro Pro Trp Arg Leu Met Pro
180 185 190
Ile Thr Ala Ala Ala Glu Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu
195 200 205
Gly Leu Glu Asn Glu Ala Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg
210 215 220
Ala Trp Ser Pro Gly Trp Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe
225 230 235 240
Ile Glu Lys Gln Leu Ile Asp Tyr Ala Lys Asn Ser Lys Lys Val Val
245 250 255
Gly Asn Ser Thr Ser Leu Leu Ser Pro Tyr Leu His Phe Gly Glu Ile
260 265 270
Ser Val Arg His Val Phe Gln Cys Ala Arg Met Lys Gln Ile Ile Trp
275 280 285
Ala Arg Asp Lys Asn Ser Glu Gly Glu Glu Ser Ala Asp Leu Phe Leu
290 295 300
Arg Gly Ile Gly Leu Arg Glu Tyr Ser Arg Tyr Ile Cys Phe Asn Phe
305 310 315 320
Pro Phe Thr His Glu Gln Ser Leu Leu Ser His Leu Arg Phe Phe Pro
325 330 335
Trp Asp Ala Asp Val Asp Lys Phe Lys Ala Trp Arg Gln Gly Arg Thr
340 345 350
Gly Tyr Pro Leu Val Asp Ala Gly Met Arg Glu Leu Trp Ala Thr Gly
355 360 365
Trp Met His Asn Arg Ile Arg Val Ile Val Ser Ser Phe Ala Val Lys
370 375 380
Phe Leu Leu Leu Pro Trp Lys Trp Gly Met Lys Tyr Phe Trp Asp Thr
385 390 395 400
Leu Leu Asp Ala Asp Leu Glu Cys Asp Ile Leu Gly Trp Gln Tyr Ile
405 410 415
Ser Gly Ser Ile Pro Asp Gly His Glu Leu Asp Arg Leu Asp Asn Pro
420 425 430
Ala Leu Gln Gly Ala Lys Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln
435 440 445
Trp Leu Pro Glu Leu Ala Arg Leu Pro Thr Glu Trp Ile His His Pro
450 455 460
Trp Asp Ala Pro Leu Thr Val Leu Lys Ala Ser Gly Val Glu Leu Gly
465 470 475 480
Thr Asn Tyr Ala Lys Pro Ile Val Asp Ile Asp Thr Ala Arg Glu Leu
485 490 495
Leu Ala Lys Ala Ile Ser Arg Thr Arg Glu Ala Gln Ile Met Ile Gly
500 505 510
Ala Ala Pro Ala Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Gly
515 520 525
Ser Gly Arg Ala Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly
530 535 540
Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala
545 550 555 560
Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp
565 570 575
Phe Asp Leu Asp Met Leu Ile Asn Ser Arg Gly Ser Gly Glu Gly Arg
580 585 590
Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Val
595 600 605
Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu Val Glu
610 615 620
Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly Glu Gly
625 630 635 640
Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr
645 650 655
Thr Gly Lys Leu Pro Val Pro Trp Pro Thr Leu Val Thr Thr Leu Thr
660 665 670
Tyr Gly Val Gln Cys Phe Ser Arg Tyr Pro Asp His Met Lys Gln His
675 680 685
Asp Phe Phe Lys Ser Ala Met Pro Glu Gly Tyr Val Gln Glu Arg Thr
690 695 700
Ile Phe Phe Lys Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys
705 710 715 720
Phe Glu Gly Asp Thr Leu Val Asn Arg Ile Glu Leu Lys Gly Ile Asp
725 730 735
Phe Lys Glu Asp Gly Asn Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr
740 745 750
Asn Ser His Asn Val Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly Ile
755 760 765
Lys Val Asn Phe Lys Ile Arg His Asn Ile Glu Asp Gly Ser Val Gln
770 775 780
Leu Ala Asp His Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro Val
785 790 795 800
Leu Leu Pro Asp Asn His Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys
805 810 815
Asp Pro Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe Val Thr
820 825 830
Ala Ala Gly Ile Thr Leu Gly Met Asp Glu Leu Tyr Lys Val
835 840 845
<210> 172
<211> 840
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 172
Met Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Lys Met Asp Lys Lys
1 5 10 15
Thr Ile Val Trp Phe Arg Arg Asp Leu Arg Ile Glu Asp Asn Pro Ala
20 25 30
Leu Ala Ala Ala Ala His Glu Gly Ser Val Phe Pro Val Phe Ile Trp
35 40 45
Cys Pro Glu Glu Glu Gly Gln Phe Tyr Pro Gly Arg Ala Ser Arg Trp
50 55 60
Trp Met Lys Gln Ser Leu Ala His Leu Ser Gln Ser Leu Lys Ala Ala
65 70 75 80
Gly Ser Asp Ala Thr Leu Ile Lys Thr His Asn Thr Ile Ser Ala Ile
85 90 95
Leu Asp Cys Ile Arg Val Thr Gly Ala Thr Lys Val Val Phe Asn His
100 105 110
Leu Tyr Asp Pro Val Ser Leu Val Arg Asp His Thr Val Lys Glu Lys
115 120 125
Leu Val Glu Arg Gly Ile Ser Val Gln Ser Tyr Asn Gly Asp Leu Leu
130 135 140
Tyr Glu Pro Trp Glu Ile Tyr Cys Glu Lys Gly Lys Pro Phe Thr Ser
145 150 155 160
Phe Asn Ser Tyr Trp Lys Lys Cys Leu Asp Met Ser Ile Glu Ser Val
165 170 175
Met Leu Pro Pro Pro Trp Arg Leu Met Pro Ile Thr Ala Ala Ala Glu
180 185 190
Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu Gly Leu Glu Asn Glu Ala
195 200 205
Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg Ala Trp Ser Pro Gly Trp
210 215 220
Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe Ile Glu Lys Gln Leu Ile
225 230 235 240
Asp Tyr Ala Lys Asn Ser Lys Lys Val Val Gly Asn Ser Thr Ser Leu
245 250 255
Leu Ser Pro Tyr Leu His Phe Gly Glu Ile Ser Val Arg His Val Phe
260 265 270
Gln Cys Ala Arg Met Lys Gln Ile Ile Trp Ala Arg Asp Lys Asn Ser
275 280 285
Glu Gly Glu Glu Ser Ala Asp Leu Phe Leu Arg Gly Ile Gly Leu Arg
290 295 300
Glu Tyr Ser Arg Tyr Ile Cys Phe Asn Phe Pro Phe Thr His Glu Gln
305 310 315 320
Ser Leu Leu Ser His Leu Arg Phe Phe Pro Trp Asp Ala Asp Val Asp
325 330 335
Lys Phe Lys Ala Trp Arg Gln Gly Arg Thr Gly Tyr Pro Leu Val Asp
340 345 350
Ala Gly Met Arg Glu Leu Trp Ala Thr Gly Trp Met His Asn Arg Ile
355 360 365
Arg Val Ile Val Ser Ser Phe Ala Val Lys Phe Leu Leu Leu Pro Trp
370 375 380
Lys Trp Gly Met Lys Tyr Phe Trp Asp Thr Leu Leu Asp Ala Asp Leu
385 390 395 400
Glu Cys Asp Ile Leu Gly Trp Gln Tyr Ile Ser Gly Ser Ile Pro Asp
405 410 415
Gly His Glu Leu Asp Arg Leu Asp Asn Pro Ala Leu Gln Gly Ala Lys
420 425 430
Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln Trp Leu Pro Glu Leu Ala
435 440 445
Arg Leu Pro Thr Glu Trp Ile His His Pro Trp Asp Ala Pro Leu Thr
450 455 460
Val Leu Lys Ala Ser Gly Val Glu Leu Gly Thr Asn Tyr Ala Lys Pro
465 470 475 480
Ile Val Asp Ile Asp Thr Ala Arg Glu Leu Leu Ala Lys Ala Ile Ser
485 490 495
Arg Thr Arg Glu Ala Gln Ile Met Ile Gly Ala Ala Pro Ala Ser Pro
500 505 510
Lys Lys Lys Arg Lys Val Glu Ala Ser Gly Ser Gly Arg Ala Asp Ala
515 520 525
Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp
530 535 540
Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu
545 550 555 560
Asp Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu
565 570 575
Ile Asn Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys
580 585 590
Gly Asp Val Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu Glu Leu
595 600 605
Phe Thr Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn
610 615 620
Gly His Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr Tyr
625 630 635 640
Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro Val
645 650 655
Pro Trp Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe
660 665 670
Ser Arg Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala
675 680 685
Met Pro Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp
690 695 700
Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu
705 710 715 720
Val Asn Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn
725 730 735
Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr
740 745 750
Ile Met Ala Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe Lys Ile
755 760 765
Arg His Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr Gln
770 775 780
Gln Asn Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His
785 790 795 800
Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg
805 810 815
Asp His Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu
820 825 830
Gly Met Asp Glu Leu Tyr Lys Val
835 840
<210> 173
<211> 841
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 173
Met Lys Met Asp Lys Lys Thr Ile Val Trp Phe Arg Arg Asp Leu Arg
1 5 10 15
Ile Glu Asp Asn Pro Ala Leu Ala Ala Ala Ala His Glu Gly Ser Val
20 25 30
Phe Pro Val Phe Ile Trp Cys Pro Glu Glu Glu Gly Gln Phe Tyr Pro
35 40 45
Gly Arg Ala Ser Arg Trp Trp Met Lys Gln Ser Leu Ala His Leu Ser
50 55 60
Gln Ser Leu Lys Ala Leu Gly Ser Asp Leu Thr Leu Ile Lys Thr His
65 70 75 80
Asn Thr Ile Ser Ala Ile Leu Asp Cys Ile Arg Val Thr Gly Ala Thr
85 90 95
Lys Val Val Phe Asn His Leu Tyr Asp Pro Val Ser Leu Val Arg Asp
100 105 110
His Thr Val Lys Glu Lys Leu Val Glu Arg Gly Ile Ser Val Gln Ser
115 120 125
Tyr Asn Gly Asp Leu Leu Tyr Glu Pro Trp Glu Ile Tyr Cys Glu Lys
130 135 140
Gly Lys Pro Phe Thr Ser Phe Asn Ser Tyr Trp Lys Lys Cys Leu Asp
145 150 155 160
Met Ser Ile Glu Ser Val Met Leu Pro Pro Pro Trp Arg Leu Met Pro
165 170 175
Ile Thr Ala Ala Ala Glu Ala Ile Trp Ala Cys Ser Ile Glu Glu Leu
180 185 190
Gly Leu Glu Asn Glu Ala Glu Lys Pro Ser Asn Ala Leu Leu Thr Arg
195 200 205
Ala Trp Ser Pro Gly Trp Ser Asn Ala Asp Lys Leu Leu Asn Glu Phe
210 215 220
Ile Glu Lys Gln Leu Ile Asp Tyr Ala Lys Asn Ser Lys Lys Val Val
225 230 235 240
Gly Asn Ser Thr Ser Leu Leu Ser Pro Tyr Leu His Phe Gly Glu Ile
245 250 255
Ser Val Arg His Val Phe Gln Cys Ala Arg Met Lys Gln Ile Ile Trp
260 265 270
Ala Arg Asp Lys Asn Ser Glu Gly Glu Glu Ser Ala Asp Leu Phe Leu
275 280 285
Arg Gly Ile Gly Leu Arg Glu Tyr Ser Arg Tyr Ile Cys Phe Asn Phe
290 295 300
Pro Phe Thr His Glu Gln Ser Leu Leu Ser His Leu Arg Phe Phe Pro
305 310 315 320
Trp Asp Ala Asp Val Asp Lys Phe Lys Ala Trp Arg Gln Gly Arg Thr
325 330 335
Gly Tyr Pro Leu Val Asp Ala Gly Met Arg Glu Leu Trp Ala Thr Gly
340 345 350
Trp Met His Asn Arg Ile Arg Val Ile Val Ser Ser Phe Ala Val Lys
355 360 365
Phe Leu Leu Leu Pro Trp Lys Trp Gly Met Lys Tyr Phe Trp Asp Thr
370 375 380
Leu Leu Asp Ala Asp Leu Glu Cys Asp Ile Leu Gly Trp Gln Tyr Ile
385 390 395 400
Ser Gly Ser Ile Pro Asp Gly His Glu Leu Asp Arg Leu Asp Asn Pro
405 410 415
Ala Leu Gln Gly Ala Lys Tyr Asp Pro Glu Gly Glu Tyr Ile Arg Gln
420 425 430
Trp Leu Pro Glu Leu Ala Arg Leu Pro Thr Glu Trp Ile His His Pro
435 440 445
Trp Asp Ala Pro Leu Thr Val Leu Lys Ala Ser Gly Val Glu Leu Gly
450 455 460
Thr Asn Tyr Ala Lys Pro Ile Val Asp Ile Asp Thr Ala Arg Glu Leu
465 470 475 480
Leu Ala Lys Ala Ile Ser Arg Thr Arg Glu Ala Gln Ile Met Ile Gly
485 490 495
Ala Ala Pro Ala Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Gly
500 505 510
Ser Gly Arg Ala Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly
515 520 525
Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala
530 535 540
Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu Asp Asp
545 550 555 560
Phe Asp Leu Asp Met Leu Ile Asn Ser Pro Lys Lys Lys Arg Lys Val
565 570 575
Glu Ala Ser Ser Arg Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr
580 585 590
Cys Gly Asp Val Glu Glu Asn Pro Gly Pro Val Ser Lys Gly Glu Glu
595 600 605
Leu Phe Thr Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val
610 615 620
Asn Gly His Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr
625 630 635 640
Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro
645 650 655
Val Pro Trp Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys
660 665 670
Phe Ser Arg Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser
675 680 685
Ala Met Pro Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp
690 695 700
Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr
705 710 715 720
Leu Val Asn Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly
725 730 735
Asn Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val
740 745 750
Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly Ile Lys Val Asn Phe Lys
755 760 765
Ile Arg His Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr
770 775 780
Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn
785 790 795 800
His Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys
805 810 815
Arg Asp His Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr
820 825 830
Leu Gly Met Asp Glu Leu Tyr Lys Val
835 840
<210> 174
<211> 954
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 174
Met Ser Arg Thr Arg Leu Pro Ser Pro Pro Ala Pro Ser Pro Ala Phe
1 5 10 15
Ser Ala Asp Ser Phe Ser Asp Leu Leu Arg Gln Phe Asp Pro Ser Leu
20 25 30
Phe Asn Thr Ser Leu Phe Asp Ser Leu Pro Pro Phe Gly Ala His His
35 40 45
Thr Glu Ala Ala Thr Gly Glu Trp Asp Glu Val Gln Ser Gly Leu Arg
50 55 60
Ala Ala Asp Ala Pro Pro Pro Thr Met Arg Val Ala Val Thr Ala Ala
65 70 75 80
Arg Pro Pro Arg Ala Lys Pro Ala Pro Arg Arg Arg Ala Ala Gln Pro
85 90 95
Ser Asp Ala Ser Pro Ala Ala Gln Val Asp Leu Arg Thr Leu Gly Tyr
100 105 110
Ser Gln Gln Gln Gln Glu Lys Ile Lys Pro Lys Val Arg Ser Thr Val
115 120 125
Ala Gln His His Glu Ala Leu Val Gly His Gly Phe Thr His Ala His
130 135 140
Ile Val Ala Leu Ser Gln His Pro Ala Ala Leu Gly Thr Val Ala Val
145 150 155 160
Lys Tyr Gln Asp Met Ile Ala Ala Leu Pro Glu Ala Thr His Glu Ala
165 170 175
Ile Val Gly Val Gly Lys Gln Trp Ser Gly Ala Arg Ala Leu Glu Ala
180 185 190
Leu Leu Thr Val Ala Gly Glu Leu Arg Gly Pro Pro Leu Gln Leu Asp
195 200 205
Thr Gly Gln Leu Leu Lys Ile Ala Lys Arg Gly Gly Val Thr Ala Val
210 215 220
Glu Ala Val His Ala Trp Arg Asn Ala Leu Thr Gly Ala Pro Leu Asn
225 230 235 240
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser His Asn Gly Gly Lys
245 250 255
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
260 265 270
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly
275 280 285
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
290 295 300
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
305 310 315 320
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
325 330 335
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
340 345 350
Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
355 360 365
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
370 375 380
Ile Ala Ser His Asn Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg
385 390 395 400
Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val
405 410 415
Val Ala Ile Ala Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val
420 425 430
Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu
435 440 445
Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys Gln Ala Leu Glu
450 455 460
Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly Leu Thr
465 470 475 480
Pro Glu Gln Val Val Ala Ile Ala Ser His Asn Gly Gly Lys Gln Ala
485 490 495
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
500 505 510
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Ile Gly Gly Lys
515 520 525
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
530 535 540
His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly
545 550 555 560
Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys
565 570 575
Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn
580 585 590
Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val
595 600 605
Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala Ile Ala
610 615 620
Ser Asn Ile Gly Gly Lys Gln Ala Leu Glu Thr Val Gln Arg Leu Leu
625 630 635 640
Pro Val Leu Cys Gln Ala His Gly Leu Thr Pro Glu Gln Val Val Ala
645 650 655
Ile Ala Ser Asn Gly Gly Gly Arg Pro Ala Leu Glu Ser Ile Val Ala
660 665 670
Gln Leu Ser Arg Pro Asp Pro Ala Leu Ala Ala Leu Thr Asn Asp His
675 680 685
Leu Val Ala Leu Ala Cys Leu Gly Gly Arg Pro Ala Leu Asp Ala Val
690 695 700
Lys Lys Gly Leu Pro His Ala Pro Ala Leu Ile Lys Arg Thr Asn Arg
705 710 715 720
Arg Ile Pro Glu Arg Thr Ser His Arg Val Ala Ala Ser Met Ala Asn
725 730 735
Ser Glu Ser Ser Ser Ser Pro Val Asn Glu Glu Glu Asn Ser Gln Arg
740 745 750
Ile Ser Thr Leu His His Gln Thr Met Pro Ser Asp Leu Thr Gln Asp
755 760 765
Glu Phe Thr Gln Leu Ser Gln Ser Ile Ala Glu Phe His Thr Tyr Gln
770 775 780
Leu Gly Asn Gly Arg Cys Ser Ser Leu Leu Ala Gln Arg Ile His Ala
785 790 795 800
Pro Pro Glu Thr Val Trp Ser Val Val Arg Arg Phe Asp Arg Pro Gln
805 810 815
Ile Tyr Lys His Phe Ile Lys Ser Cys Asn Val Ser Glu Asp Phe Glu
820 825 830
Met Arg Val Gly Cys Thr Arg Asp Val Asn Val Ile Ser Gly Leu Pro
835 840 845
Ala Asn Thr Ser Arg Glu Arg Leu Asp Leu Leu Asp Asp Asp Arg Arg
850 855 860
Val Thr Gly Phe Ser Ile Thr Gly Gly Glu His Arg Leu Arg Asn Tyr
865 870 875 880
Lys Ser Val Thr Thr Val His Arg Phe Glu Lys Glu Glu Glu Glu Glu
885 890 895
Arg Ile Trp Thr Val Val Leu Glu Ser Tyr Val Val Asp Val Pro Glu
900 905 910
Gly Asn Ser Glu Glu Asp Thr Arg Leu Phe Ala Asp Thr Val Ile Arg
915 920 925
Leu Asn Leu Gln Lys Leu Ala Ser Ile Thr Glu Ala Met Asn Arg Asn
930 935 940
Asn Asn Asn Asn Asn Ser Ser Gln Val Arg
945 950
<210> 175
<211> 379
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 175
Met Val Pro Leu Tyr Gly Phe Thr Ser Ile Cys Gly Arg Arg Pro Glu
1 5 10 15
Met Glu Ala Ala Val Ser Thr Ile Pro Arg Phe Leu Gln Ser Ser Ser
20 25 30
Gly Ser Met Leu Asp Gly Arg Phe Asp Pro Gln Ser Ala Ala His Phe
35 40 45
Phe Gly Val Tyr Asp Gly His Gly Gly Ser Gln Val Ala Asn Tyr Cys
50 55 60
Arg Glu Arg Met His Leu Ala Leu Ala Glu Glu Ile Ala Lys Glu Lys
65 70 75 80
Pro Met Leu Cys Asp Gly Asp Thr Trp Leu Glu Lys Trp Lys Lys Ala
85 90 95
Leu Phe Asn Ser Phe Leu Arg Val Asp Ser Glu Ile Glu Ser Val Ala
100 105 110
Pro Glu Thr Val Gly Ser Thr Ser Val Val Ala Val Val Phe Pro Ser
115 120 125
His Ile Phe Val Ala Asn Cys Gly Asp Ser Arg Ala Val Leu Cys Arg
130 135 140
Gly Lys Thr Ala Leu Pro Leu Ser Val Asp His Lys Pro Asp Arg Glu
145 150 155 160
Asp Glu Ala Ala Arg Ile Glu Ala Ala Gly Gly Lys Val Ile Gln Trp
165 170 175
Asn Gly Ala Arg Val Phe Gly Val Leu Ala Met Ser Arg Ser Ile Gly
180 185 190
Asp Arg Tyr Leu Lys Pro Ser Ile Ile Pro Asp Pro Glu Val Thr Ala
195 200 205
Val Lys Arg Val Lys Glu Asp Asp Cys Leu Ile Leu Ala Ser Asp Gly
210 215 220
Val Trp Asp Val Met Thr Asp Glu Glu Ala Cys Glu Met Ala Arg Lys
225 230 235 240
Arg Ile Leu Leu Trp His Lys Lys Asn Ala Val Ala Gly Asp Ala Ser
245 250 255
Leu Leu Ala Asp Glu Arg Arg Lys Glu Gly Lys Asp Pro Ala Ala Met
260 265 270
Ser Ala Ala Glu Tyr Leu Ser Lys Leu Ala Ile Gln Arg Gly Ser Lys
275 280 285
Asp Asn Ile Ser Val Val Val Val Asp Leu Lys Pro Arg Arg Lys Leu
290 295 300
Lys Ser Lys Pro Leu Asn Ala Ser Pro Lys Lys Lys Arg Lys Val Glu
305 310 315 320
Ala Ser Gly Ser Gly Arg Ala Asp Ala Leu Asp Asp Phe Asp Leu Asp
325 330 335
Met Leu Gly Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly
340 345 350
Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala
355 360 365
Leu Asp Asp Phe Asp Leu Asp Met Leu Ile Asn
370 375
<210> 176
<211> 1453
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 176
Met Asp Lys Lys Tyr Ser Ile Gly Leu Ala Ile Gly Thr Asn Ser Val
1 5 10 15
Gly Trp Ala Val Ile Thr Asp Glu Tyr Lys Val Pro Ser Lys Lys Phe
20 25 30
Lys Val Leu Gly Asn Thr Asp Arg His Ser Ile Lys Lys Asn Leu Ile
35 40 45
Gly Ala Leu Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala Thr Arg Leu
50 55 60
Lys Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn Arg Ile Cys
65 70 75 80
Tyr Leu Gln Glu Ile Phe Ser Asn Glu Met Ala Lys Val Asp Asp Ser
85 90 95
Phe Phe His Arg Leu Glu Glu Ser Phe Leu Val Glu Glu Asp Lys Lys
100 105 110
His Glu Arg His Pro Ile Phe Gly Asn Ile Val Asp Glu Val Ala Tyr
115 120 125
His Glu Lys Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys Leu Val Asp
130 135 140
Ser Thr Asp Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala Leu Ala His
145 150 155 160
Met Ile Lys Phe Arg Gly His Phe Leu Ile Glu Gly Asp Leu Asn Pro
165 170 175
Asp Asn Ser Asp Val Asp Lys Leu Phe Ile Gln Leu Val Gln Thr Tyr
180 185 190
Asn Gln Leu Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly Val Asp Ala
195 200 205
Lys Ala Ile Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg Leu Glu Asn
210 215 220
Leu Ile Ala Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu Phe Gly Asn
225 230 235 240
Leu Ile Ala Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys Ser Asn Phe
245 250 255
Asp Leu Ala Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp Thr Tyr Asp
260 265 270
Asp Asp Leu Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln Tyr Ala Asp
275 280 285
Leu Phe Leu Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu Leu Ser Asp
290 295 300
Ile Leu Arg Val Asn Thr Glu Ile Thr Lys Ala Pro Leu Ser Ala Ser
305 310 315 320
Met Ile Lys Arg Tyr Asp Glu His His Gln Asp Leu Thr Leu Leu Lys
325 330 335
Ala Leu Val Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu Ile Phe Phe
340 345 350
Asp Gln Ser Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Gly Ala Ser
355 360 365
Gln Glu Glu Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu Lys Met Asp
370 375 380
Gly Thr Glu Glu Leu Leu Val Lys Leu Asn Arg Glu Asp Leu Leu Arg
385 390 395 400
Lys Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu
405 410 415
Gly Glu Leu His Ala Ile Leu Arg Arg Gln Glu Asp Phe Tyr Pro Phe
420 425 430
Leu Lys Asp Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr Phe Arg Ile
435 440 445
Pro Tyr Tyr Val Gly Pro Leu Ala Arg Gly Asn Ser Arg Phe Ala Trp
450 455 460
Met Thr Arg Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn Phe Glu Glu
465 470 475 480
Val Val Asp Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu Arg Met Thr
485 490 495
Asn Phe Asp Lys Asn Leu Pro Asn Glu Lys Val Leu Pro Lys His Ser
500 505 510
Leu Leu Tyr Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys
515 520 525
Tyr Val Thr Glu Gly Met Arg Lys Pro Ala Phe Leu Ser Gly Glu Gln
530 535 540
Lys Lys Ala Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys Val Thr
545 550 555 560
Val Lys Gln Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys Phe Asp
565 570 575
Ser Val Glu Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser Leu Gly
580 585 590
Thr Tyr His Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe Leu Asp
595 600 605
Asn Glu Glu Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr Leu Thr
610 615 620
Leu Phe Glu Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr Tyr Ala
625 630 635 640
His Leu Phe Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg Arg Tyr
645 650 655
Thr Gly Trp Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile Arg Asp
660 665 670
Lys Gln Ser Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp Gly Phe
675 680 685
Ala Asn Arg Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu Thr Phe
690 695 700
Lys Glu Asp Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp Ser Leu
705 710 715 720
His Glu His Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys Lys Gly
725 730 735
Ile Leu Gln Thr Val Lys Val Val Asp Glu Leu Val Lys Val Met Gly
740 745 750
Arg His Lys Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln
755 760 765
Thr Thr Gln Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys Arg Ile
770 775 780
Glu Glu Gly Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu His Pro
785 790 795 800
Val Glu Asn Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu
805 810 815
Gln Asn Gly Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile Asn Arg
820 825 830
Leu Ser Asp Tyr Asp Val Asp Ala Ile Val Pro Gln Ser Phe Leu Lys
835 840 845
Asp Asp Ser Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg
850 855 860
Gly Lys Ser Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys
865 870 875 880
Asn Tyr Trp Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys
885 890 895
Phe Asp Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp
900 905 910
Lys Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr
915 920 925
Lys His Val Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr Asp
930 935 940
Glu Asn Asp Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu Lys Ser
945 950 955 960
Lys Leu Val Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys Val Arg
965 970 975
Glu Ile Asn Asn Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val
980 985 990
Val Gly Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser Glu Phe
995 1000 1005
Val Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met Ile Ala
1010 1015 1020
Lys Ser Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr Phe Phe
1025 1030 1035
Tyr Ser Asn Ile Met Asn Phe Phe Lys Thr Glu Ile Thr Leu Ala
1040 1045 1050
Asn Gly Glu Ile Arg Lys Arg Pro Leu Ile Glu Thr Asn Gly Glu
1055 1060 1065
Thr Gly Glu Ile Val Trp Asp Lys Gly Arg Asp Phe Ala Thr Val
1070 1075 1080
Arg Lys Val Leu Ser Met Pro Gln Val Asn Ile Val Lys Lys Thr
1085 1090 1095
Glu Val Gln Thr Gly Gly Phe Ser Lys Glu Ser Ile Leu Pro Lys
1100 1105 1110
Arg Asn Ser Asp Lys Leu Ile Ala Arg Lys Lys Asp Trp Asp Pro
1115 1120 1125
Lys Lys Tyr Gly Gly Phe Asp Ser Pro Thr Val Ala Tyr Ser Val
1130 1135 1140
Leu Val Val Ala Lys Val Glu Lys Gly Lys Ser Lys Lys Leu Lys
1145 1150 1155
Ser Val Lys Glu Leu Leu Gly Ile Thr Ile Met Glu Arg Ser Ser
1160 1165 1170
Phe Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala Lys Gly Tyr Lys
1175 1180 1185
Glu Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys Tyr Ser Leu
1190 1195 1200
Phe Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser Ala Gly
1205 1210 1215
Glu Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys Tyr Val
1220 1225 1230
Asn Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys Gly Ser
1235 1240 1245
Pro Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln His Lys
1250 1255 1260
His Tyr Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe Ser Lys
1265 1270 1275
Arg Val Ile Leu Ala Asp Ala Asn Leu Asp Lys Val Leu Ser Ala
1280 1285 1290
Tyr Asn Lys His Arg Asp Lys Pro Ile Arg Glu Gln Ala Glu Asn
1295 1300 1305
Ile Ile His Leu Phe Thr Leu Thr Asn Leu Gly Ala Pro Ala Ala
1310 1315 1320
Phe Lys Tyr Phe Asp Thr Thr Ile Asp Arg Lys Arg Tyr Thr Ser
1325 1330 1335
Thr Lys Glu Val Leu Asp Ala Thr Leu Ile His Gln Ser Ile Thr
1340 1345 1350
Gly Leu Tyr Glu Thr Arg Ile Asp Leu Ser Gln Leu Gly Gly Asp
1355 1360 1365
Ser Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1370 1375 1380
Gly Ser Gly Pro Lys Lys Lys Arg Lys Val Ala Ala Ala Gly Ser
1385 1390 1395
Gly Arg Ala Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly
1400 1405 1410
Ser Asp Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp
1415 1420 1425
Ala Leu Asp Asp Phe Asp Leu Asp Met Leu Gly Ser Asp Ala Leu
1430 1435 1440
Asp Asp Phe Asp Leu Asp Met Leu Ile Asn
1445 1450
<210> 177
<211> 1569
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 177
Met Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr
1 5 10 15
Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu
20 25 30
Pro Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr
35 40 45
Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu
50 55 60
Pro Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr
65 70 75 80
Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu
85 90 95
Pro Gly Ser Gly Met Asn Ile Gln Met Leu Leu Glu Ala Ala Asp Tyr
100 105 110
Leu Glu Arg Arg Glu Arg Glu Ala Glu His Gly Tyr Ala Ser Met Leu
115 120 125
Pro Ser Arg Ser Ala Gly Pro Lys Lys Lys Arg Lys Val Ala Ala Ala
130 135 140
Asp Tyr Lys Asp Asp Asp Asp Lys Ser Arg Leu Glu Pro Gly Glu Lys
145 150 155 160
Pro Tyr Lys Cys Pro Glu Cys Gly Lys Ser Phe Ser Gln Ser Gly Ala
165 170 175
Leu Thr Arg His Gln Arg Thr His Thr Arg Asp Lys Lys Tyr Ser Ile
180 185 190
Gly Leu Ala Ile Gly Thr Asn Ser Val Gly Trp Ala Val Ile Thr Asp
195 200 205
Glu Tyr Lys Val Pro Ser Lys Lys Phe Lys Val Leu Gly Asn Thr Asp
210 215 220
Arg His Ser Ile Lys Lys Asn Leu Ile Gly Ala Leu Leu Phe Asp Ser
225 230 235 240
Gly Glu Thr Ala Glu Ala Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg
245 250 255
Tyr Thr Arg Arg Lys Asn Arg Ile Cys Tyr Leu Gln Glu Ile Phe Ser
260 265 270
Asn Glu Met Ala Lys Val Asp Asp Ser Phe Phe His Arg Leu Glu Glu
275 280 285
Ser Phe Leu Val Glu Glu Asp Lys Lys His Glu Arg His Pro Ile Phe
290 295 300
Gly Asn Ile Val Asp Glu Val Ala Tyr His Glu Lys Tyr Pro Thr Ile
305 310 315 320
Tyr His Leu Arg Lys Lys Leu Val Asp Ser Thr Asp Lys Ala Asp Leu
325 330 335
Arg Leu Ile Tyr Leu Ala Leu Ala His Met Ile Lys Phe Arg Gly His
340 345 350
Phe Leu Ile Glu Gly Asp Leu Asn Pro Asp Asn Ser Asp Val Asp Lys
355 360 365
Leu Phe Ile Gln Leu Val Gln Thr Tyr Asn Gln Leu Phe Glu Glu Asn
370 375 380
Pro Ile Asn Ala Ser Gly Val Asp Ala Lys Ala Ile Leu Ser Ala Arg
385 390 395 400
Leu Ser Lys Ser Arg Arg Leu Glu Asn Leu Ile Ala Gln Leu Pro Gly
405 410 415
Glu Lys Lys Asn Gly Leu Phe Gly Asn Leu Ile Ala Leu Ser Leu Gly
420 425 430
Leu Thr Pro Asn Phe Lys Ser Asn Phe Asp Leu Ala Glu Asp Ala Lys
435 440 445
Leu Gln Leu Ser Lys Asp Thr Tyr Asp Asp Asp Leu Asp Asn Leu Leu
450 455 460
Ala Gln Ile Gly Asp Gln Tyr Ala Asp Leu Phe Leu Ala Ala Lys Asn
465 470 475 480
Leu Ser Asp Ala Ile Leu Leu Ser Asp Ile Leu Arg Val Asn Thr Glu
485 490 495
Ile Thr Lys Ala Pro Leu Ser Ala Ser Met Ile Lys Arg Tyr Asp Glu
500 505 510
His His Gln Asp Leu Thr Leu Leu Lys Ala Leu Val Arg Gln Gln Leu
515 520 525
Pro Glu Lys Tyr Lys Glu Ile Phe Phe Asp Gln Ser Lys Asn Gly Tyr
530 535 540
Ala Gly Tyr Ile Asp Gly Gly Ala Ser Gln Glu Glu Phe Tyr Lys Phe
545 550 555 560
Ile Lys Pro Ile Leu Glu Lys Met Asp Gly Thr Glu Glu Leu Leu Val
565 570 575
Lys Leu Asn Arg Glu Asp Leu Leu Arg Lys Gln Arg Thr Phe Asp Asn
580 585 590
Gly Ser Ile Pro His Gln Ile His Leu Gly Glu Leu His Ala Ile Leu
595 600 605
Arg Arg Gln Glu Asp Phe Tyr Pro Phe Leu Lys Asp Asn Arg Glu Lys
610 615 620
Ile Glu Lys Ile Leu Thr Phe Arg Ile Pro Tyr Tyr Val Gly Pro Leu
625 630 635 640
Ala Arg Gly Asn Ser Arg Phe Ala Trp Met Thr Arg Lys Ser Glu Glu
645 650 655
Thr Ile Thr Pro Trp Asn Phe Glu Glu Val Val Asp Lys Gly Ala Ser
660 665 670
Ala Gln Ser Phe Ile Glu Arg Met Thr Asn Phe Asp Lys Asn Leu Pro
675 680 685
Asn Glu Lys Val Leu Pro Lys His Ser Leu Leu Tyr Glu Tyr Phe Thr
690 695 700
Val Tyr Asn Glu Leu Thr Lys Val Lys Tyr Val Thr Glu Gly Met Arg
705 710 715 720
Lys Pro Ala Phe Leu Ser Gly Glu Gln Lys Lys Ala Ile Val Asp Leu
725 730 735
Leu Phe Lys Thr Asn Arg Lys Val Thr Val Lys Gln Leu Lys Glu Asp
740 745 750
Tyr Phe Lys Lys Ile Glu Cys Phe Asp Ser Val Glu Ile Ser Gly Val
755 760 765
Glu Asp Arg Phe Asn Ala Ser Leu Gly Thr Tyr His Asp Leu Leu Lys
770 775 780
Ile Ile Lys Asp Lys Asp Phe Leu Asp Asn Glu Glu Asn Glu Asp Ile
785 790 795 800
Leu Glu Asp Ile Val Leu Thr Leu Thr Leu Phe Glu Asp Arg Glu Met
805 810 815
Ile Glu Glu Arg Leu Lys Thr Tyr Ala His Leu Phe Asp Asp Lys Val
820 825 830
Met Lys Gln Leu Lys Arg Arg Arg Tyr Thr Gly Trp Gly Arg Leu Ser
835 840 845
Arg Lys Leu Ile Asn Gly Ile Arg Asp Lys Gln Ser Gly Lys Thr Ile
850 855 860
Leu Asp Phe Leu Lys Ser Asp Gly Phe Ala Asn Arg Asn Phe Met Gln
865 870 875 880
Leu Ile His Asp Asp Ser Leu Thr Phe Lys Glu Asp Ile Gln Lys Ala
885 890 895
Gln Val Ser Gly Gln Gly Asp Ser Leu His Glu His Ile Ala Asn Leu
900 905 910
Ala Gly Ser Pro Ala Ile Lys Lys Gly Ile Leu Gln Thr Val Lys Val
915 920 925
Val Asp Glu Leu Val Lys Val Met Gly Arg His Lys Pro Glu Asn Ile
930 935 940
Val Ile Glu Met Ala Arg Glu Asn Gln Thr Thr Gln Lys Gly Gln Lys
945 950 955 960
Asn Ser Arg Glu Arg Met Lys Arg Ile Glu Glu Gly Ile Lys Glu Leu
965 970 975
Gly Ser Gln Ile Leu Lys Glu His Pro Val Glu Asn Thr Gln Leu Gln
980 985 990
Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly Arg Asp Met Tyr
995 1000 1005
Val Asp Gln Glu Leu Asp Ile Asn Arg Leu Ser Asp Tyr Asp Val
1010 1015 1020
Asp Ala Ile Val Pro Gln Ser Phe Leu Lys Asp Asp Ser Ile Asp
1025 1030 1035
Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg Gly Lys Ser Asp
1040 1045 1050
Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys Asn Tyr Trp
1055 1060 1065
Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys Phe Asp
1070 1075 1080
Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp Lys
1085 1090 1095
Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr
1100 1105 1110
Lys His Val Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr
1115 1120 1125
Asp Glu Asn Asp Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu
1130 1135 1140
Lys Ser Lys Leu Val Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr
1145 1150 1155
Lys Val Arg Glu Ile Asn Asn Tyr His His Ala His Asp Ala Tyr
1160 1165 1170
Leu Asn Ala Val Val Gly Thr Ala Leu Ile Lys Lys Tyr Pro Lys
1175 1180 1185
Leu Glu Ser Glu Phe Val Tyr Gly Asp Tyr Lys Val Tyr Asp Val
1190 1195 1200
Arg Lys Met Ile Ala Lys Ser Glu Gln Glu Ile Gly Lys Ala Thr
1205 1210 1215
Ala Lys Tyr Phe Phe Tyr Ser Asn Ile Met Asn Phe Phe Lys Thr
1220 1225 1230
Glu Ile Thr Leu Ala Asn Gly Glu Ile Arg Lys Arg Pro Leu Ile
1235 1240 1245
Glu Thr Asn Gly Glu Thr Gly Glu Ile Val Trp Asp Lys Gly Arg
1250 1255 1260
Asp Phe Ala Thr Val Arg Lys Val Leu Ser Met Pro Gln Val Asn
1265 1270 1275
Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly Phe Ser Lys Glu
1280 1285 1290
Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile Ala Arg Lys
1295 1300 1305
Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser Pro Thr
1310 1315 1320
Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys Gly Lys
1325 1330 1335
Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile Thr Ile
1340 1345 1350
Met Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe Leu Glu
1355 1360 1365
Ala Lys Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile Lys Leu
1370 1375 1380
Pro Lys Tyr Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys Arg Met
1385 1390 1395
Leu Ala Ser Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu Ala Leu
1400 1405 1410
Pro Ser Lys Tyr Val Asn Phe Leu Tyr Leu Ala Ser His Tyr Glu
1415 1420 1425
Lys Leu Lys Gly Ser Pro Glu Asp Asn Glu Gln Lys Gln Leu Phe
1430 1435 1440
Val Glu Gln His Lys His Tyr Leu Asp Glu Ile Ile Glu Gln Ile
1445 1450 1455
Ser Glu Phe Ser Lys Arg Val Ile Leu Ala Asp Ala Asn Leu Asp
1460 1465 1470
Lys Val Leu Ser Ala Tyr Asn Lys His Arg Asp Lys Pro Ile Arg
1475 1480 1485
Glu Gln Ala Glu Asn Ile Ile His Leu Phe Thr Leu Thr Asn Leu
1490 1495 1500
Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp Thr Thr Ile Asp Arg
1505 1510 1515
Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp Ala Thr Leu Ile
1520 1525 1530
His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile Asp Leu Ser
1535 1540 1545
Gln Leu Gly Gly Asp Lys Arg Pro Ala Ala Thr Lys Lys Ala Gly
1550 1555 1560
Gln Ala Lys Lys Lys Lys
1565
<210> 178
<211> 84
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 178
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Asn Gly
1 5 10 15
Leu Met Glu Asp Pro Met Lys Val Tyr Lys Asp Arg Gln Phe Met Asn
20 25 30
Val Trp Thr Asp His Glu Lys Glu Ile Phe Lys Asp Lys Phe Ile Gln
35 40 45
His Pro Lys Asn Phe Gly Leu Ile Ala Ser Tyr Leu Glu Arg Lys Ser
50 55 60
Val Pro Asp Cys Val Leu Tyr Tyr Tyr Leu Thr Lys Lys Asn Glu Asn
65 70 75 80
Tyr Lys Glu Phe
<210> 179
<211> 288
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 179
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Gly Asn
1 5 10 15
Leu Met Ile Ser Phe Leu Lys Lys Asp Thr Gln Ser Ile Thr Leu Glu
20 25 30
Glu Leu Ala Lys Ile Ile Lys Lys Cys Lys His Val Val Ala Leu Thr
35 40 45
Gly Ser Gly Thr Ser Ala Glu Ser Asn Ile Pro Ser Phe Arg Gly Ser
50 55 60
Ser Asn Ser Ile Trp Ser Lys Tyr Asp Pro Arg Ile Tyr Gly Thr Ile
65 70 75 80
Trp Gly Phe Trp Lys Tyr Pro Glu Lys Ile Trp Glu Val Ile Arg Asp
85 90 95
Ile Ser Ser Asp Tyr Glu Ile Glu Ile Asn Asn Gly His Val Ala Leu
100 105 110
Ser Thr Leu Glu Ser Leu Gly Tyr Leu Lys Ser Val Val Thr Gln Asn
115 120 125
Val Asp Gly Leu His Glu Ala Ser Gly Asn Thr Lys Val Ile Ser Leu
130 135 140
His Gly Asn Val Phe Glu Ala Val Cys Cys Thr Cys Asn Lys Ile Val
145 150 155 160
Lys Leu Asn Lys Ile Met Leu Gln Lys Thr Ser His Phe Met His Gln
165 170 175
Leu Pro Pro Glu Cys Pro Cys Gly Gly Ile Phe Lys Pro Asn Ile Ile
180 185 190
Leu Phe Gly Glu Val Val Ser Ser Asp Leu Leu Lys Glu Ala Glu Glu
195 200 205
Glu Ile Ala Lys Cys Asp Leu Leu Leu Val Ile Gly Thr Ser Ser Thr
210 215 220
Val Ser Thr Ala Thr Asn Leu Cys His Phe Ala Cys Lys Lys Lys Lys
225 230 235 240
Lys Ile Val Glu Ile Asn Ile Ser Lys Thr Tyr Ile Thr Asn Lys Met
245 250 255
Ser Asp Tyr His Val Cys Ala Lys Phe Ser Glu Leu Thr Lys Val Ala
260 265 270
Asn Ile Leu Lys Gly Ser Ser Glu Lys Asn Lys Lys Ile Met Glu Phe
275 280 285
<210> 180
<211> 346
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 180
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Glu Lys
1 5 10 15
Ala Phe Arg Pro His Phe Phe Asn His Gly Lys Pro Asp Ala Asn Pro
20 25 30
Lys Glu Lys Lys Asn Cys His Trp Cys Gln Ile Arg Ser Phe Ala Thr
35 40 45
His Ala Gln Leu Pro Ile Ser Ile Val Asn Arg Glu Asp Asp Ala Phe
50 55 60
Leu Asn Pro Asn Phe Arg Phe Ile Asp His Ser Ile Ile Gly Lys Asn
65 70 75 80
Val Pro Val Ala Asp Gln Ser Phe Arg Val Gly Cys Ser Cys Ala Ser
85 90 95
Asp Glu Glu Cys Met Tyr Ser Thr Cys Gln Cys Leu Asp Glu Met Ala
100 105 110
Pro Asp Ser Asp Glu Glu Ala Asp Pro Tyr Thr Arg Lys Lys Arg Phe
115 120 125
Ala Tyr Tyr Ser Gln Gly Ala Lys Lys Gly Leu Leu Arg Asp Arg Val
130 135 140
Leu Gln Ser Gln Glu Pro Ile Tyr Glu Cys His Gln Gly Cys Ala Cys
145 150 155 160
Ser Lys Asp Cys Pro Asn Arg Val Val Glu Arg Gly Arg Thr Val Pro
165 170 175
Leu Gln Ile Phe Arg Thr Lys Asp Arg Gly Trp Gly Val Lys Cys Pro
180 185 190
Val Asn Ile Lys Arg Gly Gln Phe Val Asp Arg Tyr Leu Gly Glu Ile
195 200 205
Ile Thr Ser Glu Glu Ala Asp Arg Arg Arg Ala Glu Ser Thr Ile Ala
210 215 220
Arg Arg Lys Asp Val Tyr Leu Phe Ala Leu Asp Lys Phe Ser Asp Pro
225 230 235 240
Asp Ser Leu Asp Pro Leu Leu Ala Gly Gln Pro Leu Glu Val Asp Gly
245 250 255
Glu Tyr Met Ser Gly Pro Thr Arg Phe Ile Asn His Ser Cys Asp Pro
260 265 270
Asn Met Ala Ile Phe Ala Arg Val Gly Asp His Ala Asp Lys His Ile
275 280 285
His Asp Leu Ala Leu Phe Ala Ile Lys Asp Ile Pro Lys Gly Thr Glu
290 295 300
Leu Thr Phe Asp Tyr Val Asn Gly Leu Thr Gly Leu Glu Ser Asp Ala
305 310 315 320
His Asp Pro Ser Lys Ile Ser Glu Met Thr Lys Cys Leu Cys Gly Thr
325 330 335
Ala Lys Cys Arg Gly Tyr Leu Trp Glu Phe
340 345
<210> 181
<211> 306
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 181
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Thr Glu Met
1 5 10 15
Ser Val Arg Lys Ile Ala Ala His Met Lys Ser Asn Pro Asn Ala Lys
20 25 30
Val Ile Phe Met Val Gly Ala Gly Ile Ser Thr Ser Cys Gly Ile Pro
35 40 45
Asp Phe Arg Ser Pro Gly Thr Gly Leu Tyr His Asn Leu Ala Arg Leu
50 55 60
Lys Leu Pro Tyr Pro Glu Ala Val Phe Asp Val Asp Phe Phe Gln Ser
65 70 75 80
Asp Pro Leu Pro Phe Tyr Thr Leu Ala Lys Glu Leu Tyr Pro Gly Asn
85 90 95
Phe Arg Pro Ser Lys Phe His Tyr Leu Leu Lys Leu Phe Gln Asp Lys
100 105 110
Asp Val Leu Lys Arg Val Tyr Thr Gln Asn Ile Asp Thr Leu Glu Arg
115 120 125
Gln Ala Gly Val Lys Asp Asp Leu Ile Ile Glu Ala His Gly Ser Phe
130 135 140
Ala His Cys His Cys Ile Gly Cys Gly Lys Val Tyr Pro Pro Gln Val
145 150 155 160
Phe Lys Ser Lys Leu Ala Glu His Pro Ile Lys Asp Phe Val Lys Cys
165 170 175
Asp Val Cys Gly Glu Leu Val Lys Pro Ala Ile Val Phe Phe Gly Glu
180 185 190
Asp Leu Pro Asp Ser Phe Ser Glu Thr Trp Leu Asn Asp Ser Glu Trp
195 200 205
Leu Arg Glu Lys Ile Thr Thr Ser Gly Lys His Pro Gln Gln Pro Leu
210 215 220
Val Ile Val Val Gly Thr Ser Leu Ala Val Tyr Pro Phe Ala Ser Leu
225 230 235 240
Pro Glu Glu Ile Pro Arg Lys Val Lys Arg Val Leu Cys Asn Leu Glu
245 250 255
Thr Val Gly Asp Phe Lys Ala Asn Lys Arg Pro Thr Asp Leu Ile Val
260 265 270
His Gln Tyr Ser Asp Glu Phe Ala Glu Gln Leu Val Glu Glu Leu Gly
275 280 285
Trp Gln Glu Asp Phe Glu Lys Ile Leu Thr Ala Gln Gly Gly Met Gly
290 295 300
Glu Phe
305
<210> 182
<211> 272
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 182
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Val Gly
1 5 10 15
Ala Gly Ile Ser Thr Pro Ser Gly Ile Pro Asp Phe Arg Ser Pro Gly
20 25 30
Ser Gly Leu Tyr Ser Asn Leu Gln Gln Tyr Asp Leu Pro Tyr Pro Glu
35 40 45
Ala Ile Phe Glu Leu Pro Phe Phe Phe His Asn Pro Lys Pro Phe Phe
50 55 60
Thr Leu Ala Lys Glu Leu Tyr Pro Gly Asn Tyr Lys Pro Asn Val Thr
65 70 75 80
His Tyr Phe Leu Arg Leu Leu His Asp Lys Gly Leu Leu Leu Arg Leu
85 90 95
Tyr Thr Gln Asn Ile Asp Gly Leu Glu Arg Val Ser Gly Ile Pro Ala
100 105 110
Ser Lys Leu Val Glu Ala His Gly Thr Phe Ala Ser Ala Thr Cys Thr
115 120 125
Val Cys Gln Arg Pro Phe Pro Gly Glu Asp Ile Arg Ala Asp Val Met
130 135 140
Ala Asp Arg Val Pro Arg Cys Pro Val Cys Thr Gly Val Val Lys Pro
145 150 155 160
Asp Ile Val Phe Phe Gly Glu Pro Leu Pro Gln Arg Phe Leu Leu His
165 170 175
Val Val Asp Phe Pro Met Ala Asp Leu Leu Leu Ile Leu Gly Thr Ser
180 185 190
Leu Glu Val Glu Pro Phe Ala Ser Leu Thr Glu Ala Val Arg Ser Ser
195 200 205
Val Pro Arg Leu Leu Ile Asn Arg Asp Leu Val Gly Pro Leu Ala Trp
210 215 220
His Pro Arg Ser Arg Asp Val Ala Gln Leu Gly Asp Val Val His Gly
225 230 235 240
Val Glu Ser Leu Val Glu Leu Leu Gly Trp Thr Glu Glu Met Arg Asp
245 250 255
Leu Val Gln Arg Glu Thr Gly Lys Leu Asp Gly Pro Asp Lys Glu Phe
260 265 270
<210> 183
<211> 344
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 183
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Ala Ala
1 5 10 15
Ala Ala Asn Ser Gly Ser Ser Leu Pro Leu Phe Asp Cys Pro Thr Trp
20 25 30
Ala Gly Lys Pro Pro Pro Gly Leu His Leu Asp Val Val Lys Gly Asp
35 40 45
Lys Leu Ile Glu Lys Leu Ile Ile Asp Glu Lys Lys Tyr Tyr Leu Phe
50 55 60
Gly Arg Asn Pro Asp Leu Cys Asp Phe Thr Ile Asp His Gln Ser Cys
65 70 75 80
Ser Arg Val His Ala Ala Leu Val Tyr His Lys His Leu Lys Arg Val
85 90 95
Phe Leu Ile Asp Leu Asn Ser Thr His Gly Thr Phe Leu Gly His Ile
100 105 110
Arg Leu Glu Pro His Lys Pro Gln Gln Ile Pro Ile Asp Ser Thr Val
115 120 125
Ser Phe Gly Ala Ser Thr Arg Ala Tyr Thr Leu Arg Glu Lys Pro Gln
130 135 140
Thr Leu Pro Ser Ala Val Lys Gly Asp Glu Lys Met Gly Gly Glu Asp
145 150 155 160
Asp Glu Leu Lys Gly Leu Leu Gly Leu Pro Glu Glu Glu Thr Glu Leu
165 170 175
Asp Asn Leu Thr Glu Phe Asn Thr Ala His Asn Lys Arg Ile Ser Thr
180 185 190
Leu Thr Ile Glu Glu Gly Asn Leu Asp Ile Gln Arg Pro Lys Arg Lys
195 200 205
Arg Lys Asn Ser Arg Val Thr Phe Ser Glu Asp Asp Glu Ile Ile Asn
210 215 220
Pro Glu Asp Val Asp Pro Ser Val Gly Arg Phe Arg Asn Met Val Gln
225 230 235 240
Thr Ala Val Val Pro Val Lys Lys Lys Arg Val Glu Gly Pro Gly Ser
245 250 255
Leu Gly Leu Glu Glu Ser Gly Ser Arg Arg Met Gln Asn Phe Ala Phe
260 265 270
Ser Gly Gly Leu Tyr Gly Gly Leu Pro Pro Thr His Ser Glu Ala Gly
275 280 285
Ser Gln Pro His Gly Ile His Gly Thr Ala Leu Ile Gly Gly Leu Pro
290 295 300
Met Pro Tyr Pro Asn Leu Ala Pro Asp Val Asp Leu Thr Pro Val Val
305 310 315 320
Pro Ser Ala Val Asn Met Asn Pro Ala Pro Asn Pro Ala Val Tyr Asn
325 330 335
Pro Glu Ala Val Asn Glu Glu Phe
340
<210> 184
<211> 234
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 184
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Thr Thr
1 5 10 15
Asn Ser Thr Gln Asp Thr Leu Tyr Leu Ser Leu His Gly Gly Ile Asp
20 25 30
Ser Ala Ile Pro Tyr Pro Val Arg Arg Val Glu Gln Leu Leu Gln Phe
35 40 45
Ser Phe Leu Pro Glu Leu Gln Phe Gln Asn Ala Ala Val Lys Gln Arg
50 55 60
Ile Gln Arg Leu Cys Tyr Arg Glu Glu Lys Arg Leu Ala Val Ser Ser
65 70 75 80
Leu Ala Lys Trp Leu Gly Gln Leu His Lys Gln Arg Leu Arg Ala Pro
85 90 95
Lys Asn Pro Pro Val Ala Ile Cys Trp Ile Asn Ser Tyr Val Gly Tyr
100 105 110
Gly Val Phe Ala Arg Glu Ser Ile Pro Ala Trp Ser Tyr Ile Gly Glu
115 120 125
Tyr Thr Gly Ile Leu Arg Arg Arg Gln Ala Leu Trp Leu Asp Glu Asn
130 135 140
Asp Tyr Cys Phe Arg Tyr Pro Val Pro Arg Tyr Ser Phe Arg Tyr Phe
145 150 155 160
Thr Ile Asp Ser Gly Met Gln Gly Asn Val Thr Arg Phe Ile Asn His
165 170 175
Ser Asp Asn Pro Asn Leu Glu Ala Ile Gly Ala Phe Glu Asn Gly Ile
180 185 190
Phe His Ile Ile Ile Arg Ala Ile Lys Asp Ile Leu Pro Gly Glu Glu
195 200 205
Leu Cys Tyr His Tyr Gly Pro Leu Tyr Trp Lys His Arg Lys Lys Arg
210 215 220
Glu Glu Phe Val Pro Gln Glu Glu Glu Phe
225 230
<210> 185
<211> 233
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 185
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Ala Arg Tyr
1 5 10 15
Leu Gly Asn Thr Val Asp Leu Ser Ser Phe Asp Phe Arg Thr Gly Lys
20 25 30
Met Met Pro Ser Lys Leu Gln Lys Asn Lys Gln Arg Leu Arg Asn Asp
35 40 45
Pro Leu Asn Gln Asn Lys Gly Lys Pro Asp Leu Asn Thr Thr Leu Pro
50 55 60
Ile Arg Gln Thr Ala Ser Ile Phe Lys Gln Pro Val Thr Lys Val Thr
65 70 75 80
Asn His Pro Ser Asn Lys Val Lys Ser Asp Pro Gln Arg Met Asn Glu
85 90 95
Gln Pro Arg Gln Leu Phe Trp Glu Lys Arg Leu Gln Gly Leu Ser Ala
100 105 110
Ser Asp Val Thr Glu Gln Ile Ile Lys Thr Met Glu Leu Pro Lys Gly
115 120 125
Leu Gln Gly Val Gly Pro Gly Ser Asn Asp Glu Thr Leu Leu Ser Ala
130 135 140
Val Ala Ser Ala Leu His Thr Ser Ser Ala Pro Ile Thr Gly Gln Val
145 150 155 160
Ser Ala Ala Val Glu Lys Asn Pro Ala Val Trp Leu Asn Thr Ser Gln
165 170 175
Pro Leu Cys Lys Ala Phe Ile Val Thr Asp Glu Asp Ile Arg Lys Gln
180 185 190
Glu Glu Arg Val Gln Gln Val Arg Lys Ile Leu Glu Asp Ala Leu Met
195 200 205
Ala Asp Ile Leu Ser Arg Ala Ala Asp Thr Glu Glu Met Asp Ile Glu
210 215 220
Met Asp Ser Gly Asp Glu Ala Glu Phe
225 230
<210> 186
<211> 346
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 186
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Pro Ser
1 5 10 15
Leu Asp Asp Ile Leu Lys Pro Val Ala Glu Ala Val Lys Asn Gly Lys
20 25 30
Lys Val Thr Phe Phe Asn Gly Ala Gly Ile Ser Thr Gly Ala Gly Ile
35 40 45
Pro Asp Phe Arg Ser Pro Asp Thr Gly Leu Tyr Ala Asn Leu Ala Lys
50 55 60
Leu Asn Leu Pro Phe Ala Glu Ala Val Phe Asp Ile Asp Phe Phe Lys
65 70 75 80
Glu Asp Pro Lys Pro Phe Tyr Thr Leu Ala Glu Glu Leu Tyr Pro Gly
85 90 95
Asn Phe Ala Pro Thr Lys Phe His His Phe Ile Lys Leu Leu Gln Asp
100 105 110
Gln Gly Ser Leu Lys Arg Val Tyr Thr Gln Asn Ile Asp Thr Leu Glu
115 120 125
Arg Leu Ala Gly Val Glu Asp Lys Tyr Ile Val Glu Ala His Gly Ser
130 135 140
Phe Ala Ser Asn His Cys Val Asp Cys His Lys Glu Met Thr Thr Glu
145 150 155 160
Thr Leu Lys Thr Tyr Met Lys Asp Lys Lys Ile Pro Ser Cys Gln His
165 170 175
Cys Glu Gly Tyr Val Lys Pro Asp Ile Val Phe Phe Gly Glu Gly Leu
180 185 190
Pro Val Lys Phe Phe Asp Leu Trp Glu Asp Asp Cys Glu Asp Val Glu
195 200 205
Val Ala Ile Val Ala Gly Thr Ser Leu Thr Val Phe Pro Phe Ala Ser
210 215 220
Leu Pro Gly Glu Val Asn Lys Lys Cys Leu Arg Val Leu Val Asn Lys
225 230 235 240
Glu Lys Val Gly Thr Phe Lys His Glu Pro Arg Lys Ser Asp Ile Ile
245 250 255
Ala Leu His Asp Cys Asp Ile Val Ala Glu Arg Leu Cys Thr Leu Leu
260 265 270
Gly Leu Asp Asp Lys Leu Asn Glu Val Tyr Glu Lys Glu Lys Ile Lys
275 280 285
Tyr Ser Lys Ala Glu Thr Lys Glu Ile Lys Met His Glu Ile Glu Asp
290 295 300
Lys Leu Lys Glu Glu Ala His Leu Lys Glu Asp Lys His Thr Thr Lys
305 310 315 320
Val Asp Lys Lys Glu Lys Gln Asn Asp Ala Asn Asp Lys Glu Leu Glu
325 330 335
Gln Leu Ile Asp Lys Ala Lys Ala Glu Phe
340 345
<210> 187
<211> 350
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 187
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Glu Asn
1 5 10 15
Arg Ala Leu Asp Pro Gly Thr Arg Asp Ser Tyr Gly Ala Thr Ser His
20 25 30
Leu Pro Asn Lys Gly Ala Leu Ala Lys Val Lys Asn Asn Phe Lys Asp
35 40 45
Leu Met Ser Lys Leu Thr Glu Gly Gln Tyr Val Leu Cys Arg Trp Thr
50 55 60
Asp Gly Leu Tyr Tyr Leu Gly Lys Ile Lys Arg Val Ser Ser Ser Lys
65 70 75 80
Gln Ser Cys Leu Val Thr Phe Glu Asp Asn Ser Lys Tyr Trp Val Leu
85 90 95
Trp Lys Asp Ile Gln His Ala Gly Val Pro Gly Glu Glu Pro Lys Cys
100 105 110
Asn Ile Cys Leu Gly Lys Thr Ser Gly Pro Leu Asn Glu Ile Leu Ile
115 120 125
Cys Gly Lys Cys Gly Leu Gly Tyr His Gln Gln Cys His Ile Pro Ile
130 135 140
Ala Gly Ser Ala Asp Gln Pro Leu Leu Thr Pro Trp Phe Cys Arg Arg
145 150 155 160
Cys Ile Phe Ala Leu Ala Val Arg Lys Gly Gly Ala Leu Lys Lys Gly
165 170 175
Ala Ile Ala Arg Thr Leu Gln Ala Val Lys Met Val Leu Ser Tyr Gln
180 185 190
Pro Glu Glu Leu Glu Trp Asp Ser Pro His Arg Thr Asn Gln Gln Gln
195 200 205
Cys Tyr Cys Tyr Cys Gly Gly Pro Gly Glu Trp Tyr Leu Arg Met Leu
210 215 220
Gln Cys Tyr Arg Cys Arg Gln Trp Phe His Glu Ala Cys Thr Gln Cys
225 230 235 240
Leu Asn Glu Pro Met Met Phe Gly Asp Arg Phe Tyr Leu Phe Phe Cys
245 250 255
Ser Val Cys Asn Gln Gly Pro Gly Gly Ser Gly Ser Asp Ser Ser Ala
260 265 270
Glu Gly Ala Ser Val Pro Glu Arg Pro Asp Glu Gly Ile Asp Ser His
275 280 285
Thr Phe Glu Ser Ile Ser Glu Asp Asp Ser Ser Leu Ser His Leu Lys
290 295 300
Ser Ser Ile Thr Asn Tyr Phe Gly Ala Ala Gly Arg Leu Ala Cys Gly
305 310 315 320
Glu Lys Tyr Gln Val Leu Ala Arg Arg Val Thr Pro Glu Gly Lys Val
325 330 335
Gln Tyr Leu Val Glu Trp Glu Gly Thr Thr Pro Tyr Glu Phe
340 345 350
<210> 188
<211> 362
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 188
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Leu His
1 5 10 15
Thr Thr Gln Leu Tyr Gln His Val Pro Glu Thr Arg Trp Pro Ile Val
20 25 30
Tyr Ser Pro Arg Tyr Asn Ile Thr Phe Met Gly Leu Glu Lys Leu His
35 40 45
Pro Phe Asp Ala Gly Lys Trp Gly Lys Val Ile Asn Phe Leu Lys Glu
50 55 60
Glu Lys Leu Leu Ser Asp Ser Met Leu Val Glu Ala Arg Glu Ala Ser
65 70 75 80
Glu Glu Asp Leu Leu Val Val His Thr Arg Arg Tyr Leu Asn Glu Leu
85 90 95
Lys Trp Ser Phe Ala Val Ala Thr Ile Thr Glu Ile Pro Pro Val Ile
100 105 110
Phe Leu Pro Asn Phe Leu Val Gln Arg Lys Val Leu Arg Pro Leu Arg
115 120 125
Thr Gln Thr Gly Gly Thr Ile Met Ala Gly Lys Leu Ala Val Glu Arg
130 135 140
Gly Trp Ala Ile Asn Val Gly Gly Gly Phe His His Cys Ser Ser Asp
145 150 155 160
Arg Gly Gly Gly Phe Cys Ala Tyr Ala Asp Ile Thr Leu Ala Ile Lys
165 170 175
Phe Leu Phe Glu Arg Val Glu Gly Ile Ser Arg Ala Thr Ile Ile Asp
180 185 190
Leu Asp Ala His Gln Gly Asn Gly His Glu Arg Asp Phe Met Asp Asp
195 200 205
Lys Arg Val Tyr Ile Met Asp Val Tyr Asn Arg His Ile Tyr Pro Gly
210 215 220
Asp Arg Phe Ala Lys Gln Ala Ile Arg Arg Lys Val Glu Leu Glu Trp
225 230 235 240
Gly Thr Glu Asp Asp Glu Tyr Leu Asp Lys Val Glu Arg Asn Ile Lys
245 250 255
Lys Ser Leu Gln Glu His Leu Pro Asp Val Val Val Tyr Asn Ala Gly
260 265 270
Thr Asp Ile Leu Glu Gly Asp Arg Leu Gly Gly Leu Ser Ile Ser Pro
275 280 285
Ala Gly Ile Val Lys Arg Asp Glu Leu Val Phe Arg Met Val Arg Gly
290 295 300
Arg Arg Val Pro Ile Leu Met Val Thr Ser Gly Gly Tyr Gln Lys Arg
305 310 315 320
Thr Ala Arg Ile Ile Ala Asp Ser Ile Leu Asn Leu Phe Gly Leu Gly
325 330 335
Leu Ile Gly Pro Glu Ser Pro Ser Val Ser Ala Gln Asn Ser Asp Thr
340 345 350
Pro Leu Leu Pro Pro Ala Val Pro Glu Phe
355 360
<210> 189
<211> 315
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 189
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Pro Leu
1 5 10 15
Gln Ile Val His His Pro Asp Tyr Asp Ala Gly Phe Ala Thr Asn His
20 25 30
Arg Phe Pro Met Ser Lys Tyr Pro Leu Leu Met Glu Ala Leu Arg Ala
35 40 45
Arg Gly Leu Ala Ser Pro Asp Ala Leu Asn Thr Thr Glu Pro Ala Pro
50 55 60
Ala Ser Trp Leu Lys Leu Ala His Ala Ala Asp Tyr Val Asp Gln Val
65 70 75 80
Ile Ser Cys Ser Val Pro Glu Lys Ile Glu Arg Glu Ile Gly Phe Pro
85 90 95
Val Gly Pro Arg Val Ser Leu Arg Ala Gln Leu Ala Thr Gly Gly Thr
100 105 110
Ile Leu Ala Ala Arg Leu Ala Leu Arg His Gly Ile Ala Cys Asn Thr
115 120 125
Ala Gly Gly Ser His His Ala Arg Arg Ala Gln Gly Ala Gly Phe Cys
130 135 140
Thr Phe Asn Asp Val Ala Val Ala Ser Leu Val Leu Leu Asp Glu Gly
145 150 155 160
Ala Ala Gln Asn Ile Leu Val Val Asp Leu Asp Val His Gln Gly Asp
165 170 175
Gly Thr Ala Asp Ile Leu Ser Asp Glu Pro Gly Val Phe Thr Phe Ser
180 185 190
Met His Gly Glu Arg Asn Tyr Pro Val Arg Lys Ile Ala Ser Asp Leu
195 200 205
Asp Ile Ala Leu Pro Asp Gly Thr Gly Asp Ala Ala Tyr Leu Arg Arg
210 215 220
Leu Ala Thr Ile Leu Pro Glu Leu Ser Ala Arg Ala Arg Trp Asp Ile
225 230 235 240
Val Phe Tyr Asn Ala Gly Val Asp Val His Ala Glu Asp Arg Leu Gly
245 250 255
Arg Leu Ala Leu Ser Asn Gly Gly Leu Arg Ala Arg Asp Glu Met Val
260 265 270
Ile Gly His Phe Arg Ala Leu Gly Ile Pro Val Cys Gly Val Ile Gly
275 280 285
Gly Gly Tyr Ser Thr Asp Val Pro Ala Leu Ala Ser Arg His Ala Ile
290 295 300
Leu Phe Glu Val Ala Ser Thr Tyr Ala Glu Phe
305 310 315
<210> 190
<211> 134
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 190
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Phe Asn
1 5 10 15
Asp Arg Val Ile Val Lys Lys Ser Pro Leu Gly Gly Tyr Gly Val Phe
20 25 30
Ala Arg Lys Ser Phe Glu Lys Gly Glu Leu Val Glu Glu Cys Leu Cys
35 40 45
Ile Val Arg His Asn Asp Asp Trp Gly Thr Ala Leu Glu Asp Tyr Leu
50 55 60
Phe Ser Arg Lys Asn Met Ser Ala Met Ala Leu Gly Phe Gly Ala Ile
65 70 75 80
Phe Asn His Ser Lys Asp Pro Asn Ala Arg His Glu Leu Thr Ala Gly
85 90 95
Leu Lys Arg Met Arg Ile Phe Thr Ile Lys Pro Ile Ala Ile Gly Glu
100 105 110
Glu Ile Thr Ile Ser Tyr Gly Asp Asp Tyr Trp Leu Ser Arg Pro Arg
115 120 125
Leu Thr Gln Asn Glu Phe
130
<210> 191
<211> 282
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 191
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Asp Ile Ser
1 5 10 15
Gly Gly Leu Glu Phe Lys Gly Ile Pro Ala Thr Asn Arg Val Asp Asp
20 25 30
Ser Pro Val Ser Pro Thr Ser Gly Phe Thr Tyr Ile Lys Ser Leu Ile
35 40 45
Ile Glu Pro Asn Val Ile Ile Pro Lys Ser Ser Thr Gly Cys Asn Cys
50 55 60
Arg Gly Ser Cys Thr Asp Ser Lys Lys Cys Ala Cys Ala Lys Leu Asn
65 70 75 80
Gly Gly Asn Phe Pro Tyr Val Asp Leu Asn Asp Gly Arg Leu Ile Glu
85 90 95
Ser Arg Asp Val Val Phe Glu Cys Gly Pro His Cys Gly Cys Gly Pro
100 105 110
Lys Cys Val Asn Arg Thr Ser Gln Lys Arg Leu Arg Phe Asn Leu Glu
115 120 125
Val Phe Arg Ser Ala Lys Lys Gly Trp Ala Val Arg Ser Trp Glu Tyr
130 135 140
Ile Pro Ala Gly Ser Pro Val Cys Glu Tyr Ile Gly Val Val Arg Arg
145 150 155 160
Thr Ala Asp Val Asp Thr Ile Ser Asp Asn Glu Tyr Ile Phe Glu Ile
165 170 175
Asp Cys Gln Gln Thr Met Gln Gly Leu Gly Gly Arg Gln Arg Arg Leu
180 185 190
Arg Asp Val Ala Val Pro Met Asn Asn Gly Val Ser Gln Ser Ser Glu
195 200 205
Asp Glu Asn Ala Pro Glu Phe Cys Ile Asp Ala Gly Ser Thr Gly Asn
210 215 220
Phe Ala Arg Phe Ile Asn His Ser Cys Glu Pro Asn Leu Phe Val Gln
225 230 235 240
Cys Val Leu Ser Ser His Gln Asp Ile Arg Leu Ala Arg Val Val Leu
245 250 255
Phe Ala Ala Asp Asn Ile Ser Pro Met Gln Glu Leu Thr Tyr Asp Tyr
260 265 270
Gly Tyr Ala Leu Asp Ser Val His Glu Phe
275 280
<210> 192
<211> 319
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 192
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Ala Ser Arg
1 5 10 15
Arg Thr Gly Glu Phe Leu Arg Asp Ala Gln Ala Pro Ser Arg Trp Leu
20 25 30
Lys Arg Ser Lys Thr Gly Gln Asp Asp Gly Ala Phe Cys Leu Glu Thr
35 40 45
Trp Leu Ala Gly Ala Gly Asp Asp Ala Ala Gly Gly Glu Arg Gly Arg
50 55 60
Asp Arg Glu Gly Ala Ala Asp Lys Ala Lys Gln Arg Glu Glu Arg Arg
65 70 75 80
Gln Lys Glu Leu Glu Glu Arg Phe Glu Glu Met Lys Val Glu Phe Glu
85 90 95
Glu Lys Ala Gln Arg Met Ile Ala Arg Arg Ala Ala Leu Thr Gly Glu
100 105 110
Ile Tyr Ser Asp Gly Lys Gly Ser Lys Lys Pro Arg Val Pro Ser Leu
115 120 125
Pro Glu Asn Asp Asp Asp Ala Leu Ile Glu Ile Ile Ile Asp Pro Glu
130 135 140
Gln Gly Ile Leu Lys Trp Pro Leu Ser Val Met Ser Ile Arg Gln Arg
145 150 155 160
Thr Val Ile Tyr Gln Glu Cys Leu Arg Arg Asp Leu Thr Ala Cys Ile
165 170 175
His Leu Thr Lys Val Pro Gly Lys Gly Arg Ala Val Phe Ala Ala Asp
180 185 190
Thr Ile Leu Lys Asp Asp Phe Val Val Glu Tyr Lys Gly Glu Leu Cys
195 200 205
Ser Glu Arg Glu Ala Arg Glu Arg Glu Gln Arg Tyr Asn Arg Ser Lys
210 215 220
Val Pro Met Gly Ser Phe Met Phe Tyr Phe Lys Asn Gly Ser Arg Met
225 230 235 240
Met Ala Ile Asp Ala Thr Asp Glu Lys Gln Asp Phe Gly Pro Ala Arg
245 250 255
Leu Ile Asn His Ser Arg Arg Asn Pro Asn Met Thr Pro Arg Ala Ile
260 265 270
Thr Leu Gly Asp Phe Asn Ser Glu Pro Arg Leu Ile Phe Val Ala Arg
275 280 285
Arg Asn Ile Glu Lys Gly Glu Glu Leu Leu Val Asp Tyr Gly Glu Arg
290 295 300
Asp Pro Asp Val Ile Lys Glu His Pro Trp Leu Asn Ser Glu Phe
305 310 315
<210> 193
<211> 304
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 193
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Ser Val
1 5 10 15
Asn Tyr Ala Ala Gly Leu Ser Pro Tyr Ala Asp Lys Gly Lys Cys Gly
20 25 30
Leu Pro Glu Ile Phe Asp Pro Pro Glu Glu Leu Glu Arg Lys Val Trp
35 40 45
Glu Leu Ala Arg Leu Val Trp Gln Ser Ser Ser Val Val Phe His Thr
50 55 60
Gly Ala Gly Ile Ser Thr Ala Ser Gly Ile Pro Asp Phe Arg Gly Pro
65 70 75 80
His Gly Val Trp Thr Met Glu Glu Arg Gly Leu Ala Pro Lys Phe Asp
85 90 95
Thr Thr Phe Glu Ser Ala Arg Pro Thr Gln Thr His Met Ala Leu Val
100 105 110
Gln Leu Glu Arg Val Gly Leu Leu Arg Phe Leu Val Ser Gln Asn Val
115 120 125
Asp Gly Leu His Val Arg Ser Gly Phe Pro Arg Asp Lys Leu Ala Glu
130 135 140
Leu His Gly Asn Met Phe Val Glu Glu Cys Ala Lys Cys Lys Thr Gln
145 150 155 160
Tyr Val Arg Asp Thr Val Val Gly Thr Met Gly Leu Lys Ala Thr Gly
165 170 175
Arg Leu Cys Thr Val Ala Lys Ala Arg Gly Leu Arg Ala Cys Arg Gly
180 185 190
Glu Leu Arg Asp Thr Ile Leu Asp Trp Glu Asp Ser Leu Pro Asp Arg
195 200 205
Asp Leu Ala Leu Ala Asp Glu Ala Ser Arg Asn Ala Asp Leu Ser Ile
210 215 220
Thr Leu Gly Thr Ser Leu Gln Ile Arg Pro Ser Gly Asn Leu Pro Leu
225 230 235 240
Ala Thr Lys Arg Arg Gly Gly Arg Leu Val Ile Val Asn Leu Gln Pro
245 250 255
Thr Lys His Asp Arg His Ala Asp Leu Arg Ile His Gly Tyr Val Asp
260 265 270
Glu Val Met Thr Arg Leu Met Lys His Leu Gly Leu Glu Ile Pro Ala
275 280 285
Trp Asp Gly Pro Arg Val Leu Glu Arg Ala Leu Pro Pro Leu Glu Phe
290 295 300
<210> 194
<211> 257
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 194
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Lys Val
1 5 10 15
Ala Ala Lys Lys Leu Ala Thr Ser Arg Met Arg Lys Asp Arg Ala Ala
20 25 30
Ala Ala Ser Pro Ser Ser Asp Ile Glu Asn Ser Glu Asn Pro Ser Ser
35 40 45
Leu Ala Ser His Ser Ser Ser Ser Gly Arg Met Thr Pro Ser Lys Asn
50 55 60
Thr Arg Ser Arg Lys Gly Val Ser Val Lys Asp Val Ser Asn His Lys
65 70 75 80
Ile Thr Glu Phe Phe Gln Val Arg Arg Ser Asn Arg Lys Thr Ser Lys
85 90 95
Gln Ile Ser Asp Glu Ala Lys His Ala Leu Arg Asp Thr Val Leu Lys
100 105 110
Gly Thr Asn Glu Arg Leu Leu Glu Val Tyr Lys Asp Val Val Lys Gly
115 120 125
Arg Gly Ile Arg Thr Lys Val Asn Phe Glu Lys Gly Asp Phe Val Val
130 135 140
Glu Tyr Arg Gly Val Met Met Glu Tyr Ser Glu Ala Lys Val Ile Glu
145 150 155 160
Glu Gln Tyr Ser Asn Asp Glu Glu Ile Gly Ser Tyr Met Tyr Phe Phe
165 170 175
Glu His Asn Asn Lys Lys Trp Cys Ile Asp Ala Thr Lys Glu Ser Pro
180 185 190
Trp Lys Gly Arg Leu Ile Asn His Ser Val Leu Arg Pro Asn Leu Lys
195 200 205
Thr Lys Val Val Glu Ile Asp Gly Ser His His Leu Ile Leu Val Ala
210 215 220
Arg Arg Gln Ile Ala Gln Gly Glu Glu Leu Leu Tyr Asp Tyr Gly Asp
225 230 235 240
Arg Ser Ala Glu Thr Ile Ala Lys Asn Pro Trp Leu Val Asn Thr Glu
245 250 255
Phe
<210> 195
<211> 310
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 195
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Val Arg Thr
1 5 10 15
Glu Lys Ile Ile Cys Arg Asp Val Ala Arg Gly Tyr Glu Asn Val Pro
20 25 30
Ile Pro Cys Val Asn Gly Val Asp Gly Glu Pro Cys Pro Glu Asp Tyr
35 40 45
Lys Tyr Ile Ser Glu Asn Cys Glu Thr Ser Thr Met Asn Ile Asp Arg
50 55 60
Asn Ile Thr His Leu Gln His Cys Thr Cys Val Asp Asp Cys Ser Ser
65 70 75 80
Ser Asn Cys Leu Cys Gly Gln Leu Ser Ile Arg Cys Trp Tyr Asp Lys
85 90 95
Asp Gly Arg Leu Leu Gln Glu Phe Asn Lys Ile Glu Pro Pro Leu Ile
100 105 110
Phe Glu Cys Asn Gln Ala Cys Ser Cys Trp Arg Ser Cys Lys Asn Arg
115 120 125
Val Val Gln Ser Gly Ile Lys Val Arg Leu Gln Leu Tyr Arg Thr Ala
130 135 140
Lys Met Gly Trp Gly Val Arg Ala Leu Gln Thr Ile Pro Gln Gly Thr
145 150 155 160
Phe Ile Cys Glu Tyr Val Gly Glu Leu Ile Ser Asp Ala Glu Ala Asp
165 170 175
Val Arg Glu Asp Asp Ser Tyr Leu Phe Asp Leu Asp Asn Lys Asp Gly
180 185 190
Glu Val Tyr Cys Ile Asp Ala Arg Tyr Tyr Gly Asn Ile Ser Arg Phe
195 200 205
Ile Asn His Leu Cys Asp Pro Asn Ile Ile Pro Val Arg Val Phe Met
210 215 220
Leu His Gln Asp Leu Arg Phe Pro Arg Ile Ala Phe Phe Ser Ser Arg
225 230 235 240
Asp Ile Arg Thr Gly Glu Glu Leu Gly Phe Asp Tyr Gly Asp Arg Phe
245 250 255
Trp Asp Ile Lys Ser Lys Tyr Phe Thr Cys Gln Cys Gly Ser Glu Lys
260 265 270
Cys Lys His Ser Ala Glu Ala Ile Ala Leu Glu Gln Ser Arg Leu Ala
275 280 285
Arg Leu Asp Pro His Pro Glu Leu Leu Pro Asp Leu Ser Ser Leu Pro
290 295 300
Pro Ile Asn Thr Glu Phe
305 310
<210> 196
<211> 289
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 196
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Ser Ser Ser
1 5 10 15
Met Ala Asp Phe Arg Lys Phe Phe Ala Lys Ala Lys His Ile Val Ile
20 25 30
Ile Ser Gly Ala Gly Val Ser Ala Glu Ser Gly Val Pro Thr Phe Arg
35 40 45
Gly Ala Gly Gly Tyr Trp Arg Lys Trp Gln Ala Gln Asp Leu Ala Thr
50 55 60
Pro Leu Ala Phe Ala His Asn Pro Ser Arg Val Trp Glu Phe Tyr His
65 70 75 80
Tyr Arg Arg Glu Val Met Gly Ser Lys Glu Pro Asn Ala Gly His Arg
85 90 95
Ala Ile Ala Glu Cys Glu Thr Arg Leu Gly Lys Gln Gly Arg Arg Val
100 105 110
Val Val Ile Thr Gln Asn Ile Asp Glu Leu His Arg Lys Ala Gly Thr
115 120 125
Lys Asn Leu Leu Glu Ile His Gly Ser Leu Phe Lys Thr Arg Cys Thr
130 135 140
Ser Cys Gly Val Val Ala Glu Asn Tyr Lys Ser Pro Ile Cys Pro Ala
145 150 155 160
Leu Ser Gly Lys Gly Ala Pro Glu Pro Gly Thr Gln Asp Ala Ser Ile
165 170 175
Pro Val Glu Lys Leu Pro Arg Cys Glu Glu Ala Gly Cys Gly Gly Leu
180 185 190
Leu Arg Pro His Val Val Trp Phe Gly Glu Asn Leu Asp Pro Ala Ile
195 200 205
Leu Glu Glu Val Asp Arg Glu Leu Ala His Cys Asp Leu Cys Leu Val
210 215 220
Val Gly Thr Ser Ser Val Val Tyr Pro Ala Ala Met Phe Ala Pro Gln
225 230 235 240
Val Ala Ala Arg Gly Val Pro Val Ala Glu Phe Asn Thr Glu Thr Thr
245 250 255
Pro Ala Thr Asn Arg Phe Arg Phe His Phe Gln Gly Pro Cys Gly Thr
260 265 270
Thr Leu Pro Glu Ala Leu Ala Cys His Glu Asn Glu Thr Val Ser Glu
275 280 285
Phe
<210> 197
<211> 340
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 197
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Ser Arg
1 5 10 15
Val Val Lys Pro Lys Val Ala Ser Met Glu Glu Met Ala Ala Phe His
20 25 30
Thr Asp Ala Tyr Leu Gln His Leu His Lys Val Ser Glu Glu Gly Asp
35 40 45
Asn Asp Asp Pro Glu Thr Leu Glu Tyr Gly Leu Gly Tyr Asp Cys Pro
50 55 60
Ile Thr Glu Gly Ile Tyr Asp Tyr Ala Ala Ala Val Gly Gly Ala Thr
65 70 75 80
Leu Thr Ala Ala Glu Gln Leu Ile Glu Gly Lys Thr Arg Ile Ala Val
85 90 95
Asn Trp Pro Gly Gly Trp His His Ala Lys Lys Asp Glu Ala Ser Gly
100 105 110
Phe Cys Tyr Leu Asn Asp Ala Val Leu Gly Ile Leu Lys Leu Arg Glu
115 120 125
Lys Phe Asp Arg Val Leu Tyr Val Asp Met Asp Leu His His Gly Asp
130 135 140
Gly Val Glu Asp Ala Phe Ser Phe Thr Ser Lys Val Met Thr Val Ser
145 150 155 160
Leu His Lys Phe Ser Pro Gly Phe Phe Pro Gly Thr Gly Asp Val Ser
165 170 175
Asp Ile Gly Leu Gly Lys Gly Arg Tyr Tyr Ser Ile Asn Val Pro Leu
180 185 190
Gln Asp Gly Ile Gln Asp Asp Lys Tyr Tyr Gln Ile Cys Glu Gly Val
195 200 205
Leu Lys Glu Val Phe Thr Thr Phe Asn Pro Glu Ala Val Val Leu Gln
210 215 220
Leu Gly Ala Asp Thr Ile Ala Gly Asp Pro Met Cys Ser Phe Asn Met
225 230 235 240
Thr Pro Glu Gly Ile Gly Lys Cys Leu Lys Tyr Val Leu Gln Trp Gln
245 250 255
Leu Pro Thr Leu Ile Leu Gly Gly Gly Gly Tyr His Leu Pro Asn Thr
260 265 270
Ala Arg Cys Trp Thr Tyr Leu Thr Ala Leu Ile Val Gly Arg Thr Leu
275 280 285
Ser Ser Glu Ile Pro Asp His Glu Phe Phe Thr Glu Tyr Gly Pro Asp
290 295 300
Tyr Val Leu Glu Ile Thr Pro Ser Cys Arg Pro Asp Arg Asn Asp Thr
305 310 315 320
Gln Lys Val Gln Glu Ile Leu Gln Ser Ile Lys Gly Asn Leu Lys Arg
325 330 335
Val Val Glu Phe
340
<210> 198
<211> 134
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 198
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Lys Glu
1 5 10 15
Gly Glu Asn Asn Lys Pro Arg Glu Lys Ser Glu Gly Asn Lys Arg Lys
20 25 30
Ser Ser Phe Ser Asn Ser Ala Asp Asp Ile Lys Ser Lys Lys Lys Arg
35 40 45
Glu Gln Ser Asn Asp Ile Ala Arg Gly Phe Glu Arg Gly Leu Glu Pro
50 55 60
Glu Lys Ile Ile Gly Ala Thr Asp Ser Cys Gly Asp Leu Met Phe Leu
65 70 75 80
Met Lys Trp Lys Asp Thr Asp Glu Ala Asp Leu Val Leu Ala Lys Glu
85 90 95
Ala Asn Val Lys Cys Pro Gln Ile Val Ile Ala Phe Tyr Glu Glu Arg
100 105 110
Leu Thr Trp His Ala Tyr Pro Glu Asp Ala Glu Asn Lys Glu Lys Glu
115 120 125
Ser Ala Lys Ser Glu Phe
130
<210> 199
<211> 226
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 199
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Glu Phe
1 5 10 15
Trp Gly Ile Glu Val Lys Ser Gly Lys Pro Val Thr Val Thr Pro Glu
20 25 30
Glu Gly Ile Leu Ile His Val Ser Gln Ala Ser Leu Gly Glu Cys Lys
35 40 45
Asn Lys Lys Gly Glu Phe Val Pro Leu His Val Lys Val Gly Asn Gln
50 55 60
Asn Leu Val Leu Gly Thr Leu Ser Thr Glu Asn Ile Pro Gln Leu Phe
65 70 75 80
Cys Asp Leu Val Phe Asp Lys Glu Phe Glu Leu Ser His Thr Trp Gly
85 90 95
Lys Gly Ser Val Tyr Phe Val Gly Tyr Lys Thr Pro Asn Ile Glu Pro
100 105 110
Gln Gly Tyr Ser Glu Glu Glu Glu Glu Glu Glu Glu Glu Val Pro Ala
115 120 125
Gly Asn Ala Ala Lys Ala Val Ala Lys Pro Lys Ala Lys Pro Ala Glu
130 135 140
Val Lys Pro Ala Val Asp Asp Glu Glu Asp Glu Ser Asp Ser Asp Gly
145 150 155 160
Met Asp Glu Asp Asp Ser Asp Gly Glu Asp Ser Glu Glu Glu Glu Pro
165 170 175
Thr Pro Lys Lys Pro Ala Ser Ser Lys Lys Arg Ala Asn Glu Thr Thr
180 185 190
Pro Lys Ala Pro Val Ser Ala Lys Lys Ala Lys Val Ala Val Thr Pro
195 200 205
Gln Lys Thr Asp Glu Lys Lys Lys Gly Gly Lys Ala Ala Asn Gln Ser
210 215 220
Glu Phe
225
<210> 200
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 200
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Ser Arg
1 5 10 15
Arg Lys Gln Ala Lys Pro Gln His Phe Gln Ser Asp Pro Glu Val Ala
20 25 30
Ser Leu Pro Arg Arg Asp Gly Asp Thr Glu Lys Gly Gln Pro Ser Arg
35 40 45
Pro Thr Lys Ser Lys Asp Ala His Val Cys Gly Arg Cys Cys Ala Glu
50 55 60
Phe Phe Glu Leu Ser Asp Leu Leu Leu His Lys Lys Ser Cys Thr Lys
65 70 75 80
Asn Gln Leu Val Leu Ile Val Asn Glu Ser Pro Ala Ser Pro Ala Lys
85 90 95
Thr Phe Pro Pro Gly Pro Ser Leu Asn Asp Glu Phe
100 105
<210> 201
<211> 224
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 201
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Ser Cys Asp
1 5 10 15
Ser Thr Asn Ala Ala Ile Ala Lys Gln Ala Leu Lys Lys Pro Ile Lys
20 25 30
Gly Lys Gln Ala Pro Arg Lys Lys Ala Gln Gly Lys Thr Gln Gln Asn
35 40 45
Arg Lys Leu Thr Asp Phe Tyr Pro Val Arg Arg Ser Ser Arg Lys Ser
50 55 60
Lys Ala Glu Leu Gln Ser Glu Glu Arg Lys Arg Ile Asp Glu Leu Ile
65 70 75 80
Glu Ser Gly Lys Glu Glu Gly Met Lys Ile Asp Leu Ile Asp Gly Lys
85 90 95
Gly Arg Gly Val Ile Ala Thr Lys Gln Phe Ser Arg Gly Asp Phe Val
100 105 110
Val Glu Tyr His Gly Asp Leu Ile Glu Ile Thr Asp Ala Lys Lys Arg
115 120 125
Glu Ala Leu Tyr Ala Gln Asp Pro Ser Thr Gly Cys Tyr Met Tyr Tyr
130 135 140
Phe Gln Tyr Leu Ser Lys Thr Tyr Cys Val Asp Ala Thr Arg Glu Thr
145 150 155 160
Asn Arg Leu Gly Arg Leu Ile Asn His Ser Lys Cys Gly Asn Cys Gln
165 170 175
Thr Lys Leu His Asp Ile Asp Gly Val Pro His Leu Ile Leu Ile Ala
180 185 190
Ser Arg Asp Ile Ala Ala Gly Glu Glu Leu Leu Tyr Asp Tyr Gly Asp
195 200 205
Arg Ser Lys Ala Ser Ile Glu Ala Phe Pro Trp Leu Lys His Glu Phe
210 215 220
<210> 202
<211> 337
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 202
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Arg Arg Val
1 5 10 15
Ala Tyr Phe Tyr Asp Ala Asp Val Gly Asn Tyr Ala Tyr Gly Ala Gly
20 25 30
His Pro Met Lys Pro His Arg Ile Arg Met Ala His Ser Leu Ile Met
35 40 45
Asn Tyr Gly Leu Tyr Lys Lys Met Glu Ile Tyr Arg Ala Lys Pro Ala
50 55 60
Thr Lys Gln Glu Met Cys Gln Phe His Thr Asp Glu Tyr Ile Asp Phe
65 70 75 80
Leu Ser Arg Val Thr Pro Asp Asn Leu Glu Met Phe Lys Arg Glu Ser
85 90 95
Val Lys Phe Asn Val Gly Asp Asp Cys Pro Val Phe Asp Gly Leu Tyr
100 105 110
Glu Tyr Cys Ser Ile Ser Gly Gly Gly Ser Met Glu Gly Ala Ala Arg
115 120 125
Leu Asn Arg Gly Lys Cys Asp Val Ala Val Asn Tyr Ala Gly Gly Leu
130 135 140
His His Ala Lys Lys Ser Glu Ala Ser Gly Phe Cys Tyr Leu Asn Asp
145 150 155 160
Ile Val Leu Gly Ile Ile Glu Leu Leu Arg Tyr His Pro Arg Val Leu
165 170 175
Tyr Ile Asp Ile Asp Val His His Gly Asp Gly Val Glu Glu Ala Phe
180 185 190
Tyr Thr Thr Asp Arg Val Met Thr Cys Ser Phe His Lys Tyr Gly Glu
195 200 205
Phe Phe Pro Gly Thr Gly Glu Leu Arg Asp Ile Gly Val Gly Ala Gly
210 215 220
Lys Asn Tyr Ala Val Asn Val Pro Leu Arg Asp Gly Ile Asp Asp Ala
225 230 235 240
Thr Tyr Arg Ser Val Phe Glu Pro Val Ile Lys Lys Ile Met Glu Trp
245 250 255
Tyr Gln Pro Ser Ala Val Val Leu Gln Cys Gly Gly Asp Ser Leu Ser
260 265 270
Gly Asp Arg Leu Gly Cys Phe Asn Leu Ser Met Glu Gly His Ala Asn
275 280 285
Cys Val Asn Tyr Val Lys Ser Phe Gly Ile Pro Met Met Val Val Gly
290 295 300
Gly Gly Gly Tyr Thr Met Arg Asn Val Ala Arg Thr Trp Cys Phe Glu
305 310 315 320
Thr Gly Leu Leu Asn Asn Val Val Leu Asp Lys Asp Leu Pro Tyr Glu
325 330 335
Phe
<210> 203
<211> 257
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 203
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Glu Lys
1 5 10 15
Pro Arg Val Leu Val Leu Thr Gly Ala Gly Ile Ser Ala Glu Ser Gly
20 25 30
Ile Arg Thr Phe Arg Ala Ala Asp Gly Leu Trp Glu Glu His Arg Val
35 40 45
Glu Asp Val Ala Thr Pro Glu Gly Phe Asp Arg Asp Pro Glu Leu Val
50 55 60
Gln Ala Phe Tyr Asn Ala Arg Arg Arg Gln Leu Gln Gln Pro Glu Ile
65 70 75 80
Gln Pro Asn Ala Ala His Leu Ala Leu Ala Lys Leu Gln Asp Ala Leu
85 90 95
Gly Asp Arg Phe Leu Leu Val Thr Gln Asn Ile Asp Asn Leu His Glu
100 105 110
Arg Ala Gly Asn Thr Asn Val Ile His Met His Gly Glu Leu Leu Lys
115 120 125
Val Arg Cys Ser Gln Ser Gly Gln Val Leu Asp Trp Thr Gly Asp Val
130 135 140
Thr Pro Glu Asp Lys Cys His Cys Cys Gln Phe Pro Ala Pro Leu Arg
145 150 155 160
Pro His Val Val Trp Phe Gly Glu Met Pro Leu Gly Met Asp Glu Ile
165 170 175
Tyr Met Ala Leu Ser Met Ala Asp Ile Phe Ile Ala Ile Gly Thr Ser
180 185 190
Gly His Val Tyr Pro Ala Ala Gly Phe Val His Glu Ala Lys Leu His
195 200 205
Gly Ala His Thr Val Glu Leu Asn Leu Glu Pro Ser Gln Val Gly Asn
210 215 220
Glu Phe Ala Glu Lys Tyr Tyr Gly Pro Ala Ser Gln Val Val Pro Glu
225 230 235 240
Phe Val Glu Lys Leu Leu Lys Gly Leu Lys Ala Gly Ser Ile Ala Glu
245 250 255
Phe
<210> 204
<211> 349
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 204
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Asn Leu Lys
1 5 10 15
Cys Val Arg Ile Leu Lys Gln Phe His Lys Asp Leu Glu Arg Glu Leu
20 25 30
Leu Arg Arg His His Arg Ser Lys Thr Pro Arg His Leu Asp Pro Ser
35 40 45
Leu Ala Asn Tyr Leu Val Gln Lys Ala Lys Gln Arg Arg Ala Leu Arg
50 55 60
Arg Trp Glu Gln Glu Leu Asn Ala Lys Arg Ser His Leu Gly Arg Ile
65 70 75 80
Thr Val Glu Asn Glu Val Asp Leu Asp Gly Pro Pro Arg Ala Phe Val
85 90 95
Tyr Ile Asn Glu Tyr Arg Val Gly Glu Gly Ile Thr Leu Asn Gln Val
100 105 110
Ala Val Gly Cys Glu Cys Gln Asp Cys Leu Trp Ala Pro Thr Gly Gly
115 120 125
Cys Cys Pro Gly Ala Ser Leu His Lys Phe Ala Tyr Asn Asp Gln Gly
130 135 140
Gln Val Arg Leu Arg Ala Gly Leu Pro Ile Tyr Glu Cys Asn Ser Arg
145 150 155 160
Cys Arg Cys Gly Tyr Asp Cys Pro Asn Arg Val Val Gln Lys Gly Ile
165 170 175
Arg Tyr Asp Leu Cys Ile Phe Arg Thr Asp Asp Gly Arg Gly Trp Gly
180 185 190
Val Arg Thr Leu Glu Lys Ile Arg Lys Asn Ser Phe Val Met Glu Tyr
195 200 205
Val Gly Glu Ile Ile Thr Ser Glu Glu Ala Glu Arg Arg Gly Gln Ile
210 215 220
Tyr Asp Arg Gln Gly Ala Thr Tyr Leu Phe Asp Leu Asp Tyr Val Glu
225 230 235 240
Asp Val Tyr Thr Val Asp Ala Ala Tyr Tyr Gly Asn Ile Ser His Phe
245 250 255
Val Asn His Ser Cys Asp Pro Asn Leu Gln Val Tyr Asn Val Phe Ile
260 265 270
Asp Asn Leu Asp Glu Arg Leu Pro Arg Ile Ala Phe Phe Ala Thr Arg
275 280 285
Thr Ile Arg Ala Gly Glu Glu Leu Thr Phe Asp Tyr Asn Met Gln Val
290 295 300
Asp Pro Val Asp Met Glu Ser Thr Arg Met Asp Ser Asn Phe Gly Leu
305 310 315 320
Ala Gly Leu Pro Gly Ser Pro Lys Lys Arg Val Arg Ile Glu Cys Lys
325 330 335
Cys Gly Thr Glu Ser Cys Arg Lys Tyr Leu Phe Glu Phe
340 345
<210> 205
<211> 235
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 205
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Ser Asn Ser
1 5 10 15
Trp Glu Glu Gly Ser Ser Gly Ser Ser Ser Asp Glu Glu His Gly Gly
20 25 30
Gly Gly Met Arg Val Gly Pro Gln Tyr Gln Ala Val Val Pro Asp Phe
35 40 45
Asp Pro Ala Lys Leu Ala Arg Arg Ser Gln Glu Arg Asp Asn Leu Gly
50 55 60
Met Leu Val Trp Ser Pro Asn Gln Asn Leu Ser Glu Ala Lys Leu Asp
65 70 75 80
Glu Tyr Ile Ala Ile Ala Lys Glu Lys His Gly Tyr Asn Met Glu Gln
85 90 95
Ala Leu Gly Met Leu Phe Trp His Lys His Asn Ile Glu Lys Ser Leu
100 105 110
Ala Asp Leu Pro Asn Phe Thr Pro Phe Pro Asp Glu Trp Thr Val Glu
115 120 125
Asp Lys Val Leu Phe Glu Gln Ala Phe Ser Phe His Gly Lys Thr Phe
130 135 140
His Arg Ile Gln Gln Met Leu Pro Asp Lys Ser Ile Ala Ser Leu Val
145 150 155 160
Lys Phe Tyr Tyr Ser Trp Lys Lys Thr Arg Thr Lys Thr Ser Val Met
165 170 175
Asp Arg His Ala Arg Lys Gln Lys Arg Glu Arg Glu Glu Ser Glu Asp
180 185 190
Glu Leu Glu Glu Ala Asn Gly Asn Asn Pro Ile Asp Ile Glu Val Asp
195 200 205
Gln Asn Lys Glu Ser Lys Lys Glu Val Pro Pro Thr Glu Thr Val Pro
210 215 220
Gln Val Lys Lys Glu Lys His Ser Thr Glu Phe
225 230 235
<210> 206
<211> 343
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 206
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Tyr Lys Glu
1 5 10 15
Ser Val His Leu Glu Thr Tyr Pro Lys Glu Arg Ala Thr Glu Gly Ile
20 25 30
Ala Met Glu Ile Asp Tyr Ala Ser Cys Lys Arg Leu Gly Ser Ser Tyr
35 40 45
Arg Ala Leu Pro Lys Ser Tyr Gln Gln Pro Lys Cys Thr Gly Arg Thr
50 55 60
Pro Leu Cys Lys Glu Val Leu Asn Asp Thr Trp Val Ser Phe Pro Ser
65 70 75 80
Trp Ser Glu Asp Ser Thr Phe Val Ser Ser Lys Lys Thr Gln Tyr Glu
85 90 95
Glu His Ile Tyr Arg Cys Glu Asp Glu Arg Phe Glu Leu Asp Val Val
100 105 110
Leu Glu Thr Asn Leu Ala Thr Ile Arg Val Leu Glu Ala Ile Gln Lys
115 120 125
Lys Leu Ser Arg Leu Ser Ala Glu Glu Gln Ala Lys Phe Arg Leu Asp
130 135 140
Asn Thr Leu Gly Gly Thr Ser Glu Val Ile His Arg Lys Ala Leu Gln
145 150 155 160
Arg Ile Tyr Ala Asp Lys Ala Ala Asp Ile Ile Asp Gly Leu Arg Lys
165 170 175
Asn Pro Ser Ile Ala Val Pro Ile Val Leu Lys Arg Leu Lys Met Lys
180 185 190
Glu Glu Glu Trp Arg Glu Ala Gln Arg Gly Phe Asn Lys Val Trp Arg
195 200 205
Glu Gln Asn Glu Lys Tyr Tyr Leu Lys Ser Leu Asp His Gln Gly Ile
210 215 220
Asn Phe Lys Gln Asn Asp Thr Lys Val Leu Arg Ser Lys Ser Leu Leu
225 230 235 240
Asn Glu Ile Glu Ser Ile Tyr Asp Glu Arg Gln Glu Gln Ala Thr Glu
245 250 255
Glu Asn Ala Gly Val Pro Val Gly Pro His Leu Ser Leu Ala Tyr Glu
260 265 270
Asp Lys Gln Ile Leu Glu Asp Ala Ala Ala Leu Ile Ile His His Val
275 280 285
Lys Arg Gln Thr Gly Ile Gln Lys Glu Asp Lys Tyr Lys Ile Lys Gln
290 295 300
Ile Met His His Phe Ile Pro Asp Leu Leu Phe Ala Gln Arg Gly Asp
305 310 315 320
Leu Ser Asp Val Glu Glu Glu Glu Glu Glu Glu Met Asp Val Asp Glu
325 330 335
Ala Thr Gly Ala Val Glu Phe
340
<210> 207
<211> 328
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 207
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Gln Ser Ala
1 5 10 15
Tyr Leu His Val Ser Leu Ala Arg Ile Ser Asp Glu Asp Cys Cys Ala
20 25 30
Asn Cys Lys Gly Asn Cys Leu Ser Ala Asp Phe Pro Cys Thr Cys Ala
35 40 45
Arg Glu Thr Ser Gly Glu Tyr Ala Tyr Thr Lys Glu Gly Leu Leu Lys
50 55 60
Glu Lys Phe Leu Asp Thr Cys Leu Lys Met Lys Lys Glu Pro Asp Ser
65 70 75 80
Phe Pro Lys Val Tyr Cys Lys Asp Cys Pro Leu Glu Arg Asp His Asp
85 90 95
Lys Gly Thr Tyr Gly Lys Cys Asp Gly His Leu Ile Arg Lys Phe Ile
100 105 110
Lys Glu Cys Trp Arg Lys Cys Gly Cys Asp Met Gln Cys Gly Asn Arg
115 120 125
Val Val Gln Arg Gly Ile Arg Cys Gln Leu Gln Val Tyr Phe Thr Gln
130 135 140
Glu Gly Lys Gly Trp Gly Leu Arg Thr Leu Gln Asp Leu Pro Lys Gly
145 150 155 160
Thr Phe Ile Cys Glu Tyr Ile Gly Glu Ile Leu Thr Asn Thr Glu Leu
165 170 175
Tyr Asp Arg Asn Val Arg Ser Ser Ser Glu Arg His Thr Tyr Pro Val
180 185 190
Thr Leu Asp Ala Asp Trp Gly Ser Glu Lys Asp Leu Lys Asp Glu Glu
195 200 205
Ala Leu Cys Leu Asp Ala Thr Ile Cys Gly Asn Val Ala Arg Phe Ile
210 215 220
Asn His Arg Cys Glu Asp Ala Asn Met Ile Asp Ile Pro Ile Glu Ile
225 230 235 240
Glu Thr Pro Asp Arg His Tyr Tyr His Ile Ala Phe Phe Thr Leu Arg
245 250 255
Asp Val Lys Ala Met Asp Glu Leu Thr Trp Asp Tyr Met Ile Asp Phe
260 265 270
Asn Asp Lys Ser His Pro Val Lys Ala Phe Arg Cys Cys Cys Gly Ser
275 280 285
Glu Ser Cys Arg Asp Arg Lys Ile Lys Gly Ser Gln Gly Lys Ser Ile
290 295 300
Glu Arg Arg Lys Ile Val Ser Ala Lys Lys Gln Gln Gly Ser Lys Glu
305 310 315 320
Val Ser Lys Lys Arg Lys Glu Phe
325
<210> 208
<211> 301
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 208
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Val Gln Val
1 5 10 15
Lys Arg Val Leu Glu Lys Ser Pro Gly Lys Leu Leu Val Lys Met Pro
20 25 30
Phe Gln Ala Ser Pro Gly Gly Lys Gly Glu Gly Gly Gly Ala Thr Thr
35 40 45
Ser Ala Gln Val Met Val Ile Lys Arg Pro Gly Arg Lys Arg Lys Ala
50 55 60
Glu Ala Asp Pro Gln Ala Ile Pro Lys Lys Arg Gly Arg Lys Pro Gly
65 70 75 80
Ser Val Val Ala Ala Ala Ala Ala Glu Ala Lys Lys Lys Ala Val Lys
85 90 95
Glu Ser Ser Ile Arg Ser Val Gln Glu Thr Val Leu Pro Ile Lys Lys
100 105 110
Arg Lys Thr Arg Glu Thr Val Ser Ile Glu Val Lys Glu Val Val Lys
115 120 125
Pro Leu Leu Val Ser Thr Leu Gly Glu Lys Ser Gly Lys Gly Leu Lys
130 135 140
Thr Cys Lys Ser Pro Gly Arg Lys Ser Lys Glu Ser Ser Pro Lys Gly
145 150 155 160
Arg Ser Ser Ser Ala Ser Ser Pro Pro Lys Lys Glu His His His His
165 170 175
His His His Ala Glu Ser Pro Lys Ala Pro Met Pro Leu Leu Pro Pro
180 185 190
Pro Pro Pro Pro Glu Pro Gln Ser Ser Glu Asp Pro Ile Ser Pro Pro
195 200 205
Glu Pro Gln Asp Leu Ser Ser Ser Ile Cys Lys Glu Glu Lys Met Pro
210 215 220
Arg Ala Gly Ser Leu Glu Ser Asp Gly Cys Pro Lys Glu Pro Ala Lys
225 230 235 240
Thr Gln Pro Met Val Ala Ala Ala Ala Thr Thr Thr Thr Thr Thr Thr
245 250 255
Thr Thr Val Ala Glu Lys Tyr Lys His Arg Gly Glu Gly Glu Arg Lys
260 265 270
Asp Ile Val Ser Ser Ser Met Pro Arg Pro Asn Arg Glu Glu Pro Val
275 280 285
Asp Ser Arg Thr Pro Val Thr Glu Arg Val Ser Glu Phe
290 295 300
<210> 209
<211> 304
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 209
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Ala Pro
1 5 10 15
Lys Arg Gln Ser Ala Ile Leu Pro Gln Pro Lys Lys Pro Arg Pro Ala
20 25 30
Ala Ala Pro Lys Leu Glu Asp Lys Ser Ala Ser Pro Gly Leu Pro Lys
35 40 45
Gly Glu Lys Glu Gln Gln Glu Ala Ile Glu His Ile Asp Glu Val Gln
50 55 60
Asn Glu Ile Asp Arg Leu Asn Glu Gln Ala Ser Glu Glu Ile Leu Lys
65 70 75 80
Val Glu Gln Lys Tyr Asn Lys Leu Arg Gln Pro Phe Phe Gln Lys Arg
85 90 95
Ser Glu Leu Ile Ala Lys Ile Pro Asn Phe Trp Val Thr Thr Phe Val
100 105 110
Asn His Pro Gln Val Ser Ala Leu Leu Gly Glu Glu Asp Glu Glu Ala
115 120 125
Leu His Tyr Leu Thr Arg Val Glu Val Thr Glu Phe Glu Asp Ile Lys
130 135 140
Ser Gly Tyr Arg Ile Asp Phe Tyr Phe Asp Glu Asn Pro Tyr Phe Glu
145 150 155 160
Asn Lys Val Leu Ser Lys Glu Phe His Leu Asn Glu Ser Gly Asp Pro
165 170 175
Ser Ser Lys Ser Thr Glu Ile Lys Trp Lys Ser Gly Lys Asp Leu Thr
180 185 190
Lys Arg Ser Ser Gln Thr Gln Asn Lys Ala Ser Arg Lys Arg Gln His
195 200 205
Glu Glu Pro Glu Ser Phe Phe Thr Trp Phe Thr Asp His Ser Asp Ala
210 215 220
Gly Ala Asp Glu Leu Gly Glu Val Ile Lys Asp Asp Ile Trp Pro Asn
225 230 235 240
Pro Leu Gln Tyr Tyr Leu Val Pro Asp Met Asp Asp Glu Glu Gly Glu
245 250 255
Ala Glu Asp Asp Asp Asp Asp Asp Glu Glu Glu Glu Gly Leu Glu Asp
260 265 270
Ile Asp Glu Glu Gly Asp Glu Asp Glu Gly Glu Glu Asp Asp Asp Glu
275 280 285
Asp Glu Gly Glu Glu Gly Glu Glu Asp Glu Gly Glu Asp Asp Glu Phe
290 295 300
<210> 210
<211> 303
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 210
Ala Ser Ser Pro Lys Lys Lys Arg Lys Val Glu Ala Ser Met Gln Leu
1 5 10 15
His Glu Gln Ile Ala Asn Ile Ser Val Thr Phe Asn Asp Ile Pro Arg
20 25 30
Ser Asp His Ser Met Thr Pro Thr Glu Leu Cys Tyr Phe Asp Asp Phe
35 40 45
Ala Thr Thr Leu Val Val Asp Ser Val Leu Asn Phe Thr Thr His Lys
50 55 60
Met Ser Lys Lys Arg Arg Tyr Leu Tyr Gln Asp Glu Tyr Arg Thr Ala
65 70 75 80
Arg Thr Val Met Lys Thr Phe Arg Glu Gln Arg Asp Trp Thr Asn Ala
85 90 95
Ile Tyr Gly Leu Leu Thr Leu Arg Ser Val Ser His Phe Leu Ser Lys
100 105 110
Leu Pro Pro Asn Lys Leu Phe Glu Phe Arg Asp His Ile Val Arg Phe
115 120 125
Leu Asn Met Phe Ile Leu Asp Ser Gly Tyr Thr Ile Gln Glu Cys Lys
130 135 140
Arg Tyr Ser Gln Glu Gly His Gln Gly Ala Lys Leu Val Ser Thr Gly
145 150 155 160
Val Trp Ser Arg Gly Asp Lys Ile Glu Arg Leu Ser Gly Val Val Cys
165 170 175
Leu Leu Ser Ser Glu Asp Glu Asp Ser Ile Leu Ala Gln Glu Gly Ser
180 185 190
Asp Phe Ser Val Met Tyr Ser Thr Arg Lys Arg Cys Ser Thr Leu Trp
195 200 205
Leu Gly Pro Gly Ala Tyr Ile Asn His Asp Cys Arg Pro Thr Cys Glu
210 215 220
Phe Val Ser His Gly Ser Thr Ala His Ile Arg Val Leu Arg Asp Met
225 230 235 240
Val Pro Gly Asp Glu Ile Thr Cys Phe Tyr Gly Ser Glu Phe Phe Gly
245 250 255
Pro Asn Asn Ile Asp Cys Glu Cys Cys Thr Cys Glu Lys Asn Met Asn
260 265 270
Gly Ala Phe Ser Tyr Leu Arg Gly Asn Glu Asn Ala Glu Pro Ile Ile
275 280 285
Ser Glu Lys Lys Thr Lys Tyr Glu Leu Arg Ser Arg Ser Glu Phe
290 295 300
<210> 211
<211> 14164
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 211
caactttgta tagaaaagtt ggctccgaat tcgcccttca ggtccgaggt tctagacgag 60
tttactccct atcagtgata gagaacgatg tcgagtttac tccctatcag tgatagagaa 120
cgtatgtcga gtttactccc tatcagtgat agagaacgta tgtcgagttt actccctatc 180
agtgatagag aacgtatgtc gagtttatcc ctatcagtga tagagaacgt atgtcgagtt 240
tactccctat cagtgataga gaacgtatgt cgaggtaggc gtgtacggtg ggaggcctat 300
ataagcagag ctcgtttagt gaaccgtcag atcgcaaagg gcgaattcga cccaagtttg 360
tacagccacc atggactata aggaccacga cggagactac aaggatcatg atattgatta 420
caaagacgat gacgataaga tggccccaaa gaagaagcgg aaggtcggta tccacggagt 480
cccagcagcc gacaagaagt acagcatcgg cctggacatc ggcaccaact ctgtgggctg 540
ggccgtgatc accgacgagt acaaggtgcc cagcaagaaa ttcaaggtgc tgggcaacac 600
cgaccggcac agcatcaaga agaacctgat cggagccctg ctgttcgaca gcggcgaaac 660
agccgaggcc acccggctga agagaaccgc cagaagaaga tacaccagac ggaagaaccg 720
gatctgctat ctgcaagaga tcttcagcaa cgagatggcc aaggtggacg acagcttctt 780
ccacagactg gaagagtcct tcctggtgga agaggataag aagcacgagc ggcaccccat 840
cttcggcaac atcgtggacg aggtggccta ccacgagaag taccccacca tctaccacct 900
gagaaagaaa ctggtggaca gcaccgacaa ggccgacctg cggctgatct atctggccct 960
ggcccacatg atcaagttcc ggggccactt cctgatcgag ggcgacctga accccgacaa 1020
cagcgacgtg gacaagctgt tcatccagct ggtgcagacc tacaaccagc tgttcgagga 1080
aaaccccatc aacgccagcg gcgtggacgc caaggccatc ctgtctgcca gactgagcaa 1140
gagcagacgg ctggaaaatc tgatcgccca gctgcccggc gagaagaaga atggcctgtt 1200
cggcaacctg attgccctga gcctgggcct gacccccaac ttcaagagca acttcgacct 1260
ggccgaggat gccaaactgc agctgagcaa ggacacctac gacgacgacc tggacaacct 1320
gctggcccag atcggcgacc agtacgccga cctgtttctg gccgccaaga acctgtccga 1380
cgccatcctg ctgagcgaca tcctgagagt gaacaccgag atcaccaagg cccccctgag 1440
cgcctctatg atcaagagat acgacgagca ccaccaggac ctgaccctgc tgaaagctct 1500
cgtgcggcag cagctgcctg agaagtacaa agagattttc ttcgaccaga gcaagaacgg 1560
ctacgccggc tacattgacg gcggagccag ccaggaagag ttctacaagt tcatcaagcc 1620
catcctggaa aagatggacg gcaccgagga actgctcgtg aagctgaaca gagaggacct 1680
gctgcggaag cagcggacct tcgacaacgg cagcatcccc caccagatcc acctgggaga 1740
gctgcacgcc attctgcggc ggcaggaaga tttttaccca ttcctgaagg acaaccggga 1800
aaagatcgag aagatcctga ccttccgcat cccctactac gtgggccctc tggccagggg 1860
aaacagcaga ttcgcctgga tgaccagaaa gagcgaggaa accatcaccc cctggaactt 1920
cgaggaagtg gtggacaagg gcgcttccgc ccagagcttc atcgagcgga tgaccaactt 1980
cgataagaac ctgcccaacg agaaggtgct gcccaagcac agcctgctgt acgagtactt 2040
caccgtgtat aacgagctga ccaaagtgaa atacgtgacc gagggaatga gaaagcccgc 2100
cttcctgagc ggcgagcaga aaaaggccat cgtggacctg ctgttcaaga ccaaccggaa 2160
agtgaccgtg aagcagctga aagaggacta cttcaagaaa atcgagtgct tcgactccgt 2220
ggaaatctcc ggcgtggaag atcggttcaa cgcctccctg ggcacatacc acgatctgct 2280
gaaaattatc aaggacaagg acttcctgga caatgaggaa aacgaggaca ttctggaaga 2340
tatcgtgctg accctgacac tgtttgagga cagagagatg atcgaggaac ggctgaaaac 2400
ctatgcccac ctgttcgacg acaaagtgat gaagcagctg aagcggcgga gatacaccgg 2460
ctggggcagg ctgagccgga agctgatcaa cggcatccgg gacaagcagt ccggcaagac 2520
aatcctggat ttcctgaagt ccgacggctt cgccaacaga aacttcatgc agctgatcca 2580
cgacgacagc ctgaccttta aagaggacat ccagaaagcc caggtgtccg gccagggcga 2640
tagcctgcac gagcacattg ccaatctggc cggcagcccc gccattaaga agggcatcct 2700
gcagacagtg aaggtggtgg acgagctcgt gaaagtgatg ggccggcaca agcccgagaa 2760
catcgtgatc gaaatggcca gagagaacca gaccacccag aagggacaga agaacagccg 2820
cgagagaatg aagcggatcg aagagggcat caaagagctg ggcagccaga tcctgaaaga 2880
acaccccgtg gaaaacaccc agctgcagaa cgagaagctg tacctgtact acctgcagaa 2940
tgggcgggat atgtacgtgg accaggaact ggacatcaac cggctgtccg actacgatgt 3000
ggaccatatc gtgcctcaga gctttctgaa ggacgactcc atcgacaaca aggtgctgac 3060
cagaagcgac aagaaccggg gcaagagcga caacgtgccc tccgaagagg tcgtgaagaa 3120
gatgaagaac tactggcggc agctgctgaa cgccaagctg attacccaga gaaagttcga 3180
caatctgacc aaggccgaga gaggcggcct gagcgaactg gataaggccg gcttcatcaa 3240
gagacagctg gtggaaaccc ggcagatcac aaagcacgtg gcacagatcc tggactcccg 3300
gatgaacact aagtacgacg agaatgacaa gctgatccgg gaagtgaaag tgatcaccct 3360
gaagtccaag ctggtgtccg atttccggaa ggatttccag ttttacaaag tgcgcgagat 3420
caacaactac caccacgccc acgacgccta cctgaacgcc gtcgtgggaa ccgccctgat 3480
caaaaagtac cctaagctgg aaagcgagtt cgtgtacggc gactacaagg tgtacgacgt 3540
gcggaagatg atcgccaaga gcgagcagga aatcggcaag gctaccgcca agtacttctt 3600
ctacagcaac atcatgaact ttttcaagac cgagattacc ctggccaacg gcgagatccg 3660
gaagcggcct ctgatcgaga caaacggcga aaccggggag atcgtgtggg ataagggccg 3720
ggattttgcc accgtgcgga aagtgctgag catgccccaa gtgaatatcg tgaaaaagac 3780
cgaggtgcag acaggcggct tcagcaaaga gtctatcctg cccaagagga acagcgataa 3840
gctgatcgcc agaaagaagg actgggaccc taagaagtac ggcggcttcg acagccccac 3900
cgtggcctat tctgtgctgg tggtggccaa agtggaaaag ggcaagtcca agaaactgaa 3960
gagtgtgaaa gagctgctgg ggatcaccat catggaaaga agcagcttcg agaagaatcc 4020
catcgacttt ctggaagcca agggctacaa agaagtgaaa aaggacctga tcatcaagct 4080
gcctaagtac tccctgttcg agctggaaaa cggccggaag agaatgctgg cctctgccgg 4140
cgaactgcag aagggaaacg aactggccct gccctccaaa tatgtgaact tcctgtacct 4200
ggccagccac tatgagaagc tgaagggctc ccccgaggat aatgagcaga aacagctgtt 4260
tgtggaacag cacaagcact acctggacga gatcatcgag cagatcagcg agttctccaa 4320
gagagtgatc ctggccgacg ctaatctgga caaagtgctg tccgcctaca acaagcaccg 4380
ggataagccc atcagagagc aggccgagaa tatcatccac ctgtttaccc tgaccaatct 4440
gggagcccct gccgccttca agtactttga caccaccatc gaccggaaga ggtacaccag 4500
caccaaagag gtgctggacg ccaccctgat ccaccagagc atcaccggcc tgtacgagac 4560
acggatcgac ctgtctcagc tgggaggcga caaaaggccg gcggccacga aaaaggccgg 4620
ccaggcaaaa aagaaaaagg aattctctag aggcagtgga gagggcagag gaagtctgct 4680
aacatgcggt gacgtcgagg agaatcctgg cccagtgagc aagggcgagg agctgttcac 4740
cggggtggtg cccatcctgg tcgagctgga cggcgacgta aacggccaca agttcagcgt 4800
gtccggcgag ggcgagggcg atgccaccta cggcaagctg accctgaagt tcatctgcac 4860
caccggcaag ctgcccgtgc cctggcccac cctcgtgacc accctgacct acggcgtgca 4920
gtgcttcagc cgctaccccg accacatgaa gcagcacgac ttcttcaagt ccgccatgcc 4980
cgaaggctac gtccaggagc gcaccatctt cttcaaggac gacggcaact acaagacccg 5040
cgccgaggtg aagttcgagg gcgacaccct ggtgaaccgc atcgagctga agggcatcga 5100
cttcaaggag gacggcaaca tcctggggca caagctggag tacaactaca acagccacaa 5160
cgtctatatc atggccgaca agcagaagaa cggcatcaag gtgaacttca agatccgcca 5220
caacatcgag gacggcagcg tgcagctcgc cgaccactac cagcagaaca cccccatcgg 5280
cgacggcccc gtgctgctgc ccgacaacca ctacctgagc acccagtccg ccctgagcaa 5340
agaccccaac gagaagcgcg atcacatggt cctgctggag ttcgtgaccg ccgccgggat 5400
cactctcggc atggacgagc tgtacaagct cgagggaagc ggagctacta acttcagcct 5460
gctgaagcag gctggcgacg tggaggagaa ccctggacct atgtctaggc tggacaagag 5520
caaagtcata aacggagctc tggaattact caatggtgtc ggtatcgaag gcctgacgac 5580
aaggaaactc gctcaaaagc tgggagttga gcagcctacc ctgtactggc acgtgaagaa 5640
caagcgggcc ctgctcgatg ccctgccaat cgagatgctg gacaggcatc atacccactt 5700
ctgccccctg gaaggcgagt catggcaaga ctttctgcgg aacaacgcca agtcataccg 5760
ctgtgctctc ctctcacatc gcgacggggc taaagtgcat ctcggcaccc gcccaacaga 5820
gaaacagtac gaaaccctgg aaaatcagct cgcgttcctg tgtcagcaag gcttctccct 5880
ggagaacgca ctgtacgctc tgtccgccgt gggccacttt acactgggct gcgtattgga 5940
ggaacaggag catcaagtag caaaagagga aagagagaca cctaccaccg attctatgcc 6000
cccacttctg agacaagcaa ttgagctgtt cgaccggcag ggagccgaac ctgccttcct 6060
tttcggcctg gaactaatca tatgtggcct ggagaaacag ctaaagtgcg aaagcggcgg 6120
gccgaccgac gcccttgacg attttgactt agacatgctc ccagccgatg cccttgacga 6180
ttttgacctt gacatgctcc ccgggtaatg tacaaagtgg tgaattccgg caattcgata 6240
tcaagcttat cgataatcaa cctctggatt acaaaatttg tgaaagattg actggtattc 6300
ttaactatgt tgctcctttt acgctatgtg gatacgctgc tttaatgcct ttgtatcatg 6360
ctattgcttc ccgtatggct ttcattttct cctccttgta taaatcctgg ttgctgtctc 6420
tttatgagga gttgtggccc gttgtcaggc aacgtggcgt ggtgtgcact gtgtttgctg 6480
acgcaacccc cactggttgg ggcattgcca ccacctgtca gctcctttcc gggactttcg 6540
ctttccccct ccctattgcc acggcggaac tcatcgccgc ctgccttgcc cgctgctgga 6600
caggggctcg gctgttgggc actgacaatt ccgtggtgtt gtcggggaaa tcatcgtcct 6660
ttccttggct gctcgcctgt gttgccacct ggattctgcg cgggacgtcc ttctgctacg 6720
tcccttcggc cctcaatcca gcggaccttc cttcccgcgg cctgctgccg gctctgcggc 6780
ctcttccgcg tcttcgcctt cgccctcaga cgagtcggat ctccctttgg gccgcctccc 6840
cgcatcgata ccgtcgacct cgagacctag aaaaacatgg agcaatcaca agtagcaata 6900
cagcagctac caatgctgat tgtgcctggc tagaagcaca agaggaggag gaggtgggtt 6960
ttccagtcac acctcaggta cctttaagac caatgactta caaggcagct gtagatctta 7020
gccacttttt aaaagaaaag gggggactgg aagggctaat tcactcccaa cgaagacaag 7080
atatccttga tctgtggatc taccacacac aaggctactt ccctgattgg cagaactaca 7140
caccagggcc agggatcaga tatccactga cctttggatg gtgctacaag ctagtaccag 7200
ttgagcaaga gaaggtagaa gaagccaatg aaggagagaa cacccgcttg ttacaccctg 7260
tgagcctgca tgggatggat gacccggaga gagaagtatt agagtggagg tttgacagcc 7320
gcctagcatt tcatcacatg gcccgagagc tgcatccgga ctgtactggg tctctctggt 7380
tagaccagat ctgagcctgg gagctctctg gctaactagg gaacccactg cttaagcctc 7440
aataaagctt gccttgagtg cttcaagtag tgtgtgcccg tctgttgtgt gactctggta 7500
actagagatc cctcagaccc ttttagtcag tgtggaaaat ctctagcagg gcccgtttaa 7560
acccgctgat cagcctcgac tgtgccttct agttgccagc catctgttgt ttgcccctcc 7620
cccgtgcctt ccttgaccct ggaaggtgcc actcccactg tcctttccta ataaaatgag 7680
gaaattgcat cgcattgtct gagtaggtgt cattctattc tggggggtgg ggtggggcag 7740
gacagcaagg gggaggattg ggaagacaat agcaggcatg ctggggatgc ggtgggctct 7800
atggcttctg aggcggaaag aaccagctgg ggctctaggg ggtatcccca cgcgccctgt 7860
agcggcgcat taagcgcggc gggtgtggtg gttacgcgca gcgtgaccgc tacacttgcc 7920
agcgccctag cgcccgctcc tttcgctttc ttcccttcct ttctcgccac gttcgccggc 7980
tttccccgtc aagctctaaa tcgggggctc cctttagggt tccgatttag tgctttacgg 8040
cacctcgacc ccaaaaaact tgattagggt gatggttcac gtagtgggcc atcgccctga 8100
tagacggttt ttcgcccttt gacgttggag tccacgttct ttaatagtgg actcttgttc 8160
caaactggaa caacactcaa ccctatctcg gtctattctt ttgatttata agggattttg 8220
ccgatttcgg cctattggtt aaaaaatgag ctgatttaac aaaaatttaa cgcgaattaa 8280
ttctgtggaa tgtgtgtcag ttagggtgtg gaaagtcccc aggctcccca gcaggcagaa 8340
gtatgcaaag catgcatctc aattagtcag caaccaggtg tggaaagtcc ccaggctccc 8400
cagcaggcag aagtatgcaa agcatgcatc tcaattagtc agcaaccata gtcccgcccc 8460
taactccgcc catcccgccc ctaactccgc ccagttccgc ccattctccg ccccatggct 8520
gactaatttt ttttatttat gcagaggccg aggccgcctc tgcctctgag ctattccaga 8580
agtagtgagg aggctttttt ggaggcctag gcttttgcaa aaagctcccg ggagcttgta 8640
tatccatttt cggatctgat cagcacgtgt tgacaattaa tcatcggcat agtatatcgg 8700
catagtataa tacgacaagg tgaggaacta aaccatggcc aagttgacca gtgccgttcc 8760
ggtgctcacc gcgcgcgacg tcgccggagc ggtcgagttc tggaccgacc ggctcgggtt 8820
ctcccgggac ttcgtggagg acgacttcgc cggtgtggtc cgggacgacg tgaccctgtt 8880
catcagcgcg gtccaggacc aggtggtgcc ggacaacacc ctggcctggg tgtgggtgcg 8940
cggcctggac gagctgtacg ccgagtggtc ggaggtcgtg tccacgaact tccgggacgc 9000
ctccgggccg gccatgaccg agatcggcga gcagccgtgg gggcgggagt tcgccctgcg 9060
cgacccggcc ggcaactgcg tgcacttcgt ggccgaggag caggactgac acgtgctacg 9120
agatttcgat tccaccgccg ccttctatga aaggttgggc ttcggaatcg ttttccggga 9180
cgccggctgg atgatcctcc agcgcgggga tctcatgctg gagttcttcg cccaccccaa 9240
cttgtttatt gcagcttata atggttacaa ataaagcaat agcatcacaa atttcacaaa 9300
taaagcattt ttttcactgc attctagttg tggtttgtcc aaactcatca atgtatctta 9360
tcatgtctgt ataccgtcga cctctagcta gagcttggcg taatcatggt catagctgtt 9420
tcctgtgtga aattgttatc cgctcacaat tccacacaac atacgagccg gaagcataaa 9480
gtgtaaagcc tggggtgcct aatgagtgag ctaactcaca ttaattgcgt tgcgctcact 9540
gcccgctttc cagtcgggaa acctgtcgtg ccagctgcat taatgaatcg gccaacgcgc 9600
ggggagaggc ggtttgcgta ttgggcgctc ttccgcttcc tcgctcactg actcgctgcg 9660
ctcggtcgtt cggctgcggc gagcggtatc agctcactca aaggcggtaa tacggttatc 9720
cacagaatca ggggataacg caggaaagaa catgtgagca aaaggccagc aaaaggccag 9780
gaaccgtaaa aaggccgcgt tgctggcgtt tttccatagg ctccgccccc ctgacgagca 9840
tcacaaaaat cgacgctcaa gtcagaggtg gcgaaacccg acaggactat aaagatacca 9900
ggcgtttccc cctggaagct ccctcgtgcg ctctcctgtt ccgaccctgc cgcttaccgg 9960
atacctgtcc gcctttctcc cttcgggaag cgtggcgctt tctcatagct cacgctgtag 10020
gtatctcagt tcggtgtagg tcgttcgctc caagctgggc tgtgtgcacg aaccccccgt 10080
tcagcccgac cgctgcgcct tatccggtaa ctatcgtctt gagtccaacc cggtaagaca 10140
cgacttatcg ccactggcag cagccactgg taacaggatt agcagagcga ggtatgtagg 10200
cggtgctaca gagttcttga agtggtggcc taactacggc tacactagaa gaacagtatt 10260
tggtatctgc gctctgctga agccagttac cttcggaaaa agagttggta gctcttgatc 10320
cggcaaacaa accaccgctg gtagcggtgg tttttttgtt tgcaagcagc agattacgcg 10380
cagaaaaaaa ggatctcaag aagatccttt gatcttttct acggggtctg acgctcagtg 10440
gaacgaaaac tcacgttaag ggattttggt catgagatta tcaaaaagga tcttcaccta 10500
gatcctttta aattaaaaat gaagttttaa atcaatctaa agtatatatg agtaaacttg 10560
gtctgacagt taccaatgct taatcagtga ggcacctatc tcagcgatct gtctatttcg 10620
ttcatccata gttgcctgac tccccgtcgt gtagataact acgatacggg agggcttacc 10680
atctggcccc agtgctgcaa tgataccgcg agacccacgc tcaccggctc cagatttatc 10740
agcaataaac cagccagccg gaagggccga gcgcagaagt ggtcctgcaa ctttatccgc 10800
ctccatccag tctattaatt gttgccggga agctagagta agtagttcgc cagttaatag 10860
tttgcgcaac gttgttgcca ttgctacagg catcgtggtg tcacgctcgt cgtttggtat 10920
ggcttcattc agctccggtt cccaacgatc aaggcgagtt acatgatccc ccatgttgtg 10980
caaaaaagcg gttagctcct tcggtcctcc gatcgttgtc agaagtaagt tggccgcagt 11040
gttatcactc atggttatgg cagcactgca taattctctt actgtcatgc catccgtaag 11100
atgcttttct gtgactggtg agtactcaac caagtcattc tgagaatagt gtatgcggcg 11160
accgagttgc tcttgcccgg cgtcaatacg ggataatacc gcgccacata gcagaacttt 11220
aaaagtgctc atcattggaa aacgttcttc ggggcgaaaa ctctcaagga tcttaccgct 11280
gttgagatcc agttcgatgt aacccactcg tgcacccaac tgatcttcag catcttttac 11340
tttcaccagc gtttctgggt gagcaaaaac aggaaggcaa aatgccgcaa aaaagggaat 11400
aagggcgaca cggaaatgtt gaatactcat actcttcctt tttcaatatt attgaagcat 11460
ttatcagggt tattgtctca tgagcggata catatttgaa tgtatttaga aaaataaaca 11520
aataggggtt ccgcgcacat ttccccgaaa agtgccacct gacgtcgacg gatcgggaga 11580
tctcccgatc ccctatggtg cactctcagt acaatctgct ctgatgccgc atagttaagc 11640
cagtatctgc tccctgcttg tgtgttggag gtcgctgagt agtgcgcgag caaaatttaa 11700
gctacaacaa ggcaaggctt gaccgacaat tgcatgaaga atctgcttag ggttaggcgt 11760
tttgcgctgc ttcgcgatgt acgggccaga tatacgcgtt gacattgatt attgactagt 11820
tattaatagt aatcaattac ggggtcatta gttcatagcc catatatgga gttccgcgtt 11880
acataactta cggtaaatgg cccgcctggc tgaccgccca acgacccccg cccattgacg 11940
tcaataatga cgtatgttcc catagtaacg ccaataggga ctttccattg acgtcaatgg 12000
gtggagtatt tacggtaaac tgcccacttg gcagtacatc aagtgtatca tatgccaagt 12060
acgcccccta ttgacgtcaa tgacggtaaa tggcccgcct ggcattatgc ccagtacatg 12120
accttatggg actttcctac ttggcagtac atctacgtat tagtcatcgc tattaccatg 12180
gtgatgcggt tttggcagta catcaatggg cgtggatagc ggtttgactc acggggattt 12240
ccaagtctcc accccattga cgtcaatggg agtttgtttt ggcaccaaaa tcaacgggac 12300
tttccaaaat gtcgtaacaa ctccgcccca ttgacgcaaa tgggcggtag gcgtgtacgg 12360
tgggaggtct atataagcag cgcgttttgc ctgtactggg tctctctggt tagaccagat 12420
ctgagcctgg gagctctctg gctaactagg gaacccactg cttaagcctc aataaagctt 12480
gccttgagtg cttcaagtag tgtgtgcccg tctgttgtgt gactctggta actagagatc 12540
cctcagaccc ttttagtcag tgtggaaaat ctctagcagt ggcgcccgaa cagggacttg 12600
aaagcgaaag ggaaaccaga ggagctctct cgacgcagga ctcggcttgc tgaagcgcgc 12660
acggcaagag gcgaggggcg gcgactggtg agtacgccaa aaattttgac tagcggaggc 12720
tagaaggaga gagatgggtg cgagagcgtc agtattaagc gggggagaat tagatcgcga 12780
tgggaaaaaa ttcggttaag gccaggggga aagaaaaaat ataaattaaa acatatagta 12840
tgggcaagca gggagctaga acgattcgca gttaatcctg gcctgttaga aacatcagaa 12900
ggctgtagac aaatactggg acagctacaa ccatcccttc agacaggatc agaagaactt 12960
agatcattat ataatacagt agcaaccctc tattgtgtgc atcaaaggat agagataaaa 13020
gacaccaagg aagctttaga caagatagag gaagagcaaa acaaaagtaa gaccaccgca 13080
cagcaagcgg ccgctgatct tcagacctgg aggaggagat atgagggaca attggagaag 13140
tgaattatat aaatataaag tagtaaaaat tgaaccatta ggagtagcac ccaccaaggc 13200
aaagagaaga gtggtgcaga gagaaaaaag agcagtggga ataggagctt tgttccttgg 13260
gttcttggga gcagcaggaa gcactatggg cgcagcgtca atgacgctga cggtacaggc 13320
cagacaatta ttgtctggta tagtgcagca gcagaacaat ttgctgaggg ctattgaggc 13380
gcaacagcat ctgttgcaac tcacagtctg gggcatcaag cagctccagg caagaatcct 13440
ggctgtggaa agatacctaa aggatcaaca gctcctgggg atttggggtt gctctggaaa 13500
actcatttgc accactgctg tgccttggaa tgctagttgg agtaataaat ctctggaaca 13560
gatttggaat cacacgacct ggatggagtg ggacagagaa attaacaatt acacaagctt 13620
aatacactcc ttaattgaag aatcgcaaaa ccagcaagaa aagaatgaac aagaattatt 13680
ggaattagat aaatgggcaa gtttgtggaa ttggtttaac ataacaaatt ggctgtggta 13740
tataaaatta ttcataatga tagtaggagg cttggtaggt ttaagaatag tttttgctgt 13800
actttctata gtgaatagag ttaggcaggg atattcacca ttatcgtttc agacccacct 13860
cccaaccccg aggggacccg acaggcccga aggaatagaa gaagaaggtg gagagagaga 13920
cagagacaga tccattcgat tagtgaacgg atcggcactg cgtgcgccaa ttctgcagac 13980
aaatggcagt attcatccac aattttaaaa gaaaaggggg gattgggggg tacagtgcag 14040
gggaaagaat agtagacata atagcaacag acatacaaac taaagaatta caaaaacaaa 14100
ttacaaaaat tcaaaatttt cgggtttatt acagggacag cagagatcca gtttggttaa 14160
ttaa 14164
<210> 212
<211> 14
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 212
tttattccct gacc 14
<210> 213
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 213
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Asn Gly Gly Gly Lys
1 5 10 15
Gln Ala Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala
20 25 30
His Gly
<210> 214
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 214
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 215
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 215
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 216
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 216
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 217
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 217
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 218
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 218
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 219
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 219
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 220
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 220
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 221
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 221
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 222
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 222
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 223
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 223
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 224
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 224
Leu Thr Pro Asn Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 225
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 225
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 226
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 226
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 227
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 227
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 228
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 228
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 229
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 229
Leu Thr Gln Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 230
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 230
Leu Ser Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 231
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 231
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 232
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 232
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 233
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 233
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Arg Gln Ala His Gly
20 25 30
<210> 234
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 234
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Asn Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 235
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 235
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Ala Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 236
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 236
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 237
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 237
Leu Thr Leu Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 238
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 238
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Cys Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 239
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 239
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Gln Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 240
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 240
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Gly Gly Arg Pro Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 241
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 241
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 242
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 242
Leu Thr Pro Asn Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 243
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 243
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Gly Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 244
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 244
Leu Thr Leu Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 245
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 245
Leu Thr Pro Ala Gln Ala Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 246
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 246
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Asn Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 247
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 247
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 248
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 248
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 249
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 249
Leu Thr Pro Asp Gln Val Val Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 250
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 250
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Arg Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 251
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 251
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Asn Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 252
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 252
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 253
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 253
Leu Pro Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 254
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 254
Leu Thr Ser Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 255
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 255
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 256
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 256
Leu Ile Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 257
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 257
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Met Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 258
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 258
Leu Thr Arg Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 259
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 259
Leu Thr Pro Asp Gln Val Val Ala Thr Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 260
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 260
Leu Ile Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 261
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 261
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asn His Gly
20 25 30
<210> 262
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 262
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Lys Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 263
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 263
Leu Thr Pro Asp Gln Leu Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 264
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 264
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Gly His Gly
20 25 30
<210> 265
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 265
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Glu His Gly
20 25 30
<210> 266
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 266
Leu Thr Leu Asp Lys Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 267
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 267
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 268
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 268
Leu Thr Pro Asp Lys Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 269
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 269
Leu Thr Gln Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Tyr Gln Asp His Gly
20 25 30
<210> 270
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 270
Leu Thr Pro Ala Gln Val Val Ala Ile Val Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 271
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 271
Leu Thr Pro Asp Lys Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 272
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 272
Leu Thr Gln Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 273
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 273
Leu Thr Pro Asp Gln Val Met Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 274
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 274
Leu Thr Thr Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 275
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 275
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 276
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 276
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Leu Val Leu Cys Gln Ala His Gly
20 25 30
<210> 277
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 277
Leu Thr Gln Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 278
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 278
Leu Thr Pro Asp Gln Val Val Thr Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 279
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 279
Leu Ser Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys His Asp His Gly
20 25 30
<210> 280
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 280
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Met Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 281
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 281
Leu Ile Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 282
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 282
Leu Thr Pro Val Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 283
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 283
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Lys Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 284
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 284
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Met Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 285
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 285
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Phe Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 286
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 286
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Gln Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 287
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 287
Leu Thr Pro Ala Gln Val Val Ala Leu Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 288
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 288
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Arg Pro Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 289
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 289
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Ala Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 290
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 290
Leu Thr Gln Val Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 291
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 291
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Arg Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 292
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 292
Leu Pro Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 293
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 293
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 294
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 294
Leu Ser Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Ala
20 25 30
<210> 295
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 295
Leu Asn Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 296
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 296
Leu Thr Pro Asp Gln Val Met Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 297
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 297
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Arg Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 298
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 298
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Thr
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 299
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 299
Leu Thr Pro Asp Gln Val Met Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 300
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 300
Leu Thr Pro Ala Gln Val Val Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 301
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 301
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Arg Ala His Gly
20 25 30
<210> 302
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 302
Leu Ser Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 303
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 303
Leu Thr Pro Asp Gln Val Val Gly Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 304
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 304
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala Asn Gly
20 25 30
<210> 305
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 305
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 306
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 306
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Met Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 307
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 307
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Met Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 308
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 308
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Ala Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 309
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 309
Leu Thr Pro Asp Gln Val Val Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 310
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 310
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Thr Val Leu Cys Gln Asp His Gly
20 25 30
<210> 311
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 311
Met Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 312
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 312
Leu Ala Pro Asp Gln Val Val Ala Val Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 313
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 313
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Lys Thr Val Gln Gln Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 314
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 314
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Arg Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 315
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 315
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Gln Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 316
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 316
Leu Thr Pro Asp Gln Val Leu Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 317
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 317
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Arg Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 318
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 318
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Met Gln Arg Leu Leu Pro Val Leu Cys Arg Ala His Gly
20 25 30
<210> 319
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 319
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Met Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 320
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 320
Leu Thr Thr Asp Gln Val Val Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 321
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 321
Leu Thr Pro Thr Gln Val Met Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 322
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 322
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 323
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 323
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Met Leu Cys Gln Asp His Gly
20 25 30
<210> 324
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 324
Leu Thr Ser Ala Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 325
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 325
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Gln Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 326
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 326
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Ala Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 327
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 327
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Met Leu Cys Gln Ala His Gly
20 25 30
<210> 328
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 328
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala Arg Gly
20 25 30
<210> 329
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 329
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 330
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 330
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asn His Gly
20 25 30
<210> 331
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 331
Leu Thr Pro Asp Gln Val Val Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Met Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 332
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 332
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Arg Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 333
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 333
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Cys Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Arg Gln Ala His Gly
20 25 30
<210> 334
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 334
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Arg Asp His Gly
20 25 30
<210> 335
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 335
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Met Leu Cys Gln Ala His Gly
20 25 30
<210> 336
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 336
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Cys Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Arg His Ala His Gly
20 25 30
<210> 337
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 337
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln His His Gly
20 25 30
<210> 338
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 338
Leu Ile Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln His His Gly
20 25 30
<210> 339
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 339
Leu Thr Arg Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 340
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 340
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Val Gly Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 341
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 341
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 342
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 342
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Met Leu Cys Gln Asp His Gly
20 25 30
<210> 343
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 343
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Met Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 344
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 344
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Lys Gln His Gly
20 25 30
<210> 345
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 345
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 346
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 346
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Ala Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 347
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 347
Leu Thr Pro Ala Gln Val Val Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 348
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 348
Leu Thr Pro Ala Gln Val Met Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 349
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 349
Leu Thr Arg Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Arg Gln Ala His Gly
20 25 30
<210> 350
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 350
Leu Thr Leu Ala Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 351
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 351
Leu Thr Leu Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 352
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 352
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 353
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 353
Leu Ser Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 354
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 354
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln His His Gly
20 25 30
<210> 355
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 355
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Arg Gln Ala His Gly
20 25 30
<210> 356
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 356
Leu Ser Gln Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 357
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 357
Leu Pro Pro Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 358
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 358
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Ala Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 359
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 359
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Glu His Gly
20 25 30
<210> 360
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 360
Leu Thr Leu Asp Gln Val Ala Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 361
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 361
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Val Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 362
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 362
Leu Ile Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 363
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 363
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Arg Gln Ala His Gly
20 25 30
<210> 364
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 364
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 365
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 365
Leu Thr Pro Gln Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 366
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 366
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Val Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 367
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 367
Leu Ser Pro Asp Gln Val Val Thr Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Leu Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 368
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 368
Leu Thr Pro Val Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 369
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 369
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 370
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 370
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Cys Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Arg Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 371
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 371
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Phe Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 372
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 372
Leu Pro Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 373
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 373
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Phe Gln Glu His Gly
20 25 30
<210> 374
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 374
Leu Thr Pro Ala Lys Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 375
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 375
Leu Thr Pro Val Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Ala Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 376
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 376
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Gly Leu Cys Gln Asp His Gly
20 25 30
<210> 377
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 377
Leu Thr Leu Ala Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 378
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 378
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Thr Val Leu Cys Gln Asp His Gly
20 25 30
<210> 379
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 379
Leu Pro Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 380
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 380
Leu Thr Pro Ala Gln Ala Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 381
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 381
Leu Thr Pro Ala Gln Val Val Ala Ile Val Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 382
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 382
Leu Thr Pro Asp Gln Val Val Ala Val Ala Gly Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 383
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 383
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Gly Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 384
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 384
Leu Pro Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Ala His Gly
20 25 30
<210> 385
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 385
Leu Thr Thr Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 386
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 386
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Val Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 387
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 387
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Ala
20 25 30
<210> 388
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 388
Leu Thr Leu Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 389
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 389
Leu Thr Pro Asn Gln Leu Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 390
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 390
Leu Ser Pro Ala Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 391
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 391
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Val Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 392
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 392
Leu Thr Pro Asp Gln Val Met Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 393
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 393
Leu Thr Pro Glu Gln Val Val Ala Ile Ala Ser Gly Gly Arg Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 394
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 394
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Trp Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 395
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 395
Leu Thr Pro Asp Lys Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 396
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 396
Leu Thr Pro Ala Gln Val Met Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 397
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 397
Leu Thr Gln Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala Asn Gly
20 25 30
<210> 398
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 398
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Pro Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 399
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 399
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Ser Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Met Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 400
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 400
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Arg Gln Asp His Gly
20 25 30
<210> 401
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 401
Leu Thr Pro Tyr Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 402
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 402
Leu Thr Pro Tyr Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 403
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 403
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Glu His Gly
20 25 30
<210> 404
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 404
Leu Thr Leu Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Leu Val Leu Cys Gln Ala His Gly
20 25 30
<210> 405
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 405
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Arg Arg Leu Leu Gln Val Leu Cys Gln Asp His Gly
20 25 30
<210> 406
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 406
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Arg Gln Asp His Gly
20 25 30
<210> 407
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 407
Leu Thr Pro Asp Gln Val Val Ser Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 408
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 408
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Thr His Gly
20 25 30
<210> 409
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 409
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Lys Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 410
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 410
Leu Thr Thr Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 411
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 411
Leu Ile Pro Gln Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 412
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 412
Leu Thr Leu Thr Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 413
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 413
Leu Thr Pro Thr Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 414
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 414
Leu Thr Pro Thr Gln Val Met Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 415
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 415
Leu Thr Pro Asp Gln Val Val Ala Val Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 416
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 416
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 417
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 417
Leu Thr Pro Gly Gln Val Val Ala Ile Ala Ser Gly Gly Lys Arg Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 418
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 418
Leu Thr Pro Asp Gln Val Val Val Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 419
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 419
Leu Pro Pro Asp Gln Val Val Ala Ile Ala Ser Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 420
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 420
Leu Thr Pro Asp Gln Val Val Thr Ile Ala Asn Gly Ser Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 421
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 421
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Gln Val Leu Cys Gln Asp His Gly
20 25 30
<210> 422
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 422
Leu Thr Pro Asp His Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 423
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 423
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Gln Val Leu Cys Gln Asp His Gly
20 25 30
<210> 424
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 424
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Arg Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 425
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 425
Leu His Pro Gly Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 426
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 426
Leu Thr Leu Asp Gln Val Val Ser Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 427
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 427
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Ala Leu Cys Gln Asp His Gly
20 25 30
<210> 428
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 428
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Pro Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Glu Gln His Gly
20 25 30
<210> 429
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 429
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Lys Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 430
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 430
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Arg Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 431
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 431
Leu Asn Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 432
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 432
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Lys Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 433
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 433
Leu Thr Leu Asp Gln Val Val Ala Ile Ala Asn Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 434
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 434
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Arg Asp His Gly
20 25 30
<210> 435
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 435
Leu Thr Pro Ala Gln Val Leu Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Thr Val Leu Cys Gln Asp His Gly
20 25 30
<210> 436
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 436
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Met Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 437
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 437
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Gly Leu Cys Gln Ala His Gly
20 25 30
<210> 438
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 438
Leu Thr Arg Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Ala Leu Leu Pro Val Leu Arg Gln Ala His Gly
20 25 30
<210> 439
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 439
Leu Thr Pro Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Val His Gly
20 25 30
<210> 440
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 440
Leu Thr Pro Asn Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Leu Val Leu Cys Gln Asp His Gly
20 25 30
<210> 441
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 441
Leu Thr Pro Asp Gln Val Met Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Ala His Gly
20 25 30
<210> 442
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 442
Leu Thr Arg Glu Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 443
<211> 33
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 443
Leu Ser Thr Ala Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Gly Ile Gly Glu Gln Leu Leu Lys Leu Arg Thr Ala Pro Tyr
20 25 30
Gly
<210> 444
<211> 33
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 444
Leu Ser Thr Ala Gln Val Val Ala Val Ala Ser Gly Gly Lys Pro Ala
1 5 10 15
Leu Glu Ala Val Arg Ala Gln Leu Leu Ala Leu Arg Ala Ala Pro Tyr
20 25 30
Gly
<210> 445
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 445
Leu Thr Gln Val Gln Val Val Ala Ile Ala Ser Gly Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 446
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 446
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Asn Gly Lys Gln Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 447
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 447
Leu Thr Pro Asp Gln Val Val Ala Ile Ala Ser Gly Gly Lys Arg Ala
1 5 10 15
Leu Glu Thr Val Gln Arg Leu Leu Pro Val Leu Cys Gln Asp His Gly
20 25 30
<210> 448
<211> 14
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 448
tttattccct gaca 14
<210> 449
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 449
tcggcccctg ccggccca 18
<210> 450
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 450
tgcctgccct ccaggctcct 20
<210> 451
<211> 40
<212> DNA
<213> Homo sapiens
<400> 451
aaacggaagg gcctgagtcc gagcagaaga agaagtttta 40
<210> 452
<211> 40
<212> DNA
<213> Homo sapiens
<400> 452
aggaggaagg gcctgagtcc gagcagaaga agaagggctc 40
<210> 453
<211> 26
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 453
gaguccgagc agaagaagaa guuuua 26
<210> 454
<211> 39
<212> DNA
<213> Homo sapiens
<400> 454
aggaggaagg gcctgagtcc gagcagaaga gaagggctc 39
<210> 455
<211> 39
<212> DNA
<213> Homo sapiens
<400> 455
ggaggaaggg cctgagtccg agcagaagaa gaagggctc 39
<210> 456
<211> 38
<212> DNA
<213> Homo sapiens
<400> 456
ggaggaaggg cctgagtccg agcagaagag aagggctc 38
<210> 457
<211> 40
<212> DNA
<213> Homo sapiens
<400> 457
ggaggaaggg cctgagtccg agcagaagaa agaagggctc 40
<210> 458
<211> 37
<212> DNA
<213> Homo sapiens
<400> 458
ggaggaaggg cctgagtccg agcagaagga agggctc 37
<210> 459
<211> 36
<212> DNA
<213> Homo sapiens
<400> 459
ggaggaaggg cctgagtccg agcagaagaa gggctc 36
<210> 460
<211> 33
<212> DNA
<213> Homo sapiens
<400> 460
ggaggaaggg cctgagtccg agcagaaggg ctc 33
<210> 461
<211> 33
<212> DNA
<213> Homo sapiens
<400> 461
ggaggaaggg cctgagcccg agcagaaggg ctc 33
<210> 462
<211> 122
<212> DNA
<213> Homo sapiens
<400> 462
ggaggaaggg cctgagtccg agcagaagaa gaagggctcc catcacatca accggtggcg 60
cattgccacg aagcaggcca atggggagga catcgatgtc acctccaatg actagggtgg 120
gc 122
<210> 463
<211> 48
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (3)..(32)
<223> a, c, u, g, unknown or other
<400> 463
acnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnguuuuaga gcuaugcu 48
<210> 464
<211> 67
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 464
agcauagcaa guuaaaauaa ggctaguccg uuaucaacuu gaaaaagugg caccgagucg 60
gugcuuu 67
<210> 465
<211> 62
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (1)..(20)
<223> a, c, u, g, unknown or other
<400> 465
nnnnnnnnnn nnnnnnnnnn guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc 60
cg 62
<210> 466
<211> 48
<212> DNA
<213> Homo sapiens
<400> 466
ctggaggagg aagggcctga gtccgagcag aagaagaagg gctcccat 48
<210> 467
<211> 30
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 467
gaguccgagc agaagaagaa guuuuagagc 30
<210> 468
<211> 20
<212> RNA
<213> Homo sapiens
<400> 468
gaguccgagc agaagaagau 20
<210> 469
<211> 20
<212> RNA
<213> Homo sapiens
<400> 469
gaguccgagc agaagaagua 20
<210> 470
<211> 20
<212> RNA
<213> Homo sapiens
<400> 470
gaguccgagc agaagaacaa 20
<210> 471
<211> 20
<212> RNA
<213> Homo sapiens
<400> 471
gaguccgagc agaagaugaa 20
<210> 472
<211> 20
<212> RNA
<213> Homo sapiens
<400> 472
gaguccgagc agaaguagaa 20
<210> 473
<211> 20
<212> RNA
<213> Homo sapiens
<400> 473
gaguccgagc agaugaagaa 20
<210> 474
<211> 20
<212> RNA
<213> Homo sapiens
<400> 474
gaguccgagc acaagaagaa 20
<210> 475
<211> 20
<212> RNA
<213> Homo sapiens
<400> 475
gaguccgagg agaagaagaa 20
<210> 476
<211> 20
<212> RNA
<213> Homo sapiens
<400> 476
gaguccgugc agaagaagaa 20
<210> 477
<211> 20
<212> RNA
<213> Homo sapiens
<400> 477
gagucggagc agaagaagaa 20
<210> 478
<211> 20
<212> RNA
<213> Homo sapiens
<400> 478
gagaccgagc agaagaagaa 20
<210> 479
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 479
aatgacaagc ttgctagcgg tggg 24
<210> 480
<211> 39
<212> DNA
<213> Homo sapiens
<400> 480
aaaacggaag ggcctgagtc cgagcagaag aagaagttt 39
<210> 481
<211> 39
<212> DNA
<213> Homo sapiens
<400> 481
aaacaggggc cgagattggg tgttcagggc agaggtttt 39
<210> 482
<211> 38
<212> DNA
<213> Homo sapiens
<400> 482
aaaacggaag ggcctgagtc cgagcagaag aagaagtt 38
<210> 483
<211> 40
<212> DNA
<213> Homo sapiens
<400> 483
aacggaggga ggggcacaga tgagaaactc agggttttag 40
<210> 484
<211> 38
<212> DNA
<213> Homo sapiens
<400> 484
agcccttctt cttctgctcg gactcaggcc cttcctcc 38
<210> 485
<211> 40
<212> DNA
<213> Homo sapiens
<400> 485
cagggaggga ggggcacaga tgagaaactc aggaggcccc 40
<210> 486
<211> 80
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 486
ggcaatgcgc caccggttga tgtgatggga gcccttctag gaggccccca gagcagccac 60
tggggcctca acactcaggc 80
<210> 487
<211> 98
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 487
ggacgaaaca ccggaaccat tcaaaacagc atagcaagtt aaaataaggc tagtccgtta 60
tcaacttgaa aaagtggcac cgagtcggtg cttttttt 98
<210> 488
<211> 186
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 488
ggacgaaaca ccggtagtat taagtattgt tttatggctg ataaatttct ttgaatttct 60
ccttgattat ttgttataaa agttataaaa taatcttgtt ggaaccattc aaaacagcat 120
agcaagttaa aataaggcta gtccgttatc aacttgaaaa agtggcaccg agtcggtgct 180
tttttt 186
<210> 489
<211> 73
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 489
tgaatggtcc caaaacggaa gggcctgagt ccgagcagaa gaagaagttt tagagctatg 60
ctgttttgaa tgg 73
<210> 490
<211> 95
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 490
gggttttaga gctatgctgt tttgaatggt cccaaaacgg gtcttcgaga agacgtttta 60
gagctatgct gttttgaatg gtcccaaaac ttttt 95
<210> 491
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (5)..(34)
<223> a, c, t, g, unknown or other
<400> 491
aaacnnnnnn nnnnnnnnnn nnnnnnnnnn nnnngt 36
<210> 492
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (7)..(36)
<223> a, c, t, g, unknown or other
<400> 492
taaaacnnnn nnnnnnnnnn nnnnnnnnnn nnnnnn 36
<210> 493
<211> 84
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 493
gtggaaagga cgaaacaccg ggtcttcgag aagacctgtt ttagagctag aaatagcaag 60
ttaaaataag gctagtccgt tttt 84
<210> 494
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (6)..(24)
<223> a, c, t, g, unknown or other
<400> 494
caccgnnnnn nnnnnnnnnn nnnn 24
<210> 495
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (5)..(23)
<223> a, c, t, g, unknown or other
<400> 495
aaacnnnnnn nnnnnnnnnn nnnc 24
<210> 496
<211> 69
<212> DNA
<213> Homo sapiens
<400> 496
ctggtcttcc acctctctgc cctgaacacc caatctcggc ccctctcgcc accctcctgc 60
atttctgtt 69
<210> 497
<211> 138
<212> DNA
<213> Mus sp.
<400> 497
acccaagcac tgagtgccat tagctaaatg catagggtac cacccacagg tgccaggggc 60
ctttcccaaa gttcccagcc ccttctccaa cctttcctgg cccagaggct ttcccatgtg 120
tgtggctgga ccctttga 138
<210> 498
<211> 46
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (1)..(19)
<223> a, c, u, g, unknown or other
<400> 498
nnnnnnnnnn nnnnnnnnng uuauuguacu cucaagauuu auuuuu 46
<210> 499
<211> 91
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 499
guuacuuaaa ucuugcagaa gcuacaaaga uaaggcuuca ugccgaaauc aacacccugu 60
cauuuuaugg caggguguuu ucguuauuua a 91
<210> 500
<211> 70
<212> DNA
<213> Homo sapiens
<400> 500
ttttctagtg ctgagtttct gtgactcctc tacattctac ttctctgtgt ttctgtatac 60
tacctcctcc 70
<210> 501
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 501
Lys Arg Lys Phe Asp Thr Glu Thr Lys Asp Cys Asn Glu Lys Lys Lys
1 5 10 15
Lys
<210> 502
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 502
Leu Lys Ala Leu Gly Ser Asp Leu Thr Leu
1 5 10
<210> 503
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 503
Arg Arg Gly Gln
1
<210> 504
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 504
Arg Val Arg Arg Glu
1 5
<210> 505
<211> 14
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 505
tctgcaagag tagg 14
<210> 506
<211> 14
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 506
tgcactccac cttg 14
<210> 507
<211> 14
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 507
tcaagcagct tctc 14
<210> 508
<211> 23
<212> DNA
<213> Homo sapiens
<400> 508
gccccctttc atgcaaaacc cgg 23
<210> 509
<211> 20
<212> DNA
<213> Homo sapiens
<400> 509
gccccctttc atgcaaaacc 20
<210> 510
<211> 276
<212> DNA
<213> Homo sapiens
<400> 510
cgcccatggc cgcgcgcgct ccacacaact caccggagtc cgcgccttgc gccgccgacc 60
agttcgcagc tccgcgccac ggcagccagt ctctgagtgg aagagagcgc agcccggcca 120
ccggacctac ttactcgcct tgctgattgt ctatttttgc gtttacaact tttctaagaa 180
cttttgtata caaaggaact ttttaaaaaa gacgcttcca agttatattt aatccaaaga 240
agaaggatct cggccaattt ggggttttgg gttttg 276
<210> 511
<211> 151
<212> DNA
<213> Homo sapiens
<400> 511
tcccccgcgc ggccggcggc gcgggaggcc ccgccccctt tcatgcaaaa cccggcagcg 60
aggctgggct cgagtggagg agccgccgcg cgctgattgg tcgctagaaa cccatttatt 120
ccctgacagc ccccgtcaca tggatggttg t 151
<210> 512
<211> 30
<212> DNA
<213> Homo sapiens
<400> 512
gaagccgccg ggcccgcagc aaacttcggg 30
<210> 513
<211> 30
<212> DNA
<213> Homo sapiens
<400> 513
ccctacatgt tgtactacct ctgcctcgac 30
Claims (16)
- CRISPR-Cas 복합체를 포함하는 단리된 진핵세포로서, CRISPR-Cas 복합체는 하기를 포함하고:
- Cas9, 여기서 Cas9 은 둘 이상의 핵 국부화 신호를 포함하고;
- tracr mate 서열에 연결된 안내 서열; 및
- tracr mate 서열의 전부 또는 일부와 하이브리드화 가능한 tracr 서열,
상기 안내 서열은 진핵세포 내 표적 서열과 상보성을 갖도록 설계되고, 진핵세포 내 표적 서열에 대한 CRISPR 복합체의 서열-특이적 결합을 유도할 수 있는 것인, 단리된 진핵세포. - 제 1 항에 있어서, 핵 국부화 신호는 PKKKRKV, KRPAATKKAGQAKKKK, PAAKRVKLD, RQRRNELKRSP, NQSSNFGPMKGGNFGGRSSGPYGGGGQYFAKPRNQGGY, RMRIZFKNKGKDTAELRRRRVEVSVELRKAKKDEQILKRRNV, VSRKRPRP, PPKKARED, POPKKKPL, SALIKKKKKMAP, DRLRR, PKQKKRK, RKLKKKIKKL, REKKKFLKRR, KRKGDEVDGVDEVAKKKSKK, 및 RKCLQAGMNLEARKTKK 로부터 독립적으로 선택되는, 단리된 진핵세포.
- 제 1 항에 있어서, 하나 이상의 핵 국부화 신호는 PKKKRKV 인, 단리된 진핵세포.
- 제 1 항에 있어서, Cas9 는 아미노-말단에서 하나 이상의 핵 국부화 신호, 및 카르복시-말단에서 하나 이상의 핵 국부화 신호를 포함하는, 단리된 진핵세포.
- 제 1 항에 있어서, Cas9 은 표적 서열을 함유하는 표적 폴리뉴클레오티드의 양쪽 가닥의 절단을 유도하는, 단리된 진핵세포.
- 제 1 항에 있어서, Cas9 는 촉매 도메인에 하나 이상의 돌연변이를 포함하고; 바람직하게는 상기 돌연변이가 D10A, H840A, N854A 또는 N863A 로부터 선택되는, 단리된 진핵세포.
- 제 1 항에 있어서, Cas9 는 S. 피오게네스 (S.pyogenes) Cas9 인, 단리된 진핵세포.
- 제 1 항에 있어서, Cas9 는 S. 써모필루스 (S. thermophilus) Cas9 인, 단리된 진핵세포.
- 제 1 항에 있어서, tracr mate 서열은 tracr 서열에 연결되어 키메라 RNA 를 형성하고, 상기 키메라 RNA 는, 5' 에서 3' 으로, 안내 서열, tracr mate 서열 및 tracr 서열을 포함하는, 단리된 진핵세포.
- 제 1 항에 있어서, CRISPR-Cas 복합체는 단리된 진핵세포의 핵에 위치하는, 단리된 진핵세포.
- 제 1 항에 있어서, 단리된 진핵세포는 둘 이상의 상이한 안내 서열을 포함하는, 단리된 진핵세포.
- 제 1 항에 있어서, 단리된 진핵세포는 상동성-유도 복구를 위한 주형 폴리뉴클레오티드를 추가로 포함하는, 단리된 진핵세포.
- 제 1 항에 있어서, 안내 서열은 프로토스페이서 인접 모티프 (PAM) 에 인접한 표적 서열에 하이브리드화되고, tracr-mate 서열은 tracr 서열에 하이브리드화되는, 단리된 진핵세포.
- 제 1 항에 있어서, CRISPR-Cas 복합체는 표적 서열에서 절단을 일으키는, 단리된 진핵세포.
- 제 1 항에 있어서, 단리된 진핵세포는 인간 세포 또는 포유동물 세포이고, 상기 진핵세포는 인간 생식세포가 아닌, 단리된 진핵세포.
- 제 1 항 내지 제 15 항 중 어느 한 항에 있어서, 치료에 사용되는, 단리된 진핵세포.
Applications Claiming Priority (12)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201261675778P | 2012-07-25 | 2012-07-25 | |
| US61/675,778 | 2012-07-25 | ||
| US201261721283P | 2012-11-01 | 2012-11-01 | |
| US61/721,283 | 2012-11-01 | ||
| US201261736465P | 2012-12-12 | 2012-12-12 | |
| US61/736,465 | 2012-12-12 | ||
| US201361794458P | 2013-03-15 | 2013-03-15 | |
| US61/794,458 | 2013-03-15 | ||
| US201361835973P | 2013-06-17 | 2013-06-17 | |
| US61/835,973 | 2013-06-17 | ||
| KR1020157004939A KR102530118B1 (ko) | 2012-07-25 | 2013-07-21 | 유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 |
| PCT/US2013/051418 WO2014018423A2 (en) | 2012-07-25 | 2013-07-21 | Inducible dna binding proteins and genome perturbation tools and applications thereof |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020157004939A Division KR102530118B1 (ko) | 2012-07-25 | 2013-07-21 | 유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20230065381A true KR20230065381A (ko) | 2023-05-11 |
Family
ID=48914461
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020237014846A KR20230065381A (ko) | 2012-07-25 | 2013-07-21 | 유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 |
| KR1020157004939A KR102530118B1 (ko) | 2012-07-25 | 2013-07-21 | 유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020157004939A KR102530118B1 (ko) | 2012-07-25 | 2013-07-21 | 유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 |
Country Status (13)
| Country | Link |
|---|---|
| US (4) | US20150291966A1 (ko) |
| EP (3) | EP2877213B1 (ko) |
| JP (2) | JP2015527889A (ko) |
| KR (2) | KR20230065381A (ko) |
| CN (2) | CN105188767A (ko) |
| AU (3) | AU2013293270B2 (ko) |
| CA (1) | CA2879997A1 (ko) |
| DK (1) | DK3494997T3 (ko) |
| ES (1) | ES2757623T3 (ko) |
| HK (1) | HK1210965A1 (ko) |
| PL (1) | PL3494997T3 (ko) |
| PT (1) | PT3494997T (ko) |
| WO (1) | WO2014018423A2 (ko) |
Families Citing this family (444)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20160144003A1 (en) * | 2011-05-19 | 2016-05-26 | The Scripps Research Institute | Compositions and methods for treating charcot-marie-tooth diseases and related neuronal diseases |
| AU2012284365B2 (en) | 2011-07-15 | 2017-04-20 | The General Hospital Corporation | Methods of transcription activator like effector assembly |
| US10323236B2 (en) | 2011-07-22 | 2019-06-18 | President And Fellows Of Harvard College | Evaluation and improvement of nuclease cleavage specificity |
| US10465042B2 (en) | 2011-12-02 | 2019-11-05 | Yale University | Poly(amine-co-ester) nanoparticles and methods of use thereof |
| GB201122458D0 (en) | 2011-12-30 | 2012-02-08 | Univ Wageningen | Modified cascade ribonucleoproteins and uses thereof |
| US9745548B2 (en) | 2012-03-15 | 2017-08-29 | Flodesign Sonics, Inc. | Acoustic perfusion devices |
| US10704021B2 (en) | 2012-03-15 | 2020-07-07 | Flodesign Sonics, Inc. | Acoustic perfusion devices |
| US10967298B2 (en) | 2012-03-15 | 2021-04-06 | Flodesign Sonics, Inc. | Driver and control for variable impedence load |
| US10322949B2 (en) | 2012-03-15 | 2019-06-18 | Flodesign Sonics, Inc. | Transducer and reflector configurations for an acoustophoretic device |
| US9950282B2 (en) | 2012-03-15 | 2018-04-24 | Flodesign Sonics, Inc. | Electronic configuration and control for acoustic standing wave generation |
| US9752113B2 (en) | 2012-03-15 | 2017-09-05 | Flodesign Sonics, Inc. | Acoustic perfusion devices |
| US10689609B2 (en) | 2012-03-15 | 2020-06-23 | Flodesign Sonics, Inc. | Acoustic bioreactor processes |
| US9458450B2 (en) | 2012-03-15 | 2016-10-04 | Flodesign Sonics, Inc. | Acoustophoretic separation technology using multi-dimensional standing waves |
| US10737953B2 (en) | 2012-04-20 | 2020-08-11 | Flodesign Sonics, Inc. | Acoustophoretic method for use in bioreactors |
| CA2869016C (en) | 2012-04-25 | 2020-05-05 | Regeneron Pharmaceuticals, Inc. | Nuclease-mediated targeting with large targeting vectors |
| EP2841572B1 (en) | 2012-04-27 | 2019-06-19 | Duke University | Genetic correction of mutated genes |
| LT2800811T (lt) | 2012-05-25 | 2017-09-11 | The Regents Of The University Of California | Būdai ir kompozicijos, skirti tikslinės dnr modifikavimui, panaudojant adresuotą rnr, ir transkripcijos moduliavimui, panaudojant adresuotą rnr |
| US9890364B2 (en) | 2012-05-29 | 2018-02-13 | The General Hospital Corporation | TAL-Tet1 fusion proteins and methods of use thereof |
| EP2877213B1 (en) * | 2012-07-25 | 2020-12-02 | The Broad Institute, Inc. | Inducible dna binding proteins and genome perturbation tools and applications thereof |
| WO2014059255A1 (en) | 2012-10-12 | 2014-04-17 | The General Hospital Corporation | Transcription activator-like effector (tale) - lysine-specific demethylase 1 (lsd1) fusion proteins |
| KR101656237B1 (ko) | 2012-10-23 | 2016-09-12 | 주식회사 툴젠 | 표적 DNA에 특이적인 가이드 RNA 및 Cas 단백질을 암호화하는 핵산 또는 Cas 단백질을 포함하는, 표적 DNA를 절단하기 위한 조성물 및 이의 용도 |
| PL3138910T3 (pl) | 2012-12-06 | 2018-01-31 | Sigma Aldrich Co Llc | Oparta na CRISPR modyfikacja i regulacja genomu |
| US10676749B2 (en) * | 2013-02-07 | 2020-06-09 | The General Hospital Corporation | Tale transcriptional activators |
| JP2016519652A (ja) | 2013-03-14 | 2016-07-07 | カリブー・バイオサイエンシーズ・インコーポレイテッド | 核酸ターゲティング核酸の組成物および方法 |
| US10760064B2 (en) | 2013-03-15 | 2020-09-01 | The General Hospital Corporation | RNA-guided targeting of genetic and epigenomic regulatory proteins to specific genomic loci |
| WO2014144592A2 (en) | 2013-03-15 | 2014-09-18 | The General Hospital Corporation | Using truncated guide rnas (tru-grnas) to increase specificity for rna-guided genome editing |
| BR112015026197B1 (pt) | 2013-04-16 | 2022-12-06 | Regeneron Pharmaceuticals, Inc | Método para modificação marcada de um lócus genômico de interesse em uma célula de rato pluripotente |
| WO2014186435A2 (en) | 2013-05-14 | 2014-11-20 | University Of Georgia Research Foundation, Inc. | Compositions and methods for reducing neointima formation |
| US9873907B2 (en) | 2013-05-29 | 2018-01-23 | Agilent Technologies, Inc. | Method for fragmenting genomic DNA using CAS9 |
| EP3597755A1 (en) * | 2013-06-17 | 2020-01-22 | The Broad Institute, Inc. | Delivery, use and therapeutic applications of the crispr-cas systems and compositions for targeting disorders and diseases using viral components |
| US10421957B2 (en) | 2013-07-29 | 2019-09-24 | Agilent Technologies, Inc. | DNA assembly using an RNA-programmable nickase |
| US9163284B2 (en) | 2013-08-09 | 2015-10-20 | President And Fellows Of Harvard College | Methods for identifying a target site of a Cas9 nuclease |
| CN105829536A (zh) | 2013-08-22 | 2016-08-03 | 纳幕尔杜邦公司 | 用于在不掺入选择性转基因标记的情况下,在植物基因组中产生基因修饰的方法,以及用于这种方法的组合物 |
| US9359599B2 (en) | 2013-08-22 | 2016-06-07 | President And Fellows Of Harvard College | Engineered transcription activator-like effector (TALE) domains and uses thereof |
| US9340799B2 (en) | 2013-09-06 | 2016-05-17 | President And Fellows Of Harvard College | MRNA-sensing switchable gRNAs |
| US9737604B2 (en) | 2013-09-06 | 2017-08-22 | President And Fellows Of Harvard College | Use of cationic lipids to deliver CAS9 |
| US9322037B2 (en) | 2013-09-06 | 2016-04-26 | President And Fellows Of Harvard College | Cas9-FokI fusion proteins and uses thereof |
| CN105722500B (zh) | 2013-09-11 | 2019-08-30 | 伊戈尔生物药品股份有限公司 | 包含离子性液体的液体蛋白质制剂 |
| US9745569B2 (en) | 2013-09-13 | 2017-08-29 | Flodesign Sonics, Inc. | System for generating high concentration factors for low cell density suspensions |
| WO2015065964A1 (en) | 2013-10-28 | 2015-05-07 | The Broad Institute Inc. | Functional genomics using crispr-cas systems, compositions, methods, screens and applications thereof |
| LT3066201T (lt) | 2013-11-07 | 2018-08-10 | Editas Medicine, Inc. | Su crispr susiję būdai ir kompozicijos su valdančiomis grnr |
| RU2725520C2 (ru) | 2013-12-11 | 2020-07-02 | Регенерон Фармасьютикалс, Инк. | Способы и композиции для направленной модификации генома |
| US20150165054A1 (en) | 2013-12-12 | 2015-06-18 | President And Fellows Of Harvard College | Methods for correcting caspase-9 point mutations |
| SG10201804977UA (en) | 2013-12-12 | 2018-07-30 | Broad Inst Inc | Delivery, Use and Therapeutic Applications of the Crispr-Cas Systems and Compositions for Targeting Disorders and Diseases Using Particle Delivery Components |
| WO2015089364A1 (en) | 2013-12-12 | 2015-06-18 | The Broad Institute Inc. | Crystal structure of a crispr-cas system, and uses thereof |
| CA2932478A1 (en) | 2013-12-12 | 2015-06-18 | Massachusetts Institute Of Technology | Delivery, use and therapeutic applications of the crispr-cas systems and compositions for genome editing |
| EP4219699A1 (en) | 2013-12-12 | 2023-08-02 | The Broad Institute, Inc. | Engineering of systems, methods and optimized guide compositions with new architectures for sequence manipulation |
| BR112016013547A2 (pt) | 2013-12-12 | 2017-10-03 | Broad Inst Inc | Composições e métodos de uso de sistemas crispr-cas em distúrbios de repetições de nucleotídeos |
| WO2015089486A2 (en) | 2013-12-12 | 2015-06-18 | The Broad Institute Inc. | Systems, methods and compositions for sequence manipulation with optimized functional crispr-cas systems |
| EP3080261B1 (en) | 2013-12-12 | 2019-05-22 | The Broad Institute, Inc. | Delivery, use and therapeutic applications of the crispr-cas systems and compositions for hbv and viral diseases and disorders |
| WO2015105955A1 (en) | 2014-01-08 | 2015-07-16 | Flodesign Sonics, Inc. | Acoustophoresis device with dual acoustophoretic chamber |
| US10354746B2 (en) | 2014-01-27 | 2019-07-16 | Georgia Tech Research Corporation | Methods and systems for identifying CRISPR/Cas off-target sites |
| US10287590B2 (en) | 2014-02-12 | 2019-05-14 | Dna2.0, Inc. | Methods for generating libraries with co-varying regions of polynuleotides for genome modification |
| CN113215219A (zh) | 2014-02-13 | 2021-08-06 | 宝生物工程(美国) 有限公司 | 从核酸的初始集合中耗尽靶分子的方法、以及用于实践其的组合物和试剂盒 |
| WO2015126884A1 (en) * | 2014-02-18 | 2015-08-27 | Stc.Unm | Compositions and methods for controlling cellular function |
| CA3194412A1 (en) | 2014-02-27 | 2015-09-03 | Monsanto Technology Llc | Compositions and methods for site directed genomic modification |
| EP3514246B1 (en) | 2014-02-27 | 2021-11-17 | The Broad Institute, Inc. | T cell balance gene expression and methods of use thereof |
| CA2947941C (en) * | 2014-03-05 | 2021-02-23 | National University Corporation Kobe University | Genome sequence modification method for specifically converting nucleic acid bases of targeted dna sequence, and molecular complex for use in same |
| EP3114227B1 (en) | 2014-03-05 | 2021-07-21 | Editas Medicine, Inc. | Crispr/cas-related methods and compositions for treating usher syndrome and retinitis pigmentosa |
| US11141493B2 (en) | 2014-03-10 | 2021-10-12 | Editas Medicine, Inc. | Compositions and methods for treating CEP290-associated disease |
| DK3116997T3 (da) | 2014-03-10 | 2019-08-19 | Editas Medicine Inc | Crispr/cas-relaterede fremgangsmåder og sammensætninger til behandling af lebers kongenitale amaurose 10 (lca10) |
| US11339437B2 (en) | 2014-03-10 | 2022-05-24 | Editas Medicine, Inc. | Compositions and methods for treating CEP290-associated disease |
| WO2015148863A2 (en) | 2014-03-26 | 2015-10-01 | Editas Medicine, Inc. | Crispr/cas-related methods and compositions for treating sickle cell disease |
| WO2015153940A1 (en) * | 2014-04-03 | 2015-10-08 | Massachusetts Institute Of Technology | Methods and compositions for the production of guide rna |
| EP3129485B2 (en) * | 2014-04-09 | 2022-12-21 | Editas Medicine, Inc. | Crispr/cas-related methods and compositions for treating cystic fibrosis |
| CN103952424B (zh) | 2014-04-23 | 2017-01-11 | 尹熙俊 | 生产mstn双侧基因敲除的双肌性状体细胞克隆猪的方法 |
| CN104630267B (zh) * | 2014-07-17 | 2017-10-31 | 清华大学 | 利用tale转录抑制子在哺乳动物细胞中模块化构建合成基因线路的试剂盒 |
| US11918695B2 (en) | 2014-05-09 | 2024-03-05 | Yale University | Topical formulation of hyperbranched polymer-coated particles |
| US10238581B2 (en) | 2014-05-09 | 2019-03-26 | Yale University | Hyperbranched polyglycerol-coated particles and methods of making and using thereof |
| WO2015188065A1 (en) * | 2014-06-05 | 2015-12-10 | Sangamo Biosciences, Inc. | Methods and compositions for nuclease design |
| EP3155125A1 (en) | 2014-06-13 | 2017-04-19 | Illumina Cambridge Limited | Methods and compositions for preparing sequencing libraries |
| AU2015284464B2 (en) | 2014-06-30 | 2021-06-17 | Illumina, Inc. | Methods and compositions using one-sided transposition |
| US9744483B2 (en) | 2014-07-02 | 2017-08-29 | Flodesign Sonics, Inc. | Large scale acoustic separation device |
| AU2015288157A1 (en) | 2014-07-11 | 2017-01-19 | E. I. Du Pont De Nemours And Company | Compositions and methods for producing plants resistant to glyphosate herbicide |
| EP3169776A4 (en) * | 2014-07-14 | 2018-07-04 | The Regents of The University of California | Crispr/cas transcriptional modulation |
| US10077453B2 (en) | 2014-07-30 | 2018-09-18 | President And Fellows Of Harvard College | CAS9 proteins including ligand-dependent inteins |
| WO2016025759A1 (en) | 2014-08-14 | 2016-02-18 | Shen Yuelei | Dna knock-in system |
| EP3180426B1 (en) | 2014-08-17 | 2019-12-25 | The Broad Institute, Inc. | Genome editing using cas9 nickases |
| US9970030B2 (en) | 2014-08-27 | 2018-05-15 | Caribou Biosciences, Inc. | Methods for increasing CAS9-mediated engineering efficiency |
| EP3191595B1 (en) | 2014-09-12 | 2019-12-25 | E. I. du Pont de Nemours and Company | Generation of site-specific-integration sites for complex trait loci in corn and soybean, and methods of use |
| WO2016049024A2 (en) | 2014-09-24 | 2016-03-31 | The Broad Institute Inc. | Delivery, use and therapeutic applications of the crispr-cas systems and compositions for modeling competition of multiple cancer mutations in vivo |
| WO2016049251A1 (en) | 2014-09-24 | 2016-03-31 | The Broad Institute Inc. | Delivery, use and therapeutic applications of the crispr-cas systems and compositions for modeling mutations in leukocytes |
| WO2016049163A2 (en) | 2014-09-24 | 2016-03-31 | The Broad Institute Inc. | Use and production of chd8+/- transgenic animals with behavioral phenotypes characteristic of autism spectrum disorder |
| WO2016049258A2 (en) | 2014-09-25 | 2016-03-31 | The Broad Institute Inc. | Functional screening with optimized functional crispr-cas systems |
| CN107205353B (zh) | 2014-09-26 | 2021-03-19 | 谱赛科美国股份有限公司 | 用于甜菊的单核苷酸多态性(snp)标志物 |
| MX2017004306A (es) | 2014-10-01 | 2017-12-20 | Eagle Biologics Inc | Formulaciones de polisacáridos y ácido nucleico que contienen agentes de reducción de viscosidad. |
| EP3212788A2 (en) | 2014-10-27 | 2017-09-06 | The Broad Institute, Inc. | Compositions, methods and use of synthetic lethal screening |
| CA2963820A1 (en) | 2014-11-07 | 2016-05-12 | Editas Medicine, Inc. | Methods for improving crispr/cas-mediated genome-editing |
| KR20230070319A (ko) | 2014-11-21 | 2023-05-22 | 리제너론 파마슈티칼스 인코포레이티드 | 쌍 형성된 가이드 rna를 사용하는 표적화된 유전자 변형을 위한 방법 및 조성물 |
| WO2016086197A1 (en) | 2014-11-25 | 2016-06-02 | The Brigham And Women's Hospital, Inc. | Method of identifying and treating a person having a predisposition to or afflicted with a cardiometabolic disease |
| CA2969145A1 (en) | 2014-11-26 | 2016-06-02 | The Regents Of The University Of California | Therapeutic compositions comprising transcription factors and methods of making and using the same |
| GB201421096D0 (en) | 2014-11-27 | 2015-01-14 | Imp Innovations Ltd | Genome editing methods |
| CN113699116A (zh) | 2014-12-10 | 2021-11-26 | 明尼苏达大学董事会 | 用于治疗疾病的遗传修饰的细胞、组织和器官 |
| WO2016094872A1 (en) | 2014-12-12 | 2016-06-16 | The Broad Institute Inc. | Dead guides for crispr transcription factors |
| EP3230451B1 (en) | 2014-12-12 | 2021-04-07 | The Broad Institute, Inc. | Protected guide rnas (pgrnas) |
| WO2016094874A1 (en) | 2014-12-12 | 2016-06-16 | The Broad Institute Inc. | Escorted and functionalized guides for crispr-cas systems |
| WO2016094880A1 (en) | 2014-12-12 | 2016-06-16 | The Broad Institute Inc. | Delivery, use and therapeutic applications of crispr systems and compositions for genome editing as to hematopoietic stem cells (hscs) |
| WO2016100974A1 (en) | 2014-12-19 | 2016-06-23 | The Broad Institute Inc. | Unbiased identification of double-strand breaks and genomic rearrangement by genome-wide insert capture sequencing |
| US20190054117A1 (en) * | 2014-12-19 | 2019-02-21 | Novartis Ag | Dimerization switches and uses thereof |
| WO2016106236A1 (en) | 2014-12-23 | 2016-06-30 | The Broad Institute Inc. | Rna-targeting system |
| EP3702456A1 (en) | 2014-12-24 | 2020-09-02 | The Broad Institute, Inc. | Crispr having or associated with destabilization domains |
| US20170369855A1 (en) * | 2014-12-24 | 2017-12-28 | Dana-Farber Cancer Institute, Inc. | Systems and methods for genome modification and regulation |
| WO2016108926A1 (en) | 2014-12-30 | 2016-07-07 | The Broad Institute Inc. | Crispr mediated in vivo modeling and genetic screening of tumor growth and metastasis |
| KR20170126875A (ko) | 2015-01-28 | 2017-11-20 | 파이어니어 하이 부렛드 인터내쇼날 인코포레이팃드 | Crispr 하이브리드 dna/rna 폴리뉴클레오티드 및 사용 방법 |
| EP3262193A2 (en) | 2015-02-26 | 2018-01-03 | The Broad Institute Inc. | T cell balance gene expression, compositions of matters and methods of use thereof |
| US11261466B2 (en) | 2015-03-02 | 2022-03-01 | Sinai Health System | Homologous recombination factors |
| EP3279321A4 (en) * | 2015-03-16 | 2018-10-31 | Institute Of Genetics And Developmental Biology, Chinese Academy Of Sciences | Method of applying non-genetic substance to perform site-directed reform of plant genome |
| BR112017017260A2 (pt) | 2015-03-27 | 2018-04-17 | Du Pont | construções de dna, vetor, célula, plantas, semente, método de expressão de rna e método de modificação de local alvo |
| WO2016167300A1 (ja) * | 2015-04-13 | 2016-10-20 | 国立大学法人東京大学 | 光依存的に又は薬物存在下でヌクレアーゼ活性若しくはニッカーゼ活性を示す、又は標的遺伝子の発現を抑制若しくは活性化するポリペプチドのセット |
| US11180793B2 (en) | 2015-04-24 | 2021-11-23 | Editas Medicine, Inc. | Evaluation of Cas9 molecule/guide RNA molecule complexes |
| US11377651B2 (en) | 2016-10-19 | 2022-07-05 | Flodesign Sonics, Inc. | Cell therapy processes utilizing acoustophoresis |
| US11021699B2 (en) | 2015-04-29 | 2021-06-01 | FioDesign Sonics, Inc. | Separation using angled acoustic waves |
| US11708572B2 (en) | 2015-04-29 | 2023-07-25 | Flodesign Sonics, Inc. | Acoustic cell separation techniques and processes |
| NZ738068A (en) | 2015-05-06 | 2019-07-26 | Snipr Tech Ltd | Altering microbial populations & modifying microbiota |
| WO2016182893A1 (en) | 2015-05-08 | 2016-11-17 | Teh Broad Institute Inc. | Functional genomics using crispr-cas systems for saturating mutagenesis of non-coding elements, compositions, methods, libraries and applications thereof |
| WO2016182959A1 (en) | 2015-05-11 | 2016-11-17 | Editas Medicine, Inc. | Optimized crispr/cas9 systems and methods for gene editing in stem cells |
| EP3095870A1 (en) | 2015-05-19 | 2016-11-23 | Kws Saat Se | Methods for the in planta transformation of plants and manufacturing processes and products based and obtainable therefrom |
| EP3302575A4 (en) * | 2015-05-28 | 2019-01-16 | Coda Biotherapeutics | GENOME EDITION VECTORS |
| JP7396783B2 (ja) | 2015-06-09 | 2023-12-12 | エディタス・メディシン、インコーポレイテッド | 移植を改善するためのcrispr/cas関連方法および組成物 |
| WO2016205728A1 (en) | 2015-06-17 | 2016-12-22 | Massachusetts Institute Of Technology | Crispr mediated recording of cellular events |
| WO2016205745A2 (en) * | 2015-06-18 | 2016-12-22 | The Broad Institute Inc. | Cell sorting |
| EP3436575A1 (en) | 2015-06-18 | 2019-02-06 | The Broad Institute Inc. | Novel crispr enzymes and systems |
| EP3800255A3 (en) * | 2015-06-18 | 2021-06-23 | Robert D. Bowles | Rna-guided transcriptional regulation and methods of using the same for the treatment of back pain |
| US9790490B2 (en) | 2015-06-18 | 2017-10-17 | The Broad Institute Inc. | CRISPR enzymes and systems |
| EP3666895A1 (en) | 2015-06-18 | 2020-06-17 | The Broad Institute, Inc. | Novel crispr enzymes and systems |
| JP2018519811A (ja) | 2015-06-29 | 2018-07-26 | アイオーニス ファーマシューティカルズ, インコーポレーテッドIonis Pharmaceuticals,Inc. | 修飾crispr rna及び修飾単一crispr rnaならびにその使用 |
| US11459540B2 (en) | 2015-07-28 | 2022-10-04 | Flodesign Sonics, Inc. | Expanded bed affinity selection |
| US11474085B2 (en) | 2015-07-28 | 2022-10-18 | Flodesign Sonics, Inc. | Expanded bed affinity selection |
| WO2017023803A1 (en) | 2015-07-31 | 2017-02-09 | Regents Of The University Of Minnesota | Modified cells and methods of therapy |
| WO2017027910A1 (en) | 2015-08-14 | 2017-02-23 | The University Of Sydney | Connexin 45 inhibition for therapy |
| US11214800B2 (en) | 2015-08-18 | 2022-01-04 | The Broad Institute, Inc. | Methods and compositions for altering function and structure of chromatin loops and/or domains |
| KR20180038558A (ko) | 2015-08-25 | 2018-04-16 | 듀크 유니버시티 | Rna-가이드된 엔도뉴클레아제를 이용하는 게놈 조작에서 특이성을 개선하는 조성물 및 방법 |
| US9926546B2 (en) | 2015-08-28 | 2018-03-27 | The General Hospital Corporation | Engineered CRISPR-Cas9 nucleases |
| US9512446B1 (en) | 2015-08-28 | 2016-12-06 | The General Hospital Corporation | Engineered CRISPR-Cas9 nucleases |
| WO2017048316A1 (en) * | 2015-09-18 | 2017-03-23 | President And Fellows Of Harvard College | Small molecule biosensors |
| CN108350454B (zh) | 2015-09-21 | 2022-05-10 | 阿克丘勒斯治疗公司 | 等位基因选择性基因编辑及其用途 |
| EP3786294A1 (en) | 2015-09-24 | 2021-03-03 | Editas Medicine, Inc. | Use of exonucleases to improve crispr/cas-mediated genome editing |
| CN105177038B (zh) * | 2015-09-29 | 2018-08-24 | 中国科学院遗传与发育生物学研究所 | 一种高效定点编辑植物基因组的CRISPR/Cas9系统 |
| BR112018006480A2 (pt) * | 2015-10-01 | 2018-10-09 | Goleini Inc | ?vetor, método de tratamento de uma doença ou condição, composição, combinação de um vetor, kit, uso de um vetor, método para a modulação in vivo da atividade eletrofisiológica de uma célula |
| WO2017069958A2 (en) | 2015-10-09 | 2017-04-27 | The Brigham And Women's Hospital, Inc. | Modulation of novel immune checkpoint targets |
| US11970710B2 (en) | 2015-10-13 | 2024-04-30 | Duke University | Genome engineering with Type I CRISPR systems in eukaryotic cells |
| JP6936952B2 (ja) * | 2015-10-16 | 2021-09-22 | アストラゼネカ アクチボラグ | 細胞ゲノムの誘導性改変 |
| KR20180133374A (ko) | 2015-10-22 | 2018-12-14 | 더 브로드 인스티튜트, 인코퍼레이티드 | 타입 vi-b crispr 효소 및 시스템 |
| WO2017070633A2 (en) | 2015-10-23 | 2017-04-27 | President And Fellows Of Harvard College | Evolved cas9 proteins for gene editing |
| EP3368687B1 (en) | 2015-10-27 | 2021-09-29 | The Broad Institute, Inc. | Compositions and methods for targeting cancer-specific sequence variations |
| WO2017075451A1 (en) | 2015-10-28 | 2017-05-04 | The Broad Institute Inc. | Compositions and methods for evaluating and modulating immune responses by detecting and targeting pou2af1 |
| US11092607B2 (en) * | 2015-10-28 | 2021-08-17 | The Board Institute, Inc. | Multiplex analysis of single cell constituents |
| WO2017075465A1 (en) | 2015-10-28 | 2017-05-04 | The Broad Institute Inc. | Compositions and methods for evaluating and modulating immune responses by detecting and targeting gata3 |
| WO2017075294A1 (en) | 2015-10-28 | 2017-05-04 | The Board Institute Inc. | Assays for massively combinatorial perturbation profiling and cellular circuit reconstruction |
| WO2017075478A2 (en) | 2015-10-28 | 2017-05-04 | The Broad Institute Inc. | Compositions and methods for evaluating and modulating immune responses by use of immune cell gene signatures |
| US11261435B2 (en) | 2015-11-05 | 2022-03-01 | Agency For Science, Technology And Research | Chemical-inducible genome engineering technology |
| WO2017087708A1 (en) | 2015-11-19 | 2017-05-26 | The Brigham And Women's Hospital, Inc. | Lymphocyte antigen cd5-like (cd5l)-interleukin 12b (p40) heterodimers in immunity |
| WO2017106657A1 (en) | 2015-12-18 | 2017-06-22 | The Broad Institute Inc. | Novel crispr enzymes and systems |
| AU2016382512A1 (en) | 2015-12-30 | 2018-07-12 | Novartis Ag | Immune effector cell therapies with enhanced efficacy |
| IL304088A (en) * | 2016-01-11 | 2023-08-01 | Univ Leland Stanford Junior | Systems containing chimeric proteins and their uses for controlling gene expression |
| MY194127A (en) | 2016-01-11 | 2022-11-14 | Univ Leland Stanford Junior | Chimeric proteins and methods of immunotherapy |
| EP3416976A2 (en) | 2016-02-16 | 2018-12-26 | Yale University | Compositions for enhancing targeted gene editing and methods of use thereof |
| CA3014795A1 (en) | 2016-02-16 | 2017-08-24 | Yale University | Compositions and methods for treatment of cystic fibrosis |
| US20190144942A1 (en) | 2016-02-22 | 2019-05-16 | Massachusetts Institute Of Technology | Methods for identifying and modulating immune phenotypes |
| WO2017147278A1 (en) | 2016-02-25 | 2017-08-31 | The Children's Medical Center Corporation | Customized class switch of immunoglobulin genes in lymphoma and hybridoma by crispr/cas9 technology |
| WO2017161325A1 (en) | 2016-03-17 | 2017-09-21 | Massachusetts Institute Of Technology | Methods for identifying and modulating co-occurant cellular phenotypes |
| CA3018729A1 (en) | 2016-03-23 | 2017-09-28 | The Regents Of The University Of California | Methods of treating mitochondrial disorders |
| WO2017165826A1 (en) | 2016-03-25 | 2017-09-28 | Editas Medicine, Inc. | Genome editing systems comprising repair-modulating enzyme molecules and methods of their use |
| EP3433364A1 (en) | 2016-03-25 | 2019-01-30 | Editas Medicine, Inc. | Systems and methods for treating alpha 1-antitrypsin (a1at) deficiency |
| US20190117799A1 (en) | 2016-04-01 | 2019-04-25 | The Brigham And Women's Hospital, Inc. | Stimuli-responsive nanoparticles for biomedical applications |
| WO2017180694A1 (en) | 2016-04-13 | 2017-10-19 | Editas Medicine, Inc. | Cas9 fusion molecules gene editing systems, and methods of use thereof |
| EP3443088A1 (en) | 2016-04-13 | 2019-02-20 | Editas Medicine, Inc. | Grna fusion molecules, gene editing systems, and methods of use thereof |
| EP3445853A1 (en) | 2016-04-19 | 2019-02-27 | The Broad Institute, Inc. | Cpf1 complexes with reduced indel activity |
| CA3026110A1 (en) | 2016-04-19 | 2017-11-02 | The Broad Institute, Inc. | Novel crispr enzymes and systems |
| CA3026055A1 (en) | 2016-04-19 | 2017-10-26 | The Broad Institute, Inc. | Novel crispr enzymes and systems |
| EP3448997B1 (en) | 2016-04-27 | 2020-10-14 | Massachusetts Institute of Technology | Stable nanoscale nucleic acid assemblies and methods thereof |
| WO2017189914A1 (en) | 2016-04-27 | 2017-11-02 | Massachusetts Institute Of Technology | Sequence-controlled polymer random access memory storage |
| US11214789B2 (en) | 2016-05-03 | 2022-01-04 | Flodesign Sonics, Inc. | Concentration and washing of particles with acoustics |
| US11085035B2 (en) | 2016-05-03 | 2021-08-10 | Flodesign Sonics, Inc. | Therapeutic cell washing, concentration, and separation utilizing acoustophoresis |
| CN115837006A (zh) | 2016-05-06 | 2023-03-24 | 布里格姆及妇女医院股份有限公司 | 用于向软骨中受控递送被囊封的药剂的二元自组装凝胶 |
| WO2017197128A1 (en) | 2016-05-11 | 2017-11-16 | Yale University | Poly(amine-co-ester) nanoparticles and methods of use thereof |
| GB201609811D0 (en) | 2016-06-05 | 2016-07-20 | Snipr Technologies Ltd | Methods, cells, systems, arrays, RNA and kits |
| CA3028158A1 (en) | 2016-06-17 | 2017-12-21 | The Broad Institute, Inc. | Type vi crispr orthologs and systems |
| US11293021B1 (en) | 2016-06-23 | 2022-04-05 | Inscripta, Inc. | Automated cell processing methods, modules, instruments, and systems |
| WO2018005873A1 (en) | 2016-06-29 | 2018-01-04 | The Broad Institute Inc. | Crispr-cas systems having destabilization domain |
| BR112019000057A2 (pt) * | 2016-07-05 | 2019-04-02 | The Johns Hopkins University | composições e métodos com base em crispr/cas9 para o tratamento de degeneração de retina |
| US11459587B2 (en) | 2016-07-06 | 2022-10-04 | Vertex Pharmaceuticals Incorporated | Materials and methods for treatment of pain related disorders |
| AU2017292169B2 (en) | 2016-07-06 | 2021-12-23 | Vertex Pharmaceuticals Incorporated | Materials and methods for treatment of pain related disorders |
| WO2018013840A1 (en) | 2016-07-13 | 2018-01-18 | Vertex Pharmaceuticals Incorporated | Methods, compositions and kits for increasing genome editing efficiency |
| WO2018020323A2 (en) * | 2016-07-25 | 2018-02-01 | Crispr Therapeutics Ag | Materials and methods for treatment of fatty acid disorders |
| JP7200104B2 (ja) * | 2016-08-01 | 2023-01-06 | ヴァイロジン バイオテック カナダ リミテッド | 免疫系刺激分子を発現する腫瘍溶解性単純ヘルペスウイルスベクター |
| US11566263B2 (en) | 2016-08-02 | 2023-01-31 | Editas Medicine, Inc. | Compositions and methods for treating CEP290 associated disease |
| SG11201900907YA (en) | 2016-08-03 | 2019-02-27 | Harvard College | Adenosine nucleobase editors and uses thereof |
| EP3497214B1 (en) | 2016-08-09 | 2023-06-28 | President and Fellows of Harvard College | Programmable cas9-recombinase fusion proteins and uses thereof |
| WO2018035387A1 (en) | 2016-08-17 | 2018-02-22 | The Broad Institute, Inc. | Novel crispr enzymes and systems |
| US20210166783A1 (en) | 2016-08-17 | 2021-06-03 | The Broad Institute, Inc. | Methods for identifying class 2 crispr-cas systems |
| US11352647B2 (en) | 2016-08-17 | 2022-06-07 | The Broad Institute, Inc. | Crispr enzymes and systems |
| US11630103B2 (en) | 2016-08-17 | 2023-04-18 | The Broad Institute, Inc. | Product and methods useful for modulating and evaluating immune responses |
| WO2018039438A1 (en) | 2016-08-24 | 2018-03-01 | President And Fellows Of Harvard College | Incorporation of unnatural amino acids into proteins using base editing |
| WO2018044920A1 (en) | 2016-08-29 | 2018-03-08 | The Regents Of The University Of California | Topical formulations based on ionic species for skin treatment |
| WO2018049025A2 (en) | 2016-09-07 | 2018-03-15 | The Broad Institute Inc. | Compositions and methods for evaluating and modulating immune responses |
| SG11201901718VA (en) * | 2016-09-08 | 2019-03-28 | Centro De Investigaciones Energeticas Medioambientales Y Tecnologicas | Gene therapy for patients with fanconi anemia |
| WO2018067991A1 (en) | 2016-10-07 | 2018-04-12 | The Brigham And Women's Hospital, Inc. | Modulation of novel immune checkpoint targets |
| KR20190084053A (ko) | 2016-10-13 | 2019-07-15 | 주노 쎄러퓨티크스 인코퍼레이티드 | 트립토판 대사 경로 조절인자 관련 면역 치료 방법 및 조성물 |
| CA3039928A1 (en) | 2016-10-14 | 2018-04-19 | President And Fellows Of Harvard College | Aav delivery of nucleobase editors |
| CN110520530A (zh) | 2016-10-18 | 2019-11-29 | 明尼苏达大学董事会 | 肿瘤浸润性淋巴细胞和治疗方法 |
| CA3041517A1 (en) | 2016-10-19 | 2018-04-26 | Flodesign Sonics, Inc. | Affinity cell extraction by acoustics |
| WO2018081138A1 (en) | 2016-10-24 | 2018-05-03 | Yale University | Biodegradable contraceptive implants |
| WO2018083606A1 (en) | 2016-11-01 | 2018-05-11 | Novartis Ag | Methods and compositions for enhancing gene editing |
| WO2018144097A1 (en) | 2016-11-04 | 2018-08-09 | Akeagen Llc | Genetically modified non-human animals and methods for producing heavy chain-only antibodies |
| US20190359661A1 (en) * | 2016-11-25 | 2019-11-28 | Nanoscope Technologies, LLC | Method and device for pain modulation by optical activation of neurons and other cells |
| US20200085758A1 (en) | 2016-12-16 | 2020-03-19 | The Brigham And Women's Hospital, Inc. | Co-delivery of nucleic acids for simultaneous suppression and expression of target genes |
| WO2018119359A1 (en) | 2016-12-23 | 2018-06-28 | President And Fellows Of Harvard College | Editing of ccr5 receptor gene to protect against hiv infection |
| CN110139676A (zh) | 2016-12-29 | 2019-08-16 | 应用干细胞有限公司 | 使用病毒的基因编辑方法 |
| TW201839136A (zh) | 2017-02-06 | 2018-11-01 | 瑞士商諾華公司 | 治療血色素異常症之組合物及方法 |
| US11898179B2 (en) | 2017-03-09 | 2024-02-13 | President And Fellows Of Harvard College | Suppression of pain by gene editing |
| US11542496B2 (en) | 2017-03-10 | 2023-01-03 | President And Fellows Of Harvard College | Cytosine to guanine base editor |
| WO2018170184A1 (en) | 2017-03-14 | 2018-09-20 | Editas Medicine, Inc. | Systems and methods for the treatment of hemoglobinopathies |
| EP4361261A2 (en) | 2017-03-15 | 2024-05-01 | The Broad Institute Inc. | Novel cas13b orthologues crispr enzymes and systems |
| US20200115753A1 (en) | 2017-03-17 | 2020-04-16 | Massachusetts Institute Of Technology | Methods for identifying and modulating co-occurant cellular phenotypes |
| EP3595634B1 (en) | 2017-03-17 | 2024-03-27 | Rescue Hearing Inc | Gene therapy constructs and methods for treatment of hearing loss |
| IL306092A (en) | 2017-03-23 | 2023-11-01 | Harvard College | Nucleic base editors that include nucleic acid programmable DNA binding proteins |
| WO2018183908A1 (en) | 2017-03-31 | 2018-10-04 | Dana-Farber Cancer Institute, Inc. | Compositions and methods for treating ovarian tumors |
| WO2018183921A1 (en) | 2017-04-01 | 2018-10-04 | The Broad Institute, Inc. | Methods and compositions for detecting and modulating an immunotherapy resistance gene signature in cancer |
| WO2018187493A1 (en) | 2017-04-04 | 2018-10-11 | Yale University | Compositions and methods for in utero delivery |
| WO2018191553A1 (en) | 2017-04-12 | 2018-10-18 | Massachusetts Eye And Ear Infirmary | Tumor signature for metastasis, compositions of matter methods of use thereof |
| WO2018191520A1 (en) | 2017-04-12 | 2018-10-18 | The Broad Institute, Inc. | Respiratory and sweat gland ionocytes |
| US11840711B2 (en) | 2017-04-12 | 2023-12-12 | The Broad Institute, Inc. | Type VI CRISPR orthologs and systems |
| US20200405639A1 (en) | 2017-04-14 | 2020-12-31 | The Broad Institute, Inc. | Novel delivery of large payloads |
| EP3612629A1 (en) | 2017-04-18 | 2020-02-26 | The Broad Institute, Inc. | Compositions for detecting secretion and methods of use |
| US20200384115A1 (en) | 2017-04-21 | 2020-12-10 | The Broad Institute , Inc. | Targeted delivery to beta cells |
| EP3615672A1 (en) | 2017-04-28 | 2020-03-04 | Editas Medicine, Inc. | Methods and systems for analyzing guide rna molecules |
| WO2018204777A2 (en) | 2017-05-05 | 2018-11-08 | The Broad Institute, Inc. | Methods for identification and modification of lncrna associated with target genotypes and phenotypes |
| US20200124505A1 (en) * | 2017-05-09 | 2020-04-23 | Ultragenyx Pharmaceutical Inc. | Scalable method for producing transfection reagents |
| WO2018209158A2 (en) | 2017-05-10 | 2018-11-15 | Editas Medicine, Inc. | Crispr/rna-guided nuclease systems and methods |
| US11560566B2 (en) | 2017-05-12 | 2023-01-24 | President And Fellows Of Harvard College | Aptazyme-embedded guide RNAs for use with CRISPR-Cas9 in genome editing and transcriptional activation |
| EP3625343A1 (en) | 2017-05-18 | 2020-03-25 | The Broad Institute, Inc. | Systems, methods, and compositions for targeted nucleic acid editing |
| CA3102054A1 (en) | 2017-06-05 | 2018-12-13 | Fred Hutchinson Cancer Research Center | Genomic safe harbors for genetic therapies in human stem cells and engineered nanoparticles to provide targeted genetic therapies |
| JP2020524497A (ja) | 2017-06-09 | 2020-08-20 | エディタス・メディシン,インコーポレイテッド | 操作されたcas9ヌクレアーゼ |
| KR102634727B1 (ko) * | 2017-06-14 | 2024-02-07 | 위스콘신 얼럼나이 리서어치 화운데이션 | 변형된 가이드 rna, crispr-리보뉴클레오프로테인 복합체 및 사용 방법 |
| WO2018232195A1 (en) | 2017-06-14 | 2018-12-20 | The Broad Institute, Inc. | Compositions and methods targeting complement component 3 for inhibiting tumor growth |
| US10011849B1 (en) | 2017-06-23 | 2018-07-03 | Inscripta, Inc. | Nucleic acid-guided nucleases |
| US9982279B1 (en) | 2017-06-23 | 2018-05-29 | Inscripta, Inc. | Nucleic acid-guided nucleases |
| AU2018290843A1 (en) | 2017-06-26 | 2020-01-16 | Massachusetts Institute Of Technology | CRISPR/Cas-adenine deaminase based compositions, systems, and methods for targeted nucleic acid editing |
| US20200140896A1 (en) | 2017-06-30 | 2020-05-07 | Novartis Ag | Methods for the treatment of disease with gene editing systems |
| RU2021123421A (ru) | 2017-06-30 | 2021-09-10 | Инскрипта, Инк. | Автоматизированные способы обработки клеток, модули, инструменты и системы |
| WO2019006418A2 (en) | 2017-06-30 | 2019-01-03 | Intima Bioscience, Inc. | ADENO-ASSOCIATED VIRAL VECTORS FOR GENE THERAPY |
| CN109207518B (zh) * | 2017-07-07 | 2020-12-01 | 中国科学院动物研究所 | 用于基因转录激活的药物诱导型CRISPR/Cas9系统 |
| CN109207517B (zh) * | 2017-07-07 | 2020-12-01 | 中国科学院动物研究所 | 用于基因组编辑和转录调控的药物诱导型CRISPR/Cas9系统 |
| CN109207516B (zh) * | 2017-07-07 | 2020-12-01 | 中国科学院动物研究所 | 一种药物诱导的基因转录激活方法 |
| CN109206520A (zh) * | 2017-07-07 | 2019-01-15 | 中国科学院动物研究所 | 用于基因组编辑的药物诱导型融合蛋白及其编码基因与应用 |
| CN111183226A (zh) * | 2017-07-11 | 2020-05-19 | 西格马-奥尔德里奇有限责任公司 | 使用核小体相互作用蛋白结构域来增强靶向基因组修饰 |
| US11866726B2 (en) | 2017-07-14 | 2024-01-09 | Editas Medicine, Inc. | Systems and methods for targeted integration and genome editing and detection thereof using integrated priming sites |
| US11732274B2 (en) | 2017-07-28 | 2023-08-22 | President And Fellows Of Harvard College | Methods and compositions for evolving base editors using phage-assisted continuous evolution (PACE) |
| EP3662061A4 (en) * | 2017-07-31 | 2021-05-05 | Sigma-Aldrich Co. LLC | RNA SYNTHETIC GUIDE FOR CRISPR / CAS ACTIVATOR SYSTEMS |
| EP3663310A4 (en) | 2017-08-04 | 2021-08-11 | Peking University | TALE-RVD WITH SPECIFIC DETECTION OF A DNA BASE MODIFIED BY METHYLATION AND APPLICATION OF IT |
| US11624077B2 (en) | 2017-08-08 | 2023-04-11 | Peking University | Gene knockout method |
| CN111465857A (zh) * | 2017-08-08 | 2020-07-28 | 昆士兰科技大学 | 诊断早期心力衰竭的方法 |
| EP3664854A4 (en) * | 2017-08-10 | 2021-04-21 | University of Massachusetts | HUMAN ADAPTIVE TISSUE PRECURSORS FOR AUTOLOGOUS CELL THERAPY FOR LIPODYSTROPHY |
| US10738327B2 (en) | 2017-08-28 | 2020-08-11 | Inscripta, Inc. | Electroporation cuvettes for automation |
| US11319532B2 (en) | 2017-08-30 | 2022-05-03 | President And Fellows Of Harvard College | High efficiency base editors comprising Gam |
| CN109495227B (zh) * | 2017-09-11 | 2020-10-20 | 维沃移动通信有限公司 | 一种控制资源集的配置方法、网络设备及终端 |
| WO2019052577A1 (zh) | 2017-09-18 | 2019-03-21 | 博雅辑因(北京)生物科技有限公司 | 一种基因编辑t细胞及其用途 |
| EP3684172A4 (en) * | 2017-09-19 | 2021-06-23 | The State of Israel, Ministry of Agriculture & Rural Development, Agricultural Research Organization (ARO) (Volcani Center) | GENOME EDITING BIRDS |
| CA3073848A1 (en) | 2017-09-21 | 2019-03-28 | The Broad Institute, Inc. | Systems, methods, and compositions for targeted nucleic acid editing |
| US10443074B2 (en) | 2017-09-30 | 2019-10-15 | Inscripta, Inc. | Modification of cells by introduction of exogenous material |
| US20200255828A1 (en) | 2017-10-04 | 2020-08-13 | The Broad Institute, Inc. | Methods and compositions for altering function and structure of chromatin loops and/or domains |
| WO2019071240A1 (en) | 2017-10-06 | 2019-04-11 | The Research Foundation For The State University For The State Of New York | AQUEOUS AND NONAQUEOUS SELECTIVE OPTICAL DETECTION OF FREE SULPHITES |
| WO2019079347A1 (en) | 2017-10-16 | 2019-04-25 | The Broad Institute, Inc. | USES OF BASIC EDITORS ADENOSINE |
| US11680296B2 (en) | 2017-10-16 | 2023-06-20 | Massachusetts Institute Of Technology | Mycobacterium tuberculosis host-pathogen interaction |
| KR101870694B1 (ko) * | 2017-10-18 | 2018-06-25 | 주식회사 무진메디 | Cas9 단백질, SRD5A2 유전자의 발현을 억제하는 가이드 RNA 및 양이온성 폴리머의 복합체가 봉입된 나노 리포좀-마이크로버블 결합체 및 이를 함유하는 탈모 개선 또는 치료용 조성물 |
| BR112020007823A2 (pt) | 2017-10-20 | 2020-10-27 | Fred Hutchinson Cancer Research Center | sistemas e métodos para produzir células b geneticamente modificadas para expressar anticorpos selecionados |
| WO2019135816A2 (en) * | 2017-10-23 | 2019-07-11 | The Broad Institute, Inc. | Novel nucleic acid modifiers |
| CN109722415B (zh) | 2017-10-27 | 2021-01-26 | 博雅辑因(北京)生物科技有限公司 | 一种造血干细胞的培养组合物、培养基以及造血干细胞的培养方法 |
| WO2019089803A1 (en) | 2017-10-31 | 2019-05-09 | The Broad Institute, Inc. | Methods and compositions for studying cell evolution |
| EP3704245A1 (en) | 2017-11-01 | 2020-09-09 | Novartis AG | Synthetic rnas and methods of use |
| WO2019094928A1 (en) | 2017-11-10 | 2019-05-16 | Massachusetts Institute Of Technology | Microbial production of pure single stranded nucleic acids |
| EP3710583A1 (en) | 2017-11-16 | 2020-09-23 | Astrazeneca AB | Compositions and methods for improving the efficacy of cas9-based knock-in strategies |
| CN107858430B (zh) * | 2017-11-20 | 2019-01-04 | 武汉迈特维尔生物科技有限公司 | 一种用于诊断预示Her-2过表达型乳腺癌骨转移的基因诊断试剂盒 |
| EP3713644A1 (en) | 2017-11-20 | 2020-09-30 | University of Georgia Research Foundation, Inc. | Compositions and methods for modulating hif-2& x391; to improve muscle generation and repair |
| EP3717505A4 (en) * | 2017-12-01 | 2021-12-01 | Encoded Therapeutics, Inc. | GENENICALLY MODIFIED DNA-BINDING PROTEINS |
| US11332736B2 (en) | 2017-12-07 | 2022-05-17 | The Broad Institute, Inc. | Methods and compositions for multiplexing single cell and single nuclei sequencing |
| JP2021507561A (ja) | 2017-12-14 | 2021-02-22 | フロデザイン ソニックス, インク.Flodesign Sonics, Inc. | 音響トランスデューサドライバ及びコントローラ |
| WO2019126643A1 (en) * | 2017-12-22 | 2019-06-27 | Massachusetts Institute Of Technology | Compositions and methods for williams syndrome (ws) therapy |
| CN111566483A (zh) * | 2018-01-12 | 2020-08-21 | 细胞基因公司 | 筛选小脑蛋白改性化合物的方法 |
| JP7466448B2 (ja) | 2018-01-17 | 2024-04-12 | バーテックス ファーマシューティカルズ インコーポレイテッド | Dna-pk阻害剤 |
| CN111757876B (zh) | 2018-01-17 | 2024-03-22 | 沃泰克斯药物股份有限公司 | Dna-pk抑制剂 |
| CN109507429A (zh) * | 2018-02-14 | 2019-03-22 | 复旦大学 | 基于CRISPR/cas9和过氧化物酶APEX2系统识别分析特异性位点相互作用蛋白的方法 |
| US10760075B2 (en) | 2018-04-30 | 2020-09-01 | Snipr Biome Aps | Treating and preventing microbial infections |
| US10435662B1 (en) | 2018-03-29 | 2019-10-08 | Inscripta, Inc. | Automated control of cell growth rates for induction and transformation |
| CN108410909B (zh) * | 2018-04-12 | 2021-08-03 | 吉林大学 | 一种利用植物激素aba及小分子物质pyr调节细胞通路的方法 |
| US10376889B1 (en) | 2018-04-13 | 2019-08-13 | Inscripta, Inc. | Automated cell processing instruments comprising reagent cartridges |
| EP3781711A4 (en) | 2018-04-19 | 2022-01-26 | Massachusetts Institute Of Technology | SINGLE STRAND BREAK DETECTION IN DOUBLE STRAND DNA |
| US10858761B2 (en) | 2018-04-24 | 2020-12-08 | Inscripta, Inc. | Nucleic acid-guided editing of exogenous polynucleotides in heterologous cells |
| US10557216B2 (en) | 2018-04-24 | 2020-02-11 | Inscripta, Inc. | Automated instrumentation for production of T-cell receptor peptide libraries |
| US10508273B2 (en) | 2018-04-24 | 2019-12-17 | Inscripta, Inc. | Methods for identifying selective binding pairs |
| DK3560330T3 (da) | 2018-04-24 | 2022-07-11 | Kws Saat Se & Co Kgaa | Planter med forbedret fordøjelighed og markørhaplotyper |
| US11957695B2 (en) | 2018-04-26 | 2024-04-16 | The Broad Institute, Inc. | Methods and compositions targeting glucocorticoid signaling for modulating immune responses |
| US20210147831A1 (en) | 2018-04-27 | 2021-05-20 | The Broad Institute, Inc. | Sequencing-based proteomics |
| US20210386829A1 (en) | 2018-05-04 | 2021-12-16 | The Broad Institute, Inc. | Compositions and methods for modulating cgrp signaling to regulate innate lymphoid cell inflammatory responses |
| JP2021523745A (ja) | 2018-05-16 | 2021-09-09 | シンテゴ コーポレイション | ガイドrna設計および使用のための方法およびシステム |
| US20210371932A1 (en) | 2018-06-01 | 2021-12-02 | Massachusetts Institute Of Technology | Methods and compositions for detecting and modulating microenvironment gene signatures from the csf of metastasis patients |
| US10227576B1 (en) | 2018-06-13 | 2019-03-12 | Caribou Biosciences, Inc. | Engineered cascade components and cascade complexes |
| KR20210024009A (ko) | 2018-06-26 | 2021-03-04 | 더 브로드 인스티튜트, 인코퍼레이티드 | Crispr/cas 및 트랜스포사제 기반 증폭 조성물, 시스템 및 방법 |
| WO2020006036A1 (en) | 2018-06-26 | 2020-01-02 | Massachusetts Institute Of Technology | Crispr effector system based amplification methods, systems, and diagnostics |
| EP3813974A4 (en) | 2018-06-30 | 2022-08-03 | Inscripta, Inc. | INSTRUMENTS, MODULES AND METHODS FOR ENHANCED DETECTION OF EDITED SEQUENCES IN LIVING CELLS |
| EP3807399A1 (en) * | 2018-07-13 | 2021-04-21 | Lonza Ltd. | Methods for improving production of biological products by reducing the level of endogenous protein |
| US20210317412A1 (en) * | 2018-07-16 | 2021-10-14 | Agency For Science, Technology And Research | Method For Isolating A Cardiomyocyte Population |
| WO2020028555A2 (en) | 2018-07-31 | 2020-02-06 | The Broad Institute, Inc. | Novel crispr enzymes and systems |
| US11939593B2 (en) | 2018-08-01 | 2024-03-26 | University Of Georgia Research Foundation, Inc. | Compositions and methods for improving embryo development |
| AU2019318079A1 (en) | 2018-08-07 | 2021-01-28 | Massachusetts Institute Of Technology | Novel Cas12b enzymes and systems |
| US10752874B2 (en) | 2018-08-14 | 2020-08-25 | Inscripta, Inc. | Instruments, modules, and methods for improved detection of edited sequences in live cells |
| US11142740B2 (en) | 2018-08-14 | 2021-10-12 | Inscripta, Inc. | Detection of nuclease edited sequences in automated modules and instruments |
| US10532324B1 (en) | 2018-08-14 | 2020-01-14 | Inscripta, Inc. | Instruments, modules, and methods for improved detection of edited sequences in live cells |
| WO2020041380A1 (en) | 2018-08-20 | 2020-02-27 | The Broad Institute, Inc. | Methods and compositions for optochemical control of crispr-cas9 |
| WO2020041387A1 (en) | 2018-08-20 | 2020-02-27 | The Brigham And Women's Hospital, Inc. | Degradation domain modifications for spatio-temporal control of rna-guided nucleases |
| CN110857440B (zh) * | 2018-08-23 | 2021-02-19 | 武汉纽福斯生物科技有限公司 | 重组人ⅱ型线粒体动力蛋白样gtp酶基因序列及其应用 |
| AU2019363487A1 (en) | 2018-08-30 | 2021-04-15 | Inscripta, Inc. | Improved detection of nuclease edited sequences in automated modules and instruments |
| WO2020047353A1 (en) | 2018-08-31 | 2020-03-05 | Yale University | Compositions and methods for enhancing triplex and nuclease-based gene editing |
| EP3847650A1 (en) | 2018-09-06 | 2021-07-14 | The Broad Institute, Inc. | Nucleic acid assemblies for use in targeted delivery |
| CN112654702A (zh) | 2018-09-07 | 2021-04-13 | 阿斯利康(瑞典)有限公司 | 改进的核酸酶的组合物和方法 |
| US20220098613A1 (en) | 2018-09-12 | 2022-03-31 | Fred Hutchinson Cancer Research Center | Reducing cd33 expression to selectively protect therapeutic cells |
| CN113164623A (zh) | 2018-09-18 | 2021-07-23 | 维恩维纽克公司 | 基于arc的衣壳及其用途 |
| CN109408766B (zh) * | 2018-09-26 | 2023-05-16 | 国网山西省电力公司电力科学研究院 | 一种梯形图频率计算的方法 |
| CN109557315B (zh) * | 2018-09-28 | 2020-04-17 | 山东大学 | 一种光控微管示踪剂及其应用 |
| WO2020077236A1 (en) | 2018-10-12 | 2020-04-16 | The Broad Institute, Inc. | Method for extracting nuclei or whole cells from formalin-fixed paraffin-embedded tissues |
| US11851663B2 (en) | 2018-10-14 | 2023-12-26 | Snipr Biome Aps | Single-vector type I vectors |
| WO2020081730A2 (en) | 2018-10-16 | 2020-04-23 | Massachusetts Institute Of Technology | Methods and compositions for modulating microenvironment |
| US11214781B2 (en) | 2018-10-22 | 2022-01-04 | Inscripta, Inc. | Engineered enzyme |
| CN113227368B (zh) | 2018-10-22 | 2023-07-07 | 因思科瑞普特公司 | 工程化酶 |
| CN109406469B (zh) * | 2018-10-24 | 2021-04-09 | 中国医科大学 | 基于蛋白结合诱导dna双链变构的检测色氨酸的方法 |
| WO2020092057A1 (en) | 2018-10-30 | 2020-05-07 | Yale University | Compositions and methods for rapid and modular generation of chimeric antigen receptor t cells |
| JPWO2020091069A1 (ja) * | 2018-11-01 | 2021-09-30 | 国立大学法人 東京大学 | 2分割されたCpf1タンパク質 |
| US11739320B2 (en) | 2018-11-05 | 2023-08-29 | Wisconsin Alumni Research Foundation | Gene correction of Pompe disease and other autosomal recessive disorders via RNA-guided nucleases |
| CN111655042A (zh) | 2018-11-08 | 2020-09-11 | 特里同阿盖亚创新公司 | 藻类中过量产生原卟啉ix的方法及由此产生的组合物 |
| WO2020097411A1 (en) * | 2018-11-08 | 2020-05-14 | Massachusetts Institute Of Technology | Encryption and steganography of synthetic gene circuits |
| WO2020112195A1 (en) | 2018-11-30 | 2020-06-04 | Yale University | Compositions, technologies and methods of using plerixafor to enhance gene editing |
| CN113166744A (zh) | 2018-12-14 | 2021-07-23 | 先锋国际良种公司 | 用于基因组编辑的新颖crispr-cas系统 |
| US20220062394A1 (en) | 2018-12-17 | 2022-03-03 | The Broad Institute, Inc. | Methods for identifying neoantigens |
| EP3898958A1 (en) | 2018-12-17 | 2021-10-27 | The Broad Institute, Inc. | Crispr-associated transposase systems and methods of use thereof |
| US11739156B2 (en) | 2019-01-06 | 2023-08-29 | The Broad Institute, Inc. Massachusetts Institute of Technology | Methods and compositions for overcoming immunosuppression |
| CN113557036A (zh) * | 2019-01-18 | 2021-10-26 | 奥泽生物疗法公司 | 基因编辑以改善关节功能 |
| EP3914708A1 (en) | 2019-01-24 | 2021-12-01 | Massachusetts Institute Of Technology | Nucleic acid nanostructure platform for antigen presentation and vaccine formulations formed therefrom |
| WO2020178759A1 (en) | 2019-03-04 | 2020-09-10 | King Abdullah University Of Science And Technology | Compositions and methods of targeted nucleic acid enrichment by loop adapter protection and exonuclease digestion |
| WO2020178822A1 (en) | 2019-03-05 | 2020-09-10 | The State Of Israel, Ministry Of Agriculture & Rural Development, Agricultural Research Organization (Aro) (Volcani Center) | Genome-edited birds |
| US20220177863A1 (en) | 2019-03-18 | 2022-06-09 | The Broad Institute, Inc. | Type vii crispr proteins and systems |
| AU2020242032A1 (en) | 2019-03-19 | 2021-10-07 | Massachusetts Institute Of Technology | Methods and compositions for editing nucleotide sequences |
| US11001831B2 (en) | 2019-03-25 | 2021-05-11 | Inscripta, Inc. | Simultaneous multiplex genome editing in yeast |
| US10815467B2 (en) | 2019-03-25 | 2020-10-27 | Inscripta, Inc. | Simultaneous multiplex genome editing in yeast |
| WO2020206036A1 (en) | 2019-04-01 | 2020-10-08 | The Broad Institute, Inc. | Novel nucleic acid modifier |
| CN111778277B (zh) * | 2019-04-04 | 2023-02-28 | 中国科学院动物研究所 | 克氏综合征动物模型和其应用 |
| WO2020214619A1 (en) * | 2019-04-15 | 2020-10-22 | Emendobio Inc. | Crispr compositions and methods for promoting gene editing of gata2 |
| CN113766932A (zh) * | 2019-04-18 | 2021-12-07 | 米特健康有限公司 | 用于治疗呼吸性心律失常的方法和组合物 |
| CN113785059A (zh) | 2019-04-30 | 2021-12-10 | 博雅辑因(北京)生物科技有限公司 | 一种血红蛋白病治疗有效性预测方法 |
| BR112021022411A2 (pt) | 2019-05-13 | 2022-03-15 | Kws Saat Se & Co Kgaa | Tolerância à seca em milho |
| US20220220469A1 (en) | 2019-05-20 | 2022-07-14 | The Broad Institute, Inc. | Non-class i multi-component nucleic acid targeting systems |
| US20220235340A1 (en) | 2019-05-20 | 2022-07-28 | The Broad Institute, Inc. | Novel crispr-cas systems and uses thereof |
| AR118995A1 (es) | 2019-05-25 | 2021-11-17 | Kws Saat Se & Co Kgaa | Mejorador de la inducción de haploides |
| US20220243178A1 (en) | 2019-05-31 | 2022-08-04 | The Broad Institute, Inc. | Methods for treating metabolic disorders by targeting adcy5 |
| EP3953477A4 (en) | 2019-06-06 | 2022-06-22 | Inscripta, Inc. | HARDENING FOR RECURSIVE NUCLEIC ACID-DRIVEN CELL EDITING |
| CN110244048A (zh) * | 2019-06-19 | 2019-09-17 | 中国人民解放军总医院第八医学中心 | Serping1蛋白作为标志物在开发诊断活动性结核病的试剂中的应用 |
| US10907125B2 (en) | 2019-06-20 | 2021-02-02 | Inscripta, Inc. | Flow through electroporation modules and instrumentation |
| CA3139124C (en) | 2019-06-21 | 2023-01-31 | Inscripta, Inc. | Genome-wide rationally-designed mutations leading to enhanced lysine production in e. coli |
| US10927385B2 (en) | 2019-06-25 | 2021-02-23 | Inscripta, Inc. | Increased nucleic-acid guided cell editing in yeast |
| US11905532B2 (en) | 2019-06-25 | 2024-02-20 | Massachusetts Institute Of Technology | Compositions and methods for molecular memory storage and retrieval |
| CN110373459B (zh) * | 2019-06-27 | 2023-06-27 | 郑湘榕 | 基于STIP1基因rs2236647位点用于儿童哮喘病检测的试剂、试剂盒及应用 |
| BR112021026832A2 (pt) | 2019-07-02 | 2022-05-10 | Hutchinson Fred Cancer Res | Vetores ad35 recombinantes e aprimoramentos da terapia gênica relacionada |
| KR102183208B1 (ko) * | 2019-07-25 | 2020-11-25 | 한국과학기술연구원 | Ngn2(neurogenin-2)를 이용하여 척수 손상 모델에서 반응성 성상세포를 신경세포로 리프로그래밍하는 방법 |
| CN110551755A (zh) * | 2019-07-26 | 2019-12-10 | 天津大学 | 一种光控蛋白降解系统及构建方法和光调控蛋白降解的方法 |
| EP3772542A1 (en) | 2019-08-07 | 2021-02-10 | KWS SAAT SE & Co. KGaA | Modifying genetic variation in crops by modulating the pachytene checkpoint protein 2 |
| CN110563822B (zh) * | 2019-08-27 | 2021-09-24 | 上海交通大学 | 灵芝免疫调节蛋白突变体及应用 |
| WO2021041922A1 (en) | 2019-08-30 | 2021-03-04 | The Broad Institute, Inc. | Crispr-associated mu transposase systems |
| AU2020336992A1 (en) | 2019-08-30 | 2022-04-14 | Yale University | Compositions and methods for delivery of nucleic acids to cells |
| KR20220053671A (ko) | 2019-09-04 | 2022-04-29 | 에디진 인크. | 오프 타겟 평가 기반의 유전자 편집 치료의 평가 방법 |
| WO2021050974A1 (en) | 2019-09-12 | 2021-03-18 | The Broad Institute, Inc. | Engineered adeno-associated virus capsids |
| WO2021055874A1 (en) | 2019-09-20 | 2021-03-25 | The Broad Institute, Inc. | Novel type vi crispr enzymes and systems |
| JP2022169813A (ja) * | 2019-09-30 | 2022-11-10 | 国立研究開発法人産業技術総合研究所 | Cas9ヌクレアーゼのDNA切断活性を制御する方法 |
| EP4045522A1 (en) | 2019-10-17 | 2022-08-24 | KWS SAAT SE & Co. KGaA | Enhanced disease resistance of crops by downregulation of repressor genes |
| EP4048792A4 (en) * | 2019-10-21 | 2024-02-28 | Univ California | COMPOSITIONS AND METHODS OF EDITING THE CDKL5 GENE |
| WO2021102059A1 (en) | 2019-11-19 | 2021-05-27 | Inscripta, Inc. | Methods for increasing observed editing in bacteria |
| US20230031670A1 (en) * | 2019-12-02 | 2023-02-02 | Counci of Scientific & Industrial Research | Method and kit for detection of polynucleotide |
| CN114787347A (zh) | 2019-12-10 | 2022-07-22 | 因思科瑞普特公司 | 新颖的mad核酸酶 |
| US10704033B1 (en) | 2019-12-13 | 2020-07-07 | Inscripta, Inc. | Nucleic acid-guided nucleases |
| CA3157127A1 (en) | 2019-12-18 | 2021-06-24 | Aamir MIR | Cascade/dcas3 complementation assays for in vivo detection of nucleic acid-guided nuclease edited cells |
| JP2023509058A (ja) | 2019-12-30 | 2023-03-06 | 博雅▲緝▼因(北京)生物科技有限公司 | T細胞リンパ腫細胞を標的とするユニバーサルcar-tならびにその調製方法及び使用 |
| US20230054266A1 (en) | 2019-12-30 | 2023-02-23 | Edigene Biotechnology Inc. | Method for purifying ucart cell and use thereof |
| WO2021138560A2 (en) | 2020-01-02 | 2021-07-08 | The Trustees Of Columbia University In The City Of New York | Programmable and portable crispr-cas transcriptional activation in bacteria |
| US10689669B1 (en) | 2020-01-11 | 2020-06-23 | Inscripta, Inc. | Automated multi-module cell processing methods, instruments, and systems |
| WO2021154706A1 (en) | 2020-01-27 | 2021-08-05 | Inscripta, Inc. | Electroporation modules and instrumentation |
| EP3872190A1 (en) | 2020-02-26 | 2021-09-01 | Antibodies-Online GmbH | A method of using cut&run or cut&tag to validate crispr-cas targeting |
| CN111376422B (zh) * | 2020-04-18 | 2021-08-27 | 山东宇能环境工程有限公司 | 一种保温毡的自动成型生产线 |
| US20210332388A1 (en) | 2020-04-24 | 2021-10-28 | Inscripta, Inc. | Compositions, methods, modules and instruments for automated nucleic acid-guided nuclease editing in mammalian cells |
| CN111534590A (zh) * | 2020-04-30 | 2020-08-14 | 福建西陇生物技术有限公司 | 一种结直肠癌多基因甲基化联合检测试剂盒及其应用 |
| KR20230019843A (ko) | 2020-05-08 | 2023-02-09 | 더 브로드 인스티튜트, 인코퍼레이티드 | 표적 이중 가닥 뉴클레오티드 서열의 두 가닥의 동시 편집을 위한 방법 및 조성물 |
| US11787841B2 (en) | 2020-05-19 | 2023-10-17 | Inscripta, Inc. | Rationally-designed mutations to the thrA gene for enhanced lysine production in E. coli |
| UY39237A (es) | 2020-05-29 | 2021-12-31 | Kws Saat Se & Co Kgaa | Inducción de haploides en plantas |
| CN111850043A (zh) * | 2020-06-28 | 2020-10-30 | 武汉纽福斯生物科技有限公司 | Ecel1重组腺相关病毒载体及应用 |
| CN111778333B (zh) * | 2020-07-03 | 2022-11-18 | 东莞市滨海湾中心医院 | 测定edar表达水平的试剂的应用和试剂盒 |
| CA3185231A1 (en) | 2020-07-06 | 2022-01-13 | Pengfei YUAN | Improved rna editing method |
| CN111849991B (zh) * | 2020-08-05 | 2022-04-08 | 武汉纽福斯生物科技有限公司 | 一种寡核苷酸及其应用 |
| AU2021320902A1 (en) | 2020-08-07 | 2023-04-06 | Spacecraft Seven, Llc | Plakophilin-2 (PKP2) gene therapy using AAV vector |
| JP2023538303A (ja) | 2020-08-13 | 2023-09-07 | イェール ユニバーシティー | 所望の表現型を有するcar t細胞の操作および選択のための組成物および方法 |
| IL301014A (en) | 2020-08-31 | 2023-05-01 | Univ Yale | Compositions and methods for administering nucleic acids to cells |
| EP4214314A1 (en) | 2020-09-15 | 2023-07-26 | Inscripta, Inc. | Crispr editing to embed nucleic acid landing pads into genomes of live cells |
| CN112138146B (zh) * | 2020-09-25 | 2022-08-09 | 安徽医科大学 | Manf蛋白的应用 |
| US11512297B2 (en) | 2020-11-09 | 2022-11-29 | Inscripta, Inc. | Affinity tag for recombination protein recruitment |
| EP4001429A1 (en) | 2020-11-16 | 2022-05-25 | Antibodies-Online GmbH | Analysis of crispr-cas binding and cleavage sites followed by high-throughput sequencing (abc-seq) |
| WO2022109421A1 (en) * | 2020-11-20 | 2022-05-27 | Senti Biosciences, Inc. | Inducible cell death systems |
| CN112618963A (zh) * | 2020-12-12 | 2021-04-09 | 安徽省旌一农业旅游发展有限公司 | 一种用于光疗箱的新生儿保护装置 |
| US11777863B2 (en) * | 2020-12-21 | 2023-10-03 | Landis+ Gyr Innovations | Optimized route for time-critical traffic in mesh network |
| CN112516288B (zh) * | 2020-12-22 | 2023-04-18 | 西藏阿那达生物医药科技有限责任公司 | N-糖基化修饰之灵芝免疫调节蛋白的应用 |
| CA3204158A1 (en) | 2021-01-04 | 2022-07-07 | Juhan Kim | Mad nucleases |
| US11332742B1 (en) | 2021-01-07 | 2022-05-17 | Inscripta, Inc. | Mad nucleases |
| CA3207102A1 (en) | 2021-01-12 | 2022-07-21 | March Therapeutics, Inc. | Context-dependent, double-stranded dna-specific deaminases and uses thereof |
| US11884924B2 (en) | 2021-02-16 | 2024-01-30 | Inscripta, Inc. | Dual strand nucleic acid-guided nickase editing |
| CN113444730A (zh) * | 2021-03-17 | 2021-09-28 | 昆明市延安医院 | 一种原发性肝细胞klotho基因转导干细胞筛选构建方法 |
| CN113181218A (zh) * | 2021-03-22 | 2021-07-30 | 中国福利会国际和平妇幼保健院 | 人羊膜上皮细胞在制备修复子宫瘢痕细胞制剂中的应用 |
| AU2022258329A1 (en) * | 2021-04-12 | 2023-10-12 | The Regents Of The University Of California | Gene therapy for arrhythmogenic right ventricular cardiomyopathy |
| CN113124052B (zh) * | 2021-04-16 | 2022-08-23 | 中国航空发动机研究院 | 电磁轴承-转子系统不平衡振动控制方法及电子设备 |
| JP2024516168A (ja) | 2021-04-22 | 2024-04-12 | デイナ ファーバー キャンサー インスティチュート,インコーポレイテッド | がんを治療するための組成物および方法 |
| CN113178263A (zh) * | 2021-04-30 | 2021-07-27 | 上海市公共卫生临床中心 | 肺结核病变活动性标志物、试剂盒、方法及模型构建方法 |
| CN113416713A (zh) * | 2021-05-11 | 2021-09-21 | 中国农业科学院哈尔滨兽医研究所(中国动物卫生与流行病学中心哈尔滨分中心) | 一种重组腺病毒的构建及其应用 |
| EP4352226A1 (en) | 2021-06-07 | 2024-04-17 | Yale University | Peptide nucleic acids for spatiotemporal control of crispr-cas binding |
| CN113519460B (zh) * | 2021-06-30 | 2023-02-17 | 华南农业大学 | 一种诱导型子宫上皮特异性基因工程鼠的构建及应用 |
| AR126622A1 (es) | 2021-07-30 | 2023-10-25 | Kws Saat Se & Co Kgaa | Plantas con digestibilidad mejorada y haplotipos marcadores |
| WO2023018674A1 (en) * | 2021-08-09 | 2023-02-16 | Amicus Therapeutics, Inc. | Determination of gene transduction potency in neuron-like cells |
| JP7125727B1 (ja) | 2021-09-07 | 2022-08-25 | 国立大学法人千葉大学 | 核酸配列改変用組成物および核酸配列の標的部位を改変する方法 |
| CA3232376A1 (en) | 2021-09-20 | 2023-03-23 | Maria KOKKINAKI | Multitransgenic pigs comprising ten genetic modifications for xenotransplantation |
| WO2023052508A2 (en) | 2021-09-30 | 2023-04-06 | Astrazeneca Ab | Use of inhibitors to increase efficiency of crispr/cas insertions |
| WO2023064732A1 (en) | 2021-10-15 | 2023-04-20 | Georgia State University Research Foundation, Inc. | Delivery of therapeutic recombinant uricase using nanoparticles |
| WO2023070043A1 (en) | 2021-10-20 | 2023-04-27 | Yale University | Compositions and methods for targeted editing and evolution of repetitive genetic elements |
| WO2023093862A1 (en) | 2021-11-26 | 2023-06-01 | Epigenic Therapeutics Inc. | Method of modulating pcsk9 and uses thereof |
| US20230302423A1 (en) | 2022-03-28 | 2023-09-28 | Massachusetts Institute Of Technology | Rna scaffolded wireframe origami and methods thereof |
| WO2023196818A1 (en) | 2022-04-04 | 2023-10-12 | The Regents Of The University Of California | Genetic complementation compositions and methods |
| CN114632156B (zh) * | 2022-05-17 | 2022-08-16 | 中国人民解放军军事科学院军事医学研究院 | Tim-3在预防、治疗或减轻疼痛中的应用 |
| CN115350176B (zh) * | 2022-07-14 | 2023-10-13 | 深圳大学 | 一种胃癌肿瘤细胞和胃癌肿瘤干细胞治疗药物及其应用 |
| WO2024020346A2 (en) | 2022-07-18 | 2024-01-25 | Renagade Therapeutics Management Inc. | Gene editing components, systems, and methods of use |
| CN114990110B (zh) * | 2022-07-19 | 2023-07-14 | 江西农业大学 | 一种野外蝴蝶监测无损伤取样方法 |
| WO2024020597A1 (en) | 2022-07-22 | 2024-01-25 | The Johns Hopkins University | Dendrimer-enabled targeted intracellular crispr/cas system delivery and gene editing |
| WO2024023034A1 (en) * | 2022-07-25 | 2024-02-01 | Institut National de la Santé et de la Recherche Médicale | Use of apelin for the treatment of lymphedema |
| CN115177730B (zh) * | 2022-08-05 | 2024-02-27 | 华中科技大学同济医学院附属协和医院 | Ptpn22及其表达抑制剂的新用途 |
| WO2024044723A1 (en) | 2022-08-25 | 2024-02-29 | Renagade Therapeutics Management Inc. | Engineered retrons and methods of use |
| WO2024042199A1 (en) | 2022-08-26 | 2024-02-29 | KWS SAAT SE & Co. KGaA | Use of paired genes in hybrid breeding |
| WO2024047587A1 (en) | 2022-08-31 | 2024-03-07 | Regel Therapeutics, Inc. | Cas-phi compositions and methods of use |
| WO2024064824A2 (en) | 2022-09-21 | 2024-03-28 | Yale University | Compositions and methods for identification of membrane targets for enhancement of nk cell therapy |
| WO2024081736A2 (en) | 2022-10-11 | 2024-04-18 | Yale University | Compositions and methods of using cell-penetrating antibodies |
| CN115487301B (zh) * | 2022-11-08 | 2023-07-07 | 四川大学华西医院 | Il-13抑制剂在制备延缓或治疗视网膜色素变性的药物中的用途 |
| CN116519950B (zh) * | 2023-05-10 | 2023-11-07 | 首都医科大学附属北京天坛医院 | 一种预测卒中后抑郁的生物标志物及其应用 |
| CN117257958B (zh) * | 2023-11-21 | 2024-02-09 | 四川大学华西医院 | Trps1抑制剂的新用途及用于治疗和/或预防雄激素性秃发的药物 |
Family Cites Families (49)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AT395893B (de) | 1990-01-10 | 1993-03-25 | Mayreder Kraus & Co Ing | Befestigungsvorrichtung zum verbinden von bauteilen |
| NL9100084A (nl) | 1991-01-17 | 1992-08-17 | Verstraeten Funderingstech Bv | Veiligheidsbassin. |
| WO1996039154A1 (en) | 1995-06-06 | 1996-12-12 | Isis Pharmaceuticals, Inc. | Oligonucleotides having phosphorothioate linkages of high chiral purity |
| US5985662A (en) | 1995-07-13 | 1999-11-16 | Isis Pharmaceuticals Inc. | Antisense inhibition of hepatitis B virus replication |
| US5944710A (en) | 1996-06-24 | 1999-08-31 | Genetronics, Inc. | Electroporation-mediated intravascular delivery |
| JP2002512501A (ja) | 1996-07-03 | 2002-04-23 | メリアル インコーポレイテッド | 外来性dnaを含む組換えイヌアデノウィルス(cav) |
| US5869326A (en) | 1996-09-09 | 1999-02-09 | Genetronics, Inc. | Electroporation employing user-configured pulsing scheme |
| US5990091A (en) | 1997-03-12 | 1999-11-23 | Virogenetics Corporation | Vectors having enhanced expression, and methods of making and uses thereof |
| GB9710049D0 (en) | 1997-05-19 | 1997-07-09 | Nycomed Imaging As | Method |
| US6348450B1 (en) | 1997-08-13 | 2002-02-19 | The Uab Research Foundation | Noninvasive genetic immunization, expression products therefrom and uses thereof |
| US6716823B1 (en) | 1997-08-13 | 2004-04-06 | The Uab Research Foundation | Noninvasive genetic immunization, expression products therefrom, and uses thereof |
| US6706693B1 (en) | 1997-08-13 | 2004-03-16 | The Uab Research Foundation | Vaccination by topical application of genetic vectors |
| ATE421888T1 (de) | 1997-08-13 | 2009-02-15 | Uab Research Foundation | Impfung durch topische verwendung genetischer vektoren |
| AU4004699A (en) | 1998-05-15 | 1999-12-06 | Quark Biotech, Inc. | Mechanical stress induced genes, expression products therefrom, and uses thereof |
| US6534261B1 (en) * | 1999-01-12 | 2003-03-18 | Sangamo Biosciences, Inc. | Regulation of endogenous gene expression in cells using zinc finger proteins |
| US20030049799A1 (en) | 2000-10-23 | 2003-03-13 | Schwartz John Jacob | Engineered stimulus-responsive switches |
| WO2002088346A2 (en) | 2001-05-01 | 2002-11-07 | National Research Council Of Canada | A system for inducible expression in eukaryotic cells |
| US20050220796A1 (en) | 2004-03-31 | 2005-10-06 | Dynan William S | Compositions and methods for modulating DNA repair |
| BRPI0817233A2 (pt) * | 2007-09-28 | 2012-11-06 | Intrexon Corp | construções terapêuticas de gene de trca e bireatores para a expressão de moléculas bioterapêuticas, e usos das mesmas |
| US20100076057A1 (en) | 2008-09-23 | 2010-03-25 | Northwestern University | TARGET DNA INTERFERENCE WITH crRNA |
| WO2010075424A2 (en) | 2008-12-22 | 2010-07-01 | The Regents Of University Of California | Compositions and methods for downregulating prokaryotic genes |
| US20110041195A1 (en) * | 2009-08-11 | 2011-02-17 | Sangamo Biosciences, Inc. | Organisms homozygous for targeted modification |
| PL2510096T5 (pl) * | 2009-12-10 | 2018-06-29 | Regents Of The University Of Minnesota | Modyfikacja DNA zależna od efektora TAL |
| WO2011100058A1 (en) * | 2010-02-09 | 2011-08-18 | Sangamo Biosciences, Inc. | Targeted genomic modification with partially single-stranded donor molecules |
| EP2569425B1 (en) | 2010-05-10 | 2016-07-06 | The Regents of The University of California | Endoribonuclease compositions and methods of use thereof |
| AU2011256838B2 (en) | 2010-05-17 | 2014-10-09 | Sangamo Therapeutics, Inc. | Novel DNA-binding proteins and uses thereof |
| EP2714936B1 (en) * | 2011-06-01 | 2018-12-12 | Precision Biosciences, Inc. | Methods and products for producing engineered mammalian cell lines with amplified transgenes |
| WO2013020078A1 (en) * | 2011-08-04 | 2013-02-07 | University Of Medicine And Dentistry Of New Jersey | Sequence-specific mrna interferase and uses thereof |
| ES2773020T3 (es) | 2011-09-02 | 2020-07-09 | Carrier Corp | Sistema de refrigeración y procedimiento de refrigeración que proporciona la recuperación de calor |
| US8450107B1 (en) * | 2011-11-30 | 2013-05-28 | The Broad Institute Inc. | Nucleotide-specific recognition sequences for designer TAL effectors |
| US20130137173A1 (en) | 2011-11-30 | 2013-05-30 | Feng Zhang | Nucleotide-specific recognition sequences for designer tal effectors |
| US9637739B2 (en) * | 2012-03-20 | 2017-05-02 | Vilnius University | RNA-directed DNA cleavage by the Cas9-crRNA complex |
| LT2800811T (lt) | 2012-05-25 | 2017-09-11 | The Regents Of The University Of California | Būdai ir kompozicijos, skirti tikslinės dnr modifikavimui, panaudojant adresuotą rnr, ir transkripcijos moduliavimui, panaudojant adresuotą rnr |
| EP2877213B1 (en) * | 2012-07-25 | 2020-12-02 | The Broad Institute, Inc. | Inducible dna binding proteins and genome perturbation tools and applications thereof |
| KR101656237B1 (ko) | 2012-10-23 | 2016-09-12 | 주식회사 툴젠 | 표적 DNA에 특이적인 가이드 RNA 및 Cas 단백질을 암호화하는 핵산 또는 Cas 단백질을 포함하는, 표적 DNA를 절단하기 위한 조성물 및 이의 용도 |
| PL3138910T3 (pl) | 2012-12-06 | 2018-01-31 | Sigma Aldrich Co Llc | Oparta na CRISPR modyfikacja i regulacja genomu |
| US8697359B1 (en) * | 2012-12-12 | 2014-04-15 | The Broad Institute, Inc. | CRISPR-Cas systems and methods for altering expression of gene products |
| KR20150105634A (ko) * | 2012-12-12 | 2015-09-17 | 더 브로드 인스티튜트, 인코퍼레이티드 | 서열 조작을 위한 개선된 시스템, 방법 및 효소 조성물의 유전자 조작 및 최적화 |
| WO2014093694A1 (en) * | 2012-12-12 | 2014-06-19 | The Broad Institute, Inc. | Crispr-cas nickase systems, methods and compositions for sequence manipulation in eukaryotes |
| PL2931898T3 (pl) * | 2012-12-12 | 2016-09-30 | Le Cong | Projektowanie i optymalizacja systemów, sposoby i kompozycje do manipulacji sekwencją z domenami funkcjonalnymi |
| PT2784162E (pt) * | 2012-12-12 | 2015-08-27 | Broad Inst Inc | Engenharia de sistemas, métodos e composições guia otimizadas para a manipulação de sequências |
| EP3825401A1 (en) * | 2012-12-12 | 2021-05-26 | The Broad Institute, Inc. | Crispr-cas component systems, methods and compositions for sequence manipulation |
| KR20160044457A (ko) * | 2013-06-17 | 2016-04-25 | 더 브로드 인스티튜트, 인코퍼레이티드 | 서열 조작을 위한 탠덤 안내 시스템, 방법 및 조성물의 전달, 조작 및 최적화 |
| BR112016013547A2 (pt) * | 2013-12-12 | 2017-10-03 | Broad Inst Inc | Composições e métodos de uso de sistemas crispr-cas em distúrbios de repetições de nucleotídeos |
| EP3230451B1 (en) * | 2014-12-12 | 2021-04-07 | The Broad Institute, Inc. | Protected guide rnas (pgrnas) |
| WO2016205759A1 (en) * | 2015-06-18 | 2016-12-22 | The Broad Institute Inc. | Engineering and optimization of systems, methods, enzymes and guide scaffolds of cas9 orthologs and variants for sequence manipulation |
| EP3929287A3 (en) * | 2015-06-18 | 2022-04-13 | The Broad Institute, Inc. | Crispr enzyme mutations reducing off-target effects |
| US11696302B2 (en) | 2018-09-10 | 2023-07-04 | Ntt Docomo, Inc. | User terminal |
| US11696305B2 (en) | 2020-06-18 | 2023-07-04 | Qualcomm Incorporated | Scheduling uplink transmissions using relay devices |
-
2013
- 2013-07-21 EP EP13744916.1A patent/EP2877213B1/en active Active
- 2013-07-21 KR KR1020237014846A patent/KR20230065381A/ko active Application Filing
- 2013-07-21 PL PL18200462T patent/PL3494997T3/pl unknown
- 2013-07-21 PT PT182004622T patent/PT3494997T/pt unknown
- 2013-07-21 DK DK18200462T patent/DK3494997T3/da active
- 2013-07-21 JP JP2015524358A patent/JP2015527889A/ja active Pending
- 2013-07-21 WO PCT/US2013/051418 patent/WO2014018423A2/en active Application Filing
- 2013-07-21 KR KR1020157004939A patent/KR102530118B1/ko active IP Right Grant
- 2013-07-21 CN CN201380049665.5A patent/CN105188767A/zh active Pending
- 2013-07-21 AU AU2013293270A patent/AU2013293270B2/en active Active
- 2013-07-21 CA CA2879997A patent/CA2879997A1/en not_active Abandoned
- 2013-07-21 EP EP18200462.2A patent/EP3494997B1/en active Active
- 2013-07-21 CN CN202310274470.6A patent/CN116622704A/zh active Pending
- 2013-07-21 ES ES18200462T patent/ES2757623T3/es active Active
- 2013-07-21 EP EP20195578.8A patent/EP3808844A1/en active Pending
-
2015
- 2015-01-23 US US14/604,641 patent/US20150291966A1/en not_active Abandoned
- 2015-12-03 HK HK15111902.7A patent/HK1210965A1/xx unknown
-
2016
- 2016-12-22 US US15/388,248 patent/US20170166903A1/en not_active Abandoned
-
2017
- 2017-12-26 JP JP2017249186A patent/JP2018082709A/ja active Pending
-
2018
- 2018-08-03 AU AU2018211340A patent/AU2018211340A1/en not_active Abandoned
- 2018-10-12 AU AU2018247306A patent/AU2018247306B2/en active Active
-
2019
- 2019-03-08 US US16/297,560 patent/US20190203212A1/en not_active Abandoned
- 2019-08-07 US US16/535,042 patent/US20190390204A1/en active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| ES2757623T3 (es) | 2020-04-29 |
| EP2877213A2 (en) | 2015-06-03 |
| US20150291966A1 (en) | 2015-10-15 |
| EP3808844A1 (en) | 2021-04-21 |
| AU2018211340A1 (en) | 2018-08-23 |
| US20190390204A1 (en) | 2019-12-26 |
| EP2877213B1 (en) | 2020-12-02 |
| PL3494997T3 (pl) | 2020-04-30 |
| EP3494997A1 (en) | 2019-06-12 |
| AU2013293270A1 (en) | 2015-02-26 |
| HK1210965A1 (en) | 2016-05-13 |
| WO2014018423A3 (en) | 2014-04-03 |
| WO2014018423A2 (en) | 2014-01-30 |
| US20190203212A1 (en) | 2019-07-04 |
| PT3494997T (pt) | 2019-12-05 |
| DK3494997T3 (da) | 2019-12-02 |
| KR102530118B1 (ko) | 2023-05-08 |
| US20170166903A1 (en) | 2017-06-15 |
| JP2015527889A (ja) | 2015-09-24 |
| KR20150056539A (ko) | 2015-05-26 |
| CN105188767A (zh) | 2015-12-23 |
| AU2018247306B2 (en) | 2021-08-19 |
| JP2018082709A (ja) | 2018-05-31 |
| AU2018247306A1 (en) | 2018-11-08 |
| EP3494997B1 (en) | 2019-09-18 |
| WO2014018423A8 (en) | 2014-06-19 |
| CN116622704A (zh) | 2023-08-22 |
| AU2013293270B2 (en) | 2018-08-16 |
| CA2879997A1 (en) | 2014-01-30 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR102530118B1 (ko) | 유도 dna 결합 단백질 및 게놈 교란 도구 및 이의 적용 | |
| EP3237611B1 (en) | Cas9-dna targeting unit chimeras | |
| KR20210056329A (ko) | 신규 cas12b 효소 및 시스템 | |
| CN105658805B (zh) | Rna指导的基因编辑和基因调节 | |
| KR20210077732A (ko) | Nme2cas9-데아미나아제 융합 단백질에 의한 프로그램 가능한 dna 염기 편집 | |
| WO2016014837A1 (en) | Gene editing for hiv gene therapy | |
| JP2016521994A (ja) | 配列操作のための最適化されたCRISPR−Cas二重ニッカーゼ系、方法および組成物 | |
| US20230203481A1 (en) | Effector proteins and methods of use | |
| EP4349979A1 (en) | Engineered cas12i nuclease, effector protein and use thereof | |
| Fleury et al. | The APE2 nuclease is essential for DNA double-strand break repair by microhomology-mediated end joining | |
| KR20240021218A (ko) | 신규 유형 v rna 프로그래밍가능 엔도뉴클레아제 시스템 | |
| WO2022042557A1 (en) | Split cas12 systems and methods of use thereof | |
| WO2021166306A1 (en) | Production method of genome-edited cells and kit therefor | |
| CN115838719B (zh) | 特异性促进腺嘌呤碱基编辑器活性的化合物、化学调控方法及其应用 | |
| US20230323406A1 (en) | Effector proteins and methods of use | |
| Alves | Understanding the molecular pathogenesis of HIV-associated Burkitt Lymphoma–the impact of HIV-1 protein Tat on lymphoma driver genes | |
| Nguyen | Investigating the causal role of DNA methylation in transcriptional regulation | |
| WO2023091704A1 (en) | Compositions and methods for production of circular nucleic acid molecules | |
| Carusillo | Hijacking-DNA-Repair (HDR)-CRISPR promotes seamless gene editing in human primary cells | |
| KR20240021211A (ko) | Klk2-gpi 융합 단백질에 대한 핵산 코딩, 재조합 세포 및 이의 용도 | |
| Gall-Duncan | Unravelling Mechanisms of Repeat Expansions and Pathogenesis in Neurodegenerative Repeat Diseases | |
| Kounatidou | Generation of CRISPR engineered prostate cancer cell line models to study androgen receptor signalling in advanced prostate cancer | |
| WO2022221581A1 (en) | Programmable nucleases and methods of use | |
| KR20230142740A (ko) | Omni-103 crispr 뉴클레아제 | |
| WO2022241032A1 (en) | Enhanced guide nucleic acids and methods of use |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A107 | Divisional application of patent |