JP4740738B2 - ワクチン - Google Patents
ワクチン Download PDFInfo
- Publication number
- JP4740738B2 JP4740738B2 JP2005506112A JP2005506112A JP4740738B2 JP 4740738 B2 JP4740738 B2 JP 4740738B2 JP 2005506112 A JP2005506112 A JP 2005506112A JP 2005506112 A JP2005506112 A JP 2005506112A JP 4740738 B2 JP4740738 B2 JP 4740738B2
- Authority
- JP
- Japan
- Prior art keywords
- los
- bleb
- neisseria
- strain
- strains
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images





Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/095—Neisseria
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/102—Pasteurellales, e.g. Actinobacillus, Pasteurella; Haemophilus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/104—Pseudomonadales, e.g. Pseudomonas
- A61K39/1045—Moraxella
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/22—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Neisseriaceae (F)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/52—Bacterial cells; Fungal cells; Protozoal cells
- A61K2039/521—Bacterial cells; Fungal cells; Protozoal cells inactivated (killed)
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55505—Inorganic adjuvants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55516—Proteins; Peptides
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55572—Lipopolysaccharides; Lipid A; Monophosphoryl lipid A
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55577—Saponins; Quil A; QS21; ISCOMS
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/70—Multivalent vaccine
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Description
本発明は、ナイセリアワクチン組成物、それらの製造、およびかかる組成物の医薬における使用の分野に関する。より具体的には、ナイセリアの、特に髄膜炎菌の外膜小胞(すなわち、ブレブ)ワクチンの産生により適した新規の操作された髄膜炎菌株を製造する方法に関する。新規のLOSサブユニットまたは髄膜炎菌の外膜小胞(すなわち、ブレブ)ワクチンの使用に基づく有利な方法およびワクチン製品もまた記載しており、これはヒト被験体における使用のために安全性および有効性をより高めたものである。
ナイセリア・メニンジティディス(Neisseria meningitidis)(髄膜炎菌)は、しばしばヒト上気道より単離されるグラム陰性細菌である。この細菌は、菌血症および髄膜炎等の重篤な侵襲性細菌性疾患の原因である。髄膜炎菌疾患の発生数は、地理的、季節的および年次的な差異を示す(Schwartz, B., Moore, P.S., Broome, C.V.; Clin. Microbiol. Rev. 2(Supplement), S18-S24, 1989)。この菌は通常、その莢膜多糖による血清型に従って分類される。
本明細書中に記載される刊行物および特許、または特許出願の主題またはそこに開示される情報は、参照により本明細書に組み入れる。
ヒトスフィンゴ糖脂質に存在するラクト-N-ネオテトラオースオリゴ糖基(Galβ1-4GlcNAcβ1-3Galβ1-4Glcβ1-、図1)と類似した構造が存在することから、L3またはL2 LOSに対して生じた抗体の安全性が問われてきている。多くの人々が残存量のL3 LOSを含むデオキシコレート抽出小胞ワクチンで安全にワクチン接種されてきているが(G. Bjuneら, Lancet(1991), 338, 1093-1096、GVG. Sierraら, NIPH ann(1991), 14, 195-210)、LOSが本明細書に記載されるように抗原として保持される場合、LOS糖鎖構造の末端部分の欠失が、ヒト組織の表面に提示される構造との抗LOS免疫反応の交差反応の阻止において有利であることが、本発明者らによって発見された。好ましい実施形態において、lgtB遺伝子の不活性化が末端ガラクトース残基およびシアル酸が欠損した中間体LOS構造を生じる(図1および2を参照、突然変異はL2およびL3 LOSに4GlcNAcβ1-3Galβ1-4Glcβ1-構造を残存させる)。そのような中間体は、L3および/またはL2 LOS株において獲得され得る。別のおよびあまり好ましくないLOSの(短い)バージョンは、lgtE遺伝子のスイッチオフによって得られうる。さらに別のおよびあまり好ましくない型のLOSは、lgtA遺伝子のスイッチオフによって得られうる。そのようなlgtA-突然変異を選択する場合、非免疫原性L1免疫型の形成を阻止するためにlgtC発現をスイッチオフすることがまた好ましい。
本発明の上記の精製LOSまたはブレブ免疫原性組成物はまた、それらが由来する細菌性産生株における特定の遺伝子の発現の下方制御によって、毒性をより少なくすることができる。そのような無毒化は、天然のOMVによる鼻腔内免疫化には不必要であるだろうが(J.J Drabickら, Vaccine(2000), 18, 160-172)、非経口ワクチン接種における無毒化には有用であろう。好ましくは、本発明の精製LOSまたはブレブ免疫原性組成物であるLOSは、リピドA生合成に関与する遺伝子(特にリピドAへの第2アシル鎖の付加に関与する遺伝子)の突然変異/修飾/不活性化により(特にmsbBおよび/またはhtrB遺伝子由来の機能的遺伝子産物の発現の下方制御により、および好ましくは遺伝子のスイッチオフにより、最も好ましくは遺伝子のプロモーターおよび/またはオープンリーディングフレームの全体または一部の欠失により、ナイセリア産生株を遺伝的に操作することによって無毒化される。あるいはまた(またはさらに)、精製LOSまたはブレブ免疫原性組成物は、遺伝的に改変して以下の遺伝子:pmrA、pmrB、pmrEおよびpmrFのうちの1つ以上の遺伝子が(より強力なプロモーターの導入、もしくは遺伝子の余分なコピーの組み込みによって)上方制御されるようにしたナイセリア株から得ることができる。あるいはまた(またはさらに)、精製LOSまたはブレブ免疫原性組成物は、ポリミキシンBの機能的等価物である非毒性ペプチド[リピドAに対して高い親和性の有する分子]を組成物に添加することにより無毒化され得る。
本発明のさらなる態様は、本発明のナイセリア株から単離されるLOS調製物(特に上述した任意のもの)である。好ましくは、単離されたLOS(またはLOS含有ブレブ)は、L2またはL3免疫型であり、好ましくは本発明の免疫原性組成物は本発明のL2およびL3 LOS(またはブレブ)調製物を両方含む。
LOS(特に本発明のLOS)がブレブ製剤中に存在する場合、LOSは好ましくは、ブレブ調製物上にも存在する1以上の外膜タンパク質(例えば髄膜炎菌におけるPorAまたはPorB)へのLOSのコンジュゲーションを可能にする方法により、in situにおいてコンジュゲートされる。従って、本発明のさらなる態様は、その外膜中に、LOSにコンジュゲートした外膜タンパク質が組み込まれるグラム陰性細菌株由来のブレブ調製物である。LOSはコンジュゲートするためにブレブ調製物に添加してもよいが、LOSはブレブ調製物の表面に天然に存在することが好ましい。
本発明の外膜小胞(OMVまたはブレブ)は、多くの公知の技術により単離され得る(Fredriksenら, NIPH Annals(1991), 14, 67-79、Zollingerら, J. Clin Invest(1979), 63, 836-848、Saundersら, Infect Immun(1999), 67, 113-119、J.J. Drabickら, Vaccine(1999), 18, 160-172)。これらは2つの主要な群に分類される。すなわち、デオキシコレート(約0.5%)を使用して髄膜炎菌由来のブレブを抽出するために技術と、低レベルのデオキシコレート(DOC)を使用するかまたは全くデオキシコレートを使用しない技術である。DOC非含有法では、ブレブはOMV中に高レベルのLOSを維持するという興味深い特徴を有する。このことはLOSが防御抗原であるワクチンにおいて有利である。DOC抽出ブレブと比較して、DOC非含有法により得られるOMV中のL3抗原(Ag)の濃度は約10倍高い。界面活性剤を用いない(好ましくはDOCを用いない)ブレブ調製法は、この理由により本発明の方法の目的にとって好ましいが、低レベルの界面活性剤(好ましくはDOC)を含むバッファーでの抽出もまた、ブレブ内で強固に相互作用するLOSの大半を残留する一方、より毒性があり緩く保持されたLOSを除去するステップであるという点で有利であり得る。典型的には、0〜0.5%および0.02〜0.4%、0.04〜3%または0.06〜2%界面活性剤(好ましくはDOC)がブレブ抽出に使用され、より好ましくは0.08〜0.15%、および最も好ましくはおよそまたは正確に0.1%を用いて、最適な量のLOSをブレブ中に安定に存在させて得る。LOSが1以上の上記の方法により無毒化されている場合には、DOC非含有(または低DOC〜0.3%DOCもしくはそれ以下の)抽出法が特に好ましい。
本発明の免疫原性組成物は、製薬上許容される賦形剤の添加によってワクチン組成物として容易に製剤化され得る。
本発明の免疫原性組成物は、適切なアジュバントとともに製剤化することで本発明のワクチン組成物を作製することができる。
本発明の上記のブレブ組成物は、それらが由来するナイセリア株(淋菌および好ましくは髄膜炎菌、最も好ましくは髄膜炎菌Bを含む)が、ゲノム中への遺伝子のさらなるコピーを挿入することにより、または既存の遺伝子の上流により強力なプロモーターを導入することにより、または非改変株と比較して1.2、1.5、2、3、5もしくは10倍を超える抗原レベルを生じるよう改変された株を誘導することが可能なWO 01/09350に記載のあらゆる他の方法により上方制御された1以上の下記遺伝子(防御抗原をコードする)を有する場合、本発明のワクチンにおける有効性がさらに改良され得る:NspA(WO 96/29412)、Hsfまたはその末端切断物(WO 99/31132およびWO 01/55182、NhhAとしても知られる)、Hap(PCT/EP99/02766)、OMP85(WO 00/23595)、PilQ(PCT/EP99/03603)、PldA(PCT/EP99/06718)、FrpB(WO 96/31618)、TbpA(WO 92/03467、US5912336、WO 93/06861およびEP586266)、TbpB(WO 93/06861およびEP586266)、NadA(Comanducciら J. Exp. Med. 2002 195:1445-1454、NMB 1994)、FrpA/FrpC、または5以上の反復配列を含むこれらの抗原間に共通の部分(WO 92/01460; Thompsonら,(1993) J. Bacteriol. 175:811-818; Thompsonら,(1993) Infect. Immun. 61:2906-2911)、LbpA、LbpB(PCT/EP98/05117)、FhaB(WO 98/02547 配列番号38(ヌクレオチド3083-9025))、HasR(PCT/EP99/05989)、lipo02(PCT/EP99/08315)、Tbp2(WO 99/57280; NMB 0460)、MltA(WO 99/57280; NMB 0033)、TspA(WO 00/03003)、TspB(WO 00/03003)、ctrA(PCT/EP00/00135)、MafA(NMB 0652)、MafB(NMB 0643)、Omp26(NMB 0181)、アドヘシンX(NMB 0315)、アドヘシンY(NMB 0995)、アドヘシンZ(NMB 1119)およびOstA(NMB 0280)。NMB配列の例は、www.neisseria.orgのデータベースで見出すことができる。本明細書でHsfについて言及する場合、この用語はあらゆる場合においてHsf末端切断物と置換可能である(特にWO 01/55182に開示されたものにおいて)。
本発明者らは、ブレブに関わる上記組成物およびワクチンが、(同一の利点を有する)ゴーストまたは死菌全細胞調製物およびワクチンに関する方法にまで容易に拡張できることを意図している。グラム陰性株からゴースト調製物(完全なエンペロープを有する空の細胞)を製造する方法は当技術分野で周知である(例えばWO 92/01791を参照)。全細胞を殺滅して、ワクチンに使用するための不活性化細胞調製物を作製する方法もまた周知である。従って、本明細書を通して記載されるブレブを含む組成物およびワクチンは、等価物である本発明のゴーストおよび死菌全細胞調製物を含む同じ組成物またはワクチンにも当てはまることを意図している。
血清殺菌アッセイは、本発明の免疫原性組成物中で組み合わせた場合に抗原間の相乗作用的な関係を評価するための好ましい方法である。
髄膜炎菌BのB莢膜多糖産生に関わるタンパク質をコードする遺伝子の欠失、PorA遺伝子の欠失、髄膜炎菌ブレブの表面上の様々な防御外膜タンパク質の上方制御、免疫優性タンパク質または生合成酵素の下方制御、およびブレブを単離する方法を記載する例は、WO 01/09350に記載される。
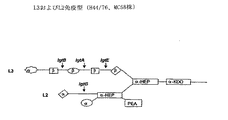
潜在的な交差防御抗原としてのLOSの役割を評価するために、H44/76野生型(WT)の髄膜炎菌B株(L3 LOSを発現する)および「galE-様LOS」(lgtE- LOSに関しては短い構造を有する)を発現する改変H44/76株を使用し、2つの異なる方法に従ってブレブを生成した。第1の方法では高レベルのLOSを含むブレブを得るために0.1%DOCを使用し、第2の方法では生じたブレブ中に低レベルのLOSを得るために0.5%DOCを使用した。
SBAは、個々の血清について異なるNmenB株を使用して行った:同種のWT H44/76株、PorA(-)H44/76株、および2つの異種の株(血清亜型に基づくもの)Cu385およびNZ124。これら4つの株はL3 LOSを発現する。5番目の株も加えられた。H44/76と比較して、この株(B16B6)はPorAに対してだけでなくLOS(これは免疫型L2株である)に対しても異種性である。
WT DOC 0.1%ブレブにより誘導される反応が主として抗LOS抗体によることを示すために、血清プールを異なる濃度の精製L3 LOSを用いて枯渇させた。枯渇後、血清を、同種WT H44/76株に対する殺菌アッセイにおいて使用した。
この結果は以下のL2 LOSについてのものと類似する。
使用したMC58髄膜炎菌誘導株はB:P1.7.16、opc-、siaD-である。この株を遺伝学的に改変して、L3(2G2株)または中間体エピトープ(2G EcoN1b-1株、2G2と同様であるがさらにlgtB-)またはLPSの短いバージョン(lgtE-であるC6株)のいずれかを発現させた。OMVを、通常の高(0.5%)DOC法またはDOC非含有法のどちらかに従って生成した。

使用したMenBブレブは、SiaD-(従って莢膜多糖を発現しない)およびPorA-であるH44/76株(LOS免疫型L3)から得た。2種の異なる株を使用した:完全なL3(B1717株、siad(-) PorA(-) 完全なL3)および末端切断型L3(B1727株、siad(-) PorA(-) lgtB(-) TrL3)。
OMV(ブレブ)は、MenB株H44/76 siaD- PorA- L3から、またはH44/76 siad- porA- TrL3から産生された。2つの異なる抽出を行い、使用したDOCの割合を0.1または0.5%のいずれかとした。2つの異なるアジュバント製剤:Al(OH)3、またはリン酸アルミニウム+3D-MPLもまた、評価した。マウス(OF1メスマウス、6〜8週齢、1群につき30匹)にIM経路によって3回(0、21および28日目)注入した(5μgブレブ/注入)。SBAを、PostII(28日)およびPostIII(42日目)血清(プールされた血清または個々の血清)について収集した。
L3およびtrL3構造が、防御抗体に関して免疫学的に非常に近い関係である場合、L3(およびL2)LOSに付随する可能性としての自己免疫の問題に関して、構造(ラクト-N-ネオテトラオース部分による)の間に何らかの差異はあるのだろうか。本発明者らは、寒冷凝集素がtrL3 LOSを認識することが可能か否かに注目することによってこの問題に取り組んできた。
ブレブ(B1820株由来[siaD(-)PorA(-)FrpB(-)であり、上方制御された末端切断型Hsfを有するH44/76から得られ、lgtB(-)突然変異株であることによりL3が末端切断されており、デスフェラル(desferral)存在下で培養され、ブレブはDOC 0.1%を用いて抽出される])は、異なる濃度のEDAC(EDACがより多く存在するほど、より多くのブレブが架橋される)を使用して架橋された。架橋は、ブレブの無菌的濾過によって示されるとおりブレブ内でなされる。
2種の製剤(Al(OH)3またはAlPO4に吸着したブレブ)を試験し、発熱試験においてウサギにヨーロッパ薬局方に記載されるようにして500ng/kgをIV経路によって投与した。
上記ブレブ(非吸着)の抗原性を評価して、この架橋がブレブの抗原性に対して何らかの影響を有するか否かを調べた。ブレブの異なる調製物(架橋されているまたはされていないもの)をマイクロプレート上にコートした(10μg/ml、4℃で一晩)。洗浄および飽和の後、MAb L379の連続希釈物またはB1820 DOC 0.1%または0.5%で免疫したマウス由来の血清を該プレートに添加した(振盪しながら室温で30分間)。コートしたブレブに対する抗体の固定は、ビオチンと結合した抗マウスIgを用い、次にストレプトアビジンペルオキシダーゼ複合体を使用し、その後OPDおよびH2O2を用いた発色によって、示した。各マイクロウエルの吸光度はマイクロプレートリーダーを用いて測定した。
この実験において、ブレブはB1727株より産生された。この株は遺伝的に改変されたH44/76株であり、これはsiaD(-)PorA(-)trL3(lgtB-)HsfでありそしてTbpAが上方制御されている。ブレブは0.5%DOCを用いて抽出した。マウスは3回(0、21および28日目)、IM経路により免疫した。1回の注入で、それらマウスにはAl(OH)3に吸着したブレブ5μgを投与した。
以下の実験を評価した:
- LNnT(ラクト-N-ネオテトラオース)と反応することができる抗体の誘導に対するTrL3(lgtB(-)L3 LOS)の影響、
- 上記構築物の殺菌抗体の誘導。
方法:マイクロプレートは、スペーサー(ADH)を介してヒト血清アルブミンにコンジュゲートしたLNnTでコートした(PBS中1ml当たり5μgのコンジュゲート、マイクロウエル当たり100μl)。4℃で一晩インキュベートした後、プレートをPBS-BSA 1%で洗浄し飽和させた(室温で40分)。洗浄の後、PBS 0.2%、BSA 0.05%、Tween20中への連続希釈液を添加した(室温で30分)。IgGのLNnTへの固定は、ペルオキシダーゼ(Jackson)に結合した抗マウスIgGによって、続いてOPDAおよびH2O2によるインキュベーションによって明らかにされた。
H44/76株に対するSBAアッセイを、3回目の注入の14日後に採取した個々の血清について実施した。以下の結果はtrL3(lgtB(-))LOSブレブがL3 LOSと同等のレベルで殺菌抗体を誘導することを明白に示す(GMTの他、>1/100のSBA力価を有するマウスの数(=SC)を参照)。
以下のデータは2つの前臨床試験の概要である。
- B1733:siaD(-) PorA(-) Tr(末端切断型)Hsfの上方制御 lgtB(-)
- B1820:siaD(-) PorA(-) TrHsfの上方制御 lgtB(-) FrpB(-)。
殺菌アッセイは3つのL3株について行った(同種の野生型株H44/76および2つの異種のL3株:NZ124およびM97250687)。この結果は、FrpB(-)(ノックアウト)(B1820)ブレブが、FrpB(+)ブレブ(B1733)よりも優れた異種交差殺菌反応(高い力価およびより優れた血清転換SC)を誘導することを明白に示す。同種反応は、FrpB欠失により低下するにも関わらず、十分なままである。
2つのNmenB株をこの評価に使用した:
- galE(-)である(従って莢膜多糖を生成できない)対照株。
lgtB(-)およびmsbB(-)突然変異を有する髄膜炎菌に由来し、かつより低い(例えば0.1%)デオキシコレート濃度で抽出されたL3およびL2ブレブを含む組成物は、髄膜炎菌Bに対する効果的で安全なワクチンに対する強力な基礎を提供する。ブレブ産生株は理想的には莢膜多糖合成欠損性であり、かつ該ブレブはブレブ内で外膜タンパク質に架橋しているLOSを有する。PorA(-)およびFrpB(-)のいずれかまたは両方が、交差殺菌効果を改良する上でさらに有用であり、Hsfおよび/またはTbpA抗原の上方制御も同様に有用である。
Claims (18)
- L2 LOS免疫型を有するナイセリア株もしくはL3 LOS免疫型を有するナイセリア株に由来し、ここで該株がlgtB-であるナイセリアブレブ調製物;または、L2 LOS免疫型を有するナイセリア株とL3 LOS免疫型を有するナイセリア株とに由来するブレブの混合物を含み、ここでその各株がlgtB-であるナイセリアブレブ調製物。
- ナイセリア株が、髄膜炎菌である、請求項1記載のナイセリアブレブ調製物。
- ナイセリア株が、血清型Bの髄膜炎菌である、請求項2記載のナイセリアブレブ調製物。
- ナイセリア株が莢膜多糖を合成できないものである、請求項1または2記載のナイセリアブレブ調製物。
- ナイセリア株が、以下の莢膜多糖遺伝子:ctrA、ctrB、ctrC、ctrD、synA、synB、synCまたはsiaDのうち1つについて、それらが由来する天然株と比較して発現が下方制御されているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株はそれぞれの株において同じ莢膜多糖遺伝子の発現が下方制御されている、請求項4記載のナイセリアブレブ調製物。
- ナイセリア株が、以下のリピドA遺伝子:msbBもしくはhtrBのいずれかまたは両方について、それらが由来する天然株と比較して発現が下方制御されているものである、請求項1〜5のいずれか1項記載のナイセリアブレブ調製物。
- ナイセリア株が、以下の外膜タンパク質遺伝子:porA、porB、opA、opC、pilCまたはfrpBのうち1以上について、それらが由来する天然株と比較して発現が下方制御されているものである、請求項1〜6のいずれか1項記載のナイセリアブレブ調製物。
- ナイセリア株が、以下の外膜タンパク質遺伝子の組み合わせ:porAおよびopA、porAおよびopC、opAおよびopC、porAおよびopAおよびopC、porAおよびfrpB、opCおよびfrpB、opAおよびfrpB、porAおよびopAおよびopCおよびfrpB、のいずれかについて、それらが由来する天然株と比較して発現が下方制御されている、請求項7記載のナイセリアブレブ調製物。
- ナイセリア株が、以下の外膜タンパク質抗原:NspA、TbpA低、TbpA高、Hsf、Hap、OMP85、PilQ、NadA、LbpA、MltAのうち1以上について、発現が上方制御されているものであり、そして、L2およびL3ブレブが両方存在する場合には、それらが由来する株はそれぞれの株において1以上の異なる外膜タンパク質抗原の発現が上方制御されている、請求項1〜8のいずれか1項記載のナイセリアブレブ調製物。
- 前記の遺伝子についての発現の下方制御が、該遺伝子の欠失である、請求項5〜8のいずれか1項記載のナイセリアブレブ調製物。
- 含まれるLOSが、Tヘルパーエピトープの供給源にコンジュゲートしている、請求項1〜10のいずれか1項記載のナイセリアブレブ調製物。
- ブレブ内架橋の工程により得られる、請求項11記載のナイセリアブレブ調製物。
- 請求項1〜8のいずれか1項記載のナイセリアブレブ調製物と、製薬上許容される賦形剤とを含む、免疫原性組成物またはワクチン。
- アジュバントをさらに含む請求項13記載のワクチン。
- アジュバントが水酸化アルミニウム、または3D-MPLおよびリン酸アルミニウムである、請求項14に記載のワクチン。
- 以下の株:髄膜炎菌血清型A、髄膜炎菌血清型C、髄膜炎菌血清型W-135、髄膜炎菌血清型Yおよびインフルエンザ菌(H. influenzae)b型、に由来する1以上のコンジュゲート化莢膜多糖またはオリゴ糖をさらに含む、請求項13または14記載のワクチン。
- 請求項1〜8のいずれか1項記載のナイセリアブレブ調製物が由来するナイセリア株を培養し、そこからブレブを単離し、L2 LOS免疫型を有するナイセリア株に由来するL2ブレブまたはL3 LOS免疫型を有するナイセリア株に由来するL3ブレブは単独で提供するかまたはそれらを組み合わせ、そしてそれらのブレブを製薬上許容される賦形剤とともに製剤化するステップを含み、ここで該ナイセリア株はL2またはL3 LOS免疫型を有し、かつlgtB - である、請求項13記載のナイセリアブレブ調製物ワクチンの製造方法。
- 単離ステップが、0.5、0.02〜0.4、0.04〜0.3、0.06〜0.2、0.08〜0.15%、または0.1%のデオキシコレートで抽出することにより行われる、請求項17記載の方法。
Applications Claiming Priority (25)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| GB0218037.0 | 2002-08-02 | ||
| GB0218036A GB0218036D0 (en) | 2002-08-02 | 2002-08-02 | Vaccine |
| GB0218036.2 | 2002-08-02 | ||
| GB0218035A GB0218035D0 (en) | 2002-08-02 | 2002-08-02 | Vaccine composition |
| GB0218037A GB0218037D0 (en) | 2002-08-02 | 2002-08-02 | Vaccine composition |
| GB0218051A GB0218051D0 (en) | 2002-08-02 | 2002-08-02 | Vaccine composition |
| GB0218035.4 | 2002-08-02 | ||
| GB0218051.1 | 2002-08-02 | ||
| GB0220197.8 | 2002-08-30 | ||
| GBGB0220199.4A GB0220199D0 (en) | 2002-08-30 | 2002-08-30 | Mutant protein and refolding method |
| GBGB0220197.8A GB0220197D0 (en) | 2002-08-30 | 2002-08-30 | Refolding method |
| GB0220199.4 | 2002-08-30 | ||
| GB0225524.8 | 2002-11-01 | ||
| GB0225524A GB0225524D0 (en) | 2002-11-01 | 2002-11-01 | Vaccine composition |
| GB0225531A GB0225531D0 (en) | 2002-11-01 | 2002-11-01 | Vaccine |
| GB0225531.3 | 2002-11-01 | ||
| GB0230170.3 | 2002-12-24 | ||
| GB0230168.7 | 2002-12-24 | ||
| GB0230164.6 | 2002-12-24 | ||
| GB0230168A GB0230168D0 (en) | 2002-12-24 | 2002-12-24 | Vaccine composition |
| GB0230164A GB0230164D0 (en) | 2002-12-24 | 2002-12-24 | Vaccine composition |
| GB0230170A GB0230170D0 (en) | 2002-12-24 | 2002-12-24 | Vaccine |
| GB0305028.3 | 2003-03-05 | ||
| GB0305028A GB0305028D0 (en) | 2003-03-05 | 2003-03-05 | Vaccine |
| PCT/EP2003/008568 WO2004014417A2 (en) | 2002-08-02 | 2003-07-31 | Vaccine compositions comprising l2 and/or l3 immunotype lipooligosaccharides from lgtb- neisseria minigitidis |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2011047056A Division JP2011142916A (ja) | 2002-08-02 | 2011-03-04 | ワクチン |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2006500962A JP2006500962A (ja) | 2006-01-12 |
| JP4740738B2 true JP4740738B2 (ja) | 2011-08-03 |
Family
ID=31722002
Family Applications (7)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005506111A Expired - Fee Related JP5409986B2 (ja) | 2002-08-02 | 2003-07-31 | グラム陰性細菌由来のトランスフェリン結合タンパク質およびHsfを含んでなるワクチン組成物 |
| JP2005506112A Expired - Fee Related JP4740738B2 (ja) | 2002-08-02 | 2003-07-31 | ワクチン |
| JP2005506113A Pending JP2006500963A (ja) | 2002-08-02 | 2003-07-31 | ワクチン組成物 |
| JP2005506114A Pending JP2006506467A (ja) | 2002-08-02 | 2003-07-31 | ワクチン組成物 |
| JP2010237357A Expired - Fee Related JP5414651B2 (ja) | 2002-08-02 | 2010-10-22 | ワクチン組成物 |
| JP2011047056A Pending JP2011142916A (ja) | 2002-08-02 | 2011-03-04 | ワクチン |
| JP2012026928A Expired - Fee Related JP5789203B2 (ja) | 2002-08-02 | 2012-02-10 | ワクチン組成物 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005506111A Expired - Fee Related JP5409986B2 (ja) | 2002-08-02 | 2003-07-31 | グラム陰性細菌由来のトランスフェリン結合タンパク質およびHsfを含んでなるワクチン組成物 |
Family Applications After (5)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005506113A Pending JP2006500963A (ja) | 2002-08-02 | 2003-07-31 | ワクチン組成物 |
| JP2005506114A Pending JP2006506467A (ja) | 2002-08-02 | 2003-07-31 | ワクチン組成物 |
| JP2010237357A Expired - Fee Related JP5414651B2 (ja) | 2002-08-02 | 2010-10-22 | ワクチン組成物 |
| JP2011047056A Pending JP2011142916A (ja) | 2002-08-02 | 2011-03-04 | ワクチン |
| JP2012026928A Expired - Fee Related JP5789203B2 (ja) | 2002-08-02 | 2012-02-10 | ワクチン組成物 |
Country Status (26)
| Country | Link |
|---|---|
| US (9) | US20060034854A1 (ja) |
| EP (11) | EP1524993B1 (ja) |
| JP (7) | JP5409986B2 (ja) |
| KR (6) | KR101239242B1 (ja) |
| CN (2) | CN1674933B (ja) |
| AU (6) | AU2003250204B8 (ja) |
| CA (4) | CA2493977A1 (ja) |
| CO (3) | CO5680456A2 (ja) |
| CY (3) | CY1114243T1 (ja) |
| DE (2) | DE20321889U1 (ja) |
| DK (2) | DK1524993T3 (ja) |
| ES (3) | ES2537737T3 (ja) |
| HK (1) | HK1077014A1 (ja) |
| HU (1) | HUE029200T2 (ja) |
| IL (3) | IL165660A0 (ja) |
| IS (3) | IS7593A (ja) |
| LU (1) | LU92262I2 (ja) |
| MX (3) | MXPA05001265A (ja) |
| MY (1) | MY149591A (ja) |
| NO (3) | NO20050010L (ja) |
| NZ (4) | NZ574530A (ja) |
| PL (5) | PL375408A1 (ja) |
| PT (2) | PT2255826E (ja) |
| SI (2) | SI1524993T1 (ja) |
| TW (1) | TWI360424B (ja) |
| WO (4) | WO2004014417A2 (ja) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2011142916A (ja) * | 2002-08-02 | 2011-07-28 | Glaxosmithkline Biologicals Sa | ワクチン |
Families Citing this family (136)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP2261347A3 (en) | 1998-05-01 | 2012-01-11 | Novartis Vaccines and Diagnostics, Inc. | Neisseria meningitidis antigens and compositions |
| GB9823978D0 (en) * | 1998-11-02 | 1998-12-30 | Microbiological Res Authority | Multicomponent meningococcal vaccine |
| US10967045B2 (en) * | 1998-11-02 | 2021-04-06 | Secretary of State for Health and Social Care | Multicomponent meningococcal vaccine |
| CN100392082C (zh) | 1999-04-30 | 2008-06-04 | 启龙股份公司 | 保守的奈瑟球菌抗原 |
| DK2270172T3 (en) | 1999-05-19 | 2016-02-29 | Glaxosmithkline Biolog Sa | Combination Compositions of Neisseria. |
| GB9918319D0 (en) * | 1999-08-03 | 1999-10-06 | Smithkline Beecham Biolog | Vaccine composition |
| ES2541830T3 (es) | 1999-10-29 | 2015-07-27 | Novartis Vaccines And Diagnositics S.R.L. | Péptidos antigénicos de Neisseria |
| GB9928196D0 (en) | 1999-11-29 | 2000-01-26 | Chiron Spa | Combinations of B, C and other antigens |
| WO2001055182A1 (en) * | 2000-01-25 | 2001-08-02 | The University Of Queensland | PROTEINS COMPRISING CONSERVED REGIONS OF NEISSERIA MENINGITIDIS SURFACE ANTIGEN NhhA |
| CA2744921C (en) | 2000-02-28 | 2014-05-13 | Chiron S.R.L. | Hybrid expression of neisserial proteins |
| MXPA03000822A (es) | 2000-07-27 | 2004-11-01 | Childrens Hosp & Res Ct Oak | Vacunas para proteccion de espectro amplio contra enfermedades causadas por neisseria meningitidis. |
| GB0118249D0 (en) | 2001-07-26 | 2001-09-19 | Chiron Spa | Histidine vaccines |
| GB0121591D0 (en) | 2001-09-06 | 2001-10-24 | Chiron Spa | Hybrid and tandem expression of neisserial proteins |
| PT2248822T (pt) | 2001-07-27 | 2017-02-14 | Glaxosmithkline Biologicals Sa | Adesinas de meningococos |
| US7214660B2 (en) | 2001-10-10 | 2007-05-08 | Neose Technologies, Inc. | Erythropoietin: remodeling and glycoconjugation of erythropoietin |
| US7173003B2 (en) | 2001-10-10 | 2007-02-06 | Neose Technologies, Inc. | Granulocyte colony stimulating factor: remodeling and glycoconjugation of G-CSF |
| GB0220194D0 (en) * | 2002-08-30 | 2002-10-09 | Chiron Spa | Improved vesicles |
| DE60335477D1 (de) * | 2002-10-11 | 2011-02-03 | Novartis Vaccines & Diagnostic | Polypeptidimpstoffe zum breiten schutz gegen hypervirulente meningokokken-linien |
| GB0227346D0 (en) | 2002-11-22 | 2002-12-31 | Chiron Spa | 741 |
| SG155777A1 (en) | 2003-04-09 | 2009-10-29 | Neose Technologies Inc | Glycopegylation methods and proteins/peptides produced by the methods |
| GB0316560D0 (en) | 2003-07-15 | 2003-08-20 | Chiron Srl | Vesicle filtration |
| WO2005012484A2 (en) | 2003-07-25 | 2005-02-10 | Neose Technologies, Inc. | Antibody-toxin conjugates |
| CA2539074C (en) | 2003-09-19 | 2017-11-07 | Epitopix, Llc | Campylobacter polypeptides and methods of use |
| CA2885040C (en) | 2003-10-02 | 2018-10-30 | Novartis Vaccines And Diagnostics S.R.L. | Liquid vaccines for multiple meningococcal serogroups |
| GB0323709D0 (en) * | 2003-10-09 | 2003-11-12 | Health Prot Agency | Modified whole cell,cell extract and omv-based vaccines |
| US20080305992A1 (en) | 2003-11-24 | 2008-12-11 | Neose Technologies, Inc. | Glycopegylated erythropoietin |
| CA2550927A1 (en) | 2003-12-23 | 2005-07-14 | Glaxosmithkline Biologicals S.A. | A gram negative bacterium with reduced lps level in the outer membrane and use thereof for treating gram negative bacterial infection |
| GB0408977D0 (en) | 2004-04-22 | 2004-05-26 | Chiron Srl | Immunising against meningococcal serogroup Y using proteins |
| MXPA06013093A (es) | 2004-05-11 | 2007-08-08 | Nederlanden Staat | Lipo-oligo-sacarido de neisseria meningitidis igtb como adyuvante. |
| WO2006010143A2 (en) | 2004-07-13 | 2006-01-26 | Neose Technologies, Inc. | Branched peg remodeling and glycosylation of glucagon-like peptide-1 [glp-1] |
| GB0419408D0 (en) * | 2004-09-01 | 2004-10-06 | Chiron Srl | 741 chimeric polypeptides |
| GB0419627D0 (en) * | 2004-09-03 | 2004-10-06 | Chiron Srl | Immunogenic bacterial vesicles with outer membrane proteins |
| EP3061461A1 (en) | 2004-10-29 | 2016-08-31 | ratiopharm GmbH | Remodeling and glycopegylation of fibroblast growth factor (fgf) |
| GB0424092D0 (en) | 2004-10-29 | 2004-12-01 | Chiron Srl | Immunogenic bacterial vesicles with outer membrane proteins |
| GB0428381D0 (en) * | 2004-12-24 | 2005-02-02 | Isis Innovation | Vaccine |
| ES2449195T3 (es) | 2005-01-10 | 2014-03-18 | Ratiopharm Gmbh | Factor estimulante de colonias de granulocitos glicopegilado |
| JP2008528512A (ja) | 2005-01-21 | 2008-07-31 | エピトピックス,リミティド ライアビリティ カンパニー | エルシニアspp.ポリペプチド及び使用方法 |
| CA2590974C (en) * | 2005-01-27 | 2017-10-03 | Children's Hospital & Research Center At Oakland | Gna1870-based vesicle vaccines for broad spectrum protection against diseases caused by neisseria meningitidis |
| WO2006121569A2 (en) | 2005-04-08 | 2006-11-16 | Neose Technologies, Inc. | Compositions and methods for the preparation of protease resistant human growth hormone glycosylation mutants |
| PE20110072A1 (es) | 2005-06-27 | 2011-02-04 | Glaxosmithkline Biolog Sa | Composicion inmunogenica |
| US20070105755A1 (en) | 2005-10-26 | 2007-05-10 | Neose Technologies, Inc. | One pot desialylation and glycopegylation of therapeutic peptides |
| US7955817B2 (en) | 2005-09-02 | 2011-06-07 | Glaxosmithkline Biologicals S.A. | Vaccine protection assay |
| DE602005024320D1 (de) * | 2005-09-05 | 2010-12-02 | Glaxosmithkline Biolog Sa | Bakterizidie-serumtest für n. meningitidis spezifische antiseren |
| WO2007056191A2 (en) | 2005-11-03 | 2007-05-18 | Neose Technologies, Inc. | Nucleotide sugar purification using membranes |
| GB0524066D0 (en) | 2005-11-25 | 2006-01-04 | Chiron Srl | 741 ii |
| CA2632434A1 (en) * | 2005-12-06 | 2007-06-14 | Universita Degli Studi Di Padova | Methods and compositions relating to adhesins as adjuvants |
| GB0607088D0 (en) | 2006-04-07 | 2006-05-17 | Glaxosmithkline Biolog Sa | Vaccine |
| CA2816182C (en) | 2005-12-22 | 2018-02-20 | Glaxosmithkline Biologicals S.A. | Pneumococcal polysaccharide conjugate vaccine |
| CU23549A1 (es) * | 2005-12-29 | 2010-07-20 | Ct Ingenieria Genetica Biotech | Composiciones farmacéuticas que contienen la proteína nma0939 |
| US20100021503A1 (en) | 2006-03-30 | 2010-01-28 | Glaxosmithkline Biologicals S.A. | Immunogenic composition |
| CA2654709A1 (en) * | 2006-06-12 | 2007-12-21 | Nathalie Devos | Neisseria meningitidis lipooligosaccharide vaccine |
| US20080274958A1 (en) | 2006-07-21 | 2008-11-06 | Neose Technologies, Inc. | Glycosylation of peptides via o-linked glycosylation sequences |
| EP2066344B2 (en) | 2006-09-07 | 2016-06-29 | GlaxoSmithKline Biologicals S.A. | Inactivated Poliovirus combination vaccine |
| US8969532B2 (en) | 2006-10-03 | 2015-03-03 | Novo Nordisk A/S | Methods for the purification of polypeptide conjugates comprising polyalkylene oxide using hydrophobic interaction chromatography |
| GB0700136D0 (en) | 2007-01-04 | 2007-02-14 | Glaxosmithkline Biolog Sa | Process for manufacturing vaccines |
| GB0700562D0 (en) | 2007-01-11 | 2007-02-21 | Novartis Vaccines & Diagnostic | Modified Saccharides |
| NZ580030A (en) | 2007-04-03 | 2012-06-29 | Biogenerix Ag | Methods of treatment using glycopegylated g-csf |
| WO2008154639A2 (en) | 2007-06-12 | 2008-12-18 | Neose Technologies, Inc. | Improved process for the production of nucleotide sugars |
| KR101579947B1 (ko) | 2007-06-26 | 2015-12-28 | 글락소스미스클라인 바이오로지칼즈 에스.에이. | 스트렙토코쿠스 뉴모니애 캡슐 다당류 컨쥬게이트를 포함하는 백신 |
| BRPI0814793A2 (pt) * | 2007-08-02 | 2015-02-03 | Glaxosmithkline Biolog Sa | Método de tipagem molecular los de uma cepa neisseria, kit, e, método de diagnóstico e classificação de uma colonização de neisseria e/ou infecção em um hospedeiro susceptível à colonização de neisseria. |
| WO2009038889A1 (en) * | 2007-08-02 | 2009-03-26 | Children's Hospital And Research Center At Oakland | Fhbp- and lpxl1-based vesicle vaccines for broad spectrum protection against diseases caused by neisseria meningitidis |
| PL2200642T3 (pl) | 2007-10-19 | 2012-09-28 | Novartis Ag | Preparaty szczepionek meningokokowych |
| ES2532946T3 (es) | 2008-02-21 | 2015-04-06 | Novartis Ag | Polipéptidos PUfH meningocócicos |
| KR101582841B1 (ko) | 2008-02-27 | 2016-01-11 | 노보 노르디스크 에이/에스 | 콘쥬게이트된 인자 viii 분자 |
| NZ588191A (en) | 2008-03-03 | 2012-06-29 | Irm Llc | Compounds and compositions as tlr activity modulators |
| MX2010012999A (es) | 2008-05-30 | 2012-03-07 | U S A As Represented By The Secretary Of The Army On Behalf Of Walter Reed Army | Vacuna de vesícula de membrana externa nativa multivalente del meningococo, método para su fabricación y uso. |
| KR101042541B1 (ko) * | 2008-07-25 | 2011-06-17 | 한국생명공학연구원 | 외막소체를 생산하는 재조합 g(-) 박테리아 및 이를이용한 다가항원이 실린 외막소체의 제조방법 |
| WO2010027729A2 (en) * | 2008-08-25 | 2010-03-11 | Creatv Microtech, Inc. | Gonococcal vaccines |
| GB0816447D0 (en) * | 2008-09-08 | 2008-10-15 | Glaxosmithkline Biolog Sa | Vaccine |
| GB0822634D0 (en) | 2008-12-11 | 2009-01-21 | Novartis Ag | Meningitis vaccines |
| CA2747340A1 (en) | 2008-12-17 | 2010-06-24 | Novartis Ag | Meningococcal vaccines including hemoglobin receptor |
| CN101759781B (zh) * | 2008-12-25 | 2013-04-03 | 上海市第六人民医院 | 一种细菌表层黏附蛋白及其用途 |
| ES2537426T3 (es) | 2008-12-25 | 2015-06-08 | The Chemo-Sero-Therapeutic Research Institute | Vacuna recombinante para la rinitis infecciosa aviar y método para producirla |
| EP2208787A1 (en) * | 2009-01-19 | 2010-07-21 | Université de Liège | A recombinant alpha-hemolysin polypeptide of Staphylococcus aureus, having a deletion in the stem domain and heterologous sequences inserted |
| NZ595234A (en) | 2009-03-24 | 2013-12-20 | Novartis Ag | Adjuvanting meningococcal factor h binding protein |
| JP5806204B2 (ja) * | 2009-03-24 | 2015-11-10 | ノバルティス アーゲー | 肺炎球菌血清型14の糖を含む組み合わせ |
| WO2010109324A1 (en) | 2009-03-24 | 2010-09-30 | Novartis Ag | Combinations of meningococcal factor h binding protein and pneumococcal saccharide conjugates |
| NZ597008A (en) * | 2009-05-14 | 2013-03-28 | Sanofi Pasteur | Meningococcal vaccine based on lipooligosaccharide (los) and neisseria meningitidis protein |
| AU2010247254A1 (en) | 2009-05-14 | 2012-01-12 | Sanofi Pasteur | Menigococcus vaccine containing lipooligosaccharide (LOS) from modified strains of L6 immunotype Neisseria meningitidis |
| WO2010130896A2 (fr) | 2009-05-14 | 2010-11-18 | Sanofi Pasteur | Procédé pour adjuver le lipopolysaccharide (lps) des bactéries à gram-négatif |
| JP5536765B2 (ja) * | 2009-05-20 | 2014-07-02 | 国立大学法人鳥取大学 | 部分糖鎖エピトープを用いた、病原性ナイセリア属細菌感染の検出方法およびそれら細菌に対するワクチン |
| JP5867952B2 (ja) | 2009-06-10 | 2016-02-24 | ノバルティス アーゲー | ベンゾナフチリジン含有ワクチン |
| GB0913681D0 (en) | 2009-08-05 | 2009-09-16 | Glaxosmithkline Biolog Sa | Immunogenic composition |
| CN104650241A (zh) | 2009-08-27 | 2015-05-27 | 诺华股份有限公司 | 包括脑膜炎球菌fHBP序列的杂交多肽 |
| CN102740882A (zh) | 2009-08-27 | 2012-10-17 | 诺华有限公司 | 含有铝、寡核苷酸和聚阳离子的佐剂 |
| WO2011027956A2 (ko) | 2009-09-04 | 2011-03-10 | 주식회사이언메딕스 | 그람 양성 박테리아에서 유래한 세포밖 소포체 및 이를 이용한 질병 모델 |
| CN102480932B (zh) | 2009-09-01 | 2015-01-14 | 阿昂梅迪克斯公司 | 源于肠道共生细菌的细胞外小泡及利用其的疫苗、备选药物筛选方法 |
| CA2772916C (en) | 2009-09-02 | 2019-01-15 | Novartis Ag | Immunogenic compositions including tlr activity modulators |
| JO3257B1 (ar) | 2009-09-02 | 2018-09-16 | Novartis Ag | مركبات وتركيبات كمعدلات لفاعلية tlr |
| GB0917002D0 (en) * | 2009-09-28 | 2009-11-11 | Novartis Vaccines Inst For Global Health Srl | Improved shigella blebs |
| GB0917003D0 (en) | 2009-09-28 | 2009-11-11 | Novartis Vaccines Inst For Global Health Srl | Purification of bacterial vesicles |
| US20130022639A1 (en) | 2009-09-30 | 2013-01-24 | Novartis Ag | Expression of meningococcal fhbp polypeptides |
| US20130022633A1 (en) | 2009-10-27 | 2013-01-24 | University Of Florence | MENINGOCOCCAL fHBP POLYPEPTIDES |
| WO2011057148A1 (en) | 2009-11-05 | 2011-05-12 | Irm Llc | Compounds and compositions as tlr-7 activity modulators |
| EP2496597A4 (en) * | 2009-11-06 | 2013-08-14 | Childrens Hosp & Res Ct Oak | T CELL STIMULATING PROTEIN B AND METHOD FOR ITS USE |
| MX339146B (es) | 2009-12-15 | 2016-05-13 | Novartis Ag | Suspension homogenea de compuestos inmunopotenciadores y usos de los mismos. |
| JP2013521770A (ja) * | 2010-03-10 | 2013-06-13 | グラクソスミスクライン バイオロジカルズ ソシエテ アノニム | ワクチン組成物 |
| BR112012022800A2 (pt) * | 2010-03-11 | 2018-05-15 | Glaxosmithkline Biologicals Sa | composição imunogênica ou vacina, cepa bacteriana gram-negativa geneticamente engenheirada, métodos para o tratamento ou prevenção de infecção ou doença, para produzir uma composição imunogênica ou uma vacina, e para preparar uma imunoglobulina, preparação da imunoglobulina, e, preparação farmacêutica |
| CN102802662A (zh) | 2010-03-18 | 2012-11-28 | 诺华有限公司 | 用于脑膜炎球菌血清组b的含佐剂疫苗 |
| WO2011119759A1 (en) | 2010-03-23 | 2011-09-29 | Irm Llc | Compounds (cystein based lipopeptides) and compositions as tlr2 agonists used for treating infections, inflammations, respiratory diseases etc. |
| AU2013202310C1 (en) * | 2010-03-29 | 2017-01-05 | Nationwide Children's Hospital, Inc. | Compositions and methods for the removal of biofilms |
| CN103097399A (zh) * | 2010-03-29 | 2013-05-08 | 南加利福尼亚大学 | 用于去除生物膜的组合物及方法 |
| EP2585106A1 (en) | 2010-06-25 | 2013-05-01 | Novartis AG | Combinations of meningococcal factor h binding proteins |
| CA2810851C (en) | 2010-09-09 | 2022-08-02 | University Of Southern California | Compositions and methods for the removal of biofilms comprising a high-mobility group-box (hmg-box) domain containing polypeptide |
| US9259462B2 (en) | 2010-09-10 | 2016-02-16 | Glaxosmithkline Biologicals Sa | Developments in meningococcal outer membrane vesicles |
| GB201015132D0 (en) | 2010-09-10 | 2010-10-27 | Univ Bristol | Vaccine composition |
| WO2012041899A1 (en) * | 2010-09-28 | 2012-04-05 | Xbrane Bioscience Ab | Fusion protein for secretory protein expression |
| WO2012054879A1 (en) * | 2010-10-22 | 2012-04-26 | Duke University | Compositions and methods for the treatment of septic arthritis, osteomyelitis, and bacteremia |
| US20150147356A1 (en) | 2011-05-12 | 2015-05-28 | Alan Kimura | Antipyretics to enhance tolerability of vesicle-based vaccines |
| PL2729167T3 (pl) * | 2011-07-07 | 2018-08-31 | De Staat Der Nederlanden, Vert. Door De Minister Van Vws | Sposób wolnego od detergentów wytwarzania pęcherzyków zewnątrzbłonowych bakterii Gram-ujemnych |
| US10596246B2 (en) | 2011-12-29 | 2020-03-24 | Glaxosmithkline Biological Sa | Adjuvanted combinations of meningococcal factor H binding proteins |
| ES2654613T3 (es) | 2012-02-02 | 2018-02-14 | Glaxosmithkline Biologicals Sa | Promotores para una expresión aumentada de proteínas en meningococos |
| JP5698693B2 (ja) * | 2012-03-13 | 2015-04-08 | グラクソスミスクライン バイオロジカルズ ソシエテ アノニム | 髄膜炎菌特異的抗血清に対する血清殺菌アッセイ |
| WO2013186753A1 (en) | 2012-06-14 | 2013-12-19 | Novartis Ag | Vaccines for serogroup x meningococcus |
| EP2892553A1 (en) | 2012-09-06 | 2015-07-15 | Novartis AG | Combination vaccines with serogroup b meningococcus and d/t/p |
| AU2013320313B2 (en) | 2012-09-18 | 2018-07-12 | Glaxosmithkline Biologicals Sa | Outer membrane vesicles |
| US20160120818A1 (en) | 2013-02-07 | 2016-05-05 | Glaxosmithline Biological Sa | Pharmaceutical compositions comprising vesicles |
| WO2014138290A1 (en) * | 2013-03-05 | 2014-09-12 | Trudeau Institute, Inc. | Compositions and methods for treating bacterial infections |
| US11274144B2 (en) | 2013-06-13 | 2022-03-15 | Research Institute At Nationwide Children's Hospital | Compositions and methods for the removal of biofilms |
| US9745366B2 (en) | 2013-09-23 | 2017-08-29 | University Of Southern California | Compositions and methods for the prevention of microbial infections |
| US10233234B2 (en) | 2014-01-13 | 2019-03-19 | Trellis Bioscience, Llc | Binding moieties for biofilm remediation |
| US11248040B2 (en) | 2013-09-26 | 2022-02-15 | Trellis Bioscience, Llc | Binding moieties for biofilm remediation |
| WO2015128480A1 (en) | 2014-02-28 | 2015-09-03 | Novartis Ag | Modified meningococcal fhbp polypeptides |
| EP3229835A1 (en) * | 2014-12-09 | 2017-10-18 | Sanofi Pasteur | Compositions comprising n. meningitidis proteins |
| US11066453B2 (en) | 2015-05-18 | 2021-07-20 | Biomvis Srl | Immunogenic compositions containing bacterial outer membrane vesicles and therapeutic uses thereof |
| US10940204B2 (en) | 2015-07-31 | 2021-03-09 | Research Institute At Nationwide Children's Hospital | Peptides and antibodies for the removal of biofilms |
| EP3365027B1 (en) | 2015-10-14 | 2022-03-30 | Research Institute at Nationwide Children's Hospital | Hu specific antibodies and their use in inhibiting biofilm |
| KR101825439B1 (ko) * | 2016-04-15 | 2018-02-05 | 배재대학교 산학협력단 | 염산 처리에 의한 그람양성 박테리아 고스트의 제조 방법 |
| US11564982B2 (en) | 2017-01-04 | 2023-01-31 | Research Institute At Nationwide Children's Hospital | DNABII vaccines and antibodies with enhanced activity |
| WO2018170178A1 (en) | 2017-03-15 | 2018-09-20 | Research Institute At Nationwide Children's Hospital | Composition and methods for disruption of bacterial biofilms without accompanying inflammation |
| EP3607967A1 (en) | 2018-08-09 | 2020-02-12 | GlaxoSmithKline Biologicals S.A. | Modified meningococcal fhbp polypeptides |
| CN112996538A (zh) * | 2018-11-06 | 2021-06-18 | 葛兰素史密丝克莱恩生物有限公司 | 免疫原性组合物 |
| US20220125908A1 (en) * | 2019-02-14 | 2022-04-28 | University Of Florida Research Foundation, Incorporated | Honeybee commensal snodgrassella alvi vaccine against pathogenic neisseriaceae |
| FR3099160B1 (fr) * | 2019-07-23 | 2022-05-06 | Univ Grenoble Alpes | Anticorps dirigé contre la protéine oprf depseudomonas aeruginosa, son utilisation en tant que médicament et composition pharmaceutique le contenant |
| WO2023097652A1 (en) * | 2021-12-03 | 2023-06-08 | National Center For Nanoscience And Technology | An engineered cell and application thereof |
| GB202203250D0 (en) | 2022-03-09 | 2022-04-20 | Glaxosmithkline Biologicals Sa | Immunogenic compositions |
Family Cites Families (131)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4239746A (en) * | 1973-06-30 | 1980-12-16 | Dezso Istvan Bartos | Complement fixation test employing reactants in a disposable package |
| DE2744721A1 (de) | 1977-10-05 | 1979-04-19 | Veba Chemie Ag | Pulverfoermige ueberzugsmittel und deren anwendung |
| DE2848965A1 (de) | 1978-11-11 | 1980-05-22 | Behringwerke Ag | Verfahren zur herstellung von membranproteinen aus neisseria meningitidis und diese enthaltende vaccine |
| EP0027888B1 (en) | 1979-09-21 | 1986-04-16 | Hitachi, Ltd. | Semiconductor switch |
| US4271147A (en) | 1980-01-10 | 1981-06-02 | Behringwerke Aktiengesellschaft | Process for the isolation of membrane proteins from Neisseria meningitidis and vaccines containing same |
| US4673574A (en) | 1981-08-31 | 1987-06-16 | Anderson Porter W | Immunogenic conjugates |
| US4695624A (en) | 1984-05-10 | 1987-09-22 | Merck & Co., Inc. | Covalently-modified polyanionic bacterial polysaccharides, stable covalent conjugates of such polysaccharides and immunogenic proteins with bigeneric spacers, and methods of preparing such polysaccharides and conjugates and of confirming covalency |
| US20020146764A1 (en) * | 1985-03-28 | 2002-10-10 | Chiron Corporation | Expression using fused genes providing for protein product |
| IT1187753B (it) | 1985-07-05 | 1987-12-23 | Sclavo Spa | Coniugati glicoproteici ad attivita' immunogenica trivalente |
| DE3622221A1 (de) * | 1986-07-02 | 1988-01-14 | Max Planck Gesellschaft | Verfahren zur gentechnologischen gewinnung von proteinen unter verwendung gramnegativer wirtszellen |
| US5173294A (en) | 1986-11-18 | 1992-12-22 | Research Foundation Of State University Of New York | Dna probe for the identification of haemophilus influenzae |
| RU2023448C1 (ru) | 1987-07-30 | 1994-11-30 | Сентро Насьональ Де Биопрепарадос | Способ получения вакцины против различных патогенных серотипов менингита нейссера группы в |
| CA2005704C (en) | 1988-12-16 | 2003-02-11 | James C. Paton | Pneumolysin mutants and pneumococcal vaccines made therefrom |
| US7118757B1 (en) * | 1988-12-19 | 2006-10-10 | Wyeth Holdings Corporation | Meningococcal class 1 outer-membrane protein vaccine |
| CU22302A1 (es) * | 1990-09-07 | 1995-01-31 | Cigb | Secuencia nucleotidica codificante para una proteina de la membrana externa de neisseria meningitidis y uso de dicha proteina en preparados vacunales |
| ES2202297T3 (es) | 1990-07-16 | 2004-04-01 | University Of North Carolina At Chapel Hill | Proteinas antigenicas de n. meningitidis reprimibles con hierro relacionadas con la familia de toxinas hemolisina. |
| DE4023721A1 (de) | 1990-07-26 | 1992-01-30 | Boehringer Mannheim Gmbh | Verfahren zur herstellung von vakzinen und ihre verwendung |
| DK0546118T3 (da) | 1990-08-23 | 2004-02-16 | Univ North Carolina | Transferrinbindende proteiner fra Neisseria gonorrhoeae og Neisseria meningtidis |
| US5912336A (en) | 1990-08-23 | 1999-06-15 | University Of North Carolina At Chapel Hill | Isolated nucleic acid molecules encoding transferrin binding proteins from Neisseria gonorrhoeae and Neisseria meningitidis |
| US5153312A (en) | 1990-09-28 | 1992-10-06 | American Cyanamid Company | Oligosaccharide conjugate vaccines |
| US5371186A (en) | 1991-02-11 | 1994-12-06 | Biosynth S.R.L. | Synthetic peptides for detoxification of bacterial endotoxins and for the prevention and treatment of septic shock |
| US5652211A (en) | 1991-02-11 | 1997-07-29 | Biosynth S.R.L. | Peptides for neutralizing the toxicity of Lipid A |
| US6592876B1 (en) | 1993-04-20 | 2003-07-15 | Uab Research Foundation | Pneumococcal genes, portions thereof, expression products therefrom, and uses of such genes, portions and products |
| US5476929A (en) | 1991-02-15 | 1995-12-19 | Uab Research Foundation | Structural gene of pneumococcal protein |
| ATE174625T1 (de) * | 1991-03-14 | 1999-01-15 | Imclone Systems Inc | Rekombinante hybride porinepitope |
| US5552146A (en) | 1991-08-15 | 1996-09-03 | Board Of Regents, The University Of Texas System | Methods and compositions relating to useful antigens of Moraxella catarrhalis |
| FR2682041B1 (fr) | 1991-10-03 | 1994-01-14 | Pasteur Merieux Serums Vaccins | Vaccin contre les infections a neisseria meningitidis. |
| CA2127871A1 (en) | 1992-01-13 | 1993-07-22 | Andreas Herman Hogt | Crosslinking of rubbers with engineering plastics |
| DE69333107T2 (de) | 1992-02-11 | 2004-01-29 | Jackson H M Found Military Med | Dualer träger für immunogene konstrukte |
| FR2692592B1 (fr) | 1992-06-19 | 1995-03-31 | Pasteur Merieux Serums Vacc | Fragments d'ADN codant pour les sous-unités du récepteur de la transferrine de Neisseria meningitidis et procédés les exprimant. |
| UA40597C2 (uk) | 1992-06-25 | 2001-08-15 | Смітклайн Бічем Байолоджікалс С.А. | Вакцинна композиція,спосіб лікування ссавців, що страждають або сприйнятливі до інфекції, спосіб лікування ссавців, що страждають на рак, спосіб одержання вакцинної композиції, композиція ад'ювантів |
| NL9201716A (nl) * | 1992-10-02 | 1994-05-02 | Nederlanden Staat | Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk buitenmembraanvesikel omvat. |
| GB9224584D0 (en) | 1992-11-23 | 1993-01-13 | Connaught Lab | Use of outer membrane protein d15 and its peptides as vaccine against haempohilus influenzae diseases |
| DE69431624T2 (de) | 1993-05-18 | 2003-07-10 | Univ Ohio State Res Found | Impfstoff gegen mittelohrentzündung |
| US5439808A (en) * | 1993-07-23 | 1995-08-08 | North American Vaccine, Inc. | Method for the high level expression, purification and refolding of the outer membrane group B porin proteins from Neisseria meningitidis |
| CA2175332C (en) | 1993-11-08 | 2009-04-07 | Sheena M. Loosmore | Haemophilus transferrin receptor genes |
| US6361779B1 (en) | 1993-11-08 | 2002-03-26 | Aventis Pasteur Limited | Transferrin receptor genes |
| GB9326253D0 (en) | 1993-12-23 | 1994-02-23 | Smithkline Beecham Biolog | Vaccines |
| CA2560114A1 (en) | 1994-07-15 | 1996-02-01 | The University Of Iowa Research Foundation | Immunomodulatory oligonucleotides |
| US5565204A (en) | 1994-08-24 | 1996-10-15 | American Cyanamid Company | Pneumococcal polysaccharide-recombinant pneumolysin conjugate vaccines for immunization against pneumococcal infections |
| US5545553A (en) | 1994-09-26 | 1996-08-13 | The Rockefeller University | Glycosyltransferases for biosynthesis of oligosaccharides, and genes encoding them |
| IL117483A (en) | 1995-03-17 | 2008-03-20 | Bernard Brodeur | MENINGITIDIS NEISSERIA shell protein is resistant to proteinase K. |
| US6287574B1 (en) * | 1995-03-17 | 2001-09-11 | Biochem Pharma Inc. | Proteinase K resistant surface protein of neisseria meningitidis |
| US6265567B1 (en) | 1995-04-07 | 2001-07-24 | University Of North Carolina At Chapel Hill | Isolated FrpB nucleic acid molecule |
| UA56132C2 (uk) | 1995-04-25 | 2003-05-15 | Смітклайн Бічем Байолоджікалс С.А. | Композиція вакцини (варіанти), спосіб стабілізації qs21 відносно гідролізу (варіанти), спосіб приготування композиції вакцини |
| US6440425B1 (en) | 1995-05-01 | 2002-08-27 | Aventis Pasteur Limited | High molecular weight major outer membrane protein of moraxella |
| US5843464A (en) | 1995-06-02 | 1998-12-01 | The Ohio State University | Synthetic chimeric fimbrin peptides |
| EA199800046A1 (ru) | 1995-06-07 | 1998-06-25 | Байокем Вэксинс Инк. | Полипептид, последовательность днк, вакцинная композиция (варианты), антитело или его фрагмент, вакцина, применения указанных полипептида, последовательности днк и антитела или его фрагмента |
| US6007838A (en) | 1995-06-07 | 1999-12-28 | The United States Of America As Represented By The Secretary Of The Army | Process for making liposome preparation |
| GB9513074D0 (en) | 1995-06-27 | 1995-08-30 | Cortecs Ltd | Novel anigen |
| CZ298460B6 (cs) * | 1995-09-18 | 2007-10-10 | United States Army Medical Research Materiel Command (Usamrmc) | Zlepšené zpusoby výroby nekovalentne komplexovaných a multivalentních proteosomových podjednotkových vakcin |
| US6290970B1 (en) | 1995-10-11 | 2001-09-18 | Aventis Pasteur Limited | Transferrin receptor protein of Moraxella |
| US6440701B1 (en) | 1996-03-08 | 2002-08-27 | Aventis Pasteur Limited | Transferrin receptor genes of Moraxella |
| US6090576A (en) | 1996-03-08 | 2000-07-18 | Connaught Laboratories Limited | DNA encoding a transferrin receptor of Moraxella |
| EP0912608B1 (en) | 1996-05-01 | 2006-04-19 | The Rockefeller University | Choline binding proteins for anti-pneumococcal vaccines |
| US7341727B1 (en) | 1996-05-03 | 2008-03-11 | Emergent Product Development Gaithersburg Inc. | M. catarrhalis outer membrane protein-106 polypeptide, methods of eliciting an immune response comprising same |
| FR2751000B1 (fr) | 1996-07-12 | 1998-10-30 | Inst Nat Sante Rech Med | Adn specifiques des bacteries de l'espece neisseria meningitidis, leurs procedes d'obtention et leurs applications biologiques |
| US5882871A (en) | 1996-09-24 | 1999-03-16 | Smithkline Beecham Corporation | Saliva binding protein |
| US5882896A (en) | 1996-09-24 | 1999-03-16 | Smithkline Beecham Corporation | M protein |
| AU6909098A (en) | 1996-10-31 | 1998-05-22 | Human Genome Sciences, Inc. | Streptococcus pneumoniae polynucleotides and sequences |
| CA2292838A1 (en) | 1997-06-03 | 1998-12-10 | Connaught Laboratories Limited | Lactoferrin receptor genes of moraxella |
| US6764686B2 (en) | 1997-07-21 | 2004-07-20 | Baxter International Inc. | Modified immunogenic pneumolysin compositions as vaccines |
| WO1999006781A1 (de) | 1997-07-31 | 1999-02-11 | Wilo Gmbh | Latentwärmespeicher für ein kraftfahrzeug |
| CN1204253C (zh) | 1997-08-15 | 2005-06-01 | 乌得勒支大学 | 奈瑟氏球菌乳铁蛋白结合蛋白 |
| GB9717953D0 (en) | 1997-08-22 | 1997-10-29 | Smithkline Beecham Biolog | Vaccine |
| US6914131B1 (en) * | 1998-10-09 | 2005-07-05 | Chiron S.R.L. | Neisserial antigens |
| GB9726398D0 (en) * | 1997-12-12 | 1998-02-11 | Isis Innovation | Polypeptide and coding sequences |
| AU1979599A (en) * | 1998-01-14 | 1999-08-02 | Chiron S.P.A. | (neisseria meningitidis) antigens |
| CA2264970A1 (en) | 1998-03-10 | 1999-09-10 | American Cyanamid Company | Antigenic conjugates of conserved lipolysaccharides of gram negative bacteria |
| CA2325340A1 (en) | 1998-04-07 | 1999-10-14 | Medimmune, Inc. | Derivatives of pneumococcal choline binding proteins for vaccines |
| GB9808734D0 (en) | 1998-04-23 | 1998-06-24 | Smithkline Beecham Biolog | Novel compounds |
| GB9808866D0 (en) * | 1998-04-24 | 1998-06-24 | Smithkline Beecham Biolog | Novel compounds |
| EP2261347A3 (en) * | 1998-05-01 | 2012-01-11 | Novartis Vaccines and Diagnostics, Inc. | Neisseria meningitidis antigens and compositions |
| US20070026021A1 (en) * | 1998-05-01 | 2007-02-01 | Chiron S.R.I. | Neisseria meningitidis antigens and compositions |
| GB9809683D0 (en) | 1998-05-06 | 1998-07-01 | Smithkline Beecham Biolog | Novel compounds |
| GB9810285D0 (en) | 1998-05-13 | 1998-07-15 | Smithkline Beecham Biolog | Novel compounds |
| GB9810276D0 (en) * | 1998-05-13 | 1998-07-15 | Smithkline Beecham Biolog | Novel compounds |
| GB9811260D0 (en) | 1998-05-26 | 1998-07-22 | Smithkline Beecham Biolog | Novel compounds |
| US6248329B1 (en) * | 1998-06-01 | 2001-06-19 | Ramaswamy Chandrashekar | Parasitic helminth cuticlin nucleic acid molecules and uses thereof |
| ATE341628T1 (de) | 1998-06-03 | 2006-10-15 | Glaxosmithkline Biolog Sa | Proteine und gene aus moraxella catarrhalis, antigene, antikörper, und verwendungen |
| GB9812163D0 (en) | 1998-06-05 | 1998-08-05 | Smithkline Beecham Biolog | Novel compounds |
| GB9812440D0 (en) | 1998-06-09 | 1998-08-05 | Smithkline Beecham Biolog | Novel compounds |
| GB9814902D0 (en) | 1998-07-10 | 1998-09-09 | Univ Nottingham | Screening of neisserial vaccine candidates against pathogenic neisseria |
| US6951652B2 (en) | 1998-07-29 | 2005-10-04 | Biosynth S.R.L. | Vaccine for prevention of gram-negative bacterial infections and endotoxin related diseases |
| GB9818004D0 (en) * | 1998-08-18 | 1998-10-14 | Smithkline Beecham Biolog | Novel compounds |
| GB9820003D0 (en) | 1998-09-14 | 1998-11-04 | Smithkline Beecham Biolog | Novel compounds |
| GB9820002D0 (en) | 1998-09-14 | 1998-11-04 | Smithkline Beecham Biolog | Novel compounds |
| MXPA01003557A (es) * | 1998-10-09 | 2004-04-05 | Chiron Corp | Secuencias genomicas de neisseria y metodos para su uso. |
| DE69926647T2 (de) | 1998-10-16 | 2006-11-09 | Fraunhofer-Gesellschaft zur Förderung der angewandten Forschung e.V. | Durch molekulares pathogenicid vermittelte krankheitsresistenz in pflanzen |
| US6610306B2 (en) * | 1998-10-22 | 2003-08-26 | The University Of Montana | OMP85 protein of neisseria meningitidis, compositions containing the same and methods of use thereof |
| WO2000023595A1 (en) | 1998-10-22 | 2000-04-27 | The University Of Montana | Omp85 proteins of neisseria gonorrhoeae and neisseria meningitidis, compositions containing same and methods of use thereof |
| GB9823978D0 (en) * | 1998-11-02 | 1998-12-30 | Microbiological Res Authority | Multicomponent meningococcal vaccine |
| US20030215469A1 (en) * | 1998-11-02 | 2003-11-20 | Microbiological Research Authority | Multicomponent meningococcal vaccine |
| ES2294821T5 (es) | 1998-11-03 | 2014-03-05 | De Staat Der Nederlanden Vertegenwoordigd Door De Minister Van Welzijn, Volksgezondheid En Cultuur | LPS con toxicidad reducida a partir de bacterias gram negativas modificadas genéticamente |
| CN100392082C (zh) * | 1999-04-30 | 2008-06-04 | 启龙股份公司 | 保守的奈瑟球菌抗原 |
| ES2323845T3 (es) * | 1999-04-30 | 2009-07-27 | Novartis Vaccines And Diagnostics, Inc. | Secuencias genomicas de neisseria y procedimientos de uso. |
| GB9911683D0 (en) * | 1999-05-19 | 1999-07-21 | Chiron Spa | Antigenic peptides |
| DK2270172T3 (en) | 1999-05-19 | 2016-02-29 | Glaxosmithkline Biolog Sa | Combination Compositions of Neisseria. |
| CA2378687A1 (en) | 1999-06-18 | 2000-12-28 | Elitra Pharmaceuticals, Inc. | Nucleotide sequences of moraxella catarrhalis genome |
| GB2351515B (en) | 1999-06-29 | 2002-09-11 | Pandrol Ltd | Adjustable railway rail fastening assembly and methods for use therewith |
| GB9917977D0 (en) | 1999-07-30 | 1999-09-29 | Smithkline Beecham Biolog | Novel compounds |
| GB9918038D0 (en) | 1999-07-30 | 1999-09-29 | Smithkline Beecham Biolog | Novel compounds |
| GB9918208D0 (en) | 1999-08-03 | 1999-10-06 | Smithkline Beecham Biolog | Novel compounds |
| GB9918319D0 (en) | 1999-08-03 | 1999-10-06 | Smithkline Beecham Biolog | Vaccine composition |
| GB9918302D0 (en) | 1999-08-03 | 1999-10-06 | Smithkline Beecham Biolog | Novel compounds |
| US6531131B1 (en) * | 1999-08-10 | 2003-03-11 | The United States Of America As Represented By The Department Of Health And Human Services | Conjugate vaccine for Neisseria meningitidis |
| AUPQ275799A0 (en) * | 1999-09-10 | 1999-10-07 | Luminis Pty Limited | Recombinant bacterium expressing an oligosaccharide receptor mimic |
| ATE460484T1 (de) | 1999-11-29 | 2010-03-15 | Novartis Vaccines & Diagnostic | 85 kda antigen von neisseria |
| PT1248647E (pt) * | 2000-01-17 | 2010-11-18 | Novartis Vaccines & Diagnostics Srl | Vacina de vesícula da membrana externa (omv) compreendendo proteínas de membrana externa de n. meningitidis do serogrupo b |
| WO2001055182A1 (en) * | 2000-01-25 | 2001-08-02 | The University Of Queensland | PROTEINS COMPRISING CONSERVED REGIONS OF NEISSERIA MENINGITIDIS SURFACE ANTIGEN NhhA |
| CA2744921C (en) * | 2000-02-28 | 2014-05-13 | Chiron S.R.L. | Hybrid expression of neisserial proteins |
| GB0007432D0 (en) * | 2000-03-27 | 2000-05-17 | Microbiological Res Authority | Proteins for use as carriers in conjugate vaccines |
| GB0011108D0 (en) * | 2000-05-08 | 2000-06-28 | Microscience Ltd | Virulence gene and protein and their use |
| MXPA03000822A (es) | 2000-07-27 | 2004-11-01 | Childrens Hosp & Res Ct Oak | Vacunas para proteccion de espectro amplio contra enfermedades causadas por neisseria meningitidis. |
| GB0103424D0 (en) * | 2001-02-12 | 2001-03-28 | Chiron Spa | Gonococcus proteins |
| GB0108024D0 (en) * | 2001-03-30 | 2001-05-23 | Chiron Spa | Bacterial toxins |
| GB0115176D0 (en) * | 2001-06-20 | 2001-08-15 | Chiron Spa | Capular polysaccharide solubilisation and combination vaccines |
| AU2002330681C1 (en) | 2001-07-26 | 2015-04-02 | Glaxosmithkline Biologicals S.A. | Vaccines comprising aluminium adjuvants and histidine |
| GB0118249D0 (en) * | 2001-07-26 | 2001-09-19 | Chiron Spa | Histidine vaccines |
| PT2248822T (pt) * | 2001-07-27 | 2017-02-14 | Glaxosmithkline Biologicals Sa | Adesinas de meningococos |
| GB0121591D0 (en) | 2001-09-06 | 2001-10-24 | Chiron Spa | Hybrid and tandem expression of neisserial proteins |
| EP1438323A4 (en) | 2001-10-03 | 2007-08-01 | Novartis Vaccines & Diagnostic | ADJUVOUS MENINGOCOCCUS COMPOSITIONS |
| US20060034854A1 (en) * | 2002-08-02 | 2006-02-16 | Berthet Francois-Xavier J | Vaccine composition comprising transferrin binding protein and hsf from gram negative bacteria |
| DE60335477D1 (de) * | 2002-10-11 | 2011-02-03 | Novartis Vaccines & Diagnostic | Polypeptidimpstoffe zum breiten schutz gegen hypervirulente meningokokken-linien |
| CN101926988B (zh) * | 2003-01-30 | 2014-06-04 | 诺华疫苗和诊断有限公司 | 抗多种脑膜炎球菌血清组的可注射性疫苗 |
| GB0315021D0 (en) * | 2003-06-26 | 2003-07-30 | Chiron Srl | Immunogenic gonococcal compositions |
| CA2885040C (en) * | 2003-10-02 | 2018-10-30 | Novartis Vaccines And Diagnostics S.R.L. | Liquid vaccines for multiple meningococcal serogroups |
| GB0323103D0 (en) * | 2003-10-02 | 2003-11-05 | Chiron Srl | De-acetylated saccharides |
| GB0408977D0 (en) * | 2004-04-22 | 2004-05-26 | Chiron Srl | Immunising against meningococcal serogroup Y using proteins |
| GB0409748D0 (en) * | 2004-04-30 | 2004-06-09 | Chiron Srl | Lactoferrin cleavage |
| RU2450019C2 (ru) * | 2006-06-29 | 2012-05-10 | Новартис Аг | Полипептиды из neisseria meningitidis |
-
2003
- 2003-07-31 US US10/523,114 patent/US20060034854A1/en not_active Abandoned
- 2003-07-31 EP EP03784153A patent/EP1524993B1/en not_active Revoked
- 2003-07-31 ES ES03784152.5T patent/ES2537737T3/es not_active Expired - Lifetime
- 2003-07-31 PT PT101778900T patent/PT2255826E/pt unknown
- 2003-07-31 ES ES03784153T patent/ES2408251T3/es not_active Expired - Lifetime
- 2003-07-31 PL PL03375408A patent/PL375408A1/xx not_active Application Discontinuation
- 2003-07-31 KR KR1020127002482A patent/KR101239242B1/ko not_active IP Right Cessation
- 2003-07-31 JP JP2005506111A patent/JP5409986B2/ja not_active Expired - Fee Related
- 2003-07-31 DK DK03784153.3T patent/DK1524993T3/da active
- 2003-07-31 PL PL399214A patent/PL220107B1/pl unknown
- 2003-07-31 EP EP10177881A patent/EP2258384A3/en not_active Withdrawn
- 2003-07-31 DK DK10177890.0T patent/DK2255826T3/en active
- 2003-07-31 JP JP2005506112A patent/JP4740738B2/ja not_active Expired - Fee Related
- 2003-07-31 EP EP10177892A patent/EP2258387A3/en not_active Withdrawn
- 2003-07-31 CA CA002493977A patent/CA2493977A1/en not_active Abandoned
- 2003-07-31 MX MXPA05001265A patent/MXPA05001265A/es active IP Right Grant
- 2003-07-31 US US10/523,055 patent/US20060057160A1/en not_active Abandoned
- 2003-07-31 TW TW092121011A patent/TWI360424B/zh not_active IP Right Cessation
- 2003-07-31 CN CN038186489A patent/CN1674933B/zh not_active Expired - Fee Related
- 2003-07-31 EP EP10177890.0A patent/EP2255826B1/en not_active Expired - Lifetime
- 2003-07-31 EP EP10177891A patent/EP2258386A3/en not_active Withdrawn
- 2003-07-31 WO PCT/EP2003/008568 patent/WO2004014417A2/en active Application Filing
- 2003-07-31 WO PCT/EP2003/008569 patent/WO2004015099A2/en active Application Filing
- 2003-07-31 EP EP08153843A patent/EP1961427A3/en not_active Withdrawn
- 2003-07-31 SI SI200332262T patent/SI1524993T1/sl unknown
- 2003-07-31 CA CA2493124A patent/CA2493124C/en not_active Expired - Fee Related
- 2003-07-31 EP EP10177887A patent/EP2258385A3/en not_active Withdrawn
- 2003-07-31 EP EP03784152.5A patent/EP1524992B1/en not_active Expired - Lifetime
- 2003-07-31 US US10/523,044 patent/US7838014B2/en not_active Expired - Fee Related
- 2003-07-31 PL PL03375382A patent/PL375382A1/xx not_active IP Right Cessation
- 2003-07-31 PL PL375407A patent/PL216662B1/pl unknown
- 2003-07-31 PL PL399492A patent/PL399492A1/pl not_active Application Discontinuation
- 2003-07-31 SI SI200332485A patent/SI2255826T1/sl unknown
- 2003-07-31 EP EP03750408A patent/EP1524990A2/en not_active Withdrawn
- 2003-07-31 AU AU2003250204A patent/AU2003250204B8/en not_active Ceased
- 2003-07-31 EP EP12157807.4A patent/EP2481419A3/en not_active Withdrawn
- 2003-07-31 KR KR1020057001941A patent/KR101140033B1/ko not_active IP Right Cessation
- 2003-07-31 WO PCT/EP2003/008571 patent/WO2004014418A2/en active Application Filing
- 2003-07-31 MX MXPA05001349A patent/MXPA05001349A/es active IP Right Grant
- 2003-07-31 DE DE20321889U patent/DE20321889U1/de not_active Expired - Lifetime
- 2003-07-31 CA CA002493092A patent/CA2493092A1/en not_active Abandoned
- 2003-07-31 JP JP2005506113A patent/JP2006500963A/ja active Pending
- 2003-07-31 CN CNA038184540A patent/CN1671413A/zh active Pending
- 2003-07-31 US US10/523,117 patent/US20060240045A1/en not_active Abandoned
- 2003-07-31 CA CA002489030A patent/CA2489030A1/en not_active Abandoned
- 2003-07-31 MX MXPA05000842A patent/MXPA05000842A/es active IP Right Grant
- 2003-07-31 WO PCT/EP2003/008567 patent/WO2004014419A1/en active Application Filing
- 2003-07-31 KR KR1020087019883A patent/KR101139976B1/ko not_active IP Right Cessation
- 2003-07-31 AU AU2003269864A patent/AU2003269864A1/en not_active Abandoned
- 2003-07-31 MY MYPI20032904A patent/MY149591A/en unknown
- 2003-07-31 NZ NZ574530A patent/NZ574530A/en not_active IP Right Cessation
- 2003-07-31 NZ NZ587398A patent/NZ587398A/en not_active IP Right Cessation
- 2003-07-31 PT PT37841533T patent/PT1524993E/pt unknown
- 2003-07-31 KR KR1020117004904A patent/KR101237329B1/ko not_active IP Right Cessation
- 2003-07-31 AU AU2003260357A patent/AU2003260357B2/en not_active Ceased
- 2003-07-31 AU AU2003253375A patent/AU2003253375B2/en not_active Ceased
- 2003-07-31 ES ES10177890.0T patent/ES2575014T3/es not_active Expired - Lifetime
- 2003-07-31 HU HUE10177890A patent/HUE029200T2/en unknown
- 2003-07-31 JP JP2005506114A patent/JP2006506467A/ja active Pending
- 2003-07-31 NZ NZ537904A patent/NZ537904A/en not_active IP Right Cessation
- 2003-07-31 NZ NZ537181A patent/NZ537181A/en not_active IP Right Cessation
- 2003-07-31 KR KR1020057001933A patent/KR20050039839A/ko not_active Application Discontinuation
- 2003-07-31 DE DE20321890U patent/DE20321890U1/de not_active Expired - Lifetime
- 2003-07-31 EP EP03784151A patent/EP1524991A1/en not_active Withdrawn
- 2003-07-31 KR KR1020057001924A patent/KR20050028051A/ko not_active Application Discontinuation
-
2004
- 2004-12-09 IL IL16566004A patent/IL165660A0/xx unknown
- 2004-12-13 IS IS7593A patent/IS7593A/is unknown
- 2004-12-16 IS IS7601A patent/IS7601A/is unknown
-
2005
- 2005-01-03 NO NO20050010A patent/NO20050010L/no not_active Application Discontinuation
- 2005-01-03 NO NO20050008A patent/NO20050008L/no not_active Application Discontinuation
- 2005-01-20 IL IL166433A patent/IL166433A/en active IP Right Grant
- 2005-01-20 IS IS7658A patent/IS7658A/is unknown
- 2005-01-25 NO NO20050421A patent/NO20050421L/no not_active Application Discontinuation
- 2005-01-28 CO CO05007268A patent/CO5680456A2/es not_active Application Discontinuation
- 2005-01-28 CO CO05007265A patent/CO5680454A2/es active IP Right Grant
- 2005-01-28 CO CO05007266A patent/CO5680455A2/es not_active Application Discontinuation
- 2005-10-13 HK HK05109073.6A patent/HK1077014A1/xx not_active IP Right Cessation
-
2008
- 2008-06-04 AU AU2008202479A patent/AU2008202479C1/en not_active Ceased
- 2008-12-12 AU AU2008255270A patent/AU2008255270A1/en not_active Abandoned
-
2010
- 2010-10-21 US US12/909,621 patent/US8221770B2/en not_active Expired - Fee Related
- 2010-10-22 JP JP2010237357A patent/JP5414651B2/ja not_active Expired - Fee Related
-
2011
- 2011-03-04 JP JP2011047056A patent/JP2011142916A/ja active Pending
- 2011-05-31 IL IL213264A patent/IL213264A0/en unknown
- 2011-09-29 US US13/248,693 patent/US20120027800A1/en not_active Abandoned
- 2011-11-17 US US13/299,007 patent/US20120064119A1/en not_active Abandoned
- 2011-11-17 US US13/299,041 patent/US20120064120A1/en not_active Abandoned
-
2012
- 2012-02-10 JP JP2012026928A patent/JP5789203B2/ja not_active Expired - Fee Related
-
2013
- 2013-05-31 CY CY20131100435T patent/CY1114243T1/el unknown
- 2013-07-31 LU LU92262C patent/LU92262I2/fr unknown
- 2013-09-05 CY CY2013036C patent/CY2013036I2/el unknown
-
2015
- 2015-09-03 US US14/844,989 patent/US20160082097A1/en not_active Abandoned
-
2016
- 2016-06-15 CY CY20161100532T patent/CY1117643T1/el unknown
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2011142916A (ja) * | 2002-08-02 | 2011-07-28 | Glaxosmithkline Biologicals Sa | ワクチン |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4740738B2 (ja) | ワクチン | |
| JP5275983B2 (ja) | ワクチン | |
| CN1688333A (zh) | 疫苗 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20060726 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20090825 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20091119 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20091127 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20100225 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20110118 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20110304 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20110426 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20110502 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20140513 Year of fee payment: 3 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| LAPS | Cancellation because of no payment of annual fees |