WO2010061890A1 - L-アミノ酸の製造法 - Google Patents
L-アミノ酸の製造法 Download PDFInfo
- Publication number
- WO2010061890A1 WO2010061890A1 PCT/JP2009/069956 JP2009069956W WO2010061890A1 WO 2010061890 A1 WO2010061890 A1 WO 2010061890A1 JP 2009069956 W JP2009069956 W JP 2009069956W WO 2010061890 A1 WO2010061890 A1 WO 2010061890A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- amino acid
- gene
- seq
- protein
- medium
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/08—Lysine; Diaminopimelic acid; Threonine; Valine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/06—Alanine; Leucine; Isoleucine; Serine; Homoserine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/12—Methionine; Cysteine; Cystine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/20—Aspartic acid; Asparagine
Landscapes
- Organic Chemistry (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Life Sciences & Earth Sciences (AREA)
- Wood Science & Technology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Microbiology (AREA)
- General Chemical & Material Sciences (AREA)
- Biotechnology (AREA)
- Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
グルタミン酸デカルボキシラーゼ活性が増強するように改変された株において、グルタミン酸/GABAアンチポーター活性を増強することによって、L-リジン、L-スレオニン及びL-トリプトファンの生産性が向上することが知られているが(特許文献6)、gltP遺伝子またはgltS遺伝子を増幅した微生物を用いたL-アミノ酸の生産の報告はない。
(1)L-アミノ酸生産能を有する腸内細菌科に属する細菌を培地で培養して、L-アミノ酸を該培地中に生成蓄積させ、該培地より前記L-アミノ酸を採取する、L-アミノ酸の製造法において、前記細菌は、gltP遺伝子及び/又はgltS遺伝子の発現が増大するように改変され、前記L-アミノ酸が、L-リジン、L-スレオニン、L-アスパラギン、L-アスパラギン酸、L-メチオニン、L-アラニン、L-イソロイシン、及びL-ホモセリンからなる群より選択されることを特徴とする方法。
(2)前記gltP遺伝子が、下記(A)又は(B)に示すタンパク質をコードする、前記方法:
(A)配列番号12に記載のアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質、
(B)配列番号12に記載のアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。
(3)前記gltP遺伝子が、下記(A1)又は(B1)に記載のタンパク質をコードする、前記方法:
(A1)配列番号2に示すアミノ酸配列を有するタンパク質、
(B1)配列番号2に示すアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。
(4)前記gltP遺伝子が、下記(a)又は(b)に記載のDNAである、前記方法:
(a)配列番号1に示す塩基配列を有するDNA、
(b)配列番号1に示す塩基配列の相補配列又は同塩基配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質をコードするDNA。
(5)前記gltS遺伝子が、下記(C)又は(D)に示すタンパク質をコードする、前記方法:
(C)配列番号13に記載のアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質、
(D)配列番号13に記載のアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。
(6)前記gltS遺伝子が、下記(C1)又は(D1)に記載のタンパク質をコードする、前記方法:
(C1)配列番号4に示すアミノ酸配列を有するタンパク質、
(D1)配列番号4に示すアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。
(7)前記gltS遺伝子が、下記(c)又は(d)に記載のDNAである、前記方法:
(c)配列番号3に示す塩基配列を有するDNA、
(d)配列番号3に示す塩基配列の相補配列又は同塩基配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質をコードするDNA。
(8)前記遺伝子の発現が、該遺伝子のコピー数を高めること、又は該遺伝子の発現調節配列を改変されたことによって増大された、前記方法。
(9)前記L-アミノ酸がL-リジンであり、前記細菌はybjE遺伝子の発現が増大した、前記方法。
(10)前記ybjE遺伝子が、下記(E)又は(F)に記載のタンパク質をコードする、前記方法:
(E)配列番号6に示すアミノ酸配列又は配列番号6のアミノ酸番号17~315のアミノ酸配列を有するタンパク質、
(F)配列番号6に示すアミノ酸配列又は配列番号6のアミノ酸番号17~315のアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-リジン排出活性を有するタンパク質。
(11)前記ybjE遺伝子が、下記(e)又は(f)に記載のDNAである、前記方法:
(e)配列番号5に示す塩基配列又は配列番号5の塩基番号49~948の塩基配列を有するDNA、
(f)配列番号5に示す塩基配列もしくは配列番号5の塩基番号49~948の塩基配列の相補配列、又はこれらの塩基配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、L-リジン排出活性を有するタンパク質をコードするDNA。
(12)前記L-アミノ酸がL-リジンであり、その生産を行う培養中の培地のpHが6.0~9.0、培養終了時の培地のpHが7.2~9.0となるように制御し、培地中の重炭酸イオン及び/又は炭酸イオンが少なくとも20mM以上存在する培養期があるようにし、前記重炭酸イオン及び/又は炭酸イオンを塩基性アミノ酸のカウンタイオンとすることを特徴とする方法。
(13)前記細菌がエシェリヒア属細菌である前記方法。
<1>本発明の細菌
本発明の細菌は、L-アミノ酸生産能を有し、かつgltP遺伝子及び/又はgltS遺伝子の発現が増大するように改変された腸内細菌科に属する細菌である。前記L-アミノ酸は、L-リジン、L-スレオニン、L-アスパラギン酸、L-アスパラギン、L-メチオニン、L-アラニン、L-イソロイシン及びL-ホモセリンからなる群より選択される。これらのL-アミノ酸を微生物を用いて発酵生産させると、L-グルタミン酸が副生することが多い。L-グルタミン酸の副生に関しては、gltP遺伝子及び/又はgltS遺伝子の発現が増大するように改変することにより、非改変株より減少していればよいが、40%以上、好ましくは50%以上、好ましくは60%以上減少していることが望ましい。
また、「遺伝子の発現の増大」とは、遺伝子の転写及び/又は翻訳の量が増大することをいう。
以下に、細菌にL-リジン、L-スレオニン、L-アスパラギン酸、L-アスパラギン、L-メチオニン、L-イソロイシン、L-アラニン、及びL-ホモセリンから選ばれるL-アミノ酸の生産能を付与する方法及び本発明で使用することのできる前記L-アミノ酸の生産能が付与された細菌を例示する。ただし、前記L-アミノ酸の生産能を有する限り、これらに制限されない。
以下、L-リジン生産菌及びその構築方法を例として示す。
例えば、L-リジン生産能を有する株としては、L-リジンアナログ耐性株又は代謝制御変異株が挙げられる。L-リジンアナログの例としては、オキサリジン、リジンヒドロキサメート、S-(2-アミノエチル)-L-システイン(AEC)、γ-メチルリジン、α-クロロカプロラクタムなどが挙げられるが、これらに限定されない。これらのリジンアナログに対して耐性を有する変異株は、腸内細菌科に属する細菌を通常の人工変異処理に付すことによって得ることができる。L-リジン生産菌として具体的には、エシェリヒア・コリAJ11442株(FERM BP-1543、NRRL B-12185;特開昭56-18596号公報及び米国特許第4346170号明細書参照)、エシェリヒア・コリ VL611株(特開2000-189180号公報)等が挙げられる。また、エシェリヒア・コリのL-リジン生産菌として、WC196株(国際公開第96/17930号パンフレット参照)を用いることも出来る。
本発明に用いられるL-スレオニン生産菌として好ましいものとしては、L-スレオニン生合成系酵素を強化した腸内細菌科に属する細菌が挙げられる。L-スレオニン生合成系酵素をコードする遺伝子としては、アスパルトキナーゼIII遺伝子(lysC)、アスパラギン酸セミアルデヒドデヒドロゲナーゼ遺伝子(asd)、thrオペロンにコードされるアスパルトキナーゼI遺伝子(thrA)、ホモセリンキナーゼ遺伝子(thrB)、スレオニンシンターゼ遺伝子(thrC)が挙げられる。これらの遺伝子は2種類以上導入してもよい。L-スレオニン生合成系遺伝子は、スレオニン分解が抑制された腸内細菌科に属する細菌に導入してもよい。スレオニン分解が抑制されたエシェリヒア属細菌としては、例えば、スレオニンデヒドロゲナーゼ活性が欠損したTDH6株(特開2001-346578号)等が挙げられる。
エシェリヒア属に属するL-スレオニン生産菌として、NZ10(thrB)株が挙げられる。この株は、公知のC600株(thrB, leuB(Appleyard R.K., Genetics, 39, 440-452 (1954))から誘導されたLeu+復帰変異株である。アスパルトキナーゼ-ホモセリンデヒドロゲナーゼIをコードするthrA遺伝子で形質転換されたNZ10形質転換株も、好適に使用できる。
細菌のrhtB遺伝子のコピー数を増加させるとL-ホモセリンに耐性になり、L-ホモセリン、L-スレオニン、L-アラニン、L-バリン、及びL-イソロイシンの生産性が向上する(EP994190A2)。また、細菌のrthC遺伝子のコピー数を増加させるとL-ホモセリン及びL-スレオニンに耐性になり、L-ホモセリン、L-スレオニン、及びL-ロイシンの生産性が向上する(EP1013765A1)。
さらに、エシェリヒア・コリ44株(ロシアン・ナショナル・コレクション・オブ・インダストリアル・マイクロオーガニズムに登録番号VKPM B-2175として寄託されている)も、使用することができる。
エシェリヒア属に属するL-メチオニン生産菌としては、エシェリヒア・コリAJ11539 (NRRL B-12399)、AJ11540 (NRRL B-12400)、AJ11541 (NRRL B-12401)、 AJ11542 (NRRL B-12402)(GB2075055)、218 (VKPM B-8125)(EP1239041)等の菌株が挙げられる。
エシェリヒア属に属するL-アスパラギン酸生産菌としては、フマル酸からL-アスパラギン酸を生成するアスパルターゼ活性が増強されたエシェリヒア・コリ菌株(特公昭38-6588号)が挙げられる。
L-アラニンは、アスパラギン酸のβ-脱炭酸により生産される。したがって、エシェリヒア属に属するL-アラニン生産菌として、アスパラギン酸β-デカルボキシラーゼが増強されたエシェリヒア・コリ菌株(特開昭2-242690号)が挙げられる。
本発明のL-イソロイシン生産菌を誘導するための親株の例としては、6-ジメチルアミノプリンに耐性を有する変異株(特開平5-304969号)、チアイソロイシン、イソロイシンヒドロキサメートなどのイソロイシンアナログに耐性を有する変異株、さらにDL-エチオニン及び/又はアルギニンヒドロキサメートに耐性を有する変異株(特開平5-130882号).が挙げられるが、これらに限定されない。さらに、スレオニンデアミナーゼ、アセトヒドロキシ酸シンターゼなどのL-イソロイシン生合成に関与するタンパク質をコードする遺伝子で形質転換された組換え株もまた親株として使用できる(特開平2-458号, FR 0356739, 及び米国特許第5,998,178号)。
L-アスパラギンは、アスパラギン酸へアミノ基を付与することにより生産される(Boehlein, S. K., Richards, N. G. J., & Schuster, S. M. (1994a) J. Biol. Chem. 269, 7450-7457.)。したがって、エシェリヒア属に属するL-アスパラギン生産菌として、L-アスパラギン酸生産菌のアスパラギンシンテターゼが増強されたエシェリヒア・コリ菌株が挙げられる。
尚、本明細書において、「相同性」(homology)」は、「同一性」(identity)を指すことがある。
また、それぞれgltP及び/又はgltSが導入される宿主で使用しやすいコドンに置換したものでもよい。同様にgltP及び/又はgltSにコードされるタンパク質は、L-グルタミン酸トランスポーター活性を有する限り、N末端側、C末端側が延長したものあるいは削られているものでもよい。例えば延長・削除する長さは、アミノ酸残基で50以下、好ましくは20以下、より好ましくは10以下、特に好ましくは5以下である。より具体的には、配列番号2のアミノ酸配列のN末端側より50アミノ酸から5アミノ酸、C末端側より50アミノ酸から5アミノ酸延長・削除したものでもよい。
本発明のL-アミノ酸の製造法は、本発明の細菌を培地で培養して、L-リジン、L-スレオニン、L-アスパラギン、L-アスパラギン酸、L-メチオニン、L-アラニン、L-イソロイシン及びL-ホモセリンから選ばれるL-アミノ酸を該培地中に生成蓄積させ、該培地又は菌体より該L-アミノ酸を回収することを特徴とする。

エシェリヒア・コリK-12株のゲノムの全塩基配列(GenBank Accession No. U00096)は既に明らかにされている(Science, 277, 1453-1474 (1997))。目的遺伝子の増幅を行うためにプラスミドベクターpMW118(ニッポンジーン社製)を用いた。本プラスミドは、任意の遺伝子をクローニングするためのマルチクローニングサイトを有しており、これらのサイトを利用して遺伝子をクローニングし、遺伝子の増幅が可能なプラスミドである。
L-リジン生産菌の親株としてとして、国際公開第2006/078039号パンフレット記載のエシェリヒア・コリWC196ΔcadAΔldcCを使用した。この菌株は、エシェリヒア・コリWC196のリジンデカルボキシラーゼ遺伝子cadA及びldcCを、Red-driven integration法(Datsenko, K. A, and Wanner, B. L. Proc. Natl. Acad. Sci. U S A. 97:6640-6645 (2000))とλファージ由来の切り出しシステム(J. Bacteriol. 184. 5200-5203 (2002))を組みあわせた方法により、破壊した株である。本菌株は、平成20年10月7日に、独立行政法人 産業技術総合研究所 特許生物寄託センター(〒305-8566 日本国茨城県つくば市東1丁目1番地1 中央第6)に、受託番号FERM BP-11027として寄託されている。
<3-1>WC196LCY株へのリジン生産用プラスミド導入
WC196LCY株をdapA、dapB及びlysC遺伝子を搭載したリジン生産用プラスミドpCABD2(EP0733710パンフレット)で常法に従い形質転換し、WC196LCY/pCABD2株を得た。pCABD2は、L-リジンによるフィードバック阻害が解除された変異を有するエシェリヒア・コリ由来のジヒドロジピコリン酸合成酵素(DDPS)をコードするDNA と、L-リジンによるフィードバック阻害が解除された変異を有するエシェリヒア・コリ由来のアスパルトキナーゼIIIをコードするDNAと、エシェリヒア・コリ由来のジヒドロジピコリン酸レダクターゼをコードするDNAと、及びブレビバクテリウム・ラクトファーメンタム由来ジアミノピメリン酸デヒドロゲナーゼをコードするDNAを有している
上記のグリセロールストックを融解し、各500μLを、25 mg/Lのストレプトマイシンと100 mg/Lのアンピシリンを含むLプレートに均一に塗布し、37℃にて24時間培養した。得られたプレートのおよそ1/8量の菌体を、500 mL容坂口フラスコの、25 mg/Lのストレプトマイシンと100 mg/Lのアンピシリンを含む以下に記載の発酵培地の20 mLに接種し、往復振とう培養装置で37℃において22時間培養した。培養後、培地中に蓄積したL-リジン及びL-グルタミン酸の量をバイオテックアナライザーAS210(サクラ精機)により測定した。培養に用いた培地組成を以下に示す。
グルコース 40 g/L
(NH4)2SO4 24 g/L
KH2PO4 1.0 g/L
MgSO4・7H2O 1.0 g/L
FeSO4・7H2O 0.01 g/L
MnSO4・7H2O 0.08 g/L
Yeast Extract 2.0 g/L
L-isoleucine 0.1 g/L
NaCl 1.0 g/L
CaCO3(日本薬局方) 50 g/L
KOHでpH7.0に調整し、115℃で10分オートクレーブを行なった。但し、グルコースとMgSO4・7H2Oは混合し、他の成分とは別に殺菌した。CaCO3は乾熱滅菌後に添加した。
培養22時間目のL-リジン収率、及びL-グルタミン酸濃度を、対照であるWC196LCY/pCABD2/pMW118の値を100とした相対値にて、表3に示す。

L-スレオニン生産菌として、エシェリヒア・コリVKPM B-3996株(米国特許第5,175,107号明細書参照)を用いた。
グルコース 100g/L
酵母エキス 1.8g/L
KH2PO4 1.0g/L
NaCl 0.6g/L
MgSO4・7H2O 0.36g/L
FeSO4・7H2O 18mg/L
MnSO4・4H2O 18mg/L
ストレプトマイシン硫酸塩 20mg/L
アンピシリン 100mg/L
グルコースとMgSO4・7H2Oは混合し、他の成分とは別に殺菌した。

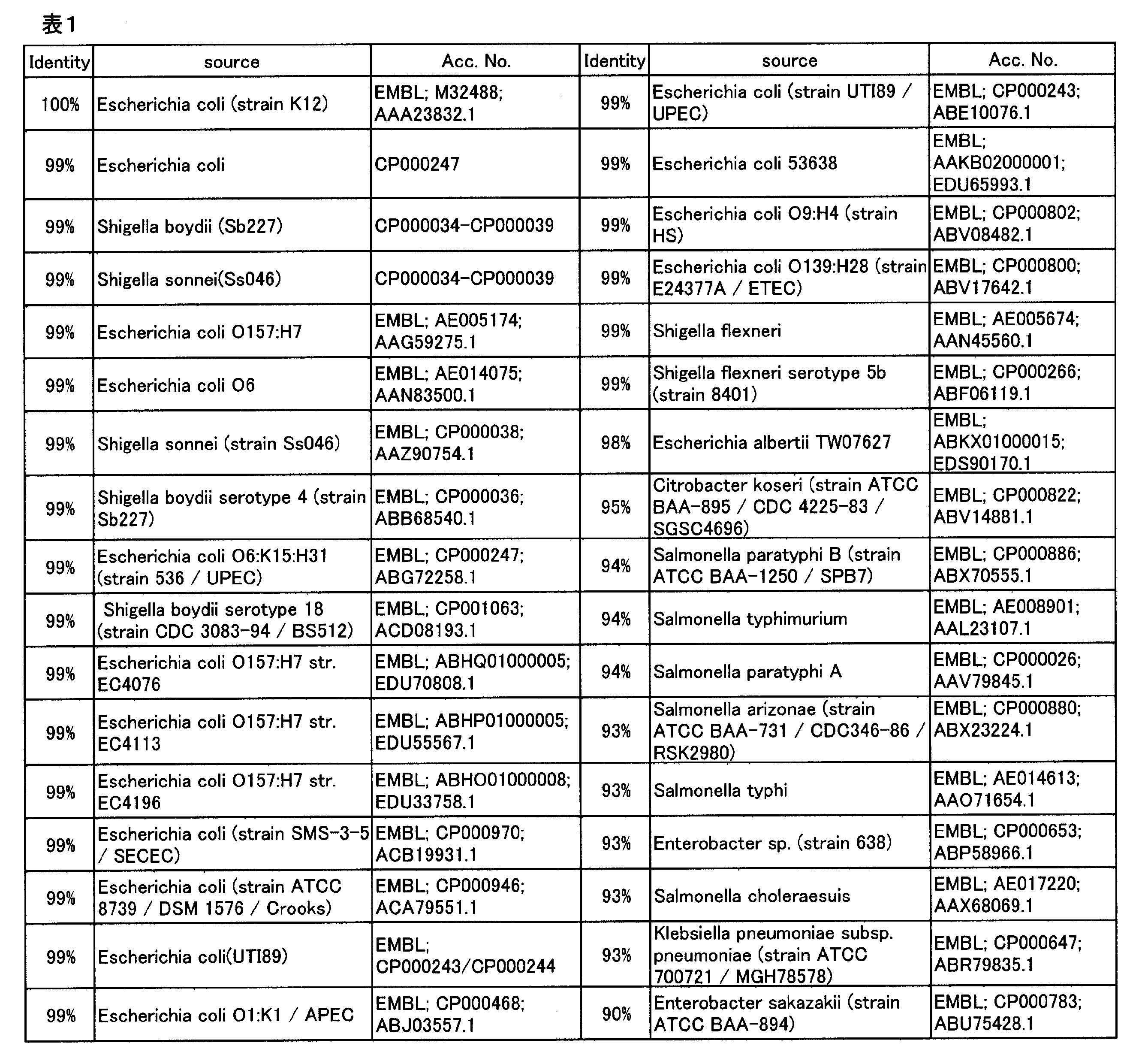
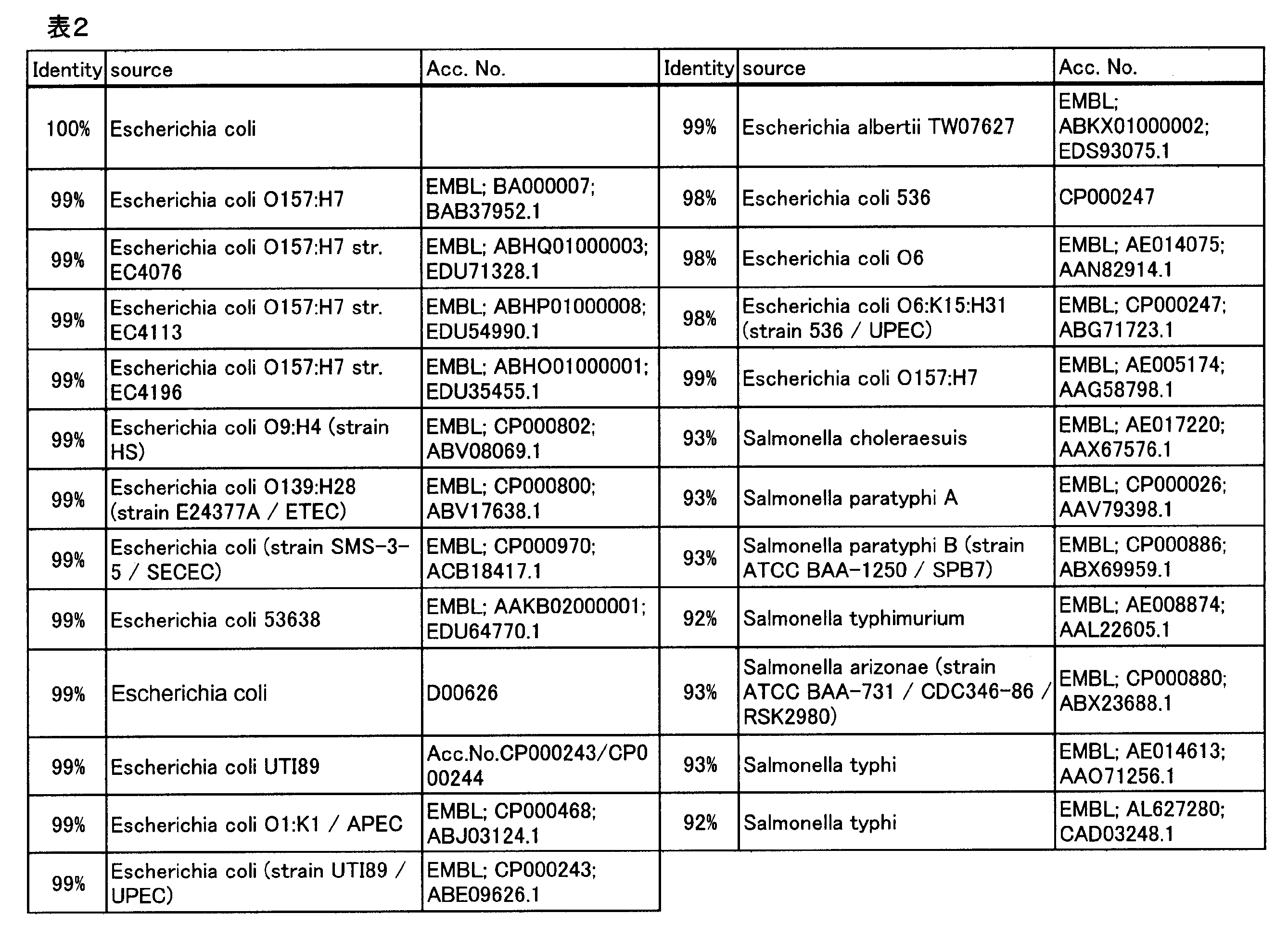
L-グルタミン酸トランスポーター活性を有するタンパク質を増幅したL-アミノ酸の製造法の先行知見として、L-グルタミン酸/GABAアンチポーター活性を有するタンパク質をコードするgadCと、グルタミン酸デカルボキシラーゼ活性を有するタンパク質をコードするgadBを同時に増幅することにより、L-リジン、L-スレオニン、及びL-トリプトファンの生産性が向上する事が知られている(WO2008/044453)。そこでgltPまたはgltS増幅によるグルタミン酸副生量の低減効果を、gadC増幅と比較した。

配列番号1:gltP遺伝子配列
配列番号2:GltPアミノ酸配列
配列番号3:gltS遺伝子配列
配列番号4:GltSアミノ酸配列
配列番号5:ybjE遺伝子配列
配列番号6:YbjEアミノ酸配列
配列番号7:ybjE上流置換配列
配列番号8:gltP増幅用プライマー(5’側)
配列番号9:gltP増幅用プライマー(3’側)
配列番号10:gltS増幅用プライマー(5’側)
配列番号11:gltS増幅用プライマー(3’側)
配列番号12:gltP保存配列
配列番号13:gltS保存配列
配列番号14:gadC増幅用プライマー(5’側)
配列番号15:gadC増幅用プライマー(3’側)
Claims (13)
- L-アミノ酸生産能を有する腸内細菌科に属する細菌を培地で培養して、L-アミノ酸を該培地中に生成蓄積させ、該培地より前記L-アミノ酸を採取する、L-アミノ酸の製造法において、
前記細菌は、gltP遺伝子及び/又はgltS遺伝子の発現が増大するように改変され、前記L-アミノ酸が、L-リジン、L-スレオニン、L-アスパラギン、L-アスパラギン酸、L-メチオニン、L-アラニン、L-イソロイシン、及びL-ホモセリンからなる群より選択されることを特徴とする方法。 - 前記gltP遺伝子が、下記(A)又は(B)に示すタンパク質をコードする、請求項1に記載の方法:
(A)配列番号12に記載のアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質、
(B)配列番号12に記載のアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。 - 前記gltP遺伝子が、下記(A1)又は(B1)に記載のタンパク質をコードする、請求項2に記載の方法:
(A1)配列番号2に示すアミノ酸配列を有するタンパク質、
(B1)配列番号2に示すアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。 - 前記gltP遺伝子が、下記(a)又は(b)に記載のDNAである、請求項1~3のいずれか一項に記載の方法:
(a)配列番号1に示す塩基配列を有するDNA、
(b)配列番号1に示す塩基配列の相補配列又は同塩基配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質をコードするDNA。 - 前記gltS遺伝子が、下記(C)又は(D)に示すタンパク質をコードする、請求項1~4のいずれか一項に記載の方法:
(C)配列番号13に記載のアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質、
(D)配列番号13に記載のアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。 - 前記gltS遺伝子が、下記(C1)又は(D1)に記載のタンパク質をコードする、請求項5に記載の方法:
(C1)配列番号4に示すアミノ酸配列を有するタンパク質、
(D1)配列番号4に示すアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質。 - 前記gltS遺伝子が、下記(c)又は(d)に記載のDNAである、請求項1~6のいずれか一項に記載の方法:
(c)配列番号3に示す塩基配列を有するDNA、
(d)配列番号3に示す塩基配列の相補配列又は同塩基配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、L-グルタミン酸トランスポーター活性を有するタンパク質をコードするDNA。 - 前記遺伝子の発現が、該遺伝子のコピー数を高めること、又は該遺伝子の発現調節配列を改変されたことによって増大された、請求項1~7のいずれか一項に記載の方法。
- 前記L-アミノ酸がL-リジンであり、前記細菌はybjE遺伝子の発現が増大した、請求項1~8のいずれか一項に記載の方法。
- 前記ybjE遺伝子が、下記(E)又は(F)に記載のタンパク質をコードする、請求項9に記載の方法:
(E)配列番号6に示すアミノ酸配列又は配列番号6のアミノ酸番号17~315のアミノ酸配列を有するタンパク質、
(F)配列番号6に示すアミノ酸配列又は配列番号6のアミノ酸番号17~315のアミノ酸配列において、1若しくは数個のアミノ酸残基の置換、欠失、挿入、付加、又は逆位を含むアミノ酸配列を有し、かつ、L-リジン排出活性を有するタンパク質。 - 前記ybjE遺伝子が、下記(e)又は(f)に記載のDNAである、請求項9又は10に記載の方法:
(e)配列番号5に示す塩基配列又は配列番号5の塩基番号49~948の塩基配列を有するDNA、
(f)配列番号5に示す塩基配列もしくは配列番号5の塩基番号49~948の塩基配列の相補配列、又はこれらの塩基配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、L-リジン排出活性を有するタンパク質をコードするDNA。 - 前記L-アミノ酸がL-リジンであり、その生産を行う培養中の培地のpHが6.0~9.0、培養終了時の培地のpHが7.2~9.0となるように制御し、培地中の重炭酸イオン及び/又は炭酸イオンが少なくとも20mM以上存在する培養期があるようにし、前記重炭酸イオン及び/又は炭酸イオンを塩基性アミノ酸のカウンタイオンとすることを特徴とする、請求項1~11のいずれか一項に記載の方法。
- 前記細菌がエシェリヒア属細菌である請求項1~12のいずれか一項に記載の方法。
Priority Applications (8)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP09829135.4A EP2360263B1 (en) | 2008-11-27 | 2009-11-26 | PROCESS FOR PRODUCING AMINO ACIDS USING A BACTERIUM HAVING INCREASED EXPRESSION OF THE GENES gltP AND/OR gltS |
| BRPI0921655-3A BRPI0921655B1 (pt) | 2008-11-27 | 2009-11-26 | Método para produzir um l-aminoácido |
| RU2011126198/10A RU2518677C2 (ru) | 2008-11-27 | 2009-11-26 | Способ получения l-аминокислоты |
| CA2744852A CA2744852C (en) | 2008-11-27 | 2009-11-26 | Method for producing l-amino acid using enterobacteriaceae comprising increased gltp and glts gene expression |
| CN200980147429.0A CN102227504B (zh) | 2008-11-27 | 2009-11-26 | 产生l-氨基酸的方法 |
| PL09829135T PL2360263T3 (pl) | 2008-11-27 | 2009-11-26 | SPOSÓB WYTWARZANIA AMINOKWASÓW Z UŻYCIEM BAKTERII O ZWIĘKSZONEJ EKSPRESJI GENÓW gltP I/LUB gltS |
| ES09829135T ES2739909T3 (es) | 2008-11-27 | 2009-11-26 | Procedimiento para la producción de aminoácidos usando una bacteria que tiene expresión aumentada de los genes gltP y/o gltS |
| US13/114,251 US8673597B2 (en) | 2008-11-27 | 2011-05-24 | Method for producing L-amino acid |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008-302521 | 2008-11-27 | ||
| JP2008302521A JP2012029565A (ja) | 2008-11-27 | 2008-11-27 | L−アミノ酸の製造法 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| US13/114,251 Continuation US8673597B2 (en) | 2008-11-27 | 2011-05-24 | Method for producing L-amino acid |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2010061890A1 true WO2010061890A1 (ja) | 2010-06-03 |
Family
ID=42225761
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2009/069956 WO2010061890A1 (ja) | 2008-11-27 | 2009-11-26 | L-アミノ酸の製造法 |
Country Status (10)
| Country | Link |
|---|---|
| US (1) | US8673597B2 (ja) |
| EP (1) | EP2360263B1 (ja) |
| JP (1) | JP2012029565A (ja) |
| CN (1) | CN102227504B (ja) |
| BR (1) | BRPI0921655B1 (ja) |
| CA (1) | CA2744852C (ja) |
| ES (1) | ES2739909T3 (ja) |
| PL (1) | PL2360263T3 (ja) |
| RU (1) | RU2518677C2 (ja) |
| WO (1) | WO2010061890A1 (ja) |
Cited By (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN102329765A (zh) * | 2010-08-31 | 2012-01-25 | 安徽华恒生物工程有限公司 | 一种高产l-丙氨酸的xz-a26菌株及构建方法与应用 |
| JP2013106588A (ja) * | 2011-11-24 | 2013-06-06 | Kyowa Hakko Bio Co Ltd | L−アスパラギンの製造法 |
| WO2015060314A1 (ja) | 2013-10-21 | 2015-04-30 | 味の素株式会社 | L-アミノ酸の製造法 |
| JP6020443B2 (ja) * | 2011-04-01 | 2016-11-02 | 味の素株式会社 | L−システインの製造法 |
| EP3385389A1 (en) | 2017-04-03 | 2018-10-10 | Ajinomoto Co., Inc. | Method for producing l-amino acid from fructose |
| WO2020071538A1 (en) | 2018-10-05 | 2020-04-09 | Ajinomoto Co., Inc. | Method for producing target substance by bacterial fermentation |
| EP3716274A1 (en) | 2019-03-29 | 2020-09-30 | Ajinomoto Co., Inc. | Control device, control method, computer program, and method for producing organic compound |
Families Citing this family (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP2818554A3 (en) | 2004-10-07 | 2015-02-11 | Ajinomoto Co., Inc. | Method for producing a basic substance |
| WO2013134625A1 (en) * | 2012-03-08 | 2013-09-12 | Novus International Inc. | Recombinant bacterium for l-homoserine production |
| RU2015114955A (ru) | 2015-04-22 | 2016-11-10 | Аджиномото Ко., Инк. | Способ получения L-изолейцина с использованием бактерии семейства Enterobacteriaceae, в которой сверхэкспрессирован ген cycA |
| JP2019062742A (ja) * | 2016-02-24 | 2019-04-25 | 味の素株式会社 | L−アミノ酸の製造法 |
| CN109666617B (zh) * | 2017-10-13 | 2021-02-02 | 四川利尔生物科技有限公司 | 一种l-高丝氨酸的生产菌株及其构建方法和应用 |
| CN111621455B (zh) * | 2020-04-28 | 2023-01-17 | 江南大学 | 一种表达天冬酰胺合成酶的重组枯草芽孢杆菌 |
| RU2748676C1 (ru) * | 2020-10-05 | 2021-05-28 | Федеральное государственное бюджетное учреждение "Государственный научно-исследовательский институт генетики и селекции промышленных микроорганизмов Национального исследовательского центра "Курчатовский институт" (НИЦ "Курчатовский институт" - ГосНИИгенетика) | Штамм Escherichia coli с инактивированным геном ydgI - продуцент L-треонина |
| CN113046398A (zh) * | 2021-05-18 | 2021-06-29 | 通辽梅花生物科技有限公司 | 一种稳定高效生产l-异亮氨酸的发酵方法及发酵稳定剂 |
| CN115490761B (zh) * | 2021-11-01 | 2023-06-09 | 中国科学院天津工业生物技术研究所 | 基于赖氨酸外排蛋白构建的重组微生物及生产赖氨酸的方法 |
Citations (50)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| FR356739A (fr) | 1904-09-20 | 1905-12-07 | Charles Glauser Perrin | Mécanisme de remontoir et de mise à l'heure |
| JPS5618596A (en) | 1979-07-23 | 1981-02-21 | Ajinomoto Co Inc | Production of l-lysine through fermentation process |
| GB2075055A (en) | 1980-04-14 | 1981-11-11 | Ajinomoto Kk | L-methionone-producing Microorganisms |
| JPS6087788A (ja) | 1983-08-29 | 1985-05-17 | Ajinomoto Co Inc | 発酵法によるl―アミノ酸の製造法 |
| JPH02458A (ja) | 1987-10-12 | 1990-01-05 | Ajinomoto Co Inc | 発酵法によるl―イソロイシンの製造法 |
| JPH02109985A (ja) | 1988-02-22 | 1990-04-23 | Eurolysine | 細菌染色体上ヘの目的遺伝子の組み込み方法及び該方法によって得られた細菌 |
| WO1990004636A1 (en) | 1988-10-25 | 1990-05-03 | Vsesojuzny Nauchno-Issledovatelsky Institut Genetiki I Selektsii Promyshlennykh Mikroorganizmov (Vniigenetika) | Strain of bacteria escherichia coli, producer of l-threonine |
| JPH02242690A (ja) | 1989-03-16 | 1990-09-27 | Mitsubishi Petrochem Co Ltd | L―アラニンの製造法 |
| JPH0386588A (ja) | 1989-08-30 | 1991-04-11 | Ricoh Co Ltd | 昇華型熱転写体 |
| FR2669935A1 (fr) | 1990-11-30 | 1992-06-05 | Ajinomoto Kk | Procede et appareil pour la regulation de la concentration de source de carbone dans la culture aerobie d'un micro-organisme. |
| US5168056A (en) | 1991-02-08 | 1992-12-01 | Purdue Research Foundation | Enhanced production of common aromatic pathway compounds |
| JPH057491A (ja) | 1990-10-15 | 1993-01-19 | Ajinomoto Co Inc | 温度感受性プラスミド |
| JPH05130882A (ja) | 1991-11-11 | 1993-05-28 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−イソロイシンの製造法 |
| JPH05276935A (ja) * | 1991-10-18 | 1993-10-26 | Degussa Ag | コリネ型細菌のアミノ酸排出性菌株の効率を増大させる方法及びアミノ酸の発酵的製造方法 |
| JPH05304969A (ja) | 1992-02-25 | 1993-11-19 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるアミノ酸の製造法 |
| EP0593792A1 (en) | 1992-10-14 | 1994-04-27 | Ajinomoto Co., Inc. | Novel L-threonine-producing microbacteria and a method for the production of L-threonine |
| WO1995006114A1 (fr) | 1993-08-24 | 1995-03-02 | Ajinomoto Co., Inc. | Allele de phosphenolpyruvate carboxylase, gene de cet allele et procede de production de l'acide amine |
| WO1995023864A1 (fr) | 1994-03-04 | 1995-09-08 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| WO1996017930A1 (fr) | 1994-12-09 | 1996-06-13 | Ajinomoto Co., Inc. | Nouveau gene de decarboxylase de lysine et procede de production de lysine l |
| EP0733712A1 (en) | 1993-10-28 | 1996-09-25 | Ajinomoto Co., Inc. | Process for producing substance |
| EP0733710A1 (en) | 1993-12-08 | 1996-09-25 | Ajinomoto Co., Inc. | Process for producing l-lysine by fermentation |
| WO1996040934A1 (fr) | 1995-06-07 | 1996-12-19 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| US5661012A (en) | 1992-11-10 | 1997-08-26 | Ajinomoto Co., Inc. | Method for the production of L-threonine by fermentation, using mutated DNA encoding aspartokinase III |
| WO1998004715A1 (en) | 1996-07-30 | 1998-02-05 | Archer-Daniels-Midland Company | Novel strains of escherichia coli, methods of preparing the same and use thereof in fermentation processes for l-threonine production |
| US5763230A (en) | 1996-03-22 | 1998-06-09 | Triple-A B.V. P/A Produkschap Voor Veevoedor | Amino acid fermentation processes |
| US5776736A (en) | 1992-12-21 | 1998-07-07 | Purdue Research Foundation | Deblocking the common pathway of aromatic amino acid synthesis |
| EP0877090A1 (en) | 1995-08-30 | 1998-11-11 | Ajinomoto Co., Inc. | Process for producing l-amino acids |
| WO1999018228A2 (de) | 1997-10-04 | 1999-04-15 | Forschungszentrum Jülich GmbH | Verfahren zur mikrobiellen herstellung von aminosäuren der aspartat- und/oder glutamatfamilie und im verfahren einsetzbare mittel |
| US5906925A (en) | 1994-09-16 | 1999-05-25 | Liao; James C. | Microorganisms and methods for overproduction of DAHP by cloned pps gene |
| US5932453A (en) | 1996-10-15 | 1999-08-03 | Ajinomoto Co., Ltd. | Process for producing L-amino acid through fermentation |
| US5998178A (en) | 1994-05-30 | 1999-12-07 | Ajinomoto Co., Ltd. | L-isoleucine-producing bacterium and method for preparing L-isoleucine through fermentation |
| WO2000018935A1 (fr) | 1998-09-25 | 2000-04-06 | Ajinomoto Co.,Inc. | Procede de construction d'une bacterie produisant des acides amines, et procede de production d'acides amines par une technique de fermentation utilisant ladite bacterie |
| EP0994190A2 (en) | 1998-10-13 | 2000-04-19 | Ajinomoto Co., Inc. | DNA conferring L-homoserine resistance to bacteria, and its use |
| JP2000157276A (ja) | 1998-11-24 | 2000-06-13 | Ajinomoto Co Inc | サーマス属細菌のl−リジン生合成系遺伝子 |
| EP1013765A1 (en) | 1998-12-23 | 2000-06-28 | Ajinomoto Co., Ltd. | Gene and method for producing L-amino acids |
| EP1016710A2 (en) | 1998-12-30 | 2000-07-05 | Ajinomoto Co., Inc. | Method for producing L-amino acids |
| EP1092776A1 (en) | 1999-10-14 | 2001-04-18 | Ajinomoto Co., Inc. | Method for producing L-amino acid by fermentation |
| US6303383B1 (en) | 1999-03-16 | 2001-10-16 | Ajinomoto Co., Inc. | Temperature sensitive plasmid for coryneform bacteria |
| JP2001346578A (ja) | 2000-04-26 | 2001-12-18 | Ajinomoto Co Inc | アミノ酸生産菌及びアミノ酸の製造法 |
| EP1170376A1 (en) | 2000-07-05 | 2002-01-09 | Ajinomoto Co., Inc. | Method for producing substances utilizing microorganisms |
| US20020025564A1 (en) | 2000-08-24 | 2002-02-28 | Ajinomoto Co., Inc. | Method for producing basic amino acid |
| WO2002026993A1 (en) | 2000-09-28 | 2002-04-04 | Archer-Daniels-Midland Company | Escherichia coli strains which over-produce l-threonine and processes for the production of l-threonine by fermentation |
| EP1239041A2 (en) | 2001-02-13 | 2002-09-11 | Ajinomoto Co., Inc. | Method for producing L-amino acid using bacteria belonging to the genus Escherichia |
| WO2005010175A1 (en) | 2003-07-29 | 2005-02-03 | Ajinomoto Co., Inc. | Method for producing l-lysine or l-threonine using escherichia bacteria having attnuated malic enzyme activity |
| WO2005049808A1 (en) | 2003-11-21 | 2005-06-02 | Ajinomoto Co., Inc. | Method for producing l-amino acid by fermentation |
| WO2005073390A2 (en) | 2004-01-30 | 2005-08-11 | Ajinomoto Co., Inc. | L-amino acid-producing microorganism and method for producing l-amino acid |
| JP2005237379A (ja) * | 2004-01-30 | 2005-09-08 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| WO2006038695A1 (ja) | 2004-10-07 | 2006-04-13 | Ajinomoto Co., Inc. | 塩基性物質の製造法 |
| WO2006078039A1 (en) | 2005-01-18 | 2006-07-27 | Ajinomoto Co., Inc. | L-amino acid producing microorganism and a method for producing l-amino acid |
| WO2008044453A1 (fr) | 2006-10-10 | 2008-04-17 | Ajinomoto Co., Inc. | Procédé de production d'un acide aminé l |
Family Cites Families (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP3473042B2 (ja) | 1992-04-28 | 2003-12-02 | 味の素株式会社 | 変異型アスパルトキナーゼ遺伝子 |
| US6132999A (en) | 1992-09-21 | 2000-10-17 | Ajinomoto Co., Inc. | L-threonine-producing microbacteria and a method for the production of L-threonine |
| JP4061769B2 (ja) | 1999-03-25 | 2008-03-19 | 味の素株式会社 | L−グルタミン酸の製造法 |
| ES2313878T3 (es) | 2000-01-21 | 2009-03-16 | Ajinomoto Co., Inc. | Procedimiento para la produccion de l-lisina. |
| JP4265093B2 (ja) | 2000-08-11 | 2009-05-20 | 味の素株式会社 | スレオニン及びイソロイシンの製造法 |
| RU2215782C2 (ru) * | 2001-02-26 | 2003-11-10 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ Escherichia coli - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) |
| RU2244007C2 (ru) | 2002-02-27 | 2005-01-10 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Способ получения l-треонина, штамм escherichia coli - продуцент треонина (варианты) |
| RU2275424C2 (ru) | 2003-12-05 | 2006-04-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) | Способ получения l-треонина с использованием бактерий, принадлежащих к роду escherichia |
| US7915018B2 (en) | 2004-10-22 | 2011-03-29 | Ajinomoto Co., Inc. | Method for producing L-amino acids using bacteria of the Enterobacteriaceae family |
| KR20070086634A (ko) * | 2004-11-26 | 2007-08-27 | 교와 핫꼬 고교 가부시끼가이샤 | 공업적으로 유용한 미생물 |
| US20070004014A1 (en) | 2005-06-29 | 2007-01-04 | Yuichiro Tsuji | Method for producing l-threonine |
| JP2007185184A (ja) | 2005-12-16 | 2007-07-26 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| WO2007086618A1 (en) | 2006-01-30 | 2007-08-02 | Ajinomoto Co., Inc. | L-amino acid producing bacterium and method of producing l-amino acid |
| JP2009060791A (ja) | 2006-03-30 | 2009-03-26 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| JP2011067095A (ja) | 2008-01-10 | 2011-04-07 | Ajinomoto Co Inc | 発酵法による目的物質の製造法 |
-
2008
- 2008-11-27 JP JP2008302521A patent/JP2012029565A/ja active Pending
-
2009
- 2009-11-26 CN CN200980147429.0A patent/CN102227504B/zh active Active
- 2009-11-26 ES ES09829135T patent/ES2739909T3/es active Active
- 2009-11-26 WO PCT/JP2009/069956 patent/WO2010061890A1/ja active Application Filing
- 2009-11-26 BR BRPI0921655-3A patent/BRPI0921655B1/pt active IP Right Grant
- 2009-11-26 PL PL09829135T patent/PL2360263T3/pl unknown
- 2009-11-26 EP EP09829135.4A patent/EP2360263B1/en active Active
- 2009-11-26 CA CA2744852A patent/CA2744852C/en active Active
- 2009-11-26 RU RU2011126198/10A patent/RU2518677C2/ru active
-
2011
- 2011-05-24 US US13/114,251 patent/US8673597B2/en active Active
Patent Citations (60)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| FR356739A (fr) | 1904-09-20 | 1905-12-07 | Charles Glauser Perrin | Mécanisme de remontoir et de mise à l'heure |
| JPS5618596A (en) | 1979-07-23 | 1981-02-21 | Ajinomoto Co Inc | Production of l-lysine through fermentation process |
| US4346170A (en) | 1979-07-23 | 1982-08-24 | Ajinomoto Company, Incorporated | Method for producing L-lysine by fermentation |
| GB2075055A (en) | 1980-04-14 | 1981-11-11 | Ajinomoto Kk | L-methionone-producing Microorganisms |
| JPS6087788A (ja) | 1983-08-29 | 1985-05-17 | Ajinomoto Co Inc | 発酵法によるl―アミノ酸の製造法 |
| JPH02458A (ja) | 1987-10-12 | 1990-01-05 | Ajinomoto Co Inc | 発酵法によるl―イソロイシンの製造法 |
| JPH02109985A (ja) | 1988-02-22 | 1990-04-23 | Eurolysine | 細菌染色体上ヘの目的遺伝子の組み込み方法及び該方法によって得られた細菌 |
| US5175107A (en) | 1988-10-25 | 1992-12-29 | Ajinomoto Co., Inc. | Bacterial strain of escherichia coli bkiim b-3996 as the producer of l-threonine |
| WO1990004636A1 (en) | 1988-10-25 | 1990-05-03 | Vsesojuzny Nauchno-Issledovatelsky Institut Genetiki I Selektsii Promyshlennykh Mikroorganizmov (Vniigenetika) | Strain of bacteria escherichia coli, producer of l-threonine |
| JPH02242690A (ja) | 1989-03-16 | 1990-09-27 | Mitsubishi Petrochem Co Ltd | L―アラニンの製造法 |
| JPH0386588A (ja) | 1989-08-30 | 1991-04-11 | Ricoh Co Ltd | 昇華型熱転写体 |
| JPH057491A (ja) | 1990-10-15 | 1993-01-19 | Ajinomoto Co Inc | 温度感受性プラスミド |
| FR2669935A1 (fr) | 1990-11-30 | 1992-06-05 | Ajinomoto Kk | Procede et appareil pour la regulation de la concentration de source de carbone dans la culture aerobie d'un micro-organisme. |
| US5912113A (en) | 1990-11-30 | 1999-06-15 | Ajinomoto Co., Inc. | Method and apparatus for controlling carbon source concentration in aerobic cultivation of a microorganism |
| US5168056A (en) | 1991-02-08 | 1992-12-01 | Purdue Research Foundation | Enhanced production of common aromatic pathway compounds |
| JPH05276935A (ja) * | 1991-10-18 | 1993-10-26 | Degussa Ag | コリネ型細菌のアミノ酸排出性菌株の効率を増大させる方法及びアミノ酸の発酵的製造方法 |
| JPH05130882A (ja) | 1991-11-11 | 1993-05-28 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるl−イソロイシンの製造法 |
| JPH05304969A (ja) | 1992-02-25 | 1993-11-19 | Kyowa Hakko Kogyo Co Ltd | 発酵法によるアミノ酸の製造法 |
| EP0593792A1 (en) | 1992-10-14 | 1994-04-27 | Ajinomoto Co., Inc. | Novel L-threonine-producing microbacteria and a method for the production of L-threonine |
| US5661012A (en) | 1992-11-10 | 1997-08-26 | Ajinomoto Co., Inc. | Method for the production of L-threonine by fermentation, using mutated DNA encoding aspartokinase III |
| US5776736A (en) | 1992-12-21 | 1998-07-07 | Purdue Research Foundation | Deblocking the common pathway of aromatic amino acid synthesis |
| WO1995006114A1 (fr) | 1993-08-24 | 1995-03-02 | Ajinomoto Co., Inc. | Allele de phosphenolpyruvate carboxylase, gene de cet allele et procede de production de l'acide amine |
| EP0733712A1 (en) | 1993-10-28 | 1996-09-25 | Ajinomoto Co., Inc. | Process for producing substance |
| US5830716A (en) | 1993-10-28 | 1998-11-03 | Ajinomoto Co., Inc. | Increased amounts of substances by modifying a microorganism to increase production of NADPH from NADH |
| EP0733710A1 (en) | 1993-12-08 | 1996-09-25 | Ajinomoto Co., Inc. | Process for producing l-lysine by fermentation |
| US6040160A (en) | 1993-12-08 | 2000-03-21 | Ajinomoto Co., Inc. | Method of producing L-lysine by fermentation |
| WO1995023864A1 (fr) | 1994-03-04 | 1995-09-08 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| US5998178A (en) | 1994-05-30 | 1999-12-07 | Ajinomoto Co., Ltd. | L-isoleucine-producing bacterium and method for preparing L-isoleucine through fermentation |
| US5906925A (en) | 1994-09-16 | 1999-05-25 | Liao; James C. | Microorganisms and methods for overproduction of DAHP by cloned pps gene |
| WO1996017930A1 (fr) | 1994-12-09 | 1996-06-13 | Ajinomoto Co., Inc. | Nouveau gene de decarboxylase de lysine et procede de production de lysine l |
| US5827698A (en) | 1994-12-09 | 1998-10-27 | Ajinomoto Co., Inc. | Lysine decarboxylase gene and method of producing l-lysine |
| WO1996040934A1 (fr) | 1995-06-07 | 1996-12-19 | Ajinomoto Co., Inc. | Procede de production de l-lysine |
| EP0877090A1 (en) | 1995-08-30 | 1998-11-11 | Ajinomoto Co., Inc. | Process for producing l-amino acids |
| US5763230A (en) | 1996-03-22 | 1998-06-09 | Triple-A B.V. P/A Produkschap Voor Veevoedor | Amino acid fermentation processes |
| WO1998004715A1 (en) | 1996-07-30 | 1998-02-05 | Archer-Daniels-Midland Company | Novel strains of escherichia coli, methods of preparing the same and use thereof in fermentation processes for l-threonine production |
| US5932453A (en) | 1996-10-15 | 1999-08-03 | Ajinomoto Co., Ltd. | Process for producing L-amino acid through fermentation |
| WO1999018228A2 (de) | 1997-10-04 | 1999-04-15 | Forschungszentrum Jülich GmbH | Verfahren zur mikrobiellen herstellung von aminosäuren der aspartat- und/oder glutamatfamilie und im verfahren einsetzbare mittel |
| WO2000018935A1 (fr) | 1998-09-25 | 2000-04-06 | Ajinomoto Co.,Inc. | Procede de construction d'une bacterie produisant des acides amines, et procede de production d'acides amines par une technique de fermentation utilisant ladite bacterie |
| EP0994190A2 (en) | 1998-10-13 | 2000-04-19 | Ajinomoto Co., Inc. | DNA conferring L-homoserine resistance to bacteria, and its use |
| JP2000157276A (ja) | 1998-11-24 | 2000-06-13 | Ajinomoto Co Inc | サーマス属細菌のl−リジン生合成系遺伝子 |
| EP1013765A1 (en) | 1998-12-23 | 2000-06-28 | Ajinomoto Co., Ltd. | Gene and method for producing L-amino acids |
| EP1016710A2 (en) | 1998-12-30 | 2000-07-05 | Ajinomoto Co., Inc. | Method for producing L-amino acids |
| JP2000189180A (ja) | 1998-12-30 | 2000-07-11 | Ajinomoto Co Inc | L―アミノ酸の製造法 |
| US6303383B1 (en) | 1999-03-16 | 2001-10-16 | Ajinomoto Co., Inc. | Temperature sensitive plasmid for coryneform bacteria |
| EP1092776A1 (en) | 1999-10-14 | 2001-04-18 | Ajinomoto Co., Inc. | Method for producing L-amino acid by fermentation |
| JP2001346578A (ja) | 2000-04-26 | 2001-12-18 | Ajinomoto Co Inc | アミノ酸生産菌及びアミノ酸の製造法 |
| EP1170376A1 (en) | 2000-07-05 | 2002-01-09 | Ajinomoto Co., Inc. | Method for producing substances utilizing microorganisms |
| US20020025564A1 (en) | 2000-08-24 | 2002-02-28 | Ajinomoto Co., Inc. | Method for producing basic amino acid |
| JP2002065287A (ja) | 2000-08-24 | 2002-03-05 | Ajinomoto Co Inc | 塩基性アミノ酸の製造方法 |
| WO2002026993A1 (en) | 2000-09-28 | 2002-04-04 | Archer-Daniels-Midland Company | Escherichia coli strains which over-produce l-threonine and processes for the production of l-threonine by fermentation |
| EP1239041A2 (en) | 2001-02-13 | 2002-09-11 | Ajinomoto Co., Inc. | Method for producing L-amino acid using bacteria belonging to the genus Escherichia |
| WO2005010175A1 (en) | 2003-07-29 | 2005-02-03 | Ajinomoto Co., Inc. | Method for producing l-lysine or l-threonine using escherichia bacteria having attnuated malic enzyme activity |
| WO2005049808A1 (en) | 2003-11-21 | 2005-06-02 | Ajinomoto Co., Inc. | Method for producing l-amino acid by fermentation |
| WO2005073390A2 (en) | 2004-01-30 | 2005-08-11 | Ajinomoto Co., Inc. | L-amino acid-producing microorganism and method for producing l-amino acid |
| JP2005237379A (ja) * | 2004-01-30 | 2005-09-08 | Ajinomoto Co Inc | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| US20060019355A1 (en) | 2004-01-30 | 2006-01-26 | Takuji Ueda | L-Amino acid-producing microorganism and method for producing L-amino acid |
| WO2006038695A1 (ja) | 2004-10-07 | 2006-04-13 | Ajinomoto Co., Inc. | 塩基性物質の製造法 |
| EP1813677A1 (en) | 2004-10-07 | 2007-08-01 | Ajinomoto Co., Inc. | Process for producing basic substance |
| WO2006078039A1 (en) | 2005-01-18 | 2006-07-27 | Ajinomoto Co., Inc. | L-amino acid producing microorganism and a method for producing l-amino acid |
| WO2008044453A1 (fr) | 2006-10-10 | 2008-04-17 | Ajinomoto Co., Inc. | Procédé de production d'un acide aminé l |
Non-Patent Citations (36)
| Title |
|---|
| "Amino Acid Fermentation", 30 May 1986, GAKKAI SHUPPAN CENTER (LTD., pages: 77 - 100 |
| "Experiment Manual for Biotechnology", 1992, BAIFUKAN CO., LTD., pages: 97 - 98 |
| "Molecular Cloning", 2001, COLD SPRING HARBOR LABORATORY PRESS |
| APPLEYARD R.K., GENETICS, vol. 39, 1954, pages 440 - 452 |
| BACKMANN B.J.: "Escherichia coli and Salmonella Cellular and Molecular Biology", 1996, AMERICAN SOCIETY FOR MICROBIOLOGY PRESS, article "Derivations and Genotypes of some mutant derivatives of Escherichia coli K-12", pages: 2460 - 2488 |
| BIBB, M.J., WARD, J.M., HOPWOOD, O.A., NATURE, vol. 274, 1978, pages 398 |
| BOEHLEIN, S.K., RICHARDS, N.G.J., SCHUSTER, S.M., J. BIOL. CHEM., vol. 269, 1994, pages 7450 - 7457 |
| CHANG, S., CHOEN, S.N., MOLEC. GEN. GENET., vol. 168, 1979, pages 111 |
| CHISTORERDOV, A.Y., TSYGANKOV, Y.D., PLASMID, vol. 16, 1986, pages 161 - 167 |
| CHO, E.H., GUMPORT, R.I., GARDNER, J.F., J. BACTERIOL., vol. 184, 2002, pages 5200 - 5203 |
| DATSENKO, K.A, WANNER, B.L., PROC. NATL. ACAD. SCI. USA, vol. 97, 2000, pages 6640 - 6645 |
| DATSENKO, K.A., WANNER B.L., PROC. NATL. ACAD. SCI. USA., vol. 97, 2000, pages 6640 - 6645 |
| DEGUCHI, Y. ET AL.: "Molecular cloning of gltS and gltP, which encode glutamate carriers of Escherichia coli B", JOURNAL OF BACTERIOLOGY, vol. 171, no. 3, March 1989 (1989-03-01), pages 1314 - 1319, XP008148835 * |
| DEGUCHI, Y. ET AL.: "Nucleotide sequence of gltS, the Na+/glutamate symport carrier gene of Escherichia coli B", THE JOURNAL OF BIOLOGICAL CHEMISTRY, vol. 265, no. 35, December 1990 (1990-12-01), pages 21704 - 21708, XP008148844 * |
| DUNCAN, C.H., WILSON, G.A., YOUNG, F.E., GENE, vol. 1, 1977, pages 153 |
| GENE, vol. 23, 1983, pages 199 - 209 |
| GOLDSTEIN ET AL.: "Prokaryotic promoters in biotechnology", BIOTECHNOL. ANNU. REV., vol. 1, 1995, pages 105 - 128, XP008038474 |
| HASEGAWA, K. ET AL.: "Isolation and purification technology in fermentation industry of amino acids", BIOSCIENCE AND INDUSTRY, vol. 66, no. 3, March 2008 (2008-03-01), pages 124 - 129, XP008148846 * |
| HINNEN, A., HICKS, J.B., FINK, G.R., PROC. NATL. ACAD. SCI. USA, vol. 75, 1978, pages 1929 |
| J. BACTERIOL., vol. 174, no. 7, April 1992 (1992-04-01), pages 2391 - 3 |
| J. BACTERIOL., vol. 184, 2002, pages 5200 - 5203 |
| J. BACTERIOL., vol. 188, no. 23, December 2006 (2006-12-01), pages 8118 - 27 |
| J. BIOL. CHEM., vol. 265, no. 35, 15 December 1990 (1990-12-15), pages 21704 - 8 |
| J. BIOL. CHEM., vol. 272, 1997, pages 8611 - 8617 |
| KARLIN, ALTSCHUL, PROC. NATL. ACAD. SCI. USA, vol. 90, 1993, pages 5873 |
| LYNN, S.P., BURTON, W.S., DONOHUE, T.J., GOULD, R.M., GUMPORT, R.L, GARDNER, J.F., J. MOL. BIOL., vol. 194, 1987, pages 59 - 69 |
| MANDEL, M., HIGA, A., J. MOL. BIOL., vol. 53, 1970, pages 159 |
| METHODS ENZYMOL., vol. 183, 1990, pages 63 |
| RES. MICROBIOL., vol. 154, 2003, pages 123 - 135 |
| SAITO H., MIURA K., BIOCHEM. BIOPHYS. ACTA, vol. 72, 1963, pages 619 |
| SCIENCE, vol. 277, 1997, pages 1453 - 1474 |
| TOLNER, B. ET AL.: "Revised nucleotide sequence of the gltP gene, which encodes the proton-glutamate-aspartate transport protein of Escherichia coli K-12", JOURNAL OF BACTERIOLOGY, vol. 174, no. 7, April 1992 (1992-04-01), pages 2391 - 2393, XP008148854 * |
| TROTSCHEL, C. ET AL.: "GltS, the sodium-coupled L-glutamate uptake system of Corynebacterium glutamicum: identification of the corresponding gene and impact on L-glutamate production", APPL. MICROBIOL. BIOTECHNOL., vol. 60, 2003, pages 738 - 742, XP008148836 * |
| WALLACE, B, YANG, YJ, HONG, JS, LUM, D, J. BACTERIOL., vol. 172, no. 6, June 1990 (1990-06-01), pages 3214 - 3220 |
| WALLACE, B. ET AL.: "Cloning and sequencing of a gene encoding a glutamate and aspartate carrier of Escherichia coli K-12", JOURNAL OF BACTERIOLOGY, vol. 172, no. 6, June 1990 (1990-06-01), pages 3214 - 3220, XP008148837 * |
| WHITE, T.J. ET AL., TRENDS GENET., vol. 5, 1989, pages 185 |
Cited By (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN102329765A (zh) * | 2010-08-31 | 2012-01-25 | 安徽华恒生物工程有限公司 | 一种高产l-丙氨酸的xz-a26菌株及构建方法与应用 |
| CN102329765B (zh) * | 2010-08-31 | 2013-08-07 | 安徽华恒生物工程有限公司 | 一种高产l-丙氨酸的xz-a26菌株及构建方法与应用 |
| JP6020443B2 (ja) * | 2011-04-01 | 2016-11-02 | 味の素株式会社 | L−システインの製造法 |
| JP2013106588A (ja) * | 2011-11-24 | 2013-06-06 | Kyowa Hakko Bio Co Ltd | L−アスパラギンの製造法 |
| WO2015060314A1 (ja) | 2013-10-21 | 2015-04-30 | 味の素株式会社 | L-アミノ酸の製造法 |
| EP3385389A1 (en) | 2017-04-03 | 2018-10-10 | Ajinomoto Co., Inc. | Method for producing l-amino acid from fructose |
| WO2020071538A1 (en) | 2018-10-05 | 2020-04-09 | Ajinomoto Co., Inc. | Method for producing target substance by bacterial fermentation |
| EP3716274A1 (en) | 2019-03-29 | 2020-09-30 | Ajinomoto Co., Inc. | Control device, control method, computer program, and method for producing organic compound |
Also Published As
| Publication number | Publication date |
|---|---|
| CA2744852A1 (en) | 2010-06-03 |
| ES2739909T3 (es) | 2020-02-04 |
| US8673597B2 (en) | 2014-03-18 |
| PL2360263T3 (pl) | 2019-10-31 |
| EP2360263A1 (en) | 2011-08-24 |
| BRPI0921655A2 (pt) | 2020-08-04 |
| US20110281311A1 (en) | 2011-11-17 |
| RU2518677C2 (ru) | 2014-06-10 |
| BRPI0921655B1 (pt) | 2021-05-25 |
| EP2360263A4 (en) | 2012-08-15 |
| CN102227504B (zh) | 2014-06-11 |
| RU2011126198A (ru) | 2013-01-10 |
| EP2360263B1 (en) | 2019-06-19 |
| JP2012029565A (ja) | 2012-02-16 |
| CA2744852C (en) | 2017-01-24 |
| CN102227504A (zh) | 2011-10-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8673597B2 (en) | Method for producing L-amino acid | |
| EP2083083B1 (en) | Method for production of l-amino acid | |
| EP1664318B1 (en) | L-amino acid-producing microorganism and method for producing l-amino acid | |
| US9644009B2 (en) | L-amino acid-producing bacterium and a method for producing an L-amino acid | |
| EP1979486B1 (en) | L-amino acid producing bacterium and method of producing l-amino acid | |
| EP1963486B1 (en) | L-amino acid producing bacterium and method of producing l-amino acid | |
| US8354255B2 (en) | L-amino acid-producing bacterium and a method for producing L-amino acids | |
| EP1928899B1 (en) | An l-amino acid-producing bacterium and a method for producing an l-amino acid | |
| EP2471942B1 (en) | Process for production of l-amino acid | |
| US7871801B2 (en) | L-amino acid-producing bacterium and a method for producing L-amino acids | |
| CN108699576B (zh) | L-氨基酸的制造方法 | |
| CN101273138A (zh) | 产生l-氨基酸的细菌和用于产生l-氨基酸的方法 | |
| JP2010187541A (ja) | L−リジンの製造法 | |
| JP2010183841A (ja) | L−アミノ酸の製造法 | |
| JP2007117076A (ja) | L−アミノ酸生産菌及びl−アミノ酸の製造法 | |
| JP2010142200A (ja) | L−リジンの製造法 | |
| JP2007117077A (ja) | L−アミノ酸生産菌及びl−アミノ酸の製造法 | |
| JP2009240161A (ja) | L−アミノ酸の製造法 | |
| CN101273139A (zh) | 产生l-氨基酸的细菌和用于产生l-氨基酸的方法 | |
| JP2008086289A (ja) | L−リジンの製造法 | |
| JP2016149983A (ja) | L−アミノ酸の製造法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WWE | Wipo information: entry into national phase |
Ref document number: 200980147429.0 Country of ref document: CN |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 09829135 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2744852 Country of ref document: CA |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2009829135 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2011126198 Country of ref document: RU |
|
| NENP | Non-entry into the national phase |
Ref country code: JP |
|
| ENP | Entry into the national phase |
Ref document number: PI0921655 Country of ref document: BR Kind code of ref document: A2 Effective date: 20110526 |