ES2906344T3 - Animal murino transgénico productor de anticuerpos - Google Patents
Animal murino transgénico productor de anticuerpos Download PDFInfo
- Publication number
- ES2906344T3 ES2906344T3 ES18183980T ES18183980T ES2906344T3 ES 2906344 T3 ES2906344 T3 ES 2906344T3 ES 18183980 T ES18183980 T ES 18183980T ES 18183980 T ES18183980 T ES 18183980T ES 2906344 T3 ES2906344 T3 ES 2906344T3
- Authority
- ES
- Spain
- Prior art keywords
- human
- light chain
- expression
- transgenic
- antibody
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 241001529936 Murinae Species 0.000 title claims abstract description 92
- 230000009261 transgenic effect Effects 0.000 title claims abstract description 75
- 210000004602 germ cell Anatomy 0.000 claims abstract description 50
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 43
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 41
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 41
- 108060003951 Immunoglobulin Proteins 0.000 claims abstract description 35
- 102000018358 immunoglobulin Human genes 0.000 claims abstract description 35
- 241001465754 Metazoa Species 0.000 claims abstract description 20
- 230000008707 rearrangement Effects 0.000 claims abstract description 17
- 101150117115 V gene Proteins 0.000 claims abstract description 14
- 101150008942 J gene Proteins 0.000 claims abstract description 7
- 229940072221 immunoglobulins Drugs 0.000 claims abstract description 6
- 230000014509 gene expression Effects 0.000 claims description 153
- 210000004027 cell Anatomy 0.000 claims description 132
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 97
- 238000000034 method Methods 0.000 claims description 85
- 101001138089 Homo sapiens Immunoglobulin kappa variable 1-39 Proteins 0.000 claims description 64
- 102100020910 Immunoglobulin kappa variable 1-39 Human genes 0.000 claims description 64
- 238000004519 manufacturing process Methods 0.000 claims description 41
- 239000000427 antigen Substances 0.000 claims description 39
- 108091007433 antigens Proteins 0.000 claims description 39
- 102000036639 antigens Human genes 0.000 claims description 39
- 238000011161 development Methods 0.000 claims description 21
- 230000030279 gene silencing Effects 0.000 claims description 15
- 230000005875 antibody response Effects 0.000 claims description 12
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 claims description 8
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 claims description 8
- 230000010354 integration Effects 0.000 claims description 7
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 5
- 238000012258 culturing Methods 0.000 claims description 4
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 claims description 3
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 claims description 3
- 101001091242 Homo sapiens Immunoglobulin kappa joining 1 Proteins 0.000 claims description 2
- 102100034892 Immunoglobulin kappa joining 1 Human genes 0.000 claims description 2
- 101100351020 Mus musculus Pax5 gene Proteins 0.000 claims description 2
- 102100037504 Paired box protein Pax-5 Human genes 0.000 claims description 2
- 101710149067 Paired box protein Pax-5 Proteins 0.000 claims description 2
- 101100351021 Xenopus laevis pax5 gene Proteins 0.000 claims description 2
- 238000003306 harvesting Methods 0.000 claims description 2
- 108090000623 proteins and genes Proteins 0.000 description 114
- 241000699666 Mus <mouse, genus> Species 0.000 description 66
- 108700019146 Transgenes Proteins 0.000 description 63
- 239000013598 vector Substances 0.000 description 59
- 241000699670 Mus sp. Species 0.000 description 53
- 230000027455 binding Effects 0.000 description 47
- 239000003623 enhancer Substances 0.000 description 37
- 108090000765 processed proteins & peptides Proteins 0.000 description 36
- 101001100327 Homo sapiens RNA-binding protein 45 Proteins 0.000 description 33
- 102100038823 RNA-binding protein 45 Human genes 0.000 description 33
- 241000700159 Rattus Species 0.000 description 31
- 108020004414 DNA Proteins 0.000 description 30
- 102000004196 processed proteins & peptides Human genes 0.000 description 28
- 102000004169 proteins and genes Human genes 0.000 description 27
- 238000002965 ELISA Methods 0.000 description 22
- 230000008685 targeting Effects 0.000 description 22
- 102000004889 Interleukin-6 Human genes 0.000 description 21
- 108090001005 Interleukin-6 Proteins 0.000 description 21
- 239000006228 supernatant Substances 0.000 description 21
- 238000011830 transgenic mouse model Methods 0.000 description 21
- 230000018109 developmental process Effects 0.000 description 20
- 238000010276 construction Methods 0.000 description 19
- 239000000047 product Substances 0.000 description 19
- 101000956885 Homo sapiens Immunoglobulin lambda variable 2-14 Proteins 0.000 description 18
- 102100038429 Immunoglobulin lambda variable 2-14 Human genes 0.000 description 18
- 239000012634 fragment Substances 0.000 description 18
- 230000008569 process Effects 0.000 description 18
- 241000699660 Mus musculus Species 0.000 description 17
- 238000002744 homologous recombination Methods 0.000 description 17
- 230000006801 homologous recombination Effects 0.000 description 17
- 239000002609 medium Substances 0.000 description 17
- 238000002823 phage display Methods 0.000 description 17
- 238000004458 analytical method Methods 0.000 description 16
- 230000035772 mutation Effects 0.000 description 16
- 102000005962 receptors Human genes 0.000 description 16
- 108020003175 receptors Proteins 0.000 description 16
- 230000000903 blocking effect Effects 0.000 description 15
- 230000003053 immunization Effects 0.000 description 15
- 230000005847 immunogenicity Effects 0.000 description 15
- 230000000392 somatic effect Effects 0.000 description 15
- 238000006243 chemical reaction Methods 0.000 description 14
- 238000002649 immunization Methods 0.000 description 14
- 230000002163 immunogen Effects 0.000 description 14
- 210000000952 spleen Anatomy 0.000 description 14
- 238000010561 standard procedure Methods 0.000 description 14
- 101100284398 Bos taurus BoLA-DQB gene Proteins 0.000 description 13
- 101150034979 DRB3 gene Proteins 0.000 description 13
- 101100278514 Oryza sativa subsp. japonica DRB2 gene Proteins 0.000 description 13
- 238000012217 deletion Methods 0.000 description 13
- 230000037430 deletion Effects 0.000 description 13
- 230000002829 reductive effect Effects 0.000 description 13
- 108700028369 Alleles Proteins 0.000 description 12
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 12
- 241000282412 Homo Species 0.000 description 12
- 238000003780 insertion Methods 0.000 description 12
- 230000037431 insertion Effects 0.000 description 12
- 210000004698 lymphocyte Anatomy 0.000 description 12
- 229920001184 polypeptide Polymers 0.000 description 12
- 239000000243 solution Substances 0.000 description 12
- 238000010186 staining Methods 0.000 description 12
- 239000011230 binding agent Substances 0.000 description 11
- 238000010367 cloning Methods 0.000 description 11
- 229960000814 tetanus toxoid Drugs 0.000 description 11
- 108700026244 Open Reading Frames Proteins 0.000 description 10
- 230000003321 amplification Effects 0.000 description 10
- 238000003199 nucleic acid amplification method Methods 0.000 description 10
- 101001076408 Homo sapiens Interleukin-6 Proteins 0.000 description 9
- 150000001413 amino acids Chemical group 0.000 description 9
- 210000001185 bone marrow Anatomy 0.000 description 9
- 239000002299 complementary DNA Substances 0.000 description 9
- 102000052611 human IL6 Human genes 0.000 description 9
- 239000002773 nucleotide Substances 0.000 description 9
- 125000003729 nucleotide group Chemical group 0.000 description 9
- 238000001262 western blot Methods 0.000 description 9
- 241000894006 Bacteria Species 0.000 description 8
- 108091026890 Coding region Proteins 0.000 description 8
- 101000854908 Homo sapiens WD repeat-containing protein 11 Proteins 0.000 description 8
- 102100020705 WD repeat-containing protein 11 Human genes 0.000 description 8
- -1 for example Proteins 0.000 description 8
- 210000004408 hybridoma Anatomy 0.000 description 8
- 238000012163 sequencing technique Methods 0.000 description 8
- 238000001890 transfection Methods 0.000 description 8
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 8
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 7
- 210000002459 blastocyst Anatomy 0.000 description 7
- 238000001514 detection method Methods 0.000 description 7
- 239000000203 mixture Substances 0.000 description 7
- 230000002441 reversible effect Effects 0.000 description 7
- 210000001519 tissue Anatomy 0.000 description 7
- 206010069754 Acquired gene mutation Diseases 0.000 description 6
- 238000011740 C57BL/6 mouse Methods 0.000 description 6
- 101150050798 IGKV1-39 gene Proteins 0.000 description 6
- 108091029795 Intergenic region Proteins 0.000 description 6
- 210000000349 chromosome Anatomy 0.000 description 6
- 230000003247 decreasing effect Effects 0.000 description 6
- 230000000694 effects Effects 0.000 description 6
- 210000002257 embryonic structure Anatomy 0.000 description 6
- 238000002474 experimental method Methods 0.000 description 6
- 239000003550 marker Substances 0.000 description 6
- 230000004048 modification Effects 0.000 description 6
- 238000012986 modification Methods 0.000 description 6
- 239000013641 positive control Substances 0.000 description 6
- 210000001948 pro-b lymphocyte Anatomy 0.000 description 6
- 230000004044 response Effects 0.000 description 6
- 230000037439 somatic mutation Effects 0.000 description 6
- 230000009466 transformation Effects 0.000 description 6
- 238000005406 washing Methods 0.000 description 6
- 102000019034 Chemokines Human genes 0.000 description 5
- 108010012236 Chemokines Proteins 0.000 description 5
- 102000010781 Interleukin-6 Receptors Human genes 0.000 description 5
- 108010038501 Interleukin-6 Receptors Proteins 0.000 description 5
- 206010035226 Plasma cell myeloma Diseases 0.000 description 5
- 101000777312 Rattus norvegicus Choline kinase alpha Proteins 0.000 description 5
- 101000777311 Rattus norvegicus Choline/ethanolamine kinase Proteins 0.000 description 5
- 101001047152 Rattus norvegicus UMP-CMP kinase Proteins 0.000 description 5
- 238000012300 Sequence Analysis Methods 0.000 description 5
- 210000001744 T-lymphocyte Anatomy 0.000 description 5
- 208000035199 Tetraploidy Diseases 0.000 description 5
- 210000004369 blood Anatomy 0.000 description 5
- 239000008280 blood Substances 0.000 description 5
- 230000011712 cell development Effects 0.000 description 5
- 230000000875 corresponding effect Effects 0.000 description 5
- 239000013604 expression vector Substances 0.000 description 5
- 238000000338 in vitro Methods 0.000 description 5
- 238000001727 in vivo Methods 0.000 description 5
- 210000003519 mature b lymphocyte Anatomy 0.000 description 5
- 201000000050 myeloid neoplasm Diseases 0.000 description 5
- 239000013642 negative control Substances 0.000 description 5
- 230000002093 peripheral effect Effects 0.000 description 5
- 230000006798 recombination Effects 0.000 description 5
- 238000005215 recombination Methods 0.000 description 5
- 238000012216 screening Methods 0.000 description 5
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 4
- 102000001301 EGF receptor Human genes 0.000 description 4
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 4
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 4
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 description 4
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 description 4
- 229920001213 Polysorbate 20 Polymers 0.000 description 4
- 238000002123 RNA extraction Methods 0.000 description 4
- 238000002105 Southern blotting Methods 0.000 description 4
- 230000004913 activation Effects 0.000 description 4
- 230000009824 affinity maturation Effects 0.000 description 4
- 230000004075 alteration Effects 0.000 description 4
- FPPNZSSZRUTDAP-UWFZAAFLSA-N carbenicillin Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)C(C(O)=O)C1=CC=CC=C1 FPPNZSSZRUTDAP-UWFZAAFLSA-N 0.000 description 4
- 229960003669 carbenicillin Drugs 0.000 description 4
- 239000003153 chemical reaction reagent Substances 0.000 description 4
- 238000009402 cross-breeding Methods 0.000 description 4
- 230000004069 differentiation Effects 0.000 description 4
- 238000004520 electroporation Methods 0.000 description 4
- 210000001671 embryonic stem cell Anatomy 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 230000002779 inactivation Effects 0.000 description 4
- 238000011534 incubation Methods 0.000 description 4
- 230000001939 inductive effect Effects 0.000 description 4
- 238000002955 isolation Methods 0.000 description 4
- 210000005210 lymphoid organ Anatomy 0.000 description 4
- 230000001404 mediated effect Effects 0.000 description 4
- 210000000066 myeloid cell Anatomy 0.000 description 4
- 210000003200 peritoneal cavity Anatomy 0.000 description 4
- 239000013612 plasmid Substances 0.000 description 4
- 230000008488 polyadenylation Effects 0.000 description 4
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 4
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 4
- 230000035755 proliferation Effects 0.000 description 4
- 230000001105 regulatory effect Effects 0.000 description 4
- 238000009877 rendering Methods 0.000 description 4
- 239000000523 sample Substances 0.000 description 4
- 235000020183 skimmed milk Nutrition 0.000 description 4
- 230000009870 specific binding Effects 0.000 description 4
- 210000004988 splenocyte Anatomy 0.000 description 4
- 230000001225 therapeutic effect Effects 0.000 description 4
- 238000013518 transcription Methods 0.000 description 4
- 230000035897 transcription Effects 0.000 description 4
- 238000013519 translation Methods 0.000 description 4
- 238000011282 treatment Methods 0.000 description 4
- 229960005486 vaccine Drugs 0.000 description 4
- 101000775252 Arabidopsis thaliana NADPH-dependent oxidoreductase 2-alkenal reductase Proteins 0.000 description 3
- 101100112922 Candida albicans CDR3 gene Proteins 0.000 description 3
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 3
- 102000004127 Cytokines Human genes 0.000 description 3
- 108090000695 Cytokines Proteins 0.000 description 3
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 3
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 3
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- 101001041031 Homo sapiens Lariat debranching enzyme Proteins 0.000 description 3
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 3
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 3
- 102100021155 Lariat debranching enzyme Human genes 0.000 description 3
- 101001077374 Oryza sativa subsp. japonica UMP-CMP kinase 3 Proteins 0.000 description 3
- 238000012408 PCR amplification Methods 0.000 description 3
- 241000700605 Viruses Species 0.000 description 3
- 238000009395 breeding Methods 0.000 description 3
- 230000001488 breeding effect Effects 0.000 description 3
- 150000001720 carbohydrates Chemical class 0.000 description 3
- 101150076615 ck gene Proteins 0.000 description 3
- 239000011248 coating agent Substances 0.000 description 3
- 238000000576 coating method Methods 0.000 description 3
- 239000003814 drug Substances 0.000 description 3
- 238000010828 elution Methods 0.000 description 3
- 238000000684 flow cytometry Methods 0.000 description 3
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 3
- 230000004927 fusion Effects 0.000 description 3
- 239000000499 gel Substances 0.000 description 3
- 230000002068 genetic effect Effects 0.000 description 3
- BRZYSWJRSDMWLG-CAXSIQPQSA-N geneticin Chemical compound O1C[C@@](O)(C)[C@H](NC)[C@@H](O)[C@H]1O[C@@H]1[C@@H](O)[C@H](O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](C(C)O)O2)N)[C@@H](N)C[C@H]1N BRZYSWJRSDMWLG-CAXSIQPQSA-N 0.000 description 3
- 239000008103 glucose Substances 0.000 description 3
- 238000009396 hybridization Methods 0.000 description 3
- 210000003297 immature b lymphocyte Anatomy 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 238000000520 microinjection Methods 0.000 description 3
- 230000003134 recirculating effect Effects 0.000 description 3
- 230000009467 reduction Effects 0.000 description 3
- 238000000926 separation method Methods 0.000 description 3
- 210000002966 serum Anatomy 0.000 description 3
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 3
- 239000000725 suspension Substances 0.000 description 3
- 238000012360 testing method Methods 0.000 description 3
- 238000011144 upstream manufacturing Methods 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- LOGFVTREOLYCPF-KXNHARMFSA-N (2s,3r)-2-[[(2r)-1-[(2s)-2,6-diaminohexanoyl]pyrrolidine-2-carbonyl]amino]-3-hydroxybutanoic acid Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H]1CCCN1C(=O)[C@@H](N)CCCCN LOGFVTREOLYCPF-KXNHARMFSA-N 0.000 description 2
- OOIBFPKQHULHSQ-UHFFFAOYSA-N (3-hydroxy-1-adamantyl) 2-methylprop-2-enoate Chemical compound C1C(C2)CC3CC2(O)CC1(OC(=O)C(=C)C)C3 OOIBFPKQHULHSQ-UHFFFAOYSA-N 0.000 description 2
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 2
- IVLXQGJVBGMLRR-UHFFFAOYSA-N 2-aminoacetic acid;hydron;chloride Chemical compound Cl.NCC(O)=O IVLXQGJVBGMLRR-UHFFFAOYSA-N 0.000 description 2
- UAIUNKRWKOVEES-UHFFFAOYSA-N 3,3',5,5'-tetramethylbenzidine Chemical compound CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 UAIUNKRWKOVEES-UHFFFAOYSA-N 0.000 description 2
- WOVKYSAHUYNSMH-RRKCRQDMSA-N 5-bromodeoxyuridine Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(Br)=C1 WOVKYSAHUYNSMH-RRKCRQDMSA-N 0.000 description 2
- 102100031585 ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Human genes 0.000 description 2
- 108010032595 Antibody Binding Sites Proteins 0.000 description 2
- 101100067974 Arabidopsis thaliana POP2 gene Proteins 0.000 description 2
- 238000011725 BALB/c mouse Methods 0.000 description 2
- 101150013553 CD40 gene Proteins 0.000 description 2
- 101100191768 Caenorhabditis elegans pbs-4 gene Proteins 0.000 description 2
- HEDRZPFGACZZDS-UHFFFAOYSA-N Chloroform Chemical compound ClC(Cl)Cl HEDRZPFGACZZDS-UHFFFAOYSA-N 0.000 description 2
- 108700010070 Codon Usage Proteins 0.000 description 2
- 108010051219 Cre recombinase Proteins 0.000 description 2
- 101150097493 D gene Proteins 0.000 description 2
- 238000000116 DAPI staining Methods 0.000 description 2
- 101150082328 DRB5 gene Proteins 0.000 description 2
- 108060006698 EGF receptor Proteins 0.000 description 2
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 2
- 108091008794 FGF receptors Proteins 0.000 description 2
- 102000044168 Fibroblast Growth Factor Receptor Human genes 0.000 description 2
- 241000724791 Filamentous phage Species 0.000 description 2
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 2
- 101710154606 Hemagglutinin Proteins 0.000 description 2
- 101000777636 Homo sapiens ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Proteins 0.000 description 2
- 101100118549 Homo sapiens EGFR gene Proteins 0.000 description 2
- 101001057504 Homo sapiens Interferon-stimulated gene 20 kDa protein Proteins 0.000 description 2
- 101001055144 Homo sapiens Interleukin-2 receptor subunit alpha Proteins 0.000 description 2
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 2
- 101000716102 Homo sapiens T-cell surface glycoprotein CD4 Proteins 0.000 description 2
- 101000851376 Homo sapiens Tumor necrosis factor receptor superfamily member 8 Proteins 0.000 description 2
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 2
- 102100023915 Insulin Human genes 0.000 description 2
- 102000003777 Interleukin-1 beta Human genes 0.000 description 2
- 108090000193 Interleukin-1 beta Proteins 0.000 description 2
- 102100026878 Interleukin-2 receptor subunit alpha Human genes 0.000 description 2
- 102000004058 Leukemia inhibitory factor Human genes 0.000 description 2
- 108090000581 Leukemia inhibitory factor Proteins 0.000 description 2
- 102000003960 Ligases Human genes 0.000 description 2
- 108090000364 Ligases Proteins 0.000 description 2
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 2
- 239000006137 Luria-Bertani broth Substances 0.000 description 2
- 241001436793 Meru Species 0.000 description 2
- 208000005647 Mumps Diseases 0.000 description 2
- 206010028980 Neoplasm Diseases 0.000 description 2
- 239000012124 Opti-MEM Substances 0.000 description 2
- 101100117565 Oryza sativa subsp. japonica DRB4 gene Proteins 0.000 description 2
- 101100117569 Oryza sativa subsp. japonica DRB6 gene Proteins 0.000 description 2
- 101710093908 Outer capsid protein VP4 Proteins 0.000 description 2
- 101710135467 Outer capsid protein sigma-1 Proteins 0.000 description 2
- 208000002606 Paramyxoviridae Infections Diseases 0.000 description 2
- 102000003992 Peroxidases Human genes 0.000 description 2
- 241000288906 Primates Species 0.000 description 2
- 101710176177 Protein A56 Proteins 0.000 description 2
- 108010076504 Protein Sorting Signals Proteins 0.000 description 2
- 108020004511 Recombinant DNA Proteins 0.000 description 2
- 108091081062 Repeated sequence (DNA) Proteins 0.000 description 2
- 101100123851 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) HER1 gene Proteins 0.000 description 2
- 102100036011 T-cell surface glycoprotein CD4 Human genes 0.000 description 2
- 239000004098 Tetracycline Substances 0.000 description 2
- 108010034949 Thyroglobulin Proteins 0.000 description 2
- 102000009843 Thyroglobulin Human genes 0.000 description 2
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 2
- 102100036857 Tumor necrosis factor receptor superfamily member 8 Human genes 0.000 description 2
- 108091008605 VEGF receptors Proteins 0.000 description 2
- 102000009484 Vascular Endothelial Growth Factor Receptors Human genes 0.000 description 2
- 230000009471 action Effects 0.000 description 2
- 230000007720 allelic exclusion Effects 0.000 description 2
- 230000002096 anti-tetanic effect Effects 0.000 description 2
- 230000009833 antibody interaction Effects 0.000 description 2
- 238000003556 assay Methods 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 230000033228 biological regulation Effects 0.000 description 2
- 230000015572 biosynthetic process Effects 0.000 description 2
- 238000010804 cDNA synthesis Methods 0.000 description 2
- 239000006285 cell suspension Substances 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 208000037976 chronic inflammation Diseases 0.000 description 2
- 238000001976 enzyme digestion Methods 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 210000002950 fibroblast Anatomy 0.000 description 2
- 239000012737 fresh medium Substances 0.000 description 2
- 239000006481 glucose medium Substances 0.000 description 2
- 239000000185 hemagglutinin Substances 0.000 description 2
- 230000028993 immune response Effects 0.000 description 2
- 208000015181 infectious disease Diseases 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- PHTQWCKDNZKARW-UHFFFAOYSA-N isoamylol Chemical compound CC(C)CCO PHTQWCKDNZKARW-UHFFFAOYSA-N 0.000 description 2
- 238000001638 lipofection Methods 0.000 description 2
- 230000013011 mating Effects 0.000 description 2
- 208000010805 mumps infectious disease Diseases 0.000 description 2
- 238000006386 neutralization reaction Methods 0.000 description 2
- 230000009871 nonspecific binding Effects 0.000 description 2
- 210000000287 oocyte Anatomy 0.000 description 2
- 238000005457 optimization Methods 0.000 description 2
- 239000002245 particle Substances 0.000 description 2
- 244000052769 pathogen Species 0.000 description 2
- 108040007629 peroxidase activity proteins Proteins 0.000 description 2
- 239000000825 pharmaceutical preparation Substances 0.000 description 2
- 230000003389 potentiating effect Effects 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 230000000644 propagated effect Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 238000011536 re-plating Methods 0.000 description 2
- 102000027426 receptor tyrosine kinases Human genes 0.000 description 2
- 108091008598 receptor tyrosine kinases Proteins 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 238000010839 reverse transcription Methods 0.000 description 2
- 210000003705 ribosome Anatomy 0.000 description 2
- 238000001542 size-exclusion chromatography Methods 0.000 description 2
- 230000010473 stable expression Effects 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 229960002180 tetracycline Drugs 0.000 description 2
- 229930101283 tetracycline Natural products 0.000 description 2
- 235000019364 tetracycline Nutrition 0.000 description 2
- 238000002560 therapeutic procedure Methods 0.000 description 2
- 229960002175 thyroglobulin Drugs 0.000 description 2
- 230000002103 transcriptional effect Effects 0.000 description 2
- ZTENZJJCFACIAK-ADWVOTLJSA-N (2r)-2-[[(1r,3s,4s)-3-[[4-(5-benzyl-2-ethylpyrazol-3-yl)piperidin-1-yl]methyl]-4-(3-fluorophenyl)cyclopentyl]-methylamino]-3-methylbutanoic acid Chemical compound C1=C(C2CCN(C[C@@H]3[C@H](C[C@H](C3)N(C)[C@H](C(C)C)C(O)=O)C=3C=C(F)C=CC=3)CC2)N(CC)N=C1CC1=CC=CC=C1 ZTENZJJCFACIAK-ADWVOTLJSA-N 0.000 description 1
- 241000588626 Acinetobacter baumannii Species 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- HJCMDXDYPOUFDY-WHFBIAKZSA-N Ala-Gln Chemical compound C[C@H](N)C(=O)N[C@H](C(O)=O)CCC(N)=O HJCMDXDYPOUFDY-WHFBIAKZSA-N 0.000 description 1
- 108091008875 B cell receptors Proteins 0.000 description 1
- 208000003950 B-cell lymphoma Diseases 0.000 description 1
- 102100038080 B-cell receptor CD22 Human genes 0.000 description 1
- WOVKYSAHUYNSMH-UHFFFAOYSA-N BROMODEOXYURIDINE Natural products C1C(O)C(CO)OC1N1C(=O)NC(=O)C(Br)=C1 WOVKYSAHUYNSMH-UHFFFAOYSA-N 0.000 description 1
- 241001518086 Bartonella henselae Species 0.000 description 1
- 241000589969 Borreliella burgdorferi Species 0.000 description 1
- 108050005711 C Chemokine Proteins 0.000 description 1
- 102000017483 C chemokine Human genes 0.000 description 1
- 102100024217 CAMPATH-1 antigen Human genes 0.000 description 1
- 102000001902 CC Chemokines Human genes 0.000 description 1
- 108010040471 CC Chemokines Proteins 0.000 description 1
- 102100027207 CD27 antigen Human genes 0.000 description 1
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 102100032937 CD40 ligand Human genes 0.000 description 1
- 108010065524 CD52 Antigen Proteins 0.000 description 1
- 102000004325 CX3C Chemokines Human genes 0.000 description 1
- 108010081635 CX3C Chemokines Proteins 0.000 description 1
- 108050006947 CXC Chemokine Proteins 0.000 description 1
- 102000019388 CXC chemokine Human genes 0.000 description 1
- 241000589875 Campylobacter jejuni Species 0.000 description 1
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 1
- 206010068051 Chimerism Diseases 0.000 description 1
- 241001647372 Chlamydia pneumoniae Species 0.000 description 1
- 108010077544 Chromatin Proteins 0.000 description 1
- 108091062157 Cis-regulatory element Proteins 0.000 description 1
- 241000193163 Clostridioides difficile Species 0.000 description 1
- 241000193155 Clostridium botulinum Species 0.000 description 1
- 102100032768 Complement receptor type 2 Human genes 0.000 description 1
- 241000711573 Coronaviridae Species 0.000 description 1
- 241000158508 Corynebacterium amycolatum Species 0.000 description 1
- 102100035298 Cytokine SCM-1 beta Human genes 0.000 description 1
- 241000701022 Cytomegalovirus Species 0.000 description 1
- 102100039498 Cytotoxic T-lymphocyte protein 4 Human genes 0.000 description 1
- 101710101803 DNA-binding protein J Proteins 0.000 description 1
- 241000725619 Dengue virus Species 0.000 description 1
- 108010043648 Discoidin Domain Receptors Proteins 0.000 description 1
- 102000002706 Discoidin Domain Receptors Human genes 0.000 description 1
- 201000011001 Ebola Hemorrhagic Fever Diseases 0.000 description 1
- 241000605310 Ehrlichia chaffeensis Species 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 206010014596 Encephalitis Japanese B Diseases 0.000 description 1
- 102100030011 Endoribonuclease Human genes 0.000 description 1
- 101710199605 Endoribonuclease Proteins 0.000 description 1
- 241000194032 Enterococcus faecalis Species 0.000 description 1
- 241000194031 Enterococcus faecium Species 0.000 description 1
- 102000018651 Epithelial Cell Adhesion Molecule Human genes 0.000 description 1
- 108010066687 Epithelial Cell Adhesion Molecule Proteins 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 241001646719 Escherichia coli O157:H7 Species 0.000 description 1
- 108010022894 Euchromatin Proteins 0.000 description 1
- 108010087819 Fc receptors Proteins 0.000 description 1
- 102000009109 Fc receptors Human genes 0.000 description 1
- 241000711950 Filoviridae Species 0.000 description 1
- 241000710831 Flavivirus Species 0.000 description 1
- 102100041003 Glutamate carboxypeptidase 2 Human genes 0.000 description 1
- 229920002527 Glycogen Polymers 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 102100030595 HLA class II histocompatibility antigen gamma chain Human genes 0.000 description 1
- 241000590002 Helicobacter pylori Species 0.000 description 1
- 208000005176 Hepatitis C Diseases 0.000 description 1
- 102000003745 Hepatocyte Growth Factor Human genes 0.000 description 1
- 108090000100 Hepatocyte Growth Factor Proteins 0.000 description 1
- 101000884305 Homo sapiens B-cell receptor CD22 Proteins 0.000 description 1
- 101000914511 Homo sapiens CD27 antigen Proteins 0.000 description 1
- 101000914324 Homo sapiens Carcinoembryonic antigen-related cell adhesion molecule 5 Proteins 0.000 description 1
- 101000914321 Homo sapiens Carcinoembryonic antigen-related cell adhesion molecule 7 Proteins 0.000 description 1
- 101000941929 Homo sapiens Complement receptor type 2 Proteins 0.000 description 1
- 101000804771 Homo sapiens Cytokine SCM-1 beta Proteins 0.000 description 1
- 101000889276 Homo sapiens Cytotoxic T-lymphocyte protein 4 Proteins 0.000 description 1
- 101000892862 Homo sapiens Glutamate carboxypeptidase 2 Proteins 0.000 description 1
- 101001082627 Homo sapiens HLA class II histocompatibility antigen gamma chain Proteins 0.000 description 1
- 101001046686 Homo sapiens Integrin alpha-M Proteins 0.000 description 1
- 101001046677 Homo sapiens Integrin alpha-V Proteins 0.000 description 1
- 101001015004 Homo sapiens Integrin beta-3 Proteins 0.000 description 1
- 101000599852 Homo sapiens Intercellular adhesion molecule 1 Proteins 0.000 description 1
- 101000599862 Homo sapiens Intercellular adhesion molecule 3 Proteins 0.000 description 1
- 101001055145 Homo sapiens Interleukin-2 receptor subunit beta Proteins 0.000 description 1
- 101000804764 Homo sapiens Lymphotactin Proteins 0.000 description 1
- 101000946889 Homo sapiens Monocyte differentiation antigen CD14 Proteins 0.000 description 1
- 101000934338 Homo sapiens Myeloid cell surface antigen CD33 Proteins 0.000 description 1
- 101000617725 Homo sapiens Pregnancy-specific beta-1-glycoprotein 2 Proteins 0.000 description 1
- 101000934341 Homo sapiens T-cell surface glycoprotein CD5 Proteins 0.000 description 1
- 101000914484 Homo sapiens T-lymphocyte activation antigen CD80 Proteins 0.000 description 1
- 101000611183 Homo sapiens Tumor necrosis factor Proteins 0.000 description 1
- 101000610604 Homo sapiens Tumor necrosis factor receptor superfamily member 10B Proteins 0.000 description 1
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 1
- 108090000144 Human Proteins Proteins 0.000 description 1
- 102000003839 Human Proteins Human genes 0.000 description 1
- 241000701085 Human alphaherpesvirus 3 Species 0.000 description 1
- 206010020429 Human ehrlichiosis Diseases 0.000 description 1
- 241000725303 Human immunodeficiency virus Species 0.000 description 1
- 108091058560 IL8 Proteins 0.000 description 1
- 241000712431 Influenza A virus Species 0.000 description 1
- 108010001127 Insulin Receptor Proteins 0.000 description 1
- 102000003746 Insulin Receptor Human genes 0.000 description 1
- 102100022338 Integrin alpha-M Human genes 0.000 description 1
- 102100022337 Integrin alpha-V Human genes 0.000 description 1
- 102100022297 Integrin alpha-X Human genes 0.000 description 1
- 102100032999 Integrin beta-3 Human genes 0.000 description 1
- 102100037877 Intercellular adhesion molecule 1 Human genes 0.000 description 1
- 102100037871 Intercellular adhesion molecule 3 Human genes 0.000 description 1
- 102000014150 Interferons Human genes 0.000 description 1
- 108010050904 Interferons Proteins 0.000 description 1
- 102000013462 Interleukin-12 Human genes 0.000 description 1
- 108010065805 Interleukin-12 Proteins 0.000 description 1
- 102000003816 Interleukin-13 Human genes 0.000 description 1
- 108090000176 Interleukin-13 Proteins 0.000 description 1
- 102000003812 Interleukin-15 Human genes 0.000 description 1
- 108090000172 Interleukin-15 Proteins 0.000 description 1
- 102000000588 Interleukin-2 Human genes 0.000 description 1
- 108010002350 Interleukin-2 Proteins 0.000 description 1
- 108010038453 Interleukin-2 Receptors Proteins 0.000 description 1
- 102000010789 Interleukin-2 Receptors Human genes 0.000 description 1
- 102100026879 Interleukin-2 receptor subunit beta Human genes 0.000 description 1
- 102100030704 Interleukin-21 Human genes 0.000 description 1
- 108010065637 Interleukin-23 Proteins 0.000 description 1
- 102000013264 Interleukin-23 Human genes 0.000 description 1
- 102000004388 Interleukin-4 Human genes 0.000 description 1
- 108090000978 Interleukin-4 Proteins 0.000 description 1
- 102000010787 Interleukin-4 Receptors Human genes 0.000 description 1
- 108010038486 Interleukin-4 Receptors Proteins 0.000 description 1
- 102000000743 Interleukin-5 Human genes 0.000 description 1
- 108010002616 Interleukin-5 Proteins 0.000 description 1
- 102000000704 Interleukin-7 Human genes 0.000 description 1
- 108010002586 Interleukin-7 Proteins 0.000 description 1
- 102000004890 Interleukin-8 Human genes 0.000 description 1
- 108090001007 Interleukin-8 Proteins 0.000 description 1
- 101150010986 J5 gene Proteins 0.000 description 1
- 201000005807 Japanese encephalitis Diseases 0.000 description 1
- 241000710843 Japanese encephalitis virus group Species 0.000 description 1
- 201000008225 Klebsiella pneumonia Diseases 0.000 description 1
- 241000588747 Klebsiella pneumoniae Species 0.000 description 1
- 108091008555 LTK receptors Proteins 0.000 description 1
- 102100025640 Lactase-like protein Human genes 0.000 description 1
- 241000589242 Legionella pneumophila Species 0.000 description 1
- 241000186779 Listeria monocytogenes Species 0.000 description 1
- 102100035304 Lymphotactin Human genes 0.000 description 1
- 241000721701 Lynx Species 0.000 description 1
- 241000701076 Macacine alphaherpesvirus 1 Species 0.000 description 1
- 201000005505 Measles Diseases 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 102100035877 Monocyte differentiation antigen CD14 Human genes 0.000 description 1
- 241000712045 Morbillivirus Species 0.000 description 1
- 108091008553 MuSK receptors Proteins 0.000 description 1
- 102100034256 Mucin-1 Human genes 0.000 description 1
- 108010008707 Mucin-1 Proteins 0.000 description 1
- 101100381525 Mus musculus Bcl6 gene Proteins 0.000 description 1
- 101000777315 Mus musculus Choline kinase alpha Proteins 0.000 description 1
- 101000777310 Mus musculus Choline/ethanolamine kinase Proteins 0.000 description 1
- 101000974705 Mus musculus UMP-CMP kinase Proteins 0.000 description 1
- 102100025243 Myeloid cell surface antigen CD33 Human genes 0.000 description 1
- 229930193140 Neomycin Natural products 0.000 description 1
- 102000005348 Neuraminidase Human genes 0.000 description 1
- 108010006232 Neuraminidase Proteins 0.000 description 1
- 108020004485 Nonsense Codon Proteins 0.000 description 1
- 102000008297 Nuclear Matrix-Associated Proteins Human genes 0.000 description 1
- 108010035916 Nuclear Matrix-Associated Proteins Proteins 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 108091008606 PDGF receptors Proteins 0.000 description 1
- 241001647379 Parachlamydia Species 0.000 description 1
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Chemical compound OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 1
- 102000011653 Platelet-Derived Growth Factor Receptors Human genes 0.000 description 1
- 206010035664 Pneumonia Diseases 0.000 description 1
- 206010035717 Pneumonia klebsiella Diseases 0.000 description 1
- 241000711902 Pneumovirus Species 0.000 description 1
- 102100022019 Pregnancy-specific beta-1-glycoprotein 2 Human genes 0.000 description 1
- 241000125945 Protoparvovirus Species 0.000 description 1
- 238000012181 QIAquick gel extraction kit Methods 0.000 description 1
- 102000014128 RANK Ligand Human genes 0.000 description 1
- 108010025832 RANK Ligand Proteins 0.000 description 1
- 108091008554 ROR receptors Proteins 0.000 description 1
- 108091008552 RYK receptors Proteins 0.000 description 1
- 206010037742 Rabies Diseases 0.000 description 1
- 241000700157 Rattus norvegicus Species 0.000 description 1
- 108010091086 Recombinases Proteins 0.000 description 1
- 101710197208 Regulatory protein cro Proteins 0.000 description 1
- 241000725643 Respiratory syncytial virus Species 0.000 description 1
- 235000011449 Rosa Nutrition 0.000 description 1
- 241001533467 Rubulavirus Species 0.000 description 1
- 241000315672 SARS coronavirus Species 0.000 description 1
- 241000293869 Salmonella enterica subsp. enterica serovar Typhimurium Species 0.000 description 1
- 101710113029 Serine/threonine-protein kinase Proteins 0.000 description 1
- 241000700584 Simplexvirus Species 0.000 description 1
- 206010041925 Staphylococcal infections Diseases 0.000 description 1
- 241000191967 Staphylococcus aureus Species 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- 241000193985 Streptococcus agalactiae Species 0.000 description 1
- 241000193996 Streptococcus pyogenes Species 0.000 description 1
- 241001505901 Streptococcus sp. 'group A' Species 0.000 description 1
- 241000193990 Streptococcus sp. 'group B' Species 0.000 description 1
- 101710172711 Structural protein Proteins 0.000 description 1
- 206010042971 T-cell lymphoma Diseases 0.000 description 1
- 208000027585 T-cell non-Hodgkin lymphoma Diseases 0.000 description 1
- 102100025244 T-cell surface glycoprotein CD5 Human genes 0.000 description 1
- 102100027222 T-lymphocyte activation antigen CD80 Human genes 0.000 description 1
- 108091005735 TGF-beta receptors Proteins 0.000 description 1
- 102000005450 TIE receptors Human genes 0.000 description 1
- 108010006830 TIE receptors Proteins 0.000 description 1
- 108700012920 TNF Proteins 0.000 description 1
- 108010055044 Tetanus Toxin Proteins 0.000 description 1
- 208000004006 Tick-borne encephalitis Diseases 0.000 description 1
- 241000710771 Tick-borne encephalitis virus Species 0.000 description 1
- 102000006747 Transforming Growth Factor alpha Human genes 0.000 description 1
- 102000004887 Transforming Growth Factor beta Human genes 0.000 description 1
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 1
- 102000016715 Transforming Growth Factor beta Receptors Human genes 0.000 description 1
- 101800004564 Transforming growth factor alpha Proteins 0.000 description 1
- 108060008683 Tumor Necrosis Factor Receptor Proteins 0.000 description 1
- 102100040247 Tumor necrosis factor Human genes 0.000 description 1
- 102100040112 Tumor necrosis factor receptor superfamily member 10B Human genes 0.000 description 1
- 206010054094 Tumour necrosis Diseases 0.000 description 1
- 102100037236 Tyrosine-protein kinase receptor UFO Human genes 0.000 description 1
- 241000700618 Vaccinia virus Species 0.000 description 1
- 241000700647 Variola virus Species 0.000 description 1
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 1
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 1
- 241000607626 Vibrio cholerae Species 0.000 description 1
- 241000607265 Vibrio vulnificus Species 0.000 description 1
- 241000710886 West Nile virus Species 0.000 description 1
- 241000120645 Yellow fever virus group Species 0.000 description 1
- 229960004150 aciclovir Drugs 0.000 description 1
- MKUXAQIIEYXACX-UHFFFAOYSA-N aciclovir Chemical compound N1C(N)=NC(=O)C2=C1N(COCCO)C=N2 MKUXAQIIEYXACX-UHFFFAOYSA-N 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 239000008186 active pharmaceutical agent Substances 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 230000007059 acute toxicity Effects 0.000 description 1
- 231100000403 acute toxicity Toxicity 0.000 description 1
- 239000002671 adjuvant Substances 0.000 description 1
- 239000011543 agarose gel Substances 0.000 description 1
- 230000002776 aggregation Effects 0.000 description 1
- 238000004220 aggregation Methods 0.000 description 1
- 238000004873 anchoring Methods 0.000 description 1
- 238000000137 annealing Methods 0.000 description 1
- 230000000259 anti-tumor effect Effects 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 238000013459 approach Methods 0.000 description 1
- 229940092524 bartonella henselae Drugs 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 230000005540 biological transmission Effects 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 210000002798 bone marrow cell Anatomy 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 235000011089 carbon dioxide Nutrition 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 230000011748 cell maturation Effects 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 238000001516 cell proliferation assay Methods 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 230000008709 cellular rearrangement Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 210000003483 chromatin Anatomy 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 230000006020 chronic inflammation Effects 0.000 description 1
- 208000037893 chronic inflammatory disorder Diseases 0.000 description 1
- 230000004186 co-expression Effects 0.000 description 1
- 238000009500 colour coating Methods 0.000 description 1
- 238000012875 competitive assay Methods 0.000 description 1
- 230000002860 competitive effect Effects 0.000 description 1
- 230000009918 complex formation Effects 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 239000013256 coordination polymer Substances 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 239000007857 degradation product Substances 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 1
- 238000002224 dissection Methods 0.000 description 1
- 238000010494 dissociation reaction Methods 0.000 description 1
- 230000005593 dissociations Effects 0.000 description 1
- 230000019975 dosage compensation by inactivation of X chromosome Effects 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 229940126534 drug product Drugs 0.000 description 1
- 229940088679 drug related substance Drugs 0.000 description 1
- 238000001962 electrophoresis Methods 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- ZMMJGEGLRURXTF-UHFFFAOYSA-N ethidium bromide Chemical compound [Br-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CC)=C1C1=CC=CC=C1 ZMMJGEGLRURXTF-UHFFFAOYSA-N 0.000 description 1
- 229960005542 ethidium bromide Drugs 0.000 description 1
- 210000000632 euchromatin Anatomy 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 238000010195 expression analysis Methods 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 239000000945 filler Substances 0.000 description 1
- 239000012467 final product Substances 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 230000037433 frameshift Effects 0.000 description 1
- 230000005714 functional activity Effects 0.000 description 1
- 238000010230 functional analysis Methods 0.000 description 1
- 229960002963 ganciclovir Drugs 0.000 description 1
- IRSCQMHQWWYFCW-UHFFFAOYSA-N ganciclovir Chemical compound O=C1NC(N)=NC2=C1N=CN2COC(CO)CO IRSCQMHQWWYFCW-UHFFFAOYSA-N 0.000 description 1
- 238000012239 gene modification Methods 0.000 description 1
- 238000012226 gene silencing method Methods 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 230000005017 genetic modification Effects 0.000 description 1
- 235000013617 genetically modified food Nutrition 0.000 description 1
- 229940096919 glycogen Drugs 0.000 description 1
- 229940037467 helicobacter pylori Drugs 0.000 description 1
- 108010036302 hemoglobin AS Proteins 0.000 description 1
- 208000005252 hepatitis A Diseases 0.000 description 1
- 208000002672 hepatitis B Diseases 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 230000004727 humoral immunity Effects 0.000 description 1
- 230000008348 humoral response Effects 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 208000037797 influenza A Diseases 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- 229940079322 interferon Drugs 0.000 description 1
- 108010074108 interleukin-21 Proteins 0.000 description 1
- 239000013067 intermediate product Substances 0.000 description 1
- 229940065638 intron a Drugs 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 238000011813 knockout mouse model Methods 0.000 description 1
- 229940115932 legionella pneumophila Drugs 0.000 description 1
- 230000003902 lesion Effects 0.000 description 1
- TYZROVQLWOKYKF-ZDUSSCGKSA-N linezolid Chemical compound O=C1O[C@@H](CNC(=O)C)CN1C(C=C1F)=CC=C1N1CCOCC1 TYZROVQLWOKYKF-ZDUSSCGKSA-N 0.000 description 1
- 229960003907 linezolid Drugs 0.000 description 1
- 210000001165 lymph node Anatomy 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 210000001161 mammalian embryo Anatomy 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 238000013411 master cell bank Methods 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 210000004379 membrane Anatomy 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 208000015688 methicillin-resistant staphylococcus aureus infectious disease Diseases 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 239000002480 mineral oil Substances 0.000 description 1
- 235000010446 mineral oil Nutrition 0.000 description 1
- 238000007479 molecular analysis Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 210000000822 natural killer cell Anatomy 0.000 description 1
- 230000010807 negative regulation of binding Effects 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 230000036963 noncompetitive effect Effects 0.000 description 1
- 238000013421 nuclear magnetic resonance imaging Methods 0.000 description 1
- 210000000299 nuclear matrix Anatomy 0.000 description 1
- 238000002515 oligonucleotide synthesis Methods 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 210000003101 oviduct Anatomy 0.000 description 1
- 230000003071 parasitic effect Effects 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 239000008188 pellet Substances 0.000 description 1
- 210000004303 peritoneum Anatomy 0.000 description 1
- 210000001986 peyer's patch Anatomy 0.000 description 1
- 239000008194 pharmaceutical composition Substances 0.000 description 1
- 229920001481 poly(stearyl methacrylate) Polymers 0.000 description 1
- 108091033319 polynucleotide Proteins 0.000 description 1
- 102000040430 polynucleotide Human genes 0.000 description 1
- 239000002157 polynucleotide Substances 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 238000012910 preclinical development Methods 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 238000004321 preservation Methods 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 239000012264 purified product Substances 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000007115 recruitment Effects 0.000 description 1
- 230000003252 repetitive effect Effects 0.000 description 1
- 206010039073 rheumatoid arthritis Diseases 0.000 description 1
- 201000005404 rubella Diseases 0.000 description 1
- 238000007790 scraping Methods 0.000 description 1
- 230000003248 secreting effect Effects 0.000 description 1
- 238000004062 sedimentation Methods 0.000 description 1
- 230000011664 signaling Effects 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 238000009987 spinning Methods 0.000 description 1
- 210000004989 spleen cell Anatomy 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 239000008223 sterile water Substances 0.000 description 1
- 230000000638 stimulation Effects 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 239000000126 substance Substances 0.000 description 1
- 230000001360 synchronised effect Effects 0.000 description 1
- 230000002123 temporal effect Effects 0.000 description 1
- 229940118376 tetanus toxin Drugs 0.000 description 1
- 150000003522 tetracyclines Chemical class 0.000 description 1
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 description 1
- 238000011287 therapeutic dose Methods 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 238000000844 transformation Methods 0.000 description 1
- 238000012250 transgenic expression Methods 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 102000003298 tumor necrosis factor receptor Human genes 0.000 description 1
- 108091052247 type I cytokine receptor family Proteins 0.000 description 1
- 102000042286 type I cytokine receptor family Human genes 0.000 description 1
- 102000042287 type II cytokine receptor family Human genes 0.000 description 1
- 108091052254 type II cytokine receptor family Proteins 0.000 description 1
- 241001529453 unidentified herpesvirus Species 0.000 description 1
- 210000000689 upper leg Anatomy 0.000 description 1
- 210000004291 uterus Anatomy 0.000 description 1
- 210000003462 vein Anatomy 0.000 description 1
- 108700026220 vif Genes Proteins 0.000 description 1
Classifications
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
- A01K67/0275—Genetically modified vertebrates, e.g. transgenic
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
- A01K67/0275—Genetically modified vertebrates, e.g. transgenic
- A01K67/0276—Knock-out vertebrates
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/8509—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells for producing genetically modified animals, e.g. transgenic
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/05—Animals comprising random inserted nucleic acids (transgenic)
- A01K2217/052—Animals comprising random inserted nucleic acids (transgenic) inducing gain of function
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/07—Animals genetically altered by homologous recombination
- A01K2217/072—Animals genetically altered by homologous recombination maintaining or altering function, i.e. knock in
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/15—Animals comprising multiple alterations of the genome, by transgenesis or homologous recombination, e.g. obtained by cross-breeding
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2227/00—Animals characterised by species
- A01K2227/10—Mammal
- A01K2227/105—Murine
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2267/00—Animals characterised by purpose
- A01K2267/01—Animal expressing industrially exogenous proteins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/24—Immunoglobulins specific features characterized by taxonomic origin containing regions, domains or residues from different species, e.g. chimeric, humanized or veneered
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2800/00—Nucleic acids vectors
- C12N2800/30—Vector systems comprising sequences for excision in presence of a recombinase, e.g. loxP or FRT
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Zoology (AREA)
- Biotechnology (AREA)
- Organic Chemistry (AREA)
- Environmental Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Veterinary Medicine (AREA)
- General Engineering & Computer Science (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Biodiversity & Conservation Biology (AREA)
- Animal Husbandry (AREA)
- Biochemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Microbiology (AREA)
- Immunology (AREA)
- Medicinal Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
Abstract
Un hospedador murino transgénico capaz de producir inmunoglobulinas que comprenden una región VL humana, comprendiendo dicho hospedador murino integrado en su línea germinal un ácido nucleico reordenado que comprende un segmento génico V de línea germinal humano y uno J de línea germinal humano que codifica una región VL humana reordenada y una región constante de cadena ligera de cualquier especie animal, en donde dicha región variable de cadena ligera es capaz de emparejarse con al menos dos cadenas pesadas diferentes codificadas por el hospedador, de modo que se conserva diversidad de especificidad de anticuerpos que comprenden dicha región VL humana mediante reordenamientos e hipermutaciones en las cadenas pesadas.
Description
DESCRIPCIÓN
Animal murino transgénico productor de anticuerpos
Campo técnico
La invención se refiere a la producción y al uso de hospedadores murinos transgénicos capaces de producir anticuerpos, o derivados de los mismos, que se expresan a partir de ácidos nucleicos al menos parcialmente exógenos (transgenes). Se describen transgenes para producir dichos hospedadores murinos transgénicos y procedimientos para producir dichos anticuerpos heterólogos; procedimientos y vectores para producir dichos hospedadores murinos transgénicos.
Antecedentes de la invención
Los linfocitos B median en la inmunidad humoral produciendo anticuerpos específicos. La subunidad estructural básica de un anticuerpo (Ab) es una molécula de inmunoglobulina (Ig). Las moléculas de Ig constan de un complejo de dos cadenas polipeptídicas ligeras (L) idénticas y dos pesadas (H) idénticas. En el extremo amino de cada cadena H y cadena L hay una región que varía en la secuencia de aminoácidos denominada región variable (V). La parte restante de las cadenas H y L es relativamente constante en la secuencia de aminoácidos y se denomina región constante (C). En una molécula de Ig, las regiones V de cadena H y L (VH y VL) se yuxtaponen para formar el posible sitio de unión a antígeno. Los genes que codifican las regiones V de cadena H y L se ensamblan somáticamente a partir de segmentos de ADN de la línea germinal durante la diferenciación de linfocitos B precursores (pre-B): segmentos génicos V, D y J para la cadena H y segmentos génicos V y J para la cadena L. Dentro de las regiones V de Ig existen tres regiones de mayor variabilidad de secuencia de aminoácidos que interaccionan para formar el sitio de reconocimiento del antígeno y por tanto se denominan regiones determinantes de complementariedad (CDR).
El segmento génico V codifica la mayor parte del dominio de la región V, incluyendo la CDR1 y CDR2. La diversidad en la CDR1 y CDR2 deriva de la heterogeneidad de secuencias entre múltiples segmentos V codificados por diferentes líneas germinales. La CDR3 se codifica por secuencias que están formadas por la unión de segmentos génicos V, D y J de la cadena H y segmentos V y J de la cadena L y por mecanismos que crean heterogeneidad de secuencia de nucleótidos donde se combinan estos segmentos. Diversidad adicional puede derivar del emparejamiento de regiones V de cadena H y L diferentes. En su conjunto estos procesos producen un repertorio primario de anticuerpos codificados por segmentos génicos de la línea germinal y expresados por linfocitos B recién formados.
Una fuente adicional de diversidad de anticuerpos está impuesta por encima de la diversidad generada por recombinación de segmentos génicos de Ig. Los linfocitos B pueden introducir mutaciones en las regiones V del anticuerpo que expresan, un proceso denominado hipermutación somática. Por tanto, cuando un animal se encuentra por primera vez con un antígeno, el antígeno se une a un linfocito B específico que sucede que lleva anticuerpos que tienen un dominio V que se une al antígeno. Esta respuesta primaria puede activar este linfocito B para que continúe segregando el anticuerpo afín. Estos linfocitos B activados también pueden dirigir ahora un proceso de mutación somática en sus segmentos génicos de anticuerpo reordenados y por tanto permitir la producción de células hijas que producen variantes de los anticuerpos de la respuesta primaria. Un proceso de selección amplifica aquellos descendientes de linfocitos T variantes que constituyen un anticuerpo de afinidad mejorada por el antígeno. En linfocitos B, las hipermutaciones somáticas se dirigen a una región genómica limitada incluyendo los dos genes VH y VL reordenados. Por tanto, la mutación somática permite la maduración por afinidad, la producción y la selección de anticuerpos de alta afinidad. Por lo tanto, la mutación somática es importante para la generación de anticuerpos de alta afinidad.
La especificidad exquisita y la alta afinidad de los anticuerpos y el descubrimiento de la tecnología de hibridoma que permite la generación de anticuerpos monoclonales (mAb) ha generado grandes expectativas para su utilización como agentes terapéuticos diana para enfermedades humanas. Los mAb son idénticos porque están producidos por un solo linfocito B y su descendencia. Los mAb se preparan fusionando los esplenocitos de un ratón que se ha inmunizado con el antígeno deseado con células de mieloma para generar hibridomas inmortalizados. Uno de los mayores impedimentos a los que se enfrenta el desarrollo de aplicaciones in vivo para los mAb en seres humanos es la inmunogenicidad intrínseca de las Ig no humanas. Los pacientes que responden a dosis terapéuticas de mAb de ratón fabricando anticuerpos contra las secuencias de Ig de ratón (Anticuerpos Humanos Anti-ratón; HAMA), que ocasionan toxicidad aguda, alteran su biodistribución y aceleran la eliminación, reduciendo de este modo la eficacia de administraciones posteriores (Mirick, et al., (2004) Q. Nucl. Med. Mol. Imaging 48, 251-257).
Para evitar la generación de HAMA, se han desarrollado procedimientos de humanización de anticuerpos en un intento para producir mAb con menor inmunogenicidad cuando se aplican a seres humanos. Estos intentos han producido diversas estrategias basadas en ADN recombinante orientadas a aumentar el contenido de secuencias de aminoácidos humanas en los mAb conservando al mismo tiempo la especificidad y afinidad del anticuerpo no humano parental. La humanización comenzó con la construcción de mAb quiméricos ratón-ser humano (Morrison, S.
L., et al., (1984). Proc. Natl. Acad. Sci. U. S. A.., 81, 6851-5), en los que las regiones C de la Ig en los mAb murinos se reemplazaron por regiones C humanas. Los mAb quiméricos contienen el 60-70 % de las secuencias de aminoácidos de ser humano y son considerablemente menos inmunogénicos que sus homólogos murinos cuando se inyectan en seres humanos, a pesar de observarse aún una respuesta de anticuerpo antiquimérico humano (Hwang, W. Y., et al. (2005). Methods, 36, 3-10).
En intentos para humanizar adicionalmente mAb murinos, se desarrolló el injerto de CDR. En el injerto de CDR, los anticuerpos murinos se humanizan injertando sus CDR sobre los armazones VL y VH de moléculas de IgG humana, conservando al mismo tiempo los restos marco conservados murinos considerados como esenciales para la especificidad y afinidad (Jones, P.T., et al., (1986). Nature, 321, 522). En conjunto, los anticuerpos con CDR injertadas constan de más del 80 % de secuencias de aminoácidos humanas (Queen, C. et al. (1989) Proc. Natl. Acad. Sci. U. S. A. 86, 10029; Carter, P. et al. (1992) Proc. Natl. Acad. Sci. U. S. A. 89, 4285). A pesar de estos esfuerzos, se observó que los anticuerpos humanizados con CDR injertadas aún suscitaban una respuesta de anticuerpo contra la región V injertada (Hwang, W. Y., et al. (2005). Methods, 36, 3).
Posterior al injerto de CDR, se han desarrollado procedimientos de humanización basados en diferentes paradigmas, tales como el rechapado (Padlan, E. A., et al., (1991). Mol. Immunol., 28, 489), superhumanización (Tan, P., D. A., et al., (2002) J. Immunol., 169, 1119), optimización del contenido de cadenas humanas (Lazar, G. A., et al., (2007). Mol. Immunol., 44, 1986) y humanización mediante modificación por ingeniería genética (humaneering) en un intento para disminuir adicionalmente el contenido de secuencias no humanas en mAb terapéuticos (Almagro, J. C., et al., (2008). Frontiers in Bioscience 13, 1619). Al igual que en las estrategias de injerto de CDR, estos procedimientos se basan en análisis de la estructura de anticuerpos y comparación de secuencias de los mAb no humanos y humanos para evaluar el impacto del proceso de humanización en la inmunogenicidad del producto final. Cuando se compara la inmunogenicidad de anticuerpos quiméricos y humanizados, la humanización de regiones variables parece disminuir adicionalmente la inmunogenicidad (Hwang, W. Y., et al. (2005). Methods, 36, 3-10).
La desinmunización es otra estrategia desarrollada para reducir la inmunogenicidad de anticuerpos quiméricos o de ratón. Esto implica la identificación de epítopos de linfocitos T lineales en el anticuerpo de interés, usando bioinformática, y su reemplazo posterior por mutagénesis dirigida a sitio por secuencias no inmunogénicas o humanas (documento WO09852976A1). Aunque los anticuerpos desinmunizados presentan inmunogenicidad reducida en primates, en comparación con sus homólogos quiméricos, se observó alguna pérdida de afinidad de unión (Jain, M., et al., (2007). Trends in Biotechnol. 25, 307).
El desarrollo de la tecnología de presentación de fagos complementó y amplió las estrategias de humanización en intentos de obtener mAb menos inmunogénicos para terapia en seres humanos. En la presentación de fagos, se expresan grandes colecciones ('bibliotecas') de regiones Vh y VL de anticuerpos humanos sobre la superficie de partículas de bacteriófagos filamentosos. A partir de estas bibliotecas, se seleccionan fagos excepcionales mediante la interacción de unión con antígeno; los fragmentos de anticuerpos solubles se expresan de bacterias infectadas y la afinidad de unión de los anticuerpos seleccionados se mejora por mutación (Winter, G., et al. (1994). Annu. Rev. Immunol. 12, 433). Este proceso imita la selección inmunitaria y usando esta estrategia se han aislado anticuerpos con muchas especificidades de unión diferentes (Hoogenboom, H. R., et al. (2005). Nat. Biotechnol., 23, 1105). Se han usado diversas fuentes de regiones V de cadena H y L para construir bibliotecas de presentación de fagos incluyendo los aislados de donantes no inmunes o inmunes. Además, se han construido bibliotecas de presentación de fagos de regiones V que contienen regiones CDR sintéticas artificialmente aleatorizadas para crear diversidad adicional. A menudo, los anticuerpos obtenidos a partir de bibliotecas de presentación de fagos se someten a maduración por afinidad in vitro para obtener anticuerpos de alta afinidad (Hoogenboom, H. R., et al. (2005). Nat. Biotechnol., 23, 1105).
La creación de cepas de ratón transgénico que producen anticuerpos humanos en ausencia de anticuerpos de ratón ha proporcionado otra plataforma de tecnología para la generación de mAb humanos específicos y de alta afinidad para aplicación en seres humanos. En estos animales transgénicos, la maquinaria de anticuerpo de ratón endógena se inactiva y se reemplaza por locus de Ig humana para reproducir sustancialmente el sistema inmunitario humoral humano en ratones (Jakobovits, A., et al. (2007). Nat. Biotechnol. 25, 1134. Lonberg, N. (2005). Nat. Biotechnol. 23, 1117). El desarrollo de linfocitos B, así como la diversificación de Ig por recombinación de segmentos génicos, se reproduce satisfactoriamente en estos ratones, lo que conduce a un repertorio diverso de linfocitos B murinos que expresan Ig humanas. Inmunizando a estos ratones con antígenos, se demostró adicionalmente que estos animales transgénicos acumulaban mutaciones somáticas en las regiones V de las cadenas tanto pesadas como ligeras para producir una amplia diversidad de mAb humanos de alta afinidad (Lonberg, N. (2005). Nat. Biotechnol. 23, 1117).
La cuestión de si los mAb “completamente humanos”, tales como los derivados de bibliotecas de presentación de fagos o de ratones transgénicos, son menos inmunogénicos que los mAb humanizados continúa sin poder responderse, porque los datos de inmunogenicidad completa solo están disponibles para dos mAb humanos. Un mAb anti-factor de necrosis tumoral, desarrollado a partir de bibliotecas humanas de presentación de fagos, indujo respuestas de anticuerpos en el 12 % de los pacientes- al extremo de la frecuencia más alta de respuestas antianticuerpo de los anticuerpos humanizados (Hwang, W. Y., et al. (2005). Methods, 36, 3-10).
La evaluación de la inmunogenicidad del primer mAb humano registrado generado mediante estrategia transgénica demostró que el tratamiento con mAb daba como resultado la generación de anticuerpos en aproximadamente el 5,5 % de pacientes con cáncer tratados (Jakobovits, A., et al. (2007). Nat. Biotechnol. 25, 1134., Lofgren, J. A., et al. (2007). J. Immunol. 178, 7467).El documento EP0814159 desvela un experimento teórico para la producción de un ratón transgénico para la producción de anticuerpos, comprendiendo dicho ratón un dominio V humano reordenado unido a un dominio constante de cadena ligera humana. No se han producido ratones transgénicos.
Por lo tanto, continúa existiendo la necesidad de un procedimiento y un medio para producir anticuerpos que sean específicos para sus dianas, pero que sean menos inmunogénicos. Como se describe en el presente documento, la reducción de la inmunogenicidad se consigue, al menos parcialmente, proporcionando un hospedador murino transgénico que comprenda, al menos en su linaje de linfocitos B, un ácido nucleico que codifique al menos una cadena ligera de inmunoglobulina, en el que la secuencia que codifica la cadena ligera está provista de un medio que la haga resistente a reordenaciones de ADN y/o hipermutaciones somáticas, preferentemente dicho hospedador murino es un ratón. El ácido nucleico codifica una región VL humana reordenada de una inmunoglobulina humana. Como se reivindica, la presente invención se refiere a un hospedador murino transgénico capaz de producir inmunoglobulinas que comprenden una región VL humana, comprendiendo dicho hospedador murino integrado en su línea germinal un ácido nucleico reordenado que comprende un segmento génico V de línea germinal humana y un J de línea germinal humana que codifica una región Vl humana reordenada y una región constante de cadena ligera de cualquier especie animal, en donde dicha región variable de cadena ligera es capaz de emparejarse con al menos dos cadenas pesadas diferentes codificadas por el hospedador, de modo que se conserva la diversidad de especificidad de anticuerpos que comprenden dicha región VL mediante reordenamientos e hipermutaciones en las cadenas pesadas.
Como se reivindica, la presente invención se refiere además a un procedimiento para producir un anticuerpo deseado que comprende exponer un hospedador según la invención a un antígeno de modo que se induce una respuesta de anticuerpos y aislar los anticuerpos específicos para el antígeno.
Como se reivindica, la presente invención se refiere además a un procedimiento para producir un anticuerpo deseado que comprende exponer un hospedador según la invención a un antígeno de modo que se induce una respuesta de anticuerpos y aislar las células que producen dichos anticuerpos, cultivar y recoger dichos anticuerpos. Como se reivindica, la presente invención se refiere además a un procedimiento para producir un anticuerpo deseado que comprende exponer un hospedador según la invención a un antígeno de modo que se induce una respuesta de anticuerpos y aislar un ácido nucleico que codifica al menos parte de dicho anticuerpo, insertar dicho ácido nucleico o una copia o un derivado del mismo en un casete de expresión y expresar dicho anticuerpo en una célula hospedadora.
En el resto de la memoria descriptiva, los ratones se usan normalmente como ejemplos de los hospedadores murinos transgénicos. Los hospedadores murinos transgénicos pueden generar una respuesta inmunitaria contra un antígeno, donde la respuesta produce anticuerpos que tienen regiones variables de cadena ligera humana. Se han usado ratones para la producción de linfocitos B para inmortalización para la producción de anticuerpos. Dado que los ratones son fáciles de manipular, pueden reproducirse en grandes cantidades, y se sabe que tienen un amplio repertorio inmunitario, los ratones serán habitualmente el animal de elección. Por lo tanto, la siguiente explicación se referirá a ratones, aunque se describe que debe entenderse que los ratones se pueden sustituir fácilmente por otros animales, particularmente mamíferos no primates, siguiendo los mismos procedimientos.
La razón de impedir reordenaciones e hipermutación es que de esta manera puede seleccionarse de antemano un polipéptido no inmunogénico sabiendo que esta cadena polipeptídica permanecerá no inmunogénica. Al menos la cadena ligera de la inmunoglobulina resultante es por tanto menos inmunogénica. El anticuerpo resultante necesita tener (normalmente) una cadena tanto ligera como pesada. La cadena no inmunogénica debe por tanto ser capaz de emparejarse con la otra cadena. La otra cadena puede ser una cadena endógena, una cadena exógena o un híbrido de ambas. Para terapia en seres humanos, la cadena no inmunogénica debe ser tan cercana a la humana como sea posible.
Naturalmente, un medio para hacer que un gen que codifica una cadena (o cadenas) de inmunoglobulina sea resistente a reordenamiento y/o mutación de ADN es la eliminación de todos los elementos genéticos responsables de dicho reordenamiento y/o mutación. Su inconveniente es que se elimina la variabilidad de las dos cadenas, mientras que la invención conserva preferentemente la variabilidad en la cadena pesada e inhibe y/o impide el reordenamiento/mutación de la cadena ligera.
Los elementos para el reordenamiento y/o hipermutación caracterizados hasta ahora se localizan en los locus de las inmunoglobulinas. Por lo tanto, los medios para hacer que la secuencia que codifica la inmunoglobulina sea resistente a reordenamiento y/o mutación de ADN es insertar el gen en un locus fuera de los locus de la inmunoglobulina.
Por tanto, de acuerdo con la invención, se proporciona un hospedador murino transgénico en donde la secuencia que codifica la cadena ligera se integra en el genoma del hospedador murino en un locus fuera de los locus de inmunoglobulina. En un aspecto de la invención, la inserción se realiza en un locus que es resistente al silenciamiento génico. De acuerdo con un aspecto de la invención, la integración se realiza en el locus Rosa.
Se prefiere proporcionar un casete de expresión que pueda insertarse en un locus Rosa o locus similar, con un medio que permita la expresión de la cadena (o cadenas) de inmunoglobulina esencialmente limitada a células de linaje de linfocitos B, preferentemente con un medio que permita la expresión del ácido nucleico que codifica la cadena ligera durante una determinada fase del desarrollo de linfocitos B. La expresión “expresión esencialmente limitada” indica que la expresión se realiza predominantemente en células del linaje de linfocitos B, pero que son posibles niveles más bajos de expresión en otras células, en comparación con el nivel de expresión en linfocitos B. En una realización preferida, la expresión “expresión esencialmente limitada” indica que la expresión está exclusivamente presente en células del linaje de linfocitos B. Dichos medios incluyen típica y preferentemente promotores específicos de linfocitos B (fase de desarrollo), tales como CD19, CD20, pHC (todos genes V), VpreBI, VpreB2, VpreB3, X5, Iga, Igp, kLC (todos genes), ALC (todos genes), BSAP (Pax5). Aunque es muy posible que dichos promotores dirijan la expresión de la cadena resistente a reordenamiento y/o mutación del a Dn , estos son relativamente débiles. Típicamente se requerirá un promotor fuerte para garantizar la expresión adecuada en la superficie del receptor de linfocitos B (constituido por la cadena H y L de la Ig unida a la membrana) y competir con la expresión y emparejamiento de cadenas endógenas (si están presentes) a través de exclusión alélica. Sin embargo, dicho promotor es normalmente no específico de tejido. Para conferir especificidad tisular, se prefiere un sistema indirecto que emplee Cre/lox o similar. La cadena deseada se pone bajo el control de un promotor fuerte inhibido por un elemento que puede eliminarse por la acción de una proteína Cre, que conduce a la activación del gen deseado que codifica la inmunoglobulina. Este sistema se describe con detalle en Wunderlich F. T. (2004), “Generation of inducible Cre systems for conditional gene inactivation in mice”, Inauguraldissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultat der Universitat zu Koln; http://deposit.ddb.de/cgi-bin/dokserv?idn= 97557230x&dok_var=d1&dok_ext=pdf&filename=97557230x.pdf.
La cadena de inmunoglobulina es una cadena ligera capaz de emparejarse cadenas pesadas diferentes codificadas por el hospedador murino. La cadena ligera será por tanto la misma (y menos inmunogénica) en todos los anticuerpos, pero se conserva la variedad en cuanto a la especificidad a través de reordenamientos e hipermutaciones en las cadenas pesadas. En este caso puede ser preferible silenciar al menos uno de los locus endógenos que codifica una cadena ligera, aunque la exclusión alélica puede hacer que esto sea innecesario.
De acuerdo con la presente realización, preferentemente el locus de cadena ligera kappa (k) endógena está silenciado funcionalmente.
Si el locus de cadena ligera k endógena está silenciado, aunque también por otras razones, se prefiere que la cadena ligera resistente sea una cadena ligera k, que tenga una secuencia de la línea germinal. De acuerdo con la invención dicha cadena ligera conduciría a un anticuerpo con inmunogenicidad reducida. La secuencia de la línea germinal preferida se basa en la IGKV1-39 (012) humana ya que esta cadena ligera se observa muy frecuentemente en el repertorio humano (de Wildt et al. 1999. J. Mol. Biol. 285(3): 895) y tiene mejor estabilidad termodinámica, rendimiento y solubilidad (Ewert et al. 2003. J. Mol. Biol. 325(3): 53l).
A continuación se presentan realizaciones más específicas del casete de expresión con el que se puede dotar al hospedador murino de acuerdo con la invención. Normalmente, esto es ventajoso para las inmunoglobulinas.
Por tanto, la invención proporciona, en una realización específica, un hospedador murino transgénico en el que el ácido nucleico que codifica la cadena ligera comprende, en dirección 5'-3': un promotor específico de linfocitos B, un líder, un gen V humano reordenado, opcionalmente un potenciador MoEkí, una región constante (k) y opcionalmente un potenciador MoEk3' (truncado). Neuberger identificó y examinó un nuevo potenciador específico de linfocitos B localizado cadena abajo de la región constante kappa (documento EP004690251). Se ha observado que este potenciador desempeña una función crucial en la expresión de genes kappa ya que la eliminación del potenciador de 808 pb redujo fuertemente la expresión. La deleción del potenciador kappa 3' también redujo fuertemente el nivel de hipermutaciones somáticas (s Hm ). En estudios transgénicos y de expresión celular, se ha revelado que los potenciadores kappa 3' reducidos, mutados o delecionados no solo disminuyeron los niveles de expresión sino que también redujeron el nivel de hipermutaciones somáticas. Actualmente, no puede determinarse si el potenciador kappa 3' está implicado en procesos de SHM, regulación de expresión o ambos (revisado en Odegard, V. H., et al. (2006). Nat. Rev. Immunol. 6, 573; Inlay, M., et al. (2002). Nat. Immunol. 3, 463.).
Estudios de expresión detallados utilizando variantes modificadas por ingeniería genética del potenciador kappa 3' indicaron que bastaba una región de 50 nucleótidos para dirigir la expresión. Sin embargo para una expresión adecuada se prefiere una secuencia reducida de 145 nucleótidos (documento EP04690251; Meyer, K. B., et al. (1990) Nucleic Acids Res. 18(19): 5609-15).
Por tanto, en un aspecto, la invención proporciona, en el contexto del hospedador murino transgénico y en el contexto del procedimiento reivindicado, el uso de un ácido nucleico para su inserción en el genoma de un
hospedador murino, que es un casete de expresión para la expresión de una molécula proteica deseada en células que se desarrollan en linfocitos B maduros durante una determinada fase de desarrollo, comprendiendo dicho casete medios para impedir el silenciamiento de la expresión de la molécula proteica deseada después de su introducción en una célula hospedadora y medios para sincronizar la expresión de la molécula proteica deseada con la fase de desarrollo deseada de la célula hospedadora.
Un casete de expresión se define como un ácido nucleico que está provisto de medios para la introducción en el genoma de una célula hospedadora, tales como secuencias que permiten la recombinación homóloga con un sitio determinado en el genoma. Normalmente el ácido nucleico será a Dn , típicamente bicatenario. Típicamente el casete de expresión se proporcionará a la célula en un vector a partir del cual este se transfiere al genoma de la célula. El casete de expresión comprende adicionalmente todos los elementos necesarios para la expresión del gen en una célula hospedadora, aunque en determinadas realizaciones algunos de dichos elementos pueden estar presentes en un segundo ácido nucleico a introducir, mediante lo cual estos elementos actúan en trans. Los elementos necesarios para la expresión en una célula hospedadora incluyen promotores, potenciadores y otros elementos reguladores. Solo son necesarios los elementos no proporcionados por la célula hospedadora.
De acuerdo con un aspecto de la invención es importante que la expresión del gen de interés no esté silenciada en el genoma de la célula hospedadora, especialmente no en la fase del desarrollo en la que se requiere la expresión. Esto puede realizarse por diversos medios, tales como inserción en el locus endógeno o proporcionando al casete elementos de ácido nucleico que impidan el silenciamiento (Kwaks et al. (2006) Trends Biotechnol. 24(3), págs.137-142). Se prefiere que el casete de expresión se inserte en un locus que no esté silenciado en las células hospedadoras (documento EP 01439234).
Dichos medios para impedir el silenciamiento comprenden secuencias Estabilizantes Anti-Represoras (secuencias STAR®) y Regiones de Unión a la Matriz (MAR). Una secuencia STAR es una secuencia de ácido nucleico que comprende una capacidad para ejercer influencia en la transcripción de genes en cis. Típicamente, aunque no necesariamente, una secuencia STAR no codifica por sí misma un elemento de proteína funcional. En una realización se utiliza un elemento STAR. Sin embargo, preferentemente, se utiliza más de un elemento STAR. En una realización particularmente preferida, se proporciona un casete de expresión para su uso en la invención con dos secuencias STAR; una secuencia STAR en el lado 5' de la secuencia codificante del gen de inmunoglobulina y otra secuencia STAR en el lado 3' de la secuencia codificante del gen de inmunoglobulina. Las MAR son secuencias de ADN que están implicadas en el anclaje del ADN/cromatina a la matriz nuclear y se han descrito en especies tanto de mamíferos como de plantas. Las MAR poseen diversas características que facilitan la apertura y conservación de la eucromatina. Las MAR pueden aumentar la expresión transgénica y limitar efectos posicionales. De acuerdo con un aspecto de la invención es importante que la expresión del casete se produzca solo durante un determinado periodo en el desarrollo de una célula, en particular un linfocito B en desarrollo, más en particular un linfocito B en un hospedador murino transgénico, en particular un ratón. En este caso particular, el periodo de desarrollo se selecciona de tal manera que la expresión de la cadena ligera no interfiera significativamente con la diferenciación y/o maduración normal de la célula y, cuando sea aplicable, permita el emparejamiento de la cadena polipeptídica producida con su homólogo.
Esto puede conseguirse proporcionando un ácido nucleico útil en el contexto de la invención, en el que dicho medio para sincronizar la expresión es un promotor en el que la actividad se limita esencialmente a la fase de desarrollo concreta. En un linfocito B en desarrollo, que, por ejemplo, después de la inmunización, madura y/o se diferencia, la expresión del gen de interés, que es la región VL humana reordenada de una inmunoglobulina humana, no debe interferir (significativamente) con dicha maduración y/o diferenciación y requiere sincronizarse de tal manera que el polipéptido resultante pueda emparejarse con sus homólogos. Por lo tanto, también se describe un ácido nucleico útil en la invención, en donde dicha fase comienza en una fase inmediatamente anterior o coincidiendo con la aparición de la expresión de moléculas de cadena ligera por dichas células en determinada fase de desarrollo en un linfocito B maduro.
Lo anterior puede conseguirse seleccionando un promotor que sea activo solo durante dicho periodo adecuado. Dicho promotor puede ser un promotor CD 19, el promotor Ig-a, el promotor Ig-p, el promotor phc (todos los genes), el promotor Vk o análogos u homólogos de los mismos.
El promotor como se desvela anteriormente puede no dirigir directamente la expresión del gen de interés. En cambio, dirige la expresión de un gen cuyo producto activa en trans la expresión del gen de interés. Dicho gen activador puede ser un gen que codifique una proteína denominada Cre recombinasa o similar a Cre. El casete de expresión para el gen de interés puede estar provisto, por ejemplo, de una secuencia que inhiba la expresión del gen de interés. Dicha secuencia puede eliminarse por la acción de la Cre recombinasa, que está bajo el control del promotor deseado (activo durante la fase de desarrollo correcta). Puede requerirse un conjunto de casetes de expresión.
Por lo tanto, se describe un conjunto de ácidos nucleicos que son casetes de expresión, en el que un ácido nucleico comprende un casete de expresión que codifica una proteína similar a Cre bajo el control de un promotor activo
durante la fase de desarrollo deseada de la célula hospedadora y el segundo ácido nucleico comprende una secuencia que codifica una molécula proteica deseada bajo el control de un promotor constitutivo que puede activarse mediante la acción de una proteína similar a Cre. Dicha activación puede realizarse eliminando una secuencia de terminación flanqueada por sitios loxP. El sistema Cre/lox se describe con detalle en Rajewsky et al. (1996) J. Clin. Invest. 98, págs. 600-603. Dichos sistemas se revisan en Wunderlich F. T. (2004), "Generation of inducible Cre systems for conditional gene inactivation in mice", Inauguraldissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultat der Universitat zu Koln; http://deposit.ddb.de/cgibin/dokserv?idn=97557230x&dok_var=d1&dok_ext=pdf&filename=97557230x.pd.
La invención proporciona un hospedador murino transgénico como se define en las reivindicaciones, al cual se le ha proporcionado un casete de expresión, en el que la molécula proteica deseada es una región VL humana reordenada de una inmunoglobulina humana, en el que la región VL está codificada por un segmento génico V de la línea germinal humana y un J de la línea germinal humana. Un polipéptido más preferido es 012, preferentemente la cadena ligera kappa de la línea germinal reordenada IGKV1- 39*0l/lGKJ1*01 (nomenclatura de acuerdo con la base de datos de IMGT, http://www.imgt.org).
Adicionalmente, se describe que la cadena polipeptídica se vuelva esencialmente incapaz de reordenamiento y/o de excluir cualquier modificación de secuencia de tal modo que opere normalmente en la Ig durante el proceso de maduración por afinidad de linfocitos B. Por lo tanto, la invención describe un hospedador murino transgénico al cual se le ha proporcionado un casete de expresión, en el que dicho reordenamiento y/o modificaciones de secuencia se impiden por la ausencia de elementos al menos parcialmente responsables de hipermutación somática, tal como, por ejemplo, el potenciador MoEkí.
Un casete de expresión comprende medios para impedir el silenciamiento. Dichos medios para impedir el silenciamiento son medios de inserción en un locus en el genoma de la célula hospedadora que es resistente al silenciamiento. Dichos medios de inserción son preferentemente medios de recombinación homóloga en dicho sitio resistente al silenciamiento. Un locus preferido cuando el de la línea germinal humana es un ratón es el locus rosa. Un casete de expresión preferido adicional comprende, en dirección 5'-3': un promotor Vk, un líder de ratón, un gen V humano, opcionalmente un potenciador MoEkí, una región constante (Ck) de rata y opcionalmente un potenciador MoEk3' (truncado).
Incluso otro casete de expresión adicional preferido comprende, en dirección 5'-3': un promotor Vk, un líder de ser humano, un gen V humano, opcionalmente un potenciador MoEkí, una región constante (Ck) de rata y opcionalmente un potenciador MoEk3' (truncado).
Por supuesto, el objetivo final de la invención, es producir anticuerpos para usar en agentes terapéuticos humanos. Por lo tanto, la invención proporciona un procedimiento para producir un anticuerpo deseado que comprende exponer a un antígeno, un hospedador murino de acuerdo con la invención, de tal manera que se induzca una respuesta de anticuerpo y aislar los anticuerpos específicos para el antígeno.
En una realización alternativa, la invención proporciona un procedimiento para producir un anticuerpo deseado que comprende exponer a un antígeno, un hospedador murino de acuerdo con la invención, de tal manera que se induzca una respuesta de anticuerpo y aislar las células que producen dichos anticuerpos, cultivar y opcionalmente inmortalizar dichas células y recoger dichos anticuerpos.
En una realización adicional, la invención proporciona un procedimiento para producir un anticuerpo deseado que comprende exponer a un antígeno, un hospedador murino de acuerdo con la invención, de tal manera que se induzca una respuesta de anticuerpo y aislar un ácido nucleico que codifique al menos parte de dicho anticuerpo, insertar dicho ácido nucleico o una copia o un derivado del mismo en un casete de expresión y expresar dicho anticuerpo en una célula hospedadora.
Un experto en la técnica conoce procedimientos para producir anticuerpos de ratones transgénicos. Particularmente preferidos son los procedimientos para la producción de mezclas de anticuerpos procedentes de una célula, mediante los cuales, los ácidos nucleicos que codifican estos anticuerpos han derivado de ratones de acuerdo con la invención.
En los documentos WO04106375 y WO05068622 se desvelan estos denominados oligoclónicos.
La presente invención proporciona hospedadores murinos transgénicos, preferentemente ratones, capaces de generar anticuerpos híbridos ratón-ser humano de alta afinidad y especificidad con regiones variables de cadena ligera (VL) de inmunoglobulina humana en configuración de la línea germinal y preferentemente regiones variables de cadena pesada (VH) de inmunoglobulina murina que pueden tener mutaciones somáticas acumuladas durante el proceso de maduración de afinidad conducida por antígenos. Se contempla que las regiones VH murinas de los anticuerpos híbridos puedan someterse a procedimientos de humanización para producir mAb que tengan inmunogenicidad reducida cuando se apliquen en seres humanos basados en regiones VL casi de la línea germinal
o de la línea germinal y en regiones VH murinas que se han humanizado.
En particular, en la presente invención se ha observado que ratones transgénicos que llevan una construcción de expresión de ADN que codifica una región VL humana reordenada bajo el control de elementos genéticos que actúan en cis que proporcionan expresión sincronizada y regulada del transgén en una proporción significativa de linfocitos B durante el desarrollo de linfocitos B, pero carecen de elementos que dirigen la maquinaria de hipermutación somática en el transgén, son capaces de generar anticuerpos híbridos ratón-ser humano de alta afinidad y específicos con cadenas L esencialmente no mutadas. Se muestra que el transgén humano reordenado es capaz de emparejarse con una diversidad de cadenas H de inmunoglobulina murina endógena para formar inmunoglobulinas híbridas ratón-ser humano expresadas sobre la superficie de linfocitos B y facilitar cumplidamente el desarrollo de linfocitos B murinos para obtener un compartimento de linfocitos B periféricos diverso y cuantioso. En una realización preferida, la construcción de la expresión transgénica lleva las secuencias codificantes de una región V de cadena L reordenada humana bajo el control de un promotor VL humano para dirigir la expresión específica de linfocitos B. Además, la construcción lleva la secuencia potenciadora 3' Ck murina para especificidad de linfocitos B y nivel de expresión inducible y elevado del transgén. Además, la construcción se diseña de manera que carece de elementos reguladores que facilitan el reclutamiento de la maquinaria de hipermutación somática al transgén, tales como el potenciador intrónico y el potenciador C-kappa 3'.
En una realización relacionada, el gen VL humano reordenado se inserta en el locus Rosa26 murino por integración específica de sitio. El locus Rosa26 es útil en el contexto de la estrategia de “transgénesis diana” para la generación eficaz de organismos transgénicos (tales como ratones) con un modelo de expresión transgénica predecible.
En una realización preferida, la región VL humana reordenada se selecciona por su capacidad para emparejarse al menos con dos genes VH murinos diferentes para garantizar la generación de una población de linfocitos B con un repertorio de genes VH diverso. Un procedimiento para obtener dichas regiones VL comprende amplificar un repertorio de genes VH reordenados a partir de los linfocitos B de ratones y un repertorio de regiones VL humanas de la línea germinal reordenadas a partir de los linfocitos B de seres humanos y clonarlos en vectores de presentación de fagémidos para preparar diversas bibliotecas de inmunoglobulinas híbridas en bacterias. Mediante análisis de secuencias de nucleótidos de colecciones de pares VH/VL seleccionados y no seleccionados por antígeno, se identifican genes VL de la línea germinal humana que se emparejan con muchos genes VH murinos diferentes. Se describe una colección de genes VL de la línea germinal humana con esta capacidad.
Se muestra que, después de la inmunización con el antígeno, los linfocitos B son capaces de generar una respuesta inmunitaria, conduciendo a la generación de linfocitos B que segregan anticuerpos híbridos con alta especificidad y afinidad. Las regiones V que codifican estos anticuerpos se caracterizan por la cadena ligera transgénica humana que no lleva, o lleva muy pocas, mutaciones y una cadena pesada murina que lleva un número variable de mutaciones introducidas por la maquinaria de hipermutación somática.
Se contemplan estrategias para obtener anticuerpos monoclonales híbridos de alta afinidad a partir de los ratones transgénicos por tecnologías de hibridoma y de presentación, así como por procedimientos para humanizar las regiones VH murinas para obtener anticuerpos menos inmunogénicos para aplicación en seres humanos.
La invención puede implicar el uso de una construcción transgénica de cadena L de inmunoglobulina que comprende secuencias de ADN que codifican una región VL de inmunoglobulina humana en combinación con una región constante de cadena ligera (CL) de una proteína de inmunoglobulina de animal, cuyas secuencias están unidas operativamente a secuencias reguladoras transcripcionales que, cuando se integran en un hospedador transgénico murino, producen un polipéptido VL-CL de Ig con una región VL humana que no está sometida o lo está marginalmente a hipermutación somática. La VL de Ig es capaz de emparejarse con polipéptidos VH-CH reordenados que se generan durante el desarrollo de linfocitos B en el hospedador transgénico murino, conservando dichos polipéptidos VH-CH la capacidad de experimentar hipermutación somática después de estimulación. La región CL puede ser de cualquier especie animal y es generalmente capaz de emparejarse con las regiones CH del hospedador transgénico murino.
También se describe el uso de una construcción transgénica, como se ha indicado anteriormente, en la producción de un hospedador murino transgénico que puede producir anticuerpos híbridos que constan de polipéptidos VL-CL y polipéptidos VH-CH en los que la región VL es de origen humano y la CL, VH y CH puede ser de cualquier especie animal, incluyendo un ser humano. Después de la humanización, estos animales transgénicos son capaces de generar anticuerpos de alta afinidad codificados por genes VH somáticamente hipermutados y genes VL esencialmente no mutados codificados por el transgén.
Otro aspecto describe un proceso para la producción de un hospedador murino transgénico que puede producir anticuerpos híbridos en respuesta a una exposición antigénica, que comprende alterar funcionalmente el locus endógeno de la cadena ligera de inmunoglobulina e insertar en el genoma del hospedador murino transgénico una construcción transgénica descrita.
La invención describe el uso de hospedadores murinos que pueden obtenerse mediante este proceso en la producción de linfocitos B que producen inmunoglobulina que tiene cadena ligera VL humana. Se describe un proceso para la producción de linfocitos B que producen inmunoglobulina que tiene VL humana y que se unen a un antígeno seleccionado, que comprende exponer un hospedador murino que puede obtenerse mediante un proceso como se ha indicado anteriormente con dicho antígeno y explorar los linfocitos B de dicho animal que se unen a dicho antígeno. Se describen además linfocitos B que pueden obtenerse mediante este proceso e hibridomas que pueden obtenerse inmortalizando dichos linfocitos B, por ejemplo, hibridomas obtenidos fusionando linfocitos B, como se ha indicado anteriormente, con células de mieloma. La invención también describe un proceso para producir anticuerpos monoclonales que comprende cultivar dicho hibridoma. También se describe el uso de los linfocitos B anteriores en la producción de un hibridoma o anticuerpo monoclonal correspondiente.
La invención describe un proceso para la producción de inmunoglobulina que tiene cadenas VL humanas y que se une a un antígeno seleccionado, que comprende exponer un hospedador murino, que puede obtenerse como se ha indicado anteriormente, a dicho antígeno y obtener inmunoglobulina del mismo.
En una estrategia, como una etapa individual, en la línea germinal de ratón se introduce una región VL reordenada codificada por segmentos génicos V y J de la línea germinal humana y una región constante de cadena ligera de cualquier especie animal, pero preferentemente una región constante murina. El ADN transgénico puede introducirse en los pronúcleos de ovocitos fecundados o en células madre embrionarias. La integración puede ser aleatoria u homóloga dependiendo de la estrategia particular que se va a emplear. Por ejemplo, el transgén VL puede introducirse mediante inserción aleatoria, lo que da lugar a ratones que portan una o múltiples copias del transgén en el genoma. Como alternativa, el transgén VL puede dirigirse a un locus genómico específico usando recombinación específica de sitio como se describe en la técnica.
En una realización preferida, el transgén VL se dirige al locus murino ROSA26 que es un sitio de integración adecuado que muestra una fuerte y predecible expresión de los transgenes insertados (documento EP 1439234). El vector dirigido permite la inserción de una sola copia de un casete de expresión génico, impidiendo así la modulación de la expresión del transgén por la disposición de copias múltiples. Seleccionando el locus Rosa26 autosómico como sitio de inserción, el patrón de expresión del transgén insertado en el hospedador murino es predecible. Además, se impide la inactivación y/o modulación del cromosoma X al azar por efectos de posición cromosómica. Esto también elimina la necesidad de generar y analizar cepas transgénicas múltiples para cualquier transgén determinado. Finalmente, el vector dirigido Rosa26 puede usarse para la integración específica de sitio para casetes de expresión génica múltiples. Por tanto, puede contemplarse que 2 o más regiones VL humanas diferentes de la línea germinal reordenadas se inserten en el locus Rosa26 para aumentar adicionalmente la diversidad del repertorio de anticuerpos híbridos o humanos.
En otra realización, una región VL humana reordenada puede dirigirse al locus murino de cadena ligera lambda o kappa de Ig para inactivar funcionalmente el locus endógeno o pueden cruzarse ratones que contengan la región VL humana reordenada con ratones que carezcan de locus Ig lambda o kappa funcionales o ambos. Por tanto, usando transformación, usando etapas repetitivas o en combinación con reproducción, pueden obtenerse hospedadores murinos transgénicos que sean capaces de producir anticuerpos que lleven el transgén VL humano en ausencia sustancial de cadenas ligeras de inmunoglobulina de huésped endógeno.
En una realización, se selecciona un transgén VL humano por su capacidad para emparejarse con una parte sustancial de regiones VH murinas para formar un repertorio diverso de anticuerpos híbridos ratón-ser humano funcionales expresados sobre la superficie de linfocitos B. Una parte sustancial de regiones VH murinas significa que la VL humana se empareja con al menos el 0,1 % de las regiones VH murinas generadas durante el desarrollo de linfocitos B, más preferentemente con al menos el 1 % y más preferentemente con al menos el 10 %. Como procedimientos para identificar genes VL humanos con esta característica se incluyen el emparejamiento al azar de un repertorio de regiones VL humanas con un repertorio de regiones VH murinas, la co-expresión de regiones VH y VL en vectores de expresión eucariotas o procariotas adecuados y la exploración de regiones VL humanas que se emparejan con una parte sustancial de regiones VH murinas. Pueden usarse vectores fagémidos para dirigir la expresión de fragmentos de anticuerpo ratón-ser humano en células bacterianas o en la superficie de fagos filamentosos y análisis de capacidad de unión de fragmentos de anticuerpo mediante procedimientos conocidos en la técnica.
En otra realización, se selecciona un transgén VL humano por su capacidad para emparejarse con una parte sustancial de regiones VH humanas para formar un repertorio diverso de anticuerpos humanos expresados sobre la superficie de linfocitos B. Una parte sustancial de regiones VH humanas se refiere a que la VL humana se empareja con al menos el 0,1 % de las regiones VH humanas generadas durante el desarrollo de linfocitos B, más preferentemente con al menos el 1 % y más preferentemente con al menos el 10 %.
En la última realización, los ratones transgénicos VL humanos se cruzan con ratones que llevan locus funcionales de inmunoglobulina de cadena H humana no reordenada o reordenada y locus endógenos de Ig de cadena H inactivados funcionalmente como se describe en la técnica. La inactivación funcional de las dos copias de cada uno de los tres locus de Ig hospedadora (cadena pesada, cadena ligera kappa y lambda), en la que el hospedador
contiene la IgH humana y el transgén VL humano reordenado, permitiría la producción de moléculas de anticuerpo puramente humanas sin la producción de anticuerpos hospedadores o quiméricos humanos hospedadores. Dicha cepa hospedadora, por inmunización con antígenos específicos, respondería mediante la producción de linfocitos B de ratón productores de anticuerpos humanos específicos, cuyos linfocitos B se fusionan posteriormente con células de mieloma de ratón o se inmortalizan de cualquier otra manera para la producción estable continua de anticuerpos monoclonales humanos. Como alternativa, dicha población de linfocitos B se usa como una fuente de regiones VH que puede obtenerse mediante la construcción de bibliotecas de ADNc o por amplificación mediante PCR usando cebadores para regiones VH humanas como se conoce en la técnica.
Un gen VL reordenado humano se reconstruye en un microorganismo eucariota o procariota apropiado y los fragmentos de ADN resultantes pueden introducirse en los pronúcleos de ovocitos de ratón fecundados o en células madre embrionarias. En la técnica se han descrito diversas construcciones que dirigen la expresión directa, específica de linfocitos B, de transgenes VL y tienen el siguiente formato general: una secuencia líder y secuencias relevantes cadena arriba para dirigir la expresión específica de linfocitos B del transgén, una secuencia codificante de un transgén VL humano, una secuencia potenciadora que dirige la expresión a alto nivel y específica de linfocitos B del transgén y un gen murino de región constante. En un formato preferido, el potenciador es el potenciador C-kappa 3' porque dirige la expresión a alto nivel en linfocitos de linaje B, pero no recluta hipermutación somática cuando se usa en construcciones transgénicas.
En una realización, los hospedadores murinos, preferentemente ratones, que comprenden una o múltiples copias del transgén en el genoma, se aíslan y analizan con respecto a la expresión estable. Se seleccionan hospedadores murinos que muestran expresión estable del transgén durante periodos de tiempo más largos, preferentemente en linfocitos B. Si es necesario, se cruzan diferentes líneas de hospedadores murinos que comprenden inserciones independientes de una o múltiples copias del transgén, preferentemente en diferentes cromosomas, para obtener hospedadores murinos con diferentes inserciones de una o múltiples copias del transgén para aumentar la expresión del transgén en hospedadores murinos, preferentemente en linfocitos B.
Se describe además, la descendencia de un hospedador murino transgénico de acuerdo con la invención, comprendiendo la descendencia, al menos en su linaje de linfocitos B, una secuencia codificante de cadena ligera junto con un medio que hace que la secuencia sea resistente a reordenamientos de ADN y/o hipermutaciones somáticas.
Se describe además la descendencia de un hospedador murino transgénico de acuerdo con la invención, comprendiendo la descendencia un casete de expresión para la expresión de una molécula proteica deseada en células durante una determinada fase del desarrollo en células que se desarrollan en linfocitos B maduros.
Adicionalmente, se describe una célula que se aísla de un hospedador murino transgénico de acuerdo con la invención, comprendiendo la célula una secuencia codificante de cadena ligera junto con un medio que hace que la secuencia sea resistente a reordenamientos de ADN y/o hipermutaciones somáticas.
Adicionalmente, se describe una célula que está aislada de un hospedador murino transgénico de acuerdo con la presente invención, comprendiendo la célula un casete de expresión para la expresión de una molécula proteica deseada en células durante una determinada fase del desarrollo en células que se desarrollan en linfocitos B maduros. Puede usarse una célula, posiblemente un linfocito B productor de anticuerpos o una célula que sea capaz de diferenciarse o madurar en un linfocito B productor de anticuerpos, para la producción in vitro de anticuerpos, como conoce un experto habitual en la técnica, por ejemplo, a partir del documento de Gascan et al. 1991. J. Exp. Med. 173: 747-750. En la técnica se conocen procedimientos para la inmortalización de una célula e incluyen la generación de hibridomas, por ejemplo, por fusión con una célula de mieloma, transformación con el virus de Epstein Barr; expresión del transductor de señal de la activación y transcripción (STAT), activación mediante c D40 y señalización de receptores de IL4, y/o expresión de Bcl6 (Shvarts et al. 2002. Genes Dev 16: 681-686).
En una etapa distinta, los locus endógenos de cadena ligera Kappa y Lambda de ratón se vuelven esencialmente no funcionales de tal manera que al menos la mayoría de los linfocitos B en los ratones transgénicos llevan receptores de Ig que contienen la región VL humana transgénica. La inactivación de los locus de inmunoglobulina de ratón endógena se realiza por alteración diana de locus apropiados por recombinación homóloga en células madre embrionarias de ratón. Dicha alteración diana comprende la alteración de la secuencia genómica de tal manera que se produce una cadena ligera Kappa y/o Lambda de inmunoglobulina de ratón endógena sustancialmente no funcional. La expresión “inmunoglobulina de ratón endógena sustancialmente no funcional” indica que los locus endógenos de cadena ligera Kappa y/o Lambda se silencian funcionalmente de tal manera que el nivel de expresión de proteína funcional de los locus endógenos de cadena ligera Kappa y/o Lambda, preferentemente el locus endógeno de cadena ligera Kappa, se reduce a aproximadamente un 20 % del nivel de expresión en un ratón de referencia, más preferentemente a aproximadamente un 10 %, más preferentemente a aproximadamente un 5 %, más preferentemente a aproximadamente un 2 % y más preferentemente a aproximadamente un 1 %. En una realización más preferida, el nivel de expresión de proteína funcional de los locus endógenos de cadena ligera Kappa y/o Lambda se reduce al 0 %. El nivel de expresión de proteína funcional puede determinarse por medios conocidos por un experto en la técnica, incluyendo transferencia de Western y emparejamiento con una cadena
pesada de ratón. Dicho ratón de referencia es un ratón en el que los locus endógenos de cadena ligera Kappa y/o Lambda no están alterados. Dicha alteración comprende mutación y/o deleción de secuencias génicas que son necesarias para la expresión funcional de genes de inmunoglobulina endógenos. Como alternativa, dicha alteración comprende la inserción de un ácido nucleico en los locus endógenos de cadena ligera Kappa y/o Lambda de inmunoglobulina de ratón de tal manera que se reduce la expresión funcional de los genes de inmunoglobulina endógenos. En un aspecto, dicho ácido nucleico comprende un elemento silenciador dando como resultado el silenciamiento transcripcional del gen de inmunoglobulina endógeno. En un aspecto adicional, o además, dicho ácido nucleico comprende una secuencia que altere el corte y empalme y/o la traducción del gen de inmunoglobulina endógeno, por ejemplo, introduciendo un exón que dé lugar a un desplazamiento de fase de lectura en la secuencia codificante, o que comprenda un codón de terminación prematuro. En cada caso se generan animales quiméricos que derivan en parte de las células madre embrionarias modificadas y que son capaces de transmitir las modificaciones genéticas a través de la línea germinal. El apareamiento de cepas de ratón con locus de inmunoglobulina humana con cepas con locus de ratón inactivados conduce a animales que producen anticuerpos que comprenden cadenas ligeras esencialmente solo humanas.
Por medios conocidos en la técnica se prepara una construcción para recombinación homóloga y se elimina cualquiera de las secuencias no deseables, por ejemplo, las secuencias procariotas. Puede emplearse cualquier técnica conveniente para introducir una construcción para recombinación homóloga en una célula diana. Estas técnicas incluyen fusión de esferoplastos, lipofección, electroporación, transferencia de ADN mediada con fosfato cálcico, o microinyección directa. Después de la transformación o transfección de las células diana, estas se seleccionan mediante marcadores positivos y/o negativos, por ejemplo, por resistencia a neomicina y/o resistencia a aciclovir y/o a ganciclovir. Estas células, que muestran el fenotipo deseado, pueden después analizarse posteriormente por análisis de restricción, electroforesis, análisis de Southern, PCR o similar. Identificando fragmentos que muestren la presencia de lesión (o lesiones) en el locus diana, se identifican células en las que se ha producido recombinación homóloga para inactivar una copia del locus diana.
Adicionalmente, se muestra que, después de la inmunización en los ratones transgénicos mencionados anteriormente, las regiones VH humanas y murinas, pero no las regiones VL, son capaces de experimentar mutaciones somáticas para generar anticuerpos de alta afinidad. Ventajosamente, se prevé que estos anticuerpos codificados por regiones VL de la línea germinal contribuyan a disminuir la inmunogenicidad cuando se apliquen en seres humanos y produzcan anticuerpos más estables que sean menos propensos a la agregación y por tanto más seguros para su uso terapéutico en seres humanos.
Todos los mAb derivados de las células o de hospedadores transgénicos murinos mencionados anteriormente, comparten las mismas regiones VL humanas idénticas. Se ha descrito que los mAb que comparten la misma región VL idéntica pueden co-expresarse en una sola célula clonal para la producción de mezclas de anticuerpos recombinantes con sitios de unión funcionales (véanse los documentos WO04106375 y WO05068622). Por tanto, la invención describe una plataforma para la generación de mAb específicos y de alta afinidad que constituye la base para mezclas de mAb producidos por células clonales.
Se prefiere que los mAb derivados de las células o de los hospedadores transgénicos murinos mencionados anteriormente se dirijan contra dianas celulares. Las dianas preferidas son proteínas o moléculas de carbohidrato humanas solubles o expresadas en la superficie. Las dianas adicionalmente preferidas son proteínas o moléculas de carbohidrato expresadas en la superficie que se expresan sobre la superficie de bacterias, virus u otros patógenos, especialmente de seres humanos.
Más específicamente, las dianas preferidas incluyen citocinas y quimiocinas, incluyendo, pero sin limitación, interleucina 1beta (IL1beta), IL2, IL4, IL5, IL7, IL8, IL12, IL13, IL15, iL18, IL21, IL23 y quimiocinas tales como, por ejemplo, quimiocinas CXC, quimiocinas CC, quimiocinas C (o quimiocinas y) tales como quimiocinas XCL1 (linfotactina-a) y XCL2 (linfotactina-p) y quimiocinas CX3C. Adicionalmente, como dianas preferidas se incluyen moléculas receptoras de las citocinas y quimiocinas, incluyendo receptores de citocina de tipo I tales como, por ejemplo, el receptor de IL-2, receptores de citocina de tipo II, tales como, por ejemplo, receptores de interferón, receptores de la superfamilia de inmunoglobulina (Ig), la familia de receptores del factor de necrosis tumoral, incluyendo receptores para CD40, CD27 y CD30, receptores de serina/treonina-proteína cinasa tales como receptores del TGF beta, receptores acoplados a proteína G, tales como CXCR1-CXCR7, y receptores de tirosina cinasa, tales como los miembros de la familia de receptores del factor de crecimiento de fibroblastos (FGFR), miembros de la familia de receptores del EGF, incluyendo erbBI (EGF-R; HERI), erbB2, (HER2), erbB3 (HeR3) y erbB4 (HER4), miembros de la familia de receptores de insulina, incluyendo IGF-R1 e IGF-RII, miembros de la familia de receptores del PDGF, miembros de la familia de receptores del factor de crecimiento de hepatocitos, incluyendo c-Met (HGF-R), miembros de la familia de receptores de Trk, miembros de la familia de receptores de AXL, miembros de la familia de receptores de LTK, miembros de la familia de receptores de TIE, miembros de la familia de receptores ROR, miembros de la familia de receptores DDR, miembros de la familia de receptores de KLG, miembros de la familia de receptores RYK, miembros de la familia de receptores de MuSK y miembros de la familia de receptores del factor de crecimiento endotelial vascular (VEGFR).
Las dianas adicionalmente preferidas son dianas que se sobreexpresan o se expresan selectivamente en tumores,
tales como, por ejemplo, VEGF, CD20, CD38, CD33, CEA, EpCAM, PSMA, CD54, Lewis Y, CD52, CD40, CD22, CD51/CD61, CD74, MUC-1, CD38, CD 19, CD262 (TRAIL-R2), RANKL, CTLA4 y CD30; dianas que están implicadas en inflamación crónica, tales como, por ejemplo, CD25, CD11a, TNF, CD4, CD80, CD23, c D3, CD14, IFNgamma, CD40L, CD50, CD122, TGFbeta y TGFalfa.
Las proteínas o moléculas de carbohidrato expresadas en superficie preferidas que se expresan sobre la superficie de bacterias, virus y otros patógenos parásitos, especialmente de seres humanos, incluyen marcadores de superficie de virus de la gripe A y B tales como hemaglutinina (HA) y neuraminidasa (NA), filovirus tales como el virus del Ébola, de la rabia, sarampión, rubeola, paperas, flavivirus tales como los tipos 1-4 del virus del dengue, virus de la encefalitis transmitido por garrapatas, virus del Nilo occidental, virus de la encefalitis japonesa, y virus de la fiebre amarilla, Paramixovirus incluyendo Paramixovirus tales como virus paragripal 1, 3, Rubulavirus tales como virus de las paperas y paragripal 2, 4, Morbilivirus y Pneumovirus, tales como el virus respiratorio sincitial, virus de la vacuna, de la viruela, coronavirus, incluyendo el virus del Síndrome Respiratorio Agudo Grave (SARS), virus de la hepatitis A, B y C, virus de la inmunodeficiencia humana, herpesvirus, incluyendo citomegalovirus, virus de Epstein Barr, virus del herpes simple y virus de la varicela zóster, parvovirus, tal como, por ejemplo, B19; Legionella pneumophila; Listeria monocytogenes; Campylobacter jejuni; Staphylococcus aureus; E. coli O157:H7 Borrelia burgdorferi; Helicobacter pylori; Ehrlichia chaffeensis; Clostridium difficile; Vibrio cholera; Salmonella enterica Serotipo Typhimurium; Bartonella henselae; Streptococcus pyogenes (estreptococo de Grupo A); Streptococcus agalactiae (estreptococo de Grupo B); S. aureus resistente a múltiples fármacos (por ejemplo, MRSA); Chlamydia pneumoniae; Clostridium botulinum; Vibrio vulnificus; Parachlamydia pneumonia; Corynebacterium amycolatum; Klebsiella pneumonia; enterococos resistentes a Linezolid (E. faecalis y E. faecium); y Acinetobacter baumannii resistente a múltiples fármacos
Las dianas más preferidas son IL-6 y su receptor, IL-6Ralfa, gp130 denominada glucoproteína, RSV, especialmente las proteínas de superficie F, G y SH y proteínas no estructurales tales como N y M, y tirosina cinasas receptoras en particular erbB1 (EGF-R; HER1), erbB2, (HER2), erbB3 (HER3), erbB4 (HER4), IGF-R1 e IGF-RII, c-Met (HGF-R). Por lo tanto, la invención describe una plataforma para la generación de mAb específicos y de alta afinidad contra las dianas mencionadas anteriormente que constituyen la base para las mezclas de los mAb producidos por células clonales. Dichos mAb específicos y de alta afinidad comprenden mAb que se dirigen contra diferentes epítopos sobre al menos una de las dianas. Dichos mAb específicos y de alta afinidad comprenden mAb que se dirigen contra diferentes dianas tales como, por ejemplo, uno o más miembros de la familia de receptores de EGF incluyendo, erbB1 (EGF-R; HER1), erbB2, (HER2), erbB3 (HER3) y erbB4 (HER4).
Salvo que se defina en contra, los términos científicos y técnicos utilizados en conexión con la presente invención tendrán los significados normalmente entendidos por los expertos habituales en la técnica. Además, salvo que el contexto lo requiera de otra manera, los términos en singular incluirán pluralidades y los términos en plural incluirán el singular. Generalmente, las nomenclaturas utilizadas en conexión con, y técnicas de, cultivo de células y tejidos, biología molecular y química de proteínas y oligo o polinucleótidos e hibridación descritas en el presente documento son las bien conocidas y normalmente utilizadas en la técnica. Se utilizan técnicas convencionales para ADN recombinante, síntesis de oligonucleótidos y cultivo y transformación de tejidos (por ejemplo, electroporación, lipofección). Las reacciones enzimáticas y técnicas de purificación se realizan de acuerdo con las especificaciones del fabricante o como normalmente se llevan a cabo en la técnica o se describe en el presente documento. Las técnicas y procedimientos anteriores se realizan generalmente de acuerdo con procedimientos convencionales bien conocidos en la técnica y se describen en diversas referencias generales y más específicas citadas y comentadas a lo largo de la presente memoria descriptiva. Véase, por ejemplo, Sambrook et al. Molecular Cloning: A Laboratory Manual (3a edición, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001)). Las nomenclaturas utilizadas en conexión con, y los procedimientos y técnicas de laboratorio de, química analítica, química orgánica sintética y química medicinal y farmacéutica descritos en el presente documento son los bien conocidos y normalmente utilizados en la técnica. Para la síntesis química, análisis químicos, preparaciones, formulaciones y administraciones farmacéuticas y tratamiento de pacientes se utilizan técnicas convencionales.
Leyenda de las figuras
Figura 1
Mapa topológico de las localizaciones de hibridación de cebadores de VH específicos de ratón y la posición de los sitios de restricción requeridos que se introducen mediante secuencias salientes en el extremo 3' de los cebadores.
Figura 2
Etapas de amplificación por PCR (amplificación, productos intermedios e introducción en sitio). La localización y nombres de los cebadores de amplificación de VH de ratón (y mezclas de cebadores) se indican por etapa. Figura 3
Topología del vector MV1043. Este vector se usa para la clonación de fragmentos VH humanos o murinos. 012 (IGKV1-39) se indica como el gen de VL. Los productos de este vector en combinación con fagos auxiliares en
células E. coli permiten la generación de fagos que presentan fragmentos Fab en la superficie de las partículas de fago como un producto de fusión con la proteína g3 y presencia del vector en el fago como el contenido genético (F1 ORI).
Figura 4
Topología del locus Ckappa de ratón cadena abajo de los segmentos J. Se indican ambos potenciadores y la región Ckappa. La flecha inferior indica la región que se elimina para silenciar el locus.
Figura 5
Topología del locus C-lambda de ratón. Se indican las tres regiones V activas (V1, V2 y V3 de Igl) como los segmentos J (Igl-J1, Igl-J2, Igl-J3, Igl-J4 y el pseudo fragmento Igl-J3p) y regiones constantes (Igl-C1, Igl-C2, Igl-C3 y Igl-C4). Las regiones que se delecionan para silenciar el locus se indican por marcadores de deleción. Estas deleciones incluyen todos los agentes V (1, 2 y 3) activos y el segmento intergénico entre V2 y V3.
Figura 6
Topología de la construcción de IGKV1-39/J-Ck con un intrón localizado en la fase de lectura abierta (ORF) líder. Figura 7
Topología de la construcción de IGLV2-14/J-Ck con un intrón localizado en la fase de lectura abierta (ORF) líder. Figura 8
Topología de la construcción de VkP-IGKV1-39/J-Ck (VkP-O12). El promotor se origina desde el gen IGKV1-39 y se coloca directamente delante de los elementos requeridos para la transcripción y traducción eficaces. Las secuencias intergénicas (incluyendo los potenciadores) derivan de ratones y se obtienen a partir de clones BAC. La secuencia C-kappa codifica la región constante kappa de rata.
Figura 9
Topología de la construcción de VkP-IGLV2-14/J-Ck (VkP-2a2). El promotor se origina del gen IGKV1-39 y se coloca directamente delante de los elementos requeridos para la transcripción y traducción eficaces. Las secuencias intergénicas (incluyendo los potenciadores) derivan de ratones y se obtienen a partir de clones BAC. La secuencia C-kappa codifica la región constante kappa de rata.
Figura 10
La topología de la construcción de VkP-IGKV1-39/J-Ck-A1 (VkP-O12-del1) es idéntica a la de VkP-IGKV1-39/J-Ck de la figura 9, excepto que la región potenciadora intrónica se elimina.
Figura 11
La topología de la construcción de VkP-IGKV1-39/J-Ck-A2 VkP-O12-del2) es idéntica a la de VkP-IGKV1-39/J-Ck-Al de la figura 10, excepto que se deleciona una gran parte de la región intergénica entre el gen Ck y el potenciador 3'. Además, el potenciador 3' se reduce de tamaño de 809 pb a 125 pb.
Figura 12
Visión general de las secuencias utilizadas o las que se hace referencia en la presente solicitud
Figura 13
Generación del alelo Rosa26-IgVk1-39 KI. (A) Dibujo esquemático del vector dirigido pCAGGS-IgVK1-39. (B) Secuencia de nucleótidos del vector dirigido pCAGGS-IgVK1-39. (C) Estrategia de direccionamiento.
Figura 14
(A) Análisis de transferencia de Southern de ADN genómico de clones ES que comprende una inserción del vector dirigido pCAGGS-IgVK1-39. El ADN genómico de 4 clones independientes se sometió a digestión con AseI y se exploró con 5e1 indicando el límite en 5' del vector dirigido. Todos los clones comprenden una inserción correcta del vector dirigido en el extremo 5'.
(B) Análisis de transferencia de Southern de ADN genómico de clones ES que comprende una inserción del vector dirigido pCAGGS-IgVK1-39. El ADN genómico de 4 clones independientes se sometió a digestión con MscI y se exploró con 3e1 indicando el límite en 3' del vector dirigido. Todos los clones comprenden una inserción correcta del vector dirigido en el extremo 3'.
(C) Análisis de transferencia de Southern de ADN genómico de clones ES que comprende una inserción del vector dirigido pCAGGS-IgVK1-39. El ADN genómico de 4 clones independientes se sometió a digestión con BamHI y se exploró con una sonda Neo interna indicando el límite en 5' del vector dirigido. Todos los clones comprenden una sola inserción correcta del vector dirigido.
Figura 15
Generación del alelo Rosa26-IgVl2-14 KI. (A) Dibujo esquemático del vector dirigido pCAGGS-IgVL2-14. (B) Secuencia de nucleótidos del vector dirigido pCAGGS-IgVL2-14 que contiene el inserto de expresión CAGGS basado en la región lambda IGLV2-14/J V de la línea germinal reordenada (IGLV2-14/J-Ck). (C) Estrategia de direccionamiento.
Figura 16
Perfil Epibase® de los restos 1-107 de IGKV1-39. La subfigura A presenta el tramo de unión para alotipos DRB1, mientras que C presenta el tramo de unión para los alotipos DRB3/4/5, DQ y DP. Los valores en la figura representan constantes de disociación (Kd) y se representan gráficamente en una escala logarítmica en el intervalo 0,01 pM - 0,1 pM (los que tienen unión muy fuerte pueden haberse salido de la gráfica). Para péptidos de unión media, solo se proporcionan valores cualitativos y los péptidos de unión débil y los que no se unen no se muestran. Los valores se representan gráficamente en el primer resto del péptido en la secuencia diana (el propio péptido se extiende por otros 9 restos). Cabe destacar que solo se muestra el receptor de unión más fuerte de cada péptido: en esta gráfica no se observan alotipos de reacción cruzada con afinidad más baja. El receptor de unión más fuerte se indica por su nombre serotípico. Finalmente, en el mapa epitópico, cualquiera de los péptidos filtrados por línea germinal se representa gráficamente con un color más claro (en este caso, no se encontraron no-auto-epítopos). La subfigura B muestra la promiscuidad de unión a HLA para cada péptido decamérico (eje Y: número de alotipos HLA que reconocen epítopos críticos en cada uno de los péptidos comenzando en el resto indicado mostrado en el eje X). La promiscuidad se mide como el número de alotipos del total de 47 para el que el péptido se une de forma crítica. Las columnas en blanco se refieren a auto-péptidos, y las columnas en negro (ausentes aquí), a no- auto- péptidos.
Figura 17
Mapa epitópico de IGKV1-39 que muestra la presencia de péptidos de unión predichos en la secuencia de IGKV1-39 por serotipo en el formato de 15 unidades. Cada oligómero de 15 unidades se numera como se indica en la parte superior de la figura. La secuencia completa de oligómero de 15 unidades correspondiente se indica en la Tabla 7. Las cajas en negrita indican la presencia de uno o más auto-epítopos críticos en el oligómero de 15 unidades para el serotipo indicado a la izquierda. Se definen epítopos críticos desde el punto de vista operativo como de unión a DRB1 fuerte o media y de unión a DRB3/4/5 o DP o DQ fuerte.
Figura 18
Desactivación (knock-out, KO) constitutiva del locus kappa de Ig. (A) Estrategia de direccionamiento. (B) Dibujo esquemático del vector dirigido a pIgKappa
Figura 19
KO constitutiva del locus lambda de Ig. (A) Primera etapa de la estrategia de direccionamiento. (B) Segunda etapa de la estrategia de direccionamiento.
Figura 20
Dibujo esquemático de vectores de direccionamiento. (A) pVkP-O12 (VkP-IGKV1-39/J-Ck); (B) pVkP-O12-del1 (VkP-IGKV1- 39/JCk-A1); (C) pVkP-O12-del2 (VkP-IGKV1-39/J-Ck-A2).
Figura 21
Estrategias de direccionamiento para la inserción de transgenes en el locus Rosa26 por transgénesis diana usando RMCE. (A) VkP-O12 (VkP-IGKV1-39/J-Ck); (B) VkP-O12-del1 (VkP-IGKV1-39/J-Ck-A1); (C) VkP-O12-del2 (VkP-IGKV1- 39/JCk-A2).
Figura 22
Topología del vector MV1057. Reemplazando el fragmento de relleno indicado con un fragmento VH se produce un vector de expresión que puede transfectarse a células eucariotas para la producción de anticuerpos IgG1 con cadenas ligeras que contienen un gen 012 (IGKV1-39).
Figura 23
Ausencia de expresión de cadena ligera Vk1 humana transgénica en poblaciones de linfocitos no B del bazo. Figura 24
La cadena ligera Vk1 humana transgénica se expresa en todas las poblaciones de linfocitos B del bazo.
Figura 25
La cadena ligera Vk1 humana transgénica se expresa en linfocitos B1 de la cavidad peritoneal.
Figura 26
La cadena ligera Vk1 humana transgénica no se expresa en linfocitos pro- y pre-B sino en los linfocitos B de poblaciones inmaduras y recirculantes en la médula ósea. (A) Selección de células de médula ósea. (B) Histogramas de expresión transgénica con solapamiento de un control de TS (Tipo Silvestre).
Figura 27
La cadena ligera Vk1 humana transgénica se correlaciona directamente con la cadena ligera endógena y la expresión de IgM en linfocitos B circulantes en la sangre.
Ejemplos
Ejemplo 1
Clones del gen V de cadena ligera humana
Este ejemplo describe el razonamiento detrás de la elección de dos genes V de cadena ligera humana, un gen de tipo kappa y un gen de tipo lambda, que se usan como prueba de hipótesis para ratones transgénicos que expresan cadenas ligeras. De Wildt et al. 1999 (de Wildt et al. (1999) J. Mol. Biol. 285(3): 895) analizaron la expresión de cadenas ligeras humanas en linfocitos B positivos a IgG periféricos. Basándose en estos datos, se seleccionaron IGKV1-39 (012) y IGLV2-14 (2a2) como cadenas ligeras ya que estaban bien representadas en el repertorio de linfocitos B. La secuencia de segmento J de las cadenas ligeras se ha seleccionado basándose en secuencias como se presentan en GenBank ABA26122 para IGKV1-39 (Rabquer, B.J., Smithson, S.L., Shriner, A.K. y Westerink, M.A.J.) y en GenBank AAF20450 para IGLV2-14 (Ignatovich, O., Tomlinson, I.M., Popov, A.V., Bruggemann, M. y Winter, G. J. Mol. Biol. 294 (2), 457-465 (1999)).
Todos los segmentos armazón se convierten en secuencias de aminoácidos de la línea germinal para proporcionar la inmunogenicidad más baja posible en posibles aplicaciones clínicas.
Ejemplo 2
Obtención de genes V de cadena pesada de ratón que se emparejan con el segmento génico IGKV1-39 humano para formar sitios de unión a anticuerpo funcionales.
Este ejemplo describe la identificación de genes V de cadena pesada de ratón que son capaces de emparejarse con una sola región IGKV1-39/J de la línea germinal humana reordenada. Un repertorio VH de bazo de ratones que se inmunizaron con el toxoide tetánico se clonó en un vector Fab de presentación de fagos con una sola cadena ligera kappa IGKV1-39-C humana y se sometió a selección contra el toxoide tetánico. Los clones obtenidos después de una sola ronda de selección se analizaron con respecto a su especificidad de unión. Los genes VH murinos que codifican fragmentos Fab específicos del toxoide tetánico se sometieron a análisis de secuencia para identificar clones únicos y asignar la utilización de VH, DH y JH.
Muchos de los protocolos descritos en el presente documento son protocolos convencionales para la construcción de bibliotecas de presentación de fagos y la selección de fagos para la unión a un antígeno de interés y se describen en Antibody Phage Display: Methods and Protocols (editor(s): Philippa M. O'Brien, Robert Aitken).
Inmunizaciones
Ratones BALB/c recibieron una inmunización con el toxoide tetánico y volvieron a vacunarse 6 semanas después con el toxoide tetánico.
Aislamiento de esplenocitos
Preparación de suspensión de células del bazo. Después de realizar la disección, el bazo se lavó con PBS y se transfirió a una placa de Petri de 60 mm con 20 ml de PBS. Se utilizó una jeringa protegida con 20 ml de PBS y una aguja G20 para aclarar el bazo repetidamente. Después de lavar las células aclaradas con PBS, las células se llevaron cuidadosamente a suspensión usando 20 ml de PBS y se dejaron en una bancada durante 5 minutos para separar los esplenocitos de los restos y agregados celulares. La suspensión de esplenocitos se transfirió a la parte superior de un tubo cargado con Ficoll-Paque™ PLUS y se procesó de acuerdo con los procedimientos del fabricante para el aislamiento de linfocitos (Amersham Biosciences).
Aislamiento de ARN y síntesis de ADNc
Después del aislamiento y sedimentación de linfocitos, las células se suspendieron en Reactivo TRIzol LS (Invitrogen) para el aislamiento de ARN total de acuerdo con el protocolo adjunto del fabricante y se sometieron a reacción de transcripción inversa usando 1 microgramo de ARN, Superscript III RT en combinación con dT20 de acuerdo con los procedimientos del fabricante (Invitrogen).
Amplificación de ADNc por PCR
El ADNc se amplificó en una reacción PCR usando combinaciones de cebadores que permiten la amplificación de aproximadamente 110 genes V murinos diferentes que pertenecen a 15 familias VH (tabla 1; RefSec NG_005838;
Thiebe et al., 1999. European Journal of Immunology 29: 2072 - 2081). En la primera ronda, se usaron las combinaciones de cebadores que se unían al extremo 5' de los genes V y al extremo 3' de las regiones J. En la segunda ronda, los productos de la PCR que se generaron con el cebador MJHRev2 se amplificaron para introducir modificaciones en la región 3' para permitir la clonación eficaz de los productos. En la última ronda de amplificación, se amplificaron todos los productos de la PCR usando cebadores que introducían un sitio de restricción SfiI en el extremo 5' y un sitio de restricción BstEII en el extremo 3' (véanse las figuras 1 y 2, y la tabla 1).
Condiciones de reacción para la 1a ronda de PCR: 4 reacciones diferentes combinando los 25 cebadores directos (MVH1 a MVH25, tabla 1 y figura 2) y 1 cebador inverso por reacción (MJH-Rev1, MJH-Rev2, MJH-Rev3 o MJH-Rev4; véase tabla 1 y figura 2). Volúmenes de PCR de 50 microlitros estaban compuestos por 2 microlitros de ADNc (de reacciones RT), 10 microlitros de tampón de polimerasa HF 5* Phusion, 40 nM de cada uno de los 25 cebadores directos (concentración total de 1 micromolar), cebador inverso 1 micromolar, 1 microlitro de reserva de dNTP 10 mM, polimerasa Phusion 1,25 unidades y agua MQ estéril. El programa del termociclador consistió en un programa de toma de contacto: 1 ciclo a 98 °C durante 30 segundos, 30 ciclos a 98 °C durante 10 segundos, 58 °C disminuyendo 0,2 °C por ciclo 10 segundos, 72 °C 20 segundos y 1 ciclo a 72 °C durante 3 minutos. El programa PCR de la segunda ronda se estableció solo para los productos de la 1a PCR que contenía el cebador MJH-Rev2: 2 reacciones diferentes combinando cualquiera de los cebadores ExtMVH-1 o ExtMVH-2 (tabla 1 y figura 2) en combinación con el cebador inverso ExtMJH-Rev2int (tabla 1 y figura 2). 50 microlitros de volúmenes pCr estaban compuestos por 50 ng de producto de PCR (de la primera ronda de PCR), 10 microlitros de tampón de polimerasa HF 5* Phusion, 500 nM de cada cebador directo, cebador directo 1 micromolar, 1 microlitro de reserva de dNTP 10 nM, polimerasa Phusion 1,25 unidades y agua MQ estéril. El programa del termociclador consistió en un programa de toma de contacto seguido de una etapa de amplificación regular: 1 ciclo a 98 °C durante 30 segundos, 10 ciclos a 98 °C durante 10 segundos, 65 °C disminuyendo 1,5 °C por ciclo 10 segundos, 72 °C 20 segundos, 10 ciclos a 98 °C durante 10 segundos, 55 °C 10 segundos, 72 °C 20 segundos y 1 ciclo a 72 °C durante 3 minutos. El programa PCR de la tercera ronda se estableció como se describe en la figura 2. Volúmenes de PCR de 50 microlitros estaban compuestos por 50 ng de producto de PCR (de las rondas de PCR anteriores, figura 2), 10 microlitros de tampón de polimerasa HF 5* Phusion, cebador directo 1 micromolar (tabla 1 y figura 2), cebador inverso 1 micromolar, 1 microlitro de reserva de dNTP 10 mM, polimerasa Phusion 1,25 unidades y agua MQ estéril. El programa consistió en un programa de toma de contacto seguido de una etapa de amplificación regular: 1 ciclo a 98 °C durante 30 segundos, 10 ciclos a 98 °C durante 10 segundos, 65 °C disminuyendo 1,5 °C por ciclo de 10 segundos, 72 °C 20 segundos, 10 ciclos a 98 °C durante 10 segundos, 55 °C 10 segundos, 72 °C 20 segundos y 1 ciclo a 72 °C durante 3 minutos. Después de las amplificaciones por PCR, todos los productos de la PCR se purificaron en gel usando Qiaex II de acuerdo con los protocolos del fabricante.
Digestiones con enzimas de restricción
Los productos purificados se sometieron a digestión con BstEII y Sfil en dos etapas. En primer lugar, se realizó la digestión de 1 microgramo de ADN en reacciones de 100 microlitros que consistían en 10 microlitros de tampón 10* NEB 3 (New England Biolabs), 1 microlitro de 100* BSA, 12,5 unidades de BstEII y agua estéril durante 6 horas a 60 °C en una estufa. Los productos se purificaron usando el kit de Purificación de PCR Qiaquick de Qiagen de acuerdo con el manual de instrucciones y se eluyeron en 40 microlitros de agua. A continuación, todos los productos se sometieron adicionalmente a digestión con Sfil en reacciones de 100 microlitros que consistían en 10 microlitros de tampón 10* NEB 2 (New England Biolabs), 1 microlitro de 100* BSA, 12,5 unidades de Sfil y agua estéril durante 12 horas a 50 °C en una estufa. Los fragmentos digeridos se purificaron usando el kit de Extracción en Gel Qiaquick seguido de separación en gel sobre un gel de bromuro de etidio más TBE agarosa 1,5 % de 20 cm a 80 V. 100 microgramos del vector aceptor (MV1043, figuras 3 y 12) se sometieron a digestión con 50 unidades de Eco91l en 600 microlitros en condiciones convencionales (tampón Tango) y después se purificó en un gel de agarosa al 0,9 %. Después de una segunda etapa de digestión en condiciones recomendadas con 400 unidades de Sfil en 500 microlitros durante 12 horas, se añadieron 100 unidades de BsrGI durante 3 horas a 50 °C.
Ligamientos
Cada producto de la PCR se ligó por separado de acuerdo con el siguiente esquema: 70 ng de productos PCR digeridos, 300 ng de vector aceptor digerido, 100 unidades de T4 Ligasa (NEB), tampón ligasa 1* en 30 microlitros durante 16 horas a 12 °C. Las reacciones de ligamiento se purificaron con extracciones de fenol/cloroformo/alcohol isoamílico seguido de precipitaciones con glucógeno (Sigma-Aldrich N.° G1767) de acuerdo con el protocolo del fabricante y finalmente se disolvieron en 25 microlitros de agua estéril.
Transformaciones y conservación de bibliotecas
Los productos de ligamiento purificados se transformaron por electroporación usando 1200 microlitros de bacterias electrocomponentes TG1 (Stratagene N.° 200123) por lote de ligamiento y se sembraron en placas de caldo LB con carbenicilina que contenía glucosa al 4 %. Las bibliotecas se recogieron raspando las bacterias en 50 ml de caldo LB con carbenicilina. Después de centrifugar a 2000 g durante 20 minutos a 4 °C, los sedimentos bacterianos se resuspendieron cuidadosamente en 2 ml de 2*TY/glicerol al 30 % enfriado con hielo en agua helada y se congelaron en hielo seco/etanol antes de conservar a -80 °C.
Amplificación de bibliotecas
Las bibliotecas se cultivaron y se recogieron de acuerdo con los procedimientos descritos por Kramer et al. 2003 (Kramer et al. 2003. Nucleic Acids Res. 31(11): e59) usando VCSM13 (Stratagene) como cepa fago auxiliar.
Selección de fagos en inmunotubos revestidos
El toxoide tetánico se disolvió en PBS en una concentración de 2 pg/ml y se revistió en tubos MaxiSorp Nunclmmuno (Nunc 444474) durante una noche a 4 °C. Después de desechar la solución de revestimiento, los tubos se bloquearon con leche desnatada (ELK) al 2 % en PBS (tampón de bloqueo) durante 1 hora a TA. En paralelo, 0,5 ml de la fagoteca se mezclaron con 1 ml de tampón de bloqueo y se incubó durante 20 minutos a temperatura ambiente. Después de bloquear los fagos, la solución de fagos se añadió a los tubos revestidos con el toxoide tetánico y se incubó durante 2 horas a TA en una plataforma rotatoria lenta para permitir la unión. A continuación, los tubos se lavaron 10 veces con PBS/Tween-20 al 0,05 % seguido por elución de fagos mediante una incubación con 1 ml de glicina-HCl 50 mM a pH 2,2 durante 10 min a TA en una rueda rotatoria y directamente seguido de neutralización del eluyente recogido con 0,5 ml de Tris-HCl 1 M a pH 7,5.
Recogida de clones de fagos. A la solución de fagos recogida se añadieron 5 ml de cultivo XL1-Blue MRF (Stratagene) a una D.O. de 0,4 y se incubó durante 30 minutos a 37 °C sin agitar para permitir la infección de los fagos. Las bacterias se sembraron en placas 2*TY de glucosa al 4 % Tetraciclina/Carbenicilina y se cultivó durante una noche a 37 °C.
Producción de fagos
Los fagos se cultivaron y procesaron como describen Kramer et al. 2003 (Kramer et al. 2003. Nucleic Acids Res.
31(11): e59) usando VCSM13 como cepa fago auxiliar.
ELISA para fagos
Se revistieron placas ELISA con 100 microlitros de IL-6 recombinante por pocillo a una concentración de 2 microgramos/ml en PBS durante una noche a 4 °C. Se usaron placas revestidas con 100 microlitros de tiroglobulina a una concentración de 2 microgramo/ml en PBS como control negativo. Los pocillos se vaciaron, se secaron golpeteando suavemente sobre una toallita de papel, se cargaron completamente con PBS-leche desnatada (ELK) al 4 % y se incubaron durante 1 hora a temperatura ambiente para bloquear los pocillos. Después de desechar la solución de bloqueo, las mini-preparaciones de fagos premezcladas con 50 pl de solución de bloqueo se añadieron y se incubaron durante 1 hora a tA. Después, 5 etapas de lavado con PBS-Tween-20 al 0,05 % retiraron los fagos no unidos. Los fagos no unidos se detectaron incubando los pocillos con 100 microlitros de anticuerpo anti-M13 conjugado con HRP (diluido a 1/5000 en tampón de bloqueo) durante 1 hora a temperatura ambiente. Se retiraron los anticuerpos libres repitiendo las etapas de lavado como se ha descrito anteriormente, seguido de incubación en sustrato TMB hasta ser visible el revelado del color. La reacción se detuvo añadiendo 100 microlitros de H2 SO4 2 M por pocillo y se analizó en un lector ELISA a una longitud de onda de emisión de 450 nm (tabla 2). Números mayores indican señales más fuertes y, por tanto, mayor incidencia de unión específica del complejo de fago-Fab.
Secuenciación
Los clones que dieron señales al menos 3 veces por encima de la señal de fondo (Tabla 2) se propagaron, se usaron para procedimientos miniprep ADN (véase el manual de procedimientos miniPrep de Qiagen) y se sometieron a análisis de secuencia de nucleótidos. La secuenciación se realizó de acuerdo con el manual adjunto al kit Big Dye 1.1 (Applied Biosystems) usando un cebador inverso (CH1_Rev1, tabla 1) que reconoce una secuencia 5' de la región CH1 de la cadena pesada IgG1 humana (presente en el vector de presentación Fab MV1043, figuras 3 y 12). Las secuencias VH de ratón de 28 clones de unión al toxoide tetánico se representan en la Tabla 3. Los resultados muestran que los genes VH murinos seleccionados pertenecen a diferentes familias de genes y diferentes miembros individuales de estas familias de genes son capaces de emparejarse con la región VH de IGKV1-39/J humana reordenada para formar sitios de unión a anticuerpo específicos del toxoide tetánico funcionales. A partir de los análisis de secuencia, se llegó a la conclusión de que las regiones VH murinas utilizan una diversidad de segmentos génicos DH y JH.
Ejemplo 3
Silenciamiento del locus de cadena ligera kappa de ratón
Este ejemplo describe el silenciamiento de los locus endógenos de cadena ligera kappa de ratón. El locus kappa endógeno se modifica por recombinación homóloga en células ES, seguido de la introducción de células Es modificadas genéticamente en embriones de ratón para obtener descendencia genéticamente adaptada.
Un vector que contiene una secuencia de nucleótidos ensamblada que consta de una parte que comprende la región J a 338 pb cadena abajo del segmento génico J5 fusionada con una secuencia 3' terminal del potenciador CK 3' se usa para recombinación homóloga en células ES. La secuencia ensamblada se usa para delecionar un fragmento de ADN genómico que abarca desde el extremo 3' de la región JK justo hasta el extremo 3' del potenciador CK 3'. Como una consecuencia de este procedimiento, se elimina el gen constante CK, el potenciador 3' y algunas regiones intergénicas (véanse las figuras 4 y 18).
Construcción del vector de direccionamiento
Un vector que recibía brazos flanqueantes de 4,5-8 kb en los extremos 3' y 5' fusionado con el segmento de deleción se usó para dirigir la recombinación homóloga en una línea de células ES. Ambos brazos se obtuvieron por medio de PCR garantizando una homología máxima. La estrategia de direccionamiento permite la generación del alelo KO constitutivo. La secuencia genómica de ratón que abarca el potenciador intrónico Igk, la región constante Igk y el potenciador 3' Igk se reemplazó con un casete PuroR, que estaba flanqueado por sitios F3 y se insertó cadena abajo de los elementos Jk. La eliminación mediada por Flp del marcador de selección dio como resultado un alelo KO constitutivo. El reemplazo de la región genómica Igk MiEk-Igk C-Igk 3'E (aproximadamente 10 kb) con un casete F3-Puro (aproximadamente 3 kb) disminuyó probamente la eficacia de la recombinación homóloga. Por lo tanto, los brazos de homología se extendieron por consiguiente y se analizaron más colonias de células ES después de la transfección para identificar clones recombinantes homólogos.
Generación de células ES que llevan el fragmento kappa delecionado
La generación de células ES modificadas genéticamente se realizó esencialmente como se ha descrito (Seibler et al. Nucleic Acids Res. 15 de febrero de 2003;31(4):e12). Véase también el ejemplo 14 para una descripción detallada. Generación de ratones ES por complementación de embriones tetraploides.
La producción de ratones por complementación de embriones tetraploides utilizando células ES modificadas genéticamente se realizó esencialmente como se ha descrito (Eggan et al., PNAS 98, 6209-6214; Seibler J, et al. Nucleic Acids Res. 15 de febrero de 2003;31 (4):e12; Hogan et al., (Summary of mouse development. Manipulating the Mouse Embryo, (1994) Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.), páginas 253-289.)). Ejemplo 4
Silenciamiento del locus de cadena ligera lambda de ratón
Este ejemplo describe el silenciamiento del locus endógeno de cadena ligera lambda de ratón. El locus endógeno lambda se modifica por recombinación homóloga en células ES seguido de la introducción de células ES modificadas genéticamente en embriones de ratón para obtener descendencia genéticamente adaptada
Dos regiones del locus lambda murino que en su conjunto contiene todas las regiones V lambda funcionales se someten a deleción.
La primera región dirigida para deleción basada en recombinación homóloga es una región que se localiza 408 pb cadena arriba del sitio de inicio del segmento génico IGLV2 y acaba 215 pb cadena abajo de segmento génico IGLV3, incluyendo el tramo de secuencia intergénica entre estos segmentos génicos IGLV. La segunda región que se somete a una deleción implica el segmento génico IGLV1 que consiste en un fragmento que abarca desde 392 pb cadena arriba a 171 pb cadena abajo del segmento génico IGLV1. Como una consecuencia de estas dos etapas de deleción, se delecionaron todos los segmentos génicos lambda V funcionales, que vuelven al locus funcionalmente inactivo (figuras 5 y 19).
Construcción de los vectores de direccionamiento
Los vectores que recibieron brazos flanqueantes de 3-9,6 kb en los extremos 3' y 5' fusionados con el segmento de deleción se usaron para recombinación homóloga diana en una línea de células ES. Ambos brazos se obtuvieron por medios PCR garantizando máxima homología. En una primera etapa, la secuencia genómica de ratón que abarcaba las regiones V2-V3 de Igl se reemplazó con un casete PuroR flanqueado por sitios F3, que produce un alelo KO constitutivo después de la retirada mediada por Flp del marcador de selección (véase la figura 19A). En una segunda etapa, la secuencia genómica de ratón que abarcaba la región V1 de Igl se reemplazó con un casete Neo en clones de células ES que ya llevaban una deleción de las regiones V2-V3 de Igl (véase la figura 19B). El marcador de selección (NeoR) se flanqueó por sitios FRT. Se obtuvo un alelo KO constitutivo después de retirada mediada por Flp de marcadores de selección.
Generación de células ES que llevan el fragmento lambda delecionado
La generación de células ES modificadas genéticamente se realizó esencialmente como se ha descrito (Seibler J,
Zevnik B, Küter-Luks B, Andreas S, Kern H, Hennek T, Rode A, Heimann C, Faust N, Kauselmann G, Schoor M, Jaenisch R, Rajewsky K, Kühn R, Schwenk F. Nucleic Acids Res. 15 feb 2003;31(4):e12). Véase también el ejemplo 14 para una descripción detallada. Para demostrar que ambos eventos de direccionamiento se producían en el mismo cromosoma se seleccionaron diversos clones doble diana para la deleción in vitro con pCMV C31deltaCpG. Los clones se expandieron bajo presión con antibióticos en una capa alimentadora mitóticamente inactivada que comprendía fibroblastos embrionarios de ratón en medio DMEM con Alto contenido en Glucosa que contenía FCS (Pa N) al 20 % y Factor Inhibidor de Leucemia 1200 u/ml (Millipore ESG 1107). 1x107 células de cada clon se sometieron a electroporación con 20 |jg de pCMV C31deltaCpG circular a 240 V y 500 jF y se sembraron en 4 placas de 10 cm cada una. 2 - 3 días después de la electroporación las células se recogieron y se analizaron por PCR. Los cebadores utilizados fueron:
2005_5: CCCTTTCCAATCTTTATGGG
2005_7: AGGTGGATTGGTGTCTTTTTCTC
2005_9: GTCATGTCGGCGACCCTACGCC
Las reacciones PCR se realizaron en mezclas que comprendían 5 j l de Tampón PCR 10x (Invitrogen), 2 j l de MgCh (50 mM), 1 j l de dNTP (10 mM), 1 j l del primer cebador (5 jM), 1 j l del segundo cebador (5 jM), 0,4 j l de Taq (5 U/ul, Invitrogen), 37,6 j l de H2 O y 2 j l de ADN. El programa usado fue a 95 °C, 5'; seguido de 35 ciclos a 95 °C 30”; 60 °C 30”; 72 °C 1'; seguido de 72 °C 10'.
Generación de ratones ES por complementación con embriones tetraploides
La producción de ratones por complementación con embriones tetraploides usando células ES modificadas genéticamente se realizó esencialmente como se ha descrito (Eggan et al., PNAS 98, 6209-6214; Seibler J, Zevnik B, Küter-Luks B, Andreas S, Kern H, Hennek T, Rode A, HeimannC, Faust N, Kauselmann G, Schoor M, Jaenisch R, Rajewsky K, Kühn R, Schwenk F. Nucleic Acids Res. 15 feb 2003;31(4):e12; Hogan et al., (Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.), páginas 253-289).
Ejemplo 5
Construcción del inserto de expresión CAGGS basado en un gen IGKV1-39/J-Ck de la línea germinal humana reordenado (IGKV1-39/J-Ck)
Este ejemplo describe la construcción de un casete de expresión CAGGS que incorpora la región IGKV1-39/J de la línea germinal humana reordenada. Este casete de expresión inserto abarca sitios de clonación, una secuencia Kozak, una secuencia líder que contiene un intrón, una fase de lectura abierta de la región IGKV1-39 reordenada, una región constante CK de rata del alelo a y una secuencia de terminación traduccional (IGKV1-39/J-Ck; figura 6). La construcción primaria consta de secuencias de origen natural y se ha analizado y optimizado eliminando elementos no deseados que actúan en cis como cajas TATA internas, señales de poliadenilación, sitios chi, sitios de entrada ribosomales, tramos de secuencia ricos en AT o ricos en GC, elementos de secuencia ARE, INS y CRS, secuencias de repetición, estructuras secundarias de ARN, sitios donantes y aceptores de corte y empalme (crípticos) y puntos de ramificación de corte y empalme (GeneArt GmbH). Además, para la expresión en ratones, se optimizó el uso de codones en las regiones de fase de lectura abierta. La secuencia intrónica es invariable y por tanto representa la secuencia idéntica a la parte codificante del intrón líder IGKV1-39 humano.
En el extremo 5' del casete de expresión, se introdujo un sitio Notl y en el extremo 3' un sitio Nhel. Ambos sitios se usan para la clonación en el módulo de expresión CAGGS. Después del ensamblaje génico de acuerdo con los procedimientos usados por GeneArt, el inserto se somete a digestión con NotI-NheI y se clona en el módulo de expresión que contiene un promotor CAGGS, una secuencia bloqueadora flanqueada por sitios LoxP ('floxed'), una secuencia señal de poliadenilación y, en el extremo 5' y 3', secuencias para facilitar la recombinación homóloga en el locus Rosa26 de las líneas de células ES de ratón. Los fragmentos promotores y/o de ADNc se amplificaron por PCR, se confirmaron por secuenciación y/o se clonaron directamente a partir de plásmidos administrados en un vector de intercambio RMCE que llevaba las características indicadas. En la figura 13A y 13B se muestra un dibujo esquemático y la secuencia confirmada del vector de direccionamiento final pCAGGS-IgVK1-39. La estrategia diana se representa en la figura 13c.
Ejemplo 6
Inserto de expresión CAGGS en base a la región lambda IGLV2-14/J V de la línea germinal reordenada (IGLV2-14/J-Ck)
Este ejemplo describe la secuencia e inserción de un casete de expresión que incorpora la región lambda V de IGLV2-14/J de la línea germinal reordenada. Este inserto abarca sitios de clonación, una secuencia Kozak, una secuencia líder que contiene un intrón, una fase de lectura abierta de la región IGLV2-14/J reordenada, una región constante CK de rata del alelo a y una secuencia de terminación traduccional (IGLV2-14/J-Ck; figura 7). La construcción primaria consta de secuencias de origen natural y se ha analizado y optimizado eliminando elementos
no deseados que actúan en cis como: cajas TATA internas, señales de poliadenilación, sitios chi, sitios de entrada ribosomales, tramos de secuencia ricos en AT o ricos en GC, elementos de secuencia ARE, INS y CRS, secuencias de repetición, estructuras secundarias de ARN, sitios donantes y aceptores de corte y empalme (crípticos) y puntos de ramificación de corte y empalme (GeneArt GmbH). Además, para la expresión en ratones, se optimizó el uso de codones en las regiones de fase de lectura abierta. La secuencia intrónica es invariable y por tanto representa la secuencia idéntica a la del intrón líder IGKV1-39 humano.
En el extremo 5' del casete de expresión, se introdujo un sitio NotI y en el extremo 3' un sitio NheI. Ambos sitios se usan para la clonación en el módulo de expresión CAGGS como describe TaconicArtemis. Después del ensamblaje génico de acuerdo con los procedimientos usados por GeneArt, el inserto se sometió a digestión con NotI-NheI y se clonó en el módulo de expresión que contenía un promotor CAGGS, una secuencia bloqueadora flanqueada por sitios LoxP ('floxed'), una secuencia señal de poliadenilación y, en el extremo 5' y 3', secuencias para facilitar la recombinación homóloga en el locus Rosa26 de las líneas de células ES de ratón. Para construir el vector dirigido RMCE ROSA26 final, se amplificaron los fragmentos promotores y/o de ADNc por PCR. Los productos amplificados se confirmaron por secuenciación y/o se clonaron directamente a partir de plásmidos administrados en un vector de intercambio RMCE que llevaba las características indicadas. En las figuras 15A y 15B se muestra un dibujo esquemático y la secuencia confirmada del vector dirigido final pCAGGS-IgVL2-14. En la figura 15C se representa la estrategia de direccionamiento.
Ejemplo 7
Expresión de IGKV1-39/J-Ck en líneas de células HEK293 (pSELECT-IGKV1-39/J-Ck)
Este ejemplo describe un procedimiento para verificar que las construcciones IGKV1-39/J-Ck descritas en el ejemplo 5 permiten la expresión y detección de la cadena L de IGKV1-39/J-Ck en células HEK293. El inserto IGKV1-39/J (figura 6) se modificó en el extremo 5' cambiando el sitio NotI en un sitio SalI. Este cambio se requiere para la clonación del producto en el plásmido de casete de expresión pSELECT-hygro (InvivoGen). El inserto de expresión CAGGS IGKV1-39/J-Ck y pSELECT-hygro se sometieron a digestión con SalI y NheI, se ligaron y se usaron para transformar células XL1-Blue competentes usando técnicas convencionales. Las colonias se seleccionaron y el ADN se purificó usando columnas Midi-prep de Qiagen de acuerdo con los procedimientos del fabricante. El vector que expresa la cadena ligera (LC) resultante denominado 0817676_pSELECT_0815426 se usó para transfectar células HEK293 con Fugene6 (Roche) de acuerdo con los protocolos del fabricante. Los sobrenadantes se exploraron para determinar la presencia de cadenas ligeras de IGKV1-39/J-Ck por ELISA y transferencia de Western usando anticuerpos anti-Ck de rata (Beckton Dickinson N.° 550336 y 553871) y los protocolos usados en la técnica.
La VH de la IgG anti-toxoide tetánico (TT) MG1494 se clonó en el vector de expresión de IgG MV1056 usando los sitios de restricción SfiI y BstEII. El clon resultante se verificó por secuenciación. Las células HEK293T se transfectaron con cinco combinaciones de vectores diferentes, como se muestra en la Tabla 4 (véase el ejemplo 8 para detalles del vector 0817678_pSELECT_0815427). Los sobrenadantes se recogieron y se determinaron las concentraciones de IgG (véase la Tabla 4). No pudo detectarse IgG para los sobrenadantes A y B que solo contenían cadena ligera como se esperaba (parte Fc de IgG reconocida por anticuerpos de detección). La concentración de IgG en los sobrenadantes C y D fue comparable a la del sobrenadante E de control positivo, indicando expresión correcta de las construcciones de cadena ligera.
La unión a TT se analizó por ELISA para comprobar la funcionalidad de los anticuerpos producidos, usando hemoglobina como antígeno de control negativo. No pudo detectarse unión específica a tT para los sobrenadantes A y B que solo contenían cadena ligera como se esperaba. La unión específica a TT para los sobrenadantes C y D fue al menos tan buena como para el sobrenadante E de control positivo, confirmando expresión correcta de las construcciones de cadena ligera y ensamblaje funcional con cadena pesada. Se detectaron los anticuerpos no solo usando un anticuerpo secundario anti-IgG humana, sino también un anticuerpo secundario anti-cadena ligera Ckappa de rata. Los resultados confirman que el anticuerpo anti-Ckappa de rata (BD Pharmingen N.° 553871, clon MRK-1) reconoce la cadena ligera expresada por los vectores pSELECT.
Los sobrenadantes se analizaron por SDS-PAGE no reductora y transferencia de Western (no mostrada). La detección usando un anticuerpo anti-cadena pesada de IgG humana, no mostró bandas para los sobrenadantes A y B que solo contenían cadena ligera, como se esperaba. Los resultados para los sobrenadantes C y D fueron comparables a los del sobrenadante E de control positivo, con una banda cerca del marcador de 170 kD como se esperaba para la IgG intacta. Se observaron bandas adicionales de peso molecular más bajo también en los sobrenadantes C, D y E, que podrían representar productos de degradación, fragmentos IgG resultantes de reducción (parcial) y/o bandas irrelevantes de proteína debido a unión no específica del anticuerpo de detección. La detección usando un anticuerpo anti-cadena ligera Ckappa de rata mostró una banda cerca del marcador de 26 kD para los sobrenadantes A y B, como se esperaba solo para cadena ligera. Esta banda era mucho más intensa para A en comparación con B, lo que indicaba que la cadena ligera IGKV1-39 libre podía expresarse mejor y/o era más estable que la cadena ligera IGLV2-14 libre. No se detectaron bandas para el sobrenadante E de control como se esperaba, dado que la IgG expresada contiene una cadena ligera Ckappa humana. Para los sobrenadantes C y
D, se observaron bandas esperadas cerca del marcador de 170 kD; también se observaron bandas de peso molecular más bajo, pero a un grado más bajo que el anterior usando el anticuerpo anti-IgG humana.
En conclusión, la transfección de las construcciones de expresión de cadena ligera combinado con la cadena pesada de la IgG anti-toxoide tetánico (TT) MG 1494 dio como resultado la producción de IgG comparable a la de la construcción de control positivo para ambas construcciones de cadena ligera lambda y kappa pSELECT. Ambas producciones de IgG produjeron señales ELISA en un ELISA TT que eran mejores que o comparables con las del control de IgG. Los análisis de SDS-PAGE y transferencia de Western confirmaron la presencia de IgG intacta. El anticuerpo anti-Ckappa de rata ensayado trabajó eficazmente en el ELISA y en la transferencia de Western. El sobrenadante de cultivo de las células transfectadas solo con construcciones de cadena ligera no dio como resultado producción detectable de IgG ni unión específica detectable a TT, mientras que la cadena ligera libre se detectó por transferencia de Western.
Ejemplo 8
Expresión de IGLV2-14/J-Ck en líneas de células HEK293 (pSELECT-IGLV2-14/J-Ck)
Este ejemplo describe un procedimiento para verificar que las construcciones de IGLV2-14/J descritas en el ejemplo 6 permiten la expresión y detección de la cadena L de IGLV2-14/J-Ck en células HEK293. El inserto IGLV2-14/J-Ck (figura 7) se modificó en el extremo 5' cambiando el sitio NotI en un sitio SalI. Este cambio se requiere para la clonación del producto en el plásmido del casete de expresión pSELECT-hygro (InvivoGen). El inserto de expresión CAGGS IGLV2-14/J-Ck y pSELECT-hygro se sometieron a digestión con Sall y NheI, se ligaron y se usaron para transformar células XL1-Blue competentes usando técnicas convencionales. Las colonias se seleccionaron y el ADN se purificó usando columnas Midi-prep de Qiagen de acuerdo con los procedimientos del fabricante. El vector que expresaba la cadena ligera (LC) resultante denominado 0817678_pSELECT_0815427 se usó para transfectar células HEK293 con Fugene6 (Roche) de acuerdo con los protocolos del fabricante. Los sobrenadantes se exploraron para determinar la presencia de cadenas ligeras IGLV2-14/J-Ck por ELISA y transferencia de Western usando anticuerpos anti-Ck de rata (Becton Dickinson N.° 550336 y 553871) y protocolos usados en la técnica. Véase el ejemplo 7 para detalles y resultados.
Ejemplo 9
Construcción de una construcción de expresión dirigida por el promotor VK que contiene un inserto IGKV1-39/J y elementos potenciadores múltiples derivados del locus CK murino (VkP-IGKV1-39/J-Ck; VkP-O12)
Este ejemplo describe la construcción de un casete de expresión que contiene elementos relevantes que permite la expresión específica de fase de linfocitos B y desarrollo/diferenciación de la región IGKV1-39 VK humana reordenada, en función de la región promotora IGKV1-39 VK, líder que contiene un intrón, gen V de línea germinal, CDR3, segmento IGKJ, región intergénica de ratón localizada entre la fase de lectura abierta Jk y Ck, y alelo a Ck de rata, y un fragmento intergénico de ratón desde el extremo 3' del gen CK de ratón acabando justo en 3' del potenciador CK 3'.
Las fases de lectura abierta optimizadas del líder, del gen IGKV1-39 reordenado y del gen a del alelo CK de rata, como se describe en el ejemplo 5, se usaron para la construcción del casete de expresión. La región promotora VK se obtuvo por procedimientos de síntesis génica (GeneArt, GmbH) y es casi idéntica a la secuencia de la región IGKV1-39 humana entre -500 pb y el ATG (sitio de inicio) del gen. La única desviación de la secuencia natural es la introducción de una secuencia Kozak GCCACCATGG en el sitio ATG (inicio) para promover la traducción. Como base para la introducción de elementos individuales se usa un fragmento genómico de un clon BAC de ratón (TaconicArtemis). Este fragmento es idéntico a la secuencia del locus VK de ratón comenzando con el sitio donante intrónico localizado directamente en 3' de la región JK5 y finalizando justo en 3' del potenciador CK 3' y cubre aproximadamente 12,5 kb.
La construcción final contiene desde el extremo 5' a 3' los siguientes elementos: promotor IGKV1-39 genómico humano (500 pb), una secuencia Kozak, una parte 1 del líder IGKV1-39 humano (optimizado), un intrón líder IGKV1-39 humano, una parte 2 del líder IGKV1-39 humano (optimizado), un gen de la línea germinal IGKV1-39 humana (optimizado), una región J humana (optimizada), una región intergénica de ratón incluyendo el elemento potenciador intrónico, una región constante kappa de rata (Rattus norvegicus) (optimizada), y una región intergénica de ratón incluyendo el potenciador kappa 3'. Los elementos de este casete de expresión se muestran en la figura 8 y se denominan VkP-IGKV1-39/J-Ck (VkP-O12). En las figuras 20A y 21A se representa un esquema del vector pVkP-O12 y la estrategia de direccionamiento. El vector se introdujo en células ES siguiendo procedimientos convencionales (véase el ejemplo 14).
Ejemplo 10
Construcción de una construcción de expresión dirigida por el promotor VK que contiene un clon IGLV2-14/J y elementos potenciadores derivados del locus CK múltiple (VkP-IGLVL2-14/J-Ck; VkP-2a2).
Este ejemplo describe la misma construcción a la descrita en el ejemplo 9, excepto que el gen IGKV1-39 y la región J se reemplazan por el gen de la línea germinal IGLV2-14 humana optimizado incluyendo una región V-J única (VkP-IGLV2-14/JCk; VkP-2a2; figura 9).
Ejemplo 11
Construcción de una construcción de expresión dirigida por el promotor VK que contiene un clon IGKV1-39 que carece del elemento potenciador intrónico de CK (VkP-IGKV1-39/J-Ck-A1; VkP-O12-del1)
La construcción descrita en el ejemplo 9 se modificó eliminando el elemento potenciador intrónico de CK, localizado en la región intergénica entre la región J humana y la región CK de rata por modificación PCR convencional y metodologías de clonación de ADN (GeneArt, GmBH). El casete de expresión resultante se muestra en la figura 10 y se denominó VkP-IGKV1-39/JCk-A1 (VkP-012-dell).
En las figuras 20B y 21B se representa un esquema del vector pVkP-O12-del1 y la estrategia de direccionamiento. El vector se introdujo en células ES siguiendo procedimientos convencionales (véase el ejemplo 14).
Ejemplo 12
Construcción de una construcción de expresión dirigida por el promotor VK que contiene un clon IGKV1-39 que carece del elemento potenciador intrónico de CK y un elemento potenciador de Ck 3' truncado (VkP-IGKV1-39/J-Ck-A2; VkP-O12-del2)
La construcción descrita en el ejemplo 11 se modificó truncando el elemento potenciador CK 3' y delecionando parte de la región intergénica 3' del gen Ck de rata, para eliminar posibles elementos inhibidores. Esto se consiguió eliminando la secuencia intergénica entre un sitio EcoRV (localizado 3' del gen Ck de rata) y el sitio NcoI presente en el potenciador 3' (5993 pb) y eliminando adicionalmente la secuencia entre el sitio BstXI potenciador 3' y el sitio Bstxi 3' del potenciador 3' (474 pb) usando procedimientos convencionales. El casete de expresión resultante se muestra en la figura 11 y se denominó VkPIGKvi-39/J-Ck-A2 (VkP-O12-del2).
En las figuras 20C y 21C se representa un esquema del vector pVkP-O12-del2 y la estrategia diana. El vector se introdujo en células ES siguiendo procedimientos convencionales (véase el ejemplo 14).
Ejemplo 13
Expresión de construcciones Vk en líneas celulares
Las construcciones descritas en los ejemplos 9-12 se sometieron a ensayo con respecto a su capacidad para producir proteínas de cadena ligera en las líneas de células de mieloma MPC11 (ATCC CCL167), linfoma de linfocitos B WEHI231 (ATCC CRL-1702), el linfoma de linfocitos T EL4 (ATCC TIB-39) y en HEK293 (ATCC CRL1573). Los elementos potenciadores y promotores en la construcción permiten la expresión en líneas de linfocitos B pero no en líneas de células derivadas de otros tejidos. Después de la transfección de las líneas celulares usando ADN linealizado purificado y Fugene6 (Roche), las células se cultivan para expresión transitoria. Las células y el sobrenadante se recogen y se someten a análisis SDS-PAGE seguido de transferencia de Western usando un anticuerpo anti-C-kappa de rata específico. Los sobrenadantes se analizan en ELISA para detectar cadenas L segregadas usando el anticuerpo anti-CK de rata (Beckton Dickinson N.° 550336).
Ejemplo 14
Generación de líneas ES transgénicas
Todas las construcciones descritas en los ejemplos 3, 4, 5, 6, 9, 10, 11 y 12 se usaron para generar líneas ES transgénicas estables individuales mediante recombinación homóloga. En la técnica se conocen procedimientos para generar líneas ES transgénicas mediante recombinación homóloga (por ejemplo, Eggan et al., PNAS 98, 6209 6214; Seibler J, Zevnik B, Küter-Luks B, Andreas S, Kern H, Hennek T, Rode A, HeimannC, Faust N, Kauselmann G, Schoor M, Jaenisch R, Rajewsky K, Kühn R, Schwenk F. Nucleic Acids Res. 15 de febrero de 2003;31(4):e12; Hogan et al. (Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY.), páginas 253-289.).
Para todas las construcciones descritas en los ejemplos 5-6, y en los ejemplos 9-12, la línea celular ES RMCE (derivada de la cepa de ratón 129S6B6F1-Gt(ROSA)26Sortm10Arte) se cultivó en una placa alimentadora mitóticamente inactivada que comprendía fibroblastos embrionarios de ratón (FER) en medio DMEM con Alto contenido de Glucosa que contenía FbS al 15 % (PAN 1302-P220821). Se añadió Factor Inhibidor de Leucemia (Chemicon ESG 1107) al medio a una concentración de 900 U/ml. Para la manipulación, 2x105 células ES se sembraron en placas de 3,5 cm con 2 ml de medio. Directamente antes de la transfección, se añadieron 2 ml de medio reciente a las células. Se mezclaron 3 ul de reactivo Fugene6 (Roche; Catálogo N.° 1814443) con 100 ul de
medio asérico (OptiMEM I con Glutamax I; Invitrogen; Catálogo N.° 51985-035) y se incubó durante 5 min. Se añadieron 100 ul de la solución Fugene/OptiMEM a 2 ug del vector circular y 2 ug de CAGGS-Flp y se incubó durante 20 min. Este complejo de transfección se añadió gota a gota a las células y se mezcló. Se añadió medio reciente a las células al día siguiente. Desde el día 2 en adelante el medio se reemplazó diariamente con medio que contenía G418250 ug/ml (Geneticin; Invitrogen; Catálogo N.° 10131-019). Siete días después de la transfección, se aislaron clones sencillos, se expandieron y se realizó su análisis molecular por transferencia de Southern de acuerdo con procedimientos convencionales.
Para cada construcción, el análisis de clones múltiples por digestión con enzimas de restricción de ADN genómico de clones sencillos seguido de hibridación con sondas 5', sondas 3' y sondas internas dio como resultado clones que comprendían una sola inserción correcta en la posición correcta en el locus Rosa26. En la figura 14 se proporciona un ejemplo.
Ejemplo 15
Generación de cepas transgénicas de ratón
Todas las líneas de células ES que se generaron y se verificaron con respecto a sus modificaciones, como se describe en el ejemplo 14, se usaron para generar ratones transgénicos estables mediante recombinación tetraploide. Los procedimientos son conocidos en la técnica. En general, después de la administración de hormonas, hembras Balb/c superovuladas se aparearon con machos Balb/c. Los blastocistos se aislaron del útero en una capa hidrófuga (“dpc, damp-proof course”) de 3,5. Para la microinyección, los blastocistos se colocaron en una gota de DMEM con FCS al 15 % con aceite mineral. Una pipeta de microinyección, accionada de forma piezoeléctrica, de punta plana con un diámetro interno de 12 - 15 micrómetros se usó para inyectar 10-15 células ES C57BL/6 N.tac diana en cada blastocisto. Después de su recuperación, los blastocistos inyectados se transfirieron a cada trompa uterina de hembras NMRI pseudopreñadas, 2,5 días post-coito. El quimerismo se midió en quimeras (G0) por contribución de recubrimiento de color de células ES en hospedador Balb/c (negro/blanco). Los ratones muy quiméricos se cruzaron con hembras de la cepa C57BL/6. Dependiendo de las necesidades del proyecto, los compañeros de apareamiento C57BL/6 son no mutantes (W) o mutantes para la presencia de un gen de recombinasa (supresor Flp o supresor Cre o supresor inducible CreER o combinación de supresor-Flp/CreER). La transmisión de la línea germinal se identificó por la presencia de descendencia de color negro, cepa C57BL/6, (G1). Por ejemplo, el clon ESC IgVK1-3926838 (véase los ejemplos 5 y 14) se inyectó en un total de 62 blastocitos en 3 experimentos independientes. Se obtuvieron 3 camadas con un total de 6 crías. Todas las crías eran quiméricas. Se obtuvieron 3 crías de descendencia heterocigota que se usaron para cruzamiento posterior.
El clon ESC Kappa 2692 A-C10 (véase los ejemplos 3 y 14) se inyectó en un total de 54 blastocistos en 3 experimentos independientes. Se obtuvieron 3 camadas con un total de 11 crías, de las cuales 10 eran quiméricas. Se obtuvieron 8 crías de descendencia heterocigota que se usaron para cruzamiento posterior.
El clon ESC Kappa 2692 B-C1 (véase los ejemplos 3 y 14) se inyectó en un total de 51 blastocistos en 3 experimentos independientes. Se obtuvieron 2 camadas con un total de 6 crías, de las cuales 4 eran quiméricas. Se obtuvieron 3 crías de descendencia heterocigota que se usaron para cruzamiento posterior.
Ejemplo 16
Cruzamiento
Este ejemplo describe el cruzamiento para obtener ratones que contienen casetes de expresión transgénicos como se describe en el ejemplo 14 y ratones genosuprimidos (“knock out”) en los que se habían silenciado los locus lambda y kappa endógenos. La localización de V-lambda en el cromosoma 16 y CD19 en el cromosoma 7 permite realizar procedimientos de cruzamiento convencionales. El cruzamiento del locus Vk y el locus Rosa26 co localizados en el cromosoma 6 con una distancia de aproximadamente 24 cM requiere especial atención durante la exploración ya que solo un porcentaje de la descendencia muestra entrecruzamiento de una manera que ambas modificaciones se integran en un cromosoma.
Los cuatro locus deben combinarse en una sola cepa de ratón que es homo o heterocigota para el transgén CD19-cre (no descrito) y Rosa26 modificado y homocigota para los otros locus. El cruzamiento se realiza mediante técnicas de cruzamiento y de exploración convencionales según sea apropiado y propuesto por compañías de cría del mercado (por ejemplo, TaconicArtemis).
Ejemplo 17
Inmunizaciones de ratones
Usando protocolos convencionales se realizan inmunizaciones primarias y de refuerzo
Para validar la expresión transgénica de cadenas ligeras de Ck de rata - Vk 012 (IGKV1-39) reordenadas humanas (véanse los ejemplos 5, 14-16) en linfocitos B de ratones CD19-HuVk1 y para evaluar su impacto sobre el tamaño del repertorio VH, diversidad de uso de familia VH y recombinación V(D)J después de inmunización, se inmunizaron los ratones transgénicos CD19-HuVk1 con la vacuna de la toxina tetánica (vacuna TT) y la diversidad de secuencia de VH de clones seleccionados al azar de ratones CD19-HuVk1 se comparan con ratones de ts inmunizados con TT y miembros de la misma camada negativos a CD19-Cre HuVkl. Se obtuvieron datos sobre la frecuencia SHM del transgén Vk 012 humano en los ratones inmunizados. Se recuperó una colección diversa de al menos 40 mAb no relacionados clonalmente, específicos de TT, que contenían la Vk 012 humana de ratones CD19-HuVk1 por presentación de fagos.
Para realizar esto, tres ratones CD19-HuVk1 adultos se vacunaron con vacuna TT usando procedimientos de inmunización convencionales. Después de la inmunización, se midieron los títulos en suero usando ELISA específico de TT (TT: Statens Serum Institute, Art. N.° 2674) y las suspensiones de bazo se sometieron a separación de células por el procedimiento FACS después de tinción con un anticuerpo monoclonal específico de Ck de rata para aislar linfocitos B transgénicos (clon RG7/9.1; BD Pharmingen N.° 553901, Lote N.° 06548). El ARN de linfocitos B positivos a Ck se extrajo y el material de ADNc resultante se usó para construir bibliotecas y realizar análisis SHM. El anticuerpo anti-CK de rata monoclonal convencional (clon RG7/9.1; BD Pharmingen N.° 553901, Lote N.° 06548) se usa en análisis FACS de linfocitos B que expresan el transgén (Meyer et al., 1996, Int. Immunol., 8: 1561). El anticuerpo RG7/9.1 del clon reacciona con un determinante de cadena kappa monotípico (común). Este anticuerpo anti-CK de rata (clon RG7/9.1 (BD Pharmingen N.° 553901, Lote N.° 06548) se marca con R-ficoeritrina (PE), usando el kit de conjugación rápida LYNX de acuerdo con las instrucciones del fabricante para análisis y separación por FACS. El anticuerpo marcado se somete en primer lugar a ensayo por citometría de flujo con respecto a la unión a proteínas de cadena ligera funcionales que contienen Ck de rata producidas en células HEK-293T transfectadas transitoriamente; el anticuerpo no conjugado sirve como un control positivo. Dos anticuerpos distintos que mostraron unirse a Ck de rata por ELISA y transferencia de Western (véase el ejemplo 7) se sometieron también a ensayo por citometría de flujo.
Se realiza la construcción de una biblioteca de presentación de fagos Fab con un conjunto de cebadores de PCR degenerados optimizados, diseñados para amplificar genes C57BL/6 VH; el tamaño mínimo de la biblioteca es de 106 clones y la frecuencia mínima de inserto es del 80 %. El vector usado, MV1043 (figuras 3 y 12), contiene el Vk 012 humano fusionado con una región Ck humana. La Ck de rata se intercambia por lo tanto por el homólogo humano en el proceso de generación de la biblioteca.
Antes de la selección, se realizó secuenciación de VH de 96 clones seleccionados al azar para validar la diversidad del repertorio de VH que se compara con la diversidad obtenida a partir de una biblioteca no seleccionada previamente generada usando los mismos procedimientos de los ratones BALB/c inmunizados con TT. Una biblioteca de ratones C57BI/6 de ts que se inmunizaron de la misma manera permite comparar la diversidad entre dos bibliotecas preseleccionadas que comparten la misma vacuna y el mismo fondo genético.
Diversas selecciones independientes se realizaron sobre TT que reviste inmunotubos. Las variables que pueden incluirse son selecciones usando antígenos biotinilados en solución o selecciones en TT de captura. Basándose en el número y diversidad de clones positivos a ELISA obtenidos en las primeras selecciones, se realizaron decisiones sobre rondas de selección adicionales. Los clones se consideran positivos cuando > 3x positivos sobre un clon control negativo. Los clones positivos se analizan por ELISA frente a un panel de antígenos de control negativos para verificar la especificidad antigénica. El objetivo es identificar al menos 40 regiones VH únicas, en función de secuencias CDR3 únicas y reordenamientos de VhDJh.
La amplificación del material de ADNc de linfocitos B clasificados positivos-CK de rata se realiza con un cebador directo de PCR específico para la secuencia líder humana y un cebador inverso de PCR específico para la secuencia Ck de rata, en una región no redundante con la secuencia Ck de ratón, como se describe en un estudio reciente (Brady et al., 2006, JIM, 315: 61). Las combinaciones de los cebadores y las temperaturas de hibridación se sometieron en primer lugar a ensayo sobre ADNc de células HEK-293T transfectadas con el vector 0817676_pSELECT_0815426 = pSELECT con el casete de ADN IGKV1-39 (véase el ejemplo 7).
Los productos de amplificación se clonan en el vector pJET-1 y después de transformación con XL1-blue, se secuencian 96 colonias para evaluar la frecuencia SHM de VL por comparación directa con la secuencia de la línea germinal Vk 012 (IGKV1-39). El procedimiento de proporción R/S, como se describe en este estudio en anticuerpos específicos de TT humanos (de Kruif et al., 2009, J. Mol. Biol., 387: 548) permite discriminar entre mutaciones al azar y mutaciones dirigidas por antígeno que se produjeron en secuencias VL.
Ejemplo 18
Análisis inmunofluorescente de poblaciones de linfocitos B en líneas de ratón transgénico
Este ejemplo describe el uso de anticuerpos y citometría de flujo para analizar poblaciones de linfocitos B en órganos linfoides primarios (médula ósea) y secundarios (bazo, cavidad peritoneal) y sangre. Se describen procedimientos y reactivos en Middendorp et al. (2002) J. Immunol. 168: 2695 y Middendorp et al. (2004) J. Immunol.
172: 1371. Para el análisis del desarrollo temprano de linfocitos B en médula ósea, se tiñó la superficie de las células con combinaciones de anticuerpos (Becton Dickinson) específicas para B220, CD19, CD25, IgM, IgD, Ckappa de ratón, Clambda de ratón y Ckappa de rata para detectar poblaciones de prolinfocitos B, prelinfocitos B, prelinfocitos B grandes, linfocitos B inmaduros tempranos y tardíos y linfocitos B recirculantes que expresen el transgén en su superficie. Se incluyó tinción con DAPi (Invitrogen) para excluir del análisis células muertas y bloqueo con FC (Becton Dickinson) para inhibir la interacción de anticuerpos con receptores de Fc en células mieloides. Para análisis de expresión transgénica de superficie sobre poblaciones de linfocitos B en órganos linfoides periféricos y sangre, las células se tiñeron con combinaciones de anticuerpos (Becton Dickinson) específicos para B220, CD5, CD19, CD21, CD23, IgM, IgD, Ckappa de ratón, Clambda de ratón y Ckappa de rata. Se incluyó tinción con DAPI para excluir del análisis las células muertas y bloqueo con FC para inhibir la interacción de anticuerpos con receptores de FC en células mieloides. Además, se incluyeron combinaciones de anticuerpos (Becton Dickinson) específicos para CD3, CD4, CD11b, CD11c y NK1.1 para determinar si se producía la expresión transgénica en los tipos de células fuera de compartimento de linfocitos B.
Se analizaron tres ratones heterocigotos para el transgén Ckappa de rata/IGKV1-39 humano y heterocigotos para el transgén CD19-Cre en un fondo C57BL6 (HuVk1/CD19-Cre). Como controles para el análisis FACS, se incluyeron tres ratones de la misma camada de tipo silvestre para el transgén IGKV1-39 humano/Ckappa de rata y heterocigotos para el transgén CD19-Cre en un fondo C57BL6 (CD19-Cre) y dos ratones C57BL6/NTac (Ts). Se permitió que todos los animales se aclimatasen en la instalación de animales durante 1 semana antes del análisis y todos los ratones eran machos y tenían 6 semanas de vida. Se aislaron linfocitos de los fémures, bazos, cavidad peritoneal y sangre de los ratones usando técnicas convencionales como se ha descrito previamente (Middendorp et al. (2002) J. Immunol. 168: 2695 y Middendorp et al. (2004) J. Immunol. 172: 1371). Los anticuerpos se precombinaron como se muestra en la Tabla 10 y la tinción se realizó en placas de 96 pocillos. La incubación con el anti-C kappa de rata conjugado con PE (descrito anteriormente) se realizó antes de la tinción con los anticuerpos anti-murinos de rata para impedir la unión no específica. Después de finalizar la tinción celular, las células marcadas se analizaron en un aparato FACS de Becton Dickinson LSR II y los datos adquiridos se analizaron con el programa informático FlowJo (v6.4.7).
Los ratones transgénicos eran similares en cuanto a peso, aspecto y actividad a los ratones de tipo silvestre. No se observaron importantes alteraciones anatómicas durante la extracción de tejidos. No se observó ninguna diferencia en cuanto al número de linfocitos B en médula ósea (MO) y bazo (Tabla 11) ni en cuanto al número de linfocitos B, linfocitos T y células mieloides en órganos periféricos entre los ratones de tipo silvestre y transgénicos. Además, la frecuencia o proporción de las células en las diferentes rutas de desarrollo de linfocitos no estaba alterada en los ratones transgénicos cuando se comparó con los ratones de tipo silvestre. Por tanto, en los ratones doble transgénicos (HuVk1/CD19-Cre) y transgénicos (CD19-Cre) el desarrollo de órganos linfoides y, lo que es más importante, de linfocitos B no podía distinguirse de los ratones de tipo silvestre.
En órganos linfoides periféricos, la tinción con el anticuerpo específico de transgén (anti-Ckappa de rata-PE) solo se observó en las poblaciones de linfocitos B. Todas las poblaciones de linfocitos T, de células mieloides y de linfocitos citóliticos (NK) fueron negativas para la expresión en superficie del transgén en el bazo (Figura 23). En cambio, en las células teñidas con los marcadores de todos los linfocitos B B220 y CD19 todas las células se desplazaron hacia la derecha en la gráfica FACS indicando expresión en la superficie celular del transgén (Figura 24). Se midió una tinción específica de transgén similar en linfocitos B1 CD5+ del peritoneo, una población distinta desde el punto de vista evolutivo de los linfocitos B (Figura 25).
La diferenciación de linfocitos B de precursores multilinaje de linfocitos B maduros se produce en la médula ósea. En los linfocitos analizados de la médula ósea, la expresión extracelular y transgénica no fue detectable en los progenitores más precoces de linfocitos B, los pro- y pre-linfocitos B, coherente con el patrón de expresión de cadena ligera normal (Figura 26). La expresión transgénica se vuelve primero detectable en linfocitos B inmaduros, la fase de desarrollo en la que la cadena ligera murina de la línea germinal experimenta reordenamiento y se expresa en la superficie celular en el contexto de la cadena pesada preseleccionada (Figura 26). Coherente con la tinción en la expresión de la cadena ligera transgénica en bazo esta también se detecta en linfocitos B recirculantes maduros (Figura 26). Por tanto, la expresión dirigida por CD19-Cre del transgén es coherente con el patrón normal de expresión de cadena ligera. La tinción con el anticuerpo específico de cadena ligera endógena es más intensa que la del anticuerpo de cadena ligera específico de transgén. Esto puede indicar un nivel de expresión más alto de la cadena ligera endógena, una tinción más sensible con el anticuerpo específico de cadena ligera endógena o una combinación de ambas cosas. Cabe destacar que la intensidad de la expresión en superficie de la cadena ligera transgénica se correlaciona con la expresión tanto de la cadena ligera endógena como con la de la superficie de IgM como se observa en la tinción de linfocitos B circulantes en la sangre (Figura 27).
Por tanto, en conjunto este análisis demuestra que la expresión del transgén IGKV1-39/Ckappa humano está restringida al compartimento de linfocitos B y la regulación temporal de su expresión es similar a la de las cadenas ligeras kappa y lambda endógenas que resultan en el desarrollo normal de todas las poblaciones de linfocitos B. El
nivel de expresión aparente más bajo del transgén podría explicarse por la fuerza del promotor en comparación con el promotor y potenciadores presentes en los genes de cadena ligera endógena o por un retraso en la expresión del transgén que otorga a las cadenas ligeras endógenas una ventaja competitiva en el emparejamiento con la cadena pesada reordenada. Esto es coherente con la observación de que a medida que los linfocitos B maduran la intensidad relativa de la tinción del transgén aumenta en comparación con las cadenas ligeras endógenas. Además, la observación de que los números de linfocitos B son normales y que cada linfocito B Ig+ de superficie co-expresa una cadena ligera endógena y transgénica refuerza la conclusión de que la región variable IGKV1-39 es capaz de emparejarse con un repertorio normal de regiones variables de cadena pesada murina diferentes. A partir de este análisis se llega a la conclusión de que la inserción del transgén IGKV1-39/Ckappa de rata dirigida por el promotor CAGGS activado por CD19-Cre en el locus Rosa facilita la expresión oportuna y específica de linfocitos B del transgén y que el transgén es capaz de emparejarse con un repertorio normal de cadenas pesadas murinas.
Ejemplo 19
Perfil epitópico de linfocitos T Epibase® para IGKV1-39.
La secuencia proteica de IGKV1-39 (figura 12, Proteína IGKV1-39/J de la línea germinal humana) se examinó para determinar la presencia de supuestos epítopos restringidos a HLA de clase II, conocidos también como epítopos Th. Para esto, se aplicó la plataforma Epibase® de Algonomics a IGKV1-39. En resumen, la plataforma analiza las especificidades de unión a HLA de todos los posibles péptidos de 10 unidades derivados de una secuencia diana (Desmet et al. Nature 1992, 356: 539-542; Desmet et al. FASEB J. 1997, 11: 164-172; Desmet et al. Proteins 2002, 48: 31-43; Desmet et al. Proteins 2005, 58: 53-69). El perfilado se realiza a nivel alotípico para 20 DRB1, 7 DRB3/4/5, 13 DQ y 7 DP, es decir, 47 receptores de HLA de clase II en total (véase la Tabla 5). Epibase® calcula una estimación cuantitativa de la energía de unión libre AGunión de un péptido para cada uno de los 47 receptores de HLA de clase II. Estos datos se procesaron adicionalmente de la siguiente manera:
• Las energías libres se convirtieron a valores Kd a través de AGunión = TR ln(Kd).
• Los péptidos se clasificaron como de unión fuerte (F), media (M) débil y sin unión (N). Se aplicaron los siguientes límites:
S: unión fuerte: Kd < 0,1 pM.
M: unión media: 0,1 pM < Kd < 0,8 pM.
N: unión débil y sin unión: 0,8 pM < Kd.
• Los péptidos correspondientes a auto-péptidos se trataron independientemente. La lista de auto-péptidos se extrajo de secuencias de la línea germinal de 293 anticuerpos. Estos se denominan péptidos 'depurados por la línea germinal'.
Los péptidos S y M se mapean sobre la secuencia diana en mapas denominados epitópicos; las afinidades de S se representaron gráficamente de modo cuantitativo; los valores de M se presentan de modo cualitativo. Como una visión general de los resultados, la Tabla 6 indica el número de aglutinantes fuertes y medios en las proteínas analizadas, para los grupos de receptores de HLA de clase II correspondientes a los genes DRB1, DQ, DP y DRB3/4/5. El recuento se realizó individualmente para los aglutinantes de afinidad fuerte y media. Los péptidos de unión a alotipos múltiples del mismo grupo se contaron como uno. Los valores entre paréntesis se refieren a péptidos depurados por la línea germinal. En la Tabla 7, la secuencia se muestra en un formato adecuado para el trabajo experimental. La secuencia se desglosa en oligómeros de 15 unidades (15 meros) consecutivos que se solapan por 12 restos. Se indica la promiscuidad (el número de alotipos de un total de 47 para el cual los 15 meros contienen un agente de unión crítico) de cada uno de los 15 meros, así como los serotipos implicados. El perfil Epibase® y los mapas epitópicos se muestran en las figuras 16 y 17.
Se llegó a la conclusión de que IGKV1-39 no contenía agentes de unión DRB1 no propios fuertes. Típicamente, de manera significativa, se encontraron más agentes de unión para DBR1 que para otros genes HLA. Esto concuerda con la confirmación experimental de que los alotipos que pertenecen al grupo DRB1 son agentes de unión peptídicos más potentes. Se espera que los epítopos de fuerza media para los alotipos de DBR1 contribuyan a la respuesta de la población y no pueden pasarse por alto. De nuevo, no se encontraron agentes de unión de DBR1 no propios en IGKV1-39.
En la respuesta humoral suscitada contra un antígeno, la activación/proliferación de linfocitos Th observada se interpreta generalmente en términos de especificidad de DRB1. Sin embargo, no puede ignorarse la posible contribución de los genes DRB3/4/5, DQ y DP. Dados los niveles de expresión más bajos de estos genes en comparación con DRB1, el enfoque fue sobre la clase de epítopos fuertes para DRB3/4/5, DQ y DP. Los 'epítopos críticos' son aquellos epítopos con unión fuerte para cualquier alotipo de d RB1, DRB3/4/5, DQ o DP o son agentes de unión medios para DRB1. IGKV1-39 no contiene agentes de unión no propios fuertes o medios para DRB3/4/5, DQ, o DP.
Diversos péptidos también están presentes en las secuencias de la línea germinal (valores entre paréntesis en la
Tabla 6). Dichos péptidos pueden unirse muy bien a HLA pero se supone que son propios y, por tanto, no inmunogénicos. En total, en IGKV1 se encontraron 6 agentes de unión DRB1 depurados por la línea germinal fuertes y 16 agentes de unión DRB1 depurados por la línea germinal medios. La región marco conservada 1 hasta la región marco conservada 3 es una concordancia exacta para el segmento V de la línea germinal VKI 2-1-(1) 012 (VBase), también conocido como IGKV1-39*01 (IMGT). La región marco conservada 4 es una concordancia exacta para el segmento-J de la línea germinal JK1 (V-base) también conocido como IGKJ1*01(IMGT). No resulta sorprendente que estos segmentos no contengan ningún epítopo no propio.
Ejemplo 20
Características de producción de IGKV1-39
Existe una gran demanda de plataformas de descubrimiento de anticuerpos que produzcan anticuerpos terapéuticos que sean termodinámicamente estables y proporcionen buenos rendimientos de expresión. Estas características son importantes para garantizar la estabilidad de la sustancia farmacológica durante la producción y después de la inyección del producto farmacológico en el paciente. Además, una buena expresión produce un impacto directamente sobre el coste de la fabricación del fármaco y, por tanto, en el precio, acceso al paciente y rentabilidad. Prácticamente todos los anticuerpos terapéuticos en uso clínico actual están compuestos de regiones constantes kappa e IgG1 humana pero usan regiones variables de cadena pesada y ligera diferentes que confieren especificidad. Los dominios de cadena ligera y pesada variable humana pueden dividirse en familias que tienen divergencia de secuencias mayor del 80 %. Cuando se combinan ejemplos reordenados de estas familias en la configuración de la línea germinal y se compara la estabilidad y producción es obvio que las familias de genes no son iguales en cuanto a propiedades biofísicas. En particular, Vh3, Vh1 y Vh5 tienen estabilidad favorable para las cadenas pesadas y Vk1 y Vk3 tienen la mejor estabilidad y producción de cadenas ligeras. Además, cuando se introducen mutaciones como parte del proceso de hipermutación somática estas pueden interferir con el emparejamiento de Vh/Vl. Para evaluar el efecto que tienen diferentes genes de cadena ligera con diferentes tasas de mutación sobre las características de producción de una cadena Vh fija, se construyó una biblioteca de presentación de fagos Fab de cadenas ligeras (kappa y lambda) de seis donantes sanos sin tratar combinada con un panel de 44 cadenas pesadas de unión a TT de donantes inmunizados. Después de una ronda de selección se aislaron clones Fab de unión a TT. Muchos de estos compartían el mismo gen Vh que el del clon TT PG1433 en combinación con diferentes cadenas ligeras. Los fragmentos de cadena ligera Fab se reclonaron en un vector de expresión kappa y se transfectaron en combinación con ADN que codificaba la cadena pesada de PG1433 en células 293 y la producción específica de IgG se midió por ELISA. Como se demuestra en la tabla 8, los clones seleccionados que contienen Vh PG1433 combinado con diferentes cadenas ligeras tenían entre 5 y 10 veces menor expresión de proteínas que Vh PG1433 combinado con IGKV1-39. Obsérvese que todas las cadenas ligeras contenían mutaciones de aminoácidos en sus regiones codificantes que podrían alterar el emparejamiento de Vh y reducir la estabilidad en producción. Por tanto, además de reducir la probabilidad de inmunogenicidad no deseada, se espera que el uso de IGKV1-39 de cadena ligera sin mutaciones contribuya a una estabilidad de producción y a rendimientos mejorados de diversos genes Vh que contribuyen con diversa especificidad. Por supuesto, clones estables por transfección de genes Vh diferentes todos emparejados con IGKV1-39 pueden realizar pases ampliamente y aún conservar características de producción fuertes como se muestra en la tabla 9.
Ejemplo 21
Generación de ratones que expresan regiones VH y VL completamente humanas.
Ratones transgénicos de acuerdo con la invención, se cruzaron con ratones que ya contenían un locus VH humano. En Taylor et al. (1992). Nucleic Acids Res 20: 6287-95; Lonberg et al. (1994). Nature 368: 856-9; Green et al. (1994). Nat Genet 7: 13-21; Dechiara et al. (2009). Methods Mol Biol 530: 311-24) se describen ejemplos de ratones apropiados que contienen un locus VH humano.
Después de cruzar y seleccionar ratones que son al menos heterocigotos para el transgén IGKV1-39 y el locus VH humano, los ratones seleccionados se inmunizan con una diana. Los genes VH se recogieron como se ha descrito anteriormente en el presente documento. Este procedimiento tiene la ventaja de que los genes VH ya son completamente humanos y por tanto no requieren humanización.
Ejemplo 22
Aislamiento, caracterización, formateo con Oligoclonics y producción de anticuerpos que se dirigen a IL6 humana para el tratamiento de enfermedades inflamatorias crónicas tales como artritis reumatoide.
Un repertorio de VH de bazo de ratones transgénicos que se inmunizan con IL6 recombinante humana se clona en un vector Fab de presentación de fagos con una sola cadena ligera kappa IGKV1-39-C humana (idéntica a la del transgén de ratón) y se sometió a selección contra la IL6 humana inmunogénica. Los clones que se obtienen después de dos a cuatro rondas de selección se analizan para determinar su especificidad de unión. Los genes VH que codifican los fragmentos Fab específicos de IL6 se someten a análisis de secuencia para identificar clones
únicos y asignar la utilización de VH, DH y JH. Los fragmentos Fab se reformatean como moléculas de IgG1 y se expresan transitoriamente. Los clones únicos se agrupan después basándose en ensayos de unión no competitiva y se someten a análisis de afinidad y funcional. Los mAb anti-IgG1 de IL6 más fuertes se expresan posteriormente como combinaciones de dos, tres, cuatro o cinco cadenas pesadas que comprenden diferentes regiones VH del formato Oligoclonics, junto con una cadena ligera kappa basada en IGKV1-39-C y se somete a ensayo in vitro para determinar la formación de complejos con IL-6. Los Oligoclonics también se someten a ensayo in vivo para la eliminación de la IL-6 humana de los ratones. Se seleccionó un Oligoclonic con la actividad de eliminación más fuerte y los genes VH murinos se humanizan de acuerdo con procedimientos convencionales. La IgG1 humanizada se transfecta en una línea de células de mamífero para generar un clon estable. Se selecciona un subclón óptimo para la generación de un banco de células maestro y la generación de material de ensayo clínico.
Muchos de los protocolos descritos en el presente documento son protocolos convencionales para la construcción de bibliotecas de presentación de fagos y la selección de fagos para la unión a un antígeno de interés y se describen, por ejemplo, en Antibody Phage Display: Methods and Protocols. 2002. Editor(es): Philippa M. O'Brien, Robert Aitken. Humana Press, Totowa, Nueva Jersey, Estados Unidos.
Inmunizaciones
Ratones transgénicos recibieron tres inmunizaciones con IL6 humana cada dos semanas usando el adyuvante Sigma titerMax de acuerdo con las instrucciones del fabricante.
Aislamiento de ARN y síntesis de ADNc
Tres días después de la última inmunización, se extirparon los bazos y los ganglios linfáticos de los ratones y se hicieron pasar a través de un filtro de 70 micrómetros en un tubo que contenía PBS pH 7,4 para generar una suspensión de células sencillas. Después de lavar y sedimentar los linfocitos, las células se suspenden en Reactivo TRIzol LS (Invitrogen) para el aislamiento de ARN total de acuerdo con el protocolo del fabricante y se someten a reacción de transcripción inversa usando 1 microgramo de ARN, Superscript III RT en comparación con dT20 de acuerdo con los procedimientos del fabricante (Invitrogen).
La generación de bibliotecas de presentación de fagos Fab se realiza como se describe en el Ejemplo 2.
Selección de fagos en inmunotubos revestidos
La IL6 recombinante humana se disuelve en PBS en una concentración de 5 pg/ml y se aplica a Tubos MaxiSorp Nunc-Immuno (Nunc 444474) durante una noche a 4 °C. Después de desechar la solución de revestimiento, los tubos se bloquean con leche desnatada (ELK) al 2 % en PBS (tampón de bloqueo) durante 1 hora a Temperatura Ambiente (TA). En paralelo, 0,5 ml de la biblioteca de fagos se mezcla con 1 ml de tampón de bloqueo y se incuba durante 20 minutos a temperatura ambiente. Después de bloquear los fagos, la solución de fagos se añade a los tubos revestidos con IL6 y se incuba durante 2 horas a TA en una plataforma rotatoria lenta para permitir la unión. A continuación, los tubos se lavan 10 veces con PBS/Tween-20 al 0,05 % seguido de elución de los fagos incubando con 1 ml de glicina-HCl 50 mM pH 2,2 durante 10 min a TA en una rueda giratoria y directamente seguido de neutralización del eluyente recogido con 0,5 ml de Tris-HCl 1 M pH 7,5.
Recogida de clones de fagos
A la solución de fagos recogida se añade un cultivo de 5 ml de XL1-Blue MRF (Stratagene) a una D.O. de 0,4 y se incuba durante 30 minutos a 37 °C sin agitación para permitir la infección de los fagos. Las bacterias se siembran en placas de 2*TY con Carbenicilina/Tetraciclina glucosa al 4 % y se cultivan durante una noche a 37 °C.
Producción de fagos.
Los fagos se cultivan y procesan como describen Kramer et al. 2003 (Kramer et al. 2003. Nucleic Acids Res. 31(11): e59) usando VCSM13 como cepa de fago auxiliar.
ELISA para fagos
Se revisten placas de ELISA con 100 microlitros de IL6 recombinante humana por pocillo a una concentración de 2,5 microgramos/ml en PBS durante una noche a 4 °C. Las placas revestidas con 100 microlitros de tiroglobulina a una concentración de 2 microgramos/ml en PBS se usan como un control negativo. Los pocillos se vacían, se secan golpeteando sobre una toallita de papel, se cargan completamente con PBS-leche desnatada al 4 % (ELK) y se incuban durante 1 hora a temperatura ambiente para bloquear los pocillos. Después de desechar la solución de bloqueo, las minipreparaciones de fagos premezcladas con 50 pl de solución de bloqueo se añaden y se incuban durante 1 hora a Ta . Los fagos no unidos se retiran posteriormente mediante 5 etapas de lavado con PBS-Tween-20 al 0,05 %. Los fagos unidos se detectan incubando los pocillos con 100 microlitros de anticuerpo anti-M13 conjugado con HRP (diluido 1/5000 en tampón de bloqueo) durante 1 hora a temperatura ambiente. Los anticuerpos libres se
retiran repitiendo las etapas de lavado como se ha descrito anteriormente, seguido por incubación con sustrato TMB hasta que fue visible el desarrollo del color. La reacción se detiene añadiendo 100 microlitros de H2SO42 M por pocillo y se analiza en un lector ELISA a una longitud de onda de emisión 450 nm.
Secuenciación
Los clones que dan señales al menos 3 veces por encima de la señal de fondo se propagan, se usan para procedimientos miniprep ADN (véase el manual de procedimientos miniPrep de Qiagen) y se someten a análisis de secuencia de nucleótidos. La secuenciación se realiza de acuerdo con el manual adjunto al kit Big Dye 1.1 (Applied Biosystems) usando un cebador inverso (CH1_Rev1, tabla 1) que reconoce una secuencia 5' de la región CH1 de la cadena pesada IgG1 humana (presente en el vector de presentación Fab MV1043, figuras 3 y 12). Las secuencias de las regiones VH murinas se analizan con respecto a la diversidad de los segmentos de los genes DH y JH.
Construcción y expresión de IgG1 quimérica
Se generó el vector MV1057 (figuras 12 y 22) por clonación del fragmento transgénico de cadena L (IGKV1-39) en un derivado del vector pcDNA3000Neo (Crucell, Leiden, Países Bajos) que contiene las regiones constantes kappa e IgG1 humana. Las regiones VH se clonan en MV1057 y las secuencias de nucleótidos de todas las construcciones se verifican de acuerdo con técnicas convencionales. Las construcciones resultantes se expresan transitoriamente en células HEK293T y usando procedimientos convencionales como se ha descrito anteriormente (Throsby, M.
2006. J Virol 80: 6982-92) se obtienen y purifican los sobrenadantes que contienen la IgG1 quimérica.
Unión a IgG1 y análisis de competencia
Los anticuerpos IgG1 se titularon en ELISA usando placas revestidas con IL6, como se ha descrito anteriormente, y un anti-IgG humano conjugado con peroxidasa. Se realizan ensayos ELISA de competencia para agrupar anticuerpos basándose en el reconocimiento epitópico, incubando fagos Fab junto con IgG1 o con anticuerpos comerciales contra IL6 (por ejemplo, Abcam cat. N.° ab9324) en placas revestidas con IL6, seguido por la detección de los fagos Fab unidos usando un anti-M13 conjugado con peroxidasa
Mediciones de afinidad de IgG1
Las afinidades de los anticuerpos contra IL6 se determinan con el protocolo cinético cuantitativo del Octet (ForteBio). Los anticuerpos se capturan sobre un biodetector de Captura Fc Anti-IgG Humano y se exponen a IL6 libre y se analizan usando un programa informático patentado para calcular el valor de la Kd de cada anticuerpo.
Actividad funcional de los anticuerpos IL6
Para ensayar la capacidad de los anticuerpos seleccionados para inhibir la unión entre IL6 y el receptor de IL6 (IL6R), se usa un ensayo basado en ELISA. Diversas concentraciones de anticuerpos se mezclan con una concentración fija (10 ng/ml) de IL6 marcada con biotina descrito por Naoko et al. 2007, Can. Res. 67: 817-875. El complejo inmunitario anticuerpo-IL6 se añade al IL6R inmovilizado. La unión de IL6 marcada con biotina a IL6R se detecta con estreptavidina conjugada con peroxidasa de rábano picante. La reducción de la señal ELISA es una medición de la inhibición. Como control positivo para la inhibición de la unión entre IL6 e IL6R se usa cualquier anticuerpo anti-IL6R (Abcam cat. N.° ab34351; clon B-R6) o anticuerpo anti-IL6 (Abcam cat. N.° ab9324).
La actividad bloqueante in vitro de los anticuerpos anti-IL6 seleccionados se mide en un ensayo de proliferación usando la línea celular 7TD1 dependiente de IL6. En resumen, las células se incuban con diferentes concentraciones de IL6 humana con o sin el anticuerpo anti-IL6. La cantidad disponible de IL6 determina el grado de proliferación. Por tanto, si un anticuerpo añadido bloquea la unión de IL6 la lectura de proliferación se reduce en comparación con un control de anticuerpo no unido. La proliferación se mide por la incorporación de 5-bromo-2'-desoxiuridina (BrdU) al ADN usando el kit de proliferación BrdU (Roche cat. N.°. 11444611001) de acuerdo con las instrucciones del fabricante.
Generación de Oligoclonics anti-IL6
Se seleccionan los anticuerpos anti-IL6 más potentes de cada grupo epitópico. Las construcciones de expresión que expresan estos anticuerpos se transfectan en células HEK293T en grupos de tres no competidores en diferentes proporciones (1:1:1; 3:1:1; 1:3:1; 1:1:3; 3:3:1; 1:3:3; 3:1:3; 10:1:1; 1:10:1; 1:1:10; 10:10:1; 1:10:10; 10:1:10; 3:10:1; 10:3:1; 1:10:3; 3:1:10; 10:1:3; 1:3:10). Los sobrenadantes que contienen anticuerpos se recogen, se purifican y se caracterizan como se ha indicado anteriormente.
Formación de complejos y eliminación in vivo de Oligoclonics anti-IL6
Para medir la capacidad de Oligoclonics anti-IL6 para formar complejos inmunitarios y analizar estos complejos se usa Cromatografía de Exclusión por Tamaño (SEC) de acuerdo con la estrategia desvelada por Min-Soo Kim et al.
(2007) JMB 374: 1374-1388 para caracterizar los complejos inmunitarios formados con diferentes anticuerpos contra TNFa. Se mezclan diferentes proporciones molares de los Oligoclonics anti-IL6 con IL6 humana y se incuban durante 20 horas a 4 °C o 25 °C. La mezcla se analiza en un sistema HPLC ajustado con una columna de exclusión por tamaño; se correlacionan tiempos de elución diferentes con peso molecular usando patrones de peso molecular. La capacidad de los anticuerpos para formar complejos con IL6 se correlaciona con su capacidad para eliminar rápidamente la citocina de la circulación in vivo. Esto se confirma midiendo la eliminación de IL6 radiomarcada de ratones. En resumen, se obtuvieron hembras Balb/c de 6 a 8 semanas de vida y 18 horas antes del experimento, a los animales se les inyecta por vía intravenosa (IV) en la vena lateral de la cola diferentes dosis de Oligoclonics anti-IL6 purificado. El día 0 los ratones reciben una inyección IV de 50 microlitros de IL6 radiomarcada (1x10E7 cpm/ml) en las mismas condiciones. Se extraen muestras de sangre (aproximadamente 50 microlitros) a diversos intervalos de tiempo y se conservan a 4 °C. Las muestras se centrifugan durante 5 minutos a 4000 x g y se determina la radioactividad del suero. Todos los experimentos farmacocinéticos se realizan simultáneamente con tres animales por cada tratamiento.
Generación de clones estables Oligoclonics anti-IL6 y desarrollo preclínico
Se selecciona un Oligoclonic anti-IL6 líder basándose en la potencia in vitro e in vivo como se ha determinado anteriormente. Los genes VH murinos se humanizan de acuerdo con procedimientos convencionales y se combinan con la cadena ligera IGKV1-39 completamente humana en un vector de expresión como se ha descrito anteriormente. Los ejemplos de procedimientos de humanización incluyen aquellos basados en paradigmas tales como rechapado (Padlan, E. A., et al., (1991). Mol. Immunol., 28, 489), superhumanización (Tan, P., D. A., et al., (2002) J. Immunol., 169, 1119) y optimización del contenido de cadenas humanas (Lazar, G. A., et al., (2007). Mol. Immunol., 44, 1986). Las tres construcciones se transfectan en células PER.C6 a la proporción óptima predeterminada (descrita anteriormente) a presión selectiva de G418 de acuerdo con procedimientos convencionales. Se selecciona un clon Oligoclonic anti-IL6 de producción altamente estable y se genera un banco de trabajo y cualificado de células maestro.
Tabla 1 Listado de cebadores

continuación


Tabla 3

Tabla 4

Tabla 5. Alotipos HLA considerados en el perfilado epitópico de Th. Se muestran los serotipos correspondientes, así como las frecuencias alotípicas en la población caucásica (Klitz et al. Tissue Antigens 2003, 62: 296-307; Gjertson y Terasake (eds) en: HLA 1997; Gjertson y Terasake (eds) en: HLA 1998; Castelli et al. J. Immunol. 2002, 169: 6928 6934). Las frecuencias pueden sumar más del 100 % dado que cada individuo tiene 2 alelos para cada gen. Si se conociesen todas las frecuencias alélicas de un solo gen, sumarían ligeramente menos del 200 % debido a individuos homocigotos.
Tipo HLA Serotipo Población % Tipo HLA Serotipo Población % DRB1*0101 DR1 17,4 DRB4*0103 DR53 21
DRB1*0102 DR1 4,9 DRB5*0101 DR51 15,8 DRB1*0301 DR17(3) 21,2 DRB5*0202 DR51 5,7
DRB1*0401 DR4 11,5 DQA1*0101/DQB1*0501 DQ5(1) 20,5 DRB1*0402 DR4 3,1 DQA1*0102/DQB1*0502 DQ5(1) 2,6
DRB1*0404 DR4 5,5 DQA1*0102/DQB1*0602 DQ6(1) 26,5 DRB1*0405 DR4 2,2 DQA1*0102/DQB1*0604 DQ6(1) 6,7
DRB1*0407 DR4 <2 DQA1*0103/DQB1*0603 DQ6(1) 11
DRB1*0701 DR7 23,4 DQA1*0104/DQB1*0503 DQ5(1) 4
(continuación)
Tipo HLA Serotipo Población % Tipo HLA Serotipo Población % DRB1*0801 DR8 3,3 DQA1*0201/DQB1*0202 DQ2 20,9
DRB1*0802 DR8 <2 DQA1*0201/DQB1*0303 DQ9(3) 7,2
DRB1*0901 DR9 <2 DQA1*0301/DQB1*0301 DQ7(3) 12,5
DRB1*1101 DR11(5) 17 DQA1*0301/DQB1*0302 DQ8(3) 18,3 DRB1*1104 DR11(5) 5,7 DQA1*0401/DQB1*0402 DQ4 4,5
DRB1*1201 DR12(5) 3,1 DQA1*0501/DQB1*0201 DQ2 24,6
DRB1*1301 DR13(6) 15,4 DQA1*0501/DQB1*0301 DQ7(3) 20,9
DRB1*1302 DR13(6) 10,8 DPA1*0103/DPB1*0201 DPw2 199
DRB1*1401 DR14(6) 4,2 DPA1*0103/DPB1*0401 DPw4 65,1
DRB1*1501 DR15(2) 13,2 DPA1*0103/DPB1*0402 DPw4 24,3
DRB1*1601 DR16(2) 5,5 DPA1*0201/DPB1*0101 DPw1 6,3
DRB3*0101 DR52 24,6 DPA1*0201/DPB1*0301 DPw3 <2
DRB3*0202 DR52 43 DPA1*0201/DPB1*0501 DPw5 <2
DRB3*0301 DR52 10 DPA1*0201/DPB1*0901 - 2,4
DRB4*0101 DR53 25,5
Tabla 6. Recuentos epitópicos de Th para IGKV1-39. Los péptidos que se unen a HLA múltiples del mismo grupo (DRB1, DRB3/4/5, DP, DQ) se cuentan como uno. Los valores entre paréntesis se refieren a péptidos depurados por ________________________________________la línea germinal.________________________________________
DRB1 DRB3/4/5 DQ DP

Tabla 7. Mapeo de predicciones Epibase® para IGKV1-39 Merus en el formato clásico de péptidos de 15 unidades (15 mero). Esta tabla muestra el recuento alotípico de epítopos críticos y serotipos implicados para cada uno de los _______________________ 15 meros que abarcan la secuencia de IGKV1-39 Merus._______________________ 15 mero Posición de inicio Secuencia de 15 mero Recuento
alotípico Serotipos implicados
1 1 DIQMTQSPSSLSASV 6 DR1, DR4, DR7, DR9
2 4 MTQSPSSLSASVGDR 5 DR1, DR4, DR9
3 7 SPSSLSASVTGDRVTI 0
4 10 SLSASVGDRVTITCR 0
5 13 ASVGDRVTITCRASQ 0
6 16 GDRVTITCPASQSIS 2 DR11(5), DR7
7 19 VTITCRASOSISSYL 4 DQ2 DR11(5), DR4, DR7
8 22 TCRASQSISSYLNWY 2 DQ2, DR4
9 25 ASQSISSYLNWYQQR 5 DR13(6), DR15(2), DR4
10 28 SISSYLNWYQQKPGK 8 DR12(5), DR13(6), DR15(2),
DR16(2), DR4, DR8 11 31 SYLNWYQQKPGKAPK 10 DR1, DR12(5), DR16(2), DR4,
DR51, DR8 12 34 NWYQQKPGKAPKLLI 9 DR1, DR15(2), DR4. DR51,
DR8
13 37 QQKPGKAPKLLIYAA 7 DQ4, DR1, DR11(5), DR15(2),
DR51, DR8 DQ4, DR1, DR11(5), DR4, DR8
14 40 PGKAPKLLIYAASSL 7 DR1, DR11(5), DR12(5),
DR13(6), DR14(6), DR15(2),
DR4, DR51, DR8, DR9 DR1, 15 43 APKLLIYAASSLQSG 15 DR11(5), DR12(5), DR13(6),
DR14(6), DR15(2), DR4,
16 46 LLIYAASSLQSGVPS 15 DR51, DR8, DR9
17 49 YAASSLQSGVPSRFS 1 DR15(2)
18 52 SSLQSGVPSRFSGSG 1 DR15(2)
19 55 QSGVPSRFSGSGSGT 0
20 58 VPSRFSGSGSGTDFT 0
21 61 RFSGSGSGTDFTLTI 0
22 64 GSGSGTDFTLTISSL 1 DR52
23 67 SGTDFTLTISSLQPE 4 DR4, DR52, DR7, DR9
24 70 DFTLTISSLQPEDFA 4 DQ2, DR4, DR7, DR9
(continuación)
15 mero Posición de inicio Secuencia de 15 mero Recuento
alotípico Serotipos implicados
25 73 LTISSLQPEDFATYY 1 DQ2
26 76 SSLQPEDFATYYCQQ 0
27 79 QPEDFATYYCQQSYS 1 DR4
28 82 DFATYYCQQSYSTPP 5 DR4, DR51, DR7
29 85 TYYCQQSYSTPPTFG 4 DR4, DR51, DR7
30 88 CQQSYSTPPTFGQGT 0
31 91 SYSTPPTFGQGTKVE 0
32 94 TPPTFGQGTKVEIK 0
Tabla 8: El gen Vh de PG1433 emparejado con diversos genes de cadena ligera con diferentes tasas de mutación de aminoácidos se comparó con respecto a los niveles de producción con el clon original que contenía el gen IGKV1-
39.

Tabla 9 Parámetros de estabilidad para clones estables que contienen el gen IGKV1-39 de línea germinal

Tabla 9, continuación

Tabla 9, continuación

Tabla 9, continuación

Tabla 9, continuación

Tabla 9, continuación
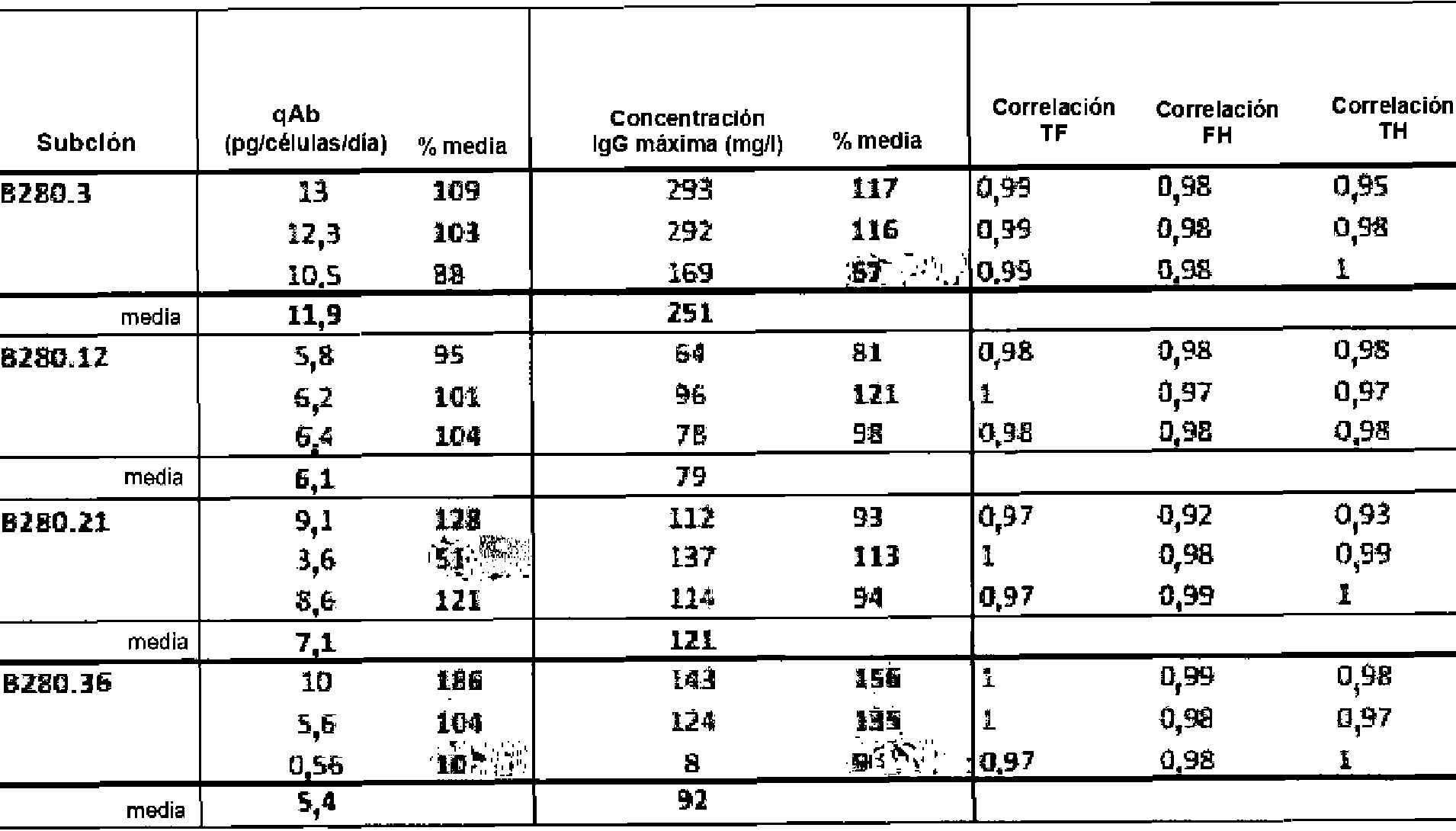
Tabla 10 Mezclas de anticuerpos usadas para la tinción de poblaciones de linfocitos. MO = médula ósea, CP = cavidad peritoneal, PP = placas de Peyer.

Tabla 10, continuación

Tabla 11. Números de linfocitos recoidos de la médula ósea de bazo de ratones de ti o silvestre trans énicos

Claims (19)
1. Un hospedador murino transgénico capaz de producir inmunoglobulinas que comprenden una región VL humana, comprendiendo dicho hospedador murino integrado en su línea germinal un ácido nucleico reordenado que comprende un segmento génico V de línea germinal humano y uno J de línea germinal humano que codifica una región VL humana reordenada y una región constante de cadena ligera de cualquier especie animal, en donde dicha región variable de cadena ligera es capaz de emparejarse con al menos dos cadenas pesadas diferentes codificadas por el hospedador, de modo que se conserva diversidad de especificidad de anticuerpos que comprenden dicha región VL humana mediante reordenamientos e hipermutaciones en las cadenas pesadas.
2. Un hospedador transgénico según la reivindicación 1, en donde el hospedador es un ratón.
3. Un hospedador transgénico según la reivindicación 1 o 2, en donde dicha región constante de cadena ligera es una región constante de cadena ligera murina.
4. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-3, en donde la integración es en un locus que es resistente al silenciamiento.
5. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-3, en donde la integración es en el locus Rosa.
6. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-5, en donde el ácido nucleico que codifica la región variable de cadena ligera está provisto de un medio que permite la expresión de dicho ácido nucleico esencialmente limitado a células del linaje de linfocitos B.
7. Un hospedador transgénico según la reivindicación 6, en donde el ácido nucleico que codifica la región variable de cadena ligera está provisto de un medio que permite la expresión del ácido nucleico que codifica la región variable de cadena ligera predominantemente durante un cierto estadio del desarrollo de linfocitos B.
8. Un hospedador transgénico según la reivindicación 7, en donde dicho medio comprende un promotor seleccionado del grupo de CD19, CD20, |jHC, VpreB1, VpreB2, VpreB3, A5, Iga, Igp, kLC, ALC y BSAP (Pax5).
9. Un hospedador transgénico según la reivindicación 7 u 8, en donde dicho medio comprende un sistema cre-lox.
10. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-9, en donde dicha región variable de cadena ligera es capaz de emparejarse con al menos dos cadenas pesadas del hospedador.
11. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-9, en donde dicha región variable de cadena ligera es capaz de emparejarse con al menos dos cadenas pesadas humanas.
12. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-11, en donde al menos uno de los locus endógenos que codifican una cadena ligera endógena está funcionalmente silenciado.
13. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-12, en donde el locus de cadena ligera k endógeno está funcionalmente silenciado.
14. Un hospedador transgénico según una cualquiera de las reivindicaciones 1-13, en donde la secuencia del ácido nucleico que codifica la región variable de cadena ligera es una secuencia vk de línea germinal humana.
15. Un hospedador transgénico según la reivindicación 14, en donde la secuencia vk de línea germinal se basa en 012.
16. Un hospedador transgénico según la reivindicación 15, en donde la secuencia vk de línea germinal es IGKV1-39*01 / IGKJ1*01.
17. Un método para producir un anticuerpo deseado que comprende exponer un hospedador según una cualquiera de las reivindicaciones 1-16 a un antígeno de modo que se induzca una respuesta de anticuerpo y aislar los anticuerpos específicos para el antígeno.
18. Un método para producir un anticuerpo deseado que comprende exponer un hospedador según una cualquiera de las reivindicaciones 1-16 a un antígeno de modo que se induzca una respuesta de anticuerpo y aislar las células que producen dichos anticuerpos, cultivar y recoger dichos anticuerpos.
19. Un método para producir un anticuerpo deseado que comprende exponer un hospedador según una cualquiera de las reivindicaciones 1-16 a un antígeno de modo que se induzca una respuesta de anticuerpo y aislar un ácido nucleico que codifica al menos parte de dicho anticuerpo, insertar dicho ácido nucleico o una copia o un derivado del
mismo en un casete de expresión y expresar dicho anticuerpo en una célula hospedadora.
Ċ
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| NL2008050430 | 2008-06-27 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2906344T3 true ES2906344T3 (es) | 2022-04-18 |
Family
ID=41280336
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES18183980T Active ES2906344T3 (es) | 2008-06-27 | 2009-06-29 | Animal murino transgénico productor de anticuerpos |
| ES09075279.1T Active ES2445193T3 (es) | 2008-06-27 | 2009-06-29 | Mamíferos no humanos productores de anticuerpos |
| ES12175544T Active ES2853226T3 (es) | 2008-06-27 | 2009-06-29 | Ratones transgénicos productores de anticuerpos |
Family Applications After (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES09075279.1T Active ES2445193T3 (es) | 2008-06-27 | 2009-06-29 | Mamíferos no humanos productores de anticuerpos |
| ES12175544T Active ES2853226T3 (es) | 2008-06-27 | 2009-06-29 | Ratones transgénicos productores de anticuerpos |
Country Status (23)
| Country | Link |
|---|---|
| EP (6) | EP3456190B1 (es) |
| JP (6) | JP5749161B2 (es) |
| KR (5) | KR101990228B1 (es) |
| CN (3) | CN105191863B (es) |
| AU (1) | AU2009263082C1 (es) |
| CA (1) | CA2729095C (es) |
| CY (2) | CY1114830T1 (es) |
| DK (3) | DK3456190T3 (es) |
| ES (3) | ES2906344T3 (es) |
| HK (1) | HK1154463A1 (es) |
| HR (2) | HRP20220205T1 (es) |
| HU (1) | HUE057994T2 (es) |
| LT (1) | LT3456190T (es) |
| MX (1) | MX2010014542A (es) |
| NZ (1) | NZ590118A (es) |
| PL (2) | PL2147594T3 (es) |
| PT (3) | PT2556747T (es) |
| RS (2) | RS53146B (es) |
| RU (3) | RU2731084C2 (es) |
| SG (3) | SG10201914027VA (es) |
| SI (2) | SI3456190T1 (es) |
| WO (1) | WO2009157771A2 (es) |
| ZA (1) | ZA201009312B (es) |
Families Citing this family (115)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| USRE47770E1 (en) | 2002-07-18 | 2019-12-17 | Merus N.V. | Recombinant production of mixtures of antibodies |
| CN101537180B (zh) | 2002-07-18 | 2016-02-10 | 莫鲁斯有限公司 | 抗体混合物的重组生产 |
| ES2408582T3 (es) | 2003-05-30 | 2013-06-21 | Merus B.V. | Biblioteca de Fab para la preparación de una mezcla de anticuerpos |
| US20100069614A1 (en) | 2008-06-27 | 2010-03-18 | Merus B.V. | Antibody producing non-human mammals |
| GB2404382B (en) | 2003-07-28 | 2008-01-30 | Oxitec Ltd | Pest control |
| GB2443186A (en) | 2006-10-25 | 2008-04-30 | Oxitec Ltd | Expression system for mediating alternative splicing |
| CN105191863B (zh) | 2008-06-27 | 2020-12-22 | 莫鲁斯股份有限公司 | 产生抗体的非人哺乳动物 |
| US9346873B2 (en) | 2008-09-30 | 2016-05-24 | Ablexis, Llc | Non-human mammals for the production of chimeric antibodies |
| PL2564695T3 (pl) * | 2009-07-08 | 2015-10-30 | Kymab Ltd | Modele zwierzęce i cząsteczki terapeutyczne |
| US9445581B2 (en) | 2012-03-28 | 2016-09-20 | Kymab Limited | Animal models and therapeutic molecules |
| US20120204278A1 (en) | 2009-07-08 | 2012-08-09 | Kymab Limited | Animal models and therapeutic molecules |
| WO2011158009A1 (en) * | 2010-06-17 | 2011-12-22 | Kymab Limited | Animal models and therapeutic molecules |
| US20130045492A1 (en) | 2010-02-08 | 2013-02-21 | Regeneron Pharmaceuticals, Inc. | Methods For Making Fully Human Bispecific Antibodies Using A Common Light Chain |
| AU2013204019B2 (en) * | 2010-02-08 | 2014-02-27 | Regeneron Pharmaceuticals, Inc. | Common light chain mouse |
| US9796788B2 (en) | 2010-02-08 | 2017-10-24 | Regeneron Pharmaceuticals, Inc. | Mice expressing a limited immunoglobulin light chain repertoire |
| ES2603559T5 (es) | 2010-02-08 | 2021-02-22 | Regeneron Pharma | Cadena ligera común de ratón |
| US20120021409A1 (en) * | 2010-02-08 | 2012-01-26 | Regeneron Pharmaceuticals, Inc. | Common Light Chain Mouse |
| EP4345163A2 (en) | 2010-03-31 | 2024-04-03 | Ablexis, LLC | Genetic engineering of non-human animals for the production of chimeric antibodies |
| US10793829B2 (en) | 2010-07-26 | 2020-10-06 | Trianni, Inc. | Transgenic mammals and methods of use thereof |
| US10662256B2 (en) | 2010-07-26 | 2020-05-26 | Trianni, Inc. | Transgenic mammals and methods of use thereof |
| CN103025884B (zh) | 2010-07-26 | 2015-11-25 | 特里安尼公司 | 转基因动物和使用方法 |
| CN113150121A (zh) * | 2010-08-02 | 2021-07-23 | 瑞泽恩制药公司 | 制造包含vl结构域的结合蛋白的小鼠 |
| SI3572517T1 (sl) * | 2011-08-05 | 2021-09-30 | Regeneron Pharmaceuticals, Inc. | Humanizirana miš z univerzalno lahko verigo |
| CA2846322A1 (en) | 2011-09-19 | 2013-03-28 | Kymab Limited | Manipulation of immunoglobulin gene diversity and multi-antibody therapeutics |
| CA2791109C (en) | 2011-09-26 | 2021-02-16 | Merus B.V. | Generation of binding molecules |
| EP2761008A1 (en) | 2011-09-26 | 2014-08-06 | Kymab Limited | Chimaeric surrogate light chains (slc) comprising human vpreb |
| US9253965B2 (en) | 2012-03-28 | 2016-02-09 | Kymab Limited | Animal models and therapeutic molecules |
| JP2015502165A (ja) | 2011-12-22 | 2015-01-22 | エフ.ホフマン−ラ ロシュ アーゲーF. Hoffmann−La Roche Aktiengesellschaft | 発現ベクター構成、新規の産生細胞生成法、およびポリペプチドの組換え産生のためのそれらの使用 |
| HUE046744T2 (hu) * | 2012-02-01 | 2020-03-30 | Regeneron Pharma | VL doméneket tartalmazó nehézláncokat expresszáló humanizált egér |
| GB2500113A (en) | 2012-03-05 | 2013-09-11 | Oxitec Ltd | Arthropod male germline gene expression system |
| CN107090471A (zh) * | 2012-03-06 | 2017-08-25 | 瑞泽恩制药公司 | 共同轻链小鼠 |
| US20140013456A1 (en) | 2012-03-16 | 2014-01-09 | Regeneron Pharmaceuticals, Inc. | Histidine Engineered Light Chain Antibodies and Genetically Modified Non-Human Animals for Generating the Same |
| SG10201607727PA (en) | 2012-03-16 | 2016-11-29 | Regeneron Pharma | Mice that produce antigen-binding proteins with ph-dependent binding characteristics |
| EP3348140B1 (en) | 2012-03-16 | 2020-12-30 | Regeneron Pharmaceuticals, Inc. | Histidine engineered light chain antibodies and genetically modified rodents for generating the same |
| SG10201700360VA (en) * | 2012-03-16 | 2017-03-30 | Regeneron Pharma | Non-human animals expressing ph-sensitive immunoglobulin sequences |
| GB2502127A (en) * | 2012-05-17 | 2013-11-20 | Kymab Ltd | Multivalent antibodies and in vivo methods for their production |
| US10251377B2 (en) | 2012-03-28 | 2019-04-09 | Kymab Limited | Transgenic non-human vertebrate for the expression of class-switched, fully human, antibodies |
| PT2838918T (pt) | 2012-04-20 | 2019-08-23 | Merus Nv | Métodos e meios para a produção de moléculas heterrodiméricas do tipo ig |
| WO2013184761A1 (en) * | 2012-06-05 | 2013-12-12 | Regeneron Pharmaceuticals, Inc. | Methods for making fully human bispecific antibodies using a common light chain |
| BR112015000167B1 (pt) | 2012-07-06 | 2021-11-23 | Genmab B.V. | Proteína dimérica, proteína, composição, kit de partes e seus usos, bem como método para aumentar a oligomerização em solução de uma proteína dimérica compreendendo um primeiro e segundo polipeptídeo, cada um compreendendo pelo menos as regiões ch2 e ch3 de uma cadeia pesada de igg1 humana e proteína dimérica variante |
| CN103571872B (zh) * | 2012-08-09 | 2016-11-23 | 山东国际生物科技园发展有限公司 | 一种能够表达人抗体的转基因动物的制备方法 |
| SI2900694T1 (sl) | 2012-09-27 | 2018-12-31 | Merus N.V. | Bispecifična IGG protitelesa kot vključitelji T-celic |
| GB201303932D0 (en) | 2013-03-05 | 2013-04-17 | Oxitec Ltd | Muscle actin promoter |
| EP3501272B1 (en) * | 2013-03-13 | 2023-03-01 | Regeneron Pharmaceuticals, Inc. | Mice expressing a limited immunoglobulin light chain repertoire |
| PL2968440T3 (pl) | 2013-03-15 | 2019-12-31 | Zymeworks Inc. | Związki cytotoksyczne i antymitotyczne oraz sposoby ich stosowania |
| US9788534B2 (en) | 2013-03-18 | 2017-10-17 | Kymab Limited | Animal models and therapeutic molecules |
| US9783618B2 (en) | 2013-05-01 | 2017-10-10 | Kymab Limited | Manipulation of immunoglobulin gene diversity and multi-antibody therapeutics |
| US11707056B2 (en) | 2013-05-02 | 2023-07-25 | Kymab Limited | Animals, repertoires and methods |
| US9783593B2 (en) | 2013-05-02 | 2017-10-10 | Kymab Limited | Antibodies, variable domains and chains tailored for human use |
| CA2925723A1 (en) | 2013-10-01 | 2015-04-09 | Kymab Limited | Animal models and therapeutic molecules |
| PL2908626T3 (pl) * | 2013-10-15 | 2017-05-31 | Regeneron Pharmaceuticals, Inc. | Zwierzęta z humanizowaną |
| EP3086815B1 (en) | 2013-12-27 | 2022-02-09 | Zymeworks Inc. | Sulfonamide-containing linkage systems for drug conjugates |
| CA2941030A1 (en) | 2014-02-28 | 2015-09-03 | Merus N.V. | Antibodies that bind egfr and erbb3 |
| CN106459212B (zh) | 2014-02-28 | 2021-07-13 | 美勒斯公司 | 结合erbb-2和erbb-3的抗体 |
| CA2942697A1 (en) | 2014-03-21 | 2015-09-24 | Lynn Macdonald | Non-human animals that make single domain binding proteins |
| CA2941514A1 (en) | 2014-03-21 | 2015-09-24 | Regeneron Pharmaceuticals, Inc. | Vl antigen binding proteins exhibiting distinct binding characteristics |
| GB2526867A (en) * | 2014-06-05 | 2015-12-09 | Oxitec Ltd | Gene expression system |
| RS62860B1 (sr) | 2014-09-17 | 2022-02-28 | Zymeworks Inc | Citotoksična i antimitotička jedinjenja, i postupci upotrebe istih |
| EP3271403A1 (en) | 2015-03-19 | 2018-01-24 | Regeneron Pharmaceuticals, Inc. | Non-human animals that select for light chain variable regions that bind antigen |
| ES2812303T3 (es) | 2015-07-10 | 2021-03-16 | Merus Nv | Anticuerpo que se une a CD3 humano |
| CA2995717A1 (en) | 2015-08-24 | 2017-03-02 | Trianni, Inc. | Enhanced production of immunoglobulins |
| HUE055222T2 (hu) | 2015-10-23 | 2021-11-29 | Merus Nv | Megkötõ molekulák, amelyek gátolják a rák növekedését |
| WO2017095939A1 (en) | 2015-12-03 | 2017-06-08 | Trianni, Inc. | Enhanced immunoglobulin diversity |
| KR20180104149A (ko) | 2016-02-04 | 2018-09-19 | 트리아니, 인코포레이티드 | 면역글로불린의 증대된 생성 |
| AU2017272337C1 (en) | 2016-06-03 | 2024-02-29 | Regeneron Pharmaceuticals, Inc. | Non-human animals expressing exogenous terminal deoxynucleotidyltransferase |
| WO2018029534A1 (en) | 2016-08-12 | 2018-02-15 | Oxitec Ltd. | A self-limiting, sex-specific gene and methods of using |
| NZ751956A (en) | 2016-09-23 | 2022-01-28 | Merus Nv | Binding molecules that modulate a biological activity expressed by a cell |
| CN110637083A (zh) * | 2017-03-20 | 2019-12-31 | 华盛顿大学 | 细胞及其使用和制备方法 |
| MX2019011660A (es) | 2017-03-31 | 2019-11-18 | Merus Nv | Anticuerpos biespecificos que se unen al receptor 2 del factor de crecimiento humano (erbb-2) y receptor 3 del factor de crecimiento humano (erbb3) para usarse en el tratamiento de celulas que tienen un gen de fusion de neuregulina-1 (nrg1). |
| BR112019020507A2 (pt) | 2017-03-31 | 2020-08-04 | Merus N.V. | agente de alvejamento de erbb-2 e um anticorpo bispecífico com locais de ligação de antígenos que ligam um epítopo em uma parte extracelular de erbb-2 e erbb-3 para tratamento de um indivíduo com um tumor positivo erbb-2, erbb-2 / erbb-3 |
| ES2951864T3 (es) | 2017-05-17 | 2023-10-25 | Merus Nv | Combinación de un anticuerpo biespecífico ERBB-2/ERBB-3 con terapia endocrina para el cáncer de mama |
| US11753470B2 (en) | 2017-07-06 | 2023-09-12 | Merus N.V. | Bispecific anti PD1-anti TIM3 antibodies |
| JP2020525533A (ja) | 2017-07-06 | 2020-08-27 | メルス ナムローゼ フェンノートシャップ | 細胞により発現される生物学的活性を調節する抗体 |
| WO2019009726A1 (en) | 2017-07-06 | 2019-01-10 | Merus N.V. | BINDING MOLECULES FOR MODULATING THE BIOLOGICAL ACTIVITY EXPRESSED BY A CELL |
| GB201710984D0 (en) * | 2017-07-07 | 2017-08-23 | Kymab Ltd | Cells, vertebrates, populations & methods |
| CN111094351A (zh) | 2017-08-09 | 2020-05-01 | 美勒斯公司 | 结合EGFR和cMET的抗体 |
| CN111432838A (zh) | 2017-12-01 | 2020-07-17 | 美勒斯公司 | 使用双特异性抗体和il-15进行联合治疗 |
| CN111432635B (zh) * | 2017-12-05 | 2022-10-21 | 瑞泽恩制药公司 | 具有经工程改造的免疫球蛋白λ轻链的非人动物以及所述非人动物的用途 |
| TWI804572B (zh) | 2018-02-09 | 2023-06-11 | 日商小野藥品工業股份有限公司 | 雙特異性抗體 |
| CN116874591A (zh) | 2018-03-24 | 2023-10-13 | 瑞泽恩制药公司 | 用于产生针对肽-mhc复合物的治疗性抗体的经过基因修饰的非人动物、其制造方法和用途 |
| CA3094318A1 (en) | 2018-03-30 | 2019-10-03 | Merus N.V. | Multivalent antibody |
| CN108624622A (zh) * | 2018-05-16 | 2018-10-09 | 湖南艾佳生物科技股份有限公司 | 一种基于CRISPR-Cas9系统构建的能分泌小鼠白细胞介素-6的基因工程细胞株 |
| EP3801010A4 (en) * | 2018-06-08 | 2022-02-23 | Crystal Bioscience Inc. | TRANSGENIC ANIMAL TO PRODUCE DIVERSIFIED ANTIBODIES WITH THE SAME LIGHT CHAIN I |
| US20190380316A1 (en) | 2018-06-14 | 2019-12-19 | Regeneron Pharmaceuticals, Inc. | Non-human animals capable of engineered dh-dh rearrangement and uses thereof |
| JP2022521036A (ja) | 2018-12-20 | 2022-04-05 | メルス ナムローゼ フェンノートシャップ | CLEC12AxCD3二重特異性抗体及び疾患の治療方法 |
| EP3897117A1 (en) | 2018-12-21 | 2021-10-27 | Compass Therapeutics LLC | Transgenic mouse expressing common human light chain |
| KR20210111767A (ko) | 2018-12-31 | 2021-09-13 | 메뤼스 엔.페. | 절단된 다가 다량체 |
| JP7443376B2 (ja) * | 2018-12-31 | 2024-03-05 | メルス ナムローゼ フェンノートシャップ | 混合結合ドメイン |
| BR112021014106A2 (pt) | 2019-01-22 | 2021-10-13 | Bristol-Myers Squibb Company | Anticorpos contra subunidade alfa de il-7r e usos dos mesmos |
| BR112021016094A2 (pt) | 2019-02-14 | 2021-10-26 | Merus N.V. | Combinações de porções de ligação que se ligam ao egfr, her2 e her3 |
| MX2021009769A (es) | 2019-02-14 | 2021-09-08 | Merus Nv | Produccion de composiciones que comprenden dos o mas anticuerpos. |
| TW202039578A (zh) | 2019-03-29 | 2020-11-01 | 荷蘭商美勒斯公司 | Cd3結合分子 |
| TW202102544A (zh) * | 2019-04-04 | 2021-01-16 | 日商小野藥品工業股份有限公司 | 雙特異性抗體 |
| JP2022534674A (ja) | 2019-05-09 | 2022-08-03 | メルス ナムローゼ フェンノートシャップ | タンパク質を多量体化する変異体ドメイン及びその分離 |
| WO2021003149A1 (en) * | 2019-07-01 | 2021-01-07 | Trianni, Inc. | Transgenic mammals and methods of use |
| WO2021006199A1 (ja) | 2019-07-05 | 2021-01-14 | 小野薬品工業株式会社 | Pd-1/cd3二重特異性タンパク質による血液がん治療 |
| TW202118790A (zh) | 2019-07-30 | 2021-05-16 | 日商小野藥品工業股份有限公司 | 雙特異性抗體 |
| EP3772518A1 (en) | 2019-08-07 | 2021-02-10 | Merus N.V. | Modified human variable domains |
| AU2020331879A1 (en) | 2019-08-19 | 2022-02-24 | Merus N.V. | Treatment of cancer with a combination of an antibody that binds LGR5 and EGFR and a topoisomerase I inhibitor. |
| GB201912008D0 (en) | 2019-08-21 | 2019-10-02 | Cambridge Entpr Ltd | Antibody |
| EP4069722A1 (en) | 2019-12-02 | 2022-10-12 | Regeneron Pharmaceuticals, Inc. | Peptide-mhc ii protein constructs and uses thereof |
| KR20220117267A (ko) | 2019-12-24 | 2022-08-23 | 메뤼스 엔.페. | Tgf-베타-rii 결합 단백질 |
| JP7480307B2 (ja) | 2020-01-29 | 2024-05-09 | メルス ナムローゼ フェンノートシャップ | 免疫細胞関与効果を調節するための手段および方法 |
| AU2021261681A1 (en) | 2020-04-24 | 2022-12-08 | Merus N.V. | Treatment of cancers with an antibody that binds LGR5 and EGFR |
| IL301137A (en) | 2020-09-11 | 2023-05-01 | Regeneron Pharma | Identification and production of antigen-specific antibodies |
| TW202237656A (zh) | 2020-12-15 | 2022-10-01 | 荷蘭商美勒斯公司 | 利用結合lgr5與egfr之抗體治療癌症 |
| KR20230147048A (ko) | 2020-12-16 | 2023-10-20 | 리제너론 파마슈티칼스 인코포레이티드 | 인간화 Fc 알파 수용체를 발현하는 마우스 |
| JP2023554422A (ja) | 2020-12-16 | 2023-12-27 | メルス ナムローゼ フェンノートシャップ | がんの治療のための多重特異性抗体 |
| EP4262867A1 (en) | 2020-12-18 | 2023-10-25 | Merus N.V. | Antibody composition |
| EP4051700A1 (en) | 2020-12-23 | 2022-09-07 | Regeneron Pharmaceuticals, Inc. | Nucleic acids encoding anchor modified antibodies and uses thereof |
| TW202321309A (zh) | 2021-10-06 | 2023-06-01 | 荷蘭商美勒斯公司 | 使用至少結合egfr之抗體治療免疫檢查點抑制劑治療之具高egfr表現的癌症 |
| WO2023146394A1 (en) | 2022-01-25 | 2023-08-03 | Merus N.V. | Combination therapy for the treatment of cancer |
| WO2023172133A1 (en) | 2022-03-07 | 2023-09-14 | Merus N.V. | Combination therapy including antibodies that bind egfr and cmet |
| WO2023172134A1 (en) | 2022-03-07 | 2023-09-14 | Merus N.V. | Treatment with an antibody that binds egfr and cmet. |
| GB202216503D0 (en) | 2022-11-05 | 2022-12-21 | Quadrucept Bio Ltd | Non-human vertebrates & cells |
Family Cites Families (25)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4531078A (en) | 1983-10-27 | 1985-07-23 | Polaroid Corporation | Control circuit for electronic flash |
| GB8909218D0 (en) | 1989-04-22 | 1989-06-07 | Medical Res Council | Improvements in or relating to enhancers |
| EP0814159B1 (en) * | 1990-08-29 | 2005-07-27 | GenPharm International, Inc. | Transgenic mice capable of producing heterologous antibodies |
| US5770429A (en) * | 1990-08-29 | 1998-06-23 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| WO1993012227A1 (en) * | 1991-12-17 | 1993-06-24 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| US7041871B1 (en) * | 1995-10-10 | 2006-05-09 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| JPH08140528A (ja) * | 1993-12-03 | 1996-06-04 | Genpharm Internatl Inc | 異種抗体を産生することができるトランスジェニック非ヒト動物 |
| US5885827A (en) * | 1996-01-23 | 1999-03-23 | The Regents Of The Universtiy Of California | Eukaryotic high rate mutagenesis system |
| WO1998050431A2 (en) | 1997-05-02 | 1998-11-12 | Genentech, Inc. | A method for making multispecific antibodies having heteromultimeric and common components |
| ES2258817T3 (es) | 1997-05-21 | 2006-09-01 | Biovation Limited | Metodo para la produccion de proteinas no inmunogenas. |
| RU2236127C2 (ru) * | 1998-03-30 | 2004-09-20 | Рисерч Дивелопмент Фаундейшн | Способ получения трансгенной мыши, не содержащей функциональный рецептор-1 рилизинг-фактора кортикотропина, способ идентификации агониста или антагониста рилизинг-фактора кортикотропина, урокортина или лиганда семейства рилизинг-фактора кортикотропина и способ скрининга соединений, которые являются аналогами или агонистами кортикостерона или кортикотропина, с использованием такой мыши |
| US20020028488A1 (en) | 2000-06-19 | 2002-03-07 | Sujay Singh | Transgenic avian species for making human and chimeric antibodies |
| US6596541B2 (en) | 2000-10-31 | 2003-07-22 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| CN101537180B (zh) * | 2002-07-18 | 2016-02-10 | 莫鲁斯有限公司 | 抗体混合物的重组生产 |
| EP1439234A1 (en) * | 2003-01-08 | 2004-07-21 | ARTEMIS Pharmaceuticals GmbH | Targeted transgenesis using the rosa26 locus |
| ES2408582T3 (es) | 2003-05-30 | 2013-06-21 | Merus B.V. | Biblioteca de Fab para la preparación de una mezcla de anticuerpos |
| DK1737971T3 (da) | 2004-01-20 | 2017-11-13 | Merus Nv | Blandinger af bindingsproteiner |
| JP4487068B2 (ja) * | 2004-10-12 | 2010-06-23 | 国立大学法人 岡山大学 | 細胞の遺伝子変異機能の制御による変異タンパク質の作製方法 |
| AU2006242854A1 (en) * | 2005-04-29 | 2006-11-09 | Inserm (Institut National De La Sante Et De La Recherche Medicale) | Transgenic animals and methods of making recombinant antibodies |
| EP2505058A1 (en) * | 2006-03-31 | 2012-10-03 | Medarex, Inc. | Transgenic animals expressing chimeric antibodies for use in preparing human antibodies |
| ES2398076T3 (es) | 2006-06-02 | 2013-03-13 | Regeneron Pharmaceuticals, Inc. | Anticuerpos de alta afinidad contra el receptor de IL-6 humano |
| LT2769992T (lt) | 2006-10-02 | 2021-04-12 | Regeneron Pharmaceuticals, Inc. | Didelio afiniškumo žmogaus antikūnai, atpažįstantys žmogaus il-4 receptorių |
| RU2448979C2 (ru) | 2006-12-14 | 2012-04-27 | Ридженерон Фармасьютикалз, Инк. | Антитела человека к дельта-подобному лиганду-4 человека |
| CN105191863B (zh) * | 2008-06-27 | 2020-12-22 | 莫鲁斯股份有限公司 | 产生抗体的非人哺乳动物 |
| ES2603559T5 (es) * | 2010-02-08 | 2021-02-22 | Regeneron Pharma | Cadena ligera común de ratón |
-
2009
- 2009-06-29 CN CN201510514640.9A patent/CN105191863B/zh active Active
- 2009-06-29 SG SG10201914027VA patent/SG10201914027VA/en unknown
- 2009-06-29 HU HUE18183980A patent/HUE057994T2/hu unknown
- 2009-06-29 RS RS20140063A patent/RS53146B/en unknown
- 2009-06-29 SG SG10201408500RA patent/SG10201408500RA/en unknown
- 2009-06-29 PT PT121755441T patent/PT2556747T/pt unknown
- 2009-06-29 RS RS20220155A patent/RS62925B1/sr unknown
- 2009-06-29 DK DK18183980.4T patent/DK3456190T3/da active
- 2009-06-29 WO PCT/NL2009/050381 patent/WO2009157771A2/en active Application Filing
- 2009-06-29 EP EP18183980.4A patent/EP3456190B1/en active Active
- 2009-06-29 ES ES18183980T patent/ES2906344T3/es active Active
- 2009-06-29 CN CN200980131844.7A patent/CN102123582B/zh active Active
- 2009-06-29 KR KR1020177015048A patent/KR101990228B1/ko active IP Right Grant
- 2009-06-29 EP EP12175544.1A patent/EP2556747B1/en active Active
- 2009-06-29 RU RU2015119670A patent/RU2731084C2/ru active
- 2009-06-29 MX MX2010014542A patent/MX2010014542A/es active IP Right Grant
- 2009-06-29 ES ES09075279.1T patent/ES2445193T3/es active Active
- 2009-06-29 KR KR1020147027471A patent/KR102261586B1/ko active IP Right Grant
- 2009-06-29 SG SG10201914019YA patent/SG10201914019YA/en unknown
- 2009-06-29 PL PL09075279T patent/PL2147594T3/pl unknown
- 2009-06-29 PT PT181839804T patent/PT3456190T/pt unknown
- 2009-06-29 RU RU2011102962/10A patent/RU2559524C2/ru active
- 2009-06-29 KR KR1020117002023A patent/KR20110029156A/ko not_active Application Discontinuation
- 2009-06-29 LT LTEP18183980.4T patent/LT3456190T/lt unknown
- 2009-06-29 JP JP2011516168A patent/JP5749161B2/ja active Active
- 2009-06-29 SI SI200932155T patent/SI3456190T1/sl unknown
- 2009-06-29 DK DK12175544.1T patent/DK2556747T3/da active
- 2009-06-29 NZ NZ590118A patent/NZ590118A/xx unknown
- 2009-06-29 EP EP18183994.5A patent/EP3456192A1/en active Pending
- 2009-06-29 CN CN202011446991.8A patent/CN112481300A/zh active Pending
- 2009-06-29 DK DK09075279.1T patent/DK2147594T3/da active
- 2009-06-29 EP EP18183986.1A patent/EP3456191A1/en active Pending
- 2009-06-29 PL PL18183980T patent/PL3456190T3/pl unknown
- 2009-06-29 ES ES12175544T patent/ES2853226T3/es active Active
- 2009-06-29 EP EP18184003.4A patent/EP3456193A1/en active Pending
- 2009-06-29 KR KR1020207013671A patent/KR102306491B1/ko active IP Right Grant
- 2009-06-29 AU AU2009263082A patent/AU2009263082C1/en active Active
- 2009-06-29 SI SI200930829T patent/SI2147594T1/sl unknown
- 2009-06-29 PT PT90752791T patent/PT2147594E/pt unknown
- 2009-06-29 KR KR1020197006239A patent/KR102112655B1/ko active IP Right Grant
- 2009-06-29 EP EP09075279.1A patent/EP2147594B1/en active Active
- 2009-06-29 CA CA2729095A patent/CA2729095C/en active Active
- 2009-06-29 HR HRP20220205TT patent/HRP20220205T1/hr unknown
-
2010
- 2010-12-23 ZA ZA2010/09312A patent/ZA201009312B/en unknown
-
2011
- 2011-08-17 HK HK11108669.0A patent/HK1154463A1/zh unknown
-
2014
- 2014-02-12 CY CY20141100105T patent/CY1114830T1/el unknown
- 2014-02-13 HR HRP20140139AT patent/HRP20140139T1/hr unknown
-
2015
- 2015-05-12 JP JP2015097258A patent/JP2015156871A/ja active Pending
-
2017
- 2017-04-12 JP JP2017078941A patent/JP6824807B2/ja active Active
-
2020
- 2020-07-13 JP JP2020119962A patent/JP7242611B2/ja active Active
- 2020-08-19 RU RU2020127564A patent/RU2020127564A/ru unknown
-
2022
- 2022-02-22 CY CY20221100149T patent/CY1125119T1/el unknown
- 2022-09-16 JP JP2022147656A patent/JP7439201B2/ja active Active
-
2024
- 2024-02-14 JP JP2024020295A patent/JP2024063038A/ja active Pending
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2906344T3 (es) | Animal murino transgénico productor de anticuerpos | |
| US20240164354A1 (en) | Antibody producing non-human animals | |
| AU2014203150B2 (en) | Antibody producing non-human mammals |