ES2779453T3 - Neuronas de dopamina (DA) del mesencéfalo para injerto - Google Patents
Neuronas de dopamina (DA) del mesencéfalo para injerto Download PDFInfo
- Publication number
- ES2779453T3 ES2779453T3 ES12846715T ES12846715T ES2779453T3 ES 2779453 T3 ES2779453 T3 ES 2779453T3 ES 12846715 T ES12846715 T ES 12846715T ES 12846715 T ES12846715 T ES 12846715T ES 2779453 T3 ES2779453 T3 ES 2779453T3
- Authority
- ES
- Spain
- Prior art keywords
- cells
- cell
- neurons
- midbrain
- signaling
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
- A61K35/30—Nerves; Brain; Eyes; Corneal cells; Cerebrospinal fluid; Neuronal stem cells; Neuronal precursor cells; Glial cells; Oligodendrocytes; Schwann cells; Astroglia; Astrocytes; Choroid plexus; Spinal cord tissue
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/14—Drugs for disorders of the nervous system for treating abnormal movements, e.g. chorea, dyskinesia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/14—Drugs for disorders of the nervous system for treating abnormal movements, e.g. chorea, dyskinesia
- A61P25/16—Anti-Parkinson drugs
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/28—Drugs for disorders of the nervous system for treating neurodegenerative disorders of the central nervous system, e.g. nootropic agents, cognition enhancers, drugs for treating Alzheimer's disease or other forms of dementia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0618—Cells of the nervous system
- C12N5/0619—Neurons
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0618—Cells of the nervous system
- C12N5/0623—Stem cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/01—Modulators of cAMP or cGMP, e.g. non-hydrolysable analogs, phosphodiesterase inhibitors, cholera toxin
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/119—Other fibroblast growth factors, e.g. FGF-4, FGF-8, FGF-10
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/13—Nerve growth factor [NGF]; Brain-derived neurotrophic factor [BDNF]; Cilliary neurotrophic factor [CNTF]; Glial-derived neurotrophic factor [GDNF]; Neurotrophins [NT]; Neuregulins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/15—Transforming growth factor beta (TGF-β)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/16—Activin; Inhibin; Mullerian inhibiting substance
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/40—Regulators of development
- C12N2501/41—Hedgehog proteins; Cyclopamine (inhibitor)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/40—Regulators of development
- C12N2501/415—Wnt; Frizzeled
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/70—Enzymes
- C12N2501/72—Transferases (EC 2.)
- C12N2501/727—Kinases (EC 2.7.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/999—Small molecules not provided for elsewhere
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2506/00—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells
- C12N2506/02—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells from embryonic cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2506/00—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells
- C12N2506/45—Differentiation of animal cells from one lineage to another; Differentiation of pluripotent cells from artificially induced pluripotent stem cells
Landscapes
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Life Sciences & Earth Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Chemical & Material Sciences (AREA)
- Neurology (AREA)
- Organic Chemistry (AREA)
- Neurosurgery (AREA)
- General Health & Medical Sciences (AREA)
- Zoology (AREA)
- Biotechnology (AREA)
- Medicinal Chemistry (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Cell Biology (AREA)
- Genetics & Genomics (AREA)
- Wood Science & Technology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Developmental Biology & Embryology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Psychology (AREA)
- Immunology (AREA)
- Virology (AREA)
- Ophthalmology & Optometry (AREA)
- Epidemiology (AREA)
- Psychiatry (AREA)
- Hospice & Palliative Care (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Macromolecular Compounds Obtained By Forming Nitrogen-Containing Linkages In General (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Nitrogen Condensed Heterocyclic Rings (AREA)
- Gastroenterology & Hepatology (AREA)
Abstract
Un método in vitro para diferenciar células pluripotentes en precursores de la placa del suelo del mesencéfalo, comprendiendo el método: exponer una pluralidad de células pluripotentes a: (a) al menos un inhibidor de la señalización de Small Mothers Against Decapentaplegic (SMAD), (b) al menos un activador de la señalización de Sonic hedgehog (SHH) y (c) al menos un activador de la señalización de wingless (Wnt), en donde: la exposición al al menos un inhibidor de la señalización de SMAD comienza el día 0; dichas células pluripotentes se exponen al al menos un activador de la señalización de Wnt 3 días después del inicio de la exposición al al menos un inhibidor de la señalización de SMAD; dichas células pluripotentes están expuestas al al menos un inhibidor de la señalización de SMAD, el al menos un activador de la señalización de SHH y el al menos un activador de la señalización de Wnt en cantidades eficaces para producir una pluralidad de células precursoras de la placa del suelo del mesencéfalo, al menos el 10 % de las cuales expresan tanto la proteína A2 de secuencia forkhead (FOXA2) como el factor de transcripción 1 alfa de homeosecuencia LIM (LMX1A); y opcionalmente someter dicha pluralidad de células precursoras de la placa del suelo del mesencéfalo diferenciadas a condiciones que favorezcan la maduración de las células precursoras de la placa del suelo del mesencéfalo en neuronas de dopamina.
Description
DESCRIPCIÓN
Neuronas de dopamina (DA) del mesencéfalo para injerto
DERECHOS DE LICENCIA GUBERNAMENTAL
La presente invención se realizó en parte con el apoyo del gobierno según la subvención NIH/NINDS NS052671 y la subvención NIH/NINDS P50 NS047085, del Instituto Nacional de Salud de los Estados Unidos (NIH) y el Instituto Nacional de Trastornos Neurológicos y Accidentes Cerebrovasculares (NINDS).
Campo de la invención
La presente invención se refiere al campo de la biología de células madre, en particular la diferenciación específica de linaje de células madre pluripotentes o multipotentes, que pueden incluir, pero sin limitación, células madre embrionarias humanas (hESC) además de células madre pluripotentes inducidas humanas no embrionarias (hiPSC), células madre somáticas, células madre de pacientes con una enfermedad o cualquier otra célula con capacidad de diferenciación específica de linaje. Se describen específicamente métodos para dirigir la diferenciación específica de linaje de hESC y/o hiPSC en células progenitoras del mesencéfalo de la placa del suelo y después en grandes poblaciones de neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas al mesencéfalo usando condiciones de cultivo novedosas. Las neuronas de dopamina (DA) destinadas a mesencéfalo FOXA2+LMX1A+TH+ preparadas usando los métodos de la presente invención se contemplan adicionalmente para diversos usos, incluyendo, pero sin limitación, uso en ensayos de descubrimiento de fármacos in vitro, investigación neurológica y como un producto terapéutico para neutralizar la enfermedad de, o el daño a, una falta de neuronas de dopamina en un paciente. Además, se proporcionan composiciones y métodos para diferenciar neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas a mesencéfalo a partir de células madre pluripotentes humanas para su uso en el modelado de enfermedades, en particular enfermedad de Parkinson. Además, las neuronas DA auténticas están enriquecidas con respecto a marcadores, tales como CD142, y células neuronales de tipo A9.
Antecedentes de la invención
Las poblaciones celulares que conservan la capacidad de diferenciarse en numerosos tipos de células especializadas son útiles para desarrollar grandes cantidades de poblaciones celulares diferenciadas específicas de linaje. Estas poblaciones celulares que conservan una capacidad de diferenciación posterior en células especializadas contienen células pluripotentes. Las células pluripotentes pueden tener origen de células madre somáticas embrionarias y/o no embrionarias.
Se contempla que estas poblaciones celulares diferenciadas específicas de linaje encuentran uso en terapias de reemplazo celular para pacientes con enfermedades que dan como resultado una pérdida de función de una población celular definida. Además de su valor terapéutico directo, las células diferenciadas específicas de linaje también son herramientas de investigación valiosas para diversos fines, incluyendo ensayos de exploración in vitro para identificar, confirmar y probar la especificación de la función o para probar el suministro de moléculas terapéuticas para tratar la enfermedad específica de linaje celular.
Anteriormente, se han usado células madre embrionarias y somáticas como productos terapéuticos y sistemas modelo para enfermedades neurodegenerativas. Se han llevado a cabo investigación y desarrollos tecnológicos relacionados con la diferenciación dirigida de células madre embrionarias y somáticas en el campo de enfermedades del sistema nervioso central (SNC), tal como para enfermedad de Huntington, enfermedad de Alzheimer, enfermedad de Parkinson y esclerosis múltiple. Sin embargo, los resultados de estos estudios mostraron poca capacidad de estas células usadas in vivo para permitir que el paciente recupere la función neuronal y, con frecuencia, provocaron el crecimiento de tumores no deseados en los pacientes.
Por lo tanto, existe la necesidad de composiciones y métodos para obtener poblaciones celulares que pueden usarse en investigación y como un producto terapéutico para tratar enfermedades que dan como resultado una pérdida de células que tienen una función particular.
Compendio de la invención
La presente invención se refiere al campo de la biología de células madre, en particular la diferenciación específica de linaje de células madre pluripotentes o multipotentes, que pueden incluir, pero sin limitación, células madre embrionarias humanas (hESC) además de células madre pluripotentes inducidas humanas no embrionarias (hiPSC), células madre somáticas, células madre de pacientes con una enfermedad o cualquier otra célula con capacidad de diferenciación específica de linaje. Se describen específicamente métodos para dirigir la diferenciación específica de linaje de hESC y/o hiPSC en células progenitoras del mesencéfalo de la placa del suelo y después en grandes poblaciones de neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas al mesencéfalo usando condiciones de cultivo novedosas. Las neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas a mesencéfalo preparadas usando los métodos de la presente invención se contemplan adicionalmente para diversos usos, incluyendo, pero sin limitación, uso en ensayos de descubrimiento de fármacos in vitro, investigación neurológica y como un producto terapéutico para neutralizar la enfermedad de, o el daño a, una falta de neuronas de dopamina en un paciente.
Además, se proporcionan composiciones y métodos para diferenciar neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas a mesencéfalo a partir de células madre pluripotentes humanas para su uso en el modelado de enfermedades, en particular enfermedad de Parkinson. Además, las neuronas DA auténticas están enriquecidas con respecto a marcadores, tales como CD142, y células neuronales de tipo A9.
Un equipo que comprende un primer inhibidor de la señalización, un segundo inhibidor de la señalización y un tercer inhibidor de la señalización, en donde dicho primer inhibidor es capaz de reducir la señalización del factor de crecimiento transformante beta (TGFp)/Activina-Nodal, dicho segundo inhibidor es capaz de reducir la señalización de Small Mothers Against Decapentaplegic (SMAD) y dicho tercer inhibidor es capaz de reducir la glucógeno sintasa cinasa 3p (GSK3p) para activación de la señalización de Wingless (Wnt). En una realización, dicho primer inhibidor es LDN-193189. En otras realizaciones, dicho primer inhibidor se selecciona del grupo que consiste en LDN-193189, derivados del mismo y mezclas de los mismos. En una realización, dicho segundo inhibidor es SB431542. En otras realizaciones, dicho segundo inhibidor se selecciona del grupo que consiste en SB431542, derivados del mismo y mezclas de los mismos. En una realización, dicho tercer inhibidor es CHIR99021. En otras realizaciones, dicho tercer inhibidor se selecciona del grupo que consiste en CHIR99021, derivados del mismo y mezclas de los mismos. En una realización, dicho equipo comprende además un activador de la señalización de Sonic hedgehog (SHH) y un activador de la señalización de la familia del receptor del factor de crecimiento de fibroblastos (FGF) 8. En una realización, dicho equipo comprende además factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial, dibutiril AMPc y factor de crecimiento transformante tipo p3. En una realización, dicho equipo comprende además anticuerpos seleccionados del grupo que consiste en anti-tirosina hidroxilasa (TH), antiproteína A2 de secuencia forkhead (FOXA2) y anti-factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A). En una realización, dicho equipo comprende además una célula seleccionada del grupo que consiste en una célula madre, célula madre embrionaria, célula madre pluripotente inducida y una célula modificada por ingeniería genética. En una realización, dicho equipo comprende además instrucciones para diferenciar células progenitoras y neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas a mesencéfalo. En una realización, dicho equipo comprende además instrucciones para obtener una célula de un paciente con enfermedad de Parkinson (EP).
Una composición, que comprende una población celular en contacto con un primer inhibidor de la señalización y un segundo inhibidor de la señalización, en donde más del 40 % de dicha población celular es positiva para la proteína A2 de secuencia forkhead (FOXA2), en donde dicha población celular entró en contacto previamente con un primer inhibidor de la señalización, un tercer inhibidor de la señalización y un activador de la señalización de Sonic hedgehog (SHH), en donde dicho primer inhibidor es capaz de reducir la señalización del factor de crecimiento transformante beta (TGFp)/Activina-Nodal, dicho segundo inhibidor es capaz de reducir la señalización de glucógeno sintasa cinasa 3p (GSK3p) para activación de la señalización de wingless (Wnt) y dicho tercer inhibidor es capaz de reducir la señalización de las Small Mothers Against Decapentaplegic (SMAD). En una realización, dicho primer inhibidor es una molécula pequeña seleccionada del grupo que consiste en LDN-193189, derivados del mismo y mezclas de los mismos. En una realización, dicho segundo inhibidor se selecciona del grupo que consiste en CHIR99021 y derivados del mismo. En una realización, dicho tercer inhibidor se selecciona del grupo que consiste en SB431542, derivados del mismo y mezclas de los mismos. En una realización, dicho activador de la señalización de Sonic hedgehog (SHH) se selecciona del grupo que consiste en C25II de Sonic hedgehog (SHH) y agonista de molécula pequeña del receptor smoothened (SMO), en donde dicho agonista es purmorfamina. En algunas realizaciones, dicha población celular había entrado en contacto previamente además con el factor de crecimiento de fibroblastos 8 (FGF8). En una realización, dicha mayoría de células que comprenden dicha población celular son células progenitoras del mesencéfalo de la placa del suelo proteína A2 de secuencia forkhead (FOXA2)+factor de transcripción 1+ de homeosecuencia LIM+, alfa (LMX1A),+ NGN2+ y DDC+. En una realización, dicha población celular se selecciona del grupo que consiste en células de roedores, células de primates y células humanas. En una realización, dichas células se obtienen de células de pacientes con enfermedad de Parkinson (EP). En una realización, dicha población celular es al menos 50 % positiva para la proteína A2 de secuencia forkhead (FOXA2). En una realización, dicha población celular es al menos 60 % positiva para la proteína A2 de secuencia forkhead. En una realización, dicha población celular es al menos 70 % positiva para la proteína A2 de secuencia forkhead. En una realización, dicha población celular es al menos 80 % positiva para la proteína A2 de secuencia forkhead. En una realización, dicha población celular es al menos 90 % positiva para la proteína A2 de secuencia forkhead. En una realización, dicha población celular es al menos 95 % hasta 100 % positiva para la proteína A2 de secuencia forkhead.
Una composición, que comprende una población celular in vitro en donde la mayoría de células que comprenden dicha población celular son neuronas de dopamina (DA) del mesencéfalo de la placa del suelo tirosina hidroxilasa (TH)+ proteína A2 de secuencia forkhead (FOXA2)+factor de transcripción 1+ de homeosecuencia LIM, alfa (LMX1A)+. En una realización, dicho más del 40 % de dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son tirosina hidroxilasa positivas (TH+). En una realización, dicha población celular es al menos 50 % tirosina hidroxilasa positiva. En una realización, dicha población celular es al menos 60 % tirosina hidroxilasa positiva. En una realización, dicha población celular es al menos 70 % tirosina hidroxilasa positiva. En una realización, dicha población celular es al menos 80 % tirosina hidroxilasa positiva. En una realización, dicha población celular es al menos 90 % tirosina hidroxilasa positiva. En una realización, dicha población celular es al menos 95 % hasta 100 % tirosina hidroxilasa positiva. En algunas realizaciones, dicha población celular comprende una mayoría de las neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo. En una realización, dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son positivas para marcadores seleccionados del grupo que consiste en el receptor nuclear
NURR1 (NR4A2), beta-tubulina de clase III específica de neurona (Tuj 1), TTF3, homeodominio de tipo paired 3 (PITX3), complejo achaete-scute (ASCL), factor de células B temprano 1 (EBF-1), factor de células B temprano 3 (EBF-3) y transtiretina (TTR). h una realización, dicha población de neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo es positiva para una molécula seleccionada del grupo que consiste en DA, ácido 3,4-dihidroxi-fenilacético (DOPAC) y ácido homo vanílico (HVA). En una realización, dicho marcador se selecciona del grupo que consiste en una proteína y un ácido nucleico. En algunas realizaciones, dicha población de neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo es capaz de injertarse in vivo en un paciente seleccionado del grupo que consiste en un paciente con enfermedad de Parkinson (EP). En una realización, dichas neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo son capaces de injertarse in vivo y proporcionar función neuronal de dopamina (DA).
En algunas realizaciones, las invenciones proporcionan una composición, que comprende una población celular en contacto con LDN-193189 y CHIR99021, en donde más del 40 % de dicha población celular es positiva para la proteína A2 de secuencia forkhead (FOXA2) y en donde dicha población celular entró en contacto previamente con LDN-193189, SB431542, un activador de la señalización de Sonic hedgehog (SHH) y CHIR99021. En una realización, dicho activador de la señalización de Sonic hedgehog (SHH) se selecciona del grupo que consiste en C25II de Sonic hedgehog (SHH) y purmorfamina. En una realización, dicho más del 10 % de dicha población celular es doblemente positivo para la proteína A2 de secuencia forkhead (FOXA2) y el factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A). En una realización, dicha mayoría de dicha población celular es una población de células progenitoras del mesencéfalo de la placa del suelo. En una realización, dicha población celular había entrado en contacto previamente con el factor de crecimiento de fibroblastos 8 (FGF8). En una realización, dicha población celular se selecciona del grupo que consiste en células de roedores, células de primates y células humanas. En una realización, dichas células humanas son células de un paciente con un síntoma neurológico de la enfermedad de Parkinson (EP). En una realización, dicha población celular procede de una célula madre pluripotente inducida (iPSC). En una realización, más del 10 % de dicha población celular se selecciona del grupo que consiste en doble positivo para la proteína A2 de secuencia forkhead (FOXA2)/factor de transcripción 1 de homeosecuencia lIM, alfa (LMX1A) y doble positivo para la proteína A2 de secuencia forkhead (FOXA2)/homeosecuencia 2 de orthodenticle (OTX2).
En una realización, las invenciones proporcionan un método para inducir la diferenciación dirigida de células en una población de células progenitoras del mesencéfalo de la placa del suelo, que comprende a) proporcionar: i) una población celular, en donde dicha población celular se selecciona del grupo que consiste en una célula madre no embrionaria, una célula madre embrionaria, una célula pluripotente no embrionaria inducida y una célula pluripotente modificada por ingeniería genética; y ii) un primer inhibidor de la señalización, un segundo inhibidor de la señalización, un activador de la señalización de Sonic hedgehog (SHH) y un tercer inhibidor de la señalización, en donde dicho primer inhibidor es capaz de reducir la señalización del factor de crecimiento transformante beta (TGFp)/Activina-Nodal, dicho segundo inhibidor es capaz de reducir la señalización de Small Mothers Against Decapentaplegic (SMAD) y dicho tercer inhibidor es capaz de reducir la señalización de glucógeno sintasa cinasa 3p (GSK3p) para activación de la señalización de Wingless (Wnt); b) poner en contacto dicha población celular con dicho primer y segundo inhibidor, c) después de contactar dicha población celular con dicho primer y dicho segundo inhibidor poner en contacto además dichas células con dicho activador de la señalización de Sonic hedgehog (SHH) en condiciones para diferenciar una población de células progenitoras del mesencéfalo de la placa del suelo; y d) después de poner en contacto dicha población celular con dicho activador de la señalización de Sonic hedgehog (SHH) poner en contacto además dichas células con dicho tercer inhibidor para diferenciar dicha población celular en una población de células progenitoras del mesencéfalo de la placa del suelo. En una realización, dicho contacto con dicho primer y dicho segundo inhibidor está en condiciones capaces de dar como resultado dicha población diferenciada de células progenitoras del mesencéfalo de la placa del suelo. En una realización, dicho contacto con dicho primer y dicho segundo inhibidor es en un periodo de 1 hora desde la siembra células en placa in vitro. En una realización, dicho contacto con dicho primer y dicho segundo inhibidor es en un periodo de 48 horas desde la siembra células en placa in vitro. En una realización, dicho contacto con dicho primer y dicho segundo inhibidor es en un periodo de 62 horas desde la siembra células en placa in vitro. En una realización, dicho contacto de dichas células con dicho activador de la señalización de Sonic hedgehog (SHH) está en condiciones capaces de dar como resultado dicha población diferenciada de células progenitoras del mesencéfalo de la placa del suelo. En una realización, dicho contacto de dichas células con dicho activador de la señalización de Sonic hedgehog (SHH) es de al menos 24 horas hasta 36 horas después de poner en contacto dicha población celular con dicho primer y dicho segundo inhibidor. En una realización, dicho contacto de dichas células con dicho activador de la señalización de Sonic hedgehog (SHH) es de hasta 144 horas. En una realización, dicho contacto de dichas células con dicho tercer inhibidor está en condiciones capaces de dar como resultado dicha población diferenciada de células progenitoras del mesencéfalo de la placa del suelo. En una realización, dicho contacto de dichas células con dicho tercer inhibidor es de al menos 24 horas hasta 36 horas después de poner en contacto dicha población celular con dicho activador de la señalización de Sonic hedgehog (sHh). En una realización, dicho contacto de dichas células con dicho tercer inhibidor es de hasta 192 horas. En una realización, dicha población celular se diferencia en dichas células progenitoras del mesencéfalo de la placa del suelo al menos el día 11 después de entrar en contacto dichas células con dicho primer y dicho segundo inhibidor. En una realización, dicho primer inhibidor es SB431542. En una realización, dicho segundo inhibidor es LDN-193189. En una realización, dicho tercer inhibidor es CHIR99021. En una realización, dicho activador de la señalización de Sonic hedgehog (SHH) se selecciona del grupo que consiste en C25II de Sonic hedgehog (SHH) y purmorfamina. En una realización, dicho método proporciona además el factor de crecimiento de fibroblastos 8 (FGF8) y poner en
contacto dicha población celular con dicho FGF8 en condiciones capaces de dar como resultado dicha población diferenciada de células progenitoras del mesencéfalo de la placa del suelo. En una realización, dicho contacto de dichas células con dicho FGF8 es de al menos 24 hasta 36 horas después de poner en contacto dicha población celular con dicho primer y dicho segundo inhibidor. En una realización, dicho contacto de dichas células con dicho FGF8 es de hasta 144 horas. En una realización, dicha población de células progenitoras del mesencéfalo de la placa del suelo comprende más de 40 % de células proteína A2 de secuencia forkhead (FOXA2)+. En una realización, dicha población de células progenitoras del mesencéfalo de la placa del suelo comprende más de 40 % de células proteína A2 de secuencia forkhead (FOXA2)+factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A)+. En una realización, dicho método comprende además la etapa e) poner en contacto dicha población de células progenitoras del mesencéfalo de la placa del suelo con medio de maduración neuronal, comprendiendo dicho medio medio N2, factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial, dibutiril AMPc (dbAMPc) y factor de crecimiento transformante tipo p3 para diferenciación de células progenitoras del mesencéfalo de la placa del suelo en neuronas de dopamina (DA) del mesencéfalo de la placa del suelo. En una realización, dicho método comprende además la etapa e) poner en contacto dicha población de células progenitoras del mesencéfalo de la placa del suelo con medio de maduración neuronal con complemento B27 para la diferenciación de células progenitoras del mesencéfalo de la placa del suelo en neuronas de dopamina (DA) del mesencéfalo de la placa del suelo. En una realización, dichas células que han entrado en contacto con medio neurobasal con complemento B27 se ponen en contacto con factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial, dibutiril AMPc (dbAMPc) y factor de crecimiento transformante tipo p3 para diferenciación de células progenitoras del mesencéfalo de la placa del suelo en neuronas de dopamina (DA) del mesencéfalo de la placa del suelo. En una realización, dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son proteína A2 de secuencia forkhead (FOXA2)+factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A)+, proteína 1 relacionada con el receptor nuclear (NURR1)+ y tirosina hidroxilasa (TH)+. En una realización, más del 40 % de dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son tirosina hidroxilasa (TH)+. En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo se diferencian al menos el día 25 después de entrar en contacto dicha población celular con dicho primer y dicho segundo inhibidor. En una realización, dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son positivas para marcadores que identifican moléculas. En una realización, dichos marcadores se seleccionan del grupo que consiste en tirosina hidroxilasa (TH), proteína A2 de secuencia forkhead (FOXA2), factor de transcripción 1 de homeosecuencia LIM, dompamina, ácido 3,4-dihidroxi-fenilacético (DOPAC) y ácido homovanílico (HVA), receptor nuclear alfa NURR1 (NR4A2), beta-tubulina de clase III específica de neurona (Tuj 1), TTF3, homeodominio de tipo paired 3 (PITX3), complejo achaete-scute (ASCL), factor de linfocitos B temprano 1 (EBF-1), factor de linfocitos B temprano 3 (EBF-3), transtiretina (TTR), sinapsina, transportador de dopamina (DAT), y canal de potasio rectificador hacia dentro y acoplado a proteína G (Kir3.2/GIRK2). En una realización, dicha molécula se selecciona del grupo que consiste en una proteína y un ácido nucleico. En una realización, dicha molécula se identifica usando un marcador seleccionado del grupo que consiste en un anticuerpo, un cebador de PCR, una secuencia de ácido nucleico y un ensayo enzimático. En una realización, dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son capaces de injertarse in vivo en un paciente con enfermedad de Parkinson (EP) para proporcionar función neuronal de dopamina (DA). En una realización, dicho método comprende además proporcionar a un paciente que lo necesite neuronas productoras de dopamina, en donde dicho paciente muestra al menos un síntoma neurológico, y la etapa de trasplantar neuronas de dopamina (DA) del mesencéfalo de la placa del suelo en dicho paciente para proporcionar función neuronal de dopamina (DA). En una realización, dichos síntomas neurológicos se seleccionan del grupo que consiste en temblor, bradicinesia (lentitud extrema del movimiento), postura flexionada, inestabilidad postural y rigidez. En una realización, dicho paciente muestra una reducción de dicho síntoma neurológico. En una realización, dicha célula se selecciona de una célula de roedor, una célula de primate y una célula humana. En una realización, dichas células humanas son células de un paciente con un síntoma neurológico de la enfermedad de Parkinson (EP).
En una realización, las invenciones proporcionan un método de injerto in vivo para tratamiento terapéutico, que comprende a) proporcionar: i) una población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo en donde más del 40 % de dicha población expresa tirosina hidroxilasa (TH); y ii) un sujeto, en donde dicho sujeto muestra al menos un síntoma neurológico; y b) trasplantar dichas neuronas de dopamina (Da) del mesencéfalo de la placa del suelo en dicho sujeto en condiciones para permitir injerto in vivo para proporcionar función neuronal de dopamina (DA). En una realización, dichos síntomas neurológicos se seleccionan del grupo que consiste en temblor, bradicinesia (lentitud extrema del movimiento), postura flexionada, inestabilidad postural y rigidez. En una realización, dicho sujeto muestra reducción de dicho síntoma neurológico. En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo procede de una población de células progenitoras del mesencéfalo de la placa del suelo tratadas según métodos de las presentes invenciones. En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo procede de una población de células progenitoras del mesencéfalo de la placa del suelo tratada según un método que comprende además una etapa de poner en contacto dicha población de células progenitoras del mesencéfalo de la placa del suelo con medio de maduración neuronal, comprendiendo dicho medio medio N2, factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial, dibutiril AMPc y factor de crecimiento transformante tipo p3 para diferenciación de células progenitoras del mesencéfalo de la placa del suelo en neuronas de dopamina (DA) del mesencéfalo de la placa del suelo. En una realización, dicha población de células progenitoras del mesencéfalo de la placa del suelo procede de una población celular tratada según un método de las presentes invenciones. En una realización, dicha población de células progenitoras del mesencéfalo de la placa del suelo procede de una población
celular tratada según un método para inducir diferenciación dirigida de células en una población de células progenitoras
del mesencéfalo de la placa del suelo, que comprende a) proporcionar: i) una población celular, en donde dicha
población celular se selecciona del grupo que consiste en una célula madre no embrionaria, una célula madre
embrionaria, una célula pluripotente no embrionaria inducida y una célula pluripotente modificada por ingeniería
genética; y ii) un primer inhibidor de la señalización, un segundo inhibidor de la señalización, un activador de la
señalización de Sonic hedgehog (SHH) y un tercer inhibidor de la señalización, en donde dicho primer inhibidor es
capaz de reducir la señalización del factor de crecimiento transformante beta (TGFp)/Activina-Nodal, dicho segundo
inhibidor es capaz de reducir la señalización de Small Mothers Against Decapentaplegic (SMAD) y dicho tercer
inhibidor es capaz de reducir la señalización de glucógeno sintasa cinasa 3p (GSK3p) para activación de la
señalización de Wingless (Wnt); b) poner en contacto dicha población celular con dicho primer y segundo inhibidor, c)
después de contactar dicha población celular con dicho primer y dicho segundo inhibidor poner en contacto además
dichas células con dicho activador de la señalización de Sonic hedgehog (SHH) en condiciones para diferenciar una
población de células progenitoras del mesencéfalo de la placa del suelo; y d) después de poner en contacto dicha
población celular con dicho activador de la señalización de Sonic hedgehog (SHH) poner en contacto además dichas
células con dicho tercer inhibidor para diferenciar dicha población celular en una población de células progenitoras del
mesencéfalo de la placa del suelo. En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo
de la placa del suelo procede de una población celular seleccionada del grupo que consiste en animales, primates y
seres humanos. En una realización, dichas células humanas son células de un paciente con un síntoma de enfermedad
de Parkinson (EP).
En una realización, las invenciones proporcionan una composición, que comprende una población celular en contacto
con LDN-193189 y CHIR99021, en donde más del 40 % de dicha población celular es positiva para la proteína A2 de
secuencia forkhead (FOXA2) y en donde dicha población celular entró en contacto previamente con LDN-193189,
SB431542, un activador de la señalización de Sonic hedgehog (SHH) y CHIR99021, en
señalización de Sonic hedgehog (SHH) se selecciona del grupo que consiste en C25II
purmorfamina. En una realización, dicha población celular es al menos 50 % positiva para
forkhead (FOXA2). En una realización, dicha población celular es al menos 60 % posi
secuencia forkhead. En una realización, dicha población celular positiva para la secuencia forkhead. En una realización, dicha población celular positiva para la secuencia forkhead. En una realización, dicha población celular positiva para la secuencia forkhead. En una realización, dicha población celular es al menos 95 % h
proteína A2 de secuencia forkhead. En una realización, más del 10 % de dicha población celular se selecciona del
grupo que consiste en doble positivo para la proteína A2 de secuencia forkhead (FOXA2)/factor de transcripción 1 de
homeosecuencia lIM, alfa (LMX1A) y doble positivo para la proteína A2 de secuencia forkhead
(FOXA2)/homeosecuencia 2 de orthodenticle (OTX2). En una realización, dicha población celular es al menos 20 %,
30 %, 40 %, 50 %, 60 %, 70 %, 80 %, 90 %, al menos 95 % hasta 100 % positiva. En una realización, al menos 20 %
de dicha población celular es positiva para un marcador seleccionado del grupo que consiste en Nurr1+, CD142,
DCSM1, Cd63 y CD99. En algunas realizaciones, dicha población celular es al menos 30 %, 40 %, 50 %, 60 %, 70 %,
80 %, 90 %, al menos 95 % hasta 100 % positiva. En algunas realizaciones, dicha población celular es al menos 50 %,
60 %, 70 %, 80 %, 90 %, al menos 95 % hasta 100 % positiva. En una realización, dicha población celular se
selecciona del grupo que consiste en células de roedores, células de primates, células humanas y células humanas
de un paciente con un síntoma neurológico de la enfermedad de Parkinson (EP).
En una realización, las invenciones proporcionan un método para inducir la diferenciación dirigida de células en una
población de células progenitoras del mesencéfalo de la placa del suelo, que comprende a) proporcionar: i) una
población celular, en donde dicha población celular se selecciona del grupo que consiste en una célula madre no
embrionaria, una célula madre embrionaria, una célula pluripotente no embrionaria inducida y una célula pluripotente
modificada por ingeniería genética; y ii) un primer inhibidor de la señalización, un segundo inhibidor de la señalización,
un activador de la señalización de Sonic hedgehog (SHH) y un tercer inhibidor de la señalización, en donde dicho
primer inhibidor es SB431542, dicho segundo inhibidor es LDN-193189, dicho activador de la señalización de Sonic
hedgehog (SHH) se selecciona del grupo que consiste en C25II de Sonic hedgehog (SHH) y una purmorfamina y dicho
tercer inhibidor es CHIR99021; b) poner en contacto dicha población celular con dicho primer y dicho segundo
inhibidor, en donde dicho contacto con dicho primer y dicho segundo inhibidor está en condiciones capaces de dar
como resultado dicha población diferenciada de células progenitoras del mesencéfalo de la placa del suelo de manera
que dicho contacto con dicho primer y dicho segundo inhibidor sea en un periodo de 48 horas desde la siembra de
células en placa in vitro, c) después de poner en contacto dicha población celular con dicho primer y segundo inhibidor
poner en contacto además dichas células con dicho activador de la señalización de Sonic hedgehog (SHH) en
condiciones para diferenciar una población de células progenitoras del mesencéfalo de la placa del suelo; y d) después
de poner en contacto dicha población celular con dicho activador de la señalización de Sonic hedgehog (SHH) poner
en contacto además dichas células con dicho tercer inhibidor para diferenciar dicha población celular en una población
de células progenitoras del mesencéfalo de la placa del suelo. En una realización, el contacto de dichas células con
dicho activador de la señalización de Sonic hedgehog (SHH) está en condiciones capaces de dar como resultado
dicha población diferenciada de células progenitoras del mesencéfalo de la placa del suelo de manera que dicho
contacto de dichas células con dicho activador de la señalización de Sonic hedgehog (SHH) sea al menos 24 horas y
hasta 36 horas después de entrar en contacto dicha población celular con dicho primer y dicho segundo inhibidor. En
una realización, el contacto de dichas células con dicho tercer inhibidor está en condiciones capaces de dar como
resultado dicha población diferenciada de células progenitoras del mesencéfalo de la placa del suelo de manera que dicho contacto de dichas células con dicho tercer inhibidor sea al menos 24 horas y hasta 36 horas después de entrar en contacto dicha población celular con dicho activador de la señalización de Sonic hedgehog (SHH). En una realización, dicha población de células progenitoras del mesencéfalo de la placa del suelo comprende más de 40 % de células proteína A2 de secuencia forkhead (FOXA2)+factor de transcripción 1+ de homeosecuencia LIM, alfa (LMX1A)+. En una realización, el método comprende además la etapa e) poner en contacto dicha población de células progenitoras del mesencéfalo de la placa del suelo con medio de maduración neuronal, comprendiendo dicho medio medio N2, factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial, dibutiril AMPc y factor de crecimiento transformante tipo p3 para diferenciación de células progenitoras del mesencéfalo de la placa del suelo en neuronas de dopamina (Da) del mesencéfalo de la placa del suelo. En una realización, dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son positivas para conjuntos de marcadores seleccionados del grupo que consiste en la proteína A2 de secuencia forkhead (FOXA2)/factor de transcripción 1 de homeosecuencia LIM alfa (LMX1A)/tirosina hidroxilasa (TH); proteína A2 de secuencia forkhead/factor de transcripción 1 de homeosecuencia LIM alfa/tirosina hidroxilasa/CD142; proteína A2 de secuencia forkhead/factor de transcripción 1 de homeosecuencia LIM alfa/tirosina hidroxilasa/proteína 1 relacionada con el receptor nuclear (NURR1); proteína A2 de secuencia forkhead/factor de transcripción 1 de homeosecuencia LIM alfa/tirosina hidroxilasa/CD142/proteína 1 relacionada con el receptor nuclear y tirosina hidroxilasa/a-sinucleína. En una realización, dicho método comprende además la etapa f) clasificar dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo con respecto a expresión de CD142 en una población de células al menos 80 % positivas para CD142. En una realización, dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo son positivas para un marcador que identifica una molécula seleccionada del grupo que consiste en tirosina hidroxilasa (TH), proteína A2 de secuencia forkhead (FOXA2), factor de transcripción 1 de homeosecuencia LIM, dompamina, ácido 3,4-dihidroxi-fenilacético (DOPAC) y ácido homovanílico (HVA), alfa, receptor nuclear NURR1 (NR4A2), betatubulina de clase III específica de neurona (Tuj 1), TTF3, homeodominio de tipo paired 3 (PITX3), complejo achaetescute (ASCL), factor de linfocitos B temprano 1 (EBF-1), factor de linfocitos B temprano 3 (EBF-3), transtiretina (TTR), sinapsina, transportador de dopamina (DAT) y canal de potasio rectificador hacia dentro acoplado a proteína G (Kir3.2/GIRK2), CD142, DCSM1, CD63 y Cd99. En una realización, el método comprende además proporcionar a un paciente que lo necesite neuronas productoras de dopamina y una etapa después de e) tratar a dicho paciente trasplantando dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo para proporcionar función neuronal de dopamina (DA). En una realización, dicho paciente comprende al menos un síntoma neurológico seleccionado del grupo que consiste en temblor, bradicinesia (lentitud extrema del movimiento), postura flexionada, inestabilidad postural y rigidez. En una realización, se observa que dicho paciente tiene al menos un síntoma neurológico seleccionado del grupo que consiste en temblor, bradicinesia (lentitud extrema del movimiento), postura flexionada, inestabilidad postural y rigidez. En una realización, dicho paciente muestra una reducción de al menos uno de dichos síntomas neurológicos.
En una realización, las invenciones proporcionan un método de injerto in vivo para tratamiento terapéutico, que comprende a) proporcionar: i) una población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo en donde más del 40 % de dicha población expresa tirosina hidroxilasa (TH); y ii) un sujeto, en donde dicho sujeto muestra al menos un síntoma neurológico, en donde dichos síntomas neurológicos se seleccionan del grupo que consiste en temblor, bradicinesia (lentitud extrema del movimiento), postura flexionada, inestabilidad postural y rigidez; y b) trasplantar dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo en dicho sujeto en condiciones para permitir injerto in vivo para proporcionar función neuronal de dopamina (DA). En una realización, dicho sujeto muestra reducción de al menos uno de dichos síntomas neurológicos. En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo procede de una población de células que comprende además una etapa de clasificar dichas neuronas de dopamina (Da) del mesencéfalo de la placa del suelo con respecto a la expresión de CD142 en una población de células al menos 80 % positivas para CD142. En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo procede de una población de células progenitoras del mesencéfalo de la placa del suelo después de una etapa de clasificar dichas neuronas de dopamina (DA) del mesencéfalo de la placa del suelo con respecto a la expresión de CD142 en una población de células al menos 80 % positivas para CD142. En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo procede de una población celular seleccionada del grupo que consiste en animales, primates, seres humanos y un paciente con un síntoma de la enfermedad de Parkinson (EP). En una realización, dicha población de neuronas de dopamina (DA) del mesencéfalo de la placa del suelo procede de una población celular aislada del grupo que consiste en animales, primates, seres humanos y un paciente con un síntoma de la enfermedad de Parkinson (EP).
Definiciones
Como se emplea en esta memoria, la expresión "modelado de enfermedad" se refiere al proceso de usar un organismo experimental o cultivos celulares in vitro para imitar signos o síntomas específicos observados en seres humanos como resultado de un trastorno. En una realización, células madre pluripotentes procedentes de un modelo animal con una mutación genética que da como resultado un trastorno neurológico, tal como enfermedad de Parkinson (EP), pueden cultivarse y diferenciarse en células neurales para identificar nuevas características de neuronas relacionadas con la EP. En una realización, células madre pluripotentes humanas procedentes de una persona con una mutación genética que da como resultado un trastorno neurológico, tal como enfermedad de Parkinson (EP) se pueden cultivar
y diferenciar en células neurales que albergan un defecto similar observado en la persona.
Como se emplea en esta memoria, el término "parkinsonismo" se refiere a un grupo de enfermedades que están todas ligadas a una insuficiencia de dopamina en los ganglios basales, que es una parte del cerebro que controla el movimiento. Los síntomas incluyen temblor, bradicinesia (lentitud extrema del movimiento), postura flexionada, inestabilidad postural y rigidez. Un diagnóstico de parkinsonismo requiere la presencia de al menos dos de estos síntomas, uno de los cuales debe ser temblor o bradicinesia. La forma más habitual de parkinsonismo es enfermedad de Parkinson (EP) idiopática o clásica, pero para una minoría significativa de diagnósticos, aproximadamente 15 por ciento del total, puede estar presente uno de los síndromes de Parkinson plus (PPS). Estos síndromes, también conocidos como parkinsonismo atípico, incluyen degeneración corticobasal, demencia con cuerpos de Lewy, atrofia sistémica múltiple y parálisis supranuclear progresiva. En general, la enfermedad de Parkinson implica la disfunción y muerte de células nerviosas vitales en el cerebro principalmente en un área del cerebro denominada sustancia negra. Muchas de estas células nerviosas vitales producen dopamina, de modo que, a medida que estas neuronas mueren, la cantidad de dopamina resultante de la diferenciación en el cerebro disminuye, dejando a una persona incapaz de controlar el movimiento de manera normal. Los intestinos también tienen células de dopamina que degeneran en pacientes con enfermedad de Parkinson y esto puede ser un factor causante importante en los síntomas gastrointestinales que son parte de la enfermedad. Un grupo de síntomas que experimenta un individuo varía entre personas. Los signos motores primarios de la enfermedad de Parkinson incluyen los siguientes: temblor de las manos, los brazos, las piernas, la mandíbula y la cara, bradicinesia o lentitud del movimiento, rigidez o agarrotamiento de las extremidades y el tronco e inestabilidad postural o deterioro del equilibrio y la coordinación.
Como se emplea en esta memoria, el término "sujeto" se refiere a un mamífero (humano y animal, es decir, animales no humanos) que va a ser el receptor de un tratamiento en particular que incluye cualquier tipo de control. Normalmente, los términos "sujeto" y "paciente" se usan indistintamente en la presente memoria en referencia a un sujeto humano.
Como se emplea en esta memoria, la expresión "animales no humanos" se refiere a todos los animales no humanos, incluyendo, pero sin limitación, vertebrados tales como roedores, primates no humanos, ovinos, bovinos, rumiantes, lagomorfos, porcinos, caprinos, equinos, caninos, felinos, aves, etc.
Como se emplea en esta memoria, el término "dopamina" se refiere a una sustancia química producida por neuronas de dopamina que envía mensajes a la parte del cerebro que contiene neuronas que controlan el movimiento y la coordinación.
Como se emplea en esta memoria, el término "LSB" se refiere a una combinación de dos compuestos LDN-193189 y SB431542 capaces de reducir o bloquear la señalización que consiste en la señalización del factor de crecimiento transformante beta (TGFp)/Activina-Nodal y la señalización de Small Mothers Against Decapentaplegic (SMAD) en una célula.
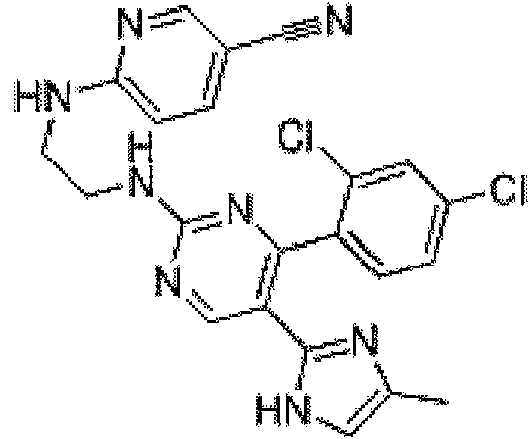
Como se emplea en esta memoria, el término "SB431542" se refiere a una molécula capaz de reducir o bloquear la señalización del factor de crecimiento transformante beta (TGFp)/Activina-Nodal con un número CAS 301836-41-9, una fórmula molecular de C22H18N4O3 y un nombre de 4-[4-( 1,3-benzodioxol-5-il)-5-(2-piridinil)-1 H-imidazol-2-il]-
benzamida, por ejemplo, véase la estructura a continuación:
En un ejemplo, SB431542 es Stemolecule™ SB431542, Stemgent, Inc. Cambridge, Massachusetts, Estados Unidos.
Como se emplea en esta memoria, el término "LDN-193189" se refiere a una molécula pequeña DM-3189, nombre de la IUPAC 4-(6-(4-(piperazin-1-il)fenil)pirazolo[1,5-a]pirimidin-3-il)quinolina, con una fórmula química de C25H22N6. LDN-193189 es capaz de actuar como un inhibidor de la señalización de SMAD. LDN-193189 también es un inhibidor de molécula pequeña muy potente de ALK2, ALK3 y ALK6, proteínas tirosina cinasas (PTK), que inhibe la señalización de miembros de las familias ALK1 y ALK3 de receptores de TGFp de tipo I, lo que da como resultado la inhibición de la transmisión de múltiples señales biológicas, incluyendo las proteínas morfogénicas óseas (BMP) BMP2, BMP4, BMP6, BMP7 y señales de citocinas activina y, posteriormente, fosforilación de SMAD de Smad1, Smad5 y Smad8 (Yu et al. (2008) Nat Med 14: 1363-1369; Cuny et al. (2008) Bioorg. Med. Chem. Lett. 18:

4388-4392, incorporado en la presente memoria por referencia).
En una realización ilustrativa, LDN-193189 es Stemolecule™ LDN-193189, Stemgent, Inc. Cambridge, Massachusetts, Estados Unidos.
Como se emplea en esta memoria, la expresión "inhibidor de glucógeno sintasa cinasa 3p" o "inhibidor de GSK3p" se refiere a un compuesto que inhibe una enzima glucógeno sintasa cinasa 3p, por ejemplo, véase, Doble, et al., J Cell Sci. 2003; 116: 1175-1186, incorporado en la presente memoria por referencia. Para los fines de las presentes invenciones, un inhibidor de GSK3p es capaz de activar una ruta de señalización de WNT, véase, por ejemplo, Cadigan, et al., J Cell Sci. 2006; 119: 395-402; Kikuchi, et al., Cell Signaling. 2007; 19: 659-671, incorporado en la presente memoria por referencia.
Como se emplea en esta memoria, el término "CHIR99021" o "CHIR" o "aminopirimidina" o "3-[3-(2-carboxietil)-4-metilpirrol-2-metilidenil]-2-indolinona" se refiere al nombre de la IUPAC 6-(2-(4-(2,4-diclorofenil)-5-(4-metil-1H-imidazol-2-il)pirimidin-2-ilamino)etilamino)nicotinonitrilo. CHIR99021 es un ejemplo de un inhibidor químico de molécula pequeña de glucógeno sintasa cinasa 3p (GSK3p) que activa una ruta de señalización de WNT y es muy selectivo, mostrando una selectividad de casi mil veces frente a un panel de cinasas relacionadas y no relacionadas, con una CI50 = 6,7 nM contra GSK3p humana y valores de CI50 nanomolares contra homólogos de GSK3p de roedores.
En una realización ilustrativa, CHIR99021 es Stemolecule™ CHIR99021, Stemgent, Inc. Cambridge, Massachusetts, Estados Unidos.
Como se emplea en esta memoria, el término "purmorfamina" se refiere a un derivado de purina, tal como número CAS: 483367-10-8, como un ejemplo, véase la estructura a continuación, que activa la ruta de Hedgehog incluyendo mediante dirección a Smoothened.

En una realización ilustrativa, la purmorfamina es Stemolecule™ purmorfamina, Stemgent, Inc. Cambridge, Massachusetts, Estados Unidos.
Como se emplea en esta memoria, el término "señalización" en referencia a una "proteína de transducción de señales" se refiere a proteínas que se activan o se ven afectadas por la unión del ligando con una proteína receptora de membrana o algún otro estímulo. Los ejemplos de la proteína de transducción de señales incluyen una SMAD, una proteína de complejo WNT, en otra realización, una proteína de complejo WNT incluyendo beta-catenina, Sonic Hedgehog (SHH), NOTCH, factor de crecimiento transformante beta (TGFp), activina, nodal, proteínas de glucógeno sintasa cinasa 3p (GSK3p) y similares. Para muchos receptores de superficie celular o proteínas receptoras internas,
las interacciones de ligando-receptor no están directamente ligadas a la respuesta de la célula. El receptor activado por ligando debe interactuar primero con otras proteínas dentro de la célula antes de que se produzca el efecto fisiológico último del ligando sobre el comportamiento de la célula. Con frecuencia, el comportamiento de una cadena de varias proteínas celulares que interactúan se altera después de la activación o inhibición del receptor. El conjunto completo de cambios celulares inducidos por activación del receptor se denomina mecanismo de transducción de señal o ruta de señalización.
Como se emplea en esta memoria, el término "LSB/S/F8/CHIR" o "LSB/SHH/FGF8/CHIR" se refiere a células en contacto con LDN-193189 y SB431542 (es decir, LSB) además de S, activador de Sonic Hedgehog, F8, FGF8 y CHIR de las presentes invenciones. A diferencia de "LSB/S/F8" o "SHH/FGF8" o "SHH/FGF", que se refieren a células en contacto con LDN-193189 y SB431542 (es decir, LSB) además de S, activador de Sonic Hedgehog, F8, FGF8 pero sin CHIR como en los métodos publicados anteriormente. En abreviaturas similares, "LDN/SB" se refiere a células en contacto con LDN, LDN-193189 y SB, SB431542.
Como se emplea en esta memoria, el término "inhibir" o "bloquear' significa una reducción en el nivel de actividad de una ruta de señalización particular de una célula tras el tratamiento con un compuesto (es decir, un inhibidor) en comparación con la actividad de dicha ruta de señalización de una célula que se deja sin tratar con dicho compuesto o se trata con un control.
Como se emplea en esta memoria, el término "activar" significa un aumento en el nivel de actividad de una ruta de señalización particular de una célula tras el tratamiento con un compuesto (es decir, un activador) en comparación con la actividad de dicha ruta de señalización de una célula que se deja sin tratar con dicho compuesto o se trata con un control. Cualquier nivel de inhibición o activación de una ruta de señalización particular se considera una realización de la invención si dicha inhibición o activación da como resultado la diferenciación dirigida de una célula madre. Como se emplea en esta memoria, la expresión "Sma Mothers Against Decapentaplegic" o "Small Mothers Against Decapentaplegic" o "SMAD" se refiere a una molécula de señalización.
Como se emplea en esta memoria, el término "WNT" o "wingless" en referencia a un ligando se refiere a un grupo de proteínas secretadas (es decir, Int1 (integración 1) en seres humanos) capaces de interactuar con un receptor de WNT, tal como un receptor en la familia de receptores Frizzled y LRPDerailed/RYK.
Como se emplea en esta memoria, el término "WNT" o "wingless" en referencia a una ruta de señalización se refiere a una ruta de señales compuesta por ligandos de la familia Wnt y receptores de la familia Wnt, tales como receptores Frizzled y LRPDerailed/RYK, mediada con o sin p-catenina. Para los fines descritos en la presente memoria, una ruta de señalización de WNT preferida incluye mediación por p-catenina, es decir, WNT/p-catenina.
Como se emplea en esta memoria, "ruta canónica" o "activación clásica" en referencia a WNT se refiere a una de las múltiples rutas de señal corriente abajo de Wnt, por ejemplo, en la ruta canónica, un efecto importante de la unión del ligando Wnt con su receptor es la estabilización de beta-catenina citoplasmática a través de la inhibición del complejo de degradación de beta-catenina. Otras rutas de Wnt no son canónicas.
Como ejemplo, la molécula pequeña CHIR afecta a una ruta corriente abajo de señalización de Wnt canónica.
Como se emplea en esta memoria, la expresión "Sonic hedgehog (SHH o Shh)" se refiere a una proteína que es una de al menos tres proteínas en la familia de rutas de señalización de mamíferos denominada hedgehog, otra es desert hedgehog (DHH) mientras que una tercera es Indian hedgehog (IHH). Shh interactúa con al menos dos proteínas transmembrana interactuando con moléculas transmembrana Patched (PTC) y Smoothened (SMO). Shh normalmente se une con PCT que después permite la activación de SMO como transductor de señal. En ausencia de SHH, PTC normalmente inhibe SMO, que a su vez activa un represor de la transcripción de modo que no se produzca transcripción de determinados genes. Cuando Shh está presente y se une con PTC, PTC no puede interferir con el funcionamiento de SMO. Con SMO desinhibido, determinadas proteínas pueden entrar en el núcleo y actuar como factores de transcripción permitiendo que determinados genes se activen (véase, Gilbert, 2000 Developmental Biology (Sunderland, Massachusetts: Sinauer Associates, Inc., editores).
Como se emplea en esta memoria, el término "activador" o "activante" se refiere a moléculas pequeñas, péptidos, proteínas y compuestos para activar moléculas que dan como resultado diferenciación dirigida de células de las presentes invenciones. Los activadores ilustrativos incluyen, pero sin limitación: CHIR, C25II de Sonic Hedgehog (SHH), un agonista de pequeña molécula Smoothened purmorfamina, factor de crecimiento de fibroblastos (FGF), etc. Como se emplea en esta memoria, la expresión "activador de la señalización de Sonic hedgehog (SHH)" se refiere a cualquier molécula o compuesto que active una ruta de señalización de SHH incluyendo una molécula o compuesto que se une con PCT o un agonista de Smoothened y similares. Son ejemplos de dichos compuestos son una proteína C25II de Sonic hedgehog (SHH) y un agonista de molécula pequeña Smoothened purmorfamina.
Como se emplea en esta memoria, la expresión "C25II de Sonic hedgehog (SHH)" se refiere a un fragmento N-terminal recombinante de una proteína sonic hedgehog murina de longitud completa capaz de unirse con el receptor de SHH para activar SHH, un ejemplo es el número de catálogo de R and D Systems: 464-SH-025/CF.
Como se emplea en esta memoria, el término "señales" se refiere a factores internos y externos que controlan cambios en la estructura y función celular. Son de naturaleza química o física.
Como se emplea en esta memoria, el término "ligando" se refiere a moléculas y proteínas que se unen con receptores (R), los ejemplos incluyen, pero sin limitación, factor de crecimiento transformante beta, activinas, nodal, proteínas morfogénicas óseas (BMP), etc.
Como se emplea en esta memoria, la expresión "inhibidor" o "inhibidor de la señalización" se refiere a la inhibición de una molécula de señalización o la ruta de una molécula de señalización, tal como un inhibidor de la señalización de SMAD, inhibidor de glucógeno sintasa cinasa 3p (GSK3p), se refiere a un compuesto o molécula (p. ej., molécula pequeña, péptido, peptidomimético, compuesto natural, proteína, ARNip, ácido nucleico antisentido, aptámero o anticuerpo) que interfiere con (es decir, reduce o suprime o elimina o bloquea) la función de señalización de la molécula o ruta. En otras palabras, un inhibidor es cualquier compuesto o molécula que cambie cualquier actividad de una proteína nombrada (molécula de señalización, cualquier molécula involucrada con la molécula de señalización nombrada, una molécula asociada nombrada, tal como una glucógeno sintasa cinasa 3p (GSK3p)) (p. ej., incluyendo, pero sin limitación, las moléculas de señalización descritas en la presente memoria), como un ejemplo, mediante el contacto directo con señalización de SMAD, contacto con ARNm de SMAD, provocando cambios conformacionales de SMAD, disminuyendo los niveles de proteína de SMAD o interfiriendo con interacciones de SMAD con compañeros de señalización (p. ej., incluyendo los descritos en la presente memoria) y afectando a la expresión de genes diana de SMAD (p. ej., los descritos en la presente memoria). Los inhibidores también incluyen moléculas que regulan indirectamente la actividad biológica de SMAD al interceptar moléculas de señalización corriente arriba. Por tanto, en una realización, un inhibidor de las presentes invenciones induce (cambia) o altera la diferenciación de un tipo celular predeterminado a uno no predeterminado, por ejemplo, uno de los métodos de las presentes invenciones que comprende LDN/SB, CHIR y un activador de SHH (que puede inhibir glucógeno sintasa cinasa 3p) diferenció células progenitoras en células progenitoras neurales no predeterminadas. En una realización preferida, un inhibidor de las presentes invenciones "altera" o "reduce" o "bloquea" la señalización predeterminada para dirigir la diferenciación celular hacia un tipo celular no predeterminado, tal como se describe en la presente memoria para diferenciar células progenitoras del mesencéfalo de la placa del suelo y neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de las presentes invenciones. Por tanto, un inhibidor de las presentes invenciones es un compuesto natural o una molécula pequeña que cambia la actividad de la molécula señal de una manera que contribuye a la diferenciación de una población celular inicial (día 0) en células progenitoras del mesencéfalo de la placa del suelo. Cuando las células progenitoras se ponen en contacto con inhibidores, estas moléculas pequeñas pueden contribuir a una diferenciación adicional en neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de las presentes invenciones. Los inhibidores se describen con respecto a inhibición competitiva (se unen con el sitio activo de manera que excluyen o reducen la unión de otro compuesto de unión conocido) e inhibición alostérica (se unen con una proteína de manera que cambia la conformación de la proteína de una manera que interfiere con la unión de un compuesto con el sitio activo de esa proteína) además de inhibición inducida por unión con y alteración de una molécula corriente arriba de la molécula de señalización nombrada lo que a su vez provoca inhibición de la molécula nombrada. En algunos casos, un inhibidor se denomina "inhibidor directo", lo que se refiere a la inhibición de una diana de señalización o una ruta de diana de señalización mediante la puesta en contacto real con la diana de señalización; por ejemplo, un inhibidor directo de una gamma secretasa es una molécula DAPT que se une con la proteína gamma secretasa.
Como se emplea en esta memoria, el término "derivado" se refiere a un compuesto químico con una estructura de núcleo similar.
Como se emplea en esta memoria, la expresión "célula progenitora del mesencéfalo de la placa del suelo" en referencia a una célula in vivo ubicada en un mesencéfalo, incluyendo durante el desarrollo embrionario de neuronas del mesencéfalo, se refiere a una célula que puede diferenciarse en una célula productora de dopamina. En algunas realizaciones, una "célula progenitora del mesencéfalo de la placa del suelo" se refiere a una célula en cultivo que se usa para producir artificialmente una célula cultivada in vitro que expresa conjuntos de marcadores solapantes o idénticos en comparación con marcadores expresados por células in vivo, es decir, la coexpresión del marcador de la placa del suelo FOXA2 y el marcador de la placa del techo LMX1A, OTX2, NGN2 y DDC, tal como en células cultivadas de las presentes invenciones alrededor del día 11 después del inicio de la diferenciación dirigida como se describe en la presente memoria. Preferiblemente, una célula progenitora del mesencéfalo de la placa del suelo es "FOXA2+LMX1A+" o "FOXA2/LMX1A+". En algunas realizaciones, bajos números de células en una población progenitora diferenciada son FOXA2/LMX1A/TH+.
Como se emplea en esta memoria, la expresión "neuronas DA procedentes de la placa del suelo" o "neuronas DA del mesencéfalo auténticas" o "neuronas de dopamina (DA) FOXA2+LMX1A+ destinadas al mesencéfalo" o "neurona de dopamina (DA) del mesencéfalo de la placa del suelo" o "neurona DA del mesencéfalo injertable" o " neurona mDA" o "FOXA2+LMX1A+TH+" o "FOXA2/LMX1A/TH" o "FOXA2+LMX1A+NURR1+TH+" o "FOXA2/LMX1A/NURR1/TH" se refiere a una población de neuronas DA del mesencéfalo injertable obtenida por métodos descritos en la presente memoria, normalmente alrededor del día 25 o antes después de iniciar diferenciación dirigida. En una realización preferida, Las "neuronas DA del mesencéfalo auténticas" son FOXA2+/LMX1A+/NURR1+/TH+. Estas neuronas se
etiquetaron como "injertables" después de experimentos de trasplante en ratones y primates que muestran la capacidad de estas neuronas para neutralizar las afecciones neurológicas de tipo Parkinson con menos interferencia del sobrecrecimiento neuronal y la formación de teratoma. Se mantuvieron las neuronas de dopamina (DA) FOXA2/LMX1A+ del mesencéfalo de las presentes invenciones in vitro durante varios meses conservando al mismo tiempo la capacidad de injerto.
Como se emplea en esta memoria, se obtienen células usadas para obtener células progenitoras del mesencéfalo de la placa del suelo y neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de diversas fuentes incluyendo fuentes embrionarias y no embrionarias, por ejemplo, hESC y hiPSC no embrionarias, células madre somáticas, células madre de la enfermedad, es decir, células pluripotentes aisladas y células madre obtenidas por ingeniería genética aisladas de pacientes con enfermedad de Parkinson, células madre cancerosas, células pluripotentes humanas o de mamífero, etc.
Como se emplea en esta memoria, la expresión "célula madre" se refiere a una célula con la capacidad de dividirse durante periodos indefinidos en cultivo y dar lugar a células especializadas. Se puede obtener una célula madre de animales y pacientes, incluyendo seres humanos; por ejemplo, una célula madre humana se refiere a una célula madre que es humana. Se puede obtener una célula madre de diversas fuentes, incluyendo embrionarias y no embrionarias, tales como células del cordón umbilical, células de niños y células de adultos. Para los fines de las presentes invenciones, las células madre de adultos en general se refieren a células que originalmente no se obtuvieron de un feto, en otras palabras, células de bebés, cordones umbilicales descartados, células placentarias descartadas, células de niños, células de adultos, etc.
Como se emplea en esta memoria, la expresión "células madre de la sangre del cordón umbilical" se refiere a células madre recogidas de un cordón umbilical en el momento del nacimiento que tienen la capacidad de producir al menos todas las células sanguíneas en el cuerpo (hematopoyéticas).
Como se emplea en esta memoria, la expresión "célula madre somática (adulta)" se refiere a una célula indiferenciada relativamente rara que se encuentra en muchos órganos y tejidos diferenciados con una capacidad limitada tanto para autorrenovación (en el laboratorio) como para diferenciación. Dichas células varían en su capacidad de diferenciación, pero están habitualmente limitadas a tipos celulares en el órgano de origen.
Como se emplea en esta memoria, la expresión "célula somática" se refiere a cualquier célula en el cuerpo distinta de gametos (óvulo o espermatozoide); en ocasiones denominadas células "adultas".
Como se emplea en esta memoria, la expresión "célula del linaje neural" se refiere a una célula que contribuye al sistema nervioso (tanto central como periférico) o a destinos de células de la cresta neural durante el desarrollo o en el adulto. El sistema nervioso incluye el cerebro, la médula espinal y el sistema nervioso periférico. Los destinos de células de la cresta neural incluyen craneal, tronco, vagal, sacro y cardíaco, dando lugar a mesectodermo, cartílago craneal, hueso craneal, timo, dientes, melanocitos, células pigmentarias del iris, ganglios craneales, ganglios de la raíz dorsal, ganglios simpáticos/parasimpáticos, células endocrinas, sistema nervioso entérico y partes del corazón. Como se emplea en esta memoria, la expresión "célula madre adulta" se refiere a una célula madre somática, como un ejemplo, una "célula madre hematopoyética" que se refiere a una célula madre en bebés, niños y adultos, que da lugar a todos los glóbulos rojos y blancos y las plaquetas.
Como se emplea en esta memoria, la expresión "célula madre embrionaria" se refiere a una célula primitiva (indiferenciada) que procede de una de varias fuentes, capaz de dividirse sin diferenciarse durante un periodo prolongado en cultivo, y se sabe que tiene la capacidad de desarrollarse a células y/o tejidos de las tres capas germinales primarias, el ectodermo, el mesodermo y el endodermo.
Como se emplea en esta memoria, el término "endodermo" se refiere a una capa de las células procedentes de la masa celular interna del blastocisto; tiene la capacidad de dar lugar a pulmones, otras estructuras respiratorias y órganos digestivos o, en general, "el intestino" "in vivo" y diversos tipos celulares in vitro.
Como se emplea en esta memoria, la expresión "línea de células madre embrionarias" se refiere a una población de células madre embrionarias que se han cultivado en condiciones in vitro que permiten la proliferación sin diferenciación durante hasta días, meses o años.
Como se emplea en esta memoria, la expresión "célula madre embrionaria humana" o "hESC" se refiere a un tipo de células madre pluripotentes procedentes de embriones humanos de etapa temprana, hasta, e incluyendo, la etapa de blastocisto, que es capaz de dividirse sin diferenciarse durante un periodo prolongado en cultivo, y se sabe que se desarrolla a células y tejidos de las tres capas germinales primarias, el ectodermo, el mesodermo y el endodermo. Como se emplea en esta memoria, la expresión "célula madre pluripotente inducida" o "iPSC" se refiere a un tipo de célula madre pluripotente, similar a una célula madre embrionaria, por la que células somáticas (adultas) se reprograman para entrar en un estado similar a células madre embrionarias al verse obligadas a expresar factores importantes para mantener la "potencialidad" de células madre embrionarias (ESC). Las iPSC de ratón se presentaron en 2006 (Takahashi y Yamanaka) y las iPSC humanas se presentaron a finales de 2007 (Takahashi et al. y Yu et al.).
Las iPSC de ratón demuestran características importantes de células madre pluripotentes, incluyendo la expresión de marcadores de células madre, la formación de tumores que contienen células de las tres capas germinales y la capacidad de contribuir a muchos tejidos diferentes cuando se inyectan en embriones de ratón en una etapa muy temprana del desarrollo. Las iPSC humanas también expresan marcadores de células madre y son capaces de generar células características de las tres capas germinales. A diferencia de una célula madre embrionaria, un iPSC se forma artificialmente por la introducción de determinados genes embrionarios (tales como un transgén OCT4, SOX2 y KLF4) (véase, por ejemplo, Takahashi y Yamanaka Cell 126, 663-676 (2006), incorporada en la presente memoria por referencia) en una célula somática, para ejemplos de líneas celulares de células inducidas, C14, C72 y similares. Otro ejemplo de una iPSC es una célula de piel humana adulta o célula fibroblástica, transformada con el uso de genes (OCT4, SOX2, NANOG, LIN28 y KLf4) clonados en un plásmido, por ejemplo, véase, Yu, et al., Science DOI: 10.1126/science. 1172482, incorporado en la presente memoria por referencia.
Como se emplea en esta memoria, el término "totipotente" se refiere a la capacidad de dar lugar a todos los tipos celulares del cuerpo más todos los tipos celulares que componen los tejidos extraembrionarios, tales como la placenta. Como se emplea en esta memoria, el término "multipotente" se refiere a la capacidad de desarrollarse en más de un tipo celular del cuerpo.
Como se emplea en esta memoria, el término "pluripotente" se refiere a una célula que tiene la capacidad de dar lugar al menos dos, pero con frecuencia numerosos tipos celulares diferentes del cuerpo. Las células pluripotentes generan con frecuencia un teratoma después de la inyección en un ratón inmunodeprimido.
Como se emplea en esta memoria, la expresión "célula madre pluripotente" se refiere a la capacidad de esta célula para desarrollarse en al menos dos tipos celulares diferentes dependiendo de factores ambientales, es decir, morfógenos, factores de crecimiento, moléculas de señalización, ya sea activadores o inhibidores, etc. En algunas realizaciones, una célula madre pluripotente se refiere a la capacidad de una célula para desarrollarse en cualquiera de las tres capas germinales del organismo, incluyendo endodermo, mesodermo y ectodermo.
Como se emplea en esta memoria, la expresión "célula especializada" se refiere a un tipo de célula que realiza una función específica en organismos multicelulares. Por ejemplo, grupos de células especializadas, tales como las neuronas, trabajan juntas para formar un sistema, tal como un sistema nervioso.
Como se emplea en esta memoria, el término "neuroectodermo" se refiere a una célula o un destino celular que se encuentra temprano en el desarrollo o durante la diferenciación de células madre pluripotentes que puede dar lugar a células del linaje neural.
Como se emplea en esta memoria, la expresión "marcadores de la proliferación celular" se refiere a la expresión de moléculas asociadas con células de ciclo rápido que normalmente no están presentes en células maduras de ciclo lento o fuera de ciclo, es decir, que se dividen activamente frente a células con tiempos de ciclo prolongados o células fuera de ciclo. Los ejemplos de dichos marcadores incluyen un marcador Ki67 de proliferación celular (Gerdes, et al., Int J Cancer 31: 13-20 (1983), incorporado en la presente memoria por referencia) y marcadores de fosfo-histona H3 de fases G2/M de mitosis (Hendzel, et al., Chromosoma 106: 348-360 (1997), incorporado en la presente memoria por referencia).
Como se emplea en esta memoria, el término "proliferación" se refiere a un aumento en el número de células.
Como se emplea en esta memoria, el término "diferenciación" se refiere a un proceso por el que una célula embrionaria no especializada adquiere las características de una célula especializada, tal como un tipo específico de neurona, célula cerebral, célula cardíaca, hepática o muscular. La diferenciación está controlada por la interacción de los genes de una célula con las condiciones físicas y químicas fuera de la célula, habitualmente a través de rutas de señalización que implican proteínas incluidas en la superficie celular.
Como se emplea en esta memoria, el término "diferenciación" como se usa con respecto a células en un sistema celular en diferenciación se refiere al proceso mediante el que las células se diferencian de un tipo celular (p. ej., una célula diferenciable multipotente, totipotente o pluripotente) a otro tipo celular tal como una célula diferenciada diana. Como se emplea en esta memoria, la expresión "diferenciación celular" se refiere a una ruta por la que una célula menos especializada (es decir, célula madre) se desarrolla o madura para poseer una forma y función más definidas (por ejemplo, una iPSC que progresa en un progenitor de la cresta neural a una célula de linaje neuronal a células progenitoras del mesencéfalo de una placa del suelo a neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de las presentes invenciones).
Como se emplea en esta memoria, el término "indiferenciado" se refiere a una célula que aún no se ha desarrollado en un tipo celular especializado.
Como se emplea en esta memoria, el término "predeterminada" o "pasiva" en referencia a una ruta de diferenciación celular se refiere a una ruta donde una célula menos especializada se convierte en un determinado tipo celular diferenciado en cultivo, cuando no se trata con determinados compuestos, es decir, condiciones normales de cultivo
celular sin contacto con al menos un morfógeno. En otras palabras, se produce una célula predeterminada cuando una célula no entra en contacto con una molécula capaz de cambiar el tipo celular diferenciado (es decir, un morfógeno), por ejemplo, cultivos tratados con LSB solo, pero no un activador de SHH o un activador de Wnt para producir una célula proteína A2 de secuencia forkhead (FOXA2)+ de las presentes invenciones, en cambio, dan como resultado la expresión de marcadores HES5, PAX6, LHX2 y EMX2. Por el contrario, "no predeterminada" en referencia a una célula se refiere a un tipo celular diferenciado que da como resultado un tipo celular que es diferente de una célula predeterminada, es decir, una célula no predeterminada es un tipo celular diferenciado resultante de condiciones no predeterminadas, tales como la célula de las presentes invenciones, incluyendo una célula neuronal proteína A2 de secuencia forkhead (FOXA2)+, una célula progenitora del mesencéfalo de la placa del suelo y neurona de dopamina (DA) FOXA2/LMX1A+ destinada al mesencéfalo de las presentes invenciones, etc. Una célula predeterminada también puede ser una célula predeterminada después de que una célula ha entrado en contacto con un morfogénico para convertirse en una célula no predeterminada sin un compuesto morfogénico posterior, tal como una célula progenitora del mesencéfalo de la placa del suelo no predeterminada que posteriormente se convierte en una célula predeterminada que no es una neurona de dopamina (DA) FOXA2/LMX1A+ destinada al mesencéfalo debido a una falta de contacto con un morfógeno tal como CHIR.
Como se emplea en esta memoria, el término "morfógeno" se refiere a un compuesto que influye en la diferenciación de una célula, es decir, determina, al menos en parte, el destino celular. Un morfógeno también puede influir en una célula para diferenciarse en un tipo celular no predeterminado.
Como se emplea en esta memoria, la expresión "diferenciación dirigida" se refiere a una manipulación de las condiciones de cultivo de células madre para inducir la diferenciación en un tipo celular particular (por ejemplo, deseado), tales como células progenitoras del mesencéfalo de la placa del suelo y neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de las presentes invenciones. En una realización, la expresión "diferenciación dirigida" en referencia a una célula se refiere al uso de moléculas pequeñas, proteínas del factor de crecimiento y otras condiciones de crecimiento para promover la transición de una célula de un estado pluripotente a un destino celular más maduro o especializado (p. ej., célula del sistema nervioso central, célula neural, célula progenitora del mesencéfalo de la placa del suelo y neurona de dopamina (DA) FOXA2/LMX1A+ destinada al mesencéfalo de las presentes invenciones, etc.). En una realización preferida, el comienzo de la diferenciación dirigida es el contacto de una célula el día 0 con LDN/SB. Una célula que se somete a diferenciación dirigida como se describe en la presente memoria da como resultado la formación de un tipo celular no predeterminado de células progenitoras del mesencéfalo de la placa del suelo y neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de las presentes invenciones.
Como se emplea en esta memoria, la expresión "inducir diferenciación" en referencia a una célula se refiere a cambiar el tipo celular predeterminado (genotipo y/o fenotipo) a un tipo celular no predeterminado (genotipo y/o fenotipo). Por tanto, "inducir la diferenciación en una célula madre" se refiere a inducir que la célula se divida en células descendientes con características que son diferentes de la célula madre, tales como cambio del genotipo (es decir, cambio en la expresión génica según lo determinado por análisis genético tal como una micromatriz) y/o fenotipo (es decir, cambio en la expresión de una proteína, tal como proteína A2 de secuencia forkhead (FOXA2) o un conjunto de proteínas, tal como proteína A2 de secuencia forkhead (FOXA2) y factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A) positivo (+) mientras que es negativo (-) para PAX6).
Como se emplea en esta memoria, el término "destino" en referencia a una célula, tal como "determinación del destino celular" en general, se refiere a una célula con un linaje genéticamente determinado cuyas células descendientes son capaces de convertirse en diversos tipos celulares o algunos tipos celulares específicos dependiendo de las condiciones de cultivo in vivo o in vitro. En otras palabras, está determinado por su entorno que el destino predeterminado de una célula se destina a una ruta de diferenciación particular de manera que una célula se convierta en un tipo celular en lugar de otro tipo celular, por ejemplo, las células descendientes de una célula madre cuyo "destino neural" es convertirse en una célula nerviosa en lugar de una célula muscular o una célula cutánea.
Como se emplea en esta memoria, la expresión "crecimiento de neuritas" o "crecimiento neuronal" se refiere a la observación de protuberancias de citoplasma alargadas, encerradas en la membrana, de las células.
A diferencia del "sobrecrecimiento neural" que se refiere al crecimiento neural no restringido indeseado, es decir, crecimiento descontrolado de neuronas, de células trasplantadas en el sitio de injerto. Como se emplea en esta memoria, el término "teratoma" se refiere a un tumor no canceroso de cualquier tipo tisular que crece a partir de células trasplantadas.
Como se emplea en esta memoria, la expresión "formación de teratoma" se refiere al crecimiento no deseado de diversos tipos tisulares en tumores no cancerosos a partir del crecimiento de células trasplantadas.
Como se emplea en esta memoria, la expresión "neurona de dopamina" o "neurona dopaminérgica" en general se refiere a una célula capaz de expresar dopamina. Las "neuronas de dopamina del mesencéfalo" o "mDA" se refieren a células que supuestamente expresan dopamina en estructuras del prosencéfalo y células que expresan dopamina en estructuras del prosencéfalo.
Como se emplea en esta memoria, la expresión "célula madre neural" se refiere a una célula madre que se encuentra en tejido neural adulto que puede dar lugar a neuronas y células gliales (de soporte). Los ejemplos de células gliales incluyen astrocitos y oligodendrocitos.
Como se emplea en esta memoria, la expresión "placa del suelo" o "PS" o "ps" se refiere a una región del tubo neural in vivo que se extiende a lo largo de toda la línea media ventral también descrita como la zona longitudinal ventral no emparejada del tubo neural o denominada centro de señalización del tubo neural. En otras palabras, el tubo neural se dividió en diferentes regiones donde las células ventrales más cercanas a la línea media constituían la placa del suelo. Como un ejemplo de identificación celular adicional, la PS del mesencéfalo de pollo se puede dividir en regiones media (PSM) y lateral (PSL) en función de la expresión génica, el modo de inducción y la función. Se encuentran células de la placa del suelo in vivo en varias áreas del embrión en desarrollo, por ejemplo,
células de la placa del suelo en el mesencéfalo, en el rombencéfalo, etc. Se contempla que células de la placa del suelo in vivo en la región del mesencéfalo dan lugar a células que son diferentes de células diferenciadas de células de la placa del suelo en otras regiones. Un marcador primario de la placa del suelo en la región del mesencéfalo es FOXA2.
Como se emplea en esta memoria, la expresión "placa del techo" se refiere a las células dorsales más cercanas a la línea media. Un marcador de la placa del techo es LMX1A. Durante el desarrollo embrionario, las células de la placa del suelo y la placa del techo están ubicadas en posiciones definidas en el SNC (ventral frente a dorsal) con requisitos de patrones diametralmente opuestos para su inducción.
Como se emplea en esta memoria, la expresión "mesencéfalo" se refiere a una región del cerebro de vertebrado en desarrollo entre el prosencéfalo (anterior) y el rombencéfalo (posterior). Las regiones del mesencéfalo dan lugar a muchas áreas del cerebro, incluyendo, pero sin limitación, formación reticular, que es parte del tegumento, una región del tronco encefálico que influye en las funciones motoras, el pie del pedúnculo cerebral, que está compuesto de fibras nerviosas que conectan los hemisferios cerebrales con el cerebelo y un núcleo pigmentado grande denominado sustancia negra. Una característica única del mesencéfalo en desarrollo es la coexpresión del marcador de la placa del suelo FOXA2 y el marcador de la placa del techo LMX1A.
Como se emplea en esta memoria, el término "neurona" se refiere a una célula nerviosa, las unidades funcionales principales del sistema nervioso. Una neurona consiste en un cuerpo celular y sus prolongaciones: un axón y una o más dendritas. Las neuronas transmiten información a otras neuronas o células mediante la liberación de neurotransmisores en las sinapsis.
Como se emplea en esta memoria, la expresión "cultivo celular" se refiere a un crecimiento de células in vitro en un medio artificial para investigación o tratamiento médico.
Como se emplea en esta memoria, la expresión "medio de cultivo" se refiere a un líquido que cubre las células en un recipiente de cultivo, tal como una placa de Petri, una placa de múltiples pocillos y similares, y contiene nutrientes para nutrir y mantener las células. El medio de cultivo también puede incluir factores de crecimiento añadidos para producir cambios deseados en las células.
Como se emplea en esta memoria, la expresión "medio de maduración neuronal" o medio "BAGCT" se refiere a un medio de cultivo que comprende medio N2, que comprende además factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial, dibutiril AMPc y factor de crecimiento transformante tipo p3 para diferenciar neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo. Como se emplea en esta memoria, la expresión "capa alimentadora" se refiere a una célula usada en cocultivo para mantener células madre pluripotentes. Para cultivo de células madre embrionarias humanas, las capas alimentadoras normales incluyen fibroblastos embrionarios de ratón (FER) o fibroblastos embrionarios humanos que se han tratado para evitar que se dividan en cultivo.
Como se emplea en esta memoria, el término "pase" en referencia a un cultivo celular, se refiere al proceso en el que las células se disocian, lavan y siembran en nuevos recipientes de cultivo después de un ciclo de crecimiento y proliferación celular. El número de pases por los que ha pasado una línea de células cultivadas es una indicación de su edad y estabilidad esperada.
Como se emplea en esta memoria, el término "expresar" en relación con un gen o proteína se refiere a producir un ARNm o una proteína que se puede observar usando ensayos tales como ensayos de micromatrices, ensayos de tinción de anticuerpos y similares.
Como se emplea en esta memoria, la expresión "gen de secuencia de paired 6" o "PAX6" se refiere a un marcador de una célula neuroprogenitora no predeterminada.
Como se emplea en esta memoria, la expresión "TUJ1" o "beta-tubulina de clase III específica de neurona" en referencia a una célula diferenciadora de las presentes invenciones se refiere a un marcador de diferenciación celular humana neural temprana, tal como células progenitoras neurales, y se encuentra expresada en neuronas del SNP y
el SNC.
Como se emplea en esta memoria, el término "homodímero" en referencia a una molécula SMAD se refiere al menos a dos moléculas de SMAD unidas entre sí, tal como por enlaces disulfuro.
Como se emplea en esta memoria, la expresión "poner en contacto" células con un compuesto las presentes invenciones se refieren a colocar el compuesto en una ubicación que le permita tocar la célula para producir (obtener) células "en contacto". El contacto puede llevarse a cabo usando cualquier método adecuado. Por ejemplo, en una realización, la puesta en contacto es mediante la adición del compuesto a un tubo de células. El contacto también se puede lograr añadiendo el compuesto a un cultivo de las células.
Como se emplea en esta memoria, la expresión "célula unida" se refiere a una célula que crece in vitro en donde la célula se adhiere al fondo o el lateral del recipiente de cultivo, una célula unida puede ponerse en contacto con el recipiente a través de moléculas de la matriz extracelular y similares y requiere el uso de una enzima para separar esta célula de la placa/recipiente de cultivo, es decir, tripsina, dispasa, etc. Una "célula unida" se diferencia de una célula en un cultivo en suspensión que no está unida y no requiere el uso de una enzima para eliminar células del recipiente de cultivo.
Como se emplea en esta memoria, la expresión "marcador" o "marcador celular" se refiere a un gen o proteína que identifica una célula o tipo celular particular. Un marcador para una célula no puede limitarse a un marcador, los marcadores pueden referirse a un "patrón" de marcadores de modo que un grupo designado de marcadores pueda identificar una célula o tipo celular de otra célula o tipo celular. Por ejemplo, las neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de las presentes invenciones expresan uno o más marcadores que distinguen una célula progenitora del mesencéfalo de la placa del suelo de una célula precursora menos diferenciada, es decir, células proteína A2 de secuencia forkhead (FOXA2) positivas y factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A) positivas frente a HES5+ y PAX6+, por ejemplo, como se muestra por patrones de expresión génica ilustrativos en las figuras 1e y 1f.
Como se emplea en esta memoria, el término "positiva" en relación con una célula, incluyendo una "célula positiva", se refiere a una célula que expresa un marcador, como un ejemplo, un anticuerpo "se tiñe" con ese marcador cuando se usa un sistema de tinción de anticuerpos (detección) o una secuencia de ácido nucleico que se hibrida con la secuencia de ácido nucleico marcador medida por una molécula indicadora, es decir, una molécula fluorescente que se une con secuencias de ácido nucleico bicatenarias, en una cantidad cuantitativa y/o cualitativa detectable por encima de una célula de control o comparativa. Por ejemplo, una célula positiva para un marcador tal como proteína A2 de secuencia forkhead (FOXA2), etc., se refiere a una célula que expresa ARNm y/o proteína FOXA2 cuando se detecta en un ensayo, tal como una matriz de genes o un anticuerpo, respectivamente. Dicha célula positiva puede denominarse FOXA2+. Cuando una célula es positiva para más de un marcador, tal como cuando se usa la notación FOXA2/LMX1A+, la célula o la mayoría de la población celular es positiva tanto para FOXA2 como para LMX1A. Como se emplea en esta memoria, el término "negativa" en relación con una célula o población celular, incluyendo una "célula negativa", se refiere a una célula o población sin señal detectable ausente para un marcador o señal a niveles de poblaciones de control. Por ejemplo, una célula que no se tiñe después del contacto con un método de detección de anticuerpos de proteína A2 de secuencia forkhead (FOXA2) o matriz génica que incluye la detección de un ARNm de FOXA2, etc., es FOXA2- o negativa para FOXA2.
Como se emplea en esta memoria, las expresiones "gen indicador" o "construcción indicadora" se refieren a construcciones genéticas que comprenden un ácido nucleico que codifica una proteína que es fácilmente detectable o fácilmente analizable, tal como una proteína coloreada, proteína fluorescente tal como GFP o una enzima tal como p-galactosidasa (gen lacZ).
Como se emplea en esta memoria, el término "GFP" se refiere a cualquier secuencia de ADN de proteína verde fluorescente capaz de producir una proteína fluorescente tras la expresión en una célula usada normalmente como un marcador de indicación para la expresión de un gen diana. Los ejemplos de GFP incluyen secuencias de GFP aisladas de celentéreos, tales como la medusa del Pacífico, Aequorea victoria, y derivados de secuencia sintética de las mismas, tales como "eGFP".
El término "muestra" se usa en su sentido más amplio. En un sentido, puede hacer referencia a una célula o un tejido. En otro sentido, se entiende que incluye una muestra de ensayo o cultivo obtenido de cualquier fuente y abarcar líquidos, sólidos y tejidos. Las muestras ambientales incluyen material ambiental tal como materia superficial, suelo, agua y muestras industriales. Estos ejemplos no deben interpretarse como limitantes de los tipos de muestra aplicables a la presente invención.
Los términos "purificada", "purificar', "purificación", "aislada", "aislar", "aislamiento", y equivalentes gramaticales de los mismos, como se emplean en esta memoria, se refieren a la reducción de la cantidad de al menos un contaminante de una muestra. Por ejemplo, un tipo celular deseado se purifica en al menos un 10 %, preferiblemente, en al menos 30 %, más preferiblemente en al menos 50 %, aún más preferiblemente en al menos 75 % y lo más preferiblemente en al menos 90 %, con una reducción correspondiente de la cantidad de tipos celulares indeseables, por ejemplo, la diferenciación dirigida de las presentes invenciones dio como resultado el aumento deseado de la pureza de células
progenitoras del mesencéfalo de la placa del suelo diferenciadas o neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo de las presentes invenciones. En otras palabras, "purificar", y sus equivalentes, se refiere a la eliminación de determinadas células (p. ej., células indeseables) de una muestra, mecánicamente, tal como por clasificación celular por citómetro de flujo o mediante diferenciación dirigida. Por ejemplo, para diferenciar una población purificada de células progenitoras proteína A2 de secuencia forkhead (FOXA2)+ factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A)+ de las presentes invenciones, las células progenitoras se purifican mediante la eliminación de células neuronales PAX6 contaminantes clasificando una población de células mixtas en células dobles positivas proteína A2 de secuencia forkhead (FOXA2)+ factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A)+ por citometría de flujo; las neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo también se purifican o "seleccionan" de no dopamina (DA) (células predeterminadas) mediante el uso de un método específico de cultivo celular que comprende composiciones y métodos de la presente invención. La eliminación o selección de las células neuronales de dopamina (DA) FOXA2/LMX1A+ no destinadas al mesencéfalo da como resultado un aumento del porcentaje de neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo deseadas en la muestra. Por tanto, la purificación de un tipo celular da como resultado un "enriquecimiento", es decir, un aumento de la cantidad, de la célula deseada, es decir, neuronas de dopamina (DA) FOXA2/LMX1A+ destinadas al mesencéfalo en la muestra.
La expresión "de origen natural" como se emplea en esta memoria cuando se aplica a un objeto (tal como célula, tejido, etc.) y/o producto químico (tal como una proteína, secuencia de aminoácidos, secuencia de ácido nucleico, codón, etc.) significa que el objeto y/o compuesto se encuentran o se han encontrado en la naturaleza. Por ejemplo, una célula de origen natural se refiere a una célula que está presente en un organismo que puede aislarse de una fuente en la naturaleza, tal como una célula embrionaria, en donde la célula no ha sido modificada intencionadamente por el ser humano en el laboratorio.
Como se emplea en esta memoria, la expresión "in vitro" se refiere a un ambiente artificial y a procesos o reacciones que se producen dentro de un ambiente artificial. Los ambientes in vitro están ilustrados, pero sin limitación, por tubos de ensayo y cultivos celulares.
Como se emplea en esta memoria, la expresión "in vivo" se refiere al ambiente natural (p. ej., un animal o una célula) y a procesos o reacciones que se producen dentro de un ambiente natural, tales como desarrollo embrionario, diferenciación celular, formación del tubo neural, etc.
La expresión "procedente de" o "establecida a partir de" o "diferenciada a partir de" cuando se usa en referencia a cualquier célula descrita en la presente memoria se refiere a una célula que se obtuvo de (p. ej., se aisló, purificó, etc.) una célula progenitora en una línea celular, tejido (tal como un embrión disociado), o fluidos usando cualquier manipulación, tal como, sin limitación, aislamiento de células individuales, cultivadas in vivo, tratamiento y/o mutagénesis. Una célula puede obtenerse de otra célula, usando, por ejemplo, tratamiento químico, radiación, induciendo expresión de nuevas proteínas, por ejemplo, mediante infección con virus, transfección con secuencias de ADN, contacto (tratamiento) con un morfógeno, etc., y selección (tal como por cultivo en serie) de cualquier tipo celular que esté contenido en células progenitoras cultivadas. Se puede seleccionar una célula derivada de una población mixta en virtud de la respuesta a un factor de crecimiento, citocina, progresión seleccionada de tratamientos con citocinas, adhesividad, falta de adhesividad, procedimiento de clasificación y similares.
Como se emplea en esta memoria, el término "célula" se refiere a una sola célula, así como a una población de (es decir, más de una) células. La población puede ser una población pura que comprende un tipo celular, tal como una población de células neuronales o una población de células embrionarias indiferenciadas. Como alternativa, la población puede comprender más de un tipo celular, por ejemplo, una población celular mixta. No se pretende que limite el número de células en una población; por ejemplo, en una realización, una población mixta de células puede comprender al menos una célula diferenciada. En las presentes invenciones, no hay límite en el número de tipos celulares que puede comprender una población celular.
Como se emplea en esta memoria, la expresión "población muy enriquecida" se refiere a una población de células, tal como una población de células en una placa de cultivo, que expresa un marcador en un porcentaje o cantidad mayor que una población de comparación, por ejemplo, el tratamiento de un cultivo celular que ha entrado en contacto con LSB el día 1 con purmorfamina y el día 3 con CHIR da como resultado una población muy enriquecida de células progenitoras del mesencéfalo de la placa del suelo en comparación con el tratamiento con LSB solo. En otros ejemplos, una población enriquecida es una población que resulta de clasificar o separar células que expresan uno o más marcadores de células que no expresan el marcador deseado, tal como una población enriquecida en CD142, una población enriquecida en A9 y similares.
La expresión "biología de la célula" o "biología celular" se refiere al estudio de una célula viva, tal como la anatomía y la función de una célula, por ejemplo, las propiedades fisiológicas, estructura, orgánulos e interacciones con su entorno de una célula, su ciclo vital, división y muerte.
La expresión "secuencia de nucleótidos de interés" se refiere a cualquier secuencia de nucleótidos (p. ej., ARN o ADN), cuya manipulación puede ser considerada deseable por cualquier motivo (p. ej., tratar una enfermedad, conferir cualidades mejoradas, expresión de una proteína de interés en una célula hospedadora, expresión de una ribozima,
etc.), por un experto en la técnica. Dichas secuencias de nucleótidos incluyen, pero sin limitación, secuencias codificantes de genes estructurales (p. ej., genes indicadores, genes marcadores de selección, oncogenes, genes de resistencia a fármacos, factores de crecimiento, etc.) y secuencias reguladoras no codificantes que no codifican un ARNm o producto proteico (p. ej., secuencia promotora, secuencia de poliadenilación, secuencia de terminación, secuencia potenciadora, etc.).
Como se emplea en esta memoria, la expresión "proteína de interés" se refiere a una proteína codificada por un ácido nucleico de interés.
El término "gen" se refiere a una secuencia de ácido nucleico (p. ej., ADN o ARN) que comprende secuencias codificantes necesarias para la producción de un polipéptido o precursor (p. ej., proinsulina). El polipéptido puede estar codificado por una secuencia codificante de longitud completa o por cualquier parte de la secuencia codificante siempre que la actividad o las propiedades funcionales deseadas (p. ej., actividad enzimática, unión a ligando, transducción de señales, etc.) de la longitud completa o el fragmento se conserven. El término también abarca la región codificante de un gen estructural e incluye secuencias ubicadas adyacentes a la región codificante en los extremos tanto 5' como 3' para una distancia de aproximadamente 1 kb o más en cualquiera de los extremos, de modo que el gen corresponda a la longitud del ARNm de longitud completa. Las secuencias que están ubicadas 5' de la región codificante y que están presentes en el ARNm se denominan secuencias no traducidas 5'. Las secuencias que están ubicadas 3' o cadena abajo de la región codificante y que están presentes en el ARNm se denominan secuencias no traducidas 3'. El término "gen" abarca las formas tanto de ADNc como genómicas de un gen. Una forma genómica o clon de un gen contiene la región codificante interrumpida con secuencias no codificantes denominadas "intrones" o "regiones intermedias" o "secuencias intermedias". Los intrones son segmentos de un gen que se transcriben en ARN nuclear (ARNnh); los intrones pueden contener elementos reguladores tales como potenciadores. Los intrones se eliminan o "se cortan y empalman" del transcrito nuclear o primario; por lo tanto, los intrones están ausentes en el transcrito de ARN mensajero (ARNm). El ARNm actúa durante la traducción para especificar la secuencia o el orden de los aminoácidos en un polipéptido naciente.
Como se emplea en esta memoria, la expresión "expresión génica" se refiere al proceso de convertir información genética codificada en un gen en ARN (p. ej., ARNm, ARNr, ARNt o ARNnp) mediante "transcripción" del gen (es decir, a través de la acción enzimática de una ARN polimerasa) y, para genes que codifican proteínas, en proteínas mediante "traducción" de ARNm. La expresión génica se puede regular en muchas etapas en el proceso. La "regulación positiva" o "activación" se refiere a regulación que aumenta la producción de productos de expresión génica (es decir, ARN o proteína), mientras que la "regulación negativa" o "represión" se refiere a regulación que disminuye la producción. Las moléculas (p. ej., factores de transcripción) que están implicadas en la regulación positiva o regulación negativa se denominan con frecuencia "activadores" y "represores", respectivamente.
Como se emplea en esta memoria, las expresiones "codificación de molécula de ácido nucleico", "codificación de secuencia de ADN", "codificación de ADN", "codificación de secuencia de ARN", y "codificación de ARN" se refieren al orden o la secuencia de desoxirribonucleótidos o ribonucleótidos a lo largo de una cadena de ácido desoxirribonucleico o ácido ribonucleico. El orden de estos desoxirribonucleótidos o ribonucleótidos determina el orden de los aminoácidos a lo largo de la cadena polipeptídica (proteína). La secuencia de ADN o ARN codifica por tanto la secuencia de aminoácidos.
El término "aislado" cuando se usa en relación con un ácido nucleico, como en "un oligonucleótido aislado" o "polinucleótido aislado" se refiere a una secuencia de ácido nucleico que se identifica y se separa de al menos un componente o contaminante con el que está asociado habitualmente en su fuente natural. El ácido nucleico aislado es el que está presente en una forma o un entorno diferente de aquel en que se encuentra en la naturaleza. Por el contrario, los ácidos nucleicos no aislados como ácidos nucleicos tales como ADN y ARN se encontraron en el estado en que existen en la naturaleza. Por ejemplo, una secuencia de ADN dada (p. ej., un gen) se encuentra en el cromosoma de la célula hospedadora próximo a genes adyacentes; las secuencias de ARN, tales como una secuencia de ARNm específica que codifica una proteína específica, se encuentran en la célula como una mezcla con numerosos ARNm adicionales que codifican una multitud de proteínas. Sin embargo, el ácido nucleico aislado que codifica una proteína dada incluye, a modo de ejemplo, dicho ácido nucleico en las células que habitualmente expresan la proteína dada donde el ácido nucleico está en una ubicación cromosómica diferente de la de células naturales o está flanqueado de otro modo por una secuencia de ácido nucleico diferente de la hallada en la naturaleza. El ácido nucleico aislado, oligonucleótido o polinucleótido pueden estar presentes en forma monocatenaria o bicatenaria. Cuando un ácido nucleico aislado, oligonucleótido o polinucleótido se va a utilizar para expresar una proteína, el oligonucleótido o polinucleótido contendrá como mínimo la cadena con sentido o codificante (es decir, el oligonucleótido o polinucleótido puede ser monocatenario), pero puede contener las cadenas tanto con sentido como antisentido (es decir, el oligonucleótido o polinucleótido puede ser bicatenario).
Como se emplea en esta memoria, el término "equipo" se refiere a cualquier sistema de suministro para suministrar materiales. En el contexto de la diferenciación celular, un equipo puede referirse a una combinación de materiales para entrar en contacto con células madre, dichos sistemas de suministro incluyen sistemas que permiten el almacenamiento, transporte o suministro de reactivos de reacción de una ubicación a otra en los recipientes adecuados (tales como tubos, etc.) y/o materiales de soporte (p. ej., tampones, instrucciones escritas para realizar diferenciación celular, etc.) (p. ej., compuestos, proteínas, agentes de detección (tales como anticuerpos que se unen con tirosina
hidroxilasa (TH), proteína A2 de secuencia forkhead (FOXA2), factor de transcripción 1 de homeosecuencia LIM, alfa (LMX1A), etc.), etc. Por ejemplo, los kits incluyen uno o más recintos (p. ej., cajas o bolsas, tubos de ensayo,
tubos Eppendorf, tubos capilares, placas de múltiples pocillos y similares) que contienen reactivos de reacción relevantes para inhibir las rutas de señalización, por ejemplo, un inhibidor para reducir la señalización del factor de crecimiento transformante beta (TGFp)/Activina-Nodal, tal como SB431542 (o reemplazo de SB431542) y similares, un inhibidor para reducir la señalización de SMAD, LDN-193189 (o reemplazo de LDN-193189) y similares, un inhibidor para reducir glucógeno sintasa cinasa 3p (GSK3p), como un ejemplo, para activación de la señalización de Wingless (Wnt o Wnts), también conocida como activador de la señalización de WNT (agonista de WNT), tal como CHIR99021 (o reemplazo de CHIR99021), etc.) y similares, un activador de la señalización de Sonic hedgehog (SHH) (tal como un agonista de molécula pequeña del receptor de smoothened (SMO)), por ejemplo, una molécula C25II de Sonic hedgehog (SHH), purmorfamina y similares, una molécula con actividad del factor de crecimiento de fibroblastos 8 (FGF8), tal como factor de crecimiento de fibroblastos 8 (FGF8), etc. y moléculas de maduración neuronal, por ejemplo, factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial, dibutiril AMPc y factor de crecimiento transformante de tipo p3, incluyendo moléculas capaces de reemplazar estos componentes y/o materiales de soporte. Los reactivos en el equipo en una realización pueden estar en solución, pueden estar congelados o pueden estar liofilizados. Los reactivos en el equipo en una realización pueden estar en recipientes individuales o proporcionarse como combinaciones específicas, tal como una combinación de LSB (LDN-193189 con SB431542), molécula C25II de Sonic hedgehog (SHH) con purmorfamina, molécula C25II de Sonic hedgehog (SHH) con purmorfamina con CHIR99021 o purmorfamina con CHIR99021, moléculas de maduración neuronal y similares.
Breve descripción de los dibujos
La figura 1 muestra inducción ilustrativa y conversión neurogénica de precursores de la placa del suelo del mesencéfalo procedentes de células ES humanas dependientes de la adición de CHIR990221. a) Análisis inmunocitoquímico el día 11 de diferenciación para expresión de FOXA2 (rojo), NESTIN (verde, paneles superiores), LMX1A (verde, paneles centrales) y OTX2 (verde, paneles inferiores). b, c) Cuantificación de los datos presentados en (a). Los datos son de tres experimentos independientes realizados cada uno por triplicado (media ± ETM). Se presentan los niveles de significación para marcadores individuales en comparación con el tratamiento solo con LSB: ANOVA; prueba de Dunnett: *** p < 0,001; ** p < 0,01; p < 0,05). d) Ilustración esquemática de las condiciones de cultivo usadas para las tres condiciones de tratamiento. e, f) Listas de genes expresados diferencialmente seleccionados el día 11 que comparan las condiciones de LSB/S/F8/CHIR con LSB (e) o LSB/S/F8 (f). g, h) Análisis de expresión génica temporal de marcadores seleccionados característicos de la identidad precursora de DA del mesencéfalo (g), prosencéfalo e identidad no precursora de DA ventral (h). Las barras de escala corresponden a 50 pm.
La figura 2 muestra un análisis inmunocitoquímico y molecular ilustrativo del destino de la neurona DA del mesencéfalo en destinos tratados con LSB/S/F8/CFIIR frente a LSB/S/F8 (ventral/hipotalámico) y LSB (prosencéfalo dorsal). a) Análisis inmunocitoquímico el día 25 para expresión de FOXA2 (azul) en combinación con Tuj1 (rojo)/LMX1A (verde) (paneles superiores) y NURRl (rojo)/TH (verde) (paneles inferiores). b) Análisis cuantitativo de coexpresión en cultivos tratados con LSB/S/F8/CHIR. Los datos son de tres experimentos independientes realizados cada uno por triplicado (media ± ETM). c, d) Se realizó análisis de expresión génica global el día 25 (muestras triplicadas para las tres condiciones). Listas seleccionadas de los genes más diferencialmente expresados que comparan el día 13 frente al día 25 en la condición de LSB/S/F8/CHIR (c) y que comparan el tratamiento con LSB/S/F8/CHIR frente a LSB (d, panel izquierdo) y LSB/S/F8 (d, panel derecho). e) Análisis de expresión génica diferencial normalizada para marcadores de neuronas DA del mesencéfalo clave. Se presentan los niveles de significación para marcadores individuales en comparación con el tratamiento solo con lSb : ANOVA; prueba de Dunnett: *** p < 0,001; ** p < 0,01; p < 0,05). Las barras de escala corresponden a 50 pm.
La figura 3-1 y 3-2 muestra una maduración in vitro ilustrativa, caracterización y evaluación funcional de neuronas DA del mesencéfalo procedentes de la placa del suelo frente a procedentes de la roseta. Figura 3-1: muestra ejemplos de: a) Análisis inmunocitoquímico el día 50 de diferenciación para TH (rojo), en combinación con LMX1A (verde, paneles izquierdos), FOXA2 (azul, paneles izquierdos) y NURR1 (verde, paneles derechos). b) Cuantificación de células TH+, FOXA2+, LMX1+ y NURR1+ de células totales en cultivos derivados de roseta frente a derivados de la placa del suelo (LSB/S/F8/CHIR). c) Cuantificación de los subtipos neuronales serotonina+ (5-HT) y GABA+ el día 50 en cultivos de neuronas DA procedentes de la placa del suelo y procedentes de la roseta. d, e) Análisis por HPLC para medir DA y metabolitos. d) Cromatograma representativo para la detección electroquímica de DA en una muestra de cultivos derivados de la placa del suelo. e) Comparación de los niveles de DA, DOPAc y HVA entre cultivos derivados de la placa del suelo frente a derivados de roseta. f) Análisis inmunocitoquímico de cultivos derivados de la placa del suelo (día 80) para TH (rojo) y sinapsina (verde). g-i) Análisis electrofisiológicos de cultivos de la placa del suelo el día 80 de diferenciación de neuronas DA. Imagen de contraste de fases de una neurona pinzada (g) y registros correspondientes (h). i) Análisis de potencia que muestra oscilaciones de potencial de membrana características de la identidad de neuronas DA (de 2 a aproximadamente 5 Hz). Los niveles de significación para marcadores individuales (paneles b, c, e) se presentan como una comparación cultivos derivados de la PS frente a derivados de la roseta: prueba de T de Student: *** p < 0,001; ** p < 0,01; p < 0,05). Las barras de escala corresponden a 50 pm en (a), 20 pm in (f, panel superior), 5 pm en (f, panel inferior) y 20 pm en (g) j: Maduración de neuronas mDA in vitro (d65). Las
neuronas TH positivas aún expresan FoxA2 y extienden fibras largas normales para neuronas mDA y k: Medición de liberación de Da por HPLC: las neuronas TH+ de edad d65 son funcionales in vitro. Figura 3-2: muestra un compendio ilustrativo de células producidas por un protocolo de neuronas DA del mesencéfalo basado en la placa del suelo como se describe en la presente memoria. a) En contraste con las estrategias pasadas (por ejemplo, Perrier, A. L. et al. Derivation of midbrain dopamine neurons from human embryonic stem cells. Proc Natl Acad Sci U S A 101, 12543-8 (2004)), el nuevo protocolo descrito en la presente memoria se basa en generar una placa del suelo del mesencéfalo positiva para LMX1A/FOXA2 (panel izquierdo) seguido de conversión neuronal (panel central) y maduración de neuronas DA (panel derecho). Las células neuronales DA maduras generadas por la placa del suelo conservan la expresión de FOXA2/LMX1A.
La figura 4 muestra una supervivencia y función in vivo ilustrativa de neuronas DA humanas procedentes de la placa del suelo en cerebro hospedador de modelo de EP de ratón, rata y mono. a-d) Trasplante de neuronas DA procedentes de la placa del suelo en ratones adultos con lesión de 6-OHDA (cepa nula para NOD-SCID IL2Rgc). a) Expresión de TH y morfología del injerto a los 4,5 meses después del trasplante. b) Expresión de marcador específico humano (hNCAM, azul), TH (verde) y FOXA2 (rojo). c) Cuantificación de células FOXA2+, TH y con doble marcador en injertos derivados de la placa del suelo (media ± ETM, n=4 a los 4,5 meses después del injerto). d) Análisis de rotación inducida por anfetaminas en injertos derivados de la placa del suelo (azul) frente a derivados de la roseta (verde). Las barras de escala corresponden a 500 pm en (a), 100 pm en (b) y 40 pm en (4c). e-p) Trasplante de neuronas DA procedentes de la placa del suelo en ratas adultas con lesión de 6-OHDA. Análisis inmunohistoquímico para coexpresión de TH (verde) y los marcadores específicos humanos (rojo) hNA. e) y hNCAM (4f). g) Cuantificación estereológica del número de células totales (hNA+), células TH+ y células TH+ que coexpresan FOXA2. El volumen promedio del injerto fue 2,6 /- 0,6 mm3). h-j) Imágenes de alta potencia que muestran la coexpresión de TH (verde) con factores de transcripción específicos del mesencéfalo FOXA2, PITX3 y NURR1 (rojo). k-m) Análisis conductual de animales tratados con injertos de neuronas DA procedentes de la placa del suelo frente a animales tratados con simulación. k) Asimetría rotacional inducida por anfetaminas. l) prueba de marcha: medición de la acinesia de las extremidades anteriores en el lado afectado frente al no afectado. m) Prueba de cilindro: medición de la preferencia de la pata ipsilateral frente a la contralateral al ponerse a dos patas. Los animales con injerto mostraron mejora significativa en al menos tres pruebas (p < 0,01 a 4,5 - 5 meses; n=4-6 cada uno). n-p) Análisis inmunohistoquímico para TH (verde) y coexpresión (rojo) con DAT (n), GIRK2 (o) y calbindina (p). Los niveles de significación (paneles d, k, 1, m) son: ** p < 0,01; p < 0,05). Las barras de escala corresponden a 200 pm en (e), 50 pm en (f), 20 pm en (h-j) y 40 pm en (n-p). q-t) Trasplante de neuronas DA procedentes de la placa del suelo en mono rhesus adulto con lesión de 1 -metil-4-fenil-1,2,3,6-tetrahidropiridina (MPTP). q) Descripción general del sitio de injerto representativo 1 mes después del trasplante marcado por la expresión de marcador de citoplasma específico humano SC-121 (verde). r) Expresión de TH en injerto con un halo circundante de fibras TH+ (flechas). s) Análisis de la coexpresión de SC-121 (rojo) con TH (verde) en el núcleo del injerto. t) Análisis de coexpresión de FOXA2 (rojo) en neuronas TH+ (verde). Las barras de escala corresponden a 2 mm para (q), 500 pm para (r), 200 pm para (s) y 50 pm para (t). u) crecimiento de fibra derivada de injerto (huNCAM+) en el cuerpo estriado del hospedador. v) Las células procedentes de injerto no se diferencian en células gliales. Sin embargo, los injertos contenían varias fibras serotoninérgicas además de la población de células TH+.
La figura 5 muestra que una sincronización ilustrativa de exposición a CHIR99021 determina la inducción de precursores de la placa del suelo del mesencéfalo FOXA2/LMX1A. a) Análisis inmunocitoquímico de FOXA2 (rojo)/lMXlA (verde) el día 11 de diferenciación después del tratamiento con LSB/S/F8 solo o en combinación con CHIR a partir de los diversos puntos temporales indicados. b) Cuantificación del porcentaje de células FOXA2+, LMX1A+ y con doble marcador el día 11 de diferenciación después del inicio diferencial de exposición a CHIR como se describe en (a). Se presentan los niveles de significación para marcadores individuales en comparación con la condición sin CHIR: ANOVA; prueba de Dunnett: *** p < 0,001; ** p < 0,01; p < 0,05). Las barras de escala corresponden a 50 pm.
La figura 6 muestra que una exposición ilustrativa a FGF8 no desempeña un papel importante en la inducción de precursores de la placa del suelo del mesencéfalo FOXA2/LMX1A. Imágenes representativas de la expresión de FOXA2 (rojo)/LMX1A (verde) mediante inmunocitoquímica el día 11 de diferenciación. Las células se expusieron a LSB/CHIR en presencia o ausencia de SHH (purmorfamina SHH C25II) y FGF8. Las barras de escala corresponden a 50 pm.
La figura 7 muestra que es necesaria una exposición ilustrativa a alta dosis de SHH y/o un agonista de molécula pequeña de smoothened (purmorfamina) para inducción eficaz de la placa del suelo del mesencéfalo en presencia de CHIR99021. Imágenes representativas de inmunocitoquímica de FOXA2 (rojo)/LMX1A (verde) el día 11 de diferenciación. Las células se trataron con LSB/F8/CHIR en presencia de diversas concentraciones de SHH (SHH-C25II) y agonista de smoothened purmorfamina. Las barras de escala corresponden a 50 pm.
La figura 8 muestra un análisis ilustrativo de genes expresados diferencialmente en cultivos tratados con LSB/S/F8/CHIR frente a cultivos tratados con LSB y LSB/S/F8 los días 11 y 25 de diferenciación. a) Agrupación jerárquica de los datos de expresión génica global obtenidos de tres condiciones los días 0, 1, 3, 5, 7, 11 y 13 (las muestras se evaluaron por triplicado para cada condición y cada día). b) Análisis de enriquecimiento génico según las clases GO usando DAVID; http://david.abcc.ncifcrf.gov. Las comparaciones el día 11 revelan el enriquecimiento de la señalización tanto de SHH como de WNT en condición de LSB/S/F8/CHIR de acuerdo con la activación mediada por CHIR99021 de la señalización de WNT canónica. Los destinos celulares alternativos tales como "desarrollo del
prosencéfalo" y "diencéfalo", así como "homeosecuencia" son otros términos GO muy enriquecidos los días 11 y 25. c) Validación en línea usando la base de datos de expresión de genes humanos Allen (http://human.brain-map.org) para marcadores candidatos enriquecidos en la condición de neuronas DA del mesencéfalo (LSB/S/F8/CHIR). TTF3, EBF1, EBF3 y TTR se expresaron en función de los datos disponibles de micromatrices específicas de la región del cerebro humano. TTR, una diana transcripcional clásica de FOXA2 solo se expresa débilmente en la sustancia negra de los adultos, lo que sugiere que su función principal puede ser durante el desarrollo o que el tratamiento con SHH puede provocar niveles de expresión de TTR artificialmente altos durante diferenciación in vitro.
La figura 9 muestra un protocolo de diferenciación ilustrativo para la inducción de la placa del suelo y el desarrollo de neuronas DA del mesencéfalo en líneas hESC e hiPSC independientes representativas. Se presentan datos de la línea de hESC H1 y las líneas de hiPSC 2C6 (de un paciente con EP esporádica) y SeV6 (basado en Sendai, sin integración). Se usó el protocolo basado en la placa del suelo descrito en la figura 1d y la figura 10, seguido de análisis de la expresión de FOXA2 (rojo) el día 11 y tH (verde)/FOXA2 (rojo) el día 25 de diferenciación.
La figura 10 muestra un compendio esquemático ilustrativo de las condiciones de diferenciación usadas para cultivos de neuronas DA procedentes de la placa del suelo y procedentes de la roseta. Ambos protocolos usaron inhibición doble de SMAD para acelerar la adquisición del destino neural. Se usó LDN para la inhibición de BMP en el protocolo de la placa del suelo, mientras que se usó la inducción de noggin tradicional para cultivos de roseta. Las abreviaturas son: LDN: LDN-193189, SB: SB431542, SHH (purmorfamina SHH C25II), FGF8: FGF8, BAGCT: BDNF ácido ascórbico GDNF dbAMPc TGFp3. SHH/FGF8 en el protocolo de roseta usó SHH C25I solo en ausencia de purmorfamina siguiendo la recomendación inicial para la distribución de cultivos de neuronas DA procedentes de la roseta. Nota: El tratamiento con purmorfamina en la etapa de roseta muestra toxicidad a una concentración adecuada para la distribución de células de la placa del suelo. BASF: BDNF ácido ascórbico SHH/FGF8.
La figura 11 muestra una maduración ilustrativa in vitro de cultivos de neuronas DA procedentes de la placa del suelo. a) Análisis inmunocitoquímico de neuronas TH procedentes de la placa del suelo (verde) para expresión de DAT (rojo; día 80). b) Se observaron tramos extensos de fibra Tuj1+ (rojo) el día 60 de diferenciación que se extendía a distancias de > 2 mm. c) En el día 80 de diferenciación, coexpresión de GIRK2 (rojo) en neuronas Th+ (verde). Las barras de escala corresponden a 20 pm en (a) 100 pm en (b) y 20 pm en (c).
La figura 12 muestra un análisis inmunohistoquímico ilustrativo de estudios de supervivencia in vivo a corto plazo (6 semanas) en el cuerpo estriado de ratón intacto (sin lesiones) adulto (cepa nula para NOD-SCID IL2Rgc). Análisis de injertos derivados de la placa del suelo (células del día 25; 150 x 103 células/animal). a) Imagen representativa del núcleo de injerto que muestra células TH+ rodeadas por fibras hospedadoras TH+. Un promedio de 6.200 células TH+ (n=3) estaban presentes en el injerto a las 6 semanas después del trasplante. b) Las células que expresan FOXA2 (rojo) solo se encontraron en el injerto pero no en el cuerpo estriado del hospedador circundante, lo que demuestra el origen del injerto y la identidad del mesencéfalo de las células. Casi todas las neuronas TH+ (verde) coexpresaron FOXA2 (rojo). Sin embargo, una proporción considerable de células FOXA2+ no coexpresó TH, lo que sugiere que estas células aún no han adquirido un fenotipo DA maduro ni representan otra población neuronal FOXA2+ negativa para marcadores de neuronas DA. Las barras de escala corresponden a 50 pm.
La figura 13 muestra un análisis histológico ilustrativo de ratones con lesión de 6-OHDA con injertos a largo plazo (4,5 meses) (cepa nula para NOD-SCID IL2Rgc) que compara el comportamiento de injertos derivados de la placa del suelo frente a derivados de la roseta. Análisis de injertos derivados de la placa del suelo. a) Ejemplo de uno de los mayores injertos derivados de la placa del suelo. El injerto conserva una citoarquitectura hNCAM+ (roja) bien localizada. b) Se observó crecimiento robusto de fibra hNCAM+ en la periferia del injerto. c) Las fibras serotoninérgicas (5-HT+) (verde) en injerto son en gran medida negativas para hNCAM (rojo), lo que sugiere origen del hospedador. d) Neuronas y fibras GABAérgicas (verde) en el injerto. Análisis de injertos derivados de la roseta: e) Sobrecrecimiento neural con compresión del tejido cerebral del hospedador. La mayoría de células fueron positivas tanto para NCAM (rojo) como para DCX (verde), lo que sugiere un destino neuronal. f) Fibras NCAM+/DCX+ extendidas al lado contralateral no trasplantado del cerebro. g) Dentro del núcleo del injerto se observaron múltiples grupos DCX+. h) Se observaron pocas células TH+ (verde) y fibras en la periferia del injerto y casi todas las células TH+ procedentes de la roseta in vivo fueron negativas para FOXA2 (azul). Análisis inmunohistoquímico de injertos derivados de la placa del suelo frente a derivados de la roseta a los 4,5 meses. Se presentan imágenes representativas para la expresión del marcador de proliferación Ki67 (rojo) y el marcador precursor neural PAX6 (verde) (i), expresión de FOXA2/DCX (j), FOXG1/hNCAM (j, recuadro) y el marcador de astrocitos GFAP (verde) (k). Las barras de escala corresponden a 500 pm en (a), 50 pm en (b-d), a 500 pm en (e), 50 pm en (g-h), 50 pm en (i), 40 pm en (j) y 20 pm en (k).
La figura 14 muestra un análisis histológico ilustrativo de ratas SD con lesión de 6-OHDA con injertos a largo plazo (5 meses). a) La gran mayoría de células hNA+ (rojas) expresaron Tuj1 (verde) lo que indica la identidad de destino neuronal. b) Las fibras GFAP+ (verde) dentro del injerto no coexpresaron hNA (rojo), lo que sugiere el origen del hospedador. c) Se observaron algunos cuerpos celulares serotoninérgicos humanos (5-HT+) (verde) mientras que la mayoría de las fibras serotoninérgicas procedieron del hospedador de manera similar a los datos en el ratón hospedador (figura 13C). d) Ejemplo adicional de neuronas TH+ (verde) dentro del injerto caracterizado por la expresión de hNCAM (rojo) para confirmar la identidad humana de células en el cerebro del hospedador. Las barras de escala corresponden a 25 pm en paneles.
La figura 15 muestra un análisis histológico ilustrativo de injertos derivados de la placa del suelo en cerebro de primate. Se injertaron neuronas DA procedentes de la placa del suelo procedentes de hESC H9 y de hESC H9-GFP (cultivos del día 25; 1,25 x 106 células/vía), 3 vías/por hemisferio, 6 vías en total) en el cuerpo estriado de monos rhesus con lesión de MPTP adultos (=2). a) Paneles superiores izquierdo y derecho: Reconstrucciones de imágenes de alta resolución de sitios de injerto representativos (1 mes después del trasplante). La citoarquitectura de injerto se ilustra mediante inmunohistoquímica para el marcador humano específico SC-121 (sin reactividad cruzada con tejidos de primates no humanos). Panel inferior izquierdo: Fibras SC-121+ que se extienden desde el núcleo del injerto. Panel inferior derecho: Imagen de mayor resolución de células SC-121+ que muestran morfologías precursoras neuronales. b) El análisis de la expresión de GFP en núcleos de injerto confirmó adicionalmente la identidad humana de las células que complementan los datos de SC-121. Nota: Para ambos animales, se injertaron en cada hemisferio células procedentes de H9 sin marcar mientras que el otro hemisferio recibió células procedentes de células H9-GFP (expresión de GFP bajo control del promotor EF1a). c) Imagen de mayor resolución del injerto GFP+ usando detección de DAB que muestra numerosas células GFP+ en la periferia del injerto que presentan morfología de neuroblastos. d) Análisis inmunohistoquímicos de áreas en el núcleo del injerto negativo para GFP y SC-121. Estas áreas contenían grandes cantidades de microglía del hospedador basadas en la expresión de Iba1 (rojo), así como pocos macrófagos ED1+ (azul), lo que sugiere inflamación persistente a pesar de la inmunosupresión de ciclosporina. Las barras de escala corresponden a 500 pm en (a, panel superior), 50 pm en (a, panel inferior izquierdo), 20 pm en (a, panel inferior derecho), 2 mm en (b), 500 pm en (c, panel izquierdo), 50 pm (c, panel superior derecho), 100 pm (c, panel inferior derecho), 100 pm (d).
La figura 16 muestra una derivación ilustrativa de células TH+ a partir de hESC usando un método basado en células alimentadoras MS5 anteriores que diferenciaba las células en células de tipo neuronal DA a través de (mediante) intermedios de células de la roseta. Parte superior) a P0 se pusieron hESCs en contacto con moléculas para comenzar la inducción neural de células Oct4+ en células de la roseta usando células alimentadoras MS5 (Perrier et al., 2004). En la etapa P1, se expandieron células de la roseta al poner en contacto células con moléculas adicionales para diferenciar las células en células en la etapa P2 con patrones de expresión específicos incluyendo células progenitoras DA Pax2+/Enl+ diferenciadas en neuronas DA TH+/En1+. Estas células se usaron para injerto en ratas con lesión de 60HDA, inmunodeprimidas a través del tratamiento con ciclosporina A. Estos estudios de trasplante mostraron muy mala viabilidad in vivo de neuronas de tipo DA, incluyendo pérdida del fenotipo TH+, y reveló preocupaciones acerca del crecimiento adicional de células no deseadas, posiblemente letales, para los animales con injertos, es decir, teratomas, y el crecimiento de células en tipos neuronales inadecuados que provocarían problemas médicos adicionales para el paciente. A: Hubo cantidades muy pequeñas de neuronas TH+ superviviente a los 4,5 meses después del trasplante (< 50 células TH+/animal) en injertos de precursores de neuronas DA procedentes de la roseta. Sin embargo, a diferencia de células TH+, las células marcadas con GFP (GFP fue impulsada por un promotor ubicuo) sobrevivieron bastante bien después del trasplante. Esto sugiere que la mayoría de las células supervivientes después del trasplante eran células neurales de identidad neuronal no DA (16B). Fig. 16C: pocas células procedentes del injerto (hNA+ (verde) coexpresan TH (rojo) lo que sugiere que la mayoría de las células humanas injertadas adoptan un fenotipo de neurona no DA. Los paneles 16 D-E muestran que D-E, a pesar de la supervivencia in vivo muy mala hubo alguna mejora (aunque muy modesta y muy variable) en algunos ensayos conductuales tales como rotaciones inducidas por anfetaminas (D), prueba de cilindro y rotaciones espontáneas (E).
La figura 17 muestra un protocolo ilustrativo para la obtención de bajas cantidades de células de la placa del suelo. Un protocolo de inhibición doble de SMAD modificado generó células de la placa del suelo (fp). Fueron necesarias altas concentraciones de SHH para la inducción de células FoxA2+ fp y que la adición de señales de distribución de caudalización tales como FGF8, Wnt1 o RA no condujeron a disminución de la expresión de FOXA2, sino a cambio en la identidad regional A) Panel izquierdo: Células el día 11 de diferenciación después del tratamiento con NSB (Noggin/SB431542); panel central izquierdo: Tratamiento con NSB+SHH (Sonic C25II); Panel central: NSB+SHH+RA; Panel central derecho: NSB+SHH+Wnt1; NSB+SHH+FGF8; Nota: El tratamiento solo con NSB no induce expresión de FoxA2. Las células de la placa del suelo FoxA2+ solo se inducen en presencia de dosis alta de SHH. La adición de RA. Wnt1 y FGF8 no inhibe la inducción de FoxA2. B) Análisis de qRT-PCR para expresión génica de FOXA2 y SIX6 que muestra el mantenimiento (o incluso el aumento) de la expresión de FOXA2 después del tratamiento con RA o FGF con una regulación negativa simultánea de la expresión de SIX6 que marca las células de tipo placa del suelo más anterior. C) Inducción de la expresión génica para precursores del mesencéfalo y marcadores de la placa del suelo del mesencéfalo en presencia de FGF8 y Wnt1.
La figura 18 muestra un protocolo ilustrativo para la obtención de células de la placa del suelo que muestra altos niveles de características del mesencéfalo en comparación con los niveles bajos o ausentes en células preparadas por los procedimientos usados en las figuras 16 y 17. A: La inducción de la placa del suelo del mesencéfalo en poblaciones celulares en contacto con altos niveles de SHH, FGF8 y CHIR dio como resultado coexpresión de FoxA2 con marcadores del mesencéfalo LMX1A y expresión de Otx2 el día 11 de diferenciación en contraste con células que han entrado en contacto con moléculas descritas en otros dos procedimientos como se muestra en las figs. 16 y 17 (N/SB y SHH/FGF8, respectivamente). B: Análisis de expresión génica global el día 11 que compara los tres grupos de células que entraron en contacto con moléculas de cada uno de los tres procedimientos, incluyendo el tercer procedimiento de las presentes invenciones (LDN/SB, SHH/FGF8, LSB/SHH/FGF8/CHIR) El diagrama B muestra específicamente los genes que son comunes entre las tres condiciones de tratamiento frente a los genes que son únicos para cada condición individual. El diagrama B es un análisis preliminar usado para los resultados de
micromatrices presentados en la figura 1.C: Los niveles de ARNm de los marcadores del mesencéfalo FoxA2, LMX1A, están muy enriquecidos en el grupo tratado con LSB/SHH/FGF8/CHIR en comparación con el grupo tratado con SHH/FGF.
La figura 19 muestra una caracterización ilustrativa in vitro de neuronas de dopamina procedentes de la región del mesencéfalo de la placa del suelo. A: Marcaje conjunto de neuronas FoxA2+ con marcadores de neuronas mDA TH, Nurr1 y LMX1A en d25 de diferenciación. B: Niveles de expresión de ARNm por QRT-PCR de marcadores de neurona mDA, así como otros tipos celulares del mesencéfalo en grupos tratados con LDN/SB SHH/FGF8 SHH/FGF8 CHIR.
La figura 20 muestra un potencial de diferenciación ilustrativo comparable hacia el destino de neurona DA del mesencéfalo de células EP-iPS mutantes PINK1 frente a células hES (o iPS) de tipo silvestre. La línea de EP-iPSC mutante PINK1 Q456X se diferenció usando los nuevos métodos de neuronas dA del mesencéfalo basadas en la placa del suelo de las presentes invenciones que producían perfiles de diferenciación del mesencéfalo comparables a los obtenidos de la línea H9. A-C) Análisis inmunocitoquímico de la línea EP-iPSC mutante PINK1 el día 11 de diferenciación (etapa precursora del mesencéfalo) para FOXA2 (rojo), LMX1A (verde) y DAPI (azul) (A), día 25 de diferenciación (etapa neuronal DA postmitótica temprana) para FOXA2 (rojo) y TH (verde) (B) y para NURR1 (rojo) y TH (verde) (C). D-F) El mismo conjunto de análisis inmunocitoquímicos realizados usando células procedentes de H9 el día 11 de diferenciación para FOXA2 (rojo), LMX1A (verde) y DAPI (azul) (D), el día 25 de diferenciación para FOXA2 (rojo) y TH (verde) (E) y para NURR1 (rojo) y TH (verde) (F).
La figura 21 muestra que una EP-iPSC mutante PINK1 ilustrativa mostró fenotipo de tipo EP de agregación de proteínas después de la diferenciación y maduración a largo plazo in vitro. El día 55 de diferenciación, la EP-iPSC mutante PINK1 mostró pruebas de expresión de a-sinucleína (un componente principal de la formación de cuerpos de Lewy en pacientes con EP) en el citosol de neuronas DA TH+. Las células también mostraron alta expresión de ubiquitina (un marcador clásico de cuerpos de Lewy). Por el contrario, las neuronas DA procedentes de la línea iPS de control mostraron expresión de expresión sináptica normal (en oposición a citosólica) de a-sinucleína y niveles muy bajos de ubiquitina. A, B) Análisis inmunocitoquímico de la línea EP-iPSC mutante PINK1 el día 55 de diferenciación para a-sinucleína (LB509, rojo), TH (verde) e imagen fusionada (A) y a-sinucleína (rojo) y ubiquitina (verde) (B). C, D) Análisis inmunocitoquímico de la línea iPSC de control el día 55 de diferenciación para a-sinucleína (rojo) y TH (verde) (C) y a-sinucleína (rojo) y ubiquitina (verde) (D).
La figura 22 muestra una expresión ilustrativa de forma agregada de a-sinucleína. En el cerebro del paciente con EP, las formas insolubles dimerizadas de a-sinucleína conducen a la agregación en el cuerpo de Lewy. La forma dimerizada de a-sinucleína muestra fosfolilación de serina 129 en a-sinucleína. Las células procedentes de EP-iPSC mutantes PINK1 mostraron fuerte expresión de a-sinucleína fosforilada Serl29 en contraste con células procedentes de iPSC de control que mostraron niveles muy bajos de expresión. A, B) Análisis inmunocitoquímico para a-sinucleína fosforilada Serl29 (verde) y DAPI (azul) en células procedentes de EP-iPSC mutantes PINK1 el día 55 de diferenciación (A) y células procedentes de iPSC de control (B).
La figura 23 muestra que se observan diferencias ilustrativas en los patrones de expresión de a-sinucleína dependiendo del protocolo de diferenciación. Los inventores muestran que las neuronas DA 'auténticas' del mesencéfalo tienen vulnerabilidad específica de EP y fenotipos correspondientes, específico in vitro. Las neuronas DA obtenidas usando el protocolo clásico de diferenciación basado en alimentador del estroma MS5 (Perrier et al., PNAS 2004, incorporado en el presente documento por referencia) puede producir grandes cantidades de neuronas TH+. Sin embargo, en función de los datos de las presentes invenciones, las células TH+ resultantes de diferenciación por el protocolo clásico de alimentación del estroma MS5 no son neuronas DA auténticas del mesencéfalo. En cultivos diferenciados a través del protocolo MS5, hubo muchas células positivas para a-sinucleína. Sin embargo, esas células no coexpresaron TH. Por otro lado, no hubo ninguna diferencia en los patrones de expresión entre EP-iPSC e iPSC de control cuando se usó la estrategia de diferenciación MS5. Estos datos indican que la a-sinucleína también se expresa en otros tipos celulares que no son DA y que dicha a-sinucleína que no es DA no cambia en células procedentes de iPSC de enfermedad frente a control, en particular cuando se usan protocolos de diferenciación MS5 convencionales. Finalmente, el nuevo protocolo de diferenciación basado en la placa del suelo de las presentes invenciones produce un gran número de células TH+ que coexpresan a-sinucleína. Esas células TH+ expresan asinucleína en un patrón de expresión citosólica. A, B) Análisis inmunocitoquímico para a-sinucleína (LB509, rojo), TH (verde) de la línea EP-iPSC mutante PINK1 el día 60 de diferenciación basada en MS5 (A) e iPSC de control (B). C) Análisis inmunocitoquímico de la línea EP-iPSC mutante PINK1 el día 55 de diferenciación basada en la placa del suelo para a-sinucleína (rojo), TH (verde).
La figura 24 muestra que las neuronas DA ilustrativas procedentes de EP-iPSC mutantes PINK1 son más vulnerables a la estimulación tóxica. Las neuronas DA TH+ procedentes de EP-iPSC obtenidas a través de los protocolos basados en la placa del suelo de las presentes invenciones fueron más vulnerables a la exposición a toxinas (valinomicina: ionóforo de mitocondrias, 5 pM, 48 h) que las células procedentes de iPSC de control. Por el contrario, las neuronas TH+ obtenidas a través del protocolo clásico basado en MS5 no mostraron vulnerabilidad diferencial entre las células procedentes de EP frente a control. A-F) Inmunocitoquímica de TH representativa el día 60 de diferenciación: Condición normal (sin tratamiento con toxinas) para células procedentes de EP-iPSC y de control obtenidas a través del protocolo basado en la placa del suelo (A, se muestran las células procedentes de EP-iPSC), degradación casi
completa de neuronas DA TH+ en EP-iPSC después del tratamiento con toxinas (B), neuronas DA TH+ parcialmente degradadas de iPSC de control (C), Condición normal de cultivos tanto procedentes de EP-iPSC como de control obtenidos a través del protocolo basado en MS5 (D, se muestran las células procedentes de EP-iPSC), Neuronas TH+ después de exposición a toxinas en cultivos procedentes de EP-iPSC (E) e iPSC de control (F) obtenidos a través del protocolo MS5. G-H) imágenes de baja potencia de inmunocitoquímica para Tuj1 (rojo) y TH (verde) mediante el protocolo basado en la placa del suelo el día 60 de diferenciación: EP-iPSC de condiciones normales (G) frente a exposición a toxina (H) e iPSC de control de condiciones normales (I) frente a exposición a toxina (J). K-N) imágenes de baja potencia de inmunocitoquímica para Tuj 1 (rojo) y TH (verde) mediante el protocolo basado en MS5 el día 60 de diferenciación: EP-iPSC de condiciones normales (K) frente a exposición a toxina (L) e iPSC de control de condiciones normales (M) frente a exposición a toxina (N).
La figura 25 muestra una cuantificación ilustrativa del ensayo de viabilidad celular-respuesta a dosis para exposición a toxina. El ensayo de viabilidad celular con azul alamar después de 48 h de tratamiento con valinomicina mostró supervivencia celular diferencial en un intervalo de dosis específico para exposición a toxinas (5 y 10 uM) al comparar EP-iPSC e iPSC de control (día 60 de diferenciación basada en la placa del suelo de las presentes invenciones). Nota: este ensayo prueba la muerte celular general mientras que los efectos más impresionantes se observaron específicamente en neuronas DA (véase figura 14). Por lo tanto, la cuantificación basada en azul de alamar probablemente subestimará el alcance del efecto diferencial observado en linajes de neuronas DA.
La figura 26 muestra que las neuronas DA humanas injertadas ilustrativas procedentes de células madre pluripotentes tienen características electrofisiológicas habituales de las observadas en la pars compacta de la sustancia negra (SNpc) de ratón. A) Parte superior-Reconstrucción de una neurona marcapasos en la región del injerto. Parte inferior-Microfotografía de un corte cerebral tomado de la rata en la que se inyectaron las neuronas procedentes de hES 9 meses antes; el injerto está delineado; se muestra una imagen de mayor aumento insertada en la parte inferior. El corte se procesó para detectar tirosina hidroxilasa, véase áreas blancas. B). Parte superior-Registro de parche unido a la célula de una posible neurona DA en el injerto; Parte inferior-Registro de célula completa de la misma célula. Se realizaron registros en presencia de glutamato y antagonistas del receptor de GABA (AP5 50 pM, CNQX 10 pM y GABAzina 10 pM) para eliminar el aporte sináptico. Estos registros demuestran que las neuronas procedentes de p S eran marcapasos autónomos con trayectorias de tensión intrasomática normales. Otra neurona registrada en el injerto tenía propiedades similares. C) A modo de comparación, se muestran registros unidos a células y de células completas de una neurona dopaminérgica en SNpc de un ratón adulto. Abreviaturas (CTx = corteza, ST r = cuerpo estriado, SNpc = pars compacta de sustancia negra, DA = dopaminérgica).
La figura 27 muestra un marcador de superficie candidato A9 ilustrativo y exploración de CD en hPSC. a) Diagrama de Venn de datos de transcriptoma de neuronas mDA procedentes de ESC de ratón purificadas con FACS. Entre los 107 genes compartidos entre PITX3 y Nurr1, la mayoría eran marcadores conocidos de neuronas DA del mesencéfalo, y además se confirmó que nuevos marcadores se expresaban en el mesencéfalo ventral: b) Uno de esos marcadores era DCSM1, un posible marcador de superficie que parece estar enriquecido dentro de la región A9, en función de datos de expresión in situ de ARNm (Allen Brain Atlas, Lein, E. S. et al., Genome-wide atlas of gene expression in the adult mouse brain, Nature 445: 168-176 (2007)). c-f) exploración de CD: c) placa representativa de 96 pocillos (1 de cada 3 x 96 placas usadas para explorar el panel de CD completo). Los pocillos oscuros marcan marcadores de CD que tienen alta expresión en neuronas DA de hESC el día 25. e) Compendio de los resultados de exploración de CD en neuronas DA procedentes de hESC. f) Un marcador ilustrativo, CD142, un marcador de superficie que se enriquece específicamente para las neuronas DA en la etapa Nurr1+ fue como se encontró después del aislamiento mediado por FACS de células CD142+ frente a células CD142- y análisis el día 7 después de la clasificación.
La figura 28 muestra CD142 ilustrativo enriquecido para la etapa de neurona DA del mesencéfalo Nurr1+ y se agota para las neuronas GABA y serotoninérgicas. a) La citometría de flujo mostró expresión representativa de CD142 el día 25 de diferenciación. b) CD142 enriquecido para la etapa Nurr1+ entre las neuronas DA del mesencéfalo FOXA2/TH en hESC (p. ej., WA09; figura 27f) y líneas hiPSC probadas (C29 y M3X representan dos líneas iPSC humanas el día 25 de diferenciación). c, d) CD142 agota los contaminantes GABAérgicos y serotoninérgicos después de la clasificación del día 25 y cultivo in vitro durante 3 semanas. e, f) CD142 agota las neuronas GABAérgicas y serotoninérgicas in vivo 3 meses después del trasplante. Las células se clasificaron con respecto a CD142 el día 25. Nota: Las células CD142 también se enriquecieron para TH y AADC.
La figura 29 muestra un uso experimental contemplado ilustrativo de PSA. Se evaluarán las neuronas DA procedentes de hESC que expresan PST y expuestas a PSTnm in vitro para determinar la influencia en el fenotipo DA y el crecimiento de fibras. Los estudios in vivo en el modelo de rata 6OHDA se analizarán para determinar si una menor cantidad de neuronas DA con expresión forzada de PSA puede igualar la recuperación conductual de los injertos convencionales y si la expresión forzada de PSA en neuronas DA procedentes de hESC es capaz de inducir la recuperación en ensayos de función motora compleja.
La figura 30 muestra un uso ejemplar de PST. La sobreexpresión (PST de ratón) dio como resultado mayores niveles de PSA-NCAM en células ES de ratón en diferenciación. (A) Cuantificación de ARNm de PST mediante qPCR en células de control (Nurr1) y en células que sobreexpresan PST (Nurr1/PST). Los datos se expresan como el factor de enriquecimiento de los niveles de PST en células Nurr1/PST frente a Nurr1. (B) La inmunotinción de PSA en cultivos de neuronas DA el día 14 de diferenciación muestra mayores niveles aumentados de PSA en células Nurr1/PST (barra
de escala: 100 pm). (C) transferencia de Western para NCAM en células diferenciadas. Las células Nurr1/PST (carril 2) muestran mayores niveles de la forma polisialilada de NCAM (mancha, llave) en comparación con el control (carril 1). El PSA se elimina de NCAM después de tratamiento con endoN (carril 3). (D) Cuantificación de la intensidad de la mancha de PSA expresada en unidades arbitrarias. (E) Análisis FACS de PSA el día 14 de diferenciación. El tratamiento de células con 20 unidades de endoN, 24 horas antes del final de la diferenciación, abolió el efecto de PST. (F) Microfotografías representativas que comparan células diferenciadas Nurr1 y Nurr1/PST con respecto a inmunofluorescencia de GFP y marcadores de DA. Las células clasificadas con respecto a GFP y que se volvieron a sembrar aún conservaban el fenotipo DA (después de la clasificación). Barras de escala: 100 pm.
La figura 31 muestra un análisis FACS ilustrativo de neuronas DA procedentes de ES. Aislamiento basado en citometría de flujo de células GFP+ y SSEA-1. Como controles negativos dobles y GFP negativos, se usaron células ES de ratón J1. Aproximadamente de 5 a 10 % de las células se clasificaron como positivas.
La figura 32 muestra que los injertos ilustrativos de Nurr1/PST fueron más eficaces para inducir la recuperación conductual en el modelo de ratón 60HDA. Las células Nurr1::GFP se diferenciaron y clasificaron el día 14 con respecto a la población GFP+/SSEA-1. Las células tratadas con endoN se cultivaron durante 12 horas antes de clasificarlas con 20 unidades de la enzima. Se injertaron 55.000 células en 1 ml de medio N2 con BDNF y AA. (A) Los animales se puntuaron para la rotación inducida por anfetaminas (rotaciones/min durante 20 min) durante 3 semanas antes del injerto, a continuación durante 7 semanas después. Las células Nurr1/PST mejoran significativamente el resultado en comparación con los controles de Nurr1 (ANOVA de 2 vías: p<0,01, con prueba posterior de Bonferroni: *p<0,05, **p<0,01, ***p<0,001; 6 animales/grupo). La eliminación de PSA por endoN elimina el efecto de PST (p = 0,26). (B) Había más células GFP+ en el injerto de PST en el punto final que en el control (p < 0,05, prueba de t). (C) Relación de inmunofluorescencia de PSA/GFP en el núcleo. **p<0,01. Prueba de t de Student (n=5/tipo de injerto). (D) Inmunofluorescencia de GFP, TH y PSA. Barra de escala: 200 pm. (E) Las células injertadas expresan marcadores de DA. Se muestran planos z individuales de micrografías confocales. Barra de escala: 20 pm. Los valores son medias /- ETM.
La figura 33 muestra un aumento de PSA ilustrativo que aumentó la inervación del cuerpo estriado del hospedador por neuronas DA procedentes de ES. La sobreexpresión de PSA-NCAM aumentó el crecimiento de la prolongación. (A) Microfotografías representativas de prolongaciones GFP/TH+, GFP/Girk2+ y GFP/sinapsina+ en controles. La tinción en muestras de Nurr1/PST fue similar. Barra de escala: 20 pm. (B) Proyecciones de apilamiento de enfoques representativas que muestran prolongaciones GFP+ que se extienden desde los injertos tanto Nurr1 como Nurr1/PST. Hay más prolongaciones GFP+ que se extienden desde el injerto Nurr1/PST (barra de escala: 50 pm). Los recuadros muestran prolongaciones GFP+/TH+ en las mismas secciones. Flecha: dirección de crecimiento. (C-E) Cuantificación de la intensidad de las prolongaciones GFP+ (C, E) y TH+ (D) a diferentes distancias del sitio del injerto normalizadas con respecto a la intensidad más cercana al sitio de inyección (las áreas se dividieron en cinco zonas separadas por ~100 pm. La normalización es relativa a la zona I; ANOVA de dos vías, p < 0,01 para datos tanto de g Fp como de TH, n=5/tipo de célula; los valores son medias /- ETM). Los injertos de Nurr1/PST hicieron crecer más neuritas en el cuerpo estriado del hospedador, que fue parcialmente suprimido (E) por el tratamiento con endoN. (F) La recuperación del animal se correlacionó con el grado de crecimiento de la prolongación. El gráfico muestra la correlación (análisis de regresión lineal) entre la intensidad de las neuritas GFP+ en la zona IV y la recuperación del animal para animales con injertos de Nurr1 y Nurr1/PST no tratados y tratados con endoN. Cada valor representa un animal (r 2 = 0,65, p<0,001, n=17).
La figura 34 muestra un uso ilustrativo de PST en métodos asociados con la lesión de la médula espinal y para la expresión en motoneuronas. En la médula espinal de control, las células de Schwann (CS) GFP injertadas están restringidas al sitio de lesión por el tejido cicatricial del hospedador (A), mientras que las CS modificadas con PST migran fácilmente distancias considerables (B). Esta ingeniería de PSA dio como resultado una mejora de la locomoción, subpuntuación de la BHE; línea superior en (C) mostrada frente a control de línea inferior) y destreza de las extremidades posteriores (prueba de ambulación en rejilla; controles de línea inferior en (D) frente a línea superior). E, H): diferenciación de ESC de ratón HB9::GFP en motoneuronas en las que la introducción de PST aumenta el brote de fibras y la migración celular (puntas de flecha) in vitro (E, F). El injerto de estas células PST en el nervio ciático da como resultado mejor inervación de la diana, como se muestra por los números de uniones neuromusculares (flechas) en el músculo EDL (G, H).
La figura 35 muestra un uso ilustrativo de la enzima PSTnm. (A) El PSA producido por PSTnm inhibe la adhesión de células de Schwann en suspensión a una monocapa de células de Schwann aún más eficazmente (línea roja-línea más baja) que el PSA producido por la expresión forzada de PST (línea verde-línea media). (B) La inmunotransferencia de PSA en motoneuronas HB9 procedentes de ESC muestra que las muestras de control tratadas con PSTnm tienen niveles indetectables de PSA. La incubación con sustrato de PSTnm CMP-ácido siálico produce una gran banda de PSA, que se elimina con tratamiento con endoN. (C, D) De manera similar a los efectos obtenidos con el gen de PST, la polisialilación de estas células por PSTnm y sustrato durante la diferenciación potencia el crecimiento de neuritas y la migración celular (puntas de flecha). (E) Inmunotinción de PSA de neuronas Da procedentes de hESC del día 30. (F) Esta tinción aumenta significativamente después del tratamiento con PSTnm y sustrato. (G) La inyección in vivo de PSTnm solo no tiene efecto, mientras que su administración conjunta con el sustrato (H) produce grandes cantidades de expresión de PSA en el cuerpo estriado de ratón.
Descripción de la invención
La presente invención se refiere al campo de la biología de células madre, en particular la diferenciación específica de linaje de células madre pluripotentes o multipotentes, que pueden incluir, pero sin limitación, células madre embrionarias humanas (hESC) además de células madre pluripotentes inducidas por seres humanos no embrionarias (hiPSC), células madre somáticas, células madre de pacientes con una enfermedad o cualquier otra célula con capacidad de diferenciación específica de linaje. Se describen específicamente métodos para dirigir la diferenciación específica de linaje de hESC y/o hiPSC en células progenitoras del mesencéfalo de la placa del suelo y después adicionalmente en grandes poblaciones de neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas al mesencéfalo usando condiciones de cultivo novedosas. Las neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas al mesencéfalo producidas usando los métodos de la presente invención se contemplan adicionalmente para diversos usos incluyendo, pero sin limitación, uso en ensayos de descubrimiento de fármacos in vitro, investigación neurológica y como un producto terapéutico para neutralizar la enfermedad de, o el daño a, una falta de neuronas de dopamina en un paciente. Además, se proporcionan composiciones y métodos para diferenciar las neuronas de dopamina (DA) FOXA2+LMX1A+TH+ destinadas al mesencéfalo de células madre pluripotentes humanas para su uso en el modelado de enfermedades, en particular enfermedad de Parkinson.
Las presentes invenciones se refieren a características de la enfermedad de Parkinson (EP), incluyendo la degradación selectiva de las neuronas de dopamina del mesencéfalo (mDA) en el cerebro de los pacientes. Debido a que los síntomas de EP se deben principalmente a la pérdida selectiva de neuronas DA en la sustancia negra del mesencéfalo ventral, la EP se considera una de las enfermedades más adecuadas para estrategias terapéuticas de reemplazo celular para el tratamiento. Por tanto, se realizaron muchos intentos para trasplantar células en el cerebro de los pacientes con el fin de reemplazar la pérdida de función de las neuronas de dopamina (mDA) del mesencéfalo. Sin embargo, estos experimentos no tuvieron éxito y actualmente los tratamientos sintomáticos usados en pacientes tienen una amplia variabilidad de éxito. Por lo tanto, se necesitan nuevos tratamientos para pacientes con EP con el fin de retrasar la pérdida de función neuronal.
Las células madre pluripotentes humanas (hPSC) son una fuente de células para aplicaciones en medicina regenerativa. Diferenciación dirigida de hPSC en células especializadas tales como motoneuronas espinales (Li, et al. Nat. Biotechnol. 23, 215-221 (2005), o neuronas de tipo dopamina (DA) del mesencéfalo resultantes de la diferenciación por métodos distintos de los métodos de la presente invención. Los inventores descubrieron, como se describe en la presente memoria, que las neuronas de dopamina (DA) anteriores (es decir, en Perrier, et al Proc Natl Acad Sci USA 101, 12543-8 (2004), a las que se hace referencia en la presente memoria como neuronas de tipo dopamina (DA), no son neuronas auténticas de dopamina (DA) del mesencéfalo de las presentes invenciones (véase figura 3, 10, 13 y 16). Por lo tanto, los inventores marcaron las neuronas productoras de dopamina procedentes de la placa del suelo producidas por métodos descritos en la presente memoria, es decir,
las neuronas productoras de dopamina de las presentes invenciones son "auténticas" porque, a diferencia de las neuronas productoras de dopamina producidas por métodos publicados, cuando las neuronas productoras de dopamina "auténticas" de las presentes invenciones se trasplantan en roedores y primates, neutralizan afecciones neurológicas de tipo Parkinson con menos interferencia de la hiperproliferación neural y la formación de teratoma. Además, a diferencia de los métodos anteriores para producir neuronas productoras de dopamina, las neuronas productoras de dopamina "auténticas" de las presentes invenciones se producen en porcentajes mayores a partir de poblaciones iniciales y conservan la capacidad de injerto durante varios meses en cultivo.
Por tanto, se descubrieron métodos para producir neuronas DA del mesencéfalo auténticas usando los métodos de diferenciación descritos en la presente memoria. Sin embargo, el uso eficaz de hPSC para terapia celular ha quedado muy rezagado con respecto a avances de cultivo celular. Aunque las neuronas DA procedentes de PSC de ratón han mostrado eficacia en modelos de enfermedad de Parkinson (EP) (Tabar, et al. Nature Med. 14, 379-381 (2008); Wemig, et al. Proc. Natl. Acad. Sci. U.S. A 105, 5856-5861 (2008), Las neuronas DA procedentes de PSC humanas presentan en general mal rendimiento in vivo (Lindvall y Kokaia, J. Clin. Invest 120, 29-40 (2010). Además de no compensar la pérdida endógena de la función neuronal, existen graves preocupaciones de seguridad cuando las neuronas procedentes de hPSC se usan para trasplante y están relacionadas con su potencial para la formación de teratomas o hiperproliferación neural (Roy, et al. Nature Med. 12, 1259-1268 (2006); Elkabetz, et al. Genes Dev. 22, 152-165 (2008).
Otra posible fuente de células para trasplante son neuronas DA procedentes de ESC humanas. Los intentos previos usando estas células como población celular inicial para producir células diferenciadas que parecían ser neuronas del mesencéfalo de tipo DA procedentes de células madre embrionarias humanas (hESC) que se trasplantaron en modelos de EP de roedores dieron como resultado mala supervivencia in vivo de los trasplantes después del trasplante. Se consideró que este fracaso probablemente se deba a una diferenciación incompleta de neuronas DA del mesencéfalo in vitro dando como resultado células que parecían ser neuronas DA del mesencéfalo pero que no eran capaces de injerto para reemplazar la función neuronal perdida. De hecho, los inventores muestran en la presente memoria que las neuronas de tipo DA producidas previamente en sus laboratorios y descritas en publicaciones no eran del mismo tipo celular ni tenían funciones o capacidades de injerto similares a las células neuronales DA del mesencéfalo de la placa del suelo de la presente invención, véase, figs. 16 y 17 para ejemplos. Por lo tanto, los inventores también descubrieron que para que las células experimenten diferenciación dirigida en el laboratorio para
producir poblaciones celulares que contengan grandes cantidades de neuronas que actúen de manera adecuada, era necesario que las células pasaran por las etapas específicas del desarrollo para convertirse en una población celular de reemplazo adecuada para terapias de reemplazo basadas en células. Los inventores también descubrieron que, al menos para obtener las neuronas DA injertables de las presentes invenciones, determinadas etapas del desarrollo deben estar presentes, tales como los intermedios FOX2A/LIM1A+ del día 11. Si dichas etapas del desarrollo no están presentes, los inventores descubrieron que las neuronas de tipo DA resultantes no tienen las mismas capacidades funcionales que las neuronas DA del mesencéfalo de las presentes invenciones procedentes de intermedios FOX2A/LIM1A+ del día 11.
En contraste con las observaciones anteriores, se describen en la presente memoria técnicas de cultivo novedosas relacionadas con estrategias basadas en la inducción de células de la placa del suelo para la obtención de neuronas DA humanas que se injertan eficazmente in vivo. Por tanto, se consideró que los fracasos anteriores en la obtención de poblaciones celulares que comprenden neuronas DA principalmente comprometidas de las presentes invenciones (es decir, neuronas DA FOXA2+ y LMX1A+ capaces de injertarse eficazmente) eran la razón por la que el injerto de neuronas de tipo DA fracasó, es decir, debido a la especificación incompleta de las células de tipo DA. La hipótesis anterior era que el fracaso del trasplante se debía a la vulnerabilidad específica de las células, es decir, las neuronas cultivadas de tipo DA no pudieron sobrevivir a la tensión del trasplante. Como se describe en la presente memoria, los precursores de la placa del suelo FOXA2+/LMX1A+ del mesencéfalo se obtuvieron de hPSC en 11 días después de la exposición a activadores de molécula pequeña de sonic hedgehog (SHH) y señalización canónica de WNT. Estas células del día 11, doblemente positivas para FOXA2+ y LMX1A+, se ponen en contacto con moléculas pequeñas adicionales para inducir diferenciación adicional en neuronas DA del mesencéfalo injertables, positivas para TH+ FOXA2+ y LMX1A+, antes del día 25. Estas neuronas DA maduras del mesencéfalo de la placa del suelo pueden mantenerse in vitro durante varios meses. El perfil molecular in vitro extenso, datos bioquímicos y electrofisiológicos definieron la progresión del desarrollo y confirmaron la identidad de las neuronas DA del mesencéfalo procedentes de hPSC. Se demostró la supervivencia y función in vivo en modelos animales de EP en tres especies hospedadoras. El injerto a largo plazo en ratones y ratas con lesiones de 6-OHDA demuestra supervivencia robusta de las neuronas DA del mesencéfalo, restauración completa del comportamiento de rotación inducido por anfetaminas y mejoras en las pruebas de uso de extremidades anteriores y acinesia. Finalmente, se demuestra la capacidad de cambio de escala mediante el trasplante en monos parkinsonianos. La excelente supervivencia, función y falta de hiperproliferación neural de neuronas DA en los tres modelos animales probados resultan prometedoras para el desarrollo de terapias basadas en células en la EP basadas en las composiciones y los métodos de las presentes invenciones.
Por lo tanto, los inventores contemplan el uso principal de sus descubrimientos como la capacidad de producir un suministro ilimitado de neuronas DA del mesencéfalo procedentes de la placa del suelo completamente funcionales adecuadas para aplicaciones terapéuticas preclínicas y clínicas. De manera específica, los inventores descubrieron un nuevo protocolo para la diferenciación eficaz de neuronas mDA de al menos poblaciones celulares pluripotentes aisladas de roedores y seres humanos (células madre embrionarias humanas (hESC) y células madre pluripotentes inducidas por seres humanos (hiPSC)). Esos estudios incluyeron líneas celulares iPS mutantes PINK1 derivadas de un paciente humano que padecía una forma genética de la enfermedad de Parkinson Seibler, et al., The Journal of Neuroscience, 2011, 31(16): 5970-5976, incorporado en la presente memoria por referencia. Las poblaciones de células madre humanas (hESC o hiPSC) se diferenciaron en un fenotipo de mesencéfalo, que después de entrar en contacto con moléculas de maduración neuronal dio lugar a más neuronas DA injertables auténticas. Este protocolo se usó para demostrar altos rendimientos de la descendencia de hESC el día 11 de diferenciación dirigida en un fenotipo neuronal DA del mesencéfalo (mDA) que incluía la expresión de factores de transcripción clave, p. ej., TH, FoxA2 y LMX1A que tras una diferenciación adicional produjeron proteínas clave adicionales, p. ej., TH. El trasplante de estas neuronas mDA procedentes de hESC en hospedadores inmunodeprimidos de roedores y primates, a diferencia de neuronas DA obtenidas in vitro anteriores, mostró buena supervivencia in vivo de las células injertadas con restauración funcional de déficits conductuales.
Las ventajas de usar los métodos de la presente invención para producir células neuronales DA con respecto a otros métodos son, en parte, evidentes a partir de la siguiente información. El uso de células madre somáticas y células madre neurales en otros métodos con el objetivo de generar neuronas DA del mesencéfalo auténticas que se injertan eficazmente in vivo no ha tenido éxito (para una revisión, véase Kriks y Studer, Protocols for generating ES cell-derived dopamine neurons in Development and engineering of dopamine neurons (eds. Pasterkamp, et al.) (Landes Biosciences, 2008, incorporado en la presente memoria por referencia). Las células madre pluripotentes, tales como células ES, se usando después como fuentes para generar células injertables. Los primeros estudios en la década de 1990 que usaban células ES de ratón demostraron la viabilidad de obtener linajes específicos de células pluripotentes in vitro, incluyendo neuronas (Okabe, et al., Mech. Dev. 59: 89-102 (1996); Bain, et al., Dev. Biol. 168v342-357 (1995). De hecho, se generaron neuronas DA del mesencéfalo usando una estrategia de diferenciación dirigida (Lee, et al., Nat. Biotechnol. 18v675-679 (2000), basada en información del desarrollo de los primeros estudios de explantes (Ye, et al., Cell 93: 755-766 (1998), incorporado en la presente memoria por referencia). Se usaron otras estrategias de diferenciación dirigida para producir otros tipos de neuronas, como las motoneuronas somáticas (Wichterle, et al., Cell 110, 385-397 (2002). Sin embargo, estos esfuerzos no dieron como resultado poblaciones celulares que contuvieran altos porcentajes de neuronas DA del mesencéfalo o células capaces de restaurar la función neuronal in vivo. De hecho, la población resultante contenía una mezcla de tipos celulares además de neuronas DA del mesencéfalo. Incluso los inventores desarrollaron otros métodos para producir neuronas DA del mesencéfalo, en parte, véase
posteriormente, sin embargo, estas poblaciones celulares también eran mezclas de tipos celulares y no pudieron restaurar la función neuronal. En particular, las células ES humanas se diferenciaron en células neuroepiteliales tempranas, denominadas rosetas neurales, seguido de la inducción de células en etapa de roseta a células que expresan precursor de neurona DA del mesencéfalo y marcadores diferenciados. Esas células presentaron posteriormente características neuronales funcionales por electrofisiología, liberación de DA in vitro y la formación de contactos sinápticos positivos para TH-inmunooro (Perrier, et al. De la portada: Derivation of midbrain dopamine neurons from human embryonic stem cells. Proc Natl Acad Sci U S A 101, 12543-8 (2004). Sin embargo, a pesar de estos prometedores datos in vitro, el trasplante de las células en hospedadores murinos con lesiones de 60HDA dio como resultado un número muy pequeño de neuronas dopaminérgicas supervivientes. Este fue un resultado negativo sorprendente dadas las pruebas sólidas de funcionalidad in vivo de las neuronas DA procedentes de ES de ratón (Barberi, et al., Nat. Biotechnol. 21: 1200-1207 (2003); Tabar, et al. Nature Med. 14: 379-381 (2008); Bjorklund, et al. Proc. Natl. Acad. Sci. U. S A. 99: 2344-2349 (2002); Kim et al. Nature 418: 50-56 (2002), características funcionales in vitro robustas de neuronas DA procedentes de ES humanas (Perrier, et al., De la portada: Derivation of midbrain dopamine neurons from human embryonic stem cells. Proc Natl Acad Sci USA 101: 12543-8 (2004) y pruebas claras de que las neuronas DA fetales humanas pueden sobrevivir como xenoinjertos del cuerpo estriado (Brundin, et al. Exp. Brain Res. 70: 192-208 (1988); Bjorklund, et al., Neuronal replacement by intracerebral neural implants in animal models of neurodegenerative disease. Raven. Press., Nueva York. 455-492 (1988).
Durante casi 8 años después de los intentos fallidos iniciales de injertar neuronas DA procedentes de células ES humanas, hubo muy poco progreso en el campo. Se observaron algunas mejoras limitadas usando fuentes de células madre pluripotentes de primates (Sanchez-Pemaute, et al. Long-term survival of dopamine neurons derived from parthenogenetic primate embryonic stem cells (Cyno1) in rat and primate striatum. Stem Cells 23: 914-922 (2005), células pretratadas con FGF20 o Wnt5A o células ES humanas diferenciadas en presencia de factores secretados por astrocitos inmortalizados del mesencéfalo (Roy, et al., Nature Med. 12: 1259-1268 (2006). Sin embargo, ninguna de las estrategias anteriores tuvo éxito en la producción de una población enriquecida de neuronas DA de las presentes invenciones para su uso en procedimientos de injerto para restaurar la función neuronal in vivo.
I. Métodos de cultivo celular para inducir células precursoras neuronales (linaje): La puesta en contacto de células madre pluripotentes humanas con SB431542 y LDN-193189 dio como resultado células de linaje neural diferenciadas.
El siguiente ejemplo describe métodos ilustrativos para proporcionar células de un linaje neural para su uso durante el desarrollo de las presentes invenciones.
La inhibición doble de SMAD se usó anteriormente como un método rápido y muy eficaz para inducir un tipo de células de linaje neural a partir de hPSC (Chambers, et al., Nat Biotechnol 27, (2009). Estas células de linaje neural inducidas por moléculas, incluyendo Noggin, tenían una ruta predeterminada que permitió el desarrollo en células del sistema nervioso central, es decir, destino de células neurales. Los estudios de seguimiento informaron del uso de una molécula pequeña de dorsomorfina (DM) en lugar de Noggin, al menos en parte, dieron como resultado células diferenciadas similares, pero no iguales, con diferencias importantes en la consistencia de los cultivos (Kim, et al., Robust enhancement of neural differentiation from human Es and iPS cells regardless of their innate difference in differentiation propensity. Stem Cell Rev 6, 270-281, (2010); Zhou, et al., High-Efficiency Induction of Neural Conversion in hESCs and hiPSCs with a Single Chemical Inhibitor of TGF-beta Superfamily Receptors. Stem Cells, 504, (2010).
Los inventores observaron que las células generadas usando Noggin, a pesar de mostrar la misma etapa del desarrollo que las células tratadas con LDN, expresan la gran mayoría de los mismos marcadores y tienen capacidad para tener un potencial de desarrollo similar para producir diversos linajes neurales, pero también mostraron diferencias, tales como ser más anteriores en un eje anterior-posterior (es decir, más prosencéfalo, más células expresan FOXG1 y similares) en comparación con las células neurales inducidas usando LDN. Por tanto, aunque se utilizó LDN en lugar de Noggin para inhibir BMP entre otras rutas de señalización, Noggin y LDN pueden tener otros tipos de actividades que son diferentes a la inhibición de BMP.
En parte debido al alto coste del uso de Noggin, los autores contemplaron que el uso de un inhibidor de BMP podría sustituir Noggin en la diferenciación de células con destino de células neurales. Por lo tanto, se usó un inhibidor de BMP de molécula pequeña, LDN-193189, (Yu, et al., Nat Med 14, 1363-1369, (2008) y se encontró durante el desarrollo de las presentes invenciones para reemplazar Noggin, en combinación con SB431542, para generar neuroectodermo primitivo de hPSC, células que tienen un destino de células neurales, es decir, células del SNC (figura 2A). Este tratamiento combinado se denominó LSB para la combinación de estos dos inhibidores LDN-193189 y SB431542.
En general, la diferenciación celular se inició mediante el tratamiento de hES o hiPS de monocapa de alta confluencia con doble inhibición de la señalización de SMAD. Una realización preferida utiliza un porcentaje de confluencia de 50 %-100 %, con una realización más preferida de 70 %-80 % de confluencia. Resultará obvio para un experto en la técnica que la densidad de siembra en placas inicial necesaria para lograr una confluencia preferida de la presente invención dependerá del tipo celular, el tamaño, la eficacia de la siembra en placas, la supervivencia, la adhesión y otros parámetros que pueden determinarse empíricamente sin experimentación excesiva por parte del experto en la
técnica. La inhibición doble de SMAD se puede lograr con diversos compuestos incluyendo Noggin, SB431542, LDN-193189, dorsomorfina u otras moléculas que bloquean TGFp, BMP y la señalización de Activina/Nodal. Una realización preferida utiliza la composición que comprende SB431542 y LDN-193189 (colectivamente, LSB) a una concentración de 0,1 pM-250 pM, o más preferiblemente 1-25 pM, o lo más preferiblemente 10 pM de SB431542 y 10-5000 nM, o lo más preferiblemente 100-500 nM de LDN-193189.
II. Obtención de neuronas DA de hESC mediante intermedios de células de la roseta y resultados de estudios de trasplante que usaron estas neuronas de tipo DA.
Los inventores usaron anteriormente otros métodos de diferenciación dirigida que dieron como resultado poblaciones celulares que contenían neuronas de tipo DA. Estas neuronas de tipo DA se usaron en estudios de trasplante que dieron como resultado preocupaciones sobre el uso posterior de estas células para aplicaciones terapéuticas. Por ejemplo, los procedimientos descritos en Perrier et al., 2004 y Fasano et al., 2010, incluyendo inducción neural de MS5, dieron como resultado la formación de células de la roseta y se usaron para producir precursores del día 11, véase figs. 2, 16 y 17 para los ejemplos, que después se usaron para obtener neuronas de tipo DA. Estas neuronas resultaron de un bajo porcentaje de las células precursoras en las poblaciones celulares del día 11 resultantes. Los estudios de trasplante que usaron estas neuronas mostraron mala viabilidad posterior al trasplante y pérdida del fenotipo neuronal de tipo DA, además de observaciones de desarrollo posterior al trasplante de tipos neurales inadecuados junto con pérdida de control del crecimiento, que condujo al desarrollo de teratomas. Véase figs. 16 y 17.
De manera específica, en P0 se pusieron en contacto hESC con moléculas para comenzar la inducción neural de células Oct4+ en células de la roseta usando células alimentadoras MS5 (Perrier et al., 2004). En la etapa P1, las células de la roseta se expandieron al poner en contacto las células con moléculas adicionales para diferenciar células en células en la etapa P2 con patrones de expresión específicos que incluyen células progenitoras DA Pax2+/En1+ y se diferenciaron adicionalmente en neuronas DA TH+/En1+. Estas células se usaron para injerto en ratas con lesión de 6OHDA, inmunodeprimidas con ciclosporina A. Esos estudios de trasplante mostraron mala viabilidad in vivo, pérdida del fenotipo TH+, preocupaciones acerca de crecimiento adicional en células no deseadas, posiblemente letales, es decir, teratomas, y crecimiento de células en tipos neuronales inadecuados que provocarían problemas médicos adicionales para el paciente.
Hubo cantidades muy pequeñas de neuronas TH+ supervivientes a los 4,5 meses después del trasplante (<50 células TH+/animal) en injertos de precursores de neuronas DA procedentes de la roseta fig 16A. Sin embargo, en contraste con las células TH+, las células marcadas con GFP (GFP fue impulsada por un promotor ubicuo) sobrevivieron bastante bien después del trasplante. Esto sugiere que la mayoría de las células supervivientes después del trasplante eran células neurales de identidad neuronal no Da (16B). Pocas células procedentes del injerto (hNA+ (verde) coexpresan TH (rojo) de nuevo, lo que sugiere que la mayoría de las células humanas injertadas adoptan un fenotipo distinto de neurona DA fig 16C.
Los paneles 16 D-E muestran que D-E, a pesar de la supervivencia muy baja in vivo hubo alguna mejora (baja y muy variable) en algunos ensayos conductuales tales como rotaciones inducidas por anfetaminas (D), prueba de cilindro y rotaciones espontáneas (E). Se llevó a cabo inducción neural sin alimentador como se ha descrito anteriormente (Chambers et al., 2009) pero se modificó adicionalmente para producir células de la placa del suelo (Fasano et al., 2010, incorporado en la presente memoria por referencia).
En el método de inhibición doble de SMAD modificado para diferenciar células pluripotentes en células de la placa del suelo, los inventores descubrieron previamente que para el día 11 eran necesarias altas concentraciones de SHH para inducción de PS. Por ejemplo, en algunas realizaciones, se añadió Sonic C25II a 200 ng/ml. En algunos experimentos, se añadieron DKK-1 (R&D; 100 ng/ml) FGF8 (R&D; 50 ng/ml), Wnt-1 (Peprotech; 50 ng/ml) y ácido retinoico (R&D; 1 mM), véase figura 17. Sin embargo, ninguna de las poblaciones celulares resultantes el día 11 usando métodos anteriores, contenía el alto porcentaje de células progenitoras de la placa del suelo del mesencéfalo FOXA2+/LMX1A+ usando métodos de las presentes invenciones.
III. Compuestos para su uso en diferenciación dirigida: La exploración de moléculas pequeñas usando células de linaje neuronal de las presentes invenciones dio como resultado compuestos que diferenciaron las células en células neuronales FOX2A+ y LIMX1A+ para el día 11 de cultivo.
El siguiente ejemplo describe el uso de células ilustrativas de la sección I para explorar compuestos candidatos de moléculas pequeñas y determinar si su uso daría como resultado diferenciación dirigida de una población celular que contiene un alto porcentaje de neuronas de la placa del suelo del mesencéfalo para el día 11 después del contacto inicial con los inhibidores dobles de SMAD. Los resultados de esta exploración mostraron inicialmente que una molécula activadora de SHH junto con la activación de FGF8 y Wnt condujo a la obtención eficaz de células de la placa del suelo del mesencéfalo positivo para FOXA2+/LMX1A+ de hESC para el día 11 de diferenciación. Los inventores muestran resultados en la presente memoria de experimentos ilustrativos que definieron qué moléculas eran necesarias y el momento óptimo de contacto para obtener la población de células positivas para FOXA2+/LMX1A+ deseada el día 11.
Estudios genéticos recientes de ratones demostraron un papel importante para el factor de transcripción FOXA2 en el
desarrollo y la supervivencia de neuronas DA del mesencéfalo. Una característica única del mesencéfalo en desarrollo es la coexpresión del marcador de la placa del suelo FOXA2 y el marcador de placa del techo LMX1A. Normalmente, las células de la placa del suelo y la placa del techo están ubicadas en posiciones distintas en el SNC (ventral frente a dorsal) con requisitos de distribución diametralmente opuestos para su inducción. Recientemente se ha descrito la obtención de precursores de la placa del suelo específicos de región a partir de hESC usando un protocolo de inhibición doble de SMAd modificado. La señalización de Wnt canónica fue importante tanto para la función de la placa del techo como para el desarrollo de neuronas DA del mesencéfalo.
A. CHIR99021 (CHIR) indujo un alto rendimiento de destino de precursor de neuronas DA del mesencéfalo para el día 11 de cultivo. Como se describe en la presente memoria, la exposición a CHIR99021 (CHIR), un potente inhibidor de GSK3p que se sabe que activa fuertemente la señalización de WNT, LMX1A inducido en precursores de la placa del suelo FOXA2+ (figura 1a). CHIR fue más potente que Wnt3A o Wnt1 recombinante en la inducción de la expresión de LMX1A. La eficacia de inducción de LMX1A dependía del momento de exposición a CHIR con un efecto máximo en los días 3-11 (figura 5). CHIR indujo la coexpresión de FOXA2/LMX1A, mientras que otros factores tales como FGF8 solo tuvieron efectos marginales (figura 6). La inducción de coexpresión de FOXA2/LMX1A requirió fuerte activación de la señalización de SHH usando purmorfamina, un agonista de molécula pequeña, solo o en combinación con SHH recombinante (figura 7).
El tratamiento con agonistas de SHH (purmorfamina SHH) y FGF8 (S/F8) en ausencia de CHIR99021 mostró expresión significativamente menor de FOXA2 para el día 11 y falta completa de expresión de LMX1A (figura 1a, b). La inhibición doble de SMAD (exposición a l Dn -193189 SB431542 = "LSB") no produjo células que expresan FOXA2, pero produjo un subconjunto de células LMX1A+ (figura 1a, b). El marcador anterior OTX2 fue inducido de manera robusta en cultivos tratados con LSB y LSB/S/F8/CHIR, pero no en condiciones de LSB/S/F8 (figura 1a, c). Se realizaron comparaciones sistemáticas de las tres condiciones de cultivo (figura 1d) usando perfiles de expresión de genes temporales globales. La agrupación jerárquica de genes expresados diferencialmente segregó las tres condiciones de tratamiento para el día 11 de diferenciación (figura 8a). FOXA1, FOXA2 y varias otras dianas de SHH corriente abajo, incluyendo PTCH1, estaban entre los transcritos con regulación más diferencial en conjuntos de tratamiento de LSB/S/F8/CHIR frente a LSB (figura 1e). La expresión de LMX1A, NGN2 y DDC indicó el establecimiento del destino precursor de neuronas DA del mesencéfalo ya para el día 11 (figura 1 e, f). Por el contrario, los cultivos de LSB se enriquecieron con respecto a marcadores precursores del prosencéfalo dorsal tales como HES5, PAX6, LHX2 y EMX2. La comparación directa del tratamiento con LSB/S/f 8/CHIR frente a LSB/S/F8 (figura 1f) confirmó el enriquecimiento selectivo con respecto a marcadores precursores de DA del mesencéfalo en el grupo LSB/S/F8/CHIR y sugirió la identidad precursora hipotalámica en cultivos tratados con LSB/S/F8 basándose en la expresión diferencial de RAX1, SIX3 y SlX6 (véase también expresión de POMC, OTP en la figura 2d a continuación). La lista completa de transcritos expresados diferencialmente tablas 1, 2 y análisis de ontología génica figura 8b (DAVID; http://david.abcc.ncifcrf.gov) confirmó el enriquecimiento de la señalización de WNT canónica tras el tratamiento con CHIR. Los datos sin procesar aún no están disponibles en GEO worldwideweb.ncbi.nlm.nih.gov/geo/ n.° de referencia: [por determinar]). El análisis temporal comparativo de la expresión génica para marcadores de precursores de DA del mesencéfalo (figura 1 g) frente a marcadores de destinos distintos de neuronas DA anteriores y ventrales (figura 1h) dividió las tres condiciones de inducción en: i) LSB: prosencéfalo dorsal; ii) LSB/S/F8: ventral/hipotalámico y iii) LSB/S/F8/CHIR: identidad precursora de DA del mesencéfalo.
Las células resultantes del protocolo obtenidas durante el desarrollo de la presente invención se compararon con células obtenidas por métodos anteriores. Véase, figs. 2 y 18 para una comparación visual ilustrativa con las células tratadas con SHH/FGF8+CHIR de las presentes invenciones. La figura 19A muestra ejemplos de células con doble marcaje FOXA2/Tuj1 después del tratamiento con LSB/S/F8/CHIR (paneles superiores) y marcaje conjunto de FOXA2 con t H, Nurr1 y LMX1A (paneles inferiores). Esas combinaciones de marcadores son diagnósticos para precursores de neuronas DA del mesencéfalo en etapa temprana. La figura 19 B muestra datos de expresión génica (para comparación con la figura 2E) para marcadores precursores de neuronas de dopamina clave.
En general, se describen materiales y métodos usados en la presente memoria: Las líneas de referencia de ESC humana (H9, H1) e iPSC (2C6 y SeV6) se sometieron a un protocolo de inducción de la placa del suelo basada en inhibición doble de SMAD modificada (Chambers, et al. Nat. Biotechnol. 27: 275-280 (2009) (Fasano, et al., Cell Stem Cell 6: 336-347 (2010). La exposición a SHH C25II, purmorfamina, FGF8 y CHIR99021 se optimizó para la placa del suelo del mesencéfalo y el rendimiento de nuevas poblaciones de neuronas DA (véase figura 1d). Después de la inducción de la placa del suelo, se llevó a cabo maduración adicional (días 11-25 o más de 25 días en cultivo hasta al menos 100 días en cultivo) en medio de diferenciación basado en Neurobasal/B27 en presencia de factores de supervivencia y maduración de neuronas DA (Perrier, et al. Proc Natl Acad Sci USA 101: 12543-8 (2004), incorporado en la presente memoria por referencia) tales como AA, BDNF, GDNF, TGFp3 y dbAMPc (véase métodos completos para más detalles). Las poblaciones de neuronas DA resultantes se sometieron a caracterización fenotípica exhaustiva mediante inmunocitoquímica, qRT-PCR, perfil de expresión génica global, análisis por HPLC para la detección de dopamina y registros electrofisiológicos in vitro. Se realizaron estudios in vivo en roedores hemiparkinsonianos (ratones o ratas con inyección de la toxina 60HDA en un lado del cerebro del animal. Los estudios se llevaron a cabo en ratones adultos NOD-SCID IL2Rgc (Jackson labs) y ratas adultas Sprague Dawley Taconic Farms, que recibieron lesiones de 6-hidroxidopamina mediante inyecciones estereotácticas de la toxina como se ha descrito anteriormente, así como dos monos rhesus adultos que fueron tratados con inyecciones carotídeas unilaterales de MPTP.
Se inyectaron neuronas DA de manera estereotáctica en el cuerpo estriado de los animales (150 x 103 células en ratones, 250 x 103 células en ratas) y un total de 7,5 x 106 células (distribuidas en 6 tramos; 3 en cada lado del cerebro) en monos. Se realizaron ensayos conductuales a intervalos mensuales después del injerto, incluyendo análisis rotacional mediado por anfetaminas, así como una prueba de acinesia focal ("prueba de marcha") y uso de las extremidades (prueba de cilindro). Se sacrificaron las ratas y los ratones a las 18-20 semanas y los primates al cabo de 1 mes después del injerto. Se realizó caracterización de los injertos mediante análisis estereológicos del número de células y los volúmenes de injerto, así como una caracterización fenotípica exhaustiva mediante inmunohistoquímica.
Cultivo de células ES humanas indiferenciadas Referencia Líneas de hESC H9 (WA-09, XX, pases 27-55 desde la fecha 10/2009), Referencia (WA-01, XY, pases 30-40 desde la fecha 6/2010) y líneas celulares de iPS 2C6 (Kim, et al. Cell Stem Cell 8: 695-706 (2011), (XY, pases 20-30) y SeV6 (XY, pases 20-30; obtenido de fibroblastos embrionarios MRC-5 usando un sistema de vector Sendai de 4 factores no integrante (Ban, et al. Proc. Natl. Acad. Sci. U.S.A. (2011) 108 (34): 14234-14239: 10.1073/pnas.l103509108, se mantuvieron en fibroblastos embrionarios de ratón a concentraciones de siembra estimadas que variaban de 0,5x103 por cm2 a 100x103 por cm2 basado en células ES humanas que tienden a formar agrupamientos celulares. (MEF, Global Stem, Rockville, MD) en un reemplazo de suero knockout óptimo al 20 % (KSR, Invitrogen, Carlsbad, California) - que contiene medio de células ES humanas (como se ha descrito anteriormente (Kim, et al. Cell Stem Cell 8: 695-706 (2011). El uso de reemplazo de suero knockout puede variar de 0 % a 40 %.
Inducción Neural. Para la inducción de neuronas de dopamina del mesencéfalo basada en la placa del suelo, se usó una versión modificada del protocolo de inhibición doble de SMAD (Chambers, et al. Nat. Biotechnol. 27: 275-280 (2009), incorporado en la presente memoria por referencia) e inducción de la placa del suelo (Fasano, et al Cell Stem Cell 6: 336-347 (2010) basándose en la exposición sincronizada a LDN-193189 (100 nM (intervalo de concentración de 0,5-50 pM, Stemgent, Cambridge, Massachusetts), SB431542 (10 pM (intervalo de concentración de 0,5-50 pM, Tocris, Ellisville, MI), SHH C25II (100 ng/ml (intervalo de concentración de 10-2000 ng/ml, R&D, Mineápolis, MN), purmorfamina (2 pM (intervalo de concentración de 10-500 ng/ml, Stemgent), FGF8 (100 ng/ml (intervalo de concentración de 10-500 ng/ml, R&D) y CHIR99021 (CHIR; 3 pM (intervalo de concentración de 0,1-10 pM, Stemgent).
Para el protocolo de inducción de la placa del suelo, el tratamiento de "SHH" se refiere a la exposición, es decir, contacto, de células a una combinación de SHH C25II 100 ng/ml purmorfamina (2 pM). Las células se sembraron en placas (35-40 x 103 células/cm2) y se cultivaron durante 11 días en matrigel o geltrex (usado tal como se adquirió) (BD, Franklin Lakes, Nueva Jersey) en medio de reemplazo de suero Knockout (KSR) que contiene DMEM, suero de reemplazo knockout al 15 %, L-glutamina 2 mM y 10 pM (intervalo de concentración de 1-25 pM de p-mercaptoetanol. El medio KSR se cambió gradualmente a medio N2 a partir del día 5 de diferenciación, mezclando en proporciones de 75 % (KSR):25 % (N2) el día 5-6, 50 % (KSR):50 % (N2) día 7-8 y 25 % (KSR):75 % (N2) el día 9-10, como se ha descrito anteriormente (Chambers, et al. Nat. Biotechnol. 27: 275-280 (2009). El día 11, los medios se cambiaron a medio neurobasal/medio B27 (dilución 1:50)/medio que contiene l-Glut (intervalos eficaces 0,2-2 mM)) (NB/B27; Invitrogen) complementado con CHIR (hasta el día 13) y con BDNF (factor neurotrófico derivado del cerebro, 20 ng/ml que varía de 5 a 100; R&D), ácido ascórbico (AA; 0,2 mM (intervalo de concentración de 0,01-1 mM), Sigma, San Luis, MI), GDNF (factor neurotrófico derivado de la línea celular glial, 20 ng/ml (intervalo de concentración de 1-200 ng/ml); R&D), TGFp3 (factor de crecimiento transformante de tipo p3, 1 ng/ml (intervalo de concentración de 0,1-25 ng/ml); R&D), dibutiril AMPc (0,5 mM (intervalo de concentración de 0,05-2 mM); Sigma) y DAPT (10 nM (intervalo de concentración de 0,5-50 nM); Tocris,) durante 9 días. El día 20, las células se disociaron usando Accutase® (Innovative Cell Technology, San Diego, California) y se volvieron a sembrar en condiciones de alta densidad celular (por ejemplo de 300-400k células/cm2) en placas previamente recubiertas con poliornitina (PO); 15 pg/ml (intervalo de concentración de 1-50 pg/ml)/Laminina (1 pg/ml) (intervalo de concentración de 0,1-10 pg/ml)/Fibronectina (2 pg/ml (intervalo de concentración de 0,1-20 pg/ml) en medio de diferenciación (NB/B27 Bd NF, AA, GDNF, dbAMPc (intervalo de concentración como se describe en la presente memoria), TGFp3 y DAPT (que varían en concentración como se describe en la presente memoria) hasta la etapa de maduración deseada para un experimento dado.
Para la inducción de neuronas DA basada en la roseta, se siguieron los protocolos descritos anteriormente en parte (Perrier, et al. Proc Natl Acad Sci U S A 101: 12543-8 (2004), con al menos una excepción donde se usó inhibición doble de SMAD para acelerar la etapa de inducción neural inicial. En resumen, se indujeron hESC hacia destino neural mediante cocultivo con células MS5 irradiadas en KSR complementado con SB431542 y Noggin (250 ng/ml (intervalo de concentración de 10-1000 ng/ml); R&D), desde el día 2-8 y SHH+FGF8 desde el día 6-11 de diferenciación. Después de 11 días en KSR, las rosetas neurales se aislaron manualmente y se cultivaron (etapa P1) en medio N2 complementado con SHH, FGF8, BDNF y AA como se ha descrito anteriormente (Perrier, et al. Proc Natl Acad Sci U S A 101: 12543-8 (2004). Después de 5-7 días en la etapa P1, las rosetas se recogieron de nuevo mecánicamente y se trituraron después de incubación en solución salina equilibrada de Hanks sin Ca2/Mg2 (HBSS) durante 1 hora y se volvieron a sembrar en placas recubiertas con poliornitina (PO)/laminina/fibronectina. La distribución con SHH/FGF8 continuó durante 7 días en la etapa P2 seguido de diferenciación final en presencia de BDNF, AA, GDNF, TGFb3 y dbAMPc como se ha descrito anteriormente hasta la etapa de maduración deseada para un experimento dado (normalmente 5-7 días para estudios de trasplante o 32 días para estudios funcionales in vitro).
Análisis de expresión génica. Se extrajo ARN total durante la diferenciación los días: 0, 1, 3, 5, 7, 9, 11, 13 y 25 de cada condición de LSB de control, LSB/SHH/FGF8 y LSB/SHH/FGF8/CHIR usando el equipo RNeasy (Qiagen,
Valencia, CA). Para análisis de micromatrices, el ARN total fue procesado por la instalación principal de genómica del MSKCC e hibridado en matrices de perlas ref-12 humana de Illumina según las especificaciones del fabricante. Se realizaron comparaciones entre cada día y condición usando el paquete LIMMA de Bioconductor (worldwideweb.bioconductor.org). Los genes que se descubrió que tenían un valor de P ajustado <0,05 y un factor de cambio mayor de dos se consideraron significativos. Algunos de los análisis y presentaciones descriptivos de datos de micromatrices se realizaron usando un paquete de software disponible en el mercado (Partek Genomics Suite (versión 6.10.0915)). Para análisis por qRT-PCR, el ARN total el día 25 de cada condición se transcribió de manera inversa (Quantitech, Qiagen) y se detectó material amplificado usando ensayos de expresión génica Taqman disponibles en el mercado (Applied Biosystems, Carlsbad, CA) con los datos normalizados con respecto a HPRT. Cada punto de datos representa 9 repeticiones técnicas de 3 muestras biológicas independientes. Aún no están disponibles datos sin procesar de estudios de micromatrices en GEO worldwideweb.ncbi.nlm.nih.gov/geo).
Cirugía animal. Se realizaron procedimientos de roedores y monos siguiendo las directrices del NIH y fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales (IACUC), el Comité Institucional de Bioseguridad (IBC) y el Comité de Investigación de Células Madre Embrionarias (ESCRO).
Ratones. Se anestesiaron ratones nulos para NOD-SCID IL2Rgc (20-35 g de peso; Jackson Laboratory, Bar Harbor, ME) con ketamina (90 mg/kg; Akom, Decatur, IL) y xilazina (4 mg/kg Fort Dodge, LA). Se inyectó 6-hidroxidopamina (10 pg (intervalo de concentración de 1-20 pg) 6-OHDA (Sigma-Aldrich) de manera estereotáctica en el cuerpo estriado en las siguientes coordenadas (en milímetros): AP, 0,5 (de bregma; una sutura de cráneo usada como referencia para cirugía estereotáctica); ML, -2,0; DV, -3,0 (de duramadre, una membrana que cubre el cerebro usada como referencia). Se seleccionaron para trasplante ratones con lesiones exitosas (un promedio de >6 rotaciones/minuto). Se inyectaron un total de 150 x 103 células en un volumen de 1,5 pl en el cuerpo estriado en las siguientes coordenadas (en mm): AP, 0,5; ML, -1,8; DV, 3,2. Los ratones se sacrificaron 18 semanas después del trasplante.
Ratas. Se anestesiaron ratas hembras adultas Sprague-Dawley (Taconic, Hudson, NY (180-230 g) con ketamina (90 mg/kg) y xilazina (4 mg/kg) durante los procedimientos quirúrgicos. Se establecieron lesiones unilaterales del haz prosencefálico medial de la ruta nigroestriatal mediante inyección estereotáctica de 6-OHDA (3,6 mg/ml en ácido ascórbico al 0,2 % y solución salina al 0,9 %, Sigma) en dos sitios (Studer, et al. Nature Neurosci. 1: 290-295 (1998). Se seleccionaron ratas para trasplante si la rotación inducida por anfetaminas superaba 6 rotaciones/min a las 6-8 semanas después de la inyección. Se trasplantaron 250 x 103 células en el cuerpo estriado de cada animal (Coordenadas: AP 1,0 mm, ML -2,5 mm y V -4,7 mm; barra de dientes ajustada a -2,5). Las ratas de control recibieron PBS en su lugar. Los procedimientos quirúrgicos se han descrito previamente (Studer, et al. Nature Neurosci. 1: 290 295 (1998). Se iniciaron inyecciones intraperitoneales diarias de ciclosporina 15 mg/kg (Bedford Labs, Bedford, OH) 24 horas antes del injerto celular y continuaron hasta el sacrificio, 20 semanas después del injerto celular. Primates. Dos monos rhesus adultos (17-18 años de edad; 10-12 kg; hembras) se hicieron hemiparkinsonianos mediante administración de MPTP carotídea seguida de administración I.V. semanal de MPTP para crear un síndrome parkinsoniano bilateral (Kordower, et al. Science 290: 767-773 (2000). Ambos animales presentaron síntomas parkinsonianos coherentes con una lesión moderadamente grave basada en análisis conductual, incluyendo postura torcida, arrastre de la pierna y síntomas de rigor (inflexibilidad de movimiento), inatención (percepción motora a estímulo lateralizado) y bradicinesia (inicio de movimiento lento). Estos parámetros se pueden evaluar en monos usando una escala de calificación clínica (CRS) de Parkinson modificada. El día de la cirugía de trasplante, los animales se tranquilizaron con ketamina (3,0 mg/kg, IM) y dexdomitor (0,02-0,04 mg/kg IM), intubado para mantener una vía respiratoria estable y anestesiado con isoflurano. Después se colocaron en un marco estereotáctico para cirugía. Ambos monos rhesus se sometieron a una única cirugía con tres inyecciones intracraneales de cultivos de DA procedentes de la placa del suelo humana basadas en coordenadas estereotácticas (Paxinos, et al. The Rhesus Monkey Brain in Stereotaxic Coordinates (Academic Press, 2000). Se realizaron inyecciones bilaterales de células (10 ul/inyección; 125.000 células/ul) en tres sitios (1-caudado posterior, 2-putamen pre-comisural y materia blanca suprayacente) para un volumen total de 30 pl por hemisferio. Se utilizó una bomba de infusión unida a un micromanipulador estereotáctico para administrar las células a una velocidad de 1 pl/min a través de una jeringa Hamilton de 50 pl con aguja de 28 G. Después de completarse las inyecciones, la aguja se dejó en su sitio durante 2 5 minutos adicionales para permitir que el infundido se difunda por la punta de la aguja antes de retraer lentamente la jeringa. Inmediatamente después de la cirugía, los animales recibieron analgésicos (buprenex, 0,01 mg/kg IM, dos veces al día durante 72 horas después de la cirugía; meloxicam, 0,1 mg/kg s.c., una vez al día durante 72 horas después de la cirugía), así como un antibiótico (cefazolina, 25 mg/kg IM, dos veces al día) hasta 72 horas después de la cirugía. Los animales recibieron ciclosporina A (Neoral, Sandimmune) por vía oral (30 mg/kg que se disminuye a 15 mg/kg) una vez al día, comenzando 48 horas antes de la cirugía hasta el sacrificio, un mes después del trasplante.
Ensayos conductuales. Se llevaron a cabo rotaciones inducidas por anfetaminas (ratones y ratas) y la prueba de marcha (rata) antes del trasplante y 4, 8, 12, 18 semanas después del trasplante. El comportamiento de rotación en ratones se registró 10 minutos después de inyección i.p. de d-anfetamina (10 mg/kg, Sigma) y se registró durante 30 minutos. El comportamiento de rotación en ratas se registró 40 minutos después de la inyección i.p. de d-anfetamina (5 mg/kg) y se evaluó automáticamente mediante el sistema TSE VideoMot2 (Alemania). Los datos se presentaron como el número promedio de rotaciones por minuto. La prueba de marcha se modificó a partir de Blume, et al. Exp. Neurol. 219: 208-211 (2009) y Crawley, et al. What's Wrong With My Mouse: Behavioral Phenotyping of Transgenic and Knockout Mice (Wiley-Liss, 2000).
En resumen, cada rata se colocó sobre una superficie plana; sus patas traseras se elevaron levantando suavemente la cola para permitir que solo las patas delanteras toquen la mesa. El experimentador tiró de la rata hacia atrás 1 metro a un ritmo constante. Se contaron los números de paso de ajuste de las patas delanteras tanto contralaterales como ipsilaterales. Los datos se presentaron como el porcentaje de pasos de ajuste contralaterales / (contralaterales ipsilaterales). La prueba del cilindro se realizó colocando cada animal en un cilindro de vidrio y contando el número de toques de patas ipsilaterales frente a contralaterales (de 20 toques) a la pared del cilindro como se ha descrito anteriormente (Tabar, et al. Nature Med. 14: 379-381 (2008). El procesamiento de tejidos para roedores y primates se describe a continuación.
Ratones y ratas: Los animales (ratones y ratas) recibieron sobredosis de pentobarbital por vía intraperitoneal (50 mg/kg) para inducir anestesia profunda y se perfundieron en paraformaldehído (PFA) al 4 %. Se extrajeron cerebros, se fijaron posteriormente en PFA al 4 % y después se empaparon en soluciones de sacarosa al 30 % durante 2-5 días. Se seccionaron en un criostato después de incluirlo en un compuesto O.C.T. (Sakura-Finetek, Torrance, California).
Primates: Los animales se sacrificaron con anestesia profunda con ketamina (10 mg/kg, intramuscular (IM)) y pentobarbital (25 mg/kg, intravenoso (IV)) a través de perfusión cardíaca con solución salina heparinizada al 0,9 % seguida de fijador de PFA frío al 4 % nuevo (pH 7,4). Inmediatamente después de la fijación primaria, los cerebros se retiraron del cráneo y se fijaron posteriormente en PFA al 4 %, en flotación libre, durante 24-36 h. Después se aclararon y se volvieron a suspender en sacarosa al 10 % en un agitador lento a 4 °C y se permitió que se "hundieran". Después se repitió el proceso en sacarosa al 20 % seguido de sacarosa al 30 %. Los cerebros completos se cortaron coronalmente en secciones en serie de 40 um en un microtomo deslizante congelado y se almacenaron en flotación libre en medio crioconservador a -20 °C.
Inmunohistoquímica: Las células se fijaron en PFA al 4 % y se bloquearon con seroalbúmina bovina (BSA) al 1 % con Triton al 0,3 %. Las secciones de tejido cerebral se lavaron en PBS frío y se procesaron de manera similar. Los anticuerpos primarios se diluyeron en BSA al 1-5 % o suero de cabra normal y se incubaron según las recomendaciones del fabricante. En la tabla 6 se proporciona una lista exhaustiva de anticuerpos y fuentes. Se usaron anticuerpos secundarios conjugados con Alexa488, Alexa555 y Alexa647 (Molecular Probes, Carlsbad, California) con contratinción nuclear de 4',6-diamidino-2-fenilindol (DAPI) (Thermo Fisher, Rockford, Illinois). Para algunos análisis, se usaron anticuerpos secundarios biotinilados seguidos de visualización mediante cromógeno DAB (3,3'-diaminobencidina).
Análisis de HPLC. HPLC de fase inversa con detección electroquímica para medir niveles de dopamina, Ácido homovanílico (HVA) y se realizó DOPAC (ácido 3,4-dihidroxi-fenilacético) como se ha descrito anteriormente (Roy, et al. Nature Med. 12: 1259-1268 (2006); Studer, et al. Brain Res. Bull. 41: 143-150 (1996). Se recogieron muestras de cultivo en ácido perclórico el día 65 de diferenciación. Para algunos experimentos, se midió DA directamente en el medio usando el mismo sistema de detección pero después de la extracción de aluminio de dopamina y sus metabolitos usando un equipo disponible en el mercado como se descrito anteriormente (Studer, et al. Brain Res. Bull.
41: 143-150 (1996).
Registros electrofisiológicos: Los cultivos se transfirieron a una cámara de registro en un microscopio vertical equipado con un objetivo de inmersión en agua 40X (Eclipse E600FN; Nikon); los cultivos se perfundieron con solución salina que contenía en mM: NaCl 125, KCl 2,5, NaHCOa 25, NaH2PO41,25, CaCl 2, MgCh 1 y glucosa 25 (34 °C; saturado con O295 %-CO25 %; pH 7,4; 298 mOsm/l). El caudal de solución salina fue de 2-3 ml/min a través de un calentador en línea (SH-27B con controlador TC-324B; Warner Instruments). Las neuronas se visualizaron mediante microscopia de vídeo con una cámara digital CCD enfriada (CoolSNAP ES2, Photometrics, Roper Scientific, T ucson, Arizona). Las células seleccionadas para registros electrofisiológicos tenían formas de tipo neuronal con neuritas ramificadas finas. Se realizaron registros de pinzamiento zonal de células somáticas completas en la configuración de pinza actual con un amplificador MultiClamp 700B (Molecular Devices). Las señales se filtraron a 1-4 kHz y se digitalizaron a 5-20 kHz con un Digidata 1440A (Molecular Devices). Los electrodos de parche de registro estaban hechos de vidrio de borosilicato filamentado (Sutter Instruments) extraído en un extractor Flaming-Brown (P-97, Sutter Instruments) y tenían resistencias de 4-6 MQ en el baño. Los electrodos se llenaron con solución interna que contenía en mM: K-MeSO4135, KCl 5, HEPES 5, EGTA 0,25, fosfocreatina-di(tris) 10, ATP-Mg 2 y GTP-Na 0,5 (pH 7,3, osmolaridad ajustada a 290-300 mOsm/l). El circuito del puente amplificador se ajustó para compensar la resistencia del electrodo y se supervisó. Se compensó la capacidad del electrodo. Cuando la resistencia en serie aumentó >20 % durante el registro, los datos se descartaron porque una mayor resistencia sugirió un fallo técnico parcial durante los registros.
Recuentos celulares y análisis estereológicos. Los porcentajes de células positivas para marcador en la placa del suelo (día 11) figura 1, precursor de neuronas de dopamina del mesencéfalo (día 25), figura 2 y etapas de neuronas DA maduras (día 50 o posterior) figura 3 y 11, se determinaron en muestras procedentes de al menos 3 experimentos independientes cada una. Las imágenes para cuantificación se seleccionaron de manera aleatoria uniforme y cada imagen se puntuó primero con respecto al número de núcleos positivos para DAPI, seguido de recuento del número de células que expresan el marcador de interés. Los datos se presentan como la media ± ETM. Se realizó cuantificación de células humanas (identificadas con anti-hNA) y neuronas TH+ dentro de los injertos en cada décima sección donde un injerto era identificable. Los recuentos celulares y el volumen del injerto se determinaron usando la sonda del fraccionador óptico y el estimador Cavalieri usando el software Stereo Investigator (MBF bioscience,
Vermont) como se ha descrito anteriormente en Tabar, et al. Nat. Biotechnol. 23: 601-606 (2005). Los datos se presentan como el número total estimado de células y el volumen total de injerto /- error típico de las medias (ETM). Las siguientes formulaciones describen medios de cultivo celular ilustrativos para su uso en el desarrollo de realizaciones de las presentes invenciones.
Medio de hESC para mantenimiento (1 litro): 800 ml de DMEM/F12, 200 ml de reemplazo de suero Knockout, 5 ml de L-glutamina 200 mM, 5 ml de pen/estrep, 10 ml de solución de 15 aminoácidos no esenciales mínima MEM 10 mM, 55 pM de 13-mercaptoetanol y bFGF (la concentración final es de 4 ng/ml).
Medio KSR para diferenciación de hESC (1 litro): 820 ml de DMEM Knockout, 150 ml de reemplazo de suero Knockout, 10 ml de L-glutamina 200 mM, 10 ml de pen/estrep, 10 ml de MEM 10 mM y 55 pM de 13-mercaptoetanol.
Medio N2 para diferenciación de hESC (1 litro): 985 ml de H2O dest. con polvo de DMEM/F12, 1,55 g de glucosa (Sigma, n.° de cat. G7021), 2,00 g de bicarbonato de sodio (Sigma, cat. n.° S5761), putrescina (alícuota de 100 ul de 1,61 g disuelto en 100 ml de agua destilada; Sigma, cat n.° P5780), progesterona (alícuota de 20 ul de 0,032 g disueltos en 100 ml de etanol al 100 %; Sigma, cat. n.° P8783), selenito de sodio (alícuota de 60 pl de solución 0,5 mM en agua destilada; Bioshop Canada, cat. n.° SEL888) y 100 mg de transferrina (Celliance/Millipore, cat. n.° 4452-01) y 25 mg de insulina (Sigma, cat. n.° 16634) en 10 ml de NaOH 5 mM.
Modificación de Dulbecco del medio de Eagle (DMEM) con FBS al 10 % para preparar PMEF (células alimentadoras de fibroblastos de embrión primario de ratón (PMEF)) (1 litro): 885 ml de DMEM, 100 ml de FBS, 10 ml de pen/estrep y 5 ml de L-glutamina.
Medio esencial mínimo alfa (MEM) con FBS al 10 % para preparar medio de células alimentadoras MS-5 (1 litro): 890 ml de MEM alfa, 100 ml de FBS, 10 ml de solución de gelatina de pen/estrep (500 ml): Disolver 0,5 g de gelatina en 500 ml de agua tibia (50-60 °C) Milli-Q. Enfriar a temperatura ambiente.
En general, a continuación hay un breve compendio de métodos ilustrativos de supervisión de la producción de neuronas DA maduras para su uso en el injerto (véase, además, condiciones mostradas en la tabla 7. El día 13 es una etapa de la placa del suelo del mesencéfalo, caracterizada por la coexpresión de FOXA2/LMX1A. Además de la expresión de FOXA2 y LMX1A, se encuentra una pérdida de expresión de OCT4 y falta de inducción de marcadores del prosencéfalo PAX6 y FOXG1. El día 25 es la etapa precursora de neuronas DA del mesencéfalo, caracterizada por la expresión continua de FOXA2/LMX1A y la expresión de los marcadores neuronales (TUJ1) y DA (NURR1, TH). Se supervisan las células Ki67+ en proliferación y el número de precursores neurales del prosencéfalo PAX6 y FOXG1, donde no se desean estos marcadores. Para cuantificación imparcial y rápida de los datos de inmunofluorescencia, se utilizó un microscopio de alto contenido Operetta (Perkin Elmer) para las mediciones. También se usaron ensayos de qRT-PCR para cada marcador para confirmar los datos de inmunofluorescencia. En algunas realizaciones, se usan para injerto líneas celulares (cultivos) que pasan estas pruebas preliminares in vitro, véase, tabla 7. En algunas realizaciones, las neuronas DA maduras se crioconservaron sin suero el día 25 (intervalo del día 20 al día 25) en medio de cultivo DMSO al 7 % (intervalo de 3 % a 12 %) hasta que se descongelaron para su uso en injerto. En algunas realizaciones, las muestras celulares se almacenan en nitrógeno líquido. En algunas realizaciones, las células se almacenan en congeladores de baja temperatura. En otras realizaciones, se contemplan crioprotectores tales como mioinositol, alcohol polivinílico, reemplazo de suero, compuestos de inhibición de caspasa, para su uso además de DMSO. Después de descongelar, las células se prueban para determinar su viabilidad, expresión de marcadores, etc., antes de su uso en injertos. En algunas realizaciones, las células descongeladas se probaron para determinar el mantenimiento de la función en ensayos a largo plazo in vitro e in vivo para supervisar las condiciones de congelación y almacenamiento.
B. Estudios para identificar factores adicionales para la generación de neuronas DA funcionales. Se contemplan experimentos adicionales de "pérdida" y "adición" para componentes de cultivo tisular para su uso en la producción de células de las presentes invenciones. Por ejemplo, se mostró que FGF8, aunque su uso dio como resultado células de las presentes invenciones, no era necesario para la producción de estas células. Estos experimentos se extenderán a reactivos adicionales, tales como los enumerados en la tabla 8, como aditivos para cultivos celulares junto con las cuatro moléculas "centrales" que dieron como resultado células neuronales DA de las presentes invenciones, es decir, i) inhibidor de Alk4/5/7 ("TGFp") (SB431542), ii) inhibidor de Alk2/3 ("BMP") (LDN-193189), iii) agonista de Smoothened ("SHH") (Purmorfamina) y iv) inhibidor de GSK3p (CHIR99021).
Como se describe en la presente memoria, el uso de SB431542 y LDN193189 mostró conversión neuronal eficaz de células madre pluripotentes, mientras que la adición de Purmorfamina y CHIR99021 a estas células demostró inducción de la placa del suelo del mesencéfalo. Otros productos químicos y proteínas recombinantes se usaron o pueden usarse para proporcionar soporte trófico a largo plazo y/o acelerar la diferenciación. Algunos de estos compuestos se usarán en pruebas adicionales para definir sus funciones en la diferenciación celular descrita en la presente memoria.
Para estos experimentos, el rendimiento de otros compuestos se comparará con los límites ilustrativos para la diferenciación de neuronas DA (tabla 7) frente al uso de los 4 factores centrales (tabla 8).
Se contempla que este tipo de experimentos definen el número mínimo de factores necesarios para producir células neuronales DA auténticas de las presentes invenciones.
C. Realizaciones para establecer curvas de respuesta a dosis para contaminantes proliferantes potenciales (hESC pluripotentes, rosetas neurales). Durante el desarrollo de las presentes invenciones, no se observaron teratomas o hiperproliferaciones excesivas dentro de los injertos hasta al menos 5 meses de supervivencia in vivo. Con el fin de supervisar la seguridad de estudios a largo plazo para reflejar la longevidad contemplada de los injertos en seres humanos, se contempla un umbral de número de células para determinar un límite de contaminación clínicamente relevante de tipos celulares problemáticos, tales como hESC indiferenciadas que pueden convertirse en teratomas o precursores neuroectodérmicos primitivos con capacidad de proliferación significativa. Por lo tanto, se contemplan algunas realizaciones para enriquecimiento adicional de neuronas dopaminérgicas en células para su uso en injertos, es decir, agotar los tipos celulares contaminantes antes del injerto, véase, tabla 7 para límites ilustrativos.
A continuación se describe un conjunto normalizado ilustrativo de ensayos adicionales para validar las estrategias de mejora. Para hESC, se usará una mezcla predeterminada de células indiferenciadas (Oct4+/Nanog+) con neuronas DA procedentes de hES para supervisar los síntomas clínicos que sugieren efecto de masa y/o muerte animal en experimentos con animales. Se realizará una respuesta a dosis de una célula hES por cada 10.000 células DA procedentes de hESC, 1/5000, 1/1000 y 1/100. Las células se inyectarán por vía intraestriatal y los animales recibirán inmunodepresión. Las ratas se supervisarán estrechamente y se sacrificarán tras la manifestación de síntomas neurológicos o en un máximo de 6 meses. Los cerebros se analizarán para determinar los volúmenes y la composición de los injertos como se describe en la presente memoria. Las relaciones celulares se ajustarán hasta que se establezca un umbral claro in vivo para la aparición de teratomas. Para determinar los niveles contaminantes de precursores neuroectodérmicos primitivos, se seguirá una estrategia similar. La presencia de precursores neurales tempranos tiene un potencial significativo para proliferación y diferenciación general en destinos del sistema nervioso central así como del sistema nervioso periférico (SNP). El análisis de injerto consistirá en IHC para células de la roseta (expresión de PLZF), su descendencia del SNC (precursores neurales que expresan Nestina/Sox2 o precursores del prosencéfalo, que expresan FoxG1), así como volúmenes de injerto y un índice de proliferación (% de Ki67+ de células supervivientes totales).
IV. Enfermedad de Parkinson.
La enfermedad de Parkinson (EP) es el segundo trastorno neurodegenerativo más habitual y se estima que afecta a 4,1-4,6 millones de pacientes en todo el mundo, un número que se prevé que aumente más del doble para 2030. Es el segundo trastorno neurodegenerativo más habitual después de la enfermedad de Alzheimer, afectando a aproximadamente 1 millón de pacientes en los Estados Unidos con 60.000 pacientes nuevos diagnosticados cada año. La enfermedad tiene un gran impacto socioeconómico que provoca morbilidad y mortalidad significativas y los costes combinados directos e indirectos de la EP, incluyendo el coste de la atención sanitaria y la pérdida de ingresos, se estima en aproximadamente 25 mil millones de dólares al año solo en los Estados Unidos. Actualmente no existe cura para la enfermedad de Parkinson (EP), un trastorno progresivo e incapacitante relacionado con la edad. La EP se caracteriza patológicamente por una pérdida selectiva de neuronas DA del mesencéfalo en la sustancia negra. Por lo tanto, una característica fundamental de la EP es la pérdida progresiva, grave e irreversible de neuronas de dopamina (DA) del mesencéfalo, lo que da como resultado una discapacidad motora incapacitante. Aunque se han desarrollado terapias farmacológicas, a base de ejercicio, génicas y quirúrgicas para la EP, ninguno de esos enfoques es capaz aún de restaurar la función adecuada de neuronas DA. El control a largo plazo de los síntomas motores en pacientes con frecuencia sigue siendo mejorable y, aunque se reconoce la importancia de los síntomas motores y no motores progresivos que no responden a dopamina, el problema fundamental del control a largo plazo de los síntomas sensible a dopamina sigue siendo un área de necesidad terapéutica crítica. La patología generalizada se reconoce en la EP, que afecta a los sistemas nerviosos central y periférico, las características fundamentales de la EP (bradicinesia, rigidez y temblor parcial) están fundamentalmente relacionadas con la pérdida de células neuronales DA y son sensibles a la dopamina. Por tanto, la EP se contempla para el tratamiento usando reemplazo de células neuronales debido a la pérdida más bien selectiva de neuronas dA del mesencéfalo que es responsable de la mayoría de los síntomas motores de la enfermedad, un cerebro humano sano alberga aproximadamente un millón de neuronas DA. Por lo tanto, en una realización, se considera que el reemplazo de neuronas DA requiere una cantidad relativamente pequeña de células supervivientes en comparación con la mayoría de los otros trastornos en el SNC.
Uno de los desafíos en el desarrollo de una terapia celular para la EP fue la identificación de una fuente celular adecuada para su uso en el reemplazo neuronal. Esta búsqueda se está realizando desde hace más de 30 años, habiéndose propuesto muchas fuentes potenciales para reemplazo de neuronas DA (Krilcs, Protocols for generating ES cell-derived dopamine neurons in Development and engineering of dopamine neurons (eds. Pasterkamp, R. J., Smidt, y Burbach) (Landes Biosciences, 2008; Fitzpatrick, et al., Antioxid. Redox. Signal. 11: 2189-2208 (2009)). En el pasado, varias de esas fuentes han progresado a ensayos clínicos de etapas tempranas que incluyen células catecolaminérgicas de la médula suprarrenal Madrazo, et al., N. Engl. J. Med. 316, 831-834 (1987), trasplantes de cuerpo carotídeo (Arjona, et al., Neurosurgery 53: 321-328 (2003)) o células epiteliales de pigmento retiniano encapsulado (Spheramine trial Bakay, et al., Front Biosci. 9, 592-602 (2004). Sin embargo, esos ensayos en su mayoría no consiguieron mostrar eficacia clínica y dieron como resultado escasa supervivencia a largo plazo y baja liberación de DA de las células injertadas. Otro enfoque fue el trasplante de neuronas DA del mesencéfalo fetal realizado en más de 300 pacientes en todo el mundo (Brundin, et al., Prog. Brain Res. 184, 265-294 (2010); Lindvall,
y Kokaia, J. Clin. Invest 120: 29-40 (2010). La terapia con tejido fetal humano en estos pacientes demostró pruebas de supervivencia de neuronas DA y liberación de Da in vivo hasta 10 o 20 años después del trasplante en algunos pacientes. Sin embargo, en muchos pacientes, el trasplante de tejido fetal no consigue reemplazar la función neuronal DA. Además, el trasplante de tejido fetal está plagado de múltiples desafíos, incluyendo la baja cantidad y calidad del tejido donante, las cuestiones éticas y prácticas que rodean a la adquisición de tejidos y la naturaleza heterogénea mal definida de las células trasplantadas, que son algunos de los factores que contribuyen a los resultados clínicos variables. Se describen ejemplos de trasplante fetal en; Mendez, et al. Nature Med.(2008)); Kordower, et al. N. Engl. J. Med. 332: 1118-1124 (1995); Piccini, et al. Nature Neuroscience 2:1137-1140 (1999). Sin embargo, los resultados clínicos se mezclaron con algunos datos positivos en estudios abiertos tempranos (Lindvall, et al. Science 247: 574 577 (1990); Widner, et al. N. Engl. J. Med. 327: 1556-1563 (1992); Brundin, et al. Brain 123: 1380-1390 (2000); Freed, et al. N. Engl. J. Med. 327: 1549-1555 (1992); Freeman, et al. Bilateral fetal nigral transplantation into the postcommissural putamen in Parkinson's disease. Ann Neurol 38: 379-388 (1995). Sin embargo, se encontraron resultados modestos en dos ensayos clínicos controlados con placebo mayores, subvencionados por el NIH, en los Estados Unidos (Freed, et al. N. Engl. J. Med. 344, 710-719 (2001); Olanow, et al. Ann. Neurol. 54: 403-414 (2003)). Existen muchas hipótesis acerca de la eficacia limitada observada en los ensayos de injertos fetales humanos, incluyendo que el injerto fetal puede no proporcionar un número suficiente de células en la etapa de desarrollo correcta para un beneficio terapéutico óptimo. Asimismo, el tejido fetal está bastante mal definido por el tipo celular y es variable con respecto a la etapa y la calidad de cada muestra tisular Bjorklund, et al. Lancet Neurol. 2, 437-445 (2003). Otro factor contribuyente puede ser una respuesta inflamatoria de bajo nivel del hospedador al injerto (Bjorklund, et al. Lancet Neurol. 2, 437-445 (2003)).
Por el contrario, se contempla que una fuente de células procedente de células madre u otro tipo celular uniforme para su uso en el suministro de células para trasplante supere muchos de los desafíos asociados con el injerto de tejido fetal y podría ofrecer una fuente ilimitada de neuronas DA en la etapa óptima para el trasplante. Después de casi 20 años de intentos de usar diversas fuentes potenciales de células madre, los inventores lograron obtener auténticas neuronas DA del mesencéfalo humano a partir de células madre pluripotentes capaces de neutralizar los defectos neurológicos en animales murinos y primates. Esta estrategia de diferenciación novedosa fue muy eficaz y condujo a injerto robusto in vivo de las células, inducción de recuperación funcional en modelos de enfermedad de la EP y falta de acontecimientos adversos tales como proliferación celular inadecuada como respaldan los datos preclínicos. La aprobación de la FDA para una estrategia basada en células ES humanas para tratar la EP se contempla porque la FDA aprobó probar otros derivados de células ES humanas en la lesión de la médula espinal (Strauss, Nat. Biotechnol.
28: 989-990 (2010) y degeneración macular (Schwartz, et al. Lancet 379: 713-720 (2012)).
Se contemplan realizaciones adicionales que incluyen estrategias de "mejora" para controlar la pureza celular que promueven el crecimiento de fibras axónicas e incluyen características novedosas de seguridad/reguladoras en estrategias de injerto. Por ejemplo, en algunas realizaciones, un método de purificación celular demostró a partir de una exploración de marcadores de superficie sencilla frente a un tipo celular de interés, (tal como CD142) una estrategia de enriquecimiento significativa para un tipo de neurona específico. En algunas realizaciones, este método se contempla para su uso en el suministro de células para su uso en seres humanos. Además, en otras realizaciones, la expresión modificada por ingeniería genética de PSA-NCAM se contempla para mejorar el crecimiento axónico para su uso en la reparación neural in vivo. Dichas aplicaciones incluyen la promoción del crecimiento axónico a larga distancia para el tratamiento de la enfermedad de motoneuronas, enfermedad de Huntington u otros trastornos que afectan principalmente a neuronas de proyección. Se contemplan realizaciones adicionales para una fuente de células pluripotentes calificadas por GMP y similares. Debido a los métodos de injerto descritos en la presente memoria, requieren una pequeña cantidad de neuronas DA y se basan en métodos relativamente sencillos, rentables, de moléculas pequeñas desarrollados para la inducción de neuronas DA, se contempla que la terapia de reemplazo de neuronas Da tendría un coste razonable por paciente.
Una posible limitación biológica del enfoque de trasplante en la EP es el hecho de que la degradación neuronal en la EP continúa afectando a muchos tipos celulares además de neuronas de dopamina del mesencéfalo, en particular en etapas posteriores de la enfermedad. De hecho, en un estudio a largo plazo, los síntomas que no responden a DA predominan en la EP tardía, lo que conduce a disfagia, caídas, demencia y otras morbilidades significativas. Sin embargo, se contempla que algunos síntomas no motores se benefician de la restauración de la función dopaminérgica. Asimismo, se contempla que el uso de neuronas DA procedentes de hESC en las primeras etapas de la enfermedad prevendría algunos de los síntomas secundarios de la Ep , incluyendo la degradación de las poblaciones que reciben dopamina del cuerpo estriado. Sin embargo, incluso en ausencia de influencia en los síntomas de la enfermedad que no responden a DA, la restauración dopaminérgica funcional a largo plazo del cuerpo estriado sería un logro importante para el tratamiento de este trastorno actualmente incurable. En el caso de la enfermedad de Parkinson, existen varias terapias alternativas disponibles incluyendo estrategias a base de fármacos y enfoques quirúrgicos tales como estimulación cerebral profunda. En algunas realizaciones, se contempla que la eficacia de recuperación es similar o superior a los niveles alcanzados con terapias alternativas. En otras realizaciones, se considera que el uso de esta terapia de injerto de células neuronales DA maduras es particularmente beneficioso para un subconjunto particular de pacientes. En otras realizaciones, se contempla el uso de esta terapia de injerto de células neuronales DA maduras para su uso además de enfoques existentes de tipo farmacológico y quirúrgico. Un beneficio importante del uso de la terapia de injerto de células neuronales DA maduras de las presentes invenciones es la naturaleza neurorrestauradora única posterior al injerto, es decir, recuperación a largo plazo de la función neuronal
que se contempla para su uso en pacientes para la eliminación progresiva de la terapia farmacológica. Se contempla que el trasplante de células afecta a un espectro diferente de síntomas relacionados con DA que los que responden a fármacos u otra terapia. Por tanto, en una realización, se contempla el trasplante de neuronas DA maduras para su uso con DBS. En otra realización, se contempla la neurona DA madura para su uso con terapia.
A) Enfermedad de Parkinson y terapias actuales. Se hicieron grandes progresos en la identificación de cambios genéticos poco habituales que contribuyen a las formas familiares de la EP. Sin embargo, para la mayoría de los casos de EP, la contribución de cualquier posible predisposición genética sigue sin estar clara. Las estrategias terapéuticas tradicionales en la EP están limitadas por el hecho de que en el momento de aparición de los síntomas clínicos, 30 70 % de todas las neuronas DA en la sustancia negra se han degradado de manera irreversible. Una opción terapéutica es el reemplazo farmacológico de la deficiencia de neuronas DA usando el precursor de DA L-Dopa. Sin embargo, a pesar de la considerable respuesta inicial de algunos pacientes con EP a la terapia con L-Dopa, el resultado clínico a largo plazo sigue siendo negativo y se producen efectos secundarios graves de la terapia con L-Dopa, incluyendo fluctuaciones motoras y discinesias, con frecuencia en etapas tardías de la enfermedad. La administración pulsátil de L-dopa tiene una función importante en el desarrollo de estas complicaciones motoras de etapa tardía, por lo tanto, un suministro "más suave" y más fisiológico de dopamina, es decir, tal como de células injertadas de las presentes invenciones, sería, por lo tanto, muy deseable. Además de estrategias farmacológicas, existen varias opciones de tratamiento quirúrgico. Estas incluyen la ablación o inactivación funcional de células dentro de los ganglios basales a través de palidotomía o estimulación cerebral profunda al dirigirse al núcleo subtalámico o a la parte interna del globo pálido. Aunque estas opciones quirúrgicas son alternativas para algunos pacientes, proporcionan alivio sintomático de la enfermedad pero no restablecen la función normal de DA. Asimismo, se ha informado de efectos secundarios quirúrgicos y no quirúrgicos, incluyendo disfunción del hardware, infecciones, ictus, hemorragia y similares. Otras opciones de tratamiento incluyen la administración de factores de crecimiento tales como GDNF o Neurturina mediante infusión intraparenquimatosa directa o expresión vírica mediante terapia génica. Aunque estudios abiertos iniciales en EP mostraron resultados prometedores para ensayos controlados posteriores de GDNF en un conjunto mayor de pacientes no consiguieron confirmar ningún beneficio y plantearon posibles problemas de seguridad debido a la producción de anticuerpos anti-GDNF en un subconjunto de pacientes. El suministro de Neurturina basado en a Av , una molécula relacionada con GDNF, tampoco consiguió mostrar ningún beneficio clínico significativo en un gran ensayo multicéntrico, controlado con placebo. Se ha informado recientemente de datos tempranos de un ensayo de fase 2 de inyecciones de ácido glutámico descarboxilasa (GAD) portada por AAV en el núcleo subtalámico (Lancet Neurology, abril de 2011), sin embargo, los beneficios clínicos fueron moderados en el mejor de los casos en este estudio. Aunque los esfuerzos en estrategias neuroprotectoras basadas en factores neurotróficos o alternativas podrían proporcionar alivio temporal a los pacientes, ninguno de ellos puede recuperar/reemplazar neuronas DA ya perdidas debido a la enfermedad, el objetivo principal es una terapia de reemplazo celular.
B) Terapia celular en la EP. Los síntomas clínicos se hacen evidentes en la EP después de perderse 70-80 % de las neuronas de dopamina estriatal y aproximadamente 50 % de neuronas de dopamina nígricas. Sin embargo, las neuronas de dopamina del mesencéfalo se desarrollaron a las 8,5 semanas después de la concepción con pocas pruebas de reemplazo de neuronas de dopamina durante el resto de la vida. Por lo tanto, las neuronas de la dopamina en el momento de inicio de la enfermedad tienen muchas décadas de antigüedad sin un mecanismo natural para reemplazar estas células, por tanto, puede ser necesario el trasplante de células para reemplazar esas células en el cerebro de los pacientes con EP. El reemplazo de neuronas DA en la EP se realizó en la década de 1980 en función del uso de células cromafines procedentes de la médula suprarrenal. Se ha mostrado que esas células productoras de hormonas cambian el fenotipo de neurotransmisores de adrenalina a DA cuando se colocan de manera ectópica en el SNC. Aunque se ha injertado en varios cientos de pacientes con EP en todo el mundo usando este enfoque, con el tiempo quedó claro que las células injertadas sobreviven muy mal con un efecto transitorio en el mejor de los casos. Por lo tanto, este enfoque fue rápidamente abandonado para su uso clínico. Por el contrario, el uso del injerto de tejido del mesencéfalo fetal se basó en estudios preclínicos más extensos en modelos de roedores que demostraron injerto robusto a largo plazo y mejora funcional en un panel de ensayos conductuales relacionados con DA. Animados por esos datos preclínicos, se realizó injerto fetal en múltiples centros a fines de la década de 1980 y principios de la década de 1990. Esos estudios mostraron pruebas claras de que el injerto funcional a largo plazo con mayor liberación de DA en el área injertada medida por Fluorodopa PET y estudios histológicos posteriores en algunos pacientes que murieron debido a causas no relacionadas. Sin embargo, el uso de injerto fetal planteó dos problemas potenciales con enfoques terapéuticos basados en células. En primer lugar, un problema inesperado de los injertos fetales fue la inducción de discinesias inducidas por injerto (GID) en aproximadamente 15 % de los pacientes. Aunque el mecanismo de GID sigue siendo debatido, pruebas recientes indicaron que las neuronas serotoninérgicas tienen capacidad de almacenamiento y liberación inadecuados de DA. Otro posible mecanismo que se ha sugerido que explica GID fue la distribución desigual de neuronas DA, es decir, que provoca puntos calientes de liberación de DA. Por el contrario, durante el desarrollo de las presentes invenciones, se descubrieron métodos para detectar y reducir neuronas serotoninérgicas que reducirían la incidencia de GID. Además, la inyección de neuronas DA maduras proporcionaría una distribución uniforme de neuronas maduras, incluyendo la extensión de terminales de fibras dopaminérgicas dentro del cuerpo estriado del hospedador. Otro problema con el tratamiento de injertos fetales fue (y es) la disponibilidad limitada de tejido del mesencéfalo fetal en la etapa de desarrollo adecuada. Se probó clínicamente una estrategia alternativa para abordar el problema del suministro limitado mediante el uso de neuronas DA procedentes de cerdo fetal. Sin embargo, la supervivencia de neuronas DA en esos xenoinjertos fue mala y se abandonó el enfoque
general. Un ensayo reciente con epitelio pigmentado retiniano tampoco logró mostrar ningún beneficio. Por el contrario, se contempla el uso de células Es humanas como fuentes de células de trasplante para proporcionar una fuente ilimitada de células para producir neuronas de dopamina para su uso en el trasplante.
V. Se descubrieron compuestos y métodos de cultivo para la diferenciación dirigida de células precursoras neuronales positivas para FOXA2+ y LMX1A+ en neuronas DA del mesencéfalo (mDA) de las presentes invenciones.
Los inventores descubrieron durante el desarrollo de las presentes invenciones que la sincronización de exposición a CHIR99021 determina la inducción de precursores de la placa del suelo del mesencéfalo FOXA2/LMX1A. Por lo tanto, los inventores probaron el análisis inmunocitoquímico de FOXA2/LMX1A el día 11 de diferenciación después de LSB/S/F8 (es decir, tratamiento de células con LSB, S, es decir, tratamiento con SHH y FGF8 (F8) solo o en combinación con CHIR comenzando en diversos puntos temporales: d(día)0-d 11, d1-d11, d3-d11, d5-d11, d7-d11 en comparación con cultivos duplicados de células sin tratamiento con CHIR. Después, se determinó la cuantificación del porcentaje de células FOXA2+, LMX1A+ y con doble marcador el día 11 de diferenciación después del inicio diferencial de exposición a CHIR como se describe en el análisis inmunocitoquímico.
A. CHIR99021 (C) es un factor para inducir células FOXA2+/LMX1A+ el día 11 de células cultivadas con LSB en contacto con un activador de Hedgehog y purmorfamina. El siguiente ejemplo describe el uso de métodos ilustrativos para probar la eficacia de cada compuesto para inducir la diferenciación neuronal dirigida de neuronas mDA.
Este ejemplo describe el descubrimiento de moléculas pequeñas y la sincronización del contacto para proporcionar la diferenciación dirigida de neuronas DA FOXA2+LMX1A+ de las presentes invenciones. A continuación, hay un breve compendio de algunos de los descubrimientos experimentales descritos en la presente memoria: El tratamiento de células con inhibición doble de SMAD con agonistas de SHH (purmorfamina SHH) y FGF8 (S/F8) en ausencia de CHIR99021 mostró expresión significativamente menor de FOXA2 para el día 11 y falta completa de expresión de LMX1A (figura 1a, b). El marcador anterior OTX2 fue inducido de manera robusta en cultivos tratados con LSB y LSB/S/F8/CHIR, pero no en condiciones de LSB/S/F8 (figura 1a, c).
Los inventores eligieron una población celular que contenía células pluripotentes para una población inicial y esta se sembró en placas el día 0. Las células se cultivan casi hasta la confluencia antes de la diferenciación (entre 60 y 100 % de confluencia). Estas células se pusieron en contacto con inhibidores dobles de SMAD (es decir, exposición a LDN-193189 SB431542 = "LSB") el día 0. Los inventores siguieron una población celular con aportes regulares que contenían LSB nuevo hasta el día 11 y descubrieron que algunas células restantes eran LMX1A+ pero no expresaban FOXA2 (figura 1a, b). Los inventores sembraron en placas poblaciones de células iniciales duplicadas y después analizaron los tipos celulares (es decir, patrones de expresión de genes/proteínas) después de entrar en contacto con mezclas que contenían cualquiera de los siguientes agonistas de SHH (purmorfamina SHH) y FGF8 (S/F8) en contacto con las células con diferentes regímenes de exposición, es decir, en contacto con las células el día 0, día 1 o día 2, etc. durante cantidades específicas de tiempo, es decir, 24 horas, 48 horas, etc. Tres condiciones de cultivo ilustrativas principales probadas fueron 1) células en contacto con LDN/SB (LSB) el día 0 y después en contacto con LSB nuevo hasta el día 5, el día 5 las células se pusieron en contacto con LDN nuevo sin SB hasta el día 11,2) células en contacto con LDN/SB (LSB) el día 0 y después en contacto con LSB nuevo hasta el día 5, el día 5 las células se pusieron en contacto con LDN nuevo sin SB hasta el día 11, mientras que durante este tiempo las células se pusieron en contacto adicionalmente con purmorfamina nueva, SHH y FGF8 hasta el día 7 y 3) células en contacto con LDN/SB (LSB) el día 0 y después en contacto con LSB nuevo hasta el día 5, el día 5 las células se pusieron en contacto con LDN nuevo sin SB hasta el día 11, mientras que durante este tiempo las células se pusieron en contacto adicionalmente con purmorfamina nueva, SHH y FGF8 hasta el día 7, mientras que se pusieron en contacto adicionalmente con CHIR nuevo a partir del día 3 de cultivo hasta el día 11 con varias variaciones de estas condiciones primarias para determinar el rendimiento óptimo de los tipos celulares.
B. Caracterización in vitro de células FOXA2+/LMX1A+ procedentes de la región del mesencéfalo de la placa del suelo en comparación con células precursoras de DA generadas con otras técnicas. Se realizaron comparaciones sistemáticas de las tres condiciones de cultivo (figura 1d) usando perfiles de expresión de genes temporales globales. La agrupación jerárquica de genes expresados diferencialmente segregó las tres condiciones de tratamiento para el día 11 de diferenciación (figura 8a). FOXA1, FOXA2 y varias otras dianas de SHH corriente abajo, incluyendo PTCH1, estaban entre los transcritos con regulación más diferencial en conjuntos de tratamiento de LSB/S/F8/CHIR frente a LSB (figura 1e). La expresión de LMX1A, NGN2 y DDC indicó el establecimiento del destino precursor de neuronas DA del mesencéfalo ya para el día 11 (figura 1e, f). Por el contrario, los cultivos de LSB se enriquecieron con respecto a marcadores precursores del prosencéfalo dorsal tales como HES5, PAX6, LHX2y EMX2. La comparación directa del tratamiento con LSB/S/F8/CHIR frente a LSB/S/F8 (figura 1f) confirmó el enriquecimiento selectivo con respecto a marcadores precursores de DA del mesencéfalo en el grupo LSB/S/F8/CHIR y sugirió la identidad precursora hipotalámica en cultivos tratados con LSB/S/F8 basándose en la expresión diferencial de RAX1, SIX3 y SIX6 (véase también expresión de POMC, OTP en la figura 2d a continuación). Se muestra una lista ilustrativa de transcritos expresados diferencialmente, es decir, las tablas 1,2 y análisis de ontología génica para el día 11, figura 8b (DAVID; http://david.abcc.ncifcrf.gov) confirmaron el enriquecimiento de la señalización de WNT canónica tras el tratamiento con CHIR. Los datos sin procesar aún no están disponibles en GEO worldwideweb.ncbi.nlm.nih.gov/geo/
n.° de referencia: [por determinar]). El análisis temporal comparativo de la expresión génica para marcadores de precursores de DA del mesencéfalo (figura 1 g) frente a marcadores de destinos distintos de neuronas DA anteriores y ventrales (figura 1h) dividió las tres condiciones de inducción en: i) LSB: prosencéfalo dorsal; ii) LSB/S/F8: ventral/hipotalámico y iii) LSB/S/F8/CHIR: identidad precursora de DA del mesencéfalo.
VI. Mayor diferenciación de células FOXA2+/LMX1A+ del día 11 en neuronas DA del mesencéfalo para el día 25 y mantenidas hasta el día 65.
Para mayor diferenciación, las células precursoras FOXA2+/LMX1A+ se mantuvieron en un medio que promovía la maduración neuronal (BAGCT, véase ejemplo I). Por comparación, se usaron otras dos técnicas para generar células precursoras neuronales DA. Se realizaron los siguientes tipos de comparaciones entre las poblaciones de células diferenciadas resultantes de métodos anteriores y métodos de las presentes invenciones: A) Análisis inmunocitoquímico el día 50 de diferenciación para TFI en combinación con LMX1A, FOXA2 y NURR1, B) Cuantificación de células TH+, FOXA2+, LMX1+ y NURR1+ de las células totales que compara cultivos procedentes de la roseta frente a procedentes de la placa del suelo (LSB/S/F8/CHIR). C) Cuantificación de los porcentajes de subtipos neuronales serotonina+ (5-HT) y GABA+ (contaminantes de neuronas distintas de DA) el día 50 en cultivos de neuronas DA procedentes de la placa del suelo y la roseta. Y D) Análisis de HPLC para medir dopamina y metabolitos: Comparación de los niveles de DA, DOPAC y HVA entre cultivos procedentes de la placa del suelo frente a la roseta.
Para el día 25, tres poblaciones de células precursoras produjeron neuronas Tuj1+ (figura 2a) y células que expresan TH, la enzima limitante de la velocidad en la síntesis de DA. Sin embargo, el tratamiento con LSB/S/F8/CHIR produjo células TH+ que coexpresaban LMX1A y FOXA2 y presentaron fuerte inducción del receptor nuclear NURR1 (NR4A2) (figura 2a, b). La comparación de la expresión génica en cultivos del día 13 frente al día 25 confirmó la inducción robusta de otros marcadores de neuronas DA postmitóticas (figura 2c). La caracterización de la identidad de neuronas DA el día 25 en comparación con cultivos tratados con LSB y LSB/S/F8 confirmó el enriquecimiento de transcritos de neuronas DA del mesencéfalo conocidos e identificó múltiples marcadores candidatos novedosos (figura 2d, tablas 3 5, figura 8b). Por ejemplo, el transcrito más enriquecido en LSB/S/F8/CHIR (grupo DA del mesencéfalo) fue TTF3, un gen no asociado anteriormente con el desarrollo de neuronas DA del mesencéfalo, pero muy expresado en la sustancia negra humana (figura 8c; Allen Brain Atlas: http://human.brain-map.org).
Se obtuvieron datos similares para EBF-1, EBF-3 (figura 8c) así como TTR, una diana transcripcional conocida de FOXA2 en el hígado. Los datos obtenidos durante el desarrollo de las presentes invenciones indicaron el enriquecimiento de varios genes PITXen células precursoras de DA del mesencéfalo. PITX3, un marcador clásico de neuronas DA del mesencéfalo, también se expresó de manera sólida el día 25 de diferenciación (figura 2e). Finalmente, la inducción de neuronas tanto de la placa del suelo del mesencéfalo como DA podría reproducirse fácilmente en líneas hESC e hiPSC independientes (figura 9). Los datos demostrados en la presente memoria mostraron que el protocolo LSB/S/F8/CHIR a diferencia de otros protocolos probados produce células que expresan un perfil de marcadores que coincide con el destino de neuronas Da del mesencéfalo.
Las propiedades in vitro e in vivo de las neuronas DA procedentes de la placa del suelo se compararon con las neuronas de tipo DA obtenidas mediante un intermedio de roseta neural (figura 10 y 16). El diseño de rosetas neurales representa la estrategia más ampliamente usada en la actualidad para obtener neuronas DA de hPSC. Los protocolos basados en la placa del suelo y la roseta fueron eficaces para generar neuronas TH+ con capacidad de supervivencia a largo plazo in vitro (día 50 de diferenciación; figura 3-1a). Sin embargo, el porcentaje de células TH+ fue significativamente mayor en cultivos procedentes de la placa del suelo (figura 3-1 b). Aunque las células TH+ en ambos protocolos presentaron coexpresión de NURR1, las neuronas DA procedentes de la placa del suelo coexpresaron FOXA2 y LMX1A (figura 3-1a, b y 3-2).
Se observaron pocas neuronas positivas para GABA y serotonina (5-HT) (figura 3-1c). DA, y sus metabolitos DOPAC y HVA, estaban presentes en cultivos generados con cualquiera de los protocolos, pero los niveles de DA fueron aproximadamente 8 veces mayores en cultivos de la placa del suelo (figura 3-1 d, e). Las neuronas DA del mesencéfalo presentaron crecimiento de fibras extenso y expresión robusta de marcadores neuronales maduros, incluyendo sinapsina, transportador de dopamina (DAT) y canal de potasio de rectificación interna, acoplado a proteína G (Kir3.2, también denominado GIRK2, expresado en neuronas DA de la pars compacta de la sustancia negra (SNpc)) (figura 3-1 f, figura 11). Las neuronas DA de SNpc in vivo presentan un fenotipo electrofisiológico que las diferencia de la mayoría de las otras neuronas en el cerebro. En particular, se disparan espontáneamente a una velocidad lenta (1 3 Hz). Por otro lado, esta activación lenta está acompañada de un potencial oscilatorio lento, por debajo del umbral. Después de 2-3 semanas in vitro, estas mismas características fisiológicas son presentadas por neuronas DA de SNpc cultivadas a partir de ratones postnatales tempranos. Las neuronas DA diferenciadas a partir de hESC de manera uniforme (4/4 pruebas) presentaron este fenotipo fisiológico distintivo (figura 3-1 g-i).
El mantenimiento de neuronas mDA in vitro en d65 mostró que las neuronas positivas para TH todavía expresan FoxA2 y extienden fibras largas habituales para neuronas mDA. Figura 3-1j. La medición de la liberación de DA por HPLC mostró que neuronas TH+ de 65 días de edad son funcionales in vitro figura 3-1 k.
En resumen, en la presente memoria se describe la conversión neurogénica de precursores de la placa del suelo del
mesencéfalo y el desarrollo de un protocolo de diferenciación de neuronas DA del mesencéfalo de la placa del suelo optimizado. Se obtuvieron neuronas DA procedentes de la placa del suelo de células ES humanas después de activación basada en moléculas pequeñas de SHH y señalización de WNT canónica durante las primeras etapas de diferenciación (figura 3-2). Estas células hES progresaron desde una etapa de la placa del suelo del mesencéfalo doble positiva para FOXA2/LMX1A, a neuronas inmaduras Tuj1+ con coexpresión de FOXA2/LMX1A y después a neuronas DA maduras (figura 3-2a, b) con liberación robusta de DA y propiedades electrofisiológicas características de neuronas DA del mesencéfalo de la pars compacta de la sustancia negra (SNpc; tipo A9), incluyendo actividad autónoma de marcapasos (figura 3-2c). De manera sorprendente, este fue un proceso muy eficaz, adoptando más de la mitad de las células en la placa de cultivo que adoptaron un perfil marcador del mesencéfalo maduro (véase figura 3-2b).
VII. Caracterización de neuronas de dopamina del mesencéfalo procedentes de la placa del suelo in vivo como neuronas injertadas.
El siguiente ejemplo describe el uso de métodos ilustrativos de las presentes invenciones para su uso en el reemplazo celular terapéutico. Un desafío importante en el campo es la capacidad de generar neuronas DA del mesencéfalo procedentes de hPSC que se injerten funcionalmente in vivo sin el riesgo de hiperproliferación neural o diferenciación inadecuada en neuronas que no son del mesencéfalo o desarrollar teratomas. Basándose en estudios de trasplante de tejido fetal, los inventores contemplaron que el momento de salida del ciclo celular, marcado por la expresión de NURR1, puede ser una etapa adecuada para el injerto (aproximadamente el día 25 de diferenciación, figura 2). Los estudios iniciales que usaron células del día 25 en ratones adultos sin lesiones mostraron supervivencia robusta de las neuronas FOXA2+/TH+ procedentes de hPSC a las 6 semanas después del trasplante (figura 12). Se probó la supervivencia de las células FOXA2+/TH+ a largo plazo en hospedadores parkinsonianos sin dar lugar a hiperproliferación neural. Con este fin, se realizaron lesiones de 6-hidroxi-dopamina (6-OHDA) (Tabar, et al. Nature Med. 14: 379-381 (2008), en ratones nulos para NOD-SCID IL2Rgc, una cepa que respalda eficazmente la supervivencia de xenoinjertos con sensibilidad particular para exponer células tumorigénicas poco habituales (Quintana, et al. Efficient tumour formation by single human melanoma cells. Nature 456: 593-598 (2008). Se injertaron cultivos de neuronas DA procedentes de la placa del suelo y la roseta (150 x 103 células/animal) sin purificación previa para revelar posibles células contaminantes con alto potencial proliferativo. Cuatro meses y medio después del trasplante, los injertos de neuronas DA procedentes de la placa del suelo mostraron un núcleo de injerto bien definido compuesto por células TH+ que coexpresan FOXA2 y el marcador específico humano hNCAM (figura 4a-c). El análisis funcional mostró un rescate completo del comportamiento de rotación inducida por anfetaminas. Por el contrario, los injertos neuronales procedentes de las rosetas mostraron pocas neuronas TH+, no produjeron una reducción significativa del comportamiento de rotación (figura 4d) y mostró hiperproliferación neural masiva (volumen del injerto > 20 mm3; figura 13). La hiperproliferación extensa de células neuronales procedentes de la roseta usadas en el injerto como se informa en la presente memoria fue similar al trabajo previo con injertos de DA procedentes de la roseta del grupo de los inventores (Kim, et al. miR-371-3 Expression Predicts Neural Differentiation Propensity in Human Pluripotent Stem Cells. Cell Stem Cell 8: 695-706 (2011), incorporado en la presente memoria por referencia) y otros (Hargus, et al. Proceedings of the National Academy of Sciences of the United States of America 107: 15921-15926 (2010). La hiperproliferación probablemente se debió a los periodos de supervivencia más largos (4,5 meses frente a 6 semanas), la falta de purificación de FACS antes del trasplante y la elección de hospedador nulo para NOD-SCID IL2Rgc. El número de células Ki67+ en proliferación fue mínimo en los injertos derivados de la placa del suelo (<1 % de las células totales), mientras que los injertos procedentes de la roseta conservaban bolsas de precursores neurales proliferantes. Se cree que la hiperproliferación neural es provocado por células neuroectodérmicas anteriores primitivas dentro del injerto (Elkabetz, et al. Genes Dev. 22: 152-165 (2008); Aubry, et al. Proc. Natl Acad. Sci. U.S.A 105: 16707-16712 (2008). Esta hipótesis fue respaldada por la expresión del marcador del prosencéfalo FOXG1 en injertos procedentes de la roseta, pero no de la placa del suelo. Un pequeño porcentaje de células astrogliales estaba presente en injertos procedentes tanto de la placa del suelo como de la roseta, aunque la mayoría de células GFAP+ fueron negativas para marcadores humanos que indican el origen del hospedador (figura 13).
Los resultados en ratones nulos para NOD-SCID IL2Rgc descritos en la presente memoria demostraron supervivencia robusta a largo plazo de las neuronas FOXA2+/TH+, inversión completa del comportamiento de rotación inducida por anfetaminas y falta de ninguna señal de hiperproliferación neural. Sin embargo, algunos de estos resultados podrían atribuirse al uso específico de ratones nulos para NOD-SCID IL2Rgc. Para ensayar esta hipótesis, se trasplantaron cultivos de neuronas DA procedentes de la placa del suelo (250 x 103 células) en ratas con lesión de 6-OHDA adultas inmunodeprimidas farmacológicamente usando ciclosporina A. Cinco meses después del trasplante, la supervivencia del injerto fue robusta (figura 4e-h) con un promedio de más de 15.000 células TH+ que coexpresan FOXA2 (figura 4 g) y antígeno nuclear humano (hNA) (figura 4e); Las fibras TH+/hNCAM+ emanaron del núcleo del injerto hacia el cuerpo estriado del hospedador circundante (figura 4f). Además de FOXA2, las células TH+ expresaron marcadores de neuronas DA del mesencéfalo PITX3 y NURR1 (figura 4h-j). Los análisis conductuales mostraron un rescate completo de la asimetría rotacional inducida por anfetaminas, en contraste con animales con injertos simulados que no mostraron mejoras (figura 4k). Los animales con injertos también mostraron mejoras en la prueba de marcha (figura 4l) que mide la acinesia de las extremidades anteriores y en la prueba del cilindro (figura 4m), ensayos que no dependen de la estimulación farmacológica del sistema DA. Se espera el inicio tardío de la recuperación (aproximadamente 3-4 meses después del trasplante) para neuronas DA humanas y depende de la tasa de maduración in vivo tal como los niveles de expresión de DAT (figura 4n). La presencia de células TH+ que expresan
canales Kir3.2 (GIRK2) o calbindina indican que están presentes neuronas DA tanto de SNpc (A9) como del área tegmental ventral (A10) en el injerto (figura 4o, p).
Como en los ratones (figura 13), las células serotoninérgicas y GABAérgicas fueron poco habituales (<1 % de las células totales) en células de rata, al igual que las células gliales GFAP+ procedentes principalmente del hospedador (7 % de las células totales; (figura 14). Aunque se detectaron pocas neuronas serotonina+ en el injerto, se observaron células hNCAM negativas que probablemente eran fibras serotoninérgicas procedentes del hospedador (figura 14).
El injerto de neuronas DA procedentes de la placa del suelo en ratones, ratas y monos demostró la sorprendente recuperación de la función neuronal en especies de roedores y primates. Los ensayos de supervivencia a corto plazo (6 semanas) se ampliaron para supervivencia a plazo sorprendentemente largo de hasta 5 meses después del trasplante en el cuerpo estriado de ratón de ratones hospedadores con lesiones de 60HDA e inmunodeprimidos. De hecho, una comparación directa de un método tradicional, basado en roseta para producir neuronas DA (Perrier, et al. Proc Natl Acad Sci USA 101, 12543-8 (2004)) en comparación con el protocolo novedoso de diferenciación de neuronas DA basado en la placa del suelo descrito en la presente memoria, mostró que las neuronas DA procedentes de la placa del suelo tenían capacidad de injerto de neuronas DA a largo plazo mientras que las neuronas basadas en roseta no (figura 4a-c).
En particular, una inducción robusta de recuperación conductual en rotaciones inducidas por anfetaminas en ratones con lesión de 6-OH en los que se trasplantaron neuronas procedentes de la placa del suelo (ps) (línea azul) y neuronas DA procedentes de la roseta (línea roja) (figura 4d) mostró que las neuronas procedentes de la ps tenían mayores tasas de recuperación. Los ratones con injertos de neuronas DA procedentes de la placa del suelo mostraron una recuperación casi completa en las puntuaciones de anfetaminas. Los animales con injertos de neuronas DA procedentes de la roseta mostraron menor mejora conductual y algunos con el tiempo volvieron a números iniciales de alta rotación.
También se encontró función robusta del injerto en el modelo de rata con lesión de 6OHDA. La rata, a diferencia del ratón con EP, permite ensayos conductuales más complejos y aborda la supervivencia de neuronas DA en un entorno de xenoinjerto después de inmunodepresión farmacológica (una terapia que imita más estrechamente los protocolos de injerto humano). La excelente supervivencia del injerto, prueba del crecimiento de fibras DA y el mantenimiento de la expresión del factor de transcripción específico del mesencéfalo confirmaron la supervivencia a largo plazo de neuronas procedentes de la placa del suelo que expresan auténticos marcadores de neuronas DA del mesencéfalo (figura 4e-j). Una serie de ensayos funcionales mostró una mejora significativa en las pruebas tanto inducidas por fármacos (rotaciones inducidas por anfetaminas) como conductuales espontáneas (prueba de cilindro y marcha).
Los resultados demostrados en la presente memoria mostraron excelente supervivencia del injerto y resultado conductual en dos modelos murinos independientes. Sin embargo, el número de neuronas DA necesarias en un cerebro de ratón o rata representa una fracción pequeña del mayor número de células necesarias para injertar en primates y seres humanos. Para probar la capacidad de cambio de escala de este protocolo, se realizaron estudios piloto de injerto en dos monos rhesus adultos con lesión de MPTP.
Además, inicialmente no se sabía si los métodos y las células descritos en la presente memoria también restaurarían la función neuronal en un primate, información que podría usarse en apoyo de la habilitación para su uso de estos métodos y células en seres humanos. Por tanto, se realizó un conjunto inicial de estudios en al menos 2 monos para evaluar a corto plazo (4-6 semanas) la supervivencia in vivo y el mantenimiento del fenotipo de neuronas DA del mesencéfalo en el cerebro de los primates. Esos estudios descritos en la presente memoria, mostraron supervivencia robusta de neuronas DA del mesencéfalo TH/FOXA2 positivas y pruebas de reinervación del cuerpo estriado del hospedador (figura 4q-t). El injerto cantidades mayores de células, similares a los estimados en el intervalo numérico necesario para futuros estudios de injertos humanos, dio como resultado supervivencia robusta de las neuronas DA del mesencéfalo. Además de estos datos a corto plazo, se realizó un conjunto de estudios a más largo plazo en monos rhesus evaluando la supervivencia de las células a los 3 meses y un régimen de inmunodepresión optimizado de CellCept, Prograf y prednisona diariamente (usados como terapia triple en combinación). De manera sorprendente, se descubrió una supervivencia robusta a los 3 meses de neuronas DA del mesencéfalo procedentes de ES en el cerebro de los primates junto con una reacción inflamatoria del hospedador muy reducida a los injertos en comparación con la fuerte respuesta de la microglía del hospedador observada en el estudio de injerto inicial (véase figura 15).
De manera específica, los métodos relacionados con los estudios de monos incluyeron lotes de 50 x 106 precursores de neuronas DA trasplantables que se obtuvieron el día 25 de diferenciación usando el protocolo basado en la placa de suelo. La dosis clásica fue de 3 mg de MPTP-HCL inyectado en la arteria carótida (intervalo de 0,5 a 5 mg). Esto se siguió de inyección sistémica de MPTP 0,2 mg/kg IV de MPTP. Se inyectaron células en tres ubicaciones (núcleo caudado posterior y putamen precomisural) en cada lado del cerebro (6 tramos en total, 1,25 x 106 células/tramo) y los animales se inmunodeprimieron con ciclosporina-A. Se inyectaron precursores de DA en un lado del cerebro de un subclón de H9 que expresa GFP, mientras que en el otro lado se injertaron células procedentes de células H9 sin marcar. Se muestran resultados que muestran el injerto de neuronas en monos rhesus con expresión continua de FOX2A y producción de TH en la figura 4q-t. Un mes después del trasplante, se observó supervivencia robusta de neuronas DA del mesencéfalo en función de la expresión de GFP (figura 15) y el marcador citoplasmático específico humano (SC-121) (figura 4q). Cada núcleo de injerto estaba rodeado por un halo de fibras TH+ que se extendía hasta
3 mm dentro del hospedador (figura 4r). Los núcleos del injerto estaban compuestos de neuronas TH+ que coexpresan SC-121 (figura 4s) y FOXA2 (figura 4t). Las áreas negativas de SC-121 y GFP dentro del injerto contenían microglía del hospedador Iba1+ (figura 15) que indica inmunodepresión incompleta.
En resumen, el injerto de una nueva población de células neuronales DA en primates, es decir, monos rhesus adultos con lesión de MPTP (3 mg de MPTP-HCL (1 -metil-4-fenil-1,2,3,6-tetrahidropiridina; que varía en concentraciones de 0,5 a 5 mg de MPTP-HCl) que contenían una pérdida grave >95 % de neuronas DA endógenas del mesencéfalo. La exposición a MPTP provocó cambios y síntomas observables similares a la enfermedad de Parkinson en seres humanos.
En resumen, se descubrió un protocolo novedoso de diferenciación de hPSC basado en la placa del suelo que recapitula fielmente el desarrollo de neuronas DA del mesencéfalo. El acceso a células con las características fundamentales de neuronas DA del mesencéfalo permitirá una amplia gama de aplicaciones biomédicas, tales como estudios básicos de desarrollo, descubrimiento de fármacos de alto rendimiento y modelado de enfermedad basado en EP-iPSC. Es importante destacar que este estudio finalmente estableció un medio para obtener una fuente de neuronas FOXA2+/TH+ que se puede cambiar de escala para el trasplante neural, una etapa importante en el camino hacia la consideración de una terapia celular para la EP.
Asimismo, la obtención de neuronas DA del mesencéfalo auténticas a partir de hESC mostró excelente rendimiento in vivo (véase figura 4) y rendimiento de neuronas DA, en el momento del injerto (aproximadamente 40 % de neuronas maduras en la población el día 25-30 de diferenciación, que excedieron los porcentajes obtenidos después de la disección del tejido del mesencéfalo ventral fetal humano (normalmente aproximadamente 10 %) (Sauer, et al., Restor. Neurol. Neurosci. 2: 123-135 (1991)).
Las siguientes son breves descripciones de materiales y métodos para su uso en la evaluación de dos parámetros principales después del injerto: duración de la supervivencia y alcance de las evaluaciones conductuales. Para estudios a corto plazo, p. ej., con el objetivo de confirmar la supervivencia celular o la composición fenotípica, se contemplan evaluaciones conductuales y los animales se analizarán aproximadamente 4-8 semanas después del injerto para determinar la supervivencia. Los estudios a largo plazo incluirán la evaluación conductual después del injerto y la supervivencia de los animales durante al menos 5 meses después del injerto. Al analizar estrategias de mejora, tales como modificaciones de PSA-NCAM, se contempla que la evaluación conductual incluya parámetros más complejos, tales como la prueba de la escalera para el uso experto de las extremidades anteriores.
Los protocolos ilustrativos para la evaluación de rendimiento in vivan tienen al menos cuatro componentes principales e incluye los siguientes: i-Inducción de lesión, ii-Análisis conductual del núcleo, iii-Injerto y iv-Análisis de tejidos. Aunque estos procedimientos se usaron en ratas, en algunas realizaciones, estos procedimientos pueden usarse en otras especies.
i- Inducción de lesión. Las inyecciones unilaterales de 6-hidroxidopamina (6-OHDA) en el haz mediano del prosencéfalo (MFB) es un enfoque convencional para la inducción de síntomas de tipo Parkinsoniano en ratas. 6-OHDA es una neurotoxina que se transporta de manera retrógrada a través de la ruta nigroestriatal en el MFB de vuelta a la sustancia negra, por lo que da como resultado la muerte de células neuronales a través del deterioro de las enzimas respiratorias mitocondriales. Las neuronas dirigidas incluyen las neuronas dopaminérgicas A9 dentro de la sustancia negra compacta (SNC) así como las neuronas A10 en el área tegmental ventral. Este modelo de lesión está ampliamente estudiado y aceptado como un excelente modelo preclínico para el estudio de las consecuencias neuroquímicas y conductuales de la EP avanzada, ya que da como resultado agotamiento unilateral extenso de dopamina dentro del complejo de caudado-putamen (CPU). Las consecuencias conductuales también están bien descritas e incluyen rotaciones espontáneas e inducidas por fármacos, así como deterioro en el uso de las extremidades (véase posteriormente). Los modelos bilaterales de parkinsonismo pueden imitar mejor la enfermedad humana, pero dan como resultado adipsia y afagia en ratas.
El procedimiento se realizó en animales anestesiados (ketamina/xilazina) mediante inyección estereotáctica en 2 sitios a lo largo del haz mediano del prosencéfalo. La eficacia de la inducción de lesión completa depende en gran medida de la experiencia del operador y, durante el desarrollo de las presentes invenciones, varía de 60 a 80 %. Se permitió que los animales se recuperaran y después se sometieron a una serie de pruebas conductuales que comenzaron 2 semanas después de la cirugía.
ii- Análisis conductual central. El análisis conductual se inicia 2 semanas después de la cirugía y continúa después del trasplante hasta que se sacrifica el animal. Su fin es 1) establecer que la lesión es estable y completa y que el animal no ha presentado inversión espontánea de los síntomas, un fenómeno que se ha mostrado en animales parcialmente lesionados, 2) demostrar la influencia del trasplante de neuronas de dopamina en los parámetros conductuales establecidos.
a- Comportamiento rotacional. Las ratas se observan para rotaciones espontáneas y para rotaciones (ipsilaterales) inducidas por D-anfetamina (10 mg/kg). Se requiere un umbral de >6 rotaciones por minuto como indicador de una lesión significativa. También se pueden analizar rotaciones inducidas por apomorfina, pero se considera que los resultados positivos requieren >80-90 % de agotamiento de la inervación de dopamina en el caudado-putamen y
son menos uniformes en las lesiones de MFB, en comparación con lesiones de CPU. Se obtienen tres conjuntos de datos a intervalos de 2 semanas y se promedian. Las ratas con una puntuación de rotación de <6 no se incluyen en los estudios.
b- Prueba de marcha. Esta es una prueba para la acinesia de las extremidades anteriores. El experimentador sujeta una rata con una mano fijando las extremidades posteriores (levantando ligeramente el torso) y con la otra mano fija las extremidades anteriores que no se van a supervisar. De esta manera, la otra pata delantera tiene que soportar el peso. La rata se mueve lentamente de lado, en posición tanto del derecho como del revés. Se cuenta el número de pasos de ajuste para ambas direcciones y ambas patas.
c- Prueba de cilindro. Esta es una prueba de asimetría de uso de las extremidades anteriores. La rata se coloca en un cilindro transparente. Durante un periodo de tiempo de 5 minutos, se puntúa el comportamiento de levantamiento sobre dos patas de la rata. El comportamiento se analiza durante el levantamiento y el aterrizaje. Se determina el porcentaje de uso simultáneo y asimétrico de las patas durante estos movimientos. Esto se puede llevar a cabo a través de un sistema de supervisión de vídeo computarizado. Se sabe que los resultados negativos en estas pruebas se correlacionaban con el agotamiento de la dopamina y se mostró que los resultados mejoraron después del injerto restaurador como se describe en la presente memoria.
i¡¡- Injerto. Los animales reciben inyecciones estereotácticas de neuronas de dopamina en el cuerpo estriado, a las 3 semanas después de las lesiones inductoras de Parkinsonismo, y si las pruebas conductuales confirman una lesión adecuada. Las coordenadas para inyección están ampliamente establecidas. Después del injerto, se realiza el mismo conjunto de pruebas conductuales bimensualmente durante duraciones variables (se necesita un promedio de 5 meses para lograr recuperación estable del comportamiento).
iv- Análisis tisular. Los animales son sacrificados y perfundidos. Los cerebros se seccionan y procesan para inmunohistoquímica y análisis estereológico. Los anticuerpos incluyen TH, FoxA2, Pitx3, Nurrl, Lmxla, Girk2, DAT para la identidad y función de neuronas de dopamina; 5-HT para identificar neuronas serotoninérgicas; NCAM humano o antígeno nuclear humano para identidad humana; Nestina, Sox2 para precursores neuronales; Ki-67 para la proliferación; Oct4, Nanog para marcadores de pluripotencia; alfafetoproteína; miosina; citoqueratina para marcadores de múltiples líneas para descartar la formación de teratomas. Los parámetros cuantitativos incluyen volúmenes de injerto (estimador de Cavalieri), recuentos de células totales y recuentos de células dopaminérgicas (usando neuronas de doble marcaje TH/FoxA2) e índice de proliferación (% Ki67+). Los anticuerpos enumerados están disponibles en el mercado y se usan durante el desarrollo de las presentes invenciones.
En algunas realizaciones, se contempla el uso de ratas Sprague-Dawley (SD) para modelar mejor la situación humana. Las ratas SD recibirán inyecciones intraperitoneales diarias de ciclosporina (15 mg/kg) comenzando un día antes del injerto hasta el sacrificio. Con las inyecciones a largo plazo de la forma acuosa de ciclosporina (Neoral, una solución oral usada en seres humanos) hubo morbilidad insignificante.
En algunas realizaciones, se proporcionan métodos ilustrativos para aumentar la escala de los cultivos de neuronas mDA, véase, tabla 9. En realizaciones particulares, dichos métodos se contemplan para su uso en la producción de cultivos a nivel de GMP para su uso clínico.
Parámetros de evaluación. Se contemplaron pruebas in vivo para su uso en métodos a corto y largo plazo de eficacia del injerto. Los resultados obtenidos durante el desarrollo de las presentes invenciones mostraron una caracterización tisular idéntica en ambos casos, con la expectativa de una mayor proporción de células TH+ en los supervivientes a largo plazo, debido a la diferenciación de precursores injertados.
En promedio, cuando se trasplantaron 15.000 neuronas TH+/FoxA2+ por cada 250.000 células, los animales sobrevivieron 5 meses después del injerto. Significativamente menos células con doble marcaje sobrevivieron y se contaron en supervivencias cortas. Los rendimientos in vivo contemplados de este modo, como se muestra en la tabla 9, son estimaciones moderadas. Se muestran parámetros ilustrativos usados para evaluar un estado aprobado/suspenso para los resultados de la composición de injerto en la tabla 10.
En injertos a largo plazo, la evaluación conductual es un componente esencial del rendimiento de los productos de la línea hES. Se describieron mejor las pautas que definen la pérdida de función y la recuperación para las rotaciones por anfetaminas, por lo que un injerto robusto debería normalizar el comportamiento y ocasionalmente provocar un movimiento contralateral debido al desequilibrio de DA. Los límites enumerados en la tabla 11 son pautas ejemplares según lo determinado durante el desarrollo de las presentes invenciones.
En algunas realizaciones, las fuentes celulares para usar en los métodos de diferenciación de las presentes invenciones incluyen, pero sin limitación, líneas celulares WA09, ACT (M09), Bio-Time y Roslin. En algunas realizaciones, las fuentes celulares para usar en los métodos de diferenciación de las presentes invenciones incluyen, pero sin limitación, líneas de uso en GMP. En algunas realizaciones, las fuentes celulares para usar en los métodos de diferenciación de las presentes invenciones incluyen, pero sin limitación, la producción de neuronas DA maduras para usar en análisis de supervivencia de injerto a corto plazo. En algunas realizaciones, las fuentes celulares para usar en los métodos de diferenciación de las presentes invenciones incluyen, pero sin limitación, la producción de neuronas DA maduras para usar en experimentos de injerto que incluyen evaluaciones conductuales. Los controles
consistirán en WA09 de uso en investigación y un grupo de solución salina simulada. Los análisis estadísticos usarán ANOVA con la prueba post-hoc de Punnett.
A. Ensayos conductuales complejos para evaluar el grado de reinervación estriatal. Las pruebas conductuales convencionales (como se describe en la presente memoria) presentaron una correlación directa con el número de neuronas de dopamina supervivientes dentro del injerto. En ratas tratadas con células neuronales DA maduras de las presentes invenciones, las pruebas conductuales muestran recuperación máxima con una estimación de aproximadamente 30 % de recuperación de neuronas DA que es necesaria para reducir las rotaciones inducidas por anfetaminas. Por tanto, una vez que se alcanza un umbral para la supervivencia de las neuronas DA en el hospedador (una estimación de 800 - 1200 neuronas DA en el modelo de rata), puede ser difícil distinguir diferencias conductuales que reflejen estrategias de mejora. Se describió un enfoque de microtrasplante que da como resultado la colocación de múltiples injertos pequeños en todo el cuerpo estriado, a diferencia de injertos grandes. Mientras se controlaba el número total de células DA trasplantadas, se observaron diferencias en los resultados conductuales al comparar 2 grupos: Las ratas con múltiples injertos pequeños presentaron recuperación conductual anterior y más extensa en las rotaciones inducidas por fármacos y las pruebas de marcha en comparación con animales con un solo injerto grande y la misma cantidad de neuronas DA y la misma cantidad de liberación de dopamina. De manera sorprendente, hubo una diferencia definida en el uso experto de las extremidades anteriores, por lo que los animales con injertos pequeños ampliamente distribuidos mostraron una mejora, mientras que las ratas que portaban injertos convencionales no lo hicieron. El uso experto de las extremidades anteriores se considera una tarea que requiere control espacial y temporal sobre la liberación de DA y, por tanto, una conectividad adecuada entre las neuronas injertadas y el cuerpo estriado del hospedador. Por tanto, en algunas realizaciones, el uso de neuronas DA maduras de las presentes invenciones en procedimientos de injerto dio como resultado la recuperación del uso de las extremidades anteriores. Por consiguiente, en algunas realizaciones, se administran neuronas DA maduras de las presentes invenciones en una ubicación del cuerpo estriado. En otras realizaciones, se administran neuronas DA maduras de las presentes invenciones en al menos 2 o más ubicaciones dentro del cuerpo estriado.
B. Prueba de uso experto de las extremidades anteriores (o la escalera). La prueba de las extremidades anteriores analiza el alcance y agarre de las extremidades anteriores y se realizó antes y después de la lesión y después del trasplante. Los animales fueron privados de alimentos durante 48 horas antes de la prueba y se probaron diariamente durante 5 días antes de la lesión y después dos veces tras la lesión a intervalos de 3 semanas. Después del injerto, la prueba se repite 2 veces más, a los 3 y 5 meses. Los animales se colocaron en una cámara de plexiglás equipada con una escalera doble. Los microgránulos de alimentos se colocan en 5 etapas bilateralmente para la prueba previa al injerto y unilateralmente (en el lado de la extremidad afectada, contralateral a la rotación) después del injerto. Los animales se prueban durante un periodo de tiempo establecido (p. ej., 10 minutos) y se calcularon los números y la proporción de gránulos que fueron consumidos (alcance exitoso) y los gránulos que se tomaron y se compararon con el rendimiento previo a la lesión del animal individual. Hubo varias variaciones de esta prueba que implicaron el número de repeticiones y el momento de la prueba.
VIII. Las células diferenciadas mediante el uso de métodos descritos en la presente memoria mostraron respuestas electrofisiológicas similares a las células DA en el sitio.
El siguiente ejemplo describe el uso de métodos ilustrativos de las presentes invenciones para determinar la capacidad funcional de las neuronas DA del mesencéfalo resultantes de la diferenciación por métodos descritos en la presente memoria. Las neuronas DA de pars compacta de la sustancia negra (SNpc) in vivo presentan un fenotipo electrofisiológico que las diferencian de la mayoría de las otras neuronas en el cerebro. En particular, se disparan espontáneamente a una velocidad lenta (1-3 Hz). Por otro lado, esta activación lenta está acompañada de un potencial oscilatorio lento, por debajo del umbral. Después de 2-3 semanas in vitro, estas mismas características fisiológicas son presentadas por neuronas DA de SNpc cultivadas a partir de ratones postnatales tempranos. Para determinar si las neuronas mDA de las presentes invenciones mostraron un fenotipo electrofisiológico comparable, las neuronas mDA de las presentes invenciones se analizaron para determinar su firma de respuesta eléctrica. Las neuronas DA del mesencéfalo de las presentes invenciones el día 80-100 de cultivo se probaron mediante registro de células individuales. Estas neuronas mDA diferenciadas de hESC presentaron de manera uniforme (pruebas 4/4) este fenotipo fisiológico distintivo al mostrar un comportamiento de activación autónomo específico y cambios del potencial de membrana oscilatorio (figura 3 g-i). Este comportamiento se conoce como actividad de marcapasos autónoma y una propiedad específica de las neuronas DA del mesencéfalo y, en particular, el subtipo de neuronas de dopamina del mesencéfalo más relevantes para la enfermedad de Parkinson (neuronas DA del mesencéfalo de tipo sustancia negra).
Se contemplan mediciones electrofisiológicas para su uso en preparaciones de corte agudo, es decir, de biopsias de áreas con injerto. En una realización, se identificarán neuronas DA procedentes de injerto de tipo A9 frente a A10 in vivo en función de pruebas para la actividad de marcapasos autónoma que es específica para las neuronas de dopamina de tipo A9 que están más afectadas en la EP. En otras palabras, las neuronas de tipo A10 no tienen actividad de marcapasos.
Se establecieron condiciones para el registro in vivo de neuronas DA procedentes de células madre pluripotentes humanas en preparaciones de corte agudo, véase, figura 26. De manera específica, se midieron las neuronas DA humanas injertadas procedentes de células madre pluripotentes y se descubrió que tienen características
electrofisiológicas típicas de las observadas en la pars compacta de sustancia negra de ratón (SNpc), La figura 26A, donde la vista superior muestra la reconstrucción de una neurona marcapasos en la región del injerto. La parte inferior muestra una microfotografía ilustrativa de un corte cerebral tomado de la rata en la que se inyectaron las neuronas procedentes de hES 9 meses antes; el injerto está delineado; una imagen de mayor aumento se muestra insertada en la parte inferior. El corte se procesó para determinar la tirosina hidroxilasa que aparece en blanco, figura 26B. Además, la vista superior muestra un parche ilustrativo unido a la célula que registra desde una supuesta neurona DA en el injerto; la parte inferior muestra un registro ilustrativo de célula completa de la misma célula. Se realizaron registros en presencia de glutamato y antagonistas del receptor de GABA (AP550 pM, CNQX 10 pM y GABAzina 10 pM) para eliminar la entrada sináptica. Estos registros demostraron que las neuronas procedentes de PS eran marcapasos autónomos con trayectorias de tensión intrasomática normales. Otra neurona registrada en una muestra de injerto tenía propiedades similares, figura 26C. A efectos de comparación, se muestran registros unidos a células y de células completas de una neurona dopaminérgica en SNpc de un ratón adulto. Abreviaturas (CTx= corteza, STr= cuerpo estriado, SNpc= pars compacta de sustancia negra, DA= dopaminérgica). Estos datos muestran estudios in vivo funcionales en el cuerpo estriado de rata con injerto meses después del trasplante. Por tanto, en algunas realizaciones, los estudios funcionales in vivo en tejido injertado demuestran la recuperación de pars compacta de sustancia negra (SNpc).
IX. La diferenciación directa de células EP-iPS genéticas mutantes PINK1 (mutación PINK1) en neuronas DA reveló anomalías de tipo Parkinson en las neuronas DA maduras.
Este ejemplo describe el descubrimiento de que grandes poblaciones de neuronas DA del mesencéfalo se desarrollaron con características de las neuronas de un paciente con EP cuando la línea celular de un paciente con EP, es decir, célula EP-iPSC mutante PINK1, obtenida de una manera que no dio como resultado la destrucción de un embrión, se usaron como la población celular para obtener neuronas DA FOXA2/L1M1XA/TH+ de las presentes invenciones.
En una realización, los inventores contemplan aislar una población celular inicial de un paciente para su uso en los métodos de fabricación de neuronas DA auténticas in vitro, donde el paciente tiene un síntoma de la enfermedad de Parkinson (EP), para la ventaja potencial de usar las células tratadas en pruebas in vitro para 1) observar la diferenciación o anomalías funcionales en comparación con las neuronas DA auténticas de seres humanos que no tienen un síntoma neurológico, después 2) usar una anomalía observada para desarrollar un tratamiento terapéutico para invertir esa anomalía y 3) tratar al paciente con el tratamiento terapéutico para reducir, es decir, invertir, un síntoma de la enfermedad de Parkinson.
En una realización, los inventores contemplan aislar una población celular inicial del mismo paciente para obtener neuronas DA auténticas para su uso en el tratamiento de trasplante, donde el paciente tiene un síntoma de la enfermedad de Parkinson (EP), por la ventaja potencial de reducir el rechazo inmunológico, es decir, rechazo de trasplante. En otras realizaciones, se contempla reducción del rechazo de trasplante mediante el uso de una fuente celular inicial aislada de un ser humano cuyo antígenos mayores de histocompatibilidad (MHC) coinciden (es decir, gemelo) o un ser humano que tenga una coincidencia tisular de MHC aceptable para el trasplante (tal como un pariente del paciente o un ser humano no relacionado que expresa moléculas de MHC superpuestas.
A. La diferenciación dirigida mostró que las células EP-iPS genéticas células PINK1 contenían la capacidad de desarrollarse en neuronas DA de tipo mesencéfalo. Véase, figuras 20-25. Se distinguió una línea de EP-iPSC mutante PINK1 Q456X usando el protocolo (método) novedoso de neuronas DA del mesencéfalo basado en la placa del suelo descrito en la presente memoria que produjo neuronas DA del mesencéfalo que expresaron perfiles de diferenciación comparables a los obtenidos de la línea H9 diferenciada por protocolo novedoso de neuronas DA del mesencéfalo basado en la placa del suelo. (Figura 20). Este ejemplo describe el descubrimiento de que grandes poblaciones de neuronas DA del mesencéfalo se desarrollaron con características de las neuronas de un paciente con EP cuando la línea celular de un paciente con EP, es decir, célula EP-iPSC mutante PINK1, obtenida de una manera que no dio como resultado la destrucción de un embrión, se usaron como la población celular para obtener neuronas Da FOXA2/L1M1XA/TH+ de las presentes invenciones.
La línea de EP-iPSC mutante PINK1 Q456X se diferenció usando el protocolo (método) novedoso de neurona DA del mesencéfalo basado en la placa del suelo de las presentes invenciones que produjo perfiles de diferenciación de mesencéfalo comparables a los obtenidos de la línea de iPSC H9. A-C) Análisis inmunocitoquímico de la línea EP-iPSC mutante PENK1 el día 11 de diferenciación (etapa precursora del mesencéfalo) para FOXA2 (rojo), LMX1A (verde) y DAPI (azul) (A), día 25 de diferenciación (etapa neuronal DA postmitótica temprana) para FOXA2 (rojo) y TH (verde) (B) y para NURR1 (rojo) y TH (verde) (C). D-F) El mismo conjunto de análisis inmunocitoquímicos realizados usando células procedentes de H9 el día 11 de diferenciación para FOXA2 (rojo), LMX1A (verde) y DAPI (azul) (D), el día 25 de diferenciación para FOXA2 (rojo) y TH (verde) (E) y para NURR1 (rojo) y TH (verde) (F).
B. EP-iPSC genética expresó un fenotipo de tipo EP de agregación de proteínas. Figuras 21-24. Los inventores descubrieron que EP-iPSC mutantes PINK1 mostraron pruebas de expresión de a-sinucleína (componente principal del cuerpo de Lewy en paciente con EP) en el citosol de neuronas Da TH+ el día 55 de diferenciación usando el protocolo novedoso de inducción de neuronas DA del mesencéfalo basado en la placa del suelo, (figura 21a-b). A, B) Análisis inmunocitoquímico de la línea EP-iPSC mutante PINK1 el día 55 de diferenciación para a-sinucleína (LB509,
rojo), TH (verde) e imagen fusionada (A) y a-sinucleína (rojo) y ubiquitina (verde) (B). Estas células positivas para asinucleína también mostraron alta expresión de ubiquitina (marcador clásico de cuerpos de Lewy). Por el contrario, las neuronas DA procedentes de la línea iPS de control mostraron expresión de a sinucleína sináptica normal (a diferencia de citosólica) y niveles muy bajos de ubiquitina (figura 21c-d). C, D) Análisis inmunocitoquímico de la línea iPSC de control el día 55 de diferenciación para a-sinucleína (rojo) y TH (verde) (C) y a-sinucleína (rojo) y ubiquitina (verde) (D).
C. Expresión de la forma agregada de a-sinucleína. En el cerebro del paciente con EP, la forma insoluble dimerizada de a-sinucleína conduce a la agregación en cuerpos de Lewy. La forma dimerizada de a-sinucleína muestra fosfolilación de serina 129 en a-sinucleína. El mismo día de diferenciación, las células procedentes de EP-iPSC mutantes PINK1 mostraron fuerte expresión de a-sinucleína fosforilada en Ser129 en contraste con las células procedentes de iPSC de control que mostraron niveles de expresión muy bajos (figura 22).
Las células procedentes de EP-iPSC mutantes PINK1 mostraron fuerte expresión de a-sinucleína fosforilada en Ser129 en contraste con las células procedentes de iPSC de control que mostraron niveles de expresión muy bajos. A, B) Análisis inmunocitoquímico para a-sinucleína fosforilada en Ser129 (verde) y DAPI (azul) en células procedentes de EP-iPSC mutantes PINK1 el día 55 de diferenciación (A) y células procedentes de iPSC de control coincidentes (B).
D. Se observan diferencias en los patrones de expresión de a-sinucleína dependiendo del protocolo de diferenciación. Los inventores contemplaron que las neuronas DA del mesencéfalo "auténticas" procedentes de la placa del suelo mostraban vulnerabilidad específica de la EP y fenotipos correspondientes, específicos, in vitro. Las neuronas DA obtenidas usando el protocolo de diferenciación basado en alimentador estromal clásico MS5 (Perrier et al., PNAS 2004, incorporado en la presente memoria por referencia) produjo grandes cantidades de neuronas TH+. Sin embargo, en función de datos obtenidos durante el desarrollo de las presentes invenciones, los inventores mostraron que las células TH+ basadas en MS5 no eran auténticas neuronas DA del mesencéfalo procedentes de la placa del suelo. En cultivos diferenciados a través del protocolo MS5, había muchas células positivas para asinucleína. Sin embargo, esas células no coexpresaron TH. Por otro lado, no hubo ninguna diferencia en los patrones de expresión entre EP-iPSC e iPSC de control cuando se usó la estrategia de diferenciación MS5 (figura 23a-b). Estos datos indican que a-sinucleína también se expresa en otros tipos celulares distintos de DA y que dicha a-sinucleína distinta de DA permanece sin cambios en las células procedentes de iPSC de enfermedad frente a control, en particular cuando se usen protocolos de diferenciación MS5 convencionales. Estas son las neuronas procedentes de la roseta de tipo DA presentadas en las publicaciones (p. ej.: Perrier PNAS 2004). Esas células TH+ basadas en MS5 (=tipo DA) se usan para comparación en las figuras 3, 10, 13 y 16. Estos datos indican que a-sinucleína también se expresa en otros tipos celulares distintos de DA y que dicha a-sinucleína distinta de DA permanece sin cambios en las células procedentes de iPSC de enfermedad frente a control, en particular cuando se usen protocolos de diferenciación MS5 convencionales. Finalmente, el nuevo protocolo de diferenciación basado en la placa del suelo descrito en la presente memoria, produce gran cantidad de células TH+ que coexpresan a-sinucleína. Esas células TH+ expresan a-sinucleína en un patrón de expresión citosólica. Figura 24A, B). Análisis inmunocitoquímico para a-sinucleína (LB509, rojo), TH (verde) de la línea EP-iPSC mutante PINK1 el día 60 de diferenciación basada en MS5 (A) e iPSC de control (B). C) Análisis inmunocitoquímico de la línea de EP-iPSC mutante PINK1 el día 55 de diferenciación basada en la placa del suelo para a-sinucleína (rojo), TH (verde).
E. Las neuronas DA procedentes de células EP-iPS genéticas son más vulnerables a la estimulación tóxica. Figura 24-25. Las neuronas DA TH+ procedentes de EP-iPSC obtenidas a través del protocolo basado en la placa del suelo fueron más vulnerables a la exposición a toxinas (valinomicina: ionóforo de mitocondrias, 5 uM (intervalo de concentración de 1-10 uM), 48 h) que las células procedentes de iPSC de control. Por el contrario, las neuronas TH+ obtenidas a través del protocolo clásico basado en MS5 no mostraron vulnerabilidad diferencial entre las células procedentes de EP frente a control (figura 24). El ensayo de viabilidad celular completa con azul alamar después de 48 h de tratamiento con valinomicina también mostró supervivencia celular diferencial en un intervalo de dosis específico para exposición a toxinas (5 y 10 pM) al comparar EP-iPSC e iPSC de control (figura 25). Condición normal de cultivos procedentes tanto de EP como de iPSC de control obtenidos a través del protocolo basado en MS5 (D, se muestran células procedentes de EP-iPSC), neuronas TH+ después de la exposición a toxinas en EP-iPSC (E) y cultivos procedentes de iPSC de control (F) obtenidos a través del protocolo MS5. G-H) imágenes de baja potencia de inmunocitoquímica para Tuj1 (rojo) y TH (verde) mediante protocolo basado en la placa del suelo el día 60 de diferenciación: EP-iPSC de condiciones normales (G), frente a exposición a toxina (H) e iPSC de control de condiciones normales (I), frente a exposición a toxina (J). K-N) imágenes de baja potencia de inmunocitoquímica para Tuj1 (rojo) y TH (verde) mediante protocolo basado en MS5 el día 60 de diferenciación: EP-iPSC de condiciones normales (K), frente a exposición a toxina (L) e iPSC de control de condiciones normales (M), frente a exposición a toxina (N).
F. Cuantificación ilustrativa de la viabilidad celular - ensayo de respuesta a dosis para exposición a toxinas.
El ensayo de viabilidad celular con azul alamar después de 48 h de tratamiento con valinomicina mostró supervivencia celular diferencial en un intervalo de dosis específico para exposición a toxinas (5 y 10 pM) al comparar EP-iPSC e iPSC de control (día 60 de diferenciación basada en la placa del suelo). Nota: este ensayo prueba la muerte celular general mientras que los efectos más drásticos se observaron específicamente en neuronas Da (véase figura 14). Por lo tanto, la cuantificación basada en azul de alamar probablemente subestimará el alcance del efecto diferencial observado en linajes de neuronas DA.
X. Cultivo contemplado a gran escala usando composiciones y métodos de las presentes invenciones para proporcionar neuronas mDA ilustrativas.
Las descripciones en la presente memoria muestran métodos y usos ilustrativos para producción a gran escala de células neuronales mDA resultantes de diferenciación por composiciones y métodos descritos en la presente memoria. Se mostró que la generación que puede cambiarse de escala (es decir, métodos contemplados como exitosos para generar células neuronales mDA a partir de cultivos que contienen una cantidad relativamente pequeña de células) produjo poblaciones celulares con capacidad de neuronas mDA trasplantables el día 25. En particular, para células iPSC PINK. Véase, tabla 9).
XI. Métodos para enriquecer con respecto a neuronas DA del mesencéfalo.
Se desarrollaron y probaron varios métodos antes y durante el desarrollo de las presentes invenciones con el objetivo de enriquecer poblaciones celulares con respecto a precursores de neuronas DA del mesencéfalo superando problemas tal como agotando poblaciones de células contaminantes, incluyendo, pero sin limitación, células madre pluripotentes contaminantes. Se realizaron estudios iniciales usando neuronas embrionarias primarias de ratón, neuronas embrionarias de rata y poblaciones procedentes de ESC de ratón. Además de tener objetivos de aumentar las poblaciones neurales para su uso en el enriquecimiento de neuronas DA, los estudios de ESC en ratones incluyeron el desarrollo de procedimientos para prevenir la formación de teratomas, lo que fue problemático en procedimientos anteriores. Estas estrategias incluyeron la selección negativa para marcadores de superficie celular expresados en células pluripotentes (tales como SSEA1) junto con selección positiva de células que expresan marcador neural (NCAM). Usando células de ratón, se propusieron varias estrategias indicadoras genéticas para su uso en la identificación del enriquecimiento de células neuronales en paradigmas de trasplante de neuronas DA (por ejemplo, identificación de células con expresión de SOX1, Corin/Lmx1a, Ngn2, TH, Pitx o DAT). Se realizaron pruebas funcionales en células primarias de ratones indicadores de Ngn2 (Thomposon et al., Exp Neurol. 198 (1): 183-98 (2006)) y de ratones indicadores de Lmx1A también clasificados con respecto a Corin (Jonsson, Exp Neurol. 219(1): 341-54 (2009). Para poblaciones procedentes de ESC de ratón, se realizaron estudios usando líneas indicadoras de ESC de ratón SOX1 (Barraud et al., Eur J Neurosci. 200522(7): 1555-69), TH (Kelly et al., Minerva Endocrinol. 1991 16(4): 203-6), Pitx3 (Hedlund et al., Stem Cells. 2008 26(6): 1526-36) y DAT (Zhou et al., Stem Cells. 200927 (12): 2952-61). Además, durante el desarrollo de las presentes invenciones, los inventores realizaron un estudio exhaustivo de trasplante comparando directamente el rendimiento in vivo de tres poblaciones purificadas procedentes de ESC de ratón que representan etapas secuenciales del desarrollo de neuronas DA: Precursores del mesencéfalo (Hes5::GFP), células postmitóticas tempranas (Nurr1::GFP) y neuronas DA maduras (Pitx3::YFP). Esos estudios identificaron el desarrollo de DA que expresan Nurr1 como particularmente adecuadas para el injerto y demostraron que las neuronas DA purificadas tienen capacidad de injerto eficaz in vivo. Asimismo, estos resultados se usaron para la selección de células el día 25 de diferenciación (inicio de la expresión de Nurr1) para su uso en el injerto de neuronas hESC-DA en modelos de ratón, rata y mono rhesus de EP.
Durante el desarrollo de las presentes invenciones, la producción de neuronas DA del mesencéfalo auténticas a partir de hESC mostró excelente rendimiento in vivo (véase figura 4) y el uso de protocolos descritos en la presente memoria dio como resultado un rendimiento de neuronas DA, en el momento del injerto de aproximadamente 40 %. Este porcentaje superó los porcentajes de neuronas DA obtenidas después de la disección del tejido del mesencéfalo ventral fetal humano (normalmente aproximadamente 10 %). Por tanto, el uso de una fuente de neuronas DA basada en hESC como se describe en la presente memoria se contempla incluso para mejoras adicionales en la pureza cuando se usan estrategias de purificación celular. Además, se desarrollaron líneas indicadoras genéticas para su uso con hESC similares a las usadas en estudios con ratones, incluyendo una línea celular humana como una línea Nurr1::GFP. Sin embargo, el uso de indicadores genéticos puede ser problemático para el uso traslacional en ser humanos porque GFP es inmunogénico en seres humanos, por tanto, no es adecuado para su uso humano. Asimismo, la clasificación de FACS puede ser problemática para establecer bancos celulares maestros de neuronas DA de uso clínico (es decir, desarrollar reservas congeladas de neuronas DA humanas para su uso en el trasplante) dada la cantidad de tiempo que se requeriría para clasificar lotes de las aproximadamente 109 células necesarias para cada trasplante, además de costes potencialmente altos y menor rendimiento celular después de la recuperación del almacenamiento.
A diferencia de los sistemas indicadores genéticos y el aislamiento celular basado en FACS, los inventores contemplaron el aislamiento de neuronas DA basadas en la expresión de marcadores de superficie usando estrategias alternativas para técnicas de separación celular contempladas para su uso en pacientes con EP. Por ejemplo, la clasificación de perlas magnéticas (p. ej., sistema CliniMACS®) fue ampliamente usada en aplicaciones aprobadas por la FDA, basadas en células y permitieron un aislamiento rápido y rentable de hasta 1010 células en condiciones compatibles con GMP, es decir, condiciones aprobadas para aislar células para su uso en seres humanos. Por tanto, en algunas realizaciones, se contempla la clasificación de perlas magnéticas para el enriquecimiento de neuronas DA maduras, por ejemplo, usando CD142 unido a perlas magnéticas para enriquecer neuronas Nurr1+ para su uso en injertos.
XII. Identificación de marcadores de superficie celular para su uso en métodos para proporcionar neuronas DA maduras.
Los datos de expresión de marcadores de superficie celular recopilados durante el desarrollo de las presentes invenciones mostraron la identificación de varios marcadores novedosos de superficie celular expresados en neuronas DA del mesencéfalo. De manera específica, se encontraron marcadores para identificar más células, tales como células específicas que madurarían en neuronas DA, neuronas DA maduras de las presentes invenciones y células A9. Se usaron dos estrategias principales para identificar dichos marcadores de superficie: en primer lugar, una exploración de expresión génica imparcial en líneas indicadoras genéticas (figura 27a) mostró varios marcadores candidatos, incluyendo CD142 y un marcador denominado DCSM1, que se expresó selectivamente en neuronas DA del mesencéfalo y parecía marcar de manera específica neuronas DA de tipo A9 (figura 27b. Una segunda estrategia fue el uso de una exploración de marcador de superficie celular CD en neuronas DA procedentes de hESC que se probaron para 242 anticuerpos disponibles en el mercado en formato de 96 pocillos (figura 27c, d). Los resultados de dicha exploración ilustrativa (figura 27e) condujeron a la identificación de al menos 5 marcadores validados enriquecidos en neuronas DA del mesencéfalo, incluyendo CD142, un marcador que marcó selectivamente una etapa de neuronas DA Nurr1+ (figura 27f), además de, Cd 63, CD99 y DCSM1.
De manera específica, como se ilustra en la figura 27, una exploración de marcadores de superficie CD para neuronas DA procedentes de WA09 el día 25 de diferenciación probadas para hasta 242 anticuerpos individuales. Estos resultados se compararon con exploraciones duplicadas de una amplia gama de otros tipos celulares neurales procedentes de w A09 (p. ej., motoneuronas HB9::GFP+ procedentes de hESC, neuronas corticales procedentes de hESC, precursores del prosencéfalo ventral Nkx2.1::GFP+ de hESC y varios otros tipos de neuronas procedentes de hESC. La base de datos resultante del perfil de expresión de marcadores de superficie se usó después para seleccionar marcadores CD candidatos enriquecidos selectivamente en cualquier subtipo dado tal como neuronas DA del mesencéfalo (figura 27). Uno de los marcadores descubiertos asociados con la diferenciación de neuronas DA hESC fue CD142. La selección de células CD142 se enriqueció específicamente con respecto a neuronas DA procedentes de hESC en la etapa Nurr1+ mientras se agotaban otros subtipos de neuronas. En algunas realizaciones, CD142 se expresa antes de Nurr1+. En algunas realizaciones, una población de células neuronales DA del mesencéfalo clasificada para CD142 tiene células Nurr1+ y Nurr1-. En algunas realizaciones, una población de células neuronales DA del mesencéfalo clasificada para CD142 tiene células Nurr1-. En algunas realizaciones, la población de células neuronales DA del mesencéfalo clasificadas Nurr1- CD142+ cultivada comienza a expresar Nurr1 (es decir, se convierte en Nurr1+) hasta dos días después de la clasificación.
Además de CD142, CD63 y CD99 fueron marcadores enriquecidos en neuronas DA procedentes de hESC. Por tanto, en algunas realizaciones, los cultivos neuronales DA se enriquecen para neuronas DA clasificando o seleccionando a partir de marcadores incluyendo, pero sin limitación, CD142, CD63, CD99, DCSM1, Nurr1+, etc. CD142 normalmente marca aproximadamente 30 % de la población celular total el día 25 de diferenciación (figura 28a). La selectividad de CD142 para la etapa de neurona DA Nurr1+ se confirmó en múltiples líneas independientes de hESC y hiPSC (figura 28b). Es importante destacar que, además de enriquecer con respecto a neuronas DA, CD142 agota selectivamente otros subtipos de neuronas tales como neuronas GABA y serotoninérgicas. (Figura 28c-f). Se realizaron estudios in vivo que demostraron la capacidad de CD142 para dar lugar a injertos de neuronas DA de alta pureza sin neuronas GABA y serotoninérgicas contaminantes detectables. Las neuronas serotoninérgicas son un tipo celular que se ha implicado en el injerto de tejido fetal humano como la fuente potencial de discinesias inducidas por injerto. Aunque los métodos de injerto descritos en la presente memoria que usan células no purificadas ya dieron como resultado pocas neuronas serotoninérgicas, el uso de CD142 debería reducir adicionalmente este riesgo.
A. Marcadores para identificar neuronas mDA maduras de tipo A9. Se descubrió que las neuronas DA procedentes de A9 frente a procedentes de A10 tenían propiedades funcionales in vitro e in vivo definidas y patrones de inervación específicos de su papel en la función mesoestriatal frente a mesolímbica. Durante el desarrollo de las presentes invenciones, los inventores descubrieron que las neuronas mDA auténticas producidas por los métodos de las presentes invenciones (figura 4) dieron lugar a neuronas con más características A9 que A10. En particular, las neuronas mDA auténticas que eran TH+ expresaron al menos en parte Girk2, un marcador usado para definir las neuronas DA de tipo A9. Además, muchas neuronas DA maduras presentaron actividad de marcapasos autónoma que es una característica funcional presente en neuronas DA de tipo A9 pero no A10. Sin embargo, algunas células TH+ generadas in vitro no eran de identidad A9. Por tanto, los inventores contemplaron procedimientos de enriquecimiento tales como los descritos en la presente memoria, para proporcionar poblaciones purificadas de neuronas mDA auténticas de tipo A9 humanas (frente a A10). Como se describe en la presente memoria, los inventores descubrieron al menos dos marcadores únicos para neuronas de tipo A9 y al menos dos marcadores únicos para al menos neuronas de tipo A10. Por tanto, en algunas realizaciones, se identifican neuronas de tipo A9 mediante marcadores (Girk2, Aldh1) frente a A10 (Calbindina, Otx2).
B. Definición de un conjunto de marcadores que mejorar el rendimiento de la neurona DA del mesencéfalo con un subtipo A9. Se contemplaron al menos dos estrategias para definir marcadores de superficie específicos de A9: Los marcadores candidatos se obtuvieron de una exploración de expresión génica, tal como se describe en la presente memoria, y los anticuerpos de CD candidatos de una exploración de marcador de superficie, como se describe en la presente memoria. Poblaciones En otro método, análisis de transcriptoma global en poblaciones purificadas de neuronas mDA procedentes de ESC de ratón en distintas etapas de diferenciación (usando tecnología transgénica BAC; véase figura 27a, b). Se descubrieron marcadores de superficie en un perfil de marcador de superficie en neuronas DA procedentes de WA09 RCB con los siguientes métodos ilustrativos. Las neuronas DA procedentes de RCB WA09 el día 25 de diferenciación se disociaron y se volvieron a sembrar en placas de 96 pocillos, seguido de
exposición a los anticuerpos CD 242 y análisis de datos usando el explorador de alto contenido Operetta. Se probó la cantidad de enriquecimiento de DA para al menos 5 anticuerpos adicionales que se unían con marcadores de CD identificados en estas exploraciones (por ejemplo, CD142, CD63 y CD99). Las células candidatas CD-positivas frente a CD-negativas se evaluaron usando los ensayos de DA QC, incluyendo la expresión de FOXA2/TH y TH/Nurr1 (véase tabla 7). En algunas realizaciones, se contemplan perfiles globales de expresión génica para comparación de células no clasificadas con CD142+.
En algunas realizaciones, se usaron células con expresión clasificadas/separadas de un marcador deseado en estudios in vivo a corto plazo y largo plazo como se describe en la presente memoria. Entre los marcadores específicos de neuronas DA identificados en estos estudios se encontraba un gen marcador de superficie que se denominó DCSM1 (marcador 1 de superficie celular DA). En función de datos de expresión in situ, expresión en el mesencéfalo ventral, pero más sorprendentemente, se descubrió que la expresión es al menos parcialmente selectiva de A9, tanto en el cerebro del ratón en desarrollo como en el adulto (figura 27) y en el cerebro humano adulto. Se observaron numerosas células que expresan expresión de DCSM1 en neuronas DA procedentes de hESC. Los ensayos in vitro para identidad de A9 frente a A10 incluyeron la diferenciación a largo plazo de células marcador+ (día 50 y día 75 de diferenciación) y el análisis de i) expresión para marcadores A9, (Girk2, Aldh1) frente a A10 (Calbindina, Otx2) en neuronas maduras, (ii) respuestas diferenciales de guía axónica a Netrina-1 y Sema3 y, (iii) las neuronas enriquecidas en A9 se evaluaron mediante pruebas de electrofisiología. Las neuronas DA A9 presentaron características funcionales específicas como se describe en la presente memoria para neuronas A9 procedentes de hESC. Se realizaron estudios in vivo para células que expresan DCSM1 y otros marcadores para confirmar i) expresión del marcador A9 (Girk2, Aldh1) frente a A10 (Calbindina, Otx2), ii) crecimiento de fibra DA injertada y iii) propiedades electrofisiológicas de A9 en la preparación de corte de las células injertadas (véase figura 26).
XIII. Uso de ácido polisiálico (PSA) y enzima polisialiltransferasa (PST).
La integración del injerto y el alcance del crecimiento de fibras DA son desafíos en los métodos de injerto, incluyendo los estudios de injerto fetal en pacientes con EP. Un problema encontrado con los tejidos y células de injerto es el crecimiento limitado de fibras de estos injertos cuando se trata a los pacientes. Este problema es particularmente crítico en seres humanos ya que la recuperación del paciente requiere reinervación estriatal extensa. En métodos anteriores, lograr reinervación adecuada después de un injerto tisular requirió múltiples inyecciones de depósitos celulares a través del cuerpo estriado. Cada inyección puede provocar daño estriatal e inflamación junto con otros riesgos quirúrgicos. Dichos riesgos incluyen la lesión de un vaso sanguíneo durante la inyección celular que podría inducir potencialmente un ictus o convulsiones en el paciente. PSA es un homopolímero de ácido siálico natural de la superficie celular (es decir, ácido siálico ligado a alfa 2,8) que se ha identificado como una modificación postraduccional (a través de la acción de la enzima polisialiltransferasa (PST)) de otras moléculas de superficie celular, tales como la molécula de adhesión de células neurales (polisialilada) (NCAM), NCAM (CD56) y similares. PSA parecía actuar en la regulación de la plasticidad de algunos comportamientos celulares que requerían cambios en las interacciones célulacélula, incluyendo migración celular y crecimiento de axones. Aunque tiene alta expresión en el embrión, PSA estaba regulado negativamente en tejidos adultos con la excepción de regiones localizadas del SNC que mantienen la plasticidad estructural y fisiológica (tales como hipocampo, núcleo supraquiasmático, SVZ). Por tanto, en algunas realizaciones, el ácido polisiálico (PSA) se contempló para su uso en la promoción del crecimiento de fibras de células injertadas. En el documento WO 2006/042105 se describen ejemplos de uso de PSA en otros tipos celulares. En algunas realizaciones, los inventores contemplan el uso de neuronas DA auténticas en combinación con PSA como se describe en la presente memoria.
A. Aumento del PSA en neuronas DA para su uso en pacientes con EP. Siguen existiendo desafíos importantes para proporcionar métodos y células basados en resultados anteriores del uso de modelos de animales pequeños, incluyendo la supervivencia limitada de células trasplantadas y la inervación escasa de fibras de tejido del hospedador. Se considera que la influencia de estas limitaciones es más grave en el cuerpo estriado humano mayor y, por tanto, es necesaria mayor supervivencia e inervación para la aplicación clínica eficaz de neuronas DA procedentes de ES. Se contempla que el crecimiento de fibras mejorado y la integración de injerto en modelos animales, incluyendo el uso de al menos una hasta varias inyecciones representan reducciones en el riesgo asociado con múltiples inyecciones o mala distribución de neuronas DA in vivo.
La regulación de las interacciones celulares por el ácido polisiálico (PSA) es uno de los factores que promovieron la distribución celular, el crecimiento del axón y la inervación diana durante el desarrollo de vertebrados, véase, por ejemplo, Rutishauser, Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat Rev Neurosci 9, 26-35 (2008). El PSA era un polímero de carbohidratos unido a la molécula de adhesión celular neural (NCAM), que atenuaba las interacciones célula-célula y, por lo tanto, promovía la plasticidad tisular. En una cicatriz glial, la expresión mejorada de PSA en el cerebro adulto promovió la migración de precursores neuronales desde la zona subventricular hacia la corteza y mejoró el crecimiento axónico (El et al. Use of polysialic acid in repair of the central nervous system. Proc Natl Acad Sci U S A 103, 16989-16994 (2006). Como se describe en la presente memoria, la expresión de PSA aumentada por ingeniería genética en neuronas DA procedentes de ES purificadas de ratón dio como resultado recuentos de células de injerto mejorados, crecimiento extenso de fibras neuronales DA en el cuerpo estriado del hospedador y, sorprendentemente, recuperación conductual mejorada en ratones parkinsonianos. Además, la ingeniería genética de neuronas DA procedentes de ESC para aumentar los niveles de PSA en la superficie celular aumentó simultáneamente la supervivencia in vivo y el crecimiento de fibras en el cuerpo
estriado del hospedador. Se muestra una realización ilustrativa en la figura 29 para usar un gen de PST de mamífero, es decir, ratón o ser humano. Otra realización ilustrativa mostrada en la figura 29 muestra el uso de PST bacteriana, es decir, PSTnm. De manera específica, como se describe en la presente memoria, el aumento de PSA en las células usadas para la terapia celular basada en neuronas DA maduras se contempla para su uso en el tratamiento de la EP.
B. Uso de PST de ratón para aumentar la expresión de PSA. Los resultados in vivo demostraron aumento de las extensiones neuríticas y una inversión significativa de las rotaciones inducidas por anfetaminas en ratones 6-OHDA que recibieron injertos modificados con células PST de ratón, mientras que los números iguales de células que no fueron modificadas por PST no lograron lo mismo (figura 33). Por otro lado, se observó mejor inervación de fibras DA para correlacionarse con resultado conductual potenciado en un modelo de ratón de EP, tal como cuando se realizaron pequeños injertos de células positivas para PSA (inyección de aproximadamente 50.000 células) proporcionaron integración de injerto y crecimiento de fibras que eran aproximadamente 70 % de las áreas cubiertas en comparación con el uso de injertos comparativos mayores (100.000 o más células). Una comparación paralela de los injertos de neuronas DA procedentes de ES potenciados con PSA frente a tratados con control mostró recuperación conductual en el grupo de PSA (cuando los injertos se basaron en el trasplante de 55.000 células cada uno) pero no se observaron en células de control. El trasplante de 100.000 neuronas DA procedentes de ES en el cerebro de ratón mostró recuperación conductual en neuronas DA procedentes de ES tanto tratadas con PSA como tratadas con control, lo que sugiere que los injertos procedentes de 100.000 células son suficientes para reinervar el cerebro de ratón sin mejora de PSA. Por tanto, en otra realización, se contempla expresión modificada por ingeniería genética de la expresión de PSA en la superficie de neuronas DA auténticas para su uso en procedimientos para el tratamiento de pacientes con EP. Cuando se indujo PSA en células neuronales de las presentes invenciones, era idéntico al polímero de PSA que se produjo de manera natural en células cerebrales, a diferencia, por tanto, del uso de otras moléculas de superficie celular para obtener por ingeniería genética tipos celulares terapéuticos, se considera que las células modificadas por ingeniería genética para expresión de PSA tienen poca antigenicidad in vivo cuando se usan en células para procedimientos de injerto en seres humanos. Por otro lado, altos niveles de PSA en células injertadas, los precursores neurales endógenos (Battista et al., J Neurosci. 30(11): 3995-4003 (2010)), células de Schwann (Ghosh et al., Glia. 60 (6); 979-92 (2012)) y neuronas DA procedentes de ES de las presentes invenciones no provocaron efectos secundarios detectables cuando se usaron en diversos modelos de roedores adultos. La expresión modificada por ingeniería genética de PSA a niveles eficaces implicó la acción de una única enzima polisialiltransferasa (PST) cuyo único producto es este glucopolímero único. De manera sorprendente, la cantidad de proteína expresada y la naturaleza del producto enzimático fue notablemente constante y se parecía mucho al PSA hallado de manera natural en tejidos embrionarios.
Como se describe en la presente memoria, la expresión de PSA modificada por ingeniería genética se contempla en células neuronales para su uso en procedimientos de injerto. En particular, se contempla PSA para su uso en la preparación de células para uso terapéutico al superar los problemas encontrados cuando se usan otros tipos de expresión de marcador de superficie celular inducida. Además, el uso de expresión de PSA fue reproducible en protocolos que cruzan especies de vertebrados (tal como usando genes de PST de ratón expresados en células humanas) debido a que sus aceptores también son uniformes en su estructura entre especies y se encuentran en la mayoría de las superficies celulares. Modificación por ingeniería genética de genes de PST en hESC para aumentar el PSA en neuronas DA. Se introdujo un gen que codificaba la polisialiltransferasa humana (hPST) en una línea hESC (WA01) usando un vector lentivírico (pLenty, Invitrogen) y como se describe en la presente memoria. Se expandieron y analizaron veinte clones seleccionados para determinar la expresión de PST. Los clones de hESC que expresan PST se diferenciaron para garantizar que PST no se silenciara en neuronas DA. La cuantificación de p SA-NCAM en diferentes etapas de diferenciación (día 0, 11, 25 y 50) se realizó usando análisis de FACS e inmunofluorescencia (Operetta). Los clones positivos se sometieron al conjunto de parámetros de QC de la neurona DA descritos en la tabla 7. Al menos 3 clones que conservan niveles uniformes altos de PSA-NCAM durante la diferenciación y rinden bien en los parámetros de QC (tabla 7) avanzarán a la evaluación del crecimiento de neuritas en neuronas DA procedentes de hESC que sobreexpresan PST. Los clones de hESC de control y que sobreexpresan PST seleccionados se diferenciaron en neuronas DA usando el protocolo convencional descrito en la presente memoria, seguido de fijación celular y análisis los días 25 y 50. El número y la longitud de las fibras positivas para TH en dichos cultivos se cuantificaron con el microscopio de alto contenido Operetta. El módulo de análisis de neuritas en el software Harmony 3.0 cuantificó el número y la longitud de las neuritas, con o sin PST y los datos se analizaron estadísticamente usando un ANOVA de dos vías. Los clones de hESC que sobreexpresan PST y de control que avanzan desde estudios in vitro anteriores, se diferenciaron de nuevo en neuronas DA y se trasplantaron a un modelo de rata de EP. Se realizaron injertos a corto plazo (4-6 semanas) para determinar la supervivencia, expresión de PSA-NCAM y crecimiento de neuritas. Para cada clon que pasó, los parámetros in vivo a corto plazo se sometieron a estudios de injerto a largo plazo. Para esos estudios, los animales recibieron la mitad o un cuarto de la dosis convencional (200 x 103) de células. Estos estudios fueron para abordar si el aumento de PSA conduce a mayor supervivencia a largo plazo después de trasplante (5 meses) y si los números menores de neuronas DA son capaces de igualar o superar la capacidad funcional de los injertos no PST trasplantados a dosis celulares convencionales (figura 27).
Además, los ensayos conductuales complejos sensibles al grado de reinervación estriatal se supervisaron para distinguir adicionalmente el potencial funcional de los injertos de neuronas DA de PST frente a control. Los animales se sacrificaron después de la finalización de los ensayos conductuales y se cuantificó el crecimiento de fibras usando anticuerpos específicos humanos NCAM y SC121 y anticuerpos contra TH (véase también figura 29). Se midió la
intensidad y propagación del injerto hNCAM+, SC121+ y TH+, así como el porcentaje de células humanas que coexpresan marcadores de neuronas DA (TH, FOXA2) y PSA. La densidad de halo NCAM/TH+ de neuritas que emanan del injerto se cuantificó a diferentes distancias. Los datos se compararon entre los grupos usando un ANOVA de dos vías con una prueba post-hoc de Bonferroni. Además, se examinaron secciones para determinar cambios cualitativos (p. ej., ramificación, espesor, distribución y forma del injerto). Además, algunos injertos se procesarán para evaluación electrofisiológica de corte (véase figura 26) en con respecto a fenotipo A9, formación de sinapsis con el cuerpo estriado del hospedador, así como inervación por aferentes endógenos.
El siguiente ejemplo muestra mejora de la expresión de ácido polisiálico que mejoró la función de los injertos de neuronas de dopamina procedentes de ES en ratones parkinsonianos.
Las células ES que expresan GFP bajo el control del promotor Nurr1 (células Nurr1::GFP ES) se transdujeron de manera estable con un vector lentivírico que expresa polisialiltransferasa (PST) de manera ubicua. Las células transducidas mostraron un aumento drástico de ARNm de PST en comparación con los controles (fig. 30A). Se observó que la expresión de PST era suficiente para la síntesis de PSA en NCAM. Por consiguiente, la expresión de PSA-NCAM aumentó en gran medida en las células modificadas con PST el día 14 de la diferenciación de neuronas DA (fig. 30B-E). El PSA de la superficie celular tanto endógena como inducida en neuronas DA procedentes de ES podría eliminarse (fig. 30E) mediante una endoneuraminidasa de fago (endoN) que escindió de manera específica los polímeros de ácido siálico ligados a alfa-2,8 únicos del PSA. De manera sorprendente, no se observó que la transducción de PST afectara a la expresión de marcadores neuronales o del mesencéfalo en las neuronas DA purificadas con GFP (fig. 30F).
Otros estudios en ratones hemiparkinsonianos con lesión de 6OHDA mostraron que es necesario el trasplante de aproximadamente 100.000 precursores de neuronas DA procedentes de ES para producir recuperación funcional robusta, medido por la prueba de rotación potenciada por anfetaminas. En los presentes estudios, se buscó injertar una cantidad mejorable de células para poder evaluar el aumento mediante expresión de PSA mejorada. Para trasplantar poblaciones de neuronas DA muy enriquecidas que están agotadas de células pluripotentes contaminantes, cultivos purificados con FACS el día 14 de diferenciación para la expresión de GFP conducida por Nurr1 y para la ausencia de expresión de SSEA-1 (figura 31). Sin sobreexpresión de PST, una reducción del tamaño de injerto mínimamente eficaz a la mitad (55.000 células DA Nurr1+) no logró producir recuperación del comportamiento detectable. Por el contrario, con expresión de PSA mejorada, el mismo número de neuronas DA Nurr1/PST dio como resultado una corrección significativa del deterioro conductual por la EP (p < 0,01; ANOVA de dos vías), con recuperación completa aproximadamente 5 semanas después de la cirugía (figura 32A). La eliminación del PSA antes del trasplante por incubación con endoN indicó la especificidad de la mejora del PSA, ya que el tratamiento con endoN invirtió parcialmente la restitución funcional obtenida con Nurr1/PST (figura 32A).
Para examinar las características de las células injertadas, los animales fueron procesados con respecto a inmunohistoquímica dos meses después del trasplante. Hubo una diferencia en el número de neuronas Nurr1 supervivientes, porque los animales en los que se injertó la línea transducida por PST tuvieron en promedio el doble de células GFP+ que los animales en los que se injertaron células de control (9.300 /- 1.400 frente a 4.230 /- 1010 células GFP+ por injerto en muestras de PST frente a control, respectivamente; figura 32B, p < 0,05, prueba de t de Student). Asimismo, los injertos de Nurr1/PST también mostraron niveles mayores de expresión de PSA in vivo (figura 32C, D). Sin embargo, las proporciones de células que expresan los marcadores DA del mesencéfalo TH y FoxA2 dentro del núcleo del injerto fueron comparables para las células Nurr1 y Nurr1/PST (TH: 62,0 % /- 8,0 frente a 51,3 % /- 7,0, p = 0,33; FoxA2: 63,2 % /- 8,6 frente a 55,4 % /- 2.0, p = 0,3, respectivamente; figura 32E).
Las prolongaciones neuronales que surgieron de las células Nurr1 y Nurr1/PST mostraron niveles similares de TH, Girk2 (canal de potasio de rectificación interna, acoplado a proteína G) y sinapsina (figura 33A). A diferencia de otros estudios con células de Schwann trasplantadas (Ghosh, et al. Glia 60, 979-992 (2012)), la expresión de PSA mejorada tuvo poco efecto sobre la migración de células DA desde el sitio de injerto. Sin embargo, hubo cambios evidentes en el crecimiento de neuritas. Como se muestra en la figura 33B, surgieron más prolongaciones neuronales DA de células Nurr1/PST en comparación con controles Nurr1+. Cuando la intensidad de la inmunofluorescencia de GFP y TH se cuantificó en cinco zonas sucesivas a 100 pm del trasplante, los injertos de Nurr1/PST presentaron una densidad relativa de prolongaciones mucho mayor (figura 33C, D; p < 0,01 para GFP y TH, ANOVA de dos vías). Al cuantificar este efecto, se normalizó la densidad relativa de las prolongaciones a la densidad observada en la zona más próxima inmediata al núcleo del injerto. Dicha normalización fue necesaria para compensar el mayor número de células supervivientes en los injertos de Nurr1/PST y para confirmar un efecto específico del PSA sobre el crecimiento de neuritas. La especificidad también se demostró cuando el PSA de la superficie celular se eliminó mediante tratamiento con endoN antes del injerto.
Por tanto, el pretratamiento con endoN redujo el crecimiento de fibras distales a niveles de control (figura 33E).
Estos descubrimientos mostraron que al menos algunos de los efectos del PSA sobre la función del injerto resultaron de la inervación de fibras mejorada del cuerpo estriado. Por consiguiente, hubo una fuerte correlación entre la función del injerto y el alcance relativo del crecimiento de fibras positivas para GFP, por ejemplo, en la zona IV (figura 33F; p < 0,001, r2 = 0,65, n = 17). De manera sorprendente, la relación de crecimiento de fibra/comportamiento fue uniforme para los grupos experimentales (control, con PSA mejorado y tratado con endoN), lo que indica que la inervación del
injerto-hospedador fue un parámetro para la recuperación del comportamiento en el modelo parkinsoniano de ratón. Varios factores contribuyeron mecánicamente al aumento del crecimiento de fibras, tales como mejora de la penetración de la zona de glía reactiva que encapsula el núcleo del injerto, mayor capacidad de brote, mejor crecimiento en el tejido hospedador circundante (p. ej., translocación de cono de crecimiento más fácil) y prevención de conexiones prematuras con tejido hospedador próximo al núcleo del injerto. Los mecanismos ilustrativos son coherentes con el papel del PSA para facilitar el crecimiento de prolongaciones durante el desarrollo normal y en el sistema nervioso adulto.
Los experimentos descritos en la presente memoria demostraron el uso de PSA modificado por ingeniería genética en el injerto de neuronas DA que proporcionó resultados superiores en comparación con injertos de otros tipos de células. Los datos indicaron claramente que la mejora de PSA proporcionó un aumento significativo de la capacidad de las neuronas DA injertadas para inervar el cuerpo estriado del hospedador y atenuar los déficits funcionales de la EP. Por lo tanto, se contempla la traducción clínica que comprende neuronas DA de las presentes invenciones para proporcionar células antes del trasplante. En algunas realizaciones, las células se manipularán genéticamente para expresar PSA. En algunas realizaciones, la PST puede administrarse directamente a las células a través de la exposición a la enzima purificada y al sustrato, in vitro, antes del trasplante. En algunas realizaciones, se contempla la estrategia de PSA para la traducción humana en el injerto de EP para minimizar la necesidad de múltiples inyecciones y, por lo tanto, reducir los riesgos quirúrgicos resultantes de estas inyecciones múltiples.
En otras realizaciones, esta tecnología se contempla para su uso en otros tipos celulares y especies, por ejemplo, aumentando la migración de células de Schwann injertadas en la creación de un puente (por ejemplo, comunicación célula-célula) para el nuevo crecimiento de axones en el sitio de la lesión de la médula espinal.
Los siguientes son materiales y métodos ilustrativos usados en este ejemplo.
Animales: Se mantuvieron ratones 129S3/SvImJ de seis semanas de edad (Jackson Laboratory) a temperatura controlada con alimentos y agua disponibles a voluntad. Los procedimientos experimentales se realizaron de acuerdo con los NIH y las pautas institucionales para el uso de animales y fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales (IACUC) y el Comité Institucional de Bioseguridad (IBC).
Inyección de 60HDA y prueba de inducción por anfetaminas: Los animales se anestesiaron con pentobarbital de sodio (10 mg/kg) y se les inyectó en el cuerpo estriado derecho con 2 pl de 60HDA (4 pg/pl en solución salina, ácido ascórbico al 0,5 %). Las inyecciones se realizaron con una jeringa Hamilton en las coordenadas: 0,5 mm posterior, 1,8 mm lateral con respecto al bregma y 2,5 mm ventral con respecto a la superficie del cerebro. Antes de la cirugía, los animales recibieron una única inyección i.p. de desipramina (25 mg/kg, Sigma). Dos semanas después de la cirugía, los animales se puntuaron en la prueba de rotación inducida por anfetaminas. Se colocaron en cilindros de plástico transparente de 30 cm de diámetro durante media hora, después de lo cual recibieron una única inyección i.p. de anfetamina (10 mg/kg, Sigma). Después de 20 min, se puntuó el número de rotaciones ipsilaterales/contralaterales durante otros 20 min. Los animales se puntuaron una vez a la semana durante siete semanas, después se anestesiaron profundamente y se perfundieron a través del corazón con PBS y paraformaldehído al 4 % en tampón de fosfato 0,1 M (PB, pH 7,4). Los cerebros se extrajeron y se fijaron durante una noche a 4 °C en paraformaldehído al 4 % y después se cortaron con vibrátomo (Pelco-101, Ted Pella) en secciones sagitales de 40 pm de grosor.
Diferenciación y trasplante celular: Se transdujo una línea celular indicadora ES de ratón BAC transgénica BAC Nurr1 ::GFP (es decir, la expresión de GFP es impulsada por el promotor Nurr1) 5 con un lentivirus (pLenti, Invitrogen) que contenía el gen de PST de ratón bajo el control del promotor de CMV. Las células ES se propagaron en MEF tratados con mitomicina C (StemCell Technologies) en DMEM (Invitrogen), FBS al 10 % (HyClone) complementado con 1.400 unidades/ml de LIF (ESGRO; Invitrogen), L-glutamina 2 mM, p-mercaptoetanol 1 mM, 100 U/ml de penicilina y 100 pg/ml de estreptomicina (Invitrogen). La diferenciación de DA se indujo según Barberi et al., Nat Biotechnol 21, 1200-1207 (2003), con modificaciones. En resumen, las células se diferenciaron en células alimentadoras MS5 en placas recubiertas con gelatina (10.000 células/placa de 10 cm) y se cultivaron durante cuatro días en medios de reemplazo de suero (SRM). El día 4, se añadieron Sonic Hedgehog (SHH, 200 ng/ml) y FGF8 (100 ng/ml). El día 7 de diferenciación, los medios se cambiaron a N2 complementado con SHH, FGF8 y bFGF (10 ng/ml). El día 11, la diferenciación terminal fue inducida por retirada de SHH, FGF8 y bFGF y la adición de ácido ascórbico (AA, 200 pM) y BDNF (20 ng/ml).
Las células se recogieron los días 14-15 con tratamiento de accutase durante 45 minutos, se lavaron una vez con N2 y se incubaron con anticuerpo anti-SSEA-1 conjugado con AlexaFluor-647 (BD Pharmingen) durante 25 minutos. Las células se lavaron una vez con N2, se resuspendieron en tampón HEPES con BSA al 0,1 %. Se añadió DAPI para evaluar la viabilidad. FACS se realizó con un clasificador de células MoFlo y la población de interés se clasificó con respecto a fluorescencia de GFP (Nurr1). La población positiva para AlexaFluor-647 (SSEA-1) se clasificó negativamente. Para el control negativo de GFP, se usaron células ES de ratón J1 sin exposición previa en la misma etapa de diferenciación.
Las células clasificadas Nurr1::GFP se analizaron para determinar su viabilidad y se resuspendieron en N2 con BDN y AA a una concentración final de 55.000 células/pl. Se inyectó un pl en el cuerpo estriado del ratón con lesión con un capilar de vidrio fino con punta de 50 pm en las coordenadas: 0,3 mm posterior, 1,5 mm lateral desde el bregma y
2,2 mm ventral con respecto a la superficie del cerebro. Se volvió a sembrar una alícuota de la suspensión celular en placas de 6 mm recubiertas con matrigel para caracterización adicional.
Para análisis de inmunofluorescencia, las células se fijaron con paraformaldehído durante 10 minutos a 4 0 C, se lavaron dos veces con PBS, se bloquearon con BSA al 5 % (Triton X-100 al 0,1 % en PBS) y se incubaron con anticuerpos primarios durante 2 horas a temperatura ambiente: anti-GFP de conejo (1:1000, Invitrogen), IgM de ratón anti-PSA (1:2000, 5A5), anti-NeuN de ratón (1:800, Chemicon), anti-TH de ratón (1:1000, Sigma), anti-FoxA2 de cabra (1:800, Santa Cruz), anti-Engrailed de cabra (1:800, Santa Cruz). Las células se incubaron después con anticuerpos secundarios conjugados con Cy (1:1000, Jackson).
Tratamiento con EndoN: Para eliminar el PSA de NCAM, la noche antes de la recogida, las células se trataron con 20 unidades de endoN, una enzima de fago que elimina específicamente PSA 7-9. Las células se recogieron e inyectaron como se ha descrito anteriormente, pero se resuspendieron en N2 con BDNF y AA y 5 unidades de endoN. Anteriormente los inventores han evaluado que la inyección de la misma cantidad de endoN solo en ratones lesionados no mejoró el comportamiento animal.
Análisis de ARNm de PST y PSA-NCAM in vitro: Para análisis de transferencia de Western, las células se trataron con tampón WB (PBS con NP40 al 1 %, NaCl 150 mM, EDTA 1 mM e inhibidores de proteasa/fosfatasa 1x añadidos inmediatamente antes de la extracción, a pH de 7,4) y se sonicaron dos veces durante 5 segundos, se centrifugaron y se resuspendieron en tampón Laemli (LB). Se guardaron alícuotas sin LB para la determinación de proteínas. Se cargaron cantidades iguales de proteína en gel de electroforesis en gel de dodecilsulfato de sodio-poliacrilamida al 6 % (BioRad). Las proteínas se transfirieron mediante electroforesis a membranas de polivinilideno (Millipore). Las membranas se bloquearon durante 1-6 horas en Triton X-100 TBS 0,1 % (TBS-T) con leche en polvo desgrasada al 5 % y se incubaron durante una noche con anticuerpos anti-NCAM (1:10.000, Santa Cruz) en TBS-T con leche al 5 %. Las transferencias se incubaron después con anticuerpo secundario conjugado con peroxidasa (1:10.000, Jackson) y se detectaron con el método de detección de ECL (Amersham Pharmacia Biotech). Los niveles de proteínas se cuantificaron usando software ImageJ.
Para análisis por qRT-PCR, se extrajo ARN total con Trizol (Sigma), se transcribió de forma inversa (Qiagen) y se amplificó con 10 pl de mezcla de reacción SYBR 2x y 0,2 pM de cebadores directo e inverso hasta un volumen final de 20 pl. Para análisis FACS de PSA-NCAM, las células se recogieron con tratamiento de accutase durante 45 minutos, se lavaron una vez y se incubaron con IgM de ratón anti-PSA (1:250, 5A5) durante 25 minutos en hielo, se lavaron una vez con medio N2 y se incubaron con IgM anti-ratón conjugada con Cy3 (1:250, Jackson) durante otros 25 minutos en hielo. Las células se lavaron una vez con N2 y se resuspendieron con BSA al 0,1 % con 7AAD y se analizaron en un clasificador de células FACS Calibur. Como control, no se añadió anticuerpo primario.
Procedimientos inmunohistológicos y estereológicos: Las secciones coronales en flotación se bloquearon en Triton X-100 al 0,1 %, suero de burro al 5 % en PBS durante 30 minutos a temperatura ambiente y se incubaron 48 horas a 4 °C con diferentes anticuerpos: anti-GFP de conejo (1:300), anti-GFP de pollo (1:200, Chemicon), anti-TH de ratón (1:200), IgM de ratón anti-PSA (1:1000), anti-NeuN de ratón (1:400), anti-FoxA2 de cabra (1:300), anti-Girk2 de conejo (1:300, Alomone Labs), anti-sinapsina de ratón (1:200, BD Transduction Laboratories). Las secciones se lavaron e incubaron con anticuerpos secundarios: anticuerpos de burro conjugados con Cy2, Cy3 y Cy5 (1:400, Jackson). Para el PSA se usó un anti-IgM de burro conjugado con Cy5 (1:500 Jackson). Las incubaciones se realizaron durante 2 horas a temperatura ambiente. Las secciones se lavaron dos veces en PBS y se montaron en Mowiol (Calbiochem). Se analizaron una de cada tres secciones coronales del cerebro para cada inmunomarcaje. Las imágenes digitales fueron recogidas por un microscopio confocal de barrido láser Zeiss LSM 510 con tres láseres (Argon 488, HeNe 543 y HeNe 633) con un objetivo 40 x c-Apochromat (de inmersión en agua). El número de células GFP+ y TH+ se contó en una de cada tres secciones que abarcan el cerebro completo con un objetivo de 40x y se estimó el número total de células/injerto. Las células con doble marcaje se analizaron en planos ópticos individuales a través del eje z completo. Para el análisis del porcentaje de células marcadas con GFP/TH+ y GFP/FoxA2+, se analizaron 100 células GFP+ para cada marcador. Para el análisis de crecimiento de prolongaciones, se realizaron exploraciones z confocales a intervalos de 0,8 pm a través del eje z completo (20-40 pm) con un agujero estenopeico de 1 pm bajo un objetivo 40x. Las secciones se exploraron desde el sitio de inyección lateralmente hasta que no se observaron prolongaciones. Se combinaron secuencialmente proyecciones tridimensionales que abarcaban toda el área explorada. Para análisis de intensidad de GFP y TH, toda el área escaneada se dividió en cinco zonas sucesivas de 100 pm de distancia desde el trasplante y las intensidades se midieron usando software ImageJ. Los datos se normalizaron con respecto a la intensidad en la zona más cercana al injerto (zona I) para controlar con respecto a cualquier diferencia potencial en el tamaño del injerto.
Análisis estadístico: Los datos se presentan como la media ± error típico de la media (ETM). Se realizaron comparaciones usando la prueba t de Student o el análisis de varianza (ANOVA) de dos vías seguido de la prueba post-hoc de Bonferroni. Se realizó análisis de regresión lineal y se cuantificó usando la correlación de Pearson. C. Sobreexpresión del gen de PST humano (polisialil transferasa).
Los grupos de animales con lesión recibieron una de 3 dosis de células de tipo silvestre o células que expresan PST
o pretratadas con enzima PST. Se procesaron para pruebas conductuales siguiendo los paradigmas analizados en la tabla 11 (rotaciones por anfetaminas, prueba de cilindro, prueba de marcha). Las dosis elegidas (por ejemplo, 200k, 100k, 50k células) se usaron con éxito en ratones de manera que las neuronas resultantes mostraron resultados como se describe en la presente memoria para genes de PST de ratón.
D. Administración directa de enzima PST purificada a células. En otras palabras, un método no transgénico de inducción de PSA. Pretratamiento de las células con la enzima PST que da como resultado polisialilación y aumento de la expresión de PSA de superficie. Aunque las PST de mamíferos son proteínas de membrana de baja abundancia que actúan en el Golgi, la a2,8-polisialiltransferasa de Neisseria meningitides (PSTnm) purificada actuó en un ambiente extracelular cuando se usó un sustrato no tóxico disponible en el mercado (es decir, CMP-ácido siálico) y produjo un polímero químicamente idéntico a la PSA de mamífero. Como ejemplo, un fragmento activo de esta enzima fue eficaz para añadir PSA a proteínas terapéuticas in vitro incrementar la farmacocinética in vivo. En presencia de CMP ácido siálico, se sintetizó PSA mediante PSTnm directamente en superficies de una amplia variedad de tipos celulares in vitro, incluidos ESC de ratón y humanas (figura 35A-E). La inyección directa de PSTnm y sustrato in vivo desencadena una mayor acumulación de PSA en las superficies celulares en regiones cerebrales adultas, incluyendo la corteza cerebral, el cuerpo estriado y la médula espinal (figura 35GH). El PSA producido por PSTnm fue degradado por endoN (figura 35B) que eliminó el PSA inducido, lo que demuestra que el PSA producido por PSTnm tiene propiedades funcionales comparables al PSA endógeno (figura 35A, C, D). Se produjo expresión de PSA a través de PSTnm en menos de una hora, lo que superó la inducción transgénica de PST lenta. La expresión persistió durante varias semanas in vivo después de lo cual se redujeron los marcadores de PSA, superando de este modo los efectos secundarios de la inducción prolongada de un transgén de PST. Por lo tanto, el uso de exposición celular in vitro al sustrato PSTnm+ se contempla como una estrategia alternativa, sencilla para desencadenar mayores niveles de PSA en neuronas DA procedentes de hESC y otros tipos celulares para su uso en procedimientos de injerto. En parte, este enfoque alternativo para traducción, es decir, uso en procedimientos de injerto humano, tiene las ventajas de ser de naturaleza no invasiva, es decir, evitar el uso de métodos transgénicos, se usan reactivos de uso en GMP en estos procedimientos para cumplir con los protocolos de uso clínico y tienen una naturaleza transitoria de la expresión de PSA generada bioquímicamente que coincide con el marco temporal esperado para que las fibras de DA abandonen el núcleo del injerto y entren en el cerebro del hospedador, necesario para evitar algunos efectos secundarios peligrosos del uso de células diseñadas para injerto.
El siguiente ejemplo muestra la modificación por ingeniería enzimática de PSA en neuronas DA procedentes de hESC usando la polisialiltransferasa bacteriana purificada, PSTnm, para mejorar la eficacia del trasplante.
Aunque es eficaz, la transfección de gen de PST requirió modificaciones genéticas de hESC con control limitado sobre la duración de la polisialilación. Este ejemplo describe el descubrimiento de que PSTnm externo indujo PSA, en lugar de suministro de genes, (véase figura 35). En la figura 35A, las células de Schwann (SC) tratadas con PST (línea verde-línea media) tenían un mayor tiempo de adhesión, mientras que el PSA producido por PSTnm inhibió la adhesión. En particular, (A) El PSA producido por PSTnm inhibe la adhesión de células de Schwann en suspensión a una monocapa de células de Schwann aún más eficazmente (línea roja-línea más baja) que el PSA producido por la expresión forzada de PST (línea verde-línea media). (B) La inmunotransferencia de PSA en motoneuronas HB9 procedentes de ESC muestra que las muestras de control tratadas con PSTnm solo tenían niveles indetectables de PSA. La incubación con sustrato de PSTnm CMP-ácido siálico produce una gran banda de PSA, que se elimina con tratamiento con endoN. (C, D) Similar a los efectos obtenidos con el gen de PST, la polisialilación de estas células mediante PSTnm y sustrato durante la diferenciación mejora el crecimiento de neuritas y la migración celular (puntas de flecha). (E) Inmunotinción de PSA de neuronas DA procedentes de hESC del día 30. (F) Esta tinción aumenta significativamente después del tratamiento con PSTnm y sustrato. (G) la inyección in vivo de PSTnm solo no tiene efecto, mientras que su administración conjunta con sustrato (H) produce grandes cantidades de expresión de PSA en el cuerpo estriado de ratón.
Por tanto, las neuronas DA maduras tratadas externamente con PSTnm se contemplan para su uso en las células productoras para injerto. Tanto PST como PSTnm de mamífero produjeron cadenas de PSA químicamente idénticas. El aumento de PSA en las neuronas DA procedentes de hESC (figura 35F) debería persistir durante varias semanas, suficiente para que las fibras DA salgan del núcleo del injerto. Debido a que PSTnm se elimina antes del injerto, la inmunogenicidad a esta enzima que contamina las células injertadas no debería tenerse en cuenta.
Se produjo PSTnm a partir de un fragmento modificado por ingeniería genética con características de solubilidad y actividad mejoradas (Willis et al., Characterization of the alpha-2,8-polysialyltransferase from Neisseria meningitidis with synthetic acceptors, and the development of a self-priming polysialyltransferase fusion enzyme. Glycobiology 18, 177-186 (2008)). Se indujo que los cultivos de hESC se diferenciaran en neuronas DA antes de la exposición a PSTnm, exposición al sustrato o ambos. Los cultivos se examinaron en diferentes puntos temporales de exposición (10 min a 6 h) mediante inmunofluorescencia cuantitativa (Operetta) y transferencia de Western para determinar la velocidad y los niveles de polisialilación. Por tanto, el día 25, las neuronas DA procedentes de hESC diferenciadas se incubarán con las concentraciones óptimas de PSTnm y sustrato usando las condiciones descritas en la presente memoria. Las neuronas mDA PSA+ se trasplantarán en ensayos a corto y largo plazo como se describe en la presente memoria y en la figura 29.
E. Aplicaciones para lesión de la médula espinal. Varios estudios dirigidos hacia el uso eventual en pacientes
humanos y las estrategias descritas en la presente memoria, mostró amplio el potencial de estos procedimientos, incluyendo la administración del gen de PST mediante la inyección de vectores lentivíricos que expresan PST directamente en el SNC para promover la regeneración de axones y la migración de progenitores endógenos. Una de esas estrategias para su uso en procedimientos humanos se dirige a la lesión de la médula espinal. En un tipo de procedimiento de regeneración de la médula espinal, los injertos celulares de Schwann se usaron como parte de la terapia para reconstruir puentes celulares para su uso en regeneración de axones in vivo. Sin embargo, la recuperación del paciente de la función locomotora, incluyendo control muscular fino, resistió cualquier intervención terapéutica. Como se muestra en la presente memoria, la mejora de la expresión de PSA en las células de Schwann usadas para el injerto dio como resultado una mejora de la migración de las células de Schwann y el crecimiento axónico, lo que dio como resultado efectos drásticos en el aumento de la función locomotora (figura 30A-D). Por tanto, en algunas realizaciones, el aumento de la expresión de PSA en células de Schwann in vitro se contempla para su uso en procedimientos de injerto en seres humanos con lesión de la médula espinal. Otra estrategia para el uso de la expresión modificada por ingeniería genética de PSA mediante el aumento de la expresión de genes de PST en procedimientos humanos implicó motoneuronas procedentes de HB9 ESC, donde la introducción del gen de PST en estas neuronas a través de la expresión génica basada en vectores lentivíricos dio como resultado un aumento drástico en el crecimiento de axones tanto en cultivo como después del injerto en ratones para reparar una lesión del nervio ciático inducida mecánicamente. Esto último dio como resultado una dirección mejorada y específica del tejido muscular (figura 30E-H). Por tanto, en otra realización, la expresión modificada por ingeniería genética de la expresión de PSA en la superficie de motoneuronas procedentes de ESC para su uso en la restauración de la función del nervio ciático.
IVX. Mayor seguridad de los injertos de neuronas DA.
Se describen a continuación realizaciones contempladas para reducir adicionalmente los riesgos para la salud para pacientes para su uso en métodos de producción de células de las presentes invenciones.
Aunque las células producidas por métodos descritos en la presente memoria demostraron características de reducción de riesgos para los pacientes cuando se usan para injerto, se contemplan realizaciones adicionales para reducir adicionalmente la posibilidad de riesgo para la salud de los pacientes que reciben células injertadas. Una de varias preocupaciones para los procedimientos de terapia celular basados en hESC es la posibilidad de introducir células contaminantes indiferenciadas que resistieron la diferenciación que, en condiciones posteriores al injerto, se convierten en células que provocan daño al paciente. En el caso de una célula pluripotente, un resultado perjudicial es la formación de teratoma que pone en peligro la vida del paciente. Se informó de la formación de teratoma usando células procedentes de hESC siguiendo protocolos de diferenciación neural a corto plazo basados en la diferenciación celular espontánea. Sin embargo, el uso por parte de los inventores de tipos celulares neurales procedentes de ES humano, a diferencia de sus homólogos procedentes de ESC de ratón, rara vez dio como resultado la formación de teratomas siguiendo estrategias de diferenciación neural adecuadas como se describe en la presente invención (es decir, cultivo en monocapa, protocolo de inhibición doble de SMAD y crecimiento en citocinas que no promueven la proliferación). De hecho, después de analizar varios cientos de animales con injertos de células humanas, usando diversas estrategias de diferenciación neural en los últimos 10 años, no se observó formación de teratoma. Asimismo, no se observaron teratomas en los procedimientos de trasplante de EP de las presentes invenciones usando células humanas para injertos. Se contempla que la diferencia entre el uso de células humanas frente a células de ratón para procedimientos de injerto para su uso en seres humanos está relacionada con la diferente etapa de pluripotencia capturada en ESC humanas frente a las de ratones, por lo que se cree que las células humanas coinciden con las propiedades de una etapa pluripotente descrita como Epi-SC, a diferencia de ESC de ratón que pueden estar en una etapa de desarrollo diferente.
Sin embargo, al menos uno de los problemas al usar células de estudios de trasplante previos fue el riesgo continuo de hiperproliferación neural que es sustancial en Perrier et al., PNAS 2004 y protocolos similares que están sorprendentemente ausentes cuando se usan neuronas DA maduras y métodos de trasplante de las presentes invenciones. Además, otro problema encontrado en pruebas de injerto en estudios previos fue la formación de estructuras neuroepiteliales procedentes de hESC (es decir, tipo roseta neural) que continúan proliferando in vivo.
Esta expansión in vivo de células neuroepiteliales injertadas se observó en diversos paradigmas de trasplante neural, incluyendo estudios de trasplante de neuronas DA procedentes de hESC en modelos de enfermedad de Parkinson y Huntington de roedores. Esas células proliferativas de "tipo roseta neural" representaban células primarias no transformadas con un alto potencial de crecimiento intrínseco que daba como resultado grandes injertos compuestos de tejido ectópico, principalmente de tipo cortical, en animales con injertos. Como se describe en la presente memoria, se usaron varias estrategias para eliminar las células pluripotentes o neuroepiteliales contaminantes en el momento del injerto (p. ej., selección para SSEA-4 (marcador pluripotente) o Forse-1 (marcador neuroepitelial). Estas estrategias fueron parcialmente exitosas cuando se usaron estrategias de diferenciación basadas en roseta y aún se observó una hiperproliferación neural en un subconjunto de injertos clasificados para Forsel o clasificados negativamente para s SeA-4. De manera sorprendente, con el desarrollo de estos procedimientos de diferenciación de neuronas DA basados en la placa del suelo en lugar de basados en la roseta, se resolvió el problema de la hiperproliferación neuroepitelial. Se observaron con poca frecuencia células proliferantes, derivadas del injerto, dentro de injertos de neuronas DA procedentes de la placa del suelo hESC funcionales.
En función de los resultados adversos de otros procedimientos de injerto, otra preocupación de seguridad para el uso de injerto neuronal DA en seres humanos son los efectos secundarios de la terapia, tales como la aparición de discinesia inducida por injerto (DIG) observada en aproximadamente 15 % de los pacientes que reciben tejido fetal en los ensayos de trasplante. Sin embargo, como se analiza en la presente memoria, la ausencia casi completa de células serotoninérgicas procedentes de injerto, junto con el uso de una fuente celular más uniforme con la posibilidad de agotar adicionalmente los tipos celulares no deseados (véase, texto asociado con la figura 28) y la posibilidad de controlar la distribución de fibras DA in vivo (véase, figura 29; es decir, prevenir "puntos calientes" de grupos neuronales que secretan L-Dopa y otros compuestos) son las principales ventajas del uso de los métodos de las presentes invenciones con respecto a otros métodos para proporcionar células para su uso en procedimientos de injerto.
Por tanto, se contempla el uso de los métodos de la presente invención para minimizar el riesgo para los pacientes.
VX. Uso de ESC humanas para la traducción clínica.
En realizaciones preferidas, se contemplan ESC humanas para su uso en métodos para fabricar y usar células para procedimientos de injerto en seres humanos, en otras palabras, terapia celular para el tratamiento de la EP. En particular, las ESC humanas tienen numerosas ventajas sobre el uso de iPSC humanas en los métodos de las presentes invenciones, tal como, como un ejemplo, para su uso en el suministro de neuronas DA del mesencéfalo injertables para su uso como terapia celular de EP. En particular, el uso de células madre pluripotentes inducidas (iPSC) como fuente celular para la obtención de neuronas DA tiene varias ventajas, tales como proporcionar una fuente de células genéticamente coincidentes para cada paciente. Sin embargo, varios estudios recientes han creado incertidumbre con respecto a la seguridad y la compatibilidad genética total de las iPSC. Se ha mostrado que las células reprogramadas albergan anomalías genéticas y epigenéticas potencialmente peligrosas que son indeseables para la utilidad clínica. Asimismo, el trabajo en iPSC de ratón mostró que las células procedentes de iPSC no son completamente inmunocompatibles, lo cual es el argumento principal para respaldar su uso en el trasplante humano. Asimismo, no existen procedimientos aprobados por la FDA que usen iPSC para injerto. Finalmente, sería poco práctico y económico prever la generación de bancos de células que cumplan con GMP y que estén totalmente controlados por QC para cada paciente individual. En comparación, se estudió intensamente la estabilidad genética de las hiPSC en comparación con las hESC. Aunque se observó que las hESC adquirían mutaciones a lo largo del tiempo en cultivo, el momento y la tasa de dichas mutaciones parecían diferir de las hiPSC, donde las hESC se consideraban en general más genéticamente estables que las iPSC, por ejemplo, véase, Hussein, et al., Nature 471, 58-62 (2011); Mayshar, et al. Cell Stem Cell 7, 521-531 (2010); Lister, et al. Nature 471, 68-73 (2011); Laurent, et al. Cell Stem Cell 8, 106-118 (2011). Además, se idearon procedimientos operativos convencionales para varias líneas de hESC que satisfacían las pruebas de seguridad rigurosas requeridas por la FDA para productos de terapia celular y genética. La FDA ha aprobado dos grupos en los Estados Unidos para avanzar las terapias celulares basadas en hESC a uso clínico. Por ejemplo, Geron Corporation entró en un ensayo de fase I con células precursoras de oligodendrocitos procedentes de hESC (GRNOPC1) y Advanced Cell Technology (ACT), Inc. tiene dos ensayos actuales de fase I/II que usan células epiteliales pigmentadas retinianas procedentes de hESC para tratar la distrofia macular de Stargardt (ensayo NCT01345006) y la degeneración macular relacionada con la edad seca avanzada (ensayo NCT01344993).
Finalmente, el hecho de que ambos de los ensayos clínicos basados en hESC, aprobados por la FDA, dirigidos a los trastornos del sistema nervioso muestran ventajas de usar métodos basados en hESC para proporcionar material de injerto para tratar enfermedades y lesiones del sistema nervioso. El sistema nervioso se considera un sitio inmunoprivilegiado ya que el tejido extraño (aloinjertos) induce respuestas inmunitarias débiles en comparación con el mismo injerto colocado en la periferia. De hecho, después de veinticinco años de trasplante de células fetales en cerebros humanos, se descubrió que algunas neuronas alogénicas sobrevivieron durante hasta 16 años en el cerebro humano con inmunodepresión transitoria. Por lo tanto, parece que la coincidencia antigénica idéntica entre una fuente celular y el receptor del injerto no es esencial. Por tanto, las hESC se consideran una fuente alogénica universal de neuronas DA para el tratamiento de la EP además de otras enfermedades, trastornos y lesiones del sistema nervioso. Se contempla que los pacientes que reciben células de la presente invención tienen un diagnóstico clínico de EP. Se contemplan injertos de neuronas DA auténticas para su uso en intervención temprana y EP de moderada a grave, incluyendo pacientes en los que hay control sintomático insuficiente con los medicamentos disponibles, tales como levodopa, medicación complementaria, etc. En algunas realizaciones, los pacientes contemplados para recibir injertos de neuronas tienen señales sutiles en la EP temprana (p. ej., mediante el uso de captura de neuroimágenes para detectar déficits dopaminérgicos, FDG-PET y señales de discinesias, etc. La discinesia medida por la escala de calificación de discinesia unificada (UDysRS) (Goetz, et al., Mov Disord. 23, 2398-2403 (2008)) se contempla para su uso en el seguimiento de pacientes antes y después del injerto. Los pacientes también pueden tener "exploraciones sin pruebas de déficits dopaminérgicos" (SWEDDS), algunos de los cuales pueden tener distonía o temblor esencial. Se realizaría IRM para identificar a los pacientes con otros factores contribuyentes (no dopa) al parkinsonismo. En algunas realizaciones, los pacientes tendrían una respuesta positiva a la levodopa. Determinación de parámetros y criterios de valoración antes y después del trasplante para la supervisión del sujeto, tal como evaluación motora, evaluación no motora, calidad de vida y también el uso de captura de neuroimágenes y otros biomarcadores. Función motora: El UPDRS y el MDS-UPDRS recientemente validado (Goetz, et al., Mov Disord. 23, 2129-2170 (2008)) son ampliamente usados para medir los síntomas motores de la EP. Sin embargo, otras pruebas, incluyendo pruebas de caminata de 10 minutos o caminata de 6 minutos, levantamiento y marcha cronometrados, evaluación funcional de la
marcha, alcance funcional y otros están contemplados para medidas de resultados más orientadas al paciente. Los pacientes se evaluarían en el estado "inactivo", así como "activo", y se incluirían escalas de calificación para el tiempo "inactivo" (por ejemplo, siguiendo las recomendaciones de la Sociedad de Trastornos del Movimiento basadas en las propiedades clinimétricas de las escalas de pérdida de efecto validadas (Antonini, et al. Mov Disord. 26, 2169-2175 (2011)) y escalas de calificación de discinesia (por ejemplo, UdysRS (Goetz, et al., Mov Disord. 23, 2398-2403 (2008)). Se contempla grabar en vídeo los exámenes normalizados de pacientes en estados tanto "activo" como "inactivo". Función no motora: Las medidas se dirigirán principalmente a resultados psiquiátricos cognitivos y disautonomía además de abordar la cognición, depresión, ansiedad, apatía, sueño, fatiga, psicosis y otros síntomas no motores antes y después del injerto. Calidad de vida: Los cuestionarios específicos de EP (EP-QUALIF) y/o escalas de calidad de vida bien validadas tales como la SF-36 se contemplan para supervisar los resultados del paciente.
Captura de neuroimágenes y otros biomarcadores: La captura de imágenes funcionales se usó ampliamente en ensayos quirúrgicos de EP. Aunque la captura de imágenes basada en dopamina (tal como FDOPA-PET) se contempla para su uso en el examen del mantenimiento del injerto, se contemplan para su uso técnicas de captura de neuroimágenes que usan otros ligandos, incluyendo marcadores basados en captura de imágenes, por ejemplo, dirigidos a inflamación, y de marcadores sistémicos sin captura de imágenes como recopilación de datos exploratorios. Las imágenes se usarían en la planificación preoperatoria, p. ej., alcance y ubicación del agotamiento de DA dentro de los ganglios basales e incorporación de datos de PET en la planificación quirúrgica a medida para cada paciente. Ubicación de la colocación del injerto y número de depósitos celulares. En algunas realizaciones, el putamen (Freed, et al. N. Engl. J. Med. 344, 710-719 (2001); Lindvall, et al. Prog. Brain Res. 82, 729-734 (1990) y putamen postcomisural (Olanow, et al. Ann. Neurol. 54, 403-414 (2003)) son sitios de injerto celular (administración). En algunas realizaciones, de contemplan múltiples ubicaciones a través de diferentes pistas quirúrgicas. Se contempla IRM con un sistema Clearpoint que proporciona imágenes en tiempo real y visualización de la trayectoria y precisión de dirección para supervisar las células injertadas. Este sistema se usa en la colocación de electrodos de estimulación cerebral profunda en pacientes con EP. Número de células y composición del injerto. Se realizaron ensayos fetales con un número esencialmente desconocido de neuronas DA, ya que usaron material de injerto fetal con numerosos tipos celulares. Según los datos proporcionados en la presente memoria, se contempla una estimación de 100.000-200.000 neuronas TH+ supervivientes para recuperación de la función neuronal DA.
Inmunodepresión. En algunas realizaciones, se contempla la inmunodepresión de pacientes injertados, de al menos 6 meses hasta la vida completa de un paciente. En algunas realizaciones, los pacientes no serán inmunodeprimidos con el fin de tener tejido injertado.
Tabla 1: Datos de matriz de expresión génica de genes significativamente regulados positivamente y regulados negativamente el día de diferenciación 11 en la población basada en la placa del suelo tratada con SHH/FGF8/Chir sobre la población tratada con LSB de control.





Tabla 2: Datos de matriz de expresión génica de genes significativamente regulados positivamente y regulados negativamente el día de diferenciación 11 en la población basada en la placa del suelo tratada con SHH/FGF8/Chir sobre la población tratada solo con SHH/f Gf8.







Tabla 3: Datos de matriz de expresión génica de genes significativamente regulados positivamente y regulados negativamente el día de diferenciación 25 sobre el día 13 en la población basada en la placa del suelo tratada con SHH/FGF8/Chir.
Columna ID de Factor de valor de p Columna ID de columna Factor de valor de p n.° cambio
n.° cambio 28590 PTPR 8,70512 3,13E-10 1209 33,085 1,60E-09 30386 SLC17 8,22904 2,02E-06 3978 28,1727 4,44E-13 31767 SYT13 8,1735 3,63E-07 31604 20,9978 1,47E-08 30390 SLC18 8,15448 6,47E-15 32159 15,2965 2,12E-08 5689 EBF1 8,05344 2,30E-06 31606 15,2102 3,72E-09 5029 DCX 8,00552 8,30E-08 7685 12,5729 5,42E-09 2470 C1QL1 7,47022 8,68E-10 4926 10,7914 4,91 E-08 12609 INA 6,92121 5,72E-07 8593 10,6527 2,94E-09 26250 NR4A 6,75723 1,27E-13 4342 10,0606 3,86E-06 29624 RTN1 6,6247 6,52E-07 12149 9,60731 3,97E-08 9374 HS.19 6,58382 1,95E-08 5691 9,28771 5,13E-11 436 ADCY 6,49154 9,75E-11 29820 9,22584 1,02E-06 1607 BEX2 6,33986 8,93E-09 12655 9,01409 3,73E-12 34098 ZCCH 6,19663 9,95E-07
27988 8,79262 1,09E-10 7056 FLJ25 6,19296 2,14E-06
8,79262 1,09E-10 7056 FLJ25 6,19296 2,14E-06
31605 STMN3 6,17605 8,77E-07 12362 ID4 4,32183 6,05E-05 Columna ID de columna Factor de valor de p 26011 NHLH 4,31928 5,91 E-05 n.° cambio 2035 C140R 4,31666 9,46E-07 7997 6,1196 1,26E-06 9428 HS.20 4,29602 1,66E-09
25922  6,11347 2,51E-04 8288
6,11347 2,51E-04 8288  GRM8
GRM8 4,17846 1,62E-06
4,17846 1,62E-06
12359 ID2 6,07844 1,32E-06  7617 GADD45G 4,17663 2,21 E-06
7617 GADD45G 4,17663 2,21 E-06






Tabla 4: Datos de matriz de expresión génica de genes significativamente regulados positivamente y regulados negativamente el día de diferenciación 25 en la población basada en la placa del suelo tratada con SHH/FGF8/Chir sobre la población tratada con LSB de control.
9428 3,88697 6,71E-09 Columna ID de Factor de valor de p 6197 3,70155 3,07E-09 n.° cambio 5036 3,61254 0,000194062 32126 TFF3 25,0585 2,47E-14 26250 3,55879 9,29E-10 1209 ASCL1 14,9328 1,97E-07 2867 3,48146 6,97E-09 7395 FOXA1 12,9897 4,15E-14 26928 3,27941 3,81 E-07 29133 RGS4 11,0181 6,65E-09 5372 3,26437 1,81E-08 27988 POST 8,20403 2,16E-10 33930 3,2414 0,000921089 7396 FOXA 7,98132 1,60E-14 27664 3,23009 1,10E-05 4342 COL3A 7,58625 2,49E-05 9819 3,14523 8,26E-08 12731 IRX3 7,42399 1,64E-06 30239 3,13411 4,23E-05 9374 HS.19 7,27196 7,36E-09 2743 3,10109 1,50E-06 3978 CHGA 7,14305 2,77E-08 623 3,00487 0,000307411 2470 C1QL1 7,1366 1,38E-09 6533 2,99243 3,45E-05
30390 SLC18 7,04751 3,52E-14 28590 2,94164 4,45E-05
SLC18 7,04751 3,52E-14 28590 2,94164 4,45E-05
22823 LOC7 6,5801 8,79E-09 1875 2,93448 1,14E-06 Columna ID de Factor de valor de p 31669 2,92397 8,02E-05 n.° cambio 5348 2,92274 8,36E-05 12456 IGFBP 6,15962 3,65E-05 30066 2,76873 0,000156193 27665 PITX2 6,10679 6,79E-14 5349 2,75425 0,000173675 12655 INSM2 5,18438 1,70E-09 3186 2,71125 2,10E-07 12944 KCNJ1 4,98753 8,89E-13 30590 2,67444 2,25E-05 436 ADCY 4,94597 2,43E-09 32000 2,66777 1,46E-06 31361 SPON 4,93795 4,69E-08 3941 2,63848 0,00120585 13857 LMO3 4,83578 7,25E-08 32769 2,62647 1,95E-06 13865 LMX1 4,63407 1,51E-06 25941 2,55992 0,000599552 2543 C20O 4,4431 3,63E-13 29509 2,53138 0,00135525
12733 IRX5 4,2929 1,45E-06  32144 2,46754 8,27E-07
32144 2,46754 8,27E-07
7988 GNB3 4,13634 1,52E-10 13689 2,40983 2,56E-05 32159 TH 4,08743 0,000370318 2850 2,4009 0,000508473 5346 DLK1 4,02203 1,73E-05 31806 2,38661 0,00154495 31775 SYT4 4,00669 0,000372793 24375 2,31488 5,04E-05 5424 DNAJ 2,30405 4,42E-07 17624 -2,0676 3,05E-06 12986 KCNS 2,28802 0,000136062 13793 -2,07092 0,00040641 27855 PMP2 2,27371 1,36E-05 33165 -2,07384 0,000187156 4003 CHN2 2,26467 0,000550787 3462 -2,07613 0,00151516 5691 EBF3 2,26453 0,000571924 4305 -2,07819 0,000485834 3187 CALC 2,11359 2,49E-06 6912 -2,07924 9,69E-05 28325 PROX 2,08192 0,000135872 5195 -2,07943 0,000935686 25218 MNX1 2,05528 3,37E-06 33905 -2,08212 0,00033792 6270 F3 2,01261 5,71E-07 12933 -2,08721 3,72E-05 2245 C18O -2,00297 0,00129421 4933 -2,09051 9,24E-05 28546 PTN -2,003 0,0003544722 5960 -2,09361 0,00111856 5920 EMID2 -2,00456 1,77E-05 25930 -2,09683 0,00144313 24405 MGC1 -2,00498 2,75E-05 3288 -2,09925 0,000130992 5949 ENKU -2,00742 0,000107398 12971 -2,09934 0,00140267
26398 NUP9
NUP9 -2,01111 0,00124009 3125
-2,01111 0,00124009 3125  -2,10277 0,00147559
-2,10277 0,00147559
10837 HS.545615 -2,01378 2,65E-05 17153 LOC399959 -2,10829 0,000816932



Tabla 5: Datos de matriz de expresión génica de genes significativamente regulados positivamente y regulados negativamente el día de diferenciación 25 en la población basada en la placa del suelo tratada con SHH/FGF8/Chir sobre la población tratada solo con SHH/f Gf8.
3941 8,71553 1,37E-08 Columna ID de columna Factor de valor de p 30390 8,37911 4,77E-15 n.° cambio 5689 8,02385 2,37E-06 32126 24,0181 3,37E-14 2470 7,76325 5,90E-10 1209 22,1637 1,69E-08 22823 7,73246 1,77E-09 9374 14,7045 1,42E-11 27665 7,32855 7,16E-15 7395 13,8748 2,30E-14 12655 7,04572 4,97E-11 13865 10,953 4,33E-10 26250 6,75415 1,28E-13 12456 9,56964 1,58E-06 8156 6,28979 1,08E-05 4342 9,49528 5,64E-06 13857 6,20624 4,64E-09 12361 9,45077 3,33E-08 33930 5,8436 6,74E-06 27988 9,10453 7,82E-11 12944 5,71955 1,37E-13 7396 8,85174 5,07E-15 27821 5,62251 8,45E-07 5372 5,61111 8,09E-12 26928 3,44485 1,91E-07 7988 5,55425 2,56E-12 27664 3,40016 5,92E-06 31669 5,51821 6,78E-08 2743 3,31658 5,91 E-07 33128 5,48234 2,88E-09 30590 3,30002 1,28E-06 2543 5,42131 2,00E-14 32144 3,28727 6,00E-09 32762 5,30583 7,23E-05 18750 3,2583 2,43E-07 436 5,20834 1,29E-09 17491 3,24609 0,000469294
26977 5,10526 6,89E-06 24375 3,24122 3,31 E-07
5,10526 6,89E-06 24375 3,24122 3,31 E-07
3978 CHGA 5,07311 7,78E-07 5346 3,21349 0,000161289 Columna ID de columna Factor de valor de p 7562 3,15302 8,07E-08 n.° cambio 3186 3,14299 1,75E-08 32000 5,05883 1,27E-10 27772 3,09727 0,000150905 4003 4,53859 9,76E-08 26335 3,08699 1,10E-05 2867 4,43629 1,97E-10 34155 3,05463 6,49E-07 32417 4,40645 3,65E-08 27855 3,03785 1,12E-07 26245 4,38488 8,32E-05 4491 3,02972 0,000317992 5036 4,37185 3,55E-05 7421 3,01784 0,000461499 6700 4,28732 4,74E-06 2850 3,00725 3,47E-05 9428 4,25031 1,93E-09 27105 2,97981 1,49E-05
8288 4,16549 1,67E-06  12358 2,94361 0,000160308
12358 2,94361 0,000160308
28590 4,13365 8,06E-07 33004 2,93637 2,29E-07 32159 4,10876 0,000355952 5691 2,92465 2,26E-05 31361 4,063 4,56E-07 1086 2,92012 2,33E-06
34166 4,05459
4,05459  0,000219198 32769
0,000219198 32769 2,88914
2,88914  4,24E-07
4,24E-07
13689 LDB2 3,9497 1,90E-08 5424 DNAJC19 2,87234 6,43E-09


Tabla 6: muestra una lista ilustrativa de anticuerpos usados como marcadores, incluyendo la concentración (es decir, la dilución) de anticuerpos usados y fuentes ilustrativas de anticuerpos. En algunas realizaciones, los anticuerpos unidos se identificaron con cualquiera de anticuerpos secundarios conjugados con Alexa488, Alexa555 y Alexa647 (Molecular Probes, Carlsbad, California). En algunas realizaciones, se usaron anticuerpos secundarios biotinilados para identificar los anticuerpos primarios unidos seguidos de visualización a través del cromógeno DAB (3,3'-diaminobencidina).


Tabla 7: Diferenciación contemplada ilustrativa en neuronas DA por límites de tipo celular.

Tabla 8. Experimentos ilustrativos para determinar la función de determinados factores para producir neuronas DA.
LDN SB PURM CHIR SHH BDNF GDNF AA Dbc TGFB3 DAPT AMP
Control
Retirada
completa
/- SHH
/- BDNF
/- GDNF
/- AA
/- dbAMPc
/- TGFB3
/- DAPT 
Tabla 9. Métodos ilustrativos para aumentar la escala del cultivo de neuronas mDA, en particular para su uso en la producción de cultivos de nivel GMP para su uso en la clínica.


Tabla 10. Evaluación in vivo ilustrativa de productos de la línea hES- Composición del injerto.

Tabla 11. Evaluación in vivo ilustrativa de productos de la línea hES- Análisis conductuales.

(*) En algunas realizaciones, las ratas que presentaban >6 rotaciones/min recibieron injertos estables.
EXPERIMENTACIÓN
Los siguientes ejemplos sirven para ilustrar determinadas realizaciones y aspectos de la presente invención y no deben interpretarse como limitantes del alcance de la misma. En las descripciones experimentales a continuación, se aplican las siguientes abreviaturas: N (normal); M (molar); mM (milimolar); pM (micromolar); mol (moles); mmol (milimoles); pmol (micromoles); nmol (nanomoles); pmol (picomoles); g (gramos); mg (miligramos); pg (microgramos); ng (nanogramos); pg (picogramos); l (litros); ml (mililitros); pl (microlitros); cm (centímetros); mm (milímetros); pm (micrómetros); nm (nanómetros); U (unidades); min (minuto); s y seg (segundo); grad (grado); pen (penicilina), estrep (estreptomicina) y °C (grados centígrados/Celsius).
EJEMPLO de referencia I.
Materiales y métodos.
Compendio de métodos: Las líneas de ESC humana (H9, H1) e iPSC (2C6 y SeV6) se sometieron a una inhibición doble de SMAD modificada (Chambers, et al. Nat. Biotechnol. 27: 275-280 (2009), protocolo inducción basada en la placa del suelo (Fasano, et al., Cell Stem Cell 6: 336-347 (2010), incorporado en la presente memoria por referencia). La exposición a SHH C25II, purmorfamina, FGF8 y CHIR99021 se optimizó para la placa del suelo del mesencéfalo y el rendimiento de nuevas poblaciones de neuronas DA (véase figura 1d). Después de la inducción de la placa del suelo, Se llevó a cabo maduración adicional (días 11-25 o más de 25 días en cultivo hasta al menos 100 días en cultivo) en medio de diferenciación basado en Neurobasal/B27 en presencia de factores de supervivencia y maduración de neuronas DA (Perrier, et al. Proc Natl Acad Sci USA 101: 12543-8 (2004), tales como AA, BDNF, GDNF, TGFp3 y dbAMPc (véase métodos completos para más detalles). La población de neuronas DA resultante se sometió a caracterización fenotípica exhaustiva mediante inmunocitoquímica, qRT-PCR, perfil de expresión génica global, análisis por HPLC para la detección de dopamina y registros electrofisiológicos in vitro. Se realizaron estudios in vivo en roedores hemiparkinsonianos (ratones o ratas con inyección de la toxina 60HDA en un lado del cerebro del animal. Los estudios se llevaron a cabo en ratones adultos NOD-SCID IL2Rgc (Jackson labs) y ratas adultas Sprague
Dawley Taconic Farms, que recibieron lesiones de 6-hidroxidopamina mediante inyecciones estereotácticas de la toxina como se ha descrito anteriormente, así como dos monos rhesus adultos que fueron tratados con inyecciones carotídeas unilaterales de MPTP. Se inyectaron neuronas DA de manera estereotáctica en el cuerpo estriado de los animales (150 x 103 células en ratones, 250 x 103 células en ratas) y un total de 7,5 x 106 células (distribuidas en 6 tramos; 3 en cada lado del cerebro) en monos. Se realizaron ensayos conductuales a intervalos mensuales después del injerto, incluyendo análisis rotacional mediado por anfetaminas, así como una prueba de acinesia focal ("prueba de marcha") y uso de las extremidades (prueba de cilindro). Se sacrificaron las ratas y los ratones a las 18-20 semanas y los primates al cabo de 1 mes después del injerto. Se realizó caracterización de los injertos mediante análisis estereológicos del número de células y los volúmenes de injerto, así como una caracterización fenotípica exhaustiva mediante inmunohistoquímica. Cultivo de células ES humanas indiferenciadas. Las líneas de hESC H9 (WA-09, XX, pases 27-55 desde la fecha 10/2009), H1 (WA-01, XY, pases 30-40 desde la fecha 6/2010) y líneas celulares de iPS 2C6 (Kim, et al. Cell Stem Cell 8: 695-706 (2011), (XY, pases 20-30) y SeV6 (XY, pases 20-30; procedente de fibroblastos embrionarios MRC-5 usando un sistema de vector Sendai de 4 factores no integrante (Ban, et al. Proc. Natl. Acad. Sci. U.S.A (2011) 108 (34): 14234-14239: 10.1073/pnas.1103509108) se mantuvieron en fibroblastos embrionarios de ratón a concentraciones de siembra en placas estimadas que varían de 0,5 x103 por cm2 a 100 x103 por cm2 basado en células ES humanas que tienden a formar agrupamientos celulares. (MEF, Global Stem, Rockville, MD) en un reemplazo de suero knockout óptimo al 20 % (KSR, Invitrogen, Carlsbad, California) que contiene medio de células ES humanas (como se ha descrito anteriormente (Kim, et al. Cell Stem Cell 8: 695-706 (2011). El uso de reemplazo de suero knockout puede variar de 0 % a 40 %.
Inducción Neural. Para la inducción de neuronas de dopamina del mesencéfalo basada en la placa del suelo, se usó una versión modificada del protocolo de inhibición doble de SMAD (Chambers, et al. Nat. Biotechnol. 27: 275-280 (2009) e inducción de la placa del suelo (Fasano, et al. Cell Stem Cell 6: 336-347 (2010) basándose en la exposición sincronizada a LDN-193189 (100 nM (intervalo de concentración de 0,5-50 pM, Stemgent, Cambridge, Massachusetts), SB431542 (10 pM (intervalo de concentración de 0,5-50 pM, Tocris, Ellisville, MI), SHH C25II (100 ng/ml (intervalo de concentración de 10-2000 ng/ml, R&D, Mineápolis, Mn ), purmorfamina (2 pM (intervalo de concentración de 10-500 ng/ml, Stemgent), FGF8 (100 ng/ml (intervalo de concentración de 10-500 ng/ml, R&D) y CHIR99021 (CHIR; 3 pM (intervalo de concentración de 0,1-10 pM, Stemgent). Nota: para el protocolo de inducción de la placa del suelo, el tratamiento de "SHH" se refiere a la exposición, es decir, contacto, de células a una combinación de SHH C25II 100 ng/ml purmorfamina (2 pM). Las células se sembraron en placas (35-40 x 103 células/cm2) y se cultivaron durante 11 días en matrigel o geltrex (usado tal como se adquirió) (BD, Franklin Lakes, Nueva Jersey) en medio de reemplazo de suero Knockout (KSR) que contiene DMEM, suero de reemplazo knockout al 15 %, L-glutamina 2 mM y 10 pM (intervalo de concentración de 1-25 pM de p-mercaptoetanol. El medio KSR se cambió gradualmente a medio N2 a partir del día 5 de diferenciación, mezclando en proporciones de 75 % (KSR):25 % (N2) el día 5-6, 50 % (KSR):50 % (N2) día 7-8 y 25 % (KSR):75 % (N2) el día 9-10, como se ha descrito anteriormente (Chambers, et al. Nat. Biotechnol. 27: 275-280 (2009), incorporado en la presente memoria por referencia). El día 11, los medios se cambiaron a medio neurobasal/medio B27 (dilución 1:50)/medio que contiene l-Glut (intervalos eficaces 0,2-2 mM)) (NB/B27; Invitrogen) complementado con CHIR (hasta el día 13) y con BDNF (factor neurotrófico derivado del cerebro, 20 ng/ml que varía de 5 a 100; R&D), ácido ascórbico (AA; 0,2 mM (intervalo de concentración de 0,01 1 mM), Sigma, San Luis, MI), GDNF (factor neurotrófico derivado de la línea celular glial, 20 ng/ml (intervalo de concentración de 1-200 ng/ml); R&D), TGFp3 (factor de crecimiento transformante de tipo p3, 1 ng/ml (intervalo de concentración de 0,1-25 ng/ml); R&D), dibutiril AMPc (0,5 mM (intervalo de concentración de _0,05-2 mM); Sigma) y DAPT (10 nM (intervalo de concentración de 0,5-50 nM); Tocris,) durante 9 días. El día 20, las células se disociaron usando Accutase® (Innovative Cell Technology, San Diego, California) y se volvieron a sembrar en condiciones de alta densidad celular (por ejemplo de 300-400k células/cm2) en placas previamente recubiertas con poliornitina (PO); 15 pg/ml (intervalo de concentración de 1-50 pg/ml)/Laminina (1 pg/ml) (intervalo de concentración de 0,1 10 pg/ml)/Fibronectina (2 pg/ml (intervalo de concentración de 0,1-20 pg/ml) en medio de diferenciación (NB/B27 BDNF, AA, GDNF, dbAMPc (intervalo de concentración como se describe en la presente memoria), TGFp3 y DAPT (que varían en concentración como se describe en la presente memoria) hasta la etapa de maduración deseada para un experimento dado.
Para la inducción de neuronas DA basada en la roseta, se siguieron los protocolos descritos anteriormente en parte (Perrier, et al. Proc Natl Acad Sci U S A 101: 12543-8 (2004) con al menos una excepción donde se usó inhibición doble de SMAD para acelerar la etapa de inducción neural inicial. En resumen, se indujeron hESC hacia destino neural mediante cocultivo con células MS5 irradiadas en KSR complementado con SB431542 y Noggin (250 ng/ml (intervalo de concentración de 10-1000 ng/ml); R&D), desde el día 2-8 y SHH+FGF8 desde el día 6-11 de diferenciación. Después de 11 días en KSR, las rosetas neurales se aislaron manualmente y se cultivaron (etapa P1) en medio N2 complementado con SHH, FGF8, BDNF y AA como se ha descrito anteriormente (Perrier, et al. Proc Natl Acad Sci U S A 101: 12543-8 (2004). Después de 5-7 días en la etapa P1, las rosetas se recogieron de nuevo mecánicamente y se trituraron después de incubación en solución salina equilibrada de Hanks sin Ca2/Mg2 (HBSS) durante 1 hora y se volvieron a sembrar en placas recubiertas con poliornitina (PO)/laminina/fibronectina. La distribución con SHH/FGF8 continuó durante 7 días en la etapa P2 seguido de diferenciación final en presencia de BDNF, AA, GDNF, TGFb3 y dbAMPc como se ha descrito anteriormente hasta la etapa de maduración deseada para un experimento dado (normalmente 5-7 días para estudios de trasplante o 32 días para estudios funcionales in vitro).
Análisis de expresión génica. Se extrajo ARN total durante la diferenciación los días: 0, 1, 3, 5, 7, 9, 11, 13 y 25 de
cada condición de LSB de control, LSB/SHH/FGF8 y LSB/SHH/FGF8/CHIR usando el equipo RNeasy (Qiagen, Valencia, CA). Para análisis de micromatrices, el ARN total fue procesado por la instalación principal de genómica del MSKCC e hibridado en matrices de perlas ref-12 humana de Illumina según las especificaciones del fabricante. Se realizaron comparaciones entre cada día y condición usando el paquete LIMMA de Bioconductor (worldwideweb.bioconductor.org). Los genes que se descubrió que tenían un valor de P ajustado <0,05 y un factor de cambio mayor de dos se consideraron significativos. Algunos de los análisis y presentaciones descriptivos de datos de micromatrices se realizaron usando un paquete de software disponible en el mercado (Partek Genomics Suite (versión 6.10.0915)). Para análisis por qRT-PCR, el ARN total el día 25 de cada condición se transcribió de manera inversa (Quantitech, Qiagen) y se detectó material amplificado usando ensayos de expresión génica Taqman disponibles en el mercado (Applied Biosystems, Carlsbad, CA) con los datos normalizados con respecto a HPRT. Cada punto de datos representa 9 repeticiones técnicas de 3 muestras biológicas independientes. Aún no están disponibles datos sin procesar de estudios de micromatrices en GEO worldwideweb.ncbi.nlm.nih.gov/geo). Cirugía animal. Se realizaron procedimientos de roedores y monos siguiendo las directrices del NIH y fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales (IACUC), el Comité Institucional de Bioseguridad (IBC) y el Comité de Investigación de Células Madre Embrionarias (ESCRO). Ratones. Ratones nulos para NOD-SCID IL2Rgc (20-35 g de peso; Jackson Laboratory, Bar Harbor, ME) fueron anestesiados con ketamina (90 mg/kg; Akorn, Decatur, IL) y xilazina (4 mg/kg Fort Dodge, IA). Se inyectó 6-hidroxidopamina (10 pg (intervalo de concentración de 0,1-20 pg) 6-OHDA (Sigma-Aldrich) de manera estereotáctica en el cuerpo estriado en las siguientes coordenadas (en milímetros): AP, 0,5 (de bregma; una sutura de cráneo usada como referencia para cirugía estereotáctica); ML, -2,0; DV, -3,0 (de duramadre, una membrana que cubre el cerebro usada como referencia). Se seleccionaron para trasplante ratones con lesiones exitosas (un promedio de >6 rotaciones/minuto). Se inyectaron un total de 150 x 103 células en un volumen de 1,5 pl en el cuerpo estriado en las siguientes coordenadas (en mm): AP, 0,5; ML, -1,8; DV, 3,2. Los ratones se sacrificaron 18 semanas después del trasplante.
Ratas. Se anestesiaron ratas hembras adultas Sprague-Dawley (Taconic, Hudson, NY (180-230 g) con ketamina (90 mg/kg) y xilazina (4 mg/kg) durante los procedimientos quirúrgicos. Se establecieron lesiones unilaterales del haz prosencefálico medial de la ruta nigroestriatal mediante inyección estereotáctica de 6-OHDA (3,6 mg/ml en ácido ascórbico al 0,2 % y solución salina al 0,9 %, Sigma) en dos sitios (Studer, et al. Nature Neurosci. 1: 290-295 (1998). Se seleccionaron ratas para trasplante si la rotación inducida por anfetaminas superaba 6 rotaciones/min a las 6-8 semanas después de la inyección. Se trasplantaron 250 x 103 células en el cuerpo estriado de cada animal (Coordenadas: AP 1,0 mm, ML -2,5 mm y V -4,7 mm; barra de dientes ajustada a -2,5). Las ratas de control recibieron PBS en su lugar. Los procedimientos quirúrgicos se han descrito previamente (Studer, et al. Nature Neurosci. 1: 290 295 (1998). Se iniciaron inyecciones intraperitoneales diarias de ciclosporina 15 mg/kg (Bedford Labs, Bedford, OH) 24 horas antes del injerto celular y continuaron hasta el sacrificio, 20 semanas después del injerto celular.
Primates no humanos: Dos monos rhesus adultos (17-18 años de edad; 10-12 kg; hembras) se volvieron hemiparkinsonianos mediante administración de MPTP carotídea seguida de I.V. semanal. Administración de MPTP para crear un síndrome parkinsoniano bilateral (Kordower, et al. Science 290:767-773 (2000). Ambos animales presentaron síntomas parkinsonianos coherentes con una lesión moderadamente grave basada en análisis conductual, incluyendo postura torcida, arrastre de la pierna y síntomas de rigor (inflexibilidad de movimiento), inatención (percepción motora a estímulo lateralizado) y bradicinesia (inicio de movimiento lento). Estos parámetros se pueden evaluar en monos usando una escala de calificación clínica (CRS) de Parkinson modificada. El día de la cirugía de trasplante, los animales se tranquilizaron con ketamina (3,0 mg/kg, IM) y dexdomitor (0,02-0,04 mg/kg IM), intubado para mantener una vía respiratoria estable y anestesiado con isoflurano. Después se colocaron en un marco estereotáctico para cirugía. Ambos monos rhesus se sometieron a una única cirugía con tres inyecciones intracraneales de cultivos de DA procedentes de la placa del suelo humana basadas en coordenadas estereotácticas (Paxinos, et al. The Rhesus Monkey Brain in Stereotaxic Coordinates (Academic Press, (2000). Se realizaron inyecciones bilaterales de células (10 ul/inyección; 125.000 células/ul) en tres sitios (1-caudado posterior, 2-putamen pre-comisural y materia blanca suprayacente) para un volumen total de 30 pl por hemisferio. Se utilizó una bomba de infusión unida a un micromanipulador estereotáctico para administrar las células a una velocidad de 1 pl/min a través de una jeringa Hamilton de 50 pl con aguja de 28 G. Después de completarse las inyecciones, la aguja se dejó en su sitio durante 2-5 minutos adicionales para permitir que el infundido se difunda por la punta de la aguja antes de retraer lentamente la jeringa. Inmediatamente después de la cirugía, los animales recibieron analgésicos (buprenex, 0,01 mg/kg IM, dos veces al día durante 72 horas después de la cirugía; meloxicam, 0,1 mg/kg s.c., una vez al día durante 72 horas después de la cirugía), así como un antibiótico (cefazolina, 25 mg/kg IM, dos veces al día) hasta 72 horas después de la cirugía. Los animales recibieron ciclosporina A (Neoral, Sandimmune) por vía oral (30 mg/kg que se disminuye a 15 mg/kg) una vez al día, comenzando 48 horas antes de la cirugía hasta el sacrificio, un mes después del trasplante. Ensayos conductuales. Se llevaron a cabo rotaciones inducidas por anfetaminas (ratones y ratas) y la prueba de marcha (rata) antes del trasplante y 4, 8, 12, 18 semanas después del trasplante. El comportamiento de rotación en ratones se registró 10 minutos después de inyección i.p. de d-anfetamina (10 mg/kg, Sigma) y se registró durante 30 minutos. El comportamiento de rotación en ratas se registró 40 minutos después de la inyección i.p. de danfetamina (5 mg/kg) y se evaluó automáticamente mediante el sistema TSE VideoMot2 (Alemania). Los datos se presentaron como el número promedio de rotaciones por minuto. La prueba de marcha se modificó a partir de Blume, et al. Exp. Neurol. 219: 208-211 (2009) y Crawley, et al. What's Wrong With My Mouse: Behavioral Phenotyping of Transgenic and Knockout Mice (Wiley-Liss, 2000).
En resumen, cada rata se colocó sobre una superficie plana; sus patas traseras se elevaron levantando suavemente la cola para permitir que solo las patas delanteras tocaran la mesa. El experimentador tiró de la rata hacia atrás 1 metro a un ritmo constante. Se contaron los números de paso de ajuste de las patas delanteras tanto contralaterales como ipsilaterales. Los datos se presentaron como el porcentaje de pasos de ajuste contralaterales / (contralaterales ipsilaterales). La prueba del cilindro se realizó colocando cada animal en un cilindro de vidrio y contando el número de toques de patas ipsilaterales frente a contralaterales (de 20 toques) a la pared del cilindro como se ha descrito anteriormente (Tabar, et al. Nature Med. 14: 379-381 (2008), incorporado en la presente memoria por referencia). Procesamiento de tejidos. Ratones y ratas: Los animales (ratones y ratas) recibieron sobredosis de pentobarbital por vía intraperitoneal (50 mg/kg) para inducir anestesia profunda y se perfundieron en paraformaldehído (PFA) al 4 %. Se extrajeron cerebros, se fijaron posteriormente en PFA al 4 % y después se empaparon en soluciones de sacarosa al 30 % durante 2-5 días. Se seccionaron en un criostato después de incluirlo en un compuesto O.C.T. (Sakura-Finetek, Torrance, California).
Primates no humanos: Los animales se sacrificaron con anestesia profunda con ketamina (10 mg/kg, intramuscular (IM)) y pentobarbital (25 mg/kg, intravenoso (IV)) a través de perfusión cardíaca con solución salina heparinizada al 0,9 % seguida de fijador de PFA frío al 4 % nuevo (pH 7,4). Inmediatamente después de la fijación primaria, los cerebros se retiraron del cráneo y se fijaron posteriormente en PFA al 4 %, en flotación libre, durante 24-36 h. Después se aclararon y se volvieron a suspender en sacarosa al 10 % en un agitador lento a 4 °C y se permitió que se "hundieran". Después se repitió el proceso en sacarosa al 20 % seguido de sacarosa al 30 %. Los cerebros completos se cortaron coronalmente en secciones en serie de 40 um en un microtomo deslizante congelado y se almacenaron en flotación libre en medio crioconservador a -20 °C.
Inmunohistoquímica: Las células se fijaron en PFA al 4 % y se bloquearon con seroalbúmina bovina (BSA) al 1 % con Triton al 0,3 %. Las secciones de tejido cerebral se lavaron en PBS frío y se procesaron de manera similar. Los anticuerpos primarios se diluyeron en BSA al 1-5 % o suero de cabra normal y se incubaron según las recomendaciones del fabricante. En la tabla 6 se proporciona una lista exhaustiva de anticuerpos y fuentes. Se usaron anticuerpos secundarios conjugados con Alexa488, Alexa555 y Alexa647 (Molecular Probes, Carlsbad, California) con contratinción nuclear de 4',6-diamidino-2-fenilindol (DAPI) (Thermo Fisher, Rockford, Illinois). Para algunos análisis, se usaron anticuerpos secundarios biotinilados seguidos de visualización mediante cromógeno DAB (3,3'-diaminobencidina). Análisis de HPLC. HPLC de fase inversa con detección electroquímica para medir niveles de dopamina, Ácido homovanílico (HVA) y se realizó DOPAC (ácido 3,4-dihidroxi-fenilacético) como se ha descrito anteriormente (Roy, et al. Nature Med. 12: 1259-1268 (2006); Studer, et al. Brain Res. Bull. 41: 143-150 (1996). Se recogieron muestras de cultivo en ácido perclórico el día 65 de diferenciación. Para algunos experimentos, se midió DA directamente en el medio usando el mismo sistema de detección pero después de la extracción de aluminio de dopamina y sus metabolitos usando un equipo disponible en el mercado como se descrito anteriormente (Studer, et al. Brain Res. Bull. 41: 143-150 (1996), incorporado en la presente memoria por referencia). Registros electrofisiológicos: Los cultivos se transfirieron a una cámara de registro en un microscopio vertical equipado con un objetivo de inmersión en agua 40X (Eclipse E600FN; Nikon); los cultivos se perfundieron con solución salina que contenía en mM: NaCl 125, KCl 2,5, NaHCO325, NaH2PO41,25, CaCl 2, MgCb 1 y glucosa 25 (34 °C; saturado con O295 %-CO25 %; pH 7,4; 298 mOsm/l). El caudal de solución salina fue de 2-3 ml/min a través de un calentador en línea (SH-27B con controlador TC-324B; Warner Instruments). Las neuronas se visualizaron mediante microscopia de vídeo con una cámara digital CCD enfriada (CoolSNAP ES2, Photometries, Roper Scientific, Tucson, Arizona). Las células seleccionadas para registros electrofisiológicos tenían formas de tipo neuronal con neuritas ramificadas finas. Se realizaron registros de pinzamiento zonal de células somáticas completas en la configuración de pinza actual con un amplificador MultiClamp 700B (Molecular Devices). Las señales se filtraron a 1-4 kHz y se digitalizaron a 5-20 kHz con un Digidata 1440A (Molecular Devices). Los electrodos de parche de registro estaban hechos de vidrio de borosilicato filamentado (Sutter Instruments) extraído en un extractor Flaming-Brown (P-97, Sutter Instruments) y tenían resistencias de 4-6 MQ en el baño. Los electrodos se llenaron con solución interna que contenía en mM: K-MeSO4135, KCl 5, HEPES 5, EGTA 0,25, fosfocreatina-di(tris) 10, ATP-Mg 2 y GTP-Na 0,5 (pH 7,3, osmolaridad ajustada a 290-300 mOsm/l). El circuito del puente amplificador se ajustó para compensar la resistencia del electrodo y se supervisó. Se compensó la capacidad del electrodo. Cuando la resistencia en serie aumentó >20 % durante el registro, los datos se descartaron porque una mayor resistencia sugirió un fallo técnico parcial durante los registros.
Recuentos celulares y análisis estereológicos. Los porcentajes de células positivas para marcador en la placa del suelo (día 11) figura 1, precursor de neuronas de dopamina del mesencéfalo (día 25), figura 2 y etapas de neuronas DA maduras (día 50 o posterior) figura 3 y 11, se determinaron en muestras procedentes de 3 experimentos independientes cada una. Las imágenes para cuantificación se seleccionaron de manera aleatoria uniforme y cada imagen se puntuó primero con respecto al número de núcleos positivos para DAPI, seguido de recuento del número de células que expresan el marcador de interés. Los datos se presentan como la media ± ETM. Se realizó cuantificación de células humanas (identificadas con anti-hNA) y neuronas TH+ dentro de los injertos en cada décima sección donde un injerto era identificable. Los recuentos celulares y el volumen del injerto se determinaron usando la sonda del fraccionador óptico y el estimador Cavalieri usando el software Stereo Investigator (MBF bioscience, Vermont) como se ha descrito anteriormente en Tabar, et al. Nat. Biotechnol. 23: 601-606 (2005). Los datos se presentan como el número total estimado de células y el volumen total de injerto /- error típico de las medias (ETM).
Las siguientes formulaciones describen medios de cultivo celular ilustrativos para su uso en el desarrollo de realizaciones de las presentes invenciones.
Medio de hESC para mantenimiento (1 litro): 800 ml de DMEM/F12, 200 ml de reemplazo de suero Knockout, 5 ml de L-glutamina 200 mM, 5 ml de pen/estrep, 10 ml de solución de 15 aminoácidos no esenciales mínima MEM 10 mM, 55 pM de 13-mercaptoetanol y bFGF (la concentración final es de 4 ng/ml).
Medio KSR para diferenciación de hESC (1 litro): 820 ml de DMEM Knockout, 150 ml de reemplazo de suero Knockout, 10 ml de L-glutamina 200 mM, 10 ml de pen/estrep, 10 ml de MEM 10 mM y 55 pM de 13-mercaptoetanol.
Medio N2 para diferenciación de hESC (1 litro): 985 ml de H2O dest. con polvo de DMEM/F12, 1,55 g de glucosa (Sigma, n.° de cat. G7021), 2,00 g de bicarbonato de sodio (Sigma, cat. n.° S5761), putrescina (alícuota de 100 ul de 1,61 g disuelto en 100 ml de agua destilada; Sigma, cat n.° P5780), progesterona (alícuota de 20 ul de 0,032 g disueltos en 100 ml de etanol al 100 %; Sigma, cat. n.° P8783), selenito de sodio (alícuota de 60 pl de solución 0,5 mM en agua destilada; Bioshop Canada, cat. n.° SEL888) y 100 mg de transferrina (Celliance/Millipore, cat. n.° 4452-01) y 25 mg de insulina (Sigma, cat. n.° 16634) en 10 ml de NaOH 5 mM.
Medio de Eagle con modificación de Dulbecco (DMEM), con FBS al 10 % para preparar PMEF (células alimentadoras (fibroblastos primarios de embrión de ratón (PMEF)) (1 litro): 885 ml de DMEM, 100 ml de FBS, 10 ml de pen/estrep y 5 ml de L-glutamina.
Medio esencial mínimo alfa (MEM) con FBS al 10 % para preparar medio de células alimentadoras MS-5 (1 litro): 890 ml de MEM alfa, 100 ml de FBS, 10 ml de solución de gelatina de pen/estrep (500 ml): Disolver 0,5 g de gelatina en 500 ml de agua tibia (50-60 °C) Milli-Q. Enfriar a temperatura ambiente.
EJEMPLO de referencia II.
Este ejemplo describe el descubrimiento de moléculas pequeñas y la sincronización del contacto para proporcionar la diferenciación dirigida de neuronas DA FOXA2+LMX1A+ de las presentes invenciones.
A continuación hay un breve compendio de algunos de los descubrimientos experimentales descritos en la presente memoria: El tratamiento de células con inhibición doble de SMAD con agonistas de SHH (purmorfamina SHH) y FGF8 (S/F8) en ausencia de CHIR99021 mostró expresión significativamente menor de FOXA2 para el día 11 y falta completa de expresión de LMX1A (figura 1a, b). El marcador anterior OTX2 fue inducido de manera robusta en cultivos tratados con lSb y LSB/S/F8/CHlR, pero no en condiciones de LSB/S/F8 (figura 1a, c).
Los inventores usaron anteriormente otros métodos de diferenciación dirigida que dieron como resultado poblaciones celulares que contenían neuronas de tipo DA. Estas neuronas de tipo DA se usaron en estudios de trasplante que dieron como resultado preocupaciones sobre el uso posterior de estas células para aplicaciones terapéuticas. Por ejemplo, los procedimientos descritos en Perrier et al., 2004 y Fasano et al., 2010, incluyendo inducción neural de MS5, dieron como resultado la formación de células de la roseta y se usaron para producir precursores del día 11, véase figs. 2, 16 y 17 para los ejemplos, que después se usaron para obtener neuronas de tipo DA. Estas neuronas resultaron de un bajo porcentaje de las células precursoras en las poblaciones celulares del día 11 resultantes. Los estudios de trasplante que usaron estas neuronas mostraron mala viabilidad posterior al trasplante y pérdida del fenotipo neuronal de tipo DA, además de observaciones de desarrollo posterior al trasplante de tipos neurales inadecuados junto con pérdida de control del crecimiento, que condujo al desarrollo de teratomas. Véase figs. 16 y 17.
De manera específica, en P0 se pusieron en contacto hESC con moléculas para comenzar la inducción neural de células Oct4+ en células de la roseta usando células alimentadoras MS5 (Perrier et al., 2004). En la etapa P1, las células de la roseta se expandieron al poner en contacto las células con moléculas adicionales para diferenciar células en células en la etapa P2 con patrones de expresión específicos que incluyen células progenitoras DA Pax2+/En1+ y se diferenciaron adicionalmente en neuronas DA TH+/En1+. Estas células se usaron para injerto en ratas con lesión de 6OHDA, inmunodeprimidas con ciclosporina A. Esos estudios de trasplante mostraron mala viabilidad in vivo, pérdida del fenotipo TH+, preocupaciones acerca de crecimiento adicional en células no deseadas, posiblemente letales, es decir, teratomas, y crecimiento de células en tipos neuronales inadecuados que provocarían problemas médicos adicionales para el paciente.
Hubo cantidades muy pequeñas de neuronas TH+ supervivientes a los 4,5 meses después del trasplante (<50 células TH+/animal) en injertos de precursores de neuronas DA procedentes de la roseta fig 16A. Sin embargo, en contraste con las células TH+, las células marcadas con GFP (GFP fue impulsada por un promotor ubicuo) sobrevivieron bastante bien después del trasplante. Esto sugiere que la mayoría de las células supervivientes después del trasplante eran células neurales de identidad neuronal no DA (16B). Pocas células procedentes del injerto (hNA+ (verde) coexpresan TH (rojo) de nuevo, lo que sugiere que la mayoría de las células humanas injertadas adoptan un fenotipo distinto de neurona Da fig 16C. Los paneles 16 D-E muestran que D-E, a pesar de la supervivencia muy baja in vivo hubo alguna mejora (baja y muy variable) en algunos ensayos conductuales tales como rotaciones inducidas por anfetaminas (D), prueba de cilindro y rotaciones espontáneas (E). Se llevó a cabo inducción neural sin alimentador como se ha descrito anteriormente (Chambers et al., 2009) pero se modificó adicionalmente para producir células de la placa del suelo (Fasano et al., 2010). En el método de inhibición doble de SMAD modificado para diferenciar células pluripotentes en células de la placa del suelo, los inventores descubrieron previamente que para el día 11 eran necesarias altas concentraciones de SHH para inducción de PS. Por ejemplo, en algunas realizaciones, se añadió Sonic C25II a 200 ng/ml. En algunos experimentos, se añadieron DKK-1 (R&D; 100 ng/ml) FGF8 (R&D; 50 ng/ml),
Wnt-1 (Peprotech; 50 ng/ml) y ácido retinoico (R&D; 1 mM), véase figura 17. Sin embargo, ninguna de las poblaciones celulares resultantes el día 11 usando métodos anteriores, contenía el alto porcentaje de células progenitoras de la placa del suelo del mesencéfalo FOXA2+/LMX1A+ usando métodos de las presentes invenciones.
Como se muestra en la presente memoria, los inventores eligieron una población celular que contenía células pluripotentes para una población inicial y esta se sembró en placas el día 0. Las células se cultivan casi hasta la confluencia antes de la diferenciación (entre 60 y 100 % de confluencia). Estas células se pusieron en contacto con inhibidores dobles de SMAD (es decir, exposición a LDN-193189 SB431542 = "LSB") el día 0. Los inventores siguieron una población celular con aportes regulares que contenían LSB nuevo hasta el día 11 y descubrieron que algunas células restantes eran LMX1A+ pero no expresaban FOXA2 (figura 1a, b). Los inventores sembraron en placas poblaciones de células iniciales duplicadas y después analizaron los tipos celulares (es decir, patrones de expresión de genes/proteínas) después de entrar en contacto con mezclas que contenían cualquiera de los siguientes agonistas de SHH (purmorfamina SHH) y FGF8 (S/F8) en contacto con las células con diferentes regímenes de exposición, es decir, en contacto con las células el día 0, día 1 o día 2, etc. durante cantidades específicas de tiempo, es decir, 24 horas, 48 horas, etc. Tres condiciones de cultivo ilustrativas principales probadas fueron 1) células en contacto con LDN/SB (LSB) el día 0 y después en contacto con LSB nuevo hasta el día 5, el día 5 las células se pusieron en contacto con LDN nuevo sin SB hasta el día 11, 2) células en contacto con LDN/SB (LSB) el día 0 y después en contacto con LSB nuevo hasta el día 5, el día 5 las células se pusieron en contacto con LDN nuevo sin SB hasta el día 11, mientras que durante este tiempo las células se pusieron en contacto adicionalmente con purmorfamina nueva, SHH y FGF8 hasta el día 7 y 3) células en contacto con LDN/SB (LSB) el día 0 y después en contacto con LSB nuevo hasta el día 5, el día 5 las células se pusieron en contacto con LDN nuevo sin SB hasta el día 11, mientras que durante este tiempo las células se pusieron en contacto adicionalmente con purmorfamina nueva, SHH y FGF8 hasta el día 7, mientras que se pusieron en contacto adicionalmente con CHIR nuevo a partir del día 3 de cultivo hasta el día 11 con varias variaciones de estas condiciones primarias para determinar el rendimiento óptimo de los tipos celulares. Se realizaron comparaciones sistemáticas de las tres condiciones de cultivo (figura 1d) usando perfiles de expresión de genes temporales globales. Véase la figura 8 ilustrativa y las tablas 1-6. La agrupación jerárquica de genes expresados diferencialmente segregó las tres condiciones de tratamiento para el día 11 de diferenciación (figura 8a). FOXA1, FOXA2 y varias otras dianas de SHH corriente abajo, incluyendo PTCH1, estaban entre los transcritos con regulación más diferencial en conjuntos de tratamiento de LSB/S/F8/CHIR frente a LSB (figura 1e). La expresión de LMX1A, NGN2 y DDC indicó el establecimiento del destino precursor de neuronas DA del mesencéfalo ya para el día 11 (figura 1e, f). Por el contrario, los cultivos de LSB para el día 11 se enriquecieron con respecto a marcadores precursores del prosencéfalo dorsal tales como HES5, PAX6, LHX2 y EMX2. La comparación directa del tratamiento con LSB/S/F8/CHIR frente a LSB/S/F8 (figura 1f) confirmó el enriquecimiento selectivo con respecto a marcadores precursores de DA del mesencéfalo en el grupo LSB/S/F8/CHIR y sugirió la identidad precursora hipotalámica en cultivos tratados con LSB/S/F8 basándose en la expresión diferencial de RAX1, SIX3 y SIX6 (véase también expresión de POMC, OTP en la figura 2d).
Se muestran listas ilustrativas de transcritos expresados diferencialmente para el día 11 en las tablas 1, 2 y el día 25 en las tablas 3-5 y el análisis de ontología génica figura 8b (DAVID; http://david.abcc.nciferf.gov) confirmó el enriquecimiento de la señalización de WNT canónica tras el tratamiento con CHIR. Los datos sin procesar aún no están disponibles en GEO worldwideweb.ncbi.nlm.nih.gov/geo/ n.° de referencia: [por determinar]). El análisis temporal comparativo de la expresión génica para marcadores precursores de DA del mesencéfalo (figura 1g) frente a marcadores de destinos neuronales anteriores y ventrales distintos de DA (figura 1h) dividió las tres condiciones de inducción en: i) LSB: prosencéfalo dorsal; ii) LSB/S/F8: ventral/hipotalámico y iii) LSB/S/F8/CHIR: identidad precursora de DA del mesencéfalo.
EJEMPLO de referencia III.
Diferenciación de neuronas DA. Para mayor diferenciación, las células precursoras se mantuvieron en un medio que promueve la maduración neuronal (BAGCT - véase material y métodos). Se realizaron los siguientes tipos de comparaciones entre las poblaciones de células diferenciadas resultantes de métodos anteriores y métodos de las presentes invenciones: A) Análisis inmunocitoquímico el día 50 de diferenciación para TH en combinación con LMX1A, FOXA2 y NURR1, B) Cuantificación de células TH+, FOXA2+, LMX1+ y NURR1+ de las células totales que compara cultivos procedentes de la roseta frente a procedentes de la placa del suelo (LSB/S/F8/CHIR). C) Cuantificación de los porcentajes de subtipos neuronales serotonina+ (5-HT) y GABA+ (contaminantes de neuronas distintas de DA) el día 50 en cultivos de neuronas DA procedentes de la placa del suelo y la roseta. Y D) Análisis de HPLC para medir dopamina y metabolitos: Comparación de los niveles de DA, DOPAC y HVA entre cultivos procedentes de la placa del suelo frente a la roseta. Para el día 25, tres poblaciones de células precursoras produjeron neuronas Tuj1+ (figura 2a) y células que expresan TH, la enzima limitante de la velocidad en la síntesis de DA. Sin embargo, el tratamiento con LSB/S/F8/CHIR produjo células TH+ que coexpresaban LMX1A y FOXA2 y presentaron fuerte inducción del receptor nuclear NURR1 (NR4A2) (figura 2a, b). La comparación de la expresión génica en cultivos del día 13 frente al día 25 confirmó la inducción robusta de otros marcadores de neuronas DA postmitóticas (figura 2c). La caracterización de la identidad de neuronas DA el día 25 en comparación con cultivos tratados con LSB y LSB/S/F8 confirmó el enriquecimiento de transcritos de neuronas DA del mesencéfalo conocidos e identificó múltiples marcadores candidatos novedosos (figura 2d, tablas 3-5, figura 8b). Por ejemplo, el transcrito más enriquecido en LSB/S/F8/CHIR (grupo DA del mesencéfalo) fue TTF3, un gen no asociado anteriormente con el desarrollo de neuronas DA del mesencéfalo, pero muy expresado en la sustancia negra humana (figura 8c; Allen Brain Atlas: http://human.brainmap.org).
Se obtuvieron datos similares para EBF-1, EBF-3 (figura 8c) así como TTR, una diana transcripcional conocida de FOXA2 en el hígado. Los datos obtenidos durante el desarrollo de las presentes invenciones indicaron el enriquecimiento de varios genes PITXen células precursoras de DA del mesencéfalo. PITX3, un marcador clásico de neuronas DA del mesencéfalo, también se expresó de manera sólida el día 25 de diferenciación (figura 2e). Finalmente, la inducción de neuronas tanto de la placa del suelo del mesencéfalo como DA podría reproducirse fácilmente en líneas hESC e hiPSC independientes (figura 9). Los datos demostrados en la presente memoria mostraron que el protocolo LSB/S/F8/CHIR a diferencia de otros protocolos probados produce células que expresan un perfil de marcadores que coincide con el destino de neuronas Da del mesencéfalo.
Las propiedades in vitro e in vivo de las neuronas DA procedentes de la placa del suelo se compararon con las neuronas de tipo DA obtenidas mediante un intermedio de roseta neural (figura 10 y 16). El diseño de rosetas neurales representa la estrategia más ampliamente usada en la actualidad para obtener neuronas DA de hPSC. Los protocolos basados en la placa del suelo y la roseta fueron eficaces para generar neuronas TH+ con capacidad de supervivencia a largo plazo in vitro (día 50 de diferenciación; Figura 3 a). Sin embargo, el porcentaje de células TH+ fue significativamente mayor en cultivos procedentes de la placa del suelo (figura 3b). Aunque las células TH+ en ambos protocolos presentaron coexpresión de NURR1, las neuronas DA procedentes de la placa del suelo coexpresaron FOXA2 y LMX1A (figura 3a,b). Se observaron pocas neuronas positivas para GABA y serotonina (5-HT) (figura 3c). DA, y sus metabolitos DOPAC y HVA, estaban presentes en cultivos generados con cualquiera de los protocolos, pero los niveles de DA fueron aproximadamente 8 veces mayores en cultivos de la placa del suelo (figura 3d,e). Las neuronas DA del mesencéfalo presentaron crecimiento de fibras extenso y expresión robusta de marcadores neuronales maduros, incluyendo sinapsina, transportador de dopamina (DAT) y canal de potasio de rectificación interna, acoplado a proteína G (Kir3.2, también denominado GIRK2, expresado en neuronas Da de pars compacta de sustancia negra (SNpc)) (figura 3f, figura 11). Las neuronas DA de SNpc in vivo presentan un fenotipo electrofisiológico que las diferencias de la mayoría de las otras neuronas en el cerebro. En particular, se disparan espontáneamente a una velocidad lenta (1-3 Hz). Por otro lado, esta activación lenta está acompañada de un potencial oscilatorio lento, por debajo del umbral. Después de 2-3 semanas in vitro, estas mismas características fisiológicas son presentadas por neuronas DA de SNpc cultivadas a partir de ratones postnatales tempranos. Las neuronas DA diferenciadas a partir de hESC de manera uniforme (4/4) presentaron este fenotipo fisiológico distintivo (figura 3 g-i).
El mantenimiento de neuronas mDA in vitro en d65 mostró que las neuronas positivas para TH todavía expresan FoxA2 y extienden fibras largas habituales para neuronas mDA. Figura 3A. La medición de la liberación de Da por HPLC mostró que neuronas TH+ de 65 días de edad son funcionales in vitro figura 3B.
EJEMPLO de referencia IV.
Injerto de una población novedosa de células neuronales DA en roedores, es decir, ratones y ratas que contienen neuronas dañadas.
Uno de los desafíos en el campo es la capacidad de generar neuronas DA del mesencéfalo procedentes de hPSC que se injerten funcionalmente in vivo sin el riesgo de hiperproliferación neural o diferenciación inadecuada en neuronas que no son del mesencéfalo o desarrollar teratomas. Basándose en estudios de trasplante de tejido fetal, los inventores contemplaron que el momento de salida del ciclo celular, marcado por la expresión de NURR1, puede ser una etapa adecuada para el injerto (aproximadamente el día 25 de diferenciación - figura 2). Los estudios iniciales que usaron células del día 25 en ratones adultos sin lesiones mostraron supervivencia robusta de las neuronas FOXA2+/TH+ procedentes de hPSC a las 6 semanas después del trasplante (figura 12). Se probó la supervivencia de las células FOXA2+/TH+ a largo plazo en hospedadores parkinsonianos sin dar lugar a hiperproliferación neural. Con este fin, se realizaron 6 lesiones de hidroxi-dopamina (6-OHDA) (Tabar, et al. Nature Med. 14: 379-381 (2008), incorporado en la presente memoria por referencia) en ratones nulos para NOD-SCID IL2Rgc, una cepa que respalda eficazmente la supervivencia de xenoinjertos con sensibilidad particular para exponer células tumorigénicas poco habituales (Quintana, et al. Efficient tumour formation by single human melanoma cells. Nature 456: 593-598 (2008). Se injertaron cultivos de neuronas DA procedentes de la placa del suelo y la roseta (150 x 103/animal) sin purificación previa para revelar posibles células contaminantes con alto potencial proliferativo. Cuatro meses y medio después del trasplante, los injertos de neuronas DA procedentes de la placa del suelo mostraron un núcleo de injerto bien definido compuesto por células TH+ que coexpresan FOXA2 y el marcador específico humano hNCAM (figura 4a-c). El análisis funcional mostró un rescate completo del comportamiento de rotación inducida por anfetaminas. Por el contrario, los injertos neuronales procedentes de la rosetas mostraron pocas neuronas TH+, no produjeron una reducción significativa del comportamiento de rotación (figura 4d) y mostró hiperproliferación neural masiva (volumen del injerto > 20 mm3; figura 13). La hiperproliferación extensa de células neuronales procedentes de la roseta usadas en el injerto como se informa en la presente memoria fue similar al trabajo previo con injertos de DA procedentes de la roseta del grupo de los inventores (Kim, et al. miR-371-3 Expression Predicts Neural Differentiation Propensity in Human Pluripotent Stem Cells. Cell Stem Cell 8: 695-706 (2011), y otros (Hargus, et al. Proceedings of the National Academy of Sciences of the United States of America 107:15921-15926 (2010). La hiperproliferación probablemente se debió a los periodos de supervivencia más largos (4,5 meses frente a 6 semanas), la falta de purificación de FACS antes del trasplante y la elección de hospedador nulo para NOD-SCID IL2Rgc. El número de células Ki67+ en proliferación fue mínimo en los injertos derivados de la placa del suelo (<1 % de las células totales), mientras que los injertos procedentes de la
roseta conservaban bolsas de precursores neurales proliferantes. Se cree que la hiperproliferación neural es provocado por células neuroectodérmicas anteriores primitivas dentro del injerto (Elkabetz, et al. Genes Dev. 22: 152 165 (2008); Aubry, et al. Proc. Natl. Acad. Sci. U. SA 105:16707-16712 (2008) Esta hipótesis fue respaldada por la expresión del marcador del prosencéfalo FOXG1 en injertos procedentes de la roseta, pero no de la placa del suelo. Un pequeño porcentaje de células astrogliales estaba presente en injertos procedentes tanto de la placa del suelo como de la roseta, aunque la mayoría de células GFAP+ fueron negativas para marcadores humanos que indican el origen del hospedador (figura 13).
Los resultados en ratones nulos para NOD-SCID IL2Rgc descritos en la presente memoria demostraron supervivencia robusta a largo plazo de las neuronas FOXA2+/TH+, inversión completa del comportamiento de rotación inducida por anfetaminas y falta de ninguna señal de hiperproliferación neural. Sin embargo, algunos de estos resultados podrían atribuirse al uso específico de ratones nulos para NOD-SCID IL2Rgc. Para ensayar esta hipótesis, se trasplantaron cultivos de neuronas DA procedentes de la placa del suelo (250 x 10 células) en ratas con lesión de 6-OHDA adultas inmunodeprimidas farmacológicamente usando ciclosporina A. Cinco meses después del trasplante, la supervivencia del injerto fue robusta (figura 4e-h) con un promedio de más de 15.000 células TH+ que coexpresan FOXA2 (figura 4 g) y antígeno nuclear humano (hNA) (figura 4e); Las fibras TH+/hNCAM+ emanaron del núcleo del injerto hacia el cuerpo estriado del hospedador circundante (figura 4f). Además de FOXA2, las células TH+ expresaron marcadores de neuronas DA del mesencéfalo PITX3 y NURR1 (figura 4h-j). Los análisis conductuales mostraron un rescate completo de la asimetría rotacional inducida por anfetaminas, en contraste con animales con injertos simulados que no mostraron mejoras (figura 4k). Los animales con injertos también mostraron mejoras en la prueba de marcha (figura 4l) que mide la acinesia de las extremidades anteriores y en la prueba del cilindro (figura 4m), ensayos que no dependen de la estimulación farmacológica del sistema DA. Se espera el inicio tardío de la recuperación (aproximadamente 3-4 meses después del trasplante) para neuronas DA humanas y depende de la tasa de maduración in vivo tal como los niveles de expresión de DAT (figura 4n). La presencia de células TFT+ que expresan canales Kir3.2 (GIRK2) o calbindina indican que están presentes neuronas DA tanto de SNpc (A9) como del área tegmental ventral (A10) en el injerto (figura 4o, p).
Como en los ratones (figura 13), las células serotoninérgicas y GABAérgicas fueron poco habituales (<1 % de las células totales) en células de rata, al igual que las células gliales GFAP+ procedentes principalmente del hospedador (7 % de las células totales; (figura 14). Aunque se detectaron pocas neuronas serotonina+ en el injerto, se observaron células hNCAM negativas que probablemente eran fibras serotoninérgicas procedentes del hospedador (figura 14).
EJEMPLO de referencia V.
Injerto de una población novedosa de células neuronales DA en primates que contienen neuronas dañadas.
Los resultados demostrados en la presente memoria mostraron excelente supervivencia del injerto y resultado conductual en dos modelos murinos independientes. Sin embargo, el número de neuronas DA necesarias en un cerebro de ratón o rata representa una fracción pequeña del mayor número de células necesarias para injertar en primates y seres humanos. Para probar la capacidad de cambio de escala de este protocolo, se realizaron estudios piloto de injerto en dos monos rhesus adultos con lesión de MPTP.
Se obtuvieron lotes de 50 x 106 precursores de neuronas DA trasplantables el día 25 de diferenciación usando el protocolo basado en la placa del suelo. La dosis clásica para inducir una condición de tipo Parkinson fue a través de 3 mg de MPTP-HCL inyectado en la arteria carótida (intervalo 0,5-5 mg). Esto se siguió de inyección sistémica de MPTP a 0,2 mg/kg IV de MPTP. Se inyectaron células en tres ubicaciones (núcleo caudado posterior y putamen precomisural) en cada lado del cerebro (6 tramos en total, 1,25 x 106 células/tramo) y los animales se inmunodeprimieron con ciclosporina-A. Se inyectaron precursores de DA en un lado del cerebro de un subclón de H9 que expresa GFP, mientras que en el otro lado se injertaron células procedentes de células H9 sin marcar. Se muestran resultados que muestran el injerto de neuronas en monos rhesus con expresión continua de FOX2A y producción de TH en la figura 4q-t. Un mes después del trasplante, se observó supervivencia robusta de neuronas DA del mesencéfalo en función de la expresión de GFP (figura 15) y el marcador citoplasmático específico humano (SC-121) (figura 4q). Cada núcleo de injerto estaba rodeado por un halo de fibras TH+ que se extendía hasta 3 mm dentro del hospedador (figura 4r). Los núcleos del injerto estaban compuestos de neuronas TH+ que coexpresan SC-121 (figura 4s) y FOXA2 (figura 4t). Las áreas negativas de SC-121 y GFP dentro del injerto contenían microglía del hospedador Iba1+ (figura 15) que indica inmunodepresión incompleta. En resumen, el injerto de una nueva población de células neuronales DA en primates, es decir, monos rhesus adultos con lesión de MPTP (3 mg de MPTP-HCL (1-metil-4-fenil-1,2,3,6-tetrahidropiridina; que varía en concentraciones de 0,5 a 5 mg de MPTP-HCl) que contenían una pérdida grave >95 % de neuronas DA endógenas del mesencéfalo. La exposición a MPTP provocó cambios y síntomas observables similares a la enfermedad de Parkinson en seres humanos.
EJEMPLO de referencia VI.
Potencial de diferenciación comparable hacia el destino de neurona DA del mesencéfalo de células EP-iPSC mutantes PINK1 frente a células hES (o iPSC) de tipo silvestre.
Este ejemplo describe el descubrimiento de que grandes poblaciones de neuronas DA del mesencéfalo se
desarrollaron con características de las neuronas de un paciente con EP cuando la línea celular de un paciente con EP, es decir, célula EP-iPSC mutante PINK1, obtenida de una manera que no dio como resultado la destrucción de un embrión, se usaron como la población celular para obtener neuronas DA FOXA2/L1M1XA/TH+ de las presentes invenciones.
La línea de EP-iPSC mutante PINK1 Q456X se diferenció usando el protocolo (método) novedoso de neurona DA del mesencéfalo basado en la placa del suelo de las presentes invenciones que produjo perfiles de diferenciación de mesencéfalo comparables a los obtenidos de la línea de iPSC H9. (Figura 20). A-C) Análisis inmunocitoquímico de la línea EP-iPSC mutante PINK1 el día 11 de diferenciación (etapa precursora del mesencéfalo) para FOXA2 (rojo), LMX1A (verde) y DAPI (azul) (A), día 25 de diferenciación (etapa neuronal DA postmitótica temprana) para FOXA2 (rojo) y TH (verde) (B) y para NURR1 (rojo) y TH (verde) (C). D-F) El mismo conjunto de análisis inmunocitoquímicos realizados usando células procedentes de H9 el día 11 de diferenciación para FOXA2 (rojo), LMX1A (verde) y DAPI (azul) (D), el día 25 de diferenciación para FOXA2 (rojo) y TH (verde) (E) y para NURR1 (rojo) y TH (verde) (F).
Las EP-iPSC mutantes PINK1 mostraron fenotipo de tipo EP de agregación de proteínas después de la diferenciación y maduración a largo plazo in vitro. Los inventores descubrieron que EP-iPSC mutantes PINK1 mostraron pruebas de expresión de a-sinucleína (componente principal del cuerpo de Lewy en paciente con EP) en el citosol de neuronas DA TH+ el día 55 de diferenciación usando el protocolo novedoso de inducción de neuronas DA del mesencéfalo basado en la placa del suelo, (figura 21a-b). A, B) Análisis inmunocitoquímico de la línea EP-iPSC mutante PINK1 el día 55 de diferenciación para a-sinucleína (LB509, rojo), TH (verde) e imagen fusionada (A) y a-sinucleína (rojo) y ubiquitina (verde) (B). Estas células positivas para a-sinucleína también mostraron alta expresión de ubiquitina (marcador clásico de cuerpos de Lewy). Por el contrario, las neuronas DA procedentes de la línea iPS de control mostraron expresión de a sinucleína sináptica normal (a diferencia de citosólica) y niveles muy bajos de ubiquitina (figura 21c-d). C, D) Análisis inmunocitoquímico de la línea iPSC de control el día 55 de diferenciación para a-sinucleína (rojo) y TH (verde) (C) y a-sinucleína (rojo) y ubiquitina (verde) (D).
Expresión de la forma agregada de a-sinucleína. En el cerebro del paciente con EP, las formas insolubles dimerizadas de a-sinucleína conducen a la agregación en cuerpos de Lewy. La forma dimerizada de a-sinucleína muestra fosfolilación de serina 129 en a-sinucleína.
El mismo día de diferenciación, las células procedentes de EP-iPSC mutantes PINK1 mostraron fuerte expresión de a-sinucleína fosforilada en Ser129 en contraste con las células procedentes de iPSC de control que mostraron niveles de expresión muy bajos (figura 22). Las células procedentes de EP-iPSC mutantes PINK1 mostraron fuerte expresión de a-sinucleína fosforilada en Ser129 en contraste con las células procedentes de iPSC de control que mostraron niveles de expresión muy bajos. A, B) Análisis inmunocitoquímico para a-sinucleína fosforilada en Ser129 (verde) y DAPI (azul) en células procedentes de EP-iPSC mutantes PINK1 el día 55 de diferenciación (A) y células procedentes de iPSC de control coincidentes (B).
Se observan diferencias en los patrones de expresión de a-sinucleína dependiendo del protocolo de diferenciación. Los inventores contemplaron que las neuronas DA del mesencéfalo "auténticas" procedentes de la placa del suelo mostraban vulnerabilidad específica de la EP y fenotipos correspondientes, específicos, in vitro. Las neuronas DA obtenidas usando el protocolo de diferenciación basado en alimentador estromal clásico MS5 (Perrier et al., PNAS 2004, incorporado en la presente memoria por referencia) produjo grandes cantidades de neuronas TH+. Sin embargo, en función de datos obtenidos durante el desarrollo de las presentes invenciones, los inventores mostraron que las células TH+ basadas en MS5 no eran auténticas neuronas DA del mesencéfalo procedentes de la placa del suelo. En cultivos diferenciados a través del protocolo MS5, había muchas células positivas para a-sinucleína. Sin embargo, esas células no coexpresaron TH. Por otro lado, no hubo ninguna diferencia en los patrones de expresión entre EP-iPSC e iPSC de control cuando se usó la estrategia de diferenciación MS5 (figura 23a-b). Estos datos indican que asinucleína también se expresa en otros tipos celulares distintos de DA y que dicha a-sinucleína distinta de DA permanece sin cambios en las células procedentes de iPSC de enfermedad frente a control, en particular cuando se usen protocolos de diferenciación MS5 convencionales. Estas son las neuronas procedentes de la roseta de tipo DA señaladas en publicaciones (p. ej., Perrier PNAS 2004). Esas células TH+ basadas en MS5 (=tipo DA) se usan para comparación en las figuras 3, 10, 13 y 16. Estos datos indican que también se expresa a-sinucleína en otros tipos celulares distintos de DA y que dicha a-sinucleína distinta de DA no cambia en las células procedentes de iPSC de enfermedad frente a control, en particular cuando se usan protocolos de diferenciación MS5 convencionales. Finalmente, el nuevo protocolo de diferenciación basado en la placa del suelo descrito en la presente memoria, produce gran cantidad de células TH+ que coexpresan a-sinucleína. Esas células TH+ expresan a-sinucleína en un patrón de expresión citosólica. Figura 24A, B) Análisis inmunocitoquímico para a-sinucleína (LB509, rojo), TH (verde) de la línea EP-iPSC mutante PINK1 el día 60 de diferenciación basada en MS5 (A) e iPSC de control (b ). C) Análisis inmunocitoquímico de la línea de EP-iPSC mutante PINK1 el día 55 de diferenciación basada en la placa del suelo para a-sinucleína (rojo), TH (verde).
Las neuronas DA ilustrativas procedentes de EP-iPSC mutante PINK1 son más vulnerables a la estimulación tóxica. Las neuronas DA TH+ procedentes de EP-iPSC obtenidas a través del protocolo basado en la placa del suelo fueron más vulnerables a la exposición a toxinas (valinomicina: ionóforo de mitocondrias, 5 uM (intervalo de concentración de 1-10 uM), 48 h) que las células procedentes de iPSC de control. Por el contrario, las neuronas TH+ obtenidas a través del protocolo clásico basado en MS5 no mostraron vulnerabilidad diferencial entre las células procedentes de
EP frente a control. (Figura 24). A-F) Inmunocitoquímica TH representativa el día 60 de diferenciación: Condición normal (sin tratamiento con toxinas) para células procedentes de iPSC tanto de EP como de control obtenidas a través del protocolo basado en la placa del suelo (A, se muestran células procedentes de EP-iPSC), degradación casi completa de neuronas DA t H+ en EP-iPSC después del tratamiento con toxinas (B), neuronas DA TH+ parcialmente degradadas de control-iPSC (C). El ensayo de viabilidad celular completa con azul alamar después de 48 h de tratamiento con valinomicina también mostró supervivencia celular diferencial en un intervalo de dosis específico para exposición a toxinas (5 y 10 pM) al comparar EP-iPSC e iPSC de control (figura 25).
Condición normal de cultivos procedentes tanto de EP como de iPSC de control obtenidos a través del protocolo basado en MS5 (D, se muestran células procedentes de EP-iPSC), neuronas TH+ después de la exposición a toxinas en EP-iPSC (E) y cultivos procedentes de iPSC de control (F) obtenidos a través del protocolo MS5. G-H) imágenes de baja potencia de inmunocitoquímica para Tuj1 (rojo) y TH (verde) mediante protocolo basado en la placa del suelo el día 60 de diferenciación: EP-iPSC de condiciones normales (G), frente a exposición a toxina (H) e iPSC de control de condiciones normales (I), frente a exposición a toxina (J). K-N) imágenes de baja potencia de inmunocitoquímica para Tuj1 (rojo) y TH (verde) mediante protocolo basado en MS5 el día 60 de diferenciación: EP-iPSC de condiciones normales (K), frente a exposición a toxina (L) e iPSC de control de condiciones normales (M), frente a exposición a toxina (N).
Cuantificación ilustrativa del ensayo de respuesta de dosis-viabilidad celular para la exposición a toxinas. El ensayo de viabilidad celular con azul alamar después de 48 h de tratamiento con valinomicina mostró supervivencia celular diferencial en un intervalo de dosis específico para exposición a toxinas (5 y 10 pM) al comparar EP-iPSC e iPSC de control (día 60 de diferenciación basada en la placa del suelo). Nota: este ensayo prueba la muerte celular general mientras que los efectos más drásticos se observaron específicamente en neuronas Da (véase figura 14). Por lo tanto, la cuantificación basada en azul de alamar probablemente subestimará el alcance del efecto diferencial observado en linajes de neuronas DA.
Bibliografía: Li, et al. Nat. Biotechnol. 23, 215-221 (2005); Perrier, et al. Proc Natl Acad Sci US 101, 12543-8 (2004); Perrier, et al. Proc Natl Acad Sci USA 101, 12543-8 (2004); Tabar, et al. Nature Med. 14, 379-381 (2008); Perrier, et al. Proc Natl Acad Sci USA 101, 12543-8 (2004); Wernig, et al. Proc. Natl. Acad. Sci. U.S. A 105, 5856-5861 (2008); Lindvall, et al. J. Clin. Invest 120, 29-40 (2010); Roy, et al. Nature Med. 12, 1259-1268 (2006); Elkabetz, et al. Genes Dev. 22, 152-165 (2008); Kittappa, et al. PLoS. Biol. 5, e325 (2007; Ferri, et al. Development 134, 2761-2769 (2007); Roelink, et al. Cell 76, 761-775 (1994); Liem, et al. Cell 82, 969-979 (1995); Fasano, et al. Cell Stem Cell 6, 336-347 (2010); Chambers, et al. Nat. Biotechnol. 27, 275-280 (2009); Muroyama, et al. Genes Dev. 16, 548-553 (2002); Joksimovic et al. Nat Neurosci 12, 125-131 (2009); Lyashenko, et al. Nat. Cell Biol. 13, 753-761 (2011); VanDunk, et al. J. Neurosci. 31, 6457-6467 (2011); Huang, et al. Nat. Protoc. 4, 44-57 (2009); Costa, et al. Mol. Cell Biol. 9, 1415 1425 (1989); Elkabetz, et al. Genes Dev. 22, 152-165 (2008); Soldner, et al. Cell 136, 964-977 (2009); Guzman, et al. J. Neurosci. 29, 11011-11019 (2009); Nedergaard, et al. J. Physiol 466, 727-747 (1993); Ferrari, et al. Eur. J. Neurosci.
24, 1885-1896 (2006); Olanow, et al. Trends Neurosci. 19, 102-109 (1996); Zetterstrom, et al. Science 276, 248-250 (1997); Quintana, et al. Nature 456, 593-598 (2008); Kim, et al. diets Neural Differentiation Propensity in Human Pluripotent Stem Cells. Cell Stem Cell 8, 695-706 (2011); Hargu, et al. Proceedings of the National Academy of Sciences of the United States of America 107, 15921-15926 (2010); Aubry, et al. Proc. Natl. Acad. Sci. U. SA 105, 16707-16712 (2008); Blume, et al., Exp. Neurol. 219, 208-211 (2009); Ban, et al., Proc. Natl. Acad. Sci. U.S. A (2011); Studer, et al., Nature Neurosci. 1, 290-295 (1998); Kordower, et al., Science 290, 767-773 (2000); Paxinos, et al., The Rhesus Monkey Brain in Stereotaxic Coordinates (Academic Press, 2000); Crawley, What's Wrong With My Mouse: Behavioral Phenotyping of Transgenic and Knockout Mice (Wiley-Liss, 2000); Studer, et al., Brain Res. Bull. 41, 143 150 (1996); Tabar, et al., Nat. Biotechnol. 23, 601-606 (2005); y Placantonakis, et al., Stem Cells 27, 521-532 (2009).
EJEMPLO de referencia VII.
Se establecieron condiciones ilustrativas para el registro in vivo de neuronas DA procedentes de células madre pluripotentes humanas en preparaciones de corte agudo; véase resultados ilustrativos mostrados en la figura 26.
Se contemplan mediciones electrofisiológicas para su uso en preparaciones de corte agudo, es decir, de biopsias de áreas con injerto. En una realización, se identificarán neuronas DA procedentes de injerto de tipo A9 frente a A10 in vivo en función de pruebas para la actividad de marcapasos autónoma que es específica para las neuronas de dopamina de tipo A9 que están más afectadas en la EP. En otras palabras, las neuronas de tipo A10 no tienen actividad de marcapasos. Se establecieron condiciones para el registro in vivo de neuronas DA procedentes de células madre pluripotentes humanas en preparaciones de corte agudo, véase, figura 26. De manera específica, se midieron las neuronas DA humanas injertadas procedentes de células madre pluripotentes y se descubrió que tienen características electrofisiológicas típicas de las observadas en la pars compacta de sustancia negra de ratón (SNpc), La figura 26A, donde la vista superior muestra la reconstrucción de una neurona marcapasos en la región del injerto. La parte inferior muestra una microfotografía ilustrativa de un corte cerebral tomado de la rata en la que se inyectaron las neuronas procedentes de hES 9 meses antes; el injerto está delineado; una imagen de mayor aumento se muestra insertada en la parte inferior. El corte se procesó para determinar la tirosina hidroxilasa que aparece en blanco, figura 26B. Además, la vista superior muestra un parche ilustrativo unido a la célula que registra desde una supuesta neurona DA en el injerto; la parte inferior muestra un registro ilustrativo de célula completa de la misma célula. Se realizaron registros en presencia de glutamato y antagonistas del receptor de GABA (AP550 pM, CNQX 10 pM y GABAzina 10 pM) para
eliminar la entrada sináptica. Estos registros demostraron que las neuronas procedentes de PS eran marcapasos autónomos con trayectorias de tensión intrasomática normales. Otra neurona registrada en una muestra de injerto tenía propiedades similares, figura 26C. A efectos de comparación, se muestran registros unidos a células y de células completas de una neurona dopaminérgica en SNpc de un ratón adulto. Abreviaturas (CTx= corteza, STr= cuerpo estriado, SNpc= pars compacta de sustancia negra, DA= dopaminérgica). Estos datos muestran estudios in vivo funcionales en el cuerpo estriado de rata con injerto meses después del trasplante. Por tanto, en algunas realizaciones, los estudios funcionales in vivo en tejido injertado demuestran la recuperación de pars compacta de sustancia negra (SNpc).
EJEMPLO de referencia VIII.
Métodos ilustrativos para identificar marcadores de superficie celular para su uso en métodos de las presentes invenciones. En particular, CD142 se identificó con estos métodos.
Dos estrategias principales para identificar marcadores de superficie candidatos: Una exploración de expresión génica imparcial en líneas indicadoras genéticas (figura 27a) que encontró varios marcadores candidatos, incluyendo un marcador, denominado DCSM1, que se expresa selectivamente en neuronas DA del mesencéfalo y parece marcar especialmente neuronas DA de tipo A9 (figura 27b). Una segunda estrategia es el uso de una exploración de marcador de superficie celular CD en neuronas DA procedentes de hESC que prueban 242 anticuerpos disponibles en el mercado en formato de 96 pocillos (figura 27c, d). Los resultados de dicha exploración (figura 27e) condujeron a la identificación de al menos 5 marcadores validados enriquecidos en neuronas dA del mesencéfalo, incluyendo CD142, un marcador que marca selectivamente una etapa de neuronas DA Nurr1+ (figura 27f). Con el uso del procedimiento celular de neuronas DA descritas en la presente memoria, CD142 normalmente marcó aproximadamente 30 % de la población celular total el día 25 de diferenciación (figura 28a). La selectividad de CD142 para una etapa de neurona DA Nurr1+ se confirmó en múltiples líneas independientes de hESC y hiPSC (figura 28b). Además de enriquecer con respecto a neuronas DA, el enriquecimiento de células positivas para CD142 dio como resultado un agotamiento selectivo de subtipos de neuronas no deseadas, tales como neuronas GABA y serotoninérgicas. (Figura 28c-f). Los estudios in vivo confirmaron la capacidad de una población de células CD142 positivas para dar lugar a injertos de neuronas DA de alta pureza que superaron los problemas de contaminar neuronas GABA y serotoninérgicas. Aunque el procedimiento de injerto que usó células sin purificar ya dio como resultado muy pocas neuronas serotoninérgicas, se contempla el uso de la selección de células precursoras basada en CD142 para reducir adicionalmente el riesgo de introducir neuronas serotoninérgicas, un tipo de célula contaminante que estaba implicado en el fracaso del injerto de tejido fetal humano como la fuente potencial de las discinesias inducidas por injerto de tejido fetal no deseadas.
EJEMPLO de referencia IX.
Este ejemplo describe métodos para la transformación de células con genes PST humanos para aumentar la expresión en superficie celular de PSA. Este ejemplo también muestra métodos ilustrativos para usar células que tienen mayor expresión en superficie celular de p Sa .
De manera específica, este ejemplo muestra genes de PST modificados por ingeniería genética en hESC para aumentar la expresión de PSA en neuronas DA. Se introdujo un gen que codificaba la polisialiltransferasa humana (hPST) en una línea hESC (WA01) usando un vector lentivírico (pLenty, Invitrogen). Se expandieron y analizaron veinte clones seleccionados para determinar la expresión de PST. Los clones de hESC que expresan PST se diferenciaron para garantizar que PST no se silenciara en neuronas DA. La cuantificación de PSA-NCAM en diferentes etapas de diferenciación (día 0, 11, 25 y 50) se realizó usando análisis de FACS e inmunofluorescencia (Operetta). Los clones positivos se sometieron al conjunto de parámetros de QC de la neurona DA descritos en la tabla 7. Al menos 3 clones que conservan niveles uniformes altos de PSA-NCAM durante la diferenciación y rinden bien en los parámetros de QC (tabla 7) avanzarán a la evaluación del crecimiento de neuritas en neuronas DA procedentes de hESC que sobreexpresan PST. Los clones de hESC de control y que sobreexpresan PST seleccionados se diferenciaron en neuronas DA usando el protocolo convencional descrito en la presente memoria, seguido de fijación celular y análisis los días 25 y 50. El número y la longitud de las fibras positivas para TH en dichos cultivos se cuantificaron con el microscopio de alto contenido Operetta. El módulo de análisis de neuritas en el software Harmony 3.0 cuantificó el número y la longitud de las neuritas, con o sin PST y los datos se analizaron estadísticamente usando un ANOVA de dos vías. Los clones de hESC que sobreexpresan PST y de control que avanzan desde estudios in vitro anteriores, se diferenciaron de nuevo en neuronas DA y se trasplantaron a un modelo de rata de EP. Se realizaron injertos a corto plazo (4-6 semanas) para determinar la supervivencia, expresión de PSA-NCAM y crecimiento de neuritas. Para cada clon que pasó, los parámetros in vivo a corto plazo se sometieron a estudios de injerto a largo plazo. Para esos estudios, los animales recibieron la mitad o un cuarto de la dosis convencional (200 x 103) de células. Estos estudios fueron para abordar si el aumento de PSA conduce a mayor supervivencia a largo plazo después de trasplante (5 meses) y si los números menores de neuronas DA son capaces de igualar o superar la capacidad funcional de los injertos no PST trasplantados a dosis celulares convencionales (no la figura 27).
Además, los ensayos conductuales complejos sensibles al grado de reinervación estriatal se supervisaron para distinguir adicionalmente el potencial funcional de los injertos de neuronas DA de PST frente a control. Los animales se sacrificaron después de la finalización de los ensayos conductuales y se cuantificó el crecimiento de fibras usando anticuerpos específicos humanos NCAM y SC121 y anticuerpos contra TH (véase también no la figura 29). Se midió la intensidad y propagación del injerto hNCAM+, SC121+ y TH+, así como el porcentaje de células humanas que
coexpresan marcadores de neuronas DA (TH, FOXA2) y PSA. La densidad de halo NCAM/TH+ de neuritas que emanan del injerto se cuantificó a diferentes distancias. Los datos se compararon entre los grupos usando un ANOVA de dos vías con una prueba post-hoc de Bonferroni. Además, se examinaron secciones para determinar cambios cualitativos (p. ej., ramificación, espesor, distribución y forma del injerto). Además, algunos injertos se procesarán para la evaluación electrofisiológica de corte con respecto al fenotipo A9, formación de sinapsis con el cuerpo estriado del hospedador, así como inervación por aferentes endógenos.
EJEMPLO de referencia X .
El siguiente ejemplo muestra mejora de la expresión de ácido polisiálico que mejoró la función de los injertos de neuronas de dopamina procedentes de ES en ratones parkinsonianos.
Las células ES que expresan GFP bajo el control del promotor Nurr1 (células Nurr1::GFP ES) se transdujeron de manera estable con un vector lentivírico que expresa polisialiltransferasa (PST) de manera ubicua. Las células transducidas mostraron un aumento drástico de ARNm de PST en comparación con los controles (figura 30A). Se observó que la expresión de PST era suficiente para la síntesis de PSA en NCAM. Por consiguiente, la expresión de PSA-NCAM aumentó en gran medida en las células modificadas con PST el día 14 de la diferenciación de neuronas DA (figura 30B-E). El PSA de la superficie celular tanto endógena como inducida en neuronas DA procedentes de ES podría eliminarse (figura 30E) mediante una endoneuraminidasa de fago (endoN) que escindió de manera específica los polímeros de ácido siálico ligados a alfa-2,8 únicos del PSA. De manera sorprendente, no se observó que la transducción de PST afectara a la expresión de marcadores neuronales o del mesencéfalo en las neuronas DA purificadas con GFP (figura 30F).
Otros estudios en ratones hemiparkinsonianos con lesión de 60HDA mostraron que es necesario el trasplante de aproximadamente 100.000 precursores de neuronas DA procedentes de ES para producir recuperación funcional robusta, medido por la prueba de rotación potenciada por anfetaminas. En los presentes estudios, se buscó injertar una cantidad mejorable de células para poder evaluar el aumento mediante expresión de PSA mejorada. Para trasplantar poblaciones de neuronas DA muy enriquecidas que están agotadas de células pluripotentes contaminantes, cultivos purificados con FACS el día 14 de diferenciación para la expresión de GFP conducida por Nurr1 y para la ausencia de expresión de SSEA-1 (figura 31). Sin sobreexpresión de PST, una reducción del tamaño de injerto mínimamente eficaz a la mitad (55.000 células DA Nurr1+) no logró producir recuperación del comportamiento detectable. Por el contrario, con expresión de PSA mejorada, el mismo número de neuronas DA Nurr1/PST dio como resultado una corrección significativa del deterioro conductual por la EP (p < 0,01; ANOVA de dos vías), con recuperación completa aproximadamente 5 semanas después de la cirugía (figura 32A). La eliminación del PSA antes del trasplante por incubación con endoN indicó la especificidad de la mejora del PSA, ya que el tratamiento con endoN invirtió parcialmente la restitución funcional obtenida con Nurr1/PST (figura 32A).
Para examinar las características de las células injertadas, los animales fueron procesados con respecto a inmunohistoquímica dos meses después del trasplante. Hubo una diferencia en el número de neuronas Nurr1 supervivientes, porque los animales en los que se injertó la línea transducida por PST tuvieron en promedio el doble de células GFP+ que los animales en los que se injertaron células de control (9.300 /- 1.400 frente a 4.230 /- 1010 células GFP+ por injerto en muestras de PST frente a control, respectivamente; figura 32B, p < 0,05, prueba de t de Student). Asimismo, los injertos de Nurr1/PST también mostraron niveles mayores de expresión de PSA in vivo (figura 32C, D). Sin embargo, las proporciones de células que expresan los marcadores DA del mesencéfalo TH y FoxA2 dentro del núcleo del injerto fueron comparables para las células Nurr1 y Nurr1/PST (TH: 62,0 % /- 8,0 frente a 51,3 % /- 7,0, p = 0,33; FoxA2: 63,2 % /- 8,6 frente a 55,4 % /- 2.0, p = 0,3, respectivamente; figura 32E).
Las prolongaciones neuronales que surgieron de las células Nurr1 y Nurr1/PST mostraron niveles similares de TH, Girk2 (canal de potasio de rectificación interna, acoplado a proteína G) y sinapsina (figura 33A). A diferencia de otros estudios con células de Schwann trasplantadas (Ghosh, M., et al. Extensive cell migration, axon regeneration, and improved cells after spinal cord injury. Glia 60, 979-992 (2012)), la expresión de PSA mejorada tuvo poco efecto sobre la migración de células DA desde el sitio de injerto. Sin embargo, hubo cambios evidentes en el crecimiento de neuritas. Como se muestra en la figura 33B, surgieron más prolongaciones neuronales DA de células Nurr1/PST en comparación con controles Nurr1+. Cuando la intensidad de la inmunofluorescencia de GFP y TH se cuantificó en cinco zonas sucesivas a 100 pm del trasplante, los injertos de Nurr1/PST presentaron una densidad relativa de prolongaciones mucho mayor (figura 33C, D; p < 0,01 para GFP y TH, ANOVA de dos vías). Al cuantificar este efecto, se normalizó la densidad relativa de las prolongaciones a la densidad observada en la zona más próxima inmediata al núcleo del injerto. Dicha normalización fue necesaria para compensar el mayor número de células supervivientes en los injertos de Nurr1/PST y para confirmar un efecto específico del PSA sobre el crecimiento de neuritas. La especificidad también se demostró cuando el PSA de la superficie celular se eliminó mediante tratamiento con endoN antes del injerto. Por tanto, el pretratamiento con endoN redujo el crecimiento de fibras distales a niveles de control (figura 33E).
Estos descubrimientos mostraron que al menos algunos de los efectos del PSA sobre la función del injerto resultaron de la inervación de fibras mejorada del cuerpo estriado. Por consiguiente, hubo una fuerte correlación entre la función del injerto y el alcance relativo del crecimiento de fibras positivas para GFP, por ejemplo, en la zona IV (figura 33F; p < 0,001, r2 = 0,65, n = 17). De manera sorprendente, la relación de crecimiento de fibra/comportamiento fue uniforme
para los grupos experimentales (control, con PSA mejorado y tratado con endoN), lo que indica que la inervación del injerto-hospedador fue un parámetro para la recuperación del comportamiento en el modelo parkinsoniano de ratón. Varios factores contribuyeron mecánicamente al aumento del crecimiento de fibras, tales como mejora de la penetración de la zona de glía reactiva que encapsula el núcleo del injerto, mayor capacidad de brote, mejor crecimiento en el tejido hospedador circundante (p. ej., translocación de cono de crecimiento más fácil) y prevención de conexiones prematuras con tejido hospedador próximo al núcleo del injerto. Los mecanismos ilustrativos son coherentes con el papel del PSA para facilitar el crecimiento de prolongaciones durante el desarrollo normal y en el sistema nervioso adulto.
Los experimentos descritos en la presente memoria demostraron el uso de PSA modificado por ingeniería genética en el injerto de neuronas DA que proporcionó resultados superiores en comparación con injertos de otros tipos de células. Los datos indicaron claramente que la mejora de PSA proporcionó un aumento significativo de la capacidad de las neuronas DA injertadas para inervar el cuerpo estriado del hospedador y atenuar los déficits funcionales de la EP. Por lo tanto, se contempla la traducción clínica que comprende neuronas DA de las presentes invenciones para proporcionar células antes del trasplante. En algunas realizaciones, las células se manipularán genéticamente para expresar PSA. En algunas realizaciones, la PST puede administrarse directamente a las células a través de la exposición a la enzima purificada y al sustrato, in vitro, antes del trasplante. En algunas realizaciones, se contempla la estrategia de PSA para la traducción humana en el injerto de EP para minimizar la necesidad de múltiples inyecciones y, por lo tanto, reducir los riesgos quirúrgicos resultantes de estas inyecciones múltiples.
En otras realizaciones, esta tecnología se contempla para su uso en otros tipos celulares y especies, por ejemplo, aumentando la migración de células de Schwann injertadas en la creación de un puente (por ejemplo, comunicación célula-célula) para el nuevo crecimiento de axones en el sitio de la lesión de la médula espinal.
Los siguientes son materiales y métodos ilustrativos usados en este ejemplo.
Animales: Se mantuvieron ratones 129S3/SvImJ de seis semanas de edad (Jackson Laboratory) a temperatura controlada con alimentos y agua disponibles a voluntad. Los procedimientos experimentales se realizaron de acuerdo con los NIH y las pautas institucionales para el uso de animales y fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales (IACUC) y el Comité Institucional de Bioseguridad (IBC).
Inyección de 6OHDA y prueba de inducción por anfetaminas: Los animales se anestesiaron con pentobarbital de sodio (10 mg/kg) y se les inyectó en el cuerpo estriado derecho con 2 pl de 6OHDA (4 pg/pl en solución salina, ácido ascórbico al 0,5 %). Las inyecciones se realizaron con una jeringa Hamilton en las coordenadas: 0,5 mm posterior, 1,8 mm lateral con respecto al bregma y 2,5 mm ventral con respecto a la superficie del cerebro. Antes de la cirugía, los animales recibieron una única inyección i.p. de desipramina (25 mg/kg, Sigma). Dos semanas después de la cirugía, los animales se puntuaron en la prueba de rotación inducida por anfetaminas. Se colocaron en cilindros de plástico transparente de 30 cm de diámetro durante media hora, después de lo cual recibieron una única inyección i.p. de anfetamina (10 mg/kg, Sigma). Después de 20 min, se puntuó el número de rotaciones ipsilaterales/contralaterales durante otros 20 min. Los animales se puntuaron una vez a la semana durante siete semanas, después se anestesiaron profundamente y se perfundieron a través del corazón con PBS y paraformaldehído al 4 % en tampón de fosfato 0,1 M (PB, pH 7,4). Los cerebros se extrajeron y se fijaron durante una noche a 4 °C en paraformaldehído al 4 % y después se cortaron con vibrátomo (Pelco-101, Ted Pella) en secciones sagitales de 40 pm de grosor.
Diferenciación y trasplante celular: Se transdujo una línea celular indicadora ES de ratón BAC transgénica BAC Nurr1 ::GFP (es decir, la expresión de GFP es impulsada por el promotor Nurr1) 5 con un lentivirus (pLenti, Invitrogen) que contenía el gen de p St de ratón bajo el control del promotor de CMV. Las células ES se propagaron en MEF tratados con mitomicina C (StemCell Technologies) en DMEM (Invitrogen), FBS al 10 % (HyClone) complementado con 1.400 unidades/ml de LIF (ESGRO; Invitrogen), L-glutamina 2 mM, p-mercaptoetanol 1 mM, 100 U/ml de penicilina y 100 pg/ml de estreptomicina (Invitrogen). La diferenciación de DA se indujo según Barberi et al., Nat Biotechnol 21, 1200-1207 (2003), con modificaciones. En resumen, las células se diferenciaron en células alimentadoras MS5 en placas recubiertas con gelatina (10.000 células/placa de 10 cm) y se cultivaron durante cuatro días en medios de reemplazo de suero (SRM). El día 4, se añadieron Sonic Hedgehog (SHH, 200 ng/ml) y FGF8 (100 ng/ml). El día 7 de diferenciación, los medios se cambiaron a N2 complementado con SHH, FGF8 y bFGF (10 ng/ml). El día 11, la diferenciación terminal fue inducida por retirada de SHH, FGF8 y bFGF y la adición de ácido ascórbico (AA, 200 pM) y BDNF (20 ng/ml).
Las células se recogieron los días 14-15 con tratamiento de accutase durante 45 minutos, se lavaron una vez con N2 y se incubaron con anticuerpo anti-SSEA-1 conjugado con AlexaFluor-647 (BD Pharmingen) durante 25 minutos. Las células se lavaron una vez con N2, se resuspendieron en tampón HEPES con BSA al 0,1 %. Se añadió DAPI para evaluar la viabilidad. FACS se realizó con un clasificador de células MoFlo y la población de interés se clasificó con respecto a fluorescencia de GFP (Nurr1). La población positiva para AlexaFluor-647 (SSEA-1) se clasificó negativamente. Para el control negativo de GFP, se usaron células ES de ratón J1 sin exposición previa en la misma etapa de diferenciación.
Las células clasificadas Nurr1::GFP se analizaron para determinar su viabilidad y se resuspendieron en N2 con BDN y AA a una concentración final de 55.000 células/pl. Se inyectó un pl en el cuerpo estriado del ratón con lesión con un
capilar de vidrio fino con punta de 50 pm en las coordenadas: 0,3 mm posterior, 1,5 mm lateral desde el bregma y 2,2 mm ventral con respecto a la superficie del cerebro. Se volvió a sembrar una alícuota de la suspensión celular en placas de 6 mm recubiertas con matrigel para caracterización adicional.
Para análisis de inmunofluorescencia, las células se fijaron con paraformaldehído durante 10 minutos a 4 0 C, se lavaron dos veces con PBS, se bloquearon con BSA al 5 % (Triton X-100 al 0,1 % en PBS) y se incubaron con anticuerpos primarios durante 2 horas a temperatura ambiente: anti-GFP de conejo (1:1000, Invitrogen), IgM de ratón anti-PSA (1:2000, 5A5), anti-NeuN de ratón (1:800, Chemicon), anti-TH de ratón (1:1000, Sigma), anti-FoxA2 de cabra (1:800, Santa Cruz), anti-Engrailed de cabra (1:800, Santa Cruz). Las células se incubaron después con anticuerpos secundarios conjugados con Cy (1:1000, Jackson).
Tratamiento con EndoN: Para eliminar el PSA de NCAM, la noche antes de la recogida, las células se trataron con 20 unidades de endoN, una enzima de fago que elimina específicamente PSA 7-9. Las células se recogieron e inyectaron como se ha descrito anteriormente, pero se resuspendieron en N2 con BDNF y AA y 5 unidades de endoN. Anteriormente los inventores han evaluado que la inyección de la misma cantidad de endoN solo en ratones lesionados no mejoró el comportamiento animal.
Análisis de ARNm de PST y PSA-NCAM in vitro: Para análisis de transferencia de Western, las células se trataron con tampón WB (PBS con NP40 al 1 %, NaCl 150 mM, EDTA 1 mM e inhibidores de proteasa/fosfatasa 1x añadidos inmediatamente antes de la extracción, a pH de 7,4) y se sonicaron dos veces durante 5 segundos, se centrifugaron y se resuspendieron en tampón Laemli (LB). Se guardaron alícuotas sin LB para la determinación de proteínas. Se cargaron cantidades iguales de proteína en gel de electroforesis en gel de dodecilsulfato de sodio-poliacrilamida al 6 % (BioRad). Las proteínas se transfirieron mediante electroforesis a membranas de polivinilideno (Millipore). Las membranas se bloquearon durante 1-6 horas en Triton X-100 TBS 0,1 % (TBS-T) con leche en polvo desgrasada al 5 % y se incubaron durante una noche con anticuerpos anti-NCAM (1:10.000, Santa Cruz) en TBS-T con leche al 5 %. Las transferencias se incubaron después con anticuerpo secundario conjugado con peroxidasa (1:10.000, Jackson) y se detectaron con el método de detección de ECL (Amersham Pharmacia Biotech). Los niveles de proteínas se cuantificaron usando software ImageJ.
Para análisis por qRT-PCR, se extrajo ARN total con Trizol (Sigma), se transcribió de forma inversa (Qiagen) y se amplificó con 10 pl de mezcla de reacción SYBR 2x y 0,2 pM de cebadores directo e inverso hasta un volumen final de 20 pl. Para análisis FACS de PSA-NCAM, las células se recogieron con tratamiento de accutase durante 45 minutos, se lavaron una vez y se incubaron con IgM de ratón anti-PSA (1:250, 5A5) durante 25 minutos en hielo, se lavaron una vez con medio N2 y se incubaron con IgM anti-ratón conjugada con Cy3 (1:250, Jackson) durante otros 25 minutos en hielo. Las células se lavaron una vez con N2 y se resuspendieron con BSA al 0,1 % con 7AAD y se analizaron en un clasificador de células FACS Calibur. Como control, no se añadió anticuerpo primario.
Procedimientos inmunohistológicos y estereológicos: Las secciones coronales en flotación se bloquearon en Triton X-100 al 0,1 %, suero de burro al 5 % en PBS durante 30 minutos a temperatura ambiente y se incubaron 48 horas a 4 °C con diferentes anticuerpos: anti-GFP de conejo (1:300), anti-GFP de pollo (1:200, Chemicon), anti-TH de ratón (1:200), IgM de ratón anti-PSA (1:1000), anti-NeuN de ratón (1:400), anti-FoxA2 de cabra (1:300), anti-Girk2 de conejo (1:300, Alomone Labs), anti-sinapsina de ratón (1:200, BD Transduction Laboratories). Las secciones se lavaron e incubaron con anticuerpos secundarios: anticuerpos de burro conjugados con Cy2, Cy3 y Cy5 (1:400, Jackson). Para el PSA se usó un anti-IgM de burro conjugado con Cy5 (1:500 Jackson). Las incubaciones se realizaron durante 2 horas a temperatura ambiente. Las secciones se lavaron dos veces en PBS y se montaron en Mowiol (Calbiochem). Se analizaron una de cada tres secciones coronales del cerebro para cada inmunomarcaje. Las imágenes digitales fueron recogidas por un microscopio confocal de barrido láser Zeiss LSM 510 con tres láseres (Argon 488, HeNe 543 y HeNe 633) con un objetivo 40 x c-Apochromat (de inmersión en agua). El número de células GFP+ y TH+ se contó en una de cada tres secciones que abarcan el cerebro completo con un objetivo de 40x y se estimó el número total de células/injerto. Las células con doble marcaje se analizaron en planos ópticos individuales a través del eje z completo.
Para el análisis del porcentaje de células marcadas con GFP/TH+ y GFP/FoxA2+, se analizaron 100 células GFP+ para cada marcador. Para el análisis de crecimiento de prolongaciones, se realizaron exploraciones z confocales a intervalos de 0,8 pm a través del eje z completo (20-40 pm) con un agujero estenopeico de 1 pm bajo un objetivo 40x. Las secciones se exploraron desde el sitio de inyección lateralmente hasta que no se observaron prolongaciones. Se combinaron secuencialmente proyecciones tridimensionales que abarcaban toda el área explorada. Para análisis de intensidad de GFP y TH, toda el área escaneada se dividió en cinco zonas sucesivas de 100 pm de distancia desde el trasplante y las intensidades se midieron usando software ImageJ. Los datos se normalizaron con respecto a la intensidad en la zona más cercana al injerto (zona I) para controlar con respecto a cualquier diferencia potencial en el tamaño del injerto.
Análisis estadístico: Los datos se presentan como la media ± error típico de la media (ETM). Se realizaron comparaciones usando la prueba t de Student o el análisis de varianza (ANOVA) de dos vías seguido de la prueba post-hoc de Bonferroni. Se realizó análisis de regresión lineal y se cuantificó usando la correlación de Pearson.
EJEMPLO de referencia XI.
El siguiente ejemplo muestra la modificación por ingeniería enzimática de PSA en neuronas DA procedentes de hESC usando la polisialiltransferasa bacteriana purificada, PSTnm, para mejorar la eficacia del trasplante.
Aunque es eficaz, la transfección de gen de PST requirió modificaciones genéticas de hESC con control limitado sobre la duración de la polisialilación. Este ejemplo describe el descubrimiento de que PSTnm externo indujo PSA, en lugar de suministro de genes, (véase figura 35). En la figura 35A, las células de Schwann (SC) tratadas con PST (línea verde-línea media) tenían un mayor tiempo de adhesión, mientras que el PSA producido por PSTnm inhibió la adhesión. En particular, (A) El PSA producido por PSTnm inhibe la adhesión de células de Schwann en suspensión a una monocapa de células de Schwann aún más eficazmente (línea roja-línea más baja) que el PSA producido por la expresión forzada de PST (línea verde-línea media). (B) La inmunotransferencia de PSA en motoneuronas HB9 procedentes de ESC muestra que las muestras de control tratadas con PSTnm solo tenían niveles indetectables de PSA. La incubación con sustrato de PSTnm CMP-ácido siálico produce una gran banda de PSA, que se elimina con tratamiento con endoN. (C, D) Similar a los efectos obtenidos con el gen de PST, la polisialilación de estas células mediante PSTnm y sustrato durante la diferenciación mejora el crecimiento de neuritas y la migración celular (puntas de flecha). (E) Inmunotinción de PSA de neuronas DA procedentes de hESC del día 30. (F) Esta tinción aumenta significativamente después del tratamiento con PSTnm y sustrato. (G) la inyección in vivo de PSTnm solo no tiene efecto, mientras que su administración conjunta con sustrato (H) produce grandes cantidades de expresión de PSA en el cuerpo estriado de ratón.
Por tanto, las neuronas DA maduras tratadas externamente con PSTnm se contemplan para su uso en las células productoras para injerto. Tanto PST como PSTnm de mamífero produjeron cadenas de PSA químicamente idénticas. El aumento de PSA en las neuronas DA procedentes de hESC (figura 35F) debería persistir durante varias semanas, suficiente para que las fibras DA salgan del núcleo del injerto. Debido a que PSTnm se elimina antes del injerto, la inmunogenicidad a esta enzima que contamina las células injertadas no debería tenerse en cuenta.
Se produjo PSTnm a partir de un fragmento modificado por ingeniería genética con características de solubilidad y actividad mejoradas (Willis et al., Characterization of the alpha-2,8-polysialyltransferase from Neisseria meningitidis with synthetic acceptors, and the development of a self-priming polysialyltransferase fusion enzyme. Glycobiology 18, 177-186 (2008)). Se indujo que los cultivos de hESC se diferenciaran en neuronas DA antes de la exposición a PSTnm, exposición al sustrato o ambos. Los cultivos se examinaron en diferentes puntos temporales de exposición (10 min a 6 h) mediante inmunofluorescencia cuantitativa (Operetta) y transferencia de Western para determinar la velocidad y los niveles de polisialilación. Por tanto, el día 25, las neuronas DA procedentes de hESC diferenciadas se incubarán con las concentraciones óptimas de PSTnm y sustrato usando las condiciones descritas en la presente memoria. Las neuronas mDA PSA+ se trasplantarán en ensayos a corto y largo plazo como se describe en la presente memoria y en la figura 29.
Claims (12)
1. Un método in vitro para diferenciar células pluripotentes en precursores de la placa del suelo del mesencéfalo, comprendiendo el método:
exponer una pluralidad de células pluripotentes a: (a) al menos un inhibidor de la señalización de Small Mothers Against Decapentaplegic (SMAD), (b) al menos un activador de la señalización de Sonic hedgehog (SHH) y (c) al menos un activador de la señalización de wingless (Wnt), en donde:
la exposición al al menos un inhibidor de la señalización de SMAD comienza el día 0;
dichas células pluripotentes se exponen al al menos un activador de la señalización de Wnt 3 días después del inicio de la exposición al al menos un inhibidor de la señalización de SMAD;
dichas células pluripotentes están expuestas al al menos un inhibidor de la señalización de SMAD, el al menos un activador de la señalización de SHH y el al menos un activador de la señalización de Wnt en cantidades eficaces para producir una pluralidad de células precursoras de la placa del suelo del mesencéfalo, al menos el 10 % de las cuales expresan tanto la proteína A2 de secuencia forkhead (FOXA2) como el factor de transcripción 1 alfa de homeosecuencia LiM (LMX1A); y
opcionalmente someter dicha pluralidad de células precursoras de la placa del suelo del mesencéfalo diferenciadas a condiciones que favorezcan la maduración de las células precursoras de la placa del suelo del mesencéfalo en neuronas de dopamina.
2. El método de la reivindicación 1, en donde dichas células pluripotentes se seleccionan del grupo que consiste en células madre no embrionarias, células madre embrionarias, células pluripotentes no embrionarias inducidas y células pluripotentes modificadas por ingeniería genética humanas, de primates no humanos o de roedores.
3. El método de la reivindicación 1, que comprende poner en contacto las células pluripotentes con al menos una de purmorfamina y/o CHIR99021.
4. El método de la reivindicación 1, en donde dichas neuronas de dopamina que expresan FOXA2 y LMX1A expresan además al menos un marcador adicional seleccionado del grupo que consiste en tirosina hidroxilasa (TH), homeosecuencia de orthodenticle 2 (OTX2), proteína 1 relacionada con el receptor nuclear (NURR1), beta-tubulina de clase III específica de neurona (Tuj1), familia de factor Trefoil 3 (TTF3), homeodominio de tipo paired 3 (PITX3), complejo achaete-scute (ASCL), factor temprano de linfocitos B 1 (EBF-1), factor temprano de linfocitos B 3 (EBF-3), transtiretina (TTR), sinapsina, transportador de dopamina (DAT) y canal de potasio rectificador hacia dentro acoplado a proteína G (Kir3.2/GIRK2), CD142, DCSM1, CD63 y CD99.
5. El método de la reivindicación 1, en donde pertenecen uno o más de los siguientes:
a. dichas células pluripotentes se diferencian en células neuronales que expresan tanto proteína A2 de secuencia forkhead (FOXA2) como factor de transcripción 1 alfa de homeosecuencia LIM (LMX1A) en los primeros 11 días después del inicio del contacto con al menos dos inhibidores de la señalización de SMAD, dicho al menos un activador de la señalización de SHH y dicho al menos un activador de la señalización de Wnt; o
b. dichas células pluripotentes se diferencian en dichas neuronas de dopamina no más tarde de 25 días después del inicio del contacto con al menos dos inhibidores de la señalización de SMAD, dicho al menos un activador de la señalización de SHH y dicho al menos un activador de la señalización de Wnt.
6. El método de la reivindicación 4, en donde al menos 20 % de dichas neuronas de dopamina expresan FOXA2 y LMX1A y expresan además al menos un marcador para las neuronas de dopamina del mesencéfalo de subtipo a 9 humano seleccionadas del grupo que consiste en Girk2, CD142 y DCSM1.
7. El método de la reivindicación 4, que comprende además seleccionar una población de neuronas de dopamina que expresan CD142 o DCM1 o ambas.
8. El método de la reivindicación 1, en donde pertenecen uno o más de los siguientes:
a. dicho al menos un inhibidor de la señalización de SMAD se selecciona del grupo que consiste en LDN-193189, SB431542, Noggin, dorsomorfina y combinaciones de dos o más de los anteriores;
b. dicho al menos un activador de la señalización de SHH se selecciona del grupo que consiste en purmorfamina, SHH recombinante, SHH purificado, agonistas de Smoothened (SAG) y combinaciones de dos o más de los anteriores;
c. dicho al menos un activador de la señalización de Wnt se selecciona del grupo que consiste en CHIR99021, Wnt3A y Wnt1 y combinaciones de dos o más de los anteriores.
9. El método de la reivindicación 1, en donde dichas células precursoras de la placa del suelo del mesencéfalo diferenciadas que expresan FOXA2 y LMX1A no expresan al menos un marcador seleccionado del grupo que consiste en PAX6, HES5, EMX2 y LHX2.
10. El método de la reivindicación 1, en donde las condiciones que favorecen la maduración de las células precursoras
de la placa del suelo del mesencéfalo en neuronas de dopamina comprenden poner en contacto las células precursoras de la placa del suelo del mesencéfalo con medio N2, factor neurotrófico derivado del cerebro (BDNF), ácido ascórbico (AA), factor neurotrófico derivado de línea celular glial (GDNF), dibutiril AMPc (dbAMPc) y factor de crecimiento transformante de tipo p3 (TGFp3).
11. El método de la reivindicación 1, en donde el al menos un inhibidor de la señalización de Small Mothers Against Decapentaplegic (SMAD) comprende al menos un inhibidor de la señalización de TGFp/Activina-Nodal y al menos un inhibidor de la señalización de la proteína morfogenética ósea (BMP).
12. El método de la reivindicación 1, en donde el al menos un activador de la señalización de wingless (Wnt) comprende un inhibidor de la señalización de glucógeno sintasa cinasa 3p (GSK3p).
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201161555828P | 2011-11-04 | 2011-11-04 | |
| PCT/US2012/063339 WO2013067362A1 (en) | 2011-11-04 | 2012-11-02 | Midbrain dopamine (da) neurons for engraftment |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2779453T3 true ES2779453T3 (es) | 2020-08-17 |
Family
ID=48192829
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES12846715T Active ES2779453T3 (es) | 2011-11-04 | 2012-11-02 | Neuronas de dopamina (DA) del mesencéfalo para injerto |
Country Status (16)
| Country | Link |
|---|---|
| US (3) | US10280398B2 (es) |
| EP (2) | EP3693455A1 (es) |
| JP (6) | JP6461602B2 (es) |
| KR (4) | KR102115273B1 (es) |
| CN (5) | CN104379731A (es) |
| AU (3) | AU2012332269B2 (es) |
| BR (2) | BR122020000182B8 (es) |
| CA (2) | CA2854578C (es) |
| DK (1) | DK2773748T3 (es) |
| EA (1) | EA201490796A1 (es) |
| ES (1) | ES2779453T3 (es) |
| HK (1) | HK1201879A1 (es) |
| IL (4) | IL286793B (es) |
| PL (1) | PL2773748T3 (es) |
| PT (1) | PT2773748T (es) |
| WO (1) | WO2013067362A1 (es) |
Families Citing this family (54)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2010096496A2 (en) | 2009-02-17 | 2010-08-26 | Memorial Sloan-Kettering Cancer Center | Methods of neural conversion of human embryonic stem cells |
| EP2694644B1 (en) * | 2011-03-30 | 2018-01-24 | Cellular Dynamics International, Inc. | Priming of pluripotent stem cells for neural differentiation |
| PT2773748T (pt) * | 2011-11-04 | 2020-03-26 | Memorial Sloan Kettering Cancer Center | Neurónios dopaminérgicos (da) do mesencéfalo para enxerto |
| EP3498824A1 (en) | 2013-04-26 | 2019-06-19 | Memorial Sloan-Kettering Cancer Center | Cortical interneurons and other neuronal cells produced by the directed differentiation of pluripotent and multipotent cells |
| CA2920287C (en) | 2013-08-06 | 2022-03-08 | Takeda Pharmaceutical Company Limited | Method for producing dopaminergic neurons |
| US11767507B2 (en) | 2013-11-08 | 2023-09-26 | The Mclean Hospital Corporation | Methods for efficient generation of GABAergic interneurons from pluripotent stem cells |
| EP3119881B1 (en) | 2014-03-21 | 2023-03-01 | FUJIFILM Cellular Dynamics, Inc. | Production of midbrain dopaminergic neurons and methods for the use thereof |
| CN105121633B (zh) * | 2014-03-25 | 2018-05-01 | 云南中科灵长类生物医学重点实验室 | 一种建立神经上皮干细胞的培养基、方法及其应用 |
| EP3183338B1 (en) | 2014-08-19 | 2020-03-04 | FUJIFILM Cellular Dynamics, Inc. | Neural networks formed from cells derived from pluripotent stem cells |
| US9730975B2 (en) | 2014-11-25 | 2017-08-15 | The Penn State Research Foundation | Chemical reprogramming of human glial cells into neurons for brain and spinal cord repair |
| EP3061809B1 (en) | 2015-02-27 | 2019-01-30 | Miltenyi Biotec GmbH | Method for generation of a cell composition of mesencephalic dopaminergic progenitor cells |
| WO2016164857A1 (en) * | 2015-04-10 | 2016-10-13 | President And Fellows Of Harvard College | Methods and devices for live cell imaging analysis |
| US10081791B2 (en) | 2015-05-07 | 2018-09-25 | William J. Freed | Modeling connections between dopaminergic neurons and the cerebral cortex |
| HRP20220482T1 (hr) | 2015-06-01 | 2022-07-08 | Memorial Sloan Kettering Cancer Center | Postupci in vitro diferencijacije neurona dopamina srednjeg mozga (mda) |
| CN108368486B (zh) | 2015-10-07 | 2023-06-27 | 纪念斯隆-凯特琳癌症中心 | 鉴定神经肌肉接头活动的调节剂的体外方法 |
| CN105567642B (zh) * | 2016-02-01 | 2019-07-12 | 中国科学院生物物理研究所 | 一种着色性干皮病人多能干细胞的制备方法 |
| EP3411048A4 (en) * | 2016-02-05 | 2019-07-31 | Memorial Sloan-Kettering Cancer Center | METHOD FOR THE DISTINCTION OF PRECURSORS OF STEM CELLS FROM THE ECOTODERMAL DESCALING LINE |
| WO2017176779A1 (en) * | 2016-04-05 | 2017-10-12 | The Regents Of The University Of California | Cellularized hydrogels and methods of using the same |
| CN105861532A (zh) * | 2016-04-08 | 2016-08-17 | 北京大学深圳医院 | 一种基于miRNA491调节DAT基因表达的方法 |
| KR101870240B1 (ko) * | 2016-04-12 | 2018-06-22 | 서울대학교산학협력단 | 지방줄기세포에서 신경줄기세포, 신경세포 및 가바성 신경세포로의 분화 유도 방법 |
| CA3021628A1 (en) * | 2016-04-22 | 2017-10-26 | Kyoto University | Method for producing dopamine-producing neural precursor cells |
| DK3500664T3 (da) * | 2016-08-16 | 2021-12-06 | Fujifilm Cellular Dynamics Inc | Fremgangsmåde til differentiering af pluripotente celler |
| FR3058892B1 (fr) * | 2016-11-23 | 2021-04-09 | Univ Bordeaux | Unite de tissu neural et utilisation d'une telle unite pour l'implantation dans le systeme nerveux d'un mammifere |
| CA3057104A1 (en) * | 2017-03-21 | 2018-09-27 | Memorial Sloan-Kettering Cancer Center | Stem cell-derived astrocytes, methods of making and methods of use |
| EP3630951A4 (en) * | 2017-05-25 | 2021-03-17 | New York Stem Cell Foundation, Inc. | METHOD AND COMPOSITION FOR GENERATING CHOLINERGIC NEURONS OF THE ANTERIOR BASAL BRAIN (BFCN) |
| CN107828728A (zh) * | 2017-10-26 | 2018-03-23 | 中山大学 | 一种制备中脑多巴胺神经元的方法 |
| KR102201417B1 (ko) * | 2018-05-02 | 2021-01-11 | (주) 에스바이오메딕스 | 도파민 신경세포의 분리방법 및 이를 이용하여 분리된 도파민 신경세포를 포함하는 파킨슨병 치료용 약제학적 조성물 |
| KR102111030B1 (ko) * | 2018-05-10 | 2020-05-14 | 한국과학기술원 | Jab1을 포함하는 신경세포 분화 탐지용 바이오마커 또는 신경세포 분화 촉진용 조성물 |
| WO2020123663A1 (en) * | 2018-12-11 | 2020-06-18 | University Of Rochester | Methods of treating schizophrenia and other neuropsychiatric disorders |
| CN112011510B (zh) * | 2019-05-30 | 2023-01-03 | 中国科学院上海有机化学研究所 | 一种由人多能干细胞制备多巴胺神经元的方法 |
| MX2022001016A (es) | 2019-07-25 | 2022-07-19 | Scripps Research Inst | Métodos para la identificación de neuronas dopaminérgicas y células progenitoras. |
| KR102183208B1 (ko) * | 2019-07-25 | 2020-11-25 | 한국과학기술연구원 | Ngn2(neurogenin-2)를 이용하여 척수 손상 모델에서 반응성 성상세포를 신경세포로 리프로그래밍하는 방법 |
| JP2023503302A (ja) * | 2019-11-22 | 2023-01-27 | ノヴォ ノルディスク アー/エス | スピン凝集した神経系マイクロスフェアおよびその応用 |
| AU2021241643A1 (en) | 2020-03-25 | 2022-11-24 | Sana Biotechnology, Inc. | Hypoimmunogenic neural cells for the treatment of neurological disorders and conditions |
| JP2023520466A (ja) * | 2020-04-02 | 2023-05-17 | メモリアル スローン ケタリング キャンサー センター | 中脳ドーパミンニューロンの生成方法、中脳ニューロンおよびその使用 |
| WO2021216622A1 (en) | 2020-04-21 | 2021-10-28 | Aspen Neuroscience, Inc. | Gene editing of gba1 in stem cells and method of use of cells differentiated therefrom |
| WO2021216623A1 (en) | 2020-04-21 | 2021-10-28 | Aspen Neuroscience, Inc. | Gene editing of lrrk2 in stem cells and method of use of cells differentiated therefrom |
| CN114292814A (zh) * | 2020-09-22 | 2022-04-08 | 上海跃赛生物科技有限公司 | 一种多能干细胞分化为中脑黒质多巴胺能神经细胞的方法 |
| KR20230165846A (ko) | 2021-04-07 | 2023-12-05 | 후지필름 셀룰러 다이내믹스, 인코포레이티드 | 도파민성 전구세포 및 사용 방법 |
| CN117500916A (zh) | 2021-04-16 | 2024-02-02 | 博恩赛尔公司 | 制备、扩增和纯化中脑多巴胺能祖细胞的方法 |
| WO2022222974A1 (zh) * | 2021-04-21 | 2022-10-27 | 上海跃赛生物科技有限公司 | 一种质控和富集人多巴胺能神经前体细胞的方法 |
| CN113388580B (zh) * | 2021-06-16 | 2022-02-18 | 中国医学科学院医学实验动物研究所 | 一种诱导脂肪干细胞分化为功能性多巴胺能神经元的方法及用途 |
| AU2022291933A1 (en) * | 2021-06-18 | 2024-01-18 | Memorial Hospital For Cancer And Allied Diseases | Methods of generating sacral neural crest lineages and uses thereof |
| CA3222505A1 (en) * | 2021-07-14 | 2023-01-19 | Sang Hun Lee | Composition for inhibiting alpha-synuclein aggregation and method for inhibiting alpha-synuclein aggregation |
| WO2023004371A1 (en) * | 2021-07-21 | 2023-01-26 | Aspen Neuroscience, Inc. | Methods of differentiating neural cells and predicting engraftment thereof and related compositions |
| WO2023004366A1 (en) | 2021-07-21 | 2023-01-26 | Aspen Neuroscience, Inc. | Transposon-based modulation of gba1 and related compositions and uses thereof |
| WO2023004370A1 (en) | 2021-07-21 | 2023-01-26 | Aspen Neuroscience, Inc. | Aav-based modulation of gba1 and related compositions and uses thereof |
| CN113481160B (zh) * | 2021-08-20 | 2022-04-29 | 北京太东生物科技有限公司 | 视网膜色素上皮细胞诱导培养基及其应用 |
| CA3231278A1 (en) | 2021-09-10 | 2023-03-16 | Deepika Rajesh | Compositions of induced pluripotent stem cell-derived cells and methods of use thereof |
| CN115354028B (zh) * | 2021-10-28 | 2023-05-23 | 中国科学院动物研究所 | 一种中脑多巴胺细胞群、其制造方法以及用途 |
| AU2022376962A1 (en) | 2021-10-29 | 2024-04-18 | FUJIFILM Cellular Dynamics, Inc. | Dopaminergic neurons comprising mutations and methods of use thereof |
| WO2023104792A1 (en) | 2021-12-07 | 2023-06-15 | Vib Vzw | Enhancers of neuronal maturation |
| WO2023177919A1 (en) * | 2022-03-18 | 2023-09-21 | Memorial Sloan-Kettering Cancer Center | Methods and compositions for improving in vivo survival of midbrain dopamine neurons |
| WO2023201361A1 (en) | 2022-04-15 | 2023-10-19 | Aspen Neuroscience, Inc. | Methods of classifying the differentiation state of cells and related compositions of differentiated cells |
Family Cites Families (77)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1990001541A1 (en) | 1988-08-04 | 1990-02-22 | Amrad Corporation Limited | In vitro propagation of embryonic stem cells |
| US5340740A (en) | 1992-05-15 | 1994-08-23 | North Carolina State University | Method of producing an avian embryonic stem cell culture and the avian embryonic stem cell culture produced by the process |
| US5589376A (en) | 1992-07-27 | 1996-12-31 | California Institute Of Technology | Mammalian neural crest stem cells |
| US5453357A (en) | 1992-10-08 | 1995-09-26 | Vanderbilt University | Pluripotential embryonic stem cells and methods of making same |
| US5523226A (en) | 1993-05-14 | 1996-06-04 | Biotechnology Research And Development Corp. | Transgenic swine compositions and methods |
| US6787356B1 (en) | 1998-07-24 | 2004-09-07 | The United States Of America As Represented By The Department Of Health And Human Services | Cell expansion system for use in neural transplantation |
| US7410798B2 (en) | 2001-01-10 | 2008-08-12 | Geron Corporation | Culture system for rapid expansion of human embryonic stem cells |
| US6667176B1 (en) | 2000-01-11 | 2003-12-23 | Geron Corporation | cDNA libraries reflecting gene expression during growth and differentiation of human pluripotent stem cells |
| US6610540B1 (en) | 1998-11-18 | 2003-08-26 | California Institute Of Technology | Low oxygen culturing of central nervous system progenitor cells |
| US20050042749A1 (en) | 2001-05-16 | 2005-02-24 | Carpenter Melissa K. | Dopaminergic neurons and proliferation-competent precursor cells for treating Parkinson's disease |
| US7005252B1 (en) | 2000-03-09 | 2006-02-28 | Wisconsin Alumni Research Foundation | Serum free cultivation of primate embryonic stem cells |
| US7011828B2 (en) | 2000-03-14 | 2006-03-14 | Es Cell International Pte. Ltd. | Implanting neural progenitor cells derived for human embryonic stem cells |
| EP1263932A4 (en) | 2000-03-14 | 2004-06-23 | Es Cell Int Pte Ltd | EMBRYONIC CELLS AND NEURONAL STEM CELLS DERIVED THEREFROM |
| US7250294B2 (en) * | 2000-05-17 | 2007-07-31 | Geron Corporation | Screening small molecule drugs using neural cells differentiated from human embryonic stem cells |
| JP5943533B2 (ja) | 2000-05-17 | 2016-07-06 | アステリアス バイオセラピューティクス インコーポレイテッド | 神経前駆細胞の集団 |
| EP2298867A3 (en) | 2000-06-20 | 2013-01-16 | ES Cell International Pte Ltd. | Method of controlling differentiation of embryonic stem (ES) cells by culturing ES cells in the presence of bmp-2 pathway antagonists |
| DE10037759A1 (de) | 2000-08-03 | 2002-07-04 | Gruenenthal Gmbh | Screeningverfahren |
| AU2001293586A1 (en) | 2000-09-29 | 2002-04-08 | Vincent Tropepe | Primitive neural stem cells and method for differentiation of stem cells to neural cells |
| JP3660601B2 (ja) | 2001-03-30 | 2005-06-15 | 独立行政法人科学技術振興機構 | 胚性幹細胞からの神経幹細胞、運動ニューロン及びgaba作動性ニューロンの製造法 |
| AU2002307455A1 (en) | 2001-04-20 | 2002-11-05 | Memorial Sloan-Kettering Cancer Center | Generation of differentiated tissue from nuclear transfer embryonic stem cells and methods of use |
| US20030211605A1 (en) | 2001-05-01 | 2003-11-13 | Lee Sang-Hun | Derivation of midbrain dopaminergic neurons from embryonic stem cells |
| US6887706B2 (en) | 2001-10-03 | 2005-05-03 | Wisconsin Alumni Research Foundation | Method of in vitro differentiation of transplantable neural precursor cells from primate embryonic stem cells |
| US20040224302A1 (en) | 2002-07-16 | 2004-11-11 | Thomas Jessel | Systems and methods for screening for modulators of neural differentiation |
| EP1546198A1 (en) | 2002-07-31 | 2005-06-29 | Stem Cell Therapeutics Inc. | Method of enhancing neural stem cell proliferation, differentiation, and survival using pituitary adenylate cyclase activating polypeptide (pacap) |
| AU2003276924A1 (en) | 2002-09-25 | 2004-04-19 | Bresagen, Inc. | Compositions and methods for enrichment of neural stem cells using ceramide analogs |
| WO2004053084A2 (en) | 2002-12-09 | 2004-06-24 | The Mclean Hospital Corporation | Dopaminergic neurons differentiated from embryonic cells for treating neurodegenerative diseases |
| US7252995B2 (en) | 2002-12-13 | 2007-08-07 | Yu-Show Fu | Method of generating neurons from stem cells and medium for culturing stem cells |
| ZA200401646B (en) * | 2003-03-12 | 2004-06-07 | Reliance Life Sciences Pvt Ltd | Derivation of terminally differentiated dopaminergic neurons from human embryonic stem cells. |
| EP1645286A1 (en) | 2003-04-18 | 2006-04-12 | Kyowa Hakko Kogyo Co., Ltd. | Drug for nerve regeneration |
| CA2457296A1 (en) | 2003-08-19 | 2005-02-19 | Takahiro Ochiya | Methods for inducing differentiation of pluripotent cells |
| EP1799272A2 (en) | 2004-10-05 | 2007-06-27 | Memorial Sloan-Kettering Cancer Center | Promotion of cell migration and axon regeneration in the cns |
| AU2006315668B2 (en) * | 2005-11-10 | 2013-06-20 | Genervon Biopharmaceuticals Llc | MNTF differentiation and growth of stem cells |
| EP1999249B8 (en) | 2006-03-30 | 2012-02-15 | The University Court Of The University of Edinburgh | Culture medium containing kinase inhibitors. and uses thereof |
| WO2008018190A1 (fr) | 2006-08-09 | 2008-02-14 | Biomaster, Inc. | Cellules de la crête neurale dérivées de tissu adipeux |
| JP2008099662A (ja) | 2006-09-22 | 2008-05-01 | Institute Of Physical & Chemical Research | 幹細胞の培養方法 |
| GB0622394D0 (en) | 2006-11-09 | 2006-12-20 | Univ Cambridge Tech | Differentiation of pluripotent cells |
| EP2101819B1 (en) | 2006-11-20 | 2013-01-09 | President and Fellows of Harvard College | Methods, compositions, and kits for treating pain and pruritis |
| SE0950586L (sv) | 2007-01-17 | 2009-08-13 | Wisconsin Alumni Res Found | Förbättrad odling av stamceller |
| EP2246414B1 (en) | 2008-02-06 | 2018-04-04 | Public University Corporation Yokohama City University | Cell culture method and screening method |
| CN101550406B (zh) | 2008-04-03 | 2016-02-10 | 北京大学 | 制备多潜能干细胞的方法,试剂盒及用途 |
| WO2009155301A2 (en) | 2008-06-17 | 2009-12-23 | The Mclean Hospital Corporation | Multipotent neural cells |
| US20110262956A1 (en) * | 2008-10-07 | 2011-10-27 | Guillermo Munoz Elias | Co-culture compositions and methods |
| DK2356218T3 (en) | 2008-12-05 | 2017-08-21 | Inserm (Institut Nat De La Santé Et De La Rech Médicale) | METHOD AND MEDIUM FOR NEURAL DIFFERENTIZATION OF PLURIPOTENT CELLS |
| WO2010096496A2 (en) * | 2009-02-17 | 2010-08-26 | Memorial Sloan-Kettering Cancer Center | Methods of neural conversion of human embryonic stem cells |
| WO2010141622A2 (en) | 2009-06-02 | 2010-12-09 | Cornell University | Method to isolate, identify, and use embryonic stem cells directed to forebrain interneuron fate |
| ES2590036T3 (es) | 2009-08-12 | 2016-11-17 | Kyoto University | Método para inducir la diferenciación de células madre pluripotentes en células precursoras neurales |
| EP2488631B1 (en) * | 2009-10-13 | 2020-01-08 | Stemcell Technologies Inc. | Manipulation of osmolality for differentiating stem cells |
| ES2539487T3 (es) | 2009-11-04 | 2015-07-01 | Cellular Dynamics International, Inc. | Reprogramación episómica con compuestos químicos |
| KR101168053B1 (ko) | 2009-11-06 | 2012-07-24 | 연세대학교 산학협력단 | 효율적이고 보편적인 전분화능 줄기세포의 신경세포 분화 유도방법 |
| JP5846608B2 (ja) | 2010-03-03 | 2016-01-20 | 国立大学法人京都大学 | iPS細胞由来の神経細胞を用いた蛋白質ミスフォールディング病の診断方法 |
| CN102191221B (zh) | 2010-03-17 | 2014-11-19 | 中国人民解放军第二军医大学东方肝胆外科医院 | 一种能无限自我更新的神经干细胞、其制备方法及其用途 |
| WO2011130675A2 (en) | 2010-04-16 | 2011-10-20 | The Mclean Hospital Corporation | Dopaminergic neurons differentiated from pluripotent stem cells and uses of thereof |
| PT2577318T (pt) | 2010-05-25 | 2019-10-14 | Memorial Sloan Kettering Cancer Center | Método de diferenciação de nocicetor de células estaminais embrionárias humanas e utilizações do mesmo |
| US9376664B2 (en) | 2010-06-14 | 2016-06-28 | The Scripps Research Institute | Reprogramming of cells to a new fate |
| US8932857B2 (en) | 2010-06-15 | 2015-01-13 | Kyoto University | Method for selecting reduced differentiation resistance human induced pluripotent stem cells |
| CN101935635A (zh) * | 2010-07-30 | 2011-01-05 | 中国人民解放军第二军医大学 | 人骨髓多能干细胞的体外扩增和向多巴胺能神经元定向分化的方法 |
| US8883502B2 (en) | 2010-09-09 | 2014-11-11 | The Regents Of The University Of California | Expandable cell source of neuronal stem cell populations and methods for obtaining and using them |
| DK2980209T3 (en) | 2011-05-20 | 2018-04-23 | Mclean Hospital Corp | NEURONAL PROGENITOR CELLS AND APPLICATIONS |
| KR101331034B1 (ko) | 2011-06-09 | 2013-11-19 | 연세대학교 산학협력단 | 미분화 줄기세포로부터 중뇌특이적 도파민 신경세포의 제조 방법 |
| US20150086481A1 (en) | 2011-07-28 | 2015-03-26 | Memorial Sloan-Kettering Cancer Center | Diagnosis and treatment of parkinson's disease |
| PT2773748T (pt) * | 2011-11-04 | 2020-03-26 | Memorial Sloan Kettering Cancer Center | Neurónios dopaminérgicos (da) do mesencéfalo para enxerto |
| US10472607B2 (en) | 2011-12-14 | 2019-11-12 | National Chung Hsing University | Culture medium and method for inducing differentiation of pluripotent stem cells into neuroepithelial cells |
| EP2614829A1 (en) | 2012-01-11 | 2013-07-17 | Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. | Mammalian neural plate border stem cells capable of forming neural tube and neural crest cell lineages including central and peripheral neurons |
| CN102604894B (zh) | 2012-02-29 | 2014-07-30 | 中国科学院广州生物医药与健康研究院 | 用于制备神经干细胞的培养基及其用途 |
| MX368315B (es) | 2012-04-24 | 2019-09-27 | Int Stem Cell Corporation | Derivación de células madre neurales y neuronas dopaminérgicas a partir de células madre pluripotentes humanas. |
| WO2014104409A1 (en) | 2012-12-28 | 2014-07-03 | Kyoto University | Method for inducing astrocytes |
| EP2961277A4 (en) | 2013-02-15 | 2016-12-28 | Int Stem Cell Corp | USE OF NEURONS FROM HUMAN PLURIPOTENT STEM CELLS FOR THE TREATMENT OF NEURODEGENERATIVE DISEASES |
| US20140248696A1 (en) | 2013-03-01 | 2014-09-04 | Wisconsin Alumni Research Foundation | Methods of maintaining, expanding, and diffrentiating neuronal subtype specific progenitors |
| JP6774333B2 (ja) | 2013-04-16 | 2020-10-21 | メモリアル スローン ケタリング キャンサー センター | 年齢改変細胞および年齢改変細胞を作製するための方法 |
| EP3498824A1 (en) | 2013-04-26 | 2019-06-19 | Memorial Sloan-Kettering Cancer Center | Cortical interneurons and other neuronal cells produced by the directed differentiation of pluripotent and multipotent cells |
| ES2721440T3 (es) | 2013-09-05 | 2019-07-31 | Univ Kyoto | Nuevo método para inducir células precursoras neurales productoras de dopamina |
| CN105814192A (zh) | 2013-10-09 | 2016-07-27 | 加利福尼亚大学董事会 | 哺乳动物视网膜干细胞产生方法和应用 |
| KR102483685B1 (ko) | 2013-11-21 | 2023-01-03 | 메모리얼 슬로안 케터링 캔서 센터 | 인간 다능성 줄기 세포로부터 기능적 두개 기원판 유도체의 전문화 |
| EP3119881B1 (en) | 2014-03-21 | 2023-03-01 | FUJIFILM Cellular Dynamics, Inc. | Production of midbrain dopaminergic neurons and methods for the use thereof |
| CN105121633B (zh) | 2014-03-25 | 2018-05-01 | 云南中科灵长类生物医学重点实验室 | 一种建立神经上皮干细胞的培养基、方法及其应用 |
| SG11201703305SA (en) | 2014-10-24 | 2017-05-30 | Sumitomo Dainippon Pharma Co Ltd | Production method for nerve tissue |
| EP3280426A2 (en) | 2015-04-09 | 2018-02-14 | Biolamina AB | Methods and compositions for producing stem cell derived dopaminergic cells for use in treatment of neurodegenerative diseases |
-
2012
- 2012-11-02 PT PT128467156T patent/PT2773748T/pt unknown
- 2012-11-02 KR KR1020147014950A patent/KR102115273B1/ko active IP Right Grant
- 2012-11-02 EP EP19215962.2A patent/EP3693455A1/en active Pending
- 2012-11-02 IL IL286793A patent/IL286793B/en unknown
- 2012-11-02 AU AU2012332269A patent/AU2012332269B2/en active Active
- 2012-11-02 CA CA2854578A patent/CA2854578C/en active Active
- 2012-11-02 CN CN201280066145.0A patent/CN104379731A/zh active Pending
- 2012-11-02 JP JP2014541125A patent/JP6461602B2/ja active Active
- 2012-11-02 DK DK12846715.6T patent/DK2773748T3/da active
- 2012-11-02 KR KR1020227025750A patent/KR102581669B1/ko active IP Right Grant
- 2012-11-02 BR BR122020000182A patent/BR122020000182B8/pt active IP Right Grant
- 2012-11-02 IL IL295556A patent/IL295556A/en unknown
- 2012-11-02 US US14/356,042 patent/US10280398B2/en active Active
- 2012-11-02 ES ES12846715T patent/ES2779453T3/es active Active
- 2012-11-02 KR KR1020237031951A patent/KR20230136702A/ko not_active Application Discontinuation
- 2012-11-02 PL PL12846715T patent/PL2773748T3/pl unknown
- 2012-11-02 CN CN201910438393.7A patent/CN113528442A/zh active Pending
- 2012-11-02 CN CN201910438392.2A patent/CN110684734A/zh active Pending
- 2012-11-02 EA EA201490796A patent/EA201490796A1/ru unknown
- 2012-11-02 WO PCT/US2012/063339 patent/WO2013067362A1/en active Application Filing
- 2012-11-02 EP EP12846715.6A patent/EP2773748B1/en active Active
- 2012-11-02 CN CN201910438266.7A patent/CN110684733A/zh active Pending
- 2012-11-02 CA CA3177246A patent/CA3177246A1/en active Pending
- 2012-11-02 CN CN201910438394.1A patent/CN113481159A/zh active Pending
- 2012-11-02 BR BR112014010776-9A patent/BR112014010776B1/pt active IP Right Grant
- 2012-11-02 KR KR1020207014325A patent/KR102426327B1/ko active IP Right Grant
-
2014
- 2014-05-04 IL IL232450A patent/IL232450B/en active IP Right Grant
-
2015
- 2015-03-10 HK HK15102423.6A patent/HK1201879A1/xx unknown
-
2017
- 2017-06-20 JP JP2017120341A patent/JP6588500B2/ja active Active
-
2018
- 2018-08-24 JP JP2018156902A patent/JP6756786B2/ja active Active
- 2018-08-30 AU AU2018222997A patent/AU2018222997B2/en active Active
-
2019
- 2019-03-14 US US16/353,546 patent/US10711243B2/en active Active
-
2020
- 2020-04-12 IL IL273910A patent/IL273910B/en unknown
- 2020-06-03 US US16/892,000 patent/US11970712B2/en active Active
- 2020-06-19 JP JP2020106370A patent/JP7225163B2/ja active Active
- 2020-09-04 JP JP2020149072A patent/JP2021000104A/ja active Pending
-
2021
- 2021-08-19 AU AU2021218116A patent/AU2021218116A1/en active Pending
-
2022
- 2022-11-14 JP JP2022181703A patent/JP2023027064A/ja active Pending
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2779453T3 (es) | Neuronas de dopamina (DA) del mesencéfalo para injerto | |
| WO2011130675A2 (en) | Dopaminergic neurons differentiated from pluripotent stem cells and uses of thereof | |
| JP2022088676A (ja) | 幹細胞由来外胚葉系統前駆体を分化する方法 | |
| US20230233617A1 (en) | Methods for differentiating stem cells into dopaminergic progenitor cells | |
| CN112368004A (zh) | 细胞组合物和其用途 | |
| EA041083B1 (ru) | Способ индукции дифференцировки плюрипотентных стволовых клеток in vitro |