WO2020256099A1 - 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 - Google Patents
外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 Download PDFInfo
- Publication number
- WO2020256099A1 WO2020256099A1 PCT/JP2020/024131 JP2020024131W WO2020256099A1 WO 2020256099 A1 WO2020256099 A1 WO 2020256099A1 JP 2020024131 W JP2020024131 W JP 2020024131W WO 2020256099 A1 WO2020256099 A1 WO 2020256099A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- gene
- virus
- zsg
- codon
- rna virus
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images






















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
- C12N15/867—Retroviral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2720/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsRNA viruses
- C12N2720/00011—Details
- C12N2720/12011—Reoviridae
- C12N2720/12311—Rotavirus, e.g. rotavirus A
- C12N2720/12321—Viruses as such, e.g. new isolates, mutants or their genomic sequences
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2720/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsRNA viruses
- C12N2720/00011—Details
- C12N2720/12011—Reoviridae
- C12N2720/12311—Rotavirus, e.g. rotavirus A
- C12N2720/12322—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2720/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsRNA viruses
- C12N2720/00011—Details
- C12N2720/12011—Reoviridae
- C12N2720/12311—Rotavirus, e.g. rotavirus A
- C12N2720/12341—Use of virus, viral particle or viral elements as a vector
- C12N2720/12343—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2800/00—Nucleic acids vectors
- C12N2800/22—Vectors comprising a coding region that has been codon optimised for expression in a respective host
Definitions
- the present invention relates to a method for producing an artificially recombinant RNA virus that stably retains a foreign gene.
- Rotavirus which belongs to the Reoviridae family, causes acute gastroenteritis in infants, and about 200,000 people die from rotavirus infection annually.
- the development of a technique for artificially producing an arbitrary virus has been delayed, but recently, the present inventors have succeeded in developing a practical artificial synthesis method for rotavirus (Patent Document 1, Patent Document 1, Non-Patent Document 1).
- Patent Document 1 Patent Document 1, Non-Patent Document 1
- Patent Document 1 Patent Document 1
- Non-Patent Document 2 Non-Patent Document 2
- Non-Patent Document 3 When expressing a foreign gene in a heterologous host, a codon optimization technique that brings the codon composition of the foreign gene closer to the codon composition of the host gene has been used for a long time for the purpose of improving the expression of the foreign gene (Non-Patent Document 3). ), Numerous codon optimization software is provided. However, it has not been known that the codon optimization technique can be applied to the stable retention of foreign genes introduced into artificially recombined RNA virus.
- An object of the present invention is to provide a method for producing an artificially recombined RNA virus that stably retains a foreign gene and a method for stably retaining a foreign gene introduced into the artificially recombined RNA virus.
- the present invention includes the following inventions in order to solve the above problems.
- a method for producing an artificially recombinant RNA virus that stably retains a foreign gene (1) A step of obtaining a foreign gene having a codon composition modified to approximate the codon composition of an RNA virus gene. It is characterized by including (2) a step of inserting the foreign gene obtained in (1) into the genome of the RNA virus, and (3) a step of artificially synthesizing an artificially recombined RNA virus using the reverse genetics method. Manufacturing method.
- RNA virus is a virus belonging to the Reoviridae family.
- virus belonging to the Reoviridae family is a virus belonging to the genus Rotavirus or the genus Ortholeovirus.
- codon composition of the foreign gene is approximated within ⁇ 30% of the codon composition of the RNA virus gene.
- RNA virus that stably retains a foreign gene for a long period of time.
- ZsG gene Green fluorescent protein gene
- Rv-ZsG gene codon-modified green fluorescent protein gene
- AsR gene red fluorescent protein gene
- Rv-ZsG gene codon-modified green fluorescent protein gene
- Is an observation image of immunostaining using MRV-specific antibody the center is an observation image of green fluorescent protein-expressing cells, and the right is a composite image of both.
- An artificially recombined mammalian leovirus having a green fluorescent protein gene (ZsG gene) and an artificially recombined mammalian leovirus having a codon-modified green fluorescent protein gene (Mrv-ZsG gene) are passaged 1 to 3 times to obtain a foreign gene. It is a figure which shows the result of having examined the stability of.
- the present invention provides a method for producing an artificially recombined RNA virus that stably retains a foreign gene (hereinafter, referred to as "method for producing the present invention”).
- the production method of the present invention may include the following steps (1), (2) and (3).
- (1) Step of acquiring a foreign gene having a codon composition modified to resemble the codon composition of an RNA virus gene (2) Step of inserting the foreign gene obtained in (1) into the RNA virus genome (3) Step of obtaining artificially recombinant RNA virus using reverse genetics method
- the RNA virus may be a double-stranded RNA virus, a single-strand plus-strand RNA virus, or a single-strand minus-strand RNA virus.
- double-stranded RNA virus examples include viruses belonging to the family Reoviridae and Birnaviridae.
- positive-strand RNA virus examples include viruses belonging to the family Coronaviridae, Picornavirus, Togaviridae, Flaviviridae, Calisivirus, Astrovirus, and the like.
- Examples of the single-stranded minus-strand RNA virus include viruses belonging to the family Paramyxoviridae, Rabdovirus, Philovirus, Orthomyxoviridae, Arenavirus, Bunyavirus, and the like.
- the RNA virus may be a virus belonging to the Reoviridae family.
- Viruses belonging to the Reoviridae family are viruses that have a straight-chain double-stranded RNA segmented into 10 to 12 lines in their genome, and their virions show a regular icosahedron structure with a diameter of 60 to 80 nm.
- Viruses belonging to the Reoviridae family include mammalian ortholeovirus (also called mammalian reoviridae), Nelsonbey ortholeovirus (also called Nelsonbeyreovirus or bat leovirus), ortholeovirus such as trileovirus.
- Virus genus (Genus Orthoreovirus), African horse epidemic virus, Orbivirus genus such as blue tongue virus (Genus Orbivirus), Rotavirus genus such as Rotavirus (Genus Rotavirus), Cortivirus genus such as Colorado tick fever virus (Genus Coltivirus) , Aquareovirus such as Aquareovirus A (GenusAquareovirus), Cypovirus such as cytoplasmic polymorphic disease virus (GenusCypovirus), Fijivirus genus such as rice southern black streak dwarf disease virus (Genus Fijivirus), rice Includes the genus Phytoreovirus such as dwarf disease virus and the genus Oryzavirus such as Rice ragged stunt virus.
- the virus belonging to the Reoviridae family may be a virus belonging to the genus Rotavirus or a virus belonging to the genus Ortholeovirus.
- the foreign gene is not particularly limited.
- the foreign gene may be an animal gene, a plant gene, a fungal gene, a bacterial gene, or a viral gene.
- the base length of the foreign gene is not particularly limited, and may be 10 bp or more, 100 bp or more, 500 bp or more, 1000 bp or more, or 1500 bp or more. , 2000 bp or more, 3000 bp or more, 4000 bp or more, 5000 bp or more.
- the base length of the foreign gene may be 500 bp or less, 1000 bp or less, 1500 bp or less, 2000 bp or less, 3000 bp or less, 4000 bp or less. It may be present, and may be 5000 bp or less.
- the protein encoded by the foreign gene is not particularly limited.
- the protein encoded by the foreign gene may be a vaccine antigen.
- the vaccine antigen include norovirus antigen, adenovirus antigen, hepatitis A antigen, sapovirus antigen, limb virus antigen, enterovirus antigen, HIV antigen, salmonella antigen, campylobacter antigen, enteritis vibrio antigen, Escherichia coli O157 antigen, and cholera.
- examples thereof include antigens, intestinal typhoid antigens, and diarrhea antigens.
- These vaccine antigens may be epitope peptides.
- a foreign gene having a codon composition modified to approximate the codon composition of the RNA virus gene is obtained.
- the codon composition of the reference RNA virus gene may be the codon composition of the gene of the RNA virus that is artificially synthesized by incorporating the foreign gene with the modified codon, or artificially by incorporating the foreign gene with the modified codon.
- the codon composition of the gene of the RNA virus different from that of the RNA virus to be synthesized may be used.
- different RNA viruses are systematically similar to RNA viruses that are artificially synthesized by incorporating foreign genes with modified codons (eg, viruses belonging to the same family). , Viruses belonging to the same genus, viruses of different species, etc.).
- the codon composition of the reference RNA virus gene may be the codon composition of all the genes of the reference RNA virus, or may be the codon composition of some genes of the reference RNA virus.
- the codon composition of some genes of the RNA virus is used as a reference, the codon composition of one gene may be used as a reference, or the codon composition of two or more genes may be used as a reference.
- the codon composition of the reference RNA virus gene can be created based on the genetic information of the RNA virus registered in a known gene database (for example, GenBank).
- Table 1 shows the genomic composition of all genes of rotavirus SA11 strain.
- Table 2 shows the genomic composition of the rotavirus SA11 strain NSP1 gene.
- Table 3 shows the genomic composition of all genes of the bat reovirus (Pteropine orthoreovirus) MB strain.
- Table 4 shows the genomic composition of all mammalian reoviridae T1L genes.
- the codon compositions shown in Tables 1 to 4 can be used as the reference codon composition in the production method of the present invention. Among them, it can be used as a reference codon composition when a foreign gene is expressed by using a virus belonging to the Reoviridae family, particularly an artificial recombinant virus belonging to the genus Rotavirus and the genus Ortholeovirus.
- Approximating the codon composition of a foreign gene to the codon composition of a reference RNA virus gene means that it frequently appears in an RNA virus gene based on a codon corresponding to the same amino acid without mutating the amino acid of the foreign gene. It means changing to a codon.
- the degree of approximation to the codon composition of the reference RNA virus gene is not particularly limited as long as the purpose of stably retaining the foreign gene can be achieved.
- the codon composition of the foreign gene may be modified to be within ⁇ 35% of the codon composition of the reference RNA virus gene, or within ⁇ 30% of the codon composition of the reference RNA virus gene.
- the codon composition of the foreign gene may be modified so as to be within ⁇ 25% of the codon composition of the reference RNA virus gene.
- the codon composition of the foreign gene may be modified to be within ⁇ 20% of the codon composition of the RNA virus gene to be used, and should be within ⁇ 15% of the codon composition of the reference RNA virus gene.
- the codon composition of the foreign gene may be modified, or the codon composition of the foreign gene may be modified so as to be within ⁇ 10% of the codon composition of the reference RNA virus gene.
- a foreign gene with a modified codon composition can be obtained by DNA synthesis based on the modified base sequence.
- a mutation can be introduced into an existing foreign gene DNA and obtained by using a site-specific mutation introduction method based on PCR.
- the foreign gene obtained in (1) is inserted into the RNA virus genome.
- the position at which the foreign gene is inserted is not particularly limited as long as it does not interfere with the artificial synthesis of the artificially recombined RNA virus using the reverse genetics method.
- a foreign gene when a foreign gene is introduced into an artificially recombined rotavirus, it may be inserted into the NSP1 gene, the NSP3 gene, or the NSP5 gene.
- a foreign gene into a recombinant mammalian reovir it may be inserted into the L1 gene or the S1, S2, and S4 genes.
- a foreign gene when introduced into an artificially recombined influenza virus, it may be inserted into the NS1 gene, the NA gene, or the PA gene.
- a foreign gene into an artificially recombinant virus of the genus Alphavirus (Sindbis virus, Chikungunya virus, etc.)
- it when introducing a foreign gene into an artificially recombinant virus of the genus Alphavirus (Sindbis virus, Chikungunya virus, etc.), it may be inserted into the NSP3 gene or the capsid gene.
- introducing a foreign gene into an artificially recombined norovirus it may be inserted into the ORF1 gene.
- a foreign gene when introduced into an artificially recombinant virus of the genus Flavivirus (dengue virus, hepatitis C virus, etc.), it may be inserted into the NS5A gene or the capsid gene.
- a foreign gene into an artificially recombinant virus of rhabdovirus rabies virus, vesicular stomatitis virus
- it when introducing a foreign gene into an artificially recombinant virus of the genus Picornavirus (poliovirus, foot-and-mouth disease virus, etc.), it may be inserted into the P1 gene.
- a foreign gene into an artificially recombined Sendai virus it may be inserted into an NP gene. Insertion of a foreign gene into the viral genome can be performed using a known gene recombination technique.
- an artificially recombinant RNA virus is artificially synthesized using the reverse genetics method.
- the method for artificially synthesizing an artificially recombined RNA virus using the reverse genetics method may be appropriately selected from known methods according to the type of RNA virus. Methods for synthesizing artificially recombinant rotavirus using the reverse genetics method are described in, for example, Patent Document 1, Non-Patent Document 1, Non-Patent Document 2, and the like. A method for synthesizing artificially recombined poliovirus using the reverse genetics method is described in, for example, Reference 1 below.
- a method for synthesizing an artificially recombined rabies virus using the reverse genetics method is described in, for example, Reference 2 below.
- a method for synthesizing an artificially recombined measles virus using the reverse genetics method is described in, for example, Reference 3 below.
- a method for synthesizing an artificially recombined bunyavirus using the reverse genetics method is described in, for example, Reference 4 below.
- a method for synthesizing an artificially recombinant infectious bursal disease virus using the reverse genetics method is described in, for example, Reference 5 below.
- a method for synthesizing artificially recombined hepatitis C virus using the reverse genetics method is described in, for example, Reference 6 below.
- a method for synthesizing an artificially recombined influenza virus using the reverse genetics method is described in, for example, Reference 7 below.
- a method for synthesizing an artificially recombined coronavirus using the reverse genetics method is described in, for example, Reference 8 below.
- a method for synthesizing an artificially recombined Ebola virus using the reverse genetics method is described in, for example, Reference 9 below.
- a method for synthesizing an artificially recombined borunavirus using the reverse genetics method is described in, for example, Reference 10 below.
- a method for synthesizing an artificially recombined arenavirus using the reverse genetics method is described in, for example, Reference 11 below.
- a method for synthesizing an artificially recombined reovir using the reverse genetics method is described in, for example, Reference 12 below.
- a method for synthesizing an artificially recombined dengue virus using the reverse genetics method is described in, for example, Reference 13 below.
- a method for synthesizing an artificially recombined norovirus using the reverse genetics method is described in, for example, Reference 14 below.
- a method for synthesizing artificially recombined Japanese encephalitis virus using the reverse genetics method is described in, for example, Reference 15 below.
- a method for synthesizing an artificially recombined bluetongue virus using the reverse genetics method is described in, for example, 16.
- a method for synthesizing an artificially recombined human immunodeficiency virus using the reverse genetics method is described in, for example, Reference 17 below.
- the artificially recombined RNA virus produced by the production method of the present invention can stably retain a foreign gene for a long period of time and stably express a foreign gene product for a long period of time. Therefore, for example, if a vaccine antigen is used as the foreign gene. , Can be suitably used as a virus vaccine.
- a vaccine antigen is used as the foreign gene.
- normal genes are used for genetic diseases of the gastrointestinal tract (colon cancer, ulcerative colitis, Crohn's disease, celiac disease, nonspecific multiple small intestinal ulcer disease, etc.) derived from (believed to be) genetic abnormalities. It can also be applied to gene therapy by delivering it to abnormal cells as a foreign gene.
- the present invention provides a method for stably retaining a foreign gene introduced into an artificially synthesized RNA virus artificially synthesized by using a reverse genetics method (hereinafter, referred to as "the stable retention method of the present invention").
- the stable retention method of the present invention is a method characterized in that the codon composition of a foreign gene is approximated to the codon composition of an RNA virus gene.
- the method for approximating the codon composition of the foreign gene to the codon composition of the reference RNA virus gene can be the same as the method described in the production method of the present invention.
- Example 1 Artificial recombinant rotavirus expressing luciferase
- Virus Sarrotavirus SA11 strain was used. We have sequenced and registered the nucleotide sequences of each of the 11 segmental RNA genomes of this virus strain. Table 5 shows the names and GenBank ACCESSION of each of the 11 segmental RNA genomes of the Sarrotavirus SA11 strain (hereinafter referred to as "SA11 strain") used in this experiment.
- a plasmid containing each segmented RNA genome expression cassette of the SA11 strain (segmented RNA genome expression vector)
- a plasmid containing the cDNA of 11 segmental RNA genomes of the SA11 strain was prepared.
- RT-PCR was performed using double-stranded RNA extracted from the virus as a template and specific primers based on the base sequence of each segmental RNA genome.
- the obtained RT-PCR product (cDNA of each segmental RNA genome) was inserted between the T7 promoter sequence and the HDV ribozyme sequence of the p3E5 plasmid to obtain a plasmid containing the expression cassette of each segmental RNA genome (see Patent Document 1). ).
- the T7 promoter sequence is adjacent to the 5'side of the cDNA of each segmental RNA genome, the hepatitis delta virus (HDV) ribozyme sequence is adjacent to the 3'side, and the T7 terminator sequence is arranged downstream thereof.
- HDV hepatitis delta virus
- the prepared plasmids were used as pT7-VP1SA11, pT7-VP2SA11, pT7-VP3SA11, pT7-VP4SA11, pT7-VP6SA11, pT7-VP7SA11, pT7-NSP1SA11, pT7-NSP2SA11, pT7-NSP3, respectively.
- NSP4SA11 and pT7-NSP5SA11 were used as NSP4SA11 and pT7-NSP5SA11.
- Luciferase gene As the luciferase gene, the NLuc gene, which is a luciferase gene derived from Oplophorus gracilirostris, was used. The NLuc coding region is located at positions 815 to 1330 (SEQ ID NO: 12) of the pNL1.1.TK [NLuc / TK] vector (Promega, GenBank ACCESSION: KM359774, 3817 bp).
- FIG. 1 shows the codon composition of each amino acid in the RVNSP1 gene, NLuc gene, and Rv-NLuc gene.
- FIG. 2 shows the nucleotide sequence of the NLuc gene (SEQ ID NO: 12) and the nucleotide sequence of the Rv-NLuc gene (SEQ ID NO: 13), respectively.
- the Rv-NLuc gene has 70 bases changed out of the total length of 516 bases of the NLuc gene.
- the GC content of the original NLuc gene was 53%, but the GC content of the Rv-NLuc gene dropped to 40%, approaching the GC content of the RV NSP1 gene (31%) (see Table 6).
- FAST protein expression vector uses the protein coding region DNA of the Nelson baileovirus p10 gene (see GenBank ACCESSION: AB908284) as the pCAGGS plasmid (5699bp, Matsuo et al., 2006, Biochem Biophys Res Commun 340). (1): It was produced by inserting it into the BglII cleavage site of 200-208) (see Patent Document 1). The obtained Nelson baileovirus p10 expression vector is referred to as pCAG-p10.
- the capping enzyme expression vector is the protein coding region DNA of the vaccinia virus D1R gene (GenBank ACCESSION: NC006998 positions 93948 to 96482) and the protein coding region DNA of the vaccinia virus D12L gene (GenBank ACCESSION: NC006998). Positions 107332 to 108195) were inserted into the BglII cleavage site of the above pCAGGS plasmid, respectively (see Patent Document 1).
- the obtained vaccinia virus mRNA capping enzyme large subunit expression vector is referred to as pCAG-D1R
- the obtained vaccinia virus mRNA capping enzyme small subunit expression vector is referred to as pCAG-D12L.
- the NSP2 protein expression vector was prepared by inserting the protein coding region DNA of the SA11 strain NSP2 gene (GenBank ACCESSION: positions 47 to 1000 of LC178571) into the BglII cleavage site of the above pCAGGS plasmid (see Patent Document 1).
- the obtained SA11 strain NSP2 protein expression vector is referred to as pCAG-NSP2.
- the NSP5 protein expression vector was prepared by inserting the protein coding region DNA of the SA11 strain NSP5 gene (GenBank ACCESSION: positions 22 to 618 of LC178574) into the BglII cleavage site of the above pCAGGS plasmid (see Patent Document 1).
- the obtained SA11 strain NSP5 protein expression vector is referred to as pCAG-NSP5.
- NLuc gene and Rv-NLuc gene are amplified by PCR, respectively, and the amplification product is inserted between positions 128 and 129 of the NSP1 gene (SEQ ID NO: 5) of pT7-NSP1SA11. Then, the NLuc gene-inserted NSP1 gene expression plasmid (referred to as pT7-NSP1SA11-NLuc) and the Rv-NLuc gene expression plasmid (referred to as pT7-NSP1SA11-Rv-NLuc) were prepared (see FIG. 3).
- RNA genome expression vectors 0.8 ⁇ g each, FAST protein expression vector (pCAG-p10) 0.015 ⁇ g, capping enzyme expression vectors (pCAG-D1R and pCAG-D12L), NSP2 protein expression vector (pCAG-NSP2), NSP5 protein An expression vector (pCAG-NSP5), 0.8 ⁇ g each, was introduced into BHK-T7 / P5 cells using a transfection reagent (TransIT-LT1 (trade name), Miras). 2 ⁇ g of transfection reagent was used per 1 ⁇ g of DNA.
- TransIT-LT1 trade name
- BHK-T7 / P5 cells were cultured in DMEM medium containing 5% FBS, 100 units / ml penicillin and 100 ⁇ g / ml streptomycin in a 37 ° C. 5% CO 2 environment. Medium and cells were harvested 48 hours after transfection. The collected medium and cells were frozen and thawed three times to prepare a cell lysate, which was subcultured into monkey MA104 cells (ATCC CRL-2378.1). Specifically, about 0.5 ml of the cell lysate was added to MA104 cells in a confluent state on a 12-well plate in the presence of 0.5 ⁇ g / ml of trypsin. FBS-free DMEM medium was used for culturing MA104 cells.
- the cells were cultured for 7 days after passage, and if cytopathic effect was observed during that period, it was judged that an artificial recombinant virus was produced.
- the virus obtained at this point is called a P1 (Passage 1) virus.
- An artificially recombined rotavirus prepared using 11 types of plasmids containing pT7-NSP1SA11-NLuc is called "rsSA11-NLuc", and an artificial set prepared using 11 types of plasmids containing pT7-NSP1SA11-Rv-NLuc.
- the replacement rotavirus is called "rsSA11-Rv-N Luc".
- rsSA11-NLuc and rsSA11-Rv-NLuc P1 viruses was added, and the cells were cultured at 37 ° C. for 5 days. After culturing for 5 days, the cells were frozen and thawed twice to obtain P2 virus. For each recombinant virus, 5 wells were passaged.
- P10 virus clones 1 to 5 of rsSA11-NLuc and P10 virus clones 1 to 5 of rsSA11-Rv-NLuc were obtained.
- Viral genomic RNA was extracted from each clone of each P10 virus obtained and subjected to SDS-PAGE together with viral genomic RNA obtained from P1 virus and wild SA11 virus.
- Example 2 Artificial recombinant rotavirus expressing green fluorescent protein
- ZsG gene the ZsGreen gene contained in the pZsGreen1-N1 vector (Clontech) was used.
- the nucleotide sequence of the ZsG gene is shown in SEQ ID NO: 14.
- the codon composition of the ZsG gene and the codon composition of the RV NSP1 gene were examined, and the codon composition of the ZsG gene was modified to be close to the codon composition of the RV NSP1 gene based on the codon compositions of both genes obtained.
- the obtained modified ZsG gene is referred to as "Rv-ZsG gene”.
- FIG. 6 shows the base sequence of the ZsG gene (SEQ ID NO: 14) and the base sequence of the Rv-ZsG gene (SEQ ID NO: 15), respectively.
- 203 bases are changed in the total length of 696 bases of the ZsG gene.
- the GC content of the original ZsG gene was 63%, but the GC content of the Rv-ZsG gene dropped to 38%, approaching the GC content of the RVNSP1 gene (31%) (see Table 7).
- the experiment was carried out in the same manner as in Example 1 except that the ZsG gene and the Rv-ZsG gene were used instead of the NLuc gene and the Rv-NLuc gene. That is, the ZsG gene and Rv-ZsG gene amplified by PCR were inserted between the 128th and 129th positions of the NSP1 gene (SEQ ID NO: 5) of pT7-NSP1SA11, respectively, and the ZsG gene-inserted NSP1 gene expression plasmid (pT7-NSP1SA11) was inserted.
- rsSA11-ZsG an Rv-ZsG gene expression plasmid
- ZsG an Rv-ZsG gene expression plasmid
- ZsG an Rv-ZsG gene expression plasmid
- rsSA11-ZsG Artificial recombinant rotavirus prepared using 11 types of plasmids containing pT7-NSP1SA11-ZsG
- rsSA11-ZsG an artificial set prepared using 11 types of plasmids containing pT7-NSP1SA11-Rv-ZsG.
- the replacement rotavirus is called "rsSA11-Rv-ZsG”.
- the obtained rsSA11-ZsG and rsSA11-RvZsG were passaged up to P10, respectively, and viral genomic RNA was extracted and subjected to SDS-PAGE.
- Example 3 Artificial recombinant rotavirus expressing red fluorescent protein
- the AsRed gene contained in the pAsRed2-N1 vector (Clontech) was used.
- the nucleotide sequence of the ZsG gene is shown in SEQ ID NO: 16.
- the codon composition of the AsR gene and the codon composition of the RV NSP1 gene were examined, and the codon composition of the AsR gene was modified to be close to the codon composition of the RV NSP1 gene based on the codon compositions of both genes obtained.
- the obtained modified AsR gene is referred to as "Rv-AsR gene”.
- FIG. 10 shows the nucleotide sequence of the AsR gene (SEQ ID NO: 16) and the nucleotide sequence of the Rv-AsR gene (SEQ ID NO: 17), respectively.
- the Rv-AsR gene has 198 bases changed in the total length of 699 bases of the AsR gene.
- the GC content of the original AsR gene was 65%, but the GC content of the Rv-AsR gene dropped to 40%, approaching the GC content of the RVNSP1 gene (31%) (see Table 8).
- the experiment was carried out in the same manner as in Example 1 except that the AsR gene and the Rv-AsR gene were used instead of the NLuc gene and the Rv-NLuc gene. That is, the AsR gene and the Rv-AsR gene amplified by PCR were inserted between the 128th and 129th positions of the NSP1 gene (SEQ ID NO: 5) of pT7-NSP1SA11, respectively, and the AsR gene-inserted NSP1 gene expression plasmid (pT7-NSP1SA11) was inserted.
- -AsR) and Rv-AsR gene expression plasmid (referred to as pT7-NSP1SA11-Rv-AsR) were prepared (see FIG.
- rsSA11-AsR Artificial recombinant rotavirus prepared using 11 types of plasmids containing pT7-NSP1SA11-AsR is called “rsSA11-AsR”, and an artificial set prepared using 11 types of plasmids containing pT7-NSP1SA11-Rv-AsR.
- the replacement rotavirus is called "rsSA11-Rv-AsR”.
- the obtained rsSA11-AsR and rsSA11-Rv-AsR were passaged up to P10, respectively, and viral genomic RNA was extracted and subjected to SDS-PAGE.
- Example 4 Expression level of modified foreign gene-derived protein
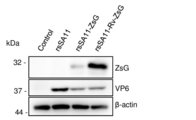
- 4-1 Materials and Methods rsSA11-ZsG and rsSA11-Rv-ZsG expressing the green fluorescent protein prepared in Example 2 and wild-type SA11 were used. Prepare MA104 cells confluent in a 12-well plate, infect each virus with MOI (Multiplicity of Infection) 0.5 PFU / cells, and observe the expression of ZsG and Rv-ZsG 24 hours after infection with a fluorescence microscope. , Quantified by Western blotting.
- MOI Multiplicity of Infection
- MA104 cells confluent in a 12-well plate were prepared, each virus was infected with MOI: 0.01 PFU / cells, and cultured in FBS-free DMEM containing 0.5 ⁇ g / ml trypsin. Twenty-four hours after infection, cells were frozen and thawed twice and virus titers in cell disruptions were measured.
- Example 1 Expression level of modified foreign gene-derived protein
- the rsSA11-AsR and rsSA11-Rv-AsR expressing the red fluorescent protein prepared in Example 3 were used.
- MA104 cells confluent in a 12-well plate were prepared, each virus was infected with MOI (Multiplicity of Infection) 0.5 PFU / cells, and AsR and Rv-AsR 24 hours after infection. The expression was observed with a fluorescence microscope.
- MOI Multiplicity of Infection
- Example 5 Artificial recombinant rotavirus expressing a foreign gene having a base length of 1.6 kbp
- 5-1 Materials and methods (1) Foreign gene As a foreign gene, the codon composition of the Akaluc gene (GenBank ACCESSION: LC320664, 1653 bp), which is a variant of the firefly luciferase (FLuc) gene, is close to that of the RV NSP1 gene.
- the codon composition of the modified Akaluc gene (hereinafter referred to as “Rv-Akaluc gene”) and the norovirus VP1 gene (GenBank ACCESSION: KM268107, 1623 bp) was modified to be close to the codon composition of the RV NSP1 gene.
- a modified norovirus VP1 gene (hereinafter referred to as "Rv-NoV VP1 gene") was used.
- the nucleotide sequence of the modified Akaluc gene is shown in SEQ ID NO: 18, and the nucleotide sequence of the modified norovirus VP1 gene is shown in SEQ ID NO: 19.
- Non-Patent Document 2 a deletion of 722 bp was added to the NSP1 gene, and the Rv-Akaluc gene-inserted NSP1 gene expression plasmid (referred to as pT7-NSP1SA11-Rv-Akaluc) and the Rv-NoV VP1 gene-inserted NSP1 A gene expression plasmid (referred to as pT7-NSP1SA11-Rv-NoV VP1) was prepared (see FIG. 18).
- the artificially recombined rotavirus produced in the above is called "rsSA11-Rv-NoV VP1".
- the obtained rsSA11-Rv-Akaluc and rsSA11-Rv-NoV VP1 were passaged up to P10, respectively, and viral genomic RNA was extracted and subjected to SDS-PAGE.
- a recombinant rotavirus that stably retains a foreign gene having a base length of 1 kbp or more is prepared by bringing the codon composition close to the codon composition of rotavirus. Shown that you can
- Virus Pteropine orthoreovirus (hereinafter referred to as "PRV") belongs to the same Reoviridae family as rotavirus, L1, L2, L3, M1, M2, M3, S1, S2. It has a 10-segment double-stranded RNA consisting of S3 and S4.
- a plasmid containing the cDNAs of the 10 segmental RNA genomes of PRV was prepared in the same manner as the preparation of the plasmid (segmented RNA genome expression vector) containing each segmental RNA genome expression cassette of Rotavirus in Example 1.
- rsMB-Rv-ZsG-2A-FAST 9 types of plasmids containing wild-type genes other than the S1 gene and plasmids containing the S1-Rv-ZsG-2A-FAST gene were transfected into BHK-T7 / P5 cells and passaged into monkey MA104 cells.
- An artificially recombinant bat leovirus (referred to as "rsMB-Rv-ZsG-2A-FAST") was obtained.
- Example 7 Artificial recombinant mammalian reovir that expresses green fluorescent protein
- Virus Mammalian orthoreovirus belongs to the same Reoviridae family as rotavirus, L1, L2, L3, M1, M2, M3, S1, S2. It has a 10-segment double-stranded RNA consisting of S3 and S4.
- a plasmid containing the cDNAs of the 10 segmental RNA genomes of MRV was prepared in the same manner as the preparation of the plasmid (segmented RNA genome expression vector) containing each segmental RNA genome expression cassette of Rotavirus in Example 1.
- (2) Green fluorescent protein gene As the green fluorescent protein, the Mrv-ZsG gene was used in which the codon composition of the ZsG gene and the ZsG gene was approximated to the codon composition of the MRV L1 gene. 23 (A) to (U) show the codon composition of each amino acid in the MRV L1 gene, ZsG gene and Mrv-ZsG gene. The GC content of the original ZsG gene was 63%, but the GC content of the Mrv-ZsG gene dropped to 43%, approaching the GC content of the MRV L1 gene (46%) (see Table 9).
- the ZsG gene and Mrv-ZsG gene were inserted into the MRV L1 gene.
- the MRV L1 gene is a gene encoding the lamda3 protein, and the 2A gene encoding a self-cleaving peptide is inserted downstream of the ZsG gene and MRV-ZsG gene to L1-ZsG-2A-Lamda3 and L1-Mrv-ZsG.
- -2A-Lamda3 was prepared (see FIG. 24).
- rsMRV-Mrv-ZsG 9 types of plasmids containing wild-type genes other than the L1 gene and plasmids containing the L1-Mrv-ZsG-2A-Lamda3 gene were transfected into BHK-T7 / P5 cells and passaged into mouse L929 cells.
- An artificially recombinant mammalian leovirus (referred to as "rsMRV-Mrv-ZsG”) was obtained.
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Organic Chemistry (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Wood Science & Technology (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Virology (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biophysics (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN202080045560.2A CN114040979B (zh) | 2019-06-21 | 2020-06-19 | 稳定地保持外源基因的人工重组rna病毒的制作方法 |
| US17/619,899 US12305192B2 (en) | 2019-06-21 | 2020-06-19 | Method for producing artificial recombinant RNA virus stably retaining foreign gene |
| EP20825608.1A EP3988650A4 (en) | 2019-06-21 | 2020-06-19 | METHOD OF PRODUCING AN ARTIFICIAL RECOMBINANT RNA VIRUS THAT KEEPS A FOREIGN GENE STABLE |
| JP2021526915A JP7517704B2 (ja) | 2019-06-21 | 2020-06-19 | 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2019115532 | 2019-06-21 | ||
| JP2019-115532 | 2019-06-21 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2020256099A1 true WO2020256099A1 (ja) | 2020-12-24 |
Family
ID=74040505
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2020/024131 Ceased WO2020256099A1 (ja) | 2019-06-21 | 2020-06-19 | 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US12305192B2 (https=) |
| EP (1) | EP3988650A4 (https=) |
| JP (1) | JP7517704B2 (https=) |
| CN (1) | CN114040979B (https=) |
| WO (1) | WO2020256099A1 (https=) |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2008507290A (ja) * | 2004-08-03 | 2008-03-13 | ジーンアート・アクチエンゲゼルシャフト | 誘導性遺伝子発現 |
| WO2018062199A1 (ja) | 2016-09-27 | 2018-04-05 | 国立大学法人大阪大学 | 人工組換えロタウイルスの作製方法 |
| WO2018060368A2 (en) * | 2016-09-28 | 2018-04-05 | Bavarian Nordic A/S | Compositions and methods for enhancing the stability of transgenes in poxviruses |
Family Cites Families (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| IL140441A0 (en) * | 1998-09-29 | 2002-02-10 | Maxygen Inc | Shuffling of codon altered genes |
| CN1934260B (zh) * | 2004-01-22 | 2013-03-13 | 株式会社载体研究所 | 利用含有巨细胞病毒增强子和鸡β-肌动蛋白启动子的杂合启动子制造负链RNA病毒载体的方法 |
| ATE453716T1 (de) | 2004-08-03 | 2010-01-15 | Geneart Ag | Verfahren zur modulation der genexpression durch änderung des cpg gehalts |
| JP2009519715A (ja) * | 2005-12-16 | 2009-05-21 | ザ ユーエービー リサーチ ファウンデーション | ウイルスベクターを使用する制御された遺伝子発現に関する組成物および方法 |
| CN101245350B (zh) * | 2007-02-17 | 2011-12-28 | 王健伟 | 密码子优化的轮状病毒蛋白的编码核苷酸序列、其重组体及其应用 |
| KR101621100B1 (ko) * | 2007-03-30 | 2016-05-13 | 더 리서치 파운데이션 오브 스테이트 유니버시티 오브 뉴욕 | 백신에 유용한 약독화 바이러스 |
| GB0800272D0 (en) * | 2008-01-08 | 2008-02-13 | Plant Bioscience Ltd | Protein expression systems |
| CN101502650B (zh) * | 2009-03-10 | 2011-12-28 | 邢益平 | 密码子优化的乙型肝炎核酸疫苗 |
| CN104862335A (zh) | 2009-07-31 | 2015-08-26 | 诺华股份有限公司 | 反向遗传系统 |
| TWI696700B (zh) * | 2015-01-16 | 2020-06-21 | 國立研究開發法人產業技術總合研究所 | 使用具有匿跡性之rna的基因表現系統及含有該rna的基因導入、表現載體 |
| CN104830874B (zh) * | 2015-04-16 | 2017-11-21 | 南京医科大学第一附属医院 | 密码子优化的严重发热伴血小板减少综合征病毒核蛋白基因及其核酸疫苗 |
| JP7054143B2 (ja) * | 2016-05-11 | 2022-04-13 | 国立大学法人大阪大学 | キメラ抗原受容体、及びその利用 |
-
2020
- 2020-06-19 CN CN202080045560.2A patent/CN114040979B/zh active Active
- 2020-06-19 US US17/619,899 patent/US12305192B2/en active Active
- 2020-06-19 EP EP20825608.1A patent/EP3988650A4/en active Pending
- 2020-06-19 WO PCT/JP2020/024131 patent/WO2020256099A1/ja not_active Ceased
- 2020-06-19 JP JP2021526915A patent/JP7517704B2/ja active Active
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2008507290A (ja) * | 2004-08-03 | 2008-03-13 | ジーンアート・アクチエンゲゼルシャフト | 誘導性遺伝子発現 |
| WO2018062199A1 (ja) | 2016-09-27 | 2018-04-05 | 国立大学法人大阪大学 | 人工組換えロタウイルスの作製方法 |
| WO2018060368A2 (en) * | 2016-09-28 | 2018-04-05 | Bavarian Nordic A/S | Compositions and methods for enhancing the stability of transgenes in poxviruses |
Non-Patent Citations (27)
| Title |
|---|
| "GenBank", Database accession no. NC006998 |
| ADACHI A ET AL.: "Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone", JOURNAL OF VIROLOGY, vol. 59, no. 2, 1986, pages 284 - 291 |
| BOYCE MCELMA CCROY P: "Development of reverse genetics systems for bluetongue virus: recovery of infectious virus from synthetic RNA transcripts", JOURNAL OF VIROLOGY, vol. 82, no. 17, 2008, pages 8339 - 8348, XP002513548, DOI: 10.1128/JVI.00808-08 |
| BRIDGEN AELLIOTT RM: "Rescue of a segmented negative-strand RNA virus entirely from cloned complementary DNAs", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 93, no. 26, 1996, pages 15400 - 15404, XP002224465, DOI: 10.1073/pnas.93.26.15400 |
| CHAUDHRY YSKINNER MAGOODFELLOW IG: "Recovery of genetically defined murine norovirus in tissue culture by using a fowlpox virus expressing T7 RNA polymerase", THE JOURNAL OF GENERAL VIROLOGY, vol. 88, 2007, pages 2091 - 2100 |
| FISCHER, M. D. ET AL.: "C odon-Optimized RPGR Improves Stability and Efficacy of AAV8 Gene Therapy in Two Mouse Models of X-Linked Retinitis Pigmentosa", MOL. THER., vol. 25, no. 8, 2017, pages 1854 - 1865, XP055399604, DOI: 10.1016/j.ymthe.2017.05.005 * |
| GUSTAFSSON ET AL., TRENDS BIOTECHNOL., vol. 22, no. 7, July 2004 (2004-07-01), pages 346 - 353 |
| KANAI ET AL., JOURNAL OF VIROLOGY, vol. 93, no. 4, 5 February 2019 (2019-02-05), pages e01774 - 18 |
| KANAI ET AL., PROC NATL ACAD SCI U S A, vol. 114, no. 9, 28 February 2017 (2017-02-28), pages 2349 - 2354 |
| KOBAYASHI T ET AL.: "A plasmid-based reverse genetics system for animal double-stranded RNA viruses", CELL HOST & MICROBE, vol. 1, no. 2, 2007, pages 147 - 157, XP002513546, DOI: 10.1016/J.CHOM.2007.03.003 |
| KOLYKHALOV AA ET AL.: "Transmission of hepatitis C by intrahepatic inoculation with transcribed RNA", SCIENCE, vol. 277, no. 5325, 1997, pages 570 - 574, XP002734276, DOI: 10.1126/science.277.5325.570 |
| LAI CJZHAO BTHORI HBRAY M: "Infectious RNA transcribed from stably cloned full-length cDNA of dengue type 4 virus", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 88, no. 12, 1991, pages 5139 - 5143, XP002073934, DOI: 10.1073/pnas.88.12.5139 |
| MADESIS, P. ET AL.: "A hepatitis C virus core polypeptide expressed in chloroplasts detects anti-core antibodies in infected human sera", J. BIOTECHNOL ., vol. 145, 2010, pages 377 - 386, XP026892462, DOI: 10.1016/j.jbiotec.2009.12.001 * |
| MATSUO ET AL., BIOCHEM BIOPHYS RES COMMUN, vol. 340, no. 1, 2006, pages 200 - 208 |
| MUNDT EVAKHARIA VN: "Synthetic transcripts of double-stranded Birnavirus genome are infectious", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 93, no. 20, 1996, pages 11131 - 11136, XP002044624, DOI: 10.1073/pnas.93.20.11131 |
| NEUMANN G ET AL.: "Generation of influenza A viruses entirely from cloned cDNAs", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 96, no. 16, 1999, pages 9345 - 9350 |
| RACANIELLO VRBALTIMORE D: "Cloned poliovirus complementary DNA is infectious in mammalian cells", SCIENCE, vol. 214, no. 4523, 1981, pages 916 - 919, XP001057398, DOI: 10.1126/science.6272391 |
| RADECKE F ET AL.: "Rescue of measles viruses from cloned DNA", EMBO J, vol. 14, no. 23, 1995, pages 5773 - 5784, XP002022952 |
| SANCHEZ ABDE LA TORRE JC: "Rescue of the prototypic Arenavirus LCMV entirely from plasmid", VIROLOGY, vol. 350, no. 2, 2006, pages 370 - 380, XP024896500, DOI: 10.1016/j.virol.2006.01.012 |
| SCHNEIDER USCHWEMMLE MSTAEHELI P: "Genome trimming: a unique strategy for replication control employed by Borna disease virus", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 102, no. 9, 2005, pages 3441 - 3446, XP008153168, DOI: 10.1073/pnas.0405965102 |
| SCHNELL MJMEBATSION TCONZELMANN KK: "Infectious Rabies Viruses from Cloned Cdna", EMBO J, vol. 13, no. 18, 1994, pages 4195 - 4203 |
| See also references of EP3988650A4 |
| T ANAKA, M. ET AL.: "Effects of codon optimization on the mRNA levels of heterologous genes in filamentous fungi", APPL. MICROBIOL. BIOTECHNOL., vol. 98, 2014, pages 3859 - 3867, XP035328382, DOI: 10.1007/s00253-014-5609-7 * |
| TOKUOKA, MASAFUMI, TANAKA MIZUKI, ONO KAZUHISA, TAKAGI SHINOBU, SHINTANI TAKAHIRO, GOMI KATSUYA: "Codon Optimization Increases Steady-State mRNA Levels in Aspergillus oryzae Heterologous Gene Expression", APPL. ENVIRON. MICROBIOL ., vol. 74, no. 21, 2008, pages 6538 - 6546, XP055773591, DOI: 10.1128/AEM.01354-08 * |
| VOLCHKOV VE ET AL.: "Recovery of infectious Ebola virus from complementary DNA: RNA editing of the GP gene and viral cytotoxicity", SCIENCE, vol. 291, no. 5510, 2001, pages 1965 - 1969, XP002404372, DOI: 10.1126/science.1057269 |
| YOUNT BCURTIS KMBARIC RS: "Strategy for systematic assembly of large RNA and DNA genomes: transmissible gastroenteritis virus model", JOURNAL OF VIROLOGY, vol. 74, no. 22, 2000, pages 10600 - 10611, XP002203456, DOI: 10.1128/JVI.74.22.10600-10611.2000 |
| YUN SIKIM SYRICE CMLEE YM: "Development and application of a reverse genetics system for Japanese encephalitis virus", JOURNAL OF VIROLOGY, vol. 77, no. 11, 2003, pages 6450 - 6465, XP002367285, DOI: 10.1128/JVI.77.11.6450-6465.2003 |
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2020256099A1 (https=) | 2020-12-24 |
| CN114040979B (zh) | 2024-06-28 |
| CN114040979A (zh) | 2022-02-11 |
| US20220356491A1 (en) | 2022-11-10 |
| EP3988650A1 (en) | 2022-04-27 |
| EP3988650A4 (en) | 2022-09-21 |
| US12305192B2 (en) | 2025-05-20 |
| JP7517704B2 (ja) | 2024-07-17 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6944213B2 (ja) | 人工組換えロタウイルスの作製方法 | |
| CN1118569C (zh) | 具有自主复制能力的负链rna病毒载体 | |
| Bukreyev et al. | Recovery of infectious respiratory syncytial virus expressing an additional, foreign gene | |
| WO1997016539A1 (en) | Recombinant sendai virus | |
| CN107190022B (zh) | 一种快速构建禽传染性支气管炎病毒反向遗传株的方法 | |
| JP7298906B2 (ja) | 光制御性のウイルスタンパク質、その遺伝子、及びその遺伝子を含むウイルスベクター | |
| CN111996174A (zh) | 猪繁殖与呼吸综合征病毒及其克隆载体和插入基因方法 | |
| CN101970645A (zh) | 呼肠弧病毒家族病毒的疫苗病毒株的制造方法 | |
| JP2019050796A (ja) | 高効率なロタウイルスの人工合成法 | |
| CN101586120A (zh) | 狂犬病病毒Flury-LEP疫苗株反向遗传操作系统及LEP绿色荧光蛋白重组病毒载体 | |
| JP7517704B2 (ja) | 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 | |
| Molouki et al. | Rescue of recombinant Newcastle disease virus: A promising vector with two decades of intensive vaccine research | |
| Enjuanes et al. | Virus-based vectors for gene expression in mammalian cells: Coronavirus | |
| CN119432748A (zh) | 拉沙病毒细胞模型及其在抗病毒药物分析、病毒感染机制及生命周期中的应用 | |
| JP3638019B2 (ja) | 組換え体センダイウイルス | |
| JP3732204B2 (ja) | 自律複製能を有する(−)鎖rnaウイルスベクター | |
| JP3991339B2 (ja) | 組換え体センダイウイルス | |
| Desselberger | A helper virus-independent, plasmid only-based reverse genetics system for rotavirus | |
| JPWO2007007921A1 (ja) | 分節ゲノム型組換えモノネガウイルスベクター | |
| CN113584063A (zh) | 一种基于转录调控序列重布的反向遗传操作方法及其应用 | |
| Theriault | The role of reverse genetics systems in the study of filovirus pathogenicity | |
| FR2841258A1 (fr) | Procede et preparation de mononegavirus recombinants et leurs applications | |
| JP2005102702A (ja) | 組換え体センダイウイルス |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 20825608 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2021526915 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2020825608 Country of ref document: EP Effective date: 20220121 |
|
| WWG | Wipo information: grant in national office |
Ref document number: 17619899 Country of ref document: US |