WO2020256099A1 - 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 - Google Patents
外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 Download PDFInfo
- Publication number
- WO2020256099A1 WO2020256099A1 PCT/JP2020/024131 JP2020024131W WO2020256099A1 WO 2020256099 A1 WO2020256099 A1 WO 2020256099A1 JP 2020024131 W JP2020024131 W JP 2020024131W WO 2020256099 A1 WO2020256099 A1 WO 2020256099A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- gene
- virus
- zsg
- codon
- rna virus
- Prior art date
Links
Images






















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
- C12N15/867—Retroviral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2720/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsRNA viruses
- C12N2720/00011—Details
- C12N2720/12011—Reoviridae
- C12N2720/12311—Rotavirus, e.g. rotavirus A
- C12N2720/12321—Viruses as such, e.g. new isolates, mutants or their genomic sequences
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2720/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsRNA viruses
- C12N2720/00011—Details
- C12N2720/12011—Reoviridae
- C12N2720/12311—Rotavirus, e.g. rotavirus A
- C12N2720/12322—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2720/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsRNA viruses
- C12N2720/00011—Details
- C12N2720/12011—Reoviridae
- C12N2720/12311—Rotavirus, e.g. rotavirus A
- C12N2720/12341—Use of virus, viral particle or viral elements as a vector
- C12N2720/12343—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2800/00—Nucleic acids vectors
- C12N2800/22—Vectors comprising a coding region that has been codon optimised for expression in a respective host
Definitions
- the present invention relates to a method for producing an artificially recombinant RNA virus that stably retains a foreign gene.
- Rotavirus which belongs to the Reoviridae family, causes acute gastroenteritis in infants, and about 200,000 people die from rotavirus infection annually.
- the development of a technique for artificially producing an arbitrary virus has been delayed, but recently, the present inventors have succeeded in developing a practical artificial synthesis method for rotavirus (Patent Document 1, Patent Document 1, Non-Patent Document 1).
- Patent Document 1 Patent Document 1, Non-Patent Document 1
- Patent Document 1 Patent Document 1
- Non-Patent Document 2 Non-Patent Document 2
- Non-Patent Document 3 When expressing a foreign gene in a heterologous host, a codon optimization technique that brings the codon composition of the foreign gene closer to the codon composition of the host gene has been used for a long time for the purpose of improving the expression of the foreign gene (Non-Patent Document 3). ), Numerous codon optimization software is provided. However, it has not been known that the codon optimization technique can be applied to the stable retention of foreign genes introduced into artificially recombined RNA virus.
- An object of the present invention is to provide a method for producing an artificially recombined RNA virus that stably retains a foreign gene and a method for stably retaining a foreign gene introduced into the artificially recombined RNA virus.
- the present invention includes the following inventions in order to solve the above problems.
- a method for producing an artificially recombinant RNA virus that stably retains a foreign gene (1) A step of obtaining a foreign gene having a codon composition modified to approximate the codon composition of an RNA virus gene. It is characterized by including (2) a step of inserting the foreign gene obtained in (1) into the genome of the RNA virus, and (3) a step of artificially synthesizing an artificially recombined RNA virus using the reverse genetics method. Manufacturing method.
- RNA virus is a virus belonging to the Reoviridae family.
- virus belonging to the Reoviridae family is a virus belonging to the genus Rotavirus or the genus Ortholeovirus.
- codon composition of the foreign gene is approximated within ⁇ 30% of the codon composition of the RNA virus gene.
- RNA virus that stably retains a foreign gene for a long period of time.
- ZsG gene Green fluorescent protein gene
- Rv-ZsG gene codon-modified green fluorescent protein gene
- AsR gene red fluorescent protein gene
- Rv-ZsG gene codon-modified green fluorescent protein gene
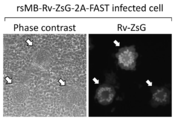
- Is an observation image of immunostaining using MRV-specific antibody the center is an observation image of green fluorescent protein-expressing cells, and the right is a composite image of both.
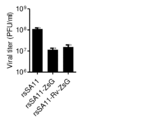
- An artificially recombined mammalian leovirus having a green fluorescent protein gene (ZsG gene) and an artificially recombined mammalian leovirus having a codon-modified green fluorescent protein gene (Mrv-ZsG gene) are passaged 1 to 3 times to obtain a foreign gene. It is a figure which shows the result of having examined the stability of.
- the present invention provides a method for producing an artificially recombined RNA virus that stably retains a foreign gene (hereinafter, referred to as "method for producing the present invention”).
- the production method of the present invention may include the following steps (1), (2) and (3).
- (1) Step of acquiring a foreign gene having a codon composition modified to resemble the codon composition of an RNA virus gene (2) Step of inserting the foreign gene obtained in (1) into the RNA virus genome (3) Step of obtaining artificially recombinant RNA virus using reverse genetics method
- the RNA virus may be a double-stranded RNA virus, a single-strand plus-strand RNA virus, or a single-strand minus-strand RNA virus.
- double-stranded RNA virus examples include viruses belonging to the family Reoviridae and Birnaviridae.
- positive-strand RNA virus examples include viruses belonging to the family Coronaviridae, Picornavirus, Togaviridae, Flaviviridae, Calisivirus, Astrovirus, and the like.
- Examples of the single-stranded minus-strand RNA virus include viruses belonging to the family Paramyxoviridae, Rabdovirus, Philovirus, Orthomyxoviridae, Arenavirus, Bunyavirus, and the like.
- the RNA virus may be a virus belonging to the Reoviridae family.
- Viruses belonging to the Reoviridae family are viruses that have a straight-chain double-stranded RNA segmented into 10 to 12 lines in their genome, and their virions show a regular icosahedron structure with a diameter of 60 to 80 nm.
- Viruses belonging to the Reoviridae family include mammalian ortholeovirus (also called mammalian reoviridae), Nelsonbey ortholeovirus (also called Nelsonbeyreovirus or bat leovirus), ortholeovirus such as trileovirus.
- Virus genus (Genus Orthoreovirus), African horse epidemic virus, Orbivirus genus such as blue tongue virus (Genus Orbivirus), Rotavirus genus such as Rotavirus (Genus Rotavirus), Cortivirus genus such as Colorado tick fever virus (Genus Coltivirus) , Aquareovirus such as Aquareovirus A (GenusAquareovirus), Cypovirus such as cytoplasmic polymorphic disease virus (GenusCypovirus), Fijivirus genus such as rice southern black streak dwarf disease virus (Genus Fijivirus), rice Includes the genus Phytoreovirus such as dwarf disease virus and the genus Oryzavirus such as Rice ragged stunt virus.
- the virus belonging to the Reoviridae family may be a virus belonging to the genus Rotavirus or a virus belonging to the genus Ortholeovirus.
- the foreign gene is not particularly limited.
- the foreign gene may be an animal gene, a plant gene, a fungal gene, a bacterial gene, or a viral gene.
- the base length of the foreign gene is not particularly limited, and may be 10 bp or more, 100 bp or more, 500 bp or more, 1000 bp or more, or 1500 bp or more. , 2000 bp or more, 3000 bp or more, 4000 bp or more, 5000 bp or more.
- the base length of the foreign gene may be 500 bp or less, 1000 bp or less, 1500 bp or less, 2000 bp or less, 3000 bp or less, 4000 bp or less. It may be present, and may be 5000 bp or less.
- the protein encoded by the foreign gene is not particularly limited.
- the protein encoded by the foreign gene may be a vaccine antigen.
- the vaccine antigen include norovirus antigen, adenovirus antigen, hepatitis A antigen, sapovirus antigen, limb virus antigen, enterovirus antigen, HIV antigen, salmonella antigen, campylobacter antigen, enteritis vibrio antigen, Escherichia coli O157 antigen, and cholera.
- examples thereof include antigens, intestinal typhoid antigens, and diarrhea antigens.
- These vaccine antigens may be epitope peptides.
- a foreign gene having a codon composition modified to approximate the codon composition of the RNA virus gene is obtained.
- the codon composition of the reference RNA virus gene may be the codon composition of the gene of the RNA virus that is artificially synthesized by incorporating the foreign gene with the modified codon, or artificially by incorporating the foreign gene with the modified codon.
- the codon composition of the gene of the RNA virus different from that of the RNA virus to be synthesized may be used.
- different RNA viruses are systematically similar to RNA viruses that are artificially synthesized by incorporating foreign genes with modified codons (eg, viruses belonging to the same family). , Viruses belonging to the same genus, viruses of different species, etc.).
- the codon composition of the reference RNA virus gene may be the codon composition of all the genes of the reference RNA virus, or may be the codon composition of some genes of the reference RNA virus.
- the codon composition of some genes of the RNA virus is used as a reference, the codon composition of one gene may be used as a reference, or the codon composition of two or more genes may be used as a reference.
- the codon composition of the reference RNA virus gene can be created based on the genetic information of the RNA virus registered in a known gene database (for example, GenBank).
- Table 1 shows the genomic composition of all genes of rotavirus SA11 strain.
- Table 2 shows the genomic composition of the rotavirus SA11 strain NSP1 gene.
- Table 3 shows the genomic composition of all genes of the bat reovirus (Pteropine orthoreovirus) MB strain.
- Table 4 shows the genomic composition of all mammalian reoviridae T1L genes.
- the codon compositions shown in Tables 1 to 4 can be used as the reference codon composition in the production method of the present invention. Among them, it can be used as a reference codon composition when a foreign gene is expressed by using a virus belonging to the Reoviridae family, particularly an artificial recombinant virus belonging to the genus Rotavirus and the genus Ortholeovirus.
- Approximating the codon composition of a foreign gene to the codon composition of a reference RNA virus gene means that it frequently appears in an RNA virus gene based on a codon corresponding to the same amino acid without mutating the amino acid of the foreign gene. It means changing to a codon.
- the degree of approximation to the codon composition of the reference RNA virus gene is not particularly limited as long as the purpose of stably retaining the foreign gene can be achieved.
- the codon composition of the foreign gene may be modified to be within ⁇ 35% of the codon composition of the reference RNA virus gene, or within ⁇ 30% of the codon composition of the reference RNA virus gene.
- the codon composition of the foreign gene may be modified so as to be within ⁇ 25% of the codon composition of the reference RNA virus gene.
- the codon composition of the foreign gene may be modified to be within ⁇ 20% of the codon composition of the RNA virus gene to be used, and should be within ⁇ 15% of the codon composition of the reference RNA virus gene.
- the codon composition of the foreign gene may be modified, or the codon composition of the foreign gene may be modified so as to be within ⁇ 10% of the codon composition of the reference RNA virus gene.
- a foreign gene with a modified codon composition can be obtained by DNA synthesis based on the modified base sequence.
- a mutation can be introduced into an existing foreign gene DNA and obtained by using a site-specific mutation introduction method based on PCR.
- the foreign gene obtained in (1) is inserted into the RNA virus genome.
- the position at which the foreign gene is inserted is not particularly limited as long as it does not interfere with the artificial synthesis of the artificially recombined RNA virus using the reverse genetics method.
- a foreign gene when a foreign gene is introduced into an artificially recombined rotavirus, it may be inserted into the NSP1 gene, the NSP3 gene, or the NSP5 gene.
- a foreign gene into a recombinant mammalian reovir it may be inserted into the L1 gene or the S1, S2, and S4 genes.
- a foreign gene when introduced into an artificially recombined influenza virus, it may be inserted into the NS1 gene, the NA gene, or the PA gene.
- a foreign gene into an artificially recombinant virus of the genus Alphavirus (Sindbis virus, Chikungunya virus, etc.)
- it when introducing a foreign gene into an artificially recombinant virus of the genus Alphavirus (Sindbis virus, Chikungunya virus, etc.), it may be inserted into the NSP3 gene or the capsid gene.
- introducing a foreign gene into an artificially recombined norovirus it may be inserted into the ORF1 gene.
- a foreign gene when introduced into an artificially recombinant virus of the genus Flavivirus (dengue virus, hepatitis C virus, etc.), it may be inserted into the NS5A gene or the capsid gene.
- a foreign gene into an artificially recombinant virus of rhabdovirus rabies virus, vesicular stomatitis virus
- it when introducing a foreign gene into an artificially recombinant virus of the genus Picornavirus (poliovirus, foot-and-mouth disease virus, etc.), it may be inserted into the P1 gene.
- a foreign gene into an artificially recombined Sendai virus it may be inserted into an NP gene. Insertion of a foreign gene into the viral genome can be performed using a known gene recombination technique.
- an artificially recombinant RNA virus is artificially synthesized using the reverse genetics method.
- the method for artificially synthesizing an artificially recombined RNA virus using the reverse genetics method may be appropriately selected from known methods according to the type of RNA virus. Methods for synthesizing artificially recombinant rotavirus using the reverse genetics method are described in, for example, Patent Document 1, Non-Patent Document 1, Non-Patent Document 2, and the like. A method for synthesizing artificially recombined poliovirus using the reverse genetics method is described in, for example, Reference 1 below.
- a method for synthesizing an artificially recombined rabies virus using the reverse genetics method is described in, for example, Reference 2 below.
- a method for synthesizing an artificially recombined measles virus using the reverse genetics method is described in, for example, Reference 3 below.
- a method for synthesizing an artificially recombined bunyavirus using the reverse genetics method is described in, for example, Reference 4 below.
- a method for synthesizing an artificially recombinant infectious bursal disease virus using the reverse genetics method is described in, for example, Reference 5 below.
- a method for synthesizing artificially recombined hepatitis C virus using the reverse genetics method is described in, for example, Reference 6 below.
- a method for synthesizing an artificially recombined influenza virus using the reverse genetics method is described in, for example, Reference 7 below.
- a method for synthesizing an artificially recombined coronavirus using the reverse genetics method is described in, for example, Reference 8 below.
- a method for synthesizing an artificially recombined Ebola virus using the reverse genetics method is described in, for example, Reference 9 below.
- a method for synthesizing an artificially recombined borunavirus using the reverse genetics method is described in, for example, Reference 10 below.
- a method for synthesizing an artificially recombined arenavirus using the reverse genetics method is described in, for example, Reference 11 below.
- a method for synthesizing an artificially recombined reovir using the reverse genetics method is described in, for example, Reference 12 below.
- a method for synthesizing an artificially recombined dengue virus using the reverse genetics method is described in, for example, Reference 13 below.
- a method for synthesizing an artificially recombined norovirus using the reverse genetics method is described in, for example, Reference 14 below.
- a method for synthesizing artificially recombined Japanese encephalitis virus using the reverse genetics method is described in, for example, Reference 15 below.
- a method for synthesizing an artificially recombined bluetongue virus using the reverse genetics method is described in, for example, 16.
- a method for synthesizing an artificially recombined human immunodeficiency virus using the reverse genetics method is described in, for example, Reference 17 below.
- the artificially recombined RNA virus produced by the production method of the present invention can stably retain a foreign gene for a long period of time and stably express a foreign gene product for a long period of time. Therefore, for example, if a vaccine antigen is used as the foreign gene. , Can be suitably used as a virus vaccine.
- a vaccine antigen is used as the foreign gene.
- normal genes are used for genetic diseases of the gastrointestinal tract (colon cancer, ulcerative colitis, Crohn's disease, celiac disease, nonspecific multiple small intestinal ulcer disease, etc.) derived from (believed to be) genetic abnormalities. It can also be applied to gene therapy by delivering it to abnormal cells as a foreign gene.
- the present invention provides a method for stably retaining a foreign gene introduced into an artificially synthesized RNA virus artificially synthesized by using a reverse genetics method (hereinafter, referred to as "the stable retention method of the present invention").
- the stable retention method of the present invention is a method characterized in that the codon composition of a foreign gene is approximated to the codon composition of an RNA virus gene.
- the method for approximating the codon composition of the foreign gene to the codon composition of the reference RNA virus gene can be the same as the method described in the production method of the present invention.
- Example 1 Artificial recombinant rotavirus expressing luciferase
- Virus Sarrotavirus SA11 strain was used. We have sequenced and registered the nucleotide sequences of each of the 11 segmental RNA genomes of this virus strain. Table 5 shows the names and GenBank ACCESSION of each of the 11 segmental RNA genomes of the Sarrotavirus SA11 strain (hereinafter referred to as "SA11 strain") used in this experiment.
- a plasmid containing each segmented RNA genome expression cassette of the SA11 strain (segmented RNA genome expression vector)
- a plasmid containing the cDNA of 11 segmental RNA genomes of the SA11 strain was prepared.
- RT-PCR was performed using double-stranded RNA extracted from the virus as a template and specific primers based on the base sequence of each segmental RNA genome.
- the obtained RT-PCR product (cDNA of each segmental RNA genome) was inserted between the T7 promoter sequence and the HDV ribozyme sequence of the p3E5 plasmid to obtain a plasmid containing the expression cassette of each segmental RNA genome (see Patent Document 1). ).
- the T7 promoter sequence is adjacent to the 5'side of the cDNA of each segmental RNA genome, the hepatitis delta virus (HDV) ribozyme sequence is adjacent to the 3'side, and the T7 terminator sequence is arranged downstream thereof.
- HDV hepatitis delta virus
- the prepared plasmids were used as pT7-VP1SA11, pT7-VP2SA11, pT7-VP3SA11, pT7-VP4SA11, pT7-VP6SA11, pT7-VP7SA11, pT7-NSP1SA11, pT7-NSP2SA11, pT7-NSP3, respectively.
- NSP4SA11 and pT7-NSP5SA11 were used as NSP4SA11 and pT7-NSP5SA11.
- Luciferase gene As the luciferase gene, the NLuc gene, which is a luciferase gene derived from Oplophorus gracilirostris, was used. The NLuc coding region is located at positions 815 to 1330 (SEQ ID NO: 12) of the pNL1.1.TK [NLuc / TK] vector (Promega, GenBank ACCESSION: KM359774, 3817 bp).
- FIG. 1 shows the codon composition of each amino acid in the RVNSP1 gene, NLuc gene, and Rv-NLuc gene.
- FIG. 2 shows the nucleotide sequence of the NLuc gene (SEQ ID NO: 12) and the nucleotide sequence of the Rv-NLuc gene (SEQ ID NO: 13), respectively.
- the Rv-NLuc gene has 70 bases changed out of the total length of 516 bases of the NLuc gene.
- the GC content of the original NLuc gene was 53%, but the GC content of the Rv-NLuc gene dropped to 40%, approaching the GC content of the RV NSP1 gene (31%) (see Table 6).
- FAST protein expression vector uses the protein coding region DNA of the Nelson baileovirus p10 gene (see GenBank ACCESSION: AB908284) as the pCAGGS plasmid (5699bp, Matsuo et al., 2006, Biochem Biophys Res Commun 340). (1): It was produced by inserting it into the BglII cleavage site of 200-208) (see Patent Document 1). The obtained Nelson baileovirus p10 expression vector is referred to as pCAG-p10.
- the capping enzyme expression vector is the protein coding region DNA of the vaccinia virus D1R gene (GenBank ACCESSION: NC006998 positions 93948 to 96482) and the protein coding region DNA of the vaccinia virus D12L gene (GenBank ACCESSION: NC006998). Positions 107332 to 108195) were inserted into the BglII cleavage site of the above pCAGGS plasmid, respectively (see Patent Document 1).
- the obtained vaccinia virus mRNA capping enzyme large subunit expression vector is referred to as pCAG-D1R
- the obtained vaccinia virus mRNA capping enzyme small subunit expression vector is referred to as pCAG-D12L.
- the NSP2 protein expression vector was prepared by inserting the protein coding region DNA of the SA11 strain NSP2 gene (GenBank ACCESSION: positions 47 to 1000 of LC178571) into the BglII cleavage site of the above pCAGGS plasmid (see Patent Document 1).
- the obtained SA11 strain NSP2 protein expression vector is referred to as pCAG-NSP2.
- the NSP5 protein expression vector was prepared by inserting the protein coding region DNA of the SA11 strain NSP5 gene (GenBank ACCESSION: positions 22 to 618 of LC178574) into the BglII cleavage site of the above pCAGGS plasmid (see Patent Document 1).
- the obtained SA11 strain NSP5 protein expression vector is referred to as pCAG-NSP5.
- NLuc gene and Rv-NLuc gene are amplified by PCR, respectively, and the amplification product is inserted between positions 128 and 129 of the NSP1 gene (SEQ ID NO: 5) of pT7-NSP1SA11. Then, the NLuc gene-inserted NSP1 gene expression plasmid (referred to as pT7-NSP1SA11-NLuc) and the Rv-NLuc gene expression plasmid (referred to as pT7-NSP1SA11-Rv-NLuc) were prepared (see FIG. 3).
- RNA genome expression vectors 0.8 ⁇ g each, FAST protein expression vector (pCAG-p10) 0.015 ⁇ g, capping enzyme expression vectors (pCAG-D1R and pCAG-D12L), NSP2 protein expression vector (pCAG-NSP2), NSP5 protein An expression vector (pCAG-NSP5), 0.8 ⁇ g each, was introduced into BHK-T7 / P5 cells using a transfection reagent (TransIT-LT1 (trade name), Miras). 2 ⁇ g of transfection reagent was used per 1 ⁇ g of DNA.
- TransIT-LT1 trade name
- BHK-T7 / P5 cells were cultured in DMEM medium containing 5% FBS, 100 units / ml penicillin and 100 ⁇ g / ml streptomycin in a 37 ° C. 5% CO 2 environment. Medium and cells were harvested 48 hours after transfection. The collected medium and cells were frozen and thawed three times to prepare a cell lysate, which was subcultured into monkey MA104 cells (ATCC CRL-2378.1). Specifically, about 0.5 ml of the cell lysate was added to MA104 cells in a confluent state on a 12-well plate in the presence of 0.5 ⁇ g / ml of trypsin. FBS-free DMEM medium was used for culturing MA104 cells.
- the cells were cultured for 7 days after passage, and if cytopathic effect was observed during that period, it was judged that an artificial recombinant virus was produced.
- the virus obtained at this point is called a P1 (Passage 1) virus.
- An artificially recombined rotavirus prepared using 11 types of plasmids containing pT7-NSP1SA11-NLuc is called "rsSA11-NLuc", and an artificial set prepared using 11 types of plasmids containing pT7-NSP1SA11-Rv-NLuc.
- the replacement rotavirus is called "rsSA11-Rv-N Luc".
- rsSA11-NLuc and rsSA11-Rv-NLuc P1 viruses was added, and the cells were cultured at 37 ° C. for 5 days. After culturing for 5 days, the cells were frozen and thawed twice to obtain P2 virus. For each recombinant virus, 5 wells were passaged.
- P10 virus clones 1 to 5 of rsSA11-NLuc and P10 virus clones 1 to 5 of rsSA11-Rv-NLuc were obtained.
- Viral genomic RNA was extracted from each clone of each P10 virus obtained and subjected to SDS-PAGE together with viral genomic RNA obtained from P1 virus and wild SA11 virus.
- Example 2 Artificial recombinant rotavirus expressing green fluorescent protein
- ZsG gene the ZsGreen gene contained in the pZsGreen1-N1 vector (Clontech) was used.
- the nucleotide sequence of the ZsG gene is shown in SEQ ID NO: 14.
- the codon composition of the ZsG gene and the codon composition of the RV NSP1 gene were examined, and the codon composition of the ZsG gene was modified to be close to the codon composition of the RV NSP1 gene based on the codon compositions of both genes obtained.
- the obtained modified ZsG gene is referred to as "Rv-ZsG gene”.
- FIG. 6 shows the base sequence of the ZsG gene (SEQ ID NO: 14) and the base sequence of the Rv-ZsG gene (SEQ ID NO: 15), respectively.
- 203 bases are changed in the total length of 696 bases of the ZsG gene.
- the GC content of the original ZsG gene was 63%, but the GC content of the Rv-ZsG gene dropped to 38%, approaching the GC content of the RVNSP1 gene (31%) (see Table 7).
- the experiment was carried out in the same manner as in Example 1 except that the ZsG gene and the Rv-ZsG gene were used instead of the NLuc gene and the Rv-NLuc gene. That is, the ZsG gene and Rv-ZsG gene amplified by PCR were inserted between the 128th and 129th positions of the NSP1 gene (SEQ ID NO: 5) of pT7-NSP1SA11, respectively, and the ZsG gene-inserted NSP1 gene expression plasmid (pT7-NSP1SA11) was inserted.
- rsSA11-ZsG an Rv-ZsG gene expression plasmid
- ZsG an Rv-ZsG gene expression plasmid
- ZsG an Rv-ZsG gene expression plasmid
- rsSA11-ZsG Artificial recombinant rotavirus prepared using 11 types of plasmids containing pT7-NSP1SA11-ZsG
- rsSA11-ZsG an artificial set prepared using 11 types of plasmids containing pT7-NSP1SA11-Rv-ZsG.
- the replacement rotavirus is called "rsSA11-Rv-ZsG”.
- the obtained rsSA11-ZsG and rsSA11-RvZsG were passaged up to P10, respectively, and viral genomic RNA was extracted and subjected to SDS-PAGE.
- Example 3 Artificial recombinant rotavirus expressing red fluorescent protein
- the AsRed gene contained in the pAsRed2-N1 vector (Clontech) was used.
- the nucleotide sequence of the ZsG gene is shown in SEQ ID NO: 16.
- the codon composition of the AsR gene and the codon composition of the RV NSP1 gene were examined, and the codon composition of the AsR gene was modified to be close to the codon composition of the RV NSP1 gene based on the codon compositions of both genes obtained.
- the obtained modified AsR gene is referred to as "Rv-AsR gene”.
- FIG. 10 shows the nucleotide sequence of the AsR gene (SEQ ID NO: 16) and the nucleotide sequence of the Rv-AsR gene (SEQ ID NO: 17), respectively.
- the Rv-AsR gene has 198 bases changed in the total length of 699 bases of the AsR gene.
- the GC content of the original AsR gene was 65%, but the GC content of the Rv-AsR gene dropped to 40%, approaching the GC content of the RVNSP1 gene (31%) (see Table 8).
- the experiment was carried out in the same manner as in Example 1 except that the AsR gene and the Rv-AsR gene were used instead of the NLuc gene and the Rv-NLuc gene. That is, the AsR gene and the Rv-AsR gene amplified by PCR were inserted between the 128th and 129th positions of the NSP1 gene (SEQ ID NO: 5) of pT7-NSP1SA11, respectively, and the AsR gene-inserted NSP1 gene expression plasmid (pT7-NSP1SA11) was inserted.
- -AsR) and Rv-AsR gene expression plasmid (referred to as pT7-NSP1SA11-Rv-AsR) were prepared (see FIG.
- rsSA11-AsR Artificial recombinant rotavirus prepared using 11 types of plasmids containing pT7-NSP1SA11-AsR is called “rsSA11-AsR”, and an artificial set prepared using 11 types of plasmids containing pT7-NSP1SA11-Rv-AsR.
- the replacement rotavirus is called "rsSA11-Rv-AsR”.
- the obtained rsSA11-AsR and rsSA11-Rv-AsR were passaged up to P10, respectively, and viral genomic RNA was extracted and subjected to SDS-PAGE.
- Example 4 Expression level of modified foreign gene-derived protein
- 4-1 Materials and Methods rsSA11-ZsG and rsSA11-Rv-ZsG expressing the green fluorescent protein prepared in Example 2 and wild-type SA11 were used. Prepare MA104 cells confluent in a 12-well plate, infect each virus with MOI (Multiplicity of Infection) 0.5 PFU / cells, and observe the expression of ZsG and Rv-ZsG 24 hours after infection with a fluorescence microscope. , Quantified by Western blotting.
- MOI Multiplicity of Infection
- MA104 cells confluent in a 12-well plate were prepared, each virus was infected with MOI: 0.01 PFU / cells, and cultured in FBS-free DMEM containing 0.5 ⁇ g / ml trypsin. Twenty-four hours after infection, cells were frozen and thawed twice and virus titers in cell disruptions were measured.
- Example 1 Expression level of modified foreign gene-derived protein
- the rsSA11-AsR and rsSA11-Rv-AsR expressing the red fluorescent protein prepared in Example 3 were used.
- MA104 cells confluent in a 12-well plate were prepared, each virus was infected with MOI (Multiplicity of Infection) 0.5 PFU / cells, and AsR and Rv-AsR 24 hours after infection. The expression was observed with a fluorescence microscope.
- MOI Multiplicity of Infection
- Example 5 Artificial recombinant rotavirus expressing a foreign gene having a base length of 1.6 kbp
- 5-1 Materials and methods (1) Foreign gene As a foreign gene, the codon composition of the Akaluc gene (GenBank ACCESSION: LC320664, 1653 bp), which is a variant of the firefly luciferase (FLuc) gene, is close to that of the RV NSP1 gene.
- the codon composition of the modified Akaluc gene (hereinafter referred to as “Rv-Akaluc gene”) and the norovirus VP1 gene (GenBank ACCESSION: KM268107, 1623 bp) was modified to be close to the codon composition of the RV NSP1 gene.
- a modified norovirus VP1 gene (hereinafter referred to as "Rv-NoV VP1 gene") was used.
- the nucleotide sequence of the modified Akaluc gene is shown in SEQ ID NO: 18, and the nucleotide sequence of the modified norovirus VP1 gene is shown in SEQ ID NO: 19.
- Non-Patent Document 2 a deletion of 722 bp was added to the NSP1 gene, and the Rv-Akaluc gene-inserted NSP1 gene expression plasmid (referred to as pT7-NSP1SA11-Rv-Akaluc) and the Rv-NoV VP1 gene-inserted NSP1 A gene expression plasmid (referred to as pT7-NSP1SA11-Rv-NoV VP1) was prepared (see FIG. 18).
- the artificially recombined rotavirus produced in the above is called "rsSA11-Rv-NoV VP1".
- the obtained rsSA11-Rv-Akaluc and rsSA11-Rv-NoV VP1 were passaged up to P10, respectively, and viral genomic RNA was extracted and subjected to SDS-PAGE.
- a recombinant rotavirus that stably retains a foreign gene having a base length of 1 kbp or more is prepared by bringing the codon composition close to the codon composition of rotavirus. Shown that you can
- Virus Pteropine orthoreovirus (hereinafter referred to as "PRV") belongs to the same Reoviridae family as rotavirus, L1, L2, L3, M1, M2, M3, S1, S2. It has a 10-segment double-stranded RNA consisting of S3 and S4.
- a plasmid containing the cDNAs of the 10 segmental RNA genomes of PRV was prepared in the same manner as the preparation of the plasmid (segmented RNA genome expression vector) containing each segmental RNA genome expression cassette of Rotavirus in Example 1.
- rsMB-Rv-ZsG-2A-FAST 9 types of plasmids containing wild-type genes other than the S1 gene and plasmids containing the S1-Rv-ZsG-2A-FAST gene were transfected into BHK-T7 / P5 cells and passaged into monkey MA104 cells.
- An artificially recombinant bat leovirus (referred to as "rsMB-Rv-ZsG-2A-FAST") was obtained.
- Example 7 Artificial recombinant mammalian reovir that expresses green fluorescent protein
- Virus Mammalian orthoreovirus belongs to the same Reoviridae family as rotavirus, L1, L2, L3, M1, M2, M3, S1, S2. It has a 10-segment double-stranded RNA consisting of S3 and S4.
- a plasmid containing the cDNAs of the 10 segmental RNA genomes of MRV was prepared in the same manner as the preparation of the plasmid (segmented RNA genome expression vector) containing each segmental RNA genome expression cassette of Rotavirus in Example 1.
- (2) Green fluorescent protein gene As the green fluorescent protein, the Mrv-ZsG gene was used in which the codon composition of the ZsG gene and the ZsG gene was approximated to the codon composition of the MRV L1 gene. 23 (A) to (U) show the codon composition of each amino acid in the MRV L1 gene, ZsG gene and Mrv-ZsG gene. The GC content of the original ZsG gene was 63%, but the GC content of the Mrv-ZsG gene dropped to 43%, approaching the GC content of the MRV L1 gene (46%) (see Table 9).
- the ZsG gene and Mrv-ZsG gene were inserted into the MRV L1 gene.
- the MRV L1 gene is a gene encoding the lamda3 protein, and the 2A gene encoding a self-cleaving peptide is inserted downstream of the ZsG gene and MRV-ZsG gene to L1-ZsG-2A-Lamda3 and L1-Mrv-ZsG.
- -2A-Lamda3 was prepared (see FIG. 24).
- rsMRV-Mrv-ZsG 9 types of plasmids containing wild-type genes other than the L1 gene and plasmids containing the L1-Mrv-ZsG-2A-Lamda3 gene were transfected into BHK-T7 / P5 cells and passaged into mouse L929 cells.
- An artificially recombinant mammalian leovirus (referred to as "rsMRV-Mrv-ZsG”) was obtained.
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Organic Chemistry (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Wood Science & Technology (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Virology (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biophysics (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
Abstract
(1)RNAウイルス遺伝子のコドン組成に近似するように改変されたコドン組成を有する外来遺伝子を取得する工程、(2)(1)で取得した外来遺伝子をRNAウイルスのゲノムに挿入する工程、および(3)リバースジェネティクス法を用いて人工組換えRNAウイルスを人工的に合成する工程を含むことを特徴とする、外来遺伝子を安定的に発現する人工組換えRNAウイルスの作製方法を提供する。
Description
本発明は、外来遺伝子を安定的に保持する人工組換えRNAウイルスの作製方法に関するものである。
レオウイルス科に属するロタウイルスは、乳幼児に急性の胃腸炎を引き起こし、年間で約20万人がロタウイルス感染により命を落としている。ロタウイルスにおいては、任意のウイルスを人工的に作製する技術の開発が遅れていたが、最近、本発明者らは、ロタウイルスの実用的な人工合成法の開発に成功した(特許文献1、非特許文献1)。この技術により、任意の変異を有する人工組換えロタウイルスや、外来遺伝子を搭載した人工組換えロタウイルスを自由自在に作製することが可能になった。その後、本発明者らは、外来遺伝子を導入した人工組換えロタウイルスにおいて、短期間で全長の外来遺伝子が消失すること、すなわち、外来遺伝子を長期間安定的に保持・発現させることが困難であることに気付き、研究を重ねて外来遺伝子を安定的に保持・発現させるための改良法の一部を報告した(非特許文献2)。
外来遺伝子を異種宿主で発現させる際に、外来遺伝子の発現を向上させる目的で、外来遺伝子のコドン組成を宿主遺伝子のコドン組成に近づけるコドン最適化技術は古くから用いられており(非特許文献3)、多数のコドン最適化ソフトウェアが提供されている。しかし、コドン最適化技術が、人工組換えRNAウイルスに導入された外来遺伝子の安定的な保持に応用できることは知られていなかった。
Kanai et al., Proc Natl Acad Sci U S A. 2017 Feb 28; 114(9):2349-2354
Kanai et al., Journal of Virology, 2019 Feb 5;93(4). pii: e01774-18
Gustafsson et al., Trends Biotechnol. 2004 Jul;22(7):346-353
本発明は、外来遺伝子を安定的に保持する人工組換えRNAウイルスの作製方法および人工組換えRNAウイルスに導入された外来遺伝子を安定的に保持させる方法を提供することを課題とする。
本発明は、上記の課題を解決するために以下の各発明を包含する。
[1]外来遺伝子を安定的に保持する人工組換えRNAウイルスの作製方法であって、
(1)RNAウイルス遺伝子のコドン組成に近似するように改変されたコドン組成を有する外来遺伝子を取得する工程、
(2)(1)で取得した外来遺伝子をRNAウイルスのゲノムに挿入する工程、および
(3)リバースジェネティクス法を用いて人工組換えRNAウイルスを人工的に合成する工程
を含むことを特徴とする作製方法。
[2]RNAウイルスがレオウイルス科に属するウイルスである前記[1]に記載の方法。
[3]レオウイルス科に属するウイルスがロタウイルス属またはオルソレオウイルス属に属するウイルスである前記[2]に記載の方法。
[4]工程(1)において、RNAウイルス遺伝子のコドン組成の±30%の範囲内になるように改変されたコドン組成を有する外来遺伝子を取得する前記[1]~[3]のいずれかに記載の方法。
[5]リバースジェネティクス法を用いて人工的に合成された人工組換えRNAウイルスに導入された外来遺伝子を安定的に保持させる方法であって、外来遺伝子のコドン組成を、RNAウイルス遺伝子のコドン組成に近似させることを特徴とする方法。
[6]RNAウイルスがレオウイルス科に属するウイルスである前記[5]に記載の方法。
[7]レオウイルス科に属するウイルスがロタウイルス属またはオルソレオウイルス属に属するウイルスである前記[6]に記載の方法。
[8]外来遺伝子のコドン組成を、RNAウイルス遺伝子のコドン組成の±30%の範囲内に近似させる前記[5]~[7]のいずれかに記載の方法。
[1]外来遺伝子を安定的に保持する人工組換えRNAウイルスの作製方法であって、
(1)RNAウイルス遺伝子のコドン組成に近似するように改変されたコドン組成を有する外来遺伝子を取得する工程、
(2)(1)で取得した外来遺伝子をRNAウイルスのゲノムに挿入する工程、および
(3)リバースジェネティクス法を用いて人工組換えRNAウイルスを人工的に合成する工程
を含むことを特徴とする作製方法。
[2]RNAウイルスがレオウイルス科に属するウイルスである前記[1]に記載の方法。
[3]レオウイルス科に属するウイルスがロタウイルス属またはオルソレオウイルス属に属するウイルスである前記[2]に記載の方法。
[4]工程(1)において、RNAウイルス遺伝子のコドン組成の±30%の範囲内になるように改変されたコドン組成を有する外来遺伝子を取得する前記[1]~[3]のいずれかに記載の方法。
[5]リバースジェネティクス法を用いて人工的に合成された人工組換えRNAウイルスに導入された外来遺伝子を安定的に保持させる方法であって、外来遺伝子のコドン組成を、RNAウイルス遺伝子のコドン組成に近似させることを特徴とする方法。
[6]RNAウイルスがレオウイルス科に属するウイルスである前記[5]に記載の方法。
[7]レオウイルス科に属するウイルスがロタウイルス属またはオルソレオウイルス属に属するウイルスである前記[6]に記載の方法。
[8]外来遺伝子のコドン組成を、RNAウイルス遺伝子のコドン組成の±30%の範囲内に近似させる前記[5]~[7]のいずれかに記載の方法。
本発明により、外来遺伝子を長期間安定的に保持する人工組換えRNAウイルスを提供することができる。
本発明は、外来遺伝子を安定的に保持する人工組換えRNAウイルスの作製方法(以下、「本発明の作製方法」と記す)を提供する。本発明の作製方法は、以下の工程(1)、(2)および(3)を含むものであればよい。
(1)RNAウイルス遺伝子のコドン組成に近似するように改変されたコドン組成を有する外来遺伝子を取得する工程
(2)(1)で取得した外来遺伝子をRNAウイルスのゲノムに挿入する工程
(3)リバースジェネティクス法を用いて人工組換えRNAウイルスを取得する工程
(1)RNAウイルス遺伝子のコドン組成に近似するように改変されたコドン組成を有する外来遺伝子を取得する工程
(2)(1)で取得した外来遺伝子をRNAウイルスのゲノムに挿入する工程
(3)リバースジェネティクス法を用いて人工組換えRNAウイルスを取得する工程
RNAウイルスは、二本鎖RNAウイルスであってもよく、一本鎖プラス鎖RNAウイルスであってもよく、一本鎖マイナス鎖RNAウイルスであってもよい。二本鎖RNAウイルスとしては、レオウイルス科、ビルナウイルス科に属するウイルスが挙げられる。一本鎖プラス鎖RNAウイルスとしては、コロナウイルス科、ピコルナウイルス科、トガウイルス科、フラビウイルス科、カリシウイルス科、アストロウイルス科等に属するウイルスが挙げられる。一本鎖マイナス鎖RNAウイルスとしては、パラミクソウイルス科、ラブドウイルス科、フィロウイルス科、オルソミクソウイルス科、アレナウイルス科、ブニャウイルス科等に属するウイルスが挙げられる。
RNAウイルスは、レオウイルス科に属するウイルスであってもよい。レオウイルス科に属するウイルスは、10~12本に分節した直鎖二本鎖RNAをゲノムに持つウイルスであり、そのビリオンは直径60~80nmの正二十面体構造を示す。レオウイルス科に属するウイルスには、哺乳類オルソレオウイルス(哺乳類レオウイルスとも称される)、ネルソンベイオルソレオウイルス(ネルソンベイレオウイルスまたはコウモリレオウイルスとも称される)、トリレオウイルス等のオルソレオウイルス属(Genus Orthoreovirus)、アフリカ馬疫ウイルス、ブルータングウイルス等のオルビウイルス属(Genus Orbivirus)、ロタウイルス等のロタウイルス属(Genus Rotavirus)、コロラドダニ熱ウイルス等のコルチウイルス属(Genus Coltivirus)、アクアレオウイルスA等のアクアレオウイルス属(Genus Aquareovirus)、細胞質多角体病ウイルス等のサイポウイルス属(Genus Cypovirus)、イネ南方黒すじ萎縮病ウイルス等のフィジウイルス属(Genus Fijivirus)、イネ萎縮病ウイルス等のフィトレオウイルス属(Genus Phytoreovirus)およびイネラギッドスタントウイルス(Rice ragged stunt virus)等のオリザウイルス属(Genus Oryzavirus)が含まれる。レオウイルス科に属するウイルスは、ロタウイルス属に属するウイルスであってもよく、オルソレオウイルス属に属するウイルスであってもよい。
外来遺伝子は特に限定されない。外来遺伝子は、動物の遺伝子であってもよく、植物の遺伝子であってもよく、菌類の遺伝子であってもよく、細菌の遺伝子であってもよく、ウイルスの遺伝子であってもよい。外来遺伝子の塩基長は特に限定されず、10bp以上であってもよく、100bp以上であってもよく、500bp以上であってもよく、1000bp以上であってもよく、1500bp以上であってもよく、2000bp以上であってもよく、3000bp以上であってもよく、4000bp以上であってもよく、5000bp以上であってもよい。外来遺伝子の塩基長は、500bp以下であってもよく、1000bp以下であってもよく、1500bp以下であってもよく、2000bp以下であってもよく、3000bp以下であってもよく、4000bp以下であってもよく、5000bp以下であってもよい。
外来遺伝子がコードするタンパク質は特に限定されない。外来遺伝子がコードするタンパク質はワクチン抗原であってもよい。ワクチン抗原としては、例えば、ノロウイルス抗原、アデノウイルス抗原、A型肝炎抗原、サポウイルス抗原、手足口病ウイルス抗原、エンテロウイルス抗原、HIV抗原、サルモネラ抗原、カンピロバクター抗原、腸炎ビブリオ抗原、大腸菌O157抗原、コレラ抗原、腸チフス抗原、赤痢抗原等が挙げられる。これらのワクチン抗原は、エピトープペプチドであってもよい。
工程(1)では、RNAウイルス遺伝子のコドン組成に近似するように改変されたコドン組成を有する外来遺伝子を取得する。基準とするRNAウイルス遺伝子のコドン組成は、コドンを改変した外来遺伝子を組み込んで人工的に合成するRNAウイルスの遺伝子のコドン組成であってもよく、コドンを改変した外来遺伝子を組み込んで人工的に合成するRNAウイルスと異なるRNAウイルスの遺伝子のコドン組成であってもよい。異なるRNAウイルスの遺伝子のコドン組成を基準とする場合、異なるRNAウイルスはコドンを改変した外来遺伝子を組み込んで人工的に合成するRNAウイルスと系統的に近似するRNAウイルス(例えば、同じ科に属するウイルス、同じ属に属するウイルス、種が異なるウイルス等)であることが好ましい。
また、基準とするRNAウイルス遺伝子のコドン組成は、基準とするRNAウイルスの全遺伝子のコドン組成であってもよく、基準とするRNAウイルスの一部の遺伝子のコドン組成であってもよい。RNAウイルスの一部の遺伝子のコドン組成を基準とする場合、1個の遺伝子のコドン組成を基準としてもよく、2個以上の遺伝子のコドン組成を基準としてもよい。
基準とするRNAウイルス遺伝子のコドン組成は、公知の遺伝子データベース(例えばGenBank等)に登録されているRNAウイルスの遺伝子情報に基づいて作成することができる。
表1にロタウイルスSA11株の全遺伝子のゲノム組成を示す。表2にロタウイルスSA11株NSP1遺伝子のゲノム組成を示す。表3にコウモリレオウイルス(Pteropine orthoreovirus)MB株の全遺伝子のゲノム組成を示す。表4に哺乳類レオウイルスT1L型の全遺伝子のゲノム組成を示す。表1~4に示したコドン組成は、本発明の作製方法における基準コドン組成として使用することができる。なかでもレオウイルス科に属するウイルス、特にロタウイルス属およびオルソレオウイルス属に属する人工組換えウイルスを用いて外来遺伝子を発現させる場合の基準コドン組成として用いることができる。




外来遺伝子のコドン組成を、基準とするRNAウイルス遺伝子のコドン組成に近似させるとは、外来遺伝子のアミノ酸を変異させることなく、同じアミノ酸に対応するコドンを基準とするRNAウイルス遺伝子において出現頻度が高いコドンに変更することをいう。基準とするRNAウイルス遺伝子のコドン組成に近似させる程度は、外来遺伝子を安定的に保持するという目的を達成できる程度であれば特に限定されない。例えば、基準とするRNAウイルス遺伝子のコドン組成の±35%の範囲内になるように外来遺伝子のコドン組成を改変してもよく、基準とするRNAウイルス遺伝子のコドン組成の±30%の範囲内になるように外来遺伝子のコドン組成を改変してもよく、基準とするRNAウイルス遺伝子のコドン組成の±25%の範囲内になるように外来遺伝子のコドン組成を改変してもよく、基準とするRNAウイルス遺伝子のコドン組成の±20%の範囲内になるように外来遺伝子のコドン組成を改変してもよく、基準とするRNAウイルス遺伝子のコドン組成の±15%の範囲内になるように外来遺伝子のコドン組成を改変してもよく、基準とするRNAウイルス遺伝子のコドン組成の±10%の範囲内になるように外来遺伝子のコドン組成を改変してもよい。
コドン組成を改変した外来遺伝子は、改変後の塩基配列に基づいてDNA合成することにより取得することができる。また、PCRをベースにした部位特異的変異導入法を用いて、既存の外来遺伝子DNAに変異を導入して取得することができる。
工程(2)では、(1)で取得した外来遺伝子をRNAウイルスのゲノムに挿入する。外来遺伝子を挿入する位置は、リバースジェネティクス法を用いて人工組換えRNAウイルスを人工的に合成することの妨げとならない限り、特に限定されない。例えば、人工組換えロタウイルスに外来遺伝子を導入する場合、NSP1遺伝子に挿入してもよく、NSP3遺伝子に挿入してもよく、NSP5遺伝子に挿入してもよい。例えば、人工組換え哺乳類レオウイルスに外来遺伝子を導入する場合、L1遺伝子に挿入してもよく、S1、S2、S4遺伝子に挿入してもよい。例えば、人工組換えインフルエンザウイルスに外来遺伝子を導入する場合、NS1遺伝子に挿入してもよく、NA遺伝子に挿入してもよく、PA遺伝子に挿入してもよい。例えば、アルファウイルス属(シンドビスウイルス、チクングニアウイルスなど)の人工組換えウイルスに外来遺伝子を導入する場合、NSP3遺伝子に挿入してもよく、カプシド遺伝子に挿入してもよい。例えば、人工組換えノロウイルスに外来遺伝子を導入する場合、ORF1遺伝子に挿入してもよい。例えば、フラビウイルス属(デングウイルス、C型肝炎ウイルスなど)の人工組換えウイルスに外来遺伝子を導入する場合、NS5A遺伝子に挿入してもよく、カプシド遺伝子に挿入してもよい。例えば、ラブドウイルス(狂犬病ウイルス、水泡性口炎ウイルス)の人工組換えウイルスに外来遺伝子を導入する場合、G蛋白質遺伝子に挿入してもよい。例えば、ピコルナウイルス属(ポリオウイルス、口蹄疫ウイルスなど)の人工組換えウイルスに外来遺伝子を導入する場合、P1遺伝子に挿入してもよい。例えば、人工組換えセンダイウイルスに外来遺伝子を導入する場合、NP遺伝子に挿入してもよい。外来遺伝子のウイルスゲノムへの挿入は、公知の遺伝子組換え技術を用いて行うことができる。
工程(3)では、リバースジェネティクス法を用いて人工組換えRNAウイルスを人工的に合成する。リバースジェネティクス法を用いて人工組換えRNAウイルスを人工的に合成する方法は、RNAウイルスの種類に応じて、公知の方法から適宜選択すればよい。リバースジェネティクス法を用いて人工組換えロタウイルスを合成する方法は、例えば、上記特許文献1、非特許文献1、非特許文献2などに記載されている。リバースジェネティクス法を用いて人工組換えポリオウイルスを合成する方法は、例えば、下記参考文献1などに記載されている。リバースジェネティクス法を用いて人工組換え狂犬病ウイルスを合成する方法は、例えば、下記参考文献2などに記載されている。リバースジェネティクス法を用いて人工組換え麻疹ウイルスを合成する方法は、例えば、下記参考文献3などに記載されている。リバースジェネティクス法を用いて人工組換えブニャウイルスを合成する方法は、例えば、下記参考文献4などに記載されている。リバースジェネティクス法を用いて人工組換え伝染性ファブリキウス嚢病ウイルスを合成する方法は、例えば、下記参考文献5などに記載されている。リバースジェネティクス法を用いて人工組換えC型肝炎ウイルスを合成する方法は、例えば、下記参考文献6などに記載されている。リバースジェネティクス法を用いて人工組換えインフルエンザウイルスを合成する方法は、例えば、下記参考文献7などに記載されている。リバースジェネティクス法を用いて人工組換えコロナウイルスを合成する方法は、例えば、下記参考文献8などに記載されている。リバースジェネティクス法を用いて人工組換えエボラウイルスを合成する方法は、例えば、下記参考文献9などに記載されている。リバースジェネティクス法を用いて人工組換えボルナウイルスを合成する方法は、例えば、下記参考文献10などに記載されている。リバースジェネティクス法を用いて人工組換えアレナウイルスを合成する方法は、例えば、下記参考文献11などに記載されている。リバースジェネティクス法を用いて人工組換えレオウイルスを合成する方法は、例えば、下記参考文献12などに記載されている。リバースジェネティクス法を用いて人工組換えデングウイルスを合成する方法は、例えば、下記参考文献13などに記載されている。リバースジェネティクス法を用いて人工組換えノロウイルスを合成する方法は、例えば、下記参考文献14などに記載されている。リバースジェネティクス法を用いて人工組換え日本脳炎ウイルスを合成する方法は、例えば、下記参考文献15などに記載されている。リバースジェネティクス法を用いて人工組換えブルータングウイルスを合成する方法は、例えば、16などに記載されている。リバースジェネティクス法を用いて人工組換えヒト免疫不全ウイルスを合成する方法は、例えば、下記参考文献17などに記載されている。
<参考文献リスト>
1.Racaniello VR & Baltimore D (1981) Cloned poliovirus complementary DNA is infectious in mammalian cells. Science 214(4523):916-919.
2.Schnell MJ, Mebatsion T, & Conzelmann KK (1994) Infectious Rabies Viruses from Cloned Cdna. Embo J 13(18):4195-4203.
3.Radecke F, et al. (1995) Rescue of measles viruses from cloned DNA. Embo J 14(23):5773-5784.
4.Bridgen A & Elliott RM (1996) Rescue of a segmented negative-strand RNA virus entirely from cloned complementary DNAs. Proceedings of the National Academy of Sciences of the United States of America 93(26):15400-15404.
5.Mundt E & Vakharia VN (1996) Synthetic transcripts of double-stranded Birnavirus genome are infectious. Proceedings of the National Academy of Sciences of the United States of America 93(20):11131-11136.
6.Kolykhalov AA, et al. (1997) Transmission of hepatitis C by intrahepatic inoculation with transcribed RNA. Science 277(5325):570-574.
7.Neumann G, et al. (1999) Generation of influenza A viruses entirely from cloned cDNAs. Proceedings of the National Academy of Sciences of the United States of America 96(16):9345-9350.
8.Yount B, Curtis KM, & Baric RS (2000) Strategy for systematic assembly of large RNA and DNA genomes: transmissible gastroenteritis virus model. Journal of virology 74(22):10600-10611.
9.olchkov VE, et al. (2001) Recovery of infectious Ebola virus from complementary DNA: RNA editing of the GP gene and viral cytotoxicity. Science 291(5510):1965-1969.
10.Schneider U, Schwemmle M, & Staeheli P (2005) Genome trimming: a unique strategy for replication control employed by Borna disease virus. Proceedings of the National Academy of Sciences of the United States of America 102(9):3441-3446.
11.Sanchez AB & de la Torre JC (2006) Rescue of the prototypic Arenavirus LCMV entirely from plasmid. Virology 350(2):370-380.
12.Kobayashi T, et al. (2007) A plasmid-based reverse genetics system for animal double-stranded RNA viruses. Cell host & microbe 1(2):147-157.
13.Lai CJ, Zhao BT, Hori H, & Bray M (1991) Infectious RNA transcribed from stably cloned full-length cDNA of dengue type 4 virus. Proceedings of the National Academy of Sciences of the United States of America 88(12):5139-5143.
14.Chaudhry Y, Skinner MA, & Goodfellow IG (2007) Recovery of genetically defined murine norovirus in tissue culture by using a fowlpox virus expressing T7 RNA polymerase. The Journal of general virology 88(Pt 8):2091-2100.
15.Yun SI, Kim SY, Rice CM, & Lee YM (2003) Development and application of a reverse genetics system for Japanese encephalitis virus. Journal of virology 77(11):6450-6465.
16.Boyce M, Celma CC, & Roy P (2008) Development of reverse genetics systems for bluetongue virus: recovery of infectious virus from synthetic RNA transcripts. Journal of virology 82(17):8339-8348.
17.Adachi A, et al. (1986) Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. Journal of virology 59(2):284-291.
1.Racaniello VR & Baltimore D (1981) Cloned poliovirus complementary DNA is infectious in mammalian cells. Science 214(4523):916-919.
2.Schnell MJ, Mebatsion T, & Conzelmann KK (1994) Infectious Rabies Viruses from Cloned Cdna. Embo J 13(18):4195-4203.
3.Radecke F, et al. (1995) Rescue of measles viruses from cloned DNA. Embo J 14(23):5773-5784.
4.Bridgen A & Elliott RM (1996) Rescue of a segmented negative-strand RNA virus entirely from cloned complementary DNAs. Proceedings of the National Academy of Sciences of the United States of America 93(26):15400-15404.
5.Mundt E & Vakharia VN (1996) Synthetic transcripts of double-stranded Birnavirus genome are infectious. Proceedings of the National Academy of Sciences of the United States of America 93(20):11131-11136.
6.Kolykhalov AA, et al. (1997) Transmission of hepatitis C by intrahepatic inoculation with transcribed RNA. Science 277(5325):570-574.
7.Neumann G, et al. (1999) Generation of influenza A viruses entirely from cloned cDNAs. Proceedings of the National Academy of Sciences of the United States of America 96(16):9345-9350.
8.Yount B, Curtis KM, & Baric RS (2000) Strategy for systematic assembly of large RNA and DNA genomes: transmissible gastroenteritis virus model. Journal of virology 74(22):10600-10611.
9.olchkov VE, et al. (2001) Recovery of infectious Ebola virus from complementary DNA: RNA editing of the GP gene and viral cytotoxicity. Science 291(5510):1965-1969.
10.Schneider U, Schwemmle M, & Staeheli P (2005) Genome trimming: a unique strategy for replication control employed by Borna disease virus. Proceedings of the National Academy of Sciences of the United States of America 102(9):3441-3446.
11.Sanchez AB & de la Torre JC (2006) Rescue of the prototypic Arenavirus LCMV entirely from plasmid. Virology 350(2):370-380.
12.Kobayashi T, et al. (2007) A plasmid-based reverse genetics system for animal double-stranded RNA viruses. Cell host & microbe 1(2):147-157.
13.Lai CJ, Zhao BT, Hori H, & Bray M (1991) Infectious RNA transcribed from stably cloned full-length cDNA of dengue type 4 virus. Proceedings of the National Academy of Sciences of the United States of America 88(12):5139-5143.
14.Chaudhry Y, Skinner MA, & Goodfellow IG (2007) Recovery of genetically defined murine norovirus in tissue culture by using a fowlpox virus expressing T7 RNA polymerase. The Journal of general virology 88(Pt 8):2091-2100.
15.Yun SI, Kim SY, Rice CM, & Lee YM (2003) Development and application of a reverse genetics system for Japanese encephalitis virus. Journal of virology 77(11):6450-6465.
16.Boyce M, Celma CC, & Roy P (2008) Development of reverse genetics systems for bluetongue virus: recovery of infectious virus from synthetic RNA transcripts. Journal of virology 82(17):8339-8348.
17.Adachi A, et al. (1986) Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. Journal of virology 59(2):284-291.
本発明の作製方法により作製された人工組換えRNAウイルスは、外来遺伝子を長期間安定に保持し、外来遺伝子産物を長期間安定に発現することができるので、例えば外来遺伝子としてワクチン抗原を用いれば、ウイルスワクチンとして好適に使用することができる。また遺伝子異常に由来する(と考えられている)消化管の遺伝病(大腸癌、潰瘍性大腸炎、クローン病、セリアック病、非特異性多発性小腸潰瘍症等)を対象とし、正常遺伝子を外来遺伝子として異常細胞に届けることで遺伝子治療にも応用できる。
本発明は、リバースジェネティクス法を用いて人工的に合成された人工組換えRNAウイルスに導入された外来遺伝子を安定的に保持させる方法(以下、「本発明の安定保持方法」と記す)を提供する。本発明の安定保持方法は、外来遺伝子のコドン組成を、RNAウイルス遺伝子のコドン組成に近似させることを特徴とする方法である。外来遺伝子のコドン組成を、基準とするRNAウイルス遺伝子のコドン組成に近似させる方法については、上記本発明の作製方法において説明した方法と同じ方法で行うことができる。
以下、実施例により本発明を詳細に説明するが、本発明はこれらに限定されるものではない。
〔実施例1:ルシフェラーゼを発現する人工組換えロタウイルス〕
1-1 材料および方法
(1)ウイルス
サルロタウイルスSA11株を使用した。本発明者らは、このウイルス株の11個の各分節RNAゲノムの塩基配列を決定し、登録している。本実験に使用したサルロタウイルスSA11株(以下「SA11株」と称する)の11個の各分節RNAゲノムの名前およびGenBank ACCESSIONを表5に示す。
1-1 材料および方法
(1)ウイルス
サルロタウイルスSA11株を使用した。本発明者らは、このウイルス株の11個の各分節RNAゲノムの塩基配列を決定し、登録している。本実験に使用したサルロタウイルスSA11株(以下「SA11株」と称する)の11個の各分節RNAゲノムの名前およびGenBank ACCESSIONを表5に示す。

(2)SA11株の各分節RNAゲノム発現カセットを含むプラスミド(分節RNAゲノム発現ベクター)
人工組換えロタウイルス作製のため、SA11株の11個の分節RNAゲノムのcDNAを含むプラスミドを作製した。ウイルスから抽出した二本鎖RNAを鋳型とし、各分節RNAゲノムの塩基配列に基づく特異的プライマーを用いてRT-PCRを行った。得られたRT-PCR産物(各分節RNAゲノムのcDNA)をp3E5プラスミドのT7プロモーター配列とHDVリボザイム配列の間に挿入し、各分節RNAゲノムの発現カセットを含むプラスミドを得た(特許文献1参照)。各分節RNAゲノムの発現カセットは、各分節RNAゲノムのcDNAの5'側にT7プロモーター配列、3'側にD型肝炎ウイルス(HDV)リボザイム配列が隣接し、その下流にT7ターミネーター配列が配置された構造を有する。作製したプラスミド(分節RNAゲノム発現ベクター)を、それぞれpT7-VP1SA11、pT7-VP2SA11、pT7-VP3SA11、pT7-VP4SA11、pT7-VP6SA11、pT7-VP7SA11、pT7-NSP1SA11、pT7-NSP2SA11、pT7-NSP3SA11、pT7-NSP4SA11およびpT7-NSP5SA11と称する。
人工組換えロタウイルス作製のため、SA11株の11個の分節RNAゲノムのcDNAを含むプラスミドを作製した。ウイルスから抽出した二本鎖RNAを鋳型とし、各分節RNAゲノムの塩基配列に基づく特異的プライマーを用いてRT-PCRを行った。得られたRT-PCR産物(各分節RNAゲノムのcDNA)をp3E5プラスミドのT7プロモーター配列とHDVリボザイム配列の間に挿入し、各分節RNAゲノムの発現カセットを含むプラスミドを得た(特許文献1参照)。各分節RNAゲノムの発現カセットは、各分節RNAゲノムのcDNAの5'側にT7プロモーター配列、3'側にD型肝炎ウイルス(HDV)リボザイム配列が隣接し、その下流にT7ターミネーター配列が配置された構造を有する。作製したプラスミド(分節RNAゲノム発現ベクター)を、それぞれpT7-VP1SA11、pT7-VP2SA11、pT7-VP3SA11、pT7-VP4SA11、pT7-VP6SA11、pT7-VP7SA11、pT7-NSP1SA11、pT7-NSP2SA11、pT7-NSP3SA11、pT7-NSP4SA11およびpT7-NSP5SA11と称する。
(3)ルシフェラーゼ遺伝子
ルシフェラーゼ遺伝子として、トゲオキヒオドシエビ(Oplophorus gracilirostris)由来のルシフェラーゼ遺伝子であるNLuc遺伝子を用いた。pNL1.1.TK[NLuc/TK]ベクター(プロメガ、GenBank ACCESSION: KM359774、3817 bp)の815位~1330位(配列番号12)がNLucコード領域である。
ルシフェラーゼ遺伝子として、トゲオキヒオドシエビ(Oplophorus gracilirostris)由来のルシフェラーゼ遺伝子であるNLuc遺伝子を用いた。pNL1.1.TK[NLuc/TK]ベクター(プロメガ、GenBank ACCESSION: KM359774、3817 bp)の815位~1330位(配列番号12)がNLucコード領域である。
(4)コドン組成を改変したルシフェラーゼ遺伝子
NLuc遺伝子のコドン組成およびロタウイルスSA11株のNSP1遺伝子(以下「RV NSP1遺伝子」と称する)のコドン組成を調べ、得られた両遺伝子のコドン組成に基づいて、NLuc遺伝子のコドン組成をRV NSP1遺伝子のコドン組成に近くなるように改変した。得られた改変NLuc遺伝子を「Rv-NLuc遺伝子」と称する。
NLuc遺伝子のコドン組成およびロタウイルスSA11株のNSP1遺伝子(以下「RV NSP1遺伝子」と称する)のコドン組成を調べ、得られた両遺伝子のコドン組成に基づいて、NLuc遺伝子のコドン組成をRV NSP1遺伝子のコドン組成に近くなるように改変した。得られた改変NLuc遺伝子を「Rv-NLuc遺伝子」と称する。
図1(A)~(U)にRV NSP1遺伝子、NLuc遺伝子およびRv-NLuc遺伝子における各アミノ酸のコドン組成を示した。また、図2にNLuc遺伝子の塩基配列(配列番号12)とRv-NLuc遺伝子の塩基配列(配列番号13)をそれぞれ示した。Rv-NLuc遺伝子は、NLuc遺伝子の全長516塩基中、70塩基が変わっている。元のNLuc遺伝子のGC含量は53%であったが、Rv-NLuc遺伝子のGC含量は40%に下がり、RV NSP1遺伝子のGC含量(31%)に近づいた(表6参照)。

(5)FASTタンパク質発現ベクター
FASTタンパク質発現ベクターは、ネルソンベイレオウイルスp10遺伝子(GenBank ACCESSION: AB908284 参照)のタンパク質コード領域DNAを、pCAGGSプラスミド(5699bp、Matsuo et al., 2006, Biochem Biophys Res Commun 340(1): 200-208)のBglII切断部位に挿入して作製した(特許文献1参照)。得られたネルソンベイレオウイルスp10発現ベクターをpCAG-p10と称する。
FASTタンパク質発現ベクターは、ネルソンベイレオウイルスp10遺伝子(GenBank ACCESSION: AB908284 参照)のタンパク質コード領域DNAを、pCAGGSプラスミド(5699bp、Matsuo et al., 2006, Biochem Biophys Res Commun 340(1): 200-208)のBglII切断部位に挿入して作製した(特許文献1参照)。得られたネルソンベイレオウイルスp10発現ベクターをpCAG-p10と称する。
(6)キャッピング酵素発現ベクター
キャッピング酵素発現ベクターは、ワクシニアウイルスD1R遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: NC006998の93948位~96482位)およびワクシニアウイルスD12L遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: NC006998の107332位~108195位)を、それぞれ上記pCAGGSプラスミドのBglII切断部位に挿入して作製した(特許文献1参照)。得られたワクシニアウイルスmRNAキャッピング酵素ラージサブユニット発現ベクターをpCAG-D1Rと称し、ワクシニアウイルスmRNAキャッピング酵素スモールサブユニット発現ベクターをpCAG-D12Lと称する。
キャッピング酵素発現ベクターは、ワクシニアウイルスD1R遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: NC006998の93948位~96482位)およびワクシニアウイルスD12L遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: NC006998の107332位~108195位)を、それぞれ上記pCAGGSプラスミドのBglII切断部位に挿入して作製した(特許文献1参照)。得られたワクシニアウイルスmRNAキャッピング酵素ラージサブユニット発現ベクターをpCAG-D1Rと称し、ワクシニアウイルスmRNAキャッピング酵素スモールサブユニット発現ベクターをpCAG-D12Lと称する。
(7)SA11株のNSP2タンパク質発現ベクター
NSP2タンパク質発現ベクターは、SA11株NSP2遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: LC178571の47位~1000位)を、上記pCAGGSプラスミドのBglII切断部位に挿入して作製した(特許文献1参照)。得られたSA11株NSP2タンパク質発現ベクターをpCAG-NSP2と称する。
NSP2タンパク質発現ベクターは、SA11株NSP2遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: LC178571の47位~1000位)を、上記pCAGGSプラスミドのBglII切断部位に挿入して作製した(特許文献1参照)。得られたSA11株NSP2タンパク質発現ベクターをpCAG-NSP2と称する。
(8)SA11株のNSP5タンパク質発現ベクター
NSP5タンパク質発現ベクターは、SA11株NSP5遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: LC178574の22位~618位)を、上記pCAGGSプラスミドのBglII切断部位に挿入して作製した(特許文献1参照)。得られたSA11株NSP5タンパク質発現ベクターをpCAG-NSP5と称する。
NSP5タンパク質発現ベクターは、SA11株NSP5遺伝子のタンパク質コード領域DNA(GenBank ACCESSION: LC178574の22位~618位)を、上記pCAGGSプラスミドのBglII切断部位に挿入して作製した(特許文献1参照)。得られたSA11株NSP5タンパク質発現ベクターをpCAG-NSP5と称する。
(9)ルシフェラーゼ発現人工組換えロタウイルスの作製
NLuc遺伝子およびRv-NLuc遺伝子をそれぞれPCRで増幅し、増幅産物をpT7-NSP1SA11のNSP1遺伝子(配列番号5)の128位と129位の間に挿入して、NLuc遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-NLucと称する)、およびRv-NLuc遺伝子発現プラスミド(pT7-NSP1SA11-Rv-NLucと称する)を作製した(図3参照)。
NLuc遺伝子およびRv-NLuc遺伝子をそれぞれPCRで増幅し、増幅産物をpT7-NSP1SA11のNSP1遺伝子(配列番号5)の128位と129位の間に挿入して、NLuc遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-NLucと称する)、およびRv-NLuc遺伝子発現プラスミド(pT7-NSP1SA11-Rv-NLucと称する)を作製した(図3参照)。
ルシフェラーゼ発現人工組換えロタウイルスの作製には、上記(2)で作製した11種類の分節RNAゲノムのcDNAを含むプラスミドからpT7-NSP1SA11を除いた10種類のプラスミドに、上記pT7-NSP1SA11-NLucまたはpT7-NSP1SA11-Rv-NLucを加えた各11種類のプラスミドを用いた。
トランスフェクションの前日に、BHK-T7/P5細胞を8×105個/ウェルで6ウェル培養プレートに播種した。11種類の分節RNAゲノム発現ベクター各0.8μg、FASTタンパク質発現ベクター(pCAG-p10)0.015μg、キャッピング酵素発現ベクター(pCAG-D1RおよびpCAG-D12L)、NSP2タンパク質発現ベクター(pCAG-NSP2)、NSP5タンパク質発現ベクター(pCAG-NSP5)、各0.8μgを、トランスフェクション試薬(TransIT-LT1(商品名)、Mirus)を用いてBHK-T7/P5細胞に導入した。DNA 1μgあたり2μgのトランスフェクション試薬を使用した。BHK-T7/P5細胞の培養には、5% FBS、100units/mlペニシリンおよび100μg/mlストレプトマイシンを含有するDMEM培地を用い、37℃5%CO2環境下で培養した。トランスフェクションの48時間後に培地および細胞を回収した。回収した培地および細胞を3回凍結融解して細胞溶解液を調製し、サルMA104細胞(ATCC CRL-2378.1)に継代した。具体的には、12ウェルプレートでコンフルエントな状態のMA104細胞に、トリプシン0.5μg/ml 存在下で前記細胞溶解液約0.5mlを添加した。MA104細胞の培養にはFBS不含DMEM培地を使用した。継代後7日間培養し、その間に細胞変性が認められた場合に人工組換えウイルスが作製されたと判断した。この時点で得られたウイルスをP1(Passage 1)ウイルスと称する。pT7-NSP1SA11-NLucを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-NLuc」と称し、pT7-NSP1SA11-Rv-NLucを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-Rv-NLuc」と称する。
(10)電気泳動によるルシフェラーゼ遺伝子の確認
rsSA11-NLucおよびrsSA11-Rv-NLucの継代後のNLuc遺伝子およびRv-NLuc遺伝子の安定性を調べるために、rsSA11-NLucおよびrsSA11-Rv-NLucをMA104細胞でP10まで継代を繰り返した。12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、ウイルス感染前に培地を除去し、0.5μg/mlのトリプシンを含有するFBS不含DMEM1mLに置換した。準備ができたMA104細胞に、rsSA11-NLucおよびrsSA11-Rv-NLucのP1ウイルスをそれぞれ1μLずつ加え37℃で5日間培養した。5日培養後2回凍結融解してP2ウイルスを得た。それぞれの組換えウイルスについて、5ウェルずつ継代を行った。
rsSA11-NLucおよびrsSA11-Rv-NLucの継代後のNLuc遺伝子およびRv-NLuc遺伝子の安定性を調べるために、rsSA11-NLucおよびrsSA11-Rv-NLucをMA104細胞でP10まで継代を繰り返した。12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、ウイルス感染前に培地を除去し、0.5μg/mlのトリプシンを含有するFBS不含DMEM1mLに置換した。準備ができたMA104細胞に、rsSA11-NLucおよびrsSA11-Rv-NLucのP1ウイルスをそれぞれ1μLずつ加え37℃で5日間培養した。5日培養後2回凍結融解してP2ウイルスを得た。それぞれの組換えウイルスについて、5ウェルずつ継代を行った。
P10まで継代を繰り返し、rsSA11-NLucのP10ウイルスクローン1~5およびrsSA11-Rv-NLucのP10ウイルスクローン1~5を得た。得られたそれぞれのP10ウイルスの各クローンからウイルスゲノムRNAを抽出し、P1ウイルス、および野生型SA11ウイルスから得られたウイルスゲノムRNAと共にSDS-PAGEに供した。
1-2 結果
結果を図4に示した。rsSA11-NLuc P1ウイルスのウイルスゲノムRNAでは、NSP1-NLuc遺伝子が認められたが、rsSA11-NLuc P10ウイルスのクローン1~5のウイルスゲノムRNAでは長いNSP1-NLuc遺伝子が消失し、短く変異したNSP1-NLuc遺伝子が観察されるクローンが認められた。一方、rsSA11-Rv-NLucでは、P1ウイルスおよびP10ウイルスのクローン1~5のいずれにも、長いNSP1-Rv-NLuc遺伝子のみが認められた。rsSA11-NLuc P10ウイルスに認められた短い変異NSP1-NLuc遺伝子をシークエンス解析したところ、NLuc遺伝子の大部分を欠失していることがわかった。この結果から、Rv-NLuc遺伝子はNLuc遺伝子と比べ、人工組換えロタウイルス中において安定的に保持されることが示された。
結果を図4に示した。rsSA11-NLuc P1ウイルスのウイルスゲノムRNAでは、NSP1-NLuc遺伝子が認められたが、rsSA11-NLuc P10ウイルスのクローン1~5のウイルスゲノムRNAでは長いNSP1-NLuc遺伝子が消失し、短く変異したNSP1-NLuc遺伝子が観察されるクローンが認められた。一方、rsSA11-Rv-NLucでは、P1ウイルスおよびP10ウイルスのクローン1~5のいずれにも、長いNSP1-Rv-NLuc遺伝子のみが認められた。rsSA11-NLuc P10ウイルスに認められた短い変異NSP1-NLuc遺伝子をシークエンス解析したところ、NLuc遺伝子の大部分を欠失していることがわかった。この結果から、Rv-NLuc遺伝子はNLuc遺伝子と比べ、人工組換えロタウイルス中において安定的に保持されることが示された。
〔実施例2:緑色蛍光タンパク質を発現する人工組換えロタウイルス〕
2-1 材料および方法
緑色蛍光タンパク質遺伝子として、pZsGreen1-N1 ベクター(クロンテック)に含まれるZsGreen遺伝子(以下「ZsG遺伝子」と称する)を使用した。ZsG遺伝子の塩基配列を配列番号14に示した。ZsG遺伝子のコドン組成およびRV NSP1遺伝子のコドン組成を調べ、得られた両遺伝子のコドン組成に基づいて、ZsG遺伝子のコドン組成をRV NSP1遺伝子のコドン組成に近くなるように改変した。得られた改変ZsG遺伝子を「Rv-ZsG遺伝子」と称する。
2-1 材料および方法
緑色蛍光タンパク質遺伝子として、pZsGreen1-N1 ベクター(クロンテック)に含まれるZsGreen遺伝子(以下「ZsG遺伝子」と称する)を使用した。ZsG遺伝子の塩基配列を配列番号14に示した。ZsG遺伝子のコドン組成およびRV NSP1遺伝子のコドン組成を調べ、得られた両遺伝子のコドン組成に基づいて、ZsG遺伝子のコドン組成をRV NSP1遺伝子のコドン組成に近くなるように改変した。得られた改変ZsG遺伝子を「Rv-ZsG遺伝子」と称する。
図5(A)~(U)にRV NSP1遺伝子、ZsG遺伝子およびRv-ZsG遺伝子における各アミノ酸のコドン組成を示した。また、図6にZsG遺伝子の塩基配列(配列番号14)とRv-ZsG遺伝子の塩基配列(配列番号15)をそれぞれ示した。Rv-ZsG遺伝子は、ZsG遺伝子の全長696塩基中、203塩基が変わっている。元のZsG遺伝子のGC含量は63%であったが、Rv-ZsG遺伝子のGC含量は38%に下がり、RV NSP1遺伝子のGC含量(31%)に近づいた(表7参照)。

NLuc遺伝子およびRv-NLuc遺伝子に代えてZsG遺伝子およびRv-ZsG遺伝子を用いた以外は、実施例1と同じ方法で実験を行った。すなわち、PCRで増幅したZsG遺伝子およびRv-ZsG遺伝子をpT7-NSP1SA11のNSP1遺伝子(配列番号5)の128位と129位の間にそれぞれ挿入して、ZsG遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-ZsGと称する)、およびRv-ZsG遺伝子発現プラスミド(pT7-NSP1SA11-Rv-ZsGと称する)を作製し(図7参照)、実施例1と同じ方法で、緑色蛍光タンパク質を発現する人工組換えロタウイルスを作製した。pT7-NSP1SA11-ZsGを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-ZsG」と称し、pT7-NSP1SA11-Rv-ZsGを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-Rv-ZsG」と称する。得られたrsSA11-ZsGおよびrsSA11-RvZsGをそれぞれP10まで継代してウイルスゲノムRNAを抽出し、SDS-PAGEに供した。
2-2 結果
結果を図8に示した。実施例1の結果と同様に、rsSA11-ZsG P10ウイルスのクローン1~5のウイルスゲノムRNAでは、短く変異したNSP1-ZsG遺伝子が観察された。一方、rsSA11-Rv-ZsGでは、10回継代後も短く変異した遺伝子は認められなかった。なお、NSP1-ZsG遺伝子およびNSP1-Rv-ZsG遺伝子(2306bp)は、VP4遺伝子(2362bp)とサイズが近いため、電気泳動で判別できなかった。
結果を図8に示した。実施例1の結果と同様に、rsSA11-ZsG P10ウイルスのクローン1~5のウイルスゲノムRNAでは、短く変異したNSP1-ZsG遺伝子が観察された。一方、rsSA11-Rv-ZsGでは、10回継代後も短く変異した遺伝子は認められなかった。なお、NSP1-ZsG遺伝子およびNSP1-Rv-ZsG遺伝子(2306bp)は、VP4遺伝子(2362bp)とサイズが近いため、電気泳動で判別できなかった。
〔実施例3:赤色蛍光タンパク質を発現する人工組換えロタウイルス〕
3-1 材料および方法
赤色蛍光タンパク質遺伝子として、pAsRed2-N1 ベクター(クロンテック)に含まれるAsRed遺伝子(以下「AsR遺伝子」と称する)を使用した。ZsG遺伝子の塩基配列を配列番号16に示した。AsR遺伝子のコドン組成およびRV NSP1遺伝子のコドン組成を調べ、得られた両遺伝子のコドン組成に基づいて、AsR遺伝子のコドン組成をRV NSP1遺伝子のコドン組成に近くなるように改変した。得られた改変AsR遺伝子を「Rv-AsR遺伝子」と称する。
3-1 材料および方法
赤色蛍光タンパク質遺伝子として、pAsRed2-N1 ベクター(クロンテック)に含まれるAsRed遺伝子(以下「AsR遺伝子」と称する)を使用した。ZsG遺伝子の塩基配列を配列番号16に示した。AsR遺伝子のコドン組成およびRV NSP1遺伝子のコドン組成を調べ、得られた両遺伝子のコドン組成に基づいて、AsR遺伝子のコドン組成をRV NSP1遺伝子のコドン組成に近くなるように改変した。得られた改変AsR遺伝子を「Rv-AsR遺伝子」と称する。
図9(A)~(U)にRV NSP1遺伝子、AsR遺伝子およびRv-AsR遺伝子における各アミノ酸のコドン組成を示した。また、図10にAsR遺伝子の塩基配列(配列番号16)とRv-AsR遺伝子の塩基配列(配列番号17)をそれぞれ示した。Rv-AsR遺伝子は、AsR遺伝子の全長699塩基中、198塩基が変わっている。元のAsR遺伝子のGC含量は65%であったが、Rv-AsR遺伝子のGC含量は40%に下がり、RV NSP1遺伝子のGC含量(31%)に近づいた(表8参照)。

NLuc遺伝子およびRv-NLuc遺伝子に代えてAsR遺伝子およびRv-AsR遺伝子を用いた以外は、実施例1と同じ方法で実験を行った。すなわち、PCRで増幅したAsR遺伝子およびRv-AsR遺伝子をpT7-NSP1SA11のNSP1遺伝子(配列番号5)の128位と129位の間にそれぞれ挿入して、AsR遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-AsRと称する)、およびRv-AsR遺伝子発現プラスミド(pT7-NSP1SA11-Rv-AsRと称する)を作製し(図11参照)、実施例1と同じ方法で、赤色蛍光タンパク質を発現する人工組換えロタウイルスを作製した。pT7-NSP1SA11-AsRを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-AsR」と称し、pT7-NSP1SA11-Rv-AsRを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-Rv-AsR」と称する。得られたrsSA11-AsRおよびrsSA11-Rv-AsRをそれぞれP10まで継代してウイルスゲノムRNAを抽出し、SDS-PAGEに供した。
3-2 結果
結果を図12に示した。実施例1および2の結果と同様に、rsSA11-AsR P10ウイルスのクローン1~5のウイルスゲノムRNAでは、短く変異したNSP1-AsR遺伝子が観察された。一方、rsSA11-Rv-AsRでは、10回継代後も短く変異した遺伝子は認められなかった。なお、NSP1-AsR遺伝子およびNSP1-Rv-AsR遺伝子(2309bp)は、VP4遺伝子(2362bp)とサイズが近いため、電気泳動で判別できなかった。
結果を図12に示した。実施例1および2の結果と同様に、rsSA11-AsR P10ウイルスのクローン1~5のウイルスゲノムRNAでは、短く変異したNSP1-AsR遺伝子が観察された。一方、rsSA11-Rv-AsRでは、10回継代後も短く変異した遺伝子は認められなかった。なお、NSP1-AsR遺伝子およびNSP1-Rv-AsR遺伝子(2309bp)は、VP4遺伝子(2362bp)とサイズが近いため、電気泳動で判別できなかった。
〔実施例4:改変外来遺伝子由来タンパク質の発現量〕
4-1 材料および方法
実施例2で作製した緑色蛍光タンパク質を発現するrsSA11-ZsGとrsSA11-Rv-ZsG、ならびに野生型SA11を使用した。12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI(Multiplicity of Infection)0.5 PFU/cellsで感染させ、感染後24時間におけるZsGおよびRv-ZsGの発現を蛍光顕微鏡で観察し、ウエスタンブロッティングで定量した。別途、12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI: 0.01 PFU/cellsで感染させ、0.5μg/mlのトリプシンを含有するFBS不含DMEMで培養した。感染24時間後に、細胞を2回凍結融解し、細胞破砕液中のウイルス力価を測定した。
4-1 材料および方法
実施例2で作製した緑色蛍光タンパク質を発現するrsSA11-ZsGとrsSA11-Rv-ZsG、ならびに野生型SA11を使用した。12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI(Multiplicity of Infection)0.5 PFU/cellsで感染させ、感染後24時間におけるZsGおよびRv-ZsGの発現を蛍光顕微鏡で観察し、ウエスタンブロッティングで定量した。別途、12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI: 0.01 PFU/cellsで感染させ、0.5μg/mlのトリプシンを含有するFBS不含DMEMで培養した。感染24時間後に、細胞を2回凍結融解し、細胞破砕液中のウイルス力価を測定した。
4-2 結果
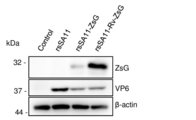
蛍光顕微鏡の観察像を図13に、ウエスタンブロッティングの結果を図14に、ウイルス力価の測定結果を図15にそれぞれ示した。蛍光顕微鏡(Nikon)で露光時間を100m秒および400m秒としてZsGおよびRv-ZsGによる緑色蛍光を観察したところ、ZsGと比較しRv-ZsGに強い発光が認められた(図13)。またZsGタンパク質特異的抗体を用いたウエスタンブロッティングによりRv-ZsGの強い発現が確認された(図14)。一方、rsSA11-ZsGとrsSA11-Rv-ZsGの増殖能は同程度であった(図15)ことから、ZsGとRv-ZsGの発現量の差はウイルス増殖能とは関係ないことが示された。
蛍光顕微鏡の観察像を図13に、ウエスタンブロッティングの結果を図14に、ウイルス力価の測定結果を図15にそれぞれ示した。蛍光顕微鏡(Nikon)で露光時間を100m秒および400m秒としてZsGおよびRv-ZsGによる緑色蛍光を観察したところ、ZsGと比較しRv-ZsGに強い発光が認められた(図13)。またZsGタンパク質特異的抗体を用いたウエスタンブロッティングによりRv-ZsGの強い発現が確認された(図14)。一方、rsSA11-ZsGとrsSA11-Rv-ZsGの増殖能は同程度であった(図15)ことから、ZsGとRv-ZsGの発現量の差はウイルス増殖能とは関係ないことが示された。
〔参考例1:改変外来遺伝子由来タンパク質の発現量〕
実施例3で作製した赤色蛍光タンパク質を発現するrsSA11-AsRおよびrsSA11-Rv-AsRを使用した。実施例4と同様に、12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI(Multiplicity of Infection)0.5 PFU/cellsで感染させ、感染後24時間におけるAsRおよびRv-AsRの発現を蛍光顕微鏡で観察した。
実施例3で作製した赤色蛍光タンパク質を発現するrsSA11-AsRおよびrsSA11-Rv-AsRを使用した。実施例4と同様に、12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI(Multiplicity of Infection)0.5 PFU/cellsで感染させ、感染後24時間におけるAsRおよびRv-AsRの発現を蛍光顕微鏡で観察した。
結果を図16に示した。蛍光顕微鏡(Nikon)で露光時間を400m秒、800m秒および1.5秒としてAsRおよびRv-AsRによる赤色蛍光を観察したところ、差は認められなかった。すなわち、AsRとRv-AsRの発現量に差がないことが示された。
〔参考例2:改変外来遺伝子由来タンパク質の発現量〕
実施例1で作製したルシフェラーゼを発現するrsSA11-NLucとrsSA11-Rv-NLuc、ならびに野生型SA11を使用した。12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI(Multiplicity of Infection)0.1 PFU/cellsで感染させ、感染後24時間におけるNLucおよびRv-NLucの発現をルシフェラーゼ活性(発光強度)として定量した。
実施例1で作製したルシフェラーゼを発現するrsSA11-NLucとrsSA11-Rv-NLuc、ならびに野生型SA11を使用した。12ウェルプレートでコンフルエントな状態にしたMA104細胞を準備し、各ウイルスをMOI(Multiplicity of Infection)0.1 PFU/cellsで感染させ、感染後24時間におけるNLucおよびRv-NLucの発現をルシフェラーゼ活性(発光強度)として定量した。
結果を図17に示した。rsSA11-NLucとrsSA11-Rv-NLucのルシフェラーゼ活性に差はなかった。すなわち、NLucとRv-NLucの発現量に差がないことが示された。
〔実施例5:1.6kbp塩基長の外来遺伝子を発現する人工組換えロタウイルス〕
1kbp以上の長い塩基長を有する外来遺伝子を発現する人工組換えロタウイルスの作製を試みた。
5-1 材料および方法
(1)外来遺伝子
外来遺伝子として、ホタルルシフェラーゼ(FLuc)遺伝子の改変体であるAkaluc遺伝子(GenBank ACCESSION: LC320664、1653 bp)のコドン組成を、RV NSP1遺伝子のコドン組成に近くなるように改変した改変Akaluc遺伝子(以下「Rv-Akaluc遺伝子」)、および、ノロウイルスVP1遺伝子(GenBank ACCESSION: KM268107、1623 bp)のコドン組成を、RV NSP1遺伝子のコドン組成に近くなるように改変した改変ノロウイルスVP1遺伝子(以下「Rv-NoV VP1遺伝子」)を使用した。改変Akaluc遺伝子の塩基配列を配列番号18に、改変ノロウイルスVP1遺伝子の塩基配列を配列番号19に示す。
1kbp以上の長い塩基長を有する外来遺伝子を発現する人工組換えロタウイルスの作製を試みた。
5-1 材料および方法
(1)外来遺伝子
外来遺伝子として、ホタルルシフェラーゼ(FLuc)遺伝子の改変体であるAkaluc遺伝子(GenBank ACCESSION: LC320664、1653 bp)のコドン組成を、RV NSP1遺伝子のコドン組成に近くなるように改変した改変Akaluc遺伝子(以下「Rv-Akaluc遺伝子」)、および、ノロウイルスVP1遺伝子(GenBank ACCESSION: KM268107、1623 bp)のコドン組成を、RV NSP1遺伝子のコドン組成に近くなるように改変した改変ノロウイルスVP1遺伝子(以下「Rv-NoV VP1遺伝子」)を使用した。改変Akaluc遺伝子の塩基配列を配列番号18に、改変ノロウイルスVP1遺伝子の塩基配列を配列番号19に示す。
(2)外来遺伝子挿入NSP1発現プラスミドの作製
PCRで増幅したRv-Akaluc遺伝子およびRv-NoV VP1遺伝子をpT7-NSP1SA11のNSP1遺伝子(配列番号5)の128位と129位の間にそれぞれ挿入した。さらに、Kanaiら(非特許文献2)に従い、NSP1遺伝子に722bpの欠失を加え、Rv-Akaluc遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-Rv-Akalucと称する)およびRv-NoV VP1遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-Rv-NoV VP1と称する)を作製した(図18参照)。
PCRで増幅したRv-Akaluc遺伝子およびRv-NoV VP1遺伝子をpT7-NSP1SA11のNSP1遺伝子(配列番号5)の128位と129位の間にそれぞれ挿入した。さらに、Kanaiら(非特許文献2)に従い、NSP1遺伝子に722bpの欠失を加え、Rv-Akaluc遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-Rv-Akalucと称する)およびRv-NoV VP1遺伝子挿入NSP1遺伝子発現プラスミド(pT7-NSP1SA11-Rv-NoV VP1と称する)を作製した(図18参照)。
(3)人工組換えウイルスの作製および外来遺伝子保持の確認
実施例1と同じ方法で、Rv-Akalucを発現する人工組換えロタウイルス、および、Rv-NoV VP1を発現する人工組換えロタウイルスを作製した。pT7-NSP1SA11-Rv-Akalucを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-Rv-Akaluc」と称し、pT7-NSP1SA11-Rv-NoV VP1を含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-Rv-NoV VP1」と称する。得られたrsSA11-Rv-AkalucおよびrsSA11-Rv-NoV VP1をそれぞれP10まで継代してウイルスゲノムRNAを抽出し、SDS-PAGEに供した。
実施例1と同じ方法で、Rv-Akalucを発現する人工組換えロタウイルス、および、Rv-NoV VP1を発現する人工組換えロタウイルスを作製した。pT7-NSP1SA11-Rv-Akalucを含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-Rv-Akaluc」と称し、pT7-NSP1SA11-Rv-NoV VP1を含む11種類のプラスミドを用いて作製した人工組換えロタウイルスを「rsSA11-Rv-NoV VP1」と称する。得られたrsSA11-Rv-AkalucおよびrsSA11-Rv-NoV VP1をそれぞれP10まで継代してウイルスゲノムRNAを抽出し、SDS-PAGEに供した。
5-2 結果
rsSA11-Rv-Akalucの結果を図19に、rsSA11-Rv-NoV VP1の結果を図20に示した。rsSA11-Rv-AkalucおよびrsSA11-Rv-NoV VP1の各ゲノムRNAのバンドは、NSP1以外すべて野生型SA11株と同じ位置に確認された。一方、rsSA11-Rv-AkalucのNSP1-Rv-Akaluc遺伝子のバンドおよびrsSA11-Rv-NoV VP1のRv-NoV VP1遺伝子のバンドは、それぞれ野生型SA11株のNSP1のバンドより高い位置に確認された。この結果から、1kbp以上の長い塩基長を有する外来遺伝子についても、コドン組成をロタウイルスのコドン組成に近づけることにより、1kbp以上の塩基長を有する外来遺伝子を安定に保持する組換えロタウイルスを作製できることが示された
rsSA11-Rv-Akalucの結果を図19に、rsSA11-Rv-NoV VP1の結果を図20に示した。rsSA11-Rv-AkalucおよびrsSA11-Rv-NoV VP1の各ゲノムRNAのバンドは、NSP1以外すべて野生型SA11株と同じ位置に確認された。一方、rsSA11-Rv-AkalucのNSP1-Rv-Akaluc遺伝子のバンドおよびrsSA11-Rv-NoV VP1のRv-NoV VP1遺伝子のバンドは、それぞれ野生型SA11株のNSP1のバンドより高い位置に確認された。この結果から、1kbp以上の長い塩基長を有する外来遺伝子についても、コドン組成をロタウイルスのコドン組成に近づけることにより、1kbp以上の塩基長を有する外来遺伝子を安定に保持する組換えロタウイルスを作製できることが示された
〔実施例6:緑色蛍光タンパク質を発現する人工組換えコウモリレオウイルス〕
6-1 材料および方法
(1)ウイルス
コウモリレオウイルス(Pteropine orthoreovirus、以下「PRV」と称する)はロタウイルスと同じレオウイルス科に属し、L1、L2、L3、M1、M2、M3、S1、S2、S3およびS4からなる10分節の2本鎖RNAを持つ。実施例1のロタウイルスの各分節RNAゲノム発現カセットを含むプラスミド(分節RNAゲノム発現ベクター)の作製と同じ方法で、PRVの10個の分節RNAゲノムのcDNAを含むプラスミドを作製した。
6-1 材料および方法
(1)ウイルス
コウモリレオウイルス(Pteropine orthoreovirus、以下「PRV」と称する)はロタウイルスと同じレオウイルス科に属し、L1、L2、L3、M1、M2、M3、S1、S2、S3およびS4からなる10分節の2本鎖RNAを持つ。実施例1のロタウイルスの各分節RNAゲノム発現カセットを含むプラスミド(分節RNAゲノム発現ベクター)の作製と同じ方法で、PRVの10個の分節RNAゲノムのcDNAを含むプラスミドを作製した。
(2)緑色蛍光タンパク質遺伝子
実施例2と同じ、ZsG遺伝子およびRv-ZsG遺伝子を使用した。ZsG遺伝子およびRv-ZsG遺伝子は、PRV S1遺伝子に挿入した。PRV S1遺伝子は、FAST、p17およびsigmaCをコードする遺伝子であるが、PRV S1遺伝子の連続したFAST-p17-sigmaCを、ZsG-2A-FASTまたはRvZsG-2A-FASTに入れかえたS1-ZsG-2A-FAST遺伝子およびS1-RvZsG-2A-FASTを作製した(図21参照)。
実施例2と同じ、ZsG遺伝子およびRv-ZsG遺伝子を使用した。ZsG遺伝子およびRv-ZsG遺伝子は、PRV S1遺伝子に挿入した。PRV S1遺伝子は、FAST、p17およびsigmaCをコードする遺伝子であるが、PRV S1遺伝子の連続したFAST-p17-sigmaCを、ZsG-2A-FASTまたはRvZsG-2A-FASTに入れかえたS1-ZsG-2A-FAST遺伝子およびS1-RvZsG-2A-FASTを作製した(図21参照)。
(3)緑色蛍光タンパク質発現人工組換えコウモリレオウイルスの作製
実施例1と同じ方法で、S1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとS1-ZsG-2A-FAST遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、サルMA104細胞に継代して人工組換えコウモリレオウイルス(「rsMB-ZsG-2A-FAST」と称する)を得た。同様に、S1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとS1-Rv-ZsG-2A-FAST遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、サルMA104細胞に継代して人工組換えコウモリレオウイルス(「rsMB-Rv-ZsG-2A-FAST」と称する)を得た。
実施例1と同じ方法で、S1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとS1-ZsG-2A-FAST遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、サルMA104細胞に継代して人工組換えコウモリレオウイルス(「rsMB-ZsG-2A-FAST」と称する)を得た。同様に、S1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとS1-Rv-ZsG-2A-FAST遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、サルMA104細胞に継代して人工組換えコウモリレオウイルス(「rsMB-Rv-ZsG-2A-FAST」と称する)を得た。
6-2 結果
プラーク法によりrsMB-ZsG-2A-FASTウイルスの緑色蛍光タンパク質(ZsG)の発現を蛍光顕微鏡で調べた結果、全ウイルス粒子中の1/50-1/100程度が緑色蛍光を発していた。しかし、ウイルスの継代を2~3回行うと緑色蛍光を発するウイルスは消失した。一方、rsMB-Rv-ZsG-2A-FASTウイルスは100%のウイルスが緑色蛍光を発しており、ウイルスの継代を2~3回行った後もウイルスは緑色蛍光を発しており(図22参照)、Rv-ZsG遺伝子が安定に保持されていることが示された。
プラーク法によりrsMB-ZsG-2A-FASTウイルスの緑色蛍光タンパク質(ZsG)の発現を蛍光顕微鏡で調べた結果、全ウイルス粒子中の1/50-1/100程度が緑色蛍光を発していた。しかし、ウイルスの継代を2~3回行うと緑色蛍光を発するウイルスは消失した。一方、rsMB-Rv-ZsG-2A-FASTウイルスは100%のウイルスが緑色蛍光を発しており、ウイルスの継代を2~3回行った後もウイルスは緑色蛍光を発しており(図22参照)、Rv-ZsG遺伝子が安定に保持されていることが示された。
〔実施例7:緑色蛍光タンパク質を発現する人工組換え哺乳類レオウイルス〕
7-1 材料および方法
(1)ウイルス
哺乳類レオウイルス(Mammalian orthoreovirus、以下「MRV」と称する)はロタウイルスと同じレオウイルス科に属し、L1、L2、L3、M1、M2、M3、S1、S2、S3およびS4からなる10分節の2本鎖RNAを持つ。実施例1のロタウイルスの各分節RNAゲノム発現カセットを含むプラスミド(分節RNAゲノム発現ベクター)の作製と同じ方法で、MRVの10個の分節RNAゲノムのcDNAを含むプラスミドを作製した。
7-1 材料および方法
(1)ウイルス
哺乳類レオウイルス(Mammalian orthoreovirus、以下「MRV」と称する)はロタウイルスと同じレオウイルス科に属し、L1、L2、L3、M1、M2、M3、S1、S2、S3およびS4からなる10分節の2本鎖RNAを持つ。実施例1のロタウイルスの各分節RNAゲノム発現カセットを含むプラスミド(分節RNAゲノム発現ベクター)の作製と同じ方法で、MRVの10個の分節RNAゲノムのcDNAを含むプラスミドを作製した。
(2)緑色蛍光タンパク質遺伝子
緑色蛍光タンパク質としてZsG遺伝子およびZsG遺伝子のコドン組成を、MRV L1遺伝子のコドン組成に近似させたMrv-ZsG遺伝子を使用した。図23(A)~(U)にMRV L1遺伝子、ZsG遺伝子およびMrv-ZsG遺伝子における各アミノ酸のコドン組成を示した。元のZsG遺伝子のGC含量は63%であったが、Mrv-ZsG遺伝子のGC含量は43%に下がり、MRV L1遺伝子のGC含量(46%)に近づいている(表9参照)。
緑色蛍光タンパク質としてZsG遺伝子およびZsG遺伝子のコドン組成を、MRV L1遺伝子のコドン組成に近似させたMrv-ZsG遺伝子を使用した。図23(A)~(U)にMRV L1遺伝子、ZsG遺伝子およびMrv-ZsG遺伝子における各アミノ酸のコドン組成を示した。元のZsG遺伝子のGC含量は63%であったが、Mrv-ZsG遺伝子のGC含量は43%に下がり、MRV L1遺伝子のGC含量(46%)に近づいている(表9参照)。

ZsG遺伝子およびMrv-ZsG遺伝子は、MRV L1遺伝子に挿入した。MRV L1遺伝子は、lamda 3タンパク質をコードする遺伝子であり、ZsG遺伝子およびMRV-ZsG遺伝子の下流に自己切断ペプチドをコードする2A遺伝子を挿入し、L1-ZsG-2A-Lamda3およびL1-Mrv-ZsG-2A-Lamda3を作製した(図24参照)。
(3)緑色蛍光タンパク質発現人工組換え哺乳類レオウイルスの作製
実施例1と同じ方法で、L1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとL1-ZsG-2A-Lamda3遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、マウスL929細胞に継代して人工組換え哺乳類レオウイルス(「rsMRV-ZsG」と称する)を得た。同様に、L1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとL1-Mrv-ZsG-2A-Lamda3遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、マウスL929細胞に継代して人工組換え哺乳類レオウイルス(「rsMRV-Mrv-ZsG」と称する)を得た。
実施例1と同じ方法で、L1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとL1-ZsG-2A-Lamda3遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、マウスL929細胞に継代して人工組換え哺乳類レオウイルス(「rsMRV-ZsG」と称する)を得た。同様に、L1遺伝子以外の野生型遺伝子を含む9種類のプラスミドとL1-Mrv-ZsG-2A-Lamda3遺伝子を含むプラスミドをBHK-T7/P5細胞にトランスフェクションし、マウスL929細胞に継代して人工組換え哺乳類レオウイルス(「rsMRV-Mrv-ZsG」と称する)を得た。
7-2 結果
MRV特異的抗体を用いた免疫染色法によりrsMRV-ZsGウイルスの緑色蛍光タンパク質(ZsG)の発現を蛍光顕微鏡で調べた結果、観察したいずれのrsMRV-ZsGウイルス感染細胞も緑色蛍光を発していなかった。一方、rsMRV-Mrv-ZsGウイルスは100%のウイルス感染細胞が緑色蛍光を発しており、ウイルスの継代を3回行った後もウイルスは緑色蛍光を発しており(図25参照)、Mrv-ZsG遺伝子が安定に保持されていることが示された。また1~3回継代した後のrsMRV-ZsGおよびrsMRV-Mrv-ZsGからウイルスゲノムRNAを抽出し、電気泳動解析を行ったところ、rsMRV-ZsGウイルスは、挿入したZsG遺伝子が1回目の継代の時点で欠失していたのに対し、rsMRV-Mrv-ZsGウイルスは、3回継代した後にもMrv-ZsG遺伝子を保持していることが確認された(図26参照)。
MRV特異的抗体を用いた免疫染色法によりrsMRV-ZsGウイルスの緑色蛍光タンパク質(ZsG)の発現を蛍光顕微鏡で調べた結果、観察したいずれのrsMRV-ZsGウイルス感染細胞も緑色蛍光を発していなかった。一方、rsMRV-Mrv-ZsGウイルスは100%のウイルス感染細胞が緑色蛍光を発しており、ウイルスの継代を3回行った後もウイルスは緑色蛍光を発しており(図25参照)、Mrv-ZsG遺伝子が安定に保持されていることが示された。また1~3回継代した後のrsMRV-ZsGおよびrsMRV-Mrv-ZsGからウイルスゲノムRNAを抽出し、電気泳動解析を行ったところ、rsMRV-ZsGウイルスは、挿入したZsG遺伝子が1回目の継代の時点で欠失していたのに対し、rsMRV-Mrv-ZsGウイルスは、3回継代した後にもMrv-ZsG遺伝子を保持していることが確認された(図26参照)。
なお本発明は上述した各実施形態および実施例に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。また、本明細書中に記載された学術文献および特許文献の全てが、本明細書中において参考として援用される。
Claims (8)
- 外来遺伝子を安定的に保持する人工組換えRNAウイルスの作製方法であって、
(1)RNAウイルス遺伝子のコドン組成に近似するように改変されたコドン組成を有する外来遺伝子を取得する工程、
(2)(1)で取得した外来遺伝子をRNAウイルスのゲノムに挿入する工程、および
(3)リバースジェネティクス法を用いて人工組換えRNAウイルスを人工的に合成する工程を含むことを特徴とする作製方法。 - RNAウイルスがレオウイルス科に属するウイルスである請求項1に記載の方法。
- レオウイルス科に属するウイルスがロタウイルス属またはオルソレオウイルス属に属するウイルスである請求項2に記載の方法。
- 工程(1)において、RNAウイルス遺伝子のコドン組成の±30%の範囲内になるように改変されたコドン組成を有する外来遺伝子を取得する請求項1~3のいずれかに記載の方法。
- リバースジェネティクス法を用いて人工的に合成された人工組換えRNAウイルスに導入された外来遺伝子を安定的に保持させる方法であって、外来遺伝子のコドン組成を、RNAウイルス遺伝子のコドン組成に近似させることを特徴とする方法。
- RNAウイルスがレオウイルス科に属するウイルスである請求項5に記載の方法。
- レオウイルス科に属するウイルスがロタウイルス属またはオルソレオウイルス属に属するウイルスである請求項6に記載の方法。
- 外来遺伝子のコドン組成を、RNAウイルス遺伝子のコドン組成の±30%の範囲内に近似させる請求項5~7のいずれかに記載の方法。
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US17/619,899 US20220356491A1 (en) | 2019-06-21 | 2020-06-19 | Method for preparing artificial recombinant rna virus that stably holds foreign gene |
| JP2021526915A JP7517704B2 (ja) | 2019-06-21 | 2020-06-19 | 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 |
| CN202080045560.2A CN114040979B (zh) | 2019-06-21 | 2020-06-19 | 稳定地保持外源基因的人工重组rna病毒的制作方法 |
| EP20825608.1A EP3988650A4 (en) | 2019-06-21 | 2020-06-19 | METHOD FOR PREPARING AN ARTIFICIAL RECOMBINANT RNA VIRUS STABLELY INTEGRATING A FOREIGN GENE |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2019-115532 | 2019-06-21 | ||
| JP2019115532 | 2019-06-21 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2020256099A1 true WO2020256099A1 (ja) | 2020-12-24 |
Family
ID=74040505
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2020/024131 WO2020256099A1 (ja) | 2019-06-21 | 2020-06-19 | 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20220356491A1 (ja) |
| EP (1) | EP3988650A4 (ja) |
| JP (1) | JP7517704B2 (ja) |
| CN (1) | CN114040979B (ja) |
| WO (1) | WO2020256099A1 (ja) |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2008507290A (ja) * | 2004-08-03 | 2008-03-13 | ジーンアート・アクチエンゲゼルシャフト | 誘導性遺伝子発現 |
| WO2018060368A2 (en) * | 2016-09-28 | 2018-04-05 | Bavarian Nordic A/S | Compositions and methods for enhancing the stability of transgenes in poxviruses |
| WO2018062199A1 (ja) | 2016-09-27 | 2018-04-05 | 国立大学法人大阪大学 | 人工組換えロタウイルスの作製方法 |
Family Cites Families (11)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2331335A1 (en) * | 1998-09-29 | 2000-04-06 | Maxygen, Inc. | Shuffling of codon altered genes |
| JP4999330B2 (ja) * | 2004-01-22 | 2012-08-15 | 株式会社ディナベック研究所 | サイトメガロウイルスエンハンサーおよびニワトリβ−アクチンプロモーターを含むハイブリッドプロモーターを利用したマイナス鎖RNAウイルスベクターの製造方法 |
| JP2009519715A (ja) * | 2005-12-16 | 2009-05-21 | ザ ユーエービー リサーチ ファウンデーション | ウイルスベクターを使用する制御された遺伝子発現に関する組成物および方法 |
| CN101245350B (zh) * | 2007-02-17 | 2011-12-28 | 王健伟 | 密码子优化的轮状病毒蛋白的编码核苷酸序列、其重组体及其应用 |
| US9476032B2 (en) * | 2007-03-30 | 2016-10-25 | The Research Foundation For The State University Of New York | Attenuated viruses useful for vaccines |
| GB0800272D0 (en) * | 2008-01-08 | 2008-02-13 | Plant Bioscience Ltd | Protein expression systems |
| CN101502650B (zh) * | 2009-03-10 | 2011-12-28 | 邢益平 | 密码子优化的乙型肝炎核酸疫苗 |
| AU2010277310B2 (en) * | 2009-07-31 | 2015-01-15 | Seqirus UK Limited | Reverse genetics systems |
| US10544431B2 (en) * | 2015-01-16 | 2020-01-28 | National Institute Of Advanced Industrial Science And Technology | Gene expression system using stealthy RNA, and gene introduction/expression vector including said RNA |
| CN104830874B (zh) * | 2015-04-16 | 2017-11-21 | 南京医科大学第一附属医院 | 密码子优化的严重发热伴血小板减少综合征病毒核蛋白基因及其核酸疫苗 |
| WO2017195749A1 (ja) * | 2016-05-11 | 2017-11-16 | 国立大学法人大阪大学 | キメラ抗原受容体、及びその利用 |
-
2020
- 2020-06-19 US US17/619,899 patent/US20220356491A1/en active Pending
- 2020-06-19 JP JP2021526915A patent/JP7517704B2/ja active Active
- 2020-06-19 EP EP20825608.1A patent/EP3988650A4/en active Pending
- 2020-06-19 WO PCT/JP2020/024131 patent/WO2020256099A1/ja unknown
- 2020-06-19 CN CN202080045560.2A patent/CN114040979B/zh active Active
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2008507290A (ja) * | 2004-08-03 | 2008-03-13 | ジーンアート・アクチエンゲゼルシャフト | 誘導性遺伝子発現 |
| WO2018062199A1 (ja) | 2016-09-27 | 2018-04-05 | 国立大学法人大阪大学 | 人工組換えロタウイルスの作製方法 |
| WO2018060368A2 (en) * | 2016-09-28 | 2018-04-05 | Bavarian Nordic A/S | Compositions and methods for enhancing the stability of transgenes in poxviruses |
Non-Patent Citations (27)
| Title |
|---|
| "GenBank", Database accession no. NC006998 |
| ADACHI A ET AL.: "Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone", JOURNAL OF VIROLOGY, vol. 59, no. 2, 1986, pages 284 - 291 |
| BOYCE MCELMA CCROY P: "Development of reverse genetics systems for bluetongue virus: recovery of infectious virus from synthetic RNA transcripts", JOURNAL OF VIROLOGY, vol. 82, no. 17, 2008, pages 8339 - 8348, XP002513548, DOI: 10.1128/JVI.00808-08 |
| BRIDGEN AELLIOTT RM: "Rescue of a segmented negative-strand RNA virus entirely from cloned complementary DNAs", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 93, no. 26, 1996, pages 15400 - 15404, XP002224465, DOI: 10.1073/pnas.93.26.15400 |
| CHAUDHRY YSKINNER MAGOODFELLOW IG: "Recovery of genetically defined murine norovirus in tissue culture by using a fowlpox virus expressing T7 RNA polymerase", THE JOURNAL OF GENERAL VIROLOGY, vol. 88, 2007, pages 2091 - 2100 |
| FISCHER, M. D. ET AL.: "C odon-Optimized RPGR Improves Stability and Efficacy of AAV8 Gene Therapy in Two Mouse Models of X-Linked Retinitis Pigmentosa", MOL. THER., vol. 25, no. 8, 2017, pages 1854 - 1865, XP055399604, DOI: 10.1016/j.ymthe.2017.05.005 * |
| GUSTAFSSON ET AL., TRENDS BIOTECHNOL., vol. 22, no. 7, July 2004 (2004-07-01), pages 346 - 353 |
| KANAI ET AL., JOURNAL OF VIROLOGY, vol. 93, no. 4, 5 February 2019 (2019-02-05), pages e01774 - 18 |
| KANAI ET AL., PROC NATL ACAD SCI U S A, vol. 114, no. 9, 28 February 2017 (2017-02-28), pages 2349 - 2354 |
| KOBAYASHI T ET AL.: "A plasmid-based reverse genetics system for animal double-stranded RNA viruses", CELL HOST & MICROBE, vol. 1, no. 2, 2007, pages 147 - 157, XP002513546, DOI: 10.1016/J.CHOM.2007.03.003 |
| KOLYKHALOV AA ET AL.: "Transmission of hepatitis C by intrahepatic inoculation with transcribed RNA", SCIENCE, vol. 277, no. 5325, 1997, pages 570 - 574, XP002734276, DOI: 10.1126/science.277.5325.570 |
| LAI CJZHAO BTHORI HBRAY M: "Infectious RNA transcribed from stably cloned full-length cDNA of dengue type 4 virus", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 88, no. 12, 1991, pages 5139 - 5143, XP002073934, DOI: 10.1073/pnas.88.12.5139 |
| MADESIS, P. ET AL.: "A hepatitis C virus core polypeptide expressed in chloroplasts detects anti-core antibodies in infected human sera", J. BIOTECHNOL ., vol. 145, 2010, pages 377 - 386, XP026892462, DOI: 10.1016/j.jbiotec.2009.12.001 * |
| MATSUO ET AL., BIOCHEM BIOPHYS RES COMMUN, vol. 340, no. 1, 2006, pages 200 - 208 |
| MUNDT EVAKHARIA VN: "Synthetic transcripts of double-stranded Birnavirus genome are infectious", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 93, no. 20, 1996, pages 11131 - 11136, XP002044624, DOI: 10.1073/pnas.93.20.11131 |
| NEUMANN G ET AL.: "Generation of influenza A viruses entirely from cloned cDNAs", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 96, no. 16, 1999, pages 9345 - 9350 |
| RACANIELLO VRBALTIMORE D: "Cloned poliovirus complementary DNA is infectious in mammalian cells", SCIENCE, vol. 214, no. 4523, 1981, pages 916 - 919, XP001057398, DOI: 10.1126/science.6272391 |
| RADECKE F ET AL.: "Rescue of measles viruses from cloned DNA", EMBO J, vol. 14, no. 23, 1995, pages 5773 - 5784, XP002022952 |
| SANCHEZ ABDE LA TORRE JC: "Rescue of the prototypic Arenavirus LCMV entirely from plasmid", VIROLOGY, vol. 350, no. 2, 2006, pages 370 - 380, XP024896500, DOI: 10.1016/j.virol.2006.01.012 |
| SCHNEIDER USCHWEMMLE MSTAEHELI P: "Genome trimming: a unique strategy for replication control employed by Borna disease virus", PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA, vol. 102, no. 9, 2005, pages 3441 - 3446, XP008153168, DOI: 10.1073/pnas.0405965102 |
| SCHNELL MJMEBATSION TCONZELMANN KK: "Infectious Rabies Viruses from Cloned Cdna", EMBO J, vol. 13, no. 18, 1994, pages 4195 - 4203 |
| See also references of EP3988650A4 |
| T ANAKA, M. ET AL.: "Effects of codon optimization on the mRNA levels of heterologous genes in filamentous fungi", APPL. MICROBIOL. BIOTECHNOL., vol. 98, 2014, pages 3859 - 3867, XP035328382, DOI: 10.1007/s00253-014-5609-7 * |
| TOKUOKA, MASAFUMI, TANAKA MIZUKI, ONO KAZUHISA, TAKAGI SHINOBU, SHINTANI TAKAHIRO, GOMI KATSUYA: "Codon Optimization Increases Steady-State mRNA Levels in Aspergillus oryzae Heterologous Gene Expression", APPL. ENVIRON. MICROBIOL ., vol. 74, no. 21, 2008, pages 6538 - 6546, XP055773591, DOI: 10.1128/AEM.01354-08 * |
| VOLCHKOV VE ET AL.: "Recovery of infectious Ebola virus from complementary DNA: RNA editing of the GP gene and viral cytotoxicity", SCIENCE, vol. 291, no. 5510, 2001, pages 1965 - 1969, XP002404372, DOI: 10.1126/science.1057269 |
| YOUNT BCURTIS KMBARIC RS: "Strategy for systematic assembly of large RNA and DNA genomes: transmissible gastroenteritis virus model", JOURNAL OF VIROLOGY, vol. 74, no. 22, 2000, pages 10600 - 10611, XP002203456, DOI: 10.1128/JVI.74.22.10600-10611.2000 |
| YUN SIKIM SYRICE CMLEE YM: "Development and application of a reverse genetics system for Japanese encephalitis virus", JOURNAL OF VIROLOGY, vol. 77, no. 11, 2003, pages 6450 - 6465, XP002367285, DOI: 10.1128/JVI.77.11.6450-6465.2003 |
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2020256099A1 (ja) | 2020-12-24 |
| US20220356491A1 (en) | 2022-11-10 |
| JP7517704B2 (ja) | 2024-07-17 |
| CN114040979B (zh) | 2024-06-28 |
| EP3988650A1 (en) | 2022-04-27 |
| CN114040979A (zh) | 2022-02-11 |
| EP3988650A4 (en) | 2022-09-21 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6944213B2 (ja) | 人工組換えロタウイルスの作製方法 | |
| Bukreyev et al. | Recovery of infectious respiratory syncytial virus expressing an additional, foreign gene | |
| KR100754091B1 (ko) | 자율 복제 능력을 갖는 (-)쇄 rna 바이러스 벡터 | |
| WO1997016539A1 (fr) | Virus sendai recombinant | |
| CN107190022B (zh) | 一种快速构建禽传染性支气管炎病毒反向遗传株的方法 | |
| JP7298906B2 (ja) | 光制御性のウイルスタンパク質、その遺伝子、及びその遺伝子を含むウイルスベクター | |
| JP5908948B2 (ja) | レオウイルス科のウイルスのワクチン用ウイルス株を製造する方法 | |
| Thiel et al. | Reverse genetics of coronaviruses using vaccinia virus vectors | |
| CN111996174A (zh) | 猪繁殖与呼吸综合征病毒及其克隆载体和插入基因方法 | |
| JP4430543B2 (ja) | 日本脳炎ウイルスの全長のゲノムRNA及び該JEVゲノムRNAに対する感染性を有するJEVcDNA及びその用途 | |
| Mebatsion et al. | Identification of a mutation in editing of defective Newcastle disease virus recombinants that modulates P-gene mRNA editing and restores virus replication and pathogenicity in chicken embryos | |
| WO2020256099A1 (ja) | 外来遺伝子を安定的に保持する人工組換えrnaウイルスの作製方法 | |
| JP2019050796A (ja) | 高効率なロタウイルスの人工合成法 | |
| JPWO2007007921A1 (ja) | 分節ゲノム型組換えモノネガウイルスベクター | |
| JP2003520573A (ja) | 分節化した(−)鎖RNAウイルスのinvitro再構築 | |
| Enjuanes et al. | Virus-based vectors for gene expression in mammalian cells: Coronavirus | |
| Desselberger | A helper virus-independent, plasmid only-based reverse genetics system for rotavirus | |
| JP3638019B2 (ja) | 組換え体センダイウイルス | |
| JP3732204B2 (ja) | 自律複製能を有する(−)鎖rnaウイルスベクター | |
| JP3991339B2 (ja) | 組換え体センダイウイルス | |
| CN113584063A (zh) | 一种基于转录调控序列重布的反向遗传操作方法及其应用 | |
| Theriault | The role of reverse genetics systems in the study of filovirus pathogenicity | |
| JP2005102702A (ja) | 組換え体センダイウイルス | |
| FR2841258A1 (fr) | Procede et preparation de mononegavirus recombinants et leurs applications |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 20825608 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2021526915 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2020825608 Country of ref document: EP Effective date: 20220121 |