CN100406562C - 与α4β7整合蛋白相作用的人化免疫球蛋白 - Google Patents
与α4β7整合蛋白相作用的人化免疫球蛋白 Download PDFInfo
- Publication number
- CN100406562C CN100406562C CNB971972214A CN97197221A CN100406562C CN 100406562 C CN100406562 C CN 100406562C CN B971972214 A CNB971972214 A CN B971972214A CN 97197221 A CN97197221 A CN 97197221A CN 100406562 C CN100406562 C CN 100406562C
- Authority
- CN
- China
- Prior art keywords
- humanized immunoglobulin
- heavy chain
- light chain
- variable region
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
Images



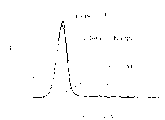














Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2839—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the integrin superfamily
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
- A61P1/04—Drugs for disorders of the alimentary tract or the digestive system for ulcers, gastritis or reflux esophagitis, e.g. antacids, inhibitors of acid secretion, mucosal protectants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/24—Immunoglobulins specific features characterized by taxonomic origin containing regions, domains or residues from different species, e.g. chimeric, humanized or veneered
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
Abstract
本发明涉及具有对α4β7整联蛋白结合特异性的人化免疫球蛋白,包括非人源的抗原结合区域(例如,啮齿类起源的)和至少一部分的人源免疫球蛋白(例如,人构架区,人恒定区)。在一个实施方案中,人化的免疫球蛋白可以鼠Act-1竞争对α4β7整合蛋白的结合。在一个优选的实施方案中,人化的免疫球蛋白的抗原结合区包括鼠Act-1抗体的重链和轻链的各自互补性决定区。本发明进一步涉及人化的免疫球蛋白的轻链和重链,包括编码本发明的人化免疫球蛋白或免疫球蛋白链的序列的核酸(例如,单链抗体),包括本发明的核酸的构建物,包括本发明的核酸的宿主细胞,用于制备人化的免疫球蛋白。人化的免疫球蛋白可以用于对人体疾病的诊断和治疗,例如,控制淋巴细胞对黏膜组织的渗入(包括征集和/或积累)。
Description
技术领域
本发明涉及人化免疫球蛋白。
背景技术
整合蛋白受体对于调节淋巴细胞循环和向炎症部位富集十分重要[Carlos,T.M.and Harlan,J.M.Blood,84:2068-2101(1994)]。人类α4β7整合蛋白有多个配体,其中一个是表达肠系膜淋巴节和派伊尔氏淋巴集结与高级内皮小静脉的黏膜血管地址素MAdCAM-1[Berlin,C.et al.Cell 74:185-195(1993);Erle,D.J.et al.J.Immunol.153:517-528(1994)])[Streeter,P.R.et al.Nature 331:41-46(1988)]。因而,α4β7整合蛋白作为一种受体,调控淋巴细胞迁向肠黏膜淋巴样组织[Schweighoffer,T.et al.J.Immunol.151:717-729(1993)]。另外,α4β7整合蛋白还同纤连蛋白和血管细胞黏附分子-1(VCAM-1)相互作用。
肠炎症疾病(IBD),诸如溃疡性结肠炎和阶段性回肠炎(Crohn’s病),都是涉及胃肠道炎症的消耗性和进行性疾病,。仅在美国就有大约200万患者,症状包括腹痛、痉挛、腹泻和直肠出血。对IBD的治疗包括消炎药物(如皮质类固醇和磺胺沙利比林),免疫抑制药物(如6-巯基嘌呤、环孢霉素和硝基咪唑硫嘌呤)以及外科手术(如结肠切除术)。参见Podolsky,New Engl.J.Med.325:928-937(1991)and Podolsky,New Engl.J.Med.325:1008-1016(1991)。
抗人α4β7整合蛋白抗体,例如鼠单克隆抗体(mAb Act-1),干扰α4β7整合蛋白结合到黏膜地址素细胞黏附分子-1(MAdCAM-1)的过程,后者出现在黏膜淋巴结的高级内皮小静脉。Act-1最早由Lazarovits,A.I.et al.J.Immunol.133:1857-1862(1984),从被人破伤风毒特异性T淋巴细胞免疫的老鼠分离出来,并被报道为是一种鼠IgG1/κ型抗体。最近的研究[Schweighoffer,T.et al.J.Immunol.151:717-729(1993)]显示,该抗体能够与特异性表达α4β7整合蛋白的人记忆CD4+T淋巴细胞的亚单位相结合。然而,利用鼠抗体来治疗人类疾病存在严重的问题,它们对于人体来讲具有高度的免疫原性,能迅速诱发人体抗鼠抗体的免疫应答(HAMA).削弱其治疗效果,也不能连续给药。HAMA反应能快速清除鼠抗体,大大限制其疗效。
因此,需要有改进的方法来治疗肠炎症疾病。
本发明的概述
本发明涉及具有α4β7整合蛋白特异结合属性的人化免疫球蛋白,该蛋白包括非人源抗原结合区(如啮齿类)和至少一部分人源免疫球蛋白(如人类框架区、人类λ型恒定区)。在一个实施方案中,这里所描述的人化免疫球蛋白将和鼠Act-1或LDP-02(见实施例4)竞争结合α4β7整合蛋白。在优选的实施方案中,人化免疫球蛋白的抗原结合区最好来源于Act-1单克隆抗体[如LDP-02,其是包含轻链可变区和重链可变区的一种免疫球蛋白,分别见图11(SEQ ID NO:19)和图12(SEQ ID NO:21)]。
例如,人化免疫球蛋白可以包括含非人源互补决定簇(CDR)的抗原结合区和从人源框架区(FR)衍生的框架区。一方面,这个有α4β7结合特性的人化免疫球蛋白包括轻链和重链,轻链包括从结合α4β7的非人源抗体衍生的CDR,和从人源轻链衍生的FR(例如,GM607’CL),重链包括从结合α4β7的非人源抗体衍生的重链和从人源重链衍生的FR(如21/28’CL)。另一方面,轻链含有三个来自于Act-1抗体轻链的CDR,重链则含有三个来自于Act-1抗体重链的CDR。
本发明也涉及人化免疫球蛋白的轻链(如Act-1抗体轻链的CDR1、CDR2、CDR3和人类轻链的FR)和人化免疫球蛋白的重链(如Act-1抗体重链的CDR1、CDR2、CDR3和人类重链的FR)。在优选的实施方案中,本发明涉及本文所描述的人化重链和轻链[例如,含有如图7所示轻链的可变区(SEQ ID NO:12)的人化轻链,含有如图9所示重链的可变区(SEQ ID NO:15)的人化重链,含有如图12所示轻链的可变区(SEQ ID NO:21)的人化轻链,含有如图11所示重链的可变区(SEQ ID NO:19)的人化重链]。本发明也包括那些含有一个或多个人化轻链和/或重链的人化免疫球蛋白。
本发明不仅涉及分离到的编码人化免疫球蛋白轻链(SEQ IDNO:20)的核酸或编码重链(SEQ ID NO:18)的核酸,而且还涉及分离到的编码人化免疫球蛋白(如单一链抗体)的核酸。例如,本发明提供了编码人化免疫球蛋白轻或重链的融合基因,这些链又由第一核酸序列和第二核酸序列组成,第一序列编码鼠源Act-1单克隆抗体的抗原结合区,第二序列至少编码人源免疫球蛋白恒定区的一部分。
本发明还涉及编码有α4β7结合特异性的人化免疫球蛋白或者它的一条链的核酸构建物。例如,本发明提供了含融合基因的表达载体,该融合基因编码人化免疫球蛋白轻链,包括对从与α4β7整合蛋白有结合特异性的非人源抗体的轻链衍生的CDR进行编码的核苷酸序列和衍生自人源轻链的框架区。这类构建物的另一个实例是这样的表达载体,其包括编码人化免疫球蛋白重链的融合基因,该融合基因包括对从与对α4β7整合蛋白有结合特异性的非人源抗体重链衍生的CDR进行编码的核苷酸序列,以及从人源重链衍生的框架区。
本发明还涉及含有本发明核酸序列的宿主细胞,其包括一个或一个以上的含有本发明核酸的构建物。在一个实施方案中,本发明涉及一种包含有编码人化免疫球蛋白轻链的第一重组核酸和编码人化免疫球蛋白重链的第二重组核酸的宿主细胞,所述的第一核酸包括编码鼠源Act-1抗体轻链CDR和人源轻链框架区的核苷酸序列,所述的第二核酸包括编码鼠源Act-1抗体重链CDR和人源重链框架区的核苷酸序列。
本发明也提供了一种制备人化免疫球蛋白的方法,该方法包括在适于表达人化免疫球蛋白的条件下培养宿主细胞,使人化免疫球蛋白链得到表达,使人源免疫球蛋白被产生。该方法还可以包括人化免疫球蛋白的分离步骤。
本发明的人化免疫球蛋白的免疫原性比它们对应的鼠源或其他非人源对应体的要小得多。因此,本文中所描述的人源免疫球蛋白能够用来治疗人体疾病,例如,用来控制淋巴细胞返回到黏膜淋巴样组织,从而减轻消化道的炎症反应。
本发明还涉及本发明的人化免疫球蛋白在诊断或治疗(包括预防)中的应用。在一个实施方案中,本发明将人化免疫球蛋白用于组织淋巴细胞渗透性疾病的治疗,例如,治疗炎症疾病,包括胃肠道淋巴细胞渗透性疾病(包括消化道内皮),其他的黏膜组织,或表达MAdCAM-1分子的组织。在特别优选的实施方案中,这种人化免疫球蛋白还可以用于治疗肠炎(IBD),例如溃疡性结肠炎和Crohn’s病。
另一方面,本发明涉及本发明的人化免疫球蛋白在制造可治疗组织淋巴细胞滤过性疾病的药物中的应用,例如,治疗炎症疾病,包括胃肠道淋巴细胞渗透性疾病,其他的黏膜组织,或者表达MAdCAM-1分子的组织。在特别优选的实施方案中,本发明涉及本发明的人化免疫球蛋白在制造用于治疗肠炎(IBD),例如溃疡性结肠炎或Crohn’s病等药物中的应用。
附图简要说明
图1说明了一致的DNA序列(SEQ ID NO:1)及其编码的氨基酸序列(SEQ ID NO:2),包括由多个独立的鼠重链可变区克隆确定的可变区。
图2说明了核酸序列(SEQ ID NO:3)及其编码的氨基酸序列(SEQ ID NO:4),包括由独立的鼠重链可变区克隆H2B#34所确定的可变区中的一部分。
图3说明了核酸序列(SEQ ID NO:5)及其编码的氨基酸序列(SEQ ID NO:6),包括由多个独立的鼠轻链可变区克隆确定的可变区。标出了为了克隆化而引入KasI位点的两突变位点。
图4A是一个荧光图,说明鼠Act-1mAb和鼠同型不相关对照抗体(MOPC21;IgG1,Kappa)对表达α4β7的HuT78细胞的染色能力。
图4B是一个荧光图,说明三种物质对表达α4β7的HuT78细胞的染色能力,(i)嵌和的Act-1抗体,(ii)人同型不相关对照抗体(IgG1,Kappa),(iii)COS-7细胞上清液。
图5排列比较了鼠Act-1轻链可变区(“Act-1,vl”)(SEQID NO:7)和人GM 607’CL轻链可变区(SEQ ID NO:8)的氨基酸序列。竖线表示相同的氨基酸,相似的氨基酸则根据相似程度分别用4或2点来表示,框起来并标记上的是CDR,每个残基按序列编号。
图6排列比较了鼠Act-1重链可变区(“Act-1,vh”)(SEQID NO:9)和人21/28’CL重链可变区(SEQ ID NO:10)的氨基酸序列。竖线表示相同的氨基酸,相似的氨基酸则根据相似程度分别用4或2点来表示,框起来并标记上的是CDR,每个残基按序列编号。
图7说明了鼠Act-1轻链上紧连于信号肽的可变区的核酸序列(SEQ ID NO:11)及其编码的氨基酸序列(SEQ ID NO:12)。
图8说明了成熟的人GM607′CL抗体Kappa轻链可变区的核酸序列(SEQ ID NO:13)及其编码的氨基酸序列(SEQ ID NO:8)。
图9说明了鼠Act-1抗体重链可变区的核酸序列及氨基酸序列。该序列连接于可编码鼠Act-1重链信号肽的核酸序列,形成一混合序列(SEQ ID NO:14和15)。(扩增重链的引物来源于退化序列,信号肽的氨基酸序列来源于引物,下游序列和别的信号肽序列。这里所示的信号肽可能与Act-1杂合体的不相同)。
图10说明了人21/28′CL抗体重链可变区的核酸序列及氨基酸序列。该序列连接于编码人HG3′CL抗体VH信号肽的核酸序列。[Rechavi,G.et al.Proc.Natl.Acad.Sci.USA 80:855-859(1983)],从而形成一混合序列(SEQ ID NOS:16和17)。
图11说明了带有重链信号肽的人化Act-1抗体(LDP-02)重链一部分的核酸序列[(SEQ ID NO:18)和氨基酸序列(SEQ ID NO:19)]。
图12说明了带有轻链的信号肽的人化Act-1抗体(LDP-02)轻链一部分的核酸序列[(SEQ ID NO:20)和氨基酸序列(SEQ IDNO:21)]。
图13说明了分别被标识为L1-L6(SEQ ID NOS:22-27)和H1-H10(SEQ ID NOS:28-37)的两组具有重叠且互补的寡聚核苷酸序列。前者编码人化Act-1免疫球蛋白(LDP-02)的轻链,后者编码人化Act-1免疫球蛋白的重链。
图14是一个荧光图,说明用构建的鼠-人Act-1免疫球蛋白,人化Act-1免疫球蛋白或一不相关的人同型对照抗体(IgG1,Kappa),对HuT-78细胞的染色效果。
图15说明了用流式细胞光度计对HuT-78细胞进行的滴定生物素化鼠Act-1和人化Act-1(LDP-02/3A9/L0T#1,实施例4)的结果。
图16是个图表,说明与对照的鼠IgG1或人IgG1相比,鼠Act-1或人化Act-1免疫球蛋白(LDP-02/3A9/L0T#1,实施例4)对生物素化鼠Act-1结合的竞争性抑制。
图17是个图表,说明分别在5种分子存在下,人周边周血单核细胞在溶胞中同位素51Cr检测的结果,(a)CAMPATH-1H,(b)CAMPATH-1G,(c)人IgG1,(d)LDP-02/3A9/Lot#1(实施例4)或(e)LDP-01(人化抗CD18,Fc突变);分别于5种浓度50,25,5,2.5,和0.5μg/ml。
图18A-18B是图表,说明鼠Act-1(图18A),鼠IgG1(图18A),LDP-02/3A9/Lot#1(图18B)或人IgG1(图18B)对α4β7携带细胞(RPMI 8866)与构建的人MAdCAM-1-Ig(免疫粘附)的吸附过程的抑制作用。
图19比较了4种方法对HuT-78细胞的染色效果,(a)LDP-02(Fc突变),(b)在轻链(MV4)有单突变,在重链有双突变(R38K,A40R)的LDP-02(Fc突变)的衍生体,(c)不相关的人同型对照抗体(IgG1,kappa)。
本发明的详细叙述
本发明涉及了对α4β7整合蛋白有结合特性的人化免疫球蛋白,它包括非人源抗原结合区和至少一部分人源免疫球蛋白。优选地,人化免疫球蛋白对α4β7整合蛋白的亲合度至少要有107M-1,以108M-1为较好,以109M-2为更好。在一个实施方案中,人化免疫球蛋白包含有结合α4β7整合蛋白的非人源抗原结合区和人源恒定区。在另一个实施方案中,与α4β7整合蛋白结合的人化免疫球蛋白,包含非人源互补决定区和一个人源可变框架区,还可以有或没有人源恒定区。例如,这种人化免疫球蛋白可以包括重链和轻链,其中的轻链包括从结合α4β7整合蛋白的非人源抗体衍生的互补决定区和人源轻链衍生的框架区,重链包括结合α4β7整合蛋白的非人源抗体衍生的互补决定区和人源重链衍生的框架区。
本发明也涉及了人化免疫球蛋白的轻链和人化免疫球蛋白的重链。在一个实施方案中,本发明涉及包含有非人源轻链CDR(一个或多个CDR)和人轻链框架区的人化轻链。在另一个实施方案中,本发明涉及包含有非人源重链CDR(一个或多个CDR)和人重链框架区的人化免疫球蛋白重链。CDR可来自于非人免疫球蛋白。
天然的免疫球蛋白都有一相同的核心结构,里面由两条相同的轻链(约24KD)和两条相同的重链(约55或70KD)形成一个四聚体。每条链的N端被认定为可变区(V),并能够同各条链其他部分更加保守的恒定区(C)相区别。轻链可变区C端部分称为J区,此外,重链可变区内,除J区,还有一D区。免疫球蛋白中绝大部分氨基酸序列变化限于V区的三个位点,称为高度可变区和互补决定区(CDR),该区直接涉及与抗原的结合。从N端数,分别被标记为CDR1、CDR2、CDR3,CDR由更保守的框架区维持位置。从N端数,框架区分别被标记为FR1、FR2、FR3和FR4。这些CDR和FR区的位置及其编号,最早由Kabatet al定义。[Kabat,E.A.et al.Sequences of Proteins ofImmunological Interest,Fifth Edition,U.S.Department of Healthand Human Services,U.S.Government Printing Office(1991);又见表3和4]。
人免疫球蛋白能被划分为几个类和几个亚类,根据重链的类型,分别为IgG、IgM、IgA、IgD和IgE。它们各自的重链分别为γ、μ、α、δ、或ε型。亚类包括IgG1、IgG2、IgG3、IgG4、IgA1和IgA2,各自的重链分别为γ1、γ2、γ3、γ4、α1和α2型。某一类或某一亚类人免疫球蛋白分子可能含有一个κ或者λ轻链。参见,Cellular and Molecular Immunology,Wonsiewicz,M.J.Ed.Chapter 45,pp.41-50,W.B.Saunders Co,Philadelphia,PA(1991);Nisonoff,A.Introduction to Melocular Immunology,2nd Ed.Chapter 4,pp.45-65,Sinauer Associates,Inc.Sunderland,MA(1984)。
这里用的术语“免疫球蛋白”包括完整的抗体和其有生物活性的片段。这种生物活性片段至少保留有相对应完整长度抗体所具有的一种抗原结合功能(如对Act-1抗体α4β7的特异性),最好保留有抑制α4β7与一个或多个它的配体(如MAdCAM-1,纤连蛋白)相互作用的能力。在特别优选的实施方案中,生物活性片段能够抑制α4β7结合到黏膜地址素(MAdCAM-1)上面。可以使用的生物活性抗体片段的例子包括能结合α4β7的片段,例如,单一链抗体,Fv、Fab、Fab’和F(ab’)2片段。这些片段能通过酶切或者重组技术来获得,例如,用木瓜蛋白酶切和胃蛋白酶切分别可制备Fab和F(ab’)2片段。抗体也能以多种截断形式被制备,即使用将一个或多个终止子导入到天然停止位置的上游的抗体基因。例如,编码F(ab’)2片段重链的嵌合基因,能被设计成包含编码CH1域和重链铰链区的DNA序列。
术语“人化免疫球蛋白”是指一种免疫球蛋白,包括不同来源的免疫球蛋白的局部,其中至少有一部分来源于人。例如,人化免疫球蛋白可以包括有必需特异性的非人源免疫球蛋白部分,如老鼠,以及人源免疫球蛋白序列(如嵌合的免疫球蛋白),运用传统技术(如合成)连接而成,或者运用遗传工程技术,将其作为一连续相邻的多肽来制备(如对编码嵌合抗体的DNA进行表达来产生出连续的多肽链)。本发明的人化免疫球蛋白的另一个实例是,包括一个或多个免疫球蛋白链,该免疫球蛋白链包括从非人源抗体衍生的CDR和由人源轻/重链衍生的框架区(例如,有或没有框架改变的CDR移植抗体)。人化免疫球蛋白这个术语也包括嵌合的或者CDR移植的单链抗体。参见,Cabilly etal.美国专利号4,816,567;Cabilly et al.欧洲专利号0,125,023B1;Boss et al.美国专利号4,816,397;Boss et al.欧洲专利号0,120,694 B1;Neuberger,M.S.et al.WO 86/01533;Neuberger,M.S.et al.欧洲专利号0,194,276 B1;Winter,美国专利号5,225,539;Winter,欧洲专利号0,239,400 B1;Padlan,E.A.et al.欧洲专利申请0,519,596 A1。又参见Ladner et al.美国专利号4,946,778;Huston,美国专利号5,476,786;及Bird,R.E.et al.Science,242:423-426(1988)等,关于单链抗体的描述。
人化免疫球蛋白(非人部分)的抗原结合区,可以来自于具有与α4β7整合蛋白特性结合的非人源免疫球蛋白(称作为供体免疫球蛋白)。例如,合适的抗原结合区可以来源于鼠Act-1单克隆抗体[Lazarovits,A.I.et al.J.Immunol.133(4):1857-1862(1984));参见实施例1-3]。其他来源包括非人源特异结合α4β7整合蛋白的抗体,例如啮齿类(小鼠、鼠)、兔、猪、山羊或非人灵长目(猴子)。其它的多克隆、单克隆抗体也能制备,例如,和Act-1抗体结合到同样或相似的抗原决定部位的抗体。[e.g.Kohler et al.Nature,256:495-497(1975);Harlow et al.1988,Antibodies:A Laboratory Manual,(Cold Spring Harbor,NY);andCurrent Protocols in Molecular Biology,Vol.2(Supplement 27,Summer’94),Ausubel et al.Eds.(John Wiley&Sons:New York,NY),Chapter 11(1991)]。
哺乳动物(例如,小鼠、大鼠、兔、羊)对一个恰当的免疫原能产生抗体。例如,α4β7携带细胞,含有α4β7的膜片段,α4β7免疫原片段,一个β7肽段共价连接上一个合适的载体,都是所述的恰当的免疫原。从接受过免疫的动物的淋巴结或脾脏都能分离到抗体生成细胞(例如淋巴细胞)。将这些细胞与某种合适的永生细胞(如骨髓瘤细胞株)进行细胞融合,形成杂合子。用选择性的培养技术能将融合细胞分离出来,用适当的检测(如ELISA)能挑选出产生所需特异性抗体的细胞。具有与α4β7特异性结合的非人源免疫球蛋白也能从抗体文库中获得(例如,具有非人源Fab分子的噬菌体文库)。
在一个实施方案中,人化免疫球蛋白的抗原结合区包括非人源CDR。在这个实施方案里,与α4β7特异性结合的人化免疫球蛋白至少带有一个非人源CDR。例如,这些CDR能来源于非人源免疫球蛋白轻链和重链的可变区,这样,人化免疫球蛋白实质上就包括一个或多个非人源免疫球蛋白的重链CDR1、CDR2和/或CDR3和/或轻链CDR1、CDR2和/或CDR3。所生成的人化免疫球蛋白因而具有与α4β7结合的特异性。特定链上全部三个CDR最好和对应的供体链的CDR实质相同,而且,每条轻链和重链各自的CDR都能与对应的供体链的实质相同则会更好。
人化免疫球蛋白的部分或人源免疫球蛋白链,可来自于任何合适的人的免疫球蛋白或免疫球蛋白链。例如,人恒定区或其片段可以来自于人类抗体的κ或λ型轻链,和/或γ(如γ1、γ2、γ3、γ4)、μ、α(如α1、α2)、δ或ε型重链,包括各自的等位变种。可以对特定的恒定区(例如IgG1)、变种或恒定区局部等进行筛选来裁选效应器功能。例如,突变的恒定区(变种)能被融入一个融合蛋白,降低其对Fc受体的结合和/或固定互补体的功能(参见,实施例3;还可以参见,Winter et al.GB2,209,757 B;Morrison et al.WO 89/07142;Morgan et al.WO94/29351,December 22,1994)。
如果存在的话,人框架区(例如轻链可变区的)最好来源于人抗体可变区,后者同抗原结合区供体的类似物或相应区域具有序列同源性。人化免疫球蛋白中人源部分的框架区的其他来源包括人可变的共有序列[参见,实施例2;又参见,Kettleborough,C.A.et al.Protein Engineering 4:773-783(1991);Carter et al.WO94/04679,1994年3月3日发表]。例如,用来获得非人源部分的抗体或可变区的序列可以与Kabat,E.A.所描述的人类序列相比较[Kabat,E.A.et al.Sequences of Proteins of ImmunologicalInterest,Fifth Edition,U.S.Department of Health and HumanServices,U.S.Government Printing Office(1991)]。在优选的实施方案中,人化免疫球蛋白链的框架区来自于拥有至少65%,最好70%的与非人供体序列同源的人类可变区(例如,小鼠Act-1)。人源部分也可来自于一种人抗体,在特定区域(如FR)和非人源供体的对应区域(如FR)相比,至少拥有65%,最好70%的序列同源。例如,在实施例2中,在鼠Act-1和人GM607’CL轻链可变区之间总体序列同源程度是71.4%,鼠Act-1和人21/28’CL重链可变区之间是68.1%。
在一个实施方案中,人化免疫球蛋白包括的框架区(FR)中至少要有一个是来自于人源抗体的单个链或多个链。因此,FR包括来自一个或一个以上的人抗体源的FR1和/或FR2和/或FR3和/或FR4。特定人化链的人源部分最好包括人源可变区的FR1、FR2、FR3和FR4(例如,来自人化免疫球蛋白链,来自一种人类共同序列)。
在本发明中使用的人源的合非人源的免疫球蛋白部分与他们各自母体的或他们的各变种的免疫球蛋白或免疫球蛋白部分具有序列同源。这些变种包括那些由于在一个或多个位点发生添加、缺失或替换而形成的各种突变体。如上述,非人源CDR实质上和其母体是相同的,最好是完全一致。如实施例2所述,人源框架区可能发生变化,在某个位点上被非人源供体的相应位点所替换。框架区可能发生一个或多个突变,包括缺失、插入和单或多氨基酸的替换。一些这样的替换模式见实施例2中人化Act-1抗体的替换模式。对于某特定的人化抗体或抗体链,框架突变可按本文所属设计。人化免疫球蛋白最好有与非人源供体相似的或更好的α4β7整合蛋白亲合性。有多种方法可以制备变种,包括非人源供体部分发生突变或者人源受体部分发生突变。
本发明的人化免疫球蛋白具有人α4β7结合特性,并且包括对异二聚体中α4和/或β7链的决定簇有结合能力的人化免疫球蛋白(包括片段)。在优选的实施方案中,本发明的人化免疫球蛋白最好具有至少一种属于鼠Act-1抗体特性的功能,例如,结合功能(例如,具有对α4β7整合蛋白的特异结合性,具有相同或相似的抗原决定部位特性),和/或抑制功能(在体内和/或体外对α4β7依赖性黏附的抑制能力,例如,抑制α4β7结合于MAdCAM-1,或者抑制α4β7携带细胞结合于它的配体)。因此,优选的人化免疫球蛋白具有鼠Act-1抗体的结合特性,抗原决定部位特性[能同鼠Act-1,一种嵌合的Act-1抗体(见实施例1),或者同人化Act-1(LDP-02)竞争结合α4β7(例如,在一个携带α4β7细胞上)],和/或抑制功能。
具有与α4β7整合蛋白特异结合性的人化免疫球蛋白的结合功能,可以通过标准免疫学实验来检测,例如,用来指示人化免疫球蛋白和α4β7形成复合物的检测[例如,包括α4β7的膜片段,α4β7的携带细胞,如人淋巴细胞(CD4+α4hi,β1lo亚单位的淋巴细胞),包括编码对α4β7进行表达的α4和/或β7的核酸的人淋巴细胞株或重组宿主细胞)。
对具有所需特性的人化免疫球蛋白的鉴别和/或分离的方法(如从一个文库中)还包括结合和/或黏附实验等等,(如,一种方法是检测α4β7携带细胞黏附于它的一个配体[例如第二个表达MAdCAM,嵌合的MAdCAM-Ig的细胞)],或者其它的适宜的方法。
本发明中用到的人源和非人源免疫球蛋白部分,包括轻链,重链和轻重链的一部分。这些免疫球蛋白部分可以取自或衍生自免疫球蛋白(如从头合成某一部分),或由对具有所需要的性质(例如,对α4β7整合蛋白的结合,序列相似性)的免疫球蛋白或其链进行编码的核酸来产生合表达而获得。具有人源和非人源特定部位(例如,抗原结合区,CDR,FR,C区)的人化免疫球蛋白,可以通过合成和/或重组核酸构建相应编码功能的基因(cDNA)来获得,为得到一条链的片段,需要将一个或多个终止子导入到预期的位点。例如,利用PCR突变技术改变现有DNA序列,使构建出的DNA能够编码新设计的人化可变区[Kamman,M.et al.Nucl.Acids Res.17:5404(1989)]。基于相同的或非常相似的人可变区,编码新CDR的PCR引物能和人化可变区的DNA模板杂交[Sato,k.et al.Cancer Research 53:851-856(1993)]。如果没有相似的DNA序列作为模板,则需要合成一段寡聚核苷酸来产生能编码可变区序列的核酸[Kolbinger,F.ProteinEngineering 8:971-980(1993)]。编码信号肽的一段序列也能掺入该核酸(例如,合成或掺入一个载体)。如果无法得到天然信号肽段,也可以用其他抗体的信号肽[Kettleborough,C.A.ProteinEngineering 4:773-783(1991)]。通过这些方法,本文中所描述的方法和其他可行的方法,都可以制备出变体(见实施例5)。在一个实施方案中,克隆的可变区可以发生突变,从中筛选出对具有所需要的特异性的变体进行编码的核酸序列(如,从一个噬菌体文库,Krebber,et al.U.S.5,514,548;Hoogenboom et al.WO93/06213,发表于1993年4月1日)。
核酸及其含有这些核酸的构建物
本发明涉及分离的和/或重组的(包括高纯度)编码人化免疫球蛋白或编码轻、重链的核酸。
这里所指“分离的”核酸是指从基因组DNA或细胞RNA中分离出来的核酸(它存在于细胞内或基因文库),包括用下述方法或其他方法获得的核酸:化学合成、联合使用生物的和化学的方法、重组核酸并被分离[参见Daugherty,B.L.et al.NucleicAcids Res.19(9):2471-2476(1991);Lewis,A.P.and J.S.Crowe,Gene,101:297-302(1991)]。
这里所指“重组的”核酸是指用重组DNA技术制备的核酸,包括依靠人为重组方法获得的核酸,例如,多聚酶链反应(PCR)和/或用限制性内切酶克隆到载体中。“重组”核酸也指细胞里以天然机理发生重组事件而获得的核酸,但需要将经设计的核酸导入细胞,以筛选出期望的重组事件。
本发明还特别涉及分离的和/或重组的编码人化Act-1免疫球蛋白(例如本发明中含有鼠Act-1单克隆抗体源非人部分的人化免疫球蛋白)及其链的核酸。在一个实施方案里,轻链包括衍生自Act-1抗体轻链源的三个互补决定区,重链包括衍生自Act-1抗体重链源的三个互补决定区。这种核酸包括,(a)编码具有人化Act-1免疫球蛋白重链可变区的氨基酸序列的多肽的核酸[例如,图11中的重链可变区(SEQ ID NO:19),图9中的重链可变区(SEQ ID NO:15)],(b)编码具有人化Act-1免疫球蛋白轻链可变区的氨基酸序列的多肽的核酸[例如,图12中的轻链可变区(SEQ ID NO:21),图7中的轻链可变区(SEQ IDNO:12)],(c)编码人化Act-1免疫球蛋白轻或重链可变区中至少一个功能部分的核酸(例如,人化免疫球蛋白中足够结合抗原的部分)。由于遗传密码的简并性,编码特定多肽的可能有多种核酸。在一个实施方案里,核酸中所包含的可变区序列被描述或实质上被描述于图11(SEQ ID NO:18),或者被描述或实质上被描述于图12(SEQ ID NO:20),包括双链或单链聚核苷酸。[尽管有多种图能够描述比可变区大得多的多肽(例如,包括信号肽编码序列或恒定区编码序列的一部分),特定图中可变区的图例说明意味着包括图示序列的可变区部分]。符合这些标准的分离的和/或重组的核酸,包括那些能够编码与上面讨论的人化Act-1抗体或它的变种的序列相等的序列的核酸。
本发明中的核酸可以用来制备特异结合α4β7整合蛋白的人化免疫球蛋白。例如,编码本发明中人化免疫球蛋白的核酸(DNA)可以被整合到一合适结构中(如一个载体),进一步操纵其序列或者在合适的宿主细胞内表达所编码的多肽。
特异结合α4β7整合蛋白的人化免疫球蛋白的制备方法
本发明的另一方面涉及特异结合α4β7的人化免疫球蛋白的制备方法。例如,在合适的宿主细胞里,对编码特异结合α4β7的人化免疫球蛋白的一个或多个重组核酸进行表达,从而获得人化免疫球蛋白。
本发明也提供了适于特异结合人化免疫球蛋白表达的构建物或表达载体。构建物可以被导入一合适的宿主细胞,产生能表达人化免疫球蛋白的细胞,并可以在培养基中生长和维持。合适的宿主细胞可以是原核生物,包括细菌,例如大肠杆菌(E.coli)、苏云金杆菌(B.subtilis)和/或其他合适的细菌,或者真核生物,例如真菌或酵母细胞(如Pichia pastoris,Aspergillus species,Saccharomyces cerevisiae,Schizosaccharomyces pombe,Neurospora crassa),或者其他低级真核细胞,以及高级真核细胞,例如昆虫细胞[如sf9昆虫细胞(WO 94/26087,O’Connor,1994年11月24日发表)]或哺乳动物细胞[如COS细胞,NSO细胞,SP2/0,中国仓鼠卵巢细胞(CHO),HuT78细胞,293细胞]。[见Ausubel,F.M.et al.eds.Current Protocols in Molecular Biology,Greene Publishing Associates and John Wiley&Sons Inc.(1993)]。
产生特异结合α4β7整合蛋白的人化免疫球蛋白的宿主细胞,可以如下述制备。例如,含有对所需的人化免疫球蛋白的全部或部分编码序列的核酸可以被插入到一核酸载体中,例如DNA载体,如质粒,病毒或其他适于表达的复制子。有多种载体可供选择,包括单拷贝或多拷贝载体,或者整合到宿主细胞染色体中的载体。
合适的表达载体可以包含许多组成部分,包括但不仅限于下述成分中的一个或几个成分:复制起点;可选择的标记基因;一个或多个表达调控因子,例如,转译控制因子(如启动子,增强子,终止子),和/或一个或多个翻译信号;用于膜靶向或膜分泌的信号序列或前导序列。在一个构建物中,信号序列可以由载体提供,也可以有其他来源,例如,免疫球蛋白的转译和/或翻译信号可以用来指导表达。
合适的宿主细胞含有用于表达的启动子,启动子可以是组成型,也可以是诱导型,例如,将启动子可工作地连接到编码人化免疫球蛋白或免疫球蛋白链的核酸上,从而指导多肽的表达。对于原核细胞(例如,适合大肠杆菌的启动子有lac、tac、T3、T7)和真核细胞(适合真核细胞的酵母乙醇脱氢酶(ADH1)、SV40、CMV)都有多种启动子可供选择。
此外,典型的表达载体包括有可供筛选的标记物,用来筛选带有载体的宿主细胞,如果是复制表达载体,则包括起源和复制区段。对具有抗生素或药物抗性的产物进行编码的基因通常用作可筛选的标记物,并且可以用于原核细胞(例如,β-内酰胺酶基因(抗青霉素),抗四环素的Tet基因)和真核细胞[例如,新霉素(G418或遗传霉素),gpt(霉菌酚酸),青霉素或潮霉素抗性基因]。二氢叶酸还原酶标记基因允许通过二氢叶酸在多种宿主细胞中进行筛选。编码营养缺陷型标记物的基因(例如LEU2,URA3,HIS3)是酵母菌常用的筛选标记。倍受关注的载体还有病毒(如杆状病毒)、噬菌体和能够整合到宿主基因组中的载体,如逆转录病毒。本发明也涉及带有这些表达载体的细胞。
例如,通过某种适合待筛选宿主细胞的方法(如转化、转染、电渗透、传染),可以将编码特异结合α4β7人化免疫球蛋白重链和轻链的核酸,或构建物(一个或多个构建物)导入合适的宿主细胞,以便这些核酸能够可工作地连接到一个或多个表达控制因子上(例如,在载体中,在细胞内的过程所创造的构建物中的核酸被整合到宿主细胞基因组中)。宿主细胞在适于表达的条件下生长(例如,存在诱导物,存在含有适当盐、生长因子、抗生素、营养补充物等的培养基),从而生产出所编码的多肽。如果需要,还可以从中(宿主细胞、培养液、奶汁)分离出所编码的蛋白(如人化Act-1抗体)。此过程包括在转基因动物宿主细胞中的表达(见WO92/03918,GenPharm International,1992年3月19日发表)。
可以制备融合蛋白,其中,人化免疫球蛋白或免疫球蛋白链在N端、C端或融合蛋白的中间位置,连接到一个非免疫蛋白部分上(天然情况下,免疫球蛋白没有此部分)。例如,一些实施例是将免疫球蛋白基因插入一合适的表达载体,如pET载体(pET-15b,Novagen),噬菌体载体(pCANTAB 5E,Pharmacia),或别的载体(pRIT2T蛋白A融合载体,Pharmacia)。所得到的构建物被导入到合适的宿主细胞中进行表达。表达后,利用恰当的亲合基质从细胞溶鲍物中分离或纯化融合蛋白{见CurrentProtocols in Molecular Biology[Ausubel,F.M.et al.eds.vol 2,Suppl.26,pp.16.4.1-16.7.8(1991)]}。
治疗方法及组合物
本发明提供了人化免疫球蛋白,它们能够(1)在体外和/或在体内结合α4β7整合蛋白;和/或(2)调节α4β7整合蛋白的活性或功能,例如(a)结合功能(α4β7结合MAdCAM-1,纤连蛋白和/或VCAM-1的能力)和/或(b)淋巴细胞渗透功能,包括富集和/或聚集于组织(抑制淋巴细胞迁向内皮黏膜组织的能力)。优选的是,人化免疫球蛋白能够在体外和/或在体内选择性结合α4β7,以及抑制α4β7介导的反应。在一个实施方案里,人化免疫球蛋白可以同α4β7整合蛋白结合,抑制了α4β7与它的配体中的一个或一个以上的结合(例如,MAdCAM-1,VCAM-1,纤连蛋白),从而抑制组织的淋巴细胞渗透(包括组织中淋巴细胞的富集和/或集中),最好具有选择性。在体外和/或在体内,此人化免疫球蛋白能够抑制在黏膜组织中α4β7携带细胞与肠内皮细胞的细胞黏附作用,这些黏膜组织包括消化道相关组织、淋巴样器官或淋巴细胞(尤其是T细胞或B细胞)。在一个最佳实施例中,人化免疫球蛋白(例如,Act-1)可以抑制α4β7与MAdCAM-1和/或纤连蛋白的反应。
本发明的人化免疫球蛋白在科研、诊断和治疗等诸多领域都是十分有价值的。例如,它们能够用来探测、分离和/或纯化α4β7整合蛋白或它的变种(例如,采用亲合纯化法或其他方法),能用来研究α4β7整合蛋白的结构(如构型)与功能。
本发明的人化免疫球蛋白也可以用于诊断(在体外)或者在治疗(包括预防)中调节α4β7的功能。
本发明的人化免疫球蛋白可用于检测和/或测量样品中α4β7整合蛋白的水平(例如,组织或体液,例如炎症渗出液、血液、血清、肠液,携带α4β7的细胞等)。例如,从个体获得样品(组织和/或体液),用合适的免疫学方法检测和/或测量α4β7的表达,这些方法包括酶联免疫吸附实验(ELISA),化学发光实验,放射免疫实验和免疫组织学等。在一种实施方案中,提供了检测样品内α4β7整合蛋白的方法,该方法包括,在适于人化免疫球蛋白特异结合α4β7的条件下,将样品与本发明的人化免疫球蛋白相接触,检测所形成的抗体-α4β7整合蛋白复合物。在应用该方法时,可以用人化免疫球蛋白来区分正常组织与炎症组织(人体的)的α4β7活性和/或表达(免疫组织学的角度),从而发现IBD或其他疾病与α4β7表达增加(受感染组织的)之间的联系。本发明的人化免疫球蛋白能用来评价正常组织和炎症组织中α4β7整合蛋白的存在情况,从而对疾病的发生、疾病进程和/或者抗α4β7整合蛋白治疗炎症的疗效等进行评价。
本发明人化免疫球蛋白也能用来调节[例如抑制(减弱或防止)]α4β7整合蛋白的结合功能和/或淋巴细胞(如淋巴细胞、单核细胞)渗透功能。例如,对于组织淋巴细胞(如淋巴细胞、单核细胞)渗透性疾病(包括组织中淋巴细胞的富集和/或积聚),尤其是表达MAdCAM分子的组织,可以用抑制α4β7与其配体(一个或多个配体)结合的人化免疫球蛋白来治疗,,为达到治疗效果,将有效剂量的本发明的人化免疫球蛋白提供给个体(例如,哺乳动物,如人或别的灵长目动物)。可用此法治疗的炎症疾病包括在下列组织的淋巴细胞渗透性疾病:胃肠道(包括肠内皮)、其他黏膜组织、或表达MAdCAM-1分子的组织(如肠相关组织,包括大小肠黏膜固有层的小静脉;和哺乳动物腺体(乳腺))。相似地,因淋巴细胞与表达MAdCAM-1细胞(如内皮细胞)结合而导致的组织淋巴渗透性疾病,也可根据本发明来治疗。
在特别优选的实施方案中,此法能用于治疗炎性肠道疾病(IBD),例如溃疡性结肠炎、Crohn’s病、回肠炎、腹腔疾病、非热带性口炎性腹泻、关节病血清反应阴性肠病、显微性或胶原性结肠炎、嗜酸性胃肠炎、直肠结肠切除及回肠吻合术后直肠瓣间小囊炎症。
胰腺炎和胰岛素依赖型糖尿病也可以用此法治疗。据报道,不仅BALB/c和SJL老鼠,而且NOD(非肥胖性糖尿病)老鼠的外分泌胰管也能表达MAdCAM-1,在胰岛素疾病早期,NOD老鼠炎症胰岛内皮上诱导出MAdCAM-1,其中以地址素为主[Hanninen,A.et al.J.Clin.Invest.92:2509-2515(1993)]。此外,表达α4β7的淋巴细胞在胰岛上聚集,α4β7介导的淋巴细胞与发炎胰岛管的结合也与MAdCAM-1有关[Hanninen,A.et al.J.Clin.Invest.92:2509-2515(1993)]。
可以用本发明的方法治疗的黏膜组织炎症疾病包括:乳腺炎(哺乳动物腺体)、胆囊炎、胆管炎或边周性胆囊炎(胆管和肝脏周围组织)、慢性支气管炎、慢性窦炎、气喘和移植排异反应(例如在胃肠道上)。象Crohn’s病一样,炎症常常扩散到黏膜表面以外,类似地,导致间隙纤维变性、过敏性肺炎、胶原疾病、肉样瘤病和其他自发疾病的慢性肺部炎症,也能用此法治疗。
人化免疫球蛋白要以能达到抑制α4β7结合其配体的有效剂量给药。在治疗中,有效剂量可以达到所需要的治疗(包括预防)效果(例如,足够减轻或防止α4β7参与的结合和/或信号的剂量,从而抑制淋巴细胞黏附和渗透和/或伴发的细胞反应)。人化免疫球蛋白可以是单剂量或者是多份剂量。剂量可以根据本领域熟知的方法来确定,并且受下列因素影响:个体年龄、敏感性、耐受性、身体状况。抗体的合适剂量可以是从0.1毫克/公斤体重到10毫克/公斤体重。
根据本方法,人化免疫球蛋白可以单独给药,或者搭配给药,可以在给其他药物之前、同时或之后给药。在一种实施方案里,给药的是多于一种的抑制α4β7结合其配体的人化免疫球蛋白。在另一实施方案里,可抑制淋巴细胞结合内皮上配体的单克隆抗体,包括抗MAdCAM-1、抗VCAM-1、抗ICAM-1抗体,与人化免疫球蛋白配合给药。在另外的一种实施方案中,一种额外的药物活性成分与人化免疫球蛋白配合给药(例如消炎化合物硫酸沙利比林、非甾体消炎药或类固醇消炎药)。
可以有多种给药途径,它们包括但并不仅限于:肠胃外给药(静脉内、动脉内、肌肉内、皮下注射),口服给药(饮食),局部给药,吸入给药(支气管内、鼻内或口腔吸入给药、鼻内滴注),或者直肠给药,具体途径视疾病和治疗条件定。肠胃外给药是最佳给药途径。
根据不同的给药途径来选择不同的药物剂型(溶液、乳剂)。适宜的含有人化抗体的组合物可以用生理学可接受的载体来制备。对于溶液和乳剂,适用的载体包括水或醇/水溶液、乳浊或悬浮液,包括盐水和缓冲液。肠胃外给药所用载体包括氯化钠溶液,Ringer’s葡萄糖,糖盐水,乳糖化Ringer’s或者固定油。静脉内给药所用载体可以包括多种添加剂,保存剂,或液体,营养剂或电解补充剂(见Remington’s Pharmaceutical Sciences,17th EditionMack Publishing Co.PA,1985)。吸入给药可以将药物溶解,再装入一种合适的喷雾器中(例如,雾化器,喷洒器,和压力气溶胶喷雾器)。
实施例
本发明由下列实施例予以进一步说明,但它们不是要限制本发明的。
如实施例1所描述,进行了鼠Act-1抗体的纯化和序列分析。编码鼠Act-1抗体轻重链可变区的cDNA被PCR扩增且测序。用蛋白质测序测得Act-1 kappa轻链可变区(VL)的氨基酸序列,发现它和VL基因DNA序列衍生出的氨基酸序列完全一致。同样,绝大部分重链可变区(VH)的氨基酸序列也已测知,并且和VH基因DNA序列衍生出的氨基酸序列一致。这些结果说明从杂交细胞株中克隆到正确的鼠Act-1可变区基因功能性嵌合Act-1抗体被产生,证实了克隆到正确的基因。特别值得注意的是,编码鼠Act-1轻链可变区和重链可变区的DNA分别与人kappa轻链恒定区DNA和人γ-1或γ-4重链恒定区DNA相连。嵌合的抗体也被用于和人化Act-1 mAb(变形的Act-1 mAb LDP-02)之间的比较分析。
为制备良好结合α4β7的人化Act-1抗体,设计出变形的人可变区(实施例2)。为方便设计,建立了一个鼠Act-1可变区的分子模型。鼠Act-1抗体中直接参与抗原结合的区域,互补决定区或CDR被移植到特定的人可变区中。在人可变区框架区(FR)上制造少量位点的氨基酸改变。这种改型的人Act-1可变区在特定人轻链可变区FR上有单个氨基酸改变,在特定人重链可变区FR上有5个氨基酸改变,每个改变都是将人型转变成鼠型。
如实施例3所描述,对改型的人Act-1可变区进行编码的DNA被构建,并连接于人恒定区DNA序列,所生成的核酸被用来制备人化Act-1免疫球蛋白。人化Act-1抗体在哺乳细胞里表达(实施例3),和鼠Act-1抗体比较,测其与人α4β7的结合(实施例4)。如表5所示,人化Act-1抗体保留了鼠Act-1也有的抗原决定部位结合特异性,而且与天然的鼠抗体相比,还表现出意外的增强的亲合性。
在设计中鉴别出几个人化Act-1抗体的变种(实施例2和5)。例如,可以在如下的一个或一个以上位点发生额外的改变:轻链突变体M4V(位点4的Met→Val突变),重链突变体R38K(位点38的Arg→Lys突变),重链突变体A40R(位点40的Ala→Arg突变)。此外,重链突变体I73T(位点73的Ile→Thr回复突变),在位点73保留原存在于人框架区同样位点的苏氨酸。既能够在单一链上制造一个或多个位点改变,又能够在多条链上制造多种位点改变。
实施例1 Act-1 V
H
和V
L
区的克隆及鼠-人Act-1嵌合
免疫球蛋白的构建和表达
Act-1 VH和VL区的克隆
根据制造商实验手册上的方法,利用TRIzol试剂(Gibco/BRL)从产Act-1单克隆抗体的杂种细胞提取RNA[Lazarovits,A.I.et al.J.Immunol.133(4):1857-1862(1984);provided by A.I.Lazarovits and R.B.Colvin)]。
根据制造商的实验手册用Ig-引物试剂盒(Novagen)对重链和轻链可变区基因进行PCR扩增,简述之,1.5μg RNA逆转录为cDNA,反应体系中包括2.0μl 5X MMLV缓冲液(5X=250mMTris-HCl,pH 8.3,25℃,375mM KCl,15mM MgCl2),1.0μl 100mMDTT(二硫苏糖醇),0.5μl 10mM dNTP混合液(dATP、dCTP、dTTP、dGTP各10mA),0.5μl寡聚dT(1μg/μl),0.25μl乙酰BSA(4mg/ml),1.0μl适用的Ig-3’引物(10pmol/μl),0.5μlMMLV逆转录酶(200单位/μl)和无RNase水,总体积为10μl。混合物于37℃温育5分钟,42℃下30分钟,99℃下5分钟,每个Ig-3’引物在单独的反应中使用。
根据制造商的实验手册,可变区在逆转录材料中被扩增,简述之,8μl的逆转录材料同下列物质混合:4μl 2.5mM dNTP,5μl 10X反应缓冲液(10X=100mM Tris-HCl,pH8.8,25℃,500mMKCl,15mM MgCl2,1%Triton X-100),2.5μl Ig-5’前导引物(10pmol/μl)(每个引物在独立的反应中使用),0.25μl(1.25单位) DNA多聚酶(Perkin-Elmer),用水补足到50μl。
DNA多聚酶(Perkin-Elmer),用水补足到50μl。
以MuIgVH5’-A,MuIgVH5’-B,MuIgkVL5’-A和MuIgkVL5’-B为引物的扩增反应,其循环参数分别为:94℃1分钟,50℃1分钟,72℃2分钟,循环35次,然后72℃延续6分钟。所有其他5’端引物的扩增反应同样适用该条件,除了退火温度上升为60℃。
不管是用MuIgGVH3’-2或MuIgMVH3’-1作3’端引物,还是用MuIgVH5’-B或MuIgVH5’-E作5’端引物,都能成功扩增重链可变区。用MuIgkVL3’-1作3’端引物和用MuIgkVL5’-G作5’端引物可以成功扩增出轻链可变区。
引物序列如下:
MuIgGVH3’-2(SEQ ID NO:56):
5’-CCC AAG CTT CCA GGG RCC ARK GGA TAR ACI GRT GG
MuIgMVH3’-1(SEQ ID NO:57):
5’-CCC AAG CTT ACG AGG GGG AAG ACA TTT GGG AA
MuIgVH5’-B(SEQ ID NO:58):
5’-GGG AAT TCA TGR AAT GSA SCT GGG TYW TYC TCT T
MuIgVH5’-E(SEQ ID NO:59):
5’-ACT AGT CGA CAT GAA GWT GTG GBT RAA CTG GRT
MuIgkVL3’-1(SEQ ID NO:60):
5’-CCC AAG CTT ACT GGA TGG TGG GAA GAT GGA
MuIgkVL5’-G(SEQ ID NO:61):
5’-ACT AGT CGA CAT GGA TTT WCA RGT GCA GAT TWT CAG CTT
用琼脂糖凝胶纯化出扩增片段,用Ig-Prime试剂盒将之共价结合到pT7Blue T载体中(Novagen),根据制造商的实验手册,用此共价连接混合物转化试剂盒中提供的NovaBlue细胞。
用T7启动子引物和U-19mer引物,对包含合适尺寸插入的白色集落进行测序。退火发生在插入位的对面,正好在pT7Blue载体多克隆位点的外面。根据制造商的实验手册,用测序酶T7DNA多聚酶试剂盒(USB/Amersham Life Science)在仪器小制备量DNA上测序。
多个独立的重链可变区克隆的共同DNA序列(SEQ ID NO:1)及其编码的氨基酸序列(SEQ ID NO:2),被示于图1。简并引物导致序列中某些简并,起始密码是核苷酸13-15编码的Met,预测的先导物多肽酶切割位点位于核苷酸67-69编码的Ser和核苷酸70-72编码的Gln之间(核苷酸13-69编码前导肽)。本图中,鼠恒定区的一部分起始于由残基433-435编码的Ala。
多个独立的轻链可变区克隆的DNA序列(SEQ ID NO:5)及氨基酸序列(SEQ ID NO:6),示于图3。不同于重链可变区,被扩增序列并不简并,可能因为所用引物并非很简并,以及可变区仅由单一的引物对扩增而来。
嵌合重链基因的构建
构建了嵌合性鼠-人重链基因,人重链恒定区的来源是一个含有野生型人γ1恒定区的克隆(由Herman Waldmann博士提供(牛津大学));被标为3818的构建物包括整合在pEE6表达载体(Celltech)的人化抗CD18重链基因,对应于人化CD18重链基因同一区域的恒定区被克隆到pEE6.hCMV中,具体如Sims,M.J.et al.J.Immunol.151(4):2296-2308(1993)andWO93/02191,1993年2月4日所述,这些文献的教导都通过在此引述而全部合并于本文。用HindIII和EcoRI酶解,可以从表达载体上释放出人化抗CD18抗体重链可变区和恒定区(野生型γ-1)的序列,回收1.421bp含有重链基因的片段,亚克隆到pCR-ScriptTM(Stratagene)的HindIII和EcoRI位点,形成一个称为pCR-CD18H的质粒。在抗-CD18重链基因可变区和恒定区之间的连接部位有一个SpeI限制性酶切点,用HindIII和SpeI酶切pCR-CD18H,释放出重链可变区,此区由如下述制备的鼠Act-1可变区替代。
合成出两个引物,使整合入新的限制性内切酶位点,引物是:
5’端引物(SEQ ID NO:41):
Hind III
5’-T[AA GCT T]CC GCC ATG GGA TGG AGC
3’端引物(SEQ ID NO:42):
Spe I
5’-GGT GAC[ACT AGT]GCC TTG ACC CCA G
黑体印刷的核苷酸说明与模板序列不同,被标为H2B#34的带有如图2中的核酸序列(SEQ ID NO:3)和氨基酸序列(SEQ IDNO:4)的独立的鼠Act-1重链克隆,可作为模板,在上述5’和3’端引物作用下,可扩增鼠可变区,同时在起始密码5’端导入一个HindIII位点,在J区3’端导入一个SpeI位点。PCR片段被直接亚克隆到pCR-ScriptTM,形成pCR-mACT1HV质粒,正确的序列得到证实。通过HindIII和SpeI酶能从pCR-mACT1HV上释放出片段,再插入pCR-CD18H的HindIII和SpeI位点,替代抗CD18可变区形成pCR-mhACT1Hchi。用HindIII和EcoRI释放出嵌合的重链基因(鼠Act-1可变区加上人γ1恒定区),再克隆回到pEE6hCMV-B载体,包括hCMV启动子,形成了一个名为pEE6mhACT1Hchi的构建物。
嵌合性轻链基因的构建
以类似重链的方式构建了嵌合性鼠-人轻链基因。然而,对于嵌合性轻链,导入了一个新的限制性酶切位点KasI到构建物中,是以一条名为KG#87的鼠Act-1轻链可变区克隆为模板对一个可变区片段进行PCR扩增,以包含有人化抗CD18 kappa轻链基因的结构体为模板读kappa轻链恒定区进行PCR扩增,从而获得此位点[由Herman Waldmann博士提供(牛津大学),3819构建物包含有存在于pEE12表达载体的人化抗CD18轻链]。恒定区对应于Sims,M.J.所描述克隆于pEE12的人化CD18轻链基因的恒定区[Sims,M.J.et al.J.Immunol.151(4):2296-2308(1993)和WO 93/02191,1993年2月4日发表]。
可变区引物为:
5’端引物(SEQ ID NO:43)
HindIII
5’-T[AA GCT T]CC GCC ATG AAG TTG CCT
3’端引物(SEQ ID NO:44)
Kas I
5’-[GGC GCC]GCA TCA GCC CGT TTT
黑体印刷的核苷酸说明与模板的序列不同,编码区内两位点的核苷酸改变,位点423T→G,位点426A→G,在图3中产生了KasI位点,它们是静息的,不改变氨基酸序列。
kappa恒定区的引物为:
5’端引物(SEQ ID NO:45)
KasI
5’-C[GG CGC C]AT CTG TCT TCA TC
3’端引物(SEQ ID NO:46)
HindIII
5’-[AAG CTT]CTA ACA CTC TCC
轻链可变区和恒定区分别在各自的模板和引物下进行扩增,PCR产物分别被亚克隆到pCR-ScriptTM中以确证序列。用HindIII和KasI酶解从载体上释放出每个片段,经凝胶纯化后共价整合到3819pEE12表达载体的HindIII位点,已经事先用HindIII酶解从该载体释放出人化抗CD18轻链基因,最后所得构建物命名为pEE12mhACT1Lchi。
嵌合性免疫球蛋白的表达
为组建包括嵌合重链和轻链基因的表达载体,用BglII和BamHI酶从pEE6表达载体上(pEE6mhACT1Hchi)释放出整个重链基因加CMV启动子,该片段再共价整合到pEE12轻链基因表达载体(pEE12mhACT1Lchi)的BamHI位点,得到了包含嵌合轻链基因和嵌合重链基因的命名为pEE12mhLHchi的单一质粒,这两条基因都分别受到CMV启动子的转译调控。
pEE6hCMV-B和pEE12表达载体和Celltech公司的谷氨酸合成酶基因扩增系统,已在前面描述过[见,WO 86/05807(Celltech),WO 87/04462(Celltech),WO 89/01036(Celltech),EP 0 323 997 B1(Celltech),和WO 89/10404(Celltech),这些有关的教导都通过在此引述而全部合并于本文]。
为短暂地表达嵌合的抗体,可以通过电渗透将20μgpEE12mhLHchi转染COS-7细胞(American Type CultureCollection,12301 Parklawn Drive,Rockville,MD,20852),用胰蛋白酶-EDTA处理从组织培养瓶中收集对数期的COS-7细胞。,对细胞用磷酸盐缓冲液(PBS)洗涤一次,用Hank’s平衡盐溶液(HBSS)洗涤一次,将细胞分散于浓度为1.5×107cells/ml的HBSS中,取0.8ml HBSS即有1.2×107个细胞与20μg质粒DNA混合,于室温孵育10分钟,将此DNA/细胞混合液转入0.4cm电渗透试管,用Bio-Rad GenePulser供以250伏特,960μF的电流,电渗透后于室温孵育10分钟,再将细胞转入20ml培养基中(Dulbecco’s改良型Eagle’s培养基(DMEM)+10%FCS),于162cm2的组织培养瓶(Costar)中培养。5天后,收集细胞培养液的上清液,测试其对表达α4β7的HuT78细胞的染色能力,HuT78细胞(一种人类T细胞株)由American Type CultureCollection提供(12301 Parklawn Drive,Rockville,MD 20852,Assession No.ATCC 71B 161)。
100μl的被瞬间转化的COS-7细胞培养上清液,不成功转染的COS-7细胞培养上清液,纯化的鼠Act-1抗体(10μg/ml),或各自纯化的鼠(鼠IgG1,kappa(MOPC21),10μg/ml,来自Sigma)和人(人IgG1,10μg/ml来自Sigma)型不相关的同型对照抗体,在冰上和1×105HuT78细胞一起孵育30分钟,用含有2%小牛血清(FCS)和0.01%叠氮化钠(FACS缓冲液)的冰冻的PBS缓冲液洗涤细胞两次,在冰上和恰当的荧光辅助抗体孵育30分钟[连有AffiniPure F(ab’)2片段山羊抗-鼠IgG(H+L)的荧光FITC或者连有AffiniPure F(ab’)2片段山羊抗-人IgG(H+L)的荧光FITC(Jackson Immno Research)]。30分钟后,用FACS缓冲液洗涤细胞两次,再溶解于300ml的同种缓冲液中,用BectonDickinson FACscan细胞流计分析细胞。图4A表示和鼠同型不相关对照抗体MOPC21(IgG1,kappa)相比,对鼠Act-1 mAb的染色。图4B表示和人同型不相关对照抗体(IgG1,kappa)以及不成功转染的COS-7细胞上清比较,嵌合的Act-1抗体对HuT78细胞的染色。因此,和鼠Act-1抗体相比,此嵌合的抗体对HuT78细胞的染色是类似的。总的来说,这些数据说明我们成功地克隆及表达了正确的鼠Act-1可变区。
氨基酸序列分析
对纯化的鼠Act-1重轻链进行氨基酸序列分析,以证实从杂合子中分离到的轻重链可变区cDNA的特性,对轻链进行的实验是这样的:
在氮气保护下,0.3M硼酸钠,0.15M氯化钠溶液,37℃下2个小时,用2mM DTT降解鼠Act-1(5mg/ml),加入碘化乙酰胺,溶液浓度调为10mM,室温下保持4小时,非变性条件下的SDS-PAGE电泳分析结果证实了此蛋白被定量降解。蛋白质溶液进一步在PBS中透析,取一部分透析干净的样品过Superdex 75柱(16/60,Pharmacia)(run 1),重链和轻链都从同一柱上通过二者洗脱,体积一样,说明两链依然相连。另一份样品用8M尿素处理,在变性条件下(6M尿素)过superdex 75柱(run2),二者还是在柱中一起洗脱,可能是由于没有打开折叠的缘故。SDS-PAGE电泳分析证实,从两种凝胶过滤洗脱下来的两样品中都有全部两条链的存在。对这些样品进行N端测序(Commonwealth Biotechnologies,Inc.),结果如下:
样品2:DVVVTQTPLSLPVSFDGQV(SEQ ID NO:47)
样品1:DVVVTQTPLSL(SEQ ID NO:48)
所得序列与从DNA推导出的成熟的轻链N端序列相一致,这里以及其他的分析重链序列的尝试,说明重链N端可能被封闭了,因此,对重链进行了内部肽段的氨基酸序列分析。
用木瓜蛋白酶切割以简化抗体F(ab)2’片段的氨基酸测序。pH3.0,0.1M柠檬酸钠,37℃下2个小时,在抗体:木瓜蛋白酶为1∶200的比例下,用木瓜蛋白酶切割鼠Act-1,通过SDS-PAGE分析来评价反应是否结束,用蛋白质G和蛋白质A柱纯化蛋白质.依上述方法将样品降解并烷基化,用制备型SDS-PAGE(15%)将重链和轻链分开,切割重链片段,并在1ml 0.1%SDS和缓冲液中电泳2小时。对样品用2ng Asp-N蛋白质内切酶切割样品30分钟,片段用SDS-PAGE(17.5%)分离。降解产物在0.1M Hepes,PH8.0,0.1%SDS中被动洗脱过夜,并进行N端测序(Commonwealth Biotechnologies,Inc.)。
从17Kda片段得到的序列为DYAIDYWG(SEQ ID NO:49),存在于重链克隆中(图1,序列AIDY对应于JH4区的起始位)。
实施例2 鼠Act-1可变区的分子模型
为帮助设计CDR-移植的可变区,建立起一个鼠Act-1可变区的分子模型,利用已建立的同源性建模方法,模型化类似于免疫球蛋白的确定特性的蛋白质结构,在UNIX操作系统下用Silicon Graphics IRIS 4D工作站建立分子模型,使用分子模型软件包QUANTA(Polygen Corp.Waltham,MA)和Brookhaven蛋白质晶体图谱数据库。第一步,依据类似的,明确了结构的免疫球蛋白可变区FRs,模型化一种新型可变区的框架区(FRs)。由于相同的氨基酸侧链被维系在原来的方位,因此就可以象在鼠Act-1抗体中那样,利用最大重叠过程以保持chi转角,从而替换突变的侧链。绝大多数新型可变区CDR的建模都是基于典型的CDR结构[Chothia,C.and A.M.Lesk,J.Mol.Biol.196:901-917(1987);Chothia,C.et al.Nature 342:877-883(1989);Tramontano.A.et al.J.Mol.Biol.215:175-182(1990);Chothia,C.et al.J.Mol.Biol.227:799-817(1992)]。而在重链可变区CDR3的情况下,却没有已知的典范结构,只能是根据存在于任何结构的蛋白质的类似环结构,来建立CDR环的模型。最后,为了缓解不需要的原子接触以及优化范德华力和静电作用,如同在QUANTA中所做的,通过CHARMm势能使模型能量最小化[Brooks,B.R.J.Comp.Chem.4:187-217(1983)]。
对于鼠Act-1可变区,是根据鼠单克隆抗体4-4-20 Fab片段FR,来建立轻链可变区FR的模型[Herron,J.N.et al.Proteins.Structure,Function and Genetics 5:271-280(1989)]。根据鼠单克隆抗体D11.15 Fab片段的FR来建立重链可变区的FR的模型[Chitarra,V.et al.Proc.Natl.Acad.Sci.USA 90:7711-7715(1993)]。建模所基于的鼠Act-1抗体和可变区之间不同的那些氨基酸侧链被替换掉,然后通过排列于残基35-39,43-47,84-88和98-102之间的空间[其定义如Kabat,E.A.et al.Sequences ofProteins of Immunological Interest,Fifth Edition,U.S.Departmentof Health and Human Services,U.S.Government PrintingOffice(1991)],Fab 4-4-20抗体的轻链被添加到D11.15轻链上面,从而将这两个异源可变区(4-4-20Kappa轻链可变区和D11.15重链可变区)安排到各自正确的方位。
Chothia,C.等人认为,mAb Act-1轻链可变区的CDR1(L1)适用于L1典范亚群4[Chothia,C.et al.Nature 342:877-883(1989)]。鼠Fab 4-4-20(见上述)的L1环与典型的亚群4,在氨基酸长度上相同,氨基酸序列上相似,二者也匹配。因此,基于Fab 4-4-20的L1环,对L1环建模。同样地,mAb Act-1轻链可变区的CDR2(L2)和CDR3(L3)既匹配各自的典范亚群1环结构,又匹配各自对应的Fab 4-4-20CDR,因此,基于Fab 4-4-20的CDR L2和L3,模型化Act-1 Kappa轻链可变区的L2和L3环。
mAb Act-1重链可变区的CDR1(H1)适合Chothia,C.定义的H1典范亚群1[Chothia,C.et al.Nature 342:877-883(1989)],相对应的鼠mAb D11.15(见上述)H1环也是这样。而且mAb D11.15CDR1环与mAb Act-1 H1在长度上相同,在氨基酸序列上十分类似。因此,如同轻链的情况,基于重链可变区CDR1环对该环建模。很难定义重链可变区(H2)的CDR2,但其似乎对应于H2典范亚群2。再者,D11.15抗体的H2环也匹配相同的典范亚群,在氨基酸序列上类似,因此可以基于D11.15的H2环,对mAb Act-1的H2环建模。
如上所讨论的,重链可变区CDR3是高度可变的,不能划分出可鉴定的结构群组。为建立H3环的模型,鉴定出有相同长度和类似氨基酸序列的环,最好是从其他抗体上找,并以此为基础去建立模型。有三种环在环大小上匹配Act-1CD3,分别来自于三种抗体,在测试全部三个环结构对模型的空间不适应之后,选出人抗体Pot的H3环[Fan,Z.C.et al.J.Mol.Biol.228:188-207(1992)]来建立mAb Act-1 H3环的模型。调节了整个模型的明显空间不适应后,按照在QUANTA上所做的,将模型能量最小化。
CDR-移植可变区的设计
设计CDR-移植可变区的第一步是选取充当人可变区基础的人轻链和重链可变区。测试并比较了两种人可变区的选择方法,一种方法是,从人可变区不同亚类的共同序列中选择人可变区[Kabat,E.A.et al.Sequences of Proteins of ImmunologicalInterest,Fifth Edition,U.S.Department of Health and HumanServices,U.S.Government Printing Office(1991)]。比较啮齿类轻重链可变区与人共同序列,最相似的人轻、重链共同序列从6个人λ轻链可变区亚群、4个人Kappa轻链可变区亚群、3个人重链可变区亚群中挑选出来[见Kettleborough,C.A.ProteinEngineering 4:773-783(1991)]。另一种方法是从所有已发表的人可变区序列中挑选人可变区[Kabat,E.A.et al.Sequences ofProteins of Immunological Interest,Fifth Edition,U.S.Departmentof Health and Human Services,U.S.Government Printing Office(1991)]。比较啮齿类与人的轻、重链可变区氨基酸序列,挑选出那些与啮齿类可变区有较高相似性的人可变区。利用来源于同一种人抗体的轻、重链可变区,以保证两可变区能够较好地相互组合[Queen,C.et al.Proc.Natl.Acad.Sci.USA 86:10029-10033(1989)]。然而这里所讲的被挑选为模板的人轻重链可变区来源于两种不同的人的抗体,这样一来,挑选出与啮齿类可变区有更高相似性的人可变区是可能的。有许多成功的CDR-移植抗体范例是基于来自两个不同人抗体的可变区。研究得最好的一个例子就是改型的人CAMPATH-1抗体[Riechmann,L.et al.Nature 332:323-327(1988)]。
为设计改型的人Act-1可变区,将鼠Act-1可变区与鼠和人可变区的所有亚群的共同序列进行比较[Kabat,E.A.et al.Sequences of Proteins of Immunological Interest,Fifth Edition,U.S.Department of Health and Human Services,U.S.GovernmentPrinting Office(1991)]。结果概括于表1和表2。
鼠ACT-1轻链可变区最类似于鼠Kappa轻链亚群II的共同序列,总体一致性是83.9%,在FR中的一致性是87.5%(表1)。对于人抗体序列,鼠ACT-1轻链可变区最类似于人Kappa轻链亚群II的共同序列,总体一致性是72.3%,在FR中的一致性是78.8%(表1)。
表1.鼠Act-1Kappa轻链可变区与鼠、人Kappa轻链可变区亚群共同序列的比较
比较鼠ACT-1 Kappa轻链可变区与鼠、人Kappa轻链可变区不同亚群的共同序列的氨基酸序列,分别都连同或不连同CDR一起比较,列出了有最高相似性鼠和人亚群之间的相同性百分比以及相似性百分比。
| 人或鼠可变区 | Kabat亚群 | 完全可变区或仅FR | 相似百分比 | 相同百分比 |
| 鼠 | II | 完全 | 91.07 | 83.93 |
| 仅FR | 95.00 | 87.50 | ||
| 人 | II | 完全 | 83.93 | 72.32 |
| 仅FR | 90.00 | 78.75 |
鼠ACT-重链可变区最类似于鼠重链亚群II B的共同序列,总体一致性是83.5%,在FR中的一致性是94.3%(表2)。对于人抗体序列,鼠ACT-1重链可变区最类似于人重链亚群I的共同序列,总体一致性是68.6%,在FR中的一致性是75.9%(表2)。结果证实,鼠ACT-1可变区是十分典型的鼠可变区,同样也说明,人可变区亚群能充当人可变区模板的良好来源和CDR-移植的受体。
表2鼠.ACT-1重链可变区与鼠、人重链可变区亚群共同序列的比较
比较鼠ACT-1重链可变区与鼠、人重链可变区不同亚群的共同序列的氨基酸序列,分别都连同或不连同CDR一起比较,列出了有最高相似性鼠和人亚群的相似百分比以及相同百分比。
| 人或鼠可变区 | Kabat亚群 | 完全可变区或仅FR | 相似百分比 | 相同百分比 |
| 鼠 | IIB | 完全 | 89.26 | 83.47 |
| 仅FR | 95.40 | 94.25 | ||
| 人 | I | 完全 | 81.82 | 68.60 |
| 仅FR | 85.06 | 75.86 |
也比较了鼠ACT-1可变区与全部已记录的鼠和人可变区范例中的个别序列[Kabat,E.A.et al.Sequences of Proteins ofImmunological Interest,Fifth Edition,U.S.Department of Healthand Human Services,U.S.Government Printing Office(1991);UWGCG package(University of Wisconsin)]。对于人抗体序列,鼠ACT-1轻链可变区十分类似于人Kappa轻链可变区的序列,后者源于人抗体GM607’CL[Klobeck,H.-G.et al,Nature,309:73-76(1984)]。图5排列出鼠ACT-1轻链可变区(SEQ ID NO:7)和人GM607’CL轻链可变区(SEQ ID NO:8)的氨基酸序列,正如我们所料,人GM607’CL轻链可变区是人Kappa轻链可变区亚群II中的一员。鼠ACT-1和人GM607’CL轻链可变区之间的总体序列相同性是71.4%。鼠ACT-1重链可变区十分类似于人抗体21/28’CL重链可变区的序列[Dersimonian,H.et al.J.Immunol.139:2496-2501(1987)]。图6排列出鼠ACT-1重链可变区(SEQ ID NO:9)和人21/28’CL重链可变区(SEQ ID NO:10)的氨基酸序列,正如我们所料,人21/28’CL重链可变区是人Kappa轻链可变区亚群II中的一员,鼠ACT-1和人GM607’CL轻链可变区之间的总体序列相同性是68.1%。根据这些比较,人GM607’CL轻链可变区被挑选为设计改型人ACT-1轻链可变区的模板,人21/28’CL重链可变区被挑选为设计改型人ACT-1重链可变区的模板。
设计过程的第二步,将啮齿类CDR插入选定的人轻、重链可变区。由Kabat定义的[Kabat,E.A.et al.Sequences of Proteinsof Immunological Interest,Fifth Edition,U.S.Department of Healthand Human Services,U.S.Government Printing Office(1991)]整个啮齿类CDR连到人FR上面,形成一个简单的CDR-移植。某些情况下,在简易CDR移植中被人化的啮齿类抗体,表现出对抗原很低或没有结合。研究人FR氨基酸序列很重要,以决定是否有氨基酸残基可能在影响同抗原的结合,或者是直接通过与抗原的作用,或间接地通过改变CDR环的位置。
第三步,决定出改变人FR中哪些氨基酸残基,来获得良好的抗原结合性。在此阶段,啮齿类可变区模型在设计中十分有用。同样有用的还有Chothia所定义的CDR典范结构[Chothia,C.et al.Nature 342:877-883(1989)]。在人化可变区中保留任何属于典范结构的啮齿类氨基酸残基是十分重要的。比较将被人化的啮齿类抗体序列与其他啮齿类抗体的相似序列,能够帮助确定在特定位点是否有异常或者稀有氨基酸。这将暗示着那个位点的啮齿类氨基酸在抗原结合中发挥重要作用。通过研究啮齿类可变区模型,可能预测特定位点上的氨基酸是否影响与抗原的结合。当个别抗体的可变区被用作设计的基础,那么我们建议,比较这个人序列与此类人可变区亚群中的共同序列,标出任何显著不寻常的氨基酸。大多数情况下,人FR中的少量氨基酸被认为将发生改变,改变成啮齿类可变区在那个位点上的氨基酸。
表3和表4概括出改型的人ACT-1可变区如何被设计出来的。表3列出了设计改型人mAb ACT-1 VL区所用的氨基酸序列,第4栏列出了将被人化的鼠ACT-1轻链可变区的氨基酸序列(SEQ ID NO:7),第5栏列出了鼠ACT-1可变区所属于的鼠可变区亚群的一致序列(SEQ ID NO:50)(鼠K-II),第6栏列出了最类似鼠ACT-1可变区的人可变区亚群的一致序列(SED ID NO:51)(人κ-II),第7栏列出了充当模板的人可变区氨基酸序列(GM607’CL)(SEQ ID NO:8),第8栏列出了所设计的改型人ACT-1可变区(SED ID NO:52)的氨基酸序列(ACT-1 RHVκ)。表4列出了设计改型人MAB ACT-1 VH区所用的氨基酸序列,第4栏列出了将被人化的鼠ACT-1重链可变区的氨基酸序列(SEQ IDNO:9),第5栏列出了鼠ACT-1可变区所属于的鼠可变区亚群的一致序列(SEQ ID NO:53)(鼠IIB),第6栏列出了最类似鼠ACT-1可变区的人可变区亚群的一致序列(SED ID NO:54)(人I型),第7栏列出了充当模板的人可变区氨基酸序列(21/28’CL)(SEQ IA NO:10),第8栏列出了所设计的改型人ACT-1可变区(SED ID NO:55)的氨基酸序列(ACT-1 RHVH)。表3和表4中倒数第二列,说明在鼠ACT-1和选定的人FR之间不同的FR残基位点(表面的或内藏的),最后一栏列出可变区此位点的有关评述。
表3所用符号:(*)不变残基(在列5和列6中),如Kabat共同序列所定义的(指Kabat亚类中有95%或更高的出现率)[Kabat,E.A.et al.Sequences of Proteins of ImmunologicalInterest,Fifth Edition,U.S.Department of Health and HumanServices,U.S.Government Printing Office(1991)],或者CDR环典范结构的一部分(在列8中),如Chothia所定义的[Chothia,c.etal.Nature 342:877-883(1989)]CDR典范结构的一部分;黑体,表示FR和CDR中人类氨基酸残基被对应的鼠残基替换的位点;下划线,表示FR中人类残基不同于鼠的位点;(△),表示人FR中的变化次数;(鼠Ab ACT-1),表示鼠ACT-1抗体VL区的氨基酸序列;(鼠κ-II),属于亚群II的鼠Kappa VL区的一致序列(Kabat,E.A,et al.Supra);(人κ-II),属于亚群II的人VL区的共同序列(Kabat,E.A,et al.通上);(GM607’CL),表示人GM607’CL抗体的氨基酸序列[Klobeck,H.-G.et al.Nature309:73-76(1984)];(表面的或内藏的),表示在全部两条抗体可变区链上氨基酸与其他残基的相对位置;(ACT-1 RHVκ),表示变型人mAb ACT-1 VL区的氨基酸序列。











表4所用符号:(*)不变残基(在列5和列6中),如由Kabat的共同序列所定义(指Kabat亚类中有95%或更高的出现率)[Kabat,E.A.et al.Sequences of Proteins of ImmunologicalInterest,Fifth Edition,U.S.Department of Health and HumanServices,U.S.Government Printing Office(1991)],或者如在列8中由Chothia定义的(Chothia,c.et al.Nature 342:877-883(1989))CDR典范结构的一部分;(黑体)表示在FR和CDR中人类氨基酸残基被对应的鼠残基替换的位点;(下划线)表示FR中人类残基不同于鼠的位点;(△)表示人FR中的变化次数;(鼠Ab ACT-1)表示鼠ACT-1抗体VL区的氨基酸序列;(鼠IIB)表示属于亚群IIB的鼠VH区的共同序列(Kabat,E.A,et al.Supra);(人I)表示属于亚群I的人VH区的共同序列(Kabat,E.A,et al.Supra);(人抗体21/28’CL的氨基酸序列(Dersimonian,H.et al.J.Immunol.139:2496-2501(1987));(表面的或内藏的)表示在全部两条抗体可变区链上氨基酸与其他残基的相对位置;(Act-1 RH VH)表示变型人mAb Act-1 VH区的氨基酸序列。











为设计改型的人ACT-1轻链可变区(表3),人FR中的1个残基被改为鼠相应位点的氨基酸,改变发生在FR1的位点2[Kabat,E.A.et al.Sequences of Proteins of Immunological Interest,Fifth Edition,U.S.Department of Health and Human Services,U.S.GovernmentPrinting Office(1991)]。具体地讲,人GM607’CL轻链可变区中的异亮氨酸被改为鼠ACT-1轻链可变区中的丙氨酸。kappa轻链可变区的该位点被视为对于维持L1环方位和结构十分关键的位点之一[Chothia,C.et al.Nature 342:877-883(1989)],也即“典范的氨基酸”之一。由于其在环构型上的重要性,这些鼠框架区残基通常保留在改型可变区中。
在FR1的位点4中,鼠型是一个丙氨酸,人型是一个甲硫氨酸,由Val变成Met都不是剧烈的改变,因为两种氨基酸都是非极性憎水的残基,因此人序列中的Met被用于改型人ACT-1可变区中。然而,模型显示,Val被埋于L1和L2环之间,当埋于旦白质时,Val的平均体积为142埃3,而Met约为171埃3,更大的Met残基可能会导致L1和L2环全部或其中之一发生构型变化。通过在改型人ACT-1轻链可变区位点4引入改变,由Met变成Val,可以提高它的抗原结合性。
为设计改型的人ACT-1重链可变区(表4),人FR中的5个残基被改为鼠相应位点的氨基酸。FR1的位点24和FR3的位点71[Kabat,E.A.et al.Sequences of Proteins of Immunological Interest,Fifth Edition,U.S.Department of Health and Human Services,U.S.Government Printing Office(1991)],存在于鼠序列的氨基酸残基被保留在改型人ACT-1重链可变区中,因为这两个氨基酸分别是H1环和H2环典范结构的一部分[Chothia,C.et al.Nature,342:877-883(1989)]。由于这些位点发生任何氨基酸改变都将破坏H1环和H2环的堆积和最终结构,因此这些关键位点的鼠型残基都被保留到人化重链可变区中。
在FR2的位点48,人序列的Met被改变成原存在于鼠ACT-1序列中的异亮氨酸。Met替换成异亮氨酸是不寻常的。更重要的是,模型显示异亮氨酸被埋在H2环的内部。结果是,在这个内藏位点发生的改变可能影响H2环的结构,进而干扰了抗原结合。
在FR3的位点69,人序列的异亮氨酸被改变成原存在于鼠Act-1序列中的亮氨酸。尽管Leu替换Ile并非是不寻常的,模型显示Leu被埋在H2环内部,因此象位点48那样,此位点的改变将影响H2环的构象,破坏抗原结合性。
最后,在FR3的位点73,人序列的苏氨酸被改变成原存在于鼠Act-1序列中的异亮氨酸。在鼠亚群IIB或人亚群I中,以前从未见过FR3上此位点是个Ile[Kabat,E.A.et al.Fifth Edition,U.S.Department of Health and Human Services,U.S.Government PrintingOffice(1991)],这意味着此位点的Ile可能在抗原结合上发挥重要作用。
模型中,位点73的Leu呈现在临近抗原结合位的表面上,根据α4β7整合蛋白上配体的大小和方位,它可能直接参与抗原结合。然而,做为一个表面残基位点,如果鼠型氨基酸未被包括在改型人抗体中,该抗体作为一个整体将有很低的免疫原性。在改型抗体的派生物中,异亮氨酸将被人型Thr替代,重新测试新形成的结构体,以确定第二次改变是否将保持类似的抗原结合水平。
除了在最初改型人ACT-1重链可变区设计中人FR的5个变化外,还有2个其他的改变,可能增强其抗原结合性,模型显示残基鼠mAb Act-1重链可变区的38Lys和40Arg被置于H2环的内部,并且堆积在CDR2中63Phe的旁边(表4的编号)。然而,这些残基也是位于重链可变区的核心部位,如果它们被用来替换对应的人型氨基酸(38Arg和40Ala),它们可能带来其他的可能是不利的效果。因此,FR2中位点38和40的改变未被包括进改型人的mAb Act-1重链可变区中。但是,这两个修改的全部或之一可以被用在改型重链上以提高抗原结合性。
结论:
建立鼠Act-1可变区的模型,主要基于已知结构的其他抗体可变区。该模型被用来设计人化Act-1可变区,特别值得注意的是,在改型的人可变区中保留了抗原结合位点。
设计了改型的人Act-1轻链可变区或改型的人Act-1重链可变区(表3和表4)。改型的人Act-1轻链可变区是基于鼠Act-1轻链可变区的CDR和人GM607’CL抗体轻链可变区的FR。在人FR的位点2有一个氨基酸改变。改型的人Act-1重链可变区是基于鼠Act-1重链可变区的CDR和人21/28’CL抗体重链可变区FR的。在人FR的位点24、48、69、71、73分别有5个氨基酸改变。
此外,在设计改型人Act-1可变区的其它版本时,Kappa轻链FR1的位点4和重链FR2的位点38、40都应当特别考虑。由于重链FR3的位点73在抗体的表面,因此也被认为参与由鼠型到人型氨基酸的回复突变。
实施例3:对改型可变区进行编码的核酸的构建
从生化方面(部分氨基酸测序)和功能方面(嵌合抗体对HuT78细胞的染色)证实克隆到正确的重链和轻链可变区之后,如上述设计了改型的氨基酸序列。接下来设计并制备编码改型抗体链的基因。人化ACT-1的设计、构建和表达
测定实施例2所述的人化抗体的一级氨基酸序列后,该序列逆翻译成简并的核酸序列,利用MacVector(Kodak,Scientific ImagingSystems)4.5.3版,分析潜在的限制性内切酶位点,选择出整合有限制性内切酶位点,而且保留一级氨基酸序列的核酸。图11列出重链核酸序列(SEQ ID NO:18)和氨基酸序列(SEQ ID NO:19),图12列出轻链核酸序列(SEQ ID NO:20)和氨基酸序列(SEQ ID NO:21),同时标出用于亚克隆的限制性内切酶位点。
依照下法,构建人化Act-1重、轻链可变区。利用Applied BiosystemDNA Synthesizer Model 392(图13)合成轻链重迭互补寡聚核苷酸L1-L6(SEQ ID NO:22-27)和重链的H1-H10(SEQ ID NO:28-37)。在55摄氏度去保护过夜,在Speed-Vac上干燥,重溶于100ml水中,在Bio-Spin6柱上脱盐(Bio-Rad)。在260mm测量吸收值来测定寡聚核苷酸的浓度,用变性聚丙烯酰胺凝胶电泳纯化核酸。
这些寡聚核苷酸各自100μg混合于2倍体积的染色液(95%甲酰胺,20mM EDTA,0.05溴酚兰,0.05%三甲苯胺FF),65摄氏度加热2分钟,250V,在1X TBE电泳3小时,用溴化乙锭染色,紫外灯下显色,将有正确长度的寡聚核苷酸切除下来,置于试管,用水和电解质透析,用苯酚/氯仿/异戊醇(25∶24∶1,V/V)(Gibco/BRL)抽提寡聚核苷酸两次,加入0.1倍体积3M醋酸钾(pH6)和2倍体积冷乙醇使沉淀,离心,用70%乙醇洗涤一次,真空干燥,溶于50μl水中。
将等摩尔的(大约100μg,50μl水中)纯化核酸与等体积的(100μl)2X退火缓冲液(2X=1M NaCl,40mM Tris-HCl,pH2.5,2mMEDTA)混合,使互补的寡聚核苷酸退火。95摄氏度10分钟,使变性,然后在65摄氏度温育8小时。照上述方法用乙醇沉淀退火寡聚核苷酸,再溶于40μl水中。
加入2μl大片段DNA多聚酶I(Klenow),5μl 2.5mM dNTP和5μl 10x缓冲液(10X=10mM Tris-HCl,10mM MgCl2,1mM DTT,pH7.9,25摄氏度),补足体积到52μl,使退火寡聚核苷酸延长。室温下温育1小时,再加入1μl dNTP和1μl Klenow,37摄氏度温育半小时。请注意重链片段A不必进行延长。
通过12%聚丙烯酰胺凝胶电泳,从单链、未退火的核酸中纯化出退火的、延长的片段,溴乙锭染色,紫外光显色,切下正确长度的片段,如上述透析回收核酸,用苯酚/氯仿/异戊醇洗涤两次,乙醇沉淀,溶解于10μl水中。
三个轻链片段(LA、LB&LC)和五个重链片段(被称为HA-HE)独立地共价连入pCR-ScriptTM,除了如以下所述之外,按照操作手册用pCR-Script试剂盒(Stratagene)转化XL-1蓝超级感受态细胞。用pCR-LA和pCR-LB片段转化DM1(Gibco/BRL)感受态细胞,以避开Dcm甲基化酶,后者可封闭限制性内切酶Msc I对DNA的降解。挑选白色集落,根据实验手册用测序酶T7 DNA多聚酶试剂盒对小制备量DNA测序。在插入位对面退火的T3和T7启动子,被用于测序。
利用合成过程中已整合到序列中的限制性位点,编排人化重链可变区和轻链可变区片段的亚克隆。重链片段HA-HD在每个序列的末端包括一个额外的Age I限制性位点,从而可以如下述进行按顺序的对这些片段的亚克隆。
用限制性内切酶Spe I和Age I降解pCR-HA和pCR-HB的小制备量DNA。DNA曾经被1%琼脂糖疑胶电泳过。从胶上回收141bp片段HB,在Spe I和Age I位点整合到pCR-HA,形成pCR-HAB,接下来,利用Xba I和Age I从pCR-HC上释放112bp片段HC,在Xba I和Age I位点整合到pCR-HAB,形成质粒pCR-AC。利用片段HD的限制性质位点Nhe I和Age I以及片段E的Best II和Age I,将片段HD(141bp)和HC(130bp)整合到一个测序质粒中,这个最后得到的包含pCR-Scipt中全部五个重链可变区片段的质粒,称为pCR-HAE。所有的降解都是利用小制备量DNA,在37摄氏度温育至少2个小时,除了对BstE II之外,它的最佳培养温度在65摄氏度。16摄氏度过夜进行共价连结,利用T4 DNA连接酶和1∶10载体,按比例整合,次日转化到DH5α亚克隆感受态细胞(Gibco/BRL)。
用Hind III和Age I降解pCR-HAE,释放出pCR-ScriptTM的Act-1人化重链可变区。用该411bp片段替换pEE6mhACT1Hchi(实施例1)的鼠可变区序列,该质粒已被Hind III和Age I降解,生成pEE6hCMV-B里的人化Act-1重链。最后得到的质粒为pEE6hACT1H,通过测序确定正确的DNA序列。
用BspE I和Msc I降解pCR-ScriptTM的轻链片段A,所得153bp片段被用于替换pCR-ScriptTM中鼠轻链可变区的BspE I到Msc I部分,这个质粒为pCR-LhAmBC。Msc I和Nru I降解的轻链片段B、Nru I和kas I降解的轻链片段C,被整合到pCR-LhAmBC的Msc I和Kas I位点,替换所剩下的鼠序列。降解、连接和转化过程如前述,除了最后一次转化之外,所有转化都用DM1感受态细胞。
用Hind III和Kas I降解pCR-ScriptTM的人化轻链可变区和质粒pEE12mhACT1Lchi(实施例1)。通过凝胶纯化360bp轻链可变区片段和315bp轻链恒定区,共价连入pEE12的Hind III位点,形成pEE12hACT1L,测序以证实正确的方位和核苷酸序列。
根据构建嵌合抗体的同样方法(见实施例1,嵌合免疫球蛋白的表达),构建一个包含人化重、轻链基因的表达载体。唯一不同的是,由于人化重链可变区有一额外的Bgl II位点,当用Bgl II切割时,只进行部份降解以生成正确的片段,含有人化重链和人化轻链基因的载体,被标为pEE12hACT1LH。
根据构建嵌合抗体的同样方法(见实施例1,嵌合免疫球蛋白的表达),临时表达全部人化抗体和细胞染色。图14显示,同不相关同型对照抗体(IgG1,Kappa)相比,用鼠-人嵌合性Act-1抗体或人化Act-1抗体对HuT78细胞的染色结果。
将pEE12hACT1LH用电穿孔法渗透到NSO细胞中,形成稳定的NSO细胞转染体,即一个骨髓瘤细胞株(Methods in Enzymol.73(B):3-46(1981);European Collection of Animal Cell Cultures,PHLSCAMR,Porton Down,Salisbury,Wiltshire SP4 OJG,U.K.ECACCNo.85110503)。
在NSO细胞中的稳定表达
用Sal I限制性内切酶降解40μg稳定转染体pEE12hACT1LH,使成线状,该酶切割构建物中细菌质粒部分。用乙醇加1/10体积的醋酸钠溶液中沉淀线性DNA,70%乙醇洗涤,干燥,重新溶解于无菌水中。
呈指数生长的NSO细胞生长于非选择性培养基Dulbecco’sModified Eagles’Medium(高葡萄糖),2mM L-谷酰胺,没有丙酮酸钠,4500mg/L葡萄糖,25mM HEPES缓冲液(GIBCO/BRL,Catalog NO.12430-021),加上10%小牛血清(Gibco/BRL,Catalog NO.16000-044)。离心收集NSO细胞,洗涤,再溶于冷PBS,以便在加入DNA后,细胞浓度为107cells/ml。将线状质粒DNA(40μg)加入到冰上电渗透管的107细胞中。细胞和DNA温和地混匀,避免产生气泡,在冰上保留10分钟,擦干管外部,用Gene Pulser(Bio-Rad)两次连续的1500V,3μf脉冲进行处理。在将管子置于冰中10分钟。
将转染细胞移到96孔盘中,50μl非选择性培养液中分别三种浓度,3×105,7.5×104和1.5×104细胞/ml,37摄氏度下温育24小时。随后在全部孔中都加入150μl选择性培养基[无谷酰胺Dulbecco’s改良Eagle’s培养基,4500mg/L葡萄糖,4mg/L的醇盐酸,110mg/L丙酮酸钠,无硝酸铁,无L-谷酰胺(JRH BioSciences,CatalogNo.51435-78P)],加上1X GS补充物(50X GS补充物由JRH Bioscience提供,Catalog No.58672-77P),加上10%透析小牛血清(Gibco/BRL,Catalog No.26300-061)。置于培养箱中,直到出现实质性的细胞死亡和出现生存集落的分离。一旦出现谷酰胺不依赖性转染体集落,便筛选单集落的孔洞,收集进行组织培养的上清液,按下述用ELISA检测人IgG的分泌情况。后面研究将要用的产抗体克隆3A9,也通过这种方法获得。类似上述方法进行第二次转染,不同的是筛选指标为L-甲硫氨酸sulphoximine(MSX,一种谷酰胺合成酶抑制剂)。
用检测人IgG分泌的ELISA普查阳性集落。将100μl浓度为2.5mg/ml pH9.5溶于碳酸盐缓冲液中的AffiniPure F(ab’)2片段驴抗人IgG(H+L)(Jackson ImmunoResearch Laboratories)涂于ELISA板上,在4摄氏度过夜,用PBS吐温20洗涤板四次,用200μl PBS、1%BSA于37摄氏度2个小时,封闭ELISA板,洗涤后,和100μl稳定转染的NSO上清液一起37摄氏度温育15分钟,溶于PBS 1%BAS的1mg/ml人IgG1 Kappa溶液用作标准品。新鲜的NSO培养基(DME+GS补充物)作为阴性对照。洗涤板,并和以0.5μl/ml溶于PBS(无Ca2+/Mg2+)的连有过氧化物酶的100μl的AffiniPure F(ab’)2片段驴抗人IgG(H+L)(Jackson ImmunoResearch Laboratories),在37摄氏度温育15分钟。5mg O一二盐酚苯二胺(OPD)药片(Sigma)溶解于12ml柠檬酸缓冲液(0.1M,pH5.0),然后加入12μl 30%过氧化氢。洗涤除去次要抗体,加入100μl溶有OPD的基质。用12.5%硫酸中止反应,在DynatechPlate Reader上于490nm检测板子。通过限制稀释度为2、1和0.5cells/孔,克隆阳性孔。当来自单一克隆的全部孔,经ELISA检测为抗体产生阳性时,可以认为克隆到了该细胞株。
用蛋白质A亲合层析[Poros A/M 4.6/100mm,5mL/min,利用Bio-Cad工作站(Perseptive Biosystems,Inc.)]从过渡的细胞培养上清或从稳定的细胞转染体培养液纯化人化Act-1抗体。柱子用PBS平衡,经0.2微米过滤器过滤的细胞培养上清液上柱,每次所给上柱量依据抗体浓度不同而不同,通常上柱抗体量不超过15mg,洗脱速度5ml/min。结合后,第一次用PBS洗柱子,至OD280nm=0。然后用至少50倍柱体积的量洗第二次,接着用0.1M醋酸钠,pH5.0洗脱柱子,最后用0.1M柠檬酸钠,pH3.5洗脱。洗脱液按每5毫升一收集,用200μl,1.5M Na2CO3 pH12调pH中性。合并含抗体部份后,通过超滤浓缩至期望的浓度(centricon,30,000KDa cut off,Amicon)。
Fc-突变的变种的构建
构建了一种无Fc结合(Fc-突变)的人化Act-1抗体。该抗体和人化Act-1抗体有相同的可变区(图11和图12),有相同的人IgG1恒定区,不同的是在IgG1重链恒定区还有两个氨基酸的替换,这样设计是为了取消FcR识别和Fc结合性能(Leu235-Ala235,Gly237-Ala237)。按如下构建Fc-突变衍生体的重链的编码核酸。称为3678的构建物(由Herman Waldmann博士提供,牛津大学)在pEE12载体中编码人化抗-CD18抗体的轻链和重链[WO 93/02191(发表于February 4,1993);Sims,M.J.et a l,J,Immunol 151(4):2296-2308(1993)],但是在IgG1重链恒定区还通过位点定向的突变导入了两个氨基酸替换(Leu235-Ala235,Gly237-Ala237),用Age I和Ecor I将之降解,释放包含有γ恒定区突变体的900bp片段。该片段被用来替换pEE6hACT1H中在Age I/EcoR I位点的重链野生型γ-1恒定区,生成pEE6hACT1H/FCmut。在用类似方法构建包含两个链的其他构建物时,要制备包含有改型轻链基因和Fc-突变改型重链基因的单结构体(pEE12hACT1LH/FCmut)。
实施例4:人化ACT-1抗体LDP-O2的特性描述
起始的特性研究是利用pEE12hACT1LH/FCmut过渡转染的COS-7细胞所产生的抗体来进行的。如上述制备和纯化此抗体,在下面被称为“1°HUM ACT-1”,后面带有组号。
利用如上述制备的(用线性pEE12hACT1LH/FCmut转染的)鼠细胞株NSO稳定的转染体所产生的抗体,进行额外的检测。该抗体制备在下面被称为“LDP-02/3A9/Lot 1”。
“LDP-02/3A9/Lot#1”抗体所用在的下述研究包括:SDS-PAGE,Western杂交分析,等电聚焦,氨基酸组成分析,品系杂交,滴定,互补介入溶胞检测,ADCC检测,结合抑制检测。“1°HUM ACT-1Lot#7”被用于亲合检测#1-2,1°HUM ACT-1 #8-9用于亲合检测#3-5和C1q结合试验。
A、物理、化学性质;
1.SDS-PAGE
为帮助确立特征,首先描述第一个制备样品的特性并评估纯度,分别在降解的和不降解的条件下进行十二烷基磺酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE),考马斯亮兰染色。
将80μl,0.82mg/ml LDP-02/3A9/Lot#1加到一个微管中,抗体溶于柠檬酸缓冲液,与160μl Tris缓冲液(0.5mM,pH 8.8)交换三次,交换后样品的最终体积为135μl,蛋白质浓度为0.486mg/ml,分别用降解的和不降解的缓冲液稀释2倍,蛋白质浓度为0.243mg/ml。将13μl的0.243mg/ml等份试样溶液内(含3.16μg蛋白质),装入SDS凝胶的样品道里,进行SDS-PAGE电泳,对照品包括Mark 12分子重量标准物(Novex.#L5677)。
在非降解条件下,LDP-02/3A9/Lot#1表现出一条主要条带,分子量略低于200,000道尔顿,几个次要组份在116,300-200,000道尔顿之间,另外三个次要组份分子量大约为97,400道尔顿、略大于55,400道尔顿的组份、和略小于31,000道尔顿的组份。用激光光密度仪扫描凝胶,定量分析各染上色的条带,计算各自百分比(表5),结果显示分子量约为200,000的主要组份占所有染色条带的84.4%。该主要组份代表的是完整的抗体分子,分子量55,000和31,000的组份则分别代表单一的重链和轻链。
在降解条件下,电泳凝胶上有两个主要组份,一个约为55,400道尔顿,代表全部条带的68.6%,第二个略低于31,000道尔顿,代表30.5%的比例(表5),这两个分子量同免疫球蛋白G的重链、轻链的相吻合,这些数据说明,样品中大约99%要么是完整的抗体,要么包含抗体的轻链或重链。除两个主要条带外,还发现有一个略小于66,300道尔顿的次要组份。
从这个分析来看,与完整的免疫球蛋白G相吻合的高分子量种类作为主要条带而存在于LDP-02/3A9/Lot#1中。其中还有多个次要条带。在降解条件里,有两个主要条带,他们的电泳特征和免疫球蛋白G的轻链、重链相同。
表5
纯化数据简表
考马斯亮蓝,非降解条件
样品 条带 面积百分比(主要成分)
LDP-02/3A9/Lot#1 5 844%
纯化数据简表
考马斯亮蓝,降解条件
低分子量面积 低分子量面积
样品 条带
百分比 百分比
LDP-02/3A9/Lot#1 9 68.6% 30.5%
2.Western杂交分析
对样品和 标准物按如上所述的SDS-PAGE进行分析。简述之,以4-20%Tris-甘氨酸凝胶分析非降解和降解的样品。Novex Mark 12分子量标准物也在胶上跑,两种样品,2.1μl和4.5μl,它们的0.2143mg/ml等份试样溶液,分别含有0.51μg和1.09μg蛋白质的样品,分别装上SDS凝胶。
SDS-PAGE电泳后,根据Novex Western Transfer Apparatus指导,将样品蛋白从凝胶转到硝酸纤维素膜上。所用转移缓冲液是1X Tris-甘氨酸溶于2%甲醇的溶液。约2小时后,从转移仪器上取出硝酸纤维素斑点,浸于DDI水中。然后,在Tris缓冲液(20mM)中37摄氏度,35分钟封闭斑点,缓冲液中含有3%明胶和0.1%吐温20。取出斑点,用Tris缓冲液洗涤两次。用20mM Tris-3%BSA溶液稀释山羊抗鼠IgG抗体溶液1000倍,生成山羊抗鼠IgG溶液,将之加到斑点上,2-8摄氏度温育过夜。然后,用Tris缓冲液洗涤斑点,每5分钟换一次溶液,共换4次。用20mM Tris-3%BSA溶液稀释连接碱性磷酸酶的抗山羊IgG 5000倍,加到斑点中,于室温保持2小时,用Tris缓冲液洗涤,每5分钟换一次,共换四次缓冲液。往斑点上一次性加入10ml BCIP/NBT(5-溴-4-氯-3’-磷酸吲哚p-甲苯胺盐/氯化氮蓝四唑),搅拌,室温下反应。将斑点浸于Tris缓冲液终止反应,用山羊抗一人IgG代替山羊抗一鼠IgG,重复上述操作。
用抗一鼠IgG试剂,在非降解和降解条件下,都能在硝酸纤维素斑点上找到0.51μg和1.09μg的IgG样品。这些条带的密度随浓度升高而升高。在非降解条件,能发现一个比20,000道尔顿标记物略快的主要条带。还有几个轻微弱的条带,其中两个跑得比主要条带慢,三个比主要条带快。在降解条件下,发现两个对应于免疫球蛋白G重链和轻链的条带。
在非降解和降解条件下,利用抗一人IgG试剂,能在硝酸纤维素斑点中清楚地找到0.51μg和1.09μg的IgG样品。这些条带的密度随浓度升高而升高。在非降解条件下,发现一个略小于200,000道尔顿的主要条带。用抗鼠IgG能检测到斑点上的几个微弱条带。当用抗人IgG检测,免疫染色的密度强于所有条带。还检测到在其他斑点不能发现的额外的条带,这可能是因为这些条带对应于IgG片段,但缺乏能被抗鼠IgG所识别的表位。在降解条件下,检测到对应于免疫球蛋白G重链的一条带。因为抗体是对人IgG的Fc部分为特异性的,所以没找到轻链。几个次要条带,用抗鼠IgG不能被发现,而用抗人IgG却能发现。这种差异可能是因为存在有缺乏与抗鼠IgG结合的表位的IgG片段。
3、等电聚焦
对LDP-02/3A9/Lot#1进行等电聚焦(IEF),用考马斯亮兰染色,将LDP-02/3A9/Lot#1的实验结果和一起聚焦的IEF标准物进行比较。
将80μl,浓度为0.82mg/ml的LDP-02/3A9/Lot#1加到微管中。含有抗体的柠檬酸缓冲液与160μl的Tris缓冲液(0.5mM,PH8.8)交换三次,最后样品体积为135μl,最终浓度计算为0.486mg/ml。用2X IEF样品缓冲液稀释溶液两倍,浓度变为0.243mg/ml。取该溶液13μl,内含3.16μg蛋白质,装入IEF胶中。对照物包括IEF标准物pI 3.6-9.3(Sigma,Cat#1-3018)。
根据标准物绘制标准图谱,8种IEF标准物的平均泳动距离对各自的pI值。对应这些数据的曲线的负性斜率为0.03459,截距8.91857,R2=0.99206。
表6包括6个IEF标准品和LDP-02/3A9/Lot#1的平均泳动距离,也包括计算出来的LDP-02/3A9/Lot/的pI值。
利用标准图的曲线参数,计算LDP-02/3A9/Lot#1的5个条带大约pI值为:7.88,7.95,8.09,8.26和8.43,以pI8.09的峰最为显著(表6),这个主要峰的pI和基于氨基酸一级序列推算的pI7.91相适应。
表6
标准 泳动距离 * pI 1
凝集素 3.3mm 8.8
凝集素 9.5mm 8.6
凝集素 17.88mm 8.2
肌红蛋白 59.3mm 6.8
碳酸酐酶I 74.0mm 6.6
碳酸酐酶II 92.5mm 5.9
β-乳球蛋白A 105.8mm 5.1
胰蛋白酶抑制物 122.0mm 4.6
样品 泳动距离 pI 1
LDP-02/3A9/Lot#1(带1) 14.0mm 8.43
LDP-02/3A9/Lot#1(带2) 19.0mm 8.26
LDP-02/3A9/Lot#1(带3) 24.0mm 8.09
LDP-02/3A9/Lot#1(带4) 28.0mm 7.95
LDP-02/3A9/Lot#1(带5) 30.0mm 7.88
*平均值
1基于标准曲线(pI对泳动距离),其中:
样品pI=截距-斜率(样品泳动距离)
4.氨基酸组成分析
进行了氨基酸组成分析,并确定了LDP-02/3A9/LOT#1的蛋白质含量、氨基酸组成、和其特性。
取出三份45μl样品进行水解,用6N HCL蒸汽于165℃水解60分钟。做为对照,在水解试管中,还包含一种标准蛋白质,其已经与LDP-02/3A9/LOT#1一起被水解。在分析LDP-02/3A9/Lot#1之前和之后,色谱分析氨基酸标准品。蛋白质标准包括以牛血清白蛋白作为蛋白质标准(Tektagen Solution Control:310:197A),而氨基酸标准品是氨基酸水解混合液(Tektagen Solution Control:310:199A)。
实验方法包括用离子交换HPLC分析再溶解的蛋白质水解液或游离氨基酸溶液,过柱后与茚三酮反应显色,双波长测定吸收值,分析氨基酸标准得到一校正表,与之对照计算在两种波长的吸收。
表7列出了氨基酸组成,LDP-02/3A9/LOT#1的蛋白质浓度为0.709mg/ml。对缺乏定量的W和C进行修正,蛋白质浓度被校正为0.740mg/ml,数据和相关计算概括于表8。
对于LDP-02/3A9/LOT#1,用6N HCL蒸汽在165℃水解一个时间点(60分钟)。从标准蛋白(BSA)得来的校正因子,被运用于蛋白质含量的确定(表8)。
在这种实验条件下,由于存在一个共同洗脱的半胱氨酸峰,因此脯氨酸的摩尔百分比,可能略有提高(表7)。脯氨酸定量的准确性具有样品依赖性,基于样品水解物中半胱氨酸的含量。对于此分析,已经利用BSA校正因子,对脯氨酸含量进行了校正(表8),校正的准确性具有样品依赖性,基于BSA(6.0%)中和样品中半胱氨酸的相对量。
以相对百分比(频度或摩尔百分比)表示的预测的LDP-02氨基酸组成,基于重链和轻链的核苷酸序列(预测的%),真实的氨基酸分析结果(真实的%)被列于表9,比较预测值与真实值,便得出很好的相关系数,除脯氨酸之外,其已经在前面有叙述,由于共同洗脱的半胱氨酸峰而使脯氨酸值偏高。
表7
| 样品 | LDP-02/3A9/LOT#1没有校正 | LDP-02/3A9/LOT#1有BSA校正系数 | LDP-02/3A9/LOT#1有W/C<sup>1</sup>和BSA校正系数 |
| AA | %摩尔 | %摩尔 | %摩尔 |
| N/D | 9.1 | 9.0 | 8.6 |
| T | 6.5 | 7.5 | 7.2 |
| S | 9.2 | 13.3 | 12.7 |
| Q/E | 11.4 | 11.3 | 10.8 |
| P | 8.2 | 9.8 | 9.4 |
| G | 7.8 | 7.4 | 7.1 |
| A | 5.9 | 5.8 | 5.6 |
| V | 10.2 | 9.5 | 9.1 |
| M | 03 | 0.7 | 0.7 |
| I | 2.5 | 2.6 | 2.5 |
| L | 82 | 7.9 | 7.6 |
| Y | 5.2 | 5.0 | 4.8 |
| F | 3.4 | 3.4 | 3.3 |
| H | 2.2 | 2.2 | 2.1 |
| K | 7.0 | 6.9 | 6.6 |
| R | 2.9 | 2.9 | 2.8 |
| 总和: | 100 |
1相关因子为0.958,基于W和C含量分别为1.8%和2.4%。
表8:蛋白质含量确定

1蛋白质含量不对半胱氨酸和胰蛋白胨校正。
2对每一个检测的氨基酸都用BSA校正因子来校正。
3总水解的ng=(总注入的ng×重新配制的体积)/注入的体积(50μl)。
表9:氨基酸组分
| 氨基酸样品 | 氨基酸 | 号 | 预测% | 实测% |
| A | Ala | 68 | 5.06 | 5.6 |
| C | Cys | 32 | 2.38 | --- |
| D | Asp | 56 | 4.17 | --- |
| E | Glu | 68 | 5.06 | --- |
| F | Phe | 40 | 2.98 | 3.3 |
| G | Gly | 90 | 6.70 | 7.1 |
| H | His | 28 | 2.08 | 2.1 |
| I | Ile | 30 | 2.23 | 2.5 |
| K | Lys | 96 | 7.14 | 6.6 |
| L | Leu | 98 | 7.29 | 7.6 |
| M | Met | 10 | 0.74 | 0.7 |
| N | Asn | 50 | 3.72 | --- |
| P | Pro | 94 | 6.99 | 9.4 |
| Q | Gln | 64 | 4.76 | --- |
| R | Arg | 36 | 2.68 | 2.8 |
| S | Ser | 170 | 12.65 | 12.7 |
| T | Thr | 100 | 7.44 | 7.2 |
| V | Val | 126 | 9.38 | 9.1 |
| W | Trp | 24 | 6.98 | --- |
| Y | Tyr | 64 | 4.76 | 4.8 |
| N/D | Asn/Asp | 106 | 7.89 | 8.6 |
| Q/E | Gln/Glu | 132 | 9.82 | 10.8 |
5.MALDI-TOF MS分析
用MALDI-TOFMS分析LDP-02/3A9/LOT#1,得到分子量。检测到一个集中于149,808Da的主要峰。集中于74,927Da的峰代表主要峰中的+2离子种类。应当注意到+2离子物质并不精确地是M+H离子的一半,这个细微偏差在±0.2%之间,可能是由于实验不准确所导致。
根据预测的抗体一级序列,预期的分子量为147,154Da,与实测的IgG分子量相差2,654Da,这最可能是由于分子内糖基化造成的,这个差异值说明分子糖基化水平约为1.8%.
B.亲合力
首先用人源HUT-78细胞在细胞流计上滴定LDP-02/3A9/LOT#1和鼠Act-1(Lot#2)。简述之,1.0×106HUT-78细胞溶于100μl生物素化鼠Act-1(Lot#2)、生物素化鼠IgG1(Lot#1,LeukoSite,Inco,)、生物素化LDP-02/3A9/LOT#1溶液,或者溶解于生物素化人IgG(JacksonImmunoResearch Avondale PA Lot 25794),4℃20分钟,此后移走抗体。除非特别声明,全部试剂稀释于0.15M PBS/1.0%FCS/0.1%叠氮化钠。两种抗体的各种浓度包括30μg/ml(仅限于鼠Act-1)、15μg/ml、7.5μg/ml,随后各自按1∶10稀释。移走主要抗体后,将细胞溶解于100μl按1∶200稀释的链霉菌素藻红蛋白的溶液(Dako CorpCarpinteria,CA)。用200μl的PBS洗涤,将细胞重新溶解于0.5ml的PBS/1%福尔马林溶液,冷藏直至进行分析。在FACScan(BectonDickinson Corp.San Jose,CA)上分析样品,用488nm激光激发藻红旦白。对每个样品,最少分析10000个细胞,计算半峰平均频道荧光(MFC),所有样品重复操作两次。
滴定研究显示,用鼠ACT-1和LDP-02/3A9/LOT#1(图15),在大约1.0μg/ml的浓度,能有最大荧光。对于能达到MFC的浓度,LDP-02比鼠ACT-1更低(生物素化鼠ACT-1 Lot#2是0.1μg/ml,LDP-02/3A9/Lot#是0.02μg/ml)。
利用细胞统计以及LDP-02、鼠ACT-1抗体对人源HUT78细胞结合的交叉竞争,相对评价亲合力(和特异性)。简述之,1.0×106HuT-78细胞溶解于100μl的0.1μg/ml带有不同浓度非连结1°HUM ACT-1或带有非连结ACT-1的生物素化鼠ACT-1(LOT#2)溶液,4℃20分钟,取出抗体。在另一个独立实验里,所用溶液是100μl的0.02μg/ml带有不同浓度非连结鼠ACT-1(LOT#2)和非连结LDP-02/3A9/LOT#1的生物素化LDP-02/3A9/LOT#1溶液。维持稳定的生物素化抗体的浓度,是在如上述相同条件下导致染色HUT-78细胞有MCF值的那个浓度。除了特别声明,所有试剂均稀释于0.15MPBS/1.0%FCS0.1%叠氮化钠。两种抗体浓度在半对数增长范围内波动,从2×10-6M到5×10-11M。移去主要抗体后,将细胞溶于100μl按1∶200稀释的链霉菌素藻红旦白溶液(Dako Corp Carpinteria,CA)。用200μl的PBS洗涤后,再溶于0.5ml的PBS/1%福尔马林,冰箱冷藏直至分析。在FACScan(Becton Dickinson Corp.San Jose,CA)上分析样品,用488nm波长激光激发藻红旦白。对每个样品最少分析10000个细胞,计算MCF。每个样品重复操作两次,IC50是生物素化同型抗体中导致MCF减半的非结合抗体的浓度。
对LDP-02(1°HUM ACT-1)和鼠ACT-1之间进行5次独立的交叉竞争实验,从而估计其亲合力。当用生物素化鼠ACT-1当作实验中保持稳定的抗体,统计出来LDP-02(5.43±0.86nM)的平均IC50值(±1SEM)比鼠ACT-1(7.94±1.17nM;P=0.02双尾t实验:两个成对样品的平均值)的要低。而不相关的人IgG1或鼠IgG1没有竞争效果(全部实验概括于表10,一个实验列于图16)。相似地,当生物素化LDP-02/3A9/LOT#1用作实验中稳定抗体,需要有比LDP-02/3A9/LOT#1更高浓度的非结合鼠Act-1,从HuT-78细胞膜竞争LDP-02(IC50=6.3nM对4.3nM)。在每一个实验里,LDP-02的IC50比鼠ACT-1的要低,这些结果说明LDP-02对鼠ACT-1所识别的配体有特异性,它的亲合性优于鼠抗体的。
表10
鼠ACT-1和人化ACT-1(LDP-02)亲和性分析
| 实验 | 抗体 | Lot# | IC<sub>50</sub>(nM) |
| # | |||
| 1 | ACT-1(鼠) | 2 | 7.57 |
| 2 | 2 | 10.95 | |
| 3 | 2 | 6.02 | |
| 4 | 2 | 4.91 | |
| 5 | 2 | 10.24 | |
| 平均值±SEM | 7.94±1.17 | ||
| 1 | LDP-02(人化) | 7 | 4.34 |
| 2 | 7 | 6.13 | |
| 3 | 8/9 | 4.71 | |
| 4 | 8/9 | 3.53 | |
| 5 | 8/9 | 8.44 | |
| 平均值±SEM | 5.43±0.86 |
P=0.02
Two-tail t-Test:Paired Two Sample for Means
C.种类交叉反应性
用细胞流计来衡量种类交叉反应性。将100μl抽自于人、狗、猫、豚鼠或老鼠的EDTA-抗凝集血加到FACS管中。移走血浆,将血细胞层重新溶于100μl生物素LDP-02/3A9/LOT#1、不相关生物素化人IgG(Jackson Immuno Research,Avondale,PA)、生物素化鼠ACT-1LOT#2、或不相关生物素化鼠IgG1(Dako Corp.Carpinteria,CA),终浓度为15μg/ml。除了特别声明,全部试剂均稀释于0.15MPBS/1%FCS/0.1%叠氮化钠。样品和抗体一起在4℃保持20分钟,洗除抗体。细胞和100μl按1∶200稀释的链霉菌素藻红旦白溶液(Southern Biotechnology Associates,Inc.Birmingham,AL)在4℃保持20min。按照实验手册用一种商业溶胞试剂FACS溶胞液(BectonDickinson,San JvseCA)溶胞红细胞。用PBS洗涤后,将细胞再溶于0.5ml PBS/1%福尔马林,冰箱冷藏直至进行分析。在FACScan(BectonDickinson Corp.San Jose,CA)上分析样品,用488nm波长激光激发藻红旦白。对每个样品最少分析10000个细胞,淋巴细胞捕获门被置于前方,90度的光散参数。
生物素化LDP-02/3A9/LOT#1以一种异源染色识别人淋巴细胞的一个亚群,该模式类似于鼠ACT-1的,不同于人或鼠同型对照物。此外,检测狗的或猫的淋巴细胞时,LDP-02/3A9/LOT#1和鼠ACT-1都有一种类似于人淋巴细胞的异源染色模式,在这种条件下,LDP-02/3A9/LOT#1或鼠ACT-1不能识别豚鼠或老鼠的淋巴细胞。
D.C1q结合
根据上述方法,用细胞流计评价LPD-02结合人补体C1q的能力[Sims,M.J.et al.J.Immunol.151:2296-2308(1993)]。根据标准Ficoll密度分离法,分离人边周血单核细胞(PBMCs)。用10%普通的兔血清/PBS封闭375000个细胞,4℃下10分钟。冲洗后,细胞与100μl下列物质温育:(a)CAMPATH-IH(Therapeutic Antibody Center,Cambridge,U.K.);(b)人IgG1(Sigma Chemical Co.St.Louis,MO),(c)LDP-01[CD-18抗体的衍生物,描述于WO 93/02191(1993年2月4日发表),以及Sims,M.J.et al.J.Immunol.151(4):2296-2308(1993),在IgG1重链恒定区有两个位点的氨基酸替换(Leu235→Ala235和Gly237→Ala237),也叫作“FcRmut CD18”,Therapeutic Antibody Center,Cambridge,U.K.],或(d)LDP-02(1℃ hum ACT-1 Lot#8/9)浓度为10μg/ml,4℃下20min。CAMPATH-1H作为一种阳性对照,LDP-01和人IgG1作为阴性对照,全部试剂均用2%BSA/PBS稀释,2%BSA/PBS也作为一种阴性对照物单独加入。洗除抗体,将细胞重溶于50μl人补体C1q(Sigma Chemical Co.St.Louis,MO),10μg/ml 4℃下30min。然后洗涤细胞,重悬浮于200μl的FITC-结合兔抗一人C1q(Dako Corp.Carpinteria,CA)抗体,浓度为20μg/ml,4℃下20min。用200μl的PBS洗涤后,再溶于0.5ml的PBS/1%福尔马林,冻箱冷藏直至分析。在FACScan(Becton Dickinson Corp.San Jose,CA)上分析样品,用488nm波长激光激发藻红旦白。对每个样品最少分析10000个细胞,记算平均频道荧光(MCF)。
人PBM与CAMPATH-1H结合的人C1q一起温育,导致MCF有一个显著漂移,而PBMC与LDP-01、BSA或人IgG一起温育所诱导的染色模式都相似,都有低背景染色。PBMC与LDP-02预先温育所形成的染色模式和阴性对照的相同,说明在这种条件下LDP-02不结合C1q。
E补体介导的溶胞
如前述方法,检测LDP-02/3A9/Lot#1参与补体介导细胞溶胞的能力(Bindon,C.I.et al.Transplantation,40:538-544(1985))。无菌收集肝素化人血,收集血浆并立即置于冰上。在一层Ficoll-Hypaque上,密度1.077g/ml,离心15分钟,分离边周血单核细胞(PBMC),用完全培养液洗涤两次,培养基包括RPMI1640/10%FCS/100U/ml青霉素/100μg/ml链霉素/2.0Mml L-谷酰胺。将2千5百万个细胞在含150μCi51铬酸钠的无菌盐水中37℃温育1小时(E.I.du Pont de Nemours&Co.Inc.Wilmington,DE)。用培养液洗涤两次,重新悬浮成106/ml。将50μl该悬浮液(5.0×104个细胞)加到U形底微滴度盘的孔中,盘中有100μl下列物质之一(a)CAMPATH-1H(Therapeutic Center,Cambridge,U.K.),(b)CAMPATH-1G(Therapeutic Center,Cambridge,U.K.),(c)人IgG1(Sigma Chemical Co.St.Louis,MO),(d)LDP-02/3A9/Lot#1,或(e)LDP-01(FcRmut CD18,Therapeutic AntibodyCenter,Cambridge,U.K.),浓度分别为50,25,5,2.5,和0.5μg/ml。CAMPATH-1抗体作为阳性对照,人IgG1和LDP-01作为阴性对照。额外的孔包含溶于含有100μl 0.1%Triton-X-100的完全培养基中的细胞。同Triton-X-100温育的细胞被用来测量总放射性,而无抗体的对照孔被用来测定自发的放射性。室温下温育15分钟后,50μl自体血浆作为补体来源,加到每个孔中,终浓度为20%。37℃下温育45分钟,100g离心2分钟,收集100μl上清液,用Cobra II gamma Counter(Packard Instruments,Downers Grove,IL)测定放射性,每个样品重复操作2次,利用公式计算51Cr的特异放射性百分比:
特异放射性=(实验-自发性)×100%/(总体-自发性)
如以前所报道[Bindon et al Transplantation 40:538-544(1985)],CAMPATH-1H和CAMPATH-1G以一种剂量依赖性方式诱导出高达35%的人PBMC补体介导溶胞。此外,人IgG1和LDP-01对照物不诱导任何可检测到的细胞溶胞,LDP-02在高于和包括25μg/ml的任何试验浓度下不参与细胞溶胞作用(图17)。
F.抗体依赖性的细胞介导细胞毒作用(ADCC)
人CD3+胚细胞被用作靶细胞,来测量LDP-02参与ADCC作用的能力。CD3+胚细胞于外包被抗CD3抗体RT66的24孔板上产生的,RT66在PBS中稀释成5μg/ml。在一层1.077g/ml Ficoll-Hypaque上离心15分钟分离PBMC细胞,洗涤,重溶于完全培养基,这些操作如前面所述。将2百万个细胞加到24孔板的每个孔中,37℃5%CO2,温育4天。将细胞转入培养瓶中,37℃5%CO2培养,培养基含有10单位/毫升的人重组IL-2(Genzyme Corp.Cambridge,MA)。三天后,10×106CD3胚细胞在含150μCi51铬酸钠的无菌盐水(E.I.duPont de Nemours&Co.Inc.Wilmington,DE;Lot#95M682)中温育于37℃45分钟。用完全培养液洗涤两次,重悬浮成2×105cells/ml,取50μl悬液(10,000个细胞)加到U形底96孔微滴定板的孔中,孔中含有50μl CAMPATH-1H(Therapeutic Antibody Center,Cambridge,UK)或LDP-02/3A9/Lot#1,终浓度为50,5,2.5,0.5,0.25μg/ml。细胞与抗体在室温下温育30分钟,效应细胞(效应器∶靶=50∶1)加到每个孔中,效应细胞是来自不同供体的0.5×106个新分离的PBMC’(ficoll-hypaque梯度,在37℃用完全培养基洗两次)。额外的孔中,加入在培养基中的100μl 5% Triton-x-100(Fisher Scientific,Fair Lawn,NJ)。用与Triton-x-100温育的细胞测量总放射性,而无抗体与效应细胞的对照被用来测量自发放射性。室温下100g离心2分钟,37℃5%CO2温育20小时,此后,将细胞转移到V形底96孔板上,室温下沉淀。收集100μl上清液,用Cobra II gamma counter(Packard Instruments,Downers Grove,IL)测定放射性。每个样品重复操作2次,利用公式计算51Cr的特异放射性百分比。
特异放射性=(实验-自发性)×100%/(总体-自发性)
如以前所证实的[Sims,M.J.et al.J.Immunol.151(4):2296-2308(1993)],CAMPATH-1H以一种剂量依赖性方式参与ADCC,在浓度≥5.0μg/ml诱导大约30%特定的51Cr放射性。在任何检测浓度的包含LDP-02的孔中未检测到任何特定放射性。
G.对粘附MAdCAM-1的抑制作用
用荧光标记的α4β7+RPMI 8866细胞(人B淋巴细胞)和嵌合的MAdCAM-1来测量LDP-02抑制α4β7粘附MAdCAM-1过程的能力,嵌合的MAdCAM-1包括人MAdCAM-1的整个胞外区域融合于人IgG1 Fc区域(来自于同一构建物的恒定区,被用于Fc-突变LDP-02的恒定区)。
1.嵌合MAdCAM-IgG的构建
人MAdCAM-1克隆pcDhuMAd4[在pCDNA3中的克隆4 cDNA;Shyjan,A.M.et al.J.Immunol.156:2851-2857(1996),该文献通过在此引述而全文合并于本文]被用作人MAdCAM-胞外区域PCR扩增的模板,该区域与人IgG1恒定区融合[国际申请号:PCT/US96/02153(WO96/24673),1996年2月12日申请,是1995年9月1日申请的美国专利申请08/523,004的后续申请,该后续申请又是1995年2月10日申请的美国专利08/386,857的后续申请]。为构建MAdCAM-IgG嵌合体,合成出包含人MAdCAM-1编码序列的5端(ATG密码子,黑体)的引物HUMADIG4/2(SEG 1D NO:62)。
HindIII
5’-GGAAGCTTCCACCATGGATTTCGGACTGGCCC-3’
这个5’端引物和3’引物HUMAD1G3结合扩增出编码人MAdCAM-1整个胞外部分的区域。3’端引物HUMAD1G3(SEQ 1DNO:63)有下列序列;
SpeI
5’-GGACTAGTGGTTTGGACGAGCCTGTTG-3’
在引物上设计有5’端的Hind III位点和3’端的Spe I位点。使用Invitrogen(San Diago,CA)的PCR试剂盒和这些引物,PCR扩增MAdCAM片段。用HindIII和Spe I降解扩增产物,生成克隆用末端,利用Glassmax DNA分离系统(Gibco,Bethesda,MD)凝胶电泳纯化。
用SpeI和EcoRI从构建物上切下包含CH1,H(铰链),CH2和CH3区的约1Kb左右的片段,该构建物编码含有Fc-突变人恒定区的人免疫球蛋白重链。该构建物中人恒定区对应于CAMPATH-1H重链的PCR扩增产物[Reichmann,L.et al.Nature,322:323-327(1988),Sims,M.J.et al.(J.Immunol.151:2296-2308(1993)and Waldmann et al.(WO 93/02191,1993年2月4日(23页),这些文献都通过在此引述而全文合并于本文]。通过寡聚核苷酸直接突变,在构建物恒定区设计了突变(Leu235-Ala235,Gly237-Ala237),减少了对人Fcγ受体的结合。因此在MAdCAM-Ig融合子中除了引入了Leu235-Ala235,Gly237-Ala237突变外,含有Spe I-EcoRI恒定区[Sims et al.J.Immunol.151:2296-2308(1993)and Waldmann et al.(WO 93/02191)]。
用GlassmaxDNA分离系统(Gibco,Bethesda,MD)凝胶电泳分离编码Fc-突变IgG1恒定区的1kb的Spe I-EcoR I片段。恒定区片段和包含MAdCAM整个胞外区的HindIII-S peI片段整合到HindIII、E coRI酶切过的pEE12载体上[Stephens,P.L.and M.LCockett,Nucl.Acids Res.17:7110(1989)and Bebbington,C.R.andC.C.G.Hentschel,1987,The use of vectors based on gene amplificationfor the expression of cloned genes in mammalian cells,(Academic Press,N.Y.)]。得到细菌DH10B株的转化子,集落生成后,用限制性内切酶图谱分析微小质粒。编码含有MAdCAM-1整个胞外区融合蛋白的构建物(构建物HuMAd Ig21)与Fc-突变IgG1恒定区融合,在整个MAdCAM-1部分进行测序,以确证各部分之间正确的融合,以及PCR过程未引入突变。这个构建体在NSO细胞里生长,通过标准蛋白质A亲合层析纯化产物。
2.粘附实验
用碳酸盐缓冲液稀释嵌合MAdCAM-1至2.5μg/ml,pH9.5,取50μl涂在一个高结合性平底96孔板(Costar)上,37℃保持1小时,在一个微板自动清洗器上(Bio-Tek Instruments,Winooski VT)用洗涤缓冲液(50mM Tris HCl,0.14M NaCl,1mM MnCl2,pH7.2)洗涤孔穴。用稀释于PBS的100μl的10%FBS封闭,37℃,1.5小时。
首先用20ml的PBS(4℃)洗涤RPMI 8866细胞[一种表达α4β7(而不是α4β1)的人B淋巴细胞(Erle,D.J,et al,J,Immunol,153:517(1994);由Erle博士赠送)],重溶于PBS浓度成为4.0×106cells/ml。BCECF(2’,7’-双-(2-羧乙基)-5-(和6)-羧基荧光素,乙酰氧基酯;Molecular Probes,Inc.Eugene,OR)溶解于DMSO成50μg/ml,再加入到细胞悬液中,最终稀释度为1∶500,37℃温育30min后,用检测缓冲液洗涤细胞(HBSS和2%小牛血清,25mMHEPES,青霉素/链毒素,PH7.2),在V形底96孔板的每一个孔中加入50,000个细胞。细胞再溶解于100μl下列物质之一:(a)鼠Act-1;(b)鼠IgG1(Sigma Chemical,Co.St,Louis,MO);(c)LDP-02/3A9/Lot#1;(d)人IgG1(Sigma Chemical Co.st,Louis,MO),浓度为15.0-0.00075μg/ml在检测缓冲液中,室温下温育10min。洗涤外覆MAdCAM-1的板,除去封闭缓冲液,将这些荧光标记的RPMI 8866细胞加到每个孔中。板置于40RPM的平板振荡器上(New Brunswick,Scientific Co.Inc,Edison,NJ),室温下30min,用铝箔包裹。未被结合的细胞通过一次洗涤便可除去,接下来,在荧光聚焦分析仪上(IDEXX Laboratories Inc.Westbrook,ME)测量荧光强度(485nm激发,535nm读数)。通过公式,根据相对荧光单位计算出每孔中结合细胞的百分比。
结合细胞%=(洗涤前RFU/洗涤后RFU)×100
LDP-02和鼠Act-1都以一种剂量依赖性方式抑制RPMI 8866细胞对人MAdCAM的粘附(图18A-18B)。半抑制粘附浓度(IC50)相对类似于鼠Act-1的(0.0018μg/ml)和LDP-02的(0.0014μg/ml),因此,LDP-02起码和鼠Act-1一样能够抑制α4β7介导的对MAdCAM-1的粘附。
实施例5额外的人化抗体
如上所述,实施例2中的改型抗体的多个变种能够提高其亲合性和/或降低抗原性。这样的构建物包括但不仅限于下列一种或一种以上的突变:轻链的M4V的突变,重链的R38K突变,重链A40R突变,重链I73T回复突变。突变可以是单个发生(例如,一条链上一个突变),也可以是联合发生。
例如,图19所示的是用改型抗体(在实施例2中设计的)或用在轻链上有一个额外突变(MV4)和在重链上有两个额外突变(R38K,A40R)的衍生体对HuT78染色的结果。这两种抗体有相似的染色模式(图19)。根据实验手册用转化子位点直接突变试剂盒(Clontech)改变核苷酸序列,从而形成突变。用克隆到pCR-ScriptTM的可变区片段制造出在重链和轻链可变区都有突变。顺式寡核苷酸ScaI/Stu I(Clontech)用于顺式寡聚核苷酸。突变寡聚核苷酸(SEQ 1DNO:38-40)的序列如下:
H/R38K(SEQ ID NO:38)
5’-C TGG CCA ACG
H/I73T(SEQ ID NO:39)
5’-CAC ATT GAC TGT AGA CAC TTC CGC TAG CAC AGC C
L/M4V(SEQ ID NO:40)
5’-CCG GAG GTG ATG TTG TGG TGA CTC
对于其他的操作,包括亚克隆到表达载体pEE6hcMV-B和pEE12,构建包含重链和轻链基因的表达质粒等,都如对主要的改型抗体的描述。瞬间转染和细胞染色也都如对主要的改型抗体的描述。
等同原则
本领域的技术人员们都知道或者都能够肯定,使用不超过常规的实验方法,都可以获得本发明的许多其他等价方案。这些等价也能够达到本项发明同样的效果,这些等价方法也被包括在下面的权利要求中。
Claims (59)
1.一种选择性地结合α4β7整合蛋白的人化免疫球蛋白或其抗原结合片段,所述的人化免疫球蛋白或抗原结合片段包括非人源互补决定区(CDRs)和由人源重链和轻链衍生而来的重链和轻链框架区。
2.根据权利要求1所述的人化免疫球蛋白或其抗原结合片段,其中,人化免疫球蛋白进一步包括人恒定区。
3.根据权利要求2所述的人化免疫球蛋白或其抗原结合片段,其中,人恒定区是γ类型的。
4.根据权利要求1的所述的人化免疫球蛋白或其抗原结合片段,其中,CDRs是啮齿类源的。
5.根据权利要求4所述的人化免疫球蛋白或其抗原结合片段,其中,CDRs来自于鼠Act-1单克隆抗体。
6.根据权利要求1所述的人化免疫球蛋白或其抗原结合片段,其中所述的免疫球蛋白或抗原结合片段包括轻链和重链,
所述轻链包括互补决定区(CDR1,CDR2和CDR3),该互补决定区具有如下所示氨基酸序列:
轻链:CDR1 SEQ ID NO:12的氨基酸44-59
CDR2 SEQ ID NO:12的氨基酸75-81
CDR3 SEQ ID NO:12的氨基酸114-122;和衍生自人源轻链可变区的框架区;
所述重链包括互补决定区(CDR1,CDR2和CDR3),该互补决定区具有如下所示氨基酸序列:
重链:CDR1 SEQ ID NO:15的氨基酸50-54
CDR2 SEQ ID NO:15的氨基酸69-85
CDR3 SEQ ID NO:15的氨基酸118-129;和衍生自人源重链可变区的框架区。
7.根据权利要求6所述的人化免疫球蛋白或其抗原结合片段,其中,所述人源轻链的可变区是SEQ ID NO:8的可变区。
8.根据权利要求6所述的人化免疫球蛋白或其抗原结合片段,其中,所述人源重链的可变区是SEQ ID NO:10的可变区。
9.根据权利要求7所述的人化免疫球蛋白或其抗原结合片段,其中,所述人源重链的可变区是SEQ ID NO:10的可变区。
10.根据权利要求6所述的人化免疫球蛋白或其抗原结合片段,其中,轻链包括SEQ ID NO:21的可变区,重链包括SEQ ID NO:19的可变区。
11.根据权利要求6所述的人化免疫球蛋白或其抗原结合片段,其中,所述的免疫球蛋白或抗原结合片段能与鼠Act-1竞争结合α4β7整合蛋白。
12.根据权利要求6所述的人化免疫球蛋白或其抗原结合片段,其中,所述免疫球蛋白或抗原结合片段具有鼠Act-1单克隆抗体的表位特异性。
13.权利要求1所述人化抗体或抗原结合片段的人化免疫球蛋白轻链或其片段,其中所述轻链或片段具有一种氨基酸序列,该氨基酸序列包括至少一种SEQ ID NO:12轻链可变区的功能部分。
14.根据权利要求13所述的人化免疫球蛋白轻链或其片段,其中,所述的轻链或片断具有包括信号肽SEQ ID NO:12的氨基酸序列。
15.一种人化免疫球蛋白轻链或其片段,其中,所述的人化免疫球蛋白轻链或片段包括鼠Act-1单克隆抗体轻链的互补决定区,CDR1,CDR2和CDR3和人源轻链框架区,所述互补决定区包括如下所示氨基酸序列,以至于包括所述轻链或其片段的抗体选择性地结合α4β7整合蛋白:
轻链:CDR1 SEQ ID NO:12的氨基酸44-59
CDR2 SEQ ID NO:12的氨基酸75-81
CDR3 SEQ ID NO:12的氨基酸114-122。
16.根据权利要求15所述的人化免疫球蛋白轻链或片段,其中,所述人框架区由SEQ ID NO:8的轻链可变区衍生而来。
17.根据权利要求15所述的人化免疫球蛋白轻链或片段,其中,所述人化免疫球蛋白轻链或片段包括SEQ ID NO:21的可变区。
18.一种被分离的核酸,包括编码权利要求13-17任意一种所述的人化免疫球蛋白轻链或片段的核苷酸序列。
19.根据权利要求18所述的被分离的核酸,其中,所述的核苷酸序列包括编码SEQ ID NO:20可变区的核苷酸序列。
20.一种表达载体,其中包括编码权利要求13-17任意一种所述的人化免疫球蛋白轻链或其片段的融合基因。
21.根据权利要求20的表达载体,其中,所述的融合基因包含SEQID NO:20的可变区编码序列。
22.权利要求1所述的人化抗体或抗原结合片段的人化免疫球蛋白重链或其片段,其中所述重链或片段具有一种氨基酸序列,该氨基酸序列包括至少一种SEQ ID NO:15重链可变区的功能部分。
23.根据权利要求22所述的人化免疫球蛋白重链或其片段,其中,所述的重链或片断具有包括信号肽SEQ ID NO:15的氨基酸序列。
24.一种人化免疫球蛋白重链或其片段,其中,所述的人化免疫球蛋白重链或片段包括鼠Act-1单克隆抗体的重链的互补决定区,CDR1,CDR2和CDR3,和人源重链框架区,所述的互补决定区包括如下所示的氨基酸序列,以至于包括所述重链或其片段的抗体选择性结合α4β7整合蛋白:
重链:CDR1 SEQ ID NO:15的氨基酸50-54
CDR2 SEQ ID NO:15的氨基酸69-85
CDR3 SEQ ID NO:15的氨基酸118-129。
25.根据权利要求24所述人化免疫球蛋白重链或片段,其中,所述的人框架区由SEQ ID NO:10可变区衍生而来。
26.根据权利要求24所述的人化免疫球蛋白重链或片段,其中,所述的人化免疫球蛋白重链或片段包含SEQ ID NO:19的可变区。
27.一种被分离的核酸,包括编码权利要求22-26任意一种所述的人化免疫球蛋白重链或片段的核苷酸序列。
28.根据权利要求27所述的被分离的核酸,其中,所述的核苷酸序列包括编码SEQ ID NO:18可变区的核苷酸序列。
29.一种表达载体,其中包括编码权利要求22-26中任意一种人化免疫球蛋白重链或其片段的融合基因。
30.根据权利要求29的表达载体,其中,所述的融合基因包含SEQID NO:18的可变区编码序列。
31.包括权利要求20所述表达载体的宿主细胞。
32.包括权利要求21所述表达载体的宿主细胞。
33.包括权利要求29所述表达载体的宿主细胞。
34.包括权利要求30所述表达载体的宿主细胞。
35.一种制备人化免疫球蛋白轻链、人化免疫球蛋白轻链片段的方法,包括,在适合人化免疫球蛋白轻链表达的条件下培养权利要求31中的宿主细胞,由此表达、制备人化免疫球蛋白轻链或人化免疫球蛋白轻链片段。
36.根据权利要求3 5所述的方法,进一步包括分离人化免疫球蛋白轻链或人化免疫球蛋白轻链片段的步骤。
37.一种制备人化免疫球蛋白轻链、人化免疫球蛋白轻链片段的方法,包括,在适合人化免疫球蛋白轻链表达的条件下培养权利要求32中的宿主细胞,由此表达、制备人化免疫球蛋白轻链或人化免疫球蛋白轻链片段。
38.根据权利要求37所述的方法,进一步包括分离人化免疫球蛋白轻链或人化免疫球蛋白轻链片段。
39.一种制备人化免疫球蛋白重链、人化免疫球蛋白重链片段的方法,包括,在适合人化免疫球蛋白重链表达的条件下培养权利要求33中的宿主细胞,由此表达、制备人化免疫球蛋白重链或人化免疫球蛋白重链片段。
40.根据权利要求39所述的方法,进一步包括分离人化免疫球蛋白重链或人化免疫球蛋白重链片段的步骤。
41.一种制备人化免疫球蛋白重链、人化免疫球蛋白重链片段的方法,包括,在适合人化免疫球蛋白重链表达的条件下培养权利要求34中的宿主细胞,由此表达、制备人化免疫球蛋白重链或人化免疫球蛋白重链片段。
42.根据权利要求41所述的方法,进一步包括分离人化免疫球蛋白重链或人化免疫球蛋白重链片段的步骤。
43.一种宿主细胞,包括编码人化免疫球蛋白轻链或其片段的第一重组核酸,和编码人化免疫球蛋白重链或其片段的第二重组核酸,其中,包含所述的轻链或其片段和所述重链或其片段的抗体或抗原结合片段选择性地结合α4β7整合蛋白,
所述第一核酸包括编码衍生自鼠Act-1单克隆抗体轻链的CDR和衍生自人源轻链框架区的核苷酸序列;以及
所述第一核酸包括编码衍生自鼠Act-1单克隆抗体重链的CDR和衍生自人源重链框架区的核苷酸序列。
44.根据权利要求43的宿主细胞,其中:
所述的第一核酸包括编码三个互补决定区,CDR1,CDR2和CDR3的核苷酸序列,具有如下所示氨基酸序列:
轻链:CDR1 SEQ ID NO:12的氨基酸44-59
CDR2 SEQ ID NO:12的氨基酸75-81
CDR3 SEQ ID NO:12的氨基酸114-122;和一种衍生自人源轻链可变区的框架区;和
所述的第二核酸包括编码三个互补决定区,CDR1,CDR2和CDR3中的至少一种的核苷酸序列,具有如下所示氨基酸序列:
重链:CDR1 SEQ ID NO:15的氨基酸50-54
CDR2 SEQ ID NO:15的氨基酸69-85
CDR3 SEQ ID NO:15的氨基酸118-129;和
一种衍生自人源重链可变区的框架区。
45.一种制备人化免疫球蛋白或其抗原结合片段的方法,包括,在适合人化免疫球蛋白或其抗原结合片段表达的条件下培养权利要求43或44所述的宿主细胞,由此表达人化免疫球蛋白链并且制备人化免疫球蛋白或抗原结合片段,和任选的分离。
46.根据权利要求45的方法,进一步包括分离所述的人化免疫球蛋白或抗原结合片段。
47.一种体外抑制携带α4β7的第一细胞和携带其配体的第二细胞的相互作用的方法,包括使足量的所述第一细胞和权利要求1-14中任意一种人化免疫球蛋白或抗原结合片段相结合,来抑制α4β7整合蛋白与其配体的结合。
48.权利要求1-12中任意一种人化免疫球蛋白或抗原结合片段在制备用于抑制黏膜组织的淋巴细胞渗透作用的药物中的应用。
49.一种药物组合物,包括权利要求1-12中任意一种人化免疫球蛋白或抗原结合片段和适宜的载体。
50.根据权利要求1-12中所述的任意一种人化免疫球蛋白或抗原结合片段在制造用于治疗炎症疾病的药物中的应用。
51.权利要求1-12中任意一种的人化免疫球蛋白或抗原结合片段在制造用于治疗与组织的淋巴细胞渗透性联系在一起的疾病的药物中的应用,其中,所述疾病是胆囊炎、胆管炎、边周性胆囊炎、乳腺炎、慢性窦炎、胰脏炎、胰岛素依赖型糖尿病。
52.权利要求1-12中任意一种的人化免疫球蛋白或抗原结合片段在制造治疗肠炎症疾病的药物中的应用。
53.根据权利要求52的应用,其中,所述的肠炎症疾病选自由溃疡性结肠炎和Crohn’s病组成的组。
54.权利要求10的人化免疫球蛋白或抗原结合片段在制造治疗肠炎症疾病的药物中的应用。
55.权利要求10的人化免疫球蛋白或抗原结合片段在制造治疗溃疡性结肠炎和Crohn’s病的药物中的应用。
56.根据权利要求50的应用,其中,所述疾病选自由慢性支气管炎、气喘和移植排异反应所组成的组。
57.根据权利要求56的应用,其中所述疾病是移植排异反应。
58.一种药物组合物,包括对包括重链和轻链的α4β7整合单位具有结合特异性的人源化免疫球蛋白或抗原结合片断,和一种药学上可接受的介质或载体,其中所述重链包括SEQ ID NO:19的可变区,所述轻链包括SEQ ID NO:21的可变区。
59.权利要求1-16任意一项所述的人化免疫球蛋白或抗原结合片段在制备用于抑制携带α4β7的第一细胞和携带其配体的第二细胞结合的药物中的应用。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US08/700,737 | 1996-08-15 | ||
| US08/700,737 US7147851B1 (en) | 1996-08-15 | 1996-08-15 | Humanized immunoglobulin reactive with α4β7 integrin |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN1227607A CN1227607A (zh) | 1999-09-01 |
| CN100406562C true CN100406562C (zh) | 2008-07-30 |
Family
ID=24814673
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CNB971972214A Expired - Lifetime CN100406562C (zh) | 1996-08-15 | 1997-08-06 | 与α4β7整合蛋白相作用的人化免疫球蛋白 |
Country Status (20)
| Country | Link |
|---|---|
| US (3) | US7147851B1 (zh) |
| EP (2) | EP1710314A1 (zh) |
| JP (3) | JP4171071B2 (zh) |
| CN (1) | CN100406562C (zh) |
| AT (1) | ATE321788T1 (zh) |
| AU (1) | AU730326C (zh) |
| BE (1) | BE2014C068I2 (zh) |
| BR (1) | BRPI9711079B1 (zh) |
| CA (1) | CA2263106C (zh) |
| DE (1) | DE69735596T3 (zh) |
| DK (1) | DK0918797T4 (zh) |
| ES (1) | ES2262186T5 (zh) |
| HK (1) | HK1021822A1 (zh) |
| IL (3) | IL128052A0 (zh) |
| LT (1) | LTC0918797I2 (zh) |
| LU (1) | LU92596I2 (zh) |
| NZ (1) | NZ334226A (zh) |
| PT (1) | PT918797E (zh) |
| SI (1) | SI0918797T2 (zh) |
| WO (1) | WO1998006248A2 (zh) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN103608071A (zh) * | 2011-05-02 | 2014-02-26 | 米伦纽姆医药公司 | 抗α4β7抗体的制剂 |
Families Citing this family (115)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7147851B1 (en) * | 1996-08-15 | 2006-12-12 | Millennium Pharmaceuticals, Inc. | Humanized immunoglobulin reactive with α4β7 integrin |
| US7122636B1 (en) | 1997-02-21 | 2006-10-17 | Genentech, Inc. | Antibody fragment-polymer conjugates and uses of same |
| US7790856B2 (en) | 1998-04-07 | 2010-09-07 | Janssen Alzheimer Immunotherapy | Humanized antibodies that recognize beta amyloid peptide |
| US7964192B1 (en) | 1997-12-02 | 2011-06-21 | Janssen Alzheimer Immunotherapy | Prevention and treatment of amyloidgenic disease |
| TWI239847B (en) | 1997-12-02 | 2005-09-21 | Elan Pharm Inc | N-terminal fragment of Abeta peptide and an adjuvant for preventing and treating amyloidogenic disease |
| US6787523B1 (en) * | 1997-12-02 | 2004-09-07 | Neuralab Limited | Prevention and treatment of amyloidogenic disease |
| US20080050367A1 (en) | 1998-04-07 | 2008-02-28 | Guriq Basi | Humanized antibodies that recognize beta amyloid peptide |
| US7090845B2 (en) * | 1998-05-13 | 2006-08-15 | Genentech, Inc. | Diagnosis and treatment of hepatic disorders |
| AU2011226857B2 (en) * | 1998-05-13 | 2013-09-19 | Genentech, Inc. | Diagnosis and treatment of hepatic disorders |
| DE69929712T2 (de) * | 1998-11-25 | 2006-10-19 | The Government Of The United States Of America As Represented By The Secretary, Department Of Health And Human Services | Antagonisten des Alpha-E-Beta-7 Integrins als therapeutische Agenzien gegen Entzündungskrankheiten |
| CA2370814C (en) * | 1999-04-22 | 2010-07-06 | Biogen, Inc. | Method for the treatment of fibrosis using an antagonist of the integrin alpha-4 subunit |
| US20010046496A1 (en) * | 2000-04-14 | 2001-11-29 | Brettman Lee R. | Method of administering an antibody |
| AU2006220426B2 (en) * | 2000-04-14 | 2008-10-30 | Genentech Inc. | Method of administering an antibody |
| US7658924B2 (en) * | 2001-10-11 | 2010-02-09 | Amgen Inc. | Angiopoietin-2 specific binding agents |
| US7521053B2 (en) * | 2001-10-11 | 2009-04-21 | Amgen Inc. | Angiopoietin-2 specific binding agents |
| CA2470643C (en) * | 2001-12-17 | 2011-10-18 | Bryan Paul Morgan | Cleavable reagents for specific delivery to disease sites |
| AU2003210802B2 (en) | 2002-02-05 | 2009-09-10 | Genentech Inc. | Protein purification |
| MY139983A (en) | 2002-03-12 | 2009-11-30 | Janssen Alzheimer Immunotherap | Humanized antibodies that recognize beta amyloid peptide |
| US20060234226A1 (en) | 2002-04-26 | 2006-10-19 | Fahner Robert L | Non-affinity purification of proteins |
| TWI379693B (en) | 2002-05-02 | 2012-12-21 | Wyeth Corp | Calicheamicin derivative-carrier conjugates |
| GB0210121D0 (en) * | 2002-05-02 | 2002-06-12 | Celltech R&D Ltd | Biological products |
| EP2368578A1 (en) * | 2003-01-09 | 2011-09-28 | Macrogenics, Inc. | Identification and engineering of antibodies with variant Fc regions and methods of using same |
| UA87453C2 (ru) * | 2003-02-01 | 2009-07-27 | Вайет | Применения фрагмента ав для лечения болезни альцгеймера |
| JP4599355B2 (ja) | 2003-07-28 | 2010-12-15 | ジェネンテック, インコーポレイテッド | プロテインaアフィニティークロマトグラフィーの間のプロテインaの浸出の低減 |
| EP3192872A1 (en) | 2003-08-26 | 2017-07-19 | The Regents of the University of Colorado, a body corporate | Inhibitors of serine protease activity and their use in methods and compositions for treatment of bacterial infections |
| AU2003300896A1 (en) | 2003-12-10 | 2005-07-14 | Millennium Pharmaceuticals, Inc. | Humanized anti-ccr2 antibodies and methods of use |
| AP2006003689A0 (en) | 2004-01-09 | 2006-08-31 | Pfizer | Antibodies to MAdCAM |
| US7655229B2 (en) | 2004-09-02 | 2010-02-02 | Chan Andrew C | Anti-FC-gamma RIIB receptor antibody and uses therefor |
| US7662926B2 (en) | 2004-09-02 | 2010-02-16 | Genentech, Inc. | Anti-Fc-gamma receptor antibodies, bispecific variants and uses therefor |
| PL1784426T3 (pl) | 2004-09-03 | 2012-04-30 | Genentech Inc | Humanizowani antagoniści anty-beta7 i ich zastosowania |
| JO3000B1 (ar) | 2004-10-20 | 2016-09-05 | Genentech Inc | مركبات أجسام مضادة . |
| JP2008523815A (ja) | 2004-12-15 | 2008-07-10 | エラン ファーマ インターナショナル リミテッド | 認知の改善における使用のためのヒト化アミロイドβ抗体 |
| MY146381A (en) * | 2004-12-22 | 2012-08-15 | Amgen Inc | Compositions and methods relating relating to anti-igf-1 receptor antibodies |
| WO2007007160A2 (en) * | 2005-07-11 | 2007-01-18 | Pfizer Limited | Anti-madcam antibodies to treat fever |
| WO2007007151A2 (en) * | 2005-07-11 | 2007-01-18 | Pfizer Limited | Use of anti-mad-cam antibody for the treatment of emphysema |
| CA2629147A1 (en) * | 2005-11-17 | 2007-05-31 | Millennium Pharmaceuticals, Inc. | Humanized immunoglobulin reactive with .alpha.4.beta.7 integrin |
| US8784810B2 (en) | 2006-04-18 | 2014-07-22 | Janssen Alzheimer Immunotherapy | Treatment of amyloidogenic diseases |
| FR2902799B1 (fr) | 2006-06-27 | 2012-10-26 | Millipore Corp | Procede et unite de preparation d'un echantillon pour l'analyse microbiologique d'un liquide |
| US20100267933A1 (en) | 2006-12-21 | 2010-10-21 | Moya Wilson | Purification of proteins |
| US8362217B2 (en) | 2006-12-21 | 2013-01-29 | Emd Millipore Corporation | Purification of proteins |
| US8569464B2 (en) | 2006-12-21 | 2013-10-29 | Emd Millipore Corporation | Purification of proteins |
| ES2426158T3 (es) | 2007-01-22 | 2013-10-21 | Genentech, Inc. | Precipitación con polielectrolito y purificación de anticuerpos |
| JP2010520204A (ja) * | 2007-03-02 | 2010-06-10 | アムジェン インコーポレイテッド | 腫瘍疾患を治療するための方法および組成物 |
| US20100297699A1 (en) * | 2007-03-20 | 2010-11-25 | Millennium Pharmaceuticals, Inc. | Nucleic Acids Encoding Humanized Immunoglobulin That Binds Alpha4Beta7 Integrin |
| US8003097B2 (en) | 2007-04-18 | 2011-08-23 | Janssen Alzheimer Immunotherapy | Treatment of cerebral amyloid angiopathy |
| CN106366192B (zh) | 2007-06-25 | 2020-03-27 | 艾斯巴技术-诺华有限责任公司 | 修饰抗体的方法和具有改善的功能性质的修饰抗体 |
| ES2751022T3 (es) | 2007-07-09 | 2020-03-30 | Genentech Inc | Prevención de la reducción de enlaces disulfuro durante la producción recombinante de polipéptidos |
| JP5889529B2 (ja) * | 2007-07-27 | 2016-03-22 | ヤンセン・サイエンシズ・アイルランド・ユーシー | アミロイド原性疾患の処置 |
| JO3076B1 (ar) | 2007-10-17 | 2017-03-15 | Janssen Alzheimer Immunotherap | نظم العلاج المناعي المعتمد على حالة apoe |
| EP3441402A1 (en) | 2007-10-30 | 2019-02-13 | Genentech, Inc. | Antibody purification by cation exchange chromatography |
| MX2010007515A (es) * | 2008-01-11 | 2010-11-12 | Chemo Sero Therapeut Res Inst | Anticuerpo antiintegrina a9 humana humanizado mejorado. |
| JO2913B1 (en) | 2008-02-20 | 2015-09-15 | امجين إنك, | Antibodies directed towards angiopoietin-1 and angiopoietin-2 proteins and their uses |
| WO2009151514A1 (en) | 2008-06-11 | 2009-12-17 | Millipore Corporation | Stirred tank bioreactor |
| DK3604324T3 (da) | 2008-08-14 | 2024-04-08 | Genentech Inc | Fremgangsmåder til at fjerne en kontaminant under anvendelse af ionbytningskromatografi med forskydning af iboende proteiner |
| EP2684570A1 (en) | 2008-09-10 | 2014-01-15 | F. Hoffmann-La Roche AG | Compositions and methods for the prevention of oxidative degradation of proteins |
| US9067981B1 (en) | 2008-10-30 | 2015-06-30 | Janssen Sciences Ireland Uc | Hybrid amyloid-beta antibodies |
| EP2358734A1 (en) | 2008-12-16 | 2011-08-24 | Millipore Corporation | Purification of proteins |
| SG171446A1 (en) | 2008-12-16 | 2011-07-28 | Millipore Corp | Stirred tank reactor and method |
| ES2547712T3 (es) | 2009-02-27 | 2015-10-08 | Genentech, Inc. | Métodos y composiciones para el marcado de proteínas |
| SG10201400798VA (en) * | 2009-03-20 | 2014-07-30 | Amgen Inc | Alpha-4-Beta-7 Heterodimer Specific Antagonist Antibody |
| AU2016208453B2 (en) * | 2009-03-20 | 2018-02-22 | Amgen Inc. | Alpha-4-beta-7 heterodimer specific antagonist antibody |
| AU2013205341B2 (en) * | 2009-03-20 | 2016-05-05 | Amgen Inc. | Alpha-4-beta-7 heterodimer specific antagonist antibody |
| NZ621170A (en) | 2009-05-13 | 2015-08-28 | Genzyme Corp | Anti-human cd52 immunoglobulins |
| LT2464725T (lt) | 2009-08-11 | 2020-06-10 | F. Hoffmann-La Roche Ag | Baltymų gamyba ląstelių mitybinėje terpėje, kurioje nėra glutamino |
| EP2473617B1 (en) | 2009-09-01 | 2020-02-26 | F.Hoffmann-La Roche Ag | Enhanced protein purification through a modified protein a elution |
| AU2010311567B2 (en) | 2009-10-26 | 2015-03-26 | Chugai Seiyaku Kabushiki Kaisha | Method for the production of a glycosylated immunoglobulin |
| KR20180000342A (ko) | 2010-03-22 | 2018-01-02 | 제넨테크, 인크. | 단백질-함유 제제의 안정화에 유용한 조성물 및 방법 |
| MX2012012743A (es) | 2010-05-03 | 2012-11-23 | Genentech Inc | Composiciones y metodos utiles para reducir la viscosidad de formulaciones que contienen proteina. |
| SG10201804385YA (en) | 2010-05-17 | 2018-06-28 | Emd Millipore Corp | Stimulus responsive polymers for the purification of biomolecules |
| MX343604B (es) | 2010-05-28 | 2016-11-11 | Genentech Inc | Disminuir nivel de lactato e incrementar producción de polipéptidos por regulación a la baja de la expresión de lactato dehidrogenasa y piruvato dehidrogenasa quinasa. |
| MY161390A (en) | 2010-06-24 | 2017-04-14 | Genentech Inc | Compositions and methods containing alkylglycosides for stabilizing protein-containing formulations |
| CN104474546A (zh) | 2010-08-13 | 2015-04-01 | 弗·哈夫曼-拉罗切有限公司 | 用于疾病治疗的针对IL-1β和IL-18的抗体 |
| WO2012030512A1 (en) | 2010-09-03 | 2012-03-08 | Percivia Llc. | Flow-through protein purification process |
| CN107998388B (zh) | 2011-05-02 | 2023-07-14 | 千禧制药公司 | 抗α4β7抗体的制剂 |
| KR102103476B1 (ko) | 2011-06-24 | 2020-04-23 | 더 리젠츠 오브 더 유니버시티 오브 콜로라도, 어 바디 코포레이트 | 알파-1 안티트립신 융합 분자용 조성물, 방법 및 용도 |
| RU2648999C2 (ru) | 2011-12-22 | 2018-03-29 | Дженентек, Инк. | Способы повышения эффективности этапов очистки белка, находящихся ниже по потоку, с использованием мембранной ионообменной хроматографии |
| EP2802653A4 (en) * | 2012-01-10 | 2015-09-02 | Univ Colorado Regents | ALPHA-1 ANTITRYPSIN FUSION MOLECULE COMPOSITIONS, METHODS AND USES THEREOF |
| EA201992881A3 (ru) * | 2012-01-12 | 2020-12-30 | Милленниум Фармасьютикалз, Инк. | Композиция анти-47 антитела |
| SG10201912877SA (en) | 2012-03-27 | 2020-02-27 | Genentech Inc | Improved harvest operations for recombinant proteins |
| CN103374073A (zh) * | 2012-04-26 | 2013-10-30 | 中国科学院上海生命科学研究院 | 识别活化形式整合素α4β7的人源化单克隆抗体 |
| SG11201407512VA (en) | 2012-05-18 | 2014-12-30 | Genentech Inc | High-concentration monoclonal antibody formulations |
| US9393327B2 (en) | 2012-12-19 | 2016-07-19 | Genentech, Inc. | Methods and compositions for radiohalogen protein labeling |
| AR095199A1 (es) | 2013-03-15 | 2015-09-30 | Genzyme Corp | Anticuerpos anti-cd52 |
| AU2015287760B2 (en) | 2014-07-09 | 2019-06-20 | Genentech, Inc. | pH adjustment to improve thaw recovery of cell banks |
| IL292655B2 (en) | 2014-10-06 | 2024-03-01 | Chemocentryx Inc | CCR9 chemokine receptor inhibitor for use in a method for treating or reducing the development of inflammatory bowel diseases |
| EP3581585A1 (en) | 2014-11-26 | 2019-12-18 | Millennium Pharmaceuticals, Inc. | Vedolizumab for the treatment of fistulizing crohn's disease |
| JP6904905B2 (ja) | 2014-12-24 | 2021-07-21 | ミレニアム ファーマシューティカルズ, インコーポレイテッドMillennium Pharmaceuticals, Inc. | 抗α4β7インテグリン抗体による治療の結果の予測 |
| MA41636A (fr) | 2015-03-06 | 2018-01-09 | Millennium Pharm Inc | Méthode de traitement de la cholangite sclérosante primitive |
| AU2016306320B2 (en) * | 2015-08-11 | 2022-04-14 | Osaka University | Antibody |
| JP6951825B2 (ja) | 2015-09-04 | 2021-10-20 | ザ スクリプス リサーチ インスティテュート | インスリン免疫グロブリン融合タンパク質 |
| WO2017117558A1 (en) | 2015-12-31 | 2017-07-06 | The Regents Of The University Of Colorado, A Body Corporate | Methods and uses for alpha-1 antitrypsin or recombinant forms thereof, on steroid-refractory graft versus host disease involving gastrointestinal complications |
| EP3430052A2 (en) * | 2016-03-14 | 2019-01-23 | Millennium Pharmaceuticals, Inc. | Method of preventing graft versus host disease |
| AU2017234010A1 (en) * | 2016-03-14 | 2018-09-27 | Millennium Pharmaceuticals, Inc. | Methods of treating or preventing graft versus host disease |
| JP7069032B2 (ja) | 2016-03-24 | 2022-05-17 | ミレニアム ファーマシューティカルズ, インコーポレイテッド | がん免疫治療における胃腸の免疫関連有害事象の治療方法 |
| TW201735949A (zh) | 2016-03-24 | 2017-10-16 | 千禧製藥公司 | 治療抗ctla4及抗pd-1組合治療中的胃腸道免疫相關不良事件之方法 |
| KR20190012145A (ko) | 2016-03-29 | 2019-02-08 | 젤터, 인코포레이티드 | 주변세포질 부피 대 세포질 부피의 비가 0.5:1 내지 10:1인 그람-음성 박테리아에서의 단백질의 발현 |
| US10918716B2 (en) | 2016-05-04 | 2021-02-16 | Millennium Pharmaceuticals, Inc. | Triple combination therapy for treating Crohn's disease |
| EP3469069A4 (en) * | 2016-06-10 | 2020-01-22 | Kota Biotherapeutics, LLC | COMPOSITIONS AND METHODS COMPRISING A B LYMPHOCYTE CELL LINE EXPRESSING A MEMBRANE IMMUNOGLOBULIN DIFFERENT FROM A SECRETED IMMUNOGLOBULIN AND CYTOLYTIC FUNCTION |
| US11389533B2 (en) | 2016-06-12 | 2022-07-19 | Takeda Pharmaceutical Company Limited | Method of treating inflammatory bowel disease |
| WO2018104893A1 (en) | 2016-12-06 | 2018-06-14 | Glaxosmithkline Intellectual Property Development Limited | Alpha4-beta7 antibodies with incrased fcrn binding and/or half-life |
| MX2019007433A (es) | 2016-12-22 | 2019-08-16 | Genentech Inc | Metodos y formulaciones para reducir el tiempo de reconstitucion de los polipeptidos liofilizados. |
| CN110612120A (zh) | 2017-04-28 | 2019-12-24 | 千禧制药公司 | 使用特异性于α4β7整联蛋白的抗体(维多珠单抗)治疗儿科病症的方法 |
| AR112336A1 (es) | 2017-07-14 | 2019-10-16 | Pfizer | ANTICUERPOS CONTRA MAdCAM |
| US11180541B2 (en) | 2017-09-28 | 2021-11-23 | Geltor, Inc. | Recombinant collagen and elastin molecules and uses thereof |
| US20210253996A1 (en) | 2018-07-03 | 2021-08-19 | Bristol-Myers Squibb Company | Methods of producing recombinant proteins |
| WO2020076776A1 (en) | 2018-10-10 | 2020-04-16 | Boehringer Ingelheim International Gmbh | Method for membrane gas transfer in high density bioreactor culture |
| WO2020113164A1 (en) * | 2018-11-30 | 2020-06-04 | Memorial Sloan Kettering Cancer Center | Heterodimeric tetravalency and specificity antibody compositions and uses thereof |
| JP2022521840A (ja) | 2019-04-01 | 2022-04-12 | ジェネンテック, インコーポレイテッド | タンパク質含有製剤を安定化するための組成物および方法 |
| WO2020210440A1 (en) | 2019-04-12 | 2020-10-15 | Geltor, Inc. | Recombinant elastin and production thereof |
| CN114206383A (zh) * | 2019-06-10 | 2022-03-18 | 武田药品工业株式会社 | 用于抗体生产的细胞培养方法和组合物 |
| WO2021021973A1 (en) | 2019-08-01 | 2021-02-04 | Bristol-Myers Squibb Company | Methods of improving protein productivity in fed-batch cell cultures |
| EP4217383A1 (en) | 2020-09-22 | 2023-08-02 | Bristol-Myers Squibb Company | Methods for producing therapeutic proteins |
| WO2023015234A1 (en) | 2021-08-05 | 2023-02-09 | Bristol-Myers Squibb Company | Cell culture methods for producing therapeutic proteins |
| WO2023049687A1 (en) | 2021-09-21 | 2023-03-30 | Bristol-Myers Squibb Company | Methods of controlling the level of dissolved oxygen (do) in a solution comprising a recombinant protein in a storage container |
| US20240076343A1 (en) | 2022-06-16 | 2024-03-07 | Cephalon Llc | Anti-pd-1 antibody-attenuated il-2 immunoconjugates and uses thereof |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1994017828A2 (en) * | 1993-02-09 | 1994-08-18 | Biogen, Inc. | Treatment for insulin dependent diabetes |
| WO1995019790A1 (en) * | 1994-01-25 | 1995-07-27 | Athena Neurosciences, Inc. | Humanized antibodies against leukocyte adhesion molecule vla-4 |
Family Cites Families (36)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB8308235D0 (en) | 1983-03-25 | 1983-05-05 | Celltech Ltd | Polypeptides |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| GB8422238D0 (en) | 1984-09-03 | 1984-10-10 | Neuberger M S | Chimeric proteins |
| US4699880A (en) * | 1984-09-25 | 1987-10-13 | Immunomedics, Inc. | Method of producing monoclonal anti-idiotype antibody |
| GB8607679D0 (en) | 1986-03-27 | 1986-04-30 | Winter G P | Recombinant dna product |
| US5225539A (en) | 1986-03-27 | 1993-07-06 | Medical Research Council | Recombinant altered antibodies and methods of making altered antibodies |
| AU600575B2 (en) | 1987-03-18 | 1990-08-16 | Sb2, Inc. | Altered antibodies |
| US5258498A (en) * | 1987-05-21 | 1993-11-02 | Creative Biomolecules, Inc. | Polypeptide linkers for production of biosynthetic proteins |
| EP0303463B1 (en) | 1987-08-11 | 1994-11-30 | The Board Of Trustees Of The Leland Stanford Junior University | Method to control leukocyte extravasation |
| US5403919A (en) | 1987-08-11 | 1995-04-04 | Board Of Trustees Of The Leland Stanford Junior University Stanford University | Method to control leukocyte extravasation |
| US5538724A (en) | 1987-08-11 | 1996-07-23 | The Board Of Trustees For The Leland Stanford Junior Univ. | Method of control leukocyte extravasation |
| US5223392A (en) * | 1988-01-25 | 1993-06-29 | Exocell, Inc. | Monoclonal antibodies against glycated albumin, hybrid cell lines producing these antibodies, and use therefore |
| JP3095168B2 (ja) | 1988-02-05 | 2000-10-03 | エル. モリソン,シェリー | ドメイン‐変性不変部を有する抗体 |
| WO1990007321A2 (en) | 1988-12-23 | 1990-07-12 | The Board Of Trustees Of The Leland Stanford Junior University | Lymphocyte receptor homing sequences and their uses |
| IL162181A (en) | 1988-12-28 | 2006-04-10 | Pdl Biopharma Inc | A method of producing humanized immunoglubulin, and polynucleotides encoding the same |
| US5530101A (en) | 1988-12-28 | 1996-06-25 | Protein Design Labs, Inc. | Humanized immunoglobulins |
| US5225538A (en) * | 1989-02-23 | 1993-07-06 | Genentech, Inc. | Lymphocyte homing receptor/immunoglobulin fusion proteins |
| IL95501A (en) | 1989-09-01 | 1997-04-15 | Hutchinson Fred Cancer Res | Inhibition by antibodies of lymphocyte adherence to vascular endothelium utilizing an extracellular matrix receptor-ligand interaction, and pharmaceutical compositions containing said antibodies |
| GB8928874D0 (en) | 1989-12-21 | 1990-02-28 | Celltech Ltd | Humanised antibodies |
| US5859205A (en) | 1989-12-21 | 1999-01-12 | Celltech Limited | Humanised antibodies |
| CZ282603B6 (cs) | 1991-03-06 | 1997-08-13 | Merck Patent Gesellschaft Mit Beschränkter Haftun G | Humanizované a chimerické monoklonální protilátky |
| US5665595A (en) * | 1991-06-07 | 1997-09-09 | Dowelanco | Immunoglobulins against insect tissue |
| LU91067I2 (fr) | 1991-06-14 | 2004-04-02 | Genentech Inc | Trastuzumab et ses variantes et dérivés immuno chimiques y compris les immotoxines |
| GB9115364D0 (en) | 1991-07-16 | 1991-08-28 | Wellcome Found | Antibody |
| US5871734A (en) | 1992-01-13 | 1999-02-16 | Biogen, Inc. | Treatment for asthma with VLA-4 blocking agents |
| WO1993015764A1 (en) | 1992-02-12 | 1993-08-19 | Biogen, Inc. | Treatment for inflammatory bowel disease |
| US5932214A (en) | 1994-08-11 | 1999-08-03 | Biogen, Inc. | Treatment for inflammatory bowel disease with VLA-4 blockers |
| WO1994012214A1 (en) | 1992-12-01 | 1994-06-09 | Protein Design Labs, Inc. | Humanized antibodies reactive with cd18 |
| WO1994013312A1 (en) | 1992-12-15 | 1994-06-23 | The Board Of Trustees Of The Leland Stanford Junior University | Mucosal vascular addressin, dna and expression |
| JPH08507680A (ja) | 1993-01-12 | 1996-08-20 | バイオジェン インコーポレイテッド | 組換え抗vla4抗体分子 |
| JPH06303990A (ja) | 1993-04-24 | 1994-11-01 | Kanebo Ltd | モノクローナル抗体、それを産生するハイブリドーマおよび該抗体の製造方法 |
| EP0714409A1 (en) | 1993-06-16 | 1996-06-05 | Celltech Therapeutics Limited | Antibodies |
| US5840299A (en) | 1994-01-25 | 1998-11-24 | Athena Neurosciences, Inc. | Humanized antibodies against leukocyte adhesion molecule VLA-4 |
| US6551593B1 (en) * | 1995-02-10 | 2003-04-22 | Millennium Pharmaceuticals, Inc. | Treatment of Inflammatory bowel disease by inhibiting binding and/or signalling through α 4 β 7 and its ligands and madcam |
| US6037324A (en) * | 1996-01-04 | 2000-03-14 | Leukosite, Inc. | Inhibitors of MAdCAM-1-mediated interactions and methods of use therefor |
| US7147851B1 (en) * | 1996-08-15 | 2006-12-12 | Millennium Pharmaceuticals, Inc. | Humanized immunoglobulin reactive with α4β7 integrin |
-
1996
- 1996-08-15 US US08/700,737 patent/US7147851B1/en not_active Expired - Lifetime
-
1997
- 1997-08-06 NZ NZ334226A patent/NZ334226A/xx not_active IP Right Cessation
- 1997-08-06 CN CNB971972214A patent/CN100406562C/zh not_active Expired - Lifetime
- 1997-08-06 SI SI9730738T patent/SI0918797T2/sl unknown
- 1997-08-06 JP JP50985398A patent/JP4171071B2/ja not_active Expired - Lifetime
- 1997-08-06 PT PT97937148T patent/PT918797E/pt unknown
- 1997-08-06 IL IL12805297A patent/IL128052A0/xx unknown
- 1997-08-06 EP EP06006108A patent/EP1710314A1/en not_active Withdrawn
- 1997-08-06 AU AU39728/97A patent/AU730326C/en not_active Expired
- 1997-08-06 EP EP97937148.1A patent/EP0918797B2/en not_active Expired - Lifetime
- 1997-08-06 CA CA2263106A patent/CA2263106C/en not_active Expired - Lifetime
- 1997-08-06 WO PCT/US1997/013884 patent/WO1998006248A2/en active IP Right Grant
- 1997-08-06 AT AT97937148T patent/ATE321788T1/de active
- 1997-08-06 BR BRPI9711079-5A patent/BRPI9711079B1/pt not_active IP Right Cessation
- 1997-08-06 DE DE69735596.9T patent/DE69735596T3/de not_active Expired - Lifetime
- 1997-08-06 ES ES97937148.1T patent/ES2262186T5/es not_active Expired - Lifetime
- 1997-08-06 DK DK97937148.1T patent/DK0918797T4/en active
-
1999
- 1999-01-14 IL IL128052A patent/IL128052A/en active Protection Beyond IP Right Term
- 1999-01-14 IL IL175570A patent/IL175570A0/en unknown
- 1999-12-02 HK HK99105648A patent/HK1021822A1/xx not_active IP Right Cessation
-
2006
- 2006-08-28 US US11/511,164 patent/US7402410B2/en not_active Expired - Fee Related
-
2008
- 2008-05-02 JP JP2008120608A patent/JP4578536B2/ja not_active Expired - Lifetime
- 2008-06-18 US US12/214,304 patent/US20090011464A1/en not_active Abandoned
-
2010
- 2010-05-24 JP JP2010118596A patent/JP5166483B2/ja not_active Expired - Lifetime
-
2014
- 2014-10-13 LT LTPA2014037C patent/LTC0918797I2/lt unknown
- 2014-11-12 BE BE2014C068C patent/BE2014C068I2/fr unknown
- 2014-11-12 LU LU92596C patent/LU92596I2/xx unknown
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1994017828A2 (en) * | 1993-02-09 | 1994-08-18 | Biogen, Inc. | Treatment for insulin dependent diabetes |
| WO1995019790A1 (en) * | 1994-01-25 | 1995-07-27 | Athena Neurosciences, Inc. | Humanized antibodies against leukocyte adhesion molecule vla-4 |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN103608071A (zh) * | 2011-05-02 | 2014-02-26 | 米伦纽姆医药公司 | 抗α4β7抗体的制剂 |
| CN108079291A (zh) * | 2011-05-02 | 2018-05-29 | 米伦纽姆医药公司 | 抗α4β7抗体的制剂 |
| CN108187042A (zh) * | 2011-05-02 | 2018-06-22 | 米伦纽姆医药公司 | 抗α4β7抗体的制剂 |
| CN108079291B (zh) * | 2011-05-02 | 2022-04-08 | 千禧制药公司 | 抗α4β7抗体的制剂 |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN100406562C (zh) | 与α4β7整合蛋白相作用的人化免疫球蛋白 | |
| KR102572091B1 (ko) | 항 gprc5d 항체, gprc5d 및 cd3에 결합하는 이중특이성 항원 결합 분자, 및 이들의 용도 | |
| US7241877B2 (en) | Humanised antibodies | |
| US6602503B1 (en) | Recombinant anti-VLA-4 antibody molecules | |
| US6750325B1 (en) | CD3 specific recombinant antibody | |
| KR101560841B1 (ko) | 신규한 화합물 | |
| US20070122404A1 (en) | Humanized immunoglobulin reactive with alpha4beta7 integrin | |
| GB2249310A (en) | Antibodies having binding affinity for cd3 antigen | |
| CA2523142A1 (en) | Humanized antibodies to interferon alpha receptor-1 (ifnar-1) | |
| AU778980B2 (en) | Humanized immunoglobulin reactive with alpha4beta7 integrin | |
| AU2005201265B2 (en) | Humanized immunoglobulin reactive with alpha4beta7 integrin | |
| GB2268745A (en) | Humanised antibodies. | |
| AU2008203508A1 (en) | Humanized immunoglobulin reactive with a4B7 integrin |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| C53 | Correction of patent of invention or patent application | ||
| CB02 | Change of applicant information |
Applicant after: Millenium Pharm Zinc Applicant before: Leukosite Inc. |
|
| COR | Change of bibliographic data |
Free format text: CORRECT: APPLICANT; FROM: LEUKOSITE, INC. TO: QIANXI MEDICINE CO.,LTD. |
|
| C14 | Grant of patent or utility model | ||
| GR01 | Patent grant | ||
| CX01 | Expiry of patent term | ||
| CX01 | Expiry of patent term |
Granted publication date: 20080730 |