WO2012001976A1 - グルコース脱水素酵素 - Google Patents
グルコース脱水素酵素 Download PDFInfo
- Publication number
- WO2012001976A1 WO2012001976A1 PCT/JP2011/003741 JP2011003741W WO2012001976A1 WO 2012001976 A1 WO2012001976 A1 WO 2012001976A1 JP 2011003741 W JP2011003741 W JP 2011003741W WO 2012001976 A1 WO2012001976 A1 WO 2012001976A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- amino acid
- glucose
- protein
- seq
- acid sequence
- Prior art date
Links
- 108010050375 Glucose 1-Dehydrogenase Proteins 0.000 title claims abstract description 64
- 230000035772 mutation Effects 0.000 claims abstract description 42
- SRBFZHDQGSBBOR-IOVATXLUSA-N D-xylopyranose Chemical compound O[C@@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-IOVATXLUSA-N 0.000 claims abstract description 37
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 claims abstract description 27
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 claims abstract description 27
- 150000001413 amino acids Chemical class 0.000 claims abstract description 19
- 241000233866 Fungi Species 0.000 claims abstract description 10
- 230000009257 reactivity Effects 0.000 claims abstract description 10
- 102000004190 Enzymes Human genes 0.000 claims description 105
- 108090000790 Enzymes Proteins 0.000 claims description 105
- 229940088598 enzyme Drugs 0.000 claims description 105
- 230000000694 effects Effects 0.000 claims description 86
- 108090000623 proteins and genes Proteins 0.000 claims description 81
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 claims description 71
- 239000008103 glucose Substances 0.000 claims description 69
- 102220093933 rs876660617 Human genes 0.000 claims description 65
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 63
- 102000004169 proteins and genes Human genes 0.000 claims description 53
- 235000018102 proteins Nutrition 0.000 claims description 50
- 125000000539 amino acid group Chemical group 0.000 claims description 43
- 102220624339 Interferon alpha-8_N176K_mutation Human genes 0.000 claims description 39
- 102220258122 rs200240922 Human genes 0.000 claims description 23
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 21
- 235000001014 amino acid Nutrition 0.000 claims description 21
- 102220370322 c.990T>A Human genes 0.000 claims description 20
- 102220344240 rs147495977 Human genes 0.000 claims description 19
- 238000004519 manufacturing process Methods 0.000 claims description 15
- 239000013598 vector Substances 0.000 claims description 12
- 108010015776 Glucose oxidase Proteins 0.000 claims description 11
- 239000004366 Glucose oxidase Substances 0.000 claims description 11
- 229940116332 glucose oxidase Drugs 0.000 claims description 11
- 235000019420 glucose oxidase Nutrition 0.000 claims description 11
- 102220554671 Transmembrane protein 199_T168P_mutation Human genes 0.000 claims description 7
- 238000003149 assay kit Methods 0.000 claims description 6
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 claims description 6
- 238000012258 culturing Methods 0.000 claims description 6
- 235000018417 cysteine Nutrition 0.000 claims description 6
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 claims description 6
- 229910052760 oxygen Inorganic materials 0.000 claims description 6
- 239000001301 oxygen Substances 0.000 claims description 6
- 102200043780 c.1065C>A Human genes 0.000 claims description 5
- 102200078805 rs118204440 Human genes 0.000 claims description 4
- 102220015927 rs117444825 Human genes 0.000 claims description 3
- 238000004458 analytical method Methods 0.000 claims description 2
- 102220600742 Calmodulin-binding transcription activator 1_G53A_mutation Human genes 0.000 claims 2
- 230000001747 exhibiting effect Effects 0.000 claims 1
- 241000588724 Escherichia coli Species 0.000 abstract description 18
- 150000002303 glucose derivatives Chemical class 0.000 abstract 2
- 108010035289 Glucose Dehydrogenases Proteins 0.000 abstract 1
- 239000002609 medium Substances 0.000 description 26
- 240000006439 Aspergillus oryzae Species 0.000 description 20
- 235000002247 Aspergillus oryzae Nutrition 0.000 description 20
- 239000000243 solution Substances 0.000 description 20
- 239000000758 substrate Substances 0.000 description 20
- 241000123649 Botryotinia Species 0.000 description 19
- 238000000034 method Methods 0.000 description 14
- 102220096727 rs876659692 Human genes 0.000 description 14
- 230000014509 gene expression Effects 0.000 description 13
- 210000004027 cell Anatomy 0.000 description 12
- 238000006243 chemical reaction Methods 0.000 description 12
- 238000005259 measurement Methods 0.000 description 12
- 239000003153 chemical reaction reagent Substances 0.000 description 11
- 102100031126 6-phosphogluconolactonase Human genes 0.000 description 10
- 108010076504 Protein Sorting Signals Proteins 0.000 description 10
- 230000002538 fungal effect Effects 0.000 description 10
- 238000002360 preparation method Methods 0.000 description 10
- 239000008280 blood Substances 0.000 description 9
- 210000004369 blood Anatomy 0.000 description 9
- CCBICDLNWJRFPO-UHFFFAOYSA-N 2,6-dichloroindophenol Chemical compound C1=CC(O)=CC=C1N=C1C=C(Cl)C(=O)C(Cl)=C1 CCBICDLNWJRFPO-UHFFFAOYSA-N 0.000 description 8
- MHAJPDPJQMAIIY-UHFFFAOYSA-N Hydrogen peroxide Chemical compound OO MHAJPDPJQMAIIY-UHFFFAOYSA-N 0.000 description 8
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 8
- LWIHDJKSTIGBAC-UHFFFAOYSA-K tripotassium phosphate Chemical compound [K+].[K+].[K+].[O-]P([O-])([O-])=O LWIHDJKSTIGBAC-UHFFFAOYSA-K 0.000 description 8
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 7
- 102100034289 Deoxynucleoside triphosphate triphosphohydrolase SAMHD1 Human genes 0.000 description 7
- 108010000445 Glycerate dehydrogenase Proteins 0.000 description 7
- 101000641031 Homo sapiens Deoxynucleoside triphosphate triphosphohydrolase SAMHD1 Proteins 0.000 description 7
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 7
- 108010006519 Molecular Chaperones Proteins 0.000 description 7
- 238000002835 absorbance Methods 0.000 description 7
- 108010029645 galactitol 2-dehydrogenase Proteins 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- RLFWWDJHLFCNIJ-UHFFFAOYSA-N 4-aminoantipyrine Chemical compound CN1C(C)=C(N)C(=O)N1C1=CC=CC=C1 RLFWWDJHLFCNIJ-UHFFFAOYSA-N 0.000 description 6
- RXGJTUSBYWCRBK-UHFFFAOYSA-M 5-methylphenazinium methyl sulfate Chemical compound COS([O-])(=O)=O.C1=CC=C2[N+](C)=C(C=CC=C3)C3=NC2=C1 RXGJTUSBYWCRBK-UHFFFAOYSA-M 0.000 description 6
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 6
- 101710088194 Dehydrogenase Proteins 0.000 description 6
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 6
- 229910052799 carbon Inorganic materials 0.000 description 6
- 230000008859 change Effects 0.000 description 6
- 239000013604 expression vector Substances 0.000 description 6
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 6
- BASFCYQUMIYNBI-UHFFFAOYSA-N platinum Chemical compound [Pt] BASFCYQUMIYNBI-UHFFFAOYSA-N 0.000 description 6
- 239000008057 potassium phosphate buffer Substances 0.000 description 6
- 238000003259 recombinant expression Methods 0.000 description 6
- 235000000346 sugar Nutrition 0.000 description 6
- 241001198387 Escherichia coli BL21(DE3) Species 0.000 description 5
- WQZGKKKJIJFFOK-PHYPRBDBSA-N alpha-D-galactose Chemical compound OC[C@H]1O[C@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-PHYPRBDBSA-N 0.000 description 5
- 238000003556 assay Methods 0.000 description 5
- GUBGYTABKSRVRQ-QUYVBRFLSA-N beta-maltose Chemical compound OC[C@H]1O[C@H](O[C@H]2[C@H](O)[C@@H](O)[C@H](O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@@H]1O GUBGYTABKSRVRQ-QUYVBRFLSA-N 0.000 description 5
- 238000005119 centrifugation Methods 0.000 description 5
- 229930182830 galactose Natural products 0.000 description 5
- 238000000746 purification Methods 0.000 description 5
- 238000012360 testing method Methods 0.000 description 5
- 108010059013 Chaperonin 10 Proteins 0.000 description 4
- 108010058432 Chaperonin 60 Proteins 0.000 description 4
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 4
- 238000012300 Sequence Analysis Methods 0.000 description 4
- 239000000872 buffer Substances 0.000 description 4
- 230000004186 co-expression Effects 0.000 description 4
- 239000005515 coenzyme Substances 0.000 description 4
- 230000003247 decreasing effect Effects 0.000 description 4
- DPXDJGUFSPAFJZ-UHFFFAOYSA-L disodium;4-[3-methyl-n-(4-sulfonatobutyl)anilino]butane-1-sulfonate Chemical compound [Na+].[Na+].CC1=CC=CC(N(CCCCS([O-])(=O)=O)CCCCS([O-])(=O)=O)=C1 DPXDJGUFSPAFJZ-UHFFFAOYSA-L 0.000 description 4
- 235000019162 flavin adenine dinucleotide Nutrition 0.000 description 4
- 239000011714 flavin adenine dinucleotide Substances 0.000 description 4
- 230000002779 inactivation Effects 0.000 description 4
- 238000011534 incubation Methods 0.000 description 4
- 230000006698 induction Effects 0.000 description 4
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 4
- 229920000642 polymer Polymers 0.000 description 4
- 229910000160 potassium phosphate Inorganic materials 0.000 description 4
- 235000011009 potassium phosphates Nutrition 0.000 description 4
- 238000006467 substitution reaction Methods 0.000 description 4
- 102100024341 10 kDa heat shock protein, mitochondrial Human genes 0.000 description 3
- 108010029731 6-phosphogluconolactonase Proteins 0.000 description 3
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 3
- 108010018962 Glucosephosphate Dehydrogenase Proteins 0.000 description 3
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 239000007993 MOPS buffer Substances 0.000 description 3
- 230000010718 Oxidation Activity Effects 0.000 description 3
- 102000003992 Peroxidases Human genes 0.000 description 3
- 230000008901 benefit Effects 0.000 description 3
- 238000003776 cleavage reaction Methods 0.000 description 3
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 3
- 238000000502 dialysis Methods 0.000 description 3
- 229930027917 kanamycin Natural products 0.000 description 3
- 229960000318 kanamycin Drugs 0.000 description 3
- 229930182823 kanamycin A Natural products 0.000 description 3
- 229930182817 methionine Natural products 0.000 description 3
- 108040007629 peroxidase activity proteins Proteins 0.000 description 3
- 229910052697 platinum Inorganic materials 0.000 description 3
- -1 potassium ferricyanide Chemical compound 0.000 description 3
- 230000007017 scission Effects 0.000 description 3
- 150000008163 sugars Chemical class 0.000 description 3
- 102100038222 60 kDa heat shock protein, mitochondrial Human genes 0.000 description 2
- 241000228212 Aspergillus Species 0.000 description 2
- 241000014175 Aspergillus carbonarius ITEM 5010 Species 0.000 description 2
- 241000228245 Aspergillus niger Species 0.000 description 2
- 241000894006 Bacteria Species 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 102000003886 Glycoproteins Human genes 0.000 description 2
- 108090000288 Glycoproteins Proteins 0.000 description 2
- 239000006142 Luria-Bertani Agar Substances 0.000 description 2
- 238000010521 absorption reaction Methods 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000008033 biological extinction Effects 0.000 description 2
- 230000015572 biosynthetic process Effects 0.000 description 2
- 238000011088 calibration curve Methods 0.000 description 2
- 239000002131 composite material Substances 0.000 description 2
- 238000007796 conventional method Methods 0.000 description 2
- 230000009849 deactivation Effects 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- KTWOOEGAPBSYNW-UHFFFAOYSA-N ferrocene Chemical compound [Fe+2].C=1C=C[CH-]C=1.C=1C=C[CH-]C=1 KTWOOEGAPBSYNW-UHFFFAOYSA-N 0.000 description 2
- VWWQXMAJTJZDQX-UYBVJOGSSA-N flavin adenine dinucleotide Chemical compound C1=NC2=C(N)N=CN=C2N1[C@@H]([C@H](O)[C@@H]1O)O[C@@H]1CO[P@](O)(=O)O[P@@](O)(=O)OC[C@@H](O)[C@@H](O)[C@@H](O)CN1C2=NC(=O)NC(=O)C2=NC2=C1C=C(C)C(C)=C2 VWWQXMAJTJZDQX-UYBVJOGSSA-N 0.000 description 2
- 229940093632 flavin-adenine dinucleotide Drugs 0.000 description 2
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 2
- 229910052737 gold Inorganic materials 0.000 description 2
- 239000010931 gold Substances 0.000 description 2
- 230000006872 improvement Effects 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- 239000013612 plasmid Substances 0.000 description 2
- 230000012846 protein folding Effects 0.000 description 2
- MMXZSJMASHPLLR-UHFFFAOYSA-N pyrroloquinoline quinone Chemical compound C12=C(C(O)=O)C=C(C(O)=O)N=C2C(=O)C(=O)C2=C1NC(C(=O)O)=C2 MMXZSJMASHPLLR-UHFFFAOYSA-N 0.000 description 2
- 238000003860 storage Methods 0.000 description 2
- 239000000725 suspension Substances 0.000 description 2
- 229930195727 α-lactose Natural products 0.000 description 2
- HMUNWXXNJPVALC-UHFFFAOYSA-N 1-[4-[2-(2,3-dihydro-1H-inden-2-ylamino)pyrimidin-5-yl]piperazin-1-yl]-2-(2,4,6,7-tetrahydrotriazolo[4,5-c]pyridin-5-yl)ethanone Chemical compound C1C(CC2=CC=CC=C12)NC1=NC=C(C=N1)N1CCN(CC1)C(CN1CC2=C(CC1)NN=N2)=O HMUNWXXNJPVALC-UHFFFAOYSA-N 0.000 description 1
- QCFYJCYNJLBDRT-UHFFFAOYSA-N Bis(2-chloro-1-methylethyl)ether Chemical compound ClCC(C)OC(C)CCl QCFYJCYNJLBDRT-UHFFFAOYSA-N 0.000 description 1
- 108700010070 Codon Usage Proteins 0.000 description 1
- PHOQVHQSTUBQQK-SQOUGZDYSA-N D-glucono-1,5-lactone Chemical compound OC[C@H]1OC(=O)[C@H](O)[C@@H](O)[C@@H]1O PHOQVHQSTUBQQK-SQOUGZDYSA-N 0.000 description 1
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 1
- 241000620209 Escherichia coli DH5[alpha] Species 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 238000012408 PCR amplification Methods 0.000 description 1
- 241000228143 Penicillium Species 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 239000012327 Ruthenium complex Substances 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 229910021607 Silver chloride Inorganic materials 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 238000001042 affinity chromatography Methods 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 229960005091 chloramphenicol Drugs 0.000 description 1
- WIIZWVCIJKGZOK-RKDXNWHRSA-N chloramphenicol Chemical compound ClC(Cl)C(=O)N[C@H](CO)[C@H](O)C1=CC=C([N+]([O-])=O)C=C1 WIIZWVCIJKGZOK-RKDXNWHRSA-N 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 229920001940 conductive polymer Polymers 0.000 description 1
- 239000003431 cross linking reagent Substances 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 206010012601 diabetes mellitus Diseases 0.000 description 1
- 238000002845 discoloration Methods 0.000 description 1
- 238000000835 electrochemical detection Methods 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 239000012634 fragment Substances 0.000 description 1
- 238000001502 gel electrophoresis Methods 0.000 description 1
- 238000002523 gelfiltration Methods 0.000 description 1
- 235000012209 glucono delta-lactone Nutrition 0.000 description 1
- 229960003681 gluconolactone Drugs 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 238000001114 immunoprecipitation Methods 0.000 description 1
- 210000003000 inclusion body Anatomy 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 239000012212 insulator Substances 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 238000001155 isoelectric focusing Methods 0.000 description 1
- 101150109249 lacI gene Proteins 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 238000004811 liquid chromatography Methods 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- VNWKTOKETHGBQD-UHFFFAOYSA-N methane Chemical compound C VNWKTOKETHGBQD-UHFFFAOYSA-N 0.000 description 1
- 239000000203 mixture Substances 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 101150093139 ompT gene Proteins 0.000 description 1
- 230000003287 optical effect Effects 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 238000000751 protein extraction Methods 0.000 description 1
- 230000009467 reduction Effects 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 102220022510 rs386833622 Human genes 0.000 description 1
- 102220218772 rs756614355 Human genes 0.000 description 1
- 102220250470 rs774802052 Human genes 0.000 description 1
- 238000005185 salting out Methods 0.000 description 1
- 238000007650 screen-printing Methods 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 238000002864 sequence alignment Methods 0.000 description 1
- HKZLPVFGJNLROG-UHFFFAOYSA-M silver monochloride Chemical compound [Cl-].[Ag+] HKZLPVFGJNLROG-UHFFFAOYSA-M 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 238000004544 sputter deposition Methods 0.000 description 1
- 238000013112 stability test Methods 0.000 description 1
- 239000012086 standard solution Substances 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 238000006257 total synthesis reaction Methods 0.000 description 1
- 238000000108 ultra-filtration Methods 0.000 description 1
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 1
- 210000005253 yeast cell Anatomy 0.000 description 1
Images







Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/001—Enzyme electrodes
- C12Q1/005—Enzyme electrodes involving specific analytes or enzymes
- C12Q1/006—Enzyme electrodes involving specific analytes or enzymes for glucose
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/26—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving oxidoreductase
- C12Q1/32—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving oxidoreductase involving dehydrogenase
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N27/00—Investigating or analysing materials by the use of electric, electrochemical, or magnetic means
- G01N27/26—Investigating or analysing materials by the use of electric, electrochemical, or magnetic means by investigating electrochemical variables; by using electrolysis or electrophoresis
Definitions
- the present invention relates to glucose dehydrogenase (FAD-GDH) with flavin adenine dinucleotide as a coenzyme, its production and its use in the determination of glucose.
- FAD-GDH glucose dehydrogenase
- Blood glucose concentration is an important marker of diabetes.
- GOD glucose oxidase
- G6PDH glucose 6-phosphate dehydrogenase
- PQQGDH glucose dehydrogenase
- pyrroloquinoline quinone pyrroloquinoline quinone
- PQQGDH has a high oxidation activity with respect to glucose and has the advantage of not requiring oxygen as an electron acceptor, but has low selectivity for glucose and high activity against maltose. It was a problem. Accordingly, there is a need for new enzymes that can be used as recognition elements for glucose sensors. Furthermore, it is desirable that the reactivity to xylose is low so that the blood glucose level can be accurately measured even during the xylose absorption test.
- An object of the present invention is to provide a new enzyme having higher productivity and / or higher thermal stability than conventional glucose dehydrogenase.
- the present inventor isolated a gene encoding a novel FAD-GDH derived from Botryothinia fuckeliana, and further replaced the specific amino acid residue, so that the productivity of recombinant expression in E. coli was remarkable. I found that it would increase. The inventor has also found that by substituting a specific amino acid residue of a fungal flavin adenine dinucleotide glucose dehydrogenase (FAD-GDH), its thermal stability is significantly increased.
- FAD-GDH fungal flavin adenine dinucleotide glucose dehydrogenase
- the present invention includes a protein comprising the amino acid sequence represented by SEQ ID NO: 2, or one or more amino acid residues deleted, substituted or inserted in the amino acid sequence represented by SEQ ID NO: 2, and In a protein having glucose dehydrogenase activity, N176K, N176R, N176E, N176S, N225K, N225E, N259K, N301K, N326K, N326E, N330K, N330S, N355K, N355E, S514G or S552C, or combinations of these amino acid mutations A protein having the same is provided.
- the protein of the present invention further has one or more mutations selected from the group consisting of G53A, E166R, T168P, N487S, S490P, N492T, A496E, D500E, V502L and A505N.
- the protein of the present invention is a protein comprising the amino acid sequence represented by SEQ ID NO: 2, N176K / S490P / D500E / S514G / S552C, N176K / A496E / D500E / S514G / S552C, N176K / S514G / S552C, A protein having an amino acid mutation selected from the group consisting of S514G / S552C and G53A / S514G / S552C.
- the protein of the present invention is a protein comprising the amino acid sequence represented by SEQ ID NO: 2, N176K / N301K / N330K / S514G / S552C, N176R / N301K / N330K / S514G / S552C, N176R / N225E / N301K / N326E / N330K / N355E / S514G / S552C, N176K / S490P / D500E / S514G / S552C, and E166R / T168P / N176R / N301K / N330K / S490P / D500E / S514G / S552C selected from the group consisting of amino acids It is.
- the present invention relates to a protein comprising the amino acid sequence represented by SEQ ID NO: 4, or one or more amino acid residues in the amino acid sequence represented by SEQ ID NO: 4 deleted, substituted or inserted. And a protein having glucose dehydrogenase activity and having amino acid mutations of V149C and G190C.
- the present invention relates to a fungus-derived glucose dehydrogenase, which is obtained by aligning the amino acid sequence with the amino acid sequence represented by SEQ ID NO: 4.
- a glucose dehydrogenase characterized in that amino acids at positions corresponding to V149 and G190 are both replaced with cysteine.
- the fungal glucose dehydrogenase of the present invention has an amino acid sequence selected from the group consisting of SEQ ID NOs: 2 and 5-9, and corresponds to V149 and G190 of the amino acid sequence represented by SEQ ID NO: 4 Both amino acids in the position are replaced with cysteine.
- the present invention relates to a protein comprising the amino acid sequence represented by SEQ ID NO: 2, or one or more amino acid residues deleted, substituted or inserted in the amino acid sequence represented by SEQ ID NO: 2. And a protein having an amino acid mutation of A150C / T192C, which is a protein having glucose dehydrogenase activity.
- the present invention provides a gene encoding the glucose dehydrogenase of the present invention, a recombinant vector containing the gene, and a transformant or transductant transformed with the recombinant vector.
- the present invention is also characterized by culturing a transformant transformed with a recombinant vector containing a gene encoding the glucose dehydrogenase of the present invention, and collecting glucose dehydrogenase from the culture, A method for producing glucose dehydrogenase is provided.
- the present invention provides a glucose analysis method characterized by measuring the glucose concentration in a sample using the glucose dehydrogenase of the present invention.
- the present invention also provides a glucose assay kit comprising the glucose dehydrogenase of the present invention.
- the present invention also provides an enzyme electrode in which the glucose dehydrogenase of the present invention is immobilized on the electrode surface, and a glucose sensor using the enzyme electrode as a working electrode.
- the present invention relates to a protein comprising the amino acid sequence represented by SEQ ID NO: 2, or one or more amino acid residues deleted, substituted or inserted in the amino acid sequence represented by SEQ ID NO: 2.
- Dissolved using glucose dehydrogenase characterized in that it has glucose oxidase activity and has glucose dehydrogenase activity, and the reactivity to xylose is 20% or less of the reactivity to glucose A biosensor for measuring glucose that is not affected by oxygen is provided.
- FIG. 1-1 shows an alignment of amino acid sequences of fungal glucose dehydrogenase.
- the 149th amino acid residue V and 190th amino acid residue G of FAD-GDH derived from Aspergillus oryzae TI strain, and amino acid residues corresponding to these amino acid residues are shown in bold.
- Each abbreviation has the following meaning.
- Aspca1_10864 Aspergillus carbonarius ITEM 5010; GenBank Aspca1_10864, Aspca1_33771: Aspergillus carbonarius ITEM 5010; GenBank Aspca1_33771, Bfu (AJ_555871): Botryothinia fuckeliana (GenBank AJ_555871), Ssc (XP_001584680): Sclenotinia sclerotiorum (GenBank XP_001584680), 40715 (XP_001394544): Aspergillus niger 40715 (GenBank XP — 001394544), 39269 (XP — 001391138): Aspergillus niger 39269 (GenBank XP — 001391138), AoT1 FADGDH: Aspergillus oryzae T1 FAD-GDH.
- FIG. 1-2 shows alignment of amino acid sequences of fungal glucose dehydrogenase.
- Fig. 1-3 shows alignment of amino acid sequences of fungal glucose dehydrogenase.
- FIG. 2 shows a culture curve of an Escherichia coli transformant having the FAD-GDH gene derived from recombinant Botryotinia fukeliana.
- FIG. 3 shows the thermal stability of wild-type and modified FAD-GDH derived from Aspergillus oryzae.
- FIG. 4 shows the enzyme activity of wild-type and modified FAD-GDH derived from Aspergillus oryzae.
- FIG. 5 shows the thermal stability of wild-type and modified FAD-GDH from Botryotinia fukeliana.
- FAD-GDH from Botryotinia fukeliana
- FAD-GDH possessed by Botryotinia fukeliana was identified. Its amino acid sequence is shown in SEQ ID NO: 1.
- SEQ ID NO: 1 Although the genome sequence of Botryotinia fukeliana has been published, there has never been reported that Botryotinia fukeliana has FAD-GDH, and there has been no gene annotated as FAD-GDH.
- the protein having the amino acid sequence represented by SEQ ID NO: 1 has been conventionally estimated to be glucose oxidase (GOD) (Rolke et al., Mol Plant Pathol. 5 (1), p.17-27, 2004).
- GOD glucose oxidase
- the amino acid sequence represented by SEQ ID NO: 1 has about 30 to 60% sequence identity with the amino acid sequence of a known fungal glucose dehydrogenase, such as glucose dehydrogenase derived from Aspergillus oryzae TI strain (GenBank ACW04779). .1) has about 57% amino acid identity.
- the predicted signal sequence of natural FAD-GDH derived from Botryotinia fukeliana represented by SEQ ID NO: 1 is removed from the N-terminal to the 17th Ser.
- An amino acid sequence added with Met was prepared.
- Modified FAD-GDH from Botryotinia fukeliana The present inventor has obtained a modified FAD-GDH having an N176K, S514G or S552C mutation of the amino acid sequence represented by SEQ ID NO: 2, or a double mutation of S514G / S552C, and a triple mutation of N176K / S514G / S552C, It has been found that modified FAD-GDH having a high productivity in Escherichia coli as compared with wild-type FAD-GDH, particularly having a triple mutation of N176K / S514G / S552C, shows high enzyme activity with high productivity.
- the position of the amino acid mutation in the amino acid sequence of Botryotinia fukeliana-derived FAD-GDH is numbered with 1 as the first Met in the amino acid sequence of SEQ ID NO: 2.
- amino acid mutation or substitution is expressed by indicating the original amino acid residue, the amino acid position, and the amino acid residue after substitution in this order. For example, “S514G” The 514th S is replaced with G. A combination of double or more mutations is represented by a “/” symbol.
- an enzyme having high productivity in Escherichia coli means a unit of enzyme obtained per unit volume of culture solution when the enzyme is isolated from the culture after recombinant expression using Escherichia coli as a host. It means an enzyme molecule characterized by high activity per volume (U / L). If productivity is high, an enzyme can be produced recombinantly at a lower cost with a smaller culture apparatus.
- the productivity of an enzyme is based on the amino acid sequence of the enzyme, such as differences in water solubility as a protein, differences in folding efficiency, differences in enzyme activity per unit protein, and differences in enzyme stability. Reflected comprehensively. Furthermore, the amino acid sequence of the enzyme is also involved in the rate of recombinant expression in the host per unit volume of the culture solution, the ease of forming inclusion bodies, the stability of the enzyme in the purification process, and the like.
- N176K, N176R, N176E, N176S, N225K, N225E, N259K, N301K, N326K, N326E, N330K, N330S, N355K, N355E, S514G or S55G It is preferred to have multiple mutations in combination with one or more mutations selected from the group consisting of E166R, T168P, N487S, S490P, N492T, A496E, D500E, V502L and A505N.
- N176K / S490P / D500E / S514G / S552C preferred are N176K / S490P / D500E / S514G / S552C, G53A / N176K / S490P / D500E / S514G / S552C, N176K / A496E / D500E / S514G / S552C, N176K / A496E / D / S514G / S552C, G53A / N176K / A496E / D500E / V502L / S514G / S552C, N176K / S514G / S552C, S514G / S552C and G53A / S514G / S552C, particularly preferred are N176K / D490P / S550P N176K / A496E / D500E / S
- N176K / N301K / N330K / S514G / S552C is particularly preferred.
- N176R / N301K / N330K / S514G / S552C is particularly preferred.
- N176R / N225E / N301K / N326E / N330K / N355E / S514G / S552C is particularly preferred.
- E166R / T168P / N176R / N301K / N330K / S490P / D500E / S514G / S552C is particularly preferred.
- Modified FAD-GDH with improved thermal stability The present inventor replaces both the 149th amino acid residue V and the 190th amino acid residue G of FAD-GDH derived from Aspergillus oryzae TI strain represented by SEQ ID NO: 4 with C, whereby wild type FAD-GDH It has been found that the thermal stability is remarkably increased as compared with the above.
- the amino acid sequence of FAD-GDH derived from Aspergillus oryzae TI strain is shown in SEQ ID NO: 3. In the present invention, for efficient recombinant expression in E.
- thermostability refers to enzyme over time when enzyme activity is measured after incubation for a predetermined time at a high temperature (eg, 45 ° C., 50 ° C., 55 ° C., 60 ° C., etc.). It means that there is little reduction in activity.
- Thermal stability is, for example, the rate of inactivation determined by the residual activity when incubated at a predetermined temperature for a predetermined time, or from the slope of a curve (inactivation curve) obtained by plotting enzyme activity against the incubation time. It can be represented by a constant and / or a half-life of enzyme activity.
- the modified FAD-GDH of the present invention may have another mutation in addition to the mutation at the 149th and 190th amino acid residues of SEQ ID NO: 4 as long as it has glucose dehydrogenase activity.
- one or more of the amino acid residues of SEQ ID NO: 4, for example 1 to 10, may be optionally replaced with other amino acid residues.
- the present inventor further determined the amino acid residues at positions corresponding to the 149th and 190th amino acid residues of FAD-GDH derived from Aspergillus oryzae TI for FAD-GDH derived from Botryotinia fukeliana and other fungi. In both cases, it was found that substitution of cysteine for the same increased thermal stability.
- FIG. 1 Alignment of amino acid sequences of several FAD-GDHs derived from fungi is shown in FIG.
- This figure shows a sequence in which a region presumed to be an N-terminal signal peptide is removed from the amino acid sequence of each enzyme and methionine is added to the N-terminus.
- the amino acid residues corresponding to the 149th amino acid residue V and the 190th amino acid residue G of Aspergillus oryzae strain FAD-GDH are shown in bold.
- the genes encoding the amino acid sequences described were fully synthesized and recombinantly expressed in E. coli, and confirmed to have GDH activity.
- a person skilled in the art will also prepare an amino acid sequence alignment according to a conventional method in FAD-GDH derived from fungi other than the fungus illustrated in FIG. 1 to obtain the 149th position of Aspergillus oryzae strain FAD-GDH.
- FAD-GDH with improved thermal stability can be obtained by understanding amino acid residues at positions corresponding to amino acid residue V and amino acid residue G at position 190 and replacing these residues with C. .
- the expression “amino acid residue at a position corresponding to...” Refers to a specific protein of the reference protein when the amino acid sequence of the target protein is aligned with the amino acid sequence of the reference protein. It means an amino acid residue present at a position corresponding to an amino acid residue.
- AlignX software Invitrogen ;, Lu, G., and Moriyama, E.N. (2004) provided by Vector NTI )
- Vector NTI a balanced all-in-one sequence analysis suite. Brief Bioinform 5, 378-88) can be used with default parameters.
- the FAD-GDH of the present invention can be produced by recombinant expression using techniques well known in the art.
- the sequences of genes encoding natural FAD-GDH derived from Botryotinia fukeliana and FAD-GDH derived from Aspergillus oryzae TI can be easily determined based on the amino acid sequences set forth in SEQ ID NOs: 2 and 4, respectively.
- the gene encoding FAD-GDH may be cloned from the genome of Botryotinia querkeriana or Aspergillus oryzae, may be produced by PCR using a series of chemically synthesized oligonucleotides, or an automated DNA synthesizer, etc. May be used for total synthesis. It is desirable that the gene sequence is appropriately designed or modified by selecting codons so that a higher expression level is achieved in the host organism to be used. The characteristics of codon usage in a particular host organism are well known in the art.
- the base sequence encoding the amino acid residue to be substituted is replaced with the base sequence encoding the desired amino acid residue.
- Various methods for such site-specific base sequence substitution are well known in the art, and can be performed, for example, by PCR using appropriately designed primers.
- a gene encoding a modified amino acid sequence may be totally synthesized.
- modified FAD-GDH preferably has at least 80% sequence identity with native FAD-GDH.
- sequence identity is at least 85%, more preferably at least 90% and even more preferably at least 95%.
- the gene thus obtained is inserted into an appropriate expression vector and transformed into an appropriate host (for example, E. coli).
- an appropriate host for example, E. coli.
- Many vectors and host systems for expressing foreign proteins are known in the art, and various hosts such as bacteria, yeast, and cultured cells can be used as the host. If it is desired to produce a glucose dehydrogenase with a sugar chain attached, eukaryotic cells are used as hosts.
- the obtained transformant is cultured according to a conventional method, and FAD-GDH can be recovered from the cells or the culture solution.
- E. coli transformants are cultured in a medium (hereinafter referred to as A medium) described as ZYP broth in F. William Studier et.al., Protein Expression and Purification (2005). . That is, in addition to 0.5% glycerol, 0.05% glucose, 0.2% alpha-lactose, 25 mM (NH 4 ) 2 SO 4 , 100 mM KH 2 PO 4 , 100 mM NaHPO 4 , 1 mM MgSO 4 the is a medium plus.
- the recombinant protein is expressed by culturing at 15 to 25 ° C., preferably about 20 ° C.
- higher productivity can be obtained than when a conventional IPTG induction method is used.
- higher productivity can be obtained by co-expressing the mutant enzyme of the present invention with chaperones GroEL and GroES, which are known to promote protein folding.
- Co-expression with a chaperone for example, an expression vector containing the gene encoding the mutant enzyme of the present invention is introduced into E. coli together with a vector that expresses the GroEL and GroES genes under the induction of arabinose (for example, the commercially available chaperone vectors pGro7 and TaKaRa) Then, after the transformant has grown to some extent, arabinose is added to induce chaperone expression.
- the recombinant FAD-GDH thus obtained can be used, for example, in gel filtration, ion exchange chromatography, affinity chromatography, liquid chromatography, filtration, ultrafiltration, salting out, solvent precipitation, immunoprecipitation, gel electrophoresis. It can be purified using any of the purification techniques known in the art such as isoelectric focusing, dialysis and the like.
- the FAD-GDH of the present invention has an action of catalyzing a reaction in which glucose is oxidized to produce gluconolactone using FAD as a coenzyme.
- the glucose dehydrogenase activity of the FAD-GDH of the present invention can quantify the amount of FAD that is reduced as the glucose is oxidized by the dehydrogenase by a color reaction of the redox dye.
- the color reagent include PMS (phenazine methosulfate), DCIP (2,6-dichlorophenolindophenol), potassium ferricyanide, ferrocene, and the like.
- the glucose oxidation activity of FAD-GDH of the present invention can be measured by quantifying hydrogen peroxide generated by the reaction between the dehydrogenase and the substrate.
- the measurement of hydrogen peroxide can be performed, for example, by measuring the change over time in the absorbance of the dye produced using peroxidase, a Trinder reagent (TODB), and 4-aminoantipyrine.
- TODB Trinder reagent
- the selectivity of the FAD-GDH for glucose of the present invention is determined by measuring enzyme activity as described above using various sugars such as mannose, galactose, xylose, lactose and maltose as substrates, and using glucose as a substrate. It can be evaluated by examining the relative activity with respect to the activity.
- the FAD-GDH of the present invention has a high selectivity for glucose, and in particular the reactivity with maltose and galactose was below the measurement limit. Therefore, the assay kit or enzyme sensor prepared using the FAD-GDH of the present invention has high selectivity for glucose measurement, and the test sample contains or may contain other sugars such as maltose. Also has the advantage that glucose can be detected with high sensitivity.
- the FAD-GDH of the present invention is characterized by low reactivity with xylose. This provides the advantage that the blood glucose level can be measured accurately even when a xylose absorption test is performed.
- the present invention also features a glucose assay kit comprising FAD-GDH according to the present invention.
- the glucose assay kit of the present invention comprises FAD-GDH according to the present invention in an amount sufficient for at least one assay.
- the kit includes the FAD-GDH of the present invention, plus buffers necessary for the assay, mediator, glucose standard solution for creating a calibration curve, and directions for use.
- the FAD-GDH according to the present invention can be provided in various forms, for example as a lyophilized reagent or as a solution in a suitable storage solution.
- the present invention also provides an enzyme electrode having FAD-GDH according to the present invention immobilized on its surface, and a glucose sensor using this enzyme electrode.
- a carbon electrode, a gold electrode, a platinum electrode or the like is used, and the enzyme of the present invention is immobilized on this electrode.
- These electrodes may be electrodes formed by screen printing or the like if they are carbon electrodes, and electrodes formed by sputtering if they are gold or platinum electrodes.
- Immobilization methods include a method using a crosslinking reagent, a method of encapsulating in a polymer matrix, a method of coating with a dialysis membrane, a photocrosslinkable polymer, a conductive polymer, a redox polymer, etc., or ferrocene or a derivative thereof. It may be fixed in a polymer or adsorbed and fixed on an electrode together with a representative electron mediator, or a combination thereof may be used.
- FAD-GDH of the present invention is immobilized on a carbon electrode using glutaraldehyde, and then treated with a reagent having an amine group to block glutaraldehyde.
- the measurement of glucose concentration can be performed as follows. Put the buffer in a thermostatic cell and add a mediator to maintain a constant temperature.
- a mediator potassium ferricyanide, phenazine methosulfate, a ruthenium complex, or the like can be used.
- An electrode on which the FAD-GDH of the present invention is immobilized is used as a working electrode, and a counter electrode (for example, a platinum electrode) and a reference electrode (for example, an Ag / AgCl electrode) are used as necessary.
- a counter electrode for example, a platinum electrode
- a reference electrode for example, an Ag / AgCl electrode
- the FAD-GDH of the present invention is particularly useful for use in a blood glucose level assay device.
- the assay device can have a structure similar to a commercially available amperometric biosensor test strip for blood glucose level measurement.
- the assay device has two electrodes (working electrode and reference electrode) mounted on an insulator, a reagent port and a sample receiving part.
- the reagent port contains the FAD-GDH and mediator of the present invention.
- an aspect represented by the expression “comprising” is an aspect represented by the expression “essentially consistent of”. And an aspect represented by the expression “consisting of”.
- Example 1 Preparation of recombinant gene for glucose dehydrogenase derived from Botryotinia quercherina (Bfu-GDH) and glucose dehydrogenase derived from Aspergillus oryzae (Ao-GDH) It is. From the N-terminal to the 17th Ser can be presumed to be a signal peptide. For example, a free access server SignalP 3.0 Server (http://www.cbs.dtu.dk/services/SignalP-3.0/) can be used as a method for predicting a signal sequence cleavage site.
- SignalP 3.0 Server http://www.cbs.dtu.dk/services/SignalP-3.0/
- This server is operated by The Center for Biological Sequence Analysis at the Technical University of Denmark, and based on the methodology described in the following paper, the possibility of the existence of a signal sequence for any amino acid sequence is confirmed. Search and predict the cleavage site. Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Henrik Nielsen, Jacob Engelbrecht, Soren Brunak and Gunnar von Heijne. Protein Engineering, 10: 1-6, 1997.
- the signal peptide from the N-terminal to the 17th Ser is predicted to be an amino acid sequence having a sequence after the 18th Thr downstream of the initiation methionine
- a gene sequence having a codon suitable for recombinant production by E. coli was designed and synthesized.
- the amino acid sequence of the protein encoded by this gene (hereinafter referred to as Bfu-GDH) is shown in SEQ ID NO: 2.
- the amino acid sequence of glucose dehydrogenase derived from Aspergillus oryzae TI strain is shown in SEQ ID NO: 3.
- the 23rd Lys from the N-terminal is presumed to be a signal peptide, so a gene encoding an amino acid sequence having a sequence after the 24th Asn downstream of the start methionine. The sequence was designed and fully synthesized.
- the amino acid sequence of the protein encoded by this gene (hereinafter referred to as Ao-GDH) is shown in SEQ ID NO: 4. E.
- Example 2 Measurement of enzyme activity
- the glucose dehydrogenase activity of the FAD-GDH of the present invention is determined by the absorbance at 600 nm of the discoloration of DCIP (2,2'-dichlorodiisopropyl ether) reduced by the reaction between the dehydrogenase and the substrate. This was done by quantifying the change over time.
- the reaction conditions were as follows unless otherwise noted. Start the reaction by adding the substrate to the reaction solution containing the enzyme solution (10 mM potassium phosphate (pH 7.0) + 0.6 mM PMS + 0.06 mM DCIP concentration is the final concentration), and measure the change in absorbance at 600 nm did.
- the final concentration of 50 mM glucose was used as the substrate, the amount of enzyme that reduced 1 ⁇ mol of DCIP was 1 Unit, and the activity value was calculated from the following formula.
- the molar extinction coefficient of DCIP at pH 7.0 was 16.3 mM ⁇ 1 cm ⁇ 1 .
- Unit / ml ⁇ ABS / min ⁇ 1 / 16.3 ⁇ 10
- the measurement of glucose oxidation activity of FAD-GDH of the present invention was carried out using hydrogen peroxide generated by the reaction of dehydrogenase and substrate using peroxidase, Trinder reagent (TODB), and 4-aminoantipyrine, and 546 nm of the generated dye. This was carried out by measuring the change in absorbance with time.
- the reaction conditions were as follows unless otherwise noted. Start the reaction by adding the substrate to the reaction solution containing enzyme solution (10 mM potassium phosphate (pH 7.0) + 1.5 mM 4-aminoantipyrine + 1.5 mM TODB + 2 U / ml peroxidase, all concentrations are final) The change in absorbance at 546 nm was measured.
- Example 3 Examination of culture conditions and preparation of crude enzyme preparation Production of Bfu-GDH using IPTG induction: Escherichia coli BL21 (DE3) was transformed with the expression vector pET30c into which the gene encoding Bfu-GDH was inserted. The obtained transformant BL21 (DE3) / pET30c (Bfu-GDH) was inoculated into 3 mL LB medium and cultured with shaking at 37 ° C. overnight. Thereafter, 1 ml of the preculture solution was inoculated into 100 mL of LB medium (kanamycin (Km) 50 ⁇ g / mL), and cultured at 37 ° C. in a conical flask with a baffle at 180 rpm.
- LB medium kanamycin (Km) 50 ⁇ g / mL
- IPTG isopropyl- ⁇ -thiogalactopyranoside; final concentration 1 mM
- the cells were cultured at 20 ° C., and the cultivation was completed 14 hours after the start of the cultivation.
- 300 ⁇ l of the culture solution was collected every few hours, 60 ⁇ l of bugbuster reagent (BugBuster Reagent) was added to the collected cells and shaken at 4 ° C. for 20 minutes to lyse the cells.
- a medium LB medium + 0.5% glycerol, 0.05% glucose, 0.2% ⁇ -lactose, 25 mM (NH 4 ) 2 SO 4 , 100 mM KH 2 PO 4 , 100 mM NaHPO 4 , 1 mM MgSO 4 (ZYP medium; F. (Modified from William Studier et.al., Protein Expression and Purification (2005)).
- Bfu-GDH was expressed as a water-soluble enzyme having GDH activity.
- the culture and enzyme production curves are shown in FIG. OD660 significantly increased 24 hours after the start of the culture, and stopped increasing at about 21 around 28 hours.
- the enzyme activity increased from around 24 hours, and after 32 hours, it reached about 2200 U per liter of culture broth, and the increase stopped.
- the protein concentration was 2.2 mg / ml after 24 hours and about 3.0 mg / ml after 36 hours, and the increase stopped.
- Arabinose (final concentration 2 mM) was added at the start of culture, 12 hours and 24 hours later, and as a result of culturing each, OD660 increased significantly 24 hours after the start of culture. The increase stopped at about 27, about 28, and about 26, respectively.
- the enzyme activity increased from around 24 hours, and after 40 hours, it became about 2500 U / L, 2600 U / L, and 1300 U / L per liter of the culture solution, respectively, and the increase stopped.
- the protein concentrations were 1.8 mg / ml, 2.0 mg / ml, and 1.9 mg / ml after 24 hours, respectively.
- Example 4 Evaluation of substrate specificity
- the substrate specificity of the enzyme obtained using the medium A of Example 3 was measured using glucose, maltose, xylose and galactose as substrates.
- the dehydrogenase activity of Bfu-GDH when glucose was 100% at a substrate concentration of 5 mM was not detectable with maltose and galactose, and 13% with xylose.
- the dehydrogenase activity of xylose was 21% when the glucose of Ao-GDH was 100%. Therefore, it was shown that Bfu-GDH has lower enzyme activity against xylose than Ao-GDH which has already been reported.
- Example 5 Mutagenesis into Bfu-GDH Site-directed mutagenesis was performed by the QuikChange (registered trademark) method.
- QuikChange registered trademark
- PCR amplification was performed using the pET30c-Bfu-GDH prepared in Example 1 as a template and primers for mutagenesis.
- DpnI was added to the sample after PCR and incubated at 37 ° C. for 60 minutes to digest only the template DNA, and Escherichia coli DH5 ⁇ was transformed using this sample.
- plasmid was extracted from an arbitrarily selected clone, and it was confirmed that the target mutation was introduced by sequence analysis.
- the obtained PCR fragment was digested with NdeI and HindIII (37 ° C., 2 hours), ligated with pET30c digested with the same restriction enzyme, and Escherichia coli BL21 (DE3) was transformed with this sample.
- plasmids were extracted from arbitrarily selected clones, and the introduction of mutations was confirmed by sequence analysis.
- transformants expressing the following mutant enzymes were obtained: Bfu-GDH (G53A), (N176K), N176R, N176E, N176S, (S514G), (S552C), (G53A / S514G), (S514G / S552C), (G53A / S514G / S552C).
- S514G, S552C, or these double mutations S514G / S552C and (N176K), N176R, N176E, N176S, N225K, N259K, N301K, N326K, N330K, N355K, N487S, T488E, V489I, S490P, 4989 , E495D, A496E, E497K, F499V, D500E, V502L, T504A, and A505N were designed in various combinations to produce transformants expressing these mutant enzymes: N176K / S490P / D500E / S514G / S552C, N176K / A496E / D500E / S514G / S552C, N176K / S514G / S552C, S514G / S552C, G53A / S514G / 552C.
- Example 6 Production of Mutant Enzyme and Measurement of Activity Transformant BL21 (DE3) expressing each mutant enzyme was pre-cultured. Using a 300 ml baffled flask, 1% of the bacterium was grown on 60 ml of medium A, and cultured with shaking at 20 ° C. for 28 hours at 125 rpm. After collecting 50 ml of the medium, BugBuster (registered trademark) protein extraction reagent (Novagen) was added to 1 g of wet cells at a rate of 5 ml, suspended, and incubated at room temperature for 15 minutes with gentle shaking.
- BugBuster registered trademark
- the resulting supernatant was dialyzed overnight at 4 ° C. against 20 mM potassium phosphate buffer (pH 6.5). After completion of dialysis, centrifugation was performed and the supernatant was used as a crude purified enzyme preparation.
- 20 mM potassium phosphate buffer (pH 6.5) was suspended at a rate of 1 m with respect to the insoluble fraction of 2 ml of the suspension.
- Transformant BL21 (DE3) expressing each mutant enzyme was precultured, and 1 vol% was inoculated into 3 ml of A medium. After shaking culture at 37 ° C for 4 hours, shaking culture was performed at 20 ° C for 20 hours. After culturing, 2 ml of the medium was collected, 400 ⁇ l of BugBuster (registered trademark) was added, and after suspension, the mixture was shaken at room temperature for 15 minutes. Thereafter, centrifugation (15000 rpm / 4 ° C./20 minutes) was performed, and the obtained supernatant was used as a crudely purified enzyme preparation.
- BugBuster registered trademark
- GDH activity was measured using a DCIP (0.3 mM) / PMS (0.6 mM) system, using 20 mM potassium phosphate buffer (pH 6.5) as the buffer, and glucose (Glc; 1, 2, 4, 10, 20 as the substrate) , 40 mM), xylose (Xyl; 4,40 mM) was used.
- the results are shown in Table 1.
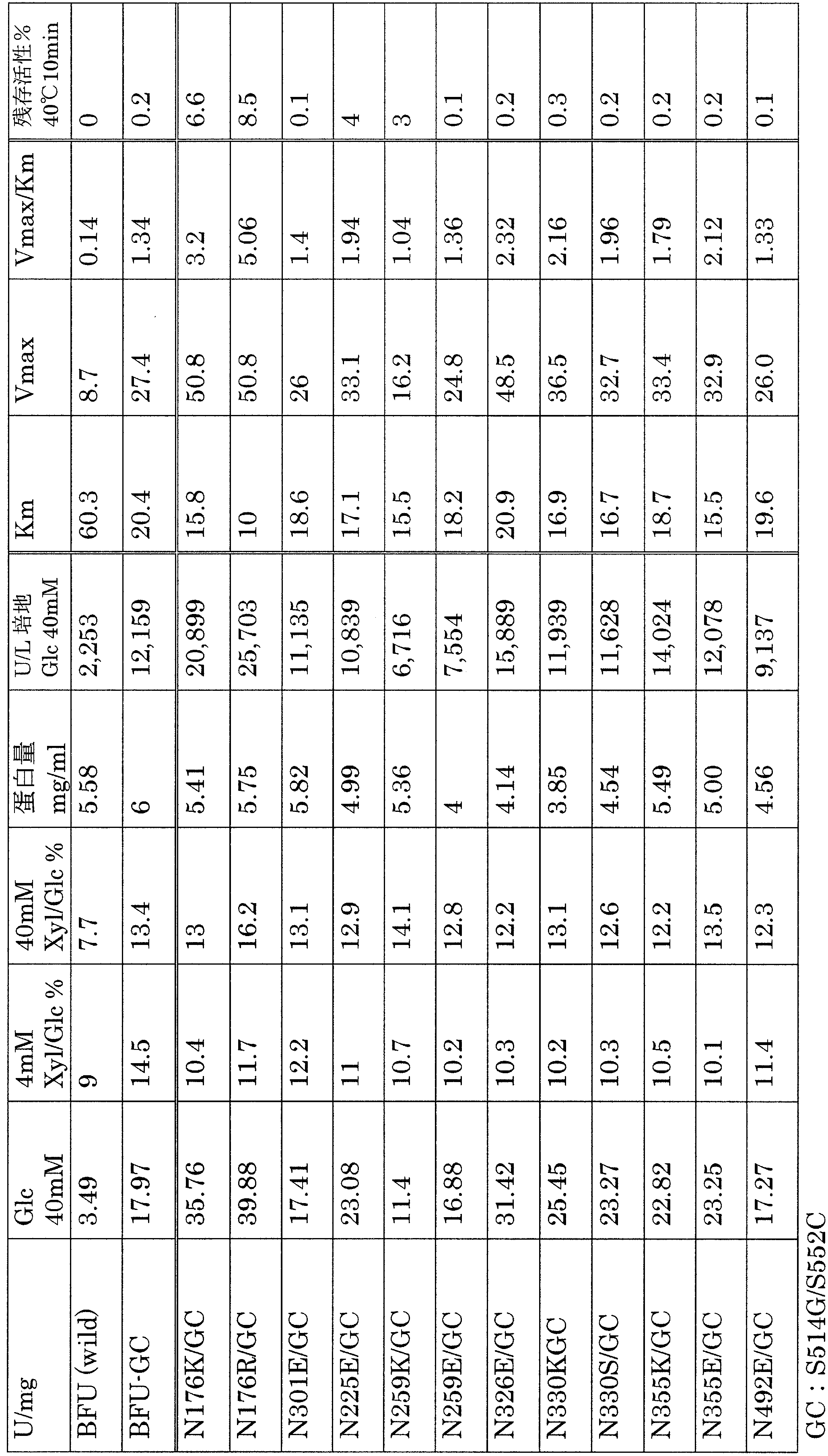
- the numerical values in the table are specific values (U / mg) and productivity (U / L), which are average values of data obtained several times under the same conditions using 40 mM glucose as a substrate.
- the correlation between the substrate (glucose; 1,2,4,10,20,40 mM) concentration and activity was observed, and Michaelis Menten obtained from the saturation curve obtained therefrom. Constants (Km value) and apparent maximum activity (Vmax) were determined.
- the activity values described in Table 1 and Table 2 of the priority application in this case had an error in the calculation of the activity value from the absorbance data. Tables 1 and 2 below show the correct values.
- N176K, N176R, N176E, N176S, S514G and S552C mutants had increased productivity and enzyme activity compared to the control (Bfu-GDH). With the S514G / S552C double mutant, higher production and enzyme activity were obtained. The N176K, N176R, N176E, N176S, and mutants obtained extremely high productivity when combined with S514G / S552C.
- the G53A mutant showed the same productivity as the control in the single mutant and the G53A / S514G double mutant, but the G53A / S514G / S552C composite mutant showed very high productivity.
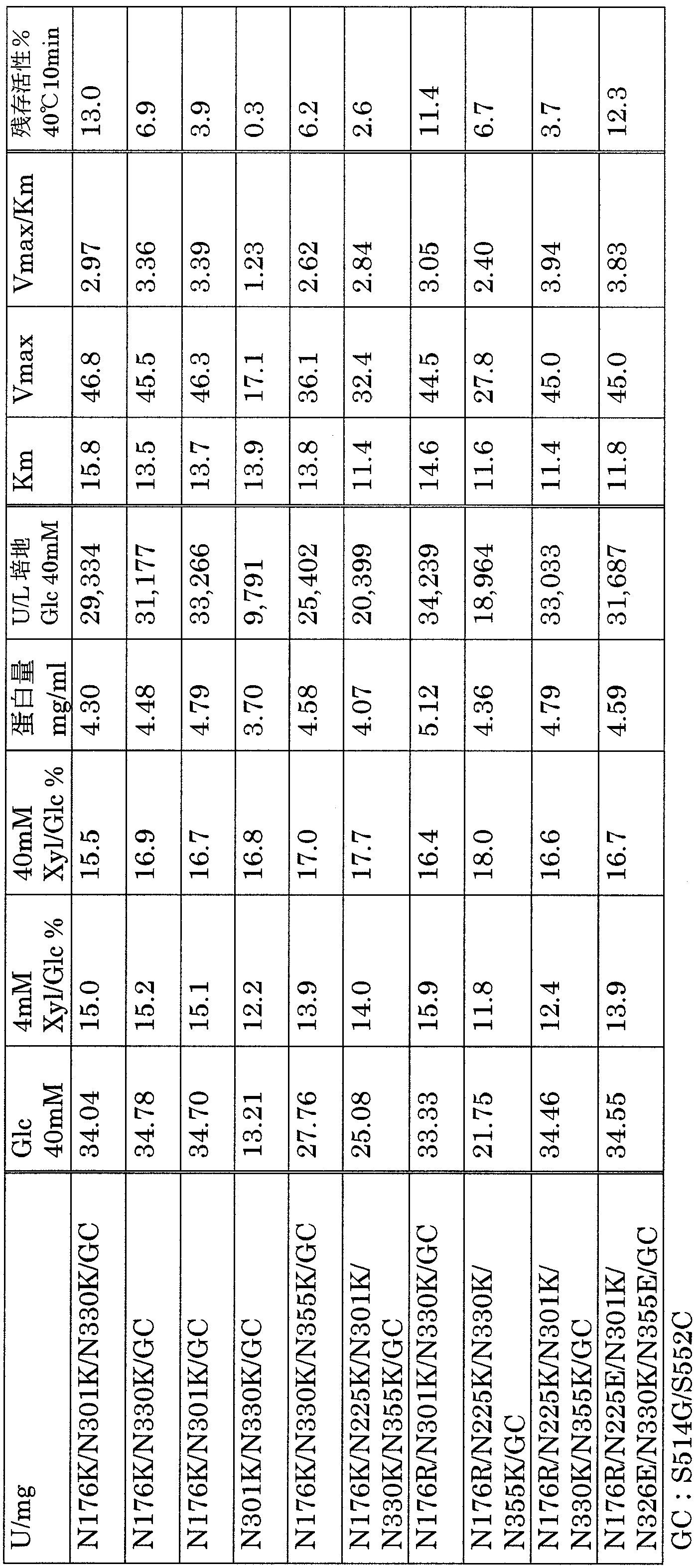
- the results for multiple mutants combining S514G and S514G / S552C with yet another mutation are shown in Table 2.
- the numerical values in the table are specific values of specific data (U / mg) and productivity (U / L) of several times of data performed under the same conditions using 40 mM glucose as a substrate. Further, using the obtained crude purified enzyme, the correlation between the substrate (glucose; 1, 2, 4, 10, 20, 40 mM) concentration and activity was observed, and Michaelis Menten obtained from the saturation curve obtained therefrom. Constant (Km value) and apparent maximum activity (Vmax) were determined.
- S514G single mutation and productivity higher than S514G / S552C were obtained in many multiple mutants.
- N176K / S490P / D500E / S514G / S552C, N176K / A496E / D500E / S514G / S552C, N176K / S514G / S552C, S514G / S552C, G53A / S514G / S552C have very high productivity and enzyme It was.
- the reactivity to xylose was 20% or less of glucose, or 15% or less. It was a high enzyme. Moreover, the activity with respect to maltose and galactose of these enzymes was not detected.
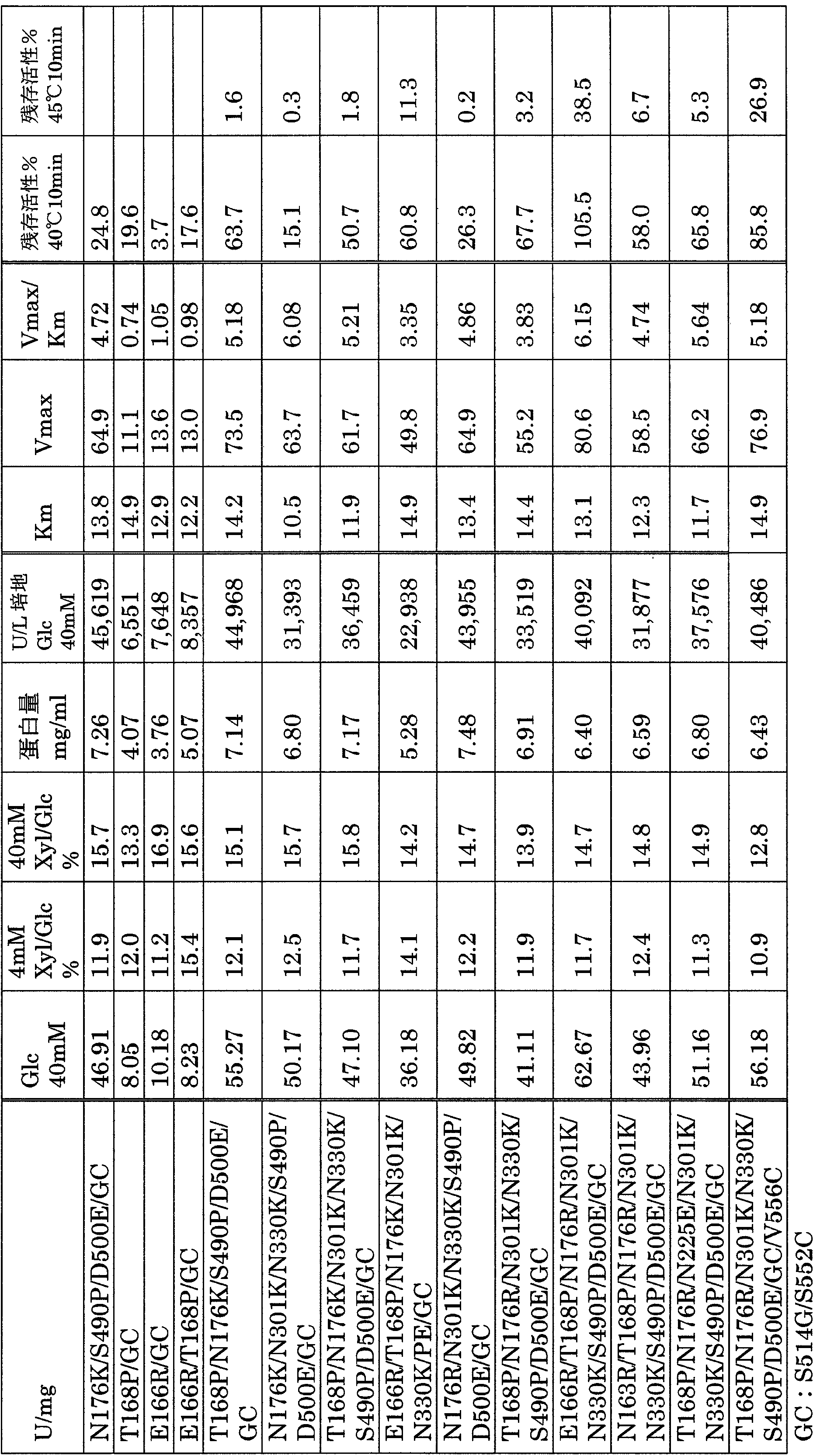
- the composite mutants shown in Table 4 below had particularly high productivity / activity and stability.
- Glu166Arg / Thr168Pro / Asn176Arg / Asn301Lys / Asn330Lys / Ser490Glu / Asp500Glu / Ser514Glys / Ser552Cys showed high Vmax / Km values and high thermal stability.
- This enzyme had a residual activity of 100% after 10 minutes incubation in a 40 ° C. solution, and no inactivation was observed. A residual activity of less than 40% was observed even after 10 minutes at 45 ° C.
- Example 7 Introduction of intramolecular SS bond into Ao-GDH
- various mutants in which various amino acids of Ao-GDH were mutated to cysteine were prepared, and the enzyme activity and thermal stability were improved. Examined. Mutation introduction and enzyme activity were measured in the same manner as in Example 5.
- the thermal stability test was conducted as follows. 200 ⁇ l of the above crude enzyme preparation was added to 800 ⁇ l of 20 mM potassium phosphate buffer (pH 6.5) at 45 ° C., and immediately mixed, and immediately 20 mM potassium phosphate buffer (pH 6. 5) 100 ⁇ l was added to 100 ⁇ l (final concentration diluted 10 times). After the addition, incubation was performed at 45 ° C. for 2, 5, 10, 15, 20, 25, or 30 minutes. After a predetermined time, it was quickly cooled in ice.
- the GDH activity was measured using a DCIP (0.3 mM) / PMS (0.6 mM) system, using 20 mM potassium phosphate buffer (pH 6.5) as a buffer and Glc (40 mM) as a substrate.
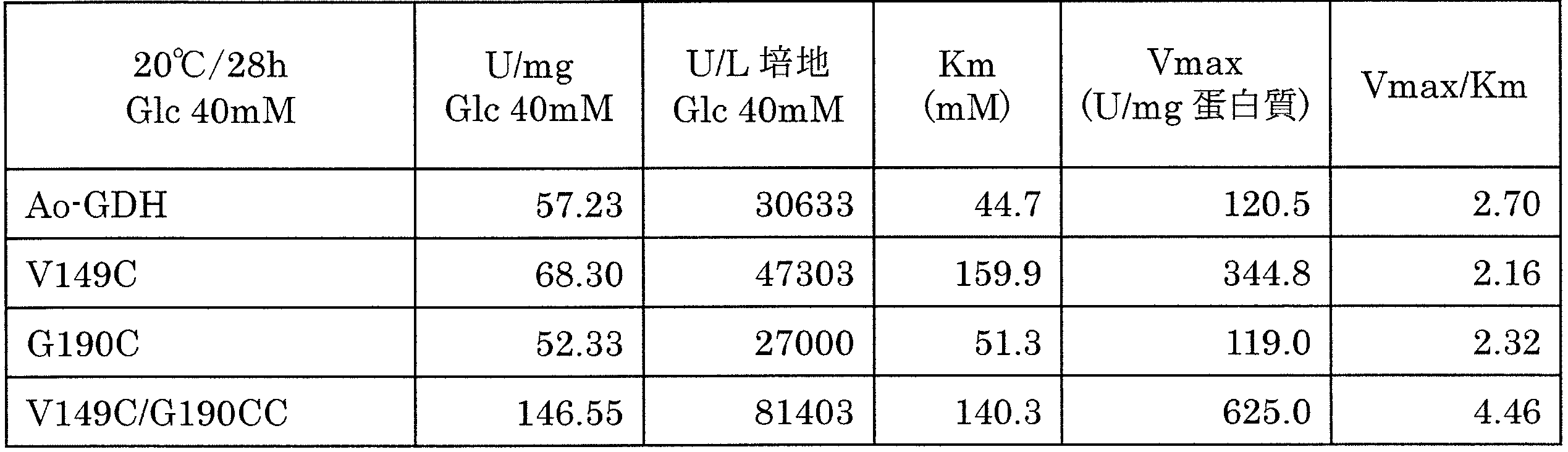
- the modified enzyme in which the 149th amino acid residue V and the 190th amino acid residue G of the Aspergillus oryzae TI-derived FAD-GDH represented by SEQ ID NO: 4 are both replaced with C is particularly high. It has been found to exhibit thermal stability as well as productivity. As shown in FIG. 3, in the case of Ao-GDH, the activity decreased with time, but V149C / G190C rapidly decreased to about 30% and then became constant at about 15%. Since a single mutation of V149C or G190C does not contribute to an increase in productivity / activity, it is presumed that this improvement in thermal stability is due to the formation of an SS bond in the molecule.
- FIG. 4 shows SV curves of the wild-type enzyme, the V149C / G190C mutant enzyme, and the G190C mutant enzyme as a reference.
- the Km value and Vmax value are shown in Table 6 below.
- V149C / G190C mutant enzyme has higher enzyme activity than the wild type. It is currently unknown whether the activity of the substrate pocket has changed due to the formation of intramolecular S—S bonds and the activity has increased, or the apparent residual activity of the enzyme has been improved by improving the thermal stability.
- Example 8 Introduction of intramolecular SS bond to Bfu-GDH and other fungal-derived GDH Based on the result of Example 7, amino acid residues A and 192 of the 150th amino acid of FAD-GDH derived from Botryotinia fukeliana represented by SEQ ID NO: 2
- a modified enzyme A150C / T192C
- both of amino acid residues T of the above were replaced with C was produced, it was found to exhibit high thermal stability.
- multiple mutants were prepared by combining such mutations and several mutations that showed high productivity in Example 6, and the enzyme activity and thermal stability were similarly evaluated for these mutants.
- Fig. 5 shows the thermal stability of the A150C / T192C mutant of Bfu-GDH.
- A150C / T192C and multiple mutations containing this mutation were introduced into Bfu-GDH, the enzyme activity was equal or decreased, but an improvement in thermal stability was observed.
- the heat treatment was performed at 45 ° C., the activity decreased immediately after that, but the rate of decrease in the activity thereafter slowed.
- the introduction of these mutations did not affect the glucose / xylose selectivity.
- genes encoding glucose dehydrogenase of Sclenotinia sclerotiorum and Aspergillus nigar 40715 having the sequence shown in FIG. 1 were synthesized (SEQ ID NOs: 7 and 8, respectively) and expressed in E. coli. It was.
- mutant enzymes into which A150C / T191C and Y150C / G191C mutations were introduced were also prepared, and the enzyme activity and thermal stability were similarly evaluated.
- Example 6 Production and Evaluation of Enzyme Sensor Enzyme electrodes were produced using the Aspergillus oryzae TI-derived V149C / G190C mutant enzyme and the Botryotinia fukeliana-derived S514G / S552C mutant enzyme. 20 mg of carbon paste was added to 5 units of the modified FAD-GDH of the present invention and lyophilized. After this was mixed well, it was filled only on the surface of the carbon paste electrode already filled with about 40 mg of carbon paste, and was polished on a filter paper.
- This electrode was treated in 10 mM MOPS buffer (pH 7.0) containing 1% glutaraldehyde for 30 minutes at room temperature, and then treated in 10 mM MOPS buffer (pH 7.0) containing 20 mM lysine for 20 minutes at room temperature. To block glutaraldehyde. This electrode was equilibrated in a 10 mM MOPS buffer (pH 7.0) at room temperature for 1 hour or longer. The electrode was stored at 4 ° C.
- the glucose concentration was measured using the prepared enzyme sensor. Using the enzyme sensor to which the modified FAD-GDH of the present invention was immobilized, glucose could be quantified in the range of 0.1 mM-5 mM.
- the present invention is useful for measuring glucose concentration, particularly for measuring blood glucose level.
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biomedical Technology (AREA)
- Physics & Mathematics (AREA)
- Immunology (AREA)
- Biophysics (AREA)
- Analytical Chemistry (AREA)
- Medicinal Chemistry (AREA)
- Emergency Medicine (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Electrochemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Plant Pathology (AREA)
- Enzymes And Modification Thereof (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Apparatus Associated With Microorganisms And Enzymes (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
Description
本出願は,日本特許出願2010-147799(2010年6月29日出願)に基づく優先権を主張しており,この内容は本明細書に参照として取り込まれる。
本発明はフラビンアデニンジヌクレオチドを補酵素とするグルコース脱水素酵素(FAD-GDH)、その製造、およびグルコースの定量におけるその使用に関する。
本発明においては、ボトリオチニア・フケリアナのもつFAD-GDHが同定された。そのアミノ酸配列は配列番号1に示される。ボトリオチニア・フケリアナのゲノム配列は公開されているが、ボトリオチニア・フケリアナがFAD-GDHを有するという報告はこれまでになく、またFAD-GDHとしてアノーテーションされた遺伝子もなかった。配列番号1で示されるアミノ酸配列を有する蛋白質は、従来はグルコースオキシダーゼ(GOD)であると推定されていたが(Rolke et al., Mol Plant Pathol. 5(1), p.17-27, 2004)、本発明者は、この蛋白質はGODではなくGDHであることを初めて明らかにした。GDHは、GODと異なり、グルコース測定において溶存酸素の影響を受けないため、グルコース計測用のバイオセンサーの認識素子として有用である。配列番号1で示されるアミノ酸配列は、既知の真菌由来グルコース脱水素酵素のアミノ酸配列と約30~60%の配列同一性を有し、例えば、アスペルギルス・オリゼTI株由来グルコース脱水素酵素(GenBank ACW04779.1)とは約57%のアミノ酸同一性を有する。
本発明者は、配列番号2で表されるアミノ酸配列のN176K,S514GまたはS552Cの変異、またはS514G/S552Cの二重変異,およびN176K/S514G/S552Cの三重変異を有する改変型FAD-GDHが、野生型FAD-GDHと比較して大腸菌における高い生産性を示し、特にN176K/S514G/S552Cの三重変異を有する改変型FAD-GDHは、高い生産性とともに高い酵素活性を示すことを見いだした。なお、本明細書においては、ボトリオチニア・フケリアナ由来FAD-GDHのアミノ酸配列中のアミノ酸変異の位置は、配列番号2のアミノ酸配列の1番目のMetを1として番号付けする。また、本明細書においては、アミノ酸の変異ないし置換は、元のアミノ酸残基とアミノ酸の位置と置換後のアミノ酸残基とをこの順序で示すことにより表記し、例えば、「S514G」とは、514番目のSがGに置き換えられていることを示す。二重またはそれ以上の変異の組み合わせは「/」の記号で表す。
本発明者は、配列番号4で示されるアスペルギルス・オリゼTI株由来FAD-GDHの149番目のアミノ酸残基Vおよび190番目のアミノ酸残基Gの両方をCで置き換えることにより、野生型FAD-GDHと比較して熱安定性が顕著に高くなることを見いだした。アスペルギルス・オリゼTI株由来FAD-GDHのアミノ酸配列は配列番号3に示される。本発明においては、大腸菌で効率よく組換え発現させるために、配列番号3で示されるアスペルギルス・オリゼ由来の天然のFAD-GDHの推定シグナル配列であるN末端から23番目のLysまでを除去してMetを付加したアミノ酸配列(配列番号4)を作製した。本明細書においては、アスペルギルス・オリゼ由来FAD-GDHのアミノ酸配列中のアミノ酸変異の位置は、配列番号4のアミノ酸配列の1番目のMetを1として番号付けする。
本発明のFAD-GDHは、当該技術分野においてよく知られる手法を用いて組換え発現により製造することができる。ボトリオチニア・フケリアナ由来の天然のFAD-GDHおよびアスペルギルス・オリゼTI由来のFAD-GDHをコードする遺伝子の配列は、それぞれ配列番号2および4に記載されるアミノ酸配列に基づいて容易に決定することができる。FAD-GDHをコードする遺伝子は、ボトリオチニア・フケリアナまたはアスペルギルス・オリゼのゲノムからクローニングしてもよく、一連の化学合成されたオリゴヌクレオチドを用いてPCRにより製造してもよく、または自動化DNA合成機などを用いて全合成してもよい。遺伝子配列は、用いる宿主生物においてより高い発現レベルが達成されるようにコドンを選択して、適宜設計または改変することが望ましい。特定の宿主生物におけるコドン使用の特徴は、当該技術分野においてよく知られている。
A培地で、15℃から25℃、好ましくは約20℃で培養して、組換え蛋白質を発現させる。このことにより、慣用のIPTG誘導法を用いた場合よりも、高い生産性を得ることができる。さらに、本発明の変異酵素を、蛋白質のフォールディングを促進することが知られているシャペロンGroELおよびGroESと共発現させることにより、さらに高い生産性を得ることができる。シャペロンとの共発現は、例えば、本発明の変異酵素をコードする遺伝子を含む発現ベクターを、GroELおよびGroES遺伝子をアラビノース誘導下に発現するベクター(例えば市販のシャペロンベクターpGro7、TaKaRa)とともに大腸菌に導入して培養し、形質転換体がある程度増殖した後にアラビノースを加えてシャペロンの発現を誘導することにより行うことができる。
本発明のFAD-GDHは、FADを補酵素として、グルコースを酸化してグルコノラクトンを生成する反応を触媒する作用を有する。本発明のFAD-GDHのグルコース脱水素酵素活性は、脱水素酵素によるグルコースの酸化に伴って還元されるFADの量を、酸化還元色素の呈色反応により定量することができる。呈色試薬としては、例えば、PMS(フェナジンメトサルフェート)、DCIP(2,6-ジクロロフェノールインドフェノール)、フェリシアン化カリウム、フェロセンなどを用いることができる。また、本発明のFAD-GDHのグルコース酸化活性測定は、脱水素酵素と基質の反応により生成する過酸化水素を定量することにより行うことができる。過酸化水素の測定は、例えば、ペルオキシダーゼ及びトリンダー試薬(TODB)、4-アミノアンチピリンを用いて、生成する色素の吸光度の経時変化を測定することにより行うことができる。
本発明のFAD-GDHのグルコースに対する選択性は、基質として、マンノース、ガラクトース、キシロース、ラクトースおよびマルトース等の各種の糖を用いて上述のように酵素活性を測定し、グルコースを基質としたときの活性に対する相対活性を調べることにより評価することができる。
本発明はまた、本発明に従うFAD-GDHを含むグルコースアッセイキットを特徴とする。本発明のグルコースアッセイキットは、本発明に従うFAD-GDHを少なくとも1回のアッセイに十分な量で含む。典型的には、キットは、本発明のFAD-GDHに加えて、アッセイに必要な緩衝液、メディエーター、キャリブレーションカーブ作製のためのグルコース標準溶液、ならびに使用の指針を含む。本発明に従うFAD-GDHは種々の形態で、例えば、凍結乾燥された試薬として、または適切な保存溶液中の溶液として提供することができる。
本発明はまた、本発明に従うFAD-GDHがその表面に固定化されている酵素電極、ならびにこの酵素電極を用いるグルコースセンサーを提供する。電極としては、カーボン電極、金電極、白金電極などを用い、この電極上に本発明の酵素を固定化する。これらの電極はカーボン電極であればスクリーンプリント印刷等によって作成される電極、および金、白金電極であればスパッタリングにより作成される電極でもよい。固定化方法としては、架橋試薬を用いる方法、高分子マトリックス中に封入する方法、透析膜で被覆する方法、光架橋性ポリマー、導電性ポリマー、酸化還元ポリマーなどがあり、あるいはフェロセンあるいはその誘導体に代表される電子メディエーターとともにポリマー中に固定あるいは電極上に吸着固定してもよく、またこれらを組み合わせて用いてもよい。典型的には、グルタルアルデヒドを用いて本発明のFAD-GDHをカーボン電極上に固定化した後、アミン基を有する試薬で処理してグルタルアルデヒドをブロッキングする。
ボトリオチニア・フケリアナ由来グルコース脱水素酵素(Bfu-GDH)およびアスペルギルス・オリゼ由来グルコース脱水素酵素(Ao-GDH)の組換え遺伝子の調製
ボトリオチニア・フケリアナ由来グルコース脱水素酵素のアミノ酸配列は配列番号1に示される。N末端から17番目のSerまではシグナルペプチドであると推定できる。例えば、シグナル配列切断部位を予測するための方法としてフリーアクセスサーバSignalP 3.0 Server(http://www.cbs.dtu.dk/services/SignalP-3.0/)を活用できる。このサーバは,The Center for Biological Sequence Analysis at the Technical University of Denmark において運営されており、以下に記す論文に記載された方法論に基づき、任意のアミノ酸配列に対して、シグナル配列の存在の可能性を検索し、その切断部位を予想する。Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Henrik Nielsen, Jacob Engelbrecht, Soren Brunak and Gunnar von Heijne. Protein Engineering, 10:1-6, 1997。SignalP 3.0 Serverを用いる予測にしたがえば、N末端から17番目のSerまではシグナルペプチドであると推定されるので、開始メチオニンの下流に18番目のThr以降の配列を有するアミノ酸配列をコードし、大腸菌による組み換え生産に適したコドンをもつ遺伝子配列を設計して、全合成した。この遺伝子によりコードされる蛋白質(以下Bfu-GDHと称する)のアミノ酸配列を配列番号2に示す。
酵素活性の測定
本発明のFAD-GDHのグルコース脱水素酵素活性は、脱水素酵素と基質の反応により還元されるDCIP(2,2’-ジクロルジイソプロピルエーテル)の退色を600 nmでの吸光度の経時変化を定量することにより行った。反応条件は断りの無い限り以下の条件で行った。酵素溶液を含む反応溶液(10 mM リン酸カリウム(pH7.0)+0.6 mM PMS+0.06 mM DCIP濃度は全て終濃度)に基質を加えることで反応を開始し、600 nmの吸光度変化を測定した。基質には終濃度50 mMグルコースを用い、1μmolのDCIPを還元させる酵素量を1Unitとし、以下の式より活性値を算出した。DCIPのpH 7.0におけるモル吸光係数は16.3 mM-1cm-1とした。
ユニット/ml = ΔABS/min × 1/ 16.3 ×10
ユニット/ml = ΔABS/min × 2 / 38 ×10
ユニット/mg = Unit/ml/蛋白質mg/ml
培養条件の検討及び粗精製酵素標品の調製
1. IPTG誘導を用いたBfu-GDHの生産:
Bfu-GDHをコードする遺伝子を挿入した発現ベクターpET30cを用いて、大腸菌BL21(DE3)を形質転換した。得られた形質転換体BL21(DE3)/pET30c(Bfu-GDH)を3 mL LB培地に植菌し、一晩37℃で振とう培養した。その後、LB培地(カナマイシン(Km)50 μg/mL)100 mLに前培養液1 mlを植菌し、37 ℃でバッフル付三角フラスコを用いて、180 rpmで振とう培養を行なった。培養液のOD660が0.6付近になった時点でIPTG(イソプロピル-β-チオガラクトピラノシド;終濃度1 mM)を添加して、Bfu-GDHの発現誘導を行った。添加後は20 ℃で培養を行い、培養開始後14時間で培養を終えた。培養中、数時間ごとに培養液300 μlを回収し、集菌した菌体に60 μlのバグバスター試薬(BugBuster Reagent)を加え、20分間、4 ℃で振盪し、菌体を溶菌させた。その後、10 mM リン酸カリウムを60 μl加え、遠心(16,000×g、4℃、20 分間)して上清を回収し、その上清を粗精製酵素標品として、酵素活性を測定した。酵素活性は菌体の濃度とともに上昇し、培養終了時には培養液1 Lあたり65 Uとなった。このとき、比活性は0.15 U/mgであり、菌体濃度はOD660=4.0であった。
A培地を用いるBfu-GDHの生産を行った。Bfu-GDHをコードする遺伝子を挿入した発現ベクターpET30cを用いて大腸菌BL21(DE3)を形質転換した。得られた形質転換体BL21(DE3)/pET30c(Bfu-GDH)を3 mL LB培地に植菌し、一晩37℃で振とう培養した。その後、下記A培地(Km 50 μg/mL)100 mLに前培養液1 mlを植菌し、20℃で坂口フラスコを用いて、120 rpmで振とう培養を行なった。
A培地: LB培地+0.5% グリセロール、0.05% グルコース、0.2% α-ラクトース、25mM (NH4)2SO4、100mM KH2PO4、100mM NaHPO4、1 mM MgSO4 (ZYP培地;F. William Studier et.al., Protein Expression and Purification (2005)を改変)。
Bfu-GDHのコドン修正遺伝子を挿入したpET30cを用いて、シャペロンベクターpGro7が既に導入されている大腸菌BL21(DE3)/pGro7を形質転換した。pGro7は、蛋白質のフォールディングを促進することが知られているシャペロンGroELおよびGroESをアラビノース誘導下に発現するベクターである。得られた形質転換体BL21(DE3)/pET30c(Bfu-GDH)・pGro7を3 mL LB培地に植菌し、一晩37℃で振とう培養した。その後、Km 50 μg/mL、クロラムフェニコール(Cm)50 μg/mLを含むA培地100 mLに前培養液1 mlを植菌し、20℃で坂口フラスコを用いて、120 rpmで振とう培養を行なった。pGro7の発現誘導はアラビノース(終濃度2 mM)の添加により行った。アラビノースの添加は培養開始時、培養開始12時間後、培養開始24時間後と添加時間を変えて検討を行った。培養中、任意の時間ごとに培養液300 μlを回収し、上記と同様の方法で粗精製酵素標品を調製した。
基質特異性の評価
上記実施例3のA培地を用いて得られた酵素について、グルコース、マルトース、キシロースおよびガラクトースを基質として基質特異性を測定した。その結果、基質濃度5 mMにおいてグルコースを100%とした時のBfu-GDHの脱水素酵素活性はマルトース、ガラクトースでは検出できず、キシロースでは13%であった。同じ条件でのAo-GDHのグルコースを100%とした時のキシロースの脱水素酵素活性は21%であった。よってBfu-GDHは既に報告されているAo-GDHよりキシロースに対する酵素活性が低いことが示された。
Bfu-GDHへの変異導入
部位特異的変異導入はQuikChange(登録商標)法により行った。QuikChange(登録商標)法では、実施例1で作製したpET30c-Bfu-GDHをテンプレートとし、変異導入用プライマーによりPCR増幅を行った。続いてPCR後の試料にDpnIを加え37℃、60分間インキュベートすることでテンプレートDNAのみを消化し、この試料を用いて大腸菌DH5αを形質転換した。LB寒天培地(50 μg/mlカナマイシン)により一晩培養後、任意に選んだクローンからプラスミドを抽出し、シークエンス解析により目的の変異が導入されていることを確認した。得られたPCR断片をNdeI、HindIII消化(37 ℃,2時間)し、同制限酵素消化したpET30cとライゲーションし、この試料を用いて大腸菌BL21(DE3)を形質転換した。LB寒天培地(50 μg/mlカナマイシン)により一晩培養後、任意に選んだクローンからプラスミドを抽出し、シークエンス解析により変異導入を確認した。
変異酵素の生産および活性測定
各変異酵素を発現する形質転換体BL21(DE3)を前培養した。300mlバッフル付きフラスコを用いA培地60ml に対し1%量殖菌し、20℃、28時間、125rpmの条件で振とう培養を行った。培地50mlを集菌後、湿菌体1gに5mlの割合でBugBuster(登録商標)蛋白質抽出試薬(Novagen)を加え懸濁し、ゆるやかに振とうしながら室温で15分間インキュベートを行った。遠心(15K rpm/4℃/20分間)により不溶性分画を除去後、得られた上清を20mM リン酸カリウム緩衝液 (pH6.5)で4℃で一晩透析した。透析終了後に遠心を行い、その上清を粗精製酵素標品とした。不溶性分画は、懸濁液2ml分の不溶性分画に対し20mM リン酸カリウム緩衝液 (pH6.5)を1mの割合で懸濁した。


次にこれらの活性向上に効果があった変異を組み合わせた変異酵素を構築し、生産性・活性ならびに安定性について検討した。結果を表4に示す。
Ao-GDHへの分子内S-S結合の導入
酵素の熱安定性の向上を目的として、Ao-GDHの種々のアミノ酸をシステインに変異させた各種変異体を作製し、その酵素活性および熱安定性を調べた。変異の導入および酵素活性の測定は実施例5と同様にして行った。
Bfu-GDHおよび他の真菌由来GDHへの分子内S-S結合の導入
実施例7の結果に基づいて、配列番号2で示されるボトリオチニア・フケリアナ由来FAD-GDHの150番目のアミノ酸残基Aおよび192番目のアミノ酸残基Tが両方ともCで置き換えられている改変型酵素(A150C/T192C)を作製したところ、高い熱安定性を示すことが見いだされた。さらに、かかる変異と、実施例6で高い生産性を示したいくつかの変異とを組み合わせた多重変異体を作製し、これらの変異体についても同様にして酵素活性および熱安定性を評価した。

酵素センサーの作製および評価
アスペルギルス・オリゼTI株由来V149C/G190C変異酵素およびボトリオチニア・フケリアナ由来S514G/S552C変異酵素を用いて酵素電極を作製した。5ユニットの本発明の改変型FAD-GDHにカーボンペースト20mgを加えて凍結乾燥させた。これをよく混合した後、既にカーボンペーストが約40mg充填されたカーボンペースト電極の表面だけに充填し、濾紙上で研磨した。この電極を1%のグルタルアルデヒドを含む10mM MOPS緩衝液(pH7.0)中で室温で30分間処理した後、20mMリジンを含む10mM MOPS緩衝液(pH7.0)中で室温で20分間処理してグルタルアルデヒドをブロッキングした。この電極を10mM MOPS緩衝液(pH7.0)中で室温で1時間以上平衡化させた。電極は4℃で保存した。
Claims (17)
- 配列番号2で表されるアミノ酸配列からなる蛋白質、または配列番号2で表されるアミノ酸配列において1またはそれ以上のアミノ酸残基が欠失、置換または挿入されており、かつグルコース脱水素酵素活性を有する蛋白質において、N176K,N176R、N176E、N176S、N225K、N225E、N259K,N301K、N326K、N326E、N330K、N330S、N355K、N355E、S514GまたはS552C、またはこれらの組み合わせのアミノ酸変異を有する蛋白質。
- さらに、G53A、E166R、T168P、N487S、S490P、N492T、A496E、D500E、V502LおよびA505Nからなる群より選択される1またはそれ以上の変異を有する、請求項1記載の蛋白質。
- 配列番号2で表されるアミノ酸配列からなる蛋白質において、N176K/S490P/D500E/S514G/S552C、N176K/A496E/D500E/S514G/S552C、N176K/S514G/S552C、S514G/S552CおよびG53A/S514G/S552Cからなる群より選択されるアミノ酸変異を有する蛋白質。
- 配列番号2で表されるアミノ酸配列からなる蛋白質において、N176K/N301K/N330K/S514G/S552C、N176R/N301K/N330K/S514G/S552C、N176R/N225E/N301K/N326E/N330K/N355E/S514G/S552C、N176K/S490P/D500E/S514G/S552C、およびE166R/T168P/N176R/N301K/N330K/S490P/D500E/S514G/S552Cからなる群より選択されるアミノ酸変異を有する蛋白質。
- 配列番号4で表されるアミノ酸配列からなる蛋白質、または配列番号4で表されるアミノ酸配列において1またはそれ以上のアミノ酸残基が欠失、置換または挿入されており、かつグルコース脱水素酵素活性を有する蛋白質において、V149CおよびG190Cのアミノ酸変異を有する蛋白質。
- 真菌由来のグルコース脱水素酵素であって、そのアミノ酸配列を配列番号4で表されるアミノ酸配列とアラインメントしたときに配列番号4で表されるアミノ酸配列のV149およびG190に相当する位置のアミノ酸が両方ともシステインで置き換えられていることを特徴とするグルコース脱水素酵素。
- 真菌由来のグルコース脱水素酵素であって、配列番号2および5-9からなる群より選択されるアミノ酸配列を有し、配列番号4で表されるアミノ酸配列のV149およびG190に相当する位置のアミノ酸が両方ともシステインで置き換えられていることを特徴とするグルコース脱水素酵素。
- 配列番号2で表されるアミノ酸配列からなる蛋白質、または配列番号2で表されるアミノ酸配列において1またはそれ以上のアミノ酸残基が欠失、置換または挿入されており、かつグルコース脱水素酵素活性を有する蛋白質において、A150C/T192Cのアミノ酸変異を有する蛋白質。
- 請求項1-8のいずれかに記載の蛋白質をコードする遺伝子。
- 請求項9に記載の遺伝子を含有する組み換えベクター。
- 請求項9に記載の組み換えベクターで形質転換した形質転換体または形質導入体。
- 請求項11に記載の形質転換体を培養して、該培養物からグルコース脱水素酵素を採取することを特徴とするグルコース脱水素酵素の製造方法。
- 請求項1-8のいずれかに記載の蛋白質を用いて試料中のグルコース濃度を測定することを特徴とするグルコース分析方法。
- 請求項1-8のいずれかに記載の蛋白質を含むことを特徴とするグルコースのアッセイキット。
- 請求項1-8のいずれかに記載の蛋白質が電極表面に固定化されている酵素電極。
- 作用極として請求項15に記載の酵素電極を用いることを特徴とするグルコースセンサー。
- 配列番号2で表されるアミノ酸配列からなる蛋白質、または配列番号2で表されるアミノ酸配列において1またはそれ以上のアミノ酸残基が欠失、置換または挿入されており、かつグルコース酸化酵素活性を示さずかつグルコース脱水素酵素活性を有し、キシロースに対する反応性がグルコースに対する反応性の20%以下であることを特徴とするグルコース脱水素酵素を用いる、溶存酸素の影響を受けないグルコース計測用のバイオセンサー。
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP11800446.4A EP2589655A4 (en) | 2010-06-29 | 2011-06-29 | GLUCOSE DEHYDROGENASE |
| JP2012522470A JP5873796B2 (ja) | 2010-06-29 | 2011-06-29 | グルコース脱水素酵素 |
| US13/807,637 US8999691B2 (en) | 2010-06-29 | 2011-06-29 | Glucose dehydrogenase |
| KR1020137002132A KR20130038914A (ko) | 2010-06-29 | 2011-06-29 | 글루코스 탈수소효소 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2010-147799 | 2010-06-29 | ||
| JP2010147799 | 2010-06-29 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2012001976A1 true WO2012001976A1 (ja) | 2012-01-05 |
Family
ID=45401715
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2011/003741 WO2012001976A1 (ja) | 2010-06-29 | 2011-06-29 | グルコース脱水素酵素 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US8999691B2 (ja) |
| EP (1) | EP2589655A4 (ja) |
| JP (1) | JP5873796B2 (ja) |
| KR (1) | KR20130038914A (ja) |
| WO (1) | WO2012001976A1 (ja) |
Cited By (20)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2013031664A1 (ja) * | 2011-08-26 | 2013-03-07 | 池田食研株式会社 | フラビン結合型グルコースデヒドロゲナーゼ |
| WO2013051704A1 (ja) * | 2011-10-05 | 2013-04-11 | 池田食研株式会社 | グルコース脱水素酵素 |
| WO2013118799A1 (ja) * | 2012-02-09 | 2013-08-15 | 東洋紡株式会社 | 新規なグルコース脱水素酵素 |
| WO2013118798A1 (ja) * | 2012-02-09 | 2013-08-15 | 東洋紡株式会社 | 新規なグルコース脱水素酵素 |
| JP2015084676A (ja) * | 2013-10-29 | 2015-05-07 | 東洋紡株式会社 | 耐熱性に優れたフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ |
| WO2015099112A1 (ja) * | 2013-12-27 | 2015-07-02 | キッコーマン株式会社 | 熱安定性が向上したフラビン結合型グルコースデヒドロゲナーゼ |
| JP2015146773A (ja) * | 2014-02-06 | 2015-08-20 | 東洋紡株式会社 | 新規なグルコースデヒドロゲナーゼ |
| JP2016007192A (ja) * | 2014-06-26 | 2016-01-18 | 株式会社村田製作所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有するタンパク質 |
| JP2016007191A (ja) * | 2014-06-26 | 2016-01-18 | 株式会社村田製作所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有するタンパク質 |
| EP2799545A4 (en) * | 2011-12-28 | 2016-01-20 | Ultizyme Int Ltd | GLUCOSE DEHYDROGENASE |
| US9260699B2 (en) | 2011-10-31 | 2016-02-16 | Toyobo Co., Ltd. | Glucose dehydrogenase |
| JP2016075553A (ja) * | 2014-10-06 | 2016-05-12 | 東洋紡株式会社 | グルコースセンサによるグルコース測定方法 |
| US9469844B2 (en) | 2010-12-02 | 2016-10-18 | Kikkoman Corporation | Flavin-bound glucose dehydrogenases, a method for producing a flavin-bound glucose dehydrogenase, and yeast transformant used for the same |
| JP2016208916A (ja) * | 2015-05-08 | 2016-12-15 | 国立研究開発法人産業技術総合研究所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有する変異型タンパク質 |
| JP2016208915A (ja) * | 2015-05-08 | 2016-12-15 | 国立研究開発法人産業技術総合研究所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有する変異型タンパク質 |
| JP2018046816A (ja) * | 2017-09-15 | 2018-03-29 | 東洋紡株式会社 | 耐熱性に優れたフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ |
| WO2018230304A1 (ja) * | 2017-06-14 | 2018-12-20 | 池田食研株式会社 | 改変型グルコース脱水素酵素 |
| WO2019172400A1 (ja) | 2018-03-08 | 2019-09-12 | 有限会社アルティザイム・インターナショナル | フラビンアデニンジヌクレオチドグルコース脱水素酵素とシトクロム分子との融合タンパク質 |
| US10913971B2 (en) | 2015-04-09 | 2021-02-09 | Toyobo Co., Ltd. | Enzyme preparation for use in measurement of glucose |
| US11072809B2 (en) | 2015-01-16 | 2021-07-27 | Toyobo Co., Ltd. | FAD-dependent glucose dehydrogenase |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP6856344B2 (ja) | 2015-10-29 | 2021-04-07 | アークレイ株式会社 | 変異グルコース脱水素酵素およびその利用 |
| US11066690B2 (en) | 2016-05-09 | 2021-07-20 | Kikkoman Corporation | Flavin-binding glucose dehydrogenase variant |
| CN111593030B (zh) * | 2020-04-17 | 2022-11-08 | 华东理工大学 | 化学耐受性的葡萄糖脱氢酶突变体及其在辅酶再生中的应用 |
| CN114107355B (zh) * | 2021-12-03 | 2024-04-23 | 浙江清肽生物科技有限公司 | 一种高效表达葡萄糖脱氢酶的工程菌的发酵方法及其应用 |
| CN114958789B (zh) * | 2022-05-23 | 2023-08-29 | 可孚医疗科技股份有限公司 | 一种葡萄糖脱氢酶突变体、其制备方法及其应用 |
Citations (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2004058958A1 (ja) | 2002-12-24 | 2004-07-15 | Ikeda Food Research Co., Ltd. | 補酵素結合型グルコース脱水素酵素 |
| WO2006101239A1 (ja) | 2005-03-25 | 2006-09-28 | Ikeda Food Research Co., Ltd. | 補酵素結合型グルコース脱水素酵素及びこれをコードするポリヌクレオチド |
| WO2007011610A2 (en) | 2005-07-18 | 2007-01-25 | Acologix, Inc. | Protein formulation for promoting hard tissue formation |
| JP2007289148A (ja) | 2006-03-31 | 2007-11-08 | Toyobo Co Ltd | アスペルギルス・オリゼ由来グルコースデヒドロゲナーゼの製造方法 |
| WO2007139013A1 (ja) | 2006-05-29 | 2007-12-06 | Amano Enzyme Inc. | フラビンアデニンジヌクレオチド結合型グルコース脱水素酵素 |
| JP2008035748A (ja) | 2006-08-03 | 2008-02-21 | Toyobo Co Ltd | アスペルギルス属菌由来新規グルコースデヒドロゲナーゼ |
| JP2008035747A (ja) | 2006-08-03 | 2008-02-21 | Toyobo Co Ltd | ペニシリウム属菌由来新規グルコースデヒドロゲナーゼ |
| JP2008154574A (ja) * | 2006-03-31 | 2008-07-10 | Toyobo Co Ltd | 糸状菌由来グルコースデヒドロゲナーゼを組換え体で高発現するための方法 |
| JP2008178380A (ja) | 2006-03-31 | 2008-08-07 | Toyobo Co Ltd | アスペルギルス・オリゼ由来グルコースデヒドロゲナーゼの製造方法 |
| JP2008237210A (ja) * | 2006-11-14 | 2008-10-09 | Toyobo Co Ltd | 改変型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ |
| JP2009273381A (ja) * | 2008-05-13 | 2009-11-26 | Toyobo Co Ltd | 熱安定性が向上したフラビンアデニン依存性グルコースデヒドロゲナーゼ改変体 |
| WO2011068050A1 (ja) * | 2009-12-05 | 2011-06-09 | 天野エンザイム株式会社 | 変異酵素及びその用途 |
| JP2011115156A (ja) * | 2009-11-06 | 2011-06-16 | Toyobo Co Ltd | フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの温度依存性を改善する方法 |
Family Cites Families (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ES2182986T5 (es) * | 1995-06-07 | 2009-03-01 | Danisco A/S | Hexosa oxidasa recombinante, procedimiento para su produccion y utilizacion de dicha enzima. |
| US7553649B2 (en) * | 2006-03-31 | 2009-06-30 | Toyo Boseki Kabushiki Kaisha | Method for producing glucose dehydrogenase from Aspergillus oryzae |
| US7494794B2 (en) * | 2006-03-31 | 2009-02-24 | Toyo Boseki Kabushiki Kaisha | Glucose dehydrogenase |
| US20080003628A1 (en) | 2006-03-31 | 2008-01-03 | Toyo Boseki Kabushiki Kaisha | Method for enhancing stability of a composition comprising soluble glucose dehydrogenase (gdh) |
| EP2083074B1 (en) | 2006-11-14 | 2011-01-19 | Toyo Boseki Kabushiki Kaisha | Modified flavine-adenine-dinucleotide-dependent glucose dehydrogenase |
| EP2223996B1 (en) * | 2007-11-28 | 2013-08-14 | Amano Enzyme Inc. | Transformant transfected with flavin adenine dinucleotide-binding glucose dehydrogenase gene and method for producing flavin adenine dinucleotide-binding glucose dehydrogenase using the same |
| EP2241621B1 (en) * | 2007-12-28 | 2012-08-15 | Ikeda Food Research Co. Ltd. | Modified glucose dehydrogenase gene |
-
2011
- 2011-06-29 EP EP11800446.4A patent/EP2589655A4/en not_active Withdrawn
- 2011-06-29 US US13/807,637 patent/US8999691B2/en active Active
- 2011-06-29 JP JP2012522470A patent/JP5873796B2/ja active Active
- 2011-06-29 KR KR1020137002132A patent/KR20130038914A/ko not_active Withdrawn
- 2011-06-29 WO PCT/JP2011/003741 patent/WO2012001976A1/ja active Application Filing
Patent Citations (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2004058958A1 (ja) | 2002-12-24 | 2004-07-15 | Ikeda Food Research Co., Ltd. | 補酵素結合型グルコース脱水素酵素 |
| WO2006101239A1 (ja) | 2005-03-25 | 2006-09-28 | Ikeda Food Research Co., Ltd. | 補酵素結合型グルコース脱水素酵素及びこれをコードするポリヌクレオチド |
| WO2007011610A2 (en) | 2005-07-18 | 2007-01-25 | Acologix, Inc. | Protein formulation for promoting hard tissue formation |
| JP2008154574A (ja) * | 2006-03-31 | 2008-07-10 | Toyobo Co Ltd | 糸状菌由来グルコースデヒドロゲナーゼを組換え体で高発現するための方法 |
| JP2007289148A (ja) | 2006-03-31 | 2007-11-08 | Toyobo Co Ltd | アスペルギルス・オリゼ由来グルコースデヒドロゲナーゼの製造方法 |
| JP2008178380A (ja) | 2006-03-31 | 2008-08-07 | Toyobo Co Ltd | アスペルギルス・オリゼ由来グルコースデヒドロゲナーゼの製造方法 |
| WO2007139013A1 (ja) | 2006-05-29 | 2007-12-06 | Amano Enzyme Inc. | フラビンアデニンジヌクレオチド結合型グルコース脱水素酵素 |
| JP2008035747A (ja) | 2006-08-03 | 2008-02-21 | Toyobo Co Ltd | ペニシリウム属菌由来新規グルコースデヒドロゲナーゼ |
| JP2008035748A (ja) | 2006-08-03 | 2008-02-21 | Toyobo Co Ltd | アスペルギルス属菌由来新規グルコースデヒドロゲナーゼ |
| JP2008237210A (ja) * | 2006-11-14 | 2008-10-09 | Toyobo Co Ltd | 改変型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ |
| JP2009273381A (ja) * | 2008-05-13 | 2009-11-26 | Toyobo Co Ltd | 熱安定性が向上したフラビンアデニン依存性グルコースデヒドロゲナーゼ改変体 |
| JP2011115156A (ja) * | 2009-11-06 | 2011-06-16 | Toyobo Co Ltd | フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼの温度依存性を改善する方法 |
| WO2011068050A1 (ja) * | 2009-12-05 | 2011-06-09 | 天野エンザイム株式会社 | 変異酵素及びその用途 |
Non-Patent Citations (8)
| Title |
|---|
| BIOCHIM. BIOPHYS. ACTA, vol. 139, no. 2, 1967, pages 265 - 276 |
| DATABASE DATABASE UNIPROT [online] 31 October 2006 (2006-10-31), ROLKE Y. ET AL.: "Definition: Glucose oxidase", XP003031796, Database accession no. Q70Q36 * |
| F. WILLIAM STUDIER ET AL.: "ZYP broth", PROTEIN EXPRESSION AND PURIFICATION, 2005 |
| F. WILLIAM; STUDIER ET AL., PROTEIN EXPRESSION AND PURIFICATION, 2005 |
| HENRIK NIELSEN; JACOB ENGELBRECHT; SOREN BRUNAK; GUNNAR VON HEIJNE: "Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites", PROTEIN ENGINEERING, vol. 10, 1997, pages 1 - 6 |
| LU, G.; MORIYAMA, E.N.: "Vector NTI, a balanced all-in-one sequence analysis suite", BRIEF BIOINFORM., vol. 5, 2004, pages 378 - 88 |
| ROLKE ET AL., MOL. PLANT PATHOL., vol. 5, no. 1, 2004, pages 17 - 27 |
| See also references of EP2589655A4 * |
Cited By (29)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US9469844B2 (en) | 2010-12-02 | 2016-10-18 | Kikkoman Corporation | Flavin-bound glucose dehydrogenases, a method for producing a flavin-bound glucose dehydrogenase, and yeast transformant used for the same |
| US8945359B2 (en) | 2011-08-26 | 2015-02-03 | Ikeda Food Research Co., Ltd. | Flavin-binding glucose dehydrogenase |
| WO2013031664A1 (ja) * | 2011-08-26 | 2013-03-07 | 池田食研株式会社 | フラビン結合型グルコースデヒドロゲナーゼ |
| WO2013051704A1 (ja) * | 2011-10-05 | 2013-04-11 | 池田食研株式会社 | グルコース脱水素酵素 |
| JP2013090620A (ja) * | 2011-10-05 | 2013-05-16 | Ikeda Shokken Kk | グルコース脱水素酵素 |
| US9260699B2 (en) | 2011-10-31 | 2016-02-16 | Toyobo Co., Ltd. | Glucose dehydrogenase |
| US9512459B2 (en) | 2011-12-28 | 2016-12-06 | Ultizyme International Ltd. | Glucose dehydrogenase |
| EP2799545A4 (en) * | 2011-12-28 | 2016-01-20 | Ultizyme Int Ltd | GLUCOSE DEHYDROGENASE |
| WO2013118798A1 (ja) * | 2012-02-09 | 2013-08-15 | 東洋紡株式会社 | 新規なグルコース脱水素酵素 |
| JP2013176363A (ja) * | 2012-02-09 | 2013-09-09 | Toyobo Co Ltd | 新規なグルコース脱水素酵素 |
| JP2013176364A (ja) * | 2012-02-09 | 2013-09-09 | Toyobo Co Ltd | 新規なグルコース脱水素酵素 |
| WO2013118799A1 (ja) * | 2012-02-09 | 2013-08-15 | 東洋紡株式会社 | 新規なグルコース脱水素酵素 |
| US9404144B2 (en) | 2012-02-09 | 2016-08-02 | Toyobo Co., Ltd. | Glucose dehydrogenase |
| JP2015084676A (ja) * | 2013-10-29 | 2015-05-07 | 東洋紡株式会社 | 耐熱性に優れたフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ |
| JPWO2015099112A1 (ja) * | 2013-12-27 | 2017-03-23 | キッコーマン株式会社 | 熱安定性が向上したフラビン結合型グルコースデヒドロゲナーゼ |
| WO2015099112A1 (ja) * | 2013-12-27 | 2015-07-02 | キッコーマン株式会社 | 熱安定性が向上したフラビン結合型グルコースデヒドロゲナーゼ |
| JP2015146773A (ja) * | 2014-02-06 | 2015-08-20 | 東洋紡株式会社 | 新規なグルコースデヒドロゲナーゼ |
| JP2016007192A (ja) * | 2014-06-26 | 2016-01-18 | 株式会社村田製作所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有するタンパク質 |
| JP2016007191A (ja) * | 2014-06-26 | 2016-01-18 | 株式会社村田製作所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有するタンパク質 |
| JP2016075553A (ja) * | 2014-10-06 | 2016-05-12 | 東洋紡株式会社 | グルコースセンサによるグルコース測定方法 |
| US11072809B2 (en) | 2015-01-16 | 2021-07-27 | Toyobo Co., Ltd. | FAD-dependent glucose dehydrogenase |
| US10913971B2 (en) | 2015-04-09 | 2021-02-09 | Toyobo Co., Ltd. | Enzyme preparation for use in measurement of glucose |
| JP2016208916A (ja) * | 2015-05-08 | 2016-12-15 | 国立研究開発法人産業技術総合研究所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有する変異型タンパク質 |
| JP2016208915A (ja) * | 2015-05-08 | 2016-12-15 | 国立研究開発法人産業技術総合研究所 | フラビンアデニンジヌクレオチド依存型グルコース脱水素酵素活性を有する変異型タンパク質 |
| WO2018230304A1 (ja) * | 2017-06-14 | 2018-12-20 | 池田食研株式会社 | 改変型グルコース脱水素酵素 |
| JP2019000020A (ja) * | 2017-06-14 | 2019-01-10 | 池田食研株式会社 | 改変型グルコース脱水素酵素 |
| US11725193B2 (en) | 2017-06-14 | 2023-08-15 | Ikeda Food Research Co., Ltd. | Modified glucose dehydrogenase |
| JP2018046816A (ja) * | 2017-09-15 | 2018-03-29 | 東洋紡株式会社 | 耐熱性に優れたフラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ |
| WO2019172400A1 (ja) | 2018-03-08 | 2019-09-12 | 有限会社アルティザイム・インターナショナル | フラビンアデニンジヌクレオチドグルコース脱水素酵素とシトクロム分子との融合タンパク質 |
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2012001976A1 (ja) | 2013-08-22 |
| EP2589655A4 (en) | 2013-12-04 |
| EP2589655A1 (en) | 2013-05-08 |
| KR20130038914A (ko) | 2013-04-18 |
| US8999691B2 (en) | 2015-04-07 |
| US20130168263A1 (en) | 2013-07-04 |
| JP5873796B2 (ja) | 2016-03-01 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5873796B2 (ja) | グルコース脱水素酵素 | |
| JP6911066B2 (ja) | フラビン結合型グルコースデヒドロゲナーゼ、フラビン結合型グルコースデヒドロゲナーゼの製造方法、およびそれを用いたグルコース測定方法 | |
| JP4348563B2 (ja) | 改変型フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ | |
| US7662600B2 (en) | Modified flavin adenine dinucleotide dependent glucose dehydrogenase | |
| JP6184326B2 (ja) | 基質特異性が向上したフラビン結合型グルコースデヒドロゲナーゼ | |
| JP6212852B2 (ja) | 新規なグルコース脱水素酵素 | |
| JP6079038B2 (ja) | 新規なグルコース脱水素酵素 | |
| JP5408125B2 (ja) | 糸状菌由来フラビンアデニンジヌクレオチド依存性グルコースデヒドロゲナーゼ(fadgdh) | |
| JP6155197B2 (ja) | グルコース脱水素酵素 | |
| JPWO2003106668A1 (ja) | グルコース脱水素酵素 | |
| JP6934720B2 (ja) | 基質特異性が向上したフラビン結合型グルコースデヒドロゲナーゼ | |
| JPWO2004005499A1 (ja) | グルコース脱水素酵素 | |
| JP2005270082A (ja) | グルコース脱水素酵素 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 11800446 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2012522470 Country of ref document: JP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 20137002132 Country of ref document: KR Kind code of ref document: A |
|
| REEP | Request for entry into the european phase |
Ref document number: 2011800446 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2011800446 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 13807637 Country of ref document: US |