RU2441663C2 - Препарат aphanizomenon flos aquae, экстракты и очищенные компоненты экстрактов для лечения неврологических, нейродегенеративных и аффективных расстройств - Google Patents
Препарат aphanizomenon flos aquae, экстракты и очищенные компоненты экстрактов для лечения неврологических, нейродегенеративных и аффективных расстройств Download PDFInfo
- Publication number
- RU2441663C2 RU2441663C2 RU2008151175/15A RU2008151175A RU2441663C2 RU 2441663 C2 RU2441663 C2 RU 2441663C2 RU 2008151175/15 A RU2008151175/15 A RU 2008151175/15A RU 2008151175 A RU2008151175 A RU 2008151175A RU 2441663 C2 RU2441663 C2 RU 2441663C2
- Authority
- RU
- Russia
- Prior art keywords
- afa
- extract
- mao
- phytochrome
- algae
- Prior art date
Links
- 0 CCC(C)CCCCCCCCCC=C(C)CCC*1*C*C*1 Chemical compound CCC(C)CCCCCCCCCC=C(C)CCC*1*C*C*1 0.000 description 1
Images














Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K36/00—Medicinal preparations of undetermined constitution containing material from algae, lichens, fungi or plants, or derivatives thereof, e.g. traditional herbal medicines
- A61K36/02—Algae
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/33—Heterocyclic compounds
- A61K31/395—Heterocyclic compounds having nitrogen as a ring hetero atom, e.g. guanethidine or rifamycins
- A61K31/40—Heterocyclic compounds having nitrogen as a ring hetero atom, e.g. guanethidine or rifamycins having five-membered rings with one nitrogen as the only ring hetero atom, e.g. sulpiride, succinimide, tolmetin, buflomedil
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/14—Drugs for disorders of the nervous system for treating abnormal movements, e.g. chorea, dyskinesia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/14—Drugs for disorders of the nervous system for treating abnormal movements, e.g. chorea, dyskinesia
- A61P25/16—Anti-Parkinson drugs
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/22—Anxiolytics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/24—Antidepressants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/28—Drugs for disorders of the nervous system for treating neurodegenerative disorders of the central nervous system, e.g. nootropic agents, cognition enhancers, drugs for treating Alzheimer's disease or other forms of dementia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Medicinal Chemistry (AREA)
- Veterinary Medicine (AREA)
- Chemical & Material Sciences (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- General Health & Medical Sciences (AREA)
- Public Health (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Neurosurgery (AREA)
- Neurology (AREA)
- Biomedical Technology (AREA)
- Organic Chemistry (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Biotechnology (AREA)
- Natural Medicines & Medicinal Plants (AREA)
- Epidemiology (AREA)
- Microbiology (AREA)
- Alternative & Traditional Medicine (AREA)
- Botany (AREA)
- Medical Informatics (AREA)
- Mycology (AREA)
- Psychiatry (AREA)
- Psychology (AREA)
- Pain & Pain Management (AREA)
- Hospice & Palliative Care (AREA)
- Medicines Containing Plant Substances (AREA)
- Acyclic And Carbocyclic Compounds In Medicinal Compositions (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
Abstract
Изобретение относится к фармацевтической промышленности, в частности к экстракту для производства композиции для профилактики, борьбы или лечения неврологического заболевания, состояния, дисфункции или расстройства. Применение экстракта микроводоросли Aphanizomenon Flos Aquae Aquae Ralfs ex Born. & Flah. Var. flos aquae (Кламатской водоросли AFA) для производства композиции для профилактики, борьбы или лечения неврологического заболевания, состояния, дисфункции или расстройства, выбранного из: болезни Альцгеймера, болезни Паркинсона, рассеянного склероза, синдрома дефицита внимания с гиперактивностью (СДВГ), аутизма, депрессии, дефицита памяти и аффективных расстройств, где экстракт получают посредством определенных стадий. Вышеописанный экстракт эффективен для профилактики, борьбы или лечения неврологического заболевания, состояния, дисфункции или расстройства. 3 з.п. ф-лы, 24 ил., 3 табл.
Description
Настоящее изобретение относится к микроводоросли Aphanizomenon Flos Aquae Aquae Ralfs ex Born. & Flah. Var. flos aquae (Кламатская водоросль AFA). Более конкретно, изобретение обеспечивает экстракты Кламатской водоросли AFA и очищенные компоненты указанных экстрактов, которые могут применяться для профилактики или лечения неврологических, нейродегенеративных и аффективных расстройств или заболеваний.
УРОВЕНЬ ТЕХНИКИ
Фенилэтиламин (ФЭА) представляет собой эндогенный амин, синтезируемый при декарбоксилировании фенилаланина в допаминергических нейронах нигростриарной системы, который может также функционировать как нейромодулятор катехоламин-опосредованной нейротрансмиссии в мозге (1). Наиболее важной функцией ФЭА является способствование нейротрансмиссии катехоламинов. Известно, что ФЭА стимулирует высвобождение ацетилхолина, а также допамина (2). Кроме того, ФЭА усиливает нейротрансмиссию норэпинефрина (НЭ) (6), а также нейротрансмиссию серотонина.
Недавно было продемонстрировано, что ФЭА может также функционировать как автономный нейромедиатор со своими специфичными нейронными рецепторами; и что ФЭА действует как истинный нейромодулятор со способностью подавлять нейротрансмиссию, если необходимо (8).
Это приводит к целому ряду эффектов: стимуляции внимания и памяти; улучшению настроения со значительной антидепрессивной активностью; стимуляции сочувствия и, таким образом, общительности, включая эмоциональное и половое поведение; подавлению чувства голода; снижению зависимости от сильнодействующих веществ и наркотической зависимости.
Связь между ФЭА и эмоциональным состоянием была подтверждена исследованиями, в соответствии с которыми значительно более низкие уровни чистого ФЭА или его метаболита ФУК (фенилуксусной кислоты) в плазме или моче были зарегистрированы у лиц с депрессивным состоянием (9).
Было отмечено, что лица, страдающие болезнью Паркинсона, имеют значительно более низкие уровни ФЭА при измерении непосредственно в плазме (12). Прогрессирующее снижение нейротрансмиссии, особенно допаминергической, у указанных пациентов связано с прогрессирующей дегенерацией допаминергических нейронов черного вещества.
Указанное снижение уровней ФЭА происходит с параллельным повышением уровней MAO-B у лиц, страдающих болезнью Паркинсона, поэтому лекарственные средства, применяемые при лечении паркинсонизма, являются ингибиторами MAO-B, такими как селегилин (14). Кроме того, ФЭА при приеме может легко проходить через гематоэнцефалический барьер и стимулировать высвобождение допамина из нигростриарной ткани даже в низких дозах. Это является важным отличительным свойством, поскольку используемое в настоящее время лекарственное средство, селегилин, ингибируя MAO-B и обратный захват допамина, не оказывает никакого действия на его высвобождение из нигростриарной ткани и таким образом не способствует повышению выработки допамина, что является серьезным ограничением в такой патологии, как болезнь Паркинсона, при которой в значительной степени нарушается именно выработка допамина.
Болезнь Альцгеймера приводит к нарушению механизма выработки и обратного захвата допамина, а также к прогрессирующему разрушению нейронов стриарной области, которое со временем приводит к снижению числа допаминергических нейронов и, следовательно, ухудшению передачи допамина (15).
Хотя каких-либо четких данных относительно того факта, что СДВГ (синдром дефицита внимания с гиперактивностью) является нейродегенеративной патологией, не существует, некоторые исследования проводились в попытке доказать, что разрушение нейронов является основной причиной СДВГ как у детей, так и у взрослых (19).
Наиболее важными являются свидетельства, согласно которым дети, пораженные СДВГ и пониженной обучаемостью, имеют значительно более низкие уровни ФЭА (21) и, как следствие, страдают снижением нейромодуляции внимания (допамин) и успокоения (серотонин). Именно поэтому препаратом выбора при СДВГ является метилфенидат, синтетическое производное ФЭА, который также стимулирует повышенную выработку ФЭА (22) и таким образом допамина и норэпинефрина, двух нейромедиаторов, непосредственно вовлеченных в этиологию СДВГ.
Хорошо известно применение амфетаминов для контроля чувства голода и, следовательно, веса. Их применение в данной области постоянно находилось под вопросом из-за побочных эффектов, которые, при наличии их переносимости, имеют тенденцию потенциально становиться крайне тяжелыми с течением времени. Это подтверждается тем фактом, что основные лекарственные средства, используемые в настоящее время для контроля чувства голода и веса, представляют собой амфетамин-подобные допаминергические антидепрессанты, такие как венлафаксин и бупроприон. Указанные вещества, как и все амфетамины, являются синтетическими производными ФЭА. Последние действуют как мощные средства подавления аппетита, поскольку предотвращено их расщепление ферментами MAO-B.
Моноаминоксидаза (MAO) A и В катализируют расщепление нейроактивных и вазоактивных аминов в ЦНС и в периферических тканях. MAO-B, в частности, с учетом ее прямого и косвенного участия в допаминергической передаче вовлечена в неврологические расстройства, в которых важную роль играет допамин, например депрессивные и аффективные расстройства, болезни Паркинсона и Альцгеймера. По этой причине ингибиторы MAO-B используются при лечении таких неврологических расстройств (26).
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на выявлении в микроводоросли Aphanizomenon Flos Aquae Aquae Ralfs ex Born. & Flah. Var. flos aquae (Кламатской водоросли AFA) веществ, которые в чистом виде или в комбинации оказывают благоприятное воздействие при различных неврологических заболеваниях, состояниях, дисфункциях или расстройствах, включая нейродегенеративные заболевания, такие как болезни Альцгеймера и Паркинсона, рассеянный склероз, гиперактивность и синдром дефицита внимания (СДВГ), аутизм, депрессию, ухудшение памяти и аффективные расстройства. В частности, было установлено, что Кламатская микроводоросль AFA, помимо фенилэтиламина, который является нейромодулятором, обладающим допаминергической и норадренергической активностью, содержит особые вещества, которые, как совершенно неожиданно оказалось, являются очень эффективными ингибиторами моноаминоксидазы В (MAO-B), а именно: a) особый AFA-фитохром; b) AFA-фикобилипротеиновый комплекс, содержащий фикобилисому, образованную C-фикоцианином (C-PC) и фикоэритроцианином (PEC, включая соответствующий хромофор фиковиолобилин или PVB) ("AFA-фикоцианины"); c) микоспорин-подобные аминокислоты или МАК. Данное открытие является очень важным, так как ФЭА, содержащийся в морских водорослях, без защиты ингибиторов MAO-B очень быстро разрушался бы ферментом MAO-B при приеме.
Те же соединения, которые действуют как избирательные ингибиторы MAO-B, также исполняют роль мощных нейропротекторов, значительно повышая, таким образом, способность экстракта содействовать неврологическому здоровью.
Таким образом, изобретение обеспечивает способ профилактики, борьбы или лечения вышеуказанных неврологических заболеваний, состояний, дисфункций или расстройств посредством введения нуждающемуся в этом субъекту препарата Кламатской водоросли AFA, в частности экстракта, обогащенного такими активными компонентами, или выделенного и очищенного компонента, выбранного из: a) AFA фитохрома, b) комплекса c-фикоцианина/фикоэритроцианинов, присутствующего в AFA или в любых других микроводорослях; c) микоспорин-подобных аминокислот, порфиры и шинорина, присутствующих в AFA или из любого другого источника, относящегося к водорослям; d) или их смеси.
Предпочтительно экстракт Кламатской водоросли AFA согласно изобретению получают посредством следующих стадий:
a) замораживание свежесобранной морской водоросли AFA и ее размораживание, или, если исходным материалом является порошок высушенной AFA, обработка ультразвуком разбавленного в воде порошка AFA для разрушения клеток;
b) центрифугирование продукта стадии a) для отделения супернатанта (содержащего в основном цитоплазматическую фракцию) от осадка (содержащего в основном фракцию обломков клеточных стенок);
c) сбор супернатанта, содержащего растворимые в воде компоненты.
Конечный продукт представляет собой экстракт (обозначенный как "Основной Экстракт"), обогащенный ФЭА, а также другими синергическими веществами, такими как AFA-фитохром, AFA-фикоцианины и МАК. Например, тогда как обычное содержание ФЭА в Кламатской микроводоросли находится в диапазоне от 2 до 4 мг/г, в Основном Экстракте указанная концентрация находится на уровне 9 - 11 мг/г (по данным анализа ВЭЖХ).
Можно дополнительно очистить экстракт, пропуская его через систему ультрафильтрации, предпочтительно через мембрану с предельной проницаемостью по молекулярной массе 30000 дальтон. Ультрафильтрационный концентрат (Экстракт A) содержит в качестве основных активных компонентов как AFA-фикоцианины (мол. масса = 121000) и AFA-фитохром (мол. масса 480000). Интересно, что хотя МАК имеют молекулярную массу значительно ниже предельной проницаемости используемой мембраны, в концентрате также возрастает концентрация МАК.
Основной Экстракт, полученный на стадиях a) - c), то есть без ультрафильтрации, в целом предпочтителен, поскольку содержит наиболее подходящие количества ФЭА, AFA-фитохрома, AFA-PC и МАК. Кроме того, указанный Основной Экстракт также включает вещества, такие как хлорофилл и каротины, хотя и в уменьшенной концентрации, что придает экстракту антиоксидантные и противовоспалительные свойства.
В альтернативном варианте активные компоненты Кламатской водоросли AFA, а именно комплекс C-фикоцианина/фикоэритроцианинов (C-PC/PEC), AFA-фитохром и МАК, могут быть выделены и очищены, как дополнительно описано ниже, и применены в способе согласно изобретению.
В предпочтительном варианте осуществления C-PC/PEC комплекс Кламатской водоросли AFA, AFA-фитохром и микоспорин-подобные аминокислоты используются как комбинированный препарат для одновременного или раздельного введения субъекту, испытывающему в этом потребность; в еще одном предпочтительном варианте осуществления такой комбинированный препарат содержит фенилэтиламин в качестве дополнительного активного компонента. Из микоспорин-подобных аминокислот наиболее предпочтительными являются шинорин и порфира-334, поскольку они содержатся в относительно более высокой концентрации в Кламатской микроводоросли AFA.
Наблюдаемое ингибирование моноаминоксидазы-B особенно актуально, поскольку позволяет усилить допаминергическую передачу и минимизировать катаболизм ФЭА. В значительной степени и фитохром, и AFA-фикоцианин ингибируют MAO-B обратимым и смешанным способом, тогда как ингибирование MAO-B под воздействием МАК является конкурентным и обратимым; таким образом, все три молекулы обеспечивают высокую эффективность в физиологических условиях при отсутствии побочных эффектов.
В дополнительном аспекте настоящее изобретение направлено на нутрицевтическую или фармацевтическую композицию, содержащую препарат Кламатской водоросли AFA, экстракт или выделенный компонент экстракта, который предпочтительно выбран из комплекса C-PC/PEC, присутствующего в AFA, или из любого другого источника, относящегося к микроводорослям, или из выделенных отдельных компонентов C-PC и PEC; AFA-фитохрома; микоспорин-подобных аминокислот, порфиры и шинорина, присутствующих в водорослях AFA, или из любого другого источника, относящегося к водорослям; или их смесей; с необязательным добавлением фенилэтиламина. В предпочтительном варианте осуществления пищевые композиции представляют собой биологически активные добавки в форме таблеток, капсул, напитков; в другом предпочтительном варианте осуществления фармацевтические композиции находятся в форме таблеток, капсул, пакетиков, сиропов, суппозиториев, драже и мазей и могут использоваться для профилактики или лечения неврологических или нейродегенеративных заболеваний или состояний, указанных выше. Жидкие экстракты Кламатской водоросли AFA согласно изобретению могут использоваться в первоначальном виде или могут быть высушены посредством таких способов, как лиофилизация, сушка распылением и т.п. Выделенные активные компоненты могут быть введены в композицию с использованием способов и последующих операций, известных любому специалисту, квалифицированному в данной области техники.
Доза активного компонента будет зависеть от предполагаемого применения композиций либо как биологически активных добавок, либо как фармацевтического препарата. Эффективное количество каждого компонента обычно находится в следующих диапазонах: ФЭА = 0,1-100 мг, предпочтительно 5-30 мг; фитохром = 0,1-1000 мг, предпочтительно 0,8-10 мг; МАК = 0,1-1000 мг, предпочтительно 10-100; фикоцианины = 1-2500 мг, предпочтительно 50-1000 мг.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Идентификация "AFA-фитохрома", уникального фитохрома, типичного для Кламатских водорослей
Фитохромы являются фоторецепторами, пигментами, которые растения используют для обнаружения света и которые являются чувствительными к свету в красном и крайней красной области видимого спектра. В растениях они выполняют много различных функций, включая регулирование цветения (посредством циркадианных ритмов), образование почек и синтез хлорофилла. Последнее особенно актуально в отношении водорослей AFA, так как присутствие данного фитохрома уникального типа в AFA можно объяснить нехваткой другого фикобилипротеина, обычно используемого другими цианобактериями в дополнение к C-фикоцианину в процессе фотосинтеза, а именно алло-фикоцианина. Так как вместо алло-фикоцианина в Кламатских водорослях присутствует фикоэритроцианин или PEC (см. ниже), вероятно, что присутствия одного PEC не достаточно, особенно учитывая то, что Кламатские водоросли живут во внетропической среде, что требует высокой эффективности светоулавливания, и таким образом морские водоросли AFA, по-видимому, объединяют свои более высокие потребности с фитохромом.
AFA-фитохром, который имеет специфическую структуру, впервые описан в настоящей заявке. За прошедшие годы в растениях были обнаружены различные типы фитохромов, которые не только кодируются различными фитохромовыми генами (3 в рисе и 6 в кукурузе, например), но в большинстве случаев они имеют различные белковые субъединицы и структуру. Фитохромами их всех делает то, что все они используют один и тот же билипротеин, называемый фитохромобилин, в качестве светоулавливающего хромофора. Указанный хромофор подобен хромофору фикоцианина, фикоцианобилину и характеризуется одной молекулой билина, состоящей из линейной цепи из четырех пиррольных колец (тетрапирролов). Более конкретно, в своем нормальном состоянии Pr указанный билипротеин поглощает свет при максимуме 650-670 нм, тогда как при активации красным светом он переходит в Pfr с максимумом поглощения 730 нм.
Первым из фитохромов цианобактерий был открыт фитохром из Synechocystis, обладающий слабым структурным подобием с фитохромами растений. Однако билипротеин Synechocystis в целом считают фитохромом, поскольку он является обратимым хромопротеином в красной/крайней красной области (48).
Очистка фитохрома AFA и определение его свойств
AFA-фитохром в качестве хромофора несет билипротеин, который поглощает свет в красном/крайнем красном спектре. Для устанавления его структуры и функций фитохром очищали согласно следующей методике:
- 1 г экстракта суспендировали в 10 мл 1 K-фосфатного буфера, pH 7,0;
- дважды встряхивали в течение 1 минуты с половиной их объема;
- клетки инкубировали в течение 35' с 2% Тритона X100;
- центрифугировали при 28000 об/мин в течение 16-18 ч;
- супернатант собирали в ступенчатом градиенте плотности сахарозы;
- градиент центрифугировали, используя колебательные роторы при 150000 g в течение 12 ч;
- хранили на -20°C.
Фитохром соответствует полосе лизата с интенсивным оранжевым цветом, который виден приблизительно при 1M сахарозы, тогда как фикобилисома остается приблизительно при 0,75М. Данное соотношение указанных двух полос также дает достоверные данные молекулярной массы фитохрома, присутствующего в водорослях, которая приблизительно в 4 раза больше, чем молекулярная масса тримерного AFA-PC: так как последняя составляет 121 кДа, можно предварительно определить молекулярную массу AFA-фитохрома как приблизительно 480 кДа (Фигура 22).
При анализе светопоглощающих свойств фитохром поглощал свет с двумя пиками при 672 нм и 694 нм, которые отвечают соответственно формам Pr (поглощению красного света) и Pfr (поглощению крайнего красного света) в состоянии равновесия (Фигура 23).
Относительно количества фитохрома, содержащегося в AFA, первая оценка дала следующий предварительный результат: 2 мг/г (или 0,2% собственного веса). Относительно экстрактов, концентрация увеличивается приблизительно до 0,5% в Основном Экстракте, и приблизительно до 1% в Экстракте B. Это довольно низкие концентрации, однако антиоксидантные и противовоспалительные свойства данного соединения настолько сильны, что даже очень малое количество может производить весьма значительные эффекты.
Антиоксидантная активность
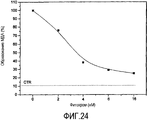
Очищенный AFA-фитохром, как было установлено, является очень мощным антиоксидантом. Более того, в абсолютных значениях самая мощная молекула на настоящий момент найдена в Кламатских водорослях. Инкубация в течение 2 ч образцов плазмы крови человека с окислителем CuCl2 при концентрации 100 мкМ приводит к образованию повышенных уровней малонового диальдегида (МДА), одного из конечных побочных продуктов перекисного окисления липидов, который определяют с помощью спектрофотометра при 535 нм после реакции с тиобарбитуровой кислотой (тест TBA). Когда плазму инкубируют в течение 2 часов при 37°C с 100 мкМ CuCl2 при повышении концентрации AFA-фитохрома (2-16 нМ), экстрагированного из водорослей AFA, наблюдается очень сильное дозозависимое снижение уровней MDA (Фигура 24). Более того, почти при полном ингибировании перекисного окисления липидов всего лишь при 16 нМ AFA-фитохрома уровни MDA были близки к контрольному значению. Что важно, значение IC50 3,6 нМ в 45 раз меньше, чем значение, полученное для PCB. Фитохром является основным соединением, ответственным за антиоксидантный и нейропротективный эффекты Основного Экстракта, которые сильнее, чем соответствующие эффекты AFA-PC.
Экстракт, очистка и количественное определение MAK
Было проанализировано присутствие МАК в цианофите Aphanizomenon flos-aquae из озера Кламат, общеизвестном как Кламатские водоросли. Насколько было известно, лишь в одной из последних статей сообщалось о присутствии МАК в некоторых видах Aphanizomenon (47); однако в указанном сообщении в качестве присутствующих МАК идентифицировали только порфиру, тогда как в настоящем исследовании показано присутствие двух МАК, и порфиры, и шинорина. С другой стороны, что касается всей литературы по водорослям, то тогда как большинство известных на сегодняшний день цианобактерий содержат шинорин в качестве основных МАК, авторы настоящего изобретения обнаружили необычное присутствие порфиры-334 в качестве основной МАК в Aphanizomenon flos-aquae в дополнение к шинорину.
| Порфира-334 |
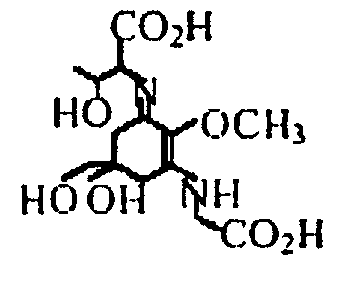 |
| Шинорин |
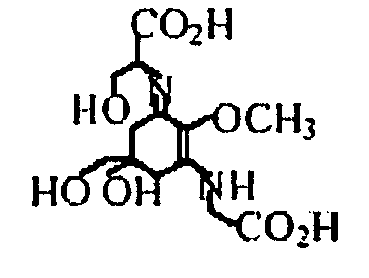 |
МАК экстрагировали, как описано ранее (29). Кратко, 20 мг порошка AFA или 20 мг водного экстракта экстрагировали 2 мл 20% (об./об.) водного раствора метанола (для ВЭЖХ) при инкубировании в водяной бане при 45°C в течение 2,5 ч. После центрифугирования (5000 g; Центрифуга GS-15R, Beckman, Пало-Альто, США) супернатант выпаривали досуха и повторно растворяли в 2 мл 100%-го метанола, встряхивали на вортексе в течение 2-3 минут и центрифугировали при 10000 g в течение 10 мин. Супернатант выпаривали, а экстракт повторно растворяли в таком же объеме 0,2% уксусной кислоты для анализа с помощью ВЭЖХ или в 200 мкл фосфатного буфера (PBS) для оценки антиоксидантных свойств. Пробы фильтровали через шприцевые фильтры с размером пор 0,2 мкм (VWR International, Милан, Италия) перед анализом с помощью ВЭЖХ или анализом антиоксидантных свойств (см. ниже).
МАК из Кламатских водорослей имеют максимум поглощения при 334 нм. Последующую очистку МАК выполняли, используя систему ВЭЖХ (Jasco Corporation, Токио, Япония), оборудованную колонкой Alltima C18 и защитной колонкой (внутр. сеч. 4,6 × 250 мм, набивка 5 мкм, Alltech, Милан, Италия), в соответствии с литературными данными (30). Длина волны для детектирования составляла 330 нм; подвижной фазой являлась 0,2% уксусная кислота при расходе 1,0 мл*мин-1. Идентификацию МАК проводили, сравнивая спектры поглощения и время удержания со стандартами, такими как Porphyra и Pterocladia sp., главным образом содержащими порфиру-334, шинорин и палитин, любезно предоставленные доктором Манфредом Клишем, университет им. Фридриха-Александра, Эрланген, Германия. Спектры поглощения проб измеряли при 200 - 800 нм на однолучевом спектрофотометре (DU 640, Beckman, Пало-Альто, США). Нерасшифрованные спектры перенесли в компьютер и обработали математически с целью анализов пиков МАК.
МАК частично очищали из образца AFA и из водного экстракта, как описано ранее. Экстракция образцов в 20%-ом метаноле при 45°C в течение 2,5 ч привела к заметному пику при 334 нм (МАК); даже если небольшие количества фотосинтетических пигментов (например, фикоцианина при 620 нм) были также экстрагированы в данной методике (см. Фигуру 1, пунктирная линия). Затем пробы МАА обрабатывали 100%-ым метанолом с целью удаления белков и солей, и в завершении 0,2% уксусной кислотой для удаления неполярных фотосинтетических пигментов. Полученные в результате частично очищенные МАК имели максимум поглощения при 334 нм (Фигура 1, сплошная линия).
Последующий анализ и очистку МАК выполняли с помощью ВЭЖХ в целях подтверждения, соответствуют ли соединения с поглощением при 334 нм одной МАК или смеси более чем одной МАК. Хроматограмма пробы (Фигура 2) показала присутствие двух МАК с временем удержания 4,2 (пик 1) и 7,6 минут (пик 2), которые были идентифицированы как шинорин и порфира-334 соответственно. Порфира-334, по-видимому, является основной МАК в AFA, так как шинорин присутствовал лишь в небольших количествах (соотношение площади пиков 1:15).
УФ-спектры очищенных МАК подтверждали соответствующий максимум поглощения при 334 нм (Фигура 3).
Принимая во внимание, что молярные коэффициенты экстинкции при 334 нм для шинорина и порфиры-334 составляют 44700 и 42300 M-1 см-1, соответственно, было вычислено:
a) для водорослей AFA, концентрация шинорина 0,49 мг г-1 собственного веса и порфиры-334 7,09 мг г-1 собственного веса; общее содержание МАК, таким образом, составляет 0,76% собственного веса водорослей;
b) для Основного Экстракта, концентрации МАК 17-21 мг (что составляет 1,7-2,1% собственного веса).
Это важные данные, поскольку цельные AFA содержат высокие уровни МАК (0,76% собственного веса), близкие к максимальной концентрации, найденной при УФ-облучении, то есть 0,84% (31). Кроме того, было обнаружено, что экстракт имеет более высокую концентрацию, чем цельные водоросли, достигая намного более высоких уровней, чем максимальная потенциальная концентрация.
МАК (шинорин и порфира-334) являются структурно простыми молекулами с молекулярной массой 300. Это позволяет указанным водорастворимым молекулам легко проходить через гематоэнцефалический барьер, что подтверждает их способность проявлять свой потенциал в ингибировании MAO-B в той области, где это наиболее необходимо, в мозге.
Фикоцианины
Фикоцианины присутствуют в экстракте в концентрации 8-10% (количественный анализ см. ниже). Фикоцианины представляют собой синие пигменты, типичные для всех цианобактерий или сине-зеленых водорослей, хотя и со специфическими свойствами для каждого конкретного вида водорослей (32). Что касается функциональных и терапевтических свойств фикоцианинов, пока исследования главным образом сфокусированы на фикоцианинах из микроводоросли Spirulina. Очищенные фикоцианины из Spirulina, как установили, обладают антиоксидантными (33) и противовоспалительными (34, 35, 36) свойствами в различных физиологических системах, таких как печень (37), дыхательная система (38) и мозг (39, 40). Такие свойства очищенного PC из Spirulina в целом могут быть также приписаны фикоцианинам других водорослей, учитывая их существенное подобие. Однако между различными фикоцианинами из различных микроводорослей могут существовать видоспецифические различия, что может привести к различной активности при проявлении вышеописанных функциональных и терапевтических свойств.
Структурное определение и специфические свойства фикобилисом Кламатских водорослей
В общем, в интактной клетке цианобактерии фикоцианины (PC) присутствуют в фикобилисоме в функциональной форме (αβ)6 (41). После вскрытия клетки белок может быть обнаружен в различных агрегированных формах (мономеры, димеры, тримеры, гексамеры) в зависимости от исследуемого организма. В случае Кламатских водорослей электрофоретический анализ PC, непосредственно содержащегося в экстракте и очищенного из экстракта, показал, что белок главным образом присутствует в форме тримера (αβ)3, с общей молекулярной массой 121000. Мономер αβ имеет массу приблизительно 40000 (18500 α субъединица + 21900 β субъединица). Большинство исследований очищенного PC из Спирулины напротив указывает, что мономерная форма αβ белка, обнаруженного в Спирулине, обладает молекулярной массой приблизительно 37500, таким образом, демонстрируя другое агрегированное состояние в отличие от очищенного PC из AFA.
Хроматографический анализ фикобилисом AFA также показал, что, как в других видах цианобактерий α субъединица PC связывает простетическую группу, тогда как β субъединица связывает две. Простетическая группа или хромофор называется фикоцианобилином (PCB) и отвечает за синий цвет белка и его антиоксидантный потенциал (42).
Фундаментальное различие между AFA и Спирулина основано на различной структуре фикобилисомы. В отличие от Спирулины, фикобилисома Кламатской водоросли AFA не содержит пигмент алло-фикоцианин, при этом только пигмент c-фикоцианин соединен со структурным компонентом, который отсутствует в Спирулине, а именно, фикоэритроцианином (PEC). PEC представляет собой фотосинтетический пигмент, который на сегодняшний день был идентифицирован лишь у ограниченного числа видов цианобактерий (43). Химическая структура PEC в значительной степени схожа со структурой PC и состоит из двух субъединиц α и β, которые соединяются с образованием мономеров и тримеров. Однако тогда как каждый мономер PC связывает 3 молекулы PCB, PEC обладает уникальной особенностью связывания двух молекул PCB с β субъединицей и одной молекулой фиковиолобилина (PVB) с α субъединицей, которая отвечает за пурпурный цвет пигмента.
Исключительно впервые фикобилисома Кламатских водорослей определена как совершенно точно состоящая из комплекса c-фикоцианина и фикоэритроцианина, причем данная различная характерная структура фикобилисомы Кламатской водоросли AFA добавляет дополнительный решающий фактор, отличающий AFA от Спирулины.
Фигура 4 подтверждает вышесказанное, сравнивая компоненты клеточного лизата AFA с компонентами лизата другой известной цианобактерии, Synechocystis PCC 6803. У обоих цианобактерий можно увидеть синюю полосу, соответствующую фикобилисоме, но в водорослях AFA фикобилисома имеет более низкую молекулярную массу, что подтверждает, в противоположность обычным микроводорослям, таким как Спирулина, что в фикобилисоме AFA присутствуют только фикоцианины, а алло-фикоцианины не присутствуют. Кроме того, Фигура 4 показывает, что в AFA также присутствует светло-пурпурная полоса (показана стрелкой), которая типична для фикоэритроцианинов, что доказывает, таким образом, их присутствие в фикобилисоме Кламатских водорослей.
В целях более интенсивного определения каждую синюю полосу дополнительно проанализировали с помощью ВЭЖХ, связанной с масс-спектрометром (ОФ-ВЭЖХ-ИЭ-МС). Благодаря различному времени удержания белки фикобилисомы были разделены и идентифицированы на основании их молекулярной массы. Полученные результаты показаны в следующих таблицах. Вначале можно увидеть, что в Synechocystis (Таблица 1) присутствуют и фикоцианин (cpcA 28,2 мин и cpcB 28,9 мин), и алло-фикоцианин (apcA 30,7 мин и apcB 31,2 мин), а в AFA (Таблица 2) только фикоцианин (cpcA 28,8 мин и cpcB 30,0 мин). Во-вторых, в AFA был идентифицирован белок с молекулярной массой 19469, который не присутствует в Synechocystis и который соответствует бета-субъединице фикоэритроцианина с двумя присоединенными билинами (pecB 25,0 мин).
| ТАБЛИЦА 1: Белки, присутствующие в фикобилисоме Synechocystis. |
||||
| Время удержания (мин) |
Измеренная молекулярная масса | Ожидаемая молекулярная масса | Белок [гомологичный организм] |
Номер NCBI |
| 14,5 | 9322 | 9322 | cpcD | gi|16329820 |
| 22,6 | 32505 32388 |
32520 30797 |
CpcC cpcC |
gi|16329821 gi|16329822 |
| 24,6 | 28770 | 27392 | cpcG | gi|16329710 |
| 24,8 | 28885 | 28522 | cpcG | gi|16332194 |
| 28,2 | 18173 | 17586 | cpcA (α субъединица фикоцианина) | gi|2493297 |
| 28,9 | 19313 | 18126 | cpcB (β субъединица фикоцианина) | gi|2493300 |
| 30,7 | 17866 | 17280 | apcA (α субъединица алло-фикоцианина) | gi|266765 |
| 31,2 | 17816 | 17215 | apcB (β субъединица алло-фикоцианина) | gi|266766 |
| ТАБЛИЦА 2: Белки, присутствующие в фикобилисоме Кламатской водоросли AFA |
||||
| Время удержания (мин) |
Измеренная молекулярная масса | Ожидаемая молекулярная масса | Белок [гомологичный организм] |
Номер NCBI |
| 15,2 | 9031 | 8925 | гипотетический белок Avar03000795 [Anabaena variabilis ATCC 29413] | gi|45510540 |
| 8895 | cpcD [Nostoc sp. PCC 7120] |
gi|131740 | ||
| 25,0 | 19469 | 18284 | pecB: бета цепь фикоэритроцианина [Nostoc sp. PCC 7120] |
gi|548504 |
| 19308 | 18370 | гипотетический белок Avar03000787 (pecB) [Anabaena variabilis ATCC 29413] | gi|45510532 | |
| 26,4 | 31044 | 32078 | срсС [Nostoc sp. PCC 7120] |
gi|20141679 |
| 32219 | гипотетический белок Avar03000794 (линкер мол. масса 32000) [Anabaena variabilis ATCC 29413] | gi|45510539 | ||
| 31295 | pecC [Nostoc sp. PCC 7120] |
gi|464511 | ||
| 31304 | гипотетический белок Avar03000789 (pecC) [Anabaena variabilis ATCC 29413] | gi|45510534 | ||
| 30124 | 29333 | гипотетический белок Avar03000801 (cpcG4) [Anabaena variabilis ATCC 29413] | gi|46135436 | |
| 26,8 | 26119 | 28637 | гипотетический белок Avar03000799 (cpcG2) [Anabaena variabilis ATCC 29413] | gi|45510544 |
| 27,8 | 10994 | 10986 | fdxH2: ferredoxin, растительный [Anabaena variabilis] | gi|1169673 |
| 28,8 | 17714 | 17457 | cpcA [Nostoc sp. PCC 7120] |
gi|9957319 |
| 30,0 | 19222 | 18332 | cpcB [Nostoc sp. PCC 7120] |
gi|38894 |
Подобная уникальная структура является важным элементом в объяснении более сильного антиоксидантного и противовоспалительного действия целого AFA-PC по отношению к его PCB. Антиоксидантные и противовоспалительные свойства становятся актуальными в данном контексте, поскольку они производят сильный нейропротективный эффект; также в отношении нейропротекции целый PC более эффективен, чем его PCB, что ясно указывает на то, что другой активный компонент помимо PCB в фикобилисоме, а именно, PEC и его специфический хромофор PVB, являются, по-видимому, наиболее активным действующим началом AFA-PC, улучшающим самочувствие. То, что очищенный AFA-PC действительно содержит не только C-PC и его хромофор PCB, но также PEC и его хромофор PVB, становится очевидным при изучении спектрометрии экстракта, полученного в результате очистки (Фигура 5). Фактически, максимум поглощения С-РС составляет 620 нм, что в спектрометрии на Фигуре 5 соответствует вершине пика. Но максимум поглощения PEC, как известно, составляет 566 нм для α-субъединицы (фиковиолобилин или PVB) и соответственно 593 нм и 639 нм для двух PCB β-субъединицы. Все три значения действительно включены в колоколообразный пик, составляющий спектральный профиль очищенного PC. Принимая во внимание сильную связь между С-РС и PEC в водорослях AFA, с большим трудом поддающуюся разрыву, это подтверждает, что помимо С-РС PEC также является обязательной частью очищенного экстракта PC. Это в свою очередь означает, что PC из AFA значительно отличен, как структурно, так и функционально, от PC из другой цианобактерии, включая PC из Спирулины, который был исследован более подробно, а также то, что указанное различие состоит в наличии лишь одной части в целом, а именно, C-PC, но не другой; с выводом, что тогда как свойства С-РС могут также быть приписаны С-РС компоненту AFA-PC, свойства всего PC из AFA, который находится в комплексе C-PC/PEC (включая его хромофоры PCB и PVB), связаны исключительно с ним (так же как и с любым комплексом C-PC/PEC, присутствующим в любых других микроводорослях).
Методики очистки (Фигура 5)
PC очищали из высушенного экстракта AFA следующим образом:
- 500 мг экстракта суспендировали в 50 мл 100 мМ Na-фосфатного буфера, pH 7,4;
- центрифугировали при 2500 об/мин в течение 10' при 4°C;
- супернатант собирали и добавляли твердый сульфат аммония до 50% насыщения;
- белки осаждали в течение 60 минут при 4°C, продолжая перемешивать пробу;
- центрифугировали при 10000 об/мин в течение 30 мин при 4°C;
- прозрачный бесцветный супернатант отбрасывали и ресуспендировали синий осадок в малом объеме 5 мМ Na-фосфатного буфера, pH 7,4;
- диализ проводили при 4°C в течение ночи против того же буфера;
- диализированный PC переносили в гидроксиапатитовую колонку, уравновешенную 5 мМ Na-фосфатным буфером, pH 7,4;
- пробу элюировали Na-фосфатным буфером, pH 7,4 с повышением ионной силы (с 5 до 150 мМ);
- фракции собирали и регистрировали поглощение при 620 нм и 280 нм с помощью спектрофотометра;
- фракции со значением Abs620/Abs280>4 (индекс чистого PC) объединяли;
- PC осаждали сульфатом аммония при 50% насыщения в течение 1 часа при 4°С;
- центрифугировали при 10000 об/мин в течение 30' при 4°C;
- супернатант отбрасывали, а PC ресуспендировали в 150 мМ Na-фосфатным буфере, рН 7,4;
- диализ проводили против того же буфера при 4°C;
- очищенный PC переносили в колбу и хранили в темноте на +4°C или -20°С.
Количественный анализ фикоцианина
Для измерения молярной концентрации чистого PC использовали его коэффициент молярной экстинкции при 620 нм, который для тримерной формы (αβ)3 равен 770000 M-1*см-1. Это означает, что 1М раствор PC при 620 нм имеет значение поглощения 770000.
Для измерения концентрации PC в экстракте использовали коэффициент удельной экстинкции E1% при 620 нм 70 л г-1 см-1. Это означает, что раствор, содержащий 1% PC (то есть 1 г/100 мл), при 620 нм поглощает с интенсивностью 70. На основе данных вычислений среднее содержание PC в экстракте равно 80-100 мг/г собственного веса (8-10% собственного веса).
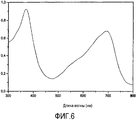
Очистка хромофора PCB (Фигура 6)
• 500 мг экстракта суспендировали в 50 мл дистиллированной H2O.
• Центрифугировали при 2500 об/мин в течение 10' при 4°C.
• Ярко-синий супернатант сливали с осадка и осаждали PC 1% трихлоруксусной кислотой.
• Инкубировали в течение 1 ч в темноте при 4°C с перемешиванием.
• Центрифугировали при 10000 об/мин в течение 30' при 4°C.
• Осадок, содержащий PC, собирали и промывали 3 раза метанолом.
• Осадок ресуспендировали в 10 мл метанола, содержащего 1 мг/мл HgCl2.
• Инкубировали в течение 20 ч при 42°C в темноте для выделения PCB из PC.
• Центрифугировали при 2500 об/мин в течение 10' для удаления белков.
• К супернатанту, содержащему PCB, добавляли β-меркаптоэтанол (1 мкл/мл) для осаждения HgCl2.
• Инкубировали при -20°C в течение 24 ч.
• Центрифугировали при 10000 об/мин в течение 30' при 4°C с целью удаления белого осадка.
• К супернатанту добавляли 10 мл метиленхлорида/бутанола (2:1, об./об.).
• Промывали 20 мл дистиллированной H2O и центрифугировали при 3000 об/мин в течение 10'.
• Верхнюю фазу удаляли, отбирали более низкую часть, содержащую PCB.
• PCB промывали 15 мл Н2О 3 раза.
• Высушили под азотом и хранили при -20°C.
ОЦЕНКА ИНГИБИРОВАНИЯ MAO-B ЭКСТРАКТОМ КЛАМАТСКОЙ ВОДОРОСЛИ AFA И ЕГО АКТИВНЫМИ КОМПОНЕНТАМИ ФОТОХРОМОМ, ФИКОЦИАНИНОМ И МАК
Была проанализирована ингибирующая активность Основного Экстракта в отношении MAO-B с использованием специфического субстрата бензиламина (1 мМ). Анализ проводили с использованием спектрофотометра при 30°C с длиной волны 250 нм, предварительно инкубируя MAO-B (2 мкг/мл) с водорастворимыми и жирорастворимыми компонентами основного экстракта при различных концентрациях, полученными на стадиях a) - c), описанных выше (начальная концентрация 10 мг/мл). Экстракт, обогащенный водорастворимыми компонентами, получали путем ресуспендирования водного экстракта в воде и сбора супернатанта после центрифугирования. Экстракт, обогащенный жирорастворимыми компонентами, получали путем ресуспендирования экстракта в ацетоне; впоследствии супернатант было высушен, а осадок ресуспендирован в ДМСО, растворителем, фармацевтически совместимым с MAO-B.
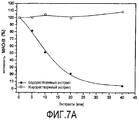
Как показано на Фигуре 7A, водорастворимая фракция ингибирует MAO-B дозозависимым образом, тогда как липофильная фракция не ингибирует фермент. Водорастворимая фракция Основного Экстракта AFA является мощным селективным ингибитором MAO-B с IC50 6,9 мкл. Селективность по отношению к MAO-B составляет 4 (IC50 MAO-B/IC50 MAO-A>4,05) (Фигура 7B).
График в координатах Лайнуивера-Берка на Фигуре 8 показывает, что такое ингибирование является обратимым смешанного типа по отношению к конкуренции, с уменьшением Vmax и увеличением константы Км Михаэлиса-Ментен. Построив график зависимости наклона от концентрации водорарастворимой фракции экстракта AFA, получили Константу ингибирования Ki при 1 мкл. По сравнению с водорарастворимой фракцией Основного Экстракта данное низкое значение Ki указывает на высокую аффинность к ферменту MAO-B.
Тот факт, что ингибирование под воздействием экстракта является обратимым, означает, что экстракт проявляет физиологическую активность без побочных эффектов. Что касается смешанной конкуренции, вероятно, это происходит из-за сложной природы экстракта, включающего молекулы с различной функциональностью, одни из которых являются конкурентными, а другие неконкурентными. Основными активными компонентами экстракта являются AFA-фитохром (0,5% собственного веса); фикоцианины (8-10% собственного веса); а также МАК или микоспорин-подобные аминокислоты (1,7-2,1% собственного веса), которые отдельно анализировали как ингибиторы MAO-B.
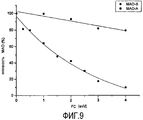
Ингибирование MAO-B под действием фикоцианинов
Тест проводили с использованием спектрофотометра при 30°C с длиной волны 250 нм, используя бензиламин в качестве субстрата, с предварительным инкубированием MAO-B с очищенным PC из AFA (0,5-4 мкМ) при различных концентрациях. Как показано на фигуре 9, AFA-PC вызывает дозозависимое снижение активности MAO-B, с IC50 1,44 мкМ. Селективность AFA-PC к MAO-B выше 3,5 (IC50 MAO-B/IC50 MAO-A > 3,5).
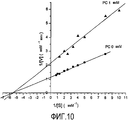
График в координатах Лайнуивера-Берка на Фигуре 10 показывает, что, как и в случае экстракта, ингибирование является обратимым смешанного типа (конкурентным и неконкурентным) с изменением Vmax и Км.
Построив график зависимости наклона от концентрации PC, получили значение константы ингибирования Ki, которая в данном случае составило 1,06 мкМ. Константа ингибирования определяет аффинность ингибитора к ферменту: высокая Ki указывает на низкую аффинность к ферменту, и наоборот. В данном случае низкое значение Ki указывает на высокую аффинность PC AFA к MAO-B.
Ингибирование MAO-B под действием МАК
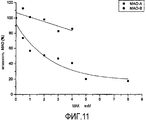
Активность MAO-B в отношении субстрата бензиламина оценивали относительно растущих концентраций МАК (0,5-8 мкМ), предварительно очищенных из Основного Экстракта с использованием 20% метанола. На Фигуре 11 показано дозозависимое ингибирование MAO-B под действием МАК с IC50 1,98 мкМ. Селективность МАК к MAO-B выше 2 (IC50 MAO-B/IC50 MAO-A > 2,02).
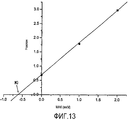
График в координатах Лайнуивера-Берка (Фигура 12) показывает, что ингибирование является обратимым и конкурентным, с ростом Км, но без изменения Vmax. Это означает, что МАК, благодаря своей химической структуре, конкурируют с субстратом за связь с активным сайтом фермента. Построив график зависимости наклона от концентрации МАК (Фигура 13), получили значение константы ингибирования Ki, которое составило 0,585 мкМ, что демонстрирует очень высокую степень аффинности к ферменту.
Ингибирование MAO-B под действием AFA-фитохрома
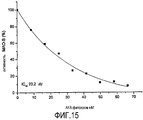
Тест проводили с использованием спектрофотометра при 30°C с длиной волны 250 нм, используя бензиламин в качестве субстрата, с предварительным инкубированием MAO-B с очищенным AFA-фитохромом при различных концентрациях (8,3 - 66,4 нМ). Как показано на Фигуре 15, AFA-фитохром вызывает дозозависимое снижение активности MAO-B, с IC50 не более 20,2 нМ.
График в координатах Лайнуивера-Берка на Фигуре 16 показывает, что, как и в случае экстракта, ингибирование является обратимым смешанного типа (конкурентным и неконкурентным) с изменением Vmax и Км.
Построив график зависимости наклона от концентрации AFA-фитохрома, получили значение константы ингибирования Ki, которое составило 10,48 нМ. Константа ингибирования определяет аффинность ингибитора к ферменту: высокая Ki указывает на низкую аффинность к ферменту, и наоборот. В данном случае, чрезвычайно низкое значение Ki указывает на очень высокую аффинность AFA-фитохрома к MAO-B.
Конкурентное и обратимое действие МАК делает указанные соединения очень мощными ингибиторами MAO-B. Фактически конкурентый и обратимый характер ингибирования MAO-B обеспечивает одновременно высокую эффективность, а также физиологическую активность без побочных эффектов. В данном смысле, МАК, содержащиеся в экстракте, также и из-за их молекулярной массы и, в результате, способности легко проходить через гематоэнцефалический барьер, составляют решающий компонент, даже in vivo, производящий терапевтические эффекты, полученные в результате ингибирования MAO-B.
Даже в большей степени, чем МАК, фитохром, как оказалось, является наиболее мощным ингибитором МАО-В из всех известных на настоящий момент веществ. Его очень высокая аффинность к ферменту MAO-B, а также его эффективное ингибирование при концентрациях немного ниже наномолярных, делает это соединение само по себе не только прекрасным терапевтическим агентом, но и фактором, который, по всей вероятности, обеспечивает наиболее весомый вклад в высокую неврологическую эффективность экстракта (экстрактов) AFA.
Следует добавить, что некоторые из факторов, касающиеся МАК и фитохрома, можно также отнести к поведению фикоцианинов in vivo. Известно, что PC оказывают нейропротективное действие на мозг in vivo, а также что они способны проходить через гематоэнцефалический барьер (44). Это означает, что они также способны проявлять in vivo свою ингибирующую активность в отношении MAO-B в мозге. Молекулярная масса хромофора действительно составляет лишь 700, что не намного больше, чем молекулярная масса МАК. То же справедливо для хромофора фитохрома - фитохромобилина - структурно подобного фикоцианобилину.
В заключение активность ингибирования MAO-B со стороны экстракта и его активных компонентов, фитохрома AFA, AFA-PC и МАК, является чрезвычайно актуальной, поскольку как указанные соединения, так и экстракт обладают наиболее высоким уровнем активности, равной или выше активности фармакологических веществ, и существенно выше любой исследованной природной молекулы, как показано в следующей таблице (45):
| Таблица 3: Сравнительные кинетические параметры (IC50 и Ki) ингибирования МАО-В под действием известных синтетических и природных ингибиторов |
|||
| Ингибиторы MAO-B | IC50 | Ki | Тип ингибирования |
| Депренил | 0,31 мкМ | 0,002 мкМ | Необратимый |
| Эпикатехин | 58,9 мкМ | 21 мкМ | Смешанный |
| Катехин | 88,6 мкМ | 74 мкМ | Смешанный |
| Алкалоид (не харман) | 6,47 мкМ | 1,12 мкМ | Смешанный |
| Пиперин | 91,3 мкМ | 79,9 мкМ | Конкурентный |
| Пеонол | 42,5 мкМ | 38,2 мкМ | Конкурентный |
| Эмодин | 35,4 мкМ | 15,1 мкМ | Смешанный |
| AFA-фикоцианин | 1,44 мкМ | 1,06 мкМ | Смешанный |
| AFA-МАК | 1,98 мкМ | 0,585 мкМ | Конкурентный |
| AFA-фитохром | 0,02 мкМ | 0,010 мкМ | Смешанный |
Как показано в таблице, только фикоцианины и МАК имеют IC50 немного выше 1 мкМ, что очень близко к IC50 Депренила (0,31 мкМ), и в десять раз ниже, чем IC50 других исследуемых соединений. AFA-фитохром, с другой стороны, имеет IC50 в 15 раз ниже, чем IC50 Депренила. То же справедливо для константы ингибирования Ki, которая определяет аффинность ингибитора к ферменту. AFA-фикоцианины имеют Ki порядка 1 мкМ, как ненаркотические (не харманового типа) алкалоиды кофе и табака (но конечно без какой-либо из проблем, связанных с двумя указанными веществами). С другой стороны, МАК и AFA-фитохром, наряду с Депренилом, являются единственными соединениями, обладающими Ki ниже 1 мкМ, и, таким образом, очень высокой аффинностью к MAO-B. Фактически, AFA-фитохром является единственной природной молекулой, помимо селегилина/Депренила, Ki которой имеет порядок нескольких наномолей. И все же существует существенное различие между селегилином/Депренилом и веществами экстракта AFA: первый является необратимым ингибитором, таким образом, отличающимися потенциальными побочными эффектами; тогда как все ингибирующие МАО-В вещества, содержащиеся в Кламатской водоросли AFA, являются обратимыми ингибиторами, отличающимися физиологической активностью, лишенной проблем, связанных с синтетическими молекулами.
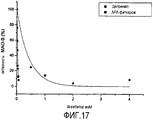
На Фигуре 14 графически показана ингибирующая активность по отношению к MAO-B трех молекул AFA в сравнении с Депренилом. Учитывая совместные действия всех трех молекул в Основном Экстракте (и других экстрактах AFA), полная ингибиторная активность Основного Экстракта в отношении MAO-B является в результате очень высокой. Также весьма существенным является высокое количество ФЭА, присутствующее в Основном Экстракте. Если сравнивать основной экстракт с Депренилом на основе содержания в нем PC, мы получим то, что Основной Экстракт достигает IC50 при концентрации PC не более 0,05 мкМ, при котором его эффективность в 7,5 раз выше, чем у Депренила (и в десятки раз выше, чем у природных веществ). Это становится понятным в свете эффективности фитохрома, содержащегося в Основном Экстракте: в среднем эффективность ингибирования PC и МАКА отличается фактически в 7,5 раз, что немного ниже по сравнению с Депренилом, а также фитохромом, который в 15 раз более эффективный (Фигура 17). Это также показывает, что более высокая эффективность экстракта по сравнению с очищенным AFA-PC главным образом обусловлена фитохромом.
Кроме того, экстракт обладает еще одним преимуществом, так как является природным физиологически активным веществом, ингибирующим MAO-B обратимо и главным образом конкурентно, в результате чего экстракт лишен побочных эффектов, потенциально связанных с необратимыми ингибиторами, такими как Депренил и другие синтетические соединения (46).
Дополнительным преимуществом экстракта является высокое содержание в нем фенилэтиламина, мощного допаминергического нейромодулятора, который работает в полной синергии с другими молекулами, при этом можно выделить следующие синергические эффекты:
- Фенилэтиламин или ФЭА обладает удвоенной допаминергической активностью, поскольку он стимулирует высвобождение допамина из нигростриарной ткани, и поскольку он ингибирует постсинаптический обратный захват самого допамина;
- Фитохром, МАК и фикоцианины, как мощные ингибиторы MAO-B, также усиливают допаминергическую передачу, поскольку сниженная активность MAO-B приводит к увеличению жизни нейроаминов, включая допамин;
- Фитохром, МАК и фикоцианины, как ингибиторы MAO-B, также продлевают жизнь и активность фенилэтиламина, который сам по себе является объектом дезаминирующей активности фермента MAO-B, с последующим созданием эффективного цикла, дополнительно поддерживающего допаминергическую передачу и активность, а также общую нейромодуляцию, производимую ФЭА.
- Наконец, мощная антиоксидантная и противовоспалительная активность фикоцианинов, вместе с их способностью или способностью их хромофора проходить через гематоэнцефалический барьер, а также чрезвычайно высокая антиокислительная активность фитохрома и менее сильная, но все же существенная антиоксидантная активность МАК, обеспечивают нейропротекцию, которая ограждает различные активные молекулы и в целом неврологический эффективный цикл, который они создают, от любого окислительного и воспалительного повреждения.
НЕЙРОПРОТЕКЦИЯ
Были исследованы нейропротективные свойства экстракта AFA, специфического AFA-PC и его хромофора PCB, а также МАК против нейротоксического эффекта глутамата.
Глутамат является главным возбуждающим нейромедиатором в центральной нервной системе млекопитающих, однако сверхстимуляция его рецептора подтипа NMDA в нейронах вызывает массивное внутриклеточное накопление Ca2+, что приводит к гибели клеток. Кроме того, внутримитохондриальное накопление
Ca2+, после стимуляции рецептора NMDA, транзиентно повышает концентрацию свободных ионов Ca2+ в цитозоле, активирующих нейронную изоформу синтазы оксида азота (NOS) (49), фермента, который синтезирует оксид азота (NO) или, главным образом в первичных нейронах, продукт его реакции с супероксидом (О2-), пероксинитрит (ONOO-).
Воздействие глутамата на нейроны проводили согласно несколько измененной методике (50): культуральную среду удаляли и однократно промывали нейроны предварительно нагретым до 37°C буфером Хэнкса (5,26 мМ KCl, 0,43 мМ KH2H2PO4, 132,4 мМ NaCl, 4,09 мМ NaHCO3, 0,33 мМ Na2HPO4, 20 мМ глюкозы, 2 мМ CaCl2, и 20 мМ HEPES, pH 7,4) и предварительно инкубировали в отсутствие или присутствии нескольких концентраций экстракта AFA (1-50 нМ), PC (10-1000 нМ), PCB (10-1000 нМ) и МАК (1-10 мкМ) в предварительно нагретом до 37°C буфере Хэнкса. Через 30 минут предварительной инкубации добавляли концентрированный раствор L-глутамата до конечной концентрации 100 мкМ плюс 10 мкМ глицина. Нейроны инкубировали при 37°C в течение 15 минут, удаляли буфер с помощью аспиратора, заменив его средой DMEM, после чего клетки инкубировали при 37°C в течение еще 24 ч в отсутствии эффекторов.
Апоптоз оценивали, окрашивая ядра клеток ДАФИ (50), флуоресцентным красителем, проходящим через мембрану, который связывает ДНК и позволяет количественно определять апоптические нейроны, то есть нейроны с фрагментированными или сжатыми ядрами. Вкратце, через 24 ч после воздействия глутамата, культуры нейронов промывали теплым PBS (37°C) и фиксировали 4% (масс./об.) раствором параформальдегида в PBS в течение 30 минут при комнатной температуре. После промывки PBS клетки обрабатывали 3 мкМ ДАФИ в течение 10 минут при комнатной температуре в темноте, а затем дважды промывали PBS. Клетки исследовали на предмет конденсации хроматина с помощью люминесцентной микроскопии, используя флюоресцеиновый фильтр (330-380 возбуждение; 30X увеличение). Подсчитывали общее количество и количество апоптических ядер. Во всех случаях приблизительно 600-1000 клеток было подсчитано оператором, неосведомленным о схеме эксперимента. Измерения отдельных культур выполняли дважды, а результаты выражали как средние значения со стандартной ошибкой среднего для указанного количества культур. Статистический анализ результатов проводили с использованием теста Крускала-Уоллиса с последующим использованием множественного критерия размаха выборки с наименьшей значимой разностью. Во всех случаях p-0,05 считали значимым.
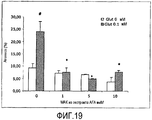
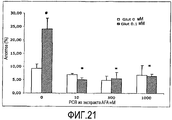
С помощью данного теста повреждения глутаматом впервые показана нейропротективная способность Основного Экстракта AFA, AFA-PC, его PCB и МАК. Как показано на Фигуре 18, добавление глутамата к культуре нейронных клеток увеличивало уровень апоптоза до 22,9% ± 3 n = 4 (p<0,05), тогда как одновременное добавление Основного Экстракта AFA обеспечивало высокую защиту от токсичности глутамата, понижая уровень апоптоза ниже контрольного уровня (6,3% ± 1 p>0,05) уже при концентрации экстракта не более 1 нМ (приведены средние значения ± СОС от 3 до 8 различных культур клеток. # Значимо различный при сравнении с группой контроля (p<0,05); * Значимо различный при сравнении с глутаматом (контроль) (p<0,05). Что касается защиты, обеспечиваемой МАК, то МАК также понижают уровень апоптоза ниже уровня контроля, при более высокой концентрации 1 мкМ (Фигура 19), приведены средние значения ± СОС от 3 до 8 различных культур клеток. # Значимо различный при сравнении с группой контроля (p <0,05); * Значимо различный при сравнении с глутаматом (контроль) (p<0,05)). В отношении AFA-PC и PCB можно видеть, что уровень их ингибирования апоптоза довольно схож: их добавление к культуре клеток понижает степень апоптоза ниже контроля при концентрации приблизительно 10 нМ (Фигуры 20 и 21 - приведены средние значения ± СОС от 3 до 8 различных культур клеток. # Значимо различный при сравнении с контрольной группой (p<0,05); * Значимо различный при сравнении с глутаматом (контроль) (p<0,05)).
Степень ингибирования AFA-PC приблизительно равна степени ингибирования PCB. Это несколько неожиданно, учитывая то, что PCB, возможно, является действующим началом AFA-PC и при очистке и, таким образом, при более высокой концентрации, должен быть значительно более эффективным, чем целая молекула, активным компонентом которой и является PCB. Тот факт, что PCB имеет фактически такую же эффективность, означает, что в целой молекуле PC присутствуют другие факторы, которые фактически могут быть еще более эффективными, чем сам PCB. Известно, что в составе молекулы PC, помимо С-РС и его хромофора PCB, присутствует PEC, который включает в качестве своих хромофоров и PCB, и PVB (фиковиолобилин). Поэтому можно предположить, что фактором, создающим существенное различие в эффективности между очищенным PCB и целым PC, является именно компонент PEC, в особенности его хромофор PVB, который, как предполагают, является очень сильным антиоксидантом.
В отношении нейропротекции, МАК, по-видимому, играют некоторую роль, однако значительно менее важную, чем PC и PCB. Впрочем, наиболее мощным нейропротектором очевидно является цельный экстракт AFA, который способен полностью ингибировать апоптоз клеток при концентрациях порядка всего лишь 1 нМ (наномоля). Это в 10 раз выше эффективности PC и PCB. Разумеется, это можно объяснить синергией многих различных антиоксидантных факторов, присутствующих в цельном экстракте AFA; и все же, так как из вышеизложенного можно видеть, что на настоящий момент AFA-фитохром возможно является наиболее мощным антиоксидантом, способным почти полностью ингибировать образование MDA (побочный продукт последних стадий перекисного окисления липидов) при концентрации всего лишь 16 наномолей, вероятно, что AFA-фитохром является наиболее важным фактором при объяснении более высокой эффективности Основного Экстракта. Таким образом, можно заключить, что AFA-фитохром, так же как и все и любые фитохромы, являются важными нейропротективными агентами.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ







Claims (4)
1. Применение экстракта микроводоросли Aphanizomenon Flos Aquae Aquae Ralfs ex Born. & Flah. Var. flos aquae (Кламатской водоросли AFA), где экстракт, содержащий следующие компоненты:
комплекс С-Фикоцианина/фикоэритроцианинов (С-РС/РЕС), и отдельных хромофоров фикоцианобилина (РСВ) и фиковиолобилина (PVB), AFA-фитохром и микоспорин-подобные аминокислоты (МАК), порфиры и шинорина, и фенилэтиламин (ФЭА)
получают посредством следующих стадий:
a) замораживания свежесобранной водоросли AFA и ее размораживания или, если исходный материал является высушенным порошком AFA, обработки ультразвуком разбавленного в воде порошка AFA для разрушения клеток;
b) центрифугирования продукта стадии а) для отделения супернатанта от осадка;
c) сбора супернатанта, содержащего водорастворимые компоненты, для производства композиции для профилактики, борьбы или лечения
неврологического заболевания, состояния дисфункции или расстройства, выбранного из: болезни Альцгеймера, болезни Паркинсона, рассеянного склероза, синдрома дефицита внимания с гиперактивностью (СДВГ), аутизма, депрессии, дефицита памяти и аффективных расстройств.
комплекс С-Фикоцианина/фикоэритроцианинов (С-РС/РЕС), и отдельных хромофоров фикоцианобилина (РСВ) и фиковиолобилина (PVB), AFA-фитохром и микоспорин-подобные аминокислоты (МАК), порфиры и шинорина, и фенилэтиламин (ФЭА)
получают посредством следующих стадий:
a) замораживания свежесобранной водоросли AFA и ее размораживания или, если исходный материал является высушенным порошком AFA, обработки ультразвуком разбавленного в воде порошка AFA для разрушения клеток;
b) центрифугирования продукта стадии а) для отделения супернатанта от осадка;
c) сбора супернатанта, содержащего водорастворимые компоненты, для производства композиции для профилактики, борьбы или лечения
неврологического заболевания, состояния дисфункции или расстройства, выбранного из: болезни Альцгеймера, болезни Паркинсона, рассеянного склероза, синдрома дефицита внимания с гиперактивностью (СДВГ), аутизма, депрессии, дефицита памяти и аффективных расстройств.
2. Применение по п.1, где экстракт Кламатской водоросли AFA дополнительно очищен посредством ультрафильтрации указанного супернатанта с использованием ультрафильтрационной мембраны с пределом пропускания по молекулярной массе 30000 Да.
3. Применение по п.1, где экстракт Кламатской водоросли AFA включен в композицию с фармацевтически приемлемыми носителями или наполнителями.
4. Применение по п.1, где указанная композиция подходит для введения человеку.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US81659306P | 2006-06-27 | 2006-06-27 | |
| US60/816,593 | 2006-06-27 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| RU2008151175A RU2008151175A (ru) | 2010-06-27 |
| RU2441663C2 true RU2441663C2 (ru) | 2012-02-10 |
Family
ID=38529410
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2008151175/15A RU2441663C2 (ru) | 2006-06-27 | 2007-06-26 | Препарат aphanizomenon flos aquae, экстракты и очищенные компоненты экстрактов для лечения неврологических, нейродегенеративных и аффективных расстройств |
Country Status (20)
| Country | Link |
|---|---|
| US (1) | US9522167B2 (ru) |
| EP (1) | EP2046354B1 (ru) |
| JP (1) | JP5259585B2 (ru) |
| KR (1) | KR20090035490A (ru) |
| CN (1) | CN101478980B (ru) |
| AT (1) | ATE519491T1 (ru) |
| AU (1) | AU2007264042B2 (ru) |
| CA (1) | CA2656154C (ru) |
| DK (1) | DK2046354T3 (ru) |
| ES (1) | ES2370533T3 (ru) |
| HR (1) | HRP20110771T1 (ru) |
| IL (1) | IL196132A (ru) |
| MY (1) | MY146594A (ru) |
| PL (1) | PL2046354T3 (ru) |
| PT (1) | PT2046354E (ru) |
| RS (1) | RS52072B (ru) |
| RU (1) | RU2441663C2 (ru) |
| SI (1) | SI2046354T1 (ru) |
| WO (1) | WO2008000430A2 (ru) |
| ZA (1) | ZA200900747B (ru) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2757424C1 (ru) * | 2018-02-23 | 2021-10-15 | СиДжей ЧеилДжеданг Корпорейшн | Микроорганизм, продуцирующий микоспорин-подобную аминокислоту, и способ получения микоспорин-подобной аминокислоты с его использованием |
Families Citing this family (17)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ITTO20070391A1 (it) * | 2007-06-05 | 2008-12-06 | Medestea Res & Production S P | Composizione di integratore alimentare,farmaco o dispositivo medicale e suo impiego per sopprimere l'appetito, migliorare tono ed umore, con attivita antidepressiva naturale e con effetto antiastenico |
| KR100969325B1 (ko) * | 2010-02-08 | 2010-07-09 | 인천대학교 산학협력단 | 홍조류로부터 비독성 자외선 차단용 추출물을 제조하는 방법 및 이를 이용한 비독성 자외선 차단제 |
| EP2568992A4 (en) * | 2010-05-11 | 2013-11-06 | Benzion Geshuri | PHARMACEUTICAL COMPOSITION COMPRISING AN ALGUE APPROPRIATE FOR INCREASING THE EFFICACY OF AN ENZYMATIC INHIBITOR |
| US8906361B2 (en) * | 2011-03-28 | 2014-12-09 | John V. Cappello | Anti-aging formulations |
| CU23963B1 (es) * | 2011-07-01 | 2013-12-11 | Ct De Ingeniería Genética Y Biotecnología | Composiciones farmacéuticas a partir de derivados de la c-ficocianina y combinaciones con interferones del tipo i para el tratamiento de enfermedades cerebrales isquémicas y neurodegenerativas |
| US20180250351A1 (en) * | 2015-09-03 | 2018-09-06 | Stefano Scoglio | Cyanobacterial microalgae phycocyanin and phycocyanobilin to beneficially inhibit the activity of the UDP-GDH enzyme while significantly increasing the absorption and circulation of curcumin |
| JP6964707B2 (ja) * | 2016-04-27 | 2021-11-10 | 長瀬産業株式会社 | 安定化されたマイコスポリン様アミノ酸を含有する溶液、およびその製造方法 |
| JP6719267B2 (ja) * | 2016-04-27 | 2020-07-08 | 長瀬産業株式会社 | 安定化されたマイコスポリン様アミノ酸を含有する溶液、およびその製造方法 |
| JP7257479B2 (ja) * | 2016-04-27 | 2023-04-13 | 長瀬産業株式会社 | 安定化されたマイコスポリン様アミノ酸を含有する溶液、およびその製造方法 |
| AU2017259736A1 (en) * | 2016-05-02 | 2018-12-20 | Stefano Scoglio | The use of phycocyanins, purified or as present in cyanobacterial microalgae or extracts thereof, as prebiotics, to enhance the viability, gastrointestinal survival, pathogen-fighting ability, and the overall health-enhancing properties of probiotic cultures and products. |
| CA3052737A1 (en) | 2017-09-06 | 2019-03-14 | Stefano Scoglio | The transdermal use of cyanobacterial algae, and/or extracts thereof, via low or high intensity transporation devices for beauty and health enhancing purposes |
| CN108484438A (zh) * | 2018-03-06 | 2018-09-04 | 佛山科学技术学院 | 一种从紫菜中提取类菌胞素氨基酸的方法 |
| CN108421031A (zh) * | 2018-04-13 | 2018-08-21 | 桂林医学院 | 藻蓝蛋白在制备防治帕金森病药物中的应用 |
| CN108752445B (zh) * | 2018-06-20 | 2021-04-30 | 广东药科大学 | 一种螺旋藻蛋白的制备工艺及其神经保护用途 |
| FR3103106B1 (fr) * | 2019-11-19 | 2022-05-27 | Algosource | Extrait liquide aqueux de Spiruline pour la prévention et/ou le traitement des neuropathies périphériques chimio-induites et leurs symptômes, composition et utilisation correspondantes. |
| CN111700903B (zh) * | 2020-08-07 | 2022-03-11 | 华侨大学 | 一种紫菜多糖的应用及紫菜多糖的制备方法 |
| KR102694496B1 (ko) * | 2021-08-18 | 2024-08-13 | 전남대학교산학협력단 | 마이코스포린-유사 아미노산을 유효성분으로 포함하는 비만 예방 또는 치료용 조성물 |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20050129709A1 (en) * | 2001-05-14 | 2005-06-16 | Jensen Gitte S. | Method for enhancing stem cell trafficking |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS5865216A (ja) | 1981-09-16 | 1983-04-18 | Dainippon Ink & Chem Inc | 抗腫瘍剤 |
| US5455276A (en) * | 1993-05-20 | 1995-10-03 | Sabelli; Hector C. | Method for treating depression |
| RU2417093C2 (ru) * | 2005-06-24 | 2011-04-27 | Дезерт Лейк Текнолоджиз | Очищенный компонент цианобактерий и способ применения |
-
2007
- 2007-06-26 PL PL07764849T patent/PL2046354T3/pl unknown
- 2007-06-26 HR HR20110771T patent/HRP20110771T1/hr unknown
- 2007-06-26 ES ES07764849T patent/ES2370533T3/es active Active
- 2007-06-26 SI SI200730753T patent/SI2046354T1/sl unknown
- 2007-06-26 MY MYPI20085257A patent/MY146594A/en unknown
- 2007-06-26 ZA ZA200900747A patent/ZA200900747B/xx unknown
- 2007-06-26 DK DK07764849.1T patent/DK2046354T3/da active
- 2007-06-26 AU AU2007264042A patent/AU2007264042B2/en not_active Ceased
- 2007-06-26 WO PCT/EP2007/005622 patent/WO2008000430A2/en not_active Ceased
- 2007-06-26 PT PT07764849T patent/PT2046354E/pt unknown
- 2007-06-26 KR KR1020087031246A patent/KR20090035490A/ko not_active Ceased
- 2007-06-26 RS RS20110480A patent/RS52072B/sr unknown
- 2007-06-26 US US12/306,483 patent/US9522167B2/en not_active Expired - Fee Related
- 2007-06-26 CN CN2007800234854A patent/CN101478980B/zh active Active
- 2007-06-26 AT AT07764849T patent/ATE519491T1/de active
- 2007-06-26 RU RU2008151175/15A patent/RU2441663C2/ru active IP Right Revival
- 2007-06-26 JP JP2009516969A patent/JP5259585B2/ja not_active Expired - Fee Related
- 2007-06-26 EP EP07764849A patent/EP2046354B1/en active Active
- 2007-06-26 CA CA2656154A patent/CA2656154C/en active Active
-
2008
- 2008-12-23 IL IL196132A patent/IL196132A/en active IP Right Grant
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20050129709A1 (en) * | 2001-05-14 | 2005-06-16 | Jensen Gitte S. | Method for enhancing stem cell trafficking |
Non-Patent Citations (1)
| Title |
|---|
| Pugh N. et al. Isolation of three high molecular weight polysaccharide preparations with potent immunostimulatory activity from Spirulina platensis, aphanizomenon flos-aquae and Chlorella pyrenoidosa. // Planta Med. 2001, 67(8):737-42. Torres A. et al. Porphyra-334, a potential natural source for UVA protective sunscreens. // Photochem Photobiol. Sci. 2006, 5(4): 432-5. * |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2757424C1 (ru) * | 2018-02-23 | 2021-10-15 | СиДжей ЧеилДжеданг Корпорейшн | Микроорганизм, продуцирующий микоспорин-подобную аминокислоту, и способ получения микоспорин-подобной аминокислоты с его использованием |
Also Published As
| Publication number | Publication date |
|---|---|
| ATE519491T1 (de) | 2011-08-15 |
| ES2370533T3 (es) | 2011-12-19 |
| JP2009541388A (ja) | 2009-11-26 |
| KR20090035490A (ko) | 2009-04-09 |
| PL2046354T3 (pl) | 2012-01-31 |
| IL196132A (en) | 2014-06-30 |
| CN101478980B (zh) | 2013-03-20 |
| ZA200900747B (en) | 2010-05-26 |
| DK2046354T3 (da) | 2011-10-31 |
| SI2046354T1 (sl) | 2011-11-30 |
| RU2008151175A (ru) | 2010-06-27 |
| CA2656154A1 (en) | 2008-01-03 |
| EP2046354B1 (en) | 2011-08-10 |
| US9522167B2 (en) | 2016-12-20 |
| AU2007264042A1 (en) | 2008-01-03 |
| CA2656154C (en) | 2017-02-28 |
| WO2008000430A3 (en) | 2008-03-27 |
| WO2008000430A2 (en) | 2008-01-03 |
| RS52072B (sr) | 2012-06-30 |
| EP2046354A2 (en) | 2009-04-15 |
| AU2007264042B2 (en) | 2012-01-12 |
| US20090311286A1 (en) | 2009-12-17 |
| MY146594A (en) | 2012-08-30 |
| HRP20110771T1 (hr) | 2011-11-30 |
| PT2046354E (pt) | 2011-11-09 |
| CN101478980A (zh) | 2009-07-08 |
| JP5259585B2 (ja) | 2013-08-07 |
| IL196132A0 (en) | 2009-09-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| RU2441663C2 (ru) | Препарат aphanizomenon flos aquae, экстракты и очищенные компоненты экстрактов для лечения неврологических, нейродегенеративных и аффективных расстройств | |
| US6210681B1 (en) | Plant proanthocyanidin extracts | |
| ES2618952T3 (es) | Composiciones que incluyen antocianidinas y métodos de uso | |
| KR101366187B1 (ko) | 프로안토시아니딘 올리고머의 제조방법 | |
| JP2006516019A (ja) | モリンダ・シトリフォリアの抗真菌効果 | |
| KR102582150B1 (ko) | 신규한 석류-산화아연 나노복합입자 및 이의 용도 | |
| WO2013172586A1 (ko) | 광 조사를 이용한 광감각제-펩타이드를 유효성분으로 포함하는 발모 개선 또는 촉진용 조성물 및 이를 이용한 방법 | |
| JP2010155840A (ja) | B型肝炎を治療するための医薬組成物およびb型肝炎ウイルス抑制効果を有する健康食品 | |
| JP2021519304A (ja) | 生理活性ミドリイガイ抽出物及びその使用 | |
| US20130109640A1 (en) | Composition for treating or preventing neurodegenerative brain diseases comprising black bean extract | |
| AU2006219286B2 (en) | Antiallergic composition | |
| Akhlaghi et al. | Improving the therapeutic performance of glycyrrhiza glabra hydroalcoholic extract using liposomal nano-carriers and their characterization | |
| US20170128503A1 (en) | Neuro-modulating preparations for treatment of neurological and mood disorders | |
| JP6831049B2 (ja) | NF−κB抑制作用を呈するグランジフロル酸誘導体の製造方法 | |
| KR101667873B1 (ko) | 한방 추출물 또는 이의 분획물을 포함하는 파킨슨 질환의 예방 또는 치료용 약학적 조성물 | |
| AQUAE et al. | THEREOF FOR THE TREATMENT OF NEUROLOGICAL, NEURODEGENERATIVE AND MOOD DISORDERS | |
| KR102926617B1 (ko) | 마이크로 및 나노버블을 활용한 초음파 보조 추출을 통해 퀘르세틴의 함량이 증가된 양파 추출물을 제조하는 방법 | |
| JP2005068081A (ja) | 発癌予防剤 | |
| HK1135315A (en) | Alphanizomenon flos aquae preparation, extracts and purified components thereof for the treatment of neurological, neurodegenerative and mood disorders | |
| TWI797279B (zh) | 球薑的花萃取物及其用途 | |
| JP2007186431A (ja) | 抗酸化組成物及び脳神経細胞保護用医薬組成物、抗酸化剤及び脳神経細胞保護剤、並びにそれらの使用方法 | |
| CN104744420B (zh) | 一种从高粱外种皮中提取原花青素四聚体的制备方法及应用 | |
| WO2024256768A1 (fr) | Extrait de punica granatum variete cybele et ses utilisations en cosmetique | |
| WO2026018050A1 (en) | Pistacia-derived bioactive peptides encapsulated in soy protein nanoparticles as an anticancer drug | |
| KR20210133603A (ko) | 굼벵이를 포함하는 치매 예방 또는 치료용 조성물 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| MM4A | The patent is invalid due to non-payment of fees |
Effective date: 20150627 |
|
| NF4A | Reinstatement of patent |
Effective date: 20170714 |