JP2008507294A - 菌株遺伝子操作による改善されたタンパク質発現のための方法 - Google Patents
菌株遺伝子操作による改善されたタンパク質発現のための方法 Download PDFInfo
- Publication number
- JP2008507294A JP2008507294A JP2007523707A JP2007523707A JP2008507294A JP 2008507294 A JP2008507294 A JP 2008507294A JP 2007523707 A JP2007523707 A JP 2007523707A JP 2007523707 A JP2007523707 A JP 2007523707A JP 2008507294 A JP2008507294 A JP 2008507294A
- Authority
- JP
- Japan
- Prior art keywords
- protein
- cell
- gene
- expression
- host cell
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 108090000623 proteins and genes Proteins 0.000 title claims abstract description 626
- 102000004169 proteins and genes Human genes 0.000 title claims abstract description 284
- 238000000034 method Methods 0.000 title claims abstract description 210
- 230000014509 gene expression Effects 0.000 title claims abstract description 179
- 230000001976 improved effect Effects 0.000 title claims description 17
- 238000010353 genetic engineering Methods 0.000 title description 5
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 claims abstract description 156
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 claims abstract description 156
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 140
- 230000002068 genetic effect Effects 0.000 claims abstract description 54
- 230000001965 increasing effect Effects 0.000 claims abstract description 35
- 210000004027 cell Anatomy 0.000 claims description 387
- 108091005804 Peptidases Proteins 0.000 claims description 140
- 102000035195 Peptidases Human genes 0.000 claims description 115
- 239000004365 Protease Substances 0.000 claims description 107
- 238000002493 microarray Methods 0.000 claims description 74
- 230000027455 binding Effects 0.000 claims description 68
- 241000588724 Escherichia coli Species 0.000 claims description 67
- 108010000521 Human Growth Hormone Proteins 0.000 claims description 55
- 102000002265 Human Growth Hormone Human genes 0.000 claims description 55
- 239000000854 Human Growth Hormone Substances 0.000 claims description 52
- 108020004999 messenger RNA Proteins 0.000 claims description 41
- 108010026552 Proteome Proteins 0.000 claims description 40
- -1 httpG Proteins 0.000 claims description 32
- 101150115543 hslU gene Proteins 0.000 claims description 31
- 241000589540 Pseudomonas fluorescens Species 0.000 claims description 27
- 230000000694 effects Effects 0.000 claims description 27
- 239000013598 vector Substances 0.000 claims description 27
- 238000013459 approach Methods 0.000 claims description 26
- 101150054715 clpY gene Proteins 0.000 claims description 26
- 108010006519 Molecular Chaperones Proteins 0.000 claims description 24
- 102000005431 Molecular Chaperones Human genes 0.000 claims description 18
- 238000003780 insertion Methods 0.000 claims description 15
- 230000037431 insertion Effects 0.000 claims description 15
- 102100037486 Reverse transcriptase/ribonuclease H Human genes 0.000 claims description 13
- 238000002744 homologous recombination Methods 0.000 claims description 13
- 230000006801 homologous recombination Effects 0.000 claims description 13
- 101150096566 clpX gene Proteins 0.000 claims description 10
- 238000000539 two dimensional gel electrophoresis Methods 0.000 claims description 9
- 101150017872 clpQ gene Proteins 0.000 claims description 8
- 101150055178 hslV gene Proteins 0.000 claims description 8
- 230000010354 integration Effects 0.000 claims description 8
- 210000004962 mammalian cell Anatomy 0.000 claims description 8
- 102000005593 Endopeptidases Human genes 0.000 claims description 7
- 108010059378 Endopeptidases Proteins 0.000 claims description 7
- 101100278084 Nostoc sp. (strain PCC 7120 / SAG 25.82 / UTEX 2576) dnaK1 gene Proteins 0.000 claims description 7
- 101100117145 Synechocystis sp. (strain PCC 6803 / Kazusa) dnaK2 gene Proteins 0.000 claims description 7
- 101150036359 clpB gene Proteins 0.000 claims description 7
- 101150052825 dnaK gene Proteins 0.000 claims description 7
- 101800000212 P1A protein Proteins 0.000 claims description 6
- 101800000862 Protease cofactor Proteins 0.000 claims description 6
- 101150033736 cbpA gene Proteins 0.000 claims description 6
- 238000010348 incorporation Methods 0.000 claims description 6
- 101100166331 Dictyostelium discoideum cbpP gene Proteins 0.000 claims description 5
- 241000238631 Hexapoda Species 0.000 claims description 5
- 101100167697 Synechococcus elongatus (strain PCC 7942 / FACHB-805) cmpA gene Proteins 0.000 claims description 5
- 101150093586 clpA gene Proteins 0.000 claims description 5
- 101150115114 dnaJ gene Proteins 0.000 claims description 4
- 238000012239 gene modification Methods 0.000 claims description 4
- 230000005017 genetic modification Effects 0.000 claims description 4
- 235000013617 genetically modified food Nutrition 0.000 claims description 4
- 241000124008 Mammalia Species 0.000 claims description 3
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 claims description 3
- 108700026220 vif Genes Proteins 0.000 claims description 3
- 229910052725 zinc Inorganic materials 0.000 claims description 3
- 239000011701 zinc Substances 0.000 claims description 3
- 102100031680 Beta-catenin-interacting protein 1 Human genes 0.000 claims description 2
- 101000993469 Homo sapiens Beta-catenin-interacting protein 1 Proteins 0.000 claims description 2
- 230000003833 cell viability Effects 0.000 claims description 2
- 230000002538 fungal effect Effects 0.000 claims description 2
- 101150077981 groEL gene Proteins 0.000 claims description 2
- 101150006844 groES gene Proteins 0.000 claims description 2
- 230000000813 microbial effect Effects 0.000 claims description 2
- 210000005253 yeast cell Anatomy 0.000 claims description 2
- 102000003850 Dipeptidase 1 Human genes 0.000 claims 2
- 108090000204 Dipeptidase 1 Proteins 0.000 claims 2
- 101710107427 Extracellular metalloprotease Proteins 0.000 claims 2
- 101710200557 Extracellular small neutral protease Proteins 0.000 claims 2
- 230000032823 cell division Effects 0.000 claims 2
- 101150054895 ftsH gene Proteins 0.000 claims 2
- 239000002243 precursor Substances 0.000 claims 2
- 101100439426 Bradyrhizobium diazoefficiens (strain JCM 10833 / BCRC 13528 / IAM 13628 / NBRC 14792 / USDA 110) groEL4 gene Proteins 0.000 claims 1
- 238000004519 manufacturing process Methods 0.000 abstract description 31
- 230000014616 translation Effects 0.000 abstract description 18
- 230000004044 response Effects 0.000 abstract description 5
- 235000018102 proteins Nutrition 0.000 description 257
- 239000000047 product Substances 0.000 description 64
- 241000192142 Proteobacteria Species 0.000 description 61
- 108020004414 DNA Proteins 0.000 description 57
- 239000000523 sample Substances 0.000 description 55
- 235000019419 proteases Nutrition 0.000 description 54
- 150000007523 nucleic acids Chemical class 0.000 description 53
- 102000039446 nucleic acids Human genes 0.000 description 52
- 108020004707 nucleic acids Proteins 0.000 description 52
- 108091034117 Oligonucleotide Proteins 0.000 description 50
- 241000589516 Pseudomonas Species 0.000 description 45
- 239000013612 plasmid Substances 0.000 description 41
- 230000006698 induction Effects 0.000 description 39
- 230000001105 regulatory effect Effects 0.000 description 37
- 102000004196 processed proteins & peptides Human genes 0.000 description 34
- 108020004635 Complementary DNA Proteins 0.000 description 28
- 238000010804 cDNA synthesis Methods 0.000 description 28
- 239000002299 complementary DNA Substances 0.000 description 28
- 238000009396 hybridization Methods 0.000 description 28
- 238000004458 analytical method Methods 0.000 description 27
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 26
- 238000002474 experimental method Methods 0.000 description 25
- 239000012634 fragment Substances 0.000 description 25
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 23
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 23
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 22
- 230000035772 mutation Effects 0.000 description 22
- 230000012010 growth Effects 0.000 description 21
- 125000003729 nucleotide group Chemical group 0.000 description 21
- 108020005187 Oligonucleotide Probes Proteins 0.000 description 20
- 230000000295 complement effect Effects 0.000 description 20
- 239000002773 nucleotide Substances 0.000 description 20
- 239000002751 oligonucleotide probe Substances 0.000 description 20
- 238000003752 polymerase chain reaction Methods 0.000 description 19
- 229920001184 polypeptide Polymers 0.000 description 19
- 239000013615 primer Substances 0.000 description 19
- 238000012217 deletion Methods 0.000 description 18
- 230000037430 deletion Effects 0.000 description 18
- 230000008569 process Effects 0.000 description 18
- 102000004190 Enzymes Human genes 0.000 description 17
- 108090000790 Enzymes Proteins 0.000 description 17
- 229940088598 enzyme Drugs 0.000 description 17
- 230000006798 recombination Effects 0.000 description 17
- 238000005215 recombination Methods 0.000 description 17
- 230000001580 bacterial effect Effects 0.000 description 16
- 230000006870 function Effects 0.000 description 16
- 239000000499 gel Substances 0.000 description 16
- 241000894006 Bacteria Species 0.000 description 15
- 241000196324 Embryophyta Species 0.000 description 15
- 230000001086 cytosolic effect Effects 0.000 description 15
- 238000003556 assay Methods 0.000 description 14
- 238000004949 mass spectrometry Methods 0.000 description 14
- 235000019833 protease Nutrition 0.000 description 14
- 238000005259 measurement Methods 0.000 description 13
- 241000589634 Xanthomonas Species 0.000 description 12
- 230000003115 biocidal effect Effects 0.000 description 12
- 230000015572 biosynthetic process Effects 0.000 description 12
- 230000002503 metabolic effect Effects 0.000 description 12
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 12
- 238000013518 transcription Methods 0.000 description 12
- 230000035897 transcription Effects 0.000 description 12
- 241000122971 Stenotrophomonas Species 0.000 description 11
- 230000015556 catabolic process Effects 0.000 description 11
- 230000002950 deficient Effects 0.000 description 11
- 238000006731 degradation reaction Methods 0.000 description 11
- 210000003000 inclusion body Anatomy 0.000 description 11
- 239000003550 marker Substances 0.000 description 11
- 241000894007 species Species 0.000 description 11
- 239000000126 substance Substances 0.000 description 11
- 229940035893 uracil Drugs 0.000 description 11
- 241001453380 Burkholderia Species 0.000 description 10
- 241000625726 Oceanimonas Species 0.000 description 10
- 241000232299 Ralstonia Species 0.000 description 10
- 150000001413 amino acids Chemical class 0.000 description 10
- 238000010276 construction Methods 0.000 description 10
- 230000035882 stress Effects 0.000 description 10
- 108700026244 Open Reading Frames Proteins 0.000 description 9
- 108020004459 Small interfering RNA Proteins 0.000 description 9
- 235000001014 amino acid Nutrition 0.000 description 9
- 229940024606 amino acid Drugs 0.000 description 9
- 239000003153 chemical reaction reagent Substances 0.000 description 9
- 210000000805 cytoplasm Anatomy 0.000 description 9
- 230000004927 fusion Effects 0.000 description 9
- 239000003112 inhibitor Substances 0.000 description 9
- 230000004048 modification Effects 0.000 description 9
- 238000012986 modification Methods 0.000 description 9
- 230000017854 proteolysis Effects 0.000 description 9
- 238000012216 screening Methods 0.000 description 9
- 238000002741 site-directed mutagenesis Methods 0.000 description 9
- 239000000758 substrate Substances 0.000 description 9
- 241000589151 Azotobacter Species 0.000 description 8
- 108091027305 Heteroduplex Proteins 0.000 description 8
- 102000005741 Metalloproteases Human genes 0.000 description 8
- 108010006035 Metalloproteases Proteins 0.000 description 8
- 108091028043 Nucleic acid sequence Proteins 0.000 description 8
- 238000006243 chemical reaction Methods 0.000 description 8
- 210000000349 chromosome Anatomy 0.000 description 8
- 239000007850 fluorescent dye Substances 0.000 description 8
- 108020001507 fusion proteins Proteins 0.000 description 8
- 238000000338 in vitro Methods 0.000 description 8
- 230000003993 interaction Effects 0.000 description 8
- 230000007246 mechanism Effects 0.000 description 8
- 239000002609 medium Substances 0.000 description 8
- 231100000219 mutagenic Toxicity 0.000 description 8
- 230000003505 mutagenic effect Effects 0.000 description 8
- 238000012545 processing Methods 0.000 description 8
- 238000011002 quantification Methods 0.000 description 8
- 241000040854 Azorhizophilus Species 0.000 description 7
- 241000131407 Brevundimonas Species 0.000 description 7
- 241000863387 Cellvibrio Species 0.000 description 7
- 102000053602 DNA Human genes 0.000 description 7
- 238000000018 DNA microarray Methods 0.000 description 7
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 7
- 102000004447 HSP40 Heat-Shock Proteins Human genes 0.000 description 7
- 108010042283 HSP40 Heat-Shock Proteins Proteins 0.000 description 7
- 241000216643 Hydrogenophaga Species 0.000 description 7
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 7
- 241000293010 Oligella Species 0.000 description 7
- 230000003321 amplification Effects 0.000 description 7
- 239000013604 expression vector Substances 0.000 description 7
- 102000037865 fusion proteins Human genes 0.000 description 7
- 239000008103 glucose Substances 0.000 description 7
- 238000001727 in vivo Methods 0.000 description 7
- 238000003199 nucleic acid amplification method Methods 0.000 description 7
- 230000028327 secretion Effects 0.000 description 7
- 239000004055 small Interfering RNA Substances 0.000 description 7
- 238000003786 synthesis reaction Methods 0.000 description 7
- 230000009466 transformation Effects 0.000 description 7
- 102000004400 Aminopeptidases Human genes 0.000 description 6
- 108090000915 Aminopeptidases Proteins 0.000 description 6
- 241000219194 Arabidopsis Species 0.000 description 6
- 241000973034 Azomonas Species 0.000 description 6
- 108090001081 Dipeptidases Proteins 0.000 description 6
- 102000004860 Dipeptidases Human genes 0.000 description 6
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 6
- 241000589345 Methylococcus Species 0.000 description 6
- 241000530467 Methylosphaera Species 0.000 description 6
- 108010076504 Protein Sorting Signals Proteins 0.000 description 6
- 241000736131 Sphingomonas Species 0.000 description 6
- 239000004098 Tetracycline Substances 0.000 description 6
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 6
- 239000004473 Threonine Substances 0.000 description 6
- 238000003491 array Methods 0.000 description 6
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 6
- 230000008859 change Effects 0.000 description 6
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 6
- 235000018417 cysteine Nutrition 0.000 description 6
- 229930027917 kanamycin Natural products 0.000 description 6
- 229960000318 kanamycin Drugs 0.000 description 6
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 6
- 229930182823 kanamycin A Natural products 0.000 description 6
- 239000000203 mixture Substances 0.000 description 6
- 230000012846 protein folding Effects 0.000 description 6
- 238000000926 separation method Methods 0.000 description 6
- 238000006467 substitution reaction Methods 0.000 description 6
- 229960002180 tetracycline Drugs 0.000 description 6
- 229930101283 tetracycline Natural products 0.000 description 6
- 235000019364 tetracycline Nutrition 0.000 description 6
- 150000003522 tetracyclines Chemical class 0.000 description 6
- 230000009261 transgenic effect Effects 0.000 description 6
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 5
- 108091023037 Aptamer Proteins 0.000 description 5
- 241001626906 Blastomonas Species 0.000 description 5
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 5
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 5
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 5
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 5
- 102100034349 Integrase Human genes 0.000 description 5
- 241000589350 Methylobacter Species 0.000 description 5
- 241000321843 Methylosarcina Species 0.000 description 5
- 108010033272 Nitrilase Proteins 0.000 description 5
- 241001135311 Pseudoalteromonas nigrifaciens Species 0.000 description 5
- 238000012228 RNA interference-mediated gene silencing Methods 0.000 description 5
- 101100273253 Rhizopus niveus RNAP gene Proteins 0.000 description 5
- 241000206217 Teredinibacter Species 0.000 description 5
- 239000002253 acid Substances 0.000 description 5
- 230000001413 cellular effect Effects 0.000 description 5
- 230000001419 dependent effect Effects 0.000 description 5
- 238000013461 design Methods 0.000 description 5
- 238000001514 detection method Methods 0.000 description 5
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 5
- 210000003527 eukaryotic cell Anatomy 0.000 description 5
- 230000009368 gene silencing by RNA Effects 0.000 description 5
- 239000005090 green fluorescent protein Substances 0.000 description 5
- 239000012528 membrane Substances 0.000 description 5
- 238000002703 mutagenesis Methods 0.000 description 5
- 231100000350 mutagenesis Toxicity 0.000 description 5
- 230000002018 overexpression Effects 0.000 description 5
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 5
- 210000001322 periplasm Anatomy 0.000 description 5
- 210000001236 prokaryotic cell Anatomy 0.000 description 5
- 238000011084 recovery Methods 0.000 description 5
- 230000010076 replication Effects 0.000 description 5
- 238000010839 reverse transcription Methods 0.000 description 5
- 239000007787 solid Substances 0.000 description 5
- 238000011144 upstream manufacturing Methods 0.000 description 5
- SEHFUALWMUWDKS-UHFFFAOYSA-N 5-fluoroorotic acid Chemical compound OC(=O)C=1NC(=O)NC(=O)C=1F SEHFUALWMUWDKS-UHFFFAOYSA-N 0.000 description 4
- 108091006112 ATPases Proteins 0.000 description 4
- 102000057290 Adenosine Triphosphatases Human genes 0.000 description 4
- 229920001817 Agar Polymers 0.000 description 4
- 241000589158 Agrobacterium Species 0.000 description 4
- 241000282693 Cercopithecidae Species 0.000 description 4
- 108090000194 Dipeptidyl-peptidases and tripeptidyl-peptidases Proteins 0.000 description 4
- 102000003779 Dipeptidyl-peptidases and tripeptidyl-peptidases Human genes 0.000 description 4
- 241000588722 Escherichia Species 0.000 description 4
- 241000206602 Eukaryota Species 0.000 description 4
- 241000589601 Francisella Species 0.000 description 4
- 102000004195 Isomerases Human genes 0.000 description 4
- 108090000769 Isomerases Proteins 0.000 description 4
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 4
- 239000004472 Lysine Substances 0.000 description 4
- 241001264650 Methylocaldum Species 0.000 description 4
- 241001533203 Methylomicrobium Species 0.000 description 4
- 241000589344 Methylomonas Species 0.000 description 4
- 108091092724 Noncoding DNA Proteins 0.000 description 4
- 102000009658 Peptidylprolyl Isomerase Human genes 0.000 description 4
- 108010020062 Peptidylprolyl Isomerase Proteins 0.000 description 4
- 108091000080 Phosphotransferase Proteins 0.000 description 4
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 4
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 4
- 238000012300 Sequence Analysis Methods 0.000 description 4
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 4
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 4
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 4
- 108091023040 Transcription factor Proteins 0.000 description 4
- 102000040945 Transcription factor Human genes 0.000 description 4
- 240000008042 Zea mays Species 0.000 description 4
- 235000005824 Zea mays ssp. parviglumis Nutrition 0.000 description 4
- 235000002017 Zea mays subsp mays Nutrition 0.000 description 4
- 239000008272 agar Substances 0.000 description 4
- 239000003242 anti bacterial agent Substances 0.000 description 4
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 4
- 229940009098 aspartate Drugs 0.000 description 4
- 239000012472 biological sample Substances 0.000 description 4
- 239000003114 blood coagulation factor Substances 0.000 description 4
- 239000000872 buffer Substances 0.000 description 4
- 229910052799 carbon Inorganic materials 0.000 description 4
- 239000003054 catalyst Substances 0.000 description 4
- 230000003197 catalytic effect Effects 0.000 description 4
- 239000013592 cell lysate Substances 0.000 description 4
- 150000001875 compounds Chemical class 0.000 description 4
- 235000005822 corn Nutrition 0.000 description 4
- 238000011161 development Methods 0.000 description 4
- 230000018109 developmental process Effects 0.000 description 4
- 238000010586 diagram Methods 0.000 description 4
- 230000029087 digestion Effects 0.000 description 4
- 239000003814 drug Substances 0.000 description 4
- 238000001962 electrophoresis Methods 0.000 description 4
- 238000004520 electroporation Methods 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 238000010195 expression analysis Methods 0.000 description 4
- 238000000855 fermentation Methods 0.000 description 4
- 230000004151 fermentation Effects 0.000 description 4
- 238000003209 gene knockout Methods 0.000 description 4
- 230000002452 interceptive effect Effects 0.000 description 4
- 239000000543 intermediate Substances 0.000 description 4
- 239000003446 ligand Substances 0.000 description 4
- 238000004811 liquid chromatography Methods 0.000 description 4
- 239000002207 metabolite Substances 0.000 description 4
- 238000002966 oligonucleotide array Methods 0.000 description 4
- 102000020233 phosphotransferase Human genes 0.000 description 4
- 102000040430 polynucleotide Human genes 0.000 description 4
- 108091033319 polynucleotide Proteins 0.000 description 4
- 239000002157 polynucleotide Substances 0.000 description 4
- 101150116440 pyrF gene Proteins 0.000 description 4
- 230000002103 transcriptional effect Effects 0.000 description 4
- 238000012085 transcriptional profiling Methods 0.000 description 4
- 241000589155 Agrobacterium tumefaciens Species 0.000 description 3
- 240000002791 Brassica napus Species 0.000 description 3
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 3
- 102000005367 Carboxypeptidases Human genes 0.000 description 3
- 108010006303 Carboxypeptidases Proteins 0.000 description 3
- 102000005600 Cathepsins Human genes 0.000 description 3
- 108010084457 Cathepsins Proteins 0.000 description 3
- 108010035563 Chloramphenicol O-acetyltransferase Proteins 0.000 description 3
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 3
- 241000305071 Enterobacterales Species 0.000 description 3
- 241001646716 Escherichia coli K-12 Species 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- 241000192128 Gammaproteobacteria Species 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- 102000001398 Granzyme Human genes 0.000 description 3
- 108060005986 Granzyme Proteins 0.000 description 3
- 102100023737 GrpE protein homolog 1, mitochondrial Human genes 0.000 description 3
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 3
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 3
- 101000829489 Homo sapiens GrpE protein homolog 1, mitochondrial Proteins 0.000 description 3
- 108090000723 Insulin-Like Growth Factor I Proteins 0.000 description 3
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 3
- 229930195725 Mannitol Natural products 0.000 description 3
- 241001465754 Metazoa Species 0.000 description 3
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 3
- 244000061176 Nicotiana tabacum Species 0.000 description 3
- 240000007594 Oryza sativa Species 0.000 description 3
- 235000007164 Oryza sativa Nutrition 0.000 description 3
- 241000288906 Primates Species 0.000 description 3
- 108090000708 Proteasome Endopeptidase Complex Proteins 0.000 description 3
- 102000004245 Proteasome Endopeptidase Complex Human genes 0.000 description 3
- 241001277679 Pseudomonas mandelii Species 0.000 description 3
- 241001312486 Pseudomonas migulae Species 0.000 description 3
- RWRDLPDLKQPQOW-UHFFFAOYSA-N Pyrrolidine Chemical compound C1CCNC1 RWRDLPDLKQPQOW-UHFFFAOYSA-N 0.000 description 3
- 238000002123 RNA extraction Methods 0.000 description 3
- 102000000574 RNA-Induced Silencing Complex Human genes 0.000 description 3
- 108010016790 RNA-Induced Silencing Complex Proteins 0.000 description 3
- 108700005075 Regulator Genes Proteins 0.000 description 3
- 241001135312 Sinorhizobium Species 0.000 description 3
- 244000061456 Solanum tuberosum Species 0.000 description 3
- 235000002595 Solanum tuberosum Nutrition 0.000 description 3
- 101710154918 Trigger factor Proteins 0.000 description 3
- 241000700605 Viruses Species 0.000 description 3
- 238000009825 accumulation Methods 0.000 description 3
- 150000007513 acids Chemical class 0.000 description 3
- 230000002776 aggregation Effects 0.000 description 3
- 238000004220 aggregation Methods 0.000 description 3
- 125000001931 aliphatic group Chemical group 0.000 description 3
- 229960000723 ampicillin Drugs 0.000 description 3
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 3
- 239000012491 analyte Substances 0.000 description 3
- 230000000692 anti-sense effect Effects 0.000 description 3
- 239000000427 antigen Substances 0.000 description 3
- 108091007433 antigens Proteins 0.000 description 3
- 102000036639 antigens Human genes 0.000 description 3
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 3
- 230000008901 benefit Effects 0.000 description 3
- 230000033228 biological regulation Effects 0.000 description 3
- 238000004113 cell culture Methods 0.000 description 3
- 238000005119 centrifugation Methods 0.000 description 3
- 238000012512 characterization method Methods 0.000 description 3
- 238000003776 cleavage reaction Methods 0.000 description 3
- 230000009089 cytolysis Effects 0.000 description 3
- 229940079593 drug Drugs 0.000 description 3
- 102000035175 foldases Human genes 0.000 description 3
- 108091005749 foldases Proteins 0.000 description 3
- 238000005462 in vivo assay Methods 0.000 description 3
- 230000000977 initiatory effect Effects 0.000 description 3
- 230000003834 intracellular effect Effects 0.000 description 3
- 102000027411 intracellular receptors Human genes 0.000 description 3
- 108091008582 intracellular receptors Proteins 0.000 description 3
- 238000001155 isoelectric focusing Methods 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 235000018977 lysine Nutrition 0.000 description 3
- 239000000594 mannitol Substances 0.000 description 3
- 235000010355 mannitol Nutrition 0.000 description 3
- 230000001404 mediated effect Effects 0.000 description 3
- 238000002705 metabolomic analysis Methods 0.000 description 3
- 230000001431 metabolomic effect Effects 0.000 description 3
- 238000012775 microarray technology Methods 0.000 description 3
- 238000000520 microinjection Methods 0.000 description 3
- 230000037361 pathway Effects 0.000 description 3
- 229920001223 polyethylene glycol Polymers 0.000 description 3
- 229920000642 polymer Polymers 0.000 description 3
- 230000009467 reduction Effects 0.000 description 3
- 230000002829 reductive effect Effects 0.000 description 3
- 238000011160 research Methods 0.000 description 3
- 235000009566 rice Nutrition 0.000 description 3
- 230000007017 scission Effects 0.000 description 3
- 150000003384 small molecules Chemical class 0.000 description 3
- 125000003396 thiol group Chemical group [H]S* 0.000 description 3
- 241000701161 unidentified adenovirus Species 0.000 description 3
- 230000003827 upregulation Effects 0.000 description 3
- 230000003612 virological effect Effects 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 2
- MXHRCPNRJAMMIM-SHYZEUOFSA-N 2'-deoxyuridine Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 MXHRCPNRJAMMIM-SHYZEUOFSA-N 0.000 description 2
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 2
- LRFVTYWOQMYALW-UHFFFAOYSA-N 9H-xanthine Chemical compound O=C1NC(=O)NC2=C1NC=N2 LRFVTYWOQMYALW-UHFFFAOYSA-N 0.000 description 2
- 241000589220 Acetobacter Species 0.000 description 2
- 241000726119 Acidovorax Species 0.000 description 2
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 2
- 229930024421 Adenine Natural products 0.000 description 2
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 2
- 241000589156 Agrobacterium rhizogenes Species 0.000 description 2
- 241000588986 Alcaligenes Species 0.000 description 2
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 2
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 2
- 241001135756 Alphaproteobacteria Species 0.000 description 2
- 241001430273 Aminobacter Species 0.000 description 2
- 102100022749 Aminopeptidase N Human genes 0.000 description 2
- 102000030431 Asparaginyl endopeptidase Human genes 0.000 description 2
- 244000063299 Bacillus subtilis Species 0.000 description 2
- 235000014469 Bacillus subtilis Nutrition 0.000 description 2
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 2
- 241001135755 Betaproteobacteria Species 0.000 description 2
- 102000015081 Blood Coagulation Factors Human genes 0.000 description 2
- 108010039209 Blood Coagulation Factors Proteins 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- 235000006008 Brassica napus var napus Nutrition 0.000 description 2
- 235000011299 Brassica oleracea var botrytis Nutrition 0.000 description 2
- 240000003259 Brassica oleracea var. botrytis Species 0.000 description 2
- 235000004977 Brassica sinapistrum Nutrition 0.000 description 2
- 108010032088 Calpain Proteins 0.000 description 2
- 102000007590 Calpain Human genes 0.000 description 2
- 101100507655 Canis lupus familiaris HSPA1 gene Proteins 0.000 description 2
- 108010080937 Carboxypeptidases A Proteins 0.000 description 2
- 102000000496 Carboxypeptidases A Human genes 0.000 description 2
- 102000014914 Carrier Proteins Human genes 0.000 description 2
- 108090000426 Caspase-1 Proteins 0.000 description 2
- 108010059081 Cathepsin A Proteins 0.000 description 2
- 102000005572 Cathepsin A Human genes 0.000 description 2
- 241001529572 Chaceon affinis Species 0.000 description 2
- 108091006146 Channels Proteins 0.000 description 2
- 241001672694 Citrus reticulata Species 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- 241000589518 Comamonas testosteroni Species 0.000 description 2
- 108020004394 Complementary RNA Proteins 0.000 description 2
- 108091035707 Consensus sequence Proteins 0.000 description 2
- 229920000742 Cotton Polymers 0.000 description 2
- 241000699800 Cricetinae Species 0.000 description 2
- 244000241257 Cucumis melo Species 0.000 description 2
- 240000008067 Cucumis sativus Species 0.000 description 2
- 235000010799 Cucumis sativus var sativus Nutrition 0.000 description 2
- 235000009854 Cucurbita moschata Nutrition 0.000 description 2
- 240000001980 Cucurbita pepo Species 0.000 description 2
- 241000192700 Cyanobacteria Species 0.000 description 2
- 102000005927 Cysteine Proteases Human genes 0.000 description 2
- 108010005843 Cysteine Proteases Proteins 0.000 description 2
- 108010071840 Cytosol nonspecific dipeptidase Proteins 0.000 description 2
- 108010066133 D-octopine dehydrogenase Proteins 0.000 description 2
- 230000004543 DNA replication Effects 0.000 description 2
- 230000006820 DNA synthesis Effects 0.000 description 2
- 101150040913 DUT gene Proteins 0.000 description 2
- 241001600125 Delftia acidovorans Species 0.000 description 2
- 102100023933 Deoxyuridine 5'-triphosphate nucleotidohydrolase, mitochondrial Human genes 0.000 description 2
- YZCKVEUIGOORGS-OUBTZVSYSA-N Deuterium Chemical compound [2H] YZCKVEUIGOORGS-OUBTZVSYSA-N 0.000 description 2
- 241000255581 Drosophila <fruit fly, genus> Species 0.000 description 2
- 108700035154 EC 3.4.15.- Proteins 0.000 description 2
- 102000051496 EC 3.4.15.- Human genes 0.000 description 2
- 108090000860 Endopeptidase Clp Proteins 0.000 description 2
- 241001528534 Ensifer Species 0.000 description 2
- 101710091045 Envelope protein Proteins 0.000 description 2
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 2
- 101000686777 Escherichia phage T7 T7 RNA polymerase Proteins 0.000 description 2
- 108700039887 Essential Genes Proteins 0.000 description 2
- VTLYFUHAOXGGBS-UHFFFAOYSA-N Fe3+ Chemical compound [Fe+3] VTLYFUHAOXGGBS-UHFFFAOYSA-N 0.000 description 2
- 241000192125 Firmicutes Species 0.000 description 2
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 2
- 102000053187 Glucuronidase Human genes 0.000 description 2
- 108010060309 Glucuronidase Proteins 0.000 description 2
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 2
- 244000068988 Glycine max Species 0.000 description 2
- 235000010469 Glycine max Nutrition 0.000 description 2
- 244000299507 Gossypium hirsutum Species 0.000 description 2
- 101150076784 HSP100 gene Proteins 0.000 description 2
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 2
- 102000003839 Human Proteins Human genes 0.000 description 2
- 108090000144 Human Proteins Proteins 0.000 description 2
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 2
- 102000018251 Hypoxanthine Phosphoribosyltransferase Human genes 0.000 description 2
- 102000004218 Insulin-Like Growth Factor I Human genes 0.000 description 2
- 101710203526 Integrase Proteins 0.000 description 2
- 108020004684 Internal Ribosome Entry Sites Proteins 0.000 description 2
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 2
- SRBFZHDQGSBBOR-HWQSCIPKSA-N L-arabinopyranose Chemical compound O[C@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-HWQSCIPKSA-N 0.000 description 2
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 2
- 102000002704 Leucyl aminopeptidase Human genes 0.000 description 2
- 241001478324 Liberibacter Species 0.000 description 2
- 101710105047 Lipoprotein B Proteins 0.000 description 2
- 239000006142 Luria-Bertani Agar Substances 0.000 description 2
- 108030000089 Metallocarboxypeptidases Proteins 0.000 description 2
- 102000006166 Metallocarboxypeptidases Human genes 0.000 description 2
- 241000699666 Mus <mouse, genus> Species 0.000 description 2
- 241000699670 Mus sp. Species 0.000 description 2
- 240000005561 Musa balbisiana Species 0.000 description 2
- 241000187482 Mycobacterium avium subsp. paratuberculosis Species 0.000 description 2
- 108090000028 Neprilysin Proteins 0.000 description 2
- 102000003729 Neprilysin Human genes 0.000 description 2
- 238000000636 Northern blotting Methods 0.000 description 2
- 238000012408 PCR amplification Methods 0.000 description 2
- 229910019142 PO4 Inorganic materials 0.000 description 2
- 108090000526 Papain Proteins 0.000 description 2
- 241001494479 Pecora Species 0.000 description 2
- 108091093037 Peptide nucleic acid Proteins 0.000 description 2
- 101710188315 Protein X Proteins 0.000 description 2
- 241000157890 Pseudoalteromonas piscicida Species 0.000 description 2
- 241000218935 Pseudomonas azotoformans Species 0.000 description 2
- 241000620655 Pseudomonas brenneri Species 0.000 description 2
- 241000180027 Pseudomonas cedrina Species 0.000 description 2
- 241001670013 Pseudomonas chlororaphis subsp. aurantiaca Species 0.000 description 2
- 241000218936 Pseudomonas corrugata Species 0.000 description 2
- 241001277052 Pseudomonas libanensis Species 0.000 description 2
- 241000204735 Pseudomonas nitroreducens Species 0.000 description 2
- 241001291513 Pseudomonas orientalis Species 0.000 description 2
- 241001291486 Pseudomonas rhodesiae Species 0.000 description 2
- 241000039935 Pseudomonas thermotolerans Species 0.000 description 2
- 241001148199 Pseudomonas tolaasii Species 0.000 description 2
- 241001291485 Pseudomonas veronii Species 0.000 description 2
- 108090000919 Pyroglutamyl-Peptidase I Proteins 0.000 description 2
- 102100031108 Pyroglutamyl-peptidase 1 Human genes 0.000 description 2
- 101150015069 RNAP gene Proteins 0.000 description 2
- 244000088415 Raphanus sativus Species 0.000 description 2
- 235000006140 Raphanus sativus var sativus Nutrition 0.000 description 2
- 108700008625 Reporter Genes Proteins 0.000 description 2
- 241000589180 Rhizobium Species 0.000 description 2
- 102000012479 Serine Proteases Human genes 0.000 description 2
- 108010022999 Serine Proteases Proteins 0.000 description 2
- 108030000574 Serine-type carboxypeptidases Proteins 0.000 description 2
- 102000034328 Serine-type carboxypeptidases Human genes 0.000 description 2
- 241000607720 Serratia Species 0.000 description 2
- 241000863432 Shewanella putrefaciens Species 0.000 description 2
- 108020004682 Single-Stranded DNA Proteins 0.000 description 2
- 241000256248 Spodoptera Species 0.000 description 2
- 108010090804 Streptavidin Proteins 0.000 description 2
- 101100502843 Streptomyces anulatus fkbP gene Proteins 0.000 description 2
- 108090000794 Streptopain Proteins 0.000 description 2
- 241000282887 Suidae Species 0.000 description 2
- 101710137500 T7 RNA polymerase Proteins 0.000 description 2
- 108090001109 Thermolysin Proteins 0.000 description 2
- 241001143310 Thermotogae <phylum> Species 0.000 description 2
- 108700009124 Transcription Initiation Site Proteins 0.000 description 2
- 102000008579 Transposases Human genes 0.000 description 2
- 108010020764 Transposases Proteins 0.000 description 2
- 235000021307 Triticum Nutrition 0.000 description 2
- 244000098338 Triticum aestivum Species 0.000 description 2
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 2
- 244000078534 Vaccinium myrtillus Species 0.000 description 2
- 208000036142 Viral infection Diseases 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 229960000643 adenine Drugs 0.000 description 2
- 238000000137 annealing Methods 0.000 description 2
- RWZYAGGXGHYGMB-UHFFFAOYSA-N anthranilic acid Chemical compound NC1=CC=CC=C1C(O)=O RWZYAGGXGHYGMB-UHFFFAOYSA-N 0.000 description 2
- 229940088710 antibiotic agent Drugs 0.000 description 2
- 108010055066 asparaginylendopeptidase Proteins 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- WPYMKLBDIGXBTP-UHFFFAOYSA-N benzoic acid Chemical compound OC(=O)C1=CC=CC=C1 WPYMKLBDIGXBTP-UHFFFAOYSA-N 0.000 description 2
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 2
- 108091008324 binding proteins Proteins 0.000 description 2
- 238000006664 bond formation reaction Methods 0.000 description 2
- 239000001506 calcium phosphate Substances 0.000 description 2
- 229910000389 calcium phosphate Inorganic materials 0.000 description 2
- 235000011010 calcium phosphates Nutrition 0.000 description 2
- 238000012219 cassette mutagenesis Methods 0.000 description 2
- 230000010261 cell growth Effects 0.000 description 2
- 108091092356 cellular DNA Proteins 0.000 description 2
- 230000010001 cellular homeostasis Effects 0.000 description 2
- 230000004637 cellular stress Effects 0.000 description 2
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 2
- 239000013611 chromosomal DNA Substances 0.000 description 2
- 230000002759 chromosomal effect Effects 0.000 description 2
- 238000010367 cloning Methods 0.000 description 2
- 108090001092 clostripain Proteins 0.000 description 2
- 238000004440 column chromatography Methods 0.000 description 2
- 239000003184 complementary RNA Substances 0.000 description 2
- 108010082025 cyan fluorescent protein Proteins 0.000 description 2
- 108010011219 dUTP pyrophosphatase Proteins 0.000 description 2
- 238000007405 data analysis Methods 0.000 description 2
- 230000007423 decrease Effects 0.000 description 2
- 239000003398 denaturant Substances 0.000 description 2
- 238000004925 denaturation Methods 0.000 description 2
- 230000036425 denaturation Effects 0.000 description 2
- MXHRCPNRJAMMIM-UHFFFAOYSA-N desoxyuridine Natural products C1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 MXHRCPNRJAMMIM-UHFFFAOYSA-N 0.000 description 2
- 239000003599 detergent Substances 0.000 description 2
- 230000009977 dual effect Effects 0.000 description 2
- 229940066758 endopeptidases Drugs 0.000 description 2
- 230000002708 enhancing effect Effects 0.000 description 2
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 2
- 238000001502 gel electrophoresis Methods 0.000 description 2
- 238000011223 gene expression profiling Methods 0.000 description 2
- 239000011521 glass Substances 0.000 description 2
- 101150053330 grpE gene Proteins 0.000 description 2
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical compound O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 2
- 238000007417 hierarchical cluster analysis Methods 0.000 description 2
- 238000004128 high performance liquid chromatography Methods 0.000 description 2
- 210000005260 human cell Anatomy 0.000 description 2
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 2
- 238000003018 immunoassay Methods 0.000 description 2
- 238000000099 in vitro assay Methods 0.000 description 2
- 230000002779 inactivation Effects 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 230000005764 inhibitory process Effects 0.000 description 2
- 238000011081 inoculation Methods 0.000 description 2
- 230000009545 invasion Effects 0.000 description 2
- 238000002955 isolation Methods 0.000 description 2
- 238000006317 isomerization reaction Methods 0.000 description 2
- 238000002372 labelling Methods 0.000 description 2
- 150000002632 lipids Chemical class 0.000 description 2
- 230000002934 lysing effect Effects 0.000 description 2
- 238000002824 mRNA display Methods 0.000 description 2
- 239000000463 material Substances 0.000 description 2
- 229910052751 metal Inorganic materials 0.000 description 2
- 239000002184 metal Substances 0.000 description 2
- 229910044991 metal oxide Inorganic materials 0.000 description 2
- 150000004706 metal oxides Chemical class 0.000 description 2
- MYWUZJCMWCOHBA-VIFPVBQESA-N methamphetamine Chemical compound CN[C@@H](C)CC1=CC=CC=C1 MYWUZJCMWCOHBA-VIFPVBQESA-N 0.000 description 2
- 238000010208 microarray analysis Methods 0.000 description 2
- 238000010369 molecular cloning Methods 0.000 description 2
- 239000013642 negative control Substances 0.000 description 2
- 108010058731 nopaline synthase Proteins 0.000 description 2
- 239000012038 nucleophile Substances 0.000 description 2
- 229940124276 oligodeoxyribonucleotide Drugs 0.000 description 2
- 238000012261 overproduction Methods 0.000 description 2
- 229940055729 papain Drugs 0.000 description 2
- 235000019834 papain Nutrition 0.000 description 2
- 239000010452 phosphate Substances 0.000 description 2
- 239000013600 plasmid vector Substances 0.000 description 2
- 229920000729 poly(L-lysine) polymer Polymers 0.000 description 2
- 229920002401 polyacrylamide Polymers 0.000 description 2
- 238000001955 polymer synthesis method Methods 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 238000007639 printing Methods 0.000 description 2
- 108010029690 procollagenase Proteins 0.000 description 2
- 230000004952 protein activity Effects 0.000 description 2
- 230000004845 protein aggregation Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- RXWNCPJZOCPEPQ-NVWDDTSBSA-N puromycin Chemical compound C1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CO RXWNCPJZOCPEPQ-NVWDDTSBSA-N 0.000 description 2
- 238000003259 recombinant expression Methods 0.000 description 2
- 108010054624 red fluorescent protein Proteins 0.000 description 2
- 230000008929 regeneration Effects 0.000 description 2
- 238000011069 regeneration method Methods 0.000 description 2
- 230000008439 repair process Effects 0.000 description 2
- 230000003252 repetitive effect Effects 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 230000001177 retroviral effect Effects 0.000 description 2
- 238000003757 reverse transcription PCR Methods 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 230000035945 sensitivity Effects 0.000 description 2
- 230000035939 shock Effects 0.000 description 2
- 239000011734 sodium Substances 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 238000010186 staining Methods 0.000 description 2
- 238000010561 standard procedure Methods 0.000 description 2
- 230000008685 targeting Effects 0.000 description 2
- 238000012360 testing method Methods 0.000 description 2
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 2
- 229940104230 thymidine Drugs 0.000 description 2
- 230000005026 transcription initiation Effects 0.000 description 2
- 238000011222 transcriptome analysis Methods 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- 230000014621 translational initiation Effects 0.000 description 2
- 108091005703 transmembrane proteins Proteins 0.000 description 2
- 102000035160 transmembrane proteins Human genes 0.000 description 2
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 2
- 101150118060 trxA gene Proteins 0.000 description 2
- 108010087967 type I signal peptidase Proteins 0.000 description 2
- 230000009385 viral infection Effects 0.000 description 2
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 2
- 238000001262 western blot Methods 0.000 description 2
- 108091005957 yellow fluorescent proteins Proteins 0.000 description 2
- DIGQNXIGRZPYDK-WKSCXVIASA-N (2R)-6-amino-2-[[2-[[(2S)-2-[[2-[[(2R)-2-[[(2S)-2-[[(2R,3S)-2-[[2-[[(2S)-2-[[2-[[(2S)-2-[[(2S)-2-[[(2R)-2-[[(2S,3S)-2-[[(2R)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[2-[[(2S)-2-[[(2R)-2-[[2-[[2-[[2-[(2-amino-1-hydroxyethylidene)amino]-3-carboxy-1-hydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1,5-dihydroxy-5-iminopentylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]hexanoic acid Chemical compound C[C@@H]([C@@H](C(=N[C@@H](CS)C(=N[C@@H](C)C(=N[C@@H](CO)C(=NCC(=N[C@@H](CCC(=N)O)C(=NC(CS)C(=N[C@H]([C@H](C)O)C(=N[C@H](CS)C(=N[C@H](CO)C(=NCC(=N[C@H](CS)C(=NCC(=N[C@H](CCCCN)C(=O)O)O)O)O)O)O)O)O)O)O)O)O)O)O)N=C([C@H](CS)N=C([C@H](CO)N=C([C@H](CO)N=C([C@H](C)N=C(CN=C([C@H](CO)N=C([C@H](CS)N=C(CN=C(C(CS)N=C(C(CC(=O)O)N=C(CN)O)O)O)O)O)O)O)O)O)O)O)O DIGQNXIGRZPYDK-WKSCXVIASA-N 0.000 description 1
- SBKVPJHMSUXZTA-MEJXFZFPSA-N (2S)-2-[[(2S)-2-[[(2S)-1-[(2S)-5-amino-2-[[2-[[(2S)-1-[(2S)-6-amino-2-[[(2S)-2-[[(2S)-5-amino-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-amino-3-(1H-indol-3-yl)propanoyl]amino]-3-(1H-imidazol-4-yl)propanoyl]amino]-3-(1H-indol-3-yl)propanoyl]amino]-4-methylpentanoyl]amino]-5-oxopentanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]pyrrolidine-2-carbonyl]amino]acetyl]amino]-5-oxopentanoyl]pyrrolidine-2-carbonyl]amino]-4-methylsulfanylbutanoyl]amino]-3-(4-hydroxyphenyl)propanoic acid Chemical compound C([C@@H](C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)NC(=O)[C@@H](N)CC=1C2=CC=CC=C2NC=1)C1=CNC=N1 SBKVPJHMSUXZTA-MEJXFZFPSA-N 0.000 description 1
- MJYQFWSXKFLTAY-OVEQLNGDSA-N (2r,3r)-2,3-bis[(4-hydroxy-3-methoxyphenyl)methyl]butane-1,4-diol;(2r,3r,4s,5s,6r)-6-(hydroxymethyl)oxane-2,3,4,5-tetrol Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O.C1=C(O)C(OC)=CC(C[C@@H](CO)[C@H](CO)CC=2C=C(OC)C(O)=CC=2)=C1 MJYQFWSXKFLTAY-OVEQLNGDSA-N 0.000 description 1
- MRXDGVXSWIXTQL-HYHFHBMOSA-N (2s)-2-[[(1s)-1-(2-amino-1,4,5,6-tetrahydropyrimidin-6-yl)-2-[[(2s)-4-methyl-1-oxo-1-[[(2s)-1-oxo-3-phenylpropan-2-yl]amino]pentan-2-yl]amino]-2-oxoethyl]carbamoylamino]-3-phenylpropanoic acid Chemical compound C([C@H](NC(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C=O)C1NC(N)=NCC1)C(O)=O)C1=CC=CC=C1 MRXDGVXSWIXTQL-HYHFHBMOSA-N 0.000 description 1
- IJWCGVPEDDQUDE-YGJAXBLXSA-N (2s)-2-[[(1s)-2-[[(2s)-5-amino-1,5-dioxo-1-[[(2s)-1-oxopropan-2-yl]amino]pentan-2-yl]amino]-1-[(6s)-2-amino-1,4,5,6-tetrahydropyrimidin-6-yl]-2-oxoethyl]carbamoylamino]-4-methylpentanoic acid Chemical compound O=C[C@H](C)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)N[C@@H](CC(C)C)C(O)=O)[C@@H]1CCN=C(N)N1 IJWCGVPEDDQUDE-YGJAXBLXSA-N 0.000 description 1
- KDXKERNSBIXSRK-RGXOIRHWSA-N (2s)-6-amino-2-(deuterioamino)hexanoic acid Chemical compound [2H]N[C@H](C(O)=O)CCCCN KDXKERNSBIXSRK-RGXOIRHWSA-N 0.000 description 1
- PMHUSCHKTSTQEP-UHFFFAOYSA-N (4-carbamimidoylphenyl)methanesulfonyl fluoride Chemical compound NC(=N)C1=CC=C(CS(F)(=O)=O)C=C1 PMHUSCHKTSTQEP-UHFFFAOYSA-N 0.000 description 1
- NDJNDUULNXNRQD-XKBRQERYSA-N 1-[(2r,4s,5s)-5-[bromo(hydroxy)methyl]-4-hydroxyoxolan-2-yl]pyrimidine-2,4-dione Chemical compound C1[C@H](O)[C@@H](C(Br)O)O[C@H]1N1C(=O)NC(=O)C=C1 NDJNDUULNXNRQD-XKBRQERYSA-N 0.000 description 1
- 102100024341 10 kDa heat shock protein, mitochondrial Human genes 0.000 description 1
- 101710122378 10 kDa heat shock protein, mitochondrial Proteins 0.000 description 1
- AXAVXPMQTGXXJZ-UHFFFAOYSA-N 2-aminoacetic acid;2-amino-2-(hydroxymethyl)propane-1,3-diol Chemical compound NCC(O)=O.OCC(N)(CO)CO AXAVXPMQTGXXJZ-UHFFFAOYSA-N 0.000 description 1
- 101710149439 20 kDa chaperonin, chloroplastic Proteins 0.000 description 1
- SUGXUUGGLDCZKB-UHFFFAOYSA-N 3,4-dichloroisocoumarin Chemical compound C1=CC=C2C(Cl)=C(Cl)OC(=O)C2=C1 SUGXUUGGLDCZKB-UHFFFAOYSA-N 0.000 description 1
- ZPZDIFSPRVHGIF-UHFFFAOYSA-N 3-aminopropylsilicon Chemical compound NCCC[Si] ZPZDIFSPRVHGIF-UHFFFAOYSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- LKDMKWNDBAVNQZ-UHFFFAOYSA-N 4-[[1-[[1-[2-[[1-(4-nitroanilino)-1-oxo-3-phenylpropan-2-yl]carbamoyl]pyrrolidin-1-yl]-1-oxopropan-2-yl]amino]-1-oxopropan-2-yl]amino]-4-oxobutanoic acid Chemical compound OC(=O)CCC(=O)NC(C)C(=O)NC(C)C(=O)N1CCCC1C(=O)NC(C(=O)NC=1C=CC(=CC=1)[N+]([O-])=O)CC1=CC=CC=C1 LKDMKWNDBAVNQZ-UHFFFAOYSA-N 0.000 description 1
- TVZGACDUOSZQKY-LBPRGKRZSA-N 4-aminofolic acid Chemical compound C1=NC2=NC(N)=NC(N)=C2N=C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 TVZGACDUOSZQKY-LBPRGKRZSA-N 0.000 description 1
- 101710144734 48 kDa protein Proteins 0.000 description 1
- 102100038222 60 kDa heat shock protein, mitochondrial Human genes 0.000 description 1
- 101710154868 60 kDa heat shock protein, mitochondrial Proteins 0.000 description 1
- 102100039602 ARF GTPase-activating protein GIT2 Human genes 0.000 description 1
- 240000004507 Abelmoschus esculentus Species 0.000 description 1
- 241000208140 Acer Species 0.000 description 1
- 244000283763 Acetobacter aceti Species 0.000 description 1
- 235000007847 Acetobacter aceti Nutrition 0.000 description 1
- 108010000700 Acetolactate synthase Proteins 0.000 description 1
- 241000580482 Acidobacteria Species 0.000 description 1
- 241001478307 Acidomonas Species 0.000 description 1
- 241000589210 Acidomonas methanolica Species 0.000 description 1
- 241001133760 Acoelorraphe Species 0.000 description 1
- 108090000107 Acrosin Proteins 0.000 description 1
- 102100026041 Acrosin Human genes 0.000 description 1
- 235000009436 Actinidia deliciosa Nutrition 0.000 description 1
- 244000298697 Actinidia deliciosa Species 0.000 description 1
- 102100032488 Acylamino-acid-releasing enzyme Human genes 0.000 description 1
- 108010061216 Acylaminoacyl-peptidase Proteins 0.000 description 1
- 241000947856 Aeromonadales Species 0.000 description 1
- 235000001674 Agaricus brunnescens Nutrition 0.000 description 1
- 102100036475 Alanine aminotransferase 1 Human genes 0.000 description 1
- 108010075409 Alanine carboxypeptidase Proteins 0.000 description 1
- 241000588813 Alcaligenes faecalis Species 0.000 description 1
- 102100034044 All-trans-retinol dehydrogenase [NAD(+)] ADH1B Human genes 0.000 description 1
- 101710193111 All-trans-retinol dehydrogenase [NAD(+)] ADH4 Proteins 0.000 description 1
- 241000234282 Allium Species 0.000 description 1
- 235000005254 Allium ampeloprasum Nutrition 0.000 description 1
- 240000006108 Allium ampeloprasum Species 0.000 description 1
- 235000002732 Allium cepa var. cepa Nutrition 0.000 description 1
- 240000002234 Allium sativum Species 0.000 description 1
- 102100033312 Alpha-2-macroglobulin Human genes 0.000 description 1
- 241000947840 Alteromonadales Species 0.000 description 1
- 102000004092 Amidohydrolases Human genes 0.000 description 1
- 108090000531 Amidohydrolases Proteins 0.000 description 1
- 108091029845 Aminoallyl nucleotide Proteins 0.000 description 1
- 241001646016 Aminobacter aminovorans Species 0.000 description 1
- 102100032126 Aminopeptidase B Human genes 0.000 description 1
- 101710082401 Aminopeptidase Ey Proteins 0.000 description 1
- 108030000961 Aminopeptidase Y Proteins 0.000 description 1
- 244000144730 Amygdalus persica Species 0.000 description 1
- FQEQMASDZFXSJI-UHFFFAOYSA-N Anagyrin Natural products C12CCCCN2CC2C3=CC=CC(=O)N3CC1C2 FQEQMASDZFXSJI-UHFFFAOYSA-N 0.000 description 1
- 108090000886 Ananain Proteins 0.000 description 1
- 244000099147 Ananas comosus Species 0.000 description 1
- 235000007119 Ananas comosus Nutrition 0.000 description 1
- 102100035765 Angiotensin-converting enzyme 2 Human genes 0.000 description 1
- 240000007087 Apium graveolens Species 0.000 description 1
- 235000015849 Apium graveolens Dulce Group Nutrition 0.000 description 1
- 235000010591 Appio Nutrition 0.000 description 1
- 108010039627 Aprotinin Proteins 0.000 description 1
- 241001142141 Aquificae <phylum> Species 0.000 description 1
- 235000017060 Arachis glabrata Nutrition 0.000 description 1
- 244000105624 Arachis hypogaea Species 0.000 description 1
- 235000010777 Arachis hypogaea Nutrition 0.000 description 1
- 235000018262 Arachis monticola Nutrition 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- 108090000101 Asclepain Proteins 0.000 description 1
- 244000003416 Asparagus officinalis Species 0.000 description 1
- 235000005340 Asparagus officinalis Nutrition 0.000 description 1
- 108010063172 Aspartate dehydrogenase Proteins 0.000 description 1
- 102000004580 Aspartic Acid Proteases Human genes 0.000 description 1
- 108010017640 Aspartic Acid Proteases Proteins 0.000 description 1
- 108090000618 Atrolysin B Proteins 0.000 description 1
- 108090000616 Atrolysin C Proteins 0.000 description 1
- 108090000664 Atrolysin E Proteins 0.000 description 1
- 108090000666 Atrolysin F Proteins 0.000 description 1
- 235000007319 Avena orientalis Nutrition 0.000 description 1
- 241000209763 Avena sativa Species 0.000 description 1
- 235000007558 Avena sp Nutrition 0.000 description 1
- 235000000832 Ayote Nutrition 0.000 description 1
- 241000217480 Azomonas agilis Species 0.000 description 1
- 241000973036 Azorhizophilus paspali Species 0.000 description 1
- 241000589154 Azotobacter group Species 0.000 description 1
- 108010007337 Azurin Proteins 0.000 description 1
- 108700003860 Bacterial Genes Proteins 0.000 description 1
- 108010066768 Bacterial leucyl aminopeptidase Proteins 0.000 description 1
- 241000605059 Bacteroidetes Species 0.000 description 1
- 101710113841 Barrierpepsin Proteins 0.000 description 1
- 241000588882 Beijerinckia Species 0.000 description 1
- 241000588883 Beijerinckia indica Species 0.000 description 1
- 235000016068 Berberis vulgaris Nutrition 0.000 description 1
- VGGGPCQERPFHOB-MCIONIFRSA-N Bestatin Chemical compound CC(C)C[C@H](C(O)=O)NC(=O)[C@@H](O)[C@H](N)CC1=CC=CC=C1 VGGGPCQERPFHOB-MCIONIFRSA-N 0.000 description 1
- VGGGPCQERPFHOB-UHFFFAOYSA-N Bestatin Natural products CC(C)CC(C(O)=O)NC(=O)C(O)C(N)CC1=CC=CC=C1 VGGGPCQERPFHOB-UHFFFAOYSA-N 0.000 description 1
- 241000335053 Beta vulgaris Species 0.000 description 1
- 241000219310 Beta vulgaris subsp. vulgaris Species 0.000 description 1
- 102100031006 Beta-Ala-His dipeptidase Human genes 0.000 description 1
- 108030004753 Beta-Ala-His dipeptidases Proteins 0.000 description 1
- 108010005785 Beta-aspartyl-peptidase Proteins 0.000 description 1
- 102100026189 Beta-galactosidase Human genes 0.000 description 1
- 108030001561 Beta-lytic metalloendopeptidases Proteins 0.000 description 1
- 241001478330 Blastomonas natatoria Species 0.000 description 1
- 108010006654 Bleomycin Proteins 0.000 description 1
- 102000004506 Blood Proteins Human genes 0.000 description 1
- 108010017384 Blood Proteins Proteins 0.000 description 1
- 108090000654 Bone morphogenetic protein 1 Proteins 0.000 description 1
- 102000004152 Bone morphogenetic protein 1 Human genes 0.000 description 1
- 241000588807 Bordetella Species 0.000 description 1
- 241000588832 Bordetella pertussis Species 0.000 description 1
- 101000984722 Bos taurus Pancreatic trypsin inhibitor Proteins 0.000 description 1
- 241000167854 Bourreria succulenta Species 0.000 description 1
- 235000014698 Brassica juncea var multisecta Nutrition 0.000 description 1
- 235000011293 Brassica napus Nutrition 0.000 description 1
- 240000000385 Brassica napus var. napus Species 0.000 description 1
- 240000007124 Brassica oleracea Species 0.000 description 1
- 235000003899 Brassica oleracea var acephala Nutrition 0.000 description 1
- 235000011301 Brassica oleracea var capitata Nutrition 0.000 description 1
- 235000004221 Brassica oleracea var gemmifera Nutrition 0.000 description 1
- 235000017647 Brassica oleracea var italica Nutrition 0.000 description 1
- 235000001169 Brassica oleracea var oleracea Nutrition 0.000 description 1
- 244000308368 Brassica oleracea var. gemmifera Species 0.000 description 1
- 235000006618 Brassica rapa subsp oleifera Nutrition 0.000 description 1
- 235000000540 Brassica rapa subsp rapa Nutrition 0.000 description 1
- 241000589539 Brevundimonas diminuta Species 0.000 description 1
- 241001598984 Bromius obscurus Species 0.000 description 1
- 235000004936 Bromus mango Nutrition 0.000 description 1
- 241000589562 Brucella Species 0.000 description 1
- 241000589567 Brucella abortus Species 0.000 description 1
- 241001148106 Brucella melitensis Species 0.000 description 1
- 241000589513 Burkholderia cepacia Species 0.000 description 1
- 108010049990 CD13 Antigens Proteins 0.000 description 1
- 102100022443 CXADR-like membrane protein Human genes 0.000 description 1
- 102100021868 Calnexin Human genes 0.000 description 1
- 108010056891 Calnexin Proteins 0.000 description 1
- 102100029968 Calreticulin Human genes 0.000 description 1
- 108090000549 Calreticulin Proteins 0.000 description 1
- 101710124576 Candidapepsin Proteins 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 244000025254 Cannabis sativa Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 235000002566 Capsicum Nutrition 0.000 description 1
- 101710167800 Capsid assembly scaffolding protein Proteins 0.000 description 1
- 101710132601 Capsid protein Proteins 0.000 description 1
- 108090000087 Carboxypeptidase B Proteins 0.000 description 1
- 102000003670 Carboxypeptidase B Human genes 0.000 description 1
- 102100032407 Carboxypeptidase D Human genes 0.000 description 1
- 108090000018 Carboxypeptidase D Proteins 0.000 description 1
- 102100032378 Carboxypeptidase E Human genes 0.000 description 1
- 108010058255 Carboxypeptidase H Proteins 0.000 description 1
- 108090000012 Carboxypeptidase T Proteins 0.000 description 1
- 235000009467 Carica papaya Nutrition 0.000 description 1
- 240000006432 Carica papaya Species 0.000 description 1
- 108010078791 Carrier Proteins Proteins 0.000 description 1
- 102100035904 Caspase-1 Human genes 0.000 description 1
- 102000053642 Catalytic RNA Human genes 0.000 description 1
- 108090000994 Catalytic RNA Proteins 0.000 description 1
- 108090000712 Cathepsin B Proteins 0.000 description 1
- 102000004225 Cathepsin B Human genes 0.000 description 1
- 102000003902 Cathepsin C Human genes 0.000 description 1
- 108090000267 Cathepsin C Proteins 0.000 description 1
- 108090000258 Cathepsin D Proteins 0.000 description 1
- 102000003908 Cathepsin D Human genes 0.000 description 1
- 102100025975 Cathepsin G Human genes 0.000 description 1
- 108090000617 Cathepsin G Proteins 0.000 description 1
- 108090000619 Cathepsin H Proteins 0.000 description 1
- 102000004175 Cathepsin H Human genes 0.000 description 1
- 108090000613 Cathepsin S Proteins 0.000 description 1
- 102100035654 Cathepsin S Human genes 0.000 description 1
- 102100026657 Cathepsin Z Human genes 0.000 description 1
- 241000218645 Cedrus Species 0.000 description 1
- 241000010977 Cellvibrio japonicus Species 0.000 description 1
- 241001532572 Cellvibrio mixtus Species 0.000 description 1
- 241000282994 Cervidae Species 0.000 description 1
- 101710163595 Chaperone protein DnaK Proteins 0.000 description 1
- 102000016078 Chaperonin Containing TCP-1 Human genes 0.000 description 1
- 108010010706 Chaperonin Containing TCP-1 Proteins 0.000 description 1
- 108050001186 Chaperonin Cpn60 Proteins 0.000 description 1
- 101710104159 Chaperonin GroEL Proteins 0.000 description 1
- 101710108115 Chaperonin GroEL, chloroplastic Proteins 0.000 description 1
- 102000052603 Chaperonins Human genes 0.000 description 1
- 102000019034 Chemokines Human genes 0.000 description 1
- 108010012236 Chemokines Proteins 0.000 description 1
- 241001185363 Chlamydiae Species 0.000 description 1
- 241000191368 Chlorobi Species 0.000 description 1
- 241000192733 Chloroflexus Species 0.000 description 1
- 241001670059 Chromohalobacter beijerinckii Species 0.000 description 1
- 206010061764 Chromosomal deletion Diseases 0.000 description 1
- 208000031639 Chromosome Deletion Diseases 0.000 description 1
- 235000007516 Chrysanthemum Nutrition 0.000 description 1
- 240000005250 Chrysanthemum indicum Species 0.000 description 1
- 241001143290 Chrysiogenetes <phylum> Species 0.000 description 1
- 102100024539 Chymase Human genes 0.000 description 1
- 108090000227 Chymases Proteins 0.000 description 1
- 108090001069 Chymopapain Proteins 0.000 description 1
- 108090000746 Chymosin Proteins 0.000 description 1
- OLVPQBGMUGIKIW-UHFFFAOYSA-N Chymostatin Natural products C=1C=CC=CC=1CC(C=O)NC(=O)C(C(C)CC)NC(=O)C(C1NC(N)=NCC1)NC(=O)NC(C(O)=O)CC1=CC=CC=C1 OLVPQBGMUGIKIW-UHFFFAOYSA-N 0.000 description 1
- 108090000317 Chymotrypsin Proteins 0.000 description 1
- 108090000205 Chymotrypsin C Proteins 0.000 description 1
- 102100039511 Chymotrypsin-C Human genes 0.000 description 1
- 102100023336 Chymotrypsin-like elastase family member 3B Human genes 0.000 description 1
- 240000006740 Cichorium endivia Species 0.000 description 1
- 235000007542 Cichorium intybus Nutrition 0.000 description 1
- 244000298479 Cichorium intybus Species 0.000 description 1
- 244000241235 Citrullus lanatus Species 0.000 description 1
- 235000012828 Citrullus lanatus var citroides Nutrition 0.000 description 1
- 241000207199 Citrus Species 0.000 description 1
- 235000008733 Citrus aurantifolia Nutrition 0.000 description 1
- 235000005979 Citrus limon Nutrition 0.000 description 1
- 244000131522 Citrus pyriformis Species 0.000 description 1
- 240000000560 Citrus x paradisi Species 0.000 description 1
- 108030007055 Classical-complement-pathway C3/C5 convertases Proteins 0.000 description 1
- 241001112695 Clostridiales Species 0.000 description 1
- 241000243321 Cnidaria Species 0.000 description 1
- 101710177832 Co-chaperonin GroES Proteins 0.000 description 1
- 101710094648 Coat protein Proteins 0.000 description 1
- 235000013162 Cocos nucifera Nutrition 0.000 description 1
- 244000060011 Cocos nucifera Species 0.000 description 1
- 108700010070 Codon Usage Proteins 0.000 description 1
- 240000007154 Coffea arabica Species 0.000 description 1
- 235000002787 Coriandrum sativum Nutrition 0.000 description 1
- 244000018436 Coriandrum sativum Species 0.000 description 1
- 101000774636 Crotalus ruber ruber Snake venom metalloproteinase HT-2 Proteins 0.000 description 1
- 240000005109 Cryptomeria japonica Species 0.000 description 1
- 235000015510 Cucumis melo subsp melo Nutrition 0.000 description 1
- 235000009847 Cucumis melo var cantalupensis Nutrition 0.000 description 1
- 235000015001 Cucumis melo var inodorus Nutrition 0.000 description 1
- 240000002495 Cucumis melo var. inodorus Species 0.000 description 1
- 235000009852 Cucurbita pepo Nutrition 0.000 description 1
- 235000009804 Cucurbita pepo subsp pepo Nutrition 0.000 description 1
- 241000219130 Cucurbita pepo subsp. pepo Species 0.000 description 1
- 235000003954 Cucurbita pepo var melopepo Nutrition 0.000 description 1
- 102000001493 Cyclophilins Human genes 0.000 description 1
- 108010068682 Cyclophilins Proteins 0.000 description 1
- 235000017788 Cydonia oblonga Nutrition 0.000 description 1
- 244000019459 Cynara cardunculus Species 0.000 description 1
- 235000019106 Cynara scolymus Nutrition 0.000 description 1
- 102000003950 Cysteine Endopeptidases Human genes 0.000 description 1
- 108090000395 Cysteine Endopeptidases Proteins 0.000 description 1
- 108030001451 Cysteine-type carboxypeptidases Proteins 0.000 description 1
- 108091000069 Cystinyl Aminopeptidase Proteins 0.000 description 1
- 102000030900 Cystinyl aminopeptidase Human genes 0.000 description 1
- 108090000695 Cytokines Proteins 0.000 description 1
- 102000004127 Cytokines Human genes 0.000 description 1
- 108030000958 Cytosol alanyl aminopeptidases Proteins 0.000 description 1
- 102100034560 Cytosol aminopeptidase Human genes 0.000 description 1
- 102000012410 DNA Ligases Human genes 0.000 description 1
- 108010061982 DNA Ligases Proteins 0.000 description 1
- 108020003215 DNA Probes Proteins 0.000 description 1
- 230000004544 DNA amplification Effects 0.000 description 1
- 239000003155 DNA primer Substances 0.000 description 1
- 239000003298 DNA probe Substances 0.000 description 1
- 238000012270 DNA recombination Methods 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 230000004568 DNA-binding Effects 0.000 description 1
- 241000252212 Danio rerio Species 0.000 description 1
- 235000002767 Daucus carota Nutrition 0.000 description 1
- 244000000626 Daucus carota Species 0.000 description 1
- 241001143296 Deferribacteres <phylum> Species 0.000 description 1
- 101710088194 Dehydrogenase Proteins 0.000 description 1
- 241000192093 Deinococcus Species 0.000 description 1
- 241001135761 Deltaproteobacteria Species 0.000 description 1
- 241001180360 Derxia Species 0.000 description 1
- 241001180351 Derxia gummosa Species 0.000 description 1
- 108030001574 Deuterolysin Proteins 0.000 description 1
- 241000970811 Dictyoglomi Species 0.000 description 1
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 1
- 240000005717 Dioscorea alata Species 0.000 description 1
- 235000002723 Dioscorea alata Nutrition 0.000 description 1
- 235000007056 Dioscorea composita Nutrition 0.000 description 1
- 235000009723 Dioscorea convolvulacea Nutrition 0.000 description 1
- 235000005362 Dioscorea floribunda Nutrition 0.000 description 1
- 235000004868 Dioscorea macrostachya Nutrition 0.000 description 1
- 235000005361 Dioscorea nummularia Nutrition 0.000 description 1
- 235000005360 Dioscorea spiculiflora Nutrition 0.000 description 1
- 102100020751 Dipeptidyl peptidase 2 Human genes 0.000 description 1
- 102100020750 Dipeptidyl peptidase 3 Human genes 0.000 description 1
- 102100025012 Dipeptidyl peptidase 4 Human genes 0.000 description 1
- 102100035966 DnaJ homolog subfamily A member 2 Human genes 0.000 description 1
- SNRUBQQJIBEYMU-UHFFFAOYSA-N Dodecane Natural products CCCCCCCCCCCC SNRUBQQJIBEYMU-UHFFFAOYSA-N 0.000 description 1
- 235000014466 Douglas bleu Nutrition 0.000 description 1
- 241000255601 Drosophila melanogaster Species 0.000 description 1
- 108700033355 EC 3.4.19.- Proteins 0.000 description 1
- 102000057898 EC 3.4.19.- Human genes 0.000 description 1
- 108700036006 EC 3.4.21.67 Proteins 0.000 description 1
- 108700036061 EC 3.4.21.88 Proteins 0.000 description 1
- 108700033921 EC 3.4.23.20 Proteins 0.000 description 1
- 108700033346 EC 3.4.24.51 Proteins 0.000 description 1
- 108700033344 EC 3.4.24.54 Proteins 0.000 description 1
- 238000002965 ELISA Methods 0.000 description 1
- 235000001950 Elaeis guineensis Nutrition 0.000 description 1
- 244000127993 Elaeis melanococca Species 0.000 description 1
- IJWCGVPEDDQUDE-UHFFFAOYSA-N Elastatinal Natural products O=CC(C)NC(=O)C(CCC(N)=O)NC(=O)C(NC(=O)NC(CC(C)C)C(O)=O)C1CCN=C(N)N1 IJWCGVPEDDQUDE-UHFFFAOYSA-N 0.000 description 1
- 108010067770 Endopeptidase K Proteins 0.000 description 1
- 102100030013 Endoribonuclease Human genes 0.000 description 1
- 108010093099 Endoribonucleases Proteins 0.000 description 1
- 108010092208 Endothia aspartic proteinase Proteins 0.000 description 1
- 244000148064 Enicostema verticillatum Species 0.000 description 1
- 241001528536 Ensifer adhaerens Species 0.000 description 1
- 241000588921 Enterobacteriaceae Species 0.000 description 1
- 102100029727 Enteropeptidase Human genes 0.000 description 1
- 108010013369 Enteropeptidase Proteins 0.000 description 1
- 239000004593 Epoxy Substances 0.000 description 1
- 241001148568 Epsilonproteobacteria Species 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 241001331845 Equus asinus x caballus Species 0.000 description 1
- 244000024675 Eruca sativa Species 0.000 description 1
- 235000014755 Eruca sativa Nutrition 0.000 description 1
- 241000588698 Erwinia Species 0.000 description 1
- 241000660147 Escherichia coli str. K-12 substr. MG1655 Species 0.000 description 1
- 241000567413 Estigmene Species 0.000 description 1
- 241000207447 Estrella Species 0.000 description 1
- 239000005977 Ethylene Substances 0.000 description 1
- 244000004281 Eucalyptus maculata Species 0.000 description 1
- 108700024394 Exon Proteins 0.000 description 1
- 108060002716 Exonuclease Proteins 0.000 description 1
- 108010088842 Fibrinolysin Proteins 0.000 description 1
- 241000923108 Fibrobacteres Species 0.000 description 1
- 108090000270 Ficain Proteins 0.000 description 1
- 240000006927 Foeniculum vulgare Species 0.000 description 1
- 235000004204 Foeniculum vulgare Nutrition 0.000 description 1
- 235000016623 Fragaria vesca Nutrition 0.000 description 1
- 240000009088 Fragaria x ananassa Species 0.000 description 1
- 235000011363 Fragaria x ananassa Nutrition 0.000 description 1
- 241000589602 Francisella tularensis Species 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 108090001126 Furin Proteins 0.000 description 1
- 102100035233 Furin Human genes 0.000 description 1
- 241001453172 Fusobacteria Species 0.000 description 1
- 230000005526 G1 to G0 transition Effects 0.000 description 1
- 102100036250 GPI mannosyltransferase 4 Human genes 0.000 description 1
- GYHNNYVSQQEPJS-UHFFFAOYSA-N Gallium Chemical compound [Ga] GYHNNYVSQQEPJS-UHFFFAOYSA-N 0.000 description 1
- 102100021023 Gamma-glutamyl hydrolase Human genes 0.000 description 1
- 108010026132 Gelatinases Proteins 0.000 description 1
- 102000013382 Gelatinases Human genes 0.000 description 1
- 241001265526 Gemmatimonadetes <phylum> Species 0.000 description 1
- 206010064571 Gene mutation Diseases 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- 101000930822 Giardia intestinalis Dipeptidyl-peptidase 4 Proteins 0.000 description 1
- 108091020100 Gingipain Cysteine Endopeptidases Proteins 0.000 description 1
- 108030005223 Glu-Glu dipeptidases Proteins 0.000 description 1
- 102000030595 Glucokinase Human genes 0.000 description 1
- 108010021582 Glucokinase Proteins 0.000 description 1
- 241000589236 Gluconobacter Species 0.000 description 1
- 241000589232 Gluconobacter oxydans Species 0.000 description 1
- 102000005731 Glucose-6-phosphate isomerase Human genes 0.000 description 1
- 108010070600 Glucose-6-phosphate isomerase Proteins 0.000 description 1
- 108010048963 Glutamate carboxypeptidase Proteins 0.000 description 1
- 108010058940 Glutamyl Aminopeptidase Proteins 0.000 description 1
- 102000006485 Glutamyl Aminopeptidase Human genes 0.000 description 1
- 108010051815 Glutamyl endopeptidase Proteins 0.000 description 1
- 102000017278 Glutaredoxin Human genes 0.000 description 1
- 108050005205 Glutaredoxin Proteins 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 1
- 102100030385 Granzyme B Human genes 0.000 description 1
- 101150051208 HSPH1 gene Proteins 0.000 description 1
- 241001670062 Halomonas utahensis Species 0.000 description 1
- 101710178376 Heat shock 70 kDa protein Proteins 0.000 description 1
- 101710152018 Heat shock cognate 70 kDa protein Proteins 0.000 description 1
- 102100031624 Heat shock protein 105 kDa Human genes 0.000 description 1
- 101710113864 Heat shock protein 90 Proteins 0.000 description 1
- 102100034051 Heat shock protein HSP 90-alpha Human genes 0.000 description 1
- 244000020551 Helianthus annuus Species 0.000 description 1
- 235000003222 Helianthus annuus Nutrition 0.000 description 1
- 241001660422 Herbaspirillum huttiense Species 0.000 description 1
- 102000005548 Hexokinase Human genes 0.000 description 1
- 108700040460 Hexokinases Proteins 0.000 description 1
- 108090000252 Histolysain Proteins 0.000 description 1
- 241001272567 Hominoidea Species 0.000 description 1
- 101000888642 Homo sapiens ARF GTPase-activating protein GIT2 Proteins 0.000 description 1
- 101000901723 Homo sapiens CXADR-like membrane protein Proteins 0.000 description 1
- 101000907951 Homo sapiens Chymotrypsin-like elastase family member 3B Proteins 0.000 description 1
- 101000931862 Homo sapiens Dipeptidyl peptidase 3 Proteins 0.000 description 1
- 101000870166 Homo sapiens DnaJ homolog subfamily C member 14 Proteins 0.000 description 1
- 101001074618 Homo sapiens GPI mannosyltransferase 4 Proteins 0.000 description 1
- 101001009603 Homo sapiens Granzyme B Proteins 0.000 description 1
- 240000005979 Hordeum vulgare Species 0.000 description 1
- 235000007340 Hordeum vulgare Nutrition 0.000 description 1
- 101150003028 Hprt1 gene Proteins 0.000 description 1
- 241000922030 Hydrogenophaga flava Species 0.000 description 1
- 108090000571 Hypodermin C Proteins 0.000 description 1
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 1
- 108010084019 IgA-specific metalloendopeptidase Proteins 0.000 description 1
- 108010002231 IgA-specific serine endopeptidase Proteins 0.000 description 1
- 102000014429 Insulin-like growth factor Human genes 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 235000002678 Ipomoea batatas Nutrition 0.000 description 1
- 244000017020 Ipomoea batatas Species 0.000 description 1
- 235000006350 Ipomoea batatas var. batatas Nutrition 0.000 description 1
- 102100033903 Isoaspartyl peptidase/L-asparaginase Human genes 0.000 description 1
- 241001148466 Janthinobacterium lividum Species 0.000 description 1
- 101710176219 Kallikrein-1 Proteins 0.000 description 1
- 101710172072 Kexin Proteins 0.000 description 1
- 241000235058 Komagataella pastoris Species 0.000 description 1
- 235000019766 L-Lysine Nutrition 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 235000003228 Lactuca sativa Nutrition 0.000 description 1
- 240000008415 Lactuca sativa Species 0.000 description 1
- 241000219732 Lathyrus cicera Species 0.000 description 1
- 241001387859 Lentisphaerae Species 0.000 description 1
- 108010004098 Leucyl aminopeptidase Proteins 0.000 description 1
- 108030007165 Leucyl endopeptidases Proteins 0.000 description 1
- 108010028275 Leukocyte Elastase Proteins 0.000 description 1
- GDBQQVLCIARPGH-UHFFFAOYSA-N Leupeptin Natural products CC(C)CC(NC(C)=O)C(=O)NC(CC(C)C)C(=O)NC(C=O)CCCN=C(N)N GDBQQVLCIARPGH-UHFFFAOYSA-N 0.000 description 1
- OJMMVQQUTAEWLP-UHFFFAOYSA-N Lincomycin Natural products CN1CC(CCC)CC1C(=O)NC(C(C)O)C1C(O)C(O)C(O)C(SC)O1 OJMMVQQUTAEWLP-UHFFFAOYSA-N 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- 235000007688 Lycopersicon esculentum Nutrition 0.000 description 1
- 102100033320 Lysosomal Pro-X carboxypeptidase Human genes 0.000 description 1
- 108010053229 Lysyl endopeptidase Proteins 0.000 description 1
- 108030001712 Macrophage elastases Proteins 0.000 description 1
- 102100027998 Macrophage metalloelastase Human genes 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- 241001670047 Malikia spinosa Species 0.000 description 1
- 235000011430 Malus pumila Nutrition 0.000 description 1
- 244000070406 Malus silvestris Species 0.000 description 1
- 235000015103 Malus silvestris Nutrition 0.000 description 1
- 235000014826 Mangifera indica Nutrition 0.000 description 1
- 240000007228 Mangifera indica Species 0.000 description 1
- 240000003183 Manihot esculenta Species 0.000 description 1
- 235000016735 Manihot esculenta subsp esculenta Nutrition 0.000 description 1
- 108010038049 Mating Factor Proteins 0.000 description 1
- 102000004318 Matrilysin Human genes 0.000 description 1
- 108090000855 Matrilysin Proteins 0.000 description 1
- 102000000380 Matrix Metalloproteinase 1 Human genes 0.000 description 1
- 108010016113 Matrix Metalloproteinase 1 Proteins 0.000 description 1
- 108010076497 Matrix Metalloproteinase 10 Proteins 0.000 description 1
- 108010016160 Matrix Metalloproteinase 3 Proteins 0.000 description 1
- 102000001776 Matrix metalloproteinase-9 Human genes 0.000 description 1
- 108010015302 Matrix metalloproteinase-9 Proteins 0.000 description 1
- 240000004658 Medicago sativa Species 0.000 description 1
- 235000017587 Medicago sativa ssp. sativa Nutrition 0.000 description 1
- 108030004769 Membrane dipeptidases Proteins 0.000 description 1
- 241000579835 Merops Species 0.000 description 1
- 101100071630 Mesocentrotus franciscanus HSP110 gene Proteins 0.000 description 1
- 102000003792 Metallothionein Human genes 0.000 description 1
- 108090000157 Metallothionein Proteins 0.000 description 1
- 108090000192 Methionyl aminopeptidases Proteins 0.000 description 1
- 102000034452 Methionyl aminopeptidases Human genes 0.000 description 1
- 241000589343 Methylobacter luteus Species 0.000 description 1
- 241001264651 Methylocaldum gracile Species 0.000 description 1
- 241000589346 Methylococcus capsulatus Species 0.000 description 1
- 241001533197 Methylomicrobium agile Species 0.000 description 1
- 241000589348 Methylomonas methanica Species 0.000 description 1
- 241000499447 Methylosphaera hansonii Species 0.000 description 1
- 108010013295 Microbial collagenase Proteins 0.000 description 1
- 241001670070 Microbulbifer elongatus Species 0.000 description 1
- 206010027626 Milia Diseases 0.000 description 1
- 108010047660 Mitochondrial intermediate peptidase Proteins 0.000 description 1
- 102100026934 Mitochondrial intermediate peptidase Human genes 0.000 description 1
- 108010042046 Mitochondrial processing peptidase Proteins 0.000 description 1
- 108010027383 Muramoylpentapeptide carboxypeptidase Proteins 0.000 description 1
- 101100451677 Mus musculus Hspa4 gene Proteins 0.000 description 1
- 235000003805 Musa ABB Group Nutrition 0.000 description 1
- 235000018290 Musa x paradisiaca Nutrition 0.000 description 1
- 102100034681 Myeloblastin Human genes 0.000 description 1
- 108090000973 Myeloblastin Proteins 0.000 description 1
- 108030001194 N-formylmethionyl-peptidases Proteins 0.000 description 1
- MQUQNUAYKLCRME-INIZCTEOSA-N N-tosyl-L-phenylalanyl chloromethyl ketone Chemical compound C1=CC(C)=CC=C1S(=O)(=O)N[C@H](C(=O)CCl)CC1=CC=CC=C1 MQUQNUAYKLCRME-INIZCTEOSA-N 0.000 description 1
- 241000244206 Nematoda Species 0.000 description 1
- 229930193140 Neomycin Natural products 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 108090000812 Neurolysin Proteins 0.000 description 1
- 102100023072 Neurolysin, mitochondrial Human genes 0.000 description 1
- 102000056189 Neutrophil collagenases Human genes 0.000 description 1
- 108030001564 Neutrophil collagenases Proteins 0.000 description 1
- 102100033174 Neutrophil elastase Human genes 0.000 description 1
- 239000000020 Nitrocellulose Substances 0.000 description 1
- JCXJVPUVTGWSNB-UHFFFAOYSA-N Nitrogen dioxide Chemical compound O=[N]=O JCXJVPUVTGWSNB-UHFFFAOYSA-N 0.000 description 1
- 241000121237 Nitrospirae Species 0.000 description 1
- 108010066503 Non-stereospecific dipeptidase Proteins 0.000 description 1
- 108091005461 Nucleic proteins Chemical class 0.000 description 1
- 101710141454 Nucleoprotein Proteins 0.000 description 1
- 108010000240 O-sialoglycoprotein endopeptidase Proteins 0.000 description 1
- 241001657389 Oceanimonas doudoroffii Species 0.000 description 1
- 240000007817 Olea europaea Species 0.000 description 1
- 241000293016 Oligella urethralis Species 0.000 description 1
- 241000237502 Ostreidae Species 0.000 description 1
- 241000283903 Ovis aries Species 0.000 description 1
- 108020002230 Pancreatic Ribonuclease Proteins 0.000 description 1
- 108010067372 Pancreatic elastase Proteins 0.000 description 1
- 102000016387 Pancreatic elastase Human genes 0.000 description 1
- 102000005891 Pancreatic ribonuclease Human genes 0.000 description 1
- 239000006002 Pepper Substances 0.000 description 1
- 108090000284 Pepsin A Proteins 0.000 description 1
- 102000057297 Pepsin A Human genes 0.000 description 1
- 108090000313 Pepsin B Proteins 0.000 description 1
- 108010033276 Peptide Fragments Proteins 0.000 description 1
- 102000007079 Peptide Fragments Human genes 0.000 description 1
- 108090000882 Peptidyl-Dipeptidase A Proteins 0.000 description 1
- 108010030544 Peptidyl-Lys metalloendopeptidase Proteins 0.000 description 1
- 101710111214 Peptidyl-prolyl cis-trans isomerase C Proteins 0.000 description 1
- 102100031653 Peptidyl-prolyl cis-trans isomerase NIMA-interacting 4 Human genes 0.000 description 1
- 244000025272 Persea americana Species 0.000 description 1
- 235000008673 Persea americana Nutrition 0.000 description 1
- 244000062780 Petroselinum sativum Species 0.000 description 1
- 235000010627 Phaseolus vulgaris Nutrition 0.000 description 1
- 244000046052 Phaseolus vulgaris Species 0.000 description 1
- 102000006335 Phosphate-Binding Proteins Human genes 0.000 description 1
- 108010058514 Phosphate-Binding Proteins Proteins 0.000 description 1
- 108010069341 Phosphofructokinases Proteins 0.000 description 1
- 102000001105 Phosphofructokinases Human genes 0.000 description 1
- ZPHBZEQOLSRPAK-UHFFFAOYSA-N Phosphoramidon Natural products C=1NC2=CC=CC=C2C=1CC(C(O)=O)NC(=O)C(CC(C)C)NP(O)(=O)OC1OC(C)C(O)C(O)C1O ZPHBZEQOLSRPAK-UHFFFAOYSA-N 0.000 description 1
- 235000008331 Pinus X rigitaeda Nutrition 0.000 description 1
- 241000018646 Pinus brutia Species 0.000 description 1
- 235000011613 Pinus brutia Nutrition 0.000 description 1
- 241001236219 Pinus echinata Species 0.000 description 1
- 235000005018 Pinus echinata Nutrition 0.000 description 1
- 235000017339 Pinus palustris Nutrition 0.000 description 1
- 235000016761 Piper aduncum Nutrition 0.000 description 1
- 240000003889 Piper guineense Species 0.000 description 1
- 235000017804 Piper guineense Nutrition 0.000 description 1
- 235000008184 Piper nigrum Nutrition 0.000 description 1
- 235000010582 Pisum sativum Nutrition 0.000 description 1
- 240000004713 Pisum sativum Species 0.000 description 1
- 108090000316 Pitrilysin Proteins 0.000 description 1
- 241001180199 Planctomycetes Species 0.000 description 1
- 108020005089 Plant RNA Proteins 0.000 description 1
- 235000015266 Plantago major Nutrition 0.000 description 1
- 108090000113 Plasma Kallikrein Proteins 0.000 description 1
- 102100034869 Plasma kallikrein Human genes 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 108010039918 Polylysine Proteins 0.000 description 1
- 101710095622 Polyporopepsin Proteins 0.000 description 1
- 241000219000 Populus Species 0.000 description 1
- 108010013381 Porins Proteins 0.000 description 1
- 108010015078 Pregnancy-Associated alpha 2-Macroglobulins Proteins 0.000 description 1
- 101710130420 Probable capsid assembly scaffolding protein Proteins 0.000 description 1
- 101710083689 Probable capsid protein Proteins 0.000 description 1
- 102100037775 Probable tRNA N6-adenosine threonylcarbamoyltransferase Human genes 0.000 description 1
- 108010064622 Procollagen N-Endopeptidase Proteins 0.000 description 1
- 102000015339 Procollagen N-endopeptidase Human genes 0.000 description 1
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 1
- 108010044159 Proprotein Convertases Proteins 0.000 description 1
- 102000006437 Proprotein Convertases Human genes 0.000 description 1
- 101800001753 Protease Proteins 0.000 description 1
- 101800001491 Protease 3C Proteins 0.000 description 1
- 101800004937 Protein C Proteins 0.000 description 1
- 102000017975 Protein C Human genes 0.000 description 1
- 235000009827 Prunus armeniaca Nutrition 0.000 description 1
- 244000018633 Prunus armeniaca Species 0.000 description 1
- 235000006029 Prunus persica var nucipersica Nutrition 0.000 description 1
- 235000006040 Prunus persica var persica Nutrition 0.000 description 1
- 244000017714 Prunus persica var. nucipersica Species 0.000 description 1
- 241000589515 Pseudoalteromonas atlantica Species 0.000 description 1
- 241000590028 Pseudoalteromonas haloplanktis Species 0.000 description 1
- 241000947836 Pseudomonadaceae Species 0.000 description 1
- 241001248479 Pseudomonadales Species 0.000 description 1
- 241000028636 Pseudomonas abietaniphila Species 0.000 description 1
- 241000589517 Pseudomonas aeruginosa Species 0.000 description 1
- 101000925883 Pseudomonas aeruginosa (strain ATCC 15692 / DSM 22644 / CIP 104116 / JCM 14847 / LMG 12228 / 1C / PRS 101 / PAO1) Elastase Proteins 0.000 description 1
- 101100084022 Pseudomonas aeruginosa (strain ATCC 15692 / DSM 22644 / CIP 104116 / JCM 14847 / LMG 12228 / 1C / PRS 101 / PAO1) lapA gene Proteins 0.000 description 1
- 241000204715 Pseudomonas agarici Species 0.000 description 1
- 241000168225 Pseudomonas alcaligenes Species 0.000 description 1
- 241001459308 Pseudomonas alcaliphila Species 0.000 description 1
- 241000218934 Pseudomonas amygdali Species 0.000 description 1
- 241001325442 Pseudomonas andersonii Species 0.000 description 1
- 241000520871 Pseudomonas asplenii Species 0.000 description 1
- 241001660019 Pseudomonas borealis Species 0.000 description 1
- 241000226031 Pseudomonas brassicacearum Species 0.000 description 1
- 241000204712 Pseudomonas caricapapayae Species 0.000 description 1
- 241001646398 Pseudomonas chlororaphis Species 0.000 description 1
- 241001508466 Pseudomonas cichorii Species 0.000 description 1
- 241000520873 Pseudomonas citronellolis Species 0.000 description 1
- 241000647960 Pseudomonas coronafaciens pv. coronafaciens Species 0.000 description 1
- 241000168053 Pseudomonas denitrificans (nomen rejiciendum) Species 0.000 description 1
- 241000946440 Pseudomonas diterpeniphila Species 0.000 description 1
- 241000429405 Pseudomonas extremorientalis Species 0.000 description 1
- 241001148192 Pseudomonas flavescens Species 0.000 description 1
- 241001358835 Pseudomonas fluorescens PF5 Species 0.000 description 1
- 241001209206 Pseudomonas fluorescens Pf0-1 Species 0.000 description 1
- 241001453326 Pseudomonas fluorescens bv. A Species 0.000 description 1
- 241000502324 Pseudomonas fluorescens bv. B Species 0.000 description 1
- 241000589641 Pseudomonas fluorescens bv. C Species 0.000 description 1
- 241000589538 Pseudomonas fragi Species 0.000 description 1
- 241001312498 Pseudomonas gessardii Species 0.000 description 1
- 241000231049 Pseudomonas gingeri Species 0.000 description 1
- 241000042121 Pseudomonas graminis Species 0.000 description 1
- 241000620589 Pseudomonas grimontii Species 0.000 description 1
- 241000520899 Pseudomonas halodenitrificans Species 0.000 description 1
- 241001531427 Pseudomonas hydrogenovora Species 0.000 description 1
- 241001300822 Pseudomonas jessenii Species 0.000 description 1
- 241000357050 Pseudomonas lini Species 0.000 description 1
- 241001670039 Pseudomonas lundensis Species 0.000 description 1
- 241000218905 Pseudomonas luteola Species 0.000 description 1
- 241000394642 Pseudomonas marginalis pv. marginalis Species 0.000 description 1
- 241000145542 Pseudomonas marginata Species 0.000 description 1
- 241000589755 Pseudomonas mendocina Species 0.000 description 1
- 241001291501 Pseudomonas monteilii Species 0.000 description 1
- 241000204709 Pseudomonas mucidolens Species 0.000 description 1
- 241000589781 Pseudomonas oleovorans Species 0.000 description 1
- 241000218904 Pseudomonas oryzihabitans Species 0.000 description 1
- 241001670066 Pseudomonas pertucinogena Species 0.000 description 1
- 241001223182 Pseudomonas plecoglossicida Species 0.000 description 1
- 241000589630 Pseudomonas pseudoalcaligenes Species 0.000 description 1
- 241000530526 Pseudomonas psychrophila Species 0.000 description 1
- 241000589776 Pseudomonas putida Species 0.000 description 1
- 101000604548 Pseudomonas sp. (strain 101) Pseudomonalisin Proteins 0.000 description 1
- 241000218901 Pseudomonas straminea Species 0.000 description 1
- 241000589614 Pseudomonas stutzeri Species 0.000 description 1
- 241000218902 Pseudomonas synxantha Species 0.000 description 1
- 241000589615 Pseudomonas syringae Species 0.000 description 1
- 241000218903 Pseudomonas taetrolens Species 0.000 description 1
- 241001478288 Pseudomonas thermocarboxydovorans Species 0.000 description 1
- 241001464820 Pseudomonas viridiflava Species 0.000 description 1
- 241000577556 Pseudomonas wisconsinensis Species 0.000 description 1
- 241000039948 Pseudomonas xiamenensis Species 0.000 description 1
- 240000001416 Pseudotsuga menziesii Species 0.000 description 1
- 235000005386 Pseudotsuga menziesii var menziesii Nutrition 0.000 description 1
- 244000294611 Punica granatum Species 0.000 description 1
- 235000014360 Punica granatum Nutrition 0.000 description 1
- 108090001002 Pyroglutamyl-peptidase II Proteins 0.000 description 1
- 102000004892 Pyroglutamyl-peptidase II Human genes 0.000 description 1
- 235000014443 Pyrus communis Nutrition 0.000 description 1
- 240000001987 Pyrus communis Species 0.000 description 1
- 108010011939 Pyruvate Decarboxylase Proteins 0.000 description 1
- 102000009572 RNA Polymerase II Human genes 0.000 description 1
- 239000013614 RNA sample Substances 0.000 description 1
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 description 1
- 241000589625 Ralstonia pickettii Species 0.000 description 1
- 241000700159 Rattus Species 0.000 description 1
- 101000658294 Rattus norvegicus Thioredoxin-like protein 1 Proteins 0.000 description 1
- 108090000783 Renin Proteins 0.000 description 1
- 102100028255 Renin Human genes 0.000 description 1
- 108020005091 Replication Origin Proteins 0.000 description 1
- 108091027981 Response element Proteins 0.000 description 1
- 241000589194 Rhizobium leguminosarum Species 0.000 description 1
- 102000006382 Ribonucleases Human genes 0.000 description 1
- 108010083644 Ribonucleases Proteins 0.000 description 1
- 240000000528 Ricinus communis Species 0.000 description 1
- 235000004443 Ricinus communis Nutrition 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 235000017848 Rubus fruticosus Nutrition 0.000 description 1
- 240000007651 Rubus glaucus Species 0.000 description 1
- 235000011034 Rubus glaucus Nutrition 0.000 description 1
- 235000009122 Rubus idaeus Nutrition 0.000 description 1
- 102100022135 S-arrestin Human genes 0.000 description 1
- 101710117586 S-arrestin Proteins 0.000 description 1
- 239000011542 SDS running buffer Substances 0.000 description 1
- 101710149263 Saccharolysin Proteins 0.000 description 1
- 101000715359 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) Carboxypeptidase S Proteins 0.000 description 1
- 101100285899 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) SSE2 gene Proteins 0.000 description 1
- 108090000077 Saccharopepsin Proteins 0.000 description 1
- 240000000111 Saccharum officinarum Species 0.000 description 1
- 235000007201 Saccharum officinarum Nutrition 0.000 description 1
- 101800001700 Saposin-D Proteins 0.000 description 1
- 101710204410 Scaffold protein Proteins 0.000 description 1
- 235000007238 Secale cereale Nutrition 0.000 description 1
- 244000082988 Secale cereale Species 0.000 description 1
- 108010004832 Serine-Type D-Ala-D-Ala Carboxypeptidase Proteins 0.000 description 1
- 102000030899 Serine-Type D-Ala-D-Ala Carboxypeptidase Human genes 0.000 description 1
- 240000003768 Solanum lycopersicum Species 0.000 description 1
- 235000002597 Solanum melongena Nutrition 0.000 description 1
- 244000061458 Solanum melongena Species 0.000 description 1
- 240000003829 Sorghum propinquum Species 0.000 description 1
- 235000011684 Sorghum saccharatum Nutrition 0.000 description 1
- 241000736110 Sphingomonas paucimobilis Species 0.000 description 1
- 235000009337 Spinacia oleracea Nutrition 0.000 description 1
- 244000300264 Spinacia oleracea Species 0.000 description 1
- 241000589970 Spirochaetales Species 0.000 description 1
- 235000009184 Spondias indica Nutrition 0.000 description 1
- 241000122973 Stenotrophomonas maltophilia Species 0.000 description 1
- 241001670040 Stenotrophomonas pictorum Species 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 102100030416 Stromelysin-1 Human genes 0.000 description 1
- 102100028848 Stromelysin-2 Human genes 0.000 description 1
- 108090000787 Subtilisin Proteins 0.000 description 1
- 235000021536 Sugar beet Nutrition 0.000 description 1
- 101150003725 TK gene Proteins 0.000 description 1
- 102000018679 Tacrolimus Binding Proteins Human genes 0.000 description 1
- 108010027179 Tacrolimus Binding Proteins Proteins 0.000 description 1
- 108010006785 Taq Polymerase Proteins 0.000 description 1
- 241001670068 Thauera butanivorans Species 0.000 description 1
- 244000269722 Thea sinensis Species 0.000 description 1
- 241000959851 Thermales Species 0.000 description 1
- 241001143138 Thermodesulfobacteria <phylum> Species 0.000 description 1
- 241001141092 Thermomicrobia Species 0.000 description 1
- 241000204667 Thermoplasma Species 0.000 description 1
- 108090000763 Thermopsin Proteins 0.000 description 1
- 241000589596 Thermus Species 0.000 description 1
- 102000002933 Thioredoxin Human genes 0.000 description 1
- 108090000190 Thrombin Proteins 0.000 description 1
- 102000006601 Thymidine Kinase Human genes 0.000 description 1
- 108020004440 Thymidine kinase Proteins 0.000 description 1
- 240000006909 Tilia x europaea Species 0.000 description 1
- 235000011941 Tilia x europaea Nutrition 0.000 description 1
- ISWQCIVKKSOKNN-UHFFFAOYSA-L Tiron Chemical compound [Na+].[Na+].OC1=CC(S([O-])(=O)=O)=CC(S([O-])(=O)=O)=C1O ISWQCIVKKSOKNN-UHFFFAOYSA-L 0.000 description 1
- 102000057032 Tissue Kallikreins Human genes 0.000 description 1
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 1
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 1
- 241000219793 Trifolium Species 0.000 description 1
- 108010070926 Tripeptide aminopeptidase Proteins 0.000 description 1
- 108010039203 Tripeptidyl-Peptidase 1 Proteins 0.000 description 1
- 102100034197 Tripeptidyl-peptidase 1 Human genes 0.000 description 1
- 102100040411 Tripeptidyl-peptidase 2 Human genes 0.000 description 1
- 108091061763 Triple-stranded DNA Proteins 0.000 description 1
- 235000019714 Triticale Nutrition 0.000 description 1
- 108090000631 Trypsin Proteins 0.000 description 1
- 102000004142 Trypsin Human genes 0.000 description 1
- 108060005989 Tryptase Proteins 0.000 description 1
- 102000001400 Tryptase Human genes 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 108030000963 Tryptophanyl aminopeptidases Proteins 0.000 description 1
- 108090000906 Tubulinyl-Tyr carboxypeptidases Proteins 0.000 description 1
- 102000004342 Tubulinyl-Tyr carboxypeptidases Human genes 0.000 description 1
- 241000397921 Turbellaria Species 0.000 description 1
- DJSISFGPUUYILV-UHFFFAOYSA-N UNPD161792 Natural products O1C(C(O)=O)C(O)C(O)C(O)C1OC(C(=C1O)O)=CC2=C1C(=O)C=C(C=1C=CC(O)=CC=1)O2 DJSISFGPUUYILV-UHFFFAOYSA-N 0.000 description 1
- 102000018390 Ubiquitin-Specific Proteases Human genes 0.000 description 1
- 108010066496 Ubiquitin-Specific Proteases Proteins 0.000 description 1
- 108010009135 Uca pugilator serine collagenase 1 Proteins 0.000 description 1
- 108010018628 Ulp1 protease Proteins 0.000 description 1
- 108090000435 Urokinase-type plasminogen activator Proteins 0.000 description 1
- 102000003990 Urokinase-type plasminogen activator Human genes 0.000 description 1
- 206010046865 Vaccinia virus infection Diseases 0.000 description 1
- 235000003095 Vaccinium corymbosum Nutrition 0.000 description 1
- 240000001717 Vaccinium macrocarpon Species 0.000 description 1
- 235000012545 Vaccinium macrocarpon Nutrition 0.000 description 1
- 235000017537 Vaccinium myrtillus Nutrition 0.000 description 1
- 235000002118 Vaccinium oxycoccus Nutrition 0.000 description 1
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 241001672648 Vieira Species 0.000 description 1
- 235000009754 Vitis X bourquina Nutrition 0.000 description 1
- 235000012333 Vitis X labruscana Nutrition 0.000 description 1
- 240000006365 Vitis vinifera Species 0.000 description 1
- 235000014787 Vitis vinifera Nutrition 0.000 description 1
- 108010038900 X-Pro aminopeptidase Proteins 0.000 description 1
- 238000004833 X-ray photoelectron spectroscopy Methods 0.000 description 1
- 102100039662 Xaa-Pro dipeptidase Human genes 0.000 description 1
- 101710171640 Xaa-Pro dipeptidase Proteins 0.000 description 1
- 108010027570 Xanthine phosphoribosyltransferase Proteins 0.000 description 1
- 241000947909 Xanthomonadales Species 0.000 description 1
- 241000589636 Xanthomonas campestris Species 0.000 description 1
- 241000589651 Zoogloea Species 0.000 description 1
- 241000589153 Zoogloea ramigera Species 0.000 description 1
- DGEZNRSVGBDHLK-UHFFFAOYSA-N [1,10]phenanthroline Chemical compound C1=CN=C2C3=NC=CC=C3C=CC2=C1 DGEZNRSVGBDHLK-UHFFFAOYSA-N 0.000 description 1
- FJJCIZWZNKZHII-UHFFFAOYSA-N [4,6-bis(cyanoamino)-1,3,5-triazin-2-yl]cyanamide Chemical compound N#CNC1=NC(NC#N)=NC(NC#N)=N1 FJJCIZWZNKZHII-UHFFFAOYSA-N 0.000 description 1
- 241001670042 [Pseudomonas] boreopolis Species 0.000 description 1
- 241001670030 [Pseudomonas] geniculata Species 0.000 description 1
- 241001670027 [Pseudomonas] hibiscicola Species 0.000 description 1
- 230000002159 abnormal effect Effects 0.000 description 1
- 229910052768 actinide Inorganic materials 0.000 description 1
- 150000001255 actinides Chemical class 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 108091006088 activator proteins Proteins 0.000 description 1
- 125000002252 acyl group Chemical group 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 229940005347 alcaligenes faecalis Drugs 0.000 description 1
- 239000013566 allergen Substances 0.000 description 1
- 102000015395 alpha 1-Antitrypsin Human genes 0.000 description 1
- 108010050122 alpha 1-Antitrypsin Proteins 0.000 description 1
- 108010052590 amastatin Proteins 0.000 description 1
- QFAADIRHLBXJJS-ZAZJUGBXSA-N amastatin Chemical compound CC(C)C[C@@H](N)[C@H](O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(O)=O QFAADIRHLBXJJS-ZAZJUGBXSA-N 0.000 description 1
- 108090000449 aminopeptidase B Proteins 0.000 description 1
- 229960003896 aminopterin Drugs 0.000 description 1
- 210000004102 animal cell Anatomy 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 108700003859 araC Genes Proteins 0.000 description 1
- 101150044616 araC gene Proteins 0.000 description 1
- 101150010487 are gene Proteins 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 235000016520 artichoke thistle Nutrition 0.000 description 1
- 235000000183 arugula Nutrition 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 238000002820 assay format Methods 0.000 description 1
- 108010070189 atroxase Proteins 0.000 description 1
- 230000002358 autolytic effect Effects 0.000 description 1
- 210000003578 bacterial chromosome Anatomy 0.000 description 1
- 244000052616 bacterial pathogen Species 0.000 description 1
- PXXJHWLDUBFPOL-UHFFFAOYSA-N benzamidine Chemical compound NC(=N)C1=CC=CC=C1 PXXJHWLDUBFPOL-UHFFFAOYSA-N 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 108010005774 beta-Galactosidase Proteins 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 230000000975 bioactive effect Effects 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 229960000074 biopharmaceutical Drugs 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 108091006004 biotinylated proteins Proteins 0.000 description 1
- 235000021029 blackberry Nutrition 0.000 description 1
- 229960001561 bleomycin Drugs 0.000 description 1
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 1
- 230000023555 blood coagulation Effects 0.000 description 1
- 229940019700 blood coagulation factors Drugs 0.000 description 1
- 235000021014 blueberries Nutrition 0.000 description 1
- 229940056450 brucella abortus Drugs 0.000 description 1
- 229940038698 brucella melitensis Drugs 0.000 description 1
- 238000004422 calculation algorithm Methods 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 229940041514 candida albicans extract Drugs 0.000 description 1
- 238000005251 capillar electrophoresis Methods 0.000 description 1
- 239000004202 carbamide Substances 0.000 description 1
- 125000000837 carbohydrate group Chemical group 0.000 description 1
- 150000001720 carbohydrates Chemical class 0.000 description 1
- 235000014633 carbohydrates Nutrition 0.000 description 1
- 150000001721 carbon Chemical group 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- NPLTVGMLNDMOQE-UHFFFAOYSA-N carthamidin Natural products C1=CC(O)=CC=C1C1OC2=CC(O)=C(O)C(O)=C2C(=O)C1 NPLTVGMLNDMOQE-UHFFFAOYSA-N 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 235000019693 cherries Nutrition 0.000 description 1
- 235000003733 chicria Nutrition 0.000 description 1
- 229960005091 chloramphenicol Drugs 0.000 description 1
- WIIZWVCIJKGZOK-RKDXNWHRSA-N chloramphenicol Chemical compound ClC(Cl)C(=O)N[C@H](CO)[C@H](O)C1=CC=C([N+]([O-])=O)C=C1 WIIZWVCIJKGZOK-RKDXNWHRSA-N 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 229960002976 chymopapain Drugs 0.000 description 1
- 229940080701 chymosin Drugs 0.000 description 1
- 108010086192 chymostatin Proteins 0.000 description 1
- 229960002376 chymotrypsin Drugs 0.000 description 1
- 235000020971 citrus fruits Nutrition 0.000 description 1
- 238000012411 cloning technique Methods 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- 108010065327 clostridial aminopeptidase Proteins 0.000 description 1
- 108010055222 clotting enzyme Proteins 0.000 description 1
- 238000007621 cluster analysis Methods 0.000 description 1
- 230000004186 co-expression Effects 0.000 description 1
- 238000000975 co-precipitation Methods 0.000 description 1
- 108010045487 coagulogen Proteins 0.000 description 1
- 235000016213 coffee Nutrition 0.000 description 1
- 235000013353 coffee beverage Nutrition 0.000 description 1
- 230000001447 compensatory effect Effects 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 230000003750 conditioning effect Effects 0.000 description 1
- 235000004634 cranberry Nutrition 0.000 description 1
- 238000004132 cross linking Methods 0.000 description 1
- 238000012866 crystallographic experiment Methods 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- NHVNXKFIZYSCEB-XLPZGREQSA-N dTTP Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)[C@@H](O)C1 NHVNXKFIZYSCEB-XLPZGREQSA-N 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 230000000368 destabilizing effect Effects 0.000 description 1
- 229910052805 deuterium Inorganic materials 0.000 description 1
- ANCLJVISBRWUTR-UHFFFAOYSA-N diaminophosphinic acid Chemical compound NP(N)(O)=O ANCLJVISBRWUTR-UHFFFAOYSA-N 0.000 description 1
- 230000009274 differential gene expression Effects 0.000 description 1
- MUCZHBLJLSDCSD-UHFFFAOYSA-N diisopropyl fluorophosphate Chemical compound CC(C)OP(F)(=O)OC(C)C MUCZHBLJLSDCSD-UHFFFAOYSA-N 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- 235000004879 dioscorea Nutrition 0.000 description 1
- 108090000212 dipeptidyl peptidase II Proteins 0.000 description 1
- 238000010494 dissociation reaction Methods 0.000 description 1
- 230000005593 dissociations Effects 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 1
- 239000000975 dye Substances 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 108010039262 elastatinal Proteins 0.000 description 1
- 239000003792 electrolyte Substances 0.000 description 1
- 108010003914 endoproteinase Asp-N Proteins 0.000 description 1
- 238000005265 energy consumption Methods 0.000 description 1
- 239000003623 enhancer Substances 0.000 description 1
- 230000002255 enzymatic effect Effects 0.000 description 1
- 238000001976 enzyme digestion Methods 0.000 description 1
- 239000003822 epoxy resin Substances 0.000 description 1
- 235000020776 essential amino acid Nutrition 0.000 description 1
- 239000003797 essential amino acid Substances 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 108010052305 exodeoxyribonuclease III Proteins 0.000 description 1
- 102000013165 exonuclease Human genes 0.000 description 1
- 235000019836 ficin Nutrition 0.000 description 1
- POTUGHMKJGOKRI-UHFFFAOYSA-N ficin Chemical compound FI=CI=N POTUGHMKJGOKRI-UHFFFAOYSA-N 0.000 description 1
- 235000004426 flaxseed Nutrition 0.000 description 1
- 108091006047 fluorescent proteins Proteins 0.000 description 1
- 102000034287 fluorescent proteins Human genes 0.000 description 1
- 230000037433 frameshift Effects 0.000 description 1
- 229940118764 francisella tularensis Drugs 0.000 description 1
- 108090000285 fruit bromelain Proteins 0.000 description 1
- 108010062699 gamma-Glutamyl Hydrolase Proteins 0.000 description 1
- 235000004611 garlic Nutrition 0.000 description 1
- 238000004817 gas chromatography Methods 0.000 description 1
- 238000012224 gene deletion Methods 0.000 description 1
- 238000010363 gene targeting Methods 0.000 description 1
- 102000034356 gene-regulatory proteins Human genes 0.000 description 1
- 108091006104 gene-regulatory proteins Proteins 0.000 description 1
- 238000011331 genomic analysis Methods 0.000 description 1
- 229960002518 gentamicin Drugs 0.000 description 1
- 230000004153 glucose metabolism Effects 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 108090000639 glutamyl endopeptidase II Proteins 0.000 description 1
- 150000004676 glycans Chemical class 0.000 description 1
- 108010092515 glycyl endopeptidase Proteins 0.000 description 1
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 1
- 229910052737 gold Inorganic materials 0.000 description 1
- 239000010931 gold Substances 0.000 description 1
- 101150106093 gpt gene Proteins 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 101150002138 grxA gene Proteins 0.000 description 1
- 101150073747 grxB gene Proteins 0.000 description 1
- 101150007702 grxC gene Proteins 0.000 description 1
- 125000001475 halogen functional group Chemical group 0.000 description 1
- 125000001072 heteroaryl group Chemical group 0.000 description 1
- 239000000833 heterodimer Substances 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 108091008039 hormone receptors Proteins 0.000 description 1
- 229910052739 hydrogen Inorganic materials 0.000 description 1
- 239000001257 hydrogen Substances 0.000 description 1
- 125000004435 hydrogen atom Chemical group [H]* 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 125000001165 hydrophobic group Chemical group 0.000 description 1
- 230000003100 immobilizing effect Effects 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 239000012535 impurity Substances 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 238000005342 ion exchange Methods 0.000 description 1
- 101150109249 lacI gene Proteins 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 238000011031 large-scale manufacturing process Methods 0.000 description 1
- GDBQQVLCIARPGH-ULQDDVLXSA-N leupeptin Chemical compound CC(C)C[C@H](NC(C)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C=O)CCCN=C(N)N GDBQQVLCIARPGH-ULQDDVLXSA-N 0.000 description 1
- 108010052968 leupeptin Proteins 0.000 description 1
- 239000004571 lime Substances 0.000 description 1
- 229960005287 lincomycin Drugs 0.000 description 1
- OJMMVQQUTAEWLP-KIDUDLJLSA-N lincomycin Chemical compound CN1C[C@H](CCC)C[C@H]1C(=O)N[C@H]([C@@H](C)O)[C@@H]1[C@H](O)[C@H](O)[C@@H](O)[C@@H](SC)O1 OJMMVQQUTAEWLP-KIDUDLJLSA-N 0.000 description 1
- 230000033001 locomotion Effects 0.000 description 1
- 238000004020 luminiscence type Methods 0.000 description 1
- 108010057284 lysosomal Pro-X carboxypeptidase Proteins 0.000 description 1
- 230000013011 mating Effects 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 238000002844 melting Methods 0.000 description 1
- 230000008018 melting Effects 0.000 description 1
- 230000004066 metabolic change Effects 0.000 description 1
- 230000037353 metabolic pathway Effects 0.000 description 1
- 230000004060 metabolic process Effects 0.000 description 1
- 230000006609 metabolic stress Effects 0.000 description 1
- 229910021645 metal ion Inorganic materials 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- HPNSFSBZBAHARI-UHFFFAOYSA-N micophenolic acid Natural products OC1=C(CC=C(C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-UHFFFAOYSA-N 0.000 description 1
- 238000012544 monitoring process Methods 0.000 description 1
- 125000004573 morpholin-4-yl group Chemical group N1(CCOCC1)* 0.000 description 1
- 230000000869 mutational effect Effects 0.000 description 1
- HPNSFSBZBAHARI-RUDMXATFSA-N mycophenolic acid Chemical compound OC1=C(C\C=C(/C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-RUDMXATFSA-N 0.000 description 1
- 229960000951 mycophenolic acid Drugs 0.000 description 1
- 108010035972 myxobacter alpha-lytic proteinase Proteins 0.000 description 1
- GNOLWGAJQVLBSM-UHFFFAOYSA-N n,n,5,7-tetramethyl-1,2,3,4-tetrahydronaphthalen-1-amine Chemical compound C1=C(C)C=C2C(N(C)C)CCCC2=C1C GNOLWGAJQVLBSM-UHFFFAOYSA-N 0.000 description 1
- YFCUZWYIPBUQBD-ZOWNYOTGSA-N n-[(3s)-7-amino-1-chloro-2-oxoheptan-3-yl]-4-methylbenzenesulfonamide;hydron;chloride Chemical compound Cl.CC1=CC=C(S(=O)(=O)N[C@@H](CCCCN)C(=O)CCl)C=C1 YFCUZWYIPBUQBD-ZOWNYOTGSA-N 0.000 description 1
- 230000037125 natural defense Effects 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 229920001220 nitrocellulos Polymers 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 238000010606 normalization Methods 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 125000003835 nucleoside group Chemical group 0.000 description 1
- 235000014571 nuts Nutrition 0.000 description 1
- 108010032563 oligopeptidase Proteins 0.000 description 1
- 108090000857 oligopeptidase B Proteins 0.000 description 1
- 229920001542 oligosaccharide Polymers 0.000 description 1
- 150000002482 oligosaccharides Chemical class 0.000 description 1
- 108010003052 omptin outer membrane protease Proteins 0.000 description 1
- 230000003287 optical effect Effects 0.000 description 1
- 235000020636 oyster Nutrition 0.000 description 1
- 238000012856 packing Methods 0.000 description 1
- 108090000155 pancreatic elastase II Proteins 0.000 description 1
- 235000020232 peanut Nutrition 0.000 description 1
- 229940066716 pepsin a Drugs 0.000 description 1
- 229950000964 pepstatin Drugs 0.000 description 1
- 108010091212 pepstatin Proteins 0.000 description 1
- FAXGPCHRFPCXOO-LXTPJMTPSA-N pepstatin A Chemical compound OC(=O)C[C@H](O)[C@H](CC(C)C)NC(=O)[C@H](C)NC(=O)C[C@H](O)[C@H](CC(C)C)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C(C)C)NC(=O)CC(C)C FAXGPCHRFPCXOO-LXTPJMTPSA-N 0.000 description 1
- 108090000756 peptidyl dipeptidase B Proteins 0.000 description 1
- 235000011197 perejil Nutrition 0.000 description 1
- 238000002823 phage display Methods 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- 239000012071 phase Substances 0.000 description 1
- 239000003016 pheromone Substances 0.000 description 1
- 101150009573 phoA gene Proteins 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 108010072906 phosphoramidon Proteins 0.000 description 1
- BWSDNRQVTFZQQD-AYVHNPTNSA-N phosphoramidon Chemical compound O([P@@](O)(=O)N[C@H](CC(C)C)C(=O)N[C@H](CC=1[C]2C=CC=CC2=NC=1)C(O)=O)[C@H]1O[C@@H](C)[C@H](O)[C@@H](O)[C@@H]1O BWSDNRQVTFZQQD-AYVHNPTNSA-N 0.000 description 1
- 230000000704 physical effect Effects 0.000 description 1
- 230000004962 physiological condition Effects 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 108010020708 plasmepsin Proteins 0.000 description 1
- 229940012957 plasmin Drugs 0.000 description 1
- 229920003023 plastic Polymers 0.000 description 1
- 239000004033 plastic Substances 0.000 description 1
- 238000007747 plating Methods 0.000 description 1
- 239000013573 pollen allergen Substances 0.000 description 1
- 229920000647 polyepoxide Polymers 0.000 description 1
- 229920000656 polylysine Polymers 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 102000007739 porin activity proteins Human genes 0.000 description 1
- 230000029279 positive regulation of transcription, DNA-dependent Effects 0.000 description 1
- 239000002244 precipitate Substances 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 108010035833 pro-ocytocin-neurophysin convertase Proteins 0.000 description 1
- 239000013587 production medium Substances 0.000 description 1
- 108010017378 prolyl aminopeptidase Proteins 0.000 description 1
- 125000001500 prolyl group Chemical group [H]N1C([H])(C(=O)[*])C([H])([H])C([H])([H])C1([H])[H] 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 238000003498 protein array Methods 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 238000001243 protein synthesis Methods 0.000 description 1
- 230000004850 protein–protein interaction Effects 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 235000015136 pumpkin Nutrition 0.000 description 1
- 229950010131 puromycin Drugs 0.000 description 1
- 238000004445 quantitative analysis Methods 0.000 description 1
- 238000011155 quantitative monitoring Methods 0.000 description 1
- 238000002708 random mutagenesis Methods 0.000 description 1
- 238000009789 rate limiting process Methods 0.000 description 1
- 230000007420 reactivation Effects 0.000 description 1
- 102000005962 receptors Human genes 0.000 description 1
- 108020003175 receptors Proteins 0.000 description 1
- 230000008263 repair mechanism Effects 0.000 description 1
- 230000003938 response to stress Effects 0.000 description 1
- 238000002702 ribosome display Methods 0.000 description 1
- 210000004708 ribosome subunit Anatomy 0.000 description 1
- 108091092562 ribozyme Proteins 0.000 description 1
- 102200027014 rs80356663 Human genes 0.000 description 1
- 239000012723 sample buffer Substances 0.000 description 1
- DJSISFGPUUYILV-ZFORQUDYSA-N scutellarin Chemical compound O1[C@H](C(O)=O)[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1OC(C(=C1O)O)=CC2=C1C(=O)C=C(C=1C=CC(O)=CC=1)O2 DJSISFGPUUYILV-ZFORQUDYSA-N 0.000 description 1
- 229930190376 scutellarin Natural products 0.000 description 1
- 230000003248 secreting effect Effects 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 238000001542 size-exclusion chromatography Methods 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 239000000243 solution Substances 0.000 description 1
- 238000001179 sorption measurement Methods 0.000 description 1
- 235000020354 squash Nutrition 0.000 description 1
- 239000003381 stabilizer Substances 0.000 description 1
- 238000012289 standard assay Methods 0.000 description 1
- 108090000346 stem bromelain Proteins 0.000 description 1
- 230000035892 strand transfer Effects 0.000 description 1
- 230000006585 stringent response Effects 0.000 description 1
- 125000000446 sulfanediyl group Chemical group *S* 0.000 description 1
- RWSOTUBLDIXVET-UHFFFAOYSA-O sulfonium Chemical compound [SH3+] RWSOTUBLDIXVET-UHFFFAOYSA-O 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 239000004094 surface-active agent Substances 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 230000009897 systematic effect Effects 0.000 description 1
- 238000004885 tandem mass spectrometry Methods 0.000 description 1
- 235000013616 tea Nutrition 0.000 description 1
- 230000002123 temporal effect Effects 0.000 description 1
- 108010031354 thermitase Proteins 0.000 description 1
- 108060008226 thioredoxin Proteins 0.000 description 1
- 229960004072 thrombin Drugs 0.000 description 1
- 230000036964 tight binding Effects 0.000 description 1
- 210000001519 tissue Anatomy 0.000 description 1
- FUSNMLFNXJSCDI-UHFFFAOYSA-N tolnaftate Chemical compound C=1C=C2C=CC=CC2=CC=1OC(=S)N(C)C1=CC=CC(C)=C1 FUSNMLFNXJSCDI-UHFFFAOYSA-N 0.000 description 1
- 231100000167 toxic agent Toxicity 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 238000006276 transfer reaction Methods 0.000 description 1
- 230000001131 transforming effect Effects 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- 230000005945 translocation Effects 0.000 description 1
- 108010039189 tripeptidyl-peptidase 2 Proteins 0.000 description 1
- 101150112435 trxC gene Proteins 0.000 description 1
- 239000012588 trypsin Substances 0.000 description 1
- 239000012137 tryptone Substances 0.000 description 1
- WFKWXMTUELFFGS-UHFFFAOYSA-N tungsten Chemical compound [W] WFKWXMTUELFFGS-UHFFFAOYSA-N 0.000 description 1
- 229910052721 tungsten Inorganic materials 0.000 description 1
- 239000010937 tungsten Substances 0.000 description 1
- 229950009811 ubenimex Drugs 0.000 description 1
- 229960005486 vaccine Drugs 0.000 description 1
- 208000007089 vaccinia Diseases 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 230000004584 weight gain Effects 0.000 description 1
- 235000019786 weight gain Nutrition 0.000 description 1
- 241000228158 x Triticosecale Species 0.000 description 1
- 229940075420 xanthine Drugs 0.000 description 1
- 108010021321 xanthomonapepsin Proteins 0.000 description 1
- 239000012138 yeast extract Substances 0.000 description 1
- 108010078692 yeast proteinase B Proteins 0.000 description 1
- 108010064359 zinc D-Ala-D-Ala carboxypeptidase Proteins 0.000 description 1
Images





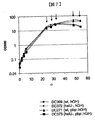




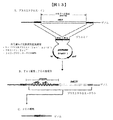


Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/06—Preparation of peptides or proteins produced by the hydrolysis of a peptide bond, e.g. hydrolysate products
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
- C12N15/78—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora for Pseudomonas
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/02—Preparation of peptides or proteins having a known sequence of two or more amino acids, e.g. glutathione
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Chemical & Material Sciences (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Microbiology (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
Description
本出願は、2004年7月26日出願の米国特許仮出願第60/591,489号の優先権を主張する。
マイクロアレイテクノロジーは、単一アッセイにおいて数多くのポリヌクレオチドの存在と発現レベルを特定するために使用することができる。例えば、全てAffymetrix,Inc.に許諾された、1995年9月15日出願の米国特許第6,040,138号、1997年6月25日出願の米国特許第6,344,316号、1999年4月15日出願の米国特許第6,261,776号、2000年10月16日出願の米国特許第6,403,957号、2000年9月27日出願の米国特許第6,451,536号、2001年8月27日出願の米国特許第6,532,462号、2001年5月9日出願の米国特許第6,551,784号、1998年2月9日出願の米国特許第6,420,108号、1998年12月14日出願の米国特許第6,410,229号、2001年1月25日出願の米国特許第6,576,424号、2000年11月2日出願の米国特許第6,687,692号、1998年4月21日出願の米国特許第6,600,031号、2001年4月16日出願の米国特許第6,567,540号参照。
組換えタンパク質の望ましくない分解は、ある種の発現系の効率的な使用に対する障害となる。外来性タンパク質の発現は、しばしば宿主細胞においてストレス応答を誘導し、それは、例えば限られた炭素源に対する天然防御であり得る。全ての細胞は、分解性タンパク質を生産することができる数多くの遺伝子を含む。いずれのプロテアーゼが特定組換えタンパク質の発現に応答して所与の宿主によって調節されるかを予測することは不可能である。例えばP.フルオレセンス細菌は、200までのプロテアーゼとプロテアーゼ関連タンパク質を含む。
宿主細胞での組換えタンパク質の生産におけるもう1つの主要な障害は、細胞がしばしば可溶性又は活性タンパク質のいずれかを生産するように適切に用意されていないことである。タンパク質の一次構造はそのアミノ酸配列によって規定されるが、二次構造はαヘリックス又はβシートの存在によって規定され、三次構造は隣接する一続きのタンパク質の間の共有結合、例えばジスルフィド結合によって規定される。組換えタンパク質を発現するとき、特に大規模生産では、タンパク質自体の二次及び三次構造が決定的に重要である。タンパク質構造の何らかの有意の変化は、機能的に不活性な分子又は有意に低い生物活性を有するタンパク質を生じ得る。多くの場合、宿主細胞は、活性組換えタンパク質の適切な生産のために必要なフォールディング調節剤(FM)を発現する。しかし、使用可能な、経済的に満足し得るバイオテクノロジー産物を生産するために一般に必要とされる高い発現レベルでは、細胞はしばしば、組換えタンパク質をプロセシングするために十分な天然フォールディング調節剤を生産することができない。
ii)前記細胞の遺伝的プロフィールを分析し、組換えタンパク質又はペプチドの発現又は過剰発現時に上方調節される1又はそれ以上の内在性遺伝子産物を特定すること;及び
iii)細胞を遺伝的に修飾することによって1又はそれ以上の特定された内在性遺伝子産物の発現を変化させること
を含む、組換えタンパク質又はペプチドの発現を改善するための方法を提供する。
i)組換え宿主細胞又は生物において組換えタンパク質又はペプチドを発現すること;
ii)組換えタンパク質を発現するように修飾されていない宿主細胞又は組換えタンパク質を発現していない組換え細胞のいずれか1つにおけるよりも高いレベルで組換え細胞において発現される補償遺伝子又は遺伝子産物を特定するために組換え細胞の遺伝的プロフィールを分析すること;及び
iii)組換えタンパク質の発現、活性又は溶解度の上昇を達成する修飾組換え細胞を提供するために、組換え細胞における特定された補償遺伝子又は遺伝子産物の発現を遺伝子修飾によって変化させること
を含む、宿主細胞又は生物における組換えタンパク質又はペプチドの発現を改善するための方法を含む。
本発明の方法の第一工程では、宿主細胞を、組換えタンパク質又はペプチドを発現する能力を有するように修飾する。宿主細胞は、当技術分野で公知の何らかの手法を用いて修飾することができる。例えば組換えタンパク質は、細胞のゲノムに内在性であり、細胞にトランスフェクト又は形質転換される発現ベクターから発現することができる。発現ベクターの構築並びにトランスフェクション又は形質転換のための手法を以下で述べる。宿主細胞はまた、以下で述べるようにゲノム挿入物から組換えタンパク質又はペプチドを発現するように修飾することもできる。組換えタンパク質又はペプチドをコードする遺伝子を、相同的又は非相同的組換えなどの手法によって宿主細胞又は生物のゲノムに挿入することができる。これらの手法を以下で述べる。

以下で述べるように、宿主細胞又は生物は、標準手法を用いて組換えタンパク質又はペプチドを発現するように工作することができる。例えば組換えタンパク質は、宿主のゲノムに挿入したベクターから又は外来性遺伝子から発現することができる。外来性タンパク質を発現するために使用できるベクターは当技術分野において周知であり、以下で述べる。組換えタンパク質又はペプチドを発現するための遺伝子はまた、以下で述べるように、相同的又は非相同的組換えなどの手法を用いてゲノムに挿入することができる。
宿主細胞は、組換えタンパク質又はペプチドを発現するように設計されてきた。これらはいかなる種及びいかなる大きさでもあり得る。しかし、ある実施形態では、組換えタンパク質又はペプチドは、治療上有用なタンパク質又はペプチドである。一部の実施形態では、タンパク質は、哺乳動物タンパク質、例えばヒトタンパク質であり得、例えば増殖因子、サイトカイン、ケモカイン又は血液タンパク質であり得る。組換えタンパク質又はペプチドは、主として宿主細胞において不活性形態で発現され得る。ある実施形態では、組換えタンパク質又はペプチドは、100kD未満、50kD未満又は30kD未満の大きさである。ある実施形態では、組換えタンパク質又はペプチドは、少なくとも5、10、15、20、30、40、50又は100アミノ酸のペプチドである。
本発明の方法は、組換えタンパク質を発現するように修飾されていない宿主細胞又は組換えタンパク質を発現しない組換え細胞よりも高いレベルで組換え細胞において発現される補償遺伝子又は遺伝子産物を特定するために、組換え細胞の遺伝的プロフィールを分析することを含む。
1つの実施形態では、分析する遺伝的プロフィールはトランスクリプトームプロフィールである。完全なトランスクリプトームとは、一度にゲノムによって生産されるmRNA転写産物の完全なセットを指す。ゲノムと異なり、トランスクリプトームは動的であり、遺伝子発現の異なるパターンの故に異なる状況においてはかなり変化する。トランスクリプトームの研究であるトランスクリプトミクスは、遺伝子発現パターンを特定する包括的な手段である。分析するトランスクリプトームは、転写される遺伝子の公知の完全なセット、すなわち宿主細胞又は宿主生物のmRNA含量又は対応するcDNAを含み得る。cDNAは、ヌクレオチドの鎖、単離ポリヌクレオチド、ヌクレオチド、核酸分子、又は組換え又は合成によって創出される何らかのフラグメント又はその相補物であり得、二本鎖又は一本鎖、コード及び/又は非コードであり得、ゲノムDNA分子のエクソン又はイントロンであり得、炭水化物、脂質、タンパク質又は無機元素又は物質と結合し得る。ヌクレオチド鎖は、少なくとも5、10、15、30、40、50、60、70、80、90又は100ヌクレオチドの長さであり得る。トランスクリプトームはまた、遺伝子転写産物の公知のセットの一部だけを含み得る。例えばトランスクリプトームは、宿主における公知の転写産物の98%、95、90、85、80、70、60又は50%未満を含み得る。トランスクリプトームはまた、遺伝子の特定セットを標的することができる。
もう1つの実施形態では、分析する遺伝的プロフィールはプロテオームプロフィールである。宿主のプロテオームは、一度にゲノムによって生産されるタンパク質の完全なセットである。プロテオームは、各々のタンパク質が合成後に化学修飾され得るので、一般にゲノム又はトランスクリプトームよりもはるかに複雑である。多くのタンパク質は、宿主細胞に依存して、生産の間に切断されたり、リン酸化、アセチル化、メチル化されたり、又はそれらに付加された炭水化物基を有する。プロテオームはまた、非常に動的である。プロテオームの研究であるプロテオミクスは、タンパク質構造、タンパク質発現及び機能の数多くの異なる態様をカバーし得る。プロテオーム分析のための手法は、トランスクリプトミクスにおいて使用されるものほど直接的ではない。しかし、プロテオミクスの利点は、細胞の機能的分子が検討されることである。
プロテオーム分析法は、多くのタンパク質の存在率と分布を同時に測定することを可能にする。しかし、プロテオームへの変化の機能的影響は間接的にしか報告されない。もう1つのアプローチは、これらの低分子又は代謝産物のレベルを測定することである。本発明の方法において分析する遺伝的プロフィールは、それ故、メタボロームプロフィールを含み得る。特定宿主のメタボロームを分析するための方法は、様々な化学的及び物理的性質に従って代謝産物を分離するための、ガスクロマトグラフィー、高圧液体クロマトグラフィー及びキャピラリー電気泳動を含む。その後、質量分析などの方法を用いて分子を特定することができる。
本発明の方法は、組換え細胞においてより高いレベルで発現される補償遺伝子又は遺伝子産物を特定するために遺伝的プロフィールを分析することを含む。一般に、この工程は、多数の遺伝子又は遺伝子産物の発現の観測(例えば発現を検出すること及び/又は定量すること)を含む。発現は一般に、上述したようなトランスクリプトーム、プロテオーム又はメタボロームプロフィールへの宿主細胞遺伝子産物の結合を検出することによって観測される。結合の分析は、組換えタンパク質又はペプチドを発現する組換え宿主細胞とナイーブ宿主細胞又は前記タンパク質又はペプチドを発現しない組換え宿主細胞の間の結合の比較を含み得る。
この工程は、多数の遺伝子又は遺伝子産物の発現の観測(例えば発現を検出すること及び/又は定量すること)を含む。発現は一般に、上述したようなトランスクリプトーム、プロテオーム又はメタボロームプロフィールへの宿主細胞遺伝子産物の結合を検出することによって観測される。典型的には、少なくとも約10個の遺伝子、又は少なくとも約100個、又は少なくとも約1000個、及び/又は少なくとも約10,000個の異なる遺伝子を一度に検定することができる。前記方法は、前記遺伝子の1又はそれ以上のRNA転写産物を含む特定された核酸又はRNA転写産物に由来する核酸のプールを提供すること;核酸のプールを、表面に固定化されたオリゴヌクレオチドプローブのアレイにハイブリダイズすること、但し、前記アレイは100個以上の異なるオリゴヌクレオチドを含み、各々の異なるオリゴヌクレオチドは前記表面のあらかじめ定められ領域に位置し、各々の異なるオリゴヌクレオチドは少なくとも1つの共有結合を通して表面に結合しており、オリゴヌクレオチドプローブはRNA転写産物又はRNA転写産物に由来する核酸に相補的である;及びアレイ内のハイブリダイズした核酸を定量すること、を含み得る。2つの試料の間の遺伝子産物の発現を観測するための1つの手法の絵画的表示を図12に示す。
特定される遺伝子は、典型的には組換えタンパク質又はペプチドを発現する宿主細胞の遺伝的プロフィールを、組換えタンパク質又はペプチドを発現しない宿主細胞の遺伝的プロフィールと比較することによって特定される。反復実施形態では、修飾すべき特定遺伝子は、修飾すべき細胞(第二細胞)の遺伝的プロフィールを、それが修飾された細胞(第一細胞)と比較することによって特定される。特定される遺伝子は、第二細胞の遺伝的プロフィールを第一細胞の遺伝的プロフィールと比較すること及び第二細胞において発現が上昇している1又はそれ以上の遺伝子を特定することによって特定される。
特定される補償遺伝子
工程ii)において特定される補償遺伝子又は遺伝子産物、又はその相同的類似体、補因子又はサブユニットは、1又はそれ以上の特定された遺伝子の発現を上昇させる、低下させる、ノックイン又はノックアウトするように細胞を遺伝的に修飾するための戦略を設計するために使用される。公的データベースにおいて特定される遺伝子配列が、上述した手法によって遺伝子の発現を調節するための戦略を設計する、特にそのための構築物を設計するために使用できる。そのような手法は周知である。
本発明の1つの実施形態では、ゲノムからの少なくとも1個のプロテアーゼの発現を低下させる、前記少なくとも1個のプロテアーゼを阻害する又は除去することによって宿主細胞を修飾する。修飾はまた、一部の実施形態では、2個以上のプロテアーゼに対してであり得る。関連実施形態では、プロテアーゼ補因子又はプロテアーゼタンパク質の発現を低下させることによって細胞を修飾する。もう1つの実施形態では、天然プロモーターであり得る、プロテアーゼ又は関連タンパク質についてのプロモーターの阻害によって宿主細胞を修飾する。遺伝子修飾は、特定された遺伝子に対して相同なタンパク質を修飾するためであり得る。
CA族は、パパイン(C1)、カルパイン(C2)、ストレプトパイン(C10)及びユビキチン特異的ペプチダーゼ(C12、C19)のファミリー、並びにウイルスシステインエンドペプチダーゼの多くのファミリーを含む。
表A
表B
表C
特定される上方調節遺伝子又は遺伝子産物は、1又はそれ以上のフォールディング調節剤であり得る。フォールディング調節剤は、例えばHSP70タンパク質、HSP110/SSEタンパク質、HSP40(DNAJ関連)タンパク質、GRPE様タンパク質、HSP90タンパク質、CPN60及びCPN10タンパク質、サイトゾルシャペロニン、HSP100タンパク質、低分子HSP、カルネキシン及びカルレティキュリン、PDI及びチオレドキシン関連タンパク質、ペプチジル−プロリルイソメラーゼ、シクロフィリンPPIアーゼ、FK−506結合タンパク質、パーブリン(Parvulin)PPIアーゼ、個々のシャペロニン、タンパク質特異的シャペロン又は細胞内シャペロンであり得る。フォールディング調節剤は、"Guidebook to Molecular Chaperones and Protein-Folding Catalysts" (1997) ed. M. Gething, Melbourne University, Australiaにおいて一般的に記述されている。
表F
工程iii)において、本発明の方法は、修飾組換え細胞を提供するために遺伝子操作によって組換え細胞内の特定された補償遺伝子又は遺伝子産物の発現を変化させることを含む。1又はそれ以上の上方調節遺伝子、タンパク質又は代謝工程の特定後に、宿主のゲノムを修飾し得る。ある種の遺伝子又は遺伝子産物は、上方調節されると特定されても、細胞に必須であるか若しくは細胞又は生物に必須であり得る他の工程に影響を及ぼすことが知られているために、調節のために使用可能でないことがある。
組換えタンパク質を発現する宿主細胞のゲノムは、挿入又は組換え、例えば相同的組換えによってであり得る、遺伝子ターゲティング事象を通して修飾することができる。相同的組換えとは、配列相同性に基づくDNA組換えの過程を指す。相同的組換えは、内在性遺伝子における部位特異的修飾を可能にし、それ故ゲノム内に新規変化を工作することができる。相同的組換えにおける1つの工程はDNA鎖交換であり、これは、ヘテロ二本鎖DNAを含む中間体組換え構造を形成するための、相補的配列を含む少なくとも1本のDNA鎖とDNA二本鎖の対合を含む(例えばRadding, C. M. (1982) Ann. Rev. Genet. 16: 405; U.S. Pat. No. 4,888,274参照)。ヘテロ二重鎖DNAは、1本の相補鎖がDNA二本鎖に侵入している三重鎖形態を有する三本鎖DNAを含む、いくつかの形態をとり得ることができ(Hsieh, et al., Genes and Development 4: 1951 (1990); Rao, et al., (1991) PNAS 88:2984))、2本の相補的DNA鎖がDNA二本鎖と対合するときは、古典的オリデイ組換え連結又はχ構造 (Holliday, R., Genet. Res. 5: 282 (1964))あるいは二重Dループ ("Diagnostic Applications of Double-D Loop Formation" U.S. Ser. No. 07/755,462, filed Sep. 4, 1991)が形成され得る。ひとたび形成されると、ヘテロ二本鎖構造は、侵入DNA鎖の全部又は一部が受容DNA二本鎖にスプライシングされて、受容DNA二本鎖のセグメントを付加する又は置換するように、鎖切断と交換によって分割することができる。あるいは、ヘテロ二本鎖構造は、侵入鎖の配列が、侵入鎖を鋳型として用いるミスマッチ塩基の修復機構によって受容DNA二本鎖に導入される、遺伝子変換をもたらし得る(Genes, 3rd Ed. (1987) Lewin, B., John Wiley, New York, N. Y.; Lopez, et al., Nucleic Acids Res. 15: 5643(1987))。切断と再連結の機構によるか又は遺伝子交換の機構によるかに関わらず、相同的に対合する連結部でのヘテロ二本鎖DNAの形成は、1個のDNA分子から別のDNA分子に遺伝子配列情報を導入するのに役立ち得る。
ゲノムはまた、特定された遺伝子、特に特定されたプロテアーゼをコードするオープンリーディングフレーム内の1又はそれ以上のヌクレオチドの突然変異によって修飾することができる。遺伝子突然変異のための手法、例えば位置指定突然変異誘発は、当技術分野において周知である。一部のアプローチは、染色体DNAにおけるランダム突然変異の生成、例えばX線及び化学物質によって誘導されるものを焦点とする。DNAの規定領域を標的する突然変異誘発は多くの手法を含み、普及度は様々である。位置指定突然変異誘発へのインビトロアプローチは一般に3つのカテゴリーに分けられる:i)DNAのフラグメントを再構築する方法、例えばカセット突然変異誘発;ii)局在化ランダム突然変異誘発;及びiii)オリゴヌクレオチド指定突然変異誘発。
別の実施形態では、特定された遺伝子、典型的にはフォールディング調節剤又はフォールディング調節剤の補因子をコードする1又はそれ以上のベクターを含めることによって宿主細胞を修飾する。もう1つの実施形態では、外来性プロモーターを宿主細胞ゲノムに付加することを含む、フォールディング調節剤又はフォールディング調節剤の補因子についてのプロモーターを増強することによって宿主細胞を修飾する。
本発明の方法は、最適には宿主細胞における組換えタンパク質又はペプチドの生産上昇を導く。生産上昇は、所与の時間での宿主タンパク質のグラム当りのタンパク質の量の上昇を含み得るか、又は宿主細胞が組換えタンパク質又はペプチドを生産している時間の長さの上昇を含み得る。生産上昇は、組換え宿主細胞の増殖のために必要条件の改善を含み得る。生産上昇は、完全長タンパク質又はペプチドの生産上昇であり得る。改善がタンパク質レベルの上昇である場合は、タンパク質又はペプチドは、宿主細胞内の1又はそれ以上の封入体において生産され得る。
組換えタンパク質の改善された発現は、タンパク質の溶解度の上昇であり得る。組換えタンパク質又はペプチドは、宿主細胞の細胞質、ペリプラズム又は細胞外媒質から生産し、回収することができる。前記タンパク質又はペプチドは、不溶性又は可溶性であり得る。タンパク質又はペプチドは、1又はそれ以上の標的配列又は精製を助けるための配列を含み得る。
ある実施形態では、タンパク質はまた、適切なシグナル分泌配列に融合している場合、ペリプラズム内に分泌され得る。1つの実施形態では、シグナル配列は、リン酸結合タンパク質、Lys−Arg−Orn結合タンパク質(LAObp又はKRObp)分泌シグナルペプチド、外膜ポーリンE(OprE))分泌シグナルペプチド、アズリン)分泌シグナルペプチド、鉄(III)結合タンパク質[Fe(III)bp])分泌シグナルペプチド、又はリポタンパク質B(LprB))分泌シグナルペプチドであり得る。
(実施例)
表1 細菌株の概略
表2 プラスミドの概略
全ての試料を200ml標準振とうフラスコ実験から収集した。試料は、図に示すような種々の時点で採取した。各々の時点で、振とうフラスコから細胞培養10mlを収集し、RNAを安定化するためにRNAlater(Ambion,Austin,TX)試薬10mlと混合した。
各々のRNA試料について、蛍光ヌクレオチドCy3−dUTP又はCy5−dUTP(Amersham Pharmacia, Piscataway, NJ)を、ランダム六量体プライマー(Amersham)を用いた逆転写(RT)反応においてcDNAに組み込んだ。2つの標識cDNAプールを一緒にし、マイクロアレイスライドに適用した。マイクロアレイスライドは、P.フルオレセンス(P. fluorescens)の各々のORFである50量体アミノ修飾オリゴデオキシリボヌクレオチド(オリゴ)を含む。各々のオリゴを、SDDC−2ロボット(Virtek,Toronto,Canada−現在はBio−Rad Laboratories,Hercules,CAを通して配給されている)及びSMP3ピン(TeleChem International Inc.,Sunnyvale,CA)を使用して異なる位置の2つのスポットに関して2回プリントした。使用した顕微鏡スライドは、効率的なDNA結合のために正に荷電したエポキシ樹脂で被覆した(MWG Inc,Alameda,CA)。プリント後、スライドをMWGの仕様書に従って後処理した。データ解析をようにするためにBioDiscovery Inc.(El Segundo,CA)からのソフトウエアパッケージを使用した。このパッケージは、CloneTracker(商標)、ImaGene(商標)、GeneSight(商標) モジュール及びGeneDirector(商標)データベースから成る。各々のハイブリダイズしたスライドを、マイクロアレイに結合したCy3及びCy5標識cDNAの蛍光を測定するためにScanArray 5000(Packard BioScience,Billerica,MA)を用いて走査した。獲得した画像をImaGene(商標)で数量化し、生データをGeneSight(商標)で処理した。データ作成の間に、各遺伝子についてのスポット強度をバックグラウンド補正した;アレイ全体についての総シグナル強度を使用して、Cy5チャネルについてのシグナルをCy3チャネルに基準化した;各遺伝子についてのCy5対Cy3の基準化した比をlog2変換し、複製を一緒にした。
IPTG誘導後様々な時点で培養アリコートを収集し、10のOD600に基準化した。細胞溶解産物を11,000gで5分間の遠心分離によって可溶性と不溶性の分画に分けた。2.5μlのアリコートを2X NuPAGE LDS試料緩衝液(Invitrogen,San Diego,CA)、50μM DTTと混合し、H2Oを加えて10μlにして、その後95℃で5分間加熱した。タンパク質を分離し、Simply Blue Safestain(Invitrogen,San Diego,CA)を用いてクマシーブルーで染色した12%Nupageゲル上で視覚化した。
緑色蛍光タンパク質(COP)とヒト成長ホルモン(hGH)の融合物の蛍光活性によってタンパク質収率も測定した。hgh::COP融合構築物を野生型又はhslU突然変異型菌株に形質転換し、ウラシルを含まないM9グルコース寒天プレートで選択した。IPTG誘導細胞培養を5のOD600に基準化した。Spectramax Geminiマイクロプレート分光蛍光計(Molecular Devices,Sunnyvale,CA)を適切な設定(Ex485、Em538530バンドパスフィルター)下で使用して相対蛍光(RF)活性を測定した。
異種タンパク質の生産の間のFM及びプロテアーゼ遺伝子発現を検討するため、P.フルオレセンス(P. fluorescens)菌株DC206、280、240及び271を初期マイクロアレイ実験において使用した。DC206は宿主菌株であり、細胞増殖についての対照として使用した;DC280はベクターだけのプラスミドであり、マイクロアレイ実験についての対照として使用した;DC240は、可溶性である細胞質ニトリラーゼ酵素をコードするプラスミドを有するDC206である;DC271は、部分的に不溶性であるペリプラズムヒト成長ホルモン(pbp::hGH)をコードするプラスミドを有するDC206である。菌株を振とうフラスコ培地200ml中で増殖させ、OD575を測定することによって細胞増殖を観測した。接種後24時間目にIPTG誘導を実施した。全ての菌株が同様に増殖し、RNA単離及びDNAマイクロアレイを用いた転写プロファイリング(TxP)のための誘導の直前(0時間目)と4時間後に培養試料を採取した(図1)。
表3 実施例1−3において実施したマイクロアレイ実験の要約
表4 定常状態mRNA比のレベルがDC240及びDC280と比較してDC271においてより高いFM及びプロテアーゼ遺伝子のリスト。列記した数値は、IPTG誘導後4時間目対0時間目の比である。
上記で得た結果を確認するため、2つの菌株DC271とDC240の直接比較によって付加的なマイクロアレイ実験を実施した(表3のスライド7−10)。誘導後4時間目の時点での2つの菌株の比較は、部分的に可溶性のpbp::hGHを発現する細胞においてほとんど同一のセットのFM及びプロテアーゼ遺伝子が上方調節されることを確認した(表5)。表5に列記する全ての遺伝子が、完全に可溶性のニトリラーゼを生産する細胞と比較したとき、部分的に不溶性のhGHを生産する菌株において有意に(すなわち≧2倍)高く発現される。DC271とDC240の直接比較において、部分的に不溶性のhGH生産の間に有意に高い遺伝子発現値を示した時点比較に比べて(表4参照)数個の付加的なタンパク質が特定された。それらの遺伝子は、ClpBをコードするrxf08347、ClpAをコードするrxf04587、及びFkbPをコードするrxf05753を含んだ。大腸菌ClpBホモログは、DnaKJ−GrpEと共に封入体の再活性化に関与する。大腸菌からのClpAはシャペロン機能を有し、ClpPと一緒になったとき、タンパク質を分解する。大腸菌において、FkbPはペプチジル−プロリルイソメラーゼとして機能する。
表5 定常状態mRNAレベルがDC240と比較してDC271においてより高いFM及びプロテアーゼ遺伝子のリスト。列記した数値は、IPTG誘導後4時間目のDC271対DC240の比である。
DC271は部分的にペリプラズムヒト成長ホルモン(pbp::hGH)を発現するので、主として不溶性の細胞質hGHを発現する菌株において類似の又は異なるFM及びプロテアーゼ遺伝子が上方調節されるかどうかを検討した。DC369をこの実験で使用した。誘導後4時間目の試料を0時間目の時点の試料と比較し、表3(スライド11及び12)に示すようにマイクロアレイ実験を実施した。やはり、類似のFM及びプロテアーゼ遺伝子が上方調節されることが認められ、特定された遺伝子がペリプラズムフォールディング及びタンパク質分解ではなく細胞質フォールディング及びタンパク質分解に関与することを示唆した(表6)。実験において特定された遺伝子の要約を、上方調節の倍数と共に図4のベン図に示す。
表6 定常状態mRNAレベルが0時間目の時点と比較して誘導後4時間目にDC369においてより高いFM及びプロテアーゼ遺伝子のリスト。列記した数値は、IPTG誘導後4時間目対0時間目(誘導の直前)の比である。
2個の遺伝子hslVUが、最も高く上方調節される特定遺伝子の中に認められた。HslUは細胞質ATPアーゼである。大腸菌における相同タンパク質は、大腸菌のエネルギー依存性タンパク質分解を促進する第二タンパク質と共同して働くことができる。HslUは、プロテアソームのαサブユニットに相同性を有するタンパク質、HslVと相互作用する。大腸菌HslVUホモログは、Missiakas, D., et al. (1996) Identification and characterization of HsIV HsIU (CIpQ CIpY) proteins involved in overall proteolysis of misfolded proteins in Escherichia coli. Embo J 15:6899-909の中で、折りたたみ異常タンパク質の全体的タンパク質分解に関与することが報告された。DNA塩基配列分析は、P.フルオレセンスhslVU遺伝子が2シストロン性オペロンの一部である可能性が高いことを示唆した(図5)。
表7 プライマー
hslU遺伝子ノックアウトの影響を検討するため、2個の外来性タンパク質発現を、親菌株DC206と新たに構築した突然変異型菌株DC370の間で比較した。pbp::hGH(pDOW1323)及びhGH(pDOW1426)をコードする遺伝子を担持するプラスミドを各々コンピテントDC370細胞に形質転換し、それぞれ菌株DC373及びDC372を生じた。4つの菌株に関して標準振とうフラスコ増殖実験を実施した。図7は、野生型と突然変異型菌株が同様の増殖速度を有することを示す。試料をSDS−PAGEゲルで泳動させた(図8及び9)。結果は、プロテアーゼサブユニットHslUの欠失の故に突然変異型がより高い量のタンパク質を生産したことを示唆する。
認められたHslUの欠如のhGHの収率への影響はSDS−PAGE分析を用いて定量することが困難であるので、COP緑色蛍光タンパク質とhGHの間の融合タンパク質の蛍光によってタンパク質生産の一時的プロフィールを観測した。hGH::COP融合物を含むプラスミドを構築し、親菌株DC206及びhslU遺伝子欠失菌株DC370に形質転換して、菌株HJ104及びHJ105を生成した(表1)。標準振とうフラスコ増殖実験を実施し、蛍光測定のために様々な時点で試料を採取した(図10)。蛍光光度計からの読み取りは、hslUプロテアーゼ突然変異型菌株が親菌株と比較して有意に高いタンパク質発現レベルを有することを明らかに示した(図11)。この所見はSDS−PAGE分析によって得られた結果を裏付ける。野生型菌株と比較して、hslU突然変異型は、誘導後24時間目に相対蛍光の33.05%を上昇させた(図11の挿入図参照)。
Hslプロテアーゼは2つのサブユニット:hslUによってコードされるATP結合サブユニット及びhslVによってコードされるプロテアーゼサブユニット、から成る。先に構築したHslプロテアーゼノックアウト菌株はhslU遺伝子の挿入不活性化である。HslVが、もう1つ別のプロテアーゼのATP結合サブユニットと結合できることによってまだプロテアーゼとして機能し得るという懸念を取り除くため、hslUとhslV遺伝子の両方が染色体から除去された欠失菌株を構築した。
hGHタンパク質を発現するhslUV欠失菌株(菌株HJ115)のSDS−PAGE分析は、hslU挿入突然変異型菌株DC372を使用して先に認められたものと同様に(データは示していない)、野生型菌株DC369よりもはるかに高いタンパク質収率を示した。
表8 IPTG誘導後4時間目対0時間目(誘導の直前)の比に基づき、その定常状態mRNAレベルが野生型菌株(DC369)と比較してhslUVプロテアーゼ欠失菌株(HJ115)においてより高いプロテアーゼ遺伝子。
図4に示す転写プロファイリングデータに基づき、フォールディング調節剤(FM)DnaK及びDnaJの発現は、対照菌株と比較して組換えタンパク質を生産する菌株において上昇した(表4及び5参照)。hGHと共にGrpE、DnaK及びDnaJを共過剰生産した菌株を生産し、これが可溶性hGHのレベル上昇の蓄積を生じさせるかどうかを特定するために試験した。
P.フルオレセンス(P. fluorescens)grpE−dnaKJ遺伝子を、MB214(DNeasy;Qiagen,Valencia,CA)から単離した染色体DNAを鋳型とし、RC199(5’−ATATACTAGTAGGAGGTAACTTATGGCTGACGAACAGACGCA−3’)及びRC200(5’−ATATTCTAGATTACAGGTCGCCGAAGAAGC−3’)をプライマーとして使用して増幅し、PfuTurbo(Stratagene,La Jolla,CA)を製造者の指示に従って使用した。生じたPCR産物(4kb)をSpeI及びXbaI(上記プライマーにおいて下線を付した制限部位)で消化し、pDOW2236に連結して、tacプロモーターの制御下にgrpE−dnaKJ遺伝子を含むpDOW2240を作製した。プラスミドpDOW2240をSpeI及びHindIIIで消化し、生じたgrpE dnaKJを含む4.0kbフラグメントを、Qiaquick(Qiagen)を用いてゲル精製し、同じくSpeIとHindIIIで消化したpDOW2247に連結した。マンニトールプロモーターの制御下にgrpE−dnaKJを含む、生じたプラスミド、pDOW3501を、250μg/ml ウラシルを添加したM9グルコースプレート上で選択することによってDC388に形質転換した。最後に、pDOW1426を上記菌株(DC462)に電気穿孔し、M9グルコースプレートで選択して、2個の誘導的プラスミド:1)PtachGHを担持するpDOW1426及び2)PmtlgrpE−dnaKJを担持するpDOW3501、を有する菌株DC463を生じた。
DC463の2つの培養を振とうフラスコで増殖させた。誘導後24時間目にhGHについては0.1mM IPTG及びGrpE−DnaKJについては0.5%マンニトールを添加してタンパク質誘導を実施した。誘導後0、4、8、24及び48時間目に試料を収集した。各々の時点で、1mLに基準化した20個のOD600細胞を採集し、EasyLyse(商標)(Epicentre,Madison,WI)を用いて溶解して、14,000rpmで30分間の遠心分離によって可溶性と不溶性の分画に分けた。等容量の試料をBioRad(Hercules,CA)2xレムリ緩衝液と混合し、95℃で5分間加熱して、30μLを、1xトリスグリシンSDSランニング緩衝液(BioRad)を用いてBioRad 15%トリスHCl Criterionゲルに負荷した。図15に示すようにSimply Blue Safestain(Invitrogen,Carlsbad,CA)でタンパク質を視覚化した。生じたクマシー染色ゲルをMolecular Devices Personal Densitometer(Molecular Devices,Sunnyvale,CA)を用いて走査し、ImageQuantとエクセルを用いて解析を行った。図15に示すように、GrpE、DnaKJの共過剰発現はhGHの溶解度を有意に上昇させ、総タンパク質収率はより低かったが、標的タンパク質のほぼ100%を可溶性分画に変換した。IPTGとマンニトールの同時添加を用いてDC463の増殖と誘導を反復する付加的な実験は、GrpE DnaKJが共過剰生産されたとき、hGH溶解度の度合は様々であったが(50−100%:データは示していない)、ここで示す結果を厳密に模倣した。これらの所見は、転写プロファイリングに基づく標的菌株遺伝子操作が、組換えタンパク質の溶解度及び/又は収率を高めるための合理的菌株設計を導き得ることをさらに明らかにする。
Claims (53)
- i)組換え宿主細胞又は生物において組換えタンパク質又はペプチドを発現すること;
ii)前記組換え細胞の遺伝的プロフィールを分析し、前記組換えタンパク質を発現するように修飾されていない宿主細胞又は前記組換えタンパク質を発現していない組換え細胞におけるよりも高いレベルで前記組換え細胞において発現される1又はそれ以上の補償遺伝子又は遺伝子産物を特定すること;及び
iii)前記組換え細胞における前記特定された補償遺伝子又は遺伝子産物の発現を遺伝子修飾によって変化させ、組換えタンパク質の発現、活性又は溶解度の上昇を達成する修飾組換え細胞を提供すること、
を含む、宿主細胞又は生物における組換えタンパク質又はペプチドの発現を改善する方法。 - 前記修飾組換え細胞において前記タンパク質又はペプチドを発現することをさらに含む、請求項1に記載の方法。
- (a)前記修飾組換え細胞において前記組換えタンパク質又はペプチドを発現すること;
(b)前記修飾組換え細胞の遺伝的プロフィールを分析し、前記修飾組換え細胞において区別して発現される少なくとも1つの第二遺伝子又は遺伝子産物を特定すること;
(c)前記修飾組換え細胞における第前記二の特定された遺伝子産物の発現を変化させ、二重修飾された細胞を提供すること;及び
(d)前記二重修飾組換え細胞において前記タンパク質又はペプチドを発現すること
をさらに含む、請求項1に記載の方法。 - 工程a)−d)を反復することをさらに含む、請求項3に記載の方法。
- 前記特定された遺伝子又は遺伝子産物の発現を変化させることによって、細胞生存率が影響を受けるまで工程a)−d)を反復することを含む、請求項4に記載の方法。
- 前記組換えタンパク質又はペプチドの発現が標的エンドポイントに達するまで工程a)−d)を反復することを含む、請求項4に記載の方法。
- 前記遺伝的プロフィールが、前記組換え細胞の遺伝的プロフィールを前記宿主細胞の第二の遺伝的プロフィールと比較することによって分析される、請求項1に記載の方法。
- 前記遺伝的プロフィールがトランスクリプトームプロフィールである、請求項1に記載の方法。
- 前記トランスクリプトームプロフィールがマイクロアレイを通して決定される、請求項8に記載の方法。
- 前記遺伝的プロフィールがプロテオームプロフィールである、請求項1に記載の方法。
- 前記プロテオームプロフィールが二次元ゲル電気泳動、ICAT又はLC/MSを通して決定される、請求項10に記載の方法。
- 前記プロテオームプロフィールがペプチドアレイを通して決定される、請求項10に記載の方法。
- 前記ペプチドアレイが抗体アレイである、請求項12に記載の方法。
- 前記特定される遺伝子産物が、プロテアーゼ、プロテアーゼのサブユニット、プロテアーゼの補因子、プロテアーゼの発現に影響を及ぼす細胞又は遺伝子調節剤である、請求項1に記載の方法。
- 前記特定される遺伝子産物がプロテアーゼである、請求項14に記載の方法。
- 前記特定される遺伝子産物がプロテアーゼのサブユニットである、請求項14に記載の方法。
- 前記特定される遺伝子産物がプロテアーゼの補因子である、請求項14に記載の方法。
- 前記特定される遺伝子産物が、プロテアーゼの発現に影響を及ぼす細胞又は遺伝子調節剤である、請求項14に記載の方法。
- 前記特定される遺伝子産物が、D−アラニル−メソ−ジアミノピメリン酸エンドペプチダーゼ、亜鉛プロテアーゼ、ミクロソームジペプチダーゼ、細胞外メタロプロテアーゼ前駆体、細胞分裂タンパク質ftsH及び、遺伝子hslV、hslU、clpX、clpA及びclpB由来の遺伝子産物から成る群より選択される、請求項14に記載の方法。
- 前記組換えタンパク質又はペプチドが前記宿主細胞において発現されるとき、特定される遺伝子産物のmRNAレベルが上方調節される、請求項14に記載の方法。
- 前記特定された遺伝子産物を宿主細胞ゲノムから除去する、請求項14に記載の方法。
- 前記特定された遺伝子産物を相同的組換えによって除去する、請求項21に記載の方法。
- 前記特定される遺伝子産物が、フォールディング調節剤、フォールディング調節剤のサブユニット、フォールディング調節剤の補因子、又はフォールディング調節剤の発現に影響を及ぼす細胞又は遺伝子調節剤である、請求項1に記載の方法。
- 前記特定される遺伝子産物がフォールディング調節剤である、請求項23に記載の方法。
- 前記特定される遺伝子産物がフォールディング調節剤のサブユニットである、請求項23に記載の方法。
- 前記特定される遺伝子産物がフォールディング調節剤の補因子である、請求項23に記載の方法。
- 前記特定される遺伝子産物が、フォールディング調節剤の発現に影響を及ぼす細胞又は遺伝子調節剤である、請求項23に記載の方法。
- 前記フォールディング調節剤がシャペロンタンパク質である、請求項23に記載の方法。
- 前記フォールディング調節剤が、遺伝子cbpA、htpG、dnaK、dnaJ、fkbP2、groES及びgroELの遺伝子産物から成る群より選択される、請求項23に記載の方法。
- 前記特定される遺伝子産物の発現が、前記特定遺伝子、特定遺伝子の補因子、又は特定遺伝子の細胞又は遺伝子調節剤の発現を上昇させることによって変化する、請求項23に記載の方法。
- 前記発現上昇が、前記特定遺伝子産物をコードするDNAの組込みによる、請求項30に記載の方法。
- 前記発現上昇が、宿主細胞ゲノムへのプロモーターの挿入による、請求項30に記載の方法。
- 前記発現上昇が、前記宿主細胞への外来性ベクターの組込みによる、請求項30に記載の方法。
- 前記宿主細胞が微生物細胞である、請求項1に記載の方法。
- 前記宿主細胞がシュードモナス菌(Pseudomonad)である、請求項1に記載の方法。
- 前記宿主細胞がP.フルオレセンス(P. fluorescence)細胞である、請求項1に記載の方法。
- 前記宿主細胞が大腸菌細胞である、請求項1に記載の方法。
- 前記宿主細胞が、昆虫細胞、哺乳動物細胞、酵母細胞、真菌細胞及び植物細胞から成る群より選択される、請求項1に記載の方法。
- 前記マイクロアレイが、前記宿主細胞のゲノムの少なくとも50%に対する結合パートナーの試料を含む、請求項9に記載の方法。
- 前記マイクロアレイ手法が、前記宿主細胞のゲノムの少なくとも80%に対する結合パートナーの試料を含む、請求項9に記載の方法。
- 前記マイクロアレイが、前記宿主細胞のゲノムの少なくとも90%に対する結合パートナーの試料を含む、請求項9に記載の方法。
- 前記マイクロアレイが、前記宿主細胞のゲノムの少なくとも95%に対する結合パートナーの試料を含む、請求項9に記載の方法。
- 前記改善された発現が、組換えタンパク質又はペプチドの量の上昇である、請求項1に記載の方法。
- 前記改善された発現が、前記組換えタンパク質又はペプチドの溶解度の上昇である、請求項1に記載の方法。
- 前記改善された発現が、前記組換えタンパク質又はペプチドの活性の上昇である、請求項1に記載の方法。
- 前記遺伝的プロフィールが、遺伝子ファミリーにおける遺伝子のプロフィールである、請求項1に記載の方法。
- 前記プロフィールが、プロテアーゼ及びフォールディング調節剤を含む、請求項1に記載の方法。
- 前記プロフィールが本質的にプロテアーゼから成る、請求項46に記載の方法。
- 少なくとも2つのプロテアーゼの発現を低下させるように遺伝的に修飾された組換えタンパク質を発現する宿主細胞又は生物。
- D−アラニル−メソ−ジアミノピメリン酸エンドペプチダーゼ、亜鉛プロテアーゼ、ミクロソームジペプチダーゼ、細胞外メタロプロテアーゼ前駆体、細胞分裂タンパク質ftsH、及び遺伝子hslV、hslU、clpX、clpA及びclpB由来の遺伝子産物から成る群より選択される少なくとも1つのプロテアーゼの発現を低下させるように遺伝的に修飾された組換えタンパク質を発現する宿主細胞又は生物。
- 少なくとも1つのプロテアーゼの発現を低下させるように遺伝的に修飾された哺乳動物由来の組換えタンパク質を発現する宿主細胞又は生物。
- 前記組換えタンパク質がヒト成長ホルモンである、請求項52に記載の宿主細胞又は生物。
- フォールディング調節剤サブユニットではない少なくとも2つのフォールディング調節剤の発現を上昇させるように遺伝的に修飾された組換えタンパク質を発現する宿主細胞又は生物。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US59148904P | 2004-07-26 | 2004-07-26 | |
| PCT/US2005/026390 WO2006014899A2 (en) | 2004-07-26 | 2005-07-26 | Process for improved protein expression by strain engineering |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2011251610A Division JP2012055318A (ja) | 2004-07-26 | 2011-11-17 | 菌株遺伝子操作による改善されたタンパク質発現のための方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2008507294A true JP2008507294A (ja) | 2008-03-13 |
| JP2008507294A5 JP2008507294A5 (ja) | 2008-09-11 |
Family
ID=35787752
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2007523707A Pending JP2008507294A (ja) | 2004-07-26 | 2005-07-26 | 菌株遺伝子操作による改善されたタンパク質発現のための方法 |
| JP2011251610A Withdrawn JP2012055318A (ja) | 2004-07-26 | 2011-11-17 | 菌株遺伝子操作による改善されたタンパク質発現のための方法 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2011251610A Withdrawn JP2012055318A (ja) | 2004-07-26 | 2011-11-17 | 菌株遺伝子操作による改善されたタンパク質発現のための方法 |
Country Status (9)
| Country | Link |
|---|---|
| US (2) | US8603824B2 (ja) |
| EP (2) | EP1774017B1 (ja) |
| JP (2) | JP2008507294A (ja) |
| KR (2) | KR20130019457A (ja) |
| CN (1) | CN101031655A (ja) |
| AU (1) | AU2005269527B2 (ja) |
| BR (1) | BRPI0513826A2 (ja) |
| CA (1) | CA2574953A1 (ja) |
| WO (1) | WO2006014899A2 (ja) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2013509181A (ja) * | 2009-10-30 | 2013-03-14 | メルク・シャープ・エンド・ドーム・コーポレイション | ジペプチジルアミノペプチダーゼ活性を欠くピチア・パストリスにおける治療用タンパク質の製造方法 |
| JP2021500892A (ja) * | 2017-10-27 | 2021-01-14 | フェネックス インク. | 組み換えE.coliアスパラギナーゼの製造のための方法 |
Families Citing this family (63)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ZA862768B (en) * | 1985-04-17 | 1986-12-30 | Zymogenetics Inc | Expression of factor vii and ix activities in mammalian cells |
| US9453251B2 (en) | 2002-10-08 | 2016-09-27 | Pfenex Inc. | Expression of mammalian proteins in Pseudomonas fluorescens |
| GB0329681D0 (en) * | 2003-12-23 | 2004-01-28 | Delta Biotechnology Ltd | Gene expression technique |
| BRPI0513826A2 (pt) | 2004-07-26 | 2010-06-22 | Dow Global Technologies Inc | processo para expressão de proteìna melhorada através de engenharia de cepa |
| AU2005317828A1 (en) * | 2004-12-23 | 2006-06-29 | Novozymes Delta Limited | Gene expression technique |
| GB0512707D0 (en) * | 2005-06-22 | 2005-07-27 | Delta Biotechnology Ltd | Gene expression technique |
| WO2007103116A2 (en) * | 2006-03-02 | 2007-09-13 | Laboratory Corporation Of America Holdings | Methods and systems for evaluating health risk factors by measurement of dna damage and dna repair |
| AU2008245696B2 (en) * | 2007-04-27 | 2013-11-07 | Pelican Technology Holdings, Inc. | Method for rapidly screening microbial hosts to identify certain strains with improved yield and/or quality in the expression of heterologous proteins |
| US9580719B2 (en) | 2007-04-27 | 2017-02-28 | Pfenex, Inc. | Method for rapidly screening microbial hosts to identify certain strains with improved yield and/or quality in the expression of heterologous proteins |
| WO2009091798A1 (en) * | 2008-01-16 | 2009-07-23 | Helicos Biosciences Corporation | Quantitative genetic analysis |
| US20090253173A1 (en) * | 2008-04-08 | 2009-10-08 | Danisco Us Inc., Genencor Division | Filamentous fungi with inactivated protease genes for altered protein production |
| WO2010008764A1 (en) * | 2008-06-23 | 2010-01-21 | Dow Global Technologies Inc. | Pseudomonas fluorescens strains for production of extracellular recombinant protein |
| US8198062B2 (en) | 2008-08-29 | 2012-06-12 | Dsm Ip Assets B.V. | Hydrolases, nucleic acids encoding them and methods for making and using them |
| UA109884C2 (uk) | 2009-10-16 | 2015-10-26 | Поліпептид, що має активність ферменту фосфатидилінозитол-специфічної фосфоліпази с, нуклеїнова кислота, що його кодує, та спосіб його виробництва і застосування | |
| UA111708C2 (uk) | 2009-10-16 | 2016-06-10 | Бандж Ойлз, Інк. | Спосіб рафінування олії |
| US8524220B1 (en) | 2010-02-09 | 2013-09-03 | David Gordon Bermudes | Protease inhibitor: protease sensitivity expression system composition and methods improving the therapeutic activity and specificity of proteins delivered by bacteria |
| US9597379B1 (en) | 2010-02-09 | 2017-03-21 | David Gordon Bermudes | Protease inhibitor combination with therapeutic proteins including antibodies |
| US8771669B1 (en) | 2010-02-09 | 2014-07-08 | David Gordon Bermudes | Immunization and/or treatment of parasites and infectious agents by live bacteria |
| EP2542574B1 (en) | 2010-03-04 | 2017-08-09 | Pfenex Inc. | Method for producing soluble recombinant interferon protein without denaturing |
| NZ602958A (en) | 2010-03-30 | 2014-07-25 | Pfenex Inc | High level expression of recombinant toxin proteins |
| AU2010201410B2 (en) * | 2010-03-30 | 2015-04-30 | Pelican Technology Holdings, Inc. | High level expression of recombinant CRM197 |
| EP2552949B1 (en) | 2010-04-01 | 2016-08-17 | Pfenex Inc. | Methods for g-csf production in a pseudomonas host cell |
| GB201101794D0 (en) | 2011-02-02 | 2011-03-16 | Fermentas Uab | Protein production |
| US20130045471A1 (en) * | 2011-02-25 | 2013-02-21 | Bio-Rad Laboratories, Inc. | Training system for investigations of bioengineered proteins |
| US20140154742A1 (en) * | 2011-04-08 | 2014-06-05 | Anthem Biosciences Pvt Ltd. | Novel expression and secretion vector systems for heterologous protein production in escherichia coli |
| US20140127749A1 (en) * | 2011-04-21 | 2014-05-08 | Arizona Borad Of Regents, A Body Corporate Of The State Of Arizona Acting For And On Behalf Of Arizo | Methods of protein production and compositions thereof |
| US9169304B2 (en) | 2012-05-01 | 2015-10-27 | Pfenex Inc. | Process for purifying recombinant Plasmodium falciparum circumsporozoite protein |
| US9737592B1 (en) | 2014-02-14 | 2017-08-22 | David Gordon Bermudes | Topical and orally administered protease inhibitors and bacterial vectors for the treatment of disorders and methods of treatment |
| CN103898150B (zh) * | 2014-04-09 | 2016-01-20 | 安徽华恒生物科技股份有限公司 | 产l-丙氨酸且耐受自来水的菌株及构建方法 |
| PL3218478T3 (pl) * | 2014-11-12 | 2020-09-07 | Lek Pharmaceuticals D.D. | Przewidywanie produktywności we wczesnym rozwoju linii komórkowej |
| PL3218514T3 (pl) * | 2014-11-12 | 2020-01-31 | Lek Pharmaceuticals D.D. | Przewidywanie genetycznie stabilnej produkcji rekombinowanego białka we wczesnym rozwoju linii komórkowej |
| ES2952405T3 (es) * | 2014-12-01 | 2023-10-31 | Pfenex Inc | Parejas de fusión para la producción de péptidos |
| CA3033143C (en) * | 2016-08-09 | 2021-05-25 | Agricultural Technology Research Institute | Composition for preventing and treating mycoplasma hyorhinis infection, and method for producing said composition |
| US11180535B1 (en) | 2016-12-07 | 2021-11-23 | David Gordon Bermudes | Saccharide binding, tumor penetration, and cytotoxic antitumor chimeric peptides from therapeutic bacteria |
| US11129906B1 (en) | 2016-12-07 | 2021-09-28 | David Gordon Bermudes | Chimeric protein toxins for expression by therapeutic bacteria |
| WO2018124189A1 (ja) * | 2016-12-28 | 2018-07-05 | 協和発酵キリン株式会社 | 細胞を選抜する方法 |
| CN106967658B (zh) * | 2017-02-23 | 2020-09-15 | 郑州大学 | 一种提高Fab抗体表达量的方法 |
| ES3011458T3 (en) | 2017-09-26 | 2025-04-07 | Bunge Global Innovation Llc | Enzymatic removal of chlorophyll substrates from triacylglycerol-based oils |
| EP3700915B1 (en) | 2017-10-27 | 2025-06-18 | Pelican Technology Holdings, Inc. | Bacterial leader sequences for periplasmic protein expression |
| CN111278852B (zh) * | 2017-10-27 | 2024-07-16 | 菲尼克斯公司 | 重组欧氏杆菌天冬酰胺酶的生产方法 |
| US10829731B2 (en) * | 2018-01-25 | 2020-11-10 | Alliance For Sustainable Energy, Llc | Biocatalysts for conversion of thermochemical waste streams |
| EP3788150A4 (en) * | 2018-05-04 | 2022-01-26 | Sigma-Aldrich Co. LLC | PRODUCTION OF RECOMBINANT PROTEINS WITH REDUCED LEVELS OF HOST CELL PROTEINS |
| JP2021521873A (ja) * | 2018-05-04 | 2021-08-30 | シグマ−アルドリッチ・カンパニー・リミテッド・ライアビリティ・カンパニーSigma−Aldrich Co. LLC | 改変された宿主細胞タンパク質プロファイルを有する操作された細胞 |
| US11541105B2 (en) | 2018-06-01 | 2023-01-03 | The Research Foundation For The State University Of New York | Compositions and methods for disrupting biofilm formation and maintenance |
| CN111518735A (zh) * | 2019-02-02 | 2020-08-11 | 江南大学 | 一种谷氨酸棒状杆菌重组菌、制备方法和应用 |
| EP3946720A1 (en) | 2019-03-27 | 2022-02-09 | Bunge Global Innovation, LLC. | Silica adsorbent treatment for removal of chlorophyll derivatives from triacylglycerol-based oils |
| US20220175848A1 (en) * | 2019-04-01 | 2022-06-09 | The Broad Institute, Inc. | Methods and compositions for cell therapy |
| CN110106128B (zh) * | 2019-04-24 | 2021-08-17 | 天津科技大学 | 一种生产重组碱性蛋白酶的基因工程菌及其构建方法 |
| WO2020237246A1 (en) * | 2019-05-23 | 2020-11-26 | University Of Maryland, Baltimore | Live attenuated non-transmissible vaccines |
| CN110484551B (zh) * | 2019-07-29 | 2021-04-02 | 因之彩生物科技(武汉)有限公司 | 金属硫蛋白表达载体及其应用 |
| US12285437B2 (en) | 2019-10-30 | 2025-04-29 | The Research Foundation For The State University Of New York | Reversing the undesirable pH-profile of doxorubicin via activation of a disubstituted maleamic acid prodrug at tumor acidity |
| CN113025601B (zh) * | 2019-12-25 | 2024-06-21 | 上海奥博生物医药股份有限公司 | 腈水解酶启动子优化表达及应用 |
| CN112941088B (zh) * | 2021-02-04 | 2023-06-16 | 中国农业科学院哈尔滨兽医研究所(中国动物卫生与流行病学中心哈尔滨分中心) | 与布氏杆菌毒力相关的基因及其在布氏杆菌毒力评价及制备弱毒布氏杆菌中的应用 |
| CN112831515A (zh) * | 2021-02-09 | 2021-05-25 | 厦门大学 | 一种变形假单胞菌HtpG抗原及其多克隆抗体的制备方法 |
| WO2022211829A1 (en) | 2021-03-30 | 2022-10-06 | Jazz Pharmaceuticals Ireland Ltd. | Dosing of recombinant l-asparaginase |
| CN113278622B (zh) * | 2021-06-09 | 2023-09-19 | 集美大学 | 一种变形假单胞菌适配体及其筛选方法 |
| KR20240021234A (ko) * | 2021-06-10 | 2024-02-16 | 펠리칸 테크놀로지 홀딩스, 인크. | 재조합 단백질 발현을 위한 박테리아 숙주 |
| CN114292800B (zh) * | 2021-12-27 | 2024-08-20 | 武汉佰乐博生物技术有限公司 | 用于igf-1基因重组表达的重组细胞和重组表达方法 |
| CN115786225B (zh) * | 2022-12-15 | 2025-06-24 | 江南大学 | 一种通过整合极端嗜酸菌分子伴侣CbpA改善E.coli耐酸性的方法 |
| WO2024186259A1 (en) * | 2023-03-07 | 2024-09-12 | Karyolaimos Alexandros | A method for enhancing recombinant production in a production host, an expression system, a library, a platform system, a production strain, an e. coli strain, a kit of parts, a method of using the platform system and a computer program, related to the method |
| CN117551678B (zh) * | 2023-12-29 | 2024-06-18 | 山东理工大学 | 一种提高丙酮酸脱羧酶SsPDC在大肠杆菌中可溶性表达的方法 |
| CN117603900B (zh) * | 2024-01-19 | 2024-04-30 | 清华大学 | 基因工程菌及其构建方法和应用 |
| CN118895293B (zh) * | 2024-10-08 | 2025-01-28 | 通化安睿特生物制药股份有限公司 | 一种产人白蛋白的重组工程菌及其构建方法和应用 |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH11510065A (ja) * | 1995-12-15 | 1999-09-07 | ユニバーシティ オブ ハワイ | 組換えタンパク質の産生のための改善されたNeurospora宿主およびそれを生成するための方法 |
Family Cites Families (308)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6696561B1 (en) | 1909-07-09 | 2004-02-24 | Basf Aktiengesellschaft | Corynebacterium glutamicum genes encoding proteins involved in membrane synthesis and membrane transport |
| GB1381306A (en) | 1972-03-03 | 1975-01-22 | Ici Ltd | Separating bacterial cells from a liquid medium |
| US3844893A (en) | 1973-02-12 | 1974-10-29 | Phillips Petroleum Co | Microbial synthesis |
| US4169010A (en) | 1976-11-18 | 1979-09-25 | Phillips Petroleum Company | Fermentation process for improved oxygen utilization |
| US4551433A (en) | 1981-05-18 | 1985-11-05 | Genentech, Inc. | Microbial hybrid promoters |
| US5399684A (en) | 1982-05-20 | 1995-03-21 | Washington Research Foundation | DNA sequences expressing mammalian alpha-1-antitrypsin |
| US4595658A (en) | 1982-09-13 | 1986-06-17 | The Rockefeller University | Method for facilitating externalization of proteins synthesized in bacteria |
| US5151350A (en) | 1982-10-27 | 1992-09-29 | Repligen Corporation | Cloned genes encoding recombinant protein a |
| US4432895A (en) | 1982-11-24 | 1984-02-21 | Hoffmann-La Roche Inc. | Monomeric interferons |
| US4511503A (en) | 1982-12-22 | 1985-04-16 | Genentech, Inc. | Purification and activity assurance of precipitated heterologous proteins |
| GB8308483D0 (en) | 1983-03-28 | 1983-05-05 | Health Lab Service Board | Secretion of gene products |
| US4755465A (en) | 1983-04-25 | 1988-07-05 | Genentech, Inc. | Secretion of correctly processed human growth hormone in E. coli and Pseudomonas |
| US4680264A (en) | 1983-07-01 | 1987-07-14 | Lubrizol Genetics, Inc. | Class II mobilizable gram-negative plasmid |
| US5281532A (en) | 1983-07-27 | 1994-01-25 | Mycogen Corporation | Pseudomas hosts transformed with bacillus endotoxin genes |
| US4637980A (en) | 1983-08-09 | 1987-01-20 | Smithkline Beckman Corporation | Externalization of products of bacteria |
| EP0155189A3 (en) | 1984-03-16 | 1987-09-16 | Genentech, Inc. | Expression of cell wall degrading proteins and host cells harboring dna encoding such protein |
| SU1364343A1 (ru) | 1984-07-13 | 1988-01-07 | Всесоюзный научно-исследовательский институт генетики и селекции промышленных микроорганизмов | Способ получени человеческого лейкоцитарного интерферона альфа-2 |
| US4963495A (en) | 1984-10-05 | 1990-10-16 | Genentech, Inc. | Secretion of heterologous proteins |
| EP0177343B1 (en) | 1984-10-05 | 1992-07-22 | Genentech, Inc. | Dna, cell cultures and methods for the secretion of heterologous proteins and periplasmic protein recovery |
| US4695462A (en) | 1985-06-28 | 1987-09-22 | Mycogen Corporation | Cellular encapsulation of biological pesticides |
| US4695455A (en) | 1985-01-22 | 1987-09-22 | Mycogen Corporation | Cellular encapsulation of pesticides produced by expression of heterologous genes |
| GB8517071D0 (en) | 1985-07-05 | 1985-08-14 | Hoffmann La Roche | Gram-positive expression control sequence |
| US4888274A (en) | 1985-09-18 | 1989-12-19 | Yale University | RecA nucleoprotein filament and methods |
| US5618920A (en) | 1985-11-01 | 1997-04-08 | Xoma Corporation | Modular assembly of antibody genes, antibodies prepared thereby and use |
| GB8529014D0 (en) | 1985-11-25 | 1986-01-02 | Biogen Nv | Enhanced secretion of heterologous proteins |
| ATE118548T1 (de) | 1986-03-24 | 1995-03-15 | Getty Scient Dev Co | Herstellungsverfahren von zuckernukleotiden mittels rekombinantverfahrens. |
| DE3751554T2 (de) | 1986-03-24 | 1996-06-13 | Texaco Development Corp | Erzeugung von xanthangummi durch rekombinant-dns. |
| US5559015A (en) | 1986-03-26 | 1996-09-24 | Getty Scientific Development Company | Recombinant-DNA mediated production of xanthan gum |
| US5232840A (en) | 1986-03-27 | 1993-08-03 | Monsanto Company | Enhanced protein production in bacteria by employing a novel ribosome binding site |
| US5165927A (en) | 1986-08-01 | 1992-11-24 | University Of Southern California | Composition with modified pertussis toxin |
| US5292507A (en) | 1986-08-01 | 1994-03-08 | Imperial Oil Limited | Method of using polysaccharides to stabilize microorganisms for inoculating plant seeds |
| US5541308A (en) | 1986-11-24 | 1996-07-30 | Gen-Probe Incorporated | Nucleic acid probes for detection and/or quantitation of non-viral organisms |
| US5084559A (en) | 1987-03-27 | 1992-01-28 | Repligen Corporation | Protein a domain mutants |
| DE3850995T2 (de) | 1987-04-23 | 1995-03-16 | Monsanto Co | Sekretion eines dem Insulin ähnlichen Wachstumsfaktors in E. coli. |
| US5591603A (en) | 1987-08-28 | 1997-01-07 | Novo Nordisk A/S | Process for preparing aprotinin and aprotinin analogs in yeast cells |
| JP2918895B2 (ja) | 1987-09-04 | 1999-07-12 | アムジエン・インコーポレーテツド | 組換え型dna由来ボルデテラ毒素サブユニット類似体 |
| GB8727489D0 (en) | 1987-11-24 | 1987-12-23 | Connaught Lab | Detoxification of pertussis toxin |
| US5128130A (en) | 1988-01-22 | 1992-07-07 | Mycogen Corporation | Hybrid Bacillus thuringiensis gene, plasmid and transformed Pseudomonas fluorescens |
| US5055294A (en) | 1988-03-03 | 1991-10-08 | Mycogen Corporation | Chimeric bacillus thuringiensis crystal protein gene comprising hd-73 and berliner 1715 toxin genes, transformed and expressed in pseudomonas fluorescens |
| JPH01257491A (ja) | 1988-04-08 | 1989-10-13 | Tosoh Corp | 不溶性融合異種蛋白質の処理方法 |
| WO1989010971A1 (en) | 1988-05-09 | 1989-11-16 | The United States Of America, As Represented By Th | Vector for secretion of proteins directly into periplasm or culture medium |
| US5169772A (en) | 1988-06-06 | 1992-12-08 | Massachusetts Institute Of Technology | Large scale method for purification of high purity heparinase from flavobacterium heparinum |
| JPH04500754A (ja) | 1988-09-30 | 1992-02-13 | アライド―シグナル・インコーポレーテッド | 異種遺伝子発現のための改良細菌株 |
| US6780613B1 (en) | 1988-10-28 | 2004-08-24 | Genentech, Inc. | Growth hormone variants |
| US5552302A (en) | 1989-04-24 | 1996-09-03 | Promega Corporation | Methods and compositions for production of human recombinant placental ribonuclease inhibitor |
| US5143854A (en) | 1989-06-07 | 1992-09-01 | Affymax Technologies N.V. | Large scale photolithographic solid phase synthesis of polypeptides and receptor binding screening thereof |
| US5744101A (en) | 1989-06-07 | 1998-04-28 | Affymax Technologies N.V. | Photolabile nucleoside protecting groups |
| US6040138A (en) | 1995-09-15 | 2000-03-21 | Affymetrix, Inc. | Expression monitoring by hybridization to high density oligonucleotide arrays |
| US6551784B2 (en) | 1989-06-07 | 2003-04-22 | Affymetrix Inc | Method of comparing nucleic acid sequences |
| DE3920358A1 (de) | 1989-06-22 | 1991-01-17 | Behringwerke Ag | Bispezifische und oligospezifische, mono- und oligovalente antikoerperkonstrukte, ihre herstellung und verwendung |
| US5683888A (en) | 1989-07-22 | 1997-11-04 | University Of Wales College Of Medicine | Modified bioluminescent proteins and their use |
| US5169760A (en) | 1989-07-27 | 1992-12-08 | Mycogen Corporation | Method, vectors, and host cells for the control of expression of heterologous genes from lac operated promoters |
| US5023171A (en) | 1989-08-10 | 1991-06-11 | Mayo Foundation For Medical Education And Research | Method for gene splicing by overlap extension using the polymerase chain reaction |
| EP0430645B1 (en) | 1989-11-28 | 1993-08-18 | The Wellcome Foundation Limited | Vaccines |
| US5292658A (en) | 1989-12-29 | 1994-03-08 | University Of Georgia Research Foundation, Inc. Boyd Graduate Studies Research Center | Cloning and expressions of Renilla luciferase |
| US5641671A (en) | 1990-07-06 | 1997-06-24 | Unilever Patent Holdings B.V. | Production of active Pseudomonas glumae lipase in homologous or heterologous hosts |
| US5639949A (en) | 1990-08-20 | 1997-06-17 | Ciba-Geigy Corporation | Genes for the synthesis of antipathogenic substances |
| US5670350A (en) | 1990-08-20 | 1997-09-23 | Novartis Finance Corporation | Genomic DNA encoding a pseudomonas global transcriptional activation element and its use in activating gene expression |
| US5662898A (en) | 1990-08-20 | 1997-09-02 | Ciba-Geigy Corporation | Genes for the synthesis of antipathogenic substances |
| US5610044A (en) | 1990-10-01 | 1997-03-11 | Lam; Stephen T. | Microorganisms with mannopine catabolizing ability |
| JP2679390B2 (ja) | 1990-10-12 | 1997-11-19 | 日本電気株式会社 | コード設定回路 |
| US5508192A (en) * | 1990-11-09 | 1996-04-16 | Board Of Regents, The University Of Texas System | Bacterial host strains for producing proteolytically sensitive polypeptides |
| US5264365A (en) | 1990-11-09 | 1993-11-23 | Board Of Regents, The University Of Texas System | Protease-deficient bacterial strains for production of proteolytically sensitive polypeptides |
| EP0834575B1 (en) | 1990-12-06 | 2001-11-28 | Affymetrix, Inc. (a Delaware Corporation) | Identification of nucleic acids in samples |
| ES2142801T3 (es) | 1991-03-11 | 2000-05-01 | Univ Georgia Res Found | Clonacion y expresion de luciferasa de renilla. |
| US6800738B1 (en) | 1991-06-14 | 2004-10-05 | Genentech, Inc. | Method for making humanized antibodies |
| US5558862A (en) | 1991-09-23 | 1996-09-24 | Monsanto Company | Method of controlling insects |
| US5348867A (en) | 1991-11-15 | 1994-09-20 | George Georgiou | Expression of proteins on bacterial surface |
| AU3178993A (en) | 1991-11-25 | 1993-06-28 | Enzon, Inc. | Multivalent antigen-binding proteins |
| IL104419A0 (en) | 1992-01-24 | 1993-05-13 | Fmc Corp | Insecticidally effective peptides |
| GB9206645D0 (en) | 1992-03-26 | 1992-05-06 | Mini Agriculture & Fisheries | Biological control of post harvest pests |
| DK154292D0 (da) | 1992-12-23 | 1992-12-23 | Novo Nordisk As | Nyt enzym |
| US5652128A (en) | 1993-01-05 | 1997-07-29 | Jarvik; Jonathan Wallace | Method for producing tagged genes, transcripts, and proteins |
| US5581476A (en) | 1993-01-28 | 1996-12-03 | Amgen Inc. | Computer-based methods and articles of manufacture for preparing G-CSF analogs |
| GB9314886D0 (en) | 1993-07-19 | 1993-09-01 | Zeneca Ltd | Production of a biological control agent |
| US5914254A (en) | 1993-08-02 | 1999-06-22 | Celtrix Pharmaceuticals, Inc. | Expression of fusion polypeptides transported out of the cytoplasm without leader sequences |
| ATE400651T1 (de) | 1993-09-10 | 2008-07-15 | Univ Columbia | Verwendung von grünem fluoreszenzprotein |
| TW300879B (ja) | 1993-11-10 | 1997-03-21 | Ibm | |
| JPH09506508A (ja) | 1993-12-03 | 1997-06-30 | メディカル リサーチ カウンシル | 組換え結合タンパク質およびペプチド |
| WO1995018634A1 (en) | 1994-01-04 | 1995-07-13 | The Scripps Research Institute | Human monoclonal antibodies to herpes simplex virus and methods therefor |
| WO1995021191A1 (en) | 1994-02-04 | 1995-08-10 | William Ward | Bioluminescent indicator based upon the expression of a gene for a modified green-fluorescent protein |
| US7294331B2 (en) | 1994-03-07 | 2007-11-13 | Immunex Corporation | Methods of using flt3-ligand in hematopoietic cell transplantation |
| AUPM516994A0 (en) | 1994-04-20 | 1994-05-12 | Gene Shears Pty. Limited | An in vivo gene expression system |
| US5527883A (en) | 1994-05-06 | 1996-06-18 | Mycogen Corporation | Delta-endotoxin expression in pseudomonas fluorescens |
| US6117670A (en) | 1994-06-08 | 2000-09-12 | Novartis Finance Corporation | Pyrrolnitrin biosynthesis genes and uses thereof |
| CA2150526C (en) | 1994-06-09 | 2005-11-15 | Andreas Kiener | Biotechnological process for the preparation of cyclic s-.alpha.-amino carboxylic acids and r-.alpha.-amino carboxamides |
| US5989808A (en) | 1994-06-14 | 1999-11-23 | American Cyanamid Company | Identification of compounds affecting specific interaction of peptide binding pairs |
| JP3565859B2 (ja) | 1994-10-28 | 2004-09-15 | ザ・トラステイーズ・オブ・ザ・ユニバーシテイ・オブ・ペンシルベニア | 改良されたアデノウイルスおよびその使用法 |
| US5625048A (en) | 1994-11-10 | 1997-04-29 | The Regents Of The University Of California | Modified green fluorescent proteins |
| US5777079A (en) | 1994-11-10 | 1998-07-07 | The Regents Of The University Of California | Modified green fluorescent proteins |
| ATE260979T1 (de) | 1994-12-09 | 2004-03-15 | Novo Nordisk As | Verfahren zur herstellung extrazellulärer proteine in bakterien |
| US5958713A (en) | 1995-01-31 | 1999-09-28 | Novo Nordisk A/S | Method of detecting biologically active substances by using green fluorescent protein |
| US5641870A (en) | 1995-04-20 | 1997-06-24 | Genentech, Inc. | Low pH hydrophobic interaction chromatography for antibody purification |
| ZA963530B (en) | 1995-05-05 | 1996-11-05 | Hoffmann La Roche | Recombinant obese (ob) proteins |
| US5711945A (en) | 1995-05-11 | 1998-01-27 | Regents Of The University Of Minnesota | Pitch degradation with Pseudomonas fluorescens |
| US6083690A (en) | 1995-06-02 | 2000-07-04 | Osteoscreen, Inc. | Methods and compositions for identifying osteogenic agents |
| FR2734842B1 (fr) | 1995-06-02 | 1998-02-27 | Rhone Poulenc Agrochimie | Sequence adn d'un gene de l'hydroxy-phenyl pyruvate dioxygenase et obtention de plantes contenant un gene de l'hydroxy-phenyl pyruvate dioxygenase, tolerantes a certains herbicides |
| US5955298A (en) | 1995-06-26 | 1999-09-21 | The United States Of America As Represented By The Secretary Of Agriculture | Sequences for production of 2,4-Diacetylphloroglucinol and methods |
| US6495357B1 (en) | 1995-07-14 | 2002-12-17 | Novozyme A/S | Lipolytic enzymes |
| US6020473A (en) | 1995-08-25 | 2000-02-01 | Genentech, Inc. | Nucleic acids encoding variants of vascular endothelial cell growth factor |
| DK0851874T3 (da) | 1995-09-22 | 2000-03-06 | Bioimage A S | Hidtil ukendte varianter af grønt fluorescerende protein, GFP |
| US5952236A (en) | 1995-10-26 | 1999-09-14 | Thompson; Richard B. | Enzyme-based fluorescence biosensor for chemical analysis |
| US5968738A (en) | 1995-12-06 | 1999-10-19 | The Board Of Trustees Of The Leland Stanford Junior University | Two-reporter FACS analysis of mammalian cells using green fluorescent proteins |
| US5874304A (en) | 1996-01-18 | 1999-02-23 | University Of Florida Research Foundation, Inc. | Humanized green fluorescent protein genes and methods |
| US6020192A (en) | 1996-01-18 | 2000-02-01 | University Of Florida | Humanized green fluorescent protein genes and methods |
| EP0880598A4 (en) | 1996-01-23 | 2005-02-23 | Affymetrix Inc | RAPID EVALUATION OF NUCLEIC ACID ABUNDANCE DIFFERENCE, WITH A HIGH-DENSITY OLIGONUCLEOTIDE SYSTEM |
| US5804387A (en) | 1996-02-01 | 1998-09-08 | The Board Of Trustees Of The Leland Stanford Junior University | FACS-optimized mutants of the green fluorescent protein (GFP) |
| US5876995A (en) | 1996-02-06 | 1999-03-02 | Bryan; Bruce | Bioluminescent novelty items |
| US6255558B1 (en) | 1996-02-14 | 2001-07-03 | Btg International Limited | Gene expression |
| KR100266924B1 (ko) | 1996-03-23 | 2000-09-15 | 무따이 마사히꼬 | 파상풍독소의 기능적 단편 항원 및 파상풍 백신 |
| US6121247A (en) | 1996-03-29 | 2000-09-19 | The Johns Hopkins University | Therapy for allergic diseases |
| US5741663A (en) | 1996-04-16 | 1998-04-21 | University Of Georgia Research Foundation, Inc. | Selective additive and media for Pseudomonas fluorescens |
| US6218185B1 (en) | 1996-04-19 | 2001-04-17 | The United States Of America As Represented By The Secretary Of Agriculture | Piggybac transposon-based genetic transformation system for insects |
| US6699838B1 (en) | 1996-05-03 | 2004-03-02 | Abbott Laboratories | Antiangiogenic peptides |
| US6096865A (en) | 1996-05-06 | 2000-08-01 | Amgen Inc. | Mutants of the green fluorescent protein having improved fluorescent properties at 37° |
| US6027881A (en) | 1996-05-08 | 2000-02-22 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Mutant Aequorea victoria fluorescent proteins having increased cellular fluorescence |
| US5948889A (en) | 1996-05-21 | 1999-09-07 | Case Western Reserve University | Compositions and methods for screening antimicrobials |
| US5912137A (en) | 1996-07-16 | 1999-06-15 | The Regents Of The University Of California | Assays for protein kinases using fluorescent |
| US5925558A (en) | 1996-07-16 | 1999-07-20 | The Regents Of The University Of California | Assays for protein kinases using fluorescent protein substrates |
| US7129061B1 (en) | 1996-08-07 | 2006-10-31 | Biogen Idec Ma Inc. | Tumor necrosis factor related ligand |
| US5948681A (en) | 1996-08-14 | 1999-09-07 | Children's Hospital Of Philadelphia | Non-viral vehicles for use in gene transfer |
| US6124128A (en) | 1996-08-16 | 2000-09-26 | The Regents Of The University Of California | Long wavelength engineered fluorescent proteins |
| US5914233A (en) | 1996-08-23 | 1999-06-22 | Osteo Screen | Screening assay for the identification of agents which alter expression of PTH-rP |
| WO1998008839A1 (en) | 1996-08-26 | 1998-03-05 | Eli Lilly And Company | Combinatorial process for preparing substituted thiophene libraries |
| US6274354B1 (en) | 1996-09-06 | 2001-08-14 | The Trustees Of The University Of Pennsylvania | Methods using cre-lox for production of recombinant adeno-associated viruses |
| US6255071B1 (en) | 1996-09-20 | 2001-07-03 | Cold Spring Harbor Laboratory | Mammalian viral vectors and their uses |
| US6025192A (en) | 1996-09-20 | 2000-02-15 | Cold Spring Harbor Laboratory | Modified retroviral vectors |
| US5976796A (en) | 1996-10-04 | 1999-11-02 | Loma Linda University | Construction and expression of renilla luciferase and green fluorescent protein fusion genes |
| US6136566A (en) | 1996-10-04 | 2000-10-24 | Lexicon Graphics Incorporated | Indexed library of cells containing genomic modifications and methods of making and utilizing the same |
| TW371617B (en) | 1996-10-09 | 1999-10-11 | Of Animal And Plant Health Inspection And Quarantine Council Of Agriculture Executive Yuan Bureau | Method to transplant GFP into autographa californica multiple nuclear polyhedrosis virus for inflicting pest in an attempt to detect and flow up it existence and to improve life span against UV |
| US5757051A (en) | 1996-11-12 | 1998-05-26 | Micron Technology, Inc. | Static memory cell and method of manufacturing a static memory cell |
| JP2001505766A (ja) | 1996-11-18 | 2001-05-08 | マクギル ユニバーシティー | アポトーシスと生長を調節するアデノウイルスベクターを有する、有糸分裂後のニューロン |
| US5891688A (en) | 1996-12-06 | 1999-04-06 | Novartis Finance Corporation | DNA encoding lemA-independent GacA and its use in activating gene expression |
| US5955348A (en) | 1997-11-25 | 1999-09-21 | Novartis Ag | Genetically modified pseudomonas strains with enhanced biocontrol activity |
| US5756087A (en) | 1996-12-06 | 1998-05-26 | Novartis Finance Corporation | Genetically modified Pseudomonas strains with enhanced biocontrol activity |
| WO1998024919A1 (en) | 1996-12-06 | 1998-06-11 | Novartis Ag | Genetically modified pseudomonas strains with enhanced biocontrol activity |
| IL129767A0 (en) | 1996-12-12 | 2000-02-29 | Prolume Ltd | Apparatus and method for detecting and identifying infectious agents |
| DE69740107D1 (de) | 1996-12-23 | 2011-03-10 | Immunex Corp | Rezeptor aktivator von nf-kappa b, rezeptor is mitglied der tnf rezeptor superfamilie |
| US6194194B1 (en) | 1996-12-23 | 2001-02-27 | Daniel Molloy | Method for controlling dreissena species |
| CA2276529A1 (en) | 1996-12-31 | 1998-07-09 | George A. Gaitanaris | Vectors and methods for the mutagenesis of mammalian genes |
| US6090919A (en) | 1997-01-31 | 2000-07-18 | The Board Of Trustees Of The Leland Stanford Junior University | FACS-optimized mutants of the green fluorescent protein (GFP) |
| US5994077A (en) | 1997-01-31 | 1999-11-30 | The Board Of Trustees Of The Leland Stanford Junior University | Flourescence-based isolation of differentially induced genes |
| US5795759A (en) | 1997-03-10 | 1998-08-18 | The University Of Iowa Research Foundation | Carboxylic acid reductase, and methods of using same |
| US6197928B1 (en) | 1997-03-14 | 2001-03-06 | The Regents Of The University Of California | Fluorescent protein sensors for detection of analytes |
| US5994071A (en) | 1997-04-04 | 1999-11-30 | Albany Medical College | Assessment of prostate cancer |
| US5952208A (en) | 1997-04-07 | 1999-09-14 | Energy Biosystems Corporation | Dsz gene expression in pseudomonas hosts |
| US6251384B1 (en) | 1997-04-28 | 2001-06-26 | Anticancer, Inc. | Metastasis models using green fluorescent protein (GFP) as a marker |
| US5993778A (en) | 1997-05-07 | 1999-11-30 | Firestein; Stuart J. | Functional expression of, and assay for, functional cellular receptors in vivo |
| US6225082B1 (en) | 1997-05-09 | 2001-05-01 | Research Corporation Technologies, Inc. | Myelin basic protein MRNA transport and translation enhancer sequences |
| US6251582B1 (en) | 1997-07-17 | 2001-06-26 | New York University | Alternative G-coupled receptors associated with retroviral entry into cells, methods of identifying the same, and diagnostic and therapeutic uses thereof |
| US6484183B1 (en) | 1997-07-25 | 2002-11-19 | Affymetrix, Inc. | Method and system for providing a polymorphism database |
| US6420108B2 (en) | 1998-02-09 | 2002-07-16 | Affymetrix, Inc. | Computer-aided display for comparative gene expression |
| IL121404A0 (en) | 1997-07-27 | 1998-01-04 | Yissum Res Dev Co | Transgenic higher plants of altered structural morphology |
| US6221612B1 (en) | 1997-08-01 | 2001-04-24 | Aurora Biosciences Corporation | Photon reducing agents for use in fluorescence assays |
| US6214563B1 (en) | 1997-08-01 | 2001-04-10 | Aurora Biosciences Corporation | Photon reducing agents for reducing undesired light emission in assays |
| US6251677B1 (en) | 1997-08-25 | 2001-06-26 | The Trustees Of The University Of Pennsylvania | Hybrid adenovirus-AAV virus and methods of use thereof |
| HUP9701446A1 (hu) | 1997-08-27 | 1999-05-28 | Phylaxia-Pharma Gyógyszer-, Oltóanyag és Agrobiológiai Készítményeket Gyártó és Forgalmazó Rt. | Eljárás a talaj mikroorganizmus populációjának előnyös kialakítására |
| US6153409A (en) | 1997-09-11 | 2000-11-28 | University Of Maryland Biotechnology Institute | Process for continuous optimized protein production in insect larvae |
| US6071738A (en) | 1997-09-19 | 2000-06-06 | Energy Biosystems Corporation | Conversion of organosulfur compounds to oxyorganosulfur compounds for desulfurization of fossil fuels |
| DK1017803T3 (da) | 1997-09-26 | 2010-08-23 | Abt Holding Co | Ekspression af endogene gener ved ikke-homolog rekombination af et vektorkonstrukt med cellulært DNA |
| US6130313A (en) | 1997-10-02 | 2000-10-10 | Clontech Laboratories, Inc. | Rapidly degrading GFP-fusion proteins |
| US6133429A (en) | 1997-10-03 | 2000-10-17 | Becton Dickinson And Company | Chromophores useful for the preparation of novel tandem conjugates |
| US6284496B1 (en) | 1997-10-03 | 2001-09-04 | University Of South Florida | DNA vector for determining the presence of out-of-reading-frame mutations |
| US6361989B1 (en) | 1997-10-13 | 2002-03-26 | Novozymes A/S | α-amylase and α-amylase variants |
| US5968773A (en) | 1997-11-14 | 1999-10-19 | Heddle; John A. | System and method for regulation of gene expression |
| US6277625B1 (en) | 1997-11-20 | 2001-08-21 | The United States Of America As Represented By The Secretary Of Agriculture | Transgenic strains of Pseudomonas for biocontrol of plant root diseases |
| US6447770B1 (en) | 1997-11-20 | 2002-09-10 | The United States Of America As Represented By The Secretary Of Agriculture | Biocontrol agents for take-all |
| US6013447A (en) | 1997-11-21 | 2000-01-11 | Innovir Laboratories, Inc. | Random intracellular method for obtaining optimally active nucleic acid molecules |
| WO1999029877A1 (en) | 1997-12-12 | 1999-06-17 | The Regents Of The University Of California | Methods for defining cell types |
| US6060594A (en) | 1997-12-18 | 2000-05-09 | Ecogen, Inc. | Nucleic acid segments encoding modified bacillus thuringiensis coleopteran-toxic crystal proteins |
| US6506559B1 (en) | 1997-12-23 | 2003-01-14 | Carnegie Institute Of Washington | Genetic inhibition by double-stranded RNA |
| US6136539A (en) | 1998-02-11 | 2000-10-24 | The Regents Of The University Of California | Compositions and methods for the inhibition of MUC-5 mucin gene expression |
| US6156552A (en) | 1998-02-18 | 2000-12-05 | Novo Nordisk A/S | Lipase variants |
| US5922576A (en) | 1998-02-27 | 1999-07-13 | The John Hopkins University | Simplified system for generating recombinant adenoviruses |
| US6210910B1 (en) | 1998-03-02 | 2001-04-03 | Trustees Of Tufts College | Optical fiber biosensor array comprising cell populations confined to microcavities |
| WO1999046592A1 (en) | 1998-03-09 | 1999-09-16 | The Regents Of The University Of California | Regulation of the cell cycle by sterols |
| AUPP249298A0 (en) | 1998-03-20 | 1998-04-23 | Ag-Gene Australia Limited | Synthetic genes and genetic constructs comprising same I |
| US6080576A (en) | 1998-03-27 | 2000-06-27 | Lexicon Genetics Incorporated | Vectors for gene trapping and gene activation |
| AU765703B2 (en) | 1998-03-27 | 2003-09-25 | Bruce J. Bryan | Luciferases, fluorescent proteins, nucleic acids encoding the luciferases and fluorescent proteins and the use thereof in diagnostics, high throughput screening and novelty items |
| US6248558B1 (en) | 1998-03-31 | 2001-06-19 | Vanderbilt University | Sequence and method for genetic engineering of proteins with cell membrane translocating activity |
| BR9909615A (pt) | 1998-04-13 | 2000-12-12 | Univ Georgia Res Found | Superexpressão de carboxilase de piruvato para a produção intensificada de produtos bioquìmicos derivados de oxaloacetato em células microbianas |
| US6037133A (en) | 1998-04-17 | 2000-03-14 | Clontech Laboratories, Inc. | IκBEGFP constructs, cell lines and methods of use |
| NZ507381A (en) | 1998-04-21 | 2003-12-19 | Micromet Ag | CD19xCD3 specific polypeptides and uses thereof |
| US6316181B1 (en) | 1998-04-24 | 2001-11-13 | Virginia Commonwealth University | Establishment of cell lines with persistent expression of a green fluorescent protein (GFP) using a pIRES/EGFP DNA vector construct |
| US6280934B1 (en) | 1998-05-13 | 2001-08-28 | Millennium Pharmaceuticals, Inc. | Assay for agents which alter G-protein coupled receptor activity |
| US20030064435A1 (en) | 1998-05-28 | 2003-04-03 | Weiner Joel Hirsch | Compositions and methods for protein secretion |
| US6140132A (en) | 1998-06-09 | 2000-10-31 | The Regents Of The University Of California | Fluorescent protein sensors for measuring the pH of a biological sample |
| US6150176A (en) | 1998-06-09 | 2000-11-21 | The Regents Of The University Of California | Fluorescent protein sensors for measuring the pH of a biological sample |
| WO1999066072A2 (en) | 1998-06-16 | 1999-12-23 | Nova Molecular, Inc. | Methods for treating a neurological disease by determining bche genotype |
| US5989837A (en) | 1998-07-13 | 1999-11-23 | Wisconsin Alumni Research Foundation | Immortalized human keratinocyte cell line |
| AUPP589798A0 (en) | 1998-09-14 | 1998-10-08 | Charles Sturt University | Novel bacterial strains and methods of controlling fungal pathogens |
| US6185561B1 (en) | 1998-09-17 | 2001-02-06 | Affymetrix, Inc. | Method and apparatus for providing and expression data mining database |
| US6323007B1 (en) | 1998-09-18 | 2001-11-27 | Novozymes A/S | 2,6-β-D-fructan hydrolase enzyme and processes for using the enzyme |
| US6180343B1 (en) | 1998-10-08 | 2001-01-30 | Rigel Pharmaceuticals, Inc. | Green fluorescent protein fusions with random peptides |
| US5985577A (en) | 1998-10-14 | 1999-11-16 | The Trustees Of Columbia University In The City Of New York | Protein conjugates containing multimers of green fluorescent protein |
| US6203986B1 (en) | 1998-10-22 | 2001-03-20 | Robert H. Singer | Visualization of RNA in living cells |
| US6268201B1 (en) | 1998-10-23 | 2001-07-31 | Albert Einstein College Of Medicine Of Yeshiva University | IniB, iniA and iniC genes of mycobacteria and methods of use |
| DK1323820T3 (da) | 1998-10-28 | 2009-03-16 | Genentech Inc | Fremgangsmåde til faciliteret isolering af heterologe proteiner fra bakterieceller |
| US6270958B1 (en) | 1998-10-29 | 2001-08-07 | Washington University | Detection of negative-strand RNA viruses |
| AU1722800A (en) | 1998-11-18 | 2000-06-05 | University Of Akron, The | Production of biological materials by simultaneous aerobic and anaerobic respiration |
| US6210922B1 (en) | 1998-11-30 | 2001-04-03 | National Research Council Of Canada | Serum free production of recombinant proteins and adenoviral vectors |
| US6136538A (en) | 1999-01-08 | 2000-10-24 | Washington University | Silent inducible virus replicons and uses thereof |
| US6015557A (en) | 1999-02-24 | 2000-01-18 | Tobinick; Edward L. | Tumor necrosis factor antagonists for the treatment of neurological disorders |
| KR100312456B1 (ko) | 1999-03-13 | 2001-11-03 | 윤덕용 | 슈도모나스 플루오레슨스 유래의 외래단백질 분비촉진유전자 |
| BR0009393A (pt) | 1999-03-29 | 2002-01-08 | Novozymes As | Polipeptìdeo, sequência de ácido nucleico isolada, construção de ácido nucleico, vetor de expressão recombinante, célula hospedeira recombinante, método para a produção do polipeptìdeo, uso de um polipeptìdeo, e, composição detergente |
| US6960652B2 (en) | 1999-03-30 | 2005-11-01 | Board Of Regents, The University Of Texas System | Compositions and methods for modifying toxic effects of proteinaceous compounds |
| US6664386B1 (en) | 1999-04-08 | 2003-12-16 | The Research Foundation Of State University Of New York | System for efficient secretion of recombinant proteins |
| AU5477500A (en) | 1999-06-09 | 2000-12-28 | General Hospital Corporation, The | Method of measuring plasma membrane targeting of glut4 |
| JP2003506088A (ja) | 1999-08-04 | 2003-02-18 | アムジェン インコーポレーテッド | TNFリガンドスーパー遺伝子ファミリーの新規のメンバーであるFhm |
| US6558939B1 (en) | 1999-08-31 | 2003-05-06 | Novozymes, A/S | Proteases and variants thereof |
| US7459540B1 (en) | 1999-09-07 | 2008-12-02 | Amgen Inc. | Fibroblast growth factor-like polypeptides |
| DE19944870A1 (de) | 1999-09-18 | 2001-03-29 | Aventis Pharma Gmbh | Signalsequenzen zur Herstellung von Leu-Hirudin über Sekretion durch E. coli in das Kulturmedium |
| BR0014758A (pt) | 1999-10-13 | 2002-07-02 | Univ Georgia Res Found | Sistemas e métodos para expressão de proteìna com alto rendimento |
| US6607885B1 (en) * | 1999-10-15 | 2003-08-19 | E. I. Du Pont De Nemours And Company | Method for high-density microarray medicated gene expression profiling |
| US6617143B1 (en) | 1999-10-20 | 2003-09-09 | Novozymes A/S | Polypeptides having glucanotransferase activity and nucleic acids encoding same |
| EP1230348A1 (en) | 1999-11-05 | 2002-08-14 | Novozymes A/S | Microtiter plate (mtp) based high throughput screening (hts) assays |
| SK8292002A3 (en) | 1999-11-12 | 2002-12-03 | Maxygen Holdings Ltd | A conjugate exhibiting interferon gamma activity, nucleotide sequence encoding for a polypeptide fraction of conjugate, an expression vector and a host cell containing nucleotide sequence, pharmaceutical composition comprising the same and use thereof |
| CA2400948A1 (en) | 2000-02-25 | 2001-08-30 | Ludwig Institute For Cancer Research | Materials and methods involving hybrid vascular endothelial growth factor dnas and proteins |
| ATE450621T2 (de) | 2000-03-30 | 2009-12-15 | Whitehead Biomedical Inst | Mediatoren von rns-interferenz, die rns- sequenzspezifisch sind |
| US6509181B1 (en) | 2000-04-14 | 2003-01-21 | Novozymes, A/S | Polypeptides having haloperoxide activity |
| JP2001299360A (ja) | 2000-04-28 | 2001-10-30 | Morihiro Matsuda | ヒト型抗破傷風毒素一本鎖組換え抗体フラグメント |
| GB0016702D0 (en) | 2000-07-08 | 2000-08-23 | Secr Defence Brit | Expression system |
| US20030108980A1 (en) | 2000-08-14 | 2003-06-12 | Sayler Gary S. | Bioluminescent methods for direct visual detection of environmental compounds |
| US20030108872A1 (en) | 2000-08-23 | 2003-06-12 | Mark Sulavik | Genomics-assisted rapid identification of targets |
| JP2002201141A (ja) | 2000-10-27 | 2002-07-16 | Hayashibara Biochem Lab Inc | 蛋白質合成抑制遺伝子の発現増強剤 |
| US6673580B2 (en) | 2000-10-27 | 2004-01-06 | Genentech, Inc. | Identification and modification of immunodominant epitopes in polypeptides |
| GB0027782D0 (en) | 2000-11-14 | 2000-12-27 | Boehringer Ingelheim Int | Methods fro large scale protein productio in prokaryotes |
| US7087412B2 (en) | 2000-11-14 | 2006-08-08 | Boehringer Ingelheim International Gmbh | Methods for large scale protein production in prokaryotes |
| JP4309656B2 (ja) | 2000-12-14 | 2009-08-05 | ジェネンテック・インコーポレーテッド | 原核生物で生成させた抗体とその用途 |
| HU230065B1 (hu) | 2000-12-14 | 2015-06-29 | Genentech, Inc. | Baktérium gazdatörzsek |
| US6979556B2 (en) | 2000-12-14 | 2005-12-27 | Genentech, Inc. | Separate-cistron contructs for secretion of aglycosylated antibodies from prokaryotes |
| EP1371377B1 (en) | 2001-01-05 | 2009-10-28 | Viromed Limited | Compositions for gene therapy of rheumatoid arthritis including a gene encoding an anti-angiogenic protein or parts thereof |
| US7202059B2 (en) | 2001-02-20 | 2007-04-10 | Sanofi-Aventis Deutschland Gmbh | Fusion proteins capable of being secreted into a fermentation medium |
| DE10108212A1 (de) | 2001-02-20 | 2002-08-22 | Aventis Pharma Gmbh | Fusionsprotein zur Sekretion von Wertprotein in bakterielle Überstände |
| US6887462B2 (en) | 2001-04-09 | 2005-05-03 | Chiron Corporation | HSA-free formulations of interferon-beta |
| US7217785B2 (en) | 2001-05-09 | 2007-05-15 | The Regents Of The University Of California | Cysteine-containing peptides having antioxidant properties |
| EP1483288A4 (en) | 2001-05-11 | 2005-09-21 | Genome Therapeutics Corp | HBM VARIANTS MODULATING BONE MASS AND LIPID LEVELS |
| CA2347657A1 (en) * | 2001-05-22 | 2002-11-22 | University Of British Columbia | Expression of heterologous peptides from caulobacter |
| JP4210737B2 (ja) | 2001-07-12 | 2009-01-21 | ユニバーシティー オブ マサチューセッツ | 遺伝子サイレンシングを仲介する低分子干渉リボ核酸のインビボにおける製造方法 |
| WO2003012052A2 (en) | 2001-07-30 | 2003-02-13 | The Government Of The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Specific inhibition of gene expression by small double stranded rnas |
| EP1424895B1 (en) | 2001-09-13 | 2009-08-12 | California Institute Of Technology | Method for expression of small antiviral rna molecules within a cell |
| US7419783B2 (en) | 2001-11-05 | 2008-09-02 | Research Development Foundation | Engineering of leader peptides for the secretion of recombinant proteins in bacteria |
| US20030114409A1 (en) | 2001-11-16 | 2003-06-19 | Mello Craig C. | Facilitation of RNA interference |
| EP1458879A2 (en) | 2001-12-21 | 2004-09-22 | Oxford Biomedica (UK) Limited | Method for producing a transgenic organism using a lentiviral expression vector such as eiav |
| US6806328B2 (en) | 2002-01-28 | 2004-10-19 | Univation Technologies, Llc | Polymerization catalyst activator complexes and their use in a polymerization process |
| WO2003064621A2 (en) | 2002-02-01 | 2003-08-07 | Ambion, Inc. | HIGH POTENCY siRNAS FOR REDUCING THE EXPRESSION OF TARGET GENES |
| WO2003068948A1 (en) | 2002-02-13 | 2003-08-21 | Dow Global Technologies Inc. | Over-expression of extremozyme genes in pseudomonads and closely related bacteria |
| EP1474433A4 (en) | 2002-02-20 | 2005-02-23 | Sirna Therapeutics Inc | VALIDATION AND IDENTIFICATION OF RNA INTERFERENCE FACILITATED TARGETS USING SHORT INTERFERENT NUCLEIC ACID (siNA) |
| WO2003079007A1 (en) | 2002-03-15 | 2003-09-25 | The Trustees Of The University Of Pennsylvania | Heterologous protein production using the twin arginine translocation pathway |
| AU2003228637B2 (en) | 2002-04-22 | 2010-08-05 | Dow Global Technologies Inc. | Low-cost production of peptides |
| NZ536475A (en) | 2002-05-24 | 2008-06-30 | Schering Corp | Neutralizing human anti-igfr antibody |
| JP2005534650A (ja) | 2002-06-11 | 2005-11-17 | ザ バーナム インスティテュート | エリスロポイエチンおよびインスリン様増殖因子の神経保護的相乗作用 |
| US7226597B2 (en) | 2002-06-17 | 2007-06-05 | The Board Of Regents Of The University Of Oklahoma | Mutants of Clostridium difficile toxin B and methods of use |
| DE60325611D1 (de) | 2002-07-03 | 2009-02-12 | Dow Global Technologies Inc | Benzoat- und antranilatinduzierbare promotoren |
| EP1539814A2 (en) | 2002-07-03 | 2005-06-15 | Maxygen Holdings Ltd. c/o Close Brothers (Cayman) Limited | Full-length interferon gamma polypeptide variants |
| AU2003243831B2 (en) | 2002-07-11 | 2008-07-24 | Sugar Research Australia Limited | Transgenic plants used as a bioreactor system |
| AU2003252111A1 (en) * | 2002-07-19 | 2004-02-09 | Diversa Corporation | Fluorescent proteins, nucleic acids encoding them and methods for making and using them |
| NO20023581D0 (no) | 2002-07-26 | 2002-07-26 | Fmc Biopolymer As | Nye mutantstammer av Pseudomonas fluorescens og varianter derav, metoder for produksjon og bruk derav til produksjon avalginat |
| US20040157289A1 (en) * | 2002-09-06 | 2004-08-12 | Salerno John C. | Protein expression system |
| RU2335535C2 (ru) | 2002-10-08 | 2008-10-10 | Дау Агросайенсиз Ллс | Измененные рекомбинантные клетки (arc), предназначенные для продуцирования и доставки антивирусных агентов, адъювантов и акселераторов вакцин |
| US9453251B2 (en) | 2002-10-08 | 2016-09-27 | Pfenex Inc. | Expression of mammalian proteins in Pseudomonas fluorescens |
| JP2006519763A (ja) | 2002-11-08 | 2006-08-31 | アブリンクス エン.ヴェー. | 治療用ポリペプチドの投与法およびそのためのポリペプチド |
| CN102584997A (zh) | 2002-11-08 | 2012-07-18 | 埃博灵克斯股份有限公司 | 针对肿瘤坏死因子-α的单结构域抗体及其用途 |
| EP1558645B1 (en) | 2002-11-08 | 2011-07-27 | Ablynx N.V. | Stabilized single domain antibodies in pharmaceutical composition adapted for inhalation |
| US20060034845A1 (en) | 2002-11-08 | 2006-02-16 | Karen Silence | Single domain antibodies directed against tumor necrosis factor alpha and uses therefor |
| DE10258885A1 (de) | 2002-12-17 | 2004-07-15 | Aventis Pharma Deutschland Gmbh | Verfahren zur Generierung eines gentechnisch veränderten Organismus |
| NZ540771A (en) | 2003-01-10 | 2009-05-31 | Ablynx Nv | Recombinant VHH single domain antibody from camelidae against von willebrand factor (vWF) or against collagen |
| EP1599499A2 (en) | 2003-02-24 | 2005-11-30 | Dow Global Technologies Inc. | Periodic antimicrobial peptides |
| JP2008501621A (ja) | 2003-05-31 | 2008-01-24 | マイクロメット アクツィエン ゲゼルシャフト | B細胞関連疾患を処置するための二重特異性抗cd3、抗cd19抗体構築物を含む薬学的組成物 |
| US7547821B2 (en) | 2003-06-17 | 2009-06-16 | Sembiosys Genetics Inc. | Methods for the production of insulin in plants |
| CA2545610C (en) | 2003-11-19 | 2014-03-25 | Dow Global Technolgies Inc. | Auxotrophic pseudomonas fluorescens bacteria for recombinant protein expression |
| EP2314602A1 (en) | 2003-11-21 | 2011-04-27 | Pfenex, Inc. | Improved expression systems with SEC-system secretion |
| US7585942B2 (en) | 2003-11-25 | 2009-09-08 | Anjin Corporation | Diphtheria toxin variant |
| CA2547511A1 (en) | 2003-12-01 | 2005-07-28 | Dow Global Technolgies Inc. | Recombinant icosahedral virus like particle production in pseudomonads |
| AU2004303783A1 (en) | 2003-12-10 | 2005-07-07 | Eli Lilly And Company | Muteins of fibroblast growth factor 21 |
| US7235641B2 (en) | 2003-12-22 | 2007-06-26 | Micromet Ag | Bispecific antibodies |
| EP1709170B1 (en) | 2004-01-16 | 2018-02-21 | Pfenex Inc. | Expression of mammalian proteins in pseudomonas fluorescens |
| DE102004014983A1 (de) | 2004-03-26 | 2005-10-20 | Univ Stuttgart | Rekombinante Polypeptide der Mitglieder der TNF Ligandenfamilie und deren Verwendung |
| US7384779B2 (en) | 2004-04-12 | 2008-06-10 | Corning Incorporated | Porous substrate plates and the use thereof |
| CA2565300A1 (en) | 2004-05-13 | 2005-12-01 | Eli Lilly And Company | Fgf-21 fusion proteins |
| EP2330120A3 (en) | 2004-06-02 | 2011-11-16 | AdAlta Pty Ltd | Binding moieties based on Shark IgNAR domains |
| BRPI0513826A2 (pt) | 2004-07-26 | 2010-06-22 | Dow Global Technologies Inc | processo para expressão de proteìna melhorada através de engenharia de cepa |
| DK1789442T3 (da) | 2004-09-02 | 2009-11-16 | Lilly Co Eli | Muteiner af fibroblastvækstfaktor 21 |
| US7563443B2 (en) | 2004-09-17 | 2009-07-21 | Domantis Limited | Monovalent anti-CD40L antibody polypeptides and compositions thereof |
| EP1814917A2 (en) | 2004-10-13 | 2007-08-08 | Ablynx N.V. | Single domain camelide anti-amyloid beta antibodies and polypeptides comprising the same for the treatment and diagnosis of degenerative neural diseases such as alzheimer's disease |
| WO2006059701A1 (ja) | 2004-12-03 | 2006-06-08 | The University Of Tokushima | 微生物または生体分子の収容容器、並びにその作成方法及び使用方法 |
| KR101263173B1 (ko) | 2004-12-17 | 2013-05-13 | 닛토덴코 가부시키가이샤 | 진핵 세포의 트랜스펙션을 위한 고정화된 분해가능한양이온성 중합체 |
| US8420102B2 (en) | 2006-03-07 | 2013-04-16 | Vaxinnate Corporation | Compositions that include hemagglutinin |
| SI1836500T1 (sl) | 2005-01-14 | 2010-11-30 | Ablynx Nv | Postopki in testi za razlikovanje med različnimioblikami bolezni in motenj ki so označene s trombocitopenijo in ali s spontano interakcijo med von Willebrandovim faktorjem vWF in krvnimi ploščicami |
| JP2008528010A (ja) | 2005-01-31 | 2008-07-31 | アブリンクス ナームローゼ フェンノートシャップ | 重鎖抗体の可変ドメイン配列を作出する方法 |
| PE20061324A1 (es) | 2005-04-29 | 2007-01-15 | Centocor Inc | Anticuerpos anti-il-6, composiciones, metodos y usos |
| EP1888640B1 (en) | 2005-05-18 | 2012-03-14 | Ablynx N.V. | Improved nanobodies against tumor necrosis factor-alpha |
| EP1907418A2 (en) | 2005-07-15 | 2008-04-09 | ZymoGenetics, Inc. | Heterodimeric four helix bundle cytokines |
| ATE518882T1 (de) | 2005-09-19 | 2011-08-15 | Allergan Inc | Mit clostridientoxin aktivierbare clostridientoxine |
| US20090252681A1 (en) | 2005-10-11 | 2009-10-08 | Ablynx N.V. | Nanobodies and Polypeptides Against EGFR and IGF-IR |
| ES2595091T3 (es) | 2005-12-21 | 2016-12-27 | Amgen Research (Munich) Gmbh | Composiciones farmacéuticas con resistencia a CEA soluble |
| AU2006328882B2 (en) | 2005-12-21 | 2011-06-16 | Amgen Research (Munich) Gmbh | Pharmaceutical antibody compositions with resistance to soluble CEA |
| US7553941B2 (en) | 2006-02-03 | 2009-06-30 | Modigene Inc | Long-acting polypeptides and methods of producing same |
| US20070269422A1 (en) | 2006-05-17 | 2007-11-22 | Ablynx N.V. | Serum albumin binding proteins with long half-lives |
| CA2695951A1 (en) | 2006-08-10 | 2008-02-14 | Plantechno S.R.L. | In-plant production of dimeric and/or oligomeric (comprising three or more units) forms of human apo a-1 protein muteins |
| ATE536369T1 (de) | 2006-10-11 | 2011-12-15 | Ablynx Nv | Im wesentlichen ph-wert-unabhängigerweise an serumproteine bindende aminosäuresequenzen, verbindungen, die diese enthalten, und deren verwendung |
| WO2008068280A1 (en) | 2006-12-05 | 2008-06-12 | Ablynx N.V. | Peptides capable of binding to serum proteins |
| JP5714230B2 (ja) | 2007-01-31 | 2015-05-07 | フェネックス インコーポレイテッド | 発現上昇のための細菌リーダー配列 |
| US9580719B2 (en) | 2007-04-27 | 2017-02-28 | Pfenex, Inc. | Method for rapidly screening microbial hosts to identify certain strains with improved yield and/or quality in the expression of heterologous proteins |
| AU2008245696B2 (en) | 2007-04-27 | 2013-11-07 | Pelican Technology Holdings, Inc. | Method for rapidly screening microbial hosts to identify certain strains with improved yield and/or quality in the expression of heterologous proteins |
| CA2695510A1 (en) | 2007-08-03 | 2009-02-12 | Thomas M. Ramseier | Translation initiation region sequences for the optimal expression of heterologous proteins |
| KR20100097719A (ko) | 2007-11-30 | 2010-09-03 | 칼로바이오스 파마슈티컬스, 아이엔씨. | 슈도모나스 아에루기노사의 PcrV 항원에 대한 항체 |
| US7755462B2 (en) | 2008-04-04 | 2010-07-13 | Cedar Ridge Research Llc | Ring magnet structure having a coded magnet pattern |
-
2005
- 2005-07-26 BR BRPI0513826-4A patent/BRPI0513826A2/pt not_active IP Right Cessation
- 2005-07-26 AU AU2005269527A patent/AU2005269527B2/en not_active Ceased
- 2005-07-26 CA CA 2574953 patent/CA2574953A1/en not_active Abandoned
- 2005-07-26 EP EP20050774619 patent/EP1774017B1/en not_active Ceased
- 2005-07-26 WO PCT/US2005/026390 patent/WO2006014899A2/en active Application Filing
- 2005-07-26 CN CNA2005800322451A patent/CN101031655A/zh active Pending
- 2005-07-26 EP EP20110173331 patent/EP2412816B1/en not_active Ceased
- 2005-07-26 US US11/189,375 patent/US8603824B2/en active Active
- 2005-07-26 JP JP2007523707A patent/JP2008507294A/ja active Pending
- 2005-07-26 KR KR1020137002343A patent/KR20130019457A/ko not_active Ceased
-
2007
- 2007-02-23 KR KR1020077004418A patent/KR101340708B1/ko not_active Expired - Fee Related
-
2011
- 2011-11-17 JP JP2011251610A patent/JP2012055318A/ja not_active Withdrawn
-
2013
- 2013-11-04 US US14/071,273 patent/US9109229B2/en active Active
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH11510065A (ja) * | 1995-12-15 | 1999-09-07 | ユニバーシティ オブ ハワイ | 組換えタンパク質の産生のための改善されたNeurospora宿主およびそれを生成するための方法 |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2013509181A (ja) * | 2009-10-30 | 2013-03-14 | メルク・シャープ・エンド・ドーム・コーポレイション | ジペプチジルアミノペプチダーゼ活性を欠くピチア・パストリスにおける治療用タンパク質の製造方法 |
| JP2021500892A (ja) * | 2017-10-27 | 2021-01-14 | フェネックス インク. | 組み換えE.coliアスパラギナーゼの製造のための方法 |
| JP7358347B2 (ja) | 2017-10-27 | 2023-10-10 | フェネックス インク. | 組み換えE.coliアスパラギナーゼの製造のための方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| AU2005269527B2 (en) | 2011-12-01 |
| JP2012055318A (ja) | 2012-03-22 |
| KR101340708B1 (ko) | 2013-12-20 |
| WO2006014899A3 (en) | 2006-09-28 |
| WO2006014899A2 (en) | 2006-02-09 |
| US9109229B2 (en) | 2015-08-18 |
| BRPI0513826A2 (pt) | 2010-06-22 |
| EP1774017A2 (en) | 2007-04-18 |
| EP2412816B1 (en) | 2014-12-03 |
| US8603824B2 (en) | 2013-12-10 |
| AU2005269527A1 (en) | 2006-02-09 |
| EP2412816A3 (en) | 2012-05-16 |
| EP1774017A4 (en) | 2009-05-06 |
| EP2412816A2 (en) | 2012-02-01 |
| US20140162279A1 (en) | 2014-06-12 |
| EP1774017B1 (en) | 2013-05-15 |
| US20060110747A1 (en) | 2006-05-25 |
| KR20130019457A (ko) | 2013-02-26 |
| KR20070041592A (ko) | 2007-04-18 |
| CA2574953A1 (en) | 2006-02-09 |
| CN101031655A (zh) | 2007-09-05 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2008507294A (ja) | 菌株遺伝子操作による改善されたタンパク質発現のための方法 | |
| US10689640B2 (en) | Method for rapidly screening microbial hosts to identify certain strains with improved yield and/or quality in the expression of heterologous proteins | |
| JP5444553B2 (ja) | 微生物宿主を迅速にスクリーニングして、異種タンパク質発現の収率および/または質が改善されている特定の株を同定する方法 | |
| EP2553102B1 (en) | High level expression of recombinant toxin proteins | |
| US20190127742A1 (en) | Method for production of recombinant erwinia asparaginase | |
| US20200248193A1 (en) | Method for production of recombinant e. coli asparaginase | |
| WO2011123139A1 (en) | High level expression of recombinant crm197 | |
| US20230100757A1 (en) | Bacterial hosts for recombinant protein expression | |
| WO2021061694A1 (en) | Method for counterselection in microorganisms | |
| MX2007000970A (es) | Proceso para expresión de proteína mejorada mediante construccion de cepa |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20080723 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20080723 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20110517 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20110811 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20110818 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20111117 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20111220 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20120420 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20120619 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20120713 |