RU2741103C2 - Рекомбинантное получение стевиол-гликозидов - Google Patents
Рекомбинантное получение стевиол-гликозидов Download PDFInfo
- Publication number
- RU2741103C2 RU2741103C2 RU2019135831A RU2019135831A RU2741103C2 RU 2741103 C2 RU2741103 C2 RU 2741103C2 RU 2019135831 A RU2019135831 A RU 2019135831A RU 2019135831 A RU2019135831 A RU 2019135831A RU 2741103 C2 RU2741103 C2 RU 2741103C2
- Authority
- RU
- Russia
- Prior art keywords
- leu
- ala
- val
- gly
- glu
- Prior art date
Links
- 235000019202 steviosides Nutrition 0.000 title claims abstract description 101
- 239000004383 Steviol glycoside Substances 0.000 title claims abstract description 65
- 229930182488 steviol glycoside Natural products 0.000 title claims abstract description 65
- 235000019411 steviol glycoside Nutrition 0.000 title claims abstract description 64
- 150000008144 steviol glycosides Chemical class 0.000 title claims abstract description 58
- 238000004519 manufacturing process Methods 0.000 title abstract description 16
- 238000000034 method Methods 0.000 claims abstract description 63
- 229930188195 rebaudioside Natural products 0.000 claims abstract description 48
- 150000001875 compounds Chemical class 0.000 claims abstract description 38
- 239000000203 mixture Substances 0.000 claims abstract description 36
- 235000003599 food sweetener Nutrition 0.000 claims abstract description 15
- 239000003765 sweetening agent Substances 0.000 claims abstract description 15
- -1 steviol-glycoside compound Chemical class 0.000 claims abstract description 14
- 239000000126 substance Substances 0.000 claims abstract description 8
- 235000013305 food Nutrition 0.000 claims abstract description 6
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 76
- RLLCWNUIHGPAJY-RYBZXKSASA-N Rebaudioside E Natural products O=C(O[C@H]1[C@H](O[C@H]2[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O2)[C@@H](O)[C@@H](O)[C@H](CO)O1)[C@]1(C)[C@@H]2[C@@](C)([C@@H]3[C@@]4(CC(=C)[C@@](O[C@@H]5[C@@H](O[C@@H]6[C@@H](O)[C@H](O)[C@@H](O)[C@H](CO)O6)[C@H](O)[C@@H](O)[C@H](CO)O5)(C4)CC3)CC2)CCC1 RLLCWNUIHGPAJY-RYBZXKSASA-N 0.000 claims description 52
- RLLCWNUIHGPAJY-SFUUMPFESA-N rebaudioside E Chemical group O([C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1O[C@]12C(=C)C[C@@]3(C1)CC[C@@H]1[C@@](C)(CCC[C@]1([C@@H]3CC2)C)C(=O)O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O RLLCWNUIHGPAJY-SFUUMPFESA-N 0.000 claims description 52
- 239000008103 glucose Substances 0.000 claims description 36
- XCCTYIAWTASOJW-XVFCMESISA-N Uridine-5'-Diphosphate Chemical compound O[C@@H]1[C@H](O)[C@@H](COP(O)(=O)OP(O)(O)=O)O[C@H]1N1C(=O)NC(=O)C=C1 XCCTYIAWTASOJW-XVFCMESISA-N 0.000 claims description 29
- 239000000758 substrate Substances 0.000 claims description 29
- 101000630755 Arabidopsis thaliana Sucrose synthase 1 Proteins 0.000 claims description 27
- 108010043934 Sucrose synthase Proteins 0.000 claims description 22
- HSCJRCZFDFQWRP-JZMIEXBBSA-N UDP-alpha-D-glucose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OP(O)(=O)OP(O)(=O)OC[C@@H]1[C@@H](O)[C@@H](O)[C@H](N2C(NC(=O)C=C2)=O)O1 HSCJRCZFDFQWRP-JZMIEXBBSA-N 0.000 claims description 22
- HSCJRCZFDFQWRP-UHFFFAOYSA-N Uridindiphosphoglukose Natural products OC1C(O)C(O)C(CO)OC1OP(O)(=O)OP(O)(=O)OCC1C(O)C(O)C(N2C(NC(=O)C=C2)=O)O1 HSCJRCZFDFQWRP-UHFFFAOYSA-N 0.000 claims description 22
- 108700023372 Glycosyltransferases Proteins 0.000 claims description 17
- 102000051366 Glycosyltransferases Human genes 0.000 claims description 15
- 229930006000 Sucrose Natural products 0.000 claims description 13
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 claims description 13
- 239000005720 sucrose Substances 0.000 claims description 13
- 241000219194 Arabidopsis Species 0.000 claims description 5
- 238000011534 incubation Methods 0.000 claims description 5
- 241000219977 Vigna Species 0.000 claims description 4
- 235000015173 baked goods and baking mixes Nutrition 0.000 claims description 3
- 235000013361 beverage Nutrition 0.000 claims description 3
- 235000015218 chewing gum Nutrition 0.000 claims description 3
- 229940112822 chewing gum Drugs 0.000 claims description 3
- 235000009508 confectionery Nutrition 0.000 claims description 3
- 125000002791 glucosyl group Chemical group C1([C@H](O)[C@@H](O)[C@H](O)[C@H](O1)CO)* 0.000 claims description 3
- 235000015895 biscuits Nutrition 0.000 claims 1
- 239000004067 bulking agent Substances 0.000 claims 1
- 239000000945 filler Substances 0.000 claims 1
- 239000003112 inhibitor Substances 0.000 claims 1
- 230000000694 effects Effects 0.000 abstract description 28
- QFVOYBUQQBFCRH-UHFFFAOYSA-N Steviol Natural products C1CC2(C3)CC(=C)C3(O)CCC2C2(C)C1C(C)(C(O)=O)CCC2 QFVOYBUQQBFCRH-UHFFFAOYSA-N 0.000 abstract description 13
- 229940032084 steviol Drugs 0.000 abstract description 13
- QFVOYBUQQBFCRH-VQSWZGCSSA-N steviol Chemical compound C([C@@]1(O)C(=C)C[C@@]2(C1)CC1)C[C@H]2[C@@]2(C)[C@H]1[C@](C)(C(O)=O)CCC2 QFVOYBUQQBFCRH-VQSWZGCSSA-N 0.000 abstract description 11
- 108090000765 processed proteins & peptides Proteins 0.000 description 151
- 102000004196 processed proteins & peptides Human genes 0.000 description 143
- 229920001184 polypeptide Polymers 0.000 description 140
- 210000004027 cell Anatomy 0.000 description 116
- RPYRMTHVSUWHSV-CUZJHZIBSA-N rebaudioside D Chemical compound O([C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)O[C@]12C(=C)C[C@@]3(C1)CC[C@@H]1[C@@](C)(CCC[C@]1([C@@H]3CC2)C)C(=O)O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O RPYRMTHVSUWHSV-CUZJHZIBSA-N 0.000 description 80
- 108090000623 proteins and genes Proteins 0.000 description 76
- 238000005516 engineering process Methods 0.000 description 71
- 150000007523 nucleic acids Chemical class 0.000 description 50
- 239000013604 expression vector Substances 0.000 description 49
- HELXLJCILKEWJH-NCGAPWICSA-N rebaudioside A Chemical compound O([C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)O[C@]12C(=C)C[C@@]3(C1)CC[C@@H]1[C@@](C)(CCC[C@]1([C@@H]3CC2)C)C(=O)O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O HELXLJCILKEWJH-NCGAPWICSA-N 0.000 description 44
- 241000196324 Embryophyta Species 0.000 description 43
- 238000006243 chemical reaction Methods 0.000 description 43
- 230000014509 gene expression Effects 0.000 description 39
- 102000004169 proteins and genes Human genes 0.000 description 38
- UEDUENGHJMELGK-HYDKPPNVSA-N Stevioside Chemical compound O([C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1O[C@]12C(=C)C[C@@]3(C1)CC[C@@H]1[C@@](C)(CCC[C@]1([C@@H]3CC2)C)C(=O)O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O UEDUENGHJMELGK-HYDKPPNVSA-N 0.000 description 37
- OHHNJQXIOPOJSC-UHFFFAOYSA-N stevioside Natural products CC1(CCCC2(C)C3(C)CCC4(CC3(CCC12C)CC4=C)OC5OC(CO)C(O)C(O)C5OC6OC(CO)C(O)C(O)C6O)C(=O)OC7OC(CO)C(O)C(O)C7O OHHNJQXIOPOJSC-UHFFFAOYSA-N 0.000 description 37
- 229940013618 stevioside Drugs 0.000 description 37
- 102000039446 nucleic acids Human genes 0.000 description 36
- 108020004707 nucleic acids Proteins 0.000 description 36
- 108091033319 polynucleotide Proteins 0.000 description 35
- 102000040430 polynucleotide Human genes 0.000 description 35
- 239000002157 polynucleotide Substances 0.000 description 35
- 239000013598 vector Substances 0.000 description 34
- 235000018102 proteins Nutrition 0.000 description 33
- 239000001512 FEMA 4601 Substances 0.000 description 30
- HELXLJCILKEWJH-SEAGSNCFSA-N Rebaudioside A Natural products O=C(O[C@H]1[C@@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1)[C@@]1(C)[C@@H]2[C@](C)([C@H]3[C@@]4(CC(=C)[C@@](O[C@H]5[C@H](O[C@H]6[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O6)[C@@H](O[C@H]6[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O6)[C@H](O)[C@@H](CO)O5)(C4)CC3)CC2)CCC1 HELXLJCILKEWJH-SEAGSNCFSA-N 0.000 description 30
- HELXLJCILKEWJH-UHFFFAOYSA-N entered according to Sigma 01432 Natural products C1CC2C3(C)CCCC(C)(C(=O)OC4C(C(O)C(O)C(CO)O4)O)C3CCC2(C2)CC(=C)C21OC(C1OC2C(C(O)C(O)C(CO)O2)O)OC(CO)C(O)C1OC1OC(CO)C(O)C(O)C1O HELXLJCILKEWJH-UHFFFAOYSA-N 0.000 description 30
- 235000019203 rebaudioside A Nutrition 0.000 description 30
- 125000003729 nucleotide group Chemical group 0.000 description 24
- 238000006206 glycosylation reaction Methods 0.000 description 23
- 239000002773 nucleotide Substances 0.000 description 23
- 101100262416 Stevia rebaudiana UGT76G1 gene Proteins 0.000 description 22
- 108020004414 DNA Proteins 0.000 description 21
- 239000000047 product Substances 0.000 description 21
- 108091026890 Coding region Proteins 0.000 description 19
- 235000001014 amino acid Nutrition 0.000 description 19
- 244000228451 Stevia rebaudiana Species 0.000 description 18
- 238000000338 in vitro Methods 0.000 description 17
- 108091028043 Nucleic acid sequence Proteins 0.000 description 16
- 150000001413 amino acids Chemical class 0.000 description 16
- 239000000284 extract Substances 0.000 description 16
- 108020004705 Codon Proteins 0.000 description 15
- 102000004190 Enzymes Human genes 0.000 description 15
- 108090000790 Enzymes Proteins 0.000 description 15
- 229940024606 amino acid Drugs 0.000 description 15
- 229940088598 enzyme Drugs 0.000 description 15
- 230000000813 microbial effect Effects 0.000 description 14
- 230000013595 glycosylation Effects 0.000 description 13
- 230000001105 regulatory effect Effects 0.000 description 13
- 238000012546 transfer Methods 0.000 description 13
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 12
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 12
- YBAFDPFAUTYYRW-UHFFFAOYSA-N N-L-alpha-glutamyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=O YBAFDPFAUTYYRW-UHFFFAOYSA-N 0.000 description 12
- 230000015572 biosynthetic process Effects 0.000 description 12
- 108010054813 diprotin B Proteins 0.000 description 12
- 239000012634 fragment Substances 0.000 description 12
- 108010073472 leucyl-prolyl-proline Proteins 0.000 description 12
- 230000009466 transformation Effects 0.000 description 12
- LRHPLDYGYMQRHN-UHFFFAOYSA-N N-Butanol Chemical compound CCCCO LRHPLDYGYMQRHN-UHFFFAOYSA-N 0.000 description 11
- 108010047857 aspartylglycine Proteins 0.000 description 11
- 239000013612 plasmid Substances 0.000 description 11
- 238000006467 substitution reaction Methods 0.000 description 11
- 241000743776 Brachypodium distachyon Species 0.000 description 10
- 240000005979 Hordeum vulgare Species 0.000 description 9
- 235000007340 Hordeum vulgare Nutrition 0.000 description 9
- XVZCXCTYGHPNEM-UHFFFAOYSA-N Leu-Leu-Pro Natural products CC(C)CC(N)C(=O)NC(CC(C)C)C(=O)N1CCCC1C(O)=O XVZCXCTYGHPNEM-UHFFFAOYSA-N 0.000 description 9
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 9
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Natural products C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 9
- 108010057821 leucylproline Proteins 0.000 description 9
- MNZHHDPWDWQJCQ-YUMQZZPRSA-N Ala-Leu-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O MNZHHDPWDWQJCQ-YUMQZZPRSA-N 0.000 description 8
- 241000880493 Leptailurus serval Species 0.000 description 8
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 8
- KZNQNBZMBZJQJO-UHFFFAOYSA-N N-glycyl-L-proline Natural products NCC(=O)N1CCCC1C(O)=O KZNQNBZMBZJQJO-UHFFFAOYSA-N 0.000 description 8
- 230000001580 bacterial effect Effects 0.000 description 8
- 239000000796 flavoring agent Substances 0.000 description 8
- 108010055341 glutamyl-glutamic acid Proteins 0.000 description 8
- 108010050848 glycylleucine Proteins 0.000 description 8
- 238000004128 high performance liquid chromatography Methods 0.000 description 8
- 238000013518 transcription Methods 0.000 description 8
- 230000035897 transcription Effects 0.000 description 8
- 241000894006 Bacteria Species 0.000 description 7
- XEKOWRVHYACXOJ-UHFFFAOYSA-N Ethyl acetate Chemical compound CCOC(C)=O XEKOWRVHYACXOJ-UHFFFAOYSA-N 0.000 description 7
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 7
- 238000005481 NMR spectroscopy Methods 0.000 description 7
- 108010086434 alanyl-seryl-glycine Proteins 0.000 description 7
- 108010005233 alanylglutamic acid Proteins 0.000 description 7
- VPZXBVLAVMBEQI-UHFFFAOYSA-N glycyl-DL-alpha-alanine Natural products OC(=O)C(C)NC(=O)CN VPZXBVLAVMBEQI-UHFFFAOYSA-N 0.000 description 7
- 238000003752 polymerase chain reaction Methods 0.000 description 7
- 210000001519 tissue Anatomy 0.000 description 7
- 238000013519 translation Methods 0.000 description 7
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 6
- QKSAZKCRVQYYGS-UWVGGRQHSA-N Arg-Gly-His Chemical compound N[C@@H](CCCN=C(N)N)C(=O)NCC(=O)N[C@@H](Cc1cnc[nH]1)C(O)=O QKSAZKCRVQYYGS-UWVGGRQHSA-N 0.000 description 6
- VHQOCWWKXIOAQI-WDSKDSINSA-N Asp-Gln-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O VHQOCWWKXIOAQI-WDSKDSINSA-N 0.000 description 6
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 6
- PBCHMHROGNUXMK-DLOVCJGASA-N Leu-Ala-His Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CN=CN1 PBCHMHROGNUXMK-DLOVCJGASA-N 0.000 description 6
- AWGBEIYZPAXXSX-RWMBFGLXSA-N Met-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCSC)N AWGBEIYZPAXXSX-RWMBFGLXSA-N 0.000 description 6
- AJHCSUXXECOXOY-UHFFFAOYSA-N N-glycyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)CN)C(O)=O)=CNC2=C1 AJHCSUXXECOXOY-UHFFFAOYSA-N 0.000 description 6
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 6
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 6
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 6
- 125000000539 amino acid group Chemical group 0.000 description 6
- 238000004458 analytical method Methods 0.000 description 6
- 230000001851 biosynthetic effect Effects 0.000 description 6
- 230000000295 complement effect Effects 0.000 description 6
- 238000001962 electrophoresis Methods 0.000 description 6
- 230000002255 enzymatic effect Effects 0.000 description 6
- 235000019634 flavors Nutrition 0.000 description 6
- 230000004927 fusion Effects 0.000 description 6
- 108010015792 glycyllysine Proteins 0.000 description 6
- 108010037850 glycylvaline Proteins 0.000 description 6
- 108020004999 messenger RNA Proteins 0.000 description 6
- QSRAJVGDWKFOGU-WBXIDTKBSA-N rebaudioside c Chemical compound O[C@@H]1[C@H](O)[C@@H](O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)[C@H](O)[C@@H](CO)O[C@H]1O[C@]1(CC[C@H]2[C@@]3(C)[C@@H]([C@](CCC3)(C)C(=O)O[C@H]3[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O3)O)CC3)C(=C)C[C@]23C1 QSRAJVGDWKFOGU-WBXIDTKBSA-N 0.000 description 6
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 6
- 230000009261 transgenic effect Effects 0.000 description 6
- XVZCXCTYGHPNEM-IHRRRGAJSA-N (2s)-1-[(2s)-2-[[(2s)-2-amino-4-methylpentanoyl]amino]-4-methylpentanoyl]pyrrolidine-2-carboxylic acid Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(O)=O XVZCXCTYGHPNEM-IHRRRGAJSA-N 0.000 description 5
- YJHKTAMKPGFJCT-NRPADANISA-N Ala-Val-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O YJHKTAMKPGFJCT-NRPADANISA-N 0.000 description 5
- 241000219195 Arabidopsis thaliana Species 0.000 description 5
- UGZUVYDKAYNCII-ULQDDVLXSA-N Arg-Phe-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O UGZUVYDKAYNCII-ULQDDVLXSA-N 0.000 description 5
- LRPXYSGPOBVBEH-IUCAKERBSA-N Glu-Gly-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O LRPXYSGPOBVBEH-IUCAKERBSA-N 0.000 description 5
- MIIVFRCYJABHTQ-ONGXEEELSA-N Gly-Leu-Val Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O MIIVFRCYJABHTQ-ONGXEEELSA-N 0.000 description 5
- JBCLFWXMTIKCCB-UHFFFAOYSA-N H-Gly-Phe-OH Natural products NCC(=O)NC(C(O)=O)CC1=CC=CC=C1 JBCLFWXMTIKCCB-UHFFFAOYSA-N 0.000 description 5
- QVFGXCVIXXBFHO-AVGNSLFASA-N Leu-Glu-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O QVFGXCVIXXBFHO-AVGNSLFASA-N 0.000 description 5
- INCJJHQRZGQLFC-KBPBESRZSA-N Leu-Phe-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)NCC(O)=O INCJJHQRZGQLFC-KBPBESRZSA-N 0.000 description 5
- DPURXCQCHSQPAN-AVGNSLFASA-N Leu-Pro-Pro Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 DPURXCQCHSQPAN-AVGNSLFASA-N 0.000 description 5
- SMVTWPOATVIXTN-NAKRPEOUSA-N Met-Ser-Ile Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O SMVTWPOATVIXTN-NAKRPEOUSA-N 0.000 description 5
- BQVUABVGYYSDCJ-UHFFFAOYSA-N Nalpha-L-Leucyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)CC(C)C)C(O)=O)=CNC2=C1 BQVUABVGYYSDCJ-UHFFFAOYSA-N 0.000 description 5
- UVKNEILZSJMKSR-FXQIFTODSA-N Pro-Asn-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H]1CCCN1 UVKNEILZSJMKSR-FXQIFTODSA-N 0.000 description 5
- OHKFXGKHSJKKAL-NRPADANISA-N Ser-Glu-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O OHKFXGKHSJKKAL-NRPADANISA-N 0.000 description 5
- 238000007792 addition Methods 0.000 description 5
- 238000000605 extraction Methods 0.000 description 5
- 108010023364 glycyl-histidyl-arginine Proteins 0.000 description 5
- 108010020688 glycylhistidine Proteins 0.000 description 5
- 108010051242 phenylalanylserine Proteins 0.000 description 5
- 239000008057 potassium phosphate buffer Substances 0.000 description 5
- 108091008146 restriction endonucleases Proteins 0.000 description 5
- YYSWCHMLFJLLBJ-ZLUOBGJFSA-N Ala-Ala-Ser Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O YYSWCHMLFJLLBJ-ZLUOBGJFSA-N 0.000 description 4
- NWVVKQZOVSTDBQ-CIUDSAMLSA-N Ala-Glu-Arg Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O NWVVKQZOVSTDBQ-CIUDSAMLSA-N 0.000 description 4
- VCSABYLVNWQYQE-UHFFFAOYSA-N Ala-Lys-Lys Natural products NCCCCC(NC(=O)C(N)C)C(=O)NC(CCCCN)C(O)=O VCSABYLVNWQYQE-UHFFFAOYSA-N 0.000 description 4
- BFMIRJBURUXDRG-DLOVCJGASA-N Ala-Phe-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=CC=C1 BFMIRJBURUXDRG-DLOVCJGASA-N 0.000 description 4
- BECXEHHOZNFFFX-IHRRRGAJSA-N Arg-Ser-Tyr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O BECXEHHOZNFFFX-IHRRRGAJSA-N 0.000 description 4
- IYVSIZAXNLOKFQ-BYULHYEWSA-N Asn-Asp-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O IYVSIZAXNLOKFQ-BYULHYEWSA-N 0.000 description 4
- 241000701489 Cauliflower mosaic virus Species 0.000 description 4
- BLGNLNRBABWDST-CIUDSAMLSA-N Cys-Leu-Asp Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CS)N BLGNLNRBABWDST-CIUDSAMLSA-N 0.000 description 4
- 241000588724 Escherichia coli Species 0.000 description 4
- 241000233866 Fungi Species 0.000 description 4
- RDPOETHPAQEGDP-ACZMJKKPSA-N Glu-Asp-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O RDPOETHPAQEGDP-ACZMJKKPSA-N 0.000 description 4
- VXQOONWNIWFOCS-HGNGGELXSA-N Glu-His-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CCC(=O)O)N VXQOONWNIWFOCS-HGNGGELXSA-N 0.000 description 4
- JWNZHMSRZXXGTM-XKBZYTNZSA-N Glu-Ser-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JWNZHMSRZXXGTM-XKBZYTNZSA-N 0.000 description 4
- BPCLDCNZBUYGOD-BPUTZDHNSA-N Glu-Trp-Glu Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCC(O)=O)N)C(=O)N[C@@H](CCC(O)=O)C(O)=O)=CNC2=C1 BPCLDCNZBUYGOD-BPUTZDHNSA-N 0.000 description 4
- JRDYDYXZKFNNRQ-XPUUQOCRSA-N Gly-Ala-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)CN JRDYDYXZKFNNRQ-XPUUQOCRSA-N 0.000 description 4
- PAWIVEIWWYGBAM-YUMQZZPRSA-N Gly-Leu-Ala Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O PAWIVEIWWYGBAM-YUMQZZPRSA-N 0.000 description 4
- WCORRBXVISTKQL-WHFBIAKZSA-N Gly-Ser-Ser Chemical compound NCC(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O WCORRBXVISTKQL-WHFBIAKZSA-N 0.000 description 4
- 244000020551 Helianthus annuus Species 0.000 description 4
- FIMNVXRZGUAGBI-AVGNSLFASA-N His-Glu-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O FIMNVXRZGUAGBI-AVGNSLFASA-N 0.000 description 4
- VCBWXASUBZIFLQ-IHRRRGAJSA-N His-Pro-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O VCBWXASUBZIFLQ-IHRRRGAJSA-N 0.000 description 4
- BQSLGJHIAGOZCD-CIUDSAMLSA-N Leu-Ala-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O BQSLGJHIAGOZCD-CIUDSAMLSA-N 0.000 description 4
- HYIFFZAQXPUEAU-QWRGUYRKSA-N Leu-Gly-Leu Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC(C)C HYIFFZAQXPUEAU-QWRGUYRKSA-N 0.000 description 4
- SGIIOQQGLUUMDQ-IHRRRGAJSA-N Leu-His-Val Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](C(C)C)C(=O)O)N SGIIOQQGLUUMDQ-IHRRRGAJSA-N 0.000 description 4
- YOKVEHGYYQEQOP-QWRGUYRKSA-N Leu-Leu-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O YOKVEHGYYQEQOP-QWRGUYRKSA-N 0.000 description 4
- BJWKOATWNQJPSK-SRVKXCTJSA-N Leu-Met-Glu Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N BJWKOATWNQJPSK-SRVKXCTJSA-N 0.000 description 4
- ZJSZPXISKMDJKQ-JYJNAYRXSA-N Lys-Phe-Glu Chemical compound NCCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CCC(O)=O)C(O)=O)CC1=CC=CC=C1 ZJSZPXISKMDJKQ-JYJNAYRXSA-N 0.000 description 4
- 108010079364 N-glycylalanine Proteins 0.000 description 4
- 240000007594 Oryza sativa Species 0.000 description 4
- 235000007164 Oryza sativa Nutrition 0.000 description 4
- CDHURCQGUDNBMA-UBHSHLNASA-N Phe-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC1=CC=CC=C1 CDHURCQGUDNBMA-UBHSHLNASA-N 0.000 description 4
- IEIFEYBAYFSRBQ-IHRRRGAJSA-N Phe-Val-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N IEIFEYBAYFSRBQ-IHRRRGAJSA-N 0.000 description 4
- YXHYJEPDKSYPSQ-AVGNSLFASA-N Pro-Leu-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 YXHYJEPDKSYPSQ-AVGNSLFASA-N 0.000 description 4
- DSGYZICNAMEJOC-AVGNSLFASA-N Ser-Glu-Phe Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O DSGYZICNAMEJOC-AVGNSLFASA-N 0.000 description 4
- CUXJENOFJXOSOZ-BIIVOSGPSA-N Ser-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)[C@H](CO)N)C(=O)O CUXJENOFJXOSOZ-BIIVOSGPSA-N 0.000 description 4
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 4
- CXPJPTFWKXNDKV-NUTKFTJISA-N Trp-Leu-Ala Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O)=CNC2=C1 CXPJPTFWKXNDKV-NUTKFTJISA-N 0.000 description 4
- RHYOAUJXSRWVJT-GVXVVHGQSA-N Val-His-Glu Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N RHYOAUJXSRWVJT-GVXVVHGQSA-N 0.000 description 4
- 238000001042 affinity chromatography Methods 0.000 description 4
- 108010024078 alanyl-glycyl-serine Proteins 0.000 description 4
- 108010018691 arginyl-threonyl-arginine Proteins 0.000 description 4
- 108010062796 arginyllysine Proteins 0.000 description 4
- 108010038633 aspartylglutamate Proteins 0.000 description 4
- 238000006555 catalytic reaction Methods 0.000 description 4
- 230000006870 function Effects 0.000 description 4
- 239000007789 gas Substances 0.000 description 4
- 108010078144 glutaminyl-glycine Proteins 0.000 description 4
- 108010075431 glycyl-alanyl-phenylalanine Proteins 0.000 description 4
- 108010027668 glycyl-alanyl-valine Proteins 0.000 description 4
- 238000003919 heteronuclear multiple bond coherence Methods 0.000 description 4
- 108010092114 histidylphenylalanine Proteins 0.000 description 4
- 230000001965 increasing effect Effects 0.000 description 4
- 230000008569 process Effects 0.000 description 4
- 108010004914 prolylarginine Proteins 0.000 description 4
- 238000000425 proton nuclear magnetic resonance spectrum Methods 0.000 description 4
- 238000001551 total correlation spectroscopy Methods 0.000 description 4
- 108010051110 tyrosyl-lysine Proteins 0.000 description 4
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 4
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 3
- BTYTYHBSJKQBQA-GCJQMDKQSA-N Ala-Asp-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](C)N)O BTYTYHBSJKQBQA-GCJQMDKQSA-N 0.000 description 3
- BGNLUHXLSAQYRQ-FXQIFTODSA-N Ala-Glu-Gln Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O BGNLUHXLSAQYRQ-FXQIFTODSA-N 0.000 description 3
- OBVSBEYOMDWLRJ-BFHQHQDPSA-N Ala-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@H](C)N OBVSBEYOMDWLRJ-BFHQHQDPSA-N 0.000 description 3
- NYDBKUNVSALYPX-NAKRPEOUSA-N Ala-Ile-Arg Chemical compound C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CCCN=C(N)N NYDBKUNVSALYPX-NAKRPEOUSA-N 0.000 description 3
- YHKANGMVQWRMAP-DCAQKATOSA-N Ala-Leu-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N YHKANGMVQWRMAP-DCAQKATOSA-N 0.000 description 3
- OYJCVIGKMXUVKB-GARJFASQSA-N Ala-Leu-Pro Chemical compound C[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N OYJCVIGKMXUVKB-GARJFASQSA-N 0.000 description 3
- 244000105624 Arachis hypogaea Species 0.000 description 3
- QJWLLRZTJFPCHA-STECZYCISA-N Arg-Tyr-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O QJWLLRZTJFPCHA-STECZYCISA-N 0.000 description 3
- CPTXATAOUQJQRO-GUBZILKMSA-N Arg-Val-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O CPTXATAOUQJQRO-GUBZILKMSA-N 0.000 description 3
- SNDBKTFJWVEVPO-WHFBIAKZSA-N Asp-Gly-Ser Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](CO)C(O)=O SNDBKTFJWVEVPO-WHFBIAKZSA-N 0.000 description 3
- VSMYBNPOHYAXSD-GUBZILKMSA-N Asp-Lys-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O VSMYBNPOHYAXSD-GUBZILKMSA-N 0.000 description 3
- 241000195493 Cryptophyta Species 0.000 description 3
- 241000192700 Cyanobacteria Species 0.000 description 3
- 102000053602 DNA Human genes 0.000 description 3
- 206010013911 Dysgeusia Diseases 0.000 description 3
- 241000588722 Escherichia Species 0.000 description 3
- 239000001776 FEMA 4720 Substances 0.000 description 3
- MZZSCEANQDPJER-ONGXEEELSA-N Gly-Ala-Phe Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 MZZSCEANQDPJER-ONGXEEELSA-N 0.000 description 3
- XUDLUKYPXQDCRX-BQBZGAKWSA-N Gly-Arg-Asn Chemical compound [H]NCC(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(O)=O XUDLUKYPXQDCRX-BQBZGAKWSA-N 0.000 description 3
- XEJTYSCIXKYSHR-WDSKDSINSA-N Gly-Asp-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)CN XEJTYSCIXKYSHR-WDSKDSINSA-N 0.000 description 3
- HQRHFUYMGCHHJS-LURJTMIESA-N Gly-Gly-Arg Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N HQRHFUYMGCHHJS-LURJTMIESA-N 0.000 description 3
- VEPBEGNDJYANCF-QWRGUYRKSA-N Gly-Lys-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCCN VEPBEGNDJYANCF-QWRGUYRKSA-N 0.000 description 3
- 235000010469 Glycine max Nutrition 0.000 description 3
- 244000068988 Glycine max Species 0.000 description 3
- WIZPFZKOFZXDQG-HTFCKZLJSA-N Ile-Ile-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O WIZPFZKOFZXDQG-HTFCKZLJSA-N 0.000 description 3
- UAELWXJFLZBKQS-WHOFXGATSA-N Ile-Phe-Gly Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](Cc1ccccc1)C(=O)NCC(O)=O UAELWXJFLZBKQS-WHOFXGATSA-N 0.000 description 3
- YJRSIJZUIUANHO-NAKRPEOUSA-N Ile-Val-Ala Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(=O)O)N YJRSIJZUIUANHO-NAKRPEOUSA-N 0.000 description 3
- SITWEMZOJNKJCH-UHFFFAOYSA-N L-alanine-L-arginine Natural products CC(N)C(=O)NC(C(O)=O)CCCNC(N)=N SITWEMZOJNKJCH-UHFFFAOYSA-N 0.000 description 3
- RVVBWTWPNFDYBE-SRVKXCTJSA-N Leu-Glu-Arg Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O RVVBWTWPNFDYBE-SRVKXCTJSA-N 0.000 description 3
- QMKFDEUJGYNFMC-AVGNSLFASA-N Leu-Pro-Arg Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCN=C(N)N)C(O)=O QMKFDEUJGYNFMC-AVGNSLFASA-N 0.000 description 3
- BMVFXOQHDQZAQU-DCAQKATOSA-N Leu-Pro-Asp Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(=O)O)C(=O)O)N BMVFXOQHDQZAQU-DCAQKATOSA-N 0.000 description 3
- KWLWZYMNUZJKMZ-IHRRRGAJSA-N Leu-Pro-Leu Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O KWLWZYMNUZJKMZ-IHRRRGAJSA-N 0.000 description 3
- FGZVGOAAROXFAB-IXOXFDKPSA-N Leu-Thr-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CC(C)C)N)O FGZVGOAAROXFAB-IXOXFDKPSA-N 0.000 description 3
- 240000006240 Linum usitatissimum Species 0.000 description 3
- WKUXWMWQTOYTFI-SRVKXCTJSA-N Lys-Met-Gln Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CCCCN)N WKUXWMWQTOYTFI-SRVKXCTJSA-N 0.000 description 3
- YSPZCHGIWAQVKQ-AVGNSLFASA-N Lys-Pro-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCCN YSPZCHGIWAQVKQ-AVGNSLFASA-N 0.000 description 3
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 3
- 244000061176 Nicotiana tabacum Species 0.000 description 3
- XEXSSIBQYNKFBX-KBPBESRZSA-N Phe-Gly-His Chemical compound C([C@H](N)C(=O)NCC(=O)N[C@@H](CC=1N=CNC=1)C(O)=O)C1=CC=CC=C1 XEXSSIBQYNKFBX-KBPBESRZSA-N 0.000 description 3
- CMHTUJQZQXFNTQ-OEAJRASXSA-N Phe-Leu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC1=CC=CC=C1)N)O CMHTUJQZQXFNTQ-OEAJRASXSA-N 0.000 description 3
- 241000235648 Pichia Species 0.000 description 3
- MCWHYUWXVNRXFV-RWMBFGLXSA-N Pro-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@@H]2CCCN2 MCWHYUWXVNRXFV-RWMBFGLXSA-N 0.000 description 3
- SNGZLPOXVRTNMB-LPEHRKFASA-N Pro-Ser-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CO)C(=O)N2CCC[C@@H]2C(=O)O SNGZLPOXVRTNMB-LPEHRKFASA-N 0.000 description 3
- 241000235070 Saccharomyces Species 0.000 description 3
- JFWDJFULOLKQFY-QWRGUYRKSA-N Ser-Gly-Phe Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O JFWDJFULOLKQFY-QWRGUYRKSA-N 0.000 description 3
- HEUVHBXOVZONPU-BJDJZHNGSA-N Ser-Leu-Ile Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O HEUVHBXOVZONPU-BJDJZHNGSA-N 0.000 description 3
- 235000006092 Stevia rebaudiana Nutrition 0.000 description 3
- HEJJDUDEHLPDAW-CUJWVEQBSA-N Thr-His-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CS)C(=O)O)N)O HEJJDUDEHLPDAW-CUJWVEQBSA-N 0.000 description 3
- UQCNIMDPYICBTR-KYNKHSRBSA-N Thr-Thr-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O UQCNIMDPYICBTR-KYNKHSRBSA-N 0.000 description 3
- OGZRZMJASKKMJZ-XIRDDKMYSA-N Trp-Leu-Asp Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N OGZRZMJASKKMJZ-XIRDDKMYSA-N 0.000 description 3
- CVUDMNSZAIZFAE-UHFFFAOYSA-N Val-Arg-Pro Natural products NC(N)=NCCCC(NC(=O)C(N)C(C)C)C(=O)N1CCCC1C(O)=O CVUDMNSZAIZFAE-UHFFFAOYSA-N 0.000 description 3
- UZDHNIJRRTUKKC-DLOVCJGASA-N Val-Gln-Val Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](C(C)C)C(=O)O)N UZDHNIJRRTUKKC-DLOVCJGASA-N 0.000 description 3
- RKIGNDAHUOOIMJ-BQFCYCMXSA-N Val-Glu-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)C(C)C)C(O)=O)=CNC2=C1 RKIGNDAHUOOIMJ-BQFCYCMXSA-N 0.000 description 3
- URIRWLJVWHYLET-ONGXEEELSA-N Val-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)C(C)C URIRWLJVWHYLET-ONGXEEELSA-N 0.000 description 3
- ZNGPROMGGGFOAA-JYJNAYRXSA-N Val-Tyr-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](C(C)C)C(O)=O)CC1=CC=C(O)C=C1 ZNGPROMGGGFOAA-JYJNAYRXSA-N 0.000 description 3
- 108010041407 alanylaspartic acid Proteins 0.000 description 3
- 108010044940 alanylglutamine Proteins 0.000 description 3
- 108010047495 alanylglycine Proteins 0.000 description 3
- 108010011559 alanylphenylalanine Proteins 0.000 description 3
- 108010087924 alanylproline Proteins 0.000 description 3
- 108010093581 aspartyl-proline Proteins 0.000 description 3
- 108010092854 aspartyllysine Proteins 0.000 description 3
- 230000033228 biological regulation Effects 0.000 description 3
- 239000000872 buffer Substances 0.000 description 3
- 238000001460 carbon-13 nuclear magnetic resonance spectrum Methods 0.000 description 3
- 238000005119 centrifugation Methods 0.000 description 3
- 239000002299 complementary DNA Substances 0.000 description 3
- 230000008878 coupling Effects 0.000 description 3
- 238000010168 coupling process Methods 0.000 description 3
- 238000005859 coupling reaction Methods 0.000 description 3
- 108010016616 cysteinylglycine Proteins 0.000 description 3
- 210000002257 embryonic structure Anatomy 0.000 description 3
- 239000006167 equilibration buffer Substances 0.000 description 3
- 235000019439 ethyl acetate Nutrition 0.000 description 3
- 108010006664 gamma-glutamyl-glycyl-glycine Proteins 0.000 description 3
- 238000010353 genetic engineering Methods 0.000 description 3
- 108010080575 glutamyl-aspartyl-alanine Proteins 0.000 description 3
- 108010049041 glutamylalanine Proteins 0.000 description 3
- 108010081551 glycylphenylalanine Proteins 0.000 description 3
- 238000003929 heteronuclear multiple quantum coherence Methods 0.000 description 3
- 108010025306 histidylleucine Proteins 0.000 description 3
- 108010018006 histidylserine Proteins 0.000 description 3
- 229910052739 hydrogen Inorganic materials 0.000 description 3
- NIKHGUQULKYIGE-UHFFFAOYSA-N kaurenoic acid Natural products C1CC2(CC3=C)CC3CCC2C2(C)C1C(C)(C(O)=O)CCC2 NIKHGUQULKYIGE-UHFFFAOYSA-N 0.000 description 3
- 108010034529 leucyl-lysine Proteins 0.000 description 3
- 108010064235 lysylglycine Proteins 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 244000005700 microbiome Species 0.000 description 3
- 230000004048 modification Effects 0.000 description 3
- 238000012986 modification Methods 0.000 description 3
- 238000010369 molecular cloning Methods 0.000 description 3
- 238000000655 nuclear magnetic resonance spectrum Methods 0.000 description 3
- 239000002245 particle Substances 0.000 description 3
- 229920000642 polymer Polymers 0.000 description 3
- 239000013615 primer Substances 0.000 description 3
- 108010090894 prolylleucine Proteins 0.000 description 3
- 108010053725 prolylvaline Proteins 0.000 description 3
- 238000000746 purification Methods 0.000 description 3
- 239000011541 reaction mixture Substances 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 238000002864 sequence alignment Methods 0.000 description 3
- 241001133760 Acoelorraphe Species 0.000 description 2
- TWCMVXMQHSVIOJ-UHFFFAOYSA-N Aglycone of yadanzioside D Natural products COC(=O)C12OCC34C(CC5C(=CC(O)C(O)C5(C)C3C(O)C1O)C)OC(=O)C(OC(=O)C)C24 TWCMVXMQHSVIOJ-UHFFFAOYSA-N 0.000 description 2
- 241000589158 Agrobacterium Species 0.000 description 2
- 241000589155 Agrobacterium tumefaciens Species 0.000 description 2
- UWQJHXKARZWDIJ-ZLUOBGJFSA-N Ala-Ala-Cys Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CS)C(O)=O UWQJHXKARZWDIJ-ZLUOBGJFSA-N 0.000 description 2
- CXRCVCURMBFFOL-FXQIFTODSA-N Ala-Ala-Pro Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O CXRCVCURMBFFOL-FXQIFTODSA-N 0.000 description 2
- YAXNATKKPOWVCP-ZLUOBGJFSA-N Ala-Asn-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(O)=O YAXNATKKPOWVCP-ZLUOBGJFSA-N 0.000 description 2
- ZIBWKCRKNFYTPT-ZKWXMUAHSA-N Ala-Asn-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O ZIBWKCRKNFYTPT-ZKWXMUAHSA-N 0.000 description 2
- WXERCAHAIKMTKX-ZLUOBGJFSA-N Ala-Asp-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O WXERCAHAIKMTKX-ZLUOBGJFSA-N 0.000 description 2
- CZPAHAKGPDUIPJ-CIUDSAMLSA-N Ala-Gln-Pro Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N1CCC[C@H]1C(O)=O CZPAHAKGPDUIPJ-CIUDSAMLSA-N 0.000 description 2
- NBTGEURICRTMGL-WHFBIAKZSA-N Ala-Gly-Ser Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O NBTGEURICRTMGL-WHFBIAKZSA-N 0.000 description 2
- CCDFBRZVTDDJNM-GUBZILKMSA-N Ala-Leu-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O CCDFBRZVTDDJNM-GUBZILKMSA-N 0.000 description 2
- MFMDKJIPHSWSBM-GUBZILKMSA-N Ala-Lys-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O MFMDKJIPHSWSBM-GUBZILKMSA-N 0.000 description 2
- NINQYGGNRIBFSC-CIUDSAMLSA-N Ala-Lys-Ser Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CO)C(O)=O NINQYGGNRIBFSC-CIUDSAMLSA-N 0.000 description 2
- NLOMBWNGESDVJU-GUBZILKMSA-N Ala-Met-Arg Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O NLOMBWNGESDVJU-GUBZILKMSA-N 0.000 description 2
- GMGWOTQMUKYZIE-UBHSHLNASA-N Ala-Pro-Phe Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 GMGWOTQMUKYZIE-UBHSHLNASA-N 0.000 description 2
- RTZCUEHYUQZIDE-WHFBIAKZSA-N Ala-Ser-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O RTZCUEHYUQZIDE-WHFBIAKZSA-N 0.000 description 2
- KUFVXLQLDHJVOG-SHGPDSBTSA-N Ala-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](C)N)O KUFVXLQLDHJVOG-SHGPDSBTSA-N 0.000 description 2
- IETUUAHKCHOQHP-KZVJFYERSA-N Ala-Thr-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@H](C)N)[C@@H](C)O)C(O)=O IETUUAHKCHOQHP-KZVJFYERSA-N 0.000 description 2
- IYKVSFNGSWTTNZ-GUBZILKMSA-N Ala-Val-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N IYKVSFNGSWTTNZ-GUBZILKMSA-N 0.000 description 2
- VHAQSYHSDKERBS-XPUUQOCRSA-N Ala-Val-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O VHAQSYHSDKERBS-XPUUQOCRSA-N 0.000 description 2
- 108010009534 Arabidopsis sucrose synthase-1 Proteins 0.000 description 2
- GIVATXIGCXFQQA-FXQIFTODSA-N Arg-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCN=C(N)N GIVATXIGCXFQQA-FXQIFTODSA-N 0.000 description 2
- HJVGMOYJDDXLMI-AVGNSLFASA-N Arg-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H](N)CCCNC(N)=N HJVGMOYJDDXLMI-AVGNSLFASA-N 0.000 description 2
- RVDVDRUZWZIBJQ-CIUDSAMLSA-N Arg-Asn-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O RVDVDRUZWZIBJQ-CIUDSAMLSA-N 0.000 description 2
- NTAZNGWBXRVEDJ-FXQIFTODSA-N Arg-Asp-Asp Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O NTAZNGWBXRVEDJ-FXQIFTODSA-N 0.000 description 2
- DGFGDPVSDQPANQ-XGEHTFHBSA-N Arg-Cys-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CCCN=C(N)N)N)O DGFGDPVSDQPANQ-XGEHTFHBSA-N 0.000 description 2
- OQCWXQJLCDPRHV-UWVGGRQHSA-N Arg-Gly-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O OQCWXQJLCDPRHV-UWVGGRQHSA-N 0.000 description 2
- JTZUZBADHGISJD-SRVKXCTJSA-N Arg-His-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(O)=O JTZUZBADHGISJD-SRVKXCTJSA-N 0.000 description 2
- BTJVOUQWFXABOI-IHRRRGAJSA-N Arg-Lys-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCNC(N)=N BTJVOUQWFXABOI-IHRRRGAJSA-N 0.000 description 2
- GRRXPUAICOGISM-RWMBFGLXSA-N Arg-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CCCN=C(N)N)N)C(=O)O GRRXPUAICOGISM-RWMBFGLXSA-N 0.000 description 2
- AUIJUTGLPVHIRT-FXQIFTODSA-N Arg-Ser-Cys Chemical compound C(C[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O)N)CN=C(N)N AUIJUTGLPVHIRT-FXQIFTODSA-N 0.000 description 2
- OQPAZKMGCWPERI-GUBZILKMSA-N Arg-Ser-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O OQPAZKMGCWPERI-GUBZILKMSA-N 0.000 description 2
- UVTGNSWSRSCPLP-UHFFFAOYSA-N Arg-Tyr Natural products NC(CCNC(=N)N)C(=O)NC(Cc1ccc(O)cc1)C(=O)O UVTGNSWSRSCPLP-UHFFFAOYSA-N 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- PFOYSEIHFVKHNF-FXQIFTODSA-N Asn-Ala-Arg Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O PFOYSEIHFVKHNF-FXQIFTODSA-N 0.000 description 2
- SLKLLQWZQHXYSV-CIUDSAMLSA-N Asn-Ala-Lys Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O SLKLLQWZQHXYSV-CIUDSAMLSA-N 0.000 description 2
- NPZJLGMWMDNQDD-GHCJXIJMSA-N Asn-Ser-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O NPZJLGMWMDNQDD-GHCJXIJMSA-N 0.000 description 2
- BFOYULZBKYOKAN-OLHMAJIHSA-N Asp-Asp-Thr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O BFOYULZBKYOKAN-OLHMAJIHSA-N 0.000 description 2
- OMMIEVATLAGRCK-BYPYZUCNSA-N Asp-Gly-Gly Chemical compound OC(=O)C[C@H](N)C(=O)NCC(=O)NCC(O)=O OMMIEVATLAGRCK-BYPYZUCNSA-N 0.000 description 2
- QCVXMEHGFUMKCO-YUMQZZPRSA-N Asp-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(O)=O QCVXMEHGFUMKCO-YUMQZZPRSA-N 0.000 description 2
- HOBNTSHITVVNBN-ZPFDUUQYSA-N Asp-Ile-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)O)NC(=O)[C@H](CC(=O)O)N HOBNTSHITVVNBN-ZPFDUUQYSA-N 0.000 description 2
- QPDUWAUSSWGJSB-NGZCFLSTSA-N Asp-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(=O)O)N QPDUWAUSSWGJSB-NGZCFLSTSA-N 0.000 description 2
- PLMKQQMDOMTZGG-UHFFFAOYSA-N Astrantiagenin E-methylester Natural products CC12CCC(O)C(C)(CO)C1CCC1(C)C2CC=C2C3CC(C)(C)CCC3(C(=O)OC)CCC21C PLMKQQMDOMTZGG-UHFFFAOYSA-N 0.000 description 2
- 244000075850 Avena orientalis Species 0.000 description 2
- 235000007319 Avena orientalis Nutrition 0.000 description 2
- 240000002791 Brassica napus Species 0.000 description 2
- 235000011299 Brassica oleracea var botrytis Nutrition 0.000 description 2
- 240000003259 Brassica oleracea var. botrytis Species 0.000 description 2
- 241001301148 Brassica rapa subsp. oleifera Species 0.000 description 2
- 241000222120 Candida <Saccharomycetales> Species 0.000 description 2
- 241000186216 Corynebacterium Species 0.000 description 2
- JDHMXPSXWMPYQZ-AAEUAGOBSA-N Cys-Gly-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CS)N JDHMXPSXWMPYQZ-AAEUAGOBSA-N 0.000 description 2
- SRBFZHDQGSBBOR-IOVATXLUSA-N D-xylopyranose Chemical group O[C@@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-IOVATXLUSA-N 0.000 description 2
- 241001057636 Dracaena deremensis Species 0.000 description 2
- CANAPGLEBDTCAF-NTIPNFSCSA-N Dulcoside A Chemical compound O[C@@H]1[C@H](O)[C@@H](O)[C@H](C)O[C@H]1O[C@H]1[C@H](O[C@]23C(C[C@]4(C2)[C@H]([C@@]2(C)[C@@H]([C@](CCC2)(C)C(=O)O[C@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)CC4)CC3)=C)O[C@H](CO)[C@@H](O)[C@@H]1O CANAPGLEBDTCAF-NTIPNFSCSA-N 0.000 description 2
- CANAPGLEBDTCAF-QHSHOEHESA-N Dulcoside A Natural products C[C@@H]1O[C@H](O[C@@H]2[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]2O[C@]34CC[C@H]5[C@]6(C)CCC[C@](C)([C@H]6CC[C@@]5(CC3=C)C4)C(=O)O[C@@H]7O[C@H](CO)[C@@H](O)[C@H](O)[C@H]7O)[C@H](O)[C@H](O)[C@H]1O CANAPGLEBDTCAF-QHSHOEHESA-N 0.000 description 2
- 241000701533 Escherichia virus T4 Species 0.000 description 2
- 108010092526 GKPV peptide Proteins 0.000 description 2
- ZQPOVSJFBBETHQ-CIUDSAMLSA-N Gln-Glu-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O ZQPOVSJFBBETHQ-CIUDSAMLSA-N 0.000 description 2
- KDXKFBSNIJYNNR-YVNDNENWSA-N Gln-Glu-Ile Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O KDXKFBSNIJYNNR-YVNDNENWSA-N 0.000 description 2
- ZBKUIQNCRIYVGH-SDDRHHMPSA-N Gln-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)N)N ZBKUIQNCRIYVGH-SDDRHHMPSA-N 0.000 description 2
- FNAJNWPDTIXYJN-CIUDSAMLSA-N Gln-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCC(N)=O FNAJNWPDTIXYJN-CIUDSAMLSA-N 0.000 description 2
- ZZLDMBMFKZFQMU-NRPADANISA-N Gln-Val-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O ZZLDMBMFKZFQMU-NRPADANISA-N 0.000 description 2
- VYOILACOFPPNQH-UMNHJUIQSA-N Gln-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)N)N VYOILACOFPPNQH-UMNHJUIQSA-N 0.000 description 2
- WOMUDRVDJMHTCV-DCAQKATOSA-N Glu-Arg-Arg Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O WOMUDRVDJMHTCV-DCAQKATOSA-N 0.000 description 2
- VTTSANCGJWLPNC-ZPFDUUQYSA-N Glu-Arg-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O VTTSANCGJWLPNC-ZPFDUUQYSA-N 0.000 description 2
- GCYFUZJHAXJKKE-KKUMJFAQSA-N Glu-Arg-Tyr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O GCYFUZJHAXJKKE-KKUMJFAQSA-N 0.000 description 2
- MUSGDMDGNGXULI-DCAQKATOSA-N Glu-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O MUSGDMDGNGXULI-DCAQKATOSA-N 0.000 description 2
- AIGROOHQXCACHL-WDSKDSINSA-N Glu-Gly-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](C)C(O)=O AIGROOHQXCACHL-WDSKDSINSA-N 0.000 description 2
- OAGVHWYIBZMWLA-YFKPBYRVSA-N Glu-Gly-Gly Chemical compound OC(=O)CC[C@H](N)C(=O)NCC(=O)NCC(O)=O OAGVHWYIBZMWLA-YFKPBYRVSA-N 0.000 description 2
- HILMIYALTUQTRC-XVKPBYJWSA-N Glu-Gly-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O HILMIYALTUQTRC-XVKPBYJWSA-N 0.000 description 2
- HVYWQYLBVXMXSV-GUBZILKMSA-N Glu-Leu-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O HVYWQYLBVXMXSV-GUBZILKMSA-N 0.000 description 2
- JDUKCSSHWNIQQZ-IHRRRGAJSA-N Glu-Phe-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O JDUKCSSHWNIQQZ-IHRRRGAJSA-N 0.000 description 2
- DXVOKNVIKORTHQ-GUBZILKMSA-N Glu-Pro-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O DXVOKNVIKORTHQ-GUBZILKMSA-N 0.000 description 2
- RMWAOBGCZZSJHE-UMNHJUIQSA-N Glu-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)O)N RMWAOBGCZZSJHE-UMNHJUIQSA-N 0.000 description 2
- MFVQGXGQRIXBPK-WDSKDSINSA-N Gly-Ala-Glu Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O MFVQGXGQRIXBPK-WDSKDSINSA-N 0.000 description 2
- UGVQELHRNUDMAA-BYPYZUCNSA-N Gly-Ala-Gly Chemical compound [NH3+]CC(=O)N[C@@H](C)C(=O)NCC([O-])=O UGVQELHRNUDMAA-BYPYZUCNSA-N 0.000 description 2
- KFMBRBPXHVMDFN-UWVGGRQHSA-N Gly-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCNC(N)=N KFMBRBPXHVMDFN-UWVGGRQHSA-N 0.000 description 2
- UHPAZODVFFYEEL-QWRGUYRKSA-N Gly-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)CN UHPAZODVFFYEEL-QWRGUYRKSA-N 0.000 description 2
- OMOZPGCHVWOXHN-BQBZGAKWSA-N Gly-Met-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)CN OMOZPGCHVWOXHN-BQBZGAKWSA-N 0.000 description 2
- WZSHYFGOLPXPLL-RYUDHWBXSA-N Gly-Phe-Glu Chemical compound NCC(=O)N[C@@H](Cc1ccccc1)C(=O)N[C@@H](CCC(O)=O)C(O)=O WZSHYFGOLPXPLL-RYUDHWBXSA-N 0.000 description 2
- SCJJPCQUJYPHRZ-BQBZGAKWSA-N Gly-Pro-Asn Chemical compound NCC(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O SCJJPCQUJYPHRZ-BQBZGAKWSA-N 0.000 description 2
- JSLVAHYTAJJEQH-QWRGUYRKSA-N Gly-Ser-Phe Chemical compound NCC(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 JSLVAHYTAJJEQH-QWRGUYRKSA-N 0.000 description 2
- LCRDMSSAKLTKBU-ZDLURKLDSA-N Gly-Ser-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)CN LCRDMSSAKLTKBU-ZDLURKLDSA-N 0.000 description 2
- YXTFLTJYLIAZQG-FJXKBIBVSA-N Gly-Thr-Arg Chemical compound NCC(=O)N[C@@H]([C@H](O)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N YXTFLTJYLIAZQG-FJXKBIBVSA-N 0.000 description 2
- YGHSQRJSHKYUJY-SCZZXKLOSA-N Gly-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)CN YGHSQRJSHKYUJY-SCZZXKLOSA-N 0.000 description 2
- SBVMXEZQJVUARN-XPUUQOCRSA-N Gly-Val-Ser Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O SBVMXEZQJVUARN-XPUUQOCRSA-N 0.000 description 2
- 235000004341 Gossypium herbaceum Nutrition 0.000 description 2
- 240000002024 Gossypium herbaceum Species 0.000 description 2
- 244000299507 Gossypium hirsutum Species 0.000 description 2
- 235000003222 Helianthus annuus Nutrition 0.000 description 2
- 241000238631 Hexapoda Species 0.000 description 2
- MWAJSVTZZOUOBU-IHRRRGAJSA-N His-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H](N)CC1=CN=CN1 MWAJSVTZZOUOBU-IHRRRGAJSA-N 0.000 description 2
- XWUIHCZETFNRPA-IHPCNDPISA-N His-His-Trp Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)C1=CN=CN1 XWUIHCZETFNRPA-IHPCNDPISA-N 0.000 description 2
- TVMNTHXFRSXZGR-IHRRRGAJSA-N His-Lys-Val Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O TVMNTHXFRSXZGR-IHRRRGAJSA-N 0.000 description 2
- UIRUVUUGUYCMBY-KCTSRDHCSA-N His-Trp-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CC3=CN=CN3)N UIRUVUUGUYCMBY-KCTSRDHCSA-N 0.000 description 2
- JRHFQUPIZOYKQP-KBIXCLLPSA-N Ile-Ala-Glu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(O)=O JRHFQUPIZOYKQP-KBIXCLLPSA-N 0.000 description 2
- WEWCEPOYKANMGZ-MMWGEVLESA-N Ile-Cys-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)N1CCC[C@@H]1C(=O)O)N WEWCEPOYKANMGZ-MMWGEVLESA-N 0.000 description 2
- MTFVYKQRLXYAQN-LAEOZQHASA-N Ile-Glu-Gly Chemical compound [H]N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O MTFVYKQRLXYAQN-LAEOZQHASA-N 0.000 description 2
- USXAYNCLFSUSBA-MGHWNKPDSA-N Ile-Phe-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N USXAYNCLFSUSBA-MGHWNKPDSA-N 0.000 description 2
- IVXJIMGDOYRLQU-XUXIUFHCSA-N Ile-Pro-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O IVXJIMGDOYRLQU-XUXIUFHCSA-N 0.000 description 2
- PELCGFMHLZXWBQ-BJDJZHNGSA-N Ile-Ser-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)O)N PELCGFMHLZXWBQ-BJDJZHNGSA-N 0.000 description 2
- PXKACEXYLPBMAD-JBDRJPRFSA-N Ile-Ser-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)O)N PXKACEXYLPBMAD-JBDRJPRFSA-N 0.000 description 2
- 108091092195 Intron Proteins 0.000 description 2
- PMGDADKJMCOXHX-UHFFFAOYSA-N L-Arginyl-L-glutamin-acetat Natural products NC(=N)NCCCC(N)C(=O)NC(CCC(N)=O)C(O)=O PMGDADKJMCOXHX-UHFFFAOYSA-N 0.000 description 2
- SENJXOPIZNYLHU-UHFFFAOYSA-N L-leucyl-L-arginine Natural products CC(C)CC(N)C(=O)NC(C(O)=O)CCCN=C(N)N SENJXOPIZNYLHU-UHFFFAOYSA-N 0.000 description 2
- UCOCBWDBHCUPQP-DCAQKATOSA-N Leu-Arg-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(O)=O UCOCBWDBHCUPQP-DCAQKATOSA-N 0.000 description 2
- DLCOFDAHNMMQPP-SRVKXCTJSA-N Leu-Asp-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O DLCOFDAHNMMQPP-SRVKXCTJSA-N 0.000 description 2
- DZQMXBALGUHGJT-GUBZILKMSA-N Leu-Glu-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O DZQMXBALGUHGJT-GUBZILKMSA-N 0.000 description 2
- WIDZHJTYKYBLSR-DCAQKATOSA-N Leu-Glu-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O WIDZHJTYKYBLSR-DCAQKATOSA-N 0.000 description 2
- VGPCJSXPPOQPBK-YUMQZZPRSA-N Leu-Gly-Ser Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O VGPCJSXPPOQPBK-YUMQZZPRSA-N 0.000 description 2
- IEWBEPKLKUXQBU-VOAKCMCISA-N Leu-Leu-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O IEWBEPKLKUXQBU-VOAKCMCISA-N 0.000 description 2
- XXXXOVFBXRERQL-ULQDDVLXSA-N Leu-Pro-Phe Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 XXXXOVFBXRERQL-ULQDDVLXSA-N 0.000 description 2
- VUBIPAHVHMZHCM-KKUMJFAQSA-N Leu-Tyr-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CO)C(O)=O)CC1=CC=C(O)C=C1 VUBIPAHVHMZHCM-KKUMJFAQSA-N 0.000 description 2
- 241000209510 Liliopsida Species 0.000 description 2
- VKCPHIOZDWUFSW-ONGXEEELSA-N Lys-Val-Gly Chemical compound OC(=O)CNC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCCN VKCPHIOZDWUFSW-ONGXEEELSA-N 0.000 description 2
- OZVXDDFYCQOPFD-XQQFMLRXSA-N Lys-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCCN)N OZVXDDFYCQOPFD-XQQFMLRXSA-N 0.000 description 2
- 239000004472 Lysine Substances 0.000 description 2
- 240000004658 Medicago sativa Species 0.000 description 2
- UZVWDRPUTHXQAM-FXQIFTODSA-N Met-Asp-Ala Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O UZVWDRPUTHXQAM-FXQIFTODSA-N 0.000 description 2
- TUSOIZOVPJCMFC-FXQIFTODSA-N Met-Asp-Asp Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O TUSOIZOVPJCMFC-FXQIFTODSA-N 0.000 description 2
- 240000007377 Petunia x hybrida Species 0.000 description 2
- XMPUYNHKEPFERE-IHRRRGAJSA-N Phe-Asp-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1 XMPUYNHKEPFERE-IHRRRGAJSA-N 0.000 description 2
- CSYVXYQDIVCQNU-QWRGUYRKSA-N Phe-Asp-Gly Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O CSYVXYQDIVCQNU-QWRGUYRKSA-N 0.000 description 2
- NAOVYENZCWFBDG-BZSNNMDCSA-N Phe-His-His Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CC=1NC=NC=1)C(O)=O)C1=CC=CC=C1 NAOVYENZCWFBDG-BZSNNMDCSA-N 0.000 description 2
- GMWNQSGWWGKTSF-LFSVMHDDSA-N Phe-Thr-Ala Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O GMWNQSGWWGKTSF-LFSVMHDDSA-N 0.000 description 2
- LXVLKXPFIDDHJG-CIUDSAMLSA-N Pro-Glu-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O LXVLKXPFIDDHJG-CIUDSAMLSA-N 0.000 description 2
- GSPPWVHVBBSPSY-FHWLQOOXSA-N Pro-His-Trp Chemical compound OC(=O)[C@H](Cc1c[nH]c2ccccc12)NC(=O)[C@H](Cc1cnc[nH]1)NC(=O)[C@@H]1CCCN1 GSPPWVHVBBSPSY-FHWLQOOXSA-N 0.000 description 2
- VZKBJNBZMZHKRC-XUXIUFHCSA-N Pro-Ile-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O VZKBJNBZMZHKRC-XUXIUFHCSA-N 0.000 description 2
- FMLRRBDLBJLJIK-DCAQKATOSA-N Pro-Leu-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 FMLRRBDLBJLJIK-DCAQKATOSA-N 0.000 description 2
- FXGIMYRVJJEIIM-UWVGGRQHSA-N Pro-Leu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 FXGIMYRVJJEIIM-UWVGGRQHSA-N 0.000 description 2
- BRJGUPWVFXKBQI-XUXIUFHCSA-N Pro-Leu-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O BRJGUPWVFXKBQI-XUXIUFHCSA-N 0.000 description 2
- SUENWIFTSTWUKD-AVGNSLFASA-N Pro-Leu-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O SUENWIFTSTWUKD-AVGNSLFASA-N 0.000 description 2
- WQUURFHRUAZQHU-VGWMRTNUSA-N Pro-Val-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N1CCC[C@H]1C(=O)O)NC(=O)[C@@H]2CCCN2 WQUURFHRUAZQHU-VGWMRTNUSA-N 0.000 description 2
- 108020004511 Recombinant DNA Proteins 0.000 description 2
- 241000607142 Salmonella Species 0.000 description 2
- DSSOYPJWSWFOLK-CIUDSAMLSA-N Ser-Cys-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(C)C)C(O)=O DSSOYPJWSWFOLK-CIUDSAMLSA-N 0.000 description 2
- RNMRYWZYFHHOEV-CIUDSAMLSA-N Ser-Gln-Arg Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O RNMRYWZYFHHOEV-CIUDSAMLSA-N 0.000 description 2
- UAJAYRMZGNQILN-BQBZGAKWSA-N Ser-Gly-Met Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CCSC)C(O)=O UAJAYRMZGNQILN-BQBZGAKWSA-N 0.000 description 2
- XXXAXOWMBOKTRN-XPUUQOCRSA-N Ser-Gly-Val Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O XXXAXOWMBOKTRN-XPUUQOCRSA-N 0.000 description 2
- UPLYXVPQLJVWMM-KKUMJFAQSA-N Ser-Phe-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O UPLYXVPQLJVWMM-KKUMJFAQSA-N 0.000 description 2
- BSXKBOUZDAZXHE-CIUDSAMLSA-N Ser-Pro-Glu Chemical compound [H]N[C@@H](CO)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O BSXKBOUZDAZXHE-CIUDSAMLSA-N 0.000 description 2
- JURQXQBJKUHGJS-UHFFFAOYSA-N Ser-Ser-Ser-Ser Chemical compound OCC(N)C(=O)NC(CO)C(=O)NC(CO)C(=O)NC(CO)C(O)=O JURQXQBJKUHGJS-UHFFFAOYSA-N 0.000 description 2
- SQHKXWODKJDZRC-LKXGYXEUSA-N Ser-Thr-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(O)=O SQHKXWODKJDZRC-LKXGYXEUSA-N 0.000 description 2
- 235000003434 Sesamum indicum Nutrition 0.000 description 2
- 244000040738 Sesamum orientale Species 0.000 description 2
- 244000062793 Sorghum vulgare Species 0.000 description 2
- IGROJMCBGRFRGI-YTLHQDLWSA-N Thr-Ala-Ala Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O IGROJMCBGRFRGI-YTLHQDLWSA-N 0.000 description 2
- TYVAWPFQYFPSBR-BFHQHQDPSA-N Thr-Ala-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(=O)NCC(O)=O TYVAWPFQYFPSBR-BFHQHQDPSA-N 0.000 description 2
- VFEHSAJCWWHDBH-RHYQMDGZSA-N Thr-Arg-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O VFEHSAJCWWHDBH-RHYQMDGZSA-N 0.000 description 2
- AMXMBCAXAZUCFA-RHYQMDGZSA-N Thr-Leu-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O AMXMBCAXAZUCFA-RHYQMDGZSA-N 0.000 description 2
- HOVLHEKTGVIKAP-WDCWCFNPSA-N Thr-Leu-Gln Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O HOVLHEKTGVIKAP-WDCWCFNPSA-N 0.000 description 2
- MUAFDCVOHYAFNG-RCWTZXSCSA-N Thr-Pro-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(O)=O MUAFDCVOHYAFNG-RCWTZXSCSA-N 0.000 description 2
- NDZYTIMDOZMECO-SHGPDSBTSA-N Thr-Thr-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O NDZYTIMDOZMECO-SHGPDSBTSA-N 0.000 description 2
- 241000723873 Tobacco mosaic virus Species 0.000 description 2
- BRBCKMMXKONBAA-KWBADKCTSA-N Trp-Ala-Ala Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O)=CNC2=C1 BRBCKMMXKONBAA-KWBADKCTSA-N 0.000 description 2
- NMCBVGFGWSIGSB-NUTKFTJISA-N Trp-Ala-Leu Chemical compound C[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N NMCBVGFGWSIGSB-NUTKFTJISA-N 0.000 description 2
- SAKLWFSRZTZQAJ-GQGQLFGLSA-N Trp-Ile-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N SAKLWFSRZTZQAJ-GQGQLFGLSA-N 0.000 description 2
- PALLCTDPFINNMM-JQHSSLGASA-N Trp-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CNC3=CC=CC=C32)N PALLCTDPFINNMM-JQHSSLGASA-N 0.000 description 2
- KSCVLGXNQXKUAR-JYJNAYRXSA-N Tyr-Leu-Glu Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O KSCVLGXNQXKUAR-JYJNAYRXSA-N 0.000 description 2
- XJPXTYLVMUZGNW-IHRRRGAJSA-N Tyr-Pro-Asp Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O XJPXTYLVMUZGNW-IHRRRGAJSA-N 0.000 description 2
- 108010064997 VPY tripeptide Proteins 0.000 description 2
- AZSHAZJLOZQYAY-FXQIFTODSA-N Val-Ala-Ser Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O AZSHAZJLOZQYAY-FXQIFTODSA-N 0.000 description 2
- QHDXUYOYTPWCSK-RCOVLWMOSA-N Val-Asp-Gly Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)NCC(=O)O)N QHDXUYOYTPWCSK-RCOVLWMOSA-N 0.000 description 2
- AGKDVLSDNSTLFA-UMNHJUIQSA-N Val-Gln-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N1CCC[C@@H]1C(=O)O)N AGKDVLSDNSTLFA-UMNHJUIQSA-N 0.000 description 2
- VVZDBPBZHLQPPB-XVKPBYJWSA-N Val-Glu-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O VVZDBPBZHLQPPB-XVKPBYJWSA-N 0.000 description 2
- XXROXFHCMVXETG-UWVGGRQHSA-N Val-Gly-Val Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O XXROXFHCMVXETG-UWVGGRQHSA-N 0.000 description 2
- PYPZMFDMCCWNST-NAKRPEOUSA-N Val-Ile-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](C(C)C)N PYPZMFDMCCWNST-NAKRPEOUSA-N 0.000 description 2
- UMPVMAYCLYMYGA-ONGXEEELSA-N Val-Leu-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O UMPVMAYCLYMYGA-ONGXEEELSA-N 0.000 description 2
- AEMPCGRFEZTWIF-IHRRRGAJSA-N Val-Leu-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(O)=O AEMPCGRFEZTWIF-IHRRRGAJSA-N 0.000 description 2
- ZHQWPWQNVRCXAX-XQQFMLRXSA-N Val-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](C(C)C)N ZHQWPWQNVRCXAX-XQQFMLRXSA-N 0.000 description 2
- RQOMPQGUGBILAG-AVGNSLFASA-N Val-Met-Leu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(O)=O RQOMPQGUGBILAG-AVGNSLFASA-N 0.000 description 2
- GQMNEJMFMCJJTD-NHCYSSNCSA-N Val-Pro-Gln Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O GQMNEJMFMCJJTD-NHCYSSNCSA-N 0.000 description 2
- NHXZRXLFOBFMDM-AVGNSLFASA-N Val-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)C(C)C NHXZRXLFOBFMDM-AVGNSLFASA-N 0.000 description 2
- QIVPZSWBBHRNBA-JYJNAYRXSA-N Val-Pro-Phe Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](Cc1ccccc1)C(O)=O QIVPZSWBBHRNBA-JYJNAYRXSA-N 0.000 description 2
- VHIZXDZMTDVFGX-DCAQKATOSA-N Val-Ser-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](C(C)C)N VHIZXDZMTDVFGX-DCAQKATOSA-N 0.000 description 2
- HWNYVQMOLCYHEA-IHRRRGAJSA-N Val-Ser-Tyr Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N HWNYVQMOLCYHEA-IHRRRGAJSA-N 0.000 description 2
- YKZVPMUGEJXEOR-JYJNAYRXSA-N Val-Val-Tyr Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N YKZVPMUGEJXEOR-JYJNAYRXSA-N 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 238000005903 acid hydrolysis reaction Methods 0.000 description 2
- 230000009471 action Effects 0.000 description 2
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- 108010013835 arginine glutamate Proteins 0.000 description 2
- 108010060035 arginylproline Proteins 0.000 description 2
- 150000001720 carbohydrates Chemical group 0.000 description 2
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 2
- 238000012217 deletion Methods 0.000 description 2
- 230000037430 deletion Effects 0.000 description 2
- 238000013461 design Methods 0.000 description 2
- 238000004520 electroporation Methods 0.000 description 2
- 239000003623 enhancer Substances 0.000 description 2
- KWVKUAKMOIEELN-UHFFFAOYSA-N ent-kaur-16-en-19-oic acid Natural products CC1(C)CCCC2(C)C1CCC34CC(=C(C3)C(=O)O)CCC24 KWVKUAKMOIEELN-UHFFFAOYSA-N 0.000 description 2
- 230000007613 environmental effect Effects 0.000 description 2
- 230000007071 enzymatic hydrolysis Effects 0.000 description 2
- 238000006047 enzymatic hydrolysis reaction Methods 0.000 description 2
- 150000002148 esters Chemical class 0.000 description 2
- 238000002474 experimental method Methods 0.000 description 2
- 239000004459 forage Substances 0.000 description 2
- 102000037865 fusion proteins Human genes 0.000 description 2
- 108020001507 fusion proteins Proteins 0.000 description 2
- 230000002068 genetic effect Effects 0.000 description 2
- 229930182478 glucoside Natural products 0.000 description 2
- 108010085059 glutamyl-arginyl-proline Proteins 0.000 description 2
- 108010090037 glycyl-alanyl-isoleucine Proteins 0.000 description 2
- 108010033719 glycyl-histidyl-glycine Proteins 0.000 description 2
- 108010074027 glycyl-seryl-phenylalanine Proteins 0.000 description 2
- 108010010147 glycylglutamine Proteins 0.000 description 2
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 2
- 239000010931 gold Substances 0.000 description 2
- 229910052737 gold Inorganic materials 0.000 description 2
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical compound O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 2
- 108010040030 histidinoalanine Proteins 0.000 description 2
- 108010036413 histidylglycine Proteins 0.000 description 2
- PFOARMALXZGCHY-UHFFFAOYSA-N homoegonol Natural products C1=C(OC)C(OC)=CC=C1C1=CC2=CC(CCCO)=CC(OC)=C2O1 PFOARMALXZGCHY-UHFFFAOYSA-N 0.000 description 2
- 229920001519 homopolymer Polymers 0.000 description 2
- 230000007062 hydrolysis Effects 0.000 description 2
- 238000006460 hydrolysis reaction Methods 0.000 description 2
- 239000012535 impurity Substances 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- 150000002500 ions Chemical class 0.000 description 2
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 2
- 108010000761 leucylarginine Proteins 0.000 description 2
- 108010009298 lysylglutamic acid Proteins 0.000 description 2
- 108010017391 lysylvaline Proteins 0.000 description 2
- 230000014759 maintenance of location Effects 0.000 description 2
- 239000003550 marker Substances 0.000 description 2
- 238000001823 molecular biology technique Methods 0.000 description 2
- 238000005457 optimization Methods 0.000 description 2
- 239000008188 pellet Substances 0.000 description 2
- 108010012581 phenylalanylglutamate Proteins 0.000 description 2
- 108010073101 phenylalanylleucine Proteins 0.000 description 2
- 239000013600 plasmid vector Substances 0.000 description 2
- 108010025826 prolyl-leucyl-arginine Proteins 0.000 description 2
- 108010031719 prolyl-serine Proteins 0.000 description 2
- 235000009566 rice Nutrition 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 238000012163 sequencing technique Methods 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 239000000243 solution Substances 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 238000010561 standard procedure Methods 0.000 description 2
- 239000000725 suspension Substances 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 2
- 230000005030 transcription termination Effects 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 108010078580 tyrosylleucine Proteins 0.000 description 2
- DRSKVOAJKLUMCL-MMUIXFKXSA-N u2n4xkx7hp Chemical compound O([C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)O[C@]12C(=C)C[C@@]3(C1)CC[C@@H]1[C@@](C)(CCC[C@]1([C@@H]3CC2)C)C(O)=O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O DRSKVOAJKLUMCL-MMUIXFKXSA-N 0.000 description 2
- IBIDRSSEHFLGSD-UHFFFAOYSA-N valinyl-arginine Natural products CC(C)C(N)C(=O)NC(C(O)=O)CCCN=C(N)N IBIDRSSEHFLGSD-UHFFFAOYSA-N 0.000 description 2
- 230000003612 virological effect Effects 0.000 description 2
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 1
- JNTMAZFVYNDPLB-PEDHHIEDSA-N (2S,3S)-2-[[[(2S)-1-[(2S,3S)-2-amino-3-methyl-1-oxopentyl]-2-pyrrolidinyl]-oxomethyl]amino]-3-methylpentanoic acid Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JNTMAZFVYNDPLB-PEDHHIEDSA-N 0.000 description 1
- FQVLRGLGWNWPSS-BXBUPLCLSA-N (4r,7s,10s,13s,16r)-16-acetamido-13-(1h-imidazol-5-ylmethyl)-10-methyl-6,9,12,15-tetraoxo-7-propan-2-yl-1,2-dithia-5,8,11,14-tetrazacycloheptadecane-4-carboxamide Chemical compound N1C(=O)[C@@H](NC(C)=O)CSSC[C@@H](C(N)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C)NC(=O)[C@@H]1CC1=CN=CN1 FQVLRGLGWNWPSS-BXBUPLCLSA-N 0.000 description 1
- HNSDLXPSAYFUHK-UHFFFAOYSA-N 1,4-bis(2-ethylhexyl) sulfosuccinate Chemical compound CCCCC(CC)COC(=O)CC(S(O)(=O)=O)C(=O)OCC(CC)CCCC HNSDLXPSAYFUHK-UHFFFAOYSA-N 0.000 description 1
- FGRBYDKOBBBPOI-UHFFFAOYSA-N 10,10-dioxo-2-[4-(N-phenylanilino)phenyl]thioxanthen-9-one Chemical compound O=C1c2ccccc2S(=O)(=O)c2ccc(cc12)-c1ccc(cc1)N(c1ccccc1)c1ccccc1 FGRBYDKOBBBPOI-UHFFFAOYSA-N 0.000 description 1
- 238000001644 13C nuclear magnetic resonance spectroscopy Methods 0.000 description 1
- 238000005160 1H NMR spectroscopy Methods 0.000 description 1
- VGONTNSXDCQUGY-RRKCRQDMSA-N 2'-deoxyinosine Chemical group C1[C@H](O)[C@@H](CO)O[C@H]1N1C(N=CNC2=O)=C2N=C1 VGONTNSXDCQUGY-RRKCRQDMSA-N 0.000 description 1
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 1
- WEZDRVHTDXTVLT-GJZGRUSLSA-N 2-[[(2s)-2-[[(2s)-2-[(2-aminoacetyl)amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]acetic acid Chemical compound OC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@@H](NC(=O)CN)CC1=CC=CC=C1 WEZDRVHTDXTVLT-GJZGRUSLSA-N 0.000 description 1
- DQVAZKGVGKHQDS-UHFFFAOYSA-N 2-[[1-[2-[(2-amino-4-methylpentanoyl)amino]-4-methylpentanoyl]pyrrolidine-2-carbonyl]amino]-4-methylpentanoic acid Chemical compound CC(C)CC(N)C(=O)NC(CC(C)C)C(=O)N1CCCC1C(=O)NC(CC(C)C)C(O)=O DQVAZKGVGKHQDS-UHFFFAOYSA-N 0.000 description 1
- 238000005084 2D-nuclear magnetic resonance Methods 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- 101710163881 5,6-dihydroxyindole-2-carboxylic acid oxidase Proteins 0.000 description 1
- WBZFUFAFFUEMEI-UHFFFAOYSA-M Acesulfame k Chemical compound [K+].CC1=CC(=O)[N-]S(=O)(=O)O1 WBZFUFAFFUEMEI-UHFFFAOYSA-M 0.000 description 1
- 241000589291 Acinetobacter Species 0.000 description 1
- 241000186361 Actinobacteria <class> Species 0.000 description 1
- 241000589156 Agrobacterium rhizogenes Species 0.000 description 1
- HHGYNJRJIINWAK-FXQIFTODSA-N Ala-Ala-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N HHGYNJRJIINWAK-FXQIFTODSA-N 0.000 description 1
- AAQGRPOPTAUUBM-ZLUOBGJFSA-N Ala-Ala-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(O)=O AAQGRPOPTAUUBM-ZLUOBGJFSA-N 0.000 description 1
- YLTKNGYYPIWKHZ-ACZMJKKPSA-N Ala-Ala-Glu Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(O)=O YLTKNGYYPIWKHZ-ACZMJKKPSA-N 0.000 description 1
- RLMISHABBKUNFO-WHFBIAKZSA-N Ala-Ala-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O RLMISHABBKUNFO-WHFBIAKZSA-N 0.000 description 1
- IMMKUCQIKKXKNP-DCAQKATOSA-N Ala-Arg-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](C)N)CCCN=C(N)N IMMKUCQIKKXKNP-DCAQKATOSA-N 0.000 description 1
- WYPUMLRSQMKIJU-BPNCWPANSA-N Ala-Arg-Tyr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O WYPUMLRSQMKIJU-BPNCWPANSA-N 0.000 description 1
- NHCPCLJZRSIDHS-ZLUOBGJFSA-N Ala-Asp-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O NHCPCLJZRSIDHS-ZLUOBGJFSA-N 0.000 description 1
- WQVYAWIMAWTGMW-ZLUOBGJFSA-N Ala-Asp-Cys Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CS)C(=O)O)N WQVYAWIMAWTGMW-ZLUOBGJFSA-N 0.000 description 1
- WDIYWDJLXOCGRW-ACZMJKKPSA-N Ala-Asp-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O WDIYWDJLXOCGRW-ACZMJKKPSA-N 0.000 description 1
- LSLIRHLIUDVNBN-CIUDSAMLSA-N Ala-Asp-Lys Chemical compound C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCCCN LSLIRHLIUDVNBN-CIUDSAMLSA-N 0.000 description 1
- CXQODNIBUNQWAS-CIUDSAMLSA-N Ala-Gln-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N CXQODNIBUNQWAS-CIUDSAMLSA-N 0.000 description 1
- FUSPCLTUKXQREV-ACZMJKKPSA-N Ala-Glu-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O FUSPCLTUKXQREV-ACZMJKKPSA-N 0.000 description 1
- WKOBSJOZRJJVRZ-FXQIFTODSA-N Ala-Glu-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O WKOBSJOZRJJVRZ-FXQIFTODSA-N 0.000 description 1
- PAIHPOGPJVUFJY-WDSKDSINSA-N Ala-Glu-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O PAIHPOGPJVUFJY-WDSKDSINSA-N 0.000 description 1
- GGNHBHYDMUDXQB-KBIXCLLPSA-N Ala-Glu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)N GGNHBHYDMUDXQB-KBIXCLLPSA-N 0.000 description 1
- HXNNRBHASOSVPG-GUBZILKMSA-N Ala-Glu-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O HXNNRBHASOSVPG-GUBZILKMSA-N 0.000 description 1
- WGDNWOMKBUXFHR-BQBZGAKWSA-N Ala-Gly-Arg Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N WGDNWOMKBUXFHR-BQBZGAKWSA-N 0.000 description 1
- QHASENCZLDHBGX-ONGXEEELSA-N Ala-Gly-Phe Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 QHASENCZLDHBGX-ONGXEEELSA-N 0.000 description 1
- NIZKGBJVCMRDKO-KWQFWETISA-N Ala-Gly-Tyr Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 NIZKGBJVCMRDKO-KWQFWETISA-N 0.000 description 1
- OKEWAFFWMHBGPT-XPUUQOCRSA-N Ala-His-Gly Chemical compound OC(=O)CNC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CN=CN1 OKEWAFFWMHBGPT-XPUUQOCRSA-N 0.000 description 1
- HJGZVLLLBJLXFC-LSJOCFKGSA-N Ala-His-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C(C)C)C(O)=O HJGZVLLLBJLXFC-LSJOCFKGSA-N 0.000 description 1
- CKLDHDOIYBVUNP-KBIXCLLPSA-N Ala-Ile-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(O)=O CKLDHDOIYBVUNP-KBIXCLLPSA-N 0.000 description 1
- DVJSJDDYCYSMFR-ZKWXMUAHSA-N Ala-Ile-Gly Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(O)=O DVJSJDDYCYSMFR-ZKWXMUAHSA-N 0.000 description 1
- SUMYEVXWCAYLLJ-GUBZILKMSA-N Ala-Leu-Gln Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O SUMYEVXWCAYLLJ-GUBZILKMSA-N 0.000 description 1
- UWIQWPWWZUHBAO-ZLIFDBKOSA-N Ala-Leu-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](NC(=O)[C@H](C)N)CC(C)C)C(O)=O)=CNC2=C1 UWIQWPWWZUHBAO-ZLIFDBKOSA-N 0.000 description 1
- VCSABYLVNWQYQE-SRVKXCTJSA-N Ala-Lys-Lys Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CCCCN)C(O)=O VCSABYLVNWQYQE-SRVKXCTJSA-N 0.000 description 1
- BLTRAARCJYVJKV-QEJZJMRPSA-N Ala-Lys-Phe Chemical compound C[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](Cc1ccccc1)C(O)=O BLTRAARCJYVJKV-QEJZJMRPSA-N 0.000 description 1
- RAAWHFXHAACDFT-FXQIFTODSA-N Ala-Met-Asn Chemical compound CSCC[C@H](NC(=O)[C@H](C)N)C(=O)N[C@@H](CC(N)=O)C(O)=O RAAWHFXHAACDFT-FXQIFTODSA-N 0.000 description 1
- DWYROCSXOOMOEU-CIUDSAMLSA-N Ala-Met-Glu Chemical compound C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N DWYROCSXOOMOEU-CIUDSAMLSA-N 0.000 description 1
- OMDNCNKNEGFOMM-BQBZGAKWSA-N Ala-Met-Gly Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCSC)C(=O)NCC(O)=O OMDNCNKNEGFOMM-BQBZGAKWSA-N 0.000 description 1
- XSTZMVAYYCJTNR-DCAQKATOSA-N Ala-Met-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(O)=O XSTZMVAYYCJTNR-DCAQKATOSA-N 0.000 description 1
- DEWWPUNXRNGMQN-LPEHRKFASA-N Ala-Met-Pro Chemical compound C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N1CCC[C@@H]1C(=O)O)N DEWWPUNXRNGMQN-LPEHRKFASA-N 0.000 description 1
- DHBKYZYFEXXUAK-ONGXEEELSA-N Ala-Phe-Gly Chemical compound OC(=O)CNC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=CC=C1 DHBKYZYFEXXUAK-ONGXEEELSA-N 0.000 description 1
- ZBLQIYPCUWZSRZ-QEJZJMRPSA-N Ala-Phe-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](C)N)CC1=CC=CC=C1 ZBLQIYPCUWZSRZ-QEJZJMRPSA-N 0.000 description 1
- IPZQNYYAYVRKKK-FXQIFTODSA-N Ala-Pro-Ala Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O IPZQNYYAYVRKKK-FXQIFTODSA-N 0.000 description 1
- DCVYRWFAMZFSDA-ZLUOBGJFSA-N Ala-Ser-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O DCVYRWFAMZFSDA-ZLUOBGJFSA-N 0.000 description 1
- AUFACLFHBAGZEN-ZLUOBGJFSA-N Ala-Ser-Cys Chemical compound N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O AUFACLFHBAGZEN-ZLUOBGJFSA-N 0.000 description 1
- MSWSRLGNLKHDEI-ACZMJKKPSA-N Ala-Ser-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O MSWSRLGNLKHDEI-ACZMJKKPSA-N 0.000 description 1
- HIIJOGIBQXHFKE-HHKYUTTNSA-N Ala-Thr-Ala-Pro Chemical compound C[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O HIIJOGIBQXHFKE-HHKYUTTNSA-N 0.000 description 1
- HCBKAOZYACJUEF-XQXXSGGOSA-N Ala-Thr-Gln Chemical compound N[C@@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CCC(N)=O)C(=O)O HCBKAOZYACJUEF-XQXXSGGOSA-N 0.000 description 1
- ISCYZXFOCXWUJU-KZVJFYERSA-N Ala-Thr-Met Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCSC)C(O)=O ISCYZXFOCXWUJU-KZVJFYERSA-N 0.000 description 1
- QRIYOHQJRDHFKF-UWJYBYFXSA-N Ala-Tyr-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=C(O)C=C1 QRIYOHQJRDHFKF-UWJYBYFXSA-N 0.000 description 1
- BVLPIIBTWIYOML-ZKWXMUAHSA-N Ala-Val-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O BVLPIIBTWIYOML-ZKWXMUAHSA-N 0.000 description 1
- LYILPUNCKACNGF-NAKRPEOUSA-N Ala-Val-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C)N LYILPUNCKACNGF-NAKRPEOUSA-N 0.000 description 1
- OMSKGWFGWCQFBD-KZVJFYERSA-N Ala-Val-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OMSKGWFGWCQFBD-KZVJFYERSA-N 0.000 description 1
- 102100034035 Alcohol dehydrogenase 1A Human genes 0.000 description 1
- 102100036826 Aldehyde oxidase Human genes 0.000 description 1
- 108020005544 Antisense RNA Proteins 0.000 description 1
- 240000007087 Apium graveolens Species 0.000 description 1
- 235000015849 Apium graveolens Dulce Group Nutrition 0.000 description 1
- 235000010591 Appio Nutrition 0.000 description 1
- DBKNLHKEVPZVQC-LPEHRKFASA-N Arg-Ala-Pro Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@@H]1C(O)=O DBKNLHKEVPZVQC-LPEHRKFASA-N 0.000 description 1
- OTOXOKCIIQLMFH-KZVJFYERSA-N Arg-Ala-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCN=C(N)N OTOXOKCIIQLMFH-KZVJFYERSA-N 0.000 description 1
- IASNWHAGGYTEKX-IUCAKERBSA-N Arg-Arg-Gly Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)NCC(O)=O IASNWHAGGYTEKX-IUCAKERBSA-N 0.000 description 1
- BHSYMWWMVRPCPA-CYDGBPFRSA-N Arg-Arg-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H](N)CCCN=C(N)N BHSYMWWMVRPCPA-CYDGBPFRSA-N 0.000 description 1
- JSLGXODUIAFWCF-WDSKDSINSA-N Arg-Asn Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(O)=O JSLGXODUIAFWCF-WDSKDSINSA-N 0.000 description 1
- NONSEUUPKITYQT-BQBZGAKWSA-N Arg-Asn-Gly Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)NCC(=O)O)N)CN=C(N)N NONSEUUPKITYQT-BQBZGAKWSA-N 0.000 description 1
- BVBKBQRPOJFCQM-DCAQKATOSA-N Arg-Asn-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O BVBKBQRPOJFCQM-DCAQKATOSA-N 0.000 description 1
- OZNSCVPYWZRQPY-CIUDSAMLSA-N Arg-Asp-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O OZNSCVPYWZRQPY-CIUDSAMLSA-N 0.000 description 1
- NAARDJBSSPUGCF-FXQIFTODSA-N Arg-Cys-Asn Chemical compound C(C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)CN=C(N)N NAARDJBSSPUGCF-FXQIFTODSA-N 0.000 description 1
- XTGGTAWGUFXJSV-NAKRPEOUSA-N Arg-Cys-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CCCN=C(N)N)N XTGGTAWGUFXJSV-NAKRPEOUSA-N 0.000 description 1
- PBSOQGZLPFVXPU-YUMQZZPRSA-N Arg-Glu-Gly Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O PBSOQGZLPFVXPU-YUMQZZPRSA-N 0.000 description 1
- NKBQZKVMKJJDLX-SRVKXCTJSA-N Arg-Glu-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O NKBQZKVMKJJDLX-SRVKXCTJSA-N 0.000 description 1
- HPSVTWMFWCHKFN-GARJFASQSA-N Arg-Glu-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CCCN=C(N)N)N)C(=O)O HPSVTWMFWCHKFN-GARJFASQSA-N 0.000 description 1
- HQIZDMIGUJOSNI-IUCAKERBSA-N Arg-Gly-Arg Chemical compound N[C@@H](CCCNC(N)=N)C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(O)=O HQIZDMIGUJOSNI-IUCAKERBSA-N 0.000 description 1
- AUFHLLPVPSMEOG-YUMQZZPRSA-N Arg-Gly-Glu Chemical compound NC(N)=NCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O AUFHLLPVPSMEOG-YUMQZZPRSA-N 0.000 description 1
- NKNILFJYKKHBKE-WPRPVWTQSA-N Arg-Gly-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O NKNILFJYKKHBKE-WPRPVWTQSA-N 0.000 description 1
- NVCIXQYNWYTLDO-IHRRRGAJSA-N Arg-His-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CCCN=C(N)N)N NVCIXQYNWYTLDO-IHRRRGAJSA-N 0.000 description 1
- OOIMKQRCPJBGPD-XUXIUFHCSA-N Arg-Ile-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O OOIMKQRCPJBGPD-XUXIUFHCSA-N 0.000 description 1
- LLUGJARLJCGLAR-CYDGBPFRSA-N Arg-Ile-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N LLUGJARLJCGLAR-CYDGBPFRSA-N 0.000 description 1
- LVMUGODRNHFGRA-AVGNSLFASA-N Arg-Leu-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O LVMUGODRNHFGRA-AVGNSLFASA-N 0.000 description 1
- OTZMRMHZCMZOJZ-SRVKXCTJSA-N Arg-Leu-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O OTZMRMHZCMZOJZ-SRVKXCTJSA-N 0.000 description 1
- WMEVEPXNCMKNGH-IHRRRGAJSA-N Arg-Leu-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N WMEVEPXNCMKNGH-IHRRRGAJSA-N 0.000 description 1
- NMRHDSAOIURTNT-RWMBFGLXSA-N Arg-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N NMRHDSAOIURTNT-RWMBFGLXSA-N 0.000 description 1
- JEOCWTUOMKEEMF-RHYQMDGZSA-N Arg-Leu-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JEOCWTUOMKEEMF-RHYQMDGZSA-N 0.000 description 1
- YVTHEZNOKSAWRW-DCAQKATOSA-N Arg-Lys-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O YVTHEZNOKSAWRW-DCAQKATOSA-N 0.000 description 1
- CLICCYPMVFGUOF-IHRRRGAJSA-N Arg-Lys-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O CLICCYPMVFGUOF-IHRRRGAJSA-N 0.000 description 1
- NPAVRDPEFVKELR-DCAQKATOSA-N Arg-Lys-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(O)=O NPAVRDPEFVKELR-DCAQKATOSA-N 0.000 description 1
- ZRNWJUAQKFUUKV-SRVKXCTJSA-N Arg-Met-Met Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCSC)C(O)=O ZRNWJUAQKFUUKV-SRVKXCTJSA-N 0.000 description 1
- DPLFNLDACGGBAK-KKUMJFAQSA-N Arg-Phe-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N DPLFNLDACGGBAK-KKUMJFAQSA-N 0.000 description 1
- BSYKSCBTTQKOJG-GUBZILKMSA-N Arg-Pro-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O BSYKSCBTTQKOJG-GUBZILKMSA-N 0.000 description 1
- HGKHPCFTRQDHCU-IUCAKERBSA-N Arg-Pro-Gly Chemical compound NC(N)=NCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O HGKHPCFTRQDHCU-IUCAKERBSA-N 0.000 description 1
- STHNZYKCJHWULY-AVGNSLFASA-N Arg-Pro-His Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CCCN=C(N)N)N)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O STHNZYKCJHWULY-AVGNSLFASA-N 0.000 description 1
- AWMAZIIEFPFHCP-RCWTZXSCSA-N Arg-Pro-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O AWMAZIIEFPFHCP-RCWTZXSCSA-N 0.000 description 1
- AMIQZQAAYGYKOP-FXQIFTODSA-N Arg-Ser-Asn Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O AMIQZQAAYGYKOP-FXQIFTODSA-N 0.000 description 1
- YHZQOSXDTFRZKU-WDSOQIARSA-N Arg-Trp-Leu Chemical compound C1=CC=C2C(C[C@@H](C(=O)N[C@@H](CC(C)C)C(O)=O)NC(=O)[C@@H](N)CCCN=C(N)N)=CNC2=C1 YHZQOSXDTFRZKU-WDSOQIARSA-N 0.000 description 1
- PJOPLXOCKACMLK-KKUMJFAQSA-N Arg-Tyr-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O PJOPLXOCKACMLK-KKUMJFAQSA-N 0.000 description 1
- CGWVCWFQGXOUSJ-ULQDDVLXSA-N Arg-Tyr-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(O)=O CGWVCWFQGXOUSJ-ULQDDVLXSA-N 0.000 description 1
- KEZVOBAKAXHMOF-GUBZILKMSA-N Arg-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCN=C(N)N KEZVOBAKAXHMOF-GUBZILKMSA-N 0.000 description 1
- PSUXEQYPYZLNER-QXEWZRGKSA-N Arg-Val-Asn Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O PSUXEQYPYZLNER-QXEWZRGKSA-N 0.000 description 1
- XEOXPCNONWHHSW-AVGNSLFASA-N Arg-Val-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N XEOXPCNONWHHSW-AVGNSLFASA-N 0.000 description 1
- WTUZDHWWGUQEKN-SRVKXCTJSA-N Arg-Val-Met Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCSC)C(O)=O WTUZDHWWGUQEKN-SRVKXCTJSA-N 0.000 description 1
- SUMJNGAMIQSNGX-TUAOUCFPSA-N Arg-Val-Pro Chemical compound CC(C)[C@H](NC(=O)[C@@H](N)CCCNC(N)=N)C(=O)N1CCC[C@@H]1C(O)=O SUMJNGAMIQSNGX-TUAOUCFPSA-N 0.000 description 1
- ANAHQDPQQBDOBM-UHFFFAOYSA-N Arg-Val-Tyr Natural products CC(C)C(NC(=O)C(N)CCNC(=N)N)C(=O)NC(Cc1ccc(O)cc1)C(=O)O ANAHQDPQQBDOBM-UHFFFAOYSA-N 0.000 description 1
- 241000722808 Arthrobotrys Species 0.000 description 1
- 235000016425 Arthrospira platensis Nutrition 0.000 description 1
- 240000002900 Arthrospira platensis Species 0.000 description 1
- XYOVHPDDWCEUDY-CIUDSAMLSA-N Asn-Ala-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O XYOVHPDDWCEUDY-CIUDSAMLSA-N 0.000 description 1
- MEFGKQUUYZOLHM-GMOBBJLQSA-N Asn-Arg-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O MEFGKQUUYZOLHM-GMOBBJLQSA-N 0.000 description 1
- KSBHCUSPLWRVEK-ZLUOBGJFSA-N Asn-Asn-Asp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O KSBHCUSPLWRVEK-ZLUOBGJFSA-N 0.000 description 1
- XSGBIBGAMKTHMY-WHFBIAKZSA-N Asn-Asp-Gly Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O XSGBIBGAMKTHMY-WHFBIAKZSA-N 0.000 description 1
- JQSWHKKUZMTOIH-QWRGUYRKSA-N Asn-Gly-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CC(=O)N)N JQSWHKKUZMTOIH-QWRGUYRKSA-N 0.000 description 1
- OOWSBIOUKIUWLO-RCOVLWMOSA-N Asn-Gly-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O OOWSBIOUKIUWLO-RCOVLWMOSA-N 0.000 description 1
- SPCONPVIDFMDJI-QSFUFRPTSA-N Asn-Ile-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O SPCONPVIDFMDJI-QSFUFRPTSA-N 0.000 description 1
- PNHQRQTVBRDIEF-CIUDSAMLSA-N Asn-Leu-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC(=O)N)N PNHQRQTVBRDIEF-CIUDSAMLSA-N 0.000 description 1
- ZMUQQMGITUJQTI-CIUDSAMLSA-N Asn-Leu-Asn Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O ZMUQQMGITUJQTI-CIUDSAMLSA-N 0.000 description 1
- UBGGJTMETLEXJD-DCAQKATOSA-N Asn-Leu-Met Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(O)=O UBGGJTMETLEXJD-DCAQKATOSA-N 0.000 description 1
- NCFJQJRLQJEECD-NHCYSSNCSA-N Asn-Leu-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O NCFJQJRLQJEECD-NHCYSSNCSA-N 0.000 description 1
- NLDNNZKUSLAYFW-NHCYSSNCSA-N Asn-Lys-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O NLDNNZKUSLAYFW-NHCYSSNCSA-N 0.000 description 1
- RTFWCVDISAMGEQ-SRVKXCTJSA-N Asn-Phe-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N RTFWCVDISAMGEQ-SRVKXCTJSA-N 0.000 description 1
- BKZFBJYIVSBXCO-KKUMJFAQSA-N Asn-Phe-His Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CNC=N1)C(O)=O BKZFBJYIVSBXCO-KKUMJFAQSA-N 0.000 description 1
- BKFXFUPYETWGGA-XVSYOHENSA-N Asn-Phe-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O BKFXFUPYETWGGA-XVSYOHENSA-N 0.000 description 1
- SNYCNNPOFYBCEK-ZLUOBGJFSA-N Asn-Ser-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O SNYCNNPOFYBCEK-ZLUOBGJFSA-N 0.000 description 1
- TZQWZQSMHDVLQL-QEJZJMRPSA-N Asn-Trp-Gln Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N TZQWZQSMHDVLQL-QEJZJMRPSA-N 0.000 description 1
- KRXIWXCXOARFNT-ZLUOBGJFSA-N Asp-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC(O)=O KRXIWXCXOARFNT-ZLUOBGJFSA-N 0.000 description 1
- WSWYMRLTJVKRCE-ZLUOBGJFSA-N Asp-Ala-Asp Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(O)=O WSWYMRLTJVKRCE-ZLUOBGJFSA-N 0.000 description 1
- VTYQAQFKMQTKQD-ACZMJKKPSA-N Asp-Ala-Gln Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(O)=O VTYQAQFKMQTKQD-ACZMJKKPSA-N 0.000 description 1
- XEDQMTWEYFBOIK-ACZMJKKPSA-N Asp-Ala-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O XEDQMTWEYFBOIK-ACZMJKKPSA-N 0.000 description 1
- XPGVTUBABLRGHY-BIIVOSGPSA-N Asp-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(=O)O)N XPGVTUBABLRGHY-BIIVOSGPSA-N 0.000 description 1
- RGKKALNPOYURGE-ZKWXMUAHSA-N Asp-Ala-Val Chemical compound N[C@@H](CC(=O)O)C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)O RGKKALNPOYURGE-ZKWXMUAHSA-N 0.000 description 1
- NYLBGYLHBDFRHL-VEVYYDQMSA-N Asp-Arg-Thr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NYLBGYLHBDFRHL-VEVYYDQMSA-N 0.000 description 1
- XYBJLTKSGFBLCS-QXEWZRGKSA-N Asp-Arg-Val Chemical compound NC(N)=NCCC[C@@H](C(=O)N[C@@H](C(C)C)C(O)=O)NC(=O)[C@@H](N)CC(O)=O XYBJLTKSGFBLCS-QXEWZRGKSA-N 0.000 description 1
- YNQIDCRRTWGHJD-ZLUOBGJFSA-N Asp-Asn-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CC(O)=O YNQIDCRRTWGHJD-ZLUOBGJFSA-N 0.000 description 1
- QRULNKJGYQQZMW-ZLUOBGJFSA-N Asp-Asn-Asp Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O QRULNKJGYQQZMW-ZLUOBGJFSA-N 0.000 description 1
- ZELQAFZSJOBEQS-ACZMJKKPSA-N Asp-Asn-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O ZELQAFZSJOBEQS-ACZMJKKPSA-N 0.000 description 1
- WCFCYFDBMNFSPA-ACZMJKKPSA-N Asp-Asp-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCC(O)=O WCFCYFDBMNFSPA-ACZMJKKPSA-N 0.000 description 1
- CELPEWWLSXMVPH-CIUDSAMLSA-N Asp-Asp-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC(O)=O CELPEWWLSXMVPH-CIUDSAMLSA-N 0.000 description 1
- WEDGJJRCJNHYSF-SRVKXCTJSA-N Asp-Cys-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC(=O)O)N WEDGJJRCJNHYSF-SRVKXCTJSA-N 0.000 description 1
- DZQKLNLLWFQONU-LKXGYXEUSA-N Asp-Cys-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC(=O)O)N)O DZQKLNLLWFQONU-LKXGYXEUSA-N 0.000 description 1
- GHODABZPVZMWCE-FXQIFTODSA-N Asp-Glu-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O GHODABZPVZMWCE-FXQIFTODSA-N 0.000 description 1
- LTXGDRFJRZSZAV-CIUDSAMLSA-N Asp-Glu-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CC(=O)O)N LTXGDRFJRZSZAV-CIUDSAMLSA-N 0.000 description 1
- DTNUIAJCPRMNBT-WHFBIAKZSA-N Asp-Gly-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](C)C(O)=O DTNUIAJCPRMNBT-WHFBIAKZSA-N 0.000 description 1
- JUWZKMBALYLZCK-WHFBIAKZSA-N Asp-Gly-Asn Chemical compound OC(=O)C[C@H](N)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O JUWZKMBALYLZCK-WHFBIAKZSA-N 0.000 description 1
- WBDWQKRLTVCDSY-WHFBIAKZSA-N Asp-Gly-Asp Chemical compound OC(=O)C[C@H](N)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(O)=O WBDWQKRLTVCDSY-WHFBIAKZSA-N 0.000 description 1
- YRBGRUOSJROZEI-NHCYSSNCSA-N Asp-His-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C(C)C)C(O)=O YRBGRUOSJROZEI-NHCYSSNCSA-N 0.000 description 1
- YFSLJHLQOALGSY-ZPFDUUQYSA-N Asp-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)O)N YFSLJHLQOALGSY-ZPFDUUQYSA-N 0.000 description 1
- PYXXJFRXIYAESU-PCBIJLKTSA-N Asp-Ile-Phe Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O PYXXJFRXIYAESU-PCBIJLKTSA-N 0.000 description 1
- RTXQQDVBACBSCW-CFMVVWHZSA-N Asp-Ile-Tyr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O RTXQQDVBACBSCW-CFMVVWHZSA-N 0.000 description 1
- JNNVNVRBYUJYGS-CIUDSAMLSA-N Asp-Leu-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O JNNVNVRBYUJYGS-CIUDSAMLSA-N 0.000 description 1
- RQHLMGCXCZUOGT-ZPFDUUQYSA-N Asp-Leu-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O RQHLMGCXCZUOGT-ZPFDUUQYSA-N 0.000 description 1
- UJGRZQYSNYTCAX-SRVKXCTJSA-N Asp-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC(O)=O UJGRZQYSNYTCAX-SRVKXCTJSA-N 0.000 description 1
- QNMKWNONJGKJJC-NHCYSSNCSA-N Asp-Leu-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O QNMKWNONJGKJJC-NHCYSSNCSA-N 0.000 description 1
- YWLDTBBUHZJQHW-KKUMJFAQSA-N Asp-Lys-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC(=O)O)N YWLDTBBUHZJQHW-KKUMJFAQSA-N 0.000 description 1
- RXBGWGRSWXOBGK-KKUMJFAQSA-N Asp-Lys-Tyr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O RXBGWGRSWXOBGK-KKUMJFAQSA-N 0.000 description 1
- IOXWDLNHXZOXQP-FXQIFTODSA-N Asp-Met-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC(=O)O)N IOXWDLNHXZOXQP-FXQIFTODSA-N 0.000 description 1
- GWIJZUVQVDJHDI-AVGNSLFASA-N Asp-Phe-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O GWIJZUVQVDJHDI-AVGNSLFASA-N 0.000 description 1
- JSHWXQIZOCVWIA-ZKWXMUAHSA-N Asp-Ser-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O JSHWXQIZOCVWIA-ZKWXMUAHSA-N 0.000 description 1
- GWWSUMLEWKQHLR-NUMRIWBASA-N Asp-Thr-Glu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CC(=O)O)N)O GWWSUMLEWKQHLR-NUMRIWBASA-N 0.000 description 1
- NAAAPCLFJPURAM-HJGDQZAQSA-N Asp-Thr-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)O)N)O NAAAPCLFJPURAM-HJGDQZAQSA-N 0.000 description 1
- GXHDGYOXPNQCKM-XVSYOHENSA-N Asp-Thr-Phe Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CC(=O)O)N)O GXHDGYOXPNQCKM-XVSYOHENSA-N 0.000 description 1
- KACWACLNYLSVCA-VHWLVUOQSA-N Asp-Trp-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O KACWACLNYLSVCA-VHWLVUOQSA-N 0.000 description 1
- YUELDQUPTAYEGM-XIRDDKMYSA-N Asp-Trp-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CC(=O)O)N YUELDQUPTAYEGM-XIRDDKMYSA-N 0.000 description 1
- XWKBWZXGNXTDKY-ZKWXMUAHSA-N Asp-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC(O)=O XWKBWZXGNXTDKY-ZKWXMUAHSA-N 0.000 description 1
- XMKXONRMGJXCJV-LAEOZQHASA-N Asp-Val-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O XMKXONRMGJXCJV-LAEOZQHASA-N 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 108010011485 Aspartame Proteins 0.000 description 1
- 241000228212 Aspergillus Species 0.000 description 1
- 241000228245 Aspergillus niger Species 0.000 description 1
- 241000193830 Bacillus <bacterium> Species 0.000 description 1
- 241000219310 Beta vulgaris subsp. vulgaris Species 0.000 description 1
- 239000002028 Biomass Substances 0.000 description 1
- 241000743774 Brachypodium Species 0.000 description 1
- 235000011331 Brassica Nutrition 0.000 description 1
- 241000219198 Brassica Species 0.000 description 1
- 244000060924 Brassica campestris Species 0.000 description 1
- 235000011293 Brassica napus Nutrition 0.000 description 1
- 240000007124 Brassica oleracea Species 0.000 description 1
- 235000003899 Brassica oleracea var acephala Nutrition 0.000 description 1
- 235000011301 Brassica oleracea var capitata Nutrition 0.000 description 1
- 235000017647 Brassica oleracea var italica Nutrition 0.000 description 1
- 235000001169 Brassica oleracea var oleracea Nutrition 0.000 description 1
- 235000004977 Brassica sinapistrum Nutrition 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- UXVMQQNJUSDDNG-UHFFFAOYSA-L Calcium chloride Chemical compound [Cl-].[Cl-].[Ca+2] UXVMQQNJUSDDNG-UHFFFAOYSA-L 0.000 description 1
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 1
- 108020004638 Circular DNA Proteins 0.000 description 1
- 241000193403 Clostridium Species 0.000 description 1
- 241000193401 Clostridium acetobutylicum Species 0.000 description 1
- 108700010070 Codon Usage Proteins 0.000 description 1
- 241000218631 Coniferophyta Species 0.000 description 1
- 229920000742 Cotton Polymers 0.000 description 1
- 235000011309 Crambe hispanica subsp abyssinica Nutrition 0.000 description 1
- 241000220247 Crambe hispanica subsp. abyssinica Species 0.000 description 1
- 244000241257 Cucumis melo Species 0.000 description 1
- 235000015510 Cucumis melo subsp melo Nutrition 0.000 description 1
- AMRLSQGGERHDHJ-FXQIFTODSA-N Cys-Ala-Arg Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O AMRLSQGGERHDHJ-FXQIFTODSA-N 0.000 description 1
- GRNOCLDFUNCIDW-ACZMJKKPSA-N Cys-Ala-Glu Chemical compound C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CS)N GRNOCLDFUNCIDW-ACZMJKKPSA-N 0.000 description 1
- DCJNIJAWIRPPBB-CIUDSAMLSA-N Cys-Ala-Lys Chemical compound C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CS)N DCJNIJAWIRPPBB-CIUDSAMLSA-N 0.000 description 1
- BGIRVSMUAJMGOK-FXQIFTODSA-N Cys-Ala-Met Chemical compound C[C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CS)N BGIRVSMUAJMGOK-FXQIFTODSA-N 0.000 description 1
- PRXCTTWKGJAPMT-ZLUOBGJFSA-N Cys-Ala-Ser Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O PRXCTTWKGJAPMT-ZLUOBGJFSA-N 0.000 description 1
- UPJGYXRAPJWIHD-CIUDSAMLSA-N Cys-Asn-Leu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O UPJGYXRAPJWIHD-CIUDSAMLSA-N 0.000 description 1
- WDQXKVCQXRNOSI-GHCJXIJMSA-N Cys-Asp-Ile Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WDQXKVCQXRNOSI-GHCJXIJMSA-N 0.000 description 1
- MBILEVLLOHJZMG-FXQIFTODSA-N Cys-Gln-Glu Chemical compound C(CC(=O)N)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CS)N MBILEVLLOHJZMG-FXQIFTODSA-N 0.000 description 1
- GCDLPNRHPWBKJJ-WDSKDSINSA-N Cys-Gly-Glu Chemical compound [H]N[C@@H](CS)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O GCDLPNRHPWBKJJ-WDSKDSINSA-N 0.000 description 1
- UXIYYUMGFNSGBK-XPUUQOCRSA-N Cys-Gly-Val Chemical compound [H]N[C@@H](CS)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O UXIYYUMGFNSGBK-XPUUQOCRSA-N 0.000 description 1
- KPENUVBHAKRDQR-GUBZILKMSA-N Cys-His-Glu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(O)=O KPENUVBHAKRDQR-GUBZILKMSA-N 0.000 description 1
- CIVXDCMSSFGWAL-YUMQZZPRSA-N Cys-Lys-Gly Chemical compound C(CCN)C[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CS)N CIVXDCMSSFGWAL-YUMQZZPRSA-N 0.000 description 1
- XBELMDARIGXDKY-GUBZILKMSA-N Cys-Pro-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CS)N XBELMDARIGXDKY-GUBZILKMSA-N 0.000 description 1
- NDNZRWUDUMTITL-FXQIFTODSA-N Cys-Ser-Val Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O NDNZRWUDUMTITL-FXQIFTODSA-N 0.000 description 1
- SAEVTQWAYDPXMU-KATARQTJSA-N Cys-Thr-Leu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O SAEVTQWAYDPXMU-KATARQTJSA-N 0.000 description 1
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Chemical group CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 1
- 108010066133 D-octopine dehydrogenase Proteins 0.000 description 1
- 108010090461 DFG peptide Proteins 0.000 description 1
- 102000012410 DNA Ligases Human genes 0.000 description 1
- 108010061982 DNA Ligases Proteins 0.000 description 1
- 102000004594 DNA Polymerase I Human genes 0.000 description 1
- 108010017826 DNA Polymerase I Proteins 0.000 description 1
- 239000003155 DNA primer Substances 0.000 description 1
- 230000004568 DNA-binding Effects 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 235000002767 Daucus carota Nutrition 0.000 description 1
- 244000000626 Daucus carota Species 0.000 description 1
- 241000235035 Debaryomyces Species 0.000 description 1
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 1
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 241000195634 Dunaliella Species 0.000 description 1
- 241001198387 Escherichia coli BL21(DE3) Species 0.000 description 1
- 108060002716 Exonuclease Proteins 0.000 description 1
- 240000009088 Fragaria x ananassa Species 0.000 description 1
- 101150094690 GAL1 gene Proteins 0.000 description 1
- 101150038242 GAL10 gene Proteins 0.000 description 1
- 102100028501 Galanin peptides Human genes 0.000 description 1
- 102100024637 Galectin-10 Human genes 0.000 description 1
- 101000892220 Geobacillus thermodenitrificans (strain NG80-2) Long-chain-alcohol dehydrogenase 1 Proteins 0.000 description 1
- YNNXQZDEOCYJJL-CIUDSAMLSA-N Gln-Arg-Asp Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CCC(=O)N)N)CN=C(N)N YNNXQZDEOCYJJL-CIUDSAMLSA-N 0.000 description 1
- MWLYSLMKFXWZPW-ZPFDUUQYSA-N Gln-Arg-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H](N)CCC(N)=O MWLYSLMKFXWZPW-ZPFDUUQYSA-N 0.000 description 1
- JESJDAAGXULQOP-CIUDSAMLSA-N Gln-Arg-Ser Chemical compound C(C[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CCC(=O)N)N)CN=C(N)N JESJDAAGXULQOP-CIUDSAMLSA-N 0.000 description 1
- INFBPLSHYFALDE-ACZMJKKPSA-N Gln-Asn-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(O)=O INFBPLSHYFALDE-ACZMJKKPSA-N 0.000 description 1
- QYTKAVBFRUGYAU-ACZMJKKPSA-N Gln-Asp-Asn Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O QYTKAVBFRUGYAU-ACZMJKKPSA-N 0.000 description 1
- COYGBRTZEVWZBW-XKBZYTNZSA-N Gln-Cys-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CS)NC(=O)[C@@H](N)CCC(N)=O COYGBRTZEVWZBW-XKBZYTNZSA-N 0.000 description 1
- CITDWMLWXNUQKD-FXQIFTODSA-N Gln-Gln-Asn Chemical compound C(CC(=O)N)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC(=O)N)C(=O)O)N CITDWMLWXNUQKD-FXQIFTODSA-N 0.000 description 1
- KVXVVDFOZNYYKZ-DCAQKATOSA-N Gln-Gln-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O KVXVVDFOZNYYKZ-DCAQKATOSA-N 0.000 description 1
- MCAVASRGVBVPMX-FXQIFTODSA-N Gln-Glu-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O MCAVASRGVBVPMX-FXQIFTODSA-N 0.000 description 1
- KCJJFESQRXGTGC-BQBZGAKWSA-N Gln-Glu-Gly Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O KCJJFESQRXGTGC-BQBZGAKWSA-N 0.000 description 1
- PNENQZWRFMUZOM-DCAQKATOSA-N Gln-Glu-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O PNENQZWRFMUZOM-DCAQKATOSA-N 0.000 description 1
- LVSYIKGMLRHKME-IUCAKERBSA-N Gln-Gly-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CCC(=O)N)N LVSYIKGMLRHKME-IUCAKERBSA-N 0.000 description 1
- KHGGWBRVRPHFMH-PEFMBERDSA-N Gln-Ile-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCC(=O)N)N KHGGWBRVRPHFMH-PEFMBERDSA-N 0.000 description 1
- ITZWDGBYBPUZRG-KBIXCLLPSA-N Gln-Ile-Ser Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O ITZWDGBYBPUZRG-KBIXCLLPSA-N 0.000 description 1
- JKGHMESJHRTHIC-SIUGBPQLSA-N Gln-Ile-Tyr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)NC(=O)[C@H](CCC(=O)N)N JKGHMESJHRTHIC-SIUGBPQLSA-N 0.000 description 1
- SHAUZYVSXAMYAZ-JYJNAYRXSA-N Gln-Leu-Phe Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCC(=O)N)N SHAUZYVSXAMYAZ-JYJNAYRXSA-N 0.000 description 1
- YPMDZWPZFOZYFG-GUBZILKMSA-N Gln-Leu-Ser Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O YPMDZWPZFOZYFG-GUBZILKMSA-N 0.000 description 1
- QMVCEWKHIUHTSD-GUBZILKMSA-N Gln-Met-Glu Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CCC(=O)N)N QMVCEWKHIUHTSD-GUBZILKMSA-N 0.000 description 1
- SFAFZYYMAWOCIC-KKUMJFAQSA-N Gln-Phe-Arg Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](CCC(=O)N)N SFAFZYYMAWOCIC-KKUMJFAQSA-N 0.000 description 1
- PBYFVIQRFLNQCO-GUBZILKMSA-N Gln-Pro-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O PBYFVIQRFLNQCO-GUBZILKMSA-N 0.000 description 1
- XQDGOJPVMSWZSO-SRVKXCTJSA-N Gln-Pro-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CCC(=O)N)N XQDGOJPVMSWZSO-SRVKXCTJSA-N 0.000 description 1
- WLRYGVYQFXRJDA-DCAQKATOSA-N Gln-Pro-Pro Chemical compound NC(=O)CC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 WLRYGVYQFXRJDA-DCAQKATOSA-N 0.000 description 1
- MKRDNSWGJWTBKZ-GVXVVHGQSA-N Gln-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)N)N MKRDNSWGJWTBKZ-GVXVVHGQSA-N 0.000 description 1
- RUFHOVYUYSNDNY-ACZMJKKPSA-N Glu-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(O)=O RUFHOVYUYSNDNY-ACZMJKKPSA-N 0.000 description 1
- CGYDXNKRIMJMLV-GUBZILKMSA-N Glu-Arg-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=O CGYDXNKRIMJMLV-GUBZILKMSA-N 0.000 description 1
- NLKVNZUFDPWPNL-YUMQZZPRSA-N Glu-Arg-Gly Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O NLKVNZUFDPWPNL-YUMQZZPRSA-N 0.000 description 1
- PBEQPAZRHDVJQI-SRVKXCTJSA-N Glu-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCC(=O)O)N PBEQPAZRHDVJQI-SRVKXCTJSA-N 0.000 description 1
- VPKBCVUDBNINAH-GARJFASQSA-N Glu-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCC(=O)O)N)C(=O)O VPKBCVUDBNINAH-GARJFASQSA-N 0.000 description 1
- SRZLHYPAOXBBSB-HJGDQZAQSA-N Glu-Arg-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SRZLHYPAOXBBSB-HJGDQZAQSA-N 0.000 description 1
- DYFJZDDQPNIPAB-NHCYSSNCSA-N Glu-Arg-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(O)=O DYFJZDDQPNIPAB-NHCYSSNCSA-N 0.000 description 1
- DSPQRJXOIXHOHK-WDSKDSINSA-N Glu-Asp-Gly Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O DSPQRJXOIXHOHK-WDSKDSINSA-N 0.000 description 1
- CKOFNWCLWRYUHK-XHNCKOQMSA-N Glu-Asp-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCC(=O)O)N)C(=O)O CKOFNWCLWRYUHK-XHNCKOQMSA-N 0.000 description 1
- OXEMJGCAJFFREE-FXQIFTODSA-N Glu-Gln-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O OXEMJGCAJFFREE-FXQIFTODSA-N 0.000 description 1
- GYCPQVFKCPPRQB-GUBZILKMSA-N Glu-Gln-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCC(=O)O)N GYCPQVFKCPPRQB-GUBZILKMSA-N 0.000 description 1
- VFZIDQZAEBORGY-GLLZPBPUSA-N Glu-Gln-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VFZIDQZAEBORGY-GLLZPBPUSA-N 0.000 description 1
- HTTSBEBKVNEDFE-AUTRQRHGSA-N Glu-Gln-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCC(=O)O)N HTTSBEBKVNEDFE-AUTRQRHGSA-N 0.000 description 1
- ILGFBUGLBSAQQB-GUBZILKMSA-N Glu-Glu-Arg Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O ILGFBUGLBSAQQB-GUBZILKMSA-N 0.000 description 1
- NKLRYVLERDYDBI-FXQIFTODSA-N Glu-Glu-Asp Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O NKLRYVLERDYDBI-FXQIFTODSA-N 0.000 description 1
- SJPMNHCEWPTRBR-BQBZGAKWSA-N Glu-Glu-Gly Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O SJPMNHCEWPTRBR-BQBZGAKWSA-N 0.000 description 1
- APHGWLWMOXGZRL-DCAQKATOSA-N Glu-Glu-His Chemical compound N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](Cc1cnc[nH]1)C(O)=O APHGWLWMOXGZRL-DCAQKATOSA-N 0.000 description 1
- AUTNXSQEVVHSJK-YVNDNENWSA-N Glu-Glu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O AUTNXSQEVVHSJK-YVNDNENWSA-N 0.000 description 1
- LGYZYFFDELZWRS-DCAQKATOSA-N Glu-Glu-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O LGYZYFFDELZWRS-DCAQKATOSA-N 0.000 description 1
- BUAKRRKDHSSIKK-IHRRRGAJSA-N Glu-Glu-Tyr Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 BUAKRRKDHSSIKK-IHRRRGAJSA-N 0.000 description 1
- QJCKNLPMTPXXEM-AUTRQRHGSA-N Glu-Glu-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O QJCKNLPMTPXXEM-AUTRQRHGSA-N 0.000 description 1
- PXXGVUVQWQGGIG-YUMQZZPRSA-N Glu-Gly-Arg Chemical compound OC(=O)CC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N PXXGVUVQWQGGIG-YUMQZZPRSA-N 0.000 description 1
- VGUYMZGLJUJRBV-YVNDNENWSA-N Glu-Ile-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(O)=O VGUYMZGLJUJRBV-YVNDNENWSA-N 0.000 description 1
- ZCOJVESMNGBGLF-GRLWGSQLSA-N Glu-Ile-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O ZCOJVESMNGBGLF-GRLWGSQLSA-N 0.000 description 1
- QXDXIXFSFHUYAX-MNXVOIDGSA-N Glu-Ile-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCC(O)=O QXDXIXFSFHUYAX-MNXVOIDGSA-N 0.000 description 1
- XTZDZAXYPDISRR-MNXVOIDGSA-N Glu-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N XTZDZAXYPDISRR-MNXVOIDGSA-N 0.000 description 1
- ZHNHJYYFCGUZNQ-KBIXCLLPSA-N Glu-Ile-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCC(O)=O ZHNHJYYFCGUZNQ-KBIXCLLPSA-N 0.000 description 1
- INGJLBQKTRJLFO-UKJIMTQDSA-N Glu-Ile-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCC(O)=O INGJLBQKTRJLFO-UKJIMTQDSA-N 0.000 description 1
- VGBSZQSKQRMLHD-MNXVOIDGSA-N Glu-Leu-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O VGBSZQSKQRMLHD-MNXVOIDGSA-N 0.000 description 1
- OQXDUSZKISQQSS-GUBZILKMSA-N Glu-Lys-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O OQXDUSZKISQQSS-GUBZILKMSA-N 0.000 description 1
- OCJRHJZKGGSPRW-IUCAKERBSA-N Glu-Lys-Gly Chemical compound NCCCC[C@@H](C(=O)NCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O OCJRHJZKGGSPRW-IUCAKERBSA-N 0.000 description 1
- SOEPMWQCTJITPZ-SRVKXCTJSA-N Glu-Met-Lys Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N SOEPMWQCTJITPZ-SRVKXCTJSA-N 0.000 description 1
- XEKAJTCACGEBOK-KKUMJFAQSA-N Glu-Met-Phe Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCC(=O)O)N XEKAJTCACGEBOK-KKUMJFAQSA-N 0.000 description 1
- JZJGEKDPWVJOLD-QEWYBTABSA-N Glu-Phe-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JZJGEKDPWVJOLD-QEWYBTABSA-N 0.000 description 1
- QNJNPKSWAHPYGI-JYJNAYRXSA-N Glu-Phe-Leu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=CC=C1 QNJNPKSWAHPYGI-JYJNAYRXSA-N 0.000 description 1
- UDEPRBFQTWGLCW-CIUDSAMLSA-N Glu-Pro-Asp Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O UDEPRBFQTWGLCW-CIUDSAMLSA-N 0.000 description 1
- DCBSZJJHOTXMHY-DCAQKATOSA-N Glu-Pro-Pro Chemical compound OC(=O)CC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 DCBSZJJHOTXMHY-DCAQKATOSA-N 0.000 description 1
- ZAPFAWQHBOHWLL-GUBZILKMSA-N Glu-Ser-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(=O)O)N ZAPFAWQHBOHWLL-GUBZILKMSA-N 0.000 description 1
- IDEODOAVGCMUQV-GUBZILKMSA-N Glu-Ser-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O IDEODOAVGCMUQV-GUBZILKMSA-N 0.000 description 1
- QOXDAWODGSIDDI-GUBZILKMSA-N Glu-Ser-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(=O)O)N QOXDAWODGSIDDI-GUBZILKMSA-N 0.000 description 1
- QCMVGXDELYMZET-GLLZPBPUSA-N Glu-Thr-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O QCMVGXDELYMZET-GLLZPBPUSA-N 0.000 description 1
- CQGBSALYGOXQPE-HTUGSXCWSA-N Glu-Thr-Phe Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)O CQGBSALYGOXQPE-HTUGSXCWSA-N 0.000 description 1
- DLISPGXMKZTWQG-IFFSRLJSSA-N Glu-Thr-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O DLISPGXMKZTWQG-IFFSRLJSSA-N 0.000 description 1
- UUTGYDAKPISJAO-JYJNAYRXSA-N Glu-Tyr-Leu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=C(O)C=C1 UUTGYDAKPISJAO-JYJNAYRXSA-N 0.000 description 1
- VIPDPMHGICREIS-GVXVVHGQSA-N Glu-Val-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O VIPDPMHGICREIS-GVXVVHGQSA-N 0.000 description 1
- PYTZFYUXZZHOAD-WHFBIAKZSA-N Gly-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)CN PYTZFYUXZZHOAD-WHFBIAKZSA-N 0.000 description 1
- YMUFWNJHVPQNQD-ZKWXMUAHSA-N Gly-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)CN YMUFWNJHVPQNQD-ZKWXMUAHSA-N 0.000 description 1
- UPOJUWHGMDJUQZ-IUCAKERBSA-N Gly-Arg-Arg Chemical compound NC(=N)NCCC[C@H](NC(=O)CN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O UPOJUWHGMDJUQZ-IUCAKERBSA-N 0.000 description 1
- KKBWDNZXYLGJEY-UHFFFAOYSA-N Gly-Arg-Pro Natural products NCC(=O)NC(CCNC(=N)N)C(=O)N1CCCC1C(=O)O KKBWDNZXYLGJEY-UHFFFAOYSA-N 0.000 description 1
- AIJAPFVDBFYNKN-WHFBIAKZSA-N Gly-Asn-Asp Chemical compound C([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)CN)C(=O)N AIJAPFVDBFYNKN-WHFBIAKZSA-N 0.000 description 1
- LURCIJSJAKFCRO-QWRGUYRKSA-N Gly-Asn-Tyr Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O LURCIJSJAKFCRO-QWRGUYRKSA-N 0.000 description 1
- MHHUEAIBJZWDBH-YUMQZZPRSA-N Gly-Asp-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)CN MHHUEAIBJZWDBH-YUMQZZPRSA-N 0.000 description 1
- YYQGVXNKAXUTJU-YUMQZZPRSA-N Gly-Cys-His Chemical compound NCC(=O)N[C@@H](CS)C(=O)N[C@@H](Cc1cnc[nH]1)C(O)=O YYQGVXNKAXUTJU-YUMQZZPRSA-N 0.000 description 1
- BYYNJRSNDARRBX-YFKPBYRVSA-N Gly-Gln-Gly Chemical compound NCC(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O BYYNJRSNDARRBX-YFKPBYRVSA-N 0.000 description 1
- MOJKRXIRAZPZLW-WDSKDSINSA-N Gly-Glu-Ala Chemical compound [H]NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O MOJKRXIRAZPZLW-WDSKDSINSA-N 0.000 description 1
- CUYLIWAAAYJKJH-RYUDHWBXSA-N Gly-Glu-Tyr Chemical compound NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 CUYLIWAAAYJKJH-RYUDHWBXSA-N 0.000 description 1
- CCQOOWAONKGYKQ-BYPYZUCNSA-N Gly-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)CN CCQOOWAONKGYKQ-BYPYZUCNSA-N 0.000 description 1
- QITBQGJOXQYMOA-ZETCQYMHSA-N Gly-Gly-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)CNC(=O)CN QITBQGJOXQYMOA-ZETCQYMHSA-N 0.000 description 1
- TVDHVLGFJSHPAX-UWVGGRQHSA-N Gly-His-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CN=CN1 TVDHVLGFJSHPAX-UWVGGRQHSA-N 0.000 description 1
- UTYGDAHJBBDPBA-BYULHYEWSA-N Gly-Ile-Asp Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)CN UTYGDAHJBBDPBA-BYULHYEWSA-N 0.000 description 1
- UESJMAMHDLEHGM-NHCYSSNCSA-N Gly-Ile-Leu Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O UESJMAMHDLEHGM-NHCYSSNCSA-N 0.000 description 1
- IUZGUFAJDBHQQV-YUMQZZPRSA-N Gly-Leu-Asn Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O IUZGUFAJDBHQQV-YUMQZZPRSA-N 0.000 description 1
- YTSVAIMKVLZUDU-YUMQZZPRSA-N Gly-Leu-Asp Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O YTSVAIMKVLZUDU-YUMQZZPRSA-N 0.000 description 1
- ULZCYBYDTUMHNF-IUCAKERBSA-N Gly-Leu-Glu Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O ULZCYBYDTUMHNF-IUCAKERBSA-N 0.000 description 1
- NNCSJUBVFBDDLC-YUMQZZPRSA-N Gly-Leu-Ser Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O NNCSJUBVFBDDLC-YUMQZZPRSA-N 0.000 description 1
- AFWYPMDMDYCKMD-KBPBESRZSA-N Gly-Leu-Tyr Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 AFWYPMDMDYCKMD-KBPBESRZSA-N 0.000 description 1
- BXICSAQLIHFDDL-YUMQZZPRSA-N Gly-Lys-Asn Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(O)=O BXICSAQLIHFDDL-YUMQZZPRSA-N 0.000 description 1
- PCPOYRCAHPJXII-UWVGGRQHSA-N Gly-Lys-Met Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCSC)C(O)=O PCPOYRCAHPJXII-UWVGGRQHSA-N 0.000 description 1
- BBTCXWTXOXUNFX-IUCAKERBSA-N Gly-Met-Arg Chemical compound CSCC[C@H](NC(=O)CN)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O BBTCXWTXOXUNFX-IUCAKERBSA-N 0.000 description 1
- FXLVSYVJDPCIHH-STQMWFEESA-N Gly-Phe-Arg Chemical compound [H]NCC(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O FXLVSYVJDPCIHH-STQMWFEESA-N 0.000 description 1
- YYXJFBMCOUSYSF-RYUDHWBXSA-N Gly-Phe-Gln Chemical compound [H]NCC(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O YYXJFBMCOUSYSF-RYUDHWBXSA-N 0.000 description 1
- IBYOLNARKHMLBG-WHOFXGATSA-N Gly-Phe-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CC=CC=C1 IBYOLNARKHMLBG-WHOFXGATSA-N 0.000 description 1
- GGLIDLCEPDHEJO-BQBZGAKWSA-N Gly-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)CN GGLIDLCEPDHEJO-BQBZGAKWSA-N 0.000 description 1
- LLWQVJNHMYBLLK-CDMKHQONSA-N Gly-Thr-Phe Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O LLWQVJNHMYBLLK-CDMKHQONSA-N 0.000 description 1
- TVTZEOHWHUVYCG-KYNKHSRBSA-N Gly-Thr-Thr Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O TVTZEOHWHUVYCG-KYNKHSRBSA-N 0.000 description 1
- JKSMZVCGQWVTBW-STQMWFEESA-N Gly-Trp-Asn Chemical compound [H]NCC(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC(N)=O)C(O)=O JKSMZVCGQWVTBW-STQMWFEESA-N 0.000 description 1
- NIOPEYHPOBWLQO-KBPBESRZSA-N Gly-Trp-Glu Chemical compound NCC(=O)N[C@@H](Cc1c[nH]c2ccccc12)C(=O)N[C@@H](CCC(O)=O)C(O)=O NIOPEYHPOBWLQO-KBPBESRZSA-N 0.000 description 1
- PYFIQROSWQERAS-LBPRGKRZSA-N Gly-Trp-Gly Chemical compound C1=CC=C2C(C[C@H](NC(=O)CN)C(=O)NCC(O)=O)=CNC2=C1 PYFIQROSWQERAS-LBPRGKRZSA-N 0.000 description 1
- MREVELMMFOLESM-HOCLYGCPSA-N Gly-Trp-Val Chemical compound [H]NCC(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](C(C)C)C(O)=O MREVELMMFOLESM-HOCLYGCPSA-N 0.000 description 1
- GWCJMBNBFYBQCV-XPUUQOCRSA-N Gly-Val-Ala Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O GWCJMBNBFYBQCV-XPUUQOCRSA-N 0.000 description 1
- DKJWUIYLMLUBDX-XPUUQOCRSA-N Gly-Val-Cys Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CS)C(=O)O DKJWUIYLMLUBDX-XPUUQOCRSA-N 0.000 description 1
- BAYQNCWLXIDLHX-ONGXEEELSA-N Gly-Val-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)CN BAYQNCWLXIDLHX-ONGXEEELSA-N 0.000 description 1
- 102100031181 Glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 description 1
- 235000009432 Gossypium hirsutum Nutrition 0.000 description 1
- RVKIPWVMZANZLI-UHFFFAOYSA-N H-Lys-Trp-OH Natural products C1=CC=C2C(CC(NC(=O)C(N)CCCCN)C(O)=O)=CNC2=C1 RVKIPWVMZANZLI-UHFFFAOYSA-N 0.000 description 1
- 239000007995 HEPES buffer Substances 0.000 description 1
- 101150009006 HIS3 gene Proteins 0.000 description 1
- 101100246753 Halobacterium salinarum (strain ATCC 700922 / JCM 11081 / NRC-1) pyrF gene Proteins 0.000 description 1
- VOEGKUNRHYKYSU-XVYDVKMFSA-N His-Asp-Ala Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O VOEGKUNRHYKYSU-XVYDVKMFSA-N 0.000 description 1
- OHOXVDFVRDGFND-YUMQZZPRSA-N His-Cys-Gly Chemical compound N[C@@H](Cc1cnc[nH]1)C(=O)N[C@@H](CS)C(=O)NCC(O)=O OHOXVDFVRDGFND-YUMQZZPRSA-N 0.000 description 1
- VHHYJBSXXMPQGZ-AVGNSLFASA-N His-Gln-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CC1=CN=CN1)N VHHYJBSXXMPQGZ-AVGNSLFASA-N 0.000 description 1
- VTMLJMNQHKBPON-QWRGUYRKSA-N His-Gly-His Chemical compound C([C@H](N)C(=O)NCC(=O)N[C@@H](CC=1NC=NC=1)C(O)=O)C1=CN=CN1 VTMLJMNQHKBPON-QWRGUYRKSA-N 0.000 description 1
- NQKRILCJYCASDV-QWRGUYRKSA-N His-Gly-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC1=CN=CN1 NQKRILCJYCASDV-QWRGUYRKSA-N 0.000 description 1
- VTZYMXGGXOFBMX-DJFWLOJKSA-N His-Ile-Asp Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O VTZYMXGGXOFBMX-DJFWLOJKSA-N 0.000 description 1
- DYKZGTLPSNOFHU-DEQVHRJGSA-N His-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CN=CN2)N DYKZGTLPSNOFHU-DEQVHRJGSA-N 0.000 description 1
- TTYKEFZRLKQTHH-MELADBBJSA-N His-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CC2=CN=CN2)N)C(=O)O TTYKEFZRLKQTHH-MELADBBJSA-N 0.000 description 1
- ZFDKSLBEWYCOCS-BZSNNMDCSA-N His-Phe-Lys Chemical compound C([C@@H](C(=O)N[C@@H](CCCCN)C(O)=O)NC(=O)[C@@H](N)CC=1NC=NC=1)C1=CC=CC=C1 ZFDKSLBEWYCOCS-BZSNNMDCSA-N 0.000 description 1
- WCHONUZTYDQMBY-PYJNHQTQSA-N His-Pro-Ile Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WCHONUZTYDQMBY-PYJNHQTQSA-N 0.000 description 1
- ZHHLTWUOWXHVQJ-YUMQZZPRSA-N His-Ser-Gly Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CO)C(=O)NCC(=O)O)N ZHHLTWUOWXHVQJ-YUMQZZPRSA-N 0.000 description 1
- XHQYFGPIRUHQIB-PBCZWWQYSA-N His-Thr-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H]([C@H](O)C)NC(=O)[C@@H](N)CC1=CN=CN1 XHQYFGPIRUHQIB-PBCZWWQYSA-N 0.000 description 1
- NBWATNYAUVSAEQ-ZEILLAHLSA-N His-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CC1=CN=CN1)N)O NBWATNYAUVSAEQ-ZEILLAHLSA-N 0.000 description 1
- MKWFGXSFLYNTKC-XIRDDKMYSA-N His-Trp-Asp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CC3=CN=CN3)N MKWFGXSFLYNTKC-XIRDDKMYSA-N 0.000 description 1
- PDLQNLSEJXOQNQ-IHPCNDPISA-N His-Trp-Lys Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCCCN)C(O)=O)C1=CN=CN1 PDLQNLSEJXOQNQ-IHPCNDPISA-N 0.000 description 1
- KDDKJKKQODQQBR-NHCYSSNCSA-N His-Val-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CC1=CN=CN1)N KDDKJKKQODQQBR-NHCYSSNCSA-N 0.000 description 1
- GBMSSORHVHAYLU-QTKMDUPCSA-N His-Val-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CC1=CN=CN1)N)O GBMSSORHVHAYLU-QTKMDUPCSA-N 0.000 description 1
- 101000780443 Homo sapiens Alcohol dehydrogenase 1A Proteins 0.000 description 1
- 101000928314 Homo sapiens Aldehyde oxidase Proteins 0.000 description 1
- 101100121078 Homo sapiens GAL gene Proteins 0.000 description 1
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 1
- 101000920618 Homo sapiens Transcription and mRNA export factor ENY2 Proteins 0.000 description 1
- 101001046426 Homo sapiens cGMP-dependent protein kinase 1 Proteins 0.000 description 1
- 108700039609 IRW peptide Proteins 0.000 description 1
- JXUGDUWBMKIJDC-NAKRPEOUSA-N Ile-Ala-Arg Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O JXUGDUWBMKIJDC-NAKRPEOUSA-N 0.000 description 1
- LQSBBHNVAVNZSX-GHCJXIJMSA-N Ile-Ala-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](CC(=O)N)C(=O)O)N LQSBBHNVAVNZSX-GHCJXIJMSA-N 0.000 description 1
- YPWHUFAAMNHMGS-QSFUFRPTSA-N Ile-Ala-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N YPWHUFAAMNHMGS-QSFUFRPTSA-N 0.000 description 1
- QICVAHODWHIWIS-HTFCKZLJSA-N Ile-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)N QICVAHODWHIWIS-HTFCKZLJSA-N 0.000 description 1
- CYHYBSGMHMHKOA-CIQUZCHMSA-N Ile-Ala-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N CYHYBSGMHMHKOA-CIQUZCHMSA-N 0.000 description 1
- ASCFJMSGKUIRDU-ZPFDUUQYSA-N Ile-Arg-Gln Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(O)=O ASCFJMSGKUIRDU-ZPFDUUQYSA-N 0.000 description 1
- FVEWRQXNISSYFO-ZPFDUUQYSA-N Ile-Arg-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N FVEWRQXNISSYFO-ZPFDUUQYSA-N 0.000 description 1
- WECYRWOMWSCWNX-XUXIUFHCSA-N Ile-Arg-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CC(C)C)C(O)=O WECYRWOMWSCWNX-XUXIUFHCSA-N 0.000 description 1
- SCHZQZPYHBWYEQ-PEFMBERDSA-N Ile-Asn-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N SCHZQZPYHBWYEQ-PEFMBERDSA-N 0.000 description 1
- UKTUOMWSJPXODT-GUDRVLHUSA-N Ile-Asn-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N1CCC[C@@H]1C(=O)O)N UKTUOMWSJPXODT-GUDRVLHUSA-N 0.000 description 1
- NKRJALPCDNXULF-BYULHYEWSA-N Ile-Asp-Gly Chemical compound [H]N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O NKRJALPCDNXULF-BYULHYEWSA-N 0.000 description 1
- LKACSKJPTFSBHR-MNXVOIDGSA-N Ile-Gln-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N LKACSKJPTFSBHR-MNXVOIDGSA-N 0.000 description 1
- PHIXPNQDGGILMP-YVNDNENWSA-N Ile-Glu-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N PHIXPNQDGGILMP-YVNDNENWSA-N 0.000 description 1
- LPXHYGGZJOCAFR-MNXVOIDGSA-N Ile-Glu-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC(C)C)C(=O)O)N LPXHYGGZJOCAFR-MNXVOIDGSA-N 0.000 description 1
- TVSPLSZTKTUYLV-ZPFDUUQYSA-N Ile-Glu-Met Chemical compound N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCSC)C(=O)O TVSPLSZTKTUYLV-ZPFDUUQYSA-N 0.000 description 1
- NZOCIWKZUVUNDW-ZKWXMUAHSA-N Ile-Gly-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O NZOCIWKZUVUNDW-ZKWXMUAHSA-N 0.000 description 1
- CDGLBYSAZFIIJO-RCOVLWMOSA-N Ile-Gly-Gly Chemical compound CC[C@H](C)[C@H]([NH3+])C(=O)NCC(=O)NCC([O-])=O CDGLBYSAZFIIJO-RCOVLWMOSA-N 0.000 description 1
- PDTMWFVVNZYWTR-NHCYSSNCSA-N Ile-Gly-Lys Chemical compound CC[C@H](C)[C@H](N)C(=O)NCC(=O)N[C@@H](CCCCN)C(O)=O PDTMWFVVNZYWTR-NHCYSSNCSA-N 0.000 description 1
- QZZIBQZLWBOOJH-PEDHHIEDSA-N Ile-Ile-Val Chemical compound N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(=O)O QZZIBQZLWBOOJH-PEDHHIEDSA-N 0.000 description 1
- YGDWPQCLFJNMOL-MNXVOIDGSA-N Ile-Leu-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N YGDWPQCLFJNMOL-MNXVOIDGSA-N 0.000 description 1
- FZWVCYCYWCLQDH-NHCYSSNCSA-N Ile-Leu-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)NCC(=O)O)N FZWVCYCYWCLQDH-NHCYSSNCSA-N 0.000 description 1
- TVYWVSJGSHQWMT-AJNGGQMLSA-N Ile-Leu-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)O)N TVYWVSJGSHQWMT-AJNGGQMLSA-N 0.000 description 1
- GVNNAHIRSDRIII-AJNGGQMLSA-N Ile-Lys-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N GVNNAHIRSDRIII-AJNGGQMLSA-N 0.000 description 1
- GLYJPWIRLBAIJH-FQUUOJAGSA-N Ile-Lys-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N1CCC[C@@H]1C(=O)O)N GLYJPWIRLBAIJH-FQUUOJAGSA-N 0.000 description 1
- GLYJPWIRLBAIJH-UHFFFAOYSA-N Ile-Lys-Pro Natural products CCC(C)C(N)C(=O)NC(CCCCN)C(=O)N1CCCC1C(O)=O GLYJPWIRLBAIJH-UHFFFAOYSA-N 0.000 description 1
- WSSGUVAKYCQSCT-XUXIUFHCSA-N Ile-Met-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(=O)O)N WSSGUVAKYCQSCT-XUXIUFHCSA-N 0.000 description 1
- LRAUKBMYHHNADU-DKIMLUQUSA-N Ile-Phe-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)[C@@H](C)CC)CC1=CC=CC=C1 LRAUKBMYHHNADU-DKIMLUQUSA-N 0.000 description 1
- IITVUURPOYGCTD-NAKRPEOUSA-N Ile-Pro-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O IITVUURPOYGCTD-NAKRPEOUSA-N 0.000 description 1
- AGGIYSLVUKVOPT-HTFCKZLJSA-N Ile-Ser-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)N AGGIYSLVUKVOPT-HTFCKZLJSA-N 0.000 description 1
- YCKPUHHMCFSUMD-IUKAMOBKSA-N Ile-Thr-Asp Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N YCKPUHHMCFSUMD-IUKAMOBKSA-N 0.000 description 1
- NAFIFZNBSPWYOO-RWRJDSDZSA-N Ile-Thr-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N NAFIFZNBSPWYOO-RWRJDSDZSA-N 0.000 description 1
- BZUOLKFQVVBTJY-SLBDDTMCSA-N Ile-Trp-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC(=O)N)C(=O)O)N BZUOLKFQVVBTJY-SLBDDTMCSA-N 0.000 description 1
- DZMWFIRHFFVBHS-ZEWNOJEFSA-N Ile-Tyr-Phe Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)O)N DZMWFIRHFFVBHS-ZEWNOJEFSA-N 0.000 description 1
- BCISUQVFDGYZBO-QSFUFRPTSA-N Ile-Val-Asp Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(O)=O BCISUQVFDGYZBO-QSFUFRPTSA-N 0.000 description 1
- DLEBSGAVWRPTIX-PEDHHIEDSA-N Ile-Val-Ile Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)[C@@H](C)CC DLEBSGAVWRPTIX-PEDHHIEDSA-N 0.000 description 1
- 108010065920 Insulin Lispro Proteins 0.000 description 1
- 241000588748 Klebsiella Species 0.000 description 1
- 241000235649 Kluyveromyces Species 0.000 description 1
- IBMVEYRWAWIOTN-UHFFFAOYSA-N L-Leucyl-L-Arginyl-L-Proline Natural products CC(C)CC(N)C(=O)NC(CCCN=C(N)N)C(=O)N1CCCC1C(O)=O IBMVEYRWAWIOTN-UHFFFAOYSA-N 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- SHZGCJCMOBCMKK-JFNONXLTSA-N L-rhamnopyranose Chemical group C[C@@H]1OC(O)[C@H](O)[C@H](O)[C@H]1O SHZGCJCMOBCMKK-JFNONXLTSA-N 0.000 description 1
- PNNNRSAQSRJVSB-UHFFFAOYSA-N L-rhamnose Chemical group CC(O)C(O)C(O)C(O)C=O PNNNRSAQSRJVSB-UHFFFAOYSA-N 0.000 description 1
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- ZRLUISBDKUWAIZ-CIUDSAMLSA-N Leu-Ala-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC(O)=O ZRLUISBDKUWAIZ-CIUDSAMLSA-N 0.000 description 1
- MJOZZTKJZQFKDK-GUBZILKMSA-N Leu-Ala-Gln Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(N)=O MJOZZTKJZQFKDK-GUBZILKMSA-N 0.000 description 1
- WNGVUZWBXZKQES-YUMQZZPRSA-N Leu-Ala-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O WNGVUZWBXZKQES-YUMQZZPRSA-N 0.000 description 1
- WSGXUIQTEZDVHJ-GARJFASQSA-N Leu-Ala-Pro Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@@H]1C(O)=O WSGXUIQTEZDVHJ-GARJFASQSA-N 0.000 description 1
- YOZCKMXHBYKOMQ-IHRRRGAJSA-N Leu-Arg-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(=O)O)N YOZCKMXHBYKOMQ-IHRRRGAJSA-N 0.000 description 1
- XYUBOFCTGPZFSA-WDSOQIARSA-N Leu-Arg-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H](N)CC(C)C)C(O)=O)=CNC2=C1 XYUBOFCTGPZFSA-WDSOQIARSA-N 0.000 description 1
- OIARJGNVARWKFP-YUMQZZPRSA-N Leu-Asn-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O OIARJGNVARWKFP-YUMQZZPRSA-N 0.000 description 1
- YKNBJXOJTURHCU-DCAQKATOSA-N Leu-Asp-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N YKNBJXOJTURHCU-DCAQKATOSA-N 0.000 description 1
- DLFAACQHIRSQGG-CIUDSAMLSA-N Leu-Asp-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O DLFAACQHIRSQGG-CIUDSAMLSA-N 0.000 description 1
- ZDSNOSQHMJBRQN-SRVKXCTJSA-N Leu-Asp-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N ZDSNOSQHMJBRQN-SRVKXCTJSA-N 0.000 description 1
- XVSJMWYYLHPDKY-DCAQKATOSA-N Leu-Asp-Met Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCSC)C(O)=O XVSJMWYYLHPDKY-DCAQKATOSA-N 0.000 description 1
- MMEDVBWCMGRKKC-GARJFASQSA-N Leu-Asp-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N MMEDVBWCMGRKKC-GARJFASQSA-N 0.000 description 1
- GBDMISNMNXVTNV-XIRDDKMYSA-N Leu-Asp-Trp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O GBDMISNMNXVTNV-XIRDDKMYSA-N 0.000 description 1
- WCTCIIAGNMFYAO-DCAQKATOSA-N Leu-Cys-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(O)=O WCTCIIAGNMFYAO-DCAQKATOSA-N 0.000 description 1
- VPKIQULSKFVCSM-SRVKXCTJSA-N Leu-Gln-Arg Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O VPKIQULSKFVCSM-SRVKXCTJSA-N 0.000 description 1
- KAFOIVJDVSZUMD-UHFFFAOYSA-N Leu-Gln-Gln Natural products CC(C)CC(N)C(=O)NC(CCC(N)=O)C(=O)NC(CCC(N)=O)C(O)=O KAFOIVJDVSZUMD-UHFFFAOYSA-N 0.000 description 1
- GLBNEGIOFRVRHO-JYJNAYRXSA-N Leu-Gln-Phe Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O GLBNEGIOFRVRHO-JYJNAYRXSA-N 0.000 description 1
- CQGSYZCULZMEDE-UHFFFAOYSA-N Leu-Gln-Pro Natural products CC(C)CC(N)C(=O)NC(CCC(N)=O)C(=O)N1CCCC1C(O)=O CQGSYZCULZMEDE-UHFFFAOYSA-N 0.000 description 1
- LLBQJYDYOLIQAI-JYJNAYRXSA-N Leu-Glu-Tyr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O LLBQJYDYOLIQAI-JYJNAYRXSA-N 0.000 description 1
- VWHGTYCRDRBSFI-ZETCQYMHSA-N Leu-Gly-Gly Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)NCC(O)=O VWHGTYCRDRBSFI-ZETCQYMHSA-N 0.000 description 1
- VBZOAGIPCULURB-QWRGUYRKSA-N Leu-Gly-His Chemical compound CC(C)C[C@@H](C(=O)NCC(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N VBZOAGIPCULURB-QWRGUYRKSA-N 0.000 description 1
- HYMLKESRWLZDBR-WEDXCCLWSA-N Leu-Gly-Thr Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(O)=O HYMLKESRWLZDBR-WEDXCCLWSA-N 0.000 description 1
- UCDHVOALNXENLC-KBPBESRZSA-N Leu-Gly-Tyr Chemical compound CC(C)C[C@H]([NH3+])C(=O)NCC(=O)N[C@H](C([O-])=O)CC1=CC=C(O)C=C1 UCDHVOALNXENLC-KBPBESRZSA-N 0.000 description 1
- POZULHZYLPGXMR-ONGXEEELSA-N Leu-Gly-Val Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O POZULHZYLPGXMR-ONGXEEELSA-N 0.000 description 1
- PBGDOSARRIJMEV-DLOVCJGASA-N Leu-His-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C)C(O)=O PBGDOSARRIJMEV-DLOVCJGASA-N 0.000 description 1
- VZBIUJURDLFFOE-IHRRRGAJSA-N Leu-His-Arg Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O VZBIUJURDLFFOE-IHRRRGAJSA-N 0.000 description 1
- KVOFSTUWVSQMDK-KKUMJFAQSA-N Leu-His-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)CC(C)C)CC1=CN=CN1 KVOFSTUWVSQMDK-KKUMJFAQSA-N 0.000 description 1
- DBSLVQBXKVKDKJ-BJDJZHNGSA-N Leu-Ile-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O DBSLVQBXKVKDKJ-BJDJZHNGSA-N 0.000 description 1
- USLNHQZCDQJBOV-ZPFDUUQYSA-N Leu-Ile-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O USLNHQZCDQJBOV-ZPFDUUQYSA-N 0.000 description 1
- SEMUSFOBZGKBGW-YTFOTSKYSA-N Leu-Ile-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O SEMUSFOBZGKBGW-YTFOTSKYSA-N 0.000 description 1
- RXGLHDWAZQECBI-SRVKXCTJSA-N Leu-Leu-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O RXGLHDWAZQECBI-SRVKXCTJSA-N 0.000 description 1
- RZXLZBIUTDQHJQ-SRVKXCTJSA-N Leu-Lys-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(O)=O RZXLZBIUTDQHJQ-SRVKXCTJSA-N 0.000 description 1
- QNTJIDXQHWUBKC-BZSNNMDCSA-N Leu-Lys-Phe Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O QNTJIDXQHWUBKC-BZSNNMDCSA-N 0.000 description 1
- JVTYXRRFZCEPPK-RHYQMDGZSA-N Leu-Met-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CC(C)C)N)O JVTYXRRFZCEPPK-RHYQMDGZSA-N 0.000 description 1
- KTOIECMYZZGVSI-BZSNNMDCSA-N Leu-Phe-His Chemical compound C([C@H](NC(=O)[C@@H](N)CC(C)C)C(=O)N[C@@H](CC=1NC=NC=1)C(O)=O)C1=CC=CC=C1 KTOIECMYZZGVSI-BZSNNMDCSA-N 0.000 description 1
- UHNQRAFSEBGZFZ-YESZJQIVSA-N Leu-Phe-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N2CCC[C@@H]2C(=O)O)N UHNQRAFSEBGZFZ-YESZJQIVSA-N 0.000 description 1
- VULJUQZPSOASBZ-SRVKXCTJSA-N Leu-Pro-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O VULJUQZPSOASBZ-SRVKXCTJSA-N 0.000 description 1
- PWPBLZXWFXJFHE-RHYQMDGZSA-N Leu-Pro-Thr Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O PWPBLZXWFXJFHE-RHYQMDGZSA-N 0.000 description 1
- JDBQSGMJBMPNFT-AVGNSLFASA-N Leu-Pro-Val Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=O JDBQSGMJBMPNFT-AVGNSLFASA-N 0.000 description 1
- IDGZVZJLYFTXSL-DCAQKATOSA-N Leu-Ser-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCN=C(N)N IDGZVZJLYFTXSL-DCAQKATOSA-N 0.000 description 1
- JIHDFWWRYHSAQB-GUBZILKMSA-N Leu-Ser-Glu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O JIHDFWWRYHSAQB-GUBZILKMSA-N 0.000 description 1
- MVHXGBZUJLWZOH-BJDJZHNGSA-N Leu-Ser-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O MVHXGBZUJLWZOH-BJDJZHNGSA-N 0.000 description 1
- ZJZNLRVCZWUONM-JXUBOQSCSA-N Leu-Thr-Ala Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O ZJZNLRVCZWUONM-JXUBOQSCSA-N 0.000 description 1
- ZDJQVSIPFLMNOX-RHYQMDGZSA-N Leu-Thr-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N ZDJQVSIPFLMNOX-RHYQMDGZSA-N 0.000 description 1
- VDIARPPNADFEAV-WEDXCCLWSA-N Leu-Thr-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O VDIARPPNADFEAV-WEDXCCLWSA-N 0.000 description 1
- LJBVRCDPWOJOEK-PPCPHDFISA-N Leu-Thr-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O LJBVRCDPWOJOEK-PPCPHDFISA-N 0.000 description 1
- CNWDWAMPKVYJJB-NUTKFTJISA-N Leu-Trp-Ala Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](N)CC(C)C)C(=O)N[C@@H](C)C(O)=O)=CNC2=C1 CNWDWAMPKVYJJB-NUTKFTJISA-N 0.000 description 1
- UCRJTSIIAYHOHE-ULQDDVLXSA-N Leu-Tyr-Arg Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N UCRJTSIIAYHOHE-ULQDDVLXSA-N 0.000 description 1
- VJGQRELPQWNURN-JYJNAYRXSA-N Leu-Tyr-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O VJGQRELPQWNURN-JYJNAYRXSA-N 0.000 description 1
- FBNPMTNBFFAMMH-UHFFFAOYSA-N Leu-Val-Arg Natural products CC(C)CC(N)C(=O)NC(C(C)C)C(=O)NC(C(O)=O)CCCN=C(N)N FBNPMTNBFFAMMH-UHFFFAOYSA-N 0.000 description 1
- AIMGJYMCTAABEN-GVXVVHGQSA-N Leu-Val-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O AIMGJYMCTAABEN-GVXVVHGQSA-N 0.000 description 1
- YQFZRHYZLARWDY-IHRRRGAJSA-N Leu-Val-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCCN YQFZRHYZLARWDY-IHRRRGAJSA-N 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 235000004431 Linum usitatissimum Nutrition 0.000 description 1
- 108010071324 Livagen Proteins 0.000 description 1
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 1
- 235000007688 Lycopersicon esculentum Nutrition 0.000 description 1
- MPGHETGWWWUHPY-CIUDSAMLSA-N Lys-Ala-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN MPGHETGWWWUHPY-CIUDSAMLSA-N 0.000 description 1
- PNPYKQFJGRFYJE-GUBZILKMSA-N Lys-Ala-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O PNPYKQFJGRFYJE-GUBZILKMSA-N 0.000 description 1
- UWKNTTJNVSYXPC-CIUDSAMLSA-N Lys-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN UWKNTTJNVSYXPC-CIUDSAMLSA-N 0.000 description 1
- VHNOAIFVYUQOOY-XUXIUFHCSA-N Lys-Arg-Ile Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O VHNOAIFVYUQOOY-XUXIUFHCSA-N 0.000 description 1
- HQVDJTYKCMIWJP-YUMQZZPRSA-N Lys-Asn-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O HQVDJTYKCMIWJP-YUMQZZPRSA-N 0.000 description 1
- QUCDKEKDPYISNX-HJGDQZAQSA-N Lys-Asn-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QUCDKEKDPYISNX-HJGDQZAQSA-N 0.000 description 1
- AAORVPFVUIHEAB-YUMQZZPRSA-N Lys-Asp-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O AAORVPFVUIHEAB-YUMQZZPRSA-N 0.000 description 1
- MWVUEPNEPWMFBD-SRVKXCTJSA-N Lys-Cys-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@H](C(O)=O)CCCCN MWVUEPNEPWMFBD-SRVKXCTJSA-N 0.000 description 1
- MRWXLRGAFDOILG-DCAQKATOSA-N Lys-Gln-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O MRWXLRGAFDOILG-DCAQKATOSA-N 0.000 description 1
- RZHLIPMZXOEJTL-AVGNSLFASA-N Lys-Gln-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCCCN)N RZHLIPMZXOEJTL-AVGNSLFASA-N 0.000 description 1
- HEWWNLVEWBJBKA-WDCWCFNPSA-N Lys-Gln-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](N)CCCCN HEWWNLVEWBJBKA-WDCWCFNPSA-N 0.000 description 1
- DRCILAJNUJKAHC-SRVKXCTJSA-N Lys-Glu-Arg Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O DRCILAJNUJKAHC-SRVKXCTJSA-N 0.000 description 1
- KZOHPCYVORJBLG-AVGNSLFASA-N Lys-Glu-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CCCCN)N KZOHPCYVORJBLG-AVGNSLFASA-N 0.000 description 1
- PAMDBWYMLWOELY-SDDRHHMPSA-N Lys-Glu-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CCCCN)N)C(=O)O PAMDBWYMLWOELY-SDDRHHMPSA-N 0.000 description 1
- WGLAORUKDGRINI-WDCWCFNPSA-N Lys-Glu-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O WGLAORUKDGRINI-WDCWCFNPSA-N 0.000 description 1
- DKTNGXVSCZULPO-YUMQZZPRSA-N Lys-Gly-Cys Chemical compound NCCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CS)C(O)=O DKTNGXVSCZULPO-YUMQZZPRSA-N 0.000 description 1
- ISHNZELVUVPCHY-ZETCQYMHSA-N Lys-Gly-Gly Chemical compound NCCCC[C@H](N)C(=O)NCC(=O)NCC(O)=O ISHNZELVUVPCHY-ZETCQYMHSA-N 0.000 description 1
- NKKFVJRLCCUJNA-QWRGUYRKSA-N Lys-Gly-Lys Chemical compound NCCCC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCCN NKKFVJRLCCUJNA-QWRGUYRKSA-N 0.000 description 1
- PBLLTSKBTAHDNA-KBPBESRZSA-N Lys-Gly-Phe Chemical compound [H]N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O PBLLTSKBTAHDNA-KBPBESRZSA-N 0.000 description 1
- OJDFAABAHBPVTH-MNXVOIDGSA-N Lys-Ile-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(N)=O)C(O)=O OJDFAABAHBPVTH-MNXVOIDGSA-N 0.000 description 1
- MYZMQWHPDAYKIE-SRVKXCTJSA-N Lys-Leu-Ala Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O MYZMQWHPDAYKIE-SRVKXCTJSA-N 0.000 description 1
- NJNRBRKHOWSGMN-SRVKXCTJSA-N Lys-Leu-Asn Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O NJNRBRKHOWSGMN-SRVKXCTJSA-N 0.000 description 1
- VMTYLUGCXIEDMV-QWRGUYRKSA-N Lys-Leu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCCCN VMTYLUGCXIEDMV-QWRGUYRKSA-N 0.000 description 1
- YPLVCBKEPJPBDQ-MELADBBJSA-N Lys-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCCN)N YPLVCBKEPJPBDQ-MELADBBJSA-N 0.000 description 1
- OIQSIMFSVLLWBX-VOAKCMCISA-N Lys-Leu-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OIQSIMFSVLLWBX-VOAKCMCISA-N 0.000 description 1
- ALGGDNMLQNFVIZ-SRVKXCTJSA-N Lys-Lys-Asp Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(=O)O)C(=O)O)N ALGGDNMLQNFVIZ-SRVKXCTJSA-N 0.000 description 1
- WBSCNDJQPKSPII-KKUMJFAQSA-N Lys-Lys-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O WBSCNDJQPKSPII-KKUMJFAQSA-N 0.000 description 1
- KJIXWRWPOCKYLD-IHRRRGAJSA-N Lys-Lys-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCCCN)N KJIXWRWPOCKYLD-IHRRRGAJSA-N 0.000 description 1
- MTBBHUKKPWKXBT-ULQDDVLXSA-N Lys-Met-Tyr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O MTBBHUKKPWKXBT-ULQDDVLXSA-N 0.000 description 1
- BPDXWKVZNCKUGG-BZSNNMDCSA-N Lys-Phe-His Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CCCCN)N BPDXWKVZNCKUGG-BZSNNMDCSA-N 0.000 description 1
- OBZHNHBAAVEWKI-DCAQKATOSA-N Lys-Pro-Asn Chemical compound NCCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O OBZHNHBAAVEWKI-DCAQKATOSA-N 0.000 description 1
- WGILOYIKJVQUPT-DCAQKATOSA-N Lys-Pro-Asp Chemical compound [H]N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O WGILOYIKJVQUPT-DCAQKATOSA-N 0.000 description 1
- LUTDBHBIHHREDC-IHRRRGAJSA-N Lys-Pro-Lys Chemical compound NCCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(O)=O LUTDBHBIHHREDC-IHRRRGAJSA-N 0.000 description 1
- YTJFXEDRUOQGSP-DCAQKATOSA-N Lys-Pro-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O YTJFXEDRUOQGSP-DCAQKATOSA-N 0.000 description 1
- LOGFVTREOLYCPF-RHYQMDGZSA-N Lys-Pro-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCCN LOGFVTREOLYCPF-RHYQMDGZSA-N 0.000 description 1
- JMNRXRPBHFGXQX-GUBZILKMSA-N Lys-Ser-Glu Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O JMNRXRPBHFGXQX-GUBZILKMSA-N 0.000 description 1
- MIFFFXHMAHFACR-KATARQTJSA-N Lys-Ser-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CCCCN MIFFFXHMAHFACR-KATARQTJSA-N 0.000 description 1
- MEQLGHAMAUPOSJ-DCAQKATOSA-N Lys-Ser-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O MEQLGHAMAUPOSJ-DCAQKATOSA-N 0.000 description 1
- QVTDVTONTRSQMF-WDCWCFNPSA-N Lys-Thr-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H]([C@H](O)C)NC(=O)[C@@H](N)CCCCN QVTDVTONTRSQMF-WDCWCFNPSA-N 0.000 description 1
- YKBSXQFZWFXFIB-VOAKCMCISA-N Lys-Thr-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CCCCN)C(O)=O YKBSXQFZWFXFIB-VOAKCMCISA-N 0.000 description 1
- ZFNYWKHYUMEZDZ-WDSOQIARSA-N Lys-Trp-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CCCCN)N ZFNYWKHYUMEZDZ-WDSOQIARSA-N 0.000 description 1
- ZVZRQKJOQQAFCF-ULQDDVLXSA-N Lys-Tyr-Arg Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O ZVZRQKJOQQAFCF-ULQDDVLXSA-N 0.000 description 1
- PSVAVKGDUAKZKU-BZSNNMDCSA-N Lys-Tyr-His Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CCCCN)N)O PSVAVKGDUAKZKU-BZSNNMDCSA-N 0.000 description 1
- NYTDJEZBAAFLLG-IHRRRGAJSA-N Lys-Val-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(O)=O NYTDJEZBAAFLLG-IHRRRGAJSA-N 0.000 description 1
- 239000007993 MOPS buffer Substances 0.000 description 1
- 239000005913 Maltodextrin Substances 0.000 description 1
- 229920002774 Maltodextrin Polymers 0.000 description 1
- 235000010624 Medicago sativa Nutrition 0.000 description 1
- 235000017587 Medicago sativa ssp. sativa Nutrition 0.000 description 1
- LMKSBGIUPVRHEH-FXQIFTODSA-N Met-Ala-Asn Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC(N)=O LMKSBGIUPVRHEH-FXQIFTODSA-N 0.000 description 1
- AHZNUGRZHMZGFL-GUBZILKMSA-N Met-Arg-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CO)C(O)=O)CCCNC(N)=N AHZNUGRZHMZGFL-GUBZILKMSA-N 0.000 description 1
- ZMYHJISLFYTQGK-FXQIFTODSA-N Met-Asp-Asn Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ZMYHJISLFYTQGK-FXQIFTODSA-N 0.000 description 1
- FJVJLMZUIGMFFU-BQBZGAKWSA-N Met-Asp-Gly Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O FJVJLMZUIGMFFU-BQBZGAKWSA-N 0.000 description 1
- RTJPUVZZCBWXSZ-BPUTZDHNSA-N Met-Cys-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CS)NC(=O)[C@@H](N)CCSC)C(O)=O)=CNC2=C1 RTJPUVZZCBWXSZ-BPUTZDHNSA-N 0.000 description 1
- YLLWCSDBVGZLOW-CIUDSAMLSA-N Met-Gln-Ala Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O YLLWCSDBVGZLOW-CIUDSAMLSA-N 0.000 description 1
- FWTBMGAKKPSTBT-GUBZILKMSA-N Met-Gln-Glu Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O FWTBMGAKKPSTBT-GUBZILKMSA-N 0.000 description 1
- NCVJJAJVWILAGI-SRVKXCTJSA-N Met-Gln-Lys Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N NCVJJAJVWILAGI-SRVKXCTJSA-N 0.000 description 1
- GPVLSVCBKUCEBI-KKUMJFAQSA-N Met-Gln-Phe Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O GPVLSVCBKUCEBI-KKUMJFAQSA-N 0.000 description 1
- AETNZPKUUYYYEK-CIUDSAMLSA-N Met-Glu-Asn Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O AETNZPKUUYYYEK-CIUDSAMLSA-N 0.000 description 1
- ORRNBLTZBBESPN-HJWJTTGWSA-N Met-Ile-Phe Chemical compound CSCC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 ORRNBLTZBBESPN-HJWJTTGWSA-N 0.000 description 1
- HWROAFGWPQUPTE-OSUNSFLBSA-N Met-Ile-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CCSC)N HWROAFGWPQUPTE-OSUNSFLBSA-N 0.000 description 1
- HGAJNEWOUHDUMZ-SRVKXCTJSA-N Met-Leu-Glu Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CCC(O)=O HGAJNEWOUHDUMZ-SRVKXCTJSA-N 0.000 description 1
- AXHNAGAYRGCDLG-UWVGGRQHSA-N Met-Lys-Gly Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)NCC(O)=O AXHNAGAYRGCDLG-UWVGGRQHSA-N 0.000 description 1
- KRLKICLNEICJGV-STQMWFEESA-N Met-Phe-Gly Chemical compound CSCC[C@H](N)C(=O)N[C@H](C(=O)NCC(O)=O)CC1=CC=CC=C1 KRLKICLNEICJGV-STQMWFEESA-N 0.000 description 1
- OIFHHODAXVWKJN-ULQDDVLXSA-N Met-Phe-Leu Chemical compound CSCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=CC=C1 OIFHHODAXVWKJN-ULQDDVLXSA-N 0.000 description 1
- RDLSEGZJMYGFNS-FXQIFTODSA-N Met-Ser-Asp Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O RDLSEGZJMYGFNS-FXQIFTODSA-N 0.000 description 1
- DSZFTPCSFVWMKP-DCAQKATOSA-N Met-Ser-Lys Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCCN DSZFTPCSFVWMKP-DCAQKATOSA-N 0.000 description 1
- GWADARYJIJDYRC-XGEHTFHBSA-N Met-Thr-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O GWADARYJIJDYRC-XGEHTFHBSA-N 0.000 description 1
- 241000589344 Methylomonas Species 0.000 description 1
- 241000589354 Methylosinus Species 0.000 description 1
- 241000235395 Mucor Species 0.000 description 1
- 108010014251 Muramidase Proteins 0.000 description 1
- 102000016943 Muramidase Human genes 0.000 description 1
- 108010062010 N-Acetylmuramoyl-L-alanine Amidase Proteins 0.000 description 1
- XZFYRXDAULDNFX-UHFFFAOYSA-N N-L-cysteinyl-L-phenylalanine Natural products SCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XZFYRXDAULDNFX-UHFFFAOYSA-N 0.000 description 1
- AUEJLPRZGVVDNU-UHFFFAOYSA-N N-L-tyrosyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CC1=CC=C(O)C=C1 AUEJLPRZGVVDNU-UHFFFAOYSA-N 0.000 description 1
- 108010002311 N-glycylglutamic acid Proteins 0.000 description 1
- 239000004384 Neotame Substances 0.000 description 1
- 101150012394 PHO5 gene Proteins 0.000 description 1
- 241000209117 Panicum Species 0.000 description 1
- 240000008114 Panicum miliaceum Species 0.000 description 1
- 235000007199 Panicum miliaceum Nutrition 0.000 description 1
- 235000006443 Panicum miliaceum subsp. miliaceum Nutrition 0.000 description 1
- 235000009037 Panicum miliaceum subsp. ruderale Nutrition 0.000 description 1
- 241000520272 Pantoea Species 0.000 description 1
- 240000004370 Pastinaca sativa Species 0.000 description 1
- 235000017769 Pastinaca sativa subsp sativa Nutrition 0.000 description 1
- 108010033276 Peptide Fragments Proteins 0.000 description 1
- 102000007079 Peptide Fragments Human genes 0.000 description 1
- 244000062780 Petroselinum sativum Species 0.000 description 1
- 235000010627 Phaseolus vulgaris Nutrition 0.000 description 1
- 244000046052 Phaseolus vulgaris Species 0.000 description 1
- FPTXMUIBLMGTQH-ONGXEEELSA-N Phe-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=CC=C1 FPTXMUIBLMGTQH-ONGXEEELSA-N 0.000 description 1
- BKWJQWJPZMUWEG-LFSVMHDDSA-N Phe-Ala-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=CC=C1 BKWJQWJPZMUWEG-LFSVMHDDSA-N 0.000 description 1
- ZWJKVFAYPLPCQB-UNQGMJICSA-N Phe-Arg-Thr Chemical compound C[C@@H](O)[C@H](NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H](N)Cc1ccccc1)C(O)=O ZWJKVFAYPLPCQB-UNQGMJICSA-N 0.000 description 1
- HXSUFWQYLPKEHF-IHRRRGAJSA-N Phe-Asn-Arg Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N HXSUFWQYLPKEHF-IHRRRGAJSA-N 0.000 description 1
- DDYIRGBOZVKRFR-AVGNSLFASA-N Phe-Asp-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N DDYIRGBOZVKRFR-AVGNSLFASA-N 0.000 description 1
- MPFGIYLYWUCSJG-AVGNSLFASA-N Phe-Glu-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1 MPFGIYLYWUCSJG-AVGNSLFASA-N 0.000 description 1
- MGECUMGTSHYHEJ-QEWYBTABSA-N Phe-Glu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1 MGECUMGTSHYHEJ-QEWYBTABSA-N 0.000 description 1
- KJJROSNFBRWPHS-JYJNAYRXSA-N Phe-Glu-Leu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O KJJROSNFBRWPHS-JYJNAYRXSA-N 0.000 description 1
- BFYHIHGIHGROAT-HTUGSXCWSA-N Phe-Glu-Thr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O BFYHIHGIHGROAT-HTUGSXCWSA-N 0.000 description 1
- VZFPYFRVHMSSNA-JURCDPSOSA-N Phe-Ile-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CC1=CC=CC=C1 VZFPYFRVHMSSNA-JURCDPSOSA-N 0.000 description 1
- MIICYIIBVYQNKE-QEWYBTABSA-N Phe-Ile-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N MIICYIIBVYQNKE-QEWYBTABSA-N 0.000 description 1
- GXDPQJUBLBZKDY-IAVJCBSLSA-N Phe-Ile-Ile Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O GXDPQJUBLBZKDY-IAVJCBSLSA-N 0.000 description 1
- DVOCGBNHAUHKHJ-DKIMLUQUSA-N Phe-Ile-Leu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O DVOCGBNHAUHKHJ-DKIMLUQUSA-N 0.000 description 1
- KBVJZCVLQWCJQN-KKUMJFAQSA-N Phe-Leu-Asn Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O KBVJZCVLQWCJQN-KKUMJFAQSA-N 0.000 description 1
- HQPWNHXERZCIHP-PMVMPFDFSA-N Phe-Leu-Trp Chemical compound C([C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)C1=CC=CC=C1 HQPWNHXERZCIHP-PMVMPFDFSA-N 0.000 description 1
- MGLBSROLWAWCKN-FCLVOEFKSA-N Phe-Phe-Thr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O MGLBSROLWAWCKN-FCLVOEFKSA-N 0.000 description 1
- IIEOLPMQYRBZCN-SRVKXCTJSA-N Phe-Ser-Cys Chemical compound N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O IIEOLPMQYRBZCN-SRVKXCTJSA-N 0.000 description 1
- BONHGTUEEPIMPM-AVGNSLFASA-N Phe-Ser-Glu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O BONHGTUEEPIMPM-AVGNSLFASA-N 0.000 description 1
- BPIMVBKDLSBKIJ-FCLVOEFKSA-N Phe-Thr-Phe Chemical compound C([C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=CC=C1 BPIMVBKDLSBKIJ-FCLVOEFKSA-N 0.000 description 1
- GNRMAQSIROFNMI-IXOXFDKPSA-N Phe-Thr-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O GNRMAQSIROFNMI-IXOXFDKPSA-N 0.000 description 1
- LKRUQZQZMXMKEQ-SFJXLCSZSA-N Phe-Trp-Thr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LKRUQZQZMXMKEQ-SFJXLCSZSA-N 0.000 description 1
- YUPRIZTWANWWHK-DZKIICNBSA-N Phe-Val-Glu Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N YUPRIZTWANWWHK-DZKIICNBSA-N 0.000 description 1
- RGMLUHANLDVMPB-ULQDDVLXSA-N Phe-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N RGMLUHANLDVMPB-ULQDDVLXSA-N 0.000 description 1
- 235000010582 Pisum sativum Nutrition 0.000 description 1
- 240000004713 Pisum sativum Species 0.000 description 1
- 241000209504 Poaceae Species 0.000 description 1
- 108010059820 Polygalacturonase Proteins 0.000 description 1
- KIZQGKLMXKGDIV-BQBZGAKWSA-N Pro-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 KIZQGKLMXKGDIV-BQBZGAKWSA-N 0.000 description 1
- XQLBWXHVZVBNJM-FXQIFTODSA-N Pro-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 XQLBWXHVZVBNJM-FXQIFTODSA-N 0.000 description 1
- LCRSGSIRKLXZMZ-BPNCWPANSA-N Pro-Ala-Tyr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O LCRSGSIRKLXZMZ-BPNCWPANSA-N 0.000 description 1
- OCSACVPBMIYNJE-GUBZILKMSA-N Pro-Arg-Asn Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(O)=O OCSACVPBMIYNJE-GUBZILKMSA-N 0.000 description 1
- SSSFPISOZOLQNP-GUBZILKMSA-N Pro-Arg-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O SSSFPISOZOLQNP-GUBZILKMSA-N 0.000 description 1
- HPXVFFIIGOAQRV-DCAQKATOSA-N Pro-Arg-Gln Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(O)=O HPXVFFIIGOAQRV-DCAQKATOSA-N 0.000 description 1
- BNBBNGZZKQUWCD-IUCAKERBSA-N Pro-Arg-Gly Chemical compound NC(N)=NCCC[C@@H](C(=O)NCC(O)=O)NC(=O)[C@@H]1CCCN1 BNBBNGZZKQUWCD-IUCAKERBSA-N 0.000 description 1
- GRIRJQGZZJVANI-CYDGBPFRSA-N Pro-Arg-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H]1CCCN1 GRIRJQGZZJVANI-CYDGBPFRSA-N 0.000 description 1
- ICTZKEXYDDZZFP-SRVKXCTJSA-N Pro-Arg-Pro Chemical compound N([C@@H](CCCN=C(N)N)C(=O)N1[C@@H](CCC1)C(O)=O)C(=O)[C@@H]1CCCN1 ICTZKEXYDDZZFP-SRVKXCTJSA-N 0.000 description 1
- FUVBEZJCRMHWEM-FXQIFTODSA-N Pro-Asn-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O FUVBEZJCRMHWEM-FXQIFTODSA-N 0.000 description 1
- UTAUEDINXUMHLG-FXQIFTODSA-N Pro-Asp-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@@H]1CCCN1 UTAUEDINXUMHLG-FXQIFTODSA-N 0.000 description 1
- ILMLVTGTUJPQFP-FXQIFTODSA-N Pro-Asp-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O ILMLVTGTUJPQFP-FXQIFTODSA-N 0.000 description 1
- KIGGUSRFHJCIEJ-DCAQKATOSA-N Pro-Asp-His Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O KIGGUSRFHJCIEJ-DCAQKATOSA-N 0.000 description 1
- YFNOUBWUIIJQHF-LPEHRKFASA-N Pro-Asp-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC(=O)O)C(=O)N2CCC[C@@H]2C(=O)O YFNOUBWUIIJQHF-LPEHRKFASA-N 0.000 description 1
- ZYBUKTMPPFQSHL-JYJNAYRXSA-N Pro-Asp-Trp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O ZYBUKTMPPFQSHL-JYJNAYRXSA-N 0.000 description 1
- LSIWVWRUTKPXDS-DCAQKATOSA-N Pro-Gln-Arg Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O LSIWVWRUTKPXDS-DCAQKATOSA-N 0.000 description 1
- PZSCUPVOJGKHEP-CIUDSAMLSA-N Pro-Gln-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O PZSCUPVOJGKHEP-CIUDSAMLSA-N 0.000 description 1
- FISHYTLIMUYTQY-GUBZILKMSA-N Pro-Gln-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H]1CCCN1 FISHYTLIMUYTQY-GUBZILKMSA-N 0.000 description 1
- DRIJZWBRGMJCDD-DCAQKATOSA-N Pro-Gln-Met Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCSC)C(O)=O DRIJZWBRGMJCDD-DCAQKATOSA-N 0.000 description 1
- QCARZLHECSFOGG-CIUDSAMLSA-N Pro-Glu-Cys Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CS)C(=O)O QCARZLHECSFOGG-CIUDSAMLSA-N 0.000 description 1
- NMELOOXSGDRBRU-YUMQZZPRSA-N Pro-Glu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CCC(=O)O)NC(=O)[C@@H]1CCCN1 NMELOOXSGDRBRU-YUMQZZPRSA-N 0.000 description 1
- LPGSNRSLPHRNBW-AVGNSLFASA-N Pro-His-Val Chemical compound C([C@@H](C(=O)N[C@@H](C(C)C)C([O-])=O)NC(=O)[C@H]1[NH2+]CCC1)C1=CN=CN1 LPGSNRSLPHRNBW-AVGNSLFASA-N 0.000 description 1
- BCNRNJWSRFDPTQ-HJWJTTGWSA-N Pro-Ile-Phe Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O BCNRNJWSRFDPTQ-HJWJTTGWSA-N 0.000 description 1
- SRBFGSGDNNQABI-FHWLQOOXSA-N Pro-Leu-Trp Chemical compound N([C@@H](CC(C)C)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)C(=O)[C@@H]1CCCN1 SRBFGSGDNNQABI-FHWLQOOXSA-N 0.000 description 1
- CDGABSWLRMECHC-IHRRRGAJSA-N Pro-Lys-His Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O CDGABSWLRMECHC-IHRRRGAJSA-N 0.000 description 1
- SMFQZMGHCODUPQ-ULQDDVLXSA-N Pro-Lys-Phe Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O SMFQZMGHCODUPQ-ULQDDVLXSA-N 0.000 description 1
- RPLMFKUKFZOTER-AVGNSLFASA-N Pro-Met-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@@H]1CCCN1 RPLMFKUKFZOTER-AVGNSLFASA-N 0.000 description 1
- VGVCNKSUVSZEIE-IHRRRGAJSA-N Pro-Phe-Asn Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(N)=O)C(O)=O VGVCNKSUVSZEIE-IHRRRGAJSA-N 0.000 description 1
- AWQGDZBKQTYNMN-IHRRRGAJSA-N Pro-Phe-Asp Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)N[C@@H](CC(=O)O)C(=O)O AWQGDZBKQTYNMN-IHRRRGAJSA-N 0.000 description 1
- BUEIYHBJHCDAMI-UFYCRDLUSA-N Pro-Phe-Phe Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O BUEIYHBJHCDAMI-UFYCRDLUSA-N 0.000 description 1
- ZVEQWRWMRFIVSD-HRCADAONSA-N Pro-Phe-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)N3CCC[C@@H]3C(=O)O ZVEQWRWMRFIVSD-HRCADAONSA-N 0.000 description 1
- KDBHVPXBQADZKY-GUBZILKMSA-N Pro-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 KDBHVPXBQADZKY-GUBZILKMSA-N 0.000 description 1
- JLMZKEQFMVORMA-SRVKXCTJSA-N Pro-Pro-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 JLMZKEQFMVORMA-SRVKXCTJSA-N 0.000 description 1
- LEIKGVHQTKHOLM-IUCAKERBSA-N Pro-Pro-Gly Chemical compound OC(=O)CNC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 LEIKGVHQTKHOLM-IUCAKERBSA-N 0.000 description 1
- DWPXHLIBFQLKLK-CYDGBPFRSA-N Pro-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 DWPXHLIBFQLKLK-CYDGBPFRSA-N 0.000 description 1
- CGSOWZUPLOKYOR-AVGNSLFASA-N Pro-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 CGSOWZUPLOKYOR-AVGNSLFASA-N 0.000 description 1
- FDMKYQQYJKYCLV-GUBZILKMSA-N Pro-Pro-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 FDMKYQQYJKYCLV-GUBZILKMSA-N 0.000 description 1
- SXJOPONICMGFCR-DCAQKATOSA-N Pro-Ser-Lys Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)O SXJOPONICMGFCR-DCAQKATOSA-N 0.000 description 1
- QKDIHFHGHBYTKB-IHRRRGAJSA-N Pro-Ser-Phe Chemical compound N([C@@H](CO)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C(=O)[C@@H]1CCCN1 QKDIHFHGHBYTKB-IHRRRGAJSA-N 0.000 description 1
- MKGIILKDUGDRRO-FXQIFTODSA-N Pro-Ser-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H]1CCCN1 MKGIILKDUGDRRO-FXQIFTODSA-N 0.000 description 1
- PRKWBYCXBBSLSK-GUBZILKMSA-N Pro-Ser-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O PRKWBYCXBBSLSK-GUBZILKMSA-N 0.000 description 1
- KIDXAAQVMNLJFQ-KZVJFYERSA-N Pro-Thr-Ala Chemical compound C[C@@H](O)[C@H](NC(=O)[C@@H]1CCCN1)C(=O)N[C@@H](C)C(O)=O KIDXAAQVMNLJFQ-KZVJFYERSA-N 0.000 description 1
- AJJDPGVVNPUZCR-RHYQMDGZSA-N Pro-Thr-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@@H]1CCCN1)O AJJDPGVVNPUZCR-RHYQMDGZSA-N 0.000 description 1
- RMJZWERKFFNNNS-XGEHTFHBSA-N Pro-Thr-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O RMJZWERKFFNNNS-XGEHTFHBSA-N 0.000 description 1
- HOJUNFDJDAPVBI-BZSNNMDCSA-N Pro-Trp-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@@H]3CCCN3 HOJUNFDJDAPVBI-BZSNNMDCSA-N 0.000 description 1
- CWZUFLWPEFHWEI-IHRRRGAJSA-N Pro-Tyr-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O CWZUFLWPEFHWEI-IHRRRGAJSA-N 0.000 description 1
- FZXSYIPVAFVYBH-KKUMJFAQSA-N Pro-Tyr-Glu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O FZXSYIPVAFVYBH-KKUMJFAQSA-N 0.000 description 1
- SHTKRJHDMNSKRM-ULQDDVLXSA-N Pro-Tyr-His Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)N[C@@H](CC3=CN=CN3)C(=O)O SHTKRJHDMNSKRM-ULQDDVLXSA-N 0.000 description 1
- QKWYXRPICJEQAJ-KJEVXHAQSA-N Pro-Tyr-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)NC(=O)[C@@H]2CCCN2)O QKWYXRPICJEQAJ-KJEVXHAQSA-N 0.000 description 1
- GIPHUOWOTCAJSR-UHFFFAOYSA-N Rebaudioside A. Natural products C1CC2C3(C)CCCC(C)(C(=O)OC4C(C(O)C(O)C(CO)O4)O)C3CCC2(C2)CC(=C)C21OC1OC(CO)C(O)C(O)C1OC(C1O)OC(CO)C(O)C1OC1OC(CO)C(O)C(O)C1O GIPHUOWOTCAJSR-UHFFFAOYSA-N 0.000 description 1
- 241000191025 Rhodobacter Species 0.000 description 1
- 241000316848 Rhodococcus <scale insect> Species 0.000 description 1
- 101100394989 Rhodopseudomonas palustris (strain ATCC BAA-98 / CGA009) hisI gene Proteins 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 108010003581 Ribulose-bisphosphate carboxylase Proteins 0.000 description 1
- 101100434411 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) ADH1 gene Proteins 0.000 description 1
- 240000000111 Saccharum officinarum Species 0.000 description 1
- 235000007201 Saccharum officinarum Nutrition 0.000 description 1
- 101001000154 Schistosoma mansoni Phosphoglycerate kinase Proteins 0.000 description 1
- 241000209056 Secale Species 0.000 description 1
- 235000007238 Secale cereale Nutrition 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- WTUJZHKANPDPIN-CIUDSAMLSA-N Ser-Ala-Lys Chemical compound C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CO)N WTUJZHKANPDPIN-CIUDSAMLSA-N 0.000 description 1
- HBZBPFLJNDXRAY-FXQIFTODSA-N Ser-Ala-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(O)=O HBZBPFLJNDXRAY-FXQIFTODSA-N 0.000 description 1
- NRCJWSGXMAPYQX-LPEHRKFASA-N Ser-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CO)N)C(=O)O NRCJWSGXMAPYQX-LPEHRKFASA-N 0.000 description 1
- VAUMZJHYZQXZBQ-WHFBIAKZSA-N Ser-Asn-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O VAUMZJHYZQXZBQ-WHFBIAKZSA-N 0.000 description 1
- FIDMVVBUOCMMJG-CIUDSAMLSA-N Ser-Asn-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CO FIDMVVBUOCMMJG-CIUDSAMLSA-N 0.000 description 1
- KNZQGAUEYZJUSQ-ZLUOBGJFSA-N Ser-Asp-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CO)N KNZQGAUEYZJUSQ-ZLUOBGJFSA-N 0.000 description 1
- OLIJLNWFEQEFDM-SRVKXCTJSA-N Ser-Asp-Phe Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 OLIJLNWFEQEFDM-SRVKXCTJSA-N 0.000 description 1
- KCFKKAQKRZBWJB-ZLUOBGJFSA-N Ser-Cys-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)N[C@@H](C)C(O)=O KCFKKAQKRZBWJB-ZLUOBGJFSA-N 0.000 description 1
- SWIQQMYVHIXPEK-FXQIFTODSA-N Ser-Cys-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(O)=O SWIQQMYVHIXPEK-FXQIFTODSA-N 0.000 description 1
- YMAWDPHQVABADW-CIUDSAMLSA-N Ser-Gln-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCSC)C(O)=O YMAWDPHQVABADW-CIUDSAMLSA-N 0.000 description 1
- YQQKYAZABFEYAF-FXQIFTODSA-N Ser-Glu-Gln Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O YQQKYAZABFEYAF-FXQIFTODSA-N 0.000 description 1
- BRGQQXQKPUCUJQ-KBIXCLLPSA-N Ser-Glu-Ile Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O BRGQQXQKPUCUJQ-KBIXCLLPSA-N 0.000 description 1
- LALNXSXEYFUUDD-GUBZILKMSA-N Ser-Glu-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O LALNXSXEYFUUDD-GUBZILKMSA-N 0.000 description 1
- WBINSDOPZHQPPM-AVGNSLFASA-N Ser-Glu-Tyr Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CO)N)O WBINSDOPZHQPPM-AVGNSLFASA-N 0.000 description 1
- UQFYNFTYDHUIMI-WHFBIAKZSA-N Ser-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)[C@@H](N)CO UQFYNFTYDHUIMI-WHFBIAKZSA-N 0.000 description 1
- ZFVFHHZBCVNLGD-GUBZILKMSA-N Ser-His-Gln Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(N)=O)C(O)=O ZFVFHHZBCVNLGD-GUBZILKMSA-N 0.000 description 1
- YMDNFPNTIPQMJP-NAKRPEOUSA-N Ser-Ile-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCSC)C(O)=O YMDNFPNTIPQMJP-NAKRPEOUSA-N 0.000 description 1
- MOINZPRHJGTCHZ-MMWGEVLESA-N Ser-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N MOINZPRHJGTCHZ-MMWGEVLESA-N 0.000 description 1
- ZOPISOXXPQNOCO-SVSWQMSJSA-N Ser-Ile-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CO)N ZOPISOXXPQNOCO-SVSWQMSJSA-N 0.000 description 1
- VZQRNAYURWAEFE-KKUMJFAQSA-N Ser-Leu-Phe Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 VZQRNAYURWAEFE-KKUMJFAQSA-N 0.000 description 1
- KCGIREHVWRXNDH-GARJFASQSA-N Ser-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N KCGIREHVWRXNDH-GARJFASQSA-N 0.000 description 1
- GVMUJUPXFQFBBZ-GUBZILKMSA-N Ser-Lys-Glu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O GVMUJUPXFQFBBZ-GUBZILKMSA-N 0.000 description 1
- CRJZZXMAADSBBQ-SRVKXCTJSA-N Ser-Lys-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CO CRJZZXMAADSBBQ-SRVKXCTJSA-N 0.000 description 1
- LRZLZIUXQBIWTB-KATARQTJSA-N Ser-Lys-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LRZLZIUXQBIWTB-KATARQTJSA-N 0.000 description 1
- UGTZYIPOBYXWRW-SRVKXCTJSA-N Ser-Phe-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O UGTZYIPOBYXWRW-SRVKXCTJSA-N 0.000 description 1
- WOJYIMBIKTWKJO-KKUMJFAQSA-N Ser-Phe-His Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CO)N WOJYIMBIKTWKJO-KKUMJFAQSA-N 0.000 description 1
- XVWDJUROVRQKAE-KKUMJFAQSA-N Ser-Phe-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)CO)CC1=CC=CC=C1 XVWDJUROVRQKAE-KKUMJFAQSA-N 0.000 description 1
- PJIQEIFXZPCWOJ-FXQIFTODSA-N Ser-Pro-Asp Chemical compound [H]N[C@@H](CO)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O PJIQEIFXZPCWOJ-FXQIFTODSA-N 0.000 description 1
- XQAPEISNMXNKGE-FXQIFTODSA-N Ser-Pro-Cys Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CO)N)C(=O)N[C@@H](CS)C(=O)O XQAPEISNMXNKGE-FXQIFTODSA-N 0.000 description 1
- RHAPJNVNWDBFQI-BQBZGAKWSA-N Ser-Pro-Gly Chemical compound OC[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O RHAPJNVNWDBFQI-BQBZGAKWSA-N 0.000 description 1
- HHJFMHQYEAAOBM-ZLUOBGJFSA-N Ser-Ser-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O HHJFMHQYEAAOBM-ZLUOBGJFSA-N 0.000 description 1
- GYDFRTRSSXOZCR-ACZMJKKPSA-N Ser-Ser-Glu Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O GYDFRTRSSXOZCR-ACZMJKKPSA-N 0.000 description 1
- PYTKULIABVRXSC-BWBBJGPYSA-N Ser-Ser-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O PYTKULIABVRXSC-BWBBJGPYSA-N 0.000 description 1
- FLMYSKVSDVHLEW-SVSWQMSJSA-N Ser-Thr-Ile Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O FLMYSKVSDVHLEW-SVSWQMSJSA-N 0.000 description 1
- DYEGLQRVMBWQLD-IXOXFDKPSA-N Ser-Thr-Phe Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CO)N)O DYEGLQRVMBWQLD-IXOXFDKPSA-N 0.000 description 1
- KIEIJCFVGZCUAS-MELADBBJSA-N Ser-Tyr-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=C(C=C2)O)NC(=O)[C@H](CO)N)C(=O)O KIEIJCFVGZCUAS-MELADBBJSA-N 0.000 description 1
- UKKROEYWYIHWBD-ZKWXMUAHSA-N Ser-Val-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O UKKROEYWYIHWBD-ZKWXMUAHSA-N 0.000 description 1
- SGZVZUCRAVSPKQ-FXQIFTODSA-N Ser-Val-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CO)N SGZVZUCRAVSPKQ-FXQIFTODSA-N 0.000 description 1
- ANOQEBQWIAYIMV-AEJSXWLSSA-N Ser-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N ANOQEBQWIAYIMV-AEJSXWLSSA-N 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- CDBYLPFSWZWCQE-UHFFFAOYSA-L Sodium Carbonate Chemical class [Na+].[Na+].[O-]C([O-])=O CDBYLPFSWZWCQE-UHFFFAOYSA-L 0.000 description 1
- 240000003768 Solanum lycopersicum Species 0.000 description 1
- 235000002595 Solanum tuberosum Nutrition 0.000 description 1
- 244000061456 Solanum tuberosum Species 0.000 description 1
- 235000007230 Sorghum bicolor Nutrition 0.000 description 1
- 235000011684 Sorghum saccharatum Nutrition 0.000 description 1
- 101100427140 Stevia rebaudiana UGT74G1 gene Proteins 0.000 description 1
- 101100048059 Stevia rebaudiana UGT85C2 gene Proteins 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 239000004376 Sucralose Substances 0.000 description 1
- 101710156775 Sucrose synthase 3 Proteins 0.000 description 1
- 235000021536 Sugar beet Nutrition 0.000 description 1
- 241000192584 Synechocystis Species 0.000 description 1
- KEGBFULVYKYJRD-LFSVMHDDSA-N Thr-Ala-Phe Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 KEGBFULVYKYJRD-LFSVMHDDSA-N 0.000 description 1
- TWLMXDWFVNEFFK-FJXKBIBVSA-N Thr-Arg-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O TWLMXDWFVNEFFK-FJXKBIBVSA-N 0.000 description 1
- TZKPNGDGUVREEB-FOHZUACHSA-N Thr-Asn-Gly Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O TZKPNGDGUVREEB-FOHZUACHSA-N 0.000 description 1
- QILPDQCTQZDHFM-HJGDQZAQSA-N Thr-Gln-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O QILPDQCTQZDHFM-HJGDQZAQSA-N 0.000 description 1
- LGNBRHZANHMZHK-NUMRIWBASA-N Thr-Glu-Asp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N)O LGNBRHZANHMZHK-NUMRIWBASA-N 0.000 description 1
- BNGDYRRHRGOPHX-IFFSRLJSSA-N Thr-Glu-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)[C@@H](C)O)C(O)=O BNGDYRRHRGOPHX-IFFSRLJSSA-N 0.000 description 1
- XPNSAQMEAVSQRD-FBCQKBJTSA-N Thr-Gly-Gly Chemical compound C[C@@H](O)[C@H](N)C(=O)NCC(=O)NCC(O)=O XPNSAQMEAVSQRD-FBCQKBJTSA-N 0.000 description 1
- FDALPRWYVKJCLL-PMVVWTBXSA-N Thr-His-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)NCC(O)=O FDALPRWYVKJCLL-PMVVWTBXSA-N 0.000 description 1
- YDWLCDQXLCILCZ-BWAGICSOSA-N Thr-His-Tyr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O YDWLCDQXLCILCZ-BWAGICSOSA-N 0.000 description 1
- WPAKPLPGQNUXGN-OSUNSFLBSA-N Thr-Ile-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O WPAKPLPGQNUXGN-OSUNSFLBSA-N 0.000 description 1
- VTVVYQOXJCZVEB-WDCWCFNPSA-N Thr-Leu-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O VTVVYQOXJCZVEB-WDCWCFNPSA-N 0.000 description 1
- NCXVJIQMWSGRHY-KXNHARMFSA-N Thr-Leu-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N)O NCXVJIQMWSGRHY-KXNHARMFSA-N 0.000 description 1
- SCSVNSNWUTYSFO-WDCWCFNPSA-N Thr-Lys-Glu Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O SCSVNSNWUTYSFO-WDCWCFNPSA-N 0.000 description 1
- QHUWWSQZTFLXPQ-FJXKBIBVSA-N Thr-Met-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCSC)C(=O)NCC(O)=O QHUWWSQZTFLXPQ-FJXKBIBVSA-N 0.000 description 1
- JMBRNXUOLJFURW-BEAPCOKYSA-N Thr-Phe-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N2CCC[C@@H]2C(=O)O)N)O JMBRNXUOLJFURW-BEAPCOKYSA-N 0.000 description 1
- NYQIZWROIMIQSL-VEVYYDQMSA-N Thr-Pro-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O NYQIZWROIMIQSL-VEVYYDQMSA-N 0.000 description 1
- DNCUODYZAMHLCV-XGEHTFHBSA-N Thr-Pro-Cys Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CS)C(=O)O)N)O DNCUODYZAMHLCV-XGEHTFHBSA-N 0.000 description 1
- FWTFAZKJORVTIR-VZFHVOOUSA-N Thr-Ser-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O FWTFAZKJORVTIR-VZFHVOOUSA-N 0.000 description 1
- STUAPCLEDMKXKL-LKXGYXEUSA-N Thr-Ser-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O STUAPCLEDMKXKL-LKXGYXEUSA-N 0.000 description 1
- MFMGPEKYBXFIRF-SUSMZKCASA-N Thr-Thr-Gln Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(O)=O MFMGPEKYBXFIRF-SUSMZKCASA-N 0.000 description 1
- LECUEEHKUFYOOV-ZJDVBMNYSA-N Thr-Thr-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@@H](N)[C@@H](C)O LECUEEHKUFYOOV-ZJDVBMNYSA-N 0.000 description 1
- LVRFMARKDGGZMX-IZPVPAKOSA-N Thr-Tyr-Thr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)O)C(O)=O)CC1=CC=C(O)C=C1 LVRFMARKDGGZMX-IZPVPAKOSA-N 0.000 description 1
- OGOYMQWIWHGTGH-KZVJFYERSA-N Thr-Val-Ala Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O OGOYMQWIWHGTGH-KZVJFYERSA-N 0.000 description 1
- XGFYGMKZKFRGAI-RCWTZXSCSA-N Thr-Val-Arg Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N XGFYGMKZKFRGAI-RCWTZXSCSA-N 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 102100031954 Transcription and mRNA export factor ENY2 Human genes 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- 241000209140 Triticum Species 0.000 description 1
- 235000021307 Triticum Nutrition 0.000 description 1
- 241000209146 Triticum sp. Species 0.000 description 1
- 229920004890 Triton X-100 Polymers 0.000 description 1
- 239000013504 Triton X-100 Substances 0.000 description 1
- UKINEYBQXPMOJO-UBHSHLNASA-N Trp-Asn-Ser Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CO)C(=O)O)N UKINEYBQXPMOJO-UBHSHLNASA-N 0.000 description 1
- HXNVJPQADLRHGR-JBACZVJFSA-N Trp-Glu-Tyr Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)O)N HXNVJPQADLRHGR-JBACZVJFSA-N 0.000 description 1
- XLVRTKPAIXJYOH-HOCLYGCPSA-N Trp-His-Gly Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CN=CN3)C(=O)NCC(=O)O)N XLVRTKPAIXJYOH-HOCLYGCPSA-N 0.000 description 1
- KULBQAVOXHQLIY-HSCHXYMDSA-N Trp-Ile-Leu Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O)=CNC2=C1 KULBQAVOXHQLIY-HSCHXYMDSA-N 0.000 description 1
- NLLARHRWSFNEMH-NUTKFTJISA-N Trp-Lys-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N NLLARHRWSFNEMH-NUTKFTJISA-N 0.000 description 1
- OWSRIUBVJOQHNY-IHPCNDPISA-N Trp-Lys-His Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC3=CN=CN3)C(=O)O)N OWSRIUBVJOQHNY-IHPCNDPISA-N 0.000 description 1
- UUIYFDAWNBSWPG-IHPCNDPISA-N Trp-Lys-Lys Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N UUIYFDAWNBSWPG-IHPCNDPISA-N 0.000 description 1
- PWPJLBWYRTVYQS-PMVMPFDFSA-N Trp-Phe-Leu Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O PWPJLBWYRTVYQS-PMVMPFDFSA-N 0.000 description 1
- BOBZBMOTRORUPT-XIRDDKMYSA-N Trp-Ser-Leu Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O)=CNC2=C1 BOBZBMOTRORUPT-XIRDDKMYSA-N 0.000 description 1
- ZKVANNIVSDOQMG-HKUYNNGSSA-N Trp-Tyr-Gly Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)NCC(=O)O)N ZKVANNIVSDOQMG-HKUYNNGSSA-N 0.000 description 1
- MXKUGFHWYYKVDV-SZMVWBNQSA-N Trp-Val-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)Cc1c[nH]c2ccccc12)C(C)C)C(O)=O MXKUGFHWYYKVDV-SZMVWBNQSA-N 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- TVOGEPLDNYTAHD-CQDKDKBSSA-N Tyr-Ala-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 TVOGEPLDNYTAHD-CQDKDKBSSA-N 0.000 description 1
- ZRPLVTZTKPPSBT-AVGNSLFASA-N Tyr-Glu-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O ZRPLVTZTKPPSBT-AVGNSLFASA-N 0.000 description 1
- NOOMDULIORCDNF-IRXDYDNUSA-N Tyr-Gly-Phe Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O NOOMDULIORCDNF-IRXDYDNUSA-N 0.000 description 1
- JJNXZIPLIXIGBX-HJPIBITLSA-N Tyr-Ile-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N JJNXZIPLIXIGBX-HJPIBITLSA-N 0.000 description 1
- GGXUDPQWAWRINY-XEGUGMAKSA-N Tyr-Ile-Gly Chemical compound OC(=O)CNC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 GGXUDPQWAWRINY-XEGUGMAKSA-N 0.000 description 1
- KHCSOLAHNLOXJR-BZSNNMDCSA-N Tyr-Leu-Leu Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O KHCSOLAHNLOXJR-BZSNNMDCSA-N 0.000 description 1
- JLKVWTICWVWGSK-JYJNAYRXSA-N Tyr-Lys-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 JLKVWTICWVWGSK-JYJNAYRXSA-N 0.000 description 1
- XDGPTBVOSHKDFT-KKUMJFAQSA-N Tyr-Met-Glu Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCC(O)=O)C(O)=O XDGPTBVOSHKDFT-KKUMJFAQSA-N 0.000 description 1
- WTTRJMAZPDHPGS-KKXDTOCCSA-N Tyr-Phe-Ala Chemical compound C[C@H](NC(=O)[C@H](Cc1ccccc1)NC(=O)[C@@H](N)Cc1ccc(O)cc1)C(O)=O WTTRJMAZPDHPGS-KKXDTOCCSA-N 0.000 description 1
- QHONGSVIVOFKAC-ULQDDVLXSA-N Tyr-Pro-His Chemical compound N[C@@H](Cc1ccc(O)cc1)C(=O)N1CCC[C@H]1C(=O)N[C@@H](Cc1cnc[nH]1)C(O)=O QHONGSVIVOFKAC-ULQDDVLXSA-N 0.000 description 1
- SYFHQHYTNCQCCN-MELADBBJSA-N Tyr-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)[C@H](CC2=CC=C(C=C2)O)N)C(=O)O SYFHQHYTNCQCCN-MELADBBJSA-N 0.000 description 1
- AKKYBQGHUAWPJR-MNSWYVGCSA-N Tyr-Thr-Trp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CC3=CC=C(C=C3)O)N)O AKKYBQGHUAWPJR-MNSWYVGCSA-N 0.000 description 1
- 101150050575 URA3 gene Proteins 0.000 description 1
- ZLFHAAGHGQBQQN-GUBZILKMSA-N Val-Ala-Pro Natural products CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O ZLFHAAGHGQBQQN-GUBZILKMSA-N 0.000 description 1
- SLLKXDSRVAOREO-KZVJFYERSA-N Val-Ala-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C)NC(=O)[C@H](C(C)C)N)O SLLKXDSRVAOREO-KZVJFYERSA-N 0.000 description 1
- COYSIHFOCOMGCF-WPRPVWTQSA-N Val-Arg-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@H](C(=O)NCC(O)=O)CCCN=C(N)N COYSIHFOCOMGCF-WPRPVWTQSA-N 0.000 description 1
- COYSIHFOCOMGCF-UHFFFAOYSA-N Val-Arg-Gly Natural products CC(C)C(N)C(=O)NC(C(=O)NCC(O)=O)CCCN=C(N)N COYSIHFOCOMGCF-UHFFFAOYSA-N 0.000 description 1
- PAPWZOJOLKZEFR-AVGNSLFASA-N Val-Arg-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(=O)O)N PAPWZOJOLKZEFR-AVGNSLFASA-N 0.000 description 1
- CVUDMNSZAIZFAE-TUAOUCFPSA-N Val-Arg-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N1CCC[C@@H]1C(=O)O)N CVUDMNSZAIZFAE-TUAOUCFPSA-N 0.000 description 1
- DNOOLPROHJWCSQ-RCWTZXSCSA-N Val-Arg-Thr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O DNOOLPROHJWCSQ-RCWTZXSCSA-N 0.000 description 1
- ZQGPWORGSNRQLN-NHCYSSNCSA-N Val-Asp-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N ZQGPWORGSNRQLN-NHCYSSNCSA-N 0.000 description 1
- XLDYBRXERHITNH-QSFUFRPTSA-N Val-Asp-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)C(C)C XLDYBRXERHITNH-QSFUFRPTSA-N 0.000 description 1
- TZVUSFMQWPWHON-NHCYSSNCSA-N Val-Asp-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](C(C)C)N TZVUSFMQWPWHON-NHCYSSNCSA-N 0.000 description 1
- XTAUQCGQFJQGEJ-NHCYSSNCSA-N Val-Gln-Arg Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N XTAUQCGQFJQGEJ-NHCYSSNCSA-N 0.000 description 1
- XGJLNBNZNMVJRS-NRPADANISA-N Val-Glu-Ala Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O XGJLNBNZNMVJRS-NRPADANISA-N 0.000 description 1
- VLDMQVZZWDOKQF-AUTRQRHGSA-N Val-Glu-Gln Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N VLDMQVZZWDOKQF-AUTRQRHGSA-N 0.000 description 1
- SZTTYWIUCGSURQ-AUTRQRHGSA-N Val-Glu-Glu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O SZTTYWIUCGSURQ-AUTRQRHGSA-N 0.000 description 1
- WDIGUPHXPBMODF-UMNHJUIQSA-N Val-Glu-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N WDIGUPHXPBMODF-UMNHJUIQSA-N 0.000 description 1
- CELJCNRXKZPTCX-XPUUQOCRSA-N Val-Gly-Ala Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O CELJCNRXKZPTCX-XPUUQOCRSA-N 0.000 description 1
- BEGDZYNDCNEGJZ-XVKPBYJWSA-N Val-Gly-Gln Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(N)=O BEGDZYNDCNEGJZ-XVKPBYJWSA-N 0.000 description 1
- PIFJAFRUVWZRKR-QMMMGPOBSA-N Val-Gly-Gly Chemical compound CC(C)[C@H]([NH3+])C(=O)NCC(=O)NCC([O-])=O PIFJAFRUVWZRKR-QMMMGPOBSA-N 0.000 description 1
- FXVDGDZRYLFQKY-WPRPVWTQSA-N Val-Gly-Met Chemical compound CSCC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)C(C)C FXVDGDZRYLFQKY-WPRPVWTQSA-N 0.000 description 1
- DJQIUOKSNRBTSV-CYDGBPFRSA-N Val-Ile-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](C(C)C)N DJQIUOKSNRBTSV-CYDGBPFRSA-N 0.000 description 1
- SYSWVVCYSXBVJG-RHYQMDGZSA-N Val-Leu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](C(C)C)N)O SYSWVVCYSXBVJG-RHYQMDGZSA-N 0.000 description 1
- WDIWOIRFNMLNKO-ULQDDVLXSA-N Val-Leu-Tyr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 WDIWOIRFNMLNKO-ULQDDVLXSA-N 0.000 description 1
- OJPRSVJGNCAKQX-SRVKXCTJSA-N Val-Met-Arg Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N OJPRSVJGNCAKQX-SRVKXCTJSA-N 0.000 description 1
- MJFSRZZJQWZHFQ-SRVKXCTJSA-N Val-Met-Val Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C(C)C)C(=O)O)N MJFSRZZJQWZHFQ-SRVKXCTJSA-N 0.000 description 1
- WMRWZYSRQUORHJ-YDHLFZDLSA-N Val-Phe-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(=O)O)C(=O)O)N WMRWZYSRQUORHJ-YDHLFZDLSA-N 0.000 description 1
- YKNOJPJWNVHORX-UNQGMJICSA-N Val-Phe-Thr Chemical compound CC(C)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)O)C(O)=O)CC1=CC=CC=C1 YKNOJPJWNVHORX-UNQGMJICSA-N 0.000 description 1
- YTNGABPUXFEOGU-SRVKXCTJSA-N Val-Pro-Arg Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCN=C(N)N)C(O)=O YTNGABPUXFEOGU-SRVKXCTJSA-N 0.000 description 1
- HPOSMQWRPMRMFO-GUBZILKMSA-N Val-Pro-Cys Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CS)C(=O)O)N HPOSMQWRPMRMFO-GUBZILKMSA-N 0.000 description 1
- KSFXWENSJABBFI-ZKWXMUAHSA-N Val-Ser-Asn Chemical compound [H]N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O KSFXWENSJABBFI-ZKWXMUAHSA-N 0.000 description 1
- UGFMVXRXULGLNO-XPUUQOCRSA-N Val-Ser-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O UGFMVXRXULGLNO-XPUUQOCRSA-N 0.000 description 1
- QZKVWWIUSQGWMY-IHRRRGAJSA-N Val-Ser-Phe Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 QZKVWWIUSQGWMY-IHRRRGAJSA-N 0.000 description 1
- GBIUHAYJGWVNLN-UHFFFAOYSA-N Val-Ser-Pro Natural products CC(C)C(N)C(=O)NC(CO)C(=O)N1CCCC1C(O)=O GBIUHAYJGWVNLN-UHFFFAOYSA-N 0.000 description 1
- UJMCYJKPDFQLHX-XGEHTFHBSA-N Val-Ser-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](C(C)C)N)O UJMCYJKPDFQLHX-XGEHTFHBSA-N 0.000 description 1
- YQYFYUSYEDNLSD-YEPSODPASA-N Val-Thr-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O YQYFYUSYEDNLSD-YEPSODPASA-N 0.000 description 1
- USXYVSTVPHELAF-RCWTZXSCSA-N Val-Thr-Met Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](C(C)C)N)O USXYVSTVPHELAF-RCWTZXSCSA-N 0.000 description 1
- AYHNXCJKBLYVOA-KSZLIROESA-N Val-Trp-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N3CCC[C@@H]3C(=O)O)N AYHNXCJKBLYVOA-KSZLIROESA-N 0.000 description 1
- VTIAEOKFUJJBTC-YDHLFZDLSA-N Val-Tyr-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N VTIAEOKFUJJBTC-YDHLFZDLSA-N 0.000 description 1
- GTACFKZDQFTVAI-STECZYCISA-N Val-Tyr-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)CC1=CC=C(O)C=C1 GTACFKZDQFTVAI-STECZYCISA-N 0.000 description 1
- NLNCNKIVJPEFBC-DLOVCJGASA-N Val-Val-Glu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCC(O)=O NLNCNKIVJPEFBC-DLOVCJGASA-N 0.000 description 1
- AEFJNECXZCODJM-UWVGGRQHSA-N Val-Val-Gly Chemical compound CC(C)[C@H]([NH3+])C(=O)N[C@@H](C(C)C)C(=O)NCC([O-])=O AEFJNECXZCODJM-UWVGGRQHSA-N 0.000 description 1
- SSKKGOWRPNIVDW-AVGNSLFASA-N Val-Val-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N SSKKGOWRPNIVDW-AVGNSLFASA-N 0.000 description 1
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 1
- 241001464837 Viridiplantae Species 0.000 description 1
- 241000219094 Vitaceae Species 0.000 description 1
- 240000008042 Zea mays Species 0.000 description 1
- 235000005824 Zea mays ssp. parviglumis Nutrition 0.000 description 1
- 235000002017 Zea mays subsp mays Nutrition 0.000 description 1
- 241000235017 Zygosaccharomyces Species 0.000 description 1
- FJJCIZWZNKZHII-UHFFFAOYSA-N [4,6-bis(cyanoamino)-1,3,5-triazin-2-yl]cyanamide Chemical compound N#CNC1=NC(NC#N)=NC(NC#N)=N1 FJJCIZWZNKZHII-UHFFFAOYSA-N 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- 235000010358 acesulfame potassium Nutrition 0.000 description 1
- 229960004998 acesulfame potassium Drugs 0.000 description 1
- 239000000619 acesulfame-K Substances 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 101150102866 adc1 gene Proteins 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 238000001261 affinity purification Methods 0.000 description 1
- 108010045350 alanyl-tyrosyl-alanine Proteins 0.000 description 1
- 108010070944 alanylhistidine Proteins 0.000 description 1
- 230000003321 amplification Effects 0.000 description 1
- 210000004102 animal cell Anatomy 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 238000013459 approach Methods 0.000 description 1
- 239000012223 aqueous fraction Substances 0.000 description 1
- 239000008346 aqueous phase Substances 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Chemical group OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 1
- 108010008355 arginyl-glutamine Proteins 0.000 description 1
- 108010009111 arginyl-glycyl-glutamic acid Proteins 0.000 description 1
- 108010068380 arginylarginine Proteins 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 239000000605 aspartame Substances 0.000 description 1
- 235000010357 aspartame Nutrition 0.000 description 1
- IAOZJIPTCAWIRG-QWRGUYRKSA-N aspartame Chemical compound OC(=O)C[C@H](N)C(=O)N[C@H](C(=O)OC)CC1=CC=CC=C1 IAOZJIPTCAWIRG-QWRGUYRKSA-N 0.000 description 1
- 229960003438 aspartame Drugs 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 108010040443 aspartyl-aspartic acid Proteins 0.000 description 1
- 108010069205 aspartyl-phenylalanine Proteins 0.000 description 1
- 238000003556 assay Methods 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Chemical group OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical group OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 230000027455 binding Effects 0.000 description 1
- 102000023732 binding proteins Human genes 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 239000002551 biofuel Substances 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 230000006696 biosynthetic metabolic pathway Effects 0.000 description 1
- 239000007853 buffer solution Substances 0.000 description 1
- 230000003139 buffering effect Effects 0.000 description 1
- 102100022422 cGMP-dependent protein kinase 1 Human genes 0.000 description 1
- 239000001110 calcium chloride Substances 0.000 description 1
- 229910001628 calcium chloride Inorganic materials 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 229910052799 carbon Inorganic materials 0.000 description 1
- 230000003197 catalytic effect Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 230000019522 cellular metabolic process Effects 0.000 description 1
- 235000013339 cereals Nutrition 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 229930002868 chlorophyll a Natural products 0.000 description 1
- ATNHDLDRLWWWCB-AENOIHSZSA-M chlorophyll a Chemical compound C1([C@@H](C(=O)OC)C(=O)C2=C3C)=C2N2C3=CC(C(CC)=C3C)=[N+]4C3=CC3=C(C=C)C(C)=C5N3[Mg-2]42[N+]2=C1[C@@H](CCC(=O)OC\C=C(/C)CCC[C@H](C)CCC[C@H](C)CCCC(C)C)[C@H](C)C2=C5 ATNHDLDRLWWWCB-AENOIHSZSA-M 0.000 description 1
- 229930002869 chlorophyll b Natural products 0.000 description 1
- NSMUHPMZFPKNMZ-VBYMZDBQSA-M chlorophyll b Chemical compound C1([C@@H](C(=O)OC)C(=O)C2=C3C)=C2N2C3=CC(C(CC)=C3C=O)=[N+]4C3=CC3=C(C=C)C(C)=C5N3[Mg-2]42[N+]2=C1[C@@H](CCC(=O)OC\C=C(/C)CCC[C@H](C)CCC[C@H](C)CCCC(C)C)[C@H](C)C2=C5 NSMUHPMZFPKNMZ-VBYMZDBQSA-M 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 239000013611 chromosomal DNA Substances 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 210000000349 chromosome Anatomy 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 238000000975 co-precipitation Methods 0.000 description 1
- 239000003184 complementary RNA Substances 0.000 description 1
- 239000000356 contaminant Substances 0.000 description 1
- 235000014510 cooky Nutrition 0.000 description 1
- 235000005822 corn Nutrition 0.000 description 1
- 108010004073 cysteinylcysteine Proteins 0.000 description 1
- 229940104302 cytosine Drugs 0.000 description 1
- 235000013365 dairy product Nutrition 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- 235000005911 diet Nutrition 0.000 description 1
- 230000000378 dietary effect Effects 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 108010054812 diprotin A Proteins 0.000 description 1
- 238000006073 displacement reaction Methods 0.000 description 1
- 238000000132 electrospray ionisation Methods 0.000 description 1
- 239000003480 eluent Substances 0.000 description 1
- 230000000408 embryogenic effect Effects 0.000 description 1
- NIKHGUQULKYIGE-SHAPNJEPSA-N ent-kaur-16-en-19-oic acid Chemical compound C([C@H]1C[C@]2(CC1=C)CC1)C[C@H]2[C@@]2(C)[C@H]1[C@](C)(C(O)=O)CCC2 NIKHGUQULKYIGE-SHAPNJEPSA-N 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- 239000002038 ethyl acetate fraction Substances 0.000 description 1
- 241001233957 eudicotyledons Species 0.000 description 1
- 102000013165 exonuclease Human genes 0.000 description 1
- 108010093305 exopolygalacturonase Proteins 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 239000012467 final product Substances 0.000 description 1
- 235000013373 food additive Nutrition 0.000 description 1
- 239000002778 food additive Substances 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 108010063718 gamma-glutamylaspartic acid Proteins 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 108010042598 glutamyl-aspartyl-glycine Proteins 0.000 description 1
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 1
- 230000001279 glycosylating effect Effects 0.000 description 1
- 108700014210 glycosyltransferase activity proteins Proteins 0.000 description 1
- 102000045442 glycosyltransferase activity proteins Human genes 0.000 description 1
- XBGGUPMXALFZOT-UHFFFAOYSA-N glycyl-L-tyrosine hemihydrate Natural products NCC(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 XBGGUPMXALFZOT-UHFFFAOYSA-N 0.000 description 1
- 108010019832 glycyl-asparaginyl-glycine Proteins 0.000 description 1
- 108010072405 glycyl-aspartyl-glycine Proteins 0.000 description 1
- 108010062266 glycyl-glycyl-argininal Proteins 0.000 description 1
- 108010050475 glycyl-leucyl-tyrosine Proteins 0.000 description 1
- 108010084389 glycyltryptophan Proteins 0.000 description 1
- 235000021021 grapes Nutrition 0.000 description 1
- 239000001257 hydrogen Substances 0.000 description 1
- 230000003301 hydrolyzing effect Effects 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 235000015243 ice cream Nutrition 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 230000001939 inductive effect Effects 0.000 description 1
- 238000001764 infiltration Methods 0.000 description 1
- 238000002347 injection Methods 0.000 description 1
- 239000007924 injection Substances 0.000 description 1
- 229940030980 inova Drugs 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 230000000968 intestinal effect Effects 0.000 description 1
- 238000010829 isocratic elution Methods 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- 108010031424 isoleucyl-prolyl-proline Proteins 0.000 description 1
- 108010027338 isoleucylcysteine Proteins 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- 108010077158 leucinyl-arginyl-tryptophan Proteins 0.000 description 1
- 108010044311 leucyl-glycyl-glycine Proteins 0.000 description 1
- 108010051673 leucyl-glycyl-phenylalanine Proteins 0.000 description 1
- 108010047926 leucyl-lysyl-tyrosine Proteins 0.000 description 1
- 239000003446 ligand Substances 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 238000009630 liquid culture Methods 0.000 description 1
- 239000012139 lysis buffer Substances 0.000 description 1
- 229960000274 lysozyme Drugs 0.000 description 1
- 239000004325 lysozyme Substances 0.000 description 1
- 235000010335 lysozyme Nutrition 0.000 description 1
- 108010044348 lysyl-glutamyl-aspartic acid Proteins 0.000 description 1
- 108010054155 lysyllysine Proteins 0.000 description 1
- 229940035034 maltodextrin Drugs 0.000 description 1
- 238000001819 mass spectrum Methods 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000001404 mediated effect Effects 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 108010063431 methionyl-aspartyl-glycine Proteins 0.000 description 1
- 108010005942 methionylglycine Proteins 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 235000021096 natural sweeteners Nutrition 0.000 description 1
- 108010070257 neotame Proteins 0.000 description 1
- 235000019412 neotame Nutrition 0.000 description 1
- HLIAVLHNDJUHFG-HOTGVXAUSA-N neotame Chemical compound CC(C)(C)CCN[C@@H](CC(O)=O)C(=O)N[C@H](C(=O)OC)CC1=CC=CC=C1 HLIAVLHNDJUHFG-HOTGVXAUSA-N 0.000 description 1
- 235000013615 non-nutritive sweetener Nutrition 0.000 description 1
- 108010058731 nopaline synthase Proteins 0.000 description 1
- 238000001208 nuclear magnetic resonance pulse sequence Methods 0.000 description 1
- 238000003199 nucleic acid amplification method Methods 0.000 description 1
- 238000000238 one-dimensional nuclear magnetic resonance spectroscopy Methods 0.000 description 1
- 230000002018 overexpression Effects 0.000 description 1
- 235000019629 palatability Nutrition 0.000 description 1
- 235000020232 peanut Nutrition 0.000 description 1
- 235000011197 perejil Nutrition 0.000 description 1
- 239000000825 pharmaceutical preparation Substances 0.000 description 1
- 229940127557 pharmaceutical product Drugs 0.000 description 1
- 230000002974 pharmacogenomic effect Effects 0.000 description 1
- 150000002989 phenols Chemical class 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 108010024607 phenylalanylalanine Proteins 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 230000029553 photosynthesis Effects 0.000 description 1
- 238000010672 photosynthesis Methods 0.000 description 1
- 230000000704 physical effect Effects 0.000 description 1
- 239000000049 pigment Substances 0.000 description 1
- 108010025488 pinealon Proteins 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 235000012015 potatoes Nutrition 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 238000003825 pressing Methods 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 108700042769 prolyl-leucyl-glycine Proteins 0.000 description 1
- 108010077112 prolyl-proline Proteins 0.000 description 1
- 108010079317 prolyl-tyrosine Proteins 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 230000006337 proteolytic cleavage Effects 0.000 description 1
- 238000010926 purge Methods 0.000 description 1
- QRGRAFPOLJOGRV-UHFFFAOYSA-N rebaudioside F Natural products CC12CCCC(C)(C1CCC34CC(=C)C(CCC23)(C4)OC5OC(CO)C(O)C(OC6OCC(O)C(O)C6O)C5OC7OC(CO)C(O)C(O)C7O)C(=O)OC8OC(CO)C(O)C(O)C8O QRGRAFPOLJOGRV-UHFFFAOYSA-N 0.000 description 1
- HYLAUKAHEAUVFE-AVBZULRRSA-N rebaudioside f Chemical compound O([C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O[C@H]1[C@@H]([C@@H](O)[C@H](O)CO1)O)O[C@]12C(=C)C[C@@]3(C1)CC[C@@H]1[C@@](C)(CCC[C@]1([C@@H]3CC2)C)C(=O)O[C@H]1[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O1)O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O HYLAUKAHEAUVFE-AVBZULRRSA-N 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 238000010992 reflux Methods 0.000 description 1
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 235000019204 saccharin Nutrition 0.000 description 1
- CVHZOJJKTDOEJC-UHFFFAOYSA-N saccharin Chemical compound C1=CC=C2C(=O)NS(=O)(=O)C2=C1 CVHZOJJKTDOEJC-UHFFFAOYSA-N 0.000 description 1
- 229940081974 saccharin Drugs 0.000 description 1
- 239000000901 saccharin and its Na,K and Ca salt Substances 0.000 description 1
- 108010069117 seryl-lysyl-aspartic acid Proteins 0.000 description 1
- 108010048397 seryl-lysyl-leucine Proteins 0.000 description 1
- 108010026333 seryl-proline Proteins 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- 239000007974 sodium acetate buffer Substances 0.000 description 1
- 235000014214 soft drink Nutrition 0.000 description 1
- 239000002689 soil Substances 0.000 description 1
- 230000003595 spectral effect Effects 0.000 description 1
- 229940082787 spirulina Drugs 0.000 description 1
- 108010005652 splenotritin Proteins 0.000 description 1
- QSIDJGUAAUSPMG-CULFPKEHSA-N steviolmonoside Chemical compound O([C@]12C(=C)C[C@@]3(C1)CC[C@@H]1[C@@](C)(CCC[C@]1([C@@H]3CC2)C)C(O)=O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O QSIDJGUAAUSPMG-CULFPKEHSA-N 0.000 description 1
- 235000021012 strawberries Nutrition 0.000 description 1
- 235000019408 sucralose Nutrition 0.000 description 1
- BAQAVOSOZGMPRM-QBMZZYIRSA-N sucralose Chemical compound O[C@@H]1[C@@H](O)[C@@H](Cl)[C@@H](CO)O[C@@H]1O[C@@]1(CCl)[C@@H](O)[C@H](O)[C@@H](CCl)O1 BAQAVOSOZGMPRM-QBMZZYIRSA-N 0.000 description 1
- 229960004793 sucrose Drugs 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 230000002459 sustained effect Effects 0.000 description 1
- 230000002194 synthesizing effect Effects 0.000 description 1
- 230000008685 targeting Effects 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 108010061238 threonyl-glycine Proteins 0.000 description 1
- 108010031491 threonyl-lysyl-glutamic acid Proteins 0.000 description 1
- 229940113082 thymine Drugs 0.000 description 1
- OQPOFZJZPYRNFF-CULFPKEHSA-N tkd5uc898q Chemical compound O=C([C@]1(C)CCC[C@@]2([C@@H]1CC[C@]13C[C@](O)(C(=C)C1)CC[C@@H]23)C)O[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O OQPOFZJZPYRNFF-CULFPKEHSA-N 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 230000001131 transforming effect Effects 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 1
- 108010080629 tryptophan-leucine Proteins 0.000 description 1
- 108010044292 tryptophyltyrosine Proteins 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 108010035534 tyrosyl-leucyl-alanine Proteins 0.000 description 1
- 238000004704 ultra performance liquid chromatography Methods 0.000 description 1
- 238000002604 ultrasonography Methods 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 108010073969 valyllysine Proteins 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 238000001262 western blot Methods 0.000 description 1
- 108010027345 wheylin-1 peptide Proteins 0.000 description 1
- 108010000998 wheylin-2 peptide Proteins 0.000 description 1
- 210000005253 yeast cell Anatomy 0.000 description 1
Images














Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/44—Preparation of O-glycosides, e.g. glucosides
- C12P19/56—Preparation of O-glycosides, e.g. glucosides having an oxygen atom of the saccharide radical directly bound to a condensed ring system having three or more carbocyclic rings, e.g. daunomycin, adriamycin
-
- A—HUMAN NECESSITIES
- A21—BAKING; EDIBLE DOUGHS
- A21D—TREATMENT OF FLOUR OR DOUGH FOR BAKING, e.g. BY ADDITION OF MATERIALS; BAKING; BAKERY PRODUCTS
- A21D13/00—Finished or partly finished bakery products
- A21D13/06—Products with modified nutritive value, e.g. with modified starch content
- A21D13/062—Products with modified nutritive value, e.g. with modified starch content with modified sugar content; Sugar-free products
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23G—COCOA; COCOA PRODUCTS, e.g. CHOCOLATE; SUBSTITUTES FOR COCOA OR COCOA PRODUCTS; CONFECTIONERY; CHEWING GUM; ICE-CREAM; PREPARATION THEREOF
- A23G3/00—Sweetmeats; Confectionery; Marzipan; Coated or filled products
- A23G3/34—Sweetmeats, confectionery or marzipan; Processes for the preparation thereof
- A23G3/36—Sweetmeats, confectionery or marzipan; Processes for the preparation thereof characterised by the composition containing organic or inorganic compounds
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23G—COCOA; COCOA PRODUCTS, e.g. CHOCOLATE; SUBSTITUTES FOR COCOA OR COCOA PRODUCTS; CONFECTIONERY; CHEWING GUM; ICE-CREAM; PREPARATION THEREOF
- A23G3/00—Sweetmeats; Confectionery; Marzipan; Coated or filled products
- A23G3/34—Sweetmeats, confectionery or marzipan; Processes for the preparation thereof
- A23G3/36—Sweetmeats, confectionery or marzipan; Processes for the preparation thereof characterised by the composition containing organic or inorganic compounds
- A23G3/38—Sucrose-free products
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23G—COCOA; COCOA PRODUCTS, e.g. CHOCOLATE; SUBSTITUTES FOR COCOA OR COCOA PRODUCTS; CONFECTIONERY; CHEWING GUM; ICE-CREAM; PREPARATION THEREOF
- A23G4/00—Chewing gum
- A23G4/06—Chewing gum characterised by the composition containing organic or inorganic compounds
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L2/00—Non-alcoholic beverages; Dry compositions or concentrates therefor; Preparation or treatment thereof
- A23L2/52—Adding ingredients
- A23L2/60—Sweeteners
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L27/00—Spices; Flavouring agents or condiments; Artificial sweetening agents; Table salts; Dietetic salt substitutes; Preparation or treatment thereof
- A23L27/30—Artificial sweetening agents
- A23L27/33—Artificial sweetening agents containing sugars or derivatives
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L27/00—Spices; Flavouring agents or condiments; Artificial sweetening agents; Table salts; Dietetic salt substitutes; Preparation or treatment thereof
- A23L27/30—Artificial sweetening agents
- A23L27/33—Artificial sweetening agents containing sugars or derivatives
- A23L27/36—Terpene glycosides
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES, NOT OTHERWISE PROVIDED FOR; PREPARATION OR TREATMENT THEREOF
- A23L33/00—Modifying nutritive qualities of foods; Dietetic products; Preparation or treatment thereof
- A23L33/10—Modifying nutritive qualities of foods; Dietetic products; Preparation or treatment thereof using additives
- A23L33/105—Plant extracts, their artificial duplicates or their derivatives
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07H—SUGARS; DERIVATIVES THEREOF; NUCLEOSIDES; NUCLEOTIDES; NUCLEIC ACIDS
- C07H15/00—Compounds containing hydrocarbon or substituted hydrocarbon radicals directly attached to hetero atoms of saccharide radicals
- C07H15/20—Carbocyclic rings
- C07H15/24—Condensed ring systems having three or more rings
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
- C12N9/1051—Hexosyltransferases (2.4.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
- C12N9/1051—Hexosyltransferases (2.4.1)
- C12N9/1062—Sucrose synthase (2.4.1.13)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y204/00—Glycosyltransferases (2.4)
- C12Y204/01—Hexosyltransferases (2.4.1)
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23V—INDEXING SCHEME RELATING TO FOODS, FOODSTUFFS OR NON-ALCOHOLIC BEVERAGES AND LACTIC OR PROPIONIC ACID BACTERIA USED IN FOODSTUFFS OR FOOD PREPARATION
- A23V2002/00—Food compositions, function of food ingredients or processes for food or foodstuffs
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y204/00—Glycosyltransferases (2.4)
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Health & Medical Sciences (AREA)
- Food Science & Technology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Polymers & Plastics (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Nutrition Science (AREA)
- Inorganic Chemistry (AREA)
- Medicinal Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Botany (AREA)
- Mycology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
- Coloring Foods And Improving Nutritive Qualities (AREA)
- Seasonings (AREA)
Abstract
Изобретение относится к области области биотехнологии. Описана группа изобретений, включающая способ получения соединения стевиол–гликозида или композиции стевиол–гликозидов, соединение ребаудиозида Z2, подсластитель, содержащий вышеуказанное соединение ребаудиозид Z2, и применение подсластителя в пищевом продукте. В одном из вариантов реализации ребаудиозид Z2 имеет структуру, представленную формулой:
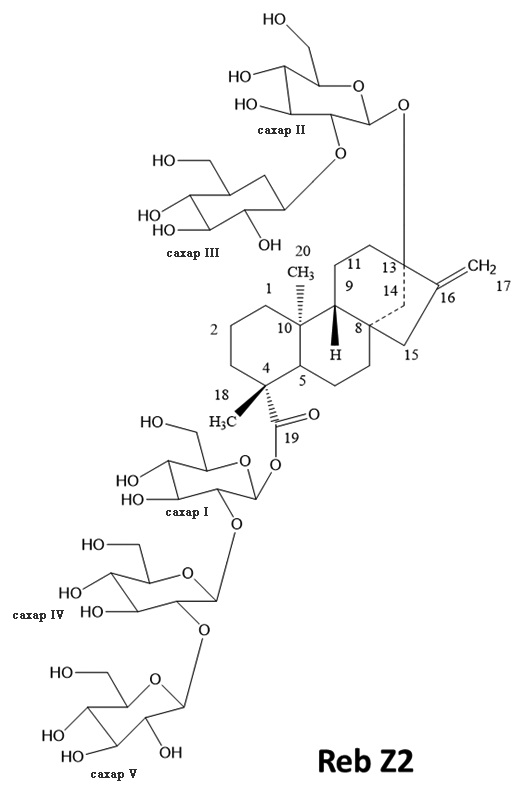
Изобретение расширяет арсенал средств стевиол–гликозидов и способов их получения для использования в пищевой промышленности. 4 н. и 8 з.п. ф-лы, 2 табл., 4 пр., 14 ил.
Description
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[1] По настоящей заявке испрашивается приоритет временной заявки на патент США 61/898,571, поданной 1 ноября 2013 года, которая настоящим полностью включена посредством отсылки.
ВКЛЮЧЕНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[2] Бумажная копия Списка последовательностей и машиночитаемая форма последовательности, содержащая файл под названием 32559-17_ST25.txt, который имеет размер 46751 байт (согласно измерению в Microsoft WINDOWS® Explorer), представлена в настоящей заявке и включена посредством отсылки. Указанный Список последовательностей состоит из SEQ ID NO: 1-12.
УРОВЕНЬ ТЕХНИКИ
[3] Настоящее описание в целом относится к биосинтезу стевиол-гликозидов. В частности, настоящее описание относится к рекомбинантному полипептиду, который катализирует продукцию стевиол-гликозидов, таких как ребаудиозид D, ребаудиозид E и новый ребаудиозид (ребаудиозид Z).
[4] Стевиол-гликозиды представляют собой природные продукты, выделенные из листьев Stevia rebaudiana, и широко применяются в качестве высокоинтенсивных, низкокалорийных подсластителей. Природные стевиол-гликозиды имеют одинаковую основную структуру (стевиол) и отличаются содержанием углеводных остатков (например, остатки глюкозы, рамнозы и ксилозы) в положениях C13 и C19. Стевиол-гликозиды с известной структурой включают стевиозид, ребаудиозид A, ребаудиозид B, ребаудиозид C, ребаудиозид D, ребаудиозид E, ребаудиозид F и дулькозид A.
[5] В пересчете на сухой вес стевиозид, ребаудиозид A, ребаудиозид C и дулькозид A составляют 9,1, 3,8, 0,6 и 0,3% от общего веса стевиол-гликозидов в листьях, соответственно, в то время как другие стевиол-глюкозиды присутствуют в намного более низком количестве. Экстракты из растения Stevia rebaudiana доступны коммерчески и обычно содержат стевиозид и ребаудиозид в качестве основных соединений. Другие стевиол-гликозиды обычно присутствуют в экстракте стевии в качестве минорных компонентов. Например, количество ребаудиозида A в коммерческих препаратах может изменяться от приблизительно 20% до более чем 90% от общего количества содержащихся стевиол-гликозидов, тогда как количество ребаудиозида B может составлять приблизительно 1-2%, количество ребаудиозида C может составлять приблизительно 7-15%, и количество ребаудиозида D может составлять приблизительно 2% от общего количества стевиол-гликозидов.
[6] Как натуральные подсластители, различные стевиол-глюкозиды имеют различные уровни сладости и послевкусие. Сладость стевиол-гликозидов значительно превышает сладость сахарозы. Например, стевиозид в 100-150 раз слаще сахарозы и имеет горькое послевкусие, тогда как ребаудиозид A и E в 250-450 раз слаще сахарозы и их послевкусие намного лучше, чем у стевиозида. Соответственно, на вкусовой профиль любого экстракта стевии существенно влияет относительное содержание стевиол-гликозидов в экстракте, на которое в свою очередь может влиять источник растения, факторы окружающей среды (такие как состав почвы и климат) и процесс экстракции. В частности, изменения условий экстракции могут привести к непостоянному содержанию стевиол-гликозидов в экстрактах стевии, в результате чего вкусовой профиль различается у разных партий продуктов экстракции. На вкусовой профиль экстрактов стевии также могут влиять содержащиеся в растении примеси (такие как пигменты, липиды, белки, фенольные соединения и сахариды), которые остаются в продукте после процесса экстракции. Такие примеси обычно имеют посторонние привкусы, нежелательные в случае применения экстракта стевии в качестве подсластителя.
[7] Большинство стевиол-гликозидов образуются в результате нескольких реакций гликозилирования стевиола, которые обычно катализируют УДФ-гликозилтрансферазы (УГТ) при использовании уридин-5’-дифосфоглюкозы (УДФ-глюкозы) в качестве донора сахарной группы. В растениях УГТ представляют крайне неоднородную группу ферментов, которые переносят остаток глюкозы от УДФ-глюкозы к стевиолу. Стевиозид является промежуточным соединением в биосинтезе ребаудиозидных соединений. Например, гликозилирование C-3’ C-13-O-глюкозы стевиозида дает ребаудиозид A; а гликозилирование C-2’ 19-O-глюкозы стевиозида дает ребаудиозид E. Дальнейшее гликозилирование ребаудиозида A (по 19-O-глюкозе) или ребаудиозида E (по C-13-O-глюкозе) дает ребаудиозид D (Фигура 1).
[8] Практический подход к улучшению вкусовых качеств экстракта стевии состоит в повышении выхода ребаудиозидных соединений в результате дальнейшего гликозилирования стевиозида. УГТ с 1,2-19-O-глюкоза гликозилирующей активностью являются важными ферментами для получения ребаудиозида D и E.
[9] Сахарозосинтазы (СС) катализируют превращение УДФ в УДФ-глюкозу в присутствии сахарозы. Таким образом, для реакции гликозилирования, использующей УДФ-глюкозу (такой как реакции, катализируемые УГТ), СС может использоваться для регенерации УДФ-глюкозы из УДФ, повышая эффективность такой реакции (Фигура 2).
[10] Таким образом, существует потребность в стевиол-гликозидах с постоянным вкусовым профилем и менее интенсивным посторонним привкусом, чем у существующих коммерческих продуктов. Как описано в настоящей заявке, в настоящем описании предложен рекомбинантный полипептид, который может применяться для получения стевиол-гликозидов (таких как ребаудиозид D и ребаудиозид E). В настоящем описании также предложен способ получения композиции стевиол-гликозида (ребаудиозида Z) с применением такого рекомбинантного полипептида.
КРАТКОЕ ОПИСАНИЕ
[11] Рассматриваемая технология в общем относится к рекомбинантным полипептидам, которые обладают активностями УДФ-гликозилтрансферазы. В частности, предложены полипептиды, обладающие 1,2-19-O-глюкоза-гликозилирующей активностью в отношении стевиол-гликозидных соединений. В одном аспекте рассматриваемая технология относится к рекомбинантному полипептиду, включающему аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. В примере осуществления аминокислотная последовательность рекомбинантного полипептида, описанного в настоящей заявке, обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с SEQ ID NO:6.
[12] В другом аспекте рассматриваемая технология относится к выделенной нуклеиновой кислоте, включающей нуклеотидную последовательность, кодирующую рекомбинантный полипептид, описанный в настоящей заявке. В другом аспекте рассматриваемая технология относится к вектору, включающему нуклеиновую кислоту, описанную в настоящей заявке, и клетке-хозяину, включающей вектор, описанный в настоящей заявке. В примере осуществления клетка-хозяин рассматриваемой технологии выбрана из группы, состоящей из клетки бактерий, дрожжей, нитчатых грибов, цианобактерий и растений.
[13] В другом аспекте рассматриваемая технология также относится к способу получения композиции стевиол-гликозида, включающему инкубирование субстрата (такого как стевиозид, ребаудиозид A, ребаудиозид E или их комбинация) с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. В примере осуществления аминокислотная последовательность рекомбинантного полипептида, применяемого в способе, описанном в настоящей заявке, обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с SEQ ID NO:6.
[14] В другом аспекте рассматриваемая технология также относится к способу получения композиции стевиол-гликозида, включающему инкубирование субстрата (такого как стевиозид и ребаудиозид E) с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:11. В примере осуществления аминокислотная последовательность рекомбинантного полипептида, применяемого в способе, описанном в настоящей заявке, обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с SEQ ID NO:11.
[15] В одном варианте осуществления способ дополнительно включает инкубирование рекомбинантной сахарозосинтазы (такой как сахарозосинтаза с аминокислотной последовательностью, обладающей по меньшей мере 80% идентичностью с аминокислотной последовательностью AtSUS1, представленной в SEQ ID NO:9) с субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. В другом варианте осуществления способ дополнительно включает инкубирование рекомбинантной УДФ-гликозилтрансферазы (такой как УДФ-гликозилтрансфераза с аминокислотной последовательностью, обладающей по меньшей мере 80% идентичностью с аминокислотной последовательностью UGT76G1, представленной в SEQ ID NO:11) с рекомбинантной сахарозосинтазой, субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. В другом варианте осуществления способ, описанный в настоящей заявке, включает инкубирование субстрата с клеткой-хозяином, экспрессирующей рекомбинантный полипептид.
[16] Рассматриваемая технология также относится к новому стевиол-гликозиду, называемому ребаудиозид Z, который отличается временем удержания приблизительно 6,68 минут на ВЭЖХ при условиях, описанных в настоящей заявке. Рассматриваемая технология также относится к способу получения ребаудиозида Z, описанного в настоящей заявке, включающему инкубирование субстрата с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. При использовании в настоящем описании, термины "ребаудиозид Z" или "Reb Z" относятся к смеси соединений, в частности смеси ребаудиозида Z1 ("Reb Z1") и ребаудиозида Z2 ("Reb Z2").
[17] В одном варианте осуществления настоящее описание также направлено на способ синтеза ребаудиозида Z из ребаудиозида E. Способ включает: получение реакционной смеси, включающей ребаудиозид E, субстрат, выбранный из группы, состоящей из сахарозы, уридиндифосфата (УДФ) и уридиндифосфат-глюкозы (УДФ-глюкозы), и HV1, и сахарозосинтазу, инкубирование реакционной смеси в течение времени, достаточного для получения ребаудиозида Z, где глюкоза ковалентно присоединяется к ребаудиозиду E с получением ребаудиозида Z, глюкоза ковалентно присоединяется к C2’-13-O-глюкозе ребаудиозида E с получением ребаудиозида Z1, и глюкоза ковалентно присоединяется к C2’-19-O-глюкозе ребаудиозида E с получением ребаудиозида Z2.
[18] В одном варианте осуществления соединение ребаудиозид Z является ребаудиозидом Z1 (Reb Z1), имеющим следующую структуру:

[19] В одном варианте осуществления соединение ребаудиозид Z является ребаудиозидом Z2 (Reb Z2), имеющим следующую структуру:

[20] Как описано в настоящей заявке, рекомбинантные полипептиды согласно настоящей технологии могут использоваться для разработки биосинтетического способа получения стевиол-гликозидов, которые обычно мало распространены в природных источниках, например, ребаудиозида D и ребаудиозида E. Таким образом, настоящая технология также обеспечивает композицию стевиол-гликозида, полученную биосинтетическим способом, описанным в настоящей заявке. Такая композиция может включать стевиол-гликозидное соединение, выбранное из группы, состоящей из ребаудиозида D, ребаудиозида E, нового ребаудиозида (указанного в настоящей заявке как "ребаудиозид Z" и "Reb Z") и их комбинаций. Кроме того, также предложен подсластитель, включающий композицию стевиол-гликозидов, описанную в настоящей заявке.
[21] В одном варианте осуществления настоящее описание направлено на подсластитель, включающий соединение, имеющее следующую химическую структуру:

[22] В другом варианте осуществления подсластитель включает соединение, имеющее следующую химическую структуру:

[23] Настоящее описание также направлено на применение подсластителей в потребительских продуктах, таких как напитки, кондитерские изделия, хлебобулочные изделия, печенье и жевательные резинки.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[24] Описание будет более понятным, и особенности, аспекты и преимущества, помимо представленных выше, станут ясны при рассмотрении их подробного описания, следующего далее. Такое подробное описание ссылается на следующие чертежи, где:
[25] Фигура 1 является схемой, на которой показаны пути биосинтеза стевиол-гликозида из стевиозида. Как описано в настоящей заявке, рекомбинантный полипептид HV1 ("HV1") содержит 1,2-19-O-глюкоза-гликозилирующую активность, которая переносит вторую сахарную группу на C-2’ 19-O-глюкозы стевиозида с получением ребаудиозида E ("Reb E") или, аналогично, с получением ребаудиозида D ("Reb D") из ребаудиозида A ("Reb A"). На Фигуре 1 также показано, что рекомбинантный фермент UGT76G1 ("UGT76G1", отличающийся от рекомбинантного полипептида HV1) катализирует реакцию, которая переносит сахарную группу на C-3’ C-13-O-глюкозы стевиозида с получением ребаудиозида A или, аналогично, с получением ребаудиозида D из ребаудиозида E.
[26] На Фигуре 2 показана примерная схема комбинированной реакционной системы УДФ-гликозилтрансферазы ("УГТ") и сахарозосинтазы ("СС"). В Реакции 1 показана катализируемая УГТ реакция, превращающая ребаудиозид A ("Reb A") в ребаудиозид D ("Reb D"), в которой УДФ-глюкоза используется в качестве донора глюкозы и приводит к получению УДФ. В Реакции 2 показана катализируемая СС реакция, превращающая УДФ в УДФ-глюкозу, в которой сахароза используется в качестве донора глюкозы. В Реакции 2 также показано, что катализируемая СС реакция может быть объединена с реакцией, катализируемой УГТ.
[27] На Фигуре 3 показано in vitro получение ребаудиозида D ("Reb D") из ребаудиозида ("Reb A"), катализируемое рекомбинантным полипептидом HV1 (SEQ ID NO:6) и рекомбинантной AtSUS1 (SEQ ID NO:9) в комбинированной реакционной системе HV1-AtSUS1, как описано в настоящей заявке. На Фигуре 3A показаны стандарты стевиозида ("Ste"), Ребаудиозида A ("Reb A") и Ребаудиозида D ("Reb D"). Результаты на 6, 9, 12 и 24 часа показаны на Фигурах 3B-E, соответственно. Результаты реакции без рекомбинантной AtSUS1 (то есть некомбинированной реакции) на 12 и 24 часа показаны на Фигурах 3F и 3G, соответственно.
[28] На Фигуре 4 показано in vitro получение ребаудиозида E ("Reb E") из стевиозида, катализируемое рекомбинантным полипептидом HV1 (SEQ ID NO:6) и рекомбинантной AtSUS1 (SEQ ID NO:9) в комбинированной реакционной системе HV1-AtSUS1, как описано в настоящей заявке. На Фигуре 4A показаны стандарты стевиозида ("Ste"), Ребаудиозида A ("Reb A") и Ребаудиозида D ("Reb D"). Результат на 20 часов показан на Фигурах 4B, который включает соединение ребаудиозид Z ("Reb Z").
[29] На Фигуре 5 показано in vitro получение ребаудиозида D ("Reb D") из стевиозида, катализируемое комбинацией рекомбинантного полипептида HV1 (SEQ ID NO:6), рекомбинантной UGT76G1 (SEQ ID NO:11) и рекомбинантной AtSUS1 (SEQ ID NO:9). На Фигуре 5A показаны стандарты стевиозида ("Ste"), Ребаудиозида A ("Reb A") и Ребаудиозида D ("Reb D"). Результаты на 6, 9, 12 и 24 часов показаны на Фигурах 5B-E, соответственно.
[30] На Фигуре 6 показан электрофоретический анализ в ДСН-ПААГ очищенного рекомбинантного полипептида HV1.
[31] На Фигуре 7 показан электрофоретический анализ в ДСН-ПААГ очищенного рекомбинантного полипептида AtSUS1.
[32] На Фигуре 8 показано in vitro получение ребаудиозида Z ("Reb Z") из ребаудиозида E ("Reb E"), катализируемое рекомбинантным полипептидом HV1 (SEQ ID NO:6) и рекомбинантной AtSUS1 (SEQ ID NO:9) в комбинированной реакционной системе HV1-AtSUS1, как описано в настоящей заявке. На Фигуре 8A показаны стандарты ребаудиозида E ("Reb E"). Результат на 24 часа показан на Фигурах 8B, который включает соединение ребаудиозид Z ("Reb Z").
[33] На Фигуре 9 показан электрофоретический анализ в ДСН-ПААГ очищенного рекомбинантного полипептида UGT76G1.
[34] Фигура 10: Структуры Ребаудиозида Z (включая Reb Z1 и Reb Z2) и Ребаудиозида E.
[35] Фигура 11: Ключевые TOCSY и HMBC корреляции Reb Z1 и Reb Z2.
[36] Фигура 12: показано in vitro получение ребаудиозида D ("Reb D") из ребаудиозида E ("Reb E"), катализируемое рекомбинантной UGT76G1 (SEQ ID NO:11). На Фигуре 12A-12B показаны стандарты Ребаудиозида E ("Reb E"), Ребаудиозида D ("Reb D"). Показаны результаты реакции без рекомбинантной AtSUS1 (Фигура 12C) (то есть некомбинированной реакции) и с рекомбинантной AtSUS1 (Фигура 12D) (то есть комбинированной УГТ-СС реакции) на 6 часов, соответственно.
[37] Хотя в отношении настоящего описания допускаются различные модификации и альтернативные формы, определенные варианты его осуществления показаны в качестве примера на чертежах и подробно описаны ниже. Впрочем, следует понимать, что описание конкретных вариантов осуществления не должно ограничивать описание и охватывает все модификации, эквиваленты и альтернативы, включенные в сущность и объем описания, как определено прилагаемой формулой изобретения.
ПОДРОБНОЕ ОПИСАНИЕ
[38] Рассматриваемая технология обеспечивает рекомбинантный полипептид, который обладает УДФ-гликозилтрансферазной активностью, такой как 1,2-19-O-глюкоза-гликозилирующая активность и 1,2-13-O-глюкоза-гликозилирующая активность, для синтеза стевиол-гликозидов. Рекомбинантный полипептид рассматриваемой технологии (который в дальнейшем может также именоваться как "рекомбинантный полипептид HV1") может применяться для биосинтеза стевиол-гликозидных соединений. В настоящем описании УДФ-гликозилтрансфераза (УГТ) относится к ферменту, который переносит остаток сахара с активированной донорной молекулы (обычно УДФ-глюкозы) на акцепторную молекулу. 1,2-19-O-глюкоза-гликозилирующая активность относится к ферментной активности, которая переносит сахарную группу на C-2’ 19-O-глюкозной группы стевиозида, ребаудиозида A или ребаудиозида E (Фигура 1 и Фигура 10). 1,2-13-O-глюкоза-гликозилирующая активность относится к ферментной активности, которая переносит сахарную группу на C-2’ 13-O-глюкозной группы ребаудиозида E (Фигура 10).
[39] Названия УГТ ферментов, используемые в настоящем описании, соответствуют системе номенклатуры, принятой Комитетом по номенклатуре УГТ (Mackenzie et al., "The UDP glycosyltransferase gene super family: recommended nomenclature updated based on evolutionary divergence", Pharmacogenetics, 1997, vol. 7, pp. 255-269), которая классифицирует гены УГТ комбинацией номера семейства, буквы, обозначающей подсемейство, и номера индивидуального гена. Например, название "UGT76G1" соответствует ферменту УГТ, кодируемому геном, который относится к семейству УГТ номер 76 (растительного происхождения), подсемейству G и гену номер 1.
[40] В растениях существует большое семейство генов УГТ. Впрочем, биологические функции большинства этих УГТ остаются неизвестными.
Определения
[41] При использовании в настоящем описании, формы единственного числа включают формы множественного числа, если из текста прямо не следует иное.
[42] В тех случаях, когда термин "включает", "содержит" или подобный используется в описании или формуле изобретения, предполагается, что такой термин является включительным так же, как термин "включать", при интерпретации "включать" при использовании в качестве переходного слова в формуле изобретения.
[43] Слово "примерный" используется в настоящем описании как "служащий в качестве примера, образца или иллюстрации". Любой вариант осуществления, описанный в настоящей заявке как "примерный", не должен обязательно рассматриваться как предпочтительный или преимущественный в отношении других вариантов осуществления.
[44] Термин "комплементарный" должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и использоваться без ограничения для описания отношения между нуклеотидными основаниями, которые способны к гибридизации друг с другом. Например, в отношении ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, рассматриваемая технология также включает выделенные фрагменты нуклеиновых кислот, которые комплементарны полным последовательностям, представленным в сопровождающем Списке последовательностей, а также те, которые по существу подобны последовательностям нуклеиновых кислот.
[45] Термины "нуклеиновая кислота" и "нуклеотид" должны иметь свои соответствующие обычные и общепринятые значения, известные среднему специалисту в данной области техники, и использоваться без ограничения для обозначения дезоксирибонуклеотидов или рибонуклеотидов и их полимеров в одно- или в двухцепочечной форме. Если нет специальных ограничений, термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые обладают подобными характеристиками связывания, как и референсная нуклеиновая кислота, и метаболизируются аналогично природным нуклеотидам. Если не указано иное, конкретная последовательность нуклеиновой кислоты также в подразумеваемой форме охватывает соответствующие консервативно модифицированные или вырожденные варианты (например, замены вырожденных кодонов) и комплементарные последовательности, а также последовательность, указанную в явной форме.
[46] Термин "выделенный" должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и при использовании в отношении выделенной нуклеиновой кислоты или выделенного полипептида должен использоваться без ограничения для обозначения нуклеиновой кислоты или полипептида, которые, благодаря человеку, существуют отдельно от своего нативного окружения и поэтому не являются продуктом природы. Выделенная нуклеиновая кислота или полипептид могут существовать в очищенной форме или могут существовать в ненативном окружении, в таком как, например, трансгенная клетка-хозяин.
[47] Термин "инкубирование", при использовании в настоящем описании, означает процесс смешивания двух или более химических или биологических единиц (таких как химическое соединение и фермент) и обеспечения их взаимодействия при условиях, благоприятных для получения композиции стевиол-гликозидов.
[48] Термин "вырожденный вариант" относится к последовательности нуклеиновой кислоты, имеющей последовательность остатков, которая отличается от референсной последовательности нуклеиновой кислоты на одну или более вырожденных замен кодонов. Вырожденные замены кодонов могут быть получены при создании последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов заменено смешанным основанием и/или остатками дезоксиинозина. Последовательность нуклеиновой кислоты и все ее вырожденные варианты будут экспрессировать ту же аминокислоту или полипептид.
[49] Термины "полипептид", "белок" и "пептид" должны иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники; эти три термина иногда используются попеременно и используются без ограничения для обозначения полимера из аминокислот или аналогов аминокислот, независимо от его размера или функции. Хотя "белок" часто используется в отношении относительно крупных полипептидов, а "пептид" часто используется в отношении малых полипептидов, применение данных терминов в уровне техники совпадает и изменяется. Термин "полипептид", при использовании в настоящем описании, относится к пептидам, полипептидам и белкам, если не отмечено иное. Термины "белок", "полипептид" и "пептид" используются в настоящем описании попеременно при обозначении продукта полинуклеотида. Таким образом, примерные полипептиды включают продукты полинуклеотидов, природные белки, гомологи, ортологи, паралоги, фрагменты и другие эквиваленты, варианты и аналоги предыдущего.
[50] Термины "полипептидный фрагмент" и "фрагмент", при использовании в отношении референсного полипептида, должны иметь свои обычные и общепринятые значения, известные среднему специалисту в данной области техники, и использоваться без ограничения для обозначения полипептида, в котором аминокислотные остатки удалены по сравнению с референсным полипептидом непосредственно, но где остальная аминокислотная последовательность обычно идентична соответствующим положениям в референсном полипептиде. Такие делеции могут происходить на N-конце или C-конце референсного полипептида или, в альтернативном варианте, на обоих концах.
[51] Термин "функциональный фрагмент" полипептида или белка относится к пептидному фрагменту, который является частью полноразмерного полипептида или белка и обладает по существу такой же биологической активностью или имеет по существу такую же функцию, что и полноразмерный полипептид или белок (например, выполняет такую же ферментативную реакцию).
[52] Термины "вариант полипептида", "модифицированная аминокислотная последовательность" или "модифицированный полипептид", которые используются попеременно, относятся к аминокислотной последовательности, которая отличается от референсного полипептида одной или более аминокислотами, например, одной или более аминокислотными заменами, делециями и/или добавлениями. В одном из аспектов вариант является "функциональным вариантом", который сохраняет некоторые или все свойства референсного полипептида.
[53] Термин "функциональный вариант" также включает консервативно замененные варианты. Термин "консервативно замененный вариант" относится к пептиду, имеющему аминокислотную последовательность, которая отличается от референсного пептида одной или более консервативными аминокислотными заменами и сохраняет некоторую часть или всю активность референсного пептида. "Консервативная аминокислотная замена" является заменой остатка аминокислоты функционально подобным остатком. Примеры консервативных замен включают замену одного неполярного (гидрофобного) остатка, такого как изолейцин, валин, лейцин или метионин, другим; замену одного заряженного или полярного (гидрофильного) остатка другим, например, между аргинином и лизином, между глутамином и аспарагином, между треонином и серином; замену одного основного остатка, такого как лизин или аргинин, другим; или замену одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота, другим; или замену одного ароматического остатка, такого как фенилаланин, тирозин или триптофан, другим. Такие замены, как ожидают, будут оказывать малое или не оказывать никакого воздействия на кажущуюся молекулярную массу или изоэлектрическую точку белка или полипептида. Фраза "консервативно замененный вариант" также включает пептиды, в которых остаток заменен химически дериватизированным остатком, при условии, что полученный в результате пептид сохраняет некоторую часть или всю активность референсного пептида, как описано в настоящей заявке.
[54] Термин "вариант", применительно к полипептидам рассматриваемой технологии, также включает функционально активный полипептид, имеющий аминокислотную последовательность, которая по меньшей мере на 75%, по меньшей мере на 76%, по меньшей мере на 77%, по меньшей мере на 78%, по меньшей мере на 79%, по меньшей мере на 80%, по меньшей мере на 81%, по меньшей мере на 82%, по меньшей мере на 83%, по меньшей мере на 84%, по меньшей мере на 85%, по меньшей мере на 86%, по меньшей мере на 87%, по меньшей мере на 88%, по меньшей мере на 89%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% и даже на 100% идентична аминокислотной последовательности референсного полипептида.
[55] Термин "гомологичный", во всех его грамматических формах и вариантах написания, относится к зависимости между полинуклеотидами или полипептидами, которые обладают "общим эволюционным происхождением", включая полинуклеотиды или полипептиды из суперсемейств, а также гомологичные полинуклеотиды или белки из различных биологических видов (Reeck и другие, Клетка 50:667, 1987). Такие полинуклеотиды или полипептиды обладают гомологией последовательности, что отражается подобием их последовательностей, либо в процентах идентичности, либо в присутствии определенных аминокислот или мотивов в консервативных положениях. Например, два гомологичных полипептида могут иметь аминокислотные последовательности, которые являются по меньшей мере на 75%, по меньшей мере на 76%, по меньшей мере на 77%, по меньшей мере на 78%, по меньшей мере на 79%, по меньшей мере на 80%, по меньшей мере на 81%, по меньшей мере на 82%, по меньшей мере на 83%, по меньшей мере на 84%, по меньшей мере на 85%, по меньшей мере на 86%, по меньшей мере на 87%, по меньшей мере на 88%, по меньшей мере на 89%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% и даже на 100% идентичными.
[56] "Процент (%) идентичности аминокислотных последовательностей" в отношении последовательностей различных полипептидов рассматриваемой технологии определяется как процент аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам референсного полипептида (такого как, например, SEQ ID NO:6), после выравнивания последовательностей и введения пропусков, при необходимости, для достижения максимального процента идентичности последовательностей, без учета каких-либо консервативных замен в качестве части идентичности последовательности.
[57] Выравнивание в целях определения процента идентичности аминокислотных последовательностей может быть выполнено различными способами, которые известны в уровне техники, например, при использовании общедоступных программ, таких как программы BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области техники сумеют определить подходящие параметры для измерения выравнивания, включая любые алгоритмы, требуемые для получения максимального выравнивания по всей длине сравниваемых последовательностей. Например, % идентичности аминокислотных последовательностей можно определить при использовании программы сравнения последовательностей NCBI-BLAST2. Программа сравнения последовательностей NCBI-BLAST2 может быть загружена с сайта ncbi.nlm.nih.gov. В NCBI BLAST2 используется несколько параметров поиска, где все параметры поиска установлены со значениями по умолчанию, включая, например, unmask yes, strand=all, expected occurrences 10, minimum low complexity length=15/5, multi-pass e-value=0.01, constant for multi-pass=25, dropoff for final gapped alignment=25 и scoring matrix=BLOSUM62. В случаях использования NCBI-BLAST2 для сравнения аминокислотных последовательностей, % идентичности аминокислотных последовательностей для данной аминокислотной последовательности A и данной аминокислотной последовательности B (что можно альтернативно сформулировать как данная аминокислотная последовательность A, которая имеет или включает некоторый % идентичности аминокислотной последовательности по отношению к данной аминокислотной последовательности B) вычисляют следующим образом: 100 умножить на частное X/Y, где X является количеством аминокислотных остатков, оцененных как идентичные совпадения программой выравнивания последовательностей NCBI-BLAST2 при выравнивании с помощью этой программы последовательностей A и B, и где Y является общим количеством аминокислотных остатков в B. Следует понимать, что если длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичности аминокислотных последовательностей A и B не будет равен % идентичности аминокислотных последовательностей B и A.
[58] В этом смысле методики определения "подобия" аминокислотных последовательностей известны в уровне техники. В общем, "подобие" означает точное сравнение аминокислоты с аминокислотой в двух или более полипептидах в подходящем положении, где аминокислоты идентичны или обладают подобными химическими и/или физическими свойствами, такими как заряд или гидрофобность. При этом между сравниваемыми полипептидными последовательностями может быть определен так называемый "процент подобия". Методики определения идентичности последовательностей нуклеиновых кислот и аминокислот также известны в уровне техники и включают определение нуклеотидной последовательности мРНК для данного гена (обычно через промежуточную кДНК) и определение аминокислотной последовательности, кодируемой в ней, и ее сравнение со второй аминокислотной последовательностью. В общем, "идентичность" относится к точному соответствию нуклеотида с нуклеотидом или аминокислоты с аминокислотой в двух полинуклеотидных или полипептидных последовательностях, соответственно. Две или более полинуклеотидных последовательности можно сравнить при определении их "процента идентичности", как и в случае двух или более аминокислотных последовательностей. Программы, доступные в пакете Wisconsin Sequence Analysis Package, Version 8 (доступном от Genetics Computer Group, Madison, Wis.), например программа GAP, позволяют вычислять идентичность между двумя полинуклеотидами и идентичность и подобие между двумя полипептидными последовательностями, соответственно. Другие программы для вычисления идентичности или подобия между последовательностями известны специалистам в данной области.
[59] Положение аминокислоты, "соответствующее" референсному положению, является положением, которое выравнивается с референсной последовательностью, как идентифицируют при выравнивании аминокислотных последовательностей. Такие выравнивания могут быть выполнены вручную или при использовании известных программ выравнивания последовательностей, таких как ClustalW2, Blast 2 и т.д.
[60] Если не определено иное, процент идентичности двух полипептидных или полинуклеотидных последовательностей именуется как процент идентичных аминокислотных или нуклеотидных остатков по всей длине более короткой из двух данных последовательностей.
[61] "Кодирующая последовательность" должна иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и использоваться без ограничения для обозначения последовательности ДНК, которая кодирует определенную аминокислотную последовательность.
[62] "Подходящие регуляторные последовательности" должны иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используются без ограничения для обозначения нуклеотидных последовательностей, которые расположены перед (5’ некодирующие последовательности), в или после (3’ некодирующие последовательности) кодирующей последовательности, и которые влияют на транскрипцию, процессинг РНК или стабильность, или трансляцию связанной кодирующей последовательности. Регуляторные последовательности могут включать промоторы, лидерные последовательности трансляции, интроны и последовательности распознавания полиаденилирования.
[63] "Промотор" должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используется без ограничения для обозначения последовательности ДНК, способной регулировать экспрессию кодирующей последовательности или функциональной РНК. В общем, кодирующая последовательность расположена 3’ относительно последовательности промотора. Промоторы могут быть получены полностью из нативного гена или собраны из различных элементов, полученных из различных промоторов, существующих в природе, или даже могут включать синтетические сегменты ДНК. Специалистам в данной области техники известно, что различные промоторы могут направлять экспрессию гена в различных тканях или типах клеток, или на различных стадиях развития, или в ответ на различные условия окружающей среды. Промоторы, которые вызывают экспрессию гена в большинстве типов клеток большую часть времени, обычно называются "конститутивными промоторами". Также известно, что поскольку в большинстве случаев точные границы регуляторных последовательностей не были полностью определены, фрагменты ДНК различной длины могут обладать идентичной промоторной активностью.
[64] Термин "функционально связанный" относится к ассоциации последовательностей нуклеиновых кислот на одном фрагменте нуклеиновой кислоты, в результате чего на функцию одной из последовательностей влияет другая. Например, промотор функционально связан с кодирующей последовательностью, когда он способен воздействовать на экспрессию данной кодирующей последовательности (то есть кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации.
[65] Термин "экспрессия", при использовании в настоящем описании, должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используется без ограничения для обозначения транскрипции и устойчивого накопления смысловой (мРНК) или антисмысловой РНК, полученной из фрагмента нуклеиновой кислоты рассматриваемой технологии. "Оверэкспрессия" относится к синтезу продукта гена в трансгенных или рекомбинантных организмах, уровни продукции которого превышают уровни продукции в нормальных или нетрансформированных организмах.
[66] "Трансформация" должна иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используется без ограничения для обозначения переноса полинуклеотида в клетку-мишень. Перенесенный полинуклеотид может быть включен в геном или хромосомную ДНК клетки-мишени, что приводит к генетически стабильному наследованию, или он может реплицироваться независимо от хромосом хозяина. Организмы-хозяева, содержащие трансформированные фрагменты нуклеиновых кислот, называются "трансгенными" или "рекомбинантными", или "трансформированными" организмами.
[67] Термины "трансформированный", "трансгенный" и "рекомбинантный", при использовании в настоящем описании в отношении клеток-хозяев, должны иметь свои соответствующие обычные и общепринятые значения, известные среднему специалисту в данной области техники, и используются без ограничения для обозначения клетки организма-хозяина, такой как растительная или микробная клетка, в которую была введена гетерологичная молекула нуклеиновой кислоты. Молекула нуклеиновой кислоты может быть стабильно интегрирована в геном клетки-хозяина, или молекула нуклеиновой кислоты может присутствовать как внехромосомная молекула. Такая внехромосомная молекула может быть самореплицирующейся. Трансформированные клетки, ткани или субъекты, как понимают, охватывают не только конечный продукт процесса трансформации, но и его трансгенное потомство.
[68] Термины "рекомбинантный", "гетерологичный" и "экзогенный", при использовании в настоящем описании в отношении полинуклеотидов, должны иметь свои обычные и общепринятые значения, известные среднему специалисту в данной области техники, и используются без ограничения для обозначения полинуклеотида (например, последовательности ДНК или гена), который происходит из источника, чужеродного для конкретной клетки-хозяина или, если он происходит из того же источника, изменен относительно своей исходной формы. Таким образом, гетерологичный ген в клетке-хозяине включает ген, который является эндогенным по отношению к конкретной клетке-хозяину, но был изменен, например, при использовании сайт-направленного мутагенеза или других рекомбинантных технологий. Указанные термины также включают не встречающиеся в природе множественные копии природной последовательности ДНК. Таким образом, термины относятся к сегменту ДНК, который является чужеродным или гетерологичным по отношению к клетке или гомологичным по отношению к клетке, но находящимся в таком положении или форме в клетке-хозяине, в котором данный элемент обычно не находится.
[69] Аналогичным образом термины "рекомбинантный", "гетерологичный" и "экзогенный", при использовании в настоящем описании в отношении полипептидной или аминокислотной последовательности, означают полипептидную или аминокислотную последовательность, которая происходит из источника, чужеродного по отношению к конкретной клетке-хозяину или, если она происходит из того же источника, изменена относительно своей исходной формы. Таким образом, рекомбинантные сегменты ДНК могут экспрессироваться в клетке-хозяине с получением рекомбинантного полипептида.
[70] Термины "плазмида", "вектор" и "кассета" должны иметь свои соответствующие обычные и общепринятые значения, известные среднему специалисту в данной области техники, и используются без ограничения для обозначения внехромосомного элемента, часто несущего гены, которые не являются частью центрального метаболизма клетки, и который обычно находится в форме кольцевых молекул двухцепочечной ДНК. Такие элементы могут быть автономно реплицирующимися последовательностями, интегрирующимися в геном последовательностями, фаговыми или нуклеотидными последовательностями, линейными или кольцевыми, одно- или двухцепочечными ДНК или РНК, полученными из любого источника, в котором несколько нуклеотидных последовательностей были соединены или рекомбинированы с получением уникальной конструкции, которая способна вводить промоторный фрагмент и последовательность ДНК для выбранного продукта гена вместе с подходящими 3’ нетранслируемыми последовательностями в клетку. "Трансформационная кассета" относится к определенному вектору, содержащему чужеродный ген и имеющему другие элементы в дополнение к чужеродному гену, которые способствуют трансформации конкретной клетки-хозяина. "Кассета экспрессии" относится к определенному вектору, содержащему чужеродный ген и имеющему другие элементы в дополнение к чужеродному гену, которые обеспечивают повышенную экспрессию данного гена в чужеродном хозяине.
[71] Стандартные методики генной инженерии и молекулярного клонирования, используемые в настоящей заявке, известны в уровне техники и описаны, например, в Sambrook, J., Fritsch, E. F. and Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, N.Y., 1989 (в дальнейшем "Maniatis"); и в Silhavy, T. J., Bennan, M. L. and Enquist, L. W. Experiments with Gene Fusions; Cold Spring Harbor Laboratory: Cold Spring Harbor, N.Y., 1984; а также в Ausubel, F. M. et al., в Current Protocols in Molecular Biology, издательство Greene Publishing and Wiley-Interscience, 1987;, которые полностью включены в настоящее описание посредством отсылки.
[72] Если не определено иное, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, под которым их обычно понимает средний специалист в области техники, к которой относится настоящее описание. Хотя любые методы и материалы, подобные или эквивалентные описанным в настоящей заявке, могут использоваться при осуществлении или тестировании настоящего описания, ниже описаны предпочтительные материалы и методы.
[73] Настоящее описание будет понято более полно при рассмотрении следующих неограничивающих Примеров. Следует понимать, что данные Примеры, притом, что в них указываются предпочтительные варианты осуществления рассматриваемой технологии, даны лишь в качестве иллюстрации. Из приведенного выше обсуждения и этих Примеров, специалист в данной области техники сумеет установить существенные характеристики рассматриваемой технологии и без отступления от ее сущности и объема сможет внести различные изменения и модификации рассматриваемой технологии, чтобы адаптировать ее к различным применениям и условиям.
Рекомбинантные полипептиды
[74] В одном аспекте настоящее описание относится к рекомбинантному полипептиду, имеющему аминокислотную последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с аминокислотной последовательностью, представленной в SEQ ID NO:6. Предпочтительно аминокислотная последовательность рекомбинантного полипептида обладает по меньшей мере 80% идентичностью с SEQ ID NO:6. Более предпочтительно аминокислотная последовательность рекомбинантного полипептида обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с SEQ ID NO:6. В примере осуществления аминокислотная последовательность рекомбинантного полипептида состоит из SEQ ID NO:6. Соответственно, рекомбинантный полипептид, описанный в настоящей заявке, включает функциональные фрагменты SEQ ID NO:6, функциональные варианты SEQ ID NO:6 и другие гомологичные полипептиды, которые обладают, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью последовательности с SEQ ID NO:6.
[75] В другом аспекте настоящее описание относится к выделенной нуклеиновой кислоте, имеющей нуклеотидную последовательность, кодирующую рекомбинантный полипептид, описанный в настоящей заявке. Например, выделенная нуклеиновая кислота может включать нуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с аминокислотной последовательностью, представленной в SEQ ID NO:6. Предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, которая по меньшей мере на 80% идентична аминокислотной последовательности, представленной в SEQ ID NO:6. Более предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, которая обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью последовательности с аминокислотной последовательностью, представленной в SEQ ID NO:6. Выделенная нуклеиновая кислота, таким образом, включает нуклеиновые кислоты, которые кодируют функциональные фрагменты SEQ ID NO:6, функциональные варианты SEQ ID NO:6 или другие гомологичные полипептиды, которые обладают, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью последовательности с SEQ ID NO:6.
[76] В варианте осуществления настоящее описание относится к выделенной нуклеиновой кислоте, имеющей нуклеотидную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с нуклеотидной последовательностью, представленной в SEQ ID NO:7. Предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, которая обладает по меньшей мере 80% идентичностью с нуклеотидной последовательностью, представленной в SEQ ID NO:7. Более предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, которая обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:7.
[77] В другом аспекте рассматриваемая технология относится к вектору, содержащему нуклеиновые кислоты, описанные в настоящей заявке, и клетке-хозяину, содержащей вектор, описанный в настоящей заявке. В некоторых вариантах осуществления настоящее описание относится к вектору экспрессии, включающему по меньшей мере один полинуклеотид рассматриваемой технологии, и где вектор экспрессии при трансфекции в клетку-хозяина способен к экспрессии по меньшей мере одного рекомбинантного полипептида HV1, описанного в настоящей заявке. В варианте осуществления вектор экспрессии включает нуклеотидную последовательность, представленную в SEQ ID NO:7, или ее вариант.
[78] Конструкция вектора экспрессии зависит от таких факторов как выбор трансформируемой клетки-хозяина, требуемый уровень экспрессии белка и т.п. Векторы экспрессии могут быть введены в клетку-хозяина для получения в результате рекомбинантного полипептида рассматриваемой технологии, такого как рекомбинантный полипептид HV1, имеющий аминокислотную последовательность SEQ ID NO:6 или ее вариант.
[79] Экспрессию белков в прокариотах чаще всего проводят в бактериальной клетке-хозяине с использованием векторов, содержащих конститутивные или индуцируемые промоторы, направляющие экспрессию слитых или неслитых белков. Слитые векторы добавляют некоторое количество аминокислот к белку, кодируемому в них, обычно на N-конец рекомбинантного белка. Такие слитые векторы обычно служат трем целям: 1) повысить экспрессию рекомбинантного белка; 2) повысить растворимость рекомбинантного белка; и 3) способствовать очистке рекомбинантного белка, действуя как лиганд при аффинной очистке. Часто в место соединения слитого фрагмента и рекомбинантного белка вводят сайт протеолитического расщепления, позволяющий отделить рекомбинантный белок от слитого фрагмента после очистки слитого белка. Такие векторы включены в объем настоящего описания.
[80] В варианте осуществления вектор экспрессии включает такие генетические элементы для экспрессии рекомбинантного полипептида в бактериальных клетках. Элементы для транскрипции и трансляции в бактериальной клетке могут включать промотор, кодирующую область белкового комплекса и терминатор транскрипции.
[81] В варианте осуществления векторы экспрессии рассматриваемой технологии включают бактериальные векторы экспрессии, например, рекомбинантную ДНК бактериофага, плазмидную ДНК или космидную ДНК, дрожжевые векторы экспрессии например, рекомбинантные дрожжевые векторы экспрессии, векторы для экспрессии в клетках насекомых, например, рекомбинантные вирусные векторы экспрессии, например бакуловирус, или векторы для экспрессии в клетках растений, например, рекомбинантные вирусные векторы экспрессии, такие как вирус мозаики цветной капусты (CaMV), вирус табачной мозаики (ВТМ), или рекомбинантные плазмидные векторы экспрессии, такие как Ti-плазмиды.
[82] В варианте осуществления вектор включает бактериальный вектор экспрессии. В другом варианте осуществления вектор экспрессии включает высококопийный вектор экспрессии; в альтернативе вектор экспрессии включает низкокопийный вектор экспрессии, например, плазмиду Mini-F.
[83] Среднему специалисту в данной области техники будут известны методики молекулярной биологии, доступные для получения векторов экспрессии. Полинуклеотид, используемый для включения в вектор экспрессии рассматриваемой технологии, как описано выше, может быть получен с помощью стандартных методик, таких как полимеразная цепная реакция (ПЦР).
[84] Было разработано много методик молекулярной биологии для функционального связывания ДНК с векторами через комплементарные липкие концы. В одном варианте осуществления дополнительные гомополимерные участки могут быть добавлены к молекуле нуклеиновой кислоты, которую предполагается вставить в векторную ДНК. Вектор и молекулу нуклеиновой кислоты затем соединяют с образованием водородной связи между дополнительными гомополимерными концевыми участками, получая рекомбинантные молекулы ДНК.
[85] В альтернативном варианте осуществления синтетические линкеры, содержащие один или более введенных сайтов рестрикции, используют для функционального связывания полинуклеотида рассматриваемой технологии с вектором экспрессии. В варианте осуществления полинуклеотид получают при обработке эндонуклеазой рестрикции. В варианте осуществления молекулу нуклеиновой кислоты обрабатывают ДНК-полимеразой бактериофага T4 или ДНК полимеразой I E. coli, ферментами, которые удаляют выступающие 3’-одноцепочечные концы с помощью своей 3’-5’-экзонуклеазной активности и заполняют углубленные 3’-концы с помощью своей полимеразной активности, с получением в результате сегментов ДНК с тупыми концами. Затем сегменты с тупыми концами инкубируют с большим молярным избытком молекул линкера в присутствии фермента, который способен катализировать лигирование молекул ДНК с тупыми концами, такого как ДНК-лигаза бактериофага T4. Таким образом, продуктом реакции является полинуклеотид, несущий полимерные линкерные последовательности на своих концах. Затем эти полинуклеотиды расщепляют подходящей рестриктазой и лигируют в вектор экспрессии, который был разрезан ферментом, который дает концы, совместимые с концами полинуклеотида.
[86] В альтернативе может использоваться вектор, имеющий сайты безлигазного клонирования (LIC). Затем требуемый ПЦР амплифицированный полинуклеотид можно клонировать в вектор LIC без рестрикции или лигирования (Aslanidis and de Jong, Nucl. Acid. Res. 18, 6069-6074, (1990), Haun, et al., Biotechniques 13, 515-518 (1992), которая включена в настоящее описание в той мере, в которой это соответствует настоящему описанию).
[87] В варианте осуществления для выделения и/или модификации целевого полинуклеотида, предназначенного для вставки в выбранную плазмиду, предпочтительно использовать ПЦР. Подходящие праймеры для использования при получении последовательности с помощью ПЦР могут быть подобраны для выделения требуемой кодирующей области молекулы нуклеиновой кислоты, добавления сайтов эндонуклеаз рестрикции или LIC, помещения кодирующей области в нужную рамку считывания.
[88] В варианте осуществления полинуклеотид для включения в вектор экспрессии рассматриваемой технологии получают при помощи ПЦР с использованием подходящих олигонуклеотидных праймеров. Кодирующую область амплифицируют, при этом сами праймеры включаются в последовательность амплифицируемого продукта. В варианте осуществления амплификационные праймеры содержат сайты узнавания эндонуклеаз рестрикции, которые позволяют клонировать последовательность амплифицированного продукта в подходящий вектор.
[89] В варианте осуществления полинуклеотид SEQ ID NO:7 или его вариант получают с помощью ПЦР и включают в вектор экспрессии при использовании расщепления эндонуклеазами рестрикции и лигирования согласно методикам, которые известны в уровне техники.
[90] Настоящее описание также относится к клетке-хозяину, включающей вектор экспрессии, описанный в настоящей заявке. Подходящие хозяева рассматриваемой технологии обычно включают микробные хозяева или растения-хозяева. Например, клетка-хозяин рассматриваемой технологии выбрана из группы, состоящей из клеток бактерий, дрожжей, нитчатых грибов, цианобактерий и растений.
[91] Микробные хозяева могут включать любой организм, способный к экспрессии полинуклеотида (такого как SEQ ID NO:7) с получением рекомбинантного полипептида HV1, описанного в настоящей заявке. Микроорганизмы, применимые в рассматриваемой технологии, включают бактерии, такие как кишечные бактерии (например, Escherichia и Salmonella), а также Bacillus, Acinetobacter, актиномицеты, такие как Streptomyces, Corynebacterium, метанотрофы, такие как Methylosinus, Methylomonas, Rhodococcus и Pseudomona; цианобактерии, такие как Rhodobacter и Synechocystis; дрожжи, такие как Saccharomyces, Zygosaccharomyces, Kluyveromyces, Candida, Hansenula, Debaryomyces, Mucor, Pichia и Torulopsis; и нитчатые грибы, такие как Aspergillus и Arthrobotrys, а также водоросли, и Escherichia, Klebsiella, Pantoea, Salmonella, Corynebacterium, Clostridium и Clostridium acetobutylicum, например. Предпочтительно, микробный хозяин является бактериями (такой как Escherichia) или дрожжами (такими как Saccharomyces). Векторы экспрессии могут быть включены в эти и другие микробные хозяева для получения большого, коммерчески применимого количества стевиол-гликозидов.
[92] В варианте осуществления рекомбинантный полипептид может быть экспрессирован в клетке-хозяине, которая является растительной клеткой. При использовании в настоящем описании, термин "растительная клетка", как подразумевают, означает любую клетку, полученную из однодольного или двудольного растения и способную формировать недифференцированные ткани, такие как каллусы, дифференцированные ткани, такие как эмбрионы, части однодольных растений, однодольные растения или семя. Термин "растение", как подразумевают, означает любой дифференцированный многоклеточный организм, способный к фотосинтезу, включая однодольные и двудольные. В некоторых вариантах осуществления растительная клетка может быть растительной клеткой Arabidopsis, растительной клеткой табака, растительной клеткой сои, растительной клеткой петунии или клеткой из другой масличной культуры, включающей, без ограничения, растительную клетку канолы, растительную клетку рапса, растительную клетку пальмы, растительную клетку подсолнечника, растительную клетку хлопка, растительную клетку кукурузы, растительную клетку арахиса, растительную клетку льна и растительную клетку кунжута.
[93] Применимые растения-хозяева могут включать любое растение, которое обеспечивает продукцию рекомбинантных полипептидов рассматриваемой технологии. Подходящие зеленые растения для использования в качестве хозяев включают, без ограничения перечисленными, сою, рапс (Brassica napus, B. campestris), подсолнечник (Helianthus annus), хлопок (Gossypium hirsutum), кукурузу, табак (Nicotiana tabacum), люцерну (Medicago sativa), пшеницу (Triticum sp.), ячмень (Hordeum vulgare), овес (Avena sativa), сорго (Sorghum bicolor), рис (Oryza sativa), Arabidopsis, крестоцветные овощи (брокколи, цветную капусту, капусту, пастернак и т.д.), дыню, морковь, сельдерей, петрушку, томаты, картофель, землянику, арахис, виноград, кормовые посевные культуры, сахарную свеклу, сахарный тростник, бобы, горох, рожь, лен, деревья лиственных пород, деревья хвойных пород и кормовые травы. Виды водорослей включают, без ограничения перечисленными, коммерчески значимых хозяев, таких как Spirulina, Haemotacoccus и Dunaliella. Подходящие растения для способа рассматриваемой технологии также включают культуры для получения биотоплива, биомассы и биоэнергии. Примеры растений включают Arabidopsis thaliana, рис (Oryza sativa), Hordeum vulgare, просо прутьевидное (Panicum vigratum), Brachypodium spp., Brassica spp. и Crambe abyssinica.
[94] В некоторых вариантах осуществления настоящее описание включает трансгенные клетки-хозяева или хозяева, которые были трансформированы одним или более векторами, раскрытыми в настоящей заявке.
[95] В альтернативе клетки-хозяева могут быть клетками-хозяевами, подходящими для продукции биосинтеза включая одноклеточные организмы, микроорганизмы, многоклеточные организмы, растения, грибы, бактерии, водоросли, сельскохозяйственные культуры, несельскохозяйственные культуры и/или подобные.
[96] Векторы экспрессии могут быть введены в растение или микробные клетки-хозяева с помощью стандартных методик трансформации или трансфекции. Трансформацию подходящих клеток вектором экспрессии рассматриваемой технологии осуществляют способами, известными в уровне техники, при этом обычно она зависит от типа вектора и клетки. Подходящие методики включают соосаждение с фосфатом кальция или хлоридом кальция, трансфекцию, опосредованную DEAE-декстраном, липофекцию, хемопорацию или электропорацию.
[97] Успешно трансформированные клетки, то есть клетки, содержащие вектор экспрессии, могут быть идентифицированы с помощью методик, известных в уровне техники. Например, клетки, трансфицированные вектором экспрессии рассматриваемой технологии, можно культивировать с получением полипептидов, описанных в настоящей заявке. Клетки можно проверить на наличие ДНК вектора экспрессии с помощью методик, известных в уровне техники.
[98] Клетки-хозяева могут содержать одну копию вектора экспрессии, описанного ранее, или, альтернативно, множество копий вектора экспрессии.
[99] В некоторых вариантах осуществления трансформированная клетка является клеткой животного, клеткой насекомого, клеткой растения, клеткой водоросли, клеткой гриба или клеткой дрожжей. В некоторых вариантах осуществления клетка является растительной клеткой, выбранной из группы, состоящей из: растительной клетки канолы, растительной клетки рапса, растительной клетки пальмы, растительной клетки подсолнечника, растительной клетки хлопка, растительной клетки кукурузы, растительной клетки арахиса, растительной клетки льна, растительной клетки кунжута, растительной клетки сои и растительной клетки петунии.
[100] Системы экспрессии на основе микробных клеток-хозяев и векторы экспрессии, содержащие регуляторные последовательности, которые направляют экспрессию с высоким уровнем чужеродных белков, известны специалистам в данной области техники. Любые из них могут применяться для создания векторов для экспрессии рекомбинантного полипептида рассматриваемой технологии в микробной клетке-хозяине. Такие векторы могут быть затем введены в подходящие микроорганизмы с помощью трансформации для обеспечения экспрессии с высоким уровнем рекомбинантного полипептида рассматриваемой технологии.
[101] Векторы или кассеты, применимые для трансформации подходящих микробных клеток-хозяев, известны в уровне техники. Как правило, вектор или кассета содержат последовательности, направляющие транскрипцию и трансляцию соответствующего полинуклеотида, селективного маркера и последовательностей, обеспечивающих автономную репликацию или хромосомную интеграцию. Подходящие векторы включают 5’ область полинуклеотида, которая содержит области регуляции инициации транскрипции и 3’ область фрагмента ДНК, которая регулирует терминацию транскрипции. Предпочтительно, чтобы обе области регуляции были получены из генов, гомологичных трансформированной клетке-хозяину, хотя следует понимать, что такие области регуляции не должны быть обязательно получены из генов, нативных по отношению к определенным видам, выбранным в качестве хозяина.
[102] Области регуляции инициации или промоторы, которые могут применяться для направления экспрессии рекомбинантного полипептида в требуемой микробной клетке-хозяине, являются многочисленными и известны специалистам в данной области техники. Фактически любой промотор, способный к направлению этих генов, подходит для рассматриваемой технологии, в том числе, без ограничения перечисленными, CYC1, HIS3, GAL1, GAL10, ADH1, PGK, PHO5, GAPDH, ADC1, TRP1, URA3, LEU2, ENO, TPI (применимый для экспрессии в Saccharomyces); AOX1 (применимый для экспрессии в Pichia); а также lac, trp, IP L , IP R , T7, tac и trc (применимые для экспрессии в Escherichia coli).
[103] Области регуляции терминации могут быть также получены из различных генов, нативных по отношению к микробным хозяевам. Необязательно может быть включен участок терминации для микробных хозяев, описанных в настоящей заявке.
[104] В растительных клетках векторы экспрессии рассматриваемой технологии могут включать кодирующую область, функционально связанную с промоторами, способными направлять экспрессию рекомбинантного полипептида рассматриваемой технологии в требуемых тканях на нужной стадии развития. По причинам удобства экспрессируемые полинуклеотиды могут включать промоторные последовательности и лидерные последовательности трансляции, полученные из одного и того же полинуклеотида. Также должны присутствовать 3’ некодирующие последовательности, кодирующие сигналы терминации транскрипции. Векторы экспрессии могут также включать один или более интронов, способствующих экспрессии полинуклеотида.
[105] В отношении растительных клеток-хозяев, любая комбинация любого промотора и любого терминатора, способная вызывать экспрессию кодирующей области, может использоваться в векторных последовательностях рассматриваемой технологии. Некоторые подходящие примеры промоторов и терминаторов включают промоторы и терминаторы генов нопалинсинтазы (nos), октопинсинтазы (ocs) и вируса мозаики цветной капусты (CaMV). Одним из типов эффективного растительного промотора, который может использоваться, является высокоэффективный растительный промотор. Такие промоторы, функционально связанные с вектором экспрессии рассматриваемой технологии, должны быть способны активировать экспрессию вектора. Высокоэффективные растительные промоторы, которые могут использоваться в рассматриваемой технологии, включают промотор малой субъединицы (ss) рибулозо-1,5-бисфосфаткарбоксилазы, например, из сои (Berry-Lowe et al., J. Molecular and App. Gen., 1:483 498 (1982), которая полностью включена в настоящее описание в той мере, в которой это соответствует настоящей заявке), и промотор хлорофилл a/b-связывающего белка. Эти два промотора, как известно, индуцирует свет в растительных клетках (см., например, Genetic Engineering of Plants, an Agricultural Perspective, A. Cashmore, Plenum, N.Y. (1983), страницы 29 38; Coruzzi, G. et al., The Journal of Biological Chemistry, 258:1399 (1983), и Dunsmuir, P. et al., Journal of Molecular and Applied Genetics, 2:285 (1983), которые полностью включены в настоящее описание в той мере, в которой это соответствует настоящей заявке).
[106] Выбор плазмидного вектора зависит от способа, который будет использоваться для трансформации растений-хозяев. Специалисту хорошо известны генетические элементы, которые должны присутствовать в плазмидном векторе для успешной трансформации, отбора и размножения клеток-хозяев, содержащих химерный полинуклеотид. Специалисту также известно, что различные независимые события трансформации приводят к различным уровням и профилям экспрессии (Jones et al., EMBO J. 4:2411 2418 (1985); De Almeida et al., Mol. Gen. Genetics 218:78 86 (1989), которые полностью включены в настоящее описание в той мере, в которой это соответствует настоящей заявке) и, следовательно, что множество событий требуется подвергнуть скринингу для получения линий, демонстрирующих требуемый уровень и профиль экспрессии. Такой скрининг может быть выполнен с помощью Саузерн-анализа ДНК-блотов, Нозерн-анализая экспрессии мРНК, Вестерн-анализа экспрессии белка или фенотипического анализа.
[107] Введение вектора экспрессии рассматриваемой технологии в растительную клетку может быть выполнено множеством способов, известных средним специалистам в данной области техники, включающих вставку целевой последовательности нуклеиновой кислоты в Ri-плазмиду Agrobacterium rhizogenes или Ti-плазмиду Agrobacterium tumefaciens, микроинъекцию, электропорацию или прямое осаждение. В качестве примера, в некоторых вариантах осуществления транзиентная экспрессия целевого полинуклеотида может быть выполнена методами агро-инфильтрации. В этом отношении, суспензия Agrobacterium tumefaciens, содержащая целевой полинуклеотид, может быть выращена в культуре и затем введена в растение с помощью шприца, конец которого прижимают к нижней стороне листа, мягко надавливая при этом на другую сторону листа. При этом раствор агробактерии поступает в воздушные полости внутри листа через устьица. Попав внутрь листа, агробактерия трансформирует целевой ген в части растительных клеток, где ген затем транзиентно экспрессируется.
[108] В качестве другого примера, трансформация целевой плазмиды в растительную клетку может быть выполнена с помощью методик бомбардировки ускоренными частицами (то есть биолистики). В этом отношении, суспензия зародышей растения может быть выращена в жидкой культуре и затем подвергнута бомбардировке плазмидами или полинуклеотидами, которые прикреплены к частицам золота, где частицы золота, связанные с целевой плазмидой или нуклеиновой кислотой, могут проникать через мембраны растительных тканей, таких как зародышевая ткань. После бомбардировки трансформированные эмбрионы могут быть отобраны при использовании подходящего антибиотика с получением новых, клонально размноженных, трансформированных эмбриогенных суспензионных культур.
[109] Клетки-хозяева могут быть немодифицированными клетками или клеточными линиями, или клеточными линиями, которые были генетически модифицированы. В некоторых вариантах осуществления клетка-хозяин является клеточной линией, которая была модифицирована с целью выращивания в требуемых условиях, например, при более низкой температуре.
[110] Стандартные методики генной инженерии могут использоваться для получения нуклеиновой кислоты, которая кодирует рекомбинантный полипептид, описанный в настоящей заявке, включения нуклеиновой кислоты в вектор экспрессии и введения вектора в клетку-хозяина, как описано в Sambrook, et al. (eds), Molecular Cloning; A Laboratory Manual, Third Edition, Cold Spring Harbor, (2001); и Ausubel, F. M. et al. (eds.) Current Protocols in Molecular Biology, John Wiley & Sons (1995). Нуклеиновая кислота, кодирующая полипептид, может быть вставлена в вектор или векторы экспрессии таким образом, чтобы нуклеиновые кислоты были функционально связаны с последовательностями регуляции транскрипции и трансляции (такими как последовательность промотора, последовательность терминации транскрипции и т.д.). Вектор экспрессии и последовательности регуляции экспрессии обычно выбирают так, чтобы они были совместимы с используемой для экспрессии клеткой-хозяином.
[111] Экспрессия полипептида в хозяине, описанном в настоящей заявке, может быть дополнительно улучшена путем оптимизации кодонов. Например, замена менее распространенного кодона более распространенным кодоном может влиять на полупериод существования мРНК или изменять ее структуру посредством введения вторичной структуры, которая препятствует трансляции мРНК. Вся или часть кодирующей области может быть оптимизирована. В некоторых случаях требуемую модуляцию экспрессии достигают путем оптимизации по существу всего гена. В других случаях требуемую модуляцию достигают при оптимизации части, а не всей последовательности гена.
[112] Использование кодонов в любой кодирующей последовательности можно регулировать для достижения требуемого свойства, например, высоких уровней экспрессии в определенном типе клеток. Исходной точкой для такой оптимизации может быть кодирующая последовательность со 100% распространенных кодонов или кодирующая последовательностью, которая содержит смесь распространенных и малораспространенных кодонов.
[113] Две или более кандидатных последовательностей, которые отличаются по своей частоте использования кодонов, могут быть получены и проверены с целью определения, обладают ли они нужным свойством. Кандидатные последовательности могут быть оценены при использовании компьютера для поиска присутствия регуляторных элементов, таких как сайленсеры или энхансеры, и поиска присутствия областей кодирующей последовательности, которая могла быть преобразована в такие регуляторные элементы при изменении использования кодонов. Дополнительные критерии могут включать обогащение специфическими нуклеотидами, например, A, C, G или U, смещение кодонов по определенной аминокислоте или присутствие или отсутствие специфической вторичной или третичной структуры мРНК. Регулирование кандидатной последовательности может быть выполнено на основе ряда таких критериев.
[114] В некоторых вариантах осуществления оптимизированная по кодонам последовательность нуклеиновой кислоты может экспрессировать свой белок на уровне, который составляет приблизительно 110%, приблизительно 150%, приблизительно 200%, приблизительно 250%, приблизительно 300%, приблизительно 350%, приблизительно 400%, приблизительно 450% или приблизительно 500% от уровня белка, экспрессируемого последовательностью нуклеиновой кислоты, которая не была оптимизирована по кодонам.
[115] В дополнение к нуклеиновой кислоте, которая кодирует рекомбинантный полипептид рассматриваемой технологии, вектор экспрессии рассматриваемой технологии может дополнительно нести регуляторные последовательности, которые регулируют экспрессию белка в клетке-хозяине, такие как промоторы, энхансеры или другие элементы регуляции экспрессии, которые регулируют транскрипцию или трансляцию нуклеиновой кислоты (кислот). Такие регуляторные последовательности известны в уровне техники. Специалистам в данной области техники будет очевидно, что конструкция вектора экспрессии, в том числе выбор регуляторных последовательностей, может зависеть от таких факторов как выбор клетки-хозяина, которую предполагается трансформировать, требуемый уровень экспрессии белка и т.д. Кроме того, рекомбинантные векторы экспрессии рассматриваемой технологии могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации) и селективные маркерные гены.
Биосинтез стевиол-гликозидов
[116] Как описано в настоящей заявке, рекомбинантные полипептиды настоящей технологии обладают УДФ-гликозилтрансферазной активностью, более конкретно включающей 1,2-19-O-глюкоза-гликозилирующую активность, и могут применяться для разработки биосинтетических способов получения стевиол-гликозидов, которые обычно мало распространены в природных источниках, таких как ребаудиозид D и ребаудиозид E. Рекомбинантные полипептиды настоящей технологии обладают УДФ-гликозилтрансферазной активностью, могут применяться для разработки биосинтетических способов получения новых стевиол-гликозидов, таких как ребаудиозид Z1 и ребаудиозид Z2.
[117] Таким образом, в одном аспекте рассматриваемая технология также относится к способу получения композиции стевиол-гликозидов, включающему инкубирование субстрата с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6.
[118] Субстрат может быть любым природным или синтетическим соединением, способным к превращению в стевиол-гликозидное соединение в реакции, катализируемой одной или более УДФ-гликозилтрансферазами. Например, субстрат может быть натуральным экстрактом стевии, стевиолом, стевиол-13-O-глюкозидом, стевиол-19-O-глюкозидом, стевиол-1,2-биозидом, рубузозидом, стевиозидом, ребаудиозидом A, ребаудиозидом G или ребаудиозидом E. Субстрат может быть чистым соединением или смесью различных соединений. Предпочтительно субстрат включает соединение, выбранное из группы, состоящей из рубузозида, стевиозида, ребаудиозида A, ребаудиозида E и их комбинаций.
[119] Способ, описанный в настоящей заявке, также обеспечивает комбинированную реакционную систему, в которой рекомбинантные пептиды, описанные в настоящей заявке, могут функционировать в комбинации с одним или более дополнительными ферментами для повышения эффективности или изменения результата общего биосинтеза стевиол-гликозидных соединений. Например, дополнительный фермент может регенерировать УДФ-глюкозу, необходимую для реакции гликозилирования, превращая УДФ, образующийся в реакции гликозилирования, обратно в УДФ-глюкозу (при использовании, например, сахарозы в качестве донора остатка глюкозы), что повышает эффективность реакции гликозилирования. В другом примере рекомбинантный полипептид рассматриваемой технологии может давать промежуточный стевиол-гликозидный продукт (например, ребаудиозид E), который затем превращается в другой стевиол-гликозид (например, ребаудиозид D) в реакции, катализируемой другой УДФ-гликозилтрансферазой, такой как UGT76G1. В другом примере рекомбинантный полипептид рассматриваемой технологии может давать промежуточный стевиол-гликозидный продукт (например, ребаудиозид E), который затем превращается в другой стевиол-гликозид (например, ребаудиозид Z1 и ребаудиозид Z2) в реакции, катализируемой УДФ-гликозилтрансферазой, такой как HV1.
[120] Таким образом, в одном варианте осуществления способ рассматриваемой технологии дополнительно включает инкубирование рекомбинантной сахарозосинтазы (СС) с субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. Рекомбинантная сахарозосинтаза превращает УДФ в УДФ-глюкозу, используя сахарозу в качестве источника глюкозы. Подходящая сахарозосинтаза включает сахарозосинтазы, полученные из генов СС Arabidopsis thaliana и Vigna radiate, или из любого гена, который кодирует функциональный гомолог сахарозосинтазы, кодируемой последовательностью SUS1 Arabidopsis thaliana и Vigna radiate, или их функциональные гомологи. Подходящими сахарозосинтазами может быть, например, сахарозосинтаза 1 Arabidopsis; сахарозосинтаза 3 Arabidopsis; и сахарозосинтаза Vigna radiate. Наиболее подходящей сахарозосинтазой может быть, например, сахарозосинтаза 1 Arabidopsis. Например, рекомбинантная СС включает аминокислотную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с аминокислотной последовательности AtSUS1, представленной в SEQ ID NO:9. Предпочтительно рекомбинантная СС рассматриваемой технологии включает аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательностью AtSUS1, представленной в SEQ ID NO:9.
[121] Рекомбинантная сахарозосинтаза рассматриваемой технологии может быть получена при экспрессии нуклеиновой кислоты, имеющей нуклеотидную последовательность, кодирующую целевую аминокислотную последовательность (например, аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательности, представленной в SEQ ID NO:9), в клетке-хозяине, как описано выше. Например, вектор, включающий нуклеотидную последовательность, представленную в SEQ ID NO:10, может быть введен в микробного хозяина (такого как E. coli) с использованием стандартных методик трансформации, с получением рекомбинантной сахарозосинтазы.
[122] В другом варианте осуществления способ рассматриваемой технологии дополнительно включает инкубирование рекомбинантной УДФ-гликозилтрансферазы с рекомбинантной сахарозосинтазой, субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. Рекомбинантная УДФ-гликозилтрансфераза может катализировать другую реакцию гликозилирования, отличную от реакции, катализируемой рекомбинантным полипептидом рассматриваемой технологии. Например, рекомбинантная УДФ-гликозилтрансфераза может катализировать реакцию, которая переносит сахарную группу на C-3’ C-13-O-глюкозы стевиозида с получением ребаудиозида A (или аналогично с получением ребаудиозида D из ребаудиозида E), тогда как рекомбинантный полипептид рассматриваемой технологии переносит вторую сахарную группу на C-2’ 19-O-глюкозы стевиозида с получением ребаудиозида E (или аналогично с получением ребаудиозида D из ребаудиозида A).
[123] Подходящая УДФ-гликозилтрансфераза включает любую УГТ, известную в уровне техники как способную катализировать одну или более реакций в биосинтезе стевиол-гликозидных соединений, таких как UGT85C2, UGT74G1, UGT76G1 или их функциональные гомологи. Например, УДФ-гликозилтрансфераза, как описано в настоящей заявке, может включать аминокислотную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с аминокислотной последовательностью UGT76G1, представленной в SEQ ID NO:11. Предпочтительно УДФ-гликозилтрансфераза включает аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательностью UGT76G1, представленной в SEQ ID NO:11.
[124] Рекомбинантная УДФ-гликозилтрансфераза может быть получена при экспрессии нуклеиновой кислоты, имеющей нуклеотидную последовательность, кодирующую целевую аминокислотную последовательность (например, аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательностью, представленной в SEQ ID NO:11), в клетке-хозяине, как описано выше. Например, вектор, включающий нуклеотидную последовательность, представленную в SEQ ID NO:12, может быть введен в микробного хозяина (такого как E. coli) с помощью стандартных методик трансформации с получением рекомбинантной УДФ-гликозилтрансферазы рассматриваемой технологии.
[125] Рассматриваемая технология охватывает как in vitro, так и in vivo получение стевиол-гликозидных соединений.
[126] Например, инкубирование может быть in vitro процессом, в котором субстрат взаимодействует с рекомбинантным полипептидом рассматриваемой технологии. Предпочтительно в in vitro процессе рекомбинантный полипептид очищают перед инкубированием с субстратом. Стандартные методики очистки полипептидов, такие как центрифугирование, лизис клеток и хроматография, включены в способы рассматриваемой технологии. Например, нуклеиновая кислота, кодирующая рекомбинантный полипептид рассматриваемой технологии, может быть клонирована в вектор экспрессии с гистидиновой меткой, в результате чего экспрессированный рекомбинантный полипептид может быть очищен с помощью аффинной колоночной хроматографии.
[127] In vitro способ рассматриваемой технологии включает любую буферную систему, которая является подходящей для получения стевиол-гликозида с применением одного или более рекомбинантных полипептидов рассматриваемой технологии. Как правило, буферная система является водным раствором, таким как буфер Трис, буфер HEPES, буфер MOPS, фосфатный буфер, с pH от приблизительно 6,0 до приблизительно 8,0. Более предпочтительно pH составляет от приблизительно 6,5 до приблизительно 7,5. Еще более предпочтительно pH составляет от приблизительно 7,0 до приблизительно 7,5.
[128] Как правило, в in vitro способе рассматриваемой технологии субстрат присутствует в буфере в концентрации от приблизительно 0,2 мг/мл до приблизительно 5 мг/мл, предпочтительно от приблизительно 0,5 мг/мл до приблизительно 2 мг/мл, более предпочтительно от приблизительно 0,7 мг/мл до приблизительно 1,5 мг/мл.
[129] Как правило, в in vitro способе рассматриваемой технологии УДФ-глюкоза включена в буфер в концентрации от приблизительно 0,2 мМ до приблизительно 5 мМ, предпочтительно от приблизительно 0,5 мМ до приблизительно 2 мМ, более предпочтительно от приблизительно 0,7 мМ до приблизительно 1,5 мМ. В варианте осуществления, в случае включения рекомбинантной сахарозосинтазы в реакцию, сахарозу также включают в буфер в концентрации от приблизительно 100 мМ до приблизительно 500 мМ, предпочтительно от приблизительно 200 мМ до приблизительно 400 мМ, более предпочтительно от приблизительно 250 мМ до приблизительно 350 мМ.
[130] Как правило, в in vitro способе рассматриваемой технологии весовое соотношение рекомбинантного полипептида к субстрату, в расчете на сухой вес, составляет от приблизительно 1:100 до приблизительно 1:5, предпочтительно от приблизительно 1:50 до приблизительно 1:10, более предпочтительно от приблизительно 1:25 до приблизительно 1:15.
[131] Как правило, температура реакции in vitro способа составляет от приблизительно 20°C до приблизительно 40°C, предпочтительно от 25°C до приблизительно 37°C, более предпочтительно от 28°C до приблизительно 32°C.
[132] В настоящем описании также предусмотрена композиция стевиол-гликозидов, полученная с применением биосинтетического способа, описанного в настоящей заявке. Точная природа композиции стевиол-гликозидов, полученной с применением способа, описанного в настоящей заявке, например, типы соединений и их процентное содержание в конечном продукте, зависит от используемого субстрата, условий инкубирования и ферментативной активности, включенных в реакционную систему. Например, в случае использования стевиозида в качестве субстрата, полученная композиция стевиол-гликозидов может включать ребаудиозид A, ребаудиозид D, ребаудиозид E, ребаудиозид Z1 и ребаудиозид Z2 и их комбинации.
[133] Рассматриваемая технология обеспечивает способ превращения преобладающих стевиол-гликозидных соединений (то есть стевиозида и ребаудиозида A) в натуральном экстракте стевии в ребаудиозид D и ребаудиозид E, которые в иных случаях мало распространены в натуральном экстракте. Таким образом, рассматриваемая технология также обеспечивает способ обогащения содержания одного или более определенных стевиол-гликозидов (таких как ребаудиозид D и ребаудиозид E), включающий инкубирование субстрата (такого как натуральный экстракт стевии) с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. Например, при использовании натурального экстракта стевии в качестве субстрата, получаемая композиция стевиол-гликозидов может быть обогащена ребаудиозидом D и/или ребаудиозидом E, которые мало распространены в натуральном экстракте стевии.
[134] Специалисту в данной области будет известно, что композиция стевиол-гликозидов, полученная способом, описанным в настоящей заявке, может быть затем очищена и смешана с другими стевиол-гликозидами, вкусоароматическими веществами или подсластителями, с получением требуемого вкуса и аромата или композиции подсластителя. Например, композиция, обогащенная ребаудиозидом D, получанная, как описано в настоящей заявке, может быть смешана с натуральным экстрактом стевии, содержащим ребаудиозид A в качестве преобладающего стевиол-гликозида, или с другими синтетическими или натуральными стевиол-гликозидными продуктами, с получением требуемой композиции подсластителя. В альтернативе по существу очищенный стевиол-гликозид (например, ребаудиозид D), полученный из композиции стевиол-гликозидов, описанной в настоящей заявке, может быть объединен с другими подсластителями, такими как сахароза, мальтодекстрин, аспартам, сукралоза, неотам, ацесульфам калия и сахарин. Количество стевиол-гликозида относительно других подсластителей можно регулировать для получения требуемого вкуса, как известно в уровне техники. Композиция стевиол-гликозидов, описанная в настоящей заявке (включающая ребаудиозид D, ребаудиозид E, ребаудиозид Z1, ребаудиозид Z2 или их комбинацию), может быть включена в пищевые продукты (такие как напитки, безалкогольные напитки, мороженое, молочные продукты, кондитерские изделия, изделия из дробленого зерна, жевательную резинку, хлебобулочные изделия и т.д.), биологически активные пищевые добавки, диетическое питание, а также фармацевтические продукты.
ПРИМЕРЫ
ПРИМЕР 1: Выбор кандидатных генов УГТ
[135] Филогенетический и белковый BLAST анализ использовали для идентификации 7 кандидатных генов, которые относятся к подсемейству UGT91, с 1,2-19-O-глюкоза-гликозилирующей активностью (Таблица 1).
| Таблица 1 Список кандидатных генов УГТ |
|||
| Название | Описание | Рег. номер | ID последовательности |
| BD1 | Предсказанный: УДФ-гликозилтрансфераза 91C1-подобный [Brachypodium distachyon] |
XP_003560664.1 | SEQ ID NO: 1 |
| BD2 | Предсказанный: УДФ-гликозилтрансфераза 91C1-подобный [Brachypodium distachyon] |
XP_003560669.1 | SEQ ID NO: 2 |
| BD3 | Предсказанный: белок низкого качества: УДФ-гликозилтрансфераза 91C1-подобный [Brachypodium distachyon] |
XP_003581636.1 | SEQ ID NO: 3 |
| BD4 | Предсказанный: УДФ-гликозилтрансфераза 91C1-подобный [Brachypodium distachyon] |
XP_003580515.1 | SEQ ID NO: 4 |
| BD5 | Предсказанный: белок низкого качества: УДФ-гликозилтрансфераза 91B1-подобный [Brachypodium distachyon] |
XP_003559500.1 | SEQ ID NO: 5 |
| HV1 | предсказанный белок [Hordeum vulgare subsp. vulgare] | BAJ98242.1 | SEQ ID NO: 6 |
| HV2 | предсказанный белок [Hordeum vulgare subsp. vulgare] | BAJ93155.1 | SEQ ID NO: 8 |
Пример 2: Скрининг ферментной активности кандидатных генов УГТ
[136] Фрагменты полноразмерной ДНК всех кандидатных генов УГТ синтезировали коммерчески. Почти все кодоны кДНК были заменены кодонами, предпочтительными для E. coli (Genscript, NJ). Синтезированную ДНК клонировали в бактериальный вектор экспрессии pETite N-His SUMO Kan Vector (Lucigen).
[137] Каждую экспрессионную конструкцию трансформировали в E. coli BL21 (DE3), которую затем выращивали в среде LB, содержащей 50 мкг/мл канамицина, при 37°C до достижения OD600 0,8-1,0. Экспрессию белка индуцировали добавлением 1 мМ изопропил-β-D-1-тиогалактопиранозида (IPTG) и выращивали культуру далее при 16°C в течение 22 ч. Клетки собирали центрифугированием (3000×g; 10 мин; 4°C). Осадки клеток собирали и использовали немедленно или хранили при -80°C.
[138] Осадки клеток обычно ресуспендировали в лизисном буфере (50 мМ калий фосфатный буфер, pH 7,2, 25 мкг/мл лизоцима, 5 мкг/мл ДНКазы I, 20 мМ имидазола, 500 мМ NaCl, 10% глицерина и 0,4% Triton X-100). Клетки разрушали ультразвуком при 4°C и удаляли клеточный дебрис центрифугированием (18000×g; 30 мин). Супернатант наносили на уравновешенную (уравновешивающий буфер: 50 мМ калий фосфатный буфер, pH 7,2, 20 мМ имидазола, 500 мМ NaCl, 10% глицерина) аффинную колонку Ni-NTA (Qiagen). После нанесения образца белка колонку промывали уравновешивающим буфером для удаления не связавшихся белковых примесей. His-меченные рекомбинантные полипептиды УГТ элюировали уравновешивающим буфером, содержащим 250 мМ имидазола.
[139] Очищенные кандидатные УГТ рекомбинантные полипептиды исследовали на наличие 1,2-19-O-глюкоза-гликозилирующей активности при использовании стевиозида или Reb A в качестве субстрата (Фигура 1). Как правило, рекомбинантный полипептид (10 мкг) тестировали в 200 мкл in vitro реакционной системе. Реакционная система содержит 50 мМ калий фосфатный буфер, pH 7,2, 3 мМ MgCl2, 1 мг/мл стевиозида или ребаудиозида A, 1 мМ УДФ-глюкозы. Реакцию проводили при 30°C и останавливали добавлением 200 мкл 1-бутанола. Образцы экстрагировали три раза 200 мкл 1-бутанола. Смешанную фракцию сушили и растворяли в 70 мкл 80% метанола для анализа с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). Экстракт стевии (Blue California, CA), содержащий 95% стевиозида, использовали в качестве стевиозид-содержащего субстрата. Ребаудиозид (чистота 99%) был также поставлен Blue California.
[140] Катализируемую УГТ реакцию гликозилирования объединяли с реакцией образования УДФ-глюкозы, катализируемой сахарозосинтазой (такой как AtSUS1 из SEQ ID NO:9). В этом способе УДФ-глюкоза образовывалась из сахарозы и УДФ (Фигура 2), при этом можно исключить добавление дополнительных количеств УДФ-глюкозы. Последовательность AtSUS1 (Bieniawska et al., Plant J. 2007, 49: 810-828) синтезировали и встраивали в бактериальный вектор экспрессии. Рекомбинантный белок AtSUS1 экспрессировали и очищали с помощью аффинной хроматографии. Очищенный рекомбинантный полипептид AtSUS1 анализировали с помощью электрофореза в ДСН-ПААГ (молекулярная масса: 106,3 кДа, Фигура 7).
[141] Таким образом, активности рекомбинантных полипептидов УГТ тестировали без AtSUS1 (50 мМ калий фосфатный буфер, pH 7,2, 3 мМ MgCl2, 1 мг/мл стевиозида или ребаудиозида A, 1 мМ УДФ) или при объединении с AtSUS (50 мМ калий фосфатный буфер, pH 7,2, 3 мМ MgCl2, 1 мг/мл стевиозида или ребаудиозида A, 1 мМ УДФ и 285 мМ сахарозы). Как правило, 10 мкг AtSUS1 использовали для 200 мкл in vitro реакции. Реакцию in vitro инкубировали при 30°C и останавливали экстракцией при использовании 1-бутанола.
[142] Затем проводили анализ ВЭЖХ при использовании системы Dionex UPLC ultimate 3000 (Sunnyvale, CA), включающей насос для четырехкомпонентных смесей, термостатируемое колоночное отделение, автодозатор и УФ-детектор. Для исследования стевиол-гликозидов использовали Phenomenex Luna NH2 с предколонкой. Ацетонитрил в воде использовали для изократического элюирования в анализе ВЭЖХ. Продукты ребаудиозид D, ребаудиозид E, ребаудиозид Z идентифицировали с помощью ЯМР-анализа.
[143] Рекомбинантный полипептид (SEQ ID NO:6), кодируемый SEQ ID NO:7, показал 1,2-19-O-глюкоза-гликозилирующую активность и был подвергнут дополнительному анализу. Ген был получен из Hordeum vulgare subsp. vulgare (в настоящей заявке сокращенно обозначен "HV1"). Очищенный рекомбинантный полипептид HV1 анализировали с помощью электрофореза в ДСН-ПААГ (Фигура 6). Как показано на Фигуре 6, рекомбинантный белок HV1 (молекулярная масса: 61,4 кДа) очищали с помощью аффинной хроматографии. Полипептиды, кодируемые другими кандидатными генами (Таблица 1), не показали обнаруживаемой активности в анализах, описанных в настоящей заявке, несмотря на то, что они обладали приблизительно 62-74% идентичностью последовательности с рекомбинантным полипептидом HV1.
[144] Как описано в настоящей заявке, рекомбинантный полипептид HV1 переносил сахарную группу на ребаудиозид A с получением ребаудиозида D при любых условиях реакции, с или без AtSUS1. Ребаудиозид A полностью превращался в ребаудиозид D под действием рекомбинантного полипептида HV1 в УГТ-СС комбинированной реакционной системе (Фигура 3B-E). Однако ребаудиозид A лишь частично превращался в ребаудиозид D через 24 часа под действием только одного рекомбинантного полипептида HV1 без объединения с AtSUS1 (Фигура 3F-G). Таким образом, рекомбинантный полипептид HV1 показал 1,2-19-O-глюкоза-гликозилирующую активность с получением ребаудиозида D из ребаудиозида A, при этом AtSUS1 повышала эффективность превращения в УГТ-СС комбинированной системе.
[145] Кроме того, рекомбинантный полипептид HV1 в комбинации с AtSUS1 (SEQ ID NO:9) превращал стевиозид в ребаудиозид E in vitro (Фигура 4). Было получено неожиданное соединение ("Reb Z"), имеющее время удержания ВЭЖХ 6,68 минуты (см. Фигуру 4), которое отличалось от ребаудиозидов D и E. Это соединение представляло собой новый стевиол-гликозид и было названо "ребаудиозид Z" ("Reb Z"). Для подтверждения превращения Reb E в Reb Z, субстрат Reb E (0,5 мг/мл) инкубировали с рекомбинантным полипептидом HV1 (20 мкг) и AtSUS1 (20 мкг) в УГТ-СС комбинированной реакционной системе (200 мкл) при условиях, аналогичных используемым в примерах выше. Как показано на Фигуре 8, Reb Z был получен с применением комбинации рекомбинантного полипептида HV1 и AtSUS1. Эти результаты показали, что HV1 может переносить глюкозную группу на Reb E с получением Reb Z.
ПРИМЕР 3: Биосинтез стевиол-гликозида с применением рекомбинантного полипептида HV1
[146] Как показано на Фигуре 1, ребаудиозид D также может образовываться при гликозилировании C-3’ C-13-O-глюкозы ребаудиозида E. Таким образом, ребаудиозид D может быть получен различными биосинтетическими методами (например, через ребаудиозид, а не ребаудиозид E), в зависимости от порядка, в котором проходят реакции гликозилирования. Например, гликозилирование по C-3’ C-13-O-глюкозы стевиозида может проходить сначала, с образованием промежуточного ребаудиозида A, с последующим гликозилированием по C-2’ 19-O-глюкозы ребаудиозида A с получением ребаудиозида D. На настоящий момент, UGT76G1 (SEQ ID NO:11) из стевии был идентифицирован как фермент, который переносит сахарный остаток на C-3’ C-13-O-глюкозы стевиозида с образованием ребаудиозида A.
[147] Кодон-оптимизированную кДНК UGT76G1 встраивали в бактериальный вектор экспрессии и рекомбинантный белок UGT76G1 экспрессировали и очищали с помощью аффинной хроматографии. Очищенный рекомбинантный полипептид UGT76G1 анализировали с помощью электрофореза в ДСН-ПААГ (молекулярная масса: 65,4 кДа, Фигура 9). Субстрат ребаудиозид E инкубировали с рекомбинантной UGT76G1, с или без AtSUS1, при условиях, аналогичных используемым в Примерах выше. Продукты анализировали с помощью ВЭЖХ. Как показано на Фигуре 12, ребаудиозид D образовывался под действием рекомбинантной UGT76G1. Добавление рекомбинантной AtSUS в реакцию повышало эффективность превращения в УГТ-СС комбинированной системе. Таким образом, рекомбинантный полипептид UGT76G1 показал 1,3-13-O-глюкоза-гликозилирующую активность с получением Reb D из Reb E.
[148] Таким образом, каталитическую активность рекомбинантного полипептида HV1 для биосинтеза стевиол-гликозидов (например, получения ребаудиозида D) также определяли в комбинации с UGT76G1. Субстрат стевиозид инкубировали с рекомбинантным полипептидом HV1 (10 мкг), UGT76G1 (10 мкг) и AtSUS1 (10 мкг) в УГТ-СС комбинированной реакционной системе (200 мкл) при условиях, аналогичных используемым в примерах выше. Продукты анализировали с помощью ВЭЖХ. Как показано на Фигуре 5, ребаудиозид D образовывался под действием комбинации рекомбинантного полипептида HV1, UGT76G1 и AtSUS1. Таким образом, рекомбинантный полипептид HV1, который показал, по меньшей мере, 1,2-19-O-глюкоза-гликозилирующую активность, может применяться в комбинации с другими УГТ ферментами (такими как UGT76G1) для комплексного, многостадийного биосинтеза стевиол-гликозидов.
Пример 4: ЯМР-анализ структуры Reb Z
[149] Материал, используемый для исследования ребаудиозида Z (Reb Z), получали при использовании ферментативного превращения ребаудиозида E и очищали с помощью ВЭЖХ.
[150] Данные МСВР были получены с использованием масс-спектрометра высокого разрешения LTQ Orbitrap Discovery, с установкой разрешения на 30000; сканирования m/z от 150 до 1500 в режиме детектирования положительно заряженных ионов с ионизацией электрораспылением. Напряжение на игле устанавливали на 4 кВ; другие исходные условия были следующими: защитный газ = 25, вспомогательный газ = 0, продувочный газ = 5 (все потоки газа указаны в условных единицах), напряжение на капилляре = 30 В, температура капилляра = 300°C и напряжение на цилиндрической линзе = 75. Образец разбавляли смесью ацетонитрила:метанола:воды 2:2:1 (такой же, как элюент для введения) и вводили 50 микролитров.
[151] Спектры ЯМР получали на спектрометрах Bruker Avance DRX 500 МГц или Varian INOVA 600 МГц при использовании стандартных последовательностей импульсов. Одномерные (1H и 13C) и двумерные (COSY, TOCSY, HMQC и HMBC) спектры ЯМР получали в C5D5N.
[152] Соединение Reb Z, показанное как смесь Reb Z1 и Reb Z2, показано на Фигуре 10. Молекулярная формула соединения Reb Z была выведена как C50H80O28 на основе его положительного масс-спектра высокого разрешения (ВР), который показал присутствие ионов-аддуктов, соответствующих [M+Na]+ при m/z 1151,4713; такой состав подтверждался данными 13C-ЯМР спектра. Данные 1H-ЯМР спектра Reb Z показали присутствие смеси двух соединений (Reb Z1 и Reb Z2) в соотношении от 60:40 до 70:30. Следовательно, данные 1H и 13C-ЯМР спектров Reb Z показали наборы пиков для каждого протона и углерода, присутствующих в его структуре. Кислотный гидролиз Reb Z 5% H2SO4 дал D-глюкозу, которую идентифицировали при непосредственном сравнении с аутентичным образцом с помощью ТСХ. Ферментативный гидролиз Reb Z дал агликон, который идентифицировали как стевиол при сравнении 1H-ЯМР и ТСХ со стандартным соединением. Значения 1H и 13C-ЯМР для соединения Reb Z были установлены на основе данных TOCSY, HMQC и HMBC. Высокие константы взаимодействия, наблюдаемые для пяти аномерных протонов глюкозных групп, указывали на их β-ориентацию, как сообщали для стевиол-гликозидов.
| Таблица 2. Данные 1H и 13C-ЯМР спектров (химические сдвиги и константы взаимодействия) ребаудиозида Z ("Reb Z") и ребаудиозида E a-c. |
||||
| Положение | Reb Z | Ребаудиозид E | ||
| 1 H ЯМР | 13 C ЯМР | 1 H ЯМР | 13 C ЯМР | |
| 1 | 0,74 т (12,7), 1,65 м | 41,2 | 0,73 т (13,2), 1,68 м | 41,0 |
| 2 | 1,45 м, 2,12 м | 20,6 | 1,46 м, 2,13 м | 20,6 |
| 3 | 1,13 м, 2,922 д (13,2)/2,79 д (12,8) |
38,2 | 1,12 м, 2,78 д (12,8) | 38,2 |
| 4 | --- | 44,9/44,8 | --- | 44,8 |
| 5 | 58,1 | 0,97 д (11,8) | 57,9 | |
| 6 | 1,87 м, 2,14 м | 22,6 | 1,85 м, 2,09 м | 22,6 |
| 7 | 1,24 м, 1,68 м | 42,2 | 1,27 м, 1,63 м | 42,1 |
| 8 | --- | 43,2/43,1 | --- | 43,0 |
| 9 | 0,88 шс | 54,5 | 0,88 шс | 54,5 |
| 10 | --- | 40,2 | --- | 40,2 |
| 11 | 21,1 | 21,1 | ||
| 12 | 1,92 м, 2,28 м | 37,8 | 1,96 м, 2,16 м | 37,8 |
| 13 | --- | 86,6 | --- | 86,6 |
| 14 | 1,72 м, 2,48 д (10,8) | 44,9/44,8 | 1,74 д (11,4), 2,54 д (11,0) | 44,8 |
| 15 | 1,92 м, 2,14 м | 48,5 | 2,04 м, 2,12 м | 48,5 |
| 16 | --- | 155,2/155,0 | --- | 154,9 |
| 17 | 5,09/5,12 с, 5,68/5,74 с | 105,4/105,2 | 5,09 с, 5,76 с | 105,4 |
| 18 | 1,43/1,49 с | 29,9 | 1,43 с | 29,8 |
| 19 | --- | 176,3/176,1 | --- | 176,2 |
| 20 | 1,09 с | 17,3 | 1,10 с | 17,2 |
| 1′ | 6,30 д (7,9)/ 6,35 д (7,8) |
93,9/93,6 | 6,30 д (7,9) | 93,9 |
| 2′ | 81,8 | 81,7 | ||
| 3′ | 78,5 | 78,4 | ||
| 4′ | 71,8 | 71,8 | ||
| 5′ | 79,6 | 79,5 | ||
| 6′ | 4,33 м, 4,46 м | 62,6 | 4,33 м, 4,43 м | 62,6 |
| 1′′ | 5,12 д (7,4)/ 5,14 д (7,4)/ |
98,4/98,2 | 5,16 д (7,5) | 98,4 |
| 2′′ | 84,9 | 84,9 | ||
| 3′′ | 78,6 | 78,5 | ||
| 4′′ | 72,1 | 72,1 | ||
| 5′′ | 78,5 | 78,2 | ||
| 6′′ | 4,32 м, 4,38 м | 62,8 | 4,26 м, 4,35 м | 62,9 |
| 1′′′ | 5,33 д (7,8)/ 5,46 д (7,8)/ |
106,7/104,5 | 5,32 д (7,5) | 107,2 |
| 2′′′ | 4,14 т (8,4) | 85,7/77,7 | 4,15 т (8,4) | 77,7 |
| 3′′′ | 78,7 | 78,6 | ||
| 4′′′ | 72,1/71,5 | 72,3 | ||
| 5′′′ | 79,1 | 79,0 | ||
| 6′′′ | 4,43 м, 4,56 м | 63,1 | 4,46 м, 4,56 м | 63,2 |
| 1′′′′ | 5,48 д (7,9)/ 5,40 д (7,6) |
106,3/104,4 | 5,48 д (7,9) | 106,2 |
| 2′′′′ | 4,04 т (7,9) | 85,6/77,3 | 4,06 т (7,9) | 76,8 |
| 3′′′′ | 78,8 | 78,7 | ||
| 4′′′′ | 71,4/71,0 | 71,2 | ||
| 5′′′′ | 79,1 | 79,1 | ||
| 6′′′′ | 4,38 м, 4,57 м | 63,4 | 4,42 м, 4,54 м | 63,4 |
| 1′′′′′ | 5,29 д (7,5)/ 5,34 д (7,5)/ |
107,1 | ||
| 2′′′′′ | 77,0 | |||
| 3′′′′′ | 78,6 | |||
| 4′′′′′ | 71,6/71,2 | |||
| 5′′′′′ | 79,1 | |||
| 6′′′′′ | 4,34 м, 4,48 м | 63,3 | ||
| a отнесения сделаны на основе TOCSY, HMQC и HMBC корреляций; b значения химических сдвигов указаны в δ (м.д.); c константы взаимодействия указаны в Гц. | ||||
[153] На основе результатов из данные ЯМР-спектров и экспериментов гидролиза Reb Z, а также тщательного сравнения значений 1H и 13C ЯМР Reb Z с ребаудиозидом E предположили, что смесь двух соединений, получаемая при ферментативном превращении, была установлена как 13-[(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопиранозил)окси]энт-каур-16-ен-19-овой кислоты 2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый эфир (Reb Z1) или 13-[(2-O-β-D-глюкопиранозил-β-D-глюкопиранозил)-окси]энт-каур-16-ен-19-овой кислоты [(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый) эфир (Reb Z2).
[154] Кислотный гидролиз соединения Reb Z. К раствору соединения Reb Z (5 мг) в MeOH (10 мл) добавляли 3 мл 5% H2SO4 и нагревали смесь с обратным холодильником в течение 24 часов. Затем реакционную смесь нейтрализовали насыщенным карбонатом натрия и экстрагировали этилацетатом (EtOAc) (2×25 мл), получив водную фракцию, содержащую сахар, и фракцию EtOAc, содержащую часть агликона. Водную фазу выпаривали и сравнивали со стандартным сахаром при использовании системы ТСХ EtOAc/н-бутанол/вода (2:7:1) и CH2Cl2/MeOH/вода (10:6:1); сахар идентифицировали как D-глюкоза.
[155] Ферментативный гидролиз соединения Reb Z. Соединение Reb Z (1 мг) растворяли в 10 мл 0,1М натрий ацетатного буфера, pH 4,5, и добавляли неочищенную пектиназу из Aspergillus niger (50 мкл, Sigma-Aldrich, P2736). Смесь перемешивали при 50°C в течение 96 ч. В ходе реакции осаждался продукт, который фильтровали и затем кристаллизовали. Полученный продукт, полученный в результате гидролиза 1, идентифицировали как стевиол при сравнении его ТСХ со стандартным соединением и данными 1H ЯМР спектра (Фигура 11).
[156] Смесь двух соединений, названную Reb Z, получили путем биоконверсии Ребаудиозида E с использованием ферментативной методики, и структуры охарактеризовали как 13-[(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопирано-зил)окси]энт-каур-16-ен-19-овой кислоты 2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый эфир (Reb Z1) или 13-[(2-O-β-D-глюкопиранозил-β-D-глюкопиранозил)окси]энт-каур-16-ен-19-овой кислоты [(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый) эфир (Reb Z2), на основе обширных исследований с помощью одно и двухмерного ЯМР, а также масс-спектральных данных высокого разрешения и гидролитических исследований.
[157] Таким образом, 1,2-19-O-глюкоза-гликозилирующая активность рекомбинантного полипептида HV1 была подтверждена его способностью переносить вторую сахарную группу на C-2' 19-O-глюкозы стевиозида с получением Ребаудиозида E. Рекомбинантный полипептид HV1 также обладает способностью переносить третью группу глюкозы на C-2’ 13-O-глюкозы или C-2’ 19-O-глюкозы ребаудиозида E с получением Reb Z1 или Reb Z2.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Conagen, Inc.
Mao, Guohong
Yu, Xiaodan
<120> RECOMBINANT PRODUCTION OF STEVIOL GLYCOSIDES
<130> 32559-17
<150> US 61/898,571
<151> 2013-11-01
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 470
<212> Белок
<213> Brachypodium distachyon
<400> 1
Met Asp Asp Ala Gly Tyr Ser Ser Ala Tyr Ser Pro Leu His Val Val
1 5 10 15
Ile Cys Pro Trp Leu Ala Phe Gly His Gln Leu Pro Cys Leu Asp Leu
20 25 30
Ala Glu Arg Leu Ala Ser Arg Gly His Arg Val Ser Phe Val Ser Thr
35 40 45
Pro Arg Ile Ile Ala Arg Leu Pro Pro Val Arg Pro Thr Ala Ala Gln
50 55 60
Leu Ile Asn Phe Val Ala Leu Pro Leu Pro Ser Val Asp Gly Leu Pro
65 70 75 80
Glu Gly Ala Glu Ser Thr Asn Asp Val Pro Phe Asp Lys Phe Glu Leu
85 90 95
His Arg Lys Ala Phe Asp Gly Leu Ala Leu Pro Phe Ser Glu Phe Leu
100 105 110
Gly Ala Ala Cys Ala Lys Gly Gln Gly His Lys Pro Asp Trp Ile Leu
115 120 125
Val Asp Ile Phe His His Trp Ala Ala Ala Ala Ala Val Glu His Lys
130 135 140
Val Pro Cys Ala Met Leu Leu Leu Gly Ala Ala Ser Phe Ile Ala Ser
145 150 155 160
Gly Ala Gly Gln Leu Phe Glu His Ala Ala Ser Gly Val Gln Val Gln
165 170 175
Glu Arg Pro Ser Ser Thr Glu Pro Pro Lys Phe Glu Ile Glu Met Arg
180 185 190
Glu Leu Ile Ile Thr Gln Arg Ala Ser Gly Met Ser Ile Ala Glu Arg
195 200 205
Val Ser Leu Thr Leu Gln Arg Ser Asn Leu Ala Ala Met Arg Ser Cys
210 215 220
Val Glu Trp Glu Pro Glu Ser Val Pro Leu Val Ala Ser Leu Gly Val
225 230 235 240
Gly Gly Lys Pro Val Val Pro Leu Gly Leu Leu Pro Pro Ser Pro Glu
245 250 255
Gly Gly Arg Gly Val Cys Lys Asp Gly Lys Lys Asp Ala Thr Val Lys
260 265 270
Trp Leu Asp Val Gln Pro Ala Lys Ser Val Val Tyr Val Ala Met Gly
275 280 285
Thr Glu Val Pro Leu Pro Ala Glu Gln Val His Glu Leu Ala Phe Gly
290 295 300
Ile Glu Leu Ala Gly Thr Arg Phe Leu Trp Ala Leu Arg Lys Pro Ser
305 310 315 320
Gly Gly Ala Pro Asp Ala Asp Ile Leu Pro Pro Gly Phe Glu Asp Arg
325 330 335
Thr Ala Gly Arg Gly Leu Val Arg Thr Gly Trp Val Pro Gln Met Ser
340 345 350
Ile Leu Gly His Asp Ala Val Gly Ala Phe Leu Thr His Cys Gly Trp
355 360 365
Asn Ser Ile Ile Glu Gly Leu Leu Phe Gly His Pro Leu Val Met Leu
370 375 380
Pro Ile Leu Gly Asp Gln Gly Pro Asn Ala Arg Leu Met Glu Gly Lys
385 390 395 400
Lys Val Gly Val Gln Val Gln Arg Asp Gly Asn Asp Gly Ser Phe Asn
405 410 415
Arg Glu Gly Val Ala Met Ala Val Arg Ala Val Met Val Glu Glu Glu
420 425 430
Ser Lys Lys Ile Phe Lys Ala Asn Ala Lys Lys Met Gln Glu Ile Val
435 440 445
Ala Asp Thr Glu Arg His Glu Arg Tyr Ile Asp Gly Phe Ile Gln Gln
450 455 460
Leu Arg Ser Tyr Lys Glu
465 470
<210> 2
<211> 460
<212> Белок
<213> Brachypodium distachyon
<400> 2
Met Asp Asn Gly Ser Ser Ser Ser Ser Ser Ser Pro Leu His Val Val
1 5 10 15
Ile Cys Pro Trp Leu Ala Phe Gly His Gln Leu Pro Cys Leu Asp Leu
20 25 30
Ala Glu Arg Leu Ala Leu Arg Gly His Arg Val Ser Phe Val Ser Thr
35 40 45
Pro Arg Ile Ile Ala Arg Leu Pro Pro Val Arg Pro Val Ala Ala Ser
50 55 60
Leu Val Asp Leu Val Ala Leu Pro Leu Pro Arg Val Asp Gly Leu Pro
65 70 75 80
Glu Gly Ala Glu Ser Thr Asn Asp Val Pro Tyr Glu Lys Phe Glu Leu
85 90 95
His Arg Lys Ala Phe Asp Gly Leu Ala Val Pro Phe Ser Glu Phe Leu
100 105 110
Arg Ala Ala Cys Ala Glu Glu Gly Lys Lys Pro Asp Trp Ile Ile Val
115 120 125
Asp Thr Phe His His Trp Ala Ala Ala Ala Ala Ile Glu His Lys Val
130 135 140
Pro Cys Ala Met Leu Met Leu Gly Ala Ala Gly Leu Ile Val Ala Trp
145 150 155 160
Ala Thr Gln Pro Ser Lys His Val Thr Ser Glu Gln Gln Glu Gln Ser
165 170 175
Ala Ala Glu Pro Pro Arg Phe Glu Thr Glu Arg Arg Lys Leu Ala Thr
180 185 190
Thr Gln Arg Ala Ser Gly Met Ser Ile Ala Glu Arg Cys Ser Val Thr
195 200 205
Leu Glu Arg Cys Asn Leu Val Ala Met Arg Ser Cys Leu Glu Trp Glu
210 215 220
Pro Glu Ser Ile Pro Leu Ala Thr Thr Ile Gly Gly Lys Gln Leu Val
225 230 235 240
Pro Leu Gly Leu Leu Pro Pro Ser Pro Glu Gly Gly Arg Gly Val Ser
245 250 255
Lys Glu Asp Ala Thr Val Arg Trp Leu Asp Ala Gln Pro Thr Lys Ser
260 265 270
Val Val Tyr Val Ala Leu Gly Ser Glu Val Pro Leu Gly Ala Lys Glu
275 280 285
Val His Glu Leu Ala Leu Gly Leu Glu Leu Ala Gly Thr Arg Phe Leu
290 295 300
Trp Ser Leu Arg Lys Pro Ser Gly Val Ser Asp Ala Asp Ile Leu Pro
305 310 315 320
Ser Gly Phe Glu Glu Arg Thr Arg Gly Arg Gly Leu Val Thr Met Gly
325 330 335
Trp Val Pro Gln Ile Ser Val Leu Ala His Gly Ala Val Gly Ala Phe
340 345 350
Leu Thr His Cys Gly Trp Asn Ser Ile Ile Glu Gly Leu Gln Phe Gly
355 360 365
His Pro Leu Val Met Leu Pro Ile Phe Gly Asp Gln Gly Pro Asn Ala
370 375 380
Arg Met Met Glu Gly Arg Lys Val Gly Val Gln Val Pro Arg Asp Glu
385 390 395 400
Ser Asn Gly Ser Phe Asp Arg Glu Gly Val Ala Thr Thr Val Arg Ala
405 410 415
Val Ala Val Glu Glu Glu Gly Asn Arg Ile Phe Thr Ala Asn Ala Lys
420 425 430
Lys Met Gln Glu Ile Val Ala Asp Lys Gly Cys His Asp Lys Tyr Val
435 440 445
Asp Lys Phe Ile Gln Lys Leu Arg Ser Tyr Met Glu
450 455 460
<210> 3
<211> 466
<212> Белок
<213> Brachypodium distachyon
<220>
<221> MISC_FEATURE
<222> (407)..(407)
<223> Неизвестный
<400> 3
Met Asp Ala Ala Gly Ser Ser Ser Ser Ser Pro Leu Arg Ile Ala Ile
1 5 10 15
Val Pro Trp Leu Ala Phe Gly His Leu Leu Pro Tyr Leu Glu Leu Ala
20 25 30
Glu Arg Leu Ala Ala Arg Gly His Arg Val Ser Phe Val Ser Thr Pro
35 40 45
Arg Asn Leu Ala Arg Leu Pro Pro Leu Arg Pro Ala Ala Ala Pro Arg
50 55 60
Val Asp Leu Val Ala Leu Pro Leu Pro Arg Val Glu Gly Leu Pro Asp
65 70 75 80
Gly Ala Glu Ser Thr Asn Asp Val Pro Asp Asp Glu Arg Glu Pro Leu
85 90 95
Trp Lys Ala Phe Asp Gly Leu Ala Ala Pro Phe Ala Gly Phe Leu Thr
100 105 110
Ala Ala Cys Ala Asp Glu Gly Thr Arg Pro His Trp Ile Ile Ala Asp
115 120 125
Ser Phe His His Trp Ala Ala Ala Ala Ala Leu Glu His Lys Val Pro
130 135 140
Cys Ala Met Leu Leu Pro Thr Ala Ala Leu Ile Ala Ala Ser Ala Gly
145 150 155 160
Ala Gly Arg Pro Ser Pro Glu Glu His Ala Glu Gln Gln Pro Gln Pro
165 170 175
Arg Tyr Glu Gln Glu Gly Arg Ala Thr Leu Leu Thr Asp Gly Asp Met
180 185 190
Ser Gly Met Ser Ile Met Gln Arg Ser Val Leu Thr Leu Glu Arg Cys
195 200 205
Lys Leu Thr Ala Ile Arg Ser Cys Val Glu Trp Glu Pro Glu Cys Leu
210 215 220
Pro Leu Val Ser Glu Phe Ile Gly Lys Pro Val Val Pro Leu Gly Leu
225 230 235 240
Leu Pro Pro Ser Pro Asp Gly Gly Arg Arg Ala Ala Asn Thr Asn Gly
245 250 255
Glu Asp Ala Thr Ile Arg Trp Leu Asp Ala Gln Pro Pro Asn Ser Val
260 265 270
Val Tyr Val Ala Leu Gly Ser Glu Val Pro Leu Pro Val Glu Gln Thr
275 280 285
His Glu Leu Ala Leu Gly Leu Glu Leu Ser Lys Thr Arg Phe Leu Trp
290 295 300
Ala Leu Arg Lys Pro Ser Gly Val Leu Asp Ala Glu Met Leu Pro Met
305 310 315 320
Gly Phe Gln Glu Arg Ile His Gly His Gly Leu Val Thr Thr Gly Trp
325 330 335
Val Pro Gln Met Ser Ile Leu Ala His Gly Ala Val Gly Ser Phe Leu
340 345 350
Thr His Cys Gly Arg Asn Ser Leu Ile Glu Gly Leu Leu Phe Gly His
355 360 365
Pro Leu Ile Met Leu Pro Ile Phe Gly Asp Gln Gly Pro Asn Ala Arg
370 375 380
Leu Met Glu Gly Lys Lys Val Gly Leu Gln Val Ala Arg Asn Glu Asn
385 390 395 400
Asp Gly Ser Phe Asp Arg Xaa Gly Val Ala Ser Ala Val Arg Ser Val
405 410 415
Met Leu Glu Glu Asp Ala Arg Lys Ser Phe Val Ala Asn Ala Leu Glu
420 425 430
Met Gln Lys Ile Val Ala Asp Lys Glu Arg His Glu Arg Tyr Ile Asp
435 440 445
Glu Phe Ile His Gln Leu Arg Ser Tyr Thr Ser Ala Pro Thr Ser Asp
450 455 460
Arg Asn
465
<210> 4
<211> 460
<212> Белок
<213> Brachypodium distachyon
<400> 4
Met Asp Ala Ala Gly Ser Ser Pro Pro Leu Arg Ile Val Ile Val Pro
1 5 10 15
Trp Leu Ala Phe Gly His Met Leu Pro Tyr Leu Glu Leu Ala Glu Arg
20 25 30
Leu Ala Ala Arg Gly His Arg Val Ser Tyr Val Ser Thr Pro Arg Asn
35 40 45
Leu Ala Arg Leu Pro Pro Leu Arg Pro Ala Ala Ala Pro Arg Val Asp
50 55 60
Leu Val Ala Leu Pro Leu Pro Arg Val Glu Gly Leu Pro Asp Gly Ala
65 70 75 80
Glu Ser Thr Asn Asp Val Pro Asp Asp Glu Arg Glu Pro Leu Trp Lys
85 90 95
Ala Phe Asp Gly Leu Ala Ala Pro Phe Arg Ser Val Pro Arg Gln Arg
100 105 110
Cys Ala Arg Asp Asp Thr Arg Pro His Trp Ile Leu Ala Asp Cys Phe
115 120 125
His His Trp Ala Val Asp Ala Ala Leu Asp His Lys Val Pro Cys Ala
130 135 140
Met Phe Leu Pro Thr Ala Ala Val Ile Ala Thr Met Pro Gln Arg Gln
145 150 155 160
Pro Asp His Ala Ala Ser Ala Pro Ala Glu His Ala Val Pro Arg His
165 170 175
Glu Ile Glu Ala Thr Ala Pro Leu Leu Ser Asp Gln Gly Val Ser Gly
180 185 190
Met Ser Ile Val Gln Arg Tyr Leu Leu Thr Lys Glu Arg Cys Thr Val
195 200 205
Gly Ala Ile Arg Ser Cys Val Glu Trp Glu Pro Asp Ser Tyr Pro Leu
210 215 220
Ala Ala Thr Ile Leu Gly Met Pro Val Val Pro Leu Gly Leu Leu Pro
225 230 235 240
Pro Ser Pro Asp Gly Gly Arg Arg Ala Pro Asp Gly Ser Glu His Ala
245 250 255
Thr Val Arg Trp Leu Asp Ala Gln Pro Pro Ser Ser Val Val Tyr Val
260 265 270
Ala Leu Gly Ser Glu Val Pro Leu Pro Val Asp His Val His Glu Leu
275 280 285
Ala Leu Gly Leu Glu Leu Ala Gly Thr Arg Phe Leu Trp Ala Leu Arg
290 295 300
Lys Pro Asn Gly Val Pro Asp Ala Asp Met Leu Pro Ala Gly Phe Gln
305 310 315 320
Asp Arg Thr Arg Gly His Gly Leu Val Thr Thr Gly Trp Val Pro Gln
325 330 335
Met Ser Ile Leu Ala His Gly Ala Val Gly Ala Phe Leu Thr His Cys
340 345 350
Gly Arg Asn Ser Leu Ile Glu Gly Leu Leu Leu Gly His Pro Leu Val
355 360 365
Met Leu Pro Ile Phe Gly Asp Gln Gly Pro Asn Ala Arg Ala Met Glu
370 375 380
Arg Lys Lys Val Gly Leu Gln Val Lys Arg Asp Asp Asn Asp Gly Ser
385 390 395 400
Phe Asn Arg Glu Gly Val Ala Asp Ala Val Arg Gly Val Met Val Asp
405 410 415
Gly Glu Ala Arg Arg Gly Phe Val Ala Asn Ala Arg Lys Met Gln Asn
420 425 430
Val Val Ala Asp Glu Glu Leu Gln Glu Arg Tyr Val Asp Gly Phe Val
435 440 445
Gln Glu Leu Arg Ser Tyr Thr Thr Ala Ala Ser Glu
450 455 460
<210> 5
<211> 471
<212> Белок
<213> Brachypodium distachyon
<220>
<221> MISC_FEATURE
<222> (323)..(323)
<223> Неизвестный
<400> 5
Met Asp Asp Gly Ser Ser Ser Ser Pro Leu His Val Val Ile Cys Pro
1 5 10 15
Trp Leu Ala Phe Gly His Leu Leu Pro Cys Leu Asp Leu Ala Gln Cys
20 25 30
Leu Ala Ser Arg Gly His Arg Val Ser Phe Val Ser Thr Pro Cys Asn
35 40 45
Ile Ala Cys Leu Leu Leu Val Pro Pro Ala Met Ala Pro Leu Val Asp
50 55 60
Phe Val Glu Leu Pro Phe Pro Phe Pro His Val Asp Gly Leu Ser Glu
65 70 75 80
Arg Lys Leu His Ile Pro Pro Pro Ile Gly Asp Lys Phe Glu Leu His
85 90 95
Arg Lys Ala Phe Asp Gly Leu Ala Met Pro Phe Ser Glu Leu Leu Gly
100 105 110
Ala Ser Cys Ala Glu Gly Gly Lys Lys Leu Asp Trp Val Ile Val Asp
115 120 125
Ile Phe His His Trp Ala Ala Ala Asp Thr Leu Glu Tyr Lys Val Pro
130 135 140
Ile Gly Ala Ala Asn Val Ile Ala Thr Trp His Gly Arg Leu Val Lys
145 150 155 160
His Thr Thr Met Ser Lys Gln Glu Gln Pro Ala Ser Lys Leu Pro Arg
165 170 175
Phe Glu Ile Glu Arg Arg Gln Leu Ser Thr Thr Gln Arg Ala Ser Gly
180 185 190
Met Ser Ile Ala Glu Arg Ile Ser Leu Thr Leu Arg Arg Cys Asn Leu
195 200 205
Val Val Met Arg Thr Arg Leu Glu Trp Glu Pro Glu Ser Val Pro Leu
210 215 220
Ala Ala Ser Leu Gly Gly Lys Pro Val Ile Ser Leu Gly Leu Leu Pro
225 230 235 240
Pro Leu Arg Lys Gly Cys Cys Gly Val Thr Glu Asp Gly Lys Asn Ile
245 250 255
Met Cys Trp Leu Asp Leu Gln Pro Ala Lys Ser Val Val Tyr Val Ala
260 265 270
Leu Gly Thr Glu Val Pro Leu Pro Val Glu Gln Met His Glu Leu Ala
275 280 285
Leu Arg Leu Glu Leu Ala Gly Met Gln Phe Leu Trp Ala Leu Arg Lys
290 295 300
Pro Arg Gly Val His Glu Ala Glu Ile Leu Pro Leu Gly Phe Glu Glu
305 310 315 320
Arg Met Xaa Gly Leu Val Thr Thr Gly Leu Ala Pro Gln Ile Asn Ile
325 330 335
Leu Ala His Gly Ala Val Gly Thr Phe Leu Thr His Cys Gly Trp Ser
340 345 350
Leu Thr Ile Glu Gly Leu Leu Phe Gly His Pro Leu Ile Met Leu Pro
355 360 365
Met Phe Gly Asp Gln Gly Pro Asn Ala Arg Leu Met Glu Gly Arg Lys
370 375 380
Val Gly Val Gln Val Pro Arg Asn Glu Gly Asp Gly Ser Phe Asp Arg
385 390 395 400
Glu Gly Val Ala Thr Thr Val Arg Ala Val Thr Val Glu Glu Glu Gly
405 410 415
Lys Arg Ile Phe Thr Ser Asn Ala Lys Lys Met Gln Glu Ile Lys Ala
420 425 430
Asp Thr Glu Cys Gln Glu Arg Tyr Ile Asn Gly Phe Ile Lys Lys Leu
435 440 445
Arg Ser Tyr Lys Glu Pro Asp Leu Ala His Pro Ile Cys Pro Val Thr
450 455 460
Ser Ser Pro Thr His Tyr Glu
465 470
<210> 6
<211> 459
<212> Белок
<213> Hordeum vulgare
<400> 6
Met Asp Gly Asn Ser Ser Ser Ser Pro Leu His Val Val Ile Cys Pro
1 5 10 15
Trp Leu Ala Leu Gly His Leu Leu Pro Cys Leu Asp Ile Ala Glu Arg
20 25 30
Leu Ala Ser Arg Gly His Arg Val Ser Phe Val Ser Thr Pro Arg Asn
35 40 45
Ile Ala Arg Leu Pro Pro Leu Arg Pro Ala Val Ala Pro Leu Val Asp
50 55 60
Phe Val Ala Leu Pro Leu Pro His Val Asp Gly Leu Pro Glu Gly Ala
65 70 75 80
Glu Ser Thr Asn Asp Val Pro Tyr Asp Lys Phe Glu Leu His Arg Lys
85 90 95
Ala Phe Asp Gly Leu Ala Ala Pro Phe Ser Glu Phe Leu Arg Ala Ala
100 105 110
Cys Ala Glu Gly Ala Gly Ser Arg Pro Asp Trp Leu Ile Val Asp Thr
115 120 125
Phe His His Trp Ala Ala Ala Ala Ala Val Glu Asn Lys Val Pro Cys
130 135 140
Val Met Leu Leu Leu Gly Ala Ala Thr Val Ile Ala Gly Phe Ala Arg
145 150 155 160
Gly Val Ser Glu His Ala Ala Ala Ala Val Gly Lys Glu Arg Pro Ala
165 170 175
Ala Glu Ala Pro Ser Phe Glu Thr Glu Arg Arg Lys Leu Met Thr Thr
180 185 190
Gln Asn Ala Ser Gly Met Thr Val Ala Glu Arg Tyr Phe Leu Thr Leu
195 200 205
Met Arg Ser Asp Leu Val Ala Ile Arg Ser Cys Ala Glu Trp Glu Pro
210 215 220
Glu Ser Val Ala Ala Leu Thr Thr Leu Ala Gly Lys Pro Val Val Pro
225 230 235 240
Leu Gly Leu Leu Pro Pro Ser Pro Glu Gly Gly Arg Gly Val Ser Lys
245 250 255
Glu Asp Ala Ala Val Arg Trp Leu Asp Ala Gln Pro Ala Lys Ser Val
260 265 270
Val Tyr Val Ala Leu Gly Ser Glu Val Pro Leu Arg Ala Glu Gln Val
275 280 285
His Glu Leu Ala Leu Gly Leu Glu Leu Ser Gly Ala Arg Phe Leu Trp
290 295 300
Ala Leu Arg Lys Pro Thr Asp Ala Pro Asp Ala Ala Val Leu Pro Pro
305 310 315 320
Gly Phe Glu Glu Arg Thr Arg Gly Arg Gly Leu Val Val Thr Gly Trp
325 330 335
Val Pro Gln Ile Gly Val Leu Ala His Gly Ala Val Ala Ala Phe Leu
340 345 350
Thr His Cys Gly Trp Asn Ser Thr Ile Glu Gly Leu Leu Phe Gly His
355 360 365
Pro Leu Ile Met Leu Pro Ile Ser Ser Asp Gln Gly Pro Asn Ala Arg
370 375 380
Leu Met Glu Gly Arg Lys Val Gly Met Gln Val Pro Arg Asp Glu Ser
385 390 395 400
Asp Gly Ser Phe Arg Arg Glu Asp Val Ala Ala Thr Val Arg Ala Val
405 410 415
Ala Val Glu Glu Asp Gly Arg Arg Val Phe Thr Ala Asn Ala Lys Lys
420 425 430
Met Gln Glu Ile Val Ala Asp Gly Ala Cys His Glu Arg Cys Ile Asp
435 440 445
Gly Phe Ile Gln Gln Leu Arg Ser Tyr Lys Ala
450 455
<210> 7
<211> 1380
<212> ДНК
<213> Hordeum vulgare
<400> 7
atggatggta actcctcctc ctcgccgctg catgtggtca tttgtccgtg gctggctctg 60
ggtcacctgc tgccgtgtct ggatattgct gaacgtctgg cgtcacgcgg ccatcgtgtc 120
agttttgtgt ccaccccgcg caacattgcc cgtctgccgc cgctgcgtcc ggctgttgca 180
ccgctggttg atttcgtcgc actgccgctg ccgcatgttg acggtctgcc ggagggtgcg 240
gaatcgacca atgatgtgcc gtatgacaaa tttgaactgc accgtaaggc gttcgatggt 300
ctggcggccc cgtttagcga atttctgcgt gcagcttgcg cagaaggtgc aggttctcgc 360
ccggattggc tgattgtgga cacctttcat cactgggcgg cggcggcggc ggtggaaaac 420
aaagtgccgt gtgttatgct gctgctgggt gcagcaacgg tgatcgctgg tttcgcgcgt 480
ggtgttagcg aacatgcggc ggcggcggtg ggtaaagaac gtccggctgc ggaagccccg 540
agttttgaaa ccgaacgtcg caagctgatg accacgcaga atgcctccgg catgaccgtg 600
gcagaacgct atttcctgac gctgatgcgt agcgatctgg ttgccatccg ctcttgcgca 660
gaatgggaac cggaaagcgt ggcagcactg accacgctgg caggtaaacc ggtggttccg 720
ctgggtctgc tgccgccgag tccggaaggc ggtcgtggcg tttccaaaga agatgctgcg 780
gtccgttggc tggacgcaca gccggcaaag tcagtcgtgt acgtcgcact gggttcggaa 840
gtgccgctgc gtgcggaaca agttcacgaa ctggcactgg gcctggaact gagcggtgct 900
cgctttctgt gggcgctgcg taaaccgacc gatgcaccgg acgccgcagt gctgccgccg 960
ggtttcgaag aacgtacccg cggccgtggt ctggttgtca cgggttgggt gccgcagatt 1020
ggcgttctgg ctcatggtgc ggtggctgcg tttctgaccc actgtggctg gaactctacg 1080
atcgaaggcc tgctgttcgg tcatccgctg attatgctgc cgatcagctc tgatcagggt 1140
ccgaatgcgc gcctgatgga aggccgtaaa gtcggtatgc aagtgccgcg tgatgaatca 1200
gacggctcgt ttcgtcgcga agatgttgcc gcaaccgtcc gcgccgtggc agttgaagaa 1260
gacggtcgtc gcgtcttcac ggctaacgcg aaaaagatgc aagaaattgt ggccgatggc 1320
gcatgccacg aacgttgtat tgacggtttt atccagcaac tgcgcagtta caaggcgtga 1380
<210> 8
<211> 488
<212> Белок
<213> Hordeum vulgare
<400> 8
Met Asp Ala Gly Ser Ser Thr Ala Gly Pro Leu Arg Ile Val Ile Cys
1 5 10 15
Pro Trp Leu Ala Phe Gly His Leu Leu Pro Tyr Leu Glu Leu Ala Glu
20 25 30
Arg Leu Ala Ser Arg Gly His Arg Val Ala Phe Val Ser Thr Pro Arg
35 40 45
Asn Leu Ala Arg Leu Pro Pro Pro Ala Ser Pro Cys Ser Val Asp Leu
50 55 60
Val Ala Leu Gln Leu Pro Arg Val Asp Gly Leu Pro Glu Gly Ala Glu
65 70 75 80
Ser Thr Asn Asp Val Pro Asp Glu Met Arg Glu Leu His Trp Lys Ala
85 90 95
Phe Asp Gly Leu Ala Ala Pro Phe Ala Asp Phe Leu Ala Ala Ala Cys
100 105 110
Ala Asp Asp Gly Arg Arg Pro His Trp Ile Ile Ala Asp Cys Phe His
115 120 125
His Trp Ala Ala Ala Ala Ala Leu Asp His Lys Val Pro Cys Ala Val
130 135 140
Leu Leu Pro Thr Ala Ala Met Leu Ala Ala Ala Pro Arg Gln Gln Pro
145 150 155 160
Leu Gly Ser Lys Pro Val Glu Ala Ala Ala Ala Ala Ser Val Leu Gly
165 170 175
Gln Ala Ala Ala Ala Val Arg Leu Ala Val Pro Ser Tyr Glu Arg Asp
180 185 190
Asp Val Ala Pro Ala Tyr Ala Asp Asp Cys Ala Ser Gly Met Ser Ile
195 200 205
Ala Gln Arg Trp Phe Leu Ala Lys Glu Arg Cys Thr Val Leu Ala Ile
210 215 220
Arg Ser Cys Val Glu Trp Glu Pro Glu Thr Phe Pro Leu Val Glu Ala
225 230 235 240
Leu Leu Gly Lys Pro Val Val Pro Leu Gly Leu Leu Pro Pro Ser Ala
245 250 255
Asp Gly Gly Arg Arg Arg Ala Ala Gly Ser Ser Glu Asp His Val Thr
260 265 270
Leu Arg Trp Leu Glu Glu Gln Pro Pro Asp Ser Val Val Tyr Ile Ala
275 280 285
Leu Gly Ser Glu Val Pro Leu Ser Ile Glu Gln Val His Glu Leu Ala
290 295 300
Leu Gly Leu Glu Leu Ala Gly Thr Arg Phe Leu Trp Ala Leu Arg Lys
305 310 315 320
Pro Ala Gly Ala Val Val Gly Asn Asn Asp Asp Asp Thr Leu Pro Pro
325 330 335
Gly Phe Arg Asp Cys Thr Arg Gly His Gly Leu Val Thr Met Gly Trp
340 345 350
Val Pro Gln Ile Ser Ile Leu Ala His Ala Ala Val Gly Ala Phe Leu
355 360 365
Thr His Cys Gly Arg Asn Ser Leu Ile Glu Gly Leu Leu Phe Gly His
370 375 380
Pro Leu Val Met Leu Pro Ile Phe Gly Asp Gln Gly Pro Asn Ala Arg
385 390 395 400
Gln Met Glu Ala Lys Lys Val Gly Leu Gln Val Ala Arg Asp Asp Asp
405 410 415
Asp Gly Ser Phe Asp Arg His Gly Val Ala Thr Ala Val Arg Ala Val
420 425 430
Met Val Asp Gly Glu Ala Arg Arg Gly Phe Val Ala Asn Ala Ile Lys
435 440 445
Met Gln Ala Ile Val Ala Asp Lys Glu Arg His Glu Arg Tyr Ile Asp
450 455 460
Gly Phe Val Gln Gln Leu Arg Ser Tyr Leu Gln Ala Thr Asp Asp Leu
465 470 475 480
Thr Thr Ala Thr Pro Asn Ser Ser
485
<210> 9
<211> 808
<212> Белок
<213> Arabidopsis thaliana
<400> 9
Met Ala Asn Ala Glu Arg Met Ile Thr Arg Val His Ser Gln Arg Glu
1 5 10 15
Arg Leu Asn Glu Thr Leu Val Ser Glu Arg Asn Glu Val Leu Ala Leu
20 25 30
Leu Ser Arg Val Glu Ala Lys Gly Lys Gly Ile Leu Gln Gln Asn Gln
35 40 45
Ile Ile Ala Glu Phe Glu Ala Leu Pro Glu Gln Thr Arg Lys Lys Leu
50 55 60
Glu Gly Gly Pro Phe Phe Asp Leu Leu Lys Ser Thr Gln Glu Ala Ile
65 70 75 80
Val Leu Pro Pro Trp Val Ala Leu Ala Val Arg Pro Arg Pro Gly Val
85 90 95
Trp Glu Tyr Leu Arg Val Asn Leu His Ala Leu Val Val Glu Glu Leu
100 105 110
Gln Pro Ala Glu Phe Leu His Phe Lys Glu Glu Leu Val Asp Gly Val
115 120 125
Lys Asn Gly Asn Phe Thr Leu Glu Leu Asp Phe Glu Pro Phe Asn Ala
130 135 140
Ser Ile Pro Arg Pro Thr Leu His Lys Tyr Ile Gly Asn Gly Val Asp
145 150 155 160
Phe Leu Asn Arg His Leu Ser Ala Lys Leu Phe His Asp Lys Glu Ser
165 170 175
Leu Leu Pro Leu Leu Lys Phe Leu Arg Leu His Ser His Gln Gly Lys
180 185 190
Asn Leu Met Leu Ser Glu Lys Ile Gln Asn Leu Asn Thr Leu Gln His
195 200 205
Thr Leu Arg Lys Ala Glu Glu Tyr Leu Ala Glu Leu Lys Ser Glu Thr
210 215 220
Leu Tyr Glu Glu Phe Glu Ala Lys Phe Glu Glu Ile Gly Leu Glu Arg
225 230 235 240
Gly Trp Gly Asp Asn Ala Glu Arg Val Leu Asp Met Ile Arg Leu Leu
245 250 255
Leu Asp Leu Leu Glu Ala Pro Asp Pro Cys Thr Leu Glu Thr Phe Leu
260 265 270
Gly Arg Val Pro Met Val Phe Asn Val Val Ile Leu Ser Pro His Gly
275 280 285
Tyr Phe Ala Gln Asp Asn Val Leu Gly Tyr Pro Asp Thr Gly Gly Gln
290 295 300
Val Val Tyr Ile Leu Asp Gln Val Arg Ala Leu Glu Ile Glu Met Leu
305 310 315 320
Gln Arg Ile Lys Gln Gln Gly Leu Asn Ile Lys Pro Arg Ile Leu Ile
325 330 335
Leu Thr Arg Leu Leu Pro Asp Ala Val Gly Thr Thr Cys Gly Glu Arg
340 345 350
Leu Glu Arg Val Tyr Asp Ser Glu Tyr Cys Asp Ile Leu Arg Val Pro
355 360 365
Phe Arg Thr Glu Lys Gly Ile Val Arg Lys Trp Ile Ser Arg Phe Glu
370 375 380
Val Trp Pro Tyr Leu Glu Thr Tyr Thr Glu Asp Ala Ala Val Glu Leu
385 390 395 400
Ser Lys Glu Leu Asn Gly Lys Pro Asp Leu Ile Ile Gly Asn Tyr Ser
405 410 415
Asp Gly Asn Leu Val Ala Ser Leu Leu Ala His Lys Leu Gly Val Thr
420 425 430
Gln Cys Thr Ile Ala His Ala Leu Glu Lys Thr Lys Tyr Pro Asp Ser
435 440 445
Asp Ile Tyr Trp Lys Lys Leu Asp Asp Lys Tyr His Phe Ser Cys Gln
450 455 460
Phe Thr Ala Asp Ile Phe Ala Met Asn His Thr Asp Phe Ile Ile Thr
465 470 475 480
Ser Thr Phe Gln Glu Ile Ala Gly Ser Lys Glu Thr Val Gly Gln Tyr
485 490 495
Glu Ser His Thr Ala Phe Thr Leu Pro Gly Leu Tyr Arg Val Val His
500 505 510
Gly Ile Asp Val Phe Asp Pro Lys Phe Asn Ile Val Ser Pro Gly Ala
515 520 525
Asp Met Ser Ile Tyr Phe Pro Tyr Thr Glu Glu Lys Arg Arg Leu Thr
530 535 540
Lys Phe His Ser Glu Ile Glu Glu Leu Leu Tyr Ser Asp Val Glu Asn
545 550 555 560
Lys Glu His Leu Cys Val Leu Lys Asp Lys Lys Lys Pro Ile Leu Phe
565 570 575
Thr Met Ala Arg Leu Asp Arg Val Lys Asn Leu Ser Gly Leu Val Glu
580 585 590
Trp Tyr Gly Lys Asn Thr Arg Leu Arg Glu Leu Ala Asn Leu Val Val
595 600 605
Val Gly Gly Asp Arg Arg Lys Glu Ser Lys Asp Asn Glu Glu Lys Ala
610 615 620
Glu Met Lys Lys Met Tyr Asp Leu Ile Glu Glu Tyr Lys Leu Asn Gly
625 630 635 640
Gln Phe Arg Trp Ile Ser Ser Gln Met Asp Arg Val Arg Asn Gly Glu
645 650 655
Leu Tyr Arg Tyr Ile Cys Asp Thr Lys Gly Ala Phe Val Gln Pro Ala
660 665 670
Leu Tyr Glu Ala Phe Gly Leu Thr Val Val Glu Ala Met Thr Cys Gly
675 680 685
Leu Pro Thr Phe Ala Thr Cys Lys Gly Gly Pro Ala Glu Ile Ile Val
690 695 700
His Gly Lys Ser Gly Phe His Ile Asp Pro Tyr His Gly Asp Gln Ala
705 710 715 720
Ala Asp Thr Leu Ala Asp Phe Phe Thr Lys Cys Lys Glu Asp Pro Ser
725 730 735
His Trp Asp Glu Ile Ser Lys Gly Gly Leu Gln Arg Ile Glu Glu Lys
740 745 750
Tyr Thr Trp Gln Ile Tyr Ser Gln Arg Leu Leu Thr Leu Thr Gly Val
755 760 765
Tyr Gly Phe Trp Lys His Val Ser Asn Leu Asp Arg Leu Glu Ala Arg
770 775 780
Arg Tyr Leu Glu Met Phe Tyr Ala Leu Lys Tyr Arg Pro Leu Ala Gln
785 790 795 800
Ala Val Pro Leu Ala Gln Asp Asp
805
<210> 10
<211> 2427
<212> ДНК
<213> Arabidopsis thaliana
<400> 10
atggcaaacg ctgaacgtat gataacgcgc gtccacagcc aacgtgagcg tttgaacgaa 60
acgcttgttt ctgagagaaa cgaagtcctt gccttgcttt ccagggttga agccaaaggt 120
aaaggtattt tacaacaaaa ccagatcatt gctgaattcg aagctttgcc tgaacaaacc 180
cggaagaaac ttgaaggtgg tcctttcttt gaccttctca aatccactca ggaagcaatt 240
gtgttgccac catgggttgc tctagctgtg aggccaaggc ctggtgtttg ggaatactta 300
cgagtcaatc tccatgctct tgtcgttgaa gaactccaac ctgctgagtt tcttcatttc 360
aaggaagaac tcgttgatgg agttaagaat ggtaatttca ctcttgagct tgatttcgag 420
ccattcaatg cgtctatccc tcgtccaaca ctccacaaat acattggaaa tggtgttgac 480
ttccttaacc gtcatttatc ggctaagctc ttccatgaca aggagagttt gcttccattg 540
cttaagttcc ttcgtcttca cagccaccag ggcaagaacc tgatgttgag cgagaagatt 600
cagaacctca acactctgca acacaccttg aggaaagcag aagagtatct agcagagctt 660
aagtccgaaa cactgtatga agagtttgag gccaagtttg aggagattgg tcttgagagg 720
ggatggggag acaatgcaga gcgtgtcctt gacatgatac gtcttctttt ggaccttctt 780
gaggcgcctg atccttgcac tcttgagact tttcttggaa gagtaccaat ggtgttcaac 840
gttgtgatcc tctctccaca tggttacttt gctcaggaca atgttcttgg ttaccctgac 900
actggtggac aggttgttta cattcttgat caagttcgtg ctctggagat agagatgctt 960
caacgtatta agcaacaagg actcaacatt aaaccaagga ttctcattct aactcgactt 1020
ctacctgatg cggtaggaac tacatgcggt gaacgtctcg agagagttta tgattctgag 1080
tactgtgata ttcttcgtgt gcccttcaga acagagaagg gtattgttcg caaatggatc 1140
tcaaggttcg aagtctggcc atatctagag acttacaccg aggatgctgc ggttgagcta 1200
tcgaaagaat tgaatggcaa gcctgacctt atcattggta actacagtga tggaaatctt 1260
gttgcttctt tattggctca caaacttggt gtcactcagt gtaccattgc tcatgctctt 1320
gagaaaacaa agtacccgga ttctgatatc tactggaaga agcttgacga caagtaccat 1380
ttctcatgcc agttcactgc ggatattttc gcaatgaacc acactgattt catcatcact 1440
agtactttcc aagaaattgc tggaagcaaa gaaactgttg ggcagtatga aagccacaca 1500
gcctttactc ttcccggatt gtatcgagtt gttcacggga ttgatgtgtt tgatcccaag 1560
ttcaacattg tctctcctgg tgctgatatg agcatctact tcccttacac agaggagaag 1620
cgtagattga ctaagttcca ctctgagatc gaggagctcc tctacagcga tgttgagaac 1680
aaagagcact tatgtgtgct caaggacaag aagaagccga ttctcttcac aatggctagg 1740
cttgatcgtg tcaagaactt gtcaggtctt gttgagtggt acgggaagaa cacccgcttg 1800
cgtgagctag ctaacttggt tgttgttgga ggagacagga ggaaagagtc aaaggacaat 1860
gaagagaaag cagagatgaa gaaaatgtat gatctcattg aggaatacaa gctaaacggt 1920
cagttcaggt ggatctcctc tcagatggac cgggtaagga acggtgagct gtaccggtac 1980
atctgtgaca ccaagggtgc ttttgtccaa cctgcattat atgaagcctt tgggttaact 2040
gttgtggagg ctatgacttg tggtttaccg actttcgcca cttgcaaagg tggtccagct 2100
gagatcattg tgcacggtaa atcgggtttc cacattgacc cttaccatgg tgatcaggct 2160
gctgatactc ttgctgattt cttcaccaag tgtaaggagg atccatctca ctgggatgag 2220
atctcaaaag gagggcttca gaggattgag gagaaataca cttggcaaat ctattcacag 2280
aggctcttga cattgactgg tgtgtatgga ttctggaagc atgtctcgaa ccttgaccgt 2340
cttgaggctc gccgttacct tgaaatgttc tatgcattga agtatcgccc attggctcag 2400
gctgttcctc ttgcacaaga tgattga 2427
<210> 11
<211> 458
<212> Белок
<213> Stevia rebaudiana
<400> 11
Met Glu Asn Lys Thr Glu Thr Thr Val Arg Arg Arg Arg Arg Ile Ile
1 5 10 15
Leu Phe Pro Val Pro Phe Gln Gly His Ile Asn Pro Ile Leu Gln Leu
20 25 30
Ala Asn Val Leu Tyr Ser Lys Gly Phe Ser Ile Thr Ile Phe His Thr
35 40 45
Asn Phe Asn Lys Pro Lys Thr Ser Asn Tyr Pro His Phe Thr Phe Arg
50 55 60
Phe Ile Leu Asp Asn Asp Pro Gln Asp Glu Arg Ile Ser Asn Leu Pro
65 70 75 80
Thr His Gly Pro Leu Ala Gly Met Arg Ile Pro Ile Ile Asn Glu His
85 90 95
Gly Ala Asp Glu Leu Arg Arg Glu Leu Glu Leu Leu Met Leu Ala Ser
100 105 110
Glu Glu Asp Glu Glu Val Ser Cys Leu Ile Thr Asp Ala Leu Trp Tyr
115 120 125
Phe Ala Gln Ser Val Ala Asp Ser Leu Asn Leu Arg Arg Leu Val Leu
130 135 140
Met Thr Ser Ser Leu Phe Asn Phe His Ala His Val Ser Leu Pro Gln
145 150 155 160
Phe Asp Glu Leu Gly Tyr Leu Asp Pro Asp Asp Lys Thr Arg Leu Glu
165 170 175
Glu Gln Ala Ser Gly Phe Pro Met Leu Lys Val Lys Asp Ile Lys Ser
180 185 190
Ala Tyr Ser Asn Trp Gln Ile Leu Lys Glu Ile Leu Gly Lys Met Ile
195 200 205
Lys Gln Thr Lys Ala Ser Ser Gly Val Ile Trp Asn Ser Phe Lys Glu
210 215 220
Leu Glu Glu Ser Glu Leu Glu Thr Val Ile Arg Glu Ile Pro Ala Pro
225 230 235 240
Ser Phe Leu Ile Pro Leu Pro Lys His Leu Thr Ala Ser Ser Ser Ser
245 250 255
Leu Leu Asp His Asp Arg Thr Val Phe Gln Trp Leu Asp Gln Gln Pro
260 265 270
Pro Ser Ser Val Leu Tyr Val Ser Phe Gly Ser Thr Ser Glu Val Asp
275 280 285
Glu Lys Asp Phe Leu Glu Ile Ala Arg Gly Leu Val Asp Ser Lys Gln
290 295 300
Ser Phe Leu Trp Val Val Arg Pro Gly Phe Val Lys Gly Ser Thr Trp
305 310 315 320
Val Glu Pro Leu Pro Asp Gly Phe Leu Gly Glu Arg Gly Arg Ile Val
325 330 335
Lys Trp Val Pro Gln Gln Glu Val Leu Ala His Gly Ala Ile Gly Ala
340 345 350
Phe Trp Thr His Ser Gly Trp Asn Ser Thr Leu Glu Ser Val Cys Glu
355 360 365
Gly Val Pro Met Ile Phe Ser Asp Phe Gly Leu Asp Gln Pro Leu Asn
370 375 380
Ala Arg Tyr Met Ser Asp Val Leu Lys Val Gly Val Tyr Leu Glu Asn
385 390 395 400
Gly Trp Glu Arg Gly Glu Ile Ala Asn Ala Ile Arg Arg Val Met Val
405 410 415
Asp Glu Glu Gly Glu Tyr Ile Arg Gln Asn Ala Arg Val Leu Lys Gln
420 425 430
Lys Ala Asp Val Ser Leu Met Lys Gly Gly Ser Ser Tyr Glu Ser Leu
435 440 445
Glu Ser Leu Val Ser Tyr Ile Ser Ser Leu
450 455
<210> 12
<211> 1377
<212> ДНК
<213> Stevia rebaudiana
<400> 12
atggagaata agacagaaac aaccgtaaga cggaggcgga ggattatctt gttccctgta 60
ccatttcagg gccatattaa tccgatcctc caattagcaa acgtcctcta ctccaaggga 120
ttttcaataa caatcttcca tactaacttt aacaagccta aaacgagtaa ttatcctcac 180
tttacattca ggttcattct agacaacgac cctcaggatg agcgtatctc aaatttacct 240
acgcatggcc ccttggcagg tatgcgaata ccaataatca atgagcatgg agccgatgaa 300
ctccgtcgcg agttagagct tctcatgctc gcaagtgagg aagacgagga agtttcgtgc 360
ctaataactg atgcgctttg gtacttcgcc caatcagtcg cagactcact gaatctacgc 420
cgtttggtcc ttatgacaag ttcattattc aactttcacg cacatgtatc actgccgcaa 480
tttgacgagt tgggttacct ggacccggat gacaaaacgc gattggagga acaagcgtcg 540
ggcttcccca tgctgaaagt caaagatatt aagagcgctt atagtaattg gcaaattctg 600
aaagaaattc tcggaaaaat gataaagcaa accaaagcgt cctctggagt aatctggaac 660
tccttcaagg agttagagga atctgaactt gaaacggtca tcagagaaat ccccgctccc 720
tcgttcttaa ttccactacc caagcacctt actgcaagta gcagttccct cctagatcat 780
gaccgaaccg tgtttcagtg gctggatcag caacccccgt cgtcagttct atatgtaagc 840
tttgggagta cttcggaagt ggatgaaaag gacttcttag agattgcgcg agggctcgtg 900
gatagcaaac agagcttcct gtgggtagtg agaccgggat tcgttaaggg ctcgacgtgg 960
gtcgagccgt tgccagatgg ttttctaggg gagagaggga gaatcgtgaa atgggttcca 1020
cagcaagagg ttttggctca cggagctata ggggcctttt ggacccactc tggttggaat 1080
tctactcttg aaagtgtctg tgaaggcgtt ccaatgatat tttctgattt tgggcttgac 1140
cagcctctaa acgctcgcta tatgtctgat gtgttgaagg ttggcgtgta cctggagaat 1200
ggttgggaaa ggggggaaat tgccaacgcc atacgccggg taatggtgga cgaggaaggt 1260
gagtacatac gtcagaacgc tcgggtttta aaacaaaaag cggacgtcag ccttatgaag 1320
ggaggtagct cctatgaatc cctagaatcc ttggtaagct atatatcttc gttataa 1377
<---
Claims (14)
1. Способ получения соединения стевиол-гликозида или композиции стевиол–гликозидов, включающий (А) инкубацию субстрата и уридиндифосфат (УДФ)-глюкозы в присутствии рекомбинантной УДФ-гликозилтрансферазы, содержащий аминокислотную последовательность, имеющую по меньшей мере 80% идентичности с SEQ ID No: 6, или (B) инкубацию субстрата, сахарозы, УДФ и УДФ-глюкозы в присутствии рекомбинантной УДФ-гликозилтрансферазы, содержащей аминокислотную последовательность, имеющую по меньшей мере 80% идентичности с SEQ ID No: 6, и сахарозосинтазы в течение времени, достаточного для получения композиции стевиол-гликозида или соединения стевиол-гликозида; причем субстратом является ребаудиозид Е.
2. Способ по п.1, в котором используют сахарозосинтазу, выбранную из группы, состоящей из сахарозосинтазы 1 Arabidopsis, сахарозосинтазы 3 Arabidopsis, сахарозосинтазы Vigna radiate и их функциональных гомологов.
3. Способ по п.2, в котором указанная сахарозосинтаза имеет по меньшей мере 80% идентичности с аминокислотной последовательностью сахарозосинтазы 1 Arabidopsis thaliana, как указано в SEQ ID No: 9.
4. Способ по п.1, в котором рекомбинантная УДФ-гликозилтрансфераза содержит аминокислотную последовательность, имеющую по меньшей мере 90% идентичности с SEQ ID NO: 6.
5. Способ по п.1, в котором рекомбинантная УДФ-гликозилтрансфераза содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности с SEQ ID NO: 6.
6. Способ по п.1, в котором рекомбинантная УДФ-гликозилтрансфераза состоит из аминокислотной последовательности гликозилтрансферазы HV1 согласно SEQ ID NO: 6.
7. Способ по п.1, в котором глюкозный фрагмент из УДФ-глюкозы переносят и ковалентно связывают с С2'-19-О-глюкозой ребаудиозида Е с получением ребаудиозида Z2.
8. Соединение ребаудиозида Z2, имеющее структуру:

9. Подсластитель, содержащий соединение, имеющее химическую структуру:
10. Подсластитель по п.9, содержащий по меньшей мере один компонент из числа наполнителя, объемообразующего вещества и ингибитора слеживания.
11. Применение подсластителя по п.9 в пищевом продукте.
12. Применение по п.11, в котором пищевой продукт выбран из группы, состоящей из напитков, кондитерских изделий, хлебобулочных изделий, печенья и жевательной резинки.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201361898571P | 2013-11-01 | 2013-11-01 | |
| US61/898,571 | 2013-11-01 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2016120635A Division RU2706789C2 (ru) | 2013-11-01 | 2014-10-03 | Рекомбинантное получение стевиол-гликозидов |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| RU2019135831A RU2019135831A (ru) | 2020-01-20 |
| RU2019135831A3 RU2019135831A3 (ru) | 2020-06-19 |
| RU2741103C2 true RU2741103C2 (ru) | 2021-01-22 |
Family
ID=53005355
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2019135831A RU2741103C2 (ru) | 2013-11-01 | 2014-10-03 | Рекомбинантное получение стевиол-гликозидов |
| RU2016120635A RU2706789C2 (ru) | 2013-11-01 | 2014-10-03 | Рекомбинантное получение стевиол-гликозидов |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| RU2016120635A RU2706789C2 (ru) | 2013-11-01 | 2014-10-03 | Рекомбинантное получение стевиол-гликозидов |
Country Status (11)
| Country | Link |
|---|---|
| US (3) | US20160251635A1 (ru) |
| EP (1) | EP3063286B1 (ru) |
| JP (1) | JP6530760B2 (ru) |
| CN (1) | CN106103729B (ru) |
| AU (2) | AU2014342939B2 (ru) |
| CA (1) | CA2928940C (ru) |
| DK (1) | DK3063286T3 (ru) |
| ES (1) | ES2709439T3 (ru) |
| MX (1) | MX367771B (ru) |
| RU (2) | RU2741103C2 (ru) |
| WO (1) | WO2015065650A2 (ru) |
Families Citing this family (28)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ES2709439T3 (es) | 2013-11-01 | 2019-04-16 | Conagen Inc | Producción recombinante de glucósidos de esteviol |
| US9522929B2 (en) | 2014-05-05 | 2016-12-20 | Conagen Inc. | Non-caloric sweetener |
| DK3200615T3 (da) | 2014-10-03 | 2019-08-19 | Conagen Inc | Kaloriefrie sødestoffer og fremgangsmåder til syntetisering |
| EP3232817A4 (en) | 2014-12-17 | 2018-10-10 | Cargill, Incorporated | Steviol glycoside compounds, compositions for oral ingestion or use, and method for enhancing steviol glycoside solubility |
| CA2979931A1 (en) | 2015-03-16 | 2016-09-22 | Dsm Ip Assets B.V. | Udp-glycosyltransferases |
| EP3718417B1 (en) | 2015-04-03 | 2025-09-03 | DSM IP Assets B.V. | Steviol glycosides |
| JP6793957B2 (ja) * | 2015-04-14 | 2020-12-02 | コナゲン インコーポレイテッド | 改変された全細胞触媒を用いたノンカロリー甘味料の生成 |
| EP3303552A4 (en) * | 2015-05-29 | 2019-01-16 | Cargill, Incorporated | FERMENTATION METHOD FOR THE PRODUCTION OF STEVIOL GLYCOSIDES WITH MULTIPHASE SUPPLY |
| CN105348337B (zh) * | 2015-11-19 | 2018-07-27 | 南京诺云生物科技有限公司 | 一种甜菊苷生物转化制备的甜菊糖衍生物、制备方法及其应用 |
| CN105380224A (zh) * | 2015-11-19 | 2016-03-09 | 南京诺云生物科技有限公司 | 一种新型甜菊糖衍生物、制备方法及其应用 |
| MX2018006599A (es) * | 2015-11-30 | 2018-09-21 | Purecircle Sdn Bhd | Proceso para producir glicosidos de esteviol de alta pureza. |
| WO2017198682A1 (en) | 2016-05-16 | 2017-11-23 | Evolva Sa | Production of steviol glycosides in recombinant hosts |
| MX2018015534A (es) | 2016-06-15 | 2019-03-14 | Codexis Inc | Beta-glucosidasas dise?adas y metodos de glucosilacion. |
| CN110191643B (zh) | 2016-10-14 | 2023-06-06 | 科纳根公司 | 甜菊醇糖苷的生物合成生产及其工艺 |
| BR112019008048A2 (pt) | 2016-10-21 | 2019-07-02 | Pepsico Inc | método para preparar rebaudiosídeo n com o uso de método enzimático |
| CA3175070A1 (en) | 2017-02-03 | 2018-08-09 | Codexis, Inc. | Engineered glycosyltransferases and steviol glycoside glucosylation methods |
| MX2019010537A (es) * | 2017-03-06 | 2019-11-21 | Conagen Inc | Produccion biosintetica del rebaudiosido d4 de glucosido de esteviol a partir del rebaudiosido e. |
| CN111093390A (zh) * | 2017-05-15 | 2020-05-01 | 谱赛科美国股份有限公司 | 高纯度甜菊醇糖苷 |
| MX2019015404A (es) | 2017-06-30 | 2020-07-20 | Conagen Inc | Hidrólisis de glucósidos de esteviol por beta-glucosidasa. |
| CN109423486B (zh) * | 2017-08-29 | 2022-02-25 | 中国科学院分子植物科学卓越创新中心 | 新型udp-糖基转移酶及其应用 |
| US11365417B2 (en) | 2017-09-12 | 2022-06-21 | Bio Capital Holdings, LLC | Biological devices and methods of use thereof to produce steviol glycosides |
| CN109868265B (zh) * | 2017-12-01 | 2023-04-25 | 中国科学院分子植物科学卓越创新中心 | 新型糖基转移酶及其应用 |
| WO2020028039A1 (en) | 2018-07-30 | 2020-02-06 | Codexis, Inc. | Engineered glycosyltransferases and steviol glycoside glucosylation methods |
| CN113302298B (zh) | 2018-11-09 | 2025-02-25 | 银杏生物制品公司 | 罗汉果甙的生物合成 |
| MX2021006986A (es) * | 2018-12-12 | 2021-08-16 | Conagen Inc | Produccion biosintetica de variantes de glucosidos de esteviol. |
| CN113388014B (zh) * | 2020-03-12 | 2023-02-03 | 中科农福(北京)生物技术有限公司 | Os02g0589400蛋白在调控植物化感作用中的应用 |
| KR102628374B1 (ko) * | 2021-10-19 | 2024-01-24 | 씨제이제일제당 주식회사 | 신규한 당전이효소 및 이의 용도 |
| WO2023183832A1 (en) | 2022-03-23 | 2023-09-28 | Sweegen, Inc. | Sweetener comprising rebaudiose m and brazzein |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2427285C2 (ru) * | 2007-03-14 | 2011-08-27 | Дзе Консентрейт Мэньюфектуринг Компани Оф Аиэлэнд | Продукты типа напитков, содержащие стевиолгликозиды и по меньшей мере одну кислоту |
| CN101701238B (zh) * | 2009-10-14 | 2011-11-16 | 南京师范大学 | 一种由甜菊糖苷制备甜茶甙的方法 |
| WO2013133689A1 (en) * | 2012-03-08 | 2013-09-12 | Purecircle Sdn Bhd | High-purity rubusoside and process for producing of the same |
| RU2599166C2 (ru) * | 2011-12-19 | 2016-10-10 | Дзе Кока-Кола Компани | Способы очистки стевиоловых гликозидов и их применение |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN105671108A (zh) * | 2010-06-02 | 2016-06-15 | 沃维公司 | 甜菊糖苷的重组生产 |
| CN108396044A (zh) * | 2011-08-08 | 2018-08-14 | 埃沃尔瓦公司 | 甜菊醇糖苷类的重组生产 |
| RU2499427C1 (ru) * | 2012-09-19 | 2013-11-27 | Нина Васильевна Мелишева | Композиция подсластителя |
| ES2709439T3 (es) | 2013-11-01 | 2019-04-16 | Conagen Inc | Producción recombinante de glucósidos de esteviol |
-
2014
- 2014-10-03 ES ES14857965T patent/ES2709439T3/es active Active
- 2014-10-03 EP EP14857965.9A patent/EP3063286B1/en active Active
- 2014-10-03 CA CA2928940A patent/CA2928940C/en active Active
- 2014-10-03 RU RU2019135831A patent/RU2741103C2/ru active
- 2014-10-03 WO PCT/US2014/059081 patent/WO2015065650A2/en not_active Ceased
- 2014-10-03 AU AU2014342939A patent/AU2014342939B2/en not_active Ceased
- 2014-10-03 CN CN201480072126.8A patent/CN106103729B/zh active Active
- 2014-10-03 MX MX2016005677A patent/MX367771B/es active IP Right Grant
- 2014-10-03 JP JP2016552419A patent/JP6530760B2/ja not_active Expired - Fee Related
- 2014-10-03 US US15/032,286 patent/US20160251635A1/en not_active Abandoned
- 2014-10-03 RU RU2016120635A patent/RU2706789C2/ru active
- 2014-10-03 DK DK14857965.9T patent/DK3063286T3/en active
-
2016
- 2016-12-01 US US15/366,209 patent/US10081826B2/en active Active
-
2017
- 2017-06-23 US US15/631,083 patent/US10253344B2/en active Active
- 2017-10-31 AU AU2017254856A patent/AU2017254856A1/en not_active Abandoned
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2427285C2 (ru) * | 2007-03-14 | 2011-08-27 | Дзе Консентрейт Мэньюфектуринг Компани Оф Аиэлэнд | Продукты типа напитков, содержащие стевиолгликозиды и по меньшей мере одну кислоту |
| CN101701238B (zh) * | 2009-10-14 | 2011-11-16 | 南京师范大学 | 一种由甜菊糖苷制备甜茶甙的方法 |
| RU2599166C2 (ru) * | 2011-12-19 | 2016-10-10 | Дзе Кока-Кола Компани | Способы очистки стевиоловых гликозидов и их применение |
| WO2013133689A1 (en) * | 2012-03-08 | 2013-09-12 | Purecircle Sdn Bhd | High-purity rubusoside and process for producing of the same |
Also Published As
| Publication number | Publication date |
|---|---|
| JP6530760B2 (ja) | 2019-06-12 |
| RU2019135831A3 (ru) | 2020-06-19 |
| MX2016005677A (es) | 2016-10-28 |
| US20160251635A1 (en) | 2016-09-01 |
| CA2928940C (en) | 2021-12-14 |
| RU2019135831A (ru) | 2020-01-20 |
| EP3063286B1 (en) | 2018-12-05 |
| AU2014342939A1 (en) | 2016-05-19 |
| US20170081691A1 (en) | 2017-03-23 |
| AU2017254856A1 (en) | 2017-11-23 |
| RU2016120635A (ru) | 2017-12-06 |
| ES2709439T3 (es) | 2019-04-16 |
| DK3063286T3 (en) | 2019-03-04 |
| AU2014342939B2 (en) | 2018-03-22 |
| MX367771B (es) | 2019-09-05 |
| CN106103729A (zh) | 2016-11-09 |
| US10253344B2 (en) | 2019-04-09 |
| EP3063286A2 (en) | 2016-09-07 |
| EP3063286A4 (en) | 2017-06-21 |
| RU2016120635A3 (ru) | 2018-05-31 |
| WO2015065650A3 (en) | 2015-11-05 |
| CA2928940A1 (en) | 2015-05-07 |
| RU2706789C2 (ru) | 2019-11-21 |
| US10081826B2 (en) | 2018-09-25 |
| JP2017500056A (ja) | 2017-01-05 |
| WO2015065650A2 (en) | 2015-05-07 |
| CN106103729B (zh) | 2020-07-07 |
| US20170298404A1 (en) | 2017-10-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| RU2741103C2 (ru) | Рекомбинантное получение стевиол-гликозидов | |
| KR102554790B1 (ko) | 레바우디오시드 e로부터 스테비올 글리코시드 레바우디오시드 d4의 생합성 제조 | |
| CN110809408B (zh) | 通过β-葡糖苷酶水解甜菊醇糖苷 | |
| JP7305213B2 (ja) | 変異体酵素を介するステビオール配糖体レバウシオシドiの生合成的産生 | |
| US20220090158A1 (en) | Biosynthetic production of udp-rhamnose | |
| US20220205007A1 (en) | Biosynthetic production of steviol glycoside rebaudioside i via variant enzymes | |
| HK1230659A1 (en) | Recombinant production of steviol glycosides | |
| HK1230659B (zh) | 甜菊醇糖苷的重组制备 | |
| HK40054956A (zh) | Udp-鼠李糖的生物合成生产 |