JP3708543B2 - ブチロベタイン/クロトノベタイン−l−カルニチン代謝のための遺伝子およびそのl−カルニチンの微生物学的生産への使用 - Google Patents
ブチロベタイン/クロトノベタイン−l−カルニチン代謝のための遺伝子およびそのl−カルニチンの微生物学的生産への使用 Download PDFInfo
- Publication number
- JP3708543B2 JP3708543B2 JP51125795A JP51125795A JP3708543B2 JP 3708543 B2 JP3708543 B2 JP 3708543B2 JP 51125795 A JP51125795 A JP 51125795A JP 51125795 A JP51125795 A JP 51125795A JP 3708543 B2 JP3708543 B2 JP 3708543B2
- Authority
- JP
- Japan
- Prior art keywords
- carnitine
- gene
- butyrobetaine
- bco
- plasmid
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images






Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/007—Carnitine; Butyrobetaine; Crotonobetaine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
- C12N15/743—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora for Agrobacterium; Rhizobium; Bradyrhizobium
Description
L−カルニチンは天然のビタミン類似物質であって、人間の代謝にきわめて重要である。脂肪酸の利用において、L−カルニチンはミトコンドリア膜を通過させる物質として、またそれによって代謝エネルギーを輸送するものとして、不可欠である。L−カルニチンが生体内で適切な量生産されなければ、その欠乏症を避けるために、食品中に添加しなければならない。とくに幼児の食品には、幼児は未だ自身でL−カルニチンを生合成することができないので、L−カルニチンは不可欠な栄養素である。
L−カルニチン製剤は、医薬製品においても活性成分として使用される。L−カルニチンの補給は、カルニチン欠乏の場合、およびその他の治療上の指示に従って、とくに心臓の不調などの場合に処方される。
高等な生物体内でのL−カルニチンの生合成は知られていて、代謝のさらなる機能および重要性が、積極的な研究活動の対象になっている。微生物とくにシュードモナス層に関して記述されている代謝経路(γ−ブチロベタインヒドロキシラーゼ触媒作用、リンデシュテットほか、Biochemistry 16,2181-2188,1977)に加えて、L−カルニチンは、ある種の微生物たとえばアグロバテリウム/リゾビウムSP.の代謝中間生産物として形成される。
EP−A−0158194は、L−カルニチンの微生物学的生産であって、たとえばγ−ブチロベタインから出発して、伝統的な微生物学的選択方法によってL−カルニチルデヒドロゲナーゼ陰性の生産変異種を得、それを使用して、相対的によいL−カルニチン収量が20〜30時間の反応時間内に得られる方法を開示している。容積/時間収率に関してこの方法をさらに最適化することは、しかし、この古典的な微生物学的方法による限り不可能である。
それゆえ、本発明の目的は、L−カルニチン生産のより経済的なバイオ技術的方法であって、L−カルニチンが顕著に短い反応時間で、かつ、むしろよりよい収率をもって得られる方法を提供することにある。
γ−ブチルロベタイン/クロトノベタイン代謝に関する研究の結果、5種の遺伝子が同定されるに至った。すなわちbcoA,bcoC,bcoD,bcoEおよびbcoTであって、これらγ−ブチロベタイン/クロトノベタイン代謝経路の酵素類をコードし、とくに、いわゆるブチロベタイン−L−カルニチンオペロン(bco)と呼ばれるオペロン中に含まれている。この関連において、略号は下記の意味を有する。
bcoA/B:γ−ブチロベタイン−CoA−シンセターゼ(bocA)/クロトノベタイン−CoA−シンセターゼ(bcoB)、すなわち、γ−ブチロベタイン−CoA−シンセターゼ活性およびクロトノベタイン−CoA−シセンターゼ活性の両方を有する酵素生産物をコードする唯一の遺伝子、
bcoC:γ−クロトノベタイン−CoAデヒドロゲナーゼ遺伝子、
bcoD:クロトノベタイン−CoAヒドラーゼ遺伝子、
bcoE:L−カルニチルデヒドロゲナーゼ遺伝子、そして
bcoT:輸送システムのポテンシャル遺伝子。
遺伝子bcoA/B,bcoCおよびbcoDの遺伝子生産物はL−カルニチンの生合成に関連するが、一方、bcoEは、代謝中間生成物L−カルニチル−CoAのベタインへの分解をひきおこすカルニチンデヒドロゲナーゼをコードし、bcoTは、ブチロベタイン代謝に割り当てられた輸送システムの輸送タンパク質をコードすると推測されている。この遺伝子は、L−カルニチン生合成に不可欠なものではない。
本発明は従って、γ−ブチロベタイン/クロトノベタイン代謝におけるL−カルニチン生合成のための酵素類をコードする遺伝子、bcoC,bcoA/BおよびbcoDの1種または2種以上からなるDNA断片およびベクターに関し、場合によっては、さらにポテンシャル輸送遺伝子bcoTにも関する。
本発明はさらに、これらDNA断片および(または)ベクターを含有する微生物にも関する。本発明はまた、本発明の微生物を使用するL−カルニチン生産のバイオ技術的方法に関する。
本明細書および請求の範囲中で使用されるbcoA/B,bcoC,bcoDおよびbcoTの表示は、定義されたように、野生種の生物のもつ遺伝子であって上記した酵素活性を有するγ−ブチロベタイン/クロトノベタイン−L−カルニチン代謝の酵素をコードするもの、とくにブチロベタイン−L−カルニチン(bco)オペロンと、その機能的に同様な遺伝子的変種すなわち、野生種の生物の遺伝子から誘導され、その遺伝子生産物が生物学的機能に関しては本質的に変化を受けていないものとの、両方から成る。機能的に同等な遺伝子的変種および突然変異種は、このようにして、たとえば遺伝子コードの既知のデジェネレーションとの関連における塩基変換、たとえば人工的な遺伝子配列を、発現を生じさせるべきある種の微生物の好ましいコドン使用に適合させるために、生産することができる。変種および突然変異種は、さらに、塩基またはコドンの削除、挿入および置換を、このようにして変性させられた遺伝子の遺伝子生産物を、生物学的機能に関しては本質的に変化させない程度に止めておいたものを包含する。ここで包含されるのは、たとえば、野生種の配列に対して高度の、たとえば70%を超える均一性を有し、野生種の配列の補体とストリンジェントハイブリダイゼーション条件下で、たとえば50〜70℃の温度および0.5〜1.5モルの塩濃度において、ハイブリッド形成可能な遺伝子配列である。
ここで使用される転写ユニットの語は、遺伝子がひとつの転写方向に配置され、ふつうの転写コントロール下に中断されない転写として転写されるようなDNA配列をするものと理解されるべきであり、遺伝子に加えてDNA配列とは、追加的に遺伝子発現に必要な遺伝子コントロール要素、たとえばプロモーターおよび染色体固定サイトを有するものである。
本発明は、下記の図面により、より詳細に説明される。
図1は、γ−ブチロベタインまたはクロトノベタイン−L−カルニチン代謝径路における酵素を示す。
図2は、bcoオペロンを有するリゾビウム/アグロバクテリウムからの10.6kbのDNA断片の制限地図を示す。
図3および図4は、プラスミドpVK1011およびpAZ101を示し、矢印はbco遺伝子およびbeu遺伝子(ベタイン利用遺伝子:EP−A−0543344)の位置および方向を示す。
図5は、recA−宿主株の生産のための出発物質と考えられる、プラスミドpCC49を示す。
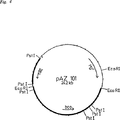
図6は、プラスミドpVK1011,pAZ101,pVK100qおよびpAZ7の構成スキームを示す。
図7は、recA−宿主株の構成スキームを示す。
γ−ブチロベタイン/クロトノベタイン−L−カルニチン代謝径路における遺伝子bcoA/B,bcoC,bcoDおよびbcoTを単離するために使用される出発物質としては、ブチロベタインおよび(または)クロトノベタインを図1に従って代謝するすべての微生物を使用することができる。適当な株は、エシェリチア、シュードモナス、アグロバクテリウム、リゾビウムおよびアグロバクテリウム/リゾビウム属の微生物であって、最後のものが好ましい。アグロバクテリウム/リゾビウム属の好ましい微生物の例は、アグロバクテリウム/リゾビウムsp.HK4(DSM2938)株であって、これに関してはEP−A−0158194に記述されている。これらの微生物はとくに好適に使用され、カルニチンデヒトロゲナーゼ陰性、すなわち、たとえばbcoE遺伝子を有しないか、または欠陥のあるbcoE遺伝子(以下、bcoE′とも記す)しか有しないものである。カルニチンデヒドロゲナーゼ陰性の好ましい微生物の例は、すでにEP−A−0158194に記述されているアグロバクテリウム/リゾビウムsp.HK13C(DSM2903)種およびHK1331b(DSM3225)種、またはEP−A−0543344に記述されているアグロバクテリウム/リゾビウムsp.HK1349(DSM3944)種である。
γ−ブチロベタイン/クロトノベタイン−L−カルニチン代謝の遺伝子bcoA/B,bcoC,bcoDおよびbcoTは、微生物の染色体中で、微生物を、トランスポゾン挿入変異処理し、それによってγ−ブチロベタイン/クロトノベタイン−L−カルニチン代謝の遺伝子を、適宜のラベルたとえばカナマイシン(Km)耐性ラベルを用いて標識づけすることによって、局在化させることができる。このようにしてラベルをつけられた、ブチロベタイン代謝の中間体としてはもはや使用できない突然変異種は、ついで単離することができる。このようにして、γ−ブチロベタイン/クロトノベタイン−L−カルニチン代謝の遺伝子をその機能に関して同定し、関連づけることができる。標識づけされた遺伝子は、ついでクローンをつくり、適当な制限酵素を使用してさらに詳細に特徴づけることができる。完全な遺伝子または本発明に従うDNA断片の単離は、ついで、対応する非変異種微生物の遺伝子バンクから出発して行なうことができ、その微生物からbco遺伝子またはその断片を単離して、既知の方法よりクローン化して得た、上記の突然変異した株の遺伝子とハイブリダイゼーションすることにより、単離して同定することができる。得られた遺伝子は、続いて所望のベクター中にクローン化し、制限酵素の助けを借りて制限地図に描くことができる。
遺伝子bcoA/B,bcoCおよびbcoDだけが、L−カルニチンの生合成に関与する。従って、これら遺伝子の存在だけがL−カルニチンの生産にとって必要である。選択した出発条件たとえば選択した出発原料または選択した生産株によって、L−カルニチン生産に使用したDNA断片およびベクターは、1種または2種以上のL−カルニチン生合成の遺伝子を含有することができる。
遺伝子bcoA/B,bcoCおよびbcoDに加えて、本発明に従うDNA断片およびベクターは、所望であれば、ポテンシャル輸送遺伝子bcoTを含むことができる。
L−カルニチルデヒドロゲナーゼ遺伝子すなわちbcoE遺伝子の存在は、望ましくない。というのは、その存在下にL−カルニチンの分解が生じるからである。しかし、欠陥のあるbcoE(bcoE′)の存在は、無害である。
好都合なことに、L−カルニチンの生産に使用するためのL−カルニチン生合成の遺伝子、すなわちbcoC,bcoA/BおよびbcoD、そして場合によってはポテンシャル輸送遺伝子bcoTは、1個のDNA断片またはベクター分子中に共に存在する。たとえば、好ましくは、遺伝子制御要素から常用の5′−3′−方向の下流において、bcoC,bcoA/BおよびbcoDの配列、またはbcoC,bcoA/B,bcoDおよびbcoTの配列であって、天然に生じるブチロベタイン−L−カルニチンオペロンの配置に対応する、上記した単一転写ユニット中に存在する。このような転写ユニット中の遺伝子bcoC,bcoA/B,bcoDおよびbcoTは、たとえば、図2の制限地図の対応するセクションによって特徴づけられる。
bco遺伝子の転写または発現は、適宜の、好ましくは協力なプロモーターのコントロールの下で好都合に起る。プロモーターの選択は所望の発現条件に依存し、たとえば、本質的なまたは誘起された発現が望ましいか否かによって、あるいは発現を起すべき微生物の種類によって異なる。適切なプロモーターは、たとえば天然のブチロベタイン−L−カルニチンオペロンのプロモーターPbcoである。L−カルニチン生合成に関与するbco遺伝子の単離を行なう場合は、たとえば、欠陥のあるbcoE(bcoE′)遺伝子を有する微生物から、有利にはたとえば全bcoオペロンを単離してbcoE′遺伝子およびこれらの微生物からの関連する遺伝子制御要素のクローンをつくることができ、ついで、適当な微生物中でL−カルニチン生産に利用することができる。変異されたbcoE遺伝子を有するこの種の転写ユニット、たとえばリゾビウム/アグロバクテリウムHK1349から単離されるようなものは、場合によっては、すなわちbcoEの欠陥が、たとえばポイント変異にだけ帰せられ、制限分解サイトには関係がないのであれば、図2に示した制限地図によって特徴づけられるものであってもよい。発現に適当なプロモーターの別の例は、たとえば、プロモーターPNm′.PS1(M.Labes et al.,Gene,89,37-46,1990)、trpプロモータ(Amann et al.,Gene,25,167-178,1983)、lacプロモーター(Amann et al.,Gene,25,167-178,1983)およびtacプロモータ、上記trpおよびlacプロモーターのハイブリッドであって、本質的な、または誘起されたプロモーター(RussellおよびBennett,Gene,20,231-243,1982)として使用することができる。
L−カルニチン生産に使用するための適当な生産株の中で、本発明に従う、前記bco遺伝子を包含するDNA断片は、好ましくは単一の転写ユニットとともに、既知の技術の助けを借りて好都合に既知の適当なベクター中に、とりわけ発現ベクターたとえばファージまたはプラスミド中に組み込まれる。使用するベクターは、自律的かつ自己複製的ベクターであってもよいし、あるいはまた、いわゆるインテグレーションベクターであってもよい。インテグレーションベクターは、この関連において、たとえば、受容株のゲノム配列に相同の配列を少なくとも1個もち、この配列に相同的な組換えによって外来の遺伝子の受容株のゲノム中への挿入を許すようなプラスミドを意味するものと解すべきである。自律的かつ自己複製的ベクターが好ましく使用される。
選択されるベクターの特性によって、L−カルニチン生合成の酵素のための遺伝子を、種々の生物体内で発現させることができる。適当なベクター特定の宿主スペクトルを有するベクターと、広い宿主スペクトル(広い宿主範囲)を有するベクターのどちらでも、また前記した集積ベクターのいずれでもよい。
使用される広い宿主範囲のベクターは、グラム陰性の細菌に適するすべてのベクターであることができる。そのような広い宿主範囲のベクターの例は、pVK100(KnaufおよびNester,Plasmid,8,45-54,1982)、pME285(HaasおよびItoh,Gene,36,27-36,1985)およびpKT240(Bagdasarian et al.,Gene,25,273-282,1983)またはこれらの誘導体である。使用されるpVK100の誘導体は、たとえば、pVK1001であることができ、使用されるpME285の誘導体は、たとえばpAZ10であることができ、使用されるpKT240の誘導体は、たとえばpLO32(すでにEp−A−0543344に記述されている)であることができる。
リゾビウム/アグロバクテリウムの場合に使用される集積ベクターは、pACYC184またはpBR322(Comai et al.,Plasmid,10,21-30,1983)にもとづくベクターであることができる。
このようにして、たとえばプラスミドpVK100q,pVK1011(図3),pAZ7,pAZ7::beu,pAZ101(図4)およびpLO41が得られる。プラスミドpVK100qは、1993年11月16日に、ドイツチェン・ザンメルグ・フュア・ミクロオルガニズメン・ウント・ツェルクルトゥーレン・Gm bH D−38124 ブラウンシュヴァイク・マシェローダヴェク1bに、リゾビウム/アグロバクテリウムHK1349中に、寄託番号DSM8726として寄託されている。
発酵のための生産株すなわちL−カルニチン生産株の生産のためにf
、本発明に従うDNA断片またはベクターを、所望であり、かつ発現に適当な宿主株の中に取り込まなければならない。この目的のために、常用されているそれ自体は既知の手法で、本発明に従うDNA断片を含有するベクターを使用して、好都合に微生物が形質転換される。そうすると微生物は、本発明に従うDNA断片を、ベクター分子上またはその染色体中に集積して含む。
適当な生産株は、L−カルニチンをクロトノベタインおよび(または)γ−ブチロベタインから生産することができ、そのL−カルニチン分解(代謝分解)の能力が完全に、または部分的に抑制されたもののすべてである。L−カルニチン代謝分解が抑制されている微生物は、たとえば、カルニチンデヒドゲナーゼ陰性の菌株、つまりその中のカルニチンデヒドロゲナーゼ遺伝子bcoEが、たとえば突然変異や削除によってスイッチオフされている菌株、および(または)L,D−カルニチンラセマーゼ陰性であってL−カルニチンデヒドロラターゼ陰性である菌株である。
適当な宿主菌株は、好ましくは高い基質および出発物質許容度をもつものであって、たとえば、エシェリチア、シュードモナス、アグロバクテリウム、リゾビウム、コマモナスおよびリゾビウム/アグロバクテリウム属の微生物であり、最後のものが好ましい。すでに記述したリゾビウム/アグロバクテリウムsp.種の微生物HX13,HK1331bおよびHK1349、ならびに、HK1349、4種(EP−A−0543344に記述されている)がとくに好適である。
L−カルニチンの収量が、組換えのための宿主菌株の能力すなわちその組換え傾向および組換え頻度が減少すると、さらに改善できることが、上記に続いて判明した。染色体相同性にもとづいてベクターとの組換えが、それによって制限される。宿主菌株の組換え能力は、たとえば既知の方法でそのrecA遺伝子を特定的に変異(recA突然変異)させることによって、減少させることができる。その組換え能力を低下させたとくに好ましい微生物は、リゾビウム/アグロバクテリウムsp.種、たとえば本発明に従って前記のようにして得られるリゾビウム/アグロバクテリウムHK1349.49株である。
適当な生産株は、このようにして、たとえばリゾビウム/アグロバクテリウムHK1349,HK1349.9およびHK1349.49種であり、いずれも、プラスミドpVK100q,pVK1001,pAZ7,pAZ::beu,pAZ101またはpLO41を含有するものである。
形質転換された宿主菌株(生産菌株)は、ベクターまたはDNA断片上に位置するマーカー遺伝子に起因して菌株がそれに対して耐性をもつ抗生物質を添加した、選択的栄養培地から単離される。EP−A−0543344に従う微生物を生産菌株として使用するとき、すなわちベタイン利用のための染色体遺伝子コーディングが変異を受け、ベタイン利用のための遺伝子コーディングを含むプラスミドと形質転換を受けている微生物もまた、ベタイン利用に関して選択使用することができる。ベタイン利用に関して選択可能な微生物の例は、すでに述べたHK1349.4およびHK1349.49であって、これらは、たとえばプラスミドpLO41,pAZ101,pAZ7::beuまたはpVK1011を含有する。
L−カルニチンのバイオ技術による生産は、本発明に従うDNA断片またはベクターを含有する微生物を利用して実施する。L−カルニチンの製造方法は、それ自体は既知の、たとえばEP−A−0158194に記述された方法で、たとえばγ−ブチロベタインから出発して、適当な炭素源および窒素源の存在下に実施する。使用できる炭素源および窒素源は、たとえば、グルタミン酸塩、酢酸塩およびベタイン、またはグリセリンおよびベタインである。バイオ技術による生産がベタイン利用に関して選択可能な微生物を手段とするならば、ベタインが唯一の窒素源として利用される。
培養およびそれに続くL−カルニチンの単離は、EP−A−0158194に記述された方法と同様な方法によって実施できる。
既存の手法において、培地中の栄養素を変更することによって、また培養条件を個々の微生物に適合させることによって、L−カルニチンの収量は、さらに改善することができる。
実施例1
トランスポゾン挿入突然変異種(Tn5)の生産およびその表現形的同定
野生種のリゾビウム/アグロバクテリウムHK4株(DSM2938,EP−B−0158194)を選択圧力により突発的に変異させ、ストレプトマイシン(Sm,1000μg/ml)耐性をもたせた。この耐性は、選択を行なわなくても50世代にわたって明らかに安定であり、選択マーカーとして利用された。
E.コリS17−1/pSUP2021(R.Simon et al.,Biotechnology,1983,1,784-790)のTsSドナー培養物0.2mlを、受容体培養物HK4の2mlと混合し、遠心分離にかけた。細胞を0.9%塩水(NaCl溶液)で洗い、0.9塩水100μl中に再懸濁させた。受容体菌株とドナー菌株との接合を、一夜、30℃において乾いた栄養寒天の上で行なった。収穫された細胞は、受容体(SmR)およびトランスポゾン(ネオマイシン耐性(NmR))のための選択媒体上に稀釈してのせた。
Tn5変異体を、栄養寒天上で、Sm(1000μg/ml)およびNn(100μg/ml)を使用して得た。突然変異種の表現形上の同定を、図1に従ってブチロベタイン代謝中間生産物を最小限培地中の炭素(C)源としての不利用を検知することによって行なった。
実施例2
Tn5で標識をつけたDNA断片のHK4ゲノムからのクローニング
Tn5変異を行なったHK4(5μg)から単離されたゲノムDNAを、EcoRI(4U/μg)で完全に消化した。pBR325(2.5μg)(Gene,1977,2,95-115)を、EcoRIによる完全な消化ののち、アルカリ性のフォスファターゼで処理した。ゲノムDNAおよびpBR325をT4−DNAリガーゼをT4−DNA400μlのリゲーション緩衝液(20mM−トリスHCl、pH7.2、10mM−DTT(ジチオスレイトール)、10mM−MgCl2、0.6mM−ATP)中のリガーゼ(0.2U/μgDNA)と混合したのち、組換えハイブリッドプラスミドが得られた。
接合混合物をいくつかに分けたものを、E.coli ED8654(Barek et al.,Mol.Gen Genet.,145,199-207,1976)の形質転換に使用した(Cohen et al.,1972,PNAS,69,2110-2114)。トランスフォーマントは、栄養培地中でアンピシリン(Ap,100μg/ml)およびカナマイシン(Km,25μg/ml)に対する耐性によって選択された。選択されたハイブリッドプラスミドはすべて、Tn5で標識されたHK4挿入部を担持していた。挿入部は、図2の制限地図に対応して、種々の制限酵素によって地図化された。制限地図の比較から、種々の表現形を有するTn5突然変異種のシリーズにおける同じゲノム断片中へのトランスポゾン挿入を確認した。
この観察結果を、同一に解裂したプラスミドDNAのサザンブロットハイブリダイゼーション(“Gentechnische Methoden”「遺伝子工学の方法」S.BertramおよびH.G.Gassen編、G.Fischer Verlag,1991,219f)、およびそれに続く電気泳動分離によって確認することができた。使用したプローブは、トランスポゾン突然変異種からクローン化したDNAのサブ断片である。
実施例3
ラムダファージ中のゲノムHK4遺伝子バンクの構成
DNAがラダムファージ中に包まれるためには、40〜52kbのサイズと「コスサイト」が必要である。ゲノムHK4遺伝子バンクをラムダファージ中に構成するために、コスミドベクターpVK100(KnaufおよびNester,1982,Plasmid,8,45-54)を使用した。これはサイズ23kbであり、従って17〜29kbの間のDNA断片のクローニングを可能にする。
pVK100をEcoRIと消化し、デフォスフォリレートしてから、部分的にEcoRIと消化したHK4DNAに接合した。HK4DNAの部分的消化のための理論的EcoRI濃度は、DNA0.58U/μgまたは反応混合物0.02U/μgとして試験消化の手段によって決定された。HK4DNAの8.5μgを反応混合物中で使用した。17kbより大きいサイズ範囲のDNA断片は、アガロース電気泳動ゲルから分離した。接合は、100ngのコスミドベクターと400ngのパッセンジャーDNAとを含有する10μlの容積の中で行なった。ついで「インビトロ・パッケージング」を、Promega Biotech社の混合物中で、製造業者の定めた手順に従って、2時間にわたり、25℃で実施した。EcoRI S17−1のトランスフェクションの後、コスミドベクターのKm耐性に関して選別を行なった。1バッチを使用して、約5500のコロニー(個々のクローン)が得られた。この遺伝子バンクのコロニーを冷凍媒体(栄養イーストブロス、NYB、オキソイドおよび50%グリセリン)中に入れた各1000クローンほどのバッチ5箇に分け、−70℃で貯蔵した。この遺伝子バンクの増殖を、各バッチの50μgを使用して、NYB10ml中で一夜培養し分画することによって行なった。
実施例4
HK4コスミド遺伝子バンクのスクリーニング、bco遺伝子担持コスミドクローンのHK突然変異種のコロニーハイブリダイゼーション、ドットハイブリダイゼーションまたは直接相補による同定
クローニングされたTn5標識をもつDNA断片を、直接ハイブリダイゼーションのプローブとして使用することが可能であった。
適切なDNA配列を有するクローンはハイブリダイゼーション信号を出し、HK4突然変異種の各場合において、欠陥のある遺伝子の相補に導いた。種々の突然変異種からのDNAのクロスハイブリダイゼーションが、10.6kbのDNA断片(図2)上におけるブチロベタイン代謝遺伝子の、若干の「クラスタリング」を裏付けた。
コロニーハイブリダイゼーションは、常法(S.BertramおよびH.G.Gassen,1991,ibid,211f)に従って実施した。ドットブロッティングも、既知の方法(S.BertranおよびH.G.Gassen,1991,ibid,217f)に従って実施した。
実施例5
HK突然変異種の相補
固定されたペプチド鎖にもとづく分子サイズを考慮に入れてブチロベタイン代謝の個々の遺伝子の正確な局在化を行なったのち、個々の突然変異種の特定の遺伝子セクションによる相補を行なうことができた。
特定の発現プラスミドpVK100::HK−DNAを、E.coli S17−1のリゾビウム/アグロバクテリウムsp.HK4株への、実施例1に従う接合を経由して一体化した。この株は、異なる代謝段階(図1参照)のための突然変異を含んでいる。ドナーのプロリン(pro)要求性およびベクターの抗生物質耐性(KmRTC「テトラサイクリンR」)にもとづいて、選別を行なった。
実施例6
6.1 HK1349(DSM3994)およびその誘導体からのbco断片のクローニング
生産菌株の遺伝子供給量効果および生産性の向上を達成するため、HK1349(DSM 3944,EP−A−054334)からのbcoオペロンのクローニングを行なった。この菌株においては、全bcoオペロンが完全な形で含有されているが、カルニチン−CoA−デヒドロゲナーゼのための第一の遺伝子bcoEが変異を受けている。bcoオペロンを有し、この株から得られたDNA断片は、このようなわけで、発現ベクターの複写数が多いという点で、生産性向上にとって理想的である。
クローニングは、E.coli S17−1において、EP−A−0543344の実施例3に対応する既知の方法に従って実施した。この目的のために、10.6kbのbcoオペロンをHK1349遺伝子バンクから単離し、pVK100中にリゲーションした。その結果、プラスミドpVK100q(図6のスキーム参照)が得られた。選別を、E.coli S17−1中で、NYB Km(25μg/ml)上で実施した。挿入の正しさは、HK突然変異種に関して、実施例5に記載した方法に従って同定した。HK1349からのEcoRI−開裂DNAを電気泳動的に分離し、サイズ範囲1.6kbの断片をゲルから単離した。単離した断片をEcoRI−開裂pVK100中にリゲーションした。
このハイブリッドプラスミド混合物を使用してE.coli S17−I(プロリン要求性)を形質転換し、栄養寒天Km(25μg/ml)上で選別した。ベクターからのハイブリッドプラスミドクローンおよびサイズ10.6kbのbcoオペロンを担持するDNA断片を、「パッチ・マッチング」法により、ブチロベタイン−CoA−シンセターゼ陰性突然変異種(HK4V4)上、C源およびN源として0.2%(w/v)のブチロベタインを含有する最小限培地上で同定した。正確なクローンは、このタイプを突然変異種中に接合的転移の後、このC源を利用して細胞を相補する能力があった。このようにして同定されたクローンpVK100qは、直接、生産プラスミドとして、またはさらなるサブクローニングのためのbcoオペロンを有するDNA断片のリザーバーとして使用された。
6.2 pAZ101,pAZ7,pVK1011およびpVK100q,pLO41およびpAZ7::beuの構成
pAZ101,pAZ7,pVK1011およびpVK100qを、図6の構成スキームに従って構成した。pLO41は、pLO32(EP−A−0543344)から出発してbco遺伝子(10.6kb EcoRI/EcoRI断片)の挿入により、またプラスミドpAZ7::beuはpAZ7(図6)から出発してbeu遺伝子(3kbサイズ、PstI/PstI−断片、EP−A−0543344)の挿入により、それぞれ構成した。対応する制限酵素は、DNA3〜5U/μgとともに、製造者の指示に従って使用した。プラスミドpVK100qはbco遺伝子をpVK100(KnaufおよびNester,ibid)中に挿入することにより得られた。出発プラスミドpVK100s1(構成スキームは図6)は、EcoRIプラスミドpVK100s(EP−A−0543344)の削除クローニングによって得た。
実施例7
recA変異のHK1349.4への導入
7.1 recAをコードする遺伝子バンククローンの同定
ゲノムHK4コスミド遺伝子バンク中に、コロニーハイブリダイゼーション法(S.BertramおよびH.G.Gassen,1991,ibid,221f)の助けを借りて、recA−エンコーディングクローンを採取した。ハイブリダイゼーションに使用したプローブは、リゾビウムレグミノサルム(leguminosarum)(W.Selbitscnka et al.,Mol.Gen.Genet.,299,1991,86-95)からのクローンされたrecA遺伝子である。標識をつけたコスミドクローンを単離し、EcoRI消化後得られたEcoRIDNA挿入断片を対象に、recA相同性配列をrecAプローブ(S.BertramおよびH.G.Gassen,1991,ibid,219f)に対するサザンブロットハイブリダイゼーションによって探索した。このようにして標識をつけられたEcoRI断片は、同じ程度まで開裂されたベクターpVK100中にリゲーションされた。得られたハイブリッドプラスミドpVK100rl,E.coli S17−1細胞を、DNA複製のために形質転換した。
7.2 染色体recA遺伝子の不活性化のための、カナマイシン耐性遺伝子のH K1349.4への導入
EcoRI断片のサイズ11.0kbのものを、pVK100rlからpACYC184(A.C.Y.ChangおよびS.N.Cohen,J.Bacteriol.,134,1978,1141-1156)のEcoRIとともに開裂させたものの中に再クローンした。
所定の制限地図に対応して、BglII−NruI開裂サブ断片で、サイズ3.1kb、recAプローブに対してサザンブロットハイブリダイゼーションで標識をつけたものを、BamHI−HindIII開裂をしたベクターpWS233中にクローニングした。pWS233は「遺伝子置換え」ベクターであって、W.Selbitschka et al.,Appl.Microbiol.Biotechnol.38,1993,615-618に記述されている。
Tn5(pSUP2021,R.Simon et al.,Biotechnol.1,1983,ibid)からの、Km耐性をコードしたXhoI−DNA断片を、得られたプラスミドpCC45のXhoI開裂サイトへクローニングした。その結果、pCC49が生じた。W.Selbitschka et al.,1993,ibidの方法と同様にして、「遺伝子の置換え」を、つぎのように実施した。
エクスポネンシャルHK1349.4培養物3mlをエクスポネンシャルドナー培養物(S17−1/pCC49)1mlと合わせ、遠心分離にかけて、0.9%NaCl溶液で洗浄した。細胞を30℃で、NYB50μl中、栄養寒天上で一夜、インキュベータ中に置いた。0.9%NaCl溶液中に再懸濁した細胞を、適宜の稀釈度で選択媒体に適用した。Sm耐性受容体HK1349.4のトランス接合体が、Sm(1000μg/ml)およびNm(150μg/ml)を使用して栄養寒天上に得られ、「置換え」ベクター上の遺伝子sacREに対応する5%シュクロース感受性が複合媒体中で認められた。
トランス接合体を栄養寒天上で選択圧力を加えることなく培養した後、二重組換えが低い頻度(10-8)で生じた。約一週間後、媒体中で5%シュクロース許容性をもち、Nm耐性は保存したがゲンタマイシン(Gm)感受性(ベクターマーカー)を再びもつに至った細胞を単離することができた。
この表現形は、対立遺伝子マーカー交換を確認するものであり、HK1349.4のrecA変異を示すものである。recA突然変異種が典型的にもつ表現形(たとえばUV感受性)を観察することができる。
このようにして得たHK1349.49株において、相同的配置(相同的組換え)をもったプラスミドの染色体組込みの傾向は、明瞭に減少した。この菌株はそれゆえ、実施例6.2に示したハイブリッドプラスミドにとって、高度に適切な宿主である。
実施例8
バイオ形質転換
バイオ形質転換を、振動フラスコ(100ml)中、Nを含まず、0.2%(w/v)のγ−ブチロベタイン(出発物質)を含有し、C源として0.4%(w/v)のグリセリンを含有する最少限媒体(MM:Kulla et al.,Arch.Microbiol.,1983,135,pp.1-7)を使用して行なった。N源および追加的なC源として、0.2%(w/v)のL−グルタミン酸塩(ベタイン利用に関して陰性の菌株に対し)、または0.2%(w/v)のベタインを添加した。
本発明に従って使用した生産株は、HK1349.4/pLO41,HK1349.4/pVK1011,HK1349.4/pAZ7::beu,HK1349.4/pAZ101,HK1349/pVK100qおよびHK1349/pAZ7である。
結果を、EP−A−0543344により既知であるHK13の誘導体HK1349(DSM 3944)およびHK1349.4と比較して、表1にまとめて示す。
γ−ブチロベタインからのL−カルニチンの生成は、DTNB(5,5′−ジチオビス(2−ニトロベンソエート))法(H.K.Bergmeyer,1994,『酵素分析の方法』“Methodeu der enzymafischen Aralyse”フェアラーク・ヒエミー,ヴァインハイム,1810fに記載)に従って行なった。

Claims (17)
- L−カルニチンの生合成のための遺伝子bcoC,bcoA/BおよびbcoDの1種または2種以上からなる分離されたDNA断片であって、γ−ブチロベタイン−CoA−デヒドロゲナーゼ(bcoC)、γ−ブチロベタイン/クロトノベタイン−CoA−シンセターゼ(bcoA/B)またはクロトノベタイン−CoA−ヒドロラーゼ(bcoD)をコードするものであり、下記から選ばれたものであるDNA断片:
a)寄託番号DSM8276として寄託されたリゾビウム/アグロバクテリウムHK1349中のプラスミドpVK100q中に挿入された10.6kbEcoRI断片、および
b)上記10.6kbEcoRI断片を混合し、γ−ブチロベタイン−CoA−デヒドロゲナーゼ、γ−ブチロベタイン/クロトノベタイン−CoA−シンセターゼおよび(または)クロトノベタイン−CoA−ヒドロラーゼの活性を有する酵素をコードする断片。 - γ−ブチロベタイン/クロトノベタイン代謝に割り当てられ、ポテンシャル輸送蛋白質をコードする遺伝子bcoTを付加的に有する請求の範囲第1項のDNA断片。
- 遺伝子bcoC,bcoA/B,bcoDおよび場合によってはbcoTが、エシエリチア、シュードモナス、アグロバクテリウム、リゾビウムおよびリゾビウム/アグロバクテリウム属の微生物から誘導されたものである請求の範囲第1項または第2項のいずれかのDNA断片。
- 遺伝子が、発現に必要な遺伝子コントロール要素と操作的に結合されている請求の範囲第1項ないし第3項のいずれかのDNA断片。
- L−カルニチン生合成の遺伝子が、bcoC,bcoA/BおよびbcoDの順に配列され、単一転写ユニットとして存在する請求の範囲第1項ないし第4項のいずれかのDNA断片。
- L−カルニチン生合成の遺伝子が、bcoC,bcoA/B,bcoDおよびbcoTの順に配列され、単一転写ユニットとして存在する請求の範囲第1項ないし第4項のいずれかのDNA断片。
- 遺伝子制御要素が、天然のbcoオペロンのプロモーターPbcoからなる請求の範囲第4項ないし第6項のいずれかのDNA断片。
- 遺伝子bcoC,bcoA/B,bcoDおよびbcoTが、下記の制限地図によって特徴づけられるものである請求の範囲第1項ないし第7項のいずれかのDNA断片。
- 請求の範囲第1項ないし第8項のいずれかによるDNA断片からなるベクター。
- 寄託番号DSM8726をもって寄託されたリゾビウム/アグロバクテリウムHK1349内に預けられたプラスミドpVK100q、第3図の制限地図によって特徴づけられるプラスミドpVK1011、および第4図の制限地図によって特徴づけられるプラスミドpAZ101のいずれかである請求の範囲第9項のベクター。
- 請求の範囲第1項ないし第10項のいずれかに従うDNA断片またはベクターを含有する組換え微生物。
- L−カルニチン代謝の能力が完全に、または部分的に抑制されたことを特徴とする請求の範囲第11項の微生物。
- 組換えの能力が低減されていることを特徴とする請求の範囲第11項および第12項のいずれかの微生物。
- エシエリチア、シュードモナス、アグロバイテリウム、リゾビウム、コマモナスおよびリゾビウム/アグロバクテリウム属の微生物からえらんだ請求の範囲第11項ないし第13項のいずれかの微生物。
- 寄託番号DSM8726をもって寄託されたリゾビウム/アグロバクテリウムHK1349であって、プラスミドpVK100qを有するもの、第3図の制限地図により特徴づけられるプラスミドpVK1011、または第4図の制限地図によって特徴づけられるプラスミドpAZ101、およびL−カルニチン生合成の能力を有する微生物から誘導された遺伝子的変種および突然変異種。
- リゾビウム/アグロバクテリウムHK1349.4であって、リゾビウム/アグロバクテリウムHK1349中に寄託番号DSM8726で寄託してあるプラスミドpVK100q、図3の制限地図によって特徴づけられるプラスミドpVK1011、または図4の制限地図によって特徴づけられるプラスミドpAZ101を含有するもの、ならびにそれらから誘導されるL−カルニチン生合成能力を有する遺伝子的変種および突然変異種。
- L−カルニチン製造のバイオ技術的方法であって、クロトノベタインおよび(または)γ−ブチロベタインを適当な炭素および窒素源の存在下に、請求の範囲11〜16のいずれかに記載の微生物を手段として発酵させ、L−カルニチンを分離回収することを特徴とする方法。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CH303693 | 1993-10-08 | ||
| CH3036/93 | 1993-10-08 | ||
| CH3694 | 1994-01-06 | ||
| CH36/94 | 1994-01-06 | ||
| PCT/EP1994/003317 WO1995010613A1 (de) | 1993-10-08 | 1994-10-07 | Gene für den butyrobetain/crotonobetain-l-carnitin-stoffwechsel und ihre verwendung zur mikrobiologischen herstellung von l-carnitin |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPH09503390A JPH09503390A (ja) | 1997-04-08 |
| JP3708543B2 true JP3708543B2 (ja) | 2005-10-19 |
Family
ID=25683317
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP51125795A Expired - Fee Related JP3708543B2 (ja) | 1993-10-08 | 1994-10-07 | ブチロベタイン/クロトノベタイン−l−カルニチン代謝のための遺伝子およびそのl−カルニチンの微生物学的生産への使用 |
Country Status (17)
| Country | Link |
|---|---|
| US (1) | US5759824A (ja) |
| EP (1) | EP0722500B1 (ja) |
| JP (1) | JP3708543B2 (ja) |
| KR (1) | KR100346293B1 (ja) |
| CN (2) | CN1058523C (ja) |
| AT (1) | ATE257515T1 (ja) |
| AU (1) | AU7854694A (ja) |
| CA (1) | CA2173115C (ja) |
| CZ (1) | CZ288247B6 (ja) |
| DE (1) | DE59410350D1 (ja) |
| FI (1) | FI118568B (ja) |
| HU (1) | HU220798B1 (ja) |
| NO (1) | NO319058B1 (ja) |
| PL (1) | PL180300B1 (ja) |
| RU (1) | RU2133773C1 (ja) |
| SK (1) | SK280580B6 (ja) |
| WO (1) | WO1995010613A1 (ja) |
Families Citing this family (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6337197B2 (en) | 1998-10-27 | 2002-01-08 | Sigma-Tau Industrie Farmaceutiche Riunite S.P.A. | Coenzymes useful for the synthesis of L-carnitine |
| DE19850426A1 (de) * | 1998-10-27 | 2000-05-04 | Sigma Tau Ind Farmaceuti | gamma-Butyrobetainyl-Coenzym A und Verfahren zu seiner Herstellung |
| ES2237954T3 (es) * | 1998-10-27 | 2005-08-01 | Sigma-Tau Industrie Farmaceutiche Riunite S.P.A. | Coenzima util para la sintesis de l-carnitina. |
| DE19850433A1 (de) * | 1998-10-27 | 2000-05-25 | Sigma Tau Ind Farmaceuti | Crotonobetainyl-Coenzym A, Verfahren zu seiner Herstellung |
| US6720168B2 (en) | 2000-08-04 | 2004-04-13 | Genencor International, Inc. | 2,5-DKG permeases |
| WO2002012481A2 (en) * | 2000-08-04 | 2002-02-14 | Genencor International, Inc. | Enhancement of industrial production by increasing substrate transport |
| CN1233832C (zh) * | 2001-01-31 | 2005-12-28 | 隆萨股份公司 | 制造l-肉毒碱的微生物学方法 |
| US7229811B2 (en) | 2001-08-03 | 2007-06-12 | Genencor International, Inc. | 2,5-diketo-D-gluconic acid (2,5-DKG) permeases |
| ATE531803T1 (de) | 2003-06-05 | 2011-11-15 | Ajinomoto Kk | Fermentierungsverfahren mittels veränderter bakterien mit einer erhöhten aufnahme von nebenprodukten |
| KR101166026B1 (ko) | 2004-07-12 | 2012-07-19 | 씨제이제일제당 (주) | 뉴로스포라 크라사 유래 l-카르니틴 생합성 관련유전자를 포함하는 엔테로박테리아세 속 미생물 및 이를이용한 l-카르니틴의 제조방법 |
| EP1901715B1 (en) | 2005-07-05 | 2012-05-02 | Lonza AG | Spray-drying process for producing a dry carnitine powder or granulate |
| US7901923B2 (en) | 2005-07-19 | 2011-03-08 | Cj Cheiljedang Corporation | Microorganism of Enterobacteriacae genus harboring genes associated with L-carnitine biosynthesis and method of producing L-carnitine using the microorganism |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS59192095A (ja) * | 1983-04-13 | 1984-10-31 | Ajinomoto Co Inc | L−カルニチンの製造法 |
| MX170707B (es) * | 1989-07-28 | 1993-09-08 | Lonza Ag | Procedimiento para la produccion discontinua de l-carnitina por transformacion microbiologica de crotonobetaina y gama-butirobetaina |
| IT1240833B (it) * | 1990-05-14 | 1993-12-17 | Sigma Tau Ind Farmaceuti | Procedimento biocatalitico per la produzione di l-(-)- carnitina da crotonilbetaina e ceppi di proteeae per l'uso in tale procedimento |
| RU2099418C1 (ru) * | 1991-11-21 | 1997-12-20 | Лонца Аг | Фрагмент днк, обеспечивающий использование бетаина, рекомбинантная плазмидная днк с фрагментом днк, обеспечивающим использование бетаина, штамм бактерий rhizobium/agrobacterium, предназначенный для стабильного хранения плазмиды с фрагментом днк, способ получения штамма бактерий |
-
1994
- 1994-10-07 CN CN94193701A patent/CN1058523C/zh not_active Expired - Fee Related
- 1994-10-07 AU AU78546/94A patent/AU7854694A/en not_active Abandoned
- 1994-10-07 CA CA002173115A patent/CA2173115C/en not_active Expired - Fee Related
- 1994-10-07 SK SK431-96A patent/SK280580B6/sk not_active IP Right Cessation
- 1994-10-07 CN CNB991207033A patent/CN1157480C/zh not_active Expired - Fee Related
- 1994-10-07 RU RU96108818A patent/RU2133773C1/ru not_active IP Right Cessation
- 1994-10-07 HU HU9600904A patent/HU220798B1/hu not_active IP Right Cessation
- 1994-10-07 WO PCT/EP1994/003317 patent/WO1995010613A1/de active IP Right Grant
- 1994-10-07 JP JP51125795A patent/JP3708543B2/ja not_active Expired - Fee Related
- 1994-10-07 EP EP94929520A patent/EP0722500B1/de not_active Expired - Lifetime
- 1994-10-07 AT AT94929520T patent/ATE257515T1/de active
- 1994-10-07 KR KR1019960701836A patent/KR100346293B1/ko not_active IP Right Cessation
- 1994-10-07 PL PL94313968A patent/PL180300B1/pl unknown
- 1994-10-07 CZ CZ19961005A patent/CZ288247B6/cs not_active IP Right Cessation
- 1994-10-07 US US08/615,191 patent/US5759824A/en not_active Expired - Lifetime
- 1994-10-07 DE DE59410350T patent/DE59410350D1/de not_active Expired - Lifetime
-
1996
- 1996-04-03 NO NO19961385A patent/NO319058B1/no not_active IP Right Cessation
- 1996-04-04 FI FI961537A patent/FI118568B/fi not_active IP Right Cessation
Also Published As
| Publication number | Publication date |
|---|---|
| PL313968A1 (en) | 1996-08-05 |
| HUT74825A (en) | 1997-02-28 |
| FI961537A (fi) | 1996-04-04 |
| DE59410350D1 (de) | 2004-02-12 |
| KR100346293B1 (ko) | 2002-11-30 |
| NO961385L (no) | 1996-06-03 |
| AU7854694A (en) | 1995-05-04 |
| HU220798B1 (hu) | 2002-05-28 |
| FI118568B (fi) | 2007-12-31 |
| CZ288247B6 (en) | 2001-05-16 |
| NO319058B1 (no) | 2005-06-13 |
| CZ100596A3 (en) | 1996-10-16 |
| SK280580B6 (sk) | 2000-04-10 |
| NO961385D0 (no) | 1996-04-03 |
| WO1995010613A1 (de) | 1995-04-20 |
| CN1282791A (zh) | 2001-02-07 |
| FI961537A0 (fi) | 1996-04-04 |
| US5759824A (en) | 1998-06-02 |
| EP0722500B1 (de) | 2004-01-07 |
| SK43196A3 (en) | 1996-10-02 |
| JPH09503390A (ja) | 1997-04-08 |
| CN1157480C (zh) | 2004-07-14 |
| CA2173115A1 (en) | 1995-04-20 |
| ATE257515T1 (de) | 2004-01-15 |
| CA2173115C (en) | 2005-07-26 |
| EP0722500A1 (de) | 1996-07-24 |
| CN1133066A (zh) | 1996-10-09 |
| PL180300B1 (pl) | 2001-01-31 |
| RU2133773C1 (ru) | 1999-07-27 |
| CN1058523C (zh) | 2000-11-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US4278765A (en) | Method for preparing strains which produce aminoacids | |
| JP2944094B2 (ja) | 細菌染色体上ヘの目的遺伝子の組み込み方法及び該方法によって得られた細菌 | |
| JP7125477B2 (ja) | グリシン生産能が増加された微生物及びこれを用いた発酵組成物の生産方法 | |
| HU224975B1 (en) | Process for producing l-amino acids by fermentation and amino acid-producing bacterium strains | |
| JP3708543B2 (ja) | ブチロベタイン/クロトノベタイン−l−カルニチン代謝のための遺伝子およびそのl−カルニチンの微生物学的生産への使用 | |
| Gibson et al. | Analysis of the cbbXYZ operon in Rhodobacter sphaeroides | |
| US7807443B2 (en) | Microorganisms providing novel gene products forming or decomposing polyamino acids | |
| Thiel et al. | [13] Conjugal transfer of plasmids to cyanobacteria | |
| Kuhlemeier et al. | Cloning of nitrate reductase genes from the cyanobacterium Anacystis nidulans | |
| KR20000060322A (ko) | 슈도모나스 플루오레슨스 유래의 외래단백질 분비촉진유전자 | |
| JPH08501694A (ja) | ビオチンの生物工学的製造方法 | |
| CN108368477A (zh) | 修饰的膜渗透性 | |
| CN111763699B (zh) | 用于发酵生产1,5-戊二胺的重组dna、菌株及其应用 | |
| US20210324391A1 (en) | Recombinant microorganism, preparation method therefor and application thereof in producing coenzyme q10 | |
| Gibson | Genetic analysis of CO 2 fixation genes | |
| Vybornaya et al. | Use of an alternative pathway for isoleucine synthesis in threonine-producing strains of Escherichia coli | |
| US20050214912A1 (en) | Method for producing an optically active amino acid | |
| JP4162383B2 (ja) | ホモグルタミン酸の生産に関与する遺伝子およびその使用 | |
| KR20230108790A (ko) | L-히스티딘 생산능이 향상된 에스케리치아 속 변이주 및 이를 이용한 l-히스티딘의 생산 방법 | |
| KR20240066510A (ko) | 호기 조건에서 카복시도써머스 하이드로게노포먼스 유래 가용성 일산화탄소 탈수소효소 대량생산 방법 | |
| KR20220126610A (ko) | L-라이신 생산능이 향상된 코리네박테리움 글루타미쿰 변이주 및 이를 이용한 l-라이신의 생산 방법 | |
| KR20240017394A (ko) | L-시트룰린 생산능이 향상된 코리네박테리움 글루타미쿰 변이주 및 이를 이용한 l-시트룰린의 생산 방법 | |
| KR20230108789A (ko) | L-히스티딘 생산능이 향상된 에스케리치아 속 변이주 및 이를 이용한 l-히스티딘의 생산 방법 | |
| KR20230165734A (ko) | L-라이신 생산능이 향상된 코리네박테리움 글루타미쿰 변이주 및 이를 이용한 l-라이신의 생산 방법 | |
| KR20220149219A (ko) | L-라이신 생산능이 향상된 코리네박테리움 글루타미쿰 변이주 및 이를 이용한 l-라이신의 생산 방법 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20040127 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20040218 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20050222 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20050523 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20050705 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20050804 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20100812 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110812 Year of fee payment: 6 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120812 Year of fee payment: 7 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130812 Year of fee payment: 8 |
|
| LAPS | Cancellation because of no payment of annual fees |