WO2009145164A1 - 改良されたポリヒドロキシアルカノエート生産微生物及びそれを用いたポリヒドロキシアルカノエートの製造方法 - Google Patents
改良されたポリヒドロキシアルカノエート生産微生物及びそれを用いたポリヒドロキシアルカノエートの製造方法 Download PDFInfo
- Publication number
- WO2009145164A1 WO2009145164A1 PCT/JP2009/059564 JP2009059564W WO2009145164A1 WO 2009145164 A1 WO2009145164 A1 WO 2009145164A1 JP 2009059564 W JP2009059564 W JP 2009059564W WO 2009145164 A1 WO2009145164 A1 WO 2009145164A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- gene
- dna
- bktb
- strain
- polyhydroxyalkanoate
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/62—Carboxylic acid esters
- C12P7/625—Polyesters of hydroxy carboxylic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1025—Acyltransferases (2.3)
- C12N9/1029—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01009—D-Xylulose reductase (1.1.1.9), i.e. xylitol dehydrogenase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01036—Acetoacetyl-CoA reductase (1.1.1.36)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y203/00—Acyltransferases (2.3)
- C12Y203/01—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
- C12Y203/01016—Acetyl-CoA C-acyltransferase (2.3.1.16)
Definitions
- the present invention relates to a microorganism that produces a polyhydroxyalkanoate copolymer having a 3-hydroxyalkanoate unit having 4 or more carbon atoms, and a method for efficiently producing the copolymer using the microorganism.
- PHA Polyhydroxyalkanoate
- PHA is a polyester-type organic molecular polymer produced by a wide range of microorganisms.
- PHA is a biodegradable thermoplastic polymer.
- PHA can also be produced from renewable resources. For these reasons, attempts have been made to industrially produce PHA as an environmentally conscious material or a biocompatible material and use it in various industries.
- P (3HB) poly-3-hydroxybutyric acid
- 3HB 3-hydroxybutyric acid
- P (3HB) was first discovered in 1925 in Bacillus megaterium .
- P (3HB) is a thermoplastic polymer and is biologically decomposed in the natural environment, and thus has attracted attention as an environmentally friendly plastic.
- P (3HB) has a hard and brittle nature because of its high crystallinity, so its practical application range is limited. In order to expand the application range, it was necessary to give flexibility to P (3HB).
- P (3HB-co-3HV) a copolymer comprising 3HB and 3-hydroxyvaleric acid
- 3HV 3-hydroxyvaleric acid
- Patent Document 1 Patent Document 2
- P (3HB-co-3HV) is a kind of PHA. Since P (3HB-co-3HV) is more flexible than P (3HB), it was considered to be applicable to a wide range of applications. However, in practice, even if the 3HV mole fraction in P (3HB-co-3HV) is increased, the accompanying change in physical properties is scarce, which is particularly required for processing into films, sheets, flexible packaging containers, etc. Since flexibility does not improve as much, it is used only in limited fields such as shampoo bottles and disposable razor handles.
- P (3HB-co-3HH) In order to increase the flexibility of P (3HB), it is sometimes referred to as a copolymer of 3HB and 3-hydroxyhexanoic acid (hereinafter sometimes referred to as 3HH) (hereinafter referred to as P (3HB-co-3HH). ) And its manufacturing method have been studied (Patent Literature 3, Patent Literature 4).
- P (3HB-co-3HH) is a kind of PHA.
- the method for producing P (3HB-co-3HH) in these reports was to fermentatively produce fatty acid such as oleic acid and palmitic acid using a wild strain of Aeromonascaviae isolated from soil as a carbon source.
- the 3HH composition ratio was 15 mol% when oleic acid was used as the carbon source, and 5 mol% when palmitic acid was used as the carbon source.
- Non-patent Document 1 Studies on the physical properties of P (3HB-co-3HH) have also been made (Non-patent Document 1).
- A. caviae is cultured using a fatty acid having 12 or more carbon atoms as a sole carbon source, and P (3HB-co-3HH) having a 3HH composition of 11 to 19 mol% is produced by fermentation.
- P (3HB-co-3HH) increases in 3HH composition, it becomes more flexible from hard and brittle properties like P (3HB), and more flexible than P (3HB-co-3HV) It was revealed that That is, P (3HB-co-3HH) can have a wide range of physical properties applicable from hard polymers to soft polymers by changing the 3HH composition ratio. Therefore, P (3HB-co-3HH) has a low 3HH composition ratio.
- Non-Patent Document 2 by using Aeromonas hydrophila and performing fed-batch culture for 43 hours using oleic acid as a carbon source, the cell mass is 95.7 g / L, the polymer content is 45.2%, and the 3HH composition ratio is 17 mol%. Of P (3HB-co-3HH) was produced. Moreover, 3HH composition ratio was 11 mol%, the amount of bacterial cells was 50 g / L, and the polymer content was 50% by cultivation using glucose and lauric acid as a carbon source using A. hydrophila (Non-patent Document 3). . However, A.
- Non-Patent Document 4 hydrophila is pathogenic to humans (Non-Patent Document 4), and therefore cannot be said to be a microorganism suitable for industrial production. Moreover, since the carbon source used in these culture productions is expensive, use of an inexpensive carbon source has been demanded from the viewpoint of manufacturing cost.
- the composition ratio of 3HH was 4 to 5 mol%, and the bacterial cell amount was 4 g / L and the polymer content was 80% (Non-patent Document 6).
- the latter production method uses inexpensive vegetable oils and fats as a carbon source and has a high polymer content, but the amount of cells is low, so the polymer productivity is low, and a 3HH composition ratio of 4 to 5 mol% is applicable to applications such as films. It was not soft.
- a P (3HB-co-3HH) producing strain using E. coli as a host was also constructed.
- a strain was constructed in which the PHA synthase gene belonging to the genus Aeromonas and the NADP-acetoacetyl-CoA reductase gene of C. necator were introduced into E. coli.
- the amount of cells was 79 g / L
- the polymer content was 27.2%
- the 3HH composition ratio was 10.8 mol% (Non-patent Document 8). Escherichia coli into which A.
- Non-patent Document 10 PHA synthase was artificially modified.
- a mutant enzyme in which the 149th amino acid asparagine is replaced with serine and a mutant enzyme in which the 171st aspartic acid is replaced with glycine are It has been shown that PHA synthase activity and 3HH composition are improved, and the mutant enzyme in which the 518th phenylalanine is replaced with isoleucine and the mutant enzyme in which the 214th valine is replaced with glycine are It has been reported that PHA synthase activity and polymer content have been improved. However, these use special E. coli as a host, and the polymer content is as low as about 13%. Therefore, further improvements for industrial production utilizing the characteristics of these mutant enzymes were necessary. .
- Patent Document 5 C.Necator by the polyester synthase gene and R- enoyl -CoA hydratase gene such pJRDEE32 and pJRDEE32d13 like was introduced (Patent Document 5, Non-Patent Document reference 5) PHA synthase expression plasmid in pJRD215 (ATCC 37533) And the PHA productivity of the transformant has been examined.
- the amount of bacterial cells of the strain was as low as 4 g / L, but the amount of bacterial cells was 45 g / L, the polymer content was 62.5%, and the 3HH composition ratio was 8.1 mol% by improving the culture conditions of the same strain using vegetable oil as a carbon source. The polymer productivity has been improved.
- Patent Document 6 an attempt was made to improve the 3HH composition ratio and polymer productivity of P (3HB-co-3HH) by a culture method
- Patent Document 6 A method of controlling the physical properties of P (3HB-co-3HH) is also disclosed (Patent Document 6).
- P (3HB-co-) By using at least two types of fats and oils and / or fatty acids having different carbon numbers as a carbon source, it becomes possible to produce a polyester having a 3HH composition ratio of 1 to 40 mol%, and P (3HB-co-) having various physical properties. 3HH) can be produced.
- this production method it is necessary to add relatively expensive hexanoic acid, octanoic acid or the like for 3HH composition control, and high concentration hexanoic acid exhibits cytotoxicity, so that the cell productivity decreases. It is the result.
- the production equipment since a multi-component carbon source is added, the production equipment may be complicated and expensive.
- a method for improving 3HH composition by culturing with C. necator as a host and fructose as a carbon source is also disclosed.
- the polyester synthase gene and the crotonyl-CoA reductase gene (hereinafter abbreviated as ccr) derived from Streptomycescinnamonensis were introduced, the 3HH composition was 0.9%, but when the R-enoyl-CoA hydratase gene and the like were further introduced.
- the 3HH composition was improved to 1.6%.
- the amount of cells at that time was about 1.5 g / L
- the polymer content was as low as about 40%, and further improvement was necessary for industrial production of high 3HH composition P (3HB-co-3HH) ( Non-patent document 7).
- P (3HB-co-3HH) having a 3HH composition ratio of 12 mol% or more is desired for application to films, sheets, flexible packaging containers, and the like.
- attempts to improve the 3HH composition ratio resulted in a decrease in the polymer content or the amount of cells.
- the object of the present invention is to produce a PHA having a high 3HH composition ratio, which is excellent in flexibility and expected to be used in a wide range of applications, with a practical productivity using vegetable oil which is an inexpensive carbon source.
- the objective is to efficiently ferment and produce P (3HB-co-3HH) having a 3HH composition ratio of 12 mol% or more while maintaining a polymer content of the cells of 70% or more and a cell amount of 150 g / L or more. .
- the present inventors have increased the amount of PHA synthase in microorganisms, decreased the ability to synthesize 3HB monomer, and created a new 3HH monomer synthesis route. It was found that PHA having an improved 3HH composition ratio can be produced and accumulated in large quantities in the microorganism by enhancing the supply system, and the present invention has been completed.
- the first of the present invention is that the phbA gene is inactivated, phaC gene and bktB ( ⁇ -ketothiolase) gene expression is enhanced, and a butyryl-CoA synthesis pathway is newly introduced by introduction of the ccr gene.
- the present invention relates to a microorganism capable of producing a 3-hydroxyhexanoic acid unit-containing polyhydroxyalkanoate with improved 3HH monomer synthesis ability.
- a microorganism having the following requirements (1) to (3); (1) The expression of the phbA gene is suppressed, or the catalytic activity of the enzyme encoded by the gene is suppressed, (2) The expression of the bktB gene is enhanced or the catalytic activity of the enzyme encoded by the gene is improved, (3) A polyhydroxyalkanoate synthase gene and / or a crotonyl-CoA reductase gene is introduced.
- the second aspect of the present invention is that the above-described microorganism is used, and the 3HH composition ratio is 12 mol% or more while maintaining a polymer content of 70% or more and a cell amount of 150 g / L or more using an inexpensive carbon source.
- the present invention relates to a method for producing 3HH unit-containing PHA.
- PHA having a 3HH composition ratio of 12 mol% or more can be produced by fermentation while maintaining a polymer content of the cells of 70% or more and a cell amount of 150 g / L or more. .
- the 3HH unit-containing PHA of the present invention is 3HH, 3HB, 3HV, 3-hydroxyoctanoic acid, and one or more 3-hydroxyalkanoic acids selected from the group consisting of 3-hydroxyalkanoic acids having a longer alkyl chain. Is a copolymerized polyester.
- the phbA gene of the present invention is a gene encoding an enzyme that catalyzes a reaction of condensing two molecules of acetyl-CoA among enzymes having ⁇ -ketothiolase activity, but hardly catalyzes ⁇ -ketovaleryl-CoA thiolysis.
- the phbA gene consisting of the base sequence shown in SEQ ID NO: 5 is exemplified. Encodes an enzyme that has a sequence identity of 85% or more with the base sequence shown in SEQ ID NO: 5 and catalyzes a reaction of condensing two molecules of acetyl-CoA, but hardly catalyzes thiolysis of ⁇ -ketovaleryl-CoA.
- the gene can be preferably used in the present invention.
- the sequence identity is more preferably 90% or more, still more preferably 95% or more, and still more preferably 98% or more.
- the bktB gene of the present invention well catalyzes both the condensation of two molecules of acetyl-CoA and the condensation of acetyl-CoA and a longer chain, such as propionyl-CoA, among enzymes having ⁇ -ketothiolase activity.
- a gene encoding an enzyme that catalyzes the thiolysis of 3-ketovaleryl-CoA for example, a gene consisting of the base sequence shown in SEQ ID NO: 29 (SLATER et al., J. Bacteriol., Vol.180, 1979) -1987, 1998).
- sequence identity with the base sequence shown in SEQ ID NO: 29, and both condensation of two molecules of acetyl-CoA and condensation of acetyl-CoA and longer chain length, for example, propionyl-CoA
- a gene encoding an enzyme that catalyzes well and catalyzes 3-ketovaleryl-CoA thiolysis can be preferably used in the present invention.
- the sequence identity is more preferably 90% or more, still more preferably 95% or more, and still more preferably 98% or more.
- Examples of a gene encoding a polymerase capable of synthesizing a 3HH unit-containing PHA include the A. caviae phaC gene or the phaC mutant gene comprising the base sequence shown in SEQ ID NO: 4, and the host results in the PHA. As long as it can be produced.
- the sequence identity is more preferably 90% or more, still more preferably 95% or more, and still more preferably 98% or more.
- the endogenous promoter of the bktB structural gene is DNA that induces transcription of the bktB structural gene, and is inherent in the microorganism originally having the phbA gene and the bktB gene used as a host.
- the microorganism originally having the phbA gene and the bktB gene in the present invention is not particularly limited as long as it is a wild strain having the phbA gene and the bktB gene, or a microorganism genetically manipulated to the wild strain, and 3HH unit-containing PHA can be synthesized. Can be used as hosts.
- the C. necator for example, C.
- necator H16 (ATCC17699), and the like.
- the strain can be obtained from the American Type Culture Collection (ATCC). Of course, even if it is a mutant obtained by artificially mutating the microorganism, or a similar strain mutated by genetic engineering techniques, it can be used in the present invention as long as it can synthesize PHA containing 3HH units. it can.
- the phbA gene present on the chromosome of the microorganism originally having the phbA gene and the bktB gene is inactivated.
- the enzyme encoded by the phbA gene catalyzes the reaction to produce acetoacetyl-CoA, a precursor of 3-hydroxybutyryl-CoA, which is one of PHA monomers, by condensation of acetyl-CoA molecules as described above.
- the enzyme encoded by the phbA gene does not catalyze the reaction of condensing acetyl-CoA and butanoyl-CoA. Therefore, inactivation of the phbA gene lowers the 3HB ratio in the polymer, and as a result, the 3HH ratio is considered to be improved.
- the method for inactivating the phbA gene is sufficient if phbA is inactivated as a result.
- a new stop codon is introduced between the start codon and the stop codon of the phbA gene, and 2) ribosome binding Introduce a mutation that decreases the ribosome binding activity into the site, 3) Introduce a mutation that decreases the catalytic activity of the enzyme inside the phbA structural gene, 4) Use RNA interference, 5) Insert a transposon, 6) phbA Examples thereof include a method of deleting part or all of the structural gene. These methods are well known to those skilled in the art.
- the enzyme encoded by the bktB gene catalyzes not only the condensation of two acetyl-CoA molecules but also the reaction of condensing acetyl-CoA and butanoyl-CoA. Condensation of acetyl-CoA and butanoyl-CoA yields 3-ketohexanoyl-CoA, a precursor of 3-hydroxyhexanoyl-CoA.
- the activity of the enzyme encoded by the bktB gene is higher for the condensation reaction of acetyl-CoA and butanoyl-CoA than for the condensation reaction of two acetyl-CoA molecules.
- the method for enhancing the enzyme encoded by the bktB gene is not particularly limited, but high expression by an expression vector and high expression by modification of an expression regulatory region are possible.
- the promoter of the gene on the chromosome of the microorganism originally having the phbA gene and the bktB gene is modified to increase transcription activity, for example, the bktB gene inherent in the microorganism originally having the phbA gene and the bktB gene If a promoter that induces the transcription of the bktB gene is further incorporated upstream of, a high expression can be achieved by increasing the transcriptional activity.
- a promoter that induces transcription of the bktB gene is preferably inserted upstream of the start codon of the bktB gene. Any promoter can be used in the present invention as long as it induces transcription of the bktB gene.
- the promoter is preferably at least one of a promoter inherent in a microorganism originally having a phbA gene and a bktB gene and a promoter of a heterologous microorganism.
- the promoter of the C. necator phbCAB operon consisting of the base sequence shown in SEQ ID NO: 1 and the promoter of the A. caviae phaPCJ operon consisting of the base sequence shown in SEQ ID NO: 2 are originally derived from the phbA gene and the bktB gene. This is suitable when C. necator is used as the microorganism.
- the DNA used as the promoter preferably has 70% or more sequence identity with the base sequences shown in SEQ ID NOS: 1 and 2, more preferably 85% or more, and still more preferably 90% or more. Even more preferably, it is 95% or more, and most preferably 98% or more. As long as it induces transcription of the bktB gene, it can be used in the present invention.
- DNA used as a promoter colony hybridization method, plaque hybridization method, Southern hybridization method or the like was performed using a nucleotide sequence complementary to the nucleotide sequences shown in SEQ ID NOS: 1 and 2 as a probe.
- DNA can be used that hybridizes under stringent conditions. As long as it induces transcription of the bktB gene, it can be used in the present invention.
- the hybridization can be performed according to the method described in Molecular Cloning, A Laboratory manual, Second edition (Cold Spring Harbor Laboratory Press, 1989) and the like.
- the DNA to hybridize is, for example, after hybridization at 65 ° C. in the presence of 0.7 to 1.0 M NaCl using a filter on which colony or plaque-derived DNA is immobilized.
- DNA that can be obtained by washing a filter under a condition of 65 ° C. using a double-concentration SSC solution (composition of a single-concentration SSC solution consists of 150 mM sodium chloride and 15 mM sodium citrate) can be mentioned. .
- it is washed with an SSC solution of 2 times concentration at 65 ° C., more preferably washed with an SSC solution of 0.2 times concentration at 65 ° C., and more preferably washed with an SSC solution of 0.1 times concentration at 65 ° C. DNA that can be obtained.
- hybridization conditions are described as described above, the conditions are not particularly limited.
- a plurality of factors such as temperature and salt concentration can be considered as factors affecting the stringency of hybridization, and those skilled in the art can realize optimum stringency by appropriately selecting these factors.
- the DNA that can hybridize under the above conditions is 70% or more, preferably 85% or more, more preferably 90% or more, and still more preferably 95%, with the DNA shown in SEQ ID NO: 1 or 2. % Or more, and most preferably 98% or more of the DNA can be mentioned and is included in the promoter as long as it induces transcription of the bktB gene.
- Crotonyl-CoA reductase encoded by the ccr gene is an enzyme that reduces crotonyl-CoA, which is an intermediate in the fatty acid ⁇ -oxidation pathway, to produce butyryl-CoA, which is a substrate for the enzyme encoded by the bktB gene.
- 3-hydroxyhexanoyl-CoA is not only supplied directly from the ⁇ -oxidation pathway but also from acetyl-CoA derived from the ⁇ -oxidation pathway.
- the ccr gene used in the present invention can be used in the present invention as long as it encodes an enzyme having the activity of reducing crotonyl-CoA to produce 3-hydroxyhexanoyl-CoA. It is preferably a gene comprising a base sequence having 70% or more sequence identity, more preferably 85% or more, still more preferably 90% or more, still more preferably 95% or more, and most preferably Is 98% or more.
- Soy DNA can also be used in the present invention as long as it encodes an enzyme having the activity of reducing crotonyl-CoA to produce butyryl-CoA.
- the polyhydroxyalkanoate synthase gene used in the present invention can be used in the present invention as long as it encodes an enzyme having polyhydroxyalkanoate synthesizing activity.
- the base sequence shown in SEQ ID NO: 4 and a sequence of 70% or more are used. It is preferably a gene comprising a base sequence having identity, more preferably 85% or more, still more preferably 90% or more, still more preferably 95% or more, and most preferably 98% or more. is there.
- a colony hybridization method, a plaque hybridization method, a Southern hybridization method, or the like is performed using a base sequence complementary to the base sequence shown in SEQ ID NO: 4 as a probe, it hybridizes under stringent conditions.
- Soy DNA can also be used in the present invention as long as it encodes an enzyme having polyhydroxyalkanoate synthesis activity.
- examples of carbon sources that can be used for the production of 3HH-containing PHA include sugars such as glucose and fructose, alcohols such as methanol, ethanol, and butanol, acetic acid, propionic acid, hexanoic acid, octanoic acid, decanoic acid, Fatty acids such as saturated and unsaturated fatty acids such as lauric acid, oleic acid, palmitic acid, linoleic acid, linolenic acid and myristic acid, fatty acid derivatives such as esters and salts of these fatty acids, organic acids such as lactic acid, Fats and oils rich in saturated / unsaturated fatty acids that are 10 or more, for example, vegetable oils and fats, especially palm oil, palm oil, palm kernel oil, palm kernel oil olein (hereinafter also referred to as PKOO), palm double olein (hereinafter referred to as POO) Also described).
- sugars such as glucose and fructose
- 3- hydroxy-hexanoyl -CoA as genes encoding high enzyme capable of converting into PHA, A.
- the method for producing a microorganism capable of producing the 3HH-containing PHA of the present invention is not particularly limited, but is exemplified below using C. necator as a host.
- a polyester synthase mutant gene encoding an enzyme having high ability to convert 3-hydroxyhexanoyl-CoA to PHA derived from A. caviae is replaced on the chromosome with the original polyester synthase gene by homologous recombination method or the like. .
- the gene disruption method is used to inactivate the phbA gene present on the chromosome, and any mode may be used as long as the activity of the protein that is the phbA gene product is reduced or eliminated.
- a stop codon may be introduced into the phbA gene, or the promoter and / or ribosome binding site upstream of the gene may be modified.
- the expression of the bktB gene is enhanced, and any mode may be used as long as the activity of the bktB gene product is increased.
- a bktB mutant gene with improved specific activity may be used, a DNA containing a heterologous or homologous promoter and ribosome binding site may be inserted upstream of the start codon of the bktB gene, Alternatively, the promoter and / or ribosome binding site may be replaced with a DNA comprising a base sequence containing a heterologous or homologous promoter and / or ribosome binding site.
- the original promoter may be completely lost from the chromosome, or a part of it may be deleted.
- a method for site-specific insertion / replacement of an arbitrary gene on the chromosome is well known to those skilled in the art, and typical methods include, but are not limited to, the following methods.
- Site-specific integration and second-stage homologous recombination caused by transposon and homologous recombination methods Ohman et al., J. Bacteriol., Vol. 162, p1068 (1985)
- homologous recombination mechanisms There is a method based on the principle of omission (Noti et al., Methods Enzymol., Vol.
- a method of inserting DNA consisting of a nucleotide sequence including a promoter and a ribosome binding site of A. caviae phaC gene immediately before the start codon of C. necator bktB gene is illustrated more specifically.
- a replacement fragment is prepared.
- DNA consisting of a base sequence including the promoter and ribosome binding site of the A. caviae phaC gene (hereinafter also referred to as phaC) is linked immediately before the start codon of the bktB gene, and then the bktB gene is linked. .
- it is a DNA fragment in which a DNA comprising a nucleotide sequence including a phaC promoter and a ribosome binding site is inserted immediately before the start codon of the gene.
- the upstream and downstream of the phaC promoter and ribosome binding site-containing DNA are sandwiched between the upstream and downstream, which are homologous sequences necessary for causing homologous recombination with DNA on the chromosome. The longer the length, the higher the frequency of recombination.
- a gene that serves as a selection marker at the time of gene replacement can be added to the replacement fragment.
- a gene serving as a selection marker for example, an antibiotic resistance gene such as kanamycin, chloramphenicol, streptomycin, ampicillin, a gene that complements various auxotrophy, and the like can be used. When C. necator is used as a host, a kanamycin resistance gene is preferred.
- a gene for facilitating selection of a microbial strain in which the region containing the selection marker gene has been eliminated by the second-stage homologous recombination can be added.
- An example of such a gene is a sacB gene derived from Bacillus subtilis. Microbial strains expressing this gene are known to be unable to grow on a medium containing sucrose, and it becomes easy to select a strain that has lost this gene by dropping off due to growth on a medium containing sucrose. .
- the replacement fragment composed of these is prepared as a plasmid for gene replacement by connecting to a vector that does not replicate in the host microorganism strain.
- vector plasmids that can be used in the genus Ralstonia and Pseudomonas include, for example, pUC vectors, pBluescript vectors, pBR322 vectors, and vectors having the same origin of replication. Furthermore, it is possible to coexist DNA sequences such as mob and oriT that enable junction transfer.
- the plasmid DNA for gene replacement prepared in such a configuration can be introduced into C. necator by a known method such as electroporation method or conjugation transfer method, and homologous recombination can be performed.
- a strain having a plasmid DNA for gene replacement inserted on the chromosome by homologous recombination is selected.
- Strains can be selected by a method based on a gene for selection coexisting with plasmid DNA for gene replacement.
- a kanamycin resistance gene is used, it can be selected from strains grown in a medium containing kanamycin.
- a strain in which the region containing the selection marker gene has been removed from the chromosome by the second homologous recombination is selected. Based on the selection gene used at the time of insertion, for example, a strain that can no longer grow on a medium containing kanamycin may be selected, but if the sacB gene is coexisting with the gene replacement plasmid, sucrose is included. It can be easily selected from strains that grow on the medium. In order to confirm whether the strain thus obtained is a strain in which a gene is substituted as desired, a known method such as a PCR method, Southern hybridization method, or determination of a DNA base sequence can be used.
- Expression vectors usually used in C. necator include an expression vector derived from pJRD215 (for example, Patent Document 5) and an expression vector derived from pBBR122 (Non-Patent Document 7). It is desirable that the expression vector be stably replicated and maintained in the host cell even under non-selective pressure conditions. Therefore, pCUP2 and the like described in International Publication WO / 2007/049716 [0041] is more preferable. pCUP2 contains a region necessary for the replication and stable maintenance of the plasmid derived from the megaplasmid pMOL28 possessed by the Cupriavidus metallidurans CH34 strain, which is closely related to C. necator .
- a plasmid in which a phaC gene expression unit derived from A. caviae and a ccr gene expression unit derived from S. cinnamonensis are inserted into pCUP2 is a pCUP2-631 vector.
- a phaP gene encoding a fasin derived from A. caviae and a phaJ gene encoding an R-specific enoyl-CoA hydratase derived therefrom may be further inserted into pCUP2-631.
- a phbP gene encoding a fasin derived from C. necator and a phaJ gene encoding an R-specific enoyl-CoA hydratase derived from A. caviae may be further inserted into pCUP2-631.
- strains obtained by introducing this expression vector into a strain in which the phbA gene on the chromosome has been disrupted and the bktB gene expression has been enhanced include strains such as the KNK-631 strain. All of these strains should be fermentatively produced PHA having a 3HH composition ratio of 12 mol% or more using an inexpensive and safe carbon source while maintaining a cell content of 70% or more and a cell amount of 150 g / L or more. Can do.
- a method for producing 3HH-containing PHA using the microorganism produced by the above-described method will be described.
- the method is not particularly limited, but can be performed as follows.
- it is preferable to culture the microorganism using a medium containing a carbon source, a nitrogen source, inorganic salts, and other organic nutrient sources.
- the carbon source saccharides, fats and oils, fatty acids and the like are preferable, vegetable oils and fats are more preferable, and palm oil and palm kernel oil are more preferable.
- the nitrogen source include ammonium salts such as ammonia, ammonium chloride, ammonium sulfate, and ammonium phosphate, peptone, meat extract, yeast extract, and the like.
- inorganic salts include potassium dihydrogen phosphate, disodium hydrogen phosphate, magnesium phosphate, magnesium sulfate, sodium chloride and the like.
- organic nutrient sources include amino acids such as glycine, alanine, serine, threonine, and proline, and vitamins such as vitamin B1, vitamin B12, and vitamin C.
- antibiotics kanamycin etc.
- the recovery of 3HH-containing PHA from the microbial cells is not particularly limited, but can be performed, for example, by the following method.
- the cells are separated from the culture solution with a centrifuge, and the cells are washed with distilled water, methanol, or the like and dried.
- 3HH-containing PHA is extracted from the dried cells using an organic solvent such as chloroform.
- Cellular components are removed from the organic solvent solution containing the 3HH-containing PHA by filtration or the like, and a poor solvent such as methanol or hexane is added to the filtrate to precipitate the 3HH-containing PHA. Further, the supernatant is removed by filtration or centrifugation, and dried to recover 3HH-containing PHA.
- Analysis of the weight average molecular weight (Mw) and 3HH composition (mol%) of the obtained 3HH-containing PHA can be performed by, for example, a gas chromatographic method, a nuclear magnetic resonance method, or the like.
- the highly flexible PHA obtained by the present invention is suitably used for films, sheets, flexible packaging containers and the like.
- Example 1 ⁇ Preparation of plasmid vector for gene insertion>
- a DNA comprising a base sequence including a promoter of phaC of A. caviae and a ribosome binding site was prepared as follows.
- PCR was performed using A. caviae genomic DNA as a template and the primers shown in SEQ ID NO: 6 and SEQ ID NO: 7.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds, and 68 ° C. for 20 seconds for 25 cycles, and the polymerase used was KOD-plus- (manufactured by TOYOBO).
- the DNA fragment obtained by PCR was terminally phosphorylated and digested with EcoRI. This DNA fragment was designated as P Ac -5P + Eco.
- the DNA insertion site is set immediately before the start codon of the bktB gene of the chromosomal DNA of the KNK-005AS strain described in JP2008-029218 [0038], and upstream of the start codon of the gene by the following procedure.
- a DNA consisting of the nucleotide sequence was prepared.
- PCR was performed using the primers shown in SEQ ID NO: 8 and SEQ ID NO: 9 to initiate the bktB gene. DNA consisting of a base sequence upstream of the codon was obtained. PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 64 ° C. for 30 seconds and 68 ° C. for 30 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was simultaneously digested with the restriction enzymes BamHI and EcoRI. This DNA fragment was designated as P bktB -Bam + Eco.
- PCR was carried out using the primers shown in SEQ ID NO: 8 and SEQ ID NO: 7 with the DNA generated in the ligation solution as the template DNA.
- PCR was (1) 98 ° C. for 2 minutes, 98 ° C. for 15 seconds, 60 ° C. for 30 seconds, and 68 ° C. for 50 seconds for 25 cycles.
- the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was terminally phosphorylated and digested with BamHI. This DNA fragment was designated as bPac-5P + Bam.
- PCR was performed using the genomic DNA of the KNK-005 strain as a template DNA source and the primers shown in SEQ ID NO: 10 and SEQ ID NO: 11 to obtain a DNA comprising a base sequence downstream from the start codon of the bktB gene. .
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 64 ° C. for 30 seconds and 68 ° C. for 30 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was terminally phosphorylated and digested with ClaI. This DNA fragment was designated ORF-5P + Cla.
- BPac-5P + Bam and ORF-5P + Cla were ligated, and PCR was performed using the primers shown in SEQ ID NO: 8 and SEQ ID NO: 11 using the DNA generated in the ligated solution as a template DNA.
- PCR (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds, and 68 ° C. for 1 minute 30 seconds were repeated 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was co-digested with BamHI and ClaI. This DNA fragment was subcloned into the site digested with the same restriction enzyme of the vector pBluescript II KS (-) (manufactured by TOYOBO).
- the obtained vector was designated as bAO / pBlu.
- the base sequence was determined using a DNA sequencer 3130xl Genetic Analyzer manufactured by APPLIED BIOSSYSTEMS, and it was confirmed that it was the same as the base sequence of the template DNA.
- a pSACKm described in JP 2008-029218 A [0037] was treated with a restriction enzyme NotI to cut out a DNA fragment of about 5.7 kb containing a kanamycin resistance gene and a sacB gene. This was inserted into a site cleaved with the same enzyme of bAO / pBlu to prepare a plasmid bAO / pBlu / SacB-Km for gene disruption / insertion.
- the obtained transformant was mixed and cultured on KNK-005AS strain and Nutrient Agar medium (manufactured by Difco) for conjugation transfer.
- Son agar medium containing 250 mg / L kanamycin (sodium citrate 2 g / L, sodium chloride 5 g / L, magnesium sulfate heptahydrate 0.2 g / L, ammonium dihydrogen phosphate 1 g / L, dihydrogen phosphate 2
- a strain that had grown on potassium 1 g / L, agar 15 g / L, pH 6.8 was selected to obtain a strain in which the plasmid was integrated on the chromosome of the KNK-005AS strain.
- This strain is cultured for 2 generations in Nutrient Broth medium (manufactured by Difco), then diluted and applied to Nutrient Agar medium containing 15% sucrose, and the grown strain is selected to perform the second recombination. The resulting stock was acquired. Furthermore, a desired gene insertion strain was isolated by PCR analysis.
- This gene insertion strain was named Pac-bktB / AS strain, its nucleotide sequence was determined using a DNA sequencer 3130xl Genetic Analyzer, and DNA comprising a phaC promoter and a ribosome binding site immediately before the start codon of the bktB gene. was confirmed to be an inserted strain.
- a DNA (hereinafter referred to as Pre) having a base sequence including a C. necator phbC promoter and a ribosome binding site was prepared as follows.
- PCR was performed using the genomic DNA of the KNK-005 strain as a template and the primers shown in SEQ ID NO: 12 and SEQ ID NO: 13.
- PCR (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 54 ° C. for 30 seconds, and 68 ° C. for 25 seconds were repeated 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was terminally phosphorylated. This DNA fragment was P re -5P.
- the DNA insertion site was set immediately before the start codon of the bktB gene of the chromosomal DNA of the KNK-005AS strain.
- ORF-5P + Cla produced in Example 1 was used as DNA consisting of a base sequence downstream from the start codon of the bktB gene.
- P re -5P and ORF-5P-Cla were ligated, and PCR was performed using the primers shown in SEQ ID NO: 12 and SEQ ID NO: 11 with the DNA produced in the ligated solution as the template DNA.
- PCR (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 54 ° C. for 30 seconds, and 68 ° C. for 50 seconds were repeated 25 cycles.
- the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was terminally phosphorylated and digested with ClaI. This DNA fragment was designated as PRO-5P + Cla.
- PCR was performed using the genomic DNA of the KNK-005 strain as a template DNA source and the primers shown in SEQ ID NO: 8 and SEQ ID NO: 14 to obtain a DNA comprising a base sequence upstream from the start codon of the bktB gene. .
- PCR (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 58 ° C. for 30 seconds, and 68 ° C. for 20 seconds were repeated 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was terminally phosphorylated and digested with BamHI. This DNA fragment was designated as P bktB ⁇ 5P + Bam.
- P bktB -5P + Bam and PRO-5P + Cla were ligated, and PCR was performed using the DNAs generated in the ligation solution as template DNAs and the primers shown in SEQ ID NO: 9 and SEQ ID NO: 11.
- PCR was performed using the DNAs generated in the ligation solution as template DNAs and the primers shown in SEQ ID NO: 9 and SEQ ID NO: 11.
- 98 ° C. for 2 minutes (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds, and 68 ° C. for 1 minute 30 seconds were repeated 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was co-digested with BamHI and ClaI. This DNA fragment was subcloned into a site obtained by digesting the vector pBluescript II KS ( ⁇ ) with the same restriction enzyme. The obtained vector was designated as bRO / pBlu.
- the base sequence
- pSACKm was treated with the restriction enzyme NotI to cut out a DNA fragment of about 5.7 kb containing the kanamycin resistance gene and the sacB gene. This was inserted into a site cleaved with the same enzyme of bRO / pBlu to prepare a gene disruption / insertion plasmid bRO / pBlu / SacB-Km.
- Pre-bktB / AS strain ⁇ Preparation of gene insertion strain Pre-bktB / AS strain>
- a nucleotide sequence including a phbC promoter and a ribosome binding site immediately before the start codon of the bktB gene A strain into which DNA consisting of was inserted was prepared.
- This DNA insertion strain was designated as Pre-bktB / AS strain.
- the nucleotide sequence was determined, and it was confirmed that the strain had a DNA having a nucleotide sequence including a phbC promoter and a ribosome binding site inserted immediately before the start codon of the bktB gene.
- Example 3 Production of BAB3 / AS strain ⁇ Production of plasmid vector for gene insertion> The DNA insertion position was set between the 91st and 92nd bases upstream of the start codon of the bktB gene of the chromosomal DNA of the KNK-005AS strain, that is, immediately after the stop codon of the ORF existing upstream of the bktB gene.
- a DNA comprising a base sequence including a promoter of phaPCJ of A. caviae and a ribosome binding site was prepared as follows.
- PCR was performed using A. caviae genomic DNA as a template and the primers shown in SEQ ID NO: 15 and SEQ ID NO: 16. PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 20 seconds for 25 cycles, and the polymerase used was KOD-plus-. The DNA fragment obtained by PCR was digested with MunI. This DNA fragment was designated as P Ac -Mun / 3.
- DNA consisting of a base sequence upstream from the insertion site was prepared according to the following procedure.
- PCR was performed using the genomic DNA of the KNK-005 strain as a template DNA source and the primers shown in SEQ ID NO: 17 and SEQ ID NO: 18 to obtain DNA consisting of the base sequence of the upstream ORF of the bktB gene.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 64 ° C. for 30 seconds and 68 ° C. for 30 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was digested with the restriction enzyme MunI. This DNA fragment was designated as miaB-Mun / 3.
- P Ac -Mun / 3 and miaB-Mun / 3 were ligated, and PCR was performed using the primers shown in SEQ ID NO: 17 and SEQ ID NO: 16 with the DNA produced in the ligation solution as the template DNA.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 60 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was digested with BamHI and ClaI. This DNA fragment was designated as BAB3-Bam + Cla.
- This DNA fragment was subcloned into a site digested with the same restriction enzyme of the vector pBluescript II KS (-).
- the obtained vector was designated as BAB3 / pBlu.
- the base sequence was determined and confirmed to be the desired base sequence.
- pSACKm was treated with the restriction enzyme NotI to cut out a DNA fragment of about 5.7 kb containing the kanamycin resistance gene and the sacB gene. This was inserted into the site of BAB3 / pBlu cleaved with the same enzyme to prepare a gene BAB3 / pBlu / SacB-Km for gene disruption / insertion.
- the nucleotide sequence was determined, and the strain was confirmed to be a strain in which a DNA comprising a nucleotide sequence including a phaPCJ promoter and a ribosome binding site was inserted between nucleotides 91 and 92 upstream of the start codon of the bktB gene.
- Example 4 Production of BAB4 / AS strain ⁇ Production of plasmid vector for gene insertion> The DNA insertion position was set between the 65th and 66th bases upstream of the start codon of the bktB gene of the chromosomal DNA of the KNK-005AS strain, that is, 26 bases downstream from the stop codon of the ORF existing upstream of the bktB gene.
- a DNA comprising a base sequence including a promoter of phaPCJ of A. caviae and a ribosome binding site was prepared as follows.
- PCR was performed using A. caviae genomic DNA as a template and the primers shown in SEQ ID NO: 15 and SEQ ID NO: 19. PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 20 seconds for 25 cycles, and the polymerase used was KOD-plus-. The DNA fragment obtained by PCR was digested with MunI. This DNA fragment was designated as P Ac -Mun / 4.
- DNA consisting of a base sequence upstream from the insertion site was prepared according to the following procedure.
- PCR was performed using the genomic DNA of the KNK-005 strain as a template DNA source and the primers shown in SEQ ID NO: 17 and SEQ ID NO: 20 to obtain DNA consisting of the base sequence of the upstream ORF of the bktB gene.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 64 ° C. for 30 seconds and 68 ° C. for 30 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was digested with the restriction enzyme MunI. This DNA fragment was designated as miaB-Mun / 4.
- P Ac -Mun / 4 and miaB-Mun / 4 were ligated, and PCR was performed using the primers shown in SEQ ID NO: 17 and SEQ ID NO: 19 with the DNA produced in the ligation solution as the template DNA.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 60 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was digested with BamHI and ClaI. This DNA fragment was designated as BAB4-Bam + Cla.
- This DNA fragment was subcloned into a site digested with the same restriction enzyme of the vector pBluescript II KS (-).
- the obtained vector was designated as BAB4 / pBlu.
- the base sequence was determined and confirmed to be the desired base sequence.
- pSACKm was treated with the restriction enzyme NotI to cut out a DNA fragment of about 5.7 kb containing the kanamycin resistance gene and the sacB gene. This was inserted into the site of BAB4 / pBlu cleaved with the same enzyme to prepare a gene BAB4 / pBlu / SacB-Km for gene disruption / insertion.
- the nucleotide sequence was determined, and the strain was confirmed to be a strain in which a DNA comprising a nucleotide sequence including a phaPCJ promoter and a ribosome binding site was inserted between the 65th and 66th bases upstream of the start codon of the bktB structural gene.
- Example 5 Production of BAB5 / AS strain ⁇ Production of plasmid vector for gene insertion> The DNA insertion position was set between the 58th and 59th bases upstream of the start codon of the bktB gene of the chromosomal DNA of the KNK-005AS strain, that is, 33 bases downstream from the stop codon of the ORF existing upstream of the bktB gene.
- a DNA comprising a base sequence including a promoter of phaPCJ of A. caviae and a ribosome binding site was prepared as follows.
- PCR was performed using A. caviae genomic DNA as a template and the primers shown in SEQ ID NO: 15 and SEQ ID NO: 21. PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 20 seconds for 25 cycles, and the polymerase used was KOD-plus-. The DNA fragment obtained by PCR was digested with MunI. This DNA fragment was designated as P Ac -Mun / 5.
- a DNA comprising a base sequence upstream from the insertion site was prepared by the following procedure.
- PCR was performed using the genomic DNA of the KNK-005 strain as a template DNA source and the primers shown in SEQ ID NO: 17 and SEQ ID NO: 22 to obtain DNA consisting of the base sequence of the upstream ORF of the bktB gene.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 64 ° C. for 30 seconds and 68 ° C. for 30 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was digested with the restriction enzyme MunI. This DNA fragment was designated as miaB-Mun / 5.
- P Ac -Mun / 5 and miaB-Mun / 5 were ligated, and PCR was performed using the primers shown in SEQ ID NO: 17 and SEQ ID NO: 21 with the DNA generated in the ligation solution as the template DNA.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 60 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was digested with BamHI and ClaI. This DNA fragment was designated as BAB5-Bam + Cla.
- This DNA fragment was subcloned into a site digested with the same restriction enzyme of the vector pBluescript II KS (-).
- the obtained vector was designated as BAB5 / pBlu.
- the nucleotide sequence was determined and confirmed to be DNA consisting of the desired nucleotide sequence.
- pSACKm was treated with the restriction enzyme NotI to cut out a DNA fragment of about 5.7 kb containing the kanamycin resistance gene and the sacB gene. This was inserted into a site where BAB5 / pBlu was cleaved with the same enzyme to prepare a gene disruption / insertion plasmid BAB5 / pBlu / SacB-Km.
- the base sequence was determined, and it was confirmed that the strain had a DNA having a base sequence including a phaPCJ promoter and a ribosome binding site inserted between the 58th and 59th bases upstream of the start codon of the bktB gene.
- a DNA consisting of a C. necator phbC promoter and a base sequence containing a ribosome binding site was prepared as follows.
- PCR was performed using C. necator genomic DNA as a template and the primers shown in SEQ ID NO: 23 and SEQ ID NO: 24.
- PCR was (1) 98 ° C. for 2 minutes, (2) 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 20 seconds for 25 cycles, and the polymerase used was KOD-plus-.
- the DNA fragment obtained by PCR was digested with the restriction enzyme MunI. This DNA fragment was designated as P Re -Mun / 5.
- P Re -Mun / 5 and miaB-Mun / 5 prepared in Example 5 were ligated, and PCR was performed using the DNA generated in the ligation solution as a template DNA and the primers shown in SEQ ID NO: 17 and SEQ ID NO: 24 It was.
- PCR was (1) 98 ° C. for 2 minutes, 98 ° C. for 15 seconds, 60 ° C. for 30 seconds and 68 ° C. for 60 seconds for 25 cycles, and KOD-plus- was used as the polymerase.
- the DNA fragment obtained by PCR was digested with BamHI and ClaI. This DNA fragment was designated as BRB5-Bam + Cla.
- This DNA fragment was subcloned into a site digested with the same restriction enzyme of the vector pBluescript II KS (-).
- the obtained vector was named BRB5 / pBlu.
- the nucleotide sequence was determined and confirmed to be DNA consisting of the desired nucleotide sequence.
- pSACKm was treated with the restriction enzyme NotI to cut out a DNA fragment of about 5.7 kb containing the kanamycin resistance gene and the sacB gene. This was inserted into a site where BRB5 / pBlu was cleaved with the same enzyme to prepare a gene disruption / insertion plasmid BRB5 / pBlu / SacB-Km.
- the base sequence was determined, and it was confirmed that the strain had a DNA having a base sequence containing a phbC promoter and a ribosome binding site inserted between the 58th and 59th bases upstream of the start codon of the bktB structural gene.
- Example 7 ⁇ Cloning of ccr gene and expression unit construction>
- the ccr gene encoding an enzyme that converts crotonyl-CoA to butyryl-CoA, a precursor of 3HH monomer, was cloned from the chromosomal DNA of the Streptomyces cinnamonensis Okami strain (DSM 1042).
- PCR was performed using the DNAs described in SEQ ID NO: 25 and SEQ ID NO: 26 as primers.
- the conditions were (1) 98 ° C. for 2 minutes, (2) 94 ° C. for 10 seconds, 55 ° C. for 20 seconds, and 68 ° C. for 90 seconds for 25 cycles.
- the polymerase used was KOD-plus-.
- the fragment amplified by PCR was purified and then cleaved with restriction enzymes BamHI and AflII. Subcloning the EE32d13 fragment (J. Bacteriol., 179, 4821 (1997)) into the EcoRI site of the pUC19 vector, cutting this plasmid with BglII and AflII, and replacing the ccr gene with the BamHI and AflII fragments.
- the ccr expression unit was constructed by
- Example 8 ⁇ Cloning of phaC gene and preparation of expression unit>
- a phaC expression unit containing SEQ ID NO: 4 was prepared as a SpeI fragment.
- PCR was performed using HG :: P Re -N149S / D171G-T / pBlu described in JP-A-2007-228894 [0031] as a template and primers shown in SEQ ID NO: 27 and SEQ ID NO: 28.
- the conditions were (1) 98 ° C. for 2 minutes, (2) 94 ° C. for 10 seconds, 55 ° C. for 30 seconds and 68 ° C. for 2 minutes for 25 cycles, and the polymerase used was KOD-plus-.
- the fragment amplified by PCR was purified and then cleaved with the restriction enzyme SpeI to prepare an expression unit.
- Expression vector pCUP2-631 was constructed as follows. As a plasmid vector for constructing an expression vector in C. necator , pCUP2 described in WO / 2007/049716 [0041] was used. First, the ccr gene expression unit constructed in Example 7 was excised by EcoRI treatment, and this fragment was ligated with pCUP2 cleaved with MunI. Next, the phaC expression unit prepared in Example 8 was prepared as a SpeI fragment and inserted into the SpeI site of pCUP2 containing the ccr gene expression unit to construct a pCUP2-631 vector.
- Example 10 Preparation of transformed cells Introduction of the pCUP2-631 vector into various cells was carried out by electrical introduction as follows.
- the gene introduction apparatus used was a gene pulser manufactured by Biorad, and the cuvette used was a gap 0.2 cm manufactured by Biorad. 400 ⁇ l of competent cells and 20 ⁇ l of expression vector were injected into a cuvette and set in a pulse device, and an electric pulse was applied under the conditions of electrostatic capacity 25 ⁇ F, voltage 1.5 kV, and resistance value 800 ⁇ .
- the bacterial solution in the cuvette was cultured with shaking on Nutrient Broth medium (manufactured by DIFCO) at 30 ° C. for 3 hours, and then on a selection plate (NutrientAgar medium (manufactured by DIFCO), kanamycin 100 mg / L) at 30 ° C.
- the cultured transformants were cultured for a day to obtain transformed transformants.
- Example 11 The produced various transformants were cultured.
- the composition of the pre-medium is 1% (w / v) Meat-extract, 1% (w / v) Bacto-Trypton, 0.2% (w / v) Yeast-extract, 0.9% (w / v) Na
- the pH was adjusted to 6.7 with 2 HPO 4 ⁇ 12H 2 O and 0.15% (w / v) KH 2 PO 4 .
- the composition of the polyester production medium is 1.1% (w / v) Na 2 HPO 4 ⁇ 12H 2 O, 0.19% (w / v) KH 2 PO 4 , 0.6% (w / v) (NH 4 ) 2 SO 4 , 0.1% (w / v) MgSO 4 .7H 2 O, 0.5% (v / v) trace metal salt solution (1.6% (w / v) FeCl in 0.1 N hydrochloric acid) 3 ⁇ 6H 2 O, 1% (w / v) CaCl 2 ⁇ 2H 2 O, 0.02% (w / v) CoCl 2 ⁇ 6H 2 O, 0.016% (w / v) CuSO 4 ⁇ 5H 2 O, 0.012% (w / v) NiCl 2 ⁇ 6H 2 O, and 0.01% (w / v) CrCl 3 ⁇ 6H 2 O. This was carried out by fed-batch culture in which PKOO (palm kernel oil
- the glycerol stock of each transformant was inoculated into the pre-culture medium and cultured for 20 hours, and 10% (v) was added to a 5 L jar fermenter (MD-500 type, manufactured by Maruhishi Bioengine) containing 2.5 L of polyester production medium. / v) Inoculated.
- the operating conditions were a culture temperature of 28 ° C., a stirring speed of 420 rpm, an aeration rate of 0.6 vvm, and a pH controlled between 6.6 and 6.8. For control, 14% aqueous ammonia was used. Incubation was carried out for up to 65 hours. After incubation, the cells were collected by centrifugation, washed with methanol, lyophilized, and analyzed for the 3HH composition ratio.
- 3HH composition analysis of the produced polyester was measured by gas chromatography as follows. To 20 mg of the dried polyester, 2 ml of a sulfuric acid-methanol mixture (15:85) and 2 ml of chloroform were added and sealed, and heated at 100 ° C. for 140 minutes to obtain a methyl ester of a polyester degradation product. After cooling, 1.5 g of sodium bicarbonate was added little by little to neutralize it, and the mixture was allowed to stand until the generation of carbon dioxide gas stopped. After adding 4 ml of diisopropyl ether and mixing well, the mixture was centrifuged and the monomer unit composition of the polyester degradation product in the supernatant was analyzed by capillary gas chromatography.
- the gas chromatograph used was Shimadzu Corporation GC-17A, and the capillary column used was GL Science's NEUTRABOND-1 (column length 25 m, column inner diameter 0.25 mm, liquid film thickness 0.4 ⁇ m). He was used as the carrier gas, the column inlet pressure was 100 kPa, and 1 ⁇ l of the sample was injected. As temperature conditions, the temperature was raised from an initial temperature of 100 to 200 ° C. at a rate of 8 ° C./min, and further from 200 to 290 ° C. at a rate of 30 ° C./min. As a result of analysis under the above conditions, it was a copolymer polyester P (3HB-co-3HH) as shown in chemical formula (1). Table 1 shows the dry cell weight, polymer content, and 3HH ratio.
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Biomedical Technology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
本発明は、ポリヒドロキシアルカノエート(PHA)を生産する微生物であって、(1)phbA遺伝子の発現が抑制されているか、該遺伝子がコードする酵素の触媒活性が抑制されており、(2)bktB遺伝子の発現が強化されているか、該遺伝子がコードする酵素の触媒活性が向上されており、及び(3)ポリヒドロキシアルカノエート合成酵素遺伝子及びクロトニル-CoA還元酵素遺伝子が導入されている微生物に関する。当該微生物を培養することにより、柔軟性にすぐれ適応用途範囲が広いPHAであるP(3HB-co-3HH)を、安価な炭素源を用いて効率よく製造することができる。
Description
本発明は、炭素数4以上の3-ヒロドキシアルカノエート単位を有するポリヒドロキシアルカノエート共重合体を生産する微生物および該微生物を用いて該共重合体を効率よく製造する方法に関する。
ポリヒドロキシアルカノエート(以下、PHAと記す)は、広範な微生物によって生成されるポリエステル型有機分子ポリマーである。PHAは生分解性を有する熱可塑性高分子である。また、PHAは再生可能資源から産生されうる。これらのことから、PHAを環境調和型素材または生体適合型素材として工業的に生産し、多様な産業へ利用する試みが行われている。
現在までに数多くの微生物において、エネルギー貯蔵物質としてポリエステルを菌体内に蓄積することが知られている。その代表例としては3-ヒドロキシ酪酸(以下、3HBと記すこともある)のホモポリマーであるポリ-3-ヒドロキシ酪酸(以下、P(3HB)と記すこともある)が挙げられる。P(3HB)は1925年にBacillusmegateriumで最初に発見された。P(3HB)は熱可塑性高分子であり、自然環境中で生物学的に分解されることから、環境にやさしいプラスチックとして注目されている。しかし、P(3HB)は結晶性が高いために硬くて脆い性質を持っていることから実用的には応用範囲が限られている。応用範囲を広げるためには、P(3HB)に柔軟性を付与することが必要であった。
そこで、3HBと3-ヒドロキシ吉草酸(以下、3HVと記す)とからなる共重合体(以下、P(3HB-co-3HV)と記す)の製造方法が開示されている(特許文献1、特許文献2)。P(3HB-co-3HV)はPHAの一種である。P(3HB-co-3HV)は、P(3HB)に比べると柔軟性に富むため、幅広い用途に応用できると考えられた。しかしながら、実際にはP(3HB-co-3HV)中の3HVモル分率を増加させてもそれに伴う物性の変化が乏しく、特にフィルムやシート、軟質系包装容器等へ加工するために要求される程には柔軟性が向上しないため、シャンプーボトルや使い捨て剃刀の取手等、硬質成型体の限られた分野にしか利用されていない。
P(3HB)の柔軟性を高めるために、3HBと3-ヒドロキシヘキサン酸(以下、3HHと記すこともある)からなる共重合体(以下、P(3HB-co-3HH)と記すこともある)及びその製造方法について研究がなされた(特許文献3、特許文献4)。P(3HB-co-3HH)もPHAの一種である。これら報告におけるP(3HB-co-3HH)の製造方法は、土壌より単離されたAeromonascaviaeの野生株を用い、オレイン酸、パルミチン酸等の脂肪酸を炭素源として発酵生産するものであった。3HH組成比はオレイン酸を炭素源としたとき15mol%、パルミチン酸を炭素源としたとき5mol%であった。
P(3HB-co-3HH)の物性に関する研究もなされている(非特許文献1)。この報告では、炭素数が12個以上の脂肪酸を唯一の炭素源としてA. caviaeを培養し、3HH組成が11~19mol%のP(3HB-co-3HH)を発酵生産している。P(3HB-co-3HH)が3HH組成の増加にしたがって、P(3HB)の様な硬くて脆い性質から次第に柔軟な性質を示すようになり、P(3HB-co-3HV)を上回る柔軟性を示すことが明らかにされた。すなわち、P(3HB-co-3HH)は3HH組成比を変えることで、硬質ポリマーから軟質ポリマーまで応用可能な幅広い物性を持たせることができるため、低3HH組成比のP(3HB-co-3HH)を用いたテレビの筐体等のように硬さを要求されるものから、高3HH組成比のP(3HB-co-3HH)を用いたフィルム等のように柔軟性を要求されるものまで、幅広い分野への応用が期待できる。しかしながら、前記製造方法では菌体生産量4g/L、菌体あたりのポリマー含量30%とポリマー生産性は低いものであったため、実用化に向けて更に高い生産性、特にポリマー含量を向上させる方法が探索されてきた。
P(3HB-co-3HH)の工業生産を目指した取組みの例として、以下の技術が挙げられる。非特許文献2では、Aeromonas hydrophilaを用い、オレイン酸を炭素源とした43時間の流加培養を行うことにより、菌体量95.7g/L、ポリマー含量45.2%、3HH組成比17mol%のP(3HB-co-3HH)が生産された。また、A.hydrophilaを用いたグルコース及びラウリン酸を炭素源とした培養により、3HH組成比が11mol%であり、菌体量50g/L、ポリマー含量50%が達成された(非特許文献3)。しかしながら、A.hydrophilaはヒトに対して病原性を有する(非特許文献4)ことから、工業生産に適した微生物とはいえなかった。また、これらの培養生産で用いられている炭素源は高価であるため、製造コストの観点より安価な炭素源の利用が求められていた。
このため、安全な宿主での生産及び生産性の向上を目指して以下のような取組みが行なわれた。A. caviaeよりクローニングされたポリヒドロキシアルカン酸(PHA)合成酵素遺伝子をCupriavidusnecator (旧分類:Ralstonia eutropha或いはAlcaligenes eutrophus)に導入した形質転換体を用い、オクタン酸を炭素源としてP(3HB-co-3HH)の生産を行った結果、3HH組成比は22mol%であり、菌体量4g/L、ポリマー含量は33重量%であった(特許文献5、非特許文献5)。更に前記形質転換体を、炭素源として植物油脂を用いて培養した結果、3HH組成比は4~5mol%であり、菌体量4g/L、ポリマー含量80%が達成された(非特許文献6)。後者の製造方法は安価な植物油脂を炭素源とし、ポリマー含量も高いものの、菌体量が低いため、ポリマー生産性が低く、且つ3HH組成比4~5mol%ではフィルム等の用途に適用できるほど柔らかいものではなかった。
大腸菌を宿主としたP(3HB-co-3HH)生産株も構築された。Aeromonas属のPHA合成酵素遺伝子、C. necatorのNADP-アセトアセチル-CoA還元酵素遺伝子等を大腸菌に導入した株が構築された。ドデカンを炭素源として該大腸菌を40.8時間培養した結果、菌体量79g/L、ポリマー含量27.2%、3HH組成比10.8mol%であった(非特許文献8)。A.caviaeのPHA合成酵素遺伝子、エノイル-CoAヒドラターゼ遺伝子及びアシル-CoAデヒドロゲナーゼ遺伝子を導入した大腸菌も構築された。ラウリン酸を含む培地で該大腸菌を培養すると、菌体生産性は1g/L、ポリマー含量約16%、3HH組成比約16mol%であった(非特許文献9)。これらの大腸菌でもポリマー生産性は低く、工業的生産への適用は困難であった。
P(3HB-co-3HH)の生産性向上並びに3HH組成制御を目指して、PHA合成酵素の人為的な改変が行なわれた(非特許文献10)。A. caviae由来のPHA合成酵素変異体のなかで、149番目のアミノ酸アスパラギンがセリンに置換された変異体酵素や、171番目のアスパラギン酸がグリシンに置換された変異体酵素は、大腸菌内でのPHA合成酵素活性や3HH組成が向上していることが示され、また、518番目のフェニルアラニンがイソロイシンに置換された変異体酵素や214番目のバリンがグリシンに置換された変異体酵素は大腸菌でのPHA合成酵素活性やポリマー含量が向上したことが報告されている。しかし、これらは宿主として特殊な大腸菌を用いており、更にポリマー含量は約13%と低いことから、これらの変異体酵素の特徴を活かした工業的生産に向けた更なる改良が必要であった。
また、pJRD215(ATCC 37533)にポリエステル合成酵素遺伝子やR-エノイル-CoAヒドラターゼ遺伝子等を導入したpJRDEE32やpJRDEE32d13等(特許文献5、非特許文献5参照)のPHA合成酵素発現プラスミドにによってC.necatorを形質転換し、該形質転換体のPHA生産性が調べられている。該菌株の菌体量は4g/Lと低かったが、植物油脂を炭素源とした同菌株の培養条件改善により菌体量45g/L、ポリマー含量62.5%、3HH組成比8.1mol%にまでポリマー生産性が向上した。このように、培養方法によってP(3HB-co-3HH)の3HH組成比やポリマー生産性を改善する試みがなされた(特許文献6)。
P(3HB-co-3HH)の物性を制御する方法も開示されている(特許文献6)。少なくとも2種類の炭素数の異なる油脂および/または脂肪酸を炭素源として用いることによって、3HH組成比が1~40mol%のポリエステルを生産することが可能となり、種々の物性を有するP(3HB-co-3HH)が生産できるようになった。しかしながら、本製造方法では、3HH組成制御のために比較的高価なヘキサン酸、オクタン酸等を添加する必要があり、また高濃度のヘキサン酸は細胞毒性を示すことから菌体生産性が低下する結果となっている。また、多成分の炭素源を添加するため、生産設備が複雑・高価になる場合がある。
C. necatorを宿主とし、果糖を炭素源とした培養で3HH組成を向上させる方法も開示されている。ポリエステル合成酵素遺伝子と放線菌Streptomycescinnamonensis由来のクロトニル-CoA還元酵素遺伝子(以下ccrと略記する)を導入すると3HH組成は0.9%であったものの、更にR-エノイル-CoAヒドラターゼ遺伝子等を導入すると3HH組成は1.6%に向上した。しかし、その時の菌体量は約1.5g/L、ポリマー含量は40%前後と低く、高3HH組成P(3HB-co-3HH)の工業的生産には一層の改良が必要であった(非特許文献7)。
フィルムやシート、軟質系包装容器等への応用には3HH組成比が12mol%以上のP(3HB-co-3HH)が望まれている。しかし、従来のP(3HB-co-3HH)生産において、3HH組成比を向上させようとすれば、ポリマー含量あるいは菌体量の低下を招いていた。工業生産において望ましいとされる70%以上のポリマー含量且つ150g/L以上の菌体量かつ3HH組成比12mol%以上を実現する方法はなく、一層の改良が必要であった。
Y. Doi, S. Kitamura, H. Abe; Macromolecules, 28, 4822-4823 (1995)
Biotechnology and Bioengineering, vol.67, 240 (2000)
Appl. Microbiol. Biotechnol., vol.57, 50 (2001)
国立感染症研究所、病原体等安全管理規定、別表1付表1(1999)
T. Fukui, Y. Doi; J. Bacteriol, 179, 15, 4821-4830 (1997)
T. Fukui等、Appl. Microbiol. Biotecnol., 49, 333 (1998)
T. Fukui等、Biomolecules, vol.3, 618 (2002)
S. Park等、Biomacromolecules, vol.2, 248 (2001)
X. Lu等、FEMS Microbiology Letters, vol.221, 97 (2003)
T. Kichise等、Appl. Environ. Microbiol., 68, 2411-2419 (2002)
本発明は、柔軟性に優れ幅広い用途が期待される高3HH組成比のPHAを、安価な炭素源である植物油を用いて、実用的な生産性で製造することを目的とする。特には、菌体のポリマー含量70%以上且つ菌体量150g/L以上を保ったまま、3HH組成比12mol%以上のP(3HB-co-3HH)を効率よく発酵生産することを目的とする。
本発明者らは上記課題を解決するために鋭意研究を重ねた結果、微生物中のPHA合成酵素量を増加させるとともに、3HBモノマー合成能を減少させ且つ新たな3HHモノマー合成経路を創出し3HHモノマー供給系を増強させることによって前記微生物中に、3HH組成比率の向上したPHAが大量に生成・蓄積できる事を見出し、本発明を完成するに至った。
即ち、本発明の第一は、phbA遺伝子が不活性化され、phaC遺伝子及びbktB(β-ケトチオラーゼ)遺伝子発現が増強されると共に新たにccr遺伝子導入でブチリル-CoA合成経路が導入されることによって3HHモノマー合成能が向上した3-ヒドロキシヘキサン酸単位含有ポリヒドロキシアルカノエートを生産可能な微生物に関する。
より詳細には、次の(1)から(3)の要件を備えた微生物に関する;
(1)phbA遺伝子の発現が抑制されているか、該遺伝子がコードする酵素の触媒活性が抑制されていること、
(2)bktB遺伝子の発現が強化されているか、該遺伝子がコードする酵素の触媒活性が向上されていること、
(3)ポリヒドロキシアルカノエート合成酵素遺伝子、および/または、クロトニル-CoA還元酵素遺伝子が導入されていること。
(1)phbA遺伝子の発現が抑制されているか、該遺伝子がコードする酵素の触媒活性が抑制されていること、
(2)bktB遺伝子の発現が強化されているか、該遺伝子がコードする酵素の触媒活性が向上されていること、
(3)ポリヒドロキシアルカノエート合成酵素遺伝子、および/または、クロトニル-CoA還元酵素遺伝子が導入されていること。
本発明の第二は、上記記載の微生物を用い、安価な炭素源を用いて菌体のポリマー含量70%以上且つ菌体量150g/L以上を保ったまま、3HH組成比12mol%以上であることを特徴とする3HH単位含有PHAの製造方法に関する。
本発明により、安価で安全な炭素源を用いて、菌体のポリマー含量70%以上且つ菌体量150g/L以上を保ったまま、3HH組成比12mol%以上のPHAを発酵生産することができる。
以下、本発明につき、さらに詳細に説明する。本発明の3HH単位含有PHAとは、3HHとともに、3HB、3HV、3-ヒドロキシオクタン酸、および、それらよりアルキル鎖の長い3-ヒドロキシアルカン酸からなる群より選らばれる1以上の3-ヒドロキシアルカン酸が共重合したポリエステルである。
本発明のphbA遺伝子とは、β-ケトチオラーゼ活性を有する酵素のうち、2分子のアセチル-CoAを縮合する反応を触媒するが、β-ケトバレリル-CoAのチオリシスを殆ど触媒しない酵素をコードする遺伝子で、例えば配列番号5に示される塩基配列からなるphbA遺伝子が例示される。配列番号5に示される塩基配列と85%以上の配列同一性を有し、2分子のアセチル-CoAを縮合する反応を触媒するが、β-ケトバレリル-CoAのチオリシスを殆ど触媒しない酵素をコードする遺伝子は本発明に好適に用いることができる。配列同一性は90%以上であればより好ましく、更に好ましくは95%以上、更により好ましくは98%以上である。
本発明のbktB遺伝子とは、β-ケトチオラーゼ活性を有する酵素のうち、2分子のアセチル-CoAの縮合およびアセチル-CoAとそれよりも鎖長の長い、例えばプロピオニル-CoAの縮合の両方をよく触媒し、且つ3-ケトバレリル-CoAのチオリシスを触媒する酵素をコードする遺伝子で、例えば配列番号29に示される塩基配列からなる遺伝子が例示される(SLATER等、J.Bacteriol., vol.180, 1979-1987, 1998)。配列番号29に示される塩基配列と85%以上の配列同一性を有し、2分子のアセチル-CoAの縮合およびアセチル-CoAとそれよりも鎖長の長い、例えばプロピオニル-CoAの縮合の両方をよく触媒し、且つ3-ケトバレリル-CoAのチオリシスを触媒する酵素をコードする遺伝子は本発明に好適に用いることができる。配列同一性は90%以上であればより好ましく、更に好ましくは95%以上、更により好ましくは98%以上である。
また、3HH単位含有PHAを合成可能なポリメラーゼをコードする遺伝子としては、A. caviaeのphaC遺伝子或いは、配列番号4に示される塩基配列からなるphaC変異遺伝子などが例示でき、宿主が結果として該PHAを産生可能であればよい。phaC遺伝子が有する塩基配列あるいは配列番号4に示される塩基配列と、85%以上の配列同一性を有する遺伝子で、3HH単位含有PHAを合成可能なポリメラーゼをコードする遺伝子は本発明に好適に用いることができる。配列同一性は90%以上であればより好ましく、更に好ましくは95%以上、更により好ましくは98%以上である。
本発明において、bktB構造遺伝子の内在のプロモーターとは、bktB構造遺伝子の転写を誘発するDNAのことであり、宿主として用いるphbA遺伝子とbktB遺伝子を元来有する微生物中に内在する。
本発明におけるphbA遺伝子とbktB遺伝子を元来有する微生物とは、phbA遺伝子とbktB遺伝子を有する野生株、或いは該野生株に遺伝子操作した微生物であれば特に制限はなく、3HH単位含有PHAを合成可能な菌株であれば、それらを宿主として使用することができる。具体的にはラルストニア(Ralstonia)属、カプリアビダス(Cupriavidus)属、ワウテルシア(Wautersia)属、アエロモナス(Aeromonas)属、エシェリキア(Escherichia)属、アルカリゲネス(Alcaligenes)属、シュードモナス(Pseudomonas)属等の細菌類を使用することが好ましい。安全性及び生産性の観点から、より好ましくはラルストニア属、カプリアビダス属、ワウテルシア属に属する細菌であり、特に好ましくはCupriavidusnecatorである。C. necatorとしては、例えば、C. necator H16(ATCC17699)等が挙げられる。当該株はAmerican Type Culture Collection (ATCC)等から入手することができる。勿論、前記微生物を人工的に突然変異処理して得られる変異株、遺伝子工学的手法により変異処理された類縁菌株であっても、3HH単位含有PHAを合成可能な菌株であれば本発明に使用できる。
そして、3HH組成比を高めるためには、phbA遺伝子とbktB遺伝子を元来有する微生物の染色体上に存在するphbA遺伝子が不活性化されていることが好ましい。phbA遺伝子がコードする酵素は、前記のようにアセチル-CoA2分子の縮合によって、PHAモノマーの一つである3-ヒドロキシブチリル-CoAの前駆体であるアセトアセチル-CoAを生成する反応を触媒する。一方、phbA遺伝子がコードする酵素はアセチル-CoAとブタノイル-CoAを縮合する反応を触媒しない。従って、phbA遺伝子の不活性化はポリマー中の3HB比率を低くするため、結果として3HH比率が向上すると考えられる。
phbA遺伝子を不活性化する方法は、結果としてphbAが不活性化されていればよく、例えば、1)phbA遺伝子の開始コドンと終止コドンの間に新たな終止コドンを導入する、2)リボソーム結合部位にリボソーム結合活性が低下する変異を導入する、3)phbA構造遺伝子内部に酵素の触媒活性を低下させる変異を導入する、4)RNA干渉を利用する、5)トランスポゾンを挿入する、6)phbA構造遺伝子の一部或いは全部を欠失させる等の方法が挙げられる。これらの方法は当業者に周知である。
また、PHAの生産性を高めつつ3HH組成比を高めるためには、bktB遺伝子がコードする酵素を増強することが好ましい。bktB遺伝子がコードする酵素は、アセチル-CoA2分子の縮合だけではなく、アセチル-CoAとブタノイル-CoAを縮合する反応も触媒する。アセチル-CoAとブタノイル-CoAが縮合すると、3-ヒドロキシヘキサノイル-CoAの前駆体である3-ケトヘキサノイル-CoAが生じる。bktB遺伝子がコードする酵素の活性は、アセチル-CoA2分子の縮合反応に対してよりもアセチル-CoAとブタノイル-CoAの縮合反応に対しての方が高い。
bktB遺伝子がコードする酵素を増強する方法は特に限定しないが、発現ベクターによる高発現化や発現調節領域の改変による高発現化が可能である。好ましくはphbA遺伝子とbktB遺伝子を元来有する微生物の染色体上にある該遺伝子のプロモーターを改変して転写活性を上げること、例えば、phbA遺伝子とbktB遺伝子を元来有する微生物に内在しているbktB遺伝子の上流に、さらにbktB遺伝子の転写を誘発するプロモーターが組み込まれていると、転写活性が上がることによって高発現化が可能である。
bktB遺伝子の転写を誘発するプロモーターは、bktB遺伝子の開始コドンの上流に挿入されることが好ましい。プロモーターは、bktB遺伝子の転写を誘発する限りはどのようなプロモーターでも本発明に用いうる。該プロモーターはphbA遺伝子とbktB遺伝子を元来有する微生物内在のプロモーター及び異種微生物のプロモーターの少なくとも何れかであることが好ましい。例えば、配列番号1に示される塩基配列からなるC.necatorのphbCABオペロンのプロモーターや、配列番号2に示される塩基配列からなるA. caviaeのphaPCJオペロンのプロモーターは、phbA遺伝子とbktB遺伝子を元来有する微生物としてC.necatorを用いる場合に好適である。
ここで、プロモーターとして用いるDNAは、配列番号1および2に示した塩基配列と70%以上の配列同一性を有することが好ましく、より好ましくは85%以上であり、更に好ましくは90%以上であり、更により好ましくは95%以上であり、最も好ましくは98%以上である。bktB遺伝子の転写を誘発する限りは本発明に用いることができる。
さらに、プロモーターとして用いるDNAとして、配列番号1および2に示した塩基配列と相補的な塩基配列をプローブとして、コロニー・ハイブリダイゼーション法、プラーク・ハイブリダイゼーション法、あるいはサザン・ハイブリダイゼーション法等を行った時に、ストリンジェントな条件でハイブリダイズするDNAを用いることができる。bktB遺伝子の転写を誘発する限りは本発明に用いることができる。
前記ハイブリダイゼーションは、Molecular Cloning, A laboratory manual, second edition(ColdSpring Harbor Laboratory Press, 1989)等に記載されている方法に準じて行うことができる。ここで、ハイブリダイズするDNAとは、例えば、コロニーあるいはプラーク由来のDNAを固定化したフィルターを用いて、0.7~1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、2倍濃度のSSC溶液(1倍濃度のSSC溶液の組成は、150mM塩化ナトリウム、15mMクエン酸ナトリウムよりなる)を用い、65℃の条件下でフィルターを洗浄することにより取得できるDNAをあげることができる。好ましくは65℃で2倍濃度のSSC溶液で洗浄、より好ましくは65℃で0.2倍濃度のSSC溶液で洗浄、更に好ましくは65℃で0.1倍濃度のSSC溶液で洗浄することにより取得できるDNAである。
以上のようにハイブリダイゼーション条件を記載したが、これらの条件に特に制限されない。ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度など複数の要素が考えられ、当業者であればこれら要素を適宜選択することで最適なストリンジェンシーを実現することが可能である。
上記の条件にてハイブリダイズ可能なDNAとしては、配列番号1、或いは2に示されるDNAと、配列同一性が70%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上、最も好ましくは98%以上のDNAをあげることができ、bktB遺伝子の転写を誘発する限り、上記プロモーターに包含される。
更に3HH組成を高める方法として、ccr遺伝子導入で3-ヒドロキシブチリル-CoA合成経路を導入することが有効である。ccr遺伝子がコードするクロトニル-CoA還元酵素は脂肪酸のβ酸化経路の中間体であるクロトニル-CoAを還元し、bktB遺伝子がコードする酵素の基質であるブチリル-CoAを生成する酵素である。この遺伝子を導入することにより、3-ヒドロキシヘキサノイル-CoAはβ酸化経路からの直接的な供給のみでなく、β酸化経路から派生するアセチル-CoAからも供給される。
本発明に用いるccr遺伝子は、クロトニル-CoAを還元し3-ヒドロキシヘキサノイル-CoAを生成する活性を有する酵素をコードする限り本発明に用いることができるが、配列番号3に示した塩基配列と70%以上の配列同一性を有する塩基配列からなる遺伝子であることが好ましく、より好ましくは85%以上であり、更に好ましくは90%以上であり、更により好ましくは95%以上であり、最も好ましくは98%以上である。
さらに、配列番号3に示した塩基配列と相補的な塩基配列をプローブとして、コロニー・ハイブリダイゼーション法、プラーク・ハイブリダイゼーション法、あるいはサザン・ハイブリダイゼーション法等を行った時に、ストリンジェントな条件でハイブリダイズするDNAも、クロトニル-CoAを還元しブチリル-CoAを生成する活性を有する酵素をコードする限り、本発明に用いることができる。
本発明に用いるポリヒドロキシアルカノエート合成酵素遺伝子は、ポリヒドロキシアルカノエート合成活性を有する酵素をコードする限り、本発明に用いることができるが、配列番号4に示した塩基配列と70%以上の配列同一性を有する塩基配列からなる遺伝子であることが好ましく、より好ましくは85%以上であり、更に好ましくは90%以上であり、更により好ましくは95%以上であり、最も好ましくは98%以上である。
さらに、配列番号4に示した塩基配列と相補的な塩基配列をプローブとして、コロニー・ハイブリダイゼーション法、プラーク・ハイブリダイゼーション法、あるいはサザン・ハイブリダイゼーション法等を行った時に、ストリンジェントな条件でハイブリダイズするDNAも、ポリヒドロキシアルカノエート合成活性を有する酵素をコードする限り、本発明に用いることができる。
本発明において、3HH含有PHAの生産に用い得る炭素源としては、例えばグルコース、フルクトース等の糖類や、メタノール、エタノール、ブタノール等のアルコール類、酢酸、プロピオン酸、ヘキサン酸、オクタン酸、デカン酸、ラウリン酸、オレイン酸、パルミチン酸、リノール酸、リノレン酸、ミリスチン酸等の飽和・不飽和脂肪酸などの脂肪酸類、あるいはこれら脂肪酸のエステルや塩等の脂肪酸誘導体、乳酸等の有機酸類、炭素数が10以上である飽和・不飽和脂肪酸を多く含む油脂、例えば植物油脂、中でも、ヤシ油、パーム油、パーム核油、パーム核油オレイン(以下、PKOOとも記載する)、パームダブルオレイン(以下、POOとも記載する)等が挙げられる。培養する際の炭素源の添加量としては、菌株が増殖し、ポリエステルを合成できる範囲であれば良い。
本発明において、宿主に用いるphbA遺伝子とbktB遺伝子を元来有する微生物の野生株が、3-ヒドロキシヘキサノイル-CoAを殆どPHAへと変換させる事ができない場合、3-ヒドロキシヘキサノイル-CoAをPHAへと変換する能力の高い酵素をコードする遺伝子を染色体上へ挿入するか、或いはプラスミドベクターなどを用いて導入する事で、細胞内にて合成された33-ヒドロキシヘキサノイル-CoAをPHAへと変換する効率を向上させる必要がある。例えば、宿主としてC.necatorを使用する場合、3-ヒドロキシヘキサノイル-CoAをPHAへと変換する能力の高い酵素をコードする遺伝子として、A. caviaeのphaC遺伝子或いは配列番号4に示したその変異体遺伝子を導入することが好ましい。
本発明の3HH含有PHAを生産可能な微生物の作製方法は、特に限定するわけではないが、C. necatorを宿主として以下に例示する。A.caviae由来で3-ヒドロキシヘキサノイル-CoAをPHAへと変換する能力の高い酵素をコードするポリエステル合成酵素変異体遺伝子を染色体上に相同組換え法などで元のポリエステル合成酵素遺伝子と置換する。そして遺伝子破壊法を用いて、染色体上に存在するphbA遺伝子を不活性化するが、その態様はphbA遺伝子産物であるタンパク質の活性が低下或いは消失すればどのような方法でも良い。例えば、phbA遺伝子の内部に終止コドンを導入しても良いし、遺伝子上流のプロモーター及び/又はリボソーム結合部位を改変しても良い。更にbktB遺伝子の発現を増強するが、その態様はbktB遺伝子産物の活性が上がればどのような方法でも良い。例えば、より比活性の向上したbktB変異体遺伝子を用いてもよく、bktB遺伝子の開始コドンの上流に異種又は同種のプロモーター及びリボソーム結合部位を含むDNAを挿入してもよいし、bktB遺伝子の本来のプロモーター及び/又はリボソーム結合部位を異種又は同種のプロモーター及び/又はリボソーム結合部位を含む塩基配列からなるDNAに置換する方法でもよい。本来のプロモーターは染色体上からすべて失われていてもよいし、一部が欠失していてもよい。
前記染色体の改変において、染色体上に任意の遺伝子を部位特異的に挿入/置換する方法は当業者に周知であり、代表的な方法としては特に限定するわけではないが以下の方法が挙げられる。トランスポゾンと相同組換えの機構を利用した方法(Ohman等、J.Bacteriol., vol.162, p1068 (1985))や相同組換えの機構によって起こる部位特異的な組み込みと第二段階の相同組換えによる脱落を原理とした方法(Noti等、MethodsEnzymol., vol.154, p197 (1987))などがあり、また、Bacillus subtilis由来のsacB遺伝子を共存させて、第二段階の相同組換えによって遺伝子が脱落した微生物株をシュークロース添加培地耐性株として容易に単離する方法(Schweizer;Mol. Microbiol., vol.6, p1195 (1992)、Lenz等; J. Bacteriol., vol.176, p4385(1994))も利用することができるが、染色体上に任意遺伝子を挿入/置換出来ればその方法は特に制限されない。
例えばC. necatorのbktB遺伝子の開始コドン直前に、A. caviaeのphaC遺伝子のプロモーター及びリボソーム結合部位を含む塩基配列からなるDNAを挿入する場合の方法を、より具体的に例示する。まず、置換フラグメントを作製する。置換フラグメントはbktB遺伝子の開始コドンの直前にA.caviaeのphaC遺伝子(以下、phaCとも記す)のプロモーター及びリボソーム結合部位を含む塩基配列からなるDNAがつながり、その後にbktB遺伝子がつながった形とする。すなわち、該遺伝子の開始コドンの直前にphaCのプロモーター及びリボソーム結合部位を含む塩基配列からなるDNAが挿入されたDNA断片である。phaCのプロモーター及びリボソーム結合部位を含む塩基配列からなるDNAを挟んで存在する上流と下流は染色体上のDNAと相同組換えを起こすために必要な相同配列であって、一般的にはその長さが長いほど組換え頻度は高くなるが、相同組換えさえ起こればよく、その長さは任意に設定できる。
置換断片には、遺伝子置換の際に選択マーカーとなる遺伝子を付加することができる。選択マーカーとなる遺伝子は、例えばカナマイシン、クロラムフェニコール、ストレプトマイシン、アンピシリン等の抗生物質の耐性遺伝子や各種の栄養要求性を相補する遺伝子等が使用できる。C.necatorを宿主とする場合にはカナマイシンの耐性遺伝子が好適である。さらにそれらに加えて、第二段階の相同組換えによって選択マーカー遺伝子を含む領域が脱落した微生物株の選択を容易にするための遺伝子が付加できる。そのような遺伝子としてはBacillussubtilis由来のsacB遺伝子が例示できる。この遺伝子が発現している微生物株はシュークロースを含む培地で生育できないことが知られており、シュークロースを含む培地での生育によりこの遺伝子を脱落によって失った株を選択することが容易となる。
これらで構成された置換フラグメントは、宿主微生物株中で複製しないベクターに接続することによって遺伝子置換用のプラスミドとして作製される。ラルストニア属やシュードモナス属等で利用できるこのようなベクタープラスミドには、例えばpUCベクター、pBluescriptベクター、pBR322ベクター、或いはそれらと同じ複製起点を持つベクター等が挙げられる。さらには、接合伝達を可能にするmob、oriTなどのDNA配列を共存させることも可能である。
このような構成で作製された遺伝子置換用のプラスミドDNAは、エレクトロポレーション法や接合伝達法など公知の方法によりC. necatorに導入することができ、相同組換えを行うことができる。
次に相同組換えによって染色体上に遺伝子置換用のプラスミドDNAが挿入された株の選択を行う。株の選択は、遺伝子置換用プラスミドDNAに共存させた選択用の遺伝子に基づいた方法によって行うことができる。カナマイシン耐性遺伝子を用いた場合には、カナマイシンを含む培地で生育してきた株から選ぶことができる。
次の段階で、第二の相同組換えによって染色体上から選択マーカー遺伝子を含む領域が脱落した株を選択する。挿入時に利用した選択用の遺伝子に基づいて、例えばカナマイシンを含む培地で生育できなくなった株を選択してもよいが、sacB遺伝子を遺伝子置換用プラスミドに共存させている場合は、シュークロースを含む培地で生育してくる株から容易に選択できる。このようにして得られた株が、所望するように遺伝子が置換された株かどうか確認するには、PCR法やサザン・ハイブリダイゼーション法、DNA塩基配列の決定など、公知の方法が使用できる。
以上のようにして、C. necatorの染色体上にあるbktB構造遺伝子の開始コドンの上流にA. caviaeのphaCのプロモーター及びリボソーム結合部位を含む塩基配列からなるDNAが挿入された株を取得することができる。
次に発現ベクターについて説明する。C. necatorで通常用いられる発現ベクターにはpJRD215由来の発現ベクター(例えば特許文献5)やpBBR122由来の発現ベクター(非特許文献7)がある。発現ベクターはその宿主細胞内において、非選択圧条件下でも安定に複製・維持されることが望ましい。従って国際公開公報WO/2007/049716号公報[0041]に記載のpCUP2等がより好適である。pCUP2にはC.necatorと近縁であるカプリアビダス・メタリデュランス(Cupriavidus metallidurans)CH34株が保有するメガプラスミドpMOL28由来のプラスミドの複製や安定維持に必要な領域が含まれている。
このpCUP2にA. caviae由来のphaC遺伝子発現ユニットとS. cinnamonensis由来のccr遺伝子発現ユニットを挿入したプラスミドがpCUP2-631ベクターである。pCUP2-631に更にA.caviae由来のフェイシンをコードするphaP遺伝子及び同由来のR-特異的エノイル-CoAヒドラターゼをコードするphaJ遺伝子を挿入しても良い。また、pCUP2-631に更にC.necator由来のフェイシンをコードするphbP遺伝子及びA. caviae由来のR-特異的エノイル-CoAヒドラターゼをコードするphaJ遺伝子を挿入しても良い。
前述した染色体上のphbA遺伝子を破壊し、且つbktB遺伝子発現を増強した株に、この発現ベクターを導入して得られる菌株としてKNK-631株等の菌株が例示できる。これらの株はいずれも安価で安全な炭素源を用いて、菌体のポリマー含量70%以上且つ菌体量150g/L以上を保ったまま、3HH組成比12mol%以上のPHAを発酵生産することができる。
前述した方法で作製した微生物を用いて3HH含有PHAを製造する方法について説明する。その方法は特に限定するわけではないが、以下のようにして行う事ができる。本発明のPHAの生産においては、炭素源、窒素源、無機塩類、そのほかの有機栄養源を含む培地を用いて、前記微生物を培養することが好ましい。
炭素源としては、糖類、油脂類、脂肪酸類などが好ましく、より好ましくは植物油脂類であり、更に好ましくはパーム油及びパーム核油である。窒素源としては、例えばアンモニア、塩化アンモニウム、硫酸アンモニウム、リン酸アンモニウム等のアンモニウム塩の他、ペプトン、肉エキス、酵母エキス等が挙げられる。無機塩類としては、例えばリン酸2水素カリウム、リン酸水素2ナトリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム等が挙げられる。
そのほかの有機栄養源としては、例えばグリシン、アラニン、セリン、スレオニン、プロリン等のアミノ酸、ビタミンB1、ビタミンB12、ビタミンC等のビタミン等が挙げられる。また、培養液中に、発現プラスミドに存在する薬剤耐性遺伝子に対応する抗生物質(カナマイシン等)を添加しても良い。
本発明において、菌体からの3HH含有PHAの回収は、特に限定はないが、例えば次のような方法により行うことができる。培養終了後、培養液から遠心分離機等で菌体を分離し、その菌体を蒸留水およびメタノール等により洗浄し、乾燥させる。この乾燥菌体から、クロロホルム等の有機溶剤を用いて3HH含有PHAを抽出する。この3HH含有PHAを含んだ有機溶剤溶液から、濾過等によって菌体成分を除去し、そのろ液にメタノールやヘキサン等の貧溶媒を加えて3HH含有PHAを沈殿させる。さらに、濾過や遠心分離によって上澄み液を除去し、乾燥させて3HH含有PHAを回収する。
得られた3HH含有PHAの重量平均分子量(Mw)や3HH組成(mol%)の分析は、例えば、ガスクロマトグラフ法や核磁気共鳴法等により行うことができる。
本発明で得られる柔軟性の高いPHAは、フィルムやシート、軟質系包装容器などに好適に用いられる。
以下に実施例を示し、本発明をより具体的に説明するが、本発明はこれらの実施例に何ら限定されるものではない。なお一般的な遺伝子操作は、MolecularCloning(Cold Spring Harbor Laboratory Press (1989))に記載されているように行うことができる。また、遺伝子操作に使用する酵素、クローニング宿主等は、市場の供給者から購入し、その取扱説明書に従い使用することができる。なお、酵素は、遺伝子操作に使用できるものであれば特に限定されない。
(実施例1)
<遺伝子挿入用プラスミドベクターの作製>
挿入用DNAとしてA. caviaeのphaCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAを次のように作製した。A.caviaeのゲノムDNAをテンプレートとして配列番号6および配列番号7で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で20秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-(TOYOBO製)を用いた。PCRで得たDNA断片を末端リン酸化およびEcoRI消化した。このDNA断片をPAc-5P+Ecoとした。
<遺伝子挿入用プラスミドベクターの作製>
挿入用DNAとしてA. caviaeのphaCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAを次のように作製した。A.caviaeのゲノムDNAをテンプレートとして配列番号6および配列番号7で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で20秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-(TOYOBO製)を用いた。PCRで得たDNA断片を末端リン酸化およびEcoRI消化した。このDNA断片をPAc-5P+Ecoとした。
次に、特開2008-029218号公報[0038]に記載のKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン直前をDNA挿入部位と設定し、以下の手順で該遺伝子の開始コドンより上流側の塩基配列からなるDNAを作製した。
特開2008-029218号公報[0036]に記載のKNK-005株のゲノムDNAを鋳型DNAの供給源として、配列番号8および配列番号9で示されるプライマーを用いてPCRを行い、bktB遺伝子の開始コドンより上流の塩基配列からなるDNAを得た。PCRは(1)98℃で2分、(2)98℃で15秒、64℃で30秒、68℃で30秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を制限酵素BamHIおよびEcoRIで2酵素同時消化した。このDNA断片をPbktB-Bam+Ecoとした。
PAc-5P+EcoおよびPbktB-Bam+Ecoをライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号8および配列番号7で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、98℃で15秒、60℃で30秒、68℃で50秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を末端リン酸化およびBamHI消化した。このDNA断片をbPac-5P+Bamとした。
次に、該遺伝子の開始コドンより下流側の塩基配列からなるDNAを作製した。KNK-005株のゲノムDNAを鋳型DNAの供給源として、配列番号10および配列番号11で示されるプライマーを用いてPCRを行い、bktB遺伝子の開始コドンより下流側の塩基配列からなるDNAを得た。PCRは(1)98℃で2分、(2)98℃で15秒、64℃で30秒、68℃で30秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を末端リン酸化およびClaI消化した。このDNA断片をORF-5P+Claとした。
bPac-5P+BamとORF-5P+Claをライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号8および配列番号11で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で1分30秒、を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をBamHIおよびClaIで2酵素同時消化した。このDNA断片を、ベクターpBluescript II KS(-)(TOYOBO製)の同制限酵素で消化した部位にサブクローニングした。得られたベクターをbAO/pBluとした。APPLIED BIOSYSTEMS社製のDNAシークエンサー3130xl Genetic Analyzerを用いて塩基配列を決定し、鋳型としたDNAの塩基配列と同一であることを確認した。
続いて、特開2008-029218号公報[0037]に記載のpSACKmを制限酵素NotIで処理することによってカナマイシン耐性遺伝子およびsacB遺伝子を含む約5.7kbのDNA断片を切り出した。これを、bAO/pBluの同酵素で切断した部位に挿入して遺伝子破壊・挿入用プラスミドbAO/pBlu/SacB-Kmを作製した。
<遺伝子挿入株Pac-bktB/AS株の作製>
次に、KNK-005AS株を親株としてbAO/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドン直前にphaCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。遺伝子挿入用プラスミドbAO/pBlu/SacB-Kmで大腸菌S17-1株(ATCC47005)を形質転換した。得られた形質転換体をKNK-005AS株とNutrient Agar培地(Difco社製)上で混合培養して接合伝達を行った。250mg/Lのカナマイシンを含むシモンズ寒天培地(クエン酸ナトリウム2g/L、塩化ナトリウム5g/L、硫酸マグネシウム・7水和物0.2g/L、リン酸二水素アンモニウム1g/L、リン酸水素二カリウム1g/L、寒天15g/L、pH6.8)上で生育してきた菌株を選択して、プラスミドがKNK-005AS株の染色体上に組み込まれた株を取得した。この株をNutrient Broth培地(Difco社製)で2世代培養した後、15%のシュークロースを含むNutrient Agar培地上に希釈して塗布し、生育してきた菌株を選択して2回目の組換えが生じた株を取得した。さらにPCRによる解析により所望の遺伝子挿入株を単離した。
次に、KNK-005AS株を親株としてbAO/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドン直前にphaCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。遺伝子挿入用プラスミドbAO/pBlu/SacB-Kmで大腸菌S17-1株(ATCC47005)を形質転換した。得られた形質転換体をKNK-005AS株とNutrient Agar培地(Difco社製)上で混合培養して接合伝達を行った。250mg/Lのカナマイシンを含むシモンズ寒天培地(クエン酸ナトリウム2g/L、塩化ナトリウム5g/L、硫酸マグネシウム・7水和物0.2g/L、リン酸二水素アンモニウム1g/L、リン酸水素二カリウム1g/L、寒天15g/L、pH6.8)上で生育してきた菌株を選択して、プラスミドがKNK-005AS株の染色体上に組み込まれた株を取得した。この株をNutrient Broth培地(Difco社製)で2世代培養した後、15%のシュークロースを含むNutrient Agar培地上に希釈して塗布し、生育してきた菌株を選択して2回目の組換えが生じた株を取得した。さらにPCRによる解析により所望の遺伝子挿入株を単離した。
この遺伝子挿入株をPac-bktB/AS株と命名し、DNAシークエンサー3130xl Genetic Analyzerを用いて塩基配列を決定し、bktB遺伝子の開始コドン直前にphaCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
(実施例2) Pre-bktB/AS株の作製
<遺伝子挿入用プラスミドベクターの作製>
挿入用DNAとしてC. necatorのphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNA(以下Preとする)を次のように作製した。KNK-005株のゲノムDNAをテンプレートとして配列番号12および配列番号13で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、54℃で30秒、68℃で25秒、を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を末端リン酸化した。このDNA断片をPre-5Pとした。
<遺伝子挿入用プラスミドベクターの作製>
挿入用DNAとしてC. necatorのphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNA(以下Preとする)を次のように作製した。KNK-005株のゲノムDNAをテンプレートとして配列番号12および配列番号13で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、54℃で30秒、68℃で25秒、を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を末端リン酸化した。このDNA断片をPre-5Pとした。
次に、KNK-005AS株の染色体DNAのbktB遺伝子の開始コドン直前をDNA挿入部位と設定した。
bktB遺伝子の開始コドンより下流側の塩基配列からなるDNAとして実施例1で作製したORF-5P+Claを用いた。
Pre-5PおよびORF-5P-Claをライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号12および配列番号11で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、54℃で30秒、68℃で50秒、を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を末端リン酸化およびClaI消化した。このDNA断片をPRO-5P+Claとした。
次に、該遺伝子の開始コドンより上流側の配列を作製した。KNK-005株のゲノムDNAを鋳型DNAの供給源として、配列番号8および配列番号14で示されるプライマーを用いてPCRを行い、bktB遺伝子の開始コドンより上流側の塩基配列からなるDNAを得た。PCRは(1)98℃で2分、(2)98℃で15秒、58℃で30秒、68℃で20秒、を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を末端リン酸化およびBamHI消化した。このDNA断片をPbktB-5P+Bamとした。
PbktB-5P+BamとPRO-5P+Claをライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号9および配列番号11で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で1分30秒、を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をBamHIおよびClaIで2酵素同時消化した。このDNA断片を、ベクターpBluescript II KS(-)を同制限酵素で消化した部位に、サブクローニングした。得られたベクターをbRO/pBluとした。塩基配列を決定し、鋳型としたDNAの塩基配列と同一であることを確認した。
続いて、pSACKmを制限酵素NotIで処理することによってカナマイシン耐性遺伝子およびsacB遺伝子を含む約5.7kbのDNA断片を切り出した。これを、bRO/pBluの同酵素で切断した部位に挿入して遺伝子破壊・挿入用プラスミドbRO/pBlu/SacB-Kmを作製した。
<遺伝子挿入株Pre-bktB/AS株の作製>
実施例1の遺伝子置換株の作製方法と同様にして、KNK-005AS株を親株としてbRO/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドン直前にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をPre-bktB/AS株と命名した。塩基配列を決定し、bktB遺伝子の開始コドン直前にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
実施例1の遺伝子置換株の作製方法と同様にして、KNK-005AS株を親株としてbRO/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドン直前にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をPre-bktB/AS株と命名した。塩基配列を決定し、bktB遺伝子の開始コドン直前にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
(実施例3) BAB3/AS株の作製
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側91塩基目と92塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドン直後と設定した。
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側91塩基目と92塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドン直後と設定した。
挿入用DNAとして、A. caviaeのphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAを次のように作製した。
A. caviaeのゲノムDNAをテンプレートとして配列番号15および配列番号16で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で20秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をMunI消化した。このDNA断片をPAc-Mun/3とした。
また以下の手順で挿入部位より上流側の塩基配列からなるDNAを作製した。
KNK-005株のゲノムDNAを鋳型DNAの供給源として、配列番号17および配列番号18で示されるプライマーを用いてPCRを行い、bktB遺伝子の上流側ORFの塩基配列からなるDNAを得た。PCRは(1)98℃で2分、(2)98℃で15秒、64℃で30秒、68℃で30秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を制限酵素MunIで消化した。このDNA断片をmiaB-Mun/3とした。
PAc-Mun/3およびmiaB-Mun/3をライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号17および配列番号16で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で60秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をBamHIおよびClaIで消化した。このDNA断片をBAB3-Bam+Claとした。
このDNA断片を、ベクターpBluescript II KS(-)の同制限酵素で消化した部位にサブクローニングした。得られたベクターをBAB3/pBluとした。塩基配列を決定し、所望の塩基配列であることを確認した。
続いて、pSACKmを制限酵素NotIで処理することによってカナマイシン耐性遺伝子およびsacB遺伝子を含む約5.7kbのDNA断片を切り出した。これを、BAB3/pBluの同酵素で切断した部位に挿入して遺伝子破壊・挿入用プラスミドBAB3/pBlu/SacB-Kmを作製した。
<遺伝子挿入株BAB3/AS株の作製>
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBAB3/pBlu/SacB-Kmを用いてbktB構造遺伝子の開始コドンの上流側にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBAB3/AS株と命名した。塩基配列を決定し、bktB遺伝子の開始コドン上流側91塩基目と92塩基目の間にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBAB3/pBlu/SacB-Kmを用いてbktB構造遺伝子の開始コドンの上流側にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBAB3/AS株と命名した。塩基配列を決定し、bktB遺伝子の開始コドン上流側91塩基目と92塩基目の間にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
(実施例4) BAB4/AS株の作製
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側65塩基目と66塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドンから26塩基下流と設定した。
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側65塩基目と66塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドンから26塩基下流と設定した。
挿入用DNAとして、A. caviaeのphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAを次のように作製した。
A. caviaeのゲノムDNAをテンプレートとして配列番号15および配列番号19で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で20秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をMunI消化した。このDNA断片をPAc-Mun/4とした。
また以下の手順で挿入部位より上流側の塩基配列からなるDNAを作製した。
KNK-005株のゲノムDNAを鋳型DNAの供給源として、配列番号17および配列番号20で示されるプライマーを用いてPCRを行い、bktB遺伝子の上流側ORFの塩基配列からなるDNAを得た。PCRは(1)98℃で2分、(2)98℃で15秒、64℃で30秒、68℃で30秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を制限酵素MunIで消化した。このDNA断片をmiaB-Mun/4とした。
PAc-Mun/4およびmiaB-Mun/4をライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号17および配列番号19で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で60秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をBamHIおよびClaIで消化した。このDNA断片をBAB4-Bam+Claとした。
このDNA断片を、ベクターpBluescript II KS(-)の同制限酵素で消化した部位にサブクローニングした。得られたベクターをBAB4/pBluとした。塩基配列を決定し、所望の塩基配列であることを確認した。
続いて、pSACKmを制限酵素NotIで処理することによってカナマイシン耐性遺伝子およびsacB遺伝子を含む約5.7kbのDNA断片を切り出した。これを、BAB4/pBluの同酵素で切断した部位に挿入して遺伝子破壊・挿入用プラスミドBAB4/pBlu/SacB-Kmを作製した。
<遺伝子挿入株BAB4/AS株の作製>
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBAB4/pBlu/SacB-Kmを用いてbktB構造遺伝子の開始コドンの上流側にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBAB4/AS株と命名した。塩基配列を決定し、bktB構造遺伝子の開始コドン上流側65塩基目と66塩基目の間にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBAB4/pBlu/SacB-Kmを用いてbktB構造遺伝子の開始コドンの上流側にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBAB4/AS株と命名した。塩基配列を決定し、bktB構造遺伝子の開始コドン上流側65塩基目と66塩基目の間にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
(実施例5) BAB5/AS株の作製
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側58塩基目と59塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドンから33塩基下流と設定した。
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側58塩基目と59塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドンから33塩基下流と設定した。
挿入用DNAとして、A. caviaeのphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAを次のように作製した。
A. caviaeのゲノムDNAをテンプレートとして配列番号15および配列番号21で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で20秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をMunI消化した。このDNA断片をPAc-Mun/5とした。
また以下の手順で挿入部位より上流側の塩基配列からなるDNAを作製した。
KNK-005株のゲノムDNAを鋳型DNAの供給源として、配列番号17および配列番号22で示されるプライマーを用いてPCRを行い、bktB遺伝子の上流側ORFの塩基配列からなるDNAを得た。PCRは(1)98℃で2分、(2)98℃で15秒、64℃で30秒、68℃で30秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を制限酵素MunIで消化した。このDNA断片をmiaB-Mun/5とした。
KNK-005株のゲノムDNAを鋳型DNAの供給源として、配列番号17および配列番号22で示されるプライマーを用いてPCRを行い、bktB遺伝子の上流側ORFの塩基配列からなるDNAを得た。PCRは(1)98℃で2分、(2)98℃で15秒、64℃で30秒、68℃で30秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を制限酵素MunIで消化した。このDNA断片をmiaB-Mun/5とした。
PAc-Mun/5およびmiaB-Mun/5をライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号17および配列番号21で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で60秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をBamHIおよびClaIで消化した。このDNA断片をBAB5-Bam+Claとした。
このDNA断片を、ベクターpBluescript II KS(-)の同制限酵素で消化した部位にサブクローニングした。得られたベクターをBAB5/pBluとした。塩基配列を決定し、所望の塩基配列からなるDNAであることを確認した。
続いて、pSACKmを制限酵素NotIで処理することによってカナマイシン耐性遺伝子およびsacB遺伝子を含む約5.7kbのDNA断片を切り出した。これを、BAB5/pBluを同酵素で切断した部位に挿入して遺伝子破壊・挿入用プラスミドBAB5/pBlu/SacB-Kmを作製した。
<遺伝子挿入株BAB5/AS株の作製>
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBAB5/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドンの上流側にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBAB5/AS株と命名した。塩基配列を決定し、bktB遺伝子の開始コドン上流側58塩基目と59塩基目の間にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBAB5/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドンの上流側にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBAB5/AS株と命名した。塩基配列を決定し、bktB遺伝子の開始コドン上流側58塩基目と59塩基目の間にphaPCJのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
(実施例6) BRB5/AS株の作製
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側58塩基目と59塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドンから33塩基下流と設定した。
<遺伝子挿入用プラスミドベクターの作製>
DNA挿入位置をKNK-005AS株の染色体DNAのbktB遺伝子の開始コドン上流側58塩基目と59塩基目の間、即ちbktB遺伝子の上流に存在するORFの停止コドンから33塩基下流と設定した。
挿入用DNAとしてC. necatorのphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAを次のように作製した。
C. necatorのゲノムDNAをテンプレートとして配列番号23および配列番号24で示されるプライマーを用いて、PCRを行った。PCRは(1)98℃で2分、(2)98℃で15秒、60℃で30秒、68℃で20秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片を制限酵素MunIで消化した。このDNA断片をPRe-Mun/5とした。
PRe-Mun/5および実施例5で調製したmiaB-Mun/5をライゲーションし、ライゲート液中に生成したDNAを鋳型DNAとして配列番号17および配列番号24で示されるプライマーを用いてPCRを行った。PCRは(1)98℃で2分、98℃で15秒、60℃で30秒、68℃で60秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで得たDNA断片をBamHIおよびClaIで消化した。このDNA断片をBRB5-Bam+Claとした。
このDNA断片を、ベクターpBluescript II KS(-)の同制限酵素で消化した部位にサブクローニングした。得られたベクターをBRB5/pBluとした。塩基配列を決定し、所望の塩基配列からなるDNAであることを確認した。
続いて、pSACKmを制限酵素NotIで処理することによってカナマイシン耐性遺伝子およびsacB遺伝子を含む約5.7kbのDNA断片を切り出した。これを、BRB5/pBluを同酵素で切断した部位に挿入して遺伝子破壊・挿入用プラスミドBRB5/pBlu/SacB-Kmを作製した。
<遺伝子挿入株BRB5/AS株の作製>
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBRB5/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドンの上流側にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBRB5/AS株と命名した。塩基配列を決定し、bktB構造遺伝子の開始コドン上流側58塩基目と59塩基目の間にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
実施例1の遺伝子挿入株の作製方法と同様にして、KNK-005AS株を親株としてBRB5/pBlu/SacB-Kmを用いてbktB遺伝子の開始コドンの上流側にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された菌株を作製した。このDNA挿入株をBRB5/AS株と命名した。塩基配列を決定し、bktB構造遺伝子の開始コドン上流側58塩基目と59塩基目の間にphbCのプロモーターおよびリボソーム結合部位を含む塩基配列からなるDNAが挿入された株であることを確認した。
(実施例7)
<ccr遺伝子のクローニング及び発現ユニット構築>
クロトニル-CoAを、3HHモノマーの前駆体であるブチリル-CoAに変換する酵素をコードするccr遺伝子を、ストレプトマイセス・シンナモネンシス(Streptomycescinnamonensis)Okami株(DSM 1042)の染色体DNAからクローニングした。配列番号25及び配列番号26記載のDNAをプライマーとしPCRを行った。その条件は(1)98℃で2分、(2)94℃で10秒、55℃で20秒、68℃で90秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで増幅した断片を精製後、制限酵素BamHI及びAflIIで切断した。EE32d13断片(J.Bacteriol., 179, 4821 (1997))を、pUC19ベクターのEcoRI部位にサブクローニングし、このプラスミドをBglIIとAflIIで切断し、BamHI及びAflII断片としたccr遺伝子と断片を置換することによってccr発現ユニットを構築した。
<ccr遺伝子のクローニング及び発現ユニット構築>
クロトニル-CoAを、3HHモノマーの前駆体であるブチリル-CoAに変換する酵素をコードするccr遺伝子を、ストレプトマイセス・シンナモネンシス(Streptomycescinnamonensis)Okami株(DSM 1042)の染色体DNAからクローニングした。配列番号25及び配列番号26記載のDNAをプライマーとしPCRを行った。その条件は(1)98℃で2分、(2)94℃で10秒、55℃で20秒、68℃で90秒を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで増幅した断片を精製後、制限酵素BamHI及びAflIIで切断した。EE32d13断片(J.Bacteriol., 179, 4821 (1997))を、pUC19ベクターのEcoRI部位にサブクローニングし、このプラスミドをBglIIとAflIIで切断し、BamHI及びAflII断片としたccr遺伝子と断片を置換することによってccr発現ユニットを構築した。
(実施例8)
<phaC遺伝子のクローニング及び発現ユニット調製>
配列番号4を含むphaC発現ユニットをSpeI断片として調製した。特開2007-228894号公報[0031]に記載のHG::PRe-N149S/D171G-T/pBluを鋳型とし、配列番号27と配列番号28に示すプライマーとしてPCRを行った。その条件は(1)98℃で2分、(2)94℃で10秒、55℃で30秒、68℃で2分を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで増幅した断片を精製後、制限酵素SpeIで切断して発現ユニットを調製した。
<phaC遺伝子のクローニング及び発現ユニット調製>
配列番号4を含むphaC発現ユニットをSpeI断片として調製した。特開2007-228894号公報[0031]に記載のHG::PRe-N149S/D171G-T/pBluを鋳型とし、配列番号27と配列番号28に示すプライマーとしてPCRを行った。その条件は(1)98℃で2分、(2)94℃で10秒、55℃で30秒、68℃で2分を25サイクル繰り返し、ポリメラーゼはKOD-plus-を用いた。PCRで増幅した断片を精製後、制限酵素SpeIで切断して発現ユニットを調製した。
(実施例9)
<発現ベクターの構築>
発現ベクターpCUP2-631は以下のようにして構築した。
C. necatorにおける発現ベクター構築用のプラスミドベクターとしては、国際公開公報WO/2007/049716号公報[0041]に記載のpCUP2を用いた。まず実施例7で構築したccr遺伝子発現ユニットをEcoRI処理により切り出し、この断片をMunIで切断したpCUP2と連結した。次に実施例8で作製したphaC発現ユニットをSpeI断片として調製し、ccr遺伝子発現ユニットを含むpCUP2のSpeI部位に挿入してpCUP2-631ベクターを構築した。
<発現ベクターの構築>
発現ベクターpCUP2-631は以下のようにして構築した。
C. necatorにおける発現ベクター構築用のプラスミドベクターとしては、国際公開公報WO/2007/049716号公報[0041]に記載のpCUP2を用いた。まず実施例7で構築したccr遺伝子発現ユニットをEcoRI処理により切り出し、この断片をMunIで切断したpCUP2と連結した。次に実施例8で作製したphaC発現ユニットをSpeI断片として調製し、ccr遺伝子発現ユニットを含むpCUP2のSpeI部位に挿入してpCUP2-631ベクターを構築した。
(実施例10)形質転換細胞の作製
pCUP2-631ベクターの種々の細胞への導入は以下のように電気導入によって行った。遺伝子導入装置はBiorad社製のジーンパルサーを用い、キュベットは同じくBiorad社製のgap0.2cmを用いた。キュベットに、コンピテント細胞400μlと発現ベクター20μlを注入してパルス装置にセットし、静電容量25μF、電圧1.5kV、抵抗値800Ωの条件で電気パルスをかけた。パルス後、キュベット内の菌液をNutrientBroth培地(DIFCO社製)で30℃、3時間振とう培養し、選択プレート(NutrientAgar培地(DIFCO社製)、カナマイシン100mg/L)で、30℃にて2日間培養して、生育してきた形質転換体を取得した。
pCUP2-631ベクターの種々の細胞への導入は以下のように電気導入によって行った。遺伝子導入装置はBiorad社製のジーンパルサーを用い、キュベットは同じくBiorad社製のgap0.2cmを用いた。キュベットに、コンピテント細胞400μlと発現ベクター20μlを注入してパルス装置にセットし、静電容量25μF、電圧1.5kV、抵抗値800Ωの条件で電気パルスをかけた。パルス後、キュベット内の菌液をNutrientBroth培地(DIFCO社製)で30℃、3時間振とう培養し、選択プレート(NutrientAgar培地(DIFCO社製)、カナマイシン100mg/L)で、30℃にて2日間培養して、生育してきた形質転換体を取得した。
(実施例11)
作製した種々の形質転換体の培養を行った。前培地の組成は1%(w/v)Meat-extract、1%(w/v)Bacto-Trypton、0.2%(w/v)Yeast-extract、0.9%(w/v)Na2HPO4・12H2O、および、0.15%(w/v)KH2PO4で、pH6.7に調整した。
作製した種々の形質転換体の培養を行った。前培地の組成は1%(w/v)Meat-extract、1%(w/v)Bacto-Trypton、0.2%(w/v)Yeast-extract、0.9%(w/v)Na2HPO4・12H2O、および、0.15%(w/v)KH2PO4で、pH6.7に調整した。
ポリエステル生産培地の組成は1.1%(w/v)Na2HPO4・12H2O、0.19%(w/v)KH2PO4、0.6%(w/v)(NH4)2SO4、0.1%(w/v)MgSO4・7H2O、0.5%(v/v)微量金属塩溶液(0.1N塩酸に1.6%(w/v)FeCl3・6H2O、1%(w/v)CaCl2・2H2O、0.02%(w/v)CoCl2・6H2O、0.016%(w/v)CuSO4・5H2O、0.012%(w/v)NiCl2・6H2O、0.01%(w/v)CrCl3・6H2Oを溶かしたもの。)とした。炭素源としてPKOO(パーム核油オレイン画分)を流加する流加培養にて行った。
それぞれの形質転換体のグリセロールストックを前培地に接種して20時間培養し、2.5Lのポリエステル生産培地を入れた5Lジャーファーメンター(丸菱バイオエンジ製MD-500型)に10%(v/v)接種した。運転条件は、培養温度28℃、攪拌速度420rpm、通気量0.6vvmとし、pHは6.6から6.8の間でコントロールした。コントロールには14%のアンモニア水を使用した。培養は65時間まで行った。培養後遠心分離によって菌体を回収し、メタノールで洗浄後、凍結乾燥し、3HH組成比率を分析した。
生産されたポリエステルの3HH組成分析は以下のようにガスクロマトグラフィーによって測定した。乾燥ポリエステルの約20mgに2mlの硫酸-メタノール混液(15:85)と2mlのクロロホルムを添加して密栓し、100℃で140分間加熱して、ポリエステル分解物のメチルエステルを得た。冷却後、これに1.5gの炭酸水素ナトリウムを少しずつ加えて中和し、炭酸ガスの発生がとまるまで放置した。4mlのジイソプロピルエーテルを添加してよく混合した後、遠心して、上清中のポリエステル分解物のモノマーユニット組成をキャピラリーガスクロマトグラフィーにより分析した。ガスクロマトグラフは島津製作所GC-17A、キャピラリーカラムはGLサイエンス社製NEUTRABOND-1(カラム長25m、カラム内径0.25mm、液膜厚0.4μm)を用いた。キャリアガスとしてHeを用い、カラム入口圧100kPaとし、サンプルは1μlを注入した。温度条件は、初発温度100から200℃まで8℃/分の速度で昇温、さらに200から290℃まで30℃/分の速度で昇温した。上記条件にて分析した結果、化学式(1)に示すような共重合体ポリエステルP(3HB-co-3HH)であった。乾燥菌体重量、ポリマー含量、3HH比率を表1に示した。
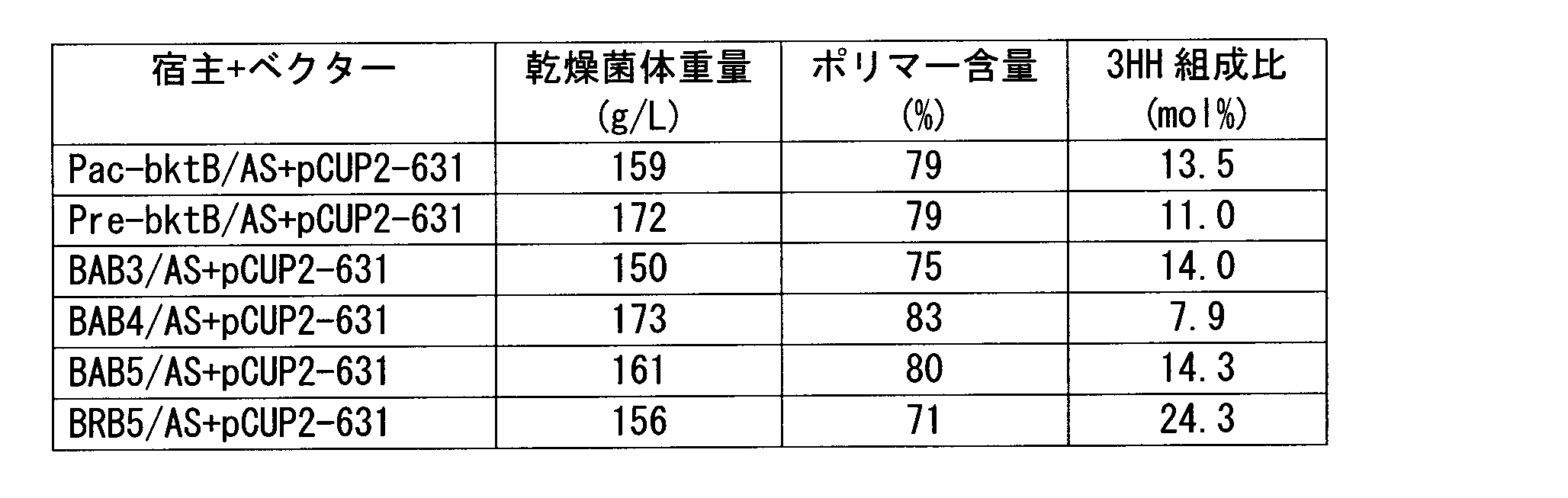
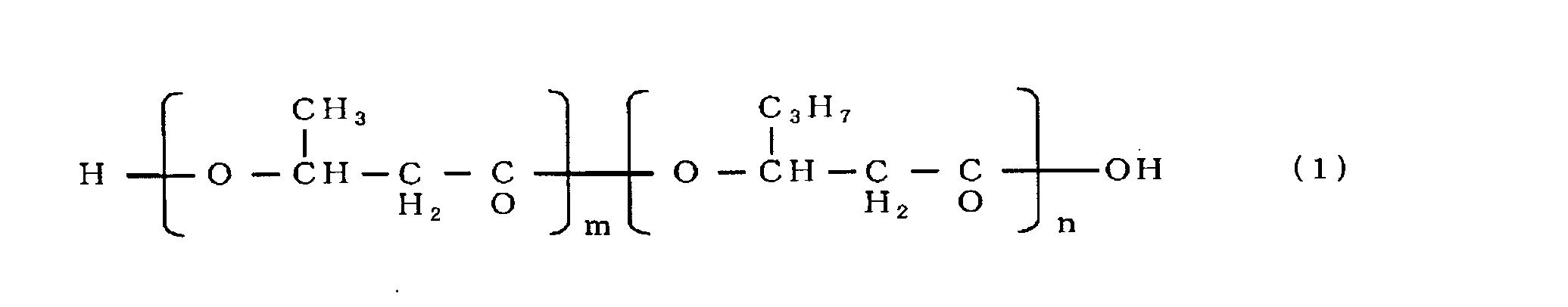
(式中、m、nは1以上の整数を表す)
Claims (12)
- 次の(1)から(3)の要件を備えた微生物;
(1)phbA遺伝子の発現が抑制されているか、該遺伝子がコードする酵素の触媒活性が抑制されていること、
(2)bktB遺伝子の発現が強化されているか、該遺伝子がコードする酵素の触媒活性が向上されていること、
(3)ポリヒドロキシアルカノエート合成酵素遺伝子、および/または、クロトニル-CoA還元酵素遺伝子が導入されていること。 - 次の(1)から(6)の少なくとも一つにより、phbA遺伝子の発現が抑制されているか、該遺伝子がコードする酵素の触媒活性が抑制されている、請求項1記載の微生物;
(1)phbA遺伝子の開始コドンと終止コドンの間に、新たな終止コドンが導入されている、
(2)phbA遺伝子のリボソーム結合部位に、リボソーム結合活性が低下する変異を導入している、
(3)phbA遺伝子内部に、phbA遺伝子がコードする酵素の触媒活性が低下する変異を導入している、
(4)RNA干渉を利用する、
(5)phbA遺伝子内部に、トランスポゾンを挿入している、
(6)phbA遺伝子の一部或いは全部を欠失している。 - bktB遺伝子の上流に、新たにbktB遺伝子の転写を誘発する活性を有するプロモーターが挿入されることにより、bktB遺伝子の発現が強化されたている、請求項1記載の微生物。
- 新たに挿入されるプロモーターが、次の(1)または(2)のDNAである、請求項3記載の微生物;
(1)bktB遺伝子の上流に、配列番号1または2に示される塩基配列からなるDNAと70%以上の配列同一性を有し、bktB遺伝子の転写を誘発する活性を有するDNA、
(2)bktB遺伝子の上流に、配列番号1または2に示される塩基配列からなるDNAと相補的なDNAとストリンジェントな条件でハイブリダイズし、bktB遺伝子の転写を誘発する活性を有するDNA。 - 次の(1)から(4)の少なくとも一つの、ポリヒドロキシアルカノエート合成酵素遺伝子、および/または、クロトニル-CoA還元酵素遺伝子が導入されている、請求項1記載の微生物;
(1)配列番号3に示される塩基配列からなるDNAと70%以上の配列同一性を有し、クロトニル-CoAを還元し、ブチリル-CoAを生成する活性を有する酵素をコードする遺伝子、
(2)配列番号3に示される塩基配列からなるDNAと相補的なDNAとストリンジェントな条件でハイブリダイズし、クロトニル-CoAを還元し、ブチリル-CoAを生成する活性を有する酵素をコードする遺伝子、
(3)配列番号4に示される塩基配列からなるDNAと70%以上の配列同一性を有し、ポリヒドロキシアルカノエート合成活性を有する酵素をコードする遺伝子、
(4)配列番号4に示される塩基配列からなるDNAと相補的なDNAとストリンジェントな条件でハイブリダイズし、ポリヒドロキシアルカノエート合成活性を有する酵素をコードする遺伝子。 - 植物油脂を炭素源として乾燥菌体量150g/L以上でポリマー含量70%以上、且つ3-ヒドロキシヘキサン酸組成比12mol%以上のポリヒドロキシアルカノエートを生産可能な微生物。
- 遺伝子組換えにより作製した請求項5記載の微生物。
- 植物油脂を炭素源として乾燥菌体量150g/L以上でポリマー含量70%以上、且つ3-ヒドロキシヘキサン酸組成比12mol%以上のポリヒドロキシアルカノエートを生産可能な、請求項1~5の何れかに記載の微生物。
- ポリヒドロキシアルカノエートが3-ヒドロキシ酪酸と3-ヒドロキシヘキサン酸を構成単位に含むポリエステルである請求項1~8の何れかに記載の微生物。
- 微生物がCupriavidus necator由来である請求項1~9の何れかに記載の微生物。
- 微生物がCupriavidus necator H16株由来である請求項10記載の微生物。
- 請求項1~11の何れかに記載の微生物を、植物油脂を炭素源として培養することを特徴とする3-ヒドロキシヘキサン酸単位含有ポリヒドロキシアルカノエートの製造方法。
Priority Applications (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP09754672.5A EP2295536B1 (en) | 2008-05-26 | 2009-05-26 | Microorganism capable of producing improved polyhydroxyalkanoate and method of producing polyhydroxyalkanoate by using the same |
| JP2010514477A JP5670728B2 (ja) | 2008-05-26 | 2009-05-26 | 改良されたポリヒドロキシアルカノエート生産微生物及びそれを用いたポリヒドロキシアルカノエートの製造方法 |
| US12/994,378 US20110091948A1 (en) | 2008-05-26 | 2009-05-26 | Microorganism capable of producing improved polyhydroxyalkanoate and method of producing polyhydroxyalkanoate by using the same |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008-136356 | 2008-05-26 | ||
| JP2008136356 | 2008-05-26 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2009145164A1 true WO2009145164A1 (ja) | 2009-12-03 |
Family
ID=41377037
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2009/059564 WO2009145164A1 (ja) | 2008-05-26 | 2009-05-26 | 改良されたポリヒドロキシアルカノエート生産微生物及びそれを用いたポリヒドロキシアルカノエートの製造方法 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US20110091948A1 (ja) |
| EP (1) | EP2295536B1 (ja) |
| JP (1) | JP5670728B2 (ja) |
| WO (1) | WO2009145164A1 (ja) |
Cited By (24)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2013009628A (ja) * | 2011-06-29 | 2013-01-17 | Kaneka Corp | ポリヒドロキシアルカノエートの製造方法 |
| US20130071892A1 (en) * | 2010-02-26 | 2013-03-21 | Tokyo Institute Of Technology | PROCESS FOR PRODUCTION OF POLYHYDROXYALKANOIC ACID USING GENETICALLY MODIFIED MICROORGANISM HAVING ENOYL-CoA HYDRATASE GENE INTRODUCED THEREIN |
| WO2013147139A1 (ja) | 2012-03-30 | 2013-10-03 | 株式会社カネカ | 生分解性ポリエステル樹脂組成物 |
| WO2014002417A1 (ja) | 2012-06-29 | 2014-01-03 | 株式会社カネカ | ポリ-3-ヒドロキシアルカノエート系樹脂組成物および成形体 |
| WO2014020838A1 (ja) | 2012-08-03 | 2014-02-06 | 株式会社カネカ | ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2014042076A1 (ja) | 2012-09-14 | 2014-03-20 | 国立大学法人東京工業大学 | 改質油脂組成物を用いたポリヒドロキシアルカノエートの製造方法 |
| WO2014054278A1 (ja) | 2012-10-05 | 2014-04-10 | 株式会社カネカ | ポリエステル樹脂組成物およびその製造方法 |
| WO2014061243A1 (ja) | 2012-10-15 | 2014-04-24 | 株式会社カネカ | ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2014068943A1 (ja) | 2012-10-29 | 2014-05-08 | 株式会社カネカ | 脂肪族ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2014188646A1 (ja) | 2013-05-20 | 2014-11-27 | 株式会社カネカ | ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2015020105A1 (ja) | 2013-08-06 | 2015-02-12 | 株式会社カネカ | 軟質熱可塑性樹脂組成物 |
| WO2015020104A1 (ja) | 2013-08-06 | 2015-02-12 | 株式会社カネカ | 軟質熱可塑性樹脂組成物 |
| WO2015052876A1 (ja) | 2013-10-11 | 2015-04-16 | 株式会社カネカ | 脂肪族ポリエステル樹脂組成物および脂肪族ポリエステル樹脂成形体 |
| WO2015098001A1 (ja) | 2013-12-24 | 2015-07-02 | 株式会社カネカ | 脂肪族ポリエステル樹脂組成物および脂肪族ポリエステル樹脂成形体 |
| WO2015114719A1 (ja) | 2014-01-30 | 2015-08-06 | 株式会社カネカ | ポリエステル樹脂組成物の製造方法およびポリエステル樹脂成形体 |
| WO2015133468A1 (ja) * | 2014-03-04 | 2015-09-11 | 国立大学法人東京工業大学 | ポリエステル顆粒結合タンパク質遺伝子座を改変した組換え株による共重合ポリエステルの製造法 |
| WO2015146195A1 (ja) * | 2014-03-28 | 2015-10-01 | 株式会社カネカ | Pha合成酵素をコードする遺伝子を複数有する微生物、およびそれを用いたphaの製造方法 |
| WO2017122679A1 (ja) | 2016-01-12 | 2017-07-20 | 国立大学法人東京工業大学 | 生分解性脂肪族ポリエステル系繊維および製造方法 |
| WO2018186278A1 (ja) | 2017-04-05 | 2018-10-11 | 株式会社カネカ | ポリヒドロキシアルカノエート粒子及びその水分散液 |
| WO2019142845A1 (ja) * | 2018-01-17 | 2019-07-25 | 株式会社カネカ | 高組成比率の3hhモノマー単位を含む共重合phaを生産する形質転換微生物およびそれによるphaの製造方法 |
| WO2020218565A1 (ja) * | 2019-04-26 | 2020-10-29 | 株式会社フューエンス | ポリヒドロキシアルカン酸及びその製造方法 |
| WO2023189102A1 (ja) | 2022-03-30 | 2023-10-05 | 株式会社カネカ | 脂肪族ポリエステル系発泡粒子の製造方法 |
| WO2023188942A1 (ja) | 2022-03-30 | 2023-10-05 | 株式会社カネカ | ポリ(3-ヒドロキシアルカノエート)系樹脂発泡粒子 |
| EP4134434A4 (en) * | 2020-04-10 | 2024-06-26 | Kaneka Corporation | METHOD FOR MANUFACTURING A COPOLYMER MIXTURE OF POLYHYDROXYALKANOIC ACID, AND TRANSFORMED MICROORGANISM |
Families Citing this family (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2010057022A1 (en) * | 2008-11-14 | 2010-05-20 | Genomatica, Inc. | Microorganisms for the production of methyl ethyl ketone and 2-butanol |
| US20160040197A1 (en) * | 2013-02-28 | 2016-02-11 | Tokyo Institute Of Technology | PRODUCTION METHOD FOR COPOLYMER POLYHYDROXYALKANOATE USING GENETICALLY MODIFIED STRAIN OF FATTY ACID ß-OXIDATION PATHWAY |
| EP3004225A1 (en) | 2013-05-30 | 2016-04-13 | Metabolix, Inc. | Recyclate blends |
| JP6663221B2 (ja) * | 2014-01-31 | 2020-03-11 | 株式会社カネカ | R体特異的エノイル−CoAヒドラターゼ遺伝子の発現が調節された微生物及びそれを用いたポリヒドロキシアルカノエート共重合体の製造方法 |
| CN106574279A (zh) * | 2014-08-04 | 2017-04-19 | 国立大学法人东京工业大学 | 自糖质原料制造共聚合聚羟基链烷酸的方法 |
| EP3741858A4 (en) * | 2018-01-16 | 2021-12-22 | Kaneka Corporation | MUTANT POLYHYDROXYALCANIC ACID SYNTHASE, GENE AND TRANSFORMER THEREOF, AND METHOD FOR THE PRODUCTION OF POLYHYDROXYALCANIC ACID |
Citations (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS57150393A (en) | 1980-11-18 | 1982-09-17 | Ici Ltd | Beta-hydroxy butyrate polymer and production thereof |
| JPS59220192A (ja) | 1983-01-18 | 1984-12-11 | インペリアル・ケミカル・インダストリ−ズ・ピ−エルシ− | ベ−タ・ヒドロキシブチレ−ト重合体の製造方法 |
| JPH0593049A (ja) | 1991-09-17 | 1993-04-16 | Kanegafuchi Chem Ind Co Ltd | 共重合体およびその製造方法 |
| JPH07265065A (ja) | 1994-03-29 | 1995-10-17 | Kanegafuchi Chem Ind Co Ltd | 共重合体の合成遺伝子による形質転換体および共重合体の製造方法 |
| JPH10108682A (ja) | 1996-08-14 | 1998-04-28 | Rikagaku Kenkyusho | ポリエステル重合酵素遺伝子及びポリエステルの製造方法 |
| JP2001340078A (ja) | 2000-06-01 | 2001-12-11 | Kanegafuchi Chem Ind Co Ltd | ポリエステルの製造方法 |
| WO2007049716A1 (ja) | 2005-10-27 | 2007-05-03 | Kaneka Corporation | 新規プラスミドベクター及びプラスミドを安定に保持する形質転換体 |
| JP2007228894A (ja) | 2006-03-01 | 2007-09-13 | Kaneka Corp | 遺伝子組み換え微生物及びそれを用いたポリエステルの製造方法 |
| JP2008029218A (ja) | 2006-07-26 | 2008-02-14 | Kaneka Corp | 酵素活性を低下させた微生物を用いる共重合ポリエステルの製造方法 |
| JP2008086238A (ja) * | 2006-09-29 | 2008-04-17 | Kaneka Corp | ポリヒドロキシアルカノエートの製造方法 |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPWO2008090873A1 (ja) * | 2007-01-24 | 2010-05-20 | 株式会社カネカ | ポリヒドロキシアルカノエートの製造方法 |
-
2009
- 2009-05-26 WO PCT/JP2009/059564 patent/WO2009145164A1/ja active Application Filing
- 2009-05-26 EP EP09754672.5A patent/EP2295536B1/en active Active
- 2009-05-26 JP JP2010514477A patent/JP5670728B2/ja active Active
- 2009-05-26 US US12/994,378 patent/US20110091948A1/en not_active Abandoned
Patent Citations (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS57150393A (en) | 1980-11-18 | 1982-09-17 | Ici Ltd | Beta-hydroxy butyrate polymer and production thereof |
| JPS59220192A (ja) | 1983-01-18 | 1984-12-11 | インペリアル・ケミカル・インダストリ−ズ・ピ−エルシ− | ベ−タ・ヒドロキシブチレ−ト重合体の製造方法 |
| JPH0593049A (ja) | 1991-09-17 | 1993-04-16 | Kanegafuchi Chem Ind Co Ltd | 共重合体およびその製造方法 |
| JPH07265065A (ja) | 1994-03-29 | 1995-10-17 | Kanegafuchi Chem Ind Co Ltd | 共重合体の合成遺伝子による形質転換体および共重合体の製造方法 |
| JPH10108682A (ja) | 1996-08-14 | 1998-04-28 | Rikagaku Kenkyusho | ポリエステル重合酵素遺伝子及びポリエステルの製造方法 |
| JP2001340078A (ja) | 2000-06-01 | 2001-12-11 | Kanegafuchi Chem Ind Co Ltd | ポリエステルの製造方法 |
| WO2007049716A1 (ja) | 2005-10-27 | 2007-05-03 | Kaneka Corporation | 新規プラスミドベクター及びプラスミドを安定に保持する形質転換体 |
| JP2007228894A (ja) | 2006-03-01 | 2007-09-13 | Kaneka Corp | 遺伝子組み換え微生物及びそれを用いたポリエステルの製造方法 |
| JP2008029218A (ja) | 2006-07-26 | 2008-02-14 | Kaneka Corp | 酵素活性を低下させた微生物を用いる共重合ポリエステルの製造方法 |
| JP2008086238A (ja) * | 2006-09-29 | 2008-04-17 | Kaneka Corp | ポリヒドロキシアルカノエートの製造方法 |
Non-Patent Citations (21)
| Title |
|---|
| "Molecular Cloning", 1989, COLD SPRING HARBOR LABORATORY PRESS |
| "Molecular Cloning, A laboratory manual", 1989, COLD SPRING HARBOR LABORATORY PRESS |
| APPL. MICROBIOL. BIOTECHNOL., vol. 57, 2001, pages 50 |
| BIOTECHNOLOGY AND BIOENGINEERING, vol. 67, 2000, pages 240 |
| FUKUI T. ET AL.: "Efficient production of polyhydroxyalkanoates from plant oils by Alcaligenes eutrophus and its recombinant strain.", APPL. MIROBIOL. BIOTECHNOL., vol. 49, 1998, pages 333 - 336, XP002979648 * |
| FUKUI TOSHIAKI ET AL.: "Cloning and Analysis of the Poly(3-Hydroxybutyrate-co-3- Hydroxyhexanoate) Biosynthesis Genes of Aeromonas caviae.", JOURNAL OF BACTERIOLOGY, vol. 179, no. 15, 1997, pages 4821 - 4830, XP002113257 * |
| FUKUI TOSHIAKI ET AL.: "Engineering of Ralstonia eutropha for Production of Poly(3- hydroxybutyrate-co-3-hydroxyhexanoate) from Fructose and Solid-State Properties of the Copolymer.", BIOMACROMOLECULES, vol. 3, 2002, pages 618 - 624, XP002538853 * |
| LENZ ET AL., J. BACTERIOL., vol. 176, 1994, pages 4385 |
| NOTI ET AL., METHODS ENZYMOL., vol. 154, 1987, pages 197 |
| OHMAN ET AL., J. BACTERIOL., vol. 162, 1985, pages 1068 |
| S. PARK ET AL., BIOMACROMOLECULES, vol. 2, 2001, pages 248 |
| SCHWEIZER, MOL. MICROBIOL., vol. 6, 1992, pages LL95 |
| See also references of EP2295536A4 |
| SLATER ET AL., J. BACTERIOL., vol. 180, 1998, pages 1979 - 1987 |
| SLATER STEVEN ET AL.: "Multiple beta-Ketothiolases Mediate Poly(beta-Hydroxyalkanoate) Copolymer Synthesis in Ralstonia eutropha.", JOURNAL OF BACTERIOLOGY, vol. 180, no. 8, 1998, pages 1979 - 1987, XP008138775 * |
| T. FUKUI ET AL., APPL. MICROBIOL. BIOTECNOL., vol. 49, 1998, pages 333 |
| T. FUKUI ET AL., BIOMOLECULES, vol. 3, 2002, pages 618 |
| T. FUKUIY. DOI, J. BACTERIOL, vol. 179, no. 15, 1997, pages 4821 - 4830 |
| T. KICHISE ET AL., APPL. ENVIRON. MICROBIOL., vol. 68, 2002, pages 2411 - 2419 |
| X. LU ET AL., FEMS MICROBIOLOGY LETTERS, vol. 221, 2003, pages 97 |
| Y. DOIS. KITAMURAH. ABE, MACROMOLECULES, vol. 28, 1995, pages 4822 - 4823 |
Cited By (38)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20130071892A1 (en) * | 2010-02-26 | 2013-03-21 | Tokyo Institute Of Technology | PROCESS FOR PRODUCTION OF POLYHYDROXYALKANOIC ACID USING GENETICALLY MODIFIED MICROORGANISM HAVING ENOYL-CoA HYDRATASE GENE INTRODUCED THEREIN |
| JP2013009628A (ja) * | 2011-06-29 | 2013-01-17 | Kaneka Corp | ポリヒドロキシアルカノエートの製造方法 |
| WO2013147139A1 (ja) | 2012-03-30 | 2013-10-03 | 株式会社カネカ | 生分解性ポリエステル樹脂組成物 |
| WO2014002417A1 (ja) | 2012-06-29 | 2014-01-03 | 株式会社カネカ | ポリ-3-ヒドロキシアルカノエート系樹脂組成物および成形体 |
| WO2014020838A1 (ja) | 2012-08-03 | 2014-02-06 | 株式会社カネカ | ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2014042076A1 (ja) | 2012-09-14 | 2014-03-20 | 国立大学法人東京工業大学 | 改質油脂組成物を用いたポリヒドロキシアルカノエートの製造方法 |
| JPWO2014042076A1 (ja) * | 2012-09-14 | 2016-08-18 | 国立大学法人東京工業大学 | 改質油脂組成物を用いたポリヒドロキシアルカノエートの製造方法 |
| WO2014054278A1 (ja) | 2012-10-05 | 2014-04-10 | 株式会社カネカ | ポリエステル樹脂組成物およびその製造方法 |
| WO2014061243A1 (ja) | 2012-10-15 | 2014-04-24 | 株式会社カネカ | ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2014068943A1 (ja) | 2012-10-29 | 2014-05-08 | 株式会社カネカ | 脂肪族ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2014188646A1 (ja) | 2013-05-20 | 2014-11-27 | 株式会社カネカ | ポリエステル樹脂組成物および該樹脂組成物を含む成形体 |
| WO2015020105A1 (ja) | 2013-08-06 | 2015-02-12 | 株式会社カネカ | 軟質熱可塑性樹脂組成物 |
| WO2015020104A1 (ja) | 2013-08-06 | 2015-02-12 | 株式会社カネカ | 軟質熱可塑性樹脂組成物 |
| WO2015052876A1 (ja) | 2013-10-11 | 2015-04-16 | 株式会社カネカ | 脂肪族ポリエステル樹脂組成物および脂肪族ポリエステル樹脂成形体 |
| WO2015098001A1 (ja) | 2013-12-24 | 2015-07-02 | 株式会社カネカ | 脂肪族ポリエステル樹脂組成物および脂肪族ポリエステル樹脂成形体 |
| WO2015114719A1 (ja) | 2014-01-30 | 2015-08-06 | 株式会社カネカ | ポリエステル樹脂組成物の製造方法およびポリエステル樹脂成形体 |
| WO2015133468A1 (ja) * | 2014-03-04 | 2015-09-11 | 国立大学法人東京工業大学 | ポリエステル顆粒結合タンパク質遺伝子座を改変した組換え株による共重合ポリエステルの製造法 |
| JPWO2015146195A1 (ja) * | 2014-03-28 | 2017-04-13 | 株式会社カネカ | Pha合成酵素をコードする遺伝子を複数有する微生物、およびそれを用いたphaの製造方法 |
| JP2020058391A (ja) * | 2014-03-28 | 2020-04-16 | 株式会社カネカ | Pha合成酵素をコードする遺伝子を複数有する微生物、およびそれを用いたphaの製造方法 |
| US11225676B2 (en) | 2014-03-28 | 2022-01-18 | Kaneka Corporation | Microorganism having multiple genes encoding PHA synthase and method for producing PHA using same |
| JP6994057B2 (ja) | 2014-03-28 | 2022-01-14 | 株式会社カネカ | Pha合成酵素をコードする遺伝子を複数有する微生物、およびそれを用いたphaの製造方法 |
| US10519473B2 (en) | 2014-03-28 | 2019-12-31 | Kaneka Corporation | Microorganism having multiple genes encoding PHA synthase and method for producing PHA using same |
| WO2015146195A1 (ja) * | 2014-03-28 | 2015-10-01 | 株式会社カネカ | Pha合成酵素をコードする遺伝子を複数有する微生物、およびそれを用いたphaの製造方法 |
| WO2017122679A1 (ja) | 2016-01-12 | 2017-07-20 | 国立大学法人東京工業大学 | 生分解性脂肪族ポリエステル系繊維および製造方法 |
| JPWO2018186278A1 (ja) * | 2017-04-05 | 2020-02-13 | 株式会社カネカ | ポリヒドロキシアルカノエート粒子及びその水分散液 |
| WO2018186278A1 (ja) | 2017-04-05 | 2018-10-11 | 株式会社カネカ | ポリヒドロキシアルカノエート粒子及びその水分散液 |
| JP7123909B2 (ja) | 2017-04-05 | 2022-08-23 | 株式会社カネカ | ポリヒドロキシアルカノエート粒子及びその水分散液 |
| JP7270556B2 (ja) | 2018-01-17 | 2023-05-10 | 株式会社カネカ | 高組成比率の3hhモノマー単位を含む共重合phaを生産する形質転換微生物およびそれによるphaの製造方法 |
| WO2019142845A1 (ja) * | 2018-01-17 | 2019-07-25 | 株式会社カネカ | 高組成比率の3hhモノマー単位を含む共重合phaを生産する形質転換微生物およびそれによるphaの製造方法 |
| JPWO2019142845A1 (ja) * | 2018-01-17 | 2021-01-07 | 株式会社カネカ | 高組成比率の3hhモノマー単位を含む共重合phaを生産する形質転換微生物およびそれによるphaの製造方法 |
| US11453896B2 (en) | 2018-01-17 | 2022-09-27 | Kaneka Corporation | Transformed microorganism for producing PHA copolymer comprising 3HH monomer unit at high composition rate and method for producing PHA using same |
| JPWO2020218565A1 (ja) * | 2019-04-26 | 2021-09-27 | 株式会社 フューエンス | ポリヒドロキシアルカン酸及びその製造方法 |
| JP7063513B2 (ja) | 2019-04-26 | 2022-05-09 | 株式会社 フューエンス | ポリヒドロキシアルカン酸及びその製造方法 |
| JP2022093402A (ja) * | 2019-04-26 | 2022-06-23 | 株式会社 フューエンス | ポリヒドロキシアルカン酸及びその製造方法 |
| WO2020218565A1 (ja) * | 2019-04-26 | 2020-10-29 | 株式会社フューエンス | ポリヒドロキシアルカン酸及びその製造方法 |
| EP4134434A4 (en) * | 2020-04-10 | 2024-06-26 | Kaneka Corporation | METHOD FOR MANUFACTURING A COPOLYMER MIXTURE OF POLYHYDROXYALKANOIC ACID, AND TRANSFORMED MICROORGANISM |
| WO2023189102A1 (ja) | 2022-03-30 | 2023-10-05 | 株式会社カネカ | 脂肪族ポリエステル系発泡粒子の製造方法 |
| WO2023188942A1 (ja) | 2022-03-30 | 2023-10-05 | 株式会社カネカ | ポリ(3-ヒドロキシアルカノエート)系樹脂発泡粒子 |
Also Published As
| Publication number | Publication date |
|---|---|
| EP2295536A4 (en) | 2013-07-24 |
| EP2295536B1 (en) | 2016-01-06 |
| JPWO2009145164A1 (ja) | 2011-10-13 |
| US20110091948A1 (en) | 2011-04-21 |
| EP2295536A1 (en) | 2011-03-16 |
| JP5670728B2 (ja) | 2015-02-18 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5670728B2 (ja) | 改良されたポリヒドロキシアルカノエート生産微生物及びそれを用いたポリヒドロキシアルカノエートの製造方法 | |
| CN111615555B (zh) | 生产包含高组成比率的3hh单体单元的共聚pha的转化微生物、以及基于该转化微生物的pha的制造方法 | |
| JP7001596B2 (ja) | 3hh単位含有共重合phaを生産する形質転換体、及び当該phaの製造方法 | |
| JP6223386B2 (ja) | ポリ(5hv)および5炭素化合物を製造するための環境に優しい方法および組成物 | |
| Insomphun et al. | Modification of β-oxidation pathway in Ralstonia eutropha for production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from soybean oil | |
| JP2007259708A (ja) | 新規生分解性ポリエステルの製造方法 | |
| WO2015115619A1 (ja) | R体特異的エノイル-CoAヒドラターゼ遺伝子の発現が調節された微生物及びそれを用いたポリヒドロキシアルカノエート共重合体の製造方法 | |
| EP2048224A1 (en) | Microorganism with replaced gene and process for producing polyester using the same | |
| JPWO2005098001A1 (ja) | 新規形質転換体 | |
| KR101273599B1 (ko) | 2-하이드록시부티레이트를 모노머로 함유하고 있는 폴리하이드록시알카노에이트의 제조방법 | |
| Kutralam-Muniasamy et al. | Genome characteristics dictate poly-R-(3)-hydroxyalkanoate production in Cupriavidus necator H16 | |
| EP2910641B1 (en) | High molecular weight pha-producing microbe and method of producing high molecular weight pha using same | |
| JP4960033B2 (ja) | 酵素活性を低下させた微生物を用いる共重合ポリエステルの製造方法 | |
| EP2818561A1 (en) | Utilization of the novel, environmental isolates Pseudomonas sp. IPB-B26 and N-128 for the efficient high yield production of mcl/lcl-PHAs | |
| JP2008086238A (ja) | ポリヒドロキシアルカノエートの製造方法 | |
| JPWO2008090873A1 (ja) | ポリヒドロキシアルカノエートの製造方法 | |
| WO2017104722A1 (ja) | スクロース資化性を有するpha生産微生物、及び該微生物を用いたphaの製造方法 | |
| US20090130731A1 (en) | Microorganism capable of accumulating ultra high molecular weight polyester | |
| Lu et al. | Molecular cloning of polyhydroxyalkanoate synthesis operon from Aeromonashydrophila and its expression in Escherichia coli | |
| WO2014133088A1 (ja) | 脂肪酸β-酸化経路改変株による共重合体ポリヒドロキシアルカン酸の製造法 | |
| JP5839854B2 (ja) | 微生物の培養方法 | |
| JP2009225775A (ja) | ポリヒドロキシアルカン酸の製造方法 | |
| JP2005333933A (ja) | 新規発現プラスミド | |
| WO2024166830A1 (ja) | 形質転換微生物、及びポリヒドロキシアルカン酸の製造方法 | |
| CN116640708A (zh) | 一种可调控聚羟基脂肪酸酯中乳酸组分含量的基因工程菌及其构建方法和应用 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 09754672 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2010514477 Country of ref document: JP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2009754672 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 12994378 Country of ref document: US |