WO2005007841A1 - 変異型セリンアセチルトランスフェラーゼ及びl−システインの製造法 - Google Patents
変異型セリンアセチルトランスフェラーゼ及びl−システインの製造法 Download PDFInfo
- Publication number
- WO2005007841A1 WO2005007841A1 PCT/JP2004/010216 JP2004010216W WO2005007841A1 WO 2005007841 A1 WO2005007841 A1 WO 2005007841A1 JP 2004010216 W JP2004010216 W JP 2004010216W WO 2005007841 A1 WO2005007841 A1 WO 2005007841A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- sat
- mutant
- amino acid
- cysteine
- serine acetyltransferase
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/12—Methionine; Cysteine; Cystine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1025—Acyltransferases (2.3)
- C12N9/1029—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Engineering & Computer Science (AREA)
- Wood Science & Technology (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
Description
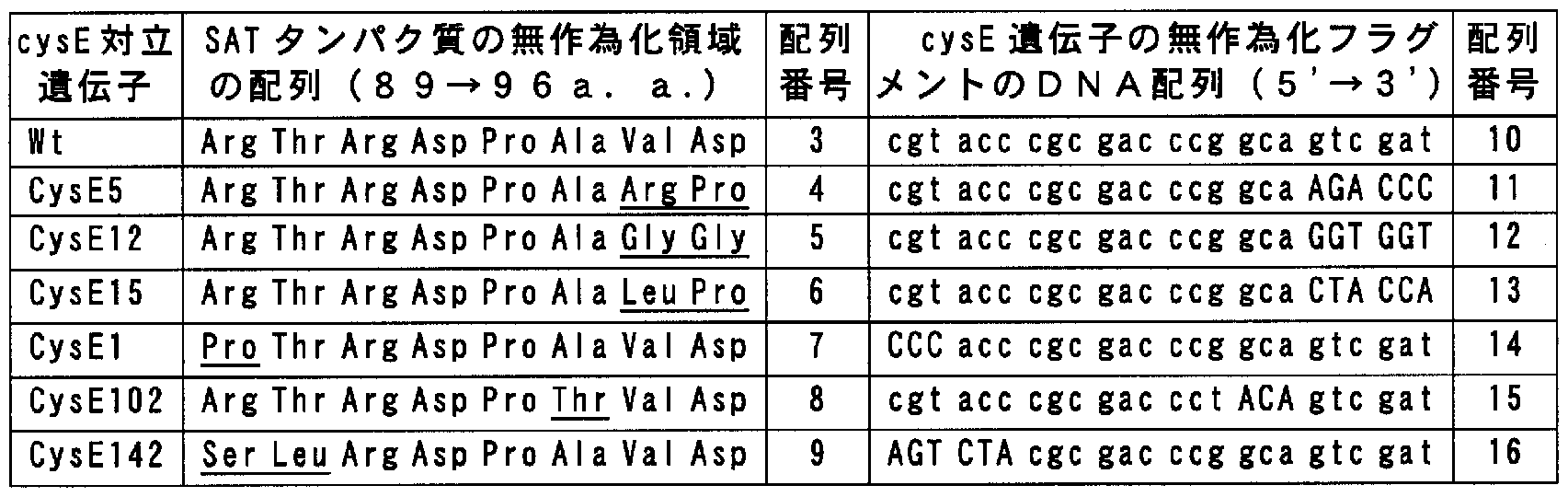


Claims
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| BRPI0412535-5A BRPI0412535A (pt) | 2003-07-16 | 2004-07-16 | serina acetiltransferase mutante, dna, bactéria, e, método para produzir l-cisteìna |
| EP04747681A EP1650296B1 (en) | 2003-07-16 | 2004-07-16 | Mutant serine acetyltransferase and process for producing l-cysteine |
| JP2005511865A JP4923573B2 (ja) | 2003-07-16 | 2004-07-16 | 変異型セリンアセチルトランスフェラーゼ及びl−システインの製造法 |
| AT04747681T ATE554162T1 (de) | 2003-07-16 | 2004-07-16 | Mutante serinacetyltransferase und vefahren zur herstellung von l-cystein |
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| RU2003121601/13A RU2003121601A (ru) | 2003-07-16 | 2003-07-16 | Мутантная серинацетилтрансфераза |
| RU2003121601 | 2003-07-16 | ||
| RU2003135291 | 2003-12-05 | ||
| RU2003135291/13A RU2279477C2 (ru) | 2003-12-05 | 2003-12-05 | Мутантная серинацетилтрансфераза, фрагмент днк, кодирующий мутантную серинацетилтрансферазу (варианты), бактерия, принадлежащая к роду escherichia, - продуцент l-цистеина, и способ продукции l-цистеина |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2005007841A1 true WO2005007841A1 (ja) | 2005-01-27 |
Family
ID=34082460
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2004/010216 WO2005007841A1 (ja) | 2003-07-16 | 2004-07-16 | 変異型セリンアセチルトランスフェラーゼ及びl−システインの製造法 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US7312058B2 (ja) |
| EP (1) | EP1650296B1 (ja) |
| JP (1) | JP4923573B2 (ja) |
| AT (1) | ATE554162T1 (ja) |
| BR (1) | BRPI0412535A (ja) |
| WO (1) | WO2005007841A1 (ja) |
Cited By (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2010027045A1 (ja) | 2008-09-08 | 2010-03-11 | 味の素株式会社 | L-アミノ酸を生産する微生物及びl-アミノ酸の製造法 |
| EP2230302A1 (en) | 2009-03-12 | 2010-09-22 | Ajinomoto Co., Inc. | An L-cysteine-producing bacterium and a method for producing L-cysteine |
| WO2011043485A1 (en) | 2009-10-05 | 2011-04-14 | Ajinomoto Co.,Inc. | A METHOD FOR PRODUCING AN L-CYSTEINE, L-CYSTINE, A DERIVATIVE OR PRECURSOR THEREOF OR A MIXTURE THEREOF USING A BACTERIUM OF Enterobacteriaceae FAMILY |
| WO2011065469A1 (ja) | 2009-11-30 | 2011-06-03 | 味の素株式会社 | L-システイン生産菌及びl-システインの製造法 |
| EP2345667A2 (en) | 2010-01-15 | 2011-07-20 | Ajinomoto Co., Inc. | A method for producing an L-amino acid using a bacterium of the enterobacteriaceae family |
| WO2012036151A1 (ja) | 2010-09-14 | 2012-03-22 | 味の素株式会社 | 含硫アミノ酸生産菌及び含硫アミノ酸の製造法 |
| WO2012137689A1 (ja) | 2011-04-01 | 2012-10-11 | 味の素株式会社 | L-システインの製造法 |
| WO2012144472A1 (ja) * | 2011-04-18 | 2012-10-26 | 味の素株式会社 | L-システインの製造法 |
| CN115135758A (zh) * | 2021-01-27 | 2022-09-30 | Cj第一制糖株式会社 | 新型半乳糖苷o-乙酰转移酶变体及使用其生产l-谷氨酸的方法 |
Families Citing this family (32)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN100529057C (zh) * | 2001-02-13 | 2009-08-19 | 味之素株式会社 | 通过埃希氏菌属细菌生产l-氨基酸的方法 |
| RU2229513C2 (ru) * | 2001-11-23 | 2004-05-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Способ получения l-аминокислот, штамм escherichia coli - продуцент l-аминокислоты (варианты) |
| JP4479283B2 (ja) * | 2004-03-04 | 2010-06-09 | 味の素株式会社 | L−システイン生産菌及びl−システインの製造法 |
| DE602006008274D1 (de) | 2005-02-18 | 2009-09-17 | Ajinomoto Kk | Verfahren zur herstellung einer l-aminosäure unter verwendung eines bakteriums der familie enterobacteriaceae |
| DE602006015302D1 (de) * | 2005-05-16 | 2010-08-19 | Ajinomoto Kk | Verfahren zur herstellung einer l-aminosäure unter verwendung eines bakteriums der familie enterobacteriaceae mit abgeschwächter expression des kefb-gens |
| WO2007086546A1 (en) * | 2006-01-26 | 2007-08-02 | Ajinomoto Co., Inc. | A method for producing an l-amino acid using a bacterium of enterobacteriaceae family with attenuated expression of the aldh gene |
| WO2007119574A2 (en) | 2006-03-23 | 2007-10-25 | Ajinomoto Co., Inc. | A method for producing an l-amino acid using bacterium of the enterobacteriaceae family with attenuated expression of a gene coding for small rna |
| WO2007119890A1 (en) * | 2006-04-18 | 2007-10-25 | Ajinomoto Co., Inc. | A METHOD FOR PRODUCING AN L-AMINO ACID USING A BACTERIUM OF THE ENTEROBACTERIACEAE FAMILY WITH ATTENUATED EXPRESSION OF THE sfmACDFH-fimZ CLUSTER OR THE fimZ GENE |
| EP2035569A1 (en) * | 2006-06-01 | 2009-03-18 | Ajinomoto Co., Inc. | A method for producing an l-amino acid using a bacterium of the enterobacteriaceae family with attenuated expression of the rcsa gene |
| RU2337961C2 (ru) * | 2006-07-04 | 2008-11-10 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) | СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ОПЕРОН rspAB |
| RU2006143864A (ru) * | 2006-12-12 | 2008-06-20 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) (RU) | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, В КОТОРОЙ ОСЛАБЛЕНА ЭКСПРЕССИЯ ГЕНОВ cynT, cynS, cynX, ИЛИ cynR, ИЛИ ИХ КОМБИНАЦИИ |
| DE102007007333A1 (de) | 2007-02-14 | 2008-08-21 | Wacker Chemie Ag | Verfahren zur Reinigung von L-Cystein |
| EP2123756B1 (en) * | 2007-02-15 | 2014-06-04 | Ajinomoto Co., Inc. | Transglutaminase having disulfide bond introduced therein |
| WO2009091045A1 (ja) * | 2008-01-18 | 2009-07-23 | Ajinomoto Co., Inc. | セリン誘導体の製造方法及びこれに用いるタンパク質 |
| JP5476545B2 (ja) * | 2008-02-21 | 2014-04-23 | 味の素株式会社 | L−システイン生産菌及びl−システインの製造法 |
| JP5332237B2 (ja) * | 2008-03-06 | 2013-11-06 | 味の素株式会社 | L−システイン生産菌及びl−システインの製造法 |
| DE602009000714D1 (de) * | 2008-03-06 | 2011-03-24 | Ajinomoto Kk | L-Zystein-produzierendes Bakterium und Verfahren zur Herstellung von L-Zystein |
| RU2008148283A (ru) * | 2008-12-09 | 2010-06-20 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) (RU) | СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, В КОТОРОЙ ОСЛАБЛЕНА ЭКСПРЕССИЯ ГЕНА artI |
| JP5521347B2 (ja) | 2009-02-16 | 2014-06-11 | 味の素株式会社 | L−アミノ酸生産菌及びl−アミノ酸の製造法 |
| JP5463528B2 (ja) * | 2009-02-25 | 2014-04-09 | 味の素株式会社 | L−システイン生産菌及びl−システインの製造法 |
| JP2014087259A (ja) | 2011-02-22 | 2014-05-15 | Ajinomoto Co Inc | L−システイン生産菌及びl−システインの製造法 |
| US9234223B2 (en) | 2011-04-01 | 2016-01-12 | Ajinomoto Co., Inc. | Method for producing L-cysteine |
| WO2014185430A1 (ja) | 2013-05-13 | 2014-11-20 | 味の素株式会社 | L-アミノ酸の製造法 |
| JP2016165225A (ja) | 2013-07-09 | 2016-09-15 | 味の素株式会社 | 有用物質の製造方法 |
| JP6459962B2 (ja) | 2013-10-21 | 2019-01-30 | 味の素株式会社 | L−アミノ酸の製造法 |
| JP7066977B2 (ja) | 2017-04-03 | 2022-05-16 | 味の素株式会社 | L-アミノ酸の製造法 |
| KR20190092951A (ko) | 2018-01-31 | 2019-08-08 | 씨제이제일제당 (주) | 연속식 크로마토그래피 공정을 이용한 천연 l-시스테인 결정의 제조 방법 |
| KR20190092950A (ko) | 2018-01-31 | 2019-08-08 | 씨제이제일제당 (주) | 연속식 크로마토그래피 공정을 이용한 천연 l-시스테인 염산염 수화물 결정의 제조 방법 |
| WO2020071538A1 (en) | 2018-10-05 | 2020-04-09 | Ajinomoto Co., Inc. | Method for producing target substance by bacterial fermentation |
| WO2020171227A1 (en) | 2019-02-22 | 2020-08-27 | Ajinomoto Co., Inc. | METHOD FOR PRODUCING L-AMINO ACIDS USING A BACTERIUM BELONGING TO THE FAMILY Enterobacteriaceae HAVING OVEREXPRESSED ydiJ GENE |
| JP2022526181A (ja) | 2019-04-05 | 2022-05-23 | 味の素株式会社 | L-アミノ酸の製造方法 |
| WO2021060438A1 (en) | 2019-09-25 | 2021-04-01 | Ajinomoto Co., Inc. | Method for producing l-amino acids by bacterial fermentation |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH11155571A (ja) * | 1997-11-25 | 1999-06-15 | Ajinomoto Co Inc | L−システインの製造法 |
| JPH11299491A (ja) * | 1997-10-08 | 1999-11-02 | Research Institute Of Innovative Technology For The Earth | 改変セリンアセチルトランスフェラーゼ、改変セリンアセチルトランスフェラーゼ遺伝子、新規な組み換え体dna及び改変セリンアセチルトランスフェラーゼの製造法 |
| JP2000504926A (ja) * | 1995-10-26 | 2000-04-25 | コンゾルティウム フュール エレクトロケミッシェ インズストリー ゲゼルシャフト ミット ベシュレンクテル ハフツング | O−アセチルセリン、l−システイン及びl−システイン−関連生成物の製法 |
Family Cites Families (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| SU875663A1 (ru) * | 1978-06-30 | 1982-09-15 | Всесоюзный научно-исследовательский институт генетики и селекции промышленных микроорганизмов | Штаммы е.coLI ВНИИГенетика VL 334 @ N6 и ВНИИГенетика VL 334 @ N7-продуценты L-треонина и способ их получени |
| JPS5618596A (en) | 1979-07-23 | 1981-02-21 | Ajinomoto Co Inc | Production of l-lysine through fermentation process |
| US5976843A (en) * | 1992-04-22 | 1999-11-02 | Ajinomoto Co., Inc. | Bacterial strain of Escherichia coli BKIIM B-3996 as the producer of L-threonine |
| ATE203769T1 (de) * | 1992-11-10 | 2001-08-15 | Ajinomoto Kk | Dna, die aspartokinase iii-mutanten kodiert und ihre verwendung zur herstellung von l-threonin durch fermentation |
| JPH07155184A (ja) * | 1993-12-08 | 1995-06-20 | Ajinomoto Co Inc | 発酵法によるl−リジンの製造法 |
| RU2229513C2 (ru) * | 2001-11-23 | 2004-05-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Способ получения l-аминокислот, штамм escherichia coli - продуцент l-аминокислоты (варианты) |
-
2004
- 2004-07-16 EP EP04747681A patent/EP1650296B1/en active Active
- 2004-07-16 BR BRPI0412535-5A patent/BRPI0412535A/pt not_active Application Discontinuation
- 2004-07-16 WO PCT/JP2004/010216 patent/WO2005007841A1/ja active Application Filing
- 2004-07-16 US US10/892,112 patent/US7312058B2/en active Active
- 2004-07-16 JP JP2005511865A patent/JP4923573B2/ja active Active
- 2004-07-16 AT AT04747681T patent/ATE554162T1/de active
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2000504926A (ja) * | 1995-10-26 | 2000-04-25 | コンゾルティウム フュール エレクトロケミッシェ インズストリー ゲゼルシャフト ミット ベシュレンクテル ハフツング | O−アセチルセリン、l−システイン及びl−システイン−関連生成物の製法 |
| US6218168B1 (en) | 1995-10-26 | 2001-04-17 | CONSORTIUM FüR ELEKTROCHEMISCHE INUDSTRIE GMBH | Process for preparing O-acetylserine, L-cysteine and L-cysteine-related products |
| JPH11299491A (ja) * | 1997-10-08 | 1999-11-02 | Research Institute Of Innovative Technology For The Earth | 改変セリンアセチルトランスフェラーゼ、改変セリンアセチルトランスフェラーゼ遺伝子、新規な組み換え体dna及び改変セリンアセチルトランスフェラーゼの製造法 |
| JPH11155571A (ja) * | 1997-11-25 | 1999-06-15 | Ajinomoto Co Inc | L−システインの製造法 |
Non-Patent Citations (4)
Cited By (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2010027045A1 (ja) | 2008-09-08 | 2010-03-11 | 味の素株式会社 | L-アミノ酸を生産する微生物及びl-アミノ酸の製造法 |
| EP2230302A1 (en) | 2009-03-12 | 2010-09-22 | Ajinomoto Co., Inc. | An L-cysteine-producing bacterium and a method for producing L-cysteine |
| WO2011043485A1 (en) | 2009-10-05 | 2011-04-14 | Ajinomoto Co.,Inc. | A METHOD FOR PRODUCING AN L-CYSTEINE, L-CYSTINE, A DERIVATIVE OR PRECURSOR THEREOF OR A MIXTURE THEREOF USING A BACTERIUM OF Enterobacteriaceae FAMILY |
| WO2011065469A1 (ja) | 2009-11-30 | 2011-06-03 | 味の素株式会社 | L-システイン生産菌及びl-システインの製造法 |
| EP2345667A2 (en) | 2010-01-15 | 2011-07-20 | Ajinomoto Co., Inc. | A method for producing an L-amino acid using a bacterium of the enterobacteriaceae family |
| WO2012036151A1 (ja) | 2010-09-14 | 2012-03-22 | 味の素株式会社 | 含硫アミノ酸生産菌及び含硫アミノ酸の製造法 |
| WO2012137689A1 (ja) | 2011-04-01 | 2012-10-11 | 味の素株式会社 | L-システインの製造法 |
| WO2012144472A1 (ja) * | 2011-04-18 | 2012-10-26 | 味の素株式会社 | L-システインの製造法 |
| CN115135758A (zh) * | 2021-01-27 | 2022-09-30 | Cj第一制糖株式会社 | 新型半乳糖苷o-乙酰转移酶变体及使用其生产l-谷氨酸的方法 |
| CN115135758B (zh) * | 2021-01-27 | 2023-10-10 | Cj第一制糖株式会社 | 新型半乳糖苷o-乙酰转移酶变体及使用其生产l-谷氨酸的方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| US7312058B2 (en) | 2007-12-25 |
| EP1650296A4 (en) | 2006-07-12 |
| ATE554162T1 (de) | 2012-05-15 |
| JP4923573B2 (ja) | 2012-04-25 |
| EP1650296A1 (en) | 2006-04-26 |
| EP1650296B1 (en) | 2012-04-18 |
| US20050112731A1 (en) | 2005-05-26 |
| BRPI0412535A (pt) | 2006-09-19 |
| JPWO2005007841A1 (ja) | 2006-11-09 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4923573B2 (ja) | 変異型セリンアセチルトランスフェラーゼ及びl−システインの製造法 | |
| ES2333055T3 (es) | Acetolactato sintasa mutante y procedimiento para producir l-aminoacidos de cadena ramificada. | |
| KR100420743B1 (ko) | 신규한리신데카복실라제유전자및l-리신의제조방법 | |
| US7211419B2 (en) | Mutant carbamoylphosphate synthetase and method for producing compounds derived from carbamoylphosphate | |
| CN100471949C (zh) | 丝氨酸乙酰转移酶突变体和生产l-半胱氨酸的方法 | |
| EP1528108B1 (en) | Method for producing l-cysteine using bacteria belonging to the genus Escherichia | |
| EP1170361B1 (en) | New mutant N-Acetylglutamate synthase and method for L-Arginine production | |
| JP4821321B2 (ja) | 腸内細菌科の細菌を用いたl−ヒスチジンの製造法 | |
| EP1801206B1 (en) | Method for producing l-arginine, l-ornithine or l-citrulline | |
| EP2486123B1 (en) | A METHOD FOR PRODUCING AN L-CYSTEINE, L-CYSTINE, A DERIVATIVE OR PRECURSOR THEREOF OR A MIXTURE THEREOF USING A BACTERIUM OF Enterobacteriaceae FAMILY | |
| US20120329105A1 (en) | Bacterium of enterobacteriaceae family producing l-aspartic acid or l-aspartic acid-derived metabolites and a method for producing l-aspartic acid or l-aspartic acid-derived metabolites | |
| JP2003164297A (ja) | 新規変異型グルタミンシンテターゼ、およびアミノ酸の生産方法 | |
| RU2279477C2 (ru) | Мутантная серинацетилтрансфераза, фрагмент днк, кодирующий мутантную серинацетилтрансферазу (варианты), бактерия, принадлежащая к роду escherichia, - продуцент l-цистеина, и способ продукции l-цистеина | |
| RU2550269C2 (ru) | СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae, СОДЕРЖАЩЕЙ N-АЦЕТИЛОРНИТИНДЕАЦЕТИЛАЗУ С НАРУШЕННОЙ АКТИВНОСТЬЮ | |
| WO2007074857A1 (ja) | L-グルタミンの製造法 | |
| JP6543734B2 (ja) | O−アセチルホモセリンを生産する微生物及びそれを用いてo−アセチルホモセリンを生産する方法 | |
| WO2008088149A1 (en) | Corynebacterium glutamicum variety producing l-arginine and method for fabricating the same | |
| WO2013154182A1 (ja) | アミノ酸の製造法 | |
| TWI832275B (zh) | 新穎YhhS變異體及使用其生產O-磷絲胺酸、半胱胺酸及半胱胺酸之衍生物的方法 | |
| KR102472558B1 (ko) | 황 함유 아미노산 또는 그 유도체 제조방법 | |
| KR102472559B1 (ko) | 황 함유 아미노산 또는 그 유도체의 제조방법 | |
| JP2003189863A (ja) | アスパラギン酸アミド並びにその誘導体の製造方法 | |
| CN117980321A (zh) | 新型MdtH变体以及使用其生产O-磷酸丝氨酸、半胱氨酸和半胱氨酸衍生物的方法 | |
| CN117897489A (zh) | 控制nadh:醌氧化还原酶表达的重组微生物,以及通过使用重组微生物生产o-磷酸丝氨酸、半胱氨酸及其衍生物的方法 | |
| JP2007135602A (ja) | L−グルタミン酸の製造法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WWE | Wipo information: entry into national phase |
Ref document number: 200480020264.8 Country of ref document: CN |
|
| AK | Designated states |
Kind code of ref document: A1 Designated state(s): AE AG AL AM AT AU AZ BA BB BG BR BW BY BZ CA CH CN CO CR CU CZ DE DK DM DZ EC EE EG ES FI GB GD GE GH GM HR HU ID IL IN IS JP KE KG KP KR KZ LC LK LR LS LT LU LV MA MD MG MK MN MW MX MZ NA NI NO NZ OM PG PH PL PT RO SC SD SE SG SK SL SY TJ TM TN TR TT TZ UA UG US UZ VC VN YU ZA ZM ZW |
|
| AL | Designated countries for regional patents |
Kind code of ref document: A1 Designated state(s): GM KE LS MW MZ NA SD SL SZ TZ UG ZM ZW AM AZ BY KG KZ MD RU TJ TM AT BE BG CH CY CZ DE DK EE ES FI FR GB GR HU IE IT LU MC NL PL PT RO SE SI SK TR BF BJ CF CG CI CM GA GN GQ GW ML MR NE SN TD TG |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application | ||
| WWE | Wipo information: entry into national phase |
Ref document number: 2005511865 Country of ref document: JP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2004747681 Country of ref document: EP |
|
| WWP | Wipo information: published in national office |
Ref document number: 2004747681 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: PI0412535 Country of ref document: BR |