JP2016190846A - インフルエンザワクチン - Google Patents
インフルエンザワクチン Download PDFInfo
- Publication number
- JP2016190846A JP2016190846A JP2016086887A JP2016086887A JP2016190846A JP 2016190846 A JP2016190846 A JP 2016190846A JP 2016086887 A JP2016086887 A JP 2016086887A JP 2016086887 A JP2016086887 A JP 2016086887A JP 2016190846 A JP2016190846 A JP 2016190846A
- Authority
- JP
- Japan
- Prior art keywords
- iscom
- domain
- ectodomain
- composition
- fraction
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images




Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/145—Orthomyxoviridae, e.g. influenza virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/39—Medicinal preparations containing antigens or antibodies characterised by the immunostimulating additives, e.g. chemical adjuvants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/16—Antivirals for RNA viruses for influenza or rhinoviruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55577—Saponins; Quil A; QS21; ISCOMS
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/70—Multivalent vaccine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16134—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Virology (AREA)
- Immunology (AREA)
- Chemical & Material Sciences (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Mycology (AREA)
- Microbiology (AREA)
- Epidemiology (AREA)
- Pulmonology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Organic Chemistry (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Peptides Or Proteins (AREA)
Abstract
【課題】インフルエンザウイルスから単離された外部ドメインを含む組成物及びキットの提供。【解決手段】少なくとも1種のISCOMマトリックス、並びに1種又は複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメイン及び少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当し、かつ前記組成物が、それぞれアジュバント画分A及び画分Cを含有する別々のISCOMマトリックス粒子を含む前記組成物。この組成物は脊椎動物、例えば鳥類および哺乳類用の、例えばインフルエンザに対する免疫刺激薬、免疫調節薬又はワクチンとして使用され得る。【選択図】図3
Description
本発明は、少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニン(HA)ドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼ(NA)ドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当する、該組成物に関する。本発明はまた、キットに関する。この組成物は脊椎動物、例えば鳥類および哺乳類用の、例えばインフルエンザに対する免疫刺激薬、免疫調節薬またはワクチンとして使用されうる。
世界的流行のブタ由来2009 A(H1N1型)インフルエンザウイルスが最近になって出現していることは、世界的規模でヒト集団において発病および死亡を引き起こすインフルエンザウイルスの可能性を強く際立たせている。世界中の200を超える国々および海外領または地域社会で、16,000超の死亡例を含む、検査室で確認された流行性ウイルスの事例が報告されている[1]。ワクチン接種はインフルエンザ疾患の罹患を防ぐまたは軽減するための第一の方法である。しかしながら、2009年の世界的流行によりかさねて例示されるように、大発生の初期段階中の迅速な対応は、時間のかかるワクチン株調製および現在使われているワクチン製造工程によって阻まれる。これは、抗原連続変異および抗原不連続変異により既存の免疫から逃避するインフルエンザウイルスの悪評高き能力と相まって、新たに発生する抗原変異体に対して迅速かつ柔軟に作製されうる新規な安全かつ好ましくは広範に有効なワクチンの必要性を強調している。
現在認可されているインフルエンザウイルスワクチンは、ウイルスエンベロープ糖タンパク質、つまりヘマグルチニン(HA)およびノイラミニダーゼ(NA)から構成されている。これらの2種の大きな糖タンパク質によって誘発される抗体は、インフルエンザウイルスに対する免疫において独特な特性を有する。HAに対する抗体は一般に、標的細胞上のシアル酸受容体へのウイルスの結合の妨害によって、または、その後、ウイルスゲノムが通過して標的細胞に侵入するウイルス膜と細胞膜との融合を防ぐことによって、ウイルス感染性を無効にする。NAに対する抗体は、NAが関連する受容体破壊酵素活性を阻害することによって、感染細胞からの子孫ウイルスの放出を不能にする。HAを介した液性免疫は最も広範囲にわたって特徴付けられており、ウイルス感染を防ぐことが示されている。疾患を防ぐことへのNA抗体の寄与は、十分に研究されていない。それらは感染した上皮の先端面からの感染性ウイルスの放出の減少によって特徴付けられる一種の許容免疫[2]を生み出し[2〜8]、ウイルスの出芽および環境への広まりの可能性を低減するように思われた。
HAおよびNAの組み合わせを用いた免疫は、インフルエンザに対する防御の強化をもたらす[5, 9, 10]。HAおよびNAは同等に免疫原性である[2]が、従来の不活化ワクチンまたはウイルス感染に対する液性免疫反応は、HAおよびNAがおよそ4:1の比率でウイルス表面に生じるため、必然的にHAに偏る[11]。さらに、無傷のウイルス粒子では、HAは、マウスにおいて示されるようにBおよびT細胞感作において、NAを免疫学的に打ち負かす[12]。この抗原競合は、HAおよびNAが別々に投与される場合は、ワクチン接種動物において観察されない[10, 13]。現在認可されている汎発性ワクチンおよび季節性三価ワクチンは、ウイルス全体から一般的に調製され、ゆえに、NA抗原よりも多くのHA抗原を含有するように偏っている。NAを優先してワクチン配合物中のHA-NA比を適合させることは、よりバランスの取れた液性免疫反応を与え、より高いNA抗体レベルおよび疾患に対する防御の増大をもたらしうる[3, 14]。
現在の不活化インフルエンザウイルスワクチンは、HAの量に対してのみ標準化されているので、NA含量は、結果としての、NAに対する抗体陽転の頻度およびレベルと同様にバラツキがあり、多くの場合、かなり不十分である[28, 29]。
インフルエンザA型ウイルスの常で、既存の免疫から逃れることが可能なある特定のウイルス亜型におけるHAおよびNAの抗原変異体がヒト集団において漸進的に選抜される。抗原連続変異のこの工程では新興の変異体に応じた季節性ワクチン組成物のほぼ毎年の調整が求められる。例えば鳥H5N1型ウイルスによって引き起こされる、将来のインフルエンザ大流行の脅威を考慮して、防御免疫を広範に誘導するワクチンが必要である。
本発明は、少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニン(HA)ドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼ(NA)ドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当する、該組成物、およびキットに関する。
ISCOMアジュバントならびにHAおよびNAまたはそれらの断片を含む組成物は、例えばWO 2008157419および[5, 9, 10]を通じて公知である。しかしながら、HAおよびNAの両方からの外部ドメインをISCOMアジュバントとともに用いることは以前には開示されていない。
今回、ISCOMまたはISCOMマトリックスを用いて増強したNAおよびHAの両方からの外部ドメインによるワクチン接種は、例えば肺力価を低下させることによって、ウイルス複製を低減し、体重減少および肺病態のような感染の臨床的影響を減らしたことが分かった。
多量体、HAおよびNA外部ドメインは、これらを大量に容易に、迅速に、柔軟にかつ安全に産生できるので、ワクチンの可能性が大いにある。インフルエンザワクチンの中にNAを含めることは、HAによる防御に大いにかつ特異的に寄与する。ワクチンにおけるその包含は必要とされるHA用量を低減する可能性が高く、防御免疫を広げる可能性が高い。
詳細な説明
本発明は、少なくとも1種のISCOM複合体、ならびにインフルエンザウイルスに由来する、少なくとも1種のHAドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のNAドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当する、該組成物に関する。
本発明は、少なくとも1種のISCOM複合体、ならびにインフルエンザウイルスに由来する、少なくとも1種のHAドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のNAドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当する、該組成物に関する。
外部ドメインは、細胞外空間に及ぶ膜タンパク質のドメインである。外部ドメインは、通常、シグナル伝達につながる表面との接触を開始する、タンパク質の一部である。組成物において、外部ドメインは抗原として機能することができ、その結果、その組成物はワクチンとして用いることができる。
単離(された)とは、外部ドメインが他のタンパク質から、ならびにインフルエンザウイルスのNAおよびHAタンパク質のそれぞれの残部から実質的に単離されることを意味する。少ない残部アミノ酸が存在してもよい。外部ドメインは、完全な外部ドメインまたは同じ酵素活性および/もしくは抗原活性を有するその一部であってよい。1つの態様によれば、外部ドメインの一部はその頭部ドメインであってよい。完全な外部ドメインまたは同じ酵素活性および/もしくは抗原活性を有するその一部であるそのような外部ドメインは、インフルエンザウイルスから単離されてもよく、または合成により産生されてもよい。それらは、相互に分離されてまたは一緒に連結されて提示されてもよい。
1つの態様によれば、外部ドメインは可溶性外部ドメインまたは可溶性頭部ドメインである。
同じまたは異なるインフルエンザウイルスの種または株に由来する、少なくとも1種のヘマグルチニンドメインからの1種または複数種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1種または複数種の外部ドメインがハイブリッドタンパク質として提示されてもよく、それは組換え体であってもよい。
組換えにより産生されたHAおよびNA抗原は、両抗原の相対量を容易に制御できるワクチンの開発を可能にする。哺乳類および昆虫の両方である真核生物発現系は、それらの、タンパク質の天然の抗原構造のより良好な保存を考慮すれば、そのような糖タンパク質の産生に好ましいプラットフォームである。実施例1において説明したように、およびGenscript http://www.genscript.com/gene_synthesis.htmlにしたがって、ハイブリッドタンパク質は産生されうる。
インフルエンザウイルスはインフルエンザの亜血清型(sub serotype)から、例えばxが1〜16でありかつyが1〜9であるHxNyから、選択されうる。したがって、xは1、2、3、4、5、6、7、8、9、10、11、12、13、14、15および16であってよく、かつyは1、2、3、4、5、6、7、8、9であってよい。インフルエンザウイルスはヒト、鳥、牛、例えばウシ種、ブタ、ヒツジ、ヤギのような任意の種由来であってよい。鳥インフルエンザウイルス、例えばH1〜6N1〜9もしくはヒトインフルエンザウイルスH1〜3N1〜2またはそのNAおよびHA由来の外部ドメインの任意の組み合わせが用いられてもよい。NA外部ドメインおよび/またはHA外部ドメインはインフルエンザウイルスの、例えばヒトおよび鳥インフルエンザウイルスの、異なる種由来であってよい。したがって、1種または複数種の、ヒトおよび鳥NA外部ドメインが、1種または複数種の、ヒトおよび鳥HA外部ドメインと組み合わされてもよく、そのためにNAおよびHAは、HxNyの異なるタイプのものであってよい。
1つの態様によれば、インフルエンザウイルスは、例えば1918 H1N1型インフルエンザウイルス(A/サウスカロライナ/1/18)から選択されるN1H1型ウイルス、ならびに/または2004 H5N1型(A/ベトナム/1203/04) H1N1型A/カリフォルニア/07/2009ウイルス、および/もしくはA/カリフォルニア/04/2009、および/もしくはA/カリフォルニア/09/2009、ならびに/またはA/ケンタッキー/UR06-0258/2007(H1N1型)、ならびに/またはA/シチメンチョウ/トルコ/1/2005(H5N1型)などのA型インフルエンザである。
NA由来およびHA由来の1種または複数種の外部ドメイン(多量体)が用いられてもよい。組成物は、少なくとも1種のヘマグルチニンドメインからの1〜5種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1〜5種の頭部ドメインを含んでもよい。
1つの態様によれば、少なくとも1種のヘマグルチニン外部ドメインはA/カリフォルニア/09/2009の三量体HA外部ドメイン(アミノ酸番号17〜522)から選択される。
1つの態様によれば、少なくとも1種のノイラミニダーゼ頭部ドメインはA/カリフォルニア/09/2009ウイルスの四量体NA頭部ドメイン(アミノ酸番号75〜469)から選択される。A/カリフォルニア/09/2009タンパク質の配列は、当技術分野において、例えば、http://www.ncbi.nlm.nih.gov/protein/227809830およびhttp://www.ncbi.nlm.nih.gov/protein/227809834を通じて公知である。
ISCOM複合体はアジュバントおよび免疫調節剤として用いられてもよい。それらはISCOMおよび/またはISCOMマトリックス複合体であってよい。
ISCOMマトリックス複合体および/またはISCOM複合体は、インフルエンザ感染に対する免疫反応を誘発するように意図された抗原の1種または複数種とともに用いられてもよい。抗原ならびにISCOMマトリックス複合体および/またはISCOM複合体は、混合物の状態で、または別々に投与されてもよい。
ISCOMマトリックス複合体は少なくとも1種のグリコシドおよび少なくとも1種の脂質を含む。グリコシドはアジュバント効果を有し、好ましくは、特にキラヤ・サポナリア・モリナ(Quillaja saponaria Molina) (Quil A)に由来するサポニンである。脂質は少なくともステロール、例えばコレステロールであり、さらに任意で、リン脂質でもよい。ISCOMマトリックス複合体は、必ずしもグリコシドではない、1種または複数種の他の免疫調節(アジュバント活性)物質を含んでもよく、EP 0 436 620 B1に記述されているように産生されてもよい。
ISCOM複合体は少なくとも1種のグリコシド、少なくとも1種の脂質、および少なくとも1種の抗原物質またはエピトープを含む。これらの抗原物質はタンパク質およびペプチド、糖タンパク質および糖ペプチド、糖質などのような異なる種類のものであってよい。グリコシドはアジュバント効果を有し、好ましくは、特にキラヤ・サポナリア・モリナ(Quil A)に由来するサポニンである。これらの複合体は、含まれる抗原の免疫原性を強化し、かつ1種または複数種の免疫調節(アジュバント活性)物質を含んでもよい。ISCOMはEP 0 109 942 B1、EP 0 242 380 B1およびEP 0 180 546 B1に記述されているように調製されてもよい。さらに、EP 9600647-3 (PCT/SE97/00289)に記述されているように、輸送および/またはパッセンジャー抗原が用いられてもよい。
外部ドメイン抗原をISCOMマトリックス複合体および/もしくはISCOM複合体と混合してもよく、ISCOMマトリックス複合体上へ結合もしくは複合化させてもよく、またはISCOMと混合してもよく、またはISCOM複合体上へカップリングさせてもよい。調製されたISCOM複合体は外部ドメイン抗原以外の1種または複数種の抗原を含んでもよい。1つの態様によれば、1種または複数種の外部ドメイン抗原はISCOM複合体に組み込まれて用いられてもよい。そのようなISCOM複合体を次いで、1種または複数種の外部ドメイン抗原と混合してもよい。1種または複数種の抗原を用いてもよく、EP 9600647-3 (PCT/SE97/00289)に記述されているように輸送およびパッセンジャー抗原を用いてもよい。
ISCOM粒子に組み込まれるためには、抗原はいくらかの疎水性部分を持つ必要があり、または、ISCOMマトリックスに静電付着される必要がある。疎水性部分を持たない抗原がそのような分子にカップリングされてもよい。疎水性分子およびカップリング方法はEP 180564に記述されている。
免疫原性複合体に含まれる脂質は、少なくともステロール、例えばコレステロールであり、さらに任意で、リン脂質でもよい。用いられる脂質は、特に、本出願人らの特許であるEP 0 109 942 B1、具体的には3頁に、および特許EP 0 436 620 B1の7頁7〜24行目に記述されているものである。特に、コレステロールなどのステロール、ならびにホスファチジル・エタノールアミンおよびホスファチジルコリンなどのリン脂質が用いられる。
ガングリオシドGM1であるコレラ毒素受容体を含む、糖脂質などの細胞結合成分に結合する脂質含有受容体、およびフコース含有血液型抗原が用いられてもよい。細胞結合成分がひいては、粘液を標的とする分子として機能し、脂質含有物質を含む複合体と単に混合するだけで脂質含有物質に結合しうる。受容体を含むISCOM複合体は、例えばWO 97/30728に記述されている。
本発明の1つの態様において、インフルエンザに対するワクチン接種で用いるための免疫原性複合体中のグリコシドは、キラヤ・サポナリア・モリナ由来のサポニン画分である。
キラヤ・サポナリア・モリナに由来するサポニンの任意の部分断片も使用されうる。さらに、キラヤ・サポナリア・モリナの部分断片の任意の組み合わせが用いられうる。したがって、2つまたはそれ以上の部分断片がそれぞれ、ISCOM複合体またはISCOMマトリックス複合体に組み込まれうる。
「キラヤ・サポナリア・モリナ由来のサポニン画分」という用語は、本明細書の全体を通しておよび特許請求の範囲においてキラヤ・サポナリアの半精製されたもしくは規定されたサポニン画分または実質的に純粋な画分の一般的記述として用いられる。この画分は、本質的に1画分を含むISCOM複合体またはISCOMマトリックス複合体の混合物が用いられる場合に得られる良好な結果に負の影響を与えるいかなる他の画分も含まないということが重要である。
1つの態様によれば、免疫原性複合体の一部として、またはそれとの混合物の状態で、キラヤ・サポナリア・モリナ由来のサポニン画分以外の少なくとも1種のさらなるアジュバント、または、ISCOMマトリックスもしくはISCOM複合体に組み込まれているもの以外の少なくとも1種のグルコシドもしくはサポニンをさらに含む、本発明にしたがって用いるための免疫原性複合体が提供される。サポニンではない有用なアジュバントの例は、さまざまな油およびAl(OH)3である。
ISCOM複合体およびISCOMマトリックス複合体にそれぞれ組み込まれうる、またはそれらと混合されうるさらなるアジュバントの例は、所望の免疫調節効果を有する天然のまたは合成の任意のアジュバント、例えば、キラヤ・サポナリア・モリナのサポニン粗抽出物に由来する天然サポニン分子、またはその誘導体、合成もしくは半合成サポニン分子であり; 例えば、Quil A由来のサポニンおよびサポニン画分、細胞壁骨格、ブロック重合体、例えば、親水性ブロック共重合体、例えばCRL-1005、TDM (トレハロースジミコレート)、リポペプチド、LPSおよびLPS誘導体、さまざまな細菌種由来の脂質Aおよびその誘導体(例えば、モノホスホリル脂質A、ムラミルジもしくはトリペプチドまたはムラミルジペプチド)、MDP誘導体(例えば脂肪酸誘導体)、置換MDP、MDPのトレオニル類似体(およびその誘導体)、CpG変種、CpGODN変種、外来性ヒトおよび動物免疫調節剤(例えばGM-CSF)、IL-2、天然または修飾毒素であるアジュバント活性細菌毒素(例えばコレラ毒素CT、およびその部分成分CTBおよびCTA1)、大腸菌(E. coli)の易熱性毒素(LT)、百日咳菌(Bordetella pertussis(BP))毒素およびBPの線維状ヘマグルチニン、DDA、硫酸デキストランのようなポリアニオンならびに(Quil A以外の)サポニンのようなリポ多糖類であり、これらの全てが参照により組み入れられる、(「Future prospects for vaccine adjuvants」, Warren, H.S. (1988) CRC Crit. Rev. Immunol. 8:2, 83-101; 「Characterisation of a non-toxic monophosphoryl lipid A」, (1987) Johnson, A.G. et al, Rev. Infect. Dis. 9:5, 5512-5516; 「Developmental status of synthetic immunomodulators」, Berendt, M.J. et al (1985), Year Immunol. 193-201; 「Immunopotentiating conjugates」, Stewart-Tull, D.E., Vaccine, 85, 3:1, 40-44)を参照されたい。
ISCOM粒子は任意のサポニンから作製されたISCOM複合体またはISCOMマトリックス複合体であってよい。アジュバント画分Aおよび他の少なくとも1種のアジュバントが異なるもしくは同じISCOM粒子もしくはISCOMマトリックス粒子上にカップリングされてもよく、またはアジュバントの1種もしくは複数種がISCOM粒子と混合されてもよい。
ISCOM粒子に組み込まれるためには、アジュバントはいくらかの疎水性分子を持つ必要がある。疎水性部分を持たないアジュバントが、そのような分子にカップリングされてもよい。疎水性分子およびカップリング方法は、EP 180564に記述されている。好ましくは、アジュバントは異なるISCOM粒子に組み込まれる。
本発明の別の態様において、Quil Aのアジュバント画分AはISCOM粒子に組み込まれるのに対し、他の少なくとも1種のアジュバントはISCOM粒子に組み込まれず、組成物において遊離形態で用いられる。
本発明の別の好ましい態様において、Quil Aのアジュバント画分はISCOM粒子またはISCOMマトリックス粒子に組み込まれるのに対し、他のアジュバントはISCOM粒子またはISCOMマトリックス粒子に組み込まれず、組成物において遊離形態で用いられる。
別の特に好ましい態様において、組成物は、ISCOM粒子またはISCOMマトリックス粒子に組み込まれているQuil Aの画分A、およびISCOM粒子またはISCOMマトリックス粒子に組み込まれていない、少なくとも1種の他のアジュバントを含む。
別の好ましい態様において、少なくとも他のアジュバントはMPLまたはコレラ毒素CTである。MPLまたはコレラ毒素は同じISCOM粒子もしくはISCOMマトリックス粒子に組み込まれてもよく、またはそれぞれ、異なるISCOM粒子もしくはISCOMマトリックス粒子に組み込まれてもよい。好ましくは、MPLまたはコレラ毒素は遊離形態である。
ISCOMマトリックス複合体、ISCOM複合体および/または少なくとも1種のさらなるアジュバントで用いるためのキラヤ・サポナリア・モリナ由来のサポニン画分は、キラヤ・サポナリア・モリナの画分A、画分B、画分C、キラヤ・サポナリア・モリナの粗画分QA 1-21から選択されうる。しかしながら、最新の強力な分離法により、60種超の異なる構造が報告および記述されている(Bankefors et al., J Chrom B Analyt Technol Biomed Life Sci in press; Bankefors et al., Rapid Commun Mass Spectrom 22:3851; Broberg et al., J Mass Spectrom 39:691; Nyberg et al., Anal Chem 75:268; Guo and Kenne Phytochemistry 55:419; Nord and Kene Carbohydr Res 329:817; Guo and Kenne Phytochemistry 54:615; Guo et al., Phytochemistry 53:861; Nyberg et al., Carbohyd. Res 323:87: Nord and Kenne Carbohyd. Res 320:70 Guo et al., Phytochemistry 48:175)。
本明細書において説明するように調製された場合、キラヤ・サポナリア・モリナの画分A、BおよびCはそれぞれ、定義できる特性を有する化学的に密接に関連している分子の群または族を表す。それらが得られるクロマトグラフィー条件は、溶出プロファイルおよび生物学的活性に関するバッチ間の再現性が高度に一致しているような条件である。
「キラヤ・サポナリア・モリナ由来の1つのサポニン画分」という用語は、本明細書の全体を通しておよび特許請求の範囲においてキラヤ・サポナリアの半精製されたもしくは規定されたサポニン画分または実質的に純粋な画分の一般的記述として用いられる。この画分は、本質的に1画分を含むISCOMまたはISCOMマトリックスの混合物が用いられる場合に得られる良好な結果に負の影響を与えるいかなる他の画分も含まないということが重要である。サポニン調製物は、所望であれば、少量の、例えば40重量%までの、例えば30重量%までの、25重量%までの、20重量%までの、15重量%までの、10重量%までの、7重量%までの、5重量%までの、2重量%までの、1重量%までの、0.5重量%までの、0.1重量%までの、他のサポニンまたは他のアジュバント材料などの他の化合物を含みうる。
本発明によるサポニン画分A、BおよびCは、WO 96/11711に記述される通りであり、B3、B4およびB4b画分は、EP 0 436 620に記述される通りであり; 画分QA1〜21はEP 0 3632 279 B2に記述される通りである。EP 0 3632 279 B2の画分QA-1-2-3-4-5-6-7-8-9-10-11-12-13-14-15-16-17-18-19-20および21、特にQA-7、17〜18が用いられてもよい。それらはEP 0 362 279 B2に、特に6頁、ならびに8頁および9頁の実施例1に記述されているように得られる。
キラヤ・サポナリア・モリナ由来のサポニンの、任意のタイプの粗画分または半精製画分が用いられてもよい。本特許の目的のために用いられるキラヤ・サポナリア・モリナの粗画分は、他の親油性の非サポニン成分からいくぶん精製された任意のサポニン組成物である。これは、キラヤ・サポナリア・モリナの精製された、しかし非分画化調製物である任意のサポニン画分であってよい。サポニンが相互に分離されていないそのような粗画分は、例えば、Desert King Chile (www.desertkingchile.cl)、Sigma-Aldrich (www.sigmaaldrich.com)、Berghausen (www.berghausen.com)、Brenntag Biosector (www.brenntaq-biosector.com)から生産および納品されうる。
WO 96/11711に記述される画分A、BおよびCは、親水性キラヤ・サポナリア・モリナ粗抽出物のクロマトグラフィー分離、および親油性画分を分離するための70%アセトニトリル水溶液を用いた溶出から得られた、親油性画分から調製される。この親油性画分を次いで、酸性水溶液中25%〜60%のアセトニトリル勾配を使用する溶出を用いた半分取HPLCにより分離する。本明細書において「画分A」または「QH-A」といわれる画分は、約39%のアセトニトリルで溶出される画分であるか、またはその画分に対応する。本明細書において「画分B」または「QH-B」といわれる画分は、約47%のアセトニトリルで溶出される画分であるか、またはその画分に対応する。本明細書において「画分C」または「QH-C」といわれる画分は、約49%のアセトニトリルで溶出される画分であるか、またはその画分に対応する。
1つの態様によれば、サポニンの粗画分が用いられる。
別の態様によれば、サポニンの粗画分が任意の他の精製されたサポニン画分、例えば上記の異なるサポニン画分とともに用いられてもよい。
本発明の1つの態様によれば、ISCOMマトリックス複合体もしくはISCOM複合体に組み込まれる、キラヤ・サポナリア・モリナ由来のサポニン画分、または、同じくISCOMもしくはISCOMマトリックス複合体に組み込まれるかもしくはそれと混合される、少なくとも1種のさらなるアジュバントは、キラヤ・サポナリア・モリナの画分A、画分B、画分Cか、キラヤ・サポナリア・モリナの半精製調製物か、キラヤ・サポナリア・モリナの精製調製物か、または精製された任意の亜画分、例えば、QA1〜21から選択される。
キラヤ・サポナリア由来の少なくとも2つのサポニン画分をそれぞれ含む、ISCOMマトリックスおよび/またはISCOM複合体が用いられてもよい。異なるサポニン画分の、任意の組み合わせの重量%が用いられてもよい。任意の2つの画分の、任意の組み合わせの重量%、例えば、それぞれキラヤ・サポナリア・モリナの、画分Aの任意の重量%と、別の画分、例えば任意の粗サポニン画分または画分Cの任意の重量%とが用いられてもよい。ISCOMマトリックスおよび/またはISCOM複合体は、ISCOMマトリックスおよび/またはISCOM複合体中のキラヤ・サポナリア・モリナの、サポニン画分の重量の合計に基づいて計算して、0,1〜99,9重量%、5〜95重量%、10〜90重量%、15〜85重量%、20〜80重量%、25〜75重量%、30〜70重量%、35〜65重量%、40〜60重量%、45〜55重量%、40〜60重量%、50〜50重量%、55〜45重量%、60〜40重量%、65〜35重量%、70〜30重量%、75〜25重量%、80〜20重量%、85〜15重量%、90〜10重量%、95〜05重量%、50〜99重量%、60〜90重量%、70〜90重量%、75〜85重量%の1つのサポニン画分、例えばキラヤ・サポナリア・モリナの画分Aと、各区間における、最大100%である残りにあたる別のサポニン、例えば任意の粗画分または任意の他の画分、例えばキラヤ・サポナリア・モリナの画分Cとを含んでもよい。
1つの態様によれば、画分Aおよび画分Cの重量に基づいて計算して、5〜99重量%の1つの画分、例えばキラヤ・サポナリア・モリナの画分Aと、重量の最大100%である残りにあたる別の画分、例えばキラヤ・サポナリア・モリナの粗サポニン画分または画分Cとを含む、本発明にしたがって用いるためのISCOMマトリックスおよび/またはISCOM複合体が提供される。
別の態様によれば、画分Aおよび画分Cの重量に基づいて計算して、40重量%〜99重量%の1つの画分、例えばキラヤ・サポナリア・モリナの画分Aと、1重量%〜60重量%の別の画分、例えばキラヤ・サポナリア・モリナの粗サポニン画分または画分Cとを含む、本発明にしたがって用いるためのISCOMマトリックスおよび/またはISCOM複合体が提供される。
さらにある態様によれば、画分Aおよび画分Cの重量に基づいて計算して、70重量%〜95重量%の1つの画分、例えばキラヤ・サポナリア・モリナの画分Aと、30重量%〜5重量%の別の画分、例えばキラヤ・サポナリア・モリナの粗サポニン画分または画分Cとを含む、本発明にしたがって用いるためのISCOMマトリックスおよび/またはISCOM複合体が提供される。
1つの態様において、キラヤ・サポナリア・モリナ由来のサポニン画分がQA1〜21の任意の1つから選択される、本発明にしたがって用いるためのISCOMマトリックスおよび/またはISCOM複合体が提供される。
本発明の1つの局面によれば、インフルエンザに対するワクチン接種で用いるための、1種もしくは複数種の外部ドメイン抗原、ならびに1つもしくは複数の薬学的に許容される賦形剤、担体および/もしくは希釈剤、ならびに/またはさらなるアジュバントとともに本発明による少なくとも1種のISCOMマトリックスおよび/またはISCOM複合体を含む組成物、例えばワクチンが提供される。
そのような組成物は、1つもしくは複数の異なるタイプのISCOMマトリックス複合体粒子および/または1つもしくは複数の異なるタイプのISCOM複合体粒子を含んでもよく、個々のタイプの複合体粒子のそれぞれがキラヤ・サポナリア・モリナ由来の1つのサポニン画分を含み、一つの複合体におけるサポニン画分は、他の複合体粒子におけるサポニン画分とは異なる。組成物はいくつかの粒子を含んでもよい。しかしながら、1つの態様によれば、ある粒子はある単一のタイプのキラヤ・サポナリア・モリナ画分を含むのみでありうる。
したがって、1つのタイプの実質的に純粋なサポニン画分または粗サポニン画分が1つのISCOMマトリックス複合体または粒子に組み込まれてもよく、別のタイプの実質的に純粋なサポニン画分または粗サポニン画分が別のISCOMマトリックス複合体または粒子に組み込まれてもよい。組成物またはワクチンは、タイプごとに、物理的に異なる粒子に組み込まれた1つのタイプのサポニンを有する、少なくとも2つのタイプの複合体または粒子を含みうる。
組成物において、キラヤ・サポナリア・モリナ由来の1つのサポニン画分およびキラヤ・サポナリア・モリナ由来の別のサポニン画分が異なるISCOMマトリックス複合体粒子および/またはISCOM複合体粒子に別々に組み込まれている、ISCOMマトリックス複合体粒子および/またはISCOM複合体粒子の混合物が用いられてもよい。1つのサポニン画分および任意の他のサポニン画分の含量に基づく、異なるISCOM複合体の、任意の組み合わせの重量%が用いられてもよく、例えば、画分Aと、別の画分、例えばそれぞれキラヤ・サポナリア・モリナの任意の粗サポニン画分または画分Cとが用いられてもよい。これらの%値は、同じ1つのISCOMマトリックス複合体粒子および/またはISCOM複合体粒子におけるサポニン画分の可能な混合物に関する上記のものと同じであってよいが、ここでは、別々のISCOMマトリックス複合体粒子および/またはISCOM複合体粒子におけるサポニン画分のものである。
さらに別の態様において、Quil Aの画分AはISCOM粒子もしくはISCOMマトリックス粒子に組み込まれ、少なくとも1種の他のアジュバントは、異なるISCOM粒子もしくはISCOMマトリックス粒子それぞれに組み込まれ、または1つもしくは複数の他のQuil A画分もしくは1つもしくは複数の他のアジュバントは、同じISCOMもしくはISCOMマトリックス粒子であるがQuil Aの画分Aが組み込まれた粒子とは異なる粒子に組み込まれ、または他の少なくとも1種のアジュバントは遊離形態である。
組成物において、画分Aは、同じまたは異なるISCOM複合体および/またはISCOMマトリックス複合体中で、キラヤ・サポナリア・モリナ由来の画分Cおよび粗サポニン画分のうちの少なくとも1つ、例えば画分Qと組み合わされてもよい。
キラヤ・サポナリア・モリナの異なる画分を含むISCOMマトリックス複合体を組み合わせることにより、動物にとって毒性の低い組成物を産生することが可能である。ゆえに、1つの態様において、本発明にしたがって用いるための組成物は、同じまたは異なるISCOM複合体および/またはISCOMマトリックス複合体中で、画分CおよびQの少なくとも1つと組み合わせて、画分Aを含む。
組成物はさらに、1つまたは複数の薬学的に許容される賦形剤、担体および/または希釈剤、ならびにさらなるアジュバントを含んでもよい。
組成物は、ISCOM複合体粒子またはISCOMマトリックス粒子に組み込まれるサポニン以外の少なくとも1種のアジュバントを含んでもよい。このさらなるアジュバントはキラヤ・サポナリア・モリナ由来のサポニン画分であってよく、これは免疫原性ISCOMマトリックス複合体粒子もしくはISCOM複合体粒子へ結合されてもよく、またはそれらと混合されてもよい。それはまた、別のタイプのサポニンまたは任意の他のタイプのアジュバントであってもよく、これらは免疫原性ISCOMマトリックス複合体粒子もしくはISCOM複合体粒子に組み込まれてもよく、それらへ結合されてもよく、またはそれらと混合されてもよい。
組成物はワクチンであってもよい。
本明細書において「ワクチン」という用語は、免疫反応を生み出すことができる材料をいう。本発明のワクチンはインフルエンザに対する免疫を生み出しうる。
組成物は、感染が起きるのを防ぐために予防的に用いられてもよく、または既存の感染を処置するために治療的であってもよく、または免疫学的試薬の産生用であってもよい。
インフルエンザウイルスに由来する外部ドメインからの抗原以外の抗原はISCOM複合体に組み込まれてもよく、ISCOM複合体もしくはISCOMマトリックス複合体へカップリングされてもよく、またはISCOM複合体もしくはISCOMマトリックス複合体と混合されてもよい。本発明はまた、処置のための混合ワクチンまたは混合獣医薬にも関する。
本発明による組成物の処方は当業者には周知である。適当な薬学的に許容される担体および/または希釈剤は、ありとあらゆる従来の溶媒、分散媒体、増量剤、固形担体、水溶液、コーティング、抗菌剤および抗真菌剤、等張剤および吸収遅延剤などを含む。薬学的に活性な物質のためのそのような媒体および薬剤の使用は、当技術分野において周知であり、例として、Remington's Pharmaceutical Sciences, 18th Edition, Mack Publishing Company, Pennsylvania, USAに記述されている。任意の従来の媒体または薬剤が活性成分と不適合でない限り、本発明の薬学的組成物において、それらの使用が企図される。追加の活性成分を組成物に組み入れることもできる。
1つの他の局面によれば、組成物は脊椎動物、例えば鳥類および哺乳類用の、例えばインフルエンザに対する免疫刺激薬、免疫調節薬またはワクチンとして使用されうる。哺乳類はヒト;ネコ、イヌ、ウマ、鳥、例えばオウムなどのコンパニオンアニマル;畜牛、例えばウシ種、ブタ、ヒツジ、ヤギまたはフェレット、ミンクなどの経済的に重要な種でありうる。
本発明はまた、1つの区画が少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部ドメインを含む組成物を含み、かつ別の区画が使用説明書を含む、少なくとも2つの区画を含むキットにも関する。
別の局面によれば、本発明は、1つの区画がISCOM複合体および/またはISCOMマトリックス複合体を含み、かつ別の区画が少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部ドメインを含む、少なくとも2つの区画を含むキットに関する。
本出願人らは、特にNA抗原の寄与に重点を置きながら、フェレットモデルにおいて同型インフルエンザウイルスに対するワクチンとして2009 A (H1N1型)インフルエンザウイルスの、組換えにより産生されたHAおよびNAサブユニットの効力にとりわけ取り組んだ。パンデミックH1N1型ウイルスの、可溶性の多量体型のHAおよびNA抗原が、哺乳類発現系において発現されている。糖タンパク質を単一段階アフィニティークロマトグラフィーによって精製し、その後、フェレットを一方の抗原または両方の抗原を用い、かつアジュバントとしてのISCOMマトリックスMを用いてまたは用いずに、免疫した。動物は両方の抗原に対して血清学的に反応したが、アジュバントとともに投与された場合に限られた。興味深いことに、ワクチンにNAを含めることで、HA抗体のレベルおよびウイルス中和活性のレベルが向上した。劇的に(5log10単位)低減したウイルス肺力価から特に判断した際の、顕著な保護が、ISCOMマトリックスMと組み合わせたHA含有ワクチンで免疫した動物での同種攻撃(homologous challenge)により観察された。興味深いことに、NAを含有するISCOMマトリックスMアジュバント配合物は、感染の臨床的影響を明らかに低減した。
本明細書において言及される全ての刊行物は、法律が許す最大の範囲内で、参照として本明細書に組み入れられる。
本発明をこれから、以下の非限定的な実施例によってさらに説明する。
実施例1. HAおよびNA抗原の調製
材料および方法
A型インフルエンザ攻撃ウイルス
A型インフルエンザウイルス/オランダ/602/2009は、11日齢の孵化鶏卵の接種により、オランダにおける検査室で確認された2009 A(H1N1型)感染の最初の事例から分離された[15]。A型インフルエンザウイルス オランダ/602/2009(H1N1型)のウイルス原液を、コンフルエントなメイディン・ダービー・イヌ腎臓(Madin-Darby Canine Kidney(MCCK))細胞に感染させることによって調製した。細胞病理学的変化が完了した後に、培養上清を低速遠心分離によって清澄化し、-70℃で貯蔵した。感染性ウイルス力価を、以前に記載のように、MCCK細胞において判定した[16]。これらのウイルスを用いた実験の全ては、生物学的安全性レベル(Bio Safety Level(BSL))-3の条件下で行った。
材料および方法
A型インフルエンザ攻撃ウイルス
A型インフルエンザウイルス/オランダ/602/2009は、11日齢の孵化鶏卵の接種により、オランダにおける検査室で確認された2009 A(H1N1型)感染の最初の事例から分離された[15]。A型インフルエンザウイルス オランダ/602/2009(H1N1型)のウイルス原液を、コンフルエントなメイディン・ダービー・イヌ腎臓(Madin-Darby Canine Kidney(MCCK))細胞に感染させることによって調製した。細胞病理学的変化が完了した後に、培養上清を低速遠心分離によって清澄化し、-70℃で貯蔵した。感染性ウイルス力価を、以前に記載のように、MCCK細胞において判定した[16]。これらのウイルスを用いた実験の全ては、生物学的安全性レベル(Bio Safety Level(BSL))-3の条件下で行った。
HAおよびNA抗原の調製
A型インフルエンザウイルス/カリフォルニア/04/2009(H1N1型)の可溶性ヘマグルチニン外部ドメイン(sHA; アミノ酸番号17〜522)およびノイラミニダーゼ頭部ドメイン(sNA; アミノ酸番号75〜469)をコードするヒトコドン最適化配列を合成し(GenScript)、HEK293T細胞における発現のために発現プラスミドpS1-lgの誘導体[17]にクローニングした。最近になって記述されているように、HA遺伝子の前にN末端CD5シグナルペプチドをコードする配列を置き、HA遺伝子の後にC末端の人工的なGCN4三量化ドメイン(GCN4-pII) [18]およびアフィニティー精製用のStrepタグ(IBA GmbH)をコードする配列を置いた[19, 20]。NA遺伝子の前に、N末端CD5シグナルペプチド、二重のStrepタグ(OneSTrEP; IBA GmbH)および人工的なGCN4四量化ドメイン(GCN4-pLI)を連続的にコードする配列を置いた[18]。
A型インフルエンザウイルス/カリフォルニア/04/2009(H1N1型)の可溶性ヘマグルチニン外部ドメイン(sHA; アミノ酸番号17〜522)およびノイラミニダーゼ頭部ドメイン(sNA; アミノ酸番号75〜469)をコードするヒトコドン最適化配列を合成し(GenScript)、HEK293T細胞における発現のために発現プラスミドpS1-lgの誘導体[17]にクローニングした。最近になって記述されているように、HA遺伝子の前にN末端CD5シグナルペプチドをコードする配列を置き、HA遺伝子の後にC末端の人工的なGCN4三量化ドメイン(GCN4-pII) [18]およびアフィニティー精製用のStrepタグ(IBA GmbH)をコードする配列を置いた[19, 20]。NA遺伝子の前に、N末端CD5シグナルペプチド、二重のStrepタグ(OneSTrEP; IBA GmbH)および人工的なGCN4四量化ドメイン(GCN4-pLI)を連続的にコードする配列を置いた[18]。
sHA 3 およびsNA 4 抗原の産生
図1Aに図示したように、2009 A(H1N1型)インフルエンザウイルスの三量体HA外部ドメイン(アミノ酸番号17〜522)および四量体NA頭部ドメイン(アミノ酸番号75〜469)を発現するように、構築体をデザインした。sHA3およびsNA4タンパク質をHEK293T細胞における発現によって産生し、アフィニティークロマトグラフィーによって培地から精製し、予想されるサイズの糖タンパク質を得た(図1B)。ゲルろ過分析により、HAおよびNAサブユニットの三量体および四量体オリゴマー性がそれぞれ示された(データ不掲載)。これらの多量体複合体はまた、そのシアル酸結合(sHA3; 投稿準備中)およびノイラミニダーゼ活性(sNA4; 以下)によって判断した場合に、生物学的に活性であり、その天然状態をさらに裏づけた。
図1Aに図示したように、2009 A(H1N1型)インフルエンザウイルスの三量体HA外部ドメイン(アミノ酸番号17〜522)および四量体NA頭部ドメイン(アミノ酸番号75〜469)を発現するように、構築体をデザインした。sHA3およびsNA4タンパク質をHEK293T細胞における発現によって産生し、アフィニティークロマトグラフィーによって培地から精製し、予想されるサイズの糖タンパク質を得た(図1B)。ゲルろ過分析により、HAおよびNAサブユニットの三量体および四量体オリゴマー性がそれぞれ示された(データ不掲載)。これらの多量体複合体はまた、そのシアル酸結合(sHA3; 投稿準備中)およびノイラミニダーゼ活性(sNA4; 以下)によって判断した場合に、生物学的に活性であり、その天然状態をさらに裏づけた。
実施例2. タンパク質発現および精製
ポリエチレンイミン(PEI)を使用して、HEK293T細胞にsHAおよびsNA発現プラスミドを、比率1:5 (μg DNA: μg PEI)でトランスフェクトした。6時間のインキュベーション時間の後、トランスフェクション培地を、重炭酸ナトリウム(3.7 g/リットル)、グルコース(2.0 g/リットル)、プリマトン(Primatone) RL-UF (3.0 g/リットル)、ペニシリン(100単位/ml)、ストレプトマイシン(100 uq/ml)、グルタMAX (Gibco)および1.5% DMSOを補充した293 SFM II発現培地(Invitrogen)に交換した。組織培養上清をトランスフェクションから5〜6日後に収集し、Strep-Tactinアフィニティークロマトグラフィー(IBA GmbH)を用いて培地からsHAおよびsNAタンパク質を精製した。sHAおよびsNAタンパク質の発現および精製を、ウエスタンブロッティングによりStrep-Tractin-HRP結合体(IBA GmbH; データ不掲載)を用いておよびSDS-PAGE分析によって確認した。タンパク質のオリゴマー形成をゲルろ過クロマトグラフィーによっておよびブルーネイティブPAGE分析によって判定した。参照基準としてBSAを用い、タンパク質量の定量化を行った。
ポリエチレンイミン(PEI)を使用して、HEK293T細胞にsHAおよびsNA発現プラスミドを、比率1:5 (μg DNA: μg PEI)でトランスフェクトした。6時間のインキュベーション時間の後、トランスフェクション培地を、重炭酸ナトリウム(3.7 g/リットル)、グルコース(2.0 g/リットル)、プリマトン(Primatone) RL-UF (3.0 g/リットル)、ペニシリン(100単位/ml)、ストレプトマイシン(100 uq/ml)、グルタMAX (Gibco)および1.5% DMSOを補充した293 SFM II発現培地(Invitrogen)に交換した。組織培養上清をトランスフェクションから5〜6日後に収集し、Strep-Tactinアフィニティークロマトグラフィー(IBA GmbH)を用いて培地からsHAおよびsNAタンパク質を精製した。sHAおよびsNAタンパク質の発現および精製を、ウエスタンブロッティングによりStrep-Tractin-HRP結合体(IBA GmbH; データ不掲載)を用いておよびSDS-PAGE分析によって確認した。タンパク質のオリゴマー形成をゲルろ過クロマトグラフィーによっておよびブルーネイティブPAGE分析によって判定した。参照基準としてBSAを用い、タンパク質量の定量化を行った。
実施例3. 免疫および感染
フェレット
健常若年成体の非近交系雌性フェレット(ムステラ・プトリウス・フロ(Mustela putorius furo); 6〜12ヶ月齢)を商業的飼育者から購入した。循環血中の季節性A/H1N1型およびA/H3N2型インフルエンザウイルスに対する抗体ならびにブタ由来A型インフルエンザ/NL/602/09ウイルスに対する抗体がないか、血球凝集阻害アッセイ法によって動物を調べた。実験の開始前に、独立した動物倫理委員会によって実験プロトコルは承認された。
フェレット
健常若年成体の非近交系雌性フェレット(ムステラ・プトリウス・フロ(Mustela putorius furo); 6〜12ヶ月齢)を商業的飼育者から購入した。循環血中の季節性A/H1N1型およびA/H3N2型インフルエンザウイルスに対する抗体ならびにブタ由来A型インフルエンザ/NL/602/09ウイルスに対する抗体がないか、血球凝集阻害アッセイ法によって動物を調べた。実験の開始前に、独立した動物倫理委員会によって実験プロトコルは承認された。
ISCOMマトリックスMアジュバントをWO2004/004762に記述されているように調製した。PBS中85%のマトリックスAおよび15%のマトリックスCを含む組成物75マイクログラムを、抗原に加えた。
免疫および感染
血清反応陰性のフェレット36匹をフェレット各6匹の6群に分け、それらに、以下の配合物を2回ワクチン接種した: リン酸緩衝生理食塩水(PBS)中3.75 μgのsHA3 + 3.75 μgのsNA4 (群1); ISCOMマトリックスM (IMM, Isconova, Uppsala, Sweden)中3.75 μgのsHA3 (群2); IMM中3.75 μgのsNA4 (群3); IMM中3.75 μgのsHA3 + 3.75 μgのsNA4 (群4); PBS (群5); IMM (群6)。ワクチン接種はケタミンによる麻酔下、20日の間隔で、全量1 mlにて後肢の大腿四頭筋に行った。フェレットは群単位で収容し、適宜、食料および水を摂取させた。最後のワクチン接種から32日後に、動物をケタミン/メデトミジンで麻酔し(アチパメゾールで打消し)、体重を量り、その後、3 ml量のPBS中1×106 TCID50のA型インフルエンザ/NL/602/09(H1N1型)で気管内から攻撃した[21, 22]。フェレットをその後、臨床兆候の発生について毎日3回モニタリングした。感染の前にならびに感染から2日および4日後、フェレットをケタミンで麻酔している間に各フェレットの鼻および咽頭スワブを回収した。接種から4日後に、動物を秤量し、その後、ケタミンおよびメデトミジンによる麻酔中に放血によって安楽死させた。標準的な手順によって剖検を行った。実験とは関係ない理由によって1回目および2回目のワクチン接種の間に群1のフェレット1匹が死亡した。
血清反応陰性のフェレット36匹をフェレット各6匹の6群に分け、それらに、以下の配合物を2回ワクチン接種した: リン酸緩衝生理食塩水(PBS)中3.75 μgのsHA3 + 3.75 μgのsNA4 (群1); ISCOMマトリックスM (IMM, Isconova, Uppsala, Sweden)中3.75 μgのsHA3 (群2); IMM中3.75 μgのsNA4 (群3); IMM中3.75 μgのsHA3 + 3.75 μgのsNA4 (群4); PBS (群5); IMM (群6)。ワクチン接種はケタミンによる麻酔下、20日の間隔で、全量1 mlにて後肢の大腿四頭筋に行った。フェレットは群単位で収容し、適宜、食料および水を摂取させた。最後のワクチン接種から32日後に、動物をケタミン/メデトミジンで麻酔し(アチパメゾールで打消し)、体重を量り、その後、3 ml量のPBS中1×106 TCID50のA型インフルエンザ/NL/602/09(H1N1型)で気管内から攻撃した[21, 22]。フェレットをその後、臨床兆候の発生について毎日3回モニタリングした。感染の前にならびに感染から2日および4日後、フェレットをケタミンで麻酔している間に各フェレットの鼻および咽頭スワブを回収した。接種から4日後に、動物を秤量し、その後、ケタミンおよびメデトミジンによる麻酔中に放血によって安楽死させた。標準的な手順によって剖検を行った。実験とは関係ない理由によって1回目および2回目のワクチン接種の間に群1のフェレット1匹が死亡した。
血清学
血清サンプルを、ワクチン接種の前、2回目のワクチン接種の日(20日目)、および攻撃の日(52日目)に回収した。血清を使用まで-20℃で貯蔵した。以前に記載のように、抗HA抗体の存在については1%シチメンチョウ赤血球を用いた血球凝集阻害アッセイ法(HIアッセイ法)を用い、ウイルス中和抗体の存在についてはマイクロウイルス中和アッセイ法(VNアッセイ法)を用いて、血清を調べた[23, 24]。血清をA型インフルエンザ/NL/602/09(H1N1型)と反応性の抗体の存在について調べた。この目的のため、逆遺伝学ウイルスを作製した。これらのウイルスで得られた力価は、野生型株に対する力価に匹敵していた(データ不掲載)。A型インフルエンザ/NL/602/09(H1N1型)に特異的な陽性対照血清を、このウイルスに感染したフェレットから得た[15]。HIアッセイ法において用いた他のH1N1型インフルエンザウイルスは、A/オランダ/386/86(NL/86)、A/オランダ/25/80(NL/80)、A/ニュージャージー/8/76(NJ/76)、A/ブタ/ショープ/1/56(Sw/56)、A/イタリア/1443/76(It/76)、A/アイオワ/15/30(lo/30)、A/プエルトリコ/8/34(Pr/34)およびA/ブリズベン/59/07(IVR-148ワクチン株; IVR/148)であった。これらのウイルスに感染したフェレットの血清サンプルをこのアッセイ法において陽性対照として用いた[25]。
血清サンプルを、ワクチン接種の前、2回目のワクチン接種の日(20日目)、および攻撃の日(52日目)に回収した。血清を使用まで-20℃で貯蔵した。以前に記載のように、抗HA抗体の存在については1%シチメンチョウ赤血球を用いた血球凝集阻害アッセイ法(HIアッセイ法)を用い、ウイルス中和抗体の存在についてはマイクロウイルス中和アッセイ法(VNアッセイ法)を用いて、血清を調べた[23, 24]。血清をA型インフルエンザ/NL/602/09(H1N1型)と反応性の抗体の存在について調べた。この目的のため、逆遺伝学ウイルスを作製した。これらのウイルスで得られた力価は、野生型株に対する力価に匹敵していた(データ不掲載)。A型インフルエンザ/NL/602/09(H1N1型)に特異的な陽性対照血清を、このウイルスに感染したフェレットから得た[15]。HIアッセイ法において用いた他のH1N1型インフルエンザウイルスは、A/オランダ/386/86(NL/86)、A/オランダ/25/80(NL/80)、A/ニュージャージー/8/76(NJ/76)、A/ブタ/ショープ/1/56(Sw/56)、A/イタリア/1443/76(It/76)、A/アイオワ/15/30(lo/30)、A/プエルトリコ/8/34(Pr/34)およびA/ブリズベン/59/07(IVR-148ワクチン株; IVR/148)であった。これらのウイルスに感染したフェレットの血清サンプルをこのアッセイ法において陽性対照として用いた[25]。
血清はまた、ノイラミニダーゼ阻害(NI)抗体の存在について、以前に記載されたフェツインに基づくアッセイ法を用いて調べた[26]。手短に言えば、96ウェルNunc MaxiSorpプレートを5 μg/mlのフェツイン100 μlにより4℃で終夜コーティングした。連続的に希釈した血清サンプル60 μl量を、sNA4含有培養上清(1.5の半最大OD450を与えるようにPBS-Ca/Mg [0.901 mM/0.493 mM]中で予め希釈された)の等量とともに37℃で30分間インキュベートし、その後、この混合物100 μlを、フェツインでコーティングされたウェルに加えた。37℃で1時間のインキュベーションの後、プレートを洗浄し、ノイラミニダーゼ活性をその後、ペルオキシダーゼ標識ピーナッツ凝集素(2.5 μg/ml; Sigma)を添加し、室温で1時間インキュベートし、プレートを洗浄し、各ウェルにペルオキシダーゼ基質(TMB) 100 μlを添加することによって、測定した。5分後、0.3 Mリン酸100 μlの添加によって反応を停止し、ELISA読取機(EL-808 [BioTEK])を用いて450 nmでOD値を測定した。交差反応NI抗体について血清を調べるため、A/カリフォルニア/04/2009(H1N1型)に関する上記のものと類似のsNA4発現構築体を、A/ケンタッキー/UR06-0258/2007(H1N1型)インフルエンザウイルスの頭部ドメイン(アミノ酸番号75〜470)およびA/シチメンチョウ/トルコ/1/2005(H5N1型)インフルエンザウイルスの頭部ドメイン(アミノ酸番号55〜449)についても作製した。これらのウイルスに感染したフェレットから得たA型インフルエンザ/NL/602/09(H1N1型)およびA/シチメンチョウ/トルコ/1/2005(H5N1型)に特異的な血清を陽性対照として用いた。
上気道および下気道におけるウイルス複製
右肺葉および副肺葉の全てのサンプルを感染フェレットから回収し、エタノールとともにドライアイス上で瞬間凍結し、さらなる処理まで-70℃で貯蔵した。肺サンプルを秤量し、その後、0.5%ラクトアルブミン、10%グリセロール、200 U/mlのペニシリン、200 μg/mlのストレプトマイシン、100 U/mlの硫酸ポリミキシンB、250 μg/mlのゲンタマイシンおよび50 U/mlのナイスタチンを含有するハンクス平衡塩類溶液(ICN Pharmaceuticals, Zoetermeer, The Netherlands)中のFastPrep-24 (MP Biomedicals, Eindhoven, The Netherlands)でホモジナイズし、手短に遠心分離した。鼻および咽頭スワブを、肺サンプルをホモジナイズするために用いたのと同じ培地中にて-70℃で直接貯蔵した。咽頭、鼻および肺サンプルの四つ組の10倍連続希釈液を用いて、以前に記載のように、MDCK細胞に感染させた[16]。接種から5日後に回収した培養上清のHA活性を、感染の指標として用いた。力価をSpearman-Karber法にしたがって計算し、肺組織の場合には1グラムあたり、またはスワブの場合には1 mlあたりのlog TCID50として表した[27]。
右肺葉および副肺葉の全てのサンプルを感染フェレットから回収し、エタノールとともにドライアイス上で瞬間凍結し、さらなる処理まで-70℃で貯蔵した。肺サンプルを秤量し、その後、0.5%ラクトアルブミン、10%グリセロール、200 U/mlのペニシリン、200 μg/mlのストレプトマイシン、100 U/mlの硫酸ポリミキシンB、250 μg/mlのゲンタマイシンおよび50 U/mlのナイスタチンを含有するハンクス平衡塩類溶液(ICN Pharmaceuticals, Zoetermeer, The Netherlands)中のFastPrep-24 (MP Biomedicals, Eindhoven, The Netherlands)でホモジナイズし、手短に遠心分離した。鼻および咽頭スワブを、肺サンプルをホモジナイズするために用いたのと同じ培地中にて-70℃で直接貯蔵した。咽頭、鼻および肺サンプルの四つ組の10倍連続希釈液を用いて、以前に記載のように、MDCK細胞に感染させた[16]。接種から5日後に回収した培養上清のHA活性を、感染の指標として用いた。力価をSpearman-Karber法にしたがって計算し、肺組織の場合には1グラムあたり、またはスワブの場合には1 mlあたりのlog TCID50として表した[27]。
病理組織診断
A型インフルエンザ/NL/602/09ウイルスの接種後日数(dpi) 4日目に、フェレットを安楽死させ、肺を肉眼的に観察し、かつ秤量し、その後右肺からのサンプルを回収してウイルス力価を判定した。その後、左肺葉を10%中性緩衝ホルマリンで膨張させた。固定およびパラフィン包埋の後、肺を4 μmの薄片にし、組織切片をヘマトキシリンおよびエオシン(HE)染色によって調べた。
A型インフルエンザ/NL/602/09ウイルスの接種後日数(dpi) 4日目に、フェレットを安楽死させ、肺を肉眼的に観察し、かつ秤量し、その後右肺からのサンプルを回収してウイルス力価を判定した。その後、左肺葉を10%中性緩衝ホルマリンで膨張させた。固定およびパラフィン包埋の後、肺を4 μmの薄片にし、組織切片をヘマトキシリンおよびエオシン(HE)染色によって調べた。
統計解析
動物群間の有意性は一元配置ANOVAおよびANOVAの後のTukey検定によって解析した。差異は、P<0.05で有意とみなした。
動物群間の有意性は一元配置ANOVAおよびANOVAの後のTukey検定によって解析した。差異は、P<0.05で有意とみなした。
結果
sHA3およびsNA4の免疫によって誘導された抗体反応
糖タンパク質を、同種ウイルス攻撃(homologous virus challenge)に対し防御免疫を誘導するその能力について試験した。フェレットを、アジュバントなしのsHA3 + sNA4 (sHA + sNA)、ISCOMマトリックスMで増強したsHA3 + sNA4 (IMM; sHA + sNA + IMM)、または同様に増強したsHA3 (sHA + IMM)もしくはsNA4 (sNA + IMM)を用いて、0日目および20日目に免疫した。血清を2回目の免疫の日および攻撃前の日(20日目および52日目)に回収し、抗体反応を、同種ウイルス(homologous virus)に対するHIおよびVNアッセイ法によって、ならびにNIアッセイ法によって測定した(図2)。これらのアッセイ法のいずれかでの反応は、PBSまたはアジュバントだけをワクチン接種した対照動物では観察されず、また、sHA3およびsNA4の非増強混合物を用いて免疫した動物においても観察されなかった。対照的に、増強したsHA3 (sHA + IMM)による免疫は、高いHI力価を誘導し、攻撃の日に91の幾何平均力価を有した。興味深いことに、sNA4をさらに含めることで(sHA + sNA + IMM)、平均HI力価が468まで著しく増加した(p<0.05; 一元配置ANOVAおよびTukey検定)。また、NI力価測定から、アジュバント依存的なNA抗体誘導が明らかとなり、これは1回の免疫後には低かった(図2)が、しかし2回の免疫後には強力にブーストされた(図2)。しかしながら、この場合、sHA3の同時投与のために、これらの力価の明らかな増大は観察されなかった。観察されたHI力価と一致して、高いVN力価がsHA + IMMワクチン接種動物およびsHA + sNA + IMMワクチン接種動物の両方において認められた(図2)。またここで、sNA4抗原とsHA + sNA + IMMワクチンとの同時投与は、平均VN力価の増加、つまりsHA + IMM群およびsHA + sNA + IMM群それぞれにおいて、1:202から1:468に及ぶ平均値をもたらした。
sHA3およびsNA4の免疫によって誘導された抗体反応
糖タンパク質を、同種ウイルス攻撃(homologous virus challenge)に対し防御免疫を誘導するその能力について試験した。フェレットを、アジュバントなしのsHA3 + sNA4 (sHA + sNA)、ISCOMマトリックスMで増強したsHA3 + sNA4 (IMM; sHA + sNA + IMM)、または同様に増強したsHA3 (sHA + IMM)もしくはsNA4 (sNA + IMM)を用いて、0日目および20日目に免疫した。血清を2回目の免疫の日および攻撃前の日(20日目および52日目)に回収し、抗体反応を、同種ウイルス(homologous virus)に対するHIおよびVNアッセイ法によって、ならびにNIアッセイ法によって測定した(図2)。これらのアッセイ法のいずれかでの反応は、PBSまたはアジュバントだけをワクチン接種した対照動物では観察されず、また、sHA3およびsNA4の非増強混合物を用いて免疫した動物においても観察されなかった。対照的に、増強したsHA3 (sHA + IMM)による免疫は、高いHI力価を誘導し、攻撃の日に91の幾何平均力価を有した。興味深いことに、sNA4をさらに含めることで(sHA + sNA + IMM)、平均HI力価が468まで著しく増加した(p<0.05; 一元配置ANOVAおよびTukey検定)。また、NI力価測定から、アジュバント依存的なNA抗体誘導が明らかとなり、これは1回の免疫後には低かった(図2)が、しかし2回の免疫後には強力にブーストされた(図2)。しかしながら、この場合、sHA3の同時投与のために、これらの力価の明らかな増大は観察されなかった。観察されたHI力価と一致して、高いVN力価がsHA + IMMワクチン接種動物およびsHA + sNA + IMMワクチン接種動物の両方において認められた(図2)。またここで、sNA4抗原とsHA + sNA + IMMワクチンとの同時投与は、平均VN力価の増加、つまりsHA + IMM群およびsHA + sNA + IMM群それぞれにおいて、1:202から1:468に及ぶ平均値をもたらした。
2009 A(Ht Nt)型インフルエンザウイルス感染後の臨床兆候に対する防御
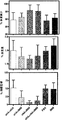
ワクチン接種したフェレットを2回目のワクチン接種から5週後に106 TCID50の2009 A(H1N1型)で攻撃した。接種後2日目以降に、臨床兆候が接種フェレットにおいて観察され、これには呼吸困難、昏睡状態、食欲減退および体重減少が含まれた。全体として、群2、3および4のフェレットでは軽度の臨床兆候しか観察されなかったが、群1、5および6のフェレットでは、より重度の症状が観察された。PBSおよびIMMをワクチン接種した対照群、およびアジュバント非添加sHA + sNAワクチン群では、体重の減少が明らかになった(図3)。興味深いことに、sHA + IMMで免疫した動物はほぼ同様の体重減少を示したが、sNA4含有配合物の両方(sNA + IMMおよびsHA + sNA + IMMの群)のワクチン接種後では、体重は大きな影響を受けなかった。程度の差はあるが一貫して、死後に確定したフェレットの肺重量は、増強したsNA4ワクチン接種動物の、一致した傾向を示し、肺硬変に起因する疾患関連の増加は最も小さかった(図3)。
ワクチン接種したフェレットを2回目のワクチン接種から5週後に106 TCID50の2009 A(H1N1型)で攻撃した。接種後2日目以降に、臨床兆候が接種フェレットにおいて観察され、これには呼吸困難、昏睡状態、食欲減退および体重減少が含まれた。全体として、群2、3および4のフェレットでは軽度の臨床兆候しか観察されなかったが、群1、5および6のフェレットでは、より重度の症状が観察された。PBSおよびIMMをワクチン接種した対照群、およびアジュバント非添加sHA + sNAワクチン群では、体重の減少が明らかになった(図3)。興味深いことに、sHA + IMMで免疫した動物はほぼ同様の体重減少を示したが、sNA4含有配合物の両方(sNA + IMMおよびsHA + sNA + IMMの群)のワクチン接種後では、体重は大きな影響を受けなかった。程度の差はあるが一貫して、死後に確定したフェレットの肺重量は、増強したsNA4ワクチン接種動物の、一致した傾向を示し、肺硬変に起因する疾患関連の増加は最も小さかった(図3)。
フェレットの肺における肉眼的病理所見および組織病理学的所見
インフルエンザウイルスの2009 A(H1N1型)型ウイルス接種から4日後、フェレットの肺を肉眼的に調べ、かつ秤量し、その後ウイルス複製および組織病理学的変化を評価するためサンプルを採取した。接種されたフェレットの肺において、赤黒く堅固に硬化した領域が肉眼的に観察された。罹患した肺組織の割合を評価したところ、群間で異なっていた。約50%である肺中罹患域平均割合が群1、5および6のフェレットでは観察されたが、群2、3および4のフェレットでは硬化の程度があまりはっきりせず、それは25%未満の罹患肺域を示した(図3)。また、相対的な肺重量は、これらの群において、群1、5および6のフェレットにおける肺重量と比べて少なかった(図3)。
インフルエンザウイルスの2009 A(H1N1型)型ウイルス接種から4日後、フェレットの肺を肉眼的に調べ、かつ秤量し、その後ウイルス複製および組織病理学的変化を評価するためサンプルを採取した。接種されたフェレットの肺において、赤黒く堅固に硬化した領域が肉眼的に観察された。罹患した肺組織の割合を評価したところ、群間で異なっていた。約50%である肺中罹患域平均割合が群1、5および6のフェレットでは観察されたが、群2、3および4のフェレットでは硬化の程度があまりはっきりせず、それは25%未満の罹患肺域を示した(図3)。また、相対的な肺重量は、これらの群において、群1、5および6のフェレットにおける肺重量と比べて少なかった(図3)。
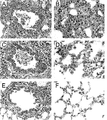
PBSもしくはアジュバントのみ(IMM)を用いてモックワクチン接種したフェレットまたは非増強sHA3 + sNA4を用いてワクチン接種したフェレットの肺において接種後4日目に観察された組織病理学的変化は、中等度から重度の壊死性の気管支間質性肺炎に特徴があった。多病巣性に、多くの好中球およびマクロファージ、ならびに不定数の赤血球、浮腫液および線維素が、これらのフェレットの肺の肺胞に存在していた。さらに、炎症性浸潤が肺胞中隔、細気管支、気管支、ならびに気管支および細気管支の壁に存在していた。組織病理学的変化の劇的な低減が増強したsNA4ワクチン接種動物(sNA + IMMおよびsHA + sNA + IMMの群)において観察されたのに対し、sHA + IMMを免疫したフェレットの病状発現の防止は部分的であった(図4)。
上気道および下気道におけるウイルス複製に対する防御
気道におけるウイルス複製に及ぼすワクチン接種の効果を測定するために、接種から4日後に肺、咽頭および鼻においてウイルス力価を判定した。図5に示されるように、攻撃ウイルス(challenge virus)は、対照フェレット(PBSおよびIMMの群)の肺において、ならびにsHA3およびsNA4の非増強混合物を免疫した動物(sHA + sNAの群)において、効率的に複製し、およそ107〜108 TCID50/グラム組織の平均ウイルス力価を有していた。これらのウイルス負荷は、アジュバント中のsHA3タンパク質で免疫した動物(sHA + IMMの群)において、ならびにアジュバント中のsHA3およびsNA4で同時免疫した動物(sHA + sNA + IMMの群)において、約5 log10単位だけ低減された。平均ウイルス負荷は、増強したsNA4抗原で免疫した動物(sNA + IMMの群)において2〜3 log10単位だけ低減された。
気道におけるウイルス複製に及ぼすワクチン接種の効果を測定するために、接種から4日後に肺、咽頭および鼻においてウイルス力価を判定した。図5に示されるように、攻撃ウイルス(challenge virus)は、対照フェレット(PBSおよびIMMの群)の肺において、ならびにsHA3およびsNA4の非増強混合物を免疫した動物(sHA + sNAの群)において、効率的に複製し、およそ107〜108 TCID50/グラム組織の平均ウイルス力価を有していた。これらのウイルス負荷は、アジュバント中のsHA3タンパク質で免疫した動物(sHA + IMMの群)において、ならびにアジュバント中のsHA3およびsNA4で同時免疫した動物(sHA + sNA + IMMの群)において、約5 log10単位だけ低減された。平均ウイルス負荷は、増強したsNA4抗原で免疫した動物(sNA + IMMの群)において2〜3 log10単位だけ低減された。
鼻における高いウイルス負荷が攻撃後4日目に対照動物(PBSおよびIMMの群; 図5)において観察された。群内の力価のバラツキが大きいために統計的に有意ではないが、これらのウイルス負荷は、増強したsHA3もしくは増強したsNA4、または非増強sHA3 + sNA4の組み合わせを用いて免疫した動物においてやや低いように思われた。鼻ウイルス力価の最大の低減は、sHA3およびsNA4抗原の増強した組み合わせを用いて免疫した動物においてみられた。咽頭におけるウイルス力価は全体として高く、sHA3およびsNA4の増強した組み合わせをワクチン接種した動物を除き、ワクチン接種による有意な影響を受けなかった。これらのフェレットは咽頭において検出可能な力価を有していなかった。
sHA 3 およびsNA 4 の免疫によって誘導される交差反応抗体の反応
sHA3およびsNA4抗原によって誘導される抗体が他のH1N1型インフルエンザウイルスと交差反応しうるかどうか調べるために、本発明者らはワクチン接種後の血清を用いてさらなるHIおよびNIアッセイ法を行った。予想通り、最大のHI力価は同種ウイルスに対して測定されたが、さまざまな程度の交差反応性が一連の他のH1株で観察された(図6A)。したがって、交差反応性はA/ブタ/ショープ/1/56、A/イタリア/1443/76、A/アイオワ/15/30、A/PR/8/34およびIVR/148について検出されなかったが、顕著な交差反応性がA/NL/25/80およびA/ニュージャージー/8/76に対して、特に、A/NL/386/86に対して測定され、多かれ少なかれ、その抗原ドメインの配列類似性と一致していた(表1参照)。
sHA3およびsNA4抗原によって誘導される抗体が他のH1N1型インフルエンザウイルスと交差反応しうるかどうか調べるために、本発明者らはワクチン接種後の血清を用いてさらなるHIおよびNIアッセイ法を行った。予想通り、最大のHI力価は同種ウイルスに対して測定されたが、さまざまな程度の交差反応性が一連の他のH1株で観察された(図6A)。したがって、交差反応性はA/ブタ/ショープ/1/56、A/イタリア/1443/76、A/アイオワ/15/30、A/PR/8/34およびIVR/148について検出されなかったが、顕著な交差反応性がA/NL/25/80およびA/ニュージャージー/8/76に対して、特に、A/NL/386/86に対して測定され、多かれ少なかれ、その抗原ドメインの配列類似性と一致していた(表1参照)。
(表1)異なるH1N1型株のHA中の抗原性領域の配列相同性
 *配列利用不可; n.a. 該当なし
*配列利用不可; n.a. 該当なし
これはsHA3 + IMMおよびsHA3 + sNA4 + IMMワクチン接種動物の両方に由来する血清にも当てはまった。同種ウイルスに対するHI活性の、初期に観察された相違(図2)と一致して、交差反応性のレベルは、sHA3 + IMMを用いて免疫した動物由来の血清よりもsHA3 + sNA4 + IMMを用いて免疫したフェレット由来の血清で著しく高く、かさねて、sNA4抗原の強化効果を裏づけた。各株に対するHI力価が同種インフルエンザA/H1N1型ウイルスに感染したフェレットの対照血清において検出された(データ不掲載)。
NA抗体の交差反応性を調べるために、本発明者らは2つの他のN1インフルエンザウイルスである、ヒトH1N1型株A/ケンタッキー/UR06-0258/2007および鳥H5N1型株A/シチメンチョウ/トルコ/1/2005の、sNA4糖タンパク質複合体を作製した。本発明者らのNIアッセイ法において試験した場合、鳥H5N1型ウイルスsNA4タンパク質に対しsNA4 + IMMおよびsHA3 + sNA4 + IMM免疫動物のプール血清で強力なノイラミニダーゼ阻害活性が認められ、その一方で季節性H1N1型ウイルスsNA4タンパク質のいくらかの阻害が観察された(図68)。注目すべきは、H5N1型ウイルスに感染したニワトリに由来する対照血清の試験結果は、両方のヒトH1N1型ウイルスsNA4タンパク質に対して陰性であった。
参考文献
多量体、HAおよびNA外部ドメインは、これらを大量に容易に、迅速に、柔軟にかつ安全に産生できるので、ワクチンの可能性が大いにある。インフルエンザワクチンの中にNAを含めることは、HAによる防御に大いにかつ特異的に寄与する。ワクチンにおけるその包含は必要とされるHA用量を低減する可能性が高く、防御免疫を広げる可能性が高い。
以下に、本発明の基本的な諸特徴および種々の態様を列挙する。
[1]
少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当する、該組成物。
[2]
少なくとも1種のヘマグルチニンドメインからの1種または複数種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1種または複数種の外部ドメインがハイブリッドタンパク質として提示される、[1]記載の組成物。
[3]
ハイブリッドタンパク質が組換え体である、[2]記載の組成物。
[4]
少なくとも1種のヘマグルチニンドメインからの1種または複数種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1種または複数種の外部ドメインが頭部ドメインである、[1]〜[3]のいずれか一項記載の組成物。
[5]
インフルエンザウイルスがインフルエンザの亜血清型(sub serotype)から、例えばxが1〜16でありかつyが1〜9であるHxNyから選択される、[1]〜[4]のいずれか一項記載の組成物。
[6]
インフルエンザウイルスが、例えば1918 H1N1型インフルエンザウイルス(A/サウスカロライナ/1/18)から選択されるN1H1型のウイルス、ならびに/または2004 H5N1型(A/ベトナム/1203/04) H1N1型A/カリフォルニア/07/2009ウイルス、および/もしくはA/カリフォルニア/04/2009、および/もしくはA/カリフォルニア/09/2009、ならびに/またはA/ケンタッキー/UR06-0258/2007(H1N1型)、ならびに/またはA/シチメンチョウ/トルコ/1/2005(H5N1型)である、[5]記載の組成物。
[7]
少なくとも1種のヘマグルチニンドメインからの1〜5種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1〜5種の頭部ドメインを含む、[1]〜[6]のいずれか一項記載の組成物。
[8]
少なくとも1種のヘマグルチニン外部ドメインがA/カリフォルニア/09/2009の三量体HA外部ドメイン(アミノ酸番号17〜522)から選択される、[7]記載の組成物。
[9]
少なくとも1種のノイラミニダーゼ頭部ドメインがA/カリフォルニア/09/2009ウイルスの四量体NA頭部ドメイン(アミノ酸番号75〜469)から選択される、[7]記載の組成物。
[10]
ISCOM複合体が、少なくとも1種のサポニン、少なくとも1種の脂質および少なくとも1種の抗原を含むISCOMである、[1]〜[9]のいずれか一項記載の組成物。
[11]
ISCOM複合体が、少なくとも1種のサポニン、および少なくとも1種の脂質を含むISCOMマトリックスである、[1]〜[9]のいずれか一項記載の組成物。
[12]
添加剤、賦形剤、さらなるアジュバントもさらに含む、[1]〜[11]のいずれか一項記載の組成物。
[13]
脊椎動物、例えば鳥類および哺乳類用の、例えばインフルエンザに対する免疫刺激薬、免疫調節薬またはワクチンとして用いるための、[1]〜[12]のいずれか一項記載の組成物。
[14]
哺乳類がヒト;ネコ、イヌ、ウマ、鳥、例えばオウムなどのコンパニオンアニマル;畜牛、例えばウシ種、ブタ、ヒツジ、ヤギまたはフェレット、ミンクなどの経済的に重要な種である、[13]記載の組成物。
[15]
1つの区画が少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部外ドメインを含んだ組成物を含み、かつ別の区画が使用説明書を含む、少なくとも2つの区画を含むキット。
以下に、本発明の基本的な諸特徴および種々の態様を列挙する。
[1]
少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当する、該組成物。
[2]
少なくとも1種のヘマグルチニンドメインからの1種または複数種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1種または複数種の外部ドメインがハイブリッドタンパク質として提示される、[1]記載の組成物。
[3]
ハイブリッドタンパク質が組換え体である、[2]記載の組成物。
[4]
少なくとも1種のヘマグルチニンドメインからの1種または複数種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1種または複数種の外部ドメインが頭部ドメインである、[1]〜[3]のいずれか一項記載の組成物。
[5]
インフルエンザウイルスがインフルエンザの亜血清型(sub serotype)から、例えばxが1〜16でありかつyが1〜9であるHxNyから選択される、[1]〜[4]のいずれか一項記載の組成物。
[6]
インフルエンザウイルスが、例えば1918 H1N1型インフルエンザウイルス(A/サウスカロライナ/1/18)から選択されるN1H1型のウイルス、ならびに/または2004 H5N1型(A/ベトナム/1203/04) H1N1型A/カリフォルニア/07/2009ウイルス、および/もしくはA/カリフォルニア/04/2009、および/もしくはA/カリフォルニア/09/2009、ならびに/またはA/ケンタッキー/UR06-0258/2007(H1N1型)、ならびに/またはA/シチメンチョウ/トルコ/1/2005(H5N1型)である、[5]記載の組成物。
[7]
少なくとも1種のヘマグルチニンドメインからの1〜5種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1〜5種の頭部ドメインを含む、[1]〜[6]のいずれか一項記載の組成物。
[8]
少なくとも1種のヘマグルチニン外部ドメインがA/カリフォルニア/09/2009の三量体HA外部ドメイン(アミノ酸番号17〜522)から選択される、[7]記載の組成物。
[9]
少なくとも1種のノイラミニダーゼ頭部ドメインがA/カリフォルニア/09/2009ウイルスの四量体NA頭部ドメイン(アミノ酸番号75〜469)から選択される、[7]記載の組成物。
[10]
ISCOM複合体が、少なくとも1種のサポニン、少なくとも1種の脂質および少なくとも1種の抗原を含むISCOMである、[1]〜[9]のいずれか一項記載の組成物。
[11]
ISCOM複合体が、少なくとも1種のサポニン、および少なくとも1種の脂質を含むISCOMマトリックスである、[1]〜[9]のいずれか一項記載の組成物。
[12]
添加剤、賦形剤、さらなるアジュバントもさらに含む、[1]〜[11]のいずれか一項記載の組成物。
[13]
脊椎動物、例えば鳥類および哺乳類用の、例えばインフルエンザに対する免疫刺激薬、免疫調節薬またはワクチンとして用いるための、[1]〜[12]のいずれか一項記載の組成物。
[14]
哺乳類がヒト;ネコ、イヌ、ウマ、鳥、例えばオウムなどのコンパニオンアニマル;畜牛、例えばウシ種、ブタ、ヒツジ、ヤギまたはフェレット、ミンクなどの経済的に重要な種である、[13]記載の組成物。
[15]
1つの区画が少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部外ドメインを含んだ組成物を含み、かつ別の区画が使用説明書を含む、少なくとも2つの区画を含むキット。
Claims (15)
- 少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部ドメインを含む組成物であって、該外部ドメインがインフルエンザウイルスから単離された外部ドメインに相当する、該組成物。
- 少なくとも1種のヘマグルチニンドメインからの1種または複数種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1種または複数種の外部ドメインがハイブリッドタンパク質として提示される、請求項1記載の組成物。
- ハイブリッドタンパク質が組換え体である、請求項2記載の組成物。
- 少なくとも1種のヘマグルチニンドメインからの1種または複数種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1種または複数種の外部ドメインが頭部ドメインである、請求項1〜3のいずれか一項記載の組成物。
- インフルエンザウイルスがインフルエンザの亜血清型(sub serotype)から、例えばxが1〜16でありかつyが1〜9であるHxNyから選択される、請求項1〜4のいずれか一項記載の組成物。
- インフルエンザウイルスが、例えば1918 H1N1型インフルエンザウイルス(A/サウスカロライナ/1/18)から選択されるN1H1型のウイルス、ならびに/または2004 H5N1型(A/ベトナム/1203/04) H1N1型A/カリフォルニア/07/2009ウイルス、および/もしくはA/カリフォルニア/04/2009、および/もしくはA/カリフォルニア/09/2009、ならびに/またはA/ケンタッキー/UR06-0258/2007(H1N1型)、ならびに/またはA/シチメンチョウ/トルコ/1/2005(H5N1型)である、請求項5記載の組成物。
- 少なくとも1種のヘマグルチニンドメインからの1〜5種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの1〜5種の頭部ドメインを含む、請求項1〜6のいずれか一項記載の組成物。
- 少なくとも1種のヘマグルチニン外部ドメインがA/カリフォルニア/09/2009の三量体HA外部ドメイン(アミノ酸番号17〜522)から選択される、請求項7記載の組成物。
- 少なくとも1種のノイラミニダーゼ頭部ドメインがA/カリフォルニア/09/2009ウイルスの四量体NA頭部ドメイン(アミノ酸番号75〜469)から選択される、請求項7記載の組成物。
- ISCOM複合体が、少なくとも1種のサポニン、少なくとも1種の脂質および少なくとも1種の抗原を含むISCOMである、請求項1〜9のいずれか一項記載の組成物。
- ISCOM複合体が、少なくとも1種のサポニン、および少なくとも1種の脂質を含むISCOMマトリックスである、請求項1〜9のいずれか一項記載の組成物。
- 添加剤、賦形剤、さらなるアジュバントもさらに含む、請求項1〜11のいずれか一項記載の組成物。
- 脊椎動物、例えば鳥類および哺乳類用の、例えばインフルエンザに対する免疫刺激薬、免疫調節薬またはワクチンとして用いるための、請求項1〜12のいずれか一項記載の組成物。
- 哺乳類がヒト;ネコ、イヌ、ウマ、鳥、例えばオウムなどのコンパニオンアニマル;畜牛、例えばウシ種、ブタ、ヒツジ、ヤギまたはフェレット、ミンクなどの経済的に重要な種である、請求項13記載の組成物。
- 1つの区画が少なくとも1種のISCOM複合体、ならびに1種または複数種のインフルエンザウイルスに由来する、少なくとも1種のヘマグルチニンドメインからの少なくとも1種の外部ドメインおよび少なくとも1種のノイラミニダーゼドメインからの少なくとも1種の外部外ドメインを含んだ組成物を含み、かつ別の区画が使用説明書を含む、少なくとも2つの区画を含むキット。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US36698310P | 2010-07-23 | 2010-07-23 | |
| US61/366,983 | 2010-07-23 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013520689A Division JP6133207B2 (ja) | 2010-07-23 | 2011-07-25 | インフルエンザワクチン |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2016190846A true JP2016190846A (ja) | 2016-11-10 |
Family
ID=45497080
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013520689A Active JP6133207B2 (ja) | 2010-07-23 | 2011-07-25 | インフルエンザワクチン |
| JP2016086887A Pending JP2016190846A (ja) | 2010-07-23 | 2016-04-25 | インフルエンザワクチン |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013520689A Active JP6133207B2 (ja) | 2010-07-23 | 2011-07-25 | インフルエンザワクチン |
Country Status (12)
| Country | Link |
|---|---|
| US (2) | US10485863B2 (ja) |
| EP (2) | EP2595653B1 (ja) |
| JP (2) | JP6133207B2 (ja) |
| KR (2) | KR20130095251A (ja) |
| CN (2) | CN103052401A (ja) |
| AU (1) | AU2011280259C1 (ja) |
| BR (1) | BR112013001514A2 (ja) |
| CA (1) | CA2805741C (ja) |
| NZ (1) | NZ606087A (ja) |
| RU (1) | RU2583297C2 (ja) |
| WO (1) | WO2012011868A1 (ja) |
| ZA (1) | ZA201300395B (ja) |
Families Citing this family (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US8927206B2 (en) | 2001-01-19 | 2015-01-06 | Vironovative B.V. | Virus causing respiratory tract illness in susceptible mammals |
| MXPA04008088A (es) | 2002-02-21 | 2005-09-30 | Medimmune Vaccines Inc | Sistema de expresion del virus de parainfluenza recombinante y vacunas que comprenden antigenos heterologos derivados del metapneumovirus. |
| CN103052401A (zh) | 2010-07-23 | 2013-04-17 | 伊斯克诺瓦公司 | 流感疫苗 |
| PL238555B1 (pl) * | 2013-12-20 | 2021-09-06 | Inst Biochemii I Biofizyki Polskiej Akademii Nauk | Sposób wytwarzania hydrofilowej domeny hemaglutyniny wirusa H5 |
| PL235555B1 (pl) | 2014-06-24 | 2020-09-07 | Inst Biotechnologii I Antybiotykow | Wyizolowany i oczyszczony polipeptyd hemaglutyniny (HA ) wirusa grypy H5N1, kompozycja zawierająca polipeptyd i jej zastosowanie, przeciwciało wiążące się specyficznie z polipeptydem oraz sposób otrzymywania tego polipeptydu |
| MX2018002728A (es) | 2015-09-03 | 2018-09-05 | Novavax Inc | Composiciones de vacuna que tienen estabilidad e inmunogenicidad mejoradas. |
| CN108103084B (zh) * | 2017-11-21 | 2020-06-26 | 浙江迪福润丝生物科技有限公司 | 区分免疫和感染动物h5亚型禽流感疫苗株及其制备方法和应用 |
| WO2022034934A1 (ko) * | 2020-08-10 | 2022-02-17 | 한국생명공학연구원 | 인플루엔자 바이러스 백신 조성물 |
| WO2023018817A1 (en) * | 2021-08-11 | 2023-02-16 | Sanofi Pasteur Inc. | Truncated influenza neuraminidase and methods of using the same |
| GB202209588D0 (en) | 2022-06-29 | 2022-08-10 | Plant Bioscience Ltd | Methods and compositions |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH10508301A (ja) * | 1994-10-12 | 1998-08-18 | イスコテック、アクチボラグ | サポニン製剤およびiscomでのその使用 |
| WO2007022425A2 (en) * | 2005-08-16 | 2007-02-22 | Hawaii Biotech, Inc. | Influenza recombinant subunit vaccine |
Family Cites Families (17)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US1048586A (en) | 1911-12-06 | 1912-12-31 | Maurice F Richardson | Spring-trap. |
| SE8205892D0 (sv) | 1982-10-18 | 1982-10-18 | Bror Morein | Immunogent membranproteinkomplex, sett for framstellning och anvendning derav som immunstimulerande medel och sasom vaccin |
| SE8405493D0 (sv) | 1984-11-01 | 1984-11-01 | Bror Morein | Immunogent komplex samt sett for framstellning derav och anvendning derav som immunstimulerande medel |
| CA1331443C (en) | 1987-05-29 | 1994-08-16 | Charlotte A. Kensil | Saponin adjuvant |
| NZ230747A (en) | 1988-09-30 | 1992-05-26 | Bror Morein | Immunomodulating matrix comprising a complex of at least one lipid and at least one saponin; certain glycosylated triterpenoid saponins derived from quillaja saponaria molina |
| SE9600647D0 (sv) | 1996-02-21 | 1996-02-21 | Bror Morein | Ny användning |
| DE60232577D1 (de) * | 2001-01-18 | 2009-07-23 | Vlaams Interuniv Inst Biotech | Oligomerische komplexe von chimären proteinen mit verbessertem immunogenen potential |
| SE0202110D0 (sv) * | 2002-07-05 | 2002-07-05 | Isconova Ab | Iscom preparation and use thereof |
| WO2005117958A1 (en) * | 2004-05-28 | 2005-12-15 | Glaxosmithkline Biologicals S.A. | Vaccine compositions comprising virosomes and a saponin adjuvant |
| EP1938835A1 (en) * | 2006-12-29 | 2008-07-02 | Pevion Biotech AG | Non-specific immunostimulating agents |
| WO2008157419A2 (en) | 2007-06-13 | 2008-12-24 | The Government Of The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Immunogenic peptides of influenza virus |
| NZ583377A (en) | 2007-10-12 | 2013-11-29 | Csl Ltd | Method of eliciting an immune response against pandemic influenza virus |
| GB0810305D0 (en) * | 2008-06-05 | 2008-07-09 | Novartis Ag | Influenza vaccination |
| CN105753948A (zh) * | 2008-07-08 | 2016-07-13 | 麦迪卡格公司 | 可溶性重组流感抗原 |
| WO2010144797A2 (en) | 2009-06-12 | 2010-12-16 | Vaccine Technologies, Incorporated | Influenza vaccines with enhanced immunogenicity and uses thereof |
| WO2011126370A1 (en) * | 2010-04-09 | 2011-10-13 | Universiteit Utrecht Holding B.V. | Recombinant multimeric influenza proteins |
| CN103052401A (zh) | 2010-07-23 | 2013-04-17 | 伊斯克诺瓦公司 | 流感疫苗 |
-
2011
- 2011-07-25 CN CN2011800356580A patent/CN103052401A/zh active Pending
- 2011-07-25 KR KR1020137001862A patent/KR20130095251A/ko active Search and Examination
- 2011-07-25 KR KR1020177035606A patent/KR102143105B1/ko active IP Right Grant
- 2011-07-25 EP EP11809955.5A patent/EP2595653B1/en active Active
- 2011-07-25 NZ NZ606087A patent/NZ606087A/en unknown
- 2011-07-25 EP EP17175284.3A patent/EP3272359A1/en not_active Withdrawn
- 2011-07-25 RU RU2013108067/10A patent/RU2583297C2/ru active
- 2011-07-25 CN CN201710126536.1A patent/CN107281478B/zh active Active
- 2011-07-25 BR BR112013001514A patent/BR112013001514A2/pt not_active Application Discontinuation
- 2011-07-25 WO PCT/SE2011/050968 patent/WO2012011868A1/en active Application Filing
- 2011-07-25 JP JP2013520689A patent/JP6133207B2/ja active Active
- 2011-07-25 US US13/811,493 patent/US10485863B2/en active Active
- 2011-07-25 CA CA2805741A patent/CA2805741C/en active Active
- 2011-07-25 AU AU2011280259A patent/AU2011280259C1/en active Active
-
2013
- 2013-01-15 ZA ZA2013/00395A patent/ZA201300395B/en unknown
-
2016
- 2016-04-25 JP JP2016086887A patent/JP2016190846A/ja active Pending
-
2019
- 2019-01-29 US US16/260,557 patent/US10736958B2/en active Active
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH10508301A (ja) * | 1994-10-12 | 1998-08-18 | イスコテック、アクチボラグ | サポニン製剤およびiscomでのその使用 |
| WO2007022425A2 (en) * | 2005-08-16 | 2007-02-22 | Hawaii Biotech, Inc. | Influenza recombinant subunit vaccine |
Non-Patent Citations (4)
| Title |
|---|
| JOURNAL OF VIROLOGY, vol. 79, no. 19, JPN7015001203, 2005, pages 12401 - 12407, ISSN: 0003503656 * |
| PROCEEDING OF THE NATIONAL ACADEMY OF SCIENCES, vol. 101, no. 5, JPN6017005544, 2004, pages 1356 - 1361, ISSN: 0003503657 * |
| PROTEINS: STRUCTURE, FUNCTION, AND GENETICS, vol. 6, JPN6017005551, 1989, pages 341 - 356, ISSN: 0003503659 * |
| 日本臨牀, vol. 55, no. 10, JPN6017005548, 1997, pages 64 - 2562, ISSN: 0003503658 * |
Also Published As
| Publication number | Publication date |
|---|---|
| AU2011280259B2 (en) | 2014-05-22 |
| EP2595653A4 (en) | 2014-03-12 |
| AU2011280259B8 (en) | 2014-09-25 |
| CN107281478A (zh) | 2017-10-24 |
| KR102143105B1 (ko) | 2020-08-11 |
| EP2595653A1 (en) | 2013-05-29 |
| BR112013001514A2 (pt) | 2016-06-07 |
| EP3272359A1 (en) | 2018-01-24 |
| NZ606087A (en) | 2014-06-27 |
| US20130129770A1 (en) | 2013-05-23 |
| KR20130095251A (ko) | 2013-08-27 |
| AU2011280259A1 (en) | 2013-02-07 |
| US10736958B2 (en) | 2020-08-11 |
| CN107281478B (zh) | 2022-04-01 |
| RU2013108067A (ru) | 2014-08-27 |
| CA2805741A1 (en) | 2012-01-26 |
| EP2595653B1 (en) | 2017-06-28 |
| CN103052401A (zh) | 2013-04-17 |
| JP2013533270A (ja) | 2013-08-22 |
| AU2011280259A8 (en) | 2013-02-28 |
| KR20170141268A (ko) | 2017-12-22 |
| WO2012011868A1 (en) | 2012-01-26 |
| US20190388534A1 (en) | 2019-12-26 |
| AU2011280259C1 (en) | 2018-06-07 |
| ZA201300395B (en) | 2013-09-25 |
| US10485863B2 (en) | 2019-11-26 |
| JP6133207B2 (ja) | 2017-05-24 |
| CA2805741C (en) | 2019-08-13 |
| RU2583297C2 (ru) | 2016-05-10 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6133207B2 (ja) | インフルエンザワクチン | |
| ES2377761T3 (es) | Vacuna contra la gripe de emulsión de aceite en agua | |
| AU2006279323B2 (en) | Influenza recombinant subunit vaccine | |
| RU2681529C2 (ru) | Иммуногенные композиции гликопротеина g вирусов hendra и nipah | |
| RU2410427C2 (ru) | Вирус собачьего гриппа, композиции, содержащие этот вирус, и способы их применения | |
| Sui et al. | Cross-protection against influenza virus infection by intranasal administration of M2-based vaccine with chitosan as an adjuvant | |
| EA018860B1 (ru) | Вакцинные композиции, содержащие сапониновый адъювант | |
| MX2012011751A (es) | Proteinas multimericas recombinantes de la influenza. | |
| CN104136456A (zh) | 以计算方式优化的h3n2、h2n2和b型流感病毒的广泛反应性抗原 | |
| JP2011524372A (ja) | インフルエンザワクチン接種のための新規ペプチドアジュバント | |
| KR20160077214A (ko) | 헨드라 및 니파 바이러스 g 당단백질 면역원성 조성물 | |
| TW202308685A (zh) | 冠狀病毒和流感病毒組合物及其使用方法 | |
| JP7107910B2 (ja) | ユニバーサルインフルエンザワクチン | |
| MX2013003451A (es) | Generacion de particulas de virosoma. | |
| RU2787820C2 (ru) | Иммуногенные композиции гликопротеина g вирусов hendra и nipah | |
| JP2023538539A (ja) | 抗コロナウイルスワクチン | |
| NZ617722B2 (en) | Hendra and nipah virus g glycoprotein immunogenic compositions |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20170222 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20170519 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20170626 |