CN1384880A - L-泛内酯水解酶和一种d-泛内酯的制备方法 - Google Patents
L-泛内酯水解酶和一种d-泛内酯的制备方法 Download PDFInfo
- Publication number
- CN1384880A CN1384880A CN00814937A CN00814937A CN1384880A CN 1384880 A CN1384880 A CN 1384880A CN 00814937 A CN00814937 A CN 00814937A CN 00814937 A CN00814937 A CN 00814937A CN 1384880 A CN1384880 A CN 1384880A
- Authority
- CN
- China
- Prior art keywords
- pantolactone
- sequence
- enzyme
- lytic enzyme
- ala
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- SERHXTVXHNVDKA-BYPYZUCNSA-N (R)-pantolactone Chemical compound CC1(C)COC(=O)[C@@H]1O SERHXTVXHNVDKA-BYPYZUCNSA-N 0.000 title claims abstract description 48
- 238000004519 manufacturing process Methods 0.000 title abstract description 4
- SERHXTVXHNVDKA-SCSAIBSYSA-N (3s)-3-hydroxy-4,4-dimethyloxolan-2-one Chemical compound CC1(C)COC(=O)[C@H]1O SERHXTVXHNVDKA-SCSAIBSYSA-N 0.000 claims abstract description 85
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 39
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 34
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 34
- 244000005700 microbiome Species 0.000 claims abstract description 17
- 238000000034 method Methods 0.000 claims description 62
- 101000925662 Enterobacteria phage PRD1 Endolysin Proteins 0.000 claims description 49
- 241000894006 Bacteria Species 0.000 claims description 34
- 229940115458 pantolactone Drugs 0.000 claims description 26
- 238000006243 chemical reaction Methods 0.000 claims description 25
- SERHXTVXHNVDKA-UHFFFAOYSA-N pantolactone Chemical compound CC1(C)COC(=O)C1O SERHXTVXHNVDKA-UHFFFAOYSA-N 0.000 claims description 24
- SIEVQTNTRMBCHO-UHFFFAOYSA-N pantolactone Natural products CC1(C)OC(=O)CC1O SIEVQTNTRMBCHO-UHFFFAOYSA-N 0.000 claims description 24
- 239000002773 nucleotide Substances 0.000 claims description 22
- 125000003729 nucleotide group Chemical group 0.000 claims description 22
- OTOIIPJYVQJATP-SCSAIBSYSA-N (2s)-2,4-dihydroxy-3,3-dimethylbutanoic acid Chemical compound OCC(C)(C)[C@H](O)C(O)=O OTOIIPJYVQJATP-SCSAIBSYSA-N 0.000 claims description 18
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 claims description 13
- 230000000694 effects Effects 0.000 claims description 10
- 150000001413 amino acids Chemical class 0.000 claims description 9
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 9
- 241000589158 Agrobacterium Species 0.000 claims description 8
- 238000000605 extraction Methods 0.000 claims description 7
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 claims description 6
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 6
- 239000002253 acid Substances 0.000 claims description 6
- 230000002068 genetic effect Effects 0.000 claims description 6
- 235000001014 amino acid Nutrition 0.000 claims description 5
- 241000190932 Rhodopseudomonas Species 0.000 claims description 4
- 229920001184 polypeptide Polymers 0.000 claims description 4
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 4
- 230000006340 racemization Effects 0.000 claims description 4
- 230000006269 (delayed) early viral mRNA transcription Effects 0.000 claims description 3
- 241000588921 Enterobacteriaceae Species 0.000 claims description 3
- 241000588722 Escherichia Species 0.000 claims description 3
- 241000947836 Pseudomonadaceae Species 0.000 claims description 3
- 241001633102 Rhizobiaceae Species 0.000 claims description 3
- 241000607142 Salmonella Species 0.000 claims description 3
- 230000003750 conditioning effect Effects 0.000 claims description 3
- 230000003287 optical effect Effects 0.000 claims description 3
- 101000924393 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) Vacuolar aminopeptidase 1 Proteins 0.000 claims description 2
- 230000005059 dormancy Effects 0.000 claims description 2
- 125000003275 alpha amino acid group Chemical group 0.000 claims 5
- 108090000623 proteins and genes Proteins 0.000 abstract description 38
- 102000004169 proteins and genes Human genes 0.000 abstract description 17
- 230000003301 hydrolyzing effect Effects 0.000 abstract description 2
- 230000002255 enzymatic effect Effects 0.000 abstract 1
- 239000013598 vector Substances 0.000 abstract 1
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 57
- 102000004190 Enzymes Human genes 0.000 description 52
- 108090000790 Enzymes Proteins 0.000 description 52
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 46
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 35
- 239000000872 buffer Substances 0.000 description 27
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 26
- 239000000243 solution Substances 0.000 description 26
- 239000007983 Tris buffer Substances 0.000 description 24
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 24
- 238000006460 hydrolysis reaction Methods 0.000 description 23
- 108020004414 DNA Proteins 0.000 description 21
- 239000000047 product Substances 0.000 description 21
- 241000588724 Escherichia coli Species 0.000 description 20
- 241001135514 Paraburkholderia caryophylli Species 0.000 description 19
- 230000007062 hydrolysis Effects 0.000 description 19
- 239000012530 fluid Substances 0.000 description 17
- OTOIIPJYVQJATP-BYPYZUCNSA-N (R)-pantoic acid Chemical compound OCC(C)(C)[C@@H](O)C(O)=O OTOIIPJYVQJATP-BYPYZUCNSA-N 0.000 description 14
- 125000000468 ketone group Chemical group 0.000 description 14
- 239000000725 suspension Substances 0.000 description 14
- 238000004448 titration Methods 0.000 description 14
- 238000013016 damping Methods 0.000 description 13
- 239000013612 plasmid Substances 0.000 description 13
- 235000018102 proteins Nutrition 0.000 description 13
- 239000011780 sodium chloride Substances 0.000 description 13
- XEKOWRVHYACXOJ-UHFFFAOYSA-N Ethyl acetate Chemical compound CCOC(C)=O XEKOWRVHYACXOJ-UHFFFAOYSA-N 0.000 description 12
- 239000000203 mixture Substances 0.000 description 12
- 239000006228 supernatant Substances 0.000 description 12
- 241000589155 Agrobacterium tumefaciens Species 0.000 description 11
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 11
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 11
- 241000233866 Fungi Species 0.000 description 10
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 10
- 229920002684 Sepharose Polymers 0.000 description 10
- 238000009396 hybridization Methods 0.000 description 10
- 238000005406 washing Methods 0.000 description 10
- CSCPPACGZOOCGX-UHFFFAOYSA-N Acetone Chemical compound CC(C)=O CSCPPACGZOOCGX-UHFFFAOYSA-N 0.000 description 9
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 9
- DBMJMQXJHONAFJ-UHFFFAOYSA-M Sodium laurylsulphate Chemical compound [Na+].CCCCCCCCCCCCOS([O-])(=O)=O DBMJMQXJHONAFJ-UHFFFAOYSA-M 0.000 description 9
- 239000004141 Sodium laurylsulphate Substances 0.000 description 9
- 230000001580 bacterial effect Effects 0.000 description 9
- 239000002585 base Substances 0.000 description 9
- 229940041514 candida albicans extract Drugs 0.000 description 9
- 230000002349 favourable effect Effects 0.000 description 9
- 238000004128 high performance liquid chromatography Methods 0.000 description 9
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 9
- 239000000523 sample Substances 0.000 description 9
- 235000019333 sodium laurylsulphate Nutrition 0.000 description 9
- 239000012138 yeast extract Substances 0.000 description 9
- 108010006519 Molecular Chaperones Proteins 0.000 description 8
- 239000001888 Peptone Substances 0.000 description 8
- 108010080698 Peptones Proteins 0.000 description 8
- 238000004458 analytical method Methods 0.000 description 8
- 238000004587 chromatography analysis Methods 0.000 description 8
- 238000002425 crystallisation Methods 0.000 description 8
- 230000008025 crystallization Effects 0.000 description 8
- 230000006870 function Effects 0.000 description 8
- JXDYKVIHCLTXOP-UHFFFAOYSA-N isatin Chemical compound C1=CC=C2C(=O)C(=O)NC2=C1 JXDYKVIHCLTXOP-UHFFFAOYSA-N 0.000 description 8
- 235000019319 peptone Nutrition 0.000 description 8
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 8
- 239000012064 sodium phosphate buffer Substances 0.000 description 8
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 7
- 239000011521 glass Substances 0.000 description 7
- -1 methane amide Chemical class 0.000 description 7
- 230000001105 regulatory effect Effects 0.000 description 7
- 239000002904 solvent Substances 0.000 description 7
- 239000000758 substrate Substances 0.000 description 7
- 239000006285 cell suspension Substances 0.000 description 6
- 150000002596 lactones Chemical class 0.000 description 6
- 238000002360 preparation method Methods 0.000 description 6
- 239000011734 sodium Substances 0.000 description 6
- 239000012137 tryptone Substances 0.000 description 6
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 5
- 102100038222 60 kDa heat shock protein, mitochondrial Human genes 0.000 description 5
- 241000193830 Bacillus <bacterium> Species 0.000 description 5
- 241000589539 Brevundimonas diminuta Species 0.000 description 5
- 108010058432 Chaperonin 60 Proteins 0.000 description 5
- 108090000604 Hydrolases Proteins 0.000 description 5
- 102000005431 Molecular Chaperones Human genes 0.000 description 5
- 241000235648 Pichia Species 0.000 description 5
- 241000589516 Pseudomonas Species 0.000 description 5
- 230000008859 change Effects 0.000 description 5
- 239000003292 glue Substances 0.000 description 5
- 238000011534 incubation Methods 0.000 description 5
- 230000002829 reductive effect Effects 0.000 description 5
- 238000003756 stirring Methods 0.000 description 5
- 239000000126 substance Substances 0.000 description 5
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 4
- 241000223679 Beauveria Species 0.000 description 4
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 4
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 4
- 241001237914 Psilocybe Species 0.000 description 4
- 241001655322 Streptomycetales Species 0.000 description 4
- QAOWNCQODCNURD-UHFFFAOYSA-L Sulfate Chemical compound [O-]S([O-])(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-L 0.000 description 4
- YXFVVABEGXRONW-UHFFFAOYSA-N Toluene Chemical compound CC1=CC=CC=C1 YXFVVABEGXRONW-UHFFFAOYSA-N 0.000 description 4
- 150000001299 aldehydes Chemical class 0.000 description 4
- 229910021529 ammonia Inorganic materials 0.000 description 4
- KLOHDWPABZXLGI-YWUHCJSESA-M ampicillin sodium Chemical compound [Na+].C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C([O-])=O)(C)C)=CC=CC=C1 KLOHDWPABZXLGI-YWUHCJSESA-M 0.000 description 4
- 239000007864 aqueous solution Substances 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 239000012153 distilled water Substances 0.000 description 4
- 238000010828 elution Methods 0.000 description 4
- 230000008014 freezing Effects 0.000 description 4
- 238000007710 freezing Methods 0.000 description 4
- 230000000968 intestinal effect Effects 0.000 description 4
- 239000002609 medium Substances 0.000 description 4
- 239000012528 membrane Substances 0.000 description 4
- 238000010369 molecular cloning Methods 0.000 description 4
- 239000012074 organic phase Substances 0.000 description 4
- 238000001556 precipitation Methods 0.000 description 4
- 230000009467 reduction Effects 0.000 description 4
- 108091008146 restriction endonucleases Proteins 0.000 description 4
- 102220023258 rs387907548 Human genes 0.000 description 4
- 229910000033 sodium borohydride Inorganic materials 0.000 description 4
- 239000012279 sodium borohydride Substances 0.000 description 4
- 230000009466 transformation Effects 0.000 description 4
- 239000003643 water by type Substances 0.000 description 4
- NWXMGUDVXFXRIG-WESIUVDSSA-N (4s,4as,5as,6s,12ar)-4-(dimethylamino)-1,6,10,11,12a-pentahydroxy-6-methyl-3,12-dioxo-4,4a,5,5a-tetrahydrotetracene-2-carboxamide Chemical compound C1=CC=C2[C@](O)(C)[C@H]3C[C@H]4[C@H](N(C)C)C(=O)C(C(N)=O)=C(O)[C@@]4(O)C(=O)C3=C(O)C2=C1O NWXMGUDVXFXRIG-WESIUVDSSA-N 0.000 description 3
- 238000005160 1H NMR spectroscopy Methods 0.000 description 3
- OPIFSICVWOWJMJ-AEOCFKNESA-N 5-bromo-4-chloro-3-indolyl beta-D-galactoside Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1OC1=CNC2=CC=C(Br)C(Cl)=C12 OPIFSICVWOWJMJ-AEOCFKNESA-N 0.000 description 3
- 241001523968 Beauveria amorpha Species 0.000 description 3
- 241001453380 Burkholderia Species 0.000 description 3
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 3
- 238000010268 HPLC based assay Methods 0.000 description 3
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 3
- 108010093096 Immobilized Enzymes Proteins 0.000 description 3
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 3
- 108090001060 Lipase Proteins 0.000 description 3
- 239000004367 Lipase Substances 0.000 description 3
- 102000004882 Lipase Human genes 0.000 description 3
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 3
- 241000187654 Nocardia Species 0.000 description 3
- 241000589776 Pseudomonas putida Species 0.000 description 3
- 241000222050 Vanrija humicola Species 0.000 description 3
- 230000004913 activation Effects 0.000 description 3
- 230000000740 bleeding effect Effects 0.000 description 3
- 230000007073 chemical hydrolysis Effects 0.000 description 3
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 3
- 239000000287 crude extract Substances 0.000 description 3
- 230000030609 dephosphorylation Effects 0.000 description 3
- 238000006209 dephosphorylation reaction Methods 0.000 description 3
- 239000000284 extract Substances 0.000 description 3
- 238000001914 filtration Methods 0.000 description 3
- 238000002290 gas chromatography-mass spectrometry Methods 0.000 description 3
- 239000000499 gel Substances 0.000 description 3
- 238000010438 heat treatment Methods 0.000 description 3
- 235000019421 lipase Nutrition 0.000 description 3
- 239000007788 liquid Substances 0.000 description 3
- BZLVMXJERCGZMT-UHFFFAOYSA-N methyl tert-butyl ether Substances COC(C)(C)C BZLVMXJERCGZMT-UHFFFAOYSA-N 0.000 description 3
- 239000002244 precipitate Substances 0.000 description 3
- 150000003839 salts Chemical class 0.000 description 3
- 229910052708 sodium Inorganic materials 0.000 description 3
- GHOKWGTUZJEAQD-ZETCQYMHSA-N (D)-(+)-Pantothenic acid Chemical compound OCC(C)(C)[C@@H](O)C(=O)NCCC(O)=O GHOKWGTUZJEAQD-ZETCQYMHSA-N 0.000 description 2
- 102100024341 10 kDa heat shock protein, mitochondrial Human genes 0.000 description 2
- 229920000936 Agarose Polymers 0.000 description 2
- PCIFXPRIFWKWLK-YUMQZZPRSA-N Ala-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@H](C)N PCIFXPRIFWKWLK-YUMQZZPRSA-N 0.000 description 2
- 241001135756 Alphaproteobacteria Species 0.000 description 2
- QGZKDVFQNNGYKY-UHFFFAOYSA-N Ammonia Chemical compound N QGZKDVFQNNGYKY-UHFFFAOYSA-N 0.000 description 2
- 241001504907 Apiotrichum Species 0.000 description 2
- 241000186063 Arthrobacter Species 0.000 description 2
- 241001135755 Betaproteobacteria Species 0.000 description 2
- BTBUEUYNUDRHOZ-UHFFFAOYSA-N Borate Chemical compound [O-]B([O-])[O-] BTBUEUYNUDRHOZ-UHFFFAOYSA-N 0.000 description 2
- 101710163595 Chaperone protein DnaK Proteins 0.000 description 2
- 108010059013 Chaperonin 10 Proteins 0.000 description 2
- 108700010070 Codon Usage Proteins 0.000 description 2
- 101710095468 Cyclase Proteins 0.000 description 2
- KIQKJXYVGSYDFS-ZLUOBGJFSA-N Cys-Asn-Asn Chemical compound SC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O KIQKJXYVGSYDFS-ZLUOBGJFSA-N 0.000 description 2
- NGOIQDYZMIKCOK-NAKRPEOUSA-N Cys-Val-Ile Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O NGOIQDYZMIKCOK-NAKRPEOUSA-N 0.000 description 2
- SNPLKNRPJHDVJA-ZETCQYMHSA-N D-panthenol Chemical compound OCC(C)(C)[C@@H](O)C(=O)NCCCO SNPLKNRPJHDVJA-ZETCQYMHSA-N 0.000 description 2
- 241000801634 Deconica coprophila Species 0.000 description 2
- YMWUJEATGCHHMB-UHFFFAOYSA-N Dichloromethane Chemical compound ClCCl YMWUJEATGCHHMB-UHFFFAOYSA-N 0.000 description 2
- 241000305071 Enterobacterales Species 0.000 description 2
- 108090000371 Esterases Proteins 0.000 description 2
- 241000206602 Eukaryota Species 0.000 description 2
- 241000282326 Felis catus Species 0.000 description 2
- 206010016825 Flushing Diseases 0.000 description 2
- 241000221778 Fusarium fujikuroi Species 0.000 description 2
- 241000192128 Gammaproteobacteria Species 0.000 description 2
- PCBBLFVHTYNQGG-LAEOZQHASA-N Glu-Asn-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CCC(=O)O)N PCBBLFVHTYNQGG-LAEOZQHASA-N 0.000 description 2
- PYTZFYUXZZHOAD-WHFBIAKZSA-N Gly-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)CN PYTZFYUXZZHOAD-WHFBIAKZSA-N 0.000 description 2
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 2
- 108010045100 HSP27 Heat-Shock Proteins Proteins 0.000 description 2
- 108010042283 HSP40 Heat-Shock Proteins Proteins 0.000 description 2
- 102000004447 HSP40 Heat-Shock Proteins Human genes 0.000 description 2
- 101710178376 Heat shock 70 kDa protein Proteins 0.000 description 2
- 101710152018 Heat shock cognate 70 kDa protein Proteins 0.000 description 2
- 102100039165 Heat shock protein beta-1 Human genes 0.000 description 2
- 102000004157 Hydrolases Human genes 0.000 description 2
- 102000004877 Insulin Human genes 0.000 description 2
- 108090001061 Insulin Proteins 0.000 description 2
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 2
- PJWOOBTYQNNRBF-BZSNNMDCSA-N Leu-Phe-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCCN)C(=O)O)N PJWOOBTYQNNRBF-BZSNNMDCSA-N 0.000 description 2
- 108010071324 Livagen Proteins 0.000 description 2
- RIJCHEVHFWMDKD-SRVKXCTJSA-N Lys-Lys-Asn Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(O)=O RIJCHEVHFWMDKD-SRVKXCTJSA-N 0.000 description 2
- DBXMFHGGHMXYHY-DCAQKATOSA-N Met-Leu-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O DBXMFHGGHMXYHY-DCAQKATOSA-N 0.000 description 2
- UIHCLUNTQKBZGK-UHFFFAOYSA-N Methyl isobutyl ketone Natural products CCC(C)C(C)=O UIHCLUNTQKBZGK-UHFFFAOYSA-N 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 108700026244 Open Reading Frames Proteins 0.000 description 2
- 101710157860 Oxydoreductase Proteins 0.000 description 2
- 238000012408 PCR amplification Methods 0.000 description 2
- 244000131316 Panax pseudoginseng Species 0.000 description 2
- 235000005035 Panax pseudoginseng ssp. pseudoginseng Nutrition 0.000 description 2
- 235000003140 Panax quinquefolius Nutrition 0.000 description 2
- 108010076039 Polyproteins Proteins 0.000 description 2
- 239000012614 Q-Sepharose Substances 0.000 description 2
- 241000316848 Rhodococcus <scale insect> Species 0.000 description 2
- 241000190946 Rhodopseudomonas sp. Species 0.000 description 2
- 241000223252 Rhodotorula Species 0.000 description 2
- WQDUMFSSJAZKTM-UHFFFAOYSA-N Sodium methoxide Chemical compound [Na+].[O-]C WQDUMFSSJAZKTM-UHFFFAOYSA-N 0.000 description 2
- 244000061458 Solanum melongena Species 0.000 description 2
- 235000002597 Solanum melongena Nutrition 0.000 description 2
- 101000693619 Starmerella bombicola Lactone esterase Proteins 0.000 description 2
- 241000063122 Streptacidiphilus griseus Species 0.000 description 2
- 241000187747 Streptomyces Species 0.000 description 2
- QAOWNCQODCNURD-UHFFFAOYSA-N Sulfuric acid Chemical compound OS(O)(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-N 0.000 description 2
- 239000012505 Superdex™ Substances 0.000 description 2
- GFDUZZACIWNMPE-KZVJFYERSA-N Thr-Ala-Met Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCSC)C(O)=O GFDUZZACIWNMPE-KZVJFYERSA-N 0.000 description 2
- XTCNBOBTROGWMW-RWRJDSDZSA-N Thr-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N XTCNBOBTROGWMW-RWRJDSDZSA-N 0.000 description 2
- 235000010724 Wisteria floribunda Nutrition 0.000 description 2
- 241000588902 Zymomonas mobilis Species 0.000 description 2
- 239000000654 additive Substances 0.000 description 2
- 239000003513 alkali Substances 0.000 description 2
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- 102220369446 c.1274G>A Human genes 0.000 description 2
- 102220369445 c.668T>C Human genes 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- RCTYPNKXASFOBE-UHFFFAOYSA-M chloromercury Chemical compound [Hg]Cl RCTYPNKXASFOBE-UHFFFAOYSA-M 0.000 description 2
- 230000004087 circulation Effects 0.000 description 2
- 238000004040 coloring Methods 0.000 description 2
- 239000012141 concentrate Substances 0.000 description 2
- NKLPQNGYXWVELD-UHFFFAOYSA-M coomassie brilliant blue Chemical compound [Na+].C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=2C=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=C1 NKLPQNGYXWVELD-UHFFFAOYSA-M 0.000 description 2
- 239000008367 deionised water Substances 0.000 description 2
- 229910021641 deionized water Inorganic materials 0.000 description 2
- 238000013461 design Methods 0.000 description 2
- 238000001514 detection method Methods 0.000 description 2
- 238000000502 dialysis Methods 0.000 description 2
- 230000008034 disappearance Effects 0.000 description 2
- 239000003814 drug Substances 0.000 description 2
- 238000004043 dyeing Methods 0.000 description 2
- 239000012636 effector Substances 0.000 description 2
- 238000001976 enzyme digestion Methods 0.000 description 2
- 238000011067 equilibration Methods 0.000 description 2
- 238000001704 evaporation Methods 0.000 description 2
- 230000008020 evaporation Effects 0.000 description 2
- 230000007717 exclusion Effects 0.000 description 2
- 238000011010 flushing procedure Methods 0.000 description 2
- 230000004927 fusion Effects 0.000 description 2
- 238000012239 gene modification Methods 0.000 description 2
- 230000005017 genetic modification Effects 0.000 description 2
- 235000013617 genetically modified food Nutrition 0.000 description 2
- 235000008434 ginseng Nutrition 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- 108010049041 glutamylalanine Proteins 0.000 description 2
- LELOWRISYMNNSU-UHFFFAOYSA-N hydrogen cyanide Chemical compound N#C LELOWRISYMNNSU-UHFFFAOYSA-N 0.000 description 2
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 2
- 230000005764 inhibitory process Effects 0.000 description 2
- 239000003999 initiator Substances 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- 108010064235 lysylglycine Proteins 0.000 description 2
- 230000002101 lytic effect Effects 0.000 description 2
- 238000013507 mapping Methods 0.000 description 2
- 239000000463 material Substances 0.000 description 2
- 230000004060 metabolic process Effects 0.000 description 2
- VNWKTOKETHGBQD-UHFFFAOYSA-N methane Natural products C VNWKTOKETHGBQD-UHFFFAOYSA-N 0.000 description 2
- 229940043265 methyl isobutyl ketone Drugs 0.000 description 2
- 230000000813 microbial effect Effects 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 230000035772 mutation Effects 0.000 description 2
- 229930027945 nicotinamide-adenine dinucleotide Natural products 0.000 description 2
- 235000015097 nutrients Nutrition 0.000 description 2
- 239000003960 organic solvent Substances 0.000 description 2
- ZNXZGRMVNNHPCA-VIFPVBQESA-N pantetheine Chemical compound OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCS ZNXZGRMVNNHPCA-VIFPVBQESA-N 0.000 description 2
- 229940101267 panthenol Drugs 0.000 description 2
- 235000020957 pantothenol Nutrition 0.000 description 2
- 239000011619 pantothenol Substances 0.000 description 2
- FDPIMTJIUBPUKL-UHFFFAOYSA-N pentan-3-one Chemical compound CCC(=O)CC FDPIMTJIUBPUKL-UHFFFAOYSA-N 0.000 description 2
- 229950000964 pepstatin Drugs 0.000 description 2
- 108010091212 pepstatin Proteins 0.000 description 2
- FAXGPCHRFPCXOO-LXTPJMTPSA-N pepstatin A Chemical compound OC(=O)C[C@H](O)[C@H](CC(C)C)NC(=O)[C@H](C)NC(=O)C[C@H](O)[C@H](CC(C)C)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C(C)C)NC(=O)CC(C)C FAXGPCHRFPCXOO-LXTPJMTPSA-N 0.000 description 2
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical compound FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 238000012545 processing Methods 0.000 description 2
- 239000011541 reaction mixture Substances 0.000 description 2
- 238000011084 recovery Methods 0.000 description 2
- 102220023257 rs387907546 Human genes 0.000 description 2
- 239000012723 sample buffer Substances 0.000 description 2
- 238000005070 sampling Methods 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 238000000926 separation method Methods 0.000 description 2
- 239000001488 sodium phosphate Substances 0.000 description 2
- 229910000162 sodium phosphate Inorganic materials 0.000 description 2
- 230000002269 spontaneous effect Effects 0.000 description 2
- 238000010561 standard procedure Methods 0.000 description 2
- LWIHDJKSTIGBAC-UHFFFAOYSA-K tripotassium phosphate Chemical compound [K+].[K+].[K+].[O-]P([O-])([O-])=O LWIHDJKSTIGBAC-UHFFFAOYSA-K 0.000 description 2
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 2
- 108010073969 valyllysine Proteins 0.000 description 2
- 239000011782 vitamin Substances 0.000 description 2
- 235000013343 vitamin Nutrition 0.000 description 2
- 229940088594 vitamin Drugs 0.000 description 2
- 229930003231 vitamin Natural products 0.000 description 2
- 150000003722 vitamin derivatives Chemical class 0.000 description 2
- RYHBNJHYFVUHQT-UHFFFAOYSA-N 1,4-Dioxane Chemical compound C1COCCO1 RYHBNJHYFVUHQT-UHFFFAOYSA-N 0.000 description 1
- UMCMPZBLKLEWAF-BCTGSCMUSA-N 3-[(3-cholamidopropyl)dimethylammonio]propane-1-sulfonate Chemical compound C([C@H]1C[C@H]2O)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(=O)NCCC[N+](C)(C)CCCS([O-])(=O)=O)C)[C@@]2(C)[C@@H](O)C1 UMCMPZBLKLEWAF-BCTGSCMUSA-N 0.000 description 1
- NLJVXZFCYKWXLH-DXTIXLATSA-N 3-[(3r,6s,9s,12s,15s,17s,20s,22r,25s,28s)-20-(2-amino-2-oxoethyl)-9-(3-aminopropyl)-3,22,25-tribenzyl-15-[(4-hydroxyphenyl)methyl]-6-(2-methylpropyl)-2,5,8,11,14,18,21,24,27-nonaoxo-12-propan-2-yl-1,4,7,10,13,16,19,23,26-nonazabicyclo[26.3.0]hentriacontan Chemical compound C([C@H]1C(=O)N[C@H](C(=O)N[C@@H](CCCN)C(=O)N[C@H](C(N[C@H](CC=2C=CC=CC=2)C(=O)N2CCC[C@H]2C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@H](CC=2C=CC=CC=2)C(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCC(O)=O)N1)=O)CC(C)C)C(C)C)C1=CC=C(O)C=C1 NLJVXZFCYKWXLH-DXTIXLATSA-N 0.000 description 1
- CXNVOWPRHWWCQR-UHFFFAOYSA-N 4-Chloro-ortho-toluidine Chemical compound CC1=CC(Cl)=CC=C1N CXNVOWPRHWWCQR-UHFFFAOYSA-N 0.000 description 1
- POUITAHNNRJWMA-UHFFFAOYSA-N 5-hydroxybenzofuran-2-one Chemical compound OC1=CC=C2OC(=O)CC2=C1 POUITAHNNRJWMA-UHFFFAOYSA-N 0.000 description 1
- HBAQYPYDRFILMT-UHFFFAOYSA-N 8-[3-(1-cyclopropylpyrazol-4-yl)-1H-pyrazolo[4,3-d]pyrimidin-5-yl]-3-methyl-3,8-diazabicyclo[3.2.1]octan-2-one Chemical class C1(CC1)N1N=CC(=C1)C1=NNC2=C1N=C(N=C2)N1C2C(N(CC1CC2)C)=O HBAQYPYDRFILMT-UHFFFAOYSA-N 0.000 description 1
- 241000186361 Actinobacteria <class> Species 0.000 description 1
- FXKNPWNXPQZLES-ZLUOBGJFSA-N Ala-Asn-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O FXKNPWNXPQZLES-ZLUOBGJFSA-N 0.000 description 1
- KXEVYGKATAMXJJ-ACZMJKKPSA-N Ala-Glu-Asp Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O KXEVYGKATAMXJJ-ACZMJKKPSA-N 0.000 description 1
- REWSWYIDQIELBE-FXQIFTODSA-N Ala-Val-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O REWSWYIDQIELBE-FXQIFTODSA-N 0.000 description 1
- 108010025188 Alcohol oxidase Proteins 0.000 description 1
- 102100036826 Aldehyde oxidase Human genes 0.000 description 1
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 1
- 244000105975 Antidesma platyphyllum Species 0.000 description 1
- 108010008184 Aryldialkylphosphatase Proteins 0.000 description 1
- 102000006996 Aryldialkylphosphatase Human genes 0.000 description 1
- DAPLJWATMAXPPZ-CIUDSAMLSA-N Asn-Asn-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CC(N)=O DAPLJWATMAXPPZ-CIUDSAMLSA-N 0.000 description 1
- HDHZCEDPLTVHFZ-GUBZILKMSA-N Asn-Leu-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O HDHZCEDPLTVHFZ-GUBZILKMSA-N 0.000 description 1
- BSYNRYMUTXBXSQ-UHFFFAOYSA-N Aspirin Chemical compound CC(=O)OC1=CC=CC=C1C(O)=O BSYNRYMUTXBXSQ-UHFFFAOYSA-N 0.000 description 1
- 235000017166 Bambusa arundinacea Nutrition 0.000 description 1
- 235000017491 Bambusa tulda Nutrition 0.000 description 1
- 239000002028 Biomass Substances 0.000 description 1
- 241000186146 Brevibacterium Species 0.000 description 1
- DKPFZGUDAPQIHT-UHFFFAOYSA-N Butyl acetate Natural products CCCCOC(C)=O DKPFZGUDAPQIHT-UHFFFAOYSA-N 0.000 description 1
- 240000001829 Catharanthus roseus Species 0.000 description 1
- GHOKWGTUZJEAQD-UHFFFAOYSA-N Chick antidermatitis factor Natural products OCC(C)(C)C(O)C(=O)NCCC(O)=O GHOKWGTUZJEAQD-UHFFFAOYSA-N 0.000 description 1
- 241000148131 Colibacter Species 0.000 description 1
- 101100321760 Comamonas testosteroni aphI gene Proteins 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- 241000186216 Corynebacterium Species 0.000 description 1
- 241000192700 Cyanobacteria Species 0.000 description 1
- PHOQVHQSTUBQQK-SQOUGZDYSA-N D-glucono-1,5-lactone Chemical compound OC[C@H]1OC(=O)[C@H](O)[C@@H](O)[C@@H]1O PHOQVHQSTUBQQK-SQOUGZDYSA-N 0.000 description 1
- 102000053602 DNA Human genes 0.000 description 1
- ZAFNJMIOTHYJRJ-UHFFFAOYSA-N Diisopropyl ether Chemical compound CC(C)OC(C)C ZAFNJMIOTHYJRJ-UHFFFAOYSA-N 0.000 description 1
- LCGLNKUTAGEVQW-UHFFFAOYSA-N Dimethyl ether Chemical group COC LCGLNKUTAGEVQW-UHFFFAOYSA-N 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 239000004593 Epoxy Substances 0.000 description 1
- 241000146391 Fusarium lichenicola Species 0.000 description 1
- 241000223221 Fusarium oxysporum Species 0.000 description 1
- 101150082479 GAL gene Proteins 0.000 description 1
- 101150094690 GAL1 gene Proteins 0.000 description 1
- 102100028501 Galanin peptides Human genes 0.000 description 1
- ITYRYNUZHPNCIK-GUBZILKMSA-N Glu-Ala-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O ITYRYNUZHPNCIK-GUBZILKMSA-N 0.000 description 1
- LRPXYSGPOBVBEH-IUCAKERBSA-N Glu-Gly-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O LRPXYSGPOBVBEH-IUCAKERBSA-N 0.000 description 1
- HKSNHPVETYYJBK-LAEOZQHASA-N Gly-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)CN HKSNHPVETYYJBK-LAEOZQHASA-N 0.000 description 1
- IUZGUFAJDBHQQV-YUMQZZPRSA-N Gly-Leu-Asn Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O IUZGUFAJDBHQQV-YUMQZZPRSA-N 0.000 description 1
- 102100031181 Glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 102100023737 GrpE protein homolog 1, mitochondrial Human genes 0.000 description 1
- 101710113864 Heat shock protein 90 Proteins 0.000 description 1
- 102100034051 Heat shock protein HSP 90-alpha Human genes 0.000 description 1
- 101710100504 Heat shock protein beta-1 Proteins 0.000 description 1
- 102100023043 Heat shock protein beta-8 Human genes 0.000 description 1
- 101000928314 Homo sapiens Aldehyde oxidase Proteins 0.000 description 1
- 101100121078 Homo sapiens GAL gene Proteins 0.000 description 1
- 101000829489 Homo sapiens GrpE protein homolog 1, mitochondrial Proteins 0.000 description 1
- 101001047341 Homo sapiens Heat shock protein beta-8 Proteins 0.000 description 1
- 101000692247 Homo sapiens Phagosome assembly factor 1 Proteins 0.000 description 1
- 101001133654 Homo sapiens Protein PALS1 Proteins 0.000 description 1
- 241000223198 Humicola Species 0.000 description 1
- 150000008575 L-amino acids Chemical group 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- CCQLQKZTXZBXTN-NHCYSSNCSA-N Leu-Gly-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H]([C@@H](C)CC)C(O)=O CCQLQKZTXZBXTN-NHCYSSNCSA-N 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 241001302042 Methanothermobacter thermautotrophicus Species 0.000 description 1
- NTIZESTWPVYFNL-UHFFFAOYSA-N Methyl isobutyl ketone Chemical compound CC(C)CC(C)=O NTIZESTWPVYFNL-UHFFFAOYSA-N 0.000 description 1
- 101100400378 Mus musculus Marveld2 gene Proteins 0.000 description 1
- 101100412856 Mus musculus Rhod gene Proteins 0.000 description 1
- MWIFXQRDVQPSNE-UHFFFAOYSA-N N1=CC=CC2=CC=C3C=CC=NC3=C12.N1=CC=CC2=CC=CC=C12 Chemical compound N1=CC=CC2=CC=C3C=CC=NC3=C12.N1=CC=CC2=CC=CC=C12 MWIFXQRDVQPSNE-UHFFFAOYSA-N 0.000 description 1
- 239000000020 Nitrocellulose Substances 0.000 description 1
- 241001655308 Nocardiaceae Species 0.000 description 1
- 108091092724 Noncoding DNA Proteins 0.000 description 1
- BPQQTUXANYXVAA-UHFFFAOYSA-N Orthosilicate Chemical compound [O-][Si]([O-])([O-])[O-] BPQQTUXANYXVAA-UHFFFAOYSA-N 0.000 description 1
- ZNXZGRMVNNHPCA-UHFFFAOYSA-N Pantetheine Natural products OCC(C)(C)C(O)C(=O)NCCC(=O)NCCS ZNXZGRMVNNHPCA-UHFFFAOYSA-N 0.000 description 1
- 102100026062 Phagosome assembly factor 1 Human genes 0.000 description 1
- 244000082204 Phyllostachys viridis Species 0.000 description 1
- 235000015334 Phyllostachys viridis Nutrition 0.000 description 1
- 108010030975 Polyketide Synthases Proteins 0.000 description 1
- 102100034054 Protein PALS1 Human genes 0.000 description 1
- 241000350481 Pterogyne nitens Species 0.000 description 1
- 108010053763 Pyruvate Carboxylase Proteins 0.000 description 1
- 102100039895 Pyruvate carboxylase, mitochondrial Human genes 0.000 description 1
- 108010091086 Recombinases Proteins 0.000 description 1
- 102000018120 Recombinases Human genes 0.000 description 1
- 229910003797 SPO1 Inorganic materials 0.000 description 1
- 229910003798 SPO2 Inorganic materials 0.000 description 1
- 241000235070 Saccharomyces Species 0.000 description 1
- 101100434411 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) ADH1 gene Proteins 0.000 description 1
- 101100150136 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) SPO1 gene Proteins 0.000 description 1
- 101100478210 Schizosaccharomyces pombe (strain 972 / ATCC 24843) spo2 gene Proteins 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical class [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 1
- 241000228389 Sporidiobolus Species 0.000 description 1
- 241000222665 Sterigmatomyces Species 0.000 description 1
- 241000134899 Streptomyces argillaceus Species 0.000 description 1
- 241000204060 Streptomycetaceae Species 0.000 description 1
- 108020005038 Terminator Codon Proteins 0.000 description 1
- 101100242191 Tetraodon nigroviridis rho gene Proteins 0.000 description 1
- 241000012469 Trimerotropis maritima Species 0.000 description 1
- 108010021006 Tyrothricin Proteins 0.000 description 1
- DPDMMXDBJGCCQC-UHFFFAOYSA-N [Na].[Cl] Chemical compound [Na].[Cl] DPDMMXDBJGCCQC-UHFFFAOYSA-N 0.000 description 1
- XJLXINKUBYWONI-DQQFMEOOSA-N [[(2r,3r,4r,5r)-5-(6-aminopurin-9-yl)-3-hydroxy-4-phosphonooxyoxolan-2-yl]methoxy-hydroxyphosphoryl] [(2s,3r,4s,5s)-5-(3-carbamoylpyridin-1-ium-1-yl)-3,4-dihydroxyoxolan-2-yl]methyl phosphate Chemical compound NC(=O)C1=CC=C[N+]([C@@H]2[C@H]([C@@H](O)[C@H](COP([O-])(=O)OP(O)(=O)OC[C@@H]3[C@H]([C@@H](OP(O)(O)=O)[C@@H](O3)N3C4=NC=NC(N)=C4N=C3)O)O2)O)=C1 XJLXINKUBYWONI-DQQFMEOOSA-N 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 238000000516 activation analysis Methods 0.000 description 1
- 101150102866 adc1 gene Proteins 0.000 description 1
- 230000000996 additive effect Effects 0.000 description 1
- 239000000853 adhesive Substances 0.000 description 1
- 230000001070 adhesive effect Effects 0.000 description 1
- 238000000246 agarose gel electrophoresis Methods 0.000 description 1
- SRBFZHDQGSBBOR-QMKXCQHVSA-N alpha-L-arabinopyranose Chemical compound O[C@H]1CO[C@@H](O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-QMKXCQHVSA-N 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 1
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 1
- 235000011130 ammonium sulphate Nutrition 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 239000012062 aqueous buffer Substances 0.000 description 1
- 239000008346 aqueous phase Substances 0.000 description 1
- 239000011425 bamboo Substances 0.000 description 1
- 238000003287 bathing Methods 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 230000003570 biosynthesizing effect Effects 0.000 description 1
- 229910021538 borax Inorganic materials 0.000 description 1
- 229940043232 butyl acetate Drugs 0.000 description 1
- 229910052799 carbon Inorganic materials 0.000 description 1
- 239000003054 catalyst Substances 0.000 description 1
- 230000003197 catalytic effect Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- XBLIFEQTVVSTIM-UHFFFAOYSA-L chembl2105392 Chemical compound [Na+].[Na+].[O-]S(=O)(=O)C1=CC2=CC(S([O-])(=O)=O)=CC=C2C(O)=C1N=NC1=CC=C([N+]([O-])=O)C=C1[N+]([O-])=O XBLIFEQTVVSTIM-UHFFFAOYSA-L 0.000 description 1
- 238000001311 chemical methods and process Methods 0.000 description 1
- 239000003795 chemical substances by application Substances 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- 101150036359 clpB gene Proteins 0.000 description 1
- 230000000295 complement effect Effects 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- MUCZHBLJLSDCSD-UHFFFAOYSA-N diisopropyl fluorophosphate Chemical compound CC(C)OP(F)(=O)OC(C)C MUCZHBLJLSDCSD-UHFFFAOYSA-N 0.000 description 1
- 230000003292 diminished effect Effects 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 239000005712 elicitor Substances 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 238000010799 enzyme reaction rate Methods 0.000 description 1
- 150000002148 esters Chemical class 0.000 description 1
- LZCLXQDLBQLTDK-BYPYZUCNSA-N ethyl (2S)-lactate Chemical compound CCOC(=O)[C@H](C)O LZCLXQDLBQLTDK-BYPYZUCNSA-N 0.000 description 1
- LZCLXQDLBQLTDK-UHFFFAOYSA-N ethyl 2-hydroxypropanoate Chemical compound CCOC(=O)C(C)O LZCLXQDLBQLTDK-UHFFFAOYSA-N 0.000 description 1
- 238000002474 experimental method Methods 0.000 description 1
- 238000000855 fermentation Methods 0.000 description 1
- 230000004151 fermentation Effects 0.000 description 1
- 239000000706 filtrate Substances 0.000 description 1
- 235000013305 food Nutrition 0.000 description 1
- 239000012634 fragment Substances 0.000 description 1
- 239000003205 fragrance Substances 0.000 description 1
- YEJRWHAVMIAJKC-UHFFFAOYSA-N gamma-butyrolactone Natural products O=C1CCCO1 YEJRWHAVMIAJKC-UHFFFAOYSA-N 0.000 description 1
- JBFHTYHTHYHCDJ-UHFFFAOYSA-N gamma-caprolactone Chemical compound CCC1CCC(=O)O1 JBFHTYHTHYHCDJ-UHFFFAOYSA-N 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 1
- 235000011187 glycerol Nutrition 0.000 description 1
- PJJJBBJSCAKJQF-UHFFFAOYSA-N guanidinium chloride Chemical compound [Cl-].NC(N)=[NH2+] PJJJBBJSCAKJQF-UHFFFAOYSA-N 0.000 description 1
- 235000009424 haa Nutrition 0.000 description 1
- FUZZWVXGSFPDMH-UHFFFAOYSA-M hexanoate Chemical compound CCCCCC([O-])=O FUZZWVXGSFPDMH-UHFFFAOYSA-M 0.000 description 1
- 239000000710 homodimer Substances 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 235000006486 human diet Nutrition 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 239000012535 impurity Substances 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 230000001939 inductive effect Effects 0.000 description 1
- 150000002500 ions Chemical class 0.000 description 1
- 229910052742 iron Inorganic materials 0.000 description 1
- 101150109249 lacI gene Proteins 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 230000000670 limiting effect Effects 0.000 description 1
- 235000019626 lipase activity Nutrition 0.000 description 1
- 238000009630 liquid culture Methods 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- BCVXHSPFUWZLGQ-UHFFFAOYSA-N mecn acetonitrile Chemical compound CC#N.CC#N BCVXHSPFUWZLGQ-UHFFFAOYSA-N 0.000 description 1
- 108020004999 messenger RNA Proteins 0.000 description 1
- 230000002906 microbiologic effect Effects 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 239000012120 mounting media Substances 0.000 description 1
- 239000013642 negative control Substances 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 238000006386 neutralization reaction Methods 0.000 description 1
- BOPGDPNILDQYTO-NNYOXOHSSA-N nicotinamide-adenine dinucleotide Chemical compound C1=CCC(C(=O)N)=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OC[C@@H]2[C@H]([C@@H](O)[C@@H](O2)N2C3=NC=NC(N)=C3N=C2)O)O1 BOPGDPNILDQYTO-NNYOXOHSSA-N 0.000 description 1
- 229920001220 nitrocellulos Polymers 0.000 description 1
- QJGQUHMNIGDVPM-UHFFFAOYSA-N nitrogen group Chemical group [N] QJGQUHMNIGDVPM-UHFFFAOYSA-N 0.000 description 1
- 238000007899 nucleic acid hybridization Methods 0.000 description 1
- 235000016709 nutrition Nutrition 0.000 description 1
- 230000035764 nutrition Effects 0.000 description 1
- 125000001477 organic nitrogen group Chemical group 0.000 description 1
- 230000003647 oxidation Effects 0.000 description 1
- 238000007254 oxidation reaction Methods 0.000 description 1
- 230000033116 oxidation-reduction process Effects 0.000 description 1
- 229940055726 pantothenic acid Drugs 0.000 description 1
- 235000019161 pantothenic acid Nutrition 0.000 description 1
- 239000011713 pantothenic acid Substances 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 1
- 230000008447 perception Effects 0.000 description 1
- 239000004848 polyfunctional curative Substances 0.000 description 1
- 229910000160 potassium phosphate Inorganic materials 0.000 description 1
- 229940093916 potassium phosphate Drugs 0.000 description 1
- 235000011009 potassium phosphates Nutrition 0.000 description 1
- 201000005484 prostate carcinoma in situ Diseases 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- 239000002994 raw material Substances 0.000 description 1
- 238000004064 recycling Methods 0.000 description 1
- 238000004153 renaturation Methods 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000000452 restraining effect Effects 0.000 description 1
- 150000003335 secondary amines Chemical class 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 239000003352 sequestering agent Substances 0.000 description 1
- 230000035939 shock Effects 0.000 description 1
- 235000011091 sodium acetates Nutrition 0.000 description 1
- 239000001509 sodium citrate Substances 0.000 description 1
- 235000010339 sodium tetraborate Nutrition 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 230000001954 sterilising effect Effects 0.000 description 1
- 238000004659 sterilization and disinfection Methods 0.000 description 1
- 238000013517 stratification Methods 0.000 description 1
- 239000003774 sulfhydryl reagent Substances 0.000 description 1
- 238000003786 synthesis reaction Methods 0.000 description 1
- 230000009897 systematic effect Effects 0.000 description 1
- 238000007669 thermal treatment Methods 0.000 description 1
- 239000011573 trace mineral Substances 0.000 description 1
- 235000013619 trace mineral Nutrition 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000001131 transforming effect Effects 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- BSVBQGMMJUBVOD-UHFFFAOYSA-N trisodium borate Chemical compound [Na+].[Na+].[Na+].[O-]B([O-])[O-] BSVBQGMMJUBVOD-UHFFFAOYSA-N 0.000 description 1
- HRXKRNGNAMMEHJ-UHFFFAOYSA-K trisodium citrate Chemical compound [Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O HRXKRNGNAMMEHJ-UHFFFAOYSA-K 0.000 description 1
- 229940038773 trisodium citrate Drugs 0.000 description 1
- GPRLSGONYQIRFK-MNYXATJNSA-N triton Chemical compound [3H+] GPRLSGONYQIRFK-MNYXATJNSA-N 0.000 description 1
- 201000008827 tuberculosis Diseases 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 229960003281 tyrothricin Drugs 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 230000029663 wound healing Effects 0.000 description 1
Images














Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P17/00—Preparation of heterocyclic carbon compounds with only O, N, S, Se or Te as ring hetero atoms
- C12P17/02—Oxygen as only ring hetero atoms
- C12P17/04—Oxygen as only ring hetero atoms containing a five-membered hetero ring, e.g. griseofulvin, vitamin C
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/18—Carboxylic ester hydrolases (3.1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P41/00—Processes using enzymes or microorganisms to separate optical isomers from a racemic mixture
- C12P41/003—Processes using enzymes or microorganisms to separate optical isomers from a racemic mixture by ester formation, lactone formation or the inverse reactions
- C12P41/005—Processes using enzymes or microorganisms to separate optical isomers from a racemic mixture by ester formation, lactone formation or the inverse reactions by esterification of carboxylic acid groups in the enantiomers or the inverse reaction
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- Biomedical Technology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Molecular Biology (AREA)
- General Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Analytical Chemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
本发明涉及具有水解L-泛内酯(L-pantolactone)酶活性的蛋白。本发明也涉及编码这些蛋白的核酸、核酸构建体、载体、遗传改造的微生物及一种D-泛内酯的制备方法。
Description
本发明涉及具有水解L-泛内酯(L-pantolactone)酶活性的蛋白。本发明也涉及编码这些蛋白的核酸、核酸构建体、载体、遗传改造的微生物及一种D-泛内酯的制备方法。
D-泛内酯是化学合成和生物合成泛酸、泛醇和泛酰硫氢乙胺及其衍生物的前体。这些物质被用来作为人类饮食、动物饲料、医药(如伤口愈合)及化妆品(如护发产品)中的维生素补充剂。因此,经济合理的合成对映体纯的D-泛内酯是十分重要的。在制备D-泛内酯方面,除了长期采用的化学方法以外,最近越来越多的生物技术方法也已被设计出来。一篇关于D-泛内酯及其化学合成的综述可以在Ullmann工业化学百科全书中找到(VCHVerlagsgesellschaft mbH,69451 Weinheim,1996年,A27卷,559-566页)。
泛内酯的生物技术遵循着各种合成策略。
Galanzert等人描述了一种使用脂肪酶或酯酶选择性地水解o-乙酰泛内酯,拆分外消旋物的方法(微生物酶学技术1988,10,689-690)。一个这种类型的外消旋物拆分方法已申请专利DE 40 05 150和EP-A-0 507278。而这种方法所达到的对映体的纯度不能达到工业使用的要求。
Degussa描述了使用醇氰醛化酶,以羟基新戊醛和氢氰酸起始,经由光学纯的羟基新戊醛氰醇制备泛内酯的方法(DE 41 26 580,EP-A-0 528 256,DE 41 39 987)。这个反应在理论上可以达到100%的收率。该反应的缺点是需要大量的酶(等摩尔的酶与底物)和产物的对映体纯度相对较低(最高82%)。
JP 47019745描述了使用节杆菌(Arthrobacter)、短杆菌(Brevibacterium)、芽孢菌(Bacillus)或棒杆菌(Corynebacterium)合成D-泛内酯的方法。在该反应中,所述的生物体通过L-泛内酯的代谢将外消旋的泛内酯转化成D-泛内酯。该方法的缺点是一半的前体被代谢并因此损失掉了。
三菱(Mitsubishi)化学公司和宇部(Ube)公司已要求了由D,L-泛内酯制备D-泛内酯的方法(JP 6067320,JP 62294092,JP 62294096,JP57152895)。这些方法描述了红酵母(Rhodotorula)、锁掷酵母(Sporidiobolus)和梗孢酵母(Sterigmatomyces)中的L-泛内酯水解酶。然而,当前的观点(参考Yamada & Shimizu,Ann.N.Y.Acad.Sci.672[1992]in Enzyme Eng.XI,Clark et al.,372-386;Chimia,47,1993:5-10,JP 62187426;JP 61293384;JP 61293386;Angew.Chem.Int.Ed.Engl.27,1988:622-642,Chemical Aspects of Enzyme Biotechnology,eds.T.O.Baldwin et al.,Plenum Press,New York,1990:151-163)认为,在这些酵母里是否存在L-泛内酯的直接水解仍值得怀疑。这一观点也为我们自己的研究所证实。该反应通过酮基泛解酸氧化还原酶将酮基泛内酯转化成L-泛内酯之说更合适一些。在我们自己进行的研究中可以检测到酮基泛内酯作为中间体存在,就是说并没有直接水解成L-泛内酯的。该反应的缺点是可被转化的泛内酯的浓度很低。
通过酮基泛内酯和酮基泛解酸氧化还原酶介导的这个反应在细菌中也有描述(例如Yamada & Shimizu,参上;Shimizu等.,生物化学杂志.1988,263,12077-12084,Kataoka等。欧洲生物化学杂志。1992,204,799-806)。虽然,由D,L-泛内酯通过酮基泛内酯和酮基泛解酸对映选择合成D-泛内酯的相关方法具有高产率的优点(理论上100%,红球菌为90.5%,见下),但由于其需要辅助因子(NADH,NADPH)、补加能量底物(葡萄糖)、低时空产率和低终浓度(18.2-72g/l D-泛内酯),其生产是很不经济的。这个反应更不利的一点是,参加反应的两个酶对于转化反应通常具有不同的最适点。这样的问题在使用单(水解)酶时是不会出现的。
富士(Fuji)与京都大学(Kyoto University)的Yamada研究组合作,已经开发了一种利用真菌D-泛内酯水解酶,拆分外消旋体的酶学方法(JP09308-497,JP 11056356,EP-B-0 436 730,EP-B-0 504 421,EP-A-0794 251,Wo 92/06182,WO 97/10341,US 5,275,949,US 5,372,940)。该酶可以从真菌(Cylindrocarpon tonkinense),藤仓赤霉(Gibberellafujikuroi)和尖镰孢(Fusarium oxysporum)中分离得到。该D-泛内酯水解酶是一种Ca2+依赖型糖基化酶,由125kDa的同型二聚体构成(Ann.N.Y.Acad.Sci.1996,799:650-658,酶工程)。该酶被Cd2+,Hg2+,Cu2+和EDTA(US 5,372,940)所抑制。Shimizu等人已对D-泛内酯水解酶的纯化进行过描述。该纯酶对许多内脂,尤其是糖内脂表现出水解活性(欧洲生物化学杂志,209,1992:383-390)。其序列与运动发酵单胞菌(Zymomonasmobilis)的葡萄糖酸内脂酶(28.9%),人和鼠的对氧磷酶(25.3%),Catharanthus roseus的strictosidin合成酶(15.9%)具有低度的同源性(EP-A-0 794 251,Kobayashi等。美国国家科学院进展1998,95,12787-12792)。Kataoko等人描述了在不同pH值下转化获得的对映体纯度依赖性很高(微生物酶学技术1996,19:307-310和应用微生物生物技术1995,44:333-338)。在接近或超过pH 7时获得的对映体浓度较低,因为在较高pH条件下,L-泛内酯的自发化学水解增加,从而使产物的对映体纯度降低。pH 5被认为是制备最高对映体纯度D-泛内酯的最适pH。然而,在此pH,酶反应速度被大大降低。为获得光学纯的产物,有必要用结晶的方法进行抽提(Yamada,H.Chimia 47,1993:5-10)。
上述方法的缺点是,其产物的光学纯度经常很低和/或时空产率不高,导致经济前景并不看好。因此,仍然非常需要一种没有上述缺点的,简单、经济的生物技术方法来制备D-泛内酯。可以说,这个方法使得以已经存在的化学合成为起始,简便地获得高收率的、不必进一步进行产物纯化的对映体纯度的D-泛内酯成为可能。
本发明的一个目标就是提供一种简单、经济地制备D-泛内酯的方法。我们已经发现,一段分离的、编码一段具有L-泛内酯水解酶活力多肽的核酸序列,可以达到此目标,该序列选自:
a)SEQ ID NO:1所述的核酸序列,
b)由于遗传密码的简并,导致从序列SEQ ID NO:1中衍生的核酸序列,
c)其编码的多肽具有SEQ ID NO:2所述氨基酸序列的SEQ ID NO:1所述核酸序列的衍生物,并且该衍生物在不明显损失该多肽酶活性的前提下,在氨基酸水平上至少应有50%的同源性,
d)(a)到(c)中所提序列的功能等同物。
这些L-泛内酯水解酶可在生物体,尤其是微生物(如细菌)中找到。此酶对于将L-泛内酯转换成L-泛解酸的水解反应具有很高的酶活力。
这些L-泛内酯水解酶不能转化D-泛内酯,因此这些生物体、抽提物或纯酶及相关的重组菌株或蛋白能被用于制备对映体纯的D-泛内酯。
本发明所述的、具有SEQ ID NO:1序列的核酸序列的衍生物是指,例如,等位基因突变体,它们在衍生的氨基酸水平上至少具有50%的同源性,优选60%以上,特别优选70%,更特别优选80%以上。同源性由Needleman & Wunsch法(分子生物学杂志1970,48:443-453)或Smith& Waterman法进行测定(应用数学进展1981,2,:482-489)。此种同源性在某些区域更高则优。由SEQ ID NO:1衍生的氨基酸序列可在SEQ ID NO:2中找到。等位基因突变体尤其含有由SEQ ID NO:1描述的序列通过核酸缺失、插入或替换所获得的功能突变体,虽然其衍生蛋白的酶活力应该有些可忽略不计的损失。活力损失可忽略不计的酶是指,酶活力至少要有20%,优选50%,特别优选75%,更特别优选90%。因此,本发明也涉及上述核酸序列组编码的氨基酸序列。本发明优选涉及到由序列SEQ ID NO:1编码的氨基酸序列。
(a)到(c)中所提序列的功能等同物是指,编码将L-泛内酯水解成相应酸的酶的核酸序列,该酶至少具有SEQ ID NO:2所示序列活力的20%,优选50%,特别优选75%,更特别优选90%,不被EDTA(1mM溶液)抑制,在pH 4和10之间稳定。除此之外,这些功能等同物的最适pH在7和8之间,优选最适温度在70℃和80℃之间。
衍生物也指SEQ ID NO:1的同系物,例如真菌或细菌的同系物,截短的序列,编码和非编码DNA序列的单链DNA或RNA。SEQ ID NO:1的同系物与SEQ ID NO:1总DNA序列相比,在DNA水平,具有至少50%的同源性,优选60%以上,特别优选70%,更特别优选80%以上。
除此之外,SEQ ID NO:1的同系物是指一些衍生物,如启动子突变体。这些在所声明核酸序列上游的启动子,可由一个或多个核苷酸的替换、插入、和/或缺失进行修饰,而不损害该启动子的功能或活力。而且,这些启动子可以通过其序列修饰提高活力,或者可以被更有效的,甚至是来自异源生物体的启动子完全取代。
衍生物也指变异体,其核苷酸序列在起始密码子前-1到-200区域或终止密码子后0到1000个碱基对处被修饰,因此基因表达和/或蛋白表达被改变,提高则更佳。
发明所述的核酸序列,原则上,可以从所有的生物体中鉴别和分离到。SEQ ID NO:1或其同系物可轻易地从真菌、酵母或细菌中分离得到。可能涉及的细菌为革兰氏阴性和阳性菌。用本领域熟练技术人员所知的方法,从革兰氏阴性细菌分离发明所述核酸,优选从α-蛋白细菌(α-proteobacteria)、β-蛋白细菌(β-proteobacteria)或γ-蛋白细菌(γ-proteobacteria)中,特别优选肠杆菌(Enterobacteriaceae)、假单胞菌(Pseudomonadaceae)或根瘤菌(Rhizobiaceae)科,更特别优选土壤杆菌(Agrobacterium)、假单胞菌(Pseudomonas)或伯克霍尔德氏菌(Burkholderia)属。可能涉及的适合的真菌优选白僵菌属(Beauveria)或裸盖菇属(Psilocybe)。优选的酵母菌可在Apiotrichum属中找到。
SEQ ID No:1或其衍生物、同系物或这些序列的一部分可以通过例如杂交或PCR技术从其它真菌或细菌中分离得到。这些DNA序列在标准条件下与本发明所述序列进行杂交。使用保守区内较短的寡核苷酸(如活性位点区)对杂交有利。这些保守区可由本领域熟练技术人员所知方法通过与D-泛内酯水解酶相比较来测定(如所谓的HTGT基元)。然而,使用更长的本发明所述的核酸序列片段,或全序列进行杂交也是可能的。这些标准条件随所使用的核酸(寡核苷酸、较长片段或全长序列)或随用来杂交的核酸类型(DNA或RNA)而改变。因此,例如,在DNA:DNA杂交混合物中所用的熔化温度就比相同长度的DNA:RNA中的要低10℃左右。
标准条件是指:例如,在0.1到5×SSC(1×SSC=0.15M氯化钠,15mM柠檬酸钠,pH 7.2)之间的水性缓冲液中,或再加50%甲酰胺,温度在20到70℃之间。这一温度依所用核酸而定。DNA:DNA杂交混合物的杂交条件在2.0×SSC及温度为20℃到70℃之间为佳,优选50℃到70℃之间。DNA:RNA杂交混合物的杂交条件在2.0×SSC及温度为20℃到60℃之间为佳,优选35℃到60℃之间。这些所述杂交温度是通过例如,以长度约1000核苷酸、G+C含量在50%之间(无甲酰胺)的核酸计算出的熔化温度。DNA杂交的实验条件在相关的遗传学书籍中有述,如Sambrook等,“分子克隆”,冷泉港实验室,1989,而且可由本领域熟练技术人员通过公式计算出来,如根据核酸长度、杂交混合液性质或G+C含量。杂交的更多信息可由本领域熟练技术人员在以下书籍中找到:Ausubel等(eds),1985,现代分子生物学方法,(John Wiley & Sons,New York);Hames和Higgins(eds),1985,核酸杂交:实用方法,(IRL Press at OxfordUniversity Press,Oxford);Brown(ed),1991,基础分子生物学:实用方法,(IRL Press at Oxford University Press,Oxford)。
本发明所述核酸构建体是指具有序列SEQ ID No:1的L-泛内酯水解酶基因,及其在功能上与一个或多个有利于增加基因表达的调节信号相连接的衍生物和同系物。这些调节序列是指,例如,一些序列,诱导子或抑制子与之结合并因此调节此核酸表达。除了这些新的调节序列以外,仍然有可能在实际的结构基因之前存在这些序列的自然调节,并且经由恰当地遗传修饰关闭自然调节,提高基因表达。然而,核酸构建体也可以有着更简单的结构,即是说,在序列SEQ ID No:1或其序列同系物前,没有插入其它的调节信号,也没有删除其用于调节的天然启动子。相反,此天然调节序列产生突变,调节作用不再发生,基因表达得到提高。对于核酸构建体来说,额外的包含一个或多个,在功能上与可提高核酸序列表达的启动子相连的所谓的增强子,也是有好处的。其它有利的序列也可以被插入到DNA的3’末端,例如其它的调节元件或终止子。本发明所述核酸可以以一个或多个拷贝的形式存在于该构建体中。此构建体也可以适当地包含其它的标记(如抗生素抗性或营养缺陷型互补基因),以供筛选之用。
对本发明所述方法有利的调节序列存在于,诸如aphII(Tn5),trc,cos,tac,trp,lacPAI,rha,tet,trp-tet,lpp,lac,lpp-lac,lacIq,T7,T5,T3,gal,trc,ara,SP6,λ-PR或λ-PL启动子中,这些启动子优选在革兰氏阴性菌中使用。更优选的调节序列存在于,如革兰氏阳性菌中的启动子,如组成型或诱导型链霉菌的aphI,ermE,melC,tipA,mcrAB,gylCAB,veg,SPO1,amy和SPO2启动子中;或酵母或真菌中的AOX1,GAL1,ADC1,MFα,AC,P-60,CYC1,GAPDH,TEF,rp28,ADH启动子中。与此相关,来自例如汉逊酵母属(Hansenula)的丙酮酸脱羧酶启动子和甲醇氧化酶启动子也非常好。也可以使用人造的启动子进行调控。
为了进行表达,核酸构建体被插入到宿主中,优选插入到可使基因在宿主中最适表达的载体中,如质粒、噬菌体或其它DNA。这些载体是本发明进一步的实施方案。适合质粒的例子有,例如大肠杆菌中的pBluescript,pBAD,pQE(His tag System),pICIC223-3,pLG338,pACYC184,pBR322,pUC18,pGEM7Z,pKK223-3,pUC19,pKC30,pRep4,pHS1,pHS2,pPLc236,pMBL24,pLG200,pUR290,pIN-III113-B1,λgt11或pBdCI,或广泛宿主范围质粒:pBBR1MCS或pRK293;链霉菌或其它放线菌中的pIJ101,pIJ364,pMVS301,pIJ702或pIJ361;芽孢杆菌中的pUB110,pC194或pBD214;棒杆菌中的pSA77或pAJ667;真菌中的pALS1,pIL2或pBB116;酵母中的2∝,pAG-1,YEp6,YEp13或pEMBLYe23;或植物中的pLGV23,pGHlac+,pBIN19,pAK2004或pDH51。所述质粒仅为可用质粒中的一小部分。更多质粒被专业人员所知,而且可以在诸如克隆载体(Eds.Pouwels P.H.et al.Elsevier,Amsterdam-New York-Oxford,1985,ISBN 0 444 904018)这样的书中找到。
为表达存在的其它基因,核酸构建体含有其它的、根据所选宿主和基因选来进行最佳表达的、并用以提高表达的,3’和/或5’终止调节序列将更加有利。
这些调节序列使特定的基因表达及蛋白表达成为可能。这可以指,例如根据宿主的不同,基因只在诱导后被表达或过表达,或是基因被立即表达或过表达。
为此目的,优选可以在被诱导基因的表达方面具有有利影响并因此增强其表达的调节序列或因子。因此,通过使用强转录信号,如启动子和/或增强子,调节元件的增强可能优选在转录水平上发生。当然,通过诸如提高mRNA稳定性的方法,也能使翻译水平得到增强。
在另一个载体的实施方案中,对于含有本发明所述核酸构建体或核酸的载体而言,优选以线形DNA的形式转入微生物,并通过异源或同源重组整合于宿主基因组。此线形DNA可以由一个线性化的载体(如质粒)或仅由本发明的核酸构建体或核酸组成。
此外,本发明涉及具有下列特征的L-泛内酯水解酶:a)将L-泛内酯转化成相应的酸,b)pH稳定性:L-泛内酯水解酶在pH 4-10范围内稳定c)最适pH:7.2到7.6d)最适温度:大约70℃到75℃e)EDTA不抑制其活性
此L-泛内酯水解酶可以作为游离酶或固定酶在发明所述的方法中使用。
为了在生物体中最适表达异源基因,按照该生物体特定的密码子用法来修饰核酸序列将更为有利。根据相应生物体其它已知基因的计算机分析,该密码子用法可被轻松建立起来。
本发明所述基因及其编码蛋白在宿主中的表达,通常都会给宿主产生一种应激。在有至少一种编码叫做应激蛋白的基因存在的情况下,或这些基因结合在一起的情况下,这些基因的同时表达使得本发明所述的核酸优选在发明所述的宿主中得到表达。应激蛋白,也叫热休克蛋白(=HSP)或分子伴侣,是一些在进化中最保守的蛋白,无论是在原核生物中还是在真核生物中。而且,这种蛋白在所有生物体中普遍存在。它们以千道尔顿的分子量来划分,如HSP60,70,90等。这些应激蛋白具有在应激条件下,如葡萄糖水平过低、热激、乙醇、紫外线、氧化剂等,诱导产生的特性,因此得名。
许多应激蛋白及相关组成型蛋白,对于蛋白的正确折叠、装配、稳定和转运都是非常重要的。本发明所述蛋白与至少一种应激蛋白的共表达,使优选表达本发明所述的核酸成为可能。这种方法也有利于防止水解酶蛋白聚合的发生。这使应激蛋白结合到蛋白的疏水端,并因此防止蛋白的错误折叠并促进其正确折叠。已经聚集或已变性的蛋白被再次解离,并被正确折叠。当这些应激蛋白执行其功能时,它们频繁地与其它称为辅助蛋白(=股蛋白)并因此有陪伴机之说的蛋白协作。这些辅助蛋白对发明所述基因的表达具有有利的影响(Frydaman等,自然1994,370:111-117)。这些陪伴机的作用可以在有ATP消耗(主陪伴机)或无ATP消耗(次分子伴侣)时发生。有利的分子伴侣或热激蛋白有,如真核基因HSP17.5,HSP22,HSP 25,HSP27,HSP60,HSP70,HSP90,TRiC,UBI1,2,3,4;或它们的原核同系物,如HtpG,DnaK,DnaJ,GroES,GroEL,HtrC,ClpB,GrpE等。优选的分子伴侣为GroES,GroEL,HtpG,DnaK,DnaJ,HSP70或HSP27。
本发明所述核酸优选在有至少一种应激蛋白存在的条件表达。在这种情况下,基因可以在一个启动子的接合控制之下,或从几个单独的启动子读出。相应地,通过在同一时间或在不同时间添加一种或多种诱导物的方法,可以诱导这些基因的表达。这些核酸可在一个载体上或在不同的载体上。也可能通过遗传操作对宿主的应激蛋白进行修饰,从而使其过表达。
为增加天然酶含量而采用的其它替代方法也可以是优选的,如在低温下培养合成本发明所述蛋白的微生物,或在本发明所述蛋白的悬浮液(添加或不加变性剂,如盐酸胍)上使用高压(优选1-2千巴),以使之复性。
适合本发明所述核酸或核酸构建体的重组宿主,原则上为所有的原核或真核生物。宿主优选使用微生物,如细菌、真菌或酵母。优选革兰氏阳性或阴性菌;更优选肠杆菌(Enterobacteriaceae)、假单胞菌(Pseudomonadaceae)、根瘤菌(Rhizobiaceae)、链霉菌(Streptomycetaceae)或诺卡氏菌(Nocardiaceae)科的细菌,酵母(如毕赤(Pichia),酵母属(Saccharomyces)或汉逊酵母属(Hansenula))或真菌(如白僵菌属(Beauveria)或裸盖菇属(Psilocybe));特别优选埃希氏菌属(Escherichia)、假单胞菌属(Pseudomonas)、链霉菌属(Streptomyces)、诺卡氏菌属(Nocardia)、伯克霍尔德氏菌属(Burkholderia)、沙门氏菌属(Salmonella)、土壤杆菌属(Agrobacterium)或红球菌属(Rhodococcus)的细菌;更特别优选大肠杆菌属和种。此外,可以在α-蛋白细菌(α-proteobacteria)、β-蛋白细菌(β-proteobacteria)或γ-蛋白细菌(γ-proteobacteria)族中找到进一步优选的细菌。
本发明所述宿主,优选含有至少一种本发明所描绘的,编码L-泛内酯水解酶的核酸序列、核酸构建体或载体。
本发明所述方法中使用的生物体,根据宿主的不同,以本领域熟练技术人员所共知的方法进行生长或培养。微生物通常在含有碳源(一般以糖的形式),氮源(一般以有机氮源如酵母抽提物,或盐如硫酸铵的形式),微量元素(如铁、锰、镁盐)及适当维生素的液体培养基中,通氧生长;培养温度为0℃到100℃之间,优选10℃到60℃之间。此外,也有可能保持培养液的pH在一固定值,就是说在培养过程中调节或不调节pH。可以进行分批的、半分批的或连续的培养。营养物可以在发酵开始时加入,也可以半连续或连续地进行补加。同样,根据所使用的启动子,可以使用不同的诱导物,如异丙基硫代半乳糖苷(IPTG),乳糖,阿拉伯糖,鼠李糖和抗生素,和/或变换温度以使发明所述基因进行表达。外消旋的泛内酯可以在培养中直接加入,或优选在培养后加入。酶可以从生物体中通过实施例中描述的方法分离得到,或以粗提物的形式用于反应。
优选含有0.5U/g DBM(=干生物量)L-泛内酯水解酶活力的宿主生物体,更优选4U/g DBM,特别优选20到150U/g DBM,更特别优选40到60U/gDBM。
本发明所述方法优选在0℃到95℃之间进行,更优选在10℃到85℃之间,特别优选在15℃到75℃之间。
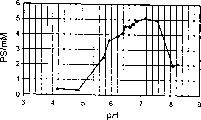
本发明所述方法中的pH优选保持在4到12之间,更优选在6到9之间,特别优选在6到8之间,更特别优选在6.5到7.5之间。
本发明所述方法中的外消旋泛内酯是指由两种对映体以50∶50的比例组成,或者由任何其它的富含两种对映体之一的混合物组成。
对映体纯或手性泛内酯(D或L对映体)在本发明所述的方法中是指,表现出富含一种对映体的对映体混合物。此方法优选达到至少70%ee对映体纯度,更优选80%ee,特别优选90%ee,更特别优选98%ee。
对于本发明所述方法,可以使用含有发明所述核酸、核酸构建体或载体的生长细胞。也可以使用休眠的或破裂的细胞。破裂的细胞是指,例如,已经通过诸如溶剂处理成为可渗透的细胞,或者通过酶处理、机械处理(如弗氏压碎器或超声波)或其它方法破损的细胞。此法获得的粗抽提物适合本发明所述的方法。纯的或部分纯化的酶也可为本法所用。同样,可在反应中优选使用的固定化微生物或酶也是适合的。
如果在本发明所述的方法中使用游离的生物体或酶,它们可以在萃取之前被方便地除去,如通过过滤或离心的方法。不必使用固定化的生物体或酶,但仍然可以使用。
本发明所述方法制备的D-泛内酯,优选从水性反应液中通过萃取或结晶(或优选萃取并结晶)的方法进行分离。这一步通过使用有机溶剂萃取水性反应液来进行。萃取可重复进行几次,以提高产率。溶液在萃取前冷冻到0℃到10℃为优选。为了使游离的酸转变成盐以使前者在反应条件下不被萃取出来,优选将水性溶液在冷冻前或冷冻后中和至大约pH6.0到7.0。被用来中和的碱是,例如,碳酸氢盐或另外一个,如NaOH或KOH。可被使用的有机溶剂原则上为所有的加入盐后可与水分层,并且可使内脂从水相中出来并进入到该相中的溶剂。优选的溶剂为仅吸收少量的水,所以仅有少量的酸进入到该溶剂的溶剂,如甲苯、二氯甲烷、乙酸丁酯、二异丙醚、苯、甲基叔丁醚、甲基异丁基酮、二乙基甲酮或乙酸乙酯。
在浓缩有机相之后,通常可以获得化学纯很高的产品,即化学纯在90%以上。萃取之后,含有产品的有机相可被部分浓缩,产品可结晶析出。优选将溶液冷冻到0℃到10℃进行操作。直接从有机相或从水溶液中进行结晶也是可能的。结晶化的产品可再次溶于相同或不同的溶剂并再次结晶。如果需要的话,可以再进行至少一次有利的结晶化操作,以进一步提高产品的对映体纯度。无论如何,对于作为目的产品的D-泛内酯来说,直接使用未进行结晶处理的有机溶液也是可以及优选的。
留在水溶液中的L对映体可以通过酸化(如硫酸)使其内脂化,然后被萃取出来,方法同上。优选将溶液加热进行内脂化。去除溶剂之后,所得的L-内脂可与碱(如NaOH,泛解酸钠或甲醇钠)的催化量(大约1到5mol%),在熔化状态下被外消旋化并被回收。优选的外消旋化处理和对多余的对映体进行回收,可使本发明所述的方法有可能达到98%的理论收率。
经过上述各类后处理之后,本发明所述方法的产品,基于用于该反应的外消旋泛内酯,可以以60%到100%的收率分离得到,优选80%到100%,特别优选90%到100%。分离到的产品具有化学纯>90%的高品质,优选>95%,特别优选>98%。除此之外,产品还有很高的对映体纯度。而且,通过必要的结晶化处理,对映体纯度还可进一步提高。
本发明所述方法可以分批、半分批或连续地进行。
此法获得的产品可作为合成泛酰醇、泛酰巯基乙胺及其衍生物的起始物。这些物质及获得的对映体纯的泛内酯可单独或彼此组合使用,用于药物、食品、动物饲料或化妆品的生产。
以下例子举例说明了本发明。
实施例:
1.石竹伯克霍尔德氏菌(Burkholderia caryophylli)Lu681的L-泛内酯水解
石竹伯克霍尔德氏菌Lu681(或表1a,1b中的其它菌株)在25ml的复合培养基(例如HFP=1%蛋白胨,1%胰化蛋白胨,0.5%酵母抽提物,0.3%NaCl)中培养1到3天,收获,洗涤,并重悬于5毫升50mM的Tris/HCl缓冲液(pH 7.0)中,与50mM D,L-泛内酯在30℃温浴3小时。去除细胞后,以GC或HPLC分析测定D,L-泛内酯,D,L-,D-和L-泛解酸的浓度(表1a)。另外,该转化反应也可以4M NaOH滴定过夜进行(4毫升细胞悬浮液,50mM D,L-泛内酯,50mM Tris/HCl pH 7.0加20毫升蒸馏水;表1b)。表1中的所有菌株均将外消旋泛内酯水解成L-泛解酸。高转化的ee(45%)和酶的对映体选择性E由Straathof和Jongejan描述的方法进行测定(酶学与微生物学技术1997,21:559-571)。
2.水解活力的确认
竹伯克霍尔德氏菌Lu681(或表1中其它菌株)在25毫升的复合培养基(例如HFP=1%蛋白胨,1%胰化蛋白胨,0.5%酵母抽提物,0.3%NaCl)中培养1到3天,收获,在Tris-HCl(50mM,pH 7.0)缓冲液中洗涤并重悬于5毫升Tris/HCl(50mM,pH 7.0)中,与50mM酮基泛内酯在30℃温浴3小时。去除细胞后,以HPLC测定酮基泛内酯,酮基泛解酸,D,L-泛内酯,D,L-,D-和L-泛解酸的浓度(表1a)。由于酮基泛内酯的自发水解,列出的除Beauveria amorpha Lu7953以外的所有菌株都能够将酮基泛解酸还原成泛解酸。既然,所有的菌株都可以形成D-泛解酸而不是L-泛解酸(如例1所述),泛内酯到L-泛解酸的转化就无法通过氧化-还原过方法(经由酮基泛内酯和酮基泛解酸)进行。在透析过的Lu681或Lu5351的粗抽提物中,及在使用来自Lu681或Lu5351的纯酶时发现,L-泛内酯的水解并不依赖于其它辅助因子。因此,该酶活性可以归于水解酶。
3.通过各种野生型菌株水解生产D-泛内酯a.石竹伯克霍尔德氏菌Lu681
石竹伯克霍尔德氏菌Lu681在200毫升复合培养基(如GYP=1% D-葡萄糖,0.5%多聚蛋白胨,0.5%酵母抽提物)(OD600=6.7,DBM=2.97g/l)中生长,然后收获并洗涤。10毫升10倍浓缩的悬浮液与50mM D,L-泛内酯(于50mM Tris/HCl pH 7.0;分批体积20毫升)在30℃温浴,以4M NaOH滴定至pH 7.0。3小时和19.5小时后,以HPLC测定转化c和ee(3h:c=45%,ee=95%;19.5 h:c=59%,ee=89%对L-泛解酸)。后者分别与ee值为73%和100%的D-泛内酯相当。b.放射形土壤杆菌(Agrobacterium radiobacter)Lu5351
放射形土壤杆菌Lu5351在200毫升复合培养基(如HFP=1%蛋白胨,1%胰化蛋白胨,0.5%酵母抽提物,0.3%NaCl)(OD600=11.5,DBM=2.90g/l)中生长,然后收获并洗涤。10毫升10倍浓缩的悬浮液与50mM D,L-泛内酯(于50mM Tris/HCl pH 7.0;分批体积20毫升)在30℃温浴,以4M NaOH滴定至pH 7.0。3小时和19.4小时后,以HPLC测定转化c和ee(3h:c=20%,ee=93%;19.4h:c=53%,ee=94%对L-泛解酸)。后者分别与ee值为21%和100%的D-泛内酯相当。
c.缺陷假单胞菌(Pseudomonas diminuta)Lu683
缺陷假单胞菌Lu683在200毫升复合培养基(如GYP=1% D-葡萄糖,0.5%多聚蛋白胨,0.5%酵母抽提物)(OD600=7.3,DBM=3.78g/l)中生长,然后收获并洗涤。10毫升10倍浓缩的悬浮液与50mM D,L-泛内酯(于50mM Tris/HCl pH 7.0;分批体积20毫升)在30℃温浴,以4M NaOH滴定至pH 7.0。3小时和19.3小时后,以HPLC测定转化c和ee(3h:c=48%,ee=97%;19.4h:c=69%,ee=79%对L-泛解酸)。后者分别与ee值为82%和100%的D-泛内酯相当。
d.Apiotrichum humicola Lu3215
Apiotrichum humicola Lu3215在200毫升复合培养基(如HFP=1%蛋白胨,1%胰化蛋白胨,0.5%酵母抽提物,0.3%NaCl)(OD600=18.5,DBM=7.34g/l)中生长,然后收获并洗涤。10毫升10倍浓缩的悬浮液与50mM D,L-泛内酯(于50mM Tris/HCl pH 7.0;分批体积20毫升)在30℃温浴,以4M NaOH滴定至pH 7.0。3小时和19.4小时后,以HPLC测定转化c和ee(3h:c=55%,ee=79%对L-泛解酸)。后者与ee值为84%的D-泛内酯相当。4.从石竹伯克霍尔德氏菌Lu681中分离L-泛内酯水解酶
石竹伯克霍尔德氏菌Lu681,于14升复合培养基(HFP=1%蛋白胨,1%胰化蛋白胨,0.5%酵母抽提物,0.3%NaCl)中,生长至OD600=10(3g/l DBM),收获并打碎。然后从粗提物中纯化出L-泛内酯水解酶(约200单位)。首先,用Ultra-Turrax shaft将细胞(1128克湿重)重悬于1.8升20mM Tris/HCl(pH 7.4)缓冲液中。终体积3升。然后,将溶液在玻璃漏斗中通过一层玻璃珠(0.1到0.2毫米,200毫升)除去粗颗粒物。此细胞悬液在z04微流化床装置上,以1500巴压力匀浆两次。再以500毫升缓冲液冲洗一次。合并后(4升),用200毫升1M MgCl2进行沉淀(终浓度为50mM)。加NaOH溶液,使其pH保持在7.0。6000rpm离心30分钟。上清(3.1升)与200毫升0.2M EDTA(pH 7.5)溶液混合。混合后,溶液pH降至5.0。所形成的沉淀再次以6000rpm(Sorvall)离心20分钟。上清(3.4升)被重新滴定至pH 7.0。
随后,加入989克硫酸氨(相当于50%饱和度),搅拌10分钟。混浊物以6000rpm,离心20分钟。所得上清(3.7升)被分开:1.2升进行苯基琼脂糖(phenyl-Sepharose)层析。
苯基琼脂糖柱(Pharmacia,直径5厘米,高25厘米,体积490毫升)以1升缓冲液A(20mM磷酸钠缓冲液,pH 7.4,40%硫酸氨)洗涤,以缓冲液B(20mM磷酸钠,pH 7.4)进行梯度洗脱。流速为10毫升/分钟,120分钟后达到100%缓冲液B;保持40分钟。收集并合并活性峰(250毫升)。
稀释到小于7mS/cm后,这3升溶液通过Q-Sepharose层析(直径5厘米,高25厘米,体积430毫升,Fast Flow,Pharmacia)进行纯化。柱子以1升缓冲液A(20mM磷酸钠缓冲液,pH 7.4,)洗涤(10毫升/分钟)。以缓冲液B(缓冲液A加1M NaCl)进行的梯度洗脱在120分钟内达到100%缓冲液B,并在100%保持40分钟。收集并合并活性峰(118毫升)。溶液被浓缩(10kD Omega膜),并对5升10mM Tris/HCl(pH 7.0)透析;终体积为21毫升。其中6毫升上Waters Q HR8柱。此柱以缓冲液A(20mMMes,pH 6.0)预平衡,然后以1%/分钟梯度从缓冲液A展开至缓冲液B(缓冲液A加0.5M NaCl)。收集活性峰(3.7毫升),并对2升10mM Tris/HCl(pH 7.0)透析两次。透析液变混,离心之(4毫升)。
然后,该原料以Mono P层析(Pharmacia,直径0.5厘米,体积5毫升)分开。所得Mono-P峰,在-20摄氏度通过丙酮沉淀浓缩至0.2毫升。
将沉淀物中加入0.005毫升无二硫苏糖醇(DTT)的十二烷基硫酸钠(SDS)样品缓冲液,然后上样至SDS凝胶(12%Tris/甘氨酸凝胶,from Novex,约2.5小时,125V,50mA,Laemmli,U.K.,1970,自然,227:680-685)。分开后,以活性染色鉴定L-泛内酯水解酶,并将其切下。此项操作先将胶于TBS缓冲液(=50mM Tris,100mM NaCl,pH 7.4)中轻轻摇动,然后和50毫升TBS+50毫升α-乙酸萘酯(Sigma N-8505,0.4g/lin 10%丙酮)溶液温浴10分钟。再加入50毫升Fast Red TR溶液(SigmaF-8764,1g/l),于室温(=大约23℃)搅动。L-泛内酯水解酶条带呈微红棕色,表观分子量约为36kDa。这块切下来的胶中的蛋白质以胰岛素进行消化,所得肽段进行测序。得到两段肽序列(SEQ ID NO:3和4)。留下的胶进行考马斯亮兰染色。5.从放射形土壤杆菌Lu5351中分离L-泛内酯水解酶
放射形土壤杆菌Lu5351,于14升复合培养基(如HFP=1%蛋白胨,1%胰化蛋白胨,0.5%酵母抽提物,0.3%NaCl)中,生长至OD600=10(3g/lDBM),收获并打碎。然后从粗提物中纯化出L-泛内酯水解酶(约60单位)(表2)。首先,用Ultra-Turrax shaft将放射形土壤杆菌(Lu 5351)细胞(400克湿重)重悬于1.8升20mM Tris/HCl(pH 7.4)缓冲液中(终体积2.2升)。然后,将溶液在玻璃漏斗中通过一层玻璃珠(0.1到0.2毫米,200毫升)除去粗颗粒物。此细胞悬液在z04微流化床装置上,以1500巴压力匀浆两次。微流化床再以500毫升缓冲液冲洗一次。合并后(2.7升),用135毫升1M MgCl2进行沉淀(终浓度为50mM)。加NaOH溶液,使其pH保持在7.0。6000rpm离心30分钟。上清(2.6升)与575ml 0.2MEDTA(pH 7.5)溶液混合并再次核查其pH。加入711克硫酸氨(相当于40%饱和度),搅拌10分钟。混浊物以6000rpm,离心30分钟。所得上清(3.3升)进行苯基琼脂糖(phenyl-Sepharose)层析。
苯基琼脂糖柱(Pharmacia,直径5厘米,高25厘米,体积490毫升)以1升缓冲液A(20mM磷酸钠缓冲液,pH 7.4,40%硫酸氨)洗涤,以缓冲液B(20mM磷酸钠,pH 7.4)进行梯度洗脱。流速为10毫升/分钟,120分钟后达到100%缓冲液B,保持40分钟。收集并合并活性峰(350毫升,20.9毫秒)。
稀释到7mS/cm后(终体积3.1升),进行Q-Sepharose层析(直径5厘米,高25厘米,体积430毫升,Fast Flow,Pharmacia)。柱子以1升缓冲液A(20mM磷酸钠缓冲液,pH 7.4,)洗涤(10毫升/分钟)。以缓冲液B(缓冲液A加1M NaCl)进行的梯度洗脱在120分钟内达到100%缓冲液B,并在100%保持40分钟。收集并合并活性峰(134毫升)。溶液被浓缩(10kD Omega膜),并对3升10mM Tris/HCl(pH 7.0)透析;(终体积为19毫升)。其中6毫升上Waters Q HR8柱。此柱以缓冲液A(20mM Mes,pH 6.0)预平衡,然后以1%/分钟梯度,从缓冲液A展开至缓冲液B(缓冲液A加0.5M NaCl)。收集活性峰(12.5毫升),并对5升10mM醋酸钠(pH 5.0)透析两次。透析液变混,离心之(4毫升)。
然后,上清以Mono P层析(Pharmacia,直径0.5厘米,体积5毫升)分开。
所得Mono-P峰在-20摄氏度通过丙酮沉淀浓缩至0.2毫升。沉淀物中加入0.005毫升无二硫苏糖醇(DTT)的十二烷基硫酸钠(SDS)样品缓冲液,然后上样至SDS凝胶。分开后,以活性染色鉴定L-泛内酯水解酶(参见例4),并将其切下。L-泛内酯水解酶带呈微红棕色,表观分子量为36kDa。切下这块切下来的胶中的蛋白质以胰岛素进行消化,所得肽段进行测序。得到两段肽序列(SEQ ID NO:5和6)。留下的胶进行考马斯亮兰染色。SEQ IDNO:5的序列表明,其第一个氨基酸不能肯定。因为此处序列不甚明确,第一位的酪氨酸也可能是一个亮氨酸。6.Lu681和Lu5351来源的,纯L-泛内酯水解酶的底物特异性
来源于Lu681或Lu5351的苯基琼脂糖层析峰段的纯酶(0.1U/ml),在150mM Pipes(pH 6.8)中与各种酯及内脂温浴。保温0、1及20小时后取样,离心(通过一个10kDa的虑膜)以终止反应。上清中的浓度及相关的酸以HPLC分析进行测定。表3a和3b所示的活力,为与以L-泛内酯为底物的酶活力相比较的结果。
Lu681的酶,在微量滴定板上进行了对脂肪酶底物1,2-O-二月桂基-rac-甘油基-3-戊二酸试卤灵酯的光学分析(Boehringer Mannheim,改进)。在45mM KH2PO4(pH 6.8)溶液中,60到482单位/升的酶,与0.18克/升试卤灵酯(resorufin ester)(2克/升,在二烷+2% SDS+10%H2O中),在室温下温浴。2分钟及82分钟后,测量572nm处E的消光。表3c显示了消光差异及由此计算出来的脂肪酶活力,其相当于大约0.05%的L-泛内酯水解酶活力。7.Lu681和Lu5351来源的,纯L-泛内酯水解酶的抑制作用与活化作用
来源于Lu681或Lu5351的苯基琼脂糖层析峰段的纯酶(0.1U/ml),与各种效应物在150mM Pipes(pH 7.0)中预温5分钟。测定以加入150mM L-泛内酯起始(30℃ 1小时),以离心(通过一个10kDa的虑膜)终止。然后,通过HPLC分析测定D,L-,D-和L-泛解酸的浓度。表4a和4b是与未加效应物样品的活力比较。总的来说,Lu681和Lu5351来源的纯酶对螯合剂、表面硬化剂(SH reagents)、蛋白酶抑制剂、去污剂及各种阳离子不敏感(残余活力>85%)(例外:Lu5351在1%SDS中残余活力为74%)。
大部分情况下,HgCl2对该酶有明显的活化作用(133/170%)。另外,检测到D-泛内酯对重组681内脂酶的竞争抑制(大肠杆菌细胞,见下面实施例8)。8.Genbank+筛选:石竹伯克霍尔德氏菌Lu681中L-泛内酯水解酶的克隆
分离到石竹伯克霍尔德氏菌Lu681的基因组DNA(Qiagen,Hilden),以EcoRI酶切,连入经EcoRI酶切并脱磷酸化的pBluescriptKS+载体中(Maniatis,T.,分子克隆:实验手册1989)。连接产物按Stratagene公司(La Jolla,Calif.)的操作说明转入大肠杆菌(E.coli)XL1Blue菌株。转化株铺于含有氨苄青霉素(100μg/ml),IPTG(=异丙基β-硫代半乳糖苷,0.2mM)和X-Gal(80mg/l)的LB平板上,于30或37℃培养过夜。在含有氨苄青霉素(100μg/ml),IPTG(=异丙基β-硫代半乳糖苷,0.2mM)和X-Gal(80mg/l)的LB平板上挑取白色菌落,并再次培养过夜。在LB-氨苄青霉素(100mg/ml)-IPTG平板上,使用灭菌的硝酸纤维素膜,以滤膜复制的方法复制样板。在此板上过夜培养后(参上),滤膜与150mML-泛内酯、0.1%硝嗪黄及10mM Tris/HCl(pH 7.0)进行活性分析(3小时-过夜,30℃)。分离得到一个黄色克隆(XL1Blue pKS+681)。9.大肠杆菌(E.coli)XL1Blue pKS+681中EcoRI插入片断的酶切图谱建立及测序
按Qiagen(Hilden)操作,从E.coli XL1Blue pKS+681中分离得到质粒DNA,并以限制酶EcoRI、BamHI、PstI和HindIII进行单或双酶切。片断化DNA以0.8%琼脂糖进行琼脂糖凝胶电泳。由得到的片断大小绘出7.5kB插入片断的限制酶切图谱,如图3。此插入片断被完全测序(Sanger等.1977),其包含核苷酸序列ID NO:1。反过来,来源于石竹伯克霍尔德氏菌Lu681和放射形土壤杆菌Lu5351的,纯的或不纯的L-泛内酯水解酶(参见例4和例5)经胰酶消化后发现,该核酸序列衍生的氨基酸序列(SEQ ID NO:2)含有肽YGIEGLNNLEAL和AKEDANSTIEAED(SEQ ID NO:3和4)。
数据库比较(Genbank,EMBL,SwissProt,1999年5月7日数据,[Sptrembel]和1999年1月5日[PIR])显示,该核酸序列及其衍生的氨基酸序列与一组假定蛋白及链霉菌中的某些四环素环化酶仅有很小的同源性(表5)。确切的说,具有共有序列HTGTHVDAP的基元在所有的蛋白中都是高度保守的。另外,与来源于恶臭假单胞菌(Pseudomonas putida)WW2(WO 94/09175)的靛红水解酶有48%的同源性(在氨基酸水平有38%的同源性),其同样含有所说的序列基元。既然未发现与其它内脂酶、酯酶或脂肪酶有同源性,所发现的L-泛内酯水解含有一个酶的新类别。序列比较表明,所说的假定蛋白、四环素环化酶、靛红水解酶及所发现的L-泛内酯水解酶属于同一个较远的系统分类相关家族。10.E.coli XL1Blue pKS+681对L-泛内酯的水解
接一环E.coli XL1Blue pKS+681菌株至含有氨苄青霉素(100μg/ml),IPTG(0.2mM)和X-Gal(80mg/l)的LB平板,于37℃培养过夜。然后重悬(OD600=2.5)于0.5毫升Tris-HCl(pH 7.0)和50mM D,L-泛内酯的溶液中。使用相应的E.coli XL1Blue pBluescriptKS+-样品(OD600=2.5)进行比较。1小时后,离下细胞。D,L-泛内酯,D,L-,D-和L-泛解酸以HPLC分析进行测定。表6a为各种样品的活力和ee值。悬液的活力≥90U/L。然而在液体培养批次中,未发现明显的活力(参考例12)。11.L-泛内酯水解酶在E.coli XL1Blue pKK223-3中的克隆表达
基于核酸序列SEQ ID NO:1,设计出寡核苷酸引物5‘-CCGGAATTCATGTGCAACAACTGC(P1)和5‘-CCCAAGCTTCAGACCAGGGCCAGAA(P2),以对L-泛内酯水解酶基因进行PCR扩增。反应条件如下:20mMTris/HCl pH 8.8,2mM MgSO4,10mM KCl,10mM(NH4)2SO4,0.1% TritonX-100,0.1mg/ml BSA,25mM/dNTP,0.96μg/ml pKS+681,P1和P2各2.2μg/ml,25U/ml Pfu聚合酶(Stratagene,LaJolla,Calif.);PCR参数如下:95℃ 1min,55℃ 1min,72℃ 2.5min,30个循环。PCR产物(0.8kb)用EcoRI和HindIII酶切,并连入经EcoRI和HindIII酶切并脱磷酸化的pKK223-3(Pharmacia,Freiburg)载体中。连接混合物转化E.coliXL1 Blue或TG1(Stratagene,LaJolla,Calif.;DSMZ,Braunschweig,DSMZ-No.6056,Inoue et al,1990,基因96:23-28)。转化体铺于LB-氨苄平板,培养过夜。在此转化平板上进行如例8的滤膜复制及接下来的活力分析,找到了约100个黄色克隆。其中十个,以质粒DNA小量制备及限制酶消化(EcoRI-HindIII,EcoRI-HindIII-BamHI)的方法进行分析(Maniatis,T.,分子克隆:实验手册,1989)。它们含有如图4所示的质粒pKK681。12.E.coli XL1Blue pKK681对L-泛内酯的水解
E.coli XL1Blue pKK681在30毫升含有氨苄青霉素(100μg/ml)和IPTG(0.5mM)的LB培养基中,37℃培养过夜。然后收获,以Tris-HCl(50mM,pH 7.0)洗涤并悬浮(3毫升50mM Tris/HCl pH 7.0)。0.25毫升悬液与150mM L-泛内酯、150mM Pipes pH 7.0加0.5毫升蒸馏水混合,30℃温浴3小时。与此平行,0.25毫升悬液与50mM D,L-泛内酯、50mM TrispH 7.0加0.5毫升蒸馏水混合,温浴3小时。另外,2毫升悬液与300mM D,L-泛内酯、50mM Tris pH 6.8加2 0毫升蒸馏水混合,以4M NaOH滴定至pH 6.8,温浴3小时。1小时和3小时后取样,去除细胞,上清进行D,L-泛内酯、D,L-、D-和L-泛解酸的分析。表6b为各个样品的活力及ee值。由此可知,此过夜培养物(1倍浓度)具有90-150U/l的活力。
在对数生长前期对E.coli XL1Blue pKK681进行诱导(OD600=0.6,+0.5mM IPTG),37℃温浴5小时后,相应对数生长后期细胞(OD600=4.1)具有大约480U/l的活力。13.L-泛内酯水解酶在E.coli TG1 pDHE19中的克隆表达
基于核酸序列SEQ ID NO:1,设计出寡核苷酸引物5‘-CAGGATGCCATATGTGCAACAACTGC(P1)和5‘-CCCAAGCTTCAGACCAGGGCCAGAA(P2),以对L-泛内酯水解酶基因进行PCR扩增。反应条件如下:20mMTris/HCl pH 8.8,2mM MgSO4,10mM KCl,10mM(NH4)2SO4,0.1% TritonX-100,0.1mg/ml BSA,25mM/dNTP,0.96μg/ml pKS+681,P1和P2各2.2μg/ml,25U/ml Pfu聚合酶(Stratagene,LaJolla,Calif.);PCR参数如下:95℃ 1min,55℃ 1min,72℃ 2.5min,30个循环。PCR产物(0.8kb)用NdeI和HindIII酶切,并连入经NdeI和HindIII酶切并脱磷酸化的pDHE19(Prof.Mattes,Stuttgart)载体中。连接混合物转化E.coli XL1Blue或TG1(Stratagene,LaJolla,Calif.;DSMZ,Braunschweig,DSMZ-No.6056,Inoue et al,1990,Gene 96:23-28)。转化株铺于LB-氨苄平板,培养过夜。在LB-氨苄(100μg/ml)/鼠李糖(2g/l)平板上(LB=Luria Broth)进行如例8的滤膜复制及接下来的活力分析,找到了约100个黄色克隆。其中十个,以质粒DNA小量制备、限制酶消化(NdeI-HindIII,NdeI-HindIII-BamHI)及序列分析的方法进行分析(Maniatis,T.,分子克隆:实验手册,1989)。它们含有如图5所示的质粒pDHE681。14.E.coli TG1 pDHE681对L-泛内酯的水解
E.coli TG1 pDHE681在14升含有40g/l甘油和2.5g/l鼠李糖的基本培养基(minimal medium)中,37℃培养6到7个小时。然后收获,以Tris-HCl(50mM,pH 7.0)洗涤并悬浮(加1.4升缓冲液)。以标准方法进行测定(150mM Pipes pH 7.0,150mM L-泛内酯,30℃,1小时),此一倍浓缩的细胞悬液具有680-2700U/l的活力或是60-160g/DBM。
10毫升10倍浓缩的悬液与50mM D,L-泛内酯,在50mM Tris/HCl pH7.0(批体积20毫升)的缓冲液中,以4M NaOH滴定至pH 7.0,30℃温浴。0.4小时和3小时后,以HPLC分析测定转化及ee值(0.4h:c=48%,ee=92%;3h:c=58%,ee=72%对L-泛解酸)。这分别与ee值为84%和100%的D-泛内酯相当。
15.通过E.coli TG1 pDHE681水解,制备D-泛内酯
3克D,L-泛内酯溶于10毫升水,并以4M NaOH滴定至pH 6.5。加5-10毫升E.coli TG1 pDHE681细胞悬液(例14),并以水补至20毫升。反应在30℃,温浴15到22小时,滴定至pH 6.8。另外,还可以再加0到2毫升的细胞悬液并继续温浴3小时,或者是加3次5毫升的细胞悬液并继续温浴90小时。图6显示了基于NaOH消耗的滴定过程。根据转化和温浴时间的不同,D-泛内酯的ee值为71%到97%。然后,22毫升批次的细胞被离下,并以5毫升50mM Tris-HCl(pH 7.0)洗涤。合并上清(20到25毫升),以1体积的乙酸乙酯抽提3次。在有机相中加入10克无水Na2SO4,室温晾干1小时。滤出沉淀并以乙酸乙酯洗涤1次,滤出液在40℃蒸发3小时。粘性残留物被称重,并以HPLC、GC、GC-MS和H-NMR进行分析(表7)。其含有纯的D-泛内酯(50%-52%的转化率,71%-87的ee)。
水相(21毫升;L-泛解酸钠)以约5毫升3M H2SO4调至pH 1,80℃加热15分钟,与8克无水Na2SO4混合。进行类似的操作:以1体积的乙酸乙酯抽提3次,以Na2SO4干燥并蒸发,得到L-泛内酯。加入NaOH及少量L-泛解酸钠后,熔化的L-泛内酯可被外消旋化(180℃,3小时),以供循环使用。16.通过E.coli菌株产生的L-泛内酯水解酶,水解制备D-泛内酯
大肠杆菌(E.coli TG1 pDHE681或共表达分子伴侣如GroEL的优选菌株)发酵液,通过破碎细胞(2×1000巴,微流化床装置)、去除细胞碎片(9000克,10℃ 20分钟)、交叉流动过滤十倍浓缩(Hmoflow F60,Fresenius,排阻限度约为10kDa的膜)及热沉淀(60℃ 20分钟,室温-10℃20分钟,离心),得到被浓缩至约3000U/g比活力的L-泛内酯水解酶。此10x匀浆的活力为63-100000U/l,蛋白浓度为20-30g/l。
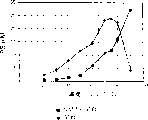
D,L-泛内酯的外消旋拆分如下:2.3M D,L-泛内酯(30% w/v)与热沉淀匀浆(16000U/l,6g/l蛋白)混合,30℃,以4到10M NaOH(pH 7.5)在弱缓冲液(6-20mM NaHCO3)中进行滴定,每批0.75到1.0升。温浴过夜后,通过交叉流动过滤(Hmoflow F60,Fresenius,排阻限度约为10kDa的膜),从含产物的溶液中分离出酶。以去离子水洗涤1到2次并浓缩。然后,酶在上述条件下被重新使用。有效期限的检测表明,6天后,对于ee>90%(D-PL),需要双倍的残留时间,从12小时到24小时(图7)。使用更大的体积(例如用F40柱,10升每批)有可能减少活力损失。
通过以1倍体积的MTBE抽提5次,逐步获得外消旋拆分的匀浆混合物(50.8%转化率,92.5%ee)。得到43%的D-泛内酯,ee为91.4%,纯度为98.2%(GCint.st.;基于100%外消旋物)。与浓硫酸(25毫升)一起加热(65℃/15分钟),并再次以MTBE抽提(5×1体积)后,得到53%的L-泛内酯,ee为62.3%,纯度为98.2%(GCint.st.)。标准方法未检测到蛋白质或DNA杂质。
*没有6天的NaOH数据。1390分钟(23.17小时),转化率50.4%,ee 89.8%。
没有7天的样品,故而没有分析数据。17.通过固定化L-泛内酯水解酶,水解制备D-泛内酯
将例16所获得的L-泛内酯水解酶结合到各种载体介质上,如商业用途的EupergitC(Rhm GmbH,Darmstadt)或Deloxan DAPIII(Degussa,Frankfurt)。
EupergitC
EupergitC是一种由环氧基活化的载体介质。因此,蛋白主要被共价结合在氨基上。
匀浆先以热处理进行沉淀。550毫升匀浆(总计1.1升)在60℃温浴30分钟,然后在冰上冷却20分钟。离心(8000rpm,GS3转头,1小时)除去变性蛋白。然后,上清以Hmoflow F40柱进行浓缩,缓冲液改为20mMHEPES,pH 7.5。每克Eupergit对结合蛋白的数量无限制。下例中,7.2克蛋白被稀释在270毫升含30毫升1M磷酸钾pH7.0的缓冲液中,然后加入17.5克固体NaCl使之盐析析出。大量的盐可促进蛋白与载体的结合。精确调节pH至6.8,然后加入15克干Eupergit,室温搅拌17小时。然后,反应混合物用玻璃漏斗抽气过滤,并用水洗涤载体。该湿介质重约60克。保存使用10mM磷酸缓冲液(pH 7.5)。
Deloxan DAPIII,还原型和非还原型
Deloxan是一种氨村修饰的硅酸盐。这些氨基可被戊二醛激活,形成席夫碱(Schiff’s base)。洗出剩余的醛后,将蛋白加到活化的载体上。蛋白质的游离氨基与戊二醛上仍游离的醛反应,形成第二个席夫碱。以这种方法固定的蛋白就可以使用了。然而,由于席夫碱易于水解,蛋白会在水溶液中从载体上慢慢地沥出来。因此,可以用硼氢化钠还原,使席夫碱转变为次级胺来加以避免。进行此项处理的前提是,酶对硼氢化钠的还原作用是稳定的。
进行匀浆的热沉淀,以固定在EupergitC上。
活化:
140克Deloxan DAPIII分别用水、1.5升0.1M磷酸钠缓冲液(pH 6)和1.5升0.1M磷酸钠缓冲液(pH 7.5)洗涤,然后重悬。加入560毫升2.5%的戊二醛溶液(在同一缓冲液中修正至pH 7.5),反应3到4小时。载体变为橙红色。然后,活化的载体用6升水洗涤并重悬于1升磷酸钠缓冲液(pH 7.5)。
例如,10000个单位(约2.7克蛋白)的L-泛内酯水解酶加到40克活化的Deloxan上。蛋白与活化的载体在室温下温浴18小时。用玻璃漏斗将载体和溶液分开。载体分别用水、1升0.1M的磷酸钠缓冲液(pH 7.2)和1M NaCl洗涤几次。移出一半的载体(未还原)。另一半再次用水洗涤,然后加入0.1M硼酸钠缓冲液(pH 7.2)。
用硼氢化钠还原:
在重悬于80毫升硼酸缓冲液的载体中加入0.4克硼氢化钠(0.5%),于室温搅拌3小时。期间,载体再次变为苍黄色。用抽气玻璃漏斗滤出载体,并用400毫升硼酸缓冲液洗涤。载体再次以1升水洗涤,然后加到20mM磷酸缓冲液中。
外消旋物拆分
用固定化酶拆分2.3M D,L-泛内酯(30%w/v),固定化酶以40毫升批次在30℃搅拌,以10M NaOH(pH 7.5,在10mM NaHCO3中)滴定至D-泛内酯的ee为90%时止(25到60小时)。该生物催化剂经过每批次的过滤后(反应混合物以HPLC洗涤液滤器通过抽气过滤),从产物中分离出来并进行回收,其间适当地洗涤3次(去离子水)。下面的图表显示了为在一个搅拌混合物中检测有效期限的NaOH分布图。
表1a
PA,泛解酸ee值指(L)-泛解酸的形成ee,对映体过量(enantiomeric excess),corr.,ee通过减去由化学水解形成的泛解酸(空白)进行修正
| LU编号 | 属 | 种 | 泛解酸形成(3h) | 直接水解(+)或氧化/还原(-) | ||||
| PA[mM] | PA[g/l] | Aspec.[U/g] | ee[%]uncorr. | ee[%]corr. | ||||
| 22 | 土壤杆菌属 | 根癌土壤杆菌种 | 5.6 | 0.8 | 1.5 | 94.6 | 97.0 | + |
| 5351 | 土壤杆菌属 | 放射形土壤杆菌种 | 4.8 | 0.7 | 1.6 | 97.3 | 100.0 | + |
| 3215 | Apiotrichum | humicola | 7.9 | 1.2 | 1.5 | 82.0 | 77.8 | + |
| 7953 | Beauveria | amorpha | 5.9 | 0.9 | 1.0 | 68.5 | 72.1 | + |
| 680 | 伯克霍尔德氏菌属 | 茄伯克霍尔德氏菌种 | 2.1 | 0.3 | 0.5 | 70.3 | 80.9 | + |
| 681 | 伯克霍尔德氏菌属 | 石竹伯克霍尔德氏菌种 | 6.7 | 1.0 | 2.0 | 96.0 | 98.7 | + |
| 683 | 假单胞菌属 | 缺陷假单胞菌种 | 6.0 | 0.9 | 1.1 | 98.0 | 100.0 | + |
| 863 | 假单胞菌属 | sp. | 7.3 | 1.1 | 1.9 | 92.0 | 94.0 | + |
| 4314 | Psilocybe | coprophila | 2.4 | 0.4 | 0.5 | 65.3 | 74.3 | + |
表1b
ee值指(L)-泛解酸的形成ee,对映体过量(enantiomeric excess),corr.,ee通过减去由化学水解形成的泛解酸(空白)进行修正
| LU编号 | 属 | 种 | 泛解酸形成 | ||||
| ee[%] | ee[%]corr. | t[min] | 转化[%] | E(corr.) | |||
| 22 | 土壤杆菌属 | 根癌土壤杆菌种 | 90.2 | 94.5 | 1352 | 6.8 | 38 |
| 5351 | 土壤杆菌属 | 放射形土壤杆菌种 | 93.8 | 96.5 | 1161 | 52.5 | >229 |
| 3215 | Apiotrichum | humicola | 79.4 | 79.1 | 91 | 54.0 | 29 |
| 7953 | Beauveria | amorpha | 31.3 | 33.8 | 288 | 20.6 | 3 |
| 680 | 伯克霍尔德氏菌属 | 茄伯克霍尔德氏菌种 | 78.1 | 81.3 | 1318 | 54.1 | 37 |
| 681 | 伯克霍尔德氏菌属 | 石竹伯克霍尔德氏菌种 | 94.7 | 97.6 | 187 | 44.6 | 198 |
| 683 | 假单胞菌属 | 缺陷假单胞菌种 | 96.3 | 98.7 | 185 | 48.5 | 524 |
| 863 | 假单胞菌属 | sp. | 89.5 | 91.9 | 1256 | 47.9 | 64 |
| 4314 | Psilocybe | coprophila | 42.6 | 49.7 | 1063 | 15.6 | 3 |
表2:Lu5351中L-泛内酯水解酶的纯化
| 样品 | 体积[升] | 活力[单位/升] | 总活力[单位] | 活力收率[%] | 蛋白量[克/升] | 总蛋白量[克] | 蛋白收率[%] | 比活力[单位/克蛋白] |
| 发酵罐 | 28.0 | 2.0 | 56.8 | - | - | - | - | - |
| 细胞破裂前 | 1.44 | 20.0 | 28.8 | 100.0 | - | - | - | - |
| 细胞破裂第一代 | 1.44 | 5.5 | 7.9 | 27.5 | - | - | - | - |
| 细胞破裂第二代 | 1.44 | 52.2 | 75.1 | 260.8 | - | - | - | - |
| 细胞破裂后的悬液 | 1.86 | 57.3 | 106.6 | 370.3 | - | - | - | - |
| 细胞破裂后的上清 | 1.82 | 84.7 | 154.1 | 535.0 | 8.52 | 15.51 | 100.0 | 9.9 |
| MnCl2处理后的上清 | 1.8 | 72.0 | 129.6 | 450.0 | 10.32 | 18.58 | 119.8 | 7.0 |
| 苯基琼脂糖加样 | 2.04 | 10.7 | 21.9 | 76.0 | 7.2 | 14.69 | 94.7 | 1.5 |
| 苯基琼脂糖峰 | 0.16 | 87.0 | 13.9 | 48.3 | 7.82 | 1.251 | 8.1 | 11.1 |
| Q-琼脂糖峰 | 0.12 | 83.5 | 10.0 | 34.8 | 0.36 | 0.043 | 0.3 | 231.9 |
| Superdex加样 | 0.012 | 605.0 | 7.3 | 25.2 | - | - | - | - |
| Superdex峰 | 0.04 | 41.3 | 1.7 | 5.7 | 0.53 | 0.021 | 0.1 | 78.0 |
表3aLu681中L-泛内脂水解酶的底物特异性
++: >50U/l+: >5U/l(+): >0.5U/l-: <0,5U/l
| 底物 | 浓度(mM) | 活力 |
| S-泛内酯 | 150 | ++ |
| D-泛内酯 | 150 | - |
| γ-丁内酯 | 150 | + |
| γ-戊内酯 | 150 | + |
| δ-戊内酯 | 50 | + |
| ε-己内酯 | 150 | ++ |
| (+/-)δ-癸内酯 | 15 | - |
| δ-壬内酯 | 75 | + |
| D(+)-乳酸乙酯 | 150 | + |
| (L)(-)-乳酸乙酯 | 150 | ++ |
| D-半乳糖酸-γ-内酯 | 150 | + |
| L-半乳糖酸-γ-内酯 | 150 | ++ |
| L-(+)-古洛糖酸-γ-内酯 | 150 | + |
| D-(-)-古洛糖酸-γ-内酯 | 150 | + |
| 1,2-O-二月桂-rac-甘油基-3-戊二酸试卤灵酯 | 0,25 | (+) |
| 5-羟基-2-香豆冉酮 | 2,5 | + |
| α-乙酸萘酯 | 2,5 | (+) |
| 靛红 | 10 | + |
表4a在标准分析中,各种添加物对石竹伯克霍尔德氏菌(Burkholderia
caryophylli)Lu681的L-泛内酯水解酶活力的影响
| 添加物 | 浓度[mM] | 相对活力 |
| 未添加 | - | 100% |
| EDTA | 1 | 97% |
| 柠檬酸pH 6.4 | 30 | 97% |
| o-菲略啉(Phenanthroline) | 1 | 95% |
| HgCl2 | 1 | 133% |
| pCMBS | 1 | 108% |
| DTT | 1 | 99% |
| PMSF | 1 | 113% |
| DIFP | 1 | 115% |
| 抑肽素(Pepstatin) | 1 | 117% |
| H2O2 | 1% | 95% |
| KCN | 1 | 102% |
| KCl | 1 | 99% |
| NH4Cl | 1 | 100% |
| MgCl2 | 1 | 99% |
| CaCl2 | 1 | 101% |
| MnCl2 | 1 | 100% |
| CoCl2 | 1 | 97% |
| FeCl2 | 1 | 104% |
| NiCl2 | 1 | 96% |
| ZnCl2 | 1 | 113% |
| SDS | 1% | 102% |
| CHAPS | 0.1% | 104% |
| Triton | 0.1% | 104% |
| 异丙醇(Isopropanol) | 10% | 93% |
| 乙腈(Acetonitrile) | 10% | 119% |
| MeOH | 10% | 100% |
表5:Lu681的L-泛内脂水解酶氨基酸序列同源性(按照gap4和bestfit5搜索进行)
4Needleman & Wunsch(1970),分子生物学杂志48,443-4535Smith & Waterman(1981),应用数学进展2,482-489
| 在pKS+681插入序列中的位置(bp) | 氨基酸(Aa) | 同系物 | 参考/接受号 | 生物体 | Gap同源性(%aa) | Gap同一性(%aa) | Bestfit同源性(%aa) | Bestfit同一性(%aa) |
| 1498-2304 | 269 | 靛红水解酶 | WO9119175 | 恶臭假单胞菌 | 47.9 | 37.4 | 48.1 | 37.6 |
| 假定蛋白 | B69206 | M.thermoautotro-phicum | 42.6 | 29.8 | 43.0 | 30.1 | ||
| D70817 | M.tuberculosis | 37.1 | 28.5 | 41.7 | 31.5 | |||
| B72430 | T.maritima | 45.4 | 31.9 | 45.9 | 32.2 | |||
| G69399 | 闪烁古生球菌 | 35.5 | 26.3 | 42.1 | 30.3 | |||
| S39964 | S.griseus | 39.6 | 29.8 | 42.0 | 32.1 | |||
| S75497 | 集胞蓝细菌 | 36.3 | 26.8 | 48.1 | 32.9 | |||
| T06135 | A.thaliana | 31.3 | 23.1 | 46.3 | 35.0 | |||
| T05418 | A.thaliana | 33.0 | 23.7 | 42.0 | 29.0 | |||
| Polyketidesynthase | O68500 | S.peuceticus | 42.7 | 32.6 | 46.6 | 37.5 | ||
| 四环素合成酶 | O86485 | S.argillaceus | 43.2 | 34.9 | 44.6 | 35.2 | ||
| 开放阅读框架(ORF) | Q54196 | S.griseus | 39.6 | 29.8 | 42.0 | 32.1 |
表6:L-泛内酯水解酶在E.coli XL1 Blue中的表达a)XL1 Blue pKS+681和pKS+(阴性对照)
| 质粒 | 分析条件 | OD600 i.a. | ΔPA/mM 1h | A/U/l | ee(L-PA) |
| pKS+681 | 300mM D,L-PL/1.1M Tris/HCl pH 7.0 | 2.5 | 5.55 | 92.5 | 79.7 |
| pKS+ | 300mM D,L-PL/1.1M Tris/HCl pH 7.0 | 2.5 | 0.21 | 3.5 | n.d. |
| pKS+681 | 300mM D,L-PL/1.1M Tris/HCl pH 7.0,1% SDS | 2.5 | 7.79 | 132.8 | n.d. |
| pKS+ | 300mM D,L-PL/1.1M Tris/HCl pH 7.0,1% SDS | 2.5 | 0.02 | 0 | n.d. |
b)XL1 Blue pKK681
n.d.,未测定
| 批次 | 分析条件 | DBM i.a./g/l | ΔPA/mM 1h | ΔPA/mM 3h | A/U/l | ee(L-PA) |
| 1 | 50mM D,L-PL,50mM Tris/HCl pH 7.0 | 8.6 | n.d. | 5.2 | 28 | 95.3 |
| 2 | 150mM L-PL,150mM Pipes pH 7.0 | 8.6 | 46 | 46 | 760 | (-) |
| 3 | 300mM D,L-PL,50mM Tris/HCl pH 6.8,Titration with4M NaOH | 1.7 | 5.6 | 16.5 | 92 | 91.3 |
表.7a
| D-泛内酯纯化 | |
| 总收率 | 69,00% |
| 纯度(GC-MS) | > 98,00% |
| 纯度(H-NMR) | > 95,00% |
| 水含量 | < 0,40% |
表.7b
| D-泛内酯纯化 | |
| 总收率 | 85,00% |
| 纯度(GC-MS)* | > 95,00% |
| 纯度(H-NMR) | > 97,00% |
| 水含量 | < 0,40% |
序列表<110>巴斯福股份公司(BASF Aktiengesellschaft)<120>L-泛内酯水解酶和一种D-泛内酯的制备方法<211>810<212>DNA<213>石竹伯克霍尔德氏菌<220><221>CDS<222>(1)..(810)<223>Lu681<400>1atg tgc aac aac tgc gtg atc gag aac gta aaa aag aac atg ctt tca 48Met Cys Asn Asn Cys Val Ile Glu Asn Val Lys Lys Asn Met Leu Ser1 5 10 15cgg cgc ctg ctg ttc aag ggc gct gcg gca ggt ttg acg gcc atg acg 96Arg Arg Leu Leu Phe Lys Gly Ala Ala Ala Gly Leu Thr Ala Met Thr
20 25 30gca ggc agt ctg gct tcc ccg gcg ctt gcg caa tcg ccc cgg cag gtc 144Ala Gly Ser Leu Ala Ser Pro Ala Leu Ala Gln Ser Pro Arg Gln Val
35 40 45gtt gat ctc act cac acc tat gat tcc gca ttt ccc acc ttc gat ggc 192Val Asp Leu Thr His Thr Tyr Asp Ser Ala Phe Pro Thr Phe Asp Gly
50 55 60aaa ccg ggc ata gaa tat gag tgg gca gcg cag atc gcc aaa gac ggc 240Lys Pro Gly Ile Glu Tyr Glu Trp Ala Ala Gln Ile Ala Lys Asp Gly65 70 75 80tat cag ctc cgc aaa ctc acc atc tac gaa cat acc ggc acc cat atc 288Tyr Gln Leu Arg Lys Leu Thr Ile Tyr Glu His Thr Gly Thr His Ile
85 90 95gat gcg cct ttc cac ttc agc gcc gat ggc gcg agc gtc gac caa ctg 336Asp Ala Pro Phe His Phe Ser Ala Asp Gly Ala Ser Val Asp Gln Leu
100 105 110gag ccg cag aaa ctt gtc gct ccg ctt gtc atc gtc gac atc acc gag 384Glu Pro Gln Lys Leu Val Ala Pro Leu Val Ile Val Asp Ile Thr Glu
115 120 125cgc gcc aaa gag gat gcc aat tcc acc att gaa gcc gaa gac atc gag 432Arg Ala Lys Glu Asp Ala Asn Ser Thr Ile Glu Ala Glu Asp Ile Glu130 135 140cgc tgg ata tct gcg aat ggc gac atc ccg aca ggt gca atc gtg gct 480Arg Trp Ile Ser Ala Asn Gly Asp Ile Pro Thr Gly Ala Ile Val Ala145 150 155 160tta cgc tcc gga tgg gca acc aaa gtg aag agt ccc tca ttc cgc aat 528Leu Arg Ser Gly Trp Ala Thr Lys Val Lys Ser Pro Ser Phe Arg Asn
165 170 175gac gaa gcc gga caa ttc gcc ttc ccc ggt ttc ggc aaa tcg gcg acc 576Asp Glu Ala Gly Gln Phe Ala Phe Pro Gly Phe Gly Lys Ser Ala Thr
180 185 190gac ctt ctg ctg aag ctc gac acc gtc gcc att ggc gtc gac aca ctt 624Asp Leu Leu Leu Lys Leu Asp Thr Val Ala Ile Gly Val Asp Thr Leu
195 200 205tct ctg gat ccg ggc aac tcc gca gat ttc gcg gtt cac aat tcc tgg 672Ser Leu Asp Pro Gly Asn Ser Ala Asp Phe Ala Val His Asn Ser Trp210 215 220ctg cca gca gga cgc tac ggt atc gaa gga ctg aac aac ctc gag gct 720Leu Pro Ala Gly Arg Tyr Gly Ile Glu Gly Leu Asn Asn Leu Glu Ala225 230 235 240ctg ccg gtc aag gga gcg acc ata atc gtc ggc gcg ccg gca cac cgc 768Leu Pro Val Lys Gly Ala Thr Ile Ile Val Gly Ala Pro Ala His Arg
245 250 255ggc gga acg ggc ggc cca gcc cgt att ctg gcc ctg gtc tga 810Gly Gly Thr Gly Gly Pro Ala Arg Ile Leu Ala Leu Val
260 265 270<210>2<211>269<212>PRT<213>石竹伯克霍尔德氏菌<400>2Met Cys Asn Asn Cys Val Ile Glu Asn Val Lys Lys Asn Met Leu Ser1 5 10 15Arg Arg Leu Leu Phe Lys Gly Ala Ala Ala Gly Leu Thr Ala Met Thr
20 25 30Ala Gly Ser Leu Ala Ser Pro Ala Leu Ala Gln Ser Pro Arg Gln Val
35 40 45Val Asp Leu Thr His Thr Tyr Asp Ser Ala Phe Pro Thr Phe Asp Gly
50 55 60Lys Pro Gly Ile Glu Tyr Glu Trp Ala Ala Gln Ile Ala Lys Asp Gly65 70 75 80Tyr Gln Leu Arg Lys Leu Thr Ile Tyr Glu His Thr Gly Thr His Ile
85 90 95Asp Ala Pro Phe His Phe Ser Ala Asp Gly Ala Ser Val Asp Gln Leu
100 105 110Glu Pro Gln Lys Leu Val Ala Pro Leu Val Ile Val Asp Ile Thr Glu
115 120 125Arg Ala Lys Glu Asp Ala Asn Ser Thr Ile Glu Ala Glu Asp Ile Glu130 135 140Arg Trp Ile Ser Ala Asn Gly Asp Ile Pro Thr Gly Ala Ile Val Ala145 150 155 160Leu Arg Ser Gly Trp Ala Thr Lys Val Lys Ser Pro Ser Phe Arg Asn
165 170 175Asp Glu Ala Gly Gln Phe Ala Phe Pro Gly Phe Gly Lys Ser Ala Thr
180 185 190Asp Leu Leu Leu Lys Leu Asp Thr Val Ala Ile Gly Val Asp Thr Leu
195 200 205Ser Leu Asp Pro Gly Asn Ser Ala Asp Phe Ala Val His Asn Ser Trp210 215 220Leu Pro Ala Gly Arg Tyr Gly Ile Glu Gly Leu Asn Asn Leu Glu Ala225 230 235 240Leu Pro Val Lys Gly Ala Thr Ile Ile Val Gly Ala Pro Ala His Arg
245 250 255Gly Gly Thr Gly Gly Pro Ala Arg Ile Leu Ala Leu Val
260 265<210>3<211>12<212>PRT<213>石竹伯克霍尔德氏菌<400>3Tyr Gly Ile Glu Gly Leu Asn Asn Leu Glu Ala Leu1 5 10<210>4<211>13<212>PRT<213>石竹伯克霍尔德氏菌<400>4Ala Lys Glu Asp Ala Asn Ser Thr Ile Glu Ala Glu Asp1 5 10<210>5<211>13<212>PRT<213>石竹伯克霍尔德氏菌<400>5Tyr Leu Gly Ile Glu Gly Leu Asn Asn Leu Glu Ala Leu1 5 10<210>6<211>10<212>PRT<213>石竹伯克霍尔德氏菌<400>6Ala Lys Glu Asp Ala Val Ser Thr Ile Glu1 5 10
Claims (20)
1.一段分离的、编码一段具有L-泛内酯水解酶活力多肽的核酸序列,选自:
a)具有SEQ ID NO:1所示序列的核酸序列,
b)由于遗传密码的简并,导致从序列SEQ ID NO:1中衍生的核酸
序列,
c)其编码的多肽具有SEQ ID NO:2所示的氨基酸序列的SEQ ID NO:
1所示核酸序列的衍生物,并且该衍生物在不明显损失该多肽酶
活性的前提下,在氨基酸水平上至少应有50%的同源性,
d)(a)到(c)中所提序列的功能等同物。
2.由权利要求1所述核酸序列编码的氨基酸序列。
3.由SEQ ID NO:1所示序列编码的,权利要求2所述的氨基酸序列。
4.含有权利要求1所述核酸序列的核酸构建体,其中该核酸序列与一或多个调节信号相连。
5.含有权利要求1所述核酸序列或权利要求4所述核酸构建体的载体。
6.含有至少一个权利要求1所述核酸序列、至少一个权利要求4所述核酸构建体或权利要求5所述载体的微生物。
7.权利要求6所述的微生物,它为革兰氏阴性菌。
8.权利要求6或7所述的微生物,它为来自α-蛋白细菌、β-蛋白细菌或γ-蛋白细菌族的细菌。
9.权利要求6到8中任何一个所述的微生物,它为来自肠杆菌科、假单胞菌科或根瘤菌科的细菌。
10.权利要求6到9中任何一个所述的微生物,它为来自土壤杆菌属、假单胞菌属、伯克霍尔德氏菌属、沙门氏菌属或埃希氏菌属的细菌。
11.具有下列性质的L-泛内酯水解酶:
a)将L-泛内酯转化成相应的酸,
b)pH稳定性:L-泛内酯水解酶在pH 4-10范围内稳定,
c)最适pH:7.2到7.6,
d)最适温度:大约70℃到75℃,
e)EDTA不抑制其活性。
12.一种制备D-泛内酯的方法,它包括下列反应步骤:
a)在权利要求11所述的L-泛内酯水解酶或在具有权利要求2所述氨基酸序列的L-泛内酯水解酶或在权利要求6所述微生物存在时,将外消旋泛内酯转化成D-泛内酯和L-泛解酸,
b)除去D-泛内酯。
13.权利要求12所述的方法,其中,在步骤(b)得到的L-泛解酸被外消旋化并被回收至步骤(a)。
14.权利要求12或13所述的方法,其中,外消旋泛内酯的转化在有固定化的、权利要求11所述的L-泛内酯水解酶或固定化的、具有权利要求2所述氨基酸序列的L-泛内酯水解酶存在时进行。
15.权利要求12或13所述的方法,其中,外消旋泛内酯的转化在有生长的、休眠的或破碎的如权利要求6所述微生物存在时进行。
16.权利要求12或15所述的方法,其中,微生物为固定化的。
17.如权利要求12到16中任何一项所述的方法,其中,该方法在pH4至12之间的水反应溶液中进行。
18.如权利要求12到17中任何一项所述的方法,其中,该方法在0℃到95℃之间进行。
19.权利要求12或13所述的方法,其中,D-泛内酯通过萃取被去除。
20.如权利要求12到18中任何一项所述的方法,其中,D-泛内酯具有至少90%ee的光学纯度。
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| DE19952501.3 | 1999-10-29 | ||
| DE19952501A DE19952501A1 (de) | 1999-10-29 | 1999-10-29 | L-Pantolacton-Hydrolase und ein Verfahren zur Herstellung von D-Pantolacton |
| DE10029194.5 | 2000-06-19 | ||
| DE10029194A DE10029194A1 (de) | 2000-06-19 | 2000-06-19 | L-Pantolacton-Hydrolase und ein Verfahren zur Herstellung von D-Pantolacton |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN1384880A true CN1384880A (zh) | 2002-12-11 |
Family
ID=26006072
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN00814937A Pending CN1384880A (zh) | 1999-10-29 | 2000-10-20 | L-泛内酯水解酶和一种d-泛内酯的制备方法 |
Country Status (17)
| Country | Link |
|---|---|
| US (1) | US6998258B1 (zh) |
| EP (1) | EP1224296A1 (zh) |
| JP (1) | JP2003530080A (zh) |
| KR (1) | KR20020043254A (zh) |
| CN (1) | CN1384880A (zh) |
| AU (1) | AU782517B2 (zh) |
| BR (1) | BR0015114A (zh) |
| CA (1) | CA2389064A1 (zh) |
| CZ (1) | CZ20021449A3 (zh) |
| EE (1) | EE200200225A (zh) |
| HU (1) | HUP0203704A3 (zh) |
| IL (1) | IL148973A0 (zh) |
| MX (1) | MXPA02003417A (zh) |
| NO (1) | NO20021931D0 (zh) |
| PL (1) | PL355487A1 (zh) |
| RU (1) | RU2002114044A (zh) |
| WO (1) | WO2001032890A1 (zh) |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101448950B (zh) * | 2006-05-16 | 2014-02-12 | 帝斯曼知识产权资产管理有限公司 | 用于生产泛醇的工艺 |
| CN107709307A (zh) * | 2015-06-19 | 2018-02-16 | 巴斯夫欧洲公司 | 泛酰内酯的制备 |
| CN112048537A (zh) * | 2019-06-07 | 2020-12-08 | 帝斯曼知识产权资产管理有限公司 | (r)-泛酸内酯乙酸酯的合成 |
Families Citing this family (83)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20030194798A1 (en) * | 2001-05-24 | 2003-10-16 | Surber Mark W. | Minicell compositions and methods |
| DE50307835D1 (de) | 2002-12-02 | 2007-09-13 | Basf Ag | L-rhamnose-induzierbare expressionssysteme |
| CN1768141B (zh) * | 2003-03-28 | 2012-10-10 | 帝斯曼知识产权资产管理有限公司 | 泛酸内酯水解酶 |
| CN101535493A (zh) | 2006-07-27 | 2009-09-16 | 惠氏公司 | 用于生产重组蛋白质的高细胞密度补料-分批发酵方法 |
| US9044600B2 (en) * | 2008-05-22 | 2015-06-02 | Vladimir Balakin | Proton tomography apparatus and method of operation therefor |
| US8129699B2 (en) | 2008-05-22 | 2012-03-06 | Vladimir Balakin | Multi-field charged particle cancer therapy method and apparatus coordinated with patient respiration |
| US8373145B2 (en) * | 2008-05-22 | 2013-02-12 | Vladimir Balakin | Charged particle cancer therapy system magnet control method and apparatus |
| US9095040B2 (en) | 2008-05-22 | 2015-07-28 | Vladimir Balakin | Charged particle beam acceleration and extraction method and apparatus used in conjunction with a charged particle cancer therapy system |
| US9974978B2 (en) | 2008-05-22 | 2018-05-22 | W. Davis Lee | Scintillation array apparatus and method of use thereof |
| US8710462B2 (en) * | 2008-05-22 | 2014-04-29 | Vladimir Balakin | Charged particle cancer therapy beam path control method and apparatus |
| US8089054B2 (en) | 2008-05-22 | 2012-01-03 | Vladimir Balakin | Charged particle beam acceleration and extraction method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8198607B2 (en) * | 2008-05-22 | 2012-06-12 | Vladimir Balakin | Tandem accelerator method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8519365B2 (en) * | 2008-05-22 | 2013-08-27 | Vladimir Balakin | Charged particle cancer therapy imaging method and apparatus |
| US9056199B2 (en) | 2008-05-22 | 2015-06-16 | Vladimir Balakin | Charged particle treatment, rapid patient positioning apparatus and method of use thereof |
| US8378311B2 (en) | 2008-05-22 | 2013-02-19 | Vladimir Balakin | Synchrotron power cycling apparatus and method of use thereof |
| US8569717B2 (en) * | 2008-05-22 | 2013-10-29 | Vladimir Balakin | Intensity modulated three-dimensional radiation scanning method and apparatus |
| US9937362B2 (en) | 2008-05-22 | 2018-04-10 | W. Davis Lee | Dynamic energy control of a charged particle imaging/treatment apparatus and method of use thereof |
| US9616252B2 (en) | 2008-05-22 | 2017-04-11 | Vladimir Balakin | Multi-field cancer therapy apparatus and method of use thereof |
| US9737272B2 (en) | 2008-05-22 | 2017-08-22 | W. Davis Lee | Charged particle cancer therapy beam state determination apparatus and method of use thereof |
| US10684380B2 (en) | 2008-05-22 | 2020-06-16 | W. Davis Lee | Multiple scintillation detector array imaging apparatus and method of use thereof |
| US8288742B2 (en) * | 2008-05-22 | 2012-10-16 | Vladimir Balakin | Charged particle cancer therapy patient positioning method and apparatus |
| US7939809B2 (en) * | 2008-05-22 | 2011-05-10 | Vladimir Balakin | Charged particle beam extraction method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8374314B2 (en) | 2008-05-22 | 2013-02-12 | Vladimir Balakin | Synchronized X-ray / breathing method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8718231B2 (en) | 2008-05-22 | 2014-05-06 | Vladimir Balakin | X-ray tomography method and apparatus used in conjunction with a charged particle cancer therapy system |
| US9177751B2 (en) | 2008-05-22 | 2015-11-03 | Vladimir Balakin | Carbon ion beam injector apparatus and method of use thereof |
| US10092776B2 (en) | 2008-05-22 | 2018-10-09 | Susan L. Michaud | Integrated translation/rotation charged particle imaging/treatment apparatus and method of use thereof |
| US8309941B2 (en) * | 2008-05-22 | 2012-11-13 | Vladimir Balakin | Charged particle cancer therapy and patient breath monitoring method and apparatus |
| US8378321B2 (en) * | 2008-05-22 | 2013-02-19 | Vladimir Balakin | Charged particle cancer therapy and patient positioning method and apparatus |
| US8637833B2 (en) | 2008-05-22 | 2014-01-28 | Vladimir Balakin | Synchrotron power supply apparatus and method of use thereof |
| US8598543B2 (en) * | 2008-05-22 | 2013-12-03 | Vladimir Balakin | Multi-axis/multi-field charged particle cancer therapy method and apparatus |
| US9782140B2 (en) | 2008-05-22 | 2017-10-10 | Susan L. Michaud | Hybrid charged particle / X-ray-imaging / treatment apparatus and method of use thereof |
| US8975600B2 (en) | 2008-05-22 | 2015-03-10 | Vladimir Balakin | Treatment delivery control system and method of operation thereof |
| US9981147B2 (en) | 2008-05-22 | 2018-05-29 | W. Davis Lee | Ion beam extraction apparatus and method of use thereof |
| US9155911B1 (en) | 2008-05-22 | 2015-10-13 | Vladimir Balakin | Ion source method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8373143B2 (en) * | 2008-05-22 | 2013-02-12 | Vladimir Balakin | Patient immobilization and repositioning method and apparatus used in conjunction with charged particle cancer therapy |
| US8969834B2 (en) | 2008-05-22 | 2015-03-03 | Vladimir Balakin | Charged particle therapy patient constraint apparatus and method of use thereof |
| US8368038B2 (en) * | 2008-05-22 | 2013-02-05 | Vladimir Balakin | Method and apparatus for intensity control of a charged particle beam extracted from a synchrotron |
| US8399866B2 (en) | 2008-05-22 | 2013-03-19 | Vladimir Balakin | Charged particle extraction apparatus and method of use thereof |
| US10143854B2 (en) | 2008-05-22 | 2018-12-04 | Susan L. Michaud | Dual rotation charged particle imaging / treatment apparatus and method of use thereof |
| US9168392B1 (en) | 2008-05-22 | 2015-10-27 | Vladimir Balakin | Charged particle cancer therapy system X-ray apparatus and method of use thereof |
| US8373146B2 (en) * | 2008-05-22 | 2013-02-12 | Vladimir Balakin | RF accelerator method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8624528B2 (en) * | 2008-05-22 | 2014-01-07 | Vladimir Balakin | Method and apparatus coordinating synchrotron acceleration periods with patient respiration periods |
| US9737734B2 (en) | 2008-05-22 | 2017-08-22 | Susan L. Michaud | Charged particle translation slide control apparatus and method of use thereof |
| US9737733B2 (en) | 2008-05-22 | 2017-08-22 | W. Davis Lee | Charged particle state determination apparatus and method of use thereof |
| US8436327B2 (en) * | 2008-05-22 | 2013-05-07 | Vladimir Balakin | Multi-field charged particle cancer therapy method and apparatus |
| US20090314960A1 (en) * | 2008-05-22 | 2009-12-24 | Vladimir Balakin | Patient positioning method and apparatus used in conjunction with a charged particle cancer therapy system |
| US9579525B2 (en) | 2008-05-22 | 2017-02-28 | Vladimir Balakin | Multi-axis charged particle cancer therapy method and apparatus |
| US8188688B2 (en) | 2008-05-22 | 2012-05-29 | Vladimir Balakin | Magnetic field control method and apparatus used in conjunction with a charged particle cancer therapy system |
| US9910166B2 (en) | 2008-05-22 | 2018-03-06 | Stephen L. Spotts | Redundant charged particle state determination apparatus and method of use thereof |
| US10548551B2 (en) | 2008-05-22 | 2020-02-04 | W. Davis Lee | Depth resolved scintillation detector array imaging apparatus and method of use thereof |
| US8907309B2 (en) | 2009-04-17 | 2014-12-09 | Stephen L. Spotts | Treatment delivery control system and method of operation thereof |
| US8178859B2 (en) * | 2008-05-22 | 2012-05-15 | Vladimir Balakin | Proton beam positioning verification method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8642978B2 (en) | 2008-05-22 | 2014-02-04 | Vladimir Balakin | Charged particle cancer therapy dose distribution method and apparatus |
| US9855444B2 (en) | 2008-05-22 | 2018-01-02 | Scott Penfold | X-ray detector for proton transit detection apparatus and method of use thereof |
| US9744380B2 (en) | 2008-05-22 | 2017-08-29 | Susan L. Michaud | Patient specific beam control assembly of a cancer therapy apparatus and method of use thereof |
| US9498649B2 (en) | 2008-05-22 | 2016-11-22 | Vladimir Balakin | Charged particle cancer therapy patient constraint apparatus and method of use thereof |
| US10070831B2 (en) | 2008-05-22 | 2018-09-11 | James P. Bennett | Integrated cancer therapy—imaging apparatus and method of use thereof |
| US10029122B2 (en) | 2008-05-22 | 2018-07-24 | Susan L. Michaud | Charged particle—patient motion control system apparatus and method of use thereof |
| US9682254B2 (en) | 2008-05-22 | 2017-06-20 | Vladimir Balakin | Cancer surface searing apparatus and method of use thereof |
| US8627822B2 (en) * | 2008-07-14 | 2014-01-14 | Vladimir Balakin | Semi-vertical positioning method and apparatus used in conjunction with a charged particle cancer therapy system |
| US8625739B2 (en) | 2008-07-14 | 2014-01-07 | Vladimir Balakin | Charged particle cancer therapy x-ray method and apparatus |
| US8229072B2 (en) * | 2008-07-14 | 2012-07-24 | Vladimir Balakin | Elongated lifetime X-ray method and apparatus used in conjunction with a charged particle cancer therapy system |
| EP3404108A1 (de) | 2009-06-05 | 2018-11-21 | Basf Se | Biokatalytische herstellung von ambroxan |
| US10179250B2 (en) | 2010-04-16 | 2019-01-15 | Nick Ruebel | Auto-updated and implemented radiation treatment plan apparatus and method of use thereof |
| US10625097B2 (en) | 2010-04-16 | 2020-04-21 | Jillian Reno | Semi-automated cancer therapy treatment apparatus and method of use thereof |
| US10188877B2 (en) | 2010-04-16 | 2019-01-29 | W. Davis Lee | Fiducial marker/cancer imaging and treatment apparatus and method of use thereof |
| US10556126B2 (en) | 2010-04-16 | 2020-02-11 | Mark R. Amato | Automated radiation treatment plan development apparatus and method of use thereof |
| US9737731B2 (en) | 2010-04-16 | 2017-08-22 | Vladimir Balakin | Synchrotron energy control apparatus and method of use thereof |
| US10376717B2 (en) | 2010-04-16 | 2019-08-13 | James P. Bennett | Intervening object compensating automated radiation treatment plan development apparatus and method of use thereof |
| US10589128B2 (en) | 2010-04-16 | 2020-03-17 | Susan L. Michaud | Treatment beam path verification in a cancer therapy apparatus and method of use thereof |
| US10349906B2 (en) | 2010-04-16 | 2019-07-16 | James P. Bennett | Multiplexed proton tomography imaging apparatus and method of use thereof |
| US11648420B2 (en) | 2010-04-16 | 2023-05-16 | Vladimir Balakin | Imaging assisted integrated tomography—cancer treatment apparatus and method of use thereof |
| US10555710B2 (en) | 2010-04-16 | 2020-02-11 | James P. Bennett | Simultaneous multi-axes imaging apparatus and method of use thereof |
| US10638988B2 (en) | 2010-04-16 | 2020-05-05 | Scott Penfold | Simultaneous/single patient position X-ray and proton imaging apparatus and method of use thereof |
| US10751551B2 (en) | 2010-04-16 | 2020-08-25 | James P. Bennett | Integrated imaging-cancer treatment apparatus and method of use thereof |
| US10518109B2 (en) | 2010-04-16 | 2019-12-31 | Jillian Reno | Transformable charged particle beam path cancer therapy apparatus and method of use thereof |
| US10086214B2 (en) | 2010-04-16 | 2018-10-02 | Vladimir Balakin | Integrated tomography—cancer treatment apparatus and method of use thereof |
| US8963112B1 (en) | 2011-05-25 | 2015-02-24 | Vladimir Balakin | Charged particle cancer therapy patient positioning method and apparatus |
| US8933651B2 (en) | 2012-11-16 | 2015-01-13 | Vladimir Balakin | Charged particle accelerator magnet apparatus and method of use thereof |
| US9907981B2 (en) | 2016-03-07 | 2018-03-06 | Susan L. Michaud | Charged particle translation slide control apparatus and method of use thereof |
| US10037863B2 (en) | 2016-05-27 | 2018-07-31 | Mark R. Amato | Continuous ion beam kinetic energy dissipater apparatus and method of use thereof |
| CN113195727A (zh) | 2018-10-18 | 2021-07-30 | 巴斯夫欧洲公司 | 用于产生丙烯酸或其盐的方法 |
| CN110452861B (zh) * | 2019-07-10 | 2021-02-09 | 杭州师范大学 | 一种基因重组工程菌及其在催化合成d-泛酰内酯中的应用 |
Family Cites Families (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS5572182A (en) * | 1978-11-28 | 1980-05-30 | Sagami Chem Res Center | Optical resolution of pantolactone |
| JPH0667320B2 (ja) * | 1986-06-13 | 1994-08-31 | 三菱化成株式会社 | D−パントラクトンの製造法 |
| JPH0655156B2 (ja) * | 1986-06-13 | 1994-07-27 | 三菱化成株式会社 | Dl−パントラクトンの光学分割法 |
| JP2844354B2 (ja) | 1989-08-03 | 1999-01-06 | 富士薬品工業株式会社 | D―パントラクトンの製造法 |
| DE4005150A1 (de) | 1990-02-17 | 1991-08-22 | Hoechst Ag | Verfahren zur enzymatischen racematspaltung von pantolacton |
| JP3011449B2 (ja) | 1990-10-05 | 2000-02-21 | 富士薬品工業株式会社 | D―パントラクトン加水分解酵素およびその製造法 |
| CA2064676A1 (en) | 1991-04-02 | 1992-10-03 | Manfred Schneider | Immobilized biocatalyst, its preparation and use for ester synthesis in a column reactor |
| DE4126580A1 (de) | 1991-08-12 | 1993-02-18 | Degussa | D-2,4-dihydroxy-3,3-dimethylbutansaeurenitril, seine herstellung und verwendung |
| KR100266037B1 (ko) | 1992-10-15 | 2000-09-15 | 에모토 간지 | 금속대의 연속침탄방법 |
| AU1810197A (en) * | 1995-09-13 | 1997-04-01 | Fuji Yakuhin Kogyo Kabushiki Kaisha | D-pantolactone hydrolase and gene encoding the same |
| US6395529B1 (en) * | 1998-11-10 | 2002-05-28 | Novozymes Biotech, Inc. | Polypeptides having lactonohydrolase activity and nucleic acids encoding same |
| WO2000028043A2 (en) * | 1998-11-10 | 2000-05-18 | Novozymes Biotech, Inc. | Polypeptides having lactonohydrolase activity and nucleic acids encoding same |
-
2000
- 2000-10-20 RU RU2002114044/13A patent/RU2002114044A/ru not_active Application Discontinuation
- 2000-10-20 US US10/111,451 patent/US6998258B1/en not_active Expired - Fee Related
- 2000-10-20 AU AU11416/01A patent/AU782517B2/en not_active Ceased
- 2000-10-20 EP EP00972817A patent/EP1224296A1/de not_active Withdrawn
- 2000-10-20 CN CN00814937A patent/CN1384880A/zh active Pending
- 2000-10-20 MX MXPA02003417A patent/MXPA02003417A/es unknown
- 2000-10-20 CZ CZ20021449A patent/CZ20021449A3/cs unknown
- 2000-10-20 JP JP2001535572A patent/JP2003530080A/ja not_active Withdrawn
- 2000-10-20 HU HU0203704A patent/HUP0203704A3/hu unknown
- 2000-10-20 WO PCT/EP2000/010320 patent/WO2001032890A1/de not_active Application Discontinuation
- 2000-10-20 CA CA002389064A patent/CA2389064A1/en not_active Abandoned
- 2000-10-20 BR BR0015114-9A patent/BR0015114A/pt not_active IP Right Cessation
- 2000-10-20 PL PL00355487A patent/PL355487A1/xx not_active Application Discontinuation
- 2000-10-20 EE EEP200200225A patent/EE200200225A/xx unknown
- 2000-10-20 KR KR1020027005466A patent/KR20020043254A/ko not_active Application Discontinuation
- 2000-10-20 IL IL14897300A patent/IL148973A0/xx unknown
-
2002
- 2002-04-24 NO NO20021931A patent/NO20021931D0/no not_active Application Discontinuation
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101448950B (zh) * | 2006-05-16 | 2014-02-12 | 帝斯曼知识产权资产管理有限公司 | 用于生产泛醇的工艺 |
| CN107709307A (zh) * | 2015-06-19 | 2018-02-16 | 巴斯夫欧洲公司 | 泛酰内酯的制备 |
| CN112048537A (zh) * | 2019-06-07 | 2020-12-08 | 帝斯曼知识产权资产管理有限公司 | (r)-泛酸内酯乙酸酯的合成 |
Also Published As
| Publication number | Publication date |
|---|---|
| HUP0203704A3 (en) | 2006-03-28 |
| PL355487A1 (en) | 2004-05-04 |
| BR0015114A (pt) | 2002-07-16 |
| HUP0203704A2 (hu) | 2003-03-28 |
| RU2002114044A (ru) | 2004-03-27 |
| NO20021931L (no) | 2002-04-24 |
| AU1141601A (en) | 2001-05-14 |
| CZ20021449A3 (cs) | 2002-10-16 |
| EP1224296A1 (de) | 2002-07-24 |
| US6998258B1 (en) | 2006-02-14 |
| NO20021931D0 (no) | 2002-04-24 |
| EE200200225A (et) | 2003-06-16 |
| JP2003530080A (ja) | 2003-10-14 |
| MXPA02003417A (es) | 2002-08-20 |
| IL148973A0 (en) | 2002-11-10 |
| WO2001032890A1 (de) | 2001-05-10 |
| CA2389064A1 (en) | 2001-05-10 |
| AU782517B2 (en) | 2005-08-04 |
| KR20020043254A (ko) | 2002-06-08 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN1384880A (zh) | L-泛内酯水解酶和一种d-泛内酯的制备方法 | |
| EP1399567B1 (en) | Method for producing alpha-hydroxy acid, glycolic acid 2-hydroxyisobutyric acid from a corresponding alpha-hydroxy nitrile using nitrilase | |
| KR100715744B1 (ko) | 니트릴라제 또는 니트릴라제의 유전자를 포함한 미생물을이용하여 니트릴로부터 키랄 카르복실산을 제조하는 방법 | |
| CN101535467B (zh) | 左旋内酯水解酶产生菌及其用于制备手性羟基酸的方法 | |
| JP6048850B2 (ja) | D−サクシニラーゼ、およびこれを用いたd−アミノ酸の製造方法 | |
| JPH11155570A (ja) | 酵素の基質特異性の改変法 | |
| CN101796192A (zh) | 使用固氮弧菌属物种(Azoarcus Sp)EBN1脱氢酶产生光学活性醇的方法 | |
| CN101824443B (zh) | 一种去除制备中性氨基酸的反应体系中的丙氨酸的方法 | |
| CN100334206C (zh) | 新型脱氢酶和编码该脱氢酶的基因 | |
| NZ292559A (en) | Preparation of racemic alpha-substituted 4-methylthiobutyric acids by enzymatic hydrolysis | |
| CN100402640C (zh) | 内酯酶的制造方法及其应用 | |
| Kesseler et al. | Development of a Novel Biocatalyst for the Resolution of rac‐Pantolactone | |
| JP4272312B2 (ja) | 新規ニトリラーゼ、および2−ヒドロキシ−4−メチルチオ酪酸の製造方法 | |
| CN101341251A (zh) | 由5-降冰片烯-2-甲腈使用芳基乙腈水解酶制备5-降冰片烯-2-甲酸的方法 | |
| JPH04320679A (ja) | L‐カルニチン‐アミダーゼを生産する微生物、微生物学的に生産されたl‐カルニチン‐アミダーゼ、その取得方法およびl‐カルニチンへのdl‐および/またはカルニチンアミドの酵素的変換方法 | |
| Naidu et al. | Optimization of tannase production from Aspergillus foetidus using statistical design methods | |
| JP4729919B2 (ja) | 微生物の培養方法及び光学活性カルボン酸の製造方法 | |
| JP4485734B2 (ja) | 5置換ヒダントインラセマーゼ、これをコードするdna、組み換えdna、形質転換された細胞および光学活性アミノ酸の製造方法 | |
| JP2009201373A (ja) | (r)−3−キヌクリジノールの製造方法 | |
| JP3917723B2 (ja) | ラクトン加水分解酵素およびその製造法 | |
| Crognale et al. | Optimization of capsaicin acylase production from Streptomyces mobaraensis in bench-top fermenter | |
| JPH1198981A (ja) | 新規エステル加水分解酵素 | |
| Crognale et al. | Optimization Of Capsaicin Acylase Production From Streptomyces Mobaraensis In Bench-Top Reactor | |
| JPH10248561A (ja) | 熱安定性耐溶媒性エステル分解酵素 | |
| JP2003319788A (ja) | 新規デヒドロゲナーゼ及びそれをコードする遺伝子 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| C02 | Deemed withdrawal of patent application after publication (patent law 2001) | ||
| WD01 | Invention patent application deemed withdrawn after publication |