JP2010142234A - 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 - Google Patents
新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 Download PDFInfo
- Publication number
- JP2010142234A JP2010142234A JP2009299177A JP2009299177A JP2010142234A JP 2010142234 A JP2010142234 A JP 2010142234A JP 2009299177 A JP2009299177 A JP 2009299177A JP 2009299177 A JP2009299177 A JP 2009299177A JP 2010142234 A JP2010142234 A JP 2010142234A
- Authority
- JP
- Japan
- Prior art keywords
- amino acid
- acid residue
- polypeptide
- glyphosate
- sequence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Withdrawn
Links
- 108090000623 proteins and genes Proteins 0.000 title claims description 118
- 108010039239 glyphosate N-acetyltransferase Proteins 0.000 title description 321
- 102000040430 polynucleotide Human genes 0.000 abstract description 250
- 108091033319 polynucleotide Proteins 0.000 abstract description 249
- 239000002157 polynucleotide Substances 0.000 abstract description 249
- XDDAORKBJWWYJS-UHFFFAOYSA-N glyphosate Chemical compound OC(=O)CNCP(O)(O)=O XDDAORKBJWWYJS-UHFFFAOYSA-N 0.000 abstract description 227
- 239000005562 Glyphosate Substances 0.000 abstract description 220
- 229940097068 glyphosate Drugs 0.000 abstract description 217
- 238000000034 method Methods 0.000 abstract description 191
- 125000003275 alpha amino acid group Chemical group 0.000 abstract description 138
- 239000002773 nucleotide Chemical group 0.000 abstract description 49
- 125000003729 nucleotide group Chemical group 0.000 abstract description 49
- 239000011159 matrix material Substances 0.000 abstract description 17
- 108020004635 Complementary DNA Proteins 0.000 abstract description 4
- 239000003153 chemical reaction reagent Substances 0.000 abstract description 4
- 125000000539 amino acid group Chemical group 0.000 description 617
- 108090000765 processed proteins & peptides Proteins 0.000 description 579
- 102000004196 processed proteins & peptides Human genes 0.000 description 567
- 229920001184 polypeptide Polymers 0.000 description 563
- 241000196324 Embryophyta Species 0.000 description 478
- 230000009261 transgenic effect Effects 0.000 description 277
- 235000001014 amino acid Nutrition 0.000 description 186
- 229940024606 amino acid Drugs 0.000 description 177
- 150000001413 amino acids Chemical class 0.000 description 176
- 150000007523 nucleic acids Chemical class 0.000 description 141
- 210000004027 cell Anatomy 0.000 description 133
- 102000039446 nucleic acids Human genes 0.000 description 120
- 108020004707 nucleic acids Proteins 0.000 description 120
- 239000004009 herbicide Substances 0.000 description 104
- 108010000700 Acetolactate synthase Proteins 0.000 description 96
- 230000002363 herbicidal effect Effects 0.000 description 92
- 230000000694 effects Effects 0.000 description 66
- 230000014509 gene expression Effects 0.000 description 59
- CAAMSDWKXXPUJR-UHFFFAOYSA-N 3,5-dihydro-4H-imidazol-4-one Chemical compound O=C1CNC=N1 CAAMSDWKXXPUJR-UHFFFAOYSA-N 0.000 description 54
- 229940124530 sulfonamide Drugs 0.000 description 53
- 150000003456 sulfonamides Chemical class 0.000 description 53
- 102000004169 proteins and genes Human genes 0.000 description 51
- 239000013598 vector Substances 0.000 description 50
- 229910052727 yttrium Inorganic materials 0.000 description 50
- 235000018102 proteins Nutrition 0.000 description 47
- 108020004705 Codon Proteins 0.000 description 42
- 238000009396 hybridization Methods 0.000 description 42
- 230000007246 mechanism Effects 0.000 description 40
- 229910052731 fluorine Inorganic materials 0.000 description 39
- 229910052739 hydrogen Inorganic materials 0.000 description 39
- 229910052700 potassium Inorganic materials 0.000 description 38
- 229910052720 vanadium Inorganic materials 0.000 description 34
- 108091028043 Nucleic acid sequence Proteins 0.000 description 33
- 108030006517 Glyphosate oxidoreductases Proteins 0.000 description 32
- 108020001991 Protoporphyrinogen Oxidase Proteins 0.000 description 30
- 102000005135 Protoporphyrinogen oxidase Human genes 0.000 description 30
- 108091026890 Coding region Proteins 0.000 description 29
- 102000016680 Dioxygenases Human genes 0.000 description 29
- 108010028143 Dioxygenases Proteins 0.000 description 29
- 238000006467 substitution reaction Methods 0.000 description 29
- 229910052721 tungsten Inorganic materials 0.000 description 29
- 108010020183 3-phosphoshikimate 1-carboxyvinyltransferase Proteins 0.000 description 28
- 229910052717 sulfur Inorganic materials 0.000 description 28
- 230000002163 immunogen Effects 0.000 description 26
- 229910052698 phosphorus Inorganic materials 0.000 description 25
- 230000000295 complement effect Effects 0.000 description 24
- 239000012634 fragment Substances 0.000 description 24
- 238000004519 manufacturing process Methods 0.000 description 24
- 238000012360 testing method Methods 0.000 description 23
- 108010082527 phosphinothricin N-acetyltransferase Proteins 0.000 description 22
- 108020004414 DNA Proteins 0.000 description 21
- 102000004190 Enzymes Human genes 0.000 description 21
- 108090000790 Enzymes Proteins 0.000 description 21
- 229940088598 enzyme Drugs 0.000 description 21
- 239000000523 sample Substances 0.000 description 21
- 230000006870 function Effects 0.000 description 20
- 238000009739 binding Methods 0.000 description 19
- 238000006640 acetylation reaction Methods 0.000 description 18
- 230000027455 binding Effects 0.000 description 18
- 241000282326 Felis catus Species 0.000 description 17
- 230000001965 increasing effect Effects 0.000 description 17
- 239000003550 marker Substances 0.000 description 17
- 229910052757 nitrogen Inorganic materials 0.000 description 17
- 210000001519 tissue Anatomy 0.000 description 17
- 229910019142 PO4 Inorganic materials 0.000 description 16
- 230000021736 acetylation Effects 0.000 description 16
- 239000000203 mixture Substances 0.000 description 16
- 239000010452 phosphate Substances 0.000 description 16
- 230000006798 recombination Effects 0.000 description 16
- 238000005215 recombination Methods 0.000 description 16
- 230000001580 bacterial effect Effects 0.000 description 15
- 229910052799 carbon Inorganic materials 0.000 description 15
- 239000013604 expression vector Substances 0.000 description 15
- IAJOBQBIJHVGMQ-UHFFFAOYSA-N 2-amino-4-[hydroxy(methyl)phosphoryl]butanoic acid Chemical group CP(O)(=O)CCC(N)C(O)=O IAJOBQBIJHVGMQ-UHFFFAOYSA-N 0.000 description 14
- ZSLZBFCDCINBPY-ZSJPKINUSA-N acetyl-CoA Chemical compound O[C@@H]1[C@H](OP(O)(O)=O)[C@@H](COP(O)(=O)OP(O)(=O)OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCSC(=O)C)O[C@H]1N1C2=NC=NC(N)=C2N=C1 ZSLZBFCDCINBPY-ZSJPKINUSA-N 0.000 description 14
- 241000588724 Escherichia coli Species 0.000 description 13
- 238000013518 transcription Methods 0.000 description 13
- 230000035897 transcription Effects 0.000 description 13
- 230000009466 transformation Effects 0.000 description 13
- 238000000338 in vitro Methods 0.000 description 12
- 238000003752 polymerase chain reaction Methods 0.000 description 12
- 238000003556 assay Methods 0.000 description 11
- 230000001276 controlling effect Effects 0.000 description 11
- 239000003112 inhibitor Substances 0.000 description 11
- 238000000746 purification Methods 0.000 description 11
- 108091034117 Oligonucleotide Proteins 0.000 description 10
- 210000003763 chloroplast Anatomy 0.000 description 10
- 230000012010 growth Effects 0.000 description 10
- 238000013519 translation Methods 0.000 description 10
- 239000005561 Glufosinate Substances 0.000 description 9
- 238000012217 deletion Methods 0.000 description 9
- 230000037430 deletion Effects 0.000 description 9
- 108020001507 fusion proteins Proteins 0.000 description 9
- 102000037865 fusion proteins Human genes 0.000 description 9
- 238000003018 immunoassay Methods 0.000 description 9
- 238000001727 in vivo Methods 0.000 description 9
- 230000004048 modification Effects 0.000 description 9
- 238000012986 modification Methods 0.000 description 9
- 239000013612 plasmid Substances 0.000 description 9
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 8
- 102000053602 DNA Human genes 0.000 description 8
- 238000007792 addition Methods 0.000 description 8
- 238000004113 cell culture Methods 0.000 description 8
- 238000001514 detection method Methods 0.000 description 8
- 230000002068 genetic effect Effects 0.000 description 8
- 230000001976 improved effect Effects 0.000 description 8
- 238000003780 insertion Methods 0.000 description 8
- 230000037431 insertion Effects 0.000 description 8
- 239000000047 product Substances 0.000 description 8
- 230000001105 regulatory effect Effects 0.000 description 8
- FSYKKLYZXJSNPZ-UHFFFAOYSA-N sarcosine Chemical compound C[NH2+]CC([O-])=O FSYKKLYZXJSNPZ-UHFFFAOYSA-N 0.000 description 8
- 239000000758 substrate Substances 0.000 description 8
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 7
- GINJFDRNADDBIN-FXQIFTODSA-N bilanafos Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCP(C)(O)=O GINJFDRNADDBIN-FXQIFTODSA-N 0.000 description 7
- 230000009260 cross reactivity Effects 0.000 description 7
- 229920000642 polymer Polymers 0.000 description 7
- 230000008569 process Effects 0.000 description 7
- 241000894007 species Species 0.000 description 7
- 239000000126 substance Substances 0.000 description 7
- 230000003612 virological effect Effects 0.000 description 7
- MGRVRXRGTBOSHW-UHFFFAOYSA-N (aminomethyl)phosphonic acid Chemical compound NCP(O)(O)=O MGRVRXRGTBOSHW-UHFFFAOYSA-N 0.000 description 6
- 108010076504 Protein Sorting Signals Proteins 0.000 description 6
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 6
- YXFVVABEGXRONW-UHFFFAOYSA-N Toluene Chemical compound CC1=CC=CC=C1 YXFVVABEGXRONW-UHFFFAOYSA-N 0.000 description 6
- 241000700605 Viruses Species 0.000 description 6
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 6
- 230000003321 amplification Effects 0.000 description 6
- 108010031100 chloroplast transit peptides Proteins 0.000 description 6
- 210000004962 mammalian cell Anatomy 0.000 description 6
- 238000003199 nucleic acid amplification method Methods 0.000 description 6
- 230000037361 pathway Effects 0.000 description 6
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 6
- BFECXRMSKVFCNB-UHFFFAOYSA-N 2-[acetyl(phosphonomethyl)amino]acetic acid Chemical compound OC(=O)CN(C(=O)C)CP(O)(O)=O BFECXRMSKVFCNB-UHFFFAOYSA-N 0.000 description 5
- 244000105975 Antidesma platyphyllum Species 0.000 description 5
- 241000701489 Cauliflower mosaic virus Species 0.000 description 5
- 241000219146 Gossypium Species 0.000 description 5
- 108700026244 Open Reading Frames Proteins 0.000 description 5
- 108091005804 Peptidases Proteins 0.000 description 5
- 108091081024 Start codon Proteins 0.000 description 5
- 108700019146 Transgenes Proteins 0.000 description 5
- 239000000427 antigen Substances 0.000 description 5
- 108091007433 antigens Proteins 0.000 description 5
- 102000036639 antigens Human genes 0.000 description 5
- 230000015572 biosynthetic process Effects 0.000 description 5
- 230000015556 catabolic process Effects 0.000 description 5
- 238000003776 cleavage reaction Methods 0.000 description 5
- 238000010367 cloning Methods 0.000 description 5
- 150000001875 compounds Chemical class 0.000 description 5
- 230000029087 digestion Effects 0.000 description 5
- 239000003623 enhancer Substances 0.000 description 5
- 230000007613 environmental effect Effects 0.000 description 5
- 230000004927 fusion Effects 0.000 description 5
- IAJOBQBIJHVGMQ-BYPYZUCNSA-N glufosinate-P Chemical compound CP(O)(=O)CC[C@H](N)C(O)=O IAJOBQBIJHVGMQ-BYPYZUCNSA-N 0.000 description 5
- 235000009424 haa Nutrition 0.000 description 5
- 230000001939 inductive effect Effects 0.000 description 5
- 230000017730 intein-mediated protein splicing Effects 0.000 description 5
- 239000000463 material Substances 0.000 description 5
- 230000035772 mutation Effects 0.000 description 5
- 238000012545 processing Methods 0.000 description 5
- 239000011541 reaction mixture Substances 0.000 description 5
- 150000003839 salts Chemical class 0.000 description 5
- 230000007017 scission Effects 0.000 description 5
- 230000003248 secreting effect Effects 0.000 description 5
- 238000003786 synthesis reaction Methods 0.000 description 5
- 241000193830 Bacillus <bacterium> Species 0.000 description 4
- 241000894006 Bacteria Species 0.000 description 4
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 4
- ZHNUHDYFZUAESO-UHFFFAOYSA-N Formamide Chemical compound NC=O ZHNUHDYFZUAESO-UHFFFAOYSA-N 0.000 description 4
- 235000010469 Glycine max Nutrition 0.000 description 4
- 244000068988 Glycine max Species 0.000 description 4
- NYHBQMYGNKIUIF-UUOKFMHZSA-N Guanosine Chemical compound C1=NC=2C(=O)NC(N)=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O NYHBQMYGNKIUIF-UUOKFMHZSA-N 0.000 description 4
- 108091092195 Intron Proteins 0.000 description 4
- 239000004365 Protease Substances 0.000 description 4
- 108010077895 Sarcosine Proteins 0.000 description 4
- XAKBSHICSHRJCL-UHFFFAOYSA-N [CH2]C(=O)C1=CC=CC=C1 Chemical group [CH2]C(=O)C1=CC=CC=C1 XAKBSHICSHRJCL-UHFFFAOYSA-N 0.000 description 4
- 239000002253 acid Substances 0.000 description 4
- 210000004102 animal cell Anatomy 0.000 description 4
- 230000008859 change Effects 0.000 description 4
- 238000006243 chemical reaction Methods 0.000 description 4
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 4
- 238000006731 degradation reaction Methods 0.000 description 4
- 238000000126 in silico method Methods 0.000 description 4
- 230000006698 induction Effects 0.000 description 4
- 238000007834 ligase chain reaction Methods 0.000 description 4
- 230000001404 mediated effect Effects 0.000 description 4
- 239000012528 membrane Substances 0.000 description 4
- 238000007899 nucleic acid hybridization Methods 0.000 description 4
- 210000000056 organ Anatomy 0.000 description 4
- 238000001742 protein purification Methods 0.000 description 4
- 230000001172 regenerating effect Effects 0.000 description 4
- 238000011160 research Methods 0.000 description 4
- 229940043230 sarcosine Drugs 0.000 description 4
- 150000003335 secondary amines Chemical class 0.000 description 4
- 230000028327 secretion Effects 0.000 description 4
- 239000002689 soil Substances 0.000 description 4
- 230000005945 translocation Effects 0.000 description 4
- 241000701161 unidentified adenovirus Species 0.000 description 4
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 3
- 108010070675 Glutathione transferase Proteins 0.000 description 3
- 102000005720 Glutathione transferase Human genes 0.000 description 3
- 235000009438 Gossypium Nutrition 0.000 description 3
- 108060003951 Immunoglobulin Proteins 0.000 description 3
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 3
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 3
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 3
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 3
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 3
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 3
- 108020004511 Recombinant DNA Proteins 0.000 description 3
- 102100037486 Reverse transcriptase/ribonuclease H Human genes 0.000 description 3
- 108091036066 Three prime untranslated region Proteins 0.000 description 3
- 230000002378 acidificating effect Effects 0.000 description 3
- 239000002671 adjuvant Substances 0.000 description 3
- 239000000872 buffer Substances 0.000 description 3
- 239000003054 catalyst Substances 0.000 description 3
- 230000003197 catalytic effect Effects 0.000 description 3
- 150000001768 cations Chemical class 0.000 description 3
- 238000005119 centrifugation Methods 0.000 description 3
- 239000003795 chemical substances by application Substances 0.000 description 3
- 238000004587 chromatography analysis Methods 0.000 description 3
- 230000002759 chromosomal effect Effects 0.000 description 3
- 230000000052 comparative effect Effects 0.000 description 3
- 244000038559 crop plants Species 0.000 description 3
- 230000003247 decreasing effect Effects 0.000 description 3
- 238000011161 development Methods 0.000 description 3
- 230000018109 developmental process Effects 0.000 description 3
- 238000005516 engineering process Methods 0.000 description 3
- 230000002255 enzymatic effect Effects 0.000 description 3
- 210000003527 eukaryotic cell Anatomy 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- RUCAXVJJQQJZGU-UHFFFAOYSA-M hydron;2-(phosphonatomethylamino)acetate;trimethylsulfanium Chemical compound C[S+](C)C.OP(O)(=O)CNCC([O-])=O RUCAXVJJQQJZGU-UHFFFAOYSA-M 0.000 description 3
- 102000018358 immunoglobulin Human genes 0.000 description 3
- 238000002955 isolation Methods 0.000 description 3
- 125000005647 linker group Chemical group 0.000 description 3
- 230000004807 localization Effects 0.000 description 3
- 239000000178 monomer Substances 0.000 description 3
- 238000010647 peptide synthesis reaction Methods 0.000 description 3
- 230000008488 polyadenylation Effects 0.000 description 3
- 235000019419 proteases Nutrition 0.000 description 3
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 3
- 238000012216 screening Methods 0.000 description 3
- 239000007790 solid phase Substances 0.000 description 3
- 238000010561 standard procedure Methods 0.000 description 3
- 230000001131 transforming effect Effects 0.000 description 3
- 238000011144 upstream manufacturing Methods 0.000 description 3
- LWTDZKXXJRRKDG-KXBFYZLASA-N (-)-phaseollin Chemical compound C1OC2=CC(O)=CC=C2[C@H]2[C@@H]1C1=CC=C3OC(C)(C)C=CC3=C1O2 LWTDZKXXJRRKDG-KXBFYZLASA-N 0.000 description 2
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 2
- 229920000936 Agarose Polymers 0.000 description 2
- QGZKDVFQNNGYKY-UHFFFAOYSA-N Ammonia Chemical compound N QGZKDVFQNNGYKY-UHFFFAOYSA-N 0.000 description 2
- 108091093088 Amplicon Proteins 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- 235000014469 Bacillus subtilis Nutrition 0.000 description 2
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 2
- 108700010070 Codon Usage Proteins 0.000 description 2
- 229920000742 Cotton Polymers 0.000 description 2
- MIKUYHXYGGJMLM-GIMIYPNGSA-N Crotonoside Natural products C1=NC2=C(N)NC(=O)N=C2N1[C@H]1O[C@@H](CO)[C@H](O)[C@@H]1O MIKUYHXYGGJMLM-GIMIYPNGSA-N 0.000 description 2
- 241000195493 Cryptophyta Species 0.000 description 2
- NYHBQMYGNKIUIF-UHFFFAOYSA-N D-guanosine Natural products C1=2NC(N)=NC(=O)C=2N=CN1C1OC(CO)C(O)C1O NYHBQMYGNKIUIF-UHFFFAOYSA-N 0.000 description 2
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 2
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 2
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 2
- 102100031780 Endonuclease Human genes 0.000 description 2
- 102100029727 Enteropeptidase Human genes 0.000 description 2
- 108010013369 Enteropeptidase Proteins 0.000 description 2
- 241000701484 Figwort mosaic virus Species 0.000 description 2
- 241000233866 Fungi Species 0.000 description 2
- 108010058683 Immobilized Proteins Proteins 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- 241001465754 Metazoa Species 0.000 description 2
- SEQKRHFRPICQDD-UHFFFAOYSA-N N-tris(hydroxymethyl)methylglycine Chemical compound OCC(CO)(CO)[NH2+]CC([O-])=O SEQKRHFRPICQDD-UHFFFAOYSA-N 0.000 description 2
- 108020004711 Nucleic Acid Probes Proteins 0.000 description 2
- 102000035195 Peptidases Human genes 0.000 description 2
- XYFCBTPGUUZFHI-UHFFFAOYSA-N Phosphine Chemical compound P XYFCBTPGUUZFHI-UHFFFAOYSA-N 0.000 description 2
- 108700001094 Plant Genes Proteins 0.000 description 2
- 108010066717 Q beta Replicase Proteins 0.000 description 2
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 description 2
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 2
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 2
- 241000714474 Rous sarcoma virus Species 0.000 description 2
- 238000002105 Southern blotting Methods 0.000 description 2
- 241000218632 Strawberry vein banding virus Species 0.000 description 2
- 108700026226 TATA Box Proteins 0.000 description 2
- 241000607479 Yersinia pestis Species 0.000 description 2
- 240000008042 Zea mays Species 0.000 description 2
- 235000005824 Zea mays ssp. parviglumis Nutrition 0.000 description 2
- 235000002017 Zea mays subsp mays Nutrition 0.000 description 2
- 230000000397 acetylating effect Effects 0.000 description 2
- 108020002494 acetyltransferase Proteins 0.000 description 2
- 102000005421 acetyltransferase Human genes 0.000 description 2
- 150000007513 acids Chemical class 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 238000001042 affinity chromatography Methods 0.000 description 2
- 238000004458 analytical method Methods 0.000 description 2
- 150000001450 anions Chemical class 0.000 description 2
- 230000000692 anti-sense effect Effects 0.000 description 2
- 230000000890 antigenic effect Effects 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- -1 aromatic amino acids Chemical class 0.000 description 2
- 210000004436 artificial bacterial chromosome Anatomy 0.000 description 2
- 210000001106 artificial yeast chromosome Anatomy 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 230000008238 biochemical pathway Effects 0.000 description 2
- 230000003115 biocidal effect Effects 0.000 description 2
- 239000011575 calcium Substances 0.000 description 2
- 238000004422 calculation algorithm Methods 0.000 description 2
- 238000004364 calculation method Methods 0.000 description 2
- 239000006143 cell culture medium Substances 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 230000009137 competitive binding Effects 0.000 description 2
- 239000002299 complementary DNA Substances 0.000 description 2
- 235000005822 corn Nutrition 0.000 description 2
- 229940104302 cytosine Drugs 0.000 description 2
- 230000001627 detrimental effect Effects 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- 239000000539 dimer Substances 0.000 description 2
- 210000002257 embryonic structure Anatomy 0.000 description 2
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 2
- 230000001747 exhibiting effect Effects 0.000 description 2
- 238000001400 expression cloning Methods 0.000 description 2
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 2
- 230000013595 glycosylation Effects 0.000 description 2
- 238000006206 glycosylation reaction Methods 0.000 description 2
- 229940029575 guanosine Drugs 0.000 description 2
- 230000005764 inhibitory process Effects 0.000 description 2
- 231100000518 lethal Toxicity 0.000 description 2
- 230000001665 lethal effect Effects 0.000 description 2
- 230000000670 limiting effect Effects 0.000 description 2
- 239000007788 liquid Substances 0.000 description 2
- 230000014759 maintenance of location Effects 0.000 description 2
- 238000004949 mass spectrometry Methods 0.000 description 2
- 239000002609 medium Substances 0.000 description 2
- 238000002844 melting Methods 0.000 description 2
- 230000008018 melting Effects 0.000 description 2
- 108020004999 messenger RNA Proteins 0.000 description 2
- 230000037353 metabolic pathway Effects 0.000 description 2
- 230000004060 metabolic process Effects 0.000 description 2
- 229910052751 metal Inorganic materials 0.000 description 2
- 239000002184 metal Substances 0.000 description 2
- 230000000813 microbial effect Effects 0.000 description 2
- 230000002906 microbiologic effect Effects 0.000 description 2
- 238000001823 molecular biology technique Methods 0.000 description 2
- 238000010369 molecular cloning Methods 0.000 description 2
- 239000002853 nucleic acid probe Substances 0.000 description 2
- 210000003463 organelle Anatomy 0.000 description 2
- 230000036961 partial effect Effects 0.000 description 2
- 150000008300 phosphoramidites Chemical class 0.000 description 2
- 229920002704 polyhistidine Polymers 0.000 description 2
- 230000001323 posttranslational effect Effects 0.000 description 2
- 150000003141 primary amines Chemical class 0.000 description 2
- 239000013615 primer Substances 0.000 description 2
- 230000000644 propagated effect Effects 0.000 description 2
- 230000009257 reactivity Effects 0.000 description 2
- 238000010188 recombinant method Methods 0.000 description 2
- 238000011084 recovery Methods 0.000 description 2
- 230000010076 replication Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 238000012552 review Methods 0.000 description 2
- 230000035945 sensitivity Effects 0.000 description 2
- 238000012163 sequencing technique Methods 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- 230000010473 stable expression Effects 0.000 description 2
- 230000008685 targeting Effects 0.000 description 2
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 2
- 230000036962 time dependent Effects 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 230000026683 transduction Effects 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 230000014621 translational initiation Effects 0.000 description 2
- 210000005253 yeast cell Anatomy 0.000 description 2
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 1
- KKADPXVIOXHVKN-UHFFFAOYSA-M 3-(4-hydroxyphenyl)pyruvate Chemical group OC1=CC=C(CC(=O)C([O-])=O)C=C1 KKADPXVIOXHVKN-UHFFFAOYSA-M 0.000 description 1
- QUTYKIXIUDQOLK-PRJMDXOYSA-N 5-O-(1-carboxyvinyl)-3-phosphoshikimic acid Chemical compound O[C@H]1[C@H](OC(=C)C(O)=O)CC(C(O)=O)=C[C@H]1OP(O)(O)=O QUTYKIXIUDQOLK-PRJMDXOYSA-N 0.000 description 1
- 241000186361 Actinobacteria <class> Species 0.000 description 1
- 229930024421 Adenine Natural products 0.000 description 1
- 241000589158 Agrobacterium Species 0.000 description 1
- 241000589155 Agrobacterium tumefaciens Species 0.000 description 1
- 102100027211 Albumin Human genes 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 108010025188 Alcohol oxidase Proteins 0.000 description 1
- 108700028369 Alleles Proteins 0.000 description 1
- 241000194110 Bacillus sp. (in: Bacteria) Species 0.000 description 1
- 244000063299 Bacillus subtilis Species 0.000 description 1
- 101100376452 Bacillus subtilis (strain 168) yitI gene Proteins 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010078791 Carrier Proteins Proteins 0.000 description 1
- 108020004638 Circular DNA Proteins 0.000 description 1
- 108091033380 Coding strand Proteins 0.000 description 1
- 241000218631 Coniferophyta Species 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- 241000557626 Corvus corax Species 0.000 description 1
- 102000018832 Cytochromes Human genes 0.000 description 1
- 108010052832 Cytochromes Proteins 0.000 description 1
- 108010066133 D-octopine dehydrogenase Proteins 0.000 description 1
- 101710177611 DNA polymerase II large subunit Proteins 0.000 description 1
- 101710184669 DNA polymerase II small subunit Proteins 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 101150070004 E8 gene Proteins 0.000 description 1
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 1
- 101000962225 Escherichia coli (strain K12) Dye-decolorizing peroxidase YfeX Proteins 0.000 description 1
- 208000000666 Fowlpox Diseases 0.000 description 1
- 101150062467 GAT gene Proteins 0.000 description 1
- 101150026259 GRP gene Proteins 0.000 description 1
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 1
- 108020000311 Glutamate Synthase Proteins 0.000 description 1
- 108010024636 Glutathione Proteins 0.000 description 1
- 108010063907 Glutathione Reductase Proteins 0.000 description 1
- 102100036442 Glutathione reductase, mitochondrial Human genes 0.000 description 1
- HVLSXIKZNLPZJJ-TXZCQADKSA-N HA peptide Chemical compound C([C@@H](C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@H]1N(CCC1)C(=O)[C@@H](N)CC=1C=CC(O)=CC=1)C1=CC=C(O)C=C1 HVLSXIKZNLPZJJ-TXZCQADKSA-N 0.000 description 1
- 101710154606 Hemagglutinin Proteins 0.000 description 1
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 229940127553 Hydroxyphenylpyruvate Dioxygenase Inhibitors Drugs 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 108020005350 Initiator Codon Proteins 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 1
- 125000000510 L-tryptophano group Chemical group [H]C1=C([H])C([H])=C2N([H])C([H])=C(C([H])([H])[C@@]([H])(C(O[H])=O)N([H])[*])C2=C1[H] 0.000 description 1
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 1
- 108090001090 Lectins Proteins 0.000 description 1
- 102000004856 Lectins Human genes 0.000 description 1
- 241000209510 Liliopsida Species 0.000 description 1
- 241000218922 Magnoliophyta Species 0.000 description 1
- 101710175625 Maltose/maltodextrin-binding periplasmic protein Proteins 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 241000736262 Microbiota Species 0.000 description 1
- 101710202061 N-acetyltransferase Proteins 0.000 description 1
- 230000004988 N-glycosylation Effects 0.000 description 1
- 108010045510 NADPH-Ferrihemoprotein Reductase Proteins 0.000 description 1
- 229930193140 Neomycin Natural products 0.000 description 1
- 101000598243 Nicotiana tabacum Probable aquaporin TIP-type RB7-18C Proteins 0.000 description 1
- 101000655028 Nicotiana tabacum Probable aquaporin TIP-type RB7-5A Proteins 0.000 description 1
- 238000000636 Northern blotting Methods 0.000 description 1
- 108010077850 Nuclear Localization Signals Proteins 0.000 description 1
- 102000015636 Oligopeptides Human genes 0.000 description 1
- 108010038807 Oligopeptides Proteins 0.000 description 1
- 101710093908 Outer capsid protein VP4 Proteins 0.000 description 1
- 101710135467 Outer capsid protein sigma-1 Proteins 0.000 description 1
- 101710091688 Patatin Proteins 0.000 description 1
- 102000057297 Pepsin A Human genes 0.000 description 1
- 108090000284 Pepsin A Proteins 0.000 description 1
- 101710163504 Phaseolin Proteins 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- 108010039918 Polylysine Proteins 0.000 description 1
- 208000020584 Polyploidy Diseases 0.000 description 1
- 241000985694 Polypodiopsida Species 0.000 description 1
- 101710176177 Protein A56 Proteins 0.000 description 1
- 101710149951 Protein Tat Proteins 0.000 description 1
- 241000589516 Pseudomonas Species 0.000 description 1
- 241000293869 Salmonella enterica subsp. enterica serovar Typhimurium Species 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 241000187391 Streptomyces hygroscopicus Species 0.000 description 1
- 108010012715 Superoxide dismutase Proteins 0.000 description 1
- UZMAPBJVXOGOFT-UHFFFAOYSA-N Syringetin Natural products COC1=C(O)C(OC)=CC(C2=C(C(=O)C3=C(O)C=C(O)C=C3O2)O)=C1 UZMAPBJVXOGOFT-UHFFFAOYSA-N 0.000 description 1
- 101150084279 TM gene Proteins 0.000 description 1
- 239000004098 Tetracycline Substances 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 108010022394 Threonine synthase Proteins 0.000 description 1
- 108700009124 Transcription Initiation Site Proteins 0.000 description 1
- 239000007997 Tricine buffer Substances 0.000 description 1
- 206010046865 Vaccinia virus infection Diseases 0.000 description 1
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 1
- 108020005202 Viral DNA Proteins 0.000 description 1
- IXKSXJFAGXLQOQ-XISFHERQSA-N WHWLQLKPGQPMY Chemical compound C([C@@H](C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)NC(=O)[C@@H](N)CC=1C2=CC=CC=C2NC=1)C1=CNC=N1 IXKSXJFAGXLQOQ-XISFHERQSA-N 0.000 description 1
- AQIBTEMVIOJQFD-NQXXGFSBSA-N [(2R,3R)-1,2-dihydroxy-4-oxo-5-phosphonooxypentan-3-yl] dihydrogen phosphate Chemical compound OC[C@@H](O)[C@@H](OP(O)(O)=O)C(=O)COP(O)(O)=O AQIBTEMVIOJQFD-NQXXGFSBSA-N 0.000 description 1
- 125000002777 acetyl group Chemical group [H]C([H])([H])C(*)=O 0.000 description 1
- JUGOREOARAHOCO-UHFFFAOYSA-M acetylcholine chloride Chemical compound [Cl-].CC(=O)OCC[N+](C)(C)C JUGOREOARAHOCO-UHFFFAOYSA-M 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 230000010933 acylation Effects 0.000 description 1
- 238000005917 acylation reaction Methods 0.000 description 1
- 229960000643 adenine Drugs 0.000 description 1
- 238000001261 affinity purification Methods 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 230000004075 alteration Effects 0.000 description 1
- 150000001408 amides Chemical class 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- IAANMKMHMYZVOC-UHFFFAOYSA-N aminomethyl dihydrogen phosphate Chemical compound NCOP(O)(O)=O IAANMKMHMYZVOC-UHFFFAOYSA-N 0.000 description 1
- 229910021529 ammonia Inorganic materials 0.000 description 1
- 238000012870 ammonium sulfate precipitation Methods 0.000 description 1
- 229960000723 ampicillin Drugs 0.000 description 1
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 1
- 230000003322 aneuploid effect Effects 0.000 description 1
- 208000036878 aneuploidy Diseases 0.000 description 1
- 238000000137 annealing Methods 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 230000003416 augmentation Effects 0.000 description 1
- XOEMATDHVZOBSG-UHFFFAOYSA-N azafenidin Chemical compound C1=C(OCC#C)C(Cl)=CC(Cl)=C1N1C(=O)N2CCCCC2=N1 XOEMATDHVZOBSG-UHFFFAOYSA-N 0.000 description 1
- 101150103518 bar gene Proteins 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 102000005936 beta-Galactosidase Human genes 0.000 description 1
- 108010005774 beta-Galactosidase Proteins 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 239000003139 biocide Substances 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 108010083912 bleomycin N-acetyltransferase Proteins 0.000 description 1
- JEDYYFXHPAIBGR-UHFFFAOYSA-N butafenacil Chemical compound O=C1N(C)C(C(F)(F)F)=CC(=O)N1C1=CC=C(Cl)C(C(=O)OC(C)(C)C(=O)OCC=C)=C1 JEDYYFXHPAIBGR-UHFFFAOYSA-N 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 239000001569 carbon dioxide Substances 0.000 description 1
- 229910002092 carbon dioxide Inorganic materials 0.000 description 1
- 230000021523 carboxylation Effects 0.000 description 1
- 238000006473 carboxylation reaction Methods 0.000 description 1
- 239000000969 carrier Substances 0.000 description 1
- 238000006555 catalytic reaction Methods 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 230000010307 cell transformation Effects 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 239000000287 crude extract Substances 0.000 description 1
- 230000001461 cytolytic effect Effects 0.000 description 1
- 210000000805 cytoplasm Anatomy 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 238000001784 detoxification Methods 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- KCFYHBSOLOXZIF-UHFFFAOYSA-N dihydrochrysin Natural products COC1=C(O)C(OC)=CC(C2OC3=CC(O)=CC(O)=C3C(=O)C2)=C1 KCFYHBSOLOXZIF-UHFFFAOYSA-N 0.000 description 1
- 102000004419 dihydrofolate reductase Human genes 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 235000013399 edible fruits Nutrition 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 238000005538 encapsulation Methods 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- 238000012869 ethanol precipitation Methods 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 238000000605 extraction Methods 0.000 description 1
- 235000013861 fat-free Nutrition 0.000 description 1
- 238000000855 fermentation Methods 0.000 description 1
- 230000004151 fermentation Effects 0.000 description 1
- 239000007850 fluorescent dye Substances 0.000 description 1
- 125000004005 formimidoyl group Chemical group [H]\N=C(/[H])* 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 125000000524 functional group Chemical group 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 229960003180 glutathione Drugs 0.000 description 1
- 239000011121 hardwood Substances 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 125000000487 histidyl group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C([H])=N1 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 239000001257 hydrogen Substances 0.000 description 1
- 238000004191 hydrophobic interaction chromatography Methods 0.000 description 1
- 238000012872 hydroxylapatite chromatography Methods 0.000 description 1
- 230000001900 immune effect Effects 0.000 description 1
- 230000003053 immunization Effects 0.000 description 1
- 238000002649 immunization Methods 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 229940072221 immunoglobulins Drugs 0.000 description 1
- 238000010324 immunological assay Methods 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 206010022000 influenza Diseases 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 229910052816 inorganic phosphate Inorganic materials 0.000 description 1
- 238000007689 inspection Methods 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 150000002500 ions Chemical class 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- 108010045069 keyhole-limpet hemocyanin Proteins 0.000 description 1
- 238000011005 laboratory method Methods 0.000 description 1
- 239000002523 lectin Substances 0.000 description 1
- 239000003446 ligand Substances 0.000 description 1
- 230000029226 lipidation Effects 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 201000001441 melanoma Diseases 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 239000002207 metabolite Substances 0.000 description 1
- 229910021645 metal ion Inorganic materials 0.000 description 1
- 150000002739 metals Chemical class 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 235000013336 milk Nutrition 0.000 description 1
- 239000008267 milk Substances 0.000 description 1
- 210000004080 milk Anatomy 0.000 description 1
- ZAHQPTJLOCWVPG-UHFFFAOYSA-N mitoxantrone dihydrochloride Chemical compound Cl.Cl.O=C1C2=C(O)C=CC(O)=C2C(=O)C2=C1C(NCCNCCO)=CC=C2NCCNCCO ZAHQPTJLOCWVPG-UHFFFAOYSA-N 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 108091027963 non-coding RNA Proteins 0.000 description 1
- 102000042567 non-coding RNA Human genes 0.000 description 1
- 108010058731 nopaline synthase Proteins 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 230000030648 nucleus localization Effects 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 238000005457 optimization Methods 0.000 description 1
- 239000003960 organic solvent Substances 0.000 description 1
- 210000001672 ovary Anatomy 0.000 description 1
- 230000002018 overexpression Effects 0.000 description 1
- KQAOIKIZSJJTII-UHFFFAOYSA-N p-mercuribenzenesulfonic acid Chemical compound OS(=O)(=O)C1=CC=C([Hg])C=C1 KQAOIKIZSJJTII-UHFFFAOYSA-N 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 244000052769 pathogen Species 0.000 description 1
- 230000001717 pathogenic effect Effects 0.000 description 1
- 229940111202 pepsin Drugs 0.000 description 1
- 210000001322 periplasm Anatomy 0.000 description 1
- 230000008823 permeabilization Effects 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- LWTDZKXXJRRKDG-UHFFFAOYSA-N phaseollin Natural products C1OC2=CC(O)=CC=C2C2C1C1=CC=C3OC(C)(C)C=CC3=C1O2 LWTDZKXXJRRKDG-UHFFFAOYSA-N 0.000 description 1
- FVZVCSNXTFCBQU-UHFFFAOYSA-N phosphanyl Chemical group [PH2] FVZVCSNXTFCBQU-UHFFFAOYSA-N 0.000 description 1
- 125000002467 phosphate group Chemical group [H]OP(=O)(O[H])O[*] 0.000 description 1
- 229940080469 phosphocellulose Drugs 0.000 description 1
- DTBNBXWJWCWCIK-UHFFFAOYSA-K phosphonatoenolpyruvate Chemical compound [O-]C(=O)C(=C)OP([O-])([O-])=O DTBNBXWJWCWCIK-UHFFFAOYSA-K 0.000 description 1
- 229910000073 phosphorus hydride Inorganic materials 0.000 description 1
- 230000026731 phosphorylation Effects 0.000 description 1
- 238000006366 phosphorylation reaction Methods 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- 238000000596 photon cross correlation spectroscopy Methods 0.000 description 1
- 210000000745 plant chromosome Anatomy 0.000 description 1
- 230000027086 plasmid maintenance Effects 0.000 description 1
- 210000002706 plastid Anatomy 0.000 description 1
- 229920000656 polylysine Polymers 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 230000001124 posttranscriptional effect Effects 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 239000002987 primer (paints) Substances 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 235000019833 protease Nutrition 0.000 description 1
- 230000016314 protein import into mitochondrial matrix Effects 0.000 description 1
- 238000001814 protein method Methods 0.000 description 1
- 230000030788 protein refolding Effects 0.000 description 1
- 208000009305 pseudorabies Diseases 0.000 description 1
- 230000002285 radioactive effect Effects 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 239000004180 red 2G Substances 0.000 description 1
- 230000009467 reduction Effects 0.000 description 1
- 230000008929 regeneration Effects 0.000 description 1
- 238000011069 regeneration method Methods 0.000 description 1
- 238000009877 rendering Methods 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 239000006152 selective media Substances 0.000 description 1
- 238000002864 sequence alignment Methods 0.000 description 1
- 210000002966 serum Anatomy 0.000 description 1
- 238000010008 shearing Methods 0.000 description 1
- 101150091813 shfl gene Proteins 0.000 description 1
- QYOJSKGCWNAKGW-HCWXCVPCSA-N shikimate-3-phosphate Chemical compound O[C@H]1CC(C(O)=O)=C[C@H](OP(O)(O)=O)[C@@H]1O QYOJSKGCWNAKGW-HCWXCVPCSA-N 0.000 description 1
- 238000004904 shortening Methods 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 244000000000 soil microbiome Species 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 239000000243 solution Substances 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 238000000527 sonication Methods 0.000 description 1
- 238000001179 sorption measurement Methods 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- BDHFUVZGWQCTTF-UHFFFAOYSA-M sulfonate Chemical compound [O-]S(=O)=O BDHFUVZGWQCTTF-UHFFFAOYSA-M 0.000 description 1
- 150000003457 sulfones Chemical class 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 239000004094 surface-active agent Substances 0.000 description 1
- 238000001308 synthesis method Methods 0.000 description 1
- 230000002194 synthesizing effect Effects 0.000 description 1
- 238000010189 synthetic method Methods 0.000 description 1
- 229960002180 tetracycline Drugs 0.000 description 1
- 229930101283 tetracycline Natural products 0.000 description 1
- 235000019364 tetracycline Nutrition 0.000 description 1
- 150000003522 tetracyclines Chemical class 0.000 description 1
- 229940113082 thymine Drugs 0.000 description 1
- 238000012090 tissue culture technique Methods 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 230000014616 translation Effects 0.000 description 1
- 230000032258 transport Effects 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 239000001226 triphosphate Substances 0.000 description 1
- 235000011178 triphosphate Nutrition 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 235000002374 tyrosine Nutrition 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 208000007089 vaccinia Diseases 0.000 description 1
- 210000003934 vacuole Anatomy 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 125000002987 valine group Chemical group [H]N([H])C([H])(C(*)=O)C([H])(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- 230000002792 vascular Effects 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 239000011782 vitamin Substances 0.000 description 1
- 235000013343 vitamin Nutrition 0.000 description 1
- 229940088594 vitamin Drugs 0.000 description 1
- 229930003231 vitamin Natural products 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
Images







Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1025—Acyltransferases (2.3)
- C12N9/1029—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8209—Selection, visualisation of transformants, reporter constructs, e.g. antibiotic resistance markers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
- C12N15/8274—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for herbicide resistance
- C12N15/8275—Glyphosate
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Organic Chemistry (AREA)
- Biotechnology (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Plant Pathology (AREA)
- Cell Biology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Medicinal Chemistry (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Enzymes And Modification Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
- Agricultural Chemicals And Associated Chemicals (AREA)
Abstract
【課題】生物体(例えば、植物)をグリホセートに対し抵抗性にするための方法および試薬を提供する。
【解決手段】単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下:(a)BLOSUM62マトリックス、11のギャップイグジステンスペナルティー、および1のギャップエクステンションペナルティーを使用して、少なくとも430の類似性スコアを生み出すために、複数の特定のアミノ酸配列からなる群から選択される配列と、最適に並べられ得るアミノ酸配列をコードするヌクレオチド配列;または(b)これらの相補鎖ヌクレオチド配列、を含む、ポリヌクレオチドからなる。
【選択図】なし
【解決手段】単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下:(a)BLOSUM62マトリックス、11のギャップイグジステンスペナルティー、および1のギャップエクステンションペナルティーを使用して、少なくとも430の類似性スコアを生み出すために、複数の特定のアミノ酸配列からなる群から選択される配列と、最適に並べられ得るアミノ酸配列をコードするヌクレオチド配列;または(b)これらの相補鎖ヌクレオチド配列、を含む、ポリヌクレオチドからなる。
【選択図】なし
Description
(関連出願の引用)
本出願は、2000年10月30日に出願された米国仮特許出願番号第60/244,385に対する優先権およびその利益を主張し、その開示は、すべての目的のために、その全体が本明細書に参考として援用される。
本出願は、2000年10月30日に出願された米国仮特許出願番号第60/244,385に対する優先権およびその利益を主張し、その開示は、すべての目的のために、その全体が本明細書に参考として援用される。
(37 C.F.R.§1.71(E)に準拠する著作権通知)
本特許文献の開示の一部は、著作権保護を受ける材料を含む。この著作権の所有者は、米国特許商標庁の特許ファイルまたは特許記録に掲載される、本特許文献および特許公開情報の何人による複製に対しても拒絶をしないが、その他の場合は、いかなるすべての著作権も保護される。
本特許文献の開示の一部は、著作権保護を受ける材料を含む。この著作権の所有者は、米国特許商標庁の特許ファイルまたは特許記録に掲載される、本特許文献および特許公開情報の何人による複製に対しても拒絶をしないが、その他の場合は、いかなるすべての著作権も保護される。
(発明の背景)
特異的な除草剤に対する農作物の選択性は、適当な除草剤代謝酵素をコードする遺伝子を農作物中の移入することによって賦与され得る。ある場合において、これらの酵素、およびそれをコードする核酸は、植物に源を発する。他の場合において、これらの酵素、およびそれをコードする核酸は、他の生物体(例えば、微生物)に由来する。例えば、Padgetteら、(1996)「New weed control oppotunities:Development of soybeans with a Round UP ReadyTM gene」、Herbicide−Resistant Crops(Duke編)、54−84ページ、CRC Press,Boca Raton;およびVasil(1996)「Phosphinothricin−resistant crops」、Herbicide−Resistant Crops(Duke編)、85−91ページを参照のこと。実際、トランスジェニック植物は、種々の生物体に由来する、種々の除草剤耐性/除草剤代謝遺伝子を発現するように操作されてきた。例えば、アセトヒドロキシ酸(acetohydroxy acid)シンターゼ(この酵素は、この酵素を発現する植物を複数の型の除草剤耐性にすることが見出されてきた)は、種々の植物に導入されてきた(例えば、Hattoriら、(1995)Mol Gen Genet 246:419、を参照のこと)。除草剤耐性を賦与する他の遺伝子として:ラットシトクロムP4507A1および酵母NADPH−シトクロムP450オキシドレダクターゼのキメラタンパク質をコードする遺伝子(Shiotaら(1994)Plant PhysiolPlant Physiol 106:17)、グルタチオンレダクターゼおよびスーパーオキシドジスムターゼについての遺伝子(Aonoら(1995)Plant Cell Physiol 36:1687)、ならびに種々のホスホトランスフェラーゼについての遺伝子(Dattaら(1992)Plant Mol Biol 20:619)が挙げられる。
特異的な除草剤に対する農作物の選択性は、適当な除草剤代謝酵素をコードする遺伝子を農作物中の移入することによって賦与され得る。ある場合において、これらの酵素、およびそれをコードする核酸は、植物に源を発する。他の場合において、これらの酵素、およびそれをコードする核酸は、他の生物体(例えば、微生物)に由来する。例えば、Padgetteら、(1996)「New weed control oppotunities:Development of soybeans with a Round UP ReadyTM gene」、Herbicide−Resistant Crops(Duke編)、54−84ページ、CRC Press,Boca Raton;およびVasil(1996)「Phosphinothricin−resistant crops」、Herbicide−Resistant Crops(Duke編)、85−91ページを参照のこと。実際、トランスジェニック植物は、種々の生物体に由来する、種々の除草剤耐性/除草剤代謝遺伝子を発現するように操作されてきた。例えば、アセトヒドロキシ酸(acetohydroxy acid)シンターゼ(この酵素は、この酵素を発現する植物を複数の型の除草剤耐性にすることが見出されてきた)は、種々の植物に導入されてきた(例えば、Hattoriら、(1995)Mol Gen Genet 246:419、を参照のこと)。除草剤耐性を賦与する他の遺伝子として:ラットシトクロムP4507A1および酵母NADPH−シトクロムP450オキシドレダクターゼのキメラタンパク質をコードする遺伝子(Shiotaら(1994)Plant PhysiolPlant Physiol 106:17)、グルタチオンレダクターゼおよびスーパーオキシドジスムターゼについての遺伝子(Aonoら(1995)Plant Cell Physiol 36:1687)、ならびに種々のホスホトランスフェラーゼについての遺伝子(Dattaら(1992)Plant Mol Biol 20:619)が挙げられる。
この点に関して多くの研究の対象であるひとつの除草剤は、N−ホスホノメチルグリシン(N−phosphonomethylglycine)(一般にはグリホセートといわれる)である。グリホセートは、2003年までに売り上げが50億ドルに達すると見積もられる、世界で一番売れている除草剤である。グリホセートは、広葉樹および芝生型植物の両方を殺す、広範なスペクトルの除草剤である。トランスジェニック植物において商業レベルのグリホセート抵抗性の成功した様式は、改変されたアグロバクテリウムCP4 5−エノールピルボイルシキミ酸(enolpyruvylshikimate)−3−リン酸シンターゼ(以下EPSPシンターゼまたはEPSPSという)遺伝子の導入による。導入遺伝子は、グリホセートの存在下においてホスホエノールピルビン酸(PEP)およびシキミ酸−3−リン酸からEPSP合成を継続し得る、葉緑体に向けられる。対照的に、ネイティブEPSPシンターゼは、グリホセートによって阻害される。導入遺伝子なしに、グリホセートを噴霧された植物は、芳香族アミノ酸、ホルモン、およびビタミンの生合成に必要とされる下流の経路を止める、EPSPシンターゼの阻害のため速やかに死ぬ。CP4グリホセート抵抗性ダイズトランスジェニック植物は、例えばMonsantoによって「Round UP ReadyTM」という名で販売されている。
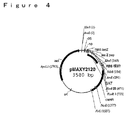
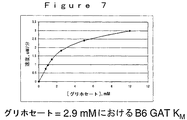
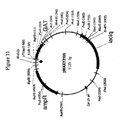
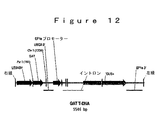
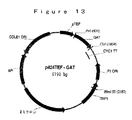
環境中において、グリホセートが分解される優勢な機構は、土壌微生物叢代謝を介する。土壌においてグリホセートの主要な代謝産物は、最終的にアンモニア、リン酸および二酸化炭素に変換される、アミノメチルリン酸(AMPA)と同定された。AMPA経路を介する土壌でのグリホセートの分解を記述する提案された代謝体系を、図8に示す。ある土壌細菌のグリホセートによる機能停止のための代替的代謝経路であるサルコシン経路は、図9に図示されるように、無機リン酸およびサルコシンを生じるC−P結合の初めの切断を経て起こる。
別の首尾よい除草剤/トランスジェニック作物パッケージは、グルホシネート(ホスフィノスリシン)およびLibertyLinkTM形質であり、これは、例えばAventisによって市販されている。グルホシネートもまた、広範なスペクトルの除草剤である。グルホシネートの標的は、葉緑体のグルタミン酸シンターゼ酵素である。抵抗性植物は、Streptomyces hygroscopicus由来のbar遺伝子を保持し、そしてグルホシネートを修正および解毒するbarのN−アセチル化活性によって抵抗性を獲得する。
AMPAの第1級アミンをアセチル化できる酵素が、PCT出願番号第WO00/29596に報告されている。この酵素が、第2級アミンを有する化合物(例えばグリホセート)をアセチル化し得るとは記載されなかった。
上記のような種々の除草剤耐性戦略が、利用され得るとはいえ、さらなる取り組みは、かなりの商業的価値を有する。本発明は、例えば除草剤耐性を賦与するための新規のポリヌクレオチドおよびポリペプチド、ならびに本開示の検討の間に明らかになるような他の多数の利点を提供する。
本願発明によって以下が提供される:
(1) 単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下:
(a)BLOSUM62マトリックス、11のギャップイグジステンスペナルティー、および1のギャップエクステンションペナルティーを使用して、少なくとも430の類似性スコアを生み出すために、配列番号300、配列番号445、および配列番号457からなる群から選択される配列と、最適に並べられ得るアミノ酸配列をコードするヌクレオチド配列;または
(b)これらの相補鎖ヌクレオチド配列、
を含む、ポリヌクレオチド。
(2) 前記ポリペプチドがグリホセートN−アセチルトランスフェラーゼ活性を有する、項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(3) 前記ポリペプチドが、グリホセートに対して、少なくとも10mM −1 min −1 のkcat/Kmでグリホセートのアセチル化を触媒する、項目2に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(4) 前記ポリペプチドがアミノメチルホスホン酸のアセチル化を触媒する、項目2に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(5) グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドをコードするヌクレオチド配列を含む、単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、該ポリペプチドが配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、少なくとも20個の連続するアミノ酸を含むアミノ酸配列を含む、ポリヌクレオチド。
(6) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、少なくとも50個の連続するアミノ酸を含むアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(7) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、少なくとも100個の連続するアミノ酸を含むアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(8) 前記ポリヌクレオチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、約140個の連続するアミノ酸を含むアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(9) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(10) 配列番号48、配列番号193および配列番号205からなる群から選択されるヌクレオチド配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(11) 親コドンが、該親コドンに対して植物中で優先的に使用される同族コドンによって置換された、項目1に記載のポリヌクレオチド。
(12) N末端葉緑体輸送ペプチドをコードするヌクレオチド配列さらにを含む、項目1に記載のポリヌクレオチド。
(13) 前記のコードされたポリペプチドの1つ以上のアミノ酸が変異された、項目1に記載のポリヌクレオチドの非ネイティブ改変体。
(14) 項目1に記載のポリヌクレオチドを含む核酸構築物。
(15) 項目1に記載のポリヌクレオチドに作動可能に連結されたプロモーターを含む、項目14に記載の核酸構築物であって、該プロモーターが、前記ポリヌクレオチドに対して異種であり、そして、前記核酸構築物を用いて形質転換された植物細胞のグリホセート耐性を強めるのに十分な前記のコードされたポリペプチドの発現を引き起こすために効果的である、核酸構築物。
(16) 項目1に記載のポリヌクレオチド配列が選択可能なマーカーとして機能する、項目14に記載の核酸構築物。
(17) 前記構築物がベクターである、項目14に記載の核酸構築物。
(18)第2のポリペプチドが効果的なレベルで発現する細胞または組織に検出可能な表現型特性を付与する、該第2ポリペプチドをコードする第2ポリヌクレオチド配列を含む、項目17に記載のベクター。
(19) 前記検出可能な表現型特性が、選択可能なマーカーとして機能する、項目18に記載のベクター。
(20) 前記検出可能な表現型特性が、除草剤耐性、害虫耐性、または可視マーカーからなる、項目19に記載のベクター。
(21) 前記ベクターが、T−DNA配列を含む、項目17に記載のベクター。
(22) 前記ポリヌクレオチドが調節配列に作動可能に連結される、項目17に記載のベクター。
(23) 前記ベクターが植物形質転換ベクターである、項目17に記載のベクター。
(24) 単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下:
(a)ストリンジェントな条件下で、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列をコードするヌクレオチド配列の実質的に全長にわたりハイブリダイズするヌクレオチド;
(b)これらの相補鎖ヌクレオチド配列;または
(c)グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドをコードする(a)または(b)のフラグメント、
を含む、ポリヌクレオチド。
(25) グリホセートN−アセチルトランスフェラーゼをコードするヌクレオチド配列を含む、項目24に記載のポリヌクレオチド。
(26) 項目1に記載の2つ以上のポリヌクレオチドを含む組成物。
(27) 項目1に記載のポリヌクレオチドを少なくとも10個含む、項目26に記載の組成物。
(28) 前記ポリヌクレオチドが細胞に対して異種である、項目1に記載のポリヌクレオチドを少なくとも1つ含む細胞。
(29) 前記ポリヌクレオチドが調節配列へ操作可能に連結される、項目28に記載の細胞。
(30) 項目17に記載のベクターによって形質導入される細胞。
(31) 前記細胞がトランスジェニック植物細胞である、項目28または項目30に記載の細胞。
(32) 前記植物細胞がグリホセートN−アセチルトランスフェラーゼ活性を有する外来ポリペプチド発現する、項目31に記載のトランスジェニック植物細胞。
(33) 項目32に記載の細胞を含む、トランスジェニック植物またはトランスジェニック植物外植片。
(34) 前記植物または前記植物外植片がグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現する、項目33に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(35) 前記トランスジェニック植物または前記植物外植片が、以下の属:Eleusine、Lollium、Bambusa、Brassica、Dactylis、Sorghum、Pennisetum、Zea、Oryza、Triticum、Secale、Avena、Hordeum、Saccharum、Coix、GlycineおよびGossypiumの中から選択される作物である、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(36) 前記トランスジェニック植物または前記植物外植片がArabidosisである、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(37) 前記トランスジェニック植物または前記トランスジェニック植物外植片がGossypiumである、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(38) 前記植物または前記植物外植片が、その同属、同株または同品種の野生型植物と比較した場合、グリホセートへの増強された耐性を示す、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(39) 項目34に記載の植物によって生成される種子。
(40) グリホセートに対して少なくとも10mM −1 min −1 のkcat/Kmを有するグリホセートN−アセチルトランスフェラーゼをコードする異種遺伝子を含むトランスジェニック植物であって、前記植物が、除草剤の適用に起因する有意な収率の減少なしに、異種遺伝子が欠如した同植物の生育を阻害するために効果的なレベルにて適用されたグリホセートに耐性を示す、トランスジェニック植物。
(41) グリホセートN−アセチルトランスフェラーゼがアミノメチルホスホン酸のアセチル化を触媒する、項目40に記載のトランスジェニック植物。
(42) BLOSUM62マトリクス、11のギャップイグジステンスペナルティー、および1のギャップエクステンションペナルティーを使用して、少なくとも430の類似性スコアを生成するよう、配列番号300、配列番号445、配列番号457からなる群から選択される配列と、最適に整列され得るアミノ酸配列を含む単離されたポリペプチドまたは組換えポリペプチドであって、該ポリペプチドがグリホセートN−アセチルトランスフェラーゼ活性を有する、ポリペプチド。
(43) 前記ポリペプチドが、グリホセートに対して少なくとも10mM −1 min −1 のkcat/Kmでグリホセートのアセチル化を触媒する、項目42に記載の単離されたポリペプチドまたは組換えポリペプチド。
(44) 前記ポリペプチドが、グリホセートに対して少なくとも100mM −1 min −1 のkcat/Kmでグリホセートのアセチル化を触媒する、項目43に記載の単離されたポリペプチドまたは組換えポリペプチド。
(45) 前記ポリペプチドがアミノメチルホスホン酸のアセチル化を触媒する、項目44に記載の単離されたポリペプチドまたは組換えポリペプチド。
(46) グリホセートN−アセチルトランスフェラーゼ活性を有する単離されたポリペプチドまたは組換えポリペプチドであって、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の少なくとも20個の連続するアミノ酸を含むアミノ酸配列を含む、ポリペプチド。
(47) 前記ポリペプチドが配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の少なくとも50個の連続したアミノ酸を含むアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(48) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の少なくとも100個の連続したアミノ酸を含むアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(49) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の約140個の連続したアミノ酸を含むアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(50) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(51) N末端葉緑体輸送ペプチドをさらに含む、項目42に記載のポリヌクレオチド配列。
(52) 前記ポリペプチドの1つ以上のアミノ酸が変異された、請求項42に記載のポリペプチドの非ネイティブ改変体。
(53) 前記ポリペプチドの1つ以上のアミノ酸が、親ポリペプチドに対して改変された、項目42に記載のポリペプチドの非ネイティブ改変体。
(54) 前記ポリペプチドが、多様な生成手順によって生成される、項目53に記載のポリペプチド。
(55) 前記多様な生成手順が、グリホセートN−アセチルトランスフェラーゼポリペプチドをコードする、少なくとも1つの親ポリヌクレオチドの変異または組換えを含む、項目54に記載のポリペプチド。
(56) 前記親ポリヌクレオチドが項目1に記載のポリヌクレオチドである、項目55に記載のポリペプチド。
(57) 分泌配列または局在化配列を含む、項目42に記載のポリペプチド。
(58) 葉緑体輸送配列を含む、項目57に記載のポリペプチド。
(59) 1つ以上の抗原に対して惹起されたポリクローナル抗血清によって特異的に結合されるポリペプチドであって、該抗原が配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列を含む、ポリペプチド。
(60) GAT活性を有するポリペプチドであって、以下:
(a)少なくとも約2mMまたはそれより少ない、グリホセートに対するkm;
(b)少なくとも約200μMまたはそれより少ない、アセチルCoAに対するkm;および
(c)少なくとも約6/分と等しいkcat、
によって特徴付けられる、ポリペプチド。
(61) グリホセート耐性トランスジェニック植物またはグリホセート耐性植物細胞を生成する方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードするポリヌクレオチドを植物または植物細胞に形質転換する工程:および
(b)該形質転換された植物細胞からトランスジェニック植物を必要に応じて再生する工程、
を包含する、方法。
(62) 前記ポリヌクレオチドが、項目1に記載のポリヌクレオチドである、項目61に記載の方法。
(63) 前記ポリヌクレオチドが細菌源由来である、項目61に記載の方法。
(64) 同属の野生型植物の生育を阻害し、形質転換された植物の生育を阻害しないグリホセート濃度において、形質転換された植物または形質転換された植物細胞を生育する工程を包含する、項目61に記載の方法。
(65) 漸増濃度のグリホセートにおいて、前記形質転換された植物もしくは前記形質転換された植物細胞、または該植物の子孫もしくは該植物細胞の子孫を生育する工程を包含する、項目64に記載の方法。
(66) 前記同属の野生型植物または野生型植物細胞に致死であるグリホセート濃度において、前記形質転換された植物または前記形質転換された植物細胞を生育する工程を包含する、項目64に記載の方法。
(67) 項目1に記載のポリヌクレオチドで形質転換した植物を増殖する工程を包含する、項目62に記載の方法。
(68) 第1の植物が、該第1の植物と第2の植物との交配によって増殖され、その結果、交配の子孫の少なくともいくつかがグリホセート耐性を示す、項目67に記載の方法。
(69) 項目1に記載のポリヌクレオチドの改変体を生成する方法であって、第2のポリヌクレオチドを用いて項目1に記載のポリヌクレオチドを再帰的に組換え、それによって改変体ポリヌクレオチドのライブラリーを形成する工程を含む、方法。
(70) グリホセートN−アセチルトランスフェラーゼ活性を基礎とする前記ライブラリーから改変体ポリヌクレオチドを選択する工程を包含する、項目69に記載の方法。
(71) 前記再帰的な組換えがインビトロで実施される、項目70に記載の方法。
(72) 前記再帰的な組換えがインビボで実施される、項目70に記載の方法。
(73) 前記再帰的な組換えがでインシリコで実施される、項目70に記載の方法。
(74) 前記再帰的な組換えがファミリーシャッフリングを包含する、項目70に記載の方法。
(75) 前記再帰的な組換えが合成シャッフリング法を包含する、項目70に記載の方法。
(76) ヌクレオチド配列中の少なくとも1つの親コドンを、該親コドンに対して植物において優先的に使用される同族コドンに置換する工程を包含する、項目70に記載の方法。
(77) 項目70に記載される方法によって生成される、改変体ポリヌクレオチドのライブラリー。
(78) 項目77に記載のライブラリーを含む、細胞集団。
(79) 項目70に記載の方法によって生成される、組換えポリヌクレオチドであって、該組換えポリヌクレオチドがグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドをコードする、ポリペプチド。
(80) 項目79に記載のポリヌクレオチドを含む、細胞。
(81) 前記細胞が植物細胞である、項目80に記載の細胞。
(82) 前記細胞がトランスジェニック植物細胞である、項目81に記載の細胞。
(83) 項目82に記載の植物によって生成される、種子。
(84) 項目79に記載のポリヌクレオチドによってコードされる、ポリペプチド。
(85) 前記ポリヌクレオチドを変異する工程を包含する、項目1に記載のポリヌクレオチドの改変体を生成する、方法。
(86) 項目85に記載の方法によって生成される、ポリヌクレオチド。
(87) 核酸構築物を含む植物または細胞を、選択する方法であって、以下:
(a)核酸構築物を含むトランスジェニック植物またはトランスジェニック細胞を提供する工程であって、該核酸構築物がグリホセートN−アセチルトラスフェラーゼをコードするヌクレオチド配列を含む、工程;
(b)該グリホセートN−アセチルトランスフェラーゼが効果的なレベルで発現される条件下、グリホセートの存在下で前記植物または前記細胞を生育する工程であって、それによって、該トランスジェニック植物または該トランスジェニック細胞が、該植物または該細胞が前記核酸構築物を含有していない場合に生育する速度よりも、明らかに速い速度にて生育する、工程、
を包含する、方法。
(88) 前記核酸構築物が、第2のヌクレオチド配列に作動可能に連結されるポリペプチドおよび調節配列をコードする、該第2のヌクレオチド配列を含む、項目87に記載の方法。
(89) 農作物を有する領域において雑草を選択的に制御する方法で、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子で形質転換された結果として、グリホセート耐性である農作物の種子または作物を該領域に植える工程;および
(b)該農作物に有意な影響を与えることなしに、該領域中の該農作物および該雑草に、該雑草を制御するために十分量のグリホセートを適用する工程、
を包含する、方法。
(90) グリホセートに対して耐性である、遺伝的に形質転換された植物を生成する方法で、以下:
(a)組換え、2本鎖DNA分子を、植物細胞のゲノムへ挿入する工程であって、該DNA分子が以下:
(i)RNA配列の生成するよう、植物細胞中で機能するプロモーター
(ii)項目42に記載のポリペプチドをコードするRNA配列を生成する、構造DNA配列
(iii)RNA配列の3’末端にポリアデニルヌクレオチドストレッチを付加するよう、植物細胞において機能する3’非翻訳領域を含み、前記プロモーターが前記構造DNA配列各々に対して異種であり、そして、該DNA分子で形質転換された植物細胞の該グリホセート耐性を強めるよう、該コードされたポリペプチドの十分な発現を引き起こすように適応されている工程、
(b)形質転換された植物細胞を獲得する工程;および
(c)該形質転換された植物細胞から、増加したグリホセート耐性を有する遺伝的に形質転換された植物を再生する工程、
を包含する、方法。
(91) 農作物を生成する方法であって、以下:
(a)農作物植物が農作物を生成する条件下で、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が形質転換された結果としてグリホセート耐性である該農作物植物を生育する工程;および
(b)該農作物植物から農作物を回収する工程
を包含する、方法。
(92) 雑草を制御するために効果的な濃度で、前記作物にグリホセートを適用する工程を包含する、項目91に記載の方法。
(93) 前記農作物が綿、トウモロコシまたは大豆である、項目92に記載の方法。
(94) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基が、少なくとも90%、以下の制限に合致し:
(a)2、4、15、19、26、28、31、45、51、54、86、90、91、97、103、105、106、114、123、129、139およ
び/または145位のアミノ酸残基がB1である;および
(b)3、5、8、10、11、14、17、18、24、27、32、37、38、47、48、49、52、57、58、61、62、63、68、69、79、80、82、83、89、92、100、101、104、119、120、124、125、126、128、131、143および/または144位のアミノ酸残基がB2である;
ここで、B1はA、I、L、M、F、W、YおよびVからなる群から選択されるアミノ酸であり、そして、B2は、R、N、D、C、Q、E、G、H、K、P、SおよびTからなる群から選択されるアミノ酸である、
ポリヌクレオチド。
(95) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下の位置に対応する前記アミノ酸配列中のアミノ酸残基が、少なくとも80%、以下の制限に合致し:
(a)2、4、15、19、26、28、51、54、86、90、91、97、103、105、106、114、129、139および/または145位のアミノ酸残基がZ1である;
(b)31および/または45位のアミノ酸残基がZ2である;
(c)8および/または89位のアミノ酸残基がZ3である;
(d)82、92、101および/または120位のアミノ酸残基がZ4である;
(e)3、11、27および/または79位のアミノ酸残基がZ5である;
(f)123位のアミノ酸残基がZ1またはZ2である;
(g)12、33、35、39、53、59、112、132、135、140および/または146位のアミノ酸残基がZ1またはZ3である;
(h)30位のアミノ酸残基がZ1またはZ4である;
(i)6位のアミノ酸残基がZ1またはZ6である;
(j)81および/または113位のアミノ酸残基がZ2またはZ3である;
(k)138および/または142位のアミノ酸残基がZ2またはZ4である;
(l)5、17、24、57、61、124および/または126位のアミノ酸残基がZ3またはZ4である;
(m)104位のアミノ酸残基がZ3またはZ5である;
(o)38、52、62および/または69位のアミノ酸残基がZ3またはZ6である;
(p)14、119および/または144位のアミノ酸残基がZ4またはZ5である;
(q)18位のアミノ酸残基がZ4またはZ6である;
(r)10、32、48、63、80および/または83位のアミノ酸残基がZ5またはZ6である;
(s)40位のアミノ酸残基がZ1、Z2またはZ3である;
(t)65および/または96位のアミノ酸残基がZ1、Z3またはZ5である;
(u)84および/または115位のアミノ酸残基がZ1、Z3またはZ4である;
(v)93位のアミノ酸残基がZ2、Z3またはZ4である;
(w)130位のアミノ酸残基がZ2、Z4またはZ6である;
(x)47および/または58位のアミノ酸残基がZ3、Z4およびZ6である;
(y)49、68、100および/または143位のアミノ酸残基がZ3、Z4およびZ5である;
(z)131位のアミノ酸残基がZ3、Z5またはZ6である;
(aa)125および/または128位のアミノ酸残基がZ4、Z5またはZ6である;
(ab)67位のアミノ酸残基がZ1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基がZ1、Z4、Z5またはZ6である;ならびに
(ad)37位のアミノ酸残基がZ3、Z4、Z5またはZ6である;
ここで、Z1はA、I、L、MおよびVからなる群から選択されるアミノ酸配列であり;Z2はF、WおよびYからなる群から選択されるアミノ酸であり;Z3はN、Q、S、およびTからなる群から選択されるアミノ酸であり;Z4はR、HおよびKからなる群から選択されるアミノ酸であり;Z5はDおよびEからなる群から選択されるアミノ酸であり;そしてZ6はC、GおよびPからなる群から選択されるアミノ酸である、
ポリヌクレオチド。
(96) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下の位置に対応する前記アミノ酸配列中のアミノ酸残基が、少なくとも90%、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位の前記アミノ酸残基がB1であり;ならびに
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2であり;
ここで、B1はA、I、L、M、F、W、YおよびVからなる群から選択されるアミノ酸であり、そして、B2はR、N、D、C、Q、E、G、H、K、P、SおよびTからなる群から選択されるアミノ酸である、
ポリヌクレオチド。
(97) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、20位、36位、42位、50位、64位、72位、75位、76位、78位、94位、98位、110位、121位および/または141位のアミノ酸残基が、Z1である;
(b)13位、46位、56位、70位、107位、117位および/または118位のアミノ酸残基が、Z2である;
(c)23位、55位、71位、77位、88位および/または109位のアミノ酸残基が、Z3である;
(d)16位、21位、41位、73位、85位、99位および/または111位のアミノ酸残基が、Z4である;
(e)34位および/または95位のアミノ酸残基が、Z5である;
(f)22位、25位、29位、43位、44位、66位、74位、87位、102位、108位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、Z6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(98) 項目94に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位,136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(99) 項目94に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位,136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(100) 項目94に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(101) 項目95に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位,136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(102) 項目に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)2位のアミノ酸残基が、IまたはLである;
(b)3位のアミノ酸残基が、EまたはDである;
(c)4位のアミノ酸残基が、V、AまたはIである;
(d)5位のアミノ酸残基が、K、RまたはNである;
(e)6位のアミノ酸残基が、PまたはLである;
(f)8位のアミノ酸残基が、N、SまたはTである;
(g)10位のアミノ酸残基が、EまたはGである;
(h)11位のアミノ酸残基が、DまたはEである;
(i)12位のアミノ酸残基が、TまたはAである;
(j)14位のアミノ酸残基が、EまたはKである;
(k)15位のアミノ酸残基が、IまたはLである;
(l)17位のアミノ酸残基が、HまたはQである;
(m)18位のアミノ酸残基が、R、CまたはKである;
(n)19位のアミノ酸残基が、IまたはVである;
(o)24位のアミノ酸残基が、QまたはRである;
(p)26位のアミノ酸残基が、LまたはIである;
(q)27位のアミノ酸残基が、EまたはDである;
(r)28位のアミノ酸残基が、AまたはVである;
(s)30位のアミノ酸残基が、K、MまたはRである;
(t)31位のアミノ酸残基が、YまたはFである;
(u)32位のアミノ酸残基が、EまたはGである;
(v)33位のアミノ酸残基が、T、AまたはSである;
(w)35位のアミノ酸残基が、L、SまたはMである;
(x)37位のアミノ酸残基が、R、G、EまたはQである;
(y)38位のアミノ酸残基が、GまたはSである;
(z)39位のアミノ酸残基が、T、AまたはSである;
(aa)40位のアミノ酸残基が、F、LまたはSである;
(ab)45位のアミノ酸残基が、YまたはFである;
(ac)47位のアミノ酸残基が、R、QまたはGである;
(ad)48位のアミノ酸残基が、GまたはDである;
(ae)49位のアミノ酸残基が、K、R、EまたはQである;
(af)51位のアミノ酸残基が、IまたはVである;
(ag)52位のアミノ酸残基が、S、CまたはGである;
(ah)53位のアミノ酸残基が、IまたはTである;
(ai)54位のアミノ酸残基が、AまたはVである;
(aj)57位のアミノ酸残基が、HまたはNである;
(ak)58位のアミノ酸残基が、Q、K、NまたはPである;
(al)59位のアミノ酸残基が、AまたはSである;
(am)60位のアミノ酸残基が、E、K、G、VまたはDである;
(an)61位のアミノ酸残基が、HまたはQである;
(ao)62位のアミノ酸残基が、P、SまたはTである;
(ap)63位のアミノ酸残基が、E、GまたはDである;
(aq)65位のアミノ酸残基が、E、D、VまたはQである;
(ar)67位のアミノ酸残基が、Q、E、R、L、HまたはKである;
(as)68位のアミノ酸残基が、K、R、EまたはNである;
(at)69位のアミノ酸残基が、QまたはPである;
(au)79位のアミノ酸残基が、EまたはDである;
(av)80位のアミノ酸残基が、GまたはEである;
(aw)81位のアミノ酸残基が、Y、NまたはFである;
(ax)82位のアミノ酸残基が、RまたはHである;
(ay)83位のアミノ酸残基が、E、GまたはDである;
(az)84位のアミノ酸残基が、Q、RまたはLである;
(ba)86位のアミノ酸残基が、AまたはVである;
(bb)89位のアミノ酸残基が、TまたはSである;
(bc)90位のアミノ酸残基が、LまたはIである;
(bd)91位のアミノ酸残基が、IまたはVである;
(be)92位のアミノ酸残基が、RまたはKである;
(bf)93位のアミノ酸残基が、H、YまたはQである;
(bg)96位のアミノ酸残基が、E、AまたはQである;
(bh)97位のアミノ酸残基が、LまたはIである;
(bi)100位のアミノ酸残基が、K、R、NまたはEである;
(bj)101位のアミノ酸残基が、KまたはRである;
(bk)103位のアミノ酸残基が、AまたはVである;
(bl)104位のアミノ酸残基が、DまたはNである;
(bm)105位のアミノ酸残基が、LまたはMである;
(bn)106位のアミノ酸残基が、LまたはIである;
(bo)112位のアミノ酸残基が、TまたはIである;
(bp)113位のアミノ酸残基が、S、TまたはFである;
(bq)114位のアミノ酸残基が、AまたはVである;
(br)115位のアミノ酸残基が、S、RまたはAである;
(bs)119位のアミノ酸残基が、K、EまたはRである;
(bt)120位のアミノ酸残基が、KまたはRである;
(bu)123位のアミノ酸残基が、FまたはLである;
(bv)124位のアミノ酸残基が、SまたはRである;
(bw)125位のアミノ酸残基が、E、K、GまたはDである;
(bx)126位のアミノ酸残基が、QまたはHである;
(by)128位のアミノ酸残基が、E、GまたはKである;
(bz)129位のアミノ酸残基が、V、IまたはAである;
(ca)130位のアミノ酸残基が、Y、H、FまたはCである;
(cb)131位のアミノ酸残基が、D、G、NまたはEである;
(cc)132位のアミノ酸残基が、I、T、A、M、VまたはLである;
(cd)135位のアミノ酸残基が、V、T、AまたはIである;
(ce)138位のアミノ酸残基が、HまたはYである;
(cf)139位のアミノ酸残基が、IまたはVである;
(cg)140位のアミノ酸残基が、LまたはSである;
(ch)142位のアミノ酸残基が、YまたはHである;
(ci)143位のアミノ酸残基が、K、TまたはEである;
(cj)144位のアミノ酸残基が、K、EまたはRである;
(ck)145位のアミノ酸残基が、LまたはIである;および
(cl)146位のアミノ酸残基が、TまたはAである。
(103) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)9位、76位、94位および110位のアミノ酸残基が、Aである;
(b)29位および108位のアミノ酸残基が、Cである;
(c)34位のアミノ酸残基が、Dである;
(d)95位のアミノ酸残基が、Eである;
(e)56位のアミノ酸残基が、Fである;
(f)43位、44位、66位、74位、87位、102位、116位、122位、127位および136位のアミノ酸残基が、Gである;
(g)41位のアミノ酸残基が、Hである;
(h)7位のアミノ酸残基が、Iである;
(i)85位のアミノ酸残基が、Kである;
(j)20位、36位、42位、50位、72位、78位、98位および121位のアミノ酸残基が、Lである;
(k)1位、75位および141位のアミノ酸残基が、Mである;
(l)23位、64位および109位のアミノ酸残基が、Nである;
(m)22位、25位、133位、134位および137位のアミノ酸残基が、Pである;
(n)71位のアミノ酸残基が、Qである;
(o)16位、21位、73位、99位および111位のアミノ酸残基が、Rである;
(p)55位および88位のアミノ酸残基が、Sである;
(q)77位のアミノ酸残基が、Tである;
(r)107位のアミノ酸残基が、Wである;および
(s)13位、46位、70位、117位および118位のアミノ酸残基が、Yである。
(104) 項目102に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(105) 項目103に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、31位、45位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、123位、129位、139位および/または145位のアミノ酸残基が、B1である;
(b)3位、5位、8位、10位、11位、14位、17位、18位、24位、27位、32位、37位、38位、47位、48位、49位、52位、57位、58位、61位、62位、63位、68位、69位、79位、80位、82位、83位、89位、92位、100位、101位、104位、119位、120位、124位、125位、126位、128位、131位、143位および/または144位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(106) 項目102に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、20位、36位、42位、50位、64位、72位、75位、76位、78位、94位、98位、110位、121位および/または141位のアミノ酸残基が、Z1である;
(b)13位、46位、56位、70位、107位、117位および/または118位のアミノ酸残基が、Z2である;
(c)23位、55位、71位、77位、88位および/または109位のアミノ酸残基が、Z3である;
(d)16位、21位、41位、73位、85位、99位および/または111位のアミノ酸残基が、Z4である;
(e)34位および/または95位のアミノ酸残基が、Z5である;
(f)22位、25位、29位、43位、44位、66位、74位、87位、102位、108位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、Z6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(107) 項目103に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、51位,54位、86位、90位、91位,97位,103位,105位,106位,114位、129位、139位および/または145位のアミノ酸残基が、Z1である;
(b)31位および/または45位のアミノ酸残基が、Z2である;
(c)8位および/または89位のアミノ酸残基が、Z3である;
(d)82位、92位、101位および/または120位のアミノ酸残基が、Z4である;
(e)3位、11位、27位および/または79位のアミノ酸残基が、Z5である;
(f)123位のアミノ酸残基が、Z1またはZ2である;
(g)12位、33位、35位、39位、53位、59位、112位、132位、135位、140位および/または146位のアミノ酸残基が、Z1またはZ3である;
(h)30位のアミノ酸残基が、Z1またはZ4である;
(i)6位のアミノ酸残基が、Z1またはZ6である;
(j)81位および/または113位のアミノ酸残基が、Z2またはZ3である;
(k)138位および/または142位のアミノ酸残基が、Z2またはZ4である;
(l)5位、17位、24位、57位、61位、124位および/または126位のアミノ酸残基が、Z3またはZ4である;
(m)104位のアミノ酸残基が、Z3またはZ5である;
(o)38位、52位、62位および/または69位のアミノ酸残基が、Z3またはZ6である;
(p)14位、119位および/または144位のアミノ酸残基が、Z4またはZ5である;
(q)18位のアミノ酸残基が、Z4またはZ6である;
(r)10位、32位、48位、63位、80位および/または83位のアミノ酸残基が、Z5またはZ6である;
(s)40位のアミノ酸残基が、Z1、Z2またはZ3である;
(t)65位および/または96位のアミノ酸残基が、Z1、Z3またはZ5である;
(u)84位および/または115位のアミノ酸残基が、Z1、Z3またはZ4である;
(v)93位のアミノ酸残基がZ2、Z3またはZ4である;
(w)130位のアミノ酸残基がZ2、Z4またはZ6である;
(x)47位および/または58位のアミノ酸残基が、Z3、Z4またはZ6である;
(y)49位、68位、100位および/または143位のアミノ酸残基が、Z3、Z4またはZ5である;
(z)131位のアミノ酸残基が、Z3、Z5またはZ6である;
(aa)125位および/または128位のアミノ酸残基が、Z4、Z5またはZ6である;
(ab)67位のアミノ酸残基が、Z1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基が、Z1、Z4、Z5またはZ6である;および
(ad)37位のアミノ酸残基が、Z3、Z4、Z5またはZ6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(108) 項目102に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)9位、76位、94位および110位のアミノ酸残基が、Aである;
(b)29位および108位のアミノ酸残基が、Cである;
(c)34位のアミノ酸残基が、Dである;
(d)95位のアミノ酸残基が、Eである;
(e)56位のアミノ酸残基が、Fである;
(f)43位、44位、66位、74位、87位、102位、116位、122位、127位および136位のアミノ酸残基が、Gである;
(g)41位のアミノ酸残基が、Hである;
(h)7位のアミノ酸残基が、Iである;
(i)85位のアミノ酸残基が、Kである;
(j)20位、36位、42位、50位、72位、78位、98位および121位のアミノ酸残基が、Lである;
(k)1位、75位および141位のアミノ酸残基が、Mである;
(l)23位、64位および109位のアミノ酸残基が、Nである;
(m)22位、25位、133位、134位および137位のアミノ酸残基が、Pである;
(n)71位のアミノ酸残基が、Qである;
(o)16位、21位、73位、99位および111位のアミノ酸残基が、Rである;
(p)55位および88位のアミノ酸残基が、Sである;
(q)77位のアミノ酸残基が、Tである;
(r)107位のアミノ酸残基が、Wである;および
(s)13位、46位、70位、117位および118位のアミノ酸残基が、Yである。
(109) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、ここで、前記アミノ酸配列中の28位に対応するアミノ酸残基が、Vである、ポリヌクレオチド。
(110) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、ここで、前記アミノ酸配列が、配列番号6〜10および263〜514からなる群から選択される、、ポリヌクレオチド。
(111) 項目42に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、31位、45位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、123位、129位、139位および/または145位のアミノ酸残基が、B1である;および
(b)3位、5位、8位、10位、11位、14位、17位、18位、24位、27位、32位、37位、38位、47位、48位、49位、52位、57位、58位、61位、62位、63位、68位、69位、79位、80位、82位、83位、89位、92位、100位、101位、104位、119位、120位、124位、125位、126位、128位、131位、143位および/または144位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(112) 項目42に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、129位、139位および/または145位のアミノ酸残基が、Z1である;
(b)31位および/または45位のアミノ酸残基が、Z2である;
(c)8位および/または89位のアミノ酸残基が、Z3である;
(d)82位、92位、101位および120位のアミノ酸残基が、Z4である;
(e)3位、11位、27位および/または79位のアミノ酸残基が、Z5である;
(f)123位のアミノ酸残基が、Z1またはZ2である;
(g)12位、33位、35位、39位、53位、59位、112位、132位、135位、140位および/または146位のアミノ酸残基が、Z1またはZ3である;
(h)30位のアミノ酸残基が、Z1またはZ4である;
(i)6位のアミノ酸残基が、Z1またはZ6である;
(j)81位および/または113位のアミノ酸残基が、Z2またはZ3である;
(k)138位および/または142位のアミノ酸残基が、Z2またはZ4である;
(l)5位、17位、24位、57位、61位、124位および/または126位のアミノ酸残基が、Z3またはZ4である;
(m)104位のアミノ酸残基が、Z3またはZ5である;
(o)38位、52位、62位および/または69位のアミノ酸残基が、Z3またはZ6である;
(p)14位、119位および/または144位のアミノ酸残基が、Z4またはZ5である;
(q)18位のアミノ酸残基が、Z4またはZ6である;
(r)10位、32位、48位、63位、80位および/または83位のアミノ酸残基が、Z5またはZ6である;
(s)40位のアミノ酸残基が、Z1、Z2またはZ3である;
(t)65位および/または96位のアミノ酸残基が、Z1、Z3またはZ5である;
(u)84位および/または115位のアミノ酸残基が、Z1、Z3またはZ4である;
(v)93位のアミノ酸残基が、Z2、Z3またはZ4である;
(w)130位のアミノ酸残基が、Z2、Z4またはZ6である;
(x)47位および/または58位のアミノ酸残基が、Z3、Z4またはZ6である;
(y)49位、68位、100位および/または143位のアミノ酸残基が、Z3、Z4またはZ5である;
(z)131位のアミノ酸残基が、Z3、Z5またはZ6である;
(aa)125位および/または128位のアミノ酸残基が、Z4、Z5またはZ6である;
(ab)67位のアミノ酸残基が、Z1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基が、Z1、Z4、Z5またはZ6である;および
(ad)37位のアミノ酸残基が、Z3、Z4、Z5またはZ6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(113) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;および
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(114) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、20、36、42、50、64、72、75、76、78、94、98、110、121および/または141位のアミノ酸残基がZ1である;
(b)13、46、56、70、107、117および/または118位のアミノ酸残基がZ2である;
(c)23、55、71、77、88および/または109位のアミノ酸残基がZ3である;
(d)16、21、41、73、85、99および/または111位のアミノ酸残基がZ4である;
(e)34および/または95位のアミノ酸残基がZ5である;
(f)22、25、29、43、44、66、74、87、102、108、116、122、127、133、134、136および/または137位のアミノ酸残基がZ6である;
ここでZ1はA、I、L、MおよびVから成る群から選択されるアミノ酸である;Z2はF、WおよびYから成る群から選択されるアミノ酸である;Z3はN、Q、SおよびTから成る群から選択されるアミノ酸である;Z4はR、HおよびKから成る群から選択されるアミノ酸である;Z5はDおよびEから成る群から選択されるアミノ酸である;そしてZ6はC、GおよびPから成る群から選択されるアミノ酸である、
ポリペプチド。
(115) 項目111に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(116) 項目111に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(117) 項目111に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136およびまたは137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(118) 項目112に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(119) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)2位のアミノ酸残基がIまたはLである;
(b)3位のアミノ酸残基がEまたはDである;
(c)4位のアミノ酸残基がV、AまたはIである;
(d)5位のアミノ酸残基がK、RまたはNである;
(e)6位のアミノ酸残基がPまたはLである;
(f)8位のアミノ酸残基がN、SまたはTである;
(g)10位のアミノ酸残基がEまたはGである;
(h)11位のアミノ酸残基がDまたはEである;
(i)12位のアミノ酸残基がTまたはAである;
(j)14位のアミノ酸残基がEまたはKである;
(k)15位のアミノ酸残基がIまたはLである;
(l)17位のアミノ酸残基がHまたはQである;
(m)18位のアミノ酸残基がR、CまたはKである;
(n)19位のアミノ酸残基がIまたはVである;
(o)24位のアミノ酸残基がQまたはRである;
(p)26位のアミノ酸残基がLまたはIである;
(q)27位のアミノ酸残基がEまたはDである;
(r)28位のアミノ酸残基がAまたはVである;
(s)30位のアミノ酸残基がK、MまたはRである;
(t)31位のアミノ酸残基がYまたはFである;
(u)32位のアミノ酸残基がEまたはGである;
(v)33位のアミノ酸残基がT、AまたはSである;
(w)35位のアミノ酸残基がL、SまたはMである;
(x)37位のアミノ酸残基がR、G、EまたはQである;
(y)38位のアミノ酸残基がGまたはSである;
(z)39位のアミノ酸残基がT、AまたはSである;
(aa)40位のアミノ酸残基がF、LまたはSである;
(ab)45位のアミノ酸残基がYまたはFである;
(ac)47位のアミノ酸残基がR、QまたはGである;
(ad)48位のアミノ酸残基がGまたはDである;
(ae)49位のアミノ酸残基がK、R、EまたはQである;
(af)51位のアミノ酸残基がIまたはVである;
(ag)52位のアミノ酸残基がS、CまたはGである;
(ah)53位のアミノ酸残基がIまたはTである;
(ai)54位のアミノ酸残基がAまたはVである;
(aj)57位のアミノ酸残基がHまたはNである;
(ak)58位のアミノ酸残基がQ、K、NまたはPである;
(al)59位のアミノ酸残基がAまたはSである;
(am)60位のアミノ酸残基がE、K、G、VまたはDである;
(an)61位のアミノ酸残基がHまたはQである;
(ao)62位のアミノ酸残基がP、SまたはTである;
(ap)63位のアミノ酸残基がE、GまたはDである;
(aq)65位のアミノ酸残基がE、D、VまたはQである;
(ar)67位のアミノ酸残基がQ、E、R、L、HまたはKである;
(as)68位のアミノ酸残基がK、R、EまたはNである;
(at)69位のアミノ酸残基がQまたはPである;
(au)79位のアミノ酸残基がEまたはDである;
(av)80位のアミノ酸残基がGまたはEである;
(aw)81位のアミノ酸残基がY、NまたはFである;
(ax)82位のアミノ酸残基がRまたはHである;
(ay)83位のアミノ酸残基がE、GまたはDである;
(az)84位のアミノ酸残基がQ、RまたはLである;
(ba)86位のアミノ酸残基がAまたはVある;
(bb)89位のアミノ酸残基がTまたはSである;
(bc)90位のアミノ酸残基がLまたはIである;
(bd)91位のアミノ酸残基がIまたはVである;
(be)92位のアミノ酸残基がRまたはKである;
(bf)93位のアミノ酸残基がH、YまたはQである;
(bg)96位のアミノ酸残基がE、AまたはQである;
(bh)97位のアミノ酸残基がLまたはIである;
(bi)100位のアミノ酸残基がK、R、NまたはEである;
(bj)101位のアミノ酸残基がKまたはRである;
(bk)103位のアミノ酸残基がAまたはVである;
(bl)104位のアミノ酸残基がDまたはNである;
(bm)105位のアミノ酸残基がLまたはMである;
(bn)106位のアミノ酸残基がLまたはIである;
(bo)112位のアミノ酸残基がTまたはIである;
(bp)113位のアミノ酸残基がS、TまたはFである;
(bq)114位のアミノ酸残基がAまたはVである;
(br)115位のアミノ酸残基がS、RまたはAである;
(bs)119位のアミノ酸残基がK、EまたはRである;
(bt)120位のアミノ酸残基がKまたはRである;
(bu)123位のアミノ酸残基がFまたはLである;
(bv)124位のアミノ酸残基がSまたはRである;
(bw)125位のアミノ酸残基がE、K、GまたはDである;
(bx)126位のアミノ酸残基がQまたはHである;
(by)128位のアミノ酸残基がE、GまたはKである;
(bz)129位のアミノ酸残基がV、IまたはAである;
(ca)130位のアミノ酸残基がY、H、FまたはCである;
(cb)131位のアミノ酸残基がD、G、NまたはEである;
(cc)132位のアミノ酸残基がI、T、A、M、VまたはLである;
(cd)135位のアミノ酸残基がV、T、AまたはIである;
(ce)138位のアミノ酸残基がHまたはYである;
(cf)139位のアミノ酸残基がIまたはVである;
(cg)140位のアミノ酸残基がLまたはSである;
(ch)142位のアミノ酸残基がYまたはHである;
(ci)143位のアミノ酸残基がK、TまたはEである;
(cj)144位のアミノ酸残基がK、EまたはRである;
(ck)145位のアミノ酸残基がLまたはIである;および
(cl)146位のアミノ酸残基がTまたはAである、
ポリペプチド。
(120) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)9、76、94および110位のアミノ酸残基がAである;
(b)29および108位のアミノ酸残基がCである;
(c)34位のアミノ酸残基がDである;
(d)95位のアミノ酸残基がEである;
(e)56位のアミノ酸残基がFである;
(f)43、44、66、74、87、102、116、122、127および136位のアミノ酸残基がGである;
(g)41位のアミノ酸残基がHである;
(h)7位のアミノ酸残基がIである;
(i)85位のアミノ酸残基がKである;
(j)20、36、42、50、72、78、98および121位のアミノ酸残基がLである;
(k)1、75および141位のアミノ酸残基がMである;
(l)23、64および109位のアミノ酸残基がNである;
(m)22、25、133、134および137位のアミノ酸残基がPである;
(n)71位のアミノ酸残基がQである;
(o)16、21、73、99および111位のアミノ酸残基がRである;
(p)55および88位のアミノ酸残基がSである;
(q)77位のアミノ酸残基がTである;
(r)107位のアミノ酸残基がWである;および
(s)13、46、70、117および118位のアミノ酸残基がYである、
ポリペプチド。
(121) 項目119に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(122) 項目120に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)2、4、15、19、26、28、31、45、51、54、86、90、91、97、103、105、106、114、123、129、139および/または145位のアミノ酸残基がB1である;
(b)3、5、8、10、11、14、17、18、24、27、32、37、38、47、48、49、52、57、58、61、62、63、68、69、79、80、82、83、89、92、100、101、104、119、120、124、125、126、128、131、143および/または144位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(123) 項目119に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、20、36、42、50、64、72、75、76、78、94、98、110、121および/または141位のアミノ酸残基がZ1である;
(b)13、46、56、70、107、117および/または118位のアミノ酸残基がZ2である;
(c)23、55、71、77、88および/または109位のアミノ酸残基がZ3である;
(d)16、21、41、73、85、99および/または111位のアミノ酸残基がZ4である;
(e)34および/または95位のアミノ酸残基がZ5である;
(f)22、25、29、43、44、66、74、87、102、108、116、122、127、133、134、136および/または137位のアミノ酸残基がZ6である;
ここでZ1はA、I、L、MおよびVから成る群から選択されるアミノ酸である;Z2はF、WおよびYから成る群から選択されるアミノ酸である;Z3はN、Q、SおよびTから成る群から選択されるアミノ酸である;Z4はR、HおよびKから成る群から選択されるアミノ酸である;Z5はDおよびEから成る群から選択されるアミノ酸である;そしてZ6はC、GおよびPから成る群から選択されるアミノ酸である、
ポリペプチド。
(124) 項目120に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)2、4、15、19、26、28、51、54、86、90、91、97、103、105、106、114、129、139および/または145位のアミノ酸残基がZ1である;
(b)31および/または45位のアミノ酸残基がZ2である;
(c)8および/または89位のアミノ酸残基がZ3である;
(d)82、92、101および/または120位のアミノ酸残基がZ4である;
(e)3、11、27および/または79位のアミノ酸残基がZ5である;
(f)123位のアミノ酸残基がZ1またはZ2である;
(g)12、33、35、39、53、59、112、132、135、140および/または146位のアミノ酸残基がZ1またはZ3である;
(h)30位のアミノ酸残基がZ1またはZ4である;
(i)6位のアミノ酸残基がZ1またはZ6である;
(j)81および/または113位のアミノ酸残基がZ2またはZ3である;
(k)138および/または142位のアミノ酸残基がZ2またはZ4である;
(l)5、17、24、57、61、124および/または126位のアミノ酸残基がZ3またはZ4である;
(m)104位のアミノ酸残基がZ3またはZ5である;
(o)38、52、62および/または69位のアミノ酸残基がZ3またはZ6である;
(p)14、119および/または144位のアミノ酸残基がZ4またはZ5である;
(q)18位のアミノ酸残基がZ4またはZ6である;
(r)10、32、48、63、80および/または83位のアミノ酸残基がZ5またはZ6である;
(s)40位のアミノ酸残基がZ1、Z2またはZ3である;
(t)65および/または96位のアミノ酸残基がZ1、Z3またはZ5である;
(u)84および/または115位のアミノ酸残基がZ1、Z3またはZ4である;
(v)93位のアミノ酸残基がZ2、Z3またはZ4である;
(w)130位のアミノ酸残基がZ2、Z4またはZ6である;
(x)47および/または58位のアミノ酸残基がZ3、Z4またはZ6である;
(y)49、68、100および/または143位のアミノ酸残基がZ3、Z4またはZ5である;
(z)131位のアミノ酸残基がZ3、Z5またはZ6である;
(aa)125および/または128位のアミノ酸残基がZ4、Z5またはZ6である;
(ab)67位のアミノ酸残基がZ1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基がZ1、Z4、Z5またはZ6である;
(ad)37位のアミノ酸残基がZ3、Z4、Z5またはZ6である;
ここでZ1はA、I、L、MおよびVから成る群から選択されるアミノ酸である;Z2はF、WおよびYから成る群から選択されるアミノ酸である;Z3はN、Q、SおよびTから成る群から選択されるアミノ酸である;Z4はR、HおよびKから成る群から選択されるアミノ酸である;Z5はDおよびEから成る群から選択されるアミノ酸である;そしてZ6はC、GおよびPから成る群から選択されるアミノ酸である、
ポリペプチド。
(125) 項目119に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)9、76、94および110位のアミノ酸残基がAである;
(b)29および108位のアミノ酸残基がCである;
(c)34位のアミノ酸残基がDである;
(d)95位のアミノ酸残基がEである;
(e)56位のアミノ酸残基がFである;
(f)43、44、66、74、87、102、116、122、127および136位のアミノ酸残基がGである;
(g)41位のアミノ酸残基がHである;
(h)7位のアミノ酸残基がIである;
(i)85位のアミノ酸残基がKである;
(j)20、36、42、50、72、78、98および121位のアミノ酸残基がLである;
(k)1、75および141位のアミノ酸残基がMである;
(l)23、64および109位のアミノ酸残基がNである;
(m)22、25、133、134および137位のアミノ酸残基がPである;
(n)71位のアミノ酸残基がQである;
(o)16、21、73、99および111位のアミノ酸残基がRである;
(p)55および88位のアミノ酸残基がSである;
(q)77位のアミノ酸残基がTである;
(r)107位のアミノ酸残基がWである;および
(s)13、46、70、117および118位のアミノ酸残基がYである、
ポリペプチド。
(126) 項目24に記載の単離ポリペプチドまたは組換えポリペプチドであって、ここで28位に対応するアミノ酸配列中のアミノ酸残基が、Vであるポリペプチド。
(127) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、ここでアミノ酸配列が、配列番号6〜10および263〜514から成る群から選択される、ポリペプチド。
(128) 増強されたグリホセート耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで植物または植物外植片が、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドおよびさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドを発現する、トランスジェニック植物またはトランスジェニック植物外植片。
(129) 項目128に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514から成る群から選択されるアミノ酸配列を含む、トランスジェニック植物またはトランスジェニック植物外植片。
(130) 項目129に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−リン酸シンターゼおよびグリホセート耐性グリホセートキシドレダクターゼから成る群から選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(131) 項目130に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−リン酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(132) 項目130に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドが、グリホセート耐性グリホセートキシドレダクターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(133) トランスジェニック植物またはトランスジェニック植物外植片であり、ここで該植物または植物外植片が、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドおよびさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドを発現する、トランスジェニック植物またはトランスジェニック植物外植片。
(134) 項目133に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514から成る群から選択されるアミノ酸配列を含む、トランスジェニック植物またはトランスジェニック植物外植片。
(135) 項目134に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、変異されたヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチル転移酵素および変異されたプロトポルフィリノゲンオキシダーゼから成る群から選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(136) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、変異されたヒドロキシフェニルピルベートジオキシゲナーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(137) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(138) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(139) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(140) 前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(141) 前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(142) 前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、請求項135に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(143) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチド、さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチド、およびさらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドを発現する、トランスジェニック植物またはトランスジェニック植物外植片。
(144) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目143に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(145) 項目144に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセートオキシドレダクターゼからなる群より選択され、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(146) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(147) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(148) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(149) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(150) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(151) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(152) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(153) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(154) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(155) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(156) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(157) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼある、トランスジェニック植物またはトランスジェニック植物外植片。
(158) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼある、トランスジェニック植物またはトランスジェニック植物外植片。
(159) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼある、トランスジェニック植物またはトランスジェニック植物外植片。
(160) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、そして少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(161) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目160に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(162) 前記少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、請求項161に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(163) 前記少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目161に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(164) トランスジェニック植物またはトランスジェニック植物外植片であって、ここで該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、そして、少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(165) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目164に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(166) 前記少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(167) 前記少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(168) 前記少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(169) 前記少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(170) 前記少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(171) 前記少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(172) 前記少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(173) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、少なくとも1つの第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択され、そして、少なくとも1つの第2ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼおよび変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(174) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目173に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(175) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(176) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(177) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、請求項174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(178) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(179) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、請求項174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(180) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、請求項174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(181) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(182) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(183) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(184) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドがスルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(185) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドがイミダゾリノン耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(186) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドがイミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(187) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(188) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが変異型プロトポルフィリノーゲンオキシダーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(189) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、そして少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼ、グリホセート耐性グリホセート酸化還元酵素、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼおよび変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(190) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目189に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(191) 前記ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(192) 前記ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(193) 前記ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(194) 前記ポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(195) 前記ポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(196) 前記ポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(197) 前記ポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(198) 前記ポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(199) 前記ポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(200) 作物を含む畑において雑草を制御するための方法であって、
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程;および、
(b)該作物への有意な影響なしで該畑における該雑草の成長を阻害するのに十分である、グリホセートの効果的な適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(201) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目200に記載の方法。
(202) 項目201に記載の方法であって、ここで、前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、方法。
(203) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目202に記載の方法。
(204) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目202に記載の方法。
(205) 作物を含む畑においてグリホセート耐性雑草の発生を防止するための方法であって、
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程;および、
(b)グリホセートの効果的な適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(206) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目205に記載の方法。
(207) 項目206に記載の方法であって、ここで、前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、方法。
(208) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目207に記載の方法。
(209) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目207に記載の方法。
(210) 作物を含む畑において選択的に雑草を制御するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)該作物への有意な影響なしで該畑における該雑草の成長を阻害するのに十分である、グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を該畑における作物および雑草に適用する工程、
を包含する、工程。
(211) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目210に記載の方法。
(212) 項目211に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(213) 項目211に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(214) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目212に記載の方法。
(215) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目212に記載の方法。
(216) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目212に記載の方法。
(217) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目212に記載の方法。
(218) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目212に記載の方法。
(219) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目212に記載の方法。
(220) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目212に記載の方法。
(221) 作物を含む畑における除草剤耐性雑草の発生を防止するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(222) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目221に記載の方法。
(223) 項目222に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(224) 項目221に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(225) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目223に記載の方法。
(226) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目223に記載の方法。
(227) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目223に記載の方法。
(228) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目223に記載の方法。
(229) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目223に記載の方法。
(230) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目223に記載の方法。
(231) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目223に記載の方法。
(232) 作物を含む畑において雑草を選択的に制御するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、さらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)該作物への有意な影響なしで該畑における該雑草の成長を阻害するのに十分である、グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を該畑における作物および雑草に適用する工程、
を包含する、方法。
(233) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目232に記載の方法。
(234) 項目233に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(235) 前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目234に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(236) 前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目234に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(237) 項目233に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(238) 項目233に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(239) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目237に記載の方法。
(240) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目237に記載の方法。
(241) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目237に記載の方法。
(242) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目237に記載の方法。
(243) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目237に記載の方法。
(244) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目237に記載の方法。
(245) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目237に記載の方法。
(246) 作物を含む畑における除草剤耐性雑草の発生を制御するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、さらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(247) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目246に記載の方法。
(248) 項目247に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(249) 項目247に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(250) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目248に記載の方法。
(251) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目248に記載の方法。
(252) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目248に記載の方法。
(253) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目248に記載の方法。
(254) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目248に記載の方法。
(255) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目248に記載の方法。
(256) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目248に記載の方法。
(本発明の要旨)
生物体(例えば、植物)をグリホセートに対し抵抗性にするための方法および試薬を提供することが、本発明の目的である。本発明のこの目的およびその他の目的は、以下に記載される一つ以上の実施形態により提供される。
(1) 単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下:
(a)BLOSUM62マトリックス、11のギャップイグジステンスペナルティー、および1のギャップエクステンションペナルティーを使用して、少なくとも430の類似性スコアを生み出すために、配列番号300、配列番号445、および配列番号457からなる群から選択される配列と、最適に並べられ得るアミノ酸配列をコードするヌクレオチド配列;または
(b)これらの相補鎖ヌクレオチド配列、
を含む、ポリヌクレオチド。
(2) 前記ポリペプチドがグリホセートN−アセチルトランスフェラーゼ活性を有する、項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(3) 前記ポリペプチドが、グリホセートに対して、少なくとも10mM −1 min −1 のkcat/Kmでグリホセートのアセチル化を触媒する、項目2に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(4) 前記ポリペプチドがアミノメチルホスホン酸のアセチル化を触媒する、項目2に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(5) グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドをコードするヌクレオチド配列を含む、単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、該ポリペプチドが配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、少なくとも20個の連続するアミノ酸を含むアミノ酸配列を含む、ポリヌクレオチド。
(6) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、少なくとも50個の連続するアミノ酸を含むアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(7) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、少なくとも100個の連続するアミノ酸を含むアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(8) 前記ポリヌクレオチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の、約140個の連続するアミノ酸を含むアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(9) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(10) 配列番号48、配列番号193および配列番号205からなる群から選択されるヌクレオチド配列を含む、項目5に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチド。
(11) 親コドンが、該親コドンに対して植物中で優先的に使用される同族コドンによって置換された、項目1に記載のポリヌクレオチド。
(12) N末端葉緑体輸送ペプチドをコードするヌクレオチド配列さらにを含む、項目1に記載のポリヌクレオチド。
(13) 前記のコードされたポリペプチドの1つ以上のアミノ酸が変異された、項目1に記載のポリヌクレオチドの非ネイティブ改変体。
(14) 項目1に記載のポリヌクレオチドを含む核酸構築物。
(15) 項目1に記載のポリヌクレオチドに作動可能に連結されたプロモーターを含む、項目14に記載の核酸構築物であって、該プロモーターが、前記ポリヌクレオチドに対して異種であり、そして、前記核酸構築物を用いて形質転換された植物細胞のグリホセート耐性を強めるのに十分な前記のコードされたポリペプチドの発現を引き起こすために効果的である、核酸構築物。
(16) 項目1に記載のポリヌクレオチド配列が選択可能なマーカーとして機能する、項目14に記載の核酸構築物。
(17) 前記構築物がベクターである、項目14に記載の核酸構築物。
(18)第2のポリペプチドが効果的なレベルで発現する細胞または組織に検出可能な表現型特性を付与する、該第2ポリペプチドをコードする第2ポリヌクレオチド配列を含む、項目17に記載のベクター。
(19) 前記検出可能な表現型特性が、選択可能なマーカーとして機能する、項目18に記載のベクター。
(20) 前記検出可能な表現型特性が、除草剤耐性、害虫耐性、または可視マーカーからなる、項目19に記載のベクター。
(21) 前記ベクターが、T−DNA配列を含む、項目17に記載のベクター。
(22) 前記ポリヌクレオチドが調節配列に作動可能に連結される、項目17に記載のベクター。
(23) 前記ベクターが植物形質転換ベクターである、項目17に記載のベクター。
(24) 単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下:
(a)ストリンジェントな条件下で、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列をコードするヌクレオチド配列の実質的に全長にわたりハイブリダイズするヌクレオチド;
(b)これらの相補鎖ヌクレオチド配列;または
(c)グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドをコードする(a)または(b)のフラグメント、
を含む、ポリヌクレオチド。
(25) グリホセートN−アセチルトランスフェラーゼをコードするヌクレオチド配列を含む、項目24に記載のポリヌクレオチド。
(26) 項目1に記載の2つ以上のポリヌクレオチドを含む組成物。
(27) 項目1に記載のポリヌクレオチドを少なくとも10個含む、項目26に記載の組成物。
(28) 前記ポリヌクレオチドが細胞に対して異種である、項目1に記載のポリヌクレオチドを少なくとも1つ含む細胞。
(29) 前記ポリヌクレオチドが調節配列へ操作可能に連結される、項目28に記載の細胞。
(30) 項目17に記載のベクターによって形質導入される細胞。
(31) 前記細胞がトランスジェニック植物細胞である、項目28または項目30に記載の細胞。
(32) 前記植物細胞がグリホセートN−アセチルトランスフェラーゼ活性を有する外来ポリペプチド発現する、項目31に記載のトランスジェニック植物細胞。
(33) 項目32に記載の細胞を含む、トランスジェニック植物またはトランスジェニック植物外植片。
(34) 前記植物または前記植物外植片がグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現する、項目33に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(35) 前記トランスジェニック植物または前記植物外植片が、以下の属:Eleusine、Lollium、Bambusa、Brassica、Dactylis、Sorghum、Pennisetum、Zea、Oryza、Triticum、Secale、Avena、Hordeum、Saccharum、Coix、GlycineおよびGossypiumの中から選択される作物である、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(36) 前記トランスジェニック植物または前記植物外植片がArabidosisである、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(37) 前記トランスジェニック植物または前記トランスジェニック植物外植片がGossypiumである、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(38) 前記植物または前記植物外植片が、その同属、同株または同品種の野生型植物と比較した場合、グリホセートへの増強された耐性を示す、項目34に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(39) 項目34に記載の植物によって生成される種子。
(40) グリホセートに対して少なくとも10mM −1 min −1 のkcat/Kmを有するグリホセートN−アセチルトランスフェラーゼをコードする異種遺伝子を含むトランスジェニック植物であって、前記植物が、除草剤の適用に起因する有意な収率の減少なしに、異種遺伝子が欠如した同植物の生育を阻害するために効果的なレベルにて適用されたグリホセートに耐性を示す、トランスジェニック植物。
(41) グリホセートN−アセチルトランスフェラーゼがアミノメチルホスホン酸のアセチル化を触媒する、項目40に記載のトランスジェニック植物。
(42) BLOSUM62マトリクス、11のギャップイグジステンスペナルティー、および1のギャップエクステンションペナルティーを使用して、少なくとも430の類似性スコアを生成するよう、配列番号300、配列番号445、配列番号457からなる群から選択される配列と、最適に整列され得るアミノ酸配列を含む単離されたポリペプチドまたは組換えポリペプチドであって、該ポリペプチドがグリホセートN−アセチルトランスフェラーゼ活性を有する、ポリペプチド。
(43) 前記ポリペプチドが、グリホセートに対して少なくとも10mM −1 min −1 のkcat/Kmでグリホセートのアセチル化を触媒する、項目42に記載の単離されたポリペプチドまたは組換えポリペプチド。
(44) 前記ポリペプチドが、グリホセートに対して少なくとも100mM −1 min −1 のkcat/Kmでグリホセートのアセチル化を触媒する、項目43に記載の単離されたポリペプチドまたは組換えポリペプチド。
(45) 前記ポリペプチドがアミノメチルホスホン酸のアセチル化を触媒する、項目44に記載の単離されたポリペプチドまたは組換えポリペプチド。
(46) グリホセートN−アセチルトランスフェラーゼ活性を有する単離されたポリペプチドまたは組換えポリペプチドであって、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の少なくとも20個の連続するアミノ酸を含むアミノ酸配列を含む、ポリペプチド。
(47) 前記ポリペプチドが配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の少なくとも50個の連続したアミノ酸を含むアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(48) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の少なくとも100個の連続したアミノ酸を含むアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(49) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列の約140個の連続したアミノ酸を含むアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(50) 前記ポリペプチドが、配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列を含む、項目46に記載の単離されたポリペプチドまたは組換えポリペプチド。
(51) N末端葉緑体輸送ペプチドをさらに含む、項目42に記載のポリヌクレオチド配列。
(52) 前記ポリペプチドの1つ以上のアミノ酸が変異された、請求項42に記載のポリペプチドの非ネイティブ改変体。
(53) 前記ポリペプチドの1つ以上のアミノ酸が、親ポリペプチドに対して改変された、項目42に記載のポリペプチドの非ネイティブ改変体。
(54) 前記ポリペプチドが、多様な生成手順によって生成される、項目53に記載のポリペプチド。
(55) 前記多様な生成手順が、グリホセートN−アセチルトランスフェラーゼポリペプチドをコードする、少なくとも1つの親ポリヌクレオチドの変異または組換えを含む、項目54に記載のポリペプチド。
(56) 前記親ポリヌクレオチドが項目1に記載のポリヌクレオチドである、項目55に記載のポリペプチド。
(57) 分泌配列または局在化配列を含む、項目42に記載のポリペプチド。
(58) 葉緑体輸送配列を含む、項目57に記載のポリペプチド。
(59) 1つ以上の抗原に対して惹起されたポリクローナル抗血清によって特異的に結合されるポリペプチドであって、該抗原が配列番号300、配列番号445および配列番号457からなる群から選択されるアミノ酸配列を含む、ポリペプチド。
(60) GAT活性を有するポリペプチドであって、以下:
(a)少なくとも約2mMまたはそれより少ない、グリホセートに対するkm;
(b)少なくとも約200μMまたはそれより少ない、アセチルCoAに対するkm;および
(c)少なくとも約6/分と等しいkcat、
によって特徴付けられる、ポリペプチド。
(61) グリホセート耐性トランスジェニック植物またはグリホセート耐性植物細胞を生成する方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードするポリヌクレオチドを植物または植物細胞に形質転換する工程:および
(b)該形質転換された植物細胞からトランスジェニック植物を必要に応じて再生する工程、
を包含する、方法。
(62) 前記ポリヌクレオチドが、項目1に記載のポリヌクレオチドである、項目61に記載の方法。
(63) 前記ポリヌクレオチドが細菌源由来である、項目61に記載の方法。
(64) 同属の野生型植物の生育を阻害し、形質転換された植物の生育を阻害しないグリホセート濃度において、形質転換された植物または形質転換された植物細胞を生育する工程を包含する、項目61に記載の方法。
(65) 漸増濃度のグリホセートにおいて、前記形質転換された植物もしくは前記形質転換された植物細胞、または該植物の子孫もしくは該植物細胞の子孫を生育する工程を包含する、項目64に記載の方法。
(66) 前記同属の野生型植物または野生型植物細胞に致死であるグリホセート濃度において、前記形質転換された植物または前記形質転換された植物細胞を生育する工程を包含する、項目64に記載の方法。
(67) 項目1に記載のポリヌクレオチドで形質転換した植物を増殖する工程を包含する、項目62に記載の方法。
(68) 第1の植物が、該第1の植物と第2の植物との交配によって増殖され、その結果、交配の子孫の少なくともいくつかがグリホセート耐性を示す、項目67に記載の方法。
(69) 項目1に記載のポリヌクレオチドの改変体を生成する方法であって、第2のポリヌクレオチドを用いて項目1に記載のポリヌクレオチドを再帰的に組換え、それによって改変体ポリヌクレオチドのライブラリーを形成する工程を含む、方法。
(70) グリホセートN−アセチルトランスフェラーゼ活性を基礎とする前記ライブラリーから改変体ポリヌクレオチドを選択する工程を包含する、項目69に記載の方法。
(71) 前記再帰的な組換えがインビトロで実施される、項目70に記載の方法。
(72) 前記再帰的な組換えがインビボで実施される、項目70に記載の方法。
(73) 前記再帰的な組換えがでインシリコで実施される、項目70に記載の方法。
(74) 前記再帰的な組換えがファミリーシャッフリングを包含する、項目70に記載の方法。
(75) 前記再帰的な組換えが合成シャッフリング法を包含する、項目70に記載の方法。
(76) ヌクレオチド配列中の少なくとも1つの親コドンを、該親コドンに対して植物において優先的に使用される同族コドンに置換する工程を包含する、項目70に記載の方法。
(77) 項目70に記載される方法によって生成される、改変体ポリヌクレオチドのライブラリー。
(78) 項目77に記載のライブラリーを含む、細胞集団。
(79) 項目70に記載の方法によって生成される、組換えポリヌクレオチドであって、該組換えポリヌクレオチドがグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドをコードする、ポリペプチド。
(80) 項目79に記載のポリヌクレオチドを含む、細胞。
(81) 前記細胞が植物細胞である、項目80に記載の細胞。
(82) 前記細胞がトランスジェニック植物細胞である、項目81に記載の細胞。
(83) 項目82に記載の植物によって生成される、種子。
(84) 項目79に記載のポリヌクレオチドによってコードされる、ポリペプチド。
(85) 前記ポリヌクレオチドを変異する工程を包含する、項目1に記載のポリヌクレオチドの改変体を生成する、方法。
(86) 項目85に記載の方法によって生成される、ポリヌクレオチド。
(87) 核酸構築物を含む植物または細胞を、選択する方法であって、以下:
(a)核酸構築物を含むトランスジェニック植物またはトランスジェニック細胞を提供する工程であって、該核酸構築物がグリホセートN−アセチルトラスフェラーゼをコードするヌクレオチド配列を含む、工程;
(b)該グリホセートN−アセチルトランスフェラーゼが効果的なレベルで発現される条件下、グリホセートの存在下で前記植物または前記細胞を生育する工程であって、それによって、該トランスジェニック植物または該トランスジェニック細胞が、該植物または該細胞が前記核酸構築物を含有していない場合に生育する速度よりも、明らかに速い速度にて生育する、工程、
を包含する、方法。
(88) 前記核酸構築物が、第2のヌクレオチド配列に作動可能に連結されるポリペプチドおよび調節配列をコードする、該第2のヌクレオチド配列を含む、項目87に記載の方法。
(89) 農作物を有する領域において雑草を選択的に制御する方法で、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子で形質転換された結果として、グリホセート耐性である農作物の種子または作物を該領域に植える工程;および
(b)該農作物に有意な影響を与えることなしに、該領域中の該農作物および該雑草に、該雑草を制御するために十分量のグリホセートを適用する工程、
を包含する、方法。
(90) グリホセートに対して耐性である、遺伝的に形質転換された植物を生成する方法で、以下:
(a)組換え、2本鎖DNA分子を、植物細胞のゲノムへ挿入する工程であって、該DNA分子が以下:
(i)RNA配列の生成するよう、植物細胞中で機能するプロモーター
(ii)項目42に記載のポリペプチドをコードするRNA配列を生成する、構造DNA配列
(iii)RNA配列の3’末端にポリアデニルヌクレオチドストレッチを付加するよう、植物細胞において機能する3’非翻訳領域を含み、前記プロモーターが前記構造DNA配列各々に対して異種であり、そして、該DNA分子で形質転換された植物細胞の該グリホセート耐性を強めるよう、該コードされたポリペプチドの十分な発現を引き起こすように適応されている工程、
(b)形質転換された植物細胞を獲得する工程;および
(c)該形質転換された植物細胞から、増加したグリホセート耐性を有する遺伝的に形質転換された植物を再生する工程、
を包含する、方法。
(91) 農作物を生成する方法であって、以下:
(a)農作物植物が農作物を生成する条件下で、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が形質転換された結果としてグリホセート耐性である該農作物植物を生育する工程;および
(b)該農作物植物から農作物を回収する工程
を包含する、方法。
(92) 雑草を制御するために効果的な濃度で、前記作物にグリホセートを適用する工程を包含する、項目91に記載の方法。
(93) 前記農作物が綿、トウモロコシまたは大豆である、項目92に記載の方法。
(94) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基が、少なくとも90%、以下の制限に合致し:
(a)2、4、15、19、26、28、31、45、51、54、86、90、91、97、103、105、106、114、123、129、139およ
び/または145位のアミノ酸残基がB1である;および
(b)3、5、8、10、11、14、17、18、24、27、32、37、38、47、48、49、52、57、58、61、62、63、68、69、79、80、82、83、89、92、100、101、104、119、120、124、125、126、128、131、143および/または144位のアミノ酸残基がB2である;
ここで、B1はA、I、L、M、F、W、YおよびVからなる群から選択されるアミノ酸であり、そして、B2は、R、N、D、C、Q、E、G、H、K、P、SおよびTからなる群から選択されるアミノ酸である、
ポリヌクレオチド。
(95) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下の位置に対応する前記アミノ酸配列中のアミノ酸残基が、少なくとも80%、以下の制限に合致し:
(a)2、4、15、19、26、28、51、54、86、90、91、97、103、105、106、114、129、139および/または145位のアミノ酸残基がZ1である;
(b)31および/または45位のアミノ酸残基がZ2である;
(c)8および/または89位のアミノ酸残基がZ3である;
(d)82、92、101および/または120位のアミノ酸残基がZ4である;
(e)3、11、27および/または79位のアミノ酸残基がZ5である;
(f)123位のアミノ酸残基がZ1またはZ2である;
(g)12、33、35、39、53、59、112、132、135、140および/または146位のアミノ酸残基がZ1またはZ3である;
(h)30位のアミノ酸残基がZ1またはZ4である;
(i)6位のアミノ酸残基がZ1またはZ6である;
(j)81および/または113位のアミノ酸残基がZ2またはZ3である;
(k)138および/または142位のアミノ酸残基がZ2またはZ4である;
(l)5、17、24、57、61、124および/または126位のアミノ酸残基がZ3またはZ4である;
(m)104位のアミノ酸残基がZ3またはZ5である;
(o)38、52、62および/または69位のアミノ酸残基がZ3またはZ6である;
(p)14、119および/または144位のアミノ酸残基がZ4またはZ5である;
(q)18位のアミノ酸残基がZ4またはZ6である;
(r)10、32、48、63、80および/または83位のアミノ酸残基がZ5またはZ6である;
(s)40位のアミノ酸残基がZ1、Z2またはZ3である;
(t)65および/または96位のアミノ酸残基がZ1、Z3またはZ5である;
(u)84および/または115位のアミノ酸残基がZ1、Z3またはZ4である;
(v)93位のアミノ酸残基がZ2、Z3またはZ4である;
(w)130位のアミノ酸残基がZ2、Z4またはZ6である;
(x)47および/または58位のアミノ酸残基がZ3、Z4およびZ6である;
(y)49、68、100および/または143位のアミノ酸残基がZ3、Z4およびZ5である;
(z)131位のアミノ酸残基がZ3、Z5またはZ6である;
(aa)125および/または128位のアミノ酸残基がZ4、Z5またはZ6である;
(ab)67位のアミノ酸残基がZ1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基がZ1、Z4、Z5またはZ6である;ならびに
(ad)37位のアミノ酸残基がZ3、Z4、Z5またはZ6である;
ここで、Z1はA、I、L、MおよびVからなる群から選択されるアミノ酸配列であり;Z2はF、WおよびYからなる群から選択されるアミノ酸であり;Z3はN、Q、S、およびTからなる群から選択されるアミノ酸であり;Z4はR、HおよびKからなる群から選択されるアミノ酸であり;Z5はDおよびEからなる群から選択されるアミノ酸であり;そしてZ6はC、GおよびPからなる群から選択されるアミノ酸である、
ポリヌクレオチド。
(96) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、以下の位置に対応する前記アミノ酸配列中のアミノ酸残基が、少なくとも90%、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位の前記アミノ酸残基がB1であり;ならびに
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2であり;
ここで、B1はA、I、L、M、F、W、YおよびVからなる群から選択されるアミノ酸であり、そして、B2はR、N、D、C、Q、E、G、H、K、P、SおよびTからなる群から選択されるアミノ酸である、
ポリヌクレオチド。
(97) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、20位、36位、42位、50位、64位、72位、75位、76位、78位、94位、98位、110位、121位および/または141位のアミノ酸残基が、Z1である;
(b)13位、46位、56位、70位、107位、117位および/または118位のアミノ酸残基が、Z2である;
(c)23位、55位、71位、77位、88位および/または109位のアミノ酸残基が、Z3である;
(d)16位、21位、41位、73位、85位、99位および/または111位のアミノ酸残基が、Z4である;
(e)34位および/または95位のアミノ酸残基が、Z5である;
(f)22位、25位、29位、43位、44位、66位、74位、87位、102位、108位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、Z6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(98) 項目94に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位,136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(99) 項目94に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位,136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(100) 項目94に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(101) 項目95に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位,136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(102) 項目に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)2位のアミノ酸残基が、IまたはLである;
(b)3位のアミノ酸残基が、EまたはDである;
(c)4位のアミノ酸残基が、V、AまたはIである;
(d)5位のアミノ酸残基が、K、RまたはNである;
(e)6位のアミノ酸残基が、PまたはLである;
(f)8位のアミノ酸残基が、N、SまたはTである;
(g)10位のアミノ酸残基が、EまたはGである;
(h)11位のアミノ酸残基が、DまたはEである;
(i)12位のアミノ酸残基が、TまたはAである;
(j)14位のアミノ酸残基が、EまたはKである;
(k)15位のアミノ酸残基が、IまたはLである;
(l)17位のアミノ酸残基が、HまたはQである;
(m)18位のアミノ酸残基が、R、CまたはKである;
(n)19位のアミノ酸残基が、IまたはVである;
(o)24位のアミノ酸残基が、QまたはRである;
(p)26位のアミノ酸残基が、LまたはIである;
(q)27位のアミノ酸残基が、EまたはDである;
(r)28位のアミノ酸残基が、AまたはVである;
(s)30位のアミノ酸残基が、K、MまたはRである;
(t)31位のアミノ酸残基が、YまたはFである;
(u)32位のアミノ酸残基が、EまたはGである;
(v)33位のアミノ酸残基が、T、AまたはSである;
(w)35位のアミノ酸残基が、L、SまたはMである;
(x)37位のアミノ酸残基が、R、G、EまたはQである;
(y)38位のアミノ酸残基が、GまたはSである;
(z)39位のアミノ酸残基が、T、AまたはSである;
(aa)40位のアミノ酸残基が、F、LまたはSである;
(ab)45位のアミノ酸残基が、YまたはFである;
(ac)47位のアミノ酸残基が、R、QまたはGである;
(ad)48位のアミノ酸残基が、GまたはDである;
(ae)49位のアミノ酸残基が、K、R、EまたはQである;
(af)51位のアミノ酸残基が、IまたはVである;
(ag)52位のアミノ酸残基が、S、CまたはGである;
(ah)53位のアミノ酸残基が、IまたはTである;
(ai)54位のアミノ酸残基が、AまたはVである;
(aj)57位のアミノ酸残基が、HまたはNである;
(ak)58位のアミノ酸残基が、Q、K、NまたはPである;
(al)59位のアミノ酸残基が、AまたはSである;
(am)60位のアミノ酸残基が、E、K、G、VまたはDである;
(an)61位のアミノ酸残基が、HまたはQである;
(ao)62位のアミノ酸残基が、P、SまたはTである;
(ap)63位のアミノ酸残基が、E、GまたはDである;
(aq)65位のアミノ酸残基が、E、D、VまたはQである;
(ar)67位のアミノ酸残基が、Q、E、R、L、HまたはKである;
(as)68位のアミノ酸残基が、K、R、EまたはNである;
(at)69位のアミノ酸残基が、QまたはPである;
(au)79位のアミノ酸残基が、EまたはDである;
(av)80位のアミノ酸残基が、GまたはEである;
(aw)81位のアミノ酸残基が、Y、NまたはFである;
(ax)82位のアミノ酸残基が、RまたはHである;
(ay)83位のアミノ酸残基が、E、GまたはDである;
(az)84位のアミノ酸残基が、Q、RまたはLである;
(ba)86位のアミノ酸残基が、AまたはVである;
(bb)89位のアミノ酸残基が、TまたはSである;
(bc)90位のアミノ酸残基が、LまたはIである;
(bd)91位のアミノ酸残基が、IまたはVである;
(be)92位のアミノ酸残基が、RまたはKである;
(bf)93位のアミノ酸残基が、H、YまたはQである;
(bg)96位のアミノ酸残基が、E、AまたはQである;
(bh)97位のアミノ酸残基が、LまたはIである;
(bi)100位のアミノ酸残基が、K、R、NまたはEである;
(bj)101位のアミノ酸残基が、KまたはRである;
(bk)103位のアミノ酸残基が、AまたはVである;
(bl)104位のアミノ酸残基が、DまたはNである;
(bm)105位のアミノ酸残基が、LまたはMである;
(bn)106位のアミノ酸残基が、LまたはIである;
(bo)112位のアミノ酸残基が、TまたはIである;
(bp)113位のアミノ酸残基が、S、TまたはFである;
(bq)114位のアミノ酸残基が、AまたはVである;
(br)115位のアミノ酸残基が、S、RまたはAである;
(bs)119位のアミノ酸残基が、K、EまたはRである;
(bt)120位のアミノ酸残基が、KまたはRである;
(bu)123位のアミノ酸残基が、FまたはLである;
(bv)124位のアミノ酸残基が、SまたはRである;
(bw)125位のアミノ酸残基が、E、K、GまたはDである;
(bx)126位のアミノ酸残基が、QまたはHである;
(by)128位のアミノ酸残基が、E、GまたはKである;
(bz)129位のアミノ酸残基が、V、IまたはAである;
(ca)130位のアミノ酸残基が、Y、H、FまたはCである;
(cb)131位のアミノ酸残基が、D、G、NまたはEである;
(cc)132位のアミノ酸残基が、I、T、A、M、VまたはLである;
(cd)135位のアミノ酸残基が、V、T、AまたはIである;
(ce)138位のアミノ酸残基が、HまたはYである;
(cf)139位のアミノ酸残基が、IまたはVである;
(cg)140位のアミノ酸残基が、LまたはSである;
(ch)142位のアミノ酸残基が、YまたはHである;
(ci)143位のアミノ酸残基が、K、TまたはEである;
(cj)144位のアミノ酸残基が、K、EまたはRである;
(ck)145位のアミノ酸残基が、LまたはIである;および
(cl)146位のアミノ酸残基が、TまたはAである。
(103) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)9位、76位、94位および110位のアミノ酸残基が、Aである;
(b)29位および108位のアミノ酸残基が、Cである;
(c)34位のアミノ酸残基が、Dである;
(d)95位のアミノ酸残基が、Eである;
(e)56位のアミノ酸残基が、Fである;
(f)43位、44位、66位、74位、87位、102位、116位、122位、127位および136位のアミノ酸残基が、Gである;
(g)41位のアミノ酸残基が、Hである;
(h)7位のアミノ酸残基が、Iである;
(i)85位のアミノ酸残基が、Kである;
(j)20位、36位、42位、50位、72位、78位、98位および121位のアミノ酸残基が、Lである;
(k)1位、75位および141位のアミノ酸残基が、Mである;
(l)23位、64位および109位のアミノ酸残基が、Nである;
(m)22位、25位、133位、134位および137位のアミノ酸残基が、Pである;
(n)71位のアミノ酸残基が、Qである;
(o)16位、21位、73位、99位および111位のアミノ酸残基が、Rである;
(p)55位および88位のアミノ酸残基が、Sである;
(q)77位のアミノ酸残基が、Tである;
(r)107位のアミノ酸残基が、Wである;および
(s)13位、46位、70位、117位および118位のアミノ酸残基が、Yである。
(104) 項目102に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位および/または141位のアミノ酸残基が、B1である;および
(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(105) 項目103に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、31位、45位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、123位、129位、139位および/または145位のアミノ酸残基が、B1である;
(b)3位、5位、8位、10位、11位、14位、17位、18位、24位、27位、32位、37位、38位、47位、48位、49位、52位、57位、58位、61位、62位、63位、68位、69位、79位、80位、82位、83位、89位、92位、100位、101位、104位、119位、120位、124位、125位、126位、128位、131位、143位および/または144位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(106) 項目102に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)1位、7位、9位、20位、36位、42位、50位、64位、72位、75位、76位、78位、94位、98位、110位、121位および/または141位のアミノ酸残基が、Z1である;
(b)13位、46位、56位、70位、107位、117位および/または118位のアミノ酸残基が、Z2である;
(c)23位、55位、71位、77位、88位および/または109位のアミノ酸残基が、Z3である;
(d)16位、21位、41位、73位、85位、99位および/または111位のアミノ酸残基が、Z4である;
(e)34位および/または95位のアミノ酸残基が、Z5である;
(f)22位、25位、29位、43位、44位、66位、74位、87位、102位、108位、116位、122位、127位、133位、134位、136位および/または137位のアミノ酸残基が、Z6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(107) 項目103に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、51位,54位、86位、90位、91位,97位,103位,105位,106位,114位、129位、139位および/または145位のアミノ酸残基が、Z1である;
(b)31位および/または45位のアミノ酸残基が、Z2である;
(c)8位および/または89位のアミノ酸残基が、Z3である;
(d)82位、92位、101位および/または120位のアミノ酸残基が、Z4である;
(e)3位、11位、27位および/または79位のアミノ酸残基が、Z5である;
(f)123位のアミノ酸残基が、Z1またはZ2である;
(g)12位、33位、35位、39位、53位、59位、112位、132位、135位、140位および/または146位のアミノ酸残基が、Z1またはZ3である;
(h)30位のアミノ酸残基が、Z1またはZ4である;
(i)6位のアミノ酸残基が、Z1またはZ6である;
(j)81位および/または113位のアミノ酸残基が、Z2またはZ3である;
(k)138位および/または142位のアミノ酸残基が、Z2またはZ4である;
(l)5位、17位、24位、57位、61位、124位および/または126位のアミノ酸残基が、Z3またはZ4である;
(m)104位のアミノ酸残基が、Z3またはZ5である;
(o)38位、52位、62位および/または69位のアミノ酸残基が、Z3またはZ6である;
(p)14位、119位および/または144位のアミノ酸残基が、Z4またはZ5である;
(q)18位のアミノ酸残基が、Z4またはZ6である;
(r)10位、32位、48位、63位、80位および/または83位のアミノ酸残基が、Z5またはZ6である;
(s)40位のアミノ酸残基が、Z1、Z2またはZ3である;
(t)65位および/または96位のアミノ酸残基が、Z1、Z3またはZ5である;
(u)84位および/または115位のアミノ酸残基が、Z1、Z3またはZ4である;
(v)93位のアミノ酸残基がZ2、Z3またはZ4である;
(w)130位のアミノ酸残基がZ2、Z4またはZ6である;
(x)47位および/または58位のアミノ酸残基が、Z3、Z4またはZ6である;
(y)49位、68位、100位および/または143位のアミノ酸残基が、Z3、Z4またはZ5である;
(z)131位のアミノ酸残基が、Z3、Z5またはZ6である;
(aa)125位および/または128位のアミノ酸残基が、Z4、Z5またはZ6である;
(ab)67位のアミノ酸残基が、Z1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基が、Z1、Z4、Z5またはZ6である;および
(ad)37位のアミノ酸残基が、Z3、Z4、Z5またはZ6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(108) 項目102に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)9位、76位、94位および110位のアミノ酸残基が、Aである;
(b)29位および108位のアミノ酸残基が、Cである;
(c)34位のアミノ酸残基が、Dである;
(d)95位のアミノ酸残基が、Eである;
(e)56位のアミノ酸残基が、Fである;
(f)43位、44位、66位、74位、87位、102位、116位、122位、127位および136位のアミノ酸残基が、Gである;
(g)41位のアミノ酸残基が、Hである;
(h)7位のアミノ酸残基が、Iである;
(i)85位のアミノ酸残基が、Kである;
(j)20位、36位、42位、50位、72位、78位、98位および121位のアミノ酸残基が、Lである;
(k)1位、75位および141位のアミノ酸残基が、Mである;
(l)23位、64位および109位のアミノ酸残基が、Nである;
(m)22位、25位、133位、134位および137位のアミノ酸残基が、Pである;
(n)71位のアミノ酸残基が、Qである;
(o)16位、21位、73位、99位および111位のアミノ酸残基が、Rである;
(p)55位および88位のアミノ酸残基が、Sである;
(q)77位のアミノ酸残基が、Tである;
(r)107位のアミノ酸残基が、Wである;および
(s)13位、46位、70位、117位および118位のアミノ酸残基が、Yである。
(109) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、ここで、前記アミノ酸配列中の28位に対応するアミノ酸残基が、Vである、ポリヌクレオチド。
(110) 項目1に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、ここで、前記アミノ酸配列が、配列番号6〜10および263〜514からなる群から選択される、、ポリヌクレオチド。
(111) 項目42に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも90%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、31位、45位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、123位、129位、139位および/または145位のアミノ酸残基が、B1である;および
(b)3位、5位、8位、10位、11位、14位、17位、18位、24位、27位、32位、37位、38位、47位、48位、49位、52位、57位、58位、61位、62位、63位、68位、69位、79位、80位、82位、83位、89位、92位、100位、101位、104位、119位、120位、124位、125位、126位、128位、131位、143位および/または144位のアミノ酸残基が、B2である;
ここで、B1は、A、I、L、M、F、W、YおよびVからなる群から選択される、アミノ酸であり;そして、B2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択される、アミノ酸である。
(112) 項目42に記載の単離されたポリヌクレオチドまたは組換えポリヌクレオチドであって、前記アミノ酸配列中の以下の位置に対応するアミノ酸残基のうち、少なくとも80%が、以下の限定に従う、ポリヌクレオチド:
(a)2位、4位、15位、19位、26位、28位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、129位、139位および/または145位のアミノ酸残基が、Z1である;
(b)31位および/または45位のアミノ酸残基が、Z2である;
(c)8位および/または89位のアミノ酸残基が、Z3である;
(d)82位、92位、101位および120位のアミノ酸残基が、Z4である;
(e)3位、11位、27位および/または79位のアミノ酸残基が、Z5である;
(f)123位のアミノ酸残基が、Z1またはZ2である;
(g)12位、33位、35位、39位、53位、59位、112位、132位、135位、140位および/または146位のアミノ酸残基が、Z1またはZ3である;
(h)30位のアミノ酸残基が、Z1またはZ4である;
(i)6位のアミノ酸残基が、Z1またはZ6である;
(j)81位および/または113位のアミノ酸残基が、Z2またはZ3である;
(k)138位および/または142位のアミノ酸残基が、Z2またはZ4である;
(l)5位、17位、24位、57位、61位、124位および/または126位のアミノ酸残基が、Z3またはZ4である;
(m)104位のアミノ酸残基が、Z3またはZ5である;
(o)38位、52位、62位および/または69位のアミノ酸残基が、Z3またはZ6である;
(p)14位、119位および/または144位のアミノ酸残基が、Z4またはZ5である;
(q)18位のアミノ酸残基が、Z4またはZ6である;
(r)10位、32位、48位、63位、80位および/または83位のアミノ酸残基が、Z5またはZ6である;
(s)40位のアミノ酸残基が、Z1、Z2またはZ3である;
(t)65位および/または96位のアミノ酸残基が、Z1、Z3またはZ5である;
(u)84位および/または115位のアミノ酸残基が、Z1、Z3またはZ4である;
(v)93位のアミノ酸残基が、Z2、Z3またはZ4である;
(w)130位のアミノ酸残基が、Z2、Z4またはZ6である;
(x)47位および/または58位のアミノ酸残基が、Z3、Z4またはZ6である;
(y)49位、68位、100位および/または143位のアミノ酸残基が、Z3、Z4またはZ5である;
(z)131位のアミノ酸残基が、Z3、Z5またはZ6である;
(aa)125位および/または128位のアミノ酸残基が、Z4、Z5またはZ6である;
(ab)67位のアミノ酸残基が、Z1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基が、Z1、Z4、Z5またはZ6である;および
(ad)37位のアミノ酸残基が、Z3、Z4、Z5またはZ6である;
ここで、Z1は、A、I、L、M、およびVからなる群から選択される、アミノ酸であり;Z2は、F、W、およびYからなる群から選択される、アミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択される、アミノ酸であり;Z4は、R、H、およびKからなる群から選択される、アミノ酸であり;Z5は、DおよびEからなる群から選択される、アミノ酸であり;そして、Z6は、C、G、およびPからなる群から選択される、アミノ酸である。
(113) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;および
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(114) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、20、36、42、50、64、72、75、76、78、94、98、110、121および/または141位のアミノ酸残基がZ1である;
(b)13、46、56、70、107、117および/または118位のアミノ酸残基がZ2である;
(c)23、55、71、77、88および/または109位のアミノ酸残基がZ3である;
(d)16、21、41、73、85、99および/または111位のアミノ酸残基がZ4である;
(e)34および/または95位のアミノ酸残基がZ5である;
(f)22、25、29、43、44、66、74、87、102、108、116、122、127、133、134、136および/または137位のアミノ酸残基がZ6である;
ここでZ1はA、I、L、MおよびVから成る群から選択されるアミノ酸である;Z2はF、WおよびYから成る群から選択されるアミノ酸である;Z3はN、Q、SおよびTから成る群から選択されるアミノ酸である;Z4はR、HおよびKから成る群から選択されるアミノ酸である;Z5はDおよびEから成る群から選択されるアミノ酸である;そしてZ6はC、GおよびPから成る群から選択されるアミノ酸である、
ポリペプチド。
(115) 項目111に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(116) 項目111に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(117) 項目111に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136およびまたは137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(118) 項目112に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(119) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)2位のアミノ酸残基がIまたはLである;
(b)3位のアミノ酸残基がEまたはDである;
(c)4位のアミノ酸残基がV、AまたはIである;
(d)5位のアミノ酸残基がK、RまたはNである;
(e)6位のアミノ酸残基がPまたはLである;
(f)8位のアミノ酸残基がN、SまたはTである;
(g)10位のアミノ酸残基がEまたはGである;
(h)11位のアミノ酸残基がDまたはEである;
(i)12位のアミノ酸残基がTまたはAである;
(j)14位のアミノ酸残基がEまたはKである;
(k)15位のアミノ酸残基がIまたはLである;
(l)17位のアミノ酸残基がHまたはQである;
(m)18位のアミノ酸残基がR、CまたはKである;
(n)19位のアミノ酸残基がIまたはVである;
(o)24位のアミノ酸残基がQまたはRである;
(p)26位のアミノ酸残基がLまたはIである;
(q)27位のアミノ酸残基がEまたはDである;
(r)28位のアミノ酸残基がAまたはVである;
(s)30位のアミノ酸残基がK、MまたはRである;
(t)31位のアミノ酸残基がYまたはFである;
(u)32位のアミノ酸残基がEまたはGである;
(v)33位のアミノ酸残基がT、AまたはSである;
(w)35位のアミノ酸残基がL、SまたはMである;
(x)37位のアミノ酸残基がR、G、EまたはQである;
(y)38位のアミノ酸残基がGまたはSである;
(z)39位のアミノ酸残基がT、AまたはSである;
(aa)40位のアミノ酸残基がF、LまたはSである;
(ab)45位のアミノ酸残基がYまたはFである;
(ac)47位のアミノ酸残基がR、QまたはGである;
(ad)48位のアミノ酸残基がGまたはDである;
(ae)49位のアミノ酸残基がK、R、EまたはQである;
(af)51位のアミノ酸残基がIまたはVである;
(ag)52位のアミノ酸残基がS、CまたはGである;
(ah)53位のアミノ酸残基がIまたはTである;
(ai)54位のアミノ酸残基がAまたはVである;
(aj)57位のアミノ酸残基がHまたはNである;
(ak)58位のアミノ酸残基がQ、K、NまたはPである;
(al)59位のアミノ酸残基がAまたはSである;
(am)60位のアミノ酸残基がE、K、G、VまたはDである;
(an)61位のアミノ酸残基がHまたはQである;
(ao)62位のアミノ酸残基がP、SまたはTである;
(ap)63位のアミノ酸残基がE、GまたはDである;
(aq)65位のアミノ酸残基がE、D、VまたはQである;
(ar)67位のアミノ酸残基がQ、E、R、L、HまたはKである;
(as)68位のアミノ酸残基がK、R、EまたはNである;
(at)69位のアミノ酸残基がQまたはPである;
(au)79位のアミノ酸残基がEまたはDである;
(av)80位のアミノ酸残基がGまたはEである;
(aw)81位のアミノ酸残基がY、NまたはFである;
(ax)82位のアミノ酸残基がRまたはHである;
(ay)83位のアミノ酸残基がE、GまたはDである;
(az)84位のアミノ酸残基がQ、RまたはLである;
(ba)86位のアミノ酸残基がAまたはVある;
(bb)89位のアミノ酸残基がTまたはSである;
(bc)90位のアミノ酸残基がLまたはIである;
(bd)91位のアミノ酸残基がIまたはVである;
(be)92位のアミノ酸残基がRまたはKである;
(bf)93位のアミノ酸残基がH、YまたはQである;
(bg)96位のアミノ酸残基がE、AまたはQである;
(bh)97位のアミノ酸残基がLまたはIである;
(bi)100位のアミノ酸残基がK、R、NまたはEである;
(bj)101位のアミノ酸残基がKまたはRである;
(bk)103位のアミノ酸残基がAまたはVである;
(bl)104位のアミノ酸残基がDまたはNである;
(bm)105位のアミノ酸残基がLまたはMである;
(bn)106位のアミノ酸残基がLまたはIである;
(bo)112位のアミノ酸残基がTまたはIである;
(bp)113位のアミノ酸残基がS、TまたはFである;
(bq)114位のアミノ酸残基がAまたはVである;
(br)115位のアミノ酸残基がS、RまたはAである;
(bs)119位のアミノ酸残基がK、EまたはRである;
(bt)120位のアミノ酸残基がKまたはRである;
(bu)123位のアミノ酸残基がFまたはLである;
(bv)124位のアミノ酸残基がSまたはRである;
(bw)125位のアミノ酸残基がE、K、GまたはDである;
(bx)126位のアミノ酸残基がQまたはHである;
(by)128位のアミノ酸残基がE、GまたはKである;
(bz)129位のアミノ酸残基がV、IまたはAである;
(ca)130位のアミノ酸残基がY、H、FまたはCである;
(cb)131位のアミノ酸残基がD、G、NまたはEである;
(cc)132位のアミノ酸残基がI、T、A、M、VまたはLである;
(cd)135位のアミノ酸残基がV、T、AまたはIである;
(ce)138位のアミノ酸残基がHまたはYである;
(cf)139位のアミノ酸残基がIまたはVである;
(cg)140位のアミノ酸残基がLまたはSである;
(ch)142位のアミノ酸残基がYまたはHである;
(ci)143位のアミノ酸残基がK、TまたはEである;
(cj)144位のアミノ酸残基がK、EまたはRである;
(ck)145位のアミノ酸残基がLまたはIである;および
(cl)146位のアミノ酸残基がTまたはAである、
ポリペプチド。
(120) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)9、76、94および110位のアミノ酸残基がAである;
(b)29および108位のアミノ酸残基がCである;
(c)34位のアミノ酸残基がDである;
(d)95位のアミノ酸残基がEである;
(e)56位のアミノ酸残基がFである;
(f)43、44、66、74、87、102、116、122、127および136位のアミノ酸残基がGである;
(g)41位のアミノ酸残基がHである;
(h)7位のアミノ酸残基がIである;
(i)85位のアミノ酸残基がKである;
(j)20、36、42、50、72、78、98および121位のアミノ酸残基がLである;
(k)1、75および141位のアミノ酸残基がMである;
(l)23、64および109位のアミノ酸残基がNである;
(m)22、25、133、134および137位のアミノ酸残基がPである;
(n)71位のアミノ酸残基がQである;
(o)16、21、73、99および111位のアミノ酸残基がRである;
(p)55および88位のアミノ酸残基がSである;
(q)77位のアミノ酸残基がTである;
(r)107位のアミノ酸残基がWである;および
(s)13、46、70、117および118位のアミノ酸残基がYである、
ポリペプチド。
(121) 項目119に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、13、20、36、42、46、50、56、64、70、72、75、76、78、94、98、107、110、117、118、121および/または141位のアミノ酸残基がB1である;
(b)16、21、22、23、25、29、34、41、43、44、55、66、71、73、74、77、85、87、88、95、99、102、108、109、111、116、122、127、133、134、136および/または137位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(122) 項目120に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)2、4、15、19、26、28、31、45、51、54、86、90、91、97、103、105、106、114、123、129、139および/または145位のアミノ酸残基がB1である;
(b)3、5、8、10、11、14、17、18、24、27、32、37、38、47、48、49、52、57、58、61、62、63、68、69、79、80、82、83、89、92、100、101、104、119、120、124、125、126、128、131、143および/または144位のアミノ酸残基がB2である;
ここでB1はA、I、L、M、F、W、Y、およびVから成る群から選択されるアミノ酸である;そしてB2はR、N、D、C、Q、E、G、H、K、P、SおよびTから成る群から選択されるアミノ酸である、
ポリペプチド。
(123) 項目119に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも90%が、以下の制限に合致し:
(a)1、7、9、20、36、42、50、64、72、75、76、78、94、98、110、121および/または141位のアミノ酸残基がZ1である;
(b)13、46、56、70、107、117および/または118位のアミノ酸残基がZ2である;
(c)23、55、71、77、88および/または109位のアミノ酸残基がZ3である;
(d)16、21、41、73、85、99および/または111位のアミノ酸残基がZ4である;
(e)34および/または95位のアミノ酸残基がZ5である;
(f)22、25、29、43、44、66、74、87、102、108、116、122、127、133、134、136および/または137位のアミノ酸残基がZ6である;
ここでZ1はA、I、L、MおよびVから成る群から選択されるアミノ酸である;Z2はF、WおよびYから成る群から選択されるアミノ酸である;Z3はN、Q、SおよびTから成る群から選択されるアミノ酸である;Z4はR、HおよびKから成る群から選択されるアミノ酸である;Z5はDおよびEから成る群から選択されるアミノ酸である;そしてZ6はC、GおよびPから成る群から選択されるアミノ酸である、
ポリペプチド。
(124) 項目120に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)2、4、15、19、26、28、51、54、86、90、91、97、103、105、106、114、129、139および/または145位のアミノ酸残基がZ1である;
(b)31および/または45位のアミノ酸残基がZ2である;
(c)8および/または89位のアミノ酸残基がZ3である;
(d)82、92、101および/または120位のアミノ酸残基がZ4である;
(e)3、11、27および/または79位のアミノ酸残基がZ5である;
(f)123位のアミノ酸残基がZ1またはZ2である;
(g)12、33、35、39、53、59、112、132、135、140および/または146位のアミノ酸残基がZ1またはZ3である;
(h)30位のアミノ酸残基がZ1またはZ4である;
(i)6位のアミノ酸残基がZ1またはZ6である;
(j)81および/または113位のアミノ酸残基がZ2またはZ3である;
(k)138および/または142位のアミノ酸残基がZ2またはZ4である;
(l)5、17、24、57、61、124および/または126位のアミノ酸残基がZ3またはZ4である;
(m)104位のアミノ酸残基がZ3またはZ5である;
(o)38、52、62および/または69位のアミノ酸残基がZ3またはZ6である;
(p)14、119および/または144位のアミノ酸残基がZ4またはZ5である;
(q)18位のアミノ酸残基がZ4またはZ6である;
(r)10、32、48、63、80および/または83位のアミノ酸残基がZ5またはZ6である;
(s)40位のアミノ酸残基がZ1、Z2またはZ3である;
(t)65および/または96位のアミノ酸残基がZ1、Z3またはZ5である;
(u)84および/または115位のアミノ酸残基がZ1、Z3またはZ4である;
(v)93位のアミノ酸残基がZ2、Z3またはZ4である;
(w)130位のアミノ酸残基がZ2、Z4またはZ6である;
(x)47および/または58位のアミノ酸残基がZ3、Z4またはZ6である;
(y)49、68、100および/または143位のアミノ酸残基がZ3、Z4またはZ5である;
(z)131位のアミノ酸残基がZ3、Z5またはZ6である;
(aa)125および/または128位のアミノ酸残基がZ4、Z5またはZ6である;
(ab)67位のアミノ酸残基がZ1、Z3、Z4またはZ5である;
(ac)60位のアミノ酸残基がZ1、Z4、Z5またはZ6である;
(ad)37位のアミノ酸残基がZ3、Z4、Z5またはZ6である;
ここでZ1はA、I、L、MおよびVから成る群から選択されるアミノ酸である;Z2はF、WおよびYから成る群から選択されるアミノ酸である;Z3はN、Q、SおよびTから成る群から選択されるアミノ酸である;Z4はR、HおよびKから成る群から選択されるアミノ酸である;Z5はDおよびEから成る群から選択されるアミノ酸である;そしてZ6はC、GおよびPから成る群から選択されるアミノ酸である、
ポリペプチド。
(125) 項目119に記載の単離ポリペプチドまたは組換えポリペプチドであって、以下の位置に対応するアミノ酸配列中のアミノ酸残基の、少なくとも80%が、以下の制限に合致し:
(a)9、76、94および110位のアミノ酸残基がAである;
(b)29および108位のアミノ酸残基がCである;
(c)34位のアミノ酸残基がDである;
(d)95位のアミノ酸残基がEである;
(e)56位のアミノ酸残基がFである;
(f)43、44、66、74、87、102、116、122、127および136位のアミノ酸残基がGである;
(g)41位のアミノ酸残基がHである;
(h)7位のアミノ酸残基がIである;
(i)85位のアミノ酸残基がKである;
(j)20、36、42、50、72、78、98および121位のアミノ酸残基がLである;
(k)1、75および141位のアミノ酸残基がMである;
(l)23、64および109位のアミノ酸残基がNである;
(m)22、25、133、134および137位のアミノ酸残基がPである;
(n)71位のアミノ酸残基がQである;
(o)16、21、73、99および111位のアミノ酸残基がRである;
(p)55および88位のアミノ酸残基がSである;
(q)77位のアミノ酸残基がTである;
(r)107位のアミノ酸残基がWである;および
(s)13、46、70、117および118位のアミノ酸残基がYである、
ポリペプチド。
(126) 項目24に記載の単離ポリペプチドまたは組換えポリペプチドであって、ここで28位に対応するアミノ酸配列中のアミノ酸残基が、Vであるポリペプチド。
(127) 項目42に記載の単離ポリペプチドまたは組換えポリペプチドであって、ここでアミノ酸配列が、配列番号6〜10および263〜514から成る群から選択される、ポリペプチド。
(128) 増強されたグリホセート耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで植物または植物外植片が、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドおよびさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドを発現する、トランスジェニック植物またはトランスジェニック植物外植片。
(129) 項目128に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514から成る群から選択されるアミノ酸配列を含む、トランスジェニック植物またはトランスジェニック植物外植片。
(130) 項目129に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−リン酸シンターゼおよびグリホセート耐性グリホセートキシドレダクターゼから成る群から選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(131) 項目130に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−リン酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(132) 項目130に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる機構によってグリホセート耐性を付与する少なくとも1つのポリペプチドが、グリホセート耐性グリホセートキシドレダクターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(133) トランスジェニック植物またはトランスジェニック植物外植片であり、ここで該植物または植物外植片が、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドおよびさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドを発現する、トランスジェニック植物またはトランスジェニック植物外植片。
(134) 項目133に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514から成る群から選択されるアミノ酸配列を含む、トランスジェニック植物またはトランスジェニック植物外植片。
(135) 項目134に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、変異されたヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチル転移酵素および変異されたプロトポルフィリノゲンオキシダーゼから成る群から選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(136) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、変異されたヒドロキシフェニルピルベートジオキシゲナーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(137) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(138) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(139) 項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここでさらなる除草剤に耐性を付与する少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(140) 前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(141) 前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目135に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(142) 前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、請求項135に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(143) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチド、さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチド、およびさらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドを発現する、トランスジェニック植物またはトランスジェニック植物外植片。
(144) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目143に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(145) 項目144に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセートオキシドレダクターゼからなる群より選択され、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(146) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(147) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(148) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(149) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(150) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(151) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(152) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(153) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(154) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(155) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(156) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、トランスジェニック植物またはトランスジェニック植物外植片。
(157) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼある、トランスジェニック植物またはトランスジェニック植物外植片。
(158) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼある、トランスジェニック植物またはトランスジェニック植物外植片。
(159) 項目145に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼある、トランスジェニック植物またはトランスジェニック植物外植片。
(160) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、そして少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(161) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目160に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(162) 前記少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、請求項161に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(163) 前記少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目161に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(164) トランスジェニック植物またはトランスジェニック植物外植片であって、ここで該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、そして、少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(165) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目164に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(166) 前記少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(167) 前記少なくとも1つのポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(168) 前記少なくとも1つのポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(169) 前記少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(170) 前記少なくとも1つのポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(171) 前記少なくとも1つのポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(172) 前記少なくとも1つのポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目165に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(173) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、少なくとも1つの第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択され、そして、少なくとも1つの第2ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼおよび変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(174) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目173に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(175) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(176) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(177) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、請求項174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(178) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(179) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、請求項174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(180) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、請求項174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(181) 前記第1ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼであり、そして前記第2ポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(182) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(183) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(184) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドがスルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(185) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドがイミダゾリノン耐性アセトラクテートシンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(186) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドがイミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(187) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(188) 前記第1ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素であり、そして前記第2ポリペプチドが変異型プロトポルフィリノーゲンオキシダーゼである、項目174に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(189) グリホセートに対する強化された耐性を有するトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、該植物または植物外植片は、グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドを発現し、そして少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼ、グリホセート耐性グリホセート酸化還元酵素、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼおよび変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(190) 前記グリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドが、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列を含む、項目189に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(191) 前記ポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(192) 前記ポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(193) 前記ポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(194) 前記ポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(195) 前記ポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(196) 前記ポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(197) 前記ポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(198) 前記ポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(199) 前記ポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目190に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(200) 作物を含む畑において雑草を制御するための方法であって、
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程;および、
(b)該作物への有意な影響なしで該畑における該雑草の成長を阻害するのに十分である、グリホセートの効果的な適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(201) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目200に記載の方法。
(202) 項目201に記載の方法であって、ここで、前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、方法。
(203) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目202に記載の方法。
(204) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目202に記載の方法。
(205) 作物を含む畑においてグリホセート耐性雑草の発生を防止するための方法であって、
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程;および、
(b)グリホセートの効果的な適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(206) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目205に記載の方法。
(207) 項目206に記載の方法であって、ここで、前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、方法。
(208) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目207に記載の方法。
(209) 前記さらなる機構によりグリホセート耐性を与えるポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目207に記載の方法。
(210) 作物を含む畑において選択的に雑草を制御するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)該作物への有意な影響なしで該畑における該雑草の成長を阻害するのに十分である、グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を該畑における作物および雑草に適用する工程、
を包含する、工程。
(211) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目210に記載の方法。
(212) 項目211に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(213) 項目211に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(214) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目212に記載の方法。
(215) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目212に記載の方法。
(216) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目212に記載の方法。
(217) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目212に記載の方法。
(218) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目212に記載の方法。
(219) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目212に記載の方法。
(220) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目212に記載の方法。
(221) 作物を含む畑における除草剤耐性雑草の発生を防止するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(222) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目221に記載の方法。
(223) 項目222に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(224) 項目221に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(225) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目223に記載の方法。
(226) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目223に記載の方法。
(227) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目223に記載の方法。
(228) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目223に記載の方法。
(229) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目223に記載の方法。
(230) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目223に記載の方法。
(231) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目223に記載の方法。
(232) 作物を含む畑において雑草を選択的に制御するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、さらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)該作物への有意な影響なしで該畑における該雑草の成長を阻害するのに十分である、グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を該畑における作物および雑草に適用する工程、
を包含する、方法。
(233) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目232に記載の方法。
(234) 項目233に記載のトランスジェニック植物またはトランスジェニック植物外植片であって、ここで、前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼおよびグリホセート耐性グリホセート酸化還元酵素からなる群より選択される、トランスジェニック植物またはトランスジェニック植物外植片。
(235) 前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性5−エノールピルビルシキメート−3−ホスフェートシンターゼである、項目234に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(236) 前記さらなる機構によりグリホセート耐性を与える少なくとも1つのポリペプチドが、グリホセート耐性グリホセート酸化還元酵素である、項目234に記載のトランスジェニック植物またはトランスジェニック植物外植片。
(237) 項目233に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(238) 項目233に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(239) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目237に記載の方法。
(240) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目237に記載の方法。
(241) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目237に記載の方法。
(242) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目237に記載の方法。
(243) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目237に記載の方法。
(244) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目237に記載の方法。
(245) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目237に記載の方法。
(246) 作物を含む畑における除草剤耐性雑草の発生を制御するための方法であって、以下:
(a)グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、さらなる機構によりグリホセート耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする少なくとも1つの遺伝子を用いて形質転換されている、作物の種または植物を該畑に植える工程、および;
(b)グリホセートおよびさらなる該除草剤の、同時適用または経時的時差適用を、該畑における作物および雑草に適用する工程、
を包含する、方法。
(247) 前記グリホセートN−アセチルトランスフェラーゼをコードする遺伝子が、配列番号1〜5および11〜262からなる群より選択されるポリヌクレオチド配列を含む、項目246に記載の方法。
(248) 項目247に記載の方法であって、ここで、前記さらなる除草剤に対する耐性を与える少なくとも1つのポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼ、スルホンアミド耐性アセトラクテートシンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセトラクテートシンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、フォスフィノスリシンアセチルトランスフェラーゼ、および変異型プロトポルフィリノーゲンオキシダーゼからなる群より選択される、方法。
(249) 項目247に記載の方法であって、ここで、前記さらなる除草剤が、ヒドロキシフェニルピルベートジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフィノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルフォシネート、およびプロトックスインヒビターからなる群より選択される、方法。
(250) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型ヒドロキシフェニルピルベートジオキシゲナーゼである、項目248に記載の方法。
(251) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトラクテートシンターゼである、項目248に記載の方法。
(252) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、スルホンアミド耐性アセトヒドロキシ酸シンターゼである、項目248に記載の方法。
(253) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトラクテートシンターゼである、項目248に記載の方法。
(254) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、イミダゾリノン耐性アセトヒドロキシ酸シンターゼである、項目248に記載の方法。
(255) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、フォスフィノスリシンアセチルトランスフェラーゼである、項目248に記載の方法。
(256) 前記さらなる除草剤に対する耐性を与えるポリペプチドが、変異型プロトポルフィリノーゲンオキシダーゼである、項目248に記載の方法。
(本発明の要旨)
生物体(例えば、植物)をグリホセートに対し抵抗性にするための方法および試薬を提供することが、本発明の目的である。本発明のこの目的およびその他の目的は、以下に記載される一つ以上の実施形態により提供される。
本発明の一つの実施形態は、本明細書中でGATポリペプチドといわれる新規のポリペプチドを提供する。GATポリペプチドは、その互いの構造の類似性(例えば、GATポリペプチドが、互いに整列させられた際の配列類似性に関して)によって特性付けられる。いくつかのGATポリペプチドは、グリホセートN−アセチルトランスフェラーゼ活性、すなわちグリホセートのアセチル化を触媒する能力を有する。いくつかのGATポリペプチドはまた、グリホセート類似物およびまたはグリホセート代謝産物(例えば、アミノメチルホスホン酸)のアセチル化を触媒し得る。
本明細書中においてGATポリヌクレオチドといわれる新規のポリヌクレオチドもまた、提供される。GATポリヌクレオチドは、それらがGATポリペプチドをコードする能力によって特性付けられる。本発明のいくつかの実施形態において、GATポリヌクレオチドは、よりよい植物の発現のため一つ以上の親コドンを、植物において親コドンと比較して優先的に使用される同義のコドンに置換することにより操作される。他の実施形態において、GATポリヌクレオチドは、N末端葉緑体輸送ペプチドをコードするヌクレオチド配列を導入することによって修正される。
GATポリペプチド、GATポリヌクレオチドおよびグリホセートN−アセチルトランスフェラーゼ活性は、以下により詳細に記載される。本発明はさらに、本明細書中に記載されるGATポリペプチドおよびGATポリヌクレオチドのある程度のフラグメントを含む。
本発明は、本明細書中に記載されるポリペプチドおよびポリヌクレオチドの非ネイティブ型改変体を含み、ここでコードされるポリペプチドの一つ以上のアミノ酸が、変異される。
本発明はさらに、本発明のポリヌクレオチドを含有する核酸構築物を提供する。本構成物は、植物形質転換ベクターのようなベクターであり得る。ある局面において、本発明に属するベクターは、T−DNA配列を含有する。本構成物は、GATポリヌクレオチドと作動可能に連結される調節配列(例えば、プロモーター)を必要に応じて含み得、ここで、本プロモーターは、ポリヌクレオチドに関して異種であり、そして核酸構築物によって形質転換された植物細胞のグリホセート耐性を強化するためにコードされたポリペプチドの、十分な発現を引き起こすために有効である。
本発明のいくつかの局面において、GATポリヌクレオチドは、選択マーカー(例えば、植物、細菌、放線菌類、酵母、藻類または他の真菌類における)として機能する。例えば、GATポリヌクレオチド選択マーカーを含むベクターによって形質転換された生物体は、グリホセート存在下でのその成長能力に基づいて選択され得る。GATマーカー遺伝子は、この遺伝子を発現する形質転換された細胞の選択またはスクリーニングのために使用され得る。
本発明はさらに、形質が重ねられたベクター、すなわちGATをコードし、かつまた第2のポリヌクレオチド配列を含むベクターを提供する。この第2のポリヌクレオチド配列は、有効なレベルの第2のポリペプチドを発現する細胞または生物体において、検出可能な表現形の形質を賦与する、第2のポリペプチド配列をコードする。この検出可能な表現形の形質は、選択マーカー(例えば、除草剤抵抗性の賦与、害虫抵抗性の賦与、またはある種の可視性マーカーを提供することにより)として機能し得る。
一つの実施形態において、本発明は、本発明の2つ以上のポリヌクレオチドを含有する組成物を供給する。
2つ以上のGATポリヌクレオチドまたはコードされたポリペプチドを含む組成物は、本発明の特徴である。いくつかの場合において、これらの組成物は、例えば、少なくとも3つ以上の上記の核酸を含む核酸のライブラリーである。デオキシリボヌクレオチド3リン酸および核酸ポリメラーゼ(例えば、熱安定性核酸ポリメラーゼ)の存在下で、本発明の核酸のインキュベートにより生成される組成物と同様に、制限エンドヌクレアーゼ、RNAseまたはDNAseによる本発明の核酸の消化、またはその核酸のその他の断片化(例えば、機械的せん断、化学的切断などによる)により生成される組成物もまた、本発明の特徴である。
本発明のベクター、またはその他の様式で本発明の核酸を組み入れるベクターにより形質導入された細胞は、本発明の一つの局面である。好ましい実施形態において、本細胞は、上記の核酸によってコードされるポリペプチドを発現する。
ある実施形態において、本発明の核酸を組み入れる細胞は、植物細胞である。本発明の核酸を組み込んでいるトランスジェニック植物、トランスジェニック植物細胞、およびトランスジェニック植物外植片もまた、本発明の特徴である。いくつかの実施形態において、トランスジェニック植物、トランスジェニック植物細胞、またはトランスジェニック植物外植片は、本発明の核酸にコードされるグリホセートN−アセチルトランスフェラーゼ活性を有する外因性のポリペプチドを発現する。本発明はまた、本発明のトランスジェニック植物により産生されたトランスジェニック種子を提供する。
本発明はさらに、グリホセートN−アセチルトランスフェラーゼ活性を持つポリペプチドおよび別の機構(例えば、グリホセート耐性5−エノールピルビルシキミ酸−3−リン酸シンターゼおよび/またはグリホセート耐性グリホセートキシドレダクターゼ)によってグリホセート耐性を与えるポリペプチドの発現のため、グリホセートに対する強化された耐性を持つトランスジェニック植物またはトランスジェニック植物外植片を提供する。さらなる実施形態において、本発明は、ポリペプチドの発現によって、さらなる除草剤に対し耐性と同様に、グリホセートに対する強化された耐性を持つトランスジェニック植物またはトランスジェニック植物外植片を提供する。さらなる実施形態において、本発明は、グリホセートに対する強化された耐性ならびにグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチド、別の機構(例えば、グリホセート耐性5−エノールピルビルシキミ酸−3−リン酸シンターゼおよび/またはグリホセート耐性グリホセートキシドレダクターゼ)によってグリホセート耐性を与えるポリペプチド、およびさらなる除草剤に対し耐性を与えるポリペプチド(例えば、変異されたヒドロキシフェニルピルビン酸ジオキシゲナーゼ、スルホンアミド耐性アセト乳酸シンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン(imidazolinone)耐性アセト乳酸シンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、ホスフィノスリシンアセチルトランスフェラーゼおよび変異されたプロトポルフィリノーゲンオキシダーゼ)の発現に起因するさらなる除草剤に対する耐性を持つトランスジェニック植物またはトランスジェニック植物外植片を提供する。
本発明はまた、グリホセートに対する強化された耐性、ならびにグリホセートN−アセチルトランスフェラーゼ活性を有するポリペプチドおよびさらなる除草剤に対し耐性を与えるポリペプチド(例えば、変異されたヒドロキシフェニルピルピン酸ジオキシゲナーゼ、スルホンアミド耐性アセト乳酸シンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセト乳酸シンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、ホスフィノスリシンアセチルトランスフェラーゼおよび変異されたプロトポルフィリノーゲンオキシダーゼ)の発現に起因するさらなる除草剤に対する耐性を持つトランスジェニック植物またはトランスジェニック植物外植片を提供する。
本発明のポリペプチドをコードする核酸を細胞中に導入し、引き続いて発現させ、そしてそれらを細胞または培地から回収することによる本発明のポリペプチドを生成する方法は、本発明の特徴である。好ましい実施形態において、本発明のポリペプチドを発現する細胞は、トランスジェニック植物細胞である。
配列番号:6〜10および263〜514に由来する抗原に対して反応するが、天然に存在する関連する配列には(例えば、GenBank登録番号CAA70664の部分配列によって表わされるペプチド)反応しないポリクローナル抗血清によって特異的に結合されるポリペプチド、ならびに配列番号:6〜10および263〜514の任意の一つ以上に由来する抗原の投与によって産生される抗体および/または前記の抗体に特異的に結合し、そしてGenBank登録番号CAA70664に対応する天然に存在するポリペプチドに対し特異的に結合しない抗体は、すべて本発明の特徴である。
本発明の別の局面は、インビトロまたはインビボにおける本発明の核酸を組換えることまたは変異させることにより新規のGATポリヌクレオチドおよびGATポリペプチドを生成するためのポリヌクレオチドの多様化の方法に関する。一つの実施形態において、組換えは、少なくとも一つの組換えGATポリヌクレオチドのライブラリーを生成する。上記のように生成されるこのライブラリーは、このライブラリーを構成する細胞と同様に、本発明の実施形態である。さらに、本発明の核酸の変異による修正されたGATポリヌクレオチドを産生する方法は、本発明の実施形態である。本発明の方法によって産生される組換えおよび変異GATポリヌクレオチドおよびポリペプチドもまた、本発明の実施形態である。
本発明のいくつかの局面において、多様化は、インビトロ、インビボ、インシリコ、またはこれらの組み合わせにおいて達成され得る、再帰的な組換えを使用して成し遂げられる。以下により詳細に記載される多様化法のいくつかの実施例は、ファミリーシャッフリング(family shuffling)法および合成シャッフリング(synthetic shuffling)法である。
本発明は、グリホセートN−アセチルトランスフェラーゼをコードするポリヌクレオチドにより植物または植物細胞を形質転換すること、および形質転換された植物細胞からトランスジェニック植物を必要に応じて再生することを包含するグリホセート抵抗性トランスジェニック植物または植物細胞を作製するための方法を提供する。ある局面において、このポリヌクレオチドは、GATポリヌクレオチドであり、必要に応じて細菌供給源に由来するGATポリヌクレオチドである。本発明のある局面において、本方法は、形質転換された植物の成長を阻害することなしに、同種の野生型植物の成長を阻害する濃度のグリホセートにより、形質転換された植物または植物細胞を増殖させることを包含し得る。本方法は、漸増する濃度のグリホセートにおいて、および/または同種の野生型植物または植物細胞に対して致死的であるグリホセート濃度中において、形質転換された植物または植物細胞あるいは植物または植物細胞の子孫を増殖させることを包含し得る。
本方法によって作製されるグリホセート抵抗性トランスジェニック植物は、例えば、その植物を第2の植物と交配することによって、繁殖させ得、その結果、少なくともいくつかのこのような交配による子孫はグリホセート耐性を示す。
本発明はさらに、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子による形質転換の結果、グリホセート耐性である農作物の種子または作物を農地に植付けること、および農作物に対する有意な影響なしに雑草を制御するために十分な量のグリホセートを農地において農作物および雑草に対して適用することを包含する、農作物を有する農地において、選択的に雑草を制御するための方法を提供する。
本発明はさらに、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子および別の機構(例えば、グリホセート耐性5−エノールピルビルシキミ酸−3−リン酸シンターゼおよび/またはグリホセート耐性グリホセートキシドレダクターゼ)によってグリホセート耐性を与えるポリペプチドをコードする遺伝子による形質転換の結果、グリホセート耐性である農作物の種子または作物を農地に植付けること、ならびに農作物に対する有意な影響なしに雑草を制御するために十分な量のグリホセートを農地において農作物および雑草に対して適用することを包含する、農地において雑草を制御し、そして農作物を含む農地においてグリホセート抵抗性の雑草の出現を予防する方法を提供する。
さらなる実施形態において、本発明は、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子、別の機構(例えば、グリホセート耐性5−エノールピルビルシキミ酸−3−リン酸シンターゼおよび/またはグリホセート耐性グリホセートキシドレダクターゼ)によってグリホセート耐性を与えるポリペプチドをコードする遺伝子、およびさらなる除草剤に対する耐性を与えるポリペプチド(例えば、変異されたヒドロキシフェニルピルビン酸ジオキシゲナーゼ、スルホンアミド耐性アセト乳酸シンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセト乳酸シンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、ホスフィノスリシンアセチルトランスフェラーゼおよび変異されたプロトポルフィリノーゲンオキシダーゼ)をコードする遺伝子による形質転換の結果、グリホセート耐性である農作物の種子または作物を用いた農地に植付けること、ならびに農作物に対する有意な影響なしに雑草を制御するための十分な量のグリホセートおよびさらなる除草剤(例えば、ヒドロキシフェニルピルビン酸ジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス(bialaphos)、フォスフォノスリシン、アザフェニジン(azafenidin)、ブタフェナシル(butafenacil)、スルフォセート(sulfosate)、グルホシネート、およびプロトポルフィリノーゲンオキシダーゼ(protox)インヒビター)を農地における農作物および雑草に適用することを包含する、農地において雑草を制御しそして農作物を含む農地においてグリホセート抵抗性の雑草の出現を予防する方法を提供する。
本発明はさらに、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子およびさらなる除草剤に対する耐性を与えるポリペプチドをコードする遺伝子(例えば、変異されたヒドロキシフェニルピルビン酸ジオキシゲナーゼ、スルホンアミド耐性アセト乳酸シンターゼ、スルホンアミド耐性アセトヒドロキシ酸シンターゼ、イミダゾリノン耐性アセト乳酸シンターゼ、イミダゾリノン耐性アセトヒドロキシ酸シンターゼ、ホスフィノスリシンアセチルトランスフェラーゼおよび変異されたプロトポルフィリノーゲンオキシダーゼ)による形質転換の結果、グリホセート耐性である農作物の種子または植物を農地に植付けること、ならびに農作物に対する有意な影響なしに雑草を制御するための十分な量のグリホセートおよびさらなる除草剤(例えば、ヒドロキシフェニルピルビン酸ジオキシゲナーゼインヒビター、スルホンアミド、イミダゾリノン、ビアラホス、フォスフォノスリシン、アザフェニジン、ブタフェナシル、スルフォセート、グルホシネート、および変異されたプロトポルフィリノーゲンオキシダーゼインヒビター)を適用することを包含する、農作物を有する農地において雑草を制御するための方法および除草剤抵抗性の雑草の出現を予防する方法を提供する。
本発明はさらに、グリホセートに対して耐性である、遺伝学的に形質転換された植物の作製のための方法を提供し、この方法は、組換え型二本鎖DNA分子を植物細胞ゲノムに挿入する工程であって、上記の組換え型二本鎖DNA分子は、(i)植物細胞においてRNA配列の生成を引き起こすため機能するプロモーター;(ii)GATをコードするRNA配列の生成を引き起こす構造DNA配列;および(iii)植物細胞においてRNA配列の3’末端にポリアデニルヌクレオチドのストレッチを付加するため、機能する3’非翻訳領域を含み;ここで、プロモーターは、構造DNA配列に関して異種であり、そしてDNA分子により形質転換される植物細胞のグリホセート耐性を強化するためにコードされたポリペプチドの十分な発現を引き起こすために適用される、工程;形質転換された植物細胞を獲得する工程;およびグリホセートに対する耐性が増加した遺伝学的に形質転換された植物を、形質転換された植物細胞から再生する工程を包含する。
本発明はさらに、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子による形質転換の結果、グリホセート耐性である作物を、作物が農作物を産出するような条件の元で生育させる工程;および作物から農作物を収穫する工程を包含する、農作物の産生のための方法を提供する。これらの方法は、しばしば作物に対する雑草の制御に有効な濃度のグリホセートの適用を含む。例示的な作物として、綿、トウモロコシ、およびダイズが挙げられる。
本発明はまた、配列番号1−514に対応する文字列を有する配列記録から構成されるデータベースを含むコンピュータ、コンピュータで読み取り可能な媒体、および集中型システムを提供する。上記の集中型システムは、互いにおよび/または任意のさらなる核酸またはアミノ酸配列と共に、配列番号1−514に対応する任意の一つ以上の文字列の選択、整列、翻訳、逆翻訳または閲覧のための一つ以上の指示セットを必要に応じて含む。
(詳細な議論)
本発明は、N−アセチルトランスフェラーゼ活性を示す新規のクラスの酵素に関する。一つの局面において、本発明は、グリホセートおよびグリホセートアナログをアセチル化できる新規のクラスの酵素(例えば、グリホセートN−トランスフェラーゼ(「GAT」)活性を有する酵素)に関する。上記の酵素は、化合物の2級アミンをアセチル化する能力によって特性付けされる。本発明のいくつかの局面において、化合物は図1に概略的に描かれるような除草剤(例えば、グリホセート)である。この化合物はまた、グリホセートアナログまたはグリホセート分解による代謝産物(例えば、アミノメチルホスホン酸)であり得る。グリホセートのアセチル化は、グリホセートの異化のための一つの代謝経路において重要な触媒ステップであるが、天然に存在する酵素、単離された酵素、または組換え型酵素によるグリホセートの酵素学的なアセチル化は、これまで記載されていない。従って、本発明の核酸およびポリペプチドは、除草剤抵抗性を移入するための新しい生化学的な経路を提供する。
本発明は、N−アセチルトランスフェラーゼ活性を示す新規のクラスの酵素に関する。一つの局面において、本発明は、グリホセートおよびグリホセートアナログをアセチル化できる新規のクラスの酵素(例えば、グリホセートN−トランスフェラーゼ(「GAT」)活性を有する酵素)に関する。上記の酵素は、化合物の2級アミンをアセチル化する能力によって特性付けされる。本発明のいくつかの局面において、化合物は図1に概略的に描かれるような除草剤(例えば、グリホセート)である。この化合物はまた、グリホセートアナログまたはグリホセート分解による代謝産物(例えば、アミノメチルホスホン酸)であり得る。グリホセートのアセチル化は、グリホセートの異化のための一つの代謝経路において重要な触媒ステップであるが、天然に存在する酵素、単離された酵素、または組換え型酵素によるグリホセートの酵素学的なアセチル化は、これまで記載されていない。従って、本発明の核酸およびポリペプチドは、除草剤抵抗性を移入するための新しい生化学的な経路を提供する。
一つの局面において、本発明は、GATポリペプチドをコードする新規の遺伝子を提供する。天然に存在するポリヌクレオチドに相当する単離されたGATポリヌクレオチドおよび組換え型GATポリヌクレオチド、ならびに組換え型および操作された(例えば、多様化された)GATポリペプチドば、本発明の特徴である。GATポリヌクレオチドは、配列番号:1〜5および11〜262によって例示される。特異的GATポリヌクレオチドおよびポリペプチド配列は、本発明の説明を助ける例示として提供され、そしてこれは、本明細書において記載されるおよび/または特許請求されるGATポリヌクレオチドおよびポリペプチドの種類の範囲を限定することを意図しない。
本発明はまた、改良されたおよび/または強化された特性(例えば、変更されたグリホセートに対するKm、増加された触媒作用速度、増加された安定性など、本明細書に記載される新しいまたは改良された活性のためのライブラリーのポリヌクレオチド構成要素の選択に基づく)を有するGATポリペプチドをコードするポリヌクレオチドを含むさらなるGATポリヌクレオチドを生成するための多様化されるライブラリーを生成するおよび選択するための方法を提供する。上記のポリヌクレオチドは、グリホセート抵抗性トランスジェニック植物の生成において特に都合よく使用される。
本発明のGATポリペプチドは、新規の酵素活性を示す。具体的には、合成除草剤のグリホセートの酵素学的アセチル化は、本発明の以前には認められていなかった。従って、本明細書に記載されるポリペプチド(例えば、配列番号:6〜10および263〜514によって例示される)は、インビボ(例えば、植物において)において機能するグリホセートの解毒のための新規の生化学的な経路を規定する。
従って、本発明の核酸およびポリペプチドは、トランスジェニック植物における除草剤選択性の設計のための、新しい核酸、ポリペプチド、および生化学的経路を提供することにより、グリホセート抵抗性植物の生成において意義深い重要性を有する。
(定義)
本発明を詳細に記載する前に、本発明は、特定の化合物または生物系に限定されず、これらは当然、変動し得るということが理解されるべきである。本明細書において使用される専門用語は、特定の実施形態を記載することのみを目的とし、そして限定することは意図しないこともまた理解されるべきである。本明細書および添付の特許請求の範囲において使用される場合に、単数形の「a」、「an」、および「the」は、その内容がそうではないと明白に示さない限り、複数の言及を含む。従って、例えば、「デバイス(a device)」との言及は、2つ以上のこのようなデバイスの組み合わせを含み、「遺伝子融合構築物(a gene fusion construct)」、との言及は、構築物の混合物を含む、などである。
本発明を詳細に記載する前に、本発明は、特定の化合物または生物系に限定されず、これらは当然、変動し得るということが理解されるべきである。本明細書において使用される専門用語は、特定の実施形態を記載することのみを目的とし、そして限定することは意図しないこともまた理解されるべきである。本明細書および添付の特許請求の範囲において使用される場合に、単数形の「a」、「an」、および「the」は、その内容がそうではないと明白に示さない限り、複数の言及を含む。従って、例えば、「デバイス(a device)」との言及は、2つ以上のこのようなデバイスの組み合わせを含み、「遺伝子融合構築物(a gene fusion construct)」、との言及は、構築物の混合物を含む、などである。
他で定義されない限り、本明細書中で使用される全ての技術用語および科学用語は、本発明が属する分野の当業者によって通常理解されているのと同じ意味を持つ。本明細書中において記載される類似または均等であるすべての方法および材料は、本発明の試験のための業務において使用され得、適当な材料および方法を使用する具体的な実施例が、本明細書中に記載される。
本発明の記載および特許請求において、下記の専門用語は、以下に提示される定義に基づいて使用される。
本発明の目的のために、用語「グリホセート」は、任意の除草のため有効な形態のN−ホスホノメチルグリシン(その任意の塩を含む)および植物(planta)におけるグルホサートアニオン生成を生じるその他の形態を含むとみなされるべきである。用語「グリホセートアナログ」は、グリホセートアナログが除草のため有効であるレベルにEPSPSを阻害するための能力を有する、グリホセート(glyphostate)の任意の構造上のアナログをいう。
本明細書中で使用される場合、用語「グリホセートN−アセチルトランスフェラーゼ活性」または「GAT活性」とは、例えば図1に図示されるように、グリホセートの2級アミン基のアセチル化を触媒する能力をいう。「グリホセートN−アセチルトランスフェラーゼ」または「GAT」は、グリホセート、グリホセートアナログ、および/またはグリホセートの主要な代謝産物(すなわち、AMPAまたはサルコシン)のアミン基のアセチル化を触媒する酵素である。本発明のいくつかの好ましい実施形態において、GATは、アセチル基をアセチルCoAからグリホセートの2級アミンおよびAMPAの1級アミンに転移し得る。本明細書中に記載される例示的なGATは、pH5〜9において活性であり、pH6.5〜8.0の範囲内で最適な活性を有する。活性は、当該分野で周知の種々の速度論的パラメーター(例えば、kcat、KM、およびkcat/KM)を用いて定量化され得る。これらの速度論的パラメーターは、実施例7において以下に記載されるように決定され得る。所定のポリヌクレオチドまたは相補的なポリヌクレオチドは、任意の特定されたヌクレオチド配列から決定され得る。
用語「ポリヌクレオチド」、「ヌクレオチド配列」および「核酸」は、ヌクレオチド(A、C、T、U、Gなど、または天然に存在するかもしくは人工的なヌクレオチドのアナログ)例えば関連する文脈に依存して、DNAまたはRNA、あるいはそれらの表現(例えば、文字列など)のポリマーをいうために、使用される。
同様に、「アミノ酸配列」とは、アミノ酸のポリマー(タンパク質、ポリペプチドなど)または、アミノ酸ポリマーを意味する文字列であり、文脈に依存する。用語「タンパク質」「ポリペプチド」および「ペプチド」は、本明細書中において交換可能に使用され得る。
ポリヌクレオチド、ポリペプチドまたは他の成分は、通常、それらが関連する成分(他のタンパク質,核酸、細胞、合成試薬など)から部分的または完全に分離される場合、「単離される」。核酸またはポリペプチドは、それが人工的もしくは操作されたタンパク質または核酸である場合、または人工的または操作されたタンパク質または核酸に由来する場合、「組換え」である。例えば、ベクターまたは他のいずれかの異種位置に(例えば、組換え生物のゲノムに)挿入され、その結果、天然に見い出されるような、そのポリヌクレオチドに通常隣接するヌクレオチド配列に関連しないポリヌクレオチドは、組換えポリヌクレオチドである。インビトロまたはインビボにおいて組換えポリヌクレオチドから発現されるタンパク質は、組換えポリペプチドの例である。同様に、天然に見られないポリヌクレオチド配列(例えば、天然に存在する遺伝子の改変体)は、組換えである。
用語「グリホセートN−アセチルトランスフェラーゼポリペプチド」および「GATポリペプチド」は、本明細書中に提供される新規のポリペプチドのファミリーのうちのいずれかをいうために互換的に用いられる。
用語「グリホセートN−アセチルトランスフェラーゼポリヌクレオチド」および「GATポリヌクレオチド」は、GATポリペプチドをコードするポリヌクレオチドをいうために互換的に用いられる。
「サブ配列」または「フラグメント」は、全配列の任意の部分である。
アミノ酸ポリマーまたはヌクレオチドポリマーの番号付けは、そのポリマーの所定のモノマー成分(アミノ酸残基、組み込まれるヌクレオチドなど)の位置が、選択される参照ポリペプチドまたはポリヌクレオチドにおいて同じ残基位置に対応するとき、選択されるアミノ酸ポリマーまたは核酸の番号付けに対応する。
ベクターは、選択される核酸による細胞形質導入または細胞における核酸の発現を促進するための組成物である。ベクターとしては、例えば、プラスミド、コスミド、ウイルス、YAC、バクテリア、ポリ−リジン、染色体組込みベクター、エピソームベクターなどが挙げられる。
「ポリヌクレオチド配列またはアミノ酸配列の実質的な全長」とは、配列の少なくとも約70%、一般的には配列の少なくとも約80%、または代表的には配列の約90%以上をいう。
本明細書中に使用される場合、「抗体」とは、イムノグロブリン遺伝子またはイムノグロブリン遺伝子フラグメントによって実質的にまたは部分的にコードされる1つ以上のポリペプチドを含むタンパク質をいう。認識されているイムノグロブリン遺伝子としては、κ、λ、α、γ、δ、εおよびμ定常領域遺伝子、ならびに無数のイムノグロブリン可変領域遺伝子と同様にが挙げられる。軽鎖は、κまたはλのいずれかとして分類される。重鎖は、γ、μ、α、δ、またはεとして分類され、これらは、それぞれ、順にイムノグロブリンクラス、IgG、IgM、IgA、IgDおよびIgEを規定する。代表的なイムノグロブリン(抗体)構造単位は、テトラマーを含む。それぞれのテトラマーは、ポリペプチド鎖の2つの同一のペア(それぞのペアは1つの「軽鎖」(約25kD)および1つの「重鎖」(約50kD〜70kD)を有する)より構成される。それぞれの鎖のN末端は、主に抗原認識を担う約100から110以上のアミノ酸の可変領域を規定する。用語、可変軽鎖(VL)および可変重鎖(VH)は、それぞれ、これらの軽鎖および重鎖をいう。抗体は、インタクトなイムノグロブリンとして、または様々なぺプチダーゼによる消化によって生成され十分特徴付けられた多々のフラグメントとして存在する。それゆえ、例えば、ペプシンは、それ自体ジスルフィド結合によってVH−CH1に結合する軽鎖であるFabのダイマー(F(ab)’2)を生成するために、ヒンジ領域におけるジスルフィド連結のもとで抗体を消化する。F(ab)’2は、温和な条件下においてヒンジ領域におけるジスルフィド連結を還元して壊し得、それによって(Fab’)2ダイマーはFab’モノマーに変換される。Fab’モノマーは、事実上、ヒンジ領域の一部を有するFabである(他の抗体フラグメントのより詳細な説明に関しては、Fundamental Immunology,第4版,W.E.Paul(編),Raven Press,N.Y.(1998)を参照)。様々な抗体フラグメントは、インタクトな抗体の消化によって定義されるが、当業者は、このようなFab’フラグメントが、化学的にまたは組換えDNA方法論のいずれかを用いることによって新規に合成され得ることを理解する。それゆえ、用語抗体はまた、本明細書中に使用される場合、全抗体の改変によって産生されるかまたは組換えDNA方法論を用いて新規に合成されるかのいずれかである、抗体フラグメントを包含する。抗体としては、一本鎖抗体(可変重鎖および可変軽鎖が、連続したポリペプチドを形成するように一緒に連結する(直接にまたはペプチドリンカーを通して)一本鎖Fv(sFv)抗体を含む)が挙げられる。
「クロロプラスト移行ペプチド(chloroplast transit peptide)」とは、タンパク質に結合して翻訳されるアミノ酸配列であり、タンパク質が生成される細胞において存在するクロロプラストまたは他の色素体型にタンパク質を方向付ける。「クロロプラスト移行配列」とは、クロロプラスト移行ペプチドをコードするヌクレオチド配列をいう。
「シグナルペプチド」とは、タンパク質に結合して翻訳されるアミノ酸配列であり、タンパク質を分泌系へ方向付ける(Chrispeels,J.J.,(1991)Ann.Rev.Plant Phys.Plant Mol.Biol.42:21−53)。タンパク質が液胞に方向付けられる場合、液胞標的化シグナル(前出)がさらに付け加えられ得、あるいは、小胞体に方向付けられれる場合、小胞体保持シグナル(前出)が付け加えられ得る。タンパク質が核に方向付けられる場合、存在するいかなるシグナルペプチドもが取り除かれ、代わりに核局在化シグナルが含まれるべきである(Raikhel,N.(1992)Plant Phys.100:1627−1632)。
ポリヌクレオチドに対して適用される用語「多様化」および「多様性」とは、親のポリヌクレオチドの複数の改変形態の生成、または複数の親のポリヌクレオチドの複数の改変形態の生成をいう。ポリヌクレオチドがポリペプチドをコードする場合、ポリヌクレオチドのヌクレオチド配列における多様性は、コードされる対応するポリペプチドに多様性(例えば、複数のポリペプチド改変体をコードするポリヌクレオチドの多様なプール)を生み出し得る。本発明のいくつかの実施形態において、この配列多様性は、望ましい機能的属性を有する改変体に対する多様化したポリヌクレオチド(例えば、増強した機能特性を有するGATポリペプチドをコードするポリヌクレオチド)のライブラリーをスクリーニング/選択することによって得られる。
用語「コードする」とは、1つ以上のアミノ酸をコードするためのヌクレオチド配列の能力をいう。この用語は、開始コドンまたは終止コドンを必要としない。アミノ酸配列は、ポリヌクレオチド配列およびその相補鎖によって提供される6つの異なるリーディングフレームのいずれか1つにおいてコードされ得る。
本明細書中に使用される場合、用語「人工的改変体」とは、改変されたGATポリヌクレオチド(例えば、配列番号1〜5および11〜262、または生物から単離された天然に存在するGATポリヌクレオチドのいずれか1つの改変された形態)によってコードされる、GAT活性を有するポリペプチドをいう。改変されたポリヌクレオチドは(これは、適切な宿主において発現される場合人工的改変体がそこから産生される)、GATポリヌクレオチドの改変による人為的介入を通して得られる。
用語「核酸構築物」または「ポリヌクレオチド構築物」は、一本鎖または二本鎖のいずれかの核酸分子を意味し、これは、天然に存在する遺伝子から単離されるか、または他に天然に存在しない様式で核酸のセグメントを含むように改変されている。用語核酸構築物は、核酸構築物が本発明のコード配列の発現に必要である制御配列を含む場合、用語「発現カセット」と同義である。
用語「制御配列」は、本明細書中において、本発明のポリペプチドの発現に必要または有利な全ての成分を含むように規定される。それぞれの制御配列は、ポリペプチドをコードするヌクレオチド配列に対してネイティブであっても外来であってもよい。このような制御配列としては、リーダー、ポリアデニル化配列、プロペプチド配列、プロモーター、シグナルペプチド配列、および転写ターミネーターが挙げられるが、これらに限定されない。制御配列は、最小でプロモーターならびに転写終止シグナルおよび翻訳終止シグナルを包含する。制御配列は、制御配列とポリペプチドをコードするヌクレオチド配列のコード領域との連結を促進する特異的制限部位を導入するためのリンカーを備え得る。
用語「作動可能に連結された」は、本明細書中において、制御配列がポリペプチドの発現を誘導するように、制御配列が、DNA配列のコード配列に対応する位置に適切に配置される立体配置として定義される。
本明細書中に使用される場合、用語「コード配列」は、そのタンパク質産物のアミノ酸配列を直接特定するヌクレオチド配列をカバーすることを意図する。コード配列の境界は、一般的に、通常ATG開始コドンから始まるオープンリーディングフレームによって決定される。コード配列は、代表的に、DNA、cDNA、および/または組換えヌクレオチド配列を包む。
本文脈において、用語「発現」は、ポリペプチドの産生に関与するいかなる工程も包含し、この工程としては、転写、転写後修飾、翻訳、翻訳後修飾、および分泌が挙げられるが、これらに限定されない。
本文脈において、用語「発現ベクター」は、本発明のポリペプチドをコードするセグメントを含む、線状DNA分子または環状DNA分子をカバーし、これは、その転写を提供するさらなるセグメントに作動可能に連結する。
本明細書中に使用される場合、用語「宿主細胞」は、核酸構築物での形質転換を受けやすい任意の細胞型を含む。
用語「植物」は、全植物、苗状の栄養器官/構造(例えば、葉、茎、および塊茎)、根、花および花器官/構造(例えば、包葉、萼片、花弁、おしべ、心皮、やく、および胚珠)、種子(胚、内乳、および種皮を含む)および果実(成熟子房)、植物組織(例えば、維管束組織、基本組織など)および細胞(例えば、孔辺細胞、卵細胞、突起様構造など)、ならびにそれらの子孫を包含する。本発明の方法において用いられ得る植物のクラスは、一般的に、被子植物(単子葉植物および双子葉植物)、裸子植物、シダ、および多細胞性藻類を含む形質転換技術の影響を受けやすい、高等植物および下等植物のクラスほど広く、異数体、倍数体、二倍体、半数体および半接合体を含む様々な倍数性レベルの植物を包含する。
本明細書中に使用される場合、用語「異種」は、一方のエレメントがもう一方のエレメントに近接して天然においては通常見い出されないことを示す2つ以上のエレメント間の関係を記載する。それゆえ、例えば、ポリヌクレオチド配列が外来種に由来する場合、または同種に由来するポリヌクレオチド配列がその元の形態から変更されている場合、そのポリヌクレオチ配列は生物または第二のポリヌクレオチド配列に対して「異種」である。例えば、異種コード配列に作動可能に連結しているプロモーターとは、プロモーターが由来する種とは異なる種に由来するコード配列をいうか、または同種に由来するコード配列である場合、そのプロモーターに天然には関連しないコード配列(例えば、遺伝子操作されたコード配列または異なる生態型もしくは変種に由来する対立遺伝子)をいう。異種のポリペプチドの例としては、トランスジェニック生物における組換えポリヌクレオチドから発現したポリペプチドが挙げられる。異種ポリヌクレオチドおよび異種ポリペプチドは、組換え分子の形態をとる。
さらなる様々な用語が、本明細書中において定義されるか、そうでなければ特徴付けられる。
(グリホセートN−アセチルトランスフェラーゼ)
1つの局面において、本発明は、単離された酵素または組換え酵素の新規ファミリー(本明細書中において「グリホセートN−アセチルトランスフェラーゼ」、「GAT」、または「GAT酵素」と称される)を提供する。GATは、GAT活性を有する酵素であり、好ましくは、GATを発現するように操作されたトランスジェニック植物に対しある程度のグリホセート耐性を与えるための十分な活性を有する酵素である。GATのいくつかの例としては、以下により詳細に記載されるGATポリペプチドが挙げられる。
(グリホセートN−アセチルトランスフェラーゼ)
1つの局面において、本発明は、単離された酵素または組換え酵素の新規ファミリー(本明細書中において「グリホセートN−アセチルトランスフェラーゼ」、「GAT」、または「GAT酵素」と称される)を提供する。GATは、GAT活性を有する酵素であり、好ましくは、GATを発現するように操作されたトランスジェニック植物に対しある程度のグリホセート耐性を与えるための十分な活性を有する酵素である。GATのいくつかの例としては、以下により詳細に記載されるGATポリペプチドが挙げられる。
当然、GATを介するグリホセート耐性は、GAT活性、トランスジェニック植物におけるGAT発現レベル、特定の植物、除草剤適用の性質およびタイミングなどの複合した機能である。当業者は、特定の状況においてグリホセート耐性をもたらすのに必要なGAT活性のレベルを過度の実験なしに決定し得る。
GAT活性は、慣習的な動的パラメータである、kcat、KM、およびkcat/KMを用いて特徴付けられ得る。kcatは、特に高基質濃度における、アセチル化の速度の基準として考えられ、KMは、その基質(例えば、アセチルCoAおよびグリホセート)に対するGATの親和性の基準であり、そしてkcat/KMは、基質親和性および触媒速度の両方を考慮に入れる触媒効果の基準である(このパラメータは、基質の濃度が少なくとも部分的に律速段階である状況において特に重要である)。一般的に、より高いkcatまたはkcat/KMを有するGATは、より低いkcatまたはkcat/KMを有する別のGATよりも効率の良い触媒である。より低いKMを有するGATは、より高いKMを有する別のGATよりも効率の良い触媒である。それゆえ、当業者は、一方のGATが別のGATより効率が良いかどうかを決定するために2つの酵素についての動的パラメータを比較し得る。kcat、kcat/KMおよびKMの相対的重要性は、GATが機能することが予想される状況(例えば、グリホセートに関するKMに対し予想されるグリホセートの有効濃度)に依存して変化する。GAT活性はまた、多くの機能特性(例えば、安定性、阻害に対する感受性、または他の分子による活性化など)のいずれかによって特徴づけられ得る。
(グリホセートN−アセチルトランスフェラーゼポリペプチド)
1つの局面において、本発明は、単離されたポリペプチドまたは組換えポリペプチドの新規ファミリー(本明細書中において「グリホセートN−アセチルトランスフェラーゼポリペプチド」または「GATポリペプチド」と称される)を提供する。GATポリペプチドは、GATの新規ファミリーに対するそれらの構造的類似性によって特徴付けられる。全てではないが多くのGATポリペプチドは、GATである。その特質は、GATポリペプチドが構造によって定義されるのに対し、GATは機能によって定義される。GATポリペプチドの一部は、好ましくは、効果的なレベルでこのタンパク質を発現するトランスジェニック植物に対してグリホセート耐性を与えるよう機能するレベルのGAT活性を有するGATポリペプチドを含む。グリホセート耐性を与える際に使用するためのいくつかの好ましいGATポリペプチドは、少なくとも1min−1のkcat、またはより好ましくは少なくとも10min−1、100min−1、または1000min−1のkcatを有する。グリホセート耐性を与える際に使用する他の好ましいGATポリペプチドは、100mM程度のKM、またはより好ましくは10mM、1mMもしくは0.1mM以下のKM、を有する。グリホセート耐性を与える際に使用するための、さらなる他の好ましいGATポリペプチドは、少なくとも1mM−1min−1以上のkcat/KM、またはより好ましくは少なくとも10mM−1min−1、100mM−1min−1、1000mM−1min−1、または10,000mM−1min−1のkcat/KMを有する。
1つの局面において、本発明は、単離されたポリペプチドまたは組換えポリペプチドの新規ファミリー(本明細書中において「グリホセートN−アセチルトランスフェラーゼポリペプチド」または「GATポリペプチド」と称される)を提供する。GATポリペプチドは、GATの新規ファミリーに対するそれらの構造的類似性によって特徴付けられる。全てではないが多くのGATポリペプチドは、GATである。その特質は、GATポリペプチドが構造によって定義されるのに対し、GATは機能によって定義される。GATポリペプチドの一部は、好ましくは、効果的なレベルでこのタンパク質を発現するトランスジェニック植物に対してグリホセート耐性を与えるよう機能するレベルのGAT活性を有するGATポリペプチドを含む。グリホセート耐性を与える際に使用するためのいくつかの好ましいGATポリペプチドは、少なくとも1min−1のkcat、またはより好ましくは少なくとも10min−1、100min−1、または1000min−1のkcatを有する。グリホセート耐性を与える際に使用する他の好ましいGATポリペプチドは、100mM程度のKM、またはより好ましくは10mM、1mMもしくは0.1mM以下のKM、を有する。グリホセート耐性を与える際に使用するための、さらなる他の好ましいGATポリペプチドは、少なくとも1mM−1min−1以上のkcat/KM、またはより好ましくは少なくとも10mM−1min−1、100mM−1min−1、1000mM−1min−1、または10,000mM−1min−1のkcat/KMを有する。
代表的なGATポリペプチドは、様々なバクテリア株から単離され、そして特徴付けられてきた。単離され、そして特徴付けられたモノマーのGATポリペプチドの1つの例は、約17kDの分子ラジウス(radius)を有する。B.licheniformisから単離された代表的なGAT酵素(配列番号7)は、グリホセートに関し約2.9mMのKmを示し、アセチルCoAに関し約2μMのKmを示す(6min−1のkcatを有す)。
用語「GATポリペプチド」とは、BLOSUM62マトリックス、11のギャップ存在ペナルティ、および1つのギャップ伸長ペナルティを用い、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列と、少なくとも430の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、任意のポリペプチドをいう。本発明のいくつかの局面は、BLOSUM62マトリックス、11のギャップ存在ペナルティ、1つのギャップ伸長ペナルティを用い、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列と、少なくとも、440、445、450、455、460、465、470、475、480、485、490、495、500、505、510、515、520、525、530、535、540、545、550、555、560、565、570、575、580、585、590、595、600、605、610、615、620、625、630、635、640、645、650、655、660、665、670、675、680、685、690、695、700、705、710、715、720、725、730、735、740、745、750、755、または760の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、GATポリペプチドに関連する。
本発明の1つの局面は、BLOSUM62マトリックス、11のギャップ存在ペナルティ、および1つのギャップ伸長ペナルティを用い、配列番号457と、少なくとも430の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、BLOSUM62マトリックス、11のギャップ存在ペナルティ、および1つのギャップ伸長ペナルティを用い、配列番号457と、少なくとも、440、445、450、455、460、465、470、475、480、485、490、495、500、505、510、515、520、525、530、535、540、545、550、555、560、565、570、575、580、585、590、595、600、605、610、615、620、625、630、635、640、645、650、655、660、665、670、675、680、685、690、695、700、705、710、715、720、725、730、735、740、745、750、755、または760の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、GATポリペプチドに関連する。
本発明の1つの局面は、BLOSUM62マトリックス、11のギャップ存在ペナルティ、および1つのギャップ伸長ペナルティを用い、配列番号445と、少なくとも430の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、BLOSUM62マトリックス、11のギャップ存在ペナルティ、および1つのギャップ伸長ペナルティを用い、配列番号445と、少なくとも、440、445、450、455、460、465、470、475、480、485、490、495、500、505、510、515、520、525、530、535、540、545、550、555、560、565、570、575、580、585、590、595、600、605、610、615、620、625、630、635、640、645、650、655、660、665、670、675、680、685、690、695、700、705、710、715、720、725、730、735、740、745、750、755、または760の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、GATポリペプチドに関連する。
本発明の1つの局面は、BLOSUM62マトリックス、11のギャップ存在ペナルティ、および1つのギャップ伸長ペナルティを用い、配列番号300と、少なくとも430の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、BLOSUM62マトリックス、11のギャップ存在ペナルティ、および1つのギャップ伸長ペナルティを用い、配列番号300と、少なくとも、440、445、450、455、460、465、470、475、480、485、490、495、500、505、510、515、520、525、530、535、540、545、550、555、560、565、570、575、580、585、590、595、600、605、610、615、620、625、630、635、640、645、650、655、660、665、670、675、680、685、690、695、700、705、710、715、720、725、730、735、740、745、750、755、または760の類似性スコアを生み出すように最適にアライメントされ得るアミノ酸配列を含む、GATポリペプチドに関連する。
2つの配列は、規定されたアミノ酸置換マトリックス(例えば、BLOSUM62)、ギャップ存在ペナルティ、およびギャップ伸長ペナルティを用い、配列のそのペアに関して可能な最も高いスコアに到達するように、類似性スコア付けに関してそれらをアライメントする場合、「最適にアライメントされる」。2つの配列間の類似性を定量する際のアミノ酸置換マトリックスおよびそれらの使用は、当該分野において周知であり、例えば、「Atlas of Protein sequence and Structure」5巻、補遺3(M.O.Dayhoff編)、345頁−352頁、Natl.Biomed.Res.Found、Washington,DCにおけるDayhoffら(1978)「A model of evolutionary change in proteins.」、およびHenikoffら(1992)Proc.Natl.Acad.Sci.USA 89:10915−10919に記載される。BLOSUM62マトリックス(図10)は、しばしば、Gapped BLAST 2.0のような配列アライメントプロトコールにおけるデフォルトスコア付け置換マトリックスとして使用される。ギャップ存在ペナルティは、アライメントされた配列の1つにおいて1つのアミノ酸ギャップの導入を課し、ギャップ伸長ペナルティは、すでに開かれたギャップに挿入されたそれぞれの付加的な空のアミノ酸位置を課す。アライメントは、アライメントの始まりから終わりまでのそれぞれの配列のアミノ酸位置によって規定され、必要に応じて、最も高い可能なスコアに到達するための1つまたは両方の配列における1つのギャップまたは複数のギャップの挿入によって規定される。最適なアライメントおよびスコア付けが手動で達成され得る一方、このプロセスは、コンピュータ実行アライメントアルゴリズム(例えば、Altschulら(1997)Nucleic Acids Res.25:3389−3402に記載され、National Center for Biotechnology Information Website(http://www.ncbi.nlm.nih.gov)にて公に利用可能なgapped BLAST 2.0)の使用によって促進される。複数のアライメントを含む最適なアライメントは、例えばhttp://www.ncbi.nlm.nih.govを通して利用可能であり、Altschulら(1997)Nucleic Acids Res.25:3389−3402に記載されているPSI−BLASTを用いて整えられ得る。
参照配列と最適にアライメントされるアミノ酸配列に関して、アミノ酸残基は、その残基がアライメントにおいてペアをなす参照配列の位置に「対応する」。「位置」は、N末端に対する位置に基づいて、参照配列において連続的にそれぞれのアミノ酸を同定する数字によって示される。例えば、配列番号300において、位置1はM、位置2はI、位置3はEなどである。あるテスト配列が配列番号300と最適にアライメントされる場合、位置3のEとアライメントするテスト配列の残基は、配列番号300の「位置3に対応する」といわれる。最適アライメントを決定する際考慮に入れられるべき欠失、挿入、短縮、融合などのために、一般的に、単純にN末端から数えられることによって決定されるテスト配列におけるアミノ酸残基番号が参照配列におけるその対応位置の番号と同じである必要はない。例えば、アライメントされたテスト配列において欠失がある場合、欠失部位において、参照配列中の位置に対応するアミノ酸は存在しない。アライメントされた参照配列において挿入がある場合、挿入は、参照配列におけるいかなるアミノ酸位置にも対応しない。短縮または融合の場合には、対応配列におけるいかなるアミノ酸にも対応しない参照配列またはアライメントされた配列のどちらかにおけるアミノ酸のストレッチが存在し得る。
用語「GATポリペプチド」とはさらに、配列番号6〜10および263〜514からなる群より選択されるアミノ酸配列に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号6〜10および263〜514からなる群より選択されるアミノ酸に関して少なくとも60%、70%、80%、90%、92%、95%、96%、97%、98%、または99%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。
本発明の1つの局面は、配列番号457に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号457に関して少なくとも60%、70%、80%、90%、92%、95%、96%、97%、98%、または99%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。
本発明の1つの局面は、配列番号445に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号445に関して少なくとも60%、70%、80%、90%、92%、95%、96%、97%、98%、または99%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。
本発明の1つの局面は、配列番号300に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号300に関して少なくとも60%、70%、80%、90%、92%、95%、96%、97%、98%、または99%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。
用語「GATポリペプチド」とはさらに、配列番号6〜10および263〜514からなる群より選択されるアミノ酸の1〜96残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む任意のポリペプチドをいう。本発明のいくつかの局面は、配列番号6〜10および263〜514からなる群より選択されるアミノ酸の1〜96残基に関して少なくとも60%、70%、80%、90%、92%、95%、96%、97%、98%、または99%の配列同一性を有するアミノ酸配列を含むGATポリペプチドに関連する。
本発明の1つの局面は、配列番号457の1〜96残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、ポリペプチドに関連する。本発明のいくつかの局面は、配列番号457の1〜96残基に関して少なくとも60%,70%,80%,90%,92%,95%,96%,97%,98%,または99%の配列同一性を有するアミノ酸配列を含むGATポリペプチドに関連する。
本発明の1つの局面は、配列番号445の1〜96残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号445の1〜96残基に関して少なくとも60%,70%,80%,90%,92%,95%,96%,97%,98%,または99%の配列同一性を有するアミノ酸配列を含むGATポリペプチドに関連する。
本発明の1つの局面は、配列番号300の1〜96残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号300の1〜96残基に関して少なくとも60%,70%,80%,90%,92%,95%,96%,97%,98%,または99%の配列同一性を有するアミノ酸配列を含むGATポリペプチドに関連する。
用語「GATポリペプチド」とはさらに、配列番号6〜10および263〜514からなる群より選択されるアミノ酸の51〜146残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む任意のポリペプチドをいう。本発明のいくつかの局面は、配列番号6〜10および263〜514からなる群より選択されるアミノ酸の51〜146残基に関して少なくとも60%、70%、80%、90%、92%、95%、96%、97%、98%、または99%の配列同一性を有するアミノ酸配列を含むポリペプチドに関連する。
本発明の1つの局面は、配列番号457の51〜146残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、ポリペプチドに関連する。本発明のいくつかの局面は、配列番号457の51〜146残基に関して少なくとも60%,70%,80%,90%,92%,95%,96%,97%,98%,または99%の配列同一性を有するアミノ酸配列を含むGATポリペプチドに関連する。
本発明の1つの局面は、配列番号445の51〜146残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号445の51〜146残基に関して少なくとも60%,70%,80%,90%,92%,95%,96%,97%,98%,または99%の配列同一性を有するアミノ酸配列を含むGATポリペプチドに関連する。
本発明の1つの局面は、配列番号300の51〜146残基に関して少なくとも40%の配列同一性を有するアミノ酸配列を含む、GATポリペプチドに関連する。本発明のいくつかの局面は、配列番号300の51〜146残基に関して少なくとも60%,70%,80%,90%,92%,95%,96%,97%,98%,または99%の配列同一性を有するアミノ酸配列を含むGATポリペプチドに関連する。
本明細書中に使用される場合、用語「同一性」または「同一性百分率」とは、アライメントされたアミノ酸配列の特定ペアに関して用いられる場合、European Bioinformatics Institute,Cambridge,UKより利用可能なClustalW解析(バージョンW1.8)(アライメントにおける同一な適合の数を数え、このような同一な適合の数を最大の(i)アライメントされた配列の長さ、および最大の(ii)96の長さによって除算し、スロウ/アキュレートペアワイズアライメントを達成するために以下のデフォルトClustalWパラメーター:ギャップオープンペナルティ:10;ギャップ伸長ペナルティー:0.10;タンパク質重量マトリックス:Gonnet series;DNA重量マトリックス:IUB;Toggle Slow/Fast ペアワイズアライメント=SLOW or FULL Alignmentを使用する)によって得られるアミノ酸配列同一性百分率をいう。
別の局面において、本発明は、配列番号6〜10および263〜514よりなる群から選択されるアミノ酸配列の、少なくとも20、あるいは50、75、100、125または140の連続したアミノ酸を含む、単離されたポリペプチドまたは組換えポリペプチドを提供する。
別の局面において、本発明は、配列番号457の、少なくとも20、あるいは50、100、または140の連続したアミノ酸を含む、単離されたポリペプチドまたは組換えポリペプチドを提供する。
別の局面において、本発明は、配列番号445の、少なくとも20、あるいは50、100、または140の連続したアミノ酸を含む、単離されたポリペプチドまたは組換えポリペプチドを提供する。
別の局面において、本発明は、配列番号300の、少なくとも20、あるいは50、100、または140の連続したアミノ酸を含む、単離されたポリペプチドまたは組換えポリペプチドを提供する。
もう一つの局面において、本発明は配列番号6〜10および配列番号263〜514からなる群より選択されるアミノ酸配列を含むポリペプチドを提供する。
いくつかの好ましい本発明のGATポリペプチドは、以下のように特徴付けられる。必要に応じて、配列番号6〜10および配列番号263〜514からなる群から選択される参照アミノ酸配列とアライメントした場合、ポリペプチド中の、以下の位置に対応するアミノ酸残基のうち少なくとも90%は、以下の制約に従う:(a)2位、4位、15位、19位、26位、28位、31位、45位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、123位、129位、139位、および/または145位で、アミノ酸残基はB1である;ならびに(b)3位、5位、8位、10位、11位、14位、17位、18位、24位、27位、32位、37位、38位、47位、48位、49位、52位、57位、58位、61位、62位、63位、68位、69位、79位、80位、82位、83位、89位、92位、100位、101位、104位、119位、120位、124位、125位、126位、128位、131位、143位、および/または144位で、アミノ酸残基はB2である;ここで、B1はA、I、L、M、F、W、Y、およびVからなる群から選択されるアミノ酸であり;B2はR、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択されるアミノ酸である。アミノ酸またはアミノ酸残基を特定するために使用するとき、一文字表記A、C、D、E、F、G、H、I、K、L、M、N、P、Q、R、S、T、V、W、およびYは、当該技術分野中で使用されるような、そして本明細書中の表2において提供されるような標準的な意味を有する。
いくつかの好ましい本発明のGATポリペプチドは、以下のように特徴付けられる。必要に応じて、配列番号6〜10および配列番号263〜514からなる群から選択される参照アミノ酸配列とアライメントした場合、ポリペプチド中の、以下の位置に対応するアミノ酸残基の少なくとも80%は、以下の制約に従う:(a)2位、4位、15位、19位、26位、28位、51位、54位、86位、90位、91位、97位、103位、105位、106位、114位、129位、139位、および/または145位で、アミノ酸残基はZ1である;(b)31位および/または45位で、アミノ酸残基はZ2である;(c)8位および/または89位で、アミノ酸残基はZ3である;(d)82位、92位、101位、および/または120位で、アミノ酸残基はZ4である;(e)3位、11位、27位、および/または79位で、アミノ酸残基はZ5である;(f)123位で、アミノ酸残基はZ1またはZ2である;(g)12位、33位、35位、39位、53位、59位、112位、132位、135位、140位、および/または146位で、アミノ酸残基はZ1またはZ3である;(h)30位で、アミノ酸残基はZ1またはZ4である;(i)6位で、アミノ酸残基はZ1またはZ6である;(j)81位および/または113位で、アミノ酸残基はZ2またはZ3である;(k)138位および/または142位で、アミノ酸残基はZ2またはZ4;(l)5位、17位、24位、57位、61位、124位、および/または126位で、アミノ酸残基はZ3またはZ4である;(m)104位で、アミノ酸残基はZ3またはZ5である;(o)38位、52位、62位、および/または69位で、アミノ酸残基はZ3またはZ6である;(p)14位、119位および/または144位で、アミノ酸残基はZ4またはZ5である;(q)18位で、アミノ酸残基はZ4またはZ6である;(r)10位、32位、48位、63位、80位および/または83位で、アミノ酸残基はZ5またはZ6である;(s)40位で、アミノ酸残基はZ1、Z2またはZ3である;(t)65位および/または96位で、アミノ酸残基はZ1、Z3またはZ5である;(u)84位および/または115位で、アミノ酸残基はZ1、Z3またはZ4である;(v)93位で、アミノ酸残基はZ2、Z3またはZ4である;(w)130位で、アミノ酸残基はZ2、Z4またはZ6である;(x)47位および/または58位で、アミノ酸残基はZ3、Z4またはZ6である;(y)49位、68位、100位および/または143位で、アミノ酸残基はZ3、Z4またはZ5である;(z)131位で、アミノ酸残基はZ3、Z5またはZ6である;(aa)125位および/または128位で、アミノ酸残基はZ4、Z5またはZ6である;(ab)67位で、アミノ酸残基はZ1、Z3、Z4またはZ5;(ac)60位で、アミノ酸残基はZ1、Z4、Z5またはZ6である;ならびに(ad)37位で、アミノ酸残基はZ3、Z4、Z5またはZ6である;ここでZ1は、A、I、L、M、およびVからなる群から選択されるアミノ酸であり;Z2は、F、W、およびYからなる群から選択されるアミノ酸であり;Z3は、N、Q、S、およびTからなる群から選択されるアミノ酸であり;Z4は、R、H、およびKからなる群から選択されるアミノ酸であり;Z5は、DおよびEからなる群から選択されるアミノ酸であり;ならびにZ6は、C、G、およびPからなる群から選択されるアミノ酸である。
いくつかの好ましい本発明のGATポリペプチドは、以下のように特徴付けられる。必要に応じて、配列番号6〜10および配列番号263〜514からなる群から選択される参照アミノ酸配列とアライメントした場合、ポリペプチド中の、以下の位置に対応するアミノ酸残基の少なくとも90%は、以下の制約に従う:(a)1位、7位、9位、13位、20位、36位、42位、46位、50位、56位、64位、70位、72位、75位、76位、78位、94位、98位、107位、110位、117位、118位、121位、および/または141位で、アミノ酸残基はB1である;ならびに(b)16位、21位、22位、23位、25位、29位、34位、41位、43位、44位、55位、66位、71位、73位、74位、77位、85位、87位、88位、95位、99位、102位、108位、109位、111位、116位、122位、127位、133位、134位、136位、および/または137位で、アミノ酸残基はB2である;ここでB1は、A、I、L、M、F、W、Y、およびVからなる群から選択されるアミノ酸残基である;そしてB2は、R、N、D、C、Q、E、G、H、K、P、S、およびTからなる群から選択されるアミノ酸残基である。
いくつかの好ましい本発明のGATポリペプチドは、以下のように特徴付けられる。必要に応じて、配列番号6〜10および配列番号263〜514からなる群から選択される参照アミノ酸配列とアライメントした場合、ポリペプチド中の、以下の位置に対応するアミノ酸残基の少なくとも90%は、以下の制約に従う:(a)1位、7位、9位、20位、36位、42位、50位、64位、72位、75位、76位、78位、94位、98位、110位、121位、および/または141位で、アミノ酸残基はZ1である;(b)13位、46位、56位、70位、107位、117位、および/または118位で、アミノ酸残基はZ2である;(c)23位、55位、71位、77位、88位、および/または109位で、アミノ酸残基はZ3である;(d)16位、21位、41位、73位、85位、99位、および/または111位で、アミノ酸残基はZ4である;(e)34位および/または95位で、アミノ酸残基はZ5である;(f)22位、25位、29位、43位、44位、66位、74位、87位、102位、108位、116位、122位、127位、133位、134位、136位、および/または137位で、アミノ酸残基はZ6である;ここでZ1は、A、I、L、M、およびVからなる群から選択されるアミノ酸である;Z2はF、W、およびYからなる群から選択されるアミノ酸である;Z3はN、Q、S、およびTからなる群から選択されるアミノ酸である;Z4はR、H、およびKからなる群から選択されるアミノ酸である;Z5はDおよびEからなる群から選択されるアミノ酸である;そしてZ6は、C、G、およびPからなる群から選択されるアミノ酸である。
いくつかの好ましい本発明のGATポリペプチドは、以下のように特徴付けられる。必要に応じて、配列番号6〜10および配列番号263〜514からなる群から選択される参照アミノ酸配列とアライメントした場合、ポリペプチド中で、以下の位置に対応するアミノ酸残基の少なくとも80%が以下の制約に従う:(a)2位で、アミノ酸残基はIまたはLである;(b)3位で、アミノ酸残基はEまたはDである;(c)4位で、アミノ酸残基はV、AまたはIである;(d)5位で、アミノ酸残基はK、RまたはNである;(e)6位で、アミノ酸残基はPまたはLである;(f)8位で、アミノ酸残基はN、SまたはTである;(g)10位で、アミノ酸残基はEまたはGである;(h)11位で、アミノ酸残基はDまたはEである;(i)12位で、アミノ酸残基はTまたはAである;(j)14位で、アミノ酸残基はEまたはKである;(k)15位で、アミノ酸残基はIまたはLである;(l)17位で、アミノ酸残基はHまたはQである;(m)18位で、アミノ酸残基はR、CまたはKである;(n)19位で、アミノ酸残基はIまたはVである;(o)24位で、アミノ酸残基はQまたはRである;(p)26位で、アミノ酸残基はLまたはIである;(q)27位で、アミノ酸残基はEまたはDである;(r)28位で、アミノ酸残基はAまたはVである;(s)30位で、アミノ酸残基はK、MまたはRである;(t)31位で、アミノ酸残基はYまたはFである;(u)32位で、アミノ酸残基はEまたはGである;(v)33位で、アミノ酸残基はT、AまたはSである;(w)35位で、アミノ酸残基はL、SまたはMである;(x)37位で、アミノ酸残基はR、G、EまたはQである;(y)38位で、アミノ酸残基はGまたはSである;(z)39位で、アミノ酸残基はT、AまたはSである;(aa)40位で、アミノ酸残基はF、LまたはSである;(ab)45位で、アミノ酸残基はYまたはFである;(ac)47位で、アミノ酸残基はR、QまたはGである;(ad)48位で、アミノ酸残基はGまたはDである;(ae)49位で、アミノ酸残基はK、R、EまたはQである;(af)51位で、アミノ酸残基はIまたはVである;(ag)52位で、アミノ酸残基はS、CまたはGである;(ah)53位で、アミノ酸残基はIまたはTである;(ai)54位で、アミノ酸残基はAまたはVである;(aj)57位で、アミノ酸残基はHまたはNである;(ak)58位で、アミノ酸残基はQ、K、NまたはPである;(al)59位で、アミノ酸残基はAまたはSである;(am)60位で、アミノ酸残基はE、K、G、VまたはDである;(an)61位で、アミノ酸残基はHまたはQである;(ao)62位で、アミノ酸残基はP、SまたはTである;(ap)63位で、アミノ酸残基はE、GまたはDである;(aq)65位で、アミノ酸残基はE、D、VまたはQである;(ar)67位で、アミノ酸残基はQ、E、R、L、HまたはKである;(as)68位で、アミノ酸残基はK、R、EまたはNである;(at)69位で、アミノ酸残基はQまたはPである;(au)79位で、アミノ酸残基はEまたはDである;(av)80位で、アミノ酸残基はGまたはEである;(aw)81位で、アミノ酸残基はY、NまたはFである;(ax)82位で、アミノ酸残基はRまたはHである;(ay)83位で、アミノ酸残基はE、GまたはDである;(az)84位で、アミノ酸残基はQ、RまたはLである;(ba)86位で、アミノ酸残基はAまたはVである;(bb)89位で、アミノ酸残基はTまたはSである;(bc)90位で、アミノ酸残基はLまたはIである;(bd)91位で、アミノ酸残基はIまたはVである;(be)92位で、アミノ酸残基はRまたはKである;(bf)93位で、アミノ酸残基はH、YまたはQである;(bg)96位で、アミノ酸残基はE、AまたはQである;(bh)97位で、アミノ酸残基はLまたはIである;(bi)100位で、アミノ酸残基はK、R、NまたはEである;(bj)101位で、アミノ酸残基はKまたはRである;(bk)103位で、アミノ酸残基はAまたはVである;(bl)104位で、アミノ酸残基はDまたはNである;(bm)105位で、アミノ酸残基はLまたはMである;(bn)106位で、アミノ酸残基はLまたはIである;(bo)112位で、アミノ酸残基はTまたはIである;(bp)113位でアミノ酸残基はS、TまたはFである;(bq)114位で、アミノ酸残基はAまたはVである;(br)115位で、アミノ酸残基はS、RまたはAである;(bs)119位で、アミノ酸残基はK、EまたはRである;(bt)120位で、アミノ酸残基はKまたはRである;(bu)123位で、アミノ酸残基はFまたはLである;(bv)124位で、アミノ酸残基はSまたはRである;(bw)125位で、アミノ酸残基はE、K、GまたはDである;(bx)126位で、アミノ酸残基はQまたはHである;(by)128位で、アミノ酸残基はE、GまたはKである;(bz)129位で、アミノ酸残基はV、IまたはAである;(ca)130位で、アミノ酸残基はY、H、FまたはCである;(cb)131位で、アミノ酸残基はD、G、NまたはEである;(cc)132位で、アミノ酸残基はI、T、A、M、VまたはLである;(cd)135位で、アミノ酸残基はV、T、AまたはIである;(ce)138位で、アミノ酸残基はHまたはYである;(cf)139位で、アミノ酸残基はIまたはVである;(cg)140位で、アミノ酸残基はLまたはSである(ch)142位で、アミノ酸残基はYまたはHである;(ci)143位で、アミノ酸残基はK、TまたはEである;(cj)144位で、アミノ酸残基はK、EまたはRである;(ck)145位で、アミノ酸残基はLまたはIである;および、(cl)146位で、アミノ酸残基はTまたはAである。
いくつかの好ましい本発明のGATポリペプチドは、以下のように特徴付けられる。必要に応じて、配列番号6〜10および配列番号263〜514からなる群から選択される参照アミノ酸とアライメントされた場合、ポリペプチド中で、以下の位置に対応するアミノ酸残基の少なくとも80%は、以下の制約に従う:(a)9位、76位、94位および110位で、アミノ酸残基はAである;(b)29位および108位で、アミノ酸残基はCである;(c)34位で、アミノ酸残基はDである;(d)95位で、アミノ酸残基はEである;(e)56位で、アミノ酸残基はFである;(f)43位、44位、66位、74位、87位、102位、116位、122位、127位および136位で、アミノ酸残基はGである;(g)41位でアミノ酸残基はHである;(h)7位で、アミノ酸残基はIである;(i)85位で、アミノ酸残基はKである;(j)20位、36位、42位、50位、72位、78位、98位および121位で、アミノ酸残基はLである;(k)1位、75位および141位で、アミノ酸残基はMである;(l)23位、64位および109位でアミノ酸残基はNである;(m)22位、25位、133位、134位および137位で、アミノ酸残基はPである;(n)71位で、アミノ酸残基はQである;(o)16位、21位、73位、99位および111位で、アミノ酸残基はRである;(p)55位および88位で、アミノ酸残基はSである;(q)77位で、アミノ酸残基はTである;(r)107位でアミノ酸残基はWである;そして(s)13位、46位、70位、117位および118位で、アミノ酸残基はYである。
いくつかの好ましい本発明のGATポリペプチドは、以下のように特徴付けられる。必要に応じて、配列番号6〜10および配列番号263〜514からなる群から選択される参照アミノ酸とアライメントした場合、28位に対応するポリペプチド中のアミノ酸残基はVまたはAである。28位のバリンは、一般的に減少したKMに関連し、一方、この位置のアラニンは、一般的に増加したkcatに関連する。他の好ましいGATポリペプチドは、I27(すなわち、27位がI)、M30、S35、R37、S39、G48、K49、N57、Q58、P62、Q65、Q67、K68、E83、S89、A96、E96、R101、T112、A114、K119、K120、E128、V129、D131、T131、V134、R144、I145、もしくはT146、またはそれらの任意の組合せを有することによって、特徴付けられる。
いくつかの好ましい本発明のGATポリペプチドは、配列番号6〜10および配列番号263〜514からなる群から選択されるアミノ酸配列を含む。
本発明は、前述のアミノ酸残基位置の制約の組合せによって特徴付けられる好ましいGATポリペプチドを、さらに提供する。
加えて、本発明は、上記の好ましいGATポリペプチドをコードするGATポリヌクレオチド、およびその相補的ヌクレオチド配列を提供する。
本発明のいくつかの局面は、本明細書中に記載されているようにGAT活性を有するGATポリペプチドの任意の上記カテゴリーのサブセットに特に関係する。これらGATポリペプチドは、例えば、植物に対してグリホセート耐性を付与するための薬剤としての使用に好ましい。所望のレベルのGAT活性の例を、本明細書中に記載する。
一つの局面において、GATポリペプチドは、組換体または天然源、例えば細菌株、から単離された天然に存在する核酸の単離型によってコードされるアミノ酸配列を含む。そのようなGATポリペプチドをコードする野生型ポリヌクレオチドは、当該技術分野で公知である標準技術によって特異的にスクリーニングされ得る。例えば、配列番号6〜配列番号10によって定義されるポリペプチドは、以下により詳細にわたって記載される、GAT活性を示すBacillus株由来の配列を発現クローニングすることによって発見された。
本発明は、配列番号1〜5および配列番号11〜262、それらの相補体、ならびに配列番号6〜10および配列番号263〜514からなる群から選択されるアミノ酸配列をコードするヌクレオチド配列(これらの相補体を含む)からなる群から選択されるヌクレオチド配列の実質的全長にわたって、ストリンジェントな条件下でハイブリダイズするヌクレオチド配列を含む、単離されたポリヌクレオチドまたは組換えポリヌクレオチドによってコードされる、単離されたポリペプチドまたは組換えポリペプチドもまた含む。
本発明は、GAT活性を有し、本明細書中に記載されている任意のGATコードポリヌクレオチドのフラグメントによってコードされる任意のポリペプチドをさらに含む。
本発明は、一緒にスプライシングされて機能性のGATポリペプチドを形成し得るGATポリペプチドのフラグメントもまた提供する。スプライシングは、インビトロまたはインビボで達成され得、シススプライシングまたはトランススプライシング(すなわち、分子内スプライシングまたは分子間スプライシング)を含み得る。このフラグメントはそれ自身、GAT活性を有し得るが、それは必要ではない。例えば、GATポリペプチドの二つ以上のセグメントはインテインによって分割され得、シススプライシングによるインテイン配列の除去によって、機能性のGATポリペプチドという結果が得られる。もう一つの例において、暗号化されたGATポリペプチドは、二つ以上の別々のフラグメントとして発現され得、これらフラグメントのトランススプラシングによって、機能性のGATポリペプチドの回復という結果が得られる。シススプライシングおよびトランススプライシングの様々な局面、遺伝子暗号化、ならびに介在配列の導入は、米国特許出願第09/517,933号および同第09/710,686号において、より詳細にわたって記述される。両者を、その全体を本明細書中に参考として援用する。
一般的に、本発明は、変異、反復配列(recursive sequence)組換え、および/または本明細書中に記載されたポリヌクレオチド配列の多様性によって誘導された改変GATポリヌクレオチドによってコードされる任意のポリペプチドを含む。本発明のいくつかの局面において、GATポリペプチドは、単一または複数のアミノ酸置換、欠失、挿入、もしくはこれらの改変型の一つ以上の組合せによって改変される。置換は、保存的でも非保存的であってもよく、機能を改変しても改変しなくてもよく、そして新規機能を付加してもよい。挿入および欠失は、例えば、配列の重要なフラグメントの短縮化の場合、あるいは内部またはN末端もしくはC末端どちらかへのさらなる配列の融合において、重要であり得る。本発明のいくつかの実施形態において、GATポリペプチドは、例えば、分泌シグナル、葉緑体輸送ペプチド、精製タグ、または当業者に明らかな任意の多くの機能性官能基のような機能付加を含む融合タンパク質の一部である。そして、そのことは本明細書中の他の箇所でより詳細に記述される。
本発明のポリペプチドは、一つ以上の改変アミノ酸を含み得る。改変アミノ酸の存在によって、例えば、(a)ポリペプチドのインビボでの半減期の増加、(b)ポリペプチドの抗原性の減少または増加、(c)ポリペプチド保存安定性の増加において有利であり得る。例えば、アミノ酸は組換え体産生中に、共翻訳修飾または翻訳後修飾され(例えば、哺乳動物細胞中での発現時の、N−X−S/TモチーフでのN−結合グリコシル化)、または合成的手段で改変される。
改変アミノ酸の非限定的な例には、グリコシル化アミノ酸、硫酸化アミノ酸、プレニル化(例えば、ファルネシル化、ゲラニルゲラニル化)アミノ酸、アセチル化アミノ酸、アシル化アミノ酸、PEG化アミノ酸、ビオチニル化アミノ酸、カルボキシル化アミノ酸、リン酸化アミノ酸、などが挙げられる。アミノ酸改変の当業者を導くのに十分な参照は、文献全体にわたって充実している。例のプロトコールは、Walker(1998)Protein Protocols on CD−ROM Human Press,Towata,NJ中に見出される。
本発明のGATポリペプチドを産生および単離する組換え方法を、本明細書中に記載する。組換え産生に加えて、ポリペプチドは固相技術を使用した直接ペプチド合成によって産生され得る。(Stewartら(1969)Solid−Phase Peptide Synthesis,WH Freeman Co,San Francisco;Merrifield J(1963)J.Am.Chem.Soc.85:2149−2154)。ペプチド合成は手動の技術の使用または自動制御によって実行され得る。例えば、自動化合成はApplied Biosystems 431Aペプチド合成器(Perkin Elmer,Foster City,Calif.)を用いて、製造業者によって提供された説明書に従って達成され得る。例えば、部分配列は別々に化学合成され、GATポリペプチドの全長を提供する化学的方法を使用することによって結合され得る。ペプチドはまた、様々な供給源から得られ得る。
本発明のもう一つの局面において、本発明のGATポリペプチドは、例えば、トランスジェニック植物の様々な組織中の、例えば、GATポリペプチドの活性、分布、および発現に関係する、例えば、診断的用途を有する抗体の産生に使用される。
抗体誘導のためのGATホモログポリペプチドは、生物学的活性を必要としないが、このポリペプチドまたはオリゴペプチドは抗原性でなければならない。特異的な抗体を誘導するために使用されるペプチドは、少なくとも10アミノ酸、好ましくは少なくとも15アミノ酸または20アミノ酸からなるアミノ酸配列を有し得る。短い一続きのGATポリペプチドは、キーホールリンペットヘモシアニンのような別のタンパク質と融合され得、キメラ分子に対する抗体が産生され得る。
ポリクローナル抗体およびモノクローナル抗体を産生する方法は、当業者に公知で、多くの抗体が入手可能である。例えば、Coligan(1991) Current Protocols in Immunology Wiley/Greene,NY;およびHarlowおよびLane(1989)Antibodies:A Laboratory Manual Cold Spring Harbor Press,NY;Stitesら(eds.)Basic and Clinical Immunology(第4版)Lange Medical Publications,Los Altos,CA、およびそれらの中で引用された文献;Goding(1986)Monoclonal Antibodies:Principles and Practice(第2版)Academic Press,New York,NY;ならびにKohlerおよびMilstein(1975)Nature 256:495−497を参照のこと。抗体の調製に適切な他の技術は、ファージまたは類似したベクター中の組換え抗体のライブラリーの選別を含む。Huseら(1989)Science 246:1275−1281;およびWardら(1989)Nature 341:544−546を参照のこと。特異的なモノクローナル抗体およびポリクローナル抗体ならびに抗血清は、通常少なくとも約0.1μMのKDで、好ましくは少なくとも約0.01μM以上のKDで、最も代表的そして最も好ましくは、0.001μM以上のKDで、結合する。
抗体の産生技術および操作技術の追加的詳細は、Borrebaeck(ed)(1995)Antibody Engineering,2nd Edition Freeman and Company,NY(Borrebaeck);McCaffertyら(1996)Antibody Engineering,A Practical Approach IRL at Oxford Press,Oxford,England(McCafferty),およびPaul(1995)Antibody Engineering Protocols Humana Press,Towata,NJ(paul)に見出され得る。
(配列のバリエーション)
本発明のGATポリペプチドは、本明細書中で配列番号6〜10および配列番号263〜514として開示される配列の保存的に改変されたバリエーションを含む。そのような保存的に改変されたバリエーションは、置換、付加または欠失を含み、それらは、配列番号6〜10および配列番号263〜514中のいずれかの、単一のアミノ酸または小さな割合のアミノ酸(代表的には約5%未満、より代表的には4%未満、2%未満、または1%未満)を改変し、付加し、または欠失する。
本発明のGATポリペプチドは、本明細書中で配列番号6〜10および配列番号263〜514として開示される配列の保存的に改変されたバリエーションを含む。そのような保存的に改変されたバリエーションは、置換、付加または欠失を含み、それらは、配列番号6〜10および配列番号263〜514中のいずれかの、単一のアミノ酸または小さな割合のアミノ酸(代表的には約5%未満、より代表的には4%未満、2%未満、または1%未満)を改変し、付加し、または欠失する。
例えば、本明細書中で配列番号6として同定された146アミノ酸ポリペプチドの保存的に改変されたバリエーション(例えば、欠失)は、少なくとも140アミノ酸長、好ましくは少なくとも141アミノ酸、より好ましくは少なくとも144アミノ酸、そしてさらにより好ましくは少なくとも146アミノ酸(これらはポリペプチド配列の約5%未満、約4%未満、約2%未満または約1%未満の欠失に対応する)を有する。
本明細書中で配列番号6として同定されたポリペプチドの保存的に改変されたバリエーションのもう一つの例(例えば「保存的に置換されたバリエーション」)は、表2(下記)で説明された六つの置換群に従って、146アミノ酸ポリペプチドの約7残基まで(すなわち、約5%未満)において「保存的な置換」を含む。
保存的に置換された配列を含む、本発明のGATポリペプチド配列ホモログは、GATポリペプチド中、(例えば、葉緑体(chloraplast)ターゲティング配列)シグナル配列を有するGAT融合体中、またはタンパク質の精製のための一つ以上のドメイン(例えば、ポリヒスチジンセグメント、FLAGタグセグメント、など)の付加において生じるような、より大きなポリペプチド配列の一部として存在し得る。後者の場合、付加的機能性ドメインは、タンパク質のGAT部分の活性に対してほとんど影響を有しないか、または全く影響しない、あるいはこの付加的ドメインは、プロテアーゼの処理によるような合成後プロセシング工程によって除去され得る。
(免疫反応性によるポリペプチドの定義)
本発明のポリペプチドは、定義された活性、すなわちグリホセートのアセチル化、を有する新規クラスの酵素を提供するので、このポリペプチドは例えば、免疫学的アッセイにおいて認識され得る新規構造的特徴もまた提供する。本発明のポリペプチドを特異的に結合する抗血清の生成は、そのような抗血清によって結合されるポリペプチドと同様に、本発明の一つの特徴である。
本発明のポリペプチドは、定義された活性、すなわちグリホセートのアセチル化、を有する新規クラスの酵素を提供するので、このポリペプチドは例えば、免疫学的アッセイにおいて認識され得る新規構造的特徴もまた提供する。本発明のポリペプチドを特異的に結合する抗血清の生成は、そのような抗血清によって結合されるポリペプチドと同様に、本発明の一つの特徴である。
本発明は、一つ以上の配列番号6〜配列番号10から選択されるアミノ酸配列を含む免疫原に対して生成された抗体または抗血清に、特異的に結合する、または特異的に免疫反応性であるGATポリペプチドを含む。他のGATホモログとの交叉反応性を除去するために、この抗体または抗血清は、本願の出願日現在入手可能なGenBankアクセッション番号に対応するタンパク質またはペプチド(それらは、CAA70664,Z99109およびY09476によって、例示される)によって示されるタンパク質のような、入手可能な関連タンパク質によって除去される。核酸に対応する登録番号の場合、核酸にコードされたポリペプチドが生成され、抗体/抗血清除去の目的のために使用される。図3は、例示のGATポリペプチドとGenbank中で入手可能な最もしっかりと関連した配列、YitIとの間の相対的同一性を表にする。ネイティブのYitIの機能は、未だに解明されていないが、この酵素は、検出可能なGAT活性を有することが示される。
一つの代表的なフォーマットにおいて、免疫アッセイは、配列番号6〜10および配列番号263〜514、またはそれらの実質的な部分配列(すなわち、提供された全長配列の少なくとも約30%)の一つ以上に対応する一つ以上の配列を含む、一つ以上のポリペプチドに対して生成されたポリクローナル抗血清を使用する。配列番号6〜10および配列番号263〜514に由来する可能なポリペプチド免疫原の全セットは、以下にひとまとめにして「免疫原性ポリペプチド」という。結果として得られる抗血清は、必要に応じて他の関連配列に対する低い交叉反応性を有するように選択され、そのようなどの交叉反応性も、免疫アッセイにおけるポリクローナル抗血清の使用前に、一つ以上の関連した配列を用いた免疫吸着によって除去される。
免疫アッセイの使用のための抗血清を産生するために、一つ以上の免疫原性ポリペプチドを、本明細書中で記載されるように、産生し、精製する。例えば、組換えタンパク質を、細菌細胞株中で産生し得る。マウスの近交系(実質的なマウスの遺伝的同一性のために、結果により再現性があるのでこのアッセイにおいて使用される)は、フロイントのアジュバンドのような標準的なアジュバンドと組合わせた免疫原性タンパク質、および標準的なマウス免疫プロトコールを用いて、免疫される(特異的な免疫反応性を決定するために使用され得る抗体産生、免疫アッセイのフォーマットおよび免疫アッセイの条件の標準的な記述については、HarlowおよびLane(1988)Antibodies,A Laboratory Manual,Cold Spring Harbor Publications,New Yorkを参照のこと)。あるいは、本明細書中に開示された配列に由来する一つ以上の合成ポリペプチドまたは組換えポリペプチドを、キャリアータンパク質と結合し、免疫原として使用する。
ポリクローナル血清が回収され、免疫アッセイ、例えば、固体支持体に固定した一つ以上の免疫原性タンパク質を用いた固相免疫アッセイ、で免疫原性ポリペプチドに対して力価測定が行われる。力価が106以上であるポリクローナル抗血清が選択され、プールされ、関連ポリペプチド、例えば、示したようにGENBANKから同定されたポリペプチド、を用いて除去されて、除去されプールされ力価測定されたポリクローナル抗血清を産生する。
この除去されプールされ力価測定されたポリクローナル抗血清は、関連したポリペプチドに対する交叉反応性について、テストされる。好ましくは、免疫原性タンパク質が特異的に結合する抗体を同定するために、少なくとも二つの免疫原性GATが、好ましくは、少なくとも二つの関連したポリペプチドとともにこの測定に使用される。
この比較アッセイにおいて、区別化するための結合条件は、関連するポリペプチドへの結合と比べて、力価測定されたポリクローナル抗血清の免疫原性GATポリペプチドへの結合に対して、少なくとも約5〜10倍高いシグナルノイズ比という結果が得られる、除去され力価測定されたポリクローナル抗血清に対して決定される。すなわち、結合反応のストリンジェンシーは、アルブミンまたは脱脂粉乳のような非特異的競合体の添加によって、もしくは塩条件、または温度などの調節によって調節される。これらの結合条件は、テストポリペプチドが、プールされ除去されたポリクローナル抗血清によって特異的に結合されるかどうか、を決定するための次のアッセイで使用される。特に、テストポリペプチドが、区別化するための結合条件下でコントロールポリペプチドに比べて、少なくとも2〜5倍高いシグナルノイズ比を示し、免疫原性ポリペプチドと比較して、少なくとも約1/2のシグナルノイズ比を示すと、公知のGATと比較して免疫原性ポリペプチドと実質的な構造類似性を共有し、従って、テストポリペプチドは本発明のポリペプチドである。
もう一つの例では、競合的結合フォーマットでの免疫アッセイを、テストポリペプチドの検出に使用する。例えば、示したように、交叉反応抗体を、コントロールGATポリペプチドとの免疫吸着により、プールされた抗血清混合物から除去する。次いで、この免疫原性ポリペプチドは、固相支持体に固定され、その支持体は、除去されプールされた抗血清にさらされる。テストタンパク質は、プールされ除去された抗血清への結合と競合するアッセイに加えられる。固定されたタンパク質と比較した、このテストタンパク質がプールされ除去された抗血清への結合について競合する能力は、このアッセイへ加えられた免疫原性ポリペプチドが結合について競合する能力と比較される(免疫原性ポリペプチドは、プールされた抗血清への結合について、固定された免疫原性ポリペプチドと効果的に競合する)。このテストタンパク質のパーセント交叉反応性を、標準の計算法を使用して、計算する。
平行アッセイにおいて、コントロールタンパク質がプールされ除去された抗血清への結合について競合する能力は、必要に応じて、免疫原性ポリペプチドが抗血清への結合について競合する能力と比べて決定される。再び、コントロールポリペプチドに対するパーセント交叉反応性を、標準の計算法を使用して、計算する。パーセント交叉反応性が、テストペプチドに対して少なくとも5〜10倍高い場合、このテストペプチドは、プールされ除去された抗血清へ特異的に結合するといわれる。
一般的に、免疫吸着された抗血清およびプールされた抗血清を、本明細書中で記載されているような競合的結合免疫アッセイにおいて使用して、任意の試験ポリペプチドを免疫原性ポリペプチドを比較し得る。この比較を行なうために、2つのポリペプチドは、広範囲の濃度でそれぞれアッセイされ、そして固定化されたタンパク質へのサブストラクトされた抗血清の結合を50%阻害するために必要とされるそれぞれのポリペプチドの量は、標準技術を使用して決定される。必要とされる試験ポリペプチドの量が、必要とされる免疫原性ポリペプチドの量の2倍より少ない場合、この試験ポリペプチドは,免疫原性タンパク質に対して産生される抗体に特異的に結合するといわれ、供給された量は、コントロールのポリペプチドに対して少なくとも約5〜10倍高い。
特異性の最終決定として、免疫吸着で使用される免疫原性ポリペプチドに対する、得られた免疫原性ポリペプチドのサブストラクトされプールされた抗血清の結合がほとんどまたは全く検出されなくなるまで、プールされた抗血清は、必要に応じて、(コントロールポリペプチドではなく)免疫原性ポリペプチドに完全に吸着される。この完全に免疫吸着された抗血清は、次に試験ポリペプチドとの反応性を試験される。反応性がほとんどないかまたは反応性がないことが観察される場合(すなわち、免疫原性ポリペプチドへの、完全に免疫吸着された抗血清の結合について観察されたノイズに対するシグナルの比が2倍しかない場合)、試験ポリペプチドは、免疫原性タンパク質によって誘発された抗血清によって特異的に結合される。
(グリホセートN−アセチル基転移酵素ポリヌクレオチド)
ひとつの局面において、本発明は、「グリホセートN−アセチル基転移酵素ポリヌクレオチド」または「GATポリヌクレオチド」のように本明細書中で参照されている、単離されたポリヌクレオチドまたは組換えポリヌクレオチドの新規ファミリーを提供する。GATポリヌクレオチド配列は、GATポリペプチドをコードする能力によって特徴付けられている。一般的に本発明は、本明細書中で記載されている新規GATポリペプチドのいずれかをコードする任意の核酸配列を含む。本発明のいくつかの局面においては、GAT活性を有するGATポリペプチドをコードするGATポリヌクレオチドが好ましい。
ひとつの局面において、本発明は、「グリホセートN−アセチル基転移酵素ポリヌクレオチド」または「GATポリヌクレオチド」のように本明細書中で参照されている、単離されたポリヌクレオチドまたは組換えポリヌクレオチドの新規ファミリーを提供する。GATポリヌクレオチド配列は、GATポリペプチドをコードする能力によって特徴付けられている。一般的に本発明は、本明細書中で記載されている新規GATポリペプチドのいずれかをコードする任意の核酸配列を含む。本発明のいくつかの局面においては、GAT活性を有するGATポリペプチドをコードするGATポリヌクレオチドが好ましい。
一つの局面において、GATポリヌクレオチドは、生物体(例えば、細菌株)から単離された天然に存在する核酸の組換え形態および単離形態を含む。例示的なGATポリヌクレオチド(例えば、配列番号1〜配列番号5)は、GAT活性を示すBacillus株に由来する配列の発現クローニングによって発見された。簡潔には、約500のBacillus株およびPseudomonas株の収集物をグリホセートをNアセチル化するネイティブな能力についてスクリーニングした。株を一晩LBで増殖させ、遠心分離によって回収し、希トルエンで透過化し、そして次に洗浄し、5mMグリホセートおよび200μMアセチルCoAの緩衝液を含む反応混合液に再懸濁した。細胞を1〜48時間の間反応混合液中でインキュベートし、その時点で同等体積のメタノールをこの反応物に添加した。細胞を、次に遠心分離によってペレット化し、そして上清を、親イオンモードの質量分析法による分析前に濾過した。反応の生成物を、反応混合液の質量分析プロフィールとN−アセチルグリホセートの標準とを図2で示すように比較することによって、N−アセチルグリホセートとして明確に同定した。生成物検出は、両方の基質(アセチルCoAおよびグリホセート)の封入に依存しており、細菌細胞を熱変性することによって消された。
個々のGATポリヌクレオチドを、次に機能的スクリーニングによって同定された株からクローン化した。ゲノムDNAを調製し、Sau3A1酵素を用いて部分的に消化した。約4Kbのフラグメントを、E.coli発現ベクターにクローン化し、そして電気的にコンピテントなE.coliに形質転換した。GAT活性を示す個々のクローンを、トルエン洗浄をPMBSを用いた透過化処理によって置換することを除いては、これまでに記載されたように、反応後の質量分析方法によって同定した。ゲノムフラグメントを配列決定し、そしてオープンリーディングフレームをコードする推定GATポリペプチドを同定した。GAT遺伝子の同定を、E.coliにおけるオープンリーディングフレームの発現および反応混合物から産生されたN−アセチルグリホセートの高レベルでの検出によって確認した。
本発明の別の局面において、GATポリヌクレオチドは、多様化することによって(例えば、一つ以上の天然に存在する単離されたGATポリヌクレオチドまたは組換えGATポリヌクレオチドを組換えおよび/または変異することによって)産生される。本明細書中の他で更に詳細に記載されているように、優れた機能的性質(例えば、多様化プロセスにおいて基質または親として使用されるGATポリヌクレオチドよりも増加した触媒機能、増加した安定性、高い発現レベル)を有するGATポリペプチドをコードする多様化されたGATポリヌクレオチドを産生することがしばしば可能である。
本発明のポリヌクレオチドは、以下の種々の用途を有する、例えば:本発明のGATポリペプチドの組換え産生(すなわち、発現);導入遺伝子として(例えば、トランスジェニック植物に除草剤耐性を与えるための);形質転換およびプラスミド維持のための選択マーカーとして;免疫原として;相補的な核酸または部分的に相補的な核酸の存在についての診断的なプローブとして(天然GATコード核酸の検出のために含まれる);さらなる多様性生成(例えば、新しいGATホモログおよび/または改良されたGATホモログなどを産生するための組換え反応または変異反応)のための基質として。
GATポリヌクレオチドのある特定の実質的かつ信頼できる有用性は、実質的なGAT活性を有するポリペプチドをコードするポリヌクレオチドを必要としないことに注目することは重要である。例えば、活性酵素をコードしないGATポリヌクレオチドは、望ましい機能的性質(例えば、高いkcatまたはkcat/Km、低いKm、熱または他の環境因子に対する高い安定性、高い転写または翻訳速度、タンパク質溶解性の切断に対する抵抗性、抗原性を減らすこと、など)を有する、GATポリヌクレオチド改変体または非GATポリヌクレオチドに到達するための多様化手順での使用のための親ポリヌクレオチドの価値ある供給源であり得る。例えば、ほとんどまたは全く検出されない活性を有するプロテアーゼ改変体をコードするヌクレオチド配列は、高活性プロテアーゼをコードする子孫を産生するためにDNAシャッフリング実験において親のポリヌクレオチドとして使用される(Nessら、(1999)Nature Biotechnology 17:893−96)。
多様性生成方法または再帰的配列組換え(「RSR])法(例えば、DNAシャッフリング)によって産生されたポリヌクレオチド配列は、本発明の特徴である。本明細書中で記載されている核酸を使用する変異方法および組換え方法は、本発明の特徴である。例えば、本発明の1つの方法は、上記および以下に記載される1つ以上の本発明のヌクレオチド配列を、1以上のさらなる配列と再帰的に組換える工程を包含する。この組換え工程は必要に応じて、インビボ、エキソビボ、インシリコまたはインビトロで実行される。前記の多様性生成または再帰的な配列組換えは、組換え改変GATポリヌクレオチドの少なくとも一つのライブラリーを生成する。このライブラリーのメンバーによってコードされるポリペプチドは、本発明に含まれる。
オリゴヌクレオチドとしてもまた本明細書中で言及されるポリヌクレオチド(代表的に少なくとも12塩基、好ましくは少なくとも15塩基、さらに好ましくは少なくとも20、30、もしくは50またはそれ以上の塩基を有する)の使用もまた企図され、これはストリンジェントまたは高ストリンジェントな条件下でGATポリヌクレオチド配列にハイブリダイズする。ポリヌクレオチドは、本明細書中に記述された方法に従って、プローブ、プライマー、センス、およびアンチセンス因子などとして使用され得る。
本発明に従って、GATポリヌクレオチド(GATポリペプチド、GATポリペプチドのフラグメント、関連する融合タンパク質、またはその機能的等価物をコードするヌクレオチド配列を含む)は、細菌細胞または植物細胞のような適切な宿主細胞でのGATポリペプチドの発現を指向する組換えDNA分子において用いられる。遺伝暗号の固有の縮重に起因して、実質的に同じアミノ酸配列または機能的に等価なアミノ酸配列をコードする他の核酸配列もまた、GATポリヌクレオチドをクローン化および発現するために使用され得る。
本発明は、続いて最終的に機能的なGATポリペプチドを産生するためにスプライシングされる転写産物および/または翻訳産物をコードする、GATポリヌクレオチドを提供する。スプライシングはインビトロまたはインビボで成し遂げられ得、そしてシススプライシングまたはトランススプライシングを含み得る。スプライシングの基質は、ポリヌクレオチド(例えば、RNA転写物)またはポリペプチドであり得る。ポリヌクレオチドのシススプライシングの例は、コード配列に挿入されたイントロンが除去され、2つの隣接したエキソン領域がスプライシングされて、GATポリペプチドコード配列を生成する場合である。トランススプライシングの例は、GATポリヌクレオチドが、別個に転写され、そして次にスプライシングされ、全長GATコード配列を形成し得る2つ以上のフラグメント内にコード配列を分けることによって暗号化される場合である。スプライシングエンハンサー配列(これは、本発明の構築物に導入され得る)の使用は、シスまたはトランスのいずれかのスプライシングを容易にし得る。ポリペプチドのシススプライシングおよびトランススプライシングは、本明細書中の他の箇所でさらに詳細に記載されている。シススプライシングおよびトランススプライシングのさらに詳細な説明は、米国特許出願第09/517,933号および同第09/710,686号に見出され得る。
したがって、いくつかのGATポリヌクレオチドは、全長GATポリペプチドを直接コードしないが、むしろGATポリペプチドのフラグメントをコードする。これらのGATポリヌクレオチドは、スプライシングに関与する機構を通して機能的なGATポリペプチドを発現するために使用され得る。ここで、スプライシングは、ポリヌクレオチド(例えば、イントロン/エキソン)および/またはポリペプチド(例えば、インテイン(intein)/イクステイン(extein))のレベルで生じ得る。スプライシングプロセスが機能的産物を産生することを可能にする実験において、すべての必要とされるフラグメントが発現する場合、機能的なGATポリペプチドが発現するだけなので、これは例えば、GAT活性の発現制御に有用であり得る。別の例では、GATポリヌクレオチドへの一つ以上の挿入配列の導入は、低い相同性のポリヌクレオチドを用いる組換えを容易にし得る;挿入配列に対するイントロンまたはインテインの使用は、介在配列の除去を容易にし、それによってコードされた改変体の機能を回復する。
当業者によって理解されるように、特定の宿主におけるその発現を増強するためのコード配列を改変する事は利点があり得る。その遺伝暗号には64の可能なコドンが縮重しているが、ほとんどの生物体は、優先的にこれらのコドンの一部を使用する。一つの種で最もよく使用されているコドンは、最適コドンと呼ばれ、それほどよく使用されないコドンは、まれなコドンまたは低使用コドンとして分類される(例えば、Zhang SPら、(1991)Gene 105: 61−72を参照のこと)。コドンは、宿主の好ましいコドン使用を反応するように置換され、このプロセスは時々「コドン最適化」または「種のコドンの傾向についての制御」と呼ばれる。
特定の原核生物または真核生物の宿主(Murray,E.ら、(1989)Nuc.Acids Res.17:477−508も参照のこと)によって好まれるコドンを含む最適化コード配列は、例えば、非最適化配列から産生された転写物と比較して、翻訳速度を増加させることまたはより長い半減期のような所望の性質を有する組換えRNA転写物を産生するために調製され得る。翻訳終止コドンはまた、宿主の優先度を反映するように改変され得る。例えば、S.cerevisiaeおよび哺乳動物に対する好ましい終止コドンは、それぞれUAAおよびUGAである。単子葉植物についての好ましい終止コドンは、UGAであり、これに対して昆虫およびE.coliは、終止コドンとしてUAAの使用を好む(Dalphin MEら、(1996)Nuc.Acids Res.24:216−218)。植物での発現のためにヌクレオチド配列を最適化するための方法論は、例えば、米国特許第6,015,891号およびその中で引用される参考文献において提供される。
本発明の一つの実施形態は、関連する宿主(例えば、トランスジェニック植物宿主)における発現のための最適コドンを有するGATポリヌクレオチドを含む。これは、細菌起源のGATポリヌクレオチドをトランスジェニック植物に導入する場合、例えば、植物にグリホセート耐性を与えるために、特に望ましい。
本発明のポリヌクレオチド配列は、種々の理由のためにGATポリヌクレオチドを変更するために設計され得、種々の理由には以下のものが挙げられるがこれらには限らない:遺伝子産物のクローニング、プロセシングおよび/または発現を改変する変更。例えば、変更は、当業者に周知である技術(例えば、部位特異的変異誘発、新しい制限部位を挿入すること、グリコシル化パターンを変更すること、コドンの優先度を変えること、スプライシング部位を導入することなど)を使用して導入され得る。
さらに詳細に本明細書中で記載されているように、本発明のポリヌクレオチドは、新規なGATポリペプチドをコードする配列、およびこのコード配列に対する相補的配列、ならびにコード配列の新規なフラグメント、およびその相補体を含む。これらのポリヌクレオチドは、RNA形態またはDNA形態であり得、そしてmRNA、cRNA、合成RNAおよび合成DNA、ゲノムDNAならびにcDNAを含む。このポリヌクレオチドは、二本鎖または一本鎖であり得、そして一本鎖の場合、コード鎖または非コード(アンチセンス、相補)鎖であり得る。このポリヌクレオチドは必要に応じて、GATポリペプチドのコード配列を以下のように含む(i)単離して、(ii)例えば、融合タンパク質、プレタンパク質、プレプロタンパク質などをコードするためのさらなるコード配列と組み合わせて、(iii)イントロンもしくはインテインのような非コード配列、プロモーター、エンハンサー、ターミネーターエレメント、または適切な宿主におけるコード配列の発現のために有効な5’および/もしくは3’非翻訳領域のような制御エレメントと組み合わせて、ならびに/あるいは(iv)GATポリヌクレオチドが異種遺伝子であるベクターまたは宿主環境。配列はまた、キャリア、緩衝液、アジュバンド、賦形剤などの存在下を含む、核酸の代表的な組成物処方と組み合わせて見出され得る。
本発明のポリヌクレオチドおよびオリゴヌクレオチドは、公知の合成法に従って、標準的な固相法によって調製され得る。代表的に、約100塩基までのフラグメントは、個々に合成され、次に実質的に任意の望ましい連続配列を形成するために結合される(例えば、酵素的連結法または化学的結合法、あるいはポリメラーゼ媒介法によって)。例えば、本発明のポリヌクレオチドおよびオリゴヌクレオチドは、例えば、Beaucageら、(1981)Tetrahedron Letters 22:1859−69によって記載される古典的なホスホラミダイト法、またはMatthesら、(1984)EMBO J. 3:801−05によって記載された方法(例えば、代表的には自動合成方法で行なわれるような方法)を使用する、化学的合成によって調製され得る。ホスホラミダイト法に従って、オリゴヌクレオチドは、例えば、自動DNA合成機で合成され、精製され、アニールされ、連結され、そして適切なベクターにクローン化される。
さらに、実質的に任意の核酸は、The Midland Certified Reagent Company(mcrc@oligos.com)、The Great American Gene Company(http://www.genco.com)、ExpressGen Inc.(www.expressgen.com)、Operon Technologies Inc.(Alameda、CA)および他多数のような任意の種々の商業的供給源から特別注文され得る。同様に、ペプチドおよび抗体は、PeptidoGenic(pkim@ccnet.com)、HTI Bio−products,Inc.(http://www.htibio.com)、BMA Biomedicals Ltd(U.K.)、Bio.Synthesis,Inc.および他多数のような任意の種々の供給源から特別注文され得る。
ポリヌクレオチドはまた、技術文献に記載されているように周知の技術によって合成され得る。例えば、Carruthersら、Cold Spring Harbor Symp.Quant.Biol.47:411−418(1982),およびAdamsら、J.Am.Chem.Soc.105:661(1983)を参照のこと。次いで、二本鎖DNAフラグメントは、相補鎖を合成し、そして適切な条件下でこれらの鎖を一緒にアニーリングすることによってか、または適切なプライマー配列を用いてDNAポリメラーゼを使用して、相補鎖を付加することによってかのいずれかで得られ得る。
変異誘発を含む本明細書中で有用な分子生物学的技術を記載する一般的なテキストとしては、以下が挙げられる:BergerおよびKimmel、Guide to Molecular Cloning Techniques,Methods in Enzymology、第152巻、Academic Press,Inc.,San Diego,CA(「Berger」);Sambookら、Molecular Cloning−A Laboratory Manual(第2版),第1〜3巻,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989(「Sambrook」);およびCurrent Prorocols in Molecular Biology,F.M.Ausubelら、編,Current Protocols,Greene Publishing Associates,Inc.とJohn Wiley&Sons,Inc.との間の合併(2000年を通じて補完された)(「Ausubel」)。ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、Qβレプリカ−ゼ増幅および他のRNAポリメラーゼ媒介技術(例えば、NASBA)を含むインビトロでの増加方法を介する、当業者を指向させるに十分な技術の例は、以下において見出される:Berger,Sambrook,およびAusubelならびにMullisら、(1987)米国特許第4,683,202号;PCR Protocols A Guide to Methods and Applications(Innisら、編)Academic Press Inc.San Diego,CA(1990);Arnheim & Levinson(1990年10月1日)Chemical and Engineering News 36−47;The Journal Of NIH Research(1991)3:81−94;Kwohら、(1989)Proc.Natl.Acad.Sci.USA 86:1173;Guatelliら、(1990)Proc.Natl.Acad.Sci.USA 87:1874;Lomellら、(1989)J.Clin.Chem.35:1826;Landegrenら、(1988)Science 241:1077−1080;Van Brunt(1990)Biotechnology 8:291−294;WuおよびWallace,(1989)Gene 4: 560;Barringerら、(1990)Gene 89:117,ならびにSooknananおよびMalek(1995)Biotechnology 13:563−564。インビトロで増幅された核酸をクローニングする改良方法は、Wallaceら、米国特許第5,426,039号に記載されている。PCRによる大きな核酸の増幅の改良方法は、Chengら、(1994)Nature 369:684−685およびその中の参考文献で要約され、ここで、40kbまでのPCRアンプリコンは生成される。当業者は、実質的な任意のRNAが、制限消化、PCR伸長ならびに逆転写酵素およびポリメラーゼを使用する配列決定に適した二本鎖DNAに変換されることを認識する。Ausbel、SambrookおよびBerger(全て上出)を参照のこと。
(配列バリエーション)
遺伝暗号の縮重に起因して、本発明のGATポリペプチドをコードするヌクレオチド配列の多数が生成され得、そのうちいくつかは、本明細書中で明白に開示された核酸配列に対する実質的な同一性を有することは、当業者によって認識される。
遺伝暗号の縮重に起因して、本発明のGATポリペプチドをコードするヌクレオチド配列の多数が生成され得、そのうちいくつかは、本明細書中で明白に開示された核酸配列に対する実質的な同一性を有することは、当業者によって認識される。
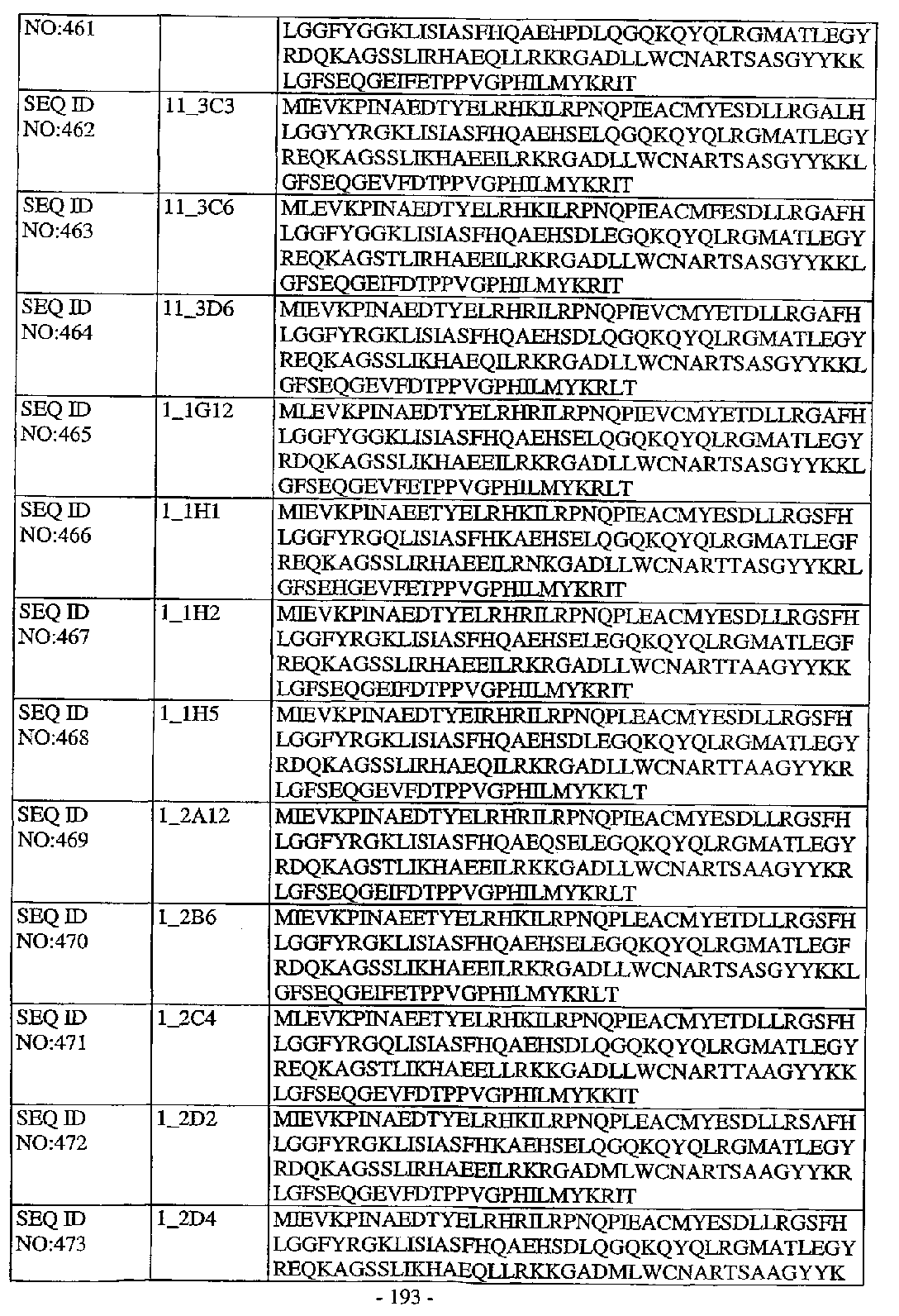
例えば、コドン表(表1)の検分は、コドンAGA、AGG、CGA、CGC、CGGおよびCGUのすべてが、アミノ酸のアルギニンをコードしていることを示す。したがって、アルギニンがコドンによって特定される本発明の核酸のすべての位置で、コドンはコードされたポリペプチドを変えずに、上記の任意の対応するコドンに変えられ得る。RNA配列のUは、DNA配列のTと対応していることは理解されている。
配列番号1(ATG ATT GAA GTC AAA)のヌクレオチド1〜15に対応する核酸配列を例として用いると、この配列のサイレントバリエーションは、AGT ATC GAG GTG AAGを含み、これらの配列は両方とも、配列番号6のアミノ酸1〜5と対応するアミノ酸配列MIEVKをコードする。
このような「サイレントバリエーション」は、「保存的改変バリエーション」の一種であり、下記で論じられている。当業者は、核酸のそれぞれのコドン(メチオニンに対する通常唯一のコドンであるAUGは除く)が標準的な技術によって改変されて、機能的に同一なポリペプチドをコードし得ることを認識する。したがって、ポリペプチドをコードする核酸のサイレントバリエーションのそれぞれは、任意の記載された配列中に潜在する。本発明は、可能なコドン選択に基づく組み合わせを選択することによって作られ得る、本発明のポリペプチドをコードする核酸配列のそれぞれおよびすべての可能なバリエーションを提供する。これらの組み合わせは、本発明のGATホモログポリペプチドをコードする核酸配列に適用される場合、標準的なトリプレット遺伝暗号(例えば、表1で示すような)に従ってなされる。本明細書中のすべての核酸のこのようなバリエーションのすべては特異的に提供され、遺伝暗号と組み合わせて配列と考慮して、記載されている。任意の改変体は、本明細書中で述べられたように生成され得る。
すべて同じアミノ酸をコードする2つ以上異なるコドンのグループは、同じ前後関係で翻訳される場合、「同義コドン」として本命書中で呼ばれる。本明細書中に記載されたように、本発明のいくつかの局面において、GATポリヌクレオチドは、例えば植物宿主のような望ましい宿主生物体内での最適化されたコドン使用のために設計される。用語「最適化された」または「最適な」は、コドンの可能な限り最良の組み合わせに限定されることを意味しないが、単純には、コード配列が全体として、これが由来する前駆体ポリヌクレオチドと比較して、コドンの改良された使用をすることを示している。したがって、一つの局面において、本発明は、親コドンと比較して、例えば、植物のような、望ましい宿主生物体で優先的に使用される同義コドンによって、ヌクレオチド配列内の少なくとも一つの親コドンを置換することによってGATポリヌクレオチド改変体を産生するための方法を提供する。
特定の核酸配列の「保存的改変バリエーション」または、単純に「保存的バリエーション」とは、同一アミノ酸配列または実質的に同一なアミノ酸配列をコードする核酸、または、核酸がアミノ酸配列をコードしない場合、実質的に同一な配列をいう。当業者は、コードされた配列内で単一アミノ酸または低%のアミノ酸(代表的に5%未満、さらに代表的には4%、2%または1%未満、あるいはそれより少ない)を変更、付加または欠失する個々の置換、欠損または付加が、「保存的改変バリエーション」であり、この変更はアミノ酸の欠失、アミノ酸の付加、または化学的に類似のアミノ酸によるアミノ酸の置換を生じるということを認識する。
機能的な類似のアミノ酸を提供する保存的置換表は、当該分野で周知である。表2は、お互いに「保存的置換」であるアミノ酸を含む6つのグループを示す。
したがって、本発明の列挙されたポリペプチドの「保存的置換バリエーション」は、同じ保存的置換グループの保存的に選択されたアミノ酸による、ポリペプチド配列のアミノ酸の低%(代表的には5%未満、さらに代表的には2%未満そしてしばしば1%未満)の置換を含む。
例えば、配列番号6として本明細書中で同定されているポリペプチドの保存的置換バリエーションは、146アミノ酸のポリペプチド内で7残基(すなわち、5%のアミノ酸)まで、上記に規定された6グループに従う「保存的置換」を含む。
さらなる例において、4つの保存的置換が配列番号6のアミノ酸21〜30とに対応する領域に局在する場合、この領域の保存的置換バリエーションの例は、 RPN QPL EAC Mであり、以下:
KPQ QPV ESC M および
KPN NPL DAC V などを含み、表2に記載された保存的置換に従う(上記例では、保存的置換に下線がひかれてある)。本明細書中でのタンパク質配列の一覧表は、上記の置換表と合わせて、すべての保存的置換タンパク質の明確な表を提供する。
KPQ QPV ESC M および
KPN NPL DAC V などを含み、表2に記載された保存的置換に従う(上記例では、保存的置換に下線がひかれてある)。本明細書中でのタンパク質配列の一覧表は、上記の置換表と合わせて、すべての保存的置換タンパク質の明確な表を提供する。
最終的に、核酸分子のコードされた活性を変えない配列の付加(例えば、非機能的配列または非コード配列の付加)は、基本核酸の保存的なバリエーションである。
当業者は、開示された核酸構築物の多数の保存的バリエーションが、機能的に同一な構築物を生じることを認識する。例えば、上記で記載されたように、遺伝暗号の縮重に起因して、「サイレント置換」(すなわち、コードされたポリペプチドでの変化を生じない核酸配列の置換)は、アミノ酸をコードするすべての核酸配列の、意味される特徴である。同様に、アミノ酸配列内での1またはいくつかのアミノ酸の「保存的アミノ酸置換」は、非常に類似した特性を有する異なるアミノ酸に置換され、また、開示された構築物に非常に類似するので容易に同定される。このような開示されたそれぞれの配列の保存的バリエーションは、本発明の特徴である。
特定の核酸の非保存的な改変は、保存的置換として特徴付けられない任意のアミノ酸を置換する改変である。例えば、6つのグループの境界を越える任意の置換は、表2に示されている。これらは、中性アミノ酸に対する塩基性または酸性のアミノ酸の置換(例えば、Val、Ile、LeuまたはMetに対するAsp、Glu、AsnまたはGln)、塩基性または酸性のアミノ酸に対する芳香族アミノ酸の置換(例えば、Asp、Asn、GluまたはGlnに対するPhe、TyrまたはTrp)あるいはアミノ酸を類似のアミノ酸で置換しない任意の他の置換、を含む。
(核酸ハイブリダイゼーション)
核酸は、それらが(代表的には溶液中で)会合する場合に、「ハイブリダイズ」する。核酸は、水素結合、溶媒排除、塩基スタッキングなどのような、よく特徴付けられた物理化学的な種々の力に起因してハイブリダイズする。核酸のハイブリダイゼーションへの広範な指針は、以下で見出される、Tijssen(1993)Laboratory Techniques in Biochemistry and Molecular Biology−−Hybridization with Nucleic Acid Probes,第I部,第2章,「Overview of principles of hybridization and the strategy of nucleic acid probe assays,」(Elsevier,New York)、ならびにAusubel,上出、HamesおよびHiggins(1995)Gene Probes 1,IRL Press、Oxford University Press,Oxford,England(HamesおよびHiggins 1)ならびにHamesおよびHiggins(1995)Gene Probes 2,IRL Press、Oxford University Press,Oxford,England(HamesおよびHiggins 2)はオリゴヌクレオチドを含むDNAおよびRNAの合成、標識、検出および定量に関する詳細を提供する。
核酸は、それらが(代表的には溶液中で)会合する場合に、「ハイブリダイズ」する。核酸は、水素結合、溶媒排除、塩基スタッキングなどのような、よく特徴付けられた物理化学的な種々の力に起因してハイブリダイズする。核酸のハイブリダイゼーションへの広範な指針は、以下で見出される、Tijssen(1993)Laboratory Techniques in Biochemistry and Molecular Biology−−Hybridization with Nucleic Acid Probes,第I部,第2章,「Overview of principles of hybridization and the strategy of nucleic acid probe assays,」(Elsevier,New York)、ならびにAusubel,上出、HamesおよびHiggins(1995)Gene Probes 1,IRL Press、Oxford University Press,Oxford,England(HamesおよびHiggins 1)ならびにHamesおよびHiggins(1995)Gene Probes 2,IRL Press、Oxford University Press,Oxford,England(HamesおよびHiggins 2)はオリゴヌクレオチドを含むDNAおよびRNAの合成、標識、検出および定量に関する詳細を提供する。
サザンハイブリダイゼーションおよびノーザンハイブリダイゼーションのような核酸のハイブリダイゼーションの実験の文脈において、「ストリンジェントなハイブリダイゼーション洗浄条件」は、配列依存性であり、そして異なる環境パラメーターの下では異なる。核酸のハイブリダイゼーションに対する広範な指針は、Tijssen(1993),上出,ならびにHamesおよびHiggins 1そしてHamesおよびHiggins 2,上出で見出される。
本発明の目的のために、一般的に「高度にストリンジェント」なハイブリダイゼーション条件および洗浄条件は、規定されたイオン強度およびpHでの特定配列についての、融解点(Tm)よりおよそ5℃以下になるように選択される(以下に記すように、高度にストリンジェントな条件とは比較の語句でも言及され得る)。Tmは、試験配列中の50%が、完全に一致したプローブとハイブリダイズする(規定されたイオン強度およびpHの下での)温度である。非常にストリンジェントな条件は、特定のプローブについてのTmと等しくなるように選択される。
核酸二重鎖のTmは、所定の条件下で二重鎖が50%変性される温度を示し、そして核酸ハイブリッドの安定性の直接的な尺度を表す。従って、Tmは、へリックスからランダムコイルへの移行の中間点に対応する温度に対応する;Tmは、長さ、ヌクレオチド組成およびヌクレオチドの長いストレッチについてのイオン強度に依存する。
ハイブリダイゼーション後、ハイブリダイズしていない核酸物質は、一連の洗浄により除去され得、洗浄のストリンジェンシーは所望される結果に依存して調整され得る。低いストリンジェンシーの洗浄条件(例えば、より高い塩濃度およびより低い温度を用いた条件)は、感度を増加するが、非特異的なハイブリダイゼーションのシグナルおよび高いバックグラウンドのシグナルを生じ得る。より高度なストリンジェンシーの条件(例えば、より低い塩濃度および、ハイブリダイゼーションの温度に近い、より高い温度を用いた条件)は、バックグラウンドシグナルを低下させ、代表的に特異的なシグナルのみが残存する。Rapley,R.およびWalker,J.M.編、Molecular Biomethods Handbook(Humana Press,Inc.1998)(本明細書中でこれより以下、「RapleyおよびWalker」)を参照のこと、この文献は本明細書中にて参考としてその全体があらゆる目的で援用される。
DNA−DNA二重鎖のTmは、以下のような式1を用いて見積もられ得る:
Tm(℃)=81.5℃+16.6(log10M)+0.41(%G+C)−0.72(%f)−500/n、
ここで、Mは1価陽イオン(普通はNa+)のモル濃度、(%G+C)はグアノシン(G)およびシトシン(C)ヌクレオチドのパーセント、(%f)はホルムアミドのパーセント、およびnはハイブリッドのヌクレオチド塩基数(すなわち、長さ)である。RapleyおよびWalker、上記を参照のこと。
Tm(℃)=81.5℃+16.6(log10M)+0.41(%G+C)−0.72(%f)−500/n、
ここで、Mは1価陽イオン(普通はNa+)のモル濃度、(%G+C)はグアノシン(G)およびシトシン(C)ヌクレオチドのパーセント、(%f)はホルムアミドのパーセント、およびnはハイブリッドのヌクレオチド塩基数(すなわち、長さ)である。RapleyおよびWalker、上記を参照のこと。
RNA−DNA二重鎖のTmは、以下のような式2を用いることで見積もられ得る:
Tm(℃)=79.8℃+18.5(log10M)+0.58(%G+C)−11.8(%G+C)2−0.56(%f)−820/n、
ここで、Mは1価陽イオン(普通は、Na+)のモル濃度、(%G+C)はグアノシン(G)およびシトシン(C)ヌクレオチドのパーセント、(%f)はホルムアミドのパーセント、およびnはハイブリッドのヌクレオチド塩基数(すなわち、長さ)である。同上。
Tm(℃)=79.8℃+18.5(log10M)+0.58(%G+C)−11.8(%G+C)2−0.56(%f)−820/n、
ここで、Mは1価陽イオン(普通は、Na+)のモル濃度、(%G+C)はグアノシン(G)およびシトシン(C)ヌクレオチドのパーセント、(%f)はホルムアミドのパーセント、およびnはハイブリッドのヌクレオチド塩基数(すなわち、長さ)である。同上。
式1および式2は典型的に約100〜200ヌクレオチドより長いハイブリッド二重鎖に対してのみ正確である。同上。
50ヌクレオチドより短い核酸配列のTmは、次のように計算され得る:
Tm(℃)=4(G+C)+2(A+T)、
ここで、A(アデニン)、C、T(チミン)、およびGは対応するヌクレオチドの数である。
Tm(℃)=4(G+C)+2(A+T)、
ここで、A(アデニン)、C、T(チミン)、およびGは対応するヌクレオチドの数である。
サザンブロットまたはノーザンブロットにおいてフィルター上の100より多いの相補的な残基を有する相補的な核酸のハイブリダイゼーションのためのストリンジェントなハイブリダイゼーション条件の例は、42℃で1mgのへパリンを補充した50%ホルマリンであり、ハイブリダイゼーションを一晩実施することである。ストリンジェントな洗浄条件の例は、65℃で15分間の0.2×SSC洗浄である(SSC緩衝液に関する記述はSambrookら、上記、を参照のこと)。しばしば、バックグラウンドのプローブシグナルを除くために、高度なストリンジェンシーの洗浄が、低いストリンジェンシーの洗浄に勝る。低いストリンジェンシーの洗浄の例は、40℃で15分間の2×SSCである。
一般的に、特定のハイブリダイゼーションアッセイにおいて無関係のプローブに見られるノイズ比シグナルよりも、2.5倍〜5倍(またはさらに高い)のノイズ比シグナルは、特異的なハイブリダイゼーションの検出を示す。本発明の状況下にある2つの配列間の少なくともストリンジェントなハイブリダイゼーションの検出は、例えば、本明細書中の配列表にて提供される、本発明の核酸に対する比較的強い構造類似性または相同性を示す。
記されるように、「高度にストリンジェント」な条件は、規定されたイオン強度およびpHでの特定の配列についての熱融点(Tm)よりも約5℃以下となるように選択される。目的のヌクレオチド配列(例えば「プローブ」)に近い関係か、または同一である標的配列は高度にストリンジェントな条件下で同定され得る。低度にストリンジェントな条件は、より相補性の低い配列に対して適切である。例えば、上記のRapleyおよびWalkerを参照のこと。
比較ハイブリダイゼーションを、本発明の核酸を同定するために使用し得、そしてこの比較ハイブリダイゼーションの方法は、本発明の核酸を区別する好ましい方法である。本発明の状況下における2つのヌクレオチド配列間での高度にストリンジェントなハイブリダイゼーションの検出は、例えば本明細書中の配列表に提供される核酸に対して、比較的強い構造類似性/構造相同性を示す。2つのヌクレオチド配列間の高度にストリンジェントなハイブリダイゼーションは、ストリンジェントなハイブリダイゼーション条件により検出されるものよりも高い程度の、構造の類似性または相同性、ヌクレオチド塩基組成の類似性または相同性、配置または順序の類似性または相同性を示す。特に、本発明の状況下における高度にストリンジェントなハイブリダイゼーションの検出は、例えば、本明細書中の配列表に提供される核酸に対する、強い構造類似性または構造相同性(例えば、核酸構造、塩基組成、塩基配置または塩基順序)を示す。例えば、ストリンジェントな条件下で本明細書中の例示的な核酸に対してハイブリダイズする試験核酸を同定することが所望される。
従って、ストリンジェントなハイブリダイゼーションの1つの尺度は、列挙された核酸(例えば、配列番号1から配列番号5の核酸配列、および配列番号11から配列番号262の核酸配列、ならびにそれらの相補的なポリヌクレオチド配列)の1つに対して高度にストリンジェントな条件(または非常にストリンジェントな条件、または超高度なストリンジェンシーのハイブリダイゼーションの条件、または超超高度なストリンジェンシーのハイブリダイゼーションの条件)下でハイブリダイズする能力である。ストリンジェントなハイブリダイゼーション(ならびに、高度にストリンジェントなハイブリダイゼーション、超高度にストリンジェントなハイブリダイゼーション、または超超高度にストリンジェントなハイブリダイゼーション)条件およびストリンジェントな洗浄条件は、任意の試験核酸について経験的に容易に決定され得る。例えば、高度にストリンジェントなハイブリダイゼーション条件および高度にストリンジェントな洗浄条件を決定する際に、ハイブリダイゼーション条件および洗浄条件は、選択された基準の組み合わせを満たすまで、(例えば、ハイブリダイゼーションまたは洗浄における温度の上昇、塩濃度の減少、界面活性剤濃度の増加および/または有機溶媒(例えば、ホルマリン)の濃度の増加により)次第に強められる。例えば、ハイブリダイゼーション条件および洗浄条件は、配列番号1〜配列番号5および配列番号11〜配列番号262より選択される1つ以上の核酸配列およびそれらの相補的なポリヌクレオチド配列を含むプローブが、完全に一致する相補的な標的(再び、配列番号1〜配列番号5および配列番号11〜配列番号262より選択される1つ以上の核酸配列およびそれらの相補的なポリヌクレオチド配列を含む、核酸)に対して、非合致標的に対するプローブのハイブリダイゼーションにて観察されるノイズ比シグナルの、少なくとも約2.5倍、場合によっては約5倍以上の強さのノイズ比シグナルで結合するまで、次第に強められる。この場合、非合致標的は、本願出願時点で公共データベース(例えばGenBankTM)に在る核酸(添付の配列表中の核酸以外)に対応する核酸である。当業者はGenBankにおいてそのような配列を同定し得る。例としては、登録番号Z99109およびY09476が挙げられる。当業者はさらなるそのような配列を、例えばGenBankにおいて、同定し得る。
試験核酸は、完全に合致する相補的な標的に対する場合の少なくとも1/2の割合でプローブとハイブリダイズする場合に、すなわち、完全に合致するプローブが、登録番号Z99109および登録番号Y09476の任意の非合致のポリヌクレオチドに対するハイブリダイゼーションに観察されるノイズ比シグナルより少なくとも約2倍〜10倍、場合によっては20倍、50倍以上であるノイズ比シグナルで、完全に合致する相補的標的と結合する条件下でのプローブの標的に対するハイブリダイゼーションの少なくとも1/2の高さのノイズ比シグナルで、プローブ核酸に対して特異的にハイブリダイズすると述べられる。
超高度なストリンジェンシーのハイブリダイゼーション条件および洗浄条件とは、ハイブリダイゼーション条件および洗浄条件のストリンジェンシーが、完全に合致する相補的な標的核酸に対するプローブの結合についてのノイズ比シグナルが、任意の非合致の標的核酸(Genbank登録番号Z99109および登録番号Y09476)に対するプローブのハイブリダイゼーションに観察されるノイズ比シグナルの、少なくとも10倍の高さになるまで強められる。完全に一致する相補的な標的核酸に対するノイズ比シグナルの少なくとも1/2のノイズ比シグナルを伴い、そのような条件下でプローブとハイブリダイズする標的核酸は、超高度なストリンジェンシーの条件下でプローブに結合すると述べられる。
同様に、ストリンジェンシーがより高度のレベルであっても、関連のハイブリダイゼーションアッセイについてのハイブリダイゼーション条件および/または洗浄条件を次第に強めることにより決定され得る。例えば、完全に一致する相補的な標的核酸へのプローブの結合に対するノイズ比シグナルが、任意の非一致の標的核酸Genbank登録番号Z99109および登録番号Y09476に対するハイブリダイゼーションに観察されるノイズ比シグナルの少なくとも10倍、20倍、50倍、100倍、または500倍以上に高度になるまで、ハイブリダイゼーション条件および洗浄条件のストリンジェンシーが強められる、ストリンジェンシーのレベルである。完全に一致する相補的な標的核酸の少なくとも1/2のノイズ比を伴う、そのような条件の下でプローブにハイブリダイズする標的核酸は、超超高度なストリンジェンシーの条件の下でプローブに対して結合すると述べられる。
高度、超高度、または超超高度なストリンジェンシーの条件の下で配列番号1〜配列番号5および配列番号11〜配列番号262に表される核酸にハイブリダイズする標的核酸は、本発明の1つの様相である。そのような核酸の例としては、所定の核酸配列と比較して1つまたは数個のサイレントな核酸置換または保存的な核酸の置換が挙げられる。
ストリンジェントな条件下でお互いにハイブリダイズしない核酸は、それらのコードするポリペプチドが実質的に同一である場合、なお実質的に同一である。このことが起こるのは、例えば、既知のヌクレオチド配列(GenBank登録番号CAA70664を含む)によりコードされるポリペプチドを使用して差し引かれた、配列番号6〜配列番号10および配列番号263〜配列番号514の遺伝子コードによって許容される最大コドン縮重を使用して核酸のコピーが生産される場合、または、既知のヌクレオチド配列(GenBank登録番号CAA70664を含む)によりコードされるポリペプチドを使用して差し引かれた、配列番号6〜配列番号10および配列番号263〜配列番号514のうちの1つ以上に対して抗血清が作製される場合、である。本発明のポリペプチドの免疫学的な同一性についてのさらなる詳細を以下に見出す。さらに、約100ヌクレオチド未満の配列を有する二重鎖の間の識別のため、当該分野の当業者に公知のTMAC1ハイブリダイゼーション手順を使用し得る。例えば、Sorg,U.ら、1 Nucleic Acids Res.(Sept.11,1991)19(17)(本明細書中に参考としてその全体が全ての目的のために援用される)を参照のこと。
1つの局面として、本発明は配列番号1〜配列番号5および配列番号11〜配列番号262より選択される核酸における独特な部分配列を含む核酸を提供する。独特な部分配列は、GenBank登録番号Z99109および登録番号Y09476のうちのいずれかに対応する核酸と比較して独特である。そのような独特な部分配列は、GenBank登録番号Z99109、登録番号Y09476または本願の出願日の時点において公共データベース中で利用可能な他の関連配列により表される、核酸の完全なセットに対して、配列番号1〜配列番号5および配列番号11〜配列番号262のうちの任意の配列を整列させることにより決定され得る。デフォルトパラメーターにセットしたBLASTアルゴリズムを使用して整列を実施し得る。任意の独特な部分配列は、例えば、本発明の核酸を同定するためのプローブとして有用である。
同様に、本発明は配列番号6〜配列番号10および配列番号263〜配列番号514から選択されるポリペプチド中の独特な部分配列を含むポリペプチドを包含する。ここでは、独特の部分配列は、GenBank登録番号CAA70664に対応するポリペプチドと比較して独特である。再度ここでは、ポリペプチドは登録番号CAA70664により表される配列に対して整列される。その配列が偽遺伝子のような非翻訳配列に対応する場合、対応するポリペプチドは単に核酸配列のアミノ酸配列へのインシリコでの翻訳により作製されることに注意するべきであり、そこでは読み取り枠が相同なGATポリヌクレオチドの読み取り枠に対応する様に選択される。
本発明はまた、配列番号6〜配列番号10および配列番号263〜配列番号514から選択されるポリペプチド中の独特な部分配列をコードする、独特なコードオリゴヌクレオチドに対して、ストリンジェントな条件下でハイブリダイズする標的核酸を提供し、ここで独特の部分配列は任意のコントロールのポリペプチドに対応するポリペプチドと比較して独特である。独特な配列は、上に記載したように決定される。
1つの例において、ストリンジェントな条件は、コードするオリゴヌクレオチドに対して完全に相補的なオリゴヌクレオチドが、任意のコントロールポリペプチドに対応するコントロール核酸に対する完全に相補的なオリゴヌクレオチドのハイブリダイゼーションよりも、少なくとも約2.5倍〜10倍高い、好ましくは少なくとも約5〜10倍高いノイズ比シグナルで、コードするオリゴヌクレオチドにハイブリダイズするように、選択される。使用される特定のアッセイにおいてより高い(例えば約15倍、約20倍、約30倍、約50倍またはそれ以上)ノイズ比シグナルが見られるような条件を選択し得る。この例では、標的核酸は、コードするオリゴヌクレオチドに対するコントロール核酸のハイブリダイゼーションと比較して、少なくとも2倍高いノイズ比シグナルを有して独特なコードオリゴヌクレオチドに対してハイブリダイズする。再び、より高いノイズ比シグナル(例えば約2.5倍、約5倍、約10倍、約20倍、約30倍、約50倍またはそれ以上)を選択し得る。特定のシグナルは関連するアッセイにおいて使用される標識(例えば、蛍光ラベル、比色ラベル、放射性ラベルなど)に依存する。
(ベクター、プロモーターおよび発現系)
本発明はまた、上で広範に記載した1つ以上の核酸配列を含む組換え構築物を包含する。その構築物は、本発明の核酸配列が順方向または逆方向にて組み込まれたベクター(例えば、プラスミド、コスミド、ファージ、ウイルス、細菌人工染色体(BAC)、酵母人工染色体(YAC)など)を含む。この実施形態の好ましい局面において、その構築物は、配列に作動可能に連結された調節配列(例えば、プロモーターを含む)をさらに含む。多くの適切なベクターおよびプロモーターは当該分野の当業者に公知であり、そして市販で利用可能である。
本発明はまた、上で広範に記載した1つ以上の核酸配列を含む組換え構築物を包含する。その構築物は、本発明の核酸配列が順方向または逆方向にて組み込まれたベクター(例えば、プラスミド、コスミド、ファージ、ウイルス、細菌人工染色体(BAC)、酵母人工染色体(YAC)など)を含む。この実施形態の好ましい局面において、その構築物は、配列に作動可能に連結された調節配列(例えば、プロモーターを含む)をさらに含む。多くの適切なベクターおよびプロモーターは当該分野の当業者に公知であり、そして市販で利用可能である。
ベクター、プロモーターおよび多くの他の関連題材の使用を含む、本明細書中で有用な分子生物学技術を記載した、一般的なテキストとして、BergerおよびKimmel、Guide to Molecular Cloning Techniques、Methods in Enzymology、152巻、Academic Press,Inc.、San Diego、CA(Berger);Sambrookら、Molecular Cloning−A Laboratory Manual(第2版)、1−3巻、Cold Spring Harbor Laboratory,Cold Spring Harbor,New York、1989(「Sambrook」)、およびCurrent Protocols in Molecular Biology、F.M.Ausubelら編、Current Protocols、Greene Publishing Associates,Inc.とJohn Wiley & Sons,Inc.の合弁事業(1999年まで追補された)(「Ausubel」)が挙げられる。例えば、本発明の相同核酸を生産するための、ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、Qβレプリカーゼ増幅、および他のRNAポリメラーゼに媒介される技術(例えば、NASBA)を含む、インビトロ増幅方法を介して、当業者を指示するのに十分なプロトコルの例は、Berger、Sambrook、およびAusubel、ならびにMullisら(1987)米国特許第4,683,202号;PCR Protocols A Guide to Methods and Applications(Innisら編)Academic Press Inc.San Diego,CA(1990)(Innis);Arnheim & Levinson(1990年10月1日)C & EN 36−47;The Journal Of NIH Research(1991)3,81−94;(Kwohら(1989)Proc.Natl.Acad.Sci.USA 86,1173;Guatelliら(1990)Proc.Natl.Acad.Sci.USA 87,1874;Lomellら(1989)J.Clin.Chem 35,1826;Landegrenら(1988)Science 241,1077−1080;Van Brunt(1990)Biotechnology 8,291−294;WuおよびWallace,(1989)Gene 4,560;Barringerら(1990)Gene 89,117、ならびにSooknananおよびMalek(1995)Biotechnology 13:563−564において、見出される。インビトロで増幅された核酸をクローニングするための改良された方法は、Wallaceら、米国特許第5,426,039号中に記載される。PCRにより大規模に核酸を増幅するために改良された方法は、Chengら(1994)Nature 369:684−685およびそこで引用される参考文献においてまとめられており、ここで、40kbにも及ぶPCRアンプリコンが生成される。当業者は、逆転写酵素およびポリメラーゼを使用して、制限消化、PCR伸長および配列決定に適した二本鎖DNAへと、実質的に任意のRNAを変換し得ることを理解する。例えば、Ausubel、Sambrook、およびBerger(すべて同上)を参照のこと。
本発明はまた、本発明のベクター(例えば、本発明のクローニングベクター、または本発明の発現ベクター)を形質導入した(形質転換した、またはトランスフェクトした)、操作された宿主細胞、ならびに組換え技術による本発明のポリペプチドの生産に関連する。そのベクターは、例えば、プラスミド、ウイルス粒子、ファージなどであり得る。プロモーター活性化、形質転換体の選択、またはGATホモログ遺伝子の増幅に対して適切であるように改変された従来の栄養培地中で、操作された宿主細胞を培養し得る。培養条件(例えば、温度、pHなど)は、発現のために選択された宿主細胞で従来から用いられる条件であり、そして当業者に明らかであり、本明細書中に記載される参考文献(例えば、Sambrook、AusubelおよびBerger、ならびに、例えば、Freshney(1994)Culture of Animal Cells、a Manual of Basic Technique、第3版、Wiley−Liss、New York、およびそれらで引用される参考文献が挙げられる)中にある。
本発明のGATポリペプチドを、非動物細胞(例えば、植物、酵母、真菌、細菌など)において生産し得る。Sambrook、BergerおよびAusubelに加えて、非動物細胞培養に関する詳細を、Payneら(1992)Plant Cell and Tissue Culture in Liquid Systems John Wiley & Sons,Inc.New York、NY;GamborgおよびPhillips(編)(1995)Plant Cell、Tissue and Organ Culture;Fundamental Methods Springer Lab Manual、Springer−Verlag(Berlin Heidelberg New York)、ならびにAtlasおよびParks(編)The Handbook of Microbiological Media(1993)CRC Press、Boca Raton、FLに見出し得る。
本発明のポリヌクレオチドを、ポリペプチドの発現に適した様々な発現ベクターのいずれかに組み込み得る。適切なベクターは、染色体、非染色体および合成DNA配列(例えば、SV40誘導体;細菌プラスミド、ファージDNA、バキュロウイルス、酵母プラスミド、ならびに、プラスミドとファージDNA、ウイルスDNA(例えば、ワクシニア、アデノウイルス、鶏痘ウイルス、仮性狂犬病、アデノウイルス、アデノ随伴ウイルス、レトロウイルスおよび多くの他のウイルス)の組み合わせに由来するベクター、が挙げられる。遺伝子物質を細胞へと形質導入させる全てのベクター、および複製が所望される場合には、関連宿主において複製可能かつ生存可能な全てのベクターが、使用され得る。
発現ベクターに組み込まれた場合、本発明のポリヌクレオチドは、mRNA合成を指示する適切な転写制御配列(プロモーター)と作動可能に連結される。特に、トランスジェニック植物における使用に適切なそのような転写制御配列の例としては、カリフラワーモザイクウイルス(CaMV)、ゴマノハグサ(figwort)モザイクウイルス(FMV)、およびイチゴベインバンディングウイルス(SVBV)プロモーターが挙げられ、米国仮特許出願番号60/245,354号に記載される。原核生物細胞、もしくは真核生物細胞、またはそれらのウイルスにおいて遺伝子発現を制御することが公知の他のプロモーター、および本発明のいくつかの実施形態において使用され得るプロモーターとしては、SV40プロモーター、E.coli lacプロモーターまたはtrpプロモーター、ファージλPLプロモーターが挙げられる。発現ベクターは、翻訳開始のためのリボソーム結合部位、および転写ターミネーターを必要に応じて含む。そのベクターはまた、発現を増幅させるための適切な配列(例えば、エンハンサー)を必要に応じて含む。さらに、本発明の発現ベクターは、形質転換した宿主細胞を選択するための表現形質を提供するために、1つ以上の選択可能なマーカー遺伝子(例えば、真核生物細胞培養用のジヒドロ葉酸レダクターゼもしくはネオマイシン耐性、または、E.coli内でのテトラサイクリン耐性もしくはアンピシリン耐性など)を必要に応じて含む。
本発明のベクターは、宿主に本発明のタンパク質またはポリペプチドを発現させるために、適切な宿主を形質転換するために利用され得る。適切な発現宿主の例としては:細菌細胞(例えば、E.coli、B.subtilis、Streptomyces、およびSalmonella typhimurium);真菌細胞(例えば、Saccharomyces cerevisiae、Pichia pastorisおよびNeurospora crassa);昆虫細胞(例えば、DrosophilaおよびSpodoptera frugiperda);哺乳動物細胞(例えば、CHO、COS、BHK、HEK 293またはBowes黒色腫;または植物細胞もしくは外植体など、が挙げられる。細胞または細胞株の全てが完全に機能するGATポリペプチドを生産する機能を有する必要はないことが理解される;例えば、GATポリペプチドの抗原性フラグメントが生産され得る。本発明は、使用される宿主細胞により制限されない。
細菌系において、多くの発現ベクターは、GATポリペプチドに意図される使用に依存して、選択され得る。例えば、GATポリペプチドまたはそのフラグメントの大部分が、商業的生産または抗体誘導に必要とされる場合、容易に精製される融合タンパク質の高レベル発現を指示するベクターが所望され得る。そのようなベクターとしては、BLUESCRIPT(Stratagene)のような多機能のE.coliクローニングベクターおよび発現ベクター(ここではGATポリペプチドのコード配列が、ハイブリッドタンパク質を発現するように、βガラクトシダーゼのアミノ末端のMetとそれに続く7残基の配列と、インフレームでベクターに連結され得る);pINベクター(Van HeekeおよびSchuster(1989)J Biol Chem 264:5503−5509);pETベクター(Novagen,Madison WI)などが挙げられるが、それらに限定されない。
同様に、酵母Saccharomyces cerevisiaeにおいて、α因子、アルコールオキシダーゼおよびPGHのような、構成的プロモーターまたは誘導性プロモーターを含む多くのベクターが、本発明のGATポリペプチド生成のために使用され得る。総説としては、Ausubelら(上記)およびGrantら(1987;Methods in Enzymology 153:516−544)を参照のこと。
哺乳動物宿主細胞において、ウイルスベースの系を含む様々な発現系が、使用され得る。アデノウイルスが発現ベクターとして使用される場合、(例えば、GATポリペプチドの)コード配列は、後期プロモーターおよび三部分リーダー配列からなるアデノウイルス転写/翻訳複合体に中に必要に応じて連結され得る。ウイルスゲノムの非必須のE1またはE3領域へのGATポリペプチドコード領域の挿入は、感染宿主細胞においてGATを発現させ得る生存ウイルスを生じさせる(LoganおよびShenk(1984)Proc Natl Acad Sci USA 81:3655−3659)。さらに、転写エンハンサー(例えば、ラウス肉腫ウイルス(RSV)エンハンサー)は、哺乳動物宿主細胞において発現を増加させるために使用され得る。
同様に、植物細胞においては、発現は植物染色体に組み込まれた導入遺伝子より、またはエピソーム核酸もしくはウイルス核酸より細胞質内で駆動され得る。安定に組み込まれた導入遺伝子の場合では、例えば、ウイルス(例えば、CaMV)または植物由来の制御配列を使用して、本発明のGATポリペプチドの構成的または誘導性の発現を駆動し得る配列を提供することがしばしば所望される。多くの植物由来の制御配列が記載されており、組織特異的な様式にて発現を指示する配列(例えば、TobRB7、パタチンB33、GRP遺伝子プロモーター、rbcS−3Aプロモーターなど)を含む。あるいは、高レベル発現は、植物ウイルスベクター(例えば、TMV、BMVなど)の外来性配列を一過的に発現させることにより達成され得る。典型的に、構成的に本発明のGATポリヌクレオチドを発現するトランスジェニック植物が好ましく、そしてGATポリペプチドの構成的な安定な発現を確実とするように、制御配列が選択される。
本発明のいくつかの実施形態において、植物細胞の形質転換に適切なGATポリヌクレオチド構築物が調製される。例えば、所望されるGATポリヌクレオチドは、植物への遺伝子導入、およびその後のコードされるポリペプチドの発現を容易にする組換え発現カセットに組み込まれ得る。発現カセットは、形質転換植物の意図される組織(例えば、植物全体、葉、種子)においてこの配列の発現を指示する、プロモーター配列ならびに他の転写および翻訳開始制御配列に作動可能に連結された、GATポリヌクレオチドまたはその機能的フラグメントを典型的に含む。
例えば、植物の全組織にてGATポリペプチドの発現を指示する、強いまたは弱い構成的な植物プロモーターが使用され得る。そのようなプロモーターは、大部分の環境条件の下、および発生または細胞分化の状態の下で活性である。構成的なプロモーターの例としては、Agrobacterium tumefaciensのT−DNAに由来する1’−または2’−プロモーター、および当業者に公知の様々な植物遺伝子由来の他の転写開始領域が挙げられる。GATポリヌクレオチドの過剰発現が植物に有害であるか、さもなくば所望されない状況においては、本開示を参照して、当業者は弱い構成的プロモーターが低レベルの発現のために使用され得ることを理解する。高レベルの発現が植物に有害ではない場合では、強力なプロモーター(例えば、t−RNAまたは他のpol IIIプロモーター、もしくは強力なpol IIプロモーター(例えば、カリフラワーモザイクウイルスプロモーター))が使用され得る。
あるいは、植物プロモーターは環境制御下であり得る。そのようなプロモーターは、本明細書で「誘導性の」プロモーターと称する。誘導性プロモーターにより転写をもたらし得る環境条件の例としては、病原体の攻撃、嫌気的な条件または光の存在が挙げられる。
本発明にて使用されるプロモーターは、「組織特異的」であり得、そして自体、ポリヌクレオチドが特定の組織(例えば、葉および種子)でのみ発現される発達制御下にあり得る。植物系に対して内在性の1つ以上の核酸配列が構築物に組み込まれる実施形態において、これら遺伝子由来の内在性プロモーター(またはその改変体)はトランスフェクトされた植物において遺伝子発現を指示するために使用され得る。組織特異的なプロモーターはまた、異種ポリヌクレオチドの発現を指示するために使用され得る。
一般的に、植物において発現カセットに使用される特定のプロモーターは、意図される用途に依存する。植物細胞において転写を指示する多くのプロモーターはどれも適切である。プロモーターは構成的かまたは誘導的のどちらかであり得る。上記のプロモーターに加え、植物において操作される微生物起源のプロモーターとして、オクトピンシンターゼプロモーター、ノパリンシンターゼプロモーターおよびネイティブのTiプラスミドに由来する他のプロモーターが挙げられる(Herrara−Estrellaら(1983)Nature 303:209−213を参照のこと)。ウイルスプロモーターとして、カリフラワーモザイクウイルスの35Sおよび19S RNAプロモーター(Odellら(1985)Nature 313:810−812)が挙げられる。他の植物プロモーターとしては、リブロース−1,3−ビスホスフェートカルボキシラーゼ小サブユニットプロモーター、およびファセオリン(phaseolin)プロモーターが挙げられる。E8遺伝子および他の遺伝子に由来するプロモーター配列がまた使用され得る。E8プロモーターの単離および配列は、DeikmanおよびFischer(1988)EMBO J.7:3315−3327に詳細に記載されている。
候補のプロモーターを同定するために、プロモーター配列に特徴的な配列について、ゲノムクローンの5’部分を解析する。例えば、プロモーター配列エレメントとして、TATAボックスのコンセンサス配列(TATAAT)が挙げられ、これは通常は転写開始部位の20塩基〜30塩基上流である。植物では、TATAボックスのさらに上流の−80位〜−100位に、3つのG(またはT)ヌクレオチドで囲まれた一連のアデニンを有する典型的なプロモーター要素があり、これは、Messingら(1983)Genetic Engineering in Plants,Kosageら(編)221−227頁に記載される。
本発明のポリヌクレオチド構築物(例えば、ベクター)の調製にあたって、プロモーター以外の配列および連結されるポリヌクレオチドが使用され得る。通常のポリペプチド発現が所望される場合、GATコード領域の3’末端にポリアデニル化領域が含まれ得る。そのポリアデニル化領域は、例えば,様々な植物遺伝子由来、またはT−DNA由来であり得る。
構築物はまた、植物細胞において選択可能な表現型を付与するマーカー遺伝子を含み得る。例えば、マーカーは、殺生剤耐性をコードし得、特に抗生物質耐性(例えば、カナマイシン耐性、G418耐性、ブレオマイシン耐性、ハイグロマイシン耐性)、または除草剤耐性(例えば、クロロスルフォロン(chlorosluforon)もしくはホスフィノトリシン(除草剤ビアラフォス(bialaphos)およびバスタ(Basta)の活性成分))をコードし得る。
特定の開始シグナルは、本発明のGATポリヌクレオチドをコードする配列の効率的な転写を補助し得る。これらのシグナルは、例えば、ATG開始コドンおよび隣接した配列を包含し得る。GATポリヌクレオチドコード配列、その開始コドンおよび上流配列が、適切な発現ベクターに挿入される場合、さらなる翻訳の制御シグナルは必要とされ得ない。しかし、コード配列(例えば、成熟タンパク質コード配列)、またはその一部のみが挿入される場合、開始コドンを含む外因性転写制御シグナルが提供されなければならない。さらに、開始コドンは、挿入物全体の転写を確実にするために正確なリーディングフレーム中に存在しなければならない。外因性の転写エレメントおよび開始コドンは、種々の起源のものであり得、天然および合成双方であり得る。発現の効率は、使用される細胞系に適切なエンハンサーを封入することにより、増強され得る(Scharf D ら(1994)Results Probl Cell Differ 20:125−62;Bittnerら(1987)Methods in Enzymol 153:516−544)。
(分泌/局在化配列)
哺乳動物細胞の所望の細胞性区画、膜もしくは細胞小器官に対してポリペプチド発現を標的化するためか、または、細胞質周辺腔もしくは細胞培養培地中にポリペプチドの分泌を指向するために、例えば、分泌/局在化配列をコードする核酸に対して、本発明のポリヌクレオチドはまた、インフレームで融合され得る。このような配列は、当業者に公知であり、そしてこれらとしては、分泌リーダーペプチド、細胞小器官標的配列(例えば、核局在化配列、ER保持シグナル、ミトコンドリア移行配列、クロロプラスト移行配列)、膜局在化/アンカー配列(例えば、終止移行配列、GPIアンカー配列)などが挙げられる。
哺乳動物細胞の所望の細胞性区画、膜もしくは細胞小器官に対してポリペプチド発現を標的化するためか、または、細胞質周辺腔もしくは細胞培養培地中にポリペプチドの分泌を指向するために、例えば、分泌/局在化配列をコードする核酸に対して、本発明のポリヌクレオチドはまた、インフレームで融合され得る。このような配列は、当業者に公知であり、そしてこれらとしては、分泌リーダーペプチド、細胞小器官標的配列(例えば、核局在化配列、ER保持シグナル、ミトコンドリア移行配列、クロロプラスト移行配列)、膜局在化/アンカー配列(例えば、終止移行配列、GPIアンカー配列)などが挙げられる。
好ましい実施形態において、本発明のポリヌクレオチドは、通常に葉緑体に標的化されるポリペプチドをコードする遺伝子由来のN末端葉緑体輸送配列(もしくは葉緑体輸送ペプチド配列)とインフレームで融合される。このような配列は、代表的には、セリンおよびスレオニンにおいて豊富であり、アスパラギン酸、グルタミン酸、およびチロシンにおいて欠失しており、一般的に正電荷のアミノ酸において豊富な中央ドメインを有する。
(発現宿主)
さらなる実施形態において、本発明は、上記の構築物を含む宿主細胞に関する。宿主細胞は真核細胞(例えば、哺乳動物細胞、酵母細胞、または植物細胞)であり得るか、あるいは、宿主細胞は原核生物(例えば、細菌細胞)であり得る。構築物の宿主細胞への導入は、リン酸カルシウムトランスフェクション、DEAE−デキストラン媒介トランスフェクション、エレクトロポレーション、または他の一般的な技術により行なわれ得る(Davis,L.,Dibner,M.およびBattey,I.(1986)Basic Methods in Molecular Biology)。
さらなる実施形態において、本発明は、上記の構築物を含む宿主細胞に関する。宿主細胞は真核細胞(例えば、哺乳動物細胞、酵母細胞、または植物細胞)であり得るか、あるいは、宿主細胞は原核生物(例えば、細菌細胞)であり得る。構築物の宿主細胞への導入は、リン酸カルシウムトランスフェクション、DEAE−デキストラン媒介トランスフェクション、エレクトロポレーション、または他の一般的な技術により行なわれ得る(Davis,L.,Dibner,M.およびBattey,I.(1986)Basic Methods in Molecular Biology)。
宿主細胞株は、必要に応じて、挿入された配列の発現を調節するかまたは所望の様式で発現されたタンパク質をプロセシングするそれらの能力について選択される。タンパク質のこのような改変は、アセチル化、カルボキシル化、グリコシル化、リン酸化、脂質化およびアシル化を含むがこれらに限定されない。タンパク質の「プレ」または「プレプロ」形態を切断する翻訳後のプロセシングはまた、正確な挿入、折り畳みおよび/または機能のために重要であり得る。E.coli、Bacillus sp.、酵母などの異なる宿主細胞もしくは、CHO、HeLa、BHK、MDCK、293、WI38などの哺乳動物細胞は、例えば、翻訳後活性について特定の細胞性機構および特徴的な機構を有し、そして導入された外来タンパク質の所望の改変およびプロセシングを確実にするように選択され得る。
組換えタンパク質の長期、高収率の生成のために、安定な発現系が使用され得る。例えば、本発明のポリペプチドを安定に発現する植物細胞、移植片もしくは組織(例えば苗条(shoot)、リーフディスク)を、ウイルスの複製起点または内因性発現エレメントおよび選択マーカー遺伝子を含む発現ベクターを使用して形質導入する。ベクターの誘導後、細胞を選択培地に切り換える前に、細胞型によって適宜(例えば、細菌細胞には1時間以上、植物細胞には1〜4日間、数種の植物移植片には2〜4週間)濃縮培地で増殖させ得る。選択マーカーの目的は、選択に対して耐性を与えることであり、そして、その存在が、導入された配列を首尾よく発現する細胞の増殖および回収を可能にする。例えば、本発明のポリペプチドを発現するトランスジェニック植物は、除草剤であるグリホセートに対する耐性について、直接選択され得る。安定して形質転換された移植片由来の耐性胚は、例えば、その細胞型に適切な組織培養技術を使用して、増殖され得る。
本発明のポリペプチドをコードする核酸配列で形質転換した宿主細胞は、必要に応じて、細胞培養物からコードされたタンパク質を発現および回収するのに適切な条件下で培養される。組換え細胞によって生成されたタンパク質またはそのフラグメントは、使用される配列および/またはベクターに依存して分泌されるか、膜結合されるか、または細胞内に含まれ得る。当業者によって理解されるように、本発明のGATポリヌクレオチドを含む発現ベクターは、原核生物膜または真核生物膜を介して成熟ポリペプチドの分泌を指向するシグナル配列を使用して設計され得る。
(付加的なポリペプチド配列)
本発明のポリヌクレオチドはまた、例えば、コードされたポリペプチドの精製を容易にするマーカー配列に対してインフレームで融合されたコード配列を含み得る。このような精製を容易にするドメインとしては、固定された金属上での精製を可能にするヒスチジン−トリプトファンモジュールのような金属キレートペプチド、グルタチオン(例えば、GST)に結合する配列、赤血球凝集素(HA)タグ(インフルエンザ赤血球凝集素タンパク質由来のエピトープに対応する;Wilsonら(1984)Cell 37:767)、マルトース結合タンパク質配列、FLAGS発現/アフィニティー精製系で利用されるFLAGエピトープ(Immunex Corp,Seattle,WA)などが挙げられるが、これらに限定されない。精製ドメインとGATホモログ配列との間のプロテアーゼ切断可能なポリペプチドリンカー配列の包含は、精製を容易にするのに有用である。本明細書中に記載される組成物および方法における使用に意図される1つの発現ベクターは、エンテロキナーゼ切断部位によって分離されるポリヒスチジン領域に融合される本発明のポリペプチドを含む融合タンパク質の発現を提供する。ヒスチジン残基は、IMIACに対する精製を容易にするが(Porathら(1992)Protein Expression and Purification 3:263−281に記載されるような、固定化された金属イオンアフィニティークロマトグラフィー)、エンテロキナーゼ切断部位は、融合タンパク質からGATホモログポリペプチドを分離するための手段を提供する。pGEXベクター(Promega;Madison,WI)はまた、グルタチオンS−トランスフェラーゼ(GST)との融合タンパク質として外来ポリペプチドを発現するために使用され得る。一般的に、このような融合タンパク質は可溶性であり、リガンド−アガロースビーズ(例えば、GST−融合の場合におけるグルタチオン−アガロース)への吸着によって溶解細胞から簡単に精製され得、その後、遊離リガンドの存在下で溶出され得る。
本発明のポリヌクレオチドはまた、例えば、コードされたポリペプチドの精製を容易にするマーカー配列に対してインフレームで融合されたコード配列を含み得る。このような精製を容易にするドメインとしては、固定された金属上での精製を可能にするヒスチジン−トリプトファンモジュールのような金属キレートペプチド、グルタチオン(例えば、GST)に結合する配列、赤血球凝集素(HA)タグ(インフルエンザ赤血球凝集素タンパク質由来のエピトープに対応する;Wilsonら(1984)Cell 37:767)、マルトース結合タンパク質配列、FLAGS発現/アフィニティー精製系で利用されるFLAGエピトープ(Immunex Corp,Seattle,WA)などが挙げられるが、これらに限定されない。精製ドメインとGATホモログ配列との間のプロテアーゼ切断可能なポリペプチドリンカー配列の包含は、精製を容易にするのに有用である。本明細書中に記載される組成物および方法における使用に意図される1つの発現ベクターは、エンテロキナーゼ切断部位によって分離されるポリヒスチジン領域に融合される本発明のポリペプチドを含む融合タンパク質の発現を提供する。ヒスチジン残基は、IMIACに対する精製を容易にするが(Porathら(1992)Protein Expression and Purification 3:263−281に記載されるような、固定化された金属イオンアフィニティークロマトグラフィー)、エンテロキナーゼ切断部位は、融合タンパク質からGATホモログポリペプチドを分離するための手段を提供する。pGEXベクター(Promega;Madison,WI)はまた、グルタチオンS−トランスフェラーゼ(GST)との融合タンパク質として外来ポリペプチドを発現するために使用され得る。一般的に、このような融合タンパク質は可溶性であり、リガンド−アガロースビーズ(例えば、GST−融合の場合におけるグルタチオン−アガロース)への吸着によって溶解細胞から簡単に精製され得、その後、遊離リガンドの存在下で溶出され得る。
(ポリペプチド生成および回収)
適切な宿主株の形質導入および適切な細胞密度に対する宿主株の増殖後、適切な手段(例えば、温度変化または化学誘導)によって、選択されたプロモーターは誘導され、そして細胞はさらなる期間培養される。細胞は、代表的には、遠心分離によって収集され、物理的または化学的手段によって破壊され、そして得られた粗抽出物をさらなる精製のために保持した。タンパク質の発現において使用される微生物細胞は、任意の簡便な方法(凍結−解凍サイクル、超音波処理、機械的破壊、もしくは細胞溶解剤の使用、または他の方法を含む)によって破壊され得、これらの方法は、当業者に周知である。
適切な宿主株の形質導入および適切な細胞密度に対する宿主株の増殖後、適切な手段(例えば、温度変化または化学誘導)によって、選択されたプロモーターは誘導され、そして細胞はさらなる期間培養される。細胞は、代表的には、遠心分離によって収集され、物理的または化学的手段によって破壊され、そして得られた粗抽出物をさらなる精製のために保持した。タンパク質の発現において使用される微生物細胞は、任意の簡便な方法(凍結−解凍サイクル、超音波処理、機械的破壊、もしくは細胞溶解剤の使用、または他の方法を含む)によって破壊され得、これらの方法は、当業者に周知である。
記載されるように、多数の参考文献が、多数の細胞(細菌、植物、動物(特に哺乳動物)および古細菌起源の細胞を含む)の培養および生成のために利用可能である。例えば、Sambrook,AusubelおよびBerger(すべて前出)、ならびに、Freshney(1994)Culture of Animal Cells,a Manual of Basic Technique、第3版、Wiely−Liss,New Yorkおよびそこに引用された参考文献;Doyle and Griffiths(1997)Mammalian Cell Culture:Essential Techniques,John Wiley and Sons,NY;Humason(1979)Animal Tissue Techniques,第4版、W.H.Freeman and Company;ならびにRicciardelliら(1989)In vitro Cell Dev.Biol.25:1016−1024を参照のこと。植物細胞培養および再分化については、Payneら(1992)Plant Cell and Tissue Culture in Liquid Systems,John Wiley&Sons,Inc.,New York,NY;GamborgおよびPhillips(編)(1995)Plant Cell,Tissue and Organ Culture;Fundamental Methods Springer Lab Manual,Springer−Verlag(Berlin Heidelberg New York);Jones(編)(1984)Plant Gene Transfer and Expression Protocols,Humana Press,Totowa,New JerseyおよびPlant Molecular Biology(1993)R.R.D.Croy編、Bios Scientific Publishers,Oxford,U.K.ISBN0 12 198370 6。一般的な細胞培養培地は、AtlasおよびParks(編)The Handbook of Microbiological Media(1993)CRC Press,Boca Raton,FLにおいて開示される。細胞培養についてのさらなる情報は、Sigma−Aldrich,Inc(StLouis,MO)からのLife Sience Research Cell Culture Catalogue(1998)(「Sigma−LSRCCC」)のような市販の文献において見出され、そして、例えば、Sigma−Aldrich,Inc(StLouis,MO)からのThe Plant Culture Catalogue and supplement(1997)(「Sigma−PCCS」)においてもまた見出される。植物細胞形質転換およびトランスジェニック植物産物に関するさらなる詳細は、以下において見出される。
本発明のポリペプチドは、当該分野で周知の多数の方法のいずれかによって、組換え細胞培養物から回収および精製され得、これらとしては、硫酸アンモニウム沈殿またはエタノール沈殿、酸性抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー(例えば、本明細書中で記載されるタギングシステムのいずれかを使用する)、ヒドロキシルアパタイトクロマトグラフィー、およびレクチンクロマトグラフィーが挙げられる。所望のように、成熟タンパク質の立体配置を完成する際、タンパク質の再折り畳み工程が使用され得る。最後に、高速液体クロマトグラフィー(HPLC)が、最終の精製工程で使用され得る。前記の参考文献に加えて、種々の精製方法が、当該分野で周知であり、Sandana(1997)Bioseparation of Preteins,Academic Press,Inc.;およびBollagら(1996)Protein Methods,第2版、Wiley−Liss,NY;Walker(1996)The Pretein Protocols Handbook,Humana Press,NJ,HarrisおよびAngal(1990)Protein Purification Applications:A Practical Approach,IRL Press at Oxford,Oxford,England;HarrisおよびAngal,Protein Purification Methods:A Practical Approach,IRL Press at Oxford,Oxford,England;Scopes(1993)Protein Purification:Principles and Practice、第3版、Springer Verlag,NY;JansonおよびRyden(1998)Protein Purification:Principles,High Resolution Methods and Applications,第2版、Wiley−VCH,NY;ならびにWalker(1998)Protein Protocols on CD−ROM,Humana Press,NJ.に開示される方法を含む。
いくつかの場合においては、工業的および/もしくは商業的適用のために適切な、本発明のGATポリペプチドを大規模に生成することが望ましい。このような場合、バルク醗酵(bulk fermentation)手順が用いられる。簡単には、GATポリヌクレオチド(例えば、配列番号1〜5および11〜262のいずれか1つを含むポリヌクレオチド、または本発明のGATポリペプチドをコードする他の核酸)は、発現ベクター中にクローン化され得る。例えば、米国特許第5,955,310号;Widnerら「METHODS FOR PRODUCING A POLYPEPTIDE IN A BACILLUS CELL」は、タンデムなプロモーターを有するベクターおよびポリペプチドコード配列に作動可能に連結された安定化配列を記載している。目的のポリペプチドをベクターに挿入したのち、ベクターは細菌性の(例えば、Bacillus subtilis PL1801IIE株(amyE、apr、npr、spoIIE::Tn917)宿主に変換される。Bacillus細胞への発現ベクターの導入は、例えば、原形質形質転換により(例えば、ChangおよびCohen(1979)Molecular General Genetics 168:111を参照)またはコンピテント細胞を用いることにより(例えば、YoungおよびSpizizin(1961)Journal of Bacteriology 81:823もしくはDubnauおよびDavidoff−Abelson(1971)Journal of Molecular Biology 56:209を参照)、またはエレクトロポレーションにより(例えば、ShigekawaおよびDower(1988)Biotechniques 6:742を参照)、または接合により(例えば、KoehlerおよびThorne(1987)Journal of Bacteriology 169:5271を参照)、また、Ausubel、SambrookおよびBerger(すべて前出)により達成される。
形質転換された細胞は、当該分野での公知な方法を用いて、ポリペプチドの産生に適した栄養培地中で培養される。例えば、細胞は、振とうフラスコ培養(shake flask cultivation)または小規模もしくは大規模な醗酵(連続、バッチ、フェドバッチ(fed−batch)、もしくは固形状態醗酵を含む)によって、実験室または適切な培地中で、かつポリペプチドが発現されおよび/もしくは単離されるのが可能な条件下で実施された工業的な醗酵槽で培養され得る。培養は、炭素源および窒素源ならびに無機塩を含む適切な栄養培地中で、当該分野で公知の手順を用いて行われる。適切な培地は、商業的な供給者から利用が可能であり、もしくは、公開された組成物に従って調製され得る(例えば、American Type Culture Collectionのカタログ中)。分泌されたポリペプチドは、培地から直接回収され得る。
得られたポリペプチドは、当該分野で公知の方法により単離され得る。例えば、このポリペプチドは、従来の手順(遠心、濾過、抽出、噴霧乾燥、エバポレーション、または沈殿などが挙げられるが、これらに限定されない)により栄養培地から単離され得る。次いで、単離されたポリペプチドを当該分野で公知の種々の方法のによって、さらに精製し得、これらの方法としては、クロマトグラフィー(例えば、イオン交換、アフィニティー、疎水性、等電点電気泳動、およびサイズ排除)、電気泳動の手順(例えば、分取用等電点電気泳動)、差次的溶解(例えば、硫酸アンモニウム沈殿)、または抽出(例えば、Bollagら(1996)Protein Methods,第2版、Wiley−Liss,NY;Walker(1996)The Protein Protocols Handbook,Humana Press,NJ;Bollagら(1996)Protein Methods,第2版、Wiley−Liss,NY;Walker(1996)The Protein Protocols Handbook,Humana Press,NJ)が挙げられるが、これらに限定されない。
無細胞転写/翻訳系もまた、本発明のDNAまたはRNAを用いてポリペプチドを産生するために使用され得る。いくつかこのような系が、市販されている。インビトロ転写および翻訳プロトコルに対する一般的なガイドは、Tymms(1995)In vitro Transcription and Translation Protocols:Methods in Molecular Biology,第37巻、Garland Publishing,NYにおいて見出される。
(配列組換えのための基質および形式)
本発明のポリヌクレオチドは、例えば、Ausubel、BergerおよびSambrookが記載するような標準的なクローニング法におけるこれらの使用、すなわち所望される特性を有するさらなるGATポリヌクレオチドおよびポリペプチドを生成するためのそれらの使用に加えて、多様な生成手順(例えば、変異、組換え、再帰的な組換え反応)のための基質として必要に応じて用いられる。多様な生成プロトコールは当該分野で利用可能であり、かつ記載されている。これらの手順はポリヌクレオチもしくはポリヌクレオチドセットの1以上の改変体、ならびにコードされるタンパク質改変体を生成するために、別々に、および/もしくは組み合わせて用いられ得る。個々におよびまとめて、これらの手順は、例えば、新規な特徴および/または改善された特徴を有する、ポリヌクレオチド、タンパク質、経路、細胞および/または生物の、技術または迅速な進化のために有用な、多様化したポリヌクレオチドおよびポリヌクレオチドのセット(例えば、ポリヌクレオチドライブラリーを含む)の、確固とした広範に適用可能な生成方法を提供する。配列を変更するプロセスは、例えば、一塩基変換、多重塩基変換、および核酸配列領域の挿入もしくは欠損をもたらし得る。
本発明のポリヌクレオチドは、例えば、Ausubel、BergerおよびSambrookが記載するような標準的なクローニング法におけるこれらの使用、すなわち所望される特性を有するさらなるGATポリヌクレオチドおよびポリペプチドを生成するためのそれらの使用に加えて、多様な生成手順(例えば、変異、組換え、再帰的な組換え反応)のための基質として必要に応じて用いられる。多様な生成プロトコールは当該分野で利用可能であり、かつ記載されている。これらの手順はポリヌクレオチもしくはポリヌクレオチドセットの1以上の改変体、ならびにコードされるタンパク質改変体を生成するために、別々に、および/もしくは組み合わせて用いられ得る。個々におよびまとめて、これらの手順は、例えば、新規な特徴および/または改善された特徴を有する、ポリヌクレオチド、タンパク質、経路、細胞および/または生物の、技術または迅速な進化のために有用な、多様化したポリヌクレオチドおよびポリヌクレオチドのセット(例えば、ポリヌクレオチドライブラリーを含む)の、確固とした広範に適用可能な生成方法を提供する。配列を変更するプロセスは、例えば、一塩基変換、多重塩基変換、および核酸配列領域の挿入もしくは欠損をもたらし得る。
明確さのために、以下の議論の中で区別および分類がなされるが、この技術は、しばしば、相互に相容れないわけではないことが認識される。実際、種々の方法が、多様な配列改変体を得るために、単独でかまたは組み合わせて、並行してかまたは連続して、使用され得る。
本明細書中に記載される任意の多様性生成手順の結果は、1つ以上のポリヌクレオチドの生成であり得、それらのポリヌクレオチドは、所望の特性を有するかまたはその特性を付与するタンパク質をコードするポリヌクレオチドについて、選択またはスクリーニングされ得る。本明細書中の方法または当業者が利用可能な方法の1つ以上による多様化に続いて、生成される任意のポリヌクレオチドが、所望される活性または特性(例えば、グリホセートの可変Km、アセチルCoAの可変Km、kcatが増大した代替的な補因子(例えばプロピオニルCoA)の使用などについて選択され得る。これは、例えば、自動化形式または自動化可能な形式で、当該分野におけるアッセイのいずれかにより検出され得る任意の活性を同定することを含み得る。例えば、増加した特異的な活性を有するGATホモログは、グリホセートのN−アセチルグリホセートへの転換をアッセイすることにより(例えば、質量分析によって)検出され得る。あるいは、グリホセートに対する抵抗を与える改善された能力は、本発明の核酸で形質転換された細菌を寒天(増加した濃度のグリホセートを含む)上で培養するか、もしくはグリホセートとともに本発明の核酸を組み込んだトランスジェニック植物をスプレーする(spraying)ことによりアッセイされ得る。種々の関連する特性が(または関連しない特性さえ)、実務家の裁量にて、連続してまたは並行して、評価され得る。除草剤の耐性のための組換えおよび選択に関するさらなる詳細は、例えば、「DNA SHUFFLING TO PRODUCE HERBICIDE RESISTANT CROPS」(1999年8月12日出願)(USSN09/373,333)内において見出され得る。
種々の多様性生成手順の詳細は、多重酵素領域をコードする改変された核酸配列を生成するためのファミリーシャッフリングおよび方法を含み、以下の出版物およびそこに引用される参考文献に見出される:Soong,N.ら(2000)「Molecular breeding of viruses」Nat Genet 25(4):436−39;Stemmerら(1999)「Molecular breeding of viruses for targeting and other clinical properties」Tumor Targeting 4:1−4;Nessら(1999)「DNA Shuffling of subgenomic sequences of subtilisin」Nature Biotechnology 17:893−896;Changら(1999)「Evolution of a cytokine using DNA family shuffling」Nature Biotechnology 17:793−797;MinshullおよびStemmer(1999)「Protein evolution by molecular breeding」Current Opinion in Chemical Biology 3:284−290;Christiansら(1999)「Directed evolution of thymidine kinase for AZT phosphorylation using DNA family shuffling」Nature Biotechnology 17:259−264;Crameriら(1998)「DNA shuffling of a family of genes from diverse species accelerates directed evolution」Nature 391:288−291;Crameriら(1997)「Molecular evolution of an arsenate detoxification pathway by DNA shuffling」Nature Biotechnology 15:436−438;Zhangら(1997)「Directed evolution of an effective fucosidase from a galactosidase by DNA shuffling and screening」Proc.Natl.Acad.Sci.USA 94:4504−4509;Pattenら(1997)「Applications of DNA Shuffling to Pharmaceuticals and Vaccines」Current Opinion in Biotechnology 8:724−733;Crameriら(1996)「Construction and evolution of antibody−phage libraries by DNA shuffling」Nature Medicine 2:100−103;Crameriら(1996)「Improved green fluorescent protein by molecular evolution using DNA shuffling」Nature Biotechnology 14:315−319;Gatesら(1996)「Affinity selective isolation of ligands from peptide libraries through display on a lac repressor「headpiece dimer」」Journal of Molecular Biology 255:373−386;Stemmer(1996)「Sexual PCR and Assembly PCR」:The Encyclopedia of Molecular Biology.VCH Publishers,New York.pp.447−457;CrameriおよびStemmer(1995)「Combinatorial multiple cassette mutagenesis creates all the permutations of mutant and wildtype cassettes」BioTechniques 18:194−195;Stemmerら(1995)「Single−step assembly of a gene and entire plasmid form large numbers of oligodeoxy−ribonucleotides」Gene,164:49−53;Stemmer(1995)「The Evolution of Molecular Computation」Science 270:1510;Stemmer(1995)「Searching Sequence Space」Bio/Technology 13:549−553;Stemmer(1994)「Rapid evolution of a protein in vitro by DNA shuffling」 Nature 370:389−391;およびStemmer(1994)「DNA shuffling by random fragmentation and reassembly:In vitro recombination for molecular evolution.」Proc.Natl Acad.Sci.USA 91:10747−10751。
多様性を生成する変異誘発方法には、例えば、以下が挙げられる:部位特異的変異誘発(Lingら(1997)「Approaches to DNA mutagenesis:an overview」Anal.Biochem.254(2):157−178;Daleら(1996)「Oligonucleotide−directed random mutagenesis using the phosphorothioate method」Methods Mol.Biol.57:369−374;Smith(1985)「In vitro mutagenesis」Ann.Rev.Genet.19:423−462;Botstein & Shortle(1985)「Strategies and applications of in vitro mutagenesis」Science 229:1193−1201;Carter(1986)「Site−directed mutagenesis」Biochem.J.237:1−7;およびKunkel(1987)「The efficiency of oligonucleotide directed mutagenesis」Nucleic Acids & Molecular Biology(Eckstein,F.およびLilley,D.M.J.編.,Springer Verlag,Berlin));テンプレートを含むウラシルを用いた変異誘発(Kunkel(1985)「Rapid and efficient site−specific mutagenesis without phenotypic selection」Proc.Natl.Acad.Sci.USA 82:488−492;Kunkelら(1987)「Rapid and efficient site−specific mutagenesis without phenotypic selection」Methods in Enzymol.154,367−382;およびBassら(1988)「Mutant Trp repressors with new DNA−binding specificities」Science 242:240−245);オリゴヌクレオチド特異的変異誘発(Methods in Enzymol.100:468−500(1983);Methods is Enzymol.154:329−350(1987);Zoller & Smith(1982)「Oligonucleotide−directed mutagenesis using M13−derived vectors:an efficient and general procedure for the production of point mutations in any DNA fragment」Nucleic Acids Res.10:6487−6500;Zoller & Smith(1983)「Oligonucleotide−directed mutagenesis of DNA fragments cloned into M13 vectors」Methods in Enzymol.100:468−500;およびZoller & Smith(1987)「Oligonucleotide−directed mutagenesis:a simple methods using two oligonucleotide primers and a single−stranded DNA templete」Methods in Enzymol.154:329−350);ホスホロチオエート修飾DNA変異誘発(Taylorら(1985)「The use of phosphorothioate−modified DNA in restriction enzyme reactions to prepare nicked DNA」Nucl.Acids Res.13:8749−8764;Taylorら(1985)「The rapid generation of oligonucleotide−directed mutations at high frequency using phosphorothioate−modified DNA」Nucl.Acids Res.13:8765−8787(1985);Nakamaye & Eckstein(1986)「Inhibition of restriction endonuclease Nci I cleavage by phosphorothioate groups and its application to oligonucleotide−directed mutagenesis」Nucl.Acids Res.14:9679−9698;Sayersら(1988)「Y−T Exonucleases in phosphorothioate−based oligonucleotide−directed mutagenesis」Nucl.Acids Res.16:791−802;およびSayersら(1988)「Strand specific cleavage of phosphorothioate−containing DNA by reaction with restriction endonucleases in the presence of ethidiurm bromide」Nucl.Acids Res.16:803−814);ギャップ二重鎖DNAを用いた変異誘発(Kramerら(1984)「The gapped duplex DNA approach to oligonucleotide−directed mutation construction」Nucl.Acids Res.12:9441−9456;Kramer & Fritz(1987)Methods in Enzymol「Oligonucleotide−directed construction of mutations via gapped duplex DNA」154.350−367;Kramerら(1988)「Improved enzymatic in vitro reactions in the gapped duplex DNA approach to oligonucleotide−directed construction of mutations」 Nucl.Acids Res.16:7207;およびFritzら(1988)「Oligonucleotide−directed construction of mutations:a gapped duplex DNA procedure without enzymatic reactions in vitro」Nucl.Acids Res.16.6987−6999)。
さらなる適切な方法としては、以下が挙げられる:点ミスマッチ修復(Kramerら、(1984)「Point Mismatch Repair」Cell 38:879−887)、修復欠陥宿主株を使用した変異誘発(Carterら(1985)「Improved oligonucleotide site−directed mutagenesis using M13 vectors」Nucl.Acids Res.13:4431−4443;およびCarter(1987)「Improved oligonucleotide−directed mutagenesis using M13 vectors」Methods in Enzymol.154:382−403)、欠失変異誘発(Eghtedarzadeh & Henikoff(1986)「Use of oligonucleotides to generate large deletions」Nucl.Acids Res.14:5115)、制限−選択および制限−精製(Wellsら(1986)「Importance of hydrogen−bond formation in stabilizing the transition state of subtilisin」Phil.Trans.R.Soc.Lond.A 317:415−423)、総遺伝子合成による変異誘発(Nambiarら(1984)「Total synthesis and cloning of a gene coding for the ribonuclease S protein」Science 223:1299−1301;SakamarおよびKhorana(1988)「Total synthesis and expression of a gene for the a−subunit of bovine rod outer segment guanine nucleotide−binding protein(transducin)」Nucl.Acids Res.14:6361−6372;Wellsら(1985)「Cassette mutagenesis:an efficient method for generation of multiple mutations at defined sites」Gene 34:315−323;およびGrundstromら(1985)「Oligonucleotide−directed mutagenesis by microscale ‘shot−gun’ gene synthesis」Nucl.Acids Res.13:3305−3316)、2本鎖切断修復(Mandecki(1986);Arnold(1993)「Protein engineering for unusual environments」Current Opinion in Biotechnology 4:450−455。「Oligonucleotide−directed double−strand break repair in plasmids of Escherichia coli:a method for site−specific mutagenesis」Proc.Natl.Acad.Sci.USA,83:7177−7181)。多くの上記方法におけるさらなる詳細は、Methods in Enzymology,第154巻に見出され得、これはまた、種々の変異誘発方法を用いたトラブルシューティング問題についての有用なコントロールを記載する。
種々の多様性生成方法に関するさらなる詳細は、以下の米国特許、PCT公開、およびEP公開に見出され得る:Stemmerに対する米国特許第5,605,793号(1997年2月25日),「Methods for In vitro Recombination;」Stemmerらに対する米国特許第5,811,238(1998年9月22日)「Methods for Generating Polynucleotides having Desired Characteristics by Iterative Selection and Recombination;」Stemmerらに対する米国特許第5,830,721号(1998年11月3日),「DNA Mutagenesis by Random Fragmentation and Reassembly」;Stemmerに対する米国特許第5,834,252号(1998年11月10日)「End−Complementary Polymerase Reaction;」Minshullに対する米国特許第5,837,458号(1998年11月17日),「Methods and Compositions for Cellular and Metabolic Engineering;」WO 95/22625,StemmerおよびCrameri,「Mutagenesis by Random Fragmentation and Reassembly;」StemmerおよびLipschutzによるWO 96/33207,「End Complementary Polymerase Chain Reaction;」StemmerおよびCrameriによるWO 97/20078,「Methods for Generating Polynucleotides having Desired Characteristics by Iterative Selection and Recombination;」MinshullおよびStemmerによるWO 97/35966,「Methods and Compositions for Cellular and Metabolic Engineering;」PunnonenらによるWO 99/41402,「Targeting of Genetic Vaccine Vectors;」PunnonenらによるWO 99/41383,「Antigen Library Immunization;」PunnonenらによるWO 99/41369,「Genetic Vaccine Vector Engineering;」PunnonenらによるWO 99/41368,「Optimization of Immunomodulatory Properties of Genetic Vaccines;」StemmerおよびCrameriによるEP 752008,「DNA Mutagenesis by Random Fragmentation and Reassembly;」StemmerによるEP 0932670,「Evolving Cellular DNA Uptake by Recursive Sequence Recombination;」StemmerらによるWO 99/23107,「Modification of Virus Tropism and Host Range by Viral Genome Shuffling;」AptらによるWO 99/21979,「Human Papillomavirus Vectors;」del CardayreらによるWO 98/31837,「Evolution of Whole Cells and Organisms by Recursive Sequence Recombination;」PattenおよびStemmerによるWO 98/27230,「Methods and Compositions for Polypeptide Engineering;」StemmerらによるWO 98/13487,「Methods for Optimization of Gene Therapy by Recursive Sequence Shuffling and Selection」WO 00/00632,「Methods for Generating Highly Diverse Libraries,」WO00/09679,「Methods for Obtaining in Vitro Recombined Polynucleotide Sequence Banks and Resulting Sequences,」ArnoldらによるWO98/42832,「Recombination of Polynucleotide Sequences Using Random or Defined Primers,」ArnoldらによるWO 99/29902,「Method for Creating Polynucleotide and Polypeptide Sequences,」VindによるWO98/41653,「An in Vitro Method for Construction of a DNA Library,」BorchertらによるWO 98/41622,「Method for Constructing a Library Using DNA Shuffling,」およびPatiおよびZarlingによるWO98/42727,「Sequence Alterations using Homologous Recombination」PattenらによるWO00/18906,「Shuffling of Codon−Altered Genes;」del CardayreらによるWO 00/04190,「Evolution of Whole Cells and Organisms by Recursive Recombination;」CrameriらによるWO00/42561,「Oligonucuclectide Mediated Nucleic Acid Recombination;」SelifonovおよびStemmerらによるWO00/42559,「Methods of Populating Data Structure for Use in Evolutionary Simulations;」SelifonovらによるWO 00/42560,「Methods for Making Character Strings,Polynucleotides & Polypeptides Having Desired Characteristics;」WelchらによるWO01/23401,「Use of Codon−Varied Oligonucleotide Synthesis for Synthetic Shuffling;」およびAffholterによるPCT/US01/06775,「Single−Stranded Nucleic Acid Template−Mediated Recombination and Nucleic Acid Fragment Isolation」。
特定の米国出願は、種々の多様性生成方法に関するさらなる詳細を提供し、これには、以下が挙げられる:「SHUFFLING OF CODON ALTERED GENES」(Pattenら、1999年9月28日出願)(USSN09/407,800);「EVOLUTION OF WHOLE CELLS AND ORGANISMS BY RECURSIVE SEQUENCE RECOMBINATION」del (Cardayreら、1998年7月15日出願)(USSN09/166,188),および1999年7月15日,(USSN09/354,922);「OLIGONUCLEOTIDE MEDIATED NUCLEIC ACID RECOMBINATION」(Crameriら,1999年9月28日出願)(USSN09/408,392);「OLIGONUCLEOTIDE MEDIATED NUCLEIC ACID RECOMBINATION」(Crameriら,2000年1月18日出願)(PCT/US00/01203);「USE OF CODON−BASED OLIGONUCLEOTIDE SYNTHESIS FOR SYNTHETIC SHUFFLING」(Welchら,1999年9月28日出願)(USSN09/408,393);「METHODS FOR MAKING CHARACTER STRINGS, POLYNUCLEOTIDES & POLYPEPTIDES HAVING DESIRED CHARACTERISTICS」(Selifonovら,2000年1月18日出願)(PCT/US00/01202);「METHODS FOR MAKING CHARACTER STRINGS,POLYNUCLEOTIDES & POLYPEPTIDES HAVING DESIRED CHARACTERISTICS」(Selifonovら,2000年7月18日出願)(USSN/09/618,579)ならびに「METHODS OF POPULATING DATA STRUCTURES FOR USE IN EVOLUTIONARY SIMULATIONS」(SelifonovおよびStemmerら、2000年1月18日出願)(PCT/US00/01138)「SINGLE−STRANDED NUCLEIC ACID TEMPLATE−MEDIATED RECOMBINATION AND NUCLEIC ACID FRAGMENT ISOLATION」Affholter(USSN 60/186,482、2000年2月2日出願)。
要約すれば、変異、組換えなどのような、いくつかの異なる配列改変方法の一般的なクラスは、本発明に適用可能であり、そして例えば上記参考文献に記載される。すなわち、成分核酸配列について、産生された改変遺伝子融合構築物への改変は、配列の結合(cojoining)の前か、または結合工程の後のいずれかで、記載される多数のプロトコルによって実施され得る。本発明の文脈において、多様性の作製のための好ましい形式の以下の例示的ないくつかの異なるタイプは、例えば、多様性作製の形式に基づいた特定の組換えを含む。
核酸は、上記参考文献において考察された種々の技術のいずれかによってインビトロで組換えられ得、例えば、組換えられる核酸のDNAse消化に続く、ライゲーションおよび/または核酸のPCR再構を含む。例えば、DNA分子のランダム(または擬似ランダム、または非ランダムでさえも)なフラグメンテーションに続く、インビトロで、異なるが関連するDNA配列を有するDNA分子間の、配列類似性に基づく組換え、次いでポリメラーゼ連鎖反応における伸長による交差の固定による、性に関するPCR変異誘発が使用され得る。このプロセスおよび多くのプロセスの変形は、上記参考文献のいくつか(例えば、Stemmer(1994)Proc.Natl.Acad.Sci.USA 91:10747〜10751)において記載される。
同様に、核酸は、例えば、細胞内の核酸の間で組換えを生じさせることによって、インビトロで再帰的に組換えられ得る。多くのこのようなインビトロでの組換えの形式は、上記に記載された参考文献に記載される。このような形式は、必要に応じて、目的の核酸の間の直接的な組換えを提供するか、または他の形式と同様に、目的の核酸を含むベクター、ウイルス、プラスミドなどの間での組換えを提供する。このような手順に関する詳細は、上記の参考文献中に見出される。
必要に応じて所望のライブラリー成分(例えば、本発明の経路に対応する遺伝子)を有するゲノムの組換え混合物のスパイキング(spiking)を含む、細胞または他の生物のゲノム全体が組換えられる、ゲノム全体の組換え方法がまた使用され得る。これらの方法は、標的遺伝子の同一性が知られていない用途を含む、多くの用途を有する。このような方法に関する詳細は、例えば、del CardayreらによるWO98/31837中の「Evolution of Whole Cells and Organisms by Recursive Sequence Recombination」;および例えば、del CardayreらによるPCT/US99/15972、同様の題名の「Evolution of Whole Cells and Organisms by Recursive Sequence Recombination」中に見出される。従って、組換え、再帰的組換え、およびゲノム全体の組換えのためのこれらのプロセスおよび技術のいずれかは、単独または組合わせて、本発明の改変された核酸配列および/または改変された遺伝子融合構築物を産生するために使用され得る。
目的の標的に対応するオリゴヌクレオチドが、2以上の親核酸に対応するオリゴヌクレオチドを含む、PCRまたはライゲーション反応において合成および再構築され、これによって新しい組換え核酸を産生する、合成的な組換え方法もまた使用され得る。オリゴヌクレオチドは、標準的なヌクレオチド付加方法によって作製され得るか、または、例えば、トリヌクレオチド合成アプローチによって作製され得る。このようなアプローチに関する詳細は、上記参考文献中に見出され、例えば、CrameriらによるWO00/42561、「Oligonucleotide(Olgonucleotide) Mediated Nucleic Acid Recombination」;WelchらによるWO01/23401、「Use of Codon−Varied Oligonucleotide Synthesis for Synthetic Shuffling」;SelifonovらによるWO00/42560、「Methods for Making Character Strings,Polynucleotides and Polypeptides Having Desired Characteristics」;ならびにSelifonovおよびStemmerによるWO00/42559「Methods of Populating Data Structures for Use in Evolutionary Simulations」を含む。
相同な(または、非相同でさえも)核酸に対応する配列のストリングを組換えるために、遺伝的アルゴリズムがコンピュータ内で使用される、インシリコでの組換えの方法が、達成され得る。その結果生じる組換えられた配列のストリングは、必要に応じて、例えば、オリゴヌクレオチド合成/遺伝子再構技術と合わせて、組換えられた配列に対応する核酸の合成によって核酸に変換される。このアプローチは、ランダム、部分的にランダムまたは設計された改変体を産生し得る。インシリコでの組換えに関する多くの詳細は、対応する核酸(および/またはタンパク質)の産生、ならびに設計された核酸および/またはタンパク質(例えば、交差位置選択に基づく)の組合わせ、ならびに設計された、擬似ランダムなまたはランダムな組換え方法と組合わせたコンピュータシステムにおける遺伝的アルゴリズム、遺伝的オペレータなどの使用を含み、SelifonovらによるWO00/42560、「Methods for Making Character Strings,Polynucleotides and Polypeptides Having Desired Characteristics」ならびにSelifonovおよびStemmerによるWO00/42559「Methods of Populating Data Structures for Use in Evolutionary Simulations」中に記載される。インシリコでの組換え方法に関する広範囲にわたる詳細は、これらの出願において見出される。この方法論は、一般的に、インシリコでの種々の代謝経路(例えば、カロチノイド生合成経路、エクトイン(ectoine)生合成経路、ポリヒドロキシアルカノエート生合成経路、芳香族ポリケチド生合成経路など)に関連するタンパク質をコードする、核酸配列および/または遺伝子融合構築物の組換えならびに/あるいは対応する核酸もしくはタンパク質の産生について提供することにおいて、本発明に適用可能である。
例えば、単鎖のテンプレートへの多種多様な核酸または核酸フラグメントのハイブリダイゼーションに続く、全長配列を再生させるための重合および/またはライゲーション、必要に応じて、テンプレートの分解および生じた改変核酸の回収によって、天然の多様性を利用する多くの方法は、同様に使用され得る。単鎖のテンプレートを利用する1つの方法において、ゲノムライブラリーに由来するフラグメントの集団は、反対側の鎖に対応するssDNAもしくはRNAの一部分、または、しばしばほぼ完全長を用いてアニールされる。次いで、この集団由来の複雑なキメラの遺伝子のアセンブリは、非ハイブリダイジングフラグメントの末端の核酸塩基の除去、このようなフラグメントとその結果生じる単鎖のライゲーションとの間のギャップを満たすための重合によって媒介される。親のポリヌクレオチド鎖は、消化(例えば、RNAまたはウラシル含有の場合)、変性条件下での磁気分離(このような分離につながる方法において標識化された場合)および他の使用可能な分離/精製方法によって除去され得る。あるいは、親鎖は、必要に応じて、キメラ鎖と一緒に精製され、そして引き続くスクリーニング工程およびプロセシング工程の間に除去される。このアプローチに関するさらなる詳細は、例えば、Affholterによる、PCT/US01/06775、「Single−Stranded Nucleic Acid Template−Mediated Recombination and Nucleic Acid Fragment Isolation」において見出される。
別のアプローチにおいて、単鎖の分子は、二本鎖のDNA(dsDNA)に変換され、そしてこのdsDNA分子は、リガンドを媒介した結合によって固体支持体に結合される。結合していないDNAの分離の後、選択されたDNA分子は、支持体から遊離され、そしてプローブにハイブリダイズするライブラリーが豊富な配列を産生するために適切な宿主細胞の中へ導入される。この方法において産生されたライブラリーは、本明細書中に記載された任意の手順を使用して、さらなる多様化のための所望の基質を提供する。
任意の前述の一般的な組換えの形式は、より多様なセットの組換え核酸を産生するために、反復様式(例えば、1以上のサイクルの変異/組換えまたは他の多様性産生方法、必要に応じて、その後に1以上の選択方法が続く)において実施され得る。
ポリヌクレオチド連鎖停止法(polynucleotide chain termination methods)を利用する変異誘発はまた、提案され(例えば、Shortによる米国特許出願第5,965,408号、「Method of DNA reassembly by interrupting synthesis」、および上記の参考文献を参照のこと)、そして本発明に適用され得る。このアプローチにおいて、配列類似性の領域を共有する1以上の遺伝子に対応する二本鎖のDNAは、その遺伝子について特異的なプライマーの存在または非存在下において、結合および変性される。次いで、単鎖のポリヌクレオチドは、ポリメラーゼおよび連鎖停止試薬(例えば、紫外線照射、γ線照射もしくはX線照射;エチジウムブロマイドもしくは他のインターカレーター;単鎖結合タンパク質、転写活性化因子、もしくはヒストンのようなDNA結合タンパク質;多環式芳香族炭化水素;三価のクロムもしくは三価のクロム塩;または急速な熱循環によって媒介される簡略された重合;など)の存在下においてアニールおよびインキュベートされ、結果として部分的に二重鎖の分子の産生を生じる。次いで、例えば、部分的に伸長された鎖を含む、部分的な二重鎖分子は、種々の程度の配列類似性を共有し、そして最初のDNA分子の集団に対して多様化されたポリヌクレオチドを生じる、複製または部分的な複製の引き続く繰り返しにおいて、変性および再アニールされる。必要に応じて、産物、または産物の部分的なプールは、このプロセスにおいて1以上の工程で増幅され得る。上記記載のような、連鎖停止法により産生されたポリヌクレオチドは、任意の他に記載された組換えの形式について適切な基質である。
多様性はまた、Ostermeierら(1999)「A combinatorial approach to hybrid enzymes independent of DNA homology」Nature Biotech 17:1205中に記載される「ハイブリッド酵素の作製のためのインクリメンタルトランケーション(incremental truncation for the creation of hybrid enzymes)」(「ITCHY」)と称される組換えの手順を使用して、核酸または核酸の集団において産生され得る。このアプローチは、必要に応じて、1以上のインビトロまたはインビボの組換え方法について基質として機能し得る改変体の初期ライブラリーを産生するために使用され得る。Ostermeierら(1999)「Combinatorial Protein Engineering by Incremental Truncation」、Proc.Natl.Acad.Sci.USA、96:3562〜67;Ostermeierら(1999)、「Incremental Truncation as a Strategy in the Engineering of Novel Biocatalysts」、Biological and Medicinal Chemistry、7:2139〜44もまた参照のこと。
個々のヌクレオチドまたは連続したヌクレオチドもしくは連続していないヌクレオチドのグループの改変を生じる変異の方法は、本発明の核酸配列および/または遺伝子融合構築物の中へヌクレオチドの多様性を導入するために好んで使用され得る。多くの変異誘発方法は、上記記載の参考文献において見出され;変異誘発方法に関するさらなる詳細は、下記において見出され得、これらはまた、本発明に適用され得る。
例えば、誤りがちなPCRは、核酸改変体を産生するために使用され得る。この技術を使用することで、DNAポリメラーゼのコピーの正確さが低下される条件下でPCRは実施され、その結果、PCR産物の全長に沿って高い率の点変異が得られる。このような技術の例は、上記の参考文献中ならびに例えば、Leungら(1989)Technique 1:11〜15およびCaldwellら(1992)PCR Methods Applic.2:28〜33中に見出される。同様に、小さなDNAフラグメントの混合物由来のPCR産物のアセンブリに関連する工程において、アセンブリPCRは、使用され得る。同じ反応混合物中で同時に、多数の異なるPCR反応は生じ得、ここで1つの反応の産物は、別の反応の産物をプライムする。
オリゴヌクレオチド指向性変異誘発は、目的の核酸配列において部位特異的な変異を誘導するために使用され得る。このような技術の例は、上記参考文献中および例えば、Reidhaar−Olsonら(1988)Science、241:53〜57中に見出される。同様に、ネイティブな配列と異なる合成オリゴヌクレオチドのカセットを使用して、二本鎖のDNA分子の小さな領域を置換する工程において、カセット式変異誘発は、使用され得る。オリゴヌクレオチドは、例えば、完全および/または部分的にランダム化されたネイティブな配列を含み得る。
再帰的アンサンブル変異誘発は、タンパク質の変異誘発についてのアルゴリズムが、表現型的に関連した変種の多種多様な集団(このメンバーはアミノ酸配列が異なる)を産生するために使用される工程である。この方法は、コンビナトリアルなカセット式変異誘発の連続的な繰り返しをモニターするために、フィードバック機構を使用する。このアプローチの例は、Arkin&Youvan(1992)Proc.Natl.Acad.Sci.USA 89:7811〜7815中に見出される。
指数的(exponential)アンサンブル変異誘発は、高い割合の独特かつ機能的な変異体を有するコンビナトリアルライブラリーを作製するために使用され得る。目的の配列における残基の小さなグループは、改変されたそれぞれの位置について、機能的なタンパク質をもたらすアミノ酸を同定するために並行してランダム化される。このような手順の例は、Delegrave&Youvan(1993)Biotechnology Research 11:1548〜1552中に見出される。
インビボ変異誘発は、例えば、1以上のDNA修復経路内に変異を保有するE.coliの株において、DNAを増やすことによって目的の任意のクローンDNA中に、ランダムな変異を産生するために使用され得る。これらの「ミューテーター」株は、野生型の親の割合よりもより高いランダム変異の割合を有する。これらの株の1つにおいてDNAを増やすことは、最終的にDNA内にランダム変異を産生する。このような手順は、上記記載の参考文献中に記載される。
ゲノム(例えば、細菌ゲノム、真菌ゲノム、動物ゲノムまたは植物ゲノム)中に多様性を導入するための他の手順は、上記記載の方法および/または参照とした方法と共に使用され得る。例えば、上記の方法に加えて、種々の種への形質転換に適切な核酸のマルチマーを産生する技術が提案されている(例えば、Schellenberger、米国特許出願第5,756,316号および上記の参考文献を参照のこと)。このようなマルチマーでの適切な宿主の形質転換は、互いに対して相違であり(例えば、天然の多様性に由来するまたは部位特異的な変異誘発、誤りがちなPCR、変異原性の細菌株にわたる継代、などの適用による)、DNAの多様化について核酸の多様性の供給源を提供する(例えば、上記に記載したようなインビボの組換え工程による)遺伝子から構築される。
あるいは、部分的な配列類似性の領域を共有する単量体のポリヌクレオチドの多重度は、宿主の種に形質転換され得、宿主細胞によってインビボで組換えされ得る。引き続く細胞分化の繰り返しは、ライブラリーを産生するために使用され得、ライブラリーのメンバーは、単一、相同な集団、または単量体ポリヌクレオチドのプールを含む。代替的に、単量体の核酸は、標準的な技術(例えば、PCRおよび/またはクローニング)によって回収され得、そして上記に記載された再帰的組換えの形式を含む任意の組換えの形式において組換えられ得る。
多種の発現ライブラリーを作製するための方法は記載されており(上記記載の参考文献に加えて、例えば、Petersonら(1998)米国特許出願第5,783,431号「METHODS FOR GENERATING AND SCREENING NOVEL METABOLIC PATHWAYS」、およびThompsonら(1998)米国特許出願第5,824,485号 METHODS FOR GENERATING AND SCREENING NOVEL METABOLIC PATHWAYSを参照のこと)、そして目的のタンパク質の活性を同定するためのそれらの使用が提案されてきた(上記記載の参考文献に加えて、Short(1999)米国特許出願第5,958,672号「PROTEIN ACTIVITY SCREENING OF CLONES HAVING DNA FROM UNCULTIVATED MICROORGANISMS」を参照のこと)。多種発現ライブラリーは、一般的に、発現カセット内で、適切な調節配列に作動可能に連結した、複数の種または株に由来するcDNA配列またはゲノム配列を包含するライブラリーを含む。cDNA配列および/またはゲノム配列は、必要に応じて、多様性をさらに強化するために、ランダムに結合される。ベクターは、宿主生物(例えば、細菌種、真核細胞)の1より多い種における形質転換および発現について適切なシャトルベクターであり得る。いくつかの場合において、ライブラリーは、目的のタンパク質をコードするか、または目的の核酸にハイブリダイズする配列を、予め選択することによって偏らせられる。任意のこのようなライブラリーは、本明細書中に記載された任意の方法について基質として提供され得る。
上記に記載された手順は、核酸の多様性および/またはコードされたタンパク質の多様性を増大させることにほとんど向けられてきた。しかし、多くの場合において、全ての多様性が有用ではなく(例えば、機能上)、そしてわずかな好ましい改変体を同定するためにスクリーニングまたは選択されねばならない改変体のバックグラウンドを単に増加することに寄与するだけである。いくつかの適用において、ライブラリー(例えば、増幅されたライブラリー、ゲノムライブラリー、cDNAライブラリー、規格化されたライブラリーなど)もしくは多様化する(例えば、組換えに基づいた変異誘発手順による)前の他の核酸基質を、予め選択もしくは予めスクリーニングするか、または、別の方法で機能的産物をコードする核酸の方向へ基質を偏らせることが所望される。例えば、抗体設計の場合において、記載された任意の方法による操作より前に、インビボでの組換え現象を利用して、機能的な抗原結合部位を有する抗体の方向へ多様性を産生する工程を偏らせることは可能である。例えば、B細胞cDNAライブラリー由来の組換えCDRは、本明細書中に記載された任意の方法に従って多様化する前に、フレームワーク領域内に増幅および構築され得る(例えば、Jirholtら(1998)「Exploiting sequence space:shuffling in vivo formed complementarity determining regions into a master framework」Gene 215:471)。
ライブラリーは、所望の酵素活性を有するタンパク質をコードする核酸の方向へ偏らせられ得る。例えば、特定の活性を示すライブラリー由来のクローンを同定した後、このクローンは、DNAの変更を導入するための任意の公知の方法を使用して変異誘発され得る。次いで、変異誘発された相同体を含むライブラリーは、最初の特定の活性と同じかまたは異なり得る、所望の活性についてスクリーニングされる。このような手順の例は、Short(1999)米国特許出願第5,939,250号「PRODUCTION OF ENZYMES HAVING DESIRED ACTIVITIES BY MUTAGENESIS」中に提案される。所望の活性は、当該分野において公知の任意の方法によって同定され得る。例えば、WO99/10539は、遺伝子ライブラリーが、遺伝子ライブラリー由来の抽出物を代謝的に豊富な細胞から入手された成分と混合し、そして所望の活性を示す組み合わせを同定することによって、スクリーニングされ得ることを提案している。所望の活性を有するクローンが、ライブラリーのサンプル中へ生物活性な基質を挿入し、そして蛍光分析器(例えば、フローサイトメトリーデバイス、CCD、蛍光測定器、または分光光度計)を使用して、所望される活性の産物に対応する生物活性な蛍光を検出することによって、同定され得ることもまた提案されてきた(例えば、WO98/58085)。
ライブラリーはまた、特定の特性(例えば、選択された核酸プローブに対するハイブリダイゼーション)を有する核酸の方向に偏らせられ得る。例えば、出願WO99/10539は、所望の活性(例えば、酵素活性(例えば:リパーゼ、エステラーゼ、プロテアーゼ、グリコシダーゼ、グリコシルトランスフェラーゼ、ホスファターゼ、キナーゼ、オキシゲナーゼ、ペルオキシダーゼ、ヒドロラーゼ、ヒドラターゼ、ニトリラーゼ、トランスアミナーゼ、アミダーゼ、またはアシラーゼ))をコードするポリヌクレオチドが、以下の方法においてゲノムDNA配列の中から同定され得ることを提案している。ゲノムDNAの集団に由来する一本鎖のDNA分子は、リガンド結合プローブにハイブリダイズされる。このゲノムDNAは、培養されたかもしくは培養されていない微生物、または環境中のサンプルに由来し得る。あるいは、ゲノムDNAは、多細胞生物、または多細胞生物由来の組織に由来し得る。第2の鎖合成は、捕捉媒体から先に放出されるか、もしくは放出されないか、または当該分野において公知の多種多様な他の方法によって、捕捉に使用されるハイブリダイゼーションプローブから直接的に実施され得る。あるいは、単離された一本鎖のゲノムDNAの集団は、さらなるクロニーングなしにフラグメント化され得、そして、上記記載のように、一本鎖のテンプレートを使用して直接的に(例えば、組換えに基づいたアプローチ)使用され得る。
核酸およびポリペプチドを産生する「非確率的な」方法は、Short「Non−Stochastic Generation of Genetic Vaccines and Enzymes」WO00/46344中に主張される。非確率的ポリヌクレオチド再構法および部位飽和(site−saturation)変異誘発法を含むこれらの方法は、本発明に同様に適用される。ドープまたは縮重したオリゴヌクレオチドを使用するランダムまたは半ランダムな変異誘発はまた、例えば、ArkinおよびYouvan(1992)「Optimizing nucleotide mixtures to encode specific subsets of amino acids for semi−random mutagenesis」Biotechnology 10:297〜300;Reidhaar−Olsonら(1991)「Random mutagenesis of protein sequences using oligonucleotide cassettes」Methods Enzymol.208:564〜86;LimおよびSauer(1991)「The role of internal packing interactions in determining the structure and stability of a protein」J.Mol.Biol.219:359〜76;BreyerおよびSauer(1989)「Mutational analysis of the fine specificity of binding of monoclonal antibody 51F to lambda repressor」J.Biol.Chem.264:13355〜60;ならびに「Walk−Through Mutagenesis」(Crea,R;米国特許出願第5,830,650号および米国特許出願第5,798,208号、ならびに欧州特許出願第0527809 B1号)中に記載される。
多様化よりも前にライブラリーを濃縮するために適切な、任意の上記に記載される技術がまた、多様性を産生する方法によって作製される、産物、または産物のライブラリーをスクリーニングするために使用され得ることは、容易に理解される。任意の上記記載の方法は、核酸(例えば、GATコードポリヌクレオチド)を変更するために、再帰的または組合わせて実施され得る。
変異誘発方法、ライブラリーの構築方法および他の多様性産生方法のためのキットもまた市販されている。例えば、キットは、例えば、Stratagene(例えば、QuickChangeTM 部位特異的突然変異誘発キット;およびChameleonTM 二本鎖部位特異的突然変異誘発キット)、Bio/Can Scientific、Bio−Rad(例えば、上述されるKunkel法を使用する)、Boehringer Mannheim Corp.、Clonetech Laboratories、DNA Technologies、Epicentre Technologies(例えば、5プライム3プライムキット);Genpak Inc、Lemargo Inc、Life Technologies(Gibco BRL)、New England Biolabs、Pharmacia Biotech、Promega Corp.、Quantum Biotechnologies、Amersham International plc(例えば、上記のEckstein法を使用する)、ならびにAnglian Biotechnology Ltd(例えば、上記のCarter/Winter法を使用する)から入手可能である。
上記の参考文献は、多くの変異の形式を提供し、変異の形式としては、以下が挙げられる:組換え、再帰的組換え、再帰的変異および他の形態の変異誘発との組み合わせまたは組換え、ならびにこれらの形式の多くの改良。使用される多様性産生形式に関わらず、本発明の核酸は、組換え(相互に、または関連配列(または関連しない配列でさえも)とともに)し、本発明の遺伝子融合構築物および改変された遺伝子融合構築物における使用のための組換え核酸の多様なセット(例えば、相同な核酸のセット、および対応するポリペプチドを含む)を産生し得る。
改変されたポリヌクレオチドを産生するための上述された方法論の多くは、親の配列の多数の多様な改変体を産生する。本発明のいくつかの好ましい実施形態において、改変技術(例えば、いくつかの形態のシャッフリング)は、改変体のライブラリーを作製するために使用され、次いで、これは、いくつかの所望される機能的特性(例えば、改良されたGAT活性)をコードする改変されたポリヌクレオチドまたは改変されたポリヌクレオチドのプールについてスクリーニングされる。スクリーニングされ得る例示的な酵素活性は、触媒速度(kcatおよびKMのような速度定数に関して慣習的に特性付けられる)、基質特異性、ならびに基質、産物もしくは他の分子(例えば、インヒビターもしくはアクチベーター)による活性化または阻害に対する感受性を含む。
所望される酵素活性についての選択の1つの例は、目的の酵素活性(例えば、GAT活性)を十分には発現しない細胞の増殖および/または生存を阻害する条件下で、宿主細胞を増殖することを伴う。このような選択工程を使用して、所望される酵素活性をコードするポリヌクレオチド以外の、全ての改変されたポリヌクレオチドを考慮から除外し得る。例えば、本発明のいくつかの実施形態において、宿主細胞は、十分なレベルのGATの非存在下で細胞の増殖または生存を阻害する条件下(例えば、GATポリヌクレオチドを欠き、これを発現しない同じ品種の野生型の植物の成長に致命的または阻害するグリホセートの濃度)で保持される。これらの条件下で、酵素活性または産物の十分なレベルの産生を触媒し得る活性をコードする改変された核酸を有する、宿主細胞のみが、生存および増殖する。本発明のいくつかの実施形態は、グリホセートまたはグリホセートアナログの濃度を増加して、複数の繰り返しのスクリーニングを使用する。
本発明のいくつかの実施形態において、質量分析法は、グリホセートまたはグリホセートアナログもしくは代謝産物のアセチル化を検出するために使用される。質量分析法の使用は、以下の実施例においてより詳細に記載される。
利便性および高処理量のために、しばしば、微生物(例えば、E.coliのような細菌)における所望される改変核酸についてのスクリーニング/選択することが所望される。他方では、植物細胞または植物におけるスクリーニングは、いくつかの場合において好まれ得る。ここで、最終的な目的は、植物系における発現のための改変核酸を産生することである。
本発明のいくつかの好ましい実施形態において、処理量は、単独かまたは遺伝子融合構築物の一部としてのいずれかで、異なる改変核酸を発現する宿主細胞のプールをスクリーニングすることによって増加される。有意な活性を示す任意のプールは、所望の活性を発現する単一のクローンを同定するために逆重畳され得る。
当業者は、関連するアッセイ、スクリーニングまたは選択方法が所望される宿主生物などに依存して変動することを認識する。ハイスループット形式において実施され得るアッセイを使用することが、通常有利である。
ハイスループットアッセイにおいて、1日に数千以下の異なる改変体をスクリーニングすることが可能である。例えば、マイクロタイタープレートの各ウェルを使用して別々のアッセイを実施し得るか、または、濃度もしくはインキュベーション時間の効果が観測される場合、5〜10ウェル毎に単一の改変体を試験し得る。
流体アプローチに加え、上述のように、単に平板培地上で細胞を増殖し、所望される酵素機能または代謝機能について選択することが可能である。このアプローチは、簡単なハイスループットスクリーニング方法を提供する。
いくつかの周知の自動装置のシステムもまた、アッセイシステムにおいて有用な溶相化学について開発されてきた。これらのシステムは、Takeda Chemical Industries,LTD.(Osaka,Japan)によって開発された自動化合成装置および科学者により実施される手動の合成操作を模倣するロボットアームを使用する多くの自動装置システム(Zymate II、Zymark Corporation、Hopkinton、MA.;Orca、Hewlett−Packard、Palo Alto、CA)のような自動化されたワークステーションを含む。任意の上記の装置は、本発明への適用について適切である。これらが、統合化されたシステムについての参考文献とともに本明細書中に記載されるように操作し得るように、これらの装置への改良の特徴および実施が(存在する場合)、当業者に理解される。
ハイスループットスクリーニングシステムは、市販されている(例えば、Zymark Corp.、Hopkinton、MA;Air Technical Industries、Mentor、OH;Beckman Instruments、Inc.Fullerton、CA;Precision Systems、Inc.、Natick、MA、などを参照のこと)。これらのシステムは、典型的に、全体の手順を自動化し、その工程としては、以下が挙げられる:全てのサンプルおよび試薬をピペッティングする工程、液体を分配する工程、指定時刻に作動するインキュベーションの工程、およびアッセイに適切な検出器におけるマイクロプレートの最終的な読み込みの工程。これらの構造化可能なシステムは、高処理量および迅速な作動ならびに高い程度の自由度および特注生産を提供する。
このようなシステムの製造業者は、種々のハイスループット装置についての詳細なプロトコルを提供する。従って、例えば、Zymark Corp.は、遺伝子転写産物の調節の検出、リガンドの結合などのためのスクリーニングシステムを記載する技術告示を提供する。試薬の操作に対する微量流体(microfluidic)のアプローチは、例えば、Caliper Technologies(Mountain View、CA)によって開発されている。
カメラまたは他の記録装置(例えば、フォトダイオードおよびデータ記憶装置)によって観察される(および、必要に応じて、記録される)光学的な画像は、必要に応じてさらに、本明細書中の任意の実施形態において(例えば、画像のデジタル化ならびに/またはコンピュータ上での画像の保存および分析をすることによって)処理される。種々の市販の周辺装置およびソフトウェアは、例えば、PC(DOSTM、OSTM WINDOWS(登録商標)、WINDOWS(登録商標) NTTMもしくはWINDOWS(登録商標) 95TMをベースにしたマシンと互換性のあるインテル x86もしくはペンティアム(登録商標)チップ)、MACINTOSHTM、またはUNIX(登録商標)ベース(例えば、SUNTMワークステーション)のコンピュータを使用して、デジタル化し、デジタル化されたビデオまたはデジタル化された光学画像の保存および分析をするために入手可能である。
1つの従来のシステムは、アッセイ装置から冷却された電荷結合素子(CCD)カメラへの光を伝達し、当該分野において通常使用される。CCDカメラは、画素(ピクセル)のアレイを含む。被験物からの光は、CCD上で画像化される。被験物の領域(例えば、生物学的ポリマーのアレイ上の個々のハイブリダイゼーション部位)に対応する特定のピクセルは、各位置について光の強度の読みを得るためにサンプリングされる。複数のピクセルは、速度の増加に応じて処理される。本発明の装置および方法は、例えば、蛍光技術または暗視野顕微鏡技術によって任意のサンプルを観測するために容易に使用される。
(他のポリヌクレオチド組成物)
本発明はまた、本発明の2つ以上のポリヌクレオチドを含む組成物(例えば、組換えのための物質のような)を含む。この組成物は、組換え核酸のライブラリーを含み得、ここでこのライブラリーは、少なくとも2、3、5、10、20、または50以上のポリヌクレオチドを含む。このポリヌクレオチドは、発現ライブラリーを提供する発現ベクター中に必要に応じてクローン化される。
本発明はまた、本発明の2つ以上のポリヌクレオチドを含む組成物(例えば、組換えのための物質のような)を含む。この組成物は、組換え核酸のライブラリーを含み得、ここでこのライブラリーは、少なくとも2、3、5、10、20、または50以上のポリヌクレオチドを含む。このポリヌクレオチドは、発現ライブラリーを提供する発現ベクター中に必要に応じてクローン化される。
本発明はまた、制限エンドヌクレアーゼであるRNAse、またはDNAseを用いて、本発明の1つ以上のポリヌクレオチドを消化すること(例えば、上記された特定の組換え様式において実施されるような)によって生成される組成物;および、機械的手段(例えば、超音波処理、ボルテックスなど)により、本発明の1つ以上のポリヌクレオチドをフラグメント化または剪断することによって生成される組成物を含み、本発明はまた、上記の方法における組換えのための物質を提供するために用いられ得る。同様に、本発明の1つより多い核酸に対応するオリゴヌクレオチドのセットを含む組成物は、組換え物質として有用であり、そして本発明の特徴である。便宜上、これらのフラグメント化された混合物、剪断された混合物、またはオリドヌクレオチド合成された混合物は、フラグメント化核酸セットといわれる。
リボヌクレオチド3リン酸またはデオキシリボヌクレオチド3リン酸および核酸ポリメラーゼの存在中で、1つ以上のフラグメント化核酸セットをインキュベートすることによって生成される組成物もまた、本発明に含まれる。この生じた組成物は、上記の多くの組換え様式のための組換え混合物を形成する。この核酸ポリメラーゼは、RNAポリメラーゼ、DNAポリメラーゼ、またはRNA指向性DNAポリメラーゼ(例えば、「逆転写酵素」)であり得;このポリメラーゼは、例えば、熱安定性DNAポリメラーゼ(例えば、VENT、TAQなど)であり得る。
(統合システム)
本発明は、例えば、本明細書中に列挙されるこれらの配列、およびこれらの種々のサイレント置換および保存的置換を含む、本明細書中のポリペプチドおよび核酸に対する本明細書中での配列情報に対応する文字列を含むコンピュータ、コンピュータ読み出し可能な媒体、および統合システムを提供する。
(統合システム)
本発明は、例えば、本明細書中に列挙されるこれらの配列、およびこれらの種々のサイレント置換および保存的置換を含む、本明細書中のポリペプチドおよび核酸に対する本明細書中での配列情報に対応する文字列を含むコンピュータ、コンピュータ読み出し可能な媒体、および統合システムを提供する。
例えば、当該分野において公知の種々の方法および遺伝子アルゴリズム(GA)は、異なる文字列の間の相同性もしくは類似性を検出するために用いられ得るか、または他の所望される機能(例えば、出力ファイルを制御すること)を実行するために用いられ得、配列などを含む情報をの提示を作成するための基礎を提供する。例としては、BLAST(前出で考察される)が挙げられる。
従って、種々のストリンジェンシーおよび長さを有する異なる型の相同性ならびに類似性は、本明細書中の統合システム中で検出および認識され得る。例えば、多くの相同性決定方法が、生体高分子の配列の比較分析、ワードプロセッシングにおけるスペルチェック、および種々のデータベースからのデータ検索のために設計されてきた。天然のポリヌクレオチドにおける主要な4つの核酸塩基間の二重らせん対形成の相補的相互作用の理解を用いて、相補的な相同ポリヌクレオチドの文字列のアニーリングをシミュレーションするモデルはまた、本明細書中の配列に対応する文字列上で代表的に実行される配列整列もしくは他の操作(例えば、ワードプロセッシング操作、配列もしくは部分配列の文字列を含む図の構築、出力表など)の基礎として用いられ得る。配列類似性を計算するためのGAを用いたソフトウェアパッケージの一例としては、BLASTがあり、これは、本明細書中の配列に対応する文字列を入力することによって本発明に適応され得る。
同様に、ワードプロセッシングソフトウェアのような標準的なデスクトップアプリケーション(例えば、Microsoft WordTMまたはCorel WordPerfectTM)およびデータベースソフトウェア(例えば、表計算ソフト(例えば、Microsoft ExcelTM、Corel Quattro ProTM)またはデータベースプログラム(例えば、Microsoft AccessTMもしくはParadoxTM))が、本発明のGATホモログ(核酸もしくはタンパク質、または両方のいずれか)に対応する文字列を入力することによって、本発明に適合され得る。例えば、この統合システムは、例えば、文字の列を操作するためにユーザーインターフェース(例えば、Windows(登録商標)システム、Macintoshシステム、もしくはLINUXシステムのような標準的な操作システムにおけるGUI)と共同して用いられる、適切な文字列情報を有する前述のソフトウェアを含み得る。記載されるように、BLASTのような専門化された整列プログラムはまた、核酸もしくはタンパク質(または対応する文字列)の整列のための本発明のシステムに組み込まれ得る。
本発明における分析のための統合システムは、代表的に、配列を整列するためのGAソフトウェアを用いたデジタルコンピュータ、および本明細書中の任意の配列を含むソフトウェアシステムに入力されるデータセットを含む。このコンピュータは、例えば、PC(Intel x86もしくはペンティアム(登録商標)のチップに適合性のDOSTM、OS2TM、WINDOWS(登録商標)、WINDOWS(登録商標)NT、WINDOWS(登録商標)95、WINDOWS(登録商標)98、LINUXに基づく機械、MACINTOSHTM、Power PCもしくはUNIX(登録商標)(例えば、SUNTMワークステーション)に基づく機械、または、当業者に公知である他の商業的に一般的なコンピュータであり得る。配列を整列するためのソフトウェア、さもなくば、配列を操作するためのソフトウェアが、利用可能であるか、または標準的なプログラミング言語(例えば、Visualbasic、Fortran、Basic、Java(登録商標)など)を用いて、当業者によって容易に構築され得る。
任意のコントローラまたはコンピュータは、必要に応じて、モニターを含み、このモニターは、しばしば、陰極線管(「CRT」)ディスプレイ、フラットパネルディスプレイ(例えば、アクティブマトリックス液晶ディスプレイ、液晶ディスプレイ)などである。コンピュータ回路は、しばしば、多数の集積回路チップ(例えば、マイクロプロセッサ、メモリー、インターフェース回路など)を含むボックス中に配置される。このボックスはまた、必要に応じて、ハードディスクドライブ、フロッピー(登録商標)ディスクドライブ、高容量の取り外し可能なドライブ(例えば、書き込み可能なCD−ROM)、および他の一般の周辺素子を含む。入力デバイス(例えば、キーボードまたはマウス)は、必要に応じて、ユーザーからの入力、および関連するコンピュータシステムにおいて比較されるか、さもなくば操作される配列のユーザー選択を提供する。
コンピュータは、代表的にセットパラメータ領域中に入力するユーザーの形態のいずれかにおいて(例えば、GUIにおいてか、または予めプログラムされた命令(例えば、種々の異なる特定の操作のために予めプログラムされた命令)の形態において)ユーザーの命令を受けるために適切なソフトウェアを含む。次いで、このソフトウェアは、所望の操作を実行するために、これらの命令を、流動方向および輸送コントローラの操作を命令するための適切な言語に変換する。
ソフトウェアはまた、(例えば、本明細書中の配列もしくは配列の整列に基づく)核酸合成を制御するための出力素子、または本明細書中の配列に対応する文字列を用いて実行される整列もしくは他の操作の下流で生じる他の操作を含み得る。従って、核酸合成装置は、本明細書中の1つ以上の統合システムにおける構成要素であり得る。
さらなる局面において、本発明は、本明細書中の方法、組成物、システムおよび装置を具体化するキットを提供する。本発明のキットは、必要に応じて次の1つ以上を含む:(1)本明細書中に記載されるような装置、システム、システムの構成要素、または装置の構成要素;(2)本明細書中に記載される方法を実行するための命令、および/または本明細書中の装置もしくは装置の構成要素を操作するための命令、および/または本明細書中の組成物を用いるための命令;(3)1つ以上のGAT組成物または構成要素;(4)構成要素または組成物を保持するための容器、ならびに(5)パッケージング物質。
さらなる局面において、本発明は、本明細書中の任意の装置、装置の構成要素、組成物、もしくはキットの使用、本明細書中の任意の方法もしくはアッセイの実行、および/または本明細書中の任意のアッセイもしくは方法を実行するための任意の装置もしくはキットの使用を提供する。
(宿主細胞および生物)
宿主細胞は、真核生物(例えば、真核細胞、植物細胞、動物細胞、プロトプラスト、または組織培養物)であり得る。宿主細胞は、必要に応じて複数(個)の細胞(例えば、生物)を含む。あるいは、宿主細胞は、細菌(すなわち、グラム陽性細菌、紅色細菌、緑色硫黄細菌、緑色非硫黄細菌、シアノバクテリア、スピロヘータ、テルモトーガ(thermatogales)、フラボバクテリア、およびバクテロイド)ならびに始原細菌(すなわち、コル古細菌、テルモプロテウス、ピロディクティウム、テルモコックス、メタン細菌、アルカエグロブスおよび高度好塩菌)を含むが、これらに限られない原核生物であり得る。
宿主細胞は、真核生物(例えば、真核細胞、植物細胞、動物細胞、プロトプラスト、または組織培養物)であり得る。宿主細胞は、必要に応じて複数(個)の細胞(例えば、生物)を含む。あるいは、宿主細胞は、細菌(すなわち、グラム陽性細菌、紅色細菌、緑色硫黄細菌、緑色非硫黄細菌、シアノバクテリア、スピロヘータ、テルモトーガ(thermatogales)、フラボバクテリア、およびバクテロイド)ならびに始原細菌(すなわち、コル古細菌、テルモプロテウス、ピロディクティウム、テルモコックス、メタン細菌、アルカエグロブスおよび高度好塩菌)を含むが、これらに限られない原核生物であり得る。
本発明のGAT核酸を組込んでいるトランスジェニック植物もしくは植物細胞、および/または本発明のGATポリペプチドを発現するトランスジェニック植物もしくは植物細胞が、本発明の特徴である。植物細胞およびプロトプラストの形質転換は、本明細書中に記載される方法を含むが、これらに限られない植物分子生物学の当業者に公知である基本的に任意の種々の方法で実行され得る。一般に、本明細書中で参考として援用されるMethods in Enzymology、Vol.153(Recombinant DNA Part D) WuおよびGrossman(編)1987、Academic Pressを参照のこと。本明細書中で使用される場合、用語「形質転換」は、核酸配列(例えば、「異種の」もしくは「外来の」核酸配列)の導入による宿主植物の遺伝子型の変化を意味する。この異種の核酸配列は、必ずしも異なる供給源に由来する必要はないが、いくつかの点で、異種核酸配列は、その導入される細胞に対して外側にある。
Berger、AusubelおよびShamblookに加えて、植物細胞のクローニング、培養および再生について有用な一般の参考として、Jones(編)(1995)Plant Gene Transfer and Expression Protocols−Methods in Molecular Biology、Volume 49 Humana Press Towata NJ;Payneら、(1992)Plant Cell and Tissue Culture in Liquid Systems John Wiley&Sons,Inc.New York,NY(Payne);ならびにGamborgおよびPhillips(編)(1995)Plant Cell,Tissue and Organ Culture;Fundamental Methods Springer Lab Manual,Springer−Verlag(Berlin Heidelberg New York)(Gamborg)が挙げられる。種々の細胞培養培地が、AtlasおよびParks(編)The Handbook of Microbiological Media(1993)CRC Press,Boca,Raton,FL(Atlas)に記載される。植物細胞培養についてのさらなる情報が、市販の文献(例えば、Sigma−Aldrich,Inc(St Louis,MO)からのLife Science Research Cell Culture Catalogue(1998)(Sigma−LSRCCC)ならびに、例えば、これもまたSigma−Aldrich,Inc(St Louis,MO)からのPlant Culture Catalogueおよび補遺(1997)(Sigma−PCCS))に見出される。植物細胞培養に関するさらなる詳細が、Croy(編)(1993)Plant Molecular Biology Bios Scientific Publishers,Oxford,U.K.)に見出される。
本発明の1つの実施形態において、植物細胞の形質転換に適した1つ以上のGATポリヌクレオチドを含む組換えベクターが、調製される。(例えば、配列番号1〜5および11〜262の中から選択される)所望のGATポリペプチドをコードするDNA配列は、所望の植物中に導入され得る組換え発現カセットを構築するために都合よく使用される。本発明の文脈において、発現カセットは、代表的に、形質転換された植物の意図される組織(例えば、植物全体、葉、根など)においてGAT配列の転写を指示するために十分な、プロモーター配列ならびに他の転写および翻訳の開始調節配列に作動可能に連結される、選択されたGATポリヌクレオチドを含む。
例えば、植物の全ての組織においてGAT核酸の発現を指向する強い構成性植物プロモーターまたは弱い構成性植物プロモーターが、都合よく利用され得る。このようなプロモーターは、ほとんどの環境条件下および発育または細胞分化の状態で活性である。構成性プロモーターの例としては、Agrobacterium tumefaciensの1’−プロモーターまたは2’−プロモーター、および当業者に公知の種々の植物遺伝子由来の他の転写開始領域が挙げられる。本発明のGATポリペプチドの過剰発現が植物に対して有害である場合、当業者は、弱い構成性プロモーターが低レベルの発現のために用いられ得ることを認識する。これらの場合において、高レベルの発現が植物に対して有害でない場合、強いプロモーター(例えば、t−RNA、または他のpol IIIプロモーター、または強いpol IIプロモーター(例えば、カリフラワーモザイクウィルスプロモーター、CaMV,35Sプロモーター))が用いられ得る。
あるいは、植物プロモーターは、環境制御下であり得る。このようなプロモーターは、「誘導性」プロモーターといわれる。誘導性プロモーターによって転写を変化し得る環境条件の例としては、病原体の襲撃、嫌気的条件、または光の存在が挙げられる。いくつかの場合において、「組織特異的」および/または発育制御下であるプロモーターを用いることが所望され、その結果、このGATポリヌクレオチドは、特定の組織もしくは発育の段階(例えば、葉、根、苗条など)においてのみ発現される。除草剤耐性および関連する表現型に関する遺伝子の内生プロモーターは、GAT核酸(例えば、P450モノオキシゲナーゼ、グルタチオン−S−トランスフェラーゼ、ホモグルタチオン−S−トランスフェラーゼ、グリホセートキシダーゼ、および5−エノールピルビルシキミ酸−2−リン酸シンターゼ)の発現の促進に対して特に有用である。
組織特異的プロモーターはまた、本明細書中に記載されるGATポリヌクレオチドを含む異種の構造遺伝子の発現を指向するために用いられ得る。従って、このプロモーターは、その発現が本発明のトランスジェニック植物において所望される任意の遺伝子(例えば、GATおよび/または除草剤抵抗性もしくは除草剤耐性を与える他の遺伝子、他の有用な特性(例えば、雑種強勢)に影響する遺伝子)の発現を駆動する組換え発現カセットにおいて用いられ得る。同様に、(例えば、5’調節配列、または異種の遺伝子のイントロンに由来する)エンハンサーエレメントもまた、GATポリヌクレオチドのような異種の構造遺伝子の発現を向上するために用いられ得る。
一般的に、植物の発現カセットにおいて用いられる特定のプロモーターは、意図される適用に依存する。植物細胞における転写を指向する任意の多数のプロモーターが、適し得る。このプロモーターは、構成性もしくは誘導性のいずれかであり得る。上記のプロモーターに加えて、植物において操作される細菌起源のプロモーターとしては、オクトピンシンターゼプロモーター、ノパリンシンターゼプロモーター、およびTiプラスミドに由来する他のプロモーターが挙げられる。Herrera−Estrellaら、(1983)Nature 303:209を参照のこと。ウィルスプロモーターとしては、CaMVの35SRNAプロモーターおよび19SRNAプロモーターが挙げられる。Odellら、(1985)Nature 313:810を参照のこと。他の植物プロモーターとしては、リブロース−1,3−二リン酸カルボキシラーゼのスモールサブユニットプロモーター、およびファゼオリンプロモーターが挙げられる。E8遺伝子(DeikmanおよびFischer(1988)EMBO J 7:3315)および他の遺伝子からのプロモーター配列がまた、都合よく用いられる。単子葉植物種に対して特異的なプロモーターもまた、考察される(McElroy D.,Brettell R.I.S. 1994. Foreign gene expression in transgenic cereals. Trends Biotech.,12:62−68)。あるいは、有用な特徴を有する新規なプロモーターが、配列分析、エンハンサートラッピングもしくはプロモータートラッピングなどを含む当該分野で公知の方法によって、任意のウィルス源、細菌源、または植物源から同定され得る。
本発明の発現ベクターの調製において、プロモーターおよびGATをコードする遺伝子以外の配列がまた、都合よく用いられる。適切なポリペプチド発現が所望される場合、ポリアデニル化領域は、天然の遺伝子、種々の他の植物遺伝子、またはT−DNAに由来し得る。(例えば、内部の細胞器官(例えば、葉緑体)への発現ポリペプチドの移動または細胞外分泌を促進する)シグナル/局在化ペプチドがまた、用いられ得る。
GATポリヌクレオチドを含むベクターはまた、植物細胞に選択可能な表現型を与えるマーカー遺伝子を含み得る。例えば、このマーカーは、殺生剤耐性、特に抗生物質耐性(例えば、カナマイシン、G418、ブレオマイシン、ハイグロマイシンに対する耐性)または除草剤耐性(例えば、クロロスルフロン、もしくはホヒノトリシン(phophinothricin)に対する耐性)をコードし得る。可視化可能な反応産物(例えば、β−グルクロニダーゼ、β−ガラクトシダーゼ、およびクロラムフェニコールアセチルトランスフェラーゼ)を介してか、または遺伝子産物自体(例えば、緑色蛍光タンパク質、GFP;Sheenら、(1995)The Plant Journal 8:777)の直接的な可視化によって、遺伝子の発現およびタンパク質の局在化をモニターするために用いられるレポーター遺伝子は、例えば、植物細胞における一過性の遺伝子の発現をモニタリングするために用いられ得る。一過性の発現系は、例えば、除草剤耐性活性について植物細胞培養物をスクリーニングする際に、植物細胞で用いられ得る。
(植物の形質転換)
(プロトプラスト)
種々の植物型からの形質転換性プロトプラストの確立についての多数のプロトコール、およびそれに続く培養されたプロトプラストの形質転換は、当該分野で利用可能であり、そして本明細書中で参考として援用される。例えば、Hashimotoら、(1990)Plant Physiol.93:857;FowkeおよびConstabel(編)(1994)Plant Protoplasts;Saundersら(1993)Applications of Plant In Vitro Technology Symposium、UPM 16−18;ならびにLyznikら(1991)BioTechniques 10:295を参照のこと、これらの各々は、本明細書中で参考として援用される。
(プロトプラスト)
種々の植物型からの形質転換性プロトプラストの確立についての多数のプロトコール、およびそれに続く培養されたプロトプラストの形質転換は、当該分野で利用可能であり、そして本明細書中で参考として援用される。例えば、Hashimotoら、(1990)Plant Physiol.93:857;FowkeおよびConstabel(編)(1994)Plant Protoplasts;Saundersら(1993)Applications of Plant In Vitro Technology Symposium、UPM 16−18;ならびにLyznikら(1991)BioTechniques 10:295を参照のこと、これらの各々は、本明細書中で参考として援用される。
(葉緑体)
葉緑体は、いくつかの除草剤耐性活性の作用の部位であり、そしていくつかの例において、このGATポリヌクレオチドは、葉緑体中への遺伝子産物の移動を容易にするために葉緑体輸送配列ペプチドに融合される。これらの場合において、GATポリヌクレオチドを植物宿主細胞の葉緑体中へ形質転換することは、好都合であり得る。葉緑体の形質転換および発現を達成するための多数の方法が、当該分野において利用可能である(例えば、Daniellら(1998)Nature Biotechnology 16:346;O’Neillら(1993)The Plant Journal 3:729;Maliga(1993)TIBTECH 11:1)。この発現構築物は、GATポリペプチドをコードするポリヌクレオチドに作動可能に連結された、植物において機能的な転写調節配列を含む。葉緑体において機能するように設計された発現カセット(例えば、GATポリヌクレオチドを含む発現カセット)は、葉緑体における発現を確実にするために必要な配列を含む。代表的に、葉緑体ゲノムを用いて相同組換えをもたらすために、葉緑色素体ゲノムに相同的な2つの領域が、コード配列に隣接し;しばしば、選択マーカー遺伝子もまた、生じる移植した(transplastonic)植物細胞における遺伝的に安定な形質転換葉緑体の選択を容易にするために、近接する色素体DNA配列内に存在する(例えば、Maliga(1993)およびDaniell(1998)、ならびにこれらに引用される参考文献を参照のこと)。
葉緑体は、いくつかの除草剤耐性活性の作用の部位であり、そしていくつかの例において、このGATポリヌクレオチドは、葉緑体中への遺伝子産物の移動を容易にするために葉緑体輸送配列ペプチドに融合される。これらの場合において、GATポリヌクレオチドを植物宿主細胞の葉緑体中へ形質転換することは、好都合であり得る。葉緑体の形質転換および発現を達成するための多数の方法が、当該分野において利用可能である(例えば、Daniellら(1998)Nature Biotechnology 16:346;O’Neillら(1993)The Plant Journal 3:729;Maliga(1993)TIBTECH 11:1)。この発現構築物は、GATポリペプチドをコードするポリヌクレオチドに作動可能に連結された、植物において機能的な転写調節配列を含む。葉緑体において機能するように設計された発現カセット(例えば、GATポリヌクレオチドを含む発現カセット)は、葉緑体における発現を確実にするために必要な配列を含む。代表的に、葉緑体ゲノムを用いて相同組換えをもたらすために、葉緑色素体ゲノムに相同的な2つの領域が、コード配列に隣接し;しばしば、選択マーカー遺伝子もまた、生じる移植した(transplastonic)植物細胞における遺伝的に安定な形質転換葉緑体の選択を容易にするために、近接する色素体DNA配列内に存在する(例えば、Maliga(1993)およびDaniell(1998)、ならびにこれらに引用される参考文献を参照のこと)。
(一般的な形質転換方法)
本発明のDNA構築物は、種々の従来の技術によって、所望される植物の宿主のゲノムに導入され得る。広範な種々の高等植物種を形質転換するための技術が、周知であり、そして技術文献および科学文献に記載される。例えば、Payne、Gamborg、Croy、Jonesら、全て前出、および例えば、Weisingら(1988)Ann.Rev.Genet.22:421を参照のこと。
本発明のDNA構築物は、種々の従来の技術によって、所望される植物の宿主のゲノムに導入され得る。広範な種々の高等植物種を形質転換するための技術が、周知であり、そして技術文献および科学文献に記載される。例えば、Payne、Gamborg、Croy、Jonesら、全て前出、および例えば、Weisingら(1988)Ann.Rev.Genet.22:421を参照のこと。
例えば、DNAは、技術(例えば、植物細胞プロトプラストのエレクトロポレーションおよびマイクロインジェクション)を用いて植物細胞のゲノムDNAに直接導入され得るか、またはDNA構築物は、DNA粒子ボンバードメントのような弾道的な方法を用いて植物組織に直接導入され得る。あるいは、DNA構築物は、適切なT−DNA近接領域と結合され得、そして従来のAgrobacterium tumefaciens宿主ベクターに導入され得る。Agrobacterium宿主の毒性機能は、植物細胞が細菌によって感染される場合、構築物および近接するマーカーの植物細胞DNAへの挿入を指向する。
マイクロインジェクション技術は、当該分野において公知であり、科学文献および特許文献に十分に記載される。ポリエチレングリコール沈殿を用いるDNA構築物の導入は、Paszkowskiら(1984)EMBO J 3:2717に記載される。エレクトロポレーション技術は、Frommら(1985)Proc Nat’l Acad Sci USA 85:5824に記載される。弾道的な形質転換技術は、Kleinら(1987)Nature 327:70;およびWeeksらPlant Physiol 102:1077に記載される。
いくつかの実施形態において、Agrobacterium媒介形質転換技術が、本発明のGAT配列をトランスジェニック植物へ移すために用いられる。Agrobacterium媒介形質転換は、双子葉植物の形質転換のために広く用いられるが、特定の単子葉植物もまた、Agrobacteriumによって形質転換され得る。例えば、コメのAgrobacterium形質転換が、Hieiら(1994)Plant J.6:271;米国特許第5,187,073号;同第5,591,616号;Liら(1991)Science in China 34:54;およびRaineriら(1990)Bio/Technology 8:33によって記載される。Agrobacterium媒介形質転換によって形質転換されたトウモロコシ、大ムギ、ライムギおよびアスパラガスもまた、記載されている(Xuら(1990)Chinese J Bot 2:81)。
Agrobacterium媒介形質転換技術は、植物細胞ゲノムに組み込み、目的の核酸を植物細胞中に同時に移すA.tumefaciensの腫瘍誘導(Ti)プラスミドの能力を利用する。代表的に、発現ベクターが生成され、ここで目的の核酸(例えば、本発明のGATポリヌクレオチド)は、T−DNA配列をまた含む自己複製プラスミドに結合される。T−DNA配列は代表的に、目的の発現カセット核酸に隣接し、そしてプラスミドの組込み配列を含む。発現カセットに加えて、T−DNAはまた代表的に、マーカー配列(例えば、抗生物質耐性遺伝子)を含む。次いで、T−DNAおよび発現カセットを有するプラスミドは、Agrobacterium細胞中にトランスフェクトされる。代表的に、植物細胞の効果的な形質転換のために、A.tumefaciens細菌はまた、プラスミド上またはその染色体に組み込まれた、必要なvir領域を所有する。Agrobacterium媒介形質転換の考察について、FiroozabadyおよびKuehnle(1995)Plant Cell Tissue and Organ Culture Fundamental Methods、GamborgおよびPhillips(編)を参照のこと。
(トランスジェニック植物の再生)
上記の技術を含む植物形質転換技術によって得られた形質転換された植物細胞は、形質転換された遺伝子型(すなわち、GATポリヌクレオチド)、従って所望の表現型(グリホセートもしくはグリホセートのアナログに対して獲得した抵抗性(すなわち、耐性))を有する植物全体を再生するために培養され得る。このような再生技術は、組織培養増殖培地中の特定の植物ホルモンの操作に依存し、代表的に、所望のヌクレオチド配列と一緒に導入されている殺生剤マーカーおよび/または除草剤マーカーに依存する。あるいは、本発明のGATポリヌクレオチドによって与えられるグリホセート抵抗性に対する選択が、実行され得る。培養されたプロトプラストからの植物再生は、Evansら(1983)Protoplasts Isolation and Culture、Handbook of Plant Cell Culture、pp 124−176、Macmillan Publishing Company、New York;およびBinding(1985)Regeneration of Plants、Plant Protoplasts pp 21−73、CRC Press、Boca Ratonに記載される。再生物はまた、植物のカルス、植物の外植片、植物の器官、もしくはこれらの一部から得られ得る。このような再生技術は、一般的にKleeら(1987)Ann Rev of Plant Phys 38:467に記載される。例えば、PayneおよびGamborgもまた参照のこと。Agrobacteriumを用いる形質転換後、この外植片は、典型的には、選択培地に移される。当業者は、選択培地が、外植片に同時にトランスフェクトされる選択マーカーに依存することを理解する。適切な長さの時間後、形質転換体は、芽を形成し始める。芽が約1〜2cmの長さになった後、この芽は、適切な根および芽の培地に移されるべきである。選択圧は、根および芽の培地中で維持されるべきである。
上記の技術を含む植物形質転換技術によって得られた形質転換された植物細胞は、形質転換された遺伝子型(すなわち、GATポリヌクレオチド)、従って所望の表現型(グリホセートもしくはグリホセートのアナログに対して獲得した抵抗性(すなわち、耐性))を有する植物全体を再生するために培養され得る。このような再生技術は、組織培養増殖培地中の特定の植物ホルモンの操作に依存し、代表的に、所望のヌクレオチド配列と一緒に導入されている殺生剤マーカーおよび/または除草剤マーカーに依存する。あるいは、本発明のGATポリヌクレオチドによって与えられるグリホセート抵抗性に対する選択が、実行され得る。培養されたプロトプラストからの植物再生は、Evansら(1983)Protoplasts Isolation and Culture、Handbook of Plant Cell Culture、pp 124−176、Macmillan Publishing Company、New York;およびBinding(1985)Regeneration of Plants、Plant Protoplasts pp 21−73、CRC Press、Boca Ratonに記載される。再生物はまた、植物のカルス、植物の外植片、植物の器官、もしくはこれらの一部から得られ得る。このような再生技術は、一般的にKleeら(1987)Ann Rev of Plant Phys 38:467に記載される。例えば、PayneおよびGamborgもまた参照のこと。Agrobacteriumを用いる形質転換後、この外植片は、典型的には、選択培地に移される。当業者は、選択培地が、外植片に同時にトランスフェクトされる選択マーカーに依存することを理解する。適切な長さの時間後、形質転換体は、芽を形成し始める。芽が約1〜2cmの長さになった後、この芽は、適切な根および芽の培地に移されるべきである。選択圧は、根および芽の培地中で維持されるべきである。
代表的に、この形質転換体は、約1〜2週間で根を発達させ、そして小植物を形成する。この小植物が約3〜5cmの高さになった後、これらは、ファイバー・ポット中の面菌下土壌に移される。当業者は、異なる順応化手順が異なる種の形質転換された植物を得るために用いられることを理解する。例えば、根および芽が発達した後、切断、および形質転換された植物の体細胞胚は、小植物の確立のための培地に移される。形質転換された植物の選択および再生の説明について、例えば、DoddsおよびRoberts(1995)Experiments in Plant Tissue Culture、第3編、Cambridge University Pressを参照のこと。
真空浸潤(Bechtold N.、Ellis J.およびPelletier G.、1993、In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants.CR Acad Sci Paris Life Sci 316:1194−1199)および顕花植物の単純な液浸(Desfeux,C.、Clough S.J.、およびBent A.F.、2000、Female reproductive tissues are the primary target of Agrobacterium−mediated transformation by the Arabidopsis floral−dip method.Plant Physiol.123:895−904)を用いるArabidopsisのAgrobacterium形質転換のための方法がまた存在する。これらの方法を用いて、トランスジェニック種子が、組織培養の必要なく生産される。
効率的なAgrobacterium媒介性形質転換プロトコルがなお開発されている植物改変体がある。例えば、形質転換された組織の再生と共役してトランスジェニック植物を生産する、首尾よい組織の形質転換は、最も商業的に関連するとともにワタの品種のいくつかについて、報告されていない。にも関わらず、これらの植物とともに使用され得るアプローチは、Agrobacterium媒介性形質転換を介して関連する植物の品種にこのポリヌクレオチドを安定に導入する工程、作動可能性を確認する工程、次いで、標準的な性交配技術もしくは戻し交配技術を用いて、この導入遺伝子を所望の商業的な系統に移す工程、を含む。例えば、ワタの場合において、Agrobacteriumを使用して、Gossypium hirustumのCoker系(例えば、Coker系310、312、5100 Deltapine61またはStoneville213)を形質転換し得、次いで、この導入遺伝子は、戻し交配によって、別のより商業的に関連するG.hirustum品種に導入され得る。
本発明のトランスジェニック植物は、本発明のGATポリヌクレオチドの存在を決定するために遺伝子型的にか、または表現型的にかのいずれかで特徴づけられ得る。遺伝子型の分析は、ゲノムDNAのPCR増幅および特異的な標識プローブを用いたゲノムDNAのハイブリダイゼーションを含む、多くの周知の技術のいずれかによって実行され得る。表現型の分析としては、例えば、グリホセートのような選択された除草剤に曝露された植物もしくは植物組織の生存が挙げられる。
本質的に、任意の植物が、本発明のGATポリヌクレオチドを用いて形質転換され得る。本発明の新規なGATポリヌクレオチドの形質転換および発現のための適切な植物としては、農学的および園芸学的に重要な種が挙げられる。このような種としては、以下の科のメンバーが挙げられるが、これらに制限されない:イネ科(トウモロコシ、ライムギ、ライコムギ、オオムギ、キビ、コメ、コムギ、カラスムギなどを含む);マメ科(エンドウマメ、マメ(bean)、レンズマメ、ピーナッツ、クズイモ、ササゲ、ハッショウマメ、ダイズ、クローバー、アルファルファ、ルビナス、ベッチ、ハス、スイートクローバー、フジ、およびスイートピーを含む);キク科(少なくとも1,000属を含む維管束植物の最大の科で、ヒマワリのような重要な商業的作物を含む)およびバラ科(ラズベリー、アンズ、アーモンド、モモ、バラなどを含む)ならびにナッツ植物(クルミ、ペカン、ヘーゼルナッツなどを含む)および森の木(Pinus、Quercus、Pseutotsuga、Sequoia、Populusなどを含む)。
本発明のGATポリヌクレオチドによる改変のためのさらなる標的、および上記で特定された標的としては、以下の属由来の植物が挙げられる:Agrostis、Allium、Antirrhinum、Apium、Arachis、Asparagus、Atropa、Avena(例えば、カラスムギ)、Bambusa、Brassica、Bromus、Browaalia、Camellia、Cannabis、Capsicum、Cicer、Chenopodium、Chichorium、Citrus、Coffea、Coix、Cucumis、Curcubita、Cynodon、Dactylis、Datura、Daucus、Digitalis、Dioscorea、Elaeis、Eleusine、Festuca、Fragaria、Geranium、Gossypium、Glycine、Helianthus、Heterocallis、Hevea、Hordeum(例えば、オオムギ)、Hyoscyamus、Ipomoea、Lactuca、Lens、Lilium、Linum、Lolium、Lotus、Lycopersicon、Majorana、Malus、Mangifera、Manihot、Medicago、Nemesia、Nicotiana、Onobrychis、Oryza(例えば、コメ)、Panicum、Pelargonium、Pennisetum(例えば、キビ)、Petunia、Pisum、Phaseolus、Phleum、Poa、Prunus、Ranunculus、Raphanus、Ribes、Ricinus、Rubus、Saccharum、Salpiglossis、Secale(例えば、ライムギ)、Senecio、Setaria、Sinapis、Solanum、Sorghum、Stenotaphrum、Theobroma、Trifolium、Trigonella、Triticum(例えば、コムギ)、Vicia、Vigna、Vitis、Zea(例えば、トウモロコシ)、ならびにOlyreae、Pharoideaeおよび他の多くの属。記載されたように、Graminae科の植物は、特に本発明の方法のための標的植物である。
本発明の標的である共通の作物植物としては、トウモロコシ、コメ、ライコムギ、ライムギ、ワタ、ダイズ、サトウキビ、小ムギ、カラスムギ、大ムギ、キビ、ヒマワリ、アブラナ、エンドウマメ、マメ(bean)、レンズマメ、ピーナッツ、クズイモ、ササゲ、ベルベットマメ、クローバー、アルファルファ、ルビナス、ベッチ、ハス、スイートクローバー、フジ、スイートピー、およびナッツ植物(例えば、クルミ、ペカンなど)が挙げられる。
1つの局面において、本発明は、作物植物が作物を産生し、そしてその作物を収集するような条件下で、グリホセートN−アセチルトランスフェラーゼをコードする遺伝子を用いて形質転換された結果としてグリホセート耐性である作物植物を生育することによって作物を産生する方法を提供する。好ましくは、グリホセートは、そのトランスジェニック作物植物が作物の生育および産生を逸すること無く、雑草を制御するのに効果的な濃度で、植物または植物の近縁に適用される。グリホセートの適用は、栽培前か、または収穫時までおよび収穫時を含む栽培後の任意の時間であり得る。グリホセートは、一回または複数回適用され得る。グリホセート適用のタイミング、適用される量、適用の様式、および他のパラメーターは、作物植物の特定の性質および生育環境に基づいて変化し、そして当業者によって容易に決定され得る。本発明はさらに、本方法によって産生される作物の増殖を提供する。
本発明は、GATポリヌクレオチド導入遺伝子を含む植物の増殖を提供する。この植物は、例えば、単子葉植物または双子葉植物であり得る。1つの局面において、増殖は、GATポリヌクレオチド導入遺伝子を含む植物と第2の植物との交雑を必然的に伴い、その結果、交雑の少なくともいくつかの子孫が、グリホセート耐性を示す。
1つの局面において、本発明は、作物が生育されている畑における雑草を選択的に制御するための方法を提供する。この方法は、GATをコードする遺伝子(例えば、GATポリヌクレオチド)を用いて形質転換された結果としてグリホセート耐性である作物の種または作物植物を栽培すること、および作物への有意の有害な影響を有さずに雑草を制御するのに十分な量のグリホセートを、作物および任意の雑草に適用することを含む。雑草阻害に由来する利点が、グリホセートまたはグリホセートアナログの、作物または作物植物上のいずれの負の影響よりも重要である限り、作物が、この除草剤に対して全く非感受性である必要は無いことに注意することが重要である。
別の局面において、本発明は、選択マーカー遺伝子としてGATポリヌクレオチドの使用を提供する。本発明のこの実施形態において、細胞中または生物中のGATポリヌクレオチドの存在は、その細胞または生物に検出可能なグリホセート耐性の表現型形質を与え、それにより、GATポリヌクレオチドに連結された目的の遺伝子で形質転換された細胞または生物について選択し得る。従って、例えば、GATポリヌクレオチドは、核酸構築物(例えば、ベクター)へと導入され得、それにより、グリホセートの存在下での宿主の増殖、ならびに核酸構築物を欠損する宿主が、生存または増殖するよりも識別可能に速い速度で生存および/または増殖する能力についての選択によって核酸構築物を含む宿主(例えば、細胞またはトランスジェニック植物)の同定を可能にする。GATポリヌクレオチドは、植物、ほとんどの細菌(E.coliを含む)、放線菌、酵母、藻類、および真菌類を含む、グリホセートに感受性である、広範な種々の種類の宿主における選択マーカーとして用いられ得る。従来の抗生物質耐性株に対して、植物においてマーカーとして除草剤耐性株を用いる1つの利点は、それが、抗生物質耐性株が環境に逸出するという、本出願の何人かの構成員の心配を、回避することである。種々の宿主系における選択マーカーとしてのGATポリヌクレオチドの使用を実証する実験からのいくつかの実験データが、本明細書の実施例の章に記載される。
(トランスジェニック植物における増強されたグリホセート耐性を与えるgatポリヌクレオチドの選択)
本明細書中に記載された方法に従い多様化されたGATコード核酸のライブラリーが、トランスジェニック植物中でグリホセートに対する耐性を与える能力について選択され得る。多様化および選択の1回以上のサイクルの後で、改変GAT遺伝子が、トランスジェニック植物の産生および評価を容易にする選択マーカー、ならびに実験植物または農学的植物に除草剤耐性を与える方法として用いられ得る。例えば、多様化されたGATポリヌクレオチドのライブラリーを産生するための、配列番号1〜配列番号5のうちの任意の1つ以上の多様化の後で、最初の機能的評価が、E.coliにおけるGATコード配列ライブラリーの発現によって実施され得る。発現したGATポリペプチドが、上述のように精製され得るか、または部分的に精製され得、そして質量分析法によって、改善された速度についてスクリーニングされ得る。多様化および選択の1つ以上の第1ラウンド(round)の後、改善されたGATポリペプチドをコードするポリヌクレオチドが、例えば、強力な構成的プロモーター(例えば、CaMV 35Sプロモーター)へと作動可能に連結された、植物発現ベクターへとクローニングされる。改変GAT核酸を含むこの発現ベクターが、代表的には、アグロバクテリウムが媒介する形質転換によって、Arabidopsis thaliana宿主植物へと形質転換される。例えば、Arabidopsis宿主は、アグロバクテリウムの溶液中に花序を浸漬することによって容易に形質転換され、そしてそれらを成長させかつ種子をつけさせ得る。数千の種子が、約6週間の間に回復する。これらの種子が、次いで、浸漬された植物から大量に収集され、そして土壌中で発芽する。この様式において、高スループット(HTP)植物形質転換形式を構成する、数千の独立に形質転換された植物を、評価のために産生し得る。大量の成長した苗木に、グリホセートを噴霧し、そしてグリホセート耐性を示す生存する苗木は、選択プロセスで生存し、一方で、非トランスジェニック植物およびあまり好ましくなく改変されたGAT核酸を組み込んだ植物は、除草剤処理によって損傷するか、または死滅する。必要に応じて、グリホセートに対する改善された耐性を与えるGATコード核酸は、例えば、ライブラリー挿入部に隣接するT−DNAプライマーを用いたPCR増幅によって回復され、そしてさらなる多様化手順においてか、または同種もしくは異種のさらなるトランスジェニック植物を産生するために用いられる。所望の場合、さらなるラウンドの多様化および選択が、各々の連続する区画においてグリホセートの濃度を増加させながら用いることで実施され得る。この様式において、畑条件において有用な濃度のグリホセートに対する耐性を与えるGATポリヌクレオチドおよびポリペプチドが得られる。
本明細書中に記載された方法に従い多様化されたGATコード核酸のライブラリーが、トランスジェニック植物中でグリホセートに対する耐性を与える能力について選択され得る。多様化および選択の1回以上のサイクルの後で、改変GAT遺伝子が、トランスジェニック植物の産生および評価を容易にする選択マーカー、ならびに実験植物または農学的植物に除草剤耐性を与える方法として用いられ得る。例えば、多様化されたGATポリヌクレオチドのライブラリーを産生するための、配列番号1〜配列番号5のうちの任意の1つ以上の多様化の後で、最初の機能的評価が、E.coliにおけるGATコード配列ライブラリーの発現によって実施され得る。発現したGATポリペプチドが、上述のように精製され得るか、または部分的に精製され得、そして質量分析法によって、改善された速度についてスクリーニングされ得る。多様化および選択の1つ以上の第1ラウンド(round)の後、改善されたGATポリペプチドをコードするポリヌクレオチドが、例えば、強力な構成的プロモーター(例えば、CaMV 35Sプロモーター)へと作動可能に連結された、植物発現ベクターへとクローニングされる。改変GAT核酸を含むこの発現ベクターが、代表的には、アグロバクテリウムが媒介する形質転換によって、Arabidopsis thaliana宿主植物へと形質転換される。例えば、Arabidopsis宿主は、アグロバクテリウムの溶液中に花序を浸漬することによって容易に形質転換され、そしてそれらを成長させかつ種子をつけさせ得る。数千の種子が、約6週間の間に回復する。これらの種子が、次いで、浸漬された植物から大量に収集され、そして土壌中で発芽する。この様式において、高スループット(HTP)植物形質転換形式を構成する、数千の独立に形質転換された植物を、評価のために産生し得る。大量の成長した苗木に、グリホセートを噴霧し、そしてグリホセート耐性を示す生存する苗木は、選択プロセスで生存し、一方で、非トランスジェニック植物およびあまり好ましくなく改変されたGAT核酸を組み込んだ植物は、除草剤処理によって損傷するか、または死滅する。必要に応じて、グリホセートに対する改善された耐性を与えるGATコード核酸は、例えば、ライブラリー挿入部に隣接するT−DNAプライマーを用いたPCR増幅によって回復され、そしてさらなる多様化手順においてか、または同種もしくは異種のさらなるトランスジェニック植物を産生するために用いられる。所望の場合、さらなるラウンドの多様化および選択が、各々の連続する区画においてグリホセートの濃度を増加させながら用いることで実施され得る。この様式において、畑条件において有用な濃度のグリホセートに対する耐性を与えるGATポリヌクレオチドおよびポリペプチドが得られる。
(除草剤耐性)
本発明のグリホセート耐性の機構が、優れたグリホセート耐性を有する植物および植物外植片を産生するための、他の当該分野において公知の様式のグリホセート耐性と組み合わせられ得る。例えば、グリホセート耐性植物が、以下によってより完全に記載されるように、高レベルの5−エノールピルビルシキメート−3−ホスフェートシンターゼ(EPSP)を産生する能力をその植物のゲノムへと挿入することによって産生され得る:米国特許第6,248,876 B1号;同第5,627,061号;同第5,804,425号;同第5,633,435号;同第5,145,783号;同第4,971,908号;同第5,312,910号;同第5,188,642号;同第4,940,835号;同第5,866,775号;同第6,225,114 B1号;同第6,130,366号;同第5,310,667号;同第4,535,060号;同第4,769,061号;同第5,633,448号;同第5,510,471号;米国特許再発行第36,449号;同第37,287 E号;および米国特許第5,491,288号;ならびに国際出願WO 97/04103号;WO 00/66746号;WO 01/66704号;およびWO 00/66747号(これらは、本発明中全体においてすべての目的のために参考として援用される。)。グリホセート耐性はまた、米国特許第5,776,760号および同第5,463,175号(これらは、すべての目的のために本明細書中全体に参考として援用される)にさらに完全に記載されるようにグリホセートキシドレダクターゼ酵素をコードする遺伝子を発現する植物にも伝達され得る。
本発明のグリホセート耐性の機構が、優れたグリホセート耐性を有する植物および植物外植片を産生するための、他の当該分野において公知の様式のグリホセート耐性と組み合わせられ得る。例えば、グリホセート耐性植物が、以下によってより完全に記載されるように、高レベルの5−エノールピルビルシキメート−3−ホスフェートシンターゼ(EPSP)を産生する能力をその植物のゲノムへと挿入することによって産生され得る:米国特許第6,248,876 B1号;同第5,627,061号;同第5,804,425号;同第5,633,435号;同第5,145,783号;同第4,971,908号;同第5,312,910号;同第5,188,642号;同第4,940,835号;同第5,866,775号;同第6,225,114 B1号;同第6,130,366号;同第5,310,667号;同第4,535,060号;同第4,769,061号;同第5,633,448号;同第5,510,471号;米国特許再発行第36,449号;同第37,287 E号;および米国特許第5,491,288号;ならびに国際出願WO 97/04103号;WO 00/66746号;WO 01/66704号;およびWO 00/66747号(これらは、本発明中全体においてすべての目的のために参考として援用される。)。グリホセート耐性はまた、米国特許第5,776,760号および同第5,463,175号(これらは、すべての目的のために本明細書中全体に参考として援用される)にさらに完全に記載されるようにグリホセートキシドレダクターゼ酵素をコードする遺伝子を発現する植物にも伝達され得る。
さらに、本発明のグリホセート耐性の機構が、グリホセートおよび1つ以上の他の除草剤に対して耐性の植物および植物外植片を産生するために、他の様式の除草剤耐性と組み合わせられ得る。例えば、ヒドロキシフェニルピルベートダイオキシゲナーゼは、パラ−ヒドロキシフェニルピルベート(HPP)が、ホモゲンチシン酸に変化する反応を触媒する酵素である。この酵素を阻害し、そしてHPPのホモゲンチシン酸への変化を阻害するためにこの酵素に結合する分子は、除草剤として有用である。特定の除草剤に対してより耐性な植物が、米国特許第6,245,968 B1号;同第6,268,549号;および6,069,115号;ならびに国際出願WO99/23886号(これらは、全ての目的のために本明細書中全体に援用される)に記載される。
スルホニル尿素およびイミダゾリノン(imidazolinone)除草剤もまた、アセト乳酸シンターゼ(ALS)またはアセトヒドロキシ酸シンターゼ(AHAS)をブロックすることによってより高い植物の成長を阻害する。スルホニル尿素およびイミダゾリノン耐性植物の産生は、米国特許第5,605,011号;同第5,013,659号;同第5,141,870号;同第5,767,361号;同第5,731,180号;同第5,304,732号;同第4,761,373号;同第5,331,107号;同第5,928,937号;および同第5,378,874号;ならびに国際出願WO 96/33270号(これらは、本明細書中全体において、全ての目的のために参考として援用される)より完全に記載される。
グルタミンシンターゼ(GS)は、ほとんどの植物細胞の発生および生存に必要な本質的酵素であることが明らかである。GSのインヒビターは、植物細胞に対して毒性である。グルフォシネート(glufosinate)除草剤が、植物中のGSの阻害に起因する毒性効果に基づき開発された。これらの除草剤は、非選択性である。これらは、存在する植物の異なる種の全ての成長を阻害し、これら全ての破壊を引き起こす。外因性ホスフィノスリシンアセチルトランスフェラーゼを含む植物の開発が、米国特許第5,969,213号;同第5,489,520号;同第5,550,318号;同第5,874,265号;同第5,919,675号;同第5,561,236号;同第5,648,477号;同第5,646,024号;同第6,177,616 B1号;および同第5,879,903号(これらは、全ての目的のために本明細書中全体に参考として援用される)に記載される。
プロトポルフィリノーゲンオキシダーゼ(protox)は、全ての植物の生存に必要な、葉緑素の産生に必要である。このprotox酵素は、種々の除草剤化合物の標的として作用する。これらの除草剤もまた、存在する植物の全ての異なる種の成長を阻害し、これら全ての破壊を引き起こす。これらの除草剤に対して耐性である変化されたprotox活性を含む植物の開発が、米国特許第6,288,306 B1号;同第6,282,837 B1号;および同第5,767,373号;ならびに国際出願WO01/12825号(これらは、全ての目的のために本明細書中全体に参考として援用される)に記載される。
(実施例)
以下の実施例は、例示的であって、限定をするものではない。当業者は、本質的に類似する結果を達成するために変化され得る、種々の非限界パラメーターを認識する。
以下の実施例は、例示的であって、限定をするものではない。当業者は、本質的に類似する結果を達成するために変化され得る、種々の非限界パラメーターを認識する。
(実施例1:新規天然GATポリヌクレオチドの単離)
5種の天然GATポリヌクレオチド(すなわち、遺伝的に改変されていない生物中に天然に存在するGATポリヌクレオチド)を、GAT活性を示すBacillus系統由来の配列の発現クローニングによって、見出した。これらのヌクレオチド配列を、決定し、そして配列番号1〜配列番号5として本明細書中に提供した。すなわち、約500種のBacillus系統およびPseudomonas系統の収集物を、N−アセチレートグリホセートに対する天然の能力についてスクリーニングした。これらの系統を、LB中で一晩増殖し、遠心分離によって収集し、希トルエン中で透過性にし、ついで洗浄し、そして緩衝液、5mMグリホセート、および200μMアセチルCoAを含む反応混合物中に再懸濁した。この細胞を、1〜48時間の間(この時間帯に、この反応物と等容量のメタノールを添加した)反応混合物中でインキュベートした。次いでこの細胞を、遠心分離によってペレットにし、そして上清を個体イオンモード質量分析法(parent ion mode mass spectrometry)による分析の前に、ろ過した。この反応生成物を、反応混合物の質量分析法プロファイルと、図2に示す標準N−アセチルグリホセートとを比較することによって、N−アセチルグリホスサートとして陽性で同定した。生成物の検出は、両方の基質(アセチルCoAおよびグリホセート)含有物に依存し、そして細菌細胞の熱変性によって破壊された。
5種の天然GATポリヌクレオチド(すなわち、遺伝的に改変されていない生物中に天然に存在するGATポリヌクレオチド)を、GAT活性を示すBacillus系統由来の配列の発現クローニングによって、見出した。これらのヌクレオチド配列を、決定し、そして配列番号1〜配列番号5として本明細書中に提供した。すなわち、約500種のBacillus系統およびPseudomonas系統の収集物を、N−アセチレートグリホセートに対する天然の能力についてスクリーニングした。これらの系統を、LB中で一晩増殖し、遠心分離によって収集し、希トルエン中で透過性にし、ついで洗浄し、そして緩衝液、5mMグリホセート、および200μMアセチルCoAを含む反応混合物中に再懸濁した。この細胞を、1〜48時間の間(この時間帯に、この反応物と等容量のメタノールを添加した)反応混合物中でインキュベートした。次いでこの細胞を、遠心分離によってペレットにし、そして上清を個体イオンモード質量分析法(parent ion mode mass spectrometry)による分析の前に、ろ過した。この反応生成物を、反応混合物の質量分析法プロファイルと、図2に示す標準N−アセチルグリホセートとを比較することによって、N−アセチルグリホスサートとして陽性で同定した。生成物の検出は、両方の基質(アセチルCoAおよびグリホセート)含有物に依存し、そして細菌細胞の熱変性によって破壊された。
次いで、個々のGATポリヌクレオチドを、機能性スクリーニングによって同定した系統からクローニングした。ゲノムDNAを、調製し、そしてSau3A1酵素で部分的に消化した。約4Kbのフラグメントを、E.coli発現ベクターへとクローニングし、そして電子受容(electrocompetent)E.coliへと形質転換した。GAT活性を示す個々のクローンを、トルエン洗浄をPMBSでの浸透に置き換えたことを除き、前に記載したような反応後の質量分析法によって同定した。ゲノムフラグメントを配列決定し、そして推定のGATポリペプチドをコードするオープンリーディングフレームを同定した。GAT遺伝子の同定を、E.coli中のオープンリーディングフレームの発現および反応混合物より産生された高レベルのN−アセチルグリホスフェートの検出によって確認した。
(実施例2:B.licheniformis系統B6から単離したGATポリペプチドの特徴づけ)
B.licheniformis系統B6からのゲノムDNAを、精製し、Sau3A1によって部分的に消化し、そして1〜10Kbのフラグメントを、E.coli発現ベクターへとクローニングした。2.5kbの挿入部を有するクローンによって、質量スペクトル分析を用いて決定したような、E.coli宿主上のグリホセートN−アセチルトランスフェラーゼ(GAT)活性を与えた。挿入部の配列決定によって、441塩基対の単一の完全なオープンリーディングフレームを明らかにした。続くこのオープンリーディングフレームのクローニングによって、それがGAT酵素をコードすることを確認した。B6のGAT酵素をコードする遺伝子を有するプラスミド(pMAXY2120、図4に示す)を、E.coli系統XL1 Blueへと形質転換した。10%の移植片(innoculum)の飽和培養物を、Luria類(broth)へと添加し、そして培養物を、37℃で、1時間インキュベートした。GATの発現を、1mMの濃度でのIPTGの添加によって誘導した。この培養物を、さらに4時間インキュベートし、その後、細胞を、遠心分離によって収集し、細胞のペレットを−80℃で保存した。
B.licheniformis系統B6からのゲノムDNAを、精製し、Sau3A1によって部分的に消化し、そして1〜10Kbのフラグメントを、E.coli発現ベクターへとクローニングした。2.5kbの挿入部を有するクローンによって、質量スペクトル分析を用いて決定したような、E.coli宿主上のグリホセートN−アセチルトランスフェラーゼ(GAT)活性を与えた。挿入部の配列決定によって、441塩基対の単一の完全なオープンリーディングフレームを明らかにした。続くこのオープンリーディングフレームのクローニングによって、それがGAT酵素をコードすることを確認した。B6のGAT酵素をコードする遺伝子を有するプラスミド(pMAXY2120、図4に示す)を、E.coli系統XL1 Blueへと形質転換した。10%の移植片(innoculum)の飽和培養物を、Luria類(broth)へと添加し、そして培養物を、37℃で、1時間インキュベートした。GATの発現を、1mMの濃度でのIPTGの添加によって誘導した。この培養物を、さらに4時間インキュベートし、その後、細胞を、遠心分離によって収集し、細胞のペレットを−80℃で保存した。
細胞の溶解を、1mlの以下の緩衝液を、0.2gの細胞に添加することによってもたらした:25mMのHEPES(pH7.3)、0.1mMのEDTAを添加した100mMのKClおよび10%メタノール(HKM)、1mMのDTT、1mg/mlのニワトリ卵リソザイム、ならびにSigmaより得、かつ製造者の指示に従い用いたプロテーゼインヒビターカクテル。室温(例えば、22〜25℃)での20分のンキュベーションの後に、溶解は、簡単な超音波処理を伴って達成された。この溶解産物を、遠心分離し、そして上清をHKMを用いて平衡化したSephadex G25を通して脱塩した。部分的精製物を、CoAアガロース(Sigma)上のアフィニティクロマトグラフィーによって得た。このカラムを、HKMを用いて平衡化し、そして清澄化された抽出物を静水圧下で通過させた。非結合タンパク質を、HKMを用いたカラムの洗浄によって取り除き、そしてGATを、1mMのCoエンザイムAを含むHKMを用いて溶出した。この手順により4倍の精製物を提供した。この段階で、粗溶解産物で充填したSDSポリアクリルアミドゲル上で観察された約65%のタンパク質染色は、GATに起因し、別の20%は、ベクターにコードされているクロラムフェニコールアセチルトランスフェラーゼに起因した。
均質性についての精製を、部分的精製されたタンパク質のSuperdex 75(Pharmacia)を介するゲルろ過によって得た。移動層は、HKMであり、ここでGAT活性が、17kDの分子半径に対応する容積で溶出された。この物質は、12%アクリルアミドゲル(厚さ1mm)上のSDSポリアクリルアミドゲル電気泳動に供される、3μgのGAT試料のクマシー染色物によって判断されるほど、均一であった。精製により、特定の活性の6倍増加を達成した。
グリホセートの見かけ上のKMを、5mMのモルホリンを含み、酢酸でpH7.7に調製された緩衝液および20%のエチレングリコールの中の、飽和(200μM)アセチルCoA、種々の濃度のグリホセート、および1μM精製GATを含む反応混合物上で決定した。反応初速度を、235nm(E=3.4 OD/mM/cm)でのアセチルCoAのチオエステル結合の加水分解を連続的にモニターすることによって決定した。飽和双曲線キネティックスを観察し(図5)、ここから2.9±0.2(SD)mMの見かけのKMを得た。
AcCoAの見かけのKMを、5mMのモルホリンを含み、酢酸でpH7.7に調製した緩衝液および50%のメタノールの中の、5mMグリホセート、種々の濃度のアセチルCoA、および0.19μM GATを含む反応混合物上で決定した。反応初速度を、N−アセチルグリホセートの質量分析法検出を用いて決定した。5μlを、この機器に繰り返し注入し、反応速度を、反応時間対積算されたピーク面積をプロットすることによって得た(図6)。飽和双曲線キネティックスが、観察され(図7)、ここから2μMの見かけのKMを導いた。既知濃度の酵素から得られたVmaxの値から、6/分のkcatを算出した。
(実施例3:質量分析法(MS)スクリーニングプロセス)
試料(5μl)を、26秒毎に1試料の速度で、96ウェルのマイクロタイタープレートから吸出し、いかなる分離もせずに、質量分析計(Micromass Quattro LC、triple quadrupole mass spectrometer)へと注入した。この試料を、流速500Ul/分の水/メタノール(50:50)の移動層によって、質量分析計へと移動させた。各々の注入された試料を、負のエレクトロスプレーイオン化プロセス(ニードルボルテージ(needle voltage)、−3.5KV;コーンボルテージ(cone voltage)、20V;供給源温度120C;非溶媒和(desolvation)温度、250C;コーン気体流量、90L/時間;および非溶媒和気体流量、600L/時間)によってイオン化した。このプロセスの間に形成されたこの分子イオン(m/z 210)を、第2の四重極(ここでの圧力を、5×10−4mBarに設定し、そして衝突エネルギーを、20 Evに調節した)における衝突誘導解離(collison induced dissociation、CID)を実施するために、第1の四重極によって選択した。第3の四重極を、親イオン(parent ion)(m/z 210)から産生された娘イオン(daughter ion)(m/z 124)の1つが、シグナル記録のための検出器へと到達し得ることのみのために設置した。第1の四重極および第3の四重極を、解像ユニットに設置した(ここで、光電子増倍機を、650 Vで作動した)。純粋の標準N−アセチルグリホセートを、比較のために用い、そして積算ピークを、濃度を見積もるため用いた。本方法により、200Nm未満のN−アセチルグリホセートが、検出可能である。
試料(5μl)を、26秒毎に1試料の速度で、96ウェルのマイクロタイタープレートから吸出し、いかなる分離もせずに、質量分析計(Micromass Quattro LC、triple quadrupole mass spectrometer)へと注入した。この試料を、流速500Ul/分の水/メタノール(50:50)の移動層によって、質量分析計へと移動させた。各々の注入された試料を、負のエレクトロスプレーイオン化プロセス(ニードルボルテージ(needle voltage)、−3.5KV;コーンボルテージ(cone voltage)、20V;供給源温度120C;非溶媒和(desolvation)温度、250C;コーン気体流量、90L/時間;および非溶媒和気体流量、600L/時間)によってイオン化した。このプロセスの間に形成されたこの分子イオン(m/z 210)を、第2の四重極(ここでの圧力を、5×10−4mBarに設定し、そして衝突エネルギーを、20 Evに調節した)における衝突誘導解離(collison induced dissociation、CID)を実施するために、第1の四重極によって選択した。第3の四重極を、親イオン(parent ion)(m/z 210)から産生された娘イオン(daughter ion)(m/z 124)の1つが、シグナル記録のための検出器へと到達し得ることのみのために設置した。第1の四重極および第3の四重極を、解像ユニットに設置した(ここで、光電子増倍機を、650 Vで作動した)。純粋の標準N−アセチルグリホセートを、比較のために用い、そして積算ピークを、濃度を見積もるため用いた。本方法により、200Nm未満のN−アセチルグリホセートが、検出可能である。
(実施例4:GAT酵素の天然または低い活性の検出)
GAT酵素の天然または低い活性は、代表的に、約1/分のKcatおよび1.5〜10MmのグリホセートについてのKMを有する。アセチルCoAについてのKMは、代表的に、25μM未満である。
GAT酵素の天然または低い活性は、代表的に、約1/分のKcatおよび1.5〜10MmのグリホセートについてのKMを有する。アセチルCoAについてのKMは、代表的に、25μM未満である。
細菌性培養物を、深い96ウェルのプレート中の栄養豊富な培地中で増殖し、そして0.5mlの静止期の細胞を遠心分離によって収集し、5mMのモルホリンアセテート(pH 8)を用いて洗浄し、そして200μMのアンモニウムアセチルCoA、5mMのアンモニウムグリホセート、および5μg/ml PMBS(Sigma)を含む0.1mlの反応混合物(5mMのモルホリンアセテート(pH 8)中)中に懸濁した。このPMBSは、細胞膜を透過性にし、細胞性内容物全てを放出することなく、基質および産生物を細胞から緩衝液へ移動させ得る。反応を、25〜37℃で1〜48時間実施した。この反応を、等容積の100%エタノールを用いて休止し、そして混合物全体を、0.45μm MAHV Multiscreenフィルタープレート(Millipore)上でろ過した。試料を、上記のように質量分析計を用いて分析し、標準の合成N−アセチルグリホセートと比較した。
(実施例5:高活性GAT酵素の検出)
高活性GAT酵素は、代表的に、400/分までのkcatおよび0.1mMグリホセート以下のKMを有する。
高活性GAT酵素は、代表的に、400/分までのkcatおよび0.1mMグリホセート以下のKMを有する。
GAT酵素をコードする遺伝子を、E.coli発現ベクター(例えば、pQE80(Qiagen))へとクローニングし、そしてE.coli系統(例えば、XL1 Blue(Stratagene))へと導入した。培養物を、浅いU底の96ウェルポリスチレンプレート中の150μlの栄養豊富な培地(例えば、50μg/mlのカルベニシリン(carbenicllin))を有するLB)中で後期対数増殖期まで増殖し、そして1mMのIPTG(USB)を含む新しい培地を用いて1:9に希釈した。4〜8時間の誘導の後で、細胞を収集し、5mMのモルホリンアセテートpH 6.8を用いて洗浄し、そして等容積の同じモルホリン緩衝液中に再懸濁した。反応を、10μlまでの洗浄した細胞を用いて実行した。より高い活性レベルでは、この細胞を、最初に1:200に希釈し、そして5μlを100μlの反応混合物へと添加した。GAT活性を測定するために、低活性について記載したのと同じ反応混合物が、用いられ得る。しかし、高活性GAT酵素を検出するために、グリホセートの濃度を0.15〜0.5mMまで下げ、pHを6.8まで下げ、そして反応を、37℃で1時間実行した。反応のワークアップおよびMS検出は、本明細書中に記載のようにした。
(実施例6:GAT酵素の精製)
酵素精製を、細胞溶解物のCoAアガロース上でのアフィニティクロマトグラフィーおよびSuperdex−75上でのゲルろ過によって達成した。10mgまでの量の精製GAT酵素を以下のように得た:50μg/mlのカルベニシリンを含むLB中で一晩増殖した、pQE80ベクター上にGATポリヌクレオチドを保有するE.coliの培養物100mlを、50μg/mlのカルベニシリンを添加した1LのLBを接種するために用いた。その後1時間、IPTGを1mMまで添加し、そして培養物をさらに6時間増殖させた。細胞を、遠心分離によって収集した。溶解を、25mMのHEPES(pH7.2)、100mMのKCl、10% メタノール(HKMと称す)、0.1mMのEDTA、1mMのDTT、Sigma−Aldrichによって供給されたプロテーゼインヒビターカクテルおよび1mg/mlのニワトリ卵リゾチーム中に細胞を懸濁することによってもたらした。その後30分、室温で、細胞を、暫時、超音波処理した。粒状の物質を、遠心分離によって取り除き、そして溶解物を、コエンザイムAアガロースの層に通した。このカラムを、層容積の数倍のHKMを用いて洗浄し、そしてGATを、1mMのアセチルコエンザイムAを含む、層容積の1.5倍のHKMを用いて溶出した。溶出物中のGATを、Centricon YM 50限外ろ過膜にわたるその保持によって、濃縮した。さらなる精製物を、そのタンパク質を、一連の0.6mlの注入を介してSuperdex 75カラムに通すことによって得た。GAT活性溶出物のピークは、17kDの分子量に対応する容積で、溶出した。この方法により、GAT酵素の85%の回復を超えるまでの均一性精製を生じた。類似の手順を用いて、0.1〜0.4mgの量の、96種までのシャッフルした改変体を同時に得た。誘導した培養物の容積を、1〜10mlまで減らし、コエンザイムA−アガロースアフィニティクロマトグラフィーを、MAHVフィルタープレート(Millipore)中に充填した0.15mlカラムにおいて実施し、そしてSuperdex 75 クロマトグラフィを、省略した。
酵素精製を、細胞溶解物のCoAアガロース上でのアフィニティクロマトグラフィーおよびSuperdex−75上でのゲルろ過によって達成した。10mgまでの量の精製GAT酵素を以下のように得た:50μg/mlのカルベニシリンを含むLB中で一晩増殖した、pQE80ベクター上にGATポリヌクレオチドを保有するE.coliの培養物100mlを、50μg/mlのカルベニシリンを添加した1LのLBを接種するために用いた。その後1時間、IPTGを1mMまで添加し、そして培養物をさらに6時間増殖させた。細胞を、遠心分離によって収集した。溶解を、25mMのHEPES(pH7.2)、100mMのKCl、10% メタノール(HKMと称す)、0.1mMのEDTA、1mMのDTT、Sigma−Aldrichによって供給されたプロテーゼインヒビターカクテルおよび1mg/mlのニワトリ卵リゾチーム中に細胞を懸濁することによってもたらした。その後30分、室温で、細胞を、暫時、超音波処理した。粒状の物質を、遠心分離によって取り除き、そして溶解物を、コエンザイムAアガロースの層に通した。このカラムを、層容積の数倍のHKMを用いて洗浄し、そしてGATを、1mMのアセチルコエンザイムAを含む、層容積の1.5倍のHKMを用いて溶出した。溶出物中のGATを、Centricon YM 50限外ろ過膜にわたるその保持によって、濃縮した。さらなる精製物を、そのタンパク質を、一連の0.6mlの注入を介してSuperdex 75カラムに通すことによって得た。GAT活性溶出物のピークは、17kDの分子量に対応する容積で、溶出した。この方法により、GAT酵素の85%の回復を超えるまでの均一性精製を生じた。類似の手順を用いて、0.1〜0.4mgの量の、96種までのシャッフルした改変体を同時に得た。誘導した培養物の容積を、1〜10mlまで減らし、コエンザイムA−アガロースアフィニティクロマトグラフィーを、MAHVフィルタープレート(Millipore)中に充填した0.15mlカラムにおいて実施し、そしてSuperdex 75 クロマトグラフィを、省略した。
(実施例7:KcatおよびKM決定のための標準プロトコール)
精製タンパク質のグリホセートについてのKcatおよびKMを、連続分光測定アッセイ(ここでAcCoAのスルホエステル結合の加水分解を、235nmで、モニターした)を用いて決定した。反応を、96ウェルアッセイプレートのウェル中で、周囲温度(約23℃)で、0.3mlの最終容積で存在する以下の成分を用いて、実施した:20mM HEPES(pH6.8)、10%エチレングリコール、0.2mMアセチルコエンザイムA、および種々の濃度のアンモニウムグルホサート。2種のGAT酵素のキネティックスの比較において、両方の酵素を、同じ条件下で(例えば、両方23℃で)アッセイするべきである。Kcatを、VmaxおよびBradfordアッセイによって決定した、酵素濃度より算出した。KMを、0.125〜10mMの範囲で変化するグリホセート濃度から得た反応初速度から、ミカエリスメンテン式のラインウェーバーバーク変換を用いて算出した。Kcat/KMを、Kcatについて決定した価を、KMについて決定した価で除することによって決定した。
精製タンパク質のグリホセートについてのKcatおよびKMを、連続分光測定アッセイ(ここでAcCoAのスルホエステル結合の加水分解を、235nmで、モニターした)を用いて決定した。反応を、96ウェルアッセイプレートのウェル中で、周囲温度(約23℃)で、0.3mlの最終容積で存在する以下の成分を用いて、実施した:20mM HEPES(pH6.8)、10%エチレングリコール、0.2mMアセチルコエンザイムA、および種々の濃度のアンモニウムグルホサート。2種のGAT酵素のキネティックスの比較において、両方の酵素を、同じ条件下で(例えば、両方23℃で)アッセイするべきである。Kcatを、VmaxおよびBradfordアッセイによって決定した、酵素濃度より算出した。KMを、0.125〜10mMの範囲で変化するグリホセート濃度から得た反応初速度から、ミカエリスメンテン式のラインウェーバーバーク変換を用いて算出した。Kcat/KMを、Kcatについて決定した価を、KMについて決定した価で除することによって決定した。
本方法論を用いて、本明細書中に例示した多くのGATポリペプチドについての速度パラメーターを、決定した。例えば、配列番号445に対応するGATポリペプチドについてのKcat、KMおよびKcat/KMを、上記のアッセイ条件を用いて各々、322/分、0.5mM、および660/mM/分であると決定した。配列番号457に対応するGATポリペプチドについてのKcat、KMおよびKcat/KMを、上記のアッセイ条件を用いて各々、118/分、0.1mM、および1184/mM/分であると決定した。配列番号300に対応するGATポリペプチドについてのKcat、KMおよびKcat/KMを、上記のアッセイ条件を用いて各々、296/分、0.65mM、および456/mM/分であると決定した。当業者は、これらの数値を用いて、GAT活性アッセイが、本明細書中で与えられる値との比較のために適した、GATについての速度パラメーターを産生することを確認し得る。例えば、GAT活性を比較するために用いられる条件は、その条件が、試験GATと本明細書中に例示されるGATポリペプチドとを比較するために使用される場合、配列番号300、445、および457について、本明細書中に報告されるこれらの値と同じ速度定数(通常の実験上の分散内で)を産み出すはずである。多くのGATポリペプチド改変体についての速度パラメーターを、本方法論に従って決定し、表3、4、5に提供した。
(表3.GATポリペプチドkcat値)
(表4.GATポリペプチド(グリホセート)KM値)
(表5.GATポリペプチドkcat/KM値)
AcCoAについてのKMを、反応にわたる繰り返しのサンプリングを伴う質量分析法を用いて測定した。アセチルコエンザイムAおよびグリホセート(アンンモニウム塩)を、50倍濃度ストック溶液として、質量分析計サンプルプレートのウェル中に配置した。反応を、モルホリンアセテートまたは炭酸アンモニウムのような揮発性緩衝液(pH6.8または7.7)中に適切に希釈した酵素の添加によって開始した。この試料を、この機器へと繰り返し注入し、そして初速度を、保持時間およびピーク面積のプロットから算出した。KMを、グリホセートについてと同様に算出した。
(実施例8:形質転換E.COLIの選択)
進化したgat遺伝子(GATコード配列の5’末端に直接的に結合された天然B.licheniformisリボソーム結合部位(AACTGAAGGAGGAATCTC;配列番号515)とのキメラ)を、EcoRI部位およびHindIII部位の間の発現ベクターpQE80(Qiagen)にクローン化し、プラスミドpMAXY2190(図11)を得た。これは、このプラスミドからHisタグドメインを排除し、そして抗生物質アンピシリンおよびカルベニシリンへの耐性を与えるB−ラクタマーゼ遺伝子を保持した。pMAXY2190を、XL1 Blue(Stratagene)E.coli細胞にエレクトロポレート(BioRad Gene Pulser)した。この細胞を、SOCリッチ培地に懸濁させ、そして1時間回復させた。次いで、この細胞を徐々にペレット化し、芳香族アミノ酸を欠くM9最小培地(12.8g/LのNa2HPO4・7H2O、3.0g/LのKH2PO4、0.5g/LのNaCl、1.0g/LのNH4Cl,0.4%のグルコース、2mMのMgSO4、0.1mMのCaCl2、10ml/Lのチアミン、10mg/Lのプロリン、30mg/Lのカルベニシリン)で1回洗浄し、20mlの同一のM9培地中に再懸濁させた。37℃での、250rpmでの終夜増殖の後、等体積の細胞を、M9培地またはM9に1mMのグリホセート(glyphosate)を加えた培地のどちらかにプレートした。gat遺伝子を有さないpQE80ベクターを、同様にE.coli細胞に導入し、そして比較のための単一コロニーにプレートした。結果を表6に要約する。そしてこの結果は、GAT活性が、バックグラウンド1%未満の形質転換E.coliの選択および増殖を可能にすることを明確に示す。IPTG誘導が、形質転換細胞の増殖を可能にするために十分なGAT活性のために必須でないことに注目されたい。グリホセートの存在下で増殖されたE.coli細胞に由来するpMAXY2190の再単離によって、形質転換を確認した。
進化したgat遺伝子(GATコード配列の5’末端に直接的に結合された天然B.licheniformisリボソーム結合部位(AACTGAAGGAGGAATCTC;配列番号515)とのキメラ)を、EcoRI部位およびHindIII部位の間の発現ベクターpQE80(Qiagen)にクローン化し、プラスミドpMAXY2190(図11)を得た。これは、このプラスミドからHisタグドメインを排除し、そして抗生物質アンピシリンおよびカルベニシリンへの耐性を与えるB−ラクタマーゼ遺伝子を保持した。pMAXY2190を、XL1 Blue(Stratagene)E.coli細胞にエレクトロポレート(BioRad Gene Pulser)した。この細胞を、SOCリッチ培地に懸濁させ、そして1時間回復させた。次いで、この細胞を徐々にペレット化し、芳香族アミノ酸を欠くM9最小培地(12.8g/LのNa2HPO4・7H2O、3.0g/LのKH2PO4、0.5g/LのNaCl、1.0g/LのNH4Cl,0.4%のグルコース、2mMのMgSO4、0.1mMのCaCl2、10ml/Lのチアミン、10mg/Lのプロリン、30mg/Lのカルベニシリン)で1回洗浄し、20mlの同一のM9培地中に再懸濁させた。37℃での、250rpmでの終夜増殖の後、等体積の細胞を、M9培地またはM9に1mMのグリホセート(glyphosate)を加えた培地のどちらかにプレートした。gat遺伝子を有さないpQE80ベクターを、同様にE.coli細胞に導入し、そして比較のための単一コロニーにプレートした。結果を表6に要約する。そしてこの結果は、GAT活性が、バックグラウンド1%未満の形質転換E.coliの選択および増殖を可能にすることを明確に示す。IPTG誘導が、形質転換細胞の増殖を可能にするために十分なGAT活性のために必須でないことに注目されたい。グリホセートの存在下で増殖されたE.coli細胞に由来するpMAXY2190の再単離によって、形質転換を確認した。
(実施例9:形質転換植物細胞の選択)
植物細胞のアグロバクテリウム(Agrobacterium)媒介形質転換は、低い有効性で生じる。非形質転換細胞の増殖を阻害しながら一方で、形質転換細胞の増殖を可能にするために、選択可能なマーカーが必要とされる。カナマイシンおよびハイグロマイシンに対する抗生物質マーカー、ならびに、遺伝子barを改変する除草剤(これは、除草性化合物ホスフィノスリシン(phosphinothricin)を解毒する)は、植物において用いられる選択可能なマーカーの例である(Methods in Molecular Biology,1995,49:9〜18)。ここで、本発明者らは、GAT活性が植物形質転換に対する有効な選択可能なマーカーとして役立つことを実証する。進化したgat遺伝子(0_5B8)を、植物プロモーター(強化イチゴ葉脈縞状ウイルス)とユビキノンターミネーターとの間でクローン化し、そして、図12に示されるように、Agrobacterium tumefaciens EHA105を介して、植物細胞の形質転換に適する二成分ベクターpMAXY3793のT−DNA領域に導入した。スクリーニング可能なGUSマーカーは、T−DNA中に存在し、形質転換の確認を可能にした。唯一の選択薬剤としてグリホセートを用いて、トランスジェニックタバコ苗条を産生した。
Nicotiana tabacum L.Xanthiの腋芽を、16時間の光(35〜42μEinsteins m−2s−1、白色蛍光灯)の下で、24℃で2〜3週間毎に、スクロース(1.5%)およびゲルライト(Gelrite)(0.3%)を含む、半強度(half−strength)MS培地で継代培養した。2週間〜3週間の継代培養の後、若葉を植物から切除し、そして3×3mmの断片に切り取った。A.tumefaciens EHA105を、LB培地に接種し、そしてA600=1.0の密度まで終夜増殖させた。4,000rpmで5分間かけて細胞をペレット化し、2mg/LのN6−ベンジルアデニン(BA)を含むMurashige and Skoog(MS)培地(pH5.2)、1%のグルコースおよび400uMのアセチシリンゴン(acetysyringone)を含む、3容量の液体共同培養培地に再懸濁させた。次いで、葉片を、100×25mmのペトリ皿中の20mlのA.tumefaciens中に30分間完全に浸し、オートクレーブ処理した濾紙を用いてブロットし、次いで固体共同培養培地(0.3%のゲルライト)に配置し、そして上記のようにインキュベートした。3日間の共同培養の後、20〜30個の断片を、2mg/LのBAを含むMS固体培地(pH5.7)、3%のスクロース、0.3%のゲルライト、0〜200uMのグリホセート、および400ug/mlのチメンチン(Timentin)を含む基礎苗条誘導(BSI)培地に移した。
3週間後、苗条は、gat遺伝子の有無に関係なく、グリホセートを含まない培地に配置された移植片上に明確に存在していた。両方の構築物からのT−DNA移入を、再生された苗条に由来する葉のGUS組織化学的染色によって確認した。20uMよりも大きなグリホセート濃度は、gat遺伝子を欠く移植片に由来する任意の苗条形成を完全に阻害した。gat構築物を有するA.tumefaciensに感染した移植片は、200uM(試験された最高レベル)までのグリホセート濃度において、苗条を再生した。GUS組織化学的染色によって、ならびに、プロモーターおよび3’領域にアニーリングするプライマーを用いるgat遺伝子のPCRフラグメント増幅によって、形質転換を確認した。結果を表7に要約する。
(実施例10:形質転換された酵母細胞のグリホセート選択)
酵母形質転換のための選択マーカーは、通常、特定のアミノ酸またはヌクレオチドを欠く培地での形質転換細胞の増殖を可能にする栄養要求性遺伝子である。Saccharomyces cerevisiaeがグリホセートに敏感であるので、GATはまた、選択可能なマーカーとして用いられ得る。このことを示すために、進化したgat遺伝子(0_6D10)を、コード領域全体を含むPstI−ClaIフラグメントとして、(実施例9に示したように)T−DNAベクターpMAXY3793からクローン化し、そして、図13に示すようなPstI−ClaI消化p424TEF(Gene,1995,156:119〜122)にライゲーションする。このプラスミドは、E.coliの複製起点、およびカルベニシリン耐性を与える遺伝子、ならびにTRP1(酵母形質転換のためのトリプトファン栄養要求体選択マーカー)を含む。
gat含有構築物を、E.coli XL1 Blue(Statagene)へと形質転換し、そしてLBカルベニシリン(50ug/ml)寒天培地にプレートする。プラスミドDNAを調製し、形質転換キット(Bio101)を用いて酵母菌株YPH499(Stratagene)を形質転換するために用いる。等量の形質転換された細胞を、全ての芳香族アミノ酸(トリプトファン、チロシン、およびフェニルアラニン)を欠いており、グリホセートを添加された、CSM−YNB−グルコース培地(Bio101)にプレートする。比較のために、gat遺伝子を欠くp424TEFもまた、YPH499に導入し、そして上記のようにプレートする。この結果は、GAT活性が有効な選択可能なマーカーとして機能することを実証する。グリホセート選択コロニーにおけるgat含有ベクターの存在は、プラスミドの再単離および制限消化分析によって確認され得る。
先述の本発明は、明確さおよび理解を目的として、いくらか詳細に記載されているが、形態または詳細の種々の変更が、本発明の真の範囲から逸脱せずになされ得ることは、当業者にとって、本開示を読めば明白である。例えば、上記の全ての技術、方法、組成物、装置および系は、種々に組み合わせて用いられ得る。本発明は、本明細書中に記載される全ての方法および試薬、ならびに全てのポリヌクレオチド、ポリペプチド、細胞、生物体、植物、穀物(これらは、これらの新規の方法および試薬の産物である)などを含むことを意図する。
本出願に引用される全ての刊行物、特許、特許出願、または他の文書は、各個の刊行物、特許、特許出願、または他の文書が全ての目的のために参考として本明細書中に援用されると個々に示されている場合と同じ程度まで、全ての目的のために参考としてその全体を本明細書中に援用される。
Claims (1)
- 本願明細書に記載されるような、遺伝子。
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US24438500P | 2000-10-30 | 2000-10-30 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002539528A Division JP2004534505A (ja) | 2000-10-30 | 2001-10-29 | 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2010142234A true JP2010142234A (ja) | 2010-07-01 |
| JP2010142234A5 JP2010142234A5 (ja) | 2010-12-24 |
Family
ID=22922516
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002539528A Pending JP2004534505A (ja) | 2000-10-30 | 2001-10-29 | 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 |
| JP2008051411A Pending JP2008206519A (ja) | 2000-10-30 | 2008-02-29 | 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 |
| JP2009299177A Withdrawn JP2010142234A (ja) | 2000-10-30 | 2009-12-29 | 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 |
Family Applications Before (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2002539528A Pending JP2004534505A (ja) | 2000-10-30 | 2001-10-29 | 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 |
| JP2008051411A Pending JP2008206519A (ja) | 2000-10-30 | 2008-02-29 | 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 |
Country Status (21)
| Country | Link |
|---|---|
| US (1) | US20030083480A1 (ja) |
| EP (1) | EP1399566A2 (ja) |
| JP (3) | JP2004534505A (ja) |
| CN (3) | CN102212534A (ja) |
| AR (3) | AR035595A1 (ja) |
| AU (2) | AU2018102A (ja) |
| BG (1) | BG107758A (ja) |
| BR (1) | BR0115046A (ja) |
| CA (1) | CA2425956C (ja) |
| CZ (1) | CZ20031120A3 (ja) |
| HR (1) | HRP20030439A2 (ja) |
| HU (1) | HUP0700153A2 (ja) |
| IL (3) | IL155599A0 (ja) |
| MX (1) | MXPA03003810A (ja) |
| NZ (1) | NZ526148A (ja) |
| PL (1) | PL366144A1 (ja) |
| RS (1) | RS32703A (ja) |
| SK (1) | SK5222003A3 (ja) |
| UA (2) | UA86918C2 (ja) |
| WO (1) | WO2002036782A2 (ja) |
| ZA (1) | ZA200303138B (ja) |
Families Citing this family (301)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20090011938A1 (en) * | 2000-10-30 | 2009-01-08 | Pioneer Hi--Bred International, Inc. | Novel glyphosate-n-acetyltransferase (gat) genes |
| US7462481B2 (en) * | 2000-10-30 | 2008-12-09 | Verdia, Inc. | Glyphosate N-acetyltransferase (GAT) genes |
| AU2009201716B2 (en) * | 2000-10-30 | 2012-05-17 | E. I. Du Pont De Nemours And Company | Novel glyphosate-N-acetyltransferase (GAT) genes |
| WO2004111245A2 (en) | 2003-03-10 | 2004-12-23 | Athenix Corporation | Gdc-2 genes conferring herbicide resistance |
| US7504561B2 (en) | 2003-03-10 | 2009-03-17 | Athenix Corporation | GDC-1 genes conferring herbicide resistance |
| US7807881B2 (en) | 2003-03-10 | 2010-10-05 | Athenix Corp. | Methods to confer herbicide resistance |
| CA2521284C (en) * | 2003-04-29 | 2014-07-08 | Pioneer Hi-Bred International, Inc. | Novel glyphosate-n-acetyltransferase (gat) genes |
| NZ575248A (en) * | 2003-04-29 | 2010-09-30 | Pioneer Hi Bred Int | Glyphosate-n-acetyltransferase (gat) genes |
| EP1664311A2 (en) * | 2003-09-25 | 2006-06-07 | Monsanto Technology LLC | Actin regulatory elements for use in plants |
| EP2868749B1 (en) | 2004-01-20 | 2017-10-04 | Monsanto Technology LLC | Chimeric promoters for use in plants |
| US20070197474A1 (en) * | 2004-03-30 | 2007-08-23 | Clinton William P | Methods for controlling plants pathogens using N-phosphonomethylglycine |
| US7405074B2 (en) | 2004-04-29 | 2008-07-29 | Pioneer Hi-Bred International, Inc. | Glyphosate-N-acetyltransferase (GAT) genes |
| EP1753866B1 (en) | 2004-06-09 | 2010-09-22 | Pioneer-Hi-Bred International, Inc. | Plastid transit peptides |
| WO2006085965A2 (en) | 2004-06-30 | 2006-08-17 | Pioneer Hi-Bred International, Inc. | Methods of protecting plants from pathogenic fungi |
| EP2230247B1 (en) | 2004-07-02 | 2013-05-29 | Pioneer-Hi-Bred International, Inc. | Antifungal polypeptides |
| WO2006023869A2 (en) | 2004-08-24 | 2006-03-02 | Monsanto Technology Llc | Adenylate translocator protein gene non-coding regulatory elements for use in plants |
| CN101442902B (zh) | 2004-12-21 | 2013-09-18 | 孟山都技术有限公司 | 具有增强的农艺性状的转基因植物的制备方法 |
| AU2005337132B2 (en) | 2004-12-21 | 2011-01-20 | Monsanto Technology, Llc | Transgenic plants with enhanced agronomic traits |
| CN101170907B (zh) | 2005-03-04 | 2012-10-03 | 孟山都技术公司 | 减轻用除草剂草甘膦制剂处理的草甘膦耐受性转基因棉花植物内的坏死 |
| AP2693A (en) | 2005-05-27 | 2013-07-16 | Monsanto Technology Llc | Soybean event MON89788 and methods for detection thereof |
| BRPI0614693A2 (pt) | 2005-07-29 | 2011-04-12 | Monsanto Technology Llc | desenvolvimento de germoplasma usando segregados de cruzamentos transgênicos |
| NZ568867A (en) | 2005-08-24 | 2010-12-24 | Pioneer Hi Bred Int | Compositions providing tolerance to multiple herbicides and methods of use thereof |
| US8993846B2 (en) | 2005-09-06 | 2015-03-31 | Monsanto Technology Llc | Vectors and methods for improved plant transformation efficiency |
| AU2006302969B2 (en) | 2005-10-13 | 2011-09-22 | Monsanto Technology, Llc | Methods for producing hybrid seed |
| CN1313614C (zh) * | 2005-10-17 | 2007-05-02 | 中国农业科学院生物技术研究所 | 草甘膦乙酰转移酶基因及其应用 |
| EP2484769A3 (en) | 2005-12-21 | 2012-09-19 | Monsanto Technology LLC | Transgenic plants with enhanced agronomic traits |
| BRPI0711376A2 (pt) * | 2006-05-12 | 2011-11-01 | Commw Scient Ind Res Org | enzimas para degradação de herbicidas |
| BRPI0711672A2 (pt) | 2006-05-16 | 2011-11-16 | Monsanto Techonology Llc | uso de espécies bacterianas não-agrobacterium para transformação de planta |
| DE602007012343D1 (de) | 2006-05-16 | 2011-03-17 | Du Pont | Antimykotische polypeptide |
| EP2308986B1 (en) | 2006-05-17 | 2014-10-08 | Pioneer Hi-Bred International Inc. | Artificial plant minichromosomes |
| BRPI0712484B1 (pt) | 2006-06-06 | 2017-06-06 | Monsanto Technology Llc | método para seleção de células transformadas |
| US7855326B2 (en) * | 2006-06-06 | 2010-12-21 | Monsanto Technology Llc | Methods for weed control using plants having dicamba-degrading enzymatic activity |
| US7968770B2 (en) | 2006-06-28 | 2011-06-28 | Pioneer Hi-Bred International, Inc. | Methods for improving yield using soybean event 3560.4.3.5 |
| US7951995B2 (en) | 2006-06-28 | 2011-05-31 | Pioneer Hi-Bred International, Inc. | Soybean event 3560.4.3.5 and compositions and methods for the identification and detection thereof |
| WO2008021543A2 (en) | 2006-08-17 | 2008-02-21 | Monsanto Technology, Llc | Transgenic plants with enhanced agronomic traits |
| US7939721B2 (en) | 2006-10-25 | 2011-05-10 | Monsanto Technology Llc | Cropping systems for managing weeds |
| US7928296B2 (en) | 2006-10-30 | 2011-04-19 | Pioneer Hi-Bred International, Inc. | Maize event DP-098140-6 and compositions and methods for the identification and/or detection thereof |
| US7897846B2 (en) | 2006-10-30 | 2011-03-01 | Pioneer Hi-Bred Int'l, Inc. | Maize event DP-098140-6 and compositions and methods for the identification and/or detection thereof |
| CL2007003744A1 (es) | 2006-12-22 | 2008-07-11 | Bayer Cropscience Ag | Composicion que comprende un derivado 2-piridilmetilbenzamida y un compuesto insecticida; y metodo para controlar de forma curativa o preventiva hongos fitopatogenos de cultivos e insectos. |
| US7838729B2 (en) * | 2007-02-26 | 2010-11-23 | Monsanto Technology Llc | Chloroplast transit peptides for efficient targeting of DMO and uses thereof |
| US8030544B2 (en) | 2007-03-09 | 2011-10-04 | Monsanto Technology Llc | Methods for plant transformation using spectinomycin selection |
| BRPI0808798A2 (pt) | 2007-03-12 | 2014-10-07 | Bayer Cropscience Ag | Fenoxifenilamidinas 3,5-dissubstituídas e seu uso como fungicidas |
| EP1969934A1 (de) | 2007-03-12 | 2008-09-17 | Bayer CropScience AG | 4-Cycloalkyl-oder 4-arylsubstituierte Phenoxyphenylamidine und deren Verwendung als Fungizide |
| JP2010520899A (ja) | 2007-03-12 | 2010-06-17 | バイエル・クロツプサイエンス・アクチエンゲゼルシヤフト | ジハロフェノキシフェニルアミジン及び殺真菌剤としてのその使用 |
| EP1969929A1 (de) | 2007-03-12 | 2008-09-17 | Bayer CropScience AG | Substituierte Phenylamidine und deren Verwendung als Fungizide |
| US8003398B2 (en) * | 2007-03-27 | 2011-08-23 | E.I. De Pont De Nemours And Company | Methods and compositions for detecting glyphosate and metabolites thereof |
| BRPI0810654B1 (pt) | 2007-04-19 | 2016-10-04 | Bayer Cropscience Ag | tiadiazoliloxifenilamidinas, seu uso e seu método de preparação, composição e método para combate de micro-organismos indesejados, semente resistente a micro-organismo indesejado, bem como método para proteger a dita semente contra micro-organismos |
| EP2543735A1 (en) | 2007-06-06 | 2013-01-09 | Monsanto Technology LLC | Genes and uses for plant enhancement |
| JP2009000046A (ja) * | 2007-06-21 | 2009-01-08 | Hitachi Zosen Corp | トチュウのメバロン酸経路の酵素をコードする遺伝子 |
| US20110265221A1 (en) | 2007-07-10 | 2011-10-27 | Monsanto Technology Llc | Transgenic plants with enhanced agronomic traits |
| DE102007045922A1 (de) | 2007-09-26 | 2009-04-02 | Bayer Cropscience Ag | Wirkstoffkombinationen mit insektiziden und akariziden Eigenschaften |
| DE102007045957A1 (de) | 2007-09-26 | 2009-04-09 | Bayer Cropscience Ag | Wirkstoffkombinationen mit insektiziden und akarziden Eigenschaften |
| DE102007045953B4 (de) | 2007-09-26 | 2018-07-05 | Bayer Intellectual Property Gmbh | Wirkstoffkombinationen mit insektiziden und akariziden Eigenschaften |
| DE102007045920B4 (de) | 2007-09-26 | 2018-07-05 | Bayer Intellectual Property Gmbh | Synergistische Wirkstoffkombinationen |
| DE102007045919B4 (de) | 2007-09-26 | 2018-07-05 | Bayer Intellectual Property Gmbh | Wirkstoffkombinationen mit insektiziden und akariziden Eigenschaften |
| DE102007045956A1 (de) | 2007-09-26 | 2009-04-09 | Bayer Cropscience Ag | Wirkstoffkombination mit insektiziden und akariziden Eigenschaften |
| DE102007045955A1 (de) | 2007-09-26 | 2009-04-09 | Bayer Cropscience Ag | Wirkstoffkombinationen mit insektiziden und akariziden Eigenschaften |
| EP2090168A1 (de) | 2008-02-12 | 2009-08-19 | Bayer CropScience AG | Methode zur Verbesserung des Pflanzenwachstums |
| US8158850B2 (en) * | 2007-12-19 | 2012-04-17 | Monsanto Technology Llc | Method to enhance yield and purity of hybrid crops |
| EP2072506A1 (de) | 2007-12-21 | 2009-06-24 | Bayer CropScience AG | Thiazolyloxyphenylamidine oder Thiadiazolyloxyphenylamidine und deren Verwendung als Fungizide |
| ES2562908T3 (es) | 2008-04-07 | 2016-03-09 | Monsanto Technology Llc | Elementos reguladores de plantas y sus usos |
| EP2537937A3 (en) | 2008-04-29 | 2013-04-10 | Monsanto Technology LLC | Genes and uses for plant enhancement |
| WO2010009353A1 (en) | 2008-07-16 | 2010-01-21 | Monsanto Technology Llc | Methods and vectors for producing transgenic plants |
| EP2168434A1 (de) | 2008-08-02 | 2010-03-31 | Bayer CropScience AG | Verwendung von Azolen zur Steigerung der Resistenz von Pflanzen oder Pflanzenteilen gegenüber abiotischem Stress |
| US9371564B2 (en) | 2008-08-08 | 2016-06-21 | Bayer Bioscience N.V. | Methods for plant fiber characterization and identification |
| US20110190365A1 (en) | 2008-08-14 | 2011-08-04 | Bayer Crop Science Ag | Insecticidal 4-phenyl-1H-pyrazoles |
| DE102008041695A1 (de) | 2008-08-29 | 2010-03-04 | Bayer Cropscience Ag | Methoden zur Verbesserung des Pflanzenwachstums |
| EP2201838A1 (de) | 2008-12-05 | 2010-06-30 | Bayer CropScience AG | Wirkstoff-Nützlings-Kombinationen mit insektiziden und akariziden Eigenschaften |
| EP2198709A1 (de) | 2008-12-19 | 2010-06-23 | Bayer CropScience AG | Verfahren zur Bekämpfung resistenter tierischer Schädlinge |
| EP2204094A1 (en) | 2008-12-29 | 2010-07-07 | Bayer CropScience AG | Method for improved utilization of the production potential of transgenic plants Introduction |
| AU2009335333B2 (en) | 2008-12-29 | 2015-04-09 | Bayer Intellectual Property Gmbh | Method for improved use of the production potential of genetically modified plants |
| EP2223602A1 (de) | 2009-02-23 | 2010-09-01 | Bayer CropScience AG | Verfahren zur verbesserten Nutzung des Produktionspotentials genetisch modifizierter Pflanzen |
| EP2039772A2 (en) | 2009-01-06 | 2009-03-25 | Bayer CropScience AG | Method for improved utilization of the production potential of transgenic plants introduction |
| EP2039770A2 (en) | 2009-01-06 | 2009-03-25 | Bayer CropScience AG | Method for improved utilization of the production potential of transgenic plants |
| EP2039771A2 (en) | 2009-01-06 | 2009-03-25 | Bayer CropScience AG | Method for improved utilization of the production potential of transgenic plants |
| CN102355820B (zh) | 2009-01-19 | 2013-10-16 | 拜尔农作物科学股份公司 | 环状二酮及其用作杀昆虫剂、杀螨剂和/或杀菌剂的用途 |
| EP2227951A1 (de) | 2009-01-23 | 2010-09-15 | Bayer CropScience AG | Verwendung von Enaminocarbonylverbindungen zur Bekämpfung von durch Insekten übertragenen Viren |
| PT2391608E (pt) | 2009-01-28 | 2013-05-13 | Bayer Cropscience Ag | Derivados de n-cicloalquil-n-biciclíco-metilenocarboxamida fungicidas |
| AR075126A1 (es) | 2009-01-29 | 2011-03-09 | Bayer Cropscience Ag | Metodo para el mejor uso del potencial de produccion de plantas transgenicas |
| CN102395277B (zh) | 2009-02-13 | 2018-08-03 | 孟山都技术公司 | 减少作物损伤的胶囊化除草剂 |
| BRPI1006006B1 (pt) | 2009-02-17 | 2018-05-22 | Bayer Intellectual Property Gmbh | Compostos, composição fungicida e método para o controle de fungos fitopatogênicos de culturas |
| EP2218717A1 (en) | 2009-02-17 | 2010-08-18 | Bayer CropScience AG | Fungicidal N-((HET)Arylethyl)thiocarboxamide derivatives |
| US20100210460A1 (en) | 2009-02-19 | 2010-08-19 | Pioneer Hi-Bred International, Inc. | Blended refuge deployment via manipulation during hybrid seed production |
| TW201031331A (en) | 2009-02-19 | 2010-09-01 | Bayer Cropscience Ag | Pesticide composition comprising a tetrazolyloxime derivative and a fungicide or an insecticide active substance |
| DE102009001469A1 (de) | 2009-03-11 | 2009-09-24 | Bayer Cropscience Ag | Verfahren zur verbesserten Nutzung des Produktionspotentials transgener Pflanzen |
| DE102009001681A1 (de) | 2009-03-20 | 2010-09-23 | Bayer Cropscience Ag | Verfahren zur verbesserten Nutzung des Produktionspotentials transgener Pflanzen |
| DE102009001728A1 (de) | 2009-03-23 | 2010-09-30 | Bayer Cropscience Ag | Verfahren zur verbesserten Nutzung des Produktionspotentials transgener Pflanzen |
| DE102009001730A1 (de) | 2009-03-23 | 2010-09-30 | Bayer Cropscience Ag | Verfahren zur verbesserten Nutzung des Produktionspotentials transgener Pflanzen |
| DE102009001732A1 (de) | 2009-03-23 | 2010-09-30 | Bayer Cropscience Ag | Verfahren zur verbesserten Nutzung des Produktionspotentials transgener Pflanzen |
| WO2010108506A1 (de) | 2009-03-25 | 2010-09-30 | Bayer Cropscience Ag | Wirkstoffkombinationen mit insektiziden und akariziden eigenschaften |
| EP2232995A1 (de) | 2009-03-25 | 2010-09-29 | Bayer CropScience AG | Verfahren zur verbesserten Nutzung des Produktionspotentials transgener Pflanzen |
| AU2009342807B2 (en) | 2009-03-25 | 2015-04-02 | Bayer Cropscience Aktiengesellschaft | Synergistic combinations of active ingredients |
| AP3073A (en) | 2009-03-25 | 2014-12-31 | Bayer Cropscience Ag | Active ingredient combinations with insecticidal and acaricidal properties |
| WO2010108505A1 (de) | 2009-03-25 | 2010-09-30 | Bayer Cropscience Ag | Wirkstoffkombinationen mit insektiziden und akariziden eigenschaften |
| BRPI0924451B1 (pt) | 2009-03-25 | 2017-12-26 | Bayer Intellectual Property Gmbh | Combinations of active substances and their uses, as well as methods for the control of animal pests and method for the manufacture of insecticides and acaricides |
| EP2239331A1 (en) | 2009-04-07 | 2010-10-13 | Bayer CropScience AG | Method for improved utilization of the production potential of transgenic plants |
| WO2010127797A2 (en) | 2009-05-06 | 2010-11-11 | Bayer Cropscience Ag | Cyclopentanedione compounds and their use as insecticides, acaricides and/or fungicides |
| AR076839A1 (es) | 2009-05-15 | 2011-07-13 | Bayer Cropscience Ag | Derivados fungicidas de pirazol carboxamidas |
| EP2251331A1 (en) | 2009-05-15 | 2010-11-17 | Bayer CropScience AG | Fungicide pyrazole carboxamides derivatives |
| WO2010135324A1 (en) | 2009-05-18 | 2010-11-25 | Monsanto Technology Llc | Use of glyphosate for disease suppression and yield enhancement in soybean |
| EP2255626A1 (de) | 2009-05-27 | 2010-12-01 | Bayer CropScience AG | Verwendung von Succinat Dehydrogenase Inhibitoren zur Steigerung der Resistenz von Pflanzen oder Pflanzenteilen gegenüber abiotischem Stress |
| UA106618C2 (uk) | 2009-06-02 | 2014-09-25 | Баєр Кропсаєнс Аг | Застосування інгібіторів сукцинатдегідрогенази для контролю підвиду sclerotinia |
| CN102803496A (zh) | 2009-06-10 | 2012-11-28 | 淡马锡生命科学研究院有限公司 | 用于棉花中基因功能性分析的病毒诱导的基因沉默(vigs) |
| EP2451947A1 (en) | 2009-07-07 | 2012-05-16 | Linda A. Castle | Crystal structure of glyphosate acetyltransferase (glyat) and methods of use |
| EP2453750A2 (de) | 2009-07-16 | 2012-05-23 | Bayer CropScience AG | Synergistische wirkstoffkombinationen mit phenyltriazolen |
| WO2011015524A2 (en) | 2009-08-03 | 2011-02-10 | Bayer Cropscience Ag | Fungicide heterocycles derivatives |
| EP2292094A1 (en) | 2009-09-02 | 2011-03-09 | Bayer CropScience AG | Active compound combinations |
| US8581046B2 (en) | 2010-11-24 | 2013-11-12 | Pioneer Hi-Bred International, Inc. | Brassica gat event DP-073496-4 and compositions and methods for the identification and/or detection thereof |
| GB0920891D0 (en) | 2009-11-27 | 2010-01-13 | Syngenta Participations Ag | Herbicidal compositions |
| EP2343280A1 (en) | 2009-12-10 | 2011-07-13 | Bayer CropScience AG | Fungicide quinoline derivatives |
| US20130012546A1 (en) | 2009-12-28 | 2013-01-10 | Christian Beier | Fungicide hydroximoyl-tetrazole derivatives |
| MX2012007539A (es) | 2009-12-28 | 2012-07-23 | Bayer Cropscience Ag | Derivados de hidroximoil-tetrazol fungicidas. |
| CN105399666A (zh) | 2009-12-28 | 2016-03-16 | 拜尔农科股份公司 | 杀真菌剂肟基-杂环衍生物 |
| CA3017321C (en) | 2010-01-14 | 2021-07-20 | Monsanto Technology Llc | Plant regulatory elements and uses thereof |
| EP2525658B1 (de) | 2010-01-22 | 2017-03-01 | Bayer Intellectual Property GmbH | Akarizide und/oder insektizide wirkstoffkombinationen |
| AR080443A1 (es) | 2010-03-04 | 2012-04-11 | Bayer Cropscience Ag | 2-amidobencimidazoles sustituidos con fluruoalquilo |
| BR112012023551A2 (pt) | 2010-03-18 | 2015-09-15 | Bayer Ip Gmbh | aril e hetaril sulfonamidas como agentes ativos contra estresse abiótico em plantas |
| JP2013523795A (ja) | 2010-04-06 | 2013-06-17 | バイエル・インテレクチユアル・プロパテイー・ゲー・エム・ベー・ハー | 植物のストレス耐性を増強させるための4−フェニル酪酸及び/又はその塩の使用 |
| AU2011237909B2 (en) | 2010-04-09 | 2015-08-20 | Bayer Intellectual Property Gmbh | Use of derivatives of the (1-cyanocyclopropyl)phenylphosphinic acid, the esters thereof and/or the salts thereof for enhancing the tolerance of plants to abiotic stress |
| JP2013525401A (ja) | 2010-04-28 | 2013-06-20 | バイエル・クロップサイエンス・アーゲー | 殺菌剤ヒドロキシモイル−複素環誘導体 |
| WO2011134911A2 (en) | 2010-04-28 | 2011-11-03 | Bayer Cropscience Ag | Fungicide hydroximoyl-tetrazole derivatives |
| BR112012027558A2 (pt) | 2010-04-28 | 2015-09-15 | Bayer Cropscience Ag | ''composto da fórmula (i), composição fungicida e método para o controle de fungos fitogênicos de colheitas'' |
| UA110703C2 (uk) | 2010-06-03 | 2016-02-10 | Байєр Кропсайнс Аг | Фунгіцидні похідні n-[(тризаміщений силіл)метил]-карбоксаміду |
| PL2576517T3 (pl) | 2010-06-03 | 2015-06-30 | Bayer Ip Gmbh | N-[(het)aryloalkilo)]pirazolo(tio)karboksyamidy i ich analogi heteropodstawione |
| ES2533026T3 (es) | 2010-06-03 | 2015-04-07 | Bayer Intellectual Property Gmbh | N-[(het)arilalquil)]pirazol (tio)carboxamidas y sus análogos heterosustituidos |
| WO2011154158A1 (en) | 2010-06-09 | 2011-12-15 | Bayer Bioscience N.V. | Methods and means to modify a plant genome at a nucleotide sequence commonly used in plant genome engineering |
| CN103119169B (zh) | 2010-06-09 | 2018-11-20 | 拜尔作物科学公司 | 植物基因组改造中常用的在核苷酸序列上修饰植物基因组的方法和工具 |
| BR122019002801B8 (pt) | 2010-06-25 | 2022-12-06 | Du Pont | Método para selecionar uma planta de milho e método para identificar uma planta de milho transformada |
| JP2013532648A (ja) | 2010-07-20 | 2013-08-19 | バイエル・クロップサイエンス・アーゲー | 抗菌剤としてのベンゾシクロアルケン類 |
| US9139842B2 (en) | 2010-08-13 | 2015-09-22 | Pioneer Hi-Bred International, Inc. | Methods and compositions for targeting sequences of interest to the chloroplast |
| EP2605646B1 (en) | 2010-08-18 | 2016-07-20 | Monsanto Technology LLC | Early applications of encapsulated acetamides for reduced injury in crops |
| US9222100B2 (en) | 2010-08-24 | 2015-12-29 | Monsanto Technology Llc | Methods and DNA constructs for autoregulating transgene silencing |
| PT2611925T (pt) | 2010-08-30 | 2018-03-02 | Dow Agrosciences Llc | Intensificador do vírus baciliforme da cana sacarina (scbv) e seu uso em genómica funcional de plantas |
| CN103228141B (zh) | 2010-09-03 | 2016-04-20 | 拜耳知识产权有限责任公司 | 取代的稠合的嘧啶酮和二氢嘧啶酮 |
| EP2460406A1 (en) | 2010-12-01 | 2012-06-06 | Bayer CropScience AG | Use of fluopyram for controlling nematodes in nematode resistant crops |
| WO2012038480A2 (en) | 2010-09-22 | 2012-03-29 | Bayer Cropscience Ag | Use of biological or chemical control agents for controlling insects and nematodes in resistant crops |
| PL2624699T3 (pl) | 2010-10-07 | 2019-05-31 | Bayer Cropscience Ag | Kompozycja grzybobójcza zawierająca pochodną tetrazolilooksymu i pochodną tiazolilopiperydyny |
| JP2013541554A (ja) | 2010-10-21 | 2013-11-14 | バイエル・インテレクチユアル・プロパテイー・ゲー・エム・ベー・ハー | N−ベンジルヘテロ環式カルボキサミド類 |
| AR083501A1 (es) | 2010-10-21 | 2013-02-27 | Bayer Cropscience Ag | 1-(heterociclo carbonil)piperidinas |
| WO2012059497A1 (en) | 2010-11-02 | 2012-05-10 | Bayer Cropscience Ag | N-hetarylmethyl pyrazolylcarboxamides |
| MX2013005258A (es) | 2010-11-15 | 2013-07-05 | Bayer Ip Gmbh | N-aril pirazol(tio)carboxamidas. |
| WO2012065945A1 (en) | 2010-11-15 | 2012-05-24 | Bayer Cropscience Ag | 5-halogenopyrazole(thio)carboxamides |
| BR112013012082A2 (pt) | 2010-11-15 | 2016-07-19 | Bayer Ip Gmbh | 5-halogenopirazolcarboxamidas |
| US8575431B2 (en) | 2010-11-24 | 2013-11-05 | Pioneer Hi-Bred International, Inc. | Brassica GAT event DP-061061-7 and compositions and methods for the identification and/or detection thereof |
| KR20180096815A (ko) | 2010-12-01 | 2018-08-29 | 바이엘 인텔렉쳐 프로퍼티 게엠베하 | 작물에서 선충류를 구제하고 수확량을 증가시키기 위한 플루오피람의 용도 |
| EP2460407A1 (de) | 2010-12-01 | 2012-06-06 | Bayer CropScience AG | Wirkstoffkombinationen umfassend Pyridylethylbenzamide und weitere Wirkstoffe |
| TWI667347B (zh) | 2010-12-15 | 2019-08-01 | 瑞士商先正達合夥公司 | 大豆品種syht0h2及偵測其之組合物及方法 |
| EP2474542A1 (en) | 2010-12-29 | 2012-07-11 | Bayer CropScience AG | Fungicide hydroximoyl-tetrazole derivatives |
| WO2012089757A1 (en) | 2010-12-29 | 2012-07-05 | Bayer Cropscience Ag | Fungicide hydroximoyl-tetrazole derivatives |
| EP2471363A1 (de) | 2010-12-30 | 2012-07-04 | Bayer CropScience AG | Verwendung von Aryl-, Heteroaryl- und Benzylsulfonamidocarbonsäuren, -carbonsäureestern, -carbonsäureamiden und -carbonitrilen oder deren Salze zur Steigerung der Stresstoleranz in Pflanzen |
| MX347631B (es) | 2011-02-01 | 2017-05-04 | Colorado Wheat Res Foundation Inc | Plantas resistentes a herbicida en acetil co-enzima a carboxilasa. |
| GB201101743D0 (en) | 2011-02-01 | 2011-03-16 | Syngenta Ltd | Herbicidal compositions |
| EP2494867A1 (de) | 2011-03-01 | 2012-09-05 | Bayer CropScience AG | Halogen-substituierte Verbindungen in Kombination mit Fungiziden |
| EP2683239A1 (en) | 2011-03-10 | 2014-01-15 | Bayer Intellectual Property GmbH | Use of lipochito-oligosaccharide compounds for safeguarding seed safety of treated seeds |
| JP2014509599A (ja) | 2011-03-14 | 2014-04-21 | バイエル・インテレクチユアル・プロパテイー・ゲー・エム・ベー・ハー | 殺菌剤ヒドロキシモイル−テトラゾール誘導体 |
| KR102003175B1 (ko) | 2011-03-25 | 2019-07-24 | 몬산토 테크놀로지 엘엘씨 | 식물 조절 요소 및 그의 용도 |
| EP2694494A1 (en) | 2011-04-08 | 2014-02-12 | Bayer Intellectual Property GmbH | Fungicide hydroximoyl-tetrazole derivatives |
| AR085568A1 (es) | 2011-04-15 | 2013-10-09 | Bayer Cropscience Ag | 5-(biciclo[4.1.0]hept-3-en-2-il)-penta-2,4-dienos y 5-(biciclo[4.1.0]hept-3-en-2-il)-pent-2-en-4-inos sustituidos como principios activos contra el estres abiotico de las plantas |
| EP2511255A1 (de) | 2011-04-15 | 2012-10-17 | Bayer CropScience AG | Substituierte Prop-2-in-1-ol- und Prop-2-en-1-ol-Derivate |
| AR085585A1 (es) | 2011-04-15 | 2013-10-09 | Bayer Cropscience Ag | Vinil- y alquinilciclohexanoles sustituidos como principios activos contra estres abiotico de plantas |
| AR090010A1 (es) | 2011-04-15 | 2014-10-15 | Bayer Cropscience Ag | 5-(ciclohex-2-en-1-il)-penta-2,4-dienos y 5-(ciclohex-2-en-1-il)-pent-2-en-4-inos sustituidos como principios activos contra el estres abiotico de las plantas, usos y metodos de tratamiento |
| ES2561296T3 (es) | 2011-04-22 | 2016-02-25 | Bayer Intellectual Property Gmbh | Combinaciones de un compuesto activo que comprenden un derivado de carboximida y un compuesto fungicida |
| US9845477B2 (en) | 2011-05-13 | 2017-12-19 | Monsanto Technology Llc | Plant regulatory elements and uses thereof |
| GB201109239D0 (en) | 2011-06-01 | 2011-07-13 | Syngenta Participations Ag | Herbicidal compositions |
| WO2012168124A1 (en) | 2011-06-06 | 2012-12-13 | Bayer Cropscience Nv | Methods and means to modify a plant genome at a preselected site |
| BR112013031723B1 (pt) | 2011-06-10 | 2020-10-13 | Temasek Life Sciences Laboratory Limited | construção de polinucleotídeo, método para transformação de uma célula fúngica e promotor isolado |
| CN103957711A (zh) | 2011-07-04 | 2014-07-30 | 拜耳知识产权有限责任公司 | 取代的异喹啉酮、异喹啉二酮、异喹啉三酮和二氢异喹啉酮或其各自的盐作为活性剂对抗植物非生物胁迫的用途 |
| WO2013020985A1 (en) | 2011-08-10 | 2013-02-14 | Bayer Intellectual Property Gmbh | Active compound combinations comprising specific tetramic acid derivatives |
| CN103717612A (zh) | 2011-08-12 | 2014-04-09 | 拜尔作物科学公司 | 转基因在棉花中的保卫细胞特异性表达 |
| CN103890181A (zh) | 2011-08-22 | 2014-06-25 | 拜尔作物科学公司 | 修饰植物基因组的方法和手段 |
| WO2013026836A1 (en) | 2011-08-22 | 2013-02-28 | Bayer Intellectual Property Gmbh | Fungicide hydroximoyl-tetrazole derivatives |
| EP2561759A1 (en) | 2011-08-26 | 2013-02-27 | Bayer Cropscience AG | Fluoroalkyl-substituted 2-amidobenzimidazoles and their effect on plant growth |
| EP2753177A1 (en) | 2011-09-09 | 2014-07-16 | Bayer Intellectual Property GmbH | Acyl-homoserine lactone derivatives for improving plant yield |
| JP6002225B2 (ja) | 2011-09-12 | 2016-10-05 | バイエル・インテレクチュアル・プロパティ・ゲゼルシャフト・ミット・ベシュレンクテル・ハフツングBayer Intellectual Property GmbH | 殺菌性4−置換−3−{フェニル[(ヘテロシクリルメトキシ)イミノ]メチル}−1,2,4−オキサジアゾール−5(4h)−オン誘導体 |
| WO2013040057A1 (en) | 2011-09-13 | 2013-03-21 | Monsanto Technology Llc | Methods and compositions for weed control |
| BR112014005951A2 (pt) | 2011-09-13 | 2017-04-04 | Monsanto Technology Llc | métodos e composições para controle de erva daninha |
| CA2848695A1 (en) | 2011-09-13 | 2013-03-21 | Monsanto Technology Llc | Methods and composition for weed control comprising inhibiting ppg oxidase |
| CA2848680C (en) | 2011-09-13 | 2020-05-19 | Monsanto Technology Llc | Methods and compositions for weed control |
| EP3434779A1 (en) | 2011-09-13 | 2019-01-30 | Monsanto Technology LLC | Methods and compositions for weed control |
| CN103958686A (zh) | 2011-09-13 | 2014-07-30 | 孟山都技术公司 | 用于杂草控制的方法和组合物 |
| CN103957697B (zh) | 2011-09-13 | 2017-10-24 | 孟山都技术公司 | 用于杂草控制的方法和组合物 |
| WO2013040021A1 (en) | 2011-09-13 | 2013-03-21 | Monsanto Technology Llc | Methods and compositions for weed control |
| EP2755484A1 (en) | 2011-09-16 | 2014-07-23 | Bayer Intellectual Property GmbH | Use of 5-phenyl- or 5-benzyl-2 isoxazoline-3 carboxylates for improving plant yield |
| EA029005B1 (ru) | 2011-09-16 | 2018-01-31 | Байер Интеллектчуал Проперти Гмбх | Применение фенилпиразолин-3-карбоксилатов для повышения урожайности растений |
| MX357299B (es) | 2011-09-16 | 2018-07-04 | Bayer Ip Gmbh | Uso de acilsulfonamidas para mejorar el rendimiento de las plantas. |
| AR087971A1 (es) | 2011-09-23 | 2014-04-30 | Bayer Ip Gmbh | Uso de derivados del acido 1-fenil-pirazol-3-carboxilico 4-sustituidos como principios activos contra estres abiotico de plantas |
| AU2012320554B2 (en) | 2011-10-04 | 2017-11-09 | Bayer Intellectual Property Gmbh | RNAi for the control of fungi and oomycetes by inhibiting saccharopine dehydrogenase gene |
| WO2013050324A1 (de) | 2011-10-06 | 2013-04-11 | Bayer Intellectual Property Gmbh | Abiotischen pflanzenstress-reduzierende kombination enthaltend 4- phenylbuttersäure (4-pba) oder eines ihrer salze (komponente (a)) und eine oder mehrere ausgewählte weitere agronomisch wirksame verbindungen (komponente(n) (b) |
| WO2013075817A1 (en) | 2011-11-21 | 2013-05-30 | Bayer Intellectual Property Gmbh | Fungicide n-[(trisubstitutedsilyl)methyl]-carboxamide derivatives |
| US9725414B2 (en) | 2011-11-30 | 2017-08-08 | Bayer Intellectual Property Gmbh | Fungicidal N-bicycloalkyl and N-tricycloalkyl pyrazole-4-(thio)carboxamide derivatives |
| US9414595B2 (en) | 2011-12-19 | 2016-08-16 | Bayer Cropscience Ag | Use of anthranilic acid diamide derivatives for pest control in transgenic crops |
| KR102028893B1 (ko) | 2011-12-29 | 2019-10-07 | 바이엘 인텔렉쳐 프로퍼티 게엠베하 | 살진균 3-[(1,3-티아졸-4-일메톡시이미노)(페닐)메틸]-2-치환-1,2,4-옥사디아졸-5(2h)-온 유도체 |
| EP2797891B1 (en) | 2011-12-29 | 2015-09-30 | Bayer Intellectual Property GmbH | Fungicidal 3-[(pyridin-2-ylmethoxyimino)(phenyl)methyl]-2-substituted-1,2,4-oxadiazol-5(2h)-one derivatives |
| WO2013102147A2 (en) | 2011-12-30 | 2013-07-04 | Butamax (Tm) Advanced Biofuels Llc | Genetic switches for butanol production |
| HUE036328T2 (hu) | 2012-02-22 | 2018-06-28 | Bayer Cropscience Ag | Fluopirám alkalmazása fabetegségek leküzdésére szõlõn |
| PE20190345A1 (es) | 2012-02-27 | 2019-03-07 | Bayer Ip Gmbh | Combinaciones de compuestos activos |
| CA2865977A1 (en) | 2012-02-29 | 2013-09-06 | Dow Agrosciences Llc | Sugarcane bacilliform viral (scbv) enhancer and its use in plant functional genomics |
| WO2013139949A1 (en) | 2012-03-23 | 2013-09-26 | Bayer Intellectual Property Gmbh | Compositions comprising a strigolactame compound for enhanced plant growth and yield |
| US9357778B2 (en) | 2012-04-12 | 2016-06-07 | Bayer Cropscience Ag | N-acyl-2-(cyclo)alkypyrrolidines and piperidines useful as fungicides |
| US20150080337A1 (en) | 2012-04-20 | 2015-03-19 | Bayer Cropscience | N-cycloalkyl-n-[(trisubstitutedsilylphenyl)methylene]-(thio)carboxamide derivatives |
| US9663793B2 (en) | 2012-04-20 | 2017-05-30 | Monsanto Technology, Llc | Plant regulatory elements and uses thereof |
| WO2013156559A1 (en) | 2012-04-20 | 2013-10-24 | Bayer Cropscience Ag | N-cycloalkyl-n-[(heterocyclylphenyl)methylene]-(thio)carboxamide derivatives |
| US11518997B2 (en) | 2012-04-23 | 2022-12-06 | BASF Agricultural Solutions Seed US LLC | Targeted genome engineering in plants |
| EP2662370A1 (en) | 2012-05-09 | 2013-11-13 | Bayer CropScience AG | 5-Halogenopyrazole benzofuranyl carboxamides |
| EP2662360A1 (en) | 2012-05-09 | 2013-11-13 | Bayer CropScience AG | 5-Halogenopyrazole indanyl carboxamides |
| EP2662361A1 (en) | 2012-05-09 | 2013-11-13 | Bayer CropScience AG | Pyrazol indanyl carboxamides |
| EP2662364A1 (en) | 2012-05-09 | 2013-11-13 | Bayer CropScience AG | Pyrazole tetrahydronaphthyl carboxamides |
| CN104768934B (zh) | 2012-05-09 | 2017-11-28 | 拜耳农作物科学股份公司 | 吡唑茚满基甲酰胺 |
| EP2662363A1 (en) | 2012-05-09 | 2013-11-13 | Bayer CropScience AG | 5-Halogenopyrazole biphenylcarboxamides |
| CN104364236B (zh) | 2012-05-09 | 2018-01-16 | 拜尔农作物科学股份公司 | 5‑卤代吡唑二氢茚基甲酰胺 |
| EP2662362A1 (en) | 2012-05-09 | 2013-11-13 | Bayer CropScience AG | Pyrazole indanyl carboxamides |
| AR091104A1 (es) | 2012-05-22 | 2015-01-14 | Bayer Cropscience Ag | Combinaciones de compuestos activos que comprenden un derivado lipo-quitooligosacarido y un compuesto nematicida, insecticida o fungicida |
| EP2871958A1 (en) | 2012-07-11 | 2015-05-20 | Bayer CropScience AG | Use of fungicidal combinations for increasing the tolerance of a plant towards abiotic stress |
| EP2892345A1 (de) | 2012-09-05 | 2015-07-15 | Bayer CropScience AG | Verwendung substituierter 2-amidobenzimidazole, 2-amidobenzoxazole und 2-amidobenzothiazole oder deren salze als wirkstoffe gegen abiotischen pflanzenstress |
| EP2895610B1 (en) | 2012-09-13 | 2019-11-06 | Indiana University Research and Technology Corporation | Compositions and systems for conferring disease resistance in plants and methods of use thereof |
| UA114937C2 (uk) | 2012-10-19 | 2017-08-28 | Байєр Кропсайнс Аг | Спосіб стимулювання росту рослин із застосуванням карбоксамідних похідних |
| WO2014060520A1 (en) | 2012-10-19 | 2014-04-24 | Bayer Cropscience Ag | Method for treating plants against fungi resistant to fungicides using carboxamide or thiocarboxamide derivatives |
| EP2908639A1 (en) | 2012-10-19 | 2015-08-26 | Bayer Cropscience AG | Active compound combinations comprising carboxamide derivatives |
| EP2908642B1 (en) | 2012-10-19 | 2022-02-23 | Bayer Cropscience AG | Method for enhancing tolerance to abiotic stress in plants by using carboxamide or thiocarboxamide derivatives |
| WO2014079957A1 (de) | 2012-11-23 | 2014-05-30 | Bayer Cropscience Ag | Selektive inhibition der ethylensignaltransduktion |
| EP2735231A1 (en) | 2012-11-23 | 2014-05-28 | Bayer CropScience AG | Active compound combinations |
| US9615578B2 (en) | 2012-11-30 | 2017-04-11 | Bayer Cropscience Ag | Binary fungicidal mixtures |
| BR112015012054A2 (pt) | 2012-11-30 | 2017-07-11 | Bayer Cropscience Ag | mistura fungicida ou pesticida binária |
| EA030235B1 (ru) | 2012-11-30 | 2018-07-31 | Байер Кропсайенс Акциенгезельшафт | Тройные фунгицидные смеси |
| US9775351B2 (en) | 2012-11-30 | 2017-10-03 | Bayer Cropscience Ag | Ternary fungicidal and pesticidal mixtures |
| WO2014083031A2 (en) | 2012-11-30 | 2014-06-05 | Bayer Cropscience Ag | Binary pesticidal and fungicidal mixtures |
| EP2740720A1 (de) | 2012-12-05 | 2014-06-11 | Bayer CropScience AG | Substituierte bicyclische- und tricyclische Pent-2-en-4-insäure -Derivate und ihre Verwendung zur Steigerung der Stresstoleranz in Pflanzen |
| EP2740356A1 (de) | 2012-12-05 | 2014-06-11 | Bayer CropScience AG | Substituierte (2Z)-5(1-Hydroxycyclohexyl)pent-2-en-4-insäure-Derivate |
| JP2016500368A (ja) | 2012-12-05 | 2016-01-12 | バイエル・クロップサイエンス・アクチェンゲゼルシャフト | 置換された1−(アリールエチニル)−、1−(ヘテロアリールエチニル)−、1−(複素環エチニル)−および1−(シクロアルケニルエチニル)−シクロヘキサノールの非生物的植物ストレスに対する活性薬剤としての使用 |
| AR093909A1 (es) | 2012-12-12 | 2015-06-24 | Bayer Cropscience Ag | Uso de ingredientes activos para controlar nematodos en cultivos resistentes a nematodos |
| US20140173781A1 (en) | 2012-12-13 | 2014-06-19 | Pioneer Hi-Bred International, Inc. | Methods and compositions for producing and selecting transgenic wheat plants |
| AR093996A1 (es) | 2012-12-18 | 2015-07-01 | Bayer Cropscience Ag | Combinaciones bactericidas y fungicidas binarias |
| CN104995174A (zh) | 2012-12-19 | 2015-10-21 | 拜耳作物科学股份公司 | 二氟甲基-烟酰-四氢萘基胺 |
| AU2013361220A1 (en) | 2012-12-21 | 2015-04-02 | Pioneer Hi-Bred International, Inc. | Compositions and methods for auxin-analog conjugation |
| EP2964614A1 (en) | 2013-03-07 | 2016-01-13 | Bayer Cropscience AG | Fungicidal 3-{phenyl[(heterocyclylmethoxy)imino]methyl}-heterocycle derivatives |
| US9273322B2 (en) | 2013-03-12 | 2016-03-01 | Pioneer Hi Bred International Inc | Root-preferred promoter and methods of use |
| US20160053277A1 (en) | 2013-03-14 | 2016-02-25 | Pioneer Hi-Bred International, Inc. | Compositions Having Dicamba Decarboxylase Activity and Methods of Use |
| EP2970935A1 (en) | 2013-03-14 | 2016-01-20 | Pioneer Hi-Bred International, Inc. | Compositions having dicamba decarboxylase activity and methods of use |
| WO2014161821A1 (en) | 2013-04-02 | 2014-10-09 | Bayer Cropscience Nv | Targeted genome engineering in eukaryotes |
| CN105308032B (zh) | 2013-04-12 | 2017-05-24 | 拜耳作物科学股份公司 | 新的三唑衍生物 |
| US9550752B2 (en) | 2013-04-12 | 2017-01-24 | Bayer Cropscience Aktiengesellschaft | Triazolinthione derivatives |
| JP2016519687A (ja) | 2013-04-19 | 2016-07-07 | バイエル・クロップサイエンス・アクチェンゲゼルシャフト | バイナリー殺虫または農薬混合物 |
| AR095867A1 (es) | 2013-04-19 | 2015-11-18 | Bayer Cropscience Ag | Método para una utilización mejorada del potencial de producción de plantas transgénicas |
| WO2014177514A1 (en) | 2013-04-30 | 2014-11-06 | Bayer Cropscience Ag | Nematicidal n-substituted phenethylcarboxamides |
| TW201507722A (zh) | 2013-04-30 | 2015-03-01 | Bayer Cropscience Ag | 做為殺線蟲劑及殺體內寄生蟲劑的n-(2-鹵素-2-苯乙基)-羧醯胺類 |
| EP3013802B1 (en) | 2013-06-26 | 2019-08-14 | Bayer Cropscience AG | N-cycloalkyl-n-[(bicyclylphenyl)methylene]-(thio)carboxamide derivatives |
| EP3019012A1 (de) | 2013-07-09 | 2016-05-18 | Bayer CropScience AG | Verwendung ausgewählter pyridoncarboxamide oder deren salzen als wirkstoffe gegen abiotischen pflanzenstress |
| EP2837287A1 (en) | 2013-08-15 | 2015-02-18 | Bayer CropScience AG | Use of prothioconazole for increasing root growth of Brassicaceae |
| US10093907B2 (en) | 2013-09-24 | 2018-10-09 | Basf Se | Hetero-transglycosylase and uses thereof |
| US20160237447A1 (en) | 2013-10-07 | 2016-08-18 | Monsanto Technology Llc | Transgenic Plants With Enhanced Traits |
| CN105874062B (zh) * | 2013-10-18 | 2021-07-20 | 先锋国际良种公司 | 草甘膦-n-乙酰转移酶(glyat)序列以及使用方法 |
| BR112016008036A2 (pt) | 2013-10-25 | 2018-01-16 | Pioneer Hi Bred Int | método de identificação de uma primeira planta, kit, polinucleotídeo isolado, planta, germoplasma ou grão de soja |
| US10071967B2 (en) | 2013-12-05 | 2018-09-11 | Bayer Cropscience Aktiengesellschaft | N-cycloalkyl-N-{[2-(1-substitutedcycloalkyl)phenyl]methylene}-(thio)carboxamide derivatives |
| US10070645B2 (en) | 2013-12-05 | 2018-09-11 | Bayer Cropscience Aktiengesellschaft | N-cycloalkyl-N-{[2-(1-substitutedcycloalkyl)phenyl]methylene}-(thio)carboxamide derivatives |
| CN103740670B (zh) * | 2014-01-13 | 2016-06-22 | 中国农业大学 | 筛选草甘膦n-乙酰转移酶抗血清的试剂盒 |
| AR099092A1 (es) | 2014-01-15 | 2016-06-29 | Monsanto Technology Llc | Métodos y composiciones para el control de malezas utilizando polinucleótidos epsps |
| JP6250826B2 (ja) | 2014-01-31 | 2017-12-20 | アグバイオーム, インコーポレイテッド | 改変された生物防除因子およびその使用 |
| US9877486B2 (en) | 2014-01-31 | 2018-01-30 | AgBiome, Inc. | Methods of growing plants using modified biological control agents |
| WO2015164457A1 (en) | 2014-04-22 | 2015-10-29 | E. I. Du Pont De Nemours And Company | Plastidic carbonic anhydrase genes for oil augmentation in seeds with increased dgat expression |
| CN103981199B (zh) * | 2014-05-15 | 2017-01-18 | 中国农业科学院生物技术研究所 | 一种含有草甘膦抗性基因的表达载体及其应用 |
| AR101214A1 (es) | 2014-07-22 | 2016-11-30 | Bayer Cropscience Ag | Ciano-cicloalquilpenta-2,4-dienos, ciano-cicloalquilpent-2-en-4-inas, ciano-heterociclilpenta-2,4-dienos y ciano-heterociclilpent-2-en-4-inas sustituidos como principios activos contra el estrés abiótico de plantas |
| WO2016064347A1 (en) | 2014-10-22 | 2016-04-28 | Temasek Life Sciences Laboratory Limited | Terpene synthases from ylang ylang (cananga odorata var. fruticosa) |
| AR103024A1 (es) | 2014-12-18 | 2017-04-12 | Bayer Cropscience Ag | Piridoncarboxamidas seleccionadas o sus sales como sustancias activas contra estrés abiótico de las plantas |
| CN108138195A (zh) | 2015-02-04 | 2018-06-08 | 孟山都技术公司 | 用于质体转化的方法 |
| EP3274462A4 (en) | 2015-03-26 | 2018-12-26 | The Texas A&M University System | Conversion of lignin into bioplastics and lipid fuels |
| EP3283476B1 (en) | 2015-04-13 | 2019-08-14 | Bayer Cropscience AG | N-cycloalkyl-n-(biheterocyclyethylene)-(thio)carboxamide derivatives |
| CN107771218B (zh) | 2015-06-17 | 2021-12-31 | 先锋国际良种公司 | 植物调节元件及其使用方法 |
| ES2773072T3 (es) | 2015-08-28 | 2020-07-09 | Pioneer Hi Bred Int | Transformación de plantas mediada por Ochrobactrum |
| WO2017059341A1 (en) | 2015-10-02 | 2017-04-06 | Monsanto Technology Llc | Recombinant maize b chromosome sequence and uses thereof |
| CA3004914A1 (en) | 2016-02-05 | 2017-08-10 | Pioneer Hi-Bred International, Inc. | Genetic loci associated with brown stem rot resistance in soybean and methods of use |
| WO2018005491A1 (en) | 2016-06-28 | 2018-01-04 | Monsanto Technology Llc | Methods and compositions for use in genome modification in plants |
| EP3490379A1 (en) | 2016-07-29 | 2019-06-05 | Bayer CropScience Aktiengesellschaft | Active compound combinations and methods to protect the propagation material of plants |
| WO2018054829A1 (en) | 2016-09-22 | 2018-03-29 | Bayer Cropscience Aktiengesellschaft | Novel triazole derivatives and their use as fungicides |
| BR112019005668A2 (pt) | 2016-09-22 | 2019-06-04 | Bayer Ag | novos derivados de triazol |
| US20190225974A1 (en) | 2016-09-23 | 2019-07-25 | BASF Agricultural Solutions Seed US LLC | Targeted genome optimization in plants |
| CA3041351A1 (en) | 2016-10-26 | 2018-05-03 | Bayer Cropscience Aktiengesellschaft | Use of pyraziflumid for controlling sclerotinia spp in seed treatment applications |
| CA3046145A1 (en) | 2016-12-08 | 2018-06-14 | Bayer Cropscience Aktiengesellschaft | Use of insecticides for controlling wireworms |
| EP3332645A1 (de) | 2016-12-12 | 2018-06-13 | Bayer Cropscience AG | Verwendung substituierter pyrimidindione oder jeweils deren salze als wirkstoffe gegen abiotischen pflanzenstress |
| WO2018108627A1 (de) | 2016-12-12 | 2018-06-21 | Bayer Cropscience Aktiengesellschaft | Verwendung substituierter indolinylmethylsulfonamide oder deren salze zur steigerung der stresstoleranz in pflanzen |
| WO2019025153A1 (de) | 2017-07-31 | 2019-02-07 | Bayer Cropscience Aktiengesellschaft | Verwendung von substituierten n-sulfonyl-n'-aryldiaminoalkanen und n-sulfonyl-n'-heteroaryldiaminoalkanen oder deren salzen zur steigerung der stresstoleranz in pflanzen |
| US20190136249A1 (en) | 2017-08-22 | 2019-05-09 | Napigen, Inc. | Organelle genome modification using polynucleotide guided endonuclease |
| EP3737747A1 (en) | 2018-01-12 | 2020-11-18 | The Texas A&M University System | Increasing plant bioproduct yield |
| CN108414768B (zh) * | 2018-02-06 | 2020-10-30 | 中国农业科学院生物技术研究所 | 一种抗草甘膦gat转基因农作物的金标检测试纸条 |
| US11905518B2 (en) | 2018-02-12 | 2024-02-20 | Curators Of The University Of Missouri | Small auxin upregulated (SAUR) gene for the improvement of root system architecture, waterlogging tolerance, drought resistance and yield in plants and methods of uses |
| WO2019233863A1 (de) | 2018-06-04 | 2019-12-12 | Bayer Aktiengesellschaft | Herbizid wirksame bizyklische benzoylpyrazole |
| UA128698C2 (uk) | 2018-07-26 | 2024-10-02 | Баєр Акціенгезельшафт | Застосування інгібітора сукцинатдегідрогенази флуопіраму для боротьби з комплексом кореневої гнилі та/або комплексом диплодіозної гнилі, спричиненим rhizoctonia solani, видом fusarium та видом pythium у рослин виду brassicaceae |
| WO2020057939A1 (en) | 2018-09-17 | 2020-03-26 | Bayer Aktiengesellschaft | Use of the fungicide isoflucypram for controlling claviceps purpurea and reducing sclerotia in cereals |
| US20220039383A1 (en) | 2018-09-17 | 2022-02-10 | Bayer Aktiengesellschaft | Use of the Succinate Dehydrogenase Inhibitor Fluopyram for Controlling Claviceps Purpurea and Reducing Sclerotia in Cereals |
| AU2020209871B2 (en) | 2019-01-14 | 2025-07-17 | Bayer Aktiengesellschaft | Herbicidal substituted n-tetrazolyl aryl carboxamides |
| AR117928A1 (es) | 2019-01-30 | 2021-09-01 | Monsanto Technology Llc | Herbicidas de acetamida microencapsulada |
| CN110218738A (zh) * | 2019-07-04 | 2019-09-10 | 安徽省农业科学院棉花研究所 | 一种抗除草剂薏苡资源获取的方法 |
| CA3189779A1 (en) | 2020-08-10 | 2022-02-17 | E. I. Du Pont De Nemours And Company | Compositions and methods for enhancing resistance to northern leaf blight in maize |
| CN115786285B (zh) * | 2020-09-23 | 2025-06-24 | 山东舜丰生物科技有限公司 | 抗除草剂乙酰辅酶a羧化酶突变体及其应用 |
| CN116064428B (zh) * | 2021-02-08 | 2025-08-26 | 山东舜丰生物科技有限公司 | 新型抗除草剂乙酰辅酶a羧化酶突变体及其应用 |
| CN118284701A (zh) | 2021-09-27 | 2024-07-02 | 孟山都技术公司 | 用于转化从单子叶植物种子切除的胚外植体的组合物和方法 |
| MX2024005691A (es) | 2021-11-12 | 2024-05-24 | Monsanto Technology Llc | Composiciones y metodos para modificar la determinacion vegetal. |
| CN116790544A (zh) * | 2022-03-15 | 2023-09-22 | 中国科学院昆明植物研究所 | 用于重楼皂苷生物合成的糖基转移酶PpUGT5 |
| CN114525292B (zh) * | 2022-04-22 | 2022-08-02 | 中国农业科学院生物技术研究所 | gat3基因及其突变体在培育抗草甘膦作物中的应用 |
| CN120005916B (zh) * | 2025-01-07 | 2026-01-06 | 浙江理工大学 | 一种薏苡油脂合成相关基因ClDGAT2及其应用 |
| CN120485126B (zh) * | 2025-07-18 | 2025-10-10 | 中国农业科学院生物技术研究所 | 一种耐除草剂蛋白杂交瘤细胞株及其产生的抗体和应用 |
| CN120485127B (zh) * | 2025-07-18 | 2025-10-28 | 中国农业科学院生物技术研究所 | 一种杂交瘤细胞株及其产生的抗体和应用 |
Family Cites Families (18)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4535060A (en) * | 1983-01-05 | 1985-08-13 | Calgene, Inc. | Inhibition resistant 5-enolpyruvyl-3-phosphoshikimate synthetase, production and use |
| DE3587548T2 (de) * | 1984-12-28 | 1993-12-23 | Bayer Ag | Rekombinante DNA, die in pflanzliche Zellen eingebracht werden kann. |
| NZ217113A (en) * | 1985-08-07 | 1988-06-30 | Monsanto Co | Production of eucaryotic plants which are glyphosate resistant, vectors (transformation and expression), chimeric gene and plant cells |
| US4940835A (en) * | 1985-10-29 | 1990-07-10 | Monsanto Company | Glyphosate-resistant plants |
| US4971908A (en) * | 1987-05-26 | 1990-11-20 | Monsanto Company | Glyphosate-tolerant 5-enolpyruvyl-3-phosphoshikimate synthase |
| US5312910A (en) * | 1987-05-26 | 1994-05-17 | Monsanto Company | Glyphosate-tolerant 5-enolpyruvyl-3-phosphoshikimate synthase |
| US5145783A (en) * | 1987-05-26 | 1992-09-08 | Monsanto Company | Glyphosate-tolerant 5-endolpyruvyl-3-phosphoshikimate synthase |
| US5310667A (en) * | 1989-07-17 | 1994-05-10 | Monsanto Company | Glyphosate-tolerant 5-enolpyruvyl-3-phosphoshikimate synthases |
| AU655197B2 (en) * | 1990-06-25 | 1994-12-08 | Monsanto Technology Llc | Glyphosate tolerant plants |
| US5633435A (en) * | 1990-08-31 | 1997-05-27 | Monsanto Company | Glyphosate-tolerant 5-enolpyruvylshikimate-3-phosphate synthases |
| US5866775A (en) * | 1990-09-28 | 1999-02-02 | Monsanto Company | Glyphosate-tolerant 5-enolpyruvyl-3-phosphoshikimate synthases |
| FR2673642B1 (fr) * | 1991-03-05 | 1994-08-12 | Rhone Poulenc Agrochimie | Gene chimere comprenant un promoteur capable de conferer a une plante une tolerance accrue au glyphosate. |
| FR2673643B1 (fr) * | 1991-03-05 | 1993-05-21 | Rhone Poulenc Agrochimie | Peptide de transit pour l'insertion d'un gene etranger dans un gene vegetal et plantes transformees en utilisant ce peptide. |
| EP1105510A2 (en) * | 1998-08-12 | 2001-06-13 | Maxygen, Inc. | Dna shuffling to produce herbicide selective crops |
| CA2351550C (en) * | 1998-11-17 | 2013-04-23 | Monsanto Company | Phosphonate metabolizing plants |
| AU2415200A (en) | 1999-01-18 | 2000-08-01 | Maxygen, Inc. | Methods of populating data structures for use in evolutionary simulations |
| US6436675B1 (en) | 1999-09-28 | 2002-08-20 | Maxygen, Inc. | Use of codon-varied oligonucleotide synthesis for synthetic shuffling |
| AU3210100A (en) | 1999-01-19 | 2000-08-01 | Maxygen, Inc. | Methods for making character strings, polynucleotides and polypeptides having desired characteristics |
-
2001
- 2001-10-29 WO PCT/US2001/046227 patent/WO2002036782A2/en not_active Ceased
- 2001-10-29 PL PL01366144A patent/PL366144A1/xx unknown
- 2001-10-29 US US10/004,357 patent/US20030083480A1/en not_active Abandoned
- 2001-10-29 NZ NZ526148A patent/NZ526148A/en not_active IP Right Cessation
- 2001-10-29 RS YU32703A patent/RS32703A/sr unknown
- 2001-10-29 SK SK522-2003A patent/SK5222003A3/sk not_active Application Discontinuation
- 2001-10-29 HU HU0700153A patent/HUP0700153A2/hu not_active Application Discontinuation
- 2001-10-29 IL IL15559901A patent/IL155599A0/xx active IP Right Grant
- 2001-10-29 JP JP2002539528A patent/JP2004534505A/ja active Pending
- 2001-10-29 EP EP01992782A patent/EP1399566A2/en not_active Withdrawn
- 2001-10-29 CN CN2011100857181A patent/CN102212534A/zh active Pending
- 2001-10-29 UA UA2003055027A patent/UA86918C2/ru unknown
- 2001-10-29 CN CN200910159780A patent/CN101684458A/zh active Pending
- 2001-10-29 AU AU2018102A patent/AU2018102A/xx active Pending
- 2001-10-29 CN CN018199755A patent/CN1531594B/zh not_active Expired - Fee Related
- 2001-10-29 BR BR0115046-4A patent/BR0115046A/pt not_active Application Discontinuation
- 2001-10-29 HR HR20030439A patent/HRP20030439A2/xx not_active Application Discontinuation
- 2001-10-29 CZ CZ20031120A patent/CZ20031120A3/cs unknown
- 2001-10-29 AU AU2002220181A patent/AU2002220181B2/en not_active Ceased
- 2001-10-29 MX MXPA03003810A patent/MXPA03003810A/es active IP Right Grant
- 2001-10-29 CA CA2425956A patent/CA2425956C/en not_active Expired - Fee Related
- 2001-10-30 AR ARP010105074A patent/AR035595A1/es not_active Application Discontinuation
-
2003
- 2003-04-23 ZA ZA2003/03138A patent/ZA200303138B/en unknown
- 2003-04-24 BG BG107758A patent/BG107758A/bg unknown
- 2003-04-28 IL IL155599A patent/IL155599A/en not_active IP Right Cessation
-
2008
- 2008-01-04 AR ARP080100038A patent/AR064756A2/es unknown
- 2008-02-29 JP JP2008051411A patent/JP2008206519A/ja active Pending
- 2008-03-05 UA UAA200802868A patent/UA94688C2/ru unknown
- 2008-06-03 IL IL191899A patent/IL191899A/en not_active IP Right Cessation
-
2009
- 2009-01-29 AR ARP090100277A patent/AR070289A2/es unknown
- 2009-12-29 JP JP2009299177A patent/JP2010142234A/ja not_active Withdrawn
Also Published As
| Publication number | Publication date |
|---|---|
| AU2002220181B2 (en) | 2007-12-20 |
| US20030083480A1 (en) | 2003-05-01 |
| IL191899A (en) | 2012-08-30 |
| BR0115046A (pt) | 2005-04-12 |
| NZ526148A (en) | 2005-09-30 |
| UA86918C2 (ru) | 2009-06-10 |
| CN102212534A (zh) | 2011-10-12 |
| SK5222003A3 (en) | 2004-12-01 |
| ZA200303138B (en) | 2005-06-29 |
| AR035595A1 (es) | 2004-06-16 |
| UA94688C2 (ru) | 2011-05-25 |
| CA2425956C (en) | 2014-12-23 |
| JP2008206519A (ja) | 2008-09-11 |
| AU2018102A (en) | 2002-05-15 |
| AR070289A2 (es) | 2010-03-25 |
| EP1399566A2 (en) | 2004-03-24 |
| CA2425956A1 (en) | 2002-05-10 |
| BG107758A (bg) | 2004-07-30 |
| RS32703A (sr) | 2006-12-15 |
| CN101684458A (zh) | 2010-03-31 |
| CN1531594A (zh) | 2004-09-22 |
| IL155599A0 (en) | 2003-11-23 |
| WO2002036782A3 (en) | 2004-01-08 |
| IL155599A (en) | 2011-09-27 |
| HUP0700153A2 (en) | 2007-08-28 |
| MXPA03003810A (es) | 2004-10-15 |
| HRP20030439A2 (en) | 2008-12-31 |
| JP2004534505A (ja) | 2004-11-18 |
| PL366144A1 (en) | 2005-01-24 |
| CZ20031120A3 (cs) | 2003-11-12 |
| AR064756A2 (es) | 2009-04-22 |
| CN1531594B (zh) | 2011-05-25 |
| WO2002036782A2 (en) | 2002-05-10 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2010142234A (ja) | 新規のグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 | |
| AU2004260931B2 (en) | Novel glyphosate-N-acetyltransferase (GAT) genes | |
| US7666644B2 (en) | Glyphosate-N-acetyltransferase (GAT) genes | |
| US7531339B2 (en) | Glyphosate-N-acetyltransferase (GAT) genes | |
| AU2002220181A1 (en) | Novel glyphosate n-acetyltransferase (gat) genes | |
| WO2003092360A2 (en) | Novel glyphosate-n-acetyltransferase (gat) genes | |
| AU2009201716B2 (en) | Novel glyphosate-N-acetyltransferase (GAT) genes | |
| JP2010227104A (ja) | 新規なグリホセートn−アセチルトランスフェラーゼ(gat)遺伝子 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A072 | Dismissal of procedure [no reply to invitation to correct request for examination] |
Free format text: JAPANESE INTERMEDIATE CODE: A073 Effective date: 20110513 |
|
| A300 | Application deemed to be withdrawn because no request for examination was validly filed |
Free format text: JAPANESE INTERMEDIATE CODE: A300 Effective date: 20110607 |