JP6784687B2 - 結合誘発型転写スイッチ及びその使用方法 - Google Patents
結合誘発型転写スイッチ及びその使用方法 Download PDFInfo
- Publication number
- JP6784687B2 JP6784687B2 JP2017544874A JP2017544874A JP6784687B2 JP 6784687 B2 JP6784687 B2 JP 6784687B2 JP 2017544874 A JP2017544874 A JP 2017544874A JP 2017544874 A JP2017544874 A JP 2017544874A JP 6784687 B2 JP6784687 B2 JP 6784687B2
- Authority
- JP
- Japan
- Prior art keywords
- cell
- domain
- amino acid
- antigen
- binding
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 230000027455 binding Effects 0.000 title claims description 364
- 238000013518 transcription Methods 0.000 title claims description 113
- 230000035897 transcription Effects 0.000 title claims description 113
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 431
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 420
- 229920001184 polypeptide Polymers 0.000 claims description 419
- 210000004027 cell Anatomy 0.000 claims description 398
- 239000000427 antigen Substances 0.000 claims description 224
- 108091007433 antigens Proteins 0.000 claims description 224
- 102000036639 antigens Human genes 0.000 claims description 224
- 230000003834 intracellular effect Effects 0.000 claims description 182
- 238000000034 method Methods 0.000 claims description 152
- 108090000623 proteins and genes Proteins 0.000 claims description 129
- 108010019670 Chimeric Antigen Receptors Proteins 0.000 claims description 103
- 108091006106 transcriptional activators Proteins 0.000 claims description 85
- 239000003446 ligand Substances 0.000 claims description 66
- 230000014509 gene expression Effects 0.000 claims description 59
- 230000006337 proteolytic cleavage Effects 0.000 claims description 57
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 52
- 102000004169 proteins and genes Human genes 0.000 claims description 52
- 102000039446 nucleic acids Human genes 0.000 claims description 47
- 108020004707 nucleic acids Proteins 0.000 claims description 47
- 150000007523 nucleic acids Chemical class 0.000 claims description 47
- 230000001105 regulatory effect Effects 0.000 claims description 37
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical group C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 claims description 35
- 238000003776 cleavage reaction Methods 0.000 claims description 32
- 230000007017 scission Effects 0.000 claims description 32
- 206010028980 Neoplasm Diseases 0.000 claims description 31
- 108091006107 transcriptional repressors Proteins 0.000 claims description 31
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 claims description 28
- 102000004127 Cytokines Human genes 0.000 claims description 27
- 108091008874 T cell receptors Proteins 0.000 claims description 27
- 230000006907 apoptotic process Effects 0.000 claims description 25
- 210000002865 immune cell Anatomy 0.000 claims description 24
- 230000001939 inductive effect Effects 0.000 claims description 24
- 102100025169 Max-binding protein MNT Human genes 0.000 claims description 23
- 102000040945 Transcription factor Human genes 0.000 claims description 23
- 108091023040 Transcription factor Proteins 0.000 claims description 23
- 239000002773 nucleotide Substances 0.000 claims description 23
- 125000003729 nucleotide group Chemical group 0.000 claims description 23
- 108090000695 Cytokines Proteins 0.000 claims description 21
- 201000011510 cancer Diseases 0.000 claims description 21
- 102400001368 Epidermal growth factor Human genes 0.000 claims description 20
- 101800003838 Epidermal growth factor Proteins 0.000 claims description 20
- 229940116977 epidermal growth factor Drugs 0.000 claims description 20
- 238000000338 in vitro Methods 0.000 claims description 20
- 210000000130 stem cell Anatomy 0.000 claims description 18
- 201000010099 disease Diseases 0.000 claims description 17
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 17
- 102000003675 cytokine receptors Human genes 0.000 claims description 16
- 108010057085 cytokine receptors Proteins 0.000 claims description 16
- 230000001225 therapeutic effect Effects 0.000 claims description 14
- 102000019034 Chemokines Human genes 0.000 claims description 12
- 108010012236 Chemokines Proteins 0.000 claims description 12
- 102000009410 Chemokine receptor Human genes 0.000 claims description 11
- 108050000299 Chemokine receptor Proteins 0.000 claims description 11
- 101710163270 Nuclease Proteins 0.000 claims description 11
- 239000000126 substance Substances 0.000 claims description 9
- 108700029229 Transcriptional Regulatory Elements Proteins 0.000 claims description 8
- 239000013604 expression vector Substances 0.000 claims description 8
- 238000011282 treatment Methods 0.000 claims description 7
- 108091026890 Coding region Proteins 0.000 claims description 5
- 101100074828 Caenorhabditis elegans lin-12 gene Proteins 0.000 claims description 4
- 239000003053 toxin Substances 0.000 claims description 4
- 231100000765 toxin Toxicity 0.000 claims description 4
- 108700012359 toxins Proteins 0.000 claims description 4
- 239000003795 chemical substances by application Substances 0.000 claims description 3
- 210000000822 natural killer cell Anatomy 0.000 claims description 3
- 238000003259 recombinant expression Methods 0.000 claims description 3
- 239000003814 drug Substances 0.000 claims description 2
- 210000003958 hematopoietic stem cell Anatomy 0.000 claims 3
- 229940124597 therapeutic agent Drugs 0.000 claims 2
- 239000012830 cancer therapeutic Substances 0.000 claims 1
- 238000012239 gene modification Methods 0.000 claims 1
- 230000005017 genetic modification Effects 0.000 claims 1
- 235000013617 genetically modified food Nutrition 0.000 claims 1
- 230000005934 immune activation Effects 0.000 claims 1
- 210000002501 natural regulatory T cell Anatomy 0.000 claims 1
- 108010070047 Notch Receptors Proteins 0.000 description 298
- 102000005650 Notch Receptors Human genes 0.000 description 298
- 125000003275 alpha amino acid group Chemical group 0.000 description 288
- 230000009870 specific binding Effects 0.000 description 245
- 150000001413 amino acids Chemical class 0.000 description 226
- 235000001014 amino acid Nutrition 0.000 description 164
- 102000005962 receptors Human genes 0.000 description 115
- 108020003175 receptors Proteins 0.000 description 115
- 239000000047 product Substances 0.000 description 75
- 230000000694 effects Effects 0.000 description 53
- 235000018102 proteins Nutrition 0.000 description 42
- 230000002401 inhibitory effect Effects 0.000 description 37
- 230000006870 function Effects 0.000 description 33
- 230000004069 differentiation Effects 0.000 description 30
- 102000010834 Extracellular Matrix Proteins Human genes 0.000 description 28
- 108010037362 Extracellular Matrix Proteins Proteins 0.000 description 28
- 210000002744 extracellular matrix Anatomy 0.000 description 28
- 230000004913 activation Effects 0.000 description 24
- 238000010586 diagram Methods 0.000 description 24
- 239000012636 effector Substances 0.000 description 24
- 238000001727 in vivo Methods 0.000 description 24
- -1 Amatsuximab Chemical compound 0.000 description 23
- 108060003951 Immunoglobulin Proteins 0.000 description 22
- 102000018358 immunoglobulin Human genes 0.000 description 22
- 230000033228 biological regulation Effects 0.000 description 16
- 239000003112 inhibitor Substances 0.000 description 16
- 230000002103 transcriptional effect Effects 0.000 description 16
- 102000016289 Cell Adhesion Molecules Human genes 0.000 description 15
- 108010067225 Cell Adhesion Molecules Proteins 0.000 description 15
- 239000003102 growth factor Substances 0.000 description 15
- 229940088597 hormone Drugs 0.000 description 15
- 239000005556 hormone Substances 0.000 description 15
- 102000004190 Enzymes Human genes 0.000 description 14
- 108090000790 Enzymes Proteins 0.000 description 14
- 230000006044 T cell activation Effects 0.000 description 14
- 230000024245 cell differentiation Effects 0.000 description 14
- 229940088598 enzyme Drugs 0.000 description 14
- 239000011159 matrix material Substances 0.000 description 14
- 230000004044 response Effects 0.000 description 14
- 108010003723 Single-Domain Antibodies Proteins 0.000 description 13
- 230000003213 activating effect Effects 0.000 description 13
- 150000003384 small molecules Chemical class 0.000 description 13
- 239000012190 activator Substances 0.000 description 12
- 230000001149 cognitive effect Effects 0.000 description 12
- 230000011664 signaling Effects 0.000 description 12
- 239000000758 substrate Substances 0.000 description 12
- 102000000905 Cadherin Human genes 0.000 description 11
- 108050007957 Cadherin Proteins 0.000 description 11
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 11
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 11
- 238000006471 dimerization reaction Methods 0.000 description 11
- 239000012634 fragment Substances 0.000 description 11
- 239000005090 green fluorescent protein Substances 0.000 description 11
- 102100037362 Fibronectin Human genes 0.000 description 10
- 108010067306 Fibronectins Proteins 0.000 description 10
- 101000716102 Homo sapiens T-cell surface glycoprotein CD4 Proteins 0.000 description 10
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 10
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 10
- 102000005755 Intercellular Signaling Peptides and Proteins Human genes 0.000 description 10
- 108010070716 Intercellular Signaling Peptides and Proteins Proteins 0.000 description 10
- 102100036011 T-cell surface glycoprotein CD4 Human genes 0.000 description 10
- 230000021164 cell adhesion Effects 0.000 description 10
- 235000018417 cysteine Nutrition 0.000 description 10
- 230000001419 dependent effect Effects 0.000 description 10
- 230000012010 growth Effects 0.000 description 10
- 230000001506 immunosuppresive effect Effects 0.000 description 10
- 230000002503 metabolic effect Effects 0.000 description 10
- 230000028327 secretion Effects 0.000 description 10
- 230000008685 targeting Effects 0.000 description 10
- 230000005909 tumor killing Effects 0.000 description 10
- 102000035195 Peptidases Human genes 0.000 description 9
- 108091005804 Peptidases Proteins 0.000 description 9
- 229950002853 bimekizumab Drugs 0.000 description 9
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 9
- 108010049777 Ankyrins Proteins 0.000 description 8
- 102000008102 Ankyrins Human genes 0.000 description 8
- 239000000411 inducer Substances 0.000 description 8
- 230000020411 cell activation Effects 0.000 description 7
- 230000001413 cellular effect Effects 0.000 description 7
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 7
- 229960000578 gemtuzumab Drugs 0.000 description 7
- 230000003308 immunostimulating effect Effects 0.000 description 7
- 230000005764 inhibitory process Effects 0.000 description 7
- 102000006495 integrins Human genes 0.000 description 7
- 108010044426 integrins Proteins 0.000 description 7
- 238000004519 manufacturing process Methods 0.000 description 7
- 230000019491 signal transduction Effects 0.000 description 7
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 6
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 6
- 102100032912 CD44 antigen Human genes 0.000 description 6
- 241000282836 Camelus dromedarius Species 0.000 description 6
- 102000001301 EGF receptor Human genes 0.000 description 6
- 108060006698 EGF receptor Proteins 0.000 description 6
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 6
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 6
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 6
- 239000004365 Protease Substances 0.000 description 6
- 230000005867 T cell response Effects 0.000 description 6
- 102100034922 T-cell surface glycoprotein CD8 alpha chain Human genes 0.000 description 6
- 108010073929 Vascular Endothelial Growth Factor A Proteins 0.000 description 6
- 102100039037 Vascular endothelial growth factor A Human genes 0.000 description 6
- 229930193936 anticarin Natural products 0.000 description 6
- 229940072221 immunoglobulins Drugs 0.000 description 6
- 210000002569 neuron Anatomy 0.000 description 6
- 238000013519 translation Methods 0.000 description 6
- 230000014616 translation Effects 0.000 description 6
- BXTJCSYMGFJEID-XMTADJHZSA-N (2s)-2-[[(2r,3r)-3-[(2s)-1-[(3r,4s,5s)-4-[[(2s)-2-[[(2s)-2-[6-[3-[(2r)-2-amino-2-carboxyethyl]sulfanyl-2,5-dioxopyrrolidin-1-yl]hexanoyl-methylamino]-3-methylbutanoyl]amino]-3-methylbutanoyl]-methylamino]-3-methoxy-5-methylheptanoyl]pyrrolidin-2-yl]-3-met Chemical compound C([C@H](NC(=O)[C@H](C)[C@@H](OC)[C@@H]1CCCN1C(=O)C[C@H]([C@H]([C@@H](C)CC)N(C)C(=O)[C@@H](NC(=O)[C@H](C(C)C)N(C)C(=O)CCCCCN1C(C(SC[C@H](N)C(O)=O)CC1=O)=O)C(C)C)OC)C(O)=O)C1=CC=CC=C1 BXTJCSYMGFJEID-XMTADJHZSA-N 0.000 description 5
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 5
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 5
- 101150029707 ERBB2 gene Proteins 0.000 description 5
- 108010087819 Fc receptors Proteins 0.000 description 5
- 102000009109 Fc receptors Human genes 0.000 description 5
- 101000851376 Homo sapiens Tumor necrosis factor receptor superfamily member 8 Proteins 0.000 description 5
- 108010088160 Staphylococcal Protein A Proteins 0.000 description 5
- 230000021541 TRAIL production Effects 0.000 description 5
- 102100036857 Tumor necrosis factor receptor superfamily member 8 Human genes 0.000 description 5
- 108010053099 Vascular Endothelial Growth Factor Receptor-2 Proteins 0.000 description 5
- 102100033177 Vascular endothelial growth factor receptor 2 Human genes 0.000 description 5
- 238000004458 analytical method Methods 0.000 description 5
- 230000000259 anti-tumor effect Effects 0.000 description 5
- 210000004899 c-terminal region Anatomy 0.000 description 5
- 230000012292 cell migration Effects 0.000 description 5
- 230000034994 death Effects 0.000 description 5
- 230000004068 intracellular signaling Effects 0.000 description 5
- 210000004962 mammalian cell Anatomy 0.000 description 5
- 210000004379 membrane Anatomy 0.000 description 5
- 239000012528 membrane Substances 0.000 description 5
- 229960003347 obinutuzumab Drugs 0.000 description 5
- 244000052769 pathogen Species 0.000 description 5
- 230000001717 pathogenic effect Effects 0.000 description 5
- 229960002621 pembrolizumab Drugs 0.000 description 5
- 239000003380 propellant Substances 0.000 description 5
- 235000019419 proteases Nutrition 0.000 description 5
- 210000001519 tissue Anatomy 0.000 description 5
- 229960003989 tocilizumab Drugs 0.000 description 5
- 230000032258 transport Effects 0.000 description 5
- 229940088872 Apoptosis inhibitor Drugs 0.000 description 4
- 108010022366 Carcinoembryonic Antigen Proteins 0.000 description 4
- 102100025475 Carcinoembryonic antigen-related cell adhesion molecule 5 Human genes 0.000 description 4
- 102000014914 Carrier Proteins Human genes 0.000 description 4
- 108010077544 Chromatin Proteins 0.000 description 4
- 102100039498 Cytotoxic T-lymphocyte protein 4 Human genes 0.000 description 4
- 108020004414 DNA Proteins 0.000 description 4
- 102100041003 Glutamate carboxypeptidase 2 Human genes 0.000 description 4
- 101000892862 Homo sapiens Glutamate carboxypeptidase 2 Proteins 0.000 description 4
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 4
- 102000019298 Lipocalin Human genes 0.000 description 4
- 108050006654 Lipocalin Proteins 0.000 description 4
- 239000004793 Polystyrene Substances 0.000 description 4
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 4
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 4
- YQMWQSMYVPLYDI-UHFFFAOYSA-N alpha-Isoaphyllin Natural products C12CCCCN2C(=O)C2C3CCCCN3CC1C2 YQMWQSMYVPLYDI-UHFFFAOYSA-N 0.000 description 4
- 239000000158 apoptosis inhibitor Substances 0.000 description 4
- 108091008324 binding proteins Proteins 0.000 description 4
- 229960000455 brentuximab vedotin Drugs 0.000 description 4
- 230000004663 cell proliferation Effects 0.000 description 4
- 210000003483 chromatin Anatomy 0.000 description 4
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 4
- 229950000932 indusatumab vedotin Drugs 0.000 description 4
- 239000000463 material Substances 0.000 description 4
- 230000005012 migration Effects 0.000 description 4
- 238000013508 migration Methods 0.000 description 4
- 229920002223 polystyrene Polymers 0.000 description 4
- 230000001323 posttranslational effect Effects 0.000 description 4
- 238000012545 processing Methods 0.000 description 4
- 230000035755 proliferation Effects 0.000 description 4
- 235000019833 protease Nutrition 0.000 description 4
- 108091007505 ADAM17 Proteins 0.000 description 3
- 102000002659 Amyloid Precursor Protein Secretases Human genes 0.000 description 3
- 108010043324 Amyloid Precursor Protein Secretases Proteins 0.000 description 3
- 101150013553 CD40 gene Proteins 0.000 description 3
- 108091033409 CRISPR Proteins 0.000 description 3
- 101100510617 Caenorhabditis elegans sel-8 gene Proteins 0.000 description 3
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 3
- 102100031111 Disintegrin and metalloproteinase domain-containing protein 17 Human genes 0.000 description 3
- 108050001049 Extracellular proteins Proteins 0.000 description 3
- 102000009465 Growth Factor Receptors Human genes 0.000 description 3
- 108010009202 Growth Factor Receptors Proteins 0.000 description 3
- 101000889276 Homo sapiens Cytotoxic T-lymphocyte protein 4 Proteins 0.000 description 3
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 3
- 108090000176 Interleukin-13 Proteins 0.000 description 3
- 102000003816 Interleukin-13 Human genes 0.000 description 3
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 102100034256 Mucin-1 Human genes 0.000 description 3
- 239000000020 Nitrocellulose Substances 0.000 description 3
- 241000283973 Oryctolagus cuniculus Species 0.000 description 3
- 101710160107 Outer membrane protein A Proteins 0.000 description 3
- 239000004698 Polyethylene Substances 0.000 description 3
- 206010060862 Prostate cancer Diseases 0.000 description 3
- 208000000236 Prostatic Neoplasms Diseases 0.000 description 3
- 241000700159 Rattus Species 0.000 description 3
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 3
- 108010091086 Recombinases Proteins 0.000 description 3
- 102000018120 Recombinases Human genes 0.000 description 3
- 102000003800 Selectins Human genes 0.000 description 3
- 108090000184 Selectins Proteins 0.000 description 3
- 102000012479 Serine Proteases Human genes 0.000 description 3
- 108010022999 Serine Proteases Proteins 0.000 description 3
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 3
- 108091008605 VEGF receptors Proteins 0.000 description 3
- 102000009484 Vascular Endothelial Growth Factor Receptors Human genes 0.000 description 3
- 229950008995 aducanumab Drugs 0.000 description 3
- 125000000539 amino acid group Chemical group 0.000 description 3
- 229950003145 apolizumab Drugs 0.000 description 3
- 230000015572 biosynthetic process Effects 0.000 description 3
- 229960002685 biotin Drugs 0.000 description 3
- 239000011616 biotin Substances 0.000 description 3
- 239000011575 calcium Substances 0.000 description 3
- 229910052791 calcium Inorganic materials 0.000 description 3
- 150000001720 carbohydrates Chemical class 0.000 description 3
- 235000014633 carbohydrates Nutrition 0.000 description 3
- 230000007348 cell dedifferentiation Effects 0.000 description 3
- 230000032823 cell division Effects 0.000 description 3
- 230000000295 complement effect Effects 0.000 description 3
- 239000013078 crystal Substances 0.000 description 3
- 150000001945 cysteines Chemical class 0.000 description 3
- 230000032459 dedifferentiation Effects 0.000 description 3
- 230000000447 dimerizing effect Effects 0.000 description 3
- 239000012645 endogenous antigen Substances 0.000 description 3
- 108020001507 fusion proteins Proteins 0.000 description 3
- 102000037865 fusion proteins Human genes 0.000 description 3
- 238000005734 heterodimerization reaction Methods 0.000 description 3
- 229910052739 hydrogen Inorganic materials 0.000 description 3
- 230000002209 hydrophobic effect Effects 0.000 description 3
- 238000009169 immunotherapy Methods 0.000 description 3
- 230000006882 induction of apoptosis Effects 0.000 description 3
- 208000027866 inflammatory disease Diseases 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 150000002632 lipids Chemical class 0.000 description 3
- 230000007246 mechanism Effects 0.000 description 3
- 230000001404 mediated effect Effects 0.000 description 3
- 201000001441 melanoma Diseases 0.000 description 3
- 102000006240 membrane receptors Human genes 0.000 description 3
- 229920001220 nitrocellulos Polymers 0.000 description 3
- 238000002823 phage display Methods 0.000 description 3
- 229920000515 polycarbonate Polymers 0.000 description 3
- 239000004417 polycarbonate Substances 0.000 description 3
- 229920000573 polyethylene Polymers 0.000 description 3
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 3
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 3
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 3
- 125000002924 primary amino group Chemical class [H]N([H])* 0.000 description 3
- AAEVYOVXGOFMJO-UHFFFAOYSA-N prometryn Chemical compound CSC1=NC(NC(C)C)=NC(NC(C)C)=N1 AAEVYOVXGOFMJO-UHFFFAOYSA-N 0.000 description 3
- 102000035160 transmembrane proteins Human genes 0.000 description 3
- 108091005703 transmembrane proteins Proteins 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- ZMEWRPBAQVSBBB-GOTSBHOMSA-N (2s)-2-[[(2s)-2-[(2-aminoacetyl)amino]-3-(4-hydroxyphenyl)propanoyl]amino]-6-[[2-[2-[2-[bis(carboxymethyl)amino]ethyl-(carboxymethyl)amino]ethyl-(carboxymethyl)amino]acetyl]amino]hexanoic acid Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CCN(CC(O)=O)CC(=O)NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CC=C(O)C=C1 ZMEWRPBAQVSBBB-GOTSBHOMSA-N 0.000 description 2
- MJZJYWCQPMNPRM-UHFFFAOYSA-N 6,6-dimethyl-1-[3-(2,4,5-trichlorophenoxy)propoxy]-1,6-dihydro-1,3,5-triazine-2,4-diamine Chemical compound CC1(C)N=C(N)N=C(N)N1OCCCOC1=CC(Cl)=C(Cl)C=C1Cl MJZJYWCQPMNPRM-UHFFFAOYSA-N 0.000 description 2
- KDCGOANMDULRCW-UHFFFAOYSA-N 7H-purine Chemical compound N1=CNC2=NC=NC2=C1 KDCGOANMDULRCW-UHFFFAOYSA-N 0.000 description 2
- 102100031585 ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Human genes 0.000 description 2
- LIWMQSWFLXEGMA-WDSKDSINSA-N Ala-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C)N LIWMQSWFLXEGMA-WDSKDSINSA-N 0.000 description 2
- 235000002198 Annona diversifolia Nutrition 0.000 description 2
- 108010039627 Aprotinin Proteins 0.000 description 2
- 208000023275 Autoimmune disease Diseases 0.000 description 2
- 102100029822 B- and T-lymphocyte attenuator Human genes 0.000 description 2
- 208000028564 B-cell non-Hodgkin lymphoma Diseases 0.000 description 2
- 102100038080 B-cell receptor CD22 Human genes 0.000 description 2
- 206010006187 Breast cancer Diseases 0.000 description 2
- 208000026310 Breast neoplasm Diseases 0.000 description 2
- 101710149863 C-C chemokine receptor type 4 Proteins 0.000 description 2
- 102100040840 C-type lectin domain family 7 member A Human genes 0.000 description 2
- 102100024217 CAMPATH-1 antigen Human genes 0.000 description 2
- 102100032976 CCR4-NOT transcription complex subunit 6 Human genes 0.000 description 2
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 2
- 108050005493 CD3 protein, epsilon/gamma/delta subunit Proteins 0.000 description 2
- 102100032937 CD40 ligand Human genes 0.000 description 2
- 108010065524 CD52 Antigen Proteins 0.000 description 2
- 241000282832 Camelidae Species 0.000 description 2
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 2
- 108010001857 Cell Surface Receptors Proteins 0.000 description 2
- 102000010970 Connexin Human genes 0.000 description 2
- 108050001175 Connexin Proteins 0.000 description 2
- 102000052510 DNA-Binding Proteins Human genes 0.000 description 2
- 101710096438 DNA-binding protein Proteins 0.000 description 2
- 108700022150 Designed Ankyrin Repeat Proteins Proteins 0.000 description 2
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 2
- 108010016626 Dipeptides Proteins 0.000 description 2
- 101100044298 Drosophila melanogaster fand gene Proteins 0.000 description 2
- 102000012545 EGF-like domains Human genes 0.000 description 2
- 108050002150 EGF-like domains Proteins 0.000 description 2
- 229940126626 Ektomab Drugs 0.000 description 2
- 102100023688 Eotaxin Human genes 0.000 description 2
- 108010008165 Etanercept Proteins 0.000 description 2
- 101150064015 FAS gene Proteins 0.000 description 2
- 229940126611 FBTA05 Drugs 0.000 description 2
- 102000006395 Globulins Human genes 0.000 description 2
- 108010044091 Globulins Proteins 0.000 description 2
- 229930186217 Glycolipid Natural products 0.000 description 2
- 102000003886 Glycoproteins Human genes 0.000 description 2
- 108090000288 Glycoproteins Proteins 0.000 description 2
- 241000282412 Homo Species 0.000 description 2
- 101000777636 Homo sapiens ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 1 Proteins 0.000 description 2
- 101000864344 Homo sapiens B- and T-lymphocyte attenuator Proteins 0.000 description 2
- 101000884305 Homo sapiens B-cell receptor CD22 Proteins 0.000 description 2
- 101000868279 Homo sapiens Leukocyte surface antigen CD47 Proteins 0.000 description 2
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 2
- 101000623901 Homo sapiens Mucin-16 Proteins 0.000 description 2
- 101000589307 Homo sapiens Natural cytotoxicity triggering receptor 3 Proteins 0.000 description 2
- 101001024605 Homo sapiens Next to BRCA1 gene 1 protein Proteins 0.000 description 2
- 101000914496 Homo sapiens T-cell antigen CD7 Proteins 0.000 description 2
- 101000831007 Homo sapiens T-cell immunoreceptor with Ig and ITIM domains Proteins 0.000 description 2
- 229940126592 Ibritsumomab Drugs 0.000 description 2
- 108010073816 IgE Receptors Proteins 0.000 description 2
- 102000009438 IgE Receptors Human genes 0.000 description 2
- 102100039688 Insulin-like growth factor 1 receptor Human genes 0.000 description 2
- 108050003558 Interleukin-17 Proteins 0.000 description 2
- 102000013691 Interleukin-17 Human genes 0.000 description 2
- 108010038453 Interleukin-2 Receptors Proteins 0.000 description 2
- 102000010789 Interleukin-2 Receptors Human genes 0.000 description 2
- 108010002616 Interleukin-5 Proteins 0.000 description 2
- 102000000743 Interleukin-5 Human genes 0.000 description 2
- 108090001005 Interleukin-6 Proteins 0.000 description 2
- 102000004889 Interleukin-6 Human genes 0.000 description 2
- 108010043610 KIR Receptors Proteins 0.000 description 2
- 125000002059 L-arginyl group Chemical group O=C([*])[C@](N([H])[H])([H])C([H])([H])C([H])([H])C([H])([H])N([H])C(=N[H])N([H])[H] 0.000 description 2
- 108010001831 LDL receptors Proteins 0.000 description 2
- 241000282842 Lama glama Species 0.000 description 2
- 108010006444 Leucine-Rich Repeat Proteins Proteins 0.000 description 2
- 102100032913 Leukocyte surface antigen CD47 Human genes 0.000 description 2
- 108090001030 Lipoproteins Proteins 0.000 description 2
- 102000004895 Lipoproteins Human genes 0.000 description 2
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 2
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 102100027159 Membrane primary amine oxidase Human genes 0.000 description 2
- 102000003735 Mesothelin Human genes 0.000 description 2
- 108090000015 Mesothelin Proteins 0.000 description 2
- 108010008707 Mucin-1 Proteins 0.000 description 2
- 102100023123 Mucin-16 Human genes 0.000 description 2
- 241000699670 Mus sp. Species 0.000 description 2
- QPCDCPDFJACHGM-UHFFFAOYSA-N N,N-bis{2-[bis(carboxymethyl)amino]ethyl}glycine Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(=O)O)CCN(CC(O)=O)CC(O)=O QPCDCPDFJACHGM-UHFFFAOYSA-N 0.000 description 2
- 102100022680 NKG2-D type II integral membrane protein Human genes 0.000 description 2
- 102100032852 Natural cytotoxicity triggering receptor 3 Human genes 0.000 description 2
- 108010069196 Neural Cell Adhesion Molecules Proteins 0.000 description 2
- 102000001068 Neural Cell Adhesion Molecules Human genes 0.000 description 2
- 206010033128 Ovarian cancer Diseases 0.000 description 2
- 206010061535 Ovarian neoplasm Diseases 0.000 description 2
- 101100335198 Pneumocystis carinii fol1 gene Proteins 0.000 description 2
- 206010041067 Small cell lung cancer Diseases 0.000 description 2
- 101000677856 Stenotrophomonas maltophilia (strain K279a) Actin-binding protein Smlt3054 Proteins 0.000 description 2
- 102100027208 T-cell antigen CD7 Human genes 0.000 description 2
- 102100024834 T-cell immunoreceptor with Ig and ITIM domains Human genes 0.000 description 2
- 102000002689 Toll-like receptor Human genes 0.000 description 2
- 108020000411 Toll-like receptor Proteins 0.000 description 2
- 108090000848 Ubiquitin Proteins 0.000 description 2
- 102000044159 Ubiquitin Human genes 0.000 description 2
- IEDXPSOJFSVCKU-HOKPPMCLSA-N [4-[[(2S)-5-(carbamoylamino)-2-[[(2S)-2-[6-(2,5-dioxopyrrolidin-1-yl)hexanoylamino]-3-methylbutanoyl]amino]pentanoyl]amino]phenyl]methyl N-[(2S)-1-[[(2S)-1-[[(3R,4S,5S)-1-[(2S)-2-[(1R,2R)-3-[[(1S,2R)-1-hydroxy-1-phenylpropan-2-yl]amino]-1-methoxy-2-methyl-3-oxopropyl]pyrrolidin-1-yl]-3-methoxy-5-methyl-1-oxoheptan-4-yl]-methylamino]-3-methyl-1-oxobutan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]-N-methylcarbamate Chemical compound CC[C@H](C)[C@@H]([C@@H](CC(=O)N1CCC[C@H]1[C@H](OC)[C@@H](C)C(=O)N[C@H](C)[C@@H](O)c1ccccc1)OC)N(C)C(=O)[C@@H](NC(=O)[C@H](C(C)C)N(C)C(=O)OCc1ccc(NC(=O)[C@H](CCCNC(N)=O)NC(=O)[C@@H](NC(=O)CCCCCN2C(=O)CCC2=O)C(C)C)cc1)C(C)C IEDXPSOJFSVCKU-HOKPPMCLSA-N 0.000 description 2
- 229950008347 abrilumab Drugs 0.000 description 2
- 229960002964 adalimumab Drugs 0.000 description 2
- 230000001640 apoptogenic effect Effects 0.000 description 2
- 210000003719 b-lymphocyte Anatomy 0.000 description 2
- 229960003270 belimumab Drugs 0.000 description 2
- 230000005540 biological transmission Effects 0.000 description 2
- 235000021152 breakfast Nutrition 0.000 description 2
- 229950001478 brontictuzumab Drugs 0.000 description 2
- 230000015556 catabolic process Effects 0.000 description 2
- 230000008568 cell cell communication Effects 0.000 description 2
- 230000030833 cell death Effects 0.000 description 2
- 229950006647 cixutumumab Drugs 0.000 description 2
- 230000002860 competitive effect Effects 0.000 description 2
- 210000000805 cytoplasm Anatomy 0.000 description 2
- 229950005026 dapirolizumab pegol Drugs 0.000 description 2
- 108010048522 dapirolizumab pegol Proteins 0.000 description 2
- 108010025838 dectin 1 Proteins 0.000 description 2
- 229950008135 dectrekumab Drugs 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- 210000004443 dendritic cell Anatomy 0.000 description 2
- 229950004079 denintuzumab mafodotin Drugs 0.000 description 2
- 229960001251 denosumab Drugs 0.000 description 2
- 229940126610 derlotuximab biotin Drugs 0.000 description 2
- 238000013461 design Methods 0.000 description 2
- 238000001514 detection method Methods 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- 239000000539 dimer Substances 0.000 description 2
- 238000010494 dissociation reaction Methods 0.000 description 2
- 230000005593 dissociations Effects 0.000 description 2
- 229950009791 durvalumab Drugs 0.000 description 2
- 229960002224 eculizumab Drugs 0.000 description 2
- 229960001776 edrecolomab Drugs 0.000 description 2
- 229960000284 efalizumab Drugs 0.000 description 2
- 229950002519 elgemtumab Drugs 0.000 description 2
- 229950004647 emactuzumab Drugs 0.000 description 2
- 210000002889 endothelial cell Anatomy 0.000 description 2
- 239000002158 endotoxin Substances 0.000 description 2
- 229950004270 enoblituzumab Drugs 0.000 description 2
- 108010087914 epidermal growth factor receptor VIII Proteins 0.000 description 2
- 210000002919 epithelial cell Anatomy 0.000 description 2
- 229950009760 epratuzumab Drugs 0.000 description 2
- 102000015694 estrogen receptors Human genes 0.000 description 2
- 108010038795 estrogen receptors Proteins 0.000 description 2
- 229960000403 etanercept Drugs 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 229960002027 evolocumab Drugs 0.000 description 2
- 102000034287 fluorescent proteins Human genes 0.000 description 2
- 108091006047 fluorescent proteins Proteins 0.000 description 2
- 229950004356 foralumab Drugs 0.000 description 2
- 229950004003 fresolimumab Drugs 0.000 description 2
- 230000004927 fusion Effects 0.000 description 2
- 229950002140 futuximab Drugs 0.000 description 2
- 229950001109 galiximab Drugs 0.000 description 2
- 229950004896 ganitumab Drugs 0.000 description 2
- 108010037850 glycylvaline Proteins 0.000 description 2
- 229950010864 guselkumab Drugs 0.000 description 2
- 239000001257 hydrogen Substances 0.000 description 2
- 229950003680 imalumab Drugs 0.000 description 2
- 230000002519 immonomodulatory effect Effects 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- 230000000968 intestinal effect Effects 0.000 description 2
- 229940121292 leronlimab Drugs 0.000 description 2
- 229950001237 lilotomab Drugs 0.000 description 2
- 229940126616 lilotomab satetraxetan Drugs 0.000 description 2
- 229950002950 lintuzumab Drugs 0.000 description 2
- 229920006008 lipopolysaccharide Polymers 0.000 description 2
- 229950000359 lokivetmab Drugs 0.000 description 2
- 229950010079 lumretuzumab Drugs 0.000 description 2
- 201000005202 lung cancer Diseases 0.000 description 2
- 208000020816 lung neoplasm Diseases 0.000 description 2
- 210000002540 macrophage Anatomy 0.000 description 2
- 229950001869 mapatumumab Drugs 0.000 description 2
- 229950008001 matuzumab Drugs 0.000 description 2
- 229960005108 mepolizumab Drugs 0.000 description 2
- 229950005674 modotuximab Drugs 0.000 description 2
- 210000001616 monocyte Anatomy 0.000 description 2
- 239000000178 monomer Substances 0.000 description 2
- 229950008897 morolimumab Drugs 0.000 description 2
- 230000035772 mutation Effects 0.000 description 2
- 229960005027 natalizumab Drugs 0.000 description 2
- 229960003301 nivolumab Drugs 0.000 description 2
- 229950003709 oxelumab Drugs 0.000 description 2
- 229950009723 ozanezumab Drugs 0.000 description 2
- 229950010626 pagibaximab Drugs 0.000 description 2
- 229960005570 pemtumomab Drugs 0.000 description 2
- 229950011098 pendetide Drugs 0.000 description 2
- 229940067082 pentetate Drugs 0.000 description 2
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 2
- 229960002087 pertuzumab Drugs 0.000 description 2
- 229950003203 pexelizumab Drugs 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 229920000642 polymer Polymers 0.000 description 2
- 238000003752 polymerase chain reaction Methods 0.000 description 2
- 230000017854 proteolysis Effects 0.000 description 2
- 230000002797 proteolythic effect Effects 0.000 description 2
- 229950009885 ralpancizumab Drugs 0.000 description 2
- 229960002633 ramucirumab Drugs 0.000 description 2
- 229960003876 ranibizumab Drugs 0.000 description 2
- 229950000987 refanezumab Drugs 0.000 description 2
- 230000010076 replication Effects 0.000 description 2
- 229950005978 rinucumab Drugs 0.000 description 2
- 229960004641 rituximab Drugs 0.000 description 2
- 229950008834 seribantumab Drugs 0.000 description 2
- 229950004951 sevirumab Drugs 0.000 description 2
- 208000000587 small cell lung carcinoma Diseases 0.000 description 2
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 230000002123 temporal effect Effects 0.000 description 2
- 238000012360 testing method Methods 0.000 description 2
- 229950008836 tosatoxumab Drugs 0.000 description 2
- 229960005267 tositumomab Drugs 0.000 description 2
- 230000009261 transgenic effect Effects 0.000 description 2
- 229960000575 trastuzumab Drugs 0.000 description 2
- 229950007217 tremelimumab Drugs 0.000 description 2
- 229950006444 trevogrumab Drugs 0.000 description 2
- 230000001960 triggered effect Effects 0.000 description 2
- 229960003824 ustekinumab Drugs 0.000 description 2
- BNJNAEJASPUJTO-DUOHOMBCSA-N vadastuximab talirine Chemical compound COc1ccc(cc1)C2=CN3[C@@H](C2)C=Nc4cc(OCCCOc5cc6N=C[C@@H]7CC(=CN7C(=O)c6cc5OC)c8ccc(NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)CCCCCN9C(=O)C[C@@H](SC[C@H](N)C(=O)O)C9=O)C(C)C)cc8)c(OC)cc4C3=O BNJNAEJASPUJTO-DUOHOMBCSA-N 0.000 description 2
- 229950001346 zolimomab aritox Drugs 0.000 description 2
- MZOFCQQQCNRIBI-VMXHOPILSA-N (3s)-4-[[(2s)-1-[[(2s)-1-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-methyl-1-oxopentan-2-yl]amino]-5-(diaminomethylideneamino)-1-oxopentan-2-yl]amino]-3-[[2-[[(2s)-2,6-diaminohexanoyl]amino]acetyl]amino]-4-oxobutanoic acid Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@@H](N)CCCCN MZOFCQQQCNRIBI-VMXHOPILSA-N 0.000 description 1
- TZCPCKNHXULUIY-RGULYWFUSA-N 1,2-distearoyl-sn-glycero-3-phosphoserine Chemical compound CCCCCCCCCCCCCCCCCC(=O)OC[C@H](COP(O)(=O)OC[C@H](N)C(O)=O)OC(=O)CCCCCCCCCCCCCCCCC TZCPCKNHXULUIY-RGULYWFUSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- 102100030310 5,6-dihydroxyindole-2-carboxylic acid oxidase Human genes 0.000 description 1
- 241000251468 Actinopterygii Species 0.000 description 1
- 208000024893 Acute lymphoblastic leukemia Diseases 0.000 description 1
- 208000014697 Acute lymphocytic leukaemia Diseases 0.000 description 1
- 108010033604 Apoptosis Inducing Factor Proteins 0.000 description 1
- 102000007272 Apoptosis Inducing Factor Human genes 0.000 description 1
- IYMAXBFPHPZYIK-BQBZGAKWSA-N Arg-Gly-Asp Chemical compound NC(N)=NCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(O)=O IYMAXBFPHPZYIK-BQBZGAKWSA-N 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- 208000002109 Argyria Diseases 0.000 description 1
- 102100022718 Atypical chemokine receptor 2 Human genes 0.000 description 1
- 208000010839 B-cell chronic lymphocytic leukemia Diseases 0.000 description 1
- 208000003950 B-cell lymphoma Diseases 0.000 description 1
- 102100032412 Basigin Human genes 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 102100036305 C-C chemokine receptor type 8 Human genes 0.000 description 1
- 102100025074 C-C chemokine receptor-like 2 Human genes 0.000 description 1
- 108090000342 C-Type Lectins Proteins 0.000 description 1
- 102000003930 C-Type Lectins Human genes 0.000 description 1
- 102100031650 C-X-C chemokine receptor type 4 Human genes 0.000 description 1
- 102000002110 C2 domains Human genes 0.000 description 1
- 108050009459 C2 domains Proteins 0.000 description 1
- 108010008629 CA-125 Antigen Proteins 0.000 description 1
- 102000007269 CA-125 Antigen Human genes 0.000 description 1
- 102100027207 CD27 antigen Human genes 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 108010021064 CTLA-4 Antigen Proteins 0.000 description 1
- 229940045513 CTLA4 antagonist Drugs 0.000 description 1
- 102100024153 Cadherin-15 Human genes 0.000 description 1
- 102100029758 Cadherin-4 Human genes 0.000 description 1
- 101100510615 Caenorhabditis elegans lag-2 gene Proteins 0.000 description 1
- 101100127656 Caenorhabditis elegans lam-2 gene Proteins 0.000 description 1
- 101100399480 Caenorhabditis elegans lmn-1 gene Proteins 0.000 description 1
- 241000282828 Camelus bactrianus Species 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 101710167800 Capsid assembly scaffolding protein Proteins 0.000 description 1
- 108010082548 Chemokine CCL11 Proteins 0.000 description 1
- 241000251730 Chondrichthyes Species 0.000 description 1
- 108020004705 Codon Proteins 0.000 description 1
- 102000008186 Collagen Human genes 0.000 description 1
- 108010035532 Collagen Proteins 0.000 description 1
- 206010009944 Colon cancer Diseases 0.000 description 1
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 1
- 235000004035 Cryptotaenia japonica Nutrition 0.000 description 1
- 230000004568 DNA-binding Effects 0.000 description 1
- 235000017274 Diospyros sandwicensis Nutrition 0.000 description 1
- 108090000204 Dipeptidase 1 Proteins 0.000 description 1
- 101100421450 Drosophila melanogaster Shark gene Proteins 0.000 description 1
- 108010024212 E-Selectin Proteins 0.000 description 1
- 102100023471 E-selectin Human genes 0.000 description 1
- 108010066687 Epithelial Cell Adhesion Molecule Proteins 0.000 description 1
- 102000018651 Epithelial Cell Adhesion Molecule Human genes 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 108010008177 Fd immunoglobulins Proteins 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 102000008946 Fibrinogen Human genes 0.000 description 1
- 108010049003 Fibrinogen Proteins 0.000 description 1
- 241001251094 Formica Species 0.000 description 1
- 102100035233 Furin Human genes 0.000 description 1
- 108090001126 Furin Proteins 0.000 description 1
- 108010001515 Galectin 4 Proteins 0.000 description 1
- 102100039556 Galectin-4 Human genes 0.000 description 1
- 229940125373 Gamma-Secretase Inhibitor Drugs 0.000 description 1
- 208000032612 Glial tumor Diseases 0.000 description 1
- 206010018338 Glioma Diseases 0.000 description 1
- 101710088083 Glomulin Proteins 0.000 description 1
- ZWZWYGMENQVNFU-UHFFFAOYSA-N Glycerophosphorylserin Natural products OC(=O)C(N)COP(O)(=O)OCC(O)CO ZWZWYGMENQVNFU-UHFFFAOYSA-N 0.000 description 1
- 102100039939 Growth/differentiation factor 8 Human genes 0.000 description 1
- 102100028976 HLA class I histocompatibility antigen, B alpha chain Human genes 0.000 description 1
- 102100030595 HLA class II histocompatibility antigen gamma chain Human genes 0.000 description 1
- 108010058607 HLA-B Antigens Proteins 0.000 description 1
- 102100034629 Hemopexin Human genes 0.000 description 1
- 108010026027 Hemopexin Proteins 0.000 description 1
- 208000017604 Hodgkin disease Diseases 0.000 description 1
- 208000021519 Hodgkin lymphoma Diseases 0.000 description 1
- 208000010747 Hodgkins lymphoma Diseases 0.000 description 1
- 101000773083 Homo sapiens 5,6-dihydroxyindole-2-carboxylic acid oxidase Proteins 0.000 description 1
- 101000678892 Homo sapiens Atypical chemokine receptor 2 Proteins 0.000 description 1
- 101000716068 Homo sapiens C-C chemokine receptor type 6 Proteins 0.000 description 1
- 101000716063 Homo sapiens C-C chemokine receptor type 8 Proteins 0.000 description 1
- 101000716070 Homo sapiens C-C chemokine receptor type 9 Proteins 0.000 description 1
- 101000922348 Homo sapiens C-X-C chemokine receptor type 4 Proteins 0.000 description 1
- 101000914511 Homo sapiens CD27 antigen Proteins 0.000 description 1
- 101000978392 Homo sapiens Eotaxin Proteins 0.000 description 1
- 101001082627 Homo sapiens HLA class II histocompatibility antigen gamma chain Proteins 0.000 description 1
- 101001103039 Homo sapiens Inactive tyrosine-protein kinase transmembrane receptor ROR1 Proteins 0.000 description 1
- 101001034652 Homo sapiens Insulin-like growth factor 1 receptor Proteins 0.000 description 1
- 101001046677 Homo sapiens Integrin alpha-V Proteins 0.000 description 1
- 101001057504 Homo sapiens Interferon-stimulated gene 20 kDa protein Proteins 0.000 description 1
- 101001055144 Homo sapiens Interleukin-2 receptor subunit alpha Proteins 0.000 description 1
- 101000697493 Homo sapiens Large proline-rich protein BAG6 Proteins 0.000 description 1
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 description 1
- 101000984626 Homo sapiens Low-density lipoprotein receptor-related protein 12 Proteins 0.000 description 1
- 101001043562 Homo sapiens Low-density lipoprotein receptor-related protein 2 Proteins 0.000 description 1
- 101001043596 Homo sapiens Low-density lipoprotein receptor-related protein 3 Proteins 0.000 description 1
- 101001043594 Homo sapiens Low-density lipoprotein receptor-related protein 5 Proteins 0.000 description 1
- 101001039199 Homo sapiens Low-density lipoprotein receptor-related protein 6 Proteins 0.000 description 1
- 101000991061 Homo sapiens MHC class I polypeptide-related sequence B Proteins 0.000 description 1
- 101001106413 Homo sapiens Macrophage-stimulating protein receptor Proteins 0.000 description 1
- 101000694615 Homo sapiens Membrane primary amine oxidase Proteins 0.000 description 1
- 101001133056 Homo sapiens Mucin-1 Proteins 0.000 description 1
- 101000934338 Homo sapiens Myeloid cell surface antigen CD33 Proteins 0.000 description 1
- 101001109501 Homo sapiens NKG2-D type II integral membrane protein Proteins 0.000 description 1
- 101000581981 Homo sapiens Neural cell adhesion molecule 1 Proteins 0.000 description 1
- 101000896414 Homo sapiens Nuclear nucleic acid-binding protein C1D Proteins 0.000 description 1
- 101001103036 Homo sapiens Nuclear receptor ROR-alpha Proteins 0.000 description 1
- 101000897042 Homo sapiens Nucleotide pyrophosphatase Proteins 0.000 description 1
- 101001098352 Homo sapiens OX-2 membrane glycoprotein Proteins 0.000 description 1
- 101001043564 Homo sapiens Prolow-density lipoprotein receptor-related protein 1 Proteins 0.000 description 1
- 101000633784 Homo sapiens SLAM family member 7 Proteins 0.000 description 1
- 101000711796 Homo sapiens Sclerostin Proteins 0.000 description 1
- 101000701411 Homo sapiens Suppressor of tumorigenicity 7 protein Proteins 0.000 description 1
- 101000874179 Homo sapiens Syndecan-1 Proteins 0.000 description 1
- 101000934346 Homo sapiens T-cell surface antigen CD2 Proteins 0.000 description 1
- 101000934341 Homo sapiens T-cell surface glycoprotein CD5 Proteins 0.000 description 1
- 101000914484 Homo sapiens T-lymphocyte activation antigen CD80 Proteins 0.000 description 1
- 101000904724 Homo sapiens Transmembrane glycoprotein NMB Proteins 0.000 description 1
- 101000610605 Homo sapiens Tumor necrosis factor receptor superfamily member 10A Proteins 0.000 description 1
- 101000610604 Homo sapiens Tumor necrosis factor receptor superfamily member 10B Proteins 0.000 description 1
- 101000851370 Homo sapiens Tumor necrosis factor receptor superfamily member 9 Proteins 0.000 description 1
- 101000666934 Homo sapiens Very low-density lipoprotein receptor Proteins 0.000 description 1
- 108010031794 IGF Type 1 Receptor Proteins 0.000 description 1
- 102000038455 IGF Type 1 Receptor Human genes 0.000 description 1
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 1
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 1
- 102000016844 Immunoglobulin-like domains Human genes 0.000 description 1
- 108050006430 Immunoglobulin-like domains Proteins 0.000 description 1
- 102100039615 Inactive tyrosine-protein kinase transmembrane receptor ROR1 Human genes 0.000 description 1
- 206010061218 Inflammation Diseases 0.000 description 1
- 108090000723 Insulin-Like Growth Factor I Proteins 0.000 description 1
- 102000004218 Insulin-Like Growth Factor I Human genes 0.000 description 1
- 102100032818 Integrin alpha-4 Human genes 0.000 description 1
- 102100022339 Integrin alpha-L Human genes 0.000 description 1
- 102100022337 Integrin alpha-V Human genes 0.000 description 1
- 108010041012 Integrin alpha4 Proteins 0.000 description 1
- 108010008212 Integrin alpha4beta1 Proteins 0.000 description 1
- 108010042918 Integrin alpha5beta1 Proteins 0.000 description 1
- 108010047852 Integrin alphaVbeta3 Proteins 0.000 description 1
- 102100037850 Interferon gamma Human genes 0.000 description 1
- 102000006992 Interferon-alpha Human genes 0.000 description 1
- 108010047761 Interferon-alpha Proteins 0.000 description 1
- 108010074328 Interferon-gamma Proteins 0.000 description 1
- 102100027268 Interferon-stimulated gene 20 kDa protein Human genes 0.000 description 1
- 108010002352 Interleukin-1 Proteins 0.000 description 1
- 102000000589 Interleukin-1 Human genes 0.000 description 1
- 108010065805 Interleukin-12 Proteins 0.000 description 1
- 108010017511 Interleukin-13 Receptors Proteins 0.000 description 1
- 102000004559 Interleukin-13 Receptors Human genes 0.000 description 1
- 102100030703 Interleukin-22 Human genes 0.000 description 1
- 108090000978 Interleukin-4 Proteins 0.000 description 1
- 102000004388 Interleukin-4 Human genes 0.000 description 1
- 102000010781 Interleukin-6 Receptors Human genes 0.000 description 1
- 108010038501 Interleukin-6 Receptors Proteins 0.000 description 1
- 230000004163 JAK-STAT signaling pathway Effects 0.000 description 1
- 108010092694 L-Selectin Proteins 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- 102100033467 L-selectin Human genes 0.000 description 1
- 102000000853 LDL receptors Human genes 0.000 description 1
- 241000282838 Lama Species 0.000 description 1
- 241000282852 Lama guanicoe Species 0.000 description 1
- 102100028047 Large proline-rich protein BAG6 Human genes 0.000 description 1
- 102000010954 Link domains Human genes 0.000 description 1
- 108050001157 Link domains Proteins 0.000 description 1
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 1
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 102100029185 Low affinity immunoglobulin gamma Fc region receptor III-B Human genes 0.000 description 1
- 102100024640 Low-density lipoprotein receptor Human genes 0.000 description 1
- 102100027120 Low-density lipoprotein receptor-related protein 12 Human genes 0.000 description 1
- 102100021922 Low-density lipoprotein receptor-related protein 2 Human genes 0.000 description 1
- 102100021917 Low-density lipoprotein receptor-related protein 3 Human genes 0.000 description 1
- 102100021926 Low-density lipoprotein receptor-related protein 5 Human genes 0.000 description 1
- 102100040704 Low-density lipoprotein receptor-related protein 6 Human genes 0.000 description 1
- 108010064548 Lymphocyte Function-Associated Antigen-1 Proteins 0.000 description 1
- 208000031422 Lymphocytic Chronic B-Cell Leukemia Diseases 0.000 description 1
- 206010025323 Lymphomas Diseases 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 108010018562 M-cadherin Proteins 0.000 description 1
- 102100030300 MHC class I polypeptide-related sequence B Human genes 0.000 description 1
- 102100021435 Macrophage-stimulating protein receptor Human genes 0.000 description 1
- 101710132836 Membrane primary amine oxidase Proteins 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- 241000699666 Mus <mouse, genus> Species 0.000 description 1
- 102100025243 Myeloid cell surface antigen CD33 Human genes 0.000 description 1
- 108010056852 Myostatin Proteins 0.000 description 1
- 108050000637 N-cadherin Proteins 0.000 description 1
- SUHQNCLNRUAGOO-UHFFFAOYSA-N N-glycoloyl-neuraminic acid Natural products OCC(O)C(O)C(O)C(NC(=O)CO)C(O)CC(=O)C(O)=O SUHQNCLNRUAGOO-UHFFFAOYSA-N 0.000 description 1
- FDJKUWYYUZCUJX-UHFFFAOYSA-N N-glycolyl-beta-neuraminic acid Natural products OCC(O)C(O)C1OC(O)(C(O)=O)CC(O)C1NC(=O)CO FDJKUWYYUZCUJX-UHFFFAOYSA-N 0.000 description 1
- FDJKUWYYUZCUJX-KVNVFURPSA-N N-glycolylneuraminic acid Chemical compound OC[C@H](O)[C@H](O)[C@@H]1O[C@](O)(C(O)=O)C[C@H](O)[C@H]1NC(=O)CO FDJKUWYYUZCUJX-KVNVFURPSA-N 0.000 description 1
- 108010001657 NK Cell Lectin-Like Receptor Subfamily K Proteins 0.000 description 1
- 102100029527 Natural cytotoxicity triggering receptor 3 ligand 1 Human genes 0.000 description 1
- 101710201161 Natural cytotoxicity triggering receptor 3 ligand 1 Proteins 0.000 description 1
- 102100027347 Neural cell adhesion molecule 1 Human genes 0.000 description 1
- 206010029260 Neuroblastoma Diseases 0.000 description 1
- 102000004108 Neurotransmitter Receptors Human genes 0.000 description 1
- 108090000590 Neurotransmitter Receptors Proteins 0.000 description 1
- 108091028043 Nucleic acid sequence Proteins 0.000 description 1
- 108091005461 Nucleic proteins Proteins 0.000 description 1
- 102100021969 Nucleotide pyrophosphatase Human genes 0.000 description 1
- 102100037589 OX-2 membrane glycoprotein Human genes 0.000 description 1
- 108010035766 P-Selectin Proteins 0.000 description 1
- 102100023472 P-selectin Human genes 0.000 description 1
- 102000000470 PDZ domains Human genes 0.000 description 1
- 108050008994 PDZ domains Proteins 0.000 description 1
- 206010061902 Pancreatic neoplasm Diseases 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 108090000284 Pepsin A Proteins 0.000 description 1
- 102000057297 Pepsin A Human genes 0.000 description 1
- 208000006664 Precursor Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 description 1
- 101710130420 Probable capsid assembly scaffolding protein Proteins 0.000 description 1
- 102100040678 Programmed cell death protein 1 Human genes 0.000 description 1
- 102100021923 Prolow-density lipoprotein receptor-related protein 1 Human genes 0.000 description 1
- 102100023832 Prolyl endopeptidase FAP Human genes 0.000 description 1
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 1
- 108010086890 R-cadherin Proteins 0.000 description 1
- 108010025832 RANK Ligand Proteins 0.000 description 1
- 230000010799 Receptor Interactions Effects 0.000 description 1
- 108091005682 Receptor kinases Proteins 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 102000014400 SH2 domains Human genes 0.000 description 1
- 108050003452 SH2 domains Proteins 0.000 description 1
- 102000000395 SH3 domains Human genes 0.000 description 1
- 108050008861 SH3 domains Proteins 0.000 description 1
- 102100029198 SLAM family member 7 Human genes 0.000 description 1
- 108050008568 SRCR domains Proteins 0.000 description 1
- 102000000185 SRCR domains Human genes 0.000 description 1
- 206010039509 Scab Diseases 0.000 description 1
- 101710204410 Scaffold protein Proteins 0.000 description 1
- 102100034201 Sclerostin Human genes 0.000 description 1
- 241000239226 Scorpiones Species 0.000 description 1
- 229940122055 Serine protease inhibitor Drugs 0.000 description 1
- 101710102218 Serine protease inhibitor Proteins 0.000 description 1
- 102000000890 Somatomedin B domains Human genes 0.000 description 1
- 108050007913 Somatomedin B domains Proteins 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- 102100035721 Syndecan-1 Human genes 0.000 description 1
- 108010092262 T-Cell Antigen Receptors Proteins 0.000 description 1
- 102100025237 T-cell surface antigen CD2 Human genes 0.000 description 1
- 102100025244 T-cell surface glycoprotein CD5 Human genes 0.000 description 1
- 102100027222 T-lymphocyte activation antigen CD80 Human genes 0.000 description 1
- 101001051488 Takifugu rubripes Neural cell adhesion molecule L1 Proteins 0.000 description 1
- 102100024554 Tetranectin Human genes 0.000 description 1
- 102000002938 Thrombospondin Human genes 0.000 description 1
- 108060008245 Thrombospondin Proteins 0.000 description 1
- 102000004887 Transforming Growth Factor beta Human genes 0.000 description 1
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 1
- 102000046299 Transforming Growth Factor beta1 Human genes 0.000 description 1
- 101800002279 Transforming growth factor beta-1 Proteins 0.000 description 1
- 102100023935 Transmembrane glycoprotein NMB Human genes 0.000 description 1
- 102000007641 Trefoil Factors Human genes 0.000 description 1
- 235000015724 Trifolium pratense Nutrition 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 1
- 102100024568 Tumor necrosis factor ligand superfamily member 11 Human genes 0.000 description 1
- 102100036922 Tumor necrosis factor ligand superfamily member 13B Human genes 0.000 description 1
- 101710181056 Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 1
- 102100040113 Tumor necrosis factor receptor superfamily member 10A Human genes 0.000 description 1
- 102100040112 Tumor necrosis factor receptor superfamily member 10B Human genes 0.000 description 1
- 102100036856 Tumor necrosis factor receptor superfamily member 9 Human genes 0.000 description 1
- 108010053096 Vascular Endothelial Growth Factor Receptor-1 Proteins 0.000 description 1
- 102100033178 Vascular endothelial growth factor receptor 1 Human genes 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 102100039066 Very low-density lipoprotein receptor Human genes 0.000 description 1
- 241001416177 Vicugna pacos Species 0.000 description 1
- 241000282840 Vicugna vicugna Species 0.000 description 1
- 102100035071 Vimentin Human genes 0.000 description 1
- 108010065472 Vimentin Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- 102100035140 Vitronectin Human genes 0.000 description 1
- 108010031318 Vitronectin Proteins 0.000 description 1
- 241000607479 Yersinia pestis Species 0.000 description 1
- XYVNHPYNSPGYLI-UUOKFMHZSA-N [(2r,3s,4r,5r)-5-(2-amino-6-oxo-3h-purin-9-yl)-4-hydroxy-2-(phosphonooxymethyl)oxolan-3-yl] dihydrogen phosphate Chemical compound C1=2NC(N)=NC(=O)C=2N=CN1[C@@H]1O[C@H](COP(O)(O)=O)[C@@H](OP(O)(O)=O)[C@H]1O XYVNHPYNSPGYLI-UUOKFMHZSA-N 0.000 description 1
- 229960003697 abatacept Drugs 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 229950004283 actoxumab Drugs 0.000 description 1
- 210000005006 adaptive immune system Anatomy 0.000 description 1
- 208000009956 adenocarcinoma Diseases 0.000 description 1
- 102000019997 adhesion receptor Human genes 0.000 description 1
- 108010013985 adhesion receptor Proteins 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 229960002459 alefacept Drugs 0.000 description 1
- 229960000548 alemtuzumab Drugs 0.000 description 1
- 108010026331 alpha-Fetoproteins Proteins 0.000 description 1
- 102000013529 alpha-Fetoproteins Human genes 0.000 description 1
- 229950009106 altumomab Drugs 0.000 description 1
- 229950001537 amatuximab Drugs 0.000 description 1
- 229950005794 anrukinzumab Drugs 0.000 description 1
- 238000013459 approach Methods 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 108010072041 arginyl-glycyl-aspartic acid Proteins 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- 229950001863 bapineuzumab Drugs 0.000 description 1
- 229960004669 basiliximab Drugs 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 229950010559 besilesomab Drugs 0.000 description 1
- 102000006635 beta-lactamase Human genes 0.000 description 1
- 229960000397 bevacizumab Drugs 0.000 description 1
- 229950008086 bezlotoxumab Drugs 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 201000000053 blastoma Diseases 0.000 description 1
- 210000001185 bone marrow Anatomy 0.000 description 1
- 210000004556 brain Anatomy 0.000 description 1
- 229950000025 brolucizumab Drugs 0.000 description 1
- 229950001178 capromab Drugs 0.000 description 1
- 229940034605 capromab pendetide Drugs 0.000 description 1
- 229910052799 carbon Inorganic materials 0.000 description 1
- 229910002092 carbon dioxide Inorganic materials 0.000 description 1
- 239000001569 carbon dioxide Substances 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 229950000771 carlumab Drugs 0.000 description 1
- 229950006754 cedelizumab Drugs 0.000 description 1
- 230000009134 cell regulation Effects 0.000 description 1
- 229960003115 certolizumab pegol Drugs 0.000 description 1
- 229960005395 cetuximab Drugs 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 238000012412 chemical coupling Methods 0.000 description 1
- 229960001231 choline Drugs 0.000 description 1
- OEYIOHPDSNJKLS-UHFFFAOYSA-N choline Chemical compound C[N+](C)(C)CCO OEYIOHPDSNJKLS-UHFFFAOYSA-N 0.000 description 1
- 208000032852 chronic lymphocytic leukemia Diseases 0.000 description 1
- 229920001436 collagen Polymers 0.000 description 1
- 238000004891 communication Methods 0.000 description 1
- 239000002299 complementary DNA Substances 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 239000000356 contaminant Substances 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000010168 coupling process Methods 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000018044 dehydration Effects 0.000 description 1
- 238000006297 dehydration reaction Methods 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 239000003398 denaturant Substances 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 1
- 125000002228 disulfide group Chemical group 0.000 description 1
- 150000002019 disulfides Chemical class 0.000 description 1
- 231100000673 dose–response relationship Toxicity 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 201000008184 embryoma Diseases 0.000 description 1
- 230000012202 endocytosis Effects 0.000 description 1
- 210000003038 endothelium Anatomy 0.000 description 1
- 229950010640 ensituximab Drugs 0.000 description 1
- 102000012803 ephrin Human genes 0.000 description 1
- 108060002566 ephrin Proteins 0.000 description 1
- 230000008472 epithelial growth Effects 0.000 description 1
- 229940011871 estrogen Drugs 0.000 description 1
- 239000000262 estrogen Substances 0.000 description 1
- 108010036236 extracellular matrix receptor Proteins 0.000 description 1
- 230000001605 fetal effect Effects 0.000 description 1
- 229940012952 fibrinogen Drugs 0.000 description 1
- 235000019688 fish Nutrition 0.000 description 1
- 238000000684 flow cytometry Methods 0.000 description 1
- 108010021843 fluorescent protein 583 Proteins 0.000 description 1
- 102000006815 folate receptor Human genes 0.000 description 1
- 108020005243 folate receptor Proteins 0.000 description 1
- 244000053095 fungal pathogen Species 0.000 description 1
- 239000003540 gamma secretase inhibitor Substances 0.000 description 1
- BTCSSZJGUNDROE-UHFFFAOYSA-N gamma-aminobutyric acid Chemical compound NCCCC(O)=O BTCSSZJGUNDROE-UHFFFAOYSA-N 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 210000002443 helper t lymphocyte Anatomy 0.000 description 1
- 210000004408 hybridoma Anatomy 0.000 description 1
- 230000036039 immunity Effects 0.000 description 1
- 230000003053 immunization Effects 0.000 description 1
- 238000002649 immunization Methods 0.000 description 1
- 239000003018 immunosuppressive agent Substances 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 230000001965 increasing effect Effects 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 229950006289 indusatumab Drugs 0.000 description 1
- 230000002757 inflammatory effect Effects 0.000 description 1
- 230000004054 inflammatory process Effects 0.000 description 1
- 108010043603 integrin alpha4beta7 Proteins 0.000 description 1
- 108010074109 interleukin-22 Proteins 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 229960005435 ixekizumab Drugs 0.000 description 1
- 210000005067 joint tissue Anatomy 0.000 description 1
- 229950010828 keliximab Drugs 0.000 description 1
- 229950000518 labetuzumab Drugs 0.000 description 1
- 229950002183 lebrikizumab Drugs 0.000 description 1
- 229950007439 lenzilumab Drugs 0.000 description 1
- 229950002884 lexatumumab Drugs 0.000 description 1
- 108020001756 ligand binding domains Proteins 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 210000001165 lymph node Anatomy 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 108020004084 membrane receptors Proteins 0.000 description 1
- 230000009401 metastasis Effects 0.000 description 1
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 1
- 229950003734 milatuzumab Drugs 0.000 description 1
- 230000003278 mimic effect Effects 0.000 description 1
- 108091005601 modified peptides Proteins 0.000 description 1
- 108091005763 multidomain proteins Proteins 0.000 description 1
- 201000006417 multiple sclerosis Diseases 0.000 description 1
- OHDXDNUPVVYWOV-UHFFFAOYSA-N n-methyl-1-(2-naphthalen-1-ylsulfanylphenyl)methanamine Chemical compound CNCC1=CC=CC=C1SC1=CC=CC2=CC=CC=C12 OHDXDNUPVVYWOV-UHFFFAOYSA-N 0.000 description 1
- 229950007708 namilumab Drugs 0.000 description 1
- 229950009793 naptumomab estafenatox Drugs 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 229960000470 omalizumab Drugs 0.000 description 1
- 201000002528 pancreatic cancer Diseases 0.000 description 1
- 208000008443 pancreatic carcinoma Diseases 0.000 description 1
- 229940126618 pankomab Drugs 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 229940111202 pepsin Drugs 0.000 description 1
- 239000000813 peptide hormone Substances 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 108010025221 plasma protein Z Proteins 0.000 description 1
- 239000013612 plasmid Substances 0.000 description 1
- 238000006116 polymerization reaction Methods 0.000 description 1
- 102000040430 polynucleotide Human genes 0.000 description 1
- 108091033319 polynucleotide Proteins 0.000 description 1
- 239000002157 polynucleotide Substances 0.000 description 1
- 230000029279 positive regulation of transcription, DNA-dependent Effects 0.000 description 1
- 230000037452 priming Effects 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 238000002818 protein evolution Methods 0.000 description 1
- 230000004850 protein–protein interaction Effects 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- 230000009257 reactivity Effects 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 210000003289 regulatory T cell Anatomy 0.000 description 1
- 230000003252 repetitive effect Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 206010039073 rheumatoid arthritis Diseases 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 238000002702 ribosome display Methods 0.000 description 1
- 108091008601 sVEGFR Proteins 0.000 description 1
- 150000003839 salts Chemical group 0.000 description 1
- 238000003118 sandwich ELISA Methods 0.000 description 1
- 238000012216 screening Methods 0.000 description 1
- 241001223796 sea lampreys Species 0.000 description 1
- 238000002864 sequence alignment Methods 0.000 description 1
- 239000003001 serine protease inhibitor Substances 0.000 description 1
- 229950003850 setoxaximab Drugs 0.000 description 1
- 210000003491 skin Anatomy 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 125000006850 spacer group Chemical group 0.000 description 1
- 238000009987 spinning Methods 0.000 description 1
- 150000003431 steroids Chemical class 0.000 description 1
- 230000000638 stimulation Effects 0.000 description 1
- 238000006467 substitution reaction Methods 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 101150047061 tag-72 gene Proteins 0.000 description 1
- 108010013645 tetranectin Proteins 0.000 description 1
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 description 1
- 125000003396 thiol group Chemical group [H]S* 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 229950005972 urelumab Drugs 0.000 description 1
- 229910052720 vanadium Inorganic materials 0.000 description 1
- 239000013598 vector Substances 0.000 description 1
- 229950010789 vesencumab Drugs 0.000 description 1
- 210000005048 vimentin Anatomy 0.000 description 1
- 108010047303 von Willebrand Factor Proteins 0.000 description 1
- 102100036537 von Willebrand factor Human genes 0.000 description 1
- 229960001134 von willebrand factor Drugs 0.000 description 1
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical group O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 1
- 229950009002 zanolimumab Drugs 0.000 description 1
- 229950007157 zolbetuximab Drugs 0.000 description 1
Images


















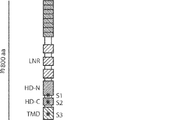









































































































































































































































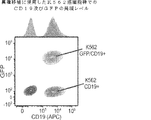




















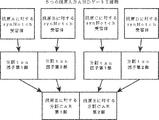








Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K19/00—Hybrid peptides, i.e. peptides covalently bound to nucleic acids, or non-covalently bound protein-protein complexes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/461—Cellular immunotherapy characterised by the cell type used
- A61K39/4611—T-cells, e.g. tumor infiltrating lymphocytes [TIL], lymphokine-activated killer cells [LAK] or regulatory T cells [Treg]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/463—Cellular immunotherapy characterised by recombinant expression
- A61K39/4631—Chimeric Antigen Receptors [CAR]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/463—Cellular immunotherapy characterised by recombinant expression
- A61K39/4632—T-cell receptors [TCR]; antibody T-cell receptor constructs
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/464—Cellular immunotherapy characterised by the antigen targeted or presented
- A61K39/4643—Vertebrate antigens
- A61K39/4644—Cancer antigens
- A61K39/464402—Receptors, cell surface antigens or cell surface determinants
- A61K39/464411—Immunoglobulin superfamily
- A61K39/464412—CD19 or B4
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/464—Cellular immunotherapy characterised by the antigen targeted or presented
- A61K39/4643—Vertebrate antigens
- A61K39/4644—Cancer antigens
- A61K39/464466—Adhesion molecules, e.g. NRCAM, EpCAM or cadherins
- A61K39/464468—Mesothelin [MSLN]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K48/00—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/475—Growth factors; Growth regulators
- C07K14/485—Epidermal growth factor [EGF] (urogastrone)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/71—Receptors; Cell surface antigens; Cell surface determinants for growth factors; for growth regulators
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/715—Receptors; Cell surface antigens; Cell surface determinants for cytokines; for lymphokines; for interferons
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/30—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants from tumour cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0636—T lymphocytes
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K39/46
- A61K2239/27—Indexing codes associated with cellular immunotherapy of group A61K39/46 characterized by targeting or presenting multiple antigens
- A61K2239/29—Multispecific CARs
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K39/46
- A61K2239/31—Indexing codes associated with cellular immunotherapy of group A61K39/46 characterized by the route of administration
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K39/46
- A61K2239/38—Indexing codes associated with cellular immunotherapy of group A61K39/46 characterised by the dose, timing or administration schedule
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
- A61K35/14—Blood; Artificial blood
- A61K35/17—Lymphocytes; B-cells; T-cells; Natural killer cells; Interferon-activated or cytokine-activated lymphocytes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/7051—T-cell receptor (TcR)-CD3 complex
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/22—Immunoglobulins specific features characterized by taxonomic origin from camelids, e.g. camel, llama or dromedary
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/569—Single domain, e.g. dAb, sdAb, VHH, VNAR or nanobody®
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/62—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising only variable region components
- C07K2317/622—Single chain antibody (scFv)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/01—Fusion polypeptide containing a localisation/targetting motif
- C07K2319/03—Fusion polypeptide containing a localisation/targetting motif containing a transmembrane segment
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/32—Fusion polypeptide fusions with soluble part of a cell surface receptor, "decoy receptors"
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/33—Fusion polypeptide fusions for targeting to specific cell types, e.g. tissue specific targeting, targeting of a bacterial subspecies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/60—Fusion polypeptide containing spectroscopic/fluorescent detection, e.g. green fluorescent protein [GFP]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/70—Fusion polypeptide containing domain for protein-protein interaction
- C07K2319/74—Fusion polypeptide containing domain for protein-protein interaction containing a fusion for binding to a cell surface receptor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2510/00—Genetically modified cells
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/566—Immunoassay; Biospecific binding assay; Materials therefor using specific carrier or receptor proteins as ligand binding reagents where possible specific carrier or receptor proteins are classified with their target compounds
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/68—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving proteins, peptides or amino acids
- G01N33/6863—Cytokines, i.e. immune system proteins modifying a biological response such as cell growth proliferation or differentiation, e.g. TNF, CNF, GM-CSF, lymphotoxin, MIF or their receptors
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/68—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving proteins, peptides or amino acids
- G01N33/6872—Intracellular protein regulatory factors and their receptors, e.g. including ion channels
Description
本出願は、2015年2月24日に出願された米国仮特許出願第62/120,256号;2015年11月18日に出願された米国仮特許出願第62/257,153号;及び2015年12月18日に出願された米国仮特許出願第62/269,758号の利益を主張し、これらの出願は、その全体が参照により本明細書に組み込まれる。
本発明は、米国国立衛生研究所により授与された助成金番号EY016546;P50 GM081879;及びR01 GM055040の下で政府支援によりなされた。政府は、本発明において一定の権利を有する。
配列表を、テキストファイル「UCSF−511WO_SeqList_ST25.txt」として本明細書と共に提供し、これは、2016年1月26日に作製され、サイズは、649KBである。テキストファイルの内容は、その全体が参照により本明細書に組み込まれる。
Notch受容体は、細胞間接触シグナル伝達を媒介し、細胞間コミュニケーション、例えば、一方の接触細胞が「受信側」細胞であり他方の接触細胞が「送信側」細胞である、2つの接触する細胞間のコミュニケーションの発達及び他の態様において中心的な役割を果たす膜貫通タンパク質である。受信側細胞において発現されるNotch受容体は、送信細胞上で発現される、それらのリガンド(タンパク質のデルタファミリー)を認識する。これらの接触細胞上でのnotchとデルタの係合は、notch受容体の2段階のタンパク質分解をもたらし、これは最終的には、膜から細胞質への受容体の細胞内部分の放出を引き起こす。この放出されたドメインが、転写調節因子として機能することにより受信側細胞の挙動を変える。Notch受容体は、発達中の様々な細胞の機能に関与し、そのために必要とされ、複数の種にわたって多数の細胞型の機能に重要である。
本開示は、結合誘発型(binding-triggered)転写スイッチポリペプチド、結合誘発型転写スイッチポリペプチドをコードするヌクレオチド配列を含む核酸、及び該核酸で遺伝子改変された宿主細胞を提供する。本開示は、本開示の結合誘発型転写スイッチポリペプチドをコードする核酸を含むトランスジェニック生物を提供する。1つまたは複数の結合誘発型転写スイッチポリペプチドを使用して細胞の活性を局所的に調節する方法、及び1つまたは複数の結合誘発型転写スイッチポリペプチドを使用する局在性細胞活性化システムを提供する。本開示の結合誘発型転写スイッチポリペプチドは、様々な用途において有用であり、それも提供する。

i)第一の特異的結合対の第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む第一の結合誘発型転写スイッチ;及び
ii)少なくとも、第二の特異的結合対の第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む第二の結合誘発型転写スイッチ
を発現し;第一の結合誘発型転写スイッチの細胞内ドメインが、第一のエフェクター機能を提供し、第二の結合誘発型転写スイッチの細胞内ドメインが、第一のエフェクター機能と異なる第二のエフェクター機能を提供し、この時、第一及び第二の特異的結合対の第一及び第二のメンバーの結合が、第一及び第二の細胞内ドメインを活性化するための結合シグナルを伝達するように結合トランスデューサを誘導する、前記方法を提供する。
[本発明1001]
N末端からC末端に向かって、共有結合状態で、
a)特異的結合対の第一のメンバーを含む細胞外ドメイン;
b)50アミノ酸〜1000アミノ酸の長さを有し、かつ1つまたは複数のリガンド誘導性タンパク質分解切断部位を含む、Notch受容体ポリペプチド;及び
c)細胞内ドメイン
を含み、
前記特異的結合対の前記第一のメンバーが、前記Notch受容体ポリペプチドに対して異種であり、前記特異的結合対の前記第一のメンバーの、前記特異的結合対の第二のメンバーへの結合が、前記Notch受容体ポリペプチドの前記1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより前記細胞内ドメインが放出される、キメラポリペプチド。
[本発明1002]
前記Notch受容体ポリペプチドが、300アミノ酸〜400アミノ酸の長さを有する、本発明1001のキメラポリペプチド。
[本発明1003]
前記細胞外ドメインと前記Notch受容体ポリペプチドの間に介在するリンカーを含む、本発明1001または本発明1002のキメラポリペプチド。
[本発明1004]
前記細胞内ドメインが、転写活性化因子である、本発明1001〜1003のいずれかのキメラポリペプチド。
[本発明1005]
前記細胞内ドメインが、転写抑制因子である、本発明1001〜1003のいずれかのキメラポリペプチド。
[本発明1006]
前記細胞内ドメインが、部位特異的ヌクレアーゼである、本発明1001〜1003のいずれかのキメラポリペプチド。
[本発明1007]
前記部位特異的ヌクレアーゼが、Cas9ポリペプチドである、本発明1006のキメラポリペプチド
[本発明1008]
前記細胞内ドメインが、リコンビナーゼである、本発明1001〜1003のいずれかのキメラポリペプチド。
[本発明1009]
前記細胞内ドメインが、抑制性免疫受容体である、本発明1001〜1003のいずれかのキメラポリペプチド。
[本発明1010]
前記細胞内ドメインが、活性化免疫受容体である、本発明1001〜1003のいずれかのキメラポリペプチド。
[本発明1011]
前記特異的結合対の前記第一のメンバーが、抗体をベースとする認識足場を含む、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1012]
前記特異的結合対の前記第一のメンバーが、抗体を含む、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1013]
前記抗体が、腫瘍特異的抗原、疾患関連抗原、病原体関連抗原、自己免疫疾患関連抗原、または細胞外マトリックス成分と特異的に結合する、本発明1012のキメラポリペプチド。
[本発明1014]
前記抗体が、細胞表面抗原、可溶性抗原、または不溶性基体上に固定された抗原と特異的に結合する、本発明1012のキメラポリペプチド。
[本発明1015]
前記抗体が、一本鎖Fvである、本発明1012のキメラポリペプチド。
[本発明1016]
前記特異的結合対の前記第一のメンバーが、ナノボディ、単一ドメイン抗体、ダイアボディ、トリアボディ、またはミニボディである、本発明1011のキメラポリペプチド。
[本発明1017]
前記特異的結合対の前記第一のメンバーが、非抗体をベースとする認識足場である、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1018]
前記非抗体をベースとする認識足場が、アビマー、DARPin、アドネクチン、アビマー、アフィボディ、アンチカリン、またはアフィリンである、本発明1017のキメラポリペプチド。
[本発明1019]
前記特異的結合対の前記第一のメンバーが、抗原である、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1020]
前記抗原が、内在性抗原である、本発明1019のキメラポリペプチド。
[本発明1021]
前記抗原が、外来性抗原である、本発明1019のキメラポリペプチド。
[本発明1022]
前記特異的結合対の前記第一のメンバーが、受容体に対するリガンドである、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1023]
前記特異的結合対の前記第一のメンバーが、受容体である、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1024]
前記特異的結合対の前記第一のメンバーが、細胞接着分子である、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1025]
前記特異的結合対の前記第一のメンバーが、第一の二量体化ドメインを含み、前記特異的結合対の前記第二のメンバーが、第二の二量体化ドメインを含む、本発明1001〜1010のいずれかのキメラポリペプチド。
[本発明1026]
前記第一の二量体化ドメインの、前記第二の二量体化ドメインへの結合が、小分子二量体化剤により誘導される、本発明1025のキメラポリペプチド。
[本発明1027]
前記第一の二量体化ドメインの、前記第二の二量体化ドメインへの結合が、光により誘導される、本発明1025のキメラポリペプチド。
[本発明1028]
前記Notch受容体ポリペプチドが、図2A〜2Gに示すアミノ酸配列のいずれか1つに対して、少なくとも75%アミノ酸配列同一性を有するアミノ酸配列を含む、本発明1001〜1027のいずれかのキメラポリペプチド。
[本発明1029]
前記1つまたは複数のリガンド誘導性タンパク質分解切断部位が、S1、S2、及びS3タンパク質分解切断部位から選択される、本発明1001〜1028のいずれかのキメラポリペプチド。
[本発明1030]
前記S1タンパク質分解切断部位が、アミノ酸配列Arg−X−(Arg/Lys)−Argを含むフューリン様プロテアーゼ切断部位であり、ここでXは任意のアミノ酸である、本発明1029のキメラポリペプチド。
[本発明1031]
前記S2タンパク質分解切断部位が、Ala−Valジペプチド配列を含むADAM−17型プロテアーゼ切断部位である、本発明1029のキメラポリペプチド。
[本発明1032]
前記S3タンパク質分解切断部位が、Gly−Valジペプチド配列を含むγ−セクレターゼ切断部位である、本発明1029のキメラポリペプチド。
[本発明1033]
本発明1001〜1032のいずれかのキメラポリペプチドをコードするヌクレオチド配列を含む、核酸。
[本発明1034]
本発明1001〜1032のいずれかのキメラポリペプチドをコードするヌクレオチド配列を含む、組換え発現ベクター。
[本発明1035]
本発明1033の核酸または本発明1034の発現ベクターで遺伝子改変された宿主細胞。
[本発明1036]
真核細胞である、本発明1035の宿主細胞。
[本発明1037]
哺乳類細胞である、本発明1036の宿主細胞。
[本発明1038]
免疫細胞、ニューロン、上皮細胞、及び内皮細胞、または幹細胞である、本発明1037の宿主細胞。
[本発明1039]
前記免疫細胞が、T細胞、B細胞、単球、ナチュラルキラー細胞、樹状細胞、マクロファージ、制御性T細胞、ヘルパーT細胞、または細胞傷害性T細胞である、本発明1038の宿主細胞。
[本発明1040]
キメラ抗原受容体(CAR)またはT細胞受容体(TCR)をコードするヌクレオチド配列を含む核酸で遺伝子改変されており、前記キメラポリペプチドの前記細胞内ドメインが、転写活性化因子である、本発明1035〜1039のいずれかの宿主細胞。
[本発明1041]
前記CARまたは前記TCRをコードする前記ヌクレオチド配列が、前記キメラポリペプチドの前記細胞内ドメインにより活性化される転写制御エレメントに作動可能に連結している、本発明1040の宿主細胞。
[本発明1042]
本発明1001〜1032のいずれかのキメラNotch受容体ポリペプチドを発現する細胞の活性を調節する方法であって、
前記細胞と前記特異的結合対の第二のメンバーを接触させること
を含み、前記特異的結合対の前記第一のメンバーの、前記特異的結合対の前記第二のメンバーへの結合が、前記Notch受容体ポリペプチドの前記1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより前記細胞内ドメインが放出され、前記細胞内ドメインの放出が、前記細胞の活性を調節する、前記方法。
[本発明1043]
前記接触が、in vivo、ex vivo、またはin vitroで実行される、本発明1042の方法。
[本発明1044]
前記特異的結合対の前記第二のメンバーが、第二の細胞の表面上にあるか、不溶性基体上に固定されているか、細胞外マトリックス中に存在するか、人工マトリックス中に存在するか、または可溶性である、本発明1042の方法。
[本発明1045]
前記細胞内ドメインの放出が、前記細胞の増殖を調節する、本発明1042の方法。
[本発明1046]
前記細胞内ドメインの放出が、前記細胞におけるアポトーシスを調節する、本発明1042の方法。
[本発明1047]
前記細胞内ドメインの放出が、アポトーシス以外の機構による細胞死を誘導する、本発明1042の方法。
[本発明1048]
前記細胞内ドメインの放出が、前記細胞における遺伝子発現を、転写調節、クロマチン制御、翻訳、トラフィッキングまたは翻訳後プロセシングを通して調節する、本発明1042の方法。
[本発明1049]
前記細胞内ドメインの放出が、前記細胞の分化を調節する、本発明1042の方法。
[本発明1050]
前記細胞内ドメインの放出が、前記細胞の遊走を調節する、本発明1042の方法。
[本発明1051]
前記細胞内ドメインの放出が、前記細胞からの分子の発現及び分泌を調節する、本発明1042の方法。
[本発明1052]
前記細胞内ドメインの放出が、前記細胞の、第二の細胞へのまたは細胞外マトリックスへの接着を調節する、本発明1042の方法。
[本発明1053]
前記細胞内ドメインの放出が、前記細胞における遺伝子産物のデノボ発現を誘導するまたは発現を調節する、本発明1042の方法。
[本発明1054]
前記遺伝子産物が、転写活性化因子、転写抑制因子、キメラ抗原受容体、第二のキメラNotch受容体ポリペプチド、翻訳調節因子、サイトカイン、ホルモン、ケモカイン、または抗体である、本発明1053の方法。
[本発明1055]
本発明1001〜1032のいずれかのキメラNotch受容体ポリペプチドを発現する細胞の活性を調節する方法であって、
前記細胞と前記特異的結合対の第二のメンバーを接触させること
を含み、ここで前記特異的結合対の前記第一のメンバーの、前記特異的結合対の前記第二のメンバーへの結合が、前記Notch受容体ポリペプチドの前記1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより前記細胞内ドメインが放出され、前記細胞内ドメインが、前記細胞の活性を調節するエフェクターポリペプチドをコードする核酸の転写を誘導する転写因子である、前記方法。
[本発明1056]
前記接触が、in vivo、ex vivo、またはin vitroで実行される、本発明1055の方法。
[本発明1057]
前記特異的結合対の前記第二のメンバーが、第二の細胞の表面上にあるか、不溶性基体上に固定されているか、細胞外マトリックス中に存在するか、人工マトリックス中に存在するか、または可溶性である、本発明1055の方法。
[本発明1058]
前記エフェクターポリペプチドが、アポトーシス誘導因子、抑制因子にあるアポトーシス、活性化免疫受容体、抑制性免疫受容体、転写活性化因子、転写抑制因子、サイトカイン、成長因子、ホルモン、受容体、抗体、部位特異的ヌクレアーゼまたはリコンビナーゼである、本発明1055の方法。
[本発明1059]
細胞の活性を調節する方法であって、
前記細胞と第一の特異的結合対の第二のメンバーを接触させること
を含み、前記細胞が、
i)第一の特異的結合対の第一のメンバーを含む、本発明1001〜1032のいずれかの第一のキメラNotch受容体ポリペプチド;及び
ii)少なくとも、第二の特異的結合対の第一のメンバーを含む本発明1001〜1032のいずれかの第二のキメラNotch受容体ポリペプチド
を発現し、
前記第一及び前記第二の特異的結合対が互いに異なり、
前記第一のキメラNotch受容体ポリペプチドの前記細胞内ドメインが、第一のエフェクター機能を提供し;前記第二のキメラNotch受容体ポリペプチドの前記細胞内ドメインが、前記第一のエフェクター機能と異なる第二のエフェクター機能を提供し、前記放出された第一及び前記第二の細胞内ドメインが、前記細胞の活性を調節する、前記方法。
[本発明1060]
前記接触が、in vivo、ex vivo、またはin vitroで実行される、本発明1059の方法。
[本発明1061]
T細胞を活性化させる方法であって、
本発明1040のT細胞と固定化抗原を接触させること
を含み、
前記キメラポリペプチドの前記細胞外ドメインが、第一の抗原に特異的な抗体を含み、
前記接触が、前記転写活性化因子の放出、及び前記細胞中の前記CARまたは前記TCRの産生をもたらし、前記CARまたはTCRが、第二の抗原の結合後に前記T細胞の活性化を提供する、前記方法。
[本発明1062]
細胞の活性を調節する方法であって、
前記細胞と第一の特異的結合対の第二のメンバーを接触させること
を含み、前記細胞が、
i)第一の特異的結合対の第一のメンバーを含む、本発明1001〜1032のいずれかの第一のキメラNotch受容体ポリペプチド;及び
ii)少なくとも、第二の特異的結合対の第一のメンバーを含む本発明1001〜1032のいずれかの第二のキメラNotch受容体ポリペプチド
を発現し、
前記第一及び前記第二の特異的結合対が互いに異なり、
前記第二のキメラNotch受容体をコードする前記ヌクレオチド配列が、前記第一のキメラNotch受容体ポリペプチドの前記細胞内ドメインにより活性化または抑制される転写制御エレメントに作動可能に連結している、前記方法。
[本発明1063]
前記接触が、in vivo、ex vivo、またはin vitroで実行される、本発明1062の方法。
[本発明1064]
T細胞を活性化させる方法であって、
本発明1040のT細胞と固定化抗原を接触させること
を含み、前記キメラポリペプチドの前記細胞外ドメインが、第一の抗原に特異的な抗体を含み、
前記接触が、前記転写活性化因子の放出、及び前記細胞における前記CARまたはTCRの産生をもたらし、前記CARまたは前記TCRが、第二の抗原の結合後に前記T細胞の活性化を提供する、前記方法。
[本発明1065]
細胞の活性を調節する方法であって、
前記細胞と表面上に固定されている抗原を接触させること
を含み、前記細胞が、本発明1001のキメラNotch受容体ポリペプチドを発現し、前記特異的結合対の前記第一のメンバーが前記抗原と結合し、
前記接触が、前記細胞内ドメインの放出及び前記細胞の活性の調節をもたらす、前記方法。
[本発明1066]
前記細胞内ドメインが、前記細胞の分化を調節する転写因子である、本発明1065の方法。
[本発明1067]
細胞の活性を局所的に調節する方法であって、
前記細胞内で、特異的結合対の第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む結合誘発型転写スイッチを発現させること;及び
前記細胞と前記特異的結合対の第二のメンバーを接触させること
を含み、前記特異的結合対の前記第一のメンバーの、前記特異的結合対の前記第二のメンバーへの結合が、前記細胞内ドメインを活性化するための結合シグナルを伝達するように前記結合トランスデューサを誘導し、それにより、活性化した細胞内ドメインが産生され、前記活性化した細胞内ドメインが、前記細胞の遺伝子産物の発現、前記細胞の増殖、前記細胞のアポトーシス、前記細胞の非アポトーシス性死、前記細胞の分化、前記細胞の脱分化、前記細胞の遊走、前記細胞からの分子の分泌及び前記細胞の細胞接着からなる群より選択される、前記細胞の活性を調節する、前記方法。
[本発明1068]
前記活性化した細胞内ドメインが、前記細胞の内在性遺伝子産物の発現を調節する、本発明1067の方法。
[本発明1069]
前記細胞の前記内在性遺伝子産物が、ケモカイン、ケモカイン受容体、サイトカイン、サイトカイン受容体、分化因子、成長因子、成長因子受容体、ホルモン、代謝酵素、増殖誘導因子、受容体、小分子セカンドメッセンジャー合成酵素、T細胞受容体、転写活性化因子、転写抑制因子、転写活性化因子、転写抑制因子、翻訳調節因子、翻訳活性化因子、翻訳抑制因子、活性化免疫受容体、抑制因子にあるアポトーシス、アポトーシス誘導因子、免疫活性化因子、免疫抑制因子及び抑制性免疫受容体からなる群より選択される、本発明1068の方法。
[本発明1070]
前記細胞の前記内在性遺伝子産物が、分泌された遺伝子産物である、本発明1068〜1069のいずれかの方法。
[本発明1071]
前記細胞の前記内在性遺伝子産物が、表面発現遺伝子産物である、本発明1068〜1069のいずれかの方法。
[本発明1072]
前記活性化した細胞内ドメインが、前記細胞の2つ以上の内在性遺伝子産物の発現を同時に調節する、本発明1068〜1071のいずれかの方法。
[本発明1073]
前記活性化した細胞内ドメインが、前記細胞の異種遺伝子産物の発現を調節する、本発明1067の方法。
[本発明1074]
前記細胞の前記異種遺伝子産物が、ケモカイン、ケモカイン受容体、キメラ抗原受容体、サイトカイン、サイトカイン受容体、分化因子、成長因子、成長因子受容体、ホルモン、代謝酵素、病原体由来タンパク質、増殖誘導因子、受容体、RNA誘導型ヌクレアーゼ、部位特異的ヌクレアーゼ、小分子セカンドメッセンジャー合成酵素、T細胞受容体、毒素由来タンパク質、転写活性化因子、転写抑制因子、転写活性化因子、転写抑制因子、翻訳調節因子、翻訳活性化因子、翻訳抑制因子、活性化免疫受容体、抗体、抑制因子にあるアポトーシス、アポトーシス誘導因子、操作T細胞受容体、免疫活性化因子、免疫抑制因子、抑制性免疫受容体、RNA誘導型DNA結合タンパク質及び第二の結合誘発型転写スイッチからなる群より選択される、本発明1073の方法。
[本発明1075]
前記細胞の前記異種遺伝子産物が、抗体である、本発明1074の方法。
[本発明1076]
前記抗体が、806、9E10、3F8、81C6、8H9、アバゴボマブ、アバタセプト、アブシキシマブ、アビツズマブ、アブリルマブ、アクトクスマブ、アダリムマブ、アデカツムマブ、アデュカヌマブ、アフェリモマブ、アフツズマブ、アラシズマブ・ペゴル、ALD518、アレファセプト、アレムツズマブ、アリロクマブ、アルツモマブ・ペンテテート、アマツキシマブ、AMG 102、アナツモマブ・マフェナトクス、アネツマブ・ラブタンシン、アニフロルマブ、アンルキンズマブ、アポリズマブ、アルシツモマブ、アスクリンバクマブ(Ascrinvacumab)、アセリズマブ、アタシセプト、アテゾリズマブ、アチヌマブ、アトリズマブ/トシリズマブ、アトロリムマブ、AVE1642、バピネオズマブ、バシリキシマブ、バビツキシマブ、ベクツモマブ、ベゲロマブ、ベリムマブ、ベンラリズマブ、ベルチリムマブ、ベシレソマブ、ベバシズマブ、ベズロトクスマブ、ビシロマブ、ビマグルマブ、ビメキズマブ(Bimekizumab)、ビバツズマブ・メルタンシン、ブリナツモマブ、ブロソズマブ、BMS−936559、ボコシズマブ、ブレンツキシマブ・ベドチン、ブリアキヌマブ、ブロダルマブ、ブロルシズマブ、ブロンチクツズマブ(Brontictuzumab)、カナキヌマブ、カンツズマブ・メルタンシン、カンツズマブ・ラブタンシン、カプラシズマブ、カプロマブ・ペンデチド、カルルマブ、カツマクソマブ、cBR96−ドキソルビシン免疫複合体、CC49、CDP791、セデリズマブ、セルトリズマブ・ペゴル、セツキシマブ、cG250、Ch.14.18、シタツズマブ・ボガトクス、シクスツムマブ、クラザキズマブ、クレノリキシマブ、クリバツズマブ・テトラキセタン、コドリツズマブ、コルツキシマブ・ラブタンシン(Coltuximab ravtansine)、コナツムマブ、コンシズマブ、CP 751871、CR6261、クレネズマブ、CS−1008、ダセツズマブ、ダクリズマブ、ダロツズマブ、ダピロリズマブ・ペゴル(Dapirolizumab pegol)、ダラツムマブ、デクトレクマブ(Dectrekumab)、デムシズマブ、デニンツズマブ・マホドチン(Denintuzumab mafodotin)、デノスマブ、デルロツキシマブ・ビオチン(Derlotuximab biotin)、デツモマブ、ジヌツキシマブ、ジリダブマブ、ドルリモマブ・アリトクス、ドロジツマブ、デュリゴツマブ、デュピルマブ、デュルバルマブ、デュシジツマブ、エクロメキシマブ、エクリズマブ、エドバコマブ、エドレコロマブ、エファリズマブ、エフングマブ、エルデルマブ、エルゲムツマブ(Elgemtumab)、エロツズマブ、エルシリモマブ、エマクツズマブ(Emactuzumab)、エミベツズマブ、エナバツズマブ、エンフォルツマブ・ベドチン、エンリモマブ・ペゴル、エノブリツズマブ(Enoblituzumab)、エノキズマブ、エノチクマブ、エンシツキシマブ、エピツモマブ・シツキセタン、エプラツズマブ、エルリズマブ、エルツマクソマブ、エタネルセプト、エタラシズマブ、エトロリズマブ、エビナクマブ、エボロクマブ、エクスビビルマブ、F19、ファノレソマブ、ファラリモマブ、ファーレツズマブ、ファシヌマブ、FBTA05、フェルビズマブ、フェザキヌマブ、フィクラツズマブ、フィジツムマブ、フィリブマブ(Firivumab)、フランボツマブ、フレチクマブ、フォントリズマブ、フォラルマブ、フォラビルマブ、フレソリムマブ、フルラヌマブ、フツキシマブ、ガリキシマブ、ガニツマブ、ガンテネルマブ、ガビリモマブ、ゲムツズマブ・オゾガマイシン、ゲボキズマブ、ギレンツキシマブ、グレムバツムマブ・ベドチン、ゴリムマブ、ゴミリキシマブ、グセルクマブ、HGS−ETR2、hu3S193、huA33、イバリズマブ、イブリツモマブ・チウキセタン、イクルクマブ、イダルシズマブ、IGN101、IgN311、イゴボマブ、IIIA4、IM−2C6、IMAB362、イマルマブ(Imalumab)、IMC−A12、イムシロマブ、イムガツズマブ、インクラクマブ、インダツキシマブ・ラブタンシン、インデュサツマブ・ベドチン(Indusatumab vedotin)、インフリキシマブ、イノリモマブ、イノツズマブ・オゾガマイシン、インテツムマブ、イピリムマブ、イラツムマブ、イサツキシマブ、イトリズマブ、イクセキズマブ、J591、KB004、ケリキシマブ、KW−2871、ラベツズマブ、ラムブロリズマブ、ラムパリズマブ、レブリキズマブ、レマレソマブ、レンジルマブ(Lenzilumab)、レルデリムマブ、レキサツムマブ、リビビルマブ、リファスツズマブ・ベドチン、リゲリズマブ、リロトマブ・サテトラキセタン(Lilotomab satetraxetan)、リンツズマブ、リリルマブ、ロデルシズマブ、ロキベトマブ(Lokivetmab)、ロルボツズマブ・メルタンシン、ルカツムマブ、ルリズマブ・ペゴル、ルミリキシマブ、ルムレツズマブ(Lumretuzumab)、マパツムマブ、マルゲツキシマブ、マスリモマブ、マツズマブ、マブリリムマブ、MEDI4736、メポリズマブ、メテリムマブ、METMAB、ミラツズマブ、ミンレツモマブ、ミルベツキシマブ・ソラブタンシン(Mirvetuximab soravtansine)、ミツモマブ、MK−0646、MK−3475、MM−121、モガムリズマブ、MORAb−003、モロリムマブ、モタビズマブ、MOv18、モキセツモマブ・パスドトクス、MPDL33280A、ムロモナブCD3、ナコロマブ・タフェナトクス、ナミルマブ、ナプツモマブ・エスタフェナトクス、ナルナツマブ、ナタリズマブ、ネバクマブ、ネシツムマブ、ネモリズマブ、ネレリモマブ、ネスバクマブ、ニモツズマブ、ニボルマブ、ノフェツモマブ・メルペンタン、オビルトキサキシマブ、オビヌツズマブ、オカラツズマブ、オクレリズマブ、オヅリモマブ、オファツムマブ、オララツマブ、オロキズマブ、オマリズマブ、オナルツズマブ、オンツキシズマブ、オピシヌマブ、オポルツズマブ・モナトクス、オレゴボマブ、オルチクマブ、オテリキシズマブ、オトレルツズマブ、オキセルマブ、オザネズマブ、オゾラリズマブ、パギバキシマブ、パリビズマブ、パニツムマブ、パンコマブ(Pankomab)、パノバクマブ、パルサツズマブ、パスコリズマブ、パソツキシズマブ、パテクリズマブ、パトリツマブ、ペムブロリズマブ、ペムツモマブ、ペラキズマブ、ペルツズマブ、ペキセリズマブ、ピジリズマブ、ピナツズマブ・ベドチン、ピンツモマブ、プラクルマブ、ポラツズマブ・ベドチン、ポネズマブ、プリリキシマブ、プリトキサキシマブ、プリツムマブ、PRO 140、キリズマブ、R1507、ラコツモマブ、ラドレツマブ、ラフィビルマブ、ラルパンシズマブ(Ralpancizumab)、ラムシルマブ、ラニビズマブ、ラキシバクマブ、レファネズマブ(Refanezumab)、レガビルマブ、レスリズマブ、リロツムマブ、リヌクマブ(Rinucumab)、リツキシマブ、ロバツムマブ、ロレヅマブ、ロモソズマブ、ロンタリズマブ、ロベリズマブ、ルプリズマブ、サシツズマブ・ゴビテカン、サマリズマブ、サリルマブ、サツモマブ・ペンデチド、SCH 900105、セクキヌマブ、セリバンツマブ、セトキサキシマブ、セビルマブ、SGN−CD19A、SGN−CD33A、シブロツズマブ、シファリムマブ、シルツキシマブ、シムツズマブ、シプリズマブ、シルクマブ、ソフィツズマブ・ベドチン、ソラネズマブ、ソリトマブ、ソネプシズマブ、ソンツズマブ、スタムルマブ、スレソマブ、スビズマブ、タバルマブ、タカツズマブ・テトラキセタン、タドシズマブ、タリズマブ、タネズマブ、タプリツモマブ・パプトクス、タレクスツマブ、テフィバズマブ、テリモマブ・アリトクス、テナツモマブ、テネリキシマブ、テプリズマブ、テプロツムマブ、テシドルマブ、テツロマブ(Tetulomab)、TGN1412、チシリムマブ/トレメリムマブ、チガツズマブ、チルドラキズマブ、TNX−650、トシリズマブ、トラリズマブ、トサトクスマブ(Tosatoxumab)、トシツモマブ、トベツマブ、トラロキヌマブ、トラスツズマブ、TRBS07、トレガリズマブ、トレメリムマブ、トレボグルマブ(Trevogrumab)、ツコツズマブ・セルモロイキン、ツビルマブ、ウブリツキシマブ、ウロクプルマブ(Ulocuplumab)、ウレルマブ、ウルトキサズマブ、ウステキヌマブ、バンドルツズマブ・ベドチン、バンチクツマブ、バヌシズマブ、バパリキシマブ、バルリルマブ、バテリズマブ、ベドリズマブ、ベルツズマブ、ベパリモマブ、ベセンクマブ、ビジリズマブ、ボロシキシマブ、ボルセツズマブ・マホドチン、ボツムマブ、ザルツムマブ、ザノリムマブ、ザツキシマブ、ジラリムマブ及びゾリモマブ・アリトクスからなる群より選択される、本発明1075の方法。
[本発明1077]
前記細胞の前記異種遺伝子産物が、分泌された遺伝子産物である、本発明1073〜1076のいずれかの方法。
[本発明1078]
前記細胞の前記異種遺伝子産物が、表面発現遺伝子産物である、本発明1073〜1076のいずれかの方法。
[本発明1079]
前記活性化した細胞内ドメインが、前記細胞の2つ以上の異種遺伝子産物の発現を同時に調節する、本発明1073〜1078のいずれかの方法。
[本発明1080]
前記接触が、in vivo、ex vivo、またはin vitroで実行される、本発明1067〜1079のいずれかの方法。
[本発明1081]
前記特異的結合対の前記第二のメンバーが、第二の細胞の表面上にあるか、不溶性基体上に固定されているか、細胞外マトリックス中に存在するか、人工マトリックス中に存在するか、または可溶性である、本発明1067〜1080のいずれかの方法。
[本発明1082]
前記細胞内ドメインが、前記細胞の分化を調節する転写因子である、本発明1067の方法。
[本発明1083]
前記転写因子が、前記細胞の分化を直接調節する、本発明1082の方法。
[本発明1084]
前記転写因子が、前記細胞の分化を、第二の転写因子の発現を調節することにより間接的に調節する、本発明1082の方法。
[本発明1085]
前記細胞が免疫細胞であり、前記細胞の前記活性が、前記免疫細胞の分化である、本発明1067の方法。
[本発明1086]
前記細胞が免疫細胞であり、前記細胞内ドメインが、前記細胞の分化を調節する転写因子であり、前記細胞の前記活性が、前記免疫細胞の分化である、本発明1067の方法。
[本発明1087]
前記転写因子が、前記免疫細胞の分化を直接調節する、本発明1086の方法。
[本発明1088]
前記転写因子が、前記免疫細胞の分化を、第二の転写因子の前記発現を調節することにより間接的に調節する、本発明1086の方法。
[本発明1089]
前記細胞が幹細胞であり、前記細胞の前記活性が、前記幹細胞の分化である、本発明1067の方法。
[本発明1090]
前記細胞が始原細胞または前駆細胞であり、前記細胞の前記活性が、前記始原細胞または前駆細胞の分化である、本発明1067の方法。
[本発明1091]
前記細胞内ドメインの活性化が、前記細胞の内在性遺伝子の発現を、転写調節、クロマチン制御、翻訳、トラフィッキングまたは翻訳後プロセシングを通して調節する、本発明1067の方法。
[本発明1092]
前記細胞内ドメインの活性化が、前記細胞の、第二の細胞へのまたは細胞外マトリックスへの細胞接着を調節する、本発明1067の方法。
[本発明1093]
前記結合トランスデューサが、リガンド誘導性タンパク質分解切断部位を含み、前記特異的結合対の前記第一のメンバーの、前記特異的結合対の前記第二のメンバーへの結合が、前記結合トランスデューサの前記リガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより、前記結合シグナルが伝達されかつ、前記細胞内ドメインが、前記細胞内ドメインをタンパク質分解的に放出することによって活性化される、本発明1067の方法。
[本発明1094]
前記細胞と、前記特異的結合対の前記第一のメンバーの前記特異的結合対の前記第二のメンバーへの結合を競合的に阻害する可溶性阻害分子とを接触させることをさらに含み、それにより、前記細胞内ドメインを活性化させるための結合シグナルを伝達するように結合トランスデューサを誘導することを防ぐ、本発明1067〜1093のいずれかの方法。
[本発明1095]
前記細胞と前記可溶性阻害分子の接触が、前記可溶性阻害分子を第一の細胞に適用または投与することを含む、本発明1094の方法。
[本発明1096]
前記細胞と前記可溶性阻害分子の接触が、前記細胞を、前記可溶性阻害分子を発現する第二の細胞の存在下に置くことを含む、本発明1094の方法。
[本発明1097]
前記第二の細胞が、前記可溶性阻害分子を構成的に発現する、本発明1096の方法。
[本発明1098]
前記第二の細胞が、前記可溶性阻害分子を条件的に発現する、本発明1096の方法。
[本発明1099]
細胞の活性を調節する方法であって、
前記細胞と第一の特異的結合対の第二のメンバー及び第二の特異的結合対の第二のメンバーを接触させること
を含み、前記細胞が、
i)前記第一の特異的結合対の第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む第一の結合誘発型転写スイッチ;及び
ii)少なくとも、第二の特異的結合対の前記第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む第二の結合誘発型転写スイッチ
を発現し;
前記第一の結合誘発型転写スイッチの前記細胞内ドメインが、第一のエフェクター機能を提供し、前記第二の結合誘発型転写スイッチの前記細胞内ドメインが、前記第一のエフェクター機能と異なる第二のエフェクター機能を提供し、この時、前記第一及び第二の特異的結合対の前記第一及び第二のメンバーの結合が、前記第一及び第二の細胞内ドメインを活性化するための結合シグナルを伝達するように前記結合トランスデューサを誘導する、前記方法。
[本発明1100]
前記第一の結合誘発型転写スイッチの前記細胞内ドメインの前記エフェクター機能が、前記細胞の遺伝子産物の発現を調節する、本発明1099の方法。
[本発明1101]
前記細胞の前記遺伝子産物が、前記細胞の内在性遺伝子産物である、本発明1100の方法。
[本発明1102]
前記細胞の前記遺伝子産物が、前記細胞の異種遺伝子産物である、本発明1100の方法。
[本発明1103]
前記細胞の前記遺伝子産物が、ケモカイン、ケモカイン受容体、サイトカイン、サイトカイン受容体、分化因子、成長因子、成長因子受容体、ホルモン、代謝酵素、増殖誘導因子、受容体、小分子セカンドメッセンジャー合成酵素、T細胞受容体、転写活性化因子、転写抑制因子、転写活性化因子、転写抑制因子、翻訳調節因子、翻訳活性化因子、翻訳抑制因子、活性化免疫受容体、抑制因子にあるアポトーシス、アポトーシス誘導因子、免疫活性化因子、免疫抑制因子及び抑制性免疫受容体からなる群より選択される、本発明1100の方法。
[本発明1104]
前記第二の結合誘発型転写スイッチの前記細胞内ドメインの前記エフェクター機能が、前記細胞の遺伝子産物の発現を調節する、本発明1100の方法。
[本発明1105]
前記細胞の前記遺伝子産物が、前記細胞の内在性遺伝子産物である、本発明1104の方法。
[本発明1106]
前記細胞の前記遺伝子産物が、前記細胞の異種遺伝子産物である、本発明1104の方法。
[本発明1107]
前記細胞の前記遺伝子産物が、ケモカイン、ケモカイン受容体、サイトカイン、サイトカイン受容体、分化因子、成長因子、成長因子受容体、ホルモン、代謝酵素、増殖誘導因子、受容体、小分子セカンドメッセンジャー合成酵素、T細胞受容体、転写活性化因子、転写抑制因子、転写活性化因子、転写抑制因子、翻訳調節因子、翻訳活性化因子、翻訳抑制因子、活性化免疫受容体、抑制因子にあるアポトーシス、アポトーシス誘導因子、免疫活性化因子、免疫抑制因子及び抑制性免疫受容体からなる群より選択される、本発明1104の方法。
[本発明1108]
前記第一及び第二の結合誘発型転写スイッチの前記結合トランスデューサのうちの少なくとも1つが、リガンド誘導性タンパク質分解切断部位を含み、それぞれの特異的結合対の前記第一及び第二のメンバーの結合が、前記結合トランスデューサの前記リガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより、前記結合シグナルが伝達されかつ、それぞれの細胞内ドメインが、前記細胞内ドメインをタンパク質分解的に放出することによって活性化される、本発明1099の方法。
[本発明1109]
前記第一及び第二の結合誘発型転写スイッチの前記結合トランスデューサが、両方ともリガンド誘導性タンパク質分解切断部位を含む、本発明1108の方法。
[本発明1110]
細胞の活性を調節する方法であって、
前記細胞と第一の特異的結合対の第二のメンバーを接触させること
を含み、前記細胞が、
i)前記第一の特異的結合対の第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む第一の結合誘発型転写スイッチ;及び
ii)少なくとも、第二の特異的結合対の前記第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む第二の結合誘発型転写スイッチ
を発現し、
前記第二の結合誘発型転写スイッチをコードするヌクレオチド配列が、前記第一の結合誘発型転写スイッチの前記細胞内ドメインにより活性化または抑制される転写制御エレメントに作動可能に連結している、前記方法。
[本発明1111]
前記接触が、in vivo、ex vivo、またはin vitroで実行される、本発明1110の方法。
[本発明1112]
前記第一の特異的結合対の前記第二のメンバーが、第二の細胞の表面上にあるか、不溶性基体上に固定されているか、細胞外マトリックス中に存在するか、人工マトリックス中に存在するか、または可溶性である、本発明1110の方法。
[本発明1113]
前記第二の結合誘発型転写スイッチの前記細胞内ドメインの活性化が、前記細胞の遺伝子産物の発現、前記細胞の増殖、前記細胞のアポトーシス、前記細胞の非アポトーシス性死、前記細胞の分化、前記細胞の脱分化、前記細胞の遊走、前記細胞からの分子の分泌及び前記細胞の細胞接着からなる群より選択される、前記細胞の活性を調節する、本発明1110の方法。
[本発明1114]
前記細胞の前記活性が、前記細胞の遺伝子産物の発現である、本発明1113の方法。
[本発明1115]
前記細胞の前記遺伝子産物が、ケモカイン、ケモカイン受容体、サイトカイン、サイトカイン受容体、分化因子、成長因子、成長因子受容体、ホルモン、代謝酵素、増殖誘導因子、受容体、小分子セカンドメッセンジャー合成酵素、T細胞受容体、転写活性化因子、転写抑制因子、転写活性化因子、転写抑制因子、翻訳調節因子、翻訳活性化因子、翻訳抑制因子、活性化免疫受容体、抑制因子にあるアポトーシス、アポトーシス誘導因子、免疫活性化因子、免疫抑制因子及び抑制性免疫受容体からなる群より選択される、前記細胞の遺伝子産物である、本発明1114の方法。
[本発明1116]
前記第一及び第二の結合誘発型転写スイッチの前記結合トランスデューサの少なくとも1つが、リガンド誘導性タンパク質分解切断部位を含み、前記それぞれの特異的結合対の前記第一及び第二のメンバーの結合が、前記結合トランスデューサの前記リガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより、前記結合シグナルが伝達されかつ、前記それぞれの細胞内ドメインが、前記細胞内ドメインをタンパク質分解的に放出することによって活性化される、本発明1110の方法。
[本発明1117]
細胞間接触を追跡する方法であって、
第一の複数の細胞の各細胞において、特異的結合対の第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む結合誘発型転写スイッチを発現させること;
第二の複数の細胞の各細胞において、前記特異的結合対の第二のメンバーを発現させること;及び
前記第一の複数の細胞と前記第二の複数の細胞を接触させること
を含み、前記特異的結合対の前記第一のメンバーの、前記特異的結合対の前記第二のメンバーへの結合が、前記結合誘発型転写スイッチの結合シグナルを伝達するように結合トランスデューサを誘導し、それにより前記細胞内ドメインが活性化され、前記細胞内ドメインの活性化が、空間的な、時間的なまたはその組み合わせにおける細胞間接触を追跡するのに十分な検出可能レポーターの発現を誘導する、前記方法。
[本発明1118]
前記第一の複数の細胞、前記第二の複数の細胞または両方が、ニューロンである、本発明1117の方法。
[本発明1119]
前記結合トランスデューサが、リガンド誘導性タンパク質分解切断部位を含み、前記特異的結合対の前記第一のメンバーの、前記特異的結合対の前記第二のメンバーへの結合が、前記結合トランスデューサの前記リガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより、前記結合シグナルが伝達されかつ、前記細胞内ドメインが、前記細胞内ドメインをタンパク質分解的に放出することによって活性化される、本発明1117の方法。
[本発明1120]
前記結合誘発型転写スイッチが、SynNotchポリペプチドである、本発明1067〜1119のいずれかの方法。
[本発明1121]
局在性細胞活性化システムであって、
第一の特異的結合対の第一のメンバーを含む細胞外ドメインと、結合トランスデューサと、細胞内ドメインとを含む発現された結合誘発型転写スイッチ;及び
前記第一の結合誘発型転写スイッチの前記細胞内ドメインにより誘導される転写制御エレメントに作動可能に連結した、第二の特異的結合対の第一のメンバーを含む結合誘発型活性化ポリペプチドをコードする核酸
を含む細胞を含み、
前記第一の特異的結合対の前記第二のメンバーとの接触の際に、前記結合誘発型活性化ポリペプチドが発現され、前記第二の特異的結合対の前記第二のメンバーとの接触の際に、前記結合誘発型活性化ポリペプチドが前記細胞を活性化させる、前記システム。
[本発明1122]
前記細胞が、免疫細胞、始原細胞または前駆細胞、幹細胞、及びニューロンからなる群より選択される、本発明1121のシステム。
[本発明1123]
前記細胞が免疫細胞であり、前記結合誘発型転写スイッチが抗原誘発型転写スイッチであり、前記結合誘発型活性化ポリペプチドが抗原誘発型活性化ポリペプチドであり、前記第一の特異的結合対の前記第二のメンバーとの接触の際に、前記抗原誘発型活性化ポリペプチドが発現され、前記第二の特異的結合対の前記第二のメンバーとの接触の際に、前記抗原誘発型活性化ポリペプチドが前記免疫細胞を活性化させて、前記第二の特異的結合対の前記第一のメンバーを発現する標的細胞を認識する、本発明1122のシステム。
[本発明1124]
前記抗原誘発型活性化ポリペプチドが、キメラ抗原受容体またはその変異体である、本発明1123のシステム。
[本発明1125]
前記抗原誘発型活性化ポリペプチドが、操作T細胞受容体またはその変異体である、本発明1123のシステム。
[本発明1126]
前記発現された結合誘発型転写スイッチが、SynNotchポリペプチドである、本発明1121〜1125のいずれかのシステム。
[本発明1127]
細胞の活性を局所的に調節する方法であって、
第一の細胞において、結合トランスデューサと、細胞内ドメインと、可溶性アダプター分子上の第一のエピトープと特異的に結合する第一のアダプター結合ドメインを含む第一の細胞外ドメインとを含む結合誘発型転写スイッチを発現させること;
前記第一の細胞と:
i)前記可溶性アダプター分子上の第二のエピトープと特異的に結合する第二のアダプター結合ドメインを含む第二の細胞外ドメインを発現する第二の細胞;及び
ii)有効濃度の前記可溶性アダプター分子
を接触させること
を含み、前記第一のアダプター結合ドメイン及び前記第二のアダプター結合ドメインの、前記アダプター分子への結合が、前記細胞内ドメインを活性化するための結合シグナルを伝達するように前記結合トランスデューサを誘導し、それにより、活性化した細胞内ドメインが産生され、前記活性化した細胞内ドメインが、前記細胞の遺伝子産物の発現、前記細胞の増殖、前記細胞のアポトーシス、前記細胞の非アポトーシス性死、前記細胞の分化、前記細胞の脱分化、前記細胞の遊走、前記細胞からの分子の分泌及び前記細胞の細胞接着からなる群より選択される前記第一の細胞の活性を調節する、前記方法。
[本発明1128]
前記接触が、前記可溶性アダプター分子を前記細胞にin vitroまたはex vivoで適用することを含む、本発明1127の方法。
[本発明1129]
前記接触が、前記可溶性アダプター分子を前記細胞にin vivoで投与することを含む、本発明1127の方法。
[本発明1130]
前記第一の細胞と有効濃度の前記可溶性アダプター分子を接触させることが、前記第一の細胞を、前記アダプター分子を発現する第三の細胞の存在下に置くことを含む、本発明1127の方法。
[本発明1131]
前記第三の細胞が、前記アダプター分子を構成的に発現する、本発明1130の方法。
[本発明1132]
前記第三の細胞が、前記アダプター分子を条件的に発現する、本発明1130の方法。
[本発明1133]
前記第一の細胞外ドメイン及び前記可溶性アダプター分子が、特異的対合対の第一及び第二のメンバーである、本発明1127〜1132のいずれかの方法。
[本発明1134]
前記第二の細胞外ドメイン及び前記可溶性アダプター分子が、特異的結合対の第一及び第二のメンバーである、本発明1127〜1132のいずれかの方法。
[本発明1135]
前記第一の細胞外ドメイン及び第二の細胞外ドメインが、抗体である、本発明1127〜1134のいずれかの方法。
[本発明1136]
前記第一の細胞外ドメイン及び第二の細胞外ドメインの一方または両方が、ナノボディである、本発明1135の方法。
[本発明1137]
前記細胞内ドメインが、転写因子である、本発明1127〜1136のいずれかの方法。
[本発明1138]
前記活性化した細胞内ドメインが、前記第一の細胞の内在性遺伝子産物の発現を調節する、本発明1127〜1137のいずれかの方法。
[本発明1139]
前記活性化した細胞内ドメインが、前記第一の細胞の異種遺伝子産物の発現を調節する、本発明1127〜1137のいずれかの方法。
[本発明1140]
前記結合トランスデューサが、リガンド誘導性タンパク質分解切断部位を含み、前記第一の細胞外ドメイン及び前記第二の細胞外ドメインの、前記可溶性アダプター分子への結合が、前記結合トランスデューサの前記リガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより、前記結合シグナルが伝達されかつ、前記細胞内ドメインが、前記細胞内ドメインをタンパク質分解的に放出することによって活性化される、本発明1127〜1139のいずれかの方法。
[本発明1141]
第一の抗原に応答する第一の結合誘発型転写スイッチをコードする核酸;
前記第一の結合誘発型転写スイッチに応答し、かつ特異的結合対の第一のメンバーに特異的に結合する細胞外ドメインを含むCARをコードする核酸に作動可能に連結している、第一のプロモーター;
第二の抗原に応答する第二の結合誘発型転写スイッチをコードする核酸;及び
前記第二の結合誘発型転写スイッチに応答し、かつ細胞内CAR阻害ドメインをコードする核酸に作動可能に連結している、第二のプロモーター
を含み、前記第二の抗原の存在下で、前記細胞内CAR阻害ドメインが発現されて前記CARによる前記細胞の活性化が阻害され、前記第一の抗原の存在下かつ前記第二の抗原の非存在下で、前記特異的結合対の前記第二のメンバーによりCARが発現されて活性化可能である、宿主細胞。
[本発明1142]
第一の抗原に応答する第一の結合誘発型転写スイッチをコードする核酸;
前記第一の結合誘発型転写スイッチに応答し、かつ特異的結合対の第一のメンバーに特異的に結合する細胞外ドメインを含むCARの第一の部分をコードする核酸に作動可能に連結している、第一のプロモーター;
第二の抗原に応答する第二の結合誘発型転写スイッチをコードする核酸;及び
前記第二の結合誘発型転写スイッチに応答し、かつ細胞内シグナル伝達ドメインを含むCARの第二の部分をコードする核酸に作動可能に連結している、第二のプロモーター
を含み、前記第一の抗原及び第二の抗原の存在下で、前記CARの前記第一及び第二の部分が発現され、前記CARが前記特異的結合対の前記第二のメンバーにより活性化可能である、宿主細胞。
[本発明1143]
第三の抗原に応答する第三の結合誘発型転写スイッチをコードする核酸;及び
前記第三の結合誘発型転写スイッチに応答し、かつ細胞内CAR阻害ドメインをコードする核酸に作動可能に連結している、第三のプロモーター
をさらに含み、前記第三の抗原の存在下で、前記細胞内CAR阻害ドメインが発現されて前記CARによる前記細胞の活性化が阻害される、本発明1142の細胞。
「ポリヌクレオチド」及び「核酸」という用語は、本明細書で互換可能に使用され、リボヌクレオチドまたはデオキシリボヌクレオチドのいずれかの、任意の長さのヌクレオチドの重合形態を指す。ゆえに、この用語には、一本鎖、二本鎖、または複数鎖DNAもしくはRNA、ゲノムDNA、cDNA、DNA−RNAハイブリッド、あるいはプリンもしくはピリミジン塩基を含むポリマー、もしくは他の天然、化学的、生物学的に修飾された、非天然もしくは誘導化されたヌクレオチド塩基を含むポリマーを含むがこれらに限定されない。
本開示は、キメラNotch受容体ポリペプチド、キメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸、及び該核酸で遺伝子改変された宿主細胞を提供する。キメラNotch受容体ポリペプチドは、様々な用途において有用であり、それも提供する。
本開示は、キメラNotch受容体ポリペプチドを提供する。本開示のキメラNotch受容体ポリペプチドは、a)特異的結合対の第一のメンバーを含む細胞外ドメイン;b)Notch受容体ポリペプチド、ここでNotch受容体ポリペプチドは、50アミノ酸〜1000アミノ酸の長さを有し、1つまたは複数のリガンド誘導性タンパク質分解切断部位を含む;及びc)細胞内ドメインを含む。特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合が、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出される。細胞内ドメインの放出は、キメラNotch受容体ポリペプチドを産生する細胞の活性を調節する。細胞外ドメインは、特異的結合対の第一のメンバーを含み;特異的結合対の第一のメンバーは、Notch受容体ポリペプチドに対して異種であるアミノ酸配列を含む。細胞内ドメインは、Notch受容体ポリペプチドに対して異種であるアミノ酸配列を含む。
上記のように、本開示のキメラNotch受容体ポリペプチドは、細胞外ドメインを含む。細胞外ドメインは、特異的結合対の第一のメンバーを含む。特異的結合対の第一のメンバーは、特異的結合対の第二のメンバーに結合し、この場合、特異的結合対の第二のメンバーは、本開示のキメラNotch受容体ポリペプチドと異なるポリペプチドの上にある。特異的結合対の第二のメンバーは、特異的結合対の第一のメンバーを含む細胞外ドメインを含むキメラNotch受容体ポリペプチドとは離れている(例えば、共有結合していない)。特異的結合対の第二のメンバーは、細胞の表面上に存在することができる。特異的結合対の第二のメンバーは、不溶性支持体上に固定されることができる。特異的結合対の第二のメンバーは、可溶性であることができる。特異的結合対の第二のメンバーは、細胞外環境(例えば、細胞外マトリックス)中に存在することができる。特異的結合対の第二のメンバーは、人工マトリックス中に存在することができる。特異的結合対の第二のメンバーは、無細胞環境中に存在することができる。
抗原−抗体特異的結合対、ここで、第一のメンバーは、抗原である第二のメンバーに特異的に結合する抗体(または抗体をベースとする認識足場)であるか、あるいはここで、第一のメンバーは抗原であり、第二のメンバーは、抗原に特異的に結合する抗体(または抗体をベースとする認識足場)である;
リガンド−受容体特異的結合対、ここで、第一のメンバーはリガンドであり、第二のメンバーはリガンドが結合する受容体であるか、あるいはここで、第一のメンバーは受容体であり、第二のメンバーは、受容体に結合するリガンドである;
非抗体をベースとする認識足場−標的特異的結合対、ここで、第一のメンバーは、非抗体をベースとする認識足場であり、第二のメンバーは、非抗体をベースとする認識足場に結合する標的であるか、あるいはここで、第一のメンバーは、標的であり、第二のメンバーは標的に結合する非抗体をベースとする認識足場である;
接着分子−細胞外マトリックス結合対;
Fc受容体−Fc結合対、ここで、第一のメンバーは、Fc受容体である第二のメンバーに結合する免疫グロブリンFcであるか、あるいはここで、第一のメンバーは、免疫グロブリンFcを含む第二のメンバーに結合するFc受容体である;及び、
受容体−共受容体結合対、ここで、第一のメンバーは、共受容体である第二のメンバーに特異的に結合する受容体であるか、あるいはここで、第一のメンバーは、受容体である第二のメンバーに特異的に結合する共受容体である。
いくつかの場合では、特異的結合対の第一のメンバーは、抗体である。抗体は、任意の抗原結合性の抗体をベースとするポリペプチドであることができ、広く様々なそれが当該分野で公知である。いくつかの例では、抗原結合ドメインは、一本鎖Fv(scFv)である。他の抗体をベースとする認識ドメイン(cAb VHH(ラクダ科抗体可変ドメイン)及びヒト化版、IgNAR VH(サメ抗体可変ドメイン)及びヒト化版、sdAb VH(単一ドメイン抗体可変ドメイン)及び「ラクダ化」抗体可変ドメインが使用に好適である。いくつかの例では、T細胞受容体(TCR)をベースとする認識ドメイン、例えば、一本鎖TCR(scTv、VαVβを含有する一本鎖2ドメインTCR)も使用に好適である。
いくつかの場合では、特異的結合対の第一のメンバーは、非抗体をベースとする認識足場である。本開示のキメラNotch受容体ポリペプチドにおける特異的結合対のメンバーが、非抗体をベースとする認識足場である場合、キメラNotch受容体ポリペプチドは、特異的結合対の第二のメンバーの存在下で活性化することができ、この場合、特異的結合対の第二のメンバーは、非抗体をベースとする認識足場に結合する標的である。
いくつかの場合では、特異的結合対の第一のメンバーは、細胞接着分子(CAM)である、すなわち、細胞外マトリックス(ECM)の成分と結合するまたは細胞表面分子と結合するポリペプチドである。例えば、いくつかの場合では、特異的結合対の第一のメンバーは、CAMの細胞外領域である。いくつかの場合では、CAMは、カルシウム非依存性接着分子であり;例えば、いくつかの場合では、CAMは、免疫グロブリンスーパーファミリーCAMである。いくつかの場合では、CAMは、カルシウム依存性接着分子であり;例えば、CAMは、インテグリン、カドヘリン、またはセレクチンである。いくつかの場合では、特異的結合対の第一のメンバーは、インテグリンである。いくつかの場合では、特異的結合対の第一のメンバーは、カドヘリン、例えば、E−カドヘリン、P−カドヘリン、N−カドヘリン、R−カドヘリン、M−カドヘリン等である。いくつかの場合では、特異的結合対の第一のメンバーは、セレクチン、例えば、E−セレクチン、L−セレクチン、またはP−セレクチンである。CAMの結合性断片は、特異的結合対の第一のメンバーとして使用することができる。
いくつかの場合では、特異的結合対の第一のメンバーは、受容体に対するリガンドである。リガンドは、ポリペプチド、核酸、糖タンパク質、小分子、炭水化物、脂質、糖脂質、リポタンパク質、リポ多糖類等を含む。いくつかの場合では、リガンドは、可溶性である。
いくつかの場合では、特異的結合対の第一のメンバーは、抗体が特異的に結合する抗原である。抗原は、任意の抗原、例えば、天然に存在する(内在性)抗原;合成(例えば、天然に存在する抗原とはもはや同一ではないように改変された;その天然の状態から改変された;等)抗原;等であることができる。
いくつかの場合では、特異的結合対の第一のメンバーは、非抗体をベースとする足場の標的である。標的には、例えば、ポリペプチド、核酸、糖タンパク質、小分子、炭水化物、脂質、糖脂質、リポタンパク質、リポ多糖類等が含まれる。
いくつかの場合では、特異的結合対の第一のメンバーは、受容体である。いくつかの場合では、受容体は、成長因子受容体である。いくつかの場合では、受容体は、サイトカイン受容体である。いくつかの場合では、受容体は、細胞上の共受容体に結合する細胞表面受容体である。いくつかの場合では、受容体は、神経伝達物質受容体である。いくつかの場合では、受容体は、細胞外マトリックス成分に結合する。いくつかの場合では、受容体は、免疫グロブリンFc受容体である。
上記のように、本開示のキメラNotch受容体ポリペプチドは、50アミノ酸〜1000アミノ酸の長さを有し、1つまたは複数のリガンド誘導性タンパク質分解切断部位を含むNotch受容体ポリペプチドを含む。
いくつかの場合では、本開示のキメラNotch受容体ポリペプチド中に存在するNotch受容体ポリペプチドは、以下のアミノ酸配列:
いくつかの場合では、本開示のキメラNotch受容体ポリペプチド中に存在するNotch受容体ポリペプチドは、N末端からC末端の順で、i)LNR−Aセグメント;ii)LNR−Bセグメント;iii)LNR−Cセグメント;iv)HD−Nセグメント、v)HD−Cセグメント;及びvi)TMドメインを含む。LNR−Aセグメント、LNR−Bセグメント、及びLNR−Cセグメントは、まとめて、「LNRセグメント」と称することができる。かかるNotch受容体ポリペプチドを図4Aに概略的に示す。
いくつかの場合では、本開示のキメラNotch受容体ポリペプチド中に存在するNotch受容体ポリペプチドは、N末端からC末端の順で、i)単一のEGFリピート;ii)LNRセグメント;iii)HD−Nセグメント、iv)HD−Cセグメント;及びv)TMドメインを含む。かかるNotch受容体ポリペプチドを図4Bに概略的に示す。
いくつかの場合では、本開示のキメラNotch受容体ポリペプチド中に存在するNotch受容体ポリペプチドは、N末端からC末端の順で、i)2〜11個のEGFリピート;ii)LNRセグメント;iii)HD−Nセグメント、iv)HD−Cセグメント;及びv)TMドメインを含む。かかるNotch受容体ポリペプチドを図4Dに概略的に示す。
いくつかの場合では、Notch受容体ポリペプチドは、N末端からC末端の順で、i)HD−Cセグメント;及びii)TMドメインを含み、ここでNotch受容体ポリペプチドは、LNRセグメントを含まない。いくつかの場合では、LNRセグメントは、異種ポリペプチドと置換される。かかるNotch受容体ポリペプチドを図4Fに概略的に示す。
上記のように、本開示のキメラNotch受容体ポリペプチドは、50アミノ酸〜1000アミノ酸の長さを有し、1つまたは複数のリガンド誘導性タンパク質分解切断部位を含むNotch受容体ポリペプチドを含む。上に考察するように、本開示のキメラNotch受容体ポリペプチドは、a)特異的結合対の第一のメンバーを含む細胞外ドメイン;b)50アミノ酸〜1000アミノ酸の長さを有し、1つまたは複数のリガンド誘導性タンパク質分解切断部位を含むNotch受容体ポリペプチド;及びc)細胞内ドメインを含み、ここで、特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合が、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出される。特異的結合対の第二のメンバー(「リガンド」)は、接触する(例えば、「送信」)細胞上に存在することができる。
上記のように、本開示のキメラNotch受容体ポリペプチドは、細胞内ドメインを含み、これは、キメラNotch受容体ポリペプチドの、特異的結合対の第二のメンバーへの結合の後に放出され、ここで、キメラNotch受容体ポリペプチドの、特異的結合対の第二のメンバーへの結合は、上述のタンパク質分解切断部位の切断を誘導する。
本開示のキメラNotchポリペプチドにおける使用に好適な細胞内シグナル伝達ドメインには、DAP10/CD28型シグナル伝達鎖が含まれる。
本開示のキメラNotchポリペプチドにおける使用に好適な細胞内シグナル伝達ドメインには、ZAP70ポリペプチド、例えば、以下のアミノ酸配列:
いくつかの場合では、細胞内ドメインは、特異的結合対の第一のメンバーの特異的結合対の第二のメンバーへの結合の際に放出されるとき、キメラNotchポリペプチドを発現する細胞において、遺伝子産物の産生を誘導する、ポリペプチドである。例えば、いくつかの場合では、本開示のキメラNotch受容体ポリペプチドの細胞内ドメインは、特異的結合対の第一のメンバーの特異的結合対の第二のメンバーへの結合の際に放出されるとき、キメラNotchポリペプチドを発現する細胞において、遺伝子産物(ポリペプチド;核酸)の産生を誘導する。いくつかの場合では、遺伝子産物は、核酸である。いくつかの場合では、遺伝子産物は、ポリペプチドである。放出された細胞内ドメインにより誘導されるポリペプチド遺伝子産物には、内在性ポリペプチド(例えば、細胞により天然にコードされるポリペプチド)及び異種ポリペプチド(例えば、細胞により天然にはコードされないポリペプチド;細胞を遺伝子改変するために使用される異種核酸によりコードされるポリペプチド)が含まれる。放出された細胞内ドメインにより誘導されるポリペプチド遺伝子産物には、分泌ポリペプチドが含まれる。放出された細胞内ドメインにより誘導されるポリペプチド遺伝子産物には、細胞表面ポリペプチドが含まれる。放出された細胞内ドメインにより誘導されるポリペプチド遺伝子産物には、細胞内ポリペプチド(通常は細胞内に存在するポリペプチド、例えば、転写因子)が含まれる。放出された細胞内ドメインにより誘導されるポリペプチド遺伝子産物には、受容体、サイトカイン、ホルモン、成長因子、ケモカイン、細胞表面ポリペプチド、転写因子(例えば、転写活性化因子;転写抑制因子)、アポトーシス誘導因子、アポトーシス抑制因子、ドミナントネガティブ変異体等が含まれる。その産生が放出された細胞内ドメインにより誘導され得るポリペプチド遺伝子産物には、転写活性化因子、転写抑制因子、キメラ抗原受容体、T細胞受容体(TCR)、第二のキメラNotchポリペプチド、CAR、翻訳調節因子、免疫抑制性受容体、免疫抑制性タンパク質、免疫活性化タンパク質、サイトカイン受容体、ケモカイン受容体、DNA結合タンパク質、エピジェネティック調節因子、RNA誘導型エンドヌクレアーゼ(例えば、Cas9ポリペプチド)、酵素的に不活性なCas9ポリペプチド、部位特異的ヌクレアーゼ、リコンビナーゼ、分化を誘導する転写因子、脱分化を誘導する転写因子等が含まれる。
a)標的細胞において本開示の第一のsynNotchポリペプチドを発現させること、ここで、第一のsynNotchポリペプチドは、可溶性アダプター分子(例えば、抗原)上の第一のエピトープと結合する抗原結合ドメイン(例えば、scFv、ナノボディ等)を含む;
b)標的細胞と、
i)本開示の第二のsynNotchポリペプチドを発現する第二の細胞、ここで、第二のsynNotchポリペプチドは、可溶性アダプター分子(例えば、抗原)上の第二のエピトープと結合する抗原結合ドメイン(例えば、scFv、ナノボディ等)を含む;及び
ii)可溶性アダプター分子(例えば、抗原)
を接触させること、ここで、第一のsynNotchポリペプチドの抗原結合ドメイン及び第二のsynNotchポリペプチドの抗原結合ドメインの、可溶性アダプター分子(例えば、抗原)への結合が、第一のsynNotchポリペプチドの細胞内ドメインの切断を誘導し、それにより、細胞内ドメインが放出され、放出された細胞内ドメインが標的細胞の活性を調節する。標的細胞の活性は、細胞の遺伝子産物の発現、細胞の増殖、細胞のアポトーシス、細胞の非アポトーシス性死、細胞の分化、細胞の脱分化、細胞の遊走、細胞からの分子の分泌及び細胞の細胞接着からなる群より選択されることができる。アダプター分子は、癌関連抗原、病原体関連抗原、抗体等であることができる。
本開示のキメラNotch受容体ポリペプチドは、1つまたは複数の追加のポリペプチドをさらに含むことができ、ここで、好適な追加のポリペプチドには、シグナル配列;エピトープタグ;親和性ドメイン;核局在化シグナル(NLS);及び検出可能シグナルを産生するポリペプチドが含まれるが、これらに限定されない。
本開示のキメラNotch受容体ポリペプチドにおける使用に好適なシグナル配列には、天然に存在するシグナル配列、合成の(例えば、人工)シグナル配列等を含む任意の真核生物シグナル配列が含まれる。
好適なエピトープタグには、
親和性ドメインは、結合パートナーと相互作用することができるペプチド配列、例えば、同定または精製に有用な、固体支持体の上に固定されたものなどを含む。複数の連続する単一のアミノ酸、例えば、ヒスチジンは、本開示のキメラNotch受容体ポリペプチドに融合したとき、ニッケルセファロースなどの樹脂カラムへの高親和性結合による組換えキメラポリペプチドのワンステップ精製のために使用されてよい。例示的な親和性ドメインには、
好適な核局在化シグナル(「NLS」;本明細書で「核局在化配列」とも称される)には、例えば、
好適な検出可能シグナル産生タンパク質には、例えば、蛍光タンパク質;検出可能シグナルを産物として生成する反応を触媒する酵素;等が含まれる。
上記の通り、本開示のキメラNotchポリペプチドの特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合は、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出される。上記の通り、特異的結合対の第二のメンバーは、様々な分子のいずれかであることができる。いくつかの場合では、キメラNotchポリペプチドは、抗原と特異的に結合する;例えば、特異的結合対の第二のメンバーは、抗原である。かかる抗原の例としては、例えば、腫瘍抗原;癌細胞関連抗原;血液学的悪性疾患抗原;固形腫瘍抗原;細胞表面抗原(例えば、T細胞受容体(TCR)により標的とされる細胞表面抗原;細胞内抗原;等が挙げられる。血液学的悪性疾患抗原の例としては、例えば、CD19(例えば、B細胞において発現する)、CD20(例えば、B細胞において発現する)、CD22(例えば、B細胞において発現する)、CD30(例えば、B細胞において発現する)、CD33(例えば、骨髄系細胞において発現する)、CD70(例えば、B細胞/T細胞において発現する)、CD123(例えば、骨髄系細胞において発現する)、カッパ(例えば、B細胞において発現する)、ルイスY(例えば、骨髄系細胞において発現する)、NKG2Dリガンド(例えば、骨髄系細胞において発現する)、ROR1(例えば、B細胞において発現する)、SLAMF7/CS1(例えば、骨髄腫細胞、ナチュラルキラー細胞、T細胞、及び大半のB細胞型において発現する)、CD138(例えば、多発性骨髄腫の悪性形質細胞において発現する)、CD56(例えば、骨髄腫細胞、神経細胞、ナチュラルキラー細胞、T細胞、及び小柱骨芽細胞(trabecular osteoblasts)において発現する)CD38(例えば、B細胞/T細胞にいて発現する)及びCD160(例えば、NK細胞/T細胞において発現する)等が挙げられる。固形腫瘍抗原の例としては、例えば、B7H3(例えば、肉腫、神経膠腫において発現する)、CAIX(例えば、腎臓において発現する)、CD44 v6/v7(例えば、子宮頸部において発現する)、CD171(例えば、神経芽細胞腫において発現する)、CEA(例えば、結腸において発現する)、EGFRvIII(例えば、神経膠腫において発現する)、EGP2(例えば、癌腫において発現する)、EGP40(例えば、結腸において発現する)、EphA2(例えば、神経膠腫、肺において発現する)、ErbB2(HER2)(例えば、乳房、肺、前立腺、神経膠腫において発現する)、ErbB受容体ファミリー(例えば、乳房、肺、前立腺、神経膠腫において発現する)、ErbB3/4(例えば、乳房、卵巣において発現する)、HLA−A1/MAGE1(例えば、メラノーマにおいて発現する)、HLA−A2/NY−ESO−1(例えば、肉腫、メラノーマにおいて発現する)、FR−a(例えば、卵巣において発現する)、FAP†(例えば、癌関連線維芽細胞において発現する)、FAR(例えば、横紋筋肉腫において発現する)、GD2(例えば、神経芽細胞腫、肉腫、メラノーマにおいて発現する)、GD3(例えば、メラノーマ、肺癌において発現する)、HMW−MAA(例えば、メラノーマにおいて発現する)、IL11Ra(例えば、骨肉腫において発現する)、IL13Ra2(例えば、神経膠腫において発現する)、ルイスY(例えば、乳房/卵巣/膵臓において発現する)、メソテリン(例えば、中皮腫、乳房、膵臓において発現する)、Muc1(例えば、卵巣、乳房、前立腺において発現する)、NCAM(例えば、神経芽細胞腫、結腸直腸において発現する)、NKG2Dリガンド(例えば、卵巣、肉腫(sacoma)において発現する)、PSCA(例えば、前立腺、膵臓において発現する)、PSMA(例えば、前立腺において発現する)、TAG72(例えば、結腸において発現する)、VEGFR−2(例えば、腫瘍血管系において発現する)、Axl(例えば、肺癌において発現する)、Met(例えば、肺癌において発現する)、α5β3(例えば、腫瘍血管系において発現する)、α5β1(例えば、腫瘍血管系において発現する)、TRAIL−R1/TRAIL−R2(例えば、固形腫瘍(結腸、肺、膵臓)及び血液学的悪性疾患において発現する)、RANKL(例えば、前立腺癌及び骨転移において発現する)、テネイシン(例えば、神経膠腫、上皮性腫瘍(乳房、前立腺)において発現する)、EpCAM(例えば、上皮性腫瘍(乳房、結腸、肺)において発現する))、CEA(例えば、上皮性腫瘍(乳房、結腸、肺)において発現する))、gpA33(例えば、大腸癌腫において発現する)、ムチン(例えば、上皮性腫瘍(乳房、結腸、肺、卵巣)において発現する))、TAG−72(例えば、上皮性腫瘍(乳房、結腸、肺)において発現する))、EphA3(例えば、肺、腎臓、メラノーマ、神経膠腫、血液学的悪性疾患において発現する)及びIGF1R(例えば、肺、乳房、頭頸部、前立腺、甲状腺、神経膠腫において発現する)が挙げられる。表面及び細胞内抗原の例としては、例えば、Her2(遺伝子記号ERBB2)、MAGE−A1(遺伝子記号MAGEA1)、MART−1(遺伝子記号MLANA)、NY−ESO(遺伝子記号CTAG1)、WT1(遺伝子記号WT1)、MUC17及びMUC13が挙げられる。他の抗原の例としては、例えば、BCMA(遺伝子記号TNFRSF17)、B7H6(遺伝子記号NCR3LG1)、CAIX(遺伝子記号CA9)、CD123(遺伝子記号IL3RA)、CD138(遺伝子記号SDC1)、CD171(遺伝子記号L1CAM)、CD19(遺伝子記号CD19)、CD20(遺伝子記号CD20)、CD22(遺伝子記号CD22)、CD30(遺伝子記号TNFRSF8)、CD33(遺伝子記号CD33)、CD38(遺伝子記号CD38)、CD44、スプライス変異体incl 7及び8(文献にてvXと示される)(遺伝子記号CD44)、CEA、CS1(遺伝子記号SLAMF7)、EGFRvIII(遺伝子記号EGFR、vIII欠失変異体)、EGP2、EGP40(遺伝子記号EPCAM)、Erbファミリーメンバー(遺伝子記号ERBB1、ERBB2、ERBB3、ERBB4)、FAP(遺伝子記号FAP)、胎児性アセチルコリン受容体(遺伝子記号AChR)、葉酸受容体アルファ(遺伝子記号FOLR1)、葉酸受容体ベータ(遺伝子記号FOLR2)、GD2、GD3、GPC3(遺伝子記号GPC3)、Her2/neu(遺伝子記号ERBB2)、IL−13Ra2(遺伝子記号IL13RA2)、カッパ軽鎖(遺伝子記号IGK)、ルイス−Y、メソテリン(遺伝子記号MSLN)、ムチン−1(遺伝子記号MUC1)、ムチン−16(遺伝子記号MUC16)、NKG2Dリガンド、前立腺特異的膜抗原(PSMA)(遺伝子記号FOLH1)、前立腺幹細胞抗原(PSCA)(遺伝子記号PSCA)、受容体チロシンキナーゼ様オーファン受容体1(遺伝子記号ROR1)、及び未分化リンパ腫受容体チロシンキナーゼ(遺伝子記号ALK)が挙げられる。
以下は、本開示のキメラNotch受容体ポリペプチドの非限定的な例である。
本開示は、本開示のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸を提供する。いくつかの場合では、本開示のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸は、発現ベクター内に含有される。ゆえに、本開示は、本開示のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸を含む組換え発現ベクターを提供する。いくつかの場合では、本開示のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列は、転写制御エレメント(例えば、プロモーター;エンハンサー;等)に作動可能に連結している。いくつかの場合では、転写制御エレメントは、誘導可能である。いくつかの場合では、転写制御エレメントは、構成的である。いくつかの場合では、プロモーターは、真核細胞において機能的である。いくつかの場合では、プロモーターは、細胞型特異的プロモーターである。いくつかの場合では、プロモーターは、組織特異的プロモーターである。
本開示は、本開示の核酸で遺伝子改変された宿主細胞、すなわち、本開示のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸で遺伝子改変された宿主細胞を提供する。本開示は、本開示のキメラNotchポリペプチドを発現する細胞の活性を調節する方法を提供する。該方法は、概して、細胞と特異的結合対の第二のメンバーを接触させることを必然的に含む。特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合は、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出される。細胞内ドメインの放出が、細胞の活性を調節する。
本開示は、本開示のキメラNotchポリペプチドをコードする核酸を含む非ヒトトランスジェニック生物を提供する。本開示のトランスジェニック非ヒト生物は、本開示のキメラNotchポリペプチドをコードするヌクレオチド配列を含む核酸を含むように遺伝子改変されているゲノムを含む。
本開示のキメラNotch受容体ポリペプチド、及び本開示の核酸(キメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸)、及び本開示の核酸を含む組換え発現ベクターが、様々な用途で有用である。本開示は、かかる用途を提供する。
本開示は、本開示のキメラNotchポリペプチドを発現する細胞の活性を調節する方法を提供する。いくつかの場合では、該方法は、細胞と特異的結合対の第二のメンバーを接触させることを含み、特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合は、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出され、細胞内ドメインの放出が、細胞の活性を調節する。細胞内ドメインは、「エフェクター機能」を提供し、ここで、エフェクター機能は、転写活性化;転写抑制;翻訳活性化;翻訳抑制;細胞小器官機能の調節;免疫細胞活性化;免疫細胞抑制;アポトーシスの誘導;アポトーシスの抑制;ヌクレアーゼ活性;分化の調節;標的核酸の置換;標的核酸の改変;等であることができる。本開示の方法を使用して調節することができる細胞の活性には、免疫細胞活性化(例えば、T細胞活性化等);アポトーシス;エフェクター分子(例えば、サイトカイン、抗体、成長因子等)の産生;標的核酸の転写;標的mRNAの翻訳;細胞小器官活性;細胞内トラフィッキング;分化;等が含まれるが、これらに限定されない。本開示の方法は、形質膜で作用するエフェクターの放出を引き起こすために使用することもでき、それにより細胞の活性の改変(例えば、免疫活性化を提供する免疫共抑制受容体モチーフの放出)がもたらされる。
i)個体から得たTリンパ球またはナチュラルキラー(NK)細胞を、本開示のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む発現ベクターで遺伝子改変すること、ここで、キメラNotch受容体ポリペプチドは個体の癌細胞上のエピトープに対して特異的であり、遺伝子改変は、ex vivoで実行される;
ii)遺伝子改変されたTリンパ球またはNK細胞を個体に導入すること、ここで、遺伝子改変されたTリンパ球またはNKが癌細胞を認識して殺傷し、それにより癌を処置する。
本開示は、本開示のキメラNotchポリペプチドを発現する細胞の活性を調節する方法を提供する。上に詳述するように、キメラNotchポリペプチドは、特異的結合対の第一のメンバーを含む細胞外ドメインを含む。キメラNotchポリペプチドは、1つまたは複数のリガンド誘導性タンパク質分解切断部位;及び細胞内ドメインを含むNotch受容体ポリペプチドも含む。いくつかの場合では、該方法は、細胞と特異的結合対の第二のメンバーを接触させることを含み、ここで、特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合は、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出され、細胞内ドメインの放出が、細胞の活性を調節する。いくつかの場合では、特異的結合対の第二のメンバーは、本開示のキメラNotchポリペプチドを発現する細胞と接触する第二の細胞の細胞表面上に存在する。いくつかの場合では、特異的結合対の第二のメンバーは、可溶性である。
本開示は、本開示のキメラNotchポリペプチドを発現する細胞の活性を調節する方法を提供する。上に詳述するように、キメラNotchポリペプチドは、特異的結合対の第一のメンバーを含む細胞外ドメインを含む。キメラNotchポリペプチドは、1つまたは複数のリガンド誘導性タンパク質分解切断部位;及び細胞内ドメインを含むNotch受容体ポリペプチドも含む。いくつかの場合では、該方法は、細胞と特異的結合対の第二のメンバーを接触させることを含み、ここで、特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合は、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出される。いくつかの場合では、特異的結合対の第二のメンバーは、本開示のキメラNotchポリペプチドを発現する細胞と接触する第二の細胞の細胞表面上に存在する。いくつかの場合では、特異的結合対の第二のメンバーは、可溶性である。
本開示のキメラNotch受容体ポリペプチドを使用して、生物学的分子、例えば、サイトカイン、成長因子、抗体、及び、遺伝子コードされている他の結合剤、アゴニスト、捕捉剤、またはブロッキング剤の発現及び分泌を制御することができる。
本開示は、i)本開示のキメラNotchポリペプチド;及びb)キメラ抗原受容体を発現する細胞の活性を調節する方法を提供する。該方法は、本開示のキメラNotch受容体ポリペプチド及びCARの両方を発現する細胞を、i)特異的結合対の第二のメンバー(ここで、特異的結合対の第二のメンバーは、特異的結合対の、キメラNotch受容体ポリペプチドに存在する、第一のメンバーに結合する);及びii)CARが結合する抗原と接触させることを必然的に含む。これらの実施形態では、細胞の活性の調節は、特異的結合対の第二のメンバー、及びCARが特異的に結合する抗原の両方が必要とされる。
本開示は、i)第一の本開示のキメラNotchポリペプチド;及びb)第二の本開示のキメラNotchポリペプチドを発現する細胞の活性を調節する方法を提供する。例えば、いくつかの場合では、細胞は、i)第一の特異的結合対の第一のメンバーを含む第一のキメラNotch受容体ポリペプチド;及びii)少なくとも、第二の特異的結合対の第一のメンバーを含む第二のキメラNotch受容体ポリペプチドを発現し、ここで、第一及び第二の特異的結合対は互いに異なり、そのため、第一の特異的結合対の第二のメンバーの第一の特異的結合対の第一のメンバーへの結合は、第二のキメラNotch受容体ポリペプチドの細胞内ドメインの放出をもたらさず、第二の特異的結合対の第二のメンバーの第二の特異的結合対の第一のメンバーへの結合は、第一の第二のキメラNotch受容体ポリペプチドの細胞内ドメインの放出をもたらさない。これらの実施形態では、第一のキメラNotch受容体ポリペプチドの細胞内ドメインは、第一のエフェクター機能を提供し;第二のキメラNotch受容体ポリペプチドの細胞内ドメインは、第一のエフェクター機能とは異なる第二のエフェクター機能を提供する。これらの実施形態の概略図を図9Aに提示する。
本開示は、a)本開示のキメラNotchポリペプチドをコードするヌクレオチド配列を含む核酸;及びb)キメラ抗原受容体をコードするヌクレオチド配列を含む核酸で遺伝子改変されている細胞の活性を調節する方法であって、キメラ抗原受容体をコードするヌクレオチド配列が誘導性プロモーターの制御下にある、前記方法を提供する。該方法は、以下を必要とする:
i)本開示のキメラNotch受容体ポリペプチドを発現する(がCARは発現しない)細胞と、特異的結合対の第二のメンバー(ここで、特異的結合対の第二のメンバーは、特異的結合対の、キメラNotch受容体ポリペプチドに存在する、第一のメンバーに結合する)を接触させること、ここで、細胞と特異的結合対の第二のメンバーの接触は、キメラNotch受容体ポリペプチドの細胞内ドメインの放出を誘導し、ここで、細胞内ドメインは、キメラ抗原受容体をコードするヌクレオチド配列を含む核酸の転写を活性化させる転写因子であり、これによりCARの発現がもたらされる;及び
ii)(i)の接触ステップの後に、細胞と、CARが結合する抗原を接触させること。二回目の接触ステップは、CARによる活性の調節をもたらす。これらの実施形態では、細胞の活性の調節は、特異的結合対の第二のメンバー、及びCARが特異的に結合する抗原の両方を必要とする。これらの実施形態の一例を図10に概略的に示す。
i)2つ以上の本開示のキメラNotch受容体ポリペプチドを発現する(がCARは発現しない)細胞と、特異的結合対の第二のメンバー(ここで、特異的結合対の第二のメンバーは、特異的結合対の、キメラNotch受容体ポリペプチドに存在する、第一のメンバーに結合する)を接触させること、ここで、細胞と特異的結合対の第二のメンバーとの接触が、2つ以上のキメラNotch受容体ポリペプチドの少なくとも1つの細胞内ドメインの放出を誘導し、ここで、細胞内ドメインは、キメラ抗原受容体をコードするヌクレオチド配列を含む核酸の転写を活性化する転写因子であり、これによりCARの発現がもたらされる;及び
ii)(i)の接触ステップの後に、細胞と、CARが結合する抗原を接触させること。二回目の接触ステップは、CARによる活性の調節をもたらす。これらの実施形態では、細胞の活性の調節は、特異的結合対の第二のメンバー、及びCARが特異的に結合する抗原の両方を必要とする。
本開示は、i)第一の本開示のキメラNotchポリペプチドをコードするヌクレオチド配列を含む核酸;及びb)第二の本開示のキメラNotchポリペプチドをコードするヌクレオチド配列を含む核酸で遺伝子改変されている細胞の活性を調節する方法を提供する。例えば、いくつかの場合では、細胞は、i)第一の特異的結合対の第一のメンバーを含む第一のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸;及びii)第二の特異的結合対の第一のメンバーを含む少なくとも第二のキメラNotch受容体ポリペプチドをコードするヌクレオチド配列を含む核酸で遺伝子改変されており、ここで第一及び第二の特異的結合対は互いに異なり、そのため、第一の特異的結合対の第二のメンバーの第一の特異的結合対の第一のメンバーへの結合は、第二のキメラNotch受容体ポリペプチドの細胞内ドメインの放出をもたらさず、第二の特異的結合対の第二のメンバーの第二の特異的結合対の第一のメンバーへの結合は、第一の第二のキメラNotch受容体ポリペプチドの細胞内ドメインの放出をもたらさない。これらの実施形態では、第一のキメラNotch受容体ポリペプチドの細胞内ドメインは、第一のエフェクター機能を提供し、ここでエフェクター機能は第二のキメラNotchポリペプチドの転写の誘導を提供し;第二のキメラNotch受容体ポリペプチドの細胞内ドメインは、第一のエフェクター機能とは異なる第二のエフェクター機能を提供する。
本開示は、第一の細胞の活性を調節するための方法であって、第一の細胞と第二の細胞を接触させることを含み、第二の細胞が、特異的結合対の第一のメンバーを含む細胞外ドメインを含むキメラNotch受容体ポリペプチドを発現し;第二の細胞が、その表面上で特異的結合対の第二のメンバーを含む分子を発現する、前記方法を提供する。いくつかの場合では、第一の細胞と第二の細胞を接触させることは、第一の細胞における活性を調節する。いくつかの場合では、第一の細胞と第二の細胞を接触させることは、第二の細胞の活性を調節する。
細胞の活性を調節するための本開示の方法は、多細胞環境において実行することができる。例えば、第一のキメラNotch受容体ポリペプチドは、第一の細胞(「第一の受信側細胞」)に存在し、近接する細胞(例えば、「送信側」細胞)上の第一のリガンドへの結合の後に放出されたとき、第二の細胞上の第二のキメラNotch受容体ポリペプチドが結合する第二のリガンドの転写を提供する、細胞内ドメインを含むことができ、ここで、第二のリガンドは、第一の細胞の表面上に発現される。第二の細胞(「第二の受信側細胞」)は、その表面上で第二のキメラNotch受容体ポリペプチドを発現し;第一の受信側細胞との相互作用の際、第二の受信側細胞における細胞内エフェクター機能は、放出され、第二の受信側細胞の活性の調節を提供する。これらの実施形態を図13に概略的に示す。このような方法は、、例えば、構造化された組織を構築すること;細胞結合性をトラッキングすること;等に有用である。
いくつかの場合では、細胞の活性を調節するための本開示の方法は、標的細胞上の2つの別々の標的分子の認識を必然的に含む。例えば、本開示のキメラNotch受容体ポリペプチドを発現する第一の細胞は、標的細胞上の第一の標的分子;及び標的細胞上の第二の標的分子と結合する受容体または他の認識分子をその細胞表面上で発現する第二の細胞を認識する。第一及び第二の細胞の、標的細胞上の第一及び第二の標的分子への結合は、第二の細胞による新しいエフェクター機能(例えば、新しいポリペプチドなどの新しい遺伝子産物)の発現をもたらす。いくつかの場合では、第一の細胞の、標的細胞上の第一の標的分子への結合は、第一の細胞において遺伝子産物(例えば、ポリペプチド)の発現を誘導し;ここで、いくつかの場合では、第一の細胞により産生される新しい遺伝子産物は、第一の細胞の表面上で発現され、第二の細胞の表面上にある受容体または他の認識分子と結合し、この結合が、第二の細胞の活性(例えば、第二の細胞が新しいエフェクター機能(例えば、新しいポリペプチドなどの新しい遺伝子産物を発現するように、核酸の転写の誘導)の調節をもたらし得る。いくつかの場合では、第一及び第二の細胞の、標的細胞上の第一及び第二の標的分子への結合は、第二の細胞による標的細胞の殺傷、または第二の細胞により発現される新しいエフェクター機能による標的細胞の殺傷をもたらす。いくつかの場合では、第一及び第二の細胞の、標的細胞上の第一及び第二の標的分子への結合は、標的細胞の活性の調節をもたらす;例えば、標的細胞の活性の調節が、第二の細胞により産生された新しいエフェクター機能により誘導される場合である。これらの実施形態を図14に概略的に示す。
本開示は、結合誘発型転写スイッチ(例えば、本開示のキメラNotchポリペプチド;MESAポリペプチド;TANGOポリペプチド;等)を発現する細胞の活性を調節する方法を提供する。細胞の活性を調節するための本開示の方法は、in vitro、ex vivo、またはin vivoで実行されることができる。細胞の活性を調節するための本開示の方法は、単一細胞において、または多細胞環境(例えば、天然に存在する組織;人工の組織;等)において実行されることができる。細胞の活性を調節するための本開示の方法は、並行してまたは一連で実行されることができる。
a)細胞において、本開示のキメラNotchポリペプチドを発現させること、ここで、キメラNotchポリペプチドは、i)特異的結合対の第一のメンバーを含む細胞外ドメイン;ii)Notch受容体ポリペプチド、ここで、Notch受容体ポリペプチドは、上記の通りであり、1つまたは複数のリガンド誘導性タンパク質分解切断部位を含む;及びiii)細胞内ドメインを含む;ならびに
b)細胞と特異的結合対の第二のメンバーを接触させること。特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合が、Notch受容体ポリペプチドの1つまたは複数のリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより細胞内ドメインが放出される。放出された細胞内ドメインは、細胞の活性を調節する。本方法を使用して調節することができる細胞の活性には、i)細胞の遺伝子産物の発現;ii)細胞の増殖;iii)細胞のアポトーシス;iv)細胞の非アポトーシス性死;v)細胞の分化;vi)細胞の脱分化;vii)細胞の遊走;viii)細胞からの分子の分泌;及びix)細胞の細胞接着が含まれるが、これらに限定されない。いくつかの場合では、接触工程は、in vivoで実行される。いくつかの場合では、接触工程は、ex vivoで実行される。いくつかの場合では、接触工程は、in vitroで実行される。
本開示の方法における使用に好適なモジュラー細胞外センサー構造(MESA)ポリペプチドは、米国特許公開第2014/0234851号に記載のMESAポリペプチドであることができる。MESAポリペプチドは、a)リガンド結合ドメイン;b)膜貫通ドメイン;c)プロテアーゼ切断部位;及びd)機能的なドメインを含む。機能的なドメインは、転写調節因子(例えば、転写活性化因子、転写抑制因子)であることができる。いくつかの場合では、MESA受容体は、2つのポリペプチド鎖を含む。いくつかの場合では、MESA受容体は、単一のポリペプチド鎖を含む。
好適なTANGOポリペプチドは、第一はタバコエッチ病ウイルス(Tev)プロテアーゼを含み、第二のポリペプチドは、転写因子に融合したTevタンパク質分解切断部位(PCS)を含む、ヘテロ二量体である。2つのポリペプチドが、互いに近接するとき、この近接は、自然なタンパク質間相互作用により媒介され、Tevは、PCSを切断して、転写因子を放出する。Barnea et al.(Proc Natl Acad Sci USA.2008 Jan.8;105(1):64−9)。
いくつかの場合では、結合誘発型スイッチは、細胞内でT細胞受容体(TCR)の発現を誘導する。本開示の方法を使用して誘導することができるTCRは、例えば、癌細胞の表面上にあるエピトープ、ウイルス感染細胞の表面上にあるエピトープ、自己抗原内に存在するエピトープ、等を含む、様々なエピトープのいずれかに対して特異的であるTCRを含む。TCRは、アルファ鎖及びベータ鎖を通常含み;主要組織適合性複合体により提示されるときに抗原を認識する。いくつかの場合では、TCRは、操作TCRである。
いくつかの場合では、結合誘発型スイッチは、細胞におけるCARの発現を誘導する。「キメラ抗原受容体」及び「CAR」という用語は、本明細書で互換可能に使用され、免疫細胞の活性化を誘発するまたは阻害することができる人工のマルチモジュール分子を指し、通常、排他的ではなく、細胞外ドメイン(例えば、リガンド/抗原結合ドメイン)、膜貫通ドメイン及び1つまたは複数の細胞内シグナル伝達ドメインを含む。CARという用語は、CAR分子に限定されず、CAR変異体も含む。CAR変異体は、分割CARを含み、CARの細胞外部分(例えば、リガンド結合部分)及び細胞内部分(例えば、細胞内シグナル伝達部分)は、2つの別々の分子上に存在する。CAR変異体は、例えば、分割CARを含む、条件的に活性化可能なCARである、オンスイッチCARも含み、分割CARの2つの部分の条件的ヘテロ二量体化は、薬理学的に制御される。CAR変異体は、一次CARの活性を増幅または阻害することができる二次CAR結合ドメインを含む、二重特異性CARも含む。CAR変異体は、阻害性キメラ抗原受容体(iCAR)も含み、これは、例えば、二重特異性CARシステムの構成要素として使用してよく、ここで、二次CAR結合ドメインの結合は、一次CAR活性化の阻害をもたらす。CAR分子及びその誘導体(すなわち、CAR変異体)は、例えば、PCT出願番号US2014/016527;Fedorov et al Sci Transl Med(2013);5(215):215ra172;Glienke et al Front Pharmacol(2015)6:21;Kakarla & Gottschalk 52 Cancer J(2014)20(2):151−5;Riddell et al Cancer J(2014)20(2):141−4;Pegram et al Cancer J(2014)20(2):127−33;Cheadle et al Immunol Rev(2014)257(1):91−106;Barrett et al Annu Rev Med(2014)65:333−47;Sadelain et al Cancer Discov(2013)3(4):388−98;Cartellieri et al.,J Biomed Biotechnol(2010)956304に記載されており;これらの開示は、それらの全体が参照により本明細書に組み込まれる。
特異的結合対の第一のメンバーは、種々の特異的結合対のいずれかの第一のメンバーであることができる。好適な特異的結合対は、上に詳述している。
特異的結合対には、例えば、抗原−抗体特異的結合対、ここで、第一のメンバーは、抗原である第二のメンバーに特異的に結合する抗体(または抗体をベースとする認識足場)であるか、または第一のメンバーは抗原であり、第二のメンバーが抗原に特異的に結合する抗体(または抗体をベースとする認識足場)である;リガンド−受容体特異的結合対、ここで、第一のメンバーはリガンドであり、第二のメンバーはリガンドが結合する受容体であるか、または第一のメンバーは受容体であり、第二のメンバーが受容体に結合するリガンドである;非抗体をベースとする認識足場−標的特異的結合対、ここで、第一のメンバーは非抗体をベースとする認識足場であり、第二のメンバーは非抗体をベースとする認識足場に結合する標的であるか、または第一のメンバーは標的であり、第二のメンバーが標的に結合する非抗体をベースとする認識足場である;接着分子−細胞外マトリックス結合対;Fc受容体−Fc結合対、ここで、第一のメンバーは、Fc受容体である第二のメンバーに結合する免疫グロブリンFcを含むか、または第一のメンバーが免疫グロブリンFcを含む第二のメンバーに結合するFc受容体である;及び受容体−共受容体結合対を含み、ここで、第一のメンバーは、共受容体である第二のメンバーに特異的に結合する受容体であるか、または第一のメンバーが受容体である第二のメンバーに特異的に結合する共受容体である。
いくつかの場合では、細胞内ドメインは、転写調節因子、例えば、転写活性化因子または転写抑制因子などの転写因子である。いくつかの場合では、転写因子は、細胞の分化を直接調節する。いくつかの場合では、転写因子は、細胞の分化を、第二の転写因子の発現を調節することにより間接的に調節する。
結合トランスデューサは、リガンド誘導性タンパク質分解切断部位を含むことができ、ここで、特異的結合対の第一のメンバーの、特異的結合対の第二のメンバーへの結合は、結合トランスデューサのリガンド誘導性タンパク質分解切断部位での切断を誘導し、それにより、結合シグナルが伝達されかつ、細胞内ドメインが、細胞内ドメインをタンパク質分解的に放出することによって活性化される。synNotchにおけるタンパク質分解切断部位の例は、上記の通りである。
カテプシンBにより切断される
本開示の方法を使用して、任意の真核細胞の活性を調節することができる。いくつかの場合では、細胞は、in vivoである。いくつかの場合では、細胞は、ex vivoである。いくつかの場合では、細胞は、in vitroである。いくつかの場合では、細胞は、哺乳類細胞である。いくつかの場合では、細胞は、ヒト細胞である。いくつかの場合では、細胞は、非ヒト霊長類細胞である。いくつかの場合では、細胞は、げっ歯類細胞である。いくつかの場合では、細胞は、マウス細胞である。いくつかの場合では、細胞は、ラット細胞である。
本開示は、細胞の活性を調節する方法を実行するためのキットを提供する。
材料及び方法
Jurkat T細胞を、レンチウイルスを介して、Tet応答エレメント(TRE)抗メソテリンCAR−EGFP/pGK mCherryデュアルベクターで安定して形質導入した。形質導入された細胞は、mCherryを構成的に発現し、抗メソテリンCAR−EGFP融合体を、Tetトランス活性化因子(tTa)の存在下で誘導性に発現した。この安定した株を、次いで、抗CD19 Notch tTA受容体で形質導入した。結果得られたJurkat細胞を、次いで、抗原CD19、メソテリン、または両方を発現する慢性骨髄性白血病K562癌細胞との共培養を介してアッセイした。T細胞を、EGFPタグ付きCARの発現及び活性化マーカーであるCD69に関してフローサイトメトリーにより24時間でアッセイした。共培養物の上清も回収し、IL−2の分泌をELISAにより決定した。
キメラNotch受容体特徴決定
キメラNotchプラットフォームを使用して、リガンド結合の際に受信側細胞において転写を誘導できることを示すために、以下のレポーター細胞株を生成した:マウスL929線維芽細胞レポーターラインを、Tet応答エレメント(TRE)−>EGFPレポーター/pGK mCherryデュアルベクター(TREレポーター)またはGal4 UAS−>EGFP/pGKピューロマイシン耐性デュアルベクター(UASレポーター)での形質導入により生成した。キメラNotch変異体を、対応するL929レポーター細胞株へと形質導入して、キメラNotchレポーター細胞を生成した。これらのキメラNotchレポーター細胞を、2つの方法により並行して刺激した。まず、キメラNotch発現細胞を、受容体の細胞外ドメインに特異的に結合する固定化抗体に暴露した。次に、細胞を、同族キメラNotchリガンドを発現するK562細胞またはL929線維芽細胞とインキュベートした。キメラNotch発現細胞を、EGFP蛍光に関して、フローサイトメトリーによりアッセイして、レポーター活性を測定した。図30〜32は、TREレポーターラインにおけるキメラNotchと抗CD19(図30A〜B)及び抗メソテリン(図31A〜B)に、及びUASレポーターラインにおけるキメラNotch抗CD19(図32A〜B)に関する結果を示す。
キメラ抗原受容体(CAR)として知られる人工のT細胞受容体を発現するように操作されたT細胞は、ある特定のB細胞癌に対する治療薬として有効である。しかしながら、CAR T細胞癌免疫療法での主な懸念事項は、オフターゲット作用であり、この場合、治療用T細胞が正常組織を破壊して、深刻な副作用、そして死亡さえももたらす。このような問題を軽減する可能性がある戦略は、治療用T細胞が、腫瘍微小環境においてのみCARを発現し、より局在化したT細胞応答を提供することである。このような戦略を実現するために、キメラNotchを治療用T細胞において用いて、最初に、腫瘍特異的細胞表面抗原と結合することにより腫瘍を検出し、腫瘍内にある第二の腫瘍特異抗原に対してのみCARの発現を開始させることができると推論された。効果的に、これは、T細胞活性に対する二重抗原制御及び腫瘍局在化応答の両方を提供する。In vitroデータでの原理証明をJurkat細胞において提示する。
図30Aに示し、実施例1に記載するような、(抗CD19 scFvを細胞外ポリペプチドとして、及びtTA転写活性化因子を細胞内ドメインとして含む)キメラNotchポリペプチドを発現し、pTet−GFP構築物を含む「受信側」細胞と、CD19をその表面上に発現する「送信側」細胞を、1mMのN−[(3,5−ジフルオロフェニル)アセチル]−L−アラニル−2−フェニル]グリシン−1,1−ジメチルエチルエステル(DAPT)の存在下または不在下で接触させた。DAPTは、ガンマセクレターゼ阻害因子である。データは、GFP+「受信側」細胞として表す。図84A及び84Bに示すように、送信側細胞及びDAPTの存在下では、GFP+「受信側」細胞は、対象、未処置レベルであり;DAPTの不在下では、送信側細胞との接触は、キメラNotchポリペプチド及びGFPの発現を強く活性化した。
最小notch膜貫通領域を、新規の細胞外及び細胞内ドメインと組み合わせて、多様なキメラ受容体を構築することができる。
Notch伝達経路のコアは、膜貫通領域にある。この設計は、多様なアレイの膜提示リガンドを検出する一連の受容体を操作するためのプラットフォームであるとして考えることができる(図85A)。細胞内側では、Notch細胞内ドメイン(NICD)を、Notchシグナル伝達活性化について報告するために、直交転写因子により置き換えることができる。細胞外では、内在性デルタ結合ドメインは、タンパク質結合ドメイン(例えば、FKBP、抗GFPナノボディ等)により置き換えることができ、切断は、同族結合パートナーが結合するときに誘導される。従って、異なる細胞内ドメインとそれぞれカップリングした異なる細胞外ドメインを有する受容体分子のライブラリーが生成された(図85B)。図86Aに示すように、合成受容体の応答は、切断依存性様式で同族リガンドを発現する送信側細胞との細胞間接触により活性化される。異なる細胞外及び細胞内ドメイン組み合わせでの結果を、図87Aに提供する。
SynNotch細胞株は、受容体活性化の際に強い活性化を示した。この結果が細胞株特異的であるか否かを検査するために、synNotch受信側細胞株を、一連の樹立細胞株:上皮性MDCK細胞、L929及びC3Hマウス線維芽細胞株、HEK293ヒト上皮性細胞株、Jurkat T細胞を操作することにより産生した。これらの全ては、synNotch受容体活性化の際に、レポーター遺伝子活性化の強い誘導を不変的に示した(図89B)。重要なことに、初代細胞も、synNotch受容体で操作することができ、こうして、これらは新規のリガンド刺激に応答するようになる。図89A(及び図90)では、synNotch及びレポーターを発現する一次海馬ニューロンを誘導して、リガンド発現送信側細胞との接触によりGFPレポーターカセットを発現することができることが示される。
SynNotch受信側細胞は、隣接細胞がリガンドを発現しているかどうかを検出することができる。上皮細胞層に配置されたsynNotch受信側細胞を、空間的に誘導してレポーター遺伝子を発現することができるかどうかを検査した。リガンド発現送信側細胞が、受信側細胞の層に低密度に播種されるとき、送信側細胞と直接接触する受信側細胞のみが、合成Notch受容体を活性化し、これは緑色送信側細胞の周囲の青色細胞(活性化受信側)の環により目視できる。一方で、離れた受信側細胞は不活性のままである(図91A)。
複数のsynNotch受容体が、クロストークすることなく、同じ細胞内で機能し得るかどうかを探求した。直交機能は、細胞が複数の入力の有無に応じて異なる出力を生成することを可能にするだろう。synNotch受容体は、シグナル伝達の機構のために、互いに直交して機能し得ると仮定された。異なる受容体の細胞内転写調節因子が異なる場合、クロストークを生じさせ得る共通の中間体(例えば、活性化キナーゼ)は存在しない。synNotchと内因性Notchとの間に何らかのクロストークがあった場合、シグナル伝達を最初に検査した。これを示すために、抗CD19−synNotchならびに異なる蛍光タンパク質を有する完全長Notchについて報告するように細胞を操作した。細胞をデルタのみで刺激したとき、完全長Notch応答のみが活性化された;同じことがCD19に対するsynNotchの応答にも当てはまる:2つの経路は独立した活性化を示す(図93A)。
複数のsynNotch受容体を同じ細胞内で使用して、多様な応答を生成することができる。組み合わせの環境の合図をインテグレートし、ある特定の基準が満たされたときにのみ応答する細胞の操作が設計された。具体的には、それらの環境上で2つの異なる抗原の存在下でのみ応答するが、それぞれに単独では応答しないだろう細胞の生成に、専念した。これを達成するために、抗CD19及び抗GFP細胞外ドメインを有する二重synNotch発現細胞(各synNotchの活性化により分割Gal4タンパク質の半分が活性化される)を操作した。このようにして、2つの入力が受信側細胞のみに提示される場合、活性化は見られない(図93C、列1〜3)。2つの受容体が活性化されたときにのみ、分割された分子が再構成され、受信側細胞における応答が誘導される(図93C、最後の列)。
細胞間コミュニケーションのための複数のsynNotch受容体を用いて、細胞間コミュニケーションの多細胞シグナル伝達システムを設計した。具体的には、上皮細胞層における自己組織化多層空間的パターニングの誘導に焦点をあてた。このために、受信側細胞は、2つのsynNotchを有し、そのうちの1つは他方のリガンドを誘導する。この特定の例において、抗GFP−synNotchは、CD19リガンド発現(及びレポータータンパク質mCherry)を誘導し;抗CD19−synNotchは、レポータータンパク質tagBFPを誘導する。次に、層のカスケードの誘導を開始するために、GFP送信側細胞を、二重synNotch受信側細胞の単層に低密度に播く。この回路は、送信側細胞島の周りに活性化の2つの同心円環の形成を可能にした:第一の隣接細胞は、抗GFP−synNotchの活性化後に赤色になり、それらはCD19送信側になる(図94A、1日目)。そして、送信側細胞から一直径離れた細胞は、第一の隣接細胞によって発現されたCD19リガンドに応答することができ、図94A、2日の二重環パターンを生成する。このようにして、2つの異なる細胞型が、空間的に制御された様式で受信側細胞の均一な集団から産生される(図94C)。ゆえに、複数のsyn−Notch受容体を使用して、自然発達過程で観察されるものと同様の、より複雑な多段階パターン形成応答を操作することができる(図94B)。
(図85A)Notchレポーター、synNotch受容体への、野生型Notchから開始した連続的な操作としての、synNotch受容体システムの概念設計。後者は、notchレポーターの細胞内直交転写因子の柔軟性を利用し、細胞外ドメインを置換することによって新規の入力に応答するプラットフォームを構築する。
(図87A)異なる細胞外及び細胞内ドメインを有する一連のsynNotch受信側細胞は、同族リガンドを用いた刺激の際に活性化を示す。細胞は、マウス線維芽細胞株L929であり、プレート結合型刺激により、またはリガンドを発現する送信側細胞を用いるかのいずれかで刺激され、これは図に示す通りである。
(図86A〜B)合成notch受容体を使用して、内因性疾患抗原を検出し、レポーター遺伝子の転写を増減させることができる。
(図88A)細胞外ドメイン上へのEGFリピートの付加は、抗メソテリンsynNotchにおける基礎活性化を低下させる。抗メソテリンScFvの前にEGFリピートを有するまたは有さない抗メソテリンsynNotch受容体を、L929線維芽細胞において誘導する。これらの受容体の活性化は、GFPレポーターを活性化する。示したデータは、2つのシナリオにおけるFACSプロットである:上、EGFリピート無し、レポーターの誘導は、リガンドの不在下であっても構成的である;下、EGFリピート有り、基礎レポーター活性化は消滅し(オフライン)、誘導によりGFP強度はオン状態になる。
(図89A)海馬ニューロン。一次海馬ニューロンを、E18ラット胚から解離し、核酸遺伝子導入(nucleofected)して、抗CD19 synNotch及びカップリングしたGFPレポーターを発現した。ニューロンは、ポリ−D−リジン及びラミニンでコーティングされたガラス底35mm培養皿上に播種する。ニューロンプレーティングの2時間後、送信側細胞(K562)を培養物に加えて共培養系を形成する。画像は、プレーティング後4日目に生細胞から取得する。右に、プレーンなK562細胞(上のパネル)またはCD19+K562送信側細胞(下のパネル)と共培養されたニューロンの代表的な画像を示す。リガンド提示送信側細胞と共培養したニューロンは、対照細胞より有意に高いGFP発現を有する。
(図90A)実験及びニューロンにおいて発現される構築物の実証。一次海馬ニューロンを、E18ラット胚から解離し、cNotch受容体ならびにTetO−GFPレポーターを発現する構築物で核酸遺伝子導入する。ニューロンを、ポリ−D−リジン及びラミニンでコーティングされたガラス底35mm培養皿上に播種する。ニューロンプレーティングの2時間後、K562送信側細胞を培養物に加えて共培養系を形成する。画像は、プレーティング後4日目に生細胞から取得する。
(図91A)上皮単層における境界検出。上皮細胞(MDCK)を以下のように操作する:送信側細胞は、膜貫通ドメインに連結した細胞外GFPを発現する;受信側細胞は、LaG17抗GFPナノボディを細胞外ドメインとして、及びGal4−VP64を細胞内ドメインとして、UAS−>BFPレポーター構築物とともに有する抗GFP synNotchを発現する。送信側及び受信側を1:50の比率で一緒に混ぜ、コンフルエントな単層のプレーティングの48時間後に共焦点画像を取得する。高倍率及び低倍率の代表的な画像を、距離に対する蛍光強度の代表的なラインとともに示す。緑色送信側細胞と接触している受信側細胞のみが青色のレポーターをオンにし、送信側細胞の周りに環を形成する。
(図92A;図91Bとの比較)C3Hマウス線維芽細胞を以下のように操作する:送信側細胞は、膜貫通ドメインに連結した細胞外CD19を、さらにtagBFPマーカーを発現する。受信側細胞は、tTA細胞内ドメインを、TRE−>myoDエフェクター構築物及び構成的mCherryマーカーとともに有する抗CD19 synNotchを発現する。送達線維芽細胞を、まずプレートの限られた領域に播種する;1時間後、送信側細胞をプレートに付着させ、受信側細胞を、ガラスプレート全体を覆うように播種する。画像は右側で10時間ごとに取得する。
下に、送信側細胞と受信側細胞の共培養(上)または受信側細胞単独(下)の実験の代表的な視野を示す。
右端の棒グラフは、様々な条件における、受信側細胞の、または親細胞のE−カドヘリン発現レベルを報告する。E−カドヘリンレベルは、同族抗原GFPを発現する送信側細胞の存在下でのみ低下する。
上パネル:L929マウス線維芽細胞を以下のように操作する:送信側細胞は、膜貫通ドメインに連結した細胞外CD19、さらにtagBFPマーカーを発現する;受信側細胞は、tTA細胞内ドメインを、TRE−>E−カドヘリン−GFPエフェクター構築物、及び構成的赤色マーカー(mCherry)とともに有する抗CD19 synNotchを発現する。顕微鏡で収集した蛍光シグナルを、送信側及び受信側細胞を有する代表的なスフェロイドについて、ならびに示すように受信側細胞のみを有するものについて、t=20時間で示す。緑色蛍光は、送信側細胞の存在下でのみ、受信側細胞において誘導される(E−カドヘリンチャネル、右端)。
下パネル:L929マウス線維芽細胞を以下のように操作する:送信側細胞は、膜貫通ドメインに連結した細胞外GFPを発現する;受信側細胞は、tTA細胞内ドメインを、TRE−>mCherryレポーター構築物、及び構成的青色マーカー(tagBFP)とともに有する抗GFP synNotchを発現する。共焦点顕微鏡で収集した蛍光シグナルを、送信側+受信側細胞を有する代表的なスフェロイドについて、及び受信側細胞のみを有するものについて、t=0 t=20時間で示す。赤色蛍光は、送信側細胞の存在下でのみ、受信側細胞において誘導される(mCherryレポーターチャネル、右端)。送信側細胞と受信側細胞の再編成は認められない。
(図93A)synNotch及び野生型Notchは、直交シグナル伝達経路を活性化する。L929マウス線維芽細胞受信側を、(i)tTA細胞内ドメイン及びTRE−>GFPレポーターを有する野生型Notch受容体、ならびに(ii)抗CD19細胞外ドメインとGal4−VP64細胞内ドメイン、及びUAS−>tagBFPレポーターを有するsynNotch受容体を発現するように操作する。右側のグラフは、異なる条件におけるBFP及びGFPレポーターに関する受信側細胞蛍光シグナルを報告する。黒色点は未処理細胞であり、青色点受信側細胞はCD19発現送信側で刺激され、橙色点はデルタ送信側で刺激した受信側細胞であり、赤色点はCD19及びデルタの両方を発現する送信側細胞で刺激された受信側細胞である。送信側細胞は、マウスL929線維芽細胞である。
上皮細胞(MDCK)を以下のように操作する:送信側細胞は、膜貫通ドメインに連結した細胞外GFPを発現する;受信側細胞は、(i)tTA細胞内ドメインを、TRE−>CD19−mCherryエフェクターカセットとともに有する抗GFP synNotch;(ii)Gal4−VP64細胞内ドメイン、及びUAS−>tagBFPレポーターを有する抗CD19受容体を発現する。
(図95A)synNotch受容体の活性化の推定上の構造機構。LNRドメインは、未結合立体構造においてプロテアーゼ切断部位をマスクする(左)。リガンドが受容体と係合すると、この部位は露出し、この事象が伝達を開始する(右)。
材料及び方法
別段の指示がない限り、以下の材料及び方法が、実施例4に記載の結果に適用される。
synNotch受容体は、CD19 scFv、LaG17(低親和性)、またはLaG16_2(高親和性)GFPナノボディをマウスNotch1(NM_008714)最小調節領域(Ile1427〜Arg1752)及びGal4VP64に融合させることにより構築した。全てのsynNotch受容体は、n末端CD8αシグナルペプチド
初代CD4+及びCD8+T細胞を、陰性選択によるアフェレーシスの後に匿名のドナー血液から単離した(STEMCELL Technologies #15062及び15063)。血液は、University Institutional Review Boardの承認を得て、Blood Centers of the Pacific(カリフォルニア州サンフランシスコ)から得た。T細胞を、20%ヒトAB血清(Valley Biomedical Inc.,#HP1022)及び10%DMSOを含むRPMI−1640(UCSF cell culture core)中で凍結保存した。解凍後、実施例4の全ての実験について、30単位/mL IL−2(NCI BRB Preclinical Repository)を補充した、X−VIVO 15(Lonza #04−418Q)、5%ヒトAB血清及び10mM中和N−アセチルL−システイン(Sigma−Aldrich #A9165)からなるヒトT細胞培地中でT細胞を培養した。
汎親和性VSV−G偽型レンチウイルスを、Fugene HD(Promega #E2312)を使用して、pHR’SIN:CSW導入遺伝子発現ベクターならびにウイルスパッケージングプラスミドpCMVdR8.91及びpMD2.Gを用いたレンチ−X 293T細胞(Clonetech #11131D)のトランスフェクションを介して産生した。初代T細胞を、同日に解凍し、培養24時間後、1:3の細胞:ビード比率でDynabeads Human T−Activator CD3/CD28(Life Technologies #11131D)で刺激した。48時間で、ウイルス上清を回収し、初代T細胞を24時間ウイルスに曝露した。T細胞刺激後4日目に、Dynabeadsを除去し、それらを静止させアッセイに使用できる9日目までT細胞を増殖させた。T細胞を、FACs ARIA IIを用いたアッセイのために選別した。
使用した癌細胞株は、K562骨髄性白血病細胞(ATCC #CCL−243)、Daudi B細胞リンパ芽球(ATCC #CCL−213)、及びHCT115結腸癌細胞(ATCC #CCL−247)とした。Daudi腫瘍と同等のレベルでヒトCD19を安定して発現させるために、K562をレンチウイルスで形質導入した。CD19レベルは、細胞をα−CD19 APC(Biolegend #302212)で染色することによって決定した。K562を、表面GFP(PDGF膜貫通ドメインに融合したGFP)を安定して発現するためにも形質導入した。全ての細胞株を導入遺伝子の発現について選別した。
全てのin vitro synNotch T細胞刺激について、2×105個のT細胞を1:1の比率で送信側細胞と共培養した。丸底96ウェル組織培養プレートでT細胞及び送信側細胞を混合した後、細胞を400xgで1分間遠心分離して、細胞の相互作用を引き起こし、培養物をレポーター発現またはカスタム遺伝子誘導の発現について、BD LSR IIを用いたフローサイトメトリーを介して24時間で分析した。全てのフローサイトメトリー分析は、FlowJoソフトウェア(TreeStar)で行った。
α−CD19 synNotch Gal4VP64受容体及びヒトIL−2またはIL−10発現のいずれかを制御する5xGal4応答エレメントを発現する初代CD4+T細胞を、K562骨髄性白血病細胞(CD19−またはCD19+)で上に記載したように刺激した。参考として、α−CD19 4−1BBζ CARを発現するCD4+T細胞は、1:3の比率でα−CD3/CD28 Dynabeadsで刺激した形質導入していないT細胞とともに刺激した。上清を24時間で回収し、製造者のプロトコルに従って、Luminex MAGPIX(Luminex Corp.)ヒトサイトカインマグネティック25−プレックスパネル(Invitrogen参照番号LHC0009M)で分析した。全てのサイトカインレベルを、xPONENTソフトウェア(Luminex Corp.)を用いて標準曲線に基づいて算出した。
IL−2産生を制御するsynNotch T細胞をアッセイして、細胞内サイトカイン染色(ICS)によって、これらがIL−2を基礎的に産生したかどうかを決定した。synNotch T細胞及び形質導入していないT細胞対照を、GolgiPlug(BD Biosciences #555029)の存在下で6時間培養した。T細胞を、次いで、α−IL−2 FITC(BD #340448)でBD Biosciences ICSキット(#555028)を用いて染色した。IL−2のレベルを、BD LSRIIを用いたフローサイトメトリーを介して分析した。
初代ヒトCD4+T細胞を、Dynabeads Human T−Activator CD3/CD28で上記のように刺激した。活性化中にT細胞をTh1サブセットへと分化するために、細胞を上記の通り、ただし、2.5ng/mLの組換えIL−12(R&Dシステム)及び12.5μg/mLのα−IL−4クローンMP4−25D2(BD Pharmigen #554481)を加えて培養した。IL−12及びα−IL−4を、少なくとも週二回加えた。並行して、初代CD4 T細胞を、レンチウイルスで形質導入して、ヒトTbet T2A mCherry(TBX21、NCBI #EAW94804.1)を発現させ、IL−2を補充したT細胞培地において正常に培養した。α−CD19 synNotch Gal4VP64受容体及びTbet T2A GFP発現を制御する5x Gal4応答エレメントを発現するCD4+T細胞を、CD19−またはCD19+K562送信側細胞の存在下でウイルス形質導入24時間後に培養した。synNotch T細胞を、K562の存在下でIL−2を補充したT細胞培地において培養した。全てのT細胞を11〜14日間培養し、次いで、細胞内サイトカイン染色(ICS)に供して、Th1 T細胞の割合(%)を決定した。
HCT116結腸癌細胞及びK562を、組換えTRAIL(1〜200ng/mL、1:2希釈シリーズ)で24時間処理した。次いで、細胞を回収し、live/dead染色、SYTOX Blue(Thermo Scientific #S34857)で染色し、死滅細胞の割合をBD LSR IIでのフローサイトメトリーによって決定した。K562により発現されるデスレセプター4(DR4)のレベルを、α−TRAIL R1(DR4)APC(Biolegend #307208)で染色することによって評価した。
動物試験を、UCSF Institutional Animal Care and Use Committeeに承認されたプロトコル下にてUCSF Preclinical Therapeutics Coreで実施した。NOD scidガンマ(NSG)(雌、8〜12週齢、Jackson Laboratory #005557)マウスを、全てのin vivoマウス実験に使用した。α−CD19 synNotch Gal4VP64受容体及びヒトIL−2 IRES mCherryを制御する5x Gal4応答エレメントを発現する初代CD4+及びCD8+T細胞を、選別し、実験において使用した。
統計学的有意性を、別段の指定がない限り、スチューデントのt検定(両側検定)によって決定した。実施例4に関する全ての統計学的分析は、Prism 6(Graphpad)を用いて行い、p値を報告した(有意差無し=p>0.05、*=p≦0.05、**=p≦0.01、***=p≦0.001、****=p≦0.0001)。全てのエラーバーは平均の標準誤差または標準偏差のいずれかを表す。
synNotch受容体は、CD4+及びCD8+ヒト初代Tリンパ球において抗原誘導型転写を推進することができる
Notch受容体は、3つの重要な要素を有する:1)リガンド結合上皮成長因子(EGF)リピート、2)活性化中に受容体の切断を制御するコア調節領域、及び3)放出され、転写を調節するNotch細胞内ドメイン(NICD)。完全にカスタマイズ可能な受容体標的化及び転写調節を可能にするsynNotch受容体プラットフォームを構築するために、リガンド依存性切断及び活性化を制御するNotchコア調節領域を最小の足場として利用したが、その後カスタマイズした入力認識及び出力転写モジュールを追加した(実施例3を参照されたい)。Notchコア調節領域には、S2切断部位のメタロプロテアーゼへのアクセス可能性を制御するLin12−Notchリピート(LNR)、ヘテロ二量体化ドメイン(HD)、及びNotch細胞内ドメイン(NICD)の放出に必要なγ−セクレターゼ切断部位を含有する膜貫通ドメイン(TMD)である。天然リガンドデルタの認識に通常関与する細胞外EGFリピートを除去し、癌抗原CD19を対象とする一本鎖可変断片(scFv)または表面提示GFPなどの直交抗原に対するナノボディで置換した(図96B〜96C)。転写調節に通常必要とされるNICDを、四量体ウイルス転写活性化因子ドメインであるVP64に融合したGal4 DNA結合ドメインで置換した(図96B〜96C)。この全体のアプローチを使用して、目的の任意の表面抗原に対するsynNotch受容体を操作し、受容体活性を、直交転写因子及びそれらの会合する応答エレメントによって制御されるカスタマイズされた細胞出力につなげることができる(実施例3を参照されたい)。ある範囲の他の細胞外及び細胞内ドメインもsynNotchと機能することが示された。
全身の免疫細胞及び組織は、サイトカインとして知られる可溶性タンパク質を分泌して、細胞挙動を情報伝達及び調節して、全体的な免疫応答を形作る。特定のサイトカインプロファイルは、病原体及び腫瘍を根絶するために重要である。多くの場合、異なるセットのサイトカインは、免疫応答を抑制する反対の効果を有する。さらに、サイトカインが癌に対する免疫を増強するか、または自己免疫設定における有害な炎症を抑制し得るある特定のシナリオでは、これらのサイトカインは存在しない。治療用T細胞がどのサイトカインを分泌するかを精密に制御することが望ましい。しかしながら、いくつかの例では、T細胞がCARまたは天然T細胞受容体を介して活性化されるとき、産生されるサイトカインに対する制御はほとんどなく、しばしばサイトカインプロファイルは、疾患及び活性化の状況ならびに受容体の特性に依存する(図98A〜98C)。多くのT細胞療法では、特定の疾患または治療の必要性に免疫応答を合わせるように特定のサイトカインの産生にバイアスをかけることが有益であり得る。
治療用T細胞の結果を精密に形作るための別の方法は、その分化を制御することである。サイトカインのようなタンパク質エフェクターを産生することを超えて、T細胞は、有効なサブタイプの免疫応答を開始するために重要な特定の分化プログラムを受ける。これらのサブタイプ分化プログラムは、通常、T細胞活性化、特定のサイトカイン、及び最終的に特定のT細胞運命を開始するマスターレギュレーター転写因子の調節の組み合わせによって決定される。Tヘルパー細胞1(Th1)またはTヘルパー細胞2(Th2)T細胞は、マスターレギュレーター転写因子であるTbet及びGATA3によりそれぞれ制御される、2つの正準のCD4+T細胞運命選択である(図99A)。Th1細胞は、病原体及び癌に対する細胞免疫のために重要であり、一方でTh2細胞は、抗体産生の刺激に関与する。多くの疾患では、局所環境は、それらが無効にされるように誤った経路に沿ってT細胞の分化を傾斜させる。これは、癌において特に当てはまり、この場合、T細胞は抑制表現型に押し込まれ、免疫反応を妨げ、腫瘍拡大がもたらされる可能性がある。
将来のT細胞治療薬の別の重要な要素は、T細胞を、それらが非未変性のものであってもカスタマイズされた治療ペイロードを送達することを可能にする、新しい能力で操作することである。天然T細胞またはCAR T細胞は、感染細胞または癌細胞を直接認識し、溶解性顆粒の送達を通してそれらを死滅させる(図100A)。しかしながら、天然のT細胞応答は、疾患を安全に根絶させるには不十分であるかまたは極端すぎることが多い。分泌された生物製剤などの、T細胞による治療用ペイロードのカスタム送達は、例えば、細胞傷害性活性を局所的に増強することにより、または免疫系により認識され殺傷されるように疾患の部位をプライミングすることにより処置困難な疾患を補助し得る。
synNotch受容体がin vitroでT細胞応答のスペクトルを精密に調節することができるため、受容体がin vivoで固形腫瘍に対して初代ヒトT細胞を選択的に標的とし、サイトカインIL−2などのカスタムペイロードの送達を誘導できるかどうかを調査した。これらの実験のために、免疫無防備状態のNOD scidガンマ(NSG)マウスにおいて、両側K562異種移植固形腫瘍モデルを確立し、ここで、非標的CD19−腫瘍及び標的CD19+腫瘍をそれぞれ左側腹部及び右側腹部に皮下移植した(図101A)。腫瘍を4日間確立させ、次にα−CD19 synNotch受容体ならびにIL−2発現及びIRES mCherryレポーターを制御する応答エレメントで操作したCD4+及びCD8+T細胞を静脈内(i.v.)注射した(図107A)。6日後、腫瘍を回収し、腫瘍浸潤T細胞をIL−2 IRES mCherryレポーターの発現について分析した(図101A〜101C)。腫瘍局在化T細胞のみが、標的CD19腫瘍においてIL−2発現に関するmCherryレポーターを発現し、レポーターレベルは、in vitroで刺激されたときに同じT細胞について観察されたレベルと同様であった(図101A〜101Cならびに図107B及び107C)。T細胞の頻度は腫瘍内で高くなかったが(CD4+では1000個の細胞中1個、CD8+T細胞では500個中1個)、T細胞の活性は標的腫瘍に対して高度に特異的であった(図101B及び101Cならびに図107D)。synNotch−>IL−2 T細胞の静脈内注射に加えて、T細胞は、非標的及び標的腫瘍にも直接注射された。次いで、腫瘍を回収し、注射2日後にフローサイトメトリーを介して分析し、同じく同様のレベルで標的腫瘍におけるIL−2レポーターの選択的発現を示して、in vitroで刺激したT細胞にマッチした(図107E)。これらの腫瘍に浸潤するsynNotch T細胞の能力はさらに改善され得るが、これらのデータは、synNotch受容体がT細胞を原発腫瘍に標的化し、局所的様式で治療物質の産生を選択的に誘導することができることを明確に示す。ゆえに、synNotch操作T細胞は、有効性を増強し、全身投与の毒性を低減する、この両方のために、局所送達から利益を得ることができる広範囲の遺伝的にコード可能な治療剤の送達に有効であると証明し得る。
(A)TCR及びCARは、T細胞応答の再形成をほとんど制御しない未変性T細胞活性化プログラムを推進するキナーゼベースのシグナル伝達カスケードを活性化する。synNotch受容体は、細胞表面抗原(例えば疾患関連抗原)を認識し、T細胞応答に対するより精密な制御を伴ってカスタム転写プログラムを直接調節する。ゆえに、synNotch受容体を用いて、アラカルト応答を操作することができる。(B)synNotch受容体は、目的の細胞表面抗原を検出するカスタムリガンド結合ドメイン(例えば、scFvまたはナノボディ)、Notchのコア調節領域、及び直交転写因子を含有する細胞質ドメイン(例えば、Gal4 VP64)を有し得る。カスタム転写プログラムを制御する直交転写因子の対応する応答エレメントは、受容体とともにT細胞へと操作されてよい。(C)癌関連抗原CD19を対象とするscFv及び直交抗原GFPに対するナノボディを有するsynNotch受容体を操作し、synNotch受容体プラットフォームの多能性を実証する。
(A)CD4+及びCD8+初代ヒトT細胞を、α−CD19 synNotch Gal4VP64受容体及びBFPレポーターの発現を制御する5x Gal4応答エレメントで操作した。(B)24時間の共培養後の、CD19−もしくはCD19+K562またはDaudi癌細胞(CD19+)を伴う送信側細胞での刺激に応答するα−CD19 synNotch受容体受信側T細胞におけるBFPレポーターの選択的誘導を示すヒストグラム(3回以上の実験の代表値)。(C)パネルBに示す複製データから算出された、送信側細胞を用いた24時間の刺激後にBFPレポーターを上方制御するα−CD19 synNotch T細胞の割合(%)(全ての条件についてn≧3、エラーバー=平均の標準誤差)。(D)CD4+及びCD8+初代ヒトT細胞を、α−GFP synNotch Gal4VP64受容体親和性変異体及びBFPレポーターで、パネルAと同様に操作した。(E)24時間の共培養後の、表面GFP−またはGFP+K562送信側細胞に応答するα−GFP synNotch受容体受信側T細胞におけるBFPレポーターの選択的誘導を示すヒストグラム(3回以上の実験の代表値、エラーバー=平均の標準誤差)。(F)パネルHに示す複製データから算出された、送信側細胞を用いた24時間の刺激後にBFPレポーターを上方制御するα−GFP synNotch T細胞の割合(%)(全ての条件についてn>3、エラーバー=平均の標準誤差)。
(A)α−CD19 synNotch Gal4VP64受容体(mycタグ付き)及びBFPの発現を制御する5xGal4応答エレメントで形質導入されたCD4+(左プロット)及びCD8+(右プロット)初代ヒトT細胞の2次元ドットプロット。応答エレメントベクターは、挿入された応答エレメントを有するT細胞を同定するためにmCherryの構成的発現を推進するPGKプロモーターも含有する。全てのBFPレポーター発現分析は、synNotch受容体及びBFP発現を制御する対応する5xGal4応答エレメントの両方を有したT細胞(右上枠の囲った集団)で行った。(B)24時間の共培養後に、CD19−K562対照細胞と比較してCD19+K562またはDaudi腫瘍細胞に応答するsynNotch受容体受信側T細胞におけるBFPレポーターの選択的誘導を示すヒストグラム(3回以上の実験の代表値)。(C)K562及びDaudi腫瘍細胞上のCD19レベルを示すヒストグラム。Daudi腫瘍は天然にCD19を発現し、K562は異所的にCD19を同様のレベルで発現する。(D)α−GFPナノボディsynNotch Gal4VP64受容体を発現するCD4(上段)及びCD8(下段)初代ヒトT細胞に関する、図103Aと同様の二次元ドットプロット。各カラムは、特定のα−GFPナノボディ親和性変異体(LaG17、LaG16−2)に関するものである。全てのBFPレポーター発現分析は、図103Aのように右上に輪郭を描いたゲート内のT細胞で行った。(E)24時間の共培養後に、表面GFP−K562と比較して表面GFP+K562癌細胞に応答するsynNotch受容体受信側T細胞におけるBFPレポーターの選択的誘導を示すヒストグラム(3回以上の実験の代表値)。(F)K562 GFP−対照と比較した、表面GFPを発現するK562癌細胞における総GFPレベルを示すヒストグラム。
(A)CAR活性化は、T細胞が多様なセットのサイトカインを産生するように推進する。(B)24時間の刺激後の、標的CD19+K562細胞(y軸)または陰性対照CD19−K562(x軸)で活性化された初代ヒトα−CD19 CAR CD4+T細胞により産生された、25種のサイトカイン(サイトカインの一覧については図98Cを参照されたい)のレベル(pg/mL)を示す散布図(n=3、エラーバー=平均の標準誤差)。(C)標的CD19+K562により刺激されたCD4+α−CD19 CAR T細胞により産生された25種のサイトカインのレベル(n=3、エラーバー=平均の標準誤差)。(D)CD4+T細胞を、α−CD19 synNotch Gal4VP64受容体及びIL−2またはIL−10のいずれかの発現を制御する対応する応答エレメントで操作した。細胞を、標的CD19+K562またはCD19−非標的K562と共培養した。(E)CD19+K562刺激に応答する単一サイトカインの産生を示すパネルBと同様の散布図(n=3、エラーバー=平均の標準誤差)。(F)CD19+K562細胞に応答してIL−2またはIL−10産生を推進するα−CD19 synNotch T細胞により産生されたサイトカインのレベル。単一のサイトカイン(IL−2またはIL−10)のみが、バックグラウンドレベルを超えて産生された(n=3、エラーバー=平均の標準誤差)。
(A)CD4+ヒト初代T細胞を、α−CD19 synNotch Gal4VP64受容体及びIL−2産生を制御する会合する5xGal4応答エレメントで操作した。(B)α−CD19 synNotch Gal4VP64受容体及び発現IL−2発現IRES mCherryを制御する5xGal4応答エレメントで形質導入されたCD4初代ヒトT細胞の2次元ドットプロット(左パネル)。応答エレメントベクターは、応答エレメントが挿入されているT細胞を同定するためにBFPの構成的発現を推進するPGKプロモーターも含有する。synNotch受容体及びIL−2 IRES mCherry発現を制御する対応する5xGal4応答エレメントの両方を有するT細胞を、選別し、図98及び図104の全ての対応するアッセイのために使用した。(右上四角)。左パネルは、形質導入していないT細胞と比較して、二重陽性T細胞においてIL−2 IRES mCherryレポーターの基礎誘導がないことを示す。(C)刺激されていないCD4+及びCD8+T細胞、IL−2産生を制御する刺激されていないα−CD19 synNotch Gal4VP64T細胞、及びPMA/イオノマイシンで6時間刺激された陽性対照T細胞について、IL−2に関する細胞内サイトカイン染色のドットプロットを示す。(D)基礎及び刺激IL−2レベルを、形質導入していないCD4+T細胞、α−CD19 4−1BBζ CAR T細胞、及びIL−2産生を制御するα−CD19 synNotch Gal4VP64 T細胞から回収された上清について提供する(n=4)。(E)α−CD3/CD28 dynabeadsで刺激された対照CD4+T細胞及びCD19−またはCD19+K562で刺激されたIL−2産生を制御するα−CD19 synNotch Gal4VP64 T細胞のCD69レベル(左カラム)及びIL−2 IRES mCherryレポーターレベル。CD69は、同族抗原での刺激の際にはsynNotch T細胞で上方制御されない。(F)CD4+ヒト初代T細胞を、α−CD19 synNotch Gal4VP64受容体及びIL−10産生を制御する会合する5xGal4応答エレメントで操作した。(G)α−CD19 synNotch Gal4VP64受容体及びIL−10 IRES mCherry発現の発現を制御する5xGal4応答エレメントで形質導入されたCD4+初代ヒトT細胞に関するパネルBに同等のデータ。(H)α−CD19 synNotch Gal4VP64受容体及び発現IL−10 IRES mCherry発現を制御する5xGal4応答エレメントで形質導入されたCD4+初代ヒトT細胞に関するパネルDに同等のデータ。(I)α−CD19 synNotch Gal4VP64受容体及び発現IL−10 IRES mCherry発現を制御する5xGal4応答エレメントで形質導入されたCD4+初代ヒトT細胞に関する図104Eに同等のデータ。
(A)天然のT細胞分化:CD4+T細胞が、抗原提示細胞上のMHC分子により提示される病原体由来ペプチドの係合を通して活性化されるとき、それらは感染に応じて特定のT細胞サブタイプへと分化する。Th1及びTh2は、異なる免疫反応を推進する正準のCD4+T細胞運命である。Th1細胞は、転写因子Tbetを発現し、IFNγを産生し、細胞免疫及び腫瘍クリアランスを助ける。Th2細胞は、B細胞による抗体産生の刺激のための重要なサイトカインであるIL−4を産生する。(B)SynNotch推進性T細胞分化:CD4+α−CD19 synNotch T細胞を、発現Tbetを、ゆえにT細胞によるTh1運命選択を調節するように操作した。synNotch T細胞を、標的CD19+または非標的CD19−K562細胞と11日間共培養して、synNotch推進性Tbet発現がCD4+T細胞をTh1運命へとCD19依存性様式で傾斜できるかどうかを決定した。(C)24時間のCD4+α−CD19 synNotch T細胞とCD19+K562の後のTbet T2A EGFPの選択的発現を示すヒストグラム(少なくとも3回の実験の代表値)。(D)CD19+またはCD19−K562のいずれかとの11日間の培養後のTbet及びIFNγに関する細胞内染色CD4+α−CD19 synNotch Gal4VP64 T細胞の2次元ドットプロット。T細胞を、染色する前にPMA/イオノマイシンで4時間にわたって刺激してサイトカイン産生を推進した(少なくとも3回の実験の代表値)。(E)11日間の指定した処理後のIFNγ+(Th1)T細胞の割合(%)(全ての処理についてn≧3、エラーバー=平均の標準誤差、有意性はスチューデントのt検定により決定、有意差無しp>0.05)。
(A)図99Cからの複製データの定量。Tbet T2A GFP発現を制御するα−CD19 synNotch Gal4VP64受容体応答エレメントを有するCD4+T細胞を、CD19+またはCD19−K562と24時間共培養した。Tbet T2A GFP発現を伴うT細胞の割合(%)を定量化し、これは、TbetはT細胞がCD19+K562に曝露されたときにのみ上方制御されたことを示す(n=3)。(B)α−CD3/CD28 dynabeadsで刺激され、Th1サイトカインIFNγについて細胞内染色された、11日間の培養後のCD4 T細胞の代表的なドットプロット。IFNγ陽性ゲートを赤色で囲った。(C)Th1分化条件(IL−12+α−IL−4)において11日間培養されたCD4+T細胞に関するパネルBと同様の代表的なドットプロット(n=4)。(D)形質導入の48時間後のTbet T2A mCherryの構成的過剰発現を伴うCD4 T細胞の代表的なヒストグラム。(E)11日間の培養後での、Tbet及びIFNγに関する、Tbet T2A mCherryの構成的過剰発現を伴うCD4+T細胞の細胞内染色の代表的なドットプロット(n=6)。二重陽性Tbet及びIFNγ T細胞を赤色で囲った。(F)対照CD19−K562と共培養したTbet T2A GFP発現を制御するα−CD19 synNotch Gal4VP64受容体応答エレメント(構成的にmCherryを発現する)を有するCD4+T細胞の代表的な顕微鏡検査。synNotch T細胞(赤色細胞)は、癌細胞と会合しないまたはTbet発現のレポーター(GFP)を上方制御しない。(G)パネルFと同様だが、T細胞をCD19+T細胞と共培養した代表的な顕微鏡検査。synNotch T細胞(赤色細胞)は、CD19+K562と会合体を形成し、Tbet T2A GFP発現を上方制御する。活性化したsynNotch T細胞は、mCherry及びGFP共発現のために黄色である。
(A)天然T細胞の細胞傷害性:CD8+細胞傷害性T細胞は、それらのTCRを介して感染細胞を認識し、パーフォリンで細胞内に孔を作製し、これによりプログラムされた細胞死を開始するグランザイムの送達を可能にし、感染した細胞を直接殺傷する。(B)SynNotchでカスタマイズしたキラーT細胞:CD4+T細胞を、表面GFPに応答してアポトーシス調節因子TRAILの発現を制御するα−GFP synNotchで操作した。(C)24時間のCD4+α−GFP synNotch T細胞と表面GFP+K562の後の表面TRAILの選択的発現を示すヒストグラム。(D)24時間後の死染色SYTOX Blueの取込みを介した表面GFP+K562細胞死を示すヒストグラム。指定したT細胞型との共培養(T細胞:標的細胞比率=1:1)。(E)パネルDに示す複製データから算出した標的細胞生存率(%)(n=4、エラーバー=平均の標準誤差)。
(A)静止させたCD4+及びCD8+T細胞を24時間にわたってα−CD3/CD28 dynabeadsで再刺激し、細胞表面TRAILについて染色した。いずれのT細胞サブセットもTRAIL発現を急性に上方制御しなかった。(B)癌細胞は、組換えTRAILが媒介するアポトーシスに対するそれらの感受性において変動する。HCT116結腸癌は、デスレセプターがTRAILに結合され、低レベルTRAIL処置に対して感受性であった。K562は、TRAILに対してデスレセプターを発現するが、TRAIL処置に対して非感受性であった。(C)表面GFP−もしくはGFP+K562またはHCT116癌細胞を、0〜200ng/mLの組換えTRAILで24時間にわたって処置し、死亡を死染色SYTOX Blueを用いた染色によりフローサイトメトリーを介してモニターした(n=4)。(D)TRAILに結合されたデスレセプター4(DR4)に関して染色したK562のヒストグラム。K562は、受容体を発現し、それらはTRAIL媒介性アポトーシスに対して耐性。(E)LaG17 α−GFPナノボディsynNotch Gal4VP64及びロイシンジッパーTRAIL(LZ−TRAIL左パネル)または完全長表面TRAIL(右パネル)の発現を制御する5xGal4応答エレメントで形質導入されたCD4+T細胞の代表的なドットプロット。TRAIL発現のmCherryレポーターのレベルを、二重陽性T細胞及び対照の形質導入していないT細胞について以下に示す。(F)LaG17 α−GFPナノボディsynNotch Gal4VP64及び発現LZ−TRAILを制御する5xGal4応答エレメントで形質導入されたCD4+T細胞を、表面GFP−またはGFP+K562と24時間共培養して、LZ−TRAILがGFP+K562に応答してのみ分泌されるかどうかを決定した。(G)表面GFP=またはGFP+K562と共培養された図106Eに示す選別されたSynNotch CD4 LZ−TRAIL T細胞におけるTRAIL産生のmCherryレポーターレベルのレベルを示すヒストグラム。レポーターは、表面GFP+K562に応答して排他的に活性化した。(H)表面GFP−またはGFP+K562のいずれかと24時間にわたって共培養された、選別されたSynNotch CD4 LZ−TRAIL T細胞からの上清のTRAIL ELISA(n=2)。
(A)NSGマウスに、CD19−非標的K562及び標的CD19+をそれぞれ左側腹部及び右側腹部に皮下注射した。IL−2 iRES mCherry発現を制御するα−CD19 synNotch T細胞を、腫瘍が確立した後にマウスに注射し、腫瘍を指定した時点で回収して、synNotch T細胞が腫瘍に浸潤し、IL−2及びmCherryレポーターの発現が誘導されたかどうかを決定した。(B)腫瘍に浸潤した静脈内注射されたCD4+及びCD8+synNotch T細胞におけるIL−2 IRES mCherryレポーターレベルのヒストグラムであり、これは、標的CD19+腫瘍におけるmCherryレポーターの選択的発現を示す(データは、3匹の複製マウスの代表値を表す)。(C)パネルBに示す複製データからIL−2発現のmCherryレポーターの発現を誘導した腫瘍浸潤T細胞の割合(%)の定量(n=3、有意性はスチューデントのt検定により決定**p<0.01)。
(A)α−CD19 synNotch Gal4VP64受容体(mycタグ付き)及びIL−2 IRES mCherryの発現を制御する5xGal4応答エレメントで形質導入されたCD4+(左プロット)及びCD8+(右プロット)初代ヒトT細胞の2次元ドットプロット。応答エレメントベクターは、応答エレメントを挿入されたT細胞を同定するためにBFPの構成的発現を推進するPGKプロモーターも含有する。右上影付き四角内のT細胞を選別し、全てのin vivo及びin vitro実験で使用した。(B)in vitroでCD19−またはCD19+K562で刺激されたIL−2発現を制御するCD4+及びCD8+synNotch T細胞の代表的なヒストグラム。IL−2 IRES mCherry発現は、CD19の存在下でのみ生じた。(C)図107Bに示す複製データからのIL−2発現のmCherryレポーターの発現を誘導したCD4+及びCD8+T細胞の割合(%)の定量(n=3、有意性はスチューデントのt検定により決定***p<0.001及び****p<0.0001)。(D)腫瘍内注射した腫瘍浸潤CD4+及びCD8+synNotch T細胞におけるIL−2 IRES mCherryレポーターレベルのヒストグラムであり、これは、標的CD19+腫瘍におけるmCherryレポーターの選択的発現を示す(データは、3匹の複製マウスの代表値を表す)。腫瘍内注射実験に関するプロトコルを示す。結果は、静脈内注射したT細胞及びin vitro刺激したT細胞で観察されるものと同様であった。(E)静脈内注射及び腫瘍内注射の後にCD19−及びCD19+腫瘍に浸潤しているCD4+及びCD8+T細胞の割合(%)(n=3、エラーバー=平均の標準誤差、有意性はスチューデントのt検定で決定、有意差無しp>0.05)。
(A)synNotch受容体は、T細胞応答の多能調節因子であり:synNotch受容体は、初代ヒトT細胞において多様な挙動を推進することができる。synNotch受容体はカスタムサイトカイン産生プロファイルを推進し、非未変性治療薬を有効に送達し、T細胞分化を制御することができ、それは全て抗原依存性でありT細胞活性化に依存しない様式である。(B)synNotchは、in vivoでT細胞を標的として治療用ペイロードを局所的に産生するのに十分である。
材料及び方法
以下の材料及び方法を、別段の指定がない限り、実施例5に記載の結果に適用する。
synNotch受容体は、CD19 scFv、LaG17(低親和性)、またはLaG16_2(高親和性)ナノボディをマウスNotch1(NM_008714)最小調節領域(Ile1427〜Arg1752)及びGal4VP64またはTetR VP64(tTa)に融合させることにより構築した。全てのsynNotch受容体は、n末端CD8αシグナルペプチド
初代CD4+及びCD8+T細胞を、陰性選択によるアフェレーシスの後に匿名のドナー血液から単離した(STEMCELL Technologies #15062及び15063)。血液は、University Institutional Review Boardの承認を得て、Blood Centers of the Pacific(カリフォルニア州サンフランシスコ)から得た。T細胞を、20%ヒトAB血清(Valley Biomedical Inc.,#HP1022)及び10%DMSOを含むRPMI−1640(UCSF cell culture core)中で凍結保存した。解凍後、全ての実験について、30単位/mL IL−2(NCI BRB Preclinical Repository)を補充した、X−VIVO 15(Lonza #04−418Q)、5%ヒトAB血清及び10mM中和N−アセチルL−システイン(Sigma−Aldrich #A9165)からなるヒトT細胞培地中でT細胞を培養した。
汎親和性VSV−G偽型レンチウイルスを、Fugene HD(Promega #E2312)を使用して、pHR’SIN:CSW導入遺伝子発現ベクターならびにウイルスパッケージングプラスミドpCMVdR8.91及びpMD2.Gを用いたレンチ−X 293T細胞(Clonetech #11131D)のトランスフェクションを介して産生した。初代T細胞を、同日に解凍し、培養24時間後、1:3の細胞:ビードの比率でDynabeads Human T−Activator CD3/CD28(Life Technologies #1113ID)で刺激した。48時間で、ウイルス上清を回収し、初代T細胞を24時間ウイルスに曝露した。T細胞刺激後4日目に、Dynabeadsを除去し、それらを静止させアッセイに使用できる9日目までT細胞を増殖させた。T細胞を、FACs ARIA IIを用いたアッセイのために選別した。
使用した癌細胞株は、K562骨髄性白血病細胞(ATCC #CCL−243)、Daudi B細胞リンパ芽球(ATCC #CCL−213)、及びHCT115結腸癌細胞(ATCC #CCL−247)とした。Daudi腫瘍と同等のレベルでヒトCD19を安定して発現させるために、K562をレンチウイルスで形質導入した。CD19レベルは、細胞をα−CD19 APC(Biolegend #302212)で染色することによって決定した。K562を、表面GFP(PDGF膜貫通ドメインに融合したGFP)を安定して発現するためにも形質導入した。全ての細胞株を導入遺伝子の発現について選別した。
全てのin vitro synNotch T細胞刺激について、2×105個のT細胞を1:1の比率で送信側細胞と共培養した。丸底96ウェル組織培養プレートでT細胞及び送信側細胞を混合した後、細胞を400xgで1分間遠心分離して、細胞の相互作用を引き起こし、培養物を、CD4+T細胞の活性化のマーカー(例えば、CD69)及びCD8+T細胞の標的腫瘍細胞の特異的溶解について、BD LSR IIを用いて、24時間で分析した。全てのフローサイトメトリー分析は、FlowJoソフトウェア(TreeStar)で行った。
α−CD19 synNotch Gal4VP64受容体及びCARを制御する5xGal4応答エレメントを発現する初代CD4+T細胞を、指定の標的癌細胞株で上に記載したように刺激した。上清を24時間で回収し、製造者のプロトコルに従って、Luminex MAGPIX(Luminex Corp.)ヒトサイトカインマグネティック25−プレックスパネル(Invitrogen参照番号LHC0009M)で分析した。全てのサイトカインレベルを、xPONENTソフトウェア(Luminex Corp.)を用いて標準曲線に基づいて算出した。
CD4+synNotch ANDゲートT細胞を、上記のように24時間にわたって指定した癌細胞株で刺激し、上清を回収した。上清におけるIL−2レベルは、IL−2 ELISA(eBiosciences #BMS2221HS)を介して決定した。T細胞も採取し、α−CD69 APC(Biolegend #310910)で染色して、それらが活性かどうかを決定した。
CD8+synNotch ANDゲートT細胞を、24時間にわたって上記のように、指定の抗原を発現する標的細胞で刺激した。標的癌細胞の特異的溶解のレベルを、形質導入していないT細胞対象を用いた処置と比べて培養物中で生存している標的細胞の割合を比較することによって決定した。細胞死を、live/dead染色SYTOX Blueの取込み、及び通常は標的細胞によって占有される正常なSSC/FSC領域から標的細胞をシフトさせることによりモニターした(Thermo Scientific #S34857)。
α−GFPナノボディ(LaG17)synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CAR IRES effluc発現を制御する対応する応答エレメントを発現するように操作した、選別されたCD4+及びCD8+初代ヒトT細胞を、GFP+またはGFP−K562細胞で24時間刺激した(2×105個のT細胞及び2×105個のK562)。efflucの産生は、ONE−gloルシフェラーゼアッセイシステム(Promega #E6110)でアッセイした。生物発光は、FlexStation 3(Molecular Devices)で測定した。
動物試験を、UCSF Institutional Animal Care and Use Committeeに承認されたプロトコル下にてUCSF Preclinical Therapeutics Coreで実施した。T細胞注射の10日前に、Daudi腫瘍及び表面GFP Daudi腫瘍をNOD scidガンマ(NSG)マウス(雌、8〜12週齢、Jackson Laboratory #005557)の左右に皮下注射した。α−GFPナノボディ(LaG17)synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CAR IRES effluc発現を制御する対応する応答エレメントを発現するように操作した、選別されたCD4+及びCD8+初代ヒトT細胞を、1:1のCD4+対CD8+T細胞割当で(1×106個の各T細胞型)、腫瘍を有するマウスに腫瘍移植の10日後に静脈内注射した。ルシフェラーゼ発現を、11日間にわたってモニターし、生物発光イメージングを、IVIS 100(Xenogen)前臨床イメージングシステムを使用して指定の時点で行った。画像は、150mg/kgのD−ルシフェリン(Gold Technology #LUCK−100)の腹腔内注射の10分後に取得した。統合した生物発光強度の定量を、ImageJ(NIH)にて定量化した。
NSGマウスに、5×106個のCD19 K562及びGFP/CD19 K562腫瘍細胞を、それぞれ左側腹部及び右側腹部で皮下移植した。腫瘍移植の4日後、α−GFP synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CAR発現を調節する対応する応答エレメントで操作した1×106個の初代ヒトCD4+及びCD8+T細胞(2×106個の総T細胞)、または形質導入していないT細胞を、マウスに静脈内注射した。腫瘍サイズは、UCSF Preclinical Therapeutics CoreのスタッフによりキャリパーによりT細胞注射後20日間にわたりモニターした。カプラン・マイヤー実験では、同一のプロトコルを使用したが、単一の腫瘍が注射された。マウスは、腫瘍サイズが安楽死基準(2000mm3)に達したときに死亡したとみなされた。
別段の記載がない限り、統計学的有意性をスチューデントのt検定(両側検定)により決定した。全ての統計学的分析及び曲線適合は、Prism 6(Graphpad)を用いて行い、p値を報告した(有意差無し=p>0.05、*=p≦0.05、**=p≦0.01、***=p≦0.001、****=p≦0.0001)。全てのエラーバーは、平均の標準誤差または標準偏差のいずれかを表す。
2つの抗原ANDゲート回路の設計:synNotch受容体は、CAR発現を誘導する。
CARの発現を制御するためにsynNotch受容体を使用するこの方法を検査するために、Jurkat T細胞の活性化に対して組み合わせ抗原制御を操作する試みがなされた。これらの実験では、2つのモデル腫瘍抗原であるCD19及びメソテリンを標的とした。Jurkat T細胞を、細胞内テトラサイクリン制御トランス活性化因子(tTa)ドメインを有するα−CD19 synNotch受容体で操作した。α−メソテリン4−1BBζ CAR遺伝子を、synNotch受容体により活性化された対応するテトラサイクリン応答エレメント(TRE)を伴うプロモーターの制御下で、挿入した(図109A及び109B)。操作したJurkatを、in vitroで、CD19、メソテリン、または両抗原一緒の異所的発現を伴う標的K562骨髄性白血病細胞と共培養した(図109A)。これらの操作したJurkat細胞が、α−CD19 synNotch刺激に応答してのみα−メソテリンCARを発現するため、T細胞は、メソテリン単独に応答して活性化するはずがない。T細胞がCD19に曝露された場合、α−メソテリンCARが発現され、T細胞は活性化のためにプライムされる(図109A及び109Bならびに図114A)。次いで、T細胞は、メソテリンを感知し、活性化することができる。
Jurkat T細胞におけるsynNotch ANDゲートの成功を考慮し、初代T細胞において同じタイプのsynNotch推進性CAR発現回路が複数の抗原を識別するように機能し得るかどうかを検査した。あるスペクトラムの異なるsynNotch Gal4VP64受容体が初代T細胞において十分に発現し、Gal4応答エレメントは初代T細胞において最小基礎活性を有することがわかった。これは、synNotch推進性CAR発現ANDゲートにとって理想的なシナリオである。それは、T細胞がsynNotch抗原を感知するまでは、CARを活性化する基礎発現がないべきであるからである。
synNotch受容体が、in vitroで初代T細胞においてCAR発現を確かにゲートするため、in vivoでT細胞がsynNotch受容体を介して腫瘍を標的とし、腫瘍微小環境にある時にのみCARを発現し得るかどうかを検査した。この実験のために、両側異種移植CD19 Daudi B細胞リンパ芽球腫瘍を、免疫無防備状態のNOD scid IL−2Rγ−/−(NSG)マウスへと注射した。野生型Daudi細胞(synNotchリガンドを含有しない)を、左側腹部に皮下注射し、一方で表面GFPを発現するDaudi腫瘍細胞を右側腹部に注射した。移植のために腫瘍に10日間を与えた後、α−GFP synNotch Gal4VP64受容体ならびにα−CD19 4−1BBζ CAR及びIRES増強ホタルルシフェラーゼ(effluc)レポーターの発現を制御する対応する応答エレメントを備えた初代CD4+及びCD8+ヒトT細胞(図116A及び116B)を注射した。ルシフェラーゼ発現を、11日間の経過にわたってCAR発現に対するレポーターとしてモニターした(図111A、図116C)。T細胞は、1日目までにGFP+Daudi腫瘍においてCARを選択的に発現し始め、二重抗原腫瘍において11日の期間にわたってCARの局所的発現を増加させ続けた(図111B及び111C)。標的腫瘍におけるルシフェラーゼシグナルの増加は、二重抗原標的腫瘍におけるsynNotch推進性CAR発現及び細胞の拡大の組み合わせである可能性が高い。(図116B)。対照GFP−腫瘍では、ルシフェラーゼの増加は観察されなかった。
α−GFP synNotch受容体がT細胞を腫瘍に対して標的化し、α−CD19 CARの局所発現を制御し得ることが示された。次いで、synNotch ANDゲートT細胞がin vivoで二重抗原腫瘍を選択的に排除し得るかどうかを検査した。これらの実験のために、同様の両側腫瘍モデルをK562腫瘍細胞で構築した(図112A及び図117A及び117B)。腫瘍を移植し、移植に4日間かけた(K562腫瘍は、Daudi細胞と比較して、より迅速に増殖し、より大きい腫瘍を確立する)。4日目に、α−GFP synNotch−>α−CD19 CAR ANDゲート回路を有するCD4+及びCD8+T細胞を注射し、腫瘍増殖を、キャリパーにより20日間モニターした(図112A)。マウスの一群を形質導入していない対照T細胞で処置して、腫瘍増殖の基準を用意した。この実験では、T細胞を、全てが同じ動物内にある、二重抗原「疾患」腫瘍を単一抗原「バイスタンダー」組織から識別するために直接攻撃する。
(A)CARまたは腫瘍特異的TCR T細胞は、通常単一抗原を標的とし、それによりしばしばオフターゲット組織損傷を引き起こす。改善された治療用T細胞は、腫瘍抗原及び組織特異的抗原の両方の組み合わせを認識する多数のセンサーを必要とするだろう。これは、細胞がそれらの環境を評価し、いつ活性化させるかをより精密に決断することを可能にする。そのような治療用細胞は、標的罹患組織を正常組織から区別するよりために備えられた方がよい。(B)抗原の組み合わせを感知し、T細胞シグナル伝達及び転写を調節する新しいタイプの受容体は、精巧な細胞の判断及びより精密な治療用T細胞応答を可能にするように組み立てなければならない。(C)synNotch受容体を、カスタム細胞外リガンド結合ドメイン、例えば、目的の抗原を対象とするscFvまたはナノボディ(例えば、腫瘍または組織特異的抗原)で操作する。synNotch受容体によるリガンド認識の際、直交転写因子(例えば、TetRVP64またはGal4VP64)は、カスタム遺伝子回路を制御する細胞質尾部から切断される。(D)synNotch ANDゲート回路の設計であり、T細胞は活性化するために2つの抗原を感知することを必要とする。このANDゲートシグナル伝達回路は、2つの連続するステップで作動する:1)synNotch受容体は、T細胞が第一の抗原Aを認識することを可能にし、2)T細胞は、第二の腫瘍抗原Bを対象とするCARを発現する。A及びBが存在する場合、T細胞は、活性化し、標的腫瘍を殺傷することができる。
(A)2つの受容体ANDゲート回路の操作:α−CD19 synNotch受容体は、α−メソテリンCAR発現を誘導する。(B)Jurkat T細胞を、α−CD19 synNotch tTa受容体及びα−メソテリンCAR発現を制御する対応する応答エレメントで操作した。Jurkat T細胞は、それらのsynNotch受容体を介して標的細胞上のCD19をまず認識して、CAR発現を開始しなければいけない。T細胞がCD19によりプライムされて活性化された後、α−メソテリンCARは、メソテリンと結合し、Jurkat細胞を活性化することができる。T細胞活性化の2つの正準のマーカーは、CD69上方制御及びIL−2産生である。synNotch ANDゲートJurkat T細胞は、CD19及びメソテリンの両方を発現する標的腫瘍細胞に曝露されたときにのみ活性化するべきである。(C)単一抗原(メソテリン単独)または二重抗原(CD19/メソテリン)K562腫瘍細胞と48時間にわたって共培養されたsynNotch ANDゲートJurkat T細胞における活性化マーカーCD69のヒストグラム。CD69は、T細胞が二重陽性K562に曝露されたときにのみ発現された(3つの独立した実験の代表値)。(D)二重抗原K562に暴露されたときにのみsynNotch AND−ゲートJurkatによるIL−2産生を示すIL−2 ELISA(n=3、エラーバーは平均の標準誤差である、有意性はスチューデントのt検定により決定、****=p≦0.0001)
(A)ヒト初代CD4+及びCD8+T細胞を、α−GFPナノボディsynNotch Gal4VP64受容体及びα−CD19 4−1BBζ CARの発現を制御する対応する応答エレメントで操作した。これらのCD4+またはCD8+synNotch ANDゲートT細胞は、まずそれらのsynNotch受容体を介して表面GFPを感知しなければならず、その後でのみ、それらはα−CD19 CARを発現し、プライミングされて活性化される。synNotch ANDゲート初代T細胞は、それらがGFPとCD19の両方を感知した場合にのみ、活性化し、サイトカインを産生するまたは標的細胞を殺傷するべきである。(B)パネルAに記載した初代CD4+synNotch ANDゲートT細胞を、CD19単独または表面GFP/CD19 K562と共培養した。α−CD19 CAR GFP受容体発現レベルのヒストグラムは、GFPが標的細胞の表面上に存在するときにのみCARが発現されることを示す(少なくとも3つの独立した実験の代表値)。(C)CD19単独またはGFP/CD19 K562のいずれかにより活性化されたCD4+synNotch ANDゲートT細胞からの上清を、Luminexを介して25種のサイトカインの存在について分析した。サイトカインは、T細胞がGFP/CD19 T細胞に曝露された時にのみ産生された(エラーバーは平均の標準誤差である、n=3)。(D)CD8+synNotch ANDゲート初代T細胞を、パネルAに記載したように操作した。CD4+T細胞と同様に、α−CD19 CAR GFP受容体発現レベルのヒストグラムは、GFPが標的細胞の表面上に存在するときにのみCARが発現されることを示す(少なくとも3つの独立した実験の代表値)。(E)CD8+synNotch ANDゲート初代T細胞とCD19単独またはGFP/CD19腫瘍細胞のいずれかとの24時間の共培養後の前方及び側方散乱フローサイトメトリープロット。T細胞は、青色ゲートの中にあり、標的K562は赤色ゲート内にある。synNotch ANDゲートT細胞は、GFP/CD19 K562のみを殺傷し、それは、K562ゲート内で減少する細胞により示される(3つの実験の代表値)。(F)パネルEに示す複製CD8+synNotch ANDゲート初代T細胞の細胞傷害性データの定量(n=3、エラーバーは平均の標準誤差である、有意性はスチューデントのt検定により決定*=p≦0.05)。
(A)初代ヒトCD4+及びCD8+T細胞を、α−GFP synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CAR IRES effluc発現を調節する対応する応答エレメントで操作し、マウスに、Daudi腫瘍(CD19単独)を左側腹部で、表面GFP Daudi(GFP/CD19)腫瘍を右側腹部で静脈内注射した。ルシフェラーゼ発現を、操作したT細胞の静脈内注射後11日間にわたってモニターした。(B)T細胞注射の7日後の、パネルAに記載のように処置されたマウスにおけるルシフェラーゼ発現の代表的な画像。ルシフェラーゼ発現は、GFP/CD19腫瘍においてのみ高く、これは、二重抗原腫瘍における局在化したCAR発現を示す(n=2マウス)。(C)左側腹部Daudi腫瘍(CD19単独)及び右側腹部の表面GFP Daudi腫瘍(GFP/CD19)におけるルシフェラーゼレベルの統合強度の定量。ルシフェラーゼ発現は、全ての時点で、二重抗原腫瘍において高い(エラーは標準偏差である、n=2)。
(A)初代ヒトCD4+及びCD8+T細胞を、α−GFP synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CAR発現を調節する対応する応答エレメントで操作し、マウスに、CD19 K562を左側腹部で、表面GFP/CD19 K562腫瘍を右側腹部で静脈内注射した。腫瘍サイズを、操作したT細胞または形質導入していないT細胞対照の静脈内注射後16日間にわたってモニターした。(B)synNotch ANDゲートT細胞(上)及び形質導入していない対照T細胞(下)で処置したマウスのCD19及びGFP/CD19腫瘍体積を示すグラフ。synNotch ANDゲートT細胞は、二重抗原腫瘍を排他的に標的とし、CD19単独腫瘍は、形質導入していない対照T細胞で処置したマウスと同一の速度で増殖した(n=5マウス、エラーバーは平均の標準誤差である、有意性はスチューデントのt検定により決定する**=p≦0.01、***=p≦0.001)。(C)synNotch ANDゲートT細胞で処置した個別のマウスに関する腫瘍体積測定。全てのマウスは、二重抗原腫瘍の選択的殺傷を示した。(D)synNotch ANDゲートT細胞がGFP/CD19腫瘍を排除し、100%のマウスが生存していることを示すカプラン・マイヤーグラフ。CD19単独腫瘍を有するマウスは、synNotch ANDゲートT細胞により排除されず、腫瘍成長は制御されない。対応する腫瘍成長曲線を、パネルDの右に提示する(n=5マウス、エラーバーは平均の標準誤差である、有意性はスチューデントのt検定により決定する**=p≦0.01)。
(A)T細胞を、腫瘍抗原を感知し、第二の抗原に対するCARの発現を上方制御するsynNotch受容体で操作した(を産生するように遺伝子改変した)。ゆえに、これらのsynNotch ANDゲートT細胞は、腫瘍微小環境における組み合わせ抗原認識に応答してのみ活性化し、単一抗原認識により媒介されるオフターゲット毒性を防ぐ。(B)単一抗原を標的とする治療用T細胞とは異なるsynNotch ANDゲートT細胞は、組み合わせ抗原標的を単一抗原バイスタンダー組織から確実に識別することができる。synNotch−CAR T細胞による組み合わせ抗原感知は、腫瘍に対するT細胞の精密な標的化を助けることができ、オフターゲット毒性を防ぐ。(C)標的可能抗原空間の拡大。腫瘍特異的抗原は、腫瘍関連抗原(正常組織において発現されるが、腫瘍上により高く発現される抗原)と比較して稀である。CARはT細胞を完全に活性化して標的組織の殺傷をもたらすため、致命的なオフターゲット毒性を減少させるために、単一CAR T細胞を腫瘍特異的抗原に標的化しなければならない(上のベン図)。synNotch受容体は、CAR発現をゲートし、T細胞が武装している場所を制御することができる。腫瘍特異的抗原の組み合わせを標的とするときには、腫瘍関連抗原を対象とするCAR受容体を使用することが可能であり得る。これは、身体の他の部分でCAR抗原を発現する組織へのオフターゲット損傷を減少させるはずである。
(A)GFPに融合したα−メソテリンCARの発現を制御するα−CD19 synNotch Jurkat T細胞を、48時間にわたってメソテリン単独またはCD19及びメソテリン+K562とインキュベートした。α−メソテリンCAR GFPレベルのヒストグラムは、CARが、JurkatがCD19に曝露されたときにのみ発現することを示す。(B)パネルA及び図2Bのヒストグラムから算出した正規化したα−メソテリンCAR GFP及びCD69レベルのプロット。CAR及びCD69の最大発現の半減期は、それぞれ6時間及び13時間であった。(C)synNotch ANDゲートJurkat T細胞を、synNotch受容体の細胞外ドメイン上のmyc−タグと結合するプレート結合型α−myc抗体で24時間刺激した。24時間後、細胞をα−myc刺激から取り除き、CAR発現を24時間モニターした。(D)synNotch刺激の除去後のα−メソテリンCAR GFP発現のヒストグラム。正規化したCARをプロットし、1フェーズ減衰にあてはめ、これは、下方制御の8時間半減期を示す。
(A)CD4+初代T細胞を、α−CD19 synNotch Gal4VP64受容体及びα−メソテリン4−1BBζ CAR EGFP発現を制御する対応する応答エレメントで操作した。次いで、T細胞を、メソテリン単独、CD19単独、またはCD19/メソテリンK562と、24時間にわたって共培養し、CD69上方制御及びIL−2産生をアッセイした。(B)パネルAに記載のように培養されたCD4+synNotch初代T細胞上のα−メソテリンCAR EGFPレベル及びCD69レベルを示すヒストグラム。α−メソテリンCARは、CD19が標的K562上にあるときにのみ発現され、T細胞は、活性化マーカーCD69を、CD19及びメソテリンの両方が標的K562上にあるときにのみ発現した(3つの実験の代表値)。(C)パネルAに記載の培養物から回収した上清からのIL−2レベル。IL−2は、T細胞がCD19及びメソテリンの両方を発現する標的細胞に曝露されたときにのみ産生された(n=3、エラーバーは平均の標準誤差である、有意性はスチューデントのt検定により決定***=p≦0.001)。(D)CD8+初代ヒトT細胞をパネルAに記載のように操作した。CD8+T細胞について、メソテリン単独、CD19単独、またはCD19/メソテリン標的K562の特異的細胞傷害性を決定した。synNotch ANDゲートCD8+T細胞は、二重陽性K562のみを殺傷するべきである。(E)パネルAに記載のように培養されたCD8+synNotch初代T細胞上のα−メソテリンCAR EGFPレベルを示すヒストグラム。α−メソテリンCARは、CD19が標的K562上にあるときにのみ発現された(3つの実験の代表値)。(F)CD19及びメソテリンの両方の発現を伴う標的K562の特異的殺傷を示す複製CD8+synNotch ANDゲート初代T細胞の細胞傷害性の定量(n=3、エラーバーは平均の標準誤差である、***=p≦0.001)。(G)CD4+初代T細胞を、α−GFPナノボディsynNotch Gal4VP64受容体及びα−メソテリン4−1BBζ CAR EGFP発現を制御する対応する応答エレメントで操作した。次いで、T細胞をメソテリン単独、GFP単独、またはGFP/メソテリンK562と24時間にわたって共培養し、CD69上方制御及びIL−2産生をアッセイした。(H)パネルGに記載のように培養されたCD4+synNotch初代T細胞上のα−メソテリンCAR EGFPレベル及びCD69レベルを示すヒストグラム。α−メソテリンCARは、GFPが標的K562上にあるときにのみ発現され、T細胞は、活性化マーカーCD69を、GFP及びメソテリンの両方が標的K562上にあるときにのみ発現した(3つの実験の代表値)。(I)パネルGに記載の培養物から回収した上清からのIL−2レベル。IL−2は、T細胞がGFP及びメソテリンの両方を発現する標的細胞に曝露されたときにのみ産生された(n=3、エラーバーは平均の標準誤差である、****=p≦0.0001)。
(A)初代CD4+及びCD8+T細胞におけるα−GFP synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CAR IRES efflucを調節する対応する応答エレメントの発現を示す代表的なドットプロット。赤色枠で囲ったT細胞を選別し、in vivo及びin vitro実験のために使用した。(B)GFP−またはGFP+K562を用いた24時間にわたる曝露後のパネルAからのsynNotch AND CD4+及びCD8+T細胞におけるルシフェラーゼ活性を示す棒グラフ。ルシフェラーゼは、GFPに応答して特異的に発現された(n=3、エラーバーは平均の標準誤差である、****=p≦0.0001)。(C)腫瘍成長曲線を、図4Cで分析したマウスについて提供する。
(A)初代ヒトCD4+及びCD8+T細胞におけるα−GFP synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CARを調節する対応する応答エレメントの発現を示す操作した代表的なドットプロット。T細胞の赤色で囲った四分の一区画を選別し、図112における実験に使用した。(B)in vitro及びin vivo実験に使用したK562におけるCD19(紫)ならびにGFP及びCD19(緑)の発現を示すフローサイトメトリープロット。(C)対照の形質導入していないCD4+及びCD8+T細胞で処置した両側CD19(左側腹部)ならびにGFP及びCD19(右側腹部)腫瘍を有する個別のマウスの腫瘍成長曲線。データは、図5B下パネルの基となる。(D)初代ヒトCD4+及びCD8+T細胞を、α−GFP synNotch Gal4VP64受容体及びα−CD19 4−1BBζ CAR発現を調節する対応する応答エレメントで操作し、マウスに、CD19 K562を左側腹部で、表面GFP K562腫瘍を右側腹部で静脈内注射して、T細胞がGFP+「プライミング腫瘍」から遊走するかどうか、及びオフターゲットCD19単独腫瘍を殺傷するかどうかを検査した。腫瘍サイズを、操作したT細胞または形質導入していないT細胞対照の静脈内注射の後16日間にわたってモニターした。(E)synNotch ANDゲートT細胞(実線)または形質導入していない対照T細胞(点線)で処置したマウスのCD19腫瘍体積を示すグラフ。CD19腫瘍は、標的とされず、これは、GFP+腫瘍からのプライムしたT細胞の遊走がないことを示す(n=5、エラーバーは平均の標準誤差である、スチューデントのt検定に基づいて有意差はいずれの時点での見られなかった、p>0.05)。
Foxp3の直接細胞内染色を使用して、抗CD19 synNotchを発現するヒトCD4 T細胞におけるFoxp3誘導を測定した。抗CD19 synNotchを発現するCD4 T細胞を、CD19+(CD19陽性)K562細胞またはCD19−(CD19陰性)K562細胞のいずれかで刺激した。Foxp3は、CD19 synNotch細胞において、CD19+K562細胞で刺激したときに発現した(図118、「cN Foxp3+刺激K562」)。比較して、CD19 synNotch細胞をCD19−K562細胞で刺激したときは、誘導は見られなかった(図118、「cN Foxp3+無関係K562」)。形質導入していない細胞は、陰性対照として使用した(図118、「形質導入していない」)。
抗体発現構築物を、ヒトT細胞synNotch誘導型抗体分泌のために設計した。全体的な抗体構築物設計を示す概略図を図119に提示する。図119では、左から右に、全体構築物のドメイン設計は、ヒトIgG重鎖シグナルペプチド、Mycタグ、VHドメイン、CHドメイン、フューリン切断部位、SGSGスペーサー、T2A配列、ヒトIgG軽鎖シグナルペプチド、HAタグ、VLドメイン及びCLドメインである。構築物は、以下の抗体のsynNotch誘導型分泌のために設計された:ペムブロリズマブ(抗PD−1)、トレメリムマブ(抗CTLA4)及び9E10(抗myc)。
分割SynNotchシグナル伝達システムを構築し、これは、GFPの異なるエピトープと結合する2つの異なる抗GFPナノボディを有し、一方のナノボディ(「LaG2」)は、「アンカー細胞」上で発現され、他方のナノボディ(「LaG17」)は、SynNotch受容体の一部分として「受信側細胞」上で発現され、GFPは可溶性アダプターとして機能する(図123を参照されたい)。LaG17ナノボディSynNotch受容体を、活性化の際に、TRE発現構築物からmCherryレポーターの発現を誘導するtTa細胞内ドメインを有するように操作した。従って、設計した分割SynNotchシステムでは、可溶性アダプター分子(すなわち、GFP)を加えることは、アンカー細胞の存在下でのみ受信側細胞においてSynNotchシグナル伝達を誘導する。
Claims (14)
- N末端からC末端に向かって、共有結合状態で、
a)抗体の抗原結合ドメインを含む細胞外ドメイン;
b)Lin−12 Notchリピート、S2タンパク質分解切断部位、および、S3タンパク質分解切断部位を含む膜貫通ドメインを含む、Notch調節領域;及び
c)該Notch調節領域に対して異種であり転写活性化因子または転写抑制因子を含む細胞内ドメインであって、該抗原結合ドメインの抗原への結合が、S2およびS3タンパク質分解切断部位での切断を誘導し、それにより該細胞内ドメインが放出される、該細胞内ドメイン
を含む、キメラNotchポリペプチド。 - 細胞外ドメイン、Notch調節領域、またはその両方が、機能的Notchリガンド結合部位を含まない、請求項1に記載のキメラNotchポリペプチド。
- 細胞内ドメインが、
(i)転写活性化因子であって、該細胞内ドメインの放出が、該転写活性化因子に、細胞における内在性遺伝子産物または異種遺伝子産物の発現を誘導させる、該転写活性化因子;または
(ii)転写抑制因子であって、該細胞内ドメインの放出が、該転写抑制因子に、細胞における内在性遺伝子産物または異種遺伝子産物の発現を抑制させる、該転写抑制因子
のいずれかを含む、請求項1または2に記載のキメラNotchポリペプチド。 - Notch調節領域が、そのN末端に、1つまたは複数の上皮成長因子(EGF)リピート、任意で2〜11個のEGFリピートをさらに含む、請求項1〜3のいずれか一項に記載のキメラNotchポリペプチド。
- 合成リンカーを、任意で前記1つまたは複数のEGFリピートと前記Lin−12 Notchリピートとの間に含む、請求項4に記載のキメラNotchポリペプチド。
- Notch調節領域が、S2タンパク質分解切断部位および/またはS1タンパク質分解切断部位を含むヘテロ二量体化ドメインをさらに含む、請求項1〜5のいずれか一項に記載のキメラNotchポリペプチド。
- 請求項1〜6のいずれか一項に記載のキメラNotchポリペプチドを産生するように遺伝子改変されたin vitroまたはex vivo細胞であって、任意で、造血性幹細胞(HSC)もしくは胚性幹(ES)細胞などの幹細胞、始原細胞、幹細胞もしくは始原細胞に由来する細胞、または、Tリンパ球、ナチュラルキラー細胞、もしくは制御性T細胞などの免疫細胞からなる群より選択される、in vitroまたはex vivo細胞。
- 請求項1〜6のいずれか一項に記載のキメラNotchポリペプチドをコードするヌクレオチド配列を含む核酸。
- 請求項8に記載の核酸を含む組換え発現ベクターであって、任意で、コード配列に作動可能に連結し転写活性化因子または転写抑制因子に応答する転写制御エレメントをさらに含む、組換え発現ベクター。
- 細胞におけるコード配列の発現を誘導または抑制する方法であって、該細胞と抗原を接触させることを含み、
該細胞が、請求項1〜6のいずれか一項に記載のキメラNotchポリペプチドを産生するように遺伝子改変され、かつ、該コード配列に作動可能に連結し該キメラNotchポリペプチドの転写活性化因子または転写抑制因子に応答する転写制御エレメントを含み、
該抗体の抗原結合ドメインへの該抗原の結合によって細胞内ドメインが放出され、それによって該コード配列の発現が誘導または抑制される、
前記方法。 - 請求項1〜6のいずれか一項に記載のキメラNotchポリペプチドをコードするヌクレオチド配列を含む発現ベクターまたはRNAでin vitroまたはex vivoにおいて細胞を遺伝子改変することを含む、請求項7に記載の細胞を作製する方法。
- a)個体から得た細胞を、
i)前記キメラNotchポリペプチドをコードするヌクレオチド配列であって、該キメラNotchポリペプチドが転写活性化因子を含み、前記抗体の抗原結合ドメインが、該個体の細胞によって発現される抗原上に存在するエピトープに対して特異的である、該ヌクレオチド配列;および
ii)治療物質をコードする配列に作動可能に連結し該転写活性化因子に応答する転写制御エレメント
を含む発現ベクターで遺伝子改変すること;ならびに
b)該遺伝子改変細胞を該個体に導入することであって、該エピトープへの該抗原結合ドメインの結合が該治療物質の発現を提供し、それにより疾患を処置する、こと
を含み、任意で、該疾患が癌であり該治療物質が癌治療物質である、個体における該疾患を処置する方法
における使用のための、請求項1〜6のいずれか一項に記載のキメラNotchポリペプチド。 - 前記治療物質が、抗体、ケモカイン、ケモカイン受容体、サイトカイン、サイトカイン受容体、キメラ抗原受容体(CAR)、T細胞受容体(TCR)、活性化免疫受容体、抑制性免疫受容体、免疫活性化因子、免疫抑制因子、アポトーシス誘導因子、アポトーシス抑制因子、毒素由来タンパク質、および部位特異的ヌクレアーゼからなる群より選択される、請求項12に記載の使用のためのキメラNotchポリペプチド。
- N末端からC末端に向かって、共有結合状態で、
a)抗体の抗原結合ドメインを含む細胞外ドメイン;
b)Lin−12 Notchリピート、S2タンパク質分解切断部位、および、S3タンパク質分解切断部位を含む膜貫通ドメインを含む、Notch調節領域;ならびに
c)該Notch調節領域に対して異種であり分割転写因子の第一の半分を含む細胞内ドメインであって、該抗原結合ドメインの抗原への結合が、S2およびS3タンパク質分解切断部位での切断を誘導し、それにより該細胞内ドメインが放出される、該細胞内ドメイン
を含む、キメラNotchポリペプチド。
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201562120256P | 2015-02-24 | 2015-02-24 | |
| US62/120,256 | 2015-02-24 | ||
| US201562257153P | 2015-11-18 | 2015-11-18 | |
| US62/257,153 | 2015-11-18 | ||
| US201562269758P | 2015-12-18 | 2015-12-18 | |
| US62/269,758 | 2015-12-18 | ||
| PCT/US2016/019188 WO2016138034A1 (en) | 2015-02-24 | 2016-02-23 | Binding-triggered transcriptional switches and methods of use thereof |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020177718A Division JP2021019625A (ja) | 2015-02-24 | 2020-10-23 | 結合誘発型転写スイッチ及びその使用方法 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2018506293A JP2018506293A (ja) | 2018-03-08 |
| JP2018506293A5 JP2018506293A5 (ja) | 2019-04-04 |
| JP6784687B2 true JP6784687B2 (ja) | 2020-11-11 |
Family
ID=56789239
Family Applications (4)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2017544874A Active JP6784687B2 (ja) | 2015-02-24 | 2016-02-23 | 結合誘発型転写スイッチ及びその使用方法 |
| JP2020177718A Pending JP2021019625A (ja) | 2015-02-24 | 2020-10-23 | 結合誘発型転写スイッチ及びその使用方法 |
| JP2022120154A Active JP7357731B2 (ja) | 2015-02-24 | 2022-07-28 | 結合誘発型転写スイッチ及びその使用方法 |
| JP2023162996A Pending JP2023165849A (ja) | 2015-02-24 | 2023-09-26 | 結合誘発型転写スイッチ及びその使用方法 |
Family Applications After (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020177718A Pending JP2021019625A (ja) | 2015-02-24 | 2020-10-23 | 結合誘発型転写スイッチ及びその使用方法 |
| JP2022120154A Active JP7357731B2 (ja) | 2015-02-24 | 2022-07-28 | 結合誘発型転写スイッチ及びその使用方法 |
| JP2023162996A Pending JP2023165849A (ja) | 2015-02-24 | 2023-09-26 | 結合誘発型転写スイッチ及びその使用方法 |
Country Status (12)
| Country | Link |
|---|---|
| US (6) | US10590182B2 (ja) |
| EP (1) | EP3262166A4 (ja) |
| JP (4) | JP6784687B2 (ja) |
| KR (2) | KR20240010752A (ja) |
| CN (1) | CN108064283B (ja) |
| AU (2) | AU2016222887B2 (ja) |
| BR (1) | BR112017017884A2 (ja) |
| CA (1) | CA2973890A1 (ja) |
| HK (1) | HK1255666A1 (ja) |
| IL (2) | IL253463B (ja) |
| MX (2) | MX2017010721A (ja) |
| WO (1) | WO2016138034A1 (ja) |
Families Citing this family (112)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2013204922B2 (en) | 2012-12-20 | 2015-05-14 | Celgene Corporation | Chimeric antigen receptors |
| CN105518018B (zh) | 2013-03-15 | 2020-04-03 | 细胞基因公司 | 修饰的t淋巴细胞 |
| US11072644B2 (en) * | 2014-11-12 | 2021-07-27 | Allogene Therapeutics, Inc. | Inhibitory chimeric antigen receptors |
| US20170151281A1 (en) | 2015-02-19 | 2017-06-01 | Batu Biologics, Inc. | Chimeric antigen receptor dendritic cell (car-dc) for treatment of cancer |
| CN108064283B (zh) * | 2015-02-24 | 2024-01-09 | 加利福尼亚大学董事会 | 结合触发的转录开关及其使用方法 |
| WO2016168594A1 (en) * | 2015-04-16 | 2016-10-20 | President And Fellows Of Harvard College | Sensor systems for target ligands and uses thereof |
| PT3298033T (pt) | 2015-05-18 | 2020-09-22 | Tcr2 Therapeutics Inc | Composições e utilizações médicas para reprogramação de tcr utilizando proteínas de fusão |
| AU2017207281B2 (en) * | 2016-01-11 | 2022-03-10 | The Board Of Trustees Of The Leland Stanford Junior University | Chimeric proteins and methods of immunotherapy |
| BR112018013679A2 (pt) | 2016-01-11 | 2019-01-22 | Univ Leland Stanford Junior | proteínas quiméricas e métodos de regulação de expressão gênica |
| WO2017180587A2 (en) | 2016-04-11 | 2017-10-19 | Obsidian Therapeutics, Inc. | Regulated biocircuit systems |
| WO2017190100A1 (en) * | 2016-04-28 | 2017-11-02 | The Trustees Of Dartmouth College | Nucleic acid constructs for co-expression of chimeric antigen receptor and transcription factor, cells containing and therapeutic use thereof |
| US11400116B2 (en) * | 2016-05-06 | 2022-08-02 | The Regents Of The University Of California | Systems and methods for targeting cancer cells |
| US10858422B2 (en) | 2016-05-31 | 2020-12-08 | Abcentra, Llc | Methods for treating systemic lupus erythematosus with an anti-apolipoprotein B antibody |
| JP2019527192A (ja) | 2016-05-31 | 2019-09-26 | カーディオバックス リミテッド ライアビリティ カンパニー | 全身性エリテマトーデスの診断及び治療方法 |
| US11242376B2 (en) | 2016-08-02 | 2022-02-08 | TCR2 Therapeutics Inc. | Compositions and methods for TCR reprogramming using fusion proteins |
| AU2017316649A1 (en) * | 2016-08-23 | 2019-03-07 | The Regents Of The University Of California | Proteolytically cleavable chimeric polypeptides and methods of use thereof |
| WO2018050851A1 (en) * | 2016-09-16 | 2018-03-22 | Nordic Nanovector Asa | Treatment of non-hodgkin lymphoma using lilotomab and 177lu-lilotomab satetraxetan |
| PL3445787T3 (pl) | 2016-10-07 | 2021-05-31 | TCR2 Therapeutics Inc. | Kompozycje i sposoby reprogramowania receptorów limfocytów t przy użyciu białek fuzyjnych |
| US11174306B2 (en) * | 2016-10-19 | 2021-11-16 | The Scripps Research Institute | Chimeric antigen receptor effector cell switches with humanized targeting moieties and/or optimized chimeric antigen receptor interacting domains and uses thereof |
| US11331380B2 (en) * | 2016-10-20 | 2022-05-17 | Celgene Corporation | Cereblon-based heterodimerizable chimeric antigen receptors |
| JP7291396B2 (ja) | 2016-11-22 | 2023-06-15 | ティーシーアール2 セラピューティクス インク. | 融合タンパク質を用いたtcrの再プログラミングのための組成物及び方法 |
| WO2018111838A1 (en) * | 2016-12-12 | 2018-06-21 | Trustees Of Boston University | Inducible dimerization of recombinases |
| WO2018119042A1 (en) * | 2016-12-20 | 2018-06-28 | Reintjes Peter B | Directed evolution through mutation rate modulation |
| AU2018219226A1 (en) | 2017-02-07 | 2019-08-15 | Seattle Children's Hospital (dba Seattle Children's Research Institute) | Phospholipid ether (PLE) CAR T cell tumor targeting (CTCT) agents |
| WO2018157000A1 (en) * | 2017-02-23 | 2018-08-30 | Olema Pharmaceuticals, Inc. | Compositions and methods for regulating immune system activity |
| CN110582288A (zh) | 2017-02-28 | 2019-12-17 | 恩多塞特公司 | 用于car t细胞疗法的组合物和方法 |
| JOP20180027A1 (ar) * | 2017-03-28 | 2019-01-30 | Cell Design Labs Inc | بوليبيبتيدات مخلطة و طرق لتغيير موضع الغشاء فيها |
| US10934336B2 (en) * | 2017-04-13 | 2021-03-02 | The Trustees Of The University Of Pennsylvania | Use of gene editing to generate universal TCR re-directed T cells for adoptive immunotherapy |
| CN110753555A (zh) | 2017-04-19 | 2020-02-04 | 得克萨斯州大学系统董事会 | 表达工程化抗原受体的免疫细胞 |
| US11851649B2 (en) * | 2017-05-17 | 2023-12-26 | Seattle Children's Hospital | Generating mammalian T cell activation inducible synthetic promoters (SYN+PRO) to improve T cell therapy |
| WO2018222880A1 (en) | 2017-05-31 | 2018-12-06 | Trustees Of Boston University | Mechano-activated control of gene expression |
| WO2018227091A1 (en) * | 2017-06-08 | 2018-12-13 | The Brigham And Women's Hospital, Inc. | Methods and compositions for identifying epitopes |
| US11325957B2 (en) * | 2017-06-19 | 2022-05-10 | Cell Design Labs, Inc. | Methods and compositions for reducing the immunogenicity of chimeric notch receptors |
| JP2020530309A (ja) * | 2017-07-12 | 2020-10-22 | レフュージ バイオテクノロジーズ, インコーポレイテッド | 遺伝子発現を条件的に調節するための方法およびシステム |
| EP3688155B1 (en) | 2017-09-28 | 2023-01-04 | Immpact-Bio Ltd. | A universal platform for preparing an inhibitory chimeric antigen receptor (icar) |
| WO2019089982A1 (en) * | 2017-11-01 | 2019-05-09 | Juno Therapeutics, Inc. | Method of assessing activity of recombinant antigen receptors |
| EP3710042A4 (en) * | 2017-11-16 | 2021-11-24 | The Regents of The University of California | CHEMERICAL POLYPEPTIDES CONTAINING A FORCE SENSOR CLIVING FIELD AND THEIR METHODS OF USE |
| WO2019125072A1 (ko) * | 2017-12-22 | 2019-06-27 | 기초과학연구원 | 자극에 의해 활성화되는 flp 재조합 효소 및 이의 용도 |
| GB201721833D0 (en) * | 2017-12-22 | 2018-02-07 | Cancer Research Tech Ltd | Fusion proteins |
| CN110055275A (zh) | 2018-01-19 | 2019-07-26 | 科济生物医药(上海)有限公司 | 一种质粒载体对、其修饰的免疫细胞及其应用 |
| BR112020014913A2 (pt) | 2018-01-22 | 2020-12-08 | Seattle Children's Hospital (dba Seattle Children's Research Institute) | Métodos para uso de células t car |
| CN110117614A (zh) * | 2018-02-05 | 2019-08-13 | 复旦大学 | 一种单输入多输出的基因回路、表达载体和宿主细胞及其应用 |
| US20200384030A1 (en) | 2018-02-21 | 2020-12-10 | Cell Design Labs, Inc. | Chimeric transmembrane receptors and uses thereof |
| US20210023139A1 (en) * | 2018-04-06 | 2021-01-28 | The Regents Of The University Of California | Methods of treating glioblastomas |
| EP3549597A1 (en) * | 2018-04-06 | 2019-10-09 | Heinrich-Heine-Universität Düsseldorf | Synthetic signalling constructs and its use |
| US20210023138A1 (en) * | 2018-04-06 | 2021-01-28 | The Regents Of The University Of California | Methods of treating egfrviii expressing glioblastomas |
| US20210032661A1 (en) * | 2018-04-09 | 2021-02-04 | The Trustees Of The University Of Pennsylvania | Methods And Compositions Comprising A Viral Vector For Expression Of A Transgene And An Effector |
| AU2019252878B2 (en) * | 2018-04-10 | 2024-02-22 | Stichting Sanquin Bloedvoorziening | Chimeric Notch receptors |
| CN108707628B (zh) * | 2018-05-28 | 2021-11-23 | 上海海洋大学 | 斑马鱼notch2基因突变体的制备方法 |
| CN112513270A (zh) | 2018-07-13 | 2021-03-16 | 加利福尼亚大学董事会 | 基于逆转录转座子的递送媒介物及其使用方法 |
| WO2020018964A1 (en) | 2018-07-20 | 2020-01-23 | Fred Hutchinson Cancer Research Center | Compositions and methods for controlled expression of antigen-specific receptors |
| CN109180805B (zh) * | 2018-08-21 | 2021-12-28 | 南方医科大学 | 新型SynNotch合成受体及其编码基因 |
| WO2020068702A1 (en) | 2018-09-24 | 2020-04-02 | Fred Hutchinson Cancer Research Center | Chimeric receptor proteins and uses thereof |
| US20220267435A1 (en) * | 2018-10-03 | 2022-08-25 | University Of Pittsburgh-Of The Commonwealth System Of Higher Education | Covalent adaptor synnotch and chimeric antigen receptors (cars) for programmable antigen-targeting |
| GB201816839D0 (en) * | 2018-10-16 | 2018-11-28 | Glaxosmithkline Ip Dev Ltd | Novel control switch |
| CN109576292B (zh) * | 2018-12-21 | 2022-11-29 | 深圳市爱思迪生物科技有限公司 | 一种抗体文库的构建方法及其应用 |
| CN111471665B (zh) * | 2019-01-23 | 2023-07-04 | 上海科技大学 | 一种dna环化分子及其用途 |
| US20220168344A1 (en) | 2019-03-06 | 2022-06-02 | Lentigen Technology, Inc. | Compositions and methods for treating cancer with self-driving chimeric antigen receptors |
| US20200345853A1 (en) | 2019-03-20 | 2020-11-05 | Massachusetts Institute Of Technology | Uses of amphiphiles in immune cell therapy and compositions therefor |
| CN111826398A (zh) * | 2019-04-16 | 2020-10-27 | 常州费洛斯药业科技有限公司 | 用于活细胞间膜蛋白展示和相互作用检测的工程质粒系统 |
| CN110093361B (zh) * | 2019-04-17 | 2023-05-09 | 拜澳泰克(沈阳)生物医学集团有限公司 | 一种增强基因表达的增强子多肽及其应用 |
| CN110082538B (zh) * | 2019-04-19 | 2021-12-03 | 中山大学附属第六医院 | CCR9+IL-17+Treg细胞在制备用于检测NEC的试剂盒中的应用 |
| CA3135531A1 (en) * | 2019-04-30 | 2020-11-05 | Daniel Getts | Engineered chimeric fusion protein compositions and methods of use thereof |
| JOP20210298A1 (ar) | 2019-05-14 | 2023-01-30 | Provention Bio Inc | طرق وتركيبات للوقاية من مرض السكري من النوع الأول |
| WO2020232366A2 (en) | 2019-05-16 | 2020-11-19 | Trustees Of Boston University | Regulated synthetic gene expression systems |
| CN111983218A (zh) * | 2019-05-24 | 2020-11-24 | 常州费洛斯药业科技有限公司 | 一种用于检测活细胞-活细胞表面受体-配体相互作用的试剂盒 |
| WO2021010951A1 (en) * | 2019-07-12 | 2021-01-21 | Children's National Medical Center | Modified natural killer cells and methods of using the same |
| EP4010697A4 (en) * | 2019-08-05 | 2022-10-12 | Charles River Laboratories, Inc. | ANTIGEN BINDING MOLECULES AND SCREENING METHODS THEREFOR |
| GB2605276A (en) | 2019-09-03 | 2022-09-28 | Myeloid Therapeutics Inc | Methods and compositions for genomic integration |
| US20230030702A1 (en) * | 2019-09-05 | 2023-02-02 | Migal Galilee Research Institute Ltd | Blocking chimeric antigen receptors for prevention of undesired activation of effector and regulatory immune cells |
| WO2021055333A1 (en) * | 2019-09-18 | 2021-03-25 | The Regents Of The University Of California | Synthetic immune cells and methods of use thereof |
| WO2021061859A1 (en) | 2019-09-24 | 2021-04-01 | The Regents Of The University Of California | Novel receptors for ligand-dependent transcriptional regulation |
| WO2021061862A1 (en) | 2019-09-24 | 2021-04-01 | The Regents Of The University Of California | Notch receptors with hinge domain |
| EP4034254A4 (en) * | 2019-09-24 | 2023-11-01 | The Regents of the University of California | HETEROLOGOUS TRANSMEMBRANE DOMAIN RECEPTORS |
| WO2021061809A1 (en) * | 2019-09-24 | 2021-04-01 | The Regents Of The University Of California | Novel receptors having a fibronectin repeat for ligand-dependent transcriptional regulation |
| US20220340637A1 (en) * | 2019-09-24 | 2022-10-27 | The Regents Of The University Of California | Notch receptors with minimal linker |
| WO2021057906A1 (zh) * | 2019-09-25 | 2021-04-01 | 科济生物医药(上海)有限公司 | 表达il-15的免疫效应细胞 |
| WO2021085580A1 (ja) * | 2019-10-31 | 2021-05-06 | 国立研究開発法人理化学研究所 | ポリヌクレオチドおよびその利用 |
| EP4054596A4 (en) * | 2019-11-07 | 2023-11-22 | The Regents of The University of California | USE OF MOG FOR PRIMING GLIOBLASTOMA TREATMENT |
| BR112022011399A2 (pt) | 2019-12-11 | 2022-08-30 | A2 Biotherapeutics Inc | Receptor de antígeno quimérico baseado em lilrb1 |
| US20230067359A1 (en) * | 2019-12-27 | 2023-03-02 | National University Corporation Kobe University | Cancer gene therapy drug |
| US20230078007A1 (en) * | 2020-02-24 | 2023-03-16 | The Regents Of The University Of California | In-series synthetic receptor and-gate circuits for expression of a therapeutic payload by engineered cells |
| IL296203A (en) * | 2020-03-09 | 2022-11-01 | Univ California | One fem protein binding agents b |
| US20210317185A1 (en) * | 2020-04-06 | 2021-10-14 | Lung Biotechnology Pbc | Modular synthetic receptors and methods of use |
| CA3179954A1 (en) * | 2020-04-09 | 2021-10-14 | The Regents Of The University Of California | Humanized notch receptors with hinge domain |
| WO2022011065A1 (en) * | 2020-07-07 | 2022-01-13 | The Nemours Foundation | Tumor-activated alloreactive and xenoreactive t cells and their use in immunotherapy against cancer |
| US11572565B2 (en) | 2020-07-21 | 2023-02-07 | Trustees Of Boston University | Inducible control of gene expression |
| CN114075294A (zh) * | 2020-08-14 | 2022-02-22 | 中国科学院分子细胞科学卓越创新中心 | 细胞间遗传标记示踪技术 |
| IL300500A (en) | 2020-08-20 | 2023-04-01 | A2 Biotherapeutics Inc | Preparations and methods for the treatment of mesothelin positive cancer |
| IL300497A (en) | 2020-08-20 | 2023-04-01 | A2 Biotherapeutics Inc | Compositions and methods for treating CEACAM-positive cancer |
| TW202214846A (zh) | 2020-08-20 | 2022-04-16 | 美商A2生物治療學股份有限公司 | 用於治療egfr陽性癌症之組合物及方法 |
| WO2022098905A2 (en) | 2020-11-04 | 2022-05-12 | Myeloid Therapeutics, Inc. | Engineered chimeric fusion protein compositions and methods of use thereof |
| AU2021376208A1 (en) * | 2020-11-04 | 2023-06-22 | Senti Biosciences, Inc. | Protein payload release |
| IL303350A (en) | 2020-12-04 | 2023-08-01 | Macrogenics Inc | Preparations containing an antibody against HER2/NEU and their use |
| WO2022125865A2 (en) * | 2020-12-11 | 2022-06-16 | President And Fellows Of Harvard College | Novel screening platform to identify immune modulatory agents |
| JP2023554441A (ja) * | 2020-12-17 | 2023-12-27 | アスガード セラピューティクス エービー | 細胞リプログラミングのための組成物、構築物及びベクター |
| WO2022140159A1 (en) | 2020-12-24 | 2022-06-30 | Kite Pharma, Inc. | Prostate cancer chimeric antigen receptors |
| WO2022204326A1 (en) * | 2021-03-24 | 2022-09-29 | The Regents Of The University Of California | Humanized synthetic notch receptors with augmented transactivation domains and uses thereof |
| CN117460741A (zh) * | 2021-06-29 | 2024-01-26 | 科济生物医药(上海)有限公司 | 调控细胞生理活动的嵌合多肽 |
| KR20240026507A (ko) | 2021-06-29 | 2024-02-28 | 플래그쉽 파이어니어링 이노베이션스 브이, 인크. | 타노트랜스미션을 촉진시키도록 엔지니어링된 면역 세포 및 이의 용도 |
| TW202317602A (zh) | 2021-07-15 | 2023-05-01 | 福瑞德哈金森腫瘤中心 | 嵌合多肽 |
| WO2023049760A1 (en) * | 2021-09-21 | 2023-03-30 | The Regents Of The University Of California | Synthetic immuno-suppressive cells and methods of use thereof |
| GB202113674D0 (en) * | 2021-09-24 | 2021-11-10 | Reflection Therapeutics Ltd | Targeted cell therapies |
| GB202113673D0 (en) * | 2021-09-24 | 2021-11-10 | Reflection Therapeutics Ltd | Targeted cell therapies |
| CN114085835A (zh) * | 2021-11-16 | 2022-02-25 | 珠海中科先进技术研究院有限公司 | 一种癌细胞分析基因电路组件及其制备方法 |
| WO2023107825A1 (en) * | 2021-12-07 | 2023-06-15 | The Regents Of The University Of California | Synthetic cytokine circuits to promote infiltration and clearance of immune excluded solid tumors by engineered immune cells |
| WO2023141480A1 (en) * | 2022-01-20 | 2023-07-27 | The Regents Of The University Of California | Synthetic targeters of ubiquitination and degradation (studs) as effectors for feedback control in mammalian cells |
| WO2023201339A1 (en) * | 2022-04-15 | 2023-10-19 | Kyverna Therapeutics, Inc. | Methods and compositions for treating autoimmune disease |
| WO2023240064A1 (en) * | 2022-06-08 | 2023-12-14 | Kyverna Therapeutics, Inc. | Methods and compositions for treating autoimmune disease |
| CN115232217B (zh) * | 2022-07-04 | 2023-06-06 | 深圳市先康达生命科学有限公司 | 一种SynNotch结构及其应用 |
| WO2024020146A2 (en) * | 2022-07-21 | 2024-01-25 | Syntax Bio, Inc. | Systems for cell programming and methods thereof |
| CN115505601A (zh) * | 2022-10-17 | 2022-12-23 | 深圳市先康达生命科学有限公司 | 一种两级SynNotch逻辑调控结构及其应用 |
Family Cites Families (48)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5906936A (en) | 1988-05-04 | 1999-05-25 | Yeda Research And Development Co. Ltd. | Endowing lymphocytes with antibody specificity |
| WO1992010591A1 (en) | 1990-12-14 | 1992-06-25 | Cell Genesys, Inc. | Chimeric chains for receptor-associated signal transduction pathways |
| IL104570A0 (en) | 1992-03-18 | 1993-05-13 | Yeda Res & Dev | Chimeric genes and cells transformed therewith |
| US5834266A (en) | 1993-02-12 | 1998-11-10 | President & Fellows Of Harvard College | Regulated apoptosis |
| US5830462A (en) | 1993-02-12 | 1998-11-03 | President & Fellows Of Harvard College | Regulated transcription of targeted genes and other biological events |
| EP1978095A1 (en) | 1993-02-12 | 2008-10-08 | The Board Of Trustees Of The Leland Stanford Junior University | Chimeric receptor proteins for controlling signal transduction and ligands binding thereto |
| US5869337A (en) | 1993-02-12 | 1999-02-09 | President And Fellows Of Harvard College | Regulated transcription of targeted genes and other biological events |
| US20030036654A1 (en) | 1994-08-18 | 2003-02-20 | Holt Dennis A. | Synthetic multimerizing agents |
| US6150527A (en) | 1994-08-18 | 2000-11-21 | Ariad Pharmaceuticals, Inc. | Synthetic multimerizing agents |
| US6133456A (en) | 1994-08-18 | 2000-10-17 | Ariad Gene Therapeutics, Inc. | Synthetic multimerizing agents |
| US5712149A (en) | 1995-02-03 | 1998-01-27 | Cell Genesys, Inc. | Chimeric receptor molecules for delivery of co-stimulatory signals |
| US6187757B1 (en) | 1995-06-07 | 2001-02-13 | Ariad Pharmaceuticals, Inc. | Regulation of biological events using novel compounds |
| US5780300A (en) * | 1995-09-29 | 1998-07-14 | Yale University | Manipulation of non-terminally differentiated cells using the notch pathway |
| GB9526131D0 (en) | 1995-12-21 | 1996-02-21 | Celltech Therapeutics Ltd | Recombinant chimeric receptors |
| AU2472400A (en) | 1998-10-20 | 2000-05-08 | City Of Hope | CD20-specific redirected T cells and their use in cellular immunotherapy of CD20+ malignancies |
| GB0025307D0 (en) | 2000-10-16 | 2000-11-29 | Celltech Chiroscience Ltd | Biological products |
| WO2002077029A2 (en) | 2000-11-07 | 2002-10-03 | City Of Hope | Cd19-specific redirected immune cells |
| GB0112818D0 (en) * | 2001-05-25 | 2001-07-18 | Lorantis Ltd | Conjugate |
| US7446190B2 (en) | 2002-05-28 | 2008-11-04 | Sloan-Kettering Institute For Cancer Research | Nucleic acids encoding chimeric T cell receptors |
| GB0220658D0 (en) | 2002-09-05 | 2002-10-16 | Lorantis Ltd | Immunotherapy |
| CA2516320C (en) | 2003-02-18 | 2015-05-26 | Kevin Slawin | Induced activation in dendritic cells |
| US7435596B2 (en) | 2004-11-04 | 2008-10-14 | St. Jude Children's Research Hospital, Inc. | Modified cell line and method for expansion of NK cell |
| US20050113564A1 (en) | 2003-11-05 | 2005-05-26 | St. Jude Children's Research Hospital | Chimeric receptors with 4-1BB stimulatory signaling domain |
| WO2005045028A1 (ja) * | 2003-11-10 | 2005-05-19 | Osaka Industrial Promotion Organization | セル・フリー・Notch切断分析方法及び薬剤スクリーニング方法 |
| US20050103332A1 (en) * | 2003-11-17 | 2005-05-19 | Bruce Gingles | Airway exchange catheter |
| EP2195658A2 (en) | 2007-09-28 | 2010-06-16 | Royal College of Surgeons in Ireland | A method of assessing colorectal cancer status in an individual |
| EP2307459B1 (en) | 2008-07-08 | 2015-01-07 | OncoMed Pharmaceuticals, Inc. | Notch1 receptor binding agents and methods of use thereof |
| RU2612788C2 (ru) | 2010-03-23 | 2017-03-13 | Интрексон Корпорейшн | Векторы, условно экспрессирующие терапевтические белки, клетки-хозяева, содержащие указанные векторы, и их применение |
| WO2011130624A2 (en) | 2010-04-16 | 2011-10-20 | Immune Disease Institute, Inc. | Sustained polypeptide expression from synthetic, modified rnas and uses thereof |
| US9089520B2 (en) | 2010-05-21 | 2015-07-28 | Baylor College Of Medicine | Methods for inducing selective apoptosis |
| SG10201510092QA (en) | 2010-12-09 | 2016-01-28 | Univ Pennsylvania | Use of chimeric antigen receptor-modified t cells to treat cancer |
| SG192010A1 (en) | 2011-01-18 | 2013-08-30 | Univ Pennsylvania | Compositions and methods for treating cancer |
| EP3530730B1 (en) | 2011-10-31 | 2022-07-20 | Riken | Method for culturing stem cell |
| KR102437522B1 (ko) | 2012-05-25 | 2022-08-26 | 셀렉티스 | 면역요법을 위한 동종이형 및 면역억제제 저항성인 t 세포의 조작 방법 |
| BR112015004522A2 (pt) | 2012-09-04 | 2017-11-21 | Cellectis | receptor de antígeno quimérico multicadeia e usos destes |
| SG11201502598SA (en) * | 2012-10-02 | 2015-05-28 | Sloan Kettering Inst Cancer | Compositions and methods for immunotherapy |
| WO2014055657A1 (en) | 2012-10-05 | 2014-04-10 | The Trustees Of The University Of Pennsylvania | Use of a trans-signaling approach in chimeric antigen receptors |
| CA2905352A1 (en) | 2013-03-14 | 2014-09-25 | Bellicum Pharmaceuticals, Inc. | Methods for controlling t cell proliferation |
| SG11201603228TA (en) * | 2013-10-31 | 2016-05-30 | Hutchinson Fred Cancer Res | Modified hematopoietic stem/progenitor and non-t effector cells, and uses thereof |
| US9730935B2 (en) | 2013-12-13 | 2017-08-15 | The Board Of Trustees Of The Leland Stanford Junior University | Targeting a non-canonical notch signaling pathway for cancer treatment |
| US20150190506A1 (en) * | 2013-12-17 | 2015-07-09 | Genentech, Inc. | Combination therapy comprising ox40 binding agonists and pd-1 axis binding antagonists |
| EP3553087A1 (en) | 2014-01-08 | 2019-10-16 | The Board of Trustees of the Leland Stanford Junior University | Targeted therapy for lung cancer |
| WO2015123527A1 (en) | 2014-02-14 | 2015-08-20 | Bellicum Pharmaceuticals, Inc. | Methods for activating t cells using an inducible chimeric polypeptide |
| CN106414748B (zh) | 2014-02-14 | 2021-05-28 | 得克萨斯州大学系统董事会 | 嵌合抗原受体及制备方法 |
| AU2015220762B2 (en) | 2014-02-21 | 2019-05-02 | Cellectis | Method for in situ inhibition of regulatory T cells |
| US20170260261A1 (en) | 2014-08-28 | 2017-09-14 | Bioatla, Llc | Conditionally Active Chimeric Antigen Receptors for Modified T-Cells |
| CN108064283B (zh) | 2015-02-24 | 2024-01-09 | 加利福尼亚大学董事会 | 结合触发的转录开关及其使用方法 |
| US11400116B2 (en) | 2016-05-06 | 2022-08-02 | The Regents Of The University Of California | Systems and methods for targeting cancer cells |
-
2016
- 2016-02-23 CN CN201680023446.3A patent/CN108064283B/zh active Active
- 2016-02-23 KR KR1020247000665A patent/KR20240010752A/ko active Application Filing
- 2016-02-23 CA CA2973890A patent/CA2973890A1/en active Pending
- 2016-02-23 KR KR1020177024941A patent/KR102624023B1/ko active IP Right Grant
- 2016-02-23 WO PCT/US2016/019188 patent/WO2016138034A1/en active Application Filing
- 2016-02-23 JP JP2017544874A patent/JP6784687B2/ja active Active
- 2016-02-23 BR BR112017017884A patent/BR112017017884A2/pt unknown
- 2016-02-23 US US15/543,220 patent/US10590182B2/en active Active
- 2016-02-23 EP EP16756215.6A patent/EP3262166A4/en active Pending
- 2016-02-23 AU AU2016222887A patent/AU2016222887B2/en active Active
- 2016-02-23 MX MX2017010721A patent/MX2017010721A/es unknown
- 2016-04-12 US US15/096,971 patent/US9670281B2/en active Active
-
2017
- 2017-05-01 US US15/583,658 patent/US9834608B2/en active Active
- 2017-07-13 IL IL253463A patent/IL253463B/en unknown
- 2017-08-21 MX MX2022011180A patent/MX2022011180A/es unknown
- 2017-12-01 US US15/829,370 patent/US10836808B2/en active Active
- 2017-12-04 US US15/831,194 patent/US10822387B2/en active Active
-
2018
- 2018-11-20 HK HK18114812.7A patent/HK1255666A1/zh unknown
-
2020
- 2020-10-13 US US17/069,717 patent/US20210107965A1/en active Pending
- 2020-10-23 JP JP2020177718A patent/JP2021019625A/ja active Pending
-
2021
- 2021-11-08 IL IL287914A patent/IL287914A/en unknown
-
2022
- 2022-07-28 JP JP2022120154A patent/JP7357731B2/ja active Active
- 2022-10-12 AU AU2022252737A patent/AU2022252737A1/en active Pending
-
2023
- 2023-09-26 JP JP2023162996A patent/JP2023165849A/ja active Pending
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7357731B2 (ja) | 結合誘発型転写スイッチ及びその使用方法 | |
| JP7108551B2 (ja) | リンパ球に形質導入を行うための方法及び組成物、並びにその制御された増加 | |
| JP2022516557A (ja) | 修飾il-2ポリペプチドを含むポリペプチド及びその使用 | |
| KR20220087454A (ko) | 힌지 도메인을 갖는 노치 수용체 | |
| JP2022105139A (ja) | キメラnotch受容体の免疫原性を低下させる方法及び組成物 | |
| WO2022046802A1 (en) | Targeted protein degradation in therapeutic cells | |
| US20220064252A1 (en) | Engineered extracellular receptor constructs and uses thereof | |
| US11701386B2 (en) | Cell | |
| CN117886949A (zh) | 结合触发的转录开关及其使用方法 | |
| US20230055337A1 (en) | Use of brain-specific antigens to home, block and deliver cell-based treatments to the brain | |
| WO2023019076A1 (en) | Use of a stromal antigen to deliver cell-based cancer therapy to a solid tumor | |
| CN116615208A (zh) | 工程化细胞粘附分子及其使用方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20170920 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20190221 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20190221 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20191129 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20200108 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20200406 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20200923 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20201023 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6784687 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |