CN103397049A - 具有增强的产量相关性状的植物及其制备方法 - Google Patents
具有增强的产量相关性状的植物及其制备方法 Download PDFInfo
- Publication number
- CN103397049A CN103397049A CN2013102981411A CN201310298141A CN103397049A CN 103397049 A CN103397049 A CN 103397049A CN 2013102981411 A CN2013102981411 A CN 2013102981411A CN 201310298141 A CN201310298141 A CN 201310298141A CN 103397049 A CN103397049 A CN 103397049A
- Authority
- CN
- China
- Prior art keywords
- plant
- nucleic acid
- sequence
- motif
- polypeptide
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 238000000034 method Methods 0.000 title claims abstract description 313
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 310
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 301
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 301
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 262
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 250
- 102000004196 processed proteins & peptides Human genes 0.000 claims abstract description 226
- 229920001184 polypeptide Polymers 0.000 claims abstract description 225
- 230000014509 gene expression Effects 0.000 claims abstract description 120
- 102000004169 proteins and genes Human genes 0.000 claims abstract description 117
- 230000002708 enhancing effect Effects 0.000 claims abstract description 28
- 230000001965 increasing effect Effects 0.000 claims abstract description 17
- 241000196324 Embryophyta Species 0.000 claims description 539
- 125000003729 nucleotide group Chemical group 0.000 claims description 145
- 239000002773 nucleotide Substances 0.000 claims description 143
- 230000009261 transgenic effect Effects 0.000 claims description 60
- 230000012010 growth Effects 0.000 claims description 53
- 230000002596 correlated effect Effects 0.000 claims description 51
- 238000004519 manufacturing process Methods 0.000 claims description 41
- 239000002028 Biomass Substances 0.000 claims description 40
- 235000007164 Oryza sativa Nutrition 0.000 claims description 37
- 235000009566 rice Nutrition 0.000 claims description 37
- 230000008676 import Effects 0.000 claims description 35
- 230000001105 regulatory effect Effects 0.000 claims description 31
- 108010048762 protochlorophyllide reductase Proteins 0.000 claims description 29
- 235000013311 vegetables Nutrition 0.000 claims description 22
- 240000008042 Zea mays Species 0.000 claims description 21
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 claims description 20
- 235000002017 Zea mays subsp mays Nutrition 0.000 claims description 19
- 235000005824 Zea mays ssp. parviglumis Nutrition 0.000 claims description 18
- 235000013339 cereals Nutrition 0.000 claims description 18
- 235000005822 corn Nutrition 0.000 claims description 18
- 101150104463 GOS2 gene Proteins 0.000 claims description 13
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 13
- 244000038559 crop plants Species 0.000 claims description 13
- 238000007899 nucleic acid hybridization Methods 0.000 claims description 13
- 235000007319 Avena orientalis Nutrition 0.000 claims description 12
- 235000021307 Triticum Nutrition 0.000 claims description 12
- 235000002566 Capsicum Nutrition 0.000 claims description 11
- 239000001390 capsicum minimum Substances 0.000 claims description 11
- 230000008635 plant growth Effects 0.000 claims description 11
- 229910052757 nitrogen Inorganic materials 0.000 claims description 10
- 241000209510 Liliopsida Species 0.000 claims description 9
- 241000208293 Capsicum Species 0.000 claims description 8
- 240000005979 Hordeum vulgare Species 0.000 claims description 8
- 235000007340 Hordeum vulgare Nutrition 0.000 claims description 8
- 235000007238 Secale cereale Nutrition 0.000 claims description 8
- 244000082988 Secale cereale Species 0.000 claims description 8
- 244000046109 Sorghum vulgare var. nervosum Species 0.000 claims description 8
- 241001233957 eudicotyledons Species 0.000 claims description 7
- 238000002360 preparation method Methods 0.000 claims description 7
- 235000019714 Triticale Nutrition 0.000 claims description 6
- 230000001737 promoting effect Effects 0.000 claims description 6
- 230000005030 transcription termination Effects 0.000 claims description 6
- 241000228158 x Triticosecale Species 0.000 claims description 6
- 230000001580 bacterial effect Effects 0.000 claims description 5
- 240000008574 Capsicum frutescens Species 0.000 claims description 4
- 241000208292 Solanaceae Species 0.000 claims description 4
- 241000209140 Triticum Species 0.000 claims description 4
- 241000589516 Pseudomonas Species 0.000 claims description 3
- 240000007594 Oryza sativa Species 0.000 claims 4
- 241000209763 Avena sativa Species 0.000 claims 2
- 108091000041 Phosphoenolpyruvate Carboxylase Proteins 0.000 abstract description 216
- 235000018102 proteins Nutrition 0.000 description 98
- 210000004027 cell Anatomy 0.000 description 74
- 238000006243 chemical reaction Methods 0.000 description 54
- 241000209094 Oryza Species 0.000 description 47
- 238000009396 hybridization Methods 0.000 description 38
- 210000001519 tissue Anatomy 0.000 description 35
- 230000002068 genetic effect Effects 0.000 description 32
- DTBNBXWJWCWCIK-UHFFFAOYSA-N phosphoenolpyruvic acid Chemical compound OC(=O)C(=C)OP(O)(O)=O DTBNBXWJWCWCIK-UHFFFAOYSA-N 0.000 description 30
- 230000000694 effects Effects 0.000 description 29
- 125000003275 alpha amino acid group Chemical group 0.000 description 28
- 108020004414 DNA Proteins 0.000 description 26
- 235000001014 amino acid Nutrition 0.000 description 25
- 239000003550 marker Substances 0.000 description 25
- 150000001413 amino acids Chemical class 0.000 description 24
- 230000000692 anti-sense effect Effects 0.000 description 24
- 108020004999 messenger RNA Proteins 0.000 description 24
- 238000005516 engineering process Methods 0.000 description 23
- 241000589158 Agrobacterium Species 0.000 description 22
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 21
- 230000008859 change Effects 0.000 description 20
- MMXZSJMASHPLLR-UHFFFAOYSA-N pyrroloquinoline quinone Chemical compound C12=C(C(O)=O)C=C(C(O)=O)N=C2C(=O)C(=O)C2=C1NC(C(=O)O)=C2 MMXZSJMASHPLLR-UHFFFAOYSA-N 0.000 description 20
- 241000894007 species Species 0.000 description 19
- 230000035882 stress Effects 0.000 description 19
- 239000000203 mixture Substances 0.000 description 18
- 241000192707 Synechococcus Species 0.000 description 17
- 244000098338 Triticum aestivum Species 0.000 description 15
- 239000003795 chemical substances by application Substances 0.000 description 15
- 239000013604 expression vector Substances 0.000 description 14
- 230000009466 transformation Effects 0.000 description 14
- 235000010469 Glycine max Nutrition 0.000 description 13
- 244000068988 Glycine max Species 0.000 description 13
- 238000003306 harvesting Methods 0.000 description 13
- 210000001161 mammalian embryo Anatomy 0.000 description 13
- 238000013507 mapping Methods 0.000 description 13
- 210000000056 organ Anatomy 0.000 description 13
- 244000075850 Avena orientalis Species 0.000 description 12
- 230000036579 abiotic stress Effects 0.000 description 12
- 239000012634 fragment Substances 0.000 description 12
- 239000000243 solution Substances 0.000 description 12
- 241000894006 Bacteria Species 0.000 description 11
- -1 Hexose phosphate Chemical class 0.000 description 11
- 206010020649 Hyperkeratosis Diseases 0.000 description 11
- 230000000295 complement effect Effects 0.000 description 11
- 230000001939 inductive effect Effects 0.000 description 11
- 230000008569 process Effects 0.000 description 11
- 230000008929 regeneration Effects 0.000 description 11
- 238000011069 regeneration method Methods 0.000 description 11
- 239000002689 soil Substances 0.000 description 11
- 230000002103 transcriptional effect Effects 0.000 description 11
- 230000001131 transforming effect Effects 0.000 description 11
- 241000190932 Rhodopseudomonas Species 0.000 description 10
- 230000004071 biological effect Effects 0.000 description 10
- 229910052799 carbon Inorganic materials 0.000 description 10
- 239000000523 sample Substances 0.000 description 10
- 102000004190 Enzymes Human genes 0.000 description 9
- 108090000790 Enzymes Proteins 0.000 description 9
- 238000009395 breeding Methods 0.000 description 9
- 230000001488 breeding effect Effects 0.000 description 9
- 230000008034 disappearance Effects 0.000 description 9
- 230000006872 improvement Effects 0.000 description 9
- 238000011534 incubation Methods 0.000 description 9
- 238000003780 insertion Methods 0.000 description 9
- 230000037431 insertion Effects 0.000 description 9
- 210000002706 plastid Anatomy 0.000 description 9
- 238000011160 research Methods 0.000 description 9
- 108091092562 ribozyme Proteins 0.000 description 9
- 238000005406 washing Methods 0.000 description 9
- 241000193830 Bacillus <bacterium> Species 0.000 description 8
- 241000219793 Trifolium Species 0.000 description 8
- 125000000539 amino acid group Chemical group 0.000 description 8
- 238000004422 calculation algorithm Methods 0.000 description 8
- 230000000875 corresponding effect Effects 0.000 description 8
- 239000011159 matrix material Substances 0.000 description 8
- 230000000442 meristematic effect Effects 0.000 description 8
- 231100000350 mutagenesis Toxicity 0.000 description 8
- 239000013612 plasmid Substances 0.000 description 8
- 230000009467 reduction Effects 0.000 description 8
- 238000009331 sowing Methods 0.000 description 8
- 230000014616 translation Effects 0.000 description 8
- 238000011144 upstream manufacturing Methods 0.000 description 8
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 8
- 241000589155 Agrobacterium tumefaciens Species 0.000 description 7
- 235000007558 Avena sp Nutrition 0.000 description 7
- 108090000994 Catalytic RNA Proteins 0.000 description 7
- 102000053642 Catalytic RNA Human genes 0.000 description 7
- 235000007688 Lycopersicon esculentum Nutrition 0.000 description 7
- 240000003768 Solanum lycopersicum Species 0.000 description 7
- 239000002253 acid Substances 0.000 description 7
- 238000004458 analytical method Methods 0.000 description 7
- 238000006555 catalytic reaction Methods 0.000 description 7
- 150000001875 compounds Chemical class 0.000 description 7
- 230000002349 favourable effect Effects 0.000 description 7
- 230000006870 function Effects 0.000 description 7
- VNWKTOKETHGBQD-UHFFFAOYSA-N methane Natural products C VNWKTOKETHGBQD-UHFFFAOYSA-N 0.000 description 7
- 238000002703 mutagenesis Methods 0.000 description 7
- KHPXUQMNIQBQEV-UHFFFAOYSA-N oxaloacetic acid Chemical compound OC(=O)CC(=O)C(O)=O KHPXUQMNIQBQEV-UHFFFAOYSA-N 0.000 description 7
- 108091033319 polynucleotide Proteins 0.000 description 7
- 102000040430 polynucleotide Human genes 0.000 description 7
- 239000002157 polynucleotide Substances 0.000 description 7
- 238000012545 processing Methods 0.000 description 7
- 238000012225 targeting induced local lesions in genomes Methods 0.000 description 7
- 238000013519 translation Methods 0.000 description 7
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 6
- 108700001094 Plant Genes Proteins 0.000 description 6
- 108010003581 Ribulose-bisphosphate carboxylase Proteins 0.000 description 6
- 239000002585 base Substances 0.000 description 6
- 238000005520 cutting process Methods 0.000 description 6
- 238000001914 filtration Methods 0.000 description 6
- 230000004927 fusion Effects 0.000 description 6
- 230000030279 gene silencing Effects 0.000 description 6
- 238000012226 gene silencing method Methods 0.000 description 6
- 238000002744 homologous recombination Methods 0.000 description 6
- 230000006801 homologous recombination Effects 0.000 description 6
- 238000005259 measurement Methods 0.000 description 6
- 239000002609 medium Substances 0.000 description 6
- 230000004048 modification Effects 0.000 description 6
- 238000012986 modification Methods 0.000 description 6
- 150000003839 salts Chemical class 0.000 description 6
- 239000000725 suspension Substances 0.000 description 6
- 238000013518 transcription Methods 0.000 description 6
- 230000035897 transcription Effects 0.000 description 6
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 5
- 241000219194 Arabidopsis Species 0.000 description 5
- 108700011259 MicroRNAs Proteins 0.000 description 5
- 108700008625 Reporter Genes Proteins 0.000 description 5
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 5
- 108090000848 Ubiquitin Proteins 0.000 description 5
- 102000044159 Ubiquitin Human genes 0.000 description 5
- 235000019519 canola oil Nutrition 0.000 description 5
- 239000000828 canola oil Substances 0.000 description 5
- 238000013461 design Methods 0.000 description 5
- 239000003623 enhancer Substances 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- BRZYSWJRSDMWLG-CAXSIQPQSA-N geneticin Chemical compound O1C[C@@](O)(C)[C@H](NC)[C@@H](O)[C@H]1O[C@@H]1[C@@H](O)[C@H](O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](C(C)O)O2)N)[C@@H](N)C[C@H]1N BRZYSWJRSDMWLG-CAXSIQPQSA-N 0.000 description 5
- 239000001963 growth medium Substances 0.000 description 5
- 238000005286 illumination Methods 0.000 description 5
- 239000002679 microRNA Substances 0.000 description 5
- 244000005700 microbiome Species 0.000 description 5
- 230000036542 oxidative stress Effects 0.000 description 5
- 238000012216 screening Methods 0.000 description 5
- 238000002741 site-directed mutagenesis Methods 0.000 description 5
- 238000012546 transfer Methods 0.000 description 5
- 241000186063 Arthrobacter Species 0.000 description 4
- 108091026821 Artificial microRNA Proteins 0.000 description 4
- 108091026890 Coding region Proteins 0.000 description 4
- 235000001950 Elaeis guineensis Nutrition 0.000 description 4
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 4
- 241001465754 Metazoa Species 0.000 description 4
- 108091092724 Noncoding DNA Proteins 0.000 description 4
- 108010038807 Oligopeptides Proteins 0.000 description 4
- 102000015636 Oligopeptides Human genes 0.000 description 4
- 108700026244 Open Reading Frames Proteins 0.000 description 4
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 4
- 235000002595 Solanum tuberosum Nutrition 0.000 description 4
- 244000061456 Solanum tuberosum Species 0.000 description 4
- 241000192584 Synechocystis Species 0.000 description 4
- 241000700605 Viruses Species 0.000 description 4
- 230000003321 amplification Effects 0.000 description 4
- 230000003115 biocidal effect Effects 0.000 description 4
- 230000001276 controlling effect Effects 0.000 description 4
- 238000001727 in vivo Methods 0.000 description 4
- 239000000463 material Substances 0.000 description 4
- 230000007246 mechanism Effects 0.000 description 4
- 238000002887 multiple sequence alignment Methods 0.000 description 4
- 230000035772 mutation Effects 0.000 description 4
- 238000003199 nucleic acid amplification method Methods 0.000 description 4
- 230000035764 nutrition Effects 0.000 description 4
- 235000016709 nutrition Nutrition 0.000 description 4
- 239000003921 oil Substances 0.000 description 4
- 235000019198 oils Nutrition 0.000 description 4
- 230000002018 overexpression Effects 0.000 description 4
- 230000006798 recombination Effects 0.000 description 4
- 238000005215 recombination Methods 0.000 description 4
- 230000021749 root development Effects 0.000 description 4
- 238000010561 standard procedure Methods 0.000 description 4
- 230000001954 sterilising effect Effects 0.000 description 4
- 238000004659 sterilization and disinfection Methods 0.000 description 4
- 239000013598 vector Substances 0.000 description 4
- CAAMSDWKXXPUJR-UHFFFAOYSA-N 3,5-dihydro-4H-imidazol-4-one Chemical compound O=C1CNC=N1 CAAMSDWKXXPUJR-UHFFFAOYSA-N 0.000 description 3
- 240000007087 Apium graveolens Species 0.000 description 3
- 240000002791 Brassica napus Species 0.000 description 3
- 235000006008 Brassica napus var napus Nutrition 0.000 description 3
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 3
- 244000025254 Cannabis sativa Species 0.000 description 3
- 241001070941 Castanea Species 0.000 description 3
- 244000241235 Citrullus lanatus Species 0.000 description 3
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 3
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 3
- 244000127993 Elaeis melanococca Species 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- 241000233866 Fungi Species 0.000 description 3
- 108700007698 Genetic Terminator Regions Proteins 0.000 description 3
- 102000053187 Glucuronidase Human genes 0.000 description 3
- 108010060309 Glucuronidase Proteins 0.000 description 3
- 235000003222 Helianthus annuus Nutrition 0.000 description 3
- 244000020551 Helianthus annuus Species 0.000 description 3
- 240000006240 Linum usitatissimum Species 0.000 description 3
- NWBJYWHLCVSVIJ-UHFFFAOYSA-N N-benzyladenine Chemical compound N=1C=NC=2NC=NC=2C=1NCC1=CC=CC=C1 NWBJYWHLCVSVIJ-UHFFFAOYSA-N 0.000 description 3
- 241000605154 Nitrobacter winogradskyi Species 0.000 description 3
- 108091034117 Oligonucleotide Proteins 0.000 description 3
- 241000219843 Pisum Species 0.000 description 3
- 108091081024 Start codon Proteins 0.000 description 3
- 108091036066 Three prime untranslated region Proteins 0.000 description 3
- 235000002041 Triticum macha Nutrition 0.000 description 3
- 244000102426 Triticum macha Species 0.000 description 3
- 102000006275 Ubiquitin-Protein Ligases Human genes 0.000 description 3
- 108010083111 Ubiquitin-Protein Ligases Proteins 0.000 description 3
- 150000007513 acids Chemical class 0.000 description 3
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 3
- 229960005305 adenosine Drugs 0.000 description 3
- 238000013459 approach Methods 0.000 description 3
- 102000005936 beta-Galactosidase Human genes 0.000 description 3
- 108010005774 beta-Galactosidase Proteins 0.000 description 3
- 230000027455 binding Effects 0.000 description 3
- 230000003570 biosynthesizing effect Effects 0.000 description 3
- 230000001413 cellular effect Effects 0.000 description 3
- 239000003086 colorant Substances 0.000 description 3
- 239000002299 complementary DNA Substances 0.000 description 3
- 230000007812 deficiency Effects 0.000 description 3
- 238000009826 distribution Methods 0.000 description 3
- 230000008641 drought stress Effects 0.000 description 3
- 238000001035 drying Methods 0.000 description 3
- 235000013399 edible fruits Nutrition 0.000 description 3
- 210000002257 embryonic structure Anatomy 0.000 description 3
- 238000011156 evaluation Methods 0.000 description 3
- 235000019688 fish Nutrition 0.000 description 3
- 230000010354 integration Effects 0.000 description 3
- 230000035800 maturation Effects 0.000 description 3
- 230000004060 metabolic process Effects 0.000 description 3
- 108091070501 miRNA Proteins 0.000 description 3
- 238000002715 modification method Methods 0.000 description 3
- 238000010369 molecular cloning Methods 0.000 description 3
- 238000012544 monitoring process Methods 0.000 description 3
- 238000003499 nucleic acid array Methods 0.000 description 3
- 230000000050 nutritive effect Effects 0.000 description 3
- 239000002245 particle Substances 0.000 description 3
- 230000026731 phosphorylation Effects 0.000 description 3
- 238000006366 phosphorylation reaction Methods 0.000 description 3
- 239000011148 porous material Substances 0.000 description 3
- 238000003753 real-time PCR Methods 0.000 description 3
- 230000002829 reductive effect Effects 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 238000000926 separation method Methods 0.000 description 3
- 230000019491 signal transduction Effects 0.000 description 3
- 238000005728 strengthening Methods 0.000 description 3
- 230000004960 subcellular localization Effects 0.000 description 3
- 238000011282 treatment Methods 0.000 description 3
- AZQWKYJCGOJGHM-UHFFFAOYSA-N 1,4-benzoquinone Chemical compound O=C1C=CC(=O)C=C1 AZQWKYJCGOJGHM-UHFFFAOYSA-N 0.000 description 2
- 239000005631 2,4-Dichlorophenoxyacetic acid Substances 0.000 description 2
- 241001075517 Abelmoschus Species 0.000 description 2
- 241000208140 Acer Species 0.000 description 2
- 241000588624 Acinetobacter calcoaceticus Species 0.000 description 2
- 241000219068 Actinidia Species 0.000 description 2
- 229920001817 Agar Polymers 0.000 description 2
- 244000198134 Agave sisalana Species 0.000 description 2
- 235000011624 Agave sisalana Nutrition 0.000 description 2
- 241000209136 Agropyron Species 0.000 description 2
- 108700028369 Alleles Proteins 0.000 description 2
- 241000234282 Allium Species 0.000 description 2
- 241000219318 Amaranthus Species 0.000 description 2
- 244000296825 Amygdalus nana Species 0.000 description 2
- 241000192542 Anabaena Species 0.000 description 2
- 244000099147 Ananas comosus Species 0.000 description 2
- 235000007119 Ananas comosus Nutrition 0.000 description 2
- 235000015849 Apium graveolens Dulce Group Nutrition 0.000 description 2
- 235000010591 Appio Nutrition 0.000 description 2
- 244000105624 Arachis hypogaea Species 0.000 description 2
- 244000018217 Artocarpus elasticus Species 0.000 description 2
- 238000012935 Averaging Methods 0.000 description 2
- 235000010082 Averrhoa carambola Nutrition 0.000 description 2
- 240000006063 Averrhoa carambola Species 0.000 description 2
- 244000036905 Benincasa cerifera Species 0.000 description 2
- 235000011274 Benincasa cerifera Nutrition 0.000 description 2
- 241000335053 Beta vulgaris Species 0.000 description 2
- 235000011293 Brassica napus Nutrition 0.000 description 2
- 240000008100 Brassica rapa Species 0.000 description 2
- 235000006618 Brassica rapa subsp oleifera Nutrition 0.000 description 2
- 241001040392 Burkholderia cenocepacia PC184 Species 0.000 description 2
- 241001040393 Burkholderia dolosa AU0158 Species 0.000 description 2
- 241000244203 Caenorhabditis elegans Species 0.000 description 2
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 2
- 235000009467 Carica papaya Nutrition 0.000 description 2
- 240000006432 Carica papaya Species 0.000 description 2
- 240000004927 Carissa macrocarpa Species 0.000 description 2
- 235000001479 Carissa macrocarpa Nutrition 0.000 description 2
- 235000003255 Carthamus tinctorius Nutrition 0.000 description 2
- 244000020518 Carthamus tinctorius Species 0.000 description 2
- 241000723418 Carya Species 0.000 description 2
- 235000014036 Castanea Nutrition 0.000 description 2
- 244000146553 Ceiba pentandra Species 0.000 description 2
- 235000003301 Ceiba pentandra Nutrition 0.000 description 2
- 108700031407 Chloroplast Genes Proteins 0.000 description 2
- 235000012828 Citrullus lanatus var citroides Nutrition 0.000 description 2
- 241000207199 Citrus Species 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 241000723377 Coffea Species 0.000 description 2
- 244000205754 Colocasia esculenta Species 0.000 description 2
- 235000006481 Colocasia esculenta Nutrition 0.000 description 2
- 108091035707 Consensus sequence Proteins 0.000 description 2
- 235000002787 Coriandrum sativum Nutrition 0.000 description 2
- 244000018436 Coriandrum sativum Species 0.000 description 2
- 241000723382 Corylus Species 0.000 description 2
- 241000186216 Corynebacterium Species 0.000 description 2
- 241000195493 Cryptophyta Species 0.000 description 2
- 244000024469 Cucumis prophetarum Species 0.000 description 2
- 241000219122 Cucurbita Species 0.000 description 2
- 108010066133 D-octopine dehydrogenase Proteins 0.000 description 2
- 240000001008 Dimocarpus longan Species 0.000 description 2
- 244000281702 Dioscorea villosa Species 0.000 description 2
- 241000192043 Echinochloa Species 0.000 description 2
- 241000512897 Elaeis Species 0.000 description 2
- 235000001942 Elaeis Nutrition 0.000 description 2
- 244000078127 Eleusine coracana Species 0.000 description 2
- 235000009008 Eriobotrya japonica Nutrition 0.000 description 2
- 244000061508 Eriobotrya japonica Species 0.000 description 2
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 2
- 235000000235 Euphoria longan Nutrition 0.000 description 2
- 240000008620 Fagopyrum esculentum Species 0.000 description 2
- 241001070947 Fagus Species 0.000 description 2
- 241000220223 Fragaria Species 0.000 description 2
- 241000187809 Frankia Species 0.000 description 2
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 2
- 235000008100 Ginkgo biloba Nutrition 0.000 description 2
- 244000194101 Ginkgo biloba Species 0.000 description 2
- 239000004471 Glycine Substances 0.000 description 2
- 244000299507 Gossypium hirsutum Species 0.000 description 2
- 241000208818 Helianthus Species 0.000 description 2
- 235000002941 Hemerocallis fulva Nutrition 0.000 description 2
- 240000009206 Hemerocallis fulva Species 0.000 description 2
- 108091027305 Heteroduplex Proteins 0.000 description 2
- 244000284380 Hibiscus rosa sinensis Species 0.000 description 2
- 101000911390 Homo sapiens Coagulation factor VIII Proteins 0.000 description 2
- 241000209219 Hordeum Species 0.000 description 2
- 244000017020 Ipomoea batatas Species 0.000 description 2
- 235000002678 Ipomoea batatas Nutrition 0.000 description 2
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 2
- 241000758789 Juglans Species 0.000 description 2
- 241000588748 Klebsiella Species 0.000 description 2
- 241000186660 Lactobacillus Species 0.000 description 2
- 235000003228 Lactuca sativa Nutrition 0.000 description 2
- 240000008415 Lactuca sativa Species 0.000 description 2
- 241000219729 Lathyrus Species 0.000 description 2
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 2
- 235000004431 Linum usitatissimum Nutrition 0.000 description 2
- 241000219745 Lupinus Species 0.000 description 2
- 241001300479 Macroptilium Species 0.000 description 2
- 241000220225 Malus Species 0.000 description 2
- 235000014826 Mangifera indica Nutrition 0.000 description 2
- 240000007228 Mangifera indica Species 0.000 description 2
- 240000003183 Manihot esculenta Species 0.000 description 2
- 244000061354 Manilkara achras Species 0.000 description 2
- 240000004658 Medicago sativa Species 0.000 description 2
- 235000017587 Medicago sativa ssp. sativa Nutrition 0.000 description 2
- 241000213996 Melilotus Species 0.000 description 2
- 241001072983 Mentha Species 0.000 description 2
- 241000218984 Momordica Species 0.000 description 2
- 240000000249 Morus alba Species 0.000 description 2
- 235000008708 Morus alba Nutrition 0.000 description 2
- 241000234295 Musa Species 0.000 description 2
- 241000186359 Mycobacterium Species 0.000 description 2
- 240000002853 Nelumbo nucifera Species 0.000 description 2
- 241000208125 Nicotiana Species 0.000 description 2
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 2
- 244000061176 Nicotiana tabacum Species 0.000 description 2
- 241000192656 Nostoc Species 0.000 description 2
- 108020004711 Nucleic Acid Probes Proteins 0.000 description 2
- 241000795633 Olea <sea slug> Species 0.000 description 2
- 240000001439 Opuntia Species 0.000 description 2
- 238000012408 PCR amplification Methods 0.000 description 2
- 241001520808 Panicum virgatum Species 0.000 description 2
- 244000288157 Passiflora edulis Species 0.000 description 2
- 240000004370 Pastinaca sativa Species 0.000 description 2
- 241000218196 Persea Species 0.000 description 2
- 241000219833 Phaseolus Species 0.000 description 2
- 235000010627 Phaseolus vulgaris Nutrition 0.000 description 2
- 244000046052 Phaseolus vulgaris Species 0.000 description 2
- 241000746983 Phleum pratense Species 0.000 description 2
- 235000014676 Phragmites communis Nutrition 0.000 description 2
- 244000064622 Physalis edulis Species 0.000 description 2
- 235000003447 Pistacia vera Nutrition 0.000 description 2
- 240000006711 Pistacia vera Species 0.000 description 2
- 241000219000 Populus Species 0.000 description 2
- 241001494501 Prosopis <angiosperm> Species 0.000 description 2
- 108010029485 Protein Isoforms Proteins 0.000 description 2
- 102000001708 Protein Isoforms Human genes 0.000 description 2
- 229940096437 Protein S Drugs 0.000 description 2
- 108010076504 Protein Sorting Signals Proteins 0.000 description 2
- 241000508269 Psidium Species 0.000 description 2
- 244000294611 Punica granatum Species 0.000 description 2
- 235000014360 Punica granatum Nutrition 0.000 description 2
- 240000001987 Pyrus communis Species 0.000 description 2
- 235000014443 Pyrus communis Nutrition 0.000 description 2
- 241000219492 Quercus Species 0.000 description 2
- 244000088415 Raphanus sativus Species 0.000 description 2
- 108020004511 Recombinant DNA Proteins 0.000 description 2
- 102000018120 Recombinases Human genes 0.000 description 2
- 108010091086 Recombinases Proteins 0.000 description 2
- 235000009411 Rheum rhabarbarum Nutrition 0.000 description 2
- 244000193032 Rheum rhaponticum Species 0.000 description 2
- 241000316848 Rhodococcus <scale insect> Species 0.000 description 2
- 108091028664 Ribonucleotide Proteins 0.000 description 2
- 240000000528 Ricinus communis Species 0.000 description 2
- 235000004443 Ricinus communis Nutrition 0.000 description 2
- 241001092459 Rubus Species 0.000 description 2
- 241000209051 Saccharum Species 0.000 description 2
- 241000607142 Salmonella Species 0.000 description 2
- 241000208829 Sambucus Species 0.000 description 2
- 108091081021 Sense strand Proteins 0.000 description 2
- 108091027967 Small hairpin RNA Proteins 0.000 description 2
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 2
- 235000002634 Solanum Nutrition 0.000 description 2
- 241000207763 Solanum Species 0.000 description 2
- 229920002472 Starch Polymers 0.000 description 2
- 229930006000 Sucrose Natural products 0.000 description 2
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 2
- 244000045719 Syzygium Species 0.000 description 2
- 241000736851 Tagetes Species 0.000 description 2
- 240000004584 Tamarindus indica Species 0.000 description 2
- 235000004298 Tamarindus indica Nutrition 0.000 description 2
- 244000269722 Thea sinensis Species 0.000 description 2
- 244000299461 Theobroma cacao Species 0.000 description 2
- 235000009470 Theobroma cacao Nutrition 0.000 description 2
- 108700019146 Transgenes Proteins 0.000 description 2
- 102000008579 Transposases Human genes 0.000 description 2
- 108010020764 Transposases Proteins 0.000 description 2
- 235000007264 Triticum durum Nutrition 0.000 description 2
- 235000007251 Triticum monococcum Nutrition 0.000 description 2
- 240000000581 Triticum monococcum Species 0.000 description 2
- 241000209143 Triticum turgidum subsp. durum Species 0.000 description 2
- 241001530121 Trollius Species 0.000 description 2
- 241000736767 Vaccinium Species 0.000 description 2
- 241000219977 Vigna Species 0.000 description 2
- 240000009038 Viola odorata Species 0.000 description 2
- 235000013487 Viola odorata Nutrition 0.000 description 2
- 241000219095 Vitis Species 0.000 description 2
- 241001247821 Ziziphus Species 0.000 description 2
- 238000009825 accumulation Methods 0.000 description 2
- DZBUGLKDJFMEHC-UHFFFAOYSA-N acridine Chemical compound C1=CC=CC2=CC3=CC=CC=C3N=C21 DZBUGLKDJFMEHC-UHFFFAOYSA-N 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 230000002411 adverse Effects 0.000 description 2
- 239000008272 agar Substances 0.000 description 2
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 2
- 239000000427 antigen Substances 0.000 description 2
- 102000036639 antigens Human genes 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 230000015572 biosynthetic process Effects 0.000 description 2
- 239000011575 calcium Substances 0.000 description 2
- 229910052791 calcium Inorganic materials 0.000 description 2
- 238000004364 calculation method Methods 0.000 description 2
- RTYJTGSCYUUYAL-YCAHSCEMSA-L carbenicillin disodium Chemical compound [Na+].[Na+].N([C@H]1[C@H]2SC([C@@H](N2C1=O)C([O-])=O)(C)C)C(=O)C(C([O-])=O)C1=CC=CC=C1 RTYJTGSCYUUYAL-YCAHSCEMSA-L 0.000 description 2
- 150000001720 carbohydrates Chemical class 0.000 description 2
- 235000014633 carbohydrates Nutrition 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- 150000001768 cations Chemical class 0.000 description 2
- 229960004261 cefotaxime Drugs 0.000 description 2
- AZZMGZXNTDTSME-JUZDKLSSSA-M cefotaxime sodium Chemical compound [Na+].N([C@@H]1C(N2C(=C(COC(C)=O)CS[C@@H]21)C([O-])=O)=O)C(=O)\C(=N/OC)C1=CSC(N)=N1 AZZMGZXNTDTSME-JUZDKLSSSA-M 0.000 description 2
- 231100000045 chemical toxicity Toxicity 0.000 description 2
- 108010031100 chloroplast transit peptides Proteins 0.000 description 2
- HZZVJAQRINQKSD-PBFISZAISA-N clavulanic acid Chemical compound OC(=O)[C@H]1C(=C/CO)/O[C@@H]2CC(=O)N21 HZZVJAQRINQKSD-PBFISZAISA-N 0.000 description 2
- 238000012937 correction Methods 0.000 description 2
- 230000006378 damage Effects 0.000 description 2
- 230000007547 defect Effects 0.000 description 2
- 238000001514 detection method Methods 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- 238000004520 electroporation Methods 0.000 description 2
- 239000012877 elongation medium Substances 0.000 description 2
- 230000000408 embryogenic effect Effects 0.000 description 2
- 238000013213 extrapolation Methods 0.000 description 2
- 235000013305 food Nutrition 0.000 description 2
- 238000012239 gene modification Methods 0.000 description 2
- 230000009368 gene silencing by RNA Effects 0.000 description 2
- 230000005017 genetic modification Effects 0.000 description 2
- 235000013617 genetically modified food Nutrition 0.000 description 2
- 230000035784 germination Effects 0.000 description 2
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 2
- 230000002363 herbicidal effect Effects 0.000 description 2
- 239000004009 herbicide Substances 0.000 description 2
- 238000003898 horticulture Methods 0.000 description 2
- 102000057593 human F8 Human genes 0.000 description 2
- 239000001257 hydrogen Substances 0.000 description 2
- 229910052739 hydrogen Inorganic materials 0.000 description 2
- 238000003384 imaging method Methods 0.000 description 2
- 230000001976 improved effect Effects 0.000 description 2
- 230000008595 infiltration Effects 0.000 description 2
- 238000001764 infiltration Methods 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 230000005764 inhibitory process Effects 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 230000003993 interaction Effects 0.000 description 2
- 150000002500 ions Chemical class 0.000 description 2
- 229940039696 lactobacillus Drugs 0.000 description 2
- 230000001404 mediated effect Effects 0.000 description 2
- 239000002207 metabolite Substances 0.000 description 2
- 231100000219 mutagenic Toxicity 0.000 description 2
- 230000003505 mutagenic effect Effects 0.000 description 2
- 108010058731 nopaline synthase Proteins 0.000 description 2
- 239000002853 nucleic acid probe Substances 0.000 description 2
- 230000008520 organization Effects 0.000 description 2
- 230000008723 osmotic stress Effects 0.000 description 2
- 239000001301 oxygen Substances 0.000 description 2
- 229910052760 oxygen Inorganic materials 0.000 description 2
- 230000005097 photorespiration Effects 0.000 description 2
- 230000029553 photosynthesis Effects 0.000 description 2
- 238000010672 photosynthesis Methods 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 229940047431 recombinate Drugs 0.000 description 2
- 238000004153 renaturation Methods 0.000 description 2
- 238000007894 restriction fragment length polymorphism technique Methods 0.000 description 2
- 239000002336 ribonucleotide Substances 0.000 description 2
- 238000002864 sequence alignment Methods 0.000 description 2
- 230000003584 silencer Effects 0.000 description 2
- 230000000392 somatic effect Effects 0.000 description 2
- 235000019698 starch Nutrition 0.000 description 2
- 239000008107 starch Substances 0.000 description 2
- 230000000638 stimulation Effects 0.000 description 2
- 238000003860 storage Methods 0.000 description 2
- 239000000126 substance Substances 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 239000005720 sucrose Substances 0.000 description 2
- 235000000346 sugar Nutrition 0.000 description 2
- 230000004083 survival effect Effects 0.000 description 2
- 238000007910 systemic administration Methods 0.000 description 2
- OHKOGUYZJXTSFX-KZFFXBSXSA-N ticarcillin Chemical compound C=1([C@@H](C(O)=O)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)C=CSC=1 OHKOGUYZJXTSFX-KZFFXBSXSA-N 0.000 description 2
- 229960004659 ticarcillin Drugs 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 239000005418 vegetable material Substances 0.000 description 2
- 230000017260 vegetative to reproductive phase transition of meristem Effects 0.000 description 2
- 230000000007 visual effect Effects 0.000 description 2
- NWXMGUDVXFXRIG-WESIUVDSSA-N (4s,4as,5as,6s,12ar)-4-(dimethylamino)-1,6,10,11,12a-pentahydroxy-6-methyl-3,12-dioxo-4,4a,5,5a-tetrahydrotetracene-2-carboxamide Chemical compound C1=CC=C2[C@](O)(C)[C@H]3C[C@H]4[C@H](N(C)C)C(=O)C(C(N)=O)=C(O)[C@@]4(O)C(=O)C3=C(O)C2=C1O NWXMGUDVXFXRIG-WESIUVDSSA-N 0.000 description 1
- MPCAJMNYNOGXPB-SLPGGIOYSA-N 1,5-anhydro-D-glucitol Chemical compound OC[C@H]1OC[C@H](O)[C@@H](O)[C@@H]1O MPCAJMNYNOGXPB-SLPGGIOYSA-N 0.000 description 1
- 108020004463 18S ribosomal RNA Proteins 0.000 description 1
- FFRBMBIXVSCUFS-UHFFFAOYSA-N 2,4-dinitro-1-naphthol Chemical compound C1=CC=C2C(O)=C([N+]([O-])=O)C=C([N+]([O-])=O)C2=C1 FFRBMBIXVSCUFS-UHFFFAOYSA-N 0.000 description 1
- KKAJSJJFBSOMGS-UHFFFAOYSA-N 3,6-diamino-10-methylacridinium chloride Chemical compound [Cl-].C1=C(N)C=C2[N+](C)=C(C=C(N)C=C3)C3=CC2=C1 KKAJSJJFBSOMGS-UHFFFAOYSA-N 0.000 description 1
- OPIFSICVWOWJMJ-AEOCFKNESA-N 5-bromo-4-chloro-3-indolyl beta-D-galactoside Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1OC1=CNC2=CC=C(Br)C(Cl)=C12 OPIFSICVWOWJMJ-AEOCFKNESA-N 0.000 description 1
- 241000726118 Acidovorax facilis Species 0.000 description 1
- 235000001674 Agaricus brunnescens Nutrition 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- 241000743339 Agrostis Species 0.000 description 1
- 240000007241 Agrostis stolonifera Species 0.000 description 1
- HOVPGJUNRLMIOZ-CIUDSAMLSA-N Ala-Ser-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@H](C)N HOVPGJUNRLMIOZ-CIUDSAMLSA-N 0.000 description 1
- 241001677738 Aleuron Species 0.000 description 1
- 241000380131 Ammophila arenaria Species 0.000 description 1
- 108091093088 Amplicon Proteins 0.000 description 1
- 235000003840 Amygdalus nana Nutrition 0.000 description 1
- 235000007755 Annona Nutrition 0.000 description 1
- 235000011518 Annona purpurea Nutrition 0.000 description 1
- 240000006199 Annona purpurea Species 0.000 description 1
- 101710117679 Anthocyanidin 3-O-glucosyltransferase Proteins 0.000 description 1
- 235000002764 Apium graveolens Nutrition 0.000 description 1
- 241001605719 Appias drusilla Species 0.000 description 1
- 101100070555 Arabidopsis thaliana HSFA4C gene Proteins 0.000 description 1
- 101100523550 Arabidopsis thaliana RABF2A gene Proteins 0.000 description 1
- 235000003911 Arachis Nutrition 0.000 description 1
- BTJVOUQWFXABOI-IHRRRGAJSA-N Arg-Lys-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCNC(N)=N BTJVOUQWFXABOI-IHRRRGAJSA-N 0.000 description 1
- 102000008682 Argonaute Proteins Human genes 0.000 description 1
- 108010088141 Argonaute Proteins Proteins 0.000 description 1
- 241001167018 Aroa Species 0.000 description 1
- 108090000328 Arrestin Proteins 0.000 description 1
- 102000003916 Arrestin Human genes 0.000 description 1
- 235000005340 Asparagus officinalis Nutrition 0.000 description 1
- 206010003694 Atrophy Diseases 0.000 description 1
- 241000972773 Aulopiformes Species 0.000 description 1
- 235000005781 Avena Nutrition 0.000 description 1
- 235000009393 Avena byzantina Nutrition 0.000 description 1
- 235000007320 Avena fatua Nutrition 0.000 description 1
- 241000209764 Avena fatua Species 0.000 description 1
- 235000018410 Avena fatua var sativa Nutrition 0.000 description 1
- 240000000372 Avena hybrida Species 0.000 description 1
- 235000009123 Avena hybrida Nutrition 0.000 description 1
- 241000209128 Bambusa Species 0.000 description 1
- 235000016068 Berberis vulgaris Nutrition 0.000 description 1
- 241000219164 Bertholletia Species 0.000 description 1
- 235000021533 Beta vulgaris Nutrition 0.000 description 1
- 241000219495 Betulaceae Species 0.000 description 1
- 108010006654 Bleomycin Proteins 0.000 description 1
- 235000004480 Bombax malabaricum Nutrition 0.000 description 1
- 241000588807 Bordetella Species 0.000 description 1
- ZOXJGFHDIHLPTG-UHFFFAOYSA-N Boron Chemical compound [B] ZOXJGFHDIHLPTG-UHFFFAOYSA-N 0.000 description 1
- 235000011331 Brassica Nutrition 0.000 description 1
- 241000219198 Brassica Species 0.000 description 1
- 244000060924 Brassica campestris Species 0.000 description 1
- 235000005637 Brassica campestris Nutrition 0.000 description 1
- 235000014698 Brassica juncea var multisecta Nutrition 0.000 description 1
- 240000000385 Brassica napus var. napus Species 0.000 description 1
- 235000011292 Brassica rapa Nutrition 0.000 description 1
- 235000000540 Brassica rapa subsp rapa Nutrition 0.000 description 1
- 235000004977 Brassica sinapistrum Nutrition 0.000 description 1
- 235000004936 Bromus mango Nutrition 0.000 description 1
- 241001453380 Burkholderia Species 0.000 description 1
- 241000933166 Burkholderia ambifaria MC40-6 Species 0.000 description 1
- 241000312517 Burkholderia cenocepacia AU 1054 Species 0.000 description 1
- 241000312512 Burkholderia cenocepacia HI2424 Species 0.000 description 1
- 241000651940 Burkholderia cenocepacia MC0-3 Species 0.000 description 1
- 241001106236 Burkholderia pseudomallei 1106a Species 0.000 description 1
- 241000312046 Burkholderia pseudomallei 1655 Species 0.000 description 1
- 241001604143 Burkholderia pseudomallei 1710a Species 0.000 description 1
- 241001604148 Burkholderia pseudomallei 668 Species 0.000 description 1
- 241000423294 Burkholderia pseudomallei K96243 Species 0.000 description 1
- 241000322329 Burkholderia pseudomallei Pasteur 52237 Species 0.000 description 1
- 235000008635 Cadaba farinosa Nutrition 0.000 description 1
- 241000628166 Cadaba farinosa Species 0.000 description 1
- 101100427201 Caenorhabditis elegans ufd-2 gene Proteins 0.000 description 1
- 235000010773 Cajanus indicus Nutrition 0.000 description 1
- 244000105627 Cajanus indicus Species 0.000 description 1
- 102000000584 Calmodulin Human genes 0.000 description 1
- 108010041952 Calmodulin Proteins 0.000 description 1
- 241001237259 Campanella <basidiomycete fungus> Species 0.000 description 1
- 244000292211 Canna coccinea Species 0.000 description 1
- 235000005273 Canna coccinea Nutrition 0.000 description 1
- 241000684239 Canna x generalis Species 0.000 description 1
- 235000008697 Cannabis sativa Nutrition 0.000 description 1
- 235000012766 Cannabis sativa ssp. sativa var. sativa Nutrition 0.000 description 1
- 235000012765 Cannabis sativa ssp. sativa var. spontanea Nutrition 0.000 description 1
- 235000002567 Capsicum annuum Nutrition 0.000 description 1
- 240000004160 Capsicum annuum Species 0.000 description 1
- 101710132601 Capsid protein Proteins 0.000 description 1
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 1
- 241000973255 Carex elata Species 0.000 description 1
- 241000701489 Cauliflower mosaic virus Species 0.000 description 1
- 108010001857 Cell Surface Receptors Proteins 0.000 description 1
- 241001148660 Cenchrus sp. Species 0.000 description 1
- 108091092236 Chimeric RNA Proteins 0.000 description 1
- 241000195628 Chlorophyta Species 0.000 description 1
- 240000006740 Cichorium endivia Species 0.000 description 1
- 235000018536 Cichorium endivia Nutrition 0.000 description 1
- 241000723347 Cinnamomum Species 0.000 description 1
- 235000009831 Citrullus lanatus Nutrition 0.000 description 1
- 244000175448 Citrus madurensis Species 0.000 description 1
- 101710094648 Coat protein Proteins 0.000 description 1
- 241001478240 Coccus Species 0.000 description 1
- 241000737241 Cocos Species 0.000 description 1
- 244000060011 Cocos nucifera Species 0.000 description 1
- 108091033380 Coding strand Proteins 0.000 description 1
- 244000228088 Cola acuminata Species 0.000 description 1
- 241000020428 Colea Species 0.000 description 1
- 241000287937 Colinus Species 0.000 description 1
- 108020004635 Complementary DNA Proteins 0.000 description 1
- 108020004394 Complementary RNA Proteins 0.000 description 1
- 235000010203 Corchorus Nutrition 0.000 description 1
- 241000332384 Corchorus Species 0.000 description 1
- 235000007706 Corchorus sp Nutrition 0.000 description 1
- 229920000742 Cotton Polymers 0.000 description 1
- 235000014493 Crataegus Nutrition 0.000 description 1
- 241001092040 Crataegus Species 0.000 description 1
- 240000000171 Crataegus monogyna Species 0.000 description 1
- 241000383377 Crocosphaera watsonii WH 8501 Species 0.000 description 1
- 235000015655 Crocus sativus Nutrition 0.000 description 1
- 244000124209 Crocus sativus Species 0.000 description 1
- 235000010071 Cucumis prophetarum Nutrition 0.000 description 1
- 241000097442 Cupriavidus metallidurans CH34 Species 0.000 description 1
- 241000846564 Cyanothece sp. CCY0110 Species 0.000 description 1
- 235000003198 Cynara Nutrition 0.000 description 1
- 241000208947 Cynara Species 0.000 description 1
- 244000019459 Cynara cardunculus Species 0.000 description 1
- 235000019106 Cynara scolymus Nutrition 0.000 description 1
- IGXWBGJHJZYPQS-SSDOTTSWSA-N D-Luciferin Chemical compound OC(=O)[C@H]1CSC(C=2SC3=CC=C(O)C=C3N=2)=N1 IGXWBGJHJZYPQS-SSDOTTSWSA-N 0.000 description 1
- 230000004544 DNA amplification Effects 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 244000000626 Daucus carota Species 0.000 description 1
- 235000002767 Daucus carota Nutrition 0.000 description 1
- CYCGRDQQIOGCKX-UHFFFAOYSA-N Dehydro-luciferin Natural products OC(=O)C1=CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 CYCGRDQQIOGCKX-UHFFFAOYSA-N 0.000 description 1
- 101710088194 Dehydrogenase Proteins 0.000 description 1
- 241000522190 Desmodium Species 0.000 description 1
- 241001163054 Dichelachne Species 0.000 description 1
- 235000000525 Dimocarpus longan Nutrition 0.000 description 1
- 235000005903 Dioscorea Nutrition 0.000 description 1
- 235000000504 Dioscorea villosa Nutrition 0.000 description 1
- 241000723267 Diospyros Species 0.000 description 1
- 241001057636 Dracaena deremensis Species 0.000 description 1
- 241000255601 Drosophila melanogaster Species 0.000 description 1
- 240000003133 Elaeis guineensis Species 0.000 description 1
- 235000007349 Eleusine coracana Nutrition 0.000 description 1
- 235000013499 Eleusine coracana subsp coracana Nutrition 0.000 description 1
- 235000014966 Eragrostis abyssinica Nutrition 0.000 description 1
- 244000140063 Eragrostis abyssinica Species 0.000 description 1
- 101000933461 Escherichia coli (strain K12) Beta-glucuronidase Proteins 0.000 description 1
- 108700039887 Essential Genes Proteins 0.000 description 1
- 244000004281 Eucalyptus maculata Species 0.000 description 1
- 244000080545 Eucalyptus sp Species 0.000 description 1
- 235000006914 Eucalyptus sp Nutrition 0.000 description 1
- 235000013420 Eugenia uniflora Nutrition 0.000 description 1
- 240000003813 Eugenia uniflora Species 0.000 description 1
- 241000206602 Eukaryota Species 0.000 description 1
- 238000001134 F-test Methods 0.000 description 1
- 235000009419 Fagopyrum esculentum Nutrition 0.000 description 1
- 241000234643 Festuca arundinacea Species 0.000 description 1
- 235000008730 Ficus carica Nutrition 0.000 description 1
- 244000025361 Ficus carica Species 0.000 description 1
- 241001412224 Firmiana Species 0.000 description 1
- BJGNCJDXODQBOB-UHFFFAOYSA-N Fivefly Luciferin Natural products OC(=O)C1CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 BJGNCJDXODQBOB-UHFFFAOYSA-N 0.000 description 1
- 235000017317 Fortunella Nutrition 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- 235000011201 Ginkgo Nutrition 0.000 description 1
- TWHDOEYLXXQYOZ-FXQIFTODSA-N Gln-Asn-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N TWHDOEYLXXQYOZ-FXQIFTODSA-N 0.000 description 1
- SDSMVVSHLAAOJL-UKJIMTQDSA-N Gln-Val-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CCC(=O)N)N SDSMVVSHLAAOJL-UKJIMTQDSA-N 0.000 description 1
- 108010024636 Glutathione Proteins 0.000 description 1
- QCTLGOYODITHPQ-WHFBIAKZSA-N Gly-Cys-Ser Chemical compound [H]NCC(=O)N[C@@H](CS)C(=O)N[C@@H](CO)C(O)=O QCTLGOYODITHPQ-WHFBIAKZSA-N 0.000 description 1
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 1
- 239000005562 Glyphosate Substances 0.000 description 1
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 1
- 241000219146 Gossypium Species 0.000 description 1
- 235000009438 Gossypium Nutrition 0.000 description 1
- 235000009429 Gossypium barbadense Nutrition 0.000 description 1
- 101150105462 HIS6 gene Proteins 0.000 description 1
- 108090001102 Hammerhead ribozyme Proteins 0.000 description 1
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 1
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 1
- 241001303799 Herpetosiphon aurantiacus DSM 785 Species 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 235000005206 Hibiscus Nutrition 0.000 description 1
- 235000007185 Hibiscus lunariifolius Nutrition 0.000 description 1
- RNAYRCNHRYEBTH-IHRRRGAJSA-N His-Met-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(O)=O RNAYRCNHRYEBTH-IHRRRGAJSA-N 0.000 description 1
- 101000808654 Homo sapiens Ubiquitin conjugation factor E4 A Proteins 0.000 description 1
- 101000809046 Homo sapiens Ubiquitin conjugation factor E4 B Proteins 0.000 description 1
- IDAHFEPYTJJZFD-PEFMBERDSA-N Ile-Asp-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N IDAHFEPYTJJZFD-PEFMBERDSA-N 0.000 description 1
- 229930010555 Inosine Natural products 0.000 description 1
- UGQMRVRMYYASKQ-KQYNXXCUSA-N Inosine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(O)=C2N=C1 UGQMRVRMYYASKQ-KQYNXXCUSA-N 0.000 description 1
- 108020005210 Integrons Proteins 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 235000013757 Juglans Nutrition 0.000 description 1
- 241001468133 Kineococcus Species 0.000 description 1
- 241000105982 Kineococcus radiotolerans SRS30216 = ATCC BAA-149 Species 0.000 description 1
- 101100288095 Klebsiella pneumoniae neo gene Proteins 0.000 description 1
- DZLNHFMRPBPULJ-VKHMYHEASA-N L-thioproline Chemical compound OC(=O)[C@@H]1CSCN1 DZLNHFMRPBPULJ-VKHMYHEASA-N 0.000 description 1
- 241000219739 Lens Species 0.000 description 1
- 240000004322 Lens culinaris Species 0.000 description 1
- 235000010666 Lens esculenta Nutrition 0.000 description 1
- KUIDCYNIEJBZBU-AJNGGQMLSA-N Leu-Ile-Leu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O KUIDCYNIEJBZBU-AJNGGQMLSA-N 0.000 description 1
- 238000003657 Likelihood-ratio test Methods 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- DDWFXDSYGUXRAY-UHFFFAOYSA-N Luciferin Natural products CCc1c(C)c(CC2NC(=O)C(=C2C=C)C)[nH]c1Cc3[nH]c4C(=C5/NC(CC(=O)O)C(C)C5CC(=O)O)CC(=O)c4c3C DDWFXDSYGUXRAY-UHFFFAOYSA-N 0.000 description 1
- 235000018780 Luffa acutangula Nutrition 0.000 description 1
- 244000280244 Luffa acutangula Species 0.000 description 1
- 108010047357 Luminescent Proteins Proteins 0.000 description 1
- 102000006830 Luminescent Proteins Human genes 0.000 description 1
- 241000605547 Luzula sylvatica Species 0.000 description 1
- 108090000856 Lyases Proteins 0.000 description 1
- 102000004317 Lyases Human genes 0.000 description 1
- 241000227653 Lycopersicon Species 0.000 description 1
- 235000002262 Lycopersicon Nutrition 0.000 description 1
- 244000276497 Lycopersicon esculentum Species 0.000 description 1
- 241000219816 Macrotyloma Species 0.000 description 1
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- 235000014791 Malpighia coccigera Nutrition 0.000 description 1
- 244000007729 Malpighia coccigera Species 0.000 description 1
- 240000003394 Malpighia glabra Species 0.000 description 1
- 235000014837 Malpighia glabra Nutrition 0.000 description 1
- 101710175625 Maltose/maltodextrin-binding periplasmic protein Proteins 0.000 description 1
- 235000000889 Mammea americana Nutrition 0.000 description 1
- 240000005984 Mammea americana Species 0.000 description 1
- 235000011339 Manilkara zapota Nutrition 0.000 description 1
- 235000000088 Maracuja Nutrition 0.000 description 1
- 235000014435 Mentha Nutrition 0.000 description 1
- 241000589308 Methylobacterium extorquens Species 0.000 description 1
- 241000589339 Methylobacterium organophilum Species 0.000 description 1
- 241000878006 Miscanthus sinensis Species 0.000 description 1
- 235000009815 Momordica Nutrition 0.000 description 1
- 101100412856 Mus musculus Rhod gene Proteins 0.000 description 1
- 101710135898 Myc proto-oncogene protein Proteins 0.000 description 1
- 102100038895 Myc proto-oncogene protein Human genes 0.000 description 1
- XMBSYZWANAQXEV-UHFFFAOYSA-N N-alpha-L-glutamyl-L-phenylalanine Natural products OC(=O)CCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XMBSYZWANAQXEV-UHFFFAOYSA-N 0.000 description 1
- KZNQNBZMBZJQJO-UHFFFAOYSA-N N-glycyl-L-proline Natural products NCC(=O)N1CCCC1C(O)=O KZNQNBZMBZJQJO-UHFFFAOYSA-N 0.000 description 1
- 241001230286 Narenga Species 0.000 description 1
- 241000588653 Neisseria Species 0.000 description 1
- 235000006508 Nelumbo nucifera Nutrition 0.000 description 1
- 235000006510 Nelumbo pentapetala Nutrition 0.000 description 1
- 244000183278 Nephelium litchi Species 0.000 description 1
- 235000015742 Nephelium litchi Nutrition 0.000 description 1
- 101100329389 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) cre-1 gene Proteins 0.000 description 1
- 101100395023 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) his-7 gene Proteins 0.000 description 1
- 101100123117 Nicotiana plumbaginifolia MSR-1 gene Proteins 0.000 description 1
- 241000605159 Nitrobacter Species 0.000 description 1
- 239000000020 Nitrocellulose Substances 0.000 description 1
- 241001649463 Nitrosococcus oceani ATCC 19707 Species 0.000 description 1
- 241000605122 Nitrosomonas Species 0.000 description 1
- 241000373063 Nitrosomonas eutropha C91 Species 0.000 description 1
- 238000000636 Northern blotting Methods 0.000 description 1
- 241000894763 Nostoc punctiforme PCC 73102 Species 0.000 description 1
- 241001478892 Nostoc sp. PCC 7120 Species 0.000 description 1
- 101710141454 Nucleoprotein Proteins 0.000 description 1
- 239000004677 Nylon Substances 0.000 description 1
- 241001446528 Ornithopus Species 0.000 description 1
- 240000001516 Oryza latifolia Species 0.000 description 1
- 108700023764 Oryza sativa OSH1 Proteins 0.000 description 1
- 101710160107 Outer membrane protein A Proteins 0.000 description 1
- 108090000417 Oxygenases Proteins 0.000 description 1
- 102000004020 Oxygenases Human genes 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 240000008114 Panicum miliaceum Species 0.000 description 1
- 235000007199 Panicum miliaceum Nutrition 0.000 description 1
- 235000000370 Passiflora edulis Nutrition 0.000 description 1
- 235000002769 Pastinaca sativa Nutrition 0.000 description 1
- 241000209046 Pennisetum Species 0.000 description 1
- 240000009164 Petroselinum crispum Species 0.000 description 1
- 235000002770 Petroselinum crispum Nutrition 0.000 description 1
- 241000745991 Phalaris Species 0.000 description 1
- 244000081757 Phalaris arundinacea Species 0.000 description 1
- RVRRHFPCEOVRKQ-KKUMJFAQSA-N Phe-His-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)N[C@@H](CC(=O)N)C(=O)O)N RVRRHFPCEOVRKQ-KKUMJFAQSA-N 0.000 description 1
- 241000233805 Phoenix Species 0.000 description 1
- IAJOBQBIJHVGMQ-UHFFFAOYSA-N Phosphinothricin Natural products CP(O)(=O)CCC(N)C(O)=O IAJOBQBIJHVGMQ-UHFFFAOYSA-N 0.000 description 1
- 244000273256 Phragmites communis Species 0.000 description 1
- 241000195888 Physcomitrella Species 0.000 description 1
- 241000218602 Pinus <genus> Species 0.000 description 1
- 241000193804 Planococcus <bacterium> Species 0.000 description 1
- 241000209048 Poa Species 0.000 description 1
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 1
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 1
- QMABBZHZMDXHKU-FKBYEOEOSA-N Pro-Tyr-Trp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O QMABBZHZMDXHKU-FKBYEOEOSA-N 0.000 description 1
- 101710083689 Probable capsid protein Proteins 0.000 description 1
- 241000192138 Prochlorococcus Species 0.000 description 1
- 241000192137 Prochlorococcus marinus Species 0.000 description 1
- 241001313098 Prochlorococcus marinus str. AS9601 Species 0.000 description 1
- 241000344823 Prochlorococcus marinus str. MIT 9211 Species 0.000 description 1
- 241000411814 Prochlorococcus marinus str. MIT 9301 Species 0.000 description 1
- 241000612222 Prochlorococcus marinus str. MIT 9303 Species 0.000 description 1
- 241001278367 Prochlorococcus marinus str. MIT 9312 Species 0.000 description 1
- 241001278366 Prochlorococcus marinus str. MIT 9313 Species 0.000 description 1
- 241000411787 Prochlorococcus marinus str. MIT 9515 Species 0.000 description 1
- 241000411816 Prochlorococcus marinus str. NATL1A Species 0.000 description 1
- 241000612207 Prochlorococcus marinus str. NATL2A Species 0.000 description 1
- 241000411779 Prochlorococcus marinus subsp. marinus str. CCMP1375 Species 0.000 description 1
- 241000611429 Prochlorococcus marinus subsp. pastoris str. CCMP1986 Species 0.000 description 1
- 235000011432 Prunus Nutrition 0.000 description 1
- 241000589540 Pseudomonas fluorescens Species 0.000 description 1
- 241000481518 Ralstonia eutropha H16 Species 0.000 description 1
- 241000079829 Ralstonia eutropha JMP134 Species 0.000 description 1
- 241000111470 Ralstonia solanacearum GMI1000 Species 0.000 description 1
- 241000696606 Ralstonia solanacearum UW551 Species 0.000 description 1
- 235000019057 Raphanus caudatus Nutrition 0.000 description 1
- 235000011380 Raphanus sativus Nutrition 0.000 description 1
- 235000006140 Raphanus sativus var sativus Nutrition 0.000 description 1
- 108700005075 Regulator Genes Proteins 0.000 description 1
- 235000011483 Ribes Nutrition 0.000 description 1
- 241000220483 Ribes Species 0.000 description 1
- 244000281247 Ribes rubrum Species 0.000 description 1
- 102000006382 Ribonucleases Human genes 0.000 description 1
- 108010083644 Ribonucleases Proteins 0.000 description 1
- 108091006597 SLC15A4 Proteins 0.000 description 1
- 240000000111 Saccharum officinarum Species 0.000 description 1
- 235000007201 Saccharum officinarum Nutrition 0.000 description 1
- 241000838182 Salinispora arenicola CNS-205 Species 0.000 description 1
- 241001505582 Salinispora tropica CNB-440 Species 0.000 description 1
- 241000124033 Salix Species 0.000 description 1
- 241000921305 Salix sp. Species 0.000 description 1
- 101100198283 Scheffersomyces stipitis (strain ATCC 58785 / CBS 6054 / NBRC 10063 / NRRL Y-11545) DHG2 gene Proteins 0.000 description 1
- 208000034189 Sclerosis Diseases 0.000 description 1
- 241000228160 Secale cereale x Triticum aestivum Species 0.000 description 1
- 108010016634 Seed Storage Proteins Proteins 0.000 description 1
- 241000125165 Selinum Species 0.000 description 1
- CWHJIJJSDGEHNS-MYLFLSLOSA-N Senegenin Chemical compound C1[C@H](O)[C@H](O)[C@@](C)(C(O)=O)[C@@H]2CC[C@@]3(C)C(CC[C@]4(CCC(C[C@H]44)(C)C)C(O)=O)=C4[C@@H](CCl)C[C@@H]3[C@]21C CWHJIJJSDGEHNS-MYLFLSLOSA-N 0.000 description 1
- 229920002684 Sepharose Polymers 0.000 description 1
- 235000009367 Sesamum alatum Nutrition 0.000 description 1
- 240000000452 Sesamum alatum Species 0.000 description 1
- 235000003434 Sesamum indicum Nutrition 0.000 description 1
- 241000220261 Sinapis Species 0.000 description 1
- 241000543826 Sinobambusa Species 0.000 description 1
- 108020004459 Small interfering RNA Proteins 0.000 description 1
- 244000244100 Solanum integrifolium Species 0.000 description 1
- 235000000099 Solanum integrifolium Nutrition 0.000 description 1
- 235000002560 Solanum lycopersicum Nutrition 0.000 description 1
- 235000002597 Solanum melongena Nutrition 0.000 description 1
- 244000061458 Solanum melongena Species 0.000 description 1
- 102100021484 Solute carrier family 15 member 4 Human genes 0.000 description 1
- 235000006745 Sonchus oleraceus Nutrition 0.000 description 1
- 244000113428 Sonchus oleraceus Species 0.000 description 1
- 244000062793 Sorghum vulgare Species 0.000 description 1
- 241000219315 Spinacia Species 0.000 description 1
- 244000300264 Spinacia oleracea Species 0.000 description 1
- 241000605008 Spirillum Species 0.000 description 1
- 235000009184 Spondias indica Nutrition 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 241001446311 Streptomyces coelicolor A3(2) Species 0.000 description 1
- 241001655322 Streptomycetales Species 0.000 description 1
- 101710172711 Structural protein Proteins 0.000 description 1
- NINIDFKCEFEMDL-UHFFFAOYSA-N Sulfur Chemical compound [S] NINIDFKCEFEMDL-UHFFFAOYSA-N 0.000 description 1
- 239000005864 Sulphur Substances 0.000 description 1
- 241000135402 Synechococcus elongatus PCC 6301 Species 0.000 description 1
- 241000192593 Synechocystis sp. PCC 6803 Species 0.000 description 1
- 108700005078 Synthetic Genes Proteins 0.000 description 1
- 235000012096 Syzygium samarangense Nutrition 0.000 description 1
- 235000012308 Tagetes Nutrition 0.000 description 1
- 108091046869 Telomeric non-coding RNA Proteins 0.000 description 1
- 108700007696 Tetrahydrofolate Dehydrogenase Proteins 0.000 description 1
- 101100242191 Tetraodon nigroviridis rho gene Proteins 0.000 description 1
- 235000006468 Thea sinensis Nutrition 0.000 description 1
- 241001504076 Thermosynechococcus elongatus BP-1 Species 0.000 description 1
- 241001136391 Thiobacillus denitrificans ATCC 25259 Species 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-N Thiophosphoric acid Chemical class OP(O)(S)=O RYYWUUFWQRZTIU-UHFFFAOYSA-N 0.000 description 1
- 244000237996 Thuarea involuta Species 0.000 description 1
- 101710150448 Transcriptional regulator Myc Proteins 0.000 description 1
- 241000970911 Trichormus variabilis ATCC 29413 Species 0.000 description 1
- 235000007218 Tripsacum dactyloides Nutrition 0.000 description 1
- 244000082267 Tripsacum dactyloides Species 0.000 description 1
- 235000007247 Triticum turgidum Nutrition 0.000 description 1
- 240000002805 Triticum turgidum Species 0.000 description 1
- 235000004424 Tropaeolum majus Nutrition 0.000 description 1
- 240000001260 Tropaeolum majus Species 0.000 description 1
- 235000018946 Tropaeolum minus Nutrition 0.000 description 1
- 240000008573 Tropaeolum minus Species 0.000 description 1
- ZKVANNIVSDOQMG-HKUYNNGSSA-N Trp-Tyr-Gly Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)NCC(=O)O)N ZKVANNIVSDOQMG-HKUYNNGSSA-N 0.000 description 1
- CRHFOYCJGVJPLE-AVGNSLFASA-N Tyr-Gln-Asn Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)O CRHFOYCJGVJPLE-AVGNSLFASA-N 0.000 description 1
- 102100038487 Ubiquitin conjugation factor E4 B Human genes 0.000 description 1
- 102000018478 Ubiquitin-Activating Enzymes Human genes 0.000 description 1
- 108010091546 Ubiquitin-Activating Enzymes Proteins 0.000 description 1
- 235000013419 Uniola paniculata Nutrition 0.000 description 1
- 235000012511 Vaccinium Nutrition 0.000 description 1
- GBESYURLQOYWLU-LAEOZQHASA-N Val-Glu-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N GBESYURLQOYWLU-LAEOZQHASA-N 0.000 description 1
- 241000847071 Verminephrobacter eiseniae EF01-2 Species 0.000 description 1
- 241000607598 Vibrio Species 0.000 description 1
- 206010047400 Vibrio infections Diseases 0.000 description 1
- 241000219873 Vicia Species 0.000 description 1
- 235000010713 Vicia narbonensis Nutrition 0.000 description 1
- 240000002570 Vicia narbonensis Species 0.000 description 1
- 244000105017 Vicia sativa Species 0.000 description 1
- 241001464837 Viridiplantae Species 0.000 description 1
- 235000018936 Vitellaria paradoxa Nutrition 0.000 description 1
- 235000009392 Vitis Nutrition 0.000 description 1
- 108010093038 XCL-2 calpain Proteins 0.000 description 1
- 241001311561 Xanthobacter autotrophicus Py2 Species 0.000 description 1
- 241000307523 Xenostegia media Species 0.000 description 1
- 108700040099 Xylose isomerases Proteins 0.000 description 1
- 235000007244 Zea mays Nutrition 0.000 description 1
- 235000016383 Zea mays subsp huehuetenangensis Nutrition 0.000 description 1
- 241001478412 Zizania palustris Species 0.000 description 1
- 210000001015 abdomen Anatomy 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 230000000996 additive effect Effects 0.000 description 1
- 244000193174 agave Species 0.000 description 1
- 230000032683 aging Effects 0.000 description 1
- 108010048916 alcohol dehydrogenase (acceptor) Proteins 0.000 description 1
- 108010050181 aleurone Proteins 0.000 description 1
- 101150099105 alien gene Proteins 0.000 description 1
- 239000003513 alkali Substances 0.000 description 1
- WQZGKKKJIJFFOK-PQMKYFCFSA-N alpha-D-mannose Chemical compound OC[C@H]1O[C@H](O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-PQMKYFCFSA-N 0.000 description 1
- SRBFZHDQGSBBOR-LECHCGJUSA-N alpha-D-xylose Chemical compound O[C@@H]1CO[C@H](O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-LECHCGJUSA-N 0.000 description 1
- KLOHDWPABZXLGI-YWUHCJSESA-M ampicillin sodium Chemical compound [Na+].C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C([O-])=O)(C)C)=CC=CC=C1 KLOHDWPABZXLGI-YWUHCJSESA-M 0.000 description 1
- 238000000540 analysis of variance Methods 0.000 description 1
- 235000019728 animal nutrition Nutrition 0.000 description 1
- 230000003064 anti-oxidating effect Effects 0.000 description 1
- 108091007433 antigens Proteins 0.000 description 1
- 239000002246 antineoplastic agent Substances 0.000 description 1
- 229940041181 antineoplastic drug Drugs 0.000 description 1
- 239000003963 antioxidant agent Substances 0.000 description 1
- 230000003078 antioxidant effect Effects 0.000 description 1
- 239000000074 antisense oligonucleotide Substances 0.000 description 1
- 238000012230 antisense oligonucleotides Methods 0.000 description 1
- 239000001387 apium graveolens Substances 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 238000003556 assay Methods 0.000 description 1
- 101150036080 at gene Proteins 0.000 description 1
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 1
- 230000037444 atrophy Effects 0.000 description 1
- 238000000211 autoradiogram Methods 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 239000002551 biofuel Substances 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 229930189065 blasticidin Natural products 0.000 description 1
- 229960001561 bleomycin Drugs 0.000 description 1
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 1
- 229910052796 boron Inorganic materials 0.000 description 1
- 229910052793 cadmium Inorganic materials 0.000 description 1
- BDOSMKKIYDKNTQ-UHFFFAOYSA-N cadmium atom Chemical compound [Cd] BDOSMKKIYDKNTQ-UHFFFAOYSA-N 0.000 description 1
- 235000009120 camo Nutrition 0.000 description 1
- 150000001722 carbon compounds Chemical class 0.000 description 1
- 235000011089 carbon dioxide Nutrition 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 230000003197 catalytic effect Effects 0.000 description 1
- 230000022131 cell cycle Effects 0.000 description 1
- 230000010001 cellular homeostasis Effects 0.000 description 1
- 230000036755 cellular response Effects 0.000 description 1
- 230000004700 cellular uptake Effects 0.000 description 1
- 235000005607 chanvre indien Nutrition 0.000 description 1
- 238000007385 chemical modification Methods 0.000 description 1
- 239000003153 chemical reaction reagent Substances 0.000 description 1
- WIIZWVCIJKGZOK-RKDXNWHRSA-N chloramphenicol Chemical compound ClC(Cl)C(=O)N[C@H](CO)[C@H](O)C1=CC=C([N+]([O-])=O)C=C1 WIIZWVCIJKGZOK-RKDXNWHRSA-N 0.000 description 1
- RCTYPNKXASFOBE-UHFFFAOYSA-M chloromercury Chemical compound [Hg]Cl RCTYPNKXASFOBE-UHFFFAOYSA-M 0.000 description 1
- 210000000349 chromosome Anatomy 0.000 description 1
- 239000001407 cinnamomum spp. Substances 0.000 description 1
- 235000020971 citrus fruits Nutrition 0.000 description 1
- 239000004927 clay Substances 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 239000003184 complementary RNA Substances 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 210000000877 corpus callosum Anatomy 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000010168 coupling process Methods 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- 230000009193 crawling Effects 0.000 description 1
- 239000012297 crystallization seed Substances 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 230000008260 defense mechanism Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 230000017858 demethylation Effects 0.000 description 1
- 238000010520 demethylation reaction Methods 0.000 description 1
- 238000010217 densitometric analysis Methods 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 102000004419 dihydrofolate reductase Human genes 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 235000004879 dioscorea Nutrition 0.000 description 1
- 238000006073 displacement reaction Methods 0.000 description 1
- 239000012153 distilled water Substances 0.000 description 1
- 238000004043 dyeing Methods 0.000 description 1
- 230000013020 embryo development Effects 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 230000006353 environmental stress Effects 0.000 description 1
- 239000003797 essential amino acid Substances 0.000 description 1
- 235000020776 essential amino acid Nutrition 0.000 description 1
- 235000008995 european elder Nutrition 0.000 description 1
- 239000003925 fat Substances 0.000 description 1
- 230000004992 fission Effects 0.000 description 1
- 235000004426 flaxseed Nutrition 0.000 description 1
- 235000013312 flour Nutrition 0.000 description 1
- 239000006260 foam Substances 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- PGBHMTALBVVCIT-VCIWKGPPSA-N framycetin Chemical compound N[C@@H]1[C@@H](O)[C@H](O)[C@H](CN)O[C@@H]1O[C@H]1[C@@H](O)[C@H](O[C@H]2[C@@H]([C@@H](N)C[C@@H](N)[C@@H]2O)O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](CN)O2)N)O[C@@H]1CO PGBHMTALBVVCIT-VCIWKGPPSA-N 0.000 description 1
- 238000011207 functional examination Methods 0.000 description 1
- 230000000799 fusogenic effect Effects 0.000 description 1
- 238000002523 gelfiltration Methods 0.000 description 1
- 238000012252 genetic analysis Methods 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 244000037671 genetically modified crops Species 0.000 description 1
- 229960002518 gentamicin Drugs 0.000 description 1
- IAJOBQBIJHVGMQ-BYPYZUCNSA-N glufosinate-P Chemical compound CP(O)(=O)CC[C@H](N)C(O)=O IAJOBQBIJHVGMQ-BYPYZUCNSA-N 0.000 description 1
- 229960003180 glutathione Drugs 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 108010077515 glycylproline Proteins 0.000 description 1
- XDDAORKBJWWYJS-UHFFFAOYSA-N glyphosate Chemical compound OC(=O)CNCP(O)(O)=O XDDAORKBJWWYJS-UHFFFAOYSA-N 0.000 description 1
- 229940097068 glyphosate Drugs 0.000 description 1
- 239000003630 growth substance Substances 0.000 description 1
- 230000002650 habitual effect Effects 0.000 description 1
- 235000015220 hamburgers Nutrition 0.000 description 1
- 239000011487 hemp Substances 0.000 description 1
- 238000013537 high throughput screening Methods 0.000 description 1
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 1
- 239000000710 homodimer Substances 0.000 description 1
- 125000004435 hydrogen atom Chemical class [H]* 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- WGCNASOHLSPBMP-UHFFFAOYSA-N hydroxyacetaldehyde Natural products OCC=O WGCNASOHLSPBMP-UHFFFAOYSA-N 0.000 description 1
- YQYJSBFKSSDGFO-FWAVGLHBSA-N hygromycin A Chemical compound O[C@H]1[C@H](O)[C@H](C(=O)C)O[C@@H]1Oc1ccc(\C=C(/C)C(=O)N[C@@H]2[C@@H]([C@H]3OCO[C@H]3[C@@H](O)[C@@H]2O)O)cc1O YQYJSBFKSSDGFO-FWAVGLHBSA-N 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 238000007901 in situ hybridization Methods 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 238000011081 inoculation Methods 0.000 description 1
- 229960003786 inosine Drugs 0.000 description 1
- 229940065638 intron a Drugs 0.000 description 1
- 230000009545 invasion Effects 0.000 description 1
- 229910052742 iron Inorganic materials 0.000 description 1
- 238000003973 irrigation Methods 0.000 description 1
- 230000002262 irrigation Effects 0.000 description 1
- 230000006122 isoprenylation Effects 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 101150066555 lacZ gene Proteins 0.000 description 1
- 125000001909 leucine group Chemical group [H]N(*)C(C(*)=O)C([H])([H])C(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 239000007788 liquid Substances 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 239000011777 magnesium Substances 0.000 description 1
- 229910052749 magnesium Inorganic materials 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 235000009973 maize Nutrition 0.000 description 1
- WPBNNNQJVZRUHP-UHFFFAOYSA-L manganese(2+);methyl n-[[2-(methoxycarbonylcarbamothioylamino)phenyl]carbamothioyl]carbamate;n-[2-(sulfidocarbothioylamino)ethyl]carbamodithioate Chemical compound [Mn+2].[S-]C(=S)NCCNC([S-])=S.COC(=O)NC(=S)NC1=CC=CC=C1NC(=S)NC(=O)OC WPBNNNQJVZRUHP-UHFFFAOYSA-L 0.000 description 1
- 235000005739 manihot Nutrition 0.000 description 1
- 230000013011 mating Effects 0.000 description 1
- 235000013372 meat Nutrition 0.000 description 1
- 235000013622 meat product Nutrition 0.000 description 1
- 238000002844 melting Methods 0.000 description 1
- 230000008018 melting Effects 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 102000006240 membrane receptors Human genes 0.000 description 1
- 210000000473 mesophyll cell Anatomy 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 1
- 238000002493 microarray Methods 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 235000013336 milk Nutrition 0.000 description 1
- 239000008267 milk Substances 0.000 description 1
- 210000004080 milk Anatomy 0.000 description 1
- 235000019713 millet Nutrition 0.000 description 1
- 108091005573 modified proteins Proteins 0.000 description 1
- 102000035118 modified proteins Human genes 0.000 description 1
- 230000003020 moisturizing effect Effects 0.000 description 1
- VYQNWZOUAUKGHI-UHFFFAOYSA-N monobenzone Chemical compound C1=CC(O)=CC=C1OCC1=CC=CC=C1 VYQNWZOUAUKGHI-UHFFFAOYSA-N 0.000 description 1
- 230000004660 morphological change Effects 0.000 description 1
- 230000007498 myristoylation Effects 0.000 description 1
- 229920001220 nitrocellulos Polymers 0.000 description 1
- 230000024121 nodulation Effects 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 239000002417 nutraceutical Substances 0.000 description 1
- 235000021436 nutraceutical agent Nutrition 0.000 description 1
- 235000021232 nutrient availability Nutrition 0.000 description 1
- 235000006286 nutrient intake Nutrition 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 229920001778 nylon Polymers 0.000 description 1
- 150000007524 organic acids Chemical class 0.000 description 1
- 229940049547 paraxin Drugs 0.000 description 1
- 244000052769 pathogen Species 0.000 description 1
- 230000001717 pathogenic effect Effects 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 239000000575 pesticide Substances 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 150000003016 phosphoric acids Chemical class 0.000 description 1
- 238000000206 photolithography Methods 0.000 description 1
- 230000000243 photosynthetic effect Effects 0.000 description 1
- 230000035479 physiological effects, processes and functions Effects 0.000 description 1
- 239000001739 pinus spp. Substances 0.000 description 1
- 238000003976 plant breeding Methods 0.000 description 1
- 239000003375 plant hormone Substances 0.000 description 1
- 230000037039 plant physiology Effects 0.000 description 1
- 238000007747 plating Methods 0.000 description 1
- 230000010152 pollination Effects 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 238000006116 polymerization reaction Methods 0.000 description 1
- 239000011591 potassium Substances 0.000 description 1
- 229910052700 potassium Inorganic materials 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 230000000750 progressive effect Effects 0.000 description 1
- 230000002062 proliferating effect Effects 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 235000014774 prunus Nutrition 0.000 description 1
- 150000003222 pyridines Chemical class 0.000 description 1
- 238000000197 pyrolysis Methods 0.000 description 1
- 230000005855 radiation Effects 0.000 description 1
- 239000010499 rapseed oil Substances 0.000 description 1
- 108020003175 receptors Proteins 0.000 description 1
- 102000005962 receptors Human genes 0.000 description 1
- 230000001172 regenerating effect Effects 0.000 description 1
- 230000008521 reorganization Effects 0.000 description 1
- 239000011347 resin Substances 0.000 description 1
- 229920005989 resin Polymers 0.000 description 1
- 230000000452 restraining effect Effects 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 238000003757 reverse transcription PCR Methods 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 101150076874 rha-1 gene Proteins 0.000 description 1
- 210000000614 rib Anatomy 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 229920002477 rna polymer Polymers 0.000 description 1
- 235000019515 salmon Nutrition 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 230000003248 secreting effect Effects 0.000 description 1
- 230000008117 seed development Effects 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 238000012772 sequence design Methods 0.000 description 1
- 210000002107 sheath cell Anatomy 0.000 description 1
- 238000004904 shortening Methods 0.000 description 1
- 230000009131 signaling function Effects 0.000 description 1
- 239000005368 silicate glass Substances 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- 239000001509 sodium citrate Substances 0.000 description 1
- 239000001488 sodium phosphate Substances 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 125000006850 spacer group Chemical group 0.000 description 1
- 229960000268 spectinomycin Drugs 0.000 description 1
- UNFWWIHTNXNPBV-WXKVUWSESA-N spectinomycin Chemical compound O([C@@H]1[C@@H](NC)[C@@H](O)[C@H]([C@@H]([C@H]1O1)O)NC)[C@]2(O)[C@H]1O[C@H](C)CC2=O UNFWWIHTNXNPBV-WXKVUWSESA-N 0.000 description 1
- 238000001228 spectrum Methods 0.000 description 1
- 238000005507 spraying Methods 0.000 description 1
- 239000010421 standard material Substances 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 238000013179 statistical model Methods 0.000 description 1
- QTENRWWVYAAPBI-YCRXJPFRSA-N streptomycin sulfate Chemical compound OS(O)(=O)=O.OS(O)(=O)=O.OS(O)(=O)=O.CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](N=C(N)N)[C@H](O)[C@@H](N=C(N)N)[C@H](O)[C@H]1O.CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](N=C(N)N)[C@H](O)[C@@H](N=C(N)N)[C@H](O)[C@H]1O QTENRWWVYAAPBI-YCRXJPFRSA-N 0.000 description 1
- 238000006467 substitution reaction Methods 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 230000002123 temporal effect Effects 0.000 description 1
- 239000009871 tenuigenin Substances 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 1
- UMGDCJDMYOKAJW-UHFFFAOYSA-N thiourea Chemical compound NC(N)=S UMGDCJDMYOKAJW-UHFFFAOYSA-N 0.000 description 1
- 108091006106 transcriptional activators Proteins 0.000 description 1
- 230000001960 triggered effect Effects 0.000 description 1
- HRXKRNGNAMMEHJ-UHFFFAOYSA-K trisodium citrate Chemical compound [Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O HRXKRNGNAMMEHJ-UHFFFAOYSA-K 0.000 description 1
- 229940038773 trisodium citrate Drugs 0.000 description 1
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 1
- 229910000406 trisodium phosphate Inorganic materials 0.000 description 1
- 235000019801 trisodium phosphate Nutrition 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
- 235000018322 upland cotton Nutrition 0.000 description 1
- 230000002792 vascular Effects 0.000 description 1
- 230000009385 viral infection Effects 0.000 description 1
- 230000028604 virus induced gene silencing Effects 0.000 description 1
- 239000002699 waste material Substances 0.000 description 1
- 238000009736 wetting Methods 0.000 description 1
- 229960003487 xylose Drugs 0.000 description 1
- 238000001086 yeast two-hybrid system Methods 0.000 description 1
Images


























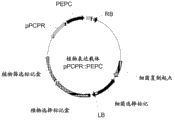


































































































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
- C12N15/8273—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for drought, cold, salt resistance
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/001—Oxidoreductases (1.) acting on the CH-CH group of donors (1.3)
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A40/00—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production
- Y02A40/10—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production in agriculture
- Y02A40/146—Genetically Modified [GMO] plants, e.g. transgenic plants
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Organic Chemistry (AREA)
- General Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Cell Biology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Medicinal Chemistry (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Peptides Or Proteins (AREA)
- Agricultural Chemicals And Associated Chemicals (AREA)
- Enzymes And Modification Thereof (AREA)
Abstract
本发明一般涉及分子生物学领域并且涉及通过调节植物中编码产量增加多肽的核酸的表达来增强产量相关性状的方法,所述多肽选自:磷酸烯醇丙酮酸羧化酶(PEPC),III类U盒蛋白和PQQC蛋白。本发明还涉及具有编码产量增加多肽的核酸的受调节表达的植物,所述植物相对于对应的野生型植物或者其他对照植物具有增强的产量相关性状,所述产量增加多肽选自:磷酸烯醇丙酮酸羧化酶(PEPC),III类U盒蛋白和PQQC蛋白。本发明还提供了可用于本发明方法的构建体。
Description
本申请是申请日为2008年7月31日、发明名称为“具有增强的产量相关性状的植物及其制备方法”的中国专利申请200880025566.2(国际申请号PCT/EP2008/060060)的分案申请。
本发明一般涉及分子生物学领域并且涉及通过调节植物中编码产量增加多肽的核酸的表达来增强产量相关性状的方法,所述多肽选自:磷酸烯醇丙酮酸羧化酶(PEPC),III类U盒蛋白和PQQC蛋白。本发明还涉及具有编码产量增加多肽的核酸的受调节表达的植物,所述植物相对于对应的野生型植物或者其他对照植物具有增强的产量相关性状。本发明还提供了可用于本发明方法的构建体。
持续增长的世界人口和农业用可耕地供应萎缩刺激了有关增加农业效率的研究。常规的作物及园艺学改良手段利用选择育种技术以鉴定具有受欢萨特性的植物。然而,此类选择育种技术具有几个缺陷,即这些技术一般耗费很多劳动并且产生经常含有异源性遗传组分的植物,所述异源性遗传组分可能不总是导致所希望性状从亲代植物中传递。分子生物学进展已经允许人类改良动物及植物的种质。植物的遗传工程使得可以分离和操作遗传物质(一般处于DNA或RNA形式)并且随后将该遗传物质导入植物中。此类技术具有产生具备多种经济学、农学或园艺学改良性状的作物或植物的能力。
具有特殊经济意义的性状是增加的产量。产量通常定义为来自作物的经济价值的可测量结果。该结果可以就数量和/或品质方面进行定义。产量直接取决于几个因素,例如器官的数目和大小、植物结构(例如枝条的数目)、种子产生、叶衰老等。根发育、养分摄入量、胁迫耐受性和早期萌发势(early vigor)也可以是决定产量的重要因素。优化前述因素因而可以对增加作物产量有贡献。
种子产量是尤其重要的性状,因为许多植物的种子对于人类和动物的营养非常重要。作物如谷物、稻、小麦、卡诺拉油菜(canola)和大豆占人类总卡路里摄取量的一半以上,不论是通过种子本身的直接消费,还是通过由加工的种子所饲养的肉类产品的消费。它们也是工业加工所用的糖类、油类和多类代谢物的来源。种子含有胚(新的芽和根的来源)和胚乳(萌发以及幼苗早期生长过程中胚生长的营养源)。种子的发育涉及许多基因,并且需要代谢物自根、叶和茎转移至正在生长的种子。特别是胚乳,吸收糖类、油类和蛋白质的代谢前体,将其合成为贮存高分子,以使谷粒饱满。
许多作物的一个重要性状是早期萌发势。提高早期萌发势是温带和热带稻栽培种中现代稻育种程序的重要目标。长根对于水中播种的稻的正确土壤锚定是重要的。当稻直接播种到被淹没的田间并且当植物必须快速出水时,较长的根与萌发势有关。当实施播种机播种时,较长的中胚轴和胚芽鞘对于好的幼苗萌出是重要的。将早期萌发势改造到植物中在农业中是重要的。例如,弱的早期萌发势局限于对于基于欧洲大西洋部玉米带种质的玉米杂种的引入产生限制。
另一重要的性状是提高的非生物胁迫耐受性。非生物胁迫是世界作物减产的主要原因,对于多数作物植物,将平均产量减少50%以上(Wang etal.,Planta(2003)218:1-14)。非生物胁迫可以由干旱、盐度、极端温度、化学毒性和氧化胁迫引起。提高植物对非生物胁迫的耐受性的能力对于世界上的农场主将具有主要的经济利益并且将允许在不利条件下和否则不可能栽培作物的领域中栽培作物。
因此,通过优化上述因素之一可以提高作物产量。
取决于最终用途,某些产量性状的修饰可以相对于其他性状是有利的。例如,对于诸如粮草或木材生产或生物燃料生产的应用,植物的营养部分的增加可以是想要的,对于诸如面粉、淀粉或油生产的应用,种子参数的增加或改善可以是尤其想要的。甚至在种子参数中,一些可以比其他有利,这取决于最终应用。多种机理可以促进提高种子产量,无论其为增加的种子大小或增加的种子数目或任何其他种子相关性状的形式。
提高植物中种子产量(种子产量和/或生物量)的一种方法是通过改变植物的内在生长机理,如涉及植物生长或防御机理的细胞周期或多种信号途径。
现在已经发现通过调节植物中编码产量增加多肽的核酸的表达可以增强植物中产量相关性状,所述多肽选自:
磷酸烯醇丙酮酸羧化酶(PEPC),
III类U盒蛋白和
PQQC蛋白。
背景
磷酸烯醇丙酮酸羧化酶(也称作PEP羧化酶,PEPC酶,或者PEPC;EC4.1.1.31)是羧基裂合酶家族的酶,其催化CO2加入到磷酸烯醇丙酮酸(PEP)以形成四碳化合物草酰乙酸:
PEP+CO2→草酰乙酸+Pi
该反应用于所称的“CAM”或“C4”植物中的碳固定,其中它在光合作用中起关键作用。除了植物外,在一些细菌中也发现此酶,但是在动物或真菌中没有发现。
通过PEP羧化酶的碳固定将可利用的CO2同化到四碳化合物(草酰乙酸,其进一步被转化为苹果酸)中,该四碳化合物可以被存储或者在植物细胞之间穿梭。这使得可以分隔通过与空气接触进行的初始CO2固定和在光合作用的光独立反应期间通过RuBisCO进行的次级CO2向糖的固定。
在适于在极干燥条件下生长的肉质CAM植物中,PEP羧化酶在夜间固定CO2,此时植物打开气孔以允许气体交换。在白天,植物关闭气孔以保留水分并释放叶子内夜间产生的来自存储化合物的CO2。这允许植物通过在白天进行光合作用而不通过开放的气孔释放水分而在干旱气候中茁壮成长。
在C4植物例如玉米中,PEP羧化酶在叶子的叶肉细胞中固定CO2并且所得的4碳化合物穿梭到维管束鞘细胞中,在这里其释放CO2用于被RuBisCO固定。从而,这两个过程在空间上分开,允许RuBisCO在低氧环境中工作以避免光呼吸作用。由于RuBisCO的固有加氧酶活性而发生光呼吸作用,其中该酶使用氧气而不是二氧化碳,不向糖中掺入碳或者产生ATP。这样,它对于植物而言是一种浪费的反应。相比较,通过PEP羧化酶进行C4碳固定更有效。
Radchuk et al.,2007(The Plant Journal51,819-839)报导了棒杆菌PEPC在Vicia narbonensis种子中过表达以改变种子代谢并且将碳引入有机酸中,导致更大的种子存储能力和增加的蛋白质含量。
U盒是~70个氨基酸的结构域,其存在于酵母到人类中。原型U盒蛋白酵母Ufd2被鉴定为泛素链装配因子(E4),其与泛素活化酶(E1)、泛素缀合酶(E2)和E3协作以催化以人工底物形成泛素链,Hatakeyama et al.,2003(Biochemical and Biophysical Research Communications302,pp635-645)。Azevedo et al.2001(Trends in Plant Science Vol.6No.8)报导,数据库研究揭示了拟南芥中37种预测的含有U盒的蛋白质,尽管据报导UFD2是酵母中唯一的U盒蛋白,并且仅仅一些在人类基因组和秀丽隐杆线虫(Caenorhabditis elegans)和黑腹果萧(Drosophila melanogaster)中被鉴定出来。基于结构域分析,发现植物U盒(PUB)蛋白质形成5个不同的亚类。发现III类由12个成员组成,具有含有~14%亮氨酸残基的结构域。与U盒结构域在其他类别成员中的位置相比,发现III类成员中的U盒结构域在N-末端区。
预测U盒蛋白为泛素化机器中的E3或E4因子。Cho et al.,2006(PlantPhysiology,December2006,Vol.142,pp.1664-1682)从水胁迫的辣椒(Capsicum annuum L.cv Pukang)中分离了推定的U盒蛋白1(CaPUB1),编码在其N末端区中含有单个U盒基序的蛋白质。体外泛素化和位点定向诱变测定法揭示CaPUB1具有E3泛素连接酶活性并且U盒基序对于其酶活性是必需的。它们在拟南芥CaPUB1中在花椰菜花叶病毒35S启动子控制下过表达。据报导转基因拟南芥植物具有比野生型显著更长的下胚轴和根并且生长更快,导致早抽苔表型。发现子叶的形态在黑暗中是相似的的。表明35S::CaPUB1的根具有增加数目的小尺寸细胞,导致在皮层、内胚层和中柱中有序的数目巨大的细胞层。转基因植物也显示出对水胁迫和轻微盐度的增加的敏感性。
在不存在任何明显辅因子的反应中,PqqC促进3a-(2-氨基-2-羧乙基)-4,5-二氧-4,5,6,7,8,9-六氢喹啉-7,9-二羧酸环化和氧化成PQQ。吡咯并喹啉醌[4,5-二氢-4,5-二氧-1H-吡咯并-[2,3-f]喹啉-2,7,9-三羧酸;PQQ]是一种芳族三环邻醌,其作为一些细菌脱氢酶的氧化还原辅因子。最著名的实例是甲醇脱氢酶和葡萄糖脱氢酶。PQQ属于醌辅因子家族,已经公知其为依赖吡啶核苷酸和黄素的辅因子之后的第三类氧化还原辅因子。尽管植物和动物自身不产生PQQ,但是PQQ因为其存在于人奶中和其抗氧化性质而引起了巨大的兴趣。
已经从乙酸钙不动杆菌(Acinetobacter calcoaceticus)、克雷伯菌属(Klebsiella)、萦光假单胞菌(Pseudomonas Fluorescens)、嗜器官甲基杆菌(Methylobacterium organophilum)和扭脱甲基杆菌(Methylobacterium extorquens)克隆了涉及PQQ合成的基因。
PqqC催化PQQ生物合成中的最后步骤。PqqC是251个残基的蛋白质(分子量28.91kDa),其在溶液中形成同型二聚体,如从凝胶过滤实验证明。PqqC折叠成紧密的七螺旋束,具有六个环形对齐的螺旋,部分包围第七个疏水螺旋。PqqC结构的分析表明这七个螺旋为活性部位腔提供了支架。该腔排列着42个最亲水的残基和芳族残基,其在来自不同细菌的PQQC蛋白内高度保守。该腔显示了不同的总的正电荷。
概述
令人惊奇地,现在已经发现调节编码产量增加多肽的核酸的表达得到了相对于对照植物具有增强的产量相关性状,尤其增加的产量的植物,所述多肽选自:
磷酸烯醇丙酮酸羧化酶(PEPC),
III类U盒蛋白和
PQQC蛋白。
尤其令人惊奇的是发现产量的增加不以更大的种子的形式表现(如已经从Radchuck et al.,2007的教导中预期),而是以一组改变的产量相关性状的形式表现,所述性状包括但不限于,相对于各自对照植物增加的地上部分面积、增加的萌发势、增加的种子总数、增加的种子重量、增加的饱满率、每圆锥花序增加的小花数、增加的种子数目、增加的收获指数、增加的高度和根厚度的增加。
根据一个实施方案,提供了相对于对照植物增强植物中产量相关性状的方法,包括调节植物中编码PEPC蛋白的核酸的表达。
根据一个实施方案,提供了相对于对照植物增强植物中产量相关性状、尤其增加的产量的方法,包括调节植物中编码III类U盒蛋白的核酸的表达。
根据一个实施方案,提供了相对于对照植物增强植物中产量相关性状、尤其增加的产量的方法,包括调节植物中编码PQQC蛋白的核酸的表达。
定义
多肽/蛋白质
术语“多肽”和“蛋白质”在本文中可互换使用并且指处于任意长度聚合形式的通过肽键连接在一起的氨基酸。
多核苷酸/核酸/核酸序列/核苷酸序列
术语“多核苷酸”、“核酸序列”、“核苷酸序列”、“核酸”、“核酸分子”在本文中可互换使用并且指处于任意长度聚合未分枝形式中的核苷酸,即核糖核苷酸或脱氧核糖核苷酸或这二者的组合。
对照植物
选择合适的对照植物是实验设置的惯用部分并且可以包括对应的野生型植物或无目的基因的对应植物。对照植物一般是与待评估植物相同的植物物种或甚至是相同的变种。对照植物也可以是待评估植物的失效合子。失效合子是通过分离而缺少所述转基因的个体。如本文中所用的“对照植物”不仅指整株植物,还指植物部分,包括种子及种子部分。
同源物
蛋白质的“同源物”包括这样的肽、寡肽、多肽、蛋白质及酶,它们相对于非修饰的上述蛋白质具有氨基酸取代、缺失和/或插入并且与所述肽、寡肽、多肽、蛋白质及酶来源的非修饰蛋白质具有相似生物学活性和功能活性。
缺失指从蛋白质中移除一个或多个氨基酸。
插入指一个或多个氨基酸残基在蛋白质中预定位点内的引入。插入可以包含单个或多个氨基酸的N端融合和/或C端融合以及序列内插入。通常,在氨基酸序列内部的插入会比N端融合或C端融合小约1-10个残基级别。N端或C端融合蛋白或融合肽的实例包括如酵母双杂交系统中所用转录激活物的结合结构域或激活结构域、噬菌体外壳蛋白、(组氨酸)-6-标签、谷胱甘肽S-转移酶-标签、A蛋白、麦芽糖结合蛋白、二氢叶酸还原酶、Tag·100表位、c-myc表位、-表位、lacZ、CMP(钙调蛋白结合肽)、HA表位、C蛋白表位和VSV表位。
取代指以具有相似特性(如相似疏水性、亲水性、抗原性、形成或破坏α-螺旋结构或β-片层结构的倾向)的其他氨基酸替换蛋白质的氨基酸。氨基酸取代一般是单个残基的,不过可以是簇集性的,这取决于置于多肽的功能性约束;插入通常会是约1-10个氨基酸残基级别。对于大的蛋白质,插入的数目甚至可以是50、100、150、200或更多级别。氨基酸取代优选地是保守性氨基酸取代。保守性取代表是本领域众所周知的(见例如Creighton(1984)Proteins.W.H.Freeman and Company(编辑)和下表1)。
表1:保守性氨基酸取代的实例
| 残基 | 保守性取代 | 残基 | 保守性取代 |
| Ala | Ser | Leu | Ile;Val |
| Arg | Lys | Lys | Arg;Gln |
| Asn | Gln;His | Met | Leu;Ile |
| Asp | Glu | Phe | Met;Leu;Tyr |
| Gln | Asn | Ser | Thr;Gly |
| Cys | Ser | Thr | Ser;Val |
| Glu | Asp | Trp | Tyr |
| Gly | Pro | Tyr | Trp;Phe |
| His | Asn;Gln | Val | Ile;Leu |
| Ile | Leu,Val |
氨基酸取代、缺失和/或插入可以使用本领域众所周知的肽合成技术如固相肽合成法等或通过重组DNA操作而轻易地进行。用于操作DNA序列以产生蛋白质的取代、插入或缺失变体的方法是本领域众所周知的。例如,用于在DNA中的预定位点处产生取代突变的技术是本领域技术人员众所周知的并且包括M13诱变法、T7-Gen体外诱变法(USB,Clevelaand,OH)、QuickChange位点定向诱变法(Stratagene,San Diego,CA)、PCR-介导的位点定向诱变或其他位点定向诱变法。
衍生物
“衍生物”包括这样的肽、寡肽、多肽,其中与天然发生形式的蛋白质(如目的蛋白质)的氨基酸序列相比,它们包含以非天然发生的氨基酸残基对氨基酸的取代或非天然发生的氨基酸残基的添加。蛋白质“衍生物”也包含这样的肽、寡肽、多肽,其中与多肽的天然发生形式的氨基酸序列相比,它们包含天然发生的改变(糖基化、酰化、异戊二烯化、磷酸化、肉豆蔻酰化、硫酸盐化等)氨基酸残基或非天然的改变氨基酸残基。与衍生物所来源的氨基酸序列相比,该衍生物可以也包含与所述氨基酸序列共价或非共价结合的一个或多个非氨基酸取代基或添加(例如报道分子或其他配体),如为促进检测该衍生物而结合的报道分子,和与天然发生的蛋白质的氨基酸序列相对比的非天然发生的氨基酸残基。此外,“衍生物”也包括天然存在形式的蛋白质与标记肽如FLAG、HIS6或硫氧还蛋白的融合物(标记肽的综述,见Terpe,Appl.Microbiol.Biotechnol.60,523-533,2003)。
直向同源物/旁系同源物
直向同源物和旁系同源物包含用来描述基因祖先关系的进化概念。旁系同源物是相同物种内起源于先祖基因复制的基因,而直向同源物是来自不同生物的起源于物种形成的基因,并且也来自共同的祖先基因。
结构域
术语”结构域"指沿进化相关蛋白质的序列比对结果而在特定位置处保守的一组氨基酸。尽管在其他位置处的氨基酸可以在同源物之间变动,然而在特定位置处的高度保守的氨基酸指示在蛋白质的结构、稳定性或功能方面可能是必需的氨基酸。结构域因通过在蛋白质同源物家族的比对序列中的高保守程度而被鉴定,它们可以用作鉴定物以确定任意的所讨论多肽是否属于先前已鉴定的多肽家族。
基序/共有序列/标签
术语”基序”或“共有序列”或“标签”指在进化相关蛋白质的序列中的短保守区。基序往往是结构域的高度保守部分,不过也可以仅包括结构域的部分,或可以位于保守结构域之外(若基序的全部氨基酸位于定义的结构域之外)。
杂交
如本文中所定义的术语”杂交"是其中基本上同源的互补核苷酸序列相互复性的过程。杂交过程可以完全在溶液中进行,即两种互补性核酸均处于溶液中。杂交过程也可以在互补性核酸之一固定到基质如磁珠、琼脂糖(Sepharose)珠或任何其他树脂的情况下发生。杂交过程也可以在互补性核酸之一固定至固相支持体如硝酸纤维素膜或尼龙膜上或通过例如照相平版印刷术固定至例如硅酸玻璃支持物(后者称作核酸阵列或微阵列或称作核酸芯片)上的情况下进行。为使杂交发生,通常将核酸分子热变性或化学变性以使双链解链成为两条单链和/或去除来自单链核酸的发夹或其他二级结构。
术语”严格性”指在其中发生杂交的条件。杂交的严格性受条件如温度、盐浓度、离子强度和杂交缓冲液组成影响。通常,将低严格性条件选择为在确定的离子强度及pH时低于特定序列热解链温度(Tm)约30℃。中等严格性条件是此时温度低于Tm约20℃并且高严格性条件是此时温度低于Tm约10℃。高严格性杂交条件一般用于分离与靶核酸序列具有高序列相似性的杂交序列。然而,核酸可以在序列上偏离并且因遗传密码子的简并性而依旧编码基本上相同的多肽。因而有时候可能需要中等严格性杂交条件以鉴定此类核酸分子。
Tm是在确定的离子强度及pH时的温度,在所述温度下50%的靶序列与完全匹配的探针杂交。Tm取决于溶液条件和探针的碱基组成及长度。例如,较长的序列在较高温度下特异性地杂交。从低于Tm约16℃直至32℃获得最大杂交速率。一价阳离子在杂交溶液中的存在降低了两条核酸链间的静电排斥,因而促进杂交体形成;这种作用对于高达0.4M的钠浓度是明显的(对于更高浓度,这种效应可以忽略)。甲酰胺降低DNA-DNA和DNA-RNA双链体的解链温度,每百分数甲酰胺降低0.6-0.7℃,并且添加50%甲酰胺允许在30-45℃进行杂交,虽然杂交速率会降低。碱基对错配降低了杂交速率及双链体的热稳定性。平均而言并且对于大的探针来说,每%碱基错配Tm下降约1℃。取决于杂交体的类型,Tm可以使用下列等式计算:
1)DNA-DNA杂交体(Meinkoth和Wahl,Anal.Biochem.,138:267-284,1984):
Tm=81.5℃+16.6xlog10[Na+]a+0.41x%[G/Cb]–500x[Lc]-1–0.61x%甲酰胺
2)DNA-RNA或RNA-RNA杂交体:
Tm=79.8+18.5(log10[Na+]a)+0.58(%G/Cbb)+11.8(%G/Cb)2-820/Lc
3)寡DNA或寡RNAd杂交体:
对于<20个核苷酸:Tm=2(ln)
对于20–35个核苷酸:Tm=22+1.46(ln)
a或对于其他一价阳离子,但是仅在0.01–0.4M范围内是精确的。
b仅对于%GC在30%-75%范围内是精确的。
c L=双链体的长度(以碱基对计)。
d Oligo,寡核苷酸;ln,=引物的有效长度=2×(G/C数)+(A/T数)。
可以以众多已知技术的任何一种控制非特异性结合,例如用含蛋白质的溶液封闭薄膜、添加异源性RNA、异源性DNA及SDS至杂交缓冲液并且用RNA酶处理。对于非同源性探针,一系列杂交可以通过改变以下条件之一进行:(i)渐进地降低复性温度(例如从68℃至42℃)或(ii)渐进地降低甲酰胺浓度(例如从50%至0%)。本领域技术人员了解杂交期间可以加以改变和将维持或改变严格性条件的多种参数。
除杂交条件之外,杂交特异性一般还取决于杂交后洗涤的功能。为除去因非特异性杂交所致的背景,样品用稀释的盐溶液洗涤。此类洗涤的关键因素包括最终洗涤溶液的离子强度及温度:盐浓度越低并且洗涤温度越高,则洗涤的严格性越高。洗涤条件一般在杂交严格性上或低于杂交严格性而进行。阳性杂交产生至少两倍于背景信号的信号。通常,用于核酸杂交分析法或基因扩增检测方法的合适严格性条件如上所述。也可以选择更严格或更不严格的条件。技术人员了解洗涤期间可以加以改变和将维持或改变严格性条件的多种参数。
例如,用于长度大于50个核苷酸的DNA杂交体的典型高严格性杂交条件包括在65℃于1×SSC中或在42℃于1×SSC和50%甲酰胺中杂交,随后在65℃于0.3×SSC中洗涤。用于长度大于50个核苷酸的DNA杂交体的中等严格性杂交条件的实例包括在50℃于4×SSC中或在40℃于6×SSC和50%甲酰胺中杂交,随后在50℃于2×SSC中洗涤。杂交体的长度是杂交核酸的预期长度。当序列已知的核酸杂交时,可以通过比对序列并鉴定本文中所述的保守区而确定杂交体长度。1×SSC是0.15M NaCl和15mM柠檬酸钠;杂交溶液和洗涤溶液可以额外地包含5×Denhardt试剂、0.5-1.0%SDS、100μg/ml变性的片段化鲑精DNA、0.5%焦磷酸钠。
为了定义严格性水平的目的,可以参考Sambrook等(2001)MolecularCloning:a laboratory manual,第三版,Cold Spring Harbor LaboratoryPress,CSH,New York或参考Current Protocols in Molecular Biology,John Wiley&Sons,N.Y.(1989和每年更新版本)。
剪接变体
如本文中所用的术语”剪接变体”包含其中已经切除、替换、移位或添加所选内含子和/或外显子或其中内含子已经缩短或加长的核酸序列的变体。此类变体将是其中基本上保留了蛋白质的生物学活性的一种变体;这可以通过选择性保留蛋白质的功能性片段而实现。此类剪接变体可以在自然界中找到或可以人工制造。用于预测和分离此类剪接变体的方法是本领域众所周知的(见例如Foissac和Schiex(2005),BMC Bioinformatics.;6:25)。
等位变体
等位基因或等位变体是给定基因的替代形式,位于相同染色体位置内。等位变体包含单核苷酸多态性(SNP)和小插入/缺失多态性(INDEL)。INDEL的尺寸通常小于100bp。SNP和INDEL形成在大部分生物的天然存在性多态性株系中序列变体的最大集合。
基因改组/定向进化
基因改组或定向进化的组成为:反复DNA改组,随后适当筛选和/或选择以产生编码具有改良生物学活性的蛋白质的核酸或其部分的变体(Castle等,(2004)Science304(5674):1151-4;美国专利5,811,238和6,395,547)。
调节元件/控制序列/启动子
术语“调节元件”、“控制序列”和“启动子”均在本文中可互换使用并且在广义上意指能够实现与之连接的序列表达的调节性核酸序列。术语”启动子”一般指位于基因转录起点上游并参与识别及结合RNA聚合酶和其他蛋白质,因而指导有效连接的核酸转录的核酸控制序列。前述术语包括从典型的真核基因组基因(包括对于精确转录启动所需的TATA框,具有或没有CCAAT框序列)中衍生的转录调节序列和应答发育刺激和/或外部刺激或以组织特异性方式改变基因表达的额外调节元件(如,上游激活序列、增强子和沉默子)。本术语还包括典型的原核基因的转录调节序列,在此情况下它可以包括–35框序列和/或–10框转录调节序列。术语“调节元件”也包含赋予、激活或增强核酸分子在细胞、组织或器官中表达的合成的融合分子或衍生物。
“植物启动子”包含介导编码序列区段在植物细胞中表达的调节元件。因此,植物启动子不需要是植物来源的,而是可以源自病毒或微生物,例如来自侵袭植物细胞的病毒。“植物启动子”也可以源自植物细胞,例如来自用待于本发明方法中表达及在本文中描述的核酸序列所转化的植物。这也适用于其他“植物”调节性信号,如“植物”终止子。用于本发明方法中的核苷酸序列上游的启动子可以由一个或多个核苷酸取代、插入和/或缺失而受到修饰,但不干扰启动子、可读框(ORF)或3′调节区如终止子或远离ORF的其他3′调节区的功能性或活性。启动子的活性还有可能因该启动子的修饰,或由更活跃的启动子、甚至来自异源生物的启动子彻底替换该启动子而增加。为在植物中表达,如上所述,核酸分子必须有效连接至或包含合适的启动子,其中所述的启动子在正确时间点上并以所需要的空间表达模式表达基因。
为鉴定功能性等效启动子,候选启动子的启动子强度和/或表达模式可以通过将该启动子与报道基因有效连接并分析该报道基因在植物多种组织的表达水平和模式而进行分析。合适的公知报道基因包括例如β-葡糖醛酸糖苷酶或β-半乳糖苷酶。启动子活性通过测量β-葡糖醛酸糖苷酶或β-半乳糖苷酶的酶活性进行分析。启动子强度和/或表达模式随后可以与参考启动子(如本发明方法中所用的一种启动子)的启动子强度和/或表达模式比较。备选地,启动子强度可以使用本领域已知方法如Northern印迹法及放射自显影图的密度计分析法、定量实时PCR或RT-PCR(Heid等,1996GenomeMethods6:986-994),通过定量mRNA水平或通过将本发明方法中所用核酸的mRNA水平与持家基因(如18S rRNA)的mRNA水平比较而分析。通常“弱启动子”意指驱动编码序列在低水平上表达的启动子。“低水平”意指在每个细胞约1/10,000转录物至约1/100,000转录物、至约1/500,0000转录物的水平上。相反,“强启动子”驱动编码序列在高水平、或在每个细胞约1/10转录物至约1/100转录物、至约1/1,000转录物上表达。
有效连接
如本文中所用的术语”有效连接”指启动子序列与目的基因之间功能性地连接,以至于启动子序列能够启动目的基因转录。
组成型启动子
“组成型启动子”指在生长和发育的大部分、但不必全部阶段期间和在大部分环境条件下在至少一个细胞、组织或器官内有转录活性的启动子。下表2a给出组成型启动子的实例。
表2a:组成型启动子的实例

遍在启动子
遍在启动子在生物基本上所有组织或细胞内有活性。
发育调节性启动子
发育调节性启动子在某个发育期期间或在经历发育变化的植物部分内有活性。
诱导性启动子
诱导性启动子在应答化学品(综述见Gatz1997,Annu.Rev.PlantPhysiol.Plant Mol.Biol.,48:89-108)、环境刺激或物理刺激时具有受诱导或增加的转录启动,或可以是“胁迫诱导性的”,即当植物暴露于多种胁迫条件时受到激活,或是“病原体诱导性的”,即当植物暴露于多种病原体时受到激活。
器官特异性/组织特异性启动子
器官特异性或组织特异性启动子是能够优先在某些器官或组织如叶、根、种子组织等内启动转录的启动子。例如,“根特异性启动子”是在植物根中优势地具有转录活性的启动子,在植物的任何其他部分内基本上无活性,尽管在植物的这些其他部分内允许任何泄露表达。能够仅在某些细胞中启动转录的启动子在本文中称作“细胞特异的”。
根特异性启动子的实例在下面的表2b中列出:
表2b:根特异性启动子的实例
| 基因来源 | 参考文献 |
| RCc3 | Plant Mol Biol.1995Jan;27(2):237-48 |
| 拟南芥磷酸转运蛋白PHT1 | Kovama et al.,2005 |
| 苜蓿磷酸转运蛋白 | Xiao et al.,2006 |
| 拟南芥Pyk10 | Nitz et al.(2001)Plant Sci161(2):337-346 |
种子特异性启动子主要在种子组织中具有转录活性,但不一定仅仅在种子组织中具有转录活性(在渗漏表达的情况下)。种子特异性启动子可以在种子发育和/或种子萌发期间有活性。种子特异性启动子可以是胚乳/糊粉粒/胚特异性的。种子特异性启动子的实例在下面的表2c、d、e和f中显示。种子特异性启动子的其他实例在Qing Qu和Takaiwa(PlantBiotechnol.J.2,113-125,2004)中给出,将其公开并入本文作为参考,就好像完整给出一样。
表2c:种子特异性启动子的实例



表2d:胚乳特异性启动子实例


表2e:胚乳特异性启动子实例:
| 基因来源 | 参考文献 |
| 稻OSH1 | Sato et al,Proc.Natl.Acad.Sci.USA,93:8117-8122,1996 |
| KNOX | Postma-Haarsma et al,Plant Mol.Biol.39:257-71,1999 |
| PRO0151 | WO2004/070039 |
| PRO0175 | WO2004/070039 |
| PRO005 | WO2004/070039 |
| PRO0095 | WO2004/070039 |
表2f:糊粉特异性启动子实例:

如本文定义的绿色组织特异性启动子是主要在绿色组织中具有转录活性的启动子,基本上排除植物的任何其他部分,而仍然允许在这些其他植物部分中的任何渗漏表达。
可以用于实施本发明方法的绿色组织特异性启动子的实例在下面的表2g中显示。
表2g:绿色组织特异性启动子实例
| 基因 | 表达 | 参考文献 |
| 玉米正磷酸二激酶 | 叶特异性 | Fukavama et al.,2001 |
| 玉米磷酸烯醇丙酮酸羧化酶 | 叶特异性 | Kausch et al.,2001 |
| 稻磷酸烯醇丙酮酸羧化酶 | 叶特异性 | Liu et al.,2003 |
| 稻小亚基Rubisco | 叶特异性 | Nomura et al.,2000 |
| 稻β扩展蛋白EXBP9 | 根特异性 | WO2004/070039 |
| 木豆小亚基Rubisco | 叶特异性 | Panguluri et al.,2005 |
| 豌豆RBCS3A | 叶特异性 |
组织特异性启动子的另一实例是分生组织特异性启动子,其主要在分生组织中具有转录活性,基本上排除植物的任何其他组织,而仍然允许在这些其他植物部分中的任何渗漏表达。
终止子
术语“终止子”包括这样的控制序列,其是在转录单位端的DNA序列,发出对初级转录物进行3’加工并聚腺苷化以及终止转录的信号。终止子可以衍生自天然基因、来自多种其他植物基因或来自T-DNA。待添加的终止子可以来自例如胭脂碱合酶或章鱼碱合酶基因或备选地来自其他植物基因或较不优选地来自任何其他真核基因。
调节
术语”调节”就表达或基因表达而言意指这样的过程,其中表达水平与对照植物相比因所述基因的表达而改变,优选地,表达水平是增加的。原先未受调节的表达可以是结构RNA(rRNA、tRNA)或mRNA的任何类型表达,随后是翻译。术语“调节活性”应当意指本发明核酸序列或所编码蛋白质的表达的任何变化,这导致植物增加的产量和/或增加的生长。
表达
术语“表达”或“基因表达”指一种或多种特定基因或者基因构建体的转录。术语“表达”或“基因表达”尤其指基因或基因构建体转录为结构RNA(rRNA,tRNA)或者mRNA,其随后翻译成蛋白质或不翻译成蛋白质。该过程包括DNA的转录和所得mRNA产物的加工。
增加的表达/过量表达
如本文中所用的术语”增加的表达”或“过量表达”意指对于原有野生型表达水平是额外的任何形式表达。
在本领域内详细记载了用于增加基因或基因产物表达的方法并且它们包括例如,由适宜启动子驱动的过量表达、使用转录增强子或翻译增强子。可以在非异源形式的多核苷酸的适宜位置(一般是上游)内导入作为启动子或增强子元件的分离核酸,以便上调编码目的多肽的核酸的表达。例如,内源性启动子可以通过突变、缺失和/或置换而在体内改变(见Kmiec,US5,565,350;Zarling等,WO9322443),或可以将分离的启动子以相对于本发明基因的正确方向及距离导入植物细胞,以便控制基因表达。
若需要多肽表达,通常希望在多核苷酸编码区的3’末端包括多聚腺苷化区。多聚腺苷化区可以来自天然基因、来自多种其它植物基因或来自T-DNA。待添加的3’末端序列可以来自例如胭脂碱合酶或章鱼碱合酶基因或备选地来自另一植物基因或更不优选来自任何其它真核基因。
内含子序列也可以添加至5′非翻译区(UTR)或部分编码性序列的编码序列上,以增加在胞浆内积累的成熟信息的量。已经证实可剪接内含子在植物表达构建体和动物表达构建体中转录单位内的包含在mRNA水平及蛋白质水平上增加基因表达至多达1000倍(Buchman和Berg(1988)Mol.Cell biol.8:4395-4405;Callis等(1987)Gens Dev1:1183-1200)。基因表达的此类内含子增强作用一般在位于转录单位5′末端附近时最强烈。使用玉米内含子Adh1-S内含子1、2和6、Bronze-1内含子是本领域已知的。对于一般信息,见:《玉米手册》,第116章,编者Freeling和Walbot,Springer,N.Y.(1994)。
内源基因
本文中提及的“内源”基因不仅仅指如在植物中以其天然形式(即没有任何人类干预)存在的所讨论基因,还指处于分离形式的随后(再)导入植物(转基因)的相同基因(或基本上同源的核酸/基因)。例如,含有这种转基因的转基因植物可以遭遇转基因表达大幅降低和/或内源基因表达的大幅降低。分离的基因可以分离自生物或者可以是人造的,例如,通过化学合成。
降低的表达
本文中提及的”降低的表达”或“降低或基本去除”表达意指内源基因表达和/或多肽水平和/或多肽活性相对于对照植物的降低。与对照植物相比,降低或基本去除以递增优选顺序是至少10%、20%、30%、40%或50%、60%、70%、80%、85%、90%,或95%、96%、97%、98%、99%或更多的降低。
为了降低或基本去除内源基因在植物中的表达,需要核酸序列的足够长度的基本上连续的核苷酸。为了开展基因沉默,这个长度可以是少至20、19、18、17、16、15、14、13、12、11、10个或更少的核苷酸,或者该长度可以多至整个基因(包括5’和/或3’UTR,部分或全体)。基本上连续的核苷酸片段可以来自编码目的蛋白的核酸(靶基因)或来自能够编码目的蛋白的直向同源物、旁系同源物或同源物的任何核酸。优选地,基本上连续的核苷酸片段能够与靶基因(有义链或反义链)形成氢键,更优选地,基本上连续的核苷酸片段以递增优选顺序与靶基因(有义链或反义链)具有50%、60%、70%、80%、85%、90%、95%、96%、97%、98%、99%、100%的序列同一性。编码(功能性)多肽的核酸序列不是本文中所讨论用于降低或基本去除内源基因表达的多种方法所需的。
表达的这种降低或基本去除可以使用常规工具和技术完成。用于降低或基本去除内源基因表达的优选方法是在植物中导入并表达这样的遗传构建体,其中核酸(在此情况下是来自目的基因或任何核酸的一段基本上连续的核苷酸序列,其中所述的任何核酸能够编码任何一种目的蛋白的直向同源物、旁系同源物或同源物)克隆至所述的遗传构建体内作为由间隔区(非编码性DNA)隔开的(部分或完全的)反向重复序列。
在这种优选的方法中,使用核酸或其部分(在此情况下是从目的基因或从任何核酸中衍生的一段基本上连续的核苷酸序列,其中所述的任何核酸能够编码目的蛋白的直向同源物、旁系同源物或同源物)的反向重复序列(优选能够形成发夹结构),通过RNA介导的沉默作用而降低或基本上去除内源基因的表达。反向重复序列在包含调控序列的表达载体中克隆。非编码性DNA核酸序列(间隔序列,例如基质附着区片段(MAR)、内含子、多接头等)位于形成反向重复序列的两个反向核酸之间。在反向重复序列转录后,形成具有(部分或完全)自身互补结构的嵌合RNA。这种双链RNA结构称作发夹RNA(hpRNA)。hpRNA由植物加工成为siRNA,其被掺入RNA诱导性沉默复合物(RISC)中。RISC进一步切开mRNA转录物,因而大幅度降低待翻译成多肽的mRNA转录物的数目。对于其它一般细节,见例如Grierson等(1998)WO98/53083;Waterhouse等(1999)WO99/53050)。
本发明方法的实施不依赖在植物中导入并表达其中克隆入核酸作为反向重复序列的遗传构建体,而是几种公知“基因沉默”方法的任何一种或多种可以用来实现相同的效果。
一种用于降低内源基因表达的如此方法是RNA介导的基因表达沉默(下调)。沉默作用在这种情况下由基本上与内源性靶基因相似的双链RNA序列(dsRNA)在植物中触发。这种dsRNA被植物进一步加工成约20到约26个核苷酸,称作短干扰性RNA(siRNA)。siRNA被掺入RNA诱导性沉默复合物(RISC)内,其中所述的RISC进一步切割内源性靶基因的mRNA转录物,因而大幅度降低待翻译成多肽的mRNA转录物的数目。优选地,双链RNA序列对应于靶基因。
RNA沉默方法的另一个实例包括以有义方向导入核酸序列或其部分(在此情况下是从目的基因或从任何核酸中衍生的一段基本上连续的核苷酸,其中所述的任何核酸能够编码目的蛋白的直向同源物、旁系同源物或同源物)至植物内。“有义方向”指与其mRNA转录物同源的DNA序列。因而将向植物中导入该核酸序列的至少一个拷贝。这种额外的核酸序列会降低内源基因表达,产生已知为共抑制作用的现象。当核酸序列的几个额外拷贝导入植物时,基因表达的降低将更明显,因为高转录物水平与共抑制作用的触发之间存在正相关。
RNA沉默方法的另一个实例包括使用反义核酸序列。"反义"核酸序列包含与编码蛋白质的"有义"核酸序列互补,即与双链cDNA分子的编码链互补,或与mRNA转录物序列互补的核苷酸序列。反义核酸序列优选地与待沉默的内源基因互补。互补性可以位于基因的“编码区”和/或”非编码区”内。术语”编码区"指包含被翻译成氨基酸残基的密码子的核苷酸序列区。术语”非编码区"指分布在编码区两侧的被转录但不翻译成氨基酸的5’和3’序列(也称作5′和3′非翻译区)。
反义核酸序列可以根据Watson和Crick碱基配对规则设计。反义核酸序列可以与整个核酸序列互补(在此情况下是从目的基因或从任何核酸中衍生的一段基本上连续的核苷酸,其中所述的任何核酸能够编码目的蛋白的直向同源物、旁系同源物或同源物),不过也可以是仅与核酸序列的一部分(包括mRNA5’和3’UTR)反义的寡核苷酸。例如,反义寡核苷酸序列可以与围绕编码多肽的mRNA转录物的翻译起点周围的区域互补。合适的反义寡核苷酸序列的长度是本领域已知的并且可以从长度约50、45、40、35、30、25、20、15或10个核苷酸或更少的核苷酸开始。本发明的反义核酸序列可以利用本领域已知方法,使用化学合成和酶连接反应而构建。例如,反义核酸序列(例如反义寡核苷酸序列)可以使用天然存在的核苷酸或多种修饰的核苷酸化学地合成,其中所述修饰的核苷酸被设计旨在增加分子的生物学稳定性或增加反义核酸序列与有义核酸序列之间所形成双链体的物理稳定性,例如,可以使用硫代磷酸酯衍生物和吖啶取代的核苷酸。可以用来产生反义核酸序列的修饰核苷酸的实例是本领域众所周知的。已知的核苷酸修饰包括甲基化、环化和′加帽′及用类似物(如肌苷)取代一个或多个天然存在的核苷酸。其它的核苷酸修饰是本领域众所周知的。
这种反义核酸序列可以使用其中核酸序列已经以反义方向加以亚克隆(即从插入的核酸中转录的RNA将与目的靶核酸呈反义方向)的表达载体以生物学方式产生。优选地,反义核酸序列在植物中的产生借助稳定整合的核酸构建体进行,其中所述的核酸构建体包含启动子、有效连接的反义寡核苷酸和终止子。
用于本发明方法中沉默作用的核酸分子(无论向植物中导入或在原位(in situ)产生)与编码多肽的mRNA转录物和/或基因组DNA杂交或结合,以便例如通过抑制转录和/或翻译而抑制蛋白质表达。杂交可以通过形成稳定双链体的常规核苷酸互补性所致,或在结合于DNA双链体的反义核酸序列的情况下,因双螺旋大沟内特异性相互作用所致。反义核酸序列可以通过转化或在特定组织部位直接注射而导入植物。备选地,反义核酸序列可以为了靶向所选择的细胞而受到修饰并且随后系统性施用。例如,对于系统性施用,反义核酸序列可以被修饰以便它们特异结合表达在所选择的细胞表面上的受体或抗原,例如通过连接反义核酸序列至与细胞表面受体或抗原结合的肽或抗体。反义核酸序列也可以使用本文中所述的载体送递至细胞内。
根据又一个方面,反义核酸序列是α-异头核酸序列。α异头核酸序列与互补性RNA形成特定双链杂交分子,其中与惯常b-单元相反,所述链相互平行(Gaultier等(1987)Nucl Ac Res15:6625-6641)。反义核酸序列也可以包含2′-o-甲基核糖核苷酸(Inoue等(1987)Nucl Ac Res15,6131-6148)或嵌合RNA-DNA类似物(Inoue等(1987)FEBS Lett.215,327-330)。
内源基因表达的降低或基本去除也可以使用核酶而进行。核酶是具有核糖核酸酶活性的催化性RNA分子,能够切割与其具有互补区域的单链核酸序列,如mRNA。因此,核酶(例如锤头核酶(在Haselhoff和Gerlach(1988)Nature334,585-591中描述)可以用来催化性地切割编码多肽的mRNA转录物,因而大幅度降低待翻译成多肽的mRNA转录物的数目。可以设计对核酸序列具特异性的核酶(见例如:Cech等美国专利号4,987,071;和Cech等美国专利号5,116,742)。备选地,对应于核酸序列的mRNA转录物可以用来从RNA分子库中选出具有特异性核糖核酸酶活性的催化性RNA(Bartel和Szostak(1993)Science261,1411-1418)。核酶用于植物中基因沉默的用途是本领域已知的(例如Atkins等(1994)WO94/00012;Lenne等(1995)WO95/03404;Lutziger等(2000)WO00/00619;Prinsen等(1997)WO97/13865和Scott等(1997)WO97/38116)。
基因沉默也可以通过插入诱变(例如T-DNA插入或转座子插入)或通过如Angell和Baulcombe((1999)Plant J.20(3):357-62)、(Amplicon VIGSWO98/36083)或Baulcombe(WO99/15682)及其它人描述的策略而实现。
当在内源基因上存在突变和/或在随后导入植物的分离的基因/核酸上存在突变时,基因沉默也可以发生。降低或基本去除可以由无功能的多肽引起。例如,多肽可以与多种相互作用性蛋白质结合;一个或多个突变和/或截短因而可以提供仍能够结合相互作用性蛋白质(如受体蛋白质)但不能表现其正常功能的多肽(如起信号作用的配体)。
另一种基因沉默的方法是靶定与基因调节区(例如启动子和/或增强子)互补的核酸序列以形成阻止基因在靶细胞中转录的三螺旋结构。见Helene,C.,Anticancer Drug Res.6,569-84,1991;Helene等,Ann.N.Y.Acad.Sci.660,27-361992和Maher,L.J.Bioassays14,807-15,1992。
其它方法,如使用针对内源性多肽的抗体以抑制此多肽在植物中的功能,或干扰所述多肽参与的信号途径,对于技术人员将是众所周知的。尤其,可以构思的是人造分子可以用于抑制靶多肽的生物学功能,或用于干扰靶多肽参与其中的信号途径。
备选地,可以建立筛选程序以鉴定在植物群体中基因的天然变体,其中所述的变体编码具有降低活性的多肽。此类天然变体也可以用于例如进行同源重组。
人工和/或天然的微RNA(miRNA)可以用来敲除基因表达和/或mRNA翻译。内源性miRNA是通常19-24个核苷酸长度的单链小RNA。它们的主要功能是调节基因表达和/或mRNA翻译。大多数的植物微RNA(miRNA)与其靶序列具有完全的或接近完全的互补性。然而,存在具有多达5个错配的天然靶标。它们由切酶家族的双链特异性RNA酶从具有特征性折回结构的较长的非编码性RNA中加工而来。在加工时,它们通过与RNA诱导性沉默复合物(RISC)的主要成分Argonaute蛋白质结合而掺入该复合体。MiRNA充当RISC的特异性成分,因此它们与细胞质中的靶核酸(大多是mRNA)碱基配对。后续的调节事件包括靶mRNA切割和破坏和/或翻译抑制。miRNA过量表达的作用因此在靶基因降低的mRNA水平上得到反映。
通常21个核苷酸长度的人工微RNA(amiRNAs)可以遗传改造以特异性地负调节单个或多个目的基因的基因表达。植物微RNA靶的选择的决定因素是本领域众所周知的。用于靶识别的经验参数已经确定并且可以用来辅助设计特定的amiRNA,Schwab等,Dev.Cell8,517–527,2005。用于设计并产生amiRNA及其前体的便利工具也是公众可获得的,Schwab等,Plant Cell18,1121-1133,2006。
为最佳性能,用于降低内源基因在植物中表达的基因沉默技术需要使用来自单子叶植物的核酸序列以转化单子叶植物,和使用来自双子叶植物的核酸序列以转化双子叶植物。优选地,将来自任何给定植物物种的核酸序列导入同一个物种内。例如,将来自稻的核酸序列转化至稻植物。然而,并非绝对要求待导入的核酸序列起源于与该核酸序列将要导入的植物相同的植物物种。只要内源性靶基因与待导入的核酸之间存在相当大的同源性就足够了。
上文描述的是用于降低或基本去除内源基因在植物中表达的多种方法的实例。本领域技术人员会轻易地能够调整前述用于沉默的方法以至于例如通过利用合适启动子而实现降低内源基因在整株植物或在其部分中的表达。
选择性标记(基因)/报道基因
“选择性标记”、“选择性标记基因”或“报道基因”包括向细胞赋予表型的任何基因,其中在所述的细胞内表达所述基因以促进鉴定和/或选择用本发明的核酸构建体所转染或转化的细胞。这些标记基因能够通过一系列不同原理鉴定核酸分子的成功转移。合适的标记可以选自赋予抗生素耐药性或除草剂抗性、引入新代谢性状或允许目视选择的标记。选择性标记基因的实例包括赋予抗生素耐药性的基因(如使新霉素和卡那霉素磷酸化的nptII或使潮霉素磷酸化的hpt或赋予对例如博来霉素、链霉素、四环素、氯霉素、氨苄青霉素、庆大霉素、遗传霉素(Geneticin)(G418)、壮观霉素或杀稻瘟素的抗性的基因)、赋予除草剂抗性的基因(例如提供 抗性的bar;提供草甘膦抗性的aroA或gox或赋予对例如咪唑啉酮、膦丝菌素或磺脲类的抗性的基因)或提供代谢性状的基因(如允许植物使用甘露糖作为唯一碳源的manA或利用木糖的木糖异构酶或抗营养标记如2-脱氧葡萄糖抗性)。目视标记基因的表达导致形成颜色(例如β-葡糖醛酸糖苷酶、GUS或β-半乳糖苷酶与其有色底物例如X-Gal)、发光(如萤光素/萤光素酶系统)或萦光(绿色萦光蛋白GFP及其衍生物)。这个名单仅代表少数的可能标记。本领域技术人员熟悉此类标记。取决于生物和选择方法,优选不同的标记。
抗性的bar;提供草甘膦抗性的aroA或gox或赋予对例如咪唑啉酮、膦丝菌素或磺脲类的抗性的基因)或提供代谢性状的基因(如允许植物使用甘露糖作为唯一碳源的manA或利用木糖的木糖异构酶或抗营养标记如2-脱氧葡萄糖抗性)。目视标记基因的表达导致形成颜色(例如β-葡糖醛酸糖苷酶、GUS或β-半乳糖苷酶与其有色底物例如X-Gal)、发光(如萤光素/萤光素酶系统)或萦光(绿色萦光蛋白GFP及其衍生物)。这个名单仅代表少数的可能标记。本领域技术人员熟悉此类标记。取决于生物和选择方法,优选不同的标记。
已知当核酸稳定或瞬时整合至植物细胞时,仅小部分的细胞摄取外来DNA并且根据需要将其整合至细胞基因组,这取决于所用表达载体和使用的转染技术。为鉴定并选择这些整合子,通常将编码选择标记(如上文所述之一)的基因连同目的基因一起导入宿主细胞。这些标记可以在其中这些基因因例如常规方法所致的缺失而无功能的突变体中使用。此外,编码选择标记的核酸分子可以导入宿主细胞中,与在包含编码本发明多肽或本发明方法中所用多肽的序列在同一载体上,或在单独的载体上。已经用导入的核酸稳定转染的细胞可以通过选择进行鉴定(例如具有整合的选择标记的细胞存活而其它细胞死亡)。
因为一旦已经成功导入了核酸,则转基因宿主细胞中不再需要或不希望有标记基因,尤其抗生素耐药性基因和除草剂抗性基因,因此用于导入核酸的本发明方法有利地使用能够去掉或切除这些标记基因的技术。一种如此方法称作共转化法。共转化法使用同时用于转化的两种载体,一种载体携带本发明的核酸而另一种载体携带标记基因。高比例的转化体接受,或在植物的情况下,包含(高达40%或更多的转化体)这两种载体。在用农杆菌(Agrobacterium)转化的情况下,转化体通常仅接受载体的一部分,即侧翼有T-DNA的序列,它通常代表表达盒。标记基因随后可以通过进行杂交而从转化的植物中去掉。在另一种方法中,整合至转座子的标记基因用来与想要的核酸一起进行转化(称作Ac/Ds技术)。转化体可以与转座酶来源植物杂交或转化体用导致转座酶表达的核酸构建体瞬时或稳定地转化。在一些情况下(大约10%),转座子在已经成功发生转化时跳出宿主细胞的基因组并丢失。在其它更多情况下,转座子跳至不同位置。在这些情况下,标记基因必须通过进行杂交而去除。在微生物学中,开发了实现或促进检测这类事件的技术。又一个有利的方法依赖于重组系统;此方法的优势在于不必通过杂交去除。该类型的最知名系统称作Cre/lox系统。Cre1是去掉位于loxP序列之间序列的重组酶。若标记基因整合于loxP序列之间,则通过重组酶表达已经成功发生转化时,标记基因被去除。其它重组系统是HIN/HIX、FLP/FRT和REP/STB系统(Tribble等,J.Biol.Chem.,275,2000:22255-22267;Velmurugan等,J.Cell Biol.,149,2000:553-566)。有可能将本发明核酸序列以位点特异性方式整合至植物基因组。这些方法自然也可以应用至微生物如酵母、真菌或细菌。
转基因的/转基因/重组
为本发明目的,“转基因的”、“转基因”或“重组”就核酸序列而言意指包含此核酸序列的表达盒、基因构建体或载体或用本发明的核酸序列、表达盒或载体转化的生物,这些构建均通过重组方法产生,其中
(a)编码用于本发明方法中的蛋白质的核酸序列,或
(b)与本发明核酸序列有效连接的遗传控制序列,例如启动子,或
(c)a)和b)。
不处于其天然遗传环境中或已经通过重组方法修饰,修饰有可能采用例如取代、添加、缺失、倒位或插入一个或多个核苷酸残基的形式。天然遗传环境理解为意指来源植物中的或在基因组文库中存在的天然基因组基因座或染色体基因座。在基因组文库的情况下,核酸序列的天然遗传环境优选地得到保留,至少部分地得以保留。该环境分布在核酸序列的至少一侧并且具有至少50bp,优选至少500bp,特别优选至少1000bp,最优选至少5000bp的序列长度。天然发生的表达盒──例如核酸序列的天然启动子与编码本发明方法中所用多肽的对应核酸序列的天然发生的组合,如上文所定义–在这种表达盒通过非天然的合成(“人工”)方法(如诱变处理)而受到修饰时,变成转基因表达盒。合适方法例如在US5,565,350或WO00/15815中描述。
为本发明目的,转基因植物因此如上理解为意指本发明方法中所用的核酸不位于所述植物基因组中(该核酸序列)的天然基因座内,所述核酸有可能同源或异源地表达。然而如所提及,转基因还意指尽管本发明核酸或在本发明方法中所用核酸处于植物基因组中该核酸序列的天然位置内,然而其序列相对于天然序列而言已经受到修饰,和/或所述天然序列的调节序列已经受到修饰。转基因优选地理解为意指本发明核酸在基因组中的非天然基因座内表达,即发生核酸的同源表达或优选异源表达。在本文中提到了优选的转基因植物。
转化
如本文中所提及的术语“引入”或“转化”包括外源性多核苷酸转移至宿主细胞内,无论用于转化的方法是什么。能够后续克隆性增殖(无论通过器官发生或胚胎发生)的植物组织可以用本发明的遗传构建体转化并且可以从中再生整株植物。选择的具体组织将取决于可用于并且最适于正进行转化的具体物种的克隆性增殖系统。示例性组织靶包括叶盘、花粉、胚、子叶、下胚轴、大配子体、胼胝体组织、现存分生组织(例如顶端分生组织、腋芽和根分生组织)和诱导的分生组织(例如子叶分生组织和下胚轴分生组织)。多核苷酸可以瞬时或稳定地引入宿主细胞并且可以非整合地维持,例如作为质粒。备选地,多核苷酸可以整合至宿主基因组内。产生的转化植物细胞随后可以用来以本领域技术人员已知的方式再生出转化植物。
外来基因转化至植物基因组内称作转化。植物物种的转化现在是相当常规的技术。有利地,几种转化方法中的任一方法可以用来将目的基因引入合适的祖先细胞。用于从植物组织或植物细胞中转化并再生出植物的方法可以用于瞬时转化或用于稳定转化。转化方法包括使用脂质体、电穿孔法、增加游离DNA摄入的化学品、DNA直接注射至植物、粒子枪轰击法、使用病毒或花粉的转化法和显微投射法(microprojection)。转化方法可以选自用于原生质体的钙/聚乙二醇法(Krens,F.A.等,(1982)Nature296,72-74;Negrutiu I等(1987)Plant Mol Biol8:363-373);原生质体的电穿孔法(Shillito R.D.等(1985)Bio/Technol3,1099-1102);对植物材料的微量注射法(Crossway A等,(1986)Mol.Gen Genet202:179-185);DNA或RNA包被的粒子轰击法(Klein TM等,(1987)Nature327:70)、(非整合性)病毒感染法等。转基因植物,包括转基因作物植物,优选地通过农杆菌介导的转化法产生。有利的转化方法是在植物中(in planta)的转化法。为此目的,例如有可能使农杆菌作用于植物种子或有可能用农杆菌接种植物的分生组织。根据本发明已经证明使转化的农杆菌混悬液作用于完整植物或至少作用于花原基是特别有利的。植物随后继续培育直至获得所处理植物的种子(Clough和Bent,Plant J.(1998)16,735–743)。用于农杆菌介导的稻转化的方法包括用于稻转化的公知方法,如在任一以下文献中描述的那些方法:欧洲专利申请EP1198985A1,Aldemita和Hodges(Planta199:612-617,1996);Chan等(Plant Mol Biol22(3):491-506,1993),Hiei等(Plant J6(2):271-282,1994),其公开内容在本文中引入作为参考,如同完全给出那样。在玉米转化的情况下,优选的方法如Ishida等(Nat.Biotechnol14(6):745-50,1996)或Frame等(Plant Physiol129(1):13-22,2002)描述,其公开内容在本文中如充分所述那样引入作为参考。所述方法通过举例方式进一步由Jenes等,Techniques for Gene Transfer,在:Transgenic Plants,第1卷,Engineering and Utilization,编者S.D.Kung和R.Wu,Academic Press(1993)128-143及在Potrykus Annu.Rev.PlantPhysiol.Plant Molec.Biol.42(1991)205-225)中描述。待表达的核酸或构建体优选地克隆至适于转化根癌农杆菌(Agrobacterium tumefaciens)的载体,例如pBin19(Bevan等,Nucl.Acids Res.12(1984)8711)。由这种载体转化的农杆菌随后可以按照已知方式用于转化植物,例如作为模型使用的植物,如拟南芥(拟南芥属于本发明的范围,不视为作物植物)或作物植物,例如烟草植物,通过在农杆菌溶液中浸泡擦伤的叶或切碎的叶并随后将它们在合适的培养基内培育。植物通过根癌农杆菌的转化例如由 和Willmitzer在Nucl.Acid Res.(1988)16,9877中描述或尤其从F.F.White,Vectors for Gene Transfer in Higher Plants;在Transgenic Plants,第1卷,Engineering and Utilization,编者S.D.Kung和R.Wu,AcademicPress,1993,第15-38页中获知。
和Willmitzer在Nucl.Acid Res.(1988)16,9877中描述或尤其从F.F.White,Vectors for Gene Transfer in Higher Plants;在Transgenic Plants,第1卷,Engineering and Utilization,编者S.D.Kung和R.Wu,AcademicPress,1993,第15-38页中获知。
除了转化体细胞(其随后必须再生成完整植物)之外,还有可能转化植物分生组织的细胞及特别转化发育成配子的那些细胞。在这种情况下,转化的配子遵循天然的植物发育过程,产生转基因植物。因此,例如拟南芥种子用农杆菌处理并且从发育植物中获得种子,其中一定比例的所述植物受到转化并且因此是转基因的[Feldman,KA和Marks MD(1987)Mol GenGenet208:274-289;Feldmann K(1992)。在:编者C Koncz,N-H Chua和J Shell,Methods in Arabidopsis Research.Word Scientific,Singapore,第274-289页]。替代性方法基于反复去掉花序并使莲座叶丛中心中的切除部位与转化的农杆菌温育,因而转化的种子同样可以在较晚的时间点获得(Chang(1994)Plant J.5:551-558;Katavic(1994)Mol Gen Genet,245:363-370)。然而,尤其有效的方法是改良的真空渗入法,如“花浸染”法。在拟南芥真空渗入法的情况下,完整植物在减压下用农杆菌混悬液处理[Bechthold,N(1993)。C R Acad Sci Paris Life Sci,316:1194-1199],而在“花浸染法”的情况下,正在发育的花组织与表面活性剂处理的农杆菌混悬液短暂温育[Clough,SJ und Bent,AF(1998).The Plant J.16,735-743]。在两种情况下收获了一定比例的转基因种子,并且这些种子可以通过在如上所述的选择条件下培育而与非转基因种子区分。此外,质体的稳定转化是有利的,因为质体在大部分作物中以母体方式遗传,降低或消除了转基因经花粉流动风险。叶绿体基因组的转化一般通过已在Klaus等,2004[Nature Biotechnology22(2),225-229]中示例性加以展示的方法实现。简而言之,待转化的序列连同选择性标记基因一起克隆至与叶绿体基因组同源的侧翼序列之间。这些同源的侧翼序列指导位点特异性整合至原质体系内。已经对众多不同植物物种描述了质体转化并且综述可以出自Bock(2001)在基础研究和植物生物技术中的转基因质体(Transgenicplastids in basic research and plant biotechnology).J Mol Biol.2001年9月21日;312(3):425-38或Maliga,P(2003)质体转化技术商业化进展(Progress towards commercialization of plastid transformationtechnology).Trends Biotechnol.21,20-28。进一步生物技术进展最近已经以无标记质体转化体的形式作了报道,所述无标记质体转化体可以通过瞬时共整合的标记基因产生(Klaus等,2004,Nature Biotechnology22(2),225-229)。
T-DNA激活标签化
T-DNA激活标签化(Hayashi等Science(1992)1350-1353)涉及在目的基因的基因组区域内或基因编码区上游或下游10kb处以如此结构插入T-DNA(通常含有启动子(也可以是翻译增强子或内含子)),使得启动子指导被靶定基因的表达。通常,由靶定基因的天然启动子对所述靶定基因表达的调节作用遭到破坏并且该基因处在新导入的启动子控制下。启动子一般嵌入在T-DNA中。这种T-DNA随机地插入植物基因组,例如通过农杆菌感染,并导致在所插入T-DNA附近的基因的改良表达。因靠近所导入启动子的基因的改良表达,产生的转基因植物表现显性表型。
TILLING
术语“TILLING”是基因组内定向诱导的局部损伤的缩写并且意指用于产生和/或鉴定核酸诱变技术,其中所述的核酸编码具有修饰表达和/或活性的蛋白质。TILLING还允许选择携带此类突变变体的植物。这些突变变体可以显示在强度方面或在位置方面或在时间方面改良的表达(例如若突变影响启动子)。这些突变变体可以显示比由处于其天然形式的基因所显出活性更高的活性。TILLING将高密度诱变与高通量筛选方法组合。一般在TILLING中遵循的步骤是:(a)EMS诱变(Redei GP和Koncz C(1992)在Methods in Arabidopsis Research,Koncz C,Chua NH,Schell J,Singapore编著,World Scientific Publishing Co,第16–82页;Feldmann等,(1994)在Meyerowitz EM,Somerville CR编著,Arabidopsis.Cold Spring HarborLaboratory Press,Cold Spring Harbor,NY,第137-172页;Lightner J和Caspar T(1998)在J Martinez-Zapater,J Salinas编者,Methods onMolecular Biology第82卷.Humana Press,Totowa,NJ,第91-104页);(b)个体的DNA制备和汇集;(c)PCR扩增目的区;(d)变性和复性以允许形成异源双链体;(e)DHPLC,其中将异源双链体是否在汇集物中的存在检测为色谱图中的一个额外峰;(f)鉴定突变个体;和(g)对突变PCR产物测序。用于TILLING的方法是本领域众所周知的(McCallum等,(2000)NatBiotechnol18:455-457;综述见Stemple(2004)Nat Rev Genet5(2):145-50)。
同源重组
同源重组允许选择的核酸在基因组中于确定的所选择位置内导入。同源重组是在生物科学中常规地用于低等生物如酵母或苔藓剑叶藓(Physcomitrella)的标准技术。用于在植物中开展同源重组的方法已经不仅对模式植物(Offringa等(1990)EMBO J9(10):3077-84)而且对作物植物例如稻(Terada等(2002)Nat Biotech20(10):1030-4;Iida和Terada(2004)Curr Opin Biotech15(2):132-8)进行了描述。
产量
术语”产量”通常意指经济价值的可测量结果,一般与指定作物、与面积并与时间段有关。单个植物部分基于它们的数目、大小和/或重量而直接对产量有贡献,或实际产量是对于某作物和一年的每英亩产量,这通过总产量(包括收获的和评估的产量)除以种植的英亩数而确定。
早期萌发势
早期萌发势(尤其在植物生长早期期间活跃、健康、良好平衡的生长)可以因植物适应性增加而产生,其原因在于例如植物更好地适应其环境(即优化能源的使用和苗与根之间的分配)。具有早期萌发势的植物也显示增加的幼苗存活和更好的作物建立,这往往导致高度均匀的田块(作物整齐地生长,即大多数植物在基本上相同的时间上达到发育的各阶段)和往往更好及更高的产量。因而,早期萌发势可以通过多种因子如千粒重、萌发百分数、出苗百分数、幼苗生长、幼苗高度、根长度、根及苗生物量和众多其它因素而确定。
增加/改善/增强
术语“增加”,”改善”或“增强”是可互换的并且应当在本申请含义上指与如本文中定义的对照植物相比至少3%、4%、5%、6%、7%、8%、9%或10%、优选至少15%或20%、更优选地25%、30%、35%或40%更多的产量和/或生长。
种子产量
增加的种子产量本身可以表现为下列一种或多种指标:a)种子生物量(种子总重量)增加,这可以基于单粒种子和/或每株植物和/或每公顷或英亩;b)每株植物增加的花数;c)增加的(饱满)种子数;d)增加的种子饱满率(其表述为饱满种子数与种子总数之间的比率);e)增加的收获指数,其表述为可收获部分(如种子)产量与总生物量的比率;和f)增加的千粒重(TKW),这从计数的饱满种子数及其总重量外推而来。增加的TKW可以因增加的种子大小和/或种子重量所致,并且也可以因胚和/或胚乳尺寸的增加所致。
种子产量的增加也可以表现为种子大小和/或种子体积的增加。此外,种子产量的增加本身也可以自我表现为种子面积和/或种子长度和/或种子宽度和/或种子周长的增加。增加的产量也可以产生改良的结构,或可以因改良的结构而出现。
绿色指数
本文所用的“绿色指数”从植物的数字图像计算而来。对于属于图像上植物物体的每个像素,计算绿色值与红色值的比值(用于编码颜色的RGB模式)。将绿色指数表示为绿色与红色比值超过给定阈值的像素百分比。在正常生长条件下,在盐胁迫生长条件下,和在降低的养分有效性生长条件下,在开花之前测量最后成像中植物的绿色指数。相比,在干旱胁迫生长条件下,测量干旱后第一次成像中植物的绿色指数。
植物
如本文中所用的术语”植物”包含整株植物、植物的祖先及后代和植物部分,包括种子、枝条、茎、叶、根(包括块茎)、花和组织及器官,其中每种所提及对象包含目的基因/核酸。术语”植物”也包含植物细胞、悬浮培养物、愈伤组织、胚、分生组织区、配子体、孢子体、花粉和小孢子,同样每种提及的对象包含目的基因/核酸。
特别用于本发明方法中的植物包括属于植物界(Viridiplantae)超家族的全部植物,尤其单子叶植物和双子叶植物,包括选自以下的饲用或饲料豆类、观赏植物、粮食作物、树或灌木:槭树属物种(Acer spp.)、猕猴桃属物种(Actinidia spp.)、秋葵属物种(Abelmoschus spp.)、剑麻(Agavesisalana)、冰草属物种(Agropyron spp.)、匍匐剪股颖(Agrostis stolonifera)、葱属物种(Allium spp.)、苋属物种(Amaranthus spp.)、欧洲海滨草(Ammophila arenaria)、凤梨(Ananas comosus)、番荔枝属物种(Annonaspp.)、芹菜(Apium graveolens)、落花生属物种(Arachis spp.)、木波罗属物种(Artocarpus spp.)、石刁柏(Asparagus officinalis)、燕麦属物种(Avenaspp.)(例如燕麦(Avena sativa)、野燕麦(Avena fatua)、比赞燕麦(Avenabyzantina)、Avena fatua var.sativa、杂种燕麦(Avena hybrida)、阳桃(Averrhoa carambola)、箣竹属(Bambusa sp.)、冬瓜(Benincasa hispida)、巴西栗(Bertholletia excelsea)、甜菜(Beta vulgaris)、芸苔属物种(Brassicaspp.)(例如欧洲油菜(Brassica napus)、芜青物种(Brassica rapa ssp.)[卡诺拉油菜、油菜(oilseed rape)、蔓青(turnip rape)])、Cadaba farinosa、茶(Camellia sinensis)、美人蕉(Canna indica)、大麻(Cannabis sativa)、辣椒属物种(Capsicum spp.)、Carex elata、番木瓜(Carica papaya)、大果假虎刺(Carissa macrocarpa)、山核桃属物种(Carya spp.)、红花(Carthamustinctorius)、栗属物种(Castanea spp.)、美洲木棉(Ceiba pentandra)、苦苣(Cichorium endivia)、樟属物种(Cinnamomum spp.)、西瓜(Citrulluslanatus)、柑橘属物种(Citrus spp.)、椰子属物种(Cocos spp.)、咖啡属物种(Coffea spp.)、芋头(Colocasia esculenta)、非洲梧桐属物种(Cola spp.)、黄麻属(Corchorus sp.)、芫荽(Coriandrum sativum)、榛属物种(Corylus spp.)、山楂属物种(Crataegus spp.)、番红花(Crocus sativus)、南瓜属物种(Cucurbita spp.)、香瓜属物种(Cucumis spp.)、菜蓟属(Cynara spp.物种)、胡萝卜(Daucus carota)、山马蝗属物种(Desmodium spp.)、龙眼(Dimocarpus longan)、薯蓣属物种(Dioscorea spp.)、柿树属物种(Diospyrosspp.)、稗属物种(Echinochloa spp.)、油棕属(Elaeis)(例如油棕(Elaeisguineensis)、美洲油棕Elaeis(oleifera))、穇子(Eleusine coracana)、Eragrostis tef、蔗茅属(Erianthus sp.)、枇杷(Eriobotrya japonica)、桉属(Eucalyptus sp.)、红仔果(Eugenia uniflora)、荞麦属物种(Fagopyrum spp.)、水青冈属物种(Fagus spp.)、苇状羊茅(Festuca arundinacea)、无花果(Ficuscarica)、金桔属物种(Fortunella spp.)、草莓属物种(Fragaria spp.)、银杏(Ginkgo biloba)、大豆属(Glycine spp.)(例如大豆、大豆(Soja hispida)或大豆(Soja max))、陆地棉(Gossypium hirstum)、向日葵属(Helianthus spp.)(例如向日葵(Helianthus annuus))、长管萱草(Hemerocallis fulva)、木槿属物种(Hibiscus spp.)、大麦属(Hordeum spp.)(例如大麦(Hordeum vulgare))、甘薯(Ipomoea batatas)、核桃属物种(Juglans spp.)、莴苣(Lactuca sativa)、山黧豆属物种(Lathyrus spp.)、兵豆(Lens culinaris)、亚麻(Linumusitatissimum)、荔枝(Litchi chinensis)、百脉根属物种(Lotus spp.)、棱角丝瓜(Luffa acutangula)、羽扇豆属物种(Lupinus spp.)、Luzula sylvatica、番茄属(Lycopersicon spp.)(例如番茄(Lycopersicon esculentum、Lycopersicon lycopersicum、Lycopersicon pyriforme))、硬皮豆属物种(Macrotyloma spp.)、苹果属物种(Malus spp.)、凹缘金虎尾(Malpighiaemarginata)、牛油果(Mammea americana)、芒果(Mangifera indica)、木薯属物种(Manihot spp.)、人心果(Manilkara zapota)、苜蓿(紫苜蓿)、草木樨属物种(Melilotus spp.)、薄荷属物种(Mentha spp.)、芒(Miscanthussinensis)、苦瓜属物种(Momordica spp.)、黑桑(Morus nigra)、芭蕉属物种(Musa spp.)、烟草属物种(Nicotiana spp.)、木犀榄属物种(Olea spp.)、仙人掌属物种(Opuntia spp.)、鸟足豆属物种(Ornithopus spp.)、稻属(Oryzaspp.)(例如稻、阔叶稻(Oryza latifolia))、稷(Panicum miliaceum)、柳枝稷(Panicum virgatum)、鸡蛋果(Passiflora edulis)、欧防风(Pastinaca sativa)、狼尾草属物种(Pennisetum sp.)、鳄梨属物种(Persea spp.)、芹菜(Petroselinum crispum)、虉草(Phalaris arundinacea)、菜豆属物种(Phaseolus spp.)、猫尾草(Phleum pratense)、刺葵属物种(Phoenix spp.)、南方芦苇(Phragmites australis)、酸浆属物种(Physalis spp.)、松属物种(Pinus spp.)、阿月浑子(Pistacia vera)、豌豆属物种(Pisum spp.)、早熟禾属物种(Poa spp.)、杨属物种(Populus spp.)、牧豆草属物种(Prosopis spp.)、李属物种(Prunus spp.)、番石榴属物种(Psidium spp.)、石榴(Punicagranatum)、西洋梨(Pyrus communis)、栎属物种(Quercus spp.)、萝卜(Raphanus sativus)、波叶大黄(Rheum rhabarbarum)、茶藨子属物种(Ribesspp.)、蓖麻(蓖麻)、悬钩子属物种(Rubus spp.)、甘蔗属物种(Saccharumspp.)、柳属物种(Salix sp.)、接骨木属物种(Sambucus spp.)、黑麦(Secalecereale)、胡麻属物种(Sesamum spp.)、白芥属物种(Sinapis sp.)、茄属(Solanum spp.)(例如马铃薯(马铃薯)、红茄(Solanum integrifolium)或番茄(Solanum lycopersicum))、两色蜀黍(两色蜀黍)、菠菜属物种(Spinaciaspp.)、蒲桃属物种(Syzygium spp.)、万寿菊属物种(Tagetes spp.)、酸豆(Tamarindus indica)、可可树(Theobroma cacao)、车轴草属物种(Trifoliumspp.)、Tripsacum dactyloides,Triticosecale rimpaui、小麦属(Triticum spp.)(例如普通小麦(普通小麦)、硬粒小麦(Triticum durum)、圆柱小麦(Triticumturgidum)、Triticum hybernum、马卡小麦(Triticum macha)、普通小麦(Triticum sativum)、一粒小麦(Triticum monococcum)或普通小麦(Triticum vulgare))、小金莲花(Tropaeolum minus)、金莲花(Tropaeolummajus)、越桔属物种(Vaccinium spp.)、野碗豆属物种(Vicia spp.)、豇豆属物种(Vigna spp.)、香堇(Viola odorata)、葡萄属物种(Vitis spp.)、玉蜀黍、Zizania palustris、枣属物种(Ziziphus spp.)等等。
发明详述
令人惊奇地,现在已经发现调节植物中编码产量增加多肽的核酸的表达得到了相对于对照植物具有增强的产量相关性状的植物,所述多肽选自:
磷酸烯醇丙酮酸羧化酶(PEPC),
III类U盒蛋白和
PQQC蛋白。
根据第一个实施方案,本发明提供了相对于对照植物增强植物中产量相关性状的方法,包括调节植物中编码PEPC蛋白质的核酸的表达。
在一个实施方案中,调节(优选增加)编码PEPC蛋白质的核酸的表达是通过在植物中导入并表达编码PEPC蛋白质的核酸。
在一个实施方案中,调节(优选增加)编码III类U盒蛋白的核酸的表达是通过在植物中导入并表达编码III类U盒蛋白的核酸。
在一个实施方案中,调节(优选增加)编码PQQC蛋白的核酸的表达是通过在植物中导入并表达编码PQQC蛋白的核酸实现。
在一个实施方案中,下文中对“用于本发明方法的蛋白质”的任何参考意指如本文定义的PEPC蛋白。下文中对“用于本发明方法的核酸”的任何参考意指能够编码这种PEPC蛋白的核酸。待导入植物的核酸(并且因此可用于实施本发明的方法)是编码现在将描述的类型的蛋白质的任何核酸,其在下文中也被称作“PEPC核酸”或“PEPC基因”。
在一个实施方案中,下文中对“用于本发明方法的蛋白质”的任何参考意指如本文定义的III类U盒蛋白。下文中对“用于本发明方法的核酸”的任何参考意指能够编码这种III类U盒蛋白的核酸。待导入植物的核酸(并且因此可用于实施本发明的方法)是编码现在将描述的类型的蛋白质的任何核酸,其在下文中也被称作“III类U盒核酸”或“III类U盒基因”。
在一个实施方案中,下文中对“用于本发明方法的蛋白质”的任何参考意指如本文定义的PQQC蛋白。下文中对“用于本发明方法的核酸”的任何参考意指能够编码这种PQQC蛋白的核酸。待导入植物的核酸(并且因此可用于实施本发明的方法)是编码现在将描述的类型的蛋白质的任何核酸,其在下文中也被称作“PQQC核酸”或“PQQC基因”。
如本文定义的“PEPC多肽”指SEQ ID NO:2表示的PEPC多肽及其直向同源物和旁系同源物。通过进行所称作的相互blast搜索可鉴定SEQID NO:2的直向同源物和旁系同源物。通常,这涉及第一次BLAST,涉及对任一序列数据库,如公众可用的NCBI数据库用查询序列(例如,使用实施例1的表A中所列的任一序列)进行BLAST。当从核苷酸序列开始时,一般使用BLASTN或TBLASTX(使用标准的默认值),当从蛋白质序列开始时,使用BLASTP或TBLASTN(使用标准的默认值)。BLAST结果可任选过滤。过滤的结果或者未过滤的结果的全长序列然后对查询序列所来源的生物(其中查询序列为SEQ ID NO:1或SEQ ID NO:2,第二次BLAST将因此是针对集胞藻属序列)的序列进行向后BLAST(第二次BLAST)。然后比较第一次和第二次BLAST的结果。如果来自第一次BLAST的高阶命中与查询序列来自相同物种,那么鉴定了旁系同源物,然后向后BLAST将理想地导致查询序列在最高命中中;如果第一次BLAST中的高阶命中与查询序列不来自相同的物种,那么鉴定了直向同源物,并且向后BLAST时优选导致查询序列在最高命中中间。
在本文实施例1的表A中给出了编码III类U盒多肽的核酸的实例。此类核酸可用于实施本发明的方法。在实施例1的表A中给出的氨基酸序列是SEQ ID NO:75表示的III类U盒多肽的直向同源物和旁系同源物的实例序列,术语“直向同源物”和“旁系同源物”在本文中定义。通过进行所称作的相互blast搜索可以容易的鉴定直向同源物和旁系同源物。通常,这涉及第一次BLAST,涉及对任一序列数据库,如公众可用的NCBI数据库用查询序列(例如,使用实施例1的表A中所列的任一序列)进行BLAST。当从核苷酸序列开始时,一般使用BLASTN或TBLASTX(使用标准的默认值),当从蛋白质序列开始时,使用BLASTP或TBLASTN(使用标准的默认值)。BLAST结果可任选过滤。过滤的结果或者未过滤的结果的全长序列然后对查询序列所来源的生物(其中查询序列为SEQ IDNO:74或SEQ ID NO:75,第二次BLAST将因此是针对辣椒属序列)的序列进行向后BLAST(第二次BLAST)。然后比较第一次和第二次BLAST的结果。如果来自第一次BLAST的高阶命中与查询序列来自相同物种,那么鉴定了旁系同源物,然后向后BLAST将理想地导致查询序列在最高命中中;如果第一次BLAST中的高阶命中与查询序列不来自相同的物种,那么鉴定了直向同源物,并且向后BLAST时优选导致查询序列在最高命中中间。
如本文定义的“PQQC多肽”指SEQ ID NO:105表示的PQQC多肽及其直向同源物和旁系同源物。通过进行所称作的相互blast搜索可鉴定SEQ ID NO:105的直向同源物和旁系同源物。通常,这涉及第一次BLAST,涉及对任一序列数据库,如公众可用的NCBI数据库用查询序列(例如,使用实施例1的表A中所列的任一序列)进行BLAST。当从核苷酸序列开始时,一般使用BLASTN或TBLASTX(使用标准的默认值),当从蛋白质序列开始时,使用BLASTP或TBLASTN(使用标准的默认值)。BLAST结果可任选过滤。过滤的结果或者未过滤的结果的全长序列然后对查询序列所来源的生物的序列进行向后BLAST(第二次BLAST)(其中查询序列为SEQ ID NO:104或SEQ ID NO:105,第二次BLAST将因此是针对假单胞菌属序列)。然后比较第一次和第二次BLAST的结果。如果来自第一次BLAST的高阶命中与查询序列来自相同物种,那么鉴定了旁系同源物,然后向后BLAST将理想地导致查询序列在最高命中中;如果第一次BLAST中的高阶命中与查询序列不来自相同的物种,那么鉴定了直向同源物,并且向后BLAST时优选导致查询序列在最高命中中间。
高阶命中是具有低E值的那些命中。E值越低,得分约显著(或者换句话说,命中偶然被发现的机会越低)。E值的计算是本领域公知的。除了E值外,还通过同一性百分比对比较打分。同一性百分比指两个比较的核酸(或者多肽)序列之间在特定长度上相同核苷酸(或氨基酸)的数目。在大家族的情况下,可以使用ClustalW,接着是邻接树,以帮助显示相关基因的聚类或者鉴定直向同源物和旁系同源物。
优选地,PEPC多肽以递增优选顺序与SEQ ID NO:2表示的多肽有至少30%、35%40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%或更高的序列同一性。
优选地,PEPC多肽具有保守的C-末端区,其以递增优选顺序与SEQID NO:2表示的多肽的C-末端区有至少70%、75%、80%、85%、90%、95%、96%、97%、98%或99%或更高的序列同一性。本领域技术人员将理解蛋白质C-末端区的程度。可以认为C-末端区为蛋白质的约最后三分之一。
PEPC多肽包含在含有PEPC的种类之间良好保守的一些基序。例如,下面的基序I到IV通常在植物和微生物PEPC中发现。
基序I:QEIMVGYSDSNKD
基序II:FHGRGGSVGRGGGPAYKAIL
基序III:LRAIPWVF
基序IV:GIAAGMRNTG。
可用于本发明方法的PEPC因此优选包含基序I到IV的任何一个或多个。
可用于本发明方法的PEPC也可以包含这样的基序,其以递增优选顺序与基序I到IV的任何一个或多个有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性。
基序I优选为:QE V/I M I/L/V GYSDS G/N/S K D/F。
基序II优选为:FHGRGG T/S V/I GRGGGP T/S/A H/Y L/I/E/D/K AIL/Q/V。
基序III优选为:LRAIPW I/L/V F。
基序IV优选为:G I/V A/S AG L/M Q/R/K N T/S G。
来自微生物的PEPC多肽也包含在微生物之间良好保守的一些基序,例如,下面的基序V。
基序V:SGFLSSNWE。
基序V优选为:S/G G F/I L/T/V S/T S N/Q/G WE。
基序V或者以递增优选顺序与基序IV有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序可以用于实施本发明的方法。
来自植物的PEPC多肽也包含在植物中良好保守的一些基序,例如,下面的基序VI。
基序VI:SYKEWPEDKR。
基序VI优选为:S/T Y S/L/K/A/R/Q E/K/D W S/P/T/D E E/D R/G/K/QR/K。
基序VI或者以递增优选顺序与基序VI有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序可以用于实施本发明的方法。
如本文定义的“III类U盒多肽”指包含蛋白质的N-末端区中的U盒结构域的任一多肽。
优选地,U盒结构域以递增优选顺序与SEQ ID NO:76的U盒结构域有至少50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性;和/或包含基序VII:GI T/S YDRE/S/D/A N/S/G/A I E/A K/R/M W L/M/V/I F/L/A和/或基序II:Q S/A/GW C/G/A I/R/A/T S/V/L/R/A H/N/R/N/V A/C/S。
最优选地,基序VII为:GITYDRENIERWIF(SEQ ID NO:77),或以递增优选顺序与基序VII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序。
基序VIII为:QSWCTLNA(SEQ ID NO:78)或以递增优选顺序与基序VIII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序。
此外,III类U盒多肽可以在其C-末端区包含一个或多个下面的基序:
基序IX:GR S/A/L A/E L/F L/V S/K/N/A H。
基序X:G/A I/L/V A A/I/V V S/C/A/G/T K/C/R R/K/A A/I/V/T M/L/TR V/L/I S/P。
基序XI:K/R L C/L F/V/M/L V/L/A V/L/A/I Q/R V/A/S D/S/G/E。
基序XII:S/A/L YP。
最优选地,基序IX为:GRAELLNH(SEQ ID NO:79)或以递增优选顺序与基序IX有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序X为:GLAIVSKKILRVS(SEQ ID NO:80)或以递增优选顺序与基序X有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序XI为:KLCLVLQVD(SEQ ID NO:81)或以递增优选顺序与基序XI有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序XII为:SYP(SEQ ID NO:82)或以递增优选顺序与基序XII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序。
III类U盒多肽通常与其他III类U盒多肽而不是与包含全长U盒蛋白质的系统树中任何其他类别的U盒蛋白质聚类。
III类U盒多肽通常以递增优选顺序与SEQ ID NO:75表示的III类U盒多肽有至少50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高的序列同一性。
优选地,PQQC多肽以递增优选顺序与SEQ ID NO:105表示的多肽有至少45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%或更高的序列同一性。
编码PQQC多肽的核酸的实例在本文实施例1的表A中给出。此类核酸可用于实施本发明方法。实施例1的表A中给出的氨基酸序列是SEQID NO:105表示的PQQC多肽的直向同源物和旁系同源物的实例,术语“直向同源物”和“旁系同源物”如本文定义。
术语“结构域”和“基序”在文中的“定义”部分定义。存在用于鉴定结构域的专业数据库,如SMART(Schultz等(1998)Proc.Natl.Acad.Sci.美国95,5857-5864;Letunic等(2002)Nucleic Acids Res30,242-244)、InterPro(Mulder等,(2003)Nucl.Acids.Res.31,315-318)、Prosite(Bucher和Bairoch(1994),A generalized profile syntax for biomolecular sequencesmotifs and its function in automatic sequence interpretation(用于生物分子序列基序的广义图谱语法及其在自动化序列判读中的功能),(在)ISMB-94;第二届分子生物学智能系统国际会议文集.Altman R.,Brutlag D.,KarpP.,Lathrop R.,Searls D.编著,第53-61页,AAAI Press,Menlo Park;Hulo等,Nucl.Acids.Res.32:D134-D137,(2004))或Pfam(Bateman等,Nucleic Acids Research30(1):276-280(2002))。用于计算机芯片上分析蛋白质序列的一组工具可在ExPASY蛋白质组学服务器上获得(瑞士生物信息学研究所(Gasteiger等,ExPASy:the proteomics server for in-depthprotein knowledge and analysis(用于深入认识和分析蛋白质的蛋白组学服务器),Nucleic Acids Res.31:3784-3788(2003))。结构域或基序也可以使用常规技术如通过序列比对而鉴定。
用于比对序列用于比较的方法是本领域众所周知的,此类方法包括GAP、BESTFIT、BLAST、FASTA和TFASTA。GAP使用Needleman和Wunsch算法((1970)J Mol Biol48:443-453)以找到使匹配数最大化并使空位数最小化的两个完整序列的全局性(即覆盖完整序列的)比对。BLAST算法(Altschul等(1990)J Mol Biol215:403-10)计算序列同一性百分数并执行对两个序列间相似性的统计分析。用于执行BLAST分析的软件是公众通过国家生物技术信息中心(NCBI)可获得的。同源物可以使用例如ClustalW多重序列比对算法(1.83版本)以默认配对比对参数和评分方法(以百分数计)而轻易地鉴定。相似性和同一性的全局百分数也可以使用在MatGAT软件包中可获得的方法之一而确定(Campanella等,BMCBioinformatics.2003年7月10日;4:29.MatGAT:用蛋白质或DNA序列而产生相似性/同一性矩阵的应用)。可以进行细微手工编著以优化保守基序之间的比对,如本领域技术人员显而易见。此外除了使用全长序列以鉴定同源物,也可以使用特定结构域。使用上文提及的程序,使用默认参数,针对整个核酸序列或氨基酸或针对选择的结构域或保守基序测定了序列同一性值。
编码PEPC多肽的核酸的实例在本文实施例1的表A中给出。此类核酸可用于实施本发明方法。实施例1的表A中给出的氨基酸序列是SEQ IDNO:2表示的PEPC多肽的直向同源物和旁系同源物的实例,术语“直向同源物”和“旁系同源物”如本文定义。
PEPC的另一优选特征是其催化CO2加入磷酸烯醇丙酮酸(PEP)以形成四碳化合物草酰乙酸:
PEP+CO2→草酰乙酸+Pi
关于PEPC活性的测量的描述,见Chen et al.,2002(Plant Cell Physiol43(2):159-169);和Chen et al.,2004(Planta219pp440-449)。
PEPC的另一特征是其落入酶分类码EC4.1.1.31。
在一个实施方案中,通过用SEQ ID NO:1表示的核酸序列转化植物来阐明本发明,所述核酸序列编码SEQ ID NO:2的多肽序列。然而,本发明的实施不局限于这些序列;本发明的方法可以使用如本文定义的任何PEPC编码核酸或PEPC多肽实施。
此外,III类U盒多肽(至少在其天然形式)通常具有E3泛素连接酶活性(见Cho et al.,2006)。
在一个实施方案中,通过用SEQ ID NO:74表示的核酸序列转化植物来阐明本发明,所述核酸序列编码SEQ ID NO:75的多肽。然而,本发明的实施不局限于这些序列;本发明的方法可以使用如本文定义的任何III类U盒编码核酸或III类U盒多肽实施。
PQQC的另一特征是它催化PQQ生物合成中的最后一步。
在一个实施方案中,通过用SEQ ID NO:104表示的核酸序列转化植物来阐明本发明,所述核酸序列编码SEQ ID NO:105的多肽序列。然而,本发明的实施不局限于这些序列;本发明的方法可以使用如本文定义的任何PQQC编码核酸或PQQC多肽实施。
核酸变体也可以用于实施本发明的方法。此类变体的实例包括编码实施例1表A中给出的任一氨基酸序列的同源物和衍生物的核酸,术语“同源物”和“衍生物”如本文定义。也可用于本发明方法的是编码实施例1表A中给出的任一氨基酸序列的直向同源物或旁系同源物的同源物和衍生物的核酸。可用于本发明方法的同源物和衍生物具有与其所来源的未修饰蛋白质基本上相同的生物和功能活性。
可用于实施本发明的方法的其他核酸变体包括编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的部分,与编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸杂交的核酸、编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的剪接变体、编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的等位变体和通过基因改组得到的编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的变体。术语杂交序列、剪接变体、等位变体和基因改组如本文所述。
编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸不必是全长核酸,因为本发明方法的实施不依赖于全长核酸序列的使用。根据本发明,提供了增强植物中产量相关性状的方法,包括在植物中导入并表达实施例1的表A中给出的任一核酸序列,或者编码实施例1的表A中给出的任一氨基酸序列的直向同源物、旁系同源物或同源物的核酸的部分。
例如,通过对核酸进行一个或多个缺失可以制备核酸的部分。所述部分可以以分离形式使用或者它们可以融合到其他编码(或非编码)序列以便例如,产生结合几种活性的蛋白质。当融合到其他编码序列时,翻译时所产生的多肽可以比为该蛋白质部分预测的更大。
在一个实施方案中,可用于本发明方法的部分编码如本文定义的PEPC多肽,并且具有与实施例1的表A中给出的氨基酸序列基本上相同的生物活性。优选地,所述部分是实施例1的表A中给出的任一核酸的部分,或者是编码实施例1的表A中给出的任一氨基酸序列的直向同源物或旁系同源物的核酸的部分。优选地,所述部分长为至少2000、2250、2500、2750、3000、3100个连续核苷酸,所述连续核苷酸为实施例1的表A中给出的任一核酸序列,或者编码实施例1的表A中给出的任一氨基酸序列的直向同源物或旁系同源物的核酸。最优选地,所述部分是SEQ ID NO:1的核酸的部分。
在一个实施方案中,可用于本发明方法的部分编码如本文定义的III类U盒多肽,并且具有与实施例1的表A中给出的氨基酸序列基本上相同的生物活性。优选地,所述部分是实施例1的表A中给出的任一核酸的部分,或者是编码实施例1的表A中给出的任一氨基酸序列的直向同源物或旁系同源物的核酸的部分。优选地,所述部分长为至少500、550、600、650、700、750、800、850、900、950、1000、1050、1100、1150、1200、1250、1300、1350个连续核苷酸,所述连续核苷酸为实施例1的表A中给出的任一核酸序列,或者编码实施例1的表A中给出的任一氨基酸序列的直向同源物或旁系同源物的核酸。最优选地,所述部分是SEQ ID NO:74的核酸的部分。
在一个实施方案中,可用于本发明方法的部分编码如本文定义的PQQC多肽,并且具有与实施例1的表A中给出的氨基酸序列基本上相同的生物活性。优选地,所述部分是实施例1的表A中给出的任一核酸的部分,或者是编码实施例1的表A中给出的任一氨基酸序列的直向同源物或旁系同源物的核酸的部分。优选地,所述部分长为至少500、550、600、650、700、750个连续核苷酸,所述连续核苷酸为实施例1的表A中给出的任一核酸序列,或者为编码实施例1的表A中给出的任一氨基酸序列的直向同源物或旁系同源物的核酸。最优选地,所述部分是SEQ ID NO:104的核酸的部分。
可用于本发明方法的另一核酸变体是这样的核酸,其在降低的严格性条件下,优选在严格条件下与编码选自如本文定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸杂交,或者与如本文定义的部分杂交。
根据本发明,提供了在植物中增强产量相关性状的方法,包括在植物中导入并表达核酸,所述核酸能够与实施例1的表A中给出的任一核酸杂交,或者包括在植物中导入并表达核酸,所述核酸能够与编码实施例1的表A中给出的任一核酸序列的直向同源物、旁系同源物或同源物的核酸杂交。
可用于本发明方法的杂交序列编码如本文定义的PEPC多肽,具有与实施例1的表A中给出的氨基酸序列相同的生物活性。优选地,所述杂交序列能够与实施例1的表A中给出的任一核酸杂交,或与任一这些序列的部分杂交,部分如生物定义,或者杂交序列能够与编码实施例1的表A中给出的任一氨基酸序列的直向同源物或旁系同源物的核酸杂交。最优选地,杂交序列能够与选自SEQ ID NO:1、SEQ ID NO:74和SEQ ID NO:104的序列表示的核酸或者与其部分杂交,部分如上文定义。
可用于本发明方法的另一核酸变体是编码选自如上文定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的剪接变体,剪接变体如本文定义。
根据本发明,提供了增强植物中产量相关性状的方法,包括在植物中导入并表达实施例1的表1中给出的任一核酸序列的剪接变体,或者编码如实施例1的表1中给出的任一氨基酸序列的直向同源物、旁系同源物或同源物的核酸的剪接变体。
在一个实施方案中,优选的剪接变体是SEQ ID NO:1表示的核酸的剪接变体,或者编码SEQ ID NO:2的直向同源物或旁系同源物的核酸的剪接变体。
在一个实施方案中,优选的剪接变体是SEQ ID NO:74表示的核酸的剪接变体,或者编码SEQ ID NO:75的直向同源物或旁系同源物的核酸的剪接变体。
在一个实施方案中,优选的剪接变体是SEQ ID NO:104表示的核酸的剪接变体,或者编码SEQ ID NO:105的直向同源物或旁系同源物的核酸的剪接变体。
可用于实施本发明方法的另一核酸变体是编码选自如上文定义的磷酸烯醇丙酮酸羧化酶(PEPC)、III类U盒蛋白和PQQC多肽的产量增加多肽的核酸的等位变体,等位变体如本文定义。
根据本发明,提供了增强植物中产量相关性状的方法,包括在植物中导入并表达实施例1的表A中给出的任一核酸的等位变体,或者包括在植物中导入并表达编码实施例1的表A中给出的任一氨基酸序列的直向同源物、旁系同源物或同源物的核酸的等位变体。
在一个实施方案中,可用于本发明方法的等位变体具有与SEQ ID NO:2的PEPC多肽或者实施例1的表A中描述的任一氨基酸基本上相同的生物活性。等位变体存在于天然中,并且本发明的方法包括使用这些天然等位基因。优选地,等位变体是SEQ ID NO:1的等位变体或者编码SEQ IDNO:2的直向同源物或旁系同源物的核酸的等位变体。
在一个实施方案中,可用于本发明方法的等位变体具有与SEQ IDNO:75的III类U盒多肽或者实施例1的表A中描述的任一氨基酸基本上相同的生物活性。等位变体存在于天然中,并且本发明的方法包括使用这些天然等位基因。优选地,等位变体是SEQ ID NO:74的等位变体或者编码SEQ ID NO:75的直向同源物或旁系同源物的核酸的等位变体。
在一个实施方案中,可用于本发明方法的等位变体具有与SEQ IDNO:105的PQQC多肽和实施例1的表A中描述的任一氨基酸基本上相同的生物活性。等位变体存在于天然中,并且本发明的方法包括使用这些天然等位基因。优选地,等位变体是SEQ ID NO:104的等位变体或者编码SEQ ID NO:105的直向同源物或旁系同源物的核酸的等位变体。
基因改组或者定向进化也可以用于产生编码选自如上文定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的变体;术语“基因改组”如本文定义。
根据本发明,提供了在植物中增强产量相关性状的方法,包括在植物中导入并表达实施例1的表A中给出的任一核酸序列的变体,或者包括在植物中导入并表达编码实施例1的表A中给出的任一氨基酸序列的直向同源物、旁系同源物或同源物的核酸的变体,所述变体核酸通过基因改组得到。
此外,通过定向诱变也可以得到核酸变体。一些方法可用于实现位点定向诱变,最常用的是基于PCR的方法(Current Protocols in MolecularBiology.Wiley Eds.)。
在一个实施方案中,编码PEPC多肽的核酸可以来自任一天然或人工来源。核酸可以通过有意的人工操作在其组成和/或基因组环境中从其天然形式修饰。优选地,PEPC多肽编码核酸是细菌的,进一步优选来自集胞藻属(Synechococcus)、倒囊藻属(Anacystis)、鱼腥藻属(Anabaena)、念珠藻属(Nostoc)、原绿球藻属(Prochlorococcus)、节杆菌属(Arthrobacter)、链霉菌属(Steptomyces)、动球菌属(Kineococcus)、硝化杆菌属(Nitobacter)、罗尔斯通氏菌属(Ralsstonia)、亚硝化单胞菌属(Nitrosomonas)、红球菌属(Rhodococcus)、红假单胞菌属(Rhodopseudomonas)、分枝杆菌属(Mycobacterium)、奈瑟菌属(Neiseria)、乳杆菌属(Lactobacillus)、棒杆菌属(Corynebacterium)的任一种,最优选来自集胞藻属。
在一个实施方案中,编码III类U盒多肽的核酸可以来自任一天然或人工来源。核酸可以通过有意的人工操作在其组成和/或基因组环境中从其天然形式修饰。优选地,编码III类U盒多肽的核酸来自植物,进一步优选来自双子叶植物,更优选来自茄科(Solanaceae),最优选地,所述核酸来自辣椒。
在一个实施方案中,编码PQQC多肽的核酸可以来自任一天然或人工来源。核酸可以通过有意的人工操作在其组成和/或基因组环境中从其天然形式修饰。优选地,PQQC多肽编码核酸是细菌的,进一步优选来自假单胞菌属。
本发明方法的实施得到了具有增强的产量相关性状的植物。具体地,本发明方法的实施得到了相对于对照植物具有增加的产量,特别是增加的种子产量的植物。术语“产量”和“种子产量”在本文的“定义”部分更详细描述。
还令人惊奇的是发现产量的增加不以更大种子的形式表现,如已经从Radchuck et al.,2007的教导预期,而是以一组改变的产量相关性状的形式表现,所述性状包括但不限于,各自相对于对照植物增加的地上部分面积、增加的萌发势、增加的种子总数、增加的种子重量、增加的饱满率、每圆锥花序增加的小花数、增加的种子数目、增加的收获指数、增加的高度和根厚度的增加。
在本文中对增强的产量相关性状的参考意指植物的一个或多个部分的生物量(重量)增加,所述的部分可以包括地上(可收获)部分和/或地下(可收获)部分。尤其,此类可收获部分是种子,并且本发明方法的实施产生相对于对照植物的种子产量具有增加的种子产量的植物。
以玉米为例,产量增加可以表现为下列一种或多种指标:每公顷或英亩生长的(establish)植物数的增加、每株植物穗数的增加、行数的增加、每行粒数、粒重、千粒重、玉米穗长度/直径、种子饱满率的增加(其中种子饱满率是饱满种子数除以种子总数并乘以100)及其他。以稻为例,产量增加本身可以表现为下列一种或多种指标的增加:每公顷或英亩植物数、每株植物圆锥花序数、每圆锥花序小穗数、每圆锥花序花(小花)数(其表达为饱满种子数对原圆锥花序数之比)、种子饱满率的增加(其中种子饱满率是饱满种子数除以种子总数并乘以100)、千粒重的增加等等。
本发明提供了相对于对照植物增加植物产量,特别是种子产量的方法,该方法包括调节植物中编码选自如本文定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的表达,优选增加其表达。
因为本发明的转基因植物具有增强的产量,所以这些植物可能显示出相对于对照植物在其生命周期的对应阶段的生长速率增加的生长速率(在至少部分其生命周期中)。
增加的生长速率可以对于植物的一个或多个部分(包括种子)是特异性的,或可以基本上遍及整株植物。具有增加的生长速率的植物可以具备较短的生活周期。植物的生活周期可以视为意指从干燥成熟种子成长至植物已经产生与起始材料相似的干燥成熟种子的阶段所需要的时间。这个生活周期可以受下列因素影响,如早期萌发势、生长速率、绿度指数、开花时间和种子成熟速度。生长速率的增加可以在植物生活周期中的一个或多个阶段上或在基本上整个植物生活周期期间发生。在植物生活周期中的早期期间增加的生长速率可以反映增强的萌发势。生长速率的增加可以改变植物的收获周期,允许植物较晚播种和/或较早收获,否则这将不可能(相似的作用可以用较早的开花时间获得)。若生长速率充分地增加,可以允许进一步播种相同植物物种的种子(例如播种并收获稻植物,随后播种并收获其他稻植物,全部过程均在一个常规生长时段内)。类似地,若生长速率足够地增加,可以允许进一步播种不同植物物种的种子(例如播种并收获玉米植物,随后例如播种并任选收获大豆、马铃薯或任何其他合适植物)。从相同的根茎中收获额外次数在一些作物植物的情况中也是可能的。改变植物的收获周期可以导致每英亩的年生物量产量的增加(因任何特定植物可以生长并收获的次数(如在一年中)增加)。生长速率的增加也可以允许比其野生型对应物在更广泛的地理区域内培育转基因植物,因为对培育作物的区域限制往往由栽种时节(早季)或在收获时期(晚季)的不利环境条件所决定。若缩短收获周期,则可以避开这类不利条件。生长速率可以通过从生长曲线中得到多种参数而确定,此类参数可以是:T-Mid(植物达到其50%最大尺寸所花费的时间)和T-90(植物达到其90%最大尺寸所花费的时间)等。
根据本发明的优选特征,本发明方法的实施产生相对于对照植物而具有增加的生长速率的植物。因而根据本发明,提供增加植物生长速率的方法,所述方法包括调节、优选增加核酸在植物中的表达,所述核酸编码选自本文中所定义的PEPC、III类U盒和PQQC多肽的产量增加多肽。
在一个实施方案中,发生产量的增加,不管与对照植物相比,植物处于非胁迫条件还是植物暴露于多种胁迫。植物一般通过生长得更慢而对暴露于胁迫作出应答。在严重胁迫条件下,植物甚至可以完全停止生长。另一方面,轻微胁迫在本文中定义为植物暴露的任何胁迫,其中所述的胁迫未导致植物完全停止生长而没有恢复生长的能力。与非胁迫条件下的对照植物相比,轻微胁迫在本发明意义中导致受胁迫植物生长降低小于40%、35%或30%,优选小于25%、20%或15%,更优选小于14%、13%、12%、11%或10%或更低。由于农业实践(灌溉、施肥、杀虫剂处理)上的进步,在栽培作物植物中并不经常遇到严重胁迫。因此,由轻微胁迫诱导的受损生长往往是农业上不希望的特征。“轻微胁迫”是植物暴露的常见生物和/或非生物(环境)胁迫。非生物胁迫可以因干旱或过多的水、缺氧胁迫、盐胁迫、化学毒性、氧化胁迫和热、冷或冰冷温度所致。非生物胁迫可以是由水胁迫(尤其因为干旱)、盐胁迫、氧化胁迫或离子胁迫引起的渗透胁迫。生物胁迫通常是病原体,如细菌、病毒、真菌和昆虫引起的那些胁迫。
具体地,本发明的方法可以在非胁迫条件或在轻微干旱条件下进行以相对于对照植物具有增加的产量。如在Wang等(Planta(2003)218:1-14)中报道,非生物胁迫导致不利地影响植物生长及生产力的一系列形态学变化、生理学变化、生物化学变化和分子变化。已知干旱、盐度、极端温度和氧化胁迫是相互联系的并可以通过相似机制而诱导生长损害及细胞损害。Rabbani等(Plant Physiol(2003)133:1755-1767)描述了干旱胁迫与高盐度胁迫间极高程度的“串话”。例如,干旱和/或盐化作用主要表现为渗透胁迫,导致细胞内稳态和离子分布的破坏。经常伴随高温或低温、盐度或干旱胁迫的氧化胁迫可以造成功能性蛋白和结构蛋白变性。因此,这些多样的环境胁迫常常激活相似的细胞信号途径和细胞应答,如产生胁迫蛋白质、上调抗氧化物质、积累相容性溶质和生长抑制。本文中所用的术语”非胁迫”条件是允许植物最佳生长的环境条件。本领域技术人员清楚对于给定地点的正常土壤条件和气候条件。
在一个实施方案中,发生产量的增加,不管植物处于非胁迫条件还是处于轻微干旱条件下。本发明方法的实施相对于在相当条件下生长的对照植物,赋予在非胁迫条件下或在轻微干旱条件下生长的植物增加的产量。因而根据本发明,提供用于在非胁迫条件下或在轻微干旱条件下生长的植物中增加产量的方法,所述方法包括增加编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸在植物中的表达。
本发明方法的实施得到了植物,其在营养缺乏,尤其氮缺乏条件下生长时与在相当条件下生长的对照植物相比具有增加的产量。因此,根据本发明,提供了增加在营养缺乏条件下生长的植物中产量的方法,该方法包括增加植物中编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的表达。营养缺乏可以由营养物的缺乏引起,所述营养物如氮、磷酸盐和其他含磷化合物、钾、钙、镉、镁、锰、铁和硼,等等。
本发明包括通过本发明的方法获得的植物或其部分(包括种子)。所述植物、或其部分包含编码选自如上文所定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸转基因。
本发明还提供遗传构建体和载体以促进在植物中引入和/或表达编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸。该基因构建体可以插入适于转化至植物内并适于在转化的细胞中表达目的基因的市售载体。本发明也提供如文中所定义的基因构建体在本发明方法中的用途。
更具体地,本发明提供构建体,其包含
(a)编码选自如上文定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸;
(b)能够驱动(a)的核酸序列表达的一个或多个控制序列;和任选地
(c)转录终止序列。
优选地,编码PEPC多肽的核酸如上文定义。
优选地,编码III类U盒多肽的核酸如上文定义。
优选地,编码PQQC多肽的核酸如上文定义。
术语“控制序列”和“终止序列”如上文所定义。
用包含上述核酸之一的载体转化植物。技术人员公知必须存在于载体上的遗传元件,以便成功转化、选择和繁殖含有目的序列的宿主细胞。目的序列有效连接一个或多个控制序列(至少启动子)。
有利地,任一类型的启动子,不管是天然还是合成的,都可以用于驱动核酸序列的表达。见本文“定义”章节中关于多种启动子类型的定义。
年轻的绿色组织特异性启动子尤其可用于方法中。
在一个实施方案中,PEPC编码核酸有效连接到如本文定义的年轻绿色组织特异性启动子。年轻绿色组织特异性启动子优选为原叶绿素酸酯还原酶(PcR)启动子,更优选为基本上类似于SEQ ID NO:71的核酸序列表示的原叶绿素酸酯还原酶启动子,最优选地,启动子如SEQ ID NO:71表示。
在一个实施方案中,III类U盒编码核酸有效连接到如本文定义的年轻绿色组织特异性启动子。年轻绿色组织特异性启动子优选为原叶绿素酸酯还原酶(PcR)启动子,更优选为基本上类似于SEQ ID NO:101的核酸序列表示的原叶绿素酸酯还原酶启动子,最优选地,启动子如SEQ ID NO:101表示。
在一个实施方案中,PQQC编码核酸有效连接到如本文定义的年轻绿色组织特异性启动子。年轻绿色组织特异性启动子优选为原叶绿素酸酯还原酶(PcR)启动子,更优选为基本上类似于SEQ ID NO:131的核酸序列表示的原叶绿素酸酯还原酶启动子,最优选地,启动子如SEQ ID NO:131表示。
此外,组成型启动子尤其可用于方法中。组成型启动子优选为GOS2启动子,优选来自稻的GOS2启动子。进一步优选地,组成型启动子由基本上类似于SEQ ID NO:227的核酸序列表示,最优选地,组成型启动子如SEQ ID NO:227所示。组成型启动子的其他实例见本文“定义”章节中表2。
应该清楚本发明的应用性不限于SEQ ID NO:1表示的PEPC编码核酸,本发明的应用性也不限于原叶绿素酸酯还原酶启动子驱动的这种PEPC编码核酸的表达。应该清楚本发明的应用性不限于SEQ ID NO:74表示的U盒编码核酸,本发明的应用性也不限于原叶绿素酸酯还原酶启动子驱动的这种U盒编码核酸的表达。应该清楚本发明的应用性不限于SEQID NO:104表示的PQQC编码核酸,本发明的应用性也不限于原叶绿素酸酯还原酶启动子驱动或GOS2启动子驱动的这种PQQC编码核酸的表达。
也可用于实施本发明方法的其他(年轻)绿色组织特异性启动子的实例在本文的表2g中显示并且也给出了其他组成型启动子的实例,见表2a。
任选地,一种或多种终止子序列可以在引入植物的构建体中使用。额外的调节元件可以包括转录增强子以及翻译增强子。本领域技术人员会知道可以在实施本发明中适用的终止子序列和增强子序列。内含子序列也可以添加至5′非翻译区(UTR)或编码序列上,以增加在胞浆内积累的成熟信息的量,如定义部分中所述。其他控制序列(除启动子、增强子、沉默子、内含子序列、3’UTR和/或5’UTR区之外)可以是蛋白质和/或RNA稳定元件。本领域技术人员会知道或可以轻易地获得此类序列。
本发明的遗传构建体还可以包括在特定细胞类型中维持和/或复制所需的复制起点序列。一个实例是当需要将遗传构建体在细菌细胞中维持为附加型遗传元件(例如质粒或粘粒分子)时。优选的复制起点包括,但不限于f1-ori和colE1。
为检测如在本发明方法中所用核酸序列的成功转移和/或选择包含这些核酸的转基因植物,使用标记基因(或报道基因)是有利的。因而,遗传构建体可以任选地包含选择性标记基因。文中的“定义”部分更加详细的描述了选择性标记。标记基因一旦不再需要可以从转基因细胞中除去或者切除。用于标记去除的技术是本领域公知的,有用的技术在上文定义部分中描述。
本发明还提供了产生相对于对照植物具有增强的产量相关性状的转基因植物的方法,包括在植物中导入并表达编码选自如上文定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的任一核酸。
更特别地,本发明提供了产生具有增强的产量相关性状,尤其增加的(种子)产量的转基因植物的方法,所述方法包括:
(i)在植物或植物细胞中导入并表达编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸;和
(ii)在促进植物生长和发育的条件下培养植物细胞。
在一个实施方案中,(i)的核酸可以是能够编码如本文定义的PEPC多肽的任一核酸。
在一个实施方案中,(i)的核酸可以是能够编码如本文定义的III类U盒多肽的任一核酸。
在一个实施方案中,(i)的核酸可以是能够编码如本文定义的PQQC多肽的任一核酸。
核酸可以直接引入植物细胞或引入植物本身(包括引入组织、器官或植物的任何其他部分)。根据本发明的优选特征,核酸优选地通过转化引入植物。文中的“定义”部分更加详细的描述了术语“转化”。
遗传修饰的植物细胞可通过本领域技术人员熟悉的所有方法再生。合适的方法可见于S.D.Kung和R.Wu、Potrykus或 和Willmitzer的上述出版物。
和Willmitzer的上述出版物。
通常在转化后,选择一种或多种标记存在的植物细胞或细胞群体,其中所述的标记由与目的基因一起共转移的植物可表达的基因编码,随后将转化的材料再生成整株植物。为了选择转化的植物,在转化中获得的植物材料原则上接受选择条件处理,以至于转化的植物可以与未转化的植物区分。例如,以上文所述方式获得的种子可以播种,在初始培育时期后,通过喷雾进行合适的选择。另一种可能性包括在使用合适选择剂的琼脂平板上生长种子(根据需要在消毒之后),使得仅转化的种子可以生长成植物。或者,转化的植物通过上述那些选择性标记的存在筛选。
在DNA转移和再生后,推测的转化植物可以例如使用Southern分析对目的基因的存在、拷贝数和/或基因组构造进行评价。备选或额外地,新引入DNA的表达水平可以使用Northern和/或Western分析进行监测,这两种技术是本领域技术人员众所周知的。
产生的转化植物可以通过多种方法加以增殖,如通过克隆增殖法或经典育种技术。例如,第一代(或T1)转化植物可以进行自交,选择纯合的第二代(或T2)转化体,并且T2植物通过经典育种技术进一步增殖。产生的转化生物可以采取多种形式。例如,它们可以是转化细胞和非转化细胞的嵌合体;克隆转化体(例如被转化以含有表达盒的全部细胞);转化组织和未转化组织的移植体(例如在植物中,与未转化嫩枝嫁接的转化的根状茎)。
本发明明确地扩展至通过文中所述的任意方法产生的任何植物细胞或植物,并扩展至全部植物部分及其繁殖体。本发明进一步扩展至包含已经通过任意前述方法产生的原代转化或转染细胞、组织、器官或整株植物的后代,唯一要求是后代表现与通过本发明方法中的亲代所产生的那些后代相同的一种或多种基因型特征和/或表型特征。
本发明也包括宿主细胞,其含有编码选自如上文所定义的PEPC、III类U盒和PQQC多肽的产量增加多肽的分离的核酸。本发明优选的宿主细胞是植物细胞。宿主植物对于本发明方法中所用核酸或载体、表达盒或构建体或载体而言原则上有利地是能够合成本发明方法中所用多肽的全部植物。
本发明方法有利地适用于任何植物。特别用于本发明方法中的植物包括属于植物界超家族的全部植物,尤其是单子叶植物和双子叶植物,包括饲用或饲料豆类、观赏植物、粮食作物、树或灌木。根据本发明优选的实施方案,植物是作物植物。作物植物的实例包括大豆、向日葵、卡诺拉油菜、苜蓿、油菜、棉花、番茄、马铃薯和烟草。还优选地,植物是单子叶植物。单子叶植物的实例包括甘蔗。更优选地,植物是谷物。谷物的实例包括稻、玉米、小麦、大麦、粟、黑麦、黑小麦、高粱和燕麦。
本发明也扩展至植物的可收获部分如,但不限于种子、叶、果实、花、茎、根、根状茎、块茎和球茎。本发明进一步涉及来自自、优选直接来自此类植物的可收获部分中的产物,如干燥颗粒或粉末、油、脂肪及脂肪酸、淀粉或蛋白质。
根据本发明优选的特征,受调控的表达是增加的表达。在本领域内详细记载了用于增加核酸或基因、或基因产物表达的方法并且实例在定义部分中提供。
如上面提到的,调节(优选增加)编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的表达的优选方法是通过在植物中导入并表达编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸;然而,实施该方法的效果,即增强产量相关性状也可以使用其他公知的技术实现,所述技术包括但不限于T-DNA活化标记、TILLING、同源重组。这些技术的描述在定义部分中提供。
在一个实施方案中,本发明还包括编码如本文所述的PEPC多肽的核酸和这些PEPC多肽用于增强植物中任一上述产量相关性状的用途。
在一个实施方案中,本发明还包括编码如本文所述的III类U盒多肽的核酸和这些III类U盒多肽用于增强植物中任一上述产量相关性状的用途。
在一个实施方案中,本发明还包括编码如本文所述的PQQC多肽的核酸和这些PQQC多肽用于增强植物中任一上述产量相关性状的用途。
编码选自如本文所述的PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸,或者选自PEPC、III类U盒和PQQC多肽的产量增加多肽自身可以用于育种程序,其中鉴定DNA标记,所述DNA标记可以与选自PEPC、III类U盒和PQQC多肽的产量增加多肽的编码基因遗传上连锁。所述核酸/基因或选自PEPC、III类U盒和PQQC多肽的产量增加多肽自身可以用于定义分子标记。该DNA或蛋白质标记然后可以用于育种程序以在本发明的方法中选择如上文定义的具有增强的产量相关性状的植物。
编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸/基因的等位变体也可以用于标记辅助的育种程序中。此类育种程序有时需要使用例如EMS诱变通过植物的诱变处理引入等位基因变异;备选地,程序可以以一组非有意引起的所谓的“天然”来源的等位变体开始。然后例如通过PCR开始等位变体的鉴定。接着是选择具有所讨论序列并且得到提高的产量的超级等位变体的步骤。一般通过监测含有所讨论序列的不同等位变体的植物的生长性能而实施选择。可以在温室中或田间监测生长性能。其他任选步骤包括将鉴定了优异等位变体的植物与另一种植物杂交。这可以用来例如产生目标表型特征的组合。
编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸也可以用作探针以便对基因进行遗传作图或物理作图,所述探针作为所述基因的一部分及与这些基因关联的性状的标记。此类信息可以用于植物育种中,以便开发具有想要表型的株系。编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸的这种用途仅需要具有至少15个核苷酸长度的核酸序列。编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸可以用作限制性片段长度多态性(RFLP)标记。限制性消化的植物基因组DNA的Southern印迹(Sambrook J,Fritsch EF和Maniatis T(1989)Molecular Cloning,A Laboratory Manual)可以用编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸探测。产生的结合图式随后可以使用计算机程序如MapMaker(Lander等(1987)Genomics1:174-181)进行遗传分析以构建遗传图。此外,该核酸可以用来探测含有经限制性内切核酸酶处理的一组个体的基因组DNA的Southern印迹,其中所述的一组个体代表具有确定的遗传杂交的亲代和后代。DNA多态性的分离被标出并用来计算编码选自PEPC、III类U盒和PQQC多肽的产量增加多肽的核酸序列在使用这个群体先前所获得的遗传图中的位置(Botstein等(1980)Am.J.Hum.Genet.32:314-331)。
在Bernatzky和Tanksley(1986)Plant Mol.Biol.Reporter4:37-41中描述了植物基因衍生的探针的产生和其在遗传作图中的用途。众多出版物描述了使用以上所提及的方法学或其改良方法对特定cDNA克隆的遗传作图。例如,F2互交群、回交群、随机交配群、邻近纯合系和其他个体群体可以用于作图。此类方法学是本领域技术人员众所周知的。
所述核酸探针也可以用于物理作图(即序列在物理图上的排列;见Hoheisel等在:Non-mammalian Genomic Analyasis:A Practical Guide,Academic press1996,第319-346页及其中引用的参考文献)。
在另一实施方案中,核酸探针可以在直接萦光原位杂交(FISH)作图法(Trask(1991)Trends Genet.7:149-154)中使用。尽管当前的FISH作图法支持使用大型克隆(几个kb至几百个kb;见Laan等(1995)Genome Res.5:13-20),然而灵敏度的改善可以允许使用更短探针进行FISH作图。
用于遗传作图及物理作图的多种基于核酸扩增的方法可以使用所述核酸而实施。实例可见于文中的“定义”部分。实例包括等位基因特异性扩增(Kazazian(1989)J.Lab.Clin.Med11:95-96)、PCR扩增片段的多态性(CAPS;Sheffield等,(1993)Genomics16:325-332)、等位基因特异性连接(Landegren等,(1988)Science241:1077-1080)、核苷酸延伸反应(Sokolov(1990)Nucleic Acid Res.18:3671)、放射杂交作图(Walter等,(1997)Nat.Genet.7:22-28)和Happy作图(Dear和Cook,(1989)Nucleic Acid Res.17:6795-6807)。为实施这些方法,使用核酸序列设计和产生用于扩增反应或引物延伸反应的引物对。这类引物的设计是本领域技术人员众所周知的。使用基于PCR的遗传作图的方法,可能需要鉴定跨越相应于本发明核酸序列区域作图的亲本之间DNA序列的差异。然而,这对作图方法通常不是必要的。
本发明方法产生具有如前文所述的增强的产量相关性状的植物。这些性状也可以与经济上有利的其他性状组合,如其他的产量增加性状,对其他非生物胁迫和生物性胁迫的耐受性,调节多种构造性特征和/或生物化学特征和/或生理学特征的性状。
用于该说明书中的术语“表A”用于说明表A1、A2和/或A3中的内容。该说明书中使用的术语“表A1”用于指表A1的内容。该说明书中使用的术语“表A2”用于指表A2的内容。该说明书中使用的术语“表A3”用于指表A3的内容。
在一个优选实施方案中,术语“表A”指表A1。在一个优选实施方案中,术语“表A”指表A2。在一个优选实施方案中,术语“表A”指表A3。
在一个实施方案中,本发明涉及如下概述的主题。
第1项.相对于对照植物在植物中增强产量相关性状的方法,包括调节植物中编码磷酸烯醇丙酮酸羧化酶(PEPC)蛋白质的核酸的表达。
第2项.根据第1项的方法,其中所述PEPC蛋白质是SEQ ID NO:2和其直向同源物和旁系同源物。
第3项.根据第1或2项的方法,其中所述PEPC蛋白质具有C-末端区,其以递增优选顺序与SEQ ID NO:2表示的多肽的C-末端区有至少70%、75%、80%、85%、90%、95%、96%、97%、98%或99%或更高序列同一性。
第4项.根据任一前面项的方法,其中所述PEPC蛋白质包含一个或多个下面的基序:
基序I:QEIMVGYSDSNKD,或以递增优选顺序与基序I有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序;
基序II:FHGRGGSVGRGGGPAYKAIL,或以递增优选顺序与基序II有至少65%70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序;
基序III:LRAIPWVF,或以递增优选顺序与基序III有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序;
基序IV:GIAAGMRNTG,或以递增优选顺序与基序IV有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序;
基序V:SGFLSSNWE,或以递增优选顺序与基序V有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序;
基序VI:SYKEWPEDKR,或以递增优选顺序与基序VI有至少65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%同一性的基序。
第5项.根据第4项的方法,其中
基序I优选为:QE V/I M I/L/V GYSDS G/N/S K D/F;
基序II优选为:FHGRGG T/S V/I GRGGGP T/S/A H/Y L/I/E/D/K AIL/Q/V;
基序III优选为:LRAIPW I/L/V F;
基序IV优选为:G I/V A/S AG L/M Q/R/K N T/S G;
基序V优选为:S/G G F/I L/T/V S/T S N/Q/G WE;
基序VI优选为:S/T Y S/L/K/A/R/Q E/K/D W S/P/T/D E E/D R/G/K/QR/K。
第6项.根据任一前面项的方法,其中通过在植物中导入并表达编码PEPC多肽的核酸实现所述受调节的表达。
第7项.根据任一前面项的方法,其中所述编码PEPC多肽的核酸编码表A中所列的任一种蛋白质或者为此类核酸的一部分,或者能够与这种核酸杂交的核酸。
第8项.根据任一前面项的方法,其中所述增强的产量相关性状包括下面的一种或多种:各自相对于对照植物增加的地上部分面积、增加的萌发势、增加的种子总数、增加的种子重量、增加的饱满率、每圆锥花序增加的小花数、增加的种子数目、增加的收获指数、增加的高度和根厚度的增加。
第9项.根据任一前面项的方法,其中在非胁迫条件和轻微干旱条件下得到所述增强的产量相关性状。
第9项.根据任一前面项的方法,其中在氮缺乏条件下得到所述增强的产量相关性状。
第10项.根据任一前面项的方法,其中所述核酸有效连接到年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
第11项.根据任一前面项的方法,其中所述编码PEPC多肽的核酸是细菌来源,优选来自集胞藻属。
第12项.通过任一前面项的方法得到的植物或其部分,包括种子,其中所述植物或其部分包含编码PEPC多肽的重组核酸。
第13项.构建体,其包含:
(i)编码如第1到5任一项定义的PEPC多肽的核酸;
(ii)能够驱动(a)的核酸序列表达的一个或多个控制序列;和任选地
(iii)转录终止序列。
第14项.根据第13项的构建体,其中所述控制序列之一是年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
第15项.根据第13或14项的构建体在用于制备植物的方法中的用途,所述植物具有下面的一种或多种:各自相对于对照植物增加的地上部分面积、增加的萌发势、增加的种子总数、增加的种子重量、增加的饱满率、每圆锥花序增加的小花数、增加的种子数目、增加的收获指数、增加的高度和根厚度的增加。
第16项.用根据第13或14项的构建体转化的植物、植物部分或植物细胞。
第17项.产生相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量的转基因植物的方法,包括:
(i)在植物中导入并表达编码如第1到5项任一项定义的PEPC多肽的核酸;和
(ii)在促进植物生长和发育的条件下培养所述植物细胞。
第18项.转基因植物,其具有下面的一种或多种:各自相对于对照植物增加的地上部分面积、增加的萌发势、增加的种子总数、增加的种子重量、增加的饱满率、每圆锥花序增加的小花数、增加的种子数目、增加的收获指数、增加的高度和根厚度的增加,所述转基因植物从编码如第1到5项任一项定义的PEPC多肽的核酸的增加的表达得到,或者来自所述转基因植物的转基因植物细胞。
第19项.根据第12、16或18项的转基因植物,或来自其的转基因植物细胞,其中所述植物是作物植物或单子叶植物或谷类,如稻、玉米、小麦、大麦、粟、黑麦、黑小麦、高粱和燕麦。
第20项.根据第19项的植物的可收获部分,其中所述可收获部分优选为苗生物量和/或种子。
第21项.来自根据第19项的植物和/或来自根据第20项的植物的可收获部分的产品。
第22项.编码PEPC多肽的核酸的用途,用于增加产量,尤其增加下面的一种或多种:各自相对于对照植物增加的地上部分面积、增加的萌发势、增加的种子总数、增加的种子重量、增加的饱满率、每圆锥花序增加的小花数、增加的种子数目、增加的收获指数、增加的高度和根厚度的增加。
在一个实施方案中,本发明涉及如下述的主题:
第23项.相对于对照植物在植物中增强产量相关性状的方法,包括调节植物中编码III类U盒蛋白的核酸的表达,所述III类U盒蛋白在其N-末端区包含U盒结构域。
第24项.根据第23项的方法,其中所述U盒结构域以递增优选顺序与SEQ ID NO:76的U盒结构域有至少50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性和/或包含基序VII:GI T/S YDR E/S/D/A N/S/G/A I E/A K/R/M WL/M/V/I F/L/A和/或基序VIII:Q S/A/G W C/G/A I/R/A/T S/V/L/R/AH/N/R/N/V A/C/S。
第25项.根据第24项的方法,其中所述基序VII是GITYDRENIERWIF(SEQ ID NO:77),或以递增优选顺序与基序VII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;并且
基序VIII是:QSWCTLNA(SEQ ID NO:78)或以递增优选顺序与基序VIII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序。
第26项.根据前面第23到25任一项的方法,其中III类U盒多肽在其C-末端区中包含一个或多个下面的基序:
基序IX:GR S/A/L A/E L/F L/V S/K/N/A H
基序X:G/A I/L/V A A/I/V V S/C/A/G/T K/C/R R/K/A A/I/V/T M/L/TR V/L/I S/P
基序XI:K/R L C/L F/V/M/L V/L/A V/L/A/I Q/R V/A/S D/S/G/E
基序XII:S/A/L YP。
第27项.根据第26项的方法,其中基序IX是:GRAELLNH(SEQ IDNO:79)或以递增优选顺序与基序IX有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序X是:GLAIVSKKILRVS(SEQ ID NO:80)或以递增优选顺序与基序X有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序XI是:KLCLVLQVD(SEQ ID NO:81)或以递增优选顺序与基序XI有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序XII是:SYP(SEQ ID NO:82)或以递增优选顺序与基序XII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序。
第28项.根据前面第23到27项任一项的方法,其中通过在植物中导入并表达编码III类U盒多肽的核酸实现所述受调节的表达。
第29项.根据前面第23到28项任一项的方法,其中所述编码III类U盒多肽的核酸编码表A2中所列的任一种蛋白质或者是这种核酸的一部分,或者能够与这种核酸杂交的核酸。
第30项.根据前面第23到29项任一项的方法,其中所述核酸序列编码表A2中给出的任一种蛋白质的直向同源物或旁系同源物。
第31项.根据前面第23到30项任一项的方法,其中所述增强的产量相关性状包括相对于对照植物增加的产量,优选增加的生物量和/或增加的种子产量。
第32项.根据前面第23到31项任一项的方法,其中在非胁迫条件下和轻微干旱条件下得到所述增强的产量相关性状。
第33项.根据前面第23到32项任一项的方法,其中在氮缺乏条件下得到所述增强的产量相关性状。
第34项.根据前面第23到33项任一项的方法,其中所述核酸有效连接到年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
第35项.根据前面第23到34项任一项的方法,其中所述编码III类多肽的核酸是植物来源,优选来自双子叶植物,进一步优选来自茄科,更优选来自辣椒属,最优选来自辣椒(Capsicum annuus)。
第36项.通过根据前面第23到35项任一项的方法得到的植物或其部分,包括种子,其中所述植物或其部分包含编码III类U盒多肽的重组核酸。
第37项.构建体,其包含:
(i)编码如第23到27任一项定义的III类U盒多肽的核酸;
(ii)能够驱动(a)的核酸序列表达的一个或多个控制序列;和任选地
(iii)转录终止序列。
第38项.根据第37项的构建体,其中所述控制序列之一是年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
第39项.根据第37或38项的构建体在用于制备植物的方法中的用途,所述植物相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量。
第40项.用根据第37或38项的构建体转化的植物、植物部分或植物细胞。
第41项.产生相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量的转基因植物的方法,包括:
(a)在植物中导入并表达编码如第23到27项任一项定义的III类U盒多肽的核酸;和
(b)在促进植物生长和发育的条件下培养所述植物细胞。
第42项.转基因植物,其相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量,所述转基因植物从编码如第23到27项任一项定义的III类U盒多肽的核酸的增加的表达得到,或者来自所述转基因植物的转基因植物细胞。
第43项.根据第36、40或42项的转基因植物,或来自其的转基因植物细胞,其中所述植物是作物植物或单子叶植物或谷类,如稻、玉米、小麦、大麦、粟、黑麦、黑小麦、高粱和燕麦。
第44项.根据第43项的植物的可收获部分,其中所述可收获部分优选为苗生物量和/或种子。
第45项.来自根据第43项的植物和/或来自根据第44项的植物的可收获部分的产品。
第46项.编码III类U盒多肽的核酸用于相对于对照植物在植物中增加产量,尤其增加种子产量和/或苗生物量的用途。
在一个实施方案中,本发明涉及如下概述的主题:
第47项.相对于对照植物在植物中增强产量相关性状的方法,包括调节植物中编码PQQC蛋白的核酸的表达。
第48项.根据第47项的方法,其中所述PQQC蛋白以递增优选顺序与SEQ ID NO:105表示的序列有至少45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性。
第49项.根据第47到48任一项的方法,其中通过在植物中导入并表达编码PQQC多肽的核酸实现所述受调节的表达。
第50项.根据前面第47到49项任一项的方法,其中所述编码PQQC多肽的核酸编码表A3中所列的任一种蛋白质或者是这种核酸的一部分,或者能够与这种核酸杂交的核酸。
第51项.根据前面第47到50项任一项的方法,其中所述增强的产量相关性状包括相对于对照植物增加的产量,优选增加的生物量和/或增加的种子产量。
第52项.根据前面第47到51项任一项的方法,其中在非胁迫条件下和轻微干旱条件下得到所述增强的产量相关性状。
第53项.根据前面第47到52项任一项的方法,其中在氮缺乏条件下得到所述增强的产量相关性状。
第54项.根据前面第47到53项任一项的方法,其中所述核酸有效连接到年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
第55项.根据前面第47到54项任一项的方法,其中所述核酸有效连接到组成型启动子,优选GOS2启动子,最优选来自稻的GOS2启动子。
第56项.根据前面第47到55项任一项的方法,其中所述编码PQQC多肽的核酸为细菌来源,优选来自假单胞菌。
第57项.通过根据前面任一项的方法得到的植物或其部分,包括种子,其中所述植物或其部分包含编码PQQC多肽的重组核酸。
第58项.构建体,其包含:
–编码如第47到48定义的PQQC多肽的核酸;
–能够驱动(a)的核酸序列表达的一个或多个控制序列;和任选地
–转录终止序列。
第59项.根据第58项的构建体,其中所述控制序列之一是年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
第60项.根据第58项的构建体,其中所述控制序列之一是组成型启动子,优选GOS2启动子,最优选来自稻的GOS2启动子。
第61项.根据第58到60任一项的构建体在用于制备植物的方法中的用途,所述植物相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量。
第62项.用根据第58到61任一项的构建体转化的植物、植物部分或植物细胞。
第63项.产生相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量的转基因植物的方法,包括:
(i)在植物中导入并表达编码如第47到48项定义的PQQC多肽的核酸;和
(ii)在促进植物生长和发育的条件下培养所述植物细胞。
第64项.转基因植物,其相对于对照植物具有增加的生物量和/或增加的种子产量,所述转基因植物从编码如第47到48项定义的PQQC多肽的核酸的增加的表达得到,或者来自所述转基因植物的转基因植物细胞。
第65项.根据第57、62或64项的转基因植物,或来自其的转基因植物细胞,其中所述植物是作物植物或单子叶植物或谷类,如稻、玉米、小麦、大麦、粟、黑麦、黑小麦、高粱和燕麦。
第66项.根据第65项的植物的可收获部分,其中所述可收获部分优选为苗生物量和/或种子。
第67项.来自根据第65项的植物和/或来自根据第66项的植物的可收获部分的产品。
第68项.编码PQQC多肽的核酸用于相对于对照植物在植物中增加产量,尤其增加种子产量和/或苗生物量的用途。
附图描述
本发明现参考以下附图描述,其中:
图1显示了集胞藻PEPC蛋白质,基序I到V突出显示。
图2显示了一组PEPC蛋白质的多序列比对。
图3表示表达载体,其用于处于原叶绿素酸酯还原酶启动子控制下的PEPC编码核酸在稻中增加的表达。
图4详述了可用于实施本发明方法的序列的实例。
图5显示了来自辣椒属、拟南芥和稻的III类U盒蛋白的氨基酸序列,其中已经指出了U盒结构域以及保守基序I到VI。U盒结构域以粗体显示;基序I为粗斜体、基序II加框,基序III加下划线,基序IV为斜体并加下划线,基序V为双下划线,基序VI为间断下划线。
图6显示了一组III类U盒蛋白的多序列表达。
图7表示双元载体,其用于处于原叶绿素酸酯还原酶启动子控制下的III类U盒编码核酸在稻中的增加的表达。
图8详述了可用于实施本发明方法的序列的实例。
图9显示了一组PQQC蛋白的多序列比对。
图10表示双元载体,其用于处于原叶绿素酸酯还原酶启动子控制下的PQQC编码核酸在稻中的增加的表达。
图11详述了可用于实施本发明方法的序列的实例。
实施例
本发明现将参考仅作为说明的下列实施例加以描述。下列实施例不意图彻底定义或限制本发明范围。
DNA操作:除非另外指出,根据(Sambrook(2001)Molecular Cloning:a laboratory manual,3rd Edition Cold Spring Harbor Laboratory Press,CSH,New York)或Ausubel et al.(1994),Current Protocols in MolecularBiology,Current Protocols的卷1和卷2中所述的标准方案进行重组DNA技术。用于植物分子工作的标准材料和方法在BIOS Scientific PublicationsLtd(UK)和Blackwell Scientific Publications(UK)出版的R.D.D.Croy的Plant Molecular Biology Labfax(1993)中描述。
实施例1:鉴定与用于本发明方法的核酸序列相关的序列
使用数据库序列搜索工具,如基本局部比对工具(BLAST)(Altschul等(1990)J.Mol.Biol.215:403-410;和Altschul等(1997)Nucleic Acids Res.25:3389-3402)在国家生物技术信息中心(NCBI)的Entrez核苷酸数据库中所维护的那些序列内鉴定到与用于本发明方法中的核酸序列相关的序列(全长cDNA、EST或基因组的)。该程序用来通过核酸序列或多肽序列与序列数据库比较并通过计算匹配的统计学显著性而找到序列间具有局部相似性的区域。例如,用于本发明方法中的核酸编码的多肽用于TBLASTN算法,采用默认设置和过滤以忽略低复杂性序列抵消。分析的结果通过配对性比较显示,并根据几率评分(E-值)排序,其中该评分反映特定比对结果因偶然而发生的概率(E-值越低,命中的显著性越高)。除了E-值外,比较还通过同一性百分数进行记分。同一性百分数指两个所比较核酸(或多肽)序列之间在特定长度范围内的相同核苷酸(或氨基酸)数目。在一些情况下,调整默认参数以调节搜索法的严格性。例如,E值可以增加以显示较低的严紧性匹配。这样,可以鉴定短的几乎精确的匹配。
表A提供了与用于本发明方法的核酸序列相关的核酸序列列表。
表A1:PEPC多肽的实例:
| 来源 | 核酸SEQ ID NO: | 蛋白质SEQ ID NO: |
| 见图4 | 见图4 | 见图4 |
表A2:III类U盒多肽的实例:

表A3:PQQC多肽的实例:
| 来源 | 核酸SEQ ID NO: | 蛋白质SEQ ID NO: |
| 见图11 | 见图11 | 见图11 |
在一些情况下,相关序列已经被试验性装配并被研究机构如基因组研究研究所(Institute for Genomic Research(TIGR))公开。Eukaryotic GeneOrthologs(EGO)数据库可用于鉴定此类相关序列,通过关键词检索或者通过使用BLAST算法用目的核酸或多肽序列进行鉴定。
下面给出了BLASTP输出,显示了与SEQ ID NO:2相关的PEPC序列并且因此可用于本发明方法。
1:NP_441713
磷酸烯醇丙酮酸羧化酶[集胞藻属PCC6803]
gi|16330985|ref|NP_441713.1|[16330985]
2:ZP_01729388
磷酸烯醇丙酮酸羧化酶[Cyanothece sp.CCY0110]
gi|126658238|ref|ZP_01729388.1|[126658238]
3:ZP_00517310
磷酸烯醇丙酮酸羧化酶[Crocosphaera watsonii WH8501]
gi|67923850|ref|ZP_00517310.1|[67923850]
4:ZP_00108354
COG2352:磷酸烯醇丙酮酸羧化酶[点形念珠藻PCC73102]
gi|23126460|ref|ZP_00108354.1|[23126460]
5:ZP_01631340
磷酸烯醇丙酮酸羧化酶[泡沫节球藻CCY9414]
gi|119512252|ref|ZP_01631340.1|[119512252]
6:YP_322649
磷酸烯醇丙酮酸羧化酶[多变鱼腥藻ATCC29413]
gi|75908353|ref|YP_322649.1|[75908353]
7:AAV90837
磷酸烯醇丙酮酸羧化酶[聚球藻属PCC7002]
gi|56417897|gb|AAV90837.1|[56417897]
8:ZP_01620914
磷酸烯醇丙酮酸羧化酶[鞘丝藻PCC8106]
gi|119487042|ref|ZP_01620914.1|[119487042]
9:P28594
磷酸烯醇丙酮酸羧化酶(PEPCase)(PEPC)
gi|25014052|sp|P28594|CAPP_ANASP[25014052]
10:YP_172556
磷酸烯醇丙酮酸羧化酶[细长聚球藻PCC6301]
gi|56751855|ref|YP_172556.1|[56751855]
11:YP_401269
磷酸烯醇丙酮酸羧化酶[细长聚球藻PCC7942]
gi|81301061|ref|YP_401269.1|[81301061]
12:YP_720722
磷酸烯醇丙酮酸羧化酶[红海束毛藻IMS101]
gi|113474661|ref|YP_720722.1|[113474661]
13:NP_682702
磷酸烯醇丙酮酸羧化酶[Thermosynechococcus elongatus BP-1]
gi|22299455|ref|NP_682702.1|[22299455]
14:NP_488901
磷酸烯醇丙酮酸羧化酶[念珠藻PCC7120]
gi|17232353|ref|NP_488901.1|[17232353]
15:YP_478060
磷酸烯醇丙酮酸羧化酶[聚球藻属JA-2-3B′a(2-13)]
gi|86609298|ref|YP_478060.1|[86609298]
16:ZP_01083776
磷酸烯醇丙酮酸羧化酶[聚球藻属WH5701]
gi|87300934|ref|ZP_01083776.1|[87300934]
17:ZP_01080788
磷酸烯醇丙酮酸羧化酶[聚球藻属RS9917]
gi|87124941|ref|ZP_01080788.1|[87124941]
18:YP_474305
磷酸烯醇丙酮酸羧化酶[聚球藻属JA-3-3Ab]
gi|86605542|ref|YP_474305.1|[86605542]
19:ZP_01469854
磷酸烯醇丙酮酸羧化酶[聚球藻属BL107]
gi|116072588|ref|ZP_01469854.1|[116072588]
20:NP_876121
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus subsp.marinus str.CCMP1375]
gi|33241179|ref|NP_876121.1|[33241179]
21:ZP_01473048
磷酸烯醇丙酮酸羧化酶[聚球藻属RS9916]
gi|116075789|ref|ZP_01473048.1|[116075789]
22:ZP_01006468
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.MIT9211]
gi|84519119|ref|ZP_01006468.1|[84519119]
23:ABE11140
磷酸烯醇丙酮酸羧化酶[未培养的Prochlorococcus marinus克隆HF10-11H11]
gi|91070220|gb|ABE11140.1|[91070220]
24:YP_001091990
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.MIT9301]
gi|126697104|ref|YP_001091990.1|[126697104]
25:YP_377934
磷酸烯醇丙酮酸羧化酶[聚球藻属CC9902]
gi|78185499|ref|YP_377934.1|[78185499]
26:YP_398164
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.MIT9312]
gi|78780052|ref|YP_398164.1|[78780052]
27:YP_292340
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.NATL2A]
gi|72382985|ref|YP_292340.1|[72382985]
28:YP_001010172
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.AS9601]
gi|123969314|ref|YP_001010172.1|[123969314]
29:NP_895540
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.MIT9313]
gi|33863980|ref|NP_895540.1|[33863980]
30:YP_001015841
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.NATL1A]
gi|124026726|ref|YP_001015841.1|[124026726]
31:YP_001018274
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.MIT9303]
gi|124023967|ref|YP_001018274.1|[124023967]
32:ZP_01123374
磷酸烯醇丙酮酸羧化酶[聚球藻属WH7805]
gi|88807863|ref|ZP_01123374.1|[88807863]
33:YP_001224177
磷酸烯醇丙酮酸羧化酶[聚球藻属WH7803]
gi|148238790|ref|YP_001224177.1|[148238790]
34:NP_898138
磷酸烯醇丙酮酸羧化酶[聚球藻属WH8102]
gi|33866579|ref|NP_898138.1|[33866579]
35:YP_380727
磷酸烯醇丙酮酸羧化酶[聚球藻属CC9605]
gi|78211948|ref|YP_380727.1|[78211948]
36:NP_893692
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus subsp.pastoris str.CCMP1986]
gi|33862131|ref|NP_893692.1|[33862131]
37:YP_001012076
磷酸烯醇丙酮酸羧化酶[Prochlorococcus marinus str.MIT9515]
gi|123966995|ref|YP_001012076.1|[123966995]
38:YP_001228346
磷酸烯醇丙酮酸羧化酶[聚球藻属RCC307]
gi|148243189|ref|YP_001228346.1|[148243189]
39:YP_729688
磷酸烯醇丙酮酸羧化酶[聚球藻属CC9311]
gi|113954224|ref|YP_729688.1|[113954224]
40:NP_923360
磷酸烯醇丙酮酸羧化酶[紫色粘杆菌PCC7421]
gi|37519983|ref|NP_923360.1|[37519983]
41:ZP_01424999
磷酸烯醇丙酮酸羧化酶[橙色滑柱菌ATCC23779]
gi|113939140|ref|ZP_01424999.1|[113939140]
42:YP_830113
磷酸烯醇丙酮酸羧化酶[节杆菌FB24]
gi|116669180|ref|YP_830113.1|[116669180]
43:YP_946565
磷酸烯醇丙酮酸羧化酶[金黄节杆菌TC1]
gi|119962288|ref|YP_946565.1|[119962288]
44:ZP_01696337
磷酸烯醇丙酮酸羧化酶[凝固芽孢杆菌36D1]
gi|124521401|ref|ZP_01696337.1|[124521401]
45:YP_342813
磷酸烯醇丙酮酸羧化酶[Nitrosococcus oceani ATCC19707]
gi|77164288|ref|YP_342813.1|[77164288]
46:NP_627344
磷酸烯醇丙酮酸羧化酶[天蓝色链霉菌A3(2)]
gi|21221565|ref|NP_627344.1|[21221565]
47:AAY89988
预测的PEP羧化酶[未培养的细菌BAC13K9BAC]
gi|68304977|gb|AAY89988.1|[68304977]
48:NP_824743
磷酸烯醇丙酮酸羧化酶[除虫链霉菌MA-4680]
gi|29830109|ref|NP_824743.1|[29830109]
49:ZP_01649489
磷酸烯醇丙酮酸羧化酶[Salinispora arenicola CNS205]
gi|119883229|ref|ZP_01649489.1|[119883229]
50:ZP_00568542
磷酸烯醇丙酮酸羧化酶[弗兰克氏菌EAN1pec]
gi|68229346|ref|ZP_00568542.1|[68229346]
51:YP_001363100
磷酸烯醇丙酮酸羧化酶[Kineococcus radiotolerans SRS30216]
gi|152967316|ref|YP_001363100.1|[152967316]
52:YP_001157738
磷酸烯醇丙酮酸羧化酶[Salinispora tropica CNB-440]
gi|145593441|ref|YP_001157738.1|[145593441]
53:YP_318884
磷酸烯醇丙酮酸羧化酶[维氏硝化杆菌Nb-255]
gi|75676463|ref|YP_318884.1|[75676463]
54:YP_294925
磷酸烯醇丙酮酸羧化酶[Ralstonia eutropha JMP134]
gi|73540405|ref|YP_294925.1|[73540405]
55:YP_577930
磷酸烯醇丙酮酸羧化酶[汉堡硝化杆菌X14]
gi|92118201|ref|YP_577930.1|[92118201]
56:ZP_02010108
磷酸烯醇丙酮酸羧化酶[皮氏罗尔斯顿氏菌12D]
gi|153888973|ref|ZP_02010108.1|[153888973]
57:NP_840673
磷酸烯醇丙酮酸羧化酶[欧洲动性球菌ATCC19718]
gi|30248603|ref|NP_840673.1|[30248603]
58:NP_947117
磷酸烯醇丙酮酸羧化酶[金红红假单胞菌CGA009]
gi|39934841|ref|NP_947117.1|[39934841]
59:YP_391787
磷酸烯醇丙酮酸羧化酶[泉生硫微螺菌XCL-2]
gi|78485862|ref|YP_391787.1|[78485862]
60:ZP_01661147
磷酸烯醇丙酮酸羧化酶[皮氏罗尔斯顿氏菌12J]
gi|121528533|ref|ZP_01661147.1|[121528533]
61:YP_001204637
磷酸烯醇丙酮酸羧化酶[短根瘤菌ORS278]
gi|146339589|ref|YP_001204637.1|[146339589]
62:YP_932496
probable磷酸烯醇丙酮酸羧化酶[固氮弧菌BH72]
gi|119897283|ref|YP_932496.1|[119897283]
63:YP_970358
磷酸烯醇丙酮酸羧化酶[类产碱西瓜亚种AAC00-1]
gi|120610680|ref|YP_970358.1|[120610680]
64:ZP_01047635
磷酸烯醇丙酮酸羧化酶[硝化杆菌Nb-311A]
gi|85716666|ref|ZP_01047635.1|[85716666]
65:YP_369991
磷酸烯醇丙酮酸羧化酶[伯克霍尔德氏菌383]
gi|78067222|ref|YP_369991.1|[78067222]
66:YP_314388
磷酸烯醇丙酮酸羧化酶[脱氮硫杆菌ATCC25259]
gi|74316648|ref|YP_314388.1|[74316648]
67:NP_520479
磷酸烯醇丙酮酸羧化酶[Ralstonia solanacearum GMI1000]
gi|17547077|ref|NP_520479.1|[17547077]
68:YP_727365
磷酸烯醇丙酮酸羧化酶[Ralstonia eutropha H16]
gi|113868876|ref|YP_727365.1|[113868876]
69:YP_001238961
磷酸烯醇丙酮酸羧化酶[慢生根瘤菌BTAi1]
gi|148254376|ref|YP_001238961.1|[148254376]
70:YP_001415233
磷酸烯醇丙酮酸羧化酶[自养黄色杆菌Py2]
gi|154244275|ref|YP_001415233.1|[154244275]
71:YP_487198
磷酸烯醇丙酮酸羧化酶[金红红假单胞菌HaA2]
gi|86750702|ref|YP_487198.1|[86750702]
72:NP_769595
磷酸烯醇丙酮酸羧化酶[大豆根瘤杆菌USDA110]
gi|27378066|ref|NP_769595.1|[27378066]
73:YP_569012
磷酸烯醇丙酮酸羧化酶[金红红假单胞菌BisB5]
gi|91976353|ref|YP_569012.1|[91976353]
74:ZP_00946208
磷酸烯醇丙酮酸羧化酶[Ralstonia solanacearum UW551]
gi|83749206|ref|ZP_00946208.1|[83749206]
75:YP_780719
磷酸烯醇丙酮酸羧化酶[金红红假单胞菌BisA53]
gi|115523808|ref|YP_780719.1|[115523808]
76:BAA21835
磷酸烯醇丙酮酸羧化酶[金红红假单胞菌]
gi|2339975|dbj|BAA21835.1|[2339975]
77:Q59757
磷酸烯醇丙酮酸羧化酶(PEPCase)(PEPC)
gi|2499464|sp|Q59757|CAPP_RHOMR[2499464]
78:YP_747257
磷酸烯醇丙酮酸羧化酶[Nitrosomonas eutropha C91]
gi|114331035|ref|YP_747257.1|[114331035]
79:ZP_01554849
磷酸烯醇丙酮酸羧化酶[Burkholderia ambifaria MC40-6]
gi|118696768|ref|ZP_01554849.1|[118696768]
80:YP_774359
磷酸烯醇丙酮酸羧化酶[洋葱伯克霍尔德氏菌AMMD]
gi|115352520|ref|YP_774359.1|[115352520]
81:EAY68193
磷酸烯醇丙酮酸羧化酶[Burkholderia dolosa AUO158]
gi|124894313|gb|EAY68193.1|[124894313]
82:YP_621690
磷酸烯醇丙酮酸羧化酶[Burkholderia cenocepacia AU1054]
gi|107023363|ref|YP_621690.1|[107023363]
83:YP_836069
磷酸烯醇丙酮酸羧化酶[Burkholderia cenocepacia HI2424]
gi|116690446|ref|YP_836069.1|[116690446]
84:ZP_00987174
COG2352:磷酸烯醇丙酮酸羧化酶[Burkholderia dolosa AUO158]
gi|84362555|ref|ZP_00987174.1|[84362555]
85:ZP_01559873
磷酸烯醇丙酮酸羧化酶[Burkholderia cenocepacia MC0-3]
gi|118707287|ref|ZP_01559873.1|[118707287]
86:YP_584892
磷酸烯醇丙酮酸羧化酶[Ralstonia metallidurans CH34]
gi|94311682|ref|YP_584892.1|[94311682]
87:YP_987106
磷酸烯醇丙酮酸羧化酶[食酸菌JS42]
gi|121595210|ref|YP_987106.1|[121595210]
88:ZP_00980295
COG2352:磷酸烯醇丙酮酸羧化酶[Burkholderia cenocepacia PC184]
gi|84355411|ref|ZP_00980295.1|[84355411]
89:EAY64002
磷酸烯醇丙酮酸羧化酶[Burkholderia cenocepacia PC184]
gi|124872286|gb|EAY64002.1|[124872286]
90:YP_531562
磷酸烯醇丙酮酸羧化酶[金红红假单胞菌BisB18]
gi|90423192|ref|YP_531562.1|[90423192]
91:YP_710875
磷酸烯醇丙酮酸羧化酶(PEPCase)(PEPC)[桤木弗兰克氏菌ACN14a]
gi|111220081|ref|YP_710875.1|[111220081]
92:ZP_01325009
hypothetical protein BpseP_03001239[类鼻疽伯克霍尔德氏菌Pasteur]
gi|100063478|ref|ZP_01325009.1|[100063478]
93:YP_995835
磷酸烯醇丙酮酸羧化酶[Verminephrobacter eiseniae EF01-2]
gi|121608028|ref|YP_995835.1|[121608028]
94:CAM76812
磷酸烯醇丙酮酸羧化酶[格利菲斯瓦尔德磁螺菌MSR-1]
gi|144899948|emb|CAM76812.1|[144899948]
95:ZP_01317342
hypothetical protein Bpse1_03003262[类鼻疽伯克霍尔德氏菌1655]
gi|99910315|ref|ZP_01317342.1|[99910315]
96:ZP_01213561
hypothetical protein Bpse17_02001265[类鼻疽伯克霍尔德氏菌1710a]
gi|90293964|ref|ZP_01213561.1|[90293964]
97:YP_001065355
磷酸烯醇丙酮酸羧化酶[类鼻疽伯克霍尔德氏菌1106a]
gi|126451534|ref|YP_001065355.1|[126451534]
98:YP_107641
磷酸烯醇丙酮酸羧化酶[类鼻疽伯克霍尔德氏菌K96243]
gi|53718655|ref|YP_107641.1|[53718655]
99:YP_707103
磷酸烯醇丙酮酸羧化酶[红球菌RHA1]
gi|111024131|ref|YP_707103.1|[111024131]
100:YP_001058119
磷酸烯醇丙酮酸羧化酶[类鼻疽伯克霍尔德氏菌668]
gi|126440608|ref|YP_001058119.1|[126440608]
实施例2:PEPC多肽序列的比对
使用来自Vector NTI(Invitrogen)的AlignX程序(其基于普遍使用的渐进比对的Clustal W算法(Thompson等,(1997)Nucleic Acids Res25:4876-4882;Chenna等,(2003)Nucleic Acids Res31:3497-3500))进行多肽序列的比对。空位开放罚分的缺省值为10,空位延伸罚分的缺省值为0.1,选择的权重矩阵是Blosum62(如果比对多肽))。进行小的手工编辑以进一步优化比对。比对见图2。
实施例3:用于实施本发明方法的多肽序列之间全局同一性百分比(global percentage identity)的计算
用于实施本发明方法的全长多肽序列之间的全局相似性和同一性百分比利用本领域可用的方法之一MatGAT(矩阵全局比对工具)软件((Campanella et al.,BMC Bioinformatics.20034:29.MatGAT:anapplication that generates similarity/identity matrices using protein orDNA sequences..Campanella JJ,Bitincka L,Smalley J;software hostedby Ledion Bitincka)来确定。MatGAT软件无需对数据进行预比对,即可产生DNA或蛋白质序列的相似性/同一性矩阵。该程序利用Myers和Miller全局比对算法(空位开放罚分为12,而空位延伸罚分为2)进行一系列的两两比对,利用例如Blosum62(对于多肽而言)计算相似性和同一性,然后将结果排列成距离矩阵。序列相似性在分界线的下半部分显示并且序列同一性在对角分界线的上半部分中显示。
比较所用的参数有:
评分矩阵:Blosum62
第一个空位:12
延伸空位:2
实施例4:鉴定在用于实施本发明方法的多肽序列内包含的结构域
蛋白质家族、结构域和位点集成资源(Integrated Resouce of ProteinFamilies,Domain and Site,InterPro)数据库是基于文本和序列的搜索的通常所用标签数据库的集成界面。InterPro数据库组合了这些数据库,所述的数据库使用不同方法学和不同程度的有关已充分表征蛋白质的生物学信息以得到蛋白质标签。合作数据库包括SWISS-PROT、PROSITE、TrEMBL、PRINTS、ProDom和Pfam、Smart和TIGRFAMs。Pfam是一组多序列比对和覆盖许多常见蛋白质结构域和家族的隐蔽马尔可夫模型。Pfam驻留于英国Sanger Institute的服务器。Interpro驻留于英国的欧洲生物信息研究所。
PEPC多肽的结果给出了INTERPRO结构域:IPR001449(磷酸烯醇丙酮酸羧化酶)和PFAM:00311。
InterPro扫描如SEQ ID NO:75所代表的多肽序列的结果下面给出。
SMART:00504
InterPro:IPR003613和IPR008938。
实施例5:用于实施本发明方法的多肽序列的拓扑学预测
TargetP1.1预测真核蛋白质的亚细胞定位。位置分配是基于任何氨基端前序列的预测的存在:叶绿体转运肽(cTP)、线粒体靶向肽(mTP)或分泌途径信号肽(SP)。作为最终预测基础的记分并不实际是概率,并且它们未必合为一体。然而,具有最高记分的位置根据TargetP是最有可能的,并且记分之间的关系(可靠性类别)可以是预测的肯定性的一个指标。可靠性类别(RC)是1-5,其中1表示最强预测。TargetP在丹麦技术大学的服务器上维护。
对于预测含有氨基端前序列的序列,也可以预测潜在的切割位点。
选择了许多参数:选择“植物”生物组,没有定义截断,并且要求了转运肽的预测长度。
众多其它算法可以用来执行此类分析,包括:
·在丹麦技术大学(Technical University of Denmar)服务器上驻留的ChloroP1.1;
·在澳大利亚布里斯班昆士兰大学分子生物科学研究所(Institute forMolecular Bioscience,University of Queensland,Brisbane,Australia)的服务器上驻留的蛋白质Prowler亚细胞定位预测程序(Protein ProwlerSubcellular Localisation Predictor)第1.2版;
·在加拿大阿尔伯塔省埃德蒙顿市阿尔伯塔大学(University of Alberta,Edmonton,Alberta,Canada)的服务器上驻留的PENCE Proteme AnalystPA-GOSUB2.5;
·在丹麦技术大学服务器上驻留的TMHMM。
实施例6:本发明多肽的功能测定法
PEPC催化CO2加入磷酸烯醇丙酮酸(PEP)以形成四碳化合物草酰乙酸:
PEP+CO2→草酰乙酸+Pi
关于测量PEPC活性的描述,见Chen et al.,2002(Plant Cell Physiol43(2):159-169);和Chen et al.,2004(Planta219pp440-449)。
III类U盒多肽显示出E3泛素连接酶活性。
PQQC催化PQQ生物合成中的最后一步。
实施例7:克隆用于本发明方法的核酸序列
通过PCR扩增用于本发明方法的编码PEPC多肽的核酸序列。使用Hifi Taq DNA聚合酶在标准条件下,用200ng模板在50μl PCR混合物中进行PCR。使用的引物为prm08777(SEQ ID NO:72;正义,起始密码子为粗体):
5-ggggacaagtttgtacaaaaaagcaggcttaaacaatgaacttggcagttcct-3’
和prm08782(SEQ ID NO:73;反向,互补的):
5’-ggggaccactttgtacaagaaagctgggttcaaccagtattacgcatt-3’,
其包括用于Gateway重组的AttB位点。也使用标准方法纯化所扩增的PCR片段。然后进行Gateway程序的第一步:BP反应,在该反应期间,PCR片段与pDONR201质粒在体内重组产生“进入克隆”(根据Gateway命名法)pPEPC。质粒pDONR201购自Invitrogen作为 技术的一部分。
技术的一部分。
包含SEQ ID NO:1的进入克隆然后用于LR反应中,使用用于稻转化的目的载体。该载体在T-DNA边界内含有作为功能元件的:植物选择标记;筛选标记表达盒;和预期用于与已经克隆在进入克隆中的目的核酸序列进行LR体内重组的Gateway盒。用于年轻绿色组织特异性表达的原叶绿素酸酯还原酶启动子(SEQ ID NO:71)位于该Gateway盒的上游。见图3。
在LR重组步骤后,根据本领域公知的方法将所得的表达载体pPCPR::PEPC(图3)转化到农杆菌菌株LBA4044中。
通过PCR用Hifi Taq DNA聚合酶在标准条件下,在50μl PCR混合物中使用200ng模板扩增编码III类U盒蛋白并且用于本发明方法中的核酸序列。使用的引物为prm9075(SEQ ID NO:102;正向,起始密码子为粗体):
5’-ggggacaagtttgtacaaaaaagcaggcttaaacaatggaagaaattcaagtacc-3’
和prm8785(SEQ ID NO:103;反向,互补的):
5’-ggggaccactttgtacaagaaagctgggtgctcttttggtcctattttcct-3’,
其包括用于Gateway重组的AttB位点。也使用标准方法纯化所扩增的PCR片段。然后进行Gateway方法的第一步:BP反应,在该反应中,PCR片段与pDONR201质粒在体内重组以产生(根据Gateway术语)“进入克隆”pU盒。质粒pDONR201购自Invitrogen,作为 技术的部分。
技术的部分。
包含SEQ ID NO:74的进入克隆然后用于LR反应中,使用用于稻转化的目的载体。该载体在T-DNA边界内含有作为功能元件的:植物选择标记;筛选标记表达盒;和预期用于与已经克隆在进入克隆中的目的核酸序列进行LR体内重组的Gateway盒。用于年轻绿色组织特异性表达的原叶绿素酸酯还原酶启动子(SEQ ID NO:101)位于该Gateway盒上游。
在LR重组步骤后,根据本领域公知的方法将所得的表达载体pPCPR::U-盒(图7)转化到农杆菌菌株LBA4044中。
通过PCR用Hifi Taq DNA聚合酶在标准条件下,在50μl PCR混合物中使用200ng模板扩增编码PQQC蛋白并且用于本发明方法中的核酸序列。使用的引物为prm08777(SEQ ID NO:228;正向,起始密码子为粗体):
5-ggggacaagtttgtacaaaaaagcaggcttaaacaatgactgacaccccaatgtc-3’
和prm08782(SEQ ID NO:229;反向,互补的):
5’-ggggaccactttgtacaagaaagctgggttcataaggtgattcctttatgc-3’,
其包括用于Gateway重组的AttB位点。也使用标准方法纯化所扩增的PCR片段。然后进行Gateway方法的第一步:BP反应,在该反应中,PCR片段与pDONR201质粒在体内重组以产生(根据Gateway术语)“进入克隆”pPQQC。质粒pDONR201购自Invitrogen,作为 技术的部分。
技术的部分。
包含SEQ ID NO:104的进入克隆然后用于LR反应中,使用用于稻转化的目的载体。该载体在T-DNA边界内含有作为功能元件的:植物选择标记;筛选标记表达盒;和预期用于与已经克隆在进入克隆中的目的核酸序列进行LR体内重组的Gateway盒。用于年轻绿色组织特异性表达的原叶绿素酸酯还原酶启动子(SEQ ID NO:226)位于该Gateway盒或GOS2启动子(SEQ ID NO227)的上游-见图9。
在LR重组步骤后,根据本领域公知的方法将所得的表达载体pGOS2∷PQQC(图9B)转化到农杆菌菌株LBA4044中。
在LR重组步骤后,根据本领域公知的方法将所得的表达载体pPCPR∷PQQC(图9A)转化到农杆菌菌株LBA4044中。
实施例8:植物转化
稻转化
含有表达载体的农杆菌用来转化稻植物。将稻的日本栽培品种Nipponbare的成熟干燥种子脱壳。通过在70%乙醇中温育一分钟,随后在2%HgCl2中30分钟,随后用无菌蒸馏水洗涤6次15分钟而实施消毒。消毒的种子随后在含有2,4-D的培养基(愈伤组织诱导培养基)上萌发。在黑暗中温育4周后,将盾片衍生的胚发生性愈伤组织切下并在同一种培养基上增殖。2周后,愈伤组织通过在同一种培养基上传代培养另外2周而繁殖或增殖。胚发生性愈伤组织片在新鲜培养基上传代培养3日,之后共培育(以增强细胞分裂活性)。
含有表达载体的农杆菌菌株LBA4404用于共培育。农杆菌接种在含有适宜抗生素的AB培养基上并在28℃培养3日。随后将细菌收集并重悬在液体共培育培养基中至密度(OD600)约1。将混悬液随后转移至培养皿并将愈伤组织在该混悬液内浸泡15分钟。愈伤组织组织随后在滤纸上吸干并转移至固化的共培育培养基上并且在黑暗中于25℃温育3日。共培育的愈伤组织在黑暗中于28℃在选择剂存在下于含有2,4-D的培养基上培育4周。在此时段期间,形成迅速生长的抗性愈伤组织岛。在这种材料转移至再生培养基并在光线下温育后,胚发生潜力释放并且苗在随后4-5周发育。将苗从愈伤组织中切下并且在含有植物生长素的培养基上温育2-3周,其中将苗从所述的培养基上转移至土壤。硬化的苗在高湿度和短日照下于温室中培育。
一个构建体产生大约35个独立T0稻转化体。将原代转化体从组织培养箱转移至温室。在定量PCR分析以验证T-DNA插入物的拷贝数后,仅保留表现选择剂耐受性的单拷贝转基因植物用于收获T1种子。种子随后在移植后3-5月收获。本方法以超过50%的比率产生单一基因座转化体(Aldemita和Hodges1996,Chan等1993,Hiei等1994)。
玉米转化
玉米的转化根据对Ishida等(1996.Nature Biotech14745-50)描述方法的改良方法进行。在玉米中的转化是基因型依赖的并且仅特定的基因型可操作用于转化和再生。近交系A188(明尼苏达大学)或以A188作为亲本的杂种是用于转化的供体材料的良好来源,但是其他基因型也可以成功地使用。玉米穗从玉米植物中在授粉后大约11日(DAP)收获,此时不成熟胚的长度是大约1至1.2mm。不成熟胚与含有表达载体的根癌农杆菌共培育并且转基因植物通过器官发生而恢复。切下的胚在愈伤组织诱导培养基上、随后在玉米再生培养基上培育,其中所述的再生培养基含有选择剂(例如咪唑啉酮,但可以使用多种选择标记)。培养板在25℃于光照下培养2-3周,或直至苗发育。将绿色苗从每个胚转移至玉米生根培养基并在25℃培养2-3周,直至根发育。将生根的苗移植至温室的土壤中。从表现选择剂耐受的并含有单拷贝的T-DNA插入物的植物中产生T1种子。
小麦转化
小麦的转化用Ishida等Ishida等(1996)Nature Biotech14(6):745-50描述的方法进行。通常在转化中使用(可从墨西哥CIMMYT获得的)栽培品种Bobwhite。不成熟胚与含有表达载体的根癌农杆菌共培育并且转基因植物通过器官发生而恢复。在与农杆菌温育后,胚在愈伤组织诱导培养基上、随后在再生培养基上体外培育,其中所述的再生培养基含有选择剂(例如咪唑啉酮,但可以使用多种选择标记)。培养平板在25℃于光照下培养2-3周,或直至苗发育。将绿色苗从每个胚转移至生根培养基并在25℃培养2-3周,直至根发育。将生根的苗移植至温室的土壤中。从表现选择剂耐受的并含有单拷贝的T-DNA插入物的植物中产生T1种子。
大豆转化
根据对Texas A&M美国专利5,164,310中所述方法的改良方法转化大豆。几个商业大豆品种对于通过这种方法的转化是可行的。栽培品种Jack(从Illinois种子基金会可获得)通常用于转化。对大豆种子消毒以便体外播种。从7日龄幼苗中切下下胚轴、胚根和一片子叶。进一步培育上胚轴和余下的子叶以发育腋生节。将这些腋生节切下并与含有表达载体的根癌农杆菌温育。在共培育处理之后,将外植体洗涤并转移至选择培养基。将再生的苗切下并置于苗伸长培养基上。将长度不超过1cm的苗置于生根培养基上直至根发育。将生根的苗移植至温室的土壤中。从表现选择剂耐受的并含有单拷贝T-DNA插入物的植物中产生T1种子。
油菜/卡诺拉油菜转化
使用5-6日龄幼苗的子叶柄和下胚轴作为用于组织培养的外植体并且根据Babic等(1998,Plant Cell Rep17:183-188)转化。商业栽培品种Westar(Agriculture Canada)是用于转化的标准品种,但是也可以使用其他品种。对卡诺拉油菜种子作表面消毒以便体外播种。从体外幼苗中切下具有附着子叶的子叶柄外植体,并以(含有表达载体的)农杆菌通过叶柄外植体的切口端浸入细菌混悬液而接种。外植体随后在含有3mg/l BAP、3%蔗糖、0.7%植物琼脂(Phytagar)的MSBAP-3培养基上在23℃,16小时光照下培养2天。在与农杆菌共培育2日后,将叶柄外植体转移至含有的3mg/l BAP、头孢噻肟、羧苄青霉素或特美汀(300mg/l)的MSBAP-3培养基上持续7日,并且随后在含头孢噻肟、羧苄青霉素或特美汀和选择剂的MSBAP-3培养基上培养,直至苗再生。当苗具有5–10mm长度时,将苗切下并转移至苗伸长培养基(含0.5mg/l BAP的MSBAP-0.5)。将长度大约2cm的苗转移至用于根诱导的生根培养基(MS0)。将生根的苗移植至温室的土壤中。从表现选择剂耐受性并含有单拷贝T-DNA插入物的植物中产生T1种子。
苜蓿转化
苜蓿的再生性克隆使用(McKersie等,1999Plant Physiol119:839–847)的方法加以转化。苜蓿的再生和转化是基因型依赖性的并且因而需要再生植物。已经描述了获得再生性植物的方法。例如,这些再生性植物可以选自栽培品种Rangelander(Agriculture Canada)或如Brown DCW与AAtanassov(1985.Plant Cell Tissue Culture4:111-112)描述的任何其他商业苜蓿品种。备选地,RA3品种(威斯康星大学)已经被选择用于组织培养中(Walker等,1978Am J Bot65:654-659)。将叶柄外植体与含有表达载体的根癌农杆菌C58C1pMP90(McKersie等,1999Plant Physiol119:839–847)或LBA4404的过夜培养物共培育。外植体在黑暗中在含有288mg/L Pro、53mg/L硫代脯氨酸、4.35g/L K2SO4和100μm乙酰丁香酮的SH诱导培养基上共培育3天.外植体在半浓缩Murashige-Skoog培养基(Murashige和Skoog,1962)中洗涤并平板接种在不含乙酰丁香酮而含有合适选择剂和合适抗生素以抑止农杆菌生长的相同SH诱导培养基上。在数周后,将体细胞胚转移至不含生长调节剂、不含抗生素而含有50g/L蔗糖的BOi2Y发育培养基中。体细胞胚随后在半浓缩Murashige-Skoog培养基上萌发。将生根的幼苗移植至花钵内并且在温室中培育。从表现选择剂耐受性并含有单拷贝T-DNA插入物的植物中产生T1种子。
实施例9:表型评估方法
9.1评估设置
产生大约35个独立的T0稻转化体。将原代转化体从组织培养室转移到温室中用于生长和收获T1种子。保留6个事件,其中T1后代对于转基因的存在/缺失以3:1分离。对于每个这些事件,通过监测可见标记表达选择约10个含有转基因的T1幼苗(杂合子和纯合子)和约10个缺少该转基因的T1幼苗(失效合子)。转基因植物和对应的失效合子在随机的位置上并排生长。温室条件是短日照(12小时光照),光照中28°C,黑暗中22°C,和70%的相对湿度。
4个T1事件在T2代中按照与T1代相同的评估步骤进一步评估,但是每个事件具有更多个体。从播种阶段到成熟阶段,植物通过数字成像箱几次。在每个时间点,从至少6个不同角度对每株植物拍下数字图像(2048x1536像素,1600万色彩数)。
干旱筛选
在正常条件下在花盆土中培养来自T2种子的植物,直到进入抽穗期。然后将其转移到“干燥”区域,停止灌溉。向随机选择的花盆中插入湿度探测仪,以监测土壤水含量(SWC)。当SWC低于一定的阈值时,自动向植物持续补水,直到再次达到正常水平。然后将植物再次重新转移到正常条件下。其余的栽培(植物成熟、种子收获)与不在非生物胁迫条件下培养的植物相同。详细记录正常条件下培养的生长和产量参数。
氮使用效率筛选
来自T2种子的稻植物在除了营养溶液之外的正常条件下生长与盆栽土壤中。花盆从移植到成熟用特定营养溶液浇灌,该溶液含有减少的氮(N)含量,通常低7到8倍。培养的剩余部分(植物成熟,种子收获)与不在非生物胁迫条件下生长的植物相同。如对正常条件下生长详述的记录生长和产量参数。
9.2统计学分析:F-检验
将双因子ANOVA(方差分析)用作植物表型特征的总体评估的统计学模型。对用本发明的基因转化的所有事件的所有植物测量的所有参数进行F-检验。进行F-检验以检查基因在所有转化事件中的效果并验证该基因的总体效果,也称作总体基因效果。F检验的真实总体基因效果的显著性阈值设为5%概率水平。显著F-检验值指向基因效果,表示并不仅仅是该基因的存在或其位置引起表型差异。
因为用重叠的事件进行了两个实验,所以进行了合并分析。这对于检查两个实验上效果的一致性是有用的,并且如果情况是这样的,积累来自两个实验的证据以便在结论中提高置信度。所用的方法是混合模型方法,其考虑了数据的多级结构(即实验-事件-分离子)。通过比较似然比检验与卡方分布得到P-值。
9.3测量的参数
生物量相关的参数测量
从播种阶段到成熟阶段,植物通过数字成像箱几次。在每个时间点,从至少6个不同角度对每株植物拍下数字图像(2048x1536像素,1600万色彩数)。通过对来自地上植物部分的数字图像上区别于背景的像素总数计数来确定植物地上部分面积(或者叶子生物量)。该值对在相同时间点从不同角度拍摄的图像平均并通过校正转化为物理表面值,其以平方mm表示。实验表明以这种方式测量的地上部分植物面积与地上植物部分的生物量相关。地上部分面积是植物达到其最大叶子生物量的时间点时测量的面积。早期萌发势是萌发后三周的植物(幼苗)地上部分面积。根生物量的增加表示为总根生物量的增加(测量为在植物寿命中观察到的根的最大生物量);或者表示为根/枝条指数的增加(测量为在根和枝条的活跃生长期中根质量和枝条质量之间的比值)。
通过对来自地上植物部分的区别于背景的像素总数计数确定早期萌发势。该值对在相同时间点从不同角度拍摄的图像平均并通过校正转化为物理表面值,以平方mm表示。下面所述的结果是萌发后三周植物的结果。
种子相关的参数测量
将成熟的总圆锥花序收获,计数,装袋,用条形码标记,然后在烘箱中37°C下干燥3天。然后将圆锥花序脱粒并收集和计数所有种子。用鼓风装置分离饱满的外壳与空壳。丢弃空壳并再次计数剩余的部分。在分析天平上对饱满外壳称重。通过计数分离步骤后剩余的饱满外壳数来确定饱满种子数。通过对从植物收获的所有饱满外壳称重来测量总种子产量。通过对从植物收获的外壳数目计数测量每株植物的总种子数。从计数的饱满种子数和它们的总重量外推千粒重(TKW)。本发明中的收获指数(HI)定义为总种子产量和地上部分面积(mm2)之间的比值,再乘以因数106。每个圆锥花序的总花数在本发明中定义为种子总数和成熟总圆锥花序数目的比值。如本文定义的种子饱满率是饱满种子数在种子(或小花)总数中占的比例(表示为%)。
实施例10:转基因植物的表型评估结果
表达处于原叶绿素酸酯还原酶启动子控制下的编码PEPC多肽的核酸(SEQ ID NO:1)的转基因稻植物在T1或T2代或者这两代中给出了下面的结果。


除了上面显示的结果外,在干旱条件下过表达PEPC的植物也显示出植物高度的增加(5株中2株,得到平均11.5%的增加);根厚度的增加(5株中4株,得到平均23.5%的增加);地上部分面积的增加(5株中2株,得到平均14%的增加);和萌发势的增加(5株中2株,得到平均47.5%的增加)。
表达处于原叶绿素酸酯还原酶启动子控制下的编码III类U盒多肽的核酸(SEQ ID NO:74)的转基因植物得到了下面的结果。
非胁迫条件下的结果:
转基因稻植物相对于对照植物显示出高度、地上部分面积、圆锥花序数、花数、TKW(千粒重)、种子总重、种子总数和饱满种子总数的增加。
(轻微)干旱条件下的结果:
转基因稻植物显示出相对于对照植物在重量、地上部分面积、种子总重和饱满种子数的增加。
表达处于原叶绿素酸酯还原酶启动子控制下的编码PQQC多肽的核酸(SEQ ID NO:104)的转基因稻植物得到了下面的结果。
非胁迫条件下的结果:
转基因稻植物相对于对照植物显示出高度、地上部分面积、圆锥花序数、花数、种子总重、种子总数和饱满种子总数的增加。
B.表达处于原叶绿素酸酯还原酶启动子控制下的编码PQQC多肽的核酸(SEQ ID NO:104)的转基因稻植物得到了下面的结果。对于表达处于GOS2启动子控制下的编码PQQC多肽的核酸(SEQ ID NO:104)的转基因稻植物得到了类似的结果。
(轻微)干旱条件下的结果:
转基因稻植物显示出相对于对照植物在重量、饱满率、TKW(千粒重)、饱满种子数和种子总重的增加。




































































































































































































































Claims (46)
1.相对于对照植物在植物中增强产量相关性状的方法,包括调节植物中编码III类U盒蛋白的核酸的表达,所述III类U盒蛋白在其N-末端区包含U盒结构域。
2.根据权利要求1的方法,其中所述U盒结构域以递增优选顺序与SEQ ID NO:76的U盒结构域有至少50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性和/或包含基序VII:GI T/S YDR E/S/D/A N/S/G/A I E/A K/R/M W L/M/V/IF/L/A和/或基序VIII:Q S/A/G W C/G/A I/R/A/T S/V/L/R/A H/N/R/N/VA/C/S。
3.根据权利要求2的方法,其中所述基序VII是GITYDRENIERWIF(SEQ ID NO:77),或以递增优选顺序与基序VII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;并且
基序VIII是:QSWCTLNA(SEQ ID NO:78)或以递增优选顺序与基序VIII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序。
4.根据权利要求1到3任一项的方法,其中III类U盒多肽在其C-末端区中包含一个或多个下面的基序:
基序IX:GR S/A/L A/E L/F L/V S/K/N/A H
基序X:G/A I/L/V A A/I/V V S/C/A/G/T K/C/R R/K/A A/I/V/T M/L/TR V/L/I S/P
基序XI:K/R L C/L F/V/M/L V/L/A V/L/A/I Q/R V/A/S D/S/G/E
基序XII:S/A/L YP。
5.根据权利要求4的方法,其中基序IX是:GRAELLNH(SEQ ID NO:79)或以递增优选顺序与基序IX有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序X是:GLAIVSKKILRVS(SEQ ID NO:80)或以递增优选顺序与基序X有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序XI是:KLCLVLQVD(SEQ ID NO:81)或以递增优选顺序与基序XI有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序;
基序XII是:SYP(SEQ ID NO:82)或以递增优选顺序与基序XII有至少60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性的基序。
6.根据权利要求1到5任一项的方法,其中通过在植物中导入并表达编码III类U盒多肽的核酸实现所述受调节的表达。
7.根据权利要求1到6任一项的方法,其中所述编码III类U盒多肽的核酸编码表A2中所列的任一种蛋白质或者是这种核酸的一部分,或者能够与这种核酸杂交的核酸。
8.根据权利要求1到7任一项的方法,其中所述核酸序列编码表A2中给出的任一种蛋白质的直向同源物或旁系同源物。
9.根据权利要求1到8任一项的方法,其中所述增强的产量相关性状包括相对于对照植物增加的产量,优选增加的生物量和/或增加的种子产量。
10.根据权利要求1到9任一项的方法,其中在非胁迫条件下和轻微干旱条件下得到所述增强的产量相关性状。
11.根据权利要求1到10任一项的方法,其中在氮缺乏条件下得到所述增强的产量相关性状。
12.根据权利要求1到11任一项的方法,其中所述核酸有效连接到年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
13.根据权利要求1到12任一项的方法,其中所述编码III类多肽的核酸是植物来源,优选来自双子叶植物,进一步优选来自茄科,更优选来自辣椒属,最优选来自辣椒(Capsicum annuus)。
14.通过权利要求1到13任一项的方法得到的植物或其部分,包括种子,其中所述植物或其部分包含编码III类U盒多肽的重组核酸。
15.构建体,其包含:
(iv)编码如权利要求1到5任一项定义的III类U盒多肽的核酸;
(v)能够驱动(a)的核酸序列表达的一个或多个控制序列;和任选地
(vi)转录终止序列。
16.根据权利要求15的构建体,其中所述控制序列之一是年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
17.根据权利要求15或16的构建体在用于制备植物的方法中的用途,所述植物相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量。
18.用根据权利要求15到16的构建体转化的植物、植物部分或植物细胞。
19.产生相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量的转基因植物的方法,包括:
(c)在植物中导入并表达编码如权利要求1到5任一项定义的III类U盒多肽的核酸;和
(d)在促进植物生长和发育的条件下培养所述植物细胞。
20.转基因植物,其相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量,所述转基因植物从编码如权利要求1到5任一项定义的III类U盒多肽的核酸的增加的表达得到,或者来自所述转基因植物的转基因植物细胞。
21.根据权利要求14、18或20的转基因植物,或来自其的转基因植物细胞,其中所述植物是作物植物或单子叶植物或谷类,如稻、玉米、小麦、大麦、粟、黑麦、黑小麦、高粱和燕麦。
22.根据权利要求21的植物的可收获部分,其中所述可收获部分优选为苗生物量和/或种子。
23.来自根据权利要求21的植物和/或来自根据权利要求22的植物的可收获部分的产品。
24.编码III类U盒多肽的核酸用于相对于对照植物在植物中增加产量,尤其增加种子产量和/或苗生物量的用途。
25.相对于对照植物在植物中增强产量相关性状的方法,包括调节植物中编码PQQC蛋白的核酸的表达。
26.根据权利要求25的方法,其中所述PQQC蛋白以递增优选顺序与SEQ ID NO:105表示的序列有至少45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或更高序列同一性。
27.根据权利要求25到26任一项的方法,其中通过在植物中导入并表达编码PQQC多肽的核酸实现所述受调节的表达。
28.根据前面权利要求25到27任一项的方法,其中所述编码PQQC多肽的核酸编码表A3中所列的任一种蛋白质或者是这种核酸的一部分,或者能够与这种核酸杂交的核酸。
29.根据前面权利要求25到28任一项的方法,其中所述增强的产量相关性状包括相对于对照植物增加的产量,优选增加的生物量和/或增加的种子产量。
30.根据前面权利要求25到29任一项的方法,其中在非胁迫条件下和轻微干旱条件下得到所述增强的产量相关性状。
31.根据前面权利要求25到30任一项的方法,其中在氮缺乏条件下得到所述增强的产量相关性状。
32.根据前面权利要求25到31任一项的方法,其中所述核酸有效连接到年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
33.根据前面权利要求25到32任一项的方法,其中所述核酸有效连接到组成型启动子,优选GOS2启动子,最优选来自稻的GOS2启动子。
34.根据前面权利要求25到33任一项的方法,其中所述编码PQQC多肽的核酸为细菌来源,优选来自假单胞菌。
35.通过根据前面权利要求任一项的方法得到的植物或其部分,包括种子,其中所述植物或其部分包含编码PQQC多肽的重组核酸。
36.构建体,其包含:
–编码如权利要求25到26定义的PQQC多肽的核酸;
–能够驱动(a)的核酸序列表达的一个或多个控制序列;和任选地
–转录终止序列。
37.根据权利要求36的构建体,其中所述控制序列之一是年轻绿色组织特异性启动子,优选原叶绿素酸酯还原酶启动子。
38.根据权利要求36的构建体,其中所述控制序列之一是组成型启动子,优选GOS2启动子,最优选来自稻的GOS2启动子。
39.根据权利要求36到38任一项的构建体在用于制备植物的方法中的用途,所述植物相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量。
40.用根据权利要求36到39任一项的构建体转化的植物、植物部分或植物细胞。
41.产生相对于对照植物具有增加的产量,尤其增加的生物量和/或增加的种子产量的转基因植物的方法,包括:
(iii)在植物中导入并表达编码如权利要求25到26定义的PQQC多肽的核酸;和
(iv)在促进植物生长和发育的条件下培养所述植物细胞。
42.转基因植物,其相对于对照植物具有增加的生物量和/或增加的种子产量,所述转基因植物从编码如权利要求25到26定义的PQQC多肽的核酸的增加的表达得到,或者来自所述转基因植物的转基因植物细胞。
43.根据权利要求35、40或42的转基因植物,或来自其的转基因植物细胞,其中所述植物是作物植物或单子叶植物或谷类,如稻、玉米、小麦、大麦、粟、黑麦、黑小麦、高粱和燕麦。
44.根据权利要求43的植物的可收获部分,其中所述可收获部分优选为苗生物量和/或种子。
45.来自根据权利要求43的植物和/或来自根据权利要求44的植物的可收获部分的产品。
46.编码PQQC多肽的核酸用于相对于对照植物在植物中增加产量,尤其增加种子产量和/或苗生物量的用途。
Applications Claiming Priority (12)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP07113571 | 2007-07-31 | ||
| EP07113570.1 | 2007-07-31 | ||
| EP07113571.9 | 2007-07-31 | ||
| EP07113570 | 2007-07-31 | ||
| US97007107P | 2007-09-05 | 2007-09-05 | |
| US97006607P | 2007-09-05 | 2007-09-05 | |
| US60/970,066 | 2007-09-05 | ||
| US60/970,071 | 2007-09-05 | ||
| EP07116620 | 2007-09-18 | ||
| EP07116620.1 | 2007-09-18 | ||
| US97587607P | 2007-09-28 | 2007-09-28 | |
| US60/975,876 | 2007-09-28 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN2008800255662A Division CN101952441B (zh) | 2007-07-31 | 2008-07-31 | 具有增强的产量相关性状的植物及其制备方法 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN103397049A true CN103397049A (zh) | 2013-11-20 |
Family
ID=39885073
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN2008800255662A Expired - Fee Related CN101952441B (zh) | 2007-07-31 | 2008-07-31 | 具有增强的产量相关性状的植物及其制备方法 |
| CN2013102981411A Pending CN103397049A (zh) | 2007-07-31 | 2008-07-31 | 具有增强的产量相关性状的植物及其制备方法 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN2008800255662A Expired - Fee Related CN101952441B (zh) | 2007-07-31 | 2008-07-31 | 具有增强的产量相关性状的植物及其制备方法 |
Country Status (12)
| Country | Link |
|---|---|
| US (1) | US20110023188A1 (zh) |
| EP (2) | EP2503000A1 (zh) |
| KR (3) | KR101269930B1 (zh) |
| CN (2) | CN101952441B (zh) |
| AR (1) | AR067766A1 (zh) |
| AU (1) | AU2008281698B2 (zh) |
| BR (1) | BRPI0813884A2 (zh) |
| CA (1) | CA2697186A1 (zh) |
| DE (1) | DE112008002020T5 (zh) |
| IN (1) | IN2010KN00318A (zh) |
| MX (1) | MX2010001128A (zh) |
| WO (1) | WO2009016232A2 (zh) |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN112442506A (zh) * | 2020-12-21 | 2021-03-05 | 浙江大学 | 一种拟南芥根肿病感病候选基因at2g35930及其应用 |
| CN112852834A (zh) * | 2021-04-06 | 2021-05-28 | 台州学院 | 水稻OsPUB23基因在调控水稻抗性中的应用 |
| CN115976067A (zh) * | 2022-12-01 | 2023-04-18 | 四川农业大学 | FaU-box E3基因家族在调控草莓果实成熟中的应用 |
Families Citing this family (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN102459614A (zh) * | 2009-04-29 | 2012-05-16 | 巴斯夫植物科学有限公司 | 具有增强的产量相关性状的植物和用于产生该植物的方法 |
| CN102811617A (zh) | 2010-01-22 | 2012-12-05 | 拜耳知识产权有限责任公司 | 杀螨和/或杀虫活性物质结合物 |
| CN103717076B (zh) | 2011-08-10 | 2016-04-13 | 拜耳知识产权股份有限公司 | 含有特定特特拉姆酸衍生物的活性化合物组合物 |
| US20140298544A1 (en) * | 2011-10-28 | 2014-10-02 | Pioneer Hi Bred International Inc | Engineered PEP carboxylase variants for improved plant productivity |
| MX2014005375A (es) * | 2011-11-03 | 2014-10-17 | Syngenta Participations Ag | Polinucleotidos, polipeptidos y metodos para mejorar la fotoasimilacion en plantas. |
| CN102559677B (zh) * | 2011-12-23 | 2013-12-18 | 北京大北农科技集团股份有限公司 | 组织特异性启动子及其用途 |
| KR102230148B1 (ko) * | 2014-10-30 | 2021-03-19 | 연세대학교 산학협력단 | 식물의 저온 스트레스 내성을 증진시키는 조성물 및 이를 이용한 형질전환 식물체 |
| AR115540A1 (es) * | 2018-06-14 | 2021-01-27 | Benson Hill Biosystems Inc | Aumento del crecimiento de las plantas y el rendimiento usando una proteína de la superfamilia de ring / u-box |
| CN108624578B (zh) * | 2018-06-25 | 2021-09-28 | 中山大学 | 花生AhPEPC5基因片段在提高微生物对渗透胁迫与盐胁迫耐受能力中的应用 |
| AU2020272047A1 (en) * | 2019-04-11 | 2021-11-25 | The Regents Of The University Of California | Engineered phosphoenolpyruvate carboxylase enzymes |
Family Cites Families (34)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4962028A (en) | 1986-07-09 | 1990-10-09 | Dna Plant Technology Corporation | Plant promotors |
| US5116742A (en) | 1986-12-03 | 1992-05-26 | University Patents, Inc. | RNA ribozyme restriction endoribonucleases and methods |
| US4987071A (en) | 1986-12-03 | 1991-01-22 | University Patents, Inc. | RNA ribozyme polymerases, dephosphorylases, restriction endoribonucleases and methods |
| AU3756889A (en) | 1988-06-01 | 1990-01-05 | The Texas A & M University System | Method for transforming plants via the shoot apex |
| AU4115693A (en) | 1992-04-24 | 1993-11-29 | Sri International | In vivo homologous sequence targeting in eukaryotic cells |
| HUT71929A (en) | 1992-06-29 | 1996-02-28 | Gene Shears Pty Ltd | Nucleic acids and methods of use thereof for controlling viral pathogens |
| WO1994012015A1 (en) | 1992-11-30 | 1994-06-09 | Chua Nam Hai | Expression motifs that confer tissue- and developmental-specific expression in plants |
| JPH09505461A (ja) | 1993-07-22 | 1997-06-03 | ジーン シェアーズ プロプライアタリー リミティド | Dnaウィルスリボザイム |
| AU687961B2 (en) | 1993-11-19 | 1998-03-05 | Biotechnology Research And Development Corporation | Chimeric regulatory regions and gene cassettes for expression of genes in plants |
| EP0733059B1 (en) | 1993-12-09 | 2000-09-13 | Thomas Jefferson University | Compounds and methods for site-directed mutations in eukaryotic cells |
| US6395547B1 (en) | 1994-02-17 | 2002-05-28 | Maxygen, Inc. | Methods for generating polynucleotides having desired characteristics by iterative selection and recombination |
| US5605793A (en) | 1994-02-17 | 1997-02-25 | Affymax Technologies N.V. | Methods for in vitro recombination |
| HUP9802535A3 (en) | 1995-10-06 | 2001-04-28 | Plant Genetic Systems Nv | Seed shattering |
| GB9607517D0 (en) | 1996-04-11 | 1996-06-12 | Gene Shears Pty Ltd | The use of DNA Sequences |
| ID21006A (id) * | 1997-02-10 | 1999-04-08 | Japan Tobacco Inc | Siklus c4 jenis pck |
| GB9703146D0 (en) | 1997-02-14 | 1997-04-02 | Innes John Centre Innov Ltd | Methods and means for gene silencing in transgenic plants |
| GB9710475D0 (en) | 1997-05-21 | 1997-07-16 | Zeneca Ltd | Gene silencing |
| GB9720148D0 (en) | 1997-09-22 | 1997-11-26 | Innes John Centre Innov Ltd | Gene silencing materials and methods |
| PT1068311E (pt) | 1998-04-08 | 2011-07-20 | Commw Scient Ind Res Org | Processos e meios de obter fenótipos modificados |
| TR200100705T2 (tr) | 1998-06-26 | 2001-10-22 | Iowa State University Research Foundation, Inc. | Bitkilerde enzim ve asetil'in değiştirilmesi için yöntemler. |
| US6555732B1 (en) | 1998-09-14 | 2003-04-29 | Pioneer Hi-Bred International, Inc. | Rac-like genes and methods of use |
| CA2366104C (en) | 1999-07-22 | 2010-07-06 | Japan As Represented By Director General Of National Institute Of Agrobiological Resources, Ministry Of Agriculture, Forestry And Fisheries | Ultra-fast transformation technique for monocotyledons |
| AU780117B2 (en) | 1999-08-26 | 2005-03-03 | Basf Plant Science Gmbh | Plant gene expression, controlled by constitutive plant V-ATpase promoters |
| CN1307104A (zh) * | 2000-01-26 | 2001-08-08 | 上海博道基因技术有限公司 | 一种新的多肽——人磷酸烯醇丙酮酸羧化酶81和编码这种多肽的多核苷酸 |
| FR2823064B1 (fr) * | 2001-04-04 | 2004-05-28 | Biogemma Fr | Procede d'obtention de plantes c4 a metabolisme carbone modifie |
| US7619146B2 (en) * | 2001-06-18 | 2009-11-17 | Frankard Valerie | Method for modifying plant morphology, biochemistry and physiology |
| CN1149917C (zh) * | 2001-12-21 | 2004-05-19 | 焦德茂 | 转入c4植物pepc基因的水稻能稳定表达的育种方法 |
| US7314974B2 (en) * | 2002-02-21 | 2008-01-01 | Monsanto Technology, Llc | Expression of microbial proteins in plants for production of plants with improved properties |
| WO2004065596A2 (en) | 2003-01-21 | 2004-08-05 | Cropdesign N.V. | Use of the regulatory sequence of the rice gos2 gene for the gene expression in dicotyledonous plants or plant cells |
| DE602004006477T2 (de) | 2003-02-04 | 2008-02-14 | Cropdesign N.V. | Promotor aus reis |
| EP2302062A1 (en) * | 2003-10-20 | 2011-03-30 | CropDesign N.V. | Identification of E2F target genes and uses thereof |
| CN1314809C (zh) * | 2004-05-31 | 2007-05-09 | 福建农林大学 | 编码甘蔗磷酸烯醇式丙酮酸羧化酶的核酸分子及其应用 |
| AR051865A1 (es) * | 2004-12-24 | 2007-02-14 | Cropdesign Nv | Plantas teniendo mayor rendimiento y metodo para producirlas |
| WO2006075807A1 (en) * | 2005-01-11 | 2006-07-20 | Seoul National University Industry Foundation | Pyrroloquinoline quinone and its biosynthetic genes for plant growth promotion and uses thereof |
-
2008
- 2008-07-31 EP EP12170712A patent/EP2503000A1/en not_active Withdrawn
- 2008-07-31 EP EP08786685A patent/EP2183372A2/en not_active Withdrawn
- 2008-07-31 KR KR1020127002696A patent/KR101269930B1/ko not_active IP Right Cessation
- 2008-07-31 CN CN2008800255662A patent/CN101952441B/zh not_active Expired - Fee Related
- 2008-07-31 AU AU2008281698A patent/AU2008281698B2/en not_active Ceased
- 2008-07-31 DE DE112008002020T patent/DE112008002020T5/de not_active Withdrawn
- 2008-07-31 WO PCT/EP2008/060060 patent/WO2009016232A2/en active Application Filing
- 2008-07-31 US US12/671,224 patent/US20110023188A1/en not_active Abandoned
- 2008-07-31 KR KR1020107004117A patent/KR101269929B1/ko not_active IP Right Cessation
- 2008-07-31 CN CN2013102981411A patent/CN103397049A/zh active Pending
- 2008-07-31 BR BRPI0813884-2A2A patent/BRPI0813884A2/pt not_active IP Right Cessation
- 2008-07-31 MX MX2010001128A patent/MX2010001128A/es not_active Application Discontinuation
- 2008-07-31 AR ARP080103335A patent/AR067766A1/es unknown
- 2008-07-31 KR KR1020127002697A patent/KR101269931B1/ko not_active IP Right Cessation
- 2008-07-31 CA CA2697186A patent/CA2697186A1/en not_active Abandoned
- 2008-07-31 IN IN318KON2010 patent/IN2010KN00318A/en unknown
Non-Patent Citations (3)
| Title |
|---|
| CHO SK,ET AL: "Heterologous Expression and Molecular and Cellular Characterization of CaPUB1 Encoding a Hot Pepper U-Box E3 Ubiquitin Ligase Homolog", 《PLANT PYSIOL》 * |
| NCBI: "ABA59556.1", 《GENBANK》 * |
| NCBI: "DQ211901.1", 《GENBANK》 * |
Cited By (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN112442506A (zh) * | 2020-12-21 | 2021-03-05 | 浙江大学 | 一种拟南芥根肿病感病候选基因at2g35930及其应用 |
| CN112852834A (zh) * | 2021-04-06 | 2021-05-28 | 台州学院 | 水稻OsPUB23基因在调控水稻抗性中的应用 |
| CN112852834B (zh) * | 2021-04-06 | 2022-06-10 | 台州学院 | 水稻OsPUB23基因在调控水稻抗性中的应用 |
| CN115976067A (zh) * | 2022-12-01 | 2023-04-18 | 四川农业大学 | FaU-box E3基因家族在调控草莓果实成熟中的应用 |
| CN115976067B (zh) * | 2022-12-01 | 2023-09-05 | 四川农业大学 | FaU-box E3基因家族在调控草莓果实成熟中的应用 |
Also Published As
| Publication number | Publication date |
|---|---|
| IN2010KN00318A (zh) | 2015-06-12 |
| EP2183372A2 (en) | 2010-05-12 |
| KR20100040939A (ko) | 2010-04-21 |
| KR101269931B1 (ko) | 2013-05-31 |
| BRPI0813884A2 (pt) | 2014-10-07 |
| MX2010001128A (es) | 2010-03-09 |
| CN101952441A (zh) | 2011-01-19 |
| KR20120096924A (ko) | 2012-08-31 |
| AR067766A1 (es) | 2009-10-21 |
| KR101269930B1 (ko) | 2013-05-31 |
| WO2009016232A3 (en) | 2009-03-26 |
| US20110023188A1 (en) | 2011-01-27 |
| CA2697186A1 (en) | 2009-02-05 |
| AU2008281698A1 (en) | 2009-02-05 |
| CN101952441B (zh) | 2013-08-14 |
| WO2009016232A8 (en) | 2010-01-21 |
| EP2503000A1 (en) | 2012-09-26 |
| WO2009016232A2 (en) | 2009-02-05 |
| KR20120034773A (ko) | 2012-04-12 |
| DE112008002020T5 (de) | 2010-10-28 |
| KR101269929B1 (ko) | 2013-06-25 |
| AU2008281698B2 (en) | 2014-02-20 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN101627125B (zh) | 具有增强的产量相关性状的植物及其制备方法 | |
| CN102131934B (zh) | 具有增强的产量相关性状的植物及其制备方法 | |
| CN101965405B (zh) | 具有增强的产量相关性状的植物和用于制备该植物的方法 | |
| CN103987848A (zh) | 具有增强的产量相关性状的植物及其制备方法 | |
| CN103249836A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN102186877A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN103773796A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN102936605A (zh) | 具有增强的产量相关性状的植物和用于制备该植物的方法 | |
| CN103397049A (zh) | 具有增强的产量相关性状的植物及其制备方法 | |
| CN102365366A (zh) | 具有增强的产量相关性状的植物及其制备方法 | |
| CN102459614A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN102066568A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN104530202A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN103834683A (zh) | 具有增强的产量相关性状的植物和产生所述植物的方法 | |
| CN104404078A (zh) | 具有增强的产率相关性状的植物及其制备方法 | |
| CN102099480A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN103119170A (zh) | 具有增强的产量相关性状的植物及其制备方法 | |
| CN103502456A (zh) | 具有增强的产量相关性状的植物和用于制备该植物的方法 | |
| CN103119167A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 | |
| CN102272309A (zh) | 具有增强的非生物胁迫耐受性和/或增强的产量相关性状的植物及其制备方法 | |
| CN103492573A (zh) | 具有增强的产量相关性状的植物和其生产方法 | |
| CN103154254A (zh) | 具有增强的产量相关性状的植物和产生该植物的方法 | |
| CN103068992A (zh) | 具有增强的产量相关性状的植物和用于制备该植物的方法 | |
| CN102892890A (zh) | 具有增强的产量相关性状的植物和用于制备该植物的方法 | |
| CN103003432A (zh) | 具有增强的产量相关性状的植物和用于产生该植物的方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| C02 | Deemed withdrawal of patent application after publication (patent law 2001) | ||
| WD01 | Invention patent application deemed withdrawn after publication |
Application publication date: 20131120 |