JP6841753B2 - ヒストンh3−リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 - Google Patents
ヒストンh3−リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 Download PDFInfo
- Publication number
- JP6841753B2 JP6841753B2 JP2017514421A JP2017514421A JP6841753B2 JP 6841753 B2 JP6841753 B2 JP 6841753B2 JP 2017514421 A JP2017514421 A JP 2017514421A JP 2017514421 A JP2017514421 A JP 2017514421A JP 6841753 B2 JP6841753 B2 JP 6841753B2
- Authority
- JP
- Japan
- Prior art keywords
- scnt
- cell
- human
- seq
- cells
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 238000010374 somatic cell nuclear transfer Methods 0.000 title claims description 323
- 238000000034 method Methods 0.000 title claims description 207
- 239000000203 mixture Substances 0.000 title claims description 71
- 230000001965 increasing effect Effects 0.000 title claims description 56
- 108010033040 Histones Proteins 0.000 title description 16
- 239000004472 Lysine Substances 0.000 title description 5
- 210000004027 cell Anatomy 0.000 claims description 326
- 150000007523 nucleic acids Chemical class 0.000 claims description 173
- 210000000287 oocyte Anatomy 0.000 claims description 161
- 210000002257 embryonic structure Anatomy 0.000 claims description 146
- 102000039446 nucleic acids Human genes 0.000 claims description 126
- 108020004707 nucleic acids Proteins 0.000 claims description 126
- 241000282414 Homo sapiens Species 0.000 claims description 123
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 123
- 239000012634 fragment Substances 0.000 claims description 108
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 106
- 210000001161 mammalian embryo Anatomy 0.000 claims description 95
- 229920001184 polypeptide Polymers 0.000 claims description 95
- 102000008157 Histone Demethylases Human genes 0.000 claims description 74
- 108010074870 Histone Demethylases Proteins 0.000 claims description 74
- 210000002459 blastocyst Anatomy 0.000 claims description 44
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 41
- 210000004962 mammalian cell Anatomy 0.000 claims description 40
- 238000004519 manufacturing process Methods 0.000 claims description 35
- 238000002347 injection Methods 0.000 claims description 26
- 239000007924 injection Substances 0.000 claims description 26
- 238000011161 development Methods 0.000 claims description 25
- 230000004913 activation Effects 0.000 claims description 21
- 238000000338 in vitro Methods 0.000 claims description 20
- 230000007159 enucleation Effects 0.000 claims description 9
- 238000012258 culturing Methods 0.000 claims description 6
- 210000000472 morula Anatomy 0.000 claims description 6
- 230000003213 activating effect Effects 0.000 claims 1
- 210000003101 oviduct Anatomy 0.000 claims 1
- 108090000623 proteins and genes Proteins 0.000 description 191
- 239000003795 chemical substances by application Substances 0.000 description 163
- 108020004459 Small interfering RNA Proteins 0.000 description 129
- 239000003112 inhibitor Substances 0.000 description 119
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 111
- 239000004055 small Interfering RNA Substances 0.000 description 111
- 230000014509 gene expression Effects 0.000 description 106
- 102000004169 proteins and genes Human genes 0.000 description 102
- 210000001082 somatic cell Anatomy 0.000 description 101
- 238000012228 RNA interference-mediated gene silencing Methods 0.000 description 97
- 230000009368 gene silencing by RNA Effects 0.000 description 97
- 125000003729 nucleotide group Chemical group 0.000 description 88
- 101150032905 Kdm4d gene Proteins 0.000 description 85
- 230000004048 modification Effects 0.000 description 85
- 238000012986 modification Methods 0.000 description 85
- 230000008672 reprogramming Effects 0.000 description 81
- 239000002773 nucleotide Substances 0.000 description 77
- 210000004940 nucleus Anatomy 0.000 description 76
- 235000018102 proteins Nutrition 0.000 description 71
- 108091034117 Oligonucleotide Proteins 0.000 description 68
- 150000001413 amino acids Chemical class 0.000 description 68
- 108060004795 Methyltransferase Proteins 0.000 description 62
- 102000016397 Methyltransferase Human genes 0.000 description 52
- -1 but not limited to Chemical class 0.000 description 49
- 108020004999 messenger RNA Proteins 0.000 description 49
- 239000012190 activator Substances 0.000 description 45
- 239000003697 methyltransferase inhibitor Substances 0.000 description 42
- 101000687346 Homo sapiens PR domain zinc finger protein 2 Proteins 0.000 description 36
- 238000012546 transfer Methods 0.000 description 36
- 230000000694 effects Effects 0.000 description 35
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 34
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical compound O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 34
- 238000006467 substitution reaction Methods 0.000 description 33
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 32
- 102100024885 PR domain zinc finger protein 2 Human genes 0.000 description 32
- 241000699666 Mus <mouse, genus> Species 0.000 description 31
- 108091027967 Small hairpin RNA Proteins 0.000 description 31
- 230000030279 gene silencing Effects 0.000 description 31
- 101001088895 Homo sapiens Lysine-specific demethylase 4D Proteins 0.000 description 30
- 102000011787 Histone Methyltransferases Human genes 0.000 description 29
- 108010036115 Histone Methyltransferases Proteins 0.000 description 29
- 229940123379 Methyltransferase inhibitor Drugs 0.000 description 29
- 239000000126 substance Substances 0.000 description 29
- 210000000130 stem cell Anatomy 0.000 description 27
- 101000613625 Homo sapiens Lysine-specific demethylase 4A Proteins 0.000 description 26
- 241001465754 Metazoa Species 0.000 description 26
- 230000027455 binding Effects 0.000 description 26
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 26
- 235000000346 sugar Nutrition 0.000 description 26
- 235000013601 eggs Nutrition 0.000 description 25
- 238000012226 gene silencing method Methods 0.000 description 25
- 238000002513 implantation Methods 0.000 description 25
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 25
- 238000007069 methylation reaction Methods 0.000 description 25
- 230000018109 developmental process Effects 0.000 description 24
- 230000002401 inhibitory effect Effects 0.000 description 24
- 230000011987 methylation Effects 0.000 description 24
- 150000003384 small molecules Chemical class 0.000 description 24
- 101000613629 Homo sapiens Lysine-specific demethylase 4B Proteins 0.000 description 23
- 125000000217 alkyl group Chemical group 0.000 description 23
- 239000000427 antigen Substances 0.000 description 23
- 108091007433 antigens Proteins 0.000 description 23
- 102000036639 antigens Human genes 0.000 description 23
- 125000002467 phosphate group Chemical group [H]OP(=O)(O[H])O[*] 0.000 description 23
- 101001088893 Homo sapiens Lysine-specific demethylase 4C Proteins 0.000 description 22
- 102100033231 Lysine-specific demethylase 4D Human genes 0.000 description 22
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 22
- 230000004888 barrier function Effects 0.000 description 22
- 241000894007 species Species 0.000 description 22
- 241000283690 Bos taurus Species 0.000 description 21
- 210000000805 cytoplasm Anatomy 0.000 description 21
- 230000002452 interceptive effect Effects 0.000 description 21
- 125000000446 sulfanediyl group Chemical group *S* 0.000 description 19
- 102100040860 Lysine-specific demethylase 4B Human genes 0.000 description 18
- 102100033230 Lysine-specific demethylase 4C Human genes 0.000 description 18
- 239000000556 agonist Substances 0.000 description 18
- 210000001671 embryonic stem cell Anatomy 0.000 description 18
- 230000005764 inhibitory process Effects 0.000 description 18
- 102100040863 Lysine-specific demethylase 4A Human genes 0.000 description 17
- 108091093037 Peptide nucleic acid Proteins 0.000 description 17
- 102000055027 Protein Methyltransferases Human genes 0.000 description 17
- 108700040121 Protein Methyltransferases Proteins 0.000 description 17
- 210000001109 blastomere Anatomy 0.000 description 17
- 239000002777 nucleoside Chemical class 0.000 description 17
- 239000013598 vector Substances 0.000 description 17
- 101000696705 Homo sapiens Histone-lysine N-methyltransferase SUV39H1 Proteins 0.000 description 16
- 230000015572 biosynthetic process Effects 0.000 description 16
- 230000000295 complement effect Effects 0.000 description 16
- 238000003786 synthesis reaction Methods 0.000 description 16
- 230000004927 fusion Effects 0.000 description 15
- 229910052760 oxygen Inorganic materials 0.000 description 15
- 230000008569 process Effects 0.000 description 15
- 230000008685 targeting Effects 0.000 description 15
- 101000696699 Homo sapiens Histone-lysine N-methyltransferase SUV39H2 Proteins 0.000 description 14
- 241000700159 Rattus Species 0.000 description 14
- 238000010367 cloning Methods 0.000 description 14
- 239000001301 oxygen Substances 0.000 description 14
- 238000002360 preparation method Methods 0.000 description 14
- 101710163270 Nuclease Proteins 0.000 description 13
- 229910019142 PO4 Inorganic materials 0.000 description 13
- 229940104302 cytosine Drugs 0.000 description 13
- 230000001973 epigenetic effect Effects 0.000 description 13
- 125000005647 linker group Chemical group 0.000 description 13
- 239000010452 phosphate Substances 0.000 description 13
- 229930024421 Adenine Natural products 0.000 description 12
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 12
- 108020004414 DNA Proteins 0.000 description 12
- 229960000643 adenine Drugs 0.000 description 12
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 12
- 230000015556 catabolic process Effects 0.000 description 12
- 150000001875 compounds Chemical class 0.000 description 12
- 238000006731 degradation reaction Methods 0.000 description 12
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 12
- 230000036961 partial effect Effects 0.000 description 12
- 229940035893 uracil Drugs 0.000 description 12
- 230000000692 anti-sense effect Effects 0.000 description 11
- 125000000623 heterocyclic group Chemical group 0.000 description 11
- 230000007246 mechanism Effects 0.000 description 11
- 150000003833 nucleoside derivatives Chemical class 0.000 description 11
- 230000009467 reduction Effects 0.000 description 11
- 108091023037 Aptamer Proteins 0.000 description 10
- 238000001353 Chip-sequencing Methods 0.000 description 10
- 125000003118 aryl group Chemical group 0.000 description 10
- 230000006870 function Effects 0.000 description 10
- 102000054786 human SUV39H1 Human genes 0.000 description 10
- 102000050832 human SUV39H2 Human genes 0.000 description 10
- 230000001404 mediated effect Effects 0.000 description 10
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 9
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 9
- 241000124008 Mammalia Species 0.000 description 9
- 101100422861 Mus musculus Suv39h1 gene Proteins 0.000 description 9
- 101100422868 Mus musculus Suv39h2 gene Proteins 0.000 description 9
- 241000283973 Oryctolagus cuniculus Species 0.000 description 9
- 108700019146 Transgenes Proteins 0.000 description 9
- 150000001412 amines Chemical class 0.000 description 9
- 125000002091 cationic group Chemical group 0.000 description 9
- 210000000349 chromosome Anatomy 0.000 description 9
- 102000045775 human KDM4A Human genes 0.000 description 9
- 102000049881 human KDM4D Human genes 0.000 description 9
- 150000003212 purines Chemical class 0.000 description 9
- 108091033409 CRISPR Proteins 0.000 description 8
- 108090000994 Catalytic RNA Proteins 0.000 description 8
- 102000053642 Catalytic RNA Human genes 0.000 description 8
- 102100028998 Histone-lysine N-methyltransferase SUV39H1 Human genes 0.000 description 8
- 108020005196 Mitochondrial DNA Proteins 0.000 description 8
- PYMYPHUHKUWMLA-LMVFSUKVSA-N Ribose Natural products OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PYMYPHUHKUWMLA-LMVFSUKVSA-N 0.000 description 8
- 125000003282 alkyl amino group Chemical group 0.000 description 8
- HMFHBZSHGGEWLO-UHFFFAOYSA-N alpha-D-Furanose-Ribose Natural products OCC1OC(O)C(O)C1O HMFHBZSHGGEWLO-UHFFFAOYSA-N 0.000 description 8
- 125000001769 aryl amino group Chemical group 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 8
- 125000004663 dialkyl amino group Chemical group 0.000 description 8
- 125000005240 diheteroarylamino group Chemical group 0.000 description 8
- 238000003197 gene knockdown Methods 0.000 description 8
- 230000002068 genetic effect Effects 0.000 description 8
- 210000004602 germ cell Anatomy 0.000 description 8
- 125000005241 heteroarylamino group Chemical group 0.000 description 8
- 239000002609 medium Substances 0.000 description 8
- 229910052698 phosphorus Inorganic materials 0.000 description 8
- 239000000047 product Substances 0.000 description 8
- 108091092562 ribozyme Proteins 0.000 description 8
- 229910052717 sulfur Inorganic materials 0.000 description 8
- HMFHBZSHGGEWLO-SOOFDHNKSA-N D-ribofuranose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H]1O HMFHBZSHGGEWLO-SOOFDHNKSA-N 0.000 description 7
- 108010053770 Deoxyribonucleases Proteins 0.000 description 7
- 102000016911 Deoxyribonucleases Human genes 0.000 description 7
- 102000006947 Histones Human genes 0.000 description 7
- 241000699670 Mus sp. Species 0.000 description 7
- 238000004458 analytical method Methods 0.000 description 7
- 125000004429 atom Chemical group 0.000 description 7
- 230000008901 benefit Effects 0.000 description 7
- 229910052799 carbon Inorganic materials 0.000 description 7
- 230000017858 demethylation Effects 0.000 description 7
- 238000010520 demethylation reaction Methods 0.000 description 7
- 238000013461 design Methods 0.000 description 7
- 125000004986 diarylamino group Chemical group 0.000 description 7
- 201000010099 disease Diseases 0.000 description 7
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 7
- 210000001654 germ layer Anatomy 0.000 description 7
- 239000003446 ligand Substances 0.000 description 7
- 239000003550 marker Substances 0.000 description 7
- 238000010373 organism cloning Methods 0.000 description 7
- 230000037361 pathway Effects 0.000 description 7
- 241000282412 Homo Species 0.000 description 6
- 239000002202 Polyethylene glycol Substances 0.000 description 6
- RWRDLPDLKQPQOW-UHFFFAOYSA-N Pyrrolidine Chemical compound C1CCNC1 RWRDLPDLKQPQOW-UHFFFAOYSA-N 0.000 description 6
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 6
- 239000000074 antisense oligonucleotide Substances 0.000 description 6
- 238000012230 antisense oligonucleotides Methods 0.000 description 6
- 210000003855 cell nucleus Anatomy 0.000 description 6
- 230000003247 decreasing effect Effects 0.000 description 6
- 230000013020 embryo development Effects 0.000 description 6
- 125000005843 halogen group Chemical group 0.000 description 6
- 229910052739 hydrogen Inorganic materials 0.000 description 6
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 6
- 238000001727 in vivo Methods 0.000 description 6
- 150000002632 lipids Chemical class 0.000 description 6
- 125000003835 nucleoside group Chemical class 0.000 description 6
- 125000004437 phosphorous atom Chemical group 0.000 description 6
- 229920001223 polyethylene glycol Polymers 0.000 description 6
- 229920000642 polymer Polymers 0.000 description 6
- 230000000392 somatic effect Effects 0.000 description 6
- 150000008163 sugars Chemical class 0.000 description 6
- 230000001225 therapeutic effect Effects 0.000 description 6
- 210000001519 tissue Anatomy 0.000 description 6
- 238000013518 transcription Methods 0.000 description 6
- 230000035897 transcription Effects 0.000 description 6
- 229940045145 uridine Drugs 0.000 description 6
- GVUOPSNMFBICMM-UHFFFAOYSA-N 5-bromo-6-morpholin-4-yl-1h-pyrimidine-2,4-dione Chemical compound OC1=NC(O)=C(Br)C(N2CCOCC2)=N1 GVUOPSNMFBICMM-UHFFFAOYSA-N 0.000 description 5
- FHYZUCKIFBIOMX-UHFFFAOYSA-N 5-bromo-6-piperidin-1-yl-1h-pyrimidine-2,4-dione Chemical compound OC1=NC(O)=C(Br)C(N2CCCCC2)=N1 FHYZUCKIFBIOMX-UHFFFAOYSA-N 0.000 description 5
- 101100422859 Bos taurus SUV39H1 gene Proteins 0.000 description 5
- 101100422864 Bos taurus SUV39H2 gene Proteins 0.000 description 5
- 108010077544 Chromatin Proteins 0.000 description 5
- 101000615488 Homo sapiens Methyl-CpG-binding domain protein 2 Proteins 0.000 description 5
- 102100021299 Methyl-CpG-binding domain protein 2 Human genes 0.000 description 5
- 108091093105 Nuclear DNA Proteins 0.000 description 5
- 102000000574 RNA-Induced Silencing Complex Human genes 0.000 description 5
- 108010016790 RNA-Induced Silencing Complex Proteins 0.000 description 5
- NINIDFKCEFEMDL-UHFFFAOYSA-N Sulfur Chemical compound [S] NINIDFKCEFEMDL-UHFFFAOYSA-N 0.000 description 5
- DRTQHJPVMGBUCF-XVFCMESISA-N Uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-XVFCMESISA-N 0.000 description 5
- 241000700605 Viruses Species 0.000 description 5
- DHKHKXVYLBGOIT-UHFFFAOYSA-N acetaldehyde Diethyl Acetal Natural products CCOC(C)OCC DHKHKXVYLBGOIT-UHFFFAOYSA-N 0.000 description 5
- 125000000304 alkynyl group Chemical group 0.000 description 5
- 238000013459 approach Methods 0.000 description 5
- 230000001413 cellular effect Effects 0.000 description 5
- 210000003483 chromatin Anatomy 0.000 description 5
- 238000003776 cleavage reaction Methods 0.000 description 5
- 230000001086 cytosolic effect Effects 0.000 description 5
- 238000010586 diagram Methods 0.000 description 5
- 230000004069 differentiation Effects 0.000 description 5
- 239000000539 dimer Substances 0.000 description 5
- 125000004119 disulfanediyl group Chemical group *SS* 0.000 description 5
- 230000001605 fetal effect Effects 0.000 description 5
- 108020001507 fusion proteins Proteins 0.000 description 5
- 102000037865 fusion proteins Human genes 0.000 description 5
- 102000052630 human KDM4B Human genes 0.000 description 5
- 102000046705 human KDM4C Human genes 0.000 description 5
- 239000001257 hydrogen Substances 0.000 description 5
- 230000006872 improvement Effects 0.000 description 5
- 239000002679 microRNA Substances 0.000 description 5
- 230000003472 neutralizing effect Effects 0.000 description 5
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 5
- 238000012545 processing Methods 0.000 description 5
- 239000002213 purine nucleotide Substances 0.000 description 5
- 150000003230 pyrimidines Chemical class 0.000 description 5
- 108700024797 rat Suv39h1 Proteins 0.000 description 5
- 230000007017 scission Effects 0.000 description 5
- 239000011593 sulfur Substances 0.000 description 5
- 238000011191 terminal modification Methods 0.000 description 5
- 230000014616 translation Effects 0.000 description 5
- 239000013603 viral vector Substances 0.000 description 5
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 4
- LRFVTYWOQMYALW-UHFFFAOYSA-N 9H-xanthine Chemical compound O=C1NC(=O)NC2=C1NC=N2 LRFVTYWOQMYALW-UHFFFAOYSA-N 0.000 description 4
- 238000010354 CRISPR gene editing Methods 0.000 description 4
- 241000282472 Canis lupus familiaris Species 0.000 description 4
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical group [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 4
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 4
- 241000283086 Equidae Species 0.000 description 4
- PIICEJLVQHRZGT-UHFFFAOYSA-N Ethylenediamine Chemical compound NCCN PIICEJLVQHRZGT-UHFFFAOYSA-N 0.000 description 4
- 241000282326 Felis catus Species 0.000 description 4
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 4
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 4
- 102100028988 Histone-lysine N-methyltransferase SUV39H2 Human genes 0.000 description 4
- 108010021466 Mutant Proteins Proteins 0.000 description 4
- 102000008300 Mutant Proteins Human genes 0.000 description 4
- 206010028980 Neoplasm Diseases 0.000 description 4
- 241000282577 Pan troglodytes Species 0.000 description 4
- 238000003559 RNA-seq method Methods 0.000 description 4
- 241000282887 Suidae Species 0.000 description 4
- 238000007792 addition Methods 0.000 description 4
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 4
- 210000004102 animal cell Anatomy 0.000 description 4
- 210000004952 blastocoel Anatomy 0.000 description 4
- 230000000903 blocking effect Effects 0.000 description 4
- 230000003197 catalytic effect Effects 0.000 description 4
- 210000000170 cell membrane Anatomy 0.000 description 4
- 239000003153 chemical reaction reagent Substances 0.000 description 4
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 4
- 230000007423 decrease Effects 0.000 description 4
- 230000007812 deficiency Effects 0.000 description 4
- 239000003814 drug Substances 0.000 description 4
- 239000000975 dye Substances 0.000 description 4
- 210000001900 endoderm Anatomy 0.000 description 4
- 210000002304 esc Anatomy 0.000 description 4
- 230000035558 fertility Effects 0.000 description 4
- 230000004720 fertilization Effects 0.000 description 4
- 239000005090 green fluorescent protein Substances 0.000 description 4
- 239000001963 growth medium Substances 0.000 description 4
- 210000005260 human cell Anatomy 0.000 description 4
- 230000001939 inductive effect Effects 0.000 description 4
- 230000003993 interaction Effects 0.000 description 4
- 238000000520 microinjection Methods 0.000 description 4
- 102000035118 modified proteins Human genes 0.000 description 4
- 108091005573 modified proteins Proteins 0.000 description 4
- 239000000178 monomer Substances 0.000 description 4
- 230000000877 morphologic effect Effects 0.000 description 4
- 229910052757 nitrogen Inorganic materials 0.000 description 4
- 210000000056 organ Anatomy 0.000 description 4
- 230000002018 overexpression Effects 0.000 description 4
- 210000001778 pluripotent stem cell Anatomy 0.000 description 4
- 230000035755 proliferation Effects 0.000 description 4
- 229940121649 protein inhibitor Drugs 0.000 description 4
- 239000012268 protein inhibitor Substances 0.000 description 4
- 239000002719 pyrimidine nucleotide Substances 0.000 description 4
- 230000002829 reductive effect Effects 0.000 description 4
- 108091069025 single-strand RNA Proteins 0.000 description 4
- 239000000243 solution Substances 0.000 description 4
- 230000000638 stimulation Effects 0.000 description 4
- 230000002194 synthesizing effect Effects 0.000 description 4
- 238000002560 therapeutic procedure Methods 0.000 description 4
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 4
- ZEMGGZBWXRYJHK-UHFFFAOYSA-N thiouracil Chemical compound O=C1C=CNC(=S)N1 ZEMGGZBWXRYJHK-UHFFFAOYSA-N 0.000 description 4
- 238000001890 transfection Methods 0.000 description 4
- 238000013519 translation Methods 0.000 description 4
- RTKIYFITIVXBLE-QEQCGCAPSA-N trichostatin A Chemical compound ONC(=O)/C=C/C(/C)=C/[C@@H](C)C(=O)C1=CC=C(N(C)C)C=C1 RTKIYFITIVXBLE-QEQCGCAPSA-N 0.000 description 4
- QOECJCJVIMVJGX-UHFFFAOYSA-N 2-cyclohexyl-6-methoxy-N-(1-propan-2-yl-4-piperidinyl)-7-[3-(1-pyrrolidinyl)propoxy]-4-quinazolinamine Chemical compound N1=C(C2CCCCC2)N=C2C=C(OCCCN3CCCC3)C(OC)=CC2=C1NC1CCN(C(C)C)CC1 QOECJCJVIMVJGX-UHFFFAOYSA-N 0.000 description 3
- BKCDJTRMYWSXMC-UHFFFAOYSA-N 5'-methoxy-6'-(3-pyrrolidin-1-ylpropoxy)spiro[cyclobutane-1,3'-indole]-2'-amine Chemical compound COC1=CC(C2(CCC2)C(N)=N2)=C2C=C1OCCCN1CCCC1 BKCDJTRMYWSXMC-UHFFFAOYSA-N 0.000 description 3
- MSSXOMSJDRHRMC-UHFFFAOYSA-N 9H-purine-2,6-diamine Chemical compound NC1=NC(N)=C2NC=NC2=N1 MSSXOMSJDRHRMC-UHFFFAOYSA-N 0.000 description 3
- 108020000948 Antisense Oligonucleotides Proteins 0.000 description 3
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 3
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 3
- 101100287812 Caenorhabditis elegans jmjd-2 gene Proteins 0.000 description 3
- KXDHJXZQYSOELW-UHFFFAOYSA-M Carbamate Chemical compound NC([O-])=O KXDHJXZQYSOELW-UHFFFAOYSA-M 0.000 description 3
- 241000255581 Drosophila <fruit fly, genus> Species 0.000 description 3
- 241000196324 Embryophyta Species 0.000 description 3
- 102000004190 Enzymes Human genes 0.000 description 3
- 108090000790 Enzymes Proteins 0.000 description 3
- NYHBQMYGNKIUIF-UUOKFMHZSA-N Guanosine Chemical group C1=NC=2C(=O)NC(N)=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O NYHBQMYGNKIUIF-UUOKFMHZSA-N 0.000 description 3
- 102100035043 Histone-lysine N-methyltransferase EHMT1 Human genes 0.000 description 3
- 101710119194 Histone-lysine N-methyltransferase SUV39H1 Proteins 0.000 description 3
- 101000907904 Homo sapiens Endoribonuclease Dicer Proteins 0.000 description 3
- 101001028703 Homo sapiens Probable JmjC domain-containing histone demethylation protein 2C Proteins 0.000 description 3
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 3
- UGQMRVRMYYASKQ-KQYNXXCUSA-N Inosine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(O)=C2N=C1 UGQMRVRMYYASKQ-KQYNXXCUSA-N 0.000 description 3
- 241000713666 Lentivirus Species 0.000 description 3
- 108060001084 Luciferase Proteins 0.000 description 3
- 241000282553 Macaca Species 0.000 description 3
- 229940122938 MicroRNA inhibitor Drugs 0.000 description 3
- 101100287809 Mus musculus Kdm4d gene Proteins 0.000 description 3
- WSDRAZIPGVLSNP-UHFFFAOYSA-N O.P(=O)(O)(O)O.O.O.P(=O)(O)(O)O Chemical compound O.P(=O)(O)(O)O.O.O.P(=O)(O)(O)O WSDRAZIPGVLSNP-UHFFFAOYSA-N 0.000 description 3
- 206010033266 Ovarian Hyperstimulation Syndrome Diseases 0.000 description 3
- 241000282320 Panthera leo Species 0.000 description 3
- 102100037169 Probable JmjC domain-containing histone demethylation protein 2C Human genes 0.000 description 3
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 3
- 101100287810 Rattus norvegicus Kdm4d gene Proteins 0.000 description 3
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 3
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 3
- 108010041897 SU(VAR)3-9 Proteins 0.000 description 3
- 229940124639 Selective inhibitor Drugs 0.000 description 3
- 108091081021 Sense strand Proteins 0.000 description 3
- 102000040945 Transcription factor Human genes 0.000 description 3
- 108091023040 Transcription factor Proteins 0.000 description 3
- 150000001241 acetals Chemical class 0.000 description 3
- 239000002253 acid Substances 0.000 description 3
- 229960005305 adenosine Drugs 0.000 description 3
- 230000002411 adverse Effects 0.000 description 3
- 125000003342 alkenyl group Chemical group 0.000 description 3
- 230000004075 alteration Effects 0.000 description 3
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 3
- 210000003719 b-lymphocyte Anatomy 0.000 description 3
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 3
- 230000010261 cell growth Effects 0.000 description 3
- 238000004590 computer program Methods 0.000 description 3
- 230000021615 conjugation Effects 0.000 description 3
- 210000001771 cumulus cell Anatomy 0.000 description 3
- 125000004122 cyclic group Chemical group 0.000 description 3
- 125000000753 cycloalkyl group Chemical group 0.000 description 3
- 238000012217 deletion Methods 0.000 description 3
- 230000037430 deletion Effects 0.000 description 3
- 230000001419 dependent effect Effects 0.000 description 3
- 238000009795 derivation Methods 0.000 description 3
- NAGJZTKCGNOGPW-UHFFFAOYSA-K dioxido-sulfanylidene-sulfido-$l^{5}-phosphane Chemical compound [O-]P([O-])([S-])=S NAGJZTKCGNOGPW-UHFFFAOYSA-K 0.000 description 3
- 238000009826 distribution Methods 0.000 description 3
- 210000002308 embryonic cell Anatomy 0.000 description 3
- 229940088598 enzyme Drugs 0.000 description 3
- 238000011156 evaluation Methods 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- 210000002950 fibroblast Anatomy 0.000 description 3
- 108091006047 fluorescent proteins Proteins 0.000 description 3
- 102000034287 fluorescent proteins Human genes 0.000 description 3
- 125000000524 functional group Chemical group 0.000 description 3
- 125000003147 glycosyl group Chemical group 0.000 description 3
- 230000012010 growth Effects 0.000 description 3
- 125000004435 hydrogen atom Chemical group [H]* 0.000 description 3
- 230000001976 improved effect Effects 0.000 description 3
- 230000006698 induction Effects 0.000 description 3
- 210000000936 intestine Anatomy 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 210000004072 lung Anatomy 0.000 description 3
- 229920002521 macromolecule Polymers 0.000 description 3
- 230000008774 maternal effect Effects 0.000 description 3
- 125000000325 methylidene group Chemical group [H]C([H])=* 0.000 description 3
- 108091070501 miRNA Proteins 0.000 description 3
- 208000012268 mitochondrial disease Diseases 0.000 description 3
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 3
- 229930014626 natural product Natural products 0.000 description 3
- 230000007935 neutral effect Effects 0.000 description 3
- 230000027758 ovulation cycle Effects 0.000 description 3
- 239000012071 phase Substances 0.000 description 3
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 3
- 150000004713 phosphodiesters Chemical class 0.000 description 3
- 239000013600 plasmid vector Substances 0.000 description 3
- 230000032361 posttranscriptional gene silencing Effects 0.000 description 3
- 239000002243 precursor Substances 0.000 description 3
- 125000002568 propynyl group Chemical group [*]C#CC([H])([H])[H] 0.000 description 3
- 230000001172 regenerating effect Effects 0.000 description 3
- 230000001105 regulatory effect Effects 0.000 description 3
- 238000011160 research Methods 0.000 description 3
- 239000000523 sample Substances 0.000 description 3
- 239000007790 solid phase Substances 0.000 description 3
- 125000001424 substituent group Chemical group 0.000 description 3
- 235000021092 sugar substitutes Nutrition 0.000 description 3
- 239000003765 sweetening agent Substances 0.000 description 3
- 230000001360 synchronised effect Effects 0.000 description 3
- 125000004001 thioalkyl group Chemical group 0.000 description 3
- 230000001052 transient effect Effects 0.000 description 3
- 239000001226 triphosphate Substances 0.000 description 3
- 241001430294 unidentified retrovirus Species 0.000 description 3
- ZEQIWKHCJWRNTH-UHFFFAOYSA-N 1h-pyrimidine-2,4-dithione Chemical compound S=C1C=CNC(=S)N1 ZEQIWKHCJWRNTH-UHFFFAOYSA-N 0.000 description 2
- SXUXMRMBWZCMEN-UHFFFAOYSA-N 2'-O-methyl uridine Natural products COC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 SXUXMRMBWZCMEN-UHFFFAOYSA-N 0.000 description 2
- SXUXMRMBWZCMEN-ZOQUXTDFSA-N 2'-O-methyluridine Chemical compound CO[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 SXUXMRMBWZCMEN-ZOQUXTDFSA-N 0.000 description 2
- IQFYYKKMVGJFEH-BIIVOSGPSA-N 2'-deoxythymidine Natural products O=C1NC(=O)C(C)=CN1[C@@H]1O[C@@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-BIIVOSGPSA-N 0.000 description 2
- 102100025425 2-oxoglutarate and iron-dependent oxygenase JMJD4 Human genes 0.000 description 2
- GJTBSTBJLVYKAU-XVFCMESISA-N 2-thiouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=S)NC(=O)C=C1 GJTBSTBJLVYKAU-XVFCMESISA-N 0.000 description 2
- OIVLITBTBDPEFK-UHFFFAOYSA-N 5,6-dihydrouracil Chemical compound O=C1CCNC(=O)N1 OIVLITBTBDPEFK-UHFFFAOYSA-N 0.000 description 2
- OSXFATOLZGZLSK-UHFFFAOYSA-N 6,7-dimethoxy-2-(4-methyl-1,4-diazepan-1-yl)-N-[1-(phenylmethyl)-4-piperidinyl]-4-quinazolinamine Chemical compound C=12C=C(OC)C(OC)=CC2=NC(N2CCN(C)CCC2)=NC=1NC(CC1)CCN1CC1=CC=CC=C1 OSXFATOLZGZLSK-UHFFFAOYSA-N 0.000 description 2
- OGHAROSJZRTIOK-KQYNXXCUSA-O 7-methylguanosine Chemical compound C1=2N=C(N)NC(=O)C=2[N+](C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OGHAROSJZRTIOK-KQYNXXCUSA-O 0.000 description 2
- 108020005544 Antisense RNA Proteins 0.000 description 2
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 2
- 102100023962 Bifunctional arginine demethylase and lysyl-hydroxylase JMJD6 Human genes 0.000 description 2
- 102100023960 Bifunctional peptidase and (3S)-lysyl hydroxylase JMJD7 Human genes 0.000 description 2
- 102100037468 Bifunctional peptidase and arginyl-hydroxylase JMJD5 Human genes 0.000 description 2
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 2
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 2
- 102000004533 Endonucleases Human genes 0.000 description 2
- 108010042407 Endonucleases Proteins 0.000 description 2
- 102100023387 Endoribonuclease Dicer Human genes 0.000 description 2
- 241000283073 Equus caballus Species 0.000 description 2
- 108060002716 Exonuclease Proteins 0.000 description 2
- 102100027727 F-box/LRR-repeat protein 19 Human genes 0.000 description 2
- 241000287828 Gallus gallus Species 0.000 description 2
- 108010034791 Heterochromatin Proteins 0.000 description 2
- 102000003964 Histone deacetylase Human genes 0.000 description 2
- 108090000353 Histone deacetylase Proteins 0.000 description 2
- 102100035864 Histone lysine demethylase PHF8 Human genes 0.000 description 2
- 102100025210 Histone-arginine methyltransferase CARM1 Human genes 0.000 description 2
- 102100023696 Histone-lysine N-methyltransferase SETDB1 Human genes 0.000 description 2
- 101000934666 Homo sapiens 2-oxoglutarate and iron-dependent oxygenase JMJD4 Proteins 0.000 description 2
- 101000975541 Homo sapiens Bifunctional arginine demethylase and lysyl-hydroxylase JMJD6 Proteins 0.000 description 2
- 101000975540 Homo sapiens Bifunctional peptidase and (3S)-lysyl hydroxylase JMJD7 Proteins 0.000 description 2
- 101001025948 Homo sapiens Bifunctional peptidase and arginyl-hydroxylase JMJD5 Proteins 0.000 description 2
- 101000862205 Homo sapiens F-box/LRR-repeat protein 19 Proteins 0.000 description 2
- 101001000378 Homo sapiens Histone lysine demethylase PHF8 Proteins 0.000 description 2
- 101000877314 Homo sapiens Histone-lysine N-methyltransferase EHMT1 Proteins 0.000 description 2
- 101000975533 Homo sapiens JmjC domain-containing protein 8 Proteins 0.000 description 2
- 101000613958 Homo sapiens Lysine-specific demethylase 2A Proteins 0.000 description 2
- 101001025945 Homo sapiens Lysine-specific demethylase 7A Proteins 0.000 description 2
- 101000692954 Homo sapiens Lysine-specific demethylase PHF2 Proteins 0.000 description 2
- 101000613960 Homo sapiens Lysine-specific histone demethylase 1B Proteins 0.000 description 2
- 101000796144 Homo sapiens Protein arginine N-methyltransferase 9 Proteins 0.000 description 2
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 2
- 102100029098 Hypoxanthine-guanine phosphoribosyltransferase Human genes 0.000 description 2
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 2
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 2
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 2
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 2
- 102100023958 JmjC domain-containing protein 8 Human genes 0.000 description 2
- 241000254158 Lampyridae Species 0.000 description 2
- 102000006830 Luminescent Proteins Human genes 0.000 description 2
- 108010047357 Luminescent Proteins Proteins 0.000 description 2
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 2
- 102100040598 Lysine-specific demethylase 2A Human genes 0.000 description 2
- 102100040581 Lysine-specific demethylase 3A Human genes 0.000 description 2
- 102100033246 Lysine-specific demethylase 5A Human genes 0.000 description 2
- 102100033247 Lysine-specific demethylase 5B Human genes 0.000 description 2
- 102100037465 Lysine-specific demethylase 7A Human genes 0.000 description 2
- 102100026395 Lysine-specific demethylase PHF2 Human genes 0.000 description 2
- 102100040596 Lysine-specific histone demethylase 1B Human genes 0.000 description 2
- 108700011259 MicroRNAs Proteins 0.000 description 2
- YNAVUWVOSKDBBP-UHFFFAOYSA-N Morpholine Chemical compound C1COCCN1 YNAVUWVOSKDBBP-UHFFFAOYSA-N 0.000 description 2
- 240000000249 Morus alba Species 0.000 description 2
- 235000008708 Morus alba Nutrition 0.000 description 2
- 101100256746 Mus musculus Setdb1 gene Proteins 0.000 description 2
- 101100489547 Mus musculus Zscan4d gene Proteins 0.000 description 2
- NWIBSHFKIJFRCO-WUDYKRTCSA-N Mytomycin Chemical compound C1N2C(C(C(C)=C(N)C3=O)=O)=C3[C@@H](COC(N)=O)[C@@]2(OC)[C@@H]2[C@H]1N2 NWIBSHFKIJFRCO-WUDYKRTCSA-N 0.000 description 2
- 108700005081 Overlapping Genes Proteins 0.000 description 2
- 208000018737 Parkinson disease Diseases 0.000 description 2
- 241001494479 Pecora Species 0.000 description 2
- 241000283080 Proboscidea <mammal> Species 0.000 description 2
- RJKFOVLPORLFTN-LEKSSAKUSA-N Progesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](C(=O)C)[C@@]1(C)CC2 RJKFOVLPORLFTN-LEKSSAKUSA-N 0.000 description 2
- 102100031369 Protein arginine N-methyltransferase 9 Human genes 0.000 description 2
- 230000004570 RNA-binding Effects 0.000 description 2
- 238000011529 RT qPCR Methods 0.000 description 2
- 108020004511 Recombinant DNA Proteins 0.000 description 2
- 241000269821 Scombridae Species 0.000 description 2
- 102000039471 Small Nuclear RNA Human genes 0.000 description 2
- RYYWUUFWQRZTIU-UHFFFAOYSA-N Thiophosphoric acid Chemical class OP(O)(S)=O RYYWUUFWQRZTIU-UHFFFAOYSA-N 0.000 description 2
- 108010022394 Threonine synthase Proteins 0.000 description 2
- 102000006601 Thymidine Kinase Human genes 0.000 description 2
- 108020004440 Thymidine kinase Proteins 0.000 description 2
- 230000002159 abnormal effect Effects 0.000 description 2
- 230000005856 abnormality Effects 0.000 description 2
- DZBUGLKDJFMEHC-UHFFFAOYSA-N acridine Chemical compound C1=CC=CC2=CC3=CC=CC=C3N=C21 DZBUGLKDJFMEHC-UHFFFAOYSA-N 0.000 description 2
- 230000009471 action Effects 0.000 description 2
- 125000005600 alkyl phosphonate group Chemical group 0.000 description 2
- 125000004103 aminoalkyl group Chemical group 0.000 description 2
- 239000005557 antagonist Substances 0.000 description 2
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 2
- 125000003710 aryl alkyl group Chemical group 0.000 description 2
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 2
- DRTQHJPVMGBUCF-PSQAKQOGSA-N beta-L-uridine Natural products O[C@H]1[C@@H](O)[C@H](CO)O[C@@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-PSQAKQOGSA-N 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 210000004556 brain Anatomy 0.000 description 2
- 201000011510 cancer Diseases 0.000 description 2
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 2
- 125000002057 carboxymethyl group Chemical group [H]OC(=O)C([H])([H])[*] 0.000 description 2
- 230000024245 cell differentiation Effects 0.000 description 2
- 230000032823 cell division Effects 0.000 description 2
- PZPPOCZWRGNKIR-PNVYSBBASA-N chaetocin Chemical compound N([C@@H]1N2C(=O)[C@]3(CO)SS[C@]2(C(N3C)=O)C2)C3=CC=CC=C3[C@]12[C@@]12C[C@]3(SS4)C(=O)N(C)[C@]4(CO)C(=O)N3[C@H]2NC2=CC=CC=C12 PZPPOCZWRGNKIR-PNVYSBBASA-N 0.000 description 2
- PZPPOCZWRGNKIR-UHFFFAOYSA-N chaetocin Natural products C1C2(C(N3C)=O)SSC3(CO)C(=O)N2C2NC3=CC=CC=C3C21C12CC3(SS4)C(=O)N(C)C4(CO)C(=O)N3C2NC2=CC=CC=C12 PZPPOCZWRGNKIR-UHFFFAOYSA-N 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 235000013330 chicken meat Nutrition 0.000 description 2
- 235000012000 cholesterol Nutrition 0.000 description 2
- 230000002759 chromosomal effect Effects 0.000 description 2
- 108010030886 coactivator-associated arginine methyltransferase 1 Proteins 0.000 description 2
- 239000003184 complementary RNA Substances 0.000 description 2
- 239000012141 concentrate Substances 0.000 description 2
- FPUGCISOLXNPPC-IOSLPCCCSA-N cordysinin B Chemical compound CO[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(N)=C2N=C1 FPUGCISOLXNPPC-IOSLPCCCSA-N 0.000 description 2
- 238000004132 cross linking Methods 0.000 description 2
- 230000002950 deficient Effects 0.000 description 2
- 230000001335 demethylating effect Effects 0.000 description 2
- 239000000412 dendrimer Substances 0.000 description 2
- 229920000736 dendritic polymer Polymers 0.000 description 2
- 239000005547 deoxyribonucleotide Substances 0.000 description 2
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 2
- 230000008021 deposition Effects 0.000 description 2
- 102000004419 dihydrofolate reductase Human genes 0.000 description 2
- 239000001177 diphosphate Substances 0.000 description 2
- NAGJZTKCGNOGPW-UHFFFAOYSA-N dithiophosphoric acid Chemical class OP(O)(S)=S NAGJZTKCGNOGPW-UHFFFAOYSA-N 0.000 description 2
- 210000004039 endoderm cell Anatomy 0.000 description 2
- 230000002255 enzymatic effect Effects 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 102000013165 exonuclease Human genes 0.000 description 2
- 238000010195 expression analysis Methods 0.000 description 2
- 239000013604 expression vector Substances 0.000 description 2
- 239000012530 fluid Substances 0.000 description 2
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 2
- 238000009472 formulation Methods 0.000 description 2
- 238000012239 gene modification Methods 0.000 description 2
- 238000010353 genetic engineering Methods 0.000 description 2
- 230000005017 genetic modification Effects 0.000 description 2
- 235000013617 genetically modified food Nutrition 0.000 description 2
- 125000001072 heteroaryl group Chemical group 0.000 description 2
- 210000004458 heterochromatin Anatomy 0.000 description 2
- 238000002744 homologous recombination Methods 0.000 description 2
- 230000006801 homologous recombination Effects 0.000 description 2
- 102000051823 human PRDM2 Human genes 0.000 description 2
- 210000004754 hybrid cell Anatomy 0.000 description 2
- 210000004408 hybridoma Anatomy 0.000 description 2
- 230000028993 immune response Effects 0.000 description 2
- 239000007943 implant Substances 0.000 description 2
- 230000003834 intracellular effect Effects 0.000 description 2
- 238000010253 intravenous injection Methods 0.000 description 2
- 239000010410 layer Substances 0.000 description 2
- 239000006166 lysate Substances 0.000 description 2
- 235000020640 mackerel Nutrition 0.000 description 2
- 238000007726 management method Methods 0.000 description 2
- 239000011159 matrix material Substances 0.000 description 2
- 210000002752 melanocyte Anatomy 0.000 description 2
- 210000004379 membrane Anatomy 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- 125000006533 methyl amino methyl group Chemical group [H]N(C([H])([H])[H])C([H])([H])* 0.000 description 2
- 125000000250 methylamino group Chemical group [H]N(*)C([H])([H])[H] 0.000 description 2
- 239000000693 micelle Substances 0.000 description 2
- 230000003278 mimic effect Effects 0.000 description 2
- 230000000394 mitotic effect Effects 0.000 description 2
- 238000002156 mixing Methods 0.000 description 2
- 230000002107 myocardial effect Effects 0.000 description 2
- 210000000633 nuclear envelope Anatomy 0.000 description 2
- 239000011824 nuclear material Substances 0.000 description 2
- 230000009437 off-target effect Effects 0.000 description 2
- 210000000496 pancreas Anatomy 0.000 description 2
- 239000002245 particle Substances 0.000 description 2
- 239000013612 plasmid Substances 0.000 description 2
- 210000004508 polar body Anatomy 0.000 description 2
- 102000040430 polynucleotide Human genes 0.000 description 2
- 108091033319 polynucleotide Proteins 0.000 description 2
- 239000002157 polynucleotide Substances 0.000 description 2
- 230000001124 posttranscriptional effect Effects 0.000 description 2
- 230000003389 potentiating effect Effects 0.000 description 2
- 108091007428 primary miRNA Proteins 0.000 description 2
- 230000002062 proliferating effect Effects 0.000 description 2
- 230000001737 promoting effect Effects 0.000 description 2
- 125000001436 propyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])[H] 0.000 description 2
- 229940048914 protamine Drugs 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- RXWNCPJZOCPEPQ-NVWDDTSBSA-N puromycin Chemical compound C1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CO RXWNCPJZOCPEPQ-NVWDDTSBSA-N 0.000 description 2
- 108010045647 puromycin N-acetyltransferase Proteins 0.000 description 2
- KIDHWZJUCRJVML-UHFFFAOYSA-N putrescine Chemical compound NCCCCN KIDHWZJUCRJVML-UHFFFAOYSA-N 0.000 description 2
- BBEAQIROQSPTKN-UHFFFAOYSA-N pyrene Chemical compound C1=CC=C2C=CC3=CC=CC4=CC=C1C2=C43 BBEAQIROQSPTKN-UHFFFAOYSA-N 0.000 description 2
- 238000003762 quantitative reverse transcription PCR Methods 0.000 description 2
- 230000006798 recombination Effects 0.000 description 2
- 238000005215 recombination Methods 0.000 description 2
- 230000008929 regeneration Effects 0.000 description 2
- 238000011069 regeneration method Methods 0.000 description 2
- 125000006853 reporter group Chemical group 0.000 description 2
- 239000011347 resin Substances 0.000 description 2
- 229920005989 resin Polymers 0.000 description 2
- 239000002342 ribonucleoside Substances 0.000 description 2
- DWRXFEITVBNRMK-JXOAFFINSA-N ribothymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 DWRXFEITVBNRMK-JXOAFFINSA-N 0.000 description 2
- 229920002477 rna polymer Polymers 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 108091029842 small nuclear ribonucleic acid Proteins 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- PFNFFQXMRSDOHW-UHFFFAOYSA-N spermine Chemical compound NCCCNCCCCNCCCN PFNFFQXMRSDOHW-UHFFFAOYSA-N 0.000 description 2
- 238000011476 stem cell transplantation Methods 0.000 description 2
- 230000004936 stimulating effect Effects 0.000 description 2
- 238000003860 storage Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 238000012360 testing method Methods 0.000 description 2
- 150000003573 thiols Chemical class 0.000 description 2
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 2
- 239000003104 tissue culture media Substances 0.000 description 2
- 230000002103 transcriptional effect Effects 0.000 description 2
- 108091006107 transcriptional repressors Proteins 0.000 description 2
- 235000011178 triphosphate Nutrition 0.000 description 2
- UNXRWKVEANCORM-UHFFFAOYSA-N triphosphoric acid Chemical compound OP(O)(=O)OP(O)(=O)OP(O)(O)=O UNXRWKVEANCORM-UHFFFAOYSA-N 0.000 description 2
- 238000009281 ultraviolet germicidal irradiation Methods 0.000 description 2
- DRTQHJPVMGBUCF-UHFFFAOYSA-N uracil arabinoside Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-UHFFFAOYSA-N 0.000 description 2
- 210000004291 uterus Anatomy 0.000 description 2
- GRYSXUXXBDSYRT-WOUKDFQISA-N (2r,3r,4r,5r)-2-(hydroxymethyl)-4-methoxy-5-[6-(methylamino)purin-9-yl]oxolan-3-ol Chemical compound C1=NC=2C(NC)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1OC GRYSXUXXBDSYRT-WOUKDFQISA-N 0.000 description 1
- YIMATHOGWXZHFX-WCTZXXKLSA-N (2r,3r,4r,5r)-5-(hydroxymethyl)-3-(2-methoxyethoxy)oxolane-2,4-diol Chemical compound COCCO[C@H]1[C@H](O)O[C@H](CO)[C@H]1O YIMATHOGWXZHFX-WCTZXXKLSA-N 0.000 description 1
- STGXGJRRAJKJRG-JDJSBBGDSA-N (3r,4r,5r)-5-(hydroxymethyl)-3-methoxyoxolane-2,4-diol Chemical compound CO[C@H]1C(O)O[C@H](CO)[C@H]1O STGXGJRRAJKJRG-JDJSBBGDSA-N 0.000 description 1
- 150000000178 1,2,4-triazoles Chemical class 0.000 description 1
- UIYWFOZZIZEEKJ-XVFCMESISA-N 1-[(2r,3r,4r,5r)-3-fluoro-4-hydroxy-5-(hydroxymethyl)oxolan-2-yl]pyrimidine-2,4-dione Chemical compound F[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 UIYWFOZZIZEEKJ-XVFCMESISA-N 0.000 description 1
- QPHRQMAYYMYWFW-FJGDRVTGSA-N 1-[(2r,3s,4r,5r)-3-fluoro-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]pyrimidine-2,4-dione Chemical compound O[C@]1(F)[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 QPHRQMAYYMYWFW-FJGDRVTGSA-N 0.000 description 1
- HBOMLICNUCNMMY-KJFJCRTCSA-N 1-[(4s,5s)-4-azido-5-(hydroxymethyl)oxolan-2-yl]-5-methylpyrimidine-2,4-dione Chemical compound O=C1NC(=O)C(C)=CN1C1O[C@H](CO)[C@@H](N=[N+]=[N-])C1 HBOMLICNUCNMMY-KJFJCRTCSA-N 0.000 description 1
- DURPTKYDGMDSBL-UHFFFAOYSA-N 1-butoxybutane Chemical compound CCCCOCCCC DURPTKYDGMDSBL-UHFFFAOYSA-N 0.000 description 1
- KAESVJOAVNADME-UHFFFAOYSA-N 1H-pyrrole Natural products C=1C=CNC=1 KAESVJOAVNADME-UHFFFAOYSA-N 0.000 description 1
- UNFCIEAAJFHALK-UHFFFAOYSA-N 1h-diazepine;quinazolin-2-amine Chemical class N1C=CC=CC=N1.C1=CC=CC2=NC(N)=NC=C21 UNFCIEAAJFHALK-UHFFFAOYSA-N 0.000 description 1
- FPUGCISOLXNPPC-UHFFFAOYSA-N 2'-O-Methyladenosine Natural products COC1C(O)C(CO)OC1N1C2=NC=NC(N)=C2N=C1 FPUGCISOLXNPPC-UHFFFAOYSA-N 0.000 description 1
- OVYNGSFVYRPRCG-UHFFFAOYSA-N 2'-O-Methylguanosine Natural products COC1C(O)C(CO)OC1N1C(NC(N)=NC2=O)=C2N=C1 OVYNGSFVYRPRCG-UHFFFAOYSA-N 0.000 description 1
- OVYNGSFVYRPRCG-KQYNXXCUSA-N 2'-O-methylguanosine Chemical compound CO[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(N=C(N)NC2=O)=C2N=C1 OVYNGSFVYRPRCG-KQYNXXCUSA-N 0.000 description 1
- MXHRCPNRJAMMIM-SHYZEUOFSA-N 2'-deoxyuridine Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 MXHRCPNRJAMMIM-SHYZEUOFSA-N 0.000 description 1
- FDZGOVDEFRJXFT-UHFFFAOYSA-N 2-(3-aminopropyl)-7h-purin-6-amine Chemical compound NCCCC1=NC(N)=C2NC=NC2=N1 FDZGOVDEFRJXFT-UHFFFAOYSA-N 0.000 description 1
- WWJMLJDSGOGNFJ-UHFFFAOYSA-N 2-amino-4-(2,4-dioxo-1h-pyrimidin-5-yl)butanoic acid Chemical compound OC(=O)C(N)CCC1=CNC(=O)NC1=O WWJMLJDSGOGNFJ-UHFFFAOYSA-N 0.000 description 1
- SCVJRXQHFJXZFZ-KVQBGUIXSA-N 2-amino-9-[(2r,4s,5r)-4-hydroxy-5-(hydroxymethyl)oxolan-2-yl]-3h-purine-6-thione Chemical compound C1=2NC(N)=NC(=S)C=2N=CN1[C@H]1C[C@H](O)[C@@H](CO)O1 SCVJRXQHFJXZFZ-KVQBGUIXSA-N 0.000 description 1
- MWBWWFOAEOYUST-UHFFFAOYSA-N 2-aminopurine Chemical compound NC1=NC=C2N=CNC2=N1 MWBWWFOAEOYUST-UHFFFAOYSA-N 0.000 description 1
- SMADWRYCYBUIKH-UHFFFAOYSA-N 2-methyl-7h-purin-6-amine Chemical compound CC1=NC(N)=C2NC=NC2=N1 SMADWRYCYBUIKH-UHFFFAOYSA-N 0.000 description 1
- 125000006325 2-propenyl amino group Chemical group [H]C([H])=C([H])C([H])([H])N([H])* 0.000 description 1
- USCCECGPGBGFOM-UHFFFAOYSA-N 2-propyl-7h-purin-6-amine Chemical compound CCCC1=NC(N)=C2NC=NC2=N1 USCCECGPGBGFOM-UHFFFAOYSA-N 0.000 description 1
- VPLZGVOSFFCKFC-UHFFFAOYSA-N 3-methyluracil Chemical compound CN1C(=O)C=CNC1=O VPLZGVOSFFCKFC-UHFFFAOYSA-N 0.000 description 1
- ASFAFOSQXBRFMV-LJQANCHMSA-N 3-n-(2-benzyl-1,3-dihydroxypropan-2-yl)-1-n-[(1r)-1-(4-fluorophenyl)ethyl]-5-[methyl(methylsulfonyl)amino]benzene-1,3-dicarboxamide Chemical compound N([C@H](C)C=1C=CC(F)=CC=1)C(=O)C(C=1)=CC(N(C)S(C)(=O)=O)=CC=1C(=O)NC(CO)(CO)CC1=CC=CC=C1 ASFAFOSQXBRFMV-LJQANCHMSA-N 0.000 description 1
- LOJNBPNACKZWAI-UHFFFAOYSA-N 3-nitro-1h-pyrrole Chemical compound [O-][N+](=O)C=1C=CNC=1 LOJNBPNACKZWAI-UHFFFAOYSA-N 0.000 description 1
- ZLOIGESWDJYCTF-UHFFFAOYSA-N 4-Thiouridine Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=S)C=C1 ZLOIGESWDJYCTF-UHFFFAOYSA-N 0.000 description 1
- 229960000549 4-dimethylaminophenol Drugs 0.000 description 1
- VHYFNPMBLIVWCW-UHFFFAOYSA-N 4-dimethylaminopyridine Substances CN(C)C1=CC=NC=C1 VHYFNPMBLIVWCW-UHFFFAOYSA-N 0.000 description 1
- PHAFOFIVSNSAPQ-UHFFFAOYSA-N 4-fluoro-6-methyl-1h-benzimidazole Chemical compound CC1=CC(F)=C2NC=NC2=C1 PHAFOFIVSNSAPQ-UHFFFAOYSA-N 0.000 description 1
- QCXGJTGMGJOYDP-UHFFFAOYSA-N 4-methyl-1h-benzimidazole Chemical compound CC1=CC=CC2=C1N=CN2 QCXGJTGMGJOYDP-UHFFFAOYSA-N 0.000 description 1
- ZLOIGESWDJYCTF-XVFCMESISA-N 4-thiouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=S)C=C1 ZLOIGESWDJYCTF-XVFCMESISA-N 0.000 description 1
- 101710169336 5'-deoxyadenosine deaminase Proteins 0.000 description 1
- 125000004032 5'-inosinyl group Chemical group 0.000 description 1
- WPQLFQWYPPALOX-UHFFFAOYSA-N 5-(2-aminopropyl)-1h-pyrimidine-2,4-dione Chemical compound CC(N)CC1=CNC(=O)NC1=O WPQLFQWYPPALOX-UHFFFAOYSA-N 0.000 description 1
- LMNPKIOZMGYQIU-UHFFFAOYSA-N 5-(trifluoromethyl)-1h-pyrimidine-2,4-dione Chemical compound FC(F)(F)C1=CNC(=O)NC1=O LMNPKIOZMGYQIU-UHFFFAOYSA-N 0.000 description 1
- DGCFDETWIXOSIF-UHFFFAOYSA-N 5-[(2,4-dioxo-1H-pyrimidin-5-yl)diazenyl]-1H-pyrimidine-2,4-dione Chemical compound N(=NC=1C(NC(NC=1)=O)=O)C=1C(NC(NC=1)=O)=O DGCFDETWIXOSIF-UHFFFAOYSA-N 0.000 description 1
- IPRQAJTUSRLECG-UHFFFAOYSA-N 5-[6-(dimethylamino)purin-9-yl]-2-(hydroxymethyl)-4-methoxyoxolan-3-ol Chemical compound COC1C(O)C(CO)OC1N1C2=NC=NC(N(C)C)=C2N=C1 IPRQAJTUSRLECG-UHFFFAOYSA-N 0.000 description 1
- LQLQRFGHAALLLE-UHFFFAOYSA-N 5-bromouracil Chemical compound BrC1=CNC(=O)NC1=O LQLQRFGHAALLLE-UHFFFAOYSA-N 0.000 description 1
- KSNXJLQDQOIRIP-UHFFFAOYSA-N 5-iodouracil Chemical compound IC1=CNC(=O)NC1=O KSNXJLQDQOIRIP-UHFFFAOYSA-N 0.000 description 1
- KELXHQACBIUYSE-UHFFFAOYSA-N 5-methoxy-1h-pyrimidine-2,4-dione Chemical compound COC1=CNC(=O)NC1=O KELXHQACBIUYSE-UHFFFAOYSA-N 0.000 description 1
- OZFPSOBLQZPIAV-UHFFFAOYSA-N 5-nitro-1h-indole Chemical compound [O-][N+](=O)C1=CC=C2NC=CC2=C1 OZFPSOBLQZPIAV-UHFFFAOYSA-N 0.000 description 1
- UJBCLAXPPIDQEE-UHFFFAOYSA-N 5-prop-1-ynyl-1h-pyrimidine-2,4-dione Chemical compound CC#CC1=CNC(=O)NC1=O UJBCLAXPPIDQEE-UHFFFAOYSA-N 0.000 description 1
- 108091027075 5S-rRNA precursor Proteins 0.000 description 1
- JTDYUFSDZATMKU-UHFFFAOYSA-N 6-(1,3-dioxo-2-benzo[de]isoquinolinyl)-N-hydroxyhexanamide Chemical compound C1=CC(C(N(CCCCCC(=O)NO)C2=O)=O)=C3C2=CC=CC3=C1 JTDYUFSDZATMKU-UHFFFAOYSA-N 0.000 description 1
- OHILKUISCGPRMQ-UHFFFAOYSA-N 6-amino-5-(trifluoromethyl)-1h-pyrimidin-2-one Chemical compound NC1=NC(=O)NC=C1C(F)(F)F OHILKUISCGPRMQ-UHFFFAOYSA-N 0.000 description 1
- CKOMXBHMKXXTNW-UHFFFAOYSA-N 6-methyladenine Chemical compound CNC1=NC=NC2=C1N=CN2 CKOMXBHMKXXTNW-UHFFFAOYSA-N 0.000 description 1
- JKUJKKGMOZDDJV-ZRNGKTOUSA-N 8-O-methylsterigmatocystin Chemical compound O([C@H]1OC=C[C@H]1C1=C2O3)C1=CC(OC)=C2C(=O)C1=C3C=CC=C1OC JKUJKKGMOZDDJV-ZRNGKTOUSA-N 0.000 description 1
- RGKBRPAAQSHTED-UHFFFAOYSA-N 8-oxoadenine Chemical compound NC1=NC=NC2=C1NC(=O)N2 RGKBRPAAQSHTED-UHFFFAOYSA-N 0.000 description 1
- DLFVBJFMPXGRIB-UHFFFAOYSA-N Acetamide Chemical compound CC(N)=O DLFVBJFMPXGRIB-UHFFFAOYSA-N 0.000 description 1
- 102220504802 Acetylcholine receptor subunit alpha_G4C_mutation Human genes 0.000 description 1
- 241000251468 Actinopterygii Species 0.000 description 1
- 102100036664 Adenosine deaminase Human genes 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- 241000282452 Ailuropoda melanoleuca Species 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 108091029845 Aminoallyl nucleotide Proteins 0.000 description 1
- 235000002198 Annona diversifolia Nutrition 0.000 description 1
- 108020004491 Antisense DNA Proteins 0.000 description 1
- 101100108891 Arabidopsis thaliana PRMT11 gene Proteins 0.000 description 1
- 208000023275 Autoimmune disease Diseases 0.000 description 1
- 108700020463 BRCA1 Proteins 0.000 description 1
- 102000036365 BRCA1 Human genes 0.000 description 1
- 108091032955 Bacterial small RNA Proteins 0.000 description 1
- 241000283726 Bison Species 0.000 description 1
- ZOXJGFHDIHLPTG-UHFFFAOYSA-N Boron Chemical compound [B] ZOXJGFHDIHLPTG-UHFFFAOYSA-N 0.000 description 1
- 241000701822 Bovine papillomavirus Species 0.000 description 1
- BVKZGUZCCUSVTD-UHFFFAOYSA-L Carbonate Chemical compound [O-]C([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-L 0.000 description 1
- 102000055007 Cartilage Oligomeric Matrix Human genes 0.000 description 1
- 101710176668 Cartilage oligomeric matrix protein Proteins 0.000 description 1
- 241000700198 Cavia Species 0.000 description 1
- 241000700199 Cavia porcellus Species 0.000 description 1
- 241000282994 Cervidae Species 0.000 description 1
- 241000221955 Chaetomium Species 0.000 description 1
- GXGJIOMUZAGVEH-UHFFFAOYSA-N Chamazulene Chemical group CCC1=CC=C(C)C2=CC=C(C)C2=C1 GXGJIOMUZAGVEH-UHFFFAOYSA-N 0.000 description 1
- 108091060290 Chromatid Proteins 0.000 description 1
- 108020004705 Codon Proteins 0.000 description 1
- 235000016795 Cola Nutrition 0.000 description 1
- 244000228088 Cola acuminata Species 0.000 description 1
- 235000011824 Cola pachycarpa Nutrition 0.000 description 1
- 241001550206 Colla Species 0.000 description 1
- 206010010099 Combined immunodeficiency Diseases 0.000 description 1
- 201000003883 Cystic fibrosis Diseases 0.000 description 1
- 125000003317 D-arabinofuranosyl group Chemical group [H]OC([H])([H])[C@@]1([H])OC([H])(*)[C@@]([H])(O[H])[C@]1([H])O[H] 0.000 description 1
- 102000053602 DNA Human genes 0.000 description 1
- 230000004543 DNA replication Effects 0.000 description 1
- 230000004568 DNA-binding Effects 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 101100447432 Danio rerio gapdh-2 gene Proteins 0.000 description 1
- 108091027757 Deoxyribozyme Proteins 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- 101000619676 Drosophila melanogaster Lipid storage droplets surface-binding protein 2 Proteins 0.000 description 1
- 101150067270 Ehmt1 gene Proteins 0.000 description 1
- 241001331845 Equus asinus x caballus Species 0.000 description 1
- 241000283070 Equus zebra Species 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 108700039887 Essential Genes Proteins 0.000 description 1
- IAYPIBMASNFSPL-UHFFFAOYSA-N Ethylene oxide Chemical compound C1CO1 IAYPIBMASNFSPL-UHFFFAOYSA-N 0.000 description 1
- 108010022894 Euchromatin Proteins 0.000 description 1
- 241000289695 Eutheria Species 0.000 description 1
- 108091060211 Expressed sequence tag Proteins 0.000 description 1
- 108091092566 Extrachromosomal DNA Proteins 0.000 description 1
- 102100038581 F-box only protein 10 Human genes 0.000 description 1
- 108090000331 Firefly luciferases Proteins 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 101150112014 Gapdh gene Proteins 0.000 description 1
- 241000282819 Giraffa Species 0.000 description 1
- 241000282818 Giraffidae Species 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 241000650678 Gorilla gorilla diehli Species 0.000 description 1
- 108010016918 Histone-Lysine N-Methyltransferase Proteins 0.000 description 1
- 102000000581 Histone-lysine N-methyltransferase Human genes 0.000 description 1
- 102100022103 Histone-lysine N-methyltransferase 2A Human genes 0.000 description 1
- 102100022102 Histone-lysine N-methyltransferase 2B Human genes 0.000 description 1
- 102100027755 Histone-lysine N-methyltransferase 2C Human genes 0.000 description 1
- 108091016366 Histone-lysine N-methyltransferase EHMT1 Proteins 0.000 description 1
- 102100035042 Histone-lysine N-methyltransferase EHMT2 Human genes 0.000 description 1
- 102100038970 Histone-lysine N-methyltransferase EZH2 Human genes 0.000 description 1
- 102100027770 Histone-lysine N-methyltransferase KMT5B Human genes 0.000 description 1
- 102100027788 Histone-lysine N-methyltransferase KMT5C Human genes 0.000 description 1
- 102100032742 Histone-lysine N-methyltransferase SETD2 Human genes 0.000 description 1
- 102100027704 Histone-lysine N-methyltransferase SETD7 Human genes 0.000 description 1
- 101710168120 Histone-lysine N-methyltransferase SETDB1 Proteins 0.000 description 1
- 102100023676 Histone-lysine N-methyltransferase SETDB2 Human genes 0.000 description 1
- 102100029239 Histone-lysine N-methyltransferase, H3 lysine-36 specific Human genes 0.000 description 1
- 102100039489 Histone-lysine N-methyltransferase, H3 lysine-79 specific Human genes 0.000 description 1
- 101710095112 Histone-lysine N-methyltransferase, H3 lysine-9 specific SUVH1 Proteins 0.000 description 1
- 101100173424 Homo sapiens FBXO10 gene Proteins 0.000 description 1
- 101001045846 Homo sapiens Histone-lysine N-methyltransferase 2A Proteins 0.000 description 1
- 101001045848 Homo sapiens Histone-lysine N-methyltransferase 2B Proteins 0.000 description 1
- 101001008892 Homo sapiens Histone-lysine N-methyltransferase 2C Proteins 0.000 description 1
- 101001008894 Homo sapiens Histone-lysine N-methyltransferase 2D Proteins 0.000 description 1
- 101000877312 Homo sapiens Histone-lysine N-methyltransferase EHMT2 Proteins 0.000 description 1
- 101000882127 Homo sapiens Histone-lysine N-methyltransferase EZH2 Proteins 0.000 description 1
- 101001008821 Homo sapiens Histone-lysine N-methyltransferase KMT5B Proteins 0.000 description 1
- 101001008824 Homo sapiens Histone-lysine N-methyltransferase KMT5C Proteins 0.000 description 1
- 101000654725 Homo sapiens Histone-lysine N-methyltransferase SETD2 Proteins 0.000 description 1
- 101000650682 Homo sapiens Histone-lysine N-methyltransferase SETD7 Proteins 0.000 description 1
- 101000684609 Homo sapiens Histone-lysine N-methyltransferase SETDB1 Proteins 0.000 description 1
- 101000684615 Homo sapiens Histone-lysine N-methyltransferase SETDB2 Proteins 0.000 description 1
- 101000634050 Homo sapiens Histone-lysine N-methyltransferase, H3 lysine-36 specific Proteins 0.000 description 1
- 101000963360 Homo sapiens Histone-lysine N-methyltransferase, H3 lysine-79 specific Proteins 0.000 description 1
- 101000614013 Homo sapiens Lysine-specific demethylase 2B Proteins 0.000 description 1
- 101000614017 Homo sapiens Lysine-specific demethylase 3A Proteins 0.000 description 1
- 101000614020 Homo sapiens Lysine-specific demethylase 3B Proteins 0.000 description 1
- 101001088892 Homo sapiens Lysine-specific demethylase 5A Proteins 0.000 description 1
- 101001088883 Homo sapiens Lysine-specific demethylase 5B Proteins 0.000 description 1
- 101001088887 Homo sapiens Lysine-specific demethylase 5C Proteins 0.000 description 1
- 101001088879 Homo sapiens Lysine-specific demethylase 5D Proteins 0.000 description 1
- 101001025971 Homo sapiens Lysine-specific demethylase 6B Proteins 0.000 description 1
- 101001050886 Homo sapiens Lysine-specific histone demethylase 1A Proteins 0.000 description 1
- 101001008816 Homo sapiens N-lysine methyltransferase KMT5A Proteins 0.000 description 1
- 101000708645 Homo sapiens N-lysine methyltransferase SMYD2 Proteins 0.000 description 1
- 101100521074 Homo sapiens PRDM2 gene Proteins 0.000 description 1
- 101001056567 Homo sapiens Protein Jumonji Proteins 0.000 description 1
- 101000757216 Homo sapiens Protein arginine N-methyltransferase 1 Proteins 0.000 description 1
- 101000757232 Homo sapiens Protein arginine N-methyltransferase 2 Proteins 0.000 description 1
- 101000924541 Homo sapiens Protein arginine N-methyltransferase 3 Proteins 0.000 description 1
- 101000775582 Homo sapiens Protein arginine N-methyltransferase 6 Proteins 0.000 description 1
- 101000693024 Homo sapiens Protein arginine N-methyltransferase 7 Proteins 0.000 description 1
- 101000796142 Homo sapiens Protein arginine N-methyltransferase 8 Proteins 0.000 description 1
- 101000755643 Homo sapiens RIMS-binding protein 2 Proteins 0.000 description 1
- 101000756365 Homo sapiens Retinol-binding protein 2 Proteins 0.000 description 1
- 108091066896 Homo sapiens miR-331 stem-loop Proteins 0.000 description 1
- 108091061778 Homo sapiens miR-615 stem-loop Proteins 0.000 description 1
- 108091068856 Homo sapiens miR-98 stem-loop Proteins 0.000 description 1
- 102000018251 Hypoxanthine Phosphoribosyltransferase Human genes 0.000 description 1
- 108060003951 Immunoglobulin Proteins 0.000 description 1
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 1
- 208000026350 Inborn Genetic disease Diseases 0.000 description 1
- 229930010555 Inosine Natural products 0.000 description 1
- 101150086476 KDM5B gene Proteins 0.000 description 1
- 101150018389 Kdm3a gene Proteins 0.000 description 1
- 101150013319 Kdm4c gene Proteins 0.000 description 1
- 150000008575 L-amino acids Chemical class 0.000 description 1
- SRBFZHDQGSBBOR-OWMBCFKOSA-N L-ribopyranose Chemical compound O[C@H]1COC(O)[C@@H](O)[C@H]1O SRBFZHDQGSBBOR-OWMBCFKOSA-N 0.000 description 1
- 241000282838 Lama Species 0.000 description 1
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 1
- 229930190887 Leptomycin Natural products 0.000 description 1
- 108090001030 Lipoproteins Proteins 0.000 description 1
- 102000004895 Lipoproteins Human genes 0.000 description 1
- 241000406668 Loxodonta cyclotis Species 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- 102100040584 Lysine-specific demethylase 2B Human genes 0.000 description 1
- 102100040582 Lysine-specific demethylase 3B Human genes 0.000 description 1
- 102100033249 Lysine-specific demethylase 5C Human genes 0.000 description 1
- 102100033143 Lysine-specific demethylase 5D Human genes 0.000 description 1
- 102100037461 Lysine-specific demethylase 6B Human genes 0.000 description 1
- 102100024985 Lysine-specific histone demethylase 1A Human genes 0.000 description 1
- 108700041567 MDR Genes Proteins 0.000 description 1
- 102000029749 Microtubule Human genes 0.000 description 1
- 108091022875 Microtubule Proteins 0.000 description 1
- 241000218213 Morus <angiosperm> Species 0.000 description 1
- 241000711408 Murine respirovirus Species 0.000 description 1
- 101100225545 Mus musculus Ehmt1 gene Proteins 0.000 description 1
- 101100517376 Mus musculus Ntm gene Proteins 0.000 description 1
- IYYIBFCJILKPCO-WOUKDFQISA-O N(2),N(2),N(7)-trimethylguanosine Chemical compound C1=2NC(N(C)C)=NC(=O)C=2N(C)C=[N+]1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O IYYIBFCJILKPCO-WOUKDFQISA-O 0.000 description 1
- ZBYRSRLCXTUFLJ-IOSLPCCCSA-O N(2),N(7)-dimethylguanosine Chemical compound CNC=1NC(C=2[N+](=CN([C@H]3[C@H](O)[C@H](O)[C@@H](CO)O3)C=2N=1)C)=O ZBYRSRLCXTUFLJ-IOSLPCCCSA-O 0.000 description 1
- BVIAOQMSVZHOJM-UHFFFAOYSA-N N(6),N(6)-dimethyladenine Chemical compound CN(C)C1=NC=NC2=C1N=CN2 BVIAOQMSVZHOJM-UHFFFAOYSA-N 0.000 description 1
- VQAYFKKCNSOZKM-IOSLPCCCSA-N N(6)-methyladenosine Chemical compound C1=NC=2C(NC)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O VQAYFKKCNSOZKM-IOSLPCCCSA-N 0.000 description 1
- 102100027771 N-lysine methyltransferase KMT5A Human genes 0.000 description 1
- 102100032806 N-lysine methyltransferase SMYD2 Human genes 0.000 description 1
- 229930193140 Neomycin Natural products 0.000 description 1
- 108091007494 Nucleic acid- binding domains Proteins 0.000 description 1
- 108091005461 Nucleic proteins Proteins 0.000 description 1
- JKUJKKGMOZDDJV-UHFFFAOYSA-N O-Methyl-sterigmatocystin Natural products O1C2=C3C4C=COC4OC3=CC(OC)=C2C(=O)C2=C1C=CC=C2OC JKUJKKGMOZDDJV-UHFFFAOYSA-N 0.000 description 1
- TTZMPOZCBFTTPR-UHFFFAOYSA-N O=P1OCO1 Chemical compound O=P1OCO1 TTZMPOZCBFTTPR-UHFFFAOYSA-N 0.000 description 1
- 208000008589 Obesity Diseases 0.000 description 1
- 108010038807 Oligopeptides Proteins 0.000 description 1
- 102000015636 Oligopeptides Human genes 0.000 description 1
- XAFYCPTXURWXGH-UHFFFAOYSA-N P(O)(O)(O)=S.P(O)(O)=O Chemical compound P(O)(O)(O)=S.P(O)(O)=O XAFYCPTXURWXGH-UHFFFAOYSA-N 0.000 description 1
- 238000012408 PCR amplification Methods 0.000 description 1
- 241000282579 Pan Species 0.000 description 1
- 241000282377 Panthera tigris sumatrae Species 0.000 description 1
- 108010079855 Peptide Aptamers Proteins 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical group OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 1
- ABLZXFCXXLZCGV-UHFFFAOYSA-N Phosphorous acid Chemical class OP(O)=O ABLZXFCXXLZCGV-UHFFFAOYSA-N 0.000 description 1
- 241000254064 Photinus pyralis Species 0.000 description 1
- 108091036407 Polyadenylation Proteins 0.000 description 1
- 241000288906 Primates Species 0.000 description 1
- 108010007568 Protamines Proteins 0.000 description 1
- 102000007327 Protamines Human genes 0.000 description 1
- 102100025733 Protein Jumonji Human genes 0.000 description 1
- 102100022985 Protein arginine N-methyltransferase 1 Human genes 0.000 description 1
- 102100022988 Protein arginine N-methyltransferase 2 Human genes 0.000 description 1
- 102100034603 Protein arginine N-methyltransferase 3 Human genes 0.000 description 1
- 102100034607 Protein arginine N-methyltransferase 5 Human genes 0.000 description 1
- 101710084427 Protein arginine N-methyltransferase 5 Proteins 0.000 description 1
- 102100032140 Protein arginine N-methyltransferase 6 Human genes 0.000 description 1
- 102100026297 Protein arginine N-methyltransferase 7 Human genes 0.000 description 1
- 102100031365 Protein arginine N-methyltransferase 8 Human genes 0.000 description 1
- 229930185560 Pseudouridine Natural products 0.000 description 1
- PTJWIQPHWPFNBW-UHFFFAOYSA-N Pseudouridine C Natural products OC1C(O)C(CO)OC1C1=CNC(=O)NC1=O PTJWIQPHWPFNBW-UHFFFAOYSA-N 0.000 description 1
- 239000005700 Putrescine Substances 0.000 description 1
- 230000006819 RNA synthesis Effects 0.000 description 1
- 241000283011 Rangifer Species 0.000 description 1
- 101100247004 Rattus norvegicus Qsox1 gene Proteins 0.000 description 1
- 108010052090 Renilla Luciferases Proteins 0.000 description 1
- 241000242743 Renilla reniformis Species 0.000 description 1
- 108700008625 Reporter Genes Proteins 0.000 description 1
- 241000282806 Rhinoceros Species 0.000 description 1
- 108010057163 Ribonuclease III Proteins 0.000 description 1
- 102000003661 Ribonuclease III Human genes 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 108020004487 Satellite DNA Proteins 0.000 description 1
- 241000555745 Sciuridae Species 0.000 description 1
- 229920002684 Sepharose Polymers 0.000 description 1
- XUIMIQQOPSSXEZ-UHFFFAOYSA-N Silicon Chemical compound [Si] XUIMIQQOPSSXEZ-UHFFFAOYSA-N 0.000 description 1
- 108091081024 Start codon Proteins 0.000 description 1
- KYGRCGGBECLWMH-UHFFFAOYSA-N Sterigmatocystin Natural products COc1cc2OC3C=COC3c2c4Oc5cccc(O)c5C(=O)c14 KYGRCGGBECLWMH-UHFFFAOYSA-N 0.000 description 1
- UTSVPXMQSFGQTM-UHFFFAOYSA-N Sterigmatrocystin Natural products O1C2=CC=CC(O)=C2C(=O)C2=C1C(C1C=COC1O1)=C1C=C2OC UTSVPXMQSFGQTM-UHFFFAOYSA-N 0.000 description 1
- 241000272534 Struthio camelus Species 0.000 description 1
- 238000000692 Student's t-test Methods 0.000 description 1
- 241000282898 Sus scrofa Species 0.000 description 1
- 210000001744 T-lymphocyte Anatomy 0.000 description 1
- 206010043276 Teratoma Diseases 0.000 description 1
- 239000004098 Tetracycline Substances 0.000 description 1
- 102000002933 Thioredoxin Human genes 0.000 description 1
- 108091036066 Three prime untranslated region Proteins 0.000 description 1
- 108020004566 Transfer RNA Proteins 0.000 description 1
- 101710188297 Trehalose synthase/amylase TreS Proteins 0.000 description 1
- GBOGMAARMMDZGR-UHFFFAOYSA-N UNPD149280 Natural products N1C(=O)C23OC(=O)C=CC(O)CCCC(C)CC=CC3C(O)C(=C)C(C)C2C1CC1=CC=CC=C1 GBOGMAARMMDZGR-UHFFFAOYSA-N 0.000 description 1
- 108091007416 X-inactive specific transcript Proteins 0.000 description 1
- 108091035715 XIST (gene) Proteins 0.000 description 1
- 230000001594 aberrant effect Effects 0.000 description 1
- 125000002777 acetyl group Chemical group [H]C([H])([H])C(*)=O 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 208000037919 acquired disease Diseases 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 125000002015 acyclic group Chemical group 0.000 description 1
- 230000002730 additional effect Effects 0.000 description 1
- 238000004115 adherent culture Methods 0.000 description 1
- 230000001464 adherent effect Effects 0.000 description 1
- 210000004504 adult stem cell Anatomy 0.000 description 1
- 239000000443 aerosol Substances 0.000 description 1
- 238000001042 affinity chromatography Methods 0.000 description 1
- PPQRONHOSHZGFQ-LMVFSUKVSA-N aldehydo-D-ribose 5-phosphate Chemical group OP(=O)(O)OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PPQRONHOSHZGFQ-LMVFSUKVSA-N 0.000 description 1
- 150000004703 alkoxides Chemical class 0.000 description 1
- 125000003545 alkoxy group Chemical group 0.000 description 1
- 150000005215 alkyl ethers Chemical group 0.000 description 1
- 125000006350 alkyl thio alkyl group Chemical group 0.000 description 1
- 125000004414 alkyl thio group Chemical group 0.000 description 1
- 150000001408 amides Chemical class 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 125000002431 aminoalkoxy group Chemical group 0.000 description 1
- HSTJJKALJLWLBR-UHFFFAOYSA-N aminophosphonic acid;2-methoxyethoxyphosphonamidic acid Chemical compound NP(O)(O)=O.COCCOP(N)(O)=O HSTJJKALJLWLBR-UHFFFAOYSA-N 0.000 description 1
- 230000002547 anomalous effect Effects 0.000 description 1
- 125000002178 anthracenyl group Chemical group C1(=CC=CC2=CC3=CC=CC=C3C=C12)* 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 239000003816 antisense DNA Substances 0.000 description 1
- 125000004104 aryloxy group Chemical group 0.000 description 1
- FIVPIPIDMRVLAY-UHFFFAOYSA-N aspergillin Natural products C1C2=CC=CC(O)C2N2C1(SS1)C(=O)N(C)C1(CO)C2=O FIVPIPIDMRVLAY-UHFFFAOYSA-N 0.000 description 1
- 238000003556 assay Methods 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- 210000000270 basal cell Anatomy 0.000 description 1
- FCPVYOBCFFNJFS-LQDWTQKMSA-M benzylpenicillin sodium Chemical compound [Na+].N([C@H]1[C@H]2SC([C@@H](N2C1=O)C([O-])=O)(C)C)C(=O)CC1=CC=CC=C1 FCPVYOBCFFNJFS-LQDWTQKMSA-M 0.000 description 1
- WGDUUQDYDIIBKT-UHFFFAOYSA-N beta-Pseudouridine Natural products OC1OC(CN2C=CC(=O)NC2=O)C(O)C1O WGDUUQDYDIIBKT-UHFFFAOYSA-N 0.000 description 1
- 239000011230 binding agent Substances 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 238000001574 biopsy Methods 0.000 description 1
- 210000004703 blastocyst inner cell mass Anatomy 0.000 description 1
- 238000006664 bond formation reaction Methods 0.000 description 1
- 229910052796 boron Inorganic materials 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 238000004422 calculation algorithm Methods 0.000 description 1
- 239000007894 caplet Substances 0.000 description 1
- 239000011203 carbon fibre reinforced carbon Substances 0.000 description 1
- 125000005587 carbonate group Chemical group 0.000 description 1
- 125000002915 carbonyl group Chemical group [*:2]C([*:1])=O 0.000 description 1
- 239000003054 catalyst Substances 0.000 description 1
- 230000022131 cell cycle Effects 0.000 description 1
- 230000011712 cell development Effects 0.000 description 1
- 230000007910 cell fusion Effects 0.000 description 1
- 239000013592 cell lysate Substances 0.000 description 1
- 238000002659 cell therapy Methods 0.000 description 1
- 230000033077 cellular process Effects 0.000 description 1
- 230000008668 cellular reprogramming Effects 0.000 description 1
- 210000002230 centromere Anatomy 0.000 description 1
- 229930191237 chaetoglobosin Natural products 0.000 description 1
- OUMWCYMRLMEZJH-VOXRAUTJSA-N chaetoglobosin A Chemical compound N([C@@H](CC=1C2=CC=CC=C2NC=1)[C@@H]1[C@@H]([C@]2(O[C@H]22)C)C)C(=O)[C@@]11[C@H]2\C=C\C[C@H](C)\C=C(C)\[C@@H](O)C(=O)\C=C\C1=O OUMWCYMRLMEZJH-VOXRAUTJSA-N 0.000 description 1
- 229930187278 chaetoglobosins Natural products 0.000 description 1
- 150000005829 chemical entities Chemical class 0.000 description 1
- 125000003636 chemical group Chemical group 0.000 description 1
- 238000007385 chemical modification Methods 0.000 description 1
- 238000006243 chemical reaction Methods 0.000 description 1
- 210000001612 chondrocyte Anatomy 0.000 description 1
- 210000004756 chromatid Anatomy 0.000 description 1
- 230000010428 chromatin condensation Effects 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 238000012411 cloning technique Methods 0.000 description 1
- 238000010835 comparative analysis Methods 0.000 description 1
- 230000000052 comparative effect Effects 0.000 description 1
- 230000001268 conjugating effect Effects 0.000 description 1
- 239000000470 constituent Substances 0.000 description 1
- 238000013270 controlled release Methods 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000010168 coupling process Methods 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- 238000012136 culture method Methods 0.000 description 1
- 108010082025 cyan fluorescent protein Proteins 0.000 description 1
- 125000004093 cyano group Chemical group *C#N 0.000 description 1
- 125000004966 cyanoalkyl group Chemical group 0.000 description 1
- GBOGMAARMMDZGR-JREHFAHYSA-N cytochalasin B Natural products C[C@H]1CCC[C@@H](O)C=CC(=O)O[C@@]23[C@H](C=CC1)[C@H](O)C(=C)[C@@H](C)[C@@H]2[C@H](Cc4ccccc4)NC3=O GBOGMAARMMDZGR-JREHFAHYSA-N 0.000 description 1
- GBOGMAARMMDZGR-TYHYBEHESA-N cytochalasin B Chemical compound C([C@H]1[C@@H]2[C@@H](C([C@@H](O)[C@@H]3/C=C/C[C@H](C)CCC[C@@H](O)/C=C/C(=O)O[C@@]23C(=O)N1)=C)C)C1=CC=CC=C1 GBOGMAARMMDZGR-TYHYBEHESA-N 0.000 description 1
- 239000000824 cytostatic agent Substances 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 239000002254 cytotoxic agent Substances 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 235000013365 dairy product Nutrition 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000007547 defect Effects 0.000 description 1
- 230000007850 degeneration Effects 0.000 description 1
- 238000002716 delivery method Methods 0.000 description 1
- MXHRCPNRJAMMIM-UHFFFAOYSA-N desoxyuridine Natural products C1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 MXHRCPNRJAMMIM-UHFFFAOYSA-N 0.000 description 1
- 230000001687 destabilization Effects 0.000 description 1
- 230000009547 development abnormality Effects 0.000 description 1
- 206010012601 diabetes mellitus Diseases 0.000 description 1
- 125000004985 dialkyl amino alkyl group Chemical group 0.000 description 1
- 238000004720 dielectrophoresis Methods 0.000 description 1
- 238000009792 diffusion process Methods 0.000 description 1
- RHGQIWVTIHZRLI-UHFFFAOYSA-N dihydrosterigmatocystin Natural products O1C2=CC=CC(O)=C2C(=O)C2=C1C(C1CCOC1O1)=C1C=C2OC RHGQIWVTIHZRLI-UHFFFAOYSA-N 0.000 description 1
- 125000000118 dimethyl group Chemical group [H]C([H])([H])* 0.000 description 1
- USIUVYZYUHIAEV-UHFFFAOYSA-N diphenyl ether Chemical compound C=1C=CC=CC=1OC1=CC=CC=C1 USIUVYZYUHIAEV-UHFFFAOYSA-N 0.000 description 1
- XPPKVPWEQAFLFU-UHFFFAOYSA-J diphosphate(4-) Chemical class [O-]P([O-])(=O)OP([O-])([O-])=O XPPKVPWEQAFLFU-UHFFFAOYSA-J 0.000 description 1
- 235000011180 diphosphates Nutrition 0.000 description 1
- POLCUAVZOMRGSN-UHFFFAOYSA-N dipropyl ether Chemical compound CCCOCCC POLCUAVZOMRGSN-UHFFFAOYSA-N 0.000 description 1
- KPUWHANPEXNPJT-UHFFFAOYSA-N disiloxane Chemical group [SiH3]O[SiH3] KPUWHANPEXNPJT-UHFFFAOYSA-N 0.000 description 1
- 239000003968 dna methyltransferase inhibitor Substances 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 239000003937 drug carrier Substances 0.000 description 1
- 238000012377 drug delivery Methods 0.000 description 1
- 238000009509 drug development Methods 0.000 description 1
- 230000004064 dysfunction Effects 0.000 description 1
- 210000004670 early embryonic cell Anatomy 0.000 description 1
- 230000005684 electric field Effects 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 238000010201 enrichment analysis Methods 0.000 description 1
- 210000001339 epidermal cell Anatomy 0.000 description 1
- 230000008995 epigenetic change Effects 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- 210000000981 epithelium Anatomy 0.000 description 1
- 210000003743 erythrocyte Anatomy 0.000 description 1
- 150000002148 esters Chemical class 0.000 description 1
- 229940011871 estrogen Drugs 0.000 description 1
- 239000000262 estrogen Substances 0.000 description 1
- 230000001158 estrous effect Effects 0.000 description 1
- 125000005745 ethoxymethyl group Chemical group [H]C([H])([H])C([H])([H])OC([H])([H])* 0.000 description 1
- 125000001495 ethyl group Chemical group [H]C([H])([H])C([H])([H])* 0.000 description 1
- 210000000632 euchromatin Anatomy 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 125000005313 fatty acid group Chemical group 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 239000000835 fiber Substances 0.000 description 1
- 235000019688 fish Nutrition 0.000 description 1
- 238000000684 flow cytometry Methods 0.000 description 1
- GVEPBJHOBDJJJI-UHFFFAOYSA-N fluoranthrene Natural products C1=CC(C2=CC=CC=C22)=C3C2=CC=CC3=C1 GVEPBJHOBDJJJI-UHFFFAOYSA-N 0.000 description 1
- 239000007850 fluorescent dye Substances 0.000 description 1
- 125000001153 fluoro group Chemical group F* 0.000 description 1
- 235000013305 food Nutrition 0.000 description 1
- 238000007306 functionalization reaction Methods 0.000 description 1
- 238000001641 gel filtration chromatography Methods 0.000 description 1
- 230000037440 gene silencing effect Effects 0.000 description 1
- 208000016361 genetic disease Diseases 0.000 description 1
- 230000007614 genetic variation Effects 0.000 description 1
- 210000004392 genitalia Anatomy 0.000 description 1
- 210000004907 gland Anatomy 0.000 description 1
- FIVPIPIDMRVLAY-RBJBARPLSA-N gliotoxin Chemical group C1C2=CC=C[C@H](O)[C@H]2N2[C@]1(SS1)C(=O)N(C)[C@@]1(CO)C2=O FIVPIPIDMRVLAY-RBJBARPLSA-N 0.000 description 1
- 229940103893 gliotoxin Drugs 0.000 description 1
- 229930190252 gliotoxin Natural products 0.000 description 1
- 150000004676 glycans Chemical class 0.000 description 1
- 150000002337 glycosamines Chemical class 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 210000002216 heart Anatomy 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 210000003494 hepatocyte Anatomy 0.000 description 1
- 125000005842 heteroatom Chemical group 0.000 description 1
- 101150113423 hisD gene Proteins 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 238000009396 hybridization Methods 0.000 description 1
- 238000004191 hydrophobic interaction chromatography Methods 0.000 description 1
- 125000004029 hydroxymethyl group Chemical group [H]OC([H])([H])* 0.000 description 1
- 210000002865 immune cell Anatomy 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000003053 immunization Effects 0.000 description 1
- 229940127121 immunoconjugate Drugs 0.000 description 1
- 102000018358 immunoglobulin Human genes 0.000 description 1
- 210000004263 induced pluripotent stem cell Anatomy 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 229960003786 inosine Drugs 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 238000009830 intercalation Methods 0.000 description 1
- 230000010468 interferon response Effects 0.000 description 1
- 239000000543 intermediate Substances 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 150000002500 ions Chemical class 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 125000001449 isopropyl group Chemical group [H]C([H])([H])C([H])(*)C([H])([H])[H] 0.000 description 1
- 210000002510 keratinocyte Anatomy 0.000 description 1
- 101150044508 key gene Proteins 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 150000002605 large molecules Chemical class 0.000 description 1
- YACHGFWEQXFSBS-RJXCBBHPSA-N leptomycin Chemical compound OC(=O)/C=C(C)/C[C@H](C)[C@@H](O)[C@H](C)C(=O)[C@H](C)/C=C(\C)/C=C/C[C@@H](C)\C=C(/CC)\C=C\[C@@H]1OC(=O)C=C[C@@H]1C YACHGFWEQXFSBS-RJXCBBHPSA-N 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 244000144972 livestock Species 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 230000000938 luteal effect Effects 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 239000003120 macrolide antibiotic agent Substances 0.000 description 1
- 229940041033 macrolides Drugs 0.000 description 1
- 210000002540 macrophage Anatomy 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 230000013011 mating Effects 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 210000003593 megakaryocyte Anatomy 0.000 description 1
- 230000021121 meiosis Effects 0.000 description 1
- 210000003716 mesoderm Anatomy 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 230000031864 metaphase Effects 0.000 description 1
- 125000004184 methoxymethyl group Chemical group [H]C([H])([H])OC([H])([H])* 0.000 description 1
- DJLUSNAYRNFVSM-UHFFFAOYSA-N methyl 2-(2,4-dioxo-1h-pyrimidin-5-yl)acetate Chemical compound COC(=O)CC1=CNC(=O)NC1=O DJLUSNAYRNFVSM-UHFFFAOYSA-N 0.000 description 1
- 108010066354 methylcobalamin-coenzyme M methyltransferase Proteins 0.000 description 1
- 125000006362 methylene amino carbonyl group Chemical group [H]N(C([*:2])=O)C([H])([H])[*:1] 0.000 description 1
- YACKEPLHDIMKIO-UHFFFAOYSA-N methylphosphonic acid Chemical compound CP(O)(O)=O YACKEPLHDIMKIO-UHFFFAOYSA-N 0.000 description 1
- 125000002816 methylsulfanyl group Chemical group [H]C([H])([H])S[*] 0.000 description 1
- 238000002406 microsurgery Methods 0.000 description 1
- 210000004688 microtubule Anatomy 0.000 description 1
- 230000002438 mitochondrial effect Effects 0.000 description 1
- 238000010852 mitochondrial transfer Methods 0.000 description 1
- 229960004857 mitomycin Drugs 0.000 description 1
- 239000003607 modifier Substances 0.000 description 1
- 238000012544 monitoring process Methods 0.000 description 1
- 210000001616 monocyte Anatomy 0.000 description 1
- 210000005087 mononuclear cell Anatomy 0.000 description 1
- 150000004712 monophosphates Chemical class 0.000 description 1
- 210000000663 muscle cell Anatomy 0.000 description 1
- 239000003471 mutagenic agent Substances 0.000 description 1
- 231100000707 mutagenic chemical Toxicity 0.000 description 1
- 230000003505 mutagenic effect Effects 0.000 description 1
- 230000035772 mutation Effects 0.000 description 1
- 239000002636 mycotoxin Substances 0.000 description 1
- BPRQFDNBWVMLPS-UHFFFAOYSA-N n-(3-methylbut-3-enyl)-7h-purin-6-amine Chemical compound CC(=C)CCNC1=NC=NC2=C1NC=N2 BPRQFDNBWVMLPS-UHFFFAOYSA-N 0.000 description 1
- FZQMZXGTZAPBAK-UHFFFAOYSA-N n-(3-methylbutyl)-7h-purin-6-amine Chemical compound CC(C)CCNC1=NC=NC2=C1NC=N2 FZQMZXGTZAPBAK-UHFFFAOYSA-N 0.000 description 1
- 239000002105 nanoparticle Substances 0.000 description 1
- 125000001624 naphthyl group Chemical group 0.000 description 1
- 239000006199 nebulizer Substances 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 230000004770 neurodegeneration Effects 0.000 description 1
- 208000015122 neurodegenerative disease Diseases 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- YCWSUKQGVSGXJO-NTUHNPAUSA-N nifuroxazide Chemical group C1=CC(O)=CC=C1C(=O)N\N=C\C1=CC=C([N+]([O-])=O)O1 YCWSUKQGVSGXJO-NTUHNPAUSA-N 0.000 description 1
- QJGQUHMNIGDVPM-UHFFFAOYSA-N nitrogen group Chemical group [N] QJGQUHMNIGDVPM-UHFFFAOYSA-N 0.000 description 1
- 230000008689 nuclear function Effects 0.000 description 1
- 238000010449 nuclear transplantation Methods 0.000 description 1
- 238000001668 nucleic acid synthesis Methods 0.000 description 1
- 230000001293 nucleolytic effect Effects 0.000 description 1
- 230000000269 nucleophilic effect Effects 0.000 description 1
- 238000011580 nude mouse model Methods 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 235000020824 obesity Nutrition 0.000 description 1
- 229940046166 oligodeoxynucleotide Drugs 0.000 description 1
- 238000002515 oligonucleotide synthesis Methods 0.000 description 1
- 230000005868 ontogenesis Effects 0.000 description 1
- 150000002894 organic compounds Chemical class 0.000 description 1
- 230000002611 ovarian Effects 0.000 description 1
- 150000002923 oximes Chemical class 0.000 description 1
- 125000004430 oxygen atom Chemical group O* 0.000 description 1
- 230000026792 palmitoylation Effects 0.000 description 1
- 230000008186 parthenogenesis Effects 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 238000002823 phage display Methods 0.000 description 1
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 1
- ACVYVLVWPXVTIT-UHFFFAOYSA-N phosphinic acid Chemical compound O[PH2]=O ACVYVLVWPXVTIT-UHFFFAOYSA-N 0.000 description 1
- PTMHPRAIXMAOOB-UHFFFAOYSA-L phosphoramidate Chemical compound NP([O-])([O-])=O PTMHPRAIXMAOOB-UHFFFAOYSA-L 0.000 description 1
- 150000008298 phosphoramidates Chemical class 0.000 description 1
- 150000008300 phosphoramidites Chemical class 0.000 description 1
- RHDXSLLGPJSSGW-VEGRVEBRSA-N phosphoric acid;(2r,3r,4r)-2,3,4,5-tetrahydroxypentanal Chemical group OP(O)(O)=O.OC[C@@H](O)[C@@H](O)[C@@H](O)C=O RHDXSLLGPJSSGW-VEGRVEBRSA-N 0.000 description 1
- 239000011574 phosphorus Substances 0.000 description 1
- LFGREXWGYUGZLY-UHFFFAOYSA-N phosphoryl Chemical class [P]=O LFGREXWGYUGZLY-UHFFFAOYSA-N 0.000 description 1
- 125000005642 phosphothioate group Chemical group 0.000 description 1
- 230000035790 physiological processes and functions Effects 0.000 description 1
- 230000006461 physiological response Effects 0.000 description 1
- 239000000049 pigment Substances 0.000 description 1
- 229920000768 polyamine Polymers 0.000 description 1
- 229920000447 polyanionic polymer Polymers 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 230000020775 positive regulation of microtubule depolymerization Effects 0.000 description 1
- 230000003334 potential effect Effects 0.000 description 1
- 244000144977 poultry Species 0.000 description 1
- 235000013594 poultry meat Nutrition 0.000 description 1
- 230000035935 pregnancy Effects 0.000 description 1
- 229960003387 progesterone Drugs 0.000 description 1
- 239000000186 progesterone Substances 0.000 description 1
- 125000006239 protecting group Chemical group 0.000 description 1
- 230000004952 protein activity Effects 0.000 description 1
- 229940076376 protein agonist Drugs 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 230000004853 protein function Effects 0.000 description 1
- 238000001243 protein synthesis Methods 0.000 description 1
- PTJWIQPHWPFNBW-GBNDHIKLSA-N pseudouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1C1=CNC(=O)NC1=O PTJWIQPHWPFNBW-GBNDHIKLSA-N 0.000 description 1
- 239000002212 purine nucleoside Substances 0.000 description 1
- 150000003834 purine nucleoside derivatives Chemical class 0.000 description 1
- 229950010131 puromycin Drugs 0.000 description 1
- 125000001725 pyrenyl group Chemical group 0.000 description 1
- UBQKCCHYAOITMY-UHFFFAOYSA-N pyridin-2-ol Chemical compound OC1=CC=CC=N1 UBQKCCHYAOITMY-UHFFFAOYSA-N 0.000 description 1
- UNZGDQIKANZIDF-UHFFFAOYSA-N pyrrolo[2,3-b]indole Chemical compound C1=CC=C2C3=CC=NC3=NC2=C1 UNZGDQIKANZIDF-UHFFFAOYSA-N 0.000 description 1
- JWVCLYRUEFBMGU-UHFFFAOYSA-N quinazoline Chemical compound N1=CN=CC2=CC=CC=C21 JWVCLYRUEFBMGU-UHFFFAOYSA-N 0.000 description 1
- 239000013074 reference sample Substances 0.000 description 1
- 210000003289 regulatory T cell Anatomy 0.000 description 1
- 230000003252 repetitive effect Effects 0.000 description 1
- 238000009256 replacement therapy Methods 0.000 description 1
- 230000001850 reproductive effect Effects 0.000 description 1
- 230000000241 respiratory effect Effects 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 230000004043 responsiveness Effects 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 108020004418 ribosomal RNA Proteins 0.000 description 1
- 210000003705 ribosome Anatomy 0.000 description 1
- 229910052594 sapphire Inorganic materials 0.000 description 1
- 239000010980 sapphire Substances 0.000 description 1
- 230000003248 secreting effect Effects 0.000 description 1
- 230000008684 selective degradation Effects 0.000 description 1
- 229910052711 selenium Inorganic materials 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- UQDJGEHQDNVPGU-UHFFFAOYSA-N serine phosphoethanolamine Chemical compound [NH3+]CCOP([O-])(=O)OCC([NH3+])C([O-])=O UQDJGEHQDNVPGU-UHFFFAOYSA-N 0.000 description 1
- 210000000717 sertoli cell Anatomy 0.000 description 1
- 210000002966 serum Anatomy 0.000 description 1
- 238000004904 shortening Methods 0.000 description 1
- 229910052710 silicon Inorganic materials 0.000 description 1
- 239000010703 silicon Substances 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 210000003491 skin Anatomy 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 125000006850 spacer group Chemical group 0.000 description 1
- 229940063675 spermine Drugs 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- UTSVPXMQSFGQTM-DCXZOGHSSA-N sterigmatocystin Chemical compound O1C2=CC=CC(O)=C2C(=O)C2=C1C([C@@H]1C=CO[C@@H]1O1)=C1C=C2OC UTSVPXMQSFGQTM-DCXZOGHSSA-N 0.000 description 1
- 150000003431 steroids Chemical class 0.000 description 1
- 210000002784 stomach Anatomy 0.000 description 1
- 230000035882 stress Effects 0.000 description 1
- 125000000547 substituted alkyl group Chemical group 0.000 description 1
- 229940124530 sulfonamide Drugs 0.000 description 1
- 150000003456 sulfonamides Chemical class 0.000 description 1
- BDHFUVZGWQCTTF-UHFFFAOYSA-M sulfonate Chemical compound [O-]S(=O)=O BDHFUVZGWQCTTF-UHFFFAOYSA-M 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 238000010189 synthetic method Methods 0.000 description 1
- 230000002123 temporal effect Effects 0.000 description 1
- 229960002180 tetracycline Drugs 0.000 description 1
- 229930101283 tetracycline Natural products 0.000 description 1
- 235000019364 tetracycline Nutrition 0.000 description 1
- 150000003522 tetracyclines Chemical class 0.000 description 1
- ABZLKHKQJHEPAX-UHFFFAOYSA-N tetramethylrhodamine Chemical compound C=12C=CC(N(C)C)=CC2=[O+]C2=CC(N(C)C)=CC=C2C=1C1=CC=CC=C1C([O-])=O ABZLKHKQJHEPAX-UHFFFAOYSA-N 0.000 description 1
- 238000010257 thawing Methods 0.000 description 1
- VOVUARRWDCVURC-UHFFFAOYSA-N thiirane Chemical compound C1CS1 VOVUARRWDCVURC-UHFFFAOYSA-N 0.000 description 1
- OFBPGACXRPVDQW-UHFFFAOYSA-N thiirane 1,1-dioxide Chemical compound O=S1(=O)CC1 OFBPGACXRPVDQW-UHFFFAOYSA-N 0.000 description 1
- 125000005309 thioalkoxy group Chemical group 0.000 description 1
- 150000003568 thioethers Chemical class 0.000 description 1
- 125000003396 thiol group Chemical class [H]S* 0.000 description 1
- 108060008226 thioredoxin Proteins 0.000 description 1
- 229940094937 thioredoxin Drugs 0.000 description 1
- 210000000115 thoracic cavity Anatomy 0.000 description 1
- 229940113082 thymine Drugs 0.000 description 1
- 210000003014 totipotent stem cell Anatomy 0.000 description 1
- 238000011222 transcriptome analysis Methods 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 210000002993 trophoblast Anatomy 0.000 description 1
- 238000010396 two-hybrid screening Methods 0.000 description 1
- 241000701161 unidentified adenovirus Species 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 210000003708 urethra Anatomy 0.000 description 1
- 210000003932 urinary bladder Anatomy 0.000 description 1
- 230000002485 urinary effect Effects 0.000 description 1
- 239000003981 vehicle Substances 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 1
- 229940075420 xanthine Drugs 0.000 description 1
- 150000003952 β-lactams Chemical group 0.000 description 1
Images



















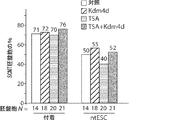

























Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/8509—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells for producing genetically modified animals, e.g. transgenic
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/873—Techniques for producing new embryos, e.g. nuclear transfer, manipulation of totipotent cells or production of chimeric embryos
- C12N15/877—Techniques for producing new mammalian cloned embryos
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
- A01K67/0273—Cloned vertebrates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
- C12N15/1137—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing against enzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/873—Techniques for producing new embryos, e.g. nuclear transfer, manipulation of totipotent cells or production of chimeric embryos
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0603—Embryonic cells ; Embryoid bodies
- C12N5/0604—Whole embryos; Culture medium therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0603—Embryonic cells ; Embryoid bodies
- C12N5/0606—Pluripotent embryonic cells, e.g. embryonic stem cells [ES]
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2227/00—Animals characterised by species
- A01K2227/10—Mammal
- A01K2227/105—Murine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/14—Type of nucleic acid interfering N.A.
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2517/00—Cells related to new breeds of animals
- C12N2517/04—Cells produced using nuclear transfer
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y201/00—Transferases transferring one-carbon groups (2.1)
- C12Y201/01—Methyltransferases (2.1.1)
- C12Y201/01043—Histone-lysine N-methyltransferase (2.1.1.43)
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Biomedical Technology (AREA)
- Zoology (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Developmental Biology & Embryology (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Environmental Sciences (AREA)
- Reproductive Health (AREA)
- Gynecology & Obstetrics (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Virology (AREA)
- Biodiversity & Conservation Biology (AREA)
- Animal Husbandry (AREA)
- Cell Biology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Agricultural Chemicals And Associated Chemicals (AREA)
Description
本願は、35 U.S.C. 119(e)の下で、2014年9月15日に出願された米国仮特許出願第62/050,308号および2014年9月22日に出願された米国仮特許出願第62/053,514号の恩典を主張し、その各内容は全体として参照により本明細書に組み入れられる。
本発明は、米国国立衛生研究所(National Institutes of Health/NIH)から授与されたNIH U01DK089565の下で政府の支援を受けてなされたものである。米国政府は、本発明に関して一定の権利を保有する。
最終分化した体細胞は、体細胞核移入(SCNT)によって除核卵母細胞内に移植されると全能状態にリプログラミングされることができる(Gurdon, 1962)。SCNTは、分化した体細胞の単一の核から動物全体の生成を可能にするので、農業、生物医学工業、および絶滅危惧種の保護に大きな潜在性を有する(Yang et al., 2007)。実際、ヒツジにおける哺乳動物クローニングが1997年に最初に成功して以来(Wilmut et al., 1997)、20種よりも多い哺乳動物種がSCNTを経てクローニングされている(Rodriguez-Osorio et al., 2012)。そのうえ、SCNTによって作製された胚盤胞から多能性胚性幹細胞を樹立することができるので(Wakayama et al., 2001)、SCNTは、ヒトの治療に大いに有望である(Hochedlinger and Jaenisch, 2003)。この有望性は、最初のヒト核移入胚性幹細胞(ntESC)の派生における最近の成功(Tachibana et al., 2013)、および高齢者またはヒト患者細胞からのヒトntESC作製(Chung et al., 2014; Yamada et al., 2014)の後に、より現実に近づいている。これらのntESCは、インビトロ疾患モデル化用の細胞の貴重な供給源として、ならびに再生療法および細胞/組織置換療法用の細胞の供給源として役立ち得る。

その基礎科学および治療用途の両方の莫大な潜在性にもかかわらず、体細胞核移入(SCNT)による哺乳動物クローニングの効率は、非常に低いままである。SCNT後の生きた幼体の出生率は、使用される種、ドナー細胞の種類、プロトコール、または技法に関わらず10%未満である。同様に、クローニングされた胚の発生率は、正常に受精した胚の発生率よりも低く、胚盤胞への発生不良および胚盤胞でのより少ない細胞数を招く。これらの欠損は、クローニングされたマウスSCNT胚からES細胞株の成功裏な樹立が滅多に起こらないことの原因にもなり、成功は、正常な胚が使用されるときの約30%の成功と比較して、マウス系統またはドナー細胞型にかかわらず約5%である。クローニングされた胚の機能不全は、主として、ドナー核における不完全な核プログラミングおよび/またはエピジェネティックなバリアが原因である。
便宜上、本出願全体に採用される特定の用語(明細書、実施例、および添付の特許請求の範囲を含む)を、ここに集める。特に定義しない限り、本明細書に使用される全ての技術用語および科学用語は、本発明が属する技術分野の技術者によって通常理解されるのと同じ意味を有する。
一局面では、本発明は、SCNTの効率を増加させる方法であって、ドナー哺乳動物細胞、またはSCNT胚、または卵母細胞の核または細胞質を、ヒストンメチル化を阻害する、特にH3K9メチル化を阻害する、特にH3H9me3トリメチル化を阻害する作用物質と接触させる段階を含む方法を提供する。いくつかの態様では、作用物質は、Kdm4ヒストンデメチラーゼ活性化剤である。
一局面では、本発明は、SCNTの効率を増加させる方法であって:ドナー哺乳動物細胞またはSCNT胚または卵母細胞の核または細胞質を、ヒストンメチル化を阻害する、特にH3K9メチル化を阻害する、特にH3H9me3トリメチル化を阻害する作用物質と接触させる段階を含む、方法を提供する。本発明の特定の態様では、作用物質は、ヒストンメチルトランスフェラーゼ活性を阻害する。本発明の特定の態様では、作用物質は、ヒストンメチルトランスフェラーゼの発現を阻害する。本発明の特定の態様では、阻害剤は、H3K9メチルトランスフェラーゼの阻害剤である。本明細書に述べられるように、本発明者らは、H3K9メチルトランスフェラーゼタンパク質の阻害をSCNTの効率を増加させるために使用することができることを発見した。いくつかの態様では、H3K9メチルトランスフェラーゼ阻害剤は、タンパク質 阻害剤であり、いくつかの態様では、阻害剤は、H3K9メチルトランスフェラーゼタンパク質の機能またはその遺伝子からのH3K9メチルトランスフェラーゼの発現を阻害する任意の作用物質である。
未修飾オリゴヌクレオチドは、一部の適用において最適未満である可能性があり、例えば未修飾オリゴヌクレオチドは、例えば細胞ヌクレアーゼによる分解を受けやすい可能性がある。ヌクレアーゼは、核酸ホスホジエステル結合を加水分解することができる。しかし、オリゴヌクレオチドの1つまたは複数のサブユニットに対する化学修飾は、改善された性質を付与することができ、例えば、オリゴヌクレオチドをヌクレアーゼに対してより安定にすることができる。
リン酸基
リン酸基は、負荷電化学種である。電荷は、2つの非架橋型酸素原子に等しく分布される。しかし、リン酸基を、酸素の一方を異なる置換基により置換することによって修飾することができる。RNAリン酸エステル骨格に対するこの修飾の1つの結果は、核酸分解性破壊に対するオリゴリボヌクレオチドの増加した耐性であることができる。したがって、理論に縛られることを望むわけではないが、非荷電リンカーまたは非対称な電荷分布を有する荷電リンカーのいずれかを生じる変更を導入することが、いくつかの態様で理想的であることができる。
リン酸基を、リン不含コネクターによって置換することができる。理論に縛られることを望むわけではないが、荷電ホスホジエステル基は、核酸分解における反応中心であるので、中性構造模倣物によるその置換は、増強したヌクレアーゼ安定性を付与するはずであると考えられる。さらに、理論に縛られることを望むわけではないが、いくつかの態様において、荷電リン酸基が中性部分によって置換される変更を導入することが理想的であることができる。
リン酸リンカーおよびリボース糖が、ヌクレアーゼ耐性のヌクレオシド代用物またはヌクレオチド代用物によって置換されている、オリゴヌクレオチド模倣足場も構築することができる。理論に縛られることを望むわけではないが、反復荷電した骨格の非存在は、ポリアニオンを認識するタンパク質(例えばヌクレアーゼ)への結合を減少させると考えられる。さらに、理論に縛られることを望むわけではないが、ある態様において、塩基が中性代用物骨格によって繋がれている変更を導入することが理想的であることができる。例には、モルホリノ、シクロブチル、ピロリジンおよびペプチド核酸(PNA)ヌクレオシド代用物が含まれる。好ましい代用物は、PNA代用物である。
オリゴヌクレオチドは、核酸の糖基の全てまたは一部の修飾を含むことができる。例えば、2'ヒドロキシル基(OH)を、いくつかの異なる「オキシ」または「デオキシ」置換基により修飾または置換することができる。理論に縛られるわけではなく、ヒドロキシルが脱プロトン化されてもはや2'-アルコキシドイオンを形成することができないので、増強した安定性が予期される。2'-アルコキシドは、リンカーのリン原子への分子内求核攻撃による分解を触媒することができる。さらに、理論に縛られることを望むわけではないが、2'位でのアルコキシド形成が不可能である変更を導入することが、いくつかの態様に理想的であることができる。
オリゴヌクレオチドの3ダッシュ(3')および5ダッシュ(5')末端を修飾することができる。そのような修飾は、分子の3'末端、5'末端または両末端にあることができる。それらは、末端リン酸全体またはリン酸基の原子の1つまたは複数の修飾または置換を含むことができる。例えば、オリゴヌクレオチドの3'および5'末端を、標識部分、例えばフルオロフォア(例えば、ピレン、TAMRA、フルオレセイン、Cy3もしくはCy5色素)または保護基(例えば、硫黄、ケイ素、ホウ素またはエステルに基づく)のような他の機能的分子実体にコンジュゲートすることができる。機能的分子実体を、リン酸基および/またはリンカーを経由して糖に結合させることができる。リンカーの末端原子は、リン酸基の結合原子または糖のC-3'もしくはC-5'のO、N、SもしくはC基と結合またはそれを置換することができる。あるいは、リンカーは、ヌクレオチド代用物(例えばPNA)の末端原子と結合またはそれを置換することができる。
アデニン、グアニン、シトシンおよびウラシルは、RNAに見出される最も一般的な塩基である。これらの塩基を、修飾または置換して、改善された性質を有するRNAを提供することができる。例えばヌクレアーゼ耐性オリゴリボヌクレオチドを、これらの塩基を用いて、または合成および天然の核酸塩基(例えば、イノシン、チミン、キサンチン、ヒポキサンチン、ヌブラリン(nubularine)、イソグアニシン(isoguanisine)、もしくはツベルシジン)および上記修飾の任意の1つを用いて調製することができる。あるいは、上記塩基および「ユニバーサル塩基」のいずれかの置換または修飾類似体を用いることができる。例には、2-(ハロ)アデニン、2-(アルキル)アデニン、2-(プロピル)アデニン、2(アミノ)アデニン、2-(アミノアルキル)アデニン、2(アミノプロピル)アデニン、2(メチルチオ)N6(イソペンテニル)アデニン、6(アルキル)アデニン、6(メチル)アデニン、7(デアザ)アデニン、8(アルケニル)アデニン、8-(アルキル)アデニン、8(アルキニル)アデニン、8(アミノ)アデニン、8-(ハロ)アデニン、8-(ヒドロキシル)アデニン、8(チオアルキル)アデニン、8-(チオール)アデニン、N6-(イソペンチル)アデニン、N6(メチル)アデニン、N6,N6(ジメチル)アデニン、2-(アルキル)グアニン、2(プロピル)グアニン、6-(アルキル)グアニン、6(メチル)グアニン、7(アルキル)グアニン、7(メチル)グアニン、7(デアザ)グアニン、8(アルキル)グアニン、8-(アルケニル)グアニン、8(アルキニル)グアニン、8-(アミノ)グアニン、8(ハロ)グアニン、8-(ヒドロキシル)グアニン、8(チオアルキル)グアニン、8-(チオール)グアニン、N(メチル)グアニン、2-(チオ)シトシン、3(デアザ)5(アザ)シトシン、3-(アルキル)シトシン、3(メチル)シトシン、5-(アルキル)シトシン、5-(アルキニル)シトシン、5(ハロ)シトシン、5(メチル)シトシン、5(プロピニル)シトシン、5(プロピニル)シトシン、5(トリフルオロメチル)シトシン、6-(アゾ)シトシン、N4(アセチル)シトシン、3(3アミノ-3カルボキシプロピル)ウラシル、2-(チオ)ウラシル、5(メチル)2(チオ)ウラシル、5(メチルアミノメチル)-2(チオ)ウラシル、4-(チオ)ウラシル、5(メチル)4(チオ)ウラシル、5(メチルアミノメチル)-4(チオ)ウラシル、5(メチル)2,4(ジチオ)ウラシル、5(メチルアミノメチル)-2,4(ジチオ)ウラシル、5(2-アミノプロピル)ウラシル、5-(アルキル)ウラシル、5-(アルキニル)ウラシル、5-(アリルアミノ)ウラシル、5(アミノアリル)ウラシル、5(アミノアルキル)ウラシル、5(グアニジニウムアルキル)ウラシル、5(1,3-ジアゾール-1-アルキル)ウラシル、5-(シアノアルキル)ウラシル、5-(ジアルキルアミノアルキル)ウラシル、5(ジメチルアミノアルキル)ウラシル、5-(ハロ)ウラシル、5-(メトキシ)ウラシル、ウラシル-5オキシ酢酸、5(メトキシカルボニルメチル)-2-(チオ)ウラシル、5(メトキシカルボニル-メチル)ウラシル、5(プロピニル)ウラシル、5(プロピニル)ウラシル、5(トリフルオロメチル)ウラシル、6(アゾ)ウラシル、ジヒドロウラシル、N3(メチル)ウラシル、5-ウラシル(すなわち、プソイドウラシル)、2(チオ)プソイドウラシル、4(チオ)プソイドウラシル、2,4-(ジチオ)プソイドウラシル、5-(アルキル)プソイドウラシル、5-(メチル)プソイドウラシル、5-(アルキル)-2-(チオ)プソイドウラシル、5-(メチル)-2-(チオ)プソイドウラシル、5-(アルキル)-4(チオ)プソイドウラシル、5-(メチル)-4(チオ)プソイドウラシル、5-(アルキル)-2,4(ジチオ)プソイドウラシル、5-(メチル)-2,4(ジチオ)プソイドウラシル、1置換プソイドウラシル、1置換2(チオ)-プソイドウラシル、1置換4(チオ)プソイドウラシル、1置換2,4-(ジチオ)プソイドウラシル、1(アミノカルボニルエチレニル)-プソイドウラシル、1(アミノカルボニルエチレニル)-2(チオ)-プソイドウラシル、1(アミノカルボニルエチレニル)-4(チオ)プソイドウラシル、1(アミノカルボニルエチレニル)-2,4-(ジチオ)プソイドウラシル、1(アミノアルキルアミノカルボニルエチレニル)-プソイドウラシル、1(アミノアルキルアミノ-カルボニルエチレニル)-2(チオ)-プソイドウラシル、1(アミノアルキルアミノカルボニルエチレニル)-4(チオ)プソイドウラシル、1(アミノアルキルアミノカルボニルエチレニル)-2,4-(ジチオ)プソイドウラシル、1,3-(ジアザ)-2-(オキソ)-フェノキサジン-1-イル、1-(アザ)-2-(チオ)-3-(アザ)-フェノキサジン-1-イル、1,3-(ジアザ)-2-(オキソ)-フェンチアジン(phenthiazin)-1-イル、1-(アザ)-2-(チオ)-3-(アザ)-フェンチアジン-1-イル、7-置換1,3-(ジアザ)-2-(オキソ)-フェノキサジン-1-イル、7-置換1-(アザ)-2-(チオ)-3-(アザ)-フェノキサジン-1-イル、7-置換1,3-(ジアザ)-2-(オキソ)-フェンチアジン-1-イル、7-置換1-(アザ)-2-(チオ)-3-(アザ)-フェンチアジン-1-イル、7-(アミノアルキルヒドロキシ)-1,3-(ジアザ)-2-(オキソ)-フェノキサジン-1-イル、7-(アミノアルキルヒドロキシ)-1-(アザ)-2-(チオ)-3-(アザ)-フェノキサジン-1-イル、7-(アミノアルキルヒドロキシ)-1,3-(ジアザ)-2-(オキソ)-フェンチアジン-1-イル、7-(アミノアルキルヒドロキシ)-1-(アザ)-2-(チオ)-3-(アザ)-フェンチアジン-1-イル、7-(グアニジニウムアルキルヒドロキシ)-1,3-(ジアザ)-2-(オキソ)-フェノキサジン-1-イル、7-(グアニジニウムアルキルヒドロキシ)-1-(アザ)-2-(チオ)-3-(アザ)-フェノキサジン-1-イル、7-(グアニジニウムアルキル-ヒドロキシ)-1,3-(ジアザ)-2-(オキソ)-フェンチアジン-1-イル、7-(グアニジニウムアルキルヒドロキシ)-1-(アザ)-2-(チオ)-3-(アザ)-フェンチアジン-1-イル、1,3,5-(トリアザ)-2,6-(ジオキサ)-ナフタレン、イノシン、キサンチン、ヒポキサンチン、ヌブラリン、ツベルシジン、イソグアニシン(isoguanisine)、イノシニル、2-アザ-イノシニル、7-デアザ-イノシニル、ニトロイミダゾリル、ニトロピラゾリル、ニトロベンゾイミダゾリル、ニトロインダゾリル、アミノインドリル、ピロールピリミジニル、3-(メチル)イソカルボスチリリル(isocarbostyrilyl)、5-(メチル)イソカルボスチリリル、3-(メチル)-7-(プロピニル)イソカルボスチリリル、7-(アザ)インドリル、6-(メチル)-7-(アザ)インドリル、イミジゾピリジニル(imidizopyridinyl)、9-(メチル)-イミジゾピリジニル、ピロールピリジニル、イソカルボスチリリル、7-(プロピニル)イソカルボスチリリル、プロピニル-7-(アザ)インドリル、2,4,5-(トリメチル)フェニル、4-(メチル)インドリル、4,6-(ジメチル)インドリル、フェニル、ナフタレニル、アントラセニル、フェナントラセニル、ピレニル、スチルベニル、テトラセニル、ペンタセニル、ジフルオロトリル、4-(フルオロ)-6-(メチル)ベンゾイミダゾール、4-(メチル)ベンゾイミダゾール、6-(アゾ)チミン、2-ピリジノン、5ニトロインドール、3ニトロピロール、6-(アザ)ピリミジン、2(アミノ)プリン、2,6-(ジアミノ)プリン、5置換ピリミジン、N2-置換プリン、N6-置換プリン、O6-置換プリン、置換1,2,4-トリアゾール、またはその任意のO-アルキル化もしくはN-アルキル化誘導体が含まれる。
オリゴヌクレオチドへの修飾は、また、リン酸エステルまたは修飾リン酸エステル骨格部分の糖、塩基、および/またはリン原子に対する1つまたは複数の陽イオン性基の結合を含むことができる。陽イオン性基は、天然、異常またはユニバーサル塩基上での置換が可能な任意の原子に結合することができる。好ましい位置は、ハイブリダイゼーションを妨害しない、すなわち、塩基対形成のために必要な水素結合形成相互作用を妨害しない位置である。陽イオン性基は、例えば糖のC2'位または環式もしくは非環式糖代用物における類似の位置を経由して結合することができる。例えば、陽イオン性基は、例えば、O-アミン(アミン=NH2;アルキルアミノ、ジアルキルアミノ、ヘテロシクリル、アリールアミノ、ジアリールアミノ、ヘテロアリールアミノ、もしくはジヘテロアリールアミノ、エチレンジアミン、ポリアミノ);アミノアルコキシ、例えば、O(CH2)nアミン、(例えば、アミン=NH2;アルキルアミノ、ジアルキルアミノ、ヘテロシクリル、アリールアミノ、ジアリールアミノ、ヘテロアリールアミノ、もしくはジヘテロアリールアミノ、エチレンジアミン、ポリアミノ);アミノ(例えばNH2;アルキルアミノ、ジアルキルアミノ、ヘテロシクリル、アリールアミノ、ジアリールアミノ、ヘテロアリールアミノ、ジヘテロアリールアミノ、もしくはアミノ酸);またはNH(CH2CH2NH)nCH2CH2-アミン(アミン=NH2;アルキルアミノ、ジアルキルアミノ、ヘテロシクリル、アリールアミノ、ジアリールアミノ、ヘテロアリールアミノ、もしくはジヘテロアリールアミノ)から得られるプロトン化アミノ基を含むことができる。
いくつかの修飾は、好ましくは、オリゴヌクレオチド上の特定の部位で、例えば鎖の内部位置で、またはオリゴヌクレオチドの5'もしくは3'末端上に含まれることができる。オリゴヌクレオチド上の修飾の好ましい部位は、作用物質に好ましい性質を付与することができる。例えば、特定の修飾の好ましい部位は、最適な遺伝子サイレンシング性質、またはエンドヌクレアーゼもしくはエキソヌクレアーゼ活性に対する増加した耐性を付与することができる。
本発明により使用されるオリゴリボヌクレオチドおよびオリゴリボヌクレオシドは、固相合成で合成することができる。例えば、"Oligonucleotide synthesis, a practical approach", Ed. M. J. Gait, IRL Press, 1984; "Oligonucleotides and Analogues, A Practical Approach", Ed. F. Eckstein, IRL Press, 1991(特にChapter 1, Modern machine-aided methods of oligodeoxyribonucleotide synthesis、Chapter 2, Oligoribonucleotide synthesis、Chapter 3, 2'-O--Methyloligoribonucleotide- s: synthesis and applications、Chapter 4, Phosphorothioate oligonucleotides、Chapter 5, Synthesis of oligonucleotide phosphorodithioates、Chapter 6, Synthesis of oligo-2'-deoxyribonucleoside methylphosphonates、およびChapter 7, Oligodeoxynucleotides containing modified basesを参照されたい。他の特に有用な合成手順、試薬、ブロッキング基および反応条件は、Martin, P., Helv. Chim. Acta, 1995, 78, 486-504; Beaucage, S. L. and Iyer, R. P., Tetrahedron, 1992, 48, 2223-2311およびBeaucage, S. L. and Iyer, R. P., Tetrahedron, 1993, 49, 6123-6194、またはその中で参照される参考文献に記載されている。国際公開公報第00/44895号、同第01/75164号、または同第02/44321号に記載されている修飾を、本明細書において使用することができる。本明細書に挙げられる全ての刊行物、特許、および公開された特許出願の開示は、参照により本明細書に組み入れられる。
ホスフィン酸オリゴリボヌクレオチドの調製は、米国特許第5,508,270号に記載されている。アルキルホスホネートオリゴリボヌクレオチドの調製は、米国特許第4,469,863号に記載されている。ホスホロアミダイトオリゴリボヌクレオチドの調製は、米国特許第5,256,775号または同第5,366,878号に記載されている。ホスホトリエステルオリゴリボヌクレオチドの調製は、米国特許第5,023,243号に記載されている。ボラノリン酸オリゴリボヌクレオチドの調製は、米国特許第5,130,302号および第5,177,198号に記載されている。3'-デオキシ-3'-アミノホスホルアミデートオリゴリボヌクレオチドの調製は、米国特許第5,476,925号に記載されている。3'-デオキシ-3'-メチレンホスホネートオリゴリボヌクレオチドは、An, H, et al. J. Org. Chem. 2001, 66, 2789-2801に記載されている。硫黄架橋ヌクレオチドの調製は、Sproat et al. Nucleosides Nucleotides 1988, 7,651およびCrosstick et al. Tetrahedron Lett. 1989, 30, 4693に記載されている。
2'修飾への修飾は、Verma, S. et al. Annu. Rev. Biochem. 1998, 67, 99-134およびその中の全ての参考文献に見出すことができる。リボースへの特異的修飾は、以下の参考文献中に見出すことができる:2'-フルオロ(Kawasaki et. al., J. Med. Chem., 1993, 36, 831-841)、2'-MOE(Martin, P. Helv. Chim. Acta 1996, 79, 1930-1938)、「LNA」(Wengel, J. Acc. Chem. Res. 1999, 32, 301-310)。
本明細書においてMMI結合型オリゴリボヌクレオシドとしても特定されるメチレンメチルイミノ結合型オリゴリボヌクレオシド、本明細書においてMDH結合型オリゴリボヌクレオシドとしても特定されるメチレンジメチルヒドラゾ結合型オリゴリボヌクレオシド、および本明細書においてアミド-3結合型オリゴリボヌクレオシドとしても特定されるメチレンカルボニルアミノ結合型オリゴヌクレオシド、および本明細書においてアミド-4結合型オリゴリボヌクレオシドとしても特定されるメチレンアミノカルボニル結合型オリゴヌクレオシド、ならびに例えば交互のMMI結合およびPOまたはPS結合として有する混合骨格化合物は、米国特許第5,378,825号、同第5,386,023号、同第5,489,677号および公開されたPCT出願PCT/US92/04294およびPCT/US92/04305(それぞれ国際公開公報第92/20822号および同第92/20823号として公開されている)に記載されているように調製することができる。ホルムアセタール結合型オリゴリボヌクレオシドおよびチオホルムアセタール結合型オリゴリボヌクレオシドは、米国特許第5,264,562号および同第5,264,564号に記載されているように調製することができる。エチレンオキシド結合型オリゴリボヌクレオシドは、米国特許第5,223,618号に記載されているように調製することができる。シロキサン置換は、Cormier,J.F. et al. Nucleic Acids Res. 1988, 16, 4583に記載されている。カルボネート置換は、Tittensor, J.R. J. Chem. Soc. C 1971, 1933に記載されている。カルボキシメチル置換は、Edge, M.D. et al. J. Chem. Soc. Perkin Trans. 1 1972, 1991に記載されている。カルバメート置換は、Stirchak, E.P. Nucleic Acids Res. 1989, 17, 6129に記載されている。
シクロブチル糖代用化合物は、米国特許第5,359,044号に記載されているように調製することができる。ピロリジン糖代用物は、米国特許第5,519,134号に記載されているように調製することができる。モルホリノ糖代用物は、米国特許第5,142,047号および同第5,235,033号および他の関連特許開示に記載されているように調製することができる。ペプチド核酸(PNA)は、それ自体が公知であり、Peptide Nucleic Acids (PNA): Synthesis, Properties and Potential Applications, Bioorganic & Medicinal Chemistry, 1996, 4, 5-23の中で参照される様々な手順のいずれかに従って調製することができる。それらは、その全体として参照により本明細書に組み入れられる米国特許第5,539,083号に従って調製することもできる。
末端修飾は、Manoharan, M. et al. Antisense and Nucleic Acid Drug Development 12, 103-128 (2002)およびその中の参考文献に記載されている。
N-2置換プリンヌクレオシドアミダイトは、米国特許第5,459,255号に記載されているように調製することができる。3-デアザプリンヌクレオシドアミダイトは、米国特許第5,457,191号に記載されているように調製することができる。5,6-置換ピリミジンヌクレオシドアミダイトは、米国特許第5,614,617号に記載されているように調製することができる。5-プロピニルピリミジンヌクレオシドアミダイトは、米国特許第5,484,908号に記載されているように調製することができる。追加的な参考文献は、塩基修飾に関する上記の節に開示されている。
本発明のオリゴヌクレオチド化合物は、溶液相または固相有機合成を使用して調製することができる。有機合成は、非天然ヌクレオチドまたは修飾ヌクレオチドを含むオリゴヌクレオチド鎖を容易に調製できるという利点を提供する。当技術分野において公知のそのような合成のための任意の他の手段を、追加的または代替的に用いることができる。ホスホロチオエート、ホスホロジチオエートおよびアルキル化誘導体のような他のオリゴヌクレオチドを調製するために類似の技法を使用することも公知である。本発明の二本鎖オリゴヌクレオチド化合物は、二段階手順を使用して調製することができる。最初に、二本鎖分子の個別の鎖は、別々に調製される。次に、構成要素の鎖がアニーリングされる。
標的細胞(例えば、基底細胞または肺および/もしくは呼吸器系の細胞または他の所望の標的細胞)にRNAi剤、例えばsiRNA、またはRNAi剤を含有するベクターを送達する方法は、当業者に周知である。いくつかの態様では、Suv39h1、Suv39h2 Setdb1、Ehmt1および/またはPRDM2の任意の1つを阻害する、RNAi剤のようなH3K9メチルトランスフェラーゼの阻害剤である、RNAi剤(例えば遺伝子サイレンシングRNAi剤)を、エアロゾル手段を介して、例えばネブライザーなどを使用して、対象に投与することができる。代替的な態様では、H3K9メチルトランスフェラーゼ阻害剤、例えば、Suv39h1、Suv39h2 Setdb1、Ehmt1および/またはPRDM2の任意の1つの阻害剤である、RNAi剤(例えば遺伝子サイレンシングRNAi剤)の投与は、例えば(i)RNA干渉剤、例えばsiRNAを含有する組成物の注射、または(ii)細胞(例えば、ドナー哺乳動物細胞、レシピエント卵母細胞、またはSCNT胚)を、RNAi剤、例えばsiRNAを含む組成物と直接接触させることを含むことができる。
いくつかの態様では、H3K9メチルトランスフェラーゼ阻害剤は、Suv39h1、Suv39h2、Setdb1、Ehmt1またはPRDM2のようなH3K9メチルトランスフェラーゼの任意の1つのタンパク質阻害剤および/またはペプチド阻害剤、例えば非限定的に、Suv39h1、Suv39h2、Setdb1、Ehmt1またはPRDM2のようなH3K9メチルトランスフェラーゼの変異型タンパク質;治療用タンパク質および組み換えタンパク質ならびにドミナントネガティブ阻害剤(例えば、H3K9メチルトランスフェラーゼの非機能的タンパク質、またはH3K9メチルトランスフェラーゼに競合的に結合するH3K9メチルトランスフェラーゼの非機能的リガンド)である。タンパク質およびペプチド阻害剤は、また、例えば変異型タンパク質、遺伝的に修飾されたタンパク質、ペプチド、合成ペプチド、組み換えタンパク質、キメラタンパク質、抗体、ヒト化タンパク質、ヒト化抗体、キメラ抗体、修飾タンパク質およびそのフラグメントを含むことができる。
いくつかの態様では、本発明の方法に有用なH3K9メチルトランスフェラーゼ阻害剤には、例えば、モノクローナル、キメラヒト化、および組み換え抗体ならびにその抗原結合性フラグメントを含めた抗体が含まれる。いくつかの態様では、中和抗体は、H3K9メチルトランスフェラーゼ阻害剤として使用することができる。抗体は、ウサギまたはマウスのような動物において、抗原を免疫処置することによって容易に産生される。免疫処置されたマウスは、ハイブリドーマを製造するためのB細胞源を提供するために特に有用であり、今度はハイブリドーマが培養されて、大量のモノクローナル抗体を産生する。Suv39h1および/またはSuv39h2の市販の抗体阻害剤が、本発明における使用のために包含され、例えばSanta Cruz biotechnologyなどから入手可能である。
上の節に述べられる適用の全ては、参照により本明細書に組み入れられる。いくつかの態様では、当業者は、H3K9メチルトランスフェラーゼ阻害剤として他の作用物質を使用することができ、例えば、抗体、デコイ抗体、またはRNAiは、本明細書開示のSCNTの効率を増加させるための方法、化合物およびキットに有効である。
本発明の目的は、SCNTの効率を増加させ、かつSCNTを使用して体細胞をより効率的にクローニングする手段を提供することである。本開示の方法は、哺乳動物をクローニングするため、全能細胞もしくは多能性細胞を得るため、または哺乳動物細胞をリプログラミングするために使用され得る。
特定の態様では、本発明の方法、キットおよび組成物に使用するためのレシピエント卵母細胞は、任意の哺乳動物種由来であり得る。特定の態様では、凍結保存した卵母細胞が、レシピエント卵母細胞として使用される。特定の態様では、レシピエント卵母細胞はヒト性である。卵母細胞の低温貯蔵および解凍は、当業者に公知である(Tucker et al., Curr Opin Obstet Gynecol. 1995 June; 7(3):188-92を参照されたい)。いくつかの態様では、レシピエント卵母細胞は、自発的な雌性ドナー、例えば卵ドナーから得られる。いくつかの態様では、卵母細胞は、卵巣刺激または卵巣の過剰刺激(すなわち排卵誘発または制御卵巣過剰刺激)を受けた雌性哺乳動物対象から得られる。制御卵巣過剰刺激の方法は、例えばその全体として参照により本明細書に組み入れられる米国特許第8,173,592号および国際特許出願である国際公開公報第2000/059542号に開示されるように、当技術分野において周知である。
本明細書開示の方法、キットおよび組成物は、ドナー哺乳動物細胞を含み、その細胞から核が除核卵母細胞内に注射されてSCNT胚が作製される。いくつかの態様では、ドナー哺乳動物細胞は、最終分化した体細胞である。いくつかの態様では、ドナー哺乳動物細胞は、胚性幹細胞または成熟幹細胞またはiPS細胞ではない。いくつかの態様では、ドナー哺乳動物細胞は、ドナー哺乳動物細胞として本明細書開示の方法に使用するためのヒトまたは動物細胞であり、その際、ドナー細胞由来の核は、除核卵母細胞内に移入される。いくつかの態様では、ドナー体細胞は、雄性哺乳動物対象、例えばXY対象から得られる。代替的な態様では、体細胞のドナーは、雌性対象、例えばXX対象から得られる。いくつかの態様では、体細胞のドナーは、XXY対象から得られる。
いくつかの態様では、ドナー哺乳動物体細胞は、H3K9メチルトランスフェラーゼ阻害剤および/またはKdm4ヒストンデメチラーゼ活性化剤で処置される、またはそれと接触される。いくつかの態様では、ドナー哺乳動物細胞の核(または核遺伝物質)は、H3K9メチルトランスフェラーゼ阻害剤および/またはKdm4ヒストンデメチラーゼ活性化剤で処置される、またはそれと接触される。いくつかの態様では、ドナー哺乳動物細胞の細胞質および/または核は、本明細書開示のH3K9メチルトランスフェラーゼ阻害剤、例えばSuv39h1、Suv39h2および/またはSetdb1の任意の1つまたは組み合わせの阻害剤で処置される、またはそれと接触される。いくつかの態様では、接触は、ドナー哺乳動物体細胞の細胞質および/または核へのH3K9メチルトランスフェラーゼ阻害剤および/またはKdm4ヒストンデメチラーゼ活性化剤のマイクロインジェクションである。
本発明の一目的は、体細胞をより効率的にクローニングする手段を提供することである。本開示の方法および組成物は、哺乳動物、例えば非ヒト哺乳動物をクローニングするために、哺乳動物(例えば、ヒトおよび非ヒト哺乳動物)多能性細胞および全能細胞を得るために、および哺乳動物細胞をリプログラミングするために、使用され得る。
卵母細胞ドナーは、以前に記載されたように同期化および過排卵させることができ(Gavin W.G., 1996)、精管切除された雄と48時間にわたり交配された。収集後、2mM L-グルタミンおよび1%ペニシリン/ストレプトマイシン(各10,000I.U./ml)を補充した10%FBSを有する平衡化M199中で卵母細胞を培養した。核移入は、インビボまたはインビトロで成熟できた卵母細胞を利用することもできる。インビボ成熟した卵母細胞を、上に説明したように派生させ、インビトロ成熟した卵母細胞を特定の細胞期までインビトロで発生させ、その後、核移入に使用するためにそれらが回収される。
卵丘細胞が付着した卵母細胞は、典型的には廃棄される。卵丘細胞のない卵母細胞を2群に分類した:停止した分裂中期II(1つの極体)および分裂終期IIのプロトコール(はっきりと視認可能な極体または部分的に突出している第二極体の存在がない)。停止した分裂中期IIのプロトコールでは卵母細胞は最初に除核される。活性化分裂終期IIのプロトコールに割り当てられた卵母細胞は、M199/10%FBS中で2〜4時間培養することによって調製された。この期間の後に、全ての活性化卵母細胞(部分的に突出した第二極体が存在)を、培養誘導カルシウム活性化分裂終期II卵母細胞(分裂終期II-Ca)として群分けし、除核した。続いて、培養期間中に活性化されなかった卵母細胞を、7%エタノール含有M199、10%FBS中で5分間インキュベートして活性化を誘導し、次に10%FBSを有するM199中で追加的に3時間培養して、分裂終期IIに到達させた(分裂終期II-EtOHプロトコール)。除核前に、全ての卵母細胞をサイトカラシン-Bで15〜30分間処置する。ガラスピペットで第一極体および極体周囲の隣接細胞質(細胞質の約30%)吸引して赤道板を除去することによって、分裂中期IIの卵母細胞を除核した。第一極体および部分的に突出している第二極体を含む周囲の細胞質(細胞質の10〜30%)を除去することによって、分裂終期II-Caおよび分裂終期II-EtOH卵母細胞を除核した。除核後、全ての卵母細胞を直ちに再構築した。
卵母細胞の除核のために使用された同じ培地中でドナー細胞の注射を行った。ガラスピペットを使用して、透明帯と卵細胞質膜との間にドナー細胞を1つ置いた。電気融合および活性化手順の前に、細胞-卵母細胞のカプレット(couplet)をM199中で30〜60分間インキュベートした。再構築された卵母細胞を、融合緩衝液(300mM マンニトール、0.05mM CaCl2、0.1mM MgSO4、1mM K2HPO4、0.1mMグルタチオン、0.1mg/ml BSA)中で2分間平衡化した。融合培地で満たされた「融合スライド」になるよう形成された2つのステンレス鋼電極(間隙500μm; BTX-Genetronics, San Diego, Calif.)を有する融合チャンバー中において、電気融合および活性化を室温で行った。
SCNT胚の培養
SCNTによって得られた胚は、胚が通常培養される条件(少なくともインビボ)以外のインビボ培養条件から利益を得る、またはそれを必要にさえし得ると示唆されている。ウシ胚の日常的な増倍において、再構築された胚(その多くが一度に)は、ヒツジ輸卵管内で5〜6日間培養されたものである(Willadsen, In Mammalian Egg Transfer (Adams, E. E., ed.) 185 CRC Press, Boca Raton, Fla. (1982)によって記載)。特定の態様では、SCNT胚は、移入前に寒天のような保護培地中に包埋され、次に一時的レシピエントから回収後に寒天から切り出され得る。保護寒天または他の培地の機能は、2つある:第一に、それは、透明帯を一緒に保持することによってSCNT胚用の構造補助物として作用し、第二に、それは、レシピエント動物の免疫系細胞に対するバリアとして作用する。このアプローチは、胚盤胞を形成する胚の比率を増加させるものの、いくつかの胚が失われ得るという欠点がある。いくつかの態様では、SCNT胚を、50μlの液滴中でフィーダー細胞、例えば初代ヤギ輸卵管上皮細胞の単層上で共培養することができる。胚培養物を、5%CO2を有する加湿39℃インキュベーター中で48時間維持し、その後、胚をレシピエント代理母に移入することができる。
全能細胞を得る段階
特定の態様では、全能細胞を得るために、SCNT胚から作製される割球は、ガラスピペットを使用して解離され得る。いくつかの態様では、解離は、0.25%トリプシンの存在下で起こり得る(Collas and Robl, 43 BIOL. REPROD. 877-84, 1992; Stice and Robl, 39 BIOL. REPROD. 657-664, 1988; Kanka et al., 43 MOL. REPROD. DEV. 135-44, 1996)。
いくつかの態様では、非ヒト哺乳動物をクローニングするためのSCNT胚の産生効率を増加させるために、該方法および組成物を使用することができる。本明細書開示の方法および組成物から得られるSCNT胚から非ヒト哺乳動物をクローニングための方法は、当技術分野において周知である。哺乳動物をクローニングするために使用される2つの主な手順は、Roslin法およびHonolulu法である。これらの手順は、1996年にスコットランドのRoslin InstituteでヒツジのDolly(Campbell, K. H. et al. (1996) Nature 380:64-66)が、および1998年にホノルルのUniversity of HawaiiでマウスのCumulina(Wakayama, T. et al. (1998) Nature 394:369-374)が作製されたことにちなんで名付けられた。
ヒツジ「Dolly」のクローニングにおけるWilmutらの発見(Wilmut, et al, Nature 385, 810 (1997)は、hESCを派生させることにおけるThomsonらの発見(Thomson et al., Science 282, 1145 (1998))と一緒になって、患者自身の核から作製されるSCNT胚またはSCNT操作された細胞塊から派生する患者特異的hESCの樹立に基づく再生細胞移植にかなりの熱狂を引き起こした。自家移植により免疫拒絶反応を避けることを目的とするこの戦略は、おそらく、SCNTについての最も強い臨床的な理論的根拠である。その証拠に、複合疾患特異的SCNT-hESCの派生は、疾患メカニズムの発見を加速し得る。細胞移植について、個別のマウス自身のSCNT由来mESCを用いたマウスSCIDおよびPDモデルの革新的な処置が有望である(Rideout et al, Cell 109, 17 (2002); Barberi, Nat. Biotechnol. 21, 1200 (2003))。最終的に、広い組織適合性を有するSCNT由来幹細胞のバンクを創出できることが、新しい卵母細胞を継続して供給する必要性を下げるであろう。
本発明の別の局面は、本明細書開示の方法によって生成されるSCNTから得られるntESC集団に関する。いくつかの態様では、ntESCは、ヒトntESC、例えば患者特異的ntESC、および/または患者特異的同質遺伝子ntESCである。いくつかの態様では、ntESCは、ntESCを全能または多能性状態に維持する培養培地のような培地中に存在する。いくつかの態様では、培養培地は、凍結保存に適した培地である。いくつかの態様では、ntESC集団は凍結保存される。低温保存は、例えば将来使用するために、例えば、治療使用のために、または他の使用、例えば研究使用のために、ntESCを貯蔵するために有用である。ntESCは増幅され得、増幅されたntESCの一部分が使用され得、別の部分が低温保存され得る。ntESCを増幅および保存できることは、かなりの柔軟性、例えば複数の患者特異的ヒトntESCの生成およびSCNT手順に使用するためのドナー体細胞の選択を可能にする。例えば、組織適合性ドナー由来の細胞が、増幅され、1つよりも多いレシピエントに使用され得る。組織バンクがntESCの低温保存を提供することができる。ntESCは、組織適合性データと共に凍結保存され得る。本明細書開示の方法を使用して生成されるntESCを、日常的な手順により凍結保存することができる。例えば、凍結保存を、適切な増殖培地、10%BSAおよび7.5%ジメチルスルホキシドを含むことができる「凍結」培地中の約100万〜1000万個の細胞に関して実施することができる。ntESCは遠心分離される。成長培地が吸引され、凍結培養培地により置換される。ntESCが球体として再懸濁される。細胞は、例えば-80℃の容器中に入れることによってゆっくりと凍結される。凍結したntESCは、37℃の浴中でかきまぜることによって解凍され、新鮮幹細胞培地中に再懸濁され、上記のように成長される。
1.
体細胞核移入(SCNT)の効率を増加させるための方法であって、
ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚を、ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚におけるH3K9me3メチル化を減少させる作用物質と接触させる段階であって、それによってSCNTの効率を増加させる、段階
を含む、SCNTの効率を増加させる方法。
2.
哺乳動物体細胞核移入(SCNT)胚を生成するための方法であって、
(a)ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物体細胞核移入(SCNT)胚の少なくとも1つを、ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚におけるH3K9me3メチル化を減少させる少なくとも1つの作用物質と接触させる段階であって、レシピエント哺乳動物卵母細胞が、有核または除核卵母細胞である、段階;
(b)レシピエント哺乳動物卵母細胞が有核である場合、レシピエント哺乳動物卵母細胞を除核する段階;
(c)ドナー哺乳動物細胞由来の核を除核卵母細胞に移入する段階;および
(d)哺乳動物SCNT胚を形成させるために十分な時間、レシピエント卵母細胞をインキュベートする段階
を含む、方法。
3.
作用物質が、Kdm4(Jmjd2)ファミリーのヒストンデメチラーゼの発現または活性を増加させる、パラグラフ1または2の方法。
4.
作用物質が、Kdm4a(Jmjd2a)、Kdm4b(Jmjd2b)、Kdm4c(Jmjd2c)、またはKdm4d(Jmjd2d)のうち少なくとも1つの発現または活性を増加させる、パラグラフ1〜3のいずれか一つの方法。
5.
作用物質が、Kdm4d(Jmjd2D)またはKdm4A(Jmjd2A)の発現または活性を増加させる、パラグラフ1〜4のいずれか一つの方法。
6.
作用物質が、SEQ ID NO:1〜8に対応する核酸配列、またはSEQ ID NO:1〜8の対応する配列と比較してSCNTの効率を同程度もしくはより大きな程度まで増加させるその生物学的に活性なフラグメントを含む、パラグラフ1〜5のいずれか一つの方法。
7.
作用物質が、SEQ ID NO:1に対応する核酸配列、またはSEQ ID NO:1の核酸配列と比較してSCNTの効率を同程度もしくはより大きな程度まで増加させるその生物学的に活性なフラグメントを含む、パラグラフ6の方法。
8.
作用物質が、H3K9メチルトランスフェラーゼの阻害剤である、パラグラフ1〜7のいずれか一つの方法。
9.
H3K9メチルトランスフェラーゼが、Suv39h1またはSuv39h2である、パラグラフ8の方法。
10.
H3K9メチルトランスフェラーゼが、Setdb1である、パラグラフ8の方法。
11.
Suv39h1、Suv39h2、およびSetdb1のうち2つ以上が阻害される、パラグラフ8の方法。
12.
H3K9メチルトランスフェラーゼを阻害する作用物質が、RNAi剤、CRISPR/Cas9、オリゴヌクレオチド、中和抗体または抗体フラグメント、アプタマー、小分子、ペプチド阻害剤、タンパク質阻害剤、アビジミル(avidimir)、およびそれらの機能的フラグメントまたは誘導体からなる群より選択される、パラグラフ8の方法。
13.
RNAi剤が、siRNAまたはshRNA分子である、パラグラフ12の方法。
14.
作用物質が、SEQ ID NO:9、11、13または15のいずれかの発現を阻害するための核酸阻害剤を含む、パラグラフ1〜13のいずれか一つの方法。
15.
作用物質が、SEQ ID NO:10、12、14または16のいずれかの発現を阻害する核酸阻害剤を含む、パラグラフ1〜13のいずれか一つの方法。
16.
RNAi剤が、SEQ ID NO:17またはSEQ ID NO:19の少なくとも一部とハイブリダイズする、パラグラフ15の方法。
17.
RNAi剤が、SEQ ID NO:18もしくはSEQ ID NO:20、またはその少なくとも10個の連続する核酸のフラグメント、またはSEQ ID NO:18もしくはSEQ ID NO:20と少なくとも80%同一の配列を有するホモログを含む、パラグラフ16の方法。
18.
レシピエント哺乳動物卵母細胞が、除核した哺乳動物卵母細胞である、パラグラフ1〜17のいずれか一つの方法。
19.
SCNT胚が、1細胞期SCNT胚、活性化の5時間後(5hpa)のSCNT胚、活性化の10〜12時間後(10〜12hpa)のSCNT胚、活性化の20〜28時間後(20〜28hpa)のSCNT胚、2細胞期SCNT胚のいずれかより選択される、パラグラフ1〜18のいずれか一つの方法。
20.
核移入前に、作用物質が、レシピエント哺乳動物卵母細胞または除核した哺乳動物卵母細胞と接触する、パラグラフ1〜19のいずれか一つの方法。
21.
活性化の前もしくは5時間後に、またはSCNT胚が1細胞期のときに、作用物質が、SCNT胚と接触する、パラグラフ1〜19のいずれか一つの方法。
22.
活性化の5時間後(5hpa)、もしくは活性化の12時間後(hpa)、もしくは活性化の20時間後(20hpa)に、またはSCNT胚が2細胞期のときに、または5hpaから28hpaの間の任意の時間に、作用物質が、SCNT胚と接触する、パラグラフ1〜19のいずれか一つの方法。
23.
レシピエント哺乳動物卵母細胞またはSCNT胚を作用物質と接触させる段階が、レシピエント哺乳動物卵母細胞またはSCNT胚の核または細胞質に作用物質を注射することを含む、パラグラフ1〜22のいずれか一つの方法。
24.
作用物質が、Kdm4ファミリーのヒストンデメチラーゼの発現または活性を増加させる、パラグラフ1〜23のいずれか一つの方法。
25.
除核した哺乳動物卵母細胞にドナー哺乳動物細胞の核を注射する前に、作用物質が、ドナー哺乳動物細胞の細胞質またはドナー哺乳動物細胞の核と接触する、パラグラフ1〜24のいずれか一つの方法。
26.
除核した哺乳動物卵母細胞にドナー哺乳動物細胞の核を注射する少なくとも24時間前に、または注射する前の少なくとも1日間、ドナー哺乳動物細胞が接触される、パラグラフ25の方法。
27.
除核した哺乳動物卵母細胞にドナー哺乳動物細胞の核を注射する前に、作用物質が、ドナー哺乳動物細胞と少なくとも24時間、または少なくとも48時間、または少なくとも3日間接触する、パラグラフ25の方法。
28.
作用物質が、H3K9メチルトランスフェラーゼを阻害する、パラグラフ25〜27のいずれか一つの方法。
29.
H3K9メチルトランスフェラーゼが、Suv39h1もしくはSuv39h2、またはSuv39h1およびSuv39h2(Suv39h1/2)である、パラグラフ25〜28のいずれか一つの方法。
30.
ドナー哺乳動物細胞が、最終分化した体細胞である、パラグラフ1〜29のいずれか一つの方法。
31.
ドナー哺乳動物細胞が、胚性幹細胞でも、人工多能性幹(iPS)細胞でも、胎児細胞でも、胚細胞でもない、パラグラフ1〜30のいずれか一つの方法。
32.
ドナー哺乳動物細胞が、卵丘細胞、上皮細胞、線維芽細胞、神経細胞、角化細胞、造血細胞、メラニン細胞、軟骨細胞、赤血球、マクロファージ、単球、筋細胞、Bリンパ球、Tリンパ球、胚性幹細胞、胚性生殖細胞、胎児細胞、胎盤細胞、および成熟細胞からなる群より選択される、パラグラフ1〜32のいずれか一つの方法。
33.
ドナー哺乳動物細胞が、線維芽細胞または卵丘細胞である、パラグラフ1〜32のいずれか一つの方法。
34.
除核したレシピエント哺乳動物卵母細胞に注射するためにドナー哺乳動物細胞から核を取り出す前に、作用物質が、ドナー哺乳動物細胞の核と接触する、パラグラフ1〜33のいずれか一つの方法。
35.
ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚が、ヒトドナー細胞、レシピエントヒト卵母細胞、またはヒトSCNT胚である、パラグラフ1〜34のいずれか一つの方法。
36.
ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚が、非ヒトドナー細胞、レシピエント非ヒト卵母細胞、または非ヒトSCNT胚である、パラグラフ1〜34のいずれか一つの方法。
37.
ドナー非ヒト哺乳動物細胞、レシピエント非ヒト哺乳動物卵母細胞、または非ヒト哺乳動物SCNT胚が、マウス、ラット、ウサギ、雌ウシ、ウマ、ブタ、ニワトリ、イヌ、ネコ、雌ウシ、マカク、チンパンジーからなる群より選択される、パラグラフ36の方法。
38.
ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚が、飼育動物または商業用動物由来である、パラグラフ36の方法。
39.
飼育動物が、アルパカ、バイソン、ラクダ、ネコ、ウシ、シカ、ゾウ、げっ歯動物、イヌ、ロバ、ガヤル、ヤギ、モルモット、ラマ、ウマ、サル、ラバ、雄ウシ、ブタ、ハト、非ヒト霊長類、ウサギ、トナカイ、ヒツジ、スイギュウ、またはヤクからなる群より選択される労役動物または競技動物または家畜動物または実験動物である、パラグラフ38の方法。
40.
ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚が、伴侶動物またはペット由来である、パラグラフ36の方法。
41.
伴侶動物が、イヌ、ネコ、雌ウシ、ハムスター、爬虫類、ウサギ、げっ歯動物、フェレット、チンチラ、鳥類ペット、モルモット、水生ペット、またはウマからなる群より選択される、パラグラフ41の方法。
42.
ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚が、絶滅に近い哺乳動物種由来である、パラグラフ36の方法。
43.
H3K9me3メチル化を減少させる作用物質の不在下で行われるSCNTと比較して、胚盤胞期へのSCNTの効率における少なくとも50%の増加を招く、パラグラフ1〜42のいずれか一つの方法。
44.
H3K9me3メチル化を減少させる作用物質の不在下で行われるSCNTと比較して、SCNTの効率における50%〜80%の増加を招く、パラグラフ1〜43のいずれか一つの方法。
45.
H3K9me3メチル化を減少させる作用物質の不在下で行われるSCNTと比較して、SCNTの効率における80%よりも大きな増加を招く、パラグラフ1〜44のいずれか一つの方法。
46.
SCNT効率における増加が、胚盤胞期へのSCNT胚の発生における増加である、パラグラフ44〜45のいずれか一つの方法。
47.
SCNT効率における増加が、SCNT胚の着床後発生における増加である、パラグラフ44〜46のいずれか一つの方法。
48.
SCNT効率における増加が、SCNT胚由来胚性幹細胞(ntESC)の派生における増加である、パラグラフ44〜47のいずれか一つの方法。
49.
ドナー哺乳動物細胞が、遺伝的に修飾されたドナー哺乳動物細胞である、パラグラフ1〜48のいずれか一つの方法。
50.
SCNT胚をインビトロ培養して胚盤胞を形成させる段階をさらに含む、パラグラフ2の方法。
51.
SCNT胚が、少なくとも1細胞期SCNT胚である、パラグラフ50の方法。
52.
SCNT胚が、少なくとも2細胞期SCNT胚である、パラグラフ50の方法。
53.
胚盤胞由来の内細胞塊から細胞を単離する段階、および
未分化状態の内細胞塊からの細胞を培養して哺乳動物胚性幹(ES)細胞を形成させる段階
をさらに含む、パラグラフ50の方法。
54.
ドナー哺乳動物細胞、レシピエント哺乳動物卵母細胞、または哺乳動物SCNT胚のうち任意の1つまたは複数が、凍結および解凍されたものである、パラグラフ1または2の方法。
55.
パラグラフ1〜54のいずれか一つの方法から生成された、哺乳動物SCNT胚由来胚性幹細胞(ntESC)集団。
56.
ntESCが、ヒトntESCである、パラグラフ55の哺乳動物ntESC集団。
57.
ntESCが、遺伝的に修飾されたntESCである、パラグラフ55の哺乳動物ntESC集団。
58.
ntESCが、多能性幹細胞である、パラグラフ55の哺乳動物ntESC集団。
59.
ntESCが、培養培地中に存在する、パラグラフ55の哺乳動物ntESC集団。
60.
培養培地が、ntESCを多能性または全能状態に維持する、パラグラフ59の哺乳動物ntESC集団。
61.
培養培地が、ntESCの凍結または凍結保存に適切な培地である、パラグラフ59の哺乳動物ntESC集団。
62.
凍結または凍結保存される、パラグラフ61の哺乳動物ntESC集団。
63.
パラグラフ1〜54の方法によって生成される、哺乳動物SCNT胚。
64.
遺伝的に修飾されている、パラグラフ63の哺乳動物SCNT胚。
65.
非ヒト哺乳動物SCNT胚である、パラグラフ63の哺乳動物SCNT胚。
66.
レシピエント哺乳動物卵母細胞由来ではないミトコンドリアDNAを含む、パラグラフ65の哺乳動物SCNT胚。
67.
培養培地中に存在する、パラグラフ63の哺乳動物SCNT胚。
68.
培養培地が、哺乳動物SCNTの凍結または凍結保存に適切な培地である、パラグラフ67の哺乳動物SCNT胚。
69.
凍結または凍結保存される、パラグラフ68の哺乳動物SCNT胚。
70.
体細胞核移入(SCNT)胚から非ヒト哺乳動物子孫を生成するための方法であって、
(a)ドナー非ヒト哺乳動物細胞、レシピエント非ヒト哺乳動物卵母細胞、または非ヒト哺乳動物体細胞核移入(SCNT)胚のうち少なくとも1つを、ドナー非ヒト哺乳動物細胞、レシピエント非ヒト哺乳動物卵母細胞、または非ヒト哺乳動物SCNT胚におけるH3K9me3メチル化を減少させる少なくとも1つの作用物質と接触させる段階であって、レシピエント非ヒト哺乳動物卵母細胞が、有核または除核卵母細胞である、段階;
(b)レシピエント非ヒト哺乳動物卵母細胞が有核である場合に、除核する段階;
(c)ドナー非ヒト哺乳動物細胞由来の核を非ヒト哺乳動物除核卵母細胞に移入し、核を除核卵母細胞と融合させ、融合した卵母細胞を活性化させる段階;
(d)非ヒト哺乳動物SCNT胚を形成させるために十分な時間、レシピエント卵母細胞をインキュベートする段階;
(e)非ヒト哺乳動物SCNT胚を非ヒト代理母の輸卵管に体内移植し、非ヒト哺乳動物SCNTの非ヒト哺乳動物子孫への発生を可能にする段階
を含む、方法。
71.
非ヒト哺乳動物SCNT胚が、胚1細胞期、胚2細胞期、胚4細胞期、桑実胚、または胚盤胞胚期に非ヒト代理母に体内移植される、パラグラフ70の方法。
72.
非ヒト代理母が、ドナー細胞またはレシピエント卵母細胞の起源ではない、パラグラフ71の方法。
73.
非ヒト代理母にSCNT胚を満期まで妊娠させる段階をさらに含む、パラグラフ70の方法。
74.
パラグラフ1〜47またはパラグラフ70〜73のいずれか一つの方法によって生成されるSCNT胚から生成される、非ヒト哺乳動物子孫。
75.
哺乳動物SCNT胚、レシピエント哺乳動物卵母細胞、または胚盤胞のうち少なくとも1つと、
(a)Kdm4ファミリーのヒストンデメチラーゼの発現もしくは活性を増加させる作用物質;または
(b)H3K9メチルトランスフェラーゼを阻害する作用物質
のうち少なくとも1つと
を含む、組成物。
76.
Kdm4(Jmjd2)ファミリーのヒストンデメチラーゼの発現または活性を増加させる作用物質が、 Kdm4a(Jmjd2a)、Kdm4b(Jmjd2b)、Kdm4c(Jmjd2c)、またはKdm4d(Jmjd2d)のうち少なくとも1つの発現または活性を増加させる、パラグラフ75の組成物。
77.
前記作用物質が、Kdm4d(Jmjd2d)またはKdm4a(Jmjd2a)の発現または活性を増加させる、パラグラフ76の組成物。
78.
前記作用物質が、SEQ ID NO:1〜8に対応する核酸、またはSEQ ID NO:1〜8の対応する配列と比較してSCNTの効率を同程度もしくはより大きな程度まで増加させるその生物学的に活性なフラグメントを含む、パラグラフ77の組成物。
79.
前記作用物質が、SEQ ID NO:1に対応する核酸、またはSEQ ID NO:1の核酸配列と比較してSCNTの効率を同程度もしくはより大きな程度まで増加させるその生物学的に活性なフラグメントを含む、パラグラフ75の組成物。
80.
H3K9メチルトランスフェラーゼの阻害剤が、Suv39h1、Suv39h2、またはSetdb1の少なくとも1つまたは任意の組み合わせを阻害する、パラグラフ75の組成物。
81.
哺乳動物SCNT胚が、1細胞期または2細胞期である、パラグラフ75の組成物。
82.
レシピエント哺乳動物卵母細胞が、除核したレシピエント哺乳動物卵母細胞である、パラグラフ75の組成物。
83.
哺乳動物SCNT胚が、最終分化した体細胞核の注射から生成され、または胚盤胞が、除核した哺乳動物卵母細胞に、最終分化した体細胞核を注射することから生成される哺乳動物SCNT胚から発生する、パラグラフ75の組成物。
84.
哺乳動物SCNT胚、レシピエント哺乳動物卵母細胞、または胚盤胞が、ヒトSCNT胚、レシピエントヒト卵母細胞、またはヒト胚盤胞である、パラグラフ75〜83のいずれか一つの組成物。
85.
哺乳動物SCNT胚、レシピエント哺乳動物卵母細胞、または胚盤胞が、非ヒト哺乳動物由来である、パラグラフ75〜84のいずれか一つの組成物。
86.
非ヒト哺乳動物が、マウス、ラット、ウサギ、雌ウシ、ウマ、ブタ、ニワトリ、イヌ、ネコ、マカク、チンパンジーからなる群より選択される、パラグラフ85の組成物。
87.
非ヒト哺乳動物が、飼育動物または商業用動物である、パラグラフ85の組成物。
88.
飼育動物または商業用動物が、アルパカ、バイソン、ラクダ、ネコ、ウシ、シカ、ゾウ、げっ歯動物、イヌ、ロバ、ガヤル、ヤギ、モルモット、ラマ、ウマ、サル、ラバ、雄ウシ、ブタ、ハト、非ヒト霊長類、ウサギ、トナカイ、ヒツジ、スイギュウ、またはヤクからなる群より選択される労役動物または競技動物または家畜動物または実験動物である、パラグラフ87の組成物。
89.
非ヒト哺乳動物が、伴侶動物またはペットである、パラグラフ85の組成物。
90.
伴侶動物が、イヌ、ネコ、雌ウシ、ハムスター、爬虫類、ウサギ、げっ歯動物、フェレット、チンチラ、鳥類ペット、モルモット、水生ペット、またはウマからなる群より選択される、パラグラフ89の組成物。
91.
非ヒト哺乳動物が、絶滅に近い哺乳動物種である、パラグラフ85の組成物。
92.
(i)Kdm4ファミリーのヒストンデメチラーゼの発現または活性を増加させる作用物質および/またはH3K9メチルトランスフェラーゼを阻害する作用物質、ならびに
(ii)哺乳動物卵母細胞
を含む、キット。
93.
哺乳動物卵母細胞が、除核した卵母細胞である、パラグラフ92のキット。
94.
哺乳動物卵母細胞が、非ヒト卵母細胞である、パラグラフ92のキット。
動物
C57BL/6J雌性マウスをDBA/2J雄と交配して、B6D2F1/J(BDF1)マウスを生成した。BDF1およびCD-1(ICR)成体雌を、それぞれレシピエント卵母細胞の収集および胚移入レシピエントのために使用した。BDF1マウスを、発生分析用のドナー体細胞の収集のために使用した。(C57BL/6×CAST/EiJ)F1マウスを、RNA-seq用のドナー細胞の収集のために使用した。GOF18デルタ-PE(Jackson Laboratory, 004654:Tg(Pou5f1-EGFP)2Mnn、C57BL/6Jバックグラウンド)を有するE13.5胚をマウス胚性線維芽細胞(MEF)の単離のために使用した。全ての動物実験は、Harvard Medical Schoolの所内動物実験委員会(Institutional Animal Care and Use Committee)によって承認された。
妊馬血清性ゴナドトロピン(PMSG; Harbor)7.5IUおよびヒト絨毛性ゴナドトロピン(hCG; Millipore)7.5IUを注射することによる過排卵を経て成体BDF1雌から卵丘細胞を収集した。hCG注射の15時間後に、卵丘-卵母細胞複合体(COC)を輸卵管から収集し、300U/mlウシ精巣ヒアルロニダーゼ(Calbiochem #385931)を含有するHepes緩衝カリウムシンプレックス最適化培地(KSOM)で短時間処置して、解離した卵丘細胞を得た。記載されたように(Matoba et al., 2011)、セルトリ細胞を3〜5日齢BDF1雄性マウスの精巣から収集した。精巣の塊を、0.1mg/mlコラゲナーゼ(Life Technologies #17104-019)を含有するPBS中に入れ、37℃で30分間インキュベートし、続いて、1mM EDTAを有する0.25%トリプシン(Life Technologies #25200-056)を用いて室温で5分処置した。3mg/mlウシ血清アルブミンを含有するPBSで4回洗浄後、解離した細胞をHepes-KSOM培地中で懸濁した。
マウスSuv39h1(Life Technologies #s74607)、Suv39h2(Life Technologies #s82300)およびSetdb1(Life Technologies #s96549)に対するsiRNAを、50μM 貯蔵液となるようにヌクレアーゼ不含水で希釈した。Lipofectamine RNAi Max(Life technologies #35050-061)を用いて、製造業者のプロトコールに従い、siRNAをMEFに導入した。簡潔には、MEF細胞1×105個を24ウェルプレート上に蒔いた(0日目;図5A参照)。24時間後に、Lipofectamine RNAi Maxを使用して5pM siRNAをMEF細胞にトランスフェクトした(1日目)。最初のトランスフェクションの24時間後に、培地を新鮮M293T培地[10%FBS、0.1mM可欠アミノ酸(Life technologies #11140-050)、2mM GlutaMAX(Life technologies #35050-079)、50U/ml ペニシリン-ストレプトマイシンおよび0.1mM 2-メルカプトエタノール(Life technologies #21985-023)を補充したDMEM]に交換した(2日目)。3日目に、MEF細胞を細胞1×105個の密度で24ウェルプレート上に再播種した。次に、トランスフェクションを上記のように1回繰り返した(4日目)。2回目のトランスフェクションの48時間後に(6日目)、MEF細胞を免疫染色、RT-qPCRまたはSCNTのために使用した。
以前に記載されたように、体細胞核移入を実施した(Matoba et al., 2011)。簡潔には、300U/ml ウシ精巣ヒアルロニダーゼ(Calbiochem)を用いた短時間処理により、過排卵成BDF1雌からレシピエントMII卵母細胞を収集した。単離されたMII卵母細胞を、7.5μg/ml サイトカラシンB(Calbiochem #250233)を含有するHepes緩衝KSOM培地中で除核した。圧電駆動マイクロマニピュレーター(Primetech #PMM-150FU)を使用して、ドナー卵丘細胞またはセルトリ細胞の核を除核卵母細胞中に注射した。不活性化センダイウイルスエンベロープ(HVJ-E; Ishihara Sangyo, Japan)によってMEF細胞を除核卵母細胞と融合した。KSOM中で1時間インキュベーション後に、再構築したSCNT卵母細胞を、5μg/ml サイトカラシンBを含有するCa不含KSOM中で1時間インキュベートすることによって活性化し、サイトカラシンBを有するKSOM中でさらに4時間培養した。活性化SCNT胚を、SrCl2処理の開始から5時間後(活性化後時間、hpa)に洗浄し、KSOM中で、5%CO2の加湿雰囲気中にて37.8℃で培養した。いくつかの実験では、圧電駆動マイクロマニピュレーター(Primetech)を使用することによって、5〜6hpaにSCNT胚に水(対照)、1800ng/μl 野生型または変異型(H189A)Kdm4d mRNA約10plを注射した。一部の実験では、培養培地にトリコスタチンA(TSA)を15nMになるよう活性化の開始から合計8時間添加した。着床前発生率は、スチューデントのT検定によって統計解析した。ドナー細胞の調製、胚移入、mRNA調製および他の手順に関するさらなる詳細は、増補の実験手順に含まれる。
完全長Kdm4d mRNAのインビトロ転写用のテンプレートプラスミドを製造するために、マウスKdm4dのオープンリーディングフレームを、ES細胞から得られたcDNAからPCRによって増幅し、In-Fusionキット(Clonetech #638909)を使用することによってpcDNA3.1-ポリ(A)83プラスミド(Inoue & Zhang, 2014)中にクローニングした。PrimeSTAR変異誘発基本キット(TAKARA # R045A)を使用して触媒欠損変異体Kdm4d(H188A)を作製した。直線化されたテンプレートプラスミドから、mMESSAGE mMACHINE T7 Ultra Kit(Life technologies # AM1345)を使用するインビトロ転写によって、製造業者の説明書に従ってmRNAを合成した。合成したmRNAを塩化リチウムで沈殿させ、ヌクレアーゼ不含水に溶解させた。NanoDrop ND-1000分光光度計(NanoDrop Technologies)によって濃度を測定した後、使用まで一定分量を-80℃で保存した。
マウスSuv39h1(Life Technologies #s74607)、Suv39h2(Life Technologies #s82300)およびSetdb1(Life Technologies #s96549)に対するsiRNAを、50μM貯蔵液となるようにヌクレアーゼ不含水で希釈した。Lipofectamine RNAi Max(Life technologies #35050-061)を用いて、製造業者のプロトコールに従ってsiRNAをMEFに導入した。簡潔には、MEF細胞1×105個を24ウェルプレート上に蒔いた(0日目;図5A参照)。24時間後に、Lipofectamine RNAi Maxを使用して5pM siRNAをMEF細胞中にトランスフェクトした(1日目)。最初のトランスフェクションの24時間後に、培地を新鮮M293T培地[10%FBS、0.1mM 可欠アミノ酸(Life technologies #11140-050)、2mM GlutaMAX(Life technologies #35050-079)、50U/ml ペニシリン-ストレプトマイシンおよび0.1mM 2-メルカプトエタノール(Life technologies #21985-023)を補充したDMEM]に交換した(2日目)。3日目に、MEF細胞を1×105個の密度で24ウェルプレート上に再播種した。次に、トランスフェクションを上記のように1回繰り返した(4日目)。2回目のトランスフェクションの48時間後に(6日目)、免疫染色、RT-qPCRまたはSCNTのためにMEF細胞を使用した。
RNeasyミニキット(Qiagen)を製造業者の説明書に従って使用して、MEF細胞から総RNAを精製した。オリゴ-dTプライマーおよびImProm-II Reverse Transcription Syetem(Promega)を用いてcDNAを合成した。Ssofast Evagreen Supermix(Bio-Rad)を使用するCFX384 Real-Time PCR検出システム(Bio-Rad)でリアルタイムPCRを行った。比較Ct法を使用して相対遺伝子発現レベルを分析し、CFX Managerソフトウェア(Bio-Rad)を使用してGapdhに対して標準化した。結果をスチューデントのT検定によって統計解析した。以下のプライマーを使用した:
SCNTまたはインビトロ受精によって作製した2細胞期胚を偽妊娠(E0.5)ICR雌の輸卵管に移入した。分娩日(E19.5)に、帝王切開によって子を回収し、泌乳ICR雌により授乳させた。
酸性タイロード処理により胚盤胞を裸化し、マイトマイシン処理MEFフィーダー細胞上で、5%FBS、10%KnockOut血清代替物(Life Technologies #10828-028)、0.1mM 可欠アミノ酸、2mM GlutaMAX、50U/ml ペニシリン-ストレプトマイシン、0.1mM 2-メルカプトエタノールおよび2000U 白血病抑制因子(LIF, Millipore #ESG1107)を補充したDMEM中に入れて、37℃および5%CO2で培養した。4〜5日後に、付着した胚からの外植片を0.25%トリプシンで解離させ、全てを新しいフィーダー細胞上に継代した。翌日、培地を、N2B27-LIF培地[0.1mM 可欠アミノ酸、2mM GlutaMAX、50U/ml ペニシリン-ストレプトマイシン、0.1mM 2-メルカプトエタノール、0.5×N2サプリメント(Life technologies #17502-048)、0.5×B27サプリメント(Life technologies #17504-044)、3μM CHIR99021(STEMGENT #04-0004)、0.5μM PD0325901(STEMGENT #04-0006)および1000U LIFを補充したDMEM/F12(Life Technologies #10565-042)]により置換した。5日後に、増大した細胞を樹立ntESCとして再播種した。
胚またはMEF細胞を、3.7%パラホルムアルデヒド(PFA)を用いて室温で20分間固定した。10mg/ml BSAを含有するPBS(PBS/BSA)で洗浄後、固定された胚または細胞を0.5%トリトン-X100と共に15分間インキュベートすることによって透過処理した。PBS/BSA中に入れて室温で1時間ブロッキング後、それらを最初の抗体の混合物中に入れ、4℃で一晩インキュベートした。抗体には、マウス抗H3K9me3(1/500: Abcam #ab71604)、ウサギ抗H3K9me3(1/500: Millipore #07-442)、ウサギ抗H3K27me3(1/500: Millipore #07-449)、ヤギ抗Oct4(1/500: Santa Cruz #SC8628)、およびマウス抗Cdx2(1/100: BioGenex #AM392-5M)が含まれる。PBS/BSAで3回洗浄後、胚または細胞を2次抗体と共に室温で1時間インキュベートした。2次抗体には、フルオレセインイソチオシアネート-コンジュゲーション型ロバ抗マウスIgG(1/400, Jackson Immuno-Research)、Alexa Flour 568ロバ抗ウサギIgG(1/400, Life technologies)、および/またはAlexa Flour 647ロバ抗ヤギIgG(1/400, Life technologies)が含まれる。最終的に、4',6-ジアミジノ-2-フェニルインドール(DAPI)を有するVectashield(Vector Laboratories #H-1200)を用いてそれらを封入した。レーザー走査共焦点顕微鏡(Zeiss LSM510)およびEM-CCDカメラ(Hamamatsu ImagEM)を使用して蛍光シグナルを観察した。
胚を直接溶解し、SMARTer Ultra Low Input RNA cDNA調製キット(Clontech)を使用するcDNA合成のために使用した。増幅後、Covaris超音波処理器(Covaris)を使用してcDNA試料をフラグメント化した。Illumina用のNEBNext Ultra DNA Library Prep Kitを使用して、製造業者(New England Biolabs)の説明書に従って、フラグメント化されたDNAを用いて配列ライブラリーを製造した。単一端50bp配列決定を、HiSeq 2500シーケンサー(Illumina)で行った。NovoalignV3.02.00を用いてシーケンシングのリードをマウスゲノム(mm9)に対してマッピングした。全てのプログラムをデフォルト設定で行った(特に規定しない限り)。続いて、一意的にマッピングされるリード(総リードの約70%)をCufflinks v2.0.2を用いて集合させ、参照アノテーション(UCSC遺伝子モデル)によってガイドされる転写物を構築した。各遺伝子の発現レベルを、標準化FPKM(エキソン1キロ塩基あたり、マッピングされるフラグメント100万個あたりのフラグメント)を用いて定量した。有意に異なる転写物の機能的アノテーションおよびエンリッチメント解析をDAVIDで行った。統計解析をRで実行した(ワールドワイドウェブwww.r-project.org/)。独立した2群Wilcoxon順位和検定を使用して、Rでのwilcox.test関数を使用して分布を比較した。デフォルトのパラメーターを用いたcor関数を使用して、Pearsonのr係数を計算した。Rでのhearmap.2関数(gplotsパッケージ)を使用して、異なる試料での全般的遺伝子発現パターンの階層的クラスタリング分析を実施した。
スライドウインドウ(サイズ100kb、ステップサイズ20kb)を使用して、1細胞期胚および2細胞期胚のゲノムワイドな発現レベルを評価した。各ウインドウについて、標準化RPM(一意的にマッピングされるリード100万個あたりのリード)を用いて発現レベルを定量した。ストリンジェントな基準(FC>5、Fisherの正確検定p値<0.01、2細胞期IVF胚においてRPM>10)で、1細胞期IVF胚と比較して2細胞期IVF胚で有意に活性化された領域を特定し、オーバーラップする領域をマージした。これらの活性化領域を、SCNTおよびIVF 2細胞期胚におけるそれらの発現差に基づき3群に分類した。
図2およびS2においてヒストン修飾およびDNaseI過感受性エンリッチメント解析を行うために、本発明者らは、以下の公表されたChIP-seqおよびDNaseI-seqデータセットを使用した:MEF細胞でのH3K9me3およびH3K96me3(Pedersen et al., 2014);MEF細胞でのH3K4me2、H3K27me3(Chang et al., 2014);MEF細胞でのH3K4me1およびH3K27ac(ENCODE/LICRプロジェクト);CH12、赤芽球、巨核(ENCODE/PSUプロジェクト)および全脳(ENCODE/LICRプロジェクト)でのH3K9me3;NIH3T3、CH12、MEL、Treg、416Bおよび全脳(ENCODE/UWプロジェクト)でのDNaseI-seq。標準化FPKMを用いてChIP-seq強度を定量した。シーケンシングリードによりゲノム位置に関するカバー率を決定し、UCSCゲノムブラウザーでのカスタムトラックとして視覚化した。
2細胞期SCNT胚における異常なZGA
インビトロ受精(IVF)およびSCNTを経て派生するマウス胚の間の最も初期の転写差を特定するために、本発明者らは、1細胞期(活性化の12時間後:hpa)および2細胞後期(28hpa)のプール胚(25〜40個/試料)を使用してRNA-seq実験を行った(図1A)。各試料について3000万個よりも多い一意的にマッピングされるリードを得、各試料の2つの生物学的反復は、高度に再現可能であった(図8Aおよび8B)。1細胞期トランスクリプトームの分析は、SCNT胚およびIVF胚がほぼ同一のトランスクリプトームを特徴とすることを明らかにした(R=0.99;図1B)。具体的には、検出された5517種の遺伝子(少なくとも1つの試料においてFPKM>5)の中で、106種の遺伝子だけがSCNT胚とIVF胚との間で3倍よりも大きな差を示した(図1B)。これは、ZGAが主としてマウス胚での最初の分割後に開始し(Schultz, 2002)、IVFまたはSCNTにかかわらず、1細胞期胚に存在する大部分の転写物が母方に保存される転写物であるという事実と一致する。したがって、本発明者らは、主要なZGAがマウス胚において明らかになる2細胞後期に本発明者らの分析の焦点を合わせた。
2細胞期SCNT胚におけるリプログラミング耐性領域(RRR)の特定
タンパク質コード遺伝子に加えて、以前の研究は、LTRクラスIIIレトロトランスポゾンおよび主要サテライト反復などの非遺伝子反復エレメントが、特に2細胞期のマウス着床前胚に高発現されることを明らかにした(Evsikov et al., 2004; Peaston et al., 2004; Probst et al., 2010)。IVF 2細胞期胚とSCNT 2細胞期胚との間のトランスクリプトームの差を包括的に特徴付けるために、本発明者らは、検出可能な転写物に関連する全てのゲノム領域を特定するためにスライドウインドウ戦略を適用した。最初に、本発明者らは、1細胞期IVF胚と比較して2細胞期IVF胚において有意に活性化された(Fisherの正確検定p値<0.01)、100〜800kbの範囲の811種のゲノム領域を特定した[図2A、FC>5、IVF 2細胞期胚におけるRPM(一意的にマッピングされるリード100万個あたりのリード)>10]。811種のゲノム領域のうち、342種の領域が、IVF胚に類似したレベルでSCNT胚において活性化されたが(IVF 2細胞期胚をSCNT 2細胞期胚と比較してFC≦2)、これらの領域を、完全リプログラミング領域(FRR)と名付けた。本発明者らは、また、IVF胚と比較してSCNT胚において部分的に活性化された(FC>2およびFC≦5)「部分的リプログラミング領域」(PRR)と名付けられた247種の領域を特定した(図2A)。興味深いことに、本明細書において「リプログラミング耐性領域」(RRR)と呼ばれる残りの222種の領域は、SCNT胚において活性化できなかった(FC>5、図2A)。特に、RRR内で産出される転写物は、13番染色体上の代表的な領域に例示されるように、主として注釈を付けられない(図9A)。実際にFRRおよびPRRを比較して、RRRは、相対的に遺伝子に乏しい領域である(図9B)。しかし、RRRは、LINEおよびLTRなどの特異的リピート配列が濃縮されているが、SINEを枯渇している(図9C)。このように、比較トランスクリプトーム解析は、本発明者らに、SCNTによって作製される2細胞期胚での転写活性化に不応性の222種のRRRを特定させた。
RRRが2細胞期SCNT胚における転写活性化に不応性であるという事実は、RRRが、SCNT介在性リプログラミングについてのバリアとして役立つ特定のエピジェネティック修飾を有し得ることを示している。SCNT胚の発育不全が、マウス胚性線維芽(MEF)細胞を含めた異なるドナー体細胞型で観察されていることを考え(Ono et al., 2001)、本発明者らは、SCNT介在性リプログラミングについてのそのようなエピジェネティックなバリアが、異なる体細胞型に共通であるかどうかを評価した。MEF細胞は、包括的ヒストン修飾データベースのデータセットを有する数少ない体細胞型の1つであるので(Bernstein et al., 2012; Chang et al., 2014; Pedersen et al., 2014)、本発明者らは、6つの主要なヒストン修飾のいずれかがRRRで特異的に濃縮されているかどうかを評価した。本発明者らは、分析される任意の他の修飾でなくH3K9me3が、RRRで特異的に濃縮されたが、一方で、FRRまたはPRRにおいていかなるヒストン修飾の明白な濃縮も観察されなかったことを発見した(図2B)。実際、7番染色体上の代表的な領域の慎重な調査から、2細胞期SCNT胚で活性化できなかったRRRは、明らかにH3K9me3マークが濃縮されており、H3K9me3濃縮領域外側の領域は、2細胞期SCNT胚で適正に活性化されたことが示された(図2C)。RRRでのH3K9me3の類似の濃縮が、ENCODEプロジェクトからのH3K9me3 ChIP-seqデータセットの分析後に(図2Dおよび9D)、4つの他の体細胞型または組織型(CH12、赤芽球、巨核球および全脳)においても観察されたので(Bernstein et al., 2012)、この観察は、MEF細胞に独特ではない。したがって、本発明者らは、体細胞においてRRRにH3K9me3が濃縮されていると結論する。
Kdm4dによるH3K9me3の除去はSCNT胚における転写リプログラミングを回復する
RRRとH3K9me3濃縮との間の相関関係を立証したので、本発明者らは、次に、H3K9me3の除去がSCNT胚におけるRRRの転写リプログラミングを促進できるかどうかに取り組もうとした。この目的を達成するために、本発明者らは、H3K9me3特異的ヒストンデメチラーゼKdm4dをコードするmRNAを合成し(Krishnan and Trievel, 2013)、5hpaに該mRNAをSCNT胚に注射した(図3A)。免疫染色は、触媒欠損変異型ではなく、野生型Kdm4d mRNAの注射が、SCNT胚におけるH3K9me3レベルを大きく低下させたことを明らかにした(図3B)。
Kdm4d注射後のSCNT胚の転写回復の生物学的結果を調査するために、本発明者らは、最初に、卵丘細胞をドナー細胞として使用し、Kdm4d mRNAを注射されたSCNT胚の発生能を分析した。対照SCNT胚において、第一分割後に発生率は減少し始め、分割した胚の26.0%だけが、96時間培養後に胚盤胞期まで成功裏に発生したが(図4Aおよび4B、ならびに表S1)、これは、以前の研究に一致する知見である(Kishigami et al., 2006)。著しくは、野生型Kdm4d mRNAを注射されたSCNT胚は、2から4細胞期および4細胞期から桑実胚期の移行の間にめったに停止せず、胚盤胞期まで高い効率で発生する(88.6%;図4Aおよび4B、ならびに表3)。対照的に、触媒欠損変異型Kdm4d mRNAの注射は、SCNT胚の発生率に有意な影響を有さず、Kdm4d注射がSCNT胚発生に及ぼす改善がその酵素活性に依存することを示している。RRRにおけるH3K9me3濃縮が、異なる体細胞型における一般的な現象に見えることを考えれば、本発明者らは、SCNT胚発生に及ぼすKdm4dのプラス効果を他のドナー体細胞型に拡張できるはずだと予測した。実際、Kdm4d mRNAの注射は、セルトリ細胞またはC57BL/6バックグラウンドのMEF細胞がドナー細胞として使用されたときに、SCNT胚の発生効率も有意に改善した(図4Aおよび4B、表3)。まとめると、これらの結果は、Kdm4d mRNAの注射によるH3K9me3の除去が、ドナー体細胞型にかかわらずSCNT胚の着床前発生を有意に改善できることを実証している。
SCNT胚の発生表現型不良の原因となる遺伝子の候補
本発明者らは、次に、H3K9me3によって抑制される遺伝子のどれがSCNT胚の発生表現型不良の原因となるかを評価した。Kdm4dの過剰発現が胚盤胞期に達しつつあるSCNT胚の率を大きく増加させることを考えれば、原因遺伝子は、野生型Kdm4d注射SCNT胚において抑制解除されていなければならない。2細胞期SCNT胚において活性化できなかった遺伝子(図1Dの群3の遺伝子)、および変異型Kdm4d注射ではなく、野生型Kdm4d注射2細胞期SCNT胚において抑制解除される遺伝子の分析により、本発明者らは49種の共通遺伝子を特定した(図5Aおよび5B、FC>5)。GO分析は、この群の遺伝子に転写およびRNA代謝過程に関与する遺伝子が濃縮されていることを示した(図5A)。着床前発生における49種の遺伝子の大部分の機能は未知であるものの、2細胞期特異的Zscan4ファミリーのメンバーであるZscan4dは、着床前発生に重要であることが示されている(Falco et al., 2007)。したがって、本発明者らは、外因性Zscan4d mRNAの補充が、SCNT胚の発生効率を高めることができるかどうかを調査した。
Suv39h1/2は体細胞におけるH3K9me3バリアの樹立の原因である
H3K9me3がSCNT介在性リプログラミングのエピジェネティックなバリアであると実証したので、本発明者らは、次に、体細胞ゲノムにおけるRRR内へのH3K9me3沈着の原因となるヒストンメチルトランスフェラーゼを特定しようとした。以前の研究は、少なくとも3つのヒストンリジンメチルトランスフェラーゼ(KMT)であるSuv39h1、Suv39h2およびSetdb1が、哺乳動物細胞におけるH3K9me3産出を触媒することができると報告している(Matsui et al., 2010; Peters et al., 2001)。最初に、本発明者らは、Suv39h1、Suv39h2およびSetdb1を標的化する低分子干渉RNA(siRNA)の混合物をトランスフェクトすることによって、MEF細胞から3つのH3K9me3メチルトランスフェラーゼを枯渇させた(図6A)。RT-qPCR分析は、トランスフェクションの48時間後に80〜60%のノックダウン効率が達成されたことを確認した(図12A〜12B)。免疫染色は、これらのsiRNAのトランスフェクション(6日の細胞培養の間に2回)がMEF細胞におけるH3K9me3レベルを大きく低下させることができたことを示した(図6Aおよび6B)が、これは、他の未特定の酵素でなく、3つのH3K9me3メチルトランスフェラーゼを体細胞におけるH3K9me3沈着の責任因子として実証するものである。ドナーとしてトリプルKD MEF細胞を使用して、本発明者らは、SCNT胚を作製し、それらの着床前発生を調査した。本発明者らは、対照SCNT胚のわずか6.7%が96時間培養後に胚盤胞期まで発生した(図6C、6D、および表3)が、一方でトリプルノックダウンMEF由来胚の65.6%が胚盤胞期まで発生したことを発見した(図6C、6Dおよび表3)。この結果は、体細胞H3K9me3がSCNT介在性リプログラミングのエピジェネティックなバリアであることを確認するばかりでなく、これらの3つの酵素がこのエピジェネティックなバリアを産出する原因となることも実証している。
ツメガエル(Xenopus)卵における体細胞核移入による動物クローニングの最初の実証以来50年超が経過した(Gurdon, 1962)。莫大な尽力にかかわらず、大部分の種でクローニング効率は比較的低いままであり、SCNT後のエピジェネティックリプログラミングの基礎をなすメカニズムは、あまり理解されていないままである。本研究において、比較トランスクリプトームおよび統合エピゲノム分析により、本発明者らは、ドナー体細胞においてSuv39h1/2によって沈着するH3K9me3が、ヒトおよびマウス卵母細胞における体細胞核リプログラミングのエピジェネティックなバリアとして機能することを発見した。IVF胚のトランスクリプトームと比較することによって、本発明者らは、RRR(リプログラミング耐性領域)と呼ばれる、SCNT胚における転写リプログラミングに耐性な222種のゲノム領域を特定した。RRRは、分析されたいくつかの体細胞型において、Suv39h1/2によって沈着されるH3K9me3の顕著な濃縮および低いDNase I到達性(どちらもヘテロクロマチンの一般的特徴である)によって特徴付けられる。Suv39h1/2のノックダウンまたは外因性Kdm4dの発現のいずれかによるH3K9me3の除去が、RRR活性化およびSCNT胚発生の有意な改善を招くので、RRR内の転写物の効率的な活性化は、SCNT胚の発生に重要に見える。したがって、本発明者らは、本明細書において、体細胞でSuv39h1/2によって沈着されるH3K9me3が、卵母細胞において発生的に重要な遺伝子の活性化についてのバリアとしての役目をし、SCNT胚の発生停止を導くモデルを実証している(図7)。SCNT後の外因性Kdm4dまたはドナー細胞におけるSuv39h1/2枯渇のいずれかによるこのエピジェネティックなバリアの除去は、発生遺伝子の発現およびSCNT胚発生の改善を可能にする(図7)。
本明細書に開示する参考文献は、その全体が参照として組み入れられる。
Claims (15)
- 体細胞核移入(SCNT)の効率を増加させるための方法であって、
ドナー非ヒト哺乳動物細胞、非ヒト除核哺乳動物卵母細胞、または非ヒト哺乳動物SCNT胚に、Kdm4ヒストンデメチラーゼまたはKdm4ヒストンデメチラーゼをコードする核酸分子を注射する段階であって、それによってSCNTの効率を増加させる、段階
を含む、SCNTの効率を増加させる方法。 - 非ヒト哺乳動物体細胞核移入(SCNT)胚を生成するための方法であって、
ドナー非ヒト哺乳動物細胞、レシピエント非ヒト哺乳動物卵母細胞、または非ヒト哺乳動物体細胞核移入(SCNT)胚の少なくとも1つに、Kdm4ヒストンデメチラーゼまたはKdm4ヒストンデメチラーゼをコードする核酸分子を注射する段階であって、レシピエント非ヒト哺乳動物卵母細胞が、有核または除核卵母細胞である、段階;
レシピエント非ヒト哺乳動物卵母細胞が有核である場合、レシピエント非ヒト哺乳動物卵母細胞を除核する段階;
ドナー非ヒト哺乳動物細胞由来の核を非ヒト除核卵母細胞に移入する段階;および
非ヒト哺乳動物SCNT胚を形成させるために十分な時間、レシピエント非ヒト卵母細胞をインキュベートする段階
を含む、方法。 - Kdm4ヒストンデメチラーゼをコードする核酸分子が、SEQ ID NO:1〜4に対応する核酸配列、またはSEQ ID NO:1〜4の対応する配列と比較してSCNTの効率を同程度もしくはより大きな程度まで増加させるその生物学的に活性なフラグメントを含む、請求項1または2記載の方法。
- 非ヒトSCNT胚が、1細胞期SCNT胚、活性化の5時間後(5hpa)のSCNT胚、活性化の10〜12時間後(10〜12hpa)のSCNT胚、活性化の20〜28時間後(20〜28hpa)のSCNT胚、2細胞期SCNT胚のいずれかより選択される、請求項1〜3のいずれか一項記載の方法。
- ドナー非ヒト哺乳動物細胞が、遺伝的に修飾されたドナー非ヒト哺乳動物細胞である、請求項1〜4のいずれか一項記載の方法。
- 非ヒトSCNT胚をインビトロ培養して非ヒト胚盤胞を形成させる段階をさらに含む、請求項2記載の方法。
- 非ヒトSCNT胚が、少なくとも1細胞期SCNT胚である、請求項2記載の方法。
- 非ヒトSCNT胚が、少なくとも2細胞期SCNT胚である、請求項2記載の方法。
- 非ヒト胚盤胞由来の内細胞塊から細胞を単離する段階、および
未分化状態の内細胞塊からの細胞を培養して非ヒト哺乳動物胚性幹(ES)細胞を形成させる段階
をさらに含む、請求項6記載の方法。 - 体細胞核移入(SCNT)胚から非ヒト哺乳動物子孫を生成するための方法であって、
ドナー非ヒト哺乳動物細胞、レシピエント非ヒト哺乳動物卵母細胞、または非ヒト哺乳動物体細胞核移入(SCNT)胚のうち少なくとも1つに、Kdm4ヒストンデメチラーゼまたはKdm4ヒストンデメチラーゼをコードする核酸分子を注射する段階であって、レシピエント非ヒト哺乳動物卵母細胞が、有核または除核卵母細胞である、段階;
レシピエント非ヒト哺乳動物卵母細胞が有核である場合に、除核する段階;
ドナー非ヒト哺乳動物細胞由来の核を非ヒト哺乳動物除核卵母細胞に移入し、核を除核卵母細胞と融合させ、融合した卵母細胞を活性化させる段階;
非ヒト哺乳動物SCNT胚を形成させるために十分な時間、レシピエント卵母細胞をインキュベートする段階;
非ヒト哺乳動物SCNT胚を非ヒト代理母の輸卵管に体内移植し、非ヒト哺乳動物SCNTの非ヒト哺乳動物子孫への発生を可能にする段階
を含む、方法。 - 非ヒト哺乳動物SCNT胚が、胚1細胞期、胚2細胞期、胚4細胞期、桑実胚、または胚盤胞胚期に非ヒト代理母に体内移植される、請求項10記載の方法。
- 非ヒト哺乳動物SCNT胚、レシピエント非ヒト哺乳動物卵母細胞、または非ヒト胚盤胞のうち少なくとも1つと、Kdm4ヒストンデメチラーゼまたはKdm4ヒストンデメチラーゼをコードする核酸分子とを含む、組成物。
- Kdm4ヒストンデメチラーゼをコードする核酸分子が、SEQ ID NO:1〜4に対応する核酸、またはSEQ ID NO:1〜4の対応する配列と比較してSCNTの効率を同程度もしくはより大きな程度まで増加させるその生物学的に活性なフラグメントを含む、請求項12記載の組成物。
- 非ヒト哺乳動物SCNT胚が、1細胞期または2細胞期である、請求項12または13記載の組成物。
- (i)Kdm4ヒストンデメチラーゼをコードする核酸分子またはKdm4ポリペプチド、および
(ii)非ヒト哺乳動物卵母細胞
を含む、キット。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201462050308P | 2014-09-15 | 2014-09-15 | |
| US62/050,308 | 2014-09-15 | ||
| US201462053514P | 2014-09-22 | 2014-09-22 | |
| US62/053,514 | 2014-09-22 | ||
| PCT/US2015/050178 WO2016044271A2 (en) | 2014-09-15 | 2015-09-15 | Methods and compositions to increase somatic cell nuclear transfer (scnt) efficiency by removing histone h3-lysine trimethylation |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021023834A Division JP7277494B2 (ja) | 2014-09-15 | 2021-02-18 | ヒストンh3-リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2017528142A JP2017528142A (ja) | 2017-09-28 |
| JP2017528142A5 JP2017528142A5 (ja) | 2018-11-01 |
| JP6841753B2 true JP6841753B2 (ja) | 2021-03-10 |
Family
ID=55534004
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2017514421A Active JP6841753B2 (ja) | 2014-09-15 | 2015-09-15 | ヒストンh3−リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 |
| JP2021023834A Active JP7277494B2 (ja) | 2014-09-15 | 2021-02-18 | ヒストンh3-リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 |
| JP2023001284A Pending JP2023026679A (ja) | 2014-09-15 | 2023-01-06 | ヒストンh3-リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 |
Family Applications After (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021023834A Active JP7277494B2 (ja) | 2014-09-15 | 2021-02-18 | ヒストンh3-リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 |
| JP2023001284A Pending JP2023026679A (ja) | 2014-09-15 | 2023-01-06 | ヒストンh3-リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 |
Country Status (5)
| Country | Link |
|---|---|
| US (3) | US11390885B2 (ja) |
| EP (1) | EP3194581A4 (ja) |
| JP (3) | JP6841753B2 (ja) |
| KR (2) | KR102473092B1 (ja) |
| WO (1) | WO2016044271A2 (ja) |
Families Citing this family (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR102473092B1 (ko) | 2014-09-15 | 2022-12-01 | 칠드런'즈 메디컬 센터 코포레이션 | 히스톤 h3-리신 트리메틸화를 제거함으로써 체세포 핵 이식(scnt) 효율을 증가시키는 방법 및 조성물 |
| CN106591374B (zh) * | 2016-12-30 | 2019-08-09 | 中国农业科学院北京畜牧兽医研究所 | 一种提高猪体细胞核移植胚胎发育效率的方法 |
| CN106676136B (zh) * | 2016-12-30 | 2019-11-29 | 中国农业科学院北京畜牧兽医研究所 | 一种提高猪体细胞核移植效率的方法 |
| CN107299113A (zh) * | 2017-06-12 | 2017-10-27 | 内蒙古大学 | H3K27me3及其去甲基化酶KDM6A/B在小鼠核移植重构胚中的应用方法 |
| CN108285906B (zh) * | 2017-12-29 | 2021-07-09 | 温氏食品集团股份有限公司 | 一种定点整合外源dna转基因猪的构建方法 |
| CN108624621B (zh) * | 2018-01-17 | 2019-04-12 | 中国科学院上海生命科学研究院 | 非人灵长类的体细胞克隆动物的制备方法 |
| CN111630176A (zh) * | 2018-01-23 | 2020-09-04 | 车医科学大学校产学协力团 | 一种包括Rad51激活剂的用于胚胎发育的组合物以及用其提高胚胎发育率的方法 |
| KR20190089782A (ko) * | 2018-01-23 | 2019-07-31 | 차의과학대학교 산학협력단 | 멜라토닌을 포함하는, 배아 발달용 조성물 및 이를 이용하여 배아 발달의 효율을 향상시키는 방법 |
| KR20200141470A (ko) * | 2018-04-06 | 2020-12-18 | 칠드런'즈 메디컬 센터 코포레이션 | 체세포 재프로그래밍 및 각인의 조정을 위한 조성물 및 방법 |
| CN110547291A (zh) * | 2019-09-27 | 2019-12-10 | 安徽医科大学 | 一种高效抗氧化的人卵母细胞冷冻保护剂 |
| CN111466367B (zh) * | 2020-02-17 | 2022-03-01 | 天津大学 | 用于保存细胞的组合物 |
| KR102520268B1 (ko) * | 2020-12-04 | 2023-04-12 | 대한민국 | microRNA-148a 직접 주입을 통한 돼지 체세포 핵치환란의 발달율 향상 방법 |
| CN113186152B (zh) * | 2021-05-19 | 2023-05-26 | 南方医科大学 | Dna甲基转移酶抑制剂在提高动物圆形精子注射胚胎发育效率中的应用 |
Family Cites Families (128)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3687808A (en) | 1969-08-14 | 1972-08-29 | Univ Leland Stanford Junior | Synthetic polynucleotides |
| US4469863A (en) | 1980-11-12 | 1984-09-04 | Ts O Paul O P | Nonionic nucleic acid alkyl and aryl phosphonates and processes for manufacture and use thereof |
| US5023243A (en) | 1981-10-23 | 1991-06-11 | Molecular Biosystems, Inc. | Oligonucleotide therapeutic agent and method of making same |
| US4441972A (en) | 1983-04-08 | 1984-04-10 | D.E.P. Systems, Inc. | Apparatus for electrofusion of biological particles |
| US4664097A (en) | 1984-05-09 | 1987-05-12 | The Wistar Institute Of Anatomy & Biology | Nuclear transplantation in the mammalian embryo by microsurgery and cell fusion |
| US4578168A (en) | 1984-07-27 | 1986-03-25 | Biotronics | Apparatus for fusing live cells with electric fields |
| IL73883A (en) | 1984-12-20 | 1990-12-23 | Yeda Res & Dev | Monoclonal antibodies against tnf-alpha,hybridomas producing them and method for the purification of tnf-alpha |
| US5235033A (en) | 1985-03-15 | 1993-08-10 | Anti-Gene Development Group | Alpha-morpholino ribonucleoside derivatives and polymers thereof |
| ATE171185T1 (de) | 1985-03-15 | 1998-10-15 | Antivirals Inc | Immunotestmittel für polynukleotid und verfahren |
| JPS6276624A (ja) * | 1985-09-30 | 1987-04-08 | Toshiba Corp | レジスト剥離装置 |
| US4994384A (en) | 1986-12-31 | 1991-02-19 | W. R. Grace & Co.-Conn. | Multiplying bovine embryos |
| US5057420A (en) | 1987-06-05 | 1991-10-15 | Granada Biosciences, Inc. | Bovine nuclear transplantation |
| US5213979A (en) | 1987-12-30 | 1993-05-25 | W. R. Grace & Co.-Conn. | In vitro culture of bovine embryos |
| JPH04500909A (ja) | 1988-09-21 | 1992-02-20 | アニマル バイオテクノロジー ケンブリッジ リミティド | 家畜からの多能性胚細胞系の誘導 |
| US4944384A (en) | 1989-01-30 | 1990-07-31 | Hay & Forage Industries | Trash discharge apparatus for crop transferring conveyor mechanism |
| US5256775A (en) | 1989-06-05 | 1993-10-26 | Gilead Sciences, Inc. | Exonuclease-resistant oligonucleotides |
| US5580774A (en) | 1989-07-31 | 1996-12-03 | Eli Lilly And Company | Chimeric antibodies directed against a human glycoprotein antigen |
| US5264564A (en) | 1989-10-24 | 1993-11-23 | Gilead Sciences | Oligonucleotide analogs with novel linkages |
| US5264562A (en) | 1989-10-24 | 1993-11-23 | Gilead Sciences, Inc. | Oligonucleotide analogs with novel linkages |
| ATE190981T1 (de) | 1989-10-24 | 2000-04-15 | Isis Pharmaceuticals Inc | 2'-modifizierte nukleotide |
| EP0747485B1 (en) | 1989-11-06 | 1998-12-02 | Cell Genesys, Inc. | Production of proteins using homologous recombination |
| US5177198A (en) | 1989-11-30 | 1993-01-05 | University Of N.C. At Chapel Hill | Process for preparing oligoribonucleoside and oligodeoxyribonucleoside boranophosphates |
| US5223168A (en) | 1989-12-12 | 1993-06-29 | Gary Holt | Surface cleaner and treatment |
| US5130302A (en) | 1989-12-20 | 1992-07-14 | Boron Bilogicals, Inc. | Boronated nucleoside, nucleotide and oligonucleotide compounds, compositions and methods for using same |
| ES2151463T5 (es) | 1989-12-22 | 2012-10-29 | Merck Serono Sa | Constructos de ADN para la activación y la modificación de la expresión de genes endógenos |
| US5272071A (en) | 1989-12-22 | 1993-12-21 | Applied Research Systems Ars Holding N.V. | Method for the modification of the expression characteristics of an endogenous gene of a given cell line |
| US5459255A (en) | 1990-01-11 | 1995-10-17 | Isis Pharmaceuticals, Inc. | N-2 substituted purines |
| US5212295A (en) | 1990-01-11 | 1993-05-18 | Isis Pharmaceuticals | Monomers for preparation of oligonucleotides having chiral phosphorus linkages |
| US5587361A (en) | 1991-10-15 | 1996-12-24 | Isis Pharmaceuticals, Inc. | Oligonucleotides having phosphorothioate linkages of high chiral purity |
| US5587470A (en) | 1990-01-11 | 1996-12-24 | Isis Pharmaceuticals, Inc. | 3-deazapurines |
| US5457191A (en) | 1990-01-11 | 1995-10-10 | Isis Pharmaceuticals, Inc. | 3-deazapurines |
| US5578718A (en) | 1990-01-11 | 1996-11-26 | Isis Pharmaceuticals, Inc. | Thiol-derivatized nucleosides |
| US5506351A (en) | 1992-07-23 | 1996-04-09 | Isis Pharmaceuticals | Process for the preparation of 2'-O-alkyl guanosine and related compounds |
| US5149797A (en) | 1990-02-15 | 1992-09-22 | The Worcester Foundation For Experimental Biology | Method of site-specific alteration of rna and production of encoded polypeptides |
| EP0651787A1 (en) | 1990-07-09 | 1995-05-10 | Amrad Corporation Limited | Enhanced implantation, development and maintenance of embryos using leukaemia inhibitory factor |
| US5096822A (en) | 1990-07-26 | 1992-03-17 | W. R. Grace & Co.- Conn. | Bovine embryo medium |
| US5608046A (en) | 1990-07-27 | 1997-03-04 | Isis Pharmaceuticals, Inc. | Conjugated 4'-desmethyl nucleoside analog compounds |
| US5138045A (en) | 1990-07-27 | 1992-08-11 | Isis Pharmaceuticals | Polyamine conjugated oligonucleotides |
| US5223618A (en) | 1990-08-13 | 1993-06-29 | Isis Pharmaceuticals, Inc. | 4'-desmethyl nucleoside analog compounds |
| US5386023A (en) | 1990-07-27 | 1995-01-31 | Isis Pharmaceuticals | Backbone modified oligonucleotide analogs and preparation thereof through reductive coupling |
| US5623070A (en) | 1990-07-27 | 1997-04-22 | Isis Pharmaceuticals, Inc. | Heteroatomic oligonucleoside linkages |
| US5541307A (en) | 1990-07-27 | 1996-07-30 | Isis Pharmaceuticals, Inc. | Backbone modified oligonucleotide analogs and solid phase synthesis thereof |
| US5489677A (en) | 1990-07-27 | 1996-02-06 | Isis Pharmaceuticals, Inc. | Oligonucleoside linkages containing adjacent oxygen and nitrogen atoms |
| US5610289A (en) | 1990-07-27 | 1997-03-11 | Isis Pharmaceuticals, Inc. | Backbone modified oligonucleotide analogues |
| US5602240A (en) | 1990-07-27 | 1997-02-11 | Ciba Geigy Ag. | Backbone modified oligonucleotide analogs |
| US5378825A (en) | 1990-07-27 | 1995-01-03 | Isis Pharmaceuticals, Inc. | Backbone modified oligonucleotide analogs |
| US5218105A (en) | 1990-07-27 | 1993-06-08 | Isis Pharmaceuticals | Polyamine conjugated oligonucleotides |
| DE69126530T2 (de) | 1990-07-27 | 1998-02-05 | Isis Pharmaceutical, Inc., Carlsbad, Calif. | Nuklease resistente, pyrimidin modifizierte oligonukleotide, die die gen-expression detektieren und modulieren |
| US6262241B1 (en) | 1990-08-13 | 2001-07-17 | Isis Pharmaceuticals, Inc. | Compound for detecting and modulating RNA activity and gene expression |
| US5177196A (en) | 1990-08-16 | 1993-01-05 | Microprobe Corporation | Oligo (α-arabinofuranosyl nucleotides) and α-arabinofuranosyl precursors thereof |
| JP3104995B2 (ja) | 1991-01-31 | 2000-10-30 | ワーナー−ランバート・コンパニー | 抗炎症剤として有用な4,6−ジ−第三ブチル−5−ヒドロキシ−1,3−ピリミジンの置換されたヘテロアリール類似体 |
| US5672697A (en) | 1991-02-08 | 1997-09-30 | Gilead Sciences, Inc. | Nucleoside 5'-methylene phosphonates |
| ES2156859T5 (es) | 1991-03-18 | 2008-03-16 | New York University | Anticuerpos monoclonales y quimericos especificos para el factor de necrosis tumoral humano. |
| AU664847B2 (en) | 1991-05-15 | 1995-12-07 | Cell Genesys, Inc. | Genomic modifications with homologous DNA targeting |
| US5539082A (en) | 1993-04-26 | 1996-07-23 | Nielsen; Peter E. | Peptide nucleic acids |
| US5173158A (en) | 1991-07-22 | 1992-12-22 | Schmukler Robert E | Apparatus and methods for electroporation and electrofusion |
| US5599797A (en) | 1991-10-15 | 1997-02-04 | Isis Pharmaceuticals, Inc. | Oligonucleotides having phosphorothioate linkages of high chiral purity |
| PT101031B (pt) | 1991-11-05 | 2002-07-31 | Transkaryotic Therapies Inc | Processo para o fornecimento de proteinas por terapia genetica |
| US5484908A (en) | 1991-11-26 | 1996-01-16 | Gilead Sciences, Inc. | Oligonucleotides containing 5-propynyl pyrimidines |
| US5359044A (en) | 1991-12-13 | 1994-10-25 | Isis Pharmaceuticals | Cyclobutyl oligonucleotide surrogates |
| AU4769893A (en) | 1992-07-17 | 1994-02-14 | Ribozyme Pharmaceuticals, Inc. | Method and reagent for treatment of animal diseases |
| US6107543A (en) | 1992-08-20 | 2000-08-22 | Infigen, Inc. | Culture of totipotent embryonic inner cells mass cells and production of bovine animals |
| TW402639B (en) | 1992-12-03 | 2000-08-21 | Transkaryotic Therapies Inc | Protein production and protein delivery |
| US5476925A (en) | 1993-02-01 | 1995-12-19 | Northwestern University | Oligodeoxyribonucleotides including 3'-aminonucleoside-phosphoramidate linkages and terminal 3'-amino groups |
| US5858988A (en) | 1993-02-24 | 1999-01-12 | Wang; Jui H. | Poly-substituted-phenyl-oligoribo nucleotides having enhanced stability and membrane permeability and methods of use |
| GB9304620D0 (en) | 1993-03-06 | 1993-04-21 | Ciba Geigy Ag | Compounds |
| GB9308271D0 (en) | 1993-04-21 | 1993-06-02 | Univ Edinburgh | Method of isolating and/or enriching and/or selectively propagating pluripotential animal cells and animals for use in said method |
| US5523226A (en) | 1993-05-14 | 1996-06-04 | Biotechnology Research And Development Corp. | Transgenic swine compositions and methods |
| US5571902A (en) | 1993-07-29 | 1996-11-05 | Isis Pharmaceuticals, Inc. | Synthesis of oligonucleotides |
| US5519134A (en) | 1994-01-11 | 1996-05-21 | Isis Pharmaceuticals, Inc. | Pyrrolidine-containing monomers and oligomers |
| US5539083A (en) | 1994-02-23 | 1996-07-23 | Isis Pharmaceuticals, Inc. | Peptide nucleic acid combinatorial libraries and improved methods of synthesis |
| US5554746A (en) | 1994-05-16 | 1996-09-10 | Isis Pharmaceuticals, Inc. | Lactam nucleic acids |
| US5843780A (en) | 1995-01-20 | 1998-12-01 | Wisconsin Alumni Research Foundation | Primate embryonic stem cells |
| US5705629A (en) | 1995-10-20 | 1998-01-06 | Hybridon, Inc. | Methods for H-phosphonate synthesis of mono- and oligonucleotides |
| JPH09260337A (ja) * | 1996-03-26 | 1997-10-03 | Nikon Corp | 薄膜構造体の製造装置 |
| US5994619A (en) | 1996-04-01 | 1999-11-30 | University Of Massachusetts, A Public Institution Of Higher Education Of The Commonwealth Of Massachusetts, As Represented By Its Amherst Campus | Production of chimeric bovine or porcine animals using cultured inner cell mass cells |
| US5945577A (en) | 1997-01-10 | 1999-08-31 | University Of Massachusetts As Represented By Its Amherst Campus | Cloning using donor nuclei from proliferating somatic cells |
| US6011197A (en) | 1997-03-06 | 2000-01-04 | Infigen, Inc. | Method of cloning bovines using reprogrammed non-embryonic bovine cells |
| US6506559B1 (en) | 1997-12-23 | 2003-01-14 | Carnegie Institute Of Washington | Genetic inhibition by double-stranded RNA |
| US6566131B1 (en) | 2000-10-04 | 2003-05-20 | Isis Pharmaceuticals, Inc. | Antisense modulation of Smad6 expression |
| JP2995179B1 (ja) * | 1998-07-31 | 1999-12-27 | 茂徳科技股▲ふん▼有限公司 | 浸漬液恒温供給システム |
| US6410323B1 (en) | 1999-08-31 | 2002-06-25 | Isis Pharmaceuticals, Inc. | Antisense modulation of human Rho family gene expression |
| US6107091A (en) | 1998-12-03 | 2000-08-22 | Isis Pharmaceuticals Inc. | Antisense inhibition of G-alpha-16 expression |
| US5981732A (en) | 1998-12-04 | 1999-11-09 | Isis Pharmaceuticals Inc. | Antisense modulation of G-alpha-13 expression |
| DE19956568A1 (de) | 1999-01-30 | 2000-08-17 | Roland Kreutzer | Verfahren und Medikament zur Hemmung der Expression eines vorgegebenen Gens |
| US8173592B1 (en) | 1999-03-31 | 2012-05-08 | Zentaris Ivf Gmbh | Method for a programmed controlled ovarian stimulation protocol |
| US6046321A (en) | 1999-04-09 | 2000-04-04 | Isis Pharmaceuticals Inc. | Antisense modulation of G-alpha-i1 expression |
| AU2001245793A1 (en) | 2000-03-16 | 2001-09-24 | Cold Spring Harbor Laboratory | Methods and compositions for rna interference |
| DK1309726T4 (en) | 2000-03-30 | 2019-01-28 | Whitehead Inst Biomedical Res | RNA Sequence-Specific Mediators of RNA Interference |
| US6365354B1 (en) | 2000-07-31 | 2002-04-02 | Isis Pharmaceuticals, Inc. | Antisense modulation of lysophospholipase I expression |
| US6566135B1 (en) | 2000-10-04 | 2003-05-20 | Isis Pharmaceuticals, Inc. | Antisense modulation of caspase 6 expression |
| CZ308053B6 (cs) | 2000-12-01 | 2019-11-27 | Max Planck Gesellschaft | Izolovaná molekula dvouřetězcové RNA, způsob její výroby a její použití |
| US7612250B2 (en) | 2002-07-29 | 2009-11-03 | Trustees Of Tufts College | Nuclear transfer embryo formation method |
| EP1534065A4 (en) | 2002-08-01 | 2005-11-09 | Gtc Biotherapeutics Inc | METHOD FOR RAPIDLY SELECTING LINES OF PRIMARY HOMOZYGOT CELLS TO PRODUCE TRANSGENIC ANIMALS BY NUCLEAR TRANSFER OF SOMATIC CELLS |
| JP2006508676A (ja) | 2002-12-10 | 2006-03-16 | ジーティーシー バイオセラピューティックス インコーポレイテッド | 追加の核移植のための細胞ドナーとして体細胞核移植胚を利用する方法および系 |
| US20040268422A1 (en) | 2003-04-09 | 2004-12-30 | Schatten Gerald P. | Methods for correcting mitotic spindle defects associated with somatic cell nuclear transfer in animals |
| US8807129B2 (en) | 2004-08-10 | 2014-08-19 | Kevin Keith Mackamul | Tracker drive system and solar energy collection system |
| EP2960328A1 (en) | 2004-11-04 | 2015-12-30 | Ocata Therapeutics, Inc. | Derivation of embryonic stem cells |
| JP2008538898A (ja) * | 2005-04-29 | 2008-11-13 | イノベイティブ デイリー プロダクツ ピーティーワイ リミティッド アズ トラスティー フォー ザ パーティシパンツ オブ ザ コーペラティブ リサーチ センター フォー イノベイティブ デイリー プ | 哺乳動物胚から幹細胞または幹細胞様細胞を作製するための方法 |
| LT2578685T (lt) | 2005-08-23 | 2019-06-10 | The Trustees Of The University Of Pennsylvania | Rnr, apimančios modifikuotus nukleozidus ir jų panaudojimo būdai |
| JP2007117081A (ja) | 2005-09-30 | 2007-05-17 | Institute Of Physical & Chemical Research | 核移植卵子の作製方法 |
| CA2597840A1 (en) | 2006-09-01 | 2008-03-01 | Government Of The United States Of America, As Represented By The Secret Ary, Department Of Health And Human Services | Methods and compositions for the treatment and prevention of cancer |
| CN101514878B (zh) * | 2008-02-18 | 2010-09-29 | 扬博科技股份有限公司 | 用于基板制程的热交换系统 |
| US20110172107A1 (en) | 2008-04-30 | 2011-07-14 | Fox Chase Cancer Center | Assay for identifying agents that modulate epigenetic silencing, and agents identified thereby |
| US20110136145A1 (en) | 2008-05-22 | 2011-06-09 | The Johns Hopkins University | Methods for promoting fusion and reprogramming of somatic cells |
| US20120034192A1 (en) | 2008-09-19 | 2012-02-09 | Young Richard A | Compositions and methods for enhancing cell reprogramming |
| DE102009039097B3 (de) | 2009-08-27 | 2010-11-25 | Siemens Aktiengesellschaft | Verfahren zum Übertragen von Daten in einem Sensornetzwerk, Sensorknoten und Zentral-Rechner |
| US20130189780A1 (en) * | 2009-12-31 | 2013-07-25 | Fate Therapeutics, Inc. | Reprogramming compositions |
| CA2796464C (en) | 2010-04-16 | 2021-08-03 | Immune Disease Institute, Inc. | Sustained polypeptide expression from synthetic, modified rnas and uses thereof |
| WO2012005769A1 (en) | 2010-07-09 | 2012-01-12 | Telecommunication Systems, Inc. | Location privacy selector |
| CN201768705U (zh) * | 2010-08-18 | 2011-03-23 | 州巧科技股份有限公司 | 节能清洗系统 |
| KR20130099071A (ko) | 2010-08-25 | 2013-09-05 | 액세스 비지니스 그룹 인터내셔날 엘엘씨 | 무선 전원 공급 시스템 및 다층 심 조립체 |
| WO2012029957A1 (ja) | 2010-09-03 | 2012-03-08 | 独立行政法人理化学研究所 | クローン動物の作出方法 |
| KR101290029B1 (ko) | 2011-01-20 | 2013-07-30 | 에스티팜 주식회사 | 시타글립틴의 중간체 제조방법 |
| JP6064231B2 (ja) | 2011-06-03 | 2017-01-25 | オンコセラピー・サイエンス株式会社 | がん治療および診断のための標的遺伝子としてのsuv39h2 |
| JP2014528959A (ja) | 2011-10-03 | 2014-10-30 | アンスティチュ ナショナル ドゥ ラ サンテ エ ドゥ ラ ルシェルシュ メディカル | Th2介在性疾患の処置のための方法および薬学的組成物 |
| US20140161785A1 (en) | 2012-12-10 | 2014-06-12 | Feiyan Liu | Verticillin A Inhibition of Histone Methyltransferases |
| JP6290931B2 (ja) | 2013-02-15 | 2018-03-07 | ソン クヮン メデイカル ファウンデーション | 体細胞核移植を利用する単為生殖幹細胞および患者特異的ヒト胚幹細胞の製造法 |
| US20160113911A1 (en) | 2013-06-06 | 2016-04-28 | The General Hospital Corporation | Methods and compositions for the treatment of cancer |
| KR102473092B1 (ko) | 2014-09-15 | 2022-12-01 | 칠드런'즈 메디컬 센터 코포레이션 | 히스톤 h3-리신 트리메틸화를 제거함으로써 체세포 핵 이식(scnt) 효율을 증가시키는 방법 및 조성물 |
| CN109641015A (zh) | 2015-10-09 | 2019-04-16 | 儿童医疗中心有限公司 | 通过移除组蛋白h3-赖氨酸三甲基化增加人类体细胞核转移(scnt)效率,以及增加人类nt-esc衍生物的方法和组合物 |
| US20200087629A1 (en) | 2016-10-19 | 2020-03-19 | Fred Hutchinson Cancer Research Center | Compositions and methods for reprogramming cells and for somatic cell nuclear transfer using duxc expression |
| CN107299113A (zh) | 2017-06-12 | 2017-10-27 | 内蒙古大学 | H3K27me3及其去甲基化酶KDM6A/B在小鼠核移植重构胚中的应用方法 |
| MA49651A (fr) | 2017-07-19 | 2021-04-28 | Childrens Medical Center | Compositions et procédés pour traiter des maladies associées à un défaut d'empreinte |
| KR20200141470A (ko) | 2018-04-06 | 2020-12-18 | 칠드런'즈 메디컬 센터 코포레이션 | 체세포 재프로그래밍 및 각인의 조정을 위한 조성물 및 방법 |
| WO2020018106A1 (en) | 2018-07-19 | 2020-01-23 | Children's Medical Center Corporation | Compositions and methods for generating physiological x chromosome inactivation |
| JP3224832U (ja) * | 2019-11-06 | 2020-01-23 | 揚▲博▼科技股▲分▼有限公司 | 基板エッチング処理装置における制御ボックスの散熱装置 |
| JP2022178121A (ja) * | 2021-05-19 | 2022-12-02 | 株式会社Screenホールディングス | 基板処理システム |
-
2015
- 2015-09-15 KR KR1020177010127A patent/KR102473092B1/ko active IP Right Grant
- 2015-09-15 JP JP2017514421A patent/JP6841753B2/ja active Active
- 2015-09-15 KR KR1020227041766A patent/KR102617137B1/ko active IP Right Grant
- 2015-09-15 EP EP15841324.5A patent/EP3194581A4/en active Pending
- 2015-09-15 US US15/511,348 patent/US11390885B2/en active Active
- 2015-09-15 WO PCT/US2015/050178 patent/WO2016044271A2/en active Application Filing
-
2018
- 2018-06-25 US US16/017,157 patent/US10266848B2/en active Active
-
2021
- 2021-02-18 JP JP2021023834A patent/JP7277494B2/ja active Active
-
2022
- 2022-06-14 US US17/840,348 patent/US20230015276A1/en active Pending
-
2023
- 2023-01-06 JP JP2023001284A patent/JP2023026679A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| US20170327846A1 (en) | 2017-11-16 |
| WO2016044271A3 (en) | 2016-05-19 |
| US10266848B2 (en) | 2019-04-23 |
| JP2023026679A (ja) | 2023-02-24 |
| JP7277494B2 (ja) | 2023-05-19 |
| WO2016044271A2 (en) | 2016-03-24 |
| KR102473092B1 (ko) | 2022-12-01 |
| US11390885B2 (en) | 2022-07-19 |
| US20180363008A1 (en) | 2018-12-20 |
| EP3194581A4 (en) | 2018-04-25 |
| KR102617137B1 (ko) | 2023-12-27 |
| JP2017528142A (ja) | 2017-09-28 |
| KR20220165807A (ko) | 2022-12-15 |
| US20230015276A1 (en) | 2023-01-19 |
| JP2021074023A (ja) | 2021-05-20 |
| KR20170120089A (ko) | 2017-10-30 |
| EP3194581A2 (en) | 2017-07-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7277494B2 (ja) | ヒストンh3-リジントリメチル化を除去することによって体細胞核移入(scnt)効率を増加させるための方法および組成物 | |
| US20200181648A1 (en) | Methods and compositions to increase human somatic cell nuclear transfer (scnt) efficiency by removing histone h3-lysine trimethylation, and derivation of human nt-esc | |
| US20240318144A1 (en) | Media for culturing naive human pluripotent stem cells | |
| ES2831424T3 (es) | Células madre pluripotentes naif aisladas y métodos para generarlas | |
| Wu et al. | Establishment of bovine embryonic stem cells after knockdown of CDX2 | |
| KR20140101390A (ko) | 반수체 세포 | |
| CN112272516B (zh) | 用于体细胞重新编程和调整印记的组合物和方法 | |
| JP2022534395A (ja) | 細胞リプログラミングのための組成物および方法 | |
| Oh | Reprogramming Pluripotent Stem Cell Towards Totipotency | |
| Wilson | MicroRNA-mediated silencing of bovine NANOG and MBD3 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20170726 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20170922 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20180831 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20180918 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20180918 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20190718 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20190719 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20191017 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20191216 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20200116 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20200528 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20200828 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20201022 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20210120 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20210218 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6841753 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |