ES2704731T3 - Proteínas de fusión de interleucina 2 y usos de las mismas - Google Patents
Proteínas de fusión de interleucina 2 y usos de las mismas Download PDFInfo
- Publication number
- ES2704731T3 ES2704731T3 ES15709419T ES15709419T ES2704731T3 ES 2704731 T3 ES2704731 T3 ES 2704731T3 ES 15709419 T ES15709419 T ES 15709419T ES 15709419 T ES15709419 T ES 15709419T ES 2704731 T3 ES2704731 T3 ES 2704731T3
- Authority
- ES
- Spain
- Prior art keywords
- igg
- lymphocytes
- dp47gs
- fusion protein
- treg
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 108010002350 Interleukin-2 Proteins 0.000 title claims abstract description 365
- 102000000588 Interleukin-2 Human genes 0.000 title claims abstract description 365
- 108020001507 fusion proteins Proteins 0.000 title claims abstract description 232
- 102000037865 fusion proteins Human genes 0.000 title claims abstract description 232
- 108060003951 Immunoglobulin Proteins 0.000 claims abstract description 185
- 102000018358 immunoglobulin Human genes 0.000 claims abstract description 185
- 150000001413 amino acids Chemical class 0.000 claims abstract description 64
- 108010038453 Interleukin-2 Receptors Proteins 0.000 claims abstract description 63
- 102000010789 Interleukin-2 Receptors Human genes 0.000 claims abstract description 63
- 239000000427 antigen Substances 0.000 claims abstract description 46
- 102000036639 antigens Human genes 0.000 claims abstract description 46
- 108091007433 antigens Proteins 0.000 claims abstract description 46
- 230000035772 mutation Effects 0.000 claims abstract description 34
- 210000004027 cell Anatomy 0.000 claims description 188
- 241000282414 Homo sapiens Species 0.000 claims description 146
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 122
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 86
- 238000000034 method Methods 0.000 claims description 86
- 230000027455 binding Effects 0.000 claims description 82
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 71
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 69
- 201000010099 disease Diseases 0.000 claims description 63
- 238000006467 substitution reaction Methods 0.000 claims description 62
- 230000004913 activation Effects 0.000 claims description 61
- 230000006698 induction Effects 0.000 claims description 60
- 108090000623 proteins and genes Proteins 0.000 claims description 56
- 210000003289 regulatory T cell Anatomy 0.000 claims description 52
- 108010087819 Fc receptors Proteins 0.000 claims description 45
- 102000009109 Fc receptors Human genes 0.000 claims description 45
- 230000014509 gene expression Effects 0.000 claims description 39
- 102000004169 proteins and genes Human genes 0.000 claims description 37
- 239000000203 mixture Substances 0.000 claims description 35
- 102000005962 receptors Human genes 0.000 claims description 35
- 108020003175 receptors Proteins 0.000 claims description 35
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 claims description 28
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 claims description 28
- 238000000338 in vitro Methods 0.000 claims description 26
- 230000004048 modification Effects 0.000 claims description 26
- 238000012986 modification Methods 0.000 claims description 26
- 239000003814 drug Substances 0.000 claims description 25
- 230000011664 signaling Effects 0.000 claims description 24
- 230000035755 proliferation Effects 0.000 claims description 19
- 238000012360 testing method Methods 0.000 claims description 15
- 230000001105 regulatory effect Effects 0.000 claims description 14
- 210000004602 germ cell Anatomy 0.000 claims description 9
- 230000003213 activating effect Effects 0.000 claims description 7
- 125000001433 C-terminal amino-acid group Chemical group 0.000 claims description 6
- 125000001429 N-terminal alpha-amino-acid group Chemical group 0.000 claims description 5
- 102100026122 High affinity immunoglobulin gamma Fc receptor I Human genes 0.000 claims description 4
- 101000913074 Homo sapiens High affinity immunoglobulin gamma Fc receptor I Proteins 0.000 claims description 4
- 101000917826 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-a Proteins 0.000 claims description 4
- 101000917824 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-b Proteins 0.000 claims description 4
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 claims description 4
- 230000001900 immune effect Effects 0.000 claims description 4
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 claims description 3
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 claims description 3
- 206010012601 diabetes mellitus Diseases 0.000 claims description 3
- 206010011878 Deafness Diseases 0.000 claims description 2
- JOCBASBOOFNAJA-UHFFFAOYSA-N N-tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid Chemical compound OCC(CO)(CO)NCCS(O)(=O)=O JOCBASBOOFNAJA-UHFFFAOYSA-N 0.000 claims 2
- 206010011224 Cough Diseases 0.000 claims 1
- 230000007123 defense Effects 0.000 claims 1
- 230000002519 immonomodulatory effect Effects 0.000 claims 1
- 238000011282 treatment Methods 0.000 description 116
- 235000001014 amino acid Nutrition 0.000 description 110
- 210000004698 lymphocyte Anatomy 0.000 description 106
- 210000004369 blood Anatomy 0.000 description 84
- 239000008280 blood Substances 0.000 description 84
- 241000282567 Macaca fascicularis Species 0.000 description 76
- 238000001727 in vivo Methods 0.000 description 66
- 229940024606 amino acid Drugs 0.000 description 65
- 210000003162 effector t lymphocyte Anatomy 0.000 description 60
- 102000040430 polynucleotide Human genes 0.000 description 57
- 108091033319 polynucleotide Proteins 0.000 description 57
- 239000002157 polynucleotide Substances 0.000 description 57
- 230000000694 effects Effects 0.000 description 56
- 230000004044 response Effects 0.000 description 54
- 241000699670 Mus sp. Species 0.000 description 51
- 101000581981 Homo sapiens Neural cell adhesion molecule 1 Proteins 0.000 description 47
- 102100027347 Neural cell adhesion molecule 1 Human genes 0.000 description 47
- 108700025316 aldesleukin Proteins 0.000 description 45
- 101000861452 Homo sapiens Forkhead box protein P3 Proteins 0.000 description 44
- 229940087463 proleukin Drugs 0.000 description 44
- 102100027581 Forkhead box protein P3 Human genes 0.000 description 41
- 102000004196 processed proteins & peptides Human genes 0.000 description 40
- 229920001184 polypeptide Polymers 0.000 description 38
- 239000012636 effector Substances 0.000 description 34
- 235000018102 proteins Nutrition 0.000 description 34
- 241000699666 Mus <mouse, genus> Species 0.000 description 33
- 230000002829 reductive effect Effects 0.000 description 33
- 230000001965 increasing effect Effects 0.000 description 30
- 101001002657 Homo sapiens Interleukin-2 Proteins 0.000 description 28
- 230000008859 change Effects 0.000 description 28
- 239000003981 vehicle Substances 0.000 description 27
- 208000023275 Autoimmune disease Diseases 0.000 description 24
- 230000037396 body weight Effects 0.000 description 24
- 231100000673 dose–response relationship Toxicity 0.000 description 24
- 230000006870 function Effects 0.000 description 24
- 102000001712 STAT5 Transcription Factor Human genes 0.000 description 23
- 108010029477 STAT5 Transcription Factor Proteins 0.000 description 23
- 238000002347 injection Methods 0.000 description 23
- 239000007924 injection Substances 0.000 description 23
- 210000002966 serum Anatomy 0.000 description 23
- 239000013598 vector Substances 0.000 description 23
- 230000003834 intracellular effect Effects 0.000 description 22
- 229940127121 immunoconjugate Drugs 0.000 description 21
- 210000000822 natural killer cell Anatomy 0.000 description 21
- 125000003729 nucleotide group Chemical group 0.000 description 21
- 230000026731 phosphorylation Effects 0.000 description 21
- 238000006366 phosphorylation reaction Methods 0.000 description 21
- 241001465754 Metazoa Species 0.000 description 20
- 239000000872 buffer Substances 0.000 description 20
- 241000282553 Macaca Species 0.000 description 19
- 206010067584 Type 1 diabetes mellitus Diseases 0.000 description 19
- 238000004519 manufacturing process Methods 0.000 description 19
- 239000002773 nucleotide Substances 0.000 description 18
- 102220274636 rs144712084 Human genes 0.000 description 18
- 102100026879 Interleukin-2 receptor subunit beta Human genes 0.000 description 17
- 229940072221 immunoglobulins Drugs 0.000 description 17
- 108040006849 interleukin-2 receptor activity proteins Proteins 0.000 description 17
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 17
- 150000007523 nucleic acids Chemical class 0.000 description 17
- 241000282693 Cercopithecidae Species 0.000 description 16
- 125000000539 amino acid group Chemical group 0.000 description 16
- 238000003556 assay Methods 0.000 description 16
- 238000010186 staining Methods 0.000 description 16
- 230000036962 time dependent Effects 0.000 description 16
- 108020004414 DNA Proteins 0.000 description 15
- 102000039446 nucleic acids Human genes 0.000 description 15
- 108020004707 nucleic acids Proteins 0.000 description 15
- 241000282412 Homo Species 0.000 description 14
- 230000004927 fusion Effects 0.000 description 14
- 201000006417 multiple sclerosis Diseases 0.000 description 14
- 239000008194 pharmaceutical composition Substances 0.000 description 14
- 238000002360 preparation method Methods 0.000 description 14
- 229940124597 therapeutic agent Drugs 0.000 description 14
- 108010076504 Protein Sorting Signals Proteins 0.000 description 13
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 13
- 239000003795 chemical substances by application Substances 0.000 description 13
- 239000012634 fragment Substances 0.000 description 13
- 102000008203 CTLA-4 Antigen Human genes 0.000 description 12
- 108010021064 CTLA-4 Antigen Proteins 0.000 description 12
- 229940045513 CTLA4 antagonist Drugs 0.000 description 12
- 108091026890 Coding region Proteins 0.000 description 12
- 239000004698 Polyethylene Substances 0.000 description 12
- 239000000090 biomarker Substances 0.000 description 12
- 230000015572 biosynthetic process Effects 0.000 description 12
- 210000005259 peripheral blood Anatomy 0.000 description 12
- 239000011886 peripheral blood Substances 0.000 description 12
- 239000002953 phosphate buffered saline Substances 0.000 description 12
- 239000000047 product Substances 0.000 description 12
- 230000000638 stimulation Effects 0.000 description 12
- 101710154942 Interleukin-2 receptor subunit beta Proteins 0.000 description 11
- 238000001514 detection method Methods 0.000 description 11
- 239000013604 expression vector Substances 0.000 description 11
- 210000003071 memory t lymphocyte Anatomy 0.000 description 11
- 239000000243 solution Substances 0.000 description 11
- 230000001225 therapeutic effect Effects 0.000 description 11
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 10
- 208000022559 Inflammatory bowel disease Diseases 0.000 description 10
- 102100037422 Receptor-type tyrosine-protein phosphatase C Human genes 0.000 description 10
- 238000001042 affinity chromatography Methods 0.000 description 10
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 10
- 238000010828 elution Methods 0.000 description 10
- 239000012467 final product Substances 0.000 description 10
- 210000004976 peripheral blood cell Anatomy 0.000 description 10
- 238000000746 purification Methods 0.000 description 10
- 238000001542 size-exclusion chromatography Methods 0.000 description 10
- 230000004936 stimulating effect Effects 0.000 description 10
- 241000124008 Mammalia Species 0.000 description 9
- 238000004458 analytical method Methods 0.000 description 9
- 230000007717 exclusion Effects 0.000 description 9
- 238000000684 flow cytometry Methods 0.000 description 9
- 230000004083 survival effect Effects 0.000 description 9
- 239000000725 suspension Substances 0.000 description 9
- 208000027930 type IV hypersensitivity disease Diseases 0.000 description 9
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 9
- 208000011231 Crohn disease Diseases 0.000 description 8
- 241000196324 Embryophyta Species 0.000 description 8
- 101000738771 Homo sapiens Receptor-type tyrosine-protein phosphatase C Proteins 0.000 description 8
- 241001529936 Murinae Species 0.000 description 8
- 108091028043 Nucleic acid sequence Proteins 0.000 description 8
- 239000004480 active ingredient Substances 0.000 description 8
- 230000017858 demethylation Effects 0.000 description 8
- 238000010520 demethylation reaction Methods 0.000 description 8
- 238000010494 dissociation reaction Methods 0.000 description 8
- 230000005593 dissociations Effects 0.000 description 8
- 239000003937 drug carrier Substances 0.000 description 8
- 230000003993 interaction Effects 0.000 description 8
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 8
- 238000012163 sequencing technique Methods 0.000 description 8
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 description 8
- 210000001519 tissue Anatomy 0.000 description 8
- 206010009900 Colitis ulcerative Diseases 0.000 description 7
- 238000002965 ELISA Methods 0.000 description 7
- 206010028980 Neoplasm Diseases 0.000 description 7
- 206010053613 Type IV hypersensitivity reaction Diseases 0.000 description 7
- 201000006704 Ulcerative Colitis Diseases 0.000 description 7
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 7
- 238000004113 cell culture Methods 0.000 description 7
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 7
- 238000010253 intravenous injection Methods 0.000 description 7
- 239000003550 marker Substances 0.000 description 7
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 7
- 150000003839 salts Chemical class 0.000 description 7
- 230000035945 sensitivity Effects 0.000 description 7
- 239000011780 sodium chloride Substances 0.000 description 7
- 210000000130 stem cell Anatomy 0.000 description 7
- 238000003786 synthesis reaction Methods 0.000 description 7
- 230000009261 transgenic effect Effects 0.000 description 7
- 230000005951 type IV hypersensitivity Effects 0.000 description 7
- 208000009329 Graft vs Host Disease Diseases 0.000 description 6
- 102100031573 Hematopoietic progenitor cell antigen CD34 Human genes 0.000 description 6
- 101000777663 Homo sapiens Hematopoietic progenitor cell antigen CD34 Proteins 0.000 description 6
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 6
- -1 PCR Proteins 0.000 description 6
- 239000012505 Superdex™ Substances 0.000 description 6
- 208000006673 asthma Diseases 0.000 description 6
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 6
- 238000004590 computer program Methods 0.000 description 6
- 230000001419 dependent effect Effects 0.000 description 6
- 208000035475 disorder Diseases 0.000 description 6
- 238000002474 experimental method Methods 0.000 description 6
- 238000009472 formulation Methods 0.000 description 6
- 208000024908 graft versus host disease Diseases 0.000 description 6
- 230000028993 immune response Effects 0.000 description 6
- 230000003053 immunization Effects 0.000 description 6
- 238000002649 immunization Methods 0.000 description 6
- 239000003018 immunosuppressive agent Substances 0.000 description 6
- 239000003446 ligand Substances 0.000 description 6
- 230000000670 limiting effect Effects 0.000 description 6
- 210000004962 mammalian cell Anatomy 0.000 description 6
- 230000001404 mediated effect Effects 0.000 description 6
- 239000000178 monomer Substances 0.000 description 6
- 230000036470 plasma concentration Effects 0.000 description 6
- 230000003389 potentiating effect Effects 0.000 description 6
- 239000003755 preservative agent Substances 0.000 description 6
- 230000009467 reduction Effects 0.000 description 6
- 102220071712 rs794728414 Human genes 0.000 description 6
- 201000000596 systemic lupus erythematosus Diseases 0.000 description 6
- 238000013518 transcription Methods 0.000 description 6
- 230000035897 transcription Effects 0.000 description 6
- 239000012099 Alexa Fluor family Substances 0.000 description 5
- 102000004127 Cytokines Human genes 0.000 description 5
- 108090000695 Cytokines Proteins 0.000 description 5
- 206010014950 Eosinophilia Diseases 0.000 description 5
- 239000004471 Glycine Substances 0.000 description 5
- 241000238631 Hexapoda Species 0.000 description 5
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 5
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 description 5
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 description 5
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 5
- 230000004989 O-glycosylation Effects 0.000 description 5
- 239000002253 acid Substances 0.000 description 5
- 238000010171 animal model Methods 0.000 description 5
- 230000000890 antigenic effect Effects 0.000 description 5
- 229960000074 biopharmaceutical Drugs 0.000 description 5
- 201000011510 cancer Diseases 0.000 description 5
- 238000005251 capillar electrophoresis Methods 0.000 description 5
- 230000001413 cellular effect Effects 0.000 description 5
- 238000004587 chromatography analysis Methods 0.000 description 5
- 210000003743 erythrocyte Anatomy 0.000 description 5
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 5
- 235000014304 histidine Nutrition 0.000 description 5
- 230000001976 improved effect Effects 0.000 description 5
- 239000013615 primer Substances 0.000 description 5
- 238000012545 processing Methods 0.000 description 5
- 230000028327 secretion Effects 0.000 description 5
- 239000001509 sodium citrate Substances 0.000 description 5
- NLJMYIDDQXHKNR-UHFFFAOYSA-K sodium citrate Chemical compound O.O.[Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O NLJMYIDDQXHKNR-UHFFFAOYSA-K 0.000 description 5
- 238000002560 therapeutic procedure Methods 0.000 description 5
- 230000001988 toxicity Effects 0.000 description 5
- 231100000419 toxicity Toxicity 0.000 description 5
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 4
- 208000004476 Acute Coronary Syndrome Diseases 0.000 description 4
- 206010027654 Allergic conditions Diseases 0.000 description 4
- 201000001320 Atherosclerosis Diseases 0.000 description 4
- 241000894006 Bacteria Species 0.000 description 4
- 208000024172 Cardiovascular disease Diseases 0.000 description 4
- 102100026234 Cytokine receptor common subunit gamma Human genes 0.000 description 4
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 4
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 4
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 4
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 4
- 241001494479 Pecora Species 0.000 description 4
- 241000288906 Primates Species 0.000 description 4
- 108010090804 Streptavidin Proteins 0.000 description 4
- 206010052779 Transplant rejections Diseases 0.000 description 4
- 230000002411 adverse Effects 0.000 description 4
- 235000004279 alanine Nutrition 0.000 description 4
- 230000004071 biological effect Effects 0.000 description 4
- 230000022131 cell cycle Effects 0.000 description 4
- 238000010276 construction Methods 0.000 description 4
- 230000008878 coupling Effects 0.000 description 4
- 238000010168 coupling process Methods 0.000 description 4
- 238000005859 coupling reaction Methods 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 210000003979 eosinophil Anatomy 0.000 description 4
- 210000003527 eukaryotic cell Anatomy 0.000 description 4
- 210000002683 foot Anatomy 0.000 description 4
- 230000002068 genetic effect Effects 0.000 description 4
- 230000013595 glycosylation Effects 0.000 description 4
- 238000006206 glycosylation reaction Methods 0.000 description 4
- 239000000833 heterodimer Substances 0.000 description 4
- 210000005260 human cell Anatomy 0.000 description 4
- 238000011577 humanized mouse model Methods 0.000 description 4
- 210000004408 hybridoma Anatomy 0.000 description 4
- 229940125721 immunosuppressive agent Drugs 0.000 description 4
- 239000004615 ingredient Substances 0.000 description 4
- 210000003292 kidney cell Anatomy 0.000 description 4
- 239000007788 liquid Substances 0.000 description 4
- 230000005923 long-lasting effect Effects 0.000 description 4
- 239000000463 material Substances 0.000 description 4
- 238000005259 measurement Methods 0.000 description 4
- 208000005069 pulmonary fibrosis Diseases 0.000 description 4
- 230000009257 reactivity Effects 0.000 description 4
- 239000003381 stabilizer Substances 0.000 description 4
- 239000000126 substance Substances 0.000 description 4
- 208000024891 symptom Diseases 0.000 description 4
- 231100000331 toxic Toxicity 0.000 description 4
- 230000002588 toxic effect Effects 0.000 description 4
- 230000014616 translation Effects 0.000 description 4
- 230000004580 weight loss Effects 0.000 description 4
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 3
- 101710117290 Aldo-keto reductase family 1 member C4 Proteins 0.000 description 3
- 108020004705 Codon Proteins 0.000 description 3
- 241000699802 Cricetulus griseus Species 0.000 description 3
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 3
- 201000004624 Dermatitis Diseases 0.000 description 3
- 208000004262 Food Hypersensitivity Diseases 0.000 description 3
- 206010016946 Food allergy Diseases 0.000 description 3
- 241000233866 Fungi Species 0.000 description 3
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 3
- 101100005713 Homo sapiens CD4 gene Proteins 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 239000004472 Lysine Substances 0.000 description 3
- 241000282560 Macaca mulatta Species 0.000 description 3
- 208000011623 Obstructive Lung disease Diseases 0.000 description 3
- 108700026244 Open Reading Frames Proteins 0.000 description 3
- 206010035226 Plasma cell myeloma Diseases 0.000 description 3
- 241000700159 Rattus Species 0.000 description 3
- 108020004511 Recombinant DNA Proteins 0.000 description 3
- 241000283984 Rodentia Species 0.000 description 3
- 239000002671 adjuvant Substances 0.000 description 3
- 239000003963 antioxidant agent Substances 0.000 description 3
- 235000006708 antioxidants Nutrition 0.000 description 3
- 238000013459 approach Methods 0.000 description 3
- 230000006472 autoimmune response Effects 0.000 description 3
- 239000002585 base Substances 0.000 description 3
- 210000004899 c-terminal region Anatomy 0.000 description 3
- 150000001720 carbohydrates Chemical group 0.000 description 3
- 239000003153 chemical reaction reagent Substances 0.000 description 3
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 3
- 238000003776 cleavage reaction Methods 0.000 description 3
- 238000010367 cloning Methods 0.000 description 3
- 230000000295 complement effect Effects 0.000 description 3
- 150000001875 compounds Chemical class 0.000 description 3
- 230000007423 decrease Effects 0.000 description 3
- 239000003085 diluting agent Substances 0.000 description 3
- 238000010790 dilution Methods 0.000 description 3
- 239000012895 dilution Substances 0.000 description 3
- 229940079593 drug Drugs 0.000 description 3
- 239000003623 enhancer Substances 0.000 description 3
- 235000020932 food allergy Nutrition 0.000 description 3
- 239000000499 gel Substances 0.000 description 3
- 238000004128 high performance liquid chromatography Methods 0.000 description 3
- 210000000987 immune system Anatomy 0.000 description 3
- 210000003734 kidney Anatomy 0.000 description 3
- 235000018977 lysine Nutrition 0.000 description 3
- 238000012423 maintenance Methods 0.000 description 3
- 239000002609 medium Substances 0.000 description 3
- 108020004999 messenger RNA Proteins 0.000 description 3
- 230000011987 methylation Effects 0.000 description 3
- 238000007069 methylation reaction Methods 0.000 description 3
- 201000000050 myeloid neoplasm Diseases 0.000 description 3
- 210000001672 ovary Anatomy 0.000 description 3
- 230000008823 permeabilization Effects 0.000 description 3
- 239000000546 pharmaceutical excipient Substances 0.000 description 3
- 239000012071 phase Substances 0.000 description 3
- 229920000642 polymer Polymers 0.000 description 3
- 239000000843 powder Substances 0.000 description 3
- 230000002335 preservative effect Effects 0.000 description 3
- 238000007639 printing Methods 0.000 description 3
- 230000002035 prolonged effect Effects 0.000 description 3
- 238000011321 prophylaxis Methods 0.000 description 3
- 238000000159 protein binding assay Methods 0.000 description 3
- 230000002441 reversible effect Effects 0.000 description 3
- 239000000523 sample Substances 0.000 description 3
- 230000007017 scission Effects 0.000 description 3
- 230000003248 secreting effect Effects 0.000 description 3
- 239000011734 sodium Substances 0.000 description 3
- 239000001488 sodium phosphate Substances 0.000 description 3
- 229910000162 sodium phosphate Inorganic materials 0.000 description 3
- 239000007787 solid Substances 0.000 description 3
- 239000002904 solvent Substances 0.000 description 3
- 230000000087 stabilizing effect Effects 0.000 description 3
- 239000006228 supernatant Substances 0.000 description 3
- 238000004448 titration Methods 0.000 description 3
- 238000013519 translation Methods 0.000 description 3
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 3
- 230000003612 virological effect Effects 0.000 description 3
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 description 2
- HZAXFHJVJLSVMW-UHFFFAOYSA-N 2-Aminoethan-1-ol Chemical compound NCCO HZAXFHJVJLSVMW-UHFFFAOYSA-N 0.000 description 2
- FPQQSJJWHUJYPU-UHFFFAOYSA-N 3-(dimethylamino)propyliminomethylidene-ethylazanium;chloride Chemical compound Cl.CCN=C=NCCCN(C)C FPQQSJJWHUJYPU-UHFFFAOYSA-N 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 2
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 2
- LSNNMFCWUKXFEE-UHFFFAOYSA-M Bisulfite Chemical compound OS([O-])=O LSNNMFCWUKXFEE-UHFFFAOYSA-M 0.000 description 2
- 101710189311 Cytokine receptor common subunit gamma Proteins 0.000 description 2
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 2
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 2
- 238000001712 DNA sequencing Methods 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 229910052693 Europium Inorganic materials 0.000 description 2
- 108010037362 Extracellular Matrix Proteins Proteins 0.000 description 2
- 102000010834 Extracellular Matrix Proteins Human genes 0.000 description 2
- 101000878602 Homo sapiens Immunoglobulin alpha Fc receptor Proteins 0.000 description 2
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 2
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical class C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 2
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 2
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 2
- 102100038005 Immunoglobulin alpha Fc receptor Human genes 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 2
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 2
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 description 2
- 241000283973 Oryctolagus cuniculus Species 0.000 description 2
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Natural products OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 2
- 239000002202 Polyethylene glycol Substances 0.000 description 2
- 229920002873 Polyethylenimine Polymers 0.000 description 2
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 2
- 201000004681 Psoriasis Diseases 0.000 description 2
- 101710138747 Receptor-type tyrosine-protein phosphatase C Proteins 0.000 description 2
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 2
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 239000004473 Threonine Substances 0.000 description 2
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 2
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- 241000700605 Viruses Species 0.000 description 2
- 230000004075 alteration Effects 0.000 description 2
- 239000012491 analyte Substances 0.000 description 2
- 230000005888 antibody-dependent cellular phagocytosis Effects 0.000 description 2
- 239000007864 aqueous solution Substances 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- 235000009697 arginine Nutrition 0.000 description 2
- 235000009582 asparagine Nutrition 0.000 description 2
- 229960001230 asparagine Drugs 0.000 description 2
- 208000010668 atopic eczema Diseases 0.000 description 2
- 230000001363 autoimmune Effects 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 239000008228 bacteriostatic water for injection Substances 0.000 description 2
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 2
- 210000000601 blood cell Anatomy 0.000 description 2
- 238000004820 blood count Methods 0.000 description 2
- 235000014633 carbohydrates Nutrition 0.000 description 2
- YCIMNLLNPGFGHC-UHFFFAOYSA-N catechol Chemical compound OC1=CC=CC=C1O YCIMNLLNPGFGHC-UHFFFAOYSA-N 0.000 description 2
- 238000006243 chemical reaction Methods 0.000 description 2
- 239000003638 chemical reducing agent Substances 0.000 description 2
- 208000017760 chronic graft versus host disease Diseases 0.000 description 2
- 238000007796 conventional method Methods 0.000 description 2
- 210000004748 cultured cell Anatomy 0.000 description 2
- 238000012258 culturing Methods 0.000 description 2
- 230000001351 cycling effect Effects 0.000 description 2
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 2
- 235000018417 cysteine Nutrition 0.000 description 2
- 125000000151 cysteine group Chemical class N[C@@H](CS)C(=O)* 0.000 description 2
- 238000002784 cytotoxicity assay Methods 0.000 description 2
- 231100000263 cytotoxicity test Toxicity 0.000 description 2
- 230000006378 damage Effects 0.000 description 2
- 230000003247 decreasing effect Effects 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- BFMYDTVEBKDAKJ-UHFFFAOYSA-L disodium;(2',7'-dibromo-3',6'-dioxido-3-oxospiro[2-benzofuran-1,9'-xanthene]-4'-yl)mercury;hydrate Chemical compound O.[Na+].[Na+].O1C(=O)C2=CC=CC=C2C21C1=CC(Br)=C([O-])C([Hg])=C1OC1=C2C=C(Br)C([O-])=C1 BFMYDTVEBKDAKJ-UHFFFAOYSA-L 0.000 description 2
- 239000002612 dispersion medium Substances 0.000 description 2
- 238000009826 distribution Methods 0.000 description 2
- 230000008030 elimination Effects 0.000 description 2
- 238000003379 elimination reaction Methods 0.000 description 2
- OGPBJKLSAFTDLK-UHFFFAOYSA-N europium atom Chemical compound [Eu] OGPBJKLSAFTDLK-UHFFFAOYSA-N 0.000 description 2
- 210000002744 extracellular matrix Anatomy 0.000 description 2
- 230000002349 favourable effect Effects 0.000 description 2
- 238000001502 gel electrophoresis Methods 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 2
- 235000004554 glutamine Nutrition 0.000 description 2
- 230000005484 gravity Effects 0.000 description 2
- 230000012010 growth Effects 0.000 description 2
- 229940093915 gynecological organic acid Drugs 0.000 description 2
- 230000036541 health Effects 0.000 description 2
- 230000002489 hematologic effect Effects 0.000 description 2
- ZFGMDIBRIDKWMY-PASTXAENSA-N heparin Chemical compound CC(O)=N[C@@H]1[C@@H](O)[C@H](O)[C@@H](COS(O)(=O)=O)O[C@@H]1O[C@@H]1[C@@H](C(O)=O)O[C@@H](O[C@H]2[C@@H]([C@@H](OS(O)(=O)=O)[C@@H](O[C@@H]3[C@@H](OC(O)[C@H](OS(O)(=O)=O)[C@H]3O)C(O)=O)O[C@@H]2O)CS(O)(=O)=O)[C@H](O)[C@H]1O ZFGMDIBRIDKWMY-PASTXAENSA-N 0.000 description 2
- 229920000669 heparin Polymers 0.000 description 2
- 238000005734 heterodimerization reaction Methods 0.000 description 2
- 210000002865 immune cell Anatomy 0.000 description 2
- 230000002163 immunogen Effects 0.000 description 2
- 230000001506 immunosuppresive effect Effects 0.000 description 2
- 229960003444 immunosuppressant agent Drugs 0.000 description 2
- 230000001861 immunosuppressant effect Effects 0.000 description 2
- 238000000099 in vitro assay Methods 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- 230000036512 infertility Effects 0.000 description 2
- 208000027866 inflammatory disease Diseases 0.000 description 2
- 238000001802 infusion Methods 0.000 description 2
- 230000005764 inhibitory process Effects 0.000 description 2
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 2
- 238000010212 intracellular staining Methods 0.000 description 2
- 238000001990 intravenous administration Methods 0.000 description 2
- 210000005229 liver cell Anatomy 0.000 description 2
- 238000011068 loading method Methods 0.000 description 2
- RLSSMJSEOOYNOY-UHFFFAOYSA-N m-cresol Chemical compound CC1=CC=CC(O)=C1 RLSSMJSEOOYNOY-UHFFFAOYSA-N 0.000 description 2
- 238000007403 mPCR Methods 0.000 description 2
- 238000013507 mapping Methods 0.000 description 2
- 229930182817 methionine Natural products 0.000 description 2
- 244000005700 microbiome Species 0.000 description 2
- 239000004005 microsphere Substances 0.000 description 2
- 230000005012 migration Effects 0.000 description 2
- 238000013508 migration Methods 0.000 description 2
- 238000002703 mutagenesis Methods 0.000 description 2
- 231100000350 mutagenesis Toxicity 0.000 description 2
- 230000007935 neutral effect Effects 0.000 description 2
- 150000007524 organic acids Chemical class 0.000 description 2
- 235000005985 organic acids Nutrition 0.000 description 2
- 230000036961 partial effect Effects 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 239000008188 pellet Substances 0.000 description 2
- AQIXEPGDORPWBJ-UHFFFAOYSA-N pentan-3-ol Chemical compound CCC(O)CC AQIXEPGDORPWBJ-UHFFFAOYSA-N 0.000 description 2
- 238000002823 phage display Methods 0.000 description 2
- 239000013612 plasmid Substances 0.000 description 2
- 229920001223 polyethylene glycol Polymers 0.000 description 2
- 239000013641 positive control Substances 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 230000000770 proinflammatory effect Effects 0.000 description 2
- QELSKZZBTMNZEB-UHFFFAOYSA-N propylparaben Chemical compound CCCOC(=O)C1=CC=C(O)C=C1 QELSKZZBTMNZEB-UHFFFAOYSA-N 0.000 description 2
- 230000005180 public health Effects 0.000 description 2
- 230000002285 radioactive effect Effects 0.000 description 2
- 230000006798 recombination Effects 0.000 description 2
- 230000008929 regeneration Effects 0.000 description 2
- 238000011069 regeneration method Methods 0.000 description 2
- GHMLBKRAJCXXBS-UHFFFAOYSA-N resorcinol Chemical compound OC1=CC=CC(O)=C1 GHMLBKRAJCXXBS-UHFFFAOYSA-N 0.000 description 2
- 238000012552 review Methods 0.000 description 2
- 206010039073 rheumatoid arthritis Diseases 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 238000002741 site-directed mutagenesis Methods 0.000 description 2
- 229910052708 sodium Inorganic materials 0.000 description 2
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 2
- 239000000600 sorbitol Substances 0.000 description 2
- 230000009870 specific binding Effects 0.000 description 2
- 238000007920 subcutaneous administration Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 230000001629 suppression Effects 0.000 description 2
- 239000004094 surface-active agent Substances 0.000 description 2
- 230000009885 systemic effect Effects 0.000 description 2
- 230000008685 targeting Effects 0.000 description 2
- 231100001274 therapeutic index Toxicity 0.000 description 2
- 235000008521 threonine Nutrition 0.000 description 2
- 229960000187 tissue plasminogen activator Drugs 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 230000001052 transient effect Effects 0.000 description 2
- 238000002054 transplantation Methods 0.000 description 2
- GETQZCLCWQTVFV-UHFFFAOYSA-N trimethylamine Chemical compound CN(C)C GETQZCLCWQTVFV-UHFFFAOYSA-N 0.000 description 2
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 2
- 235000002374 tyrosine Nutrition 0.000 description 2
- 210000005253 yeast cell Anatomy 0.000 description 2
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 description 1
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 1
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 1
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 1
- AXAVXPMQTGXXJZ-UHFFFAOYSA-N 2-aminoacetic acid;2-amino-2-(hydroxymethyl)propane-1,3-diol Chemical compound NCC(O)=O.OCC(N)(CO)CO AXAVXPMQTGXXJZ-UHFFFAOYSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- 102000007469 Actins Human genes 0.000 description 1
- 108010085238 Actins Proteins 0.000 description 1
- QGZKDVFQNNGYKY-UHFFFAOYSA-O Ammonium Chemical compound [NH4+] QGZKDVFQNNGYKY-UHFFFAOYSA-O 0.000 description 1
- 206010002091 Anaesthesia Diseases 0.000 description 1
- KWTQSFXGGICVPE-WCCKRBBISA-N Arginine hydrochloride Chemical compound Cl.OC(=O)[C@@H](N)CCCN=C(N)N KWTQSFXGGICVPE-WCCKRBBISA-N 0.000 description 1
- 108091008875 B cell receptors Proteins 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 206010006187 Breast cancer Diseases 0.000 description 1
- 102100032912 CD44 antigen Human genes 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- 241000282465 Canis Species 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 201000009030 Carcinoma Diseases 0.000 description 1
- 102000000844 Cell Surface Receptors Human genes 0.000 description 1
- 108010001857 Cell Surface Receptors Proteins 0.000 description 1
- 241000282552 Chlorocebus aethiops Species 0.000 description 1
- 208000006545 Chronic Obstructive Pulmonary Disease Diseases 0.000 description 1
- KRKNYBCHXYNGOX-UHFFFAOYSA-K Citrate Chemical compound [O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O KRKNYBCHXYNGOX-UHFFFAOYSA-K 0.000 description 1
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- 101710093674 Cyclic nucleotide-gated cation channel beta-1 Proteins 0.000 description 1
- 241000701022 Cytomegalovirus Species 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- 230000035131 DNA demethylation Effects 0.000 description 1
- 239000003155 DNA primer Substances 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 206010012438 Dermatitis atopic Diseases 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 239000004375 Dextrin Substances 0.000 description 1
- 229920001353 Dextrin Polymers 0.000 description 1
- 102100029722 Ectonucleoside triphosphate diphosphohydrolase 1 Human genes 0.000 description 1
- 102000005593 Endopeptidases Human genes 0.000 description 1
- 108010059378 Endopeptidases Proteins 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 101150027879 FOXP3 gene Proteins 0.000 description 1
- 108091006020 Fc-tagged proteins Proteins 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 108090000852 Forkhead Transcription Factors Proteins 0.000 description 1
- 230000035519 G0 Phase Effects 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 239000007995 HEPES buffer Substances 0.000 description 1
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 1
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 1
- 241000711549 Hepacivirus C Species 0.000 description 1
- 101000690301 Homo sapiens Aldo-keto reductase family 1 member C4 Proteins 0.000 description 1
- 101100220044 Homo sapiens CD34 gene Proteins 0.000 description 1
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 1
- 101000889276 Homo sapiens Cytotoxic T-lymphocyte protein 4 Proteins 0.000 description 1
- 101001012447 Homo sapiens Ectonucleoside triphosphate diphosphohydrolase 1 Proteins 0.000 description 1
- 101001018097 Homo sapiens L-selectin Proteins 0.000 description 1
- 101001131829 Homo sapiens P protein Proteins 0.000 description 1
- 101001116548 Homo sapiens Protein CBFA2T1 Proteins 0.000 description 1
- 101000997835 Homo sapiens Tyrosine-protein kinase JAK1 Proteins 0.000 description 1
- 101000934996 Homo sapiens Tyrosine-protein kinase JAK3 Proteins 0.000 description 1
- 102000012745 Immunoglobulin Subunits Human genes 0.000 description 1
- 108010079585 Immunoglobulin Subunits Proteins 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 1
- 102000004877 Insulin Human genes 0.000 description 1
- 108090001061 Insulin Proteins 0.000 description 1
- 102000000589 Interleukin-1 Human genes 0.000 description 1
- 108010002352 Interleukin-1 Proteins 0.000 description 1
- 102000019223 Interleukin-1 receptor Human genes 0.000 description 1
- 108050006617 Interleukin-1 receptor Proteins 0.000 description 1
- 108020004684 Internal Ribosome Entry Sites Proteins 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 1
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- 102100033467 L-selectin Human genes 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 241000368589 Lophotus lacepede Species 0.000 description 1
- 208000005777 Lupus Nephritis Diseases 0.000 description 1
- 239000007993 MOPS buffer Substances 0.000 description 1
- 241001503485 Mammuthus Species 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 238000000585 Mann–Whitney U test Methods 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 208000034486 Multi-organ failure Diseases 0.000 description 1
- 101100120552 Mus musculus Foxp3 gene Proteins 0.000 description 1
- NQTADLQHYWFPDB-UHFFFAOYSA-N N-Hydroxysuccinimide Chemical compound ON1C(=O)CCC1=O NQTADLQHYWFPDB-UHFFFAOYSA-N 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 101710160107 Outer membrane protein A Proteins 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- 208000025237 Polyendocrinopathy Diseases 0.000 description 1
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 1
- 108090000412 Protein-Tyrosine Kinases Proteins 0.000 description 1
- 102000004022 Protein-Tyrosine Kinases Human genes 0.000 description 1
- 241000700157 Rattus norvegicus Species 0.000 description 1
- 230000010799 Receptor Interactions Effects 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 241000220317 Rosa Species 0.000 description 1
- 241000714474 Rous sarcoma virus Species 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 108010071390 Serum Albumin Proteins 0.000 description 1
- 102000007562 Serum Albumin Human genes 0.000 description 1
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical compound [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 1
- 241000256251 Spodoptera frugiperda Species 0.000 description 1
- 235000019892 Stellar Nutrition 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 108091036066 Three prime untranslated region Proteins 0.000 description 1
- 101710120037 Toxin CcdB Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102100025946 Transforming growth factor beta activator LRRC32 Human genes 0.000 description 1
- 101710169732 Transforming growth factor beta activator LRRC32 Proteins 0.000 description 1
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 description 1
- 102100033438 Tyrosine-protein kinase JAK1 Human genes 0.000 description 1
- 102100025387 Tyrosine-protein kinase JAK3 Human genes 0.000 description 1
- 229940127174 UCHT1 Drugs 0.000 description 1
- 244000000188 Vaccinium ovalifolium Species 0.000 description 1
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 1
- 206010047115 Vasculitis Diseases 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 238000010521 absorption reaction Methods 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- DPXJVFZANSGRMM-UHFFFAOYSA-N acetic acid;2,3,4,5,6-pentahydroxyhexanal;sodium Chemical compound [Na].CC(O)=O.OCC(O)C(O)C(O)C(O)C=O DPXJVFZANSGRMM-UHFFFAOYSA-N 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 210000005006 adaptive immune system Anatomy 0.000 description 1
- 239000000443 aerosol Substances 0.000 description 1
- 230000000172 allergic effect Effects 0.000 description 1
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 230000037005 anaesthesia Effects 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 230000009830 antibody antigen interaction Effects 0.000 description 1
- 230000005875 antibody response Effects 0.000 description 1
- 229940121375 antifungal agent Drugs 0.000 description 1
- 239000003429 antifungal agent Substances 0.000 description 1
- 210000000612 antigen-presenting cell Anatomy 0.000 description 1
- 239000003125 aqueous solvent Substances 0.000 description 1
- 238000003491 array Methods 0.000 description 1
- 229960005070 ascorbic acid Drugs 0.000 description 1
- 235000010323 ascorbic acid Nutrition 0.000 description 1
- 239000011668 ascorbic acid Substances 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 239000012131 assay buffer Substances 0.000 description 1
- 201000008937 atopic dermatitis Diseases 0.000 description 1
- 208000037979 autoimmune inflammatory disease Diseases 0.000 description 1
- 230000005784 autoimmunity Effects 0.000 description 1
- 210000003719 b-lymphocyte Anatomy 0.000 description 1
- 230000008901 benefit Effects 0.000 description 1
- 229960000686 benzalkonium chloride Drugs 0.000 description 1
- UREZNYTWGJKWBI-UHFFFAOYSA-M benzethonium chloride Chemical compound [Cl-].C1=CC(C(C)(C)CC(C)(C)C)=CC=C1OCCOCC[N+](C)(C)CC1=CC=CC=C1 UREZNYTWGJKWBI-UHFFFAOYSA-M 0.000 description 1
- 229960001950 benzethonium chloride Drugs 0.000 description 1
- CADWTSSKOVRVJC-UHFFFAOYSA-N benzyl(dimethyl)azanium;chloride Chemical compound [Cl-].C[NH+](C)CC1=CC=CC=C1 CADWTSSKOVRVJC-UHFFFAOYSA-N 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 238000013357 binding ELISA Methods 0.000 description 1
- 239000011230 binding agent Substances 0.000 description 1
- 230000008033 biological extinction Effects 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- OWMVSZAMULFTJU-UHFFFAOYSA-N bis-tris Chemical compound OCCN(CCO)C(CO)(CO)CO OWMVSZAMULFTJU-UHFFFAOYSA-N 0.000 description 1
- HUTDDBSSHVOYJR-UHFFFAOYSA-H bis[(2-oxo-1,3,2$l^{5},4$l^{2}-dioxaphosphaplumbetan-2-yl)oxy]lead Chemical compound [Pb+2].[Pb+2].[Pb+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O HUTDDBSSHVOYJR-UHFFFAOYSA-H 0.000 description 1
- 238000009534 blood test Methods 0.000 description 1
- 210000001185 bone marrow Anatomy 0.000 description 1
- 108010006025 bovine growth hormone Proteins 0.000 description 1
- LRHPLDYGYMQRHN-UHFFFAOYSA-N butyl alcohol Substances CCCCO LRHPLDYGYMQRHN-UHFFFAOYSA-N 0.000 description 1
- 125000000484 butyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])C([H])([H])[H] 0.000 description 1
- 239000006227 byproduct Substances 0.000 description 1
- 239000011575 calcium Substances 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 244000309466 calf Species 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 239000001768 carboxy methyl cellulose Substances 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 230000032823 cell division Effects 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 210000003679 cervix uteri Anatomy 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 239000002738 chelating agent Substances 0.000 description 1
- 238000007385 chemical modification Methods 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 208000037976 chronic inflammation Diseases 0.000 description 1
- 208000037893 chronic inflammatory disorder Diseases 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 238000002648 combination therapy Methods 0.000 description 1
- 230000000052 comparative effect Effects 0.000 description 1
- 230000024203 complement activation Effects 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 238000011109 contamination Methods 0.000 description 1
- 239000013068 control sample Substances 0.000 description 1
- WZHCOOQXZCIUNC-UHFFFAOYSA-N cyclandelate Chemical compound C1C(C)(C)CC(C)CC1OC(=O)C(O)C1=CC=CC=C1 WZHCOOQXZCIUNC-UHFFFAOYSA-N 0.000 description 1
- HPXRVTGHNJAIIH-UHFFFAOYSA-N cyclohexanol Chemical compound OC1CCCCC1 HPXRVTGHNJAIIH-UHFFFAOYSA-N 0.000 description 1
- 102000003675 cytokine receptors Human genes 0.000 description 1
- 108010057085 cytokine receptors Proteins 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 230000001066 destructive effect Effects 0.000 description 1
- 235000019425 dextrin Nutrition 0.000 description 1
- 239000008121 dextrose Substances 0.000 description 1
- 238000003745 diagnosis Methods 0.000 description 1
- 235000014113 dietary fatty acids Nutrition 0.000 description 1
- 230000009699 differential effect Effects 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- 230000003292 diminished effect Effects 0.000 description 1
- 230000003467 diminishing effect Effects 0.000 description 1
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 1
- 150000002016 disaccharides Chemical class 0.000 description 1
- 239000002270 dispersing agent Substances 0.000 description 1
- 239000006185 dispersion Substances 0.000 description 1
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 1
- 239000002552 dosage form Substances 0.000 description 1
- 230000003828 downregulation Effects 0.000 description 1
- 239000000975 dye Substances 0.000 description 1
- 230000004064 dysfunction Effects 0.000 description 1
- QCUPYFTWJOZAOB-HYXAFXHYSA-N ectylurea Chemical compound CC\C(=C\C)C(=O)NC(N)=O QCUPYFTWJOZAOB-HYXAFXHYSA-N 0.000 description 1
- 230000002444 effect on eosinophils Effects 0.000 description 1
- 230000009982 effect on human Effects 0.000 description 1
- 230000001804 emulsifying effect Effects 0.000 description 1
- 239000000839 emulsion Substances 0.000 description 1
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 1
- 239000002158 endotoxin Substances 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 208000037902 enteropathy Diseases 0.000 description 1
- 125000001495 ethyl group Chemical group [H]C([H])([H])C([H])([H])* 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 229930195729 fatty acid Natural products 0.000 description 1
- 239000000194 fatty acid Substances 0.000 description 1
- 239000010685 fatty oil Substances 0.000 description 1
- 230000001605 fetal effect Effects 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 239000013020 final formulation Substances 0.000 description 1
- 239000003063 flame retardant Substances 0.000 description 1
- 239000000796 flavoring agent Substances 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- 235000013355 food flavoring agent Nutrition 0.000 description 1
- 235000003599 food sweetener Nutrition 0.000 description 1
- 239000012458 free base Substances 0.000 description 1
- 238000004108 freeze drying Methods 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 239000011521 glass Substances 0.000 description 1
- 238000003881 globally optimized alternating phase rectangular pulse Methods 0.000 description 1
- 102000018146 globin Human genes 0.000 description 1
- 108060003196 globin Proteins 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 108700003606 glycosylated interleukin 2 Proteins 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 230000003394 haemopoietic effect Effects 0.000 description 1
- 210000002443 helper t lymphocyte Anatomy 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 210000000548 hind-foot Anatomy 0.000 description 1
- 102000043321 human CTLA4 Human genes 0.000 description 1
- 102000047119 human OCA2 Human genes 0.000 description 1
- 102000054751 human RUNX1T1 Human genes 0.000 description 1
- 235000011167 hydrochloric acid Nutrition 0.000 description 1
- 229920001477 hydrophilic polymer Polymers 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 150000004679 hydroxides Chemical class 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 230000003832 immune regulation Effects 0.000 description 1
- 230000006058 immune tolerance Effects 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 230000008902 immunological benefit Effects 0.000 description 1
- 230000004957 immunoregulator effect Effects 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 210000002602 induced regulatory T cell Anatomy 0.000 description 1
- 239000000411 inducer Substances 0.000 description 1
- 230000002757 inflammatory effect Effects 0.000 description 1
- 230000028709 inflammatory response Effects 0.000 description 1
- 150000007529 inorganic bases Chemical class 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 229940125396 insulin Drugs 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 230000004073 interleukin-2 production Effects 0.000 description 1
- 208000028774 intestinal disease Diseases 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 239000007928 intraperitoneal injection Substances 0.000 description 1
- 238000007913 intrathecal administration Methods 0.000 description 1
- 229940065638 intron a Drugs 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 150000002500 ions Chemical class 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- FZWBNHMXJMCXLU-BLAUPYHCSA-N isomaltotriose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OC[C@@H]1[C@@H](O)[C@H](O)[C@@H](O)[C@@H](OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C=O)O1 FZWBNHMXJMCXLU-BLAUPYHCSA-N 0.000 description 1
- JJWLVOIRVHMVIS-UHFFFAOYSA-N isopropylamine Chemical compound CC(C)N JJWLVOIRVHMVIS-UHFFFAOYSA-N 0.000 description 1
- YWXYYJSYQOXTPL-SLPGGIOYSA-N isosorbide mononitrate Chemical compound [O-][N+](=O)O[C@@H]1CO[C@@H]2[C@@H](O)CO[C@@H]21 YWXYYJSYQOXTPL-SLPGGIOYSA-N 0.000 description 1
- 239000007951 isotonicity adjuster Substances 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 231100000636 lethal dose Toxicity 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 239000000314 lubricant Substances 0.000 description 1
- 210000005265 lung cell Anatomy 0.000 description 1
- 206010025135 lupus erythematosus Diseases 0.000 description 1
- 210000000207 lymphocyte subset Anatomy 0.000 description 1
- 230000000527 lymphocytic effect Effects 0.000 description 1
- 239000006166 lysate Substances 0.000 description 1
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 229920002521 macromolecule Polymers 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Chemical class 0.000 description 1
- MYWUZJCMWCOHBA-VIFPVBQESA-N methamphetamine Chemical compound CN[C@@H](C)CC1=CC=CC=C1 MYWUZJCMWCOHBA-VIFPVBQESA-N 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- 150000007522 mineralic acids Chemical class 0.000 description 1
- 230000011278 mitosis Effects 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 239000003607 modifier Substances 0.000 description 1
- 238000001823 molecular biology technique Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 150000002772 monosaccharides Chemical class 0.000 description 1
- 208000029744 multiple organ dysfunction syndrome Diseases 0.000 description 1
- AEMBWNDIEFEPTH-UHFFFAOYSA-N n-tert-butyl-n-ethylnitrous amide Chemical compound CCN(N=O)C(C)(C)C AEMBWNDIEFEPTH-UHFFFAOYSA-N 0.000 description 1
- 210000000581 natural killer T-cell Anatomy 0.000 description 1
- 108010068617 neonatal Fc receptor Proteins 0.000 description 1
- 239000002736 nonionic surfactant Substances 0.000 description 1
- 231100000956 nontoxicity Toxicity 0.000 description 1
- 239000012053 oil suspension Substances 0.000 description 1
- 238000006384 oligomerization reaction Methods 0.000 description 1
- 230000003287 optical effect Effects 0.000 description 1
- 150000007530 organic bases Chemical class 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- LXCFILQKKLGQFO-UHFFFAOYSA-N p-hydroxybenzoic acid methyl ester Natural products COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 1
- 210000000496 pancreas Anatomy 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 230000001575 pathological effect Effects 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 230000002093 peripheral effect Effects 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- 229960003742 phenol Drugs 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 235000011007 phosphoric acid Nutrition 0.000 description 1
- 150000003016 phosphoric acids Chemical class 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- 239000002504 physiological saline solution Substances 0.000 description 1
- 239000000902 placebo Substances 0.000 description 1
- 229940068196 placebo Drugs 0.000 description 1
- 239000004033 plastic Substances 0.000 description 1
- 229920003023 plastic Polymers 0.000 description 1
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 1
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 1
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 229910052700 potassium Inorganic materials 0.000 description 1
- 239000011591 potassium Substances 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000004321 preservation Methods 0.000 description 1
- 230000003449 preventive effect Effects 0.000 description 1
- MFDFERRIHVXMIY-UHFFFAOYSA-N procaine Chemical compound CCN(CC)CCOC(=O)C1=CC=C(N)C=C1 MFDFERRIHVXMIY-UHFFFAOYSA-N 0.000 description 1
- 229960004919 procaine Drugs 0.000 description 1
- 238000004393 prognosis Methods 0.000 description 1
- 125000001500 prolyl group Chemical group [H]N1C([H])(C(=O)[*])C([H])([H])C([H])([H])C1([H])[H] 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 239000004405 propyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010232 propyl p-hydroxybenzoate Nutrition 0.000 description 1
- 229960003415 propylparaben Drugs 0.000 description 1
- 230000006337 proteolytic cleavage Effects 0.000 description 1
- 239000003586 protic polar solvent Substances 0.000 description 1
- 239000002510 pyrogen Substances 0.000 description 1
- 238000003908 quality control method Methods 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 238000007420 radioactive assay Methods 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 238000004064 recycling Methods 0.000 description 1
- 230000013878 renal filtration Effects 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 230000000284 resting effect Effects 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 230000001177 retroviral effect Effects 0.000 description 1
- 238000003757 reverse transcription PCR Methods 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 238000005070 sampling Methods 0.000 description 1
- 235000004400 serine Nutrition 0.000 description 1
- 210000000717 sertoli cell Anatomy 0.000 description 1
- 239000012679 serum free medium Substances 0.000 description 1
- 239000008159 sesame oil Substances 0.000 description 1
- 235000011803 sesame oil Nutrition 0.000 description 1
- 230000019491 signal transduction Effects 0.000 description 1
- 230000007781 signaling event Effects 0.000 description 1
- 208000017520 skin disease Diseases 0.000 description 1
- 239000001632 sodium acetate Substances 0.000 description 1
- 235000017281 sodium acetate Nutrition 0.000 description 1
- 235000019812 sodium carboxymethyl cellulose Nutrition 0.000 description 1
- 229920001027 sodium carboxymethylcellulose Polymers 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 238000001228 spectrum Methods 0.000 description 1
- 210000000952 spleen Anatomy 0.000 description 1
- 230000003393 splenic effect Effects 0.000 description 1
- 210000004988 splenocyte Anatomy 0.000 description 1
- 230000010473 stable expression Effects 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- SFVFIFLLYFPGHH-UHFFFAOYSA-M stearalkonium chloride Chemical compound [Cl-].CCCCCCCCCCCCCCCCCC[N+](C)(C)CC1=CC=CC=C1 SFVFIFLLYFPGHH-UHFFFAOYSA-M 0.000 description 1
- 238000011146 sterile filtration Methods 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 239000000375 suspending agent Substances 0.000 description 1
- 239000003765 sweetening agent Substances 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 150000003626 triacylglycerols Chemical class 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 208000035408 type 1 diabetes mellitus 1 Diseases 0.000 description 1
- 125000001493 tyrosinyl group Chemical group [H]OC1=C([H])C([H])=C(C([H])=C1[H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 238000000870 ultraviolet spectroscopy Methods 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 230000003827 upregulation Effects 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 229960005486 vaccine Drugs 0.000 description 1
- 238000001291 vacuum drying Methods 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 230000009385 viral infection Effects 0.000 description 1
- 210000001835 viscera Anatomy 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 230000003442 weekly effect Effects 0.000 description 1
- 208000032620 x-linked multiple congenital anomalies-neurodevelopmental syndrome Diseases 0.000 description 1
Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- A61K38/17—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- A61K38/19—Cytokines; Lymphokines; Interferons
- A61K38/20—Interleukins [IL]
- A61K38/2013—IL-2
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
- A61K47/51—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent
- A61K47/68—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being an antibody, an immunoglobulin or a fragment thereof, e.g. an Fc-fragment
- A61K47/6801—Drug-antibody or immunoglobulin conjugates defined by the pharmacologically or therapeutically active agent
- A61K47/6803—Drugs conjugated to an antibody or immunoglobulin, e.g. cisplatin-antibody conjugates
- A61K47/6811—Drugs conjugated to an antibody or immunoglobulin, e.g. cisplatin-antibody conjugates the drug being a protein or peptide, e.g. transferrin or bleomycin
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
- A61P1/04—Drugs for disorders of the alimentary tract or the digestive system for ulcers, gastritis or reflux esophagitis, e.g. antacids, inhibitors of acid secretion, mucosal protectants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/08—Drugs for disorders of the metabolism for glucose homeostasis
- A61P3/10—Drugs for disorders of the metabolism for glucose homeostasis for hyperglycaemia, e.g. antidiabetics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/06—Immunosuppressants, e.g. drugs for graft rejection
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/52—Cytokines; Lymphokines; Interferons
- C07K14/54—Interleukins [IL]
- C07K14/55—IL-2
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/46—Hybrid immunoglobulins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K19/00—Hybrid peptides, i.e. peptides covalently bound to nucleic acids, or non-covalently bound protein-protein complexes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0636—T lymphocytes
- C12N5/0637—Immunosuppressive T lymphocytes, e.g. regulatory T cells or Treg
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/30—Non-immunoglobulin-derived peptide or protein having an immunoglobulin constant or Fc region, or a fragment thereof, attached thereto
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2302—Interleukin-2 (IL-2)
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Immunology (AREA)
- Animal Behavior & Ethology (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Pharmacology & Pharmacy (AREA)
- Zoology (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Molecular Biology (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Gastroenterology & Hepatology (AREA)
- Biophysics (AREA)
- Biomedical Technology (AREA)
- Toxicology (AREA)
- Epidemiology (AREA)
- Wood Science & Technology (AREA)
- Biotechnology (AREA)
- Diabetes (AREA)
- Hematology (AREA)
- General Engineering & Computer Science (AREA)
- Cell Biology (AREA)
- Microbiology (AREA)
- Neurology (AREA)
- Endocrinology (AREA)
- Obesity (AREA)
- Emergency Medicine (AREA)
- Neurosurgery (AREA)
- Transplantation (AREA)
- Peptides Or Proteins (AREA)
Abstract
Una proteína de fusión que comprende (i) una molécula de inmunoglobulina que no se puede unir específicamente a un antígeno y (ii) dos moléculas de interleucina 2 (IL-2) mutante que comprenden una mutación de aminoácido que reduce la afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia, en comparación con una molécula de IL-2 natural, en la que dichas moléculas de IL-2 mutante comprenden la secuencia de la SEQ ID NO: 58.
Description
DESCRIPCIÓN
Proteínas de fusión de interleucina 2 y usos de las mismas
Campo de la invención
La presente invención se refiere en general a las proteínas de fusión de inmunoglobulinas e interleucina 2 (IL-2). Más en particular, la invención aborda proteínas de fusión de inmunoglobulinas e IL-2 mutante que presentan propiedades mejoradas para su uso como agentes terapéuticos, por ejemplo, en el tratamiento de enfermedades autoinmunitarias y enfermedades inflamatorias mediadas por el sistema inmunitario. Además, la presente invención se refiere a los polinucleótidos que codifican dichas proteínas de fusión y a los vectores y células huésped que comprenden dichos polinucleótidos. La invención se refiere además a los procedimientos para producir las proteínas de fusión de la invención y a los procedimientos de uso de las mismas en el tratamiento de la enfermedad.
Antecedentes
Los linfocitos T reguladores (Treg) representan subconjuntos específicos de linfocitos T que son cruciales para el mantenimiento de la autotolerancia. Estas células CD4+CD25a con función supresora se pueden distinguir de los linfocitos T efectores mediante la expresión intracelular del factor de transcripción FOXP3, así como de otros marcadores celulares, tales como CD127baja, CTLA-4+, LAP, CD39+, PD-1+, GARP, etc. FOXP3 es crítico para la diferenciación y la función de Treg, y la carencia y las mutaciones en el gen FOXP3, tanto en ratones Scurfy como en pacientes con regulación inmunitaria incorrecta, poliendocrinopatía, enteropatía y síndrome ligado al cromosoma X (IPEX), dan como resultado la destrucción de la autotolerancia y el desarrollo de enfermedades autoinmunitarias debido a la carencia o falta de función de Treg.
Las respuestas autoinmunitarias en la diabetes de tipo 1, lupus eritematoso diseminado (LED), esclerosis múltiple y muchas otras se correlacionan con una carencia en los Treg o en las funciones de Treg. Los datos de modelos animales respaldan la hipótesis de que las respuestas autoinmunitarias se facilitan por un fallo de los Treg en el control de la respuesta inmunitaria destructora con respecto a sí mismos, en gran parte debido a los efectos de los linfocitos T efectores de memoria CD4+ autorreactivos. La diabetes de tipo 1 es una enfermedad autoinmunitaria que se produce después de la destrucción de la mayoría de las células p productoras de insulina en el páncreas. La frecuencia de la diabetes de tipo 1 es ~0,3 % de la población de los EE. UU. y su incidencia continúa incrementándose en los EE. UU., Europa y, en particular, en los países escandinavos (prácticamente un 1 %) y se espera que se duplique en los próximos veinte años.
La citocina IL-2 desempeña un papel principal en la activación y en la función tanto de los Treg como de los linfocitos T efectores (Tef). Una carencia en la producción de IL-2 o falta de reactividad da como resultado, preferentemente, una pérdida de la función de Treg y un incremento de la probabilidad de autoinmunidad. Puesto que los Treg expresan constitutivamente el receptor de IL-2 de afinidad alta en concentraciones más altas que Tef, las dosis bajas de IL-2 respaldan preferentemente el mantenimiento de los Treg en comparación con los linfocitos Tef.
Con el efecto preferente de IL-2 para activar los Treg in vitro e in vivo, el potencial del tratamiento con IL-2 de larga duración y a dosis bajas parece tener una alta probabilidad de éxito en las enfermedades autoinmunitarias. A finales de 2013, se establece el comienzo de un ensayo clínico comparativo con placebo con enmascaramiento doble sobre la diabetes de tipo 1 de 200 pacientes (Proleukin®, IL-2 humana recombinante). Los recientes ensayos clínicos con Proleukin en dosis baja diaria mejoraron algunos de los signos y síntomas de la enfermedad injerto contra huésped (EICH) crónica y vasculitis inducida por el virus de la hepatitis C (Koreth et al., New Engl J Med 365, 2055-2066 (2011), Saadoun et al., New Engl J Med 365, 2067-2077 (2011)). En ambos estudios, Proleukin® en dosis baja indujo Treg e incrementó la proporción Treg:Tef. Sin embargo, las propiedades PF deficientes de Proleukin® le hacen subóptimo para mantener concentraciones bajas y consistentes de IL-2 en el hombre. Otros procedimientos que se someten a prueba en ensayos clínicos son la expansión personalizada de Treg ex vivo seguido de reinfusión, pero este enfoque es menos que ideal y representa un arduo conjunto de problemas de control de calidad.
Por tanto, un enfoque terapéutico nuevo que restablece la tolerancia inmunitaria dominante natural mediada por los linfocitos T reguladores (Treg) y minimiza fuertemente cualquier efecto estimulador potencial sobre los linfocitos T efectores de memoria CD4+ potenciaría enormemente la capacidad para tratar pacientes con enfermedades autoinmunitarias, tales como diabetes de tipo 1 , esclerosis múltiple, lupus eritematoso diseminado, enfermedad de Crohn, así como otras enfermedades autoinmunitarias y enfermedades proinflamatorias basadas en el sistema inmunitario, tales como enfermedad injerto contra huésped crónica, asma, fibrosis pulmonar, enfermedad pulmonar obstructiva crónica, enfermedades cardiovasculares, tales como ateroesclerosis y síndrome coronario agudo, y rechazo de trasplante, tanto de víscera maciza como de médula ósea.
El documento WO 2009/135615 describe el uso de una muteína de IL-2 conocida previamente (IL-2 N88R, BAY50-4798, descrita en el documento WO 1999/60128) para el tratamiento o profilaxis de la enfermedad autoinmunitaria. Se afirma que la muteína de IL-2 tiene una actividad incrementada en los linfocitos Treg en comparación con la IL-2 natural, mientras que su efecto sobre los linfocitos T CD8+ y los linfocitos NK es pequeño. No se describe ninguna
proteína de fusión de la muteína de IL-2.
El documento WO 2010/85495 describe variantes de IL-2 que promueven selectivamente la actividad en los linfocitos Treg sobre los linfocitos T no reguladores para el tratamiento de los trastornos inflamatorios. Las variantes de IL-2 descritas comprenden una combinación de ocho o más sustituciones de aminoácido que afectan a la unión a las diferentes subunidades del receptor de IL-2.
El documento WO 2012/178137 describe proteínas de fusión inmunoglobulina-citocina. Se divulga la IL-2 como un ejemplo de citocinas. La inmunoglobulina descrita se une a un antígeno de superficie celular asociado a cáncer, y la proteína de fusión puede ser adecuada para tratar el cáncer activando una respuesta inmunitaria.
El documento WO 03/048334 describe una proteína de fusión que comprende dos variantes de IL-2. Las proteínas de fusión descritas comprenden un resto de IL-2 mutante y un resto distinto a IL-2, y son útiles para tratar el cáncer y las infecciones por virus. Se usan anticuerpos frente a diversos antígenos tumorales y víricos como un resto distinto a IL-2.
El documento WO 2012/146628 describe inmunoconjugados de IL-2 útiles para tratar el cáncer. Los inmunoconjugados descritos comprenden un resto de IL-2 única, es decir, no más de una.
Las proteínas de fusión de IL-2 de la presente invención activan preferentemente los Treg humanos y no humanos con poco o ningún efecto sobre los linfocitos T efectores de memoria CD4+ humanos, inclinando la balanza hacia una proporción Treg:Tef más alta, y reducen la respuesta autoinmunitaria. Son de larga duración, lo que permite pautas de dosificación convenientes, y están desprovistas de funciones efectoras, lo que reduce los potenciales efectos secundarios y la disminución de la eficacia.
Sumario de la invención
En un aspecto, la invención proporciona una proteína de fusión que comprende (i) una molécula de inmunoglobulina que no se puede unir específicamente a un antígeno y (ii) dos moléculas de interleucina 2 (IL-2) mutante que comprenden una mutación de aminoácido que reduce afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia en comparación con una molécula de IL-2 natural, en la que las moléculas de IL-2 mutante comprenden las sustituciones de aminoácido T3A, N88D y C125A (SEQ ID NO: 58).
En un modo de realización, dicha molécula de inmunoglobulina es una molécula de inmunoglobulina de clase IgG, en particular, una molécula de inmunoglobulina de subclase IgG1. En un modo de realización, dicha molécula de inmunoglobulina es una molécula de inmunoglobulina humana. En un modo de realización, dicha molécula de inmunoglobulina se puede unir específicamente a un antígeno. En un modo de realización, dicha molécula de inmunoglobulina es un anticuerpo monoclonal. En un modo de realización, dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena pesada basada en la secuencia de estirpe germinal de Vh3-23 humana. En un modo de realización específico, dicha molécula de inmunoglobulina comprende la secuencia de la región variable de la cadena pesada de la SEQ ID NO: 9. En un modo de realización, dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena ligera basada en la secuencia de estirpe germinal de Vk3-20 humana. En un modo de realización específico, dicha molécula de inmunoglobulina comprende la secuencia de la región variable de la cadena ligera de la SEQ ID NO: 11. En un modo de realización incluso más específico, dicha molécula de inmunoglobulina comprende la secuencia de la región variable de la cadena pesada de la SEQ ID NO: 9 y la secuencia de la región variable de la cadena ligera de la SEQ ID NO: 11. En un modo de realización, dicha molécula de inmunoglobulina no se puede unir específicamente a un antígeno, y comprende una secuencia de la región variable de la cadena pesada basada en la secuencia de estirpe germinal de Vh3-23 humana y una secuencia de la región variable de la cadena ligera basada en la secuencia de estirpe germinal de Vk3-20 humana.
En un modo de realización, dicha molécula de inmunoglobulina comprende una modificación que reduce la afinidad de unión de la molécula de inmunoglobulina con respecto a un receptor Fc en comparación con una molécula de inmunoglobulina correspondiente sin dicha modificación. En un modo de realización, dicho receptor Fc es un receptor Fcy, en particular, un receptor Fcy humano. En un modo de realización, dicho receptor Fc es un receptor Fc activador. En un modo de realización, dicho receptor Fc se selecciona del grupo de FcYRIIIa (CD16a), FcyRI (CD64), FcYRIIIa (CD32) y FcaRI (CD89). En un modo de realización específico, dicho receptor Fc es FcYRIIIa, en particular, FcYRIIIa humano. En un modo de realización, dicha modificación reduce la función efectora de la molécula de inmunoglobulina. En un modo de realización específico, dicha función efectora es la citotoxicidad celular dependiente de anticuerpos (ADCC). En un modo de realización, dicha modificación está en la región Fc, en particular, en la región CH2, de dicha molécula de inmunoglobulina. En un modo de realización, dicha molécula de inmunoglobulina comprende una sustitución de aminoácido en la posición 329 (numeración EU) de las cadenas pesadas de inmunoglobulina. En un modo de realización específico, dicha sustitución de aminoácido es P329G. En un modo de realización, dicha molécula de inmunoglobulina comprende sustituciones de aminoácido en las posiciones 234 y 235 (numeración EU) de las cadenas pesadas de inmunoglobulina. En un modo de realización específico, dichas sustituciones de aminoácido son L234A y L235A (LALA). En un modo de realización, dicha
molécula de inmunoglobulina comprende sustituciones de aminoácido en las posiciones 234, 235 y 329 (numeración EU) de las cadenas pesadas de inmunoglobulina. En un modo de realización particular, dicha molécula de inmunoglobulina comprende las sustituciones de aminoácido L234A, L235A y P329G (numeración EU) en las cadenas pesadas de inmunoglobulina.
En un modo de realización, dichas moléculas de IL-2 mutante comprenden una mutación de aminoácido en una posición correspondiente al residuo 88 de IL-2 humana (SEQ ID NO: 1); dicha mutación de aminoácido es una sustitución de aminoácido; y, dicha sustitución de aminoácido es N88D. En un modo de realización, dichas moléculas de IL-2 comprenden además una mutación de aminoácido que no altera la afinidad de unión de dichas moléculas de IL-2 con respecto a un receptor de IL-2 en comparación con la IL-2 natural. En un modo de realización, dichas moléculas de IL-2 comprenden una mutación de aminoácido en una posición correspondiente al residuo 125 de IL-2 humana; y dicha mutación de aminoácido es la sustitución de aminoácido C125A. En un modo de realización, dicha molécula de IL-2 mutante comprende además una mutación de aminoácido que elimina el sitio de O-glucosilación de IL-2 en una posición correspondiente al residuo 3 de IL-2 humana; y dicha mutación de aminoácido que elimina el sitio de O-glucosilación de IL-2 en una posición correspondiente al residuo 3 de IL-2 humana es T3A. En un modo de realización, dichas moléculas de IL-2 mutante son moléculas de IL-2 humana. En un modo de realización específico, dichas moléculas de IL-2 mutante comprenden una secuencia de la SEQ ID NO: 58. En un modo de realización, dichas moléculas de IL-2 mutante tienen una secuencia de la SEQ ID NO: 58. En un modo de realización, dichas moléculas de IL-2 mutante se fusionan cada una en su aminoácido N terminal con el aminoácido C terminal de una de las cadenas pesadas de inmunoglobulina de dicha molécula de inmunoglobulina, opcionalmente a través de un conector peptídico. En un modo de realización, dichas moléculas de IL-2 mutante se fusionan cada una con dicha molécula de inmunoglobulina a través de un conector peptídico. En un modo de realización, dicho conector peptídico comprende al menos 10, en particular, al menos 15, aminoácidos. En un modo de realización, dicho conector peptídico comprende la secuencia de aminoácidos (G4S)3 (SEQ ID NO: 66).
En un modo de realización específico, dicha proteína de fusión comprende las secuencias de polipéptido de la SEQ ID NO: 19 y la SEQ ID NO: 50. En un modo de realización específico, dicha proteína de fusión comprende una cadena ligera de inmunoglobulina de la SEQ ID NO: 19 y un polipéptido de fusión cadena pesada de inmunoglobulina-IL-2 de la SEQ ID NO: 50. En un modo de realización, dicha proteína de fusión consiste esencialmente en una molécula de inmunoglobulina, dos moléculas de interleucina 2 (IL-2) mutante que comprenden una mutación de aminoácido que reduce la afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia, en comparación con una molécula de IL-2 natural, y opcionalmente uno o más conectores peptídicos. En un modo de realización, dicha proteína de fusión consiste esencialmente en dos cadenas ligeras de inmunoglobulina de la SEQ ID NO: 19 y dos polipéptidos de fusión cadena pesada de inmunoglobulina-IL-2 de la SEQ ID NO: 50.
En un modo de realización, dicha proteína de fusión activa selectivamente los linfocitos T reguladores. En un modo de realización, dicha proteína de fusión activa selectivamente los linfocitos T reguladores sobre los linfocitos T efectores, en particular, sobre los linfocitos T CD4+ convencionales y los linfocitos T CD8+. En un modo de realización, dicha proteína de fusión activa selectivamente los linfocitos T reguladores sobre los linfocitos T de memoria CD4+ convencionales. En un modo de realización, dicha proteína de fusión activa los linfocitos T reguladores al menos 10 veces, al menos 100 veces o al menos 1000 veces más que los linfocitos T de memoria CD4+ convencionales. En un modo de realización, se determina dicha activación mediante la medición de los niveles de fosforilación de STAT intracelular, en particular, STAT5. En un modo de realización, dicha medición de los niveles de fosforilación de STAT intracelular se realiza mediante análisis por citometría de flujo.
La invención proporciona además un polinucleótido que codifica la proteína de fusión de la invención. Se proporciona además un vector, en particular, un vector de expresión, que comprende el polinucleótido de la invención. En otro aspecto, la invención proporciona una célula huésped que comprende el polinucleótido o el vector de la invención. La invención también proporciona un procedimiento para producir una proteína de fusión de la invención, que comprende las etapas de (i) cultivar la célula huésped de la invención en condiciones adecuadas para la expresión de la proteína de fusión e (i) recuperar la proteína de fusión. También se proporciona una proteína de fusión que comprende (i) una molécula de inmunoglobulina que no se puede unir específicamente a un antígeno y (ii) dos moléculas de interleucina 2 (IL-2) que comprenden una mutación de aminoácido que reduce la afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia, en comparación con una molécula de IL-2 natural, en la que las moléculas de IL-2 mutante comprenden la secuencia de la SEQ ID NO: 58, producida mediante dicho procedimiento.
En un aspecto, la invención proporciona una composición farmacéutica que comprende la proteína de fusión de la invención y un vehículo farmacéuticamente aceptable. También se proporciona la proteína de fusión o la composición farmacéutica de la invención para su uso como un medicamento y para su uso en el tratamiento o profilaxis de una enfermedad autoinmunitaria, específicamente diabetes de tipo 1, esclerosis múltiple (EM), lupus eritematoso diseminado (LED), enfermedad inflamatoria intestinal, enfermedad de Crohn o colitis ulcerosa, lo más específicamente diabetes de tipo 1, o enfermedad injerto contra huésped o rechazo de trasplante. Se proporciona además el uso de la proteína de fusión de la invención para la fabricación de un medicamento para el tratamiento de una enfermedad en un individuo que lo necesite y un procedimiento de tratamiento de una enfermedad en un
individuo, que comprende administrar a dicho individuo una cantidad terapéuticamente eficaz de una composición que comprende la proteína de fusión de la invención en una forma farmacéuticamente aceptable. En un modo de realización, dicha enfermedad es una enfermedad autoinmunitaria. En un modo de realización más específico, dicha enfermedad autoinmunitaria es diabetes de tipo 1, esclerosis múltiple (EM), lupus eritematoso diseminado (LED), enfermedad inflamatoria intestinal, enfermedad de Crohn o colitis ulcerosa. En un modo de realización incluso más específico, dicha enfermedad autoinmunitaria es diabetes de tipo 1. En otro modo de realización, dicha enfermedad es rechazo de trasplante o enfermedad injerto contra huésped. En un modo de realización, dicho individuo es un mamífero, en particular, un humano.
Se proporciona además la proteína de fusión de la invención para su uso en la activación selectiva de los linfocitos T reguladores in vitro. En un modo de realización, dicha activación comprende la inducción de la proliferación de los linfocitos T reguladores y/o la inducción de la señalización del receptor de IL-2, en particular, la fosforilación de STAT5, en los linfocitos T reguladores. En un modo de realización, dicho uso es in vitro y dicha proteína de fusión se usa a una concentración de aproximadamente 10 ng/ml o menos, en particular, aproximadamente 1 ng/ml o menos. La invención también proporciona un procedimiento para la activación selectiva de los linfocitos T reguladores in vitro, que comprende poner en contacto dichos linfocitos T reguladores con la proteína de fusión de la invención. En un modo de realización, dicha activación comprende la inducción de la proliferación de los linfocitos T reguladores y/o la inducción de la señalización del receptor de IL-2, en particular, la fosforilación de STAT5, en los linfocitos T reguladores. En un modo de realización, dicho procedimiento es in vitro y dicha proteína de fusión se usa a una concentración de aproximadamente 10 ng/ml o menos, en particular, aproximadamente 1 ng/ml o menos.
Breve descripción de los dibujos
Figura 1. Purificación de la proteína de fusión IgG DP47GS-IL-2 (véanse las SEQ ID NO 13, 15, 19). (A) Perfil de elución de la etapa de cromatografía de afinidad con proteína A. (B) Perfil de elución de la etapa de cromatografía de exclusión por tamaño. Rendimiento de 4 mg/l. (C) Electroforesis capilar analítica con SDS (Caliper) del producto final. Se observaron las siguientes bandas: no reducida - área de un 7,5 % a 111 kDa, área de un 92,5 % a 174 kDa; reducida - área de un 23,6 % a 29 kDa, área de un 23,5 % a 67 kDa, área de un 52,9 % a 82 kDa. El producto contiene aproximadamente un 7,5 % de la “mitad de IgG”. (D) Cromatografía de exclusión por tamaño analítica del producto final en una columna TSKgel G3000 SW XL (contenido en monómero de un 91 %).
Figura 2. Purificación de la proteína de fusión IgG DP47GS-(IL-2)2 (véanse las SEQ ID NO 17, 19). (A) Perfil de elución de la etapa de cromatografía de afinidad con proteína A. (B) Perfil de elución de la etapa de cromatografía de exclusión por tamaño. Rendimiento de 13 mg/l. (C) Electroforesis capilar analítica con SDS (Caliper) del producto final. Se observaron las siguientes bandas: no reducida - área de un 2,3 % a 172,5 kDa, área de un 97,7 % a 185 kDa; reducida - área de un 18,3 % a 27,3 kDa, área de un 0,6 % a 29,2 kDa, área de un 81,1 % a 78,3 kDa. (D) Cromatografía de exclusión por tamaño analítica del producto final en una columna Superdex 200 (contenido en monómero de un 100 %).
Figura 3. Purificación de la proteína de fusión IgG DP47GS-IL-2 N88D (véanse las SEQ ID NO 15, 19, 48). (A) Perfil de elución de la etapa de cromatografía de afinidad con proteína A. (B) Perfil de elución de la etapa de cromatografía de exclusión por tamaño. Rendimiento de 23,7 mg/l. Las fracciones obtenidas están dentro de un recuadro. (C) Electroforesis capilar analítica con SDS (Caliper) del producto final. Se observaron las siguientes bandas principales: no reducida - área de un 100 % a 167,0 kDa; no reducida - área de un 31,5 % a 28,6 kDa, área de un 31,7 % a 63,5 kDa, área de un 35,3 % a 77,5 kDa. (D) Cromatografía de exclusión por tamaño analítica del producto final en una columna TSKgel G3000 SW XL (contenido en monómero de un 97,3 %).
Figura 4. Purificación de la proteína de fusión IgG DP47GS-(IL-2 N88D)2 (véanse las SEQ ID NO 19 y 50). (A) Perfil de elución de la etapa de cromatografía de afinidad con proteína A. (B) Perfil de elución de la etapa de cromatografía de exclusión por tamaño. Rendimiento de 32,9 mg/l. Las fracciones obtenidas están dentro de un recuadro. (C) Electroforesis capilar analítica con SDS (Caliper) del producto final. Se observaron las siguientes bandas principales: no reducida - área de un 20,7 % a 180,4 kDa, área de un 78,0 % a 184,0 kDa; reducida - área de un 16,2 % a 27,3 kDa, área de un 82,7 % a 76,0 kDa. (D) Cromatografía de exclusión por tamaño analítica del producto final en una columna TSKgel G3000 SW XL (contenido en monómero de un 98,3 %).
Figura 5. Purificación de la proteína de fusión IgG DP47GS-(IL-2 E95A)2 (véanse las SEQ ID NO 19 y 52). (A) Perfil de elución de la etapa de cromatografía de afinidad con proteína A. (B) Perfil de elución de la etapa de cromatografía de exclusión por tamaño. Rendimiento de 8,0 mg/l. Las fracciones obtenidas están dentro de un recuadro. (C) Electroforesis capilar analítica con SDS (Caliper) del producto final. Se observaron las siguientes bandas principales: no reducida - área de un 10,3 % a 166,0 kDa, área de un 61,4 % a 175,5 kDa, área de un 28,2 % a 181,2 kDa; no reducida - área de un 16,0 % a 26,1 kDa, área de un 83,1 % a 75,0 kDa. (D) Cromatografía de exclusión por tamaño analítica del producto final en una columna TSKgel G3000 SW XL (contenido en monómero de un 100 %).
Figura 6. Expresión de CD25 (IL-2RA) y CD122 (IL-2RB) en subconjuntos de Treg CD4+, subconjuntos de linfocitos NK y linfocitos NKT. Se usaron marcadores de superficie de las células para definir subconjuntos de Treg CD4+,
linfocitos NKT y linfocitos NK. A fin de optimizar la tinción para CD25 y CD122, no se realizó ninguna tinción para FOXP3 intracelular. (A, B) Tres poblaciones de linfocitos T CD4+ reguladores (Treg): indiferenciados (CD45RA+, CD25+; línea de puntos), de memoria (CD45RA- , CD25+; línea continua) y activados (CD45RA- , CD25a; línea discontinua). (C, D) NKT (línea de puntos), linfocitos NK CD56fuerte (línea discontinua), linfocitos NK CD56intermedia (línea continua). Gris: control de isotipo (CI).
Figura 7. Expresión de CD25 (IL-2RA) y CD122 (IL-2RB) en subconjuntos de linfocitos T convencionales CD4+ y CD8+. Se usaron marcadores de superficie de las células para definir linfocitos T CD4+ convencionales, indiferenciados (CD45RA+ ; línea de puntos) y de memoria (CD45RA"; línea continua) (A, B), linfocitos T CD8+ convencionales de memoria (CD45RA"; línea continua) y linfocitos T CD8 CD45RA+ (una combinación de los subconjuntos indiferenciados y TEMRA; TEMRA se refiere a las células de memoria efectora que han vuelto a expresar CD45RA; línea de puntos) (C, D). Gris: control de isotipo (CI).
Figura 8. Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana en respuesta a IgG DP47GS-IL-2. Se evaluaron tres donantes humanos diferentes (C4 a C6) en tiempos separados para determinar los efectos de diversas dosis de IgG DP47GS-IL-2 sobre la inducción de la fosforilación de STAT5a. Los resultados se muestran para los subconjuntos de Treg CD4+: Treg activados, de memoria e indiferenciados; linfocitos T efectores de memoria CD4+ convencionales; linfocitos NK CD56fuerte; linfocitos T efectores de memoria CD8+; linfocitos T efectores indiferenciados CD4+; linfocitos NK; linfocitos NKT; y linfocitos T efectores indiferenciados CD8+ + de memoria CD45RA+ .
Figura 9. Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana en respuesta a IgG DP47GS-(IL-2)2. Se evaluaron cinco donantes humanos diferentes (N1, N2, C4-C6) en tiempos separados para determinar los efectos de diversas dosis del inmunoconjugado IgG DP47GS-(IL-2)2 sobre la inducción de la fosforilación de STAT5a. Los resultados se muestran para los subconjuntos de Treg CD4+: Treg activados, de memoria e indiferenciados; linfocitos T efectores de memoria CD4+ convencionales; linfocitos NK CD56fuerte; linfocitos T efectores de memoria CD8+ ; linfocitos T efectores indiferenciados CD4+ ; linfocitos NK; linfocitos NKT; y linfocitos T efectores indiferenciados CD8+ + de memoria CD45RA+ .
Figura 10. Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana: comparación de IgG DP47GS-IL-2 e IgG DP47GS-(IL-2)2. Los resultados para cada subconjunto de células se normalizaron al efecto máximo observado de cada subconjunto y las CE50 aproximadas para los Treg se presentan en la tabla 2. (A) Resultados normalizados para IgG DP47GS-IL-2, (B) resultados normalizados para IgG DP47GS-(IL-2)2.
Figura 11. Examen detallado de la sensibilidad del subconjunto de Treg en tres donantes comparando IgG DP47GS-IL-2 e IgG DP47GS-(IL-2)2. Los gráficos representan la media ± DE de la IMF de pSTAT5a para los tres donantes. (A) Treg CD3+CD4+FoxP3+ totales. (B) Treg activados. (C) Treg de memoria. (D) Treg indiferenciados.
Figura 12. Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana en respuesta a IgG DP47GS-(IL-2E95A)2. Se evaluaron tres donantes humanos diferentes (C4 a C6) en tiempos separados para determinar los efectos de diversas dosis de IgG DP47GS-(IL-2E95A)2 sobre la inducción de la fosforilación de STAT5a. Los resultados se muestran para los subconjuntos de Treg CD4+ : Treg activados, de memoria e indiferenciados; linfocitos T efectores de memoria CD4+ convencionales; linfocitos NK CD56fuerte ; linfocitos T efectores de memoria CD8+; linfocitos T efectores indiferenciados CD4+ ; linfocitos NK; linfocitos NKT; y linfocitos T efectores indiferenciados CD8+CD45RA+.
Figura 13. Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana en respuesta a IgG DP47GS-(IL-2N88D)2. Se evaluaron cinco donantes humanos diferentes (C4, C5, C6, N1, N2) en tiempos separados para determinar los efectos de diversas dosis de IgG DP47GS-(IL-2N88D)2 sobre la inducción de la fosforilación de STAT5a. Los resultados se muestran para los subconjuntos de Treg CD4+ : Treg activados, de memoria e indiferenciados; linfocitos T efectores de memoria CD4+ convencionales; linfocitos NK CD56fuerte ; linfocitos T efectores de memoria CD8+; linfocitos T efectores indiferenciados CD4+ ; linfocitos NK; linfocitos NKT; y linfocitos T efectores indiferenciados CD8+CD45RA+.
Figura 14. Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana: comparación de IgG DP47GS-IL-2, IgG DP47GS-(IL-2)2, IgG DP47GS-(IL-2E95A)2, IgG DP47GS-(IL-2 N88D)2 e IgG DP47GS-IL-2N88D. Los resultados se muestran para subconjuntos de Treg CD4+ (A-C): Treg activados (A), de memoria (C) e indiferenciados (B); linfocitos T efectores de memoria CD4+ convencionales (D); linfocitos NK CD56fuerte (E).
Figura 15. Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana en respuesta tanto a IgG DP47GS-(IL-2N88D)2 como a IgG DP47GS-(IL-2)2. Se evaluaron diez donantes humanos diferentes en días separados para determinar los efectos de un amplio intervalo de dosis (>6 log) de IgG DP47GS-(IL-2N88D)2 e IgG d P47GS-(iL-2)2 sobre la inducción de la fosforilación de STAT5a. Los resultados se muestran para los siguientes subconjuntos de células: (A) Treg de memoria, (B) Treg indiferenciados, (C) linfocitos NK CD56fuerte, (D) linfocitos NK totales, (E) linfocitos T CD4+ de memoria central, (F) linfocitos T CD4+ de memoria efectora, (G) linfocitos T CD4+ indiferenciados, (H) linfocitos T CD8+ de memoria central, (I) linfocitos T CD8+ de memoria efectora (J) linfocitos T
CD8+ indiferenciados, (K) linfocitos T CD8+ Temra [Tef de memoria RA+], (L) linfocitos NKT y (M) linfocitos T CD3+CD4' CD8' CD56' . Todos los resultados se muestran como la media ± EEM (n=10 donantes).
Figura 16. Comparación de los efectos in vitro de IgG DP47GS-(IL-2N88D)2 e IgG DP47GS-(IL-2)2 sobre los linfocitos NK y los linfocitos T CD8+ humanos. Se cultivaron PBMC de donantes humanos normales (n=10) a 5x106 células/pocillo de fondo en U con 50 ng/ml de IgG DP47GS-(IL-2N88D)2 (símbolos abiertos) o IgG DP47GS-(IL-2)2 (símbolos sombreados en gris) durante 6 días, momento en el que se cuantificaron números de células mediante citometría de flujo. Los efectos sobre los linfocitos NK se cuantificaron en linfocitos NK CD56debil y CD56fuerte (A). Los efectos sobre los linfocitos T CD8+ se muestran en (B). Los resultados se muestran como la mediana ± amplitud intercuartílica, las diferencias estadísticas se determinaron usando una prueba de la U de Mann-Whitney.
Figura 17. Comparación de los efectos in vivo de IgG DP47GS-(IL-2N88D)2 e IgG DP47GS-(IL-2)2 sobre ratones humanizados con células madre CD34+ . De diez a 12 semanas después del injerto de células madre CD34+ , los ratones se trataron dos veces por semana con vehículo (IgG DP47GS, sin IL-2), IgG DP47GS-(IL-2N88D)2 o IgG DP47GS-(IL-2)2. Se observaron incrementos máximos de Treg y linfocitos NK humanos en la sangre después de 3 tratamientos y los resultados se muestran en (A) para los Treg y (B) para los linfocitos NK. Los resultados se muestran como la mediana ± amplitud intercuartílica; vehículo (n=21), IgG-(IL-2N88D)2 (n=22) e IgG-(IL-2)2 (n=24).
Figura 18. Comparación de los efectos in vivo de IgG DP47GS-(IL-2N88D)2 e IgG DP47GS-(IL-2)2 sobre la supervivencia de ratones humanizados con células madre CD34+ . De diez a 12 semanas después del injerto de células madre CD34+ , los ratones se trataron dos veces por semana con vehículo (IgG DP47GS, sin IL-2), IgG DP47GS-(IL-2N88D)2 o IgG DP47GS-(IL-2)2 hasta que la gravedad de la respuesta del injerto contra huésped xenogénico humano alcanzó una etapa predeterminada (>15 % de pérdida de peso), lo que requirió su retirada del estudio. Los resultados se representan como una curva de supervivencia de Kaplan-Meier de GraphPad Prism para el vehículo (n=7), IgG DP47GS-(IL-2N88D)2 (n=12) e IgG DP47GS-(IL-2)2 (n=13).
Figura 19. Comparación de los efectos in vivo de IgG DP47GS-(IL-2N88D)2 e IgG DP47GS-IL-2)2 sobre los Treg, linfocitos NK y linfocitos T CD8+ en ratones humanizados con células madre CD34+ . De diez a 12 semanas después del injerto de células madre CD34+ , los ratones se trataron dos veces por semana con vehículo (IgG DP47GS, sin IL-2), IgG DP47GS-(IL-2N88D)2 o bien IgG DP47GS-(IL-2)2 hasta que la gravedad de la respuesta del injerto contra huésped xenogénico humano alcanzó una etapa predeterminada (>15 % de pérdida de peso), lo que requirió su retirada del estudio, momento en el que se evaluó la sangre para determinar los subconjuntos de células humanas individuales. Se muestran los Treg humanos, los linfocitos NK y las células CD8+ en la sangre como el % de células CD45+ humanas. Se muestran los resultados como la media ± el EEM para vehículo (n=6), IgG DP47GS-(IL-2N88D)2 (n=9) o IgG DP47GS-(IL-2)2 (n=9).
Figura 20. Inducción de pSTAT5a en los Treg en la sangre de macaco cangrejero en respuesta a IgG DP47GS-(IL-2)2 e IgG DP47GS-(IL-2N88D)2. Se evaluaron tres donantes sanos normales (C1-C3) al mismo tiempo para determinar los efectos de una dosis máxima (20 ng/ml) de IgG DP47GS-(IL-2)2 e IgG DP47g S-(IL-2N88D)2 sobre la inducción de la fosforilación de STAT5a en los Treg. (A) La estrategia de selección de la citometría de flujo con FOXP3 (eje y) y CD45RA (eje x) usada para identificar los subconjuntos de Treg CD4+CD25+: Treg indiferenciados (CD45RA+FOXP3+), Treg de memoria (CD45RA-FOXP3+) y activados (CD45RA-FOXP3a ). (B) La expresión de la tinción de la superficie de las células CD25+ en los subconjuntos de Treg antes de la estimulación. (C) Respuestas de pSTAT5a para los subconjuntos de Treg CD4+CD25+ : activado, de memoria e indiferenciado para cada uno de los tres macacos cangrejeros (C1-C3).
Figura 21. Activación de pSTAT5a en linfocitos T efectores de memoria CD4+ convencionales en la sangre de macaco cangrejero en respuesta a IgG DP47GS-(IL-2)2 o IgG DP47GS-(IL-2N88D)2. Usando las mismas muestras de sangre estimuladas con 20 ng/ml de los tres donantes mono descritos en la figura 11 se examinó la inducción de pSTAT5a en linfocitos T efectores de memoria CD4+ convencionales. (A) La estrategia de selección de la citometría de flujo usada para identificar linfocitos T efectores de memoria CD4+FOXP3' CD45RA' convencionales con CD45RA en el eje y c D25 en el eje x. (B) Respuestas de pSTAT5a para linfocitos T efectores de memoria CD4+FOXP3-CD45RA- totales. (C) Respuestas de pSTAT5a para linfocitos T efectores de memoria que también eran CD25-. (C) Respuestas de pSTAT5a para linfocitos T efectores de memoria que también eran CD25+.
Figura 22. Inducción de pSTAT5a en subconjuntos de linfocitos T de sangre periférica de macaco cangrejero en respuesta a IgG DP47GS-(IL-2)2 e IgG Dp47GS-(IL-2N88D)2. Se evaluaron dos donantes adultos sanos para determinar los efectos de dosis variables (0,03-300 ng/ml) de IgG DP47GS-(IL-2)2 e IgG DP47GS-(IL-2N88D)2 sobre la inducción de la fosforilación de STAT5a. Los resultados se muestran para los subconjuntos de Treg CD4+ (A-C): Treg activados (A), de memoria (C) e indiferenciados (B) y linfocitos T efectores de memoria CD4+ convencionales (D). Se obtuvieron resultados similares para ambos monos y se mostraron los resultados de un donante para ilustrar los efectos de IgG DP47GS-(IL-2)2 e IgG DP47GS-(IL-2N88D)2.
Figura 23. En ratones, IgG DP47GS-IL-2 tiene propiedades farmacocinéticas (PF) superiores en comparación con IgG DP47GS-(IL-2)2, mientras que IgG DP47g S-(iL-2N88D)2 es intermedio. (A) A ratones NOD y NOD.scid se les inyectaron por vía intravenosa (i.v.) 0,3 mg/kg de IgG DP47GS-IL-2 o 0,3 mg/kg de IgG DP47g S-(IL-2)2. (B) A
ratones NOD.sád.IL2RaI~ se les inyectaron i.v. 0,1 mg/kg de IgG DP47GS-IL-2, 0,3 mg/kg de IgG DP47GS-(IL-2)2 o 0,1 mg/kg de IgG DP47GS-(IL-2N88D)2. Se evaluó la IL-2 humana en muestras de suero en los tiempos indicados mediante un ensayo de captura basado en mAb.
Figura 24. Tanto CD25 como FoxP3 se incrementan en los Treg murinos después del tratamiento con Proleukin® , IgG DP47GS-IL-2, IgG DP47GS-(IL-2 )2 e IgG DP47GS-(IL-2N88D)2. Se trataron ratones BALB/c con Proleukin® (20.000 y 100.000 UI/ratón, n=3), IgG DP47GS-IL-2, IgG DP47GS-(IL-2 )2 o IgG DP47GS-(IL-2N88D)2 (60, 300 o 1.500 UI/ratón, n=3), los ratones tratados con vehículo se incluyeron como controles sin estimular (n=4). Veinticuatro horas después del tratamiento, se evaluaron los Treg esplénicos para determinar las concentraciones de CD25 y FOXP3. (A) Se observaron cambios dependientes de la dosis en CD25 en la superficie de las células y (B) cambios dependientes de la dosis en FoxP3 intracelular para las cuatro moléculas de IL2. Los datos se representan como la media ± DE, las barras en negro representan la dosis más alta usada y las barras en gris claro la dosis más baja.
Figura 25. Se evaluaron las propiedades PF de IgG DP47GS-IL-2 e IgG DP47GS-(IL-2)2 en macacos cangrejeros adultos sanos normales sin tratamiento previo con biofármaco. (A) Se inyectó por vía intravenosa (i.v.) IgG DP47GS-IL-2 como una corta inyección intravenosa rápida en dosis de 10, 25 y 100 pg/kg (n=2 por dosis). (B) Se inyectó i.v. IgG DP47GS-(IL-2)2 como una corta inyección intravenosa rápida en dosis de 10 y 25 pg/kg (n=2 por dosis). Se evaluó la IL-2 humana en muestras de suero en los tiempos indicados mediante un ensayo de captura basado en mAb.
Figura 26. IgG DP47GS-IL2 tiene un efecto dependiente de la dosis en macacos cangrejeros, incrementando los linfocitos T reguladores. Los cambios en los linfocitos T reguladores (Treg) CD4+ CD25+ FOXP3+ en la sangre completa en el día 7 posterior al tratamiento se muestran como (A) el número de células absoluto de Treg por mm3 de sangre completa y (B) la tasa de cambio en los Treg. Todos los datos se representan como la media ± DE. Barras abiertas: IgG DP47GS-IL-2 (n=6); barras sombreadas: vehículo (n=3).
Figura 27. Efectos del tiempo y dosis bajas de IgG DP47GS-IL-2 sobre los Treg en macacos cangrejeros. (A) Cambios dependientes del tiempo en la tasa de incremento en los Treg tras la dosificación con 2 o 6 pg/kg de IgG DP47GS-IL-2 (n=4 y 6, respectivamente). (B) Cambios dependientes del tiempo en los recuentos absolutos de Treg en la sangre después de una dosificación con 2 o 6 pg/kg de IgG DP47GS-IL-2 (n=4 y 6, respectivamente). Todos los datos se muestran como la media ± DE.
Figura 28. Proleukin en dosis única estimula una respuesta Treg dependiente de la dosis transitoria en macacos cangrejeros. (A) Cambios en los números de Treg de sangre periférica después del tratamiento de dosis única con de 3x104 a 3x105 UI/kg de Proleukin. (B) Cambios en pSTAT5a en los Treg después del tratamiento de dosis única con de 3x104 a 3x105 UI/kg de Proleukin. Los datos se muestran como la media ± DE.
Figura 29. IgG DP47GS-IL-2 en dosis baja es más eficaz que Proleukin® en dosis alta en la inducción de Treg en macacos cangrejeros. Se trataron macacos cangrejeros sanos normales (grupos de n=5) con IgG DP47GS-IL-2 en dosis baja o Proleukin en dosis alta y se sometió a prueba el cambio en los linfocitos T reguladores el día 10. En los días 0 y 7, se administró s.c. IgG DP47GS-IL-2 a una dosis de 16.800 UI/kg (12 pg/kg). Se administró s.c. un tratamiento con Proleukin (200.000 UI/kg) 3 veces por semana (LMV) durante un total de 5 dosis. Se muestran los resultados como media ± DE para (A) el cambio en los Treg totales por mm3 de sangre, (B) la tasa de incremento en los Treg y (C) el cambio en la proporción de Treg con respecto a las células CD4+ FOXP3- convencionales. Las barras abiertas representan el tratamiento con IL-2 y las barras sombreadas el control con vehículo.
Figura 30. STAT5 fosforilado de sangre completa ex vivo es un biomarcador sensible para la activación de los Treg con P47GS IgG-IL-2 in vivo. Uno y 3 días después de la administración in vivo de una dosis baja única de IgG DP47GS-IL-2 (12 pg/kg) a macacos cangrejeros sanos (n=5), se obtuvo sangre completa y se sometido a prueba inmediatamente sin estimulación para STAT5 fosforilado (pSTAT5a). Se extrajo sangre de cada mono en el día 0 antes del tratamiento y se midió la cantidad de pSTAT5a (barras sombreadas) y se usó individualmente para evaluar las tasas de cambio posteriores al tratamiento (barras abiertas). (A) Se muestra la tasa de cambio en pSTAT5a en los Treg en los días 1 y 3, (B) la tasa de cambio en pSTAT5a en los linfocitos T de memoria CD4+ CD45-convencionales y (C) la tasa de cambio en pSTAT5a en los linfocitos T indiferenciados. Los datos se muestran como la media ± DE.
Figura 31. PSTAT5a de sangre completa ex vivo como biomarcador sensible para la activación in vivo de Treg con IgG DP47GS-IL-2 en dosis baja. De uno a siete días después de la administración in vivo de una dosis baja única de IgG DP47GS-IL-2 a macacos cangrejeros sanos, se obtuvo sangre completa y se sometió a prueba inmediatamente sin estimulación para pSTAT5a. Se obtuvo sangre de cada mono en el día 0 antes del tratamiento para determinar las concentraciones sin estimular de pSTAT5a y se compararon con los cambios en pSTAT5a posteriores al tratamiento. (A) pSTAT5a en los Treg antes y después del tratamiento con 2 pg/kg de IgG DP47GS-IL-2 (n=4). (B) pSTAT5a en los Treg antes y después del tratamiento con 6 pg/kg de IgG DP47GS-IL-2 (n=6). Los datos se muestran como la media ± DE.
Figura 32. La Ki-67 de sangre completa de macaco cangrejero ex vivo sirve como un marcador para la proliferación
de linfocitos T inducida por IgG DP47GS-IL-2 in vivo. Los macacos cangrejeros tratados con 2 y 6 pg/kg de IgG DP47GS-IL-2 como se describe en la figura 14 también se supervisaron ex vivo para determinar los cambios en el marcador intracelular Ki-67 para evaluar el grado de proliferación in vivo. Se cuantificó el porcentaje de estado estable normal de células en el ciclo celular (Ki-67+) en el día 0 antes del tratamiento y, entonces, se supervisó diariamente durante los siguientes de 7 a 11 días. (A) Se muestran el % de Ki-67+ para Treg, (B) el % de Ki-67+ para linfocitos T de memoria/efectores CD4+CD45' convencionales y (C) el % de Ki-67+ para linfocitos T CD4+CD45RA+ indiferenciados. Los datos se muestran como la media ± DE.
Figura 33. Efectos del tiempo y dosis bajas de IgG DP47GS-(IL-2)2 sobre los Treg en macacos cangrejeros sanos sin tratamiento previo. (A) Cambios dependientes del tiempo en los números absolutos de Treg después de 6 pg/kg de IgG DP47-(IL-2)2. (B) Cambios dependientes del tiempo en pSTAT5a en los Treg después del tratamiento, (C) cambios dependientes del tiempo en la tasa de incremento en los Treg y (D) comparación de las tasas de cambio en los Treg de monos tratados con IgG DP47GS-IL-2 (barras abiertas, de 2 a 36 pg/kg) frente a los tratados con IgG DP47GS-(IL-2 )2 (barra sombreada, 6 pg/kg). Todos los datos se muestran como la media ± DE (n=4 a 6).
Figura 34. Efectos dependientes del tiempo y de la dosis de IgG DP47GS-(IL-2 )2 en dosis muy baja sobre los Treg en macacos cangrejeros sanos sin tratamiento previo. (A) Cambios dependientes del tiempo en las tasas de incremento de los Treg tras la administración de 0,7 y 2 pg/kg de IgG DP47GS-(IL-2)2. (B) Cambios dependientes del tiempo en pSTAT5a en los Treg medidos en el día 0 antes del tratamiento y en los días 1-4 después del tratamiento, (C) cambios dependientes del tiempo en pSTAT5a en linfocitos Tef/de mem. Todos los datos se muestran como la media ± d E (n=3 a 0,7 pg/kg y n=8 a 2 pg/kg).
Figura 35. Comparación dependiente de la dosis de IgG DP47GS-IL-2 frente a IgG DP47GS-(IL-2)2 y su capacidad para incrementar los Treg de macaco cangrejero en la sangre completa. Todos los datos se presentan como la media ± DE (n=3 a 6).
Figura 36. Se usó secuenciación de nueva generación (SNG) para analizar la desmetilación del ADN específica de subconjuntos de linfocitos T para el factor de transcripción FOXP3 y la molécula inmunodepresora CTLA-4. Se analizó la sangre de macacos cangrejeros adultos sin tratamiento previo con biofármaco antes y después del tratamiento con una dosis óptima de IgG DP47GS-IL-2 (25 pg/kg, n=4) e IgG DP47GS-(IL-2)2 (6 pg/kg, n=4). Se clasificaron subconjuntos de linfocitos T CD4+ con un BD FACSAria de PBMC de sangre completa y se usaron 100.000 células por subconjunto para la desmetilación del ADN específica de gen. Se observaron resultados similares con ambos tratamientos y los datos de los monos tratados con IgG DP47GS-(IL-2)2 se muestran para FOXP3 en el panel superior y para CTLA-4 en el panel inferior (n=4, media ± DE).
Figura 37. Se evaluaron las propiedades PF de IgG DP47GS-(IL-2N88D)2 en macacos cangrejeros sanos normales sin tratamiento previo con biofármaco. (A) Se inyectó por vía intravenosa (i.v.) IgG DP47GS-(IL-2N88D)2 en una corta inyección intravenosa rápida en dosis de 30 y 100 pg/kg (n=2 por dosis). (B) Se inyectó por vía subcutánea (s.c.) IgG DP47GS-(IL-2N88D)2 en 0,2 ml en el costado lateral en dosis de 30 y 100 pg/kg (n=2 por dosis). Se evaluó la IL-2 humana en muestras de plasma en los tiempos indicados con un ensayo de captura basado en anticuerpos monoclonales. (C) Como un biomarcador de la exposición a IL-2, se midió el CD25 soluble en plasma después de la inyección de IgG-(IL-2N88D)2 en dosis de 30 y 100 pg/kg; se midió el sCD25 de macaco cangrejero en un ensayo de captura basado en mAb para sCD25 humano que se sabe que reacciona de forma cruzada con sCD25 de macaco cangrejero. Los resultados se muestran como la media ± EEM (n=4).
Figura 38. Efectos in vivo dependientes del tiempo y de la dosis de IgG DP47GS-(IL-2N88D)2 sobre los Treg en macacos cangrejeros sanos sin tratamiento previo. Cambios dependientes del tiempo en el número absoluto (x106/ml de sangre) de Treg totales, Treg de memoria CD4+ , Treg indiferenciados CD4+ y Treg CD8+FOXP3+ tras la inyección de (A) 100 pg/kg de IgG DP47-(IL-2N88D)2 o (B) 30 pg/kg de IgG DP47-(IL-2N88D)2. Cambios dependientes del tiempo en los Treg como el % de linfocitos T CD4+ o CD8+ para Treg totales, Treg de memoria c D4+, Treg indiferenciados CD4+ y Treg CD8+FOXP3+ tras la inyección de 100 pg/kg de IgG DP47-(IL-2N88D)2 (C) o 30 pg/kg de IgG DP47-(IL-2N88D)2 (D). Los resultados se muestran como la media ± EEM (n=4).
Figura 39. Efectos in vivo dependientes del tiempo y de la dosis de IgG DP47GS-(IL-2N88D)2 sobre los linfocitos en macacos cangrejeros sanos sin tratamiento previo. Los cambios dependientes del tiempo en los linfocitos T efectores de memoria CD4+ se muestran como el % de linfocitos T CD4+ totales tras la inyección de 100 pg/kg de IgG DP47-(IL-2N88D)2 o 30 pg/kg de IgG DP47-(IL-2N88D)2 (A). Los cambios dependientes del tiempo en los linfocitos T de memoria CD4+CD25a efectores se muestran como el % de linfocitos T de memoria CD4+ efectores tras la inyección de 100 pg/kg de IgG DP47-(IL-2N88D)2 o 30 pg/kg de IgG DP47-(IL-2N88D)2 (B). Los resultados se muestran como la media ± EEM (n=4).
Figura 40. El pSTAT5a de sangre completa ex vivo es un biomarcador sensible para la activación de IL-2 in vivo. De 1 a 11 días después de la administración in vivo de una dosis única de IgG DP47GS-(IL-2N88D)2 a macacos cangrejeros, se obtuvo sangre completa y se analizó inmediatamente sin estimulación para pSTAT5a. Se extrajo sangre de cada mono en el día 0 antes del tratamiento para determinar las concentraciones sin estimular de pSTAT5a y se compararon con los cambios en pSTAT5a posteriores al tratamiento. (A) pSTAT5a (IMF, intensidad
media de fluorescencia máxima) antes y después del tratamiento con 100 pg/kg de IgG DP47GS-(IL-2N88D)2, n=4. (B) pSTAT5a antes y después del tratamiento con 30 pg/kg de IgG DP47GS-(IL-2N88D)2, n=4. Los datos se muestran como la media ± EEM para los subconjuntos de células indicados.
Figura 41. La tinción para CD25 en la superficie de las células de sangre completa ex vivo es un biomarcador sensible para la activación de IL-2 in vivo. De 1 a 14 días después de la administración in vivo de dosis únicas de IgG DP47GS-(IL-2N88D)2 a macacos cangrejeros, se obtuvo sangre completa y se sometió a prueba inmediatamente sin estimulación para CD25 de superficie en los tipos de células indicados. Se obtuvo sangre de cada mono en el día 0 antes del tratamiento para determinar las concentraciones sin estimular de CD25 y se compararon con los cambios en CD25 posteriores al tratamiento. (A) Tinción para CD25 (IMF máxima, intensidad media de fluorescencia) antes y después del tratamiento con 100 pg/kg de IgG DP47GS-(IL-2N88D)2, n=4. (B) Tinción para CD25 antes y después del tratamiento con 30 pg/kg de IgG DP47GS-(IL-2N88D)2, n=4. Los datos se muestran como la media ± EEM para los subconjuntos de células indicados.
Figura 42. La tinción in vivo para Ki-67 intracelular de sangre completa es un biomarcador sensible para la proliferación celular después de la activación de IL-2 in vivo. De 1 a 14 días después de la administración in vivo de una dosis única de IgG DP47GS-(IL-2N88D)2 a macacos cangrejeros, se obtuvo sangre completa y se sometió a prueba inmediatamente sin estimulación para determinar la Ki-67 intracelular en los tipos de células indicados. Se obtuvo sangre de cada mono en el día 0 antes del tratamiento para determinar las concentraciones sin estimular de células Ki-67+ y se compararon con los cambios en el % de células Ki-67+ posteriores al tratamiento. (A) Tinción para Ki-67 (% de células Ki-67+) antes y después del tratamiento con 100 pg/kg de IgG DP47GS-(IL-2N88d )2, n=4. (B) Tinción para Ki-67 (% de células Ki-67+) antes y después del tratamiento con 30 pg/kg de IgG Dp47GS-(IL-2N88D)2, n=4. Los resultados se muestran como la media ± e Em para los subconjuntos de células indicados.
Figura 43. Un resumen comparativo de los efectos in vivo de IL-2 sobre los Treg de macaco cangrejero. Se muestra el incremento máximo en los Treg totales como el % de linfocitos T CD4+ para Proleukin, IgG-IL-2, IgG-(IL-2)2 e IgG-(IL-2N88D)2; para la comparación, todas las dosis in vivo se convirtieron a pmol/kg. Se administró Proleukin 3 veces por semana (LMV) durante 2 semanas y se administraron las proteínas de fusión de IL-2 como una inyección única. Los resultados se muestran como la media ± EEM; Proleukin (n=5), IgG-IL-2 (n=6), IgG-(IL-2 )2 (n=6) e IgG-(IL-2N88D)2 (n=4).
Figura 44. La IL-2 en dosis alta induce eosinofilia en macacos cangrejeros. Se supervisaron los cambios en los recuentos de eosinófilos en la sangre en todas las pruebas con Proleukin o los inmunoconjugados de IL-2. Se detectó eosinofilia de 7 a 14 días después del tratamiento con Proleukin en dosis alta (2x105 UI/kg 3 veces por semana durante 2 semanas) o las dosis más altas de IgG DP47GS-IL-2 pero no con IgG DP47GS-(IL-2)2 o IgG DP47GS-(IL-2N88D)2. Los recuentos de eosinófilos en el valor de referencia son los círculos abiertos mostrados como “0” para la dosis de IL-2 y se realizaron dos veces, una vez para las pruebas con la molécula de IL-2 natural y una vez para las pruebas con IgG DP47GS-(IL-2N88D)2. Los datos se muestran como la media ± DE.
Figura 45. El tratamiento in vivo con IgG DP47GS-IL-2 suprime la hipersensibilidad de tipo retardado (DTH) de los eritrocitos de carnero murinos. Todos los datos se muestran para ratones individuales tratados con vehículo (100 % de respuesta), IgG-IL-2 (4.000 UI/ratón) o CTLA-4-Ig de ratón como control positivo (200 pg/ratón). (A) Resultados de ratones NOD y (B) ratones C57BL/6. La magnitud de las respuestas de DTH del día cuatro se muestra como el cambio en el peso de la pata en comparación con los ratones no inmunizados (A peso de la pata). Los datos se muestran como la media ± DE.
Figura 46. El tratamiento in vivo con IgG DP47GS-IL-2 (4.000 UI/ratón) suprime las respuestas de anticuerpo IgG murina con respecto a KLH en (A) ratones C57BL/621 días después de la inmunización y (B) ratones NOD siete días después de la inmunización. Los datos se muestran como la media ± DE.
Descripción detallada de la invención
Definiciones
Se usan los términos en el presente documento como se usa en general en la técnica, a menos que se defina de otro modo en lo que sigue.
Como se usa en el presente documento, el término “proteína de fusión” se refiere a una molécula de polipéptido de fusión que comprende una molécula de inmunoglobulina y una molécula de IL-2, en la que los componentes de la proteína de fusión se enlazan entre sí mediante enlaces peptídicos, directamente o bien a través de conectores peptídicos. Por claridad, las cadenas peptídicas individuales del componente de inmunoglobulina de la proteína de fusión se pueden enlazar no covalentemente, por ejemplo, mediante enlaces disulfuro.
“Fusionados” se refiere a los componentes que se enlazan mediante enlaces peptídicos, directamente o bien por medio de uno o más conectores peptídicos.
Por “unión específica” se entiende que la unión es selectiva para el antígeno y se puede discriminar de interacciones no deseadas o no específicas. Se puede medir la capacidad de una inmunoglobulina para unirse a un antígeno específico a través de un enzimoinmunoanálisis de adsorción (ELISA) o bien de otras técnicas consabidas para un experto en la técnica, por ejemplo, la técnica de resonancia de plasmón superficial (RPS) (analizada en un instrumento BIAcore) (Liljeblad et al., Glyco J 17, 323-329 (2000)) y ensayos de unión tradicionales (Heeley, Endocr Res 28, 217-229 (2002)). En un modo de realización, el grado de unión de una inmunoglobulina a una proteína no relacionada es de menos de aproximadamente un 10 % de la unión de la inmunoglobulina al antígeno como se mide, por ejemplo, mediante RPS. En determinados modos de realización, una inmunoglobulina que se une al antígeno tiene una constante de disociación (Kd ) de < 1 pM, < 100 nM, < 10 nM, < 1 nM, < 0,1 nM, < 0,01 nM o < 0,001 nM (por ejemplo, 10-8 M o menos, por ejemplo, de 10-8 M a 10-13 M, por ejemplo, de 10-9 M a 10-13 M).
“Afinidad” o “afinidad de unión” se refiere a la fuerza de la suma total de las interacciones no covalentes entre un sitio de unión único de una molécula (por ejemplo, un anticuerpo) y su compañero de unión (por ejemplo, un antígeno). A menos que se indique de otro modo, como se usa en el presente documento, “afinidad de unión” se refiere a la afinidad de unión intrínseca que refleja una interacción 1:1 entre los miembros de un par de unión (por ejemplo, anticuerpo y antígeno). La afinidad de una molécula X por su compañera Y se puede representar en general mediante la constante de disociación (KD), que es la proporción de las constantes de velocidad de disociación y asociación (kdis y kas, respectivamente). Por tanto, las afinidades equivalentes pueden comprender diferentes constantes de velocidad, siempre que la proporción de las constantes de velocidad permanezca igual. Se puede medir la afinidad mediante procedimientos comunes conocidos en la técnica, incluyendo los descritos en el presente documento. Un procedimiento particular para medir la afinidad es la resonancia de plasmón superficial (RPS).
“Unión reducida”, por ejemplo, unión reducida a un receptor Fc o a un receptor de IL-2, se refiere a una disminución de la afinidad por la interacción respectiva, como se mide, por ejemplo, mediante RPS. Por claridad, el término también incluye la reducción de la afinidad a cero (o por debajo del límite de detección del procedimiento analítico), es decir, la anulación completa de la interacción. Por el contrario, “unión incrementada” se refiere a un incremento de la afinidad de unión por la interacción respectiva.
Como se usa en el presente documento, el término “determinante antigénico” es sinónimo de “antígeno” y se refiere a un sitio (por ejemplo, un tramo contiguo de aminoácidos o una configuración conformacional compuesta de diferentes regiones de aminoácidos no contiguos) en una macromolécula polipeptídica a la que se une un anticuerpo, formando un complejo antígeno-anticuerpo. Se pueden encontrar determinantes antigénicos útiles, por ejemplo, en las superficies de las células, libres en suero sanguíneo y/o en la matriz extracelular (MEC).
Como se usa en el presente documento, el término “monocatenaria” se refiere a una molécula que comprende monómeros de aminoácido linealmente enlazados mediante enlaces peptídicos.
En el presente documento se usa el término “anticuerpo” en el sentido más amplio y engloba diversas estructuras de anticuerpo, que incluyen, pero no se limitan a, anticuerpos monoclonales, anticuerpos policlonales, anticuerpos multiespecíficos (por ejemplo, anticuerpos biespecíficos) y fragmentos de anticuerpo siempre que presenten la actividad de unión a antígeno deseada.
Un “fragmento de anticuerpo” se refiere a una molécula distinta de un anticuerpo intacto que comprende una porción de un anticuerpo intacto que se une al antígeno al que se une el anticuerpo intacto. Los ejemplos de fragmentos de anticuerpo incluyen, pero no se limitan a, Fv, Fab, Fab', Fab'-SH, F(ab')2 , diacuerpos, anticuerpos lineales, moléculas de anticuerpo monocatenario (por ejemplo, scFv) y anticuerpos de dominio único.
El término “molécula de inmunoglobulina” se refiere a una proteína que tiene la estructura de un anticuerpo natural. Por ejemplo, las inmunoglobulinas de la clase IgG son glucoproteínas heterotetraméricas de aproximadamente 150.000 dalton, compuestas por dos cadenas ligeras y dos cadenas pesadas que se unen con enlaces disulfuro. Desde el extremo N al C, cada cadena pesada de inmunoglobulina tiene una región variable (VH), también llamada dominio pesado variable o dominio variable de la cadena pesada, seguida de tres dominios constantes (CH1, CH2 y CH3), también llamados región constante de la cadena pesada. De forma similar, desde el extremo N al C, cada cadena ligera de inmunoglobulina tiene una región variable (VL), también llamada dominio ligero variable o dominio variable de la cadena ligera, seguida de un dominio ligero constante (CL), también llamado región constante de la cadena ligera. Se puede asignar la cadena pesada de una inmunoglobulina a una de cinco clases, llamadas a (IgA), ó (IgD), £ (IgE), y (IgG) o p (IgM), algunas de las cuales se pueden dividir además en subclases, por ejemplo, Y1 (IgG1), Y2 (IgG2), Y3 (IgG3), Y4 (IgG4), a1 (IgA1) y a2 (IgA2). Se puede asignar la cadena ligera de una inmunoglobulina a uno de dos tipos, llamados kappa (k) y lambda (A), en base a la secuencia de aminoácidos de su dominio constante. Una inmunoglobulina consiste esencialmente en dos moléculas Fab y un dominio Fc, enlazados por medio de la región bisagra de inmunoglobulina.
Como se usa en el presente documento, “fragmento Fab” se refiere a un fragmento de inmunoglobulina que comprende un dominio VL y un dominio constante de una cadena ligera (CL) y un dominio VH y un primer dominio constante (CH1) de una cadena pesada.
La “clase” de un anticuerpo o inmunoglobulina se refiere al tipo de dominio constante o región constante que posee su cadena pesada. Existen cinco clases principales de inmunoglobulinas: IgA, IgD, IgE, IgG e IgM, y varias de estas se pueden dividir adicionalmente en subclases (isotipos), por ejemplo, IgG1, IgG2, IgG3, IgG4, IgA1 e IgA2. Los dominios constantes de la cadena pesada que se corresponden con las diferentes clases de inmunoglobulinas se llaman a, ó, £, y y H, respectivamente.
El término “región variable” o “dominio variable” se refiere al dominio de una cadena pesada o ligera de inmunoglobulina o anticuerpo que, en general, está implicada en la unión de la inmunoglobulina o anticuerpo al antígeno. Sin embargo, la inmunoglobulina comprendida en la proteína de fusión de la presente invención puede comprender regiones variables que no confieren especificidad de unión a antígeno. Los dominios variables de la cadena pesada y de la cadena ligera (VH y VL, respectivamente) de una inmunoglobulina o anticuerpo, en general, tienen estructuras similares, comprendiendo cada dominio cuatro regiones estructurales (FR) conservadas y tres regiones hipervariables (HVR). Véase, por ejemplo, Kindt et al., Kuby Immunology, 6.a ed., W.H. Freeman y Co., página 91 (2007). Un dominio VH o VL único puede ser suficiente para conferir especificidad de unión a antígeno.
El término “región hipervariable” o “HVR”, como se usa en el presente documento, se refiere a cada una de las regiones de un dominio variable de inmunoglobulina o anticuerpo que son hipervariables en secuencia y/o forman bucles estructuralmente definidos (“bucles hipervariables”). En general, los anticuerpos tetracatenarios naturales comprenden seis HVR; tres en el VH (H1, H2, H3) y tres en el VL (L1, L2, L3). Las HVR comprenden, en general, residuos de aminoácido de los bucles hipervariables y/o de las “regiones determinantes de la complementariedad” (CDR), siendo las últimas las de variabilidad de secuencia más alta y/o estando implicadas en el reconocimiento antigénico. Los bucles hipervariables ejemplares se producen en los residuos de aminoácido 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (H2), y 96-101 (H3) (Chothia y Lesk, J. Mol. Biol. 196, 901-917 (1987)). Se producen CDR (Cd R-L1, CDR-L2, CDR-L3, c Dr -H1, c Dr -H2 y CDR-H3) ejemplares en los residuos de aminoácido 24-34 de L1, 50-56 de L2, 89-97 de L3, 31-35B de H1, 50-65 de H2 y 95-102 de H3 (Rabat et al, Sequences of Proteins of Immunological Interest, 5.a ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Con la excepción de CDR1 en VH, las CDR comprenden, en general, los residuos de aminoácido que forman los bucles hipervariables. Las CDR también comprenden “residuos determinantes de la especificidad” o “SDR”, que son los residuos que entran en contacto con el antígeno. Los SDR están contenidos en regiones de las CDR, llamadas CDR abreviadas o a-CDR. Se producen a-CDR (a-CDR-L1, a-CDR-L2, a-CDR-L3, a-CDR-H1, a-CDR-H2 y a-CDR-H3) ejemplares en los residuos de aminoácido 31-34 de L1, 50-55 de L2, 89-96 de L3, 31-35B de H1, 50-58 de H2 y 95 102 de H3 (véase Almagro y Fransson, Front. Biosci. 13, 1619-1633 (2008)). A menos que se indique de otro modo, los residuos de HVR y otros residuos en el dominio variable (por ejemplo, los residuos de FR) se numeran en el presente documento de acuerdo con Rabat et al., supra (denominado “numeración de Kabat”).
“Región estructural” o “FR” se refiere a residuos del dominio variable distintos de los residuos de la región hipervariable (HVR). La FR de un dominio variable consiste, en general, en dominios de cuatro FR: FR1, FR2, FR3 y FR4. En consecuencia, las secuencias de HVR y FR aparecen, en general, en la siguiente secuencia en VH (o VL): FR1-H1(L1 )-FR2-H2(L2)-FR3-H3(L3)-FR4.
Una “inmunoglobulina humana” es una que posee una secuencia de aminoácidos que se corresponde con la de una inmunoglobulina producida por un ser humano o una célula humana o derivada de una fuente no humana que utiliza repertorios de inmunoglobulina humana u otras secuencias que codifican inmunoglobulina humana. Esta definición de una inmunoglobulina humana excluye específicamente una inmunoglobulina humanizada que comprende residuos de unión a antígeno no humanos.
El término “anticuerpo monoclonal” como se usa en el presente documento se refiere a un anticuerpo obtenido a partir de una población de anticuerpos sustancialmente homogéneos, es decir, los anticuerpos individuales que comprenden la población son idénticos y/o se unen al mismo epítopo, excepto por posibles anticuerpos variantes, por ejemplo, que contienen mutaciones naturales o surgen durante la producción de una preparación de anticuerpos monoclonales, estando presentes, en general, dichas variantes en cantidades insignificantes. A diferencia de las preparaciones de anticuerpos policlonales, que típicamente incluyen diferentes anticuerpos dirigidos frente a diferentes determinantes (epítopos), cada anticuerpo monoclonal de una preparación de anticuerpos monoclonales se dirige frente a un determinante único en un antígeno. Por tanto, el modificador “monoclonal” indica el carácter del anticuerpo por haberse obtenido a partir de una población sustancialmente homogénea de anticuerpos, y no se debe interpretar como que requiere la producción del anticuerpo mediante ningún procedimiento particular. Por ejemplo, los anticuerpos monoclonales que se van a usar de acuerdo con la presente invención se pueden preparar mediante una variedad de técnicas, que incluyen, pero no se limitan al procedimiento de hibridoma, procedimientos de ADN recombinante, procedimientos de presentación en fagos y procedimientos que utilizan animales transgénicos que contienen todos o parte de los locus de inmunoglobulina humana; estando descritos en el presente documento dichos procedimientos y otros procedimientos ejemplares para preparar anticuerpos monoclonales.
En el presente documento se usa el término “dominio Fc” o “región Fc” para definir una región C terminal de una cadena pesada de inmunoglobulina que contiene al menos una porción de la región constante. El término incluye regiones Fc de secuencia natural y regiones Fc variantes. Una región Fc de IgG comprende un dominio CH2 de IgG
y un dominio CH3 de IgG. El “dominio CH2” de una región Fc de IgG humana se extiende normalmente desde un residuo de aminoácido en aproximadamente la posición 231 hasta un residuo de aminoácido en aproximadamente la posición 340. En un modo de realización, una cadena glucídica se une al dominio CH2. El dominio CH2 en el presente documento puede ser un dominio CH2 de secuencia natural o un dominio CH2 variante. El “dominio CH3” comprende el tramo de residuos C terminales con respecto a un dominio CH2 en una región Fc (es decir, desde un residuo de aminoácido en aproximadamente la posición 341 hasta un residuo de aminoácido en aproximadamente la posición 447 de una IgG). La región CH3 en el presente documento puede ser un dominio CH3 de secuencia natural o un dominio CH3 variante (por ejemplo, un dominio CH3 con una “protuberancia” (“botón”) introducida en una cadena del mismo y una “cavidad” (“ojal”) introducida correspondiente en la otra cadena del mismo; véase la patente de EE. UU. n.° 5.821.333). Se pueden usar dichos dominios CH3 variantes para promover la heterodimerización de dos cadenas pesadas de inmunoglobulina no idénticas como se describe en el presente documento. En un modo de realización, una región Fc de la cadena pesada de IgG humana se extiende desde Cys226, o desde Pro230, al extremo carboxílico de la cadena pesada. Sin embargo, la lisina C terminal (Lys447) de la región Fc puede estar presente o no. A menos que se especifique de otro modo en el presente documento, la numeración de los residuos de aminoácido en la región Fc o región constante es de acuerdo con el sistema de numeración EU, también llamado índice EU, como se describe en Kabat et al., Sequences of Proteins of Immunological Interest, 5.a ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
El término “funciones efectoras” se refiere a las actividades biológicas atribuibles a la región Fc de una inmunoglobulina, que varían con el isotipo de inmunoglobulina. Los ejemplos de funciones efectoras de inmunoglobulina incluyen: unión a C1q y citotoxicidad dependiente del complemento (CDC), unión al receptor Fc, citotoxicidad celular dependiente de anticuerpos (ADCC), fagocitosis celular dependiente de anticuerpos (ADCP), secreción de citocinas, captación de antígenos mediada por complejos inmunitarios mediante células presentadoras de antígeno, regulación por disminución de receptores de superficie de las células (por ejemplo, receptor de linfocitos B) y activación de linfocitos B.
Un “receptor Fc activador” es un receptor Fc que tras el acoplamiento por una región Fc de una inmunoglobulina provoca acontecimientos de señalización que estimulan que la célula que contiene el receptor realice funciones efectoras. Los receptores Fc activadores incluyen FcYRIIIa (cD16a), FcyRI (CD64), FcYRIIa (CD32) y FcaRI (CD89). Un receptor Fc activador particular es FcYRIlla humano (véase el n.° de acceso a UniProt P08637 (versión 141)).
El término “interleucina 2” o “IL-2” como se usa en el presente documento, se refiere a cualquier IL-2 natural de cualquier fuente de vertebrado, incluyendo mamíferos, tales como primates (por ejemplo, seres humanos) y roedores (por ejemplo, ratones y ratas), a menos que se indique de otro modo. El término engloba IL-2 sin procesar, así como cualquier forma de IL-2 que resulte del procesamiento en la célula. El término también engloba variantes naturales de IL-2, por ejemplo, variantes de empalme o variantes alélicas. La secuencia de aminoácidos de una IL-2 humana ejemplar se muestra en la SEQ ID NO: 1. La IL-2 humana sin procesar comprende además un péptido señal de 20 aminoácidos N terminal, que está ausente en la molécula de IL-2 madura.
Por “ IL-2 natural” se entiende una IL-2 natural. La secuencia de una molécula de IL-2 humana natural se muestra en la SEQ ID NO: 1. Para el propósito de la presente invención, el término natural también engloba formas de IL-2 que comprenden una o más mutaciones de aminoácido que no alteran la unión al receptor de IL-2 en comparación con la IL-2 natural, tal como, por ejemplo, una sustitución de cisteína en una posición correspondiente al residuo 125 de IL-2 humana con alanina. En algunos modos de realización, la IL-2 natural, para el propósito de la presente invención, comprende la sustitución de aminoácido C125A (véase la SEQ ID NO: 3).
El término “CD25” o “a del receptor de IL-2”, como se usa en el presente documento, se refiere a cualquier CD25 natural de cualquier fuente de vertebrado, incluyendo mamíferos, tales como primates (por ejemplo, seres humanos) y roedores (por ejemplo, ratones y ratas), a menos que se indique de otro modo. El término engloba CD25 sin procesar “de longitud completa”, así como cualquier forma de CD25 que resulte del procesamiento en la célula. El término también engloba variantes naturales de CD25, por ejemplo, variantes de empalme o variantes alélicas. En determinados modos de realización, CD25 es CD25 humano. La secuencia de aminoácidos de un CD25 humano ejemplar (con la secuencia señal, marca Avi y marca His) se muestra en la SEQ ID NO: 25.
El término “receptor de IL-2 de afinidad alta” como se usa en el presente documento, se refiere a la forma heterotrimérica del receptor de IL-2, que consiste en la subunidad y del receptor (también conocida como subunidad Y del receptor de citocina común, Yc, o CD132), la subunidad p del receptor (también conocida como CD122 o p70) y la subunidad a del receptor (también conocida como CD25 o p55). El término “receptor de IL-2 de afinidad intermedia” o “pY del receptor de IL-2”, en cambio, se refiere al receptor de IL-2 que incluye solo la subunidad Y y la subunidad p, sin la subunidad a (para una revisión, véase, por ejemplo, Olejniczak y Kasprzak, Med Sci Monit 14, RA179-189 (2008)). Las secuencias de aminoácidos de CD122 y CD132 humanos ejemplares (fusionadas a una región Fc con una marca His) se muestran en las SEQ ID NO 21 y 23, respectivamente.
Por “linfocito T regulador” o “linfocito Treg” se entiende un tipo especializado de linfocito T CD4+ que puede suprimir las respuestas de otros linfocitos T (linfocitos T efectores). Los linfocitos Treg se caracterizan por la expresión de CD4, la subunidad a del receptor de IL-2 (CD25) y el factor de transcripción con dominio “forkhead” P3 (FOXP3)
(Sakaguchi, Annu Rev Immunol 22, 531-62 (2004)) y desempeñan un papel crítico en la inducción y el mantenimiento de la autotolerancia periférica a antígenos, incluyendo los expresados por tumores.
Por “linfocitos T CD4+ convencionales” se entiende linfocitos T CD4+ distintos de los linfocitos T reguladores. Los linfocitos T de memoria CD4+ convencionales se caracterizan por la expresión de CD4, CD3, pero no FOXP3. Los “linfocitos T de memoria CD4+ convencionales” son un subconjunto de linfocitos T c D4+ convencionales, caracterizados además por la falta de expresión de CD45RA, a diferencia de los “linfocitos T indiferenciados CD4+ convencionales” que sí expresan CD45RA.
Por “activación selectiva de linfocitos Treg” se entiende la activación de los linfocitos Treg esencialmente sin la activación simultánea de otros subconjuntos de linfocitos T (tales como linfocitos T cooperadores CD4+, linfocitos T citotóxicos CD8+, linfocitos T NK) o linfocitos citolíticos naturales (NK). Los procedimientos para identificar y distinguir estos tipos de células se describen en los ejemplos. La activación puede incluir la inducción de la señalización del receptor de IL-2 (como se mide, por ejemplo, mediante detección de STAT5a fosforilado), la inducción de la proliferación (como se mide, por ejemplo, mediante detección de Ki-67) y/o la regulación por incremento de la expresión de marcadores de activación (tales como, por ejemplo, CD25).
El término “conector peptídico” se refiere a un péptido que comprende uno o más aminoácidos, típicamente aproximadamente 2-20 aminoácidos. Los conectores peptídicos se conocen en la técnica o se describen en el presente documento. Los péptidos de conector no inmunogénicos adecuados incluyen, por ejemplo, los conectores peptídicos (G4S)n, (SG4)n o G4(SG4)n, “n” es, en general, un número entre 1 y 10, típicamente entre 2 y 4.
El término “modificación” se refiere a cualquier manipulación del esqueleto peptídico (por ejemplo, secuencia de aminoácidos) o a las modificaciones postraduccionales (por ejemplo, glucosilación) de un polipéptido.
Una “modificación botón en ojal” se refiere a una modificación en la interfase entre dos cadenas pesadas de inmunoglobulina en el dominio CH3, en la que i) en el dominio CH3 de una cadena pesada, se reemplaza un residuo de aminoácido por un residuo de aminoácido que tiene un volumen de cadena lateral más grande, generando de este modo una protuberancia (“botón”) en la interfase en el dominio CH3 de una cadena pesada que se puede posicionar en una cavidad (“ojal”) en la interfase en el dominio CH3 de la otra cadena pesada y ii) en el dominio CH3 de la otra cadena pesada, se reemplaza un residuo de aminoácido por un residuo de aminoácido que tiene un volumen de cadena lateral más pequeño, generando de este modo una cavidad (“ojal”) en la interfase en el segundo dominio CH3 en la que se puede posicionar una protuberancia (“botón”) en la interfase en el primer dominio CH3. En un modo de realización, la “modificación botón en ojal” comprende la sustitución de aminoácido T366W y opcionalmente la sustitución de aminoácido S354C en una de las cadenas pesadas de anticuerpo y las sustituciones de aminoácido T366S, L368A, Y407V y opcionalmente Y349C en la otra de las cadenas pesadas de anticuerpo. La tecnología de botón en ojal se describe, por ejemplo, en los documentos US 5.731.168; US 7.695.936; Ridgway et al., Prot Eng 9, 617-621 (1996) y Carter, J Immunol Meth 248, 7-15 (2001). En general, el procedimiento implica introducir una protuberancia (“botón”) en la interfase de un primer polipéptido y una cavidad correspondiente (“ojal”) en la interfase de un segundo polipéptido, de tal manera que la protuberancia se pueda posicionar en la cavidad para promover la formación de heterodímeros y dificultar la formación de homodímeros. Las protuberancias se construyen reemplazando cadenas laterales de aminoácido pequeñas de la interfase del primer polipéptido por cadenas laterales más grandes (por ejemplo, tirosina o triptófano). Se crean cavidades compensadoras de tamaño idéntico o similar a las protuberancias en la interfase del segundo polipéptido reemplazando cadenas laterales de aminoácido grandes por otras más pequeñas (por ejemplo, alanina o treonina). La introducción de dos residuos de cisteína en la posición S354 e Y349, respectivamente, da como resultado la formación de un puente disulfuro entre las dos cadenas pesadas de anticuerpo en la región Fc, estabilizando además el dímero (Carter, J Immunol Methods 248, 7-15 (2001)).
Una “sustitución” de aminoácido se refiere al reemplazo en un polipéptido de un aminoácido por otro aminoácido. En un modo de realización, un aminoácido se reemplaza por otro aminoácido que tiene propiedades estructurales y/o químicas similares, por ejemplo, reemplazos de aminoácido conservadores. Se pueden realizar sustituciones de aminoácido "conservadoras", por ejemplo, en base a la similitud de polaridad, carga, solubilidad, hidrofobia, hidrofilia y/o la naturaleza anfipática de los residuos implicados. Por ejemplo, los aminoácidos no polares (hidrófobos) incluyen alanina, leucina, isoleucina, valina, prolina, fenilalanina, triptófano y metionina; los aminoácidos neutros polares incluyen glicina, serina, treonina, cisteína, tirosina, asparagina y glutamina; los aminoácidos cargados positivamente (básicos) incluyen arginina, lisina e histidina; y los aminoácidos cargados negativamente (ácidos) incluyen ácido aspártico y ácido glutámico. Las sustituciones no conservadoras conllevarán el intercambio de un miembro de una de estas clases por otra clase. Por ejemplo, las sustituciones de aminoácido también pueden dar como resultado el reemplazo de un aminoácido por otro aminoácido que tenga diferentes propiedades estructurales y/o químicas, por ejemplo, reemplazando un aminoácido de un grupo (por ejemplo, polar) por otro aminoácido de un grupo diferente (por ejemplo, básico). Se pueden generar mutaciones de aminoácido usando procedimientos genéticos o químicos bien conocidos en la técnica. Los procedimientos genéticos pueden incluir mutagénesis dirigida a sitio, PCR, síntesis génica y similares. Se contempla que también pueden ser útiles procedimientos de alteración del grupo de cadena lateral de un aminoácido mediante procedimientos distintos de genomanipulación, tales como modificación química. Se pueden usar diversas designaciones en el presente documento para indicar la
misma sustitución de aminoácido. Por ejemplo, se puede indicar una sustitución de prolina en la posición 329 de la cadena pesada de inmunoglobulina con glicina como 329G, G329, G329, P329G o Pro329Gly.
“Porcentaje (%) de identidad de secuencia de aminoácidos” con respecto a una secuencia de polipéptido de referencia se define como el porcentaje de residuos de aminoácido de una secuencia candidata que son idénticos a los residuos de aminoácido en la secuencia de polipéptido de referencia, después de alinear las secuencias e introducir huecos, si fuera necesario, para lograr el máximo porcentaje de identidad de secuencia, y sin tener en consideración ninguna sustitución conservadora como parte de la identidad de secuencia. Se puede lograr la alineación para los propósitos de determinación del porcentaje de identidad de secuencia de aminoácidos de diversas maneras que se encuentran dentro de la experiencia en la técnica, por ejemplo, usando un programa informático disponible públicamente, tal como el programa informático BLAST, BLAST-2, ALIGN o Megalign (DNASTAR). Los expertos en la técnica pueden determinar los parámetros apropiados para alinear secuencias, incluyendo cualquier algoritmo necesario para lograr una alineación máxima sobre la longitud completa de las secuencias que se comparan. Para los propósitos en el presente documento, sin embargo, se generan los valores del % de identidad de secuencia de aminoácidos usando el programa de ordenador de comparación de secuencias ALIGN-2. El programa de ordenador de comparación de secuencias ALIGN-2 se creó por Genentech, Inc., y el código fuente se ha presentado con la documentación de usuario en la Oficina de Derechos de Autor de EE. UU., Washington DC, 20559, donde se ha registrado con el n.° de registro de derechos de autor de EE. UU. TXU510087. El programa ALIGN-2 está disponible públicamente de Genentech, Inc., South San Francisco, California, o se puede compilar a partir del código fuente. El programa ALIGN-2 se debe compilar para su uso en un sistema operativo UNIX, incluyendo UNIX V4.0D digital. Todos los parámetros de comparación de secuencias se establecen por el programa ALIGN-2 y no varían. En situaciones en las que se emplea ALIGN-2 para las comparaciones de secuencias de aminoácidos, el % de identidad de secuencia de aminoácidos de una secuencia de aminoácidos dada A con respecto a, con o frente a una secuencia de aminoácidos dada B (que, de forma alternativa, se puede parafrasear como una secuencia de aminoácidos dada A que tiene o comprende un determinado % de identidad de secuencia de aminoácidos con respecto a, con o frente a una secuencia de aminoácidos dada B) se calcula como sigue:
100 veces la fracción X/Y
donde X es el número de residuos de aminoácido puntuados como emparejamientos idénticos mediante el programa de alineación de secuencias ALIGN-2 en esa alineación del programa de A y B, y donde Y es el número total de residuos de aminoácido en B. Se apreciará que si la longitud de la secuencia de aminoácidos A no es igual a la longitud de la secuencia de aminoácidos B, el % de identidad de secuencia de aminoácidos de A con respecto a B no igualará el % de identidad de secuencia de aminoácidos de B con respecto a A. A menos que se indique específicamente de otro modo, todos los valores del % de identidad de secuencia de aminoácidos usados en el presente documento se obtienen como se describe en el párrafo inmediatamente precedente usando el programa de ordenador ALIGN-2.
“Polinucleótido” o “ácido nucleico”, como se usa indistintamente en el presente documento, se refiere a polímeros de nucleótidos de cualquier longitud, e incluyen ADN y ARN. Los nucleótidos pueden ser desoxirribonucleótidos, ribonucleótidos, bases o nucleótidos modificados, y/o sus análogos, o cualquier sustrato que se pueda incorporar en un polímero mediante ADN o ARN polimerasa, o mediante una reacción sintética. Un polinucleótido puede comprender nucleótidos modificados, tales como nucleótidos metilados y sus análogos. Se puede interrumpir una secuencia de nucleótidos mediante componentes distintos a nucleótidos. Un polinucleótido puede comprender una modificación/modificaciones realizada(s) después de la síntesis, tales como su conjugación con un marcador.
Por un ácido nucleico o polinucleótido que tiene una secuencia de nucleótidos al menos, por ejemplo, un 95 % “idéntica” a una secuencia de nucleótidos de referencia de la presente invención, se pretende que la secuencia de nucleótidos del polinucleótido sea idéntica a la secuencia de referencia excepto porque la secuencia de polinucleótido pueda incluir hasta cinco mutaciones puntuales por cada 100 nucleótidos de la secuencia de nucleótidos de referencia. En otras palabras, para obtener un polinucleótido que tenga una secuencia de nucleótidos al menos un 95 % idéntica a una secuencia de nucleótidos de referencia, hasta un 5 % de los nucleótidos en la secuencia de referencia se puede delecionar o sustituir con otro nucleótido, o un número de nucleótidos hasta un 5 % de los nucleótidos totales en la secuencia de referencia se puede insertar en la secuencia de referencia. Estas alteraciones de la secuencia de referencia se pueden producir en las posiciones terminales 5' o 3' de la secuencia de nucleótidos de referencia o en cualquier parte entre esas posiciones terminales, intercaladas individualmente entre residuos en la secuencia de referencia o bien en uno o más grupos contiguos en de la secuencia de referencia. Como una cuestión práctica, si cualquier secuencia de polinucleótido particular es al menos un 80 %, 85 %, 90 %, 95 %, 96 %, 97 %, 98 % o 99 % idéntica a una secuencia de nucleótidos de la presente invención, se puede determinar convencionalmente usando programas de ordenador conocidos, tales como los analizados anteriormente para los polipéptidos (por ejemplo, ALIGN-2).
El término “vector”, como se usa en el presente documento, se refiere a una molécula de ácido nucleico que puede propagar otro ácido nucleico al que se enlaza. El término incluye el vector como una estructura de ácido nucleico autorreplicante, así como el vector incorporado en el genoma de una célula huésped en la que se ha introducido.
Determinados vectores pueden dirigir la expresión de los ácidos nucleicos a los que se enlazan funcionalmente. Dichos vectores se denominan en el presente documento “vectores de expresión”.
Los términos “célula huésped”, “línea de células huésped” y “cultivo de células huésped” se usan indistintamente y se refieren a células en las que se ha introducido ácido nucleico exógeno, incluyendo la descendencia de dichas células. Las células huésped incluyen “transformantes” y “células transformadas”, que incluyen la célula transformada primaria y la descendencia derivada de la misma independientemente del número de pases. La descendencia puede que no sea completamente idéntica en el contenido de ácido nucleico a una célula original, pero puede contener mutaciones. La descendencia mutante que tiene la misma función o actividad biológica que la cribada o seleccionada en la célula transformada originalmente se incluye en el presente documento. Una célula huésped es cualquier tipo de sistema celular que se puede usar para generar las proteínas de fusión de la presente invención. Las células huésped incluyen células cultivadas, por ejemplo, células cultivadas de mamífero, tales como células CHO, células BHK, células NS0, células SP2/0, células de mieloma YO, células de mieloma de ratón P3X63, células PER, células PER.C6 o células de hibridoma, células de levadura, células de insecto y células vegetales, por nombrar solo unas pocas, pero también células comprendidas en un animal transgénico, planta transgénica o tejido vegetal o animal cultivado.
Una “cantidad eficaz” de un agente se refiere a la cantidad que es necesaria para dar como resultado un cambio fisiológico en la célula o tejido al que se administra.
Una “cantidad terapéuticamente eficaz” de un agente, por ejemplo, una composición farmacéutica, se refiere a una cantidad eficaz, en dosificaciones y durante periodos de tiempo necesarios, para lograr el resultado terapéutico o profiláctico deseado. Una cantidad terapéuticamente eficaz de un agente, por ejemplo, elimina, disminuye, retarda, minimiza o previene los efectos adversos de una enfermedad.
Un “individuo” o “sujeto” es un mamífero. Los mamíferos incluyen, pero no se limitan a, animales domesticados (por ejemplo, vacas, ovejas, gatos, perros y caballos), primates (por ejemplo, seres humanos y primates no humanos, tales como monos), conejos y roedores (por ejemplo, ratones y ratas). En particular, el individuo o sujeto es un ser humano.
La expresión "composición farmacéutica" se refiere a una preparación que está en tal forma que permite que la actividad biológica de un ingrediente activo contenido en la misma sea eficaz, y que no contiene ningún componente adicional que sea inaceptablemente tóxico para un sujeto al que se administrará la formulación.
Un “vehículo farmacéuticamente aceptable” se refiere a un ingrediente en una composición farmacéutica, distinto de un ingrediente activo, que no sea tóxico para un sujeto. Un vehículo farmacéuticamente aceptable incluye, pero no se limita a, un tampón, excipiente, estabilizante o conservante.
Como se usa en el presente documento, “tratamiento” (y las variaciones gramaticales del mismo, tales como “tratar” o “que trata”) se refiere a la intervención clínica en un intento de alterar la evolución natural de una enfermedad en el individuo que se está tratando, y que se puede realizar por profilaxis o bien durante la evolución de una patología clínica. Los efectos deseables del tratamiento incluyen, pero no se limitan a, prevención de la aparición o recidiva de la enfermedad, alivio de los síntomas, disminución de cualquier consecuencia patológica directa o indirecta de la enfermedad, prevención de las metástasis, disminución de la tasa de progresión de la enfermedad, mejora o atenuación de la enfermedad y remisión o pronóstico mejorado. En algunos modos de realización, se usan los anticuerpos de la invención para retardar el desarrollo de una enfermedad o para ralentizar la progresión de una enfermedad.
“Enfermedad autoinmunitaria” se refiere a una enfermedad o trastorno no maligno que surge desde y se dirige frente a los tejidos de un individuo. Los ejemplos de enfermedades o trastornos autoinmunitarios incluyen, pero no se limitan a, respuestas inflamatorias, tales como dermatosis inflamatorias, incluyendo psoriasis y dermatitis (por ejemplo, dermatitis atópica); respuestas asociadas con una enfermedad inflamatoria intestinal (tal como enfermedad de Crohn y colitis ulcerosa); dermatitis; afecciones alérgicas, tales como eccema y asma; artritis reumatoide; lupus eritematoso diseminado (LED) (incluyendo, pero sin limitarse a, nefritis lúpica, lupus cutáneo); diabetes mellitus (por ejemplo, diabetes mellitus de tipo 1 o diabetes mellitus insulinodependiente); esclerosis múltiple y diabetes juvenil.
Proteínas de fusión de la invención
La invención proporciona proteínas de fusión inmunoglobulina-IL-2 novedosas con propiedades, en particular, ventajosas para el uso en procedimientos terapéuticos como se describe en el presente documento.
En un primer aspecto, la invención proporciona una proteína de fusión que comprende (i) una molécula de inmunoglobulina que no se puede unir específicamente a un antígeno y (ii) dos moléculas de interleucina 2 (IL-2) mutante que comprenden una mutación de aminoácido que reduce afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia, en comparación con una molécula de IL-2 natural, en la que las moléculas de IL-2 mutante comprenden la secuencia de la SEQ ID NO: 58.
En un modo de realización, dicha proteína de fusión consiste esencialmente en una molécula de inmunoglobulina que no se puede unir específicamente a un antígeno, dos moléculas de interleucina 2 (IL-2) mutante que comprenden una mutación de aminoácido que reduce la afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia, en comparación con una molécula de IL-2 natural, en la que las moléculas de iL-2 mutante comprenden la secuencia de la SEQ ID NO: 58, y opcionalmente uno o más conectores peptídicos.
Como se muestra en los ejemplos, una proteína de fusión que comprende dos moléculas de IL-2 proporciona sorprendentemente una eficacia y selectividad enormemente mejoradas en la activación de los linfocitos T reguladores en comparación con una proteína de fusión correspondiente que comprende una molécula de IL-2 única. Además, solo una proteína de fusión que comprende dos moléculas de IL-2 mutante (en lugar de solo una) con unión reducida al receptor de IL-2 de afinidad intermedia retiene una actividad estimuladora significativa en los linfocitos T reguladores.
En un modo de realización, dicha molécula de inmunoglobulina es una molécula de inmunoglobulina de clase IgG, en particular, una molécula de inmunoglobulina de subclase IgG1. En un modo de realización, dicha molécula de inmunoglobulina es una molécula de inmunoglobulina humana, es decir, comprende regiones variables y constantes completamente humanas. La secuencia de una región constante de IgG1 humana ejemplar se muestra en la SEQ ID NO: 8. Una molécula de inmunoglobulina de clase IgG comprende (i) dos cadenas ligeras de inmunoglobulina, comprendiendo cada una desde el extremo N al C un dominio variable de la cadena ligera (VL) y un dominio constante de la cadena ligera (CL) y (ii) dos cadenas pesadas de inmunoglobulina, comprendiendo cada una desde el extremo N al extremo C un dominio variable de la cadena pesada (VH), un dominio constante de la cadena pesada (CH)1, una región bisagra de inmunoglobulina, un dominio CH2 y un dominio CH3. Los dos últimos dominios forman parte de la región Fc de la molécula de inmunoglobulina. Las dos cadenas pesadas se dimerizan en la región Fc.
En un modo de realización de la proteína de fusión de acuerdo con la invención, dichas dos moléculas de IL-2 mutante se fusionan cada una en su aminoácido N terminal con el aminoácido C terminal de una de las cadenas pesadas de inmunoglobulina de dicha molécula de inmunoglobulina, opcionalmente a través de un conector peptídico. La fusión de dos moléculas de IL-2 (idénticas) con las cadenas pesadas de inmunoglobulina permite la producción simple de la proteína de fusión, evitando la formación de productos secundarios no deseados y obviando la necesidad de realizar modificaciones que promuevan la heterodimerización de cadenas pesadas no idénticas, tal como una modificación botón en ojal.
En determinados modos de realización de la proteína de fusión de acuerdo con la invención, dichas dos moléculas de IL-2 mutante se fusionan con dicha molécula de inmunoglobulina a través de un conector peptídico. En un modo de realización, dichas dos moléculas de IL-2 mutante se fusionan cada una con dicha molécula de inmunoglobulina a través de un conector peptídico. En un modo de realización, dichas dos moléculas de IL-2 mutante se fusionan cada una en su aminoácido N terminal con el aminoácido C terminal de una de las cadenas pesadas de inmunoglobulina de dicha molécula de inmunoglobulina, a través de un conector peptídico. En un modo de realización, cada una de dichas moléculas de IL-2 mutante se fusiona con dicha molécula de inmunoglobulina a través de un conector peptídico que tiene una secuencia de aminoácidos idéntica. En un modo de realización, dicho conector peptídico comprende al menos 10 aminoácidos. En un modo de realización particular, dicho conector peptídico comprende al menos 15 aminoácidos. Sin pretender vincularse a ninguna teoría, un conector peptídico de esta longitud puede proporcionar la flexibilidad para la unión óptima de las moléculas de IL-2 mutante al receptor de IL-2, en particular, el receptor de IL-2 de afinidad alta (heterotrimérico). En un modo de realización específico, dicho conector peptídico comprende 15 aminoácidos. En un modo de realización incluso más específico, dicho conector peptídico comprende la secuencia de aminoácidos (G4S)3 (SEQ ID NO: 66). En un modo de realización, dicho conector peptídico tiene una longitud de 15 aminoácidos. En un modo de realización, dicho conector peptídico tiene la secuencia de aminoácidos (G4S)3 (SEQ ID NO: 66). En un modo de realización, dicho conector peptídico consiste en 15 aminoácidos. En un modo de realización, dicho conector peptídico consiste en la secuencia de aminoácidos (G4S)3 (SEQ ID NO: 66).
La fusión de las moléculas de IL-2 con una molécula de inmunoglobulina proporciona propiedades farmacocinéticas favorables, incluyendo una larga semivida en suero (debido al reciclado a través de la unión a FcRn y estando el tamaño molecular muy por encima del umbral para la filtración renal) en comparación con la IL-2 libre (sin fusionar). Además, la presencia de una molécula de inmunoglobulina también posibilita la purificación simple de proteínas de fusión, por ejemplo, mediante cromatografía de afinidad con proteína A. De forma interesante, como se muestra en los ejemplos, una proteína de fusión que comprende dos moléculas de IL-2 mutante con unión reducida al receptor de IL-2 de afinidad intermedia tiene una semivida en suero más larga que una proteína de fusión correspondiente que comprende dos moléculas de IL-2 natural. La fusión con una molécula de inmunoglobulina, es decir, un tipo de molécula natural, también puede minimizar la toxicidad de la proteína de fusión a través de la formación de anticuerpos dirigidos frente a fármacos.
Mientras que la presencia de una molécula de inmunoglobulina, específicamente la región Fc de una molécula de inmunoglobulina, es favorable para las propiedades farmacocinéticas de la proteína de fusión, al mismo tiempo
puede dar lugar a un direccionamiento no deseable de la proteína de fusión hacia células que expresen receptores Fc en lugar de hacia las células que contienen el receptor de IL-2 preferentes. Además, el acoplamiento de los receptores Fc puede dar lugar a la liberación de citocinas (proinflamatorias) y la activación no deseada de diversas células inmunitarias distintas de los linfocitos T reguladores. Por lo tanto, en determinados modos de realización, dicha molécula de inmunoglobulina comprendida en la proteína de fusión de la invención comprende una modificación que reduce la afinidad de unión de la molécula de inmunoglobulina a un receptor Fc en comparación con una molécula de inmunoglobulina correspondiente sin dicha modificación. En un modo de realización específico, dicho receptor Fc es un receptor Fcy, en particular, un receptor Fcy humano. Se puede determinar fácilmente la afinidad de unión a los receptores Fc, por ejemplo, mediante ELISA, o mediante resonancia de plasmón superficial (RPS) usando instrumentación estándar, tal como un instrumento BIAcore (GE Healthcare), y se pueden obtener receptores Fc, tales como, mediante expresión recombinante. En lo que sigue se describe un modo de realización ilustrativo y ejemplar específico para medir la afinidad de unión. De acuerdo con un modo de realización, la afinidad de unión a un receptor Fc se mide mediante resonancia de plasmón superficial usando una máquina BIACORE® T100 (GE Healthcare) a 25 °C con ligando (receptor Fc) inmovilizado en chips CM5. En resumen, se activan chips de biosensor de dextrano carboximetilado (CM5, GE Healthcare) con clorhidrato de N-etil-N'-(3-dimetilaminopropil)carbodiimida (EDC) y N-hidroxisuccinimida (NHS) de acuerdo con las instrucciones del proveedor. El ligando recombinante se diluye con acetato de sodio 10 mM, pH 5,5, a 0,5-30 pg/ml antes de su inyección a un caudal de 10 pl/min para lograr aproximadamente 100-5000 unidades de respuesta (UR) de proteína acoplada. Tras la inyección del ligando, se inyecta etanolamina 1 M para bloquear los grupos sin reaccionar. Para las medidas cinéticas, se inyectan diluciones en serie de tres a cinco veces del anticuerpo (intervalo entre ~0,01 nM a 300 nM) en HBS-EP+ (GE Healthcare, HEPES 10 mM, NaCl 150 mM, EDTA 3 mM, tensioactivo P20 al 0,05 %, pH 7,4) a 25 °C a un caudal de aproximadamente 30-50 pl/min. Se calculan las velocidades de asociación (kas) y las velocidades de disociación (kd¡s) usando un simple modelo de unión uno a uno de Langmuir (programa informático de evaluación de BIACORE ® T100, versión 1.1.1) ajustando simultáneamente los sensogramas de asociación y disociación. Se calcula la constante de disociación en equilibrio (Kd ) como la proporción kdis/kas. Véase, por ejemplo, Chen et al, J Mol Biol 293, 865-881 (1999). De forma alternativa, se pueden evaluar los anticuerpos con afinidad de unión con respecto a los receptores Fc usando líneas de células que se sabe que expresan receptores Fc particulares, tales como los linfocitos Nk que expresan el receptor FcYlIIa.
En un modo de realización, la modificación comprende una o más mutaciones de aminoácido que reducen la afinidad de unión de la inmunoglobulina con respecto a un receptor Fc. En un modo de realización, la mutación de aminoácido es una sustitución de aminoácido. Típicamente, la misma o más mutaciones de aminoácido están presentes en cada una de las dos cadenas pesadas de inmunoglobulina. En un modo de realización, dicha mutación de aminoácido reduce la afinidad de unión de la inmunoglobulina con respecto al receptor Fc en al menos 2 veces, al menos 5 veces o al menos 10 veces. En modos de realización donde existe más de una mutación de aminoácido que reduce la afinidad de unión de la inmunoglobulina con respecto al receptor Fc, la combinación de estas mutaciones de aminoácido puede reducir la afinidad de unión de la inmunoglobulina con respecto al receptor Fc en al menos 10 veces, al menos 20 veces o incluso al menos 50 veces. En un modo de realización, dicha molécula de inmunoglobulina presenta menos de un 20 %, en particular, menos de un 10 %, más en particular, menos de un 5 % de la afinidad de unión con respecto a un receptor Fc en comparación con una molécula de inmunoglobulina correspondiente sin dicha modificación.
En un modo de realización, dicho receptor Fc es un receptor Fc activador. En un modo de realización específico, dicho receptor Fc se selecciona del grupo de FcYRIIIa (CD16a), FcyRI (CD64), FcYRIIa (CD32) y FcaRI (c D89). En un modo de realización específico, el receptor Fc es un receptor Fcy, más específicamente un receptor FcYRIIIa, FcyRI o FcYRIIa. Preferentemente, se reduce la afinidad de unión con respecto a cada uno de estos receptores. En un modo de realización incluso más específico, dicho receptor Fc es FcYIIIa, en particular, FcYIIIa humano. En algunos modos de realización, también se reduce la afinidad de unión con respecto a un componente del complemento, específicamente la afinidad de unión con respecto a C1q. En un modo de realización, no se reduce la afinidad de unión con respecto al receptor Fc neonatal (FcRn). La unión sustancialmente similar a FcRn, es decir, la preservación de la afinidad de unión de la molécula de inmunoglobulina con respecto a dicho receptor, se logra cuando la molécula de inmunoglobulina presenta más de aproximadamente un 70 % de la afinidad de unión de una forma sin modificar de la molécula de inmunoglobulina con respecto a FcRn. Las moléculas de inmunoglobulina comprendidas en las proteínas de fusión de la invención pueden presentar más de aproximadamente un 80 % e incluso más de aproximadamente un 90 % de dicha afinidad.
En un modo de realización, dicha modificación que reduce la afinidad de unión de la molécula de inmunoglobulina con respecto a un receptor Fc está en la región Fc, en particular, la región CH2, de la molécula de inmunoglobulina. En un modo de realización, dicha molécula de inmunoglobulina comprende una sustitución de aminoácido en la posición 329 (numeración EU) de las cadenas pesadas de inmunoglobulina. En un modo de realización más específico, la sustitución de aminoácido es P329A o P329G, en particular, P329G. En un modo de realización, dicha molécula de inmunoglobulina comprende sustituciones de aminoácido en las posiciones 234 y 235 (numeración EU) de las cadenas pesadas de inmunoglobulina. En un modo de realización específico, dichas sustituciones de aminoácido son L234A y L235A (LALA). En un modo de realización, dicha molécula de inmunoglobulina comprende una sustitución de aminoácido en la posición 329 (numeración EU) de las cadenas pesadas de anticuerpo y otra sustitución de aminoácido en una posición seleccionada de la posición 228, 233, 234, 235, 297 y 331 de las cadenas
pesadas de inmunoglobulina (numeración EU). En un modo de realización más específico, la otra sustitución de aminoácido es S228P, E233P, L234A, L235A, L235E, N297A, N297D o P331S. En un modo de realización particular, dicha molécula de inmunoglobulina comprende sustituciones de aminoácido en las posiciones P329, L234 y L235 (numeración EU) de las cadenas pesadas de inmunoglobulina. En un modo de realización más particular, dicha molécula de inmunoglobulina comprende las sustituciones de aminoácido L234A, L235A y P329g (LALA P329G; numeración EU) en las cadenas pesadas de inmunoglobulina. Esta combinación de sustituciones de aminoácido anula, en particular, eficientemente la unión al receptor Fcy de una inmunoglobulina de clase IgG humana, como se describe en la publicación PCT n.° WO 2012/130831. La publicación PCT n.° WO 2012/130831 también describe procedimientos de preparación de dicha inmunoglobulina modificada y procedimientos para determinar sus propiedades, tales como la unión al receptor Fc o las funciones efectoras.
Se pueden preparar inmunoglobulinas que comprenden modificaciones en las cadenas pesadas de inmunoglobulina mediante deleción, sustitución, inserción o modificación de aminoácido usando procedimientos genéticos o químicos bien conocidos en la técnica. Los procedimientos genéticos pueden incluir mutagénesis específica de sitio de la secuencia de ADN codificante, PCR, síntesis génica y similares. Los cambios de nucleótido correctos se pueden verificar, por ejemplo, mediante secuenciación.
Las inmunoglobulinas o anticuerpos que comprenden modificaciones que reducen la unión al receptor Fc tienen, en general, funciones efectoras reducidas, en particular, ADCC reducida en comparación con las inmunoglobulinas o anticuerpos sin modificar correspondientes. De ahí que en un modo de realización dicha modificación que reduce la afinidad de unión de la molécula de inmunoglobulina con respecto a un receptor Fc reduzca la función efectora de la molécula de inmunoglobulina. En un modo de realización específico, dicha función efectora es la citotoxicidad celular dependiente de anticuerpos (ADCC). En un modo de realización, se reduce la ADCC a menos de un 20 % de la ADCC inducida por una molécula de inmunoglobulina correspondiente sin dicha modificación. La función efectora de una inmunoglobulina o anticuerpo se puede medir mediante procedimientos conocidos en la técnica. Los ejemplos de ensayos in vitro para evaluar la actividad de ADCC de una molécula de interés se describen en la patente de EE. UU. n.° 5.500.362; Hellstrom et al. Proc Natl Acad Sci USA 83, 7059-7063 (1986) y Hellstrom et al., Proc Natl Acad Sci USA 82, 1499-1502 (1985); la patente de EE. UU. n.° 5.821.337; Bruggemann et al., J Exp Med 166, 1351 1361 (1987). De forma alternativa, se pueden emplear procedimientos de ensayos no radiactivos (véase, por ejemplo, el ensayo de citotoxicidad no radiactivo ACTI™ para citometría de flujo (CellTechnology, Inc. Mountain View, CA); y el ensayo de citotoxicidad no radiactivo CytoTox 96® (Promega, Madison, WI). Las células efectoras útiles para dichos ensayos incluyen células mononucleares de sangre periférica (PBMC) y linfocitos citolíticos naturales (NK). De forma alternativa, o adicionalmente, se puede evaluar la actividad de ADCc de la molécula de interés in vivo, por ejemplo, en un modelo animal tal como el divulgado en Clynes et al., Proc Natl Acad Sci USA 95, 652-656 (1998). En algunos modos de realización, también se reduce la unión de la molécula de inmunoglobulina a un componente del complemento, específicamente a C1q. En consecuencia, también se puede reducir la citotoxicidad dependiente del complemento (CDC). Se pueden llevar a cabo ensayos de unión a C1q para determinar si la inmunoglobulina se puede unir a C1q y, de ahí, que tenga actividad CDC. Véase, por ejemplo, ELISA de unión a C1q y C3c en los documentos WO 2006/029879 y WO 2005/100402. Para evaluar la activación del complemento se puede realizar un ensayo de CDC (véase, por ejemplo, Gazzano-Santoro et al., J Immunol Methods 202, 163 (1996); Cragg et al., Blood 101, 1045-1052 (2003); y Cragg y Glennie, Blood 103, 2738-2743 (2004)).
Además de las moléculas de inmunoglobulina descritas anteriormente en el presente documento y en la publicación PCT n.° WO 2012/130831, las inmunoglobulinas con unión al receptor Fc y/o función efectora reducidas también incluyen aquellas con sustitución de uno o más de los residuos de la región Fc 238, 265, 269, 270, 297, 327 y 329 (patente de EE. UU. n.° 6.737.056). Dichos mutantes de Fc incluyen mutantes de Fc con sustituciones en dos o más de las posiciones de aminoácido 265, 269, 270, 297 y 327, incluyendo el llamado mutante de Fc “DANA” con la sustitución de los residuos 265 y 297 a alanina (patente de EE. UU. n.° 7.332.581).
Las inmunoglobulinas de subclase IgG4 presentan una afinidad de unión reducida con respecto a los receptores Fc y funciones efectoras reducidas en comparación con las inmunoglobulinas IgG1. De ahí que en algunos modos de realización dicha molécula de inmunoglobulina comprendida en la proteína de fusión de la invención sea una inmunoglobulina de subclase IgG4, en particular, una inmunoglobulina de subclase IgG4 humana. En un modo de realización, dicha inmunoglobulina de subclase IgG4 comprende sustituciones de aminoácido en la región Fc en la posición S228 (numeración EU), específicamente la sustitución de aminoácido S228P. Para reducir además su afinidad de unión con respecto a un receptor Fc y/o su función efectora, en un modo de realización, dicha inmunoglobulina de subclase IgG4 comprende una sustitución de aminoácido en la posición L235, específicamente la sustitución de aminoácido L235E (numeración EU). En otro modo de realización, dicha inmunoglobulina de subclase IgG4 comprende una sustitución de aminoácido en la posición P329, específicamente la sustitución de aminoácido P329G (numeración EU). En un modo de realización particular, dicha inmunoglobulina de subclase IgG4 comprende las sustituciones de aminoácido en las posiciones S228, L235 y P329, específicamente las sustituciones de aminoácido S228P, L235E y P329G (numeración EU). Dichas inmunoglobulinas de subclase IgG4 modificadas y sus propiedades de unión al receptor Fcy se describen en la publicación PCT n.° WO 2012/130831.
En un modo de realización, dicha molécula de inmunoglobulina se puede unir específicamente a un antígeno. En un
modo de realización, dicha molécula de inmunoglobulina es un anticuerpo monoclonal. En un modo de realización, dicha molécula de inmunoglobulina no se puede unir específicamente a un antígeno, en particular, no se puede unir específicamente a un antígeno humano. La ausencia de unión específica de dicha molécula de inmunoglobulina a un antígeno (es decir, la ausencia de cualquier unión que se pueda discriminar de una interacción no específica) se puede determinar, por ejemplo, mediante ELISA o resonancia de plasmón superficial como se describe en el presente documento. Dicha molécula de inmunoglobulina es, en particular, útil, por ejemplo, para potenciar la semivida en suero de la proteína de fusión, cuando no se desea el direccionamiento hacia un tejido particular.
En un modo de realización, dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena pesada basada en la secuencia de estirpe germinal de Vh3-23 humana. En un modo de realización específico, dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena pesada que es al menos un 95 %, 96 %, 97 %, 98 %, 99 % o 100 % idéntica a la secuencia de la SEQ ID NO: 9. En un modo de realización, dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena ligera basada en la secuencia de estirpe germinal de Vk3-20 humana. En un modo de realización específico, dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena ligera que es al menos un 95 %, 96 %, 97 %, 98 %, 99 % o 100 % idéntica a la secuencia de la SEQ ID NO: 11. En un modo de realización incluso más específico, dicha molécula de inmunoglobulina comprende la secuencia de la región variable de la cadena pesada de la SEQ ID NO: 9 y la secuencia de la región variable de la cadena ligera de la SEQ ID NO: 11. Las moléculas de inmunoglobulina que comprenden estas secuencias de la región variable no se pueden unir específicamente a un antígeno, en particular, a un antígeno humano. Les falta la unión a tejidos normales, así como a PBMC, no tienen polirreactividad y no muestran ninguna acumulación no específica in vivo mediante formación de imágenes (datos no mostrados). Las secuencias de la región variable se basan totalmente en secuencias de estirpe germinal humana, con la excepción de la CDR3 de la cadena pesada, en la que se ha introducido una secuencia GSG para generar una inmunoglobulina que no se una.
En un modo de realización, dichas moléculas de IL-2 mutante comprenden una mutación de aminoácido en una posición correspondiente al residuo 88 de IL-2 humana (SEQ ID NO: 1); dicha mutación de aminoácido es una sustitución de aminoácido; y, dicha sustitución de aminoácido es N88D. En un modo de realización, dichas moléculas de IL-2 mutante son moléculas de IL-2 humana. En un modo de realización, dichas moléculas de IL-2 mutante comprenden solo una mutación de aminoácido que reduce la afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia en comparación con una molécula de IL-2 natural. En un modo de realización, dichas moléculas de IL-2 mutante no comprenden una mutación de aminoácido que altera la afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad alta en comparación con una molécula de IL-2 natural. En un modo de realización, dichas moléculas de IL-2 mutante comprenden solo una mutación de aminoácido única que altera la afinidad de la molécula de IL-2 mutante con respecto a un receptor de IL-2 en comparación con una molécula de IL-2 natural.
En un modo de realización, dichas moléculas de IL-2 mutante comprenden además una sustitución de aminoácido en una posición correspondiente al residuo 125 de IL-2 humana; dicha sustitución de aminoácido es C125A.
Las moléculas de IL-2 mutante comprendidas en la proteína de fusión de la invención también pueden ser moléculas de IL-2 sin glucosilar. Por ejemplo, la eliminación del sitio de O-glucosilación de la molécula de IL-2 da como resultado un producto más homogéneo cuando la proteína de fusión se expresa en células de mamífero, tales como células CHO o HEK. Por tanto, en determinados modos de realización, las moléculas de IL-2 mutante comprenden además una modificación que elimina el sitio de O-glucosilación de IL-2 en una posición correspondiente al residuo 3 de IL-2 humana; siendo dicha modificación que elimina el sitio de O-glucosilación de IL-2 en una posición correspondiente al residuo 3 de IL-2 humana una sustitución de aminoácido; y, siendo dicha sustitución de aminoácido T3A.
En un modo de realización particular, las moléculas de IL-2 mutante comprenden las sustituciones de aminoácido T3A, N88D y C125A. En un modo de realización específico, dichas moléculas de IL-2 mutante comprenden la secuencia de la SEQ ID NO: 58 (IL-2 T3A N88D C125A).
En un modo de realización, la unión de la proteína de fusión de la invención al receptor de IL-2 Py está al menos 1,5 veces, preferentemente al menos 2 veces o al menos 3 veces reducida en comparación con la unión de una proteína de fusión correspondiente que comprende dos moléculas de IL-2 natural al receptor de IL-2 Py. En un modo de realización, la proteína de fusión de la invención se une al receptor de IL-2 Py con una constante de afinidad (Kd ) al menos 2 veces más alta que la Kd de una proteína de fusión correspondiente que comprende dos moléculas de IL-2 natural, cuando se mide mediante RPS a 25 °C. En un modo de realización específico, dicho receptor de IL-2 Py es el receptor de IL-2 Py humano. En un modo de realización, la unión de la proteína de fusión de la invención al receptor de IL-2 a es aproximadamente igual a la unión de una proteína de fusión correspondiente que comprende dos moléculas de IL-2 natural al receptor de IL-2 a. En un modo de realización, la proteína de fusión de la invención se une al receptor de IL-2 a con una constante de afinidad (Kd ) aproximadamente igual a la Kd de una proteína de fusión correspondiente que comprende dos moléculas de IL-2 natural, cuando se mide mediante RPS a 25 °C. En un modo de realización específico, dicho receptor de IL-2 a es el receptor de IL-2 a humano. En el presente documento se describe un procedimiento para medir la afinidad de unión con respecto al receptor de IL-2 Py o IL-2 a mediante
RPS. De acuerdo con un modo de realización, se mide la afinidad de unión (Kd ) mediante resonancia de plasmón superficial usando una máquina BIACORE® T200 (GE Healthcare) a 25 °C con receptores de IL-2 inmovilizados en CMS o chips con estreptavidina. Se calcula la constante de afinidad (Kd ) como la proporción kdis/kas. Véase, por ejemplo, Chen et al., J Mol Biol 293, 865-881 (1999).
En un aspecto particular, la invención proporciona una proteína de fusión que comprende (i) una molécula de inmunoglobulina de subclase IgG1 que comprende las sustituciones de aminoácido L234A, L235A y P329G (numeración EU) en las cadenas pesadas de inmunoglobulina, y (ii) dos moléculas de interleucina 2 (IL-2) mutante que comprenden la sustitución de aminoácido N88D, cada una fusionada en su aminoácido N terminal con el aminoácido C terminal de una de las cadenas pesadas de inmunoglobulina a través de un conector peptídico. En un modo de realización, dicha molécula de inmunoglobulina y dichas moléculas de IL-2 son humanas. En un modo de realización específico, dicha molécula de inmunoglobulina comprende la secuencia de la región variable de la cadena pesada de la SEQ ID NO: 9 y la secuencia de la región variable de la cadena ligera de la SEQ ID NO: 11. En otro modo de realización específico, dichas moléculas de IL-2 comprenden cada una la secuencia de aminoácidos de la SEQ ID NO: 58. En otro modo de realización, dicho conector peptídico comprende la secuencia de aminoácidos (G4S)3 (SEQ ID NO: 66). En un modo de realización incluso más específico, dicha proteína de fusión comprende una secuencia de polipéptido que es al menos aproximadamente un 80 %, 85 %, 90 %, 95 %, 96 %, 97 %, 98 %, 99 % o 100 % idéntica a la secuencia de la SEQ ID NO: 50 y una secuencia de polipéptido que es al menos aproximadamente un 80 %, 85 %, 90 %, 95 %, 96 %, 97 %, 98 %, 99 % o 100 % idéntica a la secuencia de la SEQ ID NO: 19.
Como se muestra en los ejemplos, la proteína de fusión de la invención activa selectivamente los linfocitos T reguladores (es decir, esencialmente sin la activación simultánea de otros subconjuntos de linfocitos T y/o linfocitos citolíticos naturales (NK)). Por tanto, en un aspecto, la invención proporciona una proteína de fusión que comprende una molécula de inmunoglobulina y dos moléculas de IL-2 mutante, en la que dicha proteína de fusión activa selectivamente los linfocitos T reguladores sobre los linfocitos T efectores y los linfocitos NK, en particular, sobre los linfocitos T CD4+ convencionales, los linfocitos T CD8+ y los linfocitos NK. En un modo de realización, dicha proteína de fusión activa los linfocitos T reguladores al menos 10 veces, al menos 100 veces o al menos 1000 veces más que los linfocitos T efectores y los linfocitos NK. En un modo de realización, dicha proteína de fusión activa selectivamente los linfocitos T reguladores sobre los linfocitos T de memoria CD4+ convencionales. En un modo de realización, dicha proteína de fusión activa los linfocitos T reguladores al menos 10 veces, al menos 100 veces o al menos 1000 veces más que los linfocitos T de memoria CD4+ convencionales. En un modo de realización, se determina dicha activación mediante la medición de los niveles de fosforilación de STAT intracelular, en particular, STAT5. En un modo de realización, dicha medición de los niveles de fosforilación de STAT intracelular se realiza mediante análisis por citometría de flujo. En un modo de realización, el valor de CE50 de dicha proteína de fusión para la inducción de la señalización del receptor de IL-2 en los linfocitos T reguladores es al menos 10 veces, al menos 100 veces o al menos 1000 veces más bajo que su valor de CE50 para la inducción de la señalización del receptor de IL-2 en los linfocitos T efectores y en los linfocitos NK. En un modo de realización, dicha inducción de la señalización del receptor de IL-2 es la inducción de la fosforilación de STAT. En un modo de realización, el valor de CE50 de dicha proteína de fusión para la inducción de la señalización del receptor de IL-2 en los linfocitos T reguladores es al menos 10 veces, al menos 100 veces o al menos 1000 veces más bajo que su valor de CE50 para la inducción de la señalización del receptor de IL-2 en los linfocitos T de memoria CD4+ convencionales. En un modo de realización, dicha inducción de la señalización del receptor de IL-2 es la inducción de la fosforilación de STAT.
En otro aspecto, la invención proporciona, en particular, la proteína de fusión para su uso en la activación selectiva de los linfocitos T reguladores in vitro. En un modo de realización, dicho uso comprende poner en contacto linfocitos T reguladores con dicha proteína de fusión in vitro. En un modo de realización, dicho uso comprende además poner en contacto otros linfocitos T (no reguladores) con dicha proteína de fusión. En un modo de realización, dicho uso es in vitro y dicha proteína de fusión se usa a una concentración de aproximadamente 10 ng/ml o menos, en particular, aproximadamente 1 ng/ml o menos.
La invención también proporciona un procedimiento para la activación selectiva de los linfocitos T reguladores in vitro, que comprende poner en contacto dichos linfocitos T reguladores con la proteína de fusión de la invención. En un modo de realización, dicho procedimiento comprende además poner en contacto otros linfocitos T (no reguladores) con dicha proteína de fusión. En un modo de realización, dicha activación comprende la inducción de la proliferación y/o la inducción de la señalización del receptor de IL-2. En un modo de realización, dicho procedimiento es in vitro y dicha proteína de fusión se usa a una concentración de aproximadamente 10 ng/ml o menos, en particular, aproximadamente 1 ng/ml o menos.
De acuerdo con determinados modos de realización de la proteína de fusión, en el uso o procedimiento descrito en los párrafos precedentes, dicha activación comprende la inducción de la proliferación de los linfocitos T reguladores y/o la inducción de la señalización del receptor de IL-2 en los linfocitos T reguladores. Se puede medir la inducción de la proliferación, por ejemplo, mediante detección del marcador de proliferación intracelular Ki-67, como se describe en los ejemplos. En un modo de realización, la proliferación de los linfocitos T reguladores activados por la proteína de fusión de la invención se incrementa al menos aproximadamente 1,5 veces, al menos aproximadamente 2 veces o al menos aproximadamente 3 veces en comparación con la proliferación de los linfocitos T reguladores no
activados. En un modo de realización, se incrementa la proliferación de otros linfocitos T (no reguladores) y/o linfocitos NK que están en contacto con la proteína de fusión de la invención en menos de aproximadamente 1,5 veces, menos de aproximadamente 1,2 veces o menos de aproximadamente 1,1 veces en comparación con la proliferación de las células correspondientes que no están en contacto con dicha proteína de fusión. Se puede medir la inducción de la señalización del receptor de IL-2, por ejemplo, mediante detección de STAT5 fosforilado, como se describe en los ejemplos. En un modo de realización, la señalización del receptor de IL-2 en los linfocitos T reguladores activados por la proteína de fusión de la invención se incrementa al menos aproximadamente 1,5 veces, al menos aproximadamente 2 veces, al menos aproximadamente 3 veces o al menos aproximadamente 5 veces en comparación con la señalización del receptor de IL-2 en los linfocitos T reguladores no activados. En un modo de realización, se incrementa la señalización del receptor de IL-2 en otros linfocitos T (no reguladores) y/o linfocitos NK que están en contacto con la proteína de fusión de la invención en menos de aproximadamente 1,5 veces, o menos de aproximadamente 1,2 veces, o menos de aproximadamente 1,1 veces en comparación con la señalización del receptor de IL-2 en las células correspondientes que no están en contacto con dicha proteína de fusión.
Polinucleótidos
La invención proporciona además polinucleótidos que codifican una fusión como se describe en el presente documento.
Los polinucleótidos de la invención incluyen aquellos que son al menos aproximadamente un 80 %, 85 %, 90 %, 95 %, 96 %, 97 %, 98 %, 99 % o 100 % idénticos a las secuencias expuestas en las SEQ ID NO 10, 12, 20, 51 y 59, incluyendo los fragmentos funcionales o variantes de los mismos.
Los polinucleótidos que codifican las proteínas de fusión de la invención se pueden expresar como un polinucleótido único que codifica toda la proteína de fusión o como polinucleótidos múltiples (por ejemplo, dos o más) que se coexpresan. Los polipéptidos codificados por polinucleótidos que se coexpresan se pueden asociar, por ejemplo, a través de enlaces disulfuro u otros medios para formar una proteína de fusión funcional. Por ejemplo, se puede codificar la porción de la cadena ligera de una inmunoglobulina por un polinucleótido separado de la porción de la cadena pesada de inmunoglobulina. Cuando se coexpresan, los polipéptidos de cadena pesada se asociarán con los polipéptidos de cadena ligera para formar la inmunoglobulina.
En un modo de realización, la presente invención se dirige a un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia que codifica una secuencia de la región variable como se muestra en la SEQ ID NO 9 o 11. En otro modo de realización, la presente invención se dirige a un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia que codifica una secuencia de polipéptido como se muestra en la SEQ ID NO 19 o 50. En otro modo de realización, la invención se dirige además a un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia que es al menos aproximadamente un 80 %, 85 %, 90 %, 95 %, 96 %, 97 %, 98 % o 99 % idéntica a una secuencia de ácido nucleico mostrada en la SEQ ID NO 10, 12, 20, 51 o 59. En otro modo de realización, la invención se dirige a un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia de ácido nucleico mostrada en la SEQ ID NO 10, 12, 20, 51 o 59. En otro modo de realización, la invención se dirige a un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia que codifica una secuencia de la región variable que es al menos aproximadamente un 80 %, 85 %, 90 %, 95 %, 96 %, 97 %, 98 % o 99 % idéntica a la secuencia de aminoácidos de la SEQ ID NO 9 u 11. En otro modo de realización, la invención se dirige a un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia que codifica una secuencia de polipéptido que es al menos un 80 %, 85 %, 90 %, 95 %, 96 %, 97 %, 98 % o 99 % idéntica a la secuencia de aminoácidos de la SEQ ID NO 19 o 50. La invención engloba un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia que codifica las secuencias de la región variable de la SEQ ID No 9 u 11 con sustituciones de aminoácido conservadoras. La invención también engloba un polinucleótido que codifica una proteína de fusión de una molécula de inmunoglobulina y dos moléculas de IL-2, en la que el polinucleótido comprende una secuencia que codifica las secuencias de polipéptido de la SEQ ID NO 19 o 50 con sustituciones de aminoácido conservadoras.
En determinados modos de realización, el polinucleótido o ácido nucleico es ADN. En otros modos de realización, un polinucleótido de la presente invención es ARN, por ejemplo, en forma de ARN mensajero (ARNm). El ARN de la presente invención puede ser monocatenario o bicatenario.
Procedimientos recombinantes
Se pueden obtener proteínas de fusión de la invención, por ejemplo, mediante síntesis de péptidos en estado sólido (por ejemplo, síntesis en fase sólida de Merrifield) o producción recombinante. Para la producción recombinante, se aíslan e insertan uno o más polinucleótidos que codifican la proteína de fusión (fragmento), por ejemplo, como se
describe anteriormente, en uno o más vectores para su clonación y/o expresión adicionales en una célula huésped. Dicho polinucleótido se puede aislar y secuenciar fácilmente usando procedimientos convencionales. En un modo de realización, se proporciona un vector, preferentemente un vector de expresión, que comprende uno o más de los polinucleótidos de la invención. Se pueden usar los procedimientos que se conocen bien por los expertos en la técnica para construir vectores de expresión que contienen la secuencia codificante de una proteína de fusión (fragmento) junto con señales de control transcripcional/traduccional apropiadas. Estos procedimientos incluyen técnicas de ADN recombinante in vitro, técnicas sintéticas y recombinación/recombinación genética in vivo. Véanse, por ejemplo, las técnicas descritas en Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, N.Y. (1989); y Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates y Wiley Interscience, N.Y. (1989). El vector de expresión puede ser parte de un plásmido, virus o puede ser un fragmento de ácido nucleico. El vector de expresión incluye un casete de expresión en el que el polinucleótido que codifica la proteína de fusión (fragmento) (es decir, la región codificante) se clona en asociación funcional con un promotor y/u otros elementos de control de la transcripción o traducción. Como se usa en el presente documento, una “región codificante” es una porción de ácido nucleico que consiste en codones traducidos en aminoácidos. Aunque un “codón de terminación” (TAG, TGA o TAA) no se traduce en un aminoácido, se puede consider ar que es parte de una región codificante, si está presente, pero cualquier secuencia flanqueante, por ejemplo, promotores, sitios de unión a ribosoma, finalizadores de la transcripción, intrones, regiones no traducidas en 5' y 3' y similares, no son parte de una región codificante. Pueden estar presentes dos o más regiones codificantes en una construcción de polinucleótido única, por ejemplo, en un vector único, o en construcciones de polinucleótido separadas, por ejemplo, en vectores separados (diferentes). Además, cualquier vector puede contener una región codificante única o puede comprender dos o más regiones codificantes, por ejemplo, un vector de la presente invención puede codificar uno o más polipéptidos, que se separan pos o cotraduccionalmente en las proteínas finales por medio de escisión proteolítica. Además, un vector, polinucleótido o ácido nucleico de la invención puede codificar regiones codificantes heterólogas, fusionadas o bien sin fusionar con un polinucleótido que codifica la proteína de fusión (fragmento) de la invención, o variante o derivado de la misma. Las regiones codificantes heterólogas incluyen, sin limitación, elementos o motivos especializados, tales como un péptido señal secretor o un dominio funcional heterólogo. Una asociación funcional es cuando una región codificante para un producto génico, por ejemplo, un polipéptido, se asocia con una o más secuencias reguladoras de tal manera que la expresión del producto génico se encuentre bajo la influencia o el control de la(s) secuencia(s) reguladora(s). Dos fragmentos de a Dn (tales como una región codificante de polipéptidos y un promotor asociado con la misma) se “asocian funcionalmente” si la inducción de la función promotora da como resultado la transcripción de ARNm que codifica el producto génico deseado y si la naturaleza del enlace entre los dos fragmentos de ADN no interfiere con la capacidad de las secuencias reguladoras de la expresión para dirigir la expresión del producto génico o interferir con la capacidad del molde de ADN que se va a transcribir. Por tanto, una región promotora se asociaría funcionalmente con un ácido nucleico que codifica un polipéptido si el promotor pudiera efectuar la transcripción de ese ácido nucleico. El promotor puede ser un promotor específico de célula que dirige la transcripción sustancial del ADN solo en células predeterminadas. Otros elementos de control de la transcripción, además de un promotor, por ejemplo, potenciadores, operadores, represores y señales de finalización de la transcripción, se pueden asociar funcionalmente con el polinucleótido para dirigir la transcripción específica de célula. En el presente documento se divulgan promotores adecuados y otras regiones de control de la transcripción. Se conoce una variedad de regiones de control de la transcripción por los expertos en la técnica. Estas incluyen, sin limitación, regiones de control de la transcripción que funcionan en células de vertebrado, tales como, pero no limitadas a, segmentos promotores y potenciadores de citomegalovirus (por ejemplo, el promotor inmediato temprano, junto con el intrón A), virus de simio 40 (por ejemplo, el promotor temprano) y retrovirus (tales como, por ejemplo, el virus del sarcoma de Rous). Otras regiones de control de la transcripción incluyen las derivadas de genes de vertebrado, tales como actina, proteína de choque térmico, hormona de crecimiento bovina y globina a de conejo, así como otras secuencias que pueden controlar la expresión génica en células eucariotas. Las regiones de control de la transcripción adecuadas adicionales incluyen promotores y potenciadores específicos de tejido, así como promotores inducibles (por ejemplo, promotores inducibles por tetraciclinas). De forma similar, se conoce una variedad de elementos de control de la traducción por los expertos en la técnica. Estos incluyen, pero no se limitan a, sitios de unión a ribosoma, codones de inicio y finalización de la traducción y elementos derivados de sistemas víricos (en particular, un sitio interno de entrada al ribosoma o IRES, también denominado secuencia CITE). El casete de expresión también puede incluir otras características, tales como un origen de replicación y/o elementos de integración cromosómica, tales como repeticiones terminales largas (LTR) retrovíricas o repeticiones terminales invertidas (ITR) víricas adenoasociadas (AAV).
Las regiones codificantes de polinucleótidos y ácidos nucleicos de la presente invención se pueden asociar con regiones codificantes adicionales que codifican péptidos secretores o señal, que dirigen la secreción de un polipéptido codificado por un polinucleótido de la presente invención. Por ejemplo, si se desea la secreción de la fusión, el ADN que codifica una secuencia señal se puede encontrar en dirección 5' del ácido nucleico que codifica una proteína de fusión de la invención. De acuerdo con la hipótesis de las señales, las proteínas secretadas por células de mamífero tienen un péptido señal o secuencia líder secretora que se escinde desde la proteína madura una vez que se ha iniciado la exportación de la cadena de proteína en crecimiento en el retículo endoplásmico rugoso. Los expertos en la técnica saben que los polipéptidos secretados por células de vertebrado tienen, en general, un péptido señal fusionado con el extremo N del polipéptido, que se escinde del polipéptido traducido para producir una forma secretada o “madura” del polipéptido. En determinados modos de realización, se usa el péptido señal natural, por ejemplo, un péptido señal de cadena pesada o cadena ligera de inmunoglobulina, o un derivado
funcional de esa secuencia que retiene la capacidad para dirigir la secreción del polipéptido que se asocia funcionalmente con él. De forma alternativa, se puede usar un péptido señal de mamífero heterólogo o un derivado funcional del mismo. Por ejemplo, se puede sustituir la secuencia líder natural con la secuencia líder del activador tisular del plasminógeno (TPA) humano o p-glucuronidasa de ratón. Las secuencias de aminoácidos y nucleótidos de los péptidos señal secretores ejemplares se muestran en las SEQ ID NO 39-47.
El ADN que codifica una corta secuencia de proteínas que se podría usar para facilitar la purificación más tarde (por ejemplo, una marca de histidina) o ayudar en el marcaje de la proteína de fusión se puede incluir dentro de o en los extremos del polinucleótido que codifica la proteína de fusión (fragmento).
En otro modo de realización, se proporciona una célula huésped que comprende uno o más polinucleótidos de la invención. En determinados modos de realización, se proporciona una célula huésped que comprende uno o más vectores de la invención. Los polinucleótidos y vectores pueden incorporar cualquiera de las características, individualmente o en combinación, descritas en el presente documento en relación con los polinucleótidos y vectores, respectivamente. En dicho modo de realización, una célula huésped comprende (por ejemplo, se ha transformado o transfectado con) un vector que comprende un polinucleótido que codifica (parte de) una proteína de fusión de la invención. Como se usa en el presente documento, el término “célula huésped” se refiere a cualquier clase de sistema celular que se pueda genomanipular para generar las proteínas de fusión de la invención o fragmentos de las mismas. Las células huésped adecuadas para replicarse y para respaldar la expresión de las proteínas de fusión se conocen bien en la técnica. Dichas células se pueden transfectar o transducir según sea apropiado con el vector de expresión particular y se pueden cultivar grandes cantidades de células que contienen vectores para su siembra en fermentadores a gran escala para obtener cantidades suficientes de la proteína de fusión para aplicaciones clínicas. Las células huésped adecuadas incluyen microorganismos procariotas, tales como E. coli, o diversas células eucariotas, tales como células de ovario de hámster chino (CHO), células de insecto, o similares. Por ejemplo, se pueden producir polipéptidos en bacterias, en particular, cuando no se necesita glucosilación. Después de la expresión, el polipéptido se puede aislar de la pasta de células bacterianas en una fracción soluble y se puede purificar adicionalmente. Además de los procariotas, los microbios eucariotas, tales como los hongos filamentosos o las levaduras, son huéspedes de clonación o expresión adecuados para vectores que codifican polipéptidos, incluyendo cepas de hongos y levaduras cuyas vías de glucosilación se han “humanizado”, dando como resultado la producción de un polipéptido con un patrón de glucosilación parcial o completamente humano. Véanse Gerngross, Nat Biotech 22, 1409-1414 (2004), y Li et al., Nat Biotech 24, 210-215 (2006). Las células huésped adecuadas para la expresión de polipéptidos (glucosilados) también derivan de organismos multicelulares (invertebrados y vertebrados). Los ejemplos de células de invertebrado incluyen células vegetales y de insecto. Se han identificado numerosas cepas de baculovirus que se pueden usar junto con células de insecto, en particular, para la transfección de células de Spodoptera frugiperda. También se pueden utilizar cultivos de células vegetales como huéspedes. Véanse, por ejemplo, las patentes de EE. UU. n.os 5.959.177, 6.040.498, 6.420.548, 7.125.978 y 6.417.429 (que describen la tecnología PLANTIBODIES™ para producir anticuerpos en plantas transgénicas). También se pueden usar células de vertebrado como huéspedes. Por ejemplo, pueden ser útiles líneas de células de mamífero que se adapten para crecer en suspensión. Otros ejemplos de líneas de células huésped de mamífero útiles son línea CV1 de riñón de mono transformada por SV40 (COS-7); línea de riñón embrionario humano (células 293 o 293T como se describe, por ejemplo, en Graham et al., J Gen Virol 36, 59 (1977)), células de riñón de cría de hámster (BHK), células de Sertoli de ratón (células TM4 como se describe, por ejemplo, en Mather, Biol Reprod 23, 243-251 (1980)), células de riñón de mono (CV1), células de riñón de mono verde africano (VERO-76), células de carcinoma de cuello uterino humano (HELA), células de riñón canino (MDCK), células de hígado de rata búfalo (BRL 3A), células de pulmón humano (W138), células de hígado humano (Hep G2), células de tumor mamario de ratón (MMT 060562), células TRI (como se describe, por ejemplo, en Mather et al., Annals N.Y. Acad Sci 383, 44-68 (1982)), células m Rc 5 y células FS4. Otras líneas de células huésped de mamífero útiles incluyen células de ovario de hámster chino (CHO), incluyendo células CHO dhfr- (Urlaub et al., Proc Natl Acad Sci USA 77, 4216 (1980)); y líneas de células de mieloma, tales como YO, NS0, P3X63 y Sp2/0. Para una revisión de determinadas líneas de células huésped de mamífero adecuadas para la producción de proteínas, véase, por ejemplo, Yazaki y Wu, Methods in Molecular Biology, vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp.
255-268 (2003). Las células huésped incluyen células cultivadas, por ejemplo, células cultivadas de mamífero, células de levadura, células de insecto, células bacterianas y células vegetales, por nombrar solo unas pocas, pero también células comprendidas en un animal transgénico, planta transgénico o tejido animal o vegetal cultivado. En un modo de realización, la célula huésped es una célula eucariota, preferentemente una célula de mamífero, tal como una célula de ovario de hámster chino (CHO), una célula de riñón embrionario humano (HEK) o una célula linfocítica (por ejemplo, célula Y0, NS0, Sp20). Las tecnologías estándar se conocen en la técnica para expresar genes exógenos en estos sistemas. Se pueden genomanipular células que expresen un polipéptido que comprende la cadena pesada o bien la ligera de una inmunoglobulina para que también expresen la otra de las cadenas de inmunoglobulina de tal manera que el producto expresado sea una inmunoglobulina que tenga tanto una cadena pesada como una ligera.
En un modo de realización, se proporciona un procedimiento de producción de una proteína de fusión de acuerdo con la invención, en el que el procedimiento comprende cultivar una célula huésped que comprende un polinucleótido que codifica la proteína de fusión, como se proporciona en el presente documento, en condiciones adecuadas para la expresión de la proteína de fusión, y recuperar la proteína de fusión de la célula huésped (o
medio de cultivo de células huésped). En las proteínas de fusión de la invención, los componentes (molécula de inmunoglobulina y molécula de IL-2) están fusionados genéticamente entre sí. Se pueden diseñar proteínas de fusión de tal manera que sus componentes se fusionen directamente entre sí o indirectamente a través de una secuencia de conector. La composición y longitud del conector se pueden determinar de acuerdo con procedimientos bien conocidos en la técnica y se pueden someter a prueba para determinar su eficacia. Si se desea, también se pueden incluir secuencias adicionales para incorporar un sitio de escisión para separar los componentes individuales de la proteína de fusión, por ejemplo, una secuencia de reconocimiento de endopeptidasas.
En determinados modos de realización, las proteínas de fusión de la invención comprenden al menos una región variable de inmunoglobulina que se puede unir a un antígeno. Las regiones variables pueden formar parte de y derivar de anticuerpos naturales o no naturales y fragmentos de los mismos. Los procedimientos para producir anticuerpos policlonales y anticuerpos monoclonales se conocen bien en la técnica (véase, por ejemplo, Harlow y Fane, "Antibodies, a laboratory manual", Cold Spring Harbor Laboratory, 1988). Se pueden construir anticuerpos no naturales usando síntesis de péptidos en fase sólida, se pueden producir de forma recombinante (por ejemplo, como se describe en la patente de EE. UU. n.° 4.186.567) o se pueden obtener, por ejemplo, cribando colecciones combinatorias que comprenden cadenas pesadas variables y cadenas ligeras variables (véase, por ejemplo, la patente de EE. Uu . n.° 5.969.108 concedida a McCafferty).
Se puede usar cualquier especie animal de inmunoglobulina en la invención. Las inmunoglobulinas no limitantes útiles en la presente invención pueden ser de origen murino, de primate o humano. Si se pretende que la proteína de fusión sea para uso humano, se puede usar una forma quimérica de inmunoglobulina en la que las regiones constantes de la inmunoglobulina sean de un ser humano. También se puede preparar una forma humanizada o completamente humana de la inmunoglobulina de acuerdo con procedimientos bien conocidos en la técnica (véase, por ejemplo, la patente de EE. UU. n.° 5.565.332 concedida a Winter). Se puede lograr la humanización mediante diversos procedimientos incluyendo, pero sin limitarse a (a) injertar las CDR no humanas (por ejemplo, anticuerpo donante) en regiones estructurales y constantes humanas (por ejemplo anticuerpo receptor) con o sin retención de los residuos de la región estructural críticos (por ejemplo, los que son importantes para retener una buena afinidad de unión a antígeno o funciones de anticuerpo), (b) injertar solo las regiones determinantes de la especificidad no humanas (SDR o a-CDR; los residuos críticos para la interacción anticuerpo-antígeno) en regiones estructurales y constantes humanas o (c) trasplantar todos los dominios variables no humanos, pero “encubrirlos” con una sección similar a humana mediante reemplazo de los residuos de superficie. Los anticuerpos humanizados y procedimientos para prepararlos se revisan, por ejemplo, en Almagro y Fransson, Front Biosci 13, 1619-1633 (2008), y se describen además, por ejemplo, en Riechmann et al., Nature 332, 323-329 (1988); Queen et al., Proc Natl Acad Sci USA 86, 10029-10033 (1989); patentes de EE. UU. n.os 5.821.337, 7.527.791, 6.982.321 y 7.087.409; Jones et al., Nature 321, 522-525 (1986); Morrison et al., Proc Natl Acad Sci 81, 6851-6855 (1984); Morrison y Oi, Adv Immunol 44, 65 92 (1988); Verhoeyen et al., Science 239, 1534-1536 (1988); Padlan, Molec Immun 31(3), 169-217 (1994); Kashmiri et al., Methods 36, 25-34 (2005) (que describe un injerto de SDR (a-CDR)); Padlan, Mol Immunol 28, 489-498 (1991) (que describe el “rebamizado”); Dall'Acqua et al., Methods 36, 43-60 (2005) (que describe el “barajado de FR”); y Osbourn et al., Methods 36, 61-68 (2005) y Klimka et al., Br J Cancer 83, 252-260 (2000) (que describe el enfoque de “selección guiada” para el barajado de FR). Las inmunoglobulinas particulares de acuerdo con la invención son inmunoglobulinas humanas. Se pueden producir anticuerpos humanos y regiones variables humanas usando diversas técnicas conocidas en la técnica. Los anticuerpos humanos se describen, en general, en van Dijk y van de Winkel, Curr Opin Pharmacol 5, 368-74 (2001) y Lonberg, Curr Opin Immunol 20, 450-459 (2008). Las regiones variables humanas pueden formar parte de y derivar de anticuerpos monoclonales humanos preparados mediante el procedimiento de hibridoma (véase, por ejemplo, Monoclonal Antibody Production Techniques and Applications, pp.
51-63 (Marcel Dekker, Inc., Nueva York, 1987)). También se pueden preparar anticuerpos humanos y regiones variables humanas administrando un inmunógeno a un animal transgénico que se ha modificado para producir anticuerpos humanos intactos o anticuerpos intactos con regiones variables humanas en respuesta a la exposición antigénica (véase, por ejemplo, Lonberg, Nat Biotech 23, 1117-1125 (2005). También se pueden generar anticuerpos humanos y regiones variables humanas aislando secuencias de las regiones variables de clones de Fv seleccionadas de colecciones de presentación en fagos derivadas de seres humanos (véase por ejemplo, Hoogenboom et al. en Methods in Molecular Biology 178, 1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001); y McCafferty et al., Nature 348, 552-554; Clackson et al., Nature 352, 624-628 (1991)). Los fagos típicamente presentan fragmentos de anticuerpo, como fragmentos Fv monocatenarios (scFv) o bien como fragmentos Fab.
En determinados modos de realización, se genomanipulan las inmunoglobulinas comprendidas en las proteínas de fusión de la presente invención para que tengan una afinidad de unión potenciada de acuerdo con, por ejemplo, los procedimientos divulgados en la publicación PCT WO 2012/020006 (véanse los ejemplos relacionados con la maduración de la afinidad) o la pub. de sol. de pat. de EE. UU. n.° 2004/0132066. Se puede medir la capacidad de las proteínas de fusión de la invención para unirse a un determinante antigénico específico a través de un enzimoinmunoanálisis de adsorción (ELISA) o bien a través de otras técnicas consabidas para un experto en la técnica, por ejemplo, la técnica de resonancia de plasmón superficial (Liljeblad, et al., Glyco J 17, 323-329 (2000)) y ensayos de unión tradicionales (Heeley, Endocr Res 28, 217-229 (2002)). Se pueden usar ensayos de competencia para identificar un anticuerpo que compite con un anticuerpo de referencia para unirse a un antígeno particular. En determinados modos de realización, dicho anticuerpo competidor se une al mismo epítopo (por ejemplo, un epítopo lineal o uno conformacional) que se une mediante el anticuerpo de referencia. Se proporcionan procedimientos
ejemplares detallados para cartografiar un epítopo al que se une un anticuerpo en Morris (1996) “Epitope Mapping Protocols”, en Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ). En un ensayo de competencia ejemplar, se incuba el antígeno inmovilizado en una solución que comprende un primer anticuerpo marcado que se une al antígeno y un segundo anticuerpo sin marcar que se está sometiendo a prueba para determinar su capacidad para competir con el primer anticuerpo para unirse al antígeno. El segundo anticuerpo puede estar presente en un sobrenadante de hibridoma. Como control, se incuba el antígeno inmovilizado en una solución que comprende el primer anticuerpo marcado, pero no el segundo anticuerpo sin marcar. Después de su incubación en condiciones permisivas para la unión del primer anticuerpo al antígeno, se retira el exceso de anticuerpo sin unir, y se mide la cantidad de marcador asociado con el antígeno inmovilizado. Si se reduce sustancialmente la cantidad de marcador asociado con el antígeno inmovilizado en la muestra de prueba en relación con la muestra de control, entonces, eso indica que el segundo anticuerpo está compitiendo con el primer anticuerpo por su unión al antígeno. Véase Harlow y Lane (1988) Antibodies: A Laboratory Manual cap.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
Las proteínas de fusión preparadas como se describe en el presente documento se pueden purificar mediante técnicas conocidas en la técnica, tales como cromatografía de líquidos de alto rendimiento, cromatografía de intercambio iónico, electroforesis en gel, cromatografía de afinidad, cromatografía de exclusión por tamaño y similares. Las condiciones reales usadas para purificar una proteína particular dependerán, en parte, de factores tales como la carga neta, hidrofobia, hidrofilia, etc., y serán evidentes para los expertos en la técnica. Para la purificación mediante cromatografía de afinidad, se puede usar un anticuerpo, ligando, receptor o antígeno al que se una la proteína de fusión. Por ejemplo, para la purificación mediante cromatografía de afinidad de las proteínas de fusión de la invención, se puede usar una matriz con proteína A o proteína G. Se pueden usar cromatografía de afinidad secuencial con proteína A o G y cromatografía de exclusión por tamaño para aislar una proteína de fusión esencialmente como se describe en los ejemplos. Se puede determinar la pureza de la proteína de fusión mediante cualquiera de una variedad de procedimientos analíticos bien conocidos, incluyendo electroforesis en gel, cromatografía de líquidos de alta presión y similares. Por ejemplo, las proteínas de fusión expresadas como se describe en los ejemplos demostraron que estaban intactas y apropiadamente ensambladas, como se demuestra mediante SDS-PAGE reductora y no reductora (véase, por ejemplo, la figura 4).
Composiciones, formulaciones y vías de administración
En otro aspecto, la invención proporciona composiciones farmacéuticas que comprenden cualquiera de las proteínas de fusión proporcionadas en el presente documento, por ejemplo, para su uso en cualquiera de los procedimientos terapéuticos a continuación. En un modo de realización, una composición farmacéutica comprende cualquiera de las proteínas de fusión proporcionadas en el presente documento y un vehículo farmacéuticamente aceptable. En otro modo de realización, una composición farmacéutica comprende cualquiera de las proteínas de fusión proporcionadas en el presente documento y al menos un agente terapéutico adicional, por ejemplo, como se describe a continuación.
Se proporciona además un procedimiento de producción de una proteína de fusión de la invención en una forma adecuada para su administración in vivo, comprendiendo el procedimiento (a) obtener una proteína de fusión de acuerdo con la invención y (b) formular la proteína de fusión con al menos un vehículo farmacéuticamente aceptable, por lo que se formula una preparación de proteína de fusión para su administración in vivo.
Las composiciones farmacéuticas de la presente invención comprenden una cantidad terapéuticamente eficaz de una o más proteínas de fusión disueltas o dispersas en un vehículo farmacéuticamente aceptable. Las frases “farmacéuticas o farmacológicamente aceptables” se refieren a entidades moleculares y composiciones que, en general, no son tóxicas para los receptores a las dosificaciones y concentraciones empleadas, es decir, no producen una reacción adversa, alérgica u indeseable cuando se administran a un animal, tal como, por ejemplo, un ser humano, según sea apropiado. La preparación de una composición farmacéutica que contiene al menos una proteína de fusión y opcionalmente un ingrediente activo adicional se conocerá por los expertos en la técnica a la luz de la presente divulgación, como se ejemplifica en Remington's Pharmaceutical Sciences, 18.a ed. Mack Printing Company, 1990. Además, para su administración en animales (por ejemplo, un ser humano), se entenderá que las preparaciones deben cumplir las normas de esterilidad, pirogenicidad, seguridad general y pureza según se requiera por la oficina de normas biológicas de la FDA o las autoridades correspondientes en otros países. Las composiciones preferentes son formulaciones liofilizadas o soluciones acuosas. Como se usa en el presente documento, “vehículo farmacéuticamente aceptable” incluye cualquiera y todos de disolventes, tampones, medios de dispersión, recubrimientos, tensioactivos, antioxidantes, conservantes (por ejemplo, agentes antibacterianos, agentes antifúngicos), agentes isotónicos, agentes retardantes de la absorción, sales, conservantes, antioxidantes, proteínas, fármacos, estabilizantes de fármacos, polímeros, geles, aglutinantes, excipientes, agentes disgregantes, lubricantes, agentes edulcorantes, agentes saborizantes, tintes, tales como materiales y combinaciones de los mismos, como conocerá un experto en la técnica (véase, por ejemplo, Remington's Pharmaceutical Sciences, 18.a ed. Mack Printing Company, 1990, pp. 1289-1329). Excepto en la medida en que cualquier vehículo convencional sea incompatible con el ingrediente activo, se contempla su uso en las composiciones terapéuticas o farmacéuticas.
La composición puede comprender diferentes tipos de vehículos dependiendo de si se va a administrar en forma sólida, líquida o aerosol, y si se necesita que sea estéril para dichas vías de administración como inyección. Las
proteínas de fusión de la presente invención (y cualquier agente terapéutico adicional) se pueden administrar mediante cualquier procedimiento o cualquier combinación de procedimientos como se conocerían por un experto en la técnica (véase, por ejemplo, Remington's Pharmaceutical Sciences, 18.a ed. Mack Printing Company, 1990). La administración parenteral, en particular, la inyección intravenosa, se usa más comúnmente para administrar moléculas de polipéptido, tales como las proteínas de fusión de la invención.
Las composiciones parenterales incluyen las diseñadas para su administración mediante inyección, por ejemplo, inyección subcutánea, intradérmica, intralesional, intravenosa, intrarterial, intramuscular, intratecal o intraperitoneal. Para su inyección, las proteínas de fusión de la invención se pueden formular en soluciones acuosas, preferentemente en tampones fisiológicamente compatibles, tales como solución de Hanks, solución de Ringer o tampón de solución salina fisiológica. La solución puede contener agentes de formulación, tales como agentes de suspensión, estabilización y/o dispersión. De forma alternativa, las proteínas de fusión pueden estar en forma de polvo para su constitución con un vehículo adecuado, por ejemplo, agua estéril libre de pirógenos, antes de su uso. Se preparan soluciones inyectables estériles incorporando las proteínas de fusión de la invención en la cantidad requerida en el disolvente apropiado con varios de los demás ingredientes enumerados a continuación, según se requiera. Se puede conseguir fácilmente la esterilidad, por ejemplo, mediante filtración a través de membranas de filtración estériles. En general, se preparan dispersiones incorporando los diversos ingredientes activos esterilizados en un vehículo estéril que contiene el medio de dispersión básico y/o los demás ingredientes. En el caso de polvos estériles para la preparación de soluciones, suspensiones o emulsiones inyectables estériles, los procedimientos preferentes de preparación son técnicas de secado al vacío o liofilización que proporcionan un polvo del ingrediente activo más cualquier ingrediente deseado adicional de un medio líquido previamente filtrado de forma estéril del mismo. Se debe tamponar adecuadamente el medio líquido, si fuera necesario, y hacer isotónico el diluyente líquido antes de su inyección con suficiente solución salina o glucosa. La composición debe ser estable en las condiciones de fabricación y almacenamiento, y conservarse frente a la acción contaminante de microorganismos, tales como bacterias y hongos. Se apreciará que la contaminación por endotoxinas se debe minimizar hasta una concentración segura, por ejemplo, menos de 0,5 ng/mg de proteína. Los vehículos farmacéuticamente aceptables adecuados incluyen, pero no se limitan a: tampones, tales como fosfato, citrato y otros ácidos orgánicos; antioxidantes, incluyendo ácido ascórbico y metionina; conservantes (tales como cloruro de octadecildimetilbencilamonio; cloruro de hexametonio; cloruro de benzalconio; cloruro de bencetonio; fenol, alcohol butílico o bencílico; alquilparabenos, tales como metil o propilparabeno; catecol; resorcinol; ciclohexanol; 3-pentanol; y m-cresol); polipéptidos de bajo peso molecular (menos de aproximadamente 10 residuos); proteínas, tales como seroalbúmina, gelatina o inmunoglobulinas; polímeros hidrófilos, tales como polivinilpirrolidona; aminoácidos, tales como glicina, glutamina, asparagina, histidina, arginina o lisina; monosacáridos, disacáridos y otros carbohidratos, incluyendo glucosa, manosa o dextrinas; agentes quelantes, tales como EDTA; glúcidos, tales como sacarosa, manitol, trehalosa o sorbitol; contraiones formadores de sales, tales como sodio; complejos de metal (por ejemplo, complejos Znproteína); y/o tensioactivos no iónicos, tales como polietilenglicol (PEG). Las suspensiones para inyección acuosas pueden contener compuestos que incrementen la viscosidad de la suspensión, tales como carboximetilcelulosa de sodio, sorbitol, dextrano, o similares. Opcionalmente, la suspensión también puede contener estabilizantes o agentes adecuados que incrementan la solubilidad de los compuestos para permitir la preparación de soluciones altamente concentradas. Adicionalmente, se pueden preparar suspensiones de los compuestos activos como suspensiones oleosas para inyección apropiadas. Los disolventes o vehículos lipófilos adecuados incluyen aceites grasos, tales como aceite de sésamo, o ésteres de ácidos grasos sintéticos, tales como oleatos de etilo o triglicéridos, o liposomas.
Las composiciones farmacéuticas que comprenden las proteínas de fusión de la invención se pueden fabricar por medio de procedimientos convencionales de mezclado, disolución, emulsión, encapsulación, atrapamiento o liofilización. Se pueden formular las composiciones farmacéuticas de manera convencional usando uno o más vehículos, diluyentes, excipientes o coadyuvantes fisiológicamente aceptables que faciliten el procesamiento de las proteínas en preparaciones que se puedan usar farmacéuticamente. La formulación adecuada depende de la vía de administración elegida.
Las proteínas de fusión se pueden formular en una composición en forma de ácido o base libre, neutra o de sal. Las sales farmacéuticamente aceptables son sales que retienen sustancialmente la actividad biológica del ácido o base libre. Estas incluyen las sales de adición de ácido, por ejemplo, las formadas con los grupos amino libres de una composición proteica, o que se forman con ácidos inorgánicos tales como, por ejemplo, los ácidos clorhídrico o fosfórico, o dichos ácidos orgánicos como ácido acético, oxálico, tartárico o mandélico. Las sales formadas con los grupos carboxilo libres también pueden derivar de bases inorgánicas, tales como, por ejemplo, hidróxidos de sodio, potasio, amonio, calcio o férrico; o dichas bases orgánicas como isopropilamina, trimetilamina, histidina o procaína. Las sales farmacéuticas tienden a ser más solubles en disolventes acuosos y otros disolventes próticos que las formas de base libre correspondientes.
Composiciones y procedimientos terapéuticos
Se puede usar cualquiera de las proteínas de fusión proporcionadas en el presente documento en los procedimientos terapéuticos.
Para su uso en los procedimientos terapéuticos, las proteínas de fusión de la invención se formularán, dosificarán y administrarán de una manera consistente con la buena práctica médica. Los factores que se deben tener en consideración en este contexto incluyen el trastorno particular que se está tratando, el mamífero particular que se está tratando, la afección clínica del paciente individual, la causa del trastorno, el sitio de administración del agente, el procedimiento de administración, la pauta de administración y otros factores conocidos por los médicos.
En un aspecto, se proporcionan proteínas de fusión de la invención para su uso como un medicamento. En otros aspectos, se proporcionan proteínas de fusión de la invención para su uso en el tratamiento de una enfermedad. En determinados modos de realización, se proporcionan proteínas de fusión de la invención para su uso en un procedimiento de tratamiento. En un modo de realización, la invención proporciona una proteína de fusión como se describe en el presente documento para su uso en el tratamiento de una enfermedad en un individuo que lo necesite. En determinados modos de realización, la invención proporciona una proteína de fusión para su uso en un procedimiento de tratamiento a un individuo que tiene una enfermedad que comprende administrar al individuo una cantidad terapéuticamente eficaz de la proteína de fusión. En determinados modos de realización, la enfermedad que se va a tratar es una enfermedad autoinmunitaria. Las enfermedades autoinmunitarias ejemplares incluyen diabetes de tipo 1, psoriasis, asma, artritis reumatoide, enfermedad inflamatoria intestinal, enfermedad de Crohn, colitis ulcerosa, lupus eritematoso diseminado (LED) y esclerosis múltiple. En un modo de realización, la enfermedad es rechazo de trasplante o enfermedad injerto contra huésped. En un modo de realización particular, la enfermedad se selecciona del grupo de diabetes de tipo 1, enfermedad inflamatoria intestinal, enfermedad de Crohn, colitis ulcerosa, LED y esclerosis múltiple. En un modo de realización más particular, la enfermedad es diabetes de tipo 1. En otro modo de realización particular, la enfermedad es una enfermedad inflamatoria intestinal. En otro modo de realización particular, la enfermedad es esclerosis múltiple. En otros modos de realización, la enfermedad es asma, fibrosis pulmonar o enfermedad pulmonar obstructiva. En todavía otros modos de realización, la enfermedad es una enfermedad cardiovascular, en particular, ateroesclerosis y síndrome coronario agudo. En otros modos de realización, la enfermedad es una afección alérgica, en particular, una alergia alimentaria. En determinados modos de realización, el procedimiento comprende además administrar al individuo una cantidad terapéuticamente eficaz de al menos un agente terapéutico adicional, por ejemplo, un agente inmunodepresor si la enfermedad que se va a tratar es una enfermedad autoinmunitaria. Un “individuo” de acuerdo con cualquiera de los modos de realización anteriores es un mamífero, preferentemente un ser humano.
En otro aspecto, la invención proporciona el uso de una proteína de fusión de la invención en la fabricación o preparación de un medicamento para el tratamiento de una enfermedad en un individuo que lo necesite. En un modo de realización, el medicamento es para su uso en un procedimiento de tratamiento de una enfermedad que comprende administrar a un individuo que tiene la enfermedad una cantidad terapéuticamente eficaz del medicamento. En determinados modos de realización, la enfermedad que se va a tratar es una enfermedad autoinmunitaria. En un modo de realización, la enfermedad es rechazo de trasplante o enfermedad injerto contra huésped. En un modo de realización particular, la enfermedad se selecciona del grupo de diabetes de tipo 1, enfermedad inflamatoria intestinal, enfermedad de Crohn, colitis ulcerosa, LED y esclerosis múltiple. En un modo de realización más particular, la enfermedad es diabetes de tipo 1. En otro modo de realización particular, la enfermedad es una enfermedad inflamatoria intestinal. En otro modo de realización particular, la enfermedad es esclerosis múltiple. En otros modos de realización, la enfermedad es asma, fibrosis pulmonar o enfermedad pulmonar obstructiva. En todavía otros modos de realización, la enfermedad es una enfermedad cardiovascular, en particular, ateroesclerosis y síndrome coronario agudo. En otros modos de realización, la enfermedad es una afección alérgica, en particular, una alergia alimentaria. En un modo de realización, el procedimiento comprende además administrar al individuo una cantidad terapéuticamente eficaz de al menos un agente terapéutico adicional, por ejemplo, un agente inmunodepresor si la enfermedad que se va a tratar es una enfermedad autoinmunitaria. Un “individuo” de acuerdo con cualquiera de los modos de realización anteriores puede ser un mamífero, preferentemente un ser humano.
En otro aspecto, la invención proporciona un procedimiento para tratar una enfermedad en un individuo, que comprende administrar a dicho individuo una cantidad terapéuticamente eficaz de una proteína de fusión de la invención. En un modo de realización, se administra una composición a dicho individuo, que comprende una proteína de fusión de la invención en una forma farmacéuticamente aceptable. En determinados modos de realización, la enfermedad que se va a tratar es una enfermedad autoinmunitaria. En un modo de realización, la enfermedad es rechazo de trasplante o enfermedad injerto contra huésped. En un modo de realización particular, la enfermedad se selecciona del grupo de diabetes de tipo 1, enfermedad inflamatoria intestinal, enfermedad de Crohn, colitis ulcerosa, LED y esclerosis múltiple. En un modo de realización más particular, la enfermedad es diabetes de tipo 1. En otro modo de realización particular, la enfermedad es una enfermedad inflamatoria intestinal. En otro modo de realización particular, la enfermedad es esclerosis múltiple. En otros modos de realización, la enfermedad es asma, fibrosis pulmonar o enfermedad pulmonar obstructiva. En otros modos de realización, la enfermedad es una afección alérgica, en particular, una alergia alimentaria. En todavía otros modos de realización, la enfermedad es una enfermedad cardiovascular, en particular, ateroesclerosis y síndrome coronario agudo. En determinados modos de realización, el procedimiento comprende además administrar al individuo una cantidad terapéuticamente eficaz de al menos un agente terapéutico adicional, por ejemplo, un agente inmunodepresor si la enfermedad que se va a tratar es una enfermedad autoinmunitaria. Un “individuo” de acuerdo con cualquiera de los modos de realización anteriores puede ser un mamífero, preferentemente un ser humano.
En algunos modos de realización, se administra una cantidad eficaz de una proteína de fusión de la invención a una célula. En otros modos de realización, se administra una cantidad terapéuticamente eficaz de una proteína de fusión de la invención a un individuo para el tratamiento de la enfermedad.
Para la prevención o tratamiento de la enfermedad, la dosificación apropiada de una proteína de fusión de la invención (cuando se usa sola o en combinación con uno o más de otros agentes terapéuticos adicionales) dependerá del tipo de enfermedad que se va a tratar, la vía de administración, el peso corporal del paciente, el tipo de proteína de fusión, la gravedad y la evolución de la enfermedad, si la proteína de fusión se administra con propósitos preventivos o terapéuticos, las intervenciones terapéuticas previas o simultáneas, la anamnesis del paciente y la respuesta a la proteína de fusión, y el criterio del médico especialista. El médico de cabecera responsable para su administración, en cualquier caso, determinará la concentración del/de los ingrediente(s) activo(s) en una composición y la(s) dosis apropiada(s) para el sujeto individual. En el presente documento se contemplan diversas pautas de dosificación que incluyen, pero no se limitan a, administraciones únicas o múltiples durante diversos puntos temporales, administración en inyección intravenosa rápida e infusión pulsada.
La proteína de fusión se administra adecuadamente al paciente de una vez o durante una serie de tratamientos. Dependiendo del tipo y de la gravedad de la enfermedad, de aproximadamente 1 pg/kg a 15 mg/kg (por ejemplo, 0,1 mg/kg - 10 mg/kg) de proteína de fusión puede ser una dosificación inicial candidata para su administración al paciente, ya sea, por ejemplo, mediante una o más administraciones separadas o mediante infusión continua. Una dosificación diaria típica puede variar desde aproximadamente 1 pg/kg a 100 mg/kg o más, dependiendo de los factores mencionados anteriormente. Para administraciones repetidas durante varios días o más, dependiendo de la afección, el tratamiento se mantendrá, en general, hasta que se produzca una supresión deseada de los síntomas de la enfermedad. Una dosificación ejemplar de la proteína de fusión estaría en el intervalo desde aproximadamente 0,005 mg/kg a aproximadamente 10 mg/kg. En otros ejemplos no limitantes, una dosis también puede comprender desde aproximadamente 1 pg/kg de peso corporal, aproximadamente 5 pg/kg de peso corporal, aproximadamente 10 pg/kg de peso corporal, aproximadamente 50 pg/kg de peso corporal, aproximadamente 100 pg/kg de peso corporal, aproximadamente 200 pg/kg de peso corporal, aproximadamente 350 pg/kg de peso corporal, aproximadamente 500 pg/kg de peso corporal, aproximadamente 1 mg/kg de peso corporal, aproximadamente 5 mg/kg de peso corporal, aproximadamente 10 mg/kg de peso corporal, aproximadamente 50 mg/kg de peso corporal, aproximadamente 100 mg/kg de peso corporal, aproximadamente 200 mg/kg de peso corporal, aproximadamente 350 mg/kg de peso corporal, aproximadamente 500 mg/kg de peso corporal, a aproximadamente 1000 mg/kg de peso corporal o más por administración y cualquier intervalo derivable en los mismos. En ejemplos no limitantes de un intervalo derivable de los números enumerados en el presente documento, se puede administrar un intervalo de aproximadamente 5 mg/kg de peso corporal a aproximadamente 100 mg/kg de peso corporal, de aproximadamente 5 pg/kg de peso corporal a aproximadamente 500 mg/kg de peso corporal, etc., en base a los números descritos anteriormente. Por tanto, se pueden administrar una o más dosis de aproximadamente 0,5 mg/kg, 2,0 mg/kg, 5,0 mg/kg o 10 mg/kg (o cualquier combinación de las mismas) al paciente. Se pueden administrar intermitentemente dichas dosis, por ejemplo, cada semana o cada tres semanas (por ejemplo, de tal manera que el paciente reciba desde aproximadamente dos a aproximadamente veinte, o, por ejemplo, aproximadamente seis dosis de la proteína de fusión). Se puede administrar una dosis de carga más alta inicial, seguido de una o más dosis más bajas. Sin embargo, pueden ser útiles otras pautas de dosificación. La evolución de este tratamiento se supervisa fácilmente mediante técnicas y ensayos convencionales.
Las proteínas de fusión de la invención se usarán, en general, en una cantidad eficaz para lograr el propósito pretendido. Para su uso para tratar o prevenir una enfermedad, las proteínas de fusión de la invención, o las composiciones farmacéuticas de las mismas, se administran o aplican en una cantidad terapéuticamente eficaz. La determinación de una cantidad terapéuticamente eficaz está dentro de las capacidades de los expertos en la técnica, especialmente a la luz de la divulgación detallada proporcionada en el presente documento.
Para su administración sistémica, se puede estimar inicialmente una dosis terapéuticamente eficaz a partir de ensayos in vitro, tales como ensayos con cultivos de células. Entonces, se puede formular una dosis en modelos animales para lograr un intervalo de concentraciones en circulación que incluya la CI50 como se determina en un cultivo de células. Se puede usar dicha información para determinar con más exactitud dosis útiles en seres humanos.
También se pueden estimar dosificaciones iniciales a partir de datos in vivo, por ejemplo, modelos animales, usando técnicas que se conocen bien en la técnica. Un experto en la técnica podría optimizar fácilmente la administración a seres humanos en base a datos de animales.
Se pueden ajustar individualmente la cantidad de dosificación y el intervalo para proporcionar concentraciones en plasma de las proteínas de fusión que sean suficientes para mantener el efecto terapéutico. Las dosificaciones habituales para pacientes para su administración mediante inyección varían desde aproximadamente 0,1 a 50 mg/kg/día, típicamente desde aproximadamente 0,5 a 1 mg/kg/día. Se pueden lograr concentraciones en plasma terapéuticamente eficaces administrando dosis múltiples cada día. Se pueden medir las concentraciones en plasma, por ejemplo, mediante HPLC.
En casos de administración local o captación selectiva, la concentración local eficaz de la proteína de fusión puede no estar relacionada con la concentración en plasma. Un experto en la técnica podrá optimizar dosificaciones locales terapéuticamente eficaces sin demasiada experimentación.
Una dosis terapéuticamente eficaz de las proteínas de fusión descritas en el presente documento, en general, proporcionará un beneficio terapéutico sin provocar una toxicidad sustancial. Se pueden determinar la toxicidad y la eficacia terapéutica de una proteína de fusión mediante procedimientos farmacéuticos estándar en cultivos de células o en animales de experimentación. Se pueden usar ensayos con cultivos de células y estudios con animales para determinar la DL50 (la dosis letal para un 50 % de una población) y la DE50 (la dosis terapéuticamente eficaz en un 50 % de una población). La proporción de dosis entre los efectos tóxicos y terapéuticos es el índice terapéutico, que se puede expresar como la proporción DL50/DE50. Son preferentes las proteínas de fusión que presentan grandes índices terapéuticos. En un modo de realización, la proteína de fusión de acuerdo con la presente invención presenta un alto índice terapéutico. Se pueden usar los datos obtenidos de ensayos con cultivos de células y estudios con animales en la formulación de un intervalo de dosificaciones adecuadas para su uso en seres humanos. La dosificación se encuentra preferentemente en un intervalo de concentraciones en circulación que incluyen la DE50 con poca o ninguna toxicidad. La dosificación puede variar en este intervalo dependiendo de una variedad de factores, por ejemplo, la forma de dosificación empleada, la vía de administración utilizada, la afección del sujeto, y similares. La formulación exacta, la vía de administración y la dosificación se pueden elegir por el médico individual en vista de la afección del paciente (véase, por ejemplo, Fingl et al., 1975, en: The Pharmacological Basis of Therapeutics, cap. 1, p. 1).
El médico especialista para pacientes tratados con las proteínas de fusión de la invención sabrá cómo y cuándo finalizar, interrumpir o ajustar la administración debido a la toxicidad, disfunción orgánica y similares. Por el contrario, el médico especialista también sabrá ajustar el tratamiento a concentraciones mayores si la respuesta clínica no fuera adecuada (excluyendo la toxicidad). La magnitud de una dosis administrada en el tratamiento del trastorno de interés variará con la gravedad de la afección que se va a tratar, con la vía de administración, y similares. La gravedad de la afección, por ejemplo, se puede evaluar, en parte, mediante procedimientos de evaluación del pronóstico estándar. Además, la dosis y quizás la frecuencia de la dosis también variarán de acuerdo con la edad, peso corporal y respuesta del paciente individual.
Otros agentes y tratamientos
Las proteínas de fusión de la invención se pueden administrar en combinación con uno o más agentes en tratamiento. Por ejemplo, se puede coadministrar una proteína de fusión de la invención con al menos un agente terapéutico adicional. El término “agente terapéutico” engloba cualquier agente administrado para tratar un síntoma o enfermedad en un individuo que necesite dicho tratamiento. Dicho agente terapéutico adicional puede comprender cualquier ingrediente activo adecuado para la indicación particular que se está tratando, preferentemente aquellos con actividades complementarias que no se ven afectadas de manera adversa entre sí. En determinados modos de realización, un agente terapéutico adicional es un agente inmunodepresor.
Dichos otros agentes están presentes adecuadamente en combinación en cantidades que son eficaces para el propósito pretendido. La cantidad eficaz de dichos otros agentes depende de la cantidad de proteína de fusión usada, el tipo de trastorno o tratamiento y otros factores analizados anteriormente. Las proteínas de fusión se usan, en general, en las mismas dosificaciones y con las vías de administración como se describe en el presente documento, o aproximadamente desde un 1 a un 99 % de las dosificaciones descritas en el presente documento, o en cualquier dosificación y mediante cualquier vía que se determine empíricamente/clínicamente que sea apropiada.
Dichas politerapias indicadas anteriormente engloban la administración combinada (donde se incluyen dos o más agentes terapéuticos en la misma composición o en composiciones separadas) y la administración separada, en cuyo caso, la administración de la proteína de fusión de la invención se puede producir antes, simultáneamente y/o tras la administración del adyuvante y/o agente terapéutico adicional.
Artículos de fabricación
En otro aspecto de la descripción, se proporciona un artículo de fabricación que contiene materiales útiles para el tratamiento, la prevención y/o el diagnóstico de los trastornos descritos anteriormente. El artículo de fabricación comprende un recipiente y una etiqueta o prospecto del envase en o asociado con el recipiente. Los recipientes adecuados incluyen, por ejemplo, frascos, viales, jeringas, bolsas de solución i.v., etc. Los recipientes se pueden formar a partir de una variedad de materiales, tales como vidrio o plástico. El recipiente contiene una composición que es, por sí misma o combinada con otra composición, eficaz para tratar, prevenir y/o diagnosticar la afección y que puede tener un orificio de acceso estéril (por ejemplo, el recipiente puede ser una bolsa de solución intravenosa o un vial que tenga un tapón perforable mediante una aguja de inyección hipodérmica). Al menos un agente activo en la composición es una proteína de fusión de la invención. La etiqueta o prospecto del envase indica que la composición se usa para tratar la afección de elección. Además, el artículo de fabricación puede comprender (a) un primer recipiente con una composición contenida en el mismo, en el que la composición comprende una proteína de
fusión de la invención; y (b) un segundo recipiente con una composición contenida en el mismo, en el que la composición comprende un agente terapéutico adicional. El artículo de fabricación de la descripción puede comprender además un prospecto del envase que indique que las composiciones se pueden usar para tratar una afección particular. De forma alternativa, o adicionalmente, el artículo de fabricación puede comprender además un segundo (o tercer) recipiente que comprenda un tampón farmacéuticamente aceptable, tal como agua bacteriostática para inyectables (BWFI), solución salina tamponada con fosfato, solución de Ringer y solución de dextrosa. Puede incluir además otros materiales deseables desde un punto de vista comercial y del usuario, incluyendo otros tampones, diluyentes, filtros, agujas y jeringas.
Ejemplos
Lo que sigue son ejemplos de procedimientos y composiciones de la invención. Se entiende que se pueden poner en práctica otros modos de realización diversos, dada la descripción general proporcionada anteriormente.
Técnicas de ADN recombinante
Se usaron procedimientos estándar para manipular el ADN como se describe en Sambrook et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Nueva York, 1989. Se usaron los reactivos biológicos moleculares de acuerdo con las instrucciones del fabricante. La información general sobre las secuencias de nucleótidos de las cadenas ligera y pesada de inmunoglobulina humana se proporciona en: Kabat, E.A. et al., (1991) Sequences of Proteins of Immunological Interest, quinta ed., publicación del NIH n.° 91-3242.
Secuenciación de ADN
Se determinaron secuencias de ADN mediante secuenciación bicatenaria.
Síntesis génica
Cuando se requirió, se generaron segmentos génicos deseados mediante PCR usando moldes apropiados o bien se sintetizaron mediante Geneart AG (Ratisbona, Alemania) a partir de oligonucleótidos sintéticos y productos de PCR mediante síntesis génica automatizada. En los casos donde no estaba disponible ninguna secuencia génica exacta, se diseñaron cebadores de oligonucleótidos en base a las secuencias de los homólogos más próximos y se aislaron los genes mediante RT-PCR del ARN que provenía del tejido apropiado. Se clonaron los segmentos génicos flanqueados por sitios de escisión de endonucleasas de restricción singulares en vectores de clonación/secuenciación estándar. Se purificó el ADN plasmídico de bacterias transformadas y se determinó la concentración mediante espectroscopia UV. Se confirmó la secuencia de ADN de los fragmentos génicos subclonados mediante secuenciación de ADN. Se diseñaron segmentos génicos con sitios de restricción adecuados para permitir la subclonación en los vectores de expresión respectivos. Se diseñaron todas las construcciones con una secuencia de ADN en extremo 5' que codifica un péptido líder que se dirige hacia proteínas para su secreción en células eucariotas. Las SEQ ID NO 39-47 dan péptidos líder ejemplares y secuencias de polinucleótido que los codifican.
Preparación de fusiones subunidades Py de IL-2R-Fc y fusión subunidad a de IL-2R-Fc
Para estudiar la afinidad de unión con respecto al receptor de IL-2, se generó una herramienta que permitió la expresión de un receptor de IL-2 heterodimérico. La subunidad p del receptor de IL-2 se fusionó con una molécula Fc que se genomanipuló para heterodimerizar (Fc(ojal)) (véanse las SEQ ID NO 21 y 22 (humanas), las SEQ ID NO 27 y 28 (de ratón) y las SEQ ID NO 33 y 34 (de macaco cangrejero)) usando la tecnología de “botones en ojales” (Merchant et al., Nat Biotech. 16, 677-681 (1998)). Entonces, la subunidad y del receptor de IL-2 se fusionó con la variante de Fc(botón) (véanse las SEQ ID No 23 y 24 (humanas), las SEQ ID NO 29 y 30 (de ratón) y las SEQ ID NO 35 y 36 (de macaco cangrejero)), que se heterodimerizaron con Fc(ojal). Entonces, se usó esta proteína de fusión a Fc heterodimérica como sustrato para analizar la interacción IL-2/receptor de IL-2. La subunidad a de IL-2R se expresó como una cadena monomérica con un sitio de escisión de AcTev y una marca Avi His (SEQ ID NO 25 y 26 (humanas), SEQ ID NO 31 y 32 (de ratón) y SEQ ID NO 37 y 38 (de macaco cangrejero)). Se expresaron de forma transitoria subunidades de IL-2R respectivas en células HEK EBNA 293 con suero para la construcción de las subunidades pY de IL-2R y sin suero para la construcción de la subunidad a. Se purificó la construcción de las subunidades pY de IL-2R en la proteína A (GE Healthcare), seguido de cromatografía de exclusión por tamaño (GE Healthcare, Superdex 200). Se purificó la subunidad a de IL-2R por medio de marca His en una columna NiNTA (Qiagen) seguido de cromatografía de exclusión por tamaño (GE Healthcare, Superdex 75). Las secuencias de aminoácidos y nucleótidos correspondientes de diversas construcciones de receptores se dan en las SEQ ID NO 21 38.
Preparación de las proteínas de fusión
Las secuencias de ADN se generaron mediante síntesis génica y/o técnicas clásicas de biología molecular y se subclonaron en vectores de expresión en mamíferos bajo el control de un promotor MPSV y en dirección 5' de un
sitio de poliA sintética, portando cada vector una secuencia OriP de VEB. Se produjeron proteínas de fusión como se aplica en los ejemplos a continuación cotransfectando células HEK293-EBNA que crecen exponencialmente con los vectores de expresión en mamíferos usando transfección con fosfato de calcio. De forma alternativa, se transfectaron células HEK293 que crecían en suspensión con los vectores de expresión respectivos mediante polietilenimina (PEI). De forma alternativa, se usaron grupos de células CHO transfectadas de forma estable o clones de células CHO para la producción en medio libre de suero. Posteriormente, se purificaron las proteínas de fusión a partir del sobrenadante. En resumen, se purificaron proteínas de fusión mediante una etapa de afinidad con proteína A (HiTrap ProtA, GE Healthcare) equilibrada en fosfato de sodio 20 mM, citrato de sodio 20 mM, pH 7,5. Después de cargar el sobrenadante, la columna se lavó en primer lugar con fosfato de sodio 20 mM, citrato de sodio 20 mM, pH 7,5, y posteriormente se lavó con fosfato de sodio 13,3 mM, citrato de sodio 20 mM, cloruro de sodio 500 mM, pH 5,45. La proteína de fusión se eluyó con citrato de sodio 20 mM, cloruro de sodio 100 mM, glicina 100 mM, pH 3, o citrato de sodio 10 mM, pH 3,0. Las fracciones se neutralizaron con Na2HPO40,5 M, pH 8,0 (1:10), se agruparon y purificaron mediante cromatografía de exclusión por tamaño (HiLoad 16/60 Superdex 200, GE Healthcare o HiLoad 26/60 Superdex 200, GE Healthcare) en el tampón de formulación final: histidina 20 mM, NaCl 140 mM, pH 6,0. Se determinó la concentración de proteínas de las muestras de proteínas purificadas midiendo la densidad óptica (DO) a 280 nm, usando el coeficiente de extinción molar calculado en base a la secuencia de aminoácidos. También se determinó el peso molecular en base a la secuencia de aminoácidos. Se analizaron la pureza y peso molecular de las proteínas de fusión mediante SDS-PAGE en presencia y ausencia de un agente reductor (1,4-ditiotreitol 5 mM) y se tiñeron con azul de Coomassie (SimpleBlue™ SafeStain, Invitrogen). Se usó el sistema en gel NuPAGE® Pre-Cast (Invitrogen) de acuerdo con las instrucciones del fabricante (geles Tris-glicina al 4-20 % o Bis-Tris al 3-12 %). De forma alternativa, se analizaron la pureza y el peso molecular de las moléculas mediante análisis EC-SDS en presencia y ausencia de un agente reductor, usando el sistema Caliper LabChip GXII (Caliper Lifescience) de acuerdo con las instrucciones del fabricante. Se analizó el contenido total de las muestras de proteína de fusión usando una columna analítica de exclusión por tamaño Superdex 200 10/300GL (GE Healthcare) en MOPS 2 mM, NaCl 150 mM, NaN3 al 0,02 %, tampón de migración a pH 7,3 a 25 °C. De forma alternativa, se analizó el contenido total de las muestras de anticuerpo usando una columna analítica de exclusión por tamaño TSKgel G3000 SW XL (Tosoh) en K2HPO4 125 mM, NaCl 125 mM, monoclorhidrato de L-arginina 200 mM, NaN3 al 0,02 % (p/v), tampón de migración a pH 6,7 a 25 °C.
Los resultados de la purificación y caracterización de las construcciones de IgG DP47GS-IL-2, IgG DP47GS-(IL-2 )2, IgG DP47GS-IL-2 N88D, IgG DP47GS-(IL-2 N88D)2 e IgG DP47GS-(IL-2 E95A)2 se muestran en las figuras 1, 2, 3, 4 y 5, respectivamente.
Afinidad con respecto a los receptores de IL-2
La afinidad de las proteínas de fusión se determinó mediante resonancia de plasmón superficial (RPS) en un Biacore T200 (GE Healthcare) para el heterodímero Py de IL-2R humano, murino y de macaco cangrejero usando el heterodímero Py de IL-2R recombinante en las siguientes condiciones: ligandos: heterodímero botón p ojal y de IL -2R humano, murino y de macaco cangrejero biotinilado inmovilizado en un chip SA (las concentraciones de inmovilización fueron de 194, 114 y 116 UR, respectivamente), analitos: IgG DP47GS-IL-2 (véanse las SEQ ID NO 13, 15 y 19), IgG DP47GS-(IL-2)2 (véanse las SEQ ID NO 17 y 19), IgG DP47GS-IL-2 N88D (véanse las SEQ ID NO 15, 19 y 48), IgG DP47GS-(IL-2 N88D)2 (véanse las SEQ ID NO 19 y 50) e IgG DP47GS-(IL-2 E95A)2 (véanse las SEQ iD n O 19 y 52), temperatura: 25 °C, tampón: HBS-EP, concentración de analito: 100 nM hasta 1,2 nM (diluciones 1:3), caudal: 30 pl/min, asociación: 120 s, disociación: 600 s para las 2 concentraciones más altas y 120 s para las concentraciones más bajas, regeneración: 60 s MgCh 3 M, ajuste: modelo de unión de Langmuir 1:1, IR^0, Rmáx=local. Se determinaron las afinidades en base a las constantes de velocidad cinética kas y kdis. La afinidad de las proteínas de fusión también se determinó para la subunidad a de IL-2R humano, murino y de macaco cangrejero usando la subunidad a de IL-2R monomérica recombinante en las siguientes condiciones: ligandos: subunidad a de IL-2R humano, murino y de macaco cangrejero inmovilizada en un chip CM5 por medio de acoplamiento de amina (las concentraciones de inmovilización fueron de 240, 245 y 220 UR, respectivamente), analitos: IgG DP47GS-IL-2 (véanse las SEQ ID NO 13, 15 y 19), IgG DP47GS-(IL-2)2 (véanse las SEQ ID NO 17 y 19), IgG DP47GS-IL-2 N88D (véanse las SEQ ID NO 15, 19 y 48), IgG DP47GS-(IL-2 N88D)2 (véanse las SEQ ID NO 19 y 50) e IgG DP47GS-(IL-2 E95A)2 (véanse las SEQ ID NO 19 y 52), temperatura: 25 °C, tampón: HBS-EP, concentración de analito 300 nM hasta 0,41 nM (diluciones 1:3), caudal: 30 pl/min, asociación: 120 s, disociación: 180 s, regeneración: glicina 10 mM, pH 1,5, durante 60 s. Se determinaron las afinidades mediante análisis en estado estacionario.
Los resultados de las mediciones de la afinidad en base a la cinética para el heterodímero Py de IL-2R y el estado estacionario para la subunidad a de IL-2R se proporcionan en la tabla 1.
Tabla 1. Unión de proteínas de fusión a Py de IL-2R y a de IL-2R.


L a s f u s io n e s I g G D P 47 G S - I L - 2 h u m a n a s y m u t a n t e s d e la s m is m a s n o s o lo s e u n e n a la s 3 c a d e n a s d i f e r e n t e s a , p y y d e l r e c e p t o r d e I L - 2 h u m a n o , s in o t a m b ié n a la s r e s p e c t i v a s c a d e n a s d e l r e c e p t o r d e m a c a c o c a n g r e j e r o y r a tó n , a u n q u e la s a f in i d a d e s , e n p r o m e d io , t i e n d e n a s e r l i g e r a m e n t e m á s b a ja s p a r a la s ú l t i m a s d o s e s p e c ie s . L a s a f i n i d a d e s d e e s t a s f u s io n e s d e c i t o c i n a s c o n r e s p e c t o a la s c a d e n a s a h u m a n a s y d e m a c a c o c a n g r e j e r o s o n c o m p a r a b l e s , c o n r e s p e c t o a la c a d e n a a m u r i n a e x i s t e u n a d i f e r e n c i a d e a f in i d a d ~ 2 v e c e s e n c o m p a r a c ió n c o n r e s p e c t o a la c a d e n a a h u m a n a o b s e r v a d a p a r a la s m o lé c u la s q u e p o r t a n s o lo u n a p o r c ió n d e c i t o c i n a . P a r a la s m o lé c u la s c o n d o s p o r c io n e s d e c i t o c in a , lo m á s p r o b a b le m e n t e d e b id o a la a v id e z , e s t a d i f e r e n c i a n o e x i s t e . L a s a f i n i d a d e s d e I g G D P 47 G S - I L - 2 e I g G D P 47 G S - I L - 2 N 88 D c o n r e s p e c t o a la s c a d e n a s a , e n t e o r í a , d e b e r í a n s e r ig u a le s , y a q u e la m u t a c ió n N 88 D n o d e b e r í a a f e c t a r a la in t e r f a s e c o n r e s p e c t o a la c a d e n a a . N 88 s e p o s i c io n a e n la in t e r f a s e c o n r e s p e c t o a la c a d e n a p d e l r e c e p t o r d e I L - 2 y la m u t a g é n e s i s c o n r e s p e c t o a D d a l u g a r a u n a a f in i d a d d i s m i n u id a , c o m o s e p u e d e o b s e r v a r c o m p a r a n d o la u n ió n d e I g G D P 47 G S - I L - 2 e I g G D P 47 G S - I L - 2 N 88 D a p Y d e I L - 2 R d e la s t r e s e s p e c ie s ( 0 , 15 n M , 0 , 60 n M y 0 , 85 n M f r e n t e a 0 , 93 n M , 1 , 3 n M y 2 , 4 n M , r e s p e c t i v a m e n t e ) . P a r a I g G D P 47 G S - ( I L - 2 N 88 D )2 q u e p o r t a d o s c i t o c i n a s I L - 2 m u t a d a s , a l m e n o s h a c ia la s p Y d e I L - 2 R h u m a n o , e s t a d i f e r e n c i a e s m e n o s p r o n u n c ia d a , lo m á s p r o b a b le m e n t e d e b id o a la a v id e z . I g G D P 47 G S - ( I L -2 ) 2 e I g G D P 47 G S - ( I L - 2 E 95 A )2 p r e s e n t a n a v i d e c e s m u y s im i la r e s e n la u n ió n a p Y d e I L - 2 R d e la s t r e s e s p e c ie s . A u n q u e E 95 t a m b ié n s e p o s i c io n a e n la in t e r f a s e c o n r e s p e c t o a la c a d e n a p d e l r e c e p t o r d e I L - 2 , la m u t a g é n e s i s c o n r e s p e c t o a A , a l m e n o s c u a n d o d o s d e e s t o s m u t a n t e s s e f u s io n a n c o n la I g G , n o d a l u g a r a u n a d i s m i n u c ió n s ig n i f i c a t i v a d e la a v id e z .
Expresión de receptores de IL-2 en células inmunitarias
E l r e c e p t o r d e I L - 2 t r i m é r i c o d e a f in i d a d a l t a e s t á c o m p u e s t o p o r la s c a d e n a s a ( I L - 2 R A , C D 25 ) , p ( I L - 2 R B , C D 122 ) y Y (IL-2Rg , C D 1 3 2 ) y t i e n e u n a Kd d e ~ 1 0 p M . S o lo C D 2 5 t i e n e s o lo u n a a f in i d a d b a ja (Kd ~ 1 0 n M ) p a r a I L - 2 . E l d í m e r o I L - 2 R B / I L - 2 R G , q u e s e e x p r e s a e n a l g u n o s t i p o s d e c é l u la s e n a u s e n c ia d e I L - 2 R A , t a m b ié n s e u n e a I L - 2 , p e r o c o n u n a a f in i d a d in t e r m e d ia (Kd ~ 1 n M ) . L a s e ñ a l i z a c ió n p o r m e d io d e l r e c e p t o r d e I L - 2 s e m e d ia p o r la s c a d e n a s I L - 2 R B e I L - 2 R G . A p a r t i r d e lo s a n á l i s i s d e e s t r u c t u r a s c r i s t a l i n a s , I L - 2 R A n o p a r e c e e n t r a r e n c o n t a c t o c o n I L - 2 R B o b ie n c o n I L - 2 R G . S e h a p r o p u e s t o q u e la b a s e d e la c o o p e r a c ió n d e l r e c e p t o r t r i m é r i c o s e a u n a r e d u c c ió n d e e n t r o p í a c u a n d o C D 25 c a p t u r a I L - 2 e n la s u p e r f i c i e d e la s c é l u la s p a r a s u p r e s e n t a c i ó n a I L - 2 R B e I L - 2 R G , o , d e f o r m a a l t e r n a t i v a , s e p r o d u c e u n a a l t e r a c i ó n i n d u c id a p o r C D 25 e n la c o n f o r m a c ió n d e I L - 2 , e s t a b i l i z a n d o , p o r t a n t o , e l c o m p le jo . E n lo s l i n f o c i t o s T F O X P 3 C D 4 r e g u l a d o r e s e x i s t e u n g r a n e x c e s o e s t e q u i o m é t r i c o d e I L - 2 R A e n c o m p a r a c ió n c o n la s c a d e n a s p y Y d e l r e c e p t o r , lo q u e r e s p a l d a la h i p ó t e s is d e q u e lo s d í m e r o s , o in c lu s o c o m p le jo s m á s g r a n d e s , d e la c a d e n a a c o n t r ib u y e n a la u n ió n d e I L - 2. T a m b i é n e x i s t e n p r u e b a s d e q u e C D 25 e n u n a c é l u la p u e d e p r e s e n t a r I L - 2 p a r a d í m e r o s I L - 2 R B / I L - 2 R G e n o t r a c é lu la , e n u n a i n t e r a c c i ó n i n t e r c e l u la r d e a f in i d a d a l t a q u e e n f a t i z a la r e la c ió n ú n i c a e n t r e la s t r e s c a d e n a s q u e c o m p o n e n e l r e c e p t o r d e I L - 2 d e a f in i d a d a l t a .
S e d e t e r m i n ó la e x p r e s i ó n d e C D 25 ( I L - 2 R A ) y C D 122 ( I L - 2 R B ) e n s u b c o n ju n t o s d e T r e g C D 4 , s u b c o n ju n t o s d e l i n f o c i t o s N K y l i n f o c i t o s N K T , a s í c o m o e n s u b c o n ju n t o s d e l i n f o c i t o s T c o n v e n c io n a le s C D 4 y C D 8 m e d ia n t e F A C S ( f i g u r a 6 y 7 ) . S e u s a r o n m a r c a d o r e s d e s u p e r f i c i e d e la s c é l u la s p a r a d e f i n i r s u b c o n ju n t o s d e T r e g C D 4 , l i n f o c i t o s N K T y l i n f o c i t o s N K ( f i g u r a 6 ). A f in d e o p t im iz a r la t i n c i ó n p a r a C D 25 y C D 122 , n o s e r e a l i z ó n in g u n a t in c i ó n p a r a F o x P 3 i n t r a c e lu la r . E n r e s u m e n , u s a n d o 150 p l d e s a n g r e d o n a d a p o r u n v o l u n t a r i o s a n o , s e in c u b a r o n a n t i c u e r p o s f l u o r e s c e n t e s d u r a n t e 45 m in u t o s a t e m p e r a t u r a a m b ie n t e e n o s c u r id a d ( m e z c l a d a e n v ó r t e x a l c o m ie n z o y d e s p u é s d e 20 m in ) . L o s e r i t r o c i t o s s e l i s a r o n c o n t a m p ó n d e l i s is B D ( B D F A C S L y s in g S o lu t io n , 349202 ) d u r a n t e 9 m in u t o s y la s c é l u la s r e s t a n t e s s e la v a r o n ( 2 m l d e ^ P B S B S A a l 0 ,1 % ) y s e f i j a r o n ( P F A a l 1 % ) . S e a n a l i z a r o n la s c é l u la s e n u n a n a l i z a d o r d e c é l u la s L S R F o r t e s s a ™ ( B e c t o n D i c k in s o n ) y s e a n a l i z a r o n lo s d a t o s u s a n d o e l p r o g r a m a in f o r m á t i c o F lo J o ( T r e e S t a r ) . S e i d e n t i f i c a r o n s u b c o n ju n t o s d e T r e g u s a n d o a n t i c u e r p o s e s p e c í f i c o s p a r a T C R a p - F I T C ( I P 26 , B io L e g e n d ) , C D 4 - A le x a F lu o r ® 700 ( R P A - T 4 , B io L e g e n d ) , C D 127 - P E / C Y 7 ( e b io R D R 5 , E b io s c i e n c e ) , C D 4 5 R A - P a c i f i c B lu e ( H I 10 0 , B io L e g e n d ) , C D 2 5 - A P C ( 2 A 3 , M - A 25 1 , Bd B i o s c ie n c e s ) y C D 122 - P E ( T U 27 , B io L e g e n d ) . S e t i ñ e r o n l i n f o c i t o s N K y N K T e n u n t u b o s e p a r a d o c o n a n t i c u e r p o s e s p e c í f i c o s p a r a T C R a p - F I T C , C D 4 - A le x a F lu o r ® 700 , C D 8 - P E / C Y 7 ( H T T 8 a , B io L e g e n d ) , C D 56 - P a c i f i c B lu e ( H C D 56 , B io L e g e n d ) , C D 25 - A P C y C D 122 - P E . T r a s la s e l e c c ió n d e lo s l i n f o c i t o s e n b a s e a F S C / S S C , y la e x c l u s ió n d e lo s d o b le t e s , s e i d e n t i f i c a r o n lo s T r e g in d i f e r e n c ia d o s c o m o T C R a p C D 4 C D 127 'C D 25 C D 45 R A , s e i d e n t i f i c a r o n lo s T r e g d e m e m o r i a c o m o T C R a p C D 4 C D 127 'C D 25 C D 45 R A ' y s e i d e n t i f i c a r o n lo s T r e g a c t i v a d o s c o m o T C R a p C D 4 C D 127
CD25altaCD45RA' . Se identificaron los linfocitos NK como TCRap' CD56' /débil y se identificaron los linfocitos NK activados como TCRap' CD56fuerte. Se identificaron los linfocitos n Kt como TCRap+CD56+ . Se usaron controles de isotipo (CI) conjugados con APC y PE a fin de estimar la fluorescencia de fondo para CD25 y CD122, respectivamente.
De forma similar, se usaron marcadores de superficie de las celulas para definir linfocitos T CD4+ convencionales indiferenciados y de memoria (figuras 7A y 7B), linfocitos T CD8+ convencionales de memoria y linfocitos T CD8 CD45RA+ (una combinación de los subconjuntos indiferenciados y TEMRA; TEMRA se refiere a los linfocitos T de memoria efectora que han vuelto a expresar CD45RA) (figuras 7C y 7D). La tinción y el análisis se realizaron como se describe anteriormente. Usando el mismo tubo descrito anteriormente para caracterizar los Treg CD4+ , se identificaron los linfocitos T indiferenciados convencionales CD4+ como TCRap+CD4+CD127+CD25-/+CD45RA+ y se identificaron los linfocitos T de memoria convencionales CD4+ como TCRap+CD4+CD127+CD25' /+CD45RA\ Se definieron los linfocitos T CD8 usando TCRap-FITC, CD8-Alexa Fluor® 700 (HTT8a, BioLegend), CD28-PE/CY7 (CD28.2, BioLegend), CD45RA-Pacific Blue, CD25-APC y CD122-PE. Se identificaron los linfocitos T de memoria CD8+ como TCRap+CD8+CD45RA\ Se identificaron linfocitos TEMRA e indiferenciados CD8+ como TCRap+CD8+CD45RA+. No se usó CD28 para distinguir los linfocitos T indiferenciados CD8+ de los linfocitos T TEMRA CD8+ , puesto que el marcador CD28 no se incluyó en el análisis de pSTAT5a que se describe a continuación (vease la figura 8). En algunas pruebas (figura 15), se definieron los subconjuntos adicionales como sigue: linfocitos T CD4+ de memoria central (TCRap+CD4+CD56TOXP3' CD45RA' CD127+CD25' /+), linfocitos T CD4+ de memoria efectora (TCRap+CD4+CD56' FOXP3' CD45RA' CD127' CD25' /+), linfocitos T CD8+ de memoria central (TCRap+CD8+CD56"FOXP3"CD45RA"CD127+CD25"/+), linfocitos T CD8+ de memoria efectora (TCRap+CD8+CD56-FOXP3"CD45RA"CD127"CD25"/+) y linfocitos CD8+ TEMRA (TCRap+CD8+CD56TOXP3' CD45RA+CD127' CD25' /+).
En las figuras 6 y 7, se muestra la expresión específica de célula de IL-2RA e IL-2RB para subconjuntos de linfocitos T, linfocitos NK y linfocitos T NK en sangre periférica humana (IL-2RG tiene una expresión esencialmente ubicua en células hematopoyéticas, puesto que se asocia con un gran número de receptores de citocina). La concentración más alta de IL-2RA está presente en las tres poblaciones de linfocitos T CD4+ reguladores (Treg): indiferenciados (CD45RA+CD25+), de memoria (CD45RA-CD25+) y activados (CD45RA-CD25a) (figura 6A). En promedio, los linfocitos T CD4+ de memoria convencionales expresan aproximadamente 10 veces menos CD25 que los Treg (figura 7A). La expresión de CD25 en linfocitos T CD4+ indiferenciados varía significativamente entre los donantes, pero siempre es más baja que la observada en los linfocitos T CD4+ de memoria (figura 7A). La expresión de CD25 en los linfocitos T CD8, NKT y NK es muy baja o no detectable, excepto para los linfocitos NK CD56fuerte (figuras 6C y 1C). Los linfocitos NK CD56fuerte y Nk CD56+ expresan la concentración más alta de IL-2RB (figura 6D), aproximadamente 10 veces más que cualquiera de los subconjuntos de linfocitos T, incluyendo los linfocitos NKT (figuras 6B, 6D, 7B, 7D).
Inducción de pSTAT5a en subconjuntos de células de sangre periférica humana
Tras la oligomerización inducida por IL-2 del IL-2R trimérico, se activan las proteínas tirosina cinasas citoplásmicas JAK1 y JAK3, que se asocian con los dominios intracelulares de IL-2RB e IL-2RG, respectivamente. Estas cinasas fosforilan determinados residuos de tirosina de IL-2RB que actúan como sitios de acoplamiento para STAT5a y STAT5b que, a su vez, se fosforilan. La activación inducida por IL-2 de varias vías de señalización finalmente da como resultado la transcripción de genes diana que contribuyen a las diversas funciones asociadas con la vía IL-2/IL-2R. Puesto que diversos tipos de células expresan diferentes concentraciones de las moléculas IL-2RA e IL-2RB del receptor de IL-2 (figuras 6 y 7), se midieron las concentraciones de pSTAT5a en células individuales mediante citometría de flujo policromática a fin de entender la respuesta de señalización integrada a IL-2 mediada mediante diversas combinaciones de los receptores de afinidad alta e intermedia.
Se evaluaron los efectos de diversas dosis de IgG DP47GS-IL-2, IgG DP47GS-(IL-2)2 , IgG DP47GS-(IL-2E95A)2 , IgG DP47GS-IL-2N88D e IgG DP47GS-(IL-2N88D)2 sobre la inducción de la fosforilación de STAT5a en subconjuntos de Treg CD4+ humanos, linfocitos T CD4+ convencionales indiferenciados y de memoria, linfocitos T CD8+ convencionales de memoria, linfocitos T CD8 CD45RA+, linfocitos NKT y linfocitos Nk (figura 8-14 y tabla 2 y 3,). Todos los subconjuntos se caracterizaron en un tubo único para cada dosis. En resumen, se obtuvo sangre de un voluntario adulto humano sano en tubos heparinizados. Se añadieron diversas concentraciones de proteínas de fusión IgG DP47GS-IL-2 a 500 pl de sangre y se incubaron a 37 °C. Después de 30 minutos, se lisó y se fijó la sangre usando un tampón de lisado/fijación calentado previamente (Becton Dickinson n.° 558049) durante 10 minutos a 37 °C, se lavó 2 veces con PBS que contenía BSA al 0,2 % seguido de permeabilización con metanol enfriado previamente a -20 °C (Sigma, Biotech, grado 494437) durante 20 minutos en hielo. Entonces, las células se lavaron exhaustivamente 4 veces con PBS que contenía BSA al 0,2 % antes de que se realizara la tinción con FACS usando un panel de anticuerpos fluorescentes para distinguir diferentes subpoblaciones de linfocitos y linfocitos NK y el estado de pSTAT5a. Los anticuerpos utilizados fueron anti-CD4-Alexa Fluor® 700 (clon RPA-T4), CD3-PerCP/Cy5.5 (UCHT1), CD45RA-PE/Cy7 (HI100), CD8-Brilliant Violet 605 (RPA-T8), CD56-Brilliant Violet 421 (HCD56), FOXP3-PE (259D) (todos de BioLegend), CD25-APC (clones M-A251 y 2A3) y pSTAT5a-Alexa Fluor® 488 (pY694) (Becton Dickinson). Se adquirieron las muestras usando un analizador de células LSRFortessa™ (Becton Dickinson) y se analizaron los datos usando el programa informático FlowJo (FlowJo, EEC). Después de la selección de los linfocitos y la exclusión de los dobletes, se definieron los Treg como CD3+CD4+FOXP3+ y se subdividieron
como CD45" FOXP3a (Treg activados), CD3+CD4+CD45RATOXP3+ (Treg de memoria) y CD3+CD4+CD45+FOXP3+ (Treg indiferenciados). Se definieron los linfocitos T CD4+ convencionales como CD3+CD4+CD45RA+ (indiferenciados) y CD3+CD4+CD45RA' (de memoria). Se definieron los linfocitos T CD8 como CD3+CD8+CD45RA-(de memoria) y CD3+CD8+CD45RA+. Se definieron los linfocitos NKT como CD3+CD56+ y se definieron los linfocitos NK como CD3' CD56fuerte (linfocitos NK activados) o CD3' CD56+ (linfocitos NK). Se cuantificaron concentraciones de pSTAT5a intracelular en todos los subconjuntos de células a todas las dosis.
La figura 8 muestra la respuesta a la dosis del inmunoconjugado IgG DP47GS-IL-2 en los linfocitos T, linfocitos NK y linfocitos T NK en sangre periférica humana de tres donantes individuales, cada uno sometido a prueba en días diferentes. La jerarquía de la reactividad a IgG DP47GS-IL-2 fue la misma en los tres donantes y la misma que la observada cuando se usó IL-2 humana recombinante (Proleukin® ) (datos no mostrados). Las tres poblaciones de Treg, activados (CD45RA-CD25a), de memoria (CD45rA-CD25+) e indiferenciados (CD45Ra CD25+), incrementaron las concentraciones de pSTAT5a a la concentración de 0,1 ng/ml de IgG DP47GS-IL-2, mientras que otras poblaciones de células requirieron más de 1 ng/ml (linfocitos NK CD56fuerte y T CD4+ de memoria) o bien 10 100 ng/ml (linfocitos T CD8+ T de memoria, linfocitos NK CD56+, linfocitos T c D4+ indiferenciados, linfocitos NKT y linfocitos T CD8 CD45RA+) de IgG DP47-IL-2 para producir incrementos detectables de pSTAT5a. Véase también la figura 11 para obtener respuestas a la dosis más detalladas por las poblaciones de Treg que presenten su alta sensibilidad frente a IgG DP47GS-IL-2. Es notable que la expresión alta de IL-2RB en los linfocitos NK con concentraciones intermedias de CD56 (figura 6D) en comparación con la expresión de IL-2RB en los subconjuntos de linfocitos T no sea suficiente para permitir la sensibilidad de IL-2 similar a Treg. En general, los subconjuntos de Treg activados, de memoria e indiferenciados mostraron la mayor sensibilidad a IgG DP47GS-IL-2, mientras que los linfocitos NK CD56fuerte y los linfocitos T efectores de memoria convencionales CD4+ fueron 20-50 veces menos sensibles. Entre los demás subconjuntos de células analizados frente a los incrementos de pSTAT5a, los linfocitos T efectores de memoria CD8+ , los linfocitos T efectores indiferenciados CD4+, los linfocitos NKT, los linfocitos NK “en reposo” (positivos, sin tinción fuerte para CD56) y los linfocitos T efectores indiferenciados CD8+ + de memoria CD45RA+ fueron relativamente insensibles al inmunoconjugado IgG-IL-2.
La figura 9 muestra la respuesta a la dosis de IgG DP47GS-(IL-2)2 en los linfocitos T, linfocitos NK y linfocitos T NK en sangre periférica humana de cinco donantes individuales, cada uno sometido a prueba en días diferentes. Como se observó para IgG DP47GS-IL-2 (figura 8), los tres subconjuntos de Treg fueron las células más sensibles a pSTAT5a inducido por IgG DP47GS-(IL-2)2 (figuras 9 y 11) y se descubrieron respuestas a la dosis jerárquicas similares para los demás subconjuntos de células en los cinco donantes.
Para comparar más fácilmente IgG DP47GS-IL-2 e IgG DP47GS-(IL-2)2 , se normalizaron los valores de pSTAT5a (figura 10). Para normalizar los valores de IMF, se restaron los valores de IMF de pSTAT5a sin estimular específicos para cada subconjunto seleccionado de todos los valores de IMF estimulados para ese subconjunto de células. Los valores resultantes se dividieron por el valor de IMF de pSTAT5a más alto obtenido por ese subconjunto en la respuesta a la dosis. Las CE50 se estimaron en base a la cantidad de proteína de fusión de IL-2 requerida para alcanzar un 50 % de la IMF de pSTAT5a máxima observada para ese subconjunto. Como se muestra en la figura 10, el inmunoconjugado IgG DP47GS-(IL2)2 produjo una inducción más potente de pSTAT5a en células que expresaban constitutivamente CD25, potencialmente como consecuencia de una avidez incrementada del inmunoconjugado por el receptor de IL-2 de afinidad alta. Se observó que la CE50 para la activación de pSTAT5a era 5-9 veces más baja en los Treg cuando se comparaba directamente IgG DP47GS-(IL-2)2 con IgG DP47GS-IL-2. La tabla 2 resume los valores de CE50 representativos y la tasa de diferencia para la activación de pSTAT5a mediante IgG DP47GS-IL-2 frente a IgG DP47GS-(IL-2)2 en los diferentes subconjuntos de células.
Tabla 2. Valores de CE50 y tasas de diferencia para la activación de pSTAT5a mediante IgG DP47GS-IL-2 frente a IgG DP47GS-(IL-2)2 en diferentes subconjuntos de células.

Incluso a concentraciones extremadamente limitantes, IgG DP47GS-(IL-2)2 produjo concentraciones más altas de pSTAT5a en comparación con IgG DP47GS-IL-2 (figura 11). Para este experimento, se sometió a prueba individualmente la sangre de tres voluntarios sanos en el mismo día para obtener respuestas con respecto a una valoración por duplicado de IgG DP47-IL-2 e IgG DP47-(IL-2)2 a concentraciones limitantes de IL-2. Los gráficos en
la figura 11 representan la media ± DE de la IMF de pSTAT5a para los tres donantes. Además de los tres subconjuntos de Treg examinados individualmente (figura 11B-D), se aplicó una selección para evaluar pSTAT5a en los Treg CD3+CD4+FoxP3+ totales (figura 11 A). Los resultados indican claramente un incremento de 5-10 veces en la potencia para la activación por IgG DP47GS-(IL-2)2 de los de Treg a pesar de solo un incremento de 2 veces en la IL-2 por molécula de IgG. Se realizó citometría de flujo policromática como se describe anteriormente (véase la figura 8).
Los linfocitos T efectores de memoria convencionales CD4+ también respondieron a concentraciones más bajas (7 veces) de IgG DP47GS-(IL-2)2 en comparación con IgG DP47GS-IL-2. Mientras que los valores de CE50 fueron más bajos para los linfocitos NK CD56fuerte y los linfocitos NK CD56+ al comparar IgG DP47GS-(IL-2)2 con IgG DP47-IL-2, la reducción solo fue de 3 veces y 4 veces, respectivamente. Esto probablemente se debe a la dependencia de estas células del receptor de IL-2 de afinidad intermedia para la señalización mediada por IL-2. Este cambio diferencial en la DE50 para los Treg frente a los linfocitos NK incrementa la preferencia por la activación de los Treg en varias veces.
La figura 12 muestra la respuesta a la dosis del IgG DP47GS-(IL-2E95A)2 en los linfocitos T, linfocitos NK y linfocitos T NK en sangre periférica humana de tres donantes individuales, cada uno sometido a prueba en días diferentes. Se diseñó la molécula de IL-2E95A y se predijo que reduciría la actividad de unión a IL-2RpY, pero, de hecho, presentó propiedades de unión similares al receptor de IL-2RpY como lo hizo la IL-2 natural. La jerarquía de la reactividad a IgG DP47GS-(IL-2E95A)2 fue la misma en los tres donantes y la misma que la observada en los mismos donantes con el inmunoconjugado IgG DP47GS-(IL-2)2 con IL-2 natural. Las tres poblaciones de Treg, activados (CD45RA-CD25a ), de memoria (CD45RA-CD25+) e indiferenciados (CD45RA+CD25+) incrementaron las concentraciones de pSTAT5a a menos de concentraciones de 0,1 ng/ml de IgG DP47GS-(IL-2E95A)2, mientras que otras poblaciones de células requirieron 1 ng/ml (linfocitos NK CD56fuerte y linfocitos T efectores de memoria convencionales CD4+) o bien 10-100 ng/ml (linfocitos T CD8+ de memoria, linfocitos NK CD56+ , linfocitos T CD4+ indiferenciados, linfocitos NKT y linfocitos T CD45RA+CD8+) de IgG DP47-(IL-2E95A)2 para producir incrementos detectables de pSTAT5a. Es notable que la expresión alta de IL-2RB en los linfocitos NK con concentraciones intermedias de CD56 (figura 6D) en comparación con la expresión de IL-2RB en los subconjuntos de linfocitos T no sea suficiente para permitir la sensibilidad de IL-2 similar a Treg. En general, los subconjuntos de Treg activados, de memoria e indiferenciados mostraron la mayor sensibilidad a IgG DP47GS-(IL-2E95A)2 , mientras que los linfocitos NK CD56fuerte y los linfocitos T efectores de memoria convencionales CD4+ fueron 20-50 veces menos sensibles. Entre los demás subconjuntos de células analizados frente a los incrementos de pSTAT5a, los linfocitos T efectores de memoria CD8+, los linfocitos T efectores indiferenciados CD4+, los linfocitos NKT, los linfocitos NK “en reposo” (positivos, sin tinción fuerte para CD56) y los linfocitos T efectores indiferenciados CD8+ CD45RA+ fueron relativamente insensibles al inmunoconjugado IgG-(IL-2E95A)2.
La figura 13 muestra la respuesta a la dosis de IgG DP47GS-(IL-2N88D)2 en los linfocitos T, linfocitos NK y linfocitos T NK en sangre periférica humana de cinco donantes individuales, cada uno sometido a prueba en días diferentes. Se diseñó la molécula de IL-2N88D para reducir la actividad de unión a IL2RpY y, a diferencia de la mutación puntual E95A, tenía 6,2 veces menos afinidad de unión a IL2RpY mediante análisis con Biacore (tabla 1, Kd 0,15 nM frente a 0,93 nM). Como se observó para todos los inmunoconjugados de IL-2 hasta la fecha, los tres subconjuntos de Treg fueron las células más sensibles a la activación de pSTAT5a mediante IgG DP47GS-(IL-2N88D)2 y se descubrieron respuestas a la dosis jerárquicas similares para los demás subconjuntos de células en los cinco donantes. Cabe destacar la falta de reactividad de los linfocitos T efectores de memoria CD4+ cuando se estimulan con IgG DP47GS-(IL-2N88D)2 ; incluso a dosis de 1000 ng/ml, tuvo poco efecto sobre la estimulación de esta importante población de células autoinmunitarias.
En este ejemplo, una mutación puntual única en IL-2 diseñada de manera única, N88D, redujo drásticamente la actividad estimuladora en la población de células deseada, los linfocitos T efectores de memoria CD4+ , todo mientras mantenía los efectos estimuladores de Treg requeridos para una entidad terapéutica novedosa y mejorada.
Para comparar IgG DP47GS-(IL-2)2 para IL-2 natural con los inmunoconjugados con las mutaciones puntuales en IL-2 E95A y N88D, se representaron los valores de pSTAT5a frente a sí mismos para cada uno de los tipos de células que se están sometiendo a prueba (figura 14). IgG DPS47GS-(IL-2E95A)2 mostró una actividad idéntica a su homólogo natural IgG DP47GS-(IL-2)2 estimulando cada una de las poblaciones de células humanas con curvas de respuesta a la dosis superponibles. En cambio, las moléculas de unión reducida a IL2RpY, IgG DP47GS-IL-2N88D e IgG DP47GS-(IL-2N88D2)2 , tuvieron poco efecto sobre los linfocitos T efectores de memoria CD4+ y solo el conjugado bivalente IgG DP47GS-(IL-2N88D)2 retuvo una actividad estimuladora significativa en las diferentes poblaciones de Treg.
Los resultados en la figura 15 muestran las respuestas de pSTAT5a de sangre completa obtenidas de diez donantes de sangre humana adicionales. En días separados, se sometió a prueba cada donante comparando un amplio intervalo de valoración (>6 log) de IgG DP47GS-(IL-2)2 e IgG DP47GS-(IL-2N88D)2. Como se observó previamente con IgG DP47GS-(IL-2)2 (figura 9), los Treg de memoria y los Treg indiferenciados fueron las células más sensibles a pSTAT5a inducido por IgG DP47GS-(IL-2)2 e IgG Dp47GS-(IL-2N88D)2 (figura 15A, B); mientras que cada proteína de fusión estimuló las respuestas de pSTAT5a máximas, diferían en las concentraciones necesarias para el
punto de inflexión de la activación, así como la dosis máxima para la activación. Los linfocitos NK CD56fuerte se presentaron como un subconjunto de células menos sensible que Treg a IgG DP47GS-(IL-2N88D)2 en comparación con las proteínas de fusión de IL-2 natural (figura 15C); el punto de inflexión de la activación fue más alto y, en este caso, nunca logró un efecto máximo incluso a la dosis más alta sometida a prueba (5000 ng/ml). A diferencia de IgG DP47GS-(IL-2)2, los linfocitos NK fueron no reactivos a IgG DP47GS-(IL-2N88D)2 (figura 15D). Tanto los linfocitos T CD4+ de memoria central (figura 15E) como los linfocitos T CD4+ de memoria efectora (figura 15F) fueron relativamente no reactivos a IgG DP47GS-(IL-2N88D)2 con un punto de inflexión de activación 1000 veces más alto que IgG DP47GS-(IL-2)2 e induciendo solo respuestas parciales a la dosis más alta sometida a prueba (5000 ng/ml). Mientras que IgG DP47GS-(IL-2)2 indujo diversos grados de activación, IgG DP47GS-(IL-2N88D)2 no tuvo ningún efecto sobre los siguientes subconjuntos celulares: linfocitos T CD4+ indiferenciados (figura 15G), linfocitos T CD8+ de memoria central (figura 15H), linfocitos T CD8+ de memoria efectora (figura 15I), linfocitos T CD8+ indiferenciados (figura 15J), linfocitos T CD8+ CD45RA+ de memoria efectora (figura 15K), linfocitos NKT (figura 15L) y linfocitos T CD3+CD4"CD8"CD56" (figura 15M).
Los valores de CE50 se muestran en la tabla 3 y se basaron en la cantidad de proteínas de fusión de IL-2 requeridas para alcanzar un 50 % de la respuesta de pSTAT5a máxima observada para ese subconjunto de células. IgG DP47GS-(IL-2E95A)2 , como su homólogo IgG DP47GS-(IL-2)2 para IL-2 natural, produjo la inducción más potente de pSTAT5a en los Treg, las células que expresan constitutivamente el receptor de afinidad alta CD25 (IL2Ra), potencialmente como consecuencia de su avidez incrementada por el receptor de IL-2 de afinidad alta. Las respuestas a la dosis a IgG DP47GS-(IL-2E95A)2 e IgG DP47GS-(IL-2)2 fueron superponibles. La CE50 para la activación de pSTAT5a en los Treg fue de 6 a 7 veces más baja con IgG DP47GS-(iL-2E95A)2 e IgG DP47GS-(IL-2)2 en comparación con IgG DP47GS-IL-2 monovalente. Asimismo, la CE50 para la activación de los Treg fue de 7 a 10 veces más baja con IgG DP47GS-IL-2 monovalente en comparación con IgG DP47GS-(IL-2N88D)2 bivalente. Y por último, la CE50 para la activación de los Treg fue de 36 a 61 veces más baja con IgG DP47GS-(IL-2)2 bivalente en comparación con IgG DP47GS-(IL-2N88D)2 bivalente.
La ventaja inmunológica más significativa que tiene IgG DP47GS-(IL-2N88D)2 para tratar los trastornos autoinmunitarios se ilustra mejor en la tabla 3, que demuestra el drástico margen de especificidad que tiene IgG DP47GS-(IL-2N88D)2 para la activación de los Treg frente a los linfocitos T efectores de memoria c D4+ (de >320 veces a >500 veces). En comparación, el margen de especificidad para los Treg frente a los linfocitos T efectores de memoria CD4+ es sustancialmente más bajo para IgG DP47GS-(IL-2)2 (de 13 a 14 veces) e IgG DP47GS-IL-2 (de 10 a 16 veces).
Tabla 3. Valores de CE50 y diferencias en las tasas de especificidad para la activación de pSTAT5a mediante IgG DP47GS-(IL-2E95A)2 , IgG DP47GS-IL-2 e IgG DP47GS-(IL-2N88D)2 en diferentes subconjuntos de células.

Efectos diferenciales de las proteínas de fusión de IL-2 sobre los linfocitos NK y los linfocitos T CD8+ humanos in vitro Se cultivaron in vitro células mononucleares de sangre periférica (PBMC) humana recién obtenida sin otra estimulación distinta de IL-2, condiciones conocidas que favorecen la supervivencia y el crecimiento de los linfocitos NK y de los linfocitos T CD8+ . Los linfocitos INK humanos se pueden subdividir en aquellos que expresan poco o ningún CD56 detectable (denominado CD56' /debil) y un subconjunto “activado” que expresa concentraciones altas de CD56 (denominado CD56fuerte); la mayoría de los linfocitos n K en la sangre son CD56-/débil con una minoría clasificada como CD56fuerte . Se cree que los linfocitos NK “activados” que son CD56fuerte son precursores de los linfocitos NK a los que les falta la expresión de CD56 (es decir, CD56-/débil). Después de seis días in vitro con IgG DP47GS-(IL-2)2 e IgG DP47GS-(IL-2N88D)2 (cada una a^ 0, 5, 50 y 500 ng/ml), se detectaron diferencias en el crecimiento y la supervivencia de los linfocitos NK CD56-/débil, los linfocitos NK CD56fuerte y los linfocitos T CD8+ . Se observaron diferencias significativas en los números de células entre los estímulos con IgG DP47GS-(IL-2)2 que estimulaban los mayores incrementos de los linfocitos CD56fuerte (una mediana de 101x103 frente a 38x103 , p<0,005) (figura 16A) y también de los linfocitos T CD8+ (una mediana de 13,6x103 células frente a 2,5x103 células, p<0,0001) (figura 16b ).
Efectos de las proteínas de fusión de IL-2 en ratones humanizados
Se construyeron ratones humanizados usando ratones NOD.Prkdcscid ¡L2rganulados (NSG) recién nacidos ligeramente irradiados como huésped inmunodeficiente e injertándoles células madre CD34+ de hígado humano fetal inyectadas i.p. A las de 8 a 10 semanas de edad, se sometió a prueba la sangre de cada ratón para asegurar que se había producido el injerto humano. Comparando Proleukin e IgG DP47GS-(IL-2)2 , se descubrió que las células primarias afectadas in vivo eran los linfocitos NK y Treg; tras el tratamiento con IgG DP47GS-(IL-2)2 hubo un incremento sustancial tanto en los linfocitos NK como en los Treg, respondiendo los linfocitos NK proporcionalmente mejor. Los efectos in vivo de Proleukin fueron similares, pero los efectos sobre los linfocitos NK y Treg en un grupo fueron menos consistentes y el grado de los incrementos se redujo en comparación con IgG DP47GS-(IL-2)2. Los ratones humanizados también pudieron diferenciar los efectos in vivo de las proteínas de fusión de IL-2. IgG DP47GS-(IL-2)2 natural incrementó tanto los Treg (figura 17A) como los linfocitos NK (figura 17B), mientras que IgG DP47GS-(IL-2N88D)2 no tuvo ningún efecto sobre los linfocitos NK (figura 17B), pero incrementó los Treg a un grado significativamente mayor (figura 17A) que la IgG-(IL-2)2 natural.
Una consecuencia a largo plazo de la humanización de los ratones NSG es su tiempo de supervivencia más corto debido a una respuesta del injerto contra huésped xenogénico humano que da como resultado una pérdida de peso y una insuficiencia multiorgánica. La administración de vehículo dos veces por semana no tuvo ningún efecto y dio como resultado una mediana de supervivencia de 36 días después del inicio del tratamiento (figura 18). De forma similar al vehículo, el tratamiento con IgG DP47GS-(IL-2N88D)2 dio como resultado una mediana de supervivencia de 37 días. En cambio, el tratamiento dos veces por semana con IgG DP47GS-(IL-2)2 acortó la mediana del tiempo de supervivencia de este grupo a 21 días (figura 18, p<0,0037 en comparación con IgG DP47GS-(IL-2N88D)2).
Cuando una gravedad predeterminada de la respuesta del injerto contra huésped dictó el final de la porción en vida para cada ratón (pérdida de peso >15 %), se evaluó la composición de células humanas en la sangre. Con una respuesta del injerto contra huésped potencialmente mortal a la vista, se observaron diferencias sorprendentes en las ramas de tratamiento en la composición final de los Treg, linfocitos NK y linfocitos T CD8+ en la sangre (figura 19). En los ratones tratados con vehículo, los Treg humanos representaron menos de un 1 % del total de células CD45+ humanas y los linfocitos NK y T CD8+ comprendieron ~3 % de las células CD45+ humanas en la sangre. El tratamiento con IgG DP47GS-(IL-2N88D)2 incrementó los linfocitos NK y T CD8+ a un 2 % y un 5 %, respectivamente, mientras que los Treg se incrementaron a un 6 %. En cambio, el tratamiento con IgG DP47GS-(IL-2)2 natural incrementó la fracción de los linfocitos NK y los linfocitos T CD8+ a un 30 % del total de células CD45+ humanas y los Treg a un 3,5 %, quizás explicando la reducción significativa de los tiempos de supervivencia en este grupo en comparación con los ratones tratados con IgG DP47GS-(IL-2N88D)2.
Inducción de pSTAT5a en subconjuntos de células de sangre periférica de macaco cangrejero
Como se observó en la sangre periférica humana, existe una inducción de pSTAT5a dependiente de la dosis preferente y similar en los subconjuntos de Treg en la sangre periférica de macaco cangrejero estimulada con IL-2 (Proleukin) (datos no mostrados). En una comparación directa de la capacidad de IgG DP47GS-(IL-2)2 e IgG DP47GS-IL-2 para inducir pSTAT5a en los tres subconjuntos de Treg, se requirió de 2 a 8 veces menos IgG DP47GS-(IL-2)2 para alcanzar un 50 % de la concentración máxima de pSTAT5a que para IgG DP47GS-IL-2. La tabla 4 resume los valores de CE50 para la activación de pSTAT5a mediante IgG DP47GS-IL-2 frente a IgG DP47GS-(IL-2)2 en los diferentes subconjuntos de Treg de macaco cangrejero, resultados que son sorprendentemente similares a los observados con sangre periférica humana (tabla 3).
De forma similar a la sangre completa humana, se usaron marcadores intracelulares y de superficie de las células para identificar subconjuntos de linfocitos T reguladores y linfocitos T convencionales en la sangre completa de macacos cangrejeros sanos normales. Se obtuvieron muestras de sangre en el mismo día de tres macacos cangrejeros sanos en tubos con heparina de sodio y se añadieron diversas concentraciones de IgG DP47GS-IL-2 o IgG Dp47GS-(IL-2)2 a 500 pl de sangre y se incubaron a 37 °C. Después de 10 minutos a 37 °C, las muestras se lisaron y se fijaron con tampón BD Lyse/Fix calentado previamente (BD Biosciences). Después del lavado, las células se permeabilizaron con 1 ml de metanol durante 30 min en hielo. Las muestras se lavaron 3 veces y se tiñeron con un panel de FOXP3-Alexa Fluor® 647 (clon: 259D, BioLegend), CD4-V500 (clon: L200, BD Biosciences), CD45RA-V450 (clon: 5H9, BD Biosciences), CD25-PE (clon: 4E3, eBioscience), pSTAT5a-Alexa Fluor® 488 (clon: 47, BD Biosciences) y Ki-67-PerCP-Cy5.5 (clon: B56, BD Biosciences) durante 1 hora a 4 °C. Todas las muestras se adquirieron mediante un analizador de células LSRFortessa™ (Becton Dickinson) y, entonces, se analizaron con el programa informático FlowJo (FlowJo, LLC).
Tabla 4. Inducción de pSTAT5a en subconjuntos de Treg de sangre periférica de macaco cangrejero en respuesta a IgG DP47GS-IL-2 e IgG DP47GS-(IL-2)2.

En otro experimento, similar a los ensayos de pSTAT5a con sangre humana, se estimuló in vitro la sangre heparinizada recién obtenida de macacos cangrejeros adultos sanos normales con PBS como control con vehículo, IgG DP47GS-(IL-2)2 e IgG DP47GS-(IL-2N88D)2 y se examinaron los efectos sobre la fosforilación de STAT5a en subconjuntos de Treg y linfocitos T efectores de memoria CD4+ convencionales. Se eligió una dosis de estimulación de 20 ng/ml basada en las respuestas máximas de Treg humanos a < 1 ng/ml y en las respuestas cuasimáximas de los linfocitos T efectores de memoria CD4+ convencionales en ese intervalo de dosis (resultados en la figura 9).
Las condiciones experimentales fueron como se describe anteriormente. Se obtuvieron muestras de sangre en el mismo día de tres macacos cangrejeros sanos (donantes C1, C2, C3) en tubos con heparina de sodio y se añadieron 20 ng/ml de IgG DP47GS-(IL-2)2 o IgG DP47GS-(IL-2N88D)2 a 500 pl de sangre y se incubó a 37 °C durante 20 minutos antes de su lisis y análisis.
La estrategia de selección de la citometría de flujo para dividir Treg CD4+CD25+ de macaco cangrejero en subconjuntos indiferenciados (FOXP3+CD45RA+), de memoria (FOXP3+CD45RA- ) y activados (FOXP3aCD45RA-) para su análisis se muestra en la figura 20A. Como en los seres humanos, los tres subconjuntos de Treg CD4+CD25+FOXP3+ de cada donante expresaron concentraciones altas del receptor de IL-2 de afinidad alta CD25 (figura 20B). Después de la estimulación in vitro con 20 ng/ml tanto de IgG DP47GS-(IL-2)2 como de IgG DP47GS-(IL-2N88D)2 , los Treg de los tres donantes produjeron una fosforilación de STAT5a significativa (figura 20C). De forma similar a los humanos, los Treg activados y de memoria de macaco cangrejero mostraron un mayor potencial para la inducción de pSTAT5a en comparación con los Treg indiferenciados.
Los linfocitos T efectores de memoria CD4+ convencionales humanos, aunque no sean tan sensibles como los Treg, se pueden estimular con IL-2 con una DE90 estimada de 20 ng/ml para IgG DP47GS-(IL-2)2 usando pSTAT5a como el biomarcador de activación (figura 14). Los linfocitos T efectores de memoria CD4+ convencionales de macaco cangrejero se pueden identificar de forma similar en la sangre como células CD4+FOXP3-CD45RA- , siendo la mayoría de las células CD25- y siendo un subconjunto más pequeño CD25+ (figura 21A). Cuando se tomaron como una población completa, los linfocitos Tef de memoria de macaco cangrejero de los tres donantes respondieron a 20 ng/ml de IgG Dp47GS-(IL-2)2 con incrementos de pSTAT5A mientras que 20 ng/ml de IgG DP47GS-(IL-2N88D)2 tuvieron poco (mc 2) o ningún efecto (mc 1 y 3) sobre pSTAT5a (figura 21b ). Cuando los linfocitos Tef de memoria se subdividieron además en los subconjuntos CD25" y CD25+ , se descubrió que la respuesta a IgG DP47GS-(IL-2)2
principalmente residía en el subconjunto CD25+ (figura 21D) con poca respuesta en el subconjunto CD25' (figura 21C). En cambio, IgG DP47GS-(IL-2N88D)2 no tuvo efecto sobre los linfocitos Tef de memoria CD25- (figura 21C) y tuvo un 70-80 % menos de actividad estimuladora sobre el subconjunto CD25+ (figura 21D).
Los estudios adicionales en la sangre de macaco cangrejero normal recién obtenida compararon directamente concentraciones variables de IgG DP47GS-(IL-2)2 e IgG d P47GS-(IL-2N88D)2 de una manera con respuesta a la dosis para determinar su capacidad para estimular pSTAT5a en diferentes subconjuntos de linfocitos T, es decir, los Treg activados, indiferenciados y de memoria y los linfocitos T efectores de memoria CD4+ . Tanto IgG DP47GS-(IL-2)2 como IgG DP47GS-(IL-2N88D)2 estimularon los Treg de una manera dependiente de la dosis con la proteína de fusión de IL-2 bivalente natural ~10 veces más potente que la proteína de fusión de IL-2N88D bivalente en cada uno de los subconjuntos de Treg (figura 22A-C). En cambio, se observó una drástica diferencia entre las dos moléculas cuando se evaluaron los linfocitos T efectores de memoria CD4+ para determinar la inducción de pSTAT5a (figura 22D). Se presentó IgG DP47GS-(IL-2)2 con una ~CE90 a 300 ng/ml, mientras que IgG DP47g S-(IL-2N88D)2 virtualmente no tuvo efecto a esa dosis alta. Los valores de CE50 para la estimulación de pSTAT5a de macaco cangrejero se muestran en la tabla 5, así como las tasas de especificidad relativas para estimular los Treg frente a los linfocitos T efectores de memoria CD4+. Estos resultados en la sangre completa de macaco cangrejero son bastante similares a los resultados que usan sangre completa humana (figura 14 y tabla 3) y sugieren que los estudios en el macaco cangrejero pueden tener un valor predictivo alto para los ensayos clínicos en seres humanos.
Tabla 5. Valores de CE50 y diferencias en las tasas de especificidad para la activación de pSTAT5a mediante IgG DP47GS-(IL-2)2 e IgG DP47GS-(IL-2N88D)2 en subconjuntos de linfocitos T de macaco cangrejero.
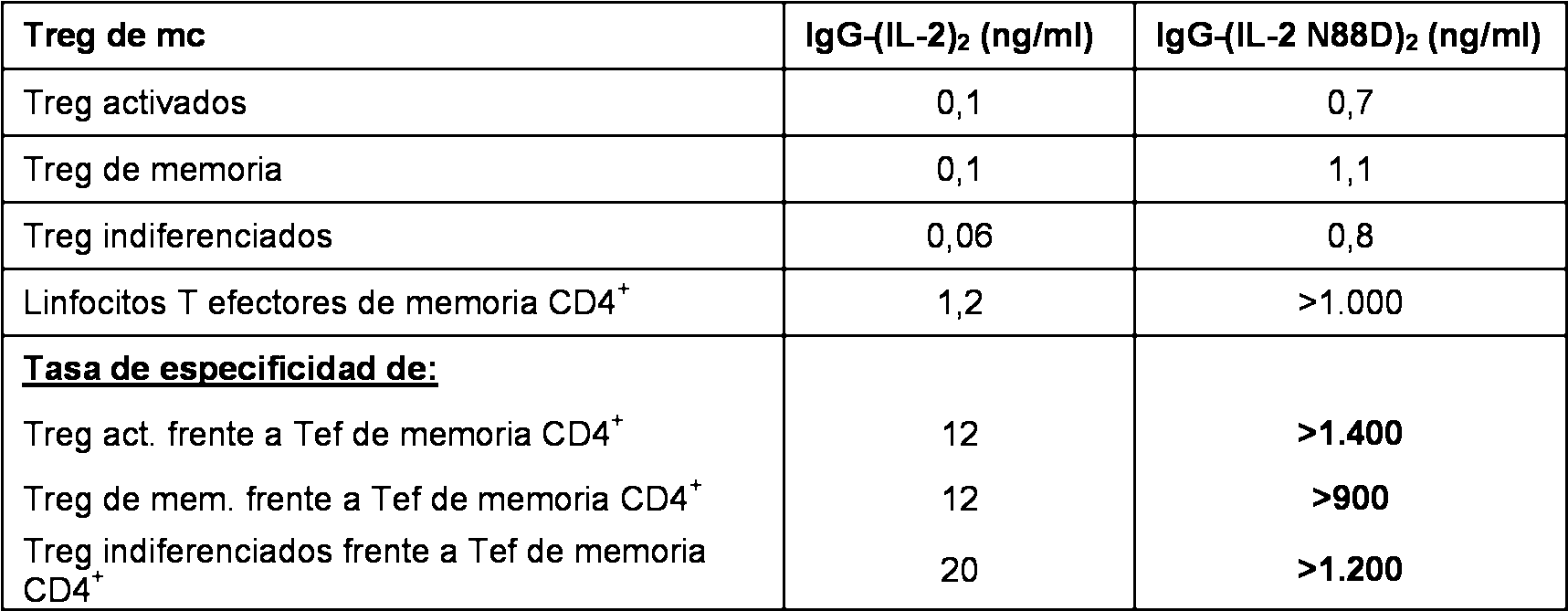
Propiedades farmacocinéticas en ratones
Antes de realizar los estudios funcionales en ratones, se evaluaron las propiedades farmacocinéticas (PF) de los inmunoconjugados de IL-2 monovalentes y bivalentes naturales y con N88D en ratones NOD, NOD.scid y NOD.scid.//2ra/_ (figura 23).
A ratones NOD y NOD.scid (n=3) se les inyectaron por vía intravenosa (i.v.) 0,3 mg/kg por ratón de IgG DP47GS-IL-2 o IgG DP47GS-(IL-2)2 en PBS que contenía suero de ratón al 0,5 % y se les extrajo sangre en diversos tiempos después de la inyección, variando desde 2 minutos a 168 horas (figura 23A). La IL-2 humana se evaluó en las muestras de suero de ratón usando mAb anti-IL-2 humana de ratón (BD Pharmingen, n.° 555051, clon 5344.111) para recubrir placas de 96 pocillos a fin de capturar la IL-2. Entonces, se detectó IL-2 humana usando mAb biotinilado anti-IL-2 humana de ratón (BD Pharmingen, n.° 555040, clon B33-2). La unión a IL-2 se visualizó y cuantificó usando estreptavidina conjugada con europio. Durante las primeras 24 horas, IgG DP47GS-(IL-2)2 se aclaró más rápidamente que IgG DP47GS-IL-2 tanto en ratones nOd como en NOD.scid. En 48 horas, las concentraciones en suero de cada uno estaban a concentraciones similares y todos estaban por debajo del límite de detección a las 72 horas. El límite de detección de IL-2 fue de 0,05 ng/ml y se indica mediante la línea de puntos. El aclaramiento sorprendentemente rápido de los inmunoconjugados IgG-IL-2 tanto monovalentes como bivalentes en ratones sin un sistema inmunitario adaptativo sugirió que existe un compartimento no hematopoyético con IL-2 o “sumidero” en ratones y podría estar promoviendo el aclaramiento rápido in vivo de los inmunoconjugados de IL-2.
Para someter a prueba esta hipótesis, se realizaron estudios PF en ratones NOD. scid que también carecían del receptor de IL-2 de afinidad alta, IL2Ra (CD25), es decir, ratones NOD.sdd.//2ra~A (figura 23B). En los ratones NOD.sdd.//2ra-/', IgG DP47GS-(IL-2)2 , a pesar de que se administra a una dosis tres veces más alta (0,3 mg/kg), se aclaró más rápidamente durante las primeras 24 a 48 horas que 0,1 mg/kg de IgG DP47GS-IL-2 y 0,1 mg/kg de IgG DP47GS-(IL-2N88D)2. Sin embargo, después de 48 horas, las concentraciones en la sangre de cada conjugado presentaron una fase de eliminación lenta y constante con IL-2 fácilmente detectable en la sangre hasta 6 días. De forma interesante, en estos ratones carentes del receptor de IL-2 de afinidad alta, la molécula IgG DP47GS-(IL2N88D)2 tuvo una fase de distribución inicial de 24 horas ligeramente más rápida que IgG-IL-2, pero luego presentó una fase de eliminación similar a la del inmunoconjugado monovalente natural hasta seis días, la última vez que se sometieron a prueba. Estos datos para IgG DP47GS-(IL-2N88D)2 son, en particular, pertinentes, puesto que los resultados PF en los ratones NOD.sdd.Il2ra~A parecen predecir estrechamente los resultados PF en primates no humanos.
Incremento de FOXP3 y CD25 en los Treg de ratón después del tratamiento con IL-2
Para comparar las capacidades de diferentes formas y configuraciones moleculares de IL-2 humana para estimular los Treg FoxP3+ in vivo, a los ratones se les inyectó por vía subcutánea un vehículo, Proleukin (IL-2 humana recombinante), IgG DP47GS-IL-2, IgG DP47GS-(IL-2)2 o IgG DP47GS-(IL-2N88D)2 y se evaluaron los Treg para determinar los cambios en la expresión de CD25 en la superficie de las células y FOXP3 intracelular en un tiempo óptimo determinado previamente de 24 horas (figura 24). A ratones BALB/c jóvenes y sanos (n=3/grupo de tratamiento, n=4/cohortes con vehículo) se les inyectó por vía subcutánea Proleukin (20.000 o 100.000 UI/ratón), IgG DP47GS-IL-2 (60, 300 o 1500 UI/ratón), IgG DP47GS-(IL-2)2 (60, 300 o 1500 UI/ratón) o IgG DP47GS-(IL-2N88D)2 (60, 300 o 1500 UI/ratón). Las dosis se administraron en un vehículo que comprendía 100 pl de PBS estéril, pH 7,2, que contenía un 0,5 % de suero de ratón filtrado de forma estéril. Después de 24 horas, se sacrificaron los ratones y se extirparon los bazos. Se generó una suspensión de células sueltas de esplenocitos en 1 ml de medio L-15 y se almacenó en hielo, hasta su procesamiento adicional. Se transfirió una alícuota filtrada de la suspensión de células sueltas, 40 pl, a tubos de FACs y se lavó con 2 ml de tampón de FACS (600 x g, 5 min). Entonces, las muestras se incubaron con anticuerpos conjugados con fluorocromo dirigidos frente a los antígenos de superficie celular: CD4 (clon RM4-5, fluorocromo A700), CD25 (eBio7D4, Af488), CD44 (IM7, e605), CD62L (MEL-14, PE), ICOS (C398.4A, PE/Cy7), CD103 (2E7, APC). La tinción se realizó durante 30 min a 4 °C en 100 pl de tampón de FACS (PBS pH 7,2 BSA al 0,2 %). Tras la tinción de la superficie de las células, las muestras se lavaron con 4 ml de tampón de FACS (600 x g, 5 min) antes de la tinción intracelular (de acuerdo con el protocolo de tinción intracelular de eBioscience). En resumen, las muestras se resuspendieron en 200 pl de tampón de fijación/peririeabiNzación (eBioscience n.° 00 5521) y se incubaron durante 1 h, 4 °C. Se añadió 1 ml de 1 x de tampón de permeabilización (eBiosciencen n.° 00 8333) a las muestras antes de 3 ml de tampón de FACS y lavado (600 x g, 5 min). Se tiñeron los antígenos intracelulares Ki-67 (B56, PerCP Cy5.5) y fOx P3 (FJK-16S, e450) en 100 pl 1x de tampón de permeabilización durante 1 h, 4 °C. Las muestras se lavaron con 4 ml de tampón de FACS (600 x g, 5 min, dos veces) y los datos se adquirieron en un analizador BD Fortessa™ y se analizaron usando el programa informático FlowJo (FlowJo, LLC). Los Treg se definieron como CD4+FOXP3+ de singletes en la selección de los linfocitos; a partir de esta población, se calcularon las intensidades medias de fluorescencia (IMF) de CD25 y FOXP3 para todas las muestras.
Como se muestra en la figura 24, los tres inmunoconjugados de IL-2 fueron igualmente potentes en la estimulación de la expresión de CD25 (24A) y FoxP3 (24B) en los Treg de ratón y mostraron respuestas dependientes de la dosis consistentes en el intervalo de 60, 300 y 1500 UI/dosis de ratón. En todos los casos, 1500 UI de cada inmunoconjugado (barras en negro) fueron significativamente más eficaces que 100.000 UI de Proleukin (barras en negro). En la mayoría de los casos, 300 UI de los inmunoconjugados (barras en gris oscuro) fueron equivalentes a 100.000 UI de Proleukin, y 60 UI de los inmunoconjugados (barras en gris claro) fueron equivalentes a 20.000 UI de Proleukin (barras en gris claro). En todos los grupos de tratamiento, los datos se muestran como la media ± DE.
Propiedades farmacocinéticas en macacos cangrejeros
Antes de realizar estudios funcionales en primates no humanos, se evaluaron las propiedades farmacocinéticas (PF) de los inmunoconjugados de IL-2 en macacos cangrejeros adultos sanos sin tratamiento previo con biofármaco (figura 25). A los monos (n=2/dosis) se les inyectó por vía intravenosa (i.v.) una corta inyección intravenosa rápida de IgG DP47GS-IL-2 o IgG DP47GS-(IL-2)2 estéril en PBS que contenía suero de macaco cangrejero al 0,5 % y se les extrajo sangre en diversos tiempos después de la inyección, variando desde 30 minutos a 72 horas. La IL-2 humana se evaluó en las muestras de suero de macaco cangrejero usando mAb anti-IL-2 humana de ratón (BD Pharmingen, n.° de catálogo 555051, clon 5344.111) para recubrir placas de 96 pocillos a fin de capturar la IL-2 humana. Entonces, se detectó IL-2 humana usando mAb biotinilado anti-IL-2 humana de ratón (BD Pharmingen, n.° de catálogo 555040, clon B33-2). La unión a IL-2 se visualizó y cuantificó usando estreptavidina conjugada con europio. La concentración de detección usando suero de macaco cangrejero en el ensayo fue de 0,05 ng/ml de IL-2 (línea de puntos en las figuras). El suero de macaco cangrejero tomado antes del tratamiento no tenía ninguna IL-2 detectable (por lo tanto, < 0,05 ng/ml).
Se detectó IL-2 en suero de macaco cangrejero desde 30 minutos a 48 horas después de la inyección i.v. de todas las dosis de IgG DP47GS-IL-2 (figura 25A) e IgG DP47GS-(IL-2)2 (figura 25B). En 72 horas, las concentraciones en suero de IL-2 estaban por debajo del límite de detección (0,05 ng/ml, línea de puntos) para todos los animales tratados con 10 y 25 pg/kg de IgG DP47GS-IL-2 o IgG DP47g S-(IL-2)2. Después de las dosis de 100 pg/kg de IgG DP47GS-IL-2, la IL-2 en suero detectable todavía estaba presente a las 72 horas.
Inducción del número de Treg en macacos cangrejeros
Los animales macacos cangrejeros tratados in vivo con IgG DP47GS-IL-2 tuvieron incrementos dependientes de la
dosis en el número absoluto de Treg, así como la tasa de incremento por encima del valor de referencia 7 días posteriores a la dosificación (figura 26A y 26B, respectivamente). En todas las pruebas se usaron macacos cangrejeros sanos normales de ambos sexos en edades que variaban desde los 3 a los 6 años y no se usó ningún animal más de una vez. Durante la anestesia, se inyectaron s.c. diversas dosis de IgG DP47GS-IL-2 (n=4-6) o vehículo (n=3) en el costado lateral. Las dosis individuales de IgG DP47GS-IL-2 se basaron en el peso corporal y se formularon para su inyección en un vehículo de PBS estéril, pH 7,2, que contenía suero de macaco cangrejero normal estéril al 0,5 %. Se obtuvieron muestras de sangre en diversos tiempos posteriores al tratamiento y se sometieron a prueba para determinar los cambios hematológicos (hemograma y diferencial) con un analizador automático para hematología Advia, así como los marcadores intracelulares y de superficie de las células detallados anteriormente (procedimientos experimentales para la tabla 4). Los cambios en los linfocitos T reguladores CD4+CD25+FOXP3+ en la sangre completa en el día 7 posterior al tratamiento se muestran en la figura 26 como el número de células absoluto por mm3 de sangre completa (figura 26A) y la tasa de cambio en los Treg (figura 26B); todos los datos se representan como la media ± DE. En las dosis más altas de 25 pg/kg y 36 pg/kg de IgG DP47GS-IL-2, se observaron incrementos en los Treg promedio de prácticamente 3 veces (intervalo de 111-255 %, n=6) y 4 veces (intervalo de 110-470 %, n=6), respectivamente. La reducción adicional de la dosis s.c. de IgG DP47GS-IL-2 (12, 6 y 2 pg/kg) continuó produciendo cambios reducidos de forma correspondiente en el número de Treg y en la tasa de cambio. Sin pretender vincularse a ninguna teoría, el incremento ~2 veces en los Treg (que varía desde un 67-133 %, n=6) con la dosis de 12 pg/kg de IgG DP47GS-IL2 puede representar el incremento deseable en los Treg para algunas enfermedades autoinmunitarias e inflamatorias. Existe un gran intervalo en los números de Treg en los seres humanos (20-90 Treg por mm3 de sangre; de un 4 a un 10 % de linfocitos T CD4+) y es razonable suponer que un incremento de los Treg inducidos por IL-2 en un individuo dará como resultado un incremento general de la supresión funcional.
Cuando se evaluaron las dosis más bajas de IgG DP47GS-IL-2 (2-6 pg/kg), se obtuvieron muestras de sangre más frecuentes para identificar los tiempos óptimos para detectar los cambios en los Treg posteriores a la administración de IgG DP47GS-IL-2 (figura 27). Mientras que estuvieron presentes cambios en los Treg a los 7 días, la estimulación máxima se produjo en el día 4, antes de lo que se midió con las dosis más altas de IgG DP47GS-IL-2 (día 7, figura 26).
Proleukin e IgG DP47GS-IL-2 son comparables in vitro en ensayos de sangre completa humana y de macaco cangrejero al comparar directamente mediante las unidades de actividad de IL-2 usadas para estimular pSTAT5a en los Treg y otras células reactivas a IL-2 (datos no mostrados). Con la corta semivida conocida de Proleukin en los seres humanos, se realizó un estudio de dosis única en macacos cangrejeros para evaluar in vivo la inducción y activación de Treg. Proleukin produjo un cambio dependiente de la dosis y del tiempo en los Treg medido mediante la tasa de cambio en los números absolutos, así como en la activación de pSTAT5a. Mientras que el incremento en los números de Treg fue nominal (figura 28A: un 10-40 %), el incremento inducido por Proleukin® en pSTAT5a en los Treg fue sustancial y dependiente de la dosis un día después del tratamiento (figura 28B), pero de corta duración in vivo, regresando a la normalidad del día 2 al 3.
En la figura 29 se muestra una comparación de la capacidad de IgG DP47GS-IL-2 para inducir un incremento en los Treg in vivo en macacos cangrejeros con la de Proleukin. Se trataron macacos cangrejeros sanos normales (grupos de n=5) con dosis bajas de IgG DP47GS-IL-2 o dosis altas de Proleukin y se sometió a prueba el cambio en los linfocitos T reguladores el día 10. En los días 0 y 7, se administró s.c. IgG DP47-IL-2 a una dosis de 16.800 UI/kg (la dosis de 12 pg/kg mostrada en la figura 26). En base al trabajo publicado donde se administró Proleukin a pacientes con diabetes de tipo 1 (4,5x106 UI/persona, 3 veces/semana), que demostró que incrementaba los Treg, y los datos de dosis única en la figura 28, se administró s.c. Proleukin 3 veces por semana (L/M/V) en un total de 5 dosis cada una a 200.000 UI/kg (el equivalente para macaco cangrejero de 4,5x106 UI/persona). Los resultados se muestran en la figura 29 como la media ± DE para el cambio en los Treg totales por mm3 de sangre (figura 29A), la tasa de incremento en el número de Treg (figura 29B) y el cambio en la proporción de Treg con respecto a las células CD4+ FOXP3- convencionales (figura 29C).
Aunque se administró IgG DP47GS-IL-2 prácticamente 30 veces menos durante el periodo de 10 días, IgG DP47GS-IL-2 indujo un incremento más grande en el número de Treg que Proleukin (figura 29A, p=0,06). La tasa de incremento de los números de Treg por encima del valor de referencia (figura 29B) y el incremento de los Treg en relación con los linfocitos T CD4 convencionales (figura 29C) también fueron más grandes (p=0,0011 y p=0,016, respectivamente) en los monos a los que se les dosificó IgG DP47GS-IL-2 en comparación con Proleukin. En los seres humanos, se usa a menudo la proporción de linfocitos T reguladores CD4+ (normalmente definidos como FOXP3+ y una combinación de marcadores de superficie) con respecto a los linfocitos T CD4 no reguladores (denominados células convencionales o efectoras) para definir las concentraciones funcionales de Treg en los pacientes a través del tiempo.
Respuesta in vivo de subconjuntos de células de sangre periférica de macaco cangrejero al tratamiento con IgG DP47GS-IL-2 en dosis baja
La especificidad celular in vivo del tratamiento con IL-2 en dosis baja es un parámetro crítico. Se ha determinado que se puede supervisar con sensibilidad la activación de las células in vivo inducida por IgG DP47GS-IL-2 o Proleukin
midiendo las concentraciones de pSTAT5a ex vivo en la sangre extraída en diversos tiempos después de su dosificación en macacos cangrejeros o ratones. La respuesta in vivo de todas las poblaciones de células que se pueden supervisar in vitro (figuras 8-10) también se puede examinar ex vivo.
Uno y 3 días después de la administración in vivo de una dosis baja única de IgG DP47GS-IL-2 (12 pg/kg) a macacos cangrejeros sanos (n=5), se obtuvo sangre completa y se sometió a prueba para determinar la fosforilación de STAT5a como se describe anteriormente (procedimientos experimentales con respecto a la tabla 4). Se extrajo sangre de cada mono en el día 0 antes del tratamiento y se midió la cantidad de fosforilación de STAT5a y se usó individualmente para evaluar las tasas de cambio posteriores al tratamiento. La tasa de cambio en pSTAT5a en los Treg en los días 1 y 3 se muestra en la figura 30A, la tasa de cambio en pSTAT5a en los linfocitos T efectores de memoria CD4+CD45RA" convencionales en la figura 30B y la tasa de cambio en pSTAT5a en los linfocitos T efectores indiferenciados CD4+CD45RA+ convencionales están en la figura 30C. Los resultados muestran claramente que la sangre de macaco cangrejero obtenida uno y tres días después de una dosis baja única (12 pg/kg) de IgG DP47GS-IL-2 mostró incrementos de pSTAT5a preferentes en los linfocitos Treg en comparación con los linfocitos T CD4+ de memoria e indiferenciados (figura 30A-C). IgG DP47GS-IL-2 en dosis baja única fue claramente superior a Proleukin en dosis alta única (figura 28B) en la producción de un estado de activación de Treg mantenido in vivo.
Los estudios iniciales que usaron la fosforilación de STAT5a como un biomarcador in vivo de la activación de los Treg experimentaron un pSTAT5a máximo solo el día 1 con Proleukin en dosis única (figura 28B) y 1 y 3 días con IgG DP47GS-IL-2 en dosis baja (12 pg/kg) (figura 30A). La valoración adicional de IgG DP47GS-IL-2 combinada con tiempos de muestreo adicionales mostró que las señales de pSTAT5a in vivo se podrían mantener durante 4 días antes de reducirse a la normalidad a los 7 días (figuras 31A, 31B). La IMF (intensidad media de fluorescencia) de las señales de pSTAT5a, tanto para la dosis de 2 pg/kg (31A, n=4) como de 6 pg/kg (31B, n=6), sugirió que se producía una señalización de pSTAT5a más baja que la máxima (véase la figura 33B). Incluso estas dosis muy bajas de IgG DP47GS-IL-2 proporcionaron una señalización de Treg mantenida in vivo y fueron superiores a Proleukin (figura 28B). Estos resultados respaldan la hipótesis de que concentraciones muy bajas de IL-2 de larga duración, es decir, IgG DP47GS-IL-2, pero no Proleukin, pueden estimular los Treg durante periodos prolongados de tiempo in vivo, reduciendo, de este modo, la frecuencia de tratamiento necesaria para restablecer la autotolerancia dominante y mejorar la evolución de la enfermedad. El incremento en los linfocitos Treg en la sangre periférica después del tratamiento con IL-2 en dosis baja podría reflejar un cambio en la distribución de las células en el cuerpo en lugar de un incremento real de las células. Para justificar que los incrementos en los Treg in vivo se deben, al menos en parte, a la inducción de la división de las células mediante el tratamiento con IL-2, se evaluó el marcador de proliferación intracelular Ki-67. Ki-67 es una proteína que se puede detectar en el núcleo durante G1, S, G2 y mitosis, pero está ausente en las células en reposo que están en la fase G0 del ciclo celular. También se supervisaron los macacos cangrejeros tratados con 2 y 6 pg/kg de IgG DP47GS-IL-2 descritos anteriormente (figura 31) para determinar cambios ex vivo en el marcador intracelular Ki-67 como se describe (procedimientos experimentales con respecto a la tabla 4) para evaluar el grado de proliferación in vivo. Se comparó el porcentaje de células que estaban en el ciclo celular (Ki-67+) en el día 0 con el porcentaje de células Ki-67+ a los 1 a 11 días posteriores al tratamiento. Los Treg Ki-67+ se muestran en la figura 32A, los linfocitos T efectores de memoria CD4+CDRA45- convencionales se muestran en la figura 32B y los linfocitos T efectores de indiferenciados CD4+CD45RA+ convencionales se muestran en la figura 32C. Las células sanguíneas de macaco cangrejero obtenidas de uno a siete días después de una dosis baja única de IgG DP47GS-IL-2 (6 pg/kg) mostraron incrementos de Ki-67 preferentes en los Treg en comparación con los linfocitos T efectores indiferenciados CD4+ convencionales (figura 32A frente a 32C). A diferencia de los linfocitos T efectores indiferenciados, IgG DP47GS-IL-2 (6 pg/kg) pudo estimular la proliferación de linfocitos T efectores CD4+ de memoria convencionales (figura 32B). En la dosis más baja de IgG DP47GS-IL-2 (2 pg/kg), hubo una activación mínima en el ciclo celular y una proliferación en comparación con la dosis de 6 pg/kg.
En los ensayos de sangre completa humana y de macaco cangrejero la actividad in vitro de IgG DP47GS-(IL-2)2 bivalente fue, en conjunto, aproximadamente 6 veces más potente en los Treg que con IgG DP47GS-IL-2 monovalente (tablas 2 y 4). De ahí que la primera dosis en los macacos cangrejeros fuera 6 veces más baja que la dosis más alta de IgG DP47GS-IL-2 sometida a prueba (36 pg/kg). IgG DP47GS-(IL-2)2 administrada a 6 pg/kg (n=4) produjo grandes incrementos en los Treg en circulación (figura 33A) que permanecieron muy por encima de la normalidad 14 días después del tratamiento, pSTAT5a significativo y prolongado en los Treg que regresaron a la normalidad en 1 semana (figura 33B) y prácticamente un incremento de 3 veces en el número de Treg (figura 33C). Las tasas de cambio inducidas mediante IgG DP47GS-IL-2 (de la figura 26, barras abiertas) se compararon con la dosis de 6 pg/kg de IgG DP47GS-(IL-2)2 bivalente (barra sombreada) (figura 33D). De forma similar a los ensayos de sangre completa humana y de macaco cangrejero, IgG DP47GS-(IL-2)2 presentó una potencia in vivo potenciada más allá de su incremento de 2 veces en IL-2 por molécula de IgG.
Se administraron dosis adicionales de IgG DP47GS-(IL-2)2 bivalente a macacos cangrejeros para evaluar sus efectos in vivo a dosis muy bajas (2 y 0,7 pg/kg) (figura 34). Mientras que 2 pg/kg produjeron tasas de cambio moderadas (media de un 46 %, variando desde un 20 a un 70 %, n=8), la dosis de 0,7 pg/kg tuvo poco efecto sobre los números de linfocitos T (media de incremento de un 16 %, n=3) (figura 34A). Ambas dosis bajas estimularon incrementos de pSTAT5a en los Treg (figura 34B, n=3 cada uno), mientras que tuvieron poco efecto sobre pSTAT5a en linfocitos Tef/de mem. (figura 34C, n=3 cada uno).
Los efectos generales del tratamiento con la proteína de fusión de IL-2 natural monovalente y bivalente y su capacidad para incrementar los Treg en macacos cangrejeros se presentan en la figura 35. Ambas formas de IgG-IL-2 de larga duración son potentes inductores de los Treg que se pueden seguir in vivo con pSTAT5a como biomarcador de activación y en esta figura como la tasa de incremento en los números de Treg en circulación. IgG DP47GS-(IL-2)2 con su potencia potenciada lograda duplicando las moléculas de IL-2 en el extremo C de la IgG tiene una superioridad dependiente de la dosis sobre IgG DP47GS-IL-2.
Desmetilación del ADN de FOXP3 y CTLA-4 en subconjuntos de linfocitos T de macaco cangrejero
Los Treg funcionalmente activos pueden producir las citocinas inmunodepresoras CTLA-4, IL-10 y TGFp y requieren la expresión estable del factor de transcripción FOXP3. La expresión de FOXP3 en los Treg humanos y de macaco cangrejero depende de la desmetilación de los diez sitios de metilación en el ADN con CpG en la región desmetilada específica de Treg (TSDR) en el locus de FOXP3. Asimismo, la expresión y producción de CTLA-4 depende de la desmetilación de los siete sitios de metilación en el ADN con CpG en el locus de CTLA-4 humano y de macaco cangrejero. En consecuencia, se evaluó el estado de desmetilación tanto de FOXP3 como de CTLA-4 en diversos subconjuntos de linfocitos T CD4+ antes y después del tratamiento con IgG DP47GS-IL-2 (n=4, 25 pg/kg s.c.) e IgG DP47GS-(IL-2)2 (n=4, 6 pg/kg s.c.) administradas a dosis y tiempos mostrados previamente para incrementar significativamente el número de Treg en la sangre. En el estudio, solo se usaron macacos cangrejeros adultos macho sin tratamiento previo con biofármaco y sus pesos variaban desde 9,1 a 10,9 kg. Se administraron los tratamientos 3 semanas después de la obtención de sangre en el valor de referencia inicial y se obtuvo sangre posterior al tratamiento 4-5 días más tarde. Se clasificaron PBMC de 30 ml de sangre heparinizada usando un FACSAria™ (Becton Dickinson) en los subconjuntos CD4+ pertinentes incluyendo los Treg (CD4+CD25+CD127baja), los linfocitos T efectores de memoria (CD4+CD45RA-), los linfocitos T efectores indiferenciados (CD4+CD45RA+) y las células CD4+CD25' CD127‘ .
Se congelaron los subconjuntos de células clasificados en alícuotas de 100.000 células y se almacenaron como sedimentos secos a -80 °C para permitir el procesamiento de todas las muestras juntas. Los sedimentos celulares se descongelaron y el ADN se extrajo y se trató con bisulfito en una etapa única usando el kit Epitect Fast Lyse All (Qiagen) siguiendo las instrucciones del fabricante. Se usaron 5 ng (en 2 pl) de ADN tratado con bisulfito en una primera tanda de PCR doble de 20 pl que contenía 10 pl del kit Multiplex PCR (Qiagen), 6 pl de agua, 0,5 pl del cebador directo de FOXP3 tgtaaaacgacggccagtTTTAGAAGTTGTATGGGGGATGTT (SEQ ID NO: 54), 0,5 pl del cebador inverso de FOXP3 caggaaacagctatgaccAAAATATCTACCCTCTTCTCTTCCTC (SEQ ID NO: 55), 0,5 pl del cebador directo de CTLA-4 tgtaaaacgacggccagtGGGTTTGGTTATGAAGGAGTATGA (SEQ ID NO: 56) y 0,5 pl del cebador inverso de CTLA-4 caggaaacagctatgaccTTCACTTAATTTCCACTAAAAATACCC (SEQ ID NO: 57). El ciclado de la primera tanda de PCR fue 95 °C durante 15 minutos, seguido de 20 ciclos de 95 °C durante 30 segundos, 60 °C durante 90 segundos y 72 °C durante 60 segundos, seguido de 10 minutos a 72 °C. El producto de PCR se purificó usando microesferas de AMPure XP (Becton Coulter) de acuerdo con las instrucciones del fabricante, y se eluyó en 20 pl de agua. Se realizó una segunda tanda de PCR que añadió una secuencia índice única al comienzo de cada muestra, a través de una PCR de 15 pl que contenía 7,5 pl del kit Multiplex PCR (Qiagen), 6,5 pl del producto de PCR de la primera tanda y 1 pl de cebador índice. Las condiciones de ciclado de la segunda tanda de PCR fueron 95 °C durante 15 minutos seguido de 7 ciclos de 95 °C durante 30 segundos, 54 °C durante 90 segundos y 72 °C durante 60 segundos, seguido de 5 minutos a 72 °C. Los productos de PCR se purificaron usando microesferas de AMPure XP y se eluyeron en 15 pl de agua, y se usaron 4 pl para cuantificar la cantidad de producto de PCR usando un Shimadzu MultiNA. Se agrupó una concentración molar equivalente de cada muestra para crear la colección de secuenciación que se cuantificó usando el kit de cuantificación de colecciones de Kapa Illumina. La colección se secuenció usando un Illumina MiSeq con reactivos v3 y secuenciación de ambos extremos de 2 x 300 pb. Los datos secuenciados se desmultiplexaron usando un archivo de instrucciones de Python hecho a medida y se usó el programa Cutadapt para retirar los adaptadores de secuenciación. Se combinaron las lecturas inversas y directas usando FLASH y se extrajo la secuencia de cada sitio de metilación.
El tratamiento de los macacos cangrejeros tanto con IgG DP47GS-IL-2 como con IgG DP47GS-(IL-2)2 indujo números sustancialmente incrementados de los Treg en la sangre en todos los animales tratados, similar a lo que se observó antes usando el mismo protocolo de tratamiento (figuras 33 y 35). Es importante destacar que el incremento rápido y drástico en los Treg tras el tratamiento con tanto IgG DP47GS-IL-2 como con IgG DP47GS-(IL-2)2 no afectó de manera adversa ni redujo el estado de desmetilación de FOXP3 (figura 36, panel superior) o CTLA-4 (figura 36, panel inferior). Se observaron resultados similares para ambas moléculas y los datos con IgG DP47GS-(IL-2)2 se muestran en la figura 36. El hecho de que el estado de desmetilación tanto de FOXP3 como de CTLA-4 no se viera afectado y no se redujera mediante el tratamiento indica que todos los Treg mantuvieron su fenotipo inmunodepresor funcional maduro natural a pesar de los incrementos sustanciales en los números absolutos. Estos resultados de desmetilación de Treg FOXP3 y CTLA-4 en primates no humanos sugieren que el tratamiento con dosis bajas de moléculas similares a IgG-(IL-2)2 de larga duración en el hombre debe poder incrementar los números de Treg maduros completamente funcionales que, a su vez, corregirá los desequilibrios inmunorreguladores presentes en las enfermedades autoinmunitarias humanas, así como otras enfermedades inflamatorias crónicas mediadas por el sistema inmunitario.
Farmacocinética de IgG DP47GS-(IL-2N88D)
2
en macacos cangrejeros
Se evaluaron las propiedades PF de IgG DP47GS-(IL-2N88D)2 en macacos cangrejeros sin tratamiento previo con biofármaco (figura 37). A los monos (n=2/dosis) se les inyectaron por vía intravenosa (i.v.) y por vía subcutánea (s.c.) 30 o 100 pg/kg de IgG DP47GS-(IL-2N88D)2 estéril en PBS que contenía suero de macaco cangrejero al 0,5 % y se les extrajo sangre en diversos tiempos después de la inyección, variando desde 30 minutos a los 14 días. Se evaluó IL-2 humana en las muestras de plasma de macaco cangrejero como se describe previamente. La concentración de detección usando plasma de macaco cangrejero en el ensayo fue de 0,05 ng/ml de IL-2. El plasma de macaco cangrejero tomado antes del tratamiento no tenía ninguna IL-2 detectable (por lo tanto, < 0,05 ng/ml).
A diferencia de la PFFC de las proteínas de fusión naturales (figura 25) donde las concentraciones en sangre solo se podían detectar hasta de las 48 a las 72 horas, se detectó IgG DP47GS-(IL-2N88D)2 en el plasma de macaco cangrejero de todos los animales en todos los puntos temporales hasta 14 días con las vías de administración i.v. (figura 37A) y s.c. (figura 37B). Las concentraciones en plasma fueron proporcionales a la dosis durante los primeros dos a tres días y presentaron una semivida prolongada en comparación con las moléculas de fusión de IL-2 natural. Como biomarcador de la exposición a IL-2 in vivo, se midieron las concentraciones en plasma de CD25 soluble (sCD25, IL-2RA) (figura 37C). Se usaron reactivos de anticuerpos monoclonales para sCD25 humanos conocidos por reaccionar de forma cruzada con sCD25 de macaco cangrejero en un inmunoensayo de tipo sándwich usando anticuerpos de captura (MAB623, R&D Systems) y de detección biotinilados (BAF223, R&D Systems) usando estreptavidina conjugada con Eu++ para detectar el sCD25 unido. Los incrementos de sCD25 después de la administración de IgG DP47GS-(IL-2N88D)2 fueron dependientes de la dosis y estuvieron presentes entre 1 a 7 días, regresando a las concentraciones normales en el día 8.
La inducción de Treg en macacos cangrejeros con IgG DP47GS-(IL-2N88D)
2
Las pruebas descritas previamente en macacos cangrejeros tratados in vivo con IgG DP47GS-IL-2 natural e IgG DP47GS-(IL-2)2 presentaron incrementos dependientes de la dosis de 2 a 4 veces en el número absoluto de Treg (figura 26 y 33, respectivamente) que reflejaban sus actividades in vitro en los análisis de sangre completa humana, es decir, diferencias in vitro de 6 a 9 veces (tablas 2 y 3). Con su pérdida de potencia in vitro en la sangre completa humana (tabla 3), se eligieron dosis in vivo de 30 y 100 pg/kg para IgG DP47GS-(IL-2N88D)2. De forma similar a las pruebas previas, se inyectó IgG DP47GS-(IL-2N88d )2 mediante una vía i.v. o s.c. con dosis individuales basadas en el peso corporal y se formuló para su inyección en un vehículo de PBS estéril, pH 7,2, que contenía suero de macaco cangrejero normal estéril al 0,5 % (n=2 por dosis y vía de inyección). Se obtuvieron muestras de sangre antes del tratamiento (días 4 y 5), inmediatamente antes del tratamiento (día 0) y en diversos tiempos posteriores al tratamiento (días 1-14) y se sometieron a prueba para determinar los cambios hematológicos (hemograma y diferencial), así como los marcadores intracelulares y de superficie de las células descritos previamente.
Los incrementos en los Treg CD4+ totales, Treg de memoria CD4+, Treg indiferenciados CD4+ y Treg CD8+ FOXP3+ se muestran como respuestas dependientes del tiempo como el número absoluto de células/ml de sangre (figura 38A y 38B) o bien como el % de linfocitos T CD4+ o c D8+ totales (figura 38C y 38D).
Los Treg CD4+ totales se incrementaron en número después de 100 pg/kg de 90.000/ml a 810.000/ml (9 veces) y desde un 4,2 % a un 26 % de los linfocitos T CD4+ totales (6,2 veces) y todavía estaban elevados a los 14 días. A 30 pg/kg, los Treg CD4+ totales se incrementaron desde 62.000/ml a 447.000/ml (7,2 veces) y desde un 4,5 % a un 16,7 % de las células CD4+ totales (3,7 veces).
Los Treg de memoria se incrementaron en número a 100 pg/kg desde 42.000/ml a 536.000/ml (12,8 veces) y desde un 2 % a un 17,4 % de los linfocitos T CD4+ totales (8,7 veces). A 30 pg/kg, los Treg de memoria se incrementaron desde 51.000/ml a 280.000/ml (5,5 veces) y desde un 2,3 % a un 10,5 % de los linfocitos T CD4+ totales (4,6 veces). Los Treg indiferenciados se incrementaron en número a 100 pg/kg desde 35.000/ml a 272.000/ml (7,8 veces) y desde un 2 % a un 8,6 % de los linfocitos T CD4+ totales (4,3 veces). A 30 pg/kg, los Treg indiferenciados se incrementaron desde 46.000/ml a 166.000/ml (3,6 veces) y desde un 2,2 % a un 6,2 % de los linfocitos T CD4+ totales (2,8 veces).
Después de la administración de 100 pg/kg de IgG DP47GS-(IL-2N88D)2 , los Treg CD8+FOXP3 + se incrementaron de un tipo de células escaso a 16.000/ml a 183.000/ml (11,4 veces) y desde un 0,7 % a un 7,8 % de los linfocitos T CD8+ totales (11,1 veces). A la dosis de 30 pg/kg de IgG d P47GS-(IL-2N88D)2 , los Treg CD8+ totales se incrementaron desde 17.000/ml a 101.000/ml (5,9 veces) y desde un 0,5 % a un 4,3 % de los linfocitos T CD8+ totales (8,6 veces). No existió ningún incremento relacionado con el tratamiento en los linfocitos Tef de memoria CD4+ después de 100 pg/kg o 30 pg/kg de IgG DP47GS-(IL-2N88D)2 (figura 39A); de hecho, los porcentajes disminuyeron en el día 4 probablemente debido al gran incremento en los Treg CD4+ en ese momento. Los linfocitos Tef de memoria CD4+CD25a se mantuvieron sin cambios después de 30 pg/kg de IgG DP47GS-(IL-2N88D)2 , pero se elevaron 1,6 veces en el día 4 con un regreso al valor de referencia a los 7-10 días (figura 39B).
Inducción de pSTAT5a, CD25 y Ki-67 en macacos cangrejeros con IgG DP47GS-(IL-2N88D) 2 Como se describe anteriormente, la especificidad celular in vivo del tratamiento con IL-2 es un parámetro crítico y se demuestra que la activación de los subconjuntos de células in vivo se podría supervisar midiendo pSTAT5a ex vivo, CD25 en la superficie de las células y Ki-67 intracelular en la sangre tomada en diversos tiempos después de la dosificación.
Los resultados en la figura 40A y 40B muestran la inducción de pSTAT5a preferente y las respuestas de señalización de pSTAT5a prolongado de los Treg CD4+ y CD8+ tras el tratamiento con IgG DP47GS-(IL-2N88D)2; las respuestas máximas se mantuvieron desde el día 1 al día 4 tanto en 100 pg/kg como en 30 pg/kg, regresando al valor de referencia a los 7 días. Los Tef de memoria CD4+CD25a respondieron con respuestas -semimáximas en el día 1 y regresaron a la normalidad en el día 4. Todos los demás tipos de células sometidos a prueba mostraron poca o ninguna inducción de pSTAT5a después del tratamiento.
Un incremento de CD25 en la superficie de las células es una consecuencia de la activación de IL-2 y es un biomarcador sensible de la activación in vivo. Poco después del tratamiento tanto con 100 pg/kg como con 3o pg/kg de IgG DP47GS-(IL-2N88D)2, se incrementó preferentemente CD25 en los Treg de memoria e indiferenciados CD4+, así como en los Treg CD8+ FOXP3+ (figura 41A, 41B). Las respuestas de CD25 de los Treg de memoria e indiferenciados CD4+ alcanzaron su punto máximo en el día 4 y regresaron al valor de referencia en 7 días, mientras que las elevaciones de CD25 en los Treg CD8+ FOXP3+ se mantuvieron a través de los 10-14 días.
En las pruebas con proteínas de fusión de IL-2 natural, se demostró que Ki-67 era un marcador intracelular sensible para las células que habían comenzado a proliferar in vivo y era un biomarcador sensible para la activación inducida por IL-2 en macacos cangrejeros. Las respuestas a 100 pg/kg de IgG DP47GS-(IL-2N88D)2 mostraron que un 80 90 % de los Treg CD4+ y CD8+ se convirtieron en Ki-67+ (figura 42A) mientras que un 60-90 % se convirtieron en Ki-67+ después de 30 pg/kg (figura 42B). Como se muestra, los demás subconjuntos de células sometidos a prueba fueron menos reactivos en comparación con los Treg.
Para capturar los penúltimos efectos in vivo de los números de Treg en incremento con las moléculas de IL-2 genomanipuladas novedosas, se convirtieron todas las dosis de macaco cangrejero a pmol/kg, lo que permitió las comparaciones directas independientemente del peso molecular; se compararon directamente las respuestas dependientes de la dosis como los incrementos máximos en los Treg CD4+ como el % de linfocitos T CD4+ totales (figura 43). A pesar de su pérdida de potencia in vitro en comparación con las proteínas de fusión naturales, IgG DP47GS-(IL-2N88D)2 indujo incrementos inesperadamente grandes en los Treg de macaco cangrejero, quizás en parte debido a su incremento de la exposición sistémica tras la administración tanto i.v. como s.c.
Eosinofilia como consecuencia de la administración de IL-2 in vivo
Proleukin en humanos estimula una eosinofilia cuando se administra diariamente en dosis de 1x106 a 4,5x106 UI/persona. En macacos cangrejeros se observaron incrementos similares en los eosinófilos en un 100 % de los animales después de un tratamiento repetido con Proleukin en dosis alta o dosis únicas de 226 pmol/kg de IgG DP47GS-IL-2 (figura 44). Los resultados en la figura 44 representan todos los animales sometidos a prueba (se tomaron valores de referencia de las colonias para los animales antes de las pruebas: n=44 y n=30) y para los diversos grupos de tratamiento con IL-2 (n=5-9) independientemente de si los números de eosinófilos eran normales o elevados. Sorprendente, a diferencia de los efectos de Proleukin e IgG DP47GS-IL-2, fue la virtual falta de efecto sobre los eosinófilos con IgG DP47GS-(IL-2N88D)2 en dosis in vivo de 170 y 570 pmol/kg (30 y 100 pg/kg, respectivamente). Esta falta de inducción de una eosinofilia sistémica por IgG DP47GS-(IL-2N88D)2 es sorprendente a la luz del grado de expansión de los Treg inducida por estos diferentes tratamientos (en comparación con la figura 43).
El tratamiento in vivo con IgG DP47GS-IL-2 suprime las respuestas inmunitarias en ratones
En estudios preliminares, se observó que una dosis de 4000 UI de IgG DP47GS-IL-2 activó los linfocitos T reguladores FOXP3+ de ratón in vivo; entonces, se usó esta dosis para evaluar su capacidad para suprimir las clásicas respuestas inmunitarias dependientes de linfocitos T en ratones, es decir, hipersensibilidad de tipo retardado (figura 45) y una respuesta de IgG anti-KLH (figura 46).
Se inmunizaron i.v. ratones NOD y ratones C57BL/6 (n=7) con eritrocitos de carnero (srbc) y se expusieron 3 días más tarde a una inyección intravenosa rápida de srbc en una pata trasera única para inducir una respuesta de hipersensibilidad de tipo retardado (DTH). Un día después de la exposición, se sacrificaron los ratones con CO2 y se extirparon y pesaron las patas. La magnitud de la respuesta de DTH se muestra como el cambio en el peso de la pata en comparación con los ratones no inmunizados (A peso de la pata). Se administró s.c. IgG DP47GS-IL-2 a 4000 UI por ratón 3 días antes y en el día de la inmunización con srbc y el vehículo era PBS estéril, pH 7,2. La significación estadística derivó de la prueba de Mann Whitney en GraphPad Prism.
La dosificación de IgG DP47GS-IL-2 tres días antes y en el día de la inmunización con eritrocitos de carnero
suprimió la respuesta de hipersensibilidad de tipo retardado posterior con respecto a una exposición a eritrocitos de carnero en un 51 % en ratones NOD (figura 45A; p=0,0023) y en un 38 % en ratones C57BL/6 (figura 45B; p=0,002). CTLA-4-Ig de ratón como inmunodepresor de control positivo inhibió la respuesta de DTH a una concentración similar a IgG DP47GS-IL-2.
IgG DP47GS-IL-2 también pudo suprimir las respuestas de IgG específicas para KLH en ratones C57BL/6 (inhibición de un 78 %, p=0,0007, figura 46A) y NOD (inhibición de un 67 %, p=0,004, figura 46B). Para este experimento, se inmunizaron i.p. crías de ratones C57BL/6 (n=7-10) y ratones NOD (n=13-14) sanos con 100 pg de KLH de calidad de vacuna humana sin adyuvante como se recomienda por el fabricante (Stellar). El tratamiento con IgG DP47GS-IL-2 consistió en 1 (NOD) o 2 (C57BL/6) tratamientos s.c. semanales con 4000 UI por ratón iniciado el día de la inmunización. Siete días (NOD) y 21 días (C57BL/6) después de la inmunización, se obtuvo sangre y se midieron las respuestas de IgG específicas para KLH en suero mediante ELISA.
La capacidad de IgG DP47GS-IL-2 para suprimir las respuestas inmunitarias in vivo respalda la hipótesis de que la activación de los linfocitos T reguladores inducida por IL-2 en dosis baja produce linfocitos T reguladores funcionales que median una reducción en la respuesta inmunitaria.
Aunque la invención anterior se ha descrito con algo de detalle a modo de ilustración y ejemplo para propósitos de claridad de entendimiento, no se deben interpretar las descripciones y ejemplos como limitantes del alcance de la invención.
Claims (37)
1. Una proteína de fusión que comprende (i) una molécula de inmunoglobulina que no se puede unir específicamente a un antígeno y (ii) dos moléculas de interleucina 2 (IL-2) mutante que comprenden una mutación de aminoácido que reduce la afinidad de la molécula de IL-2 mutante con respecto al receptor de IL-2 de afinidad intermedia, en comparación con una molécula de IL-2 natural, en la que dichas moléculas de IL-2 mutante comprenden la secuencia de la SEQ ID NO: 58.
2. La proteína de fusión de la reivindicación 1, en la que dicha molécula de inmunoglobulina es una molécula de inmunoglobulina de clase IgG, en particular, una molécula de inmunoglobulina de subclase IgG1.
3. La proteína de fusión de la reivindicación 1 o 2, en la que dicha molécula de inmunoglobulina es una molécula de inmunoglobulina humana.
4. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena pesada basada en la secuencia de estirpe germinal de Vh3-23 humana.
5. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende la secuencia de la región variable de la cadena pesada de la SEQ ID NO: 9.
6. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende una secuencia de la región variable de la cadena ligera basada en la secuencia de estirpe germinal de Vk3-20 humana.
7. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende la secuencia de la región variable de la cadena ligera de la SEQ ID NO: 11.
8. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende una modificación que reduce la afinidad de unión de la molécula de inmunoglobulina con respecto a un receptor Fc en comparación con una molécula de inmunoglobulina correspondiente sin dicha modificación.
9. La proteína de fusión de la reivindicación 8, en la que dicho receptor Fc es un receptor Fcy, en particular, un receptor Fcy humano.
10. La proteína de fusión de la reivindicación 8 o 9, en la que dicho receptor Fc es un receptor Fc activador.
11. La proteína de fusión de una cualquiera de las reivindicaciones 8 a 10, en la que dicho receptor Fc se selecciona del grupo de FcYRIIIa (CD16a), FcyRI (CD64), FcYRIIa (CD32) y FcaRI (Cd89).
12. La proteína de fusión de una cualquiera de las reivindicaciones 8 a 11, en la que dicho receptor Fc es FcYRIIIa, en particular, FcYRIIIa humano.
13. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende una sustitución de aminoácido en la posición 329 (numeración EU) de las cadenas pesadas de inmunoglobulina.
14. La proteína de fusión de la reivindicación 13, en la que dicha sustitución de aminoácido es P329G.
15. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende sustituciones de aminoácido en las posiciones 234 y 235 (numeración EU) de las cadenas pesadas de inmunoglobulina.
16. La proteína de fusión de la reivindicación 15, en la que dichas sustituciones de aminoácido son L234A y L235A (LALA).
17. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dicha molécula de inmunoglobulina comprende las sustituciones de aminoácido L234A, L235A y P329G (numeración EU) en las cadenas pesadas de inmunoglobulina.
18. La proteína de fusión de una cualquiera de las reivindicaciones precedentes, en la que dichas moléculas de IL-2 mutante se fusionan cada una en su aminoácido N terminal con el aminoácido C terminal de una de las cadenas pesadas de inmunoglobulina de dicha molécula de inmunoglobulina, opcionalmente a través de un conector peptídico.
19. L a p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s p r e c e d e n t e s , e n la q u e d i c h a s m o lé c u la s d e I L - 2 m u t a n t e s e f u s io n a n c a d a u n a c o n d i c h a m o lé c u la d e in m u n o g lo b u l i n a a t r a v é s d e u n c o n e c t o r p e p t í d ic o .
20. L a p r o t e í n a d e f u s ió n d e la r e iv i n d ic a c ió n 19 , e n la q u e d i c h o c o n e c t o r p e p t í d ic o c o m p r e n d e a l m e n o s 10 , e n p a r t i c u la r , a l m e n o s 15 , a m in o á c id o s .
21. L a p r o t e í n a d e f u s ió n d e la r e iv i n d ic a c ió n 19 o 20 , e n la q u e d i c h o c o n e c t o r p e p t í d ic o c o m p r e n d e la s e c u e n c ia d e a m in o á c id o s ( G 4S )3 ( S E Q ID N O : 66 ) .
22. L a p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s p r e c e d e n t e s , e n la q u e la p r o t e í n a d e f u s ió n c o m p r e n d e la s s e c u e n c ia s d e p o l ip é p t i d o d e la S E Q ID N O : 19 y la S E Q ID N O : 50.
23. L a p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s p r e c e d e n t e s , e n la q u e d i c h a p r o t e í n a d e f u s ió n a c t i v a s e l e c t i v a m e n t e lo s l i n f o c i t o s T r e g u l a d o r e s .
24. U n p o l i n u c le ó t i d o q u e c o d i f i c a la p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s p r e c e d e n t e s .
25. U n v e c t o r , e n p a r t i c u la r , u n v e c t o r d e e x p r e s i ó n , q u e c o m p r e n d e e l p o l i n u c le ó t i d o d e la r e iv i n d ic a c ió n 24.
26. U n a c é lu la h u é s p e d q u e c o m p r e n d e e l p o l i n u c le ó t i d o d e la r e iv i n d ic a c ió n 24 o e l v e c t o r d e la r e iv i n d ic a c ió n 25.
27. U n p r o c e d i m i e n t o p a r a p r o d u c i r u n a p r o t e í n a d e f u s ió n q u e c o m p r e n d e u n a m o lé c u la d e i n m u n o g lo b u l i n a y d o s m o lé c u la s d e I L - 2 m u t a n t e q u e c o m p r e n d e n u n a m u t a c ió n d e a m in o á c id o q u e r e d u c e la a f in i d a d d e la m o lé c u la d e I L - 2 m u t a n t e c o n r e s p e c t o a l r e c e p t o r d e I L - 2 d e a f in i d a d i n t e r m e d ia e n c o m p a r a c ió n c o n u n a m o lé c u la d e I L - 2 n a t u r a l , q u e c o m p r e n d e la s e t a p a s d e ( i ) c u l t i v a r la c é lu la h u é s p e d d e la r e iv i n d ic a c ió n 26 e n c o n d i c io n e s a d e c u a d a s p a r a la e x p r e s i ó n d e la p r o t e í n a d e f u s ió n y ( i i ) r e c u p e r a r la p r o t e í n a d e f u s ió n .
28. U n a p r o t e í n a d e f u s ió n p r o d u c i d a m e d ia n t e e l p r o c e d i m i e n t o d e la r e iv i n d ic a c ió n 27.
29. U n a c o m p o s i c ió n f a r m a c é u t i c a q u e c o m p r e n d e la p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s 1 - 23 o 28 y u n v e h í c u l o f a r m a c é u t i c a m e n t e a c e p t a b le .
30. L a p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s 1 - 23 o 28 , o la c o m p o s i c ió n f a r m a c é u t i c a d e la r e i v i n d i c a c ió n 29 , p a r a s u u s o c o m o u n m e d ic a m e n t o .
31. L a p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s 1 - 23 o 28 , o la c o m p o s i c ió n f a r m a c é u t i c a d e la r e i v i n d i c a c ió n 29 , p a r a s u u s o e n e l t r a t a m ie n t o o p r o f i l a x is d e u n a e n f e r m e d a d a u t o i n m u n i t a r i a .
32. L a p r o t e í n a d e f u s ió n o la c o m p o s i c ió n f a r m a c é u t i c a p a r a s u u s o d e la r e iv i n d ic a c ió n 31 , e n la q u e d i c h a e n f e r m e d a d a u t o i n m u n i t a r i a s e s e l e c c io n a d e l g r u p o d e d i a b e t e s d e t ip o 1 , lu p u s e r i t e m a t o s o d i s e m i n a d o , e n f e r m e d a d in f l a m a t o r i a in t e s t in a l , e n f e r m e d a d d e C r o h n , c o l i t i s u l c e r o s a y e s c l e r o s is m ú l t ip l e .
33. L a p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s 1 - 23 o 28 , o la c o m p o s i c ió n f a r m a c é u t i c a d e la r e i v i n d i c a c ió n 29 , p a r a s u u s o e n e l t r a t a m ie n t o o p r o f i l a x is d e r e c h a z o d e t r a s p la n t e o e n f e r m e d a d in je r t o c o n t r a h u é s p e d .
34. L a p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s 1 - 23 o 28 p a r a s u u s o e n la a c t i v a c i ó n s e l e c t i v a d e lo s l i n f o c i t o s T r e g u l a d o r e s in vitro.
35. L a p r o t e í n a d e f u s ió n d e la r e iv i n d ic a c ió n 34 , e n la q u e d i c h a a c t i v a c i ó n c o m p r e n d e la in d u c c ió n d e la p r o l i f e r a c i ó n d e l i n f o c i t o s T r e g u l a d o r e s y / o la in d u c c ió n d e la s e ñ a l i z a c ió n d e l r e c e p t o r d e I L - 2 e n lo s l i n f o c i t o s T r e g u la d o r e s .
36 . U n p r o c e d i m i e n t o p a r a la a c t i v a c i ó n s e l e c t i v a d e lo s l i n f o c i t o s T r e g u l a d o r e s in vitro, q u e c o m p r e n d e p o n e r e n c o n t a c t o d i c h o s l i n f o c i t o s T r e g u l a d o r e s c o n la p r o t e í n a d e f u s ió n d e u n a c u a l q u i e r a d e la s r e i v i n d i c a c io n e s 1 23 o 28.
37. E l p r o c e d i m i e n t o d e la r e iv i n d ic a c ió n 36 , e n e l q u e d i c h a a c t i v a c i ó n c o m p r e n d e la in d u c c ió n d e la p r o l i f e r a c i ó n d e lo s l i n f o c i t o s T r e g u l a d o r e s y / o la in d u c c ió n d e la s e ñ a l i z a c ió n d e l r e c e p t o r d e I L - 2 e n lo s l i n f o c i t o s T r e g u la d o r e s .
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201461936564P | 2014-02-06 | 2014-02-06 | |
| PCT/EP2015/052312 WO2015118016A1 (en) | 2014-02-06 | 2015-02-04 | Interleukin-2 fusion proteins and uses thereof |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2704731T3 true ES2704731T3 (es) | 2019-03-19 |
Family
ID=52669578
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES15709419T Active ES2704731T3 (es) | 2014-02-06 | 2015-02-04 | Proteínas de fusión de interleucina 2 y usos de las mismas |
Country Status (32)
| Country | Link |
|---|---|
| US (1) | US11098099B2 (es) |
| EP (2) | EP3508496A1 (es) |
| JP (2) | JP6306201B2 (es) |
| KR (2) | KR102042246B1 (es) |
| CN (2) | CN105980410B (es) |
| AR (1) | AR099289A1 (es) |
| AU (2) | AU2015215001B2 (es) |
| BR (1) | BR112016018288A2 (es) |
| CA (1) | CA2931114A1 (es) |
| CL (1) | CL2016001968A1 (es) |
| CR (1) | CR20160333A (es) |
| DK (1) | DK3102595T3 (es) |
| EA (2) | EA034350B1 (es) |
| ES (1) | ES2704731T3 (es) |
| HR (1) | HRP20190028T1 (es) |
| HU (1) | HUE041530T2 (es) |
| IL (1) | IL245484B (es) |
| LT (1) | LT3102595T (es) |
| MA (1) | MA39250B1 (es) |
| MX (2) | MX370283B (es) |
| NZ (2) | NZ760289A (es) |
| PE (1) | PE20161324A1 (es) |
| PH (1) | PH12016501406A1 (es) |
| PL (1) | PL3102595T3 (es) |
| PT (1) | PT3102595T (es) |
| RS (1) | RS58149B1 (es) |
| SG (1) | SG11201606369XA (es) |
| SI (1) | SI3102595T1 (es) |
| TW (1) | TWI671318B (es) |
| UA (1) | UA120847C2 (es) |
| WO (1) | WO2015118016A1 (es) |
| ZA (1) | ZA201603262B (es) |
Families Citing this family (48)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EA201892619A1 (ru) | 2011-04-29 | 2019-04-30 | Роше Гликарт Аг | Иммуноконъюгаты, содержащие мутантные полипептиды интерлейкина-2 |
| US20140044675A1 (en) | 2012-08-10 | 2014-02-13 | Roche Glycart Ag | Interleukin-2 fusion proteins and uses thereof |
| MA39250B1 (fr) | 2014-02-06 | 2018-09-28 | Hoffmann La Roche | Protéines hybrides de l'interleukine-2 et leurs utilisations |
| GB201403775D0 (en) | 2014-03-04 | 2014-04-16 | Kymab Ltd | Antibodies, uses & methods |
| SG11201700629TA (en) | 2014-08-11 | 2017-02-27 | Delinia Inc | Modified il-2 variants that selectively activate regulatory t cells for the treatment of autoimmune diseases |
| MD3226843T2 (ro) | 2014-12-05 | 2021-10-31 | Aragon Pharmaceuticals Inc | Compoziții anticancer |
| SG11201804259UA (en) | 2015-12-04 | 2018-06-28 | Novartis Ag | Antibody cytokine engrafted compositions and methods of use for immunoregulation |
| EP3402499B1 (en) * | 2016-01-14 | 2021-07-21 | Intrexon Actobiotics NV | Compositions and methods for the treatment of type 1 diabetes |
| US20170204154A1 (en) | 2016-01-20 | 2017-07-20 | Delinia, Inc. | Molecules that selectively activate regulatory t cells for the treatment of autoimmune diseases |
| US9567399B1 (en) | 2016-06-20 | 2017-02-14 | Kymab Limited | Antibodies and immunocytokines |
| US10858428B2 (en) | 2016-09-28 | 2020-12-08 | Xoma (Us) Llc | Antibodies that bind interleukin-2 and uses thereof |
| WO2018073365A1 (en) | 2016-10-19 | 2018-04-26 | F. Hoffmann-La Roche Ag | Method for producing an immunoconjugate |
| WO2018083248A1 (en) | 2016-11-03 | 2018-05-11 | Kymab Limited | Antibodies, combinations comprising antibodies, biomarkers, uses & methods |
| US11077172B2 (en) * | 2016-11-08 | 2021-08-03 | Delinia, Inc. | IL-2 variants for the treatment of psoriasis |
| BR112019011799B1 (pt) * | 2016-12-13 | 2021-12-21 | Delinia, Inc | Proteína de fusão, proteína dimérica e composição farmacêutica |
| CA3056630A1 (en) | 2017-03-15 | 2018-09-20 | Pandion Therapeutics, Inc. | Targeted immunotolerance |
| JOP20190271A1 (ar) | 2017-05-24 | 2019-11-21 | Novartis Ag | بروتينات مطعّمة بسيتوكين- الجسم المضاد وطرق الاستخدام للاضطرابات المتعلقة بالمناعة |
| CA3064435A1 (en) | 2017-05-24 | 2018-11-29 | Pandion Therapeutics, Inc. | Targeted immunotolerance |
| US20200299349A1 (en) * | 2017-11-21 | 2020-09-24 | The Board Of Trustees Of The Leland Stanford Junior University | Partial agonists of interleukin-2 |
| US10946068B2 (en) | 2017-12-06 | 2021-03-16 | Pandion Operations, Inc. | IL-2 muteins and uses thereof |
| US10174091B1 (en) | 2017-12-06 | 2019-01-08 | Pandion Therapeutics, Inc. | IL-2 muteins |
| EP3746103A4 (en) * | 2017-12-06 | 2022-03-30 | The Board of Trustees of the Leland Stanford Junior University | PROTEINS ENGINEERED TO INCREASE A CELL'S SENSITIVITY TO IL-2 |
| JP2021506291A (ja) * | 2017-12-19 | 2021-02-22 | ゼンコア インコーポレイテッド | 改変されたil−2 fc融合タンパク質 |
| EP3752178A1 (en) | 2018-02-16 | 2020-12-23 | Iltoo Pharma | Use of interleukin 2 for treating sjögren's syndrome |
| MX2020009975A (es) | 2018-03-28 | 2020-10-12 | Bristol Myers Squibb Co | Proteinas de fusion interleucina-2/receptor alfa de interleucina-2 y metodos de uso. |
| WO2020007937A1 (en) | 2018-07-03 | 2020-01-09 | Iltoo Pharma | Use of interleukin-2 for treating systemic sclerosis |
| EP3818083A2 (en) | 2018-07-03 | 2021-05-12 | Elstar Therapeutics, Inc. | Anti-tcr antibody molecules and uses thereof |
| WO2020035482A1 (en) | 2018-08-13 | 2020-02-20 | Iltoo Pharma | Combination of interleukin-2 with an interleukin 1 inhibitor, conjugates and therapeutic uses thereof |
| EP3908596A1 (en) * | 2019-01-07 | 2021-11-17 | Inhibrx, Inc. | Polypeptides comprising modified il-2 polypeptides and uses thereof |
| CN111944036A (zh) * | 2019-05-14 | 2020-11-17 | 上海盖浦生物科技有限公司 | 一种增殖免疫细胞的突变体蛋白 |
| BR112021023345A2 (pt) | 2019-05-20 | 2022-02-01 | Pandion Operations Inc | Imunotolerância com alvo em madcam |
| JP2022536347A (ja) * | 2019-06-14 | 2022-08-15 | キュージーン インコーポレイテッド | 新規インターロイキン-2バリアントおよびその二官能性融合分子 |
| EP3997115A4 (en) * | 2019-07-12 | 2023-11-22 | Proviva Therapeutics (Hong Kong) Limited | IL-2 COMPOSITIONS AND METHODS OF USE THEREOF |
| US11633488B2 (en) | 2020-01-10 | 2023-04-25 | Bright Peak Therapeutics Ag | Modified IL-2 polypeptides and uses thereof |
| KR102653906B1 (ko) | 2020-01-14 | 2024-04-03 | 신테카인, 인크. | 편향된 il2 뮤테인 방법 및 조성물 |
| BR112022015920A2 (pt) | 2020-02-13 | 2022-10-04 | Hoffmann La Roche | Método para determinar a razão de partículas de aav e uso do tempo de relaxação |
| EP4122951A1 (en) * | 2020-03-19 | 2023-01-25 | Innovent Biologics (Suzhou) Co., Ltd. | Interleukin-2 mutant and use thereof |
| WO2021231773A1 (en) | 2020-05-13 | 2021-11-18 | Good Therapeutics, Inc. | Compositions of protein complexes and methods of use thereof |
| CN113912735A (zh) * | 2020-07-08 | 2022-01-11 | 南京师范大学 | 多肽二聚体及其用途 |
| KR20230097094A (ko) | 2020-10-29 | 2023-06-30 | 브리스톨-마이어스 스큅 컴퍼니 | 질환의 치료를 위한 융합 단백질 |
| CN112724259B (zh) * | 2020-11-16 | 2022-12-20 | 天津林达生物科技有限公司 | 人血清白蛋白与白介素2的融合蛋白及其用途 |
| EP4255923A2 (en) * | 2020-12-04 | 2023-10-11 | F. Hoffmann-La Roche AG | Ph-dependent mutant interleukin-2 polypeptides |
| KR20230148413A (ko) | 2021-02-24 | 2023-10-24 | 에프. 호프만-라 로슈 아게 | 심층 필터의 재생 및 다회 사용 |
| WO2023057588A1 (en) * | 2021-10-06 | 2023-04-13 | Iltoo Pharma | Interleukin 2 chimeric constructs with targeting specificy to inflamed tissues |
| GB202115122D0 (en) * | 2021-10-21 | 2021-12-08 | Dualyx Nv | Binding molecules targeting IL-2 receptor |
| WO2024056154A1 (en) | 2022-09-12 | 2024-03-21 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Interleukin-2 for use in treating autism spectrum disorder |
| CN116041539B (zh) * | 2022-10-31 | 2023-07-21 | 山东博安生物技术股份有限公司 | Il-2突变体免疫缀合物 |
| CN116554356B (zh) * | 2023-05-16 | 2024-01-23 | 武汉大学 | 一种hyperIL-15与sCD4及Fc的融合蛋白及其应用 |
Family Cites Families (46)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| FR2388385B1 (fr) | 1977-04-18 | 1982-01-08 | Hitachi Metals Ltd | Piece d'ornement fixee par des aimants permanents |
| US4518584A (en) | 1983-04-15 | 1985-05-21 | Cetus Corporation | Human recombinant interleukin-2 muteins |
| WO1985000817A1 (en) | 1983-08-10 | 1985-02-28 | Amgen | Microbial expression of interleukin ii |
| US5206344A (en) | 1985-06-26 | 1993-04-27 | Cetus Oncology Corporation | Interleukin-2 muteins and polymer conjugation thereof |
| US6548640B1 (en) | 1986-03-27 | 2003-04-15 | Btg International Limited | Altered antibodies |
| IL85035A0 (en) | 1987-01-08 | 1988-06-30 | Int Genetic Eng | Polynucleotide molecule,a chimeric antibody with specificity for human b cell surface antigen,a process for the preparation and methods utilizing the same |
| US5959177A (en) | 1989-10-27 | 1999-09-28 | The Scripps Research Institute | Transgenic plants expressing assembled secretory antibodies |
| GB9015198D0 (en) | 1990-07-10 | 1990-08-29 | Brien Caroline J O | Binding substance |
| US5650150A (en) | 1990-11-09 | 1997-07-22 | Gillies; Stephen D. | Recombinant antibody cytokine fusion proteins |
| JP4124480B2 (ja) | 1991-06-14 | 2008-07-23 | ジェネンテック・インコーポレーテッド | 免疫グロブリン変異体 |
| US5565332A (en) | 1991-09-23 | 1996-10-15 | Medical Research Council | Production of chimeric antibodies - a combinatorial approach |
| US5731168A (en) | 1995-03-01 | 1998-03-24 | Genentech, Inc. | Method for making heteromultimeric polypeptides |
| US6040498A (en) | 1998-08-11 | 2000-03-21 | North Caroline State University | Genetically engineered duckweed |
| BR9813365A (pt) | 1997-12-05 | 2004-06-15 | Scripps Research Inst | Método para produção e humanização de um anticorpo monoclonal de rato |
| BR9908226A (pt) | 1998-02-25 | 2000-10-24 | Lexigen Pharm Corp | Melhoramento da meia vida de circulação de proteìnas de fusão com base em anticorpo |
| US6955807B1 (en) * | 1998-05-15 | 2005-10-18 | Bayer Pharmaceuticals Corporation | IL-2 selective agonists and antagonists |
| DZ2788A1 (fr) | 1998-05-15 | 2003-12-01 | Bayer Ag | Agonistes et antagonistes selectifs à IL-2. |
| US6737056B1 (en) | 1999-01-15 | 2004-05-18 | Genentech, Inc. | Polypeptide variants with altered effector function |
| US7125978B1 (en) | 1999-10-04 | 2006-10-24 | Medicago Inc. | Promoter for regulating expression of foreign genes |
| WO2001025454A2 (en) | 1999-10-04 | 2001-04-12 | Medicago Inc. | Method for regulating transcription of foreign genes in the presence of nitrogen |
| US6962696B1 (en) | 1999-10-04 | 2005-11-08 | Vion Pharmaceuticals Inc. | Compositions and methods for tumor-targeted delivery of effector molecules |
| PT1454138E (pt) * | 2001-12-04 | 2012-03-28 | Merck Patent Gmbh | Imunocitoquinas com seletividade modulada |
| US7432063B2 (en) | 2002-02-14 | 2008-10-07 | Kalobios Pharmaceuticals, Inc. | Methods for affinity maturation |
| WO2005007121A2 (en) | 2003-07-18 | 2005-01-27 | Massachusetts Institute Of Technology | Mutant interleukin-2(il-2) polypeptides |
| RU2006135112A (ru) * | 2004-03-05 | 2008-04-10 | Чирон Корпорейшн (Us) | Тест-система in vitro для прогнозирования устойчивости пациента к терапевтическим средствам |
| RU2386638C2 (ru) | 2004-03-31 | 2010-04-20 | Дженентек, Инк. | Гуманизированные анти-тфр-бета-антитела |
| CA2885854C (en) | 2004-04-13 | 2017-02-21 | F. Hoffmann-La Roche Ag | Anti-p-selectin antibodies |
| TWI380996B (zh) | 2004-09-17 | 2013-01-01 | Hoffmann La Roche | 抗ox40l抗體 |
| CA2587572A1 (en) | 2004-11-24 | 2006-06-01 | Schering Corporation | Dap-10 and uses thereof |
| EP2038417A2 (en) | 2006-07-06 | 2009-03-25 | Merck Patent GmbH | Compositions and methods for enhancing the efficacy of il-2 mediated immune responses |
| AU2008278803A1 (en) | 2007-07-25 | 2009-01-29 | Astrazeneca Ab | Targeted binging agents directed to KDR and uses thereof - 035 |
| WO2009061853A2 (en) | 2007-11-05 | 2009-05-14 | Massachusetts Institute Of Technology | Mutant interleukin-2 (il-2) polypeptides |
| WO2009085462A1 (en) | 2007-12-19 | 2009-07-09 | Centocor, Inc. | Design and generation of human de novo pix phage display libraries via fusion to pix or pvii, vectors, antibodies and methods |
| DE102008023820A1 (de) | 2008-05-08 | 2009-11-12 | Aicuris Gmbh & Co. Kg | Mittel zur Behandlung und/oder Prophylaxe einer Autoimmunerkrankung und zur Bildung von Regulatorischen T-Zellen |
| CA2749539C (en) * | 2009-01-21 | 2022-07-19 | Amgen Inc. | Compositions and methods comprising interleukin-2 mutants for treating inflammatory and autoimmune diseases |
| HUE036077T2 (hu) | 2010-08-13 | 2018-06-28 | Roche Glycart Ag | Anti-FAP ellenanyagok és alkalmazásukra szolgáló eljárások |
| CU23923B1 (es) | 2010-11-12 | 2013-07-31 | Ct De Inmunología Molecular | Polipéptidos derivados de la il-2 con actividad agonista |
| DK3489255T3 (da) | 2011-02-10 | 2021-08-23 | Roche Glycart Ag | Muterede interleukin-2-polypeptider |
| EP2683395B1 (en) | 2011-03-11 | 2018-08-01 | Assistance Publique - Hôpitaux de Paris | Use of low dose il-2 for treating type 1 diabetes |
| EP3590965A1 (en) * | 2011-03-29 | 2020-01-08 | Roche Glycart AG | Antibody fc variants |
| EA201892619A1 (ru) | 2011-04-29 | 2019-04-30 | Роше Гликарт Аг | Иммуноконъюгаты, содержащие мутантные полипептиды интерлейкина-2 |
| WO2012178137A1 (en) * | 2011-06-24 | 2012-12-27 | Gillies Stephen D | Light chain immunoglobulin fusion proteins and methods of use thereof |
| US20140044675A1 (en) | 2012-08-10 | 2014-02-13 | Roche Glycart Ag | Interleukin-2 fusion proteins and uses thereof |
| US9546203B2 (en) | 2013-03-14 | 2017-01-17 | Amgen Inc. | Aglycosylated Fc-containing polypeptides with cysteine substitutions |
| MA39250B1 (fr) | 2014-02-06 | 2018-09-28 | Hoffmann La Roche | Protéines hybrides de l'interleukine-2 et leurs utilisations |
| MA40094B1 (fr) | 2014-08-06 | 2022-05-31 | Univ Miami | Protéines de fusion de l'interleukine-2/récepteur alpha de l'interleukine-2 et procédés d'utilisation |
-
2015
- 2015-02-04 MA MA39250A patent/MA39250B1/fr unknown
- 2015-02-04 US US14/613,831 patent/US11098099B2/en active Active
- 2015-02-04 LT LTEP15709419.4T patent/LT3102595T/lt unknown
- 2015-02-04 EP EP18201416.7A patent/EP3508496A1/en not_active Withdrawn
- 2015-02-04 EP EP15709419.4A patent/EP3102595B1/en active Active
- 2015-02-04 CN CN201580007400.8A patent/CN105980410B/zh active Active
- 2015-02-04 EA EA201691576A patent/EA034350B1/ru not_active IP Right Cessation
- 2015-02-04 ES ES15709419T patent/ES2704731T3/es active Active
- 2015-02-04 DK DK15709419.4T patent/DK3102595T3/en active
- 2015-02-04 CA CA2931114A patent/CA2931114A1/en active Pending
- 2015-02-04 BR BR112016018288A patent/BR112016018288A2/pt not_active Application Discontinuation
- 2015-02-04 EA EA201992609A patent/EA201992609A1/ru unknown
- 2015-02-04 WO PCT/EP2015/052312 patent/WO2015118016A1/en active Application Filing
- 2015-02-04 PE PE2016001255A patent/PE20161324A1/es unknown
- 2015-02-04 UA UAA201609263A patent/UA120847C2/uk unknown
- 2015-02-04 MX MX2016009687A patent/MX370283B/es active IP Right Grant
- 2015-02-04 RS RS20190003A patent/RS58149B1/sr unknown
- 2015-02-04 KR KR1020167021255A patent/KR102042246B1/ko active IP Right Grant
- 2015-02-04 JP JP2016550531A patent/JP6306201B2/ja active Active
- 2015-02-04 AU AU2015215001A patent/AU2015215001B2/en active Active
- 2015-02-04 NZ NZ760289A patent/NZ760289A/en unknown
- 2015-02-04 SG SG11201606369XA patent/SG11201606369XA/en unknown
- 2015-02-04 PL PL15709419T patent/PL3102595T3/pl unknown
- 2015-02-04 NZ NZ719654A patent/NZ719654A/en unknown
- 2015-02-04 SI SI201530545T patent/SI3102595T1/sl unknown
- 2015-02-04 CN CN202010081233.4A patent/CN111423513A/zh active Pending
- 2015-02-04 KR KR1020197032546A patent/KR20190126205A/ko not_active Application Discontinuation
- 2015-02-04 PT PT15709419T patent/PT3102595T/pt unknown
- 2015-02-04 CR CR20160333A patent/CR20160333A/es unknown
- 2015-02-04 HU HUE15709419A patent/HUE041530T2/hu unknown
- 2015-02-05 TW TW104103962A patent/TWI671318B/zh active
- 2015-02-05 AR ARP150100325A patent/AR099289A1/es unknown
-
2016
- 2016-05-04 IL IL24548416A patent/IL245484B/en active IP Right Grant
- 2016-05-13 ZA ZA201603262A patent/ZA201603262B/en unknown
- 2016-07-15 PH PH12016501406A patent/PH12016501406A1/en unknown
- 2016-07-26 MX MX2019014695A patent/MX2019014695A/es unknown
- 2016-08-04 CL CL2016001968A patent/CL2016001968A1/es unknown
-
2018
- 2018-03-07 JP JP2018040501A patent/JP2018102312A/ja active Pending
-
2019
- 2019-01-03 HR HRP20190028TT patent/HRP20190028T1/hr unknown
- 2019-11-13 AU AU2019264579A patent/AU2019264579A1/en not_active Abandoned
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2704731T3 (es) | Proteínas de fusión de interleucina 2 y usos de las mismas | |
| US20200172591A1 (en) | Interleukin-2 fusion proteins and uses thereof | |
| ES2720433T3 (es) | Anticuerpos biespecíficos específicos para FAP y DR5, anticuerpos específicos para DR5 y procedimientos de uso | |
| JP2023159115A (ja) | Gprc5dに結合する抗体 | |
| ES2777778T3 (es) | Variantes de CTLA-4 | |
| KR20220028035A (ko) | Gprc5d에 결합하는 항체 | |
| EP3180361A1 (en) | Recombinant antibody molecule and its use for target cell restricted t cell activation | |
| JP2022541163A (ja) | Nkg2dに結合する抗体 | |
| EP3601346A1 (en) | Bispecific antigen binding molecule for a costimulatory tnf receptor | |
| KR20230025673A (ko) | CD3 및 FolR1에 결합하는 항체 | |
| KR20230025665A (ko) | Cd3에 결합하는 항체 | |
| JP2023538716A (ja) | Cd3及びcd19に結合する抗体 | |
| JP7436365B2 (ja) | 抗vegf抗体及び使用の方法 |