EP0184576A1 - Promotoren für die Expression von fremden Genen in Hefe, Plasmide, die diese Promotoren enthalten, sowie deren Verwendung zur Herstellung von Polypeptiden - Google Patents
Promotoren für die Expression von fremden Genen in Hefe, Plasmide, die diese Promotoren enthalten, sowie deren Verwendung zur Herstellung von Polypeptiden Download PDFInfo
- Publication number
- EP0184576A1 EP0184576A1 EP85870171A EP85870171A EP0184576A1 EP 0184576 A1 EP0184576 A1 EP 0184576A1 EP 85870171 A EP85870171 A EP 85870171A EP 85870171 A EP85870171 A EP 85870171A EP 0184576 A1 EP0184576 A1 EP 0184576A1
- Authority
- EP
- European Patent Office
- Prior art keywords
- plasmid
- yeast
- promoters
- promoter
- site
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
- 239000013612 plasmid Substances 0.000 title claims abstract description 102
- 240000004808 Saccharomyces cerevisiae Species 0.000 title claims abstract description 76
- 108090000623 proteins and genes Proteins 0.000 title claims abstract description 51
- 229920001184 polypeptide Polymers 0.000 title claims abstract description 23
- 108090000765 processed proteins & peptides Proteins 0.000 title claims abstract description 23
- 102000004196 processed proteins & peptides Human genes 0.000 title claims abstract description 23
- 238000004519 manufacturing process Methods 0.000 title claims abstract description 6
- 239000012634 fragment Substances 0.000 claims abstract description 49
- 239000013598 vector Substances 0.000 claims abstract description 22
- 239000002773 nucleotide Substances 0.000 claims abstract description 5
- 125000003729 nucleotide group Chemical group 0.000 claims abstract description 5
- 108020004414 DNA Proteins 0.000 claims description 13
- 230000010076 replication Effects 0.000 claims description 10
- 239000003550 marker Substances 0.000 claims description 9
- 108020005091 Replication Origin Proteins 0.000 claims description 7
- 230000000717 retained effect Effects 0.000 claims description 5
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 claims 8
- 108050000633 Lysozyme C Proteins 0.000 claims 1
- 238000010276 construction Methods 0.000 description 24
- 238000000034 method Methods 0.000 description 23
- 108010014251 Muramidase Proteins 0.000 description 22
- 108010062010 N-Acetylmuramoyl-L-alanine Amidase Proteins 0.000 description 22
- 210000004027 cell Anatomy 0.000 description 22
- 102000016943 Muramidase Human genes 0.000 description 18
- 229960000274 lysozyme Drugs 0.000 description 18
- 235000010335 lysozyme Nutrition 0.000 description 18
- 239000004325 lysozyme Substances 0.000 description 18
- 241000588724 Escherichia coli Species 0.000 description 14
- 230000000694 effects Effects 0.000 description 13
- 101150066555 lacZ gene Proteins 0.000 description 11
- 108010005774 beta-Galactosidase Proteins 0.000 description 9
- 238000011144 upstream manufacturing Methods 0.000 description 9
- 241000287828 Gallus gallus Species 0.000 description 8
- 108091081024 Start codon Proteins 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 8
- 102100026189 Beta-galactosidase Human genes 0.000 description 7
- 238000012217 deletion Methods 0.000 description 7
- 230000037430 deletion Effects 0.000 description 7
- 241000894007 species Species 0.000 description 7
- 108020004705 Codon Proteins 0.000 description 6
- 239000002609 medium Substances 0.000 description 6
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 5
- 230000001580 bacterial effect Effects 0.000 description 5
- 239000013604 expression vector Substances 0.000 description 5
- 238000002955 isolation Methods 0.000 description 5
- 239000002207 metabolite Substances 0.000 description 5
- 102000004190 Enzymes Human genes 0.000 description 4
- 108090000790 Enzymes Proteins 0.000 description 4
- 230000008901 benefit Effects 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 229940088598 enzyme Drugs 0.000 description 4
- 239000000284 extract Substances 0.000 description 4
- 102000004169 proteins and genes Human genes 0.000 description 4
- 238000013518 transcription Methods 0.000 description 4
- 230000035897 transcription Effects 0.000 description 4
- OPIFSICVWOWJMJ-AEOCFKNESA-N 5-bromo-4-chloro-3-indolyl beta-D-galactoside Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1OC1=CNC2=CC=C(Br)C(Cl)=C12 OPIFSICVWOWJMJ-AEOCFKNESA-N 0.000 description 3
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 3
- 241000191938 Micrococcus luteus Species 0.000 description 3
- 230000009471 action Effects 0.000 description 3
- 238000010353 genetic engineering Methods 0.000 description 3
- 239000001963 growth medium Substances 0.000 description 3
- 239000006166 lysate Substances 0.000 description 3
- 108020004999 messenger RNA Proteins 0.000 description 3
- 230000003287 optical effect Effects 0.000 description 3
- 239000000523 sample Substances 0.000 description 3
- 210000005253 yeast cell Anatomy 0.000 description 3
- 101710163881 5,6-dihydroxyindole-2-carboxylic acid oxidase Proteins 0.000 description 2
- 108090000204 Dipeptidase 1 Proteins 0.000 description 2
- 108010076504 Protein Sorting Signals Proteins 0.000 description 2
- 150000001413 amino acids Chemical class 0.000 description 2
- 238000003556 assay Methods 0.000 description 2
- 239000011324 bead Substances 0.000 description 2
- 238000005119 centrifugation Methods 0.000 description 2
- 238000003776 cleavage reaction Methods 0.000 description 2
- 238000010367 cloning Methods 0.000 description 2
- 230000009089 cytolysis Effects 0.000 description 2
- 238000002474 experimental method Methods 0.000 description 2
- 239000011521 glass Substances 0.000 description 2
- 125000001475 halogen functional group Chemical group 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- 238000012423 maintenance Methods 0.000 description 2
- 230000035772 mutation Effects 0.000 description 2
- 229930014626 natural product Natural products 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 230000007017 scission Effects 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 230000009466 transformation Effects 0.000 description 2
- 230000001131 transforming effect Effects 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 1
- XMTQQYYKAHVGBJ-UHFFFAOYSA-N 3-(3,4-DICHLOROPHENYL)-1,1-DIMETHYLUREA Chemical compound CN(C)C(=O)NC1=CC=C(Cl)C(Cl)=C1 XMTQQYYKAHVGBJ-UHFFFAOYSA-N 0.000 description 1
- 101100325906 Bacillus subtilis (strain 168) ganA gene Proteins 0.000 description 1
- 108010072454 CTGCAG-specific type II deoxyribonucleases Proteins 0.000 description 1
- 230000004544 DNA amplification Effects 0.000 description 1
- 239000005510 Diuron Substances 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 241000206602 Eukaryota Species 0.000 description 1
- 108010058643 Fungal Proteins Proteins 0.000 description 1
- 230000005526 G1 to G0 transition Effects 0.000 description 1
- 108020005350 Initiator Codon Proteins 0.000 description 1
- 102000014150 Interferons Human genes 0.000 description 1
- 108010050904 Interferons Proteins 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 241001138401 Kluyveromyces lactis Species 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- 101150007280 LEU2 gene Proteins 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- 108091028043 Nucleic acid sequence Proteins 0.000 description 1
- 102000009572 RNA Polymerase II Human genes 0.000 description 1
- 108010009460 RNA Polymerase II Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 241000235070 Saccharomyces Species 0.000 description 1
- 241000235347 Schizosaccharomyces pombe Species 0.000 description 1
- 108700026226 TATA Box Proteins 0.000 description 1
- 101100111413 Thermoanaerobacter pseudethanolicus (strain ATCC 33223 / 39E) lacZ gene Proteins 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 239000006035 Tryptophane Substances 0.000 description 1
- 241000235015 Yarrowia lipolytica Species 0.000 description 1
- 238000004458 analytical method Methods 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 239000003795 chemical substances by application Substances 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 230000000295 complement effect Effects 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- 238000001952 enzyme assay Methods 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 230000004927 fusion Effects 0.000 description 1
- 239000003966 growth inhibitor Substances 0.000 description 1
- 230000002363 herbicidal effect Effects 0.000 description 1
- 239000004009 herbicide Substances 0.000 description 1
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 1
- 230000003301 hydrolyzing effect Effects 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 238000009655 industrial fermentation Methods 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 229940047124 interferons Drugs 0.000 description 1
- 101150086432 lacA gene Proteins 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 230000002934 lysing effect Effects 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- MYWUZJCMWCOHBA-VIFPVBQESA-N methamphetamine Chemical compound CN[C@@H](C)CC1=CC=CC=C1 MYWUZJCMWCOHBA-VIFPVBQESA-N 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 230000000063 preceeding effect Effects 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 239000000047 product Substances 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 238000001179 sorption measurement Methods 0.000 description 1
- 239000006228 supernatant Substances 0.000 description 1
- 239000000725 suspension Substances 0.000 description 1
- 238000000844 transformation Methods 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- 229960004799 tryptophan Drugs 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 238000012800 visualization Methods 0.000 description 1
Images








Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
- C12N9/2405—Glucanases
- C12N9/2408—Glucanases acting on alpha -1,4-glucosidic bonds
- C12N9/2411—Amylases
- C12N9/2414—Alpha-amylase (3.2.1.1.)
- C12N9/2417—Alpha-amylase (3.2.1.1.) from microbiological source
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/80—Vectors or expression systems specially adapted for eukaryotic hosts for fungi
- C12N15/81—Vectors or expression systems specially adapted for eukaryotic hosts for fungi for yeasts
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
- C12N9/2462—Lysozyme (3.2.1.17)
Definitions
- the present invention relates to fragments of yeast DNA comprising a promoter capable of ensuring the expression in yeast of genes coding for polypeptides, generally for heterologous polypeptides. It also relates to vector plasmids comprising said fragments and to yeasts transformed by said plasmids.
- New genetic engineering techniques permit the expression in various organisms of genes coding for foreign proteins.
- One example of this, among many others, is the synthesis of human interferons by the yeast Socchoromyces cerevisiae (R.A. Hitzman et al., Science 219, 1983, 620).
- yeast promoter which is recognized by the yeast RNA polymerase II and causes the synthesis of the corresponding RNA messenger.
- An object of the present invention is to provide such promoters. Another object is to provide vector plasmids wherein said promoters are fused with heterologous genes so as to ensure the expression in yeast of said heterologous genes into the corresponding polypeptides.
- the yeasts transformed by said plasmids are a further object of the invention, as are the new polypeptides produced by said yeasts. Still another object is to provide a process for producing polypeptides by growing said transformed yeasts.
- the isolation of the base promoter of the invention hereinafter called p415, used known methods. It relied on the observation (L. Guarente, Meth. Enzymol. 101, 1983, 181; M.J.Casadaban et al., ibid. 100, 183, 293) that the gene lacZ coding for the ⁇ -golactosidase of E. coli can, under certain conditions, be expressed in the yeast S. cerevisiae and that this expression can be monitored by the blue colour of the yeast colonies when grown on the chromogenic indicator X-gal. This blue colour is due to hydrolytic cleavage of the indicator by ⁇ -galactosidase, and the rate of this cleavage depends on the amount of ⁇ -galactosidase synthesized.
- the blue colour of the colony depends on the efficiency of ⁇ -galactosidase expression, which in turn depends, among other things, on the force of the yeast promoter used.
- the strongest may give the blue colour more rapidly, and this can be verified quantitatively by enzyme assay of ⁇ -galactosidase activity.
- a prerequisite for the isolation of yeast promoters according to the above method is a vector capable of replication and selection in yeast, that contains the lacZ gene of E. coli but does not express this in yeast due to the absence of a suitable promoter.
- Said vector further has to fulfill the following conditions. It should be capable of replicating in the yeast by means of a sequence recognized by the replication machinery of the yeast. It should be capable of selection and maintenance in yeast due to the presence of a marker gene. It-should be potentially capable of expressing the lacZ gene in yeast when given a suitable promoter. It should comprise a unique restriction site upstream of this gene, for the insertion of small fragments of yeast DNA likely to comprise a promoter.
- Isolation of the promoter of the present invention with the hereabove described promoter probe vector was carried out as follows. DNA fragments resulting from a partial digest, with the Mbol restriction enzyme of total DNA of S. cerevisiae KL14-4A (G. Foge et al., J. Mol. Biol. 99, 1975, 203) were inserted into the BamHI site of the hereabove described plasmid YEpZ36, and the recombinant DNA thus obtained was used to transform the leu2 strain of S. cerevisiae GRF18.
- the transformed yeast colonies were screened on X-gal medium for ⁇ -galactosidase production. Several clones appeared positive upon transfer to X-gal medium, and the most active one, as determined by f-galactosidase assays, was selected for the isolation of the promoter of the invention. 1
- Plasmid DNA from this clone isolated after transfer to E. coli showed a 980 bp insert,that contained an EcoRI site 630 bp before the /3-galactosidase gene.
- the promoter ensuring transcription of.the ⁇ -galactosidase was found to be associated to this 630 bp fragment, as replacement of the 350 bp fragment preceding the EcoRI site did not affect f-galactosidase activity.
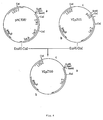
- the method of construction whereby the 350 bp fragment was deleted is detailed on Figure 2, where the initial 980 bp fragment is referred to as p414.
- both ends were flanked with unique restriction sites in current use. In this way, it can be moved from one plasmid to another, according to the application envisioned, and downstream insertion of a foreign gene for expression from the promoter is facilitated.
- promoter p415 as it is included into plasmid YEpZ101 is advantageous for the expression of a gene, such as the lacZ gene of pMC1587 or YEpZ100 ( Figure 4) that lacks an ATG initiator codon, since this is provided by the promoter itself just upstream of the BamHI site.
- promoter p415 of plasmid YEpZ101 cannot be used as such. It is indeed known that in eucaryotes, yeast included, the first ATG codon of the messenger RNA is normally used.
- a second ATG introduced would have one of two effects, according to whether it would be in phase or out of phase with the ATG codon of the promoter.
- a polypeptide would be produced which would be totally different from that coded by the gene.
- the amino acids corresponding to the codons between the first and the second ATG's would be added to the polypeptide at the amino end.
- a polypeptide differing from that normally coded by the gene, though perhaps not inactive would be obtained; this is unfavorable since substitutes as close as possible to the natural products are sought in most genetic engineering applications. This situation is especially unfavorable when it is also attempted to make use, as in the examples, of the signal sequence of a natural product to ensure its export outside of the transformed yeast cell.
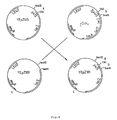
- promoter p415 it was necessary to have variants of promoter p415 not comprising the ATG initiator codon, while retaining the immediately downstream restriction site.
- the following examples describe such deletions from plasmid YEpZ101, starting from site BamHI.
- variants of promoter p415 were obtained which comprise a BamHI site at various distances from the HindIII end:p415 ⁇ 2, p415 ⁇ 4 and p415 ⁇ 5. It is obvious that the BamHI and HindIII ends in these variants can be replaced by other restriction sites just as in the case of the original p415 promoter.
- the promoters according to the invention can be used to express foreign genes in yeast if they are correctly associated to the gene to be expressed in a vector plasmid capable of replication into an as high as possible number of copies.
- This vector should therefore comprise a replication origin recognized by the host cell, and also a marker gene allowing visualisation and selection of the cells which have effectively been transformed by the plasmid.
- a great number of expression vectors comprising these different elements have been constructed, especially for the transformation of yeasts of the S.cerevisiae species. They generally comprise either the replication origin off the 2-micron plasmid present in most strains of this species, or even an autonomous ARS replication segment of chromosomal origin.
- a gene .coding for an enzyme involved in the biosynthesis of on essential metabolite, e.g. an amino acid, is generally used as marker.
- the host cell is then used in a yeast strain which has become by mutation auxotrophic for said metabolite. When growing this strain on a medium free of said metabolite, only those cells transformed by a plasmid carrying the missing gene will be able to grow.
- Typical examples of such markers are the LEU2 or TRP1 genes coding for on enzyme involved in the biosynthesis of leucine and tryptophane respectively.
- These expression vectors should also comprise one or preferably several restriction sites where to insert the coding, part of interest and the various elements required for optimizing the expression thereof: promoters, terminators and other control elements.
- these plasmids often comprise bacterial sequences capable of ensuring their replication and selection in an intermediate bacterial host, e.g. E.coli.
- shuttle plasmids it may be cited YEp13 (J.R.Broach et al., Gene 8, 1979, 121) pFLI-4 (M.R. Chevallier et al., Gene 11, 1980, 11), and pJDB207 (J.D.Beggs, Alfred Benson Symposium N°16, Munksgaard, Copenhagen 1981, p.383).
- the plasmid used comprises at least the REP functions of the 2-micron endogenous plasmid sequence. These functions generally give a better stability to the plosmid, especially if the host cell has previously been cured of its 2-micron plasmids (C.P. Hollenberg, Curr.Top.Microbiol. Immunol. 96, 1982, 119; R.M. Walmsley et al., Mol.Gen.Genet., 1983, 261).
- Classical examples of such vectors are plasmids pJDB219 and pJDB248 (J.D.Beggs, Nature 275, 1978, 104). Another vector of this type is described in the following examples.
- the transformed cells obtained in these various cases are also part of the invention.
- the invention also comprises the cells obtained by transforming other species and - genera of yeast with expression vectors of the type hereabove described.
- other yeasts it may be cited Saccharomycopsis lipolytica, Schizosaccharomyces pombe, Kluyveromyces lactis, etc....
- transformable host cells from the Saccharomyces genes and preferably from the S.cerevisiae species will be used.
- transformable strains from this species it may be cited AH22 and GRF18 amongst many others.
- the DNA fragments which can be expressed in yeast according to this invention may have various origins. They may be procaryotic genes, as the E.coli / 3-galactosidase gene whose expression is described in one of the following examples.
- the DNA fragment may also be an eucaryotic giene as for as it does not comprise introns as yeast is unable to ensure maturation of the messenger RNA from the transcription of fragmented genes (J.C.Beggs et al., Nature 283, 1980,835). In this case, it is however possible to express the corresponding cDNA, as described herebelow in another example showing chicken lysozyme expressed in yeast. Many other genes can similarly be expressed, whether they are of bacterial, vegetable, animal or human origin. New polypeptides that would thus be expressed are obviously also within the scope of the invention.
- a yeast strain When a yeast strain has been induced according to the present invention to produce one or another of these polypeptides, it is necessary to multiply it under the conditions most favourable to its growth in order to take advantage of this new property.
- One skilled in the art will easily determine these conditions according to the characteristics peculiar to the yeast strain used as host. As transformed yeasts have in most cases a more or less important tendency to lose artificially-constructed plasmids, it is advantageous to use a culture medium such as to exert a positive selection pressure on them.
- the strain is an auxotrophic mutant for one or another essential metabolite and when the vector plasmid used comprises a marker gene capable of restoring the strain prototrophy, e.g.
- this selection pressure may be exerted by omitting said metabolite from the culture medium.
- the plasmid comprises as marker a gene capable of conferring to the yeast a more or less marked resistance to a growth inhibitor, e.g. an antibiotic such as G418 (J.Jimenez and J.Davies, Nature 287, 1980, 869) or an herbicide such as diuron (Belgian Patent 899,607 to the Applicant), the selection pressure in favour of the transformed yeast can be applied by growing it in a medium supplemented with this inhibitor.
- a growth inhibitor e.g. an antibiotic such as G418 (J.Jimenez and J.Davies, Nature 287, 1980, 869) or an herbicide such as diuron (Belgian Patent 899,607 to the Applicant
- the selection pressure in favour of the transformed yeast can be applied by growing it in a medium supplemented with this inhibitor.
- Other means exist to obtain the same result and may also be used to practice the invention
- the gene expressed upon intervention of promoter p415 or one of the variants thereof will preferably be equipped with a leader sequence coding for a signal peptide capable of ensuring the transport of the product through the plasmid membrane of the transformed cell.
- the separation of the polypeptide formed will indeed be considerably easier, whether it is liberated into the medium from where it will be recovered by classical methods, including adsorption and/or precipitation, or it remains associated with the yeast wall from where it will have to be separated by other methods.
- Plasmid YEpZ415 the construction of which contributed, as hereabove described, to the isolation of promoter p415 according to the invention, was used to transform S.cerevisiae GRF18(Leu - ,His - ) yeast strain with selection of the clones prototrophic for leucine.
- promoter p415 according to the invention was flanked by two restriction sites commonly used in genetic engineering (HindIII and BamHI), and used in this form to express the lacZ gene of E.coli in a plasmid called YEpZ101. This operation was carried out in several steps which required the construction of two intermediate plasmids: YEpZ100 and pJ04.
- YEpZ100 (Figure 4) was constructed from pMC1587 (M.J.Casadaban et al., Methods in Enzymology 100, 1983, 293) and YEpZ415 of which it combines the advantages: it has the beginning of the lacZ gene and the group of restriction sites EcoRI, Smal and BamHI of pMC1587; it has the end of the lacZ gene of YEpZ415, whereby are eliminated the lacy and lacA genes of pMC1587 which would uselessly increase the size of the plasmid; it has the further advantage of keeping the high number of copies characteristic to the plasmids derived from pJDB207, e.g. . YEpZ415 (E.Erhart and C.P.Hollenberg, J.Bacteriol. 156, 1983, 625).
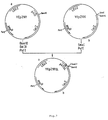
- 2.3 YEpZ101 was then constructed by combining DNA fragments from three of the plasmids described above, by a process involving three ligation events ( Figure 6).
- the lacZ gene, the amp R gene, the replication origin in E.coli, the LEU2 gene, and the replication origin in S.cerevisiae all derive from YEpZ100.
- the promoter p415 is reconstituted intact from two parts : (a) the 5' terminal half from the HindIII-ClaI fragment of YEpZ415, and (b) the 3' proximal half from the ClaI-BomHI fragment from pJ04.
- the next result of these operations is that the promoter p415 is now bounded by a unique HindIII site upstream and a unique BamHI site downstream, the latter being able to serve for the introduction of foreign genes as described herebelow.
- plasmid plys ⁇ 9 was available which comprises the complete full cDNA of chicken lysozyme with its own ATG codon as described in Belgian patent 901,223.
- This cDNA had still to be appropriately combined with a yeast promoter as described for the ⁇ -galactosidase gene in the preceding example.
- a plasmid such as YEpZ101 comprising promoter p415 which itself comprises an ATG codon.
- a variant of this plasmid free from this ATG codon was therefore required.
- the plasmids constructed as described hereabove were then transformed into GRF18 strain (Leu , His ) of the yeast S.cerevisice, followed by selection for clones prototrophic for leucine (Leu + ).
- lysozyme was evidenced around transformed colonies growing on Petri dishes covered with a lawn of M. lysodeikticus.
- lysozyme expression was visualised by a transparent halo of bacterial lysis around the colonies.
- the halo of lysis was greatest with plys ⁇ 49 indicating that this clone was the best producer of lysozyme.
- This result also showed that the lysozyme is exported from the yeast cell since it must of necessity be extracellular in order to lyse the bacterial indicator.
- Figure 10 also shows that by operating in the same way with the plys ⁇ 29 clone, no cell lysis could be observed.
- the sequences responsible for the promoter function should therefore not only be present upstream of the gene to express, but also be suitably positioned with respect to this gene.
- the vector pJDB207 upon which the lysozyme expression plasmids plys ⁇ 29, plys ⁇ 49 and plys ⁇ 59 (described hereabove) are based does not contain the entire 2-micron yeast plasmid and is consequently dependant on the presence of the endogenous 2-micron plasmid (found in most strains of S.cerevisiae) for its continued maintainance. This leads to such an unstable situation that the pJDB207-type plasmids are frequently lost from the cell (E. Erhaert & C.P.Hollenberg, J. Bacteriol. 156, 1983, 625, and M.Jarayam et al., Cell 34, 1983, 95).
- the natural 2-micron plasmid is stably inherited. It is indeed known that plasmids constructed in such a way that they contain the entire 2-micron plasmid pJDB219, are more stable than those of the pJDB207-type (C.P. Hollenberg, Curr. Top. Microbiol. Immunol. 96, 1982, 119; R.M. Walmsley et al., Mol. Gen. Genet. 1983, 361). Such plasmids are therefore more useful for long term growth, as for example in industrial fermentations.
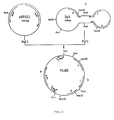
- plasmid YEpB2 ( Figure 11) was used, which was previously made by cloning the entire 2-micron plasmid into pBR322 at their mutually unique PstI sites. Two fragments from YEpB2 and two fragments from plys ⁇ 49 were then combined to give plys50 ( Figure 12).
- the three 2-micron genes A, B and C are intact and the lysozyme gene is expressed from the p415A4 promoter as in plys ⁇ 49 described in the preceeding example.
- Lysozyme activity in the supernatant and the lysate from both culture was determined by their ability to lyse M.lysodeikticus cells according to the method of D.Shugar (Biochem. Biophys. Acta, 8, 1952, 302). No activity was detected in a homogenate of the strain transformed by plasmid pJDB207, whereas a lysozyme activity of 35 units per ml of culture could be shown for that of strain GRF18(plys50).
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- General Health & Medical Sciences (AREA)
- Mycology (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Enzymes And Modification Thereof (AREA)
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AT85870171T ATE54167T1 (de) | 1984-12-06 | 1985-12-05 | Promotoren fuer die expression von fremden genen in hefe, plasmide, die diese promotoren enthalten, sowie deren verwendung zur herstellung von polypeptiden. |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| BE214123 | 1984-12-06 | ||
| BE0/214123A BE901222A (fr) | 1984-12-06 | 1984-12-06 | Promoteurs assurant l'expression de genes etrangers chez la levure, plasmides comportant ces promoteurs et leurs utilisations pour la production de polypeptides. |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| EP0184576A1 true EP0184576A1 (de) | 1986-06-11 |
| EP0184576B1 EP0184576B1 (de) | 1990-06-27 |
Family
ID=3843826
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| EP85870171A Expired - Lifetime EP0184576B1 (de) | 1984-12-06 | 1985-12-05 | Promotoren für die Expression von fremden Genen in Hefe, Plasmide, die diese Promotoren enthalten, sowie deren Verwendung zur Herstellung von Polypeptiden |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US4990446A (de) |
| EP (1) | EP0184576B1 (de) |
| JP (1) | JPH06104065B2 (de) |
| AT (1) | ATE54167T1 (de) |
| CA (1) | CA1280080C (de) |
| DE (1) | DE3578435D1 (de) |
Families Citing this family (48)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU633370B2 (en) * | 1989-10-16 | 1993-01-28 | Amgen, Inc. | N-acetylmuramidase m1 |
| WO1998016644A1 (en) | 1996-10-16 | 1998-04-23 | Zymogenetics, Inc. | Fibroblast growth factor homologs |
| BR9913942A (pt) | 1998-09-23 | 2001-10-16 | Zymogenetics Inc | Polinucleotìdeo e polipeptìdeo isolado, vetor de expressão, célula cultivada, constructo de dna codificador de uma proteìna de fusão, processos de produção de uma proteìna, de fusão de um polipeptìdeo e de um anticorpo para o polipeptìdeo e de detecção, em uma amostra de teste, da presença de um modulador da atividade da proteìna zalpha11 e de um ligante de receptor zalpha11 dentro de uma amostra de teste, e, anticorpo |
| CA2354325C (en) | 1998-12-07 | 2011-08-16 | Zymogenetics, Inc. | Growth factor homolog zvegf3 |
| US7833529B1 (en) | 1999-01-07 | 2010-11-16 | Zymogenetics, Inc. | Methods for inhibiting B lymphocyte proliferation with soluble ztnf4 receptor |
| PL393286A1 (pl) | 1999-01-07 | 2011-06-06 | Zymogenetics, Inc. | Rozpuszczalny receptor BR43x2 i sposoby jego zastosowania |
| CN101575377B (zh) | 1999-03-09 | 2012-12-26 | 津莫吉尼蒂克斯公司 | 作为zalpha受体之配体的人细胞因子及其用途 |
| ATE508193T1 (de) | 1999-07-07 | 2011-05-15 | Zymogenetics Inc | Menschliches rezeptor für zytokine |
| CA2395369C (en) | 1999-12-23 | 2014-07-08 | Zymogenetics, Inc. | Cytokine zcyto18 |
| WO2001087933A2 (en) | 2000-05-11 | 2001-11-22 | Zymogenetics, Inc. | Zsig33-like peptides |
| CA2412239C (en) | 2000-06-26 | 2013-05-28 | Zymogenetics, Inc. | Cytokine receptor zcytor17 |
| CN100448993C (zh) | 2000-06-30 | 2009-01-07 | 津莫吉尼蒂克斯公司 | 干扰素样蛋白质Zcyto21 |
| US7009045B2 (en) * | 2000-07-14 | 2006-03-07 | Archer-Daniels-Midland Company | Transformation systems for flavinogenic yeast |
| EP1736545A3 (de) | 2000-08-08 | 2007-03-28 | ZymoGenetics, Inc. | Lösliche Zcyctor 11 Cytokinrezeptoren |
| JP2004533997A (ja) | 2001-02-20 | 2004-11-11 | ザイモジェネティクス,インコーポレイティド | Bcma及びtaciの両者を結合する抗体 |
| EA007275B1 (ru) | 2001-05-24 | 2006-08-25 | Займодженетикс, Инк. | Слитые белки taci-иммуноглобулина |
| US6929932B2 (en) | 2001-11-05 | 2005-08-16 | Zymogenetics, Inc. | IL-21 antagonists |
| US7064186B2 (en) | 2002-01-18 | 2006-06-20 | Zymogenetics, Inc. | Cytokine zcytor17 ligand |
| EP1576112B1 (de) | 2002-01-18 | 2012-02-29 | ZymoGenetics, Inc. | Multimere des cytokinrezeptors zcytor17 |
| ES2356307T3 (es) | 2002-04-19 | 2011-04-06 | Zymogenetics, L.L.C. | Procedimientos para detectar o modular la interacción de un receptor de citocina con su ligando. |
| ES2303132T3 (es) | 2003-08-07 | 2008-08-01 | Zymogenetics, Inc. | Preparaciones homogeneas de il-29. |
| US7381794B2 (en) | 2004-03-08 | 2008-06-03 | Zymogenetics, Inc. | Dimeric fusion proteins and materials and methods for producing them |
| JP2008508310A (ja) | 2004-07-29 | 2008-03-21 | ザイモジェネティクス, インコーポレイテッド | がんおよび自己免疫障害を治療するためのil−28およびil−29の使用法 |
| EP2163635B1 (de) | 2004-08-02 | 2013-05-22 | BASF Plant Science GmbH | Verfahren zur Isolierung von Transkriptionsabschlusssequenzen |
| EP1856156A2 (de) | 2005-02-08 | 2007-11-21 | ZymoGenetics, Inc. | Anti-il-20-, anti-il-22- und anti-il-22ra-antikörper und bindungspartner sowie verwendungsverfahren bei entzündungen |
| ES2772674T3 (es) | 2005-05-12 | 2020-07-08 | Zymogenetics Inc | Composiciones y métodos para modular respuestas inmunitarias |
| WO2007019573A2 (en) | 2005-08-09 | 2007-02-15 | Zymogenetics, Inc. | Methods for the treatment and prevention of abnormal cell proliferation using taci-fusion molecules |
| KR20080056714A (ko) * | 2005-08-09 | 2008-06-23 | 지모제넥틱스, 인코포레이티드 | TACI―Ig 융합 분자를 사용하여 B―세포 악성종양을치료하는 방법 |
| BRPI0711823A2 (pt) * | 2006-05-15 | 2012-01-17 | Ares Trading Sa | métodos para tratamento de doenças auto-imunes com uma molécula de fusão taci-ig |
| US20100240597A1 (en) | 2007-06-15 | 2010-09-23 | Arkansas State University | Methods of delivery of molecules to cells using a ricin subunit and compositions relating to same |
| AU2008328371B2 (en) | 2007-11-21 | 2011-08-11 | Roskilde Universitet | Polypeptides comprising an ice-binding activity |
| US20110027337A1 (en) * | 2007-12-21 | 2011-02-03 | Ifxa A/S | Protease inhibitor |
| DK2245052T3 (en) * | 2008-01-25 | 2017-09-11 | Univ Aarhus | SELECTIVE EXOSITE INHIBITATION OF PAPP-A ACTIVITY AGAINST IGFBP-4 |
| ES2456296T3 (es) | 2008-03-27 | 2014-04-21 | Zymogenetics, Inc. | Composiciones y procedimientos para inhibir PDGFR beta y VEGF-A |
| US8658766B2 (en) | 2008-06-27 | 2014-02-25 | Zymogenetics, Inc. | Soluble hybrid Fcγ receptors and related methods |
| US8822642B2 (en) | 2010-06-09 | 2014-09-02 | Zymogenetics, Inc. | Dimeric fusion proteins and related compositions and methods |
| SG10201606285YA (en) | 2012-02-01 | 2016-09-29 | Synthetic Genomics Inc | Materials and methods for the synthesis of error-minimized nucleic acid molecules |
| WO2014202089A2 (en) | 2013-06-18 | 2014-12-24 | Roskilde Universitet | Variants of anti-freeze polypeptides |
| KR102440820B1 (ko) | 2015-09-08 | 2022-09-05 | 테리피온, 인크. | ApoA-1 융합 폴리펩티드 및 관련 조성물 및 방법 |
| WO2018136163A2 (en) | 2016-12-09 | 2018-07-26 | Theripion, Inc. | Tandem apoa-1 fusion polypeptides |
| BR112020001180A2 (pt) | 2017-07-20 | 2020-09-08 | Aptevo Research And Development Llc | proteínas de ligação a antígenos que se ligam a 5t4 e 4-1bb e composições e métodos relacionados |
| DK3710484T3 (da) | 2017-12-20 | 2023-11-27 | Harbour Biomed Shanghai Co Ltd | Ctla-4-bindende antistoffer og anvendelser deraf |
| US10150801B1 (en) | 2017-12-27 | 2018-12-11 | Imunami Laboratories Pte. Ltd. | Recombinant polypeptides and methods of use thereof |
| EP3732190A1 (de) | 2017-12-27 | 2020-11-04 | Imunami Laboratories Pte. Ltd. | Rekombinante polypeptide und verfahren zur verwendung davon |
| US10675332B1 (en) | 2019-08-26 | 2020-06-09 | Imunami Laboratories Pte. Ltd. | Recombinant polypeptides and methods of use thereof |
| EP4021512A1 (de) | 2019-08-26 | 2022-07-06 | Imunami Laboratories Pte. Ltd. | Rekombinante polypeptide und verfahren zur verwendung davon |
| JP2023538476A (ja) | 2020-06-22 | 2023-09-08 | イミュナミ ラボラトリーズ プライベート リミティド | 癌治療における使用のための組換えポリペプチド及び組合せ |
| CA3207134A1 (en) | 2021-02-19 | 2022-08-25 | Jeffrey A. Ledbetter | Dnase fusion polypeptides and related compositions and methods |
Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0011562A1 (de) * | 1978-11-14 | 1980-05-28 | ANVAR Agence Nationale de Valorisation de la Recherche | Mit einem Plasmid transformierte Saccharomyces cer. |
| EP0073635A2 (de) * | 1981-08-25 | 1983-03-09 | Celltech Limited | Expressionsvektoren |
| EP0109560A2 (de) * | 1982-10-20 | 1984-05-30 | SUNTORY KABUSHIKI KAISHA, also known as SUNTORY LTD. | Hefeplasmid-Vektor fähig zur induzierten Expression und Verfahren zur Verwendung dieses Plasmids |
| EP0127304A1 (de) * | 1983-04-25 | 1984-12-05 | Genentech, Inc. | Verfahren zur Herstellung von heterologem Protein in Hefe, Expressionsbildungsträger hierfür und damit transformierte Hefe |
| EP0123811B1 (de) * | 1983-02-28 | 1991-06-12 | Collaborative Research Inc. | Die Verwendung des GAL 1 Hefe-Promotors |
-
1985
- 1985-12-05 CA CA000496939A patent/CA1280080C/en not_active Expired - Lifetime
- 1985-12-05 EP EP85870171A patent/EP0184576B1/de not_active Expired - Lifetime
- 1985-12-05 AT AT85870171T patent/ATE54167T1/de not_active IP Right Cessation
- 1985-12-05 US US06/804,928 patent/US4990446A/en not_active Expired - Lifetime
- 1985-12-05 DE DE8585870171T patent/DE3578435D1/de not_active Expired - Lifetime
- 1985-12-06 JP JP60273531A patent/JPH06104065B2/ja not_active Expired - Lifetime
Patent Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0011562A1 (de) * | 1978-11-14 | 1980-05-28 | ANVAR Agence Nationale de Valorisation de la Recherche | Mit einem Plasmid transformierte Saccharomyces cer. |
| EP0073635A2 (de) * | 1981-08-25 | 1983-03-09 | Celltech Limited | Expressionsvektoren |
| EP0109560A2 (de) * | 1982-10-20 | 1984-05-30 | SUNTORY KABUSHIKI KAISHA, also known as SUNTORY LTD. | Hefeplasmid-Vektor fähig zur induzierten Expression und Verfahren zur Verwendung dieses Plasmids |
| EP0123811B1 (de) * | 1983-02-28 | 1991-06-12 | Collaborative Research Inc. | Die Verwendung des GAL 1 Hefe-Promotors |
| EP0127304A1 (de) * | 1983-04-25 | 1984-12-05 | Genentech, Inc. | Verfahren zur Herstellung von heterologem Protein in Hefe, Expressionsbildungsträger hierfür und damit transformierte Hefe |
Also Published As
| Publication number | Publication date |
|---|---|
| JPH06104065B2 (ja) | 1994-12-21 |
| EP0184576B1 (de) | 1990-06-27 |
| US4990446A (en) | 1991-02-05 |
| CA1280080C (en) | 1991-02-12 |
| ATE54167T1 (de) | 1990-07-15 |
| DE3578435D1 (de) | 1990-08-02 |
| JPS61185190A (ja) | 1986-08-18 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP0184576B1 (de) | Promotoren für die Expression von fremden Genen in Hefe, Plasmide, die diese Promotoren enthalten, sowie deren Verwendung zur Herstellung von Polypeptiden | |
| CA1224735A (en) | Cloning system for kluyveromyces species | |
| JP3437850B2 (ja) | ピキア・パストリスの酸ホスファターゼ遺伝子 | |
| USRE37767E1 (en) | Highly stable recombinant yeasts for the production of recombinant proteins | |
| US4885242A (en) | Genes from pichia histidine pathway and uses thereof | |
| US4943529A (en) | Kluyveromyces as a host strain | |
| US4882279A (en) | Site selective genomic modification of yeast of the genus pichia | |
| Lambert et al. | Genetic analysis of regulatory mutants affecting synthesis of extracellular proteinases in the yeast Yarrowia lipolytica: identification of a RIM101/pacC homolog | |
| EP0184575B1 (de) | Transformierte Hefen die Lysozym produzieren, für diese Transformation zu verwendende Plasmide und deren Verwendung | |
| EP0243338A2 (de) | Für eine spezifische Lipase kodierendes DNS-Segment, Vektoren zur Expression desselben, mit diesen Vektoren transformierte Mikroorganismen, sowie deren Anwendung zur Herstellung dieser Lipase | |
| US5068185A (en) | Trains of yeast for the expression of heterologous genes | |
| US5679544A (en) | Modified Kluyveromyces yeasts, their preparation and use | |
| EP0362183B1 (de) | Verfahren zur Herstellung von menschlichem Lysozym | |
| JPH08500014A (ja) | K.ラクチス(K.lactis)トランスアルドラーゼ遺伝子のプロモーターおよびその使用 | |
| US7132522B1 (en) | Regulatory sequences and expression cassettes for yeasts, especially for kluyveromyces | |
| EP0173668A1 (de) | Rekombinanter Mikroorganismus der Art Saccharomyces | |
| IE922007A1 (en) | Cloning and/or expression vectors, preparation and use | |
| US5541084A (en) | Hybrid yeast promoter comprising an ENO2 UAS and TATA region and an additional UAS located between said ENO2 UAS and TATA region | |
| JPH0775551B2 (ja) | アルフア−1−アンチトリプシンの発現のための安定なdna構造物 | |
| HU196454B (en) | Process for producing yeast vector of much reproduces |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PUAI | Public reference made under article 153(3) epc to a published international application that has entered the european phase |
Free format text: ORIGINAL CODE: 0009012 |
|
| AK | Designated contracting states |
Kind code of ref document: A1 Designated state(s): AT BE CH DE FR GB IT LI LU NL SE |
|
| 17P | Request for examination filed |
Effective date: 19861030 |
|
| 17Q | First examination report despatched |
Effective date: 19880803 |
|
| RAP1 | Party data changed (applicant data changed or rights of an application transferred) |
Owner name: FINA RESEARCH S.A. |
|
| GRAA | (expected) grant |
Free format text: ORIGINAL CODE: 0009210 |
|
| AK | Designated contracting states |
Kind code of ref document: B1 Designated state(s): AT BE CH DE FR GB IT LI LU NL SE |
|
| REF | Corresponds to: |
Ref document number: 54167 Country of ref document: AT Date of ref document: 19900715 Kind code of ref document: T |
|
| ET | Fr: translation filed | ||
| REF | Corresponds to: |
Ref document number: 3578435 Country of ref document: DE Date of ref document: 19900802 |
|
| ITF | It: translation for a ep patent filed |
Owner name: UFFICIO BREVETTI RICCARDI & C. |
|
| PLBE | No opposition filed within time limit |
Free format text: ORIGINAL CODE: 0009261 |
|
| STAA | Information on the status of an ep patent application or granted ep patent |
Free format text: STATUS: NO OPPOSITION FILED WITHIN TIME LIMIT |
|
| 26N | No opposition filed | ||
| ITTA | It: last paid annual fee | ||
| EPTA | Lu: last paid annual fee | ||
| EAL | Se: european patent in force in sweden |
Ref document number: 85870171.7 |
|
| REG | Reference to a national code |
Ref country code: GB Ref legal event code: IF02 |
|
| PGFP | Annual fee paid to national office [announced via postgrant information from national office to epo] |
Ref country code: GB Payment date: 20041122 Year of fee payment: 20 |
|
| PGFP | Annual fee paid to national office [announced via postgrant information from national office to epo] |
Ref country code: SE Payment date: 20041209 Year of fee payment: 20 |
|
| PGFP | Annual fee paid to national office [announced via postgrant information from national office to epo] |
Ref country code: NL Payment date: 20041210 Year of fee payment: 20 Ref country code: LU Payment date: 20041210 Year of fee payment: 20 Ref country code: CH Payment date: 20041210 Year of fee payment: 20 |
|
| PGFP | Annual fee paid to national office [announced via postgrant information from national office to epo] |
Ref country code: FR Payment date: 20041213 Year of fee payment: 20 Ref country code: DE Payment date: 20041213 Year of fee payment: 20 Ref country code: AT Payment date: 20041213 Year of fee payment: 20 |
|
| PGFP | Annual fee paid to national office [announced via postgrant information from national office to epo] |
Ref country code: BE Payment date: 20041227 Year of fee payment: 20 |
|
| REG | Reference to a national code |
Ref country code: CH Ref legal event code: PFA Owner name: TOTAL PETROCHEMICALS RESEARCH FELUY Free format text: FINA RESEARCH S.A.#ZONE INDUSTRIELLE C#SENEFFE (BE) -TRANSFER TO- TOTAL PETROCHEMICALS RESEARCH FELUY#ZONE INDUSTRIELLE C#7181 SENEFFE (FELUY) (BE) Ref country code: CH Ref legal event code: NV Representative=s name: LUCHS & PARTNER PATENTANWAELTE |
|
| NLT1 | Nl: modifications of names registered in virtue of documents presented to the patent office pursuant to art. 16 a, paragraph 1 |
Owner name: TOTAL PETROCHEMICALS RESEARCH FELUY Owner name: ATOFINA RESEARCH |
|
| PG25 | Lapsed in a contracting state [announced via postgrant information from national office to epo] |
Ref country code: GB Free format text: LAPSE BECAUSE OF EXPIRATION OF PROTECTION Effective date: 20051204 |
|
| PG25 | Lapsed in a contracting state [announced via postgrant information from national office to epo] |
Ref country code: NL Free format text: LAPSE BECAUSE OF EXPIRATION OF PROTECTION Effective date: 20051205 |
|
| REG | Reference to a national code |
Ref country code: GB Ref legal event code: PE20 |
|
| REG | Reference to a national code |
Ref country code: CH Ref legal event code: PL |
|
| NLV7 | Nl: ceased due to reaching the maximum lifetime of a patent |
Effective date: 20051205 |
|
| EUG | Se: european patent has lapsed | ||
| BE20 | Be: patent expired |
Owner name: S.A. *FINA RESEARCH Effective date: 20051205 |