JP7010902B2 - 限られた免疫グロブリン軽鎖レパートリーを発現するマウス - Google Patents
限られた免疫グロブリン軽鎖レパートリーを発現するマウス Download PDFInfo
- Publication number
- JP7010902B2 JP7010902B2 JP2019148806A JP2019148806A JP7010902B2 JP 7010902 B2 JP7010902 B2 JP 7010902B2 JP 2019148806 A JP2019148806 A JP 2019148806A JP 2019148806 A JP2019148806 A JP 2019148806A JP 7010902 B2 JP7010902 B2 JP 7010902B2
- Authority
- JP
- Japan
- Prior art keywords
- human
- light chain
- mouse
- gene segment
- immunoglobulin
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 title claims description 85
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 title claims description 81
- 241000699670 Mus sp. Species 0.000 title description 409
- 241000282414 Homo sapiens Species 0.000 claims description 1220
- 108090000623 proteins and genes Proteins 0.000 claims description 784
- 210000004027 cell Anatomy 0.000 claims description 354
- 108060003951 Immunoglobulin Proteins 0.000 claims description 175
- 102000018358 immunoglobulin Human genes 0.000 claims description 175
- 238000000034 method Methods 0.000 claims description 117
- 210000004602 germ cell Anatomy 0.000 claims description 108
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 claims description 51
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 claims description 49
- 238000004519 manufacturing process Methods 0.000 claims description 20
- 101100370002 Mus musculus Tnfsf14 gene Proteins 0.000 claims description 17
- 102100029567 Immunoglobulin kappa light chain Human genes 0.000 claims description 10
- 101710189008 Immunoglobulin kappa light chain Proteins 0.000 claims description 10
- 241000282412 Homo Species 0.000 claims description 9
- 101000998953 Homo sapiens Immunoglobulin heavy variable 1-2 Proteins 0.000 claims description 9
- 102100036887 Immunoglobulin heavy variable 1-2 Human genes 0.000 claims description 8
- 210000001161 mammalian embryo Anatomy 0.000 claims description 6
- 238000012239 gene modification Methods 0.000 claims description 3
- 230000005017 genetic modification Effects 0.000 claims description 3
- 235000013617 genetically modified food Nutrition 0.000 claims description 3
- 241000699666 Mus <mouse, genus> Species 0.000 description 522
- 239000000427 antigen Substances 0.000 description 223
- 108091007433 antigens Proteins 0.000 description 206
- 102000036639 antigens Human genes 0.000 description 206
- 210000003719 b-lymphocyte Anatomy 0.000 description 206
- 230000035772 mutation Effects 0.000 description 133
- 150000007523 nucleic acids Chemical group 0.000 description 102
- 230000014509 gene expression Effects 0.000 description 97
- 230000027455 binding Effects 0.000 description 81
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 71
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 71
- 210000001185 bone marrow Anatomy 0.000 description 58
- 108091028043 Nucleic acid sequence Proteins 0.000 description 55
- 101001094887 Ambrosia artemisiifolia Pectate lyase 1 Proteins 0.000 description 53
- 101001123576 Ambrosia artemisiifolia Pectate lyase 2 Proteins 0.000 description 53
- 101001123572 Ambrosia artemisiifolia Pectate lyase 3 Proteins 0.000 description 53
- 101000573177 Ambrosia artemisiifolia Pectate lyase 5 Proteins 0.000 description 53
- 102000004169 proteins and genes Human genes 0.000 description 53
- 235000018102 proteins Nutrition 0.000 description 52
- 210000000952 spleen Anatomy 0.000 description 51
- 230000000392 somatic effect Effects 0.000 description 48
- 230000008707 rearrangement Effects 0.000 description 43
- 239000000523 sample Substances 0.000 description 40
- 230000003393 splenic effect Effects 0.000 description 40
- 102100035360 Cerebellar degeneration-related antigen 1 Human genes 0.000 description 38
- 102000014914 Carrier Proteins Human genes 0.000 description 34
- 108091008324 binding proteins Proteins 0.000 description 34
- 230000004048 modification Effects 0.000 description 33
- 238000012986 modification Methods 0.000 description 33
- 108020004414 DNA Proteins 0.000 description 30
- 210000003519 mature b lymphocyte Anatomy 0.000 description 30
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 29
- 102000004196 processed proteins & peptides Human genes 0.000 description 29
- 108090000765 processed proteins & peptides Proteins 0.000 description 29
- 230000002441 reversible effect Effects 0.000 description 29
- 239000011324 bead Substances 0.000 description 28
- 229920001184 polypeptide Polymers 0.000 description 28
- 102000039446 nucleic acids Human genes 0.000 description 27
- 108020004707 nucleic acids Proteins 0.000 description 27
- 239000002773 nucleotide Substances 0.000 description 27
- 125000003729 nucleotide group Chemical group 0.000 description 27
- 241000700159 Rattus Species 0.000 description 26
- 125000003275 alpha amino acid group Chemical group 0.000 description 26
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 24
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 24
- 235000001014 amino acid Nutrition 0.000 description 24
- 238000010586 diagram Methods 0.000 description 24
- 102100025877 Complement component C1q receptor Human genes 0.000 description 23
- 102100032768 Complement receptor type 2 Human genes 0.000 description 23
- 101000933665 Homo sapiens Complement component C1q receptor Proteins 0.000 description 23
- 101000941929 Homo sapiens Complement receptor type 2 Proteins 0.000 description 23
- 229930193140 Neomycin Natural products 0.000 description 23
- 229960004927 neomycin Drugs 0.000 description 23
- 230000000903 blocking effect Effects 0.000 description 22
- 239000003446 ligand Substances 0.000 description 22
- 239000013598 vector Substances 0.000 description 21
- 150000001413 amino acids Chemical class 0.000 description 20
- 238000003556 assay Methods 0.000 description 20
- 239000006228 supernatant Substances 0.000 description 19
- 230000008685 targeting Effects 0.000 description 19
- 210000003297 immature b lymphocyte Anatomy 0.000 description 18
- 241000894007 species Species 0.000 description 18
- 229940024606 amino acid Drugs 0.000 description 17
- 239000003623 enhancer Substances 0.000 description 17
- 230000003325 follicular Effects 0.000 description 17
- 239000000203 mixture Substances 0.000 description 17
- 210000001948 pro-b lymphocyte Anatomy 0.000 description 17
- 230000004044 response Effects 0.000 description 16
- 239000000306 component Substances 0.000 description 15
- 239000012634 fragment Substances 0.000 description 15
- 238000003752 polymerase chain reaction Methods 0.000 description 15
- 238000006467 substitution reaction Methods 0.000 description 15
- 206010069754 Acquired gene mutation Diseases 0.000 description 14
- 230000002163 immunogen Effects 0.000 description 14
- 230000037439 somatic mutation Effects 0.000 description 14
- 239000013604 expression vector Substances 0.000 description 13
- 210000004408 hybridoma Anatomy 0.000 description 13
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 12
- 210000004988 splenocyte Anatomy 0.000 description 12
- 101150117115 V gene Proteins 0.000 description 11
- 210000004698 lymphocyte Anatomy 0.000 description 11
- 102000000844 Cell Surface Receptors Human genes 0.000 description 10
- 108010001857 Cell Surface Receptors Proteins 0.000 description 10
- 101150008942 J gene Proteins 0.000 description 10
- 102000025171 antigen binding proteins Human genes 0.000 description 10
- 108091000831 antigen binding proteins Proteins 0.000 description 10
- 210000002966 serum Anatomy 0.000 description 10
- 101000608935 Homo sapiens Leukosialin Proteins 0.000 description 9
- 102100039564 Leukosialin Human genes 0.000 description 9
- 108700026244 Open Reading Frames Proteins 0.000 description 9
- 238000010494 dissociation reaction Methods 0.000 description 9
- 230000005593 dissociations Effects 0.000 description 9
- 210000003826 marginal zone b cell Anatomy 0.000 description 9
- 101001008255 Homo sapiens Immunoglobulin kappa variable 1D-8 Proteins 0.000 description 8
- 101001047628 Homo sapiens Immunoglobulin kappa variable 2-29 Proteins 0.000 description 8
- 101001008321 Homo sapiens Immunoglobulin kappa variable 2D-26 Proteins 0.000 description 8
- 101001047619 Homo sapiens Immunoglobulin kappa variable 3-20 Proteins 0.000 description 8
- 101001008263 Homo sapiens Immunoglobulin kappa variable 3D-15 Proteins 0.000 description 8
- 241001465754 Metazoa Species 0.000 description 8
- 102000016971 Proto-Oncogene Proteins c-kit Human genes 0.000 description 8
- 108010014608 Proto-Oncogene Proteins c-kit Proteins 0.000 description 8
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 8
- 230000015572 biosynthetic process Effects 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 8
- 238000012217 deletion Methods 0.000 description 8
- 230000037430 deletion Effects 0.000 description 8
- 230000028993 immune response Effects 0.000 description 8
- 238000000338 in vitro Methods 0.000 description 8
- 108020004999 messenger RNA Proteins 0.000 description 8
- 239000013642 negative control Substances 0.000 description 8
- 230000002093 peripheral effect Effects 0.000 description 8
- 230000001225 therapeutic effect Effects 0.000 description 8
- 206010028980 Neoplasm Diseases 0.000 description 7
- 241000283984 Rodentia Species 0.000 description 7
- 102000005936 beta-Galactosidase Human genes 0.000 description 7
- 108010005774 beta-Galactosidase Proteins 0.000 description 7
- 210000004899 c-terminal region Anatomy 0.000 description 7
- 230000011712 cell development Effects 0.000 description 7
- 238000010367 cloning Methods 0.000 description 7
- 238000002474 experimental method Methods 0.000 description 7
- 210000005260 human cell Anatomy 0.000 description 7
- 230000003053 immunization Effects 0.000 description 7
- 230000008569 process Effects 0.000 description 7
- 238000003753 real-time PCR Methods 0.000 description 7
- 102000005962 receptors Human genes 0.000 description 7
- 108020003175 receptors Proteins 0.000 description 7
- 230000002207 retinal effect Effects 0.000 description 7
- 241000699800 Cricetinae Species 0.000 description 6
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 6
- 210000001744 T-lymphocyte Anatomy 0.000 description 6
- 239000000872 buffer Substances 0.000 description 6
- 238000004113 cell culture Methods 0.000 description 6
- 239000003814 drug Substances 0.000 description 6
- 238000000684 flow cytometry Methods 0.000 description 6
- 229940072221 immunoglobulins Drugs 0.000 description 6
- 230000000717 retained effect Effects 0.000 description 6
- 102220117530 rs112626848 Human genes 0.000 description 6
- 102220238658 rs1468529365 Human genes 0.000 description 6
- 102220268018 rs201210997 Human genes 0.000 description 6
- 238000010561 standard procedure Methods 0.000 description 6
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 5
- 108700024394 Exon Proteins 0.000 description 5
- 241000238631 Hexapoda Species 0.000 description 5
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 5
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 5
- 206010035226 Plasma cell myeloma Diseases 0.000 description 5
- 230000036039 immunity Effects 0.000 description 5
- 201000000050 myeloid neoplasm Diseases 0.000 description 5
- 210000005259 peripheral blood Anatomy 0.000 description 5
- 239000011886 peripheral blood Substances 0.000 description 5
- 238000012216 screening Methods 0.000 description 5
- 210000001082 somatic cell Anatomy 0.000 description 5
- 210000004881 tumor cell Anatomy 0.000 description 5
- 238000012286 ELISA Assay Methods 0.000 description 4
- 108010046276 FLP recombinase Proteins 0.000 description 4
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 4
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 4
- 108010076504 Protein Sorting Signals Proteins 0.000 description 4
- QAOWNCQODCNURD-UHFFFAOYSA-N Sulfuric acid Chemical compound OS(O)(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-N 0.000 description 4
- 108700005077 Viral Genes Proteins 0.000 description 4
- 235000004279 alanine Nutrition 0.000 description 4
- 210000004102 animal cell Anatomy 0.000 description 4
- 238000013459 approach Methods 0.000 description 4
- 230000008901 benefit Effects 0.000 description 4
- 210000004369 blood Anatomy 0.000 description 4
- 239000008280 blood Substances 0.000 description 4
- 230000000295 complement effect Effects 0.000 description 4
- 238000012258 culturing Methods 0.000 description 4
- 238000001514 detection method Methods 0.000 description 4
- 238000011161 development Methods 0.000 description 4
- 230000018109 developmental process Effects 0.000 description 4
- 210000003527 eukaryotic cell Anatomy 0.000 description 4
- 239000003102 growth factor Substances 0.000 description 4
- 239000000833 heterodimer Substances 0.000 description 4
- 239000000710 homodimer Substances 0.000 description 4
- 238000001727 in vivo Methods 0.000 description 4
- 238000002347 injection Methods 0.000 description 4
- 239000007924 injection Substances 0.000 description 4
- 239000003550 marker Substances 0.000 description 4
- 239000013612 plasmid Substances 0.000 description 4
- 238000002360 preparation method Methods 0.000 description 4
- 230000006798 recombination Effects 0.000 description 4
- 238000005215 recombination Methods 0.000 description 4
- 230000001105 regulatory effect Effects 0.000 description 4
- 230000001850 reproductive effect Effects 0.000 description 4
- 238000003757 reverse transcription PCR Methods 0.000 description 4
- 102200148758 rs116840795 Human genes 0.000 description 4
- 239000011780 sodium chloride Substances 0.000 description 4
- 229940124597 therapeutic agent Drugs 0.000 description 4
- 238000013518 transcription Methods 0.000 description 4
- 230000035897 transcription Effects 0.000 description 4
- 238000012546 transfer Methods 0.000 description 4
- 238000011144 upstream manufacturing Methods 0.000 description 4
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 description 3
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 3
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 3
- 241000282693 Cercopithecidae Species 0.000 description 3
- 238000002965 ELISA Methods 0.000 description 3
- 241000196324 Embryophyta Species 0.000 description 3
- 108010087819 Fc receptors Proteins 0.000 description 3
- 102000009109 Fc receptors Human genes 0.000 description 3
- 101000854886 Homo sapiens Immunoglobulin iota chain Proteins 0.000 description 3
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 3
- 102100020744 Immunoglobulin iota chain Human genes 0.000 description 3
- 102100022964 Immunoglobulin kappa variable 3-20 Human genes 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 3
- 125000000174 L-prolyl group Chemical group [H]N1C([H])([H])C([H])([H])C([H])([H])[C@@]1([H])C(*)=O 0.000 description 3
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 3
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 3
- 241000699729 Muridae Species 0.000 description 3
- 229920001213 Polysorbate 20 Polymers 0.000 description 3
- 101100240365 Streptomyces fradiae neoK gene Proteins 0.000 description 3
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 3
- 238000007792 addition Methods 0.000 description 3
- 230000009824 affinity maturation Effects 0.000 description 3
- 150000001412 amines Chemical class 0.000 description 3
- 235000003704 aspartic acid Nutrition 0.000 description 3
- 230000001580 bacterial effect Effects 0.000 description 3
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 3
- 102220349284 c.287A>T Human genes 0.000 description 3
- 239000006285 cell suspension Substances 0.000 description 3
- 238000006243 chemical reaction Methods 0.000 description 3
- 239000003599 detergent Substances 0.000 description 3
- 238000005516 engineering process Methods 0.000 description 3
- 230000001747 exhibiting effect Effects 0.000 description 3
- 230000006870 function Effects 0.000 description 3
- 230000002538 fungal effect Effects 0.000 description 3
- 230000004927 fusion Effects 0.000 description 3
- 238000003018 immunoassay Methods 0.000 description 3
- 230000005847 immunogenicity Effects 0.000 description 3
- 238000001802 infusion Methods 0.000 description 3
- 238000003780 insertion Methods 0.000 description 3
- 230000037431 insertion Effects 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 239000011159 matrix material Substances 0.000 description 3
- 239000004005 microsphere Substances 0.000 description 3
- 238000002823 phage display Methods 0.000 description 3
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 3
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 3
- 108091008146 restriction endonucleases Proteins 0.000 description 3
- 102220142694 rs192332456 Human genes 0.000 description 3
- 239000012146 running buffer Substances 0.000 description 3
- 238000012360 testing method Methods 0.000 description 3
- 230000010474 transient expression Effects 0.000 description 3
- 239000004474 valine Substances 0.000 description 3
- 210000005253 yeast cell Anatomy 0.000 description 3
- 241000699725 Acomys Species 0.000 description 2
- 108700028369 Alleles Proteins 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- 241000193830 Bacillus <bacterium> Species 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 2
- 102100030886 Complement receptor type 1 Human genes 0.000 description 2
- 241000398985 Cricetidae Species 0.000 description 2
- 230000006820 DNA synthesis Effects 0.000 description 2
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- -1 FcγRI Proteins 0.000 description 2
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 2
- 102000006395 Globulins Human genes 0.000 description 2
- 108010044091 Globulins Proteins 0.000 description 2
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 2
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 2
- 239000004471 Glycine Substances 0.000 description 2
- 239000007995 HEPES buffer Substances 0.000 description 2
- 101000727061 Homo sapiens Complement receptor type 1 Proteins 0.000 description 2
- 101000917826 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-a Proteins 0.000 description 2
- 101000917824 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-b Proteins 0.000 description 2
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 2
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 description 2
- 108010073807 IgG Receptors Proteins 0.000 description 2
- 241000235058 Komagataella pastoris Species 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 2
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 2
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 2
- 239000004472 Lysine Substances 0.000 description 2
- 239000007987 MES buffer Substances 0.000 description 2
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 241001452677 Ogataea methanolica Species 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 229910019142 PO4 Inorganic materials 0.000 description 2
- 108020005067 RNA Splice Sites Proteins 0.000 description 2
- 239000012980 RPMI-1640 medium Substances 0.000 description 2
- 241000121210 Sigmodontinae Species 0.000 description 2
- 108010090804 Streptavidin Proteins 0.000 description 2
- 241000187747 Streptomyces Species 0.000 description 2
- 108091008874 T cell receptors Proteins 0.000 description 2
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 2
- 239000007983 Tris buffer Substances 0.000 description 2
- 238000002835 absorbance Methods 0.000 description 2
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 2
- 239000002671 adjuvant Substances 0.000 description 2
- 125000000539 amino acid group Chemical group 0.000 description 2
- 230000003321 amplification Effects 0.000 description 2
- 239000012491 analyte Substances 0.000 description 2
- 238000004458 analytical method Methods 0.000 description 2
- 230000033115 angiogenesis Effects 0.000 description 2
- 230000007503 antigenic stimulation Effects 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- 210000000988 bone and bone Anatomy 0.000 description 2
- 239000011575 calcium Substances 0.000 description 2
- 229910052791 calcium Inorganic materials 0.000 description 2
- 230000007910 cell fusion Effects 0.000 description 2
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 2
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 2
- 238000010276 construction Methods 0.000 description 2
- 201000010099 disease Diseases 0.000 description 2
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 2
- 230000002349 favourable effect Effects 0.000 description 2
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 2
- 238000003205 genotyping method Methods 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- 235000013922 glutamic acid Nutrition 0.000 description 2
- 239000004220 glutamic acid Substances 0.000 description 2
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Chemical group OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 2
- 239000001963 growth medium Substances 0.000 description 2
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 2
- 230000006801 homologous recombination Effects 0.000 description 2
- 238000002744 homologous recombination Methods 0.000 description 2
- 239000005556 hormone Substances 0.000 description 2
- 229940088597 hormone Drugs 0.000 description 2
- 238000002649 immunization Methods 0.000 description 2
- 230000001771 impaired effect Effects 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 2
- 230000003993 interaction Effects 0.000 description 2
- 229960000310 isoleucine Drugs 0.000 description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 2
- 210000003734 kidney Anatomy 0.000 description 2
- 239000011777 magnesium Substances 0.000 description 2
- 229910052749 magnesium Inorganic materials 0.000 description 2
- 210000004962 mammalian cell Anatomy 0.000 description 2
- 239000002609 medium Substances 0.000 description 2
- 239000012533 medium component Substances 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- AEMBWNDIEFEPTH-UHFFFAOYSA-N n-tert-butyl-n-ethylnitrous amide Chemical compound CCN(N=O)C(C)(C)C AEMBWNDIEFEPTH-UHFFFAOYSA-N 0.000 description 2
- 238000003199 nucleic acid amplification method Methods 0.000 description 2
- 210000004940 nucleus Anatomy 0.000 description 2
- 230000036961 partial effect Effects 0.000 description 2
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 2
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 2
- 239000010452 phosphate Substances 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 239000000047 product Substances 0.000 description 2
- 210000001236 prokaryotic cell Anatomy 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 230000010076 replication Effects 0.000 description 2
- 239000011435 rock Substances 0.000 description 2
- 102220210869 rs1057524586 Human genes 0.000 description 2
- 102220220520 rs1060503090 Human genes 0.000 description 2
- 102220206698 rs142514490 Human genes 0.000 description 2
- 102220325921 rs1555376589 Human genes 0.000 description 2
- 102200164344 rs63751661 Human genes 0.000 description 2
- 230000003248 secreting effect Effects 0.000 description 2
- 238000000926 separation method Methods 0.000 description 2
- 210000000717 sertoli cell Anatomy 0.000 description 2
- 239000000243 solution Substances 0.000 description 2
- 238000012453 sprague-dawley rat model Methods 0.000 description 2
- 238000010186 staining Methods 0.000 description 2
- 210000000130 stem cell Anatomy 0.000 description 2
- 239000000126 substance Substances 0.000 description 2
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 210000001519 tissue Anatomy 0.000 description 2
- 239000003053 toxin Substances 0.000 description 2
- 231100000765 toxin Toxicity 0.000 description 2
- 108700012359 toxins Proteins 0.000 description 2
- 230000007704 transition Effects 0.000 description 2
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 2
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 2
- 210000003501 vero cell Anatomy 0.000 description 2
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 1
- UAIUNKRWKOVEES-UHFFFAOYSA-N 3,3',5,5'-tetramethylbenzidine Chemical compound CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 UAIUNKRWKOVEES-UHFFFAOYSA-N 0.000 description 1
- YRNWIFYIFSBPAU-UHFFFAOYSA-N 4-[4-(dimethylamino)phenyl]-n,n-dimethylaniline Chemical compound C1=CC(N(C)C)=CC=C1C1=CC=C(N(C)C)C=C1 YRNWIFYIFSBPAU-UHFFFAOYSA-N 0.000 description 1
- 102100034540 Adenomatous polyposis coli protein Human genes 0.000 description 1
- 101001073212 Arabidopsis thaliana Peroxidase 33 Proteins 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 108091008875 B cell receptors Proteins 0.000 description 1
- 102100025218 B-cell differentiation antigen CD72 Human genes 0.000 description 1
- 102100038080 B-cell receptor CD22 Human genes 0.000 description 1
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 1
- 235000017166 Bambusa arundinacea Nutrition 0.000 description 1
- 235000017491 Bambusa tulda Nutrition 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- GDOPTJXRTPNYNR-UHFFFAOYSA-N CC1CCCC1 Chemical compound CC1CCCC1 GDOPTJXRTPNYNR-UHFFFAOYSA-N 0.000 description 1
- 241000700193 Calomyscus Species 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 241000282994 Cervidae Species 0.000 description 1
- 101710098119 Chaperonin GroEL 2 Proteins 0.000 description 1
- 108091062157 Cis-regulatory element Proteins 0.000 description 1
- 102000016911 Deoxyribonucleases Human genes 0.000 description 1
- 108010053770 Deoxyribonucleases Proteins 0.000 description 1
- 241001095404 Dipodoidea Species 0.000 description 1
- 108010021468 Fc gamma receptor IIA Proteins 0.000 description 1
- 108010021472 Fc gamma receptor IIB Proteins 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 241000287828 Gallus gallus Species 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- 241000699694 Gerbillinae Species 0.000 description 1
- 229920000209 Hexadimethrine bromide Polymers 0.000 description 1
- 241001272567 Hominoidea Species 0.000 description 1
- 101000934359 Homo sapiens B-cell differentiation antigen CD72 Proteins 0.000 description 1
- 101000884305 Homo sapiens B-cell receptor CD22 Proteins 0.000 description 1
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 1
- 101001128634 Homo sapiens NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 2, mitochondrial Proteins 0.000 description 1
- 101001123325 Homo sapiens Peroxisome proliferator-activated receptor gamma coactivator 1-beta Proteins 0.000 description 1
- 101000934341 Homo sapiens T-cell surface glycoprotein CD5 Proteins 0.000 description 1
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 1
- 108090000144 Human Proteins Proteins 0.000 description 1
- 102000003839 Human Proteins Human genes 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- 102000009786 Immunoglobulin Constant Regions Human genes 0.000 description 1
- 108010009817 Immunoglobulin Constant Regions Proteins 0.000 description 1
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 1
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 1
- 108700029227 Immunoglobulin Light Chain Genes Proteins 0.000 description 1
- 108090001061 Insulin Proteins 0.000 description 1
- 102000004877 Insulin Human genes 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical group OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical group OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 1
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- 102100029205 Low affinity immunoglobulin gamma Fc region receptor II-b Human genes 0.000 description 1
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 241000282560 Macaca mulatta Species 0.000 description 1
- 102000012750 Membrane Glycoproteins Human genes 0.000 description 1
- 108010090054 Membrane Glycoproteins Proteins 0.000 description 1
- 241000398750 Muroidea Species 0.000 description 1
- 241000282339 Mustela Species 0.000 description 1
- 101000894412 Mycolicibacterium paratuberculosis (strain ATCC BAA-968 / K-10) Bacterioferritin Proteins 0.000 description 1
- 102100032194 NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 2, mitochondrial Human genes 0.000 description 1
- 241000398990 Nesomyidae Species 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 240000007594 Oryza sativa Species 0.000 description 1
- 235000007164 Oryza sativa Nutrition 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 102100028961 Peroxisome proliferator-activated receptor gamma coactivator 1-beta Human genes 0.000 description 1
- 244000082204 Phyllostachys viridis Species 0.000 description 1
- 235000015334 Phyllostachys viridis Nutrition 0.000 description 1
- 241001338313 Platacanthomyidae Species 0.000 description 1
- 241000276498 Pollachius virens Species 0.000 description 1
- 241000288906 Primates Species 0.000 description 1
- 238000011530 RNeasy Mini Kit Methods 0.000 description 1
- 241000700157 Rattus norvegicus Species 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 108010084592 Saporins Proteins 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 108010052160 Site-specific recombinase Proteins 0.000 description 1
- 241000398956 Spalacidae Species 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- NINIDFKCEFEMDL-UHFFFAOYSA-N Sulfur Chemical compound [S] NINIDFKCEFEMDL-UHFFFAOYSA-N 0.000 description 1
- 102100025244 T-cell surface glycoprotein CD5 Human genes 0.000 description 1
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 1
- 101000588258 Taenia solium Paramyosin Proteins 0.000 description 1
- 108091036066 Three prime untranslated region Proteins 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 1
- 102000004357 Transferases Human genes 0.000 description 1
- 108090000992 Transferases Proteins 0.000 description 1
- 108700019146 Transgenes Proteins 0.000 description 1
- 241000255993 Trichoplusia ni Species 0.000 description 1
- 229920004890 Triton X-100 Polymers 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 108020005202 Viral DNA Proteins 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 1
- 238000012867 alanine scanning Methods 0.000 description 1
- 125000001931 aliphatic group Chemical group 0.000 description 1
- 229930013930 alkaloid Natural products 0.000 description 1
- 230000007815 allergy Effects 0.000 description 1
- ILRRQNADMUWWFW-UHFFFAOYSA-K aluminium phosphate Chemical compound O1[Al]2OP1(=O)O2 ILRRQNADMUWWFW-UHFFFAOYSA-K 0.000 description 1
- 150000001408 amides Chemical class 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 230000009833 antibody interaction Effects 0.000 description 1
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 1
- 210000004436 artificial bacterial chromosome Anatomy 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 208000006673 asthma Diseases 0.000 description 1
- 239000011425 bamboo Substances 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 244000309464 bull Species 0.000 description 1
- 238000010805 cDNA synthesis kit Methods 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 239000013592 cell lysate Substances 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 235000013330 chicken meat Nutrition 0.000 description 1
- 150000003841 chloride salts Chemical class 0.000 description 1
- 235000012000 cholesterol Nutrition 0.000 description 1
- 230000009194 climbing Effects 0.000 description 1
- 238000012875 competitive assay Methods 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000010168 coupling process Methods 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- 102000003675 cytokine receptors Human genes 0.000 description 1
- 108010057085 cytokine receptors Proteins 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- 230000007423 decrease Effects 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- 230000003292 diminished effect Effects 0.000 description 1
- 210000001840 diploid cell Anatomy 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 210000002257 embryonic structure Anatomy 0.000 description 1
- 238000001976 enzyme digestion Methods 0.000 description 1
- 230000008472 epithelial growth Effects 0.000 description 1
- 210000003743 erythrocyte Anatomy 0.000 description 1
- 238000010195 expression analysis Methods 0.000 description 1
- 239000012091 fetal bovine serum Substances 0.000 description 1
- 239000003527 fibrinolytic agent Substances 0.000 description 1
- 210000002950 fibroblast Anatomy 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 238000011010 flushing procedure Methods 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 235000003869 genetically modified organism Nutrition 0.000 description 1
- 210000002980 germ line cell Anatomy 0.000 description 1
- 239000005337 ground glass Substances 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 238000001114 immunoprecipitation Methods 0.000 description 1
- 230000002637 immunotoxin Effects 0.000 description 1
- 239000002596 immunotoxin Substances 0.000 description 1
- 231100000608 immunotoxin Toxicity 0.000 description 1
- 229940051026 immunotoxin Drugs 0.000 description 1
- 238000000126 in silico method Methods 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 150000002484 inorganic compounds Chemical class 0.000 description 1
- 229910010272 inorganic material Inorganic materials 0.000 description 1
- 229940125396 insulin Drugs 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 230000000670 limiting effect Effects 0.000 description 1
- 210000001165 lymph node Anatomy 0.000 description 1
- 239000012139 lysis buffer Substances 0.000 description 1
- 241001515942 marmosets Species 0.000 description 1
- 230000013011 mating Effects 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 230000005012 migration Effects 0.000 description 1
- 238000013508 migration Methods 0.000 description 1
- 238000001823 molecular biology technique Methods 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 238000007857 nested PCR Methods 0.000 description 1
- 238000010449 nuclear transplantation Methods 0.000 description 1
- VYNDHICBIRRPFP-UHFFFAOYSA-N pacific blue Chemical compound FC1=C(O)C(F)=C2OC(=O)C(C(=O)O)=CC2=C1 VYNDHICBIRRPFP-UHFFFAOYSA-N 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 239000008188 pellet Substances 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 239000000651 prodrug Substances 0.000 description 1
- 229940002612 prodrug Drugs 0.000 description 1
- 230000009465 prokaryotic expression Effects 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 230000020978 protein processing Effects 0.000 description 1
- 210000001938 protoplast Anatomy 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000002829 reductive effect Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 235000009566 rice Nutrition 0.000 description 1
- 230000005070 ripening Effects 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 239000012723 sample buffer Substances 0.000 description 1
- 229920006395 saturated elastomer Polymers 0.000 description 1
- 230000028327 secretion Effects 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 238000013207 serial dilution Methods 0.000 description 1
- 230000009870 specific binding Effects 0.000 description 1
- 210000004989 spleen cell Anatomy 0.000 description 1
- 230000000638 stimulation Effects 0.000 description 1
- 238000010254 subcutaneous injection Methods 0.000 description 1
- 239000007929 subcutaneous injection Substances 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- 239000011593 sulfur Substances 0.000 description 1
- 229910052717 sulfur Inorganic materials 0.000 description 1
- 239000013595 supernatant sample Substances 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 229960000103 thrombolytic agent Drugs 0.000 description 1
- 235000013619 trace mineral Nutrition 0.000 description 1
- 239000011573 trace mineral Substances 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 238000001890 transfection Methods 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 210000000689 upper leg Anatomy 0.000 description 1
- 239000012646 vaccine adjuvant Substances 0.000 description 1
- 229940124931 vaccine adjuvant Drugs 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- 108700026220 vif Genes Proteins 0.000 description 1
- 230000003612 virological effect Effects 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
Images




























Classifications
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
- A01K67/0275—Genetically modified vertebrates, e.g. transgenic
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/46—Hybrid immunoglobulins
- C07K16/468—Immunoglobulins having two or more different antigen binding sites, e.g. multifunctional antibodies
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/8509—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells for producing genetically modified animals, e.g. transgenic
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2227/00—Animals characterised by species
- A01K2227/10—Mammal
- A01K2227/105—Murine
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2267/00—Animals characterised by purpose
- A01K2267/01—Animal expressing industrially exogenous proteins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/24—Immunoglobulins specific features characterized by taxonomic origin containing regions, domains or residues from different species, e.g. chimeric, humanized or veneered
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/31—Immunoglobulins specific features characterized by aspects of specificity or valency multispecific
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Immunology (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Zoology (AREA)
- Biotechnology (AREA)
- Environmental Sciences (AREA)
- Veterinary Medicine (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Medicinal Chemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Biodiversity & Conservation Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Animal Husbandry (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Microbiology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Description
本出願は、2013年3月13日に出願された米国特許出願第13/798,455号の利益を、米国特許法§119(e)の下で請求し、この出願は、その全体が参照によって本明細書により組み込まれる。
本出願では、2014年3月13日に「2010794-0550_ST25」という名称のascii.txtファイルとして電子形式で提出された配列表に言及する。この.txtファイルは2014年3月11日に作成されたものであり、サイズは33kbである。
Protein Sequence Database、Science 256巻:1443~45頁)に開示されているPAM250対数尤度行列において正の値を有するように成される。一部の実施形態では、置換は、置換がPAM250対数尤度行列において負ではない値を有する、中程度に保存的な置換である。
有用な多重特異性エピトープ結合タンパク質、例えば二重特異性抗体を作製するための以前の試みは、共通のパラダイムが頻繁に共有される様々な問題:ヘテロ二量体の二重特異性ヒト免疫グロブリンを対合させるための適切な形式を合理的に操作するため、または試行錯誤を通じて操作するための配列のin vitroにおける選択または処理によって妨げられている。残念ながら、in vitroにおける操作手法(engineering approach)の全てではないにしろ大多数では、全ての場合、個々の分子に適するほとんどその場限りの調整(fix)がもたらされる。他方では、複雑な生物体を使用して、ヒト治療薬をもたらすことができる適切な対合を選択するためのin vivoにおける方法は実現されていない。
本明細書に記載の組成物および方法を使用して、2つ以上のエピトープに高親和性で結合する結合タンパク質、例えば二重特異性抗体を作製することができる。本発明の利点としては、重鎖免疫グロブリン鎖であって、そのそれぞれが単一の軽鎖と結合する、適切に高い結合性(例えば、親和性成熟した)の重鎖免疫グロブリン鎖を選択できることが挙げられる。
4種の異なる抗原に対して生じた親和性成熟した抗体由来の種々のヒト可変領域を、それらのコグネイト軽鎖またはヒトVκ1-39/Jκ5、ヒトVκ3-20/Jκ1またはヒトVpreB/Jλ5から選択されるヒト軽鎖の少なくとも1つのいずれかと共に発現させた(実施例1を参照されたい)。抗原のそれぞれに対する抗体について、異なる遺伝子ファミリー由来の体細胞変異した高親和性重鎖は再構成されたヒト生殖系列Vκ1-39Jκ5領域およびVκ3-20Jκ1領域と首尾よく対合し、重鎖および軽鎖を発現している細胞から分泌された。Vκ1-39Jκ5およびVκ3-20Jκ1については、以下のヒトVH遺伝子ファミリーに由来するVHドメインが好都合に発現した:1-2、1-8、1-24、2-5、3-7、3-9、3-11、3-13、3-15、3-20、3-23、3-30、3-33、3-48、4-31、4-39、4-59、5-51、および6-1。したがって、Vκ1-39Jκ5およびVκ3-20Jκ1の一方または両方由来のヒトVLドメインの限定されたレパートリーを発現するように操作されたマウスは、マウスVH遺伝子セグメントをヒトVH遺伝子セグメントで置き換えるように改変されたVH遺伝子座から体細胞変異したヒトVHドメインの多様な集団を生成する。
種々の実施形態における第1のステップとして、それぞれヒト重鎖可変ドメイン(および二重特異性抗体を形成する任意のさらなる核酸配列)をコードする第1の核酸配列および第2の核酸配列を、例えば、異なるエピトープに結合できること(図7Aおよび図7Bを参照されたい)、異なる親和性を有することなどのような所望の特性を有する親モノクローナル抗体から選択する。通常、ヒト重鎖可変ドメインをコードする核酸配列は本明細書に記載の通り免疫したマウスから単離し、ヒトへの投与に適するようにヒト重鎖定常領域との融合を可能にする。二重特異性抗体にさらなる機能性を付加する変異を導入することによって行うことができる配列(複数可)へのさらなる改変を実現することができ、その改変としては、例えば、血清半減期を増加させること(例えば、U.S.7,217,797を参照されたい)および/または抗体依存性細胞媒介性細胞傷害性を高めること(例えば、U.S.6,737,056を参照されたい)が挙げられる。抗体の定常領域への変異の導入は当技術分野で公知である。さらに、二重特異性抗体の一部は組換えによって細胞培養物中で作製させることができ、当該分子の他の部分(複数可)は上記の技法によって作製することができる。
選択されたヒトVLと結合するヒトVH領域の同定
単一の再構成されたヒト生殖系列軽鎖を抗原特異的ヒト抗体由来のヒト重鎖と共発現させることができるかどうかを決定するためにin vitro発現系を構築した。


再構成されたヒト生殖系列軽鎖遺伝子座の生成
VELOCIGENE(登録商標)技術(例えば、米国特許第6,586,251号およびValenzuelaら(2003年)High-throughput engineering of the mouse genome coupled with high-resolution expression analysis、Nature Biotech.21巻(6号):652~659頁を参照されたい)を使用して種々の再構成されたヒト生殖系列軽鎖ターゲティングベクターを作製して、マウスゲノム細菌人工染色体(BAC)クローン302g12および254m04(Invitrogen)を改変した。これらの2種のBACクローンを使用して、ゲノム構築物を、単一の再構成されたヒト生殖系列軽鎖領域を含有するように操作し、内因性κ可変遺伝子セグメントおよび内因性連結遺伝子セグメントが欠失するように予め改変した内因性κ軽鎖遺伝子座に挿入した。
当技術分野において認められている標準の分子生物学技法を使用して3種の異なる再構成されたヒト生殖系列軽鎖領域を作製した。これらの3種の領域を構築するために使用したヒト可変遺伝子セグメントは、再構成されたヒトVκ1-39Jκ5配列、再構成されたヒトVκ3-20Jκ1配列および再構成されたヒトVpreBJλ5配列を含むものであった。
Technologies)によって作製した。5’非翻訳領域の一部から天然に存在するBlpI制限酵素部位までを含めた。ヒトVκ1-39遺伝子およびヒトVκ3-20遺伝子のエクソンをヒトゲノムBACライブラリーからPCR増幅した。フォワードプライマーはマウスVκ3-7遺伝子のイントロン1のスプライスアクセプター部位を含有する5’伸長を有した。ヒトVκ1-39配列のPCRのために使用したリバースプライマーはヒトJκ5をコードする伸長を含むものであり、ヒトVκ3-20配列のPCRのために使用したリバースプライマーはヒトJκ1をコードする伸長を含むものであった。ヒトVpreBJλ5配列をデノボDNA合成(Integrated DNA Technologies)によって作製した。スプライスドナー部位を含むヒトJκ-Cκイントロンの一部をプラスミドpBS-296-HA18-PISceIからPCR増幅した。フォワードPCRプライマーはヒトJκ5配列、Jκ1配列またはJλ5配列のいずれかの一部をコードする伸長を含むものであった。リバースプライマーはイントロンに予め操作されたPI-SceI部位を含むものであった。
操作された軽鎖挿入物の5’末端および3’末端に、ターゲティングベクターへのクローニングのための制限酵素部位を導入した:5’末端にAscI部位および3’末端にPI-SceI部位。5’AscI部位から3’PI-SceI部位内で、ターゲティング構築物は、5’から3’へ、マウスBACクローン302g12から得た内因性マウスκ軽鎖遺伝子座に対して5’側の5’相同性アーム含有配列、FRTedネオマイシン耐性遺伝子、ヒトVκ3-15プロモーターを含むゲノム配列、マウスVκ3-7可変遺伝子セグメントのリーダー配列、マウスVκ3-7可変遺伝子セグメントのイントロン配列、再構成されたヒト生殖系列Vκ1-39Jκ5領域のオープンリーディングフレーム、ヒトJκ-Cκイントロンの一部を含有するゲノム配列、およびマウスBACクローン254m04から得た内因性マウスJκ5遺伝子セグメントの3’側の3’相同性アーム含有配列を含んだ(図1、中央)。内因性マウスκ軽鎖遺伝子座の上流および最も3’側のJκ遺伝子セグメント(例えば、内因性3’エンハンサー)の下流の遺伝子および/または配列はターゲティング構築物によって改変しなかった(図1参照)。操作されたヒトVκ1-39Jκ5遺伝子座の配列が配列番号1に示されている。
A;配列番号4)および1633-h2R(TGCAAACTGG ATGCAGCATA G;配列番号5)を用いて確認した。ネオマイシンカセットを、プライマーneoF(GGTGGAGAGG CTATTCGGC;配列番号6)およびneoR(GAACACGGCG GCATCAG;配列番号7)を用いて確認した。次いで、標的BAC
DNAを使用して、マウスES細胞に電気穿孔し、再構成されたヒト生殖系列Vκ1-39Jκ5領域を発現するキメラマウスを生成するための改変ES細胞を作製した。
同様に、再構成されたヒト生殖系列Vκ3-20Jκ1領域を発現する、操作された軽鎖遺伝子座を、5’から3’へ、マウスBACクローン302g12から得た内因性マウスκ軽鎖遺伝子座に対して5’側の5’相同性アーム含有配列、FRTedネオマイシン耐性遺伝子、ヒトVκ3-15プロモーターを含むゲノム配列、マウスVκ3-7可変遺伝子セグメントのリーダー配列、マウスVκ3-7可変遺伝子セグメントのイントロン配列、再構成されたヒト生殖系列Vκ3-20Jκ1領域のオープンリーディングフレーム、ヒトJκ-Cκイントロンの一部を含有するゲノム配列、およびマウスBACクローン254m04から得た内因性マウスJκ5遺伝子セグメントの3’側の3’相同性アーム含有配列を含むターゲティング構築物を使用して作製した(図2、中央)。操作されたヒトVκ3-20Jκ1遺伝子座の配列が配列番号11に示されている。
同様に、再構成されたヒト生殖系列VpreBJλ5領域を発現する、操作された軽鎖遺伝子座を、5’から3’へ、マウスBACクローン302g12から得た内因性マウスκ軽鎖遺伝子座に対して5’側の5’相同性アーム含有配列、FRTedネオマイシン耐性遺伝子、ヒトVκ3-15プロモーターを含むゲノム配列、マウスVκ3-7可変遺伝子セグメントのリーダー配列、マウスVκ3-7可変遺伝子セグメントのイントロン配列、再構成されたヒト生殖系列VpreBJλ5領域のオープンリーディングフレーム、ヒトJκ-Cκイントロンの一部を含有するゲノム配列、およびマウスBACクローン254m04から得た内因性マウスJκ5遺伝子セグメントの3’側の3’相同性アーム含有配列を含むターゲティング構築物を使用して作製した(図3、中央)。操作されたヒトVpreBJλ5遺伝子座の配列が配列番号15に示されている。
単一の再構成されたヒト軽鎖を発現するマウスの生成
上記の標的ES細胞をドナーES細胞として使用し、VELOCIMOUSE(登録商標)法(例えば、米国特許第7,294,754号、およびPoueymirouら(2007年)F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech.25巻(1号):91~99頁を参照されたい)によって8細胞期マウス胚に導入した。操作されたヒト生殖系列Vκ1-39Jκ5軽鎖領域、Vκ3-20Jκ1軽鎖領域またはVpreBJλ5軽鎖領域をそれぞれ独立に有するVELOCIMICE(登録商標)を、対立遺伝子アッセイ(Valenzuelaら、上記)の改変を使用した遺伝子型決定によって同定し、それにより独特の再構成されたヒト生殖系列軽鎖領域の存在を検出する。
共通軽鎖マウスの正常な抗体レパートリーにおける再構成されたヒト軽鎖領域の発現を、共通軽鎖マウスの脾細胞および末梢血における免疫グロブリンκおよびλ発現を分析することにより検証した。野生型マウス(n=5)、Vκ1-39Jκ5共通軽鎖ヘテロ接合体マウス(n=3)、Vκ1-39Jκ5共通軽鎖ホモ接合体マウス(n=3)、Vκ3-20Jκ1共通軽鎖ヘテロ接合体マウス(n=2)、およびVκ3-20Jκ1共通軽鎖ホモ接合体マウス(n=2)から採取した脾臓および末梢血から、標準の方法を使用して細胞懸濁物を作製し、蛍光標識された抗体(BD Pharmigen)を使用してCD19+、Igλ+およびIgκ+で染色した。

各共通軽鎖(Vκ1-39Jκ5およびVκ3-20Jκ1)の発現を、ヘテロ接合性マウスおよびホモ接合性マウスにおいて定量的PCRアッセイ(例えば、TAQMAN(商標))を使用して解析した。
Sequence Detection System(Applied Biosystems)を使用して実施した。表4には、このアッセイに使用したプライマーおよびプローブの配列が示されている。相対的な発現をマウスCκ領域の発現に対して正規化した。結果が図5A、図5Bおよび図5Cに示されている。

内因性マウスκ軽鎖遺伝子座にVκ1-39Jκ5共通軽鎖またはVκ3-20Jκ1共通軽鎖のいずれかを有する共通軽鎖マウスを、β-ガラクトシダーゼを用いて免疫し、抗体価を測定した。
単一の再構成されたヒト生殖系列軽鎖を発現するマウスの掛け合わせ
この実施例では、本明細書に記載の共通軽鎖マウスの任意の1つと掛け合わせて、複数の遺伝子改変免疫グロブリン遺伝子座を有する複数の遺伝子改変マウス系統を作製することができるいくつかの他の遺伝子改変マウス系統が記載されている。
操作された軽鎖遺伝子座の使用を最適化するために、再構成されたヒト生殖系列軽鎖領域のうちの1つを有するマウスを、内因性λ軽鎖遺伝子座に欠失を含有する別のマウスと掛け合わせる。このように、実施例2に記載の通り、得られた後代は、それらの唯一の軽鎖として、再構成されたヒト生殖系列軽鎖領域を発現する。掛け合わせは、当技術分野において認められている標準の技法によって実施する、あるいは、商業的なブリーダー(例えば、Jackson Laboratory)によって実施される。操作された、軽鎖遺伝子座および内因性λ軽鎖遺伝子座の欠失を有するマウス系統を、独特の軽鎖領域の存在および内因性マウスλ軽鎖の非存在についてスクリーニングする。
操作された、ヒト生殖系列軽鎖遺伝子座を有するマウスを、内因性マウス重鎖可変遺伝子遺伝子座の、ヒト重鎖可変遺伝子遺伝子座での置き換えを含有するマウス(US6,596,541を参照されたい;VELOCIMMUNE(登録商標)マウス、Regeneron Pharmaceuticals,Inc.)と掛け合わせる。VELOCIMMUNE(登録商標)マウスは、内因性マウス定常領域遺伝子座に作動可能に連結したヒト重鎖可変領域を含むゲノムを含み、その結果、マウスは、抗原による刺激に応答して、ヒト重鎖可変領域およびマウス重鎖定常領域を含む抗体を産生する。抗体の重鎖の可変領域をコードするDNAを単離し、ヒト重鎖定常領域をコードするDNAに作動可能に連結する。次いで、DNAを、抗体の完全ヒト重鎖を発現することができる細胞において発現させる。
ヒト重鎖および再構成されたヒト生殖系列軽鎖領域を発現するマウスからの抗体の生成
操作されたヒト軽鎖領域を含有するマウスと他の内因性Ig遺伝子座の改変および欠失を含有する種々の所望の系統と掛け合わせた後(実施例4に記載の通り)、選択されたマウスを、目的の抗原を用いて免疫することができる。
抗原特異的共通軽鎖抗体における重鎖遺伝子セグメントの使用
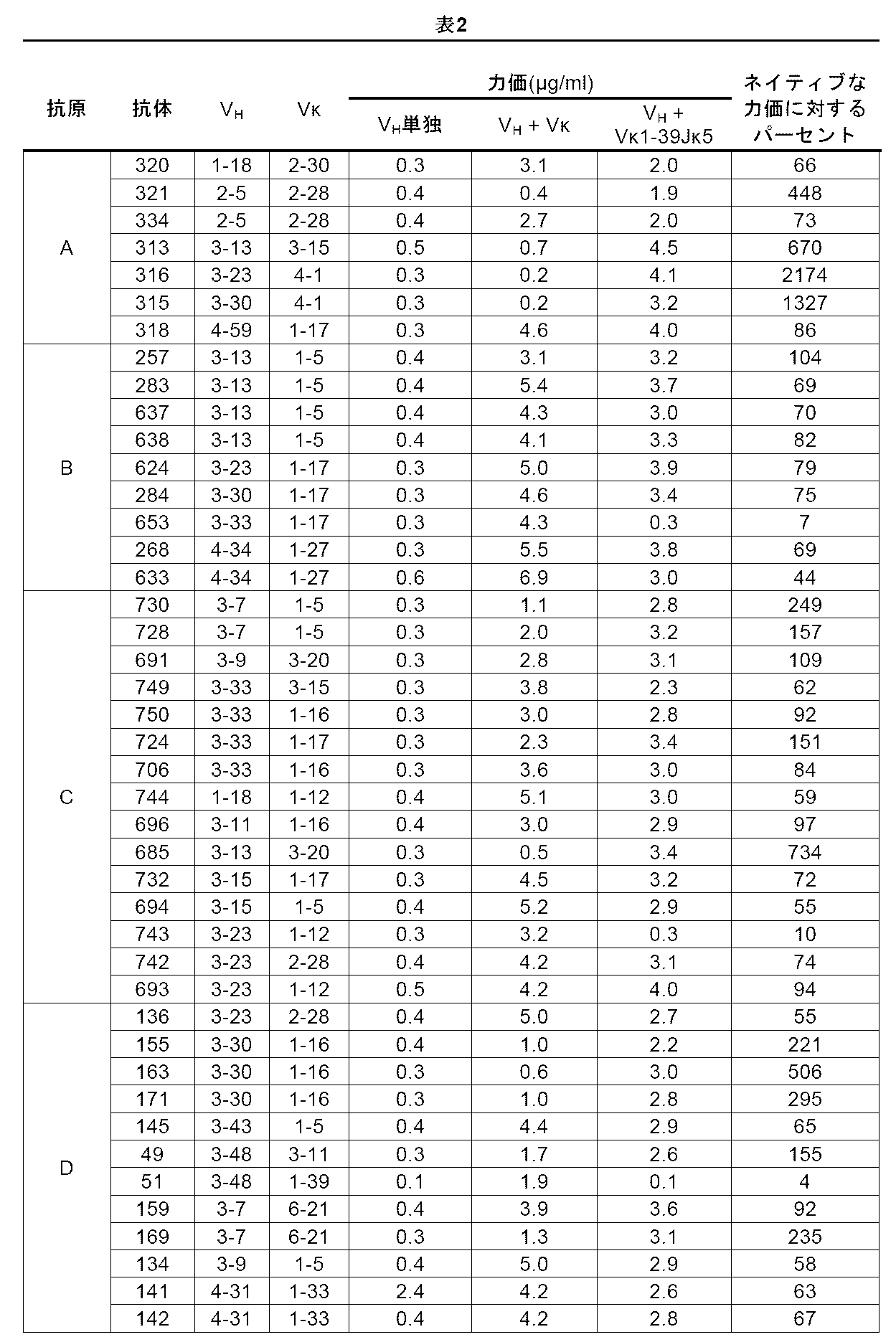
産生されるヒト抗抗原E共通軽鎖抗体の構造を解析するために、重鎖抗体可変領域をコードする核酸をクローニングし、配列決定した。抗体の核酸配列および予測されるアミノ酸配列から、操作されたヒトVκ1-39Jκ5軽鎖または操作されたヒトVκ3-20Jκ1軽鎖領域のいずれかを含有する免疫したVELOCIMMUNE(登録商標)マウスから得た、選択された共通軽鎖抗体の重鎖可変領域(HCVR)について遺伝子使用を同定した。結果が表5および表6に示されており、これにより、ヒトVκ1-39由来の軽鎖またはヒトVκ3-20由来の軽鎖のいずれかのみから軽鎖を発現するマウスを使用した場合、本発明によるマウスが、種々の再構成に起因して種々のヒト重鎖遺伝子セグメントから抗原特異的共通軽鎖抗体を生成することが実証される。2、3、4、および5ファミリーのヒトVH遺伝子セグメントを種々のヒトDHセグメントおよびヒトJHセグメントと共に再構成して抗原特異的抗体をもたらす。


LUMINEX(商標)アッセイによる抗原特異的共通軽鎖抗体の遮断能の決定
抗原Eに対して生じたヒト共通軽鎖抗体98種を、抗原Eの天然のリガンド(リガンドY)の,抗原Eへの結合を遮断するそれらの能力についてビーズに基づくアッセイで試験した。



ELISAによる抗原特異的共通軽鎖抗体の遮断能の決定
抗原Eに対して生じたヒト共通軽鎖抗体を、リガンドYをコーティングした表面への抗原Eの結合を遮断するそれらの能力についてELISAアッセイにおいて試験した。


抗原特異的共通軽鎖抗体についてのBIACORE(商標)親和性決定
選択された抗体上清についての平衡解離定数(KD)を、BIACORE(商標)T100機器(GE Healthcare)を使用したSPR(表面プラズモン共鳴)によって決定した。HBS-EP(10mMのHepes、150mMのNaCl、0.3mMのEDTA、0.05%界面活性剤P20、pH7.4)をランニング緩衝液および試料緩衝液の両方として使用して、25℃で全てのデータを得た。抗体を、粗上清試料から、標準のアミンカップリング化学を使用して高密度の抗ヒトFc抗体で予め誘導体化したCM5センサーチップ表面に捕捉した。捕捉ステップの間に、上清を抗ヒトFc表面にわたって毎分3μLの流速で合計3分間注入した。捕捉ステップの後に、ランニング緩衝液または濃度100nMの分析物のいずれかを、毎分35μLの流速で2分間注入した。捕捉された抗体からの抗原の解離を6分間モニターした。10mMのグリシン、pH1.5を簡単に注入することによって捕捉された抗体を取り出した。全てのセンサーグラムを、分析物センサーグラムから緩衝液の注入由来のセンサーグラムを引き、それにより、捕捉表面からの抗体の解離によって引き起こされるアーチファクトを除くことによって2重参照した。BIAcore T100 Evaluation software v2.1を使用した質量伝達を用いて各抗体についての結合データを1:1結合モデルに対してフィッティングした。結果が表11および表12に示されている。


LUMINEX(商標)アッセイによる抗原特異的共通軽鎖抗体の結合特異性の決定
選択された抗抗原E共通軽鎖抗体を、ヒトタンパク質とはアミノ酸残基のおよそ10%が異なるカニクイザル(cynomolgous monkey)オルソログを含めた抗原EのECDおよび抗原E ECDバリアント(Mf抗原E);ECDのC末端から最後の10アミノ酸を欠く抗原Eの欠失変異体(抗原E-ΔCT);ならびにリガンドYとの相互作用が推測される位置にアラニン置換を含有する2つの変異体(抗原E-Ala1および抗原E-Ala2)に結合するそれらの能力について試験した。抗原Eタンパク質をCHO細胞において産生させ、それぞれがmyc-myc-His C末端タグを含有した。



共通軽鎖抗体における軽鎖シャッフリング
選択された抗原特異的共通軽鎖抗体の重鎖を、重鎖を生殖系列Vκ1-39Jκ5操作軽鎖または生殖系列Vκ3-20Jκ1操作軽鎖のいずれかと再対合させた(実施例1に記載の通り)後の抗原Eへの結合について試験した。

共通軽鎖抗体における重鎖遺伝子使用および体細胞超変異の発生頻度
VELCOIMMUNE(登録商標)マウス(例えば、US6,596,541およびUS7,105,348)において生じた抗体の重鎖配列および軽鎖配列(>6000)を、抗体鎖の重鎖遺伝子セグメントの使用および体細胞超変異の発生頻度を比較するために操作された軽鎖マウス(上記)を使用する多抗原免疫スキームによって得られる共通軽鎖抗体の重鎖配列および軽鎖配列(>600)と共にまとめた。
ヒト細胞表面受容体(抗原E)、2種のヒト細胞表面糖タンパク質のヘテロ二量体(抗原F)、ヒトサイトカイン受容体(抗原G)およびヒト腫瘍分化抗原(抗原H)を用いて免疫した、内因性マウス重鎖遺伝子座のヒトVH、DH、およびJH遺伝子セグメントでの置き換えおよび内因性マウスκ軽鎖遺伝子座の操作された生殖系列Vκ1-39Jκ5ヒト軽鎖領域または操作された生殖系列Vκ3-20Jκ1ヒト軽鎖領域のいずれかでの置き換えを含有するVELOCIMMUNE(登録商標)マウス(実施例2に記載の通り)から得た重鎖配列および軽鎖配列を、重鎖遺伝子セグメントの使用について分析し、VHおよびJH遺伝子セグメントを記録した。結果が表16-18に示されている。表16-18中の百分率は丸めた値を表し、足し合わせたときに100%と等しくならない場合があり得る。
VELCOIMMUNE(登録商標)マウスおよびに操作された軽鎖マウス(上記)において生じた抗体由来の重鎖および軽鎖を、各重鎖および/または軽鎖について実証された重鎖および軽鎖の遺伝子使用に応じて生殖系列配列に対してアライメントした。各配列の重鎖と軽鎖の両方についての各フレームワーク領域(FW)および相補性決定領域(CDR)のアミノ酸の変化を算出した。結果が表19-22に示されている。表21-24中の百分率は丸めた値を表し、足し合わせたときに100%と等しくならない場合があり得る。







ユニバーサル軽鎖を有する二重特異性抗体の結合親和性
選択された単一特異性抗抗原E共通軽鎖抗体(実施例5に記載)のクローニングされたヒト重鎖可変領域から当技術分野で公知の標準の組換えDNA技法を使用して完全ヒト二重特異性抗体を構築した。表23には、選択された親単一特異性抗体由来のヒト重鎖(HC-1およびHC-2)の対合が示されており、各対を、各二重特異性抗体を構築するために生殖系列再構成されたヒトVκ1-39/Jκ1軽鎖と共に使用した。


2つのヒト軽鎖を発現するマウスの生成および分析
実施例2に記載されている方法を使用して、2つのヒトVκ遺伝子セグメント(例えば、ヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメント)を含有する2つのさらなる操作された軽鎖遺伝子座を構築した(図9)。1つの操作された軽鎖遺伝子座は、2つのヒトVκ遺伝子セグメントおよび5つのヒトJκ遺伝子セグメントを、再構成されていない立体配置(DLC-5J)で含有した。第2の操作された軽鎖遺伝子座は、2つのヒトVκ遺伝子セグメントおよび1つのヒトJκ遺伝子セグメントを、再構成されていない立体配置(DLC-1J)で含有した。2つのさらなる操作された軽鎖遺伝子座のそれぞれについて、ヒト遺伝子セグメントは3’に組換えシグナル配列が隣接し、B細胞においてヒト遺伝子セグメントのin vivo再構成が可能になった。
DLCマウスにおけるB細胞集団およびB細胞発生を脾細胞調製物および骨髄調製物のフローサイトメトリー分析によって検証した。2つのヒトVκ遺伝子セグメントおよび5つのヒトJκ遺伝子セグメントについてホモ接合性のマウス(n=4)、2つのヒトVκ遺伝子セグメントおよび1つのヒトJκ遺伝子セグメントについてホモ接合性のマウス(n=4)、および野生型マウス(n=4)からの細胞懸濁物を、標準の方法(上記)を使用して作製し、蛍光標識された抗体で染色した(実施例3に記載の通り)。
ホモ接合性のマウスにおいて両方のヒトVκ遺伝子セグメントの発現を実施例3に従って定量的PCRアッセイを使用して解析した。簡単に述べると、CD19+B細胞を野生型マウス、マウス重鎖およびκ軽鎖可変遺伝子座の、対応するヒト重鎖およびκ軽鎖可変領域遺伝子座(Hκ)での置き換えについてホモ接合性のマウス、ならびに2つのヒトVκ遺伝子セグメントおよび5つのヒトJκ遺伝子セグメント(DLC-5J)または1つのヒトJκ遺伝子セグメント(DLC-1J)のいずれかを含有する、操作されたκ軽鎖遺伝子座についてホモ接合性のマウスの骨髄および脾臓全体から精製した。相対的な発現をマウスCκ領域の発現に対して正規化した(マウス群当たりn=3~5)。結果が図17および図18に示されている。
2つの再構成されていないヒトVκ遺伝子セグメントおよび5つの再構成されていないヒトJκ遺伝子セグメントについてホモ接合性のマウス(DLC-5J)を、脾性B細胞におけるヒトVκ/Jκ遺伝子セグメントの使用について逆転写酵素ポリメラーゼ連鎖反応(RT-PCR)によって分析した。


マウスであって、
マウスまたはラット軽鎖定常領域に作動可能に連結した2つ以下のヒトVL遺伝子セグメントおよび2つまたはそれ超のヒトJL遺伝子セグメント;ならびに
非ヒト定常領域に作動可能に連結した1つまたは複数のヒトVH遺伝子セグメント、1つまたは複数のヒトDH遺伝子セグメント、および1つまたは複数のヒトJH遺伝子セグメント
を含み、
該ヒト遺伝子セグメントが、再構成することが可能であって、抗体のヒト可変ドメインをコードしており、
さらに、該マウスが、免疫グロブリン軽鎖が形成されるように再構成することが可能な内因性VL遺伝子セグメントを含まない、マウス。
(項目2)
前記軽鎖定常領域がラットCκ領域である、項目1に記載のマウス。
(項目3)
前記軽鎖定常領域がマウスCκ領域である、項目1に記載のマウス。
(項目4)
2つのヒトVL遺伝子セグメントが前記マウスまたはラット軽鎖定常領域に作動可能に連結している、項目1から3のいずれか一項に記載のマウス。
(項目5)
2つから5つまでのヒトJL遺伝子セグメントが前記マウスまたはラット軽鎖定常領域に作動可能に連結している、前記項目のいずれか一項に記載のマウス。
(項目6)
5つのヒトJL遺伝子セグメントを含む、前記項目のいずれか一項に記載のマウス。
(項目7)
前記5つのヒトJL遺伝子セグメントが、ヒトJκ1、Jκ2、Jκ3、Jκ4およびJκ5である、項目6に記載のマウス。
(項目8)
前記2つ以下のヒトVL遺伝子セグメントが、ヒトVκ1-39遺伝子セグメント、ヒトVκ3-20遺伝子セグメント、またはこれらの組合せから選択される、前記項目のいずれか一項に記載のマウス。
(項目9)
前記2つ以下のヒトVL遺伝子セグメントが、ヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメントである、前記項目のいずれか一項に記載のマウス。
(項目10)
前記2つ以下のヒトVL遺伝子セグメントおよび2つまたはそれ超のヒトJL遺伝子セグメントが、内因性軽鎖遺伝子座に存在する、前記項目のいずれか一項に記載のマウス。
(項目11)
前記1つまたは複数のヒトVH遺伝子セグメント、1つまたは複数のヒトDH遺伝子セグメント、および1つまたは複数のヒトJH遺伝子セグメントが、マウス定常領域に作動可能に連結している、前記項目のいずれか一項に記載のマウス。
(項目12)
機能的なλ軽鎖遺伝子座を含む、前記項目のいずれか一項に記載のマウス。
(項目13)
非機能的なλ軽鎖遺伝子座を含む、項目1から11のいずれか一項に記載のマウス。
(項目14)
マウスまたはラット軽鎖定常領域に作動可能に連結した前記2つ以下のヒトVL遺伝子セグメントおよび2つまたはそれ超のヒトJL遺伝子セグメントが、ヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメントである2つのヒトVκセグメント、ならびにヒトJκ1、Jκ2Jκ3、Jκ4およびJκ5を順番に含む遺伝子座に存在する、前記項目のいずれか一項に記載のマウス。
(項目15)
図9のDLCJ-5J遺伝子座を含む、前記項目のいずれか一項に記載のマウス。
(項目16)
骨髄中に、CD19+、CD43+、c-kit+発現によって特徴付けられる細胞約2.5×104個~約1.5×105個の範囲内のプロB細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目17)
骨髄中に、CD19+、CD43-、c-kit-発現によって特徴付けられる細胞約1×106個~約2×106個の範囲内のプレB細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目18)
骨髄中に、IgM+、B220int発現によって特徴付けられる細胞約5×105個~約7×105個の範囲内の未成熟B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目19)
骨髄中に、IgM+、B220hi発現によって特徴付けられる細胞約3×104個~約1.5×105個の範囲内の成熟B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目20)
骨髄中に、細胞約1×106個~約3×106個の範囲内の総CD19+B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目21)
細胞約2×106個~約7×106個の範囲内のCD19+脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目22)
細胞約1×106個~約4×106個の範囲内のCD19+、IgDhi、IgMlo脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目23)
細胞約9×105個~約2×106個の範囲内のCD19+、IgDlo、IgMhi脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目24)
CD93+B220+IgMhiCD23-発現によって特徴付けられる細胞約2×106個~約7×106個の範囲内の移行T1脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目25)
CD93+B220+IgMhiCD23+発現によって特徴付けられる細胞約1×106個~約7×106個の範囲内の移行T2脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目26)
CD93+B220+IgMloCD23+発現によって特徴付けられる細胞約1×106個~約4×106個の範囲内の移行T3脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目27)
CD93-B220+IgMhiCD21/35hiCD23-発現によって特徴付けられる細胞約1×106個~約3×106個の範囲内の辺縁帯脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目28)
CD93-B220+CD21/35intIgMloIgDhi発現によって特徴付けられる細胞約3×106個~約1.5×107個の範囲内の濾胞1型(FO-I)脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目29)
CD93-B220+CD21/35intIgMintIgDhi発現によって特徴付けられる細胞約1×106個~約2×106個の範囲内の濾胞2型(FO-II)脾性B細胞集団を含む、前記項目のいずれか一項に記載のマウス。
(項目30)
前記項目のいずれか一項に記載のマウスの単離された細胞。
(項目31)
胚性幹(ES)細胞である、項目30に記載の細胞。
(項目32)
項目31に記載のES細胞を含むマウス胚。
(項目33)
B細胞である、項目30に記載の細胞。
(項目34)
項目33に記載のB細胞を用いて作製されたハイブリドーマ。
(項目35)
抗体を生成するため、またはヒト重鎖可変領域をコードする核酸配列を同定するための、項目1から29のいずれか一項に記載のマウスの使用。
(項目36)
目的の抗原に結合する抗体を作製するための方法であって、
項目1から29のいずれか一項に記載のマウスを、目的の抗原を用いて免疫するステップ、
該マウスから免疫グロブリン可変領域遺伝子配列を得るステップ、および
該免疫グロブリン可変領域遺伝子配列を使用して、該抗原に結合する抗体を産生させるステップ
を含む方法。
(項目37)
単一細胞において、
(a)免疫された項目1から29のいずれか一項に記載のマウスの第1のVH遺伝子配列であって、ヒトCH遺伝子配列と融合した第1のVH遺伝子配列;および
(b)ヒトCL遺伝子配列と融合したVL遺伝子配列
を発現させるステップ、
該細胞を、完全ヒト抗体を発現するのに十分な条件下で維持するステップ;ならびに
該抗体を該細胞から単離するステップ
をさらに含む、項目36に記載の方法。
(項目38)
前記細胞が免疫された項目1から29のいずれか一項に記載のマウスの第2のVH遺伝子配列を含み、該VH遺伝子配列がヒトCH遺伝子配列と融合しており、前記第1のVH遺伝子配列が第1のエピトープを認識するVHドメインをコードし、該第2のVH遺伝子配列が第2のエピトープを認識するVHドメインをコードし、該第1のエピトープおよび該第2のエピトープが同一ではなく、該第1のVHドメインおよび該第2のVHドメインが前記VL遺伝子配列によりコードされるVLドメインと相互作用する、項目37に記載の方法。
(項目39)
ヒト二重特異性抗体の作製における、項目1から29のいずれか一項に記載のマウスの使用。
(項目40)
ヒト二重特異性抗体を作製するための方法であって、項目1から29のいずれか一項に記載のマウスを免疫するステップ、および該マウス由来のB細胞のヒト可変領域遺伝子配列を使用して該二重特異性抗体を作製するステップを含む方法。
(項目41)
(a)項目1から29のいずれか一項に記載のマウスのクローン選択されたリンパ球を同定するステップであって、該マウスが免疫され、目的の抗原に対する免疫応答を生じるようにされており、該リンパ球が該目的の抗原に特異的に結合する抗体を発現するステップ;
(b)該リンパ球または該抗体から、該目的の抗原に特異的に結合するヒト重鎖可変領域をコードするヌクレオチド配列を得るステップ;および
(c)(c)のヌクレオチド配列を二重特異性抗体の作製に使用するステップ
を含む、項目40に記載の方法。
(項目42)
1回目に第1のヒト重鎖可変領域配列を生成するために目的の第1の抗原に対してステップ(a)から(c)までを実施し、2回目に第2のヒト重鎖可変領域配列を生成するために目的の第2の抗原に対してステップ(a)から(c)までを実施し、該第1のヒト重鎖可変領域配列が第1のヒト重鎖定常領域と融合して発現して第1のヒト重鎖を形成し、該第2のヒト重鎖可変領域配列が第2のヒト重鎖定常領域と融合して発現して第2のヒト重鎖を形成し、該第1のヒト重鎖および該第2のヒト重鎖がVκ1-39遺伝子セグメントまたはVκ3-20遺伝子セグメントから発現した単一のヒト軽鎖の存在下で発現する、項目41に記載の方法。
(項目43)
(a)
-第1の免疫原を用いて免疫された項目1から29のいずれか一項に記載のマウス;および
-第2の免疫原を用いて免疫された同じマウス、または遺伝学的には同じである異なるマウス
由来のB細胞から可変重鎖領域をクローニングするステップ、
(b)細胞において、同じ前記重鎖定常領域および同じ前記軽鎖とともに(a)の可変重鎖領域を発現させて二重特異性抗体を作製するステップ
を含む、項目42に記載の方法。
(項目44)
前記第1のヒト重鎖が、前記第1のヒト重鎖のプロテインAに対する親和性を消失させるまたは実質的に低下させる改変を含み、前記第2のヒト重鎖がプロテインAに結合する能力を保持し、該第1のヒト重鎖のプロテインAに対する親和性を消失させるまたは実質的に低下させる改変が、95R(EU435R)、96F(EU436F)、およびこれらの組合せから選択される、項目43に記載の方法。
(項目45)
ヒト免疫グロブリン重鎖可変ドメイン遺伝子セグメントを選択するための、項目1から29のいずれか一項に記載のマウスの使用。
(項目46)
免疫されている、項目1から29のいずれか一項に記載のマウス。
(項目47)
抗体を産生する方法であって、項目1から29のいずれか一項に記載のマウスを、抗原を用いて免疫するステップ、および結果として生ずる、該マウスにより該抗原に対して産生された抗体から重鎖可変領域アミノ酸配列またはコードするヌクレオチド配列を得るステップ、および該重鎖可変領域アミノ酸配列または核酸配列を抗体の作製に利用するステップを含む方法。
(項目48)
(a)前記マウスを免疫した前記抗原の異なるエピトープに対する2種の異なる抗体から前記重鎖可変領域アミノ酸配列またはコードするヌクレオチド配列を同定するステップ;または
(b)項目1から29のいずれか一項に記載のマウスと同じマウス、またはさらなるマウスを、異なる抗原を用いて免疫し、次いで、該第2の抗原に特異的である、該マウスによって産生された抗体から重鎖可変領域配列アミノ酸配列またはコードするヌクレオチド配列を同定するステップを含み、
該2種の異なる重鎖可変領域を使用して二重特異性抗体を生成するステップをさらに含む、項目47に記載の方法。
(項目49)
前記抗体または2つの抗体から軽鎖可変領域配列も同定するステップをさらに含む、項目47または48に記載の方法。
(項目50)
項目49に記載の方法であって、該方法によって産生された抗体において同定された1つまたは複数の軽鎖重鎖可変領域も使用することを含む、方法。
(項目A1)
マウスであって、
免疫グロブリン軽鎖定常領域に作動可能に連結した2つのヒト免疫グロブリンVL遺伝子セグメントおよび2つまたはそれ超のヒト免疫グロブリンJL遺伝子セグメントであって、ここで、該2つのヒト免疫グロブリンVL遺伝子セグメントが、ヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメントであり、該2つまたはそれ超のヒト免疫グロブリンJL遺伝子セグメントが、ヒトJκ1、Jκ2、Jκ3、Jκ4、およびJκ5からなる群から選択される、2つのヒト免疫グロブリンVL遺伝子セグメントおよび2つまたはそれ超のヒト免疫グロブリンJL遺伝子セグメント;ならびに
非ヒト免疫グロブリン定常領域に作動可能に連結した1つまたは複数のヒト免疫グロブリンVH遺伝子セグメント、1つまたは複数のヒト免疫グロブリンDH遺伝子セグメント、および1つまたは複数のヒト免疫グロブリンJH遺伝子セグメント
を含み、
該ヒト遺伝子セグメントが、再構成することが可能であって、抗体のヒト可変ドメインをコードしており、さらに、該マウスが、免疫グロブリン軽鎖が形成されるように再構成することが可能な内因性免疫グロブリンVL遺伝子セグメントを含まない、マウス。
(項目A2)
前記2つのヒトVL遺伝子セグメントおよび2つまたはそれ超のヒトJL遺伝子セグメントが、マウスまたはラット免疫グロブリン軽鎖定常領域に作動可能に連結される、項目A1に記載のマウス
(項目A3)
前記軽鎖定常領域がラットCκ領域である、項目A2に記載のマウス。
(項目A4)
前記軽鎖定常領域がマウスCκ領域である、項目A2に記載のマウス。
(項目A5)
5つのヒトJL遺伝子セグメントを含む、前記項目Aのいずれか一項に記載のマウス。
(項目A6)
前記5つのヒトJL遺伝子セグメントが、ヒトJκ1、Jκ2、Jκ3、Jκ4およびJκ5である、項目A5に記載のマウス。
(項目A7)
前記2つのヒトVL遺伝子セグメントおよび2つまたはそれ超のヒトJL遺伝子セグメントが、内因性軽鎖遺伝子座に存在する、前記項目Aのいずれか一項に記載のマウス。
(項目A8)
前記1つまたは複数のヒトVH遺伝子セグメント、1つまたは複数のヒトDH遺伝子セグメント、および1つまたは複数のヒトJH遺伝子セグメントが、マウス定常領域に作動可能に連結している、前記項目Aのいずれか一項に記載のマウス。
(項目A9)
機能的なλ軽鎖遺伝子座を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A10)
非機能的なλ軽鎖遺伝子座を含む、項目A1からA8のいずれか一項に記載のマウス。
(項目A11)
マウスまたはラット軽鎖定常領域に作動可能に連結した前記2つのヒトVL遺伝子セグメントおよび2つまたはそれ超のヒトJL遺伝子セグメントが、ヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメントである2つのヒトVκセグメント、ならびにヒトJκ1、Jκ2Jκ3、Jκ4およびJκ5を順番に含む遺伝子座に存在する、前記項目Aのいずれか一項に記載のマウス。
(項目A12)
図9のDLCJ-5J遺伝子座を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A13)
骨髄中に、CD19+、CD43+、c-kit+発現によって特徴付けられる細胞約2.5×104個~約1.5×105個の範囲内のプロB細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A14)
骨髄中に、CD19+、CD43-、c-kit-発現によって特徴付けられる細胞約1×106個~約2×106個の範囲内のプレB細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A15)
骨髄中に、IgM+、B220int発現によって特徴付けられる細胞約5×105個~約7×105個の範囲内の未成熟B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A16)
骨髄中に、IgM+、B220hi発現によって特徴付けられる細胞約3×104個~約1.5×105個の範囲内の成熟B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A17)
骨髄中に、細胞約1×106個~約3×106個の範囲内の総CD19+B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A18)
細胞約2×106個~約7×106個の範囲内のCD19+脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A19)
細胞約1×106個~約4×106個の範囲内のCD19+、IgDhi、IgMlo脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A20)
細胞約9×105個~約2×106個の範囲内のCD19+、IgDlo、IgMhi脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A21)
CD93+B220+IgMhiCD23-発現によって特徴付けられる細胞約2×106個~約7×106個の範囲内の移行T1脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A22)
CD93+B220+IgMhiCD23+発現によって特徴付けられる細胞約1×106個~約7×106個の範囲内の移行T2脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A23)
CD93+B220+IgMloCD23+発現によって特徴付けられる細胞約1×106個~約4×106個の範囲内の移行T3脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A24)
CD93-B220+IgMhiCD21/35hiCD23-発現によって特徴付けられる細胞約1×106個~約3×106個の範囲内の辺縁帯脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A25)
CD93-B220+CD21/35intIgMloIgDhi発現によって特徴付けられる細胞約3×106個~約1.5×107個の範囲内の濾胞1型(FO-I)脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A26)
CD93-B220+CD21/35intIgMintIgDhi発現によって特徴付けられる細胞約1×106個~約2×106個の範囲内の濾胞2型(FO-II)脾性B細胞集団を含む、前記項目Aのいずれか一項に記載のマウス。
(項目A27)
前記項目Aのいずれか一項に記載のマウスの単離された細胞。
(項目A28)
胚性幹(ES)細胞である、項目A27に記載の細胞。
(項目A29)
項目A28に記載のES細胞を含むマウス胚。
(項目A30)
B細胞である、項目A27に記載の細胞。
(項目A31)
項目A30に記載のB細胞を用いて作製されたハイブリドーマ。
(項目A32)
抗体を生成するため、またはヒト免疫グロブリン重鎖可変領域をコードする核酸配列を同定するための、項目A1からA26のいずれか一項に記載のマウスの使用。
(項目A33)
目的の抗原に結合する抗体を作製するための方法であって、
項目A1からA26のいずれか一項に記載のマウスを、目的の抗原を用いて免疫するステップ、
該マウスから免疫グロブリン可変領域遺伝子配列を得るステップ、および
該免疫グロブリン可変領域遺伝子配列を使用して、該抗原に結合する抗体を産生させるステップ
を含む方法。
(項目A34)
単一細胞において、
(a)免疫された項目A1からA26のいずれか一項に記載のマウスの第1のVH遺伝子配列であって、ヒトCH遺伝子配列と融合した第1のVH遺伝子配列;および
(b)ヒトCL遺伝子配列と融合したVL遺伝子配列
を発現させるステップ、
該細胞を、完全ヒト抗体を発現するのに十分な条件下で維持するステップ;ならびに
該抗体を該細胞から単離するステップ
をさらに含む、項目A33に記載の方法。
(項目A35)
前記細胞が免疫された項目A1からA26のいずれか一項に記載のマウスの第2のVH遺伝子配列を含み、該VH遺伝子配列がヒトCH遺伝子配列と融合しており、前記第1のVH遺伝子配列が第1のエピトープを認識するVHドメインをコードし、該第2のVH遺伝子配列が第2のエピトープを認識するVHドメインをコードし、該第1のエピトープおよび該第2のエピトープが同一ではなく、該第1のVHドメインおよび該第2のVHドメインが前記VL遺伝子配列によりコードされるVLドメインと相互作用する、項目A34に記載の方法。
(項目A36)
ヒト二重特異性抗体の作製における、項目A1からA26のいずれか一項に記載のマウスの使用。
(項目A37)
ヒト二重特異性抗体を作製するための方法であって、項目A1からA26のいずれか一項に記載のマウスを免疫するステップ、および該マウス由来のB細胞のヒト可変領域遺伝子配列を使用して該二重特異性抗体を作製するステップを含む方法。
(項目A38)
(a)項目A1からA26のいずれか一項に記載のマウスのクローン選択されたリンパ球を同定するステップであって、該マウスが免疫され、目的の抗原に対する免疫応答を生じるようにされており、該リンパ球が該目的の抗原に特異的に結合する抗体を発現するステップ;
(b)該リンパ球または該抗体から、該目的の抗原に特異的に結合するヒト重鎖可変領域をコードするヌクレオチド配列を得るステップ;および
(c)(c)のヌクレオチド配列を二重特異性抗体の作製に使用するステップ
を含む、項目A37に記載の方法。
(項目A39)
1回目に第1のヒト重鎖可変領域配列を生成するために目的の第1の抗原に対してステップ(a)から(c)までを実施し、2回目に第2のヒト重鎖可変領域配列を生成するために目的の第2の抗原に対してステップ(a)から(c)までを実施し、該第1のヒト重鎖可変領域配列が第1のヒト重鎖定常領域と融合して発現して第1のヒト重鎖を形成し、該第2のヒト重鎖可変領域配列が第2のヒト重鎖定常領域と融合して発現して第2のヒト重鎖を形成し、該第1のヒト重鎖および該第2のヒト重鎖がVκ1-39遺伝子セグメントまたはVκ3-20遺伝子セグメントから発現した単一のヒト軽鎖の存在下で発現する、項目A38に記載の方法。
(項目A40)
(a)
-第1の免疫原を用いて免疫された項目A1からA26のいずれか一項に記載のマウス;および
-第2の免疫原を用いて免疫された同じマウス、または遺伝学的には同じである異なるマウス
由来のB細胞から可変重鎖領域をクローニングするステップ、
(b)細胞において、同じ前記重鎖定常領域および同じ前記軽鎖とともに(a)の可変重鎖領域を発現させて二重特異性抗体を作製するステップ
を含む、項目A39に記載の方法。
(項目A41)
前記第1のヒト重鎖が、前記第1のヒト重鎖のプロテインAに対する親和性を消失させるまたは実質的に低下させる改変を含み、前記第2のヒト重鎖がプロテインAに結合する能力を保持し、該第1のヒト重鎖のプロテインAに対する親和性を消失させるまたは実質的に低下させる改変が、95R(EU435R)、96F(EU436F)、およびこれらの組合せから選択される、項目A40に記載の方法。
(項目A42)
ヒト免疫グロブリン重鎖可変ドメイン遺伝子セグメントを選択するための、項目A1からA26のいずれか一項に記載のマウスの使用。
(項目A43)
免疫されている、項目A1からA26のいずれか一項に記載のマウス。
(項目A44)
抗体を産生する方法であって、項目A1からA26のいずれか一項に記載のマウスを、抗原を用いて免疫するステップ、および結果として生ずる、該マウスにより該抗原に対して産生された抗体から重鎖可変領域アミノ酸配列またはコードするヌクレオチド配列を得るステップ、および該重鎖可変領域アミノ酸配列または核酸配列を抗体の作製に利用するステップを含む方法。
(項目A45)
(a)前記マウスを免疫した前記抗原の異なるエピトープに対する2種の異なる抗体から前記重鎖可変領域アミノ酸配列またはコードするヌクレオチド配列を同定するステップ;または
(b)項目A1からA26のいずれか一項に記載のマウスと同じマウス、またはさらなるマウスを、異なる抗原を用いて免疫し、次いで、該第2の抗原に特異的である、該マウスによって産生された抗体から重鎖可変領域配列アミノ酸配列またはコードするヌクレオチド配列を同定するステップを含み、
該2種の異なる重鎖可変領域を使用して二重特異性抗体を生成するステップをさらに含む、項目A44に記載の方法。
(項目A46)
前記抗体または2つの抗体から軽鎖可変領域配列も同定するステップをさらに含む、項目A44またはA45に記載の方法。
(項目A47)
項目A46に記載の方法であって、該方法によって産生された抗体において同定された1つまたは複数の軽鎖重鎖可変領域も使用することを含む、方法。
Claims (32)
- そのゲノム内に、
マウス免疫グロブリン軽鎖定常領域配列に作動可能に連結された、ちょうど2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントと、5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントと
を含むように遺伝子改変されたマウス胚性幹(ES)細胞であって、該2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントがヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメントであり、該5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントが、ヒトJκ1遺伝子セグメント、ヒトJκ2遺伝子セグメント、ヒトJκ3遺伝子セグメント、ヒトJκ4遺伝子セグメントおよびヒトJκ5遺伝子セグメントである、遺伝子改変されたマウスES細胞。 - そのゲノム内に、
内因性重鎖遺伝子座においてマウス免疫グロブリン重鎖定常領域配列に作動可能に連結された、1つまたは複数の再構成されていないヒト免疫グロブリンVH遺伝子セグメントと、1つまたは複数の再構成されていないヒト免疫グロブリンDH遺伝子セグメントと、1つまたは複数の再構成されていないヒト免疫グロブリンJH遺伝子セグメントと
をさらに含む、請求項1に記載の遺伝子改変されたマウスES細胞。 - 前記ES細胞の前記ゲノムが、再構成して免疫グロブリン重鎖可変領域を形成できる内因性マウス免疫グロブリンVH遺伝子セグメントを含まない、請求項2に記載の遺伝子改変されたES細胞。
- 前記2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントと、5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントが、内因性免疫グロブリンκ軽鎖遺伝子座に存在する、請求項1~3のいずれか一項に記載の遺伝子改変されたES細胞。
- 前記遺伝子改変されたマウスES細胞の前記ゲノムが、機能的なλ軽鎖遺伝子座を含まない、請求項1または請求項2に記載の遺伝子改変されたES細胞。
- 前記マウス軽鎖定常領域がマウスCκ定常領域である、請求項1または請求項2に記載の遺伝子改変されたマウスES細胞。
- 前記マウスCκ定常領域が内因性マウスCκ定常領域である、請求項6に記載の遺伝子改変されたマウスES細胞。
- 前記遺伝子改変されたマウスES細胞のゲノム内の前記ヒトVκ1-39遺伝子セグメントがヒト生殖系列Vκ1-39遺伝子セグメントであり、前記遺伝子改変されたマウスES細胞の前記ゲノム内の前記ヒトVκ3-20遺伝子セグメントがヒト生殖系列Vκ3-20遺伝子セグメントである、請求項1または請求項2に記載の遺伝子改変されたマウスES細胞。
- 前記遺伝子改変されたマウスES細胞の前記ゲノムが、前記ヒトVκ1-39遺伝子セグメント、前記ヒトVκ3-20遺伝子セグメント、前記ヒトJκ1遺伝子セグメント、前記ヒトJκ2遺伝子セグメント、前記ヒトJκ3遺伝子セグメント、前記ヒトJκ4遺伝子セグメント、および、前記ヒトJκ5遺伝子セグメントを順に含む、請求項1または請求項2に記載の遺伝子改変されたマウスES細胞。
- 前記遺伝子改変されたマウスES細胞の前記ゲノムが、DLC-5J遺伝子座を含み、該DLC-5J遺伝子座が、配列番号38~49より選択されるhVκ/hJκ/mCκ接合部配列を含む、請求項1~9のいずれか一項に記載の遺伝子改変されたマウスES細胞。
- 請求項1または請求項2に記載の遺伝子改変されたES細胞に由来するマウス胚。
- 代理母において請求項11に記載のマウス胚を懐胎させ、該代理母に、全体的にまたは部分的に前記遺伝子改変されたES細胞に由来する子孫を出産させることを含む、マウスを作製する方法。
- 遺伝子改変されたマウスES細胞を作製する方法であって、ES細胞を、そのゲノム内に、
マウス免疫グロブリン軽鎖定常領域配列に作動可能に連結された、ちょうど2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントと、5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントと
を含むように遺伝子改変することを含み、ここで、該2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントがヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメントであり、該5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントが、ヒトJκ1遺伝子セグメント、ヒトJκ2遺伝子セグメント、ヒトJκ3遺伝子セグメント、ヒトJκ4遺伝子セグメントおよびヒトJκ5遺伝子セグメントである、方法。 - 前記遺伝子改変されたマウスES細胞が、そのゲノム内に、
内因性重鎖遺伝子座においてマウス免疫グロブリン重鎖定常領域配列に作動可能に連結された、1つまたは複数の再構成されていないヒト免疫グロブリンVH遺伝子セグメントと、1つまたは複数の再構成されていないヒト免疫グロブリンDH遺伝子セグメントと、1つまたは複数の再構成されていないヒト免疫グロブリンJH遺伝子セグメントと
をさらに含む、請求項13に記載の方法。 - 前記マウス軽鎖定常領域がマウスCκ定常領域である、請求項13または請求項14に記載の方法。
- 前記マウス軽鎖定常領域が内因性マウスCκ定常領域である、請求項15に記載の方法。
- 前記遺伝子改変されたES細胞の前記ゲノムが、再構成して免疫グロブリン重鎖可変領域を形成できる内因性マウス免疫グロブリンVH遺伝子セグメントを含まない、請求項14に記載の方法。
- 前記2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントと、5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントが、内因性免疫グロブリンκ軽鎖遺伝子座に存在する、請求項13~17のいずれか一項に記載の方法。
- 前記遺伝子改変されたマウスES細胞の前記ゲノムが機能的なλ軽鎖遺伝子座を含まない、請求項13または請求項14に記載の方法。
- 前記遺伝子改変されたマウスES細胞の前記ゲノム内の前記ヒトVκ1-39遺伝子セグメントがヒト生殖系列Vκ1-39遺伝子セグメントであり、前記遺伝子改変されたマウスES細胞の前記ゲノム内の前記ヒトVκ3-20遺伝子セグメントがヒト生殖系列Vκ3-20遺伝子セグメントである、請求項13または請求項14に記載の方法。
- 前記遺伝子改変されたマウスES細胞の前記ゲノムが、前記ヒトVκ1-39遺伝子セグメント、前記ヒトVκ3-20遺伝子セグメント、前記ヒトJκ1遺伝子セグメント、前記ヒトJκ2遺伝子セグメント、前記ヒトJκ3遺伝子セグメント、前記ヒトJκ4遺伝子セグメント、および、前記ヒトJκ5遺伝子セグメントを順に含む、請求項13または請求項14に記載の方法。
- 前記遺伝子改変されたマウスES細胞が、DLC-5J遺伝子座を含み、該DLC-5J遺伝子座が、配列番号38~49より選択されるhVκ/hJκ/mCκ接合部配列を含む、請求項13~21のいずれか一項に記載の方法。
- 遺伝子改変されたマウスを作製する方法であって、マウスの生殖系列ゲノムを、
マウス免疫グロブリン軽鎖定常領域配列に作動可能に連結された、ちょうど2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントと、5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントと
を含むように遺伝子改変することを含み、ここで、該2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントがヒトVκ1-39遺伝子セグメントおよびヒトVκ3-20遺伝子セグメントであり、該5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントが、ヒトJκ1遺伝子セグメント、ヒトJκ2遺伝子セグメント、ヒトJκ3遺伝子セグメント、ヒトJκ4遺伝子セグメントおよびヒトJκ5遺伝子セグメントであり、
該遺伝子改変されたマウスが、そのゲノム内に、
内因性重鎖遺伝子座においてマウス免疫グロブリン重鎖定常領域配列に作動可能に連結された、1つまたは複数の再構成されていないヒト免疫グロブリンV H 遺伝子セグメントと、1つまたは複数の再構成されていないヒト免疫グロブリンD H 遺伝子セグメントと、1つまたは複数の再構成されていないヒト免疫グロブリンJ H 遺伝子セグメントと
をさらに含む、方法。 - 前記マウス軽鎖定常領域がマウスCκ定常領域である、請求項23に記載の方法。
- 前記マウス軽鎖定常領域が内因性マウスCκ定常領域である、請求項24に記載の方法。
- 前記遺伝子改変されたマウスの前記ゲノムが、再構成して免疫グロブリン重鎖可変領域を形成できる内因性マウス免疫グロブリンVH遺伝子セグメントを含まない、請求項23に記載の方法。
- 前記2つの再構成されていないヒト免疫グロブリンVκ遺伝子セグメントと、5つの再構成されていないヒト免疫グロブリンJκ遺伝子セグメントが、内因性免疫グロブリンκ軽鎖遺伝子座に存在する、請求項23~26のいずれか一項に記載の方法。
- 前記遺伝子改変されたマウスの前記ゲノムが非機能的な免疫グロブリンλ軽鎖遺伝子座を含む、請求項23に記載の方法。
- 前記遺伝子改変されたマウスの前記ゲノムが機能的なλ軽鎖遺伝子座を含まない、請求項23に記載の方法。
- 前記遺伝子改変されたマウスの前記ゲノム内の前記ヒトVκ1-39遺伝子セグメントがヒト生殖系列Vκ1-39遺伝子セグメントであり、前記遺伝子改変されたマウスの前記ゲノム内の前記ヒトVκ3-20遺伝子セグメントがヒト生殖系列Vκ3-20遺伝子セグメントである、請求項23に記載の方法。
- 前記遺伝子改変されたマウスの前記ゲノムが、前記ヒトVκ1-39遺伝子セグメント、前記ヒトVκ3-20遺伝子セグメント、前記ヒトJκ1遺伝子セグメント、前記ヒトJκ2遺伝子セグメント、前記ヒトJκ3遺伝子セグメント、前記ヒトJκ4遺伝子セグメント、および、前記ヒトJκ5遺伝子セグメントを順に含む、請求項23に記載の方法。
- 前記遺伝子改変されたマウスが、DLC-5J遺伝子座を含み、該DLC-5J遺伝子座が、配列番号38~49より選択されるhVκ/hJκ/mCκ接合部配列を含む、請求項23~29のいずれか一項に記載の方法。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US13/798,455 US9796788B2 (en) | 2010-02-08 | 2013-03-13 | Mice expressing a limited immunoglobulin light chain repertoire |
| US13/798,455 | 2013-03-13 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019006738A Division JP6790136B2 (ja) | 2013-03-13 | 2019-01-18 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020128111A Division JP2020182490A (ja) | 2013-03-13 | 2020-07-29 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2019213549A JP2019213549A (ja) | 2019-12-19 |
| JP2019213549A5 JP2019213549A5 (ja) | 2020-09-10 |
| JP7010902B2 true JP7010902B2 (ja) | 2022-02-10 |
Family
ID=50483561
Family Applications (4)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2016502035A Active JP6529957B2 (ja) | 2013-03-13 | 2014-03-13 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
| JP2019006738A Active JP6790136B2 (ja) | 2013-03-13 | 2019-01-18 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
| JP2019148806A Active JP7010902B2 (ja) | 2013-03-13 | 2019-08-14 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
| JP2020128111A Pending JP2020182490A (ja) | 2013-03-13 | 2020-07-29 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
Family Applications Before (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2016502035A Active JP6529957B2 (ja) | 2013-03-13 | 2014-03-13 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
| JP2019006738A Active JP6790136B2 (ja) | 2013-03-13 | 2019-01-18 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020128111A Pending JP2020182490A (ja) | 2013-03-13 | 2020-07-29 | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス |
Country Status (26)
| Country | Link |
|---|---|
| EP (3) | EP3501272B1 (ja) |
| JP (4) | JP6529957B2 (ja) |
| KR (2) | KR102424756B1 (ja) |
| CN (2) | CN105188357B (ja) |
| AU (3) | AU2014244020B2 (ja) |
| BR (1) | BR112015022034B1 (ja) |
| CA (1) | CA2903698C (ja) |
| CY (1) | CY1121545T1 (ja) |
| DK (2) | DK3501272T5 (ja) |
| ES (2) | ES2942413T3 (ja) |
| FI (1) | FI3501272T3 (ja) |
| HK (1) | HK1218493A1 (ja) |
| HR (2) | HRP20230490T1 (ja) |
| HU (2) | HUE061977T2 (ja) |
| IL (2) | IL240563B (ja) |
| LT (2) | LT2967013T (ja) |
| MX (2) | MX357308B (ja) |
| PL (2) | PL2967013T3 (ja) |
| PT (2) | PT2967013T (ja) |
| RS (2) | RS58669B1 (ja) |
| RU (1) | RU2689664C2 (ja) |
| SG (2) | SG11201506278RA (ja) |
| SI (2) | SI2967013T1 (ja) |
| TR (1) | TR201901668T4 (ja) |
| WO (1) | WO2014160202A1 (ja) |
| ZA (2) | ZA201505835B (ja) |
Families Citing this family (21)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US9796788B2 (en) | 2010-02-08 | 2017-10-24 | Regeneron Pharmaceuticals, Inc. | Mice expressing a limited immunoglobulin light chain repertoire |
| US20130045492A1 (en) | 2010-02-08 | 2013-02-21 | Regeneron Pharmaceuticals, Inc. | Methods For Making Fully Human Bispecific Antibodies Using A Common Light Chain |
| RU2724663C2 (ru) | 2010-02-08 | 2020-06-25 | Ридженерон Фармасьютикалз, Инк. | Мышь с общей легкой цепью |
| MY172718A (en) | 2011-08-05 | 2019-12-11 | Regeneron Pharma | Humanized universal light chain mice |
| SG10202010120XA (en) | 2011-10-17 | 2020-11-27 | Regeneron Pharma | Restricted immunoglobulin heavy chain mice |
| KR102436654B1 (ko) | 2012-06-12 | 2022-08-26 | 리제너론 파마슈티칼스 인코포레이티드 | 제한된 면역글로불린 중쇄 유전자좌를 가지는 인간화된 비-인간 동물 |
| LT2840892T (lt) * | 2013-02-20 | 2018-07-25 | Regeneron Pharmaceuticals, Inc. | Nežmogaus tipo gyvūnai su modifikuotomis imunoglobulino sunkiųjų grandinių sekomis |
| KR102601491B1 (ko) | 2014-03-21 | 2023-11-13 | 리제너론 파마슈티칼스 인코포레이티드 | 단일 도메인 결합 단백질을 생산하는 비-인간 동물 |
| CN107438622A (zh) | 2015-03-19 | 2017-12-05 | 瑞泽恩制药公司 | 选择结合抗原的轻链可变区的非人动物 |
| CA3011359A1 (en) | 2016-01-13 | 2017-07-20 | Regeneron Pharmaceuticals, Inc. | Rodents having an engineered heavy chain diversity region |
| BR112020004977A2 (pt) * | 2017-09-29 | 2020-10-06 | Regeneron Pharmaceuticals, Inc. | moléculas de ligação de antígenos biespecíficas que ligam antígeno alvo de staphyocococcus e componente de complemento e usos dos mesmos |
| IL305557A (en) | 2018-06-14 | 2023-10-01 | Regeneron Pharma | Non-human animals with transgenic DH–DH rearrangement capacity, and their uses |
| US20220090125A1 (en) * | 2018-12-21 | 2022-03-24 | Compass Therapeutics Llc | Transgenic mouse expressing common human light chain |
| CA3125380A1 (en) | 2019-02-18 | 2020-08-27 | Biocytogen Pharmaceuticals (Beijing) Co., Ltd. | Genetically modified non-human animals with humanized immunoglobulin locus |
| BR112021016173A2 (pt) | 2019-02-22 | 2021-11-03 | Regeneron Pharma | Roedor geneticamente modificado, métodos de produção de um roedor geneticamente modificado e de produção de um anticorpo anti-nav1.7, célula ou tecido isolado de roedor, linhagem celular imortalizada, embrião de roedor, construto de ácido nucleico de direcionamento, e, hibridoma |
| KR20210095781A (ko) | 2020-01-24 | 2021-08-03 | 주식회사 에이프릴바이오 | 항원결합 단편 및 생리활성 이펙터 모이어티로 구성된 융합 컨스트럭트를 포함하는 다중결합항체 및 이를 포함하는 약학조성물 |
| EP4158034A4 (en) | 2020-06-02 | 2024-07-03 | Biocytogen Pharmaceuticals Beijing Co Ltd | GENETICALLY MODIFIED NON-HUMAN ANIMALS HAVING A LIGHT-CHAIN IMMUNOGLOBULIN LOCUS |
| BR112023011689A2 (pt) | 2020-12-23 | 2023-10-31 | Regeneron Pharma | Método para obter uma célula produtora de anticorpos, anticorpo, e, célula hospedeira de mamífero |
| US20240317849A1 (en) | 2020-12-23 | 2024-09-26 | Regeneron Pharmaceuticals, Inc. | Nucleic acids encoding anchor modified antibodies and uses thereof |
| CN114736927A (zh) * | 2022-05-19 | 2022-07-12 | 东北大学 | 一种单抗克隆与表达载体的构建及应用 |
| US20240263190A1 (en) * | 2023-01-18 | 2024-08-08 | Gilead Sciences, Inc. | Human immunoglobulin binary light chain transgene constructs and uses thereof |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2012148873A2 (en) | 2011-04-25 | 2012-11-01 | Regeneron Pharmaceuticals, Inc. | Non-human animals expressing antibodies having a common light chain |
Family Cites Families (22)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7041871B1 (en) * | 1995-10-10 | 2006-05-09 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| US6774279B2 (en) | 1997-05-30 | 2004-08-10 | Carnegie Institution Of Washington | Use of FLP recombinase in mice |
| US6737056B1 (en) | 1999-01-15 | 2004-05-18 | Genentech, Inc. | Polypeptide variants with altered effector function |
| US7105348B2 (en) | 2000-10-31 | 2006-09-12 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| US6596541B2 (en) * | 2000-10-31 | 2003-07-22 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| US6586251B2 (en) | 2000-10-31 | 2003-07-01 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| US7217797B2 (en) | 2002-10-15 | 2007-05-15 | Pdl Biopharma, Inc. | Alteration of FcRn binding affinities or serum half-lives of antibodies by mutagenesis |
| MXPA06000562A (es) * | 2003-07-15 | 2006-03-30 | Therapeutic Human Polyclonals | Loci de inmunoglobulina humanizada. |
| ES2463476T3 (es) | 2004-10-19 | 2014-05-28 | Regeneron Pharmaceuticals, Inc. | Método para generar un ratón homocigótico para una modificación genética |
| EP2505058A1 (en) * | 2006-03-31 | 2012-10-03 | Medarex, Inc. | Transgenic animals expressing chimeric antibodies for use in preparing human antibodies |
| PL2041177T3 (pl) | 2006-06-02 | 2012-09-28 | Regeneron Pharma | Przeciwciała o wysokim powinowactwie przeciw ludzkiemu receptorowi IL 6 |
| ITMI20071522A1 (it) * | 2007-07-27 | 2009-01-28 | Areta Internat S R L | Vaccino idiotipico |
| ES2906344T3 (es) * | 2008-06-27 | 2022-04-18 | Merus Nv | Animal murino transgénico productor de anticuerpos |
| RU2011129459A (ru) * | 2008-12-18 | 2013-01-27 | Эрасмус Юниверсити Медикал Сентр Роттердам | Трансгенные животные (не человек), экспрессирующие гуманизированные антитела, и их применение |
| EP3916011A1 (en) | 2009-06-26 | 2021-12-01 | Regeneron Pharmaceuticals, Inc. | Readily isolated bispecific antibodies with native immunoglobulin format |
| RU2724663C2 (ru) * | 2010-02-08 | 2020-06-25 | Ридженерон Фармасьютикалз, Инк. | Мышь с общей легкой цепью |
| US20130045492A1 (en) * | 2010-02-08 | 2013-02-21 | Regeneron Pharmaceuticals, Inc. | Methods For Making Fully Human Bispecific Antibodies Using A Common Light Chain |
| JP6009441B2 (ja) * | 2010-06-22 | 2016-10-19 | リジェネロン・ファーマシューティカルズ・インコーポレイテッドRegeneron Pharmaceuticals, Inc. | ハイブリッド軽鎖マウス |
| NZ606824A (en) * | 2010-08-02 | 2015-05-29 | Regeneron Pharma | Mice that make binding proteins comprising vl domains |
| RU2460287C1 (ru) * | 2011-03-29 | 2012-09-10 | Государственное образовательное учреждение высшего профессионального образования "Российский государственный медицинский университет Федерального агентства по здравоохранению и социальному развитию" (ГОУ ВПО РГМУ Росздрава) | Трансгенная мышь, предназначенная для моделирования состояний, угрожающих развитием синдрома внезапной сердечной смерти при дистрофин-дефицитной кардиомиопатии |
| MY172718A (en) * | 2011-08-05 | 2019-12-11 | Regeneron Pharma | Humanized universal light chain mice |
| GB201122047D0 (en) * | 2011-12-21 | 2012-02-01 | Kymab Ltd | Transgenic animals |
-
2014
- 2014-03-13 RU RU2015143110A patent/RU2689664C2/ru active
- 2014-03-13 FI FIEP18210518.9T patent/FI3501272T3/fi active
- 2014-03-13 BR BR112015022034-7A patent/BR112015022034B1/pt active IP Right Grant
- 2014-03-13 CN CN201480013894.6A patent/CN105188357B/zh active Active
- 2014-03-13 ES ES18210518T patent/ES2942413T3/es active Active
- 2014-03-13 RS RS20190467A patent/RS58669B1/sr unknown
- 2014-03-13 LT LTEP14717331.4T patent/LT2967013T/lt unknown
- 2014-03-13 SG SG11201506278RA patent/SG11201506278RA/en unknown
- 2014-03-13 HR HRP20230490TT patent/HRP20230490T1/hr unknown
- 2014-03-13 DK DK18210518.9T patent/DK3501272T5/da active
- 2014-03-13 EP EP18210518.9A patent/EP3501272B1/en active Active
- 2014-03-13 HU HUE18210518A patent/HUE061977T2/hu unknown
- 2014-03-13 LT LTEP18210518.9T patent/LT3501272T/lt unknown
- 2014-03-13 MX MX2015012539A patent/MX357308B/es active IP Right Grant
- 2014-03-13 PL PL14717331T patent/PL2967013T3/pl unknown
- 2014-03-13 PL PL18210518.9T patent/PL3501272T3/pl unknown
- 2014-03-13 DK DK14717331.4T patent/DK2967013T3/en active
- 2014-03-13 PT PT14717331T patent/PT2967013T/pt unknown
- 2014-03-13 EP EP14717331.4A patent/EP2967013B1/en active Active
- 2014-03-13 MX MX2020010837A patent/MX2020010837A/es unknown
- 2014-03-13 SG SG10201801101QA patent/SG10201801101QA/en unknown
- 2014-03-13 AU AU2014244020A patent/AU2014244020B2/en active Active
- 2014-03-13 EP EP23151398.7A patent/EP4218409A3/en active Pending
- 2014-03-13 TR TR2019/01668T patent/TR201901668T4/tr unknown
- 2014-03-13 RS RS20230350A patent/RS64185B1/sr unknown
- 2014-03-13 CA CA2903698A patent/CA2903698C/en active Active
- 2014-03-13 HU HUE14717331A patent/HUE044003T2/hu unknown
- 2014-03-13 WO PCT/US2014/026040 patent/WO2014160202A1/en active Application Filing
- 2014-03-13 CN CN201810439928.8A patent/CN108611369B/zh active Active
- 2014-03-13 KR KR1020217027031A patent/KR102424756B1/ko active IP Right Grant
- 2014-03-13 KR KR1020157026152A patent/KR102295842B1/ko active IP Right Grant
- 2014-03-13 PT PT182105189T patent/PT3501272T/pt unknown
- 2014-03-13 ES ES14717331T patent/ES2712207T3/es active Active
- 2014-03-13 SI SI201431079T patent/SI2967013T1/sl unknown
- 2014-03-13 JP JP2016502035A patent/JP6529957B2/ja active Active
- 2014-03-13 SI SI201432022T patent/SI3501272T1/sl unknown
-
2015
- 2015-08-13 IL IL240563A patent/IL240563B/en active IP Right Grant
- 2015-08-13 ZA ZA2015/05835A patent/ZA201505835B/en unknown
-
2016
- 2016-06-06 HK HK16106453.9A patent/HK1218493A1/zh unknown
-
2018
- 2018-08-13 ZA ZA2018/05369A patent/ZA201805369B/en unknown
-
2019
- 2019-01-18 JP JP2019006738A patent/JP6790136B2/ja active Active
- 2019-04-05 CY CY20191100383T patent/CY1121545T1/el unknown
- 2019-04-16 HR HRP20190706TT patent/HRP20190706T1/hr unknown
- 2019-08-14 JP JP2019148806A patent/JP7010902B2/ja active Active
-
2020
- 2020-05-19 AU AU2020203238A patent/AU2020203238B2/en active Active
- 2020-07-29 JP JP2020128111A patent/JP2020182490A/ja active Pending
-
2021
- 2021-02-15 IL IL280892A patent/IL280892B2/en unknown
-
2023
- 2023-05-04 AU AU2023202783A patent/AU2023202783A1/en active Pending
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2012148873A2 (en) | 2011-04-25 | 2012-11-01 | Regeneron Pharmaceuticals, Inc. | Non-human animals expressing antibodies having a common light chain |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7010902B2 (ja) | 限られた免疫グロブリン軽鎖レパートリーを発現するマウス | |
| US11026407B2 (en) | Mice expressing a limited immunoglobulin light chain repertoire | |
| US20240156070A1 (en) | Common light chain mouse | |
| US20190071519A1 (en) | Methods for making fully human bispecific antibodies using a common light chain | |
| AU2012249953B2 (en) | Non-human animals expressing antibodies having a common light chain | |
| JP2018138047A (ja) | 共通の軽鎖のマウス | |
| JP2015509380A (ja) | 共通の軽鎖のマウス | |
| JP2016519568A (ja) | 共通軽鎖のマウス | |
| RU2812814C2 (ru) | Мыши, экспрессирующие ограниченный репертуар легких цепей иммуноглобулина | |
| NZ711776B2 (en) | Mice expressing a limited immunoglobulin light chain repertoire |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20190912 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20200729 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20201006 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210105 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20210419 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20210716 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210917 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20211221 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20220113 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7010902 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |