本明細書で提供される発明は、特に、ホスホケトラーゼポリペプチドを発現するよう遺伝子操作を施した組換え細胞においてメバロン酸、イソプレノイド前駆体分子、イソプレン及び/又はイソプレノイドを産生させるための組成物及び方法を開示するものである。本発明のホスホケトラーゼ酵素は、以下に更に詳細に記載される通りの多様な基質を利用することができる。特定の実施形態では、本発明は、キシルロース5−リン酸のグリセルアルデヒド3−リン酸及びアセチルリン酸への変換を触媒することのできるホスホケトラーゼポリペプチドを発現するよう遺伝子操作を施した組換え細胞において、メバロン酸、イソプレノイド前駆体分子、イソプレン及び/又はイソプレノイドを産生させるための組成物及び方法を提供する。他の実施形態では、本発明は、フルクトース6−リン酸のエリトロース4−リン酸及びアセチルリン酸への変換を触媒することのできるホスホケトラーゼポリペプチドを発現するよう遺伝子操作を施した組換え細胞において、メバロン酸、イソプレノイド前駆体分子、イソプレン及び/又はイソプレノイドを産生させるための組成物及び方法を提供する。更に他の実施形態では、本発明は、セドヘプツロース−7−リン酸のリボース−5−リン酸及びアセチルリン酸への変換を触媒することのできるホスホケトラーゼポリペプチドを発現するよう遺伝子操作を施した組換え細胞において、メバロン酸、イソプレノイド前駆体分子、イソプレン及び/又はイソプレノイドを産生させるための組成物及び方法を提供する。更に他の実施形態では、本発明は、キシルロース5−リン酸のグリセルアルデヒド3−リン酸及びアセチルリン酸への変換、並びに/あるいはフルクトース6−リン酸のエリトロース4−リン酸及びアセチルリン酸への変換、並びに/あるいはセドヘプツロース−7−リン酸のリボース−5−リン酸及びアセチルリン酸への変換を触媒することのできる、ホスホケトラーゼポリペプチドを発現するよう遺伝子操作を施した組換え細胞において、メバロン酸、イソプレノイド前駆体分子、イソプレン及び/又はイソプレノイドを産生させるための組成物及び方法を提供する。
宿主細胞において操作された代謝経路には、組換え発現させたホスホケトラーゼが使用される。米国特許第7,785,858号を参照されたい。Sonderegger et al.(Applied and Environmental Microbiology,2004,70:5,2892〜97)は、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)においてエタノールを過剰産生させる際のホスホケトラーゼの使用を記載する。Fleige et al.(Appl Microbial Biotechnol.,2011,91:3,769〜76)は、生育用の唯一の炭素源としてフルクトースを利用する能力を回復させたラルストニア・ユートロファ(Ralstonia eutropha)改変株における、ビフィドバクテリウム属(bifidobacterium)のホスホケトラーゼ遺伝子(上掲Meile et al.)の発現を報告している。しかしながら、これまでにも、メバロン酸経路への炭素の取り込みを増加させるためのホスホケトラーゼについての記載はなされている。
メバロン酸依存型生合成経路は、メバロン酸、及びその他のイソプレノイド前駆体分子、例えばジメチルアリル二リン酸(DMAPP)並びにイソペンテニルピロリン酸(IPP)の産生に特に重要である。メバロン酸経路上流の酵素は、グルコースから生成されたアセチルCoAを、3つの酵素反応によりメバロン酸へと変換する。理論に束縛されるものではないが、ホスホケトラーゼポリペプチドを用いアセチルCoAの生合成を増加させることで、上流メバロン酸経路に依存する生合成経路の産生性が向上し、メバロン酸の生合成が大幅に増加し、結果として、DMAPP及びIPPなどの下流のイソプレノイド前駆体分子の生合成を増加させることができるものと考えられる。したがって、これらの代替経路によるメバロン酸の産生収率の増加は、商業用途に有利なものである。
理論上は、平衡反応においては、1分子のグルコースから3分子のアセチルCoAを誘導することができる。しかしながら、生物は、通常、2分子のアセチルCoAのみを産生し、残部の質量はCO2として失われてしまう。CO2の放出は、ピルビン酸デヒドロゲナーゼにより触媒される、ピルビン酸からのアセチルCoAの生成反応中に生じる。炭素原子が1分子失われることにより、メバロン酸、イソプレノイド前駆体、イソプレン、及びイソプレノイド分子の産生収率が低下することになる。この反応時の損失を排除したものがWood−Ljungdahl経路であり、この経路は、嫌気的酢酸産生において、一酸化炭素デヒドロゲナーゼ及びアセチルCoAシンターゼにより二酸化炭素を還元してアセチルCoAを生成する。
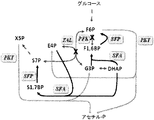
本発明は、Wood−Ljungdahl経路の酵素を利用しない経路により、1分子のグルコースから3分子のアセチル−CoAを生成し得る、代替的な代謝過程を提供する。その代わりに、特定の生物、特にビフィドバクテリウム(Bifidobacteria)属において見出されるホスホケトラーゼを使用する[例えば、Biology of the Prokaryotes(ed.Lengeler,Drews and Schlegel);Blackwell Science,New York,1999,p.299〜301;Meile et al.,J.of Bacteriology,2001,183:9,2929〜36;Jeong et al.,J.Microbiol.Biotechnol.,2007,17:5,822〜829を参照されたい]。ホスホケトラーゼ酵素は、典型的な代謝で用いられるピルビン酸の酸化以外の経路により、キシルロース5−リン酸又はフルクトース6−リン酸からのアセチル−CoA(リン酸アセチルを介する)の生成を行うことができる。
これまでに、ホスホケトラーゼは、基質選好性に基づき2系統、すなわち、X5Pのみに作用するキシルロース−5−リン酸(X5P)ホスホケトラーゼ、並びにX5P及びF6Pの両方に作用することのできるX5P/フルクトース−6−リン酸(F6P)ホスホケトラーゼに分類されてきた(Suzuki et al.,Acta Cryst.F66,2010,66:8,941〜43)。ホスホケトラーゼは、無機リン酸(Pi)を利用して、X5P又はF6Pを開裂させることにより、アセチルリン酸(アセチル−P)、H2O及びグリセルアルデヒド3−リン酸又はエリトロース4−リン酸の生成を触媒する。続いて、この経路では、酢酸キナーゼにより、高エネルギー代謝産物のアセチル−Pが酢酸へと変換され、ADPからATPが生成される。アセチルリン酸に加えて、代謝経路には最高収率、すなわちグルコース1分子毎にアセチルCoA3分子の生成が得られるよう操作を加え、酵素反応により産生されたグリセルアルデヒド3−リン酸を再利用することができる。有意に、ピルビン酸デヒドロゲナーゼが介在する反応で観察される通り、ホスホケトラーゼによりアセチルCoAを産生させることにより、炭素(例えば、CO2)の損失は排除される。
本明細書において更に詳述される通り、ホスホケトラーゼは、セドヘプツロース−7−リン酸に対しても作用し、これをリボース−5−リン酸及びアセチルリン酸に変換する。このようなホスホケトラーゼの非限定例はビフィドバクテリウム・ロングム(Bifidobacterium longum)ホスホケトラーゼであり、この酵素はセドヘプツロース−7−リン酸に対し触媒活性を有する。
本発明は、メバロン酸、イソプレノイド前駆体、イソプレン及び/又はイソプレノイドの産生においてホスホケトラーゼ酵素を使用して、産生収率を増大させることを目的とする。具体的には、イソプレンの理論収率は、次式に表されるとおりに増大する(生物が、1molのグルコースを6molのCO2に完全酸化して、ATPを産生できるものと仮定する):
MVA経路のみ
1.5個グルコース+2.00個O2→1.00個イソプレン+4.00個CO2+5.00個H2O
理論収率−イソプレン0.252(g)/グルコース(g)
DXP経路
1.25個グルコース+0.50個O2→1.00個イソプレン+2.50個CO2+3.50個H2O
理論収率−イソプレン0.302(g)/グルコース(g)
MVA+ホスホケトラーゼ経路
1.22個グルコース+0.33個O2→1.00個イソプレン+2.33個CO2+3.32個H2O
理論収率−イソプレン0.309(g)/グルコース(g)
適宜に、特定の態様では、本発明は、メバロン酸の産生を増強させることのできる組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸、及びMVA経路上流の1種以上のポリペプチドをコードしている1種以上の核酸を含み、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まない細胞と比較して多量にメバロン酸を産生する。
他の態様では、本発明はイソプレノイド前駆体の産生を増強させることのできる組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸、及び完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸を含み、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まない細胞と比較して多量にイソプレノイド前駆体を産生する。
更に他の態様では、本発明はイソプレンを産生することのできる組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸、並びに(i)完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸及び(ii)イソプレンシンターゼポリペプチドをコードしている異種核酸を含み、細胞は、回収可能な量でイソプレンを産生することができる。特定の実施形態では、本発明は、イソプレンの産生を増強させることのできる組換え細胞であって、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸と、(i)完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸、及び(ii)イソプレンシンターゼポリペプチドをコードしている異種核酸と、を含む、細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まないイソプレン産生細胞と比較して多量にイソプレンを産生する。
更に別の態様では、本発明は、イソプレノイドを産生することのできる組換え細胞であって、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸と、(i)完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸、及び(ii)ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸と、を含む、細胞を提供し、細胞は、回収可能な量でイソプレノイドを産生することができる。特定の実施形態では、本発明はイソプレノイドの産生を増強させることのできる組換え細胞であって、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸と、(i)完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸、及び(ii)ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸とを含む、細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まないイソプレノイド産生細胞と比較して多量のイソプレノイドを産生する。
本開示の任意の態様では、本発明は組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含み、更に遺伝子組換えを施して、次の遺伝子:リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(talB)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)、グルコース−6−リン酸デヒドロゲナーゼ(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトース二リン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)のうち1つ以上の活性を調節して、ホスホケトラーゼ経路に取り込まれる炭素を増加させることができる。
一部の実施形態では、本発明はイソプレンを産生することのできる組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸、並びに(i)完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸、(ii)イソプレンシンターゼポリペプチドをコードしている異種核酸を含み、かつ(iii)ホスホケトラーゼ経路に取り込まれる炭素を増加させるよう1つ以上の遺伝子の活性を調節するため、更に遺伝子操作を施され、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まないイソプレン産生細胞と比較して多量のイソプレンを産生する。
一部の実施形態では、本発明は、イソプレノイドを産生することのできる組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸、並びに(i)完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸、(ii)イソプレンシンターゼポリペプチドをコードしている異種核酸を含み、かつ(iii)ホスホケトラーゼ経路に取り込まれる炭素を増加させるよう1つ以上の遺伝子の活性を調節するため、更に遺伝子操作を施され、及び(iv)ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸を含み、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まないイソプレノイド産生細胞と比較して多量のイソプレノイドを産生する。
一般的技術
本発明の実施においては、特に断らないかぎりにおいて、当業者の技能の範囲内に含まれる従来の分子生物学(組換え技術を含む)、微生物学、細胞生物学、生化学、及び免疫学の技術を用いる。これらの手法は、次の文献:「分子クローニング:実験マニュアル(Molecular Cloning:A Laboratory Manual)」第2版(Sambrook et al.、1989年);「オリゴヌクレオチド合成(Oligonucleotide Synthesis)」(M.J.Gait,ed.,1984);「動物細胞の培養(Animal Cell Culture)」(R.I.Freshney,ed.、1987年);「酵素学的実験法(Methods in Enzymology)」(Academic Press,Inc.);「分子生物学標準プロトコル(Current Protocols in Molecular Biology)」(F.M.Ausubel et al.,eds.、1987年、定期的に改定);「PCR:ポリメラーゼ連鎖反応(PCR:The Polymerase Chain Reaction)」(Mullis et al.,eds.,1994年).Singleton et al.、「微生物学及び分子生物学辞典(Dictionary of Microbiology and Molecular Biology)第2版」、J.Wiley & Sons(New York,N.Y.1994年)、及びMarch「有機化学反応、機序及び構造第4版(Advanced Organic Chemistry Reactions,Mechanisms and Structure 4th ed.)」,John Wiley & Sons(New York,N.Y.1992)中に十分に説明されており、これらの文献は、本出願で使用される数多くの用語の一般的な指針を当業者に提供する。
用語の定義
用語「イソプレン」は、2−メチル−1,3−ブタジエン(CAS# 78−79−5)を指す。3,3−ジメチルアリル二リン酸(DMAPP)からピロリン酸を除去することで、揮発性のC5炭化水素を、直接的に及び最終的に生成することができる。IPP分子をDMAPP分子に結合又は重合させることは包含しない場合がある。用語「イソプレン」は、概して、本明細書に別途記載のない限り、産生方法を限定されることを意図しない。
本明細書で使用するとき、用語「ポリペプチド」には、ポリペプチド、タンパク質、ペプチド、ポリペプチド断片、及び融合ポリペプチドが含まれる。
本明細書で使用するとき、「単離ポリペプチド」は、2、5、10、20、又は50個以上の異なるポリペプチドのライブラリなどといった、ポリペプチドのライブラリの一部を意味するものではなく、天然に生じる少なくとも1つの成分から分離されたポリペプチドを意味する。例えば、ポリペプチドをコードしている組換え核酸を発現させることで単離ポリペプチドを得ることができる。
「異種ポリペプチド」は、宿主細胞と異なる生物、種、又は株由来の核酸配列によりコードされるポリペプチドを意味する。一部の実施形態では、異種ポリペプチドは、同様の宿主細胞に天然に見られる野生型ポリペプチドと同一ではない。
本明細書で使用するとき、「核酸」は、共有結合により単鎖又は二本鎖のいずれかの形態で連結している、2つ以上のデオキシリボヌクレオチド及び/又はリボヌクレオチドを指す。
「組換え核酸」とは、対象とする核酸が由来する生物において自然に存在するゲノムにおいて、対象とする核酸に隣接する1種以上の核酸(例えば遺伝子)を含まない、対象とする核酸のことを意味する。したがって、この用語には、例えば、ベクターに組み込まれた、プラスミド又はウィルスに自己複製的に組み込まれた、原核生物若しくは真核生物のゲノムDNAに組み込まれた、又は他の配列とは独立して別個の分子(例えば、cDNA、ゲノムDNA断片、又はPCRにより産生された若しくは制限エンドヌクレアーゼによる消化により産生されたcDNA断片)として存在する、組換えDNAが包含される。
「異種核酸」は、宿主細胞と異なる生物、種又は株由来の核酸配列を意味する。一部の実施形態では、異種核酸は、同様の宿主細胞に天然に見られる野生型核酸と同一ではない。例えば、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロングム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)に由来するホスホケトラーゼ遺伝子によりコードされ、大腸菌(E. coli)の形質転換に使用される核酸は、異種核酸である。
本明細書で使用するとき、用語「ホスホケトラーゼ」、「ホスホケトラーゼ酵素」又は「ホスホケトラーゼポリペプチド」は互換的に使用され、5−リン酸をグリセルアルデヒド3−リン酸及びアセチルリン酸へと変換し、並びに/又はフルクトース6−リン酸をエリトロース4−リン酸及びアセチルリン酸へと変換するポリペプチドを指す。一般的に、ホスホケトラーゼはケトンに作用する。特定の実施形態では、ホスホケトラーゼポリペプチドは、キシルロース5−リン酸のグリセルアルデヒド3−リン酸及びアセチルリン酸への変換を触媒する。他の実施形態では、ホスホケトラーゼポリペプチドは、フルクトース6−リン酸のエリトロース4−リン酸及びアセチルリン酸への変換を触媒する。他の実施形態では、ホスホケトラーゼポリペプチドは、セドヘプツロース−7−リン酸の生成物(例えば、リボース−5−リン酸)及びアセチルリン酸への変換を触媒する。
本明細書において使用するとき、「発現制御配列」とは、対象とする核酸の転写を指示する核酸配列のことを意味する。発現制御配列は、構成型若しくは誘導型のプロモータ、又はエンハンサなどのプロモータであり得る。発現制御配列は「ネイティブ」な配列又は異種性の配列であり得る。ネイティブな発現制御配列は、遺伝子を発現させる生物、種又は株と同様の生物、種又は株に由来する配列である。異種性の発現制御配列は、遺伝子を発現させる生物、種又は株とは異なる生物、種又は株に由来する配列である。「誘導型プロモータ」は、環境下、又は発育制限下で活性であるプロモータである。
「調節可能に連結された」は、核酸発現制御配列(プロモータなど)及び第2の核酸配列間の機能的連結を意味し、発現制御配列は、第2の配列に対応する核酸の転写を指示する。
本明細書で使用するとき、用語「最少培地(「minimal medium」又は「minimal media」)は、細胞の生育に必要とされる最低限の栄養素を含有し、概してアミノ酸は存在していない増殖培地を指す。最少培地は、典型的には:(1)細菌を生育させるための炭素源;(2)細菌種及び生育条件によって異なる様々な塩類;並びに(3)水;を含有する。炭素源は、グルコースなどの単糖から、本明細書で以下により詳細に記載されるような、その他のバイオマスの、より複雑な加水分解物、例えば、酵母エキスなどといった多様なものであり得る。塩は、概してマグネシウム、窒素、リン及びイオウなどの必須元素を提供し、細胞がタンパク質及び核酸を合成できるようにする。また、最少培地には、特定のプラスミド及び同様物を維持すべく選別するために、抗菌剤などの選択剤を添加することもできる。例えば、微生物が、例えばアンピシリン又はテトラサイクリンなどの特定の抗菌剤に耐性である場合、耐性を欠く細胞の増殖を阻害する目的で培地に抗菌剤を添加することができる。培地には、所望される生理学的又は生化学的特性について選別するのに必要とされる、例えば特定のアミノ酸などといった他の化合物を添加することができる。
本明細書で使用するとき、用語「イソプレノイド」は、2つ以上の炭化水素単位からなり、各単位は、特定の様式で配置された5つの炭素原子からなる、天然に生じる有機化合物類の、広範にわたるかつ多様な部類を指す。本明細書で使用するとき、「イソプレン」は、明らかに「イソプレノイド」の定義から除外される。
本明細書で使用するとき、用語「テルペノイド」は、多様な様式で組み立てられ及び修飾され、構成員として使用されたイソプレノイド単位の数に基づき分類される、炭素数5のイソプレノイド単位に由来する有機分子の、広範にわたるかつ多様な部類を指す。ヘミテルペノイドは、イソプレノイド単位を1つ有する。モノテルペノイドは、イソプレノイド単位を2つ有する。セスキテルペノイドは、イソプレノイド単位を3つ有する。ジテルペノイドは、イソプレン単位を4つ有する。セステルテルペノイドは、イソプレノイド単位を5つ有する。トリテルペノイドは、イソプレノイド単位を6つ有する。テトラテルペノイドは、イソプレノイド単位を8つ有する。ポリテルペノイドは、イソプレノイド単位を8つ超有する。
本明細書で使用するとき、「イソプレノイド前駆体」は、テルペノイド又はイソプレノイドの生合成時に生物により使用される任意の分子を指す。イソプレノイド前駆体分子の非限定例としては、例えば、メバロン酸塩(例えば、メバロン酸(MVA))、イソペンテニルピロリン酸(IPP)及びジメチルアリル二リン酸(DMAPP)が挙げられる。
本明細書で使用するとき、用語「質量収率」は、組換え細胞により産生された生成物の質量を、組換え細胞により消費されたグルコースの質量により除したものを指し、割合(%)として示す。
「比産生量」は、組換え細胞により産生される産生物の質量を、産生物の産生にかかった時間、細胞密度及び培養体積により除したものを意味する。
「力価」は、組換え細胞により産生される産生物の質量を、培養体積により除したものを意味する。
本明細書で使用するとき、用語「細胞産生性指数(cell productivity index)(CPI)」は、組換え細胞により産生される産生物の質量を、培養により産生された組換え細胞の質量により除したものを指す。
本明細書において別途定義されない限り、本明細書で使用されるすべての技術及び科学用語は、本発明の属する技術分野の当業者により一般に理解される意味と同様の意味を持つ。
本明細書で使用するとき、単数形「a」、「an」、及び「the」には、文脈に明示されない限り、対象物が複数ある場合をも包含する。
本明細書を通じて与えられるあらゆる最大数の限定は、あらゆるより小さい数値の限定を、あたかもそのようなより小さい数値の限定が明確に書かれているかのように包含するものと理解されることが意図される。本明細書の全体を通じて与えられるすべての最小の数値的限定には、これよりも大きいすべての数値的限定が、あたかもこうしたより大きい数値的限定が本明細書に明確に記載されているものと同様に含まれる。本明細書の全体を通じて与えられるすべての数値的範囲には、これよりも狭い数値的範囲が、あたかもこうしたより狭い数値的範囲がすべて本明細書に明確に記載されているものと同様に含まれる。
ホスホケトラーゼポリペプチド及びMVA経路の1種以上のポリペプチドを発現している組換え細胞
メバロン酸依存性生合成経路(MVA経路)は、全ての高等真核生物及び特定種の細菌に存在する、重要な代謝経路である。メバロン酸経路は、タンパク質のプレニル化、細胞膜の維持、タンパク質の固定及びN−グリコシル化などの、多様な工程において使用される分子の産生に重要であり、並びにテルペン、テルペノイド、イソプレノイド、及びイソプレンの生合成時の主成分として機能するイソプレノイド前駆体分子のDMAPP及びIPPの主要な供給源を提供する。
完全なMVA経路は、上流及び下流経路の2群に細分することができる。MVA経路の上流では、細胞内代謝により産生されたアセチルCo−Aを、(a)(i)チオラーゼ活性又は(ii)アセトアセチル−CoAシンターゼ活性、(b)HMG−CoA還元酵素、及び(c)HMG−CoAシンターゼ活性のいずれかの活性をもつポリペプチドの作用によりメバロン酸に変換する。最初に、チオラーゼ又はアセトアセチル−CoAシンターゼ(アセチル−CoA及びマロニル−CoAを利用する)の作用によりアセチルCo−AをアセトアセチルCoAに変換する。次に、アセトアセチル−CoAは、HMG−CoAシンターゼの酵素作用により、3−ヒドロキシ−3−メチルグルタリル−CoA(HMG−CoA)へと変換される。このCo−A誘導体をHMG−CoA還元酵素により還元してメバロン酸を生成する。この反応は、メバロン酸経路によるイソプレノイド産生の律速段階となる。次に、MVA経路下流では、メバロン酸は、メバロン酸キナーゼの作用により5−ホスホメバロン酸に変換され、5−ホスホメバロン酸は、続いてホスホメバロン酸キナーゼの酵素活性により、5−ジホスホメバロン酸に変換される。最後に、酵素の5−ジホスホメバロン酸デカルボキシラーゼの活性により、5−ジホスホメバロン酸からIPPが形成される。
したがって、特定の実施形態では、本発明の組換え細胞は、MVA経路によるメバロン酸、イソプレノイド前駆体、イソプレン又はイソプレノイド産生能を有する組換え細胞であって、組換え細胞は、(i)キシルロース5−リン酸からグリセルアルデヒド3−リン酸及びアセチルリン酸を合成することのできるホスホケトラーゼをコードしている異種遺伝子、(ii)MVAポリペプチドを1種以上コードしている1種以上の異種遺伝子、及び(iii)宿主細胞において、アセトアセチルCoAからのメバロン酸、イソプレノイド前駆体、イソプレン又はイソプレノイドの合成を可能にする、メバロン酸、イソプレノイド前駆体、又はイソプレン又はイソプレノイド生合成に関与する1種以上の異種遺伝子を含む。他の実施形態では、本発明の組換え細胞は、メバロン酸、イソプレノイド前駆体、イソプレン又はイソプレノイド産生能を有する組換え細胞であって、組換え細胞は、(i)フルクトース6−リン酸からエリトロース4−リン酸及びアセチルリン酸を合成することのできるホスホケトラーゼをコードしている異種遺伝子、(ii)MVAポリペプチド1種以上をコードしている1種以上の異種遺伝子及び(iii)宿主細胞において、アセトアセチルCoAからメバロン酸、イソプレノイド前駆体、イソプレン又はイソプレノイドを合成することのできる、メバロン酸、イソプレノイド前駆体、イソプレン又はイソプレノイド生合成に関与する1種以上の異種遺伝子を含む。
ホスホケトラーゼポリぺプチド及び核酸例
ホスホケトラーゼ酵素は、キシルロース5−リン酸のグリセルアルデヒド3−リン酸及びアセチルリン酸への変換、並びに/又はフルクトース6−リン酸のエリトロース4−リン酸及びアセチルリン酸への変換を触媒する。特定の実施形態では、ホスホケトラーゼ酵素は、キシルロース5−リン酸のグリセルアルデヒド3−リン酸及びアセチルリン酸への変換を触媒する。他の実施形態では、ホスホケトラーゼ酵素は、フルクトース6−リン酸のエリトロース4−リン酸及びアセチルリン酸への変換を触媒する。他の実施形態では、ホスホケトラーゼポリペプチドは、セドヘプツロース−7−リン酸の生成物(例えば、リボース−5−リン酸)及びアセチルリン酸への変換を触媒する。したがって、理論に束縛されるものではないが、本明細書で説明されるとおりホスホケトラーゼを発現させることにより、炭水化物資源から生成されるアセチルリン酸量を増加させることができる。このアセチルリン酸をアセチルCoAへと変換させ、更にこれを利用して、MVA経路に関係する酵素活性により、メバロン酸、イソプレノイド前駆体分子、イソプレン及び/又はイソプレノイドを生成させることができる。したがって、炭水化物基質より生成されるこれらの化合物量を増加させることができる。あるいは、細胞内濃度の上昇という形式で生成性の向上が反映されずとも、アセチル−P及びAcCoAの生成量を増加させることができる。特定の実施形態では、細胞内アセチル−P又はアセチルCoA濃度は、ホスホケトラーゼによる反応が生じた場合でさえ変化せず一定であり、又は減少する場合すらある。
ホスホケトラーゼの核酸の例としては、ホスホケトラーゼポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又はポリペプチド融合物をコードしている核酸が挙げられる。ホスホケトラーゼのポリペプチド及び核酸の例としては、本明細書に記載の任意の生物資源から天然に生じるポリペプチド及び核酸、並びに本明細書に記載の任意の生物資源から誘導されるポリペプチド変異体及び核酸変異体が挙げられる(例については図4を参照されたい)。加えて、表1は、本発明の実施形態に利用することのできるある種のホスホケトラーゼ例に関する、種及び生化学的特徴の非限定的な一欄を提供する。
ホスホケトラーゼ例の生化学的特徴には、限定するものではないが、タンパク質発現、タンパク質溶解度、及び活性が包含される。ホスホケトラーゼは、限定するものではないが、異なる種類の生物間(例えば、グラム陽性菌、シアノバクテリア、放線菌)の多様性、所望の種(例えば、大腸菌(E. coli))に比較的近い、通性低温好気性菌(facultative low temperature aerobe)、及び熱耐性などのその他の特徴に基づき選択することもできる。
本明細書で提供するとき、ホスホケトラーゼ活性は、イソプレノイド前駆体(例えば、メバロン酸)、イソプレン、及び/又はイソプレノイドの産生を向上し得る。本明細書では、ホスホケトラーゼを含む組換え宿主が提供され、細胞は、イソプレノイド前駆体(例えば、メバロン酸)、イソプレン、及び/又はイソプレノイドの産生を増加させる、対象とする少なくとも1つの特性を示す。一部の態様では、対象とする少なくとも1つの特性は、限定するものではないが、比産生性、収率、力価、及び細胞性能指数からなる群から選択される。
特定の実施形態では、本明細書に用いるのに好適なホスホケトラーゼには、可溶性ホスホケトラーゼが包含される。タンパク質の溶解度を測定するための手法は、当該技術分野において周知である。タンパク質溶解度を測定する手法としては、本明細書において実施例で記載されるものが挙げられる。一部の実施形態では、本明細書に用いるためのホスホケトラーゼは、少なくとも20%の溶解度を有するものを包含する。一部の実施形態では、ホスホケトラーゼの溶解度は、約5%〜約100%、約10%〜約100%、約15%〜約100%、約20%〜約100%、約25%〜約100%、約30%〜約100%、約35%〜約100%、約40%〜約100%、約45%〜約100%、約50%〜約100%、約55%〜約100%、約60%〜約100%、約65%〜約100%、約70%〜約100%、約75%〜約100%、約80%〜約100%、約85%〜約100%、又は約90%〜約100%のうちのいずれかである。一部の実施形態では、ホスホケトラーゼの溶解度は、約5%〜約100%である。一部の実施形態では、溶解度は5%〜100%である。一部の実施形態では、ホスホケトラーゼの溶解度は約100、90、80、70、60、50、40、30、20、又は10%未満のいずれかであるが、但し約5%未満ではない。一部の実施形態では、溶解度は、約5、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、又は95%超のいずれかである。
所望の動的特性を有するホスホケトラーゼは、イソプレンの産生を向上させる。動的特性としては、限定するものではないが、非活性、Kcat、Ki、及びKmが挙げられる。一部の態様では、kcatは、少なくとも約0.2、0.4、0.6、0.8、1.0、1.2、1.4、1.6、1.8、2.0、2.2、2.4、2.6、2.8、3.0、3.2、3.4、3.6、3.8、4.0、4.2、4.4、4.6、4.8、5.0、5.2、5.4、5.6、5.8、6.0、6.2、6.4、6.6、6.8、7.0、7.2、7.4、7.6、7.8、8.0、8.1、8.2、8.4、8.6、8.8、9.0、9.2、9.4、9.6、9.8、10.0、10.2、10.4、10.6、10.8、11.0、11.2、11.4、11.6、11.8、12.0、12.2、12.4、12.6、12.8、13.0、13.2、13.4、13.6、13.8、14.0、14.2、14.4、14.6、14.8、15.0、15.2、15.4、15.6、15.8、16.0、16.2、16.4、16.6、16.8、17.0、17.2、17.4、17.6、17.8、18.0、18.2、18.4、18.6、18.8、19.0、19.2、19.4、19.6、19.8、20.0、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、55、60、65、70、75、80、85、90、95、100、200、300、400、500、600、700、又は800である。他の態様では、kcatは、少なくとも約0.2、0.4、0.6、0.8、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9 2.0、2.1、2.2、2.3、2.4、2.5、2.6、2.7、2.8、2.9、3.0、3.2、3.4、3.6、3.8、4.0、4.2、4.4、4.6、4.8、5.0、5.2、5.4、5.6、5.8、6.0、6.2、6.4、6.6、6.8、7.0、7.2、7.4、7.6、7.8、8.0、8.1、8.2、8.4、8.6、8.8、9.0、9.2、9.4、9.6、9.8、10.0、10.2、10.4、10.6、10.8、11.0、11.2、11.4、11.6、11.8、12.0、12.2、12.4、12.6、12.8、13.0、13.2、13.4、13.6、13.8、14.0、14.2、14.4、14.6、14.8、15.0、15.2、15.4、15.6、15.8、16.0、16.2、16.4、又は16.6である。
一部の態様では、Kmは、少なくとも約1、1.5、2、2.5、3、3.5、4、4.5、5、5.5、6、6.5、7、7.5、8、8.5、9、9.5、10、10.5、11、11.5、12、12.5、13、13.5、14、14.5、15、16、17、17.5、18、18.5、19、19.5、20、20.5、21、21.5、22、22.5、23、23.5、24、24.5、25、25.5、26、26.5、27、27.5、28、28.5、29、29.5、30、30.5、31、31.5、32、32.5、33、33.5、34、34.5、35、35.5、36、36.5、37、37.5、38、38.5、39、39.5、40、40.5、41、41.5、42、42.5、43、43.5、44、44.5、45、45.5、46、46.5、47、47.5、48、48.5、49、49.5、50、50.5、51、51.5、52、52.5、53、53.5、54、54.5、55、55.5、又は56である。他の態様では、kmは、少なくとも約2.5、3、3.5、4、4.5、5、5.5、6、6.5、7、7.5、8、8.5、9、9.5、10、10.5、11、11.5、12、12.5、13、13.5、14、14.5、15、16、17、17.5、18、18.5、19、19.5、20、20.5、21、21.5、又は22である。
対象とする特性としては、限定するものではないが、ホスホケトラーゼポリペプチドを含まない組換え細胞と比較して、細胞内活性、比産生性、収率、及び細胞性能指数が向上していることが挙げられる。一部の実施形態では、比産生性は、少なくとも約0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、3、4、5、6 7、8、9、又は10倍以上増加する。一実施形態では、比産生性は約40mg/L/OD/hである。一部の実施形態では、収率は、少なくとも約0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、3、4、又は5倍以上向上する。他の実施形態では、MVA収率は、少なくとも約0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、3、4、又は5倍以上向上する。他の実施形態では、イソプレン収率は、少なくとも約0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、3、4、又は5倍以上向上する。他の実施形態では、細胞性能指数は、少なくとも約0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、3、4、又は5倍以上向上する。他の実施形態では、細胞内活性は、少なくとも約0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、3、4、5、6、7、8、9、又は10倍以上向上する。
使用することのできる他のホスホケトラーゼとしては、限定するものではないが、ビフィドバクテリウム・ロングム(B. longum)、ラクトバチルス・プランタルム(L. plantarum)、クロストリジウム・アセトブチリカム(C. acetobutylicum)、ラクトバチルス・ロイテリ(L. reuteri)、ラクトバチルス・パラプランタラム(L. paraplantarum)、ロードシュードモナス・パルストリス(R. palustris)、ノストック・パンクチホルム(Nostoc punctiforme)、ビフィドバクテリウム・アニマリス(B. animalis)、ビフィドバクテリウム・ブレビ(B. breve)、ガードネレラ・バギナリス(G. vaginalis)、エンテロコッカス・ガリナラム(E. gallinarum)、ムチラギニバクター・パルジス(M. paludis)、パントエア(Panteoa sp.)、ラナンキュラス・アクアティリス(R. aquatilis)、ノストック・パンクチホルム(N. punctiforme)、ストレプトマイセス・エバミティリス(S. avermetilis)、及びサーモビフィダ・フスカ(T. fusca)が挙げられる。
標準法を使用し、ペプチドがD−フルクトース6−リン酸又はD−キシルロース5−リン酸をアセチル−Pへと変換する能力を測定することで、ポリペプチドがホスホケトラーゼペプチド活性を有するかを判断できる。次に、アセチル−Pはフェリルアセチルヒドロキサム酸(ferryl acetyl hydroxamate)へと変換され得る。この変換は、分光測定により検出可能である(Meile et al.,J.Bact.183:2929〜2936,2001)。本発明での使用には、本明細書に記載されるとおりホスホケトラーゼペプチド活性を有するとして判定された任意のポリペプチドが好適である。一部の実施形態では、ホスホケトラーゼポリペプチドは、キシルロース5−リン酸のグリセルアルデヒド3−リン酸及びアセチルリン酸への変換を触媒する。他の実施形態では、ホスホケトラーゼポリペプチドは、フルクトース6−リン酸のエリトロース4−リン酸及びアセチルリン酸への変換を触媒する。更に他の実施形態では、ホスホケトラーゼポリペプチドは、セドヘプツロース−7−リン酸のリブロース−5−リン酸及びアセチルリン酸への変換を触媒することができる。更に他の実施形態では、ホスホケトラーゼポリペプチドは、キシルロース5−リン酸のグリセルアルデヒド3−リン酸及びアセチルリン酸への変換、並びに/あるいはフルクトース6−リン酸のエリトロース4−リン酸及びアセチルリン酸への変換、並びに/あるいはセドヘプツロース−7−リン酸のリブロース−5−リン酸及びアセチルリン酸への変換を触媒する。
本明細書においては、微生物から単離されたホスホケトラーゼも提供される。一部の態様では、ホスホケトラーゼは、グラム陽性菌、グラム陰性菌、好気性菌、嫌気性菌、光熱性菌、好冷性菌、好塩菌、又はシアノバクテリアからなる群から単離される。一部の態様では、ホスホケトラーゼは真菌から単離される。他の態様では、ホスホケトラーゼの核酸の例としては、限定するものではないが、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)から単離したホスホケトラーゼが挙げられる。他の態様では、ホスホケトラーゼの核酸の例としては、例えば、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea sp.)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼが挙げられる。他の態様では、ホスホケトラーゼの核酸例としては、例えば、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼが挙げられる。本明細書に記載の任意の態様では、ホスホケトラーゼの核酸は、本明細書に記載の任意のホスホケトラーゼの核酸配列に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ラクトバチルス・ロイテリ(Lactobacillus reuteri)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号1に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ビフィドバクテリウム・ロングム(Bifidobacterium longum)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号3に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。エンテロコッカス・ガリナラム(Enterococcus gallinarum)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号17に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ノストック・パンクチホルム(Nostoc punctiforme)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号18に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号19に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。パントエア(Pantoea sp.)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号20に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ムチラギニバクター・パルジス(Mucilaginibacter paludis)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号21に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。サーモビフィダ・フスカ(Thermobifida fusca)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号22に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ビフィドバクテリウム・ブレビ(Bifidobacterium breve)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号23に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ラーネラ・アクアティリス(Rahnella aquatilis)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号24に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号25に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ガードネレラ・バギナリス(Gardnerella vaginalis)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号26に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号27に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号28に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。ラクトバチルス・パラプランタラム(Lactobacillus paraplantarum)のホスホケトラーゼ遺伝子によりコードされているホスホケトラーゼの核酸は、配列番号29に対して少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有する。
一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号1の、ラクトバチルス・ロイテリ(Lactobacillus reuteri)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号3の、ビフィドバクテリウム・ロングム(Bifidobacterium longum)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号17の、エンテロコッカス・ガリナラム(Enterococcus gallinarum)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号18の、ノストック・パンクチホルム(Nostoc punctiforme)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号19の、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)ホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号20の、パントエア(Pantoea sp.)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号21の、ムチラギニバクター・パルジス(Mucilaginibacter paludis)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号22の、サーモビフィダ・フスカ(Thermobifida fusca)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号23の、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号24の、ラーネラ・アクアティリス(Rahnella aquatilis)おホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号25の、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号26の、ガードネレラ・バギナリス(Gardnerella vaginalis)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号27の、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号28の、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。一部の実施形態では、ホスホケトラーゼポリペプチドは、配列番号29の、ラクトバチルス・パラプランタラム(Lactobacillus paraplantarum)のホスホケトラーゼの核酸配列によりコードされるホスホケトラーゼポリペプチドに対して、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。本明細書に記載の任意の態様では、ホスホケトラーゼポリペプチドは、本明細書に記載の任意のホスホケトラーゼの核酸配列によりコードされる任意のホスホケトラーゼポリペプチドに対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る。
本明細書に使用することのできるホスホケトラーゼ酵素の追加例は、米国特許第7,785,858号及び国際公開第2011/159853号に記載されており、これらの特許文献は、特に、ホスホケトラーゼ酵素に対する開示に関し、参照により本明細書に組み込まれる。
MVA経路上流に関係するポリペプチド
MVA経路の上流は、細胞内代謝により産生されるアセチルCo−Aを、(a)(i)チオラーゼ活性又は(ii)アセトアセチル−CoA活性、(b)HMG−CoA還元酵素、及び(c)HMG−CoAシンターゼ活性のいずれかの活性をもつポリペプチドの作用によりメバロン酸に変換する際の開始基質として利用する。最初に、チオラーゼ又はアセトアセチル−CoAシンターゼ(アセチル−CoA及びマロニル−CoAを利用する)の作用によりアセチルCo−AをアセトアセチルCoAに変換する。次に、アセトアセチル−CoAは、HMG−CoAシンターゼの酵素作用により、3−ヒドロキシ−3−メチルグルタリル−CoA(HMG−CoA)へと変換される。このCo−A誘導体をHMG−CoA還元酵素により還元してメバロン酸を生成する。この反応は、メバロン酸経路によるイソプレノイド産生の律速段階となる。
MVA経路上流に関係するポリペプチドの非限定例としては、アセチル−CoAアセチルトランスフェラーゼ(AA−CoAチオラーゼ)ポリペプチド、アセトアセチル−CoAシンターゼポリペプチド、3−ヒドロキシ−3−メチルグルタリル−CoAシンターゼ(HMG−CoAシンターゼ)ポリペプチド、3−ヒドロキシ−3−メチルグルタリル−CoA還元酵素(HMG−CoA還元酵素)ポリペプチドが挙げられる。MVA経路上流のポリペプチドには、MVA経路上流のポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド及び融合ポリペプチドが含まれる。MVA経路上流に関係する代表的な核酸としては、MVA経路上流に関係するポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又は融合ポリペプチドをコードしている核酸が挙げられる。MVA経路に含まれる代表的なポリペプチド及び核酸としては、本明細書に記載されるような任意の生物資源から天然に生じるポリペプチド及び核酸が挙げられる。したがって、本開示では、MVA経路上流関連性ポリペプチドをコードしている任意の遺伝子は、本発明において使用できることを企図する。
特定の実施形態では、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カセリフラブス(E. casseliflavus)及び/又はエンテロコッカス・フェカリス(E. faecalis)由来の様々なmvaE及びmvaS遺伝子を、単独で、あるいはMVA経路上流関連性タンパク質をコードしている1つ以上の他のmvaE及びmvaS遺伝子と組み合わせて選択することが、本発明の範囲内のものとして企図される。他の実施形態では、アセトアセチル−CoAシンターゼの遺伝子は、(i)3−ヒドロキシ−3−メチルグルタリル−CoAシンターゼ(HMG−CoAシンターゼ)ポリペプチド及び3−ヒドロキシ−3−メチルグルタリル−CoA還元酵素(HMG−CoA還元酵素)ポリペプチドをコードしている1つ以上の他の遺伝子との組み合わせで本発明の範囲内のものとして企図される。したがって、特定の態様では、企図される遺伝子の任意の組み合わせを、本開示に記載される任意の手法により、組換え細胞で発現させることができる。
国際公開出願第2009/076676号、同第2010/003007号、及び同第2010/148150号に記載の、MVA経路上流に関係するポリペプチドのその他の非限定例を、本開示に使用できる。
mvaE及びmvaSポリペプチドをコードしている遺伝子
特定の実施形態では、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カセリフラブス(E. casseliflavus)及び/又はエンテロコッカス・フェカリス(E. faecalis)由来の様々なmvaE及びmvaS遺伝子を、単独で、あるいはMVA経路上流関連性タンパク質をコードしている1つ以上の他のmvaE及びmvaS遺伝子と組み合わせて選択することが、本発明の範囲内のものとして企図される。リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カセリフラブス(E. casseliflavus)、及びエンテロコッカス・フェカリス(E. faecalis)では、mvaE遺伝子は、チオラーゼ及びHMG−CoA還元酵素の活性のいずれをも保有しているポリペプチドをコードする。実際に、mvaE遺伝子産物は、真性細菌で見られる、IPP生合成に関係する最初の二機能性酵素となるものであり、HMG−CoA還元酵素の第一例は、天然において他のタンパク質と融合していた(Hedl,et al.,J Bacteriol.2002 April;184(8):2116〜2122)。それに対しmvaS遺伝子は、HMG−CoAシンターゼ活性を有するポリペプチドをコードする。
したがって、組換え細胞(例えば、大腸菌)は、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カセリフラブス(E. casseliflavus)及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaS遺伝子を1つ以上発現してメバロン酸を生成するよう設計することができる。1つ以上のmvaE及びmvaS遺伝子を多コピープラスミドで発現させることもできる。プラスミドは高コピー数プラスミド、低コピー数プラスミド又は中程度コピー数プラスミドであってよい。あるいは、1つ以上のmvaE及びmvaS遺伝子を宿主細胞の染色体に組み込むことができる。プラスミド上の、あるいは宿主細胞染色体の一部に組み込まれた、1つ以上のmvaE及びmvaS遺伝子のいずれもの異種発現に際し、遺伝子発現は、誘導型プロモータ又は構成的に(恒常的に)発現するプロモータのいずれかにより駆動できる。プロモータは、1つ以上のmvaE及びmvaS遺伝子の発現を強力に駆動することができ、弱く駆動することができ、あるいは中程度に駆動することができる。
mvaEポリペプチド及び核酸例
mvaE遺伝子は、チオラーゼ活性及びHMG−CoA還元酵素活性を両方保有しているポリペプチドをコードする。mvaE遺伝子によりコードされているポリペプチドのチオラーゼ活性は、アセチルCo−AをアセトアセチルCoAに変換するのに対し、HMG−CoA還元酵素ポリペプチドの酵素活性は、3−ヒドロキシ−3−メチルグルタリル−CoAをメバロン酸に変換する。mvaEポリペプチド及び核酸の例としては、本明細書に記載の任意の生物資源から天然に生じるポリペプチド及び核酸、並びにmvaEポリペプチドの活性を少なくとも1つ有する、本明細書に記載の任意の生物資源から誘導されるポリペプチド変異体及び核酸変異体が挙げられる。
変異型mvaEポリペプチドとしては、1つ以上のアミノ酸残基がアミノ酸置換を受けており、かつmvaEポリペプチド活性(すなわち、アセチルCo−AをアセトアセチルCoAに変換する能力並びに3−ヒドロキシ−3−メチルグルタリル−CoAをメバロン酸に変換する能力)を保持しているものが挙げられる。アミノ酸置換は、保存的なものであっても非保存的なものであってもよく、アミノ酸残基の置換は遺伝子暗号によりコードされていてもコードされていなくてもよい。標準的な20個のアミノ酸「記号(alphabet)」を、側鎖の類似性に基づき化学的なファミリーに分けた。これらのファミリーとしては、塩基性側鎖(例えば、リシン、アルギニン、ヒスチジン)、酸性側鎖(例えば、アスパラギン酸、グルタミン酸)、極性無電荷側鎖(例えば、グリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン)、非極性側鎖(例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、β分岐側鎖(例えば、トレオニン、バリン、イソロイシン)、及び芳香族側鎖(例えば、チロシン、フェニルアラニン、トリプトファン、ヒスチジン)を持つアミノ酸が挙げられる。「保存的アミノ酸置換」では、アミノ酸残基は、化学的に類似の側鎖を有するアミノ酸残基で置換される(すなわち、塩基性側鎖を有するアミノ酸は、塩基性側鎖を有する別のアミノ酸により置換される)。「非保存的アミノ酸置換」では、アミノ酸残基は、化学的に異なる側鎖を有するアミノ酸残基で置換される(すなわち、塩基性側鎖を有するアミノ酸は、芳香族側鎖を有する別のアミノ酸により置換される)。
mvaEポリペプチド中にアミノ酸置換を導入することで、分子の官能性を改良することができる。例えば、mvaEポリペプチドの、基質に対する結合親和性を増加させるアミノ酸置換、あるいはアセチルCo−AをアセトアセチルCoAに変換する能力及び/又は3−ヒドロキシ−3−メチルグルタリル−CoAをメバロン酸に変換する能力を改良するアミノ酸置換を、mvaEポリペプチドに導入することができる。一部の態様では、mvaEポリペプチド変異体は、1つ以上の保存的なアミノ酸置換を含有する。
一態様では、メバロン酸、イソプレン、イソプレノイド前駆体、イソプレン及び/又はイソプレノイドを産生する際には、分解されていない又は分解しにくいmvaEタンパク質を使用することができる。使用することのできる、分解されていない又は分解しにくいmvaE遺伝子産物の例としては、限定するものではないが、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カセリフラブス(E. casseliflavus)、エンテロコッカス・フェカリス(E. faecalis)及びリステリア・グレイ(L. grayi)に由来するものが挙げられる。当業者は、大腸菌(E. coli)BL21(DE3)においてmvaEタンパク質を発現させ、任意の標準的な分子生物学手法により断片が存在していないか調べることができる。例えば、Hisタグを利用して精製した後、すなわちメバロン酸、イソプレン又イソプレノイドを産生する大腸菌(E. coli)BL21において発現させた後、本明細書に記載の検出法を使用して、SDS−PAGEゲルをSafestainで染色することにより、断片が存在していないことを確認することができる。
Hedl et al.,(J Bacteriol.2002,April;184(8):2116〜2122)に記載のものなどの標準法を使用して、アセトアセチルCoAチオラーゼ活性並びにHMG−CoA還元酵素活性を測定することにより、ポリペプチドがmvaE活性を有しているか評価することができる。代表的な解析では、アセトアセチルCoAチオラーゼ活性は、アセトアセチルCoAの形成又はリオリシスに伴う吸光度の変化を分光光度計により302nmにて監視することで測定できる。アセトアセチルCoA合成を判定するための各反応に関する標準的な解析条件は、1mMのアセチルCoA、10mMのMgCl2、50mMのトリス(pH 10.5)というものであり、反応は酵素の添加により開始される。解析は最終用量200μLで実施できる。解析に関し、1酵素単位(eu)は、1分間で1μmolのアセトアセチルCoAを合成又はチオリシスすることを意味する。他の代表的な解析では、HMG−CoA還元酵素活性は、分光光度計により、340nmでのNADP(H)の出現又は消失をもとに監視することもできる。HMG−CoAのメバロン酸への還元的脱アシル化について測定した各反応の標準的な解析条件は、0.4mMのNADPH、1.0mMの(R,S)−HMG−CoA、100mMのKCl及び100mMのKxPO4(pH 6.5)というものである。解析は最終用量200μLで実施する。反応は酵素の添加により開始される。解析に関し、1euは、1分間で1μmolのNADP(H)が代謝回転されることを意味する。これは、0.5μmolのHMG−CoA又はメバロン酸が代謝回転することに相当する。
あるいは、組換え細胞におけるメバロン酸の産生量は、限定するものではないが、ガスクロマトグラフィー(米国特許出願公開番号第2005/0287655(A1)号を参照されたい)又はHPLC(米国特許出願第公開2011/0159557(A1)号を参照されたい)で測定することができる。代表的な解析の際には、1種以上の抗生物質を添加したLBブロスを入れた振とうチューブに培養物を接種し、34℃にて、250rpmで14時間インキュベートする。次に、1%グルコース、0.1%酵母エキス及び200μMのIPTGを添加したTM3培地を入れたウェルプレート中で、最終的なODが0.2になるよう培養物を希釈する。次に、プレートを、Breath Easier membrane(Diversified Biotech)でシールし、振とう/恒温器で、34℃にて、600rpmで24時間インキュベートした。次に各培養物1mLを3,000×gで5分遠心分離する。次に、上清に20%硫酸を添加し、氷上で5分インキュベートする。次に、混合物を3000×gで5分遠心分離し、HPLC解析のため上清を回収する。メバロン酸(Sigma)の標準曲線と比較して、試料中のメバロン酸濃度を測定する。更に、当該技術分野において既知の任意の手法によりグルコースオキシダーゼ解析を実施し、グルコース濃度を測定する。HPLCを使用し、各試料により得られた屈折率を、濃度既知の各種メバロン酸溶液のHPLCをもとに作成した検量線に対し比較して、メバロン酸濃度を定量することができる。
mvaE核酸の例としては、mvaEポリペプチド活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又はポリペプチド融合物をコードしている核酸が挙げられる。mvaEポリペプチド及び核酸の例としては、本明細書に記載の任意の生物資源から天然に生じるポリペプチド及び核酸、並びに本明細書に記載の任意の生物資源から誘導されるポリペプチド変異体及び核酸変異体が挙げられる。mvaE核酸の例としては、例えば、リステリア・グレイ(Listeria grayi)DSM 20601、エンテロコッカス・フェシウム(Enterococcus faecium)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)EG2、エンテロコッカス・フェカリス(Enterococcus faecalis)、及び/又はエンテロコッカス・カセリフラブス(Enterococcus casseliflavus)から単離したmvaE核酸が挙げられる。リステリア・グレイ(Listeria grayi)DSM 20601のmvaE遺伝子によりコードされるmvaE核酸は、配列番号6に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%配列同一性を有し得る。エンテロコッカス・フェシウム(Enterococcus faecium)のmvaE遺伝子によりコードされるmvaE核酸は、配列番号7に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%又は85%の配列同一性を有し得る。エンテロコッカス・ガリナラム(Enterococcus gallinarum)EG2のmvaE遺伝子によりコードされるmvaE核酸は、配列番号8に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%又は85%の配列同一性を有し得る。エンテロコッカス・カセリフラブス(Enterococcus casseliflavus)のmvaE遺伝子によりコードされるmvaE核酸は、配列番号9に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%又は85%の配列同一性を有し得る。エンテロコッカス・フェカリス(Enterococcus faecalis)のmvaE遺伝子によりコードされているmvaE核酸は、これまでにメバロン酸を産生するための大腸菌(E. coli)において開示されているmvaE遺伝子に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る(米国特許第2005/0287655 A1号;Tabata,K.and Hashimoto,S.−I.Biotechnology Letters 26:1487〜1491,2004を参照されたい)。
mvaE核酸核酸は、組換え細胞において、多コピープラスミド上で発現させることができる。プラスミドは高コピー数プラスミド、低コピー数プラスミド又は中程度コピー数プラスミドであってよい。あるいは、mvaE核酸は、宿主細胞の染色体に組み込むこともできる。プラスミド上の、あるいは宿主細胞染色体の一部に組み込まれた、mvaE核酸の異種発現に際し、核酸の発現は、誘導型プロモータ又は構成的に(恒常的に)発現するプロモータのいずれかにより駆動できる。プロモータは、mvaE核酸の発現を強力に駆動することができ、弱く駆動することができ、あるいは中程度に駆動することができる。
mvaSポリペプチド及び核酸例
mvaS遺伝子は、HMG−CoAシンターゼ活性を保有するポリペプチドをコードする。このポリペプチドは、アセトアセチルCoAを、3−ヒドロキシ−3−メチルグルタリル−CoA(HMG−CoA)を変換させることができる。mvaSポリペプチド及び核酸の例としては、本明細書に記載の任意の生物資源から天然に生じるポリペプチド及び核酸、並びにmvaSポリペプチドの活性を少なくとも1つ有する、本明細書に記載の任意の生物資源から誘導されるポリペプチド変異体及び核酸変異体が挙げられる。
変異型mvaSポリペプチドとしては、1つ以上のアミノ酸残基がアミノ酸置換を受けており、かつmvaSポリペプチド活性(すなわち、アセトアセチルCoAを3−ヒドロキシ−3−メチルグルタリル−CoAをメバロン酸に変換する能力)を保持しているものが挙げられる。mvaSポリペプチド中にアミノ酸置換を導入することで、分子の官能性を改良することができる。例えば、mvaSポリペプチドの、基質に対する結合親和性を増加させるアミノ酸置換、あるいはアセトアセチルCoAを3−ヒドロキシ−3−メチルグルタリル−CoAに変換する能力を改良するアミノ酸置換を、mvaSポリペプチドに導入することができる。一部の態様では、mvaSポリペプチド変異体は、1つ以上の保存的なアミノ酸置換を含有する。
Quant et al.(Biochem J.,1989,262:159〜164)に記載のものなどの標準法を使用して、HMG−CoAシンターゼの活性を測定することにより、ポリペプチドがmvaS活性を有するか評価することもできる。代表的な解析では、HMG−CoAシンターゼの活性は、303nmでの吸光度の変化を監視することにより、エノール形態のアセトアセチルCoAの消失を、分光光度を元に評価することで、検定できる。30℃下で、50mMのTris/HCl(pH 8.0)、10mMのMgCl2及び0.2mMのジチオスレイトールを含有する標準的な1mLの解析系に、5μLのアセチルリン酸、10,M−アセトアセチル−CoA及び5ulの抽出試料を加え、続いてアセチルCoA(100μM)及び10ユニットのPTAを同時に加えることができる。次に、HMG−CoAシンターゼの活性を、アセチルCoA添加前及び添加後の速度(rate)の差として測定する。使用した条件下(pH 8.0、10mM−MgCl2)での、アセトアセチルCoAの吸収係数は、12.2×103M−1cm−1である。定義によると、1ユニットの酵素活性により、1分あたり1μmolのアセトアセチルCoAが変換されることになる。
あるいは、組換え細胞におけるメバロン酸の産生量は、限定するものではないが、ガスクロマトグラフィー(米国特許出願公開番号第2005/0287655(A1)号を参照されたい)又はHPLC(米国特許出願第公開2011/0159557(A1)号を参照されたい)で測定することができる。代表的な解析の際には、1種以上の抗生物質を添加したLBブロスを入れた振とうチューブに培養物を接種し、34℃にて、250rpmで14時間インキュベートする。次に、1%グルコース、0.1%酵母エキス及び200μMのIPTGを添加したTM3培地を入れたウェルプレート中で、最終的なODが0.2になるよう培養物を希釈する。次に、プレートを、Breath Easier membrane(Diversified Biotech)でシールし、振とう/恒温器で、34℃にて、600rpmで24時間インキュベートした。次に各培養物1mLを3,000×gで5分遠心分離する。次に、上清に20%硫酸を添加し、氷上で5分インキュベートする。次に、混合物を3000×gで5分遠心分離し、HPLC解析のため上清を回収する。メバロン酸(Sigma)の標準曲線と比較して、試料中のメバロン酸濃度を測定する。更に、当該技術分野において既知の任意の手法によりグルコースオキシダーゼ解析を実施し、グルコース濃度を測定する。HPLCを使用し、各試料により得られた屈折率を、濃度既知の各種メバロン酸溶液のHPLCをもとに作成した検量線に対し比較して、メバロン酸濃度を定量することができる。
mvaS核酸の例としては、mvaSポリペプチド活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又はポリペプチド融合物をコードしている核酸が挙げられる。mvaSポリペプチド及び核酸の例としては、本明細書に記載の任意の生物資源から天然に生じるポリペプチド及び核酸、並びに本明細書に記載の任意の生物資源から誘導されるポリペプチド変異体及び核酸変異体が挙げられる。mvaS核酸の例としては、例えば、リステリア・グレイ(Listeria grayi)DSM 20601、エンテロコッカス・フェシウム(Enterococcus faecium)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)EG2、エンテロコッカス・フェカリス(Enterococcus faecalis)、及び/又はエンテロコッカス・カセリフラブス(Enterococcus casseliflavus)から単離したmvaS核酸が挙げられる。リステリア・グレイ(Listeria grayi)DSM 20601のmvaS遺伝子によりコードされるmvaS核酸は、配列番号10に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%又は85%の配列同一性を有し得る。エンテロコッカス・フェシウム(Enterococcus faecium)のmvaS遺伝子によりコードされるmvaS核酸は、配列番号11に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%又は85%の配列同一性を有し得る。エンテロコッカス・ガリナラム(Enterococcus gallinarum)EG2のmvaS遺伝子によりコードされるmvaS核酸は、配列番号12に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%又は85%の配列同一性を有し得る。エンテロコッカス・カセリフラブス(Enterococcus casseliflavus)のmvaS遺伝子によりコードされるmvaS核酸は、配列番号13に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%又は85%の配列同一性を有し得る。エンテロコッカス・フェカリス(Enterococcus faecalis)mvaS遺伝子によりコードされているmvaS核酸は、これまでにメバロン酸を産生するための大腸菌(E. coli)において開示されているmvaE遺伝子に対し、少なくとも約99%、98%、97%、96%、95%、95%、93%、92%、91%、90%、89%、88%、87%、86%、又は85%の配列同一性を有し得る(米国特許第2005/0287655 A1号;Tabata,K.and Hashimoto,S.−I.Biotechnology Letters 26:1487〜1491,2004を参照されたい)。
mvaS核酸核酸は、組換え細胞において、マルチコピープラスミド上で発現させることができる。プラスミドは高コピー数プラスミド、低コピー数プラスミド又は中程度コピー数プラスミドであってよい。あるいは、mvaS核酸は、宿主細胞の染色体に組み込むこともできる。プラスミド上の、あるいは宿主細胞染色体の一部に組み込まれた、mvaS核酸の異種発現に際し、核酸の発現は、誘導型プロモータ又は構成的に(恒常的に)発現するプロモータのいずれかにより駆動できる。プロモータは、mvaS核酸の発現を強力に駆動することができ、弱く駆動することができ、あるいは中程度に駆動することができる。
アセトアセチル−CoAシンターゼ遺伝子
アセトアセチルCoAシンターゼ遺伝子(nphT7としても知られる)は、マロニルCoA及びアセチルCoAからアセトアセチルCoAを合成する活性を有し、かつ2分子のアセチルCoAからアセトアセチルCoAを合成する活性は最小限である(例えば、非活性である)酵素をコードしている遺伝子である。例えば、Okamura et al.,PNAS Vol 107,No.25,pp.11265〜11270(2010)を参照されたい。この文献中のnphT7に関する教示は、本開示に明確に組み込まれる。日本国特許公開第2008−61506(A)号及び米国特許出願公開第2010/0285549号には、放線菌のストレプトマイセス属CL190株のアセトアセチル−CoAシンターゼ遺伝子が記載されている。アセトアセチル−CoAシンターゼも、アセチルCoA:マロニルCoAアシル基転移酵素として参照され得る。使用することのできる代表的なアセトアセチル−CoAシンターゼ(又はアセチルCoA:マロニルCoAアシル基転移酵素)としては、Genbank AB540131.1が挙げられる。
本開示に記載の任意の態様又は実施例では、マロニル−CoA及びアセチル−CoAからアセトアセチル−CoAを合成する能力を有する酵素を使用できる。このような酵素の非限定例を本明細書に記載する。本開示に記載の特定の実施形態では、マロニルCoA及びアセチルCoAからアセトアセチルCoAを合成する能力を有する放線菌のストレプトマイセス(Streptomyces)属から誘導されるアセトアセチルCoAシンターゼ遺伝子を使用できる。このようなアセトアセチルCoAシンターゼの遺伝子の一例としては、アミノを有するタンパク質をコードしている遺伝子が挙げられる。このような、配列番号5のアミノ酸配列を有するタンパク質は、マロニルCoA及びアセチルCoAからアセトアセチルCoAを合成する活性を有し、2分子のアセチルCoAからアセトアセチルCoAを合成する活性は有さないアセトアセチルCoAシンターゼに相当する。
一実施形態では、配列番号5のアミノ酸配列を有するタンパク質をコードしている遺伝子は、テンプレートとして、放線菌のストレプトマイセス(Streptomyces sp.)CL190株から得られるゲノムDNAと、日本国特許公開第2008−61506(A)号を参照して設計することのできるプレマー対と、を使用して核酸増幅法(例えば、PCR)により得ることができる。
本明細書に記載するとき、本発明での使用に関し、アセトアセチルCoAシンターゼ遺伝子は、放線菌のストレプトマイセス(Streptomyces sp.)CL190株に由来する配列番号5のアミノ酸配列を有するタンパク質をコードしている遺伝子に限定されない。マロニルCoA及びアセチルCoAからアセトアセチルCoAを合成することができかつ2分子のアセチルCoAからアセトアセチルCoAを合成することはできないタンパク質をコードしている任意の遺伝子を本発明に記載の方法に使用することができる。特定の実施形態では、アセトアセチルCoAシンターゼ遺伝子は、配列番号5のアミノ酸配列と類似性が高く又は実質的に同一であるアミノ酸配列を有しかつマロニルCoA及びアセチルCoAからアセトアセチルCoAを合成する機能を有するタンパク質をコードしている遺伝子であり得る。表現「類似性の高い」又は「実質的に同一」は、例えば、少なくとも約80%同一性、少なくとも約85%、少なくとも約90%、少なくとも約91%、少なくとも約92%、少なくとも約93%、少なくとも約94%、少なくとも約95%、少なくとも約96%、少なくとも約97%、少なくとも約98%、及び少なくとも約99%同一であることを意味する。上記で使用した通り、同一度の値は、異なるアミノ酸配列及び配列番号5のアミノ酸配列に含まれるアミノ酸残基間の同一度(%)に相当するものであり、配列類似性について検索するためのプログラムを使用し、配列番号5のアミノ酸配列と、異なるアミノ酸配列とを整列させることで算出される。
他の実施形態では、アセトアセチルCoAシンターゼ遺伝子は、1つ以上のアミノ酸の置換、欠失、付加又は挿入により、配列番号5のアミノ酸配列由来のアミノ酸配列を有しかつマロニルCoA及びアセチルCoAからアセトアセチルCoAを合成する機能を有するタンパク質をコードしている遺伝子であってよい。本明細書において、表現「1つ以上のアミノ酸(more amino acids)」は、例えば、2〜30個のアミノ酸、好ましくは2〜20個のアミノ酸、より好ましくは2〜10個のアミノ酸及び最も好ましくは2〜5個のアミノ酸を指す。
更に他の実施形態では、アセトアセチルCoAシンターゼ遺伝子は、厳密な条件下で配列番号5のアミノ酸配列をコードしているヌクレオチド配列と相補的であり、ヌクレオチド配列を有するポリヌクレオチドと一部又は完全にハイブリダイゼーションすることのできる、並びにマロニルCoA及びアセチルCoAからアセトアセチルCoAを合成する機能を有するタンパク質をコードすることのできる、ポリヌクレオチドからなる場合がある。本明細書において、厳密な条件下でのハイブリダイゼーションは、60℃で2回SSCで洗浄するという条件下で結合を維持させることに相当する。ハイブリダイゼーションは、J.Sambrook et al.「分子クローニング−実験室マニュアル第3版(Molecular Cloning,A Laboratory Manual,3rd Ed.)」,Cold Spring Harbor Laboratory(2001年)に記載の手法などの、既知の従来法により実施することができる。
本明細書に記載されるとおり、配列番号5のアミノ酸配列とは異なるアミノ酸配列を有するアセトアセチルCoAシンターゼをコードしている遺伝子は、任意の生物から単離できる可能性があり、例えば、ストレプトマイセス(Streptomyces sp.)CL190株ではない放線菌から得ることもできる。加えて、本明細書で用いるアセトアセチルCoAシンターゼ遺伝子は、配列番号5のアミノ酸配列をコードしているポリヌクレオチドを、当該技術分野において既知の手法により改変することで得ることもできる。ヌクレオチド配列への変異導入は、Kunkel法又はgapped duplex法あるいはこれらのいずれかに類似の手法などの既知の手法により実施することができる。例えば、変異導入は、部位特異的変異導入については、変異導入キット(例えば、製品名;Mutant−K及びMutant−G(タカラバイオ))及び製品名LA PCR in vitro Mutagenesisシリーズのキット(タカラバイオ)などを使用することで実施できる。
配列番号5のアミノ酸配列とは異なるアミノ酸配列を有するアセトアセチルCoAシンターゼの活性は、以下に記載のとおりに評価することができる。具体的には、評価されることになるタンパク質をコードしている遺伝子は、遺伝子を細胞中で発現させ、続いてクロマトグラフィーなどの手法によりタンパク質を精製することができるような様式で、まずは宿主細胞に導入される。得られた評価すべきタンパク質を含有させた緩衝液に、基質としてマロニルCoA及びアセチルCoAを加え、続いて例えば、所望の温度(例えば、10℃〜60℃)でインキュベートする。反応の完了後、基質の減少量及び/又は生成物量(アセトアセチルCoA)を測定する。したがって、試験したタンパク質がマロニルCoA及びアセチルCoAからアセトアセチルCoAを合成する機能を有するか評価すること、並びに合成度を評価することができる。このような場合では、このタンパク質が、2分子のアセチルCoAからアセトアセチルCoAを合成する活性を有しているか否かは、得られた評価すべきタンパク質を含有させた緩衝液に、アセチルCoAのみを基質として加え、基質の減少量及び/又は同様の方法において産生された産生物量を測定することにより、試験することができる。
メバロン酸の産生量を増加させることのできる組換え細胞
本明細書に記載の組換え細胞(例えば、組換え細菌細胞)は、本明細書に記載の各種酵素経路に対し何ら操作を施していない同様の細胞を上回る量及び/又は濃度でメバロン酸を産生することができる。したがって、本明細書に記載の各種経路において調節がなされるよう遺伝子操作された組換え細胞(例えば、細菌細胞)は、メバロン酸の産生を増強させるのに有用である。
適宜に、特定の態様では、本発明は、メバロン酸の産生を増強させることのできる組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸、及びMVA経路上流の1種以上のポリペプチドをコードしている1種以上の核酸を含み、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まない細胞と比較して多量にメバロン酸を産生する。
特定の態様では、本明細書に記載の組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の態様では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の態様では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
一態様では、組換え細胞は、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)、及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaSポリペプチドをコードしている異種核酸の1種以上のコピーを更に含む。別の実施形態では、組換え細胞は、アセトアセチルCoAシンターゼと、MVA経路上流の1種以上のポリペプチドをコードしている1種以上の核酸とを更に含む。
一態様では、組換え細胞には、次の、リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(tal B)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)からなる群から選択される遺伝子のうちの1つ以上の活性を増加させるよう、遺伝子操作を施すことができる。別の実施形態では、組換え細胞は、次の、グルコース−6−リン酸脱水素酵素(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトースビスリン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸脱水素酵素(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)といった遺伝子のうち、1つ以上の遺伝子の活性を減少させるよう更に遺伝子操作を施すことができる。
一態様では、本明細書に記載の組換え細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して、高比産生性でメバロン酸を産生し得る。特定の実施形態では、組換え細胞は、2.00g/L/h超のメバロン酸を産生し得る。あるいは、組換え細胞は、包括的に、約1.0g/L/h、1.2g/L/h、1.4g/L/h、1.6g/L/h、1.8g/L/h、2.0g/L/h、2.2g/L/h、2.4g/L/h、2.6g/L/h、2.8g/L/h、3.0g/L/h、3.2g/L/h、3.4g/L/h、3.6g/L/h、3.8g/L/h、4.0g/L/h、4.2g/L/h、4.4g/L/h、4.6g/L/h、4.8g/L/h、5.0g/L/h、5.2g/L/h、5.4g/L/h、5.6g/L/h、5.8g/L/h、6.0g/L/h超並びにこれらの数字の間の任意の数値でメバロン酸を産生することができる。
一態様では、本明細書に記載の組換え細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して、高力価でメバロン酸を産生し得る。組換え細胞は、48時間にわたる発酵後に、約100g/L超のピーク力価でメバロン酸を産生し得る。あるいは、組換え細胞は、48時間にわたる発酵後に、約50g/L、60g/L、70g/L、80g/L、90g/L、100g/L、110g/L、120g/L、130g/L、140g/L、150g/L、160g/L、170g/L、180g/L、190g/L、200g/L、210g/L、220g/L、230g/L、240g/L、250g/L、260g/L、270g/L、280g/L、290g/L、300g/L超並びにこれらの数字の間の任意の数値でメバロン酸を産生することができる。
他の実施形態では、本明細書に記載の組換え細胞は、MVA経路に取り込まれる炭素を増加させ、かつ同様の遺伝子操作をなされていない細胞と比較して高力価のメバロン酸を産生させ得る、1つ以上の変異を更に含む。このような実施形態では、本明細書に記載の組換え細胞は、ホスホケトラーゼ活性を有するホスホケトラーゼポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して、高ピーク力価でメバロン酸を産生し得る。一態様では、組換え細胞には、次の、リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(tal B)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)からなる群から選択される遺伝子のうちの1つ以上の活性を増加させるよう、更に遺伝子操作を施すことができる。別の実施形態では、組換え細胞は、次の、グルコース−6−リン酸脱水素酵素(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトースビスリン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸脱水素酵素(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)といった遺伝子のうち、1つ以上の遺伝子の活性を減少させるよう、更に遺伝子操作を施すことができる。
一態様では、本明細書に記載の組換え細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して、高細胞産生指数(CPI)でメバロン酸を産生し得る。組換え細胞は、メバロン酸に関し、少なくとも約3.0(g/g)のCPIを有し得る。あるいは、組換え細胞は、メバロン酸に関し、包括的に、少なくとも約1(g/g)、2(g/g)、3(g/g)、4(g/g)、5(g/g)、6(g/g)、7(g/g)、8(g/g)、9(g/g)、10(g/g)、11(g/g)、12(g/g)、13(g/g)、14(g/g)、15(g/g)、20(g/g)、25(g/g)又は30(g/g)のCPI並びにこれらの数値の間の任意の数値のCPIを有し得る。
特定の実施形態では、本明細書に記載の組換え細胞には、MVA経路に取り込まれる炭素を増加させ、かつ同様の遺伝子操作をなされていない細胞と比較して、メバロン酸に関しより高い細胞産生性指数(CPI)をもたらす1つ以上の変異(mutation)を更に含む。加えて、本明細書に記載の組換え細胞は、ホスホケトラーゼ活性を有するホスホケトラーゼポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して高CPIを有する。一態様では、組換え細胞には、次の、リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(tal B)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)からなる群から選択される遺伝子のうちの1つ以上の活性を増加させるよう、更に遺伝子操作を施すことができる。別の実施形態では、これらの組換え細胞には、次の、グルコース−6−リン酸脱水素酵素(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトースビスリン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸脱水素酵素(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)といった遺伝子のうち、1種以上の遺伝子の活性を減少させるよう更に遺伝子操作を施すことができる。
加えて、本明細書に記載の組換え細胞は、ホスホケトラーゼ活性を有するホスホケトラーゼポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して、高質量収率でグルコースからメバロン酸を産生する。組換え細胞は、少なくとも約28%の質量収率でグルコースからメバロン酸を産生できる。あるいは、組換え細胞は、少なくとも約25%、26%、27%、28%、29%、30%、31%、32%、33%、34%、35%、36%、37%、38%、39%、40%、41%、42%、43%、44%、45%、46%、47%、48%、49%、50%又は55%並びに包括的にこれらの数値の間の任意の数値の質量収率でグルコースからメバロン酸を産生できる。
特定の実施形態では、本明細書に記載の組換え細胞には、MVA経路に取り込まれる炭素量を増加させかつ同様の遺伝子操作をなされていない細胞と比較して、メバロン酸の質量収率が高くなる1つ以上の変異(mutation)を更に含む。加えて、本明細書に記載の組換え細胞は、ホスホケトラーゼ活性を有するホスホケトラーゼポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して、メバロン酸の高質量収率を有する。一態様では、組換え細胞には、次の、リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(tal B)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)からなる群から選択される遺伝子のうちの1つ以上の活性を増加させるよう、遺伝子操作を施すことができる。別の実施形態では、これらの組換え細胞には、次の、グルコース−6−リン酸脱水素酵素(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトースビスリン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸脱水素酵素(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)といった遺伝子のうち、1種以上の遺伝子の活性を減少させるよう更に遺伝子操作を施すことができる。
一態様では、本明細書に記載の組換え細胞はメバロン酸を産生し、かつホスホケトラーゼ活性を有するポリペプチドをコードしている異種核酸の1種以上のコピーを含まない同様の細胞と比較して発酵ブロスへの酢酸の蓄積が少ない。組換え細胞は、多量のメバロン酸を産生し、かつ48時間の発酵後の発酵ブロスへの酢酸の蓄積が4.5g/L未満であり得る。あるいは、組換え細胞は、メバロン酸を多量に産生することができ、かつ48時間の発酵後の発酵ブロスへの酢酸の蓄積は約8.0g/L、7.5g/L、7.0g/L、6.5g/L、6.0g/L、5.5g/L、5.0g/L、4.5g/L、4.0g/L、3.5g/L、3.0g/L、2.5g/L、2.0g/L、又は1.5g/L未満、並びにこれらの数値間の任意の数値である。特定の実施形態では、発酵ブロスへの酢酸の蓄積を減少させることにより、発酵中の細胞生存率を向上させることができる。
特定の実施形態では、本明細書に記載の組換え細胞は、結果として多量のメバロン酸が得られるようMVA経路への炭素の取り込みを増加させ、かつ同様の遺伝子操作をなされていない細胞と比較して、発酵ブロスへの酢酸の蓄積を減少させる1つ以上の変異を更に含む。特定の実施形態では、発酵ブロスへの酢酸の蓄積を減少させることにより、発酵中の細胞生存率を向上させることができる。
組換え細胞を使用して多量にメバロン酸を産生する方法
本明細書では、メバロン酸の産生方法も提供する。一部の態様では、メバロン酸の産生方法は、(a)本明細書に記載のとおりに炭素の取り込み量が増加するよう遺伝子操作された組換え細胞(上記の任意の組換え細胞)、又はメバロン酸を産生できるそれらの子孫細胞を含む、組成物を培養する工程並びに(b)メバロン酸を産生する工程、を包含する。一部の態様では、メバロン酸を産生させる方法は、メバロン酸の産生に好適な条件下で、本明細書に記載の任意の組換え細胞を培養する工程、並びに組換え細胞にメバロン酸を産生させる工程、を含む。一部の態様では、メバロン酸の産生方法は、メバロン酸を回収する工程を更に含む。
本明細書に記載するとき、メバロン酸の産生方法は、(a)ホスホケトラーゼポリペプチドを内因的に発現しない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞など)を培養する工程、細胞は、MVA経路の1種以上のペプチドを発現する1種以上の異種核酸とともに、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上のコピーを異種発現する;並びに(b)メバロン酸を産生する工程、を含む。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)由来のものである。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。加えて、組換え細胞を最少培地で培養した場合、組換え細胞は、MVA経路の1種以上のペプチドを発現する1種以上の異種核酸とともに、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)由来のホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上の異種コピーを含まない同様の細胞と比較して、高濃度でメバロン酸を産生し得る。特定の実施形態では、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)由来のホスホケトラーゼポリペプチドをコードしている異種核酸の1種以上のコピーは、宿主細胞の染色体に組み込まれた異種核酸である。
特定の実施形態では、ホスホケトラーゼポリペプチドは、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)に由来する。他の実施形態では、ホスホケトラーゼポリペプチドは、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)に由来する。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。加えて、組換え細胞を最少培地で培養した場合、組換え細胞は、MVA経路の1種以上のペプチドを発現する1種以上の異種核酸とともに、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上の異種コピーを含まない同様の細胞と比較して、高濃度のメバロン酸を産生し得る。特定の実施形態では、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている異種核酸の1種以上のコピーは、宿主細胞の染色体に組み込まれた異種核酸である。
メバロン酸産生に関係する本方法は、2.00g/L/h超のメバロン酸の体積産生性を有する細胞を使用してメバロン酸を産生させることができる。あるいは、組換え細胞は、包括的に、約1.0g/L/h、1.2g/L/h、1.4g/L/h、1.6g/L/h、1.8g/L/h、2.0g/L/h、2.2g/L/h、2.4g/L/h、2.6g/L/h、2.8g/L/h、3.0g/L/h、3.2g/L/h、3.4g/L/h、3.6g/L/h、3.8g/L/h、4.0g/L/h、4.2g/L/h、4.4g/L/h、4.6g/L/h、4.8g/L/h、5.0g/L/h、5.2g/L/h、5.4g/L/h、5.6g/L/h、5.8g/L/h、6.0g/L/h超並びにこれらの数字の間の任意の数値でメバロン酸を産生することができる。一部の態様では、メバロン酸の産生方法は、メバロン酸を回収する工程を更に含む。
他の実施形態では、メバロン酸の産生方法は、(a)ホスホケトラーゼポリペプチドを内因的に発現しない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞など)を培養する工程、細胞は、MVA経路の1種以上のペプチドを発現する1種以上の異種核酸とともに、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上のコピーを異種発現する;並びに(b)メバロン酸を産生させる工程、組換え細胞は、48時間の発酵後に、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上の異種コピーを含まない同様の細胞と比較して、高いピーク力価を有する、を包含し得る。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
メバロン酸を産生するための本発明の方法は、発酵開始から48時間後に、約100g/L超のピーク力価でメバロン酸を産生し得る細胞を使用し、メバロン酸を産生し得る。あるいは、組換え細胞は、48時間にわたる発酵後に、約50g/L、60g/L、70g/L、80g/L、90g/L、100g/L、110g/L、120g/L、130g/L、140g/L、150g/L、160g/L、170g/L、180g/L、190g/L、200g/L、210g/L、220g/L、230g/L、240g/L、250g/L、260g/L、270g/L、280g/L、290g/L、300g/L超並びにこれらの数字の間の任意の数値でメバロン酸を産生することができる。一部の態様では、メバロン酸の産生方法は、メバロン酸を回収する工程を更に含む。
他の実施形態では、メバロン酸の産生方法は、(a)ホスホケトラーゼポリペプチドを内因的に発現しない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞など)を培養する工程、細胞は、MVA経路の1種以上のペプチドを発現する1種以上の異種核酸とともに、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上のコピーを異種発現する;並びに(b)メバロン酸を産生させる工程、組換え細胞は、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上の異種コピーを含まない同様の細胞と比較して高CPIを有する、を包含し得る。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
メバロン酸産生に関する本方法は、メバロン酸のCPIが少なくとも約3.0(g/g)である細胞を使用することによりメバロン酸を産生することができる。あるいは、組換え細胞は、メバロン酸に関し、包括的に、少なくとも約1(g/g)、2(g/g)、3(g/g)、4(g/g)、5(g/g)、6(g/g)、7(g/g)、8(g/g)、9(g/g)、10(g/g)、11(g/g)、12(g/g)、13(g/g)、14(g/g)、15(g/g)、20(g/g)、25(g/g)又は30(g/g)のCPI並びにこれらの数値の間の任意の数値のCPIを有し得る。一部の態様では、メバロン酸の産生方法は、メバロン酸を回収する工程を更に含む。
特定の実施形態では、メバロン酸の産生方法は、(a)ホスホケトラーゼポリペプチドを内因的に発現しない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞など)を培養する工程、細胞は、MVA経路の1種以上のペプチドを発現する1種以上の異種核酸とともに、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上のコピーを異種発現する;並びに(b)メバロン酸を産生させる工程、組換え細胞は、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上の異種コピーを含まない同様の細胞と比較して減少している酸素の取り込み速度(OUR)を示す、を包含し得る。特定の実施形態では、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上の異種コピーを発現する組換え細胞は、ホスホケトラーゼを発現しない組換え細胞と比較した場合に、OURの、最大で1倍、2倍、3倍、4倍、5倍、6倍又は7倍の減少を示す。
本明細書では、メバロン酸の産生量を増加させるために上記の任意の細胞を使用する方法を提供する。細胞によるメバロン酸産生は、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を発現させることにより増加させることができる。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
メバロン酸の産生は、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を発現しないメバロン酸産生細胞によるメバロン酸産生と比較して、約1,000,000倍(例えば、約1〜約500,000倍、約1〜約50,000倍、約1〜約5,000倍、約1〜約1,000倍、約1〜約500倍、約1〜約100倍、約1〜約50倍、約5〜約100,000倍、約5〜約10,000倍、約5〜約1,000倍、約5〜約500倍、約5〜約100倍、約10〜約50,000倍、約50〜約10,000倍、約100〜約5,000倍、約200〜約1,000倍、約50〜約500倍、又は約50〜約200倍)だけ増加させることもできる。本明細書に記載の特定の実施形態では、宿主細胞には、MVA産生に取り込まれる炭素を増加させるよう更に遺伝子操作することができる。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
他の態様では、本明細書に記載の方法は、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を発現しないメバロン酸産生細胞によるメバロン酸の産生と比較して、少なくとも約5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、1倍、2倍、5倍、10倍、20倍、50倍、100倍、200倍、500倍、1000倍、2000倍、5000倍、10,000倍、20,000倍、50,000倍、100,000倍、200,000倍、500,000倍、又は1,000,000倍だけメバロン酸を増産することができるよう提供され得る。本明細書に記載の特定の実施形態では、宿主細胞には、MVA産生に取り込まれる炭素を増加させるよう更に遺伝子操作することができる。
加えて、より具体的な細胞培養条件を使用して、本明細書に記載の方法により細胞を培養することができる。例えば、一部の態様では、メバロン酸の産生方法は、(a)ホスホケトラーゼ遺伝子を内因的に有さない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞)を、34℃下で最少培地にて培養する工程であって、組換え細胞は、低〜通常コピー数のプラスミドにおいて、高発現型プロモータの調節下で、ホスホケトラーゼポリペプチドをコードしている異種遺伝子の1種以上のコピーを異種発現する、培養する工程と、(b)メバロン酸を産生させる工程と、を含む。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一部の態様では、メバロン酸の産生方法は、メバロン酸を回収する工程を更に含む。
イソプレン産生能を持つ組換え細胞
イソプレン(2−メチル−1,3−ブタジエン)は、多様な用途で使用される重要な有機化合物である。例えば、イソプレンは、合成ゴムの製造時など、数多くの化学組成物及びポリマーの合成時に、中間体又は出発物質として使用される。イソプレンはまた、多くの植物及び動物により天然に合成される重要な生体物質でもある。
イソプレンは、イソプレンシンターゼの触媒作用によりDMAPPから製造される。したがって、理論に束縛されるものではないが、上記の任意の組成物及び方法により、組換え細胞(細菌細胞など)において細胞によるメバロン酸産生量を増加させることは、より多量のイソプレンを産生させるのと同様であると考えられる。グルコースからのメバロン酸産生量のモル収率を上昇させ、適切な酵素活性濃度で、メバロン酸キナーゼ、ホスホメバロン酸キナーゼ、ジホスホメバロン酸デカルボキシラーゼ、イソペンテニル二リン酸イソメラーゼ(例えば、MVA経路下流)並びにイソプレン及びイソプレノイド産生に好適な他の酵素と組み合わせると、イソプレノイド前駆体、イソプレン分子及び/又はイソプレノイド(イソプレンを含む)のモル収率の上昇につながる。
本明細書に記載される通り、本発明は、イソプレンを産生することのできる組換え細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸と、(i)完全なMVA経路(すなわち、MVA経路上流、及びMVA経路下流)の1種以上のポリペプチドをコードしている1種以上の核酸及び(ii)イソプレンシンターゼポリペプチドをコードしている異種核酸と、を含み、細胞は、回収可能な量でイソプレンを産生することができる。特定の実施形態では、本発明は、イソプレンの産生を増強させることのできる組換え細胞であって、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸と、(i)完全なMVA経路の1種以上のポリペプチドをコードしている1種以上の核酸、及び(ii)イソプレンシンターゼポリペプチドをコードしている異種核酸と、を含む、細胞を提供し、細胞は、ホスホケトラーゼ活性を有するポリペプチドをコードしている1種以上の異種核酸を含まないイソプレン産生細胞と比較して多量にイソプレンを産生する。
イソプレン産生は、1つ以上の酵素経路を更に操作された、本明細書に記載の任意の組換え宿主細胞を使用することによりなすこともでき、この場合、酵素活性は、メバロン酸産生に取り込まれる炭素を増加させるよう調節される。本明細書に記載の、メバロン酸産生に取り込まれる炭素量が増加するよう操作された各種酵素経路を有する組換え細胞を、イソプレン産生に使用することができる。一態様では、組換え細胞には、次の、リブロース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(tal B)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)からなる群から選択される遺伝子のうちの1つ以上の活性を増加させるよう、遺伝子操作を施すことができる。別の実施形態では、これらの組換え細胞には、次の、グルコース−6−リン酸脱水素酵素(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトースビスリン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸脱水素酵素(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)といった遺伝子のうち、1種以上の遺伝子の活性を減少させるよう更に遺伝子操作を施すことができる。
MVA経路下流のポリペプチドをコードしている核酸
本発明の一部の態様では、本明細書に記載の組成物又は方法の任意のものに記載の細胞は、メバロン酸(MVA)経路下流のポリペプチドをコードしている核酸を1つ以上更に含む。一部の態様では、MVA経路下流のポリペプチドは内在性ポリペプチドである。一部の態様では、MVA経路下流のポリペプチドをコードしている内在性核酸は、調節可能なように構成型プロモータに連結される。一部の態様では、MVA経路下流のポリペプチドをコードしている内在性核酸は、調節可能なように誘導型プロモータに連結される。一部の態様では、MVA経路下流のポリペプチドをコードしている内在性核酸は、調節可能なように高発現型プロモータに連結される。特定の態様では、細胞は、野生型細胞と比較して、内在性MVA経路下流のポリペプチドが過剰発現するよう設計する。一部の態様では、MVA経路下流のポリペプチドをコードしている内在性核酸は、調節可能なように低発現型プロモータに連結される。
メバロン酸生合成経路の下流は、メバロン酸キナーゼ(MVK)、ホスホメバロン酸キナーゼ(PMK)及びジホスホメバロン酸デカルボキシラーゼ(MVD)を含む。一部の態様では、MVA経路の下流は、イソペンテニル二リン酸イソメラーゼ(IDI)を更に含む。本明細書に提供される細胞は、イソプレンシンターゼ、MVA経路上流の1種以上のポリペプチド及び/又はMVA経路下流の1種以上のポリペプチドをコードしている核酸を含み得る。MVA経路下流のポリペプチドは、(a)メバロン酸を5−ホスホメバロン酸ヘとリン酸化する酵素、(b)5−ホスホメバロン酸を5−ジホスホメバロン酸へと変換する酵素、(c)5−ジホスホメバロン酸をイソペンテニルピロリン酸に変換する酵素、のうちの任意の酵素であり得る。より具体的には、メバロン酸を5−ホスホメバロン酸にリン酸化する酵素は、メタノサルシナ・マゼイ(M. mazei)メバロン酸キナーゼ、ラクトバチルス(Lactobacillus)メバロン酸キナーゼポリペプチド、ラクトバチルス・サケイ(Lactobacillus sakei)メバロン酸キナーゼポリペプチド、酵母メバロン酸キナーゼポリペプチド、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)メバロン酸キナーゼポリペプチド、ストレプトコッカス(Streptococcus)メバロン酸キナーゼポリペプチド、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)メバロン酸キナーゼポリペプチド、ストレプトマイセス(Streptomyces)メバロン酸キナーゼポリペプチド、ストレプトマイセス(Streptomyces)CL190メバロン酸キナーゼポリペプチド、及びメタノコッコイド・バートニィ(M. Burtonii)メバロン酸キナーゼポリペプチドからなる群であってもよい。他の態様では、メバロン酸をメバロン酸5−リン酸へとリン酸化する酵素はメタノサルシナ・マゼイ(M. mazei)・メバロン酸キナーゼである。
一部の態様では、MVA経路下流のポリペプチドは異種ポリペプチドである。一部の態様では、細胞は、MVA経路下流のポリペプチドをコードしている異種核酸のコピーを1つ以上含む。一部の態様では、MVA経路下流のポリペプチドをコードしている異種核酸は、調節可能なように構成型プロモータに連結される。一部の態様では、MVA経路下流のポリペプチドをコードしている異種核酸は、調節可能なように誘導型プロモータに連結される。一部の態様では、MVA経路下流のポリペプチドをコードしている異種核酸は、調節可能なように高発現型プロモータに連結される。一部の態様では、MVA経路下流のポリペプチドをコードしている異種核酸は、調節可能なように低発現型プロモータに連結される。一部の態様では、異種MVA経路下流のポリペプチドは、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)、エンテロコッカス・フェカリス(Enterococcus faecalis)又はメタノサルシナ・マゼイ(Methanosarcina mazei)由来のポリペプチドである。
MVA経路下流のポリペプチドをコードしている核酸は、細胞のゲノムに組み込むことができ、あるいは細胞で安定的に発現させることができる。MVA経路下流のポリペプチドをコードしている核酸は更に、ベクター上に存在させてもよい。
MVA経路下流のポリペプチドの例としては、次の(i)メバロン酸キナーゼ(MVK);(ii)ホスホメバロン酸キナーゼ(PMK);(iii)ジホスホメバロン酸デカルボキシラーゼ(MVD);及び(iv)イソペンテニル二リン酸イソメラーゼ(IDI)も挙げられる。詳細には、下流のMVKポリペプチドは、メタノサルシナ(Methanosarcina)属由来のものであってよく、更に詳細には、下流のMVKポリペプチドは、メタノサルシナ・マゼイ(Methanosarcina mazei)由来のものであってよい。一部の実施形態では、下流のMVKポリペプチドは、メタノコッカシド・バートニイ(M. Burtonii)由来のものであってよい。MVA経路下流のポリぺプチドのその他の例は、米国特許出願公開第2010/0086978号に見出すことができ、この内容は、MVK経路下流のポリペプチド及びMVK経路下流のポリペプチド変異体に関し参照によりその全体が明示的に本明細書に組み込まれる。
特に、MVA経路下流のポリペプチドとしては、MVA経路下流のポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド及び融合ポリペプチドが挙げられる。MVA経路下流の核酸の例としては、MVA経路下流のポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又は融合ポリペプチドをコードしている核酸が挙げられる。MVA経路下流のポリペプチド及び核酸の例としては、本明細書に記載されるような任意の生物資源から天然に生じるポリペプチド及び核酸が挙げられる。更に、イソプレンの産生量を増加させるような、MVA経路下流のポリペプチド変異体も、良好に使用することができる。
一部の態様では、MVA経路下流のポリペプチドは、サッカロマイセス・セレヴィシエ(Saccharomyces cerevisiae)、エンテロコッカス・フェカリス(Enterococcus faecalis)又はメタノサルシナ・マゼイ(Methanosarcina mazei)由来のポリペプチドである。一部の態様では、MVKポリペプチドは、ラクトバチルス(Lactobacillus)メバロン酸キナーゼポリペプチド、ラクトバチルス・サケイ(Lactobacillus sakei)メバロン酸キナーゼポリペプチド、酵母メバロン酸キナーゼポリペプチド、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)メバロン酸キナーゼポリペプチド、ストレプトコッカス(Streptococcus)メバロン酸キナーゼポリペプチド、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)メバロン酸キナーゼポリペプチド、ストレプトマイセス(Streptomyces)メバロン酸キナーゼポリペプチド、ストレプトマイセス(Streptomyces)CL190メバロン酸キナーゼポリペプチド、メタノサルシナ・マゼイ(Methanosarcina mazei)メバロン酸キナーゼポリペプチド、及びメタノコッカシド・バートニイ(M. Burtonii)メバロン酸キナーゼポリペプチドからなる群から選択される。本開示のプロモータのうちのいずれか(例えば、本開示に記載され、本開示の実施例において識別される、誘導型プロモータ及び構成型プロモータなどのプロモータ)を使用して、本開示のいずれかのMVAポリペプチドの発現を駆動させることができる。
本明細書に記載の細胞のうち任意のものは、IDI核酸(例えば、IDIをコードしている内在性又は異種核酸)を含み得る。イソペンテニルジホスフェートイソメラーゼポリペプチド(イソペンテニル−ジホスフェートδ−イソメラーゼ又はIDI)は、イソペンテニルジホスフェート(IPP)及びジメチルアリル二リン酸(DMAPP)の相互変換を触媒する(例えば、IPPをDMAPPへと変換し及び/又はDMAPPをIPPヘと変換する)。例示的なIDIポリペプチドとしては、IDIポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、及び融合ポリペプチドが挙げられる。標準法(本明細書に記載されるものなど)を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがIPP及びDMAPPを相互変換する能力を測定することで、ポリペプチドがIDIポリペプチド活性を有するか否かを判定することができる。IDI核酸の例としては、IDIポリペプチド活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又はポリペプチド融合物をコードしている核酸が挙げられる。IDIポリペプチド及び核酸の例としては、本明細書に記載の任意の生物資源から天然に生じるポリペプチド及び核酸、並びに本明細書に記載の任意の生物資源から誘導されるポリペプチド変異体及び核酸変異体が挙げられる。
イソプレンシンターゼポリペプチドをコードしている核酸
本発明の一部の態様では、本明細書に記載の任意の組成物又は方法に記載の細胞(本明細書に記載されるとおりにMVA経路への炭素の取り込みを増加させるよう遺伝子操作された宿主細胞など)は、イソプレンシンターゼポリペプチド又はイソプレンシンターゼ活性を有するポリペプチドをコードしている1種以上の核酸を更に含む。一部の態様では、イソプレンシンターゼポリペプチドは内在性ポリペプチドである。一部の態様では、イソプレンシンターゼポリペプチドをコードしている内在性核酸は、構成型プロモータに調節可能なように連結される。一部の態様では、イソプレンシンターゼポリペプチドをコードしている内在性核酸は、誘導型プロモータに調節可能なように連結される。一部の態様では、イソプレンシンターゼポリペプチドをコードしている内在性核酸は、高発現型プロモータに調節可能なように連結される。特定の態様では、細胞は、野生型細胞と比較して、経路の内在性イソプレンシンターゼポリペプチドを過剰発現するよう遺伝子操作する。一部の態様では、イソプレンシンターゼポリペプチドをコードしている内在性核酸は、低発現型プロモータに調節可能なように連結される。一部の態様では、イソプレンシンターゼポリペプチドは、クズ(Pueraria)又はハコヤナギ(Populus)、又はウラジロハコヤナギとヤマナラシの交雑種(Populus alba x Populus tremula)のような交雑種に由来のポリペプチドである。
一部の態様では、イソプレンシンターゼポリペプチドは異種ポリペプチドである。一部の態様では、細胞は、イソプレンシンターゼポリペプチドをコードしている異種核酸のコピーを1つ以上含む。一部の態様では、イソプレンシンターゼポリペプチドをコードしている異種核酸は、構成型プロモータに調節可能なように連結される。一部の態様では、イソプレンシンターゼポリペプチドをコードしている異種核酸は、誘導型プロモータに調節可能なように連結される。一部の態様では、イソプレンシンターゼポリペプチドをコードしている異種核酸は、高発現型プロモータに調節可能なように連結される。一部の態様では、イソプレンシンターゼポリペプチドをコードしている異種核酸は、低発現型プロモータに調節可能なように連結される。
イソプレンシンターゼポリペプチドをコードしている核酸は、宿主細胞のゲノムに組み込むことができ、あるいは細胞で安定的に発現させることができる。イソプレンシンターゼポリペプチドをコードしている核酸は、更にベクターに組み込むこともできる。
代表的なイソプレンシンターゼの核酸としては、イソプレンシンターゼポリペプチドの活性を少なくとも1種有するポリペプチド、ポリペプチド断片、ペプチド、又は融合ポリペプチドをコードする核酸が挙げられる。イソプレンシンターゼポリペプチドは、ジメチルアリールジホスフェート(DMAPP)をイソプレンに変換する。代表的なイソプレンシンターゼポリペプチドとしては、イソプレンシンターゼポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、並びに融合ポリペプチドが挙げられる。イソプレンシンターゼポリペプチド及び核酸の例としては、本明細書に記載されるような任意の生物資源から天然に生じるポリペプチド及び核酸が挙げられる。加えて、イソプレンシンターゼ変異体は、酵素活性が向上しているなどして、活性が向上されていてよい。一部の態様では、イソプレンシンターゼ変異体は、安定性(例えば、熱安定性)が改良されている、及び/又は溶解性が改良されているなどして、その他の特性が改良されている。
インビトロで、細胞抽出物中で又はインビボで、ポリペプチドがDMAPPをイソプレンへと変換する能力を測定して、ポリペプチドがイソプレンシンターゼポリペプチド活性を有するか否かを判定する際には、標準法を使用できる。細胞抽出物中のイソプレンシンターゼポリペプチド活性は、例えば、Silver et al.,J.Biol.Chem.270:13010〜13016,1995に記載のとおりに測定することができる。代表的な解析では、DMAPP(Sigma)は窒素流下で濃縮させ、乾燥させ、100mMリン酸カリウム緩衝液(pH 8.2)を用い100mMに再水和し、−20℃で保存する。解析を実施するために、金属製スクリューキャップと、テフロンコーティングのなされたシリコン製隔壁(Agilent Technologies)とを取り付けた20mLヘッドスペースバイアル内で、5μLの1M MgCl2、1mM(250μg/mL)DMAPP、65μLの植物抽出緩衝液(PEB)(50mM Tris−HCl(pH 8.0)、20mM MgCl2、5%グリセロール、及び2mM DTT)に、25μL細胞抽出物を加え、振盪させながら37℃で15分間培養した。200μLの250mM EDTAを加えて反応をクエンチさせ、GC/MSにより定量することができる。
一部の態様では、イソプレンシンターゼポリペプチドは、植物のイソプレンシンターゼポリペプチド又はそれらの変異体である。一部の態様では、イソプレンシンターゼポリペプチドは、プエラリア(Pueraria)に由来するイソプレンシンターゼ又はそれらの変異体である。一部の態様では、イソプレンシンターゼポリペプチドは、ハコヤナギ属(Populus)由来のイソプレンシンターゼ又はそれらの変異体である。一部の態様では、イソプレンシンターゼポリペプチドは、ポプラ(poplar)のイソプレンシンターゼポリペプチド又はそれらの変異体である。一部の態様では、イソプレンシンターゼポリペプチドはクズのイソプレンシンターゼポリペプチド又はそれらの変異体である。一部の態様では、イソプレンシンターゼポリペプチドは、クズ属(Pueraria)又はハコヤナギ属(Populus)、ウラジロハコヤナギとヤマナラシの交雑種(Populus alba x Populus tremula)、又はこれらの(変異体(variant)由来のポリペプチドである。
一部の態様では、イソプレンシンターゼポリペプチド又は核酸は、マメ科、例えば、マメ亜科(Faboideae)由来のものである。特定の態様では、イソプレンシンターゼポリペプチド又は核酸は、プエラリア・モンタナ(Pueraria montana)(クズ)(Sharkey et al.,、Plant Physiology 137:700〜712,2005)、プエラリア・ロバタ(Pueraria lobata)、ポプラ(ウラジロハコヤナギ(Populus alba)、セイヨウハコヤナギ(Populus nigra)、コットンウッド(Populus trichocarpa)、又はウラジロハコヤナギ(Populus alba)xトレムラ(tremula)(CAC35696)Milleret al.、Planta 213:483〜487,2001)、アスペン(aspen)(ヤマナラシなど)Silver et al.,JBC 270(22):13010〜1316,1995)、又はヨーロッパナラ(Quercus robur)(Zimmeret al.,国際特許出願公開第98/02550号)に由来するポリペプチド又は核酸である。一部の態様では、イソプレンシンターゼポリペプチドは、プエラリア・モンタナ(Pueraria montana)、プエラリア・ロバタ(Pueraria lobata)、ポプラ・トレムロイド(Populus tremuloides)、ウラジロハコヤナギ(Populus alba)、ポプラ・ニグラ(Populus nigra)又はポプラ・トリコカルパ(Populus trichocarpa)由来のイソプレンシンターゼ、又はこれらの変異体である。一部の態様では、イソプレンシンターゼポリペプチドは、ウラジロハコヤナギ(Populus alba)由来のイソプレンシンターゼであるか、又はこれらの変異体である。一部の態様では、イソプレンシンターゼ(例えば、ウラジロハコヤナギ(Populus alba)又はこれらの変異体由来のイソプレンシンターゼ)をコードしている核酸は、コドンを最適化されている。
一部の態様では、イソプレンシンターゼ核酸又はポリペプチドは、天然に生じるポリペプチド又は核酸である(例えば、ハコヤナギ属から天然に生じるポリペプチド又は核酸)。一部の態様では、イソプレンシンターゼ核酸又はポリペプチドは、野生型又は天然に生じるポリペプチド又は核酸ではない。一部の態様では、イソプレンシンターゼ核酸又はポリペプチドは、野生型又は天然に生じるポリペプチド又は核酸の変異体である(例えば、ハコヤナギ属の野生型又は天然に生じるポリペプチド又は核酸の変異体)。
一部の態様では、イソプレンシンターゼポリペプチドは変異体である。一部の態様では、イソプレンシンターゼポリペプチドは、野生型又は天然型イソプレンシンターゼの変異体である。一部の態様では、変異体は、野生型又は天然型イソプレンシンターゼと比較して触媒活性が向上しているなど、活性が向上している。活性(例えば、触媒活性)の向上は、少なくとも約10%、20%、30%、40%、50%、60%、70%、80%、90%、又は95%のうちのいずれかの程度であり得る。一部の態様では、触媒活性などの活性の向上は、少なくとも約1倍、2倍、5倍、10倍、20倍、30倍、40倍、50倍、75倍、又は100倍のいずれかである。一部の態様では、触媒活性などの活性の向上は、約10%〜約100倍である(例えば、約20%〜約100倍、約50%〜約50倍、約1倍〜約25倍、約2倍〜約20倍、又は約5倍〜約20倍)。一部の態様では、変異体は、野生型又は天然型イソプレンシンターゼと比較して可溶性が向上している。可溶性の向上は、少なくとも約10%、20%、30%、40%、50%、60%、70%、80%、90%、又は95%のうちのいずれかの程度であり得る。可溶性の向上は、少なくとも約1倍、2倍、5倍、10倍、20倍、30倍、40倍、50倍、75倍、又は100倍のうちのいずれかの程度であり得る。一部の態様では、溶解性の向上は、約10%〜約100倍のうちのいずれかの程度であり得る(例えば、約20%〜約100倍、約50%〜約50倍、約1倍〜約25倍、約2倍〜約20倍、又は約5倍〜約20倍)。一部の態様では、イソプレンシンターゼポリペプチドは、天然型イソプレンシンターゼ変異体であり、かつ天然型イソプレンシンターゼと比較して安定性が向上している(熱安定性など)。
一部の態様では、変異体は、野生型又は天然型イソプレンシンターゼの活性の、少なくとも約10%、少なくとも約20%、少なくとも約30%、少なくとも約40%、少なくとも約50%、少なくとも約60%、少なくとも約70%、少なくとも約80%、少なくとも約90%、少なくとも約100%、少なくとも約110%、少なくとも約120%、少なくとも約130%、少なくとも約140%、少なくとも約150%、少なくとも約160%、少なくとも約170%、少なくとも約180%、少なくとも約190%、少なくとも約200%の活性を有する。変異体は、野生型又は天然型イソプレンシンターゼとの配列類似性を保有し得る。一部の態様では、野生型又は天然型イソプレンシンターゼの変異体は、野生型又は天然型イソプレンシンターゼのアミノ酸配列と、少なくとも約40%、50%、60%、70%、75%、80%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、又は99.9%のうちのいずれかの程度で配列相同性を有する。一部の態様では、野生型又は天然型イソプレンシンターゼの変異体は、野生型又は天然型イソプレンシンターゼのアミノ酸配列と、約70%〜約99.9%、約75%〜約99%、約80%〜約98%、約85%〜約97%、又は約90%〜約95%のうちのいずれかの程度で配列相同性を有する。
一部の態様では、変異体は、野生型又は天然型イソプレンシンターゼ内に変異を持つ。一部の態様では、変異体は、少なくとも1箇所にアミノ酸置換、少なくとも1箇所にアミノ酸挿入、及び/又は少なくとも1箇所にアミノ酸欠失を有する。一部の態様では、変異体は、少なくとも1箇所にアミノ酸置換を有する。一部の態様では、変異体と、野生型又は天然型イソプレンシンターゼとの間で異なっているアミノ酸残基の数は、1以上であってよく、例えば、1、2、3、4、5、10、15、20、30、40、又は50以上のアミノ酸残基が異なっていてよい。天然型イソプレンシンターゼとしては、植物由来の任意のイソプレンシンターゼを挙げることができ、例えば、クズ(kudzu)イソプレンシンターゼ、ポプラ(poplar)イソプレンシンターゼ、ヨーロッパナラ(English oak)イソプレンシンターゼ、及びヤナギ(willow)イソプレンシンターゼが挙げられる。一部の態様では、変異体は、ウラジロハコヤナギ(Populus alba)由来のイソプレンシンターゼの変異体である。一部の態様では、ウラジロハコヤナギ(Populus alba)由来のイソプレンシンターゼ変異体は、少なくとも一箇所にアミノ酸置換、少なくとも1箇所にアミノ酸挿入、及び/又は少なくとも1箇所にアミノ酸欠失を有する。一部の態様では、変異体は、トランケート型のウラジロハコヤナギ(Populus alba)イソプレンシンターゼである。一部の態様では、変異体(例えば、ウラジロハコヤナギ(Populus alba)由来のイソプレンシンターゼの変異体)をコードしている核酸は、コドンを最適化させたものである(例えば、異種イソプレンシンターゼを発現させる宿主細胞に基づきコドンを最適化している)。
本開示のイソプレンシンターゼポリペプチドは、国際公開第2009/132220号、同第2010/124146号、米国特許出願第2010/0086978号に記載のいずれかのイソプレンシンターゼ又はイソプレンシンターゼ変異体であってよい。これらの文献に記載される、イソプレンシンターゼ及びイソプレンシンターゼ変異体に関係するすべての内容は、参照により本開示に明示的に組み込む。
本開示のプロモータのうちのいずれか(例えば、本開示に記載され、本開示の実施例において識別される、誘導型プロモータ及び構成型プロモータなどのプロモータ)を使用して、本開示のいずれかのイソプレンシンターゼの発現を駆動させることができる。
好適なイソプレンシンターゼとしては、限定するものではないが、Genbank受託番号AY341431、AY316691、AY279379、AJ457070、及びAY182241として識別されるものが挙げられる。本開示のイソプレンシンターゼをコードしている細胞の製造法を含む組成物又は方法のうちのいずれか1つに使用することのできるイソプレンシンターゼの種類は、国際公開第2009/076676号、同第2010/003007号、同第2009/132220号、同第2010/031062号、同第2010/031068号、同第2010/031076号、同第2010/013077号、同第2010/031079号、同第2010/148150号、同第2010/124146号、同第2010/078457号、同第2010/148256号、同第2012/058494号、及び米国特許第8,173,410号にも記載されている。
DXP経路のポリペプチドをコードしている核酸
本発明の一部の態様では、本明細書に記載の任意の組成物又は方法に記載の細胞(本明細書に記載されるとおりにMVA経路への炭素の取り込みを増加させるよう遺伝子操作された宿主細胞など)は、DXSポリペプチド又はDXP経路の他のポリペプチドをコードしている1種以上の異種核酸を更に含む。一部の態様では、細胞は更に、DXSポリペプチド又は他のDXP経路ポリペプチドをコードしている内在性の核酸の染色体コピーを含む。一部の態様では、大腸菌(E. coli)細胞は更に、IDIポリペプチド及びDXSポリペプチド又は他のDXP経路ポリペプチドをコードしている核酸を1つ以上含む。一部の態様では、核酸は、イソプレンシンターゼ、IDIポリペプチド及びDXSポリペプチド又は他のDXP経路ポリペプチドをコードしている。一部の態様では、1つのプラスミドは、イソプレンシンターゼ、IDIポリペプチド及びDXSポリペプチド又は他のDXP経路ポリペプチドをコードしている。一部の態様では、複数のプラスミド(multiple plasmids)は、イソプレンシンターゼ、IDIポリペプチド及びDXSポリペプチド又は他のDXP経路ポリペプチドをコードしている。
例示的なDXSポリペプチドとしては、DXSポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、及び融合ポリペプチドが挙げられる。標準法(本明細書に記載されるものなど)を用い、ポリペプチドが、インビトロで、細胞抽出物中で又はインビボでピルビン酸及びD−グリセルアルデヒド−3−リン酸を1−デオキシ−D−キシルロース−5−リン酸ヘと変換する能力を測定することで、ポリペプチドがDXSポリペプチド活性を有するか否かを判定することができる。DXSポリぺプチド及び核酸、並びにDXS活性の測定方法の例は、国際公開出願第2009/076676号、国際公開第2010/003007号、同第2009/132220号、及び米国特許出願公開第2009/0203102号、同第2010/0003716号、及び同第2010/0048964号においてより詳細に記載される。
DXP経路に含まれるポリペプチドの例としては、限定するものではないが、次の任意のポリペプチド:DXSポリペプチド、DXRポリペプチド、MCTポリペプチド、CMKポリペプチド、MCSポリペプチド、HDSポリペプチド、HDRポリペプチド、及びDXP経路のポリペプチドの活性を1つ、又は2つ以上有するDXP経路ポリペプチド(例えば、融合ポリペプチド)、が挙げられる。特に、DXP経路ポリペプチドとしては、DXP経路のポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド及び融合ポリペプチドが挙げられる。例示的なDXP経路に関する核酸としては、DXP経路のポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又は融合ポリペプチドをコードしている核酸が挙げられる。DXP経路に含まれる代表的なポリペプチド及び核酸としては、本明細書に記載の任意の生物資源から天然に生じるポリペプチド及び核酸、並びに本明細書に記載の任意の生物資源から誘導されるポリペプチド変異体及び核酸変異体が挙げられる。DXP経路のポリペプチド及び核酸の例としては、並びにDXP経路のポリペプチド活性を測定する方法については、国際公開第2010/148150号に、より詳細に記載されている。
例示的なDXSポリペプチドとしては、DXSポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、及び融合ポリペプチドが挙げられる。標準法(本明細書に記載されるものなど)を用い、ポリペプチドが、インビトロで、細胞抽出物中で又はインビボでピルビン酸及びD−グリセルアルデヒド−3−リン酸を1−デオキシ−D−キシルロース−5−リン酸ヘと変換する能力を測定することで、ポリペプチドがDXSポリペプチド活性を有するか否かを判定することができる。DXSポリぺプチド及び核酸、並びにDXS活性の測定方法の例は、国際公開出願第2009/076676号、国際公開第2010/003007号、同第2009/132220号、及び米国特許出願公開第2009/0203102号、同第2010/0003716号、及び同第2010/0048964号においてより詳細に記載される。
特に、DXSポリペプチドは、ピルビン酸及びD−グリセルアルデヒド3−リン酸を1−デオキシ−d−キシルロース5−リン酸(DXP)ヘと変換する。標準法を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがピルビン酸及びD−グリセルアルデヒド−3−リン酸を変換する能力を測定することで、ポリペプチドがDXSポリペプチド活性を有するか否かを判定することができる。
DXRポリペプチドは、1−デオキシ−D−キシルロース5−リン酸(DXP)を2−C−メチル−D−エリスリトール4−リン酸(MEP)ヘと変換する。標準法を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがDXPを変換する能力を測定することで、ポリペプチドがDXRポリペプチド活性を有するか否かを判定することができる。
MCTポリペプチドは、2−C−メチル−D−エリスリトール4−リン酸(MEP)を4−(シチジン5’−ジホスホ)−2−メチル−D−エリスリトール(CDP−ME)ヘと変換する。標準法を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがMEPを変換する能力を測定することで、ポリペプチドがMCTポリペプチド活性を有するか否かを判定することができる。
CMKポリペプチドは、4−(シチジン5’−ジホスホ)−2−C−メチル−D−エリスリトール(CDP−ME)を2−ホスホ−4−(シチジン5’−ジホスホ)−2−C−メチル−D−エリスリトール(CDP−MEP)ヘと変換する。標準法を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがCDP−MEを変換する能力を測定することで、ポリペプチドがCMKポリペプチド活性を有するか否かを判定することができる。
MCSポリペプチドは、2−ホスホ−4−(シチジン5’−ジホスホ)−2−C−メチル−D−エリスリトール(CDP−MEP)を2−C−メチル−D−エリスリトール2,4−シクロジホスフェート(ME−CPP又はcMEPP)ヘと変換する。標準法を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがCDP−MEPを変換する能力を測定することで、ポリペプチドがMCSポリペプチド活性を有するか否かを判定することができる。
HDSポリペプチドは、2−C−メチル−D−エリスリトール2,4−シクロジホスフェートを(E)−4−ヒドロキシ−3−メチルブタ−2−エン−1−イルジホスフェート(HMBPP又はHDMAPP)ヘと変換する。標準法を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがME−CPPを変換する能力を測定することで、ポリペプチドがHDSポリペプチド活性を有するか否かを判定することができる。
HDRポリペプチドは、(E)−4−ヒドロキシ−3−メチルブタ−2−エン−1−イルジホスフェートをイソペンテニルジホスフェート(IPP)及びジメチルアリールジホスフェート(DMAPP)ヘと変換する。標準法を用い、インビトロで、細胞抽出物中で又はインビボでポリペプチドがHMBPPを変換する能力を測定することで、ポリペプチドがHDRポリペプチド活性を有するか否かを判定することができる。
MVA経路下流、イソプレンシンターゼ、IDI及びDXP経路のポリペプチドに関する生物資源
イソプレンシンターゼ、IDI、DXP経路及び/又はMVA経路下流の核酸(及びそれらにコードされるポリペプチド)は、イソプレンシンターゼ、IDI、DXP経路及び/又はMVA経路下流の核酸を天然に含有する任意の生物から得ることができる。イソプレンは、細菌、酵母、植物及び動物などの様々な生物により天然に生成される。一部の生物は、イソプレンの産生に関係するMVA経路を含有する。イソプレンシンターゼの核酸は、例えば、イソプレンシンターゼを含有する任意の生物から得ることができる。したがってMVA経路の核酸は、例えば、MVA経路を含有する任意の生物から得ることができる。IDI及びDXP経路の核酸は、例えば、IDI及びDXP経路を含有する任意の生物から得ることができる。
イソプレンシンターゼ、DXP経路、IDI、及び/又はMVA経路の核酸の核酸配列は、細菌、真菌、植物、藻類、又はシアノバクテリアから単離することができる。例示的な生物資源としては、例えば、酵母、例えばサッカロマイセス(Saccharomyces)種(例えば、サッカロマイセス・セレヴィシエ(S. cerevisiae))、細菌、例えば、エシェリキア(Escherichia)種(例えば、大腸菌)、又はメタノサルシナ(Methanosarcina)種(例えば、メタノサルシナ・マゼイ(Methanosarcina mazei))、植物、例えば葛又はポプラ(例えば、ウラジロハコヤナギ(Populus alba)又はウラジロハコヤナギxトレムラCAC35696(Populus alba x tremula CAC35696))又はヤマナラシ(例えば、アメリカヤマナラシ(Populus tremuloides))が挙げられる。イソプレンシンターゼ、IDI及び/又はMVA経路ポリペプチドに関係し、使用することのできる資源例は、同様に、国際公開第2009/076676号、同第2010/003007号、同第2009/132220号、同第2010/031062号、同第2010/031068号、同第2010/031076号、同第2010/013077号、同第2010/031079号、同第2010/148150号、同第2010/078457号、及び同第2010/148256号に記載されている。
一部の態様では、生物資源は酵母であり、例えば、酵母菌(Saccharomyces sp.)、シゾサッカロミセス(Schizosaccharomyces sp.)、ピキア(Pichia sp)又はカンジダ(Candida sp.)である。
一部の態様では、宿主細胞は、バチルス・リケノフォルミス(B. lichenoformis)又はバチルス・サブチリス(B. subtilis)などのバチルス株、シュードアルテロモナス・シトレア(P. citrea)などのパントエア(Pantoea)株、シュードモナス・アルカリゲネス(P. alcaligenes)などのシュードモナス属(Pseudomonas)株、ストレプトマイセス・リビダンス(S. lividans)又はストレプトマイセス・ルビギノーサス(S. rubiginosus)などのストレプトマイセス(Streptomyces)株、大腸菌(E. coli)などのエシェリキア(Escherichia)株、エンテロバクター株(Enterobacter)、ストレプトコッカス(Streptococcus)株、又はメタノサルシナ・マゼイ(Methanosarcina mazei)などの古細菌株である。
本明細書で使用するとき、「バチルス(Bacillus)属」としては、当業者に既知の「バチルス(Bacillus)」属のすべての種を包含し、限定するものではないが、例えば、バチルス・サブチリス(B. subtilis)、バチルス・リケニフォルミス(B. licheniformis)、バチルス・レンタス(B. lentus)、バチルス・ブレビス(B. brevis)、バチルス・ステアロサーモフィルス(B. stearothermophilus)、バチルス・アルカロフィルス(B. alkalophilus)、バチルス・アミロリケファシエンス(B. amyloliquefaciens)、バチルス・クラウシイ(B. clausii)、バチルス・ハロデュランス(B. halodurans)、バチルス・メガテリウム(B. megaterium)、バチルス・コアギュランス(B. coagulans)、バチルス・サークランス(B. circulans)、バチルス・ロータス(B. lautus)、及びバチルス・チューリンゲンシス(B. thuringiensis)が挙げられる。バチルス属は分類上の再編成を受け続けるものと認識される。したがって、この属は、再分類された種を包含することを意図し、例えば、限定するものではないが、現在は「ゲオバチルス・ステアロサーモフィルス」と命名されたバチルス・ステアロサーモフィルス(B. stearothermophilus)のような生物を包含する。酸素存在下での耐性内生胞子の生成はバチルス属の決定的特徴と考えられるが、この特徴は最近命名されたアリシクロバチルス(Alicyclobacillus)、アムピバチルス(Amphibacillus)、アネウリニバチルス(Aneurinibacillus)、アノキシバチルス(Anoxybacillus)、ブレビバチルス(Brevibacillus)、フィロバチルス(Filobacillus)、グラシリバチルス(Gracilibacillus)、ハロバチルス(Halobacillus)、パエニバチルス(Paenibacillus)、サリバチルス(Salibacillus)、サーモバチルス(Thermobacillus)、ウレイバチルス(Ureibacillus)、及びバルジバチルス(Virgibacillus)にも当てはまる。
一部の態様では、生物資源はグラム陽性細菌である。非限定的な例としては、ストレプトマイセス(treptomyces)株(例えば、ストレプトマイセス・リビダンス(S. lividans)、ストレプトマイセス・コエリカラー(S. coelicolor)、又はストレプトマイセス・グリセウス(S. griseus))及びバチルス株が挙げられる。一部の態様では、生物資源は、大腸菌(E. coli)又はシュードモナス属(Pseudomonas sp.)などのグラム陰性細菌である。
一部の態様では、微生物源は植物であり、例えば、マメ科(Fabaceae)、例えばマメ亜科(Faboideae)などの植物である。一部の態様では、生物資源はクズ、ポプラ(例えば、ウラジロハコヤナギ(Populus alba)×トレムラ(tremula)CAC 35696など)、アスペン(例えば、アメリカヤマナラシ(Populus tremuloides))、又はヨーロッパナラ(Quercus robur)である。
一部の態様では、微生物源は、藻類、例えば緑藻、紅藻、灰色藻、クロララクニオン藻、ミドリムシ目、クロミスタ、又は渦鞭毛藻類である。
一部の態様では、生物資源は、形態に基づき次の群:クロオコッカス(Chroococcales)、プレウロカプサ(Pleurocapsales)、ユレモ(Oscillatoriales)、ネンジュモ(Nostocales)、又はスティゴネマ(Stigonematales)のいずれかに分類される、シアノバクテリアである。
イソプレンの産生量を増大させることのできる組換え細胞
本明細書に記載の組換え細胞(本明細書に記載されるとおりに炭素の取り込みを増加させるよう遺伝子操作された宿主細胞など)は、同一条件下で培養した場合に、ホスホケトラーゼポリペプチドの異種核酸の1種以上のコピー、MVA経路のポリペプチドをコードしている異種核酸の1種以上のコピー、及びイソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸を含まない同様の細胞と比較して、高濃度でイソプレンを産生することができる。細胞には、IDIポリペプチドをコードしている1種以上の異種核酸を更に含ませてもよい。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
一部の態様では、ホスホケトラーゼをコードしている異種核酸の1種以上のコピー、MVA経路のポリペプチドをコードしている異種核酸の1種以上のコピー、及びイソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸は、宿主細胞の染色体のヌクレオチド配列に組み込まれた異種核酸である。他の態様では、1種以上の異種核酸はプラスミドに組み込まれる。更に他の態様では、1種以上の異種核酸のうち少なくとも1つは、細胞の染色体のヌクレオチド配列に組み込まれ、かつ1種以上の異種核酸配列のうち少なくとも1つはプラスミドに組み込まれる。組換え細胞は、ホスホケトラーゼポリペプチドを含まないイソプレン産生細胞と比較して、少なくとも5%多量にイソプレンを産生することができる。あるいは、組換え細胞は、約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、又は15超並びにこれらの数字の間の任意の数値でイソプレンを産生することができる。
本明細書において、本発明の一態様では、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸、メバロン酸(MVA)経路のポリペプチドをコードしている1種以上の異種核酸、DXP経路のポリペプチドをコードしている1種以上の異種核酸、並びにイソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸、を含む、組換え細胞が提供される。細胞には、IDIポリペプチドをコードしている1種以上の異種核酸を更に含ませてもよい。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。任意の1種以上の異種核酸を、調節可能なように構成型プロモータに連結させることができ、調節可能なように誘導型プロモータに連結させることができ、あるいは調節可能なように誘導型及び構成型プロモータと組み合わせて連結させることができる。1種以上の異種核酸は、更に、調節可能なように高発現型プロモータ、低発現型プロモータ及び/又は強度が発現が中程度のプロモータに連結させることができる。ホスホケトラーゼ、メバロン酸(MVA)経路ポリペプチド、DXP経路ポリペプチド、及びイソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸は、宿主細胞のゲノムに組み込むことができ、あるいは細胞において安定的に発現させることができる。加えて、1種以上の異種核酸は、ベクター上に存在させることもできる。
本明細書に記載の本明細書に記載の任意の組成物又は方法に従う細胞によるイソプレンの産生は、増強させることができる(例えば、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸、イソプレンシンターゼポリペプチド、MVA経路のポリペプチド、及び/又はDXP経路のポリペプチドの発現を増加させることにより)。本明細書で使用するとき、イソプレン産生の「増強」は、本明細書に記載の任意の組成物及び方法により表される細胞による、イソプレンに関する細胞産生性指数(CPI)、イソプレンの力価、イソプレンの質量収率及び/又はイソプレンの比産生量が、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を有さない細胞と比較して増加していることを意味する。本明細書に記載の特定の実施形態では、宿主細胞には、MVA産生に取り込まれる炭素を増加させるよう更に遺伝子操作することができる。
本明細書に記載の組換え細胞によるイソプレンの産生は、約5%〜約1,000,000倍だけ増強させることができる。特定の態様では、イソプレン産生は、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現していない細胞によるイソプレン産生と比較して、約10%〜約1,000,000倍(例えば、約1〜約500,000倍、約1〜約50,000倍、約1〜約5,000倍、約1〜約1,000倍、約1〜約500倍、約1〜約100倍、約1〜約50倍、約5〜約100,000倍、約5〜約10,000倍、約5〜約1,000倍、約5〜約500倍、約5〜約100倍、約10〜約50,000倍、約50〜約10,000倍、約100〜約5,000倍、約200〜約1,000倍、約50〜約500倍、又は約50〜約200倍)だけ増強させることができる。本明細書に記載の特定の実施形態では、宿主細胞には、MVA産生への炭素の取り込みを増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素を増加させるよう遺伝子操作されていない細胞によるイソプレン産生と比較して、イソプレン産生を増強させる遺伝子操作を更に行うことができる。
他の態様では、本明細書に記載の組換え細胞によるイソプレンの産生は、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現していない細胞によるイソプレンの産生量と比較して、少なくとも約5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、1倍、2倍、5倍、10倍、20倍、50倍、100倍、200倍、500倍、1000倍、2000倍、5000倍、10,000倍、20,000倍、50,000倍、100,000倍、200,000倍、500,000倍、又は1,000,000倍だけ増強させることもできる。本明細書に記載の特定の実施形態では、宿主細胞には、MVA産生への炭素の取り込みを増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素を増加させるよう遺伝子操作されていない細胞によるイソプレン産生と比較して、イソプレン産生を増強させる遺伝子操作を更に行うことができる。
組換え細胞を使用してイソプレンを産生する方法
本開示では、本明細書に記載の任意の組換え細胞を培養する工程を含む、イソプレンの産生方法も提供される。一態様では、イソプレンは、ホスホケトラーゼポリペプチド、MVA経路のポリペプチドを1種以上、及びイソプレンシンターゼポリペプチドをコードしている、1種以上の異種核酸を含む、組換え細胞を培養することにより産生することができる。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
別の態様では、イソプレンは、本明細書に記載のいずれかの酵素経路に調節がなされており、かつホスホケトラーゼペプチド、MVA経路のポリペプチド、及びイソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸を含む、組換え細胞を培養することにより産生され得る。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。イソプレンは、本開示の任意の方法に従い、本明細書に記載の任意の細胞から産生することができる。限定するものではないが、六炭糖(グルコースなど)などの炭水化物からイソプレンを産生する目的に際し、任意の細胞を使用することができる。他の実施形態では、ホスホケトラーゼポリペプチドは、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)に由来する。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
したがって、本明細書では、ホスホケトラーゼポリペプチド及びイソプレンシンターゼをコードしている1種以上の異種核酸を含む細胞を、イソプレンの産生に好適な条件下で培養する工程と、(b)イソプレンを産生させる工程と、を含む、イソプレンの産生方法が提供される。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
細胞は、MVA経路のポリペプチド(例えば、完全なMVA経路)及び上記の任意のイソプレンシンターゼポリペプチド(例えば、プエラリア(Pueraria)のイソプレンシンターゼ)をコードしている1種以上の核酸分子を更に含み得る。一部の態様では、組換え細胞は、本明細書に記載の細胞のうちのいずれか1つであり得る。本明細書に記載の任意のイソプレンシンターゼ又はそれらの変異体、本明細書に記載の任意の宿主細胞株、本明細書に記載の任意のプロモータ及び/又は本明細書に記載の任意のベクターも、本明細書に記載の方法において使用することができる、本明細書に記載の任意のエネルギー源(例えば、グルコース又は任意のその他の六炭糖)を使用するイソプレンの産生に使用することができる。一部の態様では、イソプレンの産生方法は更に、イソプレンを回収する工程を含む。他の実施形態では、ホスホケトラーゼポリペプチドは、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)に由来する。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。
特定の態様では、本明細書では、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチド、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)、及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaSポリペプチドをコードしている1種以上の異種核酸を含む組換え細胞を、イソプレンの産生に好適な条件下で培養する工程と、並びに(b)イソプレンを産生させる工程と、を含む、イソプレンの産生方法が提供される。細胞には、上記のMVA経路下流のポリペプチド(例えば、MVK、PMK、MVD及び/又はIDI)及び上記の任意のイソプレンシンターゼポリペプチドをコードしている1種以上の核酸分子を更に含ませることができる。一部の態様では、組換え細胞は、本明細書に記載の任意の細胞であってよい。
特定の態様では、本明細書では、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を含む組換え細胞を、イソプレンの産生に好適な条件下で培養する工程と、並びに(b)イソプレンを産生させる工程と、を含む、イソプレンの産生方法が提供される。細胞には、上記のMVA経路下流のポリペプチド(例えば、MVK、PMK、MVD及び/又はIDI)及び上記の任意のイソプレンシンターゼポリペプチドをコードしている1種以上の核酸分子を更に含ませることができる。一部の態様では、組換え細胞は、本明細書に記載の任意の細胞であってよい。本明細書に記載の、メバロン酸産生に取り込まれる炭素量が増加するよう操作された各種酵素経路を有する組換え細胞を、イソプレン産生に使用することができる。一部の態様では、組換え細胞には、次の、リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(tal B)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)からなる群から選択される遺伝子のうちの1つ以上の活性を増加させるよう、更に遺伝子操作を施すことができる。別の実施形態では、これらの組換え細胞には、次の、グルコース−6−リン酸脱水素酵素(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトースビスリン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸脱水素酵素(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)といった遺伝子のうち、1種以上の遺伝子の活性を減少させるよう更に遺伝子操作を施すことができる。
特定の態様では、本明細書では、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)由来のホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を含む組換え細胞を、イソプレンの産生に好適な条件下で培養する工程と、(b)イソプレンを産生させる工程と、を含む、イソプレンの産生方法が提供される。細胞には、上記のMVA経路下流のポリペプチド(例えば、MVK、PMK、MVD及び/又はIDI)及び上記の任意のイソプレンシンターゼポリペプチドをコードしている1種以上の核酸分子を更に含ませることができる。一部の態様では、組換え細胞は、本明細書に記載の任意の細胞であってよい。本明細書に記載の、メバロン酸産生に取り込まれる炭素量が増加するよう操作された各種酵素経路を有する組換え細胞を、イソプレン産生に使用することができる。一部の態様では、組換え細胞は、次の、rpiA、rpe、tktA、tal B、pta、及び/又はeutDからなる群から選択される1種以上の遺伝子の活性を増加させるよう、更に遺伝子操作を施すことができる。別の態様では、これらの株は、次の、zwf、pfkA、fba、gapA、ackA、gltA及び/又はptsを含む遺伝子のうち1つ以上の遺伝子の活性を減少させるよう、更に遺伝子操作を施すことができる。
一部の態様では、産生されるイソプレン量は、純産生量(absolute productivity)が最大となった時点で測定されたものである。一部の態様では、細胞に関し純産生量が最大であるとは、本明細書で開示される任意のおよそのイソプレン量を指していうものである。一部の態様では、産生されたイソプレンの量は、比産生量が最大となった時点で測定されたものである。一部の態様では、細胞に関し比産生量が最大であるとは、本明細書で開示される、細胞あたりの任意のおよそのイソプレン量を指していうものである。一部の態様では、産生されたイソプレンの総量は累積値として測定される。一部の態様では、細胞の累積産生量は、本明細書で開示される任意のおよそのイソプレン量を指していうものである。
一部の態様では、本明細書に記載の、任意の細胞(例えば、培養物中の細胞)は、約1、10、25、50、100、150、200、250、300、400、500、600、700、800、900、1,000、1,250、1,500、1,750、2,000、2,500、3,000、4,000又は5,000以上のうちのいずれかのイソプレン(nmol)/細胞湿潤重量(g)/時間(nmol/gwcm/h)値でイソプレンを産生する。一部の態様では、イソプレンの量は約2〜約5,000nmol/gwcm/hであり、例えば、約2〜約100nmol/gwcm/h、約100〜約500nmol/gwcm/h、約150〜約500nmol/gwcm/h、約500〜約1,000nmol/gwcm/h、約1,000〜約2,000nmol/gwcm/h又は約2,000〜約5,000nmol/gwcm/hである。一部の態様では、イソプレンの量は約20〜約5,000nmol/gwcm/h、約100〜約5,000nmol/gwcm/h、約200〜約2,000nmol/gwcm/h、約200〜約1,000nmol/gwcm/h、約300〜約1,000nmol/gwcm/h又は約400〜約1,000nmol/gwcm/hである。
一部の態様では、細胞は、培養時に、約1、10、25、50、100、150、200、250、300、400、500、600、700、800、900、1,000、1,250、1,500、1,750、2,000、2,500、3,000、4,000、5,000、10,000又は100,000以上のうちのいずれかのイソプレン(nmol)/細胞湿潤重量(g)/時間(ng/gwcm/h)値でイソプレンを産生する。一部の態様では、イソプレンの量は、約2〜約5,000ng/gwcm/hであり、例えば、約2〜約100ng/gwcm/h、約100〜約500ng/gwcm/h、約500〜約1,000ng/gwcm/h、約1,000〜約2,000ng/gwcm/h又は約2,000〜約5,000ng/gwcm/hである。一部の態様では、イソプレンの量は、約20〜約5,000ng/gwcm/h、約100〜約5,000ng/gwcm/h、約200〜約2,000ng/gwcm/h、約200〜約1,000ng/gwcm/h、約300〜約1,000ng/gwcm/h又は約400〜約1,000ng/gwcm/hである。
一部の態様では、細胞は、培養時に、約1、10、25、50、100、150、200、250、300、400、500、600、700、800、900、1,000、1,250、1,500、1,750、2,000、2,500、3,000、4,000、5,000、10,000、50,000、100,000以上のうちのいずれかのイソプレン(mg)/ブロス(L)(mg/Lブロス、ブロス体積には、細胞及び細胞培地の体積を含む)値の累積力価(総量)でイソプレンを産生する。一部の態様では、イソプレンの量は、約2〜約5,000mg/Lブロスであり、例えば、約2〜約100mg/Lブロス、約100〜約500mg/Lブロス、約500〜約1,000mg/Lブロス、約1,000〜約2,000mg/Lブロス、又は約2,000〜約5,000mg/Lブロスなどである。一部の態様では、イソプレンの量は、約20〜約5,000mg/Lブロス、約100〜約5,000mg/Lブロス、約200〜約2,000mg/Lブロス、約200〜約1,000mg/Lブロス、約300〜約1,000mg/Lブロス、又は約400〜約1,000mg/Lブロスである。
一部の態様では、細胞により、培養時に産生されるイソプレンは、発酵時排出気体(fermentation offgas)の体積の少なくとも約1、2、5、10、15、20又は25%を構成する。一部の態様では、イソプレンは、排出気体の体積の約1〜約25%を構成し、例えば、約5〜約15%、約15〜約25%、約10〜約20%又は約1〜約10%を構成する。
特定の実施形態では、イソプレンの産生方法は、(a)ホスホケトラーゼポリペプチドを内因的に発現しない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞など)を培養する工程、細胞は、(i)MVA経路の1種以上のペプチドを発現する1種以上の異種核酸、及び(ii)イソプレンシンターゼとともに、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上のコピーを異種発現する、並びに(b)イソプレンを産生させる工程、を含んでよく、組換え細胞は、ホスホケトラーゼポリペプチドをコードしている遺伝子の1つ以上の異種コピーを含まない同様の細胞と比較して、減少している酸素の取り込み速度(OUR)を示す特定の実施形態では、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上の異種コピーを発現する組換え細胞は、ホスホケトラーゼを発現しない組換え細胞と比較した場合に、OURの、最大で1倍、2倍、3倍、4倍、5倍、6倍又は7倍の減少を示す。
同様に、本明細書では、増強されたイソプレン産生能を有する細胞を含むイソプレン産生法を提供する。本明細書に記載の細胞によるイソプレンの産生は、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸、完全なMVA経路の1種以上のポリペプチドをコードしている異種核酸の1種以上のコピー、及びイソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸をコードしている1種以上の異種核酸を発現させることにより増加させることができる。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。本明細書で使用するとき、イソプレン産生量の「増大」は、本明細書に記載の任意の組成物及び方法により表される細胞による、イソプレンに関する細胞産生性指数(CPI)、イソプレンの力価、イソプレンの質量収率及び/又はイソプレンの比産生量が、ホスホケトラーゼポリペプチド、MVA経路のポリペプチド、及びイソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸を有さない細胞と比較して増加していることを意味する。イソプレンの産生量は、約5%〜約1,000,000倍増加させることができる。イソプレン産生量は、ホスホケトラーゼ酵素を内因的に発現しないイソプレン産生細胞によるイソプレンの産生量と比較して、約10%〜約1,000,000倍(例えば、約50%〜約1,000,000倍、約1〜約500,000倍、約1〜約50,000倍、約1〜約5,000倍、約1〜約1,000倍、約1〜約500倍、約1〜約100倍、約1〜約50倍、約5〜約100,000倍、約5〜約10,000倍、約5〜約1,000倍、約5〜約500倍、約5〜約100倍、約10〜約50,000倍、約50〜約10,000倍、約100〜約5,000倍、約200〜約1,000倍、約50〜約500倍、又は約50〜約200倍)だけ増加させることができる。本明細書に記載の特定の実施形態では、本明細書に記載の方法は、MVA産生への炭素の取り込みを増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素を増加させるよう遺伝子操作されていないイソプレン産生細胞によるイソプレン産生と比較して、イソプレン産生を増強させる遺伝子操作を更に行った宿主細胞を含む。特定の実施形態では、ホスホケトラーゼポリペプチドは、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)に由来する。
他の態様では、本明細書に記載の方法は、本明細書に記載の細胞によるイソプレン産生の増強(例えば、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を発現させることによる増強)を目的とする特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。イソプレンの産生量は、約5%〜約1,000,000倍増加させることができる。イソプレン産生は、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を発現しないイソプレン産生細胞によるイソプレンの産生と比較して、約10%〜約1,000,000倍(例えば、約50%〜約1,000,000倍、約1〜約500,000倍、約1〜約50,000倍、約1〜約5,000倍、約1〜約1,000倍、約1〜約500倍、約1〜約100倍、約1〜約50倍、約5〜約100,000倍、約5〜約10,000倍、約5〜約1,000倍、約5〜約500倍、約5〜約100倍、約10〜約50,000倍、約50〜約10,000倍、約100〜約5,000倍、約200〜約1,000倍、約50〜約500倍、又は約50〜約200倍)だけ増強させることができる。イソプレンの産生は、ホスホケトラーゼをコードしている1種以上の異種核酸を発現しないイソプレン産生細胞によるイソプレン産生と比較して、少なくとも約5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、1倍、2倍、5倍、10倍、20倍、50倍、100倍、200倍、500倍、1000倍、2000倍、5000倍、10,000倍、20,000倍、50,000倍、100,000倍、200,000倍、500,000倍、又は1,000,000倍だけ増加させることができる。本明細書に記載の特定の実施形態では、本明細書に記載の方法は、MVA産生への炭素の取り込みを増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素を増加させるよう遺伝子操作されていない細胞によるイソプレン産生と比較して、イソプレン産生を増強させる遺伝子操作を更に行った宿主細胞を含む。特定の実施形態では、ホスホケトラーゼポリペプチドは、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)に由来する。
加えて、より具体的な細胞培養条件を使用して、本明細書に記載の方法により細胞を培養することができる。例えば、一部の態様では、イソプレンの産生方法は、(a)ホスホケトラーゼ遺伝子を内因的に有さない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞)を、34℃下で、最少培地にて、培養する工程であって、組換え細胞が、(i)低〜通常コピー数のプラスミドにおいて、高発現型プロモータの調節下で、ホスホケトラーゼポリペプチドをコードしている異種遺伝子の1種以上のコピー、(ii)MVA経路のポリペプチド(MVA経路上流及びMVA経路下流)の1種以上のポリペプチドをコードしている異種核酸の1種以上のコピー、及び(iii)イソプレンシンターゼポリペプチドをコードしている1種以上の異種核酸を異種発現する、培養する工程;並びに(b)イソプレンを産生させる工程、を含む。特定の実施形態では、ホスホケトラーゼポリペプチドは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)に由来する。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一部の態様では、イソプレンの産生方法は更に、イソプレンを回収する工程を含む。
イソプレノイド前駆体及び/又はイソプレノイドの産生量を増加させることのできる組換え細胞
イソプレノイドは、多くの生物において、MVA経路の最終産物であるイソプレノイド前駆体分子の合成によりとができる。上記のように、イソプレノイドは、重要な部類の化合物として存在し、例えば、食品及び飼料用サプリメント、風味及び臭気化合物、並びに抗癌、抗マラリア、抗真菌及び抗菌化合物を含む。
分子の部類の際には、イソプレノイドは、化合物を構成しているイソプレン単位の数に基づき分類される。モノテルペンは10個の炭素原子又は2個のイソプレン単位を含み、セスキテルペンは15個の炭素原子又は3個のイソプレン単位を含み、ジテルペンは20個の炭素原子又は4個のイソプレン単位を含み、セステルテルペンは25個の炭素原子又は5個のイソプレン単位を含む。ステロイド(一般的に、炭素原子を約27個含む)は、イソプレノイドを分解又は再構成した生成物である。
イソプレノイドは、イソプレノイド前駆体分子のIPP及びDMAPPから生成することができる。これらの多様な化合物は、これらのより単純で一般的な前駆体から誘導され、及び保存されているポリプレニルピロリン酸シンターゼ群により合成される(Hsieh et al.,Plant Physiol.2011 Mar;155(3):1079〜90)。多様な鎖長を有するこれらの直鎖プレニルピロリン酸はそれぞれに特有の生理機能を示し、及びポリプレニルピロリン酸シンターゼの非常に発達した活性部位により、一般的にアリル基(ジメチルアリル二リン酸(C5−DMAPP)、ゲラニルピロリン酸(C10−GPP)、ファルネシルピロリン酸(C15−FPP)、ゲラニルゲラニルピロリン酸(C20−GGPP))と、相当する数のイソペンテニルピロリン酸との縮合反応を介し、認識される(C5−IPP)(Hsieh et al.,Plant Physiol.2011 Mar;155(3):1079〜90)。
イソプレノイド前駆体及び/又はイソプレノイドの産生は、イソプレノイド前駆体及び/又はイソプレノイドの産生を増加させるため、ホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む、任意の組換え宿主細胞を使用することにより行うことができる。一部の態様では、これらの細胞は、上記のように、MVA経路、IDI及び/又はDXP経路のポリペプチドをコードしている1種以上の異種核酸、並びにポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸を更に含む。理論に束縛されるものではないが、上記の任意の組成物及び方法により、組換え細胞におけるメバロン酸の産生量を増加させることで、同様に、イソプレノイド前駆体分子及び/又はイソプレノイドなどのより高分子量の生成物の産生量も増加するものと考えられる。グルコースからのメバロン酸産生量のモル収率を上昇させ、適切な酵素活性濃度で、メバロン酸キナーゼ、ホスホメバロン酸キナーゼ、ジホスホメバロン酸デカルボキシラーゼ、イソペンテニル二リン酸イソメラーゼ並びにイソプレン及びイソプレノイド産生に好適な他の酵素と組み合わせると、イソプレノイド前駆体分子及び/又はイソプレノイド(イソプレンを含む)のモル収率の上昇につながる。本明細書に記載の、メバロン酸産生に取り込まれる炭素量が増加するよう遺伝子操作された各種酵素経路を有する組換え細胞を、イソプレノイド前駆体及び/又はイソプレノイドの産生に使用することができる。一部の態様では、組換え細胞は、次の、rpiA、rpe、tktA、tal B、pta、及び/又はeutDからなる群から選択される1種以上の遺伝子の活性を増加させるよう、更に遺伝子操作を施すことができる。別の態様では、これらの株は、次の、zwf、pfkA、fba、gapA、ackA、gltA及び/又はptsを含む遺伝子のうち1つ以上の遺伝子の活性を減少させるよう、更に遺伝子操作を施すことができる。
イソプレノイドの種類
本発明の組換え細胞は、イソプレノイド及びイソプレノイド前駆体分子DMAPP及びIPPの産生量を増加させる能力がある。イソプレノイドの例としては、限定するものではないが、ヘミテルペノイド、モノテルペノイド、セスキテルペノイド、ジテルペノイド、セステルテルペノイド、トリテルペノイド、テトラテルペノイド、及びより高分子量のポリテルペノイドが挙げられる。一部の態様では、ヘミテルペノイドは、プレノール(すなわち、3−メチル−2−ブテン−1−オール)、イソプレノール(すなわち、3−メチル−3−ブテン−1−オール)、2−メチル−3−ブテン−2−オール、又はイソ吉草酸である。一部の態様では、モノテルペノイドは、限定するものではないが、ゲラニルピロリン酸、オイカリプトール、リモネン、又はピネンであってよい。一部の態様では、セスキテルペノイドはファルネシルピロリン酸、アルテミシニン、又はビサボロールである。一部の態様では、ジテルペノイドは、限定するものではないが、ゲラニルゲラニルピロリン酸、レチノール、レチナール、フィトール、タキソール、フォルスコリン、又はアフィジコリンであってよい。一部の態様では、トリテルペノイドは、スクアレン又はラノステロールであってよい。イソプレノイドは、アビエタジエン、アモルファジエン、カレン、α−ファルネセン(α-famesene)、β−ファルネセン、ファルネソール、ゲラニオール、ゲラニルゲラニオール、リナロール、リモネン、ミルセン、ネロリドール、オシメン、パチョロール、β−ピネン、サビネン、γ−テルピネン、テルピンデン(terpindene)及びバレンセンからなる群から選択することもできる。
一部の態様では、テトラテルペノイドは、リコピン又はカロチン(カロチノイド)である。本明細書で使用するとき、用語「カロテノイド」は、クロロプラスト中で、並びに植物、何らかの他の光合成生物、例えば藻類、ある種の真菌、ある種の細菌、のクロロプラスト中で産生される、天然に生じる有機色素群を指す。カロテノイドとしては、酸素を含有しているキサントフィル及び酸素を含有していないカロテンが挙げられる。一部の態様では、カロテノイドはキサントフィル及びカロテンからなる群から選択される。一部の態様では、キサントフィルはルテイン又はゼアキサンチンである。一部の態様では、カロチノイドは、α−カロチン、β−カロチン、γ−カロチン、β−クリプトキサンチン又はリコピンである。
ポリプレニルピロリン酸シンターゼのポリペプチドをコードしている異種核酸
本発明の一部の態様では、本明細書において任意の組成物又は方法に記載されるように、細胞は、更に、上記の通りにメバロン酸(MVA)経路のポリペプチドを1種以上コードしている核酸、並びにポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の核酸を更に含む。ポリプレニルピロリン酸シンターゼポリペプチドは、内在性ポリペプチドであってよい。ポリプレニルピロリン酸シンターゼポリペプチドをコードしている内在性核酸は、構成型プロモータに調節可能なように連結させてよく、あるいは同様に、調節可能なように誘導型プロモータに連結させてもよい。ポリプレニルピロリン酸シンターゼポリペプチドをコードしている内在性核酸は、更に、調節可能なように高発現型プロモータと連結させてもよい。あるいは、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている内在性の核酸は、調節可能なように低発現型プロモータと連結させてもよい。詳細には、細胞は、野生型細胞と比較して、内在性のポリプレニルピロリン酸シンターゼポリペプチドを過剰発現するよう遺伝子操作することができる。
一部の態様では、ポリプレニルピロリン酸シンターゼポリペプチドは、異種ポリペプチドである。本発明の細胞は、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸の1種以上のコピーを含む。一部の態様では、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸は、調節可能なように構成型プロモータに連結されている。一部の態様では、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸は、調節可能なように誘導型プロモータに連結されている。一部の態様では、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸は、調節可能なように高発現型プロモータに連結されている。一部の態様では、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸は、調節可能なように低発現型プロモータに連結されている。
ポリプレニルピロリン酸シンターゼポリペプチドをコードしている核酸は、宿主細胞のゲノムに組み込むことができ、あるいは細胞で安定的に発現させることができる。ポリプレニルピロリン酸シンターゼポリペプチドをコードしている核酸は、更にベクター上に存在させてもよい。
ポリプレニルピロリン酸シンターゼの核酸の例としては、ポリプレニルピロリン酸シンターゼの活性を少なくとも1種有するポリペプチド、ポリペプチド断片、ペプチド、又は融合ポリペプチドをコードする核酸が挙げられる。ポリプレニルピロリン酸シンターゼポリペプチドは、イソプレノイド前駆体分子をより複雑なイソプレノイド化合物に変換する。ポリプレニルピロリン酸シンターゼポリペプチドの例としては、イソプレンシンターゼポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、並びに融合ポリペプチドが挙げられる。ポリプレニルピロリン酸シンターゼのポリペプチド及び核酸の例としては、本明細書に記載されるような任意の生物資源から天然に生じるポリペプチド及び核酸が挙げられる。加えて、ポリプレニルピロリン酸シンターゼ変異体は、酵素活性が向上しているなどして、活性が向上されていてよい。一部の態様では、ポリプレニルピロリン酸シンターゼ変異体は、安定性(例えば、熱安定性)が改良されている、及び/又は溶解性が改良されているなどして、その他の特性が改良されている。ポリプレニルピロリン酸シンターゼの核酸の例としては、限定するものではないが、ゲラニルジホスフェート(geranyl diphosposphate)(GPP)シンターゼ、ファルネシルピロリン酸(FPP)シンターゼ、及びゲラニルゲラニルピロリン酸(GGPP)シンターゼ、又はその他の任意の既知のポリプレニルピロリン酸シンターゼポリペプチドなど、ポリプレニルピロリン酸シンターゼのポリペプチドをコードしている核酸を挙げることができる。
本発明の一部の態様では、本明細書において任意の組成物又は方法に記載されるように、細胞は、ファルネシルピロリン酸(FPP)シンターゼをコードしている1種以上の核酸を更に含む。FPPシンターゼポリペプチドは、内在性遺伝子によりコードされている内在性ポリペプチドであってもよい。一部の態様では、FPPシンターゼポリペプチドは、大腸菌(E. coli)において内在性ispA遺伝子によりコードされている。FPPシンターゼポリペプチドをコードしている内在性の核酸は、調節可能なように構成型プロモータに連結させることができ、あるいは同様に調節可能なように誘導型プロモータに連結させることができる。FPPシンターゼポリペプチドをコードしている内在性の核酸は、更に、調節可能なように高発現型プロモータに連結させることができる。特に、細胞は、野生型細胞と比較して、内在性FPPシンターゼポリペプチドを過剰発現するよう遺伝子操作することができる。
一部の態様では、FPPシンターゼポリペプチドは異種ポリペプチドである。本発明の細胞は、FPPシンターゼポリペプチドをコードしている異種核酸の1種以上のコピーを含み得る。一部の態様では、FPPシンターゼポリペプチドをコードしている異種核酸は、構成型プロモータに調節可能なように連結される。一部の態様では、FPPシンターゼポリペプチドをコードしている異種核酸は、誘導型プロモータに調節可能なように連結される。一部の態様では、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている異種核酸は、調節可能なように高発現型プロモータに連結されている。
FPPシンターゼポリペプチドをコードしている核酸は、宿主細胞のゲノムに組み込むことができ、あるいは細胞で安定的に発現させることができる。FPPシンターゼポリペプチドをコードしている核酸は、更にベクターに組み込むこともできる。
インビトロで、細胞抽出物中で又はインビボで、ポリペプチドがIPPをイソプレノイドへと変換する能力を測定して、ポリペプチドがポリプレニルピロリン酸シンターゼポリペプチド活性を有するか否かを判定する際には、標準法を使用できる。これらの手法は当該技術分野において周知であり、例えば、米国特許第7,915,026号;Hsieh et al.,Plant Physiol.2011 Mar;155(3):1079〜90;Danner et al.,Phytochemistry.2011 Apr 12[Epub ahead of print];Jones et al.,J Biol Chem.2011 Mar 24[Epub ahead of print];Keeling et al.,BMC Plant Biol.2011 Mar 7;11:43;Martin et al.,BMC Plant Biol.2010 Oct 21;10:226;Kumeta & Ito,Plant Physiol.2010 Dec;154(4):1998〜2007;and Kollner & Boland,J Org Chem.2010 Aug 20;75(16):5590〜600に記載されている。
イソプレノイド前駆体及び/又はイソプレノイドの産生を増強させることのできる組換え細胞
本明細書に記載の組換え細胞(例えば、組換え細菌細胞)は、同一条件下で培養した場合に、ホスホケトラーゼをコードしている異種核酸の1種以上のコピー、MVA経路のポリペプチドをコードしている異種核酸の1種以上のコピー、及びポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸を含まない同様の細胞と比較して、多量に及び/又は高濃度にイソプレノイド前駆体及び/又はイソプレノイドを産生することができる。特定の実施形態では、ホスホケトラーゼペプチドをコードしている異種核酸の1種以上のコピーは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロングム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)に由来するものである。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一部の態様では、ホスホケトラーゼをコードしている異種核酸の1種以上のコピー、MVA経路のポリペプチドをコードしている異種核酸の1種以上のコピー、及びポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸は、宿主細胞の染色体に組み込まれた異種核酸である。組換え細胞は、ホスホケトラーゼポリペプチドを含まないイソプレノイド及び/又はイソプレノイド前駆体産生組換え細胞と比較した場合に、少なくとも5%多量のイソプレノイド前駆体及び/又はイソプレノイドを産生することができる。あるいは、組換え細胞は、ホスホケトラーゼをコードしている1種以上の異種核酸を発現しないイソプレノイド及び/又はイソプレノイド前駆体産生細胞によるイソプレノイド及び/又はイソプレノイド前駆体の産生と比較して、1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、又は15%超並びにこれらの数字の間の任意の数値だけ多量にイソプレノイド前駆体及び/又はイソプレノイドを産生することができる。本明細書に記載の特定の実施形態では、本明細書に記載の方法は、MVA産生に取り込まれる炭素を増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素を増加させるよう遺伝子操作されていないイソプレノイド及び/又はイソプレノイド前駆体産生細胞によるイソプレノイド及び/又はイソプレノイド前駆体の産生と比較して、イソプレノイド及び/又はイソプレノイド前駆体の産生を増強させる遺伝子操作を更に行った宿主細胞を含む。
本発明の一態様では、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸、完全なMVA経路のポリペプチド(すなわち、MVA経路上流及びMVA経路下流)をコードしている1種以上の異種核酸、ポリプレニルピロリン酸シンターゼをコードしている1種以上の異種核酸、及び/又はDXP経路のポリペプチドをコードしている1種以上の異種核酸を含む組換え細胞を提供する。細胞には、IDIポリペプチドをコードしている1種以上の異種核酸を更に含ませてもよい。更に、ポリプレニルピロリン酸シンターゼポリペプチドは、FPPシンターゼポリペプチドであってよい。特定の実施形態では、ホスホケトラーゼペプチドをコードしている異種核酸の1種以上のコピーは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロングム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)に由来するものである。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。1種以上の異種核酸は、調節可能なように構成型プロモータに連結させることができ、調節可能なように誘導型プロモータに連結させることができ、あるいは調節可能なように誘導型及び構成型プロモータと組み合わせて連結させることができる。1種以上の異種核酸は、更に、調節可能なように高発現型プロモータ、低発現型プロモータ及び/又は強度が発現が中程度のプロモータに連結させることができる。ホスホケトラーゼポリペプチド、完全なMVA経路(すなわち、MVA経路上流及びMVA経路下流)のポリペプチドを1つ以上、ポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸、及び/又はDXP経路のポリペプチドをコードしている1種以上の異種核酸は、宿主細胞のゲノムに組み込むことができ、あるいは細胞において安定的に発現させることができる。特定の実施形態では、ホスホケトラーゼペプチドをコードしている異種核酸の1種以上のコピーは、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)に由来する。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。1種以上の異種核酸は、更に1種以上のベクターに組み込むことができる。
本明細書では、イソプレノイド前駆体及び/又はイソプレノイドの産生を増強させ得る組換え細胞が提供される。細胞によるイソプレノイド前駆体及び/又はイソプレノイドの産生は、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸、完全なMVA経路(すなわち、MVA経路上流及びMVA経路下流)の1種以上のポリペプチドをコードしている1種以上の異種核酸、及びポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸を発現させることにより増強させることができる。特定の実施形態では、ホスホケトラーゼペプチドをコードしている異種核酸の1種以上のコピーは、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロングム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)に由来するものである。一実施形態では、組換え細胞は、ラクトバチルス・ロイテリ(Lactobacillus reuteri)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、フェリモナス・バレアリカ(Ferrimonas balearica)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、組換え細胞は、ペドバクター・サルタンス(Pedobactor saltans)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ストレプトマイセス・グリセウス(Streptomyces griseus)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、ノカルジオプシス・ダッソンビエイ(Nocardiopsis dassonvillei)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に他の実施形態では、本明細書に記載の組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。一態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。別の実施形態では、組換え細胞は、ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。更に別の実施形態では、組換え細胞は、エンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼをコードしている異種核酸の1種以上のコピーを含む。本明細書で使用するとき、イソプレノイド前駆体及び/又はイソプレノイドの産生の「増強」は、ホスホケトラーゼ、完全なMVA経路の1種以上のポリペプチド、及びポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸を含まない細胞と比較して、記載の細胞による、イソプレノイド前駆体及び/又はイソプレノイド産生に関係する細胞産生性指数(CPI)、イソプレノイド前駆体及び/又はイソプレノイドの力価、イソプレノイド前駆体及び/又はイソプレノイドの質量収率、並びに/あるいはイソプレノイド前駆体及び/又はイソプレノイドの比産生性が増加していることを指す。イソプレノイド前駆体及び/又はイソプレノイドの産生量は、約5%〜約1,000,000倍増加させることができる。イソプレノイド前駆体及び/又はイソプレノイドの産生量は、ホスホケトラーゼをコードしている1種以上の異種核酸を発現しない細胞によるイソプレノイド及び/又はイソプレノイド前駆体の産生と比較して約10%〜約1,000,000倍(例えば、約1〜約500,000倍、約1〜約50,000倍、約1〜約5,000倍、約1〜約1,000倍、約1〜約500倍、約1〜約100倍、約1〜約50倍、約5〜約100,000倍、約5〜約10,000倍、約5〜約1,000倍、約5〜約500倍、約5〜約100倍、約10〜約50,000倍、約50〜約10,000倍、約100〜約5,000倍、約200〜約1,000倍、約50〜約500倍、又は約50〜約200倍)だけ増強させることができる。本明細書に記載の特定の実施形態では、組換え宿主細胞は、MVA産生に取り込まれる炭素を増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素を増加させるよう遺伝子操作されていないイソプレノイド及び/又はイソプレノイド前駆体産生細胞によるイソプレノイド及び/又はイソプレノイド前駆体の産生と比較して、イソプレノイド及び/又はイソプレノイド前駆体の産生を増強させる遺伝子操作を更に行われている。
本明細書に記載の細胞によるイソプレノイド前駆体及び/又はイソプレノイドの産生は増強させることができる(例えば、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸、MVA経路下流のポリペプチドをコードしている1種以上の異種核酸、並びにポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸を発現させることにより増強させることができる)。イソプレノイド前駆体及び/又はイソプレノイドの産生量は、約5%〜約1,000,000倍増加させることができる。イソプレノイド前駆体及び/又はイソプレノイドの産生は、天然に生じる細胞(例えば、MVA経路の1種以上のペプチドを発現する1つ以上の異種核酸とともに、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を発現しておらず、かつメバロン酸産生に取り込まれる炭素が増加するよう遺伝子操作されていない細胞)によるイソプレノイド前駆体及び/又はイソプレノイドの産生と比較して、約10%〜約1,000,000倍(例えば、約1〜約500,000倍、約1〜約50,000倍、約1〜約5,000倍、約1〜約1,000倍、約1〜約500倍、約1〜約100倍、約1〜約50倍、約5〜約100,000倍、約5〜約10,000倍、約5〜約1,000倍、約5〜約500倍、約5〜約100倍、約10〜約50,000倍、約50〜約10,000倍、約100〜約5,000倍、約200〜約1,000倍、約50〜約500倍、又は約50〜約200倍)だけ増強させることができる。
他の実施形態では、イソプレノイド前駆体及び/又はイソプレノイドの産生を提供することのできる、本明細書に記載の組換え細胞は、ホスホケトラーゼポリペプチドをコードしている1つ以上の異種核酸を発現しないイソプレノイド前駆体及び/又はイソプレノイドを産生する組換え細胞によるイソプレノイド前駆体及び/又はイソプレノイドの産生と比較して、少なくとも約5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、1倍、2倍、5倍、10倍、20倍、50倍、100倍、200倍、500倍、1000倍、2000倍、5000倍、10,000倍、20,000倍、50,000倍、100,000倍、200,000倍、500,000倍、又は1,000,000倍だけイソプレノイド前駆体及び/又はイソプレノイドの産生を増強させることもできる。
組換え細胞を使用してイソプレノイド及び/又はイソプレノイド前駆体分子を産生する方法
本明細書では、イソプレノイド前駆体分子及び/又はイソプレノイドの産生方法も提供され、方法は、ホスホケトラーゼ及びポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸を含む組換え細胞(例えば、組換え細菌細胞)を培養することを含む。特定の実施形態では、組換え細胞は、MVA経路上流のポリペプチド及びMVA経路下流のポリペプチドをコードしている1種以上の異種核酸を更に含む。イソプレノイド前駆体分子及び/又はイソプレノイドは、本明細書に記載の任意の細胞から、及び本開示の任意の方法に従って、産生することができる。グルコースなどの六炭糖といった炭水化物から、イソプレノイド前駆体分子及び/又はイソプレノイドを産生する目的で、任意の細胞を使用することができる。
特定の態様では、本明細書では、イソプレノイド前駆体分子及び/又はイソプレノイドの製造方法が提供され、方法は、イソプレノイド前駆体分子及び/又はイソプレノイドの産生に好適な条件下で、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)由来のホスホケトラーゼポリペプチドをコードしている1つ以上の異種核酸、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)、及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaSポリペプチドを含む組換え細胞を培養すること、及び(b)イソプレノイド前駆体分子及び/又はイソプレノイドを産生させること、を含む。細胞には、上記のMVA経路下流のポリペプチド(例えば、MVK、PMK、MVD及び/又はIDI)及び上記の任意のポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の核酸分子を更に含ませることができる。一部の態様では、組換え細胞は、本明細書に記載の任意の細胞であってよい。本明細書に記載の任意のポリプレニルピロリン酸シンターゼ又はそれらの変異体、本明細書に記載の任意の宿主細胞株、本明細書に記載の任意のプロモータ及び/又は本明細書に記載の任意のベクターを使用し、本明細書に記載の任意のエネルギー源(例えば、グルコース又は任意の他の六炭糖)を利用して、イソプレノイド前駆体分子及び/又はイソプレノイドを産生することもできる。一部の態様では、イソプレノイド前駆体分子及び/又はイソプレノイドの産生方法は、イソプレノイド前駆体分子及び/又はイソプレノイドを回収する工程を更に含む。
特定の態様では、本明細書では、イソプレノイド前駆体分子及び/又はイソプレノイドの製造方法が提供され、方法は、イソプレノイド前駆体分子及び/又はイソプレノイドの産生に好適な条件下で、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)、及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaSのポリペプチドをコードしている1つ以上の異種核酸を含む組換え細胞を培養すること、及び(b)イソプレノイド前駆体分子及び/又はイソプレノイドを産生させることを含む。細胞には、上記のMVA経路下流のポリペプチド(例えば、MVK、PMK、MVD及び/又はIDI)及び上記の任意のポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の核酸分子を更に含ませることができる。一部の態様では、組換え細胞は、本明細書に記載の任意の細胞であってよい。本明細書に記載の任意のポリプレニルピロリン酸シンターゼ又はそれらの変異体、本明細書に記載の任意の宿主細胞株、本明細書に記載の任意のプロモータ及び/又は本明細書に記載の任意のベクターを使用し、本明細書に記載の任意のエネルギー源(例えば、グルコース又は任意の他の六炭糖)を利用して、イソプレノイド前駆体分子及び/又はイソプレノイドを産生することもできる。一部の態様では、イソプレノイド前駆体分子及び/又はイソプレノイドの産生方法は、イソプレノイド前駆体分子及び/又はイソプレノイドを回収する工程を更に含む。
イソプレノイド前駆体分子及び/又はイソプレノイドを産生する方法には、同様に、(a)ホスホケトラーゼを内在的に有さない組換え細胞(限定するものではないが、大腸菌(E. coli)細胞)を培養する工程、ここで、組換え細胞は、ホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上のコピーを異種発現する;並びに(b)イソプレノイド前駆体分子及び/又はイソプレノイドを産生させる工程、を含ませることができ、組換え細胞は、ホスホケトラーゼポリペプチドを含まないイソプレノイド及び/又はイソプレノイド前駆体産生細胞と比較して多量にイソプレノイド前駆体及び/又はイソプレノイドを産生する。
イソプレノイド前駆体分子及び/又はイソプレノイドの産生に関係する本方法は、イソプレノイド及び/又はイソプレノイド前駆体を産生する、ホスホケトラーゼポリペプチドを含まない組換え細胞と比較して、少なくとも5%多量にイソプレノイド前駆体及び/又はイソプレノイドを産生することができる。あるいは、組換え細胞は、約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%又は15%超、並びに包括的にこれらの数値の間の任意の数値だけイソプレノイド前駆体及び/又はイソプレノイドを産生することができる。一部の態様では、イソプレノイド前駆体分子及び/又はイソプレノイドの産生方法は、イソプレノイド前駆体分子及び/又はイソプレノイドを回収する工程を更に含む。
本開示では、イソプレノイド及び/又はイソプレノイド前駆体分子の産生を増強させるために、上記の細胞のいずれかを使用する方法を提供する。細胞によるイソプレノイド前駆体分子及び/又はイソプレノイドの産生は、ホスホケトラーゼ並びに/又はリステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)、及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaSポリペプチドをコードしている1種以上の異種核酸、MVA経路下流のポリペプチドをコードしている1種以上の異種核酸、及びポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸を発現させることにより増強させることができる。本明細書で使用するとき、イソプレノイド前駆体及び/又はイソプレノイドの産生の「増強」は、本明細書に記載の任意の組成物及び方法により、細胞により、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)由来のホスホケトラーゼ、ポリプレニルピロリン酸シンターゼポリペプチド、MVA経路下流のポリペプチド、mvaE及びmvaSポリペプチドをコードしている1種以上の異種核酸を含まない細胞と比較して、イソプレノイド前駆体及び/又はイソプレノイドの産生に関する細胞産生性指数(CPI)、イソプレノイド前駆体及び/又はイソプレノイド力価、イソプレノイド前駆体及び/又はイソプレノイドの質量収率、並びに/あるいはイソプレノイド前駆体及び/又はイソプレノイドの比産生性が増加することを意味する。イソプレノイド前駆体分子及び/又はイソプレノイドの産生量は、約5%〜約1,000,000倍増加させることができる。イソプレノイド前駆体分子及び/又はイソプレノイドの産生は、ホスホケトラーゼポリペプチドをコードしている1種以上の異種核酸を発現しない細胞によるイソプレノイド前駆体分子及び/又はイソプレノイドの産生と比較して、約10%〜約1,000,000倍(例えば、約1〜約500,000倍、約1〜約50,000倍、約1〜約5,000倍、約1〜約1,000倍、約1〜約500倍、約1〜約100倍、約1〜約50倍、約5〜約100,000倍、約5〜約10,000倍、約5〜約1,000倍、約5〜約500倍、約5〜約100倍、約10〜約50,000倍、約50〜約10,000倍、約100〜約5,000倍、約200〜約1,000倍、約50〜約500倍、又は約50〜約200倍)だけ増強させることができる。本明細書に記載の特定の実施形態では、方法は、MVA産生に取り込まれる炭素を増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素が増加するよう遺伝子操作されていないイソプレノイド及び/又はイソプレノイド前駆体産生細胞によるイソプレノイド及び/又はイソプレノイド前駆体の産生と比較して、イソプレノイド及び/又はイソプレノイド前駆体の産生を増強させる遺伝子操作を更に行った組換え宿主細胞を含む。
イソプレノイド前駆体分子及び/又はイソプレノイドの産生は、本明細書に記載の方法により、イソプレノイド前駆体及び/又はイソプレノイドを産生し、ホスホケトラーゼポリペプチドをコードしている1つ以上の異種核酸を発現しない細胞による、イソプレノイド前駆体分子及び/又はイソプレノイドの産生と比較して、少なくとも約10%、20%、30%、40%、50%、60%、70%、80%、90%、1倍、2倍、5倍、10倍、20倍、50倍、100倍、200倍、500倍、1000倍、2000倍、5000倍、10,000倍、20,000倍、50,000倍、100,000倍、200,000倍、500,000倍、又は1,000,000倍だけ増強させることができる。本明細書に記載の特定の実施形態では、方法は、MVA産生に取り込まれる炭素を増加させることにより、ホスホケトラーゼペプチドをコードしている1種以上の異種核酸を発現せずかつメバロン酸産生に取り込まれる炭素が増加するよう遺伝子操作されていないイソプレノイド及び/又はイソプレノイド前駆体産生細胞によるイソプレノイド及び/又はイソプレノイド前駆体の産生と比較して、イソプレノイド及び/又はイソプレノイド前駆体の産生を増強させる遺伝子操作を更に行った組換え宿主細胞を含む。
加えて、より具体的な細胞培養条件を使用して、本明細書に記載の方法により細胞を培養することができる。例えば、一部の態様では、イソプレノイド前駆体分子及び/又はイソプレノイドの産生方法は、(a)34℃にて、最少培地においてホスホケトラーゼポリペプチドをコードしている異種核酸を含み、かつリステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)、及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE遺伝子及びmvaS遺伝子を内在的に有さない組換え細胞(例えば、限定するものではないが、大腸菌(E. coli)細胞など)を培養する工程、ここで、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アバーミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている遺伝子の1種以上のコピーを異種発現する;並びに(b)イソプレノイド前駆体分子及び/又はイソプレノイドを産生させる工程と、を含む。一部の態様では、イソプレノイド前駆分子及び/又はイソプレノイドを回収する工程を更に含む。一部の態様では、組換え細胞は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)由来のホスホケトラーゼポリペプチドをコードしている遺伝子の1つ以上のコピーを異種発現する。
ベクター
本明細書に記載の任意の組成物及び方法には好適なベクターを使用することができる。例えば、適切なベクターを使用して、ホスホケトラーゼ、MVA経路上流のポリペプチド(限定するものではないが、mvaE及びmvaSポリペプチド)、MVA経路下流のポリペプチド、イソプレンシンターゼ、又はポリプレニルピロリン酸シンターゼをコードしている遺伝子の1つ以上のコピーの、特定の宿主細胞(例えば、大腸菌(E. coli))における発現を最適化することができる。一部の態様では、ベクターには選択マーカーを含有させる。選択可能なマーカーの例としては、これらに限定されるものではないが、抗生物質耐性核酸(例えば、カナマイシン、アンピシリン、カルベニシリン、ゲンタマイシン、ヒグロマイシン、フェロマイシン、ブレオマイシン、ネオマイシン又はクロラムフェニコール)、及び/又はホスト細胞に栄養的な利点などの代謝的な利点を与える核酸が挙げられる。一部の態様では、選択マーカーは用いずに、ホスホケトラーゼ、MVA経路上流のポリペプチド(限定するものではないが、mvaE及びmvaSポリペプチド)、MVA経路下流のポリペプチド、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)、及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaS核酸、イソプレンシンターゼ、又はポリプレニルピロリン酸シンターゼの核酸の1つ以上のコピーを宿主細胞のゲノムに組み込むことができる。
本明細書において特徴付けられ、本開示の実施例において使用される任意のベクターを本発明に使用することができる。
形質転換法
好適な技術を使用して、ホスホケトラーゼ、MVA経路上流のポリペプチド(限定するものではないが、mvaE及びmvaSポリペプチドなど)、MVA経路下流のポリペプチド、及び/又はMVA経路下流のポリペプチドの1つ以上のコピーをコードしている核酸を細胞に導入することができる。更に、イソプレンシンターゼ、IDI、DXP経路及び/又はポリプレニルピロリン酸シンターゼの核酸、又はこれらを含むベクターは、形質転換、電気穿孔法、核マイクロインジェクション、形質導入、形質移入(例えばリポフェクション介在型若しくはDEAE−デキストラン介在型トランスフェクション、又は組換えファージウイルスを使用した形質移入)、リン酸カルシウムDNA沈殿物とのインキュベーション、DNAコーティングした微粒子を用いる高速遺伝子銃照射、及びプロトプラスト融合などの、宿主細胞内にDNAコンストラクト又はベクターを導入するための標準的な技術を使用して宿主細胞(例えば本明細書において述べられるような植物細胞、真菌細胞、酵母細胞、又は細菌細胞)内に挿入することができる。一般的な形質転換法は、当該技術分野で既知である(例えば、分子生物学領域の現行のプロトコル(F.M.Ausubel et al.(eds.)Chapter 9,1987;Sambrook et al.,「分子クローニング(Molecular Cloning)」:「実験室マニュアル(A Laboratory Manual)第2版」,Cold Spring Harbor,1989;及びCampbell et al.,Curr.Genet.16:53〜56,1989を参照されたい)。導入された核酸は、染色体DNAに組み込むことができ、又は染色体外の複製配列として維持することができる。形質転換体は、当該技術分野において既知の任意の方法により選別することができる。形質転換体の選別に好適な方法は、国際公開第2009/076676号、米国特許出願公開第2009/0203102号、国際公開第2010/003007号、米国特許出願公開第2010/0048964号、国際公開第2009/132220号、及び米国特許出願公開第2010/0003716に記載される。
例示的な宿主細胞
当業者であれば、具体的な宿主株における遺伝子発現を最適化する特定の配列を含有するよう、発現ベクターが設計されることを認識するであろう。このような最適化配列としては、限定するものではないが、複製起点、プロモータ、及びエンハンサが挙げられる。本明細書で参照するベクター及び配列は例示目的で記載され、本発明の範囲を狭めることを意味するものではない。
遺伝子を異種発現するために使用することのできる任意の細胞又はそれらの子孫細胞を使用して、MVA経路の1種以上のペプチド、イソプレンシンターゼ、IDI、DXP経路ポリペプチド、及び/又はポリプレニルピロリン酸シンターゼポリペプチドを発現する1種以上の異種核酸とともに、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、フェリモナス・バレアリカ(Ferrimonas balearica)、ペドバクター・サルタンス(Pedobactor saltans)、ストレプトマイセス・グリセウス(Streptomyces griseus)、及び/又はノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)から単離された1種以上のホスホケトラーゼを発現させることができる。一部の実施形態では、宿主細胞は、グラム陽性細菌である。非限定的な例としては、ストレプトマイセス株(例えば、ストレプトマイセス・リビダンス、ストレプトマイセス・コエリカラー、又はストレプトマイセス・グリセウス)、バチルス株、リステリア株(例えば、L.モノサイトゲネス)又はラクトバチルス株(例えば、L.spp)が挙げられる。一部の実施形態では、生物資源は、大腸菌、シュードモナス属、又はH.ピロリなどのグラム陰性細菌である。
グラム陽性又はグラム陰性細菌などの細菌細胞を使用して、上記の異種遺伝子のうち任意のものを発現させることができる。詳細には、mvaE及びmvaS遺伝子は、シュードアルテロモナス・シトレア(P. citrea)、バチルス・サブチリス(B. subtilis)、バチルス・リケニフォルミス(B. licheniformis)、バチルス・レンタス(B. lentus)、バチルス・ブレビス(B. brevis)、バチルス・ステアロサーモフィルス(B. stearothermophilus)、バチルス・アルカロフィルス(B. alkalophilus)、バチルス・アミロリケファシエンス(B. amyloliquefaciens)、バチルス・クラウシイ(B. clausii)、バチルス・ハロドュランス(B. halodurans)、バチルス・メガテリウム(B. megaterium)、バチルス・コアギュランス(B. coagulans)、バチルス・サーキュランス(B. circulans)、バチルス・ロータス(B. lautus)、バチルス・チューリンゲンシス(B. thuringiensis)、ストレプトマイセス・アルバス(S. albus)、ストレプトマイセス・リビダンス(S. lividans)、ストレプトマイセス・コエリカラー(S. coelicolor)、ストレプトマイセス・グリセウス(S. griseus)、シュードモナス種(Pseudomonas sp.)及びシュードモナス・アルカリゲネス(P. alcaligenes)細胞のいずれかにおいて発現させることができる。
本発明の組成物及び方法には、数多くの種類の嫌気性細胞を宿主細胞として使用することができる。本発明の一態様では、本明細書に記載の任意の組成物又は方法に関し記載される細胞は、絶対嫌気性細胞及びその子孫細胞である。絶対嫌気性菌は、典型的には、条件下に酸素が存在する場合には良好に増殖しない。絶対嫌気性菌が低濃度の酸素に対しある程度の耐性を示す場合には、少量の酸素が存在してもよいことは理解されるであろう。一態様では、メバロン酸、イソプレノイド前駆体、イソプレン、及びイソプレノイドを産生するよう遺伝子操作を行った絶対嫌気性菌を、本明細書に記載のいずれかの方法及び/又は組成物用の宿主細胞として提供し、実質的に無酸素条件下で増殖させることができる。この場合、存在する酸素量は嫌気性菌の増殖、維持、及び/又は発酵に有害なものではない。
本発明の他の態様では、本明細書に記載の組成物又は方法のいずれかにおいて記載され及び/又は使用される宿主細胞は、通性嫌気性細胞及びその子孫細胞である。酸素が存在する場合、通性嫌気性菌は、好気呼吸により(例えば、TCAサイクルを利用するなどして)細胞ATPを生成し得る。しかしながら、通性嫌気性菌も、酸素の非存在下で増殖させることができる。絶対嫌気性菌とは対照的に、通性嫌気性菌はより多量の酸素の存在下で死滅し、又は増殖性が乏しくなる。したがって、一態様では、通性嫌気性菌は、本明細書で提供される組成物及び/又は方法のいずれかにおいて、宿主細胞として使用でき、メバロン酸、イソプレノイド前駆体、イソプレン、及びイソプレノイドを生成するよう遺伝子操作を行うことができる。通性嫌気性宿主細胞は、実質的に無酸素(存在する酸素量が、増殖、嫌気性菌の維持、及び/又は発酵に有害なものではないことを意味する)条件下で増殖させることができ、あるいは酸素がより多量に存在している場合にも増殖することができる。
宿主細胞は、糸状真菌細胞及びその子孫細胞であってもよい。(例えば、Berka & Barnett,Biotechnology Advances,(1989),7(2):127〜154を参照されたい)。一部の態様では、糸状菌細胞はトリコデルマ・ロンギブラキアタム(Trichoderma longibrachiatum)、トリコデルマ・ビリデ(T. VIRIDE)、トリコデルマ・コニンギ(T. KONINGII)、トリコデルマ・ハルジアナム(T. harzianum)、ペニシリウム(Penicillium sp.)、ヒュミコラ・インソレンス(Humicola insolens)、ヒュミコラ・ラヌギノス(H. lanuginose)、ヒュミコラ・グリセア(H. grisea)、クリソスポリウム(Chrysosporium sp.)、クリソスポリウム・ラックノウエンス(C. lucknowense)、グリオクラジウム(Gliocladium sp.)、アスペルギルス(Aspergillus sp.)、例えば、アスペルギルス・オリゼ(A. oryzae)、アスペルギルス・ニガー(A. niger)、ショウユコウジカビ(A sojae)、アスペルギルス・ジャポニクス(A. japonicus)、アスペルギルス・ニデュランス(A. nidulans)、又はアワモリコウジカビ(A. awamori)、フザリウム(Fusarium sp.)、例えばフザリウム・ロゼウム(F. roseum)、フザリウム・グラミニウム(F. graminum)、フザリウム・セレアリス(F. cerealis)、フザリウム・オキシスポラム(F. oxysporuim)、又はフザリウム・ベネナタム(F. venenatum)、ニューロスポラ(Neurospora sp.)、例えば、ニューロスポラ・クラッサ(N. crassa)、ヒポクレア(Hypocrea sp.)、ムコール(Mucor sp.,)、例えば、ムコール・ミエヘイ(M. miehei)、リゾプス(Rhizopus sp.)又はエメリセラ(Emericella sp.)のいずれか由来のものであってよい。一部の態様では、真菌は、アスペルギルス・ニデュランス(A. nidulans)、アワモリコウジカビ(A. awamori)、アスペルギルス・オリゼ(A. oryzae)、アスペルギルス・アクレタス(A. aculeatus)、アスペルギルス・ニガー(A. niger)、アスペルギルス・ジャポニクス(A. japonicus)、トリコデルマ・リーゼイ(T. reesei)、トリコデルマ・ビリデ(T. VIRIDE)(T. viride)、フザリウム・オキシスポラム(F. oxysporum)又はフザリウム・ソラニ(F. solani)である。特定の実施形態では、本明細書で使用するためのプラスミド又はプラスミド配列には、米国特許出願公開第2011/0045563号に記載のものが包含される。
宿主細胞は、サッカロマイセス(Saccharomyces sp.)、シゾサッカロマイセス(Schizosaccharomyces sp.)、ピキア(Pichia sp.)、又はカンジダ(Candida sp.)などの酵母であってもよい。一部の態様では、サッカロマイセス(Saccharomyces sp.)は、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)である(例えば、Romanos et al.,Yeast,(1992),8(6):423〜488を参照されたい)。特定の実施形態では、本明細書で使用するプラスミド又はプラスミド配列には、米国特許第7,659,097号、及び米国特許出願公開第2011/0045563号に記載のものが包含される。
宿主細胞は、緑藻類、紅藻類、灰色藻類、クロララクニオン藻類、ミドリムシ類、クロミスタ類、又は渦鞭毛藻類などの藻類であってもよい。(例えば、Saunders及びWarmbrodt、「藻類及び酵母などの菌類における遺伝子発現(Gene Expression in Algae and Fungi, Including Yeast)」(1993年),National Agricultural Library,Beltsville,MDを参照されたい)。特定の実施形態では、本明細書で使用するためのプラスミド又はプラスミド配列には、米国特許出願公開第2011/0045563号に記載のものが包含される。一部の態様では、宿主細胞は、形態学に基づき次の群:クロオコッカス目(Chroococcales)、プレウロカプサ目(Pleurocapsales)、ユレモ目(Oscillatoriales)、ネンジュモ目(Nostocales)、又はスティゴネマ目(Stigonematales)(例えば、Lindberg et al.,Metab.Eng.,(2010)12(1):70〜79を参照されたい)のいずれかに分類されるシアノバクテリアなどの、シアノバクテリアである。特定の実施形態では、本明細書で使用するプラスミド又はプラスミド配列には、米国特許出願公開第2010/0297749号、同第2009/0282545号、及び国際公開第2011/034863号に記載のものが包含される。
大腸菌(E. coli)宿主細胞を使用して、MVA経路の1つ以上ポリペプチド、イソプレンシンターゼ、IDI、DXP経路のポリペプチド(e)、及び/又はポリプレニルピロリン酸シンターゼポリペプチドをコードしている1種以上の異種核酸とともに、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来の1つ以上のホスホケトラーゼ酵素を発現させることができる。一態様では、宿主細胞は、MVA経路の1種以上のペプチドを発現する1つ以上の異種核酸とともに、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼをコードしている1種以上の核酸を発現し、メバロン酸を産生することのできる大腸菌(E. coli)株の組換え細胞又はこれらの子孫細胞である。大腸菌(E. coli)宿主細胞は、MVA経路の1種以上のペプチドを発現する1つ以上の異種核酸とともに、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている1つ以上の異種核酸を発現しない同様の細胞と比較して多量に、高ピーク力価で、及び高細胞産生性で、メバロン酸を産生することができる。加えて、大腸菌(E. coli)において、MVA経路の1種以上のペプチドを発現する1つ以上の異種核酸とともに、異種発現される、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ラクトバチルス・ロイテリ(Lactobacillus reuteri)、ラクトバチルス・プランタラム(Lactobacillus plantarum)、ラクトバチルス・パラプランタルム(Lactobacillus paraplantarum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)、ビフィドバクテリウム・ブレビ(Bifidobacterium breve)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、ガードネレラ・バギナリス(Gardnerella vaginalis)、フェリモナス・バレアリカ(Ferrimonas balearica)、ムチラギニバクター・パルジス(Mucilaginibacter paludis)、ノストック・パンクチホルム(Nostoc punctiforme)、ノストック・パンクチホルム(Nostoc punctiforme)PCC 73102、パントエア(Pantoea)、ペドバクター・サルタンス(Pedobactor saltans)、ラーネラ・アクアティリス(Rahnella aquatilis)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、ストレプトマイセス・グリセウス(Streptomyces griseus)、ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)、ノカルジオプシス・ダッソンビレイ(Nocardiopsis dassonvillei)、及び/又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼポリペプチドをコードしている1種以上の核酸は、染色体のコピー(例えば、大腸菌(E. coli)染色体に組み込まれたコピー)であってよい。他の態様では、大腸菌(E. coli)細胞は培養物である。一部の態様では、1つ以上のホスホケトラーゼ酵素は、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、及び/又はエンテロコッカス・ガリナラム(Enterococcus gallinarum)に由来する。任意の態様では、1つ以上のホスホケトラーゼ酵素は、本明細書に記載の通りの任意のホスホケトラーゼである。
宿主細胞の改変例
クエン酸シンターゼによる経路
クエン酸シンターゼは、オキサロ酢酸とアセチルCoAを縮合させることによるクエン酸(トリカルボン酸(TCA)回路の代謝生成物)の生成を触媒する(Ner,S.et al.1983.Biochemistry 22,:5243〜5249;Bhayana,V.and Duckworth,H.1984.Biochemistry 23:2900〜2905)。大腸菌(E. coli)では、gltAによりコードされたこの酵素は、二量体サブユニットからなる三量体様の挙動を示す。六量体の形成により、酵素はNADHによりアロステリックに制御されるようになる。この酵素は、これまでに広く研究されている(Wiegand,G.,and Remington,S.1986.Annual Rev.Biophysics Biophys.Chem.15:97〜117;Duckworth et al.1987.Biochem Soc Symp.54:83〜92;Stockell,D.et al.2003.J.Biol.Chem.278:35435〜43;Maurus,R.et al.2003.Biochemistry.42:5555〜5565)。NADHによるアロステリック阻害を回避するにあたって、これまでに、バチルス・サブチリス(Bacillus subtilis)NADH非感受性クエン酸シンターゼによる置き換え、又はこの酵素の添加が検討されている(Underwood et al.2002.Appl.Environ.Microbiol.68:1071〜1081;Sanchez et al.2005.Met.Eng.7:229〜239)。
クエン酸シンターゼによる触媒反応は、メバロン酸経路の第一工程を触媒し、同様にアセチルCoAを基質として使用するチオラーゼと直接的に競合する(Hedl et al.2002.J.Bact.184:2116〜2122)。したがって、当業者は、クエン酸シンターゼの発現を調節して(例えば、酵素活性を減少させて)、より多量の炭素がメバロン酸経路に取り込まれるようにすることで、メバロン酸、イソプレン及びイソプレノイドの最終的な産生量を増加させることができる。クエン酸シンターゼ活性を低下させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で低下させることができる。一部の例では、酵素活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%又は100%低下させる。一部の態様では、クエン酸シンターゼに関係する内在性遺伝子の活性を減少させることにより、クエン酸シンターゼの活性を調節する。この調節は、NADH非感受性クエン酸シンターゼをコードしている導入遺伝子により、あるいは枯草菌から誘導したNADH非感受性クエン酸シンターゼをコードしている導入遺伝子により、内在性クエン酸シンターゼ遺伝子の染色体を置換することで実施できる。クエン酸シンターゼの活性は、内在性クエン酸シンターゼ遺伝子のプロモータを、常時低発現型の合成プロモータと置き換えることによっても調節(例えば、低下させる)できる。クエン酸シンターゼをコードしている遺伝子を欠失させることもできる。クエン酸シンターゼの活性を低下させることで、メバロン酸依存性の生合成経路への炭素の取り込みを、クエン酸シンターゼの発現を低下させていない細胞と比較して増加させることができる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつクエン酸シンターゼ(gltA)の活性が減少するよう更に遺伝子操作されている。クエン酸シンターゼイソ酵素の活性の調節(例えば、低下)も本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつクエン酸シンターゼイソ酵素の活性が減少するよう更に遺伝子操作されている。
ホスホトランスアセチラーゼ及び/又は酢酸キナーゼの関与する経路
ホスホトランスアセチラーゼ((大腸菌(E. coli)において(i)pta(Shimizu et al.1969.Biochim.Biophys.Acta 191:550〜558又は(ii)eutD(Bologna et al.2010.J of Microbiology.48:629〜636によりコードされている)は、アセチル−CoA及びアセチルリン酸(アセチル−P)の可逆的な変換を触媒するのに対し、酢酸キナーゼ(大腸菌(E. coli)においてackAによりコードされる)(Kakuda,H.et al.1994.J.Biochem.11:916〜922)はアセチル−Pを使用して酢酸を生成する。これらの遺伝子は、大腸菌においてオペロンとして転写され得る。これらの遺伝子は共に、ATPの放出により酢酸の異化を触媒する。したがって、ホスホトランスアセチラーゼの活性を増強させることにより、アセチル−CoAに変換されるアセチル−Pの量を増加させることができる。特定の実施形態では、増強は、染色体中の遺伝子上流に上方調節されたプロモータを配置するか、又はプラスミドにおいて適切なプロモータの下流に遺伝子のコピーを配置するかによりなすことができる。酢酸に変換されるアセチルCoAの量を減少させる目的で、酢酸キナーゼ遺伝子(例えば、内在性酢酸キナーゼ遺伝子)の活性を減少又は減弱させることができる。特定の実施形態では、減弱は、酢酸キナーゼ(ackA)を欠失させることにより実施される。このような欠失は、遺伝子をクロラムフェニコールカセットにより置き換えた後、カセットを除去させることにより実施される。一部の態様では、酢酸キナーゼの活性は、内在性酢酸キナーゼの活性を減少させることにより調節される。このような調節は、内在性酢酸キナーゼ遺伝子のプロモータを常時低発現型合成プロモータと置き換えることにより行うことができる。特定の実施形態では、このような酢酸キナーゼ遺伝子の減弱は、ホスホトランスアセチラーゼ(pta)遺伝子の発現を干渉することによりなされるべきである。酢酸は多様な理由により大腸菌(E. coli)により生成される(Wolfe,A.2005.Microb.Mol.Biol.Rev.69:12〜50)。理論に束縛されるものではないが、ackAを欠失させることにより、炭素の酢酸の産生への転用が減少し(ackAはアセチル−CoAを利用するため)、メバロン酸、イソプレノイド前駆体、イソプレン及び/又はイソプレノイドの収率が増加することになる。
一部の態様では、本明細書に記載の組換え細胞は、内在性の酢酸キナーゼ遺伝子の発現を減弱させていない、あるいはホスホトランスアセチラーゼを増強させていない細胞と比較して減少した量で酢酸を産生する。酢酸生成量の減少は、当業者に既知の所定の解析法により測定できる。酢酸の減少量は、分子的操作を行わずに内在性酢酸キナーゼ遺伝子の発現又はホスホトランスアセチラーゼ遺伝子発現の発現を実施した場合と比較して、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%である。
ホスホトランスアセチラーゼ(pta及び/又はeutD)の活性は、酵素に関係する他の分子的操作により増加させることができる。酵素活性を増加させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で増加させることができる。一部の例では、酵素活性の増加は、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%だけ増加させることができる。一実施形態では、ptaの活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつホスホトランスアセチラーゼ(pta及び/又はeutD)の活性が増加するよう更に遺伝子操作されている。ホスホトランスアセチラーゼイソ酵素の活性を調節(例えば、増加)することも、本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつホスホトランスアセチラーゼ(pta及び/又はeutD)イソ酵素の活性が増加するよう更に遺伝子操作されている。
酢酸キナーゼ(ackA)の活性は、酵素に関係する他の分子的操作によっても低下させることができる。酵素活性を低下させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で低下させることができる。一部の例では、酵素活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%低下させる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつ酢酸キナーゼ(ackA)の活性が減少するよう更に遺伝子操作されている。酢酸キナーゼイソ酵素の活性の調節(例えば、低下)も本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつ酢酸キナーゼイソ酵素の活性が減少するよう更に遺伝子操作されている。
一部の場合では、内在性酢酸キナーゼ遺伝子の活性を減弱させることで、内在性酢酸キナーゼ遺伝子の発現を減弱させていない細胞と比較して、メバロン酸依存性生合成経路への炭素原子の取り込みが増加する。
乳酸脱水素酵素の関与する経路
大腸菌(E. coli)では、D−乳酸は、乳酸デヒドロゲナーゼ(ldhAによりコード:図1)により、ピルビン酸から生成される(Bunch,P.et al.1997.Microbiol.143:187〜195)。乳酸の生成はNADHの酸化によりなされるため、乳酸は、酸素制限下で、かつ還元当量のすべてを収容できない場合に生成されることになる。したがって、乳酸の生成は、炭素消費の原因となり得る。そのため、メバロン酸産生(並びに必要に応じてイソプレン、イソプレノイド前駆体及びイソプレノイドの産生)への炭素の取り込みを向上させるため、当業者は、酵素活性を低下させるなどして乳酸デヒドロゲナーゼの活性を調節することができる。
したがって、一態様では、乳酸脱水素酵素の活性は、内在性乳酸脱水素酵素遺伝子の活性を減弱させることで調節できる。このような減弱は、内在性乳酸脱水素酵素遺伝子の欠失により実施できる。当業者に知られている、乳酸脱水素酵素遺伝子の活性を減弱させる他の手法も使用できる。乳酸脱水素酵素の関与する経路を操作することで、組換え細胞の生成する乳酸量は、内在性乳酸脱水素酵素遺伝子の発現を減弱させていない細胞と比較して、減少することになる。乳酸の生成量の減少は、当業者に知られている所定の解析法により測定できる。乳酸の生成量は、分子的な操作を行なっていない場合と比較して、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%減少する。
乳酸脱水素酵素の活性は、酵素に関係するその他の分子操作により低下させることもできる。酵素活性を低下させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で低下させることができる。一部の例では、酵素活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%低下させる。
したがって、一部の場合では、内在性乳酸デヒドロゲナーゼ遺伝子の活性を減弱させることで、内在性乳酸デヒドロゲナーゼの遺伝子発現を減弱させていない細胞と比較して、メバロン酸依存性の生合成経路への炭素の取り込みが増加する。
グリセルアルデヒド3−リン酸を包含する経路
グリセルアルデヒド3−リン酸脱水素酵素(gapA及び/又はgapB)は、解糖に重要な酵素であり、グリセルアルデヒド3−リン酸の1,3−ビホスホ−D−グリセリン酸への変換を触媒する(Branlant G.and Branlant C.1985.Eur.J.Biochem.150:61〜66)。
ホスホケトラーゼ酵素を炭素に直接作用させるために、グリセルアルデヒド3−リン酸脱水素酵素の発現を調節(例えば、酵素活性を低下させるなど)して、より多量の炭素をフルクトース6−リン酸及びキシルロース5−リン酸へと変換させ、以降に続くメバロン酸、イソプレン及びイソプレノイドの産生量を増加させることもできる。グリセルアルデヒド3−リン酸脱水素酵素の活性を低下させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で低下させることができる。一部の例では、酵素活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%又は100%低下させる。一部の態様では、グリセルアルデヒド3−リン酸脱水素酵素は、内在性グリセルアルデヒド3−リン酸脱水素酵素の活性を減少させることにより調節される。このような調節は、内在性グリセルアルデヒド3−リン酸脱水素酵素遺伝子プロモータを常時低発現型合成プロモータにより置き換えることで実施できる。グリセルアルデヒド3−リン酸脱水素酵素をコードしている遺伝子を欠失させることもできる。グリセルアルデヒド3−リン酸脱水素酵素をコードしている遺伝子は、同様の反応を触媒するもののNADPHよりはNADHを産生するバチルス(Bacillus)の酵素と置き換えることもできる。グリセルアルデヒド3−リン酸脱水素酵素の活性を減少させることで、グリセルアルデヒド3−リン酸脱水素酵素の発現を減少させていない細胞と比較して、より多量にメバロン酸依存性生合成経路に炭素を取り込ませることができる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、異種発現される、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている核酸を1つ以上含み、かつグリセルアルデヒド3−リン酸脱水素酵素(gapA及び/又はgapB)の活性が減少するよう更に遺伝子操作されている。グリセルアルデヒド3−リン酸脱水素酵素イソ酵素の活性を調節(例えば、減少)させることも本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、異種発現される、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている核酸を1つ以上含み、かつグリセルアルデヒド3−リン酸脱水素酵素(gapA及び/又はgapB)イソ酵素の活性が減少するよう更に遺伝子操作されている。
エントナー・ドゥドロフ経路に関与する経路
エントナー・ドゥドロフ(ED)経路は、エムデン・マイヤーホフ・パルナス(EMP−解糖)経路とは異なる経路である。大腸菌(E. coli)などの一部の生物がED経路及びEMP経路の両方を内包するのに対し、その他の生物はいずれか1つの経路のみを有する。バチルス・サブチリス(Bacillus subtilis)は、EMP経路のみを有するのに対し、ザイモモナス・モビリス(Zymomonas mobilis)はED経路のみを有する(Peekhaus and Conway.1998.J.Bact.180:3495〜3502;Stulke and Hillen.2000.Annu.Rev.Microbiol.54,849〜880;Dawes et al.1966.Biochem.J.98:795〜803)。フルクトース二リン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)はエントナー・ドゥドロフ経路と相互作用し、フルクトース1,6−2リン酸のジヒドロキシアセトンリン酸(DHAP)及びグリセルアルデヒド3−リン酸(GAP)への変換を可逆的に触媒する(Baldwin S.A.,et.al.,Biochem J.(1978)169(3):633〜41)。
ホスホグルコン酸デヒドラターゼ(edd)は、6−ホスホ−D−グルコネートから一分子のH2Oを除去して2−デヒドロ−3−デオキシ−D−グルコネート6−リン酸を生成するのに対し、2−ケト−3−デオキシグルコネート6−リン酸アルドラーゼ(eda)はアルドール開裂を触媒する(Egan et al.1992.J.Bact.174:4638〜4646)。2つの遺伝子はオペロンの関係である。
ホスホケトラーゼ経路に指向する代謝経路をED経路に迂回させることもできる。ED経路に取り込まれる代謝産物が損失することを回避するため、ホスホグルコン酸デヒドラターゼ遺伝子(例えば、内在性ホスホグルコン酸デヒドラターゼ遺伝子)及び/又は2−ケト−3−デオキシグルコネート6−リン酸アルドラーゼ遺伝子(例えば、内在性2−ケト−3−デオキシグルコネート6−リン酸アルドラーゼ遺伝子)活性を減弱させる。1つの手法としては、ホスホグルコン酸デヒドラターゼ(edd)及び/又は2−ケト−3−デオキシグルコネート6−リン酸アルドラーゼ(eda)を欠失させることで減弱を行うことができる。この欠失は、クロラムフェニコール又はカナマイシンカセットにより片方又はいずれもの遺伝子を置き換え、その後カセットを除去することで導入される。これらの酵素活性を存在させずとも、ホスホケトラーゼ酵素によってより多くの炭素を取り込むことができ、ひいてはメバロン酸、イソプレン、又はイソプレノイドの収率を増加させることができる。
ホスホグルコン酸デヒドラターゼ(edd)及び/又は2−ケト−3−デオキシグルコネート6−リン酸アルドラーゼ(eda)の活性は、酵素に関係する他の分子操作により減少させることもできる。酵素活性を低下させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で低下させることができる。一部の例では、酵素活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%低下させる。
一部の場合では、内在性ホスホグルコン酸デヒドラターゼ遺伝子及び/又は内在性2−ケト−3−デオキシグルコネート6−リン酸アルドラーゼ遺伝子の活性を減弱させることにより、内在性ホスホグルコン酸デヒドラターゼ遺伝子及び/又は内在性酢酸キナーゼ2−ケト−3−デオキシグルコネート6−リン酸アルドラーゼ遺伝子の発現を減弱させていない細胞と比較して、多量の炭素がメバロン酸依存性生合成経路に取り込まれることになる。
ホスホケトラーゼ経路に指向する代謝経路をED経路又はEMP経路に迂回させることもできる。代謝産物の損失を回避し、フルクトース−6−リン酸(F6P)濃度を増加させるため、フルクトース二リン酸アルドラーゼ(例えば、内在性フルクトース二リン酸アルドラーゼ)活性を減弱させる。場合によっては、内在性フルクトース二リン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)遺伝子の活性を減弱させることで、内在性のフルクトース二リン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)遺伝子の発現を減弱させていない細胞と比較して、より多量にメバロン酸依存性生合成経路に炭素が取り込まれるようになる。一部の態様では、減弱は、フルクトース二リン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)を欠失させることにより実施される。この欠失は、クロラムフェニコール又はカナマイシンカセットにより遺伝子を置き換え、その後カセットを除去することで導入される。一部の態様では、フルクトース二リン酸アルドラーゼの活性は、内在性フルクトース二リン酸アルドラーゼの活性を減少させることにより調節される。このような調節は、内在性フルクトース二リン酸アルドラーゼ遺伝子プロモータを常時低発現型合成プロモータにより置き換えることで実施できる。これらの酵素活性を存在させずとも、ホスホケトラーゼ酵素によってより多くの炭素を取り込むことができ、ひいてはメバロン酸、イソプレン、又はイソプレノイドの収率を増加させることができる。フルクトース二リン酸アルドラーゼの活性は、酵素に関係する他の分子的操作によっても減少させることができる。酵素活性を低下させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で低下させることができる。一部の例では、酵素活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%低下させる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつフルクトース二リン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)の活性が減少するよう更に遺伝子操作されている。フルクトース二リン酸アルドラーゼのイソ酵素の活性の調節(例えば、減少)も、本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつフルクトース二リン酸アルドラーゼイソ酵素の活性が減少するよう更に遺伝子操作されている。
ペントースリン酸経路の酸化経路に関与する経路
大腸菌(E. coli)は、ペントースリン酸経路を使用してヘキソース及びペントースを分解し、各種代謝経路に関係する中間体を細胞に提供する。この経路は、NADPHの主要な生成経路でもある。ペントースリン酸経路は、酸化経路(グルコース6−リン酸1−脱水素酵素(zwf)、6−ホスホグルコノラクトナーゼ(pgl)又は6−ホスホグルコネート脱水素酵素(gnd)などの酵素による)及び非酸化経路(トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼ(talA又はtalB)、リボース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼ、リブロース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)及び/又はリブロース−5−リン酸3−エピメラーゼ(rpe)などの酵素による)から構成される(Sprenger.1995.Arch.Microbiol.164:324〜330)。
炭素にホスホケトラーゼ酵素を作用させるために、ペントースリン酸経路の非酸化経路(トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リブロース−5−リン酸エピメラーゼ、リブロース−5−リン酸イソメラーゼA、リブロース−5−リン酸イソメラーゼB、及び/又はリブロース−5−リン酸3−エピメラーゼ)の発現を調節して(例えば、酵素活性を増加させて)、より多量の炭素をフルクトース6−リン酸及びキシルロース5−リン酸へと変換させることにより、最終的にメバロン酸、イソプレン及びイソプレノイドの産生を増加させることができる。トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼ活性を増加させることで、活性を操作しなかった場合と比較して比活性又は総活性を任意の量で増加させることができる。一部の例では、酵素活性は少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%又は100%上昇する。一部の態様では、トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼの活性は、内在性トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼの活性を増強させることにより調節される。このような上昇は、内在性トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼ遺伝子のプロモータを常時高発現型の合成プロモータにより置き換えることで得られる。トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼをコードしている遺伝子を、プラスミド上の適切なプロモータの後にクローニングさせてもよい。トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼの活性を増大させることにより、トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼの発現を増加させていない細胞と比較して、多量の炭素をメバロン酸依存型の生合成経路に取り込ませることができる。
本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつトランスケトラーゼ(tktA及び/又はtktB)の活性が増加するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつトランスケトラーゼ(tktA及び/又はtktB)の活性が減少するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつトランスアルドラーゼ(talA又はtalB)の活性が増加するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつリボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)の活性が増加するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつリブロース−5−リン酸3−エピメラーゼ(rpe)の活性が増加するよう更に遺伝子操作されている。グルコース6−リン酸1−脱水素酵素(zwf)、6−ホスホグルコノラクトナーゼ(PGL)、6−ホスホグルコネート脱水素酵素(gnd)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼ(talA又はtalB)、リブロース−5−リン酸−エピメラーゼ、リボース−5−リン酸エピメラーゼ、リブロース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)及び/又はリブロース−5−リン酸3−エピメラーゼ(rpe)イソメラーゼの活性の調節(例えば、減少又は増加)も本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつグルコース6−リン酸1−脱水素酵素(zwf)イソ酵素の活性が増加するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつトランスケトラーゼ(tktA及び/又はtktB)イソ酵素の活性が増加するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつトランスケトラーゼ(tktA及び/又はtktB)イソ酵素の活性が減少するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつトランスアルドラーゼ(talA又はtalB)イソ酵素の活性が増加するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつリボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)イソ酵素の活性が増加するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつリブロース−5−リン酸3−エピメラーゼ(rpe)イソ酵素の活性が増加するよう更に遺伝子操作されている。
炭素に対しホスホケトラーゼ酵素により触媒反応させるために、グルコース6−リン酸1−脱水素酵素を調節する(例えば、酵素活性を減少させる)ことができる。一部の態様では、グルコース6−リン酸1−脱水素酵素(zwf)(例えば、内在性グルコース6−リン酸1−脱水素酵素遺伝子)の活性を減少又は減弱させることができる。特定の実施形態では、減弱は、グルコース6−リン酸1−脱水素酵素を欠失させることによってなされる。一部の態様では、グルコース6−リン酸1−脱水素酵素の活性は、内在性グルコース6−リン酸1−脱水素酵素の活性を減少させることにより調節される。このような調節は、内在性グルコース6−リン酸1−脱水素酵素遺伝子プロモータを常時低発現型合成プロモータにより置き換えることで実施できる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつグルコース6−リン酸1−脱水素酵素(zwf)の活性が減少するよう更に遺伝子操作されている。グルコース6−リン酸1−脱水素酵素イソ酵素の活性の調節(例えば、減少)も、本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつグルコース6−リン酸1−脱水素酵素イソ酵素の活性が減少するよう更に遺伝子操作されている。
ホスホフルクトキナーゼに関与する経路
ホスホフルクトキナーゼは、解糖系でフルクトース6−リン酸のリン酸化を触媒する重要な酵素である。大腸菌(E. coli)はpfkA及びpfkBによりコードされる2種のイソ酵素を有する。細胞におけるホスホフルクトキナーゼ活性の大部分はpfkAによるものである(Kotlarz et al.1975,Biochim.Biophys.Acta,381:257〜268)。
ホスホケトラーゼ酵素を炭素に直接作用させるために、ホスホフルクトキナーゼの発現を調節(例えば、酵素活性を低下させるなど)して、より多量の炭素をフルクトース6−リン酸及びキシルロース5−リン酸へと変換させ、以降に続くメバロン酸、イソプレン、及びイソプレノイドの産生量を増加させることもできる。ホスホフルクトキナーゼ活性を低下させることで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の量で低下させることができる。一部の例では、酵素活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%又は100%低下させる。一部の態様では、内在性ホスホフルクトキナーゼの活性を低下させることにより、ホスホフルクトキナーゼの活性を調節する。このような調節は、内在性ホスホフルクトキナーゼ遺伝子プロモータを常時低発現型合成プロモータと置き換えることにより行うことができる。ホスホフルクトキナーゼをコードしている遺伝子を欠失させてもよい。ホスホフルクトキナーゼの活性を低下させることで、メバロン酸依存性の生合成経路への炭素の取り込みを、ホスホフルクトキナーゼの発現を低下させていない細胞と比較して、増大させることができる。
本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつフルクトース6−リン酸(pfkA及び/又はpfkB)の活性が減少するよう更に遺伝子操作されている。フルクトース6−リン酸イソ酵素の活性の調節(例えば、減少)も、本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつフルクトース6−リン酸イソ酵素の活性が減少するよう更に遺伝子操作されている。
ピルビン酸脱水素酵素複合体の関与する経路
ピルビン酸のアセチル−CoAへの脱炭酸を触媒するピルビン酸脱水素酵素複合体は、遺伝子aceE、aceF、及びlpdAによりコードされるタンパク質から構成される。これらの遺伝子の転写は、複数の制御因子により制御される。したがって、当業者は、ピルビン酸脱水素酵素複合体の活性を調節して、アセチル−CoAを増加することができる。調節により、ピルビン酸脱水素酵素複合体の活性及び/又は発現(例えば、恒常的発現)を増加させることができる。このような調節は、異なる手法により、例えば、PL.6(aattcatataaaaaacatacagataaccatctgcggtgataaattatctctggcggtgttgacataaataccactggcggtgatactgagcacatcagcaggacgcactgaccaccatgaaggtg−ラムダプロモーター、GenBank NC_001416(配列番号14))などの強力な構成型プロモータをオペロンの前に配置することにより、あるいは構成的に(恒常的に)発現する合成プロモータを1種以上用いることにより行うことができる。
したがって、一態様では、ピルビン酸デヒドロゲナーゼの活性は、(a)ピルビン酸デヒドロゲナーゼ(E1)、(b)ジヒドロリポイルトランスアセチラーゼ、及び(c)ジヒドロリポイルデヒドロゲナーゼから構成されるピルビン酸デヒドロゲナーゼ複合体に関係する、1種以上の酵素の活性を増加させることで調節される。これらの酵素をコードしている任意の1、2、又は3つの遺伝子を調節して、ピルビン酸脱水素酵素の活性を増加させることができることは理解される。他の態様では、以降に更に説明するように、ピルビン酸脱水素酵素複合体の活性は、内在性ピルビン酸脱水素酵素複合体のリプレッサーの活性を減弱させることで調節できる。内在性ピルビン酸デヒドロゲナーゼ複合体のリプレッサーの活性は、内在性ピルビン酸デヒドロゲナーゼ複合体リプレッサー遺伝子を欠失させることにより減弱させることができる。
一部の場合では、ピルビン酸脱水素酵素複合体をコードする遺伝子の1つ以上は内在性遺伝子である。ピルビン酸デヒドロゲナーゼ複合体の活性を増加させるその他の方法としては、細胞に、(a)ピルビン酸デヒドロゲナーゼ(E1)、(b)ジヒドロリポイルトランスアセチラーゼ、及び(c)ジヒドロリポイルデヒドロゲナーゼからなる群に由来するポリペプチを1つ以上コードしている異種核酸を1つ以上導入することによるものが挙げられる。
これらの方法のうち任意のものを用いることで、ピルビン酸デヒドロゲナーゼ活性の調節されていない細胞と比較して、組換え細胞によるアセチルCo−Aの産生量を増加させることができる。ピルビン酸脱水素酵素の活性を調節することで、ピルビン酸脱水素酵素の発現を調節していない細胞と比較して、メバロン酸依存性の生合成経路への炭素原子の取り込みを増加させることができる。
ホスホトランスフェラーゼ系を包含する経路
ホスホエノールピルビン酸依存型ホスホトランスフェラーゼ系(PTS)は、工程において、膜を介した糖基質の輸送及びリン酸化を同時に行う多成分系であり、解糖中間体のホスホエノールピルビン酸(PEP)によるエネルギー供給に依存する系である。PTSを調節する遺伝子の大部分はオペロンとしてクラスター化している。例えば、大腸菌(E. coli)のptsオペロン(ptsHIcrr)は、ホスホエノールピルビン酸依存型ホスホトランスフェラーゼ系(PTS)に中心的な3種類のタンパク質、すなわちHPr(ptsH)、酵素I(ptsI)及びEIIIGlc(crr)タンパク質をコードしているptsH、ptsI、及びcrr遺伝子から構成される。これら3つの遺伝子は複合型オペロンとして構成され、末端遺伝子crrの発現の大部分は、ptsI内のプロモータ領域から開始される。ptsオペロンの遺伝子の他、ptsGは、ホスホトランスフェラーゼ系のグルコース特異的な輸送体ptsGをコードしている。このプロモータ領域から開始される転写は、カタボライト活性化タンパク質(CAP)−環状AMP(cAMP)の正の調節下にあり、グルコース(PTS基質)存在下での生育中に増強される。更に、ppsA遺伝子は、ホスホトランスフェラーゼ系(PTS)の活性に必要とされるホスホエノールピルビン酸(PEP)の産生に関係するホスホエノールピルビン酸シンターゼをコードする。ピルビン酸脱水素酵素経路又はPTS経路を介し、ホスホエノールピルビン酸シンターゼにより炭素の取り込みを調整する。Postma,P.W.,et al.,Microbiol Rev.(1993),57(3):543〜94)を参照されたい。この非特許文献は、参照によりその全体が本明細書に組み込まれる。
本明細書に記載の特定の実施形態では、ptsオペロンを下方調節(例えば、減弱)することで、宿主細胞による酢酸の利用を増強させることができる。PTSオペロンの活性を下方調節することで、比活性又は総活性を、活性を操作しなかった場合と比較して任意の程度で減少させることができる。一部の例では、複合体の活性を、少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%だけ減少させる。特定の実施形態では、減弱は、ptsオペロンの欠失により実施される。一部の態様では、PTS系の活性は、内在性ptsオペロンの活性を減少させることにより調節される。活性の減少は、ptsオペロン内の内在性プロモータを、常時低発現型合成プロモータにより置き換えることで実施できる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつptsオペロンの活性が減少するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつEI(ptsI)の活性が減少するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつEIICBGlc(ptsG)の活性が減少するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつEIIAGlc(crr)の活性が減少するよう更に遺伝子操作されている。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつHPr(ptsH)の活性が減少するよう更に遺伝子操作されている。ピルビン酸脱水素酵素による炭素の損失を減少させ、かつグルコースの取り込みに関係するPEP類を増加させるため、ホスホエノールピルビン酸シンターゼ(ppsA)の活性を向上させることができる。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつホスホエノールピルビン酸シンターゼ(ppsA)の活性が増加するよう更に遺伝子操作されている。本発明の任意の更なる態様では、PTSは下方制御され、グルコース輸送経路は上方調節される。グルコース輸送系は、限定するものではないが、ガラクトース(galP)及びグルコキナーゼ(glk)を包含する。一部の実施形態では、ptsオペロンは、下方調節され、ガラクトース(galP)遺伝子は上方調節され、かつグルコキナーゼ(glk)遺伝子は上方調節される。PTSのイソ酵素の活性の調節(例えば、減少)も、本明細書において企図される。本明細書において提供される、本発明の任意の態様では、組換え細胞は、本明細書で開示されるホスホケトラーゼポリペプチドをコードしている1つ以上の異種発現される核酸を含み、かつPTSイソ酵素の活性が減少するよう更に遺伝子操作されている。
キシロース利用を包含する経路
本明細書に記載の特定の実施形態では、植物バイオマス由来の糖を、メバロン酸など、イソプレノイド前駆体、イソプレン及び/又はイソプレノイドなどの所望の産物へと変換するにあたって、キシロースを利用することが望ましい。一部の生物では、キシロースを利用するにあたり、代謝用のフルクトース−6−リン酸に変換する際に、ペントースリン酸経路を使用する必要がある。生物は、グルコースによる異化代謝抑制を不活性化させることにより、あるいは他の生物において見られるキシロースオペロン由来の遺伝子を異種発現させることにより、キシロースの利用効率を増強させるよう遺伝子操作することができる。以下に記載されるとおりにキシロース経路を遺伝子操作することで、ホスホケトラーゼ経路によるメバロン酸、イソプレノイド前駆体、イソプレン及び/又はイソプレノイドの産生を増強させることができる。
キシロースの取り込み及びキシルロース−5−リン酸への変換後の、ホスホケトラーゼ経路への直接的な取り込みを増強することは効果的である。理論に束縛されるものではないが、これにより、炭素の取り込みをペントースリン酸経路へと迂回させることができる(ただし、一部のグリセルアルデヒド−3−リン酸は、必要に応じてPPPに回収される)。キシルロキナーゼ(xyulokinase)の発現増強を用い、キシルロース−5−リン酸の総生産量を増加させることができる。キシルロキナーゼの発現及び活性の最適化を用い、ホスホケトラーゼ経路を有する株におけるキシロースの利用を増強させることができる。所望されるキシルロキナーゼ(xyulokinase)は、宿主に内在する酵素又は宿主に適合性の任意の異種キシルロキナーゼ(xyulokinase)のいずれかであり得る。一態様では、効果を増加させるためキシロースオペロン以外の成分(例えば、キシロースイソメラーゼ)を過剰発現させることができる。別の実施形態では、他のキシロース経路酵素(例えば、キシロース還元酵素)を減弱させる(例えば、活性を減少又は欠失させる)必要がある場合もある。
適宜に、明細書に記載されるとおりのホスホケトラーゼ酵素を有するよう遺伝子操作した宿主細胞を、内在性又は異種のいずれかの形態のキシルロースイソメラーゼ及び/又はキシルロキナーゼ(xyulokinase)のいずれかを過剰発現するよう更に遺伝子操作して、総収率並びにメバロン酸、イソプレノイド前駆体、イソプレン及び/又はイソプレノイド産生を向上させることができる。
ペントースリン酸経路に関与するトランスアルドラーゼ及びトランスケトラーゼ酵素
嫌気的に又はヘテロ発酵を行い生育することのできる一部の微生物は、解糖経路の代わりに、あるいは解糖経路に加えて、ホスホケトラーゼ経路を包含する。この経路は、ペントースリン酸経路の酵素、トランスアルドラーゼ及びトランスケトラーゼの活性に依存する。したがって、本明細書に記載のとおりにホスホケトラーゼ酵素を有するよう遺伝子操作した宿主細胞を、内在性又は異種のいずれかの形態のトランスケトラーゼ及びトランスアルドラーゼを過剰発現して、経路の取り込みを増加させ、毒性中間体の濃度を減少させ、非産生的経路への中間体の迂回を減少させ、並びに総収率並びにメバロン酸、イソプレノイド前駆体、イソプレン及び/又はイソプレノイド産生を向上させるよう更に遺伝子操作することができる。
変異の組み合わせ
本明細書に記載の任意の酵素及び/又は酵素経路、本明細書に記載の任意の組み合わせ(2、3、4、5、6、7、8、9、10、11、12、13、又は14)の酵素及び/又は酵素経路を調節する分子操作が明らかに企図されることは理解されたい。組み合わせての記載を容易にするため、クエン酸シンターゼ(gltA)はAとして表記し、ホスホトランスアセチラーゼ(pta)はBとして表記し、酢酸キナーゼ(ackA)はCとして表記し、乳酸脱水素酵素(ldhA)はDとして表記し、グリセルアルデヒド3−リン酸脱水素酵素(gap)はEとして表記し、ピルビン酸脱炭酸酵素(aceE、aceF、及び/又はlpdA)はFとして表記し、ホスホグルコン酸デヒドラターゼ(edd)はGとして表記し、2−ケト−3−デオキシグルコン酸6−リン酸アルドラーゼ(eda)はHとして表記し、ホスホフルクトキナーゼはIとして表記し、トランスアルドラーゼはJとして表記し、トランスケトラーゼはKとして表記し、リブロース−5−リン酸−エピメラーゼはLとして表記し、リボース−5−リン酸エピメラーゼはMとして表記し、キシルキナーゼ(xylukinase)はNとして表記し、キシロースイソメラーゼはOとして表記し、及びキシリトール還元酵素はPとして表記し、リボース−5−リン酸イソメラーゼ(rpi)はQとして表記し、D−リブロース−5−リン酸3−エピメラーゼ(rpe)はRとして表記し、ホスホエノールピルビン酸シンターゼ(pps)はSとして表記し、フルクトースビスリン酸アルドラーゼ(fba)はTとして表記し、EI(ptsI)はUとして表記し、EIICBGlc(ptsG)はVとして表記し、EIIAGlc(crr)はWとして表記し、HPr(ptsH)はXとして表記し、ガラクトース(galP)はYとして表記し、グルコキナーゼ(glk)はZとして表記し、グルコース−6−リン酸脱水素酵素(zwf)はAAとして表記する。上記の通り、ピルビン酸脱炭酸酵素複合体のaceE、aceF、及び/又はlpdA酵素を単独で使用して、又は3つの酵素のうち2つを、又は3つ酵素のうち3つを使用して、ピルビン酸脱炭酸酵素の活性を増加させることができる。したがって、任意の及びすべての酵素の組み合わせは、本明細書においてA−Mとして表記されることが明示的に企図され、並びに任意の及びすべての酵素の組み合わせはA−AAとして表記される。更に、上記の任意の組み合わせを、本明細書に記載の任意の酵素及び/又は酵素経路(例えば、ホスホケトラーゼ、MVA経路ポリペプチド、イソプレンシンターゼ、DXP経路ポリペプチド)と組み合わせて使用することができる。
産生性増加に関係する他の制御因子及び要素
他の分子的操作を使用して、メバロン酸産生に指向する炭素取り込みを増加させることができる。このような方法のうちの1つは、メバロン酸経路に合流する経路に対する負の制御効果を減少させ、低下させるか、又は除去するものである。例えば、一部の場合では、遺伝子aceEF−lpdAはオペロンであり、pdhRの上流に遺伝子を4つ有する。遺伝子pdhRは、このオペロンの転写に対する負の制御因子である。pdhRは、ピルビン酸の非存在下で標的プロモータに結合し、転写を抑制する。この制御因子は同様の手法でndh及びcyoABCDも制御する(Ogasawara,H.et al.2007.J.Bact.189:5534〜5541)。一態様では、pdhR制御因子を欠失させることにより、ピルビン酸の供給及びそれに伴うメバロン酸、イソプレノイド前駆体、及びイソプレノイドの産生を向上させることができる。
他の実施形態では、上記で得られる任意の株を更に遺伝子操作して、エントナー・ドゥドロフ経路の活性を調節することもできる。ホスホグルコン酸デヒドラターゼ又はアルドラーゼをコードしている遺伝子を減弱又は欠失させることができる。他の実施形態では、上記で得られる任意の株は、酢酸キナーゼ又はクエン酸シンターゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、得られる株のうち任意の株は、ホスホフルクトキナーゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、上記で得られる任意の株は、グリセルアルデヒド−3−リン酸脱水素酵素の活性を調節するよう遺伝子操作することもできる。グリセルアルデヒド−3−リン酸脱水素酵素の活性は、活性を減少させることにより調節することができる。他の実施形態では、ペントースリン酸経路の非酸価経路の、例えば、トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼなどの酵素を過剰発現させることができる。
他の態様では、セドヘプツロース−1,7−ビスホスファターゼ/フルクトース−1,6−ビスホスフェートアルドラーゼ及びセドヘプツロース−1,7−ビスホスファターゼ/フルクトース−1,6−ビスホスフェートホスファターゼをコードしている異種核酸を導入することにより、細胞内アセチル−リン酸濃度を増加させるよう宿主細胞を更に遺伝子操作することができる。特定の実施形態では、これらの分子操作を有する宿主細胞には、トランスアルドラーゼ(talB)及びホスホフルクトキナーゼ(pfkA及び/又はpfkB)遺伝子の減弱又は欠失を組み合わせることにより、エリトロース4−リン酸、ジヒドロキシアセトンリン酸、及びグリセルアルデヒド3−リン酸を、セドヘプツロース7−リン酸及びフルクトース1−リン酸へと迅速に変換させることができる(図5を参照されたい)。
他の態様では、メバロン酸、イソプレン、イソプレノイド及びイソプレノイド前駆体の生産を改良させるために、PGLを欠損している細胞(各種大腸菌(E. coli)株など)に6−ホスホグルコノラクトナーゼ(PGL)を導入し、使用することができる。染色体の組み込み又はプラスミドなどの染色体外媒体を用い、PGLをコードしている遺伝子を導入することにより、PGLを導入することもできる。
本明細書に記載の各種酵素経路を調節して、メバロン酸産生に取り込まれる炭素を増加させるための宿主細胞(例えば、細菌宿主細胞変異)の他、本明細書に記載の宿主細胞は、ホスホケトラーゼポリペプチド、並びに限定するものではないが、mvaE及びmvaS遺伝子産物などの上流及びMVA経路下流由来のその他の酵素をコードしている遺伝子を含む。MVA経路に含まれる代表的なポリペプチドの非限定例としては、アセチル−CoAアセチルトランスフェラーゼ(AA−CoAチオラーゼ)ポリペプチド、3−ヒドロキシ−3−メチルグルタリル−CoAシンターゼ(HMG−CoAシンターゼ)ポリペプチド、3−ヒドロキシ−3−メチルグルタリル−CoAレダクターゼ(HMG−CoAレダクターゼ)ポリペプチド、メバロン酸キナーゼ(MVK)ポリペプチド、ホスホメバロン酸キナーゼ(PMK)ポリペプチド、ジホスホメバロン酸デカルボキシラーゼ(MVD)ポリペプチド、ホスホメバロン酸デカルボキシラーゼ(PMDC)ポリペプチド、イソペンテニルリン酸キナーゼ(IPK)ポリペプチド、IDIポリペプチド、及びMVA経路ポリペプチドの活性を2つ以上有するポリペプチド(例えば、融合ポリペプチド)が挙げられる。MVA経路のポリペプチドには、MVA経路のポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド及び融合ポリペプチドが含まれる。MVA経路の代表的な核酸としては、MVA経路に含まれるポリペプチドの活性を少なくとも1つ有するポリペプチド、ポリペプチド断片、ペプチド、又は融合ポリペプチドをコードしている核酸が挙げられる。MVA経路に含まれる代表的なポリペプチド及び核酸としては、本明細書に記載されるような任意の生物資源から天然に生じるポリペプチド及び核酸が挙げられる。
国際公開出願第2009/076676号、同第2010/003007号、及び同第2010/148150号に記載の、MVA経路に関係するポリペプチドの非限定例を使用できる。
例示的な細胞培養培地
本明細書で使用するとき、用語「最少培地(minimal medium又はminimal media)」は、概して細胞増殖に必要とされる最低限の栄養素を含有している増殖培地を指すが、常に1種以上のアミノ酸(例えば、1、2、3、4、5、6、7、8、9又は10種以上のアミノ酸)が存在していないわけではない。最少培地は、典型的には:(1)細菌を増殖させるための炭素源;(2)細菌種及び増殖条件によって異なる様々な塩;並びに(3)水;を含有する。炭素源は、グルコースなどの単糖から、本明細書で以下により詳細に記載されるような、他のバイオマスのより複雑な加水分解物、例えば、酵母エキスなどといった多様なものであり得る。塩は、概してマグネシウム、窒素、リン及びイオウなどの必須元素を提供し、細胞がタンパク質及び核酸を合成できるようにする。また、最少培地には、特定のプラスミド及び同様物を維持すべく選別するために、抗菌剤などの選択剤を添加することもできる。例えば、微生物が、例えばアンピシリン又はテトラサイクリンなどの特定の抗菌剤に耐性である場合、耐性を欠く細胞の増殖を阻害する目的で培地に抗菌剤を添加することができる。培地には、所望される生理学的又は生化学的特性について選別するのに必要とされる、例えば特定のアミノ酸などといった他の化合物を添加することができる。
宿主細胞を培養するにあたり、任意の最少培地処方を使用することができる。例示的な最少培地処方としては、例えば、M9最少培地及びTM3最少培地が挙げられる。M9最少培地は、1Lにつき(1)200mLの滅菌M9塩類(1L当たり64gのNa2HPO4−7H2O、15gのKH2PO4、2.5gのNaCl及び5.0gのNH4Cl);(2)2mLの1MのMgSO4(滅菌);(3)20mLの20%(w/v)グルコース(又は他の炭素源);及び(4)100μLの1MのCaCl2(滅菌)を含有する。TM3最少培地は、1L中に(1)K2HPO4(13.6g);(2)KH2PO4(13.6g);(3)MgSO4 *7H2O(2g);(4)クエン酸一水和物(2g);(5)クエン酸鉄アンモニウム(0.3g);(6)(NH4)2SO4(3.2g);(7)酵母エキス(0.2g);及び(8)1000X微量元素溶液(1mL)を含有する。pHは約6.8に調整し、溶液は濾過滅菌する。1000X微量元素は、1L中に(1)クエン酸一水和物(40g);(2)MnSO4 *H2O(30g);(3)NaCl(10g);(4)FeSO4 *7H2O(1g);(4)CoCl2 *6H2O(1g);(5)ZnSO4 *7H2O(1g);(6)CuSO4 *5H2O(100mg);(7)H3BO3(100mg);及び(8)NaMoO4 *2H2O(100mg)を含有する。pHは約3.0に調節する。
その他の最小培地の例は、(1)リン酸カリウムK2HPO4、(2)硫酸マグネシウムMgSO4 *7H2O、(3)クエン酸一水和物C6H8O7 *H2O、(4)クエン酸鉄アンモニウムNH4FeC6H5O7、(5)酵母エキス(biospringer)、(6)1000X改変微量金属溶液、(7)硫酸50%(w/v)、(8)foamblast 882(Emerald Performance Materials)及び(9)微量塩溶液を3.36mL含有する。すべての成分を同時に加え、脱イオン水に溶解させた後、加熱滅菌した。続いて室温に冷却し、水酸化アンモニウム(28%)によりpHを7.0に調節し、用量に調整する。滅菌後にビタミン溶液及びスペクチノマイシンを加え、pHを調整する。
宿主細胞を培養するにあたり任意の炭素源を使用することができる。用語「炭素源」は、宿主細胞又は微生物により代謝させることのできる、1つ以上の炭素を含有している化合物を指す。例えば、宿主細胞を培養するにあたり使用される細胞培地には、宿主細胞の生存能を維持させる又は宿主細胞を増殖させるのに好適な任意の炭素源を包含させることができる。一部の態様では、炭素源は炭水化物(例えば、単糖、二糖、オリゴ糖、又は多糖など)、又は転化糖(例えば、酵素により処理したスクロースシロップ)である。
一部の態様では、炭素源としては、酵母エキス又は酵母エキスの1つ以上の成分が挙げられる。一部の態様では、酵母エキスの濃度は、酵母エキスの0.1%(w/v)、0.09%(w/v)、0.08%(w/v)、0.07%(w/v)、0.06%(w/v)、0.05%(w/v)、0.04%(w/v)、0.03%(w/v)、0.02%(w/v)、又は0.01%(w/v)である。一部の態様では、炭素源には、酵母エキス(又は酵母エキスの1つ以上の成分)及び他の炭素源、例えばグルコースの両方を含む。
単糖の例としては、グルコース及びフルクトースが挙げられ、オリゴ糖の一例としては、ラクトース及びスクロースが挙げられ、並びに多糖の例としては、デンプン及びセルロースが挙げられる。例示的な炭水化物としては、C6糖(例えば、フルクトース、マンノース、ガラクトース、又はグルコース)及びC5糖(例えば、キシロース又はアラビノース)が挙げられる。
一部の態様では、本明細書に記載の細胞は、エネルギー源及び/又は炭素源として合成ガスを利用することができる。一部の実施形態では、合成ガスは少なくとも一酸化炭素及び水素を含有する。一部の実施形態では、合成ガスは、二酸化炭素、水、又は窒素のうちの1つ以上を更に含む。一部の実施形態では、合成ガス中の水素対一酸化炭素のモル比は、0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2.0、3.0、4.0、5.0、又は10.0である。一部の実施形態では、合成ガスは、10、20、30、40、50、60、70、80、又は90体積%の一酸化炭素を含む。一部の実施形態では、合成ガスは、10、20、30、40、50、60、70、80、又は90体積%の水素を含む。一部の実施形態では、合成ガスは、10、20、30、40、50、60、70、80、又は90体積%の二酸化炭素を含む。一部の実施形態では、合成ガスは、10、20、30、40、50、60、70、80、又は90体積%の水を含む。一部の実施形態では、合成ガスは、10、20、30、40、50、60、70、80、又は90体積%の窒素を含む。
合成ガスは、天然又は合成資源に由来するものであってよい。合成ガスの由来する資源は「原料」と呼ばれる。一部の実施形態では、合成ガスはバイオマス(例えば、木材、スイッチグラス、農業廃棄物、都市ごみ)又は糖質(例えば、糖類)に由来する。他の実施形態では、合成ガスは、石炭、石油、ケロゲン、タールサンド、オイルシェール、又は天然ガスに由来する。他の実施形態では、合成ガスは、ゴムタイヤなどのゴムに由来する。
合成ガスは、メタン改質、石炭液化、混焼、発酵反応、酵素反応、及びバイオマスの気化などの各種加工により原材料から生成することができる。バイオマスの気化は、バイオマスを、当量未満の酸素の存在下、約700℃以上の温度にて、反応器内で部分酸化することにより、を実施する。酸素は、気体、純酸素又は蒸気の形態でバイオリアクタに導入される。気化は主に3つの工程:1)バイオマス中に存在する任意の湿分を完全に乾燥させるよう最初に加熱する工程;2)酸化剤の非存在下でバイオマスを300〜500℃に加熱して気体、タール類、油類及び固体チャー残渣を生成する熱分解工程;並び3)固体チャー、タール類及び気体を気化して合成ガスの主成分を生成する工程において生じ得る。混焼は、石炭/バイオマス混合の気化により実施される。合成ガスの組成、例えば、合成ガスの成分の同一性及びモル比などは、合成ガスの由来する原材料及び原材料を合成ガスへと変換する方法によって異なる。
合成ガスは不純物を含有し得るものであり、性質及び量は、産生に使用した原料及び加工法のいずれもに応じて変化する。発酵は一部の不純物に寛容性を示すものの、合成ガス成分から、発酵槽及び関連する装置を汚す恐れのあるタール類及び粒子を除去する必要はある。イソプレン生成物を汚染し得る、揮発性有機化合物、酸性ガス、メタン、ベンゼン、トルエン、エチルベンゼン、キシレン、H2S、COS、CS2、HCl、O3、有機硫黄化合物、アンモニア、酸化窒素、窒素含有有機化合物、及び重金属蒸気などの化合物を除去することも望ましい。合成ガスからの不純物の除去は、ガス洗浄、固相吸着剤による処理、及び気体透過膜を用いる精製などのいくつかの手法により実施することができる。
例示的な細胞培養条件
本発明の組換え細胞を維持し及び増殖させるのに好適な材料及び方法は、以下、例えば、実施例の節に記載される。微生物培養物の維持及び増殖に好適な他の材料及び方法は当該技術分野において周知である。例示的な手法としては、国際公開第2009/076676号、米国特許出願第第2009/0203102号、国際公開第2010/003007号、米国特許出願第2010/0048964号、国際公開第2009/132220号、米国特許出願第2010/0003716号、Gerhardt et al.,編の一般細菌学に関係する手法についてのマニュアル、American Society for Microbiology,Washington,D.C.(1994)又はBrock in Biotechnology:テキスト「工業微生物学(Industrial Microbiology)」第2版(1989)Sinauer Associates,Inc.(Sunderland,MA)が挙げられる。一部の態様では、細胞は、宿主細胞に挿入された核酸によりコードされている、ホスホケトラーゼポリペプチド、並びに限定するものではないが、mvaE及びmvaS遺伝子産物、イソプレンシンターゼ、DXP経路(例えば、DXS)、IDI、又はPGLポリペプチドなどの、上流及びMVA経路下流由来のその他の酵素の発現を可能にする条件下で培養培地中で培養される。
細胞を培養するにあたり、標準的な細胞培養条件を使用することができる(例えば、国際公開第2004/033646号及び該当特許中に引用される参考文献を参照されたい)。一部の態様では、細胞は適切な温度、気体混合物、及びpH(例えば、約20℃〜約37℃、約6%〜約84%のCO2、及び約5〜約9のpH)にて増殖及び維持される。一部の態様では、細胞は適切な細胞培地中で35℃で増殖する。一部の態様では、発酵の際のpH範囲は約pH 5.0〜約pH 9.0(例えば、約pH 6.0〜約pH 8.0、又は約6.5〜約7.0)である。細胞は、宿主細胞に必要とされる条件に基づき、好気性、無酸素性、又は嫌気性条件下で生育させることができる。加えて、細胞を培養するにあたり、より特異的な細胞培養条件を使用することができる。例えば一部の実施形態では、組換え細胞(大腸菌(E. coli)細胞など)は、低〜通常コピー数のプラスミド中、高発現型プロモータの調節下で、ホスホケトラーゼポリペプチド並びに限定するものではないが、リステリア・グレイ(L. grayi)、エンテロコッカス・フェシウム(E. faecium)、エンテロコッカス・ガリナラム(E. gallinarum)、エンテロコッカス・カッセリフラバス(E. casseliflavus)及び/又はエンテロコッカス・フェカリス(E. faecalis)由来のmvaE及びmvaS遺伝子産物のmvaE及びmvaSポリペプチド酵素などの上流由来の酵素をコードしている1種以上の異種核酸を含み、34℃にて培養される。
使用することのできる標準的な培養条件、並びに回分式、流加式培養、又は連続式発酵などの発酵様式は、国際公開第2009/076676号、米国特許出願公開第2009/0203102号、国際公開第2010/003007号、米国特許出願公開第2010/0048964号、国際公開第2009/132220号、米国特許出願公開第2010/0003716号に記載されている。回分式発酵及び流加式発酵は一般的かつ当該技術分野では周知のものであり、その例は、Brock,Biotechnology:テキスト「工業微生物学(Industrial Microbiology)」第2版(1989)Sinauer Associates,Inc.に見ることができる。
一部の態様では、細胞はグルコース制限条件下で培養される。「グルコース制限条件」は、添加されるグルコースの量が、細胞により消費されるグルコース量の約105%以下(例えば、約100%、90%、80%、70%、60%、50%、40%、30%、20%、又は10%)であることを意味する。特定の態様では、培養培地に添加されるグルコース量は、特定の期間中に細胞により消費されるグルコース量とほぼ同様である。一部の態様では、細胞の増殖速度は、細胞培地中のグルコースの量により維持することのできる速度で細胞が増殖するよう、添加するグルコース量を制限することで制御される。一部の態様では、グルコースは細胞培養時に蓄積しない。様々な態様で、細胞はグルコース制限条件下で、約1、2、3、5、10、15、20、25、30、35、40、50、60、又は70時間以上培養される。様々な態様で、細胞は、細胞を培養する合計時間の長さの約5、10、15、20、25、30、35、40、50、60、70、80、90、95又は100%以上の時間にわたって、グルコース制限条件下で培養される。任意の特定の理論に束縛されることを意図するものではないが、グルコース制限条件は、細胞をより都合よく制御し得るものであると考えられる。
一部の態様では、組換え細胞は回分式培養で増殖させる。組換え細胞は、流加式培養又は連続式培養により生育させることもできる。加えて、組換え細胞は、限定するものではないが、上記のいずれかの最少培地などの最少培地で培養することができる。最少培地には、更に1.0%(w/v)グルコース又は任意の他の6単糖以下の糖などを添加してもよい。具体的には、最少培地には1%(w/v)、0.9%(w/v)、0.8%(w/v)、0.7%(w/v)、0.6%(w/v)、0.5%(w/v)、0.4%(w/v)、0.3%(w/v)、0.2%(w/v)、又は0.1%(w/v)のグルコースが添加される。加えて、最少培地には0.1%(w/v)以下の酵母エキスを添加してもよい。具体的には、最少培地には0.1%(w/v)、0.09%(w/v)、0.08%(w/v)、0.07%(w/v)、0.06%(w/v)、0.05%(w/v)、0.04%(w/v)、0.03%(w/v)、0.02%(w/v)又は0.01%(w/v)の酵母エキスを添加してもよい。あるいは、最少培地には1%(w/v)、0.9%(w/v)、0.8%(w/v)、0.7%(w/v)、0.6%(w/v)、0.5%(w/v)、0.4%(w/v)、0.3%(w/v)、0.2%(w/v)又は0.1%(w/v)のグルコース及び0.1%(w/v)、0.09%(w/v)、0.08%(w/v)、0.07%(w/v)、0.06%(w/v)、0.05%(w/v)、0.04%(w/v)、0.03%(w/v)、0.02%(w/v)又は0.01%(w/v)の酵母エキスを添加してもよい。
例示的な精製方法
一部の態様では、本明細書に記載の任意の方法は更に、産生された化合物を回収する工程を包含する。一部の態様では、本明細書に記載の任意の方法は更に、イソプレンを回収する工程を包含する。一部の態様では、イソプレンは吸着ストリッピングにより回収される(例えば、米国特許第2011/0178261号を参照されたい)。一部の態様では、本明細書に記載の任意の方法は更に異種ポリペプチドを回収する工程を包含する。一部の態様では、本明細書に記載の任意の方法は更に、テルペノイド又はカロテノイドを回収する工程を含む。
好適な精製方法は、米国特許出願公開第2010/0196977(A1)号により詳細に記載されている。
本明細書を通じ、各種特許、特許出願及び他の種類の刊行物(例えば、学術論文)を参照する。本明細書に引用する、本開示に関係する全ての特許、特許出願及び刊行物は、すべての目的に関し、参照によりその全文が本明細書に組み込まれる。
本発明は、実例として提供され、制限することを意味するものではない以降の実施例を参照することにより更に理解することができる。
実施例1:ビフィドバクテリウム・インファンティス(Bifidobacterium infantis)由来のホスホケトラーゼ酵素をコードしている遺伝子のクローニング
ビフィドバクテリウム・インファンティス(Bifidobacterium infantis)の染色体DNAをATCCから得た(ATCC# 15697D−5,ATCC(Manassas,VA))。プライマーCMP283:5’−ctgtatTCATGAcgagtcctgttattggcacc−3’(列番号30)及びCMP284:5’−ctctatGAATTCTCACTCGTTGTCGCCAGCG−3’(配列番号31)、テンプレートとして100ngのDNA、並びに製造元に従ってポリメラーゼHerculase II Fusion(Agilent,Santa Clara,CA)を使用して、ホスホケトラーゼ(PKL)酵素をコードしている遺伝子を増幅させた。精製後、消化によりBspHI及びEcoRIにより2798bpの断片を切り出し、NcoI/EcoRIにより消化したpTrcHis2B(Invitrogen,Carlsbad,CA)と連結して、プラスミドpCMP1090を生成した(配列番号15−図6)。
実施例2:ラクトバチルス・ロイテリ(Lactobacillus reuteri)株F275由来のホスホケトラーゼ酵素のクローニング
ラクトバチルス・ロイテリ(Lactobacillus reuteri)株F275の染色体DNAをATCC から得た(ATCC# 23272D−5,ATCC(Manassas,VA))。プライマーCMP34:5’−taaggaggaataaacATGGCAGTAGATTACGATTCCAAG−3’(配列番号32)及びCMP335:5’−ttctagaaagcttcgttacttaagacccttccaagtccag−3’(配列番号33)、テンプレートとして100ngのDNA、並びに製造元に従ってポリメラーゼHerculase II Fusion(Agilent(Santa Clara,CA))を使用して、ホスホケトラーゼ(PKL)酵素をコードしている遺伝子を増幅させた。精製後、2442bpの断片を、NcoI/EcoRIにより消化したpTrcHis2B(Invitrogen(Carlsbad,CA))にGENEARTシームレス・クローニング・キット(Invitrogen(Carlsbad,CA))を使用して組み込み、プラスミドpCMP1029を生成した(配列番号16−図7)。
実施例3:株CMP451、CMP674、CMP1015及びCMP1047の構築(BL21 GI1.2gltA ldhA)
大腸菌(E. coli)株BL21(Novagen)中のクエン酸シンターゼ遺伝子(gltA)の前に位置するプロモータを、常時低発現型プロモータ、すなわちGI1.2(米国特許第7,371,558号)によりあらかじめ置き換えた。2つの野生型プロモータはgltAに関し記載されている(Wilde,R,and J.Guest.1986.J.Gen.Microbiol.132:3239〜3251)。末端プロモータの−35領域に直後に合成プロモータを挿入した。プライマーUpgltACm−F(5’−TATTTAATTTTTAATCATCTAATTTGACAATCATTCAACAAAGTTGTTACAATTAACCCTCACTAAAGGGCGG−3’(配列番号34))及びDngltA1.xgiCm−R(5’−TCAACAGCTGTATCCCCGTTGAGGGTGAGTTTTGCTTTTGTATCAGCCATATATTCCACCAGCTATTTGTTAGTGAATAAAAGTGGTTGAATTATTTGCTCAGGATGTGGCATHGTCAAGGGCTAATACGACTCACTATAGGGCTCG−3’(配列番号35))、及びテンプレートとしてGene Bridges(Heidelberg,Germany)プラスミドFRT−gb2−Cm−FRTを使用し、PCR産物を得た。PCR産物を精製し、λredを用いる組み換えに製造元(Gene Bridges,Heidelberg,Germany)による記載の通りに使用した。更なる評価にあたって、複数種のコロニーを選別した。プロモータ領域は、プライマーgltAPromSeqF:5’−GGCAGTATAGGCTGTTCACAAAATC−3’(配列番号36)及びgltApromSeqR:5’−CTTGACCCAGCGTGCCTTTCAGC−3’(配列番号37)と、テンプレートとしてコロニーから抽出したDNA(コロニーを30μLのH2Oに再懸濁し、95℃で4分加熱し、スピンダウンしたもの。50μLのPCR反応には、この溶液のうち2μLをテンプレートとして使用する)とを使用しPCR増幅させた。得られたPCR産物の配列決定結果を観察した後、GI1.2プロモータ(米国特許第7,371,558号)を保有しているコロニーを、CMP141と命名した。
CMP258(米国特許出願第/号を参照されたい)にMCM521株(米国特許出願第/号を参照されたい)のP1可溶化液を形質導入し、MD09−313株を構築し、20μg/mLのカナマイシンを含有させたルリア・ベルターニ培地のプレートにてコロニーを選択した。Ausubel,et al.,Current Protocols in Molecular Biology,John Wiley and Sons,Incに記載の方法に従ってP1可溶化液を調製した。製造元により推奨されるプロトコルを用い(Gene Bridges,Heidelberg,Germany)カナマイシンマーカーを除去し、MD09−314株を作製する。
株CMP141からP1可溶化液を作製し、株MD09−314に形質導入してCMP440を作成するのに使用した。製造元により推奨されるプロトコルを用い(Gene Bridges,Heidelberg,Germany)クロラムフェニコールマーカーを除去して株CMP451を作製した。
テンプレートとしてプラスミドpKD3(Datsenko,K.,and Wanner,B.2000.PNAS 97:6640〜6645)を使用し、プライマーCMP171(5’−AAAATTTTCATTCTGTGACAGAGAAAAAGTAGCCGAAGATGACGGTTTGTCACATGGAGTTGGCAGGATGTTTGATTACATGGGAATTAGCCATGGTCC−3’(配列番号38))及びCMP172(5’−GACCAGCCGCGTAACCTGGCAAAATCGGTTACGGTTGAGTAATAAATGGATGCCCTGCGTAAGCGGGGCATTTTTCTTGGTGTAGGCTGGAGCTGCTTCG−3’(配列番号39))を使用して、DNAホモログによりλ挿入部位attBの上流及び下流部位とに隣接しているクロラムフェニコールマーカーを含有しているDNA断片をPCRにより増幅した。得られたPCR産物を、製造元(Gene Bridges,Heidelberg,Germany)の推奨する通りにBL21(Novagen)における組み換え反応に使用し、λ挿入部位attBに組み入れた。これにより生成されたCMP646株を、LB+5μg/mLのクロラムフェニコールにて選択した。CMP646のP1可溶化液を調製し、株CMP451に対する形質導入反応に使用し、この株の染色体からメバロン酸経路下流の遺伝子(メバロン酸キナーゼ、ホスホメバロン酸キナーゼ、ジホスホメバロン酸デカルボキシラーゼ、及びイソペンテニル二リン酸イソメラーゼをコードしている)を除去した。形質導入物をLB+クロラムフェニコール5μg/mLに播種し、各形質導入からコロニーを1つずつ選択し、CMP674と名づけた。
テンプレートとしてKeio collection(Baba et al.2006.Mol.Syst.Biol.2:2006.0008)のJW1375株を使用し、プライマーはldhAseqR(5’−GGCTTACCGTTTACGCTTTCCAGC−3’)(配列番号40)及びldhAseqF2(5’−CTAATGCAATACGTGTCCCGAGC−3’)(配列番号41)と、を使用し、カナマイシンマーカーにより破壊されているldhA遺伝子を含有しているDNA断片をPCRにより増幅させた。得られたPCR産物を、製造元(Gene Bridges,Heidelberg,Germany)の推奨する通りに組み換え反応に使用し、CMP674株のldhA座に組み入れた。この株をCMP1015と命名した。株に電気穿孔法によりpCP20を導入して、クロラムフェニコール及びカナマイシンマーカーを同時に除去し(Datsenko and Wanner.2000.PNAS 97:6640〜6645)30℃下でLB+50ug/mLのカルベニシリンで2つのコロニーを選択し、次に、これらのコロニーを、42℃下でLBプレートに再画線した。これらのプレートからCmS及びKanSコロニーを選択し、CMP1047と名づけた。
実施例4:株CMP1036、1038及び1040の構築
プラスミドpTrcHis2B、pCMP1090(ビフィドバクテリウム・インファンティス(Bifidobacterium infantis)由来のPKL)及びpCMP1029(ラクトバチルス・ロイテリ(Lactobacillus reuteri)由来のPKL)の存在下で、株CMP674に電気穿孔法を行った。LB+カルベニシリン50μg/mLで、コロニーを単離した各形質転換からコロニーを1つ選択し、それぞれCMP1036、CMP1038及びCMP1040と命名した。
実施例5:株CMP1036、1038及び1040におかるアセチルリン酸の測定
(i)材料
TM3培地の組成(発酵培地1L当たり):
K2HPO4を13.6g、KH2PO4を13.6g、MgSO4 *7H2Oを2g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、酵母エキスを0.2g、1000X微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。0.22μmフィルタで培地を濾過滅菌する。pHを調整及び滅菌後にグルコース10.0g及び抗菌剤を加えた。
1000X微量金属溶液(発酵培地1L当たり):
クエン酸*H2O 40g、MnSO4 *H2O 30g、NaCl 10g、FeSO4*7H2O 1g、CoCl2*6H2O 1g、ZnSO4*7H2O 1g、CuSO4*5H2O 100mg、H3BO3 100mg、NaMoO4*2H2O 100mg。各成分を1成分ずつ脱イオン水に溶解する。pHをHCl/NaOHで3.0に調整した後、溶液を用量に調整し、0.22μmフィルタでフィルタ滅菌する。
(ii)実験手順
ルリア・ベルターニブロス+抗生物質で細胞を一晩生育させた。翌日、OD600が0.05になるよう、50ug/mLのカルベニシリンを含有させた20mLのTM3培地(250mLのバッフル付き三角フラスコ)により細胞を希釈し、34℃かつ200rpmでインキュベートする。2時間生育させた後、OD600を測定し、200uMのIPTGを加える。3.5時間以上経過後、1.5mLのサンプルを遠心分離し、上清を廃棄し、ペレットを100μLの無水冷メタノールに再懸濁した。
(iii)細胞内アセチル−リン酸の測定
アセチル−リン酸を抽出するため、OD 0.57〜2.26へと生育させた大腸菌(E. coli)細胞1.5mLを遠心分離によりスピンダウンし、100μLの無水冷メタノールをペレットに加えた。メタノールにより急冷したサンプルを−20℃で数日間保存した。以降のサンプル加工には、細胞を穏やかに再懸濁し、−9℃で5分間遠心分離し、上清を清潔なバイアル瓶に吸引する工程を包含した。2%酢酸を含有させた75μLの水により、ペレットを2回再抽出した。各抽出後、−9℃にて遠心分離することにより細胞片をペレット化し、全3つの抽出物由来の上清を一緒に集め、1μLのトリブチルアミンを添加した。Thermo Finnigan TSQシステム(Thermo Electron Corporation(San Jose,CA))を使用し、LCMSによるアセチルリン酸の質量分析を実施した。システムの調節、データ収集、及び質量スペクトルデータの評価は、ソフトウェアXCalibur及びLCQuan(Thermo Electron Corp)を使用して行った。移動相を勾配させて、流速0.4mL/分にてSynergi MAX−RP 5μM HPLCカラム(150×2mm,Phenomenex)に加えた。勾配プロファイルは、溶媒Aは15mMトリブチルアミン/10mM酢酸水溶液、溶媒Bはメタノール、及び溶媒Cは水として、99% A及び1% Bをt=0〜1分;80% A及び20% Bをt=11分;75% B及び25% Cをt=12〜14分;99% A及び1% Bをt=15〜16分のように適用した。エレクトロスプレーイオン化質量分析(ESI−MS/MS)ネガティブモード(ESIスプレー電位2.5〜3.0kV、イオントランスファーチューブ温度390℃)でを用い、アセチルリン酸の質量検出を実施したところ、前駆体イオンのm/z値は138.9であった。PO3 −プロダクトイオン(m/z=79.0,衝突エネルギー20V,衝突ガス圧0.22kPa(1.7mTorr),Rt=13.2分)により生成されたピークの積分強度をもとに、アセチルリン酸濃度を求めた。標準アセチルリン酸(Sigma−Aldrich)を注入することにより得られた検量線を使用して、細胞抽出物中の代謝産物濃度を算出した。OD=200の培養液1mLにおける消費に基づき、アセチルリン酸の細胞内濃度を求めた。すべての細胞の総体積は50Mlである(図8)。
(iv)結果
ホスホケトラーゼを発現している株では、ホスホケトラーゼを発現しない対照株よりも高濃度のアセチルリン酸細胞内濃度を有した(CMP1040及びCMP1038)(図8)。
実施例6:CMP1053、1055及び1057の構築
プラスミドpMCM82(発現ベクターMCM82(米国特許出願公開番号第2010/0196977号)とともに、プラスミドpTrcHis2B(Invitrogen(Carlsbad,CA))、pCMP1090(ビフィドバクテリウム・インファンティス(Bifidobacterium infantis)由来のPKL)又はpCMP1029(ラクトバチルス・ロイテリ(Lactobacillus reuteri)PKL)を使用して、CMP1047を形質転換させた。宿主CMP1047を対数増殖期の中期まで34℃にてLBで生育させ、電気穿孔に備え、培養物の当量の氷冷ddH2Oにより2回洗浄し、培養物の1/10当量の氷冷ddH2Oに再懸濁した。100μLの細胞懸濁液を1μLの各プラスミドDNAと組み合わせて、2mmの電気穿孔法キュベットに入れ、25uFD、200オーム、2.5kVで電気穿孔し、直ちに1mLのLBにより冷却した。細胞を回収し、34℃にて1時間振盪し、次に50μg/mLのスペクチノマイシン+50μg/mLのカルベニシリンを添加したLBプレートにより34℃にて一晩形質転換体させた。各形質転換用にコロニーを1つ選択し、それぞれCMP1053、1055及び1057と命名した。
他の実施形態では、上記で得られる任意の株を更に遺伝子操作して、エントナー・ドゥドロフ経路の活性を調節することもできる。ホスホグルコン酸デヒドラターゼ又はアルドラーゼをコードしている遺伝子を減弱又は欠失させることができる。他の実施形態では、上記で得られる任意の株は、酢酸キナーゼ又はクエン酸シンターゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、得られる株のうち任意の株は、ホスホフルクトキナーゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、上記で得られる任意の株は、グリセルアルデヒド−3−リン酸脱水素酵素の活性を調節するよう遺伝子操作することもできる。グリセルアルデヒド3−リン酸脱水素酵素の活性は、活性を減少させることにより調節することができる。他の実施形態では、ペントースリン酸経路の非酸価経路の、例えば、トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼなどの酵素を過剰発現させることができる。
実施例7:株CMP1053、1055及び1057によるメバロン酸産生
(i)材料
TM3培地の組成(発酵培地1L当たり):
K2HPO4を13.6g、KH2PO4を13.6g、MgSO4 *7H2Oを2g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、酵母エキスを0.2g、1000X微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。0.22μmフィルタで培地を濾過滅菌する。pH調整及び滅菌後にグルコース10.0g及び抗菌剤を添加する。
1000X微量金属溶液(発酵培地1L当たり):
クエン酸*H2Oを40g、MnSO4 *H2Oを30g、NaClを10g、FeSO4*7H2Oを1g、CoCl2*6H2Oを1g、ZnSO4*7H2Oを1g、CuSO4*5H2Oを100mg、H3BO3を100mg、NaMoO4*2H2Oを100mg。各成分を1成分ずつ脱イオン水に溶解する。pHをHCl/NaOHで3.0に調整した後、溶液を用量に調整し、0.22μmフィルタでフィルタ滅菌する。
(ii)実験手順
ルリア・ベルターニブロス+抗生物質で細胞を一晩生育させる。翌日、OD600が0.05になるよう、250mLのバッフル付き三角フラスコにおいて細胞を20mLのTM3培地(50μg/mLのスペクチノマイシン及び50μg/mLのカルベニシリンを含有)により希釈し、34℃かつ200rpmでインキュベートする。2時間生育させた後、OD600を測定し、200uMのIPTGを加える。発酵工程の間、規則的に試料を採取する。各時点でOD600を測定する。24時間後、メバロン酸をHPLCにより解析した。次の様式でHPLC解析を実施した:300μLのブロスに54μLの10%(w/v)H2SO4を加え、混合物を氷上で5分インキュベートした。次に、試料を14,000×gで5分遠心分離し、HPLC解析のため上清を回収し、次の条件でHPLC解析を実施した:(1)BioRad−AminexHPX−87Hイオン排除カラム(300mm×7.8mm)(カタログ番号125−0140)(BioRad,Hercules,California);(2)カラム温度=50℃;(3)BioRad−マイクロガードCarbo−H詰め替えカートリッジ(30mm×4.6mm)(カタログ番号125−0129)(BioRad);(4)移動相=0.01N H2SO4;(5)流速=0.6mL/分;(6)およその圧力=約6550.0kPa(950psi);(7)装填量=100μL;(8)実施時間=26分。
(iii)結果:
ホスホケトラーゼを発現している株は、対照株よりも生育が遅かった(図9)。CMP1057(ラクトバチルス・ロイテリ(L. reuteri)ホスホケトラーゼ遺伝子を発現している)は、対照の空プラスミド又はビフィドバクテリウム・インファンティス(B. infantis)由来のホスホケトラーゼを含有している株と比較して、多量にメバロン酸を産生した(図10)。
実施例8:イソプレンを産生し、ホスホケトラーゼを発現している株の構築
MCM521可溶化液を使用し、CMP674にメバロン酸経路の下流を形質導入する(表2を参照されたい)。製造元に従いカナマイシンマーカーを除去する(Gene Bridges、Heidelberg、Germany)。MCM521の下流の経路は、別の遺伝子を使用することにより各遺伝子の前に位置するrbsを改変し、オペロン上流のプロモータを変更することで改変することができる。lacI、イソプレンシンターゼ及びメタノサルシナ・マゼイ(M. mazei)メバロン酸キナーゼを発現している発現プラスミドの、プラスミドpMCM82(発現ベクターMCM82(米国特許出願公開番号第2010/0196977号))及びプラスミドpCMP1090(ビフィドバクテリウム・インファンティス(Bifidobacterium infantis)由来のPKL)、pCMP1029(ラクトバチルス・ロイテリ(Lactobacillus reuteri)由来のPKL)又はpTrcHis2BをCMP1047に電気穿孔した(2段階で)。LB+スペクチノマイシン50μg/mL+カルベニシリン50μg/mL+クロラムフェニコール25μg/mLでコロニーを選別する。
他の実施形態では、上記で得られる任意の株を更に遺伝子操作して、エントナー・ドゥドロフ経路の活性を調節することもできる。ホスホグルコン酸デヒドラターゼ又はアルドラーゼをコードしている遺伝子を減弱又は欠失させることができる。他の実施形態では、上記で得られる任意の株は、酢酸キナーゼ又はクエン酸シンターゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、得られる株のうち任意の株は、ホスホフルクトキナーゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、上記で得られる任意の株は、グリセルアルデヒド−3−リン酸脱水素酵素の活性を調節するよう遺伝子操作することもできる。グリセルアルデヒド−3−リン酸脱水素酵素の活性は、活性を減少させることにより調節することができる。他の実施形態では、ペントースリン酸経路の非酸価経路の、例えば、トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リボース−5−リン酸エピメラーゼなどの酵素を過剰発現させることができる。
実施例9:メバロン酸経路上流を発現しているプラスミド、lacI、イソプレンシンターゼ及びメバロン酸キナーゼを発現しているプラスミド、及びホスホケトラーゼを発現しているプラスミドを保有している株によるイソプレンの産生と、メバロン酸経路上流を発現しているプラスミド、lacI、イソプレンシンターゼ及びメバロン酸キナーゼを発現しているプラスミド、並びに空プラスミド(pTrcHis2B)を保有している細胞によるイソプレンの産生との比較
(i)材料
TM3培地の組成(発酵培地1L当たり):
K2HPO4を13.6g、KH2PO4を13.6g、MgSO4 *7H2Oを2g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、酵母エキスを0.2g、1000X微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。0.22μmフィルタで培地を濾過滅菌する。pH調整及び滅菌後にグルコース10.0g及び抗菌剤を添加する。
1000X微量金属溶液(発酵培地1L当たり):
クエン酸*H2Oを40g、MnSO4 *H2Oを30g、NaClを10g、FeSO4*7H2Oを1g、CoCl2*6H2Oを1g、ZnSO4*7H2Oを1g、CuSO4 *5H2Oを100mg、H3BO3を100mg、NaMoO4 *2H2Oを100mg。各成分を1成分ずつ脱イオン水に溶解する。pHをHCl/NaOHで3.0に調整した後、溶液を用量に調整し、0.22μmフィルタでフィルタ滅菌する。
(ii)実験手順
ルリア・ベルターニブロス+抗生物質で細胞を一晩生育させる。翌日、OD600が0.1になるよう、250mLのバッフル付き三角フラスコにおいて細胞を20mLのTM3培地(50μg/mLのスペクチノマイシン、25μg/mLのクロラムフェニコール、及び50μg/mLカルベニシリンを含有)により希釈し、34℃かつ200rpmでインキュベートする。2時間生育させた後、OD600を測定し、200uMのIPTGを加える。発酵工程の間、規則的に試料を採取する。各時点でOD600を測定する。また、ガスクロマトグラフ−マススペクトロメータ(GC−MS)(Agilent)によるヘッドスペース解析によりイソプレンのオフガス分析を行う。100μLの全ブロスをGCバイアルに入れ密閉し、34℃かつ200rpmで30分間インキュベートする。70℃で7分のインキュベーションからなる熱殺菌工程後、試料をGCに装填する。記録される比産生量は、イソプレン量のGCによる読み取り値(μg/L)を、インキュベート時間(30分)及びOD600測定値で除算した値である。
(iii)結果:
ホスホケトラーゼを発現している株は、ホスホケトラーゼを発現しない対照株よりも生育が遅かった。ホスホケトラーゼポリペプチドを発現している株において、イソプレンの比産生性、収率、CPI、及び/又は力価の増加が観察されたことから、ホスホケトラーゼポリペプチドを発現している株は、対照の空プラスミド(すなわち、ホスホケトラーゼを発現しない株)を含有している株と比較して、イソプレンの産生の増強を示す。
実施例10:ホスホケトラーゼを発現しかつアモルファジエン又はファルネセンを産生している株の構築
MCM521可溶化液を使用し、CMP674にメバロン酸経路の下流を形質導入する(表2を参照されたい)。製造元に従いカナマイシンマーカーを除去する(Gene Bridges、Heidelberg、Germany)。MCM521の下流の経路は、別の遺伝子を使用することにより各遺伝子の前に位置するrbsを改変し、オペロン上流のプロモータを変更することで改変することができる。染色体上のプロモータ及び/又はrbsを変更させるか、又はプラスミドから発現させるかのいずれかにより、ファルネシル二リン酸シンターゼ(ispA)を過剰発現させる。
実施例8の、lacI、イソプレンシンターゼ、及びメタノサルシナ・マゼイ(M. mazei)メバロン酸キナーゼを発現している発現プラスミドを改変して、イソプレンシンターゼをコードしている遺伝子を、ファルネセンシンターゼ又はアモルファジエンシンターゼをコードしているコドン最適化した遺伝子により置き換えた。次の発現プラスミドをコンピテントな宿主細胞に電気穿孔法により導入した(2段階で):(i)lacI、ファルネセンシンターゼ又はアモルファジエンシンターゼ、及びメタノサルシナ・マゼイ(M. mazei)メバロン酸キナーゼを有するプラスミド、(ii)pMCM82(発現ベクターMCM82(米国特許出願公開番号第2010/0196977号)、並びに(iii)pCMP1090(ビフィドバクテリウム・インファンティス(Bifidobacterium infantis)由来のPKL)、又はpCMP1029(ラクトバチルス・ロイテリ(Lactobacillus reuteri)由来のPKL)又はpTrcHis2B。LB+スペクチノマイシン50μg/mL+カルベニシリン50μg/mL+クロラムフェニコール25μg/mLでコロニーを選別する。
他の実施形態では、上記で得られる任意の株を更に遺伝子操作して、エントナー・ドゥドロフ経路の活性を調節することもできる。ホスホグルコン酸デヒドラターゼ又はアルドラーゼをコードしている遺伝子を減弱又は欠失させることができる。他の実施形態では、上記で得られる任意の株は、酢酸キナーゼ又はクエン酸シンターゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、得られる株のうち任意の株は、ホスホフルクトキナーゼの活性を減少又は除去するよう遺伝子操作することもできる。他の実施形態では、上記で得られる任意の株は、グリセルアルデヒド−3−リン酸脱水素酵素の活性を調節するよう遺伝子操作することもできる。グリセルアルデヒド−3−リン酸脱水素酵素の活性は、活性を減少させることにより調節することができる。他の実施形態では、ペントースリン酸経路の非酸価経路の、例えば、トランスケトラーゼ、トランスアルドラーゼ、リブロース−5−リン酸−エピメラーゼ及び(又は)リブロース−5−リン酸エピメラーゼなどの酵素を過剰発現させることができる。
実施例11:ホスホケトラーゼを発現しているプラスミドを含有している株におけるアモルファジエン又はファルネセンの産生と対照株におけるアモルファジエン又はファルネセンの産生との比較
(i)材料
TM3培地の組成(発酵培地1L当たり):
K2HPO4を13.6g、KH2PO4を13.6g、MgSO4 *7H2Oを2g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、酵母エキスを0.2g、1000X微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。次に、0.22μmのフィルタを用い培地を濾過滅菌した。滅菌及びpH調整後にグルコース10.0g及び抗菌剤を添加する。
1000X微量金属溶液(発酵培地1Lあたり):
クエン酸*H2Oを40g、MnSO4 *H2Oを30g、NaClを10g、FeSO4*7H2Oを1g、CoCl2*6H2Oを1g、ZnSO4*7H2Oを1g、CuSO4 *5H2Oを100mg、H3BO3を100mg、NaMoO4 *2H2Oを100mg。各成分を1成分ずつ脱イオン水に溶解する。pHをHCl/NaOHで3.0に調整した後、溶液を用量に調整し、0.22μmフィルタでフィルタ滅菌する。
(ii)実験手順
ルリア・ベルターニブロス+抗生物質で細胞を一晩生育させる。翌日、OD600が0.05になるよう、250mLのバッフル付き三角フラスコにおいて細胞を20mLのTM3培地(50μg/mLのスペクチノマイシン、25μg/mLのクロラムフェニコール、及び50μg/mLカルベニシリンを含有)により希釈し、34℃かつ200rpmでインキュベートする。接種前に、各培養フラスコには20%(v/v)ドデカン(Sigma−Aldrich)を積層し、これまでに記載のとおり、揮発性セスキテルペン産物を捕捉する(Newman et.al.,2006)。
2時間生育させた後、OD600を測定し、0.05〜0.40mMのイソプロピルβ−D−1−チオガラクトピラノシド(IPTG)を加えた。発酵工程の間、規則的に試料を採取する。各時点でOD600を測定する。また、積層したドデカンを酢酸エチルに希釈し、有機層中のアモルファジエン又はファルネセン濃度を分析する。ドデカン/エチルアセテート抽出物を、従来記載されている通りの(Martin et.al.,Nat.Biotechnol.2003,21:96〜802)GC−MS法により、アモルファジエンの分子イオン(204m/z)及び189m/zフラグメントイオン又はファルネセンの分子イオン(204m/z)の観察により解析する。濃度既知のアモルファジエン又はファルネセン試料を装填して、それぞれアモルファジエン又はファルネセンの標準曲線を生成する。試料中のアモルファジエン又はファルネセンの量は、それぞれアモルファジエン又はファルネセンの標準曲線を使用し算出する。
(iii)結果
ホスホケトラーゼポリペプチドを発現している株を、同様のバックグラウンドを持ち、空プラスミド(すなわち、ホスホケトラーゼポリペプチドを不含有である)を含有している株と比較する。ホスホケトラーゼポリペプチドを発現している株において、アモルファジエン又はファルネセンの比産生性、収率、CPI、及び/又は力価の増加が観察されたことから、ホスホケトラーゼポリペプチドを発現している株は、対照の空プラスミド(すなわち、ホスホケトラーゼを発現しない株)を含有している株と比較して、アモルファジエン又はファルネセンの産生の増強を示す。
(iv)引用文献:
Newman,J.D.,Marshal,J.L.,Chang,M.C.Y.,Nowroozi,F.,Paradise,E.M.,Pitera,D.J.,Newman,K.L.,Keasling,J.D.,2006.High−level production of amorpha−4,11−diene in a two−phase partitioning bioreactor of metabolically engineered E.coli.Biotechnol.Bioeng.95,684〜691。
Martin,V.J.,Pitera,D.J.,Withers,S.T.,Newman,J.D.,Keasling,J.D.,2003.Engineering a mevalonate pathway in E.coli for production of terpenoids.Nat.Biotechnol.21,796〜802。
実施例12:ホスホケトラーゼを発現している組み換え宿主細胞における15L規模でのメバロン酸(MVA)産生
ホスホケトラーゼポリペプチドをコードしている異種遺伝子並びにメバロン酸経路由来の遺伝子を発現している大腸菌(E. coli)においてメバロン酸産生を評価し、15L規模で流加式培養により生育させた。
MVA産生株SHG0863(CMP1053−HMB GI1.2gltA attB ldhA,pTrcHis2B,pCLPtrcUpperEfaecalis)に、標準的なMVA産生法においてMVAを産生させた。性能指数(グルコースに対するMVA産生性及びMVA収率)は、同一条件下で発現誘導し、ホスホケトラーゼポリペプチドの発現により収率が改善し得るのかを決定した試験株SHG0864(CMP1057−HMB GI1.2gltA attB ldhA,pTrcPKL_Lreuteri,pCLPtrcUpperEfaecalis)を本明細書において比較するためのものである。
方法:
培地組成(発酵培地1Lあたり):
K2HPO4(7.5g)、MgSO4*7H2O(2g)、クエン酸一水和物(2g)、クエン酸鉄アンモニウム(0.3g)、酵母エキス(0.5g)、50%硫酸(1.6mL)、1000X改変微量金属溶液(1mL)。すべての成分を共に加え、脱イオン水に溶解させた。この溶液を加熱滅菌した(123℃で20分)。水酸化アンモニウム(28%)によりpHを7.0に調整し、容量にメスアップした。滅菌及びpH調製後にグルコース10g、ビタミン溶液8mL、及び抗生物質を加えた。
1000X改変微量金属溶液(1Lあたり):
クエン酸*H2O(40g)、MnSO4*H2O(30g)、NaCl(10g)、FeSO4*7H2O(1g)、CoCl2*6H2O(1g)、ZnSO*7H2O(1g)、CuSO4*5H2O(100mg)、H3BO3(100mg)、NaMoO4*2H2O(100mg)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、0.22μmのフィルタを用いろ過滅菌した。
ビタミン溶液(1Lあたり):
チアミン塩酸塩(1.0g)、D−(+)−ビオチン(1.0g)、ニコチン酸(1.0g)、塩酸ピリドキシン(4.0g)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、孔径0.22μmのフィルタを用いろ過滅菌した。
供給溶液(1kgあたり):
グルコース(0.590kg)、脱イオン水(0.393kg)、K2HPO4(7.4g)及び100% Foamblast882(8.9g)。すべての成分を合わせて混合し、オートクレーブ処理した。
マクロ塩溶液(1Lあたり):
MgSO4*7H2O(296g)、クエン酸水和物(296g)、クエン酸鉄アンモニウム(49.6g)。すべての成分を水に溶解させ、用量に調整し、0.22μmのフィルタを用い、ろ過滅菌した。更なる添加物は加えずに、このうち16.8mLを培養槽に直接加えた後に滅菌した。
本実験は、所望の発酵pH 7.0及び温度34℃でのグルコースからのメバロン酸発酵をモニターするために実施した。大腸菌(E. coli)株を凍結したバイアルを解凍し、トリプトン酵母エキス及び適切な抗生物質を添加した培地を入れたフラスコに接種した。550nm(OD550)での吸光度が1.0になるまで接種材料を生育させた後、培養物のうち500mLを15Lのバイオリアクタに接種し、初期槽容量を5Lに調整した。
各バッチの培地には、グルコースを9.7g/L含有させた。誘導は、イソプロピル−β−D−1−チオガラクトピラノシド(IPTG)を添加し実施した。細胞のOD550が6になったなら、250μMの濃度になるようIPTGを反応槽に添加した。細胞のOD550が100になったなら、500μMの濃度になるよう2回めのIPTGを反応槽に添加した。培養によるグルコースの消費がpHの上昇により示されたなら、代謝に必要とされる量に足りるよう、10g/分以下の速度でグルコース供給溶液を供給した。グルコースに対するメバロン酸の最大質量収率を測定するのに十分な時間、すなわち発酵の開始から合計48時間にわたって発酵を行った。
分析:
4時間間隔でブロス試料を回収し、発酵ブロス中のメバロン酸濃度をHPLC分析により測定した。屈折計の測定値と、予め作成した検量線とを比較して、ブロス試料中のメバロン酸濃度を測定した。
HPLC情報:
システム:Waters Alliance 2695
カラム:BioRad−AminexHPX−87Hイオン排除カラム300mm×7.8mm(カタログ番号125−0140
カラム温度:50℃
ガードカラム:詰め替え用BioRad−Microguard Cation H(30mM×4.6mM)カタログ番号125−0129
ランニング緩衝液:0.01NのH2SO4;
ランニング緩衝液流速:0.6mL/分
最適流圧:約7.5〜8.2MPa(約1100〜1200psi)
注入容量:20μL
検出器:屈折率(Knauer K−2301)
実行時間:26分
結果:
ホスホケトラーゼを発現している株(CMP1057)により発酵させた場合、グルコースに対するメバロン酸収率は、空プラスミド対照株(CMP1053)よりも高かった。下記図11〜13及び表3を参照されたい。加えて、ホスホケトラーゼを発現している株(CMP1057)を有する発酵ブロスでは、空プラスミドの対照株(CMP1053)と比較して、発酵ブロス中の酢酸の蓄積は少なかった。図14を参照されたい。
実施例13−ホスホケトラーゼを発現しているサッカロミセス・セレビシエ(Saccharomyces cerevisiae)によるイソプレン産生
Life Technologiesの酵母発現系(コンピテントな野生型サッカロミセス・セレビシエ(S. cerevisiae)(INVSc1(カタログ番号C810−00))及びプラスミドpYES2/CT(カタログ番号V8251−20)を使用して、イソプレンシンターゼ、MVA経路上流、MVA経路下流及びホスホケトラーゼを発現させるための上記の各種遺伝子コンストラクトを調製する。
イソプレンシンターゼ、MVA経路上流、MVA経路下流、及びホスホケトラーゼをコードしている遺伝子を最初にpYES2/CTベクターにサブクローニングする。次に、プラスミドを大腸菌(E. coli)細胞中で増殖させる。次に、精製したプラスミドDNAにより、サッカロマイセス・セレヴィシエ(S. cerevisiae)(INVSc1)を形質転換させる。プラスミドを含有している酵母株を、2%グルコースと指定の選択マーカーを添加したSC最少培地で選択及び維持する。プラスミドを含有している単離コロニーを以降の実験に備え選別する。
遺伝子操作した酵母株からのイソプレンの比生産性を測定する。プラスミドのコードする遺伝子の発現を誘導するため、選択マーカーを添加した液体SC最少培地で培養物を一晩生育させた。次に培養物のOD600が約0.2になるよう希釈し、2〜3時間生育させる。100μLのブロスサンプルを2mLヘッドスペースバイアルで34℃で30分インキュベートし、続いて70℃で12分熱殺菌する。ヘッドスペース中のイソプレン濃度を、例えば、ガスクロマトグラフ(Model G1562A,Agilent Technologies)(Mergen et al.,LC GC North America,28(7):540〜543,2010)を連結した水素炎イオン化検出器により測定する。
実施例14:多様な各種細菌からのホスホケトラーゼ酵素のクローニング
ATCC15697株、ビフィドバクテリウム・ロングム(Bifidobacterium longum)亜種のインファンティス(infantis)の染色体DNAをATCC(Manassas,VA)から得た。プライマーCMP283:5’−ctgtatTCATGAcgagtcctgttattggcacc−3’(配列番号42)及びCMP284:5’−ctctatGAATTCTCACTCGTTGTCGCCAGCG−3’(配列番号43)、並びに製造元のプロトコルに従ってポリメラーゼHerculase(Life Technologies(Carlsbad,CA))を使用して、ビフィドバクテリウム・ロンガム(B. longhum)PKL(配列番号3)をコードしている遺伝子をポリメラーゼ連鎖反応(PCR)により染色体DNAから増幅させた。EcoRI及びBspHIによりPCR産物を消化した後、精製した。精製後、約2500bpの断片を、EcoRI/NcoIにより消化したpTrcHis2B(Invitrogen(Carlsbad,CA))にGENEARTシームレス・クローニング・キット(Invitrogen(Carlsbad,CA))を使用して組み込み、プラスミドpCMP1090を生成した(配列番号15−図6)。
ラクトバチルス・ロイテリ(Lactobacillus reuteri)株F275の染色体DNAをATCCから得た(ATCC# 23272D−5,ATCC(Manassas,VA))。プライマーCMP34:5’−taaggaggaataaacATGGCAGTAGATTACGATTCCAAG−3’(配列番号44)及びCMP335:5’−ttctagaaagcttcgttacttaagacccttccaagtccag−3’(配列番号45)、テンプレートとして100ngのDNA、並びに製造元に従ってポリメラーゼHerculase II Fusion(Agilent(Santa Clara,CA))を使用して、ラクトバチルス・ロイテリ(Lactobacillus reuteri)(PKL)(配列番号1)をコードしている遺伝子を増幅させた。精製後、2442bpの断片を、NcoI/EcoRIにより消化したpTrcHis2B(Invitrogen(Carlsbad,CA))にGENEARTシームレス・クローニング・キット(Invitrogen(Carlsbad,CA))を使用して組み込み、プラスミドpCMP1029を生成した(配列番号16、図7)。
エンテロコッカス・ガリナラム(Enterococcus gallinarum)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、エンテロコッカス・ガリナラム(E. gallinarum)PKL遺伝子(配列番号17)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1321を作成した。
ATCC27893株ノストック・パンクチホルム(Nostoc punctiforme)の染色体DNAをATCCから得た(Manassas,VA)。プライマーNostocpTrcHis2BF:5’−taaggaggaataaaccatgacattagccagtcctctacaaac−3’(配列番号46)及びNostocpTrcHis2BR:5’−TTCTAGAAAGCTTCGTTAATAGGGCCACTTCCAGTCACG−3’(配列番号47)、並びに製造元のプロトコルに従ってポリメラーゼHerculase(Life Technologies(Carlsbad,CA))を使用して、ノストック・パンクチホルム(N. punctiforme)PKL(配列番号18)をコードしている遺伝子をポリメラーゼ連鎖反応(PCR)により染色体DNAから増幅させた。EcoRI及びBspHIによりPCR産物を消化した後、精製した。精製後、約2500bpの断片を、EcoRI/NcoIにより消化したpTrcHis2B(Invitrogen(Carlsbad,CA))にGENEARTシームレス・クローニング・キット(Invitrogen(Carlsbad,CA))を使用して組み込み、プラスミドpCMP1305を生成した。
ATCC BAA−98株ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)の染色体DNAをATCC(Manassas,VA)から得た。プライマーRpalpTrcHis2BF:5’−taaggaggaataaaccatgtccgacgtgttgtccaacgatc−3’(配列番号48)及びRpalpTrcHis2BR:5’TTCTAGAAAGCTTCGTCAGGCCGACCAGCGCCAG−3’(配列番号49)、並びに製造元のプロトコルに従ってポリメラーゼHerculase(Life Technologies,Carlsbad,CA)を使用して、ロードシュードモナス・パルストリス(R. palustris)PKL(配列番号19)をコードしている遺伝子をポリメラーゼ連鎖反応(PCR)により染色体DNAから増幅させた。EcoRI及びBspHIによりPCR産物を消化した後、精製した。精製後、約2500bpの断片を、EcoRI/NcoIにより消化したpTrcHis2B(Invitrogen(Carlsbad,CA))にGENEARTシームレス・クローニング・キット(Invitrogen(Carlsbad,CA))を使用して組み込み、プラスミドpCMP1306を生成した。
パントエア(Pantoea sp.)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、パントエア(Pantoea sp.)PKLPKL遺伝子(配列番号20)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1324を作成した。
ムチラギニバクター・パルジス(Mucilaginibacter paludis)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、ムチラギニバクター・パルジス(M. paludis)PKL遺伝子(配列番号21)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1323を作成した。
サーモビフィダ・フスカ(Thermobifida fusca)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、サーモビフィダ・フスカ(T. fusca)PKL遺伝子(配列番号22)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1326を作成した。
ビフィドバクテリウム・ブレビ(Bfidobacterium breve)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、ビフィドバクテリウム・ブレビ(B. breve)PKL遺伝子(配列番号23)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1322を作成した。
ラーネラ・アクアティリス(Rahnella aquatilis)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、ラーネラ・アクアティリス(R. aquatilis)PKL遺伝子(配列番号24)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1325を作成した。
ビフィドバクテリウム・アニマリス(Bifidobacterium animalis)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、ビフィドバクテリウム・アニマリス(B. animalis)PKL遺伝子(配列番号25)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1320を作成した。
ガードネレラ・バギナリス(Gardnerella vaginalis)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、ガードネレラ・バギナリス(G. vaginalis)PKL遺伝子(配列番号26)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpCMP1309を作成した。
ストレプトマイセス・アベルミティリス(Streptomyces avermitilis)PKLアミノ酸配列をGeneBankから得て、大腸菌(E. coli)での発現を最適化するために、GeneArtの最適化ソフトウェアで加工した。PKL遺伝子の前に2塩基対を加えてNcoI認識部位及びSacI認識部位を作製し、終止コドンの直後に挿入した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、ストレプトマイセス・アベルミティリス(S. avermitilis)PKL遺伝子(配列番号27)を、BspHI/SacIにより消化したpTrcHis2Bベクター(Life Technologies,Carlsbad,CA)にサブクローニングして、プラスミドpEWL1362を作成した。
ATCC BAA−98株クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)の染色体DNAをATCC(Manassas,VA)から得た。プライマーCacetpTrcHisBF:5’−taaggaggaataaaccatgcaaagtataataggaaaacataaggatgaagg−3’(配列番号50)及びCacetpTrcHisBR:5’−ttctagaaagcttcgttatacatgccactgccaattagttatttc−3’(配列番号51)、並びに製造元のプロトコルに従ってポリメラーゼHerculase(Life Technologies(Carlsbad,CA))を使用して、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)PKL(配列番号28)をコードしている遺伝子をポリメラーゼ連鎖反応(PCR)により染色体DNAから増幅させた。EcoRI及びBspHIによりPCR産物を消化した後、精製した。精製後、約2500bpの断片を、EcoRI/NcoIにより消化したpTrcHis2B(Invitrogen(Carlsbad,CA))にGENEARTシームレス・クローニング・キット(Invitrogen(Carlsbad,CA))を使用して組み込み、プラスミドpCMP1364を生成した。
ラクトバチルス・パラプランタラム(Lactobacillus paraplantarum)PKLのアミノ酸配列をGeneBank及びJeong et al.,(J.Microbiol.Biotechnol.2007,17:822〜829)から得て、大腸菌(E. coli)における発現を最適化するために最適化ソフトウェアGeneArtにおいて加工した。合成したPKL遺伝子をGeneArtカナマイシン耐性クローニングプラスミドにクローニングした。次に、プライマーSML_NcoI_PhosphokLplantF(taaggaggaataaacatgaccaccgattatagcagtcc)及びv2SML_EcoRI_PhosphokLplantR(ttctagaaagcttcgTTA TTT CAG ACC TTT CCA CTG CC)、並びに製造元のプロトコルに従ってポリメラーゼとしてHerculase(Life Technologies(Carlsbad,CA))を使用して、ラクトバチルス・パラプランタラム(L. paraplantarum)PKL遺伝子(配列番号29)を増幅した。次に、得られたPCR産物を、GeneArt(登録商標)シームレス・クローニング・アンド・アセンブリ・キット(Life Technologies(Carlsbad,CA))を使用して、EcoRI/NcoIにより消化したpTrcHis2Bプラスミド(Life Technologies(Carlsbad,CA))と連結させてプラスミドpCMP1184を作製した。
実施例15:CMP1183、CMP1328、CMP 1366、CMP1182、CMP1308、CMP1309、CMP1331、CMP1330、CMP1333、CMP1329、CMP1184、及びCMP1332株の構築
CMP1133株(BL21,Δpgl PL.2mKKDyl,GI1.2gltA,yhfSFRTPyddVIspAyhfS,thiFRTtruncIspA)を形質転換してPKL発現株を構築し、20μg/mLのカナマイシンを含有させたルリア・ベルターニプレートでコロニーを選別した。製造元により推奨されるプロトコルを用い、カナマイシンマーカーを除去して(Gene Bridges,Heidelberg,Germany)記載の株を作製した(表4)。
実施例16:多様な各種細菌から単離されたホスホケトラーゼの発現、溶解度及び酵素活性の比較
TrcPKLビフィドバクテリウム・ロングム(B. longum)(CMP1183株)、pTrcPKLエンテロコッカス・ガリナラム(E. gallinarum)(CMP1328株)、pTrcPKLクロストリジウム・アセトブチリカム(C. acetobutylicum)(CMP1328株)、pTrcPKLラクトバチルス・ロイテリ(L. reuteri)(CMP1182株)、pTrcPKLのストック・パンクチホルム(N. punctiforme)(CMP1308株)、pTrcPKLロードシュードモナス・パルストリス(R. palustris)(CMP1309)、pTrcPKLパントエア(Pantaoea)(CMP1331)、pTrcPKLムチラギニバクター・パルジス(M. paludis)(CMP1330)、及びpTrcPKLサーモビフィダ・フスカ(T. fusca)(CMP1333)発現株をLB培地で生育し、OD600が約0.5の時点で200μMのIPTGにより発現誘導し、並びに30℃又は34℃の温度で4時間発現誘導した。4mLの培養ブロスを3000rpmにて10分間遠心分離して細胞を回収した。細胞ペレットを、0.1% DNAase及び0.5mMのAEBSFを添加した2mLの50mM MES/50mM NaCl(pH 6.0)に再懸濁した。96.5MPa(14,000psi)にて、フレンチ・プレス・セル(American Instrument Company)により細胞懸濁液を溶解した。次に、Eppendorf 5804R遠心機により、可溶化液を4℃にて15,000RPMで10分遠心分離した。上清及びペレットを分離した。ペレットを溶解液(50mM MES/50mM NaCl(pH 6.0)緩衝液)に再懸濁した。上清及びペレットサンプルを4〜12% SDS−PAGEゲル電気泳動により分析した。ホスホケトラーゼの可溶性画分対ペレット(不溶性)画分を比較することにより溶解度を評価した。
結果として、ビフィドバクテリウム・ロングム(B. longum)PKL、エンテロコッカス・ガリナラム(E. gallinarum)PKL、クロストリジウム・アセトブチリカム(C. acetobutylicum)PKL、ラクトバチルス・ロイテリ(L. reuteri)PKL、のストック・パンクチホルム(N. punctiforme)PKL、ロードシュードモナス・パルストリス(R. palustris)PKL、及びサーモビフィダ・フスカ(T. fusca)PKLが、30℃の温度にて70%超の溶解度を有していたことが示された。パントエア(Pantaoea)PKL及びムチラギニバクター・パルジス(M. paludis)PKLは、34℃の温度にて約50%増加した(表5)。
実施例17:ビフィドバクテリウム・ロングム(Bifidobacterium longum)及びエンテロコッカス・ガリナラム(Enterococcus gallinarum)から単離されたホスホケトラーゼの動態解析
以降の動態試験において使用するため、ビフィドバクテリウム・ロングム(B. longum)由来のホスホケトラーゼ(PKL)を精製した。ビフィドバクテリウム・ロングム(B. longum)由来のPKLを、pTrc His2Bプラスミドを入れたBL21株(CMP1183)において発現させた。細胞を、50μg/mLカルベニシリンを添加したルリア・ベルターニ培地にて34℃にて生育させた後、誘導を行った。200μM IPTGによる誘導後、培地を室温の振盪器に移し、5時間置いた。4℃、10,000rpmにて10分間遠心分離して細胞を回収した。精製前に−80℃にて細胞ペレットを保存した。精製に際し、ビフィドバクテリウム・ロングム(B. longum)PKL細胞ペレットを20mM HEPES(pH 7.0)、60mM NaCl、0.5mM AEBSF、0.5mM MgCl2、0.1mg/mL DNaseIに再懸濁した。フレンチ・プレス・セルを繰り返し通過させて細胞を溶解し、50,000rpmで30分間、超遠心により清澄化させた。ビフィドバクテリウム・ロングム(B. longum)由来のPKLを含有している清澄化させた溶解液を、まずは、50mM Tris/50mM NaCl(pH 7)により平衡化させたMonoQ 10/100GLカラム(GE Healthcare)に装填し、50mM Tris/1 M NaCl(pH 7)までび勾配を用い溶出した。得られた画分をSDS−PAGEにより分析した。ビフィドバクテリウム・ロングム(B. longum)PKLを、50mM Tris/50mM NaCl(pH 7)により平衡化したSuperdex 200 10/300GL及びMonoQ 10/300GL(pH 6.0)を使用し、50mM MES/50mM NaCl〜1M NaClの勾配を用い、更に精製した。A280を用いビフィドバクテリウム・ロングム(B. longum)PKLを定量し、149550(ベクターNTIにより測定した)のモル吸光係数を求め、及びまたゲルデンシトメトリー法によっても求めた。イオン交換及びゲル濾過クロマトグラフィーを用い精製し、見かけの均一性>95%でPKLを調製した。ヒドロキサム酸第二鉄による分析を用いPKL活性を解析するのに使用するため、ビフィドバクテリウム・ロングム(B. longum)PKLを含有している画分をプールした。
以降の動態試験において使用するため、エンテロコッカス・ガリナラム(E. gallinarum)由来のホスホケトラーゼ(PKL)を精製した。エンテロコッカス・ガリナラム(E. gallinarum)由来のPKLを、pTrc His2Bプラスミドを入れたBL21株(CMP1328)において発現させた。細胞を、50μg/mLカルベニシリンを添加したLB培地にて37℃にて生育させた後、誘導を行った。200μM IPTGによる誘導後、培地を30℃の振盪器に移し、5時間置いた。4℃、10,000rpmにて10分間遠心分離して細胞を回収した。精製前に−80℃にて細胞ペレットを保存した。精製のため、エンテロコッカス・ガリナラム(E. gallinarum)PKLの細胞ペレットを50mM MES(pH 6.0)、50mM NaCL、0.5mM AEBSF、0.1mg/mL DNaseIに再懸濁した。フレンチ・プレス・セルを繰り返し通過させて細胞を溶解し、50,000rpmで60分間、超遠心により清澄化させた。エンテロコッカス・ガリナラム(E. gallinarum)由来のPKLを含有している清澄化させた溶解液を、50mM MES/50mM NaCl(pH 6)により平衡化したDEAE HiTrap FFカラムに装填し、50mM MES/1M NaCl(pH 6)までの勾配で溶出した。得られた画分をSDS−PAGEにより分析した。PKLを含有している画分をプールし、G25脱塩カラムを用い50mM MES/50mM NaCL(pH 6.0)により脱塩した。50mM MES、50mM NaCL(pH 6)により平衡化したMonoQ 10/100GLカラムを使用し、1M NaClまでの塩勾配を用い、さらなる精製を実施した。PKLを含有している画分をプールしSDS PAGEにより分析し、ベクターNTIにより求められたA280モル吸光係数136980を用い定量化を行った。イオン交換及びゲル濾過クロマトグラフィーを用い精製し、見かけの均一性>95%でPKLを調製した。ヒドロキサム酸第二鉄による分析を用いPKL活性を解析するのに使用するため、エンテロコッカス・ガリナラム(E. gallinarum)PKLを含有している画分をプールした。
ビフィドバクテリウム・ロングム(B. longum)PKL又はエンテロコッカス・ガリナラム(E. gallinarum)PKLを含有しているプールした画分を、第二鉄ヒドロキサム酸解析を用いPKL活性について解析した。L.Meile et.al.,Bacteriol.,2001,183:2929〜2936 and Frey et.al.,Bioorganic Chem.,2008,36:121〜127に記載の、スケールダウンさせたヒドロキサム酸解析を使用し、PKLの触媒活性を測定した。この非特許文献は、参照によりその全体が本明細書に組み込まれる。96ウェルプレート(Costarカタログ番号9017)フォーマットにて、37℃下で解析を行った。各300μLの反応液には、1mM TPP、10mMリン酸カリウム(pH 6.0)、50mM MES(pH 6)、10mM MgCl2、5mM F6P及び濃度250nMのPKLを含有させた。時点を各間隔にて設定した。反応を停止させるため、60μLの反応混合物と、60μLの2Mヒドロキシルアミン(pH 6.5)とを混合し、室温で10分間インキュベートした。40μLの15% TCA、40μLの4M HCl、及び40μLの5% FeCl3の0.1M HCl水溶液を使用してタンパク質を沈殿させ、AcP検出できるようにした。次にこのサンプルを、3000rpmで10分遠心分離させた。上清の200μLのサンプルをマイクロタイタープレートに移し、プレートリーダーを使用してA505を測定した。定量化のため、12.5〜0.2mMの範囲のAcP標準曲線を作成した。飽和濃度Pi(20mM)にて0.3mM〜20mMの範囲のF6P濃度を用い、ビフィドバクテリウム・ロングム(B. longum)及びエンテロコッカス・ガリナラム(E. gallinarum)由来のPKLに関し、ミカエリス定数、KMを求めた。250nM(7μg)の精製したビフィドバクテリウム・ロングム(B. longum)又は500nM(4μg)のエンテロコッカス・ガリナラム(E. gallinarum)のPKLを添加し、反応を開始した。形成されたAcP濃度と相関した吸光度変化を505nmにてモニターし、時間に対してプロットしてPKLの反応速度を求めた。
飽和濃度のPiにてF6Pに対してビフィドバクテリウム・ロングム(B. longum)及びエンテロコッカス・ガリナラム(E. gallinarum)のPKLの、並びに飽和濃度のPiにてX5Pに対してエンテロコッカス・ガリナラム(E. gallinarum)のPKLの動態パラメータを評価した。速度定数を算出するため、反応速度をミカエリス・メンテン式に代入した(表6、図43及び図44)。エンテロコッカス・ガリナラム(E. gallinarum)のPKLは、F6P基質に対しては2.86mM及びX5P基質に対しては5.81mMのKMを有した一方、ビフィドバクテリウム・ロングム(B. longum)のPKLは、F6P基質に対し21.16mMと、より大きなKMを有した。ビフィドバクテリウム・ロングム(B. longum)のPKLのkcatは16.6s−1であり、及びエンテロコッカス・ガリナラム(E. gallinarum)のPKLのkcatはF6Pに対しては1.4s−1であり、X5Pに対しては4.4s−1であった。
実施例18:ビフィドバクテリウム・ロングム(Bifidobacterium longum)から単離されたホスホケトラーゼはセドヘプツロース−7−リン酸触媒活性を有する。
フルクトース6−リン酸(陽性対照)、リボース5−リン酸(陰性対照)、又はセドヘプツロース7−リン酸を単独で、又はビフィドバクテリウム・ロングム(B. longum)ホスホケトラーゼとともに発現している細胞を生育させ、LC−MS検出により、代謝産物の産生について解析した。
LC−MSにより代謝産物を検出する解析から、フルクトース6−リン酸(F6P)、リボース5−リン酸(R5P)、又はセドヘプツロース7−リン酸(S7P)のみを発現している細胞においては、それぞれ代謝生成物F6P、R5P、又はS7Pが主に検出されたことが示された(図45)。R5Pをビフィドバクテリウム・ロングム(B. longum)のホスホケトラーゼと共発現している細胞は、主に、一部のAcP産生に関してはR5Pとして保持していたことが示された。対照的に、F6Pをビフィドバクテリウム・ロングム(B. longum)のホスホケトラーゼと共発現している細胞は、F6Pの検出が消失し、AcPの生成が検出されることが示された(図45)。同様に、S7Pをビフィドバクテリウム・ロングム(B. longum)のホスホケトラーゼと共発現している細胞は、S7Pの検出が消失し、AcPの生成が検出されることが示された(図45)。
実施例19:ホスホケトラーゼを発現している組み換え宿主細胞による小規模におけるメバロン酸(MVA)産生
チオラーゼ、HMG−CoAシンターゼ、及びHMG−CoA還元酵素(表7)をコードする遺伝子に加え、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、クロストリジウム・アセトブチリカム(Clostridium acetubutylicum)、ノストック(Nostoc)、ロードシュードモナス・パルストリス(Rhodopseudomonas palustris)、パントエア(Pantoea)、又はサーモビフィダ・フスカ(Thermobifida fusca)由来のホスホケトラーゼを発現させることにより、メバロン酸(MVA)を産生している大腸菌(E. coli)株を作製した。ホスホケトラーゼを発現しないMVA産生株を対照として使用した(表7)。小規模実験において、グルコース存在下で生育させた場合のホスホケトラーゼ発現及びメバロン酸収率を、ホスホケトラーゼを発現しない対照株と比較して、ホスホケトラーゼ発現株を選別した。
(i)材料
酵母エキス及びMgSO4を不含有の改変TM3培地の組成(発酵培地1Lあたり):
K2HPO4を13.6g、KH2PO4を13.6g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、1000X微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。0.22μmフィルタで培地を濾過滅菌する。pHを調整及び滅菌後にグルコース10.0g及び抗菌剤を加えた。
1000X改変微量金属溶液(1Lあたり):
クエン酸*H2O(40g)、MnSO4*H2O(30g)、NaCl(10g)、FeSO4*7H2O(1g)、CoCl2*6H2O(1g)、ZnSO*7H2O(1g)、CuSO4*5H2O(100mg)、H3BO3(100mg)、NaMoO4*2H2O(100mg)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、0.22μmのフィルタを用いろ過滅菌した。
(ii)実験手順
増殖速度の測定
必要に応じて抗生物質を添加したLB培地を3mL入れた振盪チューブに、培養物のグリセロールストックを接種した。培養物を、30℃にて約15時間、220rpmでインキュベートした。TM3培地(MgSO4及び酵母エキスを不含有)、1%グルコース、8mM MgSO4、0.02%酵母エキス及び必要に応じて抗生物質を組み合わせて、追加のTM3培地を調製した。最終的なOD600が0.2になるよう、48ウェル滅菌ブロックの各ウェルに追加のTM3を2mL加えた。ブロックをBreathe Easierメンブレンによりシールし、34℃にて2時間、600rpmでインキュベートした。2時間生育させた後、マイクロタイタープレートにおいてOD600を測定し、各種濃度のIPTGにより細胞に発現誘導した。IPTGによる発現誘導から4時間にわたって、1時間毎にOD600の読み取りを行った。OD600の測定は、SpectraMax Plus190(Molecular Devices)を使用して行った。細胞を一晩生育させ、OD600を測定した。
グルコース測定
300μLの細胞培養物を96ウェルコニカルプレートにおいて4℃にて3000rpmで10分間遠心分離し、グルコースサンプルを回収した。上清を脱イオン水で10倍希釈し、Pointe Scientificから購入したグルコースオキシダーゼ解析キットを用いグルコース濃度測定した。
メバロン酸測定
34μLの10%硫酸と300μLの細胞培養物とを組み合わせ、氷上で10分間インキュベートし、メバロン酸サンプルを加工した。4℃にて10分後、混合物を4℃、3000rpmにて10分間遠心分離した。250μLの上清を96ウェルコニカルプレートに回収し、HPLCによるメバロン酸測定のため、Zone−Free(商標)Filmsプレートシーラーによりシールした。グルコース消費量に対し総メバロン酸量を算出することによりメバロン酸収率を求めた。
タンパク質発現解析
発現解析のため、発現誘導から4時間の時点の細胞培養全ブロスのうち50μLのサンプルを、50μLの2X SDSサンプル緩衝液とともに95℃で5分間ボイルし、10μLのサンプルを4〜12% Bis−Trisゲルに装填した。精製したホスホケトラーゼ酵素及び前染色した標準を各ゲルに加えた。GelをSimplyBlue Coomassie(登録商標)G−250染色により染色し、脱イオン水で脱染色した。
ホスホケトラーゼの発現及び溶解度特性解析
4mLの培養ブロスを3000rpmにて10分間遠心分離して細胞を回収した。細胞ペレットを、0.1% DNAase及び0.5mM AEBSFを添加した2mLの100mM Tris、100mM NaCl(pH 7.6)に再懸濁した。96.5MPa(14,000psi)にて、フレンチ・プレス・セル(American Instrument Company)により細胞懸濁液を溶解した。次に、Eppendorf 5804R遠心機により、可溶化液を4℃にて15,000RPMで10分遠心分離した。上清及びペレットを分離した。ペレットを溶解緩衝液に再懸濁した。ペレット及び上清の両方を調製するためにゲルサンプルを調製し、電気泳動を実施した。iBlot(登録商標)ドライ・ブロッティング・システムを使用し、タンパク質をニトロセルロース膜に転写した後、1:10,000希釈したウサギ抗ビフィドバクテリウム・ロンガム(Bifido longum)ホスホケトラーゼ血清を一次抗体とし、2μg/mL濃度のAlexa Fluor 488ヤギ抗ウサギIgGを検出抗体として、製造元のプロトコルに従って免疫検出した。Storm 860分子イメージャ(Molecular Dynamics)を使用して、青色蛍光スキャナスクリーンを用い、タンパク質のバンドを検出し、ソフトウェアImageQuantを使用してタンパク質濃度を算出した。
(iii)結果
ビフィドバクテリウム・ロングム(B. longum)のPKL(EWL1319)、エンテロコッカス・ガリナラム(E. gallinarum)のPKL(EWL1341)、ノストック(Nostoc)のPKL(EWL1344)、ロードシュードモナス・パルストリス(R. palustris)PKL(EWL1347)、パントエア(Pantoea)PKL(EWL1350)、又はサーモビフィダ・フスカ(T. fusca)PKL(EWL1353)を発現するよう遺伝子操作された大腸菌(E. coli)株によるMVA産生の解析により、発現誘導に使用するIPTGの増加と、メバロン酸収率の増加とが相関することが実証された(図46)。PKLを発現するよう遺伝子操作された株から調製した全細胞溶解液におけるタンパク質発現の解析により、IPTGによりタンパク質の発現が誘導されたことが示された(図47A〜D)。これらの所見により、メバロン酸収率の増加は、ホスホケトラーゼタンパク質の発現が増加した結果であることが示される。PKLを発現しないMVA産生対照株(CHL875)と比較した場合のメバロン酸収率の増加は、ビフィドバクテリウム・ロングム(B. longum)PKL(EWL1319)、エンテロコッカス・ガリナラム(E. gallinarum)PKL(EWL1341)、又はクロストリジウム・アセトブチリカム(C. acetobutylicum)PKL(EWL1359)発現株においても観察された。ビフィドバクテリウム・ロングム(B. longum)PKL(図48A)、ガリナラム(gallinarum)PKL(図49A)、又はクロストリジウム・アセトブチリカム(C. acetobutylicum)PKL(図50A)発現株におけるメバロン酸収率の増加は、IPTGの発現誘導濃度の増加と相関した。ビフィドバクテリウム・ロングム(B. longum)PKL、エンテロコッカス・ガリナラム(E. gallinarum)PKL、及びクロストリジウム・アセトブチリカム(C. acetobutylicum)のホスホケトラーゼ発現株のメバロン酸収率は、対照株CHL875により産生される最大メバロン酸収率と比較して増加していた。PKLを発現している遺伝子操作株から調製した細胞溶解液におけるタンパク質解析により、ビフィドバクテリウム・ロングム(B. longum)PKL(図48B及びC)、ガリナラム(gallinarum)PKL(図49B)、又はクロストリジウム・アセトブチリカム(C. acetobutylicum)のPKL(図50B)は、IPTGによる発現誘導により発現されることが確認された。ビフィドバクテリウム・ロングム(B. longum)のPKL発現株から単離された上清画分及びペレット画分の更なる解析により、ホスホケトラーゼは、不溶性画分(図48C)と比較して、主に可溶性画分(図48B)に存在することが示された。これらの結果は、メバロン酸収率の増加は、ホスホケトラーゼ発現が増加したことによる結果だという結論と一致する。
実施例20:ホスホケトラーゼを発現している組み換え宿主細胞における15L規模でのメバロン酸(MVA)産生
エンテロコッカス・ガリナラム(Enterococcus gallinarum)(EWL1341株)及びクロストリジウム・アセトブチリカム(Clostridium acetubutylicum)(EWL1359株)由来のホスホケトラーゼを発現しているメバロン酸(MVA)産生株を、ホスホケトラーゼを発現しないMVA産生株(CHL875株)と、15規模の実験において比較した。グルコースに対する累積MVA収率、グルコースに対する瞬間収率、MVAの体積産生性、MVAの比産生性及び細胞性能指数(CPI)を測定し、解析した。
(i)材料
培地組成(発酵培地1Lあたり):
K2HPO4(7.5g)、MgSO4*7H2O(2g)、クエン酸一水和物(2g)、クエン酸鉄アンモニウム(0.3g)、酵母エキス(0.5g)、50%硫酸(1.6mL)、1000X改変微量金属溶液(1mL)。すべての成分を共に加え、脱イオン水に溶解させた。この溶液を加熱滅菌した(123℃で20分)。水酸化アンモニウム(28%)によりpHを7.0に調整し、容量にメスアップした。滅菌及びpH調製後にグルコース10g、ビタミン溶液8mL、及び抗生物質を加えた。
1000X改変微量金属溶液(1Lあたり):
クエン酸*H2O(40g)、MnSO4*H2O(30g)、NaCl(10g)、FeSO4*7H2O(1g)、CoCl2*6H2O(1g)、ZnSO*7H2O(1g)、CuSO4*5H2O(100mg)、H3BO3(100mg)、NaMoO4*2H2O(100mg)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、0.22μmのフィルタを用いろ過滅菌した。
ビタミン溶液(1Lあたり):
チアミン塩酸塩(1.0g)、D−(+)−ビオチン(1.0g)、ニコチン酸(1.0g)、塩酸ピリドキシン(4.0g)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、孔径0.22μmのフィルタを用いろ過滅菌した。
マクロ塩溶液(1Lあたり):
MgSO4*7H2O(296g)、クエン酸水和物(296g)、クエン酸鉄アンモニウム(49.6g)。すべての成分を水に溶解させ、用量に調整し、0.22μmのフィルタを用い、ろ過滅菌した。
供給溶液(1kgあたり):
グルコース(0.590kg)、脱イオン水(0.393kg)、K2HPO4(7.4g)及び100% Foamblast882(8.9g)。すべての成分を合わせて混合し、オートクレーブ処理した。供給溶液をオートクレーブにより処理した後、滅菌フード中で、供給ボトルに栄養塩類を添加する。滅菌後に供給溶液に添加するものは(供給溶液1kg当たり)マクロ塩溶液5.54mL、ビタミン溶液6.55mL、1000X改変微量金属溶液0.82mLである。
(ii)実験手順
導入されたメバロン酸経路由来の遺伝子及びエンテロコッカス・ガリナラム(E. gllinarum)(EWL1341株)又はクロストリジウム・アセトブチリカム(C. acetobutylicum)(EWL1359株)のいずれかから単離されたPKLを発現している改変大腸菌(E. coli)(BL21)宿主(CMP1133)によるメバロン酸(MVA)産生を、この株を15L規模の流加式培養において生育させることにより評価した(表9)。導入されたメバロン酸経路由来の遺伝子を発現しているもののPKL(CHL875株)は発現しない改変大腸菌(E. coli)(BL21)宿主(CMP1133)と比較して、MVA産生を比較し、ホスホケトラーゼの使用により収率が増加するかを決定した。
所望の発酵pH(7.0)及び温度(34℃)にて、グルコースからのメバロン酸(MVA)生成を求めた。各実験の開始時に、大腸菌(E. coli)(BL21)株の適切な凍結バイアルを解凍し、トリプトン酵母エキス(LB)培地及び適切な抗生物質を入れたフラスコに接種した。550nm(OD550)での吸光度が約1.0になるまで接種材料を生育させた後、培養物のうち500mLを15Lのバイオリアクタに接種し、初期槽容量を5Lに調整した。
複数種の産生条件において、メバロン酸産生株に産生誘導を行った(表10)。各バッチの培地には、グルコースを9.7g/L含有させた。誘導は、イソプロピル−β−D−1−チオガラクトピラノシド(IPTG)を添加し実施した。細胞のOD550が6になったなら、記載の濃度になるよう発現誘導用のIPTGを反応槽に添加した。培養によるグルコースの消費がpHの上昇により示されたなら、代謝に必要とされる量に足りるよう、10g/分以下の速度でグルコース供給溶液を供給した。グルコースに対するメバロン酸の累積最大質量収率を測定するのに十分な時間、すなわち発酵の開始から合計60〜64時間にわたって発酵を行った。
対照株(CHL875)の性能指数を、実験株EWL1341、EWL1359のものと比較した。関連する性能指数は、グルコースに対する累積MVA収率、グルコースに対するMVA瞬間収率、MVAの体積産生性、比MVA産生性、及び細胞性能指数であった。実験株(ホスホケトラーゼ発現株)に、対照(ホスホケトラーゼ非発現株)と同一の条件下で発現誘導を行い、ホスホケトラーゼ酵素の使用により何らかの収率改善が生じ得るかを決定した。
全体的な収率は、次式を用い算出した:
グルコースに対する収率(重量%)=総MVA(t)/[(供給重量(0)−供給重量(t)+83.5)*0.59)],
式中、0.59グルコース供給溶液中のグルコースの割合(重量%)であり、及び83.5は、t=0の時点で発酵槽に供給したバッチのg重量である。各供給量は、独立して重量%として測定した。
CPIは、次式を用い算出した:
CPI=総MVA量(g)/総乾燥細胞重量
2種の質量分析器iSCAN(ハミルトン・サンドストランド)、及びHiden HPR20(Hiden Analytical)により、排出気体中の、酸素、窒素、及び二酸化炭素濃度を別個に測定した。発酵ブロス中の溶存酸素は、Hamilton Companyにより提供された光学センサーを備えた衛生的であり滅菌可能なプローブにより測定した。4時間間隔でブロス試料にHPLC解析を行い、発酵ブロス中のクエン酸、グルコース、酢酸及びメバロン酸濃度を測定した。屈折計による測定値と、濃度既知の標準を使用して予め作成した検量線とを比較して、ブロス試料中の濃度を測定した。
HPLC情報:
システム:Waters Alliance 2695
カラム:BioRad−Aminex HPX−87Hイオン排除カラム300mm×7.8mm(カタログ番号125−0140
カラム温度:50℃
ガードカラム:詰め替え用BioRad−Microguard Cation H(30mm×4.6mm)カタログ番号125−0129
ランニング緩衝液:0.01NのH2SO4;
ランニング緩衝液流速:0.6mL/分
最適流圧:約7.5〜8.2MPa(約1100〜1200psi)
注入容量:20μL検出器:屈折率(Knauer K−2301)
実行時間:26分
(iii)結果
クロストリジウム・アセトブチリカム(C. acetobutylicum)ホスホケトラーゼの発現株(EWL1359、実施番号20121056、201201057、20121058)は、ホスホケトラーゼ非発現株よりも(CHL875、実施番号20120821、20120976、20121059)グルコースに対するMVAの累積収率(%)が高かった(表11及び図51)。少なくとも100μMのIPTGを槽に加えた場合に(20121057及び20121058)、MVA収率の増加が顕著であった。
エンテロコッカス・ガリナラム(E. gallinarum)ホスホケトラーゼ(EWL1341、実施番号20120977、201200978、20120979)の発現株は、ホスホケトラーゼ非発現株よりも(CHL875、実施番号20120821、20120976、20121059)グルコースに対するMVAの累積収率(%)が高かった(表11及び図52)。少なくとも50μMのIPTGを槽に加えた場合に(20120978及び20120979)、MVA収率の増加が顕著であった。
すべての場合において、MVA収率は、添加したIPTG量と相関した(図53)。直接比較するため、実施に際し100μM IPTGを添加したところ、ホスホケトラーゼ(20120979及び20121057)発現株は、ホスホケトラーゼ非発現株(CHL875、実施番号201201059)と比較して、グルコースに対するMVAの累積収率が著しく高かった(表12及び図53)。各実施において、累積MVA収率(いずれの場合もホスホケトラーゼを発現していた)はホスホケトラーゼ活性と良好に相関していた(表12及び図54)。
クロストリジウム・アセトブチリカム(C. acetobutylicum)ホスホケトラーゼの発現株(EWL1359、実施番号20121057、100μM IPTGにより発現誘導)は、ホスホケトラーゼ非発現株(CHL875、実施番号20121059、100μM IPTGにより発現誘導)と比較して、細胞性能指数(MVA(g)/乾燥細胞重量(g))がわずかに高かった。400μM IPTG(EWL1359、実施番号20121058)により発現誘導した場合、CPIは対照(CHL875、実施番号20121059、100μM IPTGにより発現誘導)よりも高かった(図55)。
エンテロコッカス・ガリナラム(E. gallinarum)ホスホケトラーゼ(EWL1341、実施番号20120978及び20120979、それぞれ50μM及び100μM IPTGにより発現誘導)の発現株は、ホスホケトラーゼ非発現株(CHL875、実施番号20120976、IPTG未処理)と比較して、細胞性能指数(MVA(g)/乾燥細胞重量(g))が高かった。IPTGを添加しなかった場合(EWL1341、実施番号20120977)、CPIは対照(CHL875、実施番号20120976、IPTG未処理)と比較してほんのわずかに高かった(図56)。
実施例21:ホスホケトラーゼを発現している組み換え宿主細胞による小規模におけるイソプレン産生
ビフィドバクテリウム・ロンガム(Bifidobacterium longum)、エンテロコッカス・ガリナラム(Enterococcus gallinarum)、又はクロストリジウム・アセトブチリカム(Clostridium acetobutylicum)由来のホスホケトラーゼを発現させて、イソプレン産生大腸菌(E. coli)株を作製した。ホスホケトラーゼを発現していないイソプレン産生株を対照として使用した(表13)。小規模実験において、グルコース存在下で生育させた場合のホスホケトラーゼ発現及びイソプレン収率を、ホスホケトラーゼを発現しない対照株と比較して、ホスホケトラーゼ発現株を選別した。
ビフィドバクテリウム・ロングム(B. longum)PKLを発現している改変大腸菌(E. coli)(BL21)宿主(MCM2065)によりイソプレン産生株を作製し、プラスミドpEWL1418(図57)を導入することにより、EWL1427株を作製した(表13)。エンテロコッカス・ガリナラム(E. gallinarum)PKLの発現株を作製するため、プラスミドpEWL1421(図58)又はpEWL1438(図59)を改変大腸菌(E. coli)(BL21)宿主(MCM2065)に導入し、それぞれEWL1430株及びEWL1449株を生成した(表13)。クロストリジウム・アセトブチリカム(C. acetobutylicum)PKL発現株を作製するために、プラスミドpEWL1436(図60)及びプラスミドpEWL1440(図61)を使用して、それぞれEWL1446株及びEWL1452株を作製した(表13)。
bMVKはメタノコッカシド・バートニイ(M. Burtonii)メバロン酸キナーゼを意味する
mMVKはメタノサルシナ.マゼイ(M. mazei)メバロン酸キナーゼを意味する
(i)材料
TM3培地組成(発酵培地1L当たり):
K2HPO4を13.6g、KH2PO4を13.6g、MgSO4 *7H2Oを2g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、酵母エキスを0.2g、1000X微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。0.22μmフィルタで培地を濾過滅菌する。pHを調整及び滅菌後にグルコース10.0g及び抗菌剤を加えた。
酵母エキス及びMgSO4を不含有の改変TM3培地の組成(発酵培地1Lあたり):
K2HPO4を13.6g、KH2PO4を13.6g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、1000X微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。0.22μmフィルタで培地を濾過滅菌する。pHを調整及び滅菌後にグルコース10.0g及び抗菌剤を加えた。
1000X改変微量金属溶液(1Lあたり):
クエン酸*H2O(40g)、MnSO4*H2O(30g)、NaCl(10g)、FeSO4*7H2O(1g)、CoCl2*6H2O(1g)、ZnSO*7H2O(1g)、CuSO4*5H2O(100mg)、H3BO3(100mg)、NaMoO4*2H2O(100mg)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、0.22μmのフィルタを用いろ過滅菌した。
(ii)実験手順
増殖速度の測定
必要に応じて抗生物質を添加したLB培地を3mL入れた振盪チューブに、培養物のグリセロールストックを接種した。培養物を、30℃にて約15時間、220rpmでインキュベートした。TM3培地(MgSO4及び酵母エキスを不含有)、1%グルコース、8mM MgSO4、0.02%酵母エキス及び必要に応じて抗生物質を組み合わせて、追加のTM3培地を調製した。最終的なOD600が0.2になるよう、48ウェル滅菌ブロックの各ウェルに追加のTM3を2mL加えた。ブロックをBreathe Easierメンブレンによりシールし、34℃にて2時間、600rpmでインキュベートした。2時間生育させた後、マイクロタイタープレートにおいてOD600を測定し、各種濃度のIPTGにより細胞に発現誘導した。IPTGによる発現誘導から4時間にわたって、1時間毎にOD600の読み取りを行った。OD600の測定は、SpectraMax Plus190(Molecular Devices)を使用して行った。細胞を一晩生育させ、OD600を測定した。
グルコース測定
300μLの細胞培養物を96ウェルコニカルプレートにおいて4℃にて3000rpmで10分間遠心分離し、グルコースサンプルを回収した。上清を脱イオン水で10倍希釈し、Pointe Scientificから購入したグルコースオキシダーゼ解析キットを用いグルコース濃度測定した。
イソプレン比産生性測定
IPTGによる発現誘導後、4時間にわたって1時間毎に100μLのイソプレンサンプルを96ウェルガラスブロックに回収した。アルミホイルによりガラスブロックをシールし、Thermomixerを使用し、34℃にて、450rpmで振盪しながら30分間インキュベートした。30分後、ブロックを70℃の水浴に2時間保持し、ガスクロマトグラフィー質量分析法を使用し、ヘッドスペース測定法において、イソプレン濃度を決定した。
タンパク質発現解析
発現解析のため、発現誘導から4時間の時点の細胞培養全ブロスのうち50μLのサンプルを、50μLの2X SDSサンプル緩衝液とともに95℃で5分間ボイルし、10μLのサンプルを4〜12% Bis−Trisゲルに装填した。精製したホスホケトラーゼ酵素及び前染色した標準を各ゲルに加えた。GelをSimplyBlue Coomassie(登録商標)G−250染色により染色し、脱イオン水で脱染色した。
(iii)結果
メタノコッカシド・バートニイ(M. Burtonii)のメバロン酸キナーゼの発現存在下でビフィドバクテリウム・ロングム(B. longum)PKLを(EWL1427株)、メタノコッカシド・バートニイ(M. Burtonii)メバロン酸キナーゼの発現存在下でエンテロコッカス・ガリナラム(E. gallinarum)PKLを(EWL1430株)、又はメタノコッカシド・バートニイ(M. Burtonii)メバロン酸キナーゼの発現存在下でクロストリジウム・アセトブチリカム(C. acetobutylicum)PKLを(EWL1446株)、発現するよう遺伝子操作された大腸菌(E. coli)株により、グルコースから産生されたイソプレンを、PKLを発現しない対照株(MCM2158株)と比較して分析したところ、発現誘導に使用するIPTGの増加と、イソプレン収率の増加とが相関することが実証された(図62A及びB)。PKLを発現するよう遺伝子操作された株から調製した全細胞溶解液におけるタンパク質発現の解析により、IPTGによりタンパク質の発現が誘導されたことが示された(図63A及びB)。これらの所見により、イソプレン収率の増加は、ホスホケトラーゼタンパク質の発現が増加した結果であることが示される。
メタノサルシナ・マゼイ(M. mazei)のメバロン酸キナーゼ発現存在下でエンテロコッカス・ガリナラム(E. gallinarum)PKLを(EWL1449株)、又はメタノサルシナ・マゼイ(M. mazei)メバロン酸キナーゼの発現存在下でクロストリジウム・アセトブチリカム(C. acetobutylicum)を(EWL1452株)、発現するよう遺伝子操作された大腸菌(E. coli)株により、グルコースから産生されたイソプレンを、PKLを発現しない対照株(DW719株)と比較して分析したところ、発現誘導に使用するIPTGの増加と、イソプレン収率の増加とが相関することが実証された(図64A)。PKLを発現するよう遺伝子操作された株から調製した全細胞溶解液におけるタンパク質発現の解析により、IPTGによりタンパク質の発現が誘導されたことが示された(図64B)。これらの所見により、イソプレン収率の増加は、ホスホケトラーゼタンパク質の発現が増加した結果であることが示される。
総じてこれらの結果から、異なるメバロン酸キナーゼ遺伝子の存在下で、ビフィドバクテリウム・ロングム(B. longum)、エンテロコッカス・ガリナラム(E. gallinarum)、及びクロストリジウム・アセトブチリカム(C. acetobutylicum)などの異なる細菌から単離されたPKL遺伝子を発現する株において、グルコースからイソプレンが産生されることが実証された。
実施例22:ホスホケトラーゼを発現している組み換え宿主細胞による15L規模におけるイソプレン産生
ビフィドバクテリウム・ロングム(B. longum)(EWL1427株)又はエンテロコッカス・ガリナラム(E. gallinarum)(EWL1430株)由来のホスホケトラーゼを発現するイソプレン産生株を、ホスホケトラーゼを発現しないイソプレン産生株(MCM2158株)と、15L規模の実験において、イソプレン産生について比較した。グルコースに対する累積イソプレン収率、グルコースに対するイソプレン瞬間収率及び細胞性能指数(CPI)を測定し、解析した。
(i)材料
培地組成(発酵培地1Lあたり):
K2HPO4(7.5g)、MgSO4*7H2O(2g)、クエン酸一水和物(2g)、クエン酸鉄アンモニウム(0.3g)、酵母エキス(0.5g)、50%硫酸(1.6mL)、1000X改変微量金属溶液(1mL)。すべての成分を共に加え、脱イオン水に溶解させた。この溶液を加熱滅菌した(123℃で20分)。水酸化アンモニウム(28%)によりpHを7.0に調整し、容量にメスアップした。滅菌及びpH調製後にグルコース10g、ビタミン溶液8mL、及び抗生物質を加えた。
1000X改変微量金属溶液(1Lあたり):
クエン酸*H2O(40g)、MnSO4*H2O(30g)、NaCl(10g)、FeSO4*7H2O(1g)、CoCl2*6H2O(1g)、ZnSO*7H2O(1g)、CuSO4*5H2O(100mg)、H3BO3(100mg)、NaMoO4*2H2O(100mg)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、0.22μmのフィルタを用いろ過滅菌した。
ビタミン溶液(1Lあたり):
チアミン塩酸塩(1.0g)、D−(+)−ビオチン(1.0g)、ニコチン酸(1.0g)、塩酸ピリドキシン(4.0g)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、孔径0.22μmのフィルタを用いろ過滅菌した。
マクロ塩溶液(1Lあたり):
MgSO4*7H2O(296g)、クエン酸水和物(296g)、クエン酸鉄アンモニウム(49.6g)。すべての成分を水に溶解させ、用量に調整し、0.22μmのフィルタを用い、ろ過滅菌した。
供給溶液(1kgあたり):
グルコース(0.590kg)、脱イオン水(0.393kg)、K2HPO4(7.4g)及び100% Foamblast882(8.9g)。すべての成分を合わせて混合し、オートクレーブ処理した。供給溶液をオートクレーブにより処理した後、滅菌フード中で、供給ボトルに栄養塩類を添加する。滅菌後に供給溶液に添加するものは(供給溶液1kg当たり)マクロ塩溶液5.54mL、ビタミン溶液6.55mL、1000X改変微量金属溶液0.82mLである。
(ii)実験手順
メバロン酸経路由来の導入遺伝子及びビフィドバクテリウム・ロングム(B. longum)(EWL1427株)又はエンテロコッカス・ガリナラム(E. gallinarum)(EWL1430株)のいずれかから単離されたPKLを発現している改変大腸菌(E. coli)(BL21)宿主(MCM2065)によるイソプレン生産を、株を15L規模の流加式培養において生育させることにより評価した(表14)。導入されたメバロン酸経路由来の遺伝子を発現しているもののPKL(MCM2158株)は発現しない改変大腸菌(E. coli)(BL21)宿主(MCM2065)と、イソプレン産生を比較し、ホスホケトラーゼの使用により収率が増加するかを決定した。
bMVKはメタノコッカシド・バートニイ(M. Burtonii)メバロン酸キナーゼを意味する
イソプレン産生株に、標準的な産生工程において発現誘導した(表15)。発酵pH 7.0及び温度34℃にて、グルコースからのイソプレン産生をモニターした。実験の開始時に、大腸菌(E. coli)(BL21)株の凍結バイアルを解凍し、トリプトン酵母エキス(LB)培地及び適切な抗生物質を入れたフラスコに接種した。550nm(OD550)での吸光度が約1.0になるまで接種材料を生育させた後、培養物のうち500mLを15Lのバイオリアクタに接種し、初期槽容量を5Lに調整した。各バッチの培地には、グルコースを9.7g/L含有させた。誘導は、イソプロピル−β−D−1−チオガラクトピラノシド(IPTG)を添加し実施した。細胞のOD550が6になったなら、記載の濃度になるよう発現誘導用のIPTGを反応槽に添加した。培養によるグルコースの消費がpHの上昇により示されたなら、代謝に必要とされる量に足りるよう、10g/分以下の速度でグルコース供給溶液を供給した。グルコースに対するイソプレンの累積最大質量収率を測定するのに十分な時間、すなわち発酵の開始から合計60〜64時間にわたって発酵を行った。
対照株(MCM2158)の性能指数を、実験株EWL1427及びEWL1430のものと比較した。性能指数は、グルコースに対する累積イソプレン収率、グルコースに対するイソプレン瞬間収率及び細胞性能指数(CPI)であった。実験株(ホスホケトラーゼ発現株)に、対照(ホスホケトラーゼ非発現株)と同一の条件下で発現誘導した。
次式を用い、総イソプレン収率を算出した:
グルコースに対する収率(重量%)=総イソプレン(t)/[(供給重量(0)−供給重量(t)+83.5)*0.59)],
式中、0.59グルコース供給溶液中のグルコースの割合(重量%)であり、及び83.5は、t=0の時点で発酵槽に供給したバッチのg重量である。各供給量は、独立して重量%として測定した。
次式を用い、イソプレン瞬間収率を算出した:
イソプレン瞬間収率(g/g%)=産生されたイソプレン(t1−t0)/消費されたグルコース(t1−t0)*100
CPIは、次式を用い算出した:
CPI=総イソプレン量(g)/総乾燥細胞重量
2種の質量分析器iSCAN(ハミルトン・サンドストランド)、及びHiden HPR20(Hiden Analytical)により、排出気体中の、イソプレン、酸素、窒素、及び二酸化炭素濃度を別個に測定する。発酵ブロス中の溶存酸素は、Hamilton Companyにより提供された光学センサーを備えた衛生的であり滅菌可能なプローブにより測定した。4時間間隔でブロス試料にHPLC解析を行い、発酵ブロス中のクエン酸、グルコース、酢酸及びメバロン酸濃度を測定した。屈折計による測定値と、濃度既知の標準を使用して予め作成した検量線とを比較して、ブロス試料中の濃度を測定した。
HPLC情報:
システム:Waters Alliance 2695
カラム:BioRad−AminexHPX−87Hイオン排除カラム300mm×7.8mm(カタログ番号125−0140
カラム温度:50℃
ガードカラム:詰め替え用BioRad−Microguard Cation H(30mm×4.6mm)カタログ番号125−0129
ランニング緩衝液:0.01NのH2SO4;
ランニング緩衝液流速:0.6mL/分
最適流圧:約7.5〜8.2MPa(約1100〜1200psi)
注入容量:20μL
検出器:屈折率(Knauer K−2301)
実行時間:26分
(iii)結果
エンテロコッカス・ガリナラム(E. gallinarum)ホスホケトラーゼ(EWL1340、20121136)の発現株は、ホスホケトラーゼ非発現株よりも(MCM2158、20121134)グルコースに対するイソプレンの累積収率(%)が高かった(表16及び図65)。エンテロコッカス・ガリナラム(E. gallinarum)ホスホケトラーゼ(EWL1430、20121136)発現株は、ホスホケトラーゼ非発現株(MCM2158、20121134)よりもイソプレン瞬間収率(%)が高く(表16及び図66)、及びより長期間にわたって高い累積収率を維持したことから累積収率が高くなった(表16及び図65)。ビフィドバクテリウム・ロンガム(B. longum)ホスホケトラーゼ発現株(EWL1427、20121135)は、誘導実験においてより高い瞬間収率を有していたものの(図66)、この株は最高収率に達するまでにより長時間を要し、かつ終了時のグルコースに対するイソプレンの累積収率は、対照株(MCM2158、20121134)のものとおおよそ同一であった(表16及び図65)。
エンテロコッカス・ガリナラム(E. gallinarum)ホスホケトラーゼ(EWL1430、20121136)は、ホスホケトラーゼ非発現株(MCM2158、20121134)と比較して、わずかにCPIが高かった(図67)。ビフィドバクテリウム・ロンガム(B. longum)ホスホケトラーゼ(EWL1427、20121135)発現株は、ホスホケトラーゼ非発現株(MCM2158、20121134)と比較してわずかに低いCPIを有した(図67)。最高累積収率(60時間EFT)は、CPIと比較した時間であった。
実施例23:14L規模で生育させた組換え発現細胞におけるホスホケトラーゼのIn vitro特異的活性解析
エンテロコッカス・ガリナラム(E. gallinarum)(表19及び表21)、クロストリジウム・アセトブチリカム(C. acetobutylicum)(表20)、又はビフィドバクテリウム・ロンガム(B. longum)(表21)由来のホスホケトラーゼ(PKL)発現株を動態試験により解析した。試料を調製する際、細胞ペレットは、14Lの培養工程中に1mL培養物から得て−80℃で保管した。1mg/mLのリゾチーム、0.2mg/mLのDNaseI、及び0.5mMのAEBSFを添加した50mM MES(pH 6)にペレット試料を再懸濁し、OD(600)=20に揃えた。ODを揃えた細胞ペレットを、培養液の割合に対し4.8MPa(700psi)に設定したフレンチプレスMini Cellに繰り返し通過させて溶解した。次に、溶解した試料を、4℃にて14,000rpmで10分間遠心分離して清澄化させた。溶解物の可溶性画分の活性解析及びタンパク質測定を実施した。0.5〜0.05mg/mLの範囲の濃度でのBSA滴定をもとに作成した標準曲線を使用し、Bradford法(バイオラッド)により総タンパク質を測定した。
L.Meile et.al.,Bacteriol.,2001,183:2929〜2936 and Frey et.al.,Bioorganic Chem.,2008,36:121〜127に記載の、スケールダウンさせたヒドロキサム酸解析を使用し、PKLの触媒活性を測定した。この非特許文献は、参照によりその全体が本明細書に組み込まれる。96ウェルプレート(Costarカタログ番号9017)フォーマットにて、37℃下で解析を行った。標準解析において、反応混合物は、5mM F6P、1mM TPP、10mMリン酸カリウム、50mM MES緩衝液(pH 6)、及び10mM MgCl2、20mMフッ化ナトリウム、8mMヨードアセトアミド、及び1mM DTTにより構成した。F6Pを加えて反応を開始し、インキュベート開始から30分後に反応を停止させた。総反応容量は、通常300μLである(必要であればより少量を用いる)。15〜0.23mMの範囲の濃度に関し、検量線としてAcPを使用した。反応を停止させるため、60μLの反応混合物と、60μLの2Mヒドロキシルアミン(pH 6.5)とを混合し、室温で10分間インキュベートした。40μLの15% TCA、40μLの4M HCl、及び40μLの5% FeCl3の0.1M HCl水溶液を使用してタンパク質を沈殿させ、AcP検出できるようにした。次にこのサンプルを、3000rpmで10分遠心分離させた。吸光度505nmにてAcP生成率を測定するため、200μLの上清をマイクロタイタープレートに移した。
エンテロコッカス・ガリナラム(E. gallinarum)PKLを保有しているEWL1341株によるAcP生成に関し、インビトロでの比産生性は、誘導濃度に応じ0.75〜1.77mM/L/時/ODの範囲であった。(表22及び図68A)。更に、エンテロコッカス・ガリナラム(E. gallinarum)PKLを保有しているEWL1341株によるAcP生成に関し、インビトロでの比活性は、誘導濃度に応じ、7.1E−02及び1.1E−01μM/mg/分の範囲であった(表22及び図68B)。
クロストリジウム・アセトブチリカム(C. acetobutylicum)PKLを保有しているEWL1359株によるAcP生成に関し、インビトロでの比産生性は、誘導濃度に応じ、7.1E−02及び0.12〜0.74mM/L/時/ODの範囲であった(表23及び図69A)。更に、クロストリジウム・アセトブチリカム(C. acetobutylicum)PKLを保有しているEWL1359株によるAcP生成に関し、インビトロでの比活性は、誘導濃度に応じ、1.1E−02及び7.6E−02μM/mg/分の範囲であった(表23及び図69B)。
ビフィドバクテリウム・ロンガム(B. longum)PKLを保有しているEWL1427株によるAcP生成に関し、インビトロでの比産生性は、誘導濃度に応じ、7.30〜10.01mM/L/時/ODの範囲であった。(表23及び図70)。エンテロコッカス・ガリナラム(E. gallinarum)PKLを保有しているEWL1430株によるAcP生成に関し、インビトロでの比産生性は、100μM IPTGで誘導した場合には0.98〜1.13mM/L/時/ODの範囲であり、200μM IPTGで誘導した場合には1.30〜1.38mM/L/時/ODの範囲であった(表24及び図70)。
ビフィドバクテリウム・ロンガム(B. longum)PKLを保有しているEWL1427株によるAcP生成に関し、インビトロでの比活性は、100μM IPTGで誘導した場合には7.1E−01〜8.7E−01μM/mg/分の範囲であった(表23及び図71)。エンテロコッカス・ガリナラム(E. gallinarum)PKLを保有しているEWL1430株によるAcP生成に関し、インビトロでの比産生性は、100μM IPTGで誘導した場合には8.0E−02〜1.0E−01μM/mg/分の範囲であり、200μM IPTGで誘導した場合には1.1E−01〜1.2E−01μM/mg/分の範囲であった(表24及び図71)。
総じて、これらの所見は、すべての株においてホスホケトラーゼ活性が存在していたことを意味する。
実施例24:イソプレンの産生に関係する宿主由来の変異を有するホスホケトラーゼ発現株の構築
上記の通りに活性ホスホケトラーゼポリペプチドを含むイソプレン産生株を更に遺伝子操作して、次の遺伝子:リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)、D−リブロース−5−リン酸3−エピメラーゼ(rpe)、トランスケトラーゼ(tktA及び/又はtktB)、トランスアルドラーゼB(tal B)、ホスホエノールピルビン酸シンターゼ(ppsA)、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)のうちの1つ以上の活性を向上させて、ホスホケトラーゼ経路に取り込まれる炭素量を増加させることができる(図72)。特定の態様では、次の遺伝子、rpiA、rpiB、rpe、tktA、tktB、talB、ppsA、eutD、及び/又はptaの活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。一実施形態では、リボース−5−リン酸イソメラーゼ(rpiA及び/又はrpiB)の活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。他の実施形態では、D−リブロース−5−リン酸3−エピメラーゼ(rpe)の活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。他の実施形態では、トランスケトラーゼ(tktA及び/又はtktB)の活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。更に別の実施形態では、トランスアルドラーゼB(tal B)の活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。他の実施形態では、ホスホエノールピルビン酸シンターゼ(ppsA)の活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。更に他の実施形態では、リン酸アセチルトランスフェラーゼ(pta及び/又はeutD)の活性は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。特定の態様では、次の遺伝子、rpiA、rpiB、rpe、tktA、tktB、tal B、ppsA、eutD、及び/又はptaのイソ酵素は、染色体上のプロモータ及び/又はrbsを変更することにより増加させることができ、又はプラスミドに発現させることにより増加させることができる。
これらの株には、次の遺伝子、グルコース−6−リン酸脱水素酵素(zwf)、6−ホスホフルクトキナーゼ−1(pfkA及び/又はpfkB)、フルクトースビスリン酸アルドラーゼ(fba、fbaA、fbaB、及び/又はfbaC)、グリセルアルデヒド−3−リン酸脱水素酵素(gapA及び/又はgapB)、酢酸キナーゼ(ackA)、クエン酸シンターゼ(gltA)、トランスケトラーゼ(tktA及び/又はtktB)、EI(ptsI)、EIICBGlc(ptsG)、EIIAGlc(crr)、及び/又はHPr(ptsH)のうちの1つ以上の活性が低下するよう遺伝子操作を行い、ホスホケトラーゼ経路への炭素の取り込みを増加させることができる(図73)。一態様では、グルコース−6−リン酸脱水素酵素をコードしているzwf遺伝子は、下方調節される。別の実施形態では、6−ホスホフルクトキナーゼ−1AをコードしているpfkA遺伝子は、下方調節される。別の実施形態では、グリセルアルデヒド−3−リン酸脱水素酵素AをコードしているgapA遺伝子は、下方調節される。別の実施形態では、フルクトースビスリン酸アルドラーゼをコードしているfba遺伝子は、下方調節される。更に別の実施形態では、クエン酸シンターゼをコードしているgltA遺伝子は、下方調節される。一実施形態では、酢酸キナーゼをコードしているackA遺伝子は、下方調節される。別の実施形態では、EIをコードしているptsI遺伝子は、下方調節される。一実施形態では、HPrをコードしているptsH遺伝子は、下方調節される。別の実施形態では、EIICBGlcをコードしているptsG遺伝子は、下方調節される。更に別の実施形態では、EIIAGlcをコードしているcrr遺伝子は、下方調節される。ptsオペロンは、ホスホトランスフェラーゼ系の遺伝子をコードする。一部の実施形態では、株は、ホスホトランスフェラーゼ系(PTS)の活性が低下するよう遺伝子操作して、ホスホケトラーゼ経路への炭素の取り込みを増加させることができる。一部の実施形態では、PTSは、ptsオペロンを下方調節することにより下方調節される。特定の態様では、PTSは、下方調節され、かつグルコース輸送経路は上方調節される。グルコース輸送系は、限定するものではないが、ガラクトース(galP)及びグルコキナーゼ(glk)遺伝を包含する。一部の実施形態では、ptsオペロンは、下方調節され、ガラクトース(galP)遺伝子は上方調節され、かつグルコキナーゼ(glk)遺伝子は上方調節される。特定の態様では、次の遺伝子zwf、pfkA、fba、gapA、ackA、gltA、tktA、ptsG、ptsH、ptsI、及び/又はcrrによりコードされるタンパク質のイソ酵素を下方調節することで、ホスホケトラーゼ経路への炭素の取り込みを増加させることができる。一部の実施形態では、pfkB遺伝子は下方調節される。一部の実施形態では、グリセルアルデヒド−3−リン酸脱水素酵素B(gapB)遺伝子は下方調節される。一部の実施形態では、トランスケトラーゼB(tktB)遺伝子は下方調節される。
実施例25:宿主由来の変異を有するホスホケトラーゼ発現株による小規模でのイソプレン産生
(i)材料
TM3培地の組成(発酵培地1L当たり):
K2HPO4を13.6g、KH2PO4を13.6g、MgSO4 *7H2Oを2g、クエン酸一水和物を2g、クエン酸鉄アンモニウムを0.3g、(NH4)2SO4を3.2g、酵母エキスを0.2g、1000×微量金属溶液を1mL。すべての成分を共に加え、脱イオン水に溶解させる。水酸化アンモニウム(30%)によりpHを6.8に調整し、及び容量を調整する。0.22μmフィルタで培地を濾過滅菌する。pH調整及び滅菌後にグルコース10.0g及び抗菌剤を添加する。
1000X微量金属溶液(発酵培地1L当たり):
クエン酸*H2O 40g、MnSO4 *H2O 30g、NaCl 10g、FeSO4 *7H2O 1g、CoCl2 *6H2O 1g、ZnSO4 *7H2O 1g、CuSO4 *5H2O 100mg、H3BO3 100mg、NaMoO4 *2H2O 100mg。各成分を1成分ずつ脱イオン水に溶解する。pHをHCl/NaOHで3.0に調整した後、溶液を用量に調整し、0.22μmフィルタでフィルタ滅菌する。
(ii)実験手順
ルリア・ベルターニブロス+抗生物質で細胞を一晩生育させる。翌日、OD600が0.1になるよう、250mLのバッフル付き三角フラスコにおいて細胞を20mLのTM3培地(50μg/mLのスペクチノマイシン、25μg/mLのクロラムフェニコール、及び50μg/mLカルベニシリンを含有)により希釈し、34℃かつ200rpmでインキュベートする。2時間生育させた後、OD600を測定し、200uMのIPTGを加える。発酵工程の間、規則的に試料を採取する。各時点でOD600を測定する。また、ガスクロマトグラフ−マススペクトロメータ(GC−MS)(Agilent)によるヘッドスペース解析によりイソプレンのオフガス分析を行う。100μLの全ブロスをGCバイアルに入れ密閉し、34℃かつ200rpmで30分間インキュベートする。70℃で7分のインキュベーションからなる熱殺菌工程後、試料をGCに装填する。記録される比産生量は、イソプレン量のGCによる読み取り値(μg/L)を、インキュベート時間(30分)及びOD600測定値で除算した値である。
実施例26:宿主由来の変異を有するホスホケトラーゼ発現株による15L規模でのイソプレンの産生
(i)材料
培地組成(発酵培地1Lあたり):
K2HPO4(7.5g)、MgSO4*7H2O(2g)、クエン酸一水和物(2g)、クエン酸鉄アンモニウム(0.3g)、酵母エキス(0.5g)、50%硫酸(1.6mL)、1000X改変微量金属溶液(1mL)。すべての成分を共に加え、脱イオン水に溶解させた。この溶液を加熱滅菌した(123℃で20分)。水酸化アンモニウム(28%)によりpHを7.0に調整し、容量にメスアップした。滅菌及びpH調製後にグルコース10g、ビタミン溶液8mL、及び抗生物質を加えた。
1000X改変微量金属溶液(1Lあたり):
クエン酸*H2O(40g)、MnSO4*H2O(30g)、NaCl(10g)、FeSO4*7H2O(1g)、CoCl2*6H2O(1g)、ZnSO*7H2O(1g)、CuSO4*5H2O(100mg)、H3BO3(100mg)、NaMoO4*2H2O(100mg)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、0.22μmのフィルタを用いろ過滅菌した。
ビタミン溶液(1Lあたり):
チアミン塩酸塩(1.0g)、D−(+)−ビオチン(1.0g)、ニコチン酸(1.0g)、塩酸ピリドキシン(4.0g)。各成分を1つずつ脱イオン水に溶解させ、HCl/NaOHによりpHを3.0に調整し、次に溶液を用量に調整し、孔径0.22μmのフィルタを用いろ過滅菌した。
マクロ塩溶液(1Lあたり):
MgSO4*7H2O(296g)、クエン酸水和物(296g)、クエン酸鉄アンモニウム(49.6g)。すべての成分を水に溶解させ、用量に調整し、0.22μmのフィルタを用い、ろ過滅菌した。
供給溶液(1kgあたり):
グルコース(0.590kg)、脱イオン水(0.393kg)、K2HPO4(7.4g)及び100% Foamblast882(8.9g)。すべての成分を合わせて混合し、オートクレーブ処理した。供給溶液をオートクレーブにより処理した後、滅菌フード中で、供給ボトルに栄養塩類を添加する。滅菌後に供給溶液に添加するものは(供給溶液1kg当たり)マクロ塩溶液5.54mL、ビタミン溶液6.55mL、1000X改変微量金属溶液0.82mLである。
(ii)解析
2種の質量分析器iSCAN(ハミルトン・サンドストランド)、及びHiden HPR20(Hiden Analytical)により、排出気体中の、イソプレン、酸素、窒素、及び二酸化炭素濃度を別個に測定した。
発酵ブロス中の溶存酸素は、Hamilton Companyにより提供された光学センサーを備えた衛生的であり滅菌可能なプローブにより測定した。
4時間間隔でブロス試料にHPLC解析を行い、発酵ブロス中のクエン酸、グルコース、酢酸及びメバロン酸濃度を測定する。屈折計による測定値と、濃度既知の標準を使用して予め作成した検量線とを比較して、ブロス試料中の濃度を測定する。