JP4138848B2 - 免疫不全組換えポックスウイルス - Google Patents
免疫不全組換えポックスウイルス Download PDFInfo
- Publication number
- JP4138848B2 JP4138848B2 JP2007162393A JP2007162393A JP4138848B2 JP 4138848 B2 JP4138848 B2 JP 4138848B2 JP 2007162393 A JP2007162393 A JP 2007162393A JP 2007162393 A JP2007162393 A JP 2007162393A JP 4138848 B2 JP4138848 B2 JP 4138848B2
- Authority
- JP
- Japan
- Prior art keywords
- virus
- plasmid
- fragment
- gene
- alvac
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
- 241000713772 Human immunodeficiency virus 1 Species 0.000 claims description 173
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 5
- 108090000623 proteins and genes Proteins 0.000 abstract description 263
- 241000700605 Viruses Species 0.000 abstract description 226
- 229940023860 canarypox virus HIV vaccine Drugs 0.000 abstract description 97
- 238000000034 method Methods 0.000 abstract description 62
- 230000014509 gene expression Effects 0.000 abstract description 46
- 239000000427 antigen Substances 0.000 abstract description 38
- 108091007433 antigens Proteins 0.000 abstract description 38
- 102000036639 antigens Human genes 0.000 abstract description 38
- 230000003053 immunization Effects 0.000 abstract description 32
- 238000002649 immunization Methods 0.000 abstract description 32
- 230000002238 attenuated effect Effects 0.000 abstract description 26
- 239000000523 sample Substances 0.000 abstract description 26
- 101100462138 Brassica napus OlnB1 gene Proteins 0.000 abstract description 21
- 101100224481 Dictyostelium discoideum pole gene Proteins 0.000 abstract description 21
- 206010061598 Immunodeficiency Diseases 0.000 abstract description 21
- 101150110488 POL2 gene Proteins 0.000 abstract description 21
- 208000029462 Immunodeficiency disease Diseases 0.000 abstract description 20
- 230000007813 immunodeficiency Effects 0.000 abstract description 20
- 101100117436 Thermus aquaticus polA gene Proteins 0.000 abstract description 19
- 239000000203 mixture Substances 0.000 abstract description 19
- 102100035954 Choline transporter-like protein 2 Human genes 0.000 abstract description 13
- 101000948115 Homo sapiens Choline transporter-like protein 2 Proteins 0.000 abstract description 13
- 102100031699 Choline transporter-like protein 1 Human genes 0.000 abstract description 12
- 101000940912 Homo sapiens Choline transporter-like protein 1 Proteins 0.000 abstract description 12
- 230000001225 therapeutic effect Effects 0.000 abstract description 2
- 230000003449 preventive effect Effects 0.000 abstract 1
- 210000004027 cell Anatomy 0.000 description 254
- 239000012634 fragment Substances 0.000 description 243
- 239000013612 plasmid Substances 0.000 description 230
- 239000002773 nucleotide Substances 0.000 description 113
- 125000003729 nucleotide group Chemical group 0.000 description 113
- 238000011081 inoculation Methods 0.000 description 111
- 241000700618 Vaccinia virus Species 0.000 description 104
- 206010046865 Vaccinia virus infection Diseases 0.000 description 102
- 208000007089 vaccinia Diseases 0.000 description 102
- 241000699670 Mus sp. Species 0.000 description 95
- 238000003752 polymerase chain reaction Methods 0.000 description 92
- 108020004414 DNA Proteins 0.000 description 90
- 102000004169 proteins and genes Human genes 0.000 description 86
- 241000282693 Cercopithecidae Species 0.000 description 85
- 241000725303 Human immunodeficiency virus Species 0.000 description 69
- 108091034117 Oligonucleotide Proteins 0.000 description 66
- 206010037742 Rabies Diseases 0.000 description 65
- 229960005486 vaccine Drugs 0.000 description 61
- 239000013598 vector Substances 0.000 description 61
- 239000000047 product Substances 0.000 description 58
- 108700026244 Open Reading Frames Proteins 0.000 description 57
- 238000012217 deletion Methods 0.000 description 57
- 230000037430 deletion Effects 0.000 description 57
- 210000002966 serum Anatomy 0.000 description 57
- 235000018102 proteins Nutrition 0.000 description 54
- 208000014674 injury Diseases 0.000 description 47
- 241000283973 Oryctolagus cuniculus Species 0.000 description 46
- 230000008733 trauma Effects 0.000 description 45
- 230000004044 response Effects 0.000 description 44
- 238000012360 testing method Methods 0.000 description 44
- 241001465754 Metazoa Species 0.000 description 43
- 230000028993 immune response Effects 0.000 description 42
- 108090000288 Glycoproteins Proteins 0.000 description 40
- 102000003886 Glycoproteins Human genes 0.000 description 38
- 230000002163 immunogen Effects 0.000 description 38
- 238000004458 analytical method Methods 0.000 description 37
- 230000006798 recombination Effects 0.000 description 37
- 241000588724 Escherichia coli Species 0.000 description 36
- 108700004025 env Genes Proteins 0.000 description 36
- 101150030339 env gene Proteins 0.000 description 36
- 238000005215 recombination Methods 0.000 description 34
- 241000287828 Gallus gallus Species 0.000 description 33
- 101710154606 Hemagglutinin Proteins 0.000 description 33
- 101710093908 Outer capsid protein VP4 Proteins 0.000 description 33
- 101710135467 Outer capsid protein sigma-1 Proteins 0.000 description 33
- 101710176177 Protein A56 Proteins 0.000 description 33
- 235000013330 chicken meat Nutrition 0.000 description 33
- 238000000338 in vitro Methods 0.000 description 32
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 31
- 238000002965 ELISA Methods 0.000 description 30
- 238000010276 construction Methods 0.000 description 30
- 108090000765 processed proteins & peptides Proteins 0.000 description 30
- 238000002347 injection Methods 0.000 description 29
- 239000007924 injection Substances 0.000 description 29
- 230000000694 effects Effects 0.000 description 28
- 241000271566 Aves Species 0.000 description 27
- 208000015181 infectious disease Diseases 0.000 description 27
- 230000003472 neutralizing effect Effects 0.000 description 27
- 241000178270 Canarypox virus Species 0.000 description 26
- 239000000185 hemagglutinin Substances 0.000 description 25
- 108010005774 beta-Galactosidase Proteins 0.000 description 24
- 102100034349 Integrase Human genes 0.000 description 23
- 108091028043 Nucleic acid sequence Proteins 0.000 description 23
- 238000003780 insertion Methods 0.000 description 23
- 230000037431 insertion Effects 0.000 description 23
- 239000000872 buffer Substances 0.000 description 22
- 238000001114 immunoprecipitation Methods 0.000 description 22
- 239000013615 primer Substances 0.000 description 22
- 239000000126 substance Substances 0.000 description 22
- 101800001690 Transmembrane protein gp41 Proteins 0.000 description 21
- 238000002474 experimental method Methods 0.000 description 21
- 239000000902 placebo Substances 0.000 description 21
- 229940068196 placebo Drugs 0.000 description 21
- 238000002255 vaccination Methods 0.000 description 21
- 230000003612 virological effect Effects 0.000 description 21
- 101710121417 Envelope glycoprotein Proteins 0.000 description 20
- 108020004440 Thymidine kinase Proteins 0.000 description 20
- 102000005936 beta-Galactosidase Human genes 0.000 description 19
- 210000004369 blood Anatomy 0.000 description 19
- 239000008280 blood Substances 0.000 description 19
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 19
- 239000002953 phosphate buffered saline Substances 0.000 description 19
- 230000010076 replication Effects 0.000 description 19
- 102100022563 Tubulin polymerization-promoting protein Human genes 0.000 description 18
- 238000004820 blood count Methods 0.000 description 18
- 238000010790 dilution Methods 0.000 description 18
- 239000012895 dilution Substances 0.000 description 18
- 241000711404 Avian avulavirus 1 Species 0.000 description 17
- 101710147327 Calcineurin B homologous protein 1 Proteins 0.000 description 17
- 101710205625 Capsid protein p24 Proteins 0.000 description 17
- 101710177166 Phosphoprotein Proteins 0.000 description 17
- 101710149279 Small delta antigen Proteins 0.000 description 17
- 230000005875 antibody response Effects 0.000 description 17
- 239000002609 medium Substances 0.000 description 17
- 231100000419 toxicity Toxicity 0.000 description 17
- 230000001988 toxicity Effects 0.000 description 17
- 241000699666 Mus <mouse, genus> Species 0.000 description 16
- 150000001413 amino acids Chemical group 0.000 description 16
- DDRJAANPRJIHGJ-UHFFFAOYSA-N creatinine Chemical compound CN1CC(=O)NC1=N DDRJAANPRJIHGJ-UHFFFAOYSA-N 0.000 description 16
- 238000002360 preparation method Methods 0.000 description 16
- 241000282472 Canis lupus familiaris Species 0.000 description 15
- 108010017826 DNA Polymerase I Proteins 0.000 description 15
- 102000004594 DNA Polymerase I Human genes 0.000 description 15
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 15
- 102000006601 Thymidine Kinase Human genes 0.000 description 15
- 230000027455 binding Effects 0.000 description 15
- 230000002829 reductive effect Effects 0.000 description 15
- 238000011144 upstream manufacturing Methods 0.000 description 15
- 241000282326 Felis catus Species 0.000 description 14
- 238000010367 cloning Methods 0.000 description 14
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 14
- 230000001681 protective effect Effects 0.000 description 14
- 210000001519 tissue Anatomy 0.000 description 14
- 101150082239 G gene Proteins 0.000 description 13
- 241000282412 Homo Species 0.000 description 13
- 101150060895 I4L gene Proteins 0.000 description 13
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 13
- 241000711798 Rabies lyssavirus Species 0.000 description 13
- 229920002684 Sepharose Polymers 0.000 description 13
- 238000009825 accumulation Methods 0.000 description 13
- 235000001014 amino acid Nutrition 0.000 description 13
- 229940024606 amino acid Drugs 0.000 description 13
- 238000011161 development Methods 0.000 description 13
- 230000005847 immunogenicity Effects 0.000 description 13
- 229960004452 methionine Drugs 0.000 description 13
- 239000000725 suspension Substances 0.000 description 13
- 241000700198 Cavia Species 0.000 description 12
- 108091026890 Coding region Proteins 0.000 description 12
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 12
- 230000000890 antigenic effect Effects 0.000 description 12
- 108010029566 avian influenza A virus hemagglutinin Proteins 0.000 description 12
- 238000006243 chemical reaction Methods 0.000 description 12
- 230000012010 growth Effects 0.000 description 12
- 210000005260 human cell Anatomy 0.000 description 12
- CMSMOCZEIVJLDB-UHFFFAOYSA-N Cyclophosphamide Chemical compound ClCCN(CCCl)P1(=O)NCCCO1 CMSMOCZEIVJLDB-UHFFFAOYSA-N 0.000 description 11
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 11
- 230000000875 corresponding effect Effects 0.000 description 11
- 229960004397 cyclophosphamide Drugs 0.000 description 11
- 230000034994 death Effects 0.000 description 11
- 231100000517 death Toxicity 0.000 description 11
- 230000029087 digestion Effects 0.000 description 11
- 210000002950 fibroblast Anatomy 0.000 description 11
- 230000006870 function Effects 0.000 description 11
- 108700004026 gag Genes Proteins 0.000 description 11
- 230000036541 health Effects 0.000 description 11
- 230000036039 immunity Effects 0.000 description 11
- OPIFSICVWOWJMJ-AEOCFKNESA-N 5-bromo-4-chloro-3-indolyl beta-D-galactoside Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1OC1=CNC2=CC=C(Br)C(Cl)=C12 OPIFSICVWOWJMJ-AEOCFKNESA-N 0.000 description 10
- 101150039660 HA gene Proteins 0.000 description 10
- 241000282579 Pan Species 0.000 description 10
- PXIPVTKHYLBLMZ-UHFFFAOYSA-N Sodium azide Chemical compound [Na+].[N-]=[N+]=[N-] PXIPVTKHYLBLMZ-UHFFFAOYSA-N 0.000 description 10
- 101150084279 TM gene Proteins 0.000 description 10
- 108020005202 Viral DNA Proteins 0.000 description 10
- 238000003556 assay Methods 0.000 description 10
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 10
- 238000003776 cleavage reaction Methods 0.000 description 10
- 239000003814 drug Substances 0.000 description 10
- 101150098622 gag gene Proteins 0.000 description 10
- 238000009396 hybridization Methods 0.000 description 10
- 230000001965 increasing effect Effects 0.000 description 10
- 238000007917 intracranial administration Methods 0.000 description 10
- 230000007017 scission Effects 0.000 description 10
- 238000004448 titration Methods 0.000 description 10
- 210000003501 vero cell Anatomy 0.000 description 10
- 241000713311 Simian immunodeficiency virus Species 0.000 description 9
- 239000007983 Tris buffer Substances 0.000 description 9
- 238000007918 intramuscular administration Methods 0.000 description 9
- 210000000265 leukocyte Anatomy 0.000 description 9
- 210000001161 mammalian embryo Anatomy 0.000 description 9
- 229930182817 methionine Natural products 0.000 description 9
- 238000011580 nude mouse model Methods 0.000 description 9
- 230000036961 partial effect Effects 0.000 description 9
- 238000000746 purification Methods 0.000 description 9
- 238000007920 subcutaneous administration Methods 0.000 description 9
- 231100000331 toxic Toxicity 0.000 description 9
- 230000002588 toxic effect Effects 0.000 description 9
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 9
- 108010082126 Alanine transaminase Proteins 0.000 description 8
- 108010003415 Aspartate Aminotransferases Proteins 0.000 description 8
- 102000004625 Aspartate Aminotransferases Human genes 0.000 description 8
- 101150039990 B13R gene Proteins 0.000 description 8
- BPYKTIZUTYGOLE-IFADSCNNSA-N Bilirubin Chemical compound N1C(=O)C(C)=C(C=C)\C1=C\C1=C(C)C(CCC(O)=O)=C(CC2=C(C(C)=C(\C=C/3C(=C(C=C)C(=O)N\3)C)N2)CCC(O)=O)N1 BPYKTIZUTYGOLE-IFADSCNNSA-N 0.000 description 8
- 241000283707 Capra Species 0.000 description 8
- 201000006082 Chickenpox Diseases 0.000 description 8
- 239000003155 DNA primer Substances 0.000 description 8
- 230000004543 DNA replication Effects 0.000 description 8
- 241000282553 Macaca Species 0.000 description 8
- 229920001213 Polysorbate 20 Polymers 0.000 description 8
- 102000000505 Ribonucleotide Reductases Human genes 0.000 description 8
- 108010041388 Ribonucleotide Reductases Proteins 0.000 description 8
- 101150071286 SPI-2 gene Proteins 0.000 description 8
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 8
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 8
- 206010046980 Varicella Diseases 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 8
- 229940109239 creatinine Drugs 0.000 description 8
- 238000010166 immunofluorescence Methods 0.000 description 8
- 230000002779 inactivation Effects 0.000 description 8
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical compound FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 8
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 8
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 8
- 108091008146 restriction endonucleases Proteins 0.000 description 8
- 230000000405 serological effect Effects 0.000 description 8
- 239000000758 substrate Substances 0.000 description 8
- 241000712461 unidentified influenza virus Species 0.000 description 8
- 230000029812 viral genome replication Effects 0.000 description 8
- 230000001018 virulence Effects 0.000 description 8
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 8
- -1 C7L-K1L Proteins 0.000 description 7
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 7
- 241000282567 Macaca fascicularis Species 0.000 description 7
- 241000699660 Mus musculus Species 0.000 description 7
- 101900083372 Rabies virus Glycoprotein Proteins 0.000 description 7
- 230000036436 anti-hiv Effects 0.000 description 7
- 239000013553 cell monolayer Substances 0.000 description 7
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 7
- 238000001962 electrophoresis Methods 0.000 description 7
- 239000000499 gel Substances 0.000 description 7
- 239000008103 glucose Substances 0.000 description 7
- 230000002458 infectious effect Effects 0.000 description 7
- 239000002054 inoculum Substances 0.000 description 7
- 231100000252 nontoxic Toxicity 0.000 description 7
- 230000003000 nontoxic effect Effects 0.000 description 7
- 244000052769 pathogen Species 0.000 description 7
- 108700004029 pol Genes Proteins 0.000 description 7
- 238000001179 sorption measurement Methods 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- 210000003371 toe Anatomy 0.000 description 7
- 230000005030 transcription termination Effects 0.000 description 7
- 241000700199 Cavia porcellus Species 0.000 description 6
- 241000282695 Saimiri Species 0.000 description 6
- 238000012300 Sequence Analysis Methods 0.000 description 6
- 108091081024 Start codon Proteins 0.000 description 6
- 208000025865 Ulcer Diseases 0.000 description 6
- 230000008901 benefit Effects 0.000 description 6
- 230000037396 body weight Effects 0.000 description 6
- 230000008859 change Effects 0.000 description 6
- 201000010099 disease Diseases 0.000 description 6
- 235000013601 eggs Nutrition 0.000 description 6
- 230000002068 genetic effect Effects 0.000 description 6
- 230000002489 hematologic effect Effects 0.000 description 6
- 230000002008 hemorrhagic effect Effects 0.000 description 6
- 210000000987 immune system Anatomy 0.000 description 6
- 238000011534 incubation Methods 0.000 description 6
- KWGKDLIKAYFUFQ-UHFFFAOYSA-M lithium chloride Chemical compound [Li+].[Cl-] KWGKDLIKAYFUFQ-UHFFFAOYSA-M 0.000 description 6
- 210000004698 lymphocyte Anatomy 0.000 description 6
- 239000006166 lysate Substances 0.000 description 6
- 238000004519 manufacturing process Methods 0.000 description 6
- 239000012528 membrane Substances 0.000 description 6
- 230000001717 pathogenic effect Effects 0.000 description 6
- 101150088264 pol gene Proteins 0.000 description 6
- 102000004196 processed proteins & peptides Human genes 0.000 description 6
- 238000005070 sampling Methods 0.000 description 6
- 235000020183 skimmed milk Nutrition 0.000 description 6
- 239000000243 solution Substances 0.000 description 6
- 241000894007 species Species 0.000 description 6
- 238000013519 translation Methods 0.000 description 6
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 5
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 5
- 206010002091 Anaesthesia Diseases 0.000 description 5
- 241000700663 Avipoxvirus Species 0.000 description 5
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 5
- 108020004705 Codon Proteins 0.000 description 5
- UHDGCWIWMRVCDJ-CCXZUQQUSA-N Cytarabine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@@H](O)[C@H](O)[C@@H](CO)O1 UHDGCWIWMRVCDJ-CCXZUQQUSA-N 0.000 description 5
- 238000009007 Diagnostic Kit Methods 0.000 description 5
- 102000001554 Hemoglobins Human genes 0.000 description 5
- 108010054147 Hemoglobins Proteins 0.000 description 5
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 5
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 5
- 239000000020 Nitrocellulose Substances 0.000 description 5
- 206010058461 Orchitis noninfective Diseases 0.000 description 5
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 5
- 102100027378 Prothrombin Human genes 0.000 description 5
- 108010094028 Prothrombin Proteins 0.000 description 5
- ABBQHOQBGMUPJH-UHFFFAOYSA-M Sodium salicylate Chemical compound [Na+].OC1=CC=CC=C1C([O-])=O ABBQHOQBGMUPJH-UHFFFAOYSA-M 0.000 description 5
- 230000037005 anaesthesia Effects 0.000 description 5
- 230000005540 biological transmission Effects 0.000 description 5
- 210000000170 cell membrane Anatomy 0.000 description 5
- 239000003153 chemical reaction reagent Substances 0.000 description 5
- 235000012000 cholesterol Nutrition 0.000 description 5
- 230000003247 decreasing effect Effects 0.000 description 5
- 210000003743 erythrocyte Anatomy 0.000 description 5
- 239000012894 fetal calf serum Substances 0.000 description 5
- 238000005534 hematocrit Methods 0.000 description 5
- 230000005764 inhibitory process Effects 0.000 description 5
- 210000003292 kidney cell Anatomy 0.000 description 5
- 229920001220 nitrocellulos Polymers 0.000 description 5
- 230000007918 pathogenicity Effects 0.000 description 5
- 239000008188 pellet Substances 0.000 description 5
- 239000011591 potassium Substances 0.000 description 5
- 229910052700 potassium Inorganic materials 0.000 description 5
- 229940039716 prothrombin Drugs 0.000 description 5
- 239000011734 sodium Substances 0.000 description 5
- 229910052708 sodium Inorganic materials 0.000 description 5
- 229960004025 sodium salicylate Drugs 0.000 description 5
- 229940124597 therapeutic agent Drugs 0.000 description 5
- 230000036269 ulceration Effects 0.000 description 5
- LGGRPYXPOUIMKG-OJICBBQQSA-N (2S)-2-[[(2S)-2-[[(2S,3S)-2-[[(2S,3R)-2-[[2-[[(2S,3S)-2-[[(2S,3S)-2-[[(2S)-4-amino-2-[[(2S)-6-amino-2-[[(2S,3R)-2-[[(2S,3R)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[2-[[(2S)-1-[2-[[(2S,3S)-2-[[(2S)-2-[[(2S,3S)-2-[[(2S)-2-[[(2S)-6-amino-2-[[(2S)-2-[[(2S)-6-amino-2-[[(2S)-4-amino-2-[[(2S)-2-[[(2S)-4-amino-2-[[(2S)-1-[(2S)-2-[[(2S,3R)-2-amino-3-hydroxybutanoyl]amino]-5-(diaminomethylideneamino)pentanoyl]pyrrolidine-2-carbonyl]amino]-4-oxobutanoyl]amino]-3-(4-hydroxyphenyl)propanoyl]amino]-4-oxobutanoyl]amino]hexanoyl]amino]-5-(diaminomethylideneamino)pentanoyl]amino]hexanoyl]amino]-5-(diaminomethylideneamino)pentanoyl]amino]-3-methylpentanoyl]amino]-3-(1H-imidazol-4-yl)propanoyl]amino]-3-methylpentanoyl]amino]acetyl]pyrrolidine-2-carbonyl]amino]acetyl]amino]-5-(diaminomethylideneamino)pentanoyl]amino]propanoyl]amino]-3-phenylpropanoyl]amino]-3-(4-hydroxyphenyl)propanoyl]amino]-3-hydroxybutanoyl]amino]-3-hydroxybutanoyl]amino]hexanoyl]amino]-4-oxobutanoyl]amino]-3-methylpentanoyl]amino]-3-methylpentanoyl]amino]acetyl]amino]-3-hydroxybutanoyl]amino]-3-methylpentanoyl]amino]-5-(diaminomethylideneamino)pentanoyl]amino]-N-[(2S)-1-[[(2S)-1-amino-3-(1H-imidazol-4-yl)-1-oxopropan-2-yl]amino]-1-oxopropan-2-yl]pentanediamide Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N2CCC[C@H]2C(=O)NCC(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC3=CC=CC=C3)C(=O)N[C@@H](CC4=CC=C(C=C4)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC5=CNC=N5)C(=O)N)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC6=CC=C(C=C6)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@@H]7CCCN7C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H]([C@@H](C)O)N LGGRPYXPOUIMKG-OJICBBQQSA-N 0.000 description 4
- 101710172503 Chemokine-binding protein Proteins 0.000 description 4
- 241000700626 Cowpox virus Species 0.000 description 4
- 102000053602 DNA Human genes 0.000 description 4
- 239000006145 Eagle's minimal essential medium Substances 0.000 description 4
- 102000004190 Enzymes Human genes 0.000 description 4
- 108090000790 Enzymes Proteins 0.000 description 4
- 101150105849 H5 gene Proteins 0.000 description 4
- 208000031886 HIV Infections Diseases 0.000 description 4
- 229940033332 HIV-1 vaccine Drugs 0.000 description 4
- 208000002979 Influenza in Birds Diseases 0.000 description 4
- 108091005804 Peptidases Proteins 0.000 description 4
- 102000003992 Peroxidases Human genes 0.000 description 4
- 101710138270 PspA protein Proteins 0.000 description 4
- 241000700584 Simplexvirus Species 0.000 description 4
- 108010046722 Thrombospondin 1 Proteins 0.000 description 4
- 101100534084 Vaccinia virus (strain Copenhagen) B14R gene Proteins 0.000 description 4
- 101100004092 Vaccinia virus (strain Western Reserve) VACWR196 gene Proteins 0.000 description 4
- 241000700622 Vaccinia virus Copenhagen Species 0.000 description 4
- 239000002671 adjuvant Substances 0.000 description 4
- 238000010171 animal model Methods 0.000 description 4
- 206010064097 avian influenza Diseases 0.000 description 4
- 238000012742 biochemical analysis Methods 0.000 description 4
- 210000000601 blood cell Anatomy 0.000 description 4
- 239000004202 carbamide Substances 0.000 description 4
- 230000000295 complement effect Effects 0.000 description 4
- 238000001514 detection method Methods 0.000 description 4
- 239000010432 diamond Substances 0.000 description 4
- 239000013024 dilution buffer Substances 0.000 description 4
- 229940079593 drug Drugs 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 4
- 230000001976 improved effect Effects 0.000 description 4
- 210000003000 inclusion body Anatomy 0.000 description 4
- 238000001802 infusion Methods 0.000 description 4
- 210000004072 lung Anatomy 0.000 description 4
- 239000000463 material Substances 0.000 description 4
- 238000005259 measurement Methods 0.000 description 4
- 230000035772 mutation Effects 0.000 description 4
- 238000006386 neutralization reaction Methods 0.000 description 4
- 108040007629 peroxidase activity proteins Proteins 0.000 description 4
- 229920002401 polyacrylamide Polymers 0.000 description 4
- 238000001556 precipitation Methods 0.000 description 4
- 239000002243 precursor Substances 0.000 description 4
- 230000000644 propagated effect Effects 0.000 description 4
- 230000009257 reactivity Effects 0.000 description 4
- 238000011084 recovery Methods 0.000 description 4
- 210000001995 reticulocyte Anatomy 0.000 description 4
- 238000012216 screening Methods 0.000 description 4
- 239000011780 sodium chloride Substances 0.000 description 4
- 229940031626 subunit vaccine Drugs 0.000 description 4
- 230000035897 transcription Effects 0.000 description 4
- 238000013518 transcription Methods 0.000 description 4
- 229920001817 Agar Polymers 0.000 description 3
- 229920000936 Agarose Polymers 0.000 description 3
- 241000282552 Chlorocebus aethiops Species 0.000 description 3
- 108091029865 Exogenous DNA Proteins 0.000 description 3
- 108060002716 Exonuclease Proteins 0.000 description 3
- 101710177291 Gag polyprotein Proteins 0.000 description 3
- YQEZLKZALYSWHR-UHFFFAOYSA-N Ketamine Chemical compound C=1C=CC=C(Cl)C=1C1(NC)CCCCC1=O YQEZLKZALYSWHR-UHFFFAOYSA-N 0.000 description 3
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 3
- 206010024769 Local reaction Diseases 0.000 description 3
- 206010067482 No adverse event Diseases 0.000 description 3
- 102220480756 Nucleotide pyrophosphatase_I13L_mutation Human genes 0.000 description 3
- 235000014676 Phragmites communis Nutrition 0.000 description 3
- 239000004743 Polypropylene Substances 0.000 description 3
- 108020004511 Recombinant DNA Proteins 0.000 description 3
- 241000283984 Rodentia Species 0.000 description 3
- 206010039509 Scab Diseases 0.000 description 3
- 241000287231 Serinus Species 0.000 description 3
- 108010008038 Synthetic Vaccines Proteins 0.000 description 3
- 210000001744 T-lymphocyte Anatomy 0.000 description 3
- 108020005038 Terminator Codon Proteins 0.000 description 3
- 102100036034 Thrombospondin-1 Human genes 0.000 description 3
- 108700019146 Transgenes Proteins 0.000 description 3
- 241000700647 Variola virus Species 0.000 description 3
- 238000010521 absorption reaction Methods 0.000 description 3
- 239000008272 agar Substances 0.000 description 3
- 230000003321 amplification Effects 0.000 description 3
- 238000000137 annealing Methods 0.000 description 3
- 210000004899 c-terminal region Anatomy 0.000 description 3
- 239000001506 calcium phosphate Substances 0.000 description 3
- 229910000389 calcium phosphate Inorganic materials 0.000 description 3
- 235000011010 calcium phosphates Nutrition 0.000 description 3
- 239000003795 chemical substances by application Substances 0.000 description 3
- 238000004587 chromatography analysis Methods 0.000 description 3
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 3
- 235000018417 cysteine Nutrition 0.000 description 3
- 230000001461 cytolytic effect Effects 0.000 description 3
- 230000007423 decrease Effects 0.000 description 3
- 230000001627 detrimental effect Effects 0.000 description 3
- 238000002405 diagnostic procedure Methods 0.000 description 3
- 238000011156 evaluation Methods 0.000 description 3
- 102000013165 exonuclease Human genes 0.000 description 3
- 210000002683 foot Anatomy 0.000 description 3
- 230000004927 fusion Effects 0.000 description 3
- 108091008053 gene clusters Proteins 0.000 description 3
- 238000010353 genetic engineering Methods 0.000 description 3
- 208000002672 hepatitis B Diseases 0.000 description 3
- 238000003384 imaging method Methods 0.000 description 3
- 230000001506 immunosuppresive effect Effects 0.000 description 3
- 238000007901 in situ hybridization Methods 0.000 description 3
- 238000001727 in vivo Methods 0.000 description 3
- 230000000977 initiatory effect Effects 0.000 description 3
- 238000001990 intravenous administration Methods 0.000 description 3
- 229960003299 ketamine Drugs 0.000 description 3
- 231100001231 less toxic Toxicity 0.000 description 3
- 231100000636 lethal dose Toxicity 0.000 description 3
- 239000012139 lysis buffer Substances 0.000 description 3
- 210000004379 membrane Anatomy 0.000 description 3
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 3
- 238000003199 nucleic acid amplification method Methods 0.000 description 3
- 230000003287 optical effect Effects 0.000 description 3
- 239000002245 particle Substances 0.000 description 3
- 229920001184 polypeptide Polymers 0.000 description 3
- 229920001155 polypropylene Polymers 0.000 description 3
- 239000013641 positive control Substances 0.000 description 3
- 229960003127 rabies vaccine Drugs 0.000 description 3
- 229940124551 recombinant vaccine Drugs 0.000 description 3
- 238000011160 research Methods 0.000 description 3
- 230000004936 stimulating effect Effects 0.000 description 3
- 230000000638 stimulation Effects 0.000 description 3
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 3
- 229940124954 vaccinia virus vaccine Drugs 0.000 description 3
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 2
- LHEJDBBHZGISGW-UHFFFAOYSA-N 5-fluoro-3-(3-oxo-1h-2-benzofuran-1-yl)-1h-pyrimidine-2,4-dione Chemical compound O=C1C(F)=CNC(=O)N1C1C2=CC=CC=C2C(=O)O1 LHEJDBBHZGISGW-UHFFFAOYSA-N 0.000 description 2
- 208000030507 AIDS Diseases 0.000 description 2
- 241000272517 Anseriformes Species 0.000 description 2
- 108010039627 Aprotinin Proteins 0.000 description 2
- 101150068401 BSL1 gene Proteins 0.000 description 2
- 101150011571 BSL2 gene Proteins 0.000 description 2
- 102100037084 C4b-binding protein alpha chain Human genes 0.000 description 2
- 101710159767 C4b-binding protein alpha chain Proteins 0.000 description 2
- BVKZGUZCCUSVTD-UHFFFAOYSA-L Carbonate Chemical compound [O-]C([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-L 0.000 description 2
- 102000004420 Creatine Kinase Human genes 0.000 description 2
- 108010042126 Creatine kinase Proteins 0.000 description 2
- 239000003298 DNA probe Substances 0.000 description 2
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 2
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 2
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 2
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 2
- 201000010374 Down Syndrome Diseases 0.000 description 2
- 102100038132 Endogenous retrovirus group K member 6 Pro protein Human genes 0.000 description 2
- 206010015150 Erythema Diseases 0.000 description 2
- 108700039887 Essential Genes Proteins 0.000 description 2
- 241000711950 Filoviridae Species 0.000 description 2
- 241000713340 Human immunodeficiency virus 2 Species 0.000 description 2
- 206010020751 Hypersensitivity Diseases 0.000 description 2
- 108060003951 Immunoglobulin Proteins 0.000 description 2
- 206010062016 Immunosuppression Diseases 0.000 description 2
- 108010002350 Interleukin-2 Proteins 0.000 description 2
- 108010002586 Interleukin-7 Proteins 0.000 description 2
- 241000282560 Macaca mulatta Species 0.000 description 2
- 101710125418 Major capsid protein Proteins 0.000 description 2
- 241000712079 Measles morbillivirus Species 0.000 description 2
- 102000012750 Membrane Glycoproteins Human genes 0.000 description 2
- 108010090054 Membrane Glycoproteins Proteins 0.000 description 2
- 206010029350 Neurotoxicity Diseases 0.000 description 2
- 241000609499 Palicourea Species 0.000 description 2
- 206010033733 Papule Diseases 0.000 description 2
- 208000004210 Pressure Ulcer Diseases 0.000 description 2
- 241000288906 Primates Species 0.000 description 2
- 239000004365 Protease Substances 0.000 description 2
- 101710192141 Protein Nef Proteins 0.000 description 2
- 241000287219 Serinus canaria Species 0.000 description 2
- 229930006000 Sucrose Natural products 0.000 description 2
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 2
- 241000282887 Suidae Species 0.000 description 2
- 230000024932 T cell mediated immunity Effects 0.000 description 2
- 206010044221 Toxic encephalopathy Diseases 0.000 description 2
- 241000881059 Vaccinia virus Wyeth Species 0.000 description 2
- 208000027418 Wounds and injury Diseases 0.000 description 2
- 230000002411 adverse Effects 0.000 description 2
- 239000011543 agarose gel Substances 0.000 description 2
- 229960004405 aprotinin Drugs 0.000 description 2
- 229940031567 attenuated vaccine Drugs 0.000 description 2
- 230000006399 behavior Effects 0.000 description 2
- RIIWUGSYXOBDMC-UHFFFAOYSA-N benzene-1,2-diamine;hydron;dichloride Chemical compound Cl.Cl.NC1=CC=CC=C1N RIIWUGSYXOBDMC-UHFFFAOYSA-N 0.000 description 2
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 2
- 230000015572 biosynthetic process Effects 0.000 description 2
- 238000009835 boiling Methods 0.000 description 2
- 210000004556 brain Anatomy 0.000 description 2
- 238000004364 calculation method Methods 0.000 description 2
- 239000002775 capsule Substances 0.000 description 2
- 239000006285 cell suspension Substances 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 210000003837 chick embryo Anatomy 0.000 description 2
- 230000000052 comparative effect Effects 0.000 description 2
- 238000012790 confirmation Methods 0.000 description 2
- 201000003740 cowpox Diseases 0.000 description 2
- 210000004748 cultured cell Anatomy 0.000 description 2
- 238000005520 cutting process Methods 0.000 description 2
- 230000006378 damage Effects 0.000 description 2
- 238000010908 decantation Methods 0.000 description 2
- 230000002950 deficient Effects 0.000 description 2
- 210000000852 deltoid muscle Anatomy 0.000 description 2
- 238000000432 density-gradient centrifugation Methods 0.000 description 2
- 230000010460 detection of virus Effects 0.000 description 2
- 238000003745 diagnosis Methods 0.000 description 2
- 238000007865 diluting Methods 0.000 description 2
- 210000002257 embryonic structure Anatomy 0.000 description 2
- 238000009585 enzyme analysis Methods 0.000 description 2
- 239000013613 expression plasmid Substances 0.000 description 2
- 239000013604 expression vector Substances 0.000 description 2
- 230000001605 fetal effect Effects 0.000 description 2
- 125000001153 fluoro group Chemical group F* 0.000 description 2
- 235000013305 food Nutrition 0.000 description 2
- 210000003953 foreskin Anatomy 0.000 description 2
- 238000009472 formulation Methods 0.000 description 2
- 210000004408 hybridoma Anatomy 0.000 description 2
- 230000000521 hyperimmunizing effect Effects 0.000 description 2
- 102000018358 immunoglobulin Human genes 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- ZPNFWUPYTFPOJU-LPYSRVMUSA-N iniprol Chemical compound C([C@H]1C(=O)NCC(=O)NCC(=O)N[C@H]2CSSC[C@H]3C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@H](C(N[C@H](C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC=4C=CC=CC=4)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC=4C=CC=CC=4)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C)NC(=O)[C@H](CCCNC(N)=N)NC2=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC=2C=CC=CC=2)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H]2N(CCC2)C(=O)[C@@H](N)CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N2[C@@H](CCC2)C(=O)N2[C@@H](CCC2)C(=O)N[C@@H](CC=2C=CC(O)=CC=2)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N2[C@@H](CCC2)C(=O)N3)C(=O)NCC(=O)NCC(=O)N[C@@H](C)C(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@H](C(=O)N1)C(C)C)[C@@H](C)O)[C@@H](C)CC)=O)[C@@H](C)CC)C1=CC=C(O)C=C1 ZPNFWUPYTFPOJU-LPYSRVMUSA-N 0.000 description 2
- 238000010255 intramuscular injection Methods 0.000 description 2
- 239000007927 intramuscular injection Substances 0.000 description 2
- 230000003907 kidney function Effects 0.000 description 2
- 239000007788 liquid Substances 0.000 description 2
- 210000004185 liver Anatomy 0.000 description 2
- 230000003908 liver function Effects 0.000 description 2
- 210000002751 lymph Anatomy 0.000 description 2
- 239000011159 matrix material Substances 0.000 description 2
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000010172 mouse model Methods 0.000 description 2
- 239000013642 negative control Substances 0.000 description 2
- 231100000189 neurotoxic Toxicity 0.000 description 2
- 230000002887 neurotoxic effect Effects 0.000 description 2
- 231100000228 neurotoxicity Toxicity 0.000 description 2
- 230000007135 neurotoxicity Effects 0.000 description 2
- 108020004707 nucleic acids Proteins 0.000 description 2
- 102000039446 nucleic acids Human genes 0.000 description 2
- 150000007523 nucleic acids Chemical class 0.000 description 2
- 230000002688 persistence Effects 0.000 description 2
- 244000144977 poultry Species 0.000 description 2
- 235000013594 poultry meat Nutrition 0.000 description 2
- 239000002244 precipitate Substances 0.000 description 2
- 230000035935 pregnancy Effects 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 229940049155 rabies serum Drugs 0.000 description 2
- 238000012770 revaccination Methods 0.000 description 2
- 238000004062 sedimentation Methods 0.000 description 2
- 239000012679 serum free medium Substances 0.000 description 2
- 239000002356 single layer Substances 0.000 description 2
- PUZPDOWCWNUUKD-UHFFFAOYSA-M sodium fluoride Chemical compound [F-].[Na+] PUZPDOWCWNUUKD-UHFFFAOYSA-M 0.000 description 2
- 229960001922 sodium perborate Drugs 0.000 description 2
- YKLJGMBLPUQQOI-UHFFFAOYSA-M sodium;oxidooxy(oxo)borane Chemical compound [Na+].[O-]OB=O YKLJGMBLPUQQOI-UHFFFAOYSA-M 0.000 description 2
- 230000007480 spreading Effects 0.000 description 2
- 238000003892 spreading Methods 0.000 description 2
- 239000003381 stabilizer Substances 0.000 description 2
- 238000005728 strengthening Methods 0.000 description 2
- 239000005720 sucrose Substances 0.000 description 2
- 230000004083 survival effect Effects 0.000 description 2
- 208000024891 symptom Diseases 0.000 description 2
- 230000009885 systemic effect Effects 0.000 description 2
- 201000008827 tuberculosis Diseases 0.000 description 2
- 210000000689 upper leg Anatomy 0.000 description 2
- 230000009385 viral infection Effects 0.000 description 2
- 230000017613 viral reproduction Effects 0.000 description 2
- 238000005406 washing Methods 0.000 description 2
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 2
- OSBLTNPMIGYQGY-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;2-[2-[bis(carboxymethyl)amino]ethyl-(carboxymethyl)amino]acetic acid;boric acid Chemical compound OB(O)O.OCC(N)(CO)CO.OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O OSBLTNPMIGYQGY-UHFFFAOYSA-N 0.000 description 1
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 1
- 238000010600 3H thymidine incorporation assay Methods 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- 108010068327 4-hydroxyphenylpyruvate dioxygenase Proteins 0.000 description 1
- 102100036475 Alanine aminotransferase 1 Human genes 0.000 description 1
- 102100033312 Alpha-2-macroglobulin Human genes 0.000 description 1
- 241001244729 Apalis Species 0.000 description 1
- BTBUEUYNUDRHOZ-UHFFFAOYSA-N Borate Chemical compound [O-]B([O-])[O-] BTBUEUYNUDRHOZ-UHFFFAOYSA-N 0.000 description 1
- 102100021933 C-C motif chemokine 25 Human genes 0.000 description 1
- 208000005623 Carcinogenesis Diseases 0.000 description 1
- 241000867607 Chlorocebus sabaeus Species 0.000 description 1
- 102000006912 Complement C4b-Binding Protein Human genes 0.000 description 1
- 108010047548 Complement C4b-Binding Protein Proteins 0.000 description 1
- 108020004635 Complementary DNA Proteins 0.000 description 1
- 208000034656 Contusions Diseases 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- 102000012410 DNA Ligases Human genes 0.000 description 1
- 108010061982 DNA Ligases Proteins 0.000 description 1
- 108020003215 DNA Probes Proteins 0.000 description 1
- 108010041986 DNA Vaccines Proteins 0.000 description 1
- 230000004544 DNA amplification Effects 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 230000006820 DNA synthesis Effects 0.000 description 1
- 229940122029 DNA synthesis inhibitor Drugs 0.000 description 1
- 229940021995 DNA vaccine Drugs 0.000 description 1
- 206010011906 Death Diseases 0.000 description 1
- 101710170658 Endogenous retrovirus group K member 10 Gag polyprotein Proteins 0.000 description 1
- 101710186314 Endogenous retrovirus group K member 21 Gag polyprotein Proteins 0.000 description 1
- 101710162093 Endogenous retrovirus group K member 24 Gag polyprotein Proteins 0.000 description 1
- 102100039715 Endogenous retrovirus group K member 6 Gag polyprotein Human genes 0.000 description 1
- 101710094596 Endogenous retrovirus group K member 8 Gag polyprotein Proteins 0.000 description 1
- 101710177443 Endogenous retrovirus group K member 9 Gag polyprotein Proteins 0.000 description 1
- 108010067770 Endopeptidase K Proteins 0.000 description 1
- 101000621107 Enterobacteria phage N4 Probable portal protein Proteins 0.000 description 1
- 101710091045 Envelope protein Proteins 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 241000701533 Escherichia virus T4 Species 0.000 description 1
- 101900068883 Feline leukemia virus Envelope glycoprotein Proteins 0.000 description 1
- 108010093031 Galactosidases Proteins 0.000 description 1
- 108700010908 HIV-1 proteins Proteins 0.000 description 1
- 108700004031 HN Proteins 0.000 description 1
- 101710121925 Hemagglutinin glycoprotein Proteins 0.000 description 1
- 208000009889 Herpes Simplex Diseases 0.000 description 1
- 108010027412 Histocompatibility Antigens Class II Proteins 0.000 description 1
- 102000018713 Histocompatibility Antigens Class II Human genes 0.000 description 1
- 101000897486 Homo sapiens C-C motif chemokine 25 Proteins 0.000 description 1
- 241000701074 Human alphaherpesvirus 2 Species 0.000 description 1
- 108010016183 Human immunodeficiency virus 1 p16 protease Proteins 0.000 description 1
- XQFRJNBWHJMXHO-RRKCRQDMSA-N IDUR Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(I)=C1 XQFRJNBWHJMXHO-RRKCRQDMSA-N 0.000 description 1
- 101710203526 Integrase Proteins 0.000 description 1
- 241000710842 Japanese encephalitis virus Species 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 1
- 208000032420 Latent Infection Diseases 0.000 description 1
- 206010025323 Lymphomas Diseases 0.000 description 1
- 241000282341 Mustela putorius furo Species 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 108091092724 Noncoding DNA Proteins 0.000 description 1
- 101710163270 Nuclease Proteins 0.000 description 1
- 206010030113 Oedema Diseases 0.000 description 1
- 102000004316 Oxidoreductases Human genes 0.000 description 1
- 108090000854 Oxidoreductases Proteins 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 241000286209 Phasianidae Species 0.000 description 1
- 208000013544 Platelet disease Diseases 0.000 description 1
- 208000000474 Poliomyelitis Diseases 0.000 description 1
- 241000700625 Poxviridae Species 0.000 description 1
- 208000005585 Poxviridae Infections Diseases 0.000 description 1
- 108010015078 Pregnancy-Associated alpha 2-Macroglobulins Proteins 0.000 description 1
- 101710098398 Probable alanine aminotransferase, mitochondrial Proteins 0.000 description 1
- 108010076504 Protein Sorting Signals Proteins 0.000 description 1
- 101710188315 Protein X Proteins 0.000 description 1
- 101000955367 Pseudomonas putida Transcriptional regulatory protein XylR Proteins 0.000 description 1
- 206010037660 Pyrexia Diseases 0.000 description 1
- 108010092799 RNA-directed DNA polymerase Proteins 0.000 description 1
- 102000006382 Ribonucleases Human genes 0.000 description 1
- 108010083644 Ribonucleases Proteins 0.000 description 1
- 108050008465 Ribonucleoside-diphosphate reductase large subunit Proteins 0.000 description 1
- 102100036320 Ribonucleoside-diphosphate reductase large subunit Human genes 0.000 description 1
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 1
- 101100277251 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) HEM12 gene Proteins 0.000 description 1
- 241000288960 Saguinus oedipus Species 0.000 description 1
- 241000282696 Saimiri sciureus Species 0.000 description 1
- 206010040914 Skin reaction Diseases 0.000 description 1
- 238000002105 Southern blotting Methods 0.000 description 1
- 241000701093 Suid alphaherpesvirus 1 Species 0.000 description 1
- 239000008051 TBE buffer Substances 0.000 description 1
- 101100388071 Thermococcus sp. (strain GE8) pol gene Proteins 0.000 description 1
- 108090000340 Transaminases Proteins 0.000 description 1
- 102000003929 Transaminases Human genes 0.000 description 1
- 206010044688 Trisomy 21 Diseases 0.000 description 1
- 229920004890 Triton X-100 Polymers 0.000 description 1
- 108090000631 Trypsin Proteins 0.000 description 1
- 102000004142 Trypsin Human genes 0.000 description 1
- 101100268528 Vaccinia virus (strain Copenhagen) A26L gene Proteins 0.000 description 1
- 101100268529 Vaccinia virus (strain Western Reserve) VACWR149 gene Proteins 0.000 description 1
- 101900209501 Vaccinia virus Thymidine kinase Proteins 0.000 description 1
- 241000700646 Vaccinia virus WR Species 0.000 description 1
- 108010003533 Viral Envelope Proteins Proteins 0.000 description 1
- 108700005077 Viral Genes Proteins 0.000 description 1
- 238000002679 ablation Methods 0.000 description 1
- 206010000210 abortion Diseases 0.000 description 1
- 231100000176 abortion Toxicity 0.000 description 1
- 230000004913 activation Effects 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 208000002552 acute disseminated encephalomyelitis Diseases 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 230000006978 adaptation Effects 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 208000026935 allergic disease Diseases 0.000 description 1
- 230000007815 allergy Effects 0.000 description 1
- 230000000735 allogeneic effect Effects 0.000 description 1
- 210000001691 amnion Anatomy 0.000 description 1
- 208000007502 anemia Diseases 0.000 description 1
- 230000007416 antiviral immune response Effects 0.000 description 1
- 210000000436 anus Anatomy 0.000 description 1
- 230000001174 ascending effect Effects 0.000 description 1
- 238000003149 assay kit Methods 0.000 description 1
- 108010058966 bacteriophage T7 induced DNA polymerase Proteins 0.000 description 1
- 210000003651 basophil Anatomy 0.000 description 1
- 230000008512 biological response Effects 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 230000023555 blood coagulation Effects 0.000 description 1
- 238000010241 blood sampling Methods 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 230000036952 cancer formation Effects 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 231100000504 carcinogenesis Toxicity 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 108091092356 cellular DNA Proteins 0.000 description 1
- 108091092328 cellular RNA Proteins 0.000 description 1
- 230000007969 cellular immunity Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 210000004720 cerebrum Anatomy 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 210000003711 chorioallantoic membrane Anatomy 0.000 description 1
- 239000003593 chromogenic compound Substances 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 230000024203 complement activation Effects 0.000 description 1
- 239000000470 constituent Substances 0.000 description 1
- 238000011109 contamination Methods 0.000 description 1
- 238000010924 continuous production Methods 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- UQHKFADEQIVWID-UHFFFAOYSA-N cytokinin Natural products C1=NC=2C(NCC=C(CO)C)=NC=NC=2N1C1CC(O)C(CO)O1 UQHKFADEQIVWID-UHFFFAOYSA-N 0.000 description 1
- 239000004062 cytokinin Substances 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 210000000805 cytoplasm Anatomy 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000007812 deficiency Effects 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 238000013461 design Methods 0.000 description 1
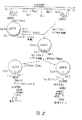
- 238000010586 diagram Methods 0.000 description 1
- 239000005546 dideoxynucleotide Substances 0.000 description 1
- 235000005911 diet Nutrition 0.000 description 1
- 230000037213 diet Effects 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- 210000001840 diploid cell Anatomy 0.000 description 1
- IRXRGVFLQOSHOH-UHFFFAOYSA-L dipotassium;oxalate Chemical compound [K+].[K+].[O-]C(=O)C([O-])=O IRXRGVFLQOSHOH-UHFFFAOYSA-L 0.000 description 1
- 230000008034 disappearance Effects 0.000 description 1
- 208000037771 disease arising from reactivation of latent virus Diseases 0.000 description 1
- 231100000676 disease causative agent Toxicity 0.000 description 1
- 239000012153 distilled water Substances 0.000 description 1
- 231100000673 dose–response relationship Toxicity 0.000 description 1
- 235000020188 drinking water Nutrition 0.000 description 1
- 239000003651 drinking water Substances 0.000 description 1
- 235000013399 edible fruits Nutrition 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 239000000839 emulsion Substances 0.000 description 1
- 230000007515 enzymatic degradation Effects 0.000 description 1
- 238000001976 enzyme digestion Methods 0.000 description 1
- 210000003979 eosinophil Anatomy 0.000 description 1
- 230000008029 eradication Effects 0.000 description 1
- 231100000321 erythema Toxicity 0.000 description 1
- ZMMJGEGLRURXTF-UHFFFAOYSA-N ethidium bromide Chemical compound [Br-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CC)=C1C1=CC=CC=C1 ZMMJGEGLRURXTF-UHFFFAOYSA-N 0.000 description 1
- 229960005542 ethidium bromide Drugs 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 108010052305 exodeoxyribonuclease III Proteins 0.000 description 1
- 210000003191 femoral vein Anatomy 0.000 description 1
- 238000011049 filling Methods 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 108020001507 fusion proteins Proteins 0.000 description 1
- 102000034356 gene-regulatory proteins Human genes 0.000 description 1
- 108091006104 gene-regulatory proteins Proteins 0.000 description 1
- 125000002791 glucosyl group Chemical group C1([C@H](O)[C@@H](O)[C@H](O)[C@H](O1)CO)* 0.000 description 1
- 229930195712 glutamate Natural products 0.000 description 1
- 210000002288 golgi apparatus Anatomy 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 230000009931 harmful effect Effects 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- 230000035876 healing Effects 0.000 description 1
- 230000003862 health status Effects 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 230000009215 host defense mechanism Effects 0.000 description 1
- 230000028996 humoral immune response Effects 0.000 description 1
- 230000004727 humoral immunity Effects 0.000 description 1
- 230000008348 humoral response Effects 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 230000009610 hypersensitivity Effects 0.000 description 1
- 230000001900 immune effect Effects 0.000 description 1
- 230000008105 immune reaction Effects 0.000 description 1
- 229940042743 immune sera Drugs 0.000 description 1
- 210000004201 immune sera Anatomy 0.000 description 1
- 238000011575 immunodeficient mouse model Methods 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 229940072221 immunoglobulins Drugs 0.000 description 1
- 239000002955 immunomodulating agent Substances 0.000 description 1
- 239000012133 immunoprecipitate Substances 0.000 description 1
- 238000009169 immunotherapy Methods 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 230000000415 inactivating effect Effects 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 230000028709 inflammatory response Effects 0.000 description 1
- 206010022000 influenza Diseases 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 230000002452 interceptive effect Effects 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000010253 intravenous injection Methods 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 238000000021 kinase assay Methods 0.000 description 1
- 231100000518 lethal Toxicity 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 229940124590 live attenuated vaccine Drugs 0.000 description 1
- 229940023012 live-attenuated vaccine Drugs 0.000 description 1
- 210000005265 lung cell Anatomy 0.000 description 1
- 210000001165 lymph node Anatomy 0.000 description 1
- 230000001589 lymphoproliferative effect Effects 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 210000004779 membrane envelope Anatomy 0.000 description 1
- 238000012544 monitoring process Methods 0.000 description 1
- 210000001616 monocyte Anatomy 0.000 description 1
- 210000005087 mononuclear cell Anatomy 0.000 description 1
- 210000000214 mouth Anatomy 0.000 description 1
- 210000003205 muscle Anatomy 0.000 description 1
- 230000007524 negative regulation of DNA replication Effects 0.000 description 1
- 210000000440 neutrophil Anatomy 0.000 description 1
- 238000011587 new zealand white rabbit Methods 0.000 description 1
- HYIMSNHJOBLJNT-UHFFFAOYSA-N nifedipine Chemical compound COC(=O)C1=C(C)NC(C)=C(C(=O)OC)C1C1=CC=CC=C1[N+]([O-])=O HYIMSNHJOBLJNT-UHFFFAOYSA-N 0.000 description 1
- 231100000957 no side effect Toxicity 0.000 description 1
- 210000001331 nose Anatomy 0.000 description 1
- 230000037360 nucleotide metabolism Effects 0.000 description 1
- 229940124276 oligodeoxyribonucleotide Drugs 0.000 description 1
- 101150029123 osa gene Proteins 0.000 description 1
- KHPXUQMNIQBQEV-UHFFFAOYSA-N oxaloacetic acid Chemical compound OC(=O)CC(=O)C(O)=O KHPXUQMNIQBQEV-UHFFFAOYSA-N 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 239000007981 phosphate-citrate buffer Substances 0.000 description 1
- 239000002504 physiological saline solution Substances 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- 229920000729 poly(L-lysine) polymer Polymers 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 230000001323 posttranslational effect Effects 0.000 description 1
- 229940024231 poxvirus vaccine Drugs 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 230000009465 prokaryotic expression Effects 0.000 description 1
- 230000035755 proliferation Effects 0.000 description 1
- 235000019419 proteases Nutrition 0.000 description 1
- 239000003223 protective agent Substances 0.000 description 1
- 230000009993 protective function Effects 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 230000007026 protein scission Effects 0.000 description 1
- 238000003908 quality control method Methods 0.000 description 1
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 1
- 231100000916 relative toxicity Toxicity 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 210000003935 rough endoplasmic reticulum Anatomy 0.000 description 1
- 229940016590 sarkosyl Drugs 0.000 description 1
- 108700004121 sarkosyl Proteins 0.000 description 1
- 230000018040 scab formation Effects 0.000 description 1
- 238000013341 scale-up Methods 0.000 description 1
- 231100000241 scar Toxicity 0.000 description 1
- 238000007790 scraping Methods 0.000 description 1
- 238000011896 sensitive detection Methods 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 239000003001 serine protease inhibitor Substances 0.000 description 1
- 230000001568 sexual effect Effects 0.000 description 1
- 238000007390 skin biopsy Methods 0.000 description 1
- 230000035483 skin reaction Effects 0.000 description 1
- 231100000430 skin reaction Toxicity 0.000 description 1
- 238000010583 slow cooling Methods 0.000 description 1
- 229940083538 smallpox vaccine Drugs 0.000 description 1
- 239000001509 sodium citrate Substances 0.000 description 1
- NLJMYIDDQXHKNR-UHFFFAOYSA-K sodium citrate Chemical compound O.O.[Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O NLJMYIDDQXHKNR-UHFFFAOYSA-K 0.000 description 1
- 239000011775 sodium fluoride Substances 0.000 description 1
- 235000013024 sodium fluoride Nutrition 0.000 description 1
- KSAVQLQVUXSOCR-UHFFFAOYSA-M sodium lauroyl sarcosinate Chemical compound [Na+].CCCCCCCCCCCC(=O)N(C)CC([O-])=O KSAVQLQVUXSOCR-UHFFFAOYSA-M 0.000 description 1
- 238000000527 sonication Methods 0.000 description 1
- 238000001228 spectrum Methods 0.000 description 1
- 210000000278 spinal cord Anatomy 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 238000001308 synthesis method Methods 0.000 description 1
- 238000003786 synthesis reaction Methods 0.000 description 1
- 239000006188 syrup Substances 0.000 description 1
- 235000020357 syrup Nutrition 0.000 description 1
- 239000008399 tap water Substances 0.000 description 1
- 235000020679 tap water Nutrition 0.000 description 1
- 238000002723 toxicity assay Methods 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- 230000000472 traumatic effect Effects 0.000 description 1
- 210000000427 trigeminal ganglion Anatomy 0.000 description 1
- 239000012588 trypsin Substances 0.000 description 1
- 238000002604 ultrasonography Methods 0.000 description 1
- 241001529453 unidentified herpesvirus Species 0.000 description 1
- 229940125575 vaccine candidate Drugs 0.000 description 1
- 210000001215 vagina Anatomy 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- 230000007442 viral DNA synthesis Effects 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 230000007923 virulence factor Effects 0.000 description 1
- 239000000304 virulence factor Substances 0.000 description 1
- 239000005723 virus inoculator Substances 0.000 description 1
Images
































































































































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/18—Antivirals for RNA viruses for HIV
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/33—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Clostridium (G)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5254—Virus avirulent or attenuated
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/16011—Herpesviridae
- C12N2710/16211—Lymphocryptovirus, e.g. human herpesvirus 4, Epstein-Barr Virus
- C12N2710/16222—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/16011—Herpesviridae
- C12N2710/16611—Simplexvirus, e.g. human herpesvirus 1, 2
- C12N2710/16622—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/16011—Herpesviridae
- C12N2710/16711—Varicellovirus, e.g. human herpesvirus 3, Varicella Zoster, pseudorabies
- C12N2710/16722—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/24011—Poxviridae
- C12N2710/24022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/24011—Poxviridae
- C12N2710/24041—Use of virus, viral particle or viral elements as a vector
- C12N2710/24043—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/24011—Poxviridae
- C12N2710/24111—Orthopoxvirus, e.g. vaccinia virus, variola
- C12N2710/24122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/24011—Poxviridae
- C12N2710/24111—Orthopoxvirus, e.g. vaccinia virus, variola
- C12N2710/24141—Use of virus, viral particle or viral elements as a vector
- C12N2710/24143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2730/00—Reverse transcribing DNA viruses
- C12N2730/00011—Details
- C12N2730/10011—Hepadnaviridae
- C12N2730/10111—Orthohepadnavirus, e.g. hepatitis B virus
- C12N2730/10122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/13011—Gammaretrovirus, e.g. murine leukeamia virus
- C12N2740/13022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/15011—Lentivirus, not HIV, e.g. FIV, SIV
- C12N2740/15022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/16011—Human Immunodeficiency Virus, HIV
- C12N2740/16111—Human Immunodeficiency Virus, HIV concerning HIV env
- C12N2740/16122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/16011—Human Immunodeficiency Virus, HIV
- C12N2740/16211—Human Immunodeficiency Virus, HIV concerning HIV gagpol
- C12N2740/16222—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/16011—Human Immunodeficiency Virus, HIV
- C12N2740/16311—Human Immunodeficiency Virus, HIV concerning HIV regulatory proteins
- C12N2740/16322—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14311—Parvovirus, e.g. minute virus of mice
- C12N2750/14322—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/12011—Bunyaviridae
- C12N2760/12022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/18011—Paramyxoviridae
- C12N2760/18111—Avulavirus, e.g. Newcastle disease virus
- C12N2760/18122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/18011—Paramyxoviridae
- C12N2760/18711—Rubulavirus, e.g. mumps virus, parainfluenza 2,4
- C12N2760/18722—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/20011—Rhabdoviridae
- C12N2760/20022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/20011—Rhabdoviridae
- C12N2760/20111—Lyssavirus, e.g. rabies virus
- C12N2760/20122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/24011—Flaviviridae
- C12N2770/24022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/24011—Flaviviridae
- C12N2770/24111—Flavivirus, e.g. yellow fever virus, dengue, JEV
- C12N2770/24122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/24011—Flaviviridae
- C12N2770/24111—Flavivirus, e.g. yellow fever virus, dengue, JEV
- C12N2770/24141—Use of virus, viral particle or viral elements as a vector
- C12N2770/24143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Virology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Immunology (AREA)
- General Engineering & Computer Science (AREA)
- Veterinary Medicine (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Biomedical Technology (AREA)
- Gastroenterology & Hepatology (AREA)
- Pharmacology & Pharmacy (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biotechnology (AREA)
- Oncology (AREA)
- Physics & Mathematics (AREA)
- Communicable Diseases (AREA)
- Plant Pathology (AREA)
- Microbiology (AREA)
- Tropical Medicine & Parasitology (AREA)
- AIDS & HIV (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Description
下記には、本発明を、例を挙げて詳しく説明してあるが、本発明はここで挙げた実施例に限定されるものではない。下記に示した図面を参照することで、本発明の理解が深まるであろう。
(2)出血性領域(u;B13R +B14R) vP553
(3)A型細胞封入体(ATI;A26L) vP618
(4)赤血球凝集素遺伝子(HA; A56R) vP723
(5)宿主域遺伝子部位(C7L-K1L) vP804
(6)リボヌクレオチド還元酵素ラージサブユニット(I4L)vP866(NYVAC)
NYVACは、遺伝子操作技術により、18個の毒性遺伝子をコードしているオープンリーディングフレーム(ORF)と宿主域領域を欠失させることで作製したワクシニアウイルスである。このNYVACは多くの基準によって高度に弱毒化されている。弱毒化させる場合の基準としては、(i)新生マウスの大脳内に接種した後の減少された毒性、(ii)遺伝的(NU+/NU+)免疫不全マウスまたは化学的(シクロホスファミド)免疫不全マウスにも接種可能であること、(iii)免疫不全マウスに接種しても播種感染を引き起こさないものであること、(iv)重大なウサギの皮膚の硬化や腫瘍化を起こさないこと、(v)接種部位から速やかに消失すること、(vi)ヒトを含む組織培養細胞における複製能力が減少されていることがある。このように弱毒化されたものであるにもかかわらず、NYVACをベースにしたベクターは、外来性の免疫原に優れた免疫応答を示し防御免疫を提供している。
従って、本発明の組み換えポックスウイルスには、種々の使用法がある。例えば血清陰性者に対して投与する抗原、免疫原、ワクチン組成物として使える。血清陽性者には、治療剤として、免疫不全ウイルスに対する免疫系の強化のために使える。本発明によれば、in vitroで、発明の抗原、免疫原、改変gp120またはgp160を作製可能であり、その場合でも、これらの作製したものは、抗原、免疫原、ワクチン組成物または治療剤として使用可能である。本発明の組み換えポックスウイルスを直接に投与するか、このウイルスの発現物質を投与して得られた抗体は、例えば血清等のサンプル中に抗原があるかないか、例えば、血清中の免疫不全ウイルスまたはCTLの有無、ウイルスや特定の抗原に免疫応答を示したかどうかを探知するための診断キット、試験キットに使用され、さらに、免疫吸着クロマトグラフィに使用される。(本発明にかかる免疫原および改変gp120またはgp160は、利用価値のある抗体を産生させるために使用可能である。)ハイブリダイゼーション・プローブとして使うDNAやPCRプライマーの作製、DNA免疫にも使用(再注入)できる。本発明の組み換えポックスウイルス、それからの発現物質、本発明の免疫原、改変gp120またはgp160は、刺激細胞を作製するのに使用でき、その刺激細胞は免疫応答(抗原反応、免疫反応、能動免疫)を刺激し、あるいは、免疫系を高めて強化(例えば、免疫不全患者あるいは血清反応陽性者の免疫系を強化)するのに使用可能である。本発明の実施例としてまだ他の使用法はある。
プラスミドpSD460の構築
今、図1を参照しているが、プラスミドpSD406は、pUC8中にクローン化されたワクシニアHindIII J(位置83359−88377)を含んでいる。pSD406はHindIIIおよびPvuIIで切断され、HindIII Jの左側からの1.7kb断片が、HindIII/SmaIで消化されたpUC8中にクローン化され、pSD447となった。pSD447はJ2R(位置83855−84385)全体の遺伝子を有している。開始コドンはNlaIII部位内部に含まれ、終止コドンはSspI部位内部に含まれている。転写方向は図1中に矢印で示されている。

プラスミドpSD486の構築
今、図2を参照しているが、プラスミドpSD419は、pUC8中にクローン化されたワクシニアSalI G(位置160744−173351)を有している。pSD422は、右側に近接したワクシニアSalI断片、pUC8中にクローン化されたSalI J(位置173351−182746)を有している。出血性領域、u、B13R−B14R(位置172549−173552)を欠失させたプラスミドを構築するために、pSD419を左側隣接アームの源として、そしてpSD422を右側の隣接アームの源として用いた。領域uの転写方向は図2において矢印で示されている。
プラスミドpMP494Δの構築
今、図3を参照しているが、pSD414はpUC8中にクローン化されたSalI Bを含んでいる。A26L領域の左側の所望でないDNA配列を取り除くため、pSD414を、XbaIでワクシニア配列の内部(位置137079)で、HindIIIでpUC/ワクシニア結合部分で切断し、続いてE.coliポリメラーゼクレノー断片で平滑化しライゲーションし、プラスミドpSD483とした。A26L領域の右側の所望でないワクシニア配列を取り除くため、pSD483をEcoRIで切断し(位置140665およびpUC/ワクシニア結合部分)、ライゲーションし、プラスミドpSD484とした。A26Lコード領域を除去するため、pSD484はNdeIで(部分的に)A26LORFの僅かに上流を(位置139004)、HpaIでA26LORFの僅かに下流を(位置137889)切断した。5.2kbpのベクター断片を単離し、アニーリングさせた人工合成オリゴヌクレオチドATI3/ATI4(配列番号12/配列番号13)
(5’ AAAATGGGCGTGGATTGTTAACTTTATATAACTTATTTTTTGAATATAC 3’)
を用いた突然変異により欠失させた(Mandecki,1986)。生成されたプラスミド、pMP494Δにおいては、A26L ORFの全体を含む位置[137889−138937]を取り巻くワクシニアウイルスDNAが欠失していた。pMP494Δと、ベータガラクトシダーゼを含むワクシニア組換体vP581との間での組換えにより、ワクシニア欠失変異体vP618が、X−gal存在下での透明プラークとして単離され、得られた。
プラスミドpSD467の構築
今、図4を参照しているが、ワクシニアSalI G制限酵素断片(位置160744−173351)はHindIII A/B結合部分(位置162539)で交差している。pSD419は、pUC8中にクローン化されたワクシニアSalI G断片を有している。血球凝集素(HA)遺伝子の転写方向は図13中に矢印で示されている。HindIII Bから派生したワクシニア配列は、pSD419をHindIIIでワクシニア配列内部とpUC/ワクシニア結合部分を消化し続いてライゲーションすることにより取り除いた。生成されたプラスミド、pSD456は、HA遺伝子A56Rを、左に0.4kbpのワクシニア配列、右に0.4kbpのワクシニア配列と隣接した状態で有している。A56Rコード配列を、pSD456をRsaIで(部分的;位置161090)A56Rコード配列の上流を、またEaqIで(位置162054)遺伝子の末端ちかくを切断することにより除去した。3.6kbpRsaI/EaqIベクター断片を単離し、アニーリングさせた合成ヌクレオチドMPSYN59(配列番号15)、MPSYN62(配列番号16)、MPSYN60(配列番号17)およびMPSYN61(配列番号18)
欠失のためのプラスミドpMPCSK1Δの構築
今は、図5を参照しているが、以下のワクシニアクローンがpMCSK1Δの構築のために利用された。pSD420はpUC8中にクローン化されたSalI Hである。pSD435は、pUC18中にクローン化されたKpnI Fである。pSD435はSphIで切断され、再び連結され、pSD451となった。pSD451中では、HindIII M中のSphI部位(位置27416)の左側のDNA配列は取り除かれている(Perkus et al.,1990)。pSD409はpUC8中にクローン化されたHindIII Mである。
5’-TGTCATTTAACACTATACTCATATTAATAAAAATAATATTTATT-3’
を用いた突然変異にかけた。その結果生成したプラスミド、pMCSK1Δは、12のワクシニアオープンリーディングフレーム[C7L−K1L]を含む位置18805−29108のワクシニア配列を欠失していた。pMCSK1Δと、ベータガラクトシダーゼを含むワクシニア組換体vP784との組換えにより、ワクシニア欠失変異体、vP804が生成し、X−gal存在下で透明プラークとして単離された。
欠失のためのプラスミドpSD548の構築
今、図6を参照しているが、プラスミドpSD405は、pUC8中にクローン化されたワクシニアHindIII I(位置63875−70367)を有している。pSD405をEcoRVでワクシニア配列内部(位置67933)を、およびSmaIでpUC/ワクシニア結合部分を消化し、連結させ、プラスミドpSD518とした。pSD518はpSD548の構築において用いられた、全てのワクシニア制限断片の供給源として用いられた。
ワクシニアH6プロモーター(Taylor et al.,1988a,b)の支配下で狂犬病糖タンパク質 Gをコードしている遺伝子を、TK欠失プラスミドpSD513中に挿入した。pSD513は、ポリリンカー領域の存在を除いてプラスミドpSD460(図1)と同一である。
融合および血球凝集素ノイラミニダーゼ糖タンパク質を発現する
TROVAC−NDVの構築
本実施例では、ニワトリポックスウイルスベクターTROVAC、およびTROVAC−NDVと命名されたニワトリポックスニューカッスル病ウイルス組換体の開発、およびその安全性並びに効率を記述する。毒性NDV株テキサスのFおよびHN遺伝子を両方発現するニワトリポックスウイルス(FPV)ベクターを構築した。創出された組換体はTROVAC−NDVと命名された。TROVAC−NDVは実際にプロセシングされたNDV糖タンパク質を、組換えウイルスを感染させた鳥類細胞中で発現し、生後1日のニワトリへの接種により、それに続く毒性NDV投与に対して防御する。
CE75:
CGATATCCGTTAAGTTTGTATCGTAATGGGCTCCAGATCTTCTACCAGGATCCCGGTAC
CE76:
CGGGATCCTGGTAGAAGATCTGGAGCCCATTACGATACAAACTTAACGGATATCG
NDV−Fの3’末端非コード領域を取り除くため、pCE13由来のSmaIからPstI断片をpUC18のSmaIおよびPstI部位に挿入し、pCE23とした。非コード領域をpCE23のSacI、BamHI、エキソヌクレアーゼIII、SIヌクレアーゼおよびEcoRIによる連続的消化により除去した。アニーリングさせキナーゼ処理したオリゴヌクレオチドCE42(配列番号29)およびCE43(配列番号30)を続いて挿入しpCE29とした。
CE43:CCCGGGGAGCTCG
NDV−F配列の3’末端を続いて、既にpCE29からのPstI−SacI断片をpCE20のPstI−SacI部位に挿入することによりNDV−Fの5’末端を有しているプラスミドpCE20に挿入しpCE32とした。pCE20の開発は以前にTaylor et al.,1990中で記述された。
CE166:CTTTTTATAAAAAGTTAACTACGTAG
CE167:GATCCTACGTAGTTAACTTTTTATAAAAAGAGCT
NDV−F完全3’末端を、pCE54をテンプレートとして、オリゴヌクレオチドCE182(配列番号33)およびCE183(配列番号34)をプライマーとしたポリメラーゼ連鎖反応(PCR)により得た。
CE182:CTTAACTCAGCTGACTATCC
CE183:TACGTAGTTAACTTTTTATAAAAATCATATTTTTGTAGTGGTC
PCR断片はPvuIIおよびHpaIにより消化させ、HpaIおよび部分的にPvuIIにより消化したpCE58中へクローン化した。その結果生成されたプラスミドはpCE64と命名された。pCE64由来の完全H6プロモーターおよびFコード配列を含むHindIII−HpaI断片を、pRW846のHindIII−HpaI部位にクローン化することにより、翻訳終結シグナルを挿入し、最終NDV−FカセットであるpCE71を生成した。プラスミドpRW846は実質的にpJCA002(以下に記述)と同一ではあるが、H6プロモーターおよび転写並びに翻訳終結シグナルを有している。pRW846をHindIIIおよびHpaIで消化することによって、H6プロモーターは除かれるが停止シグナルは手つかずで残る。
CE162:
AATTCAGGATCGTTCCTTTACTAGTTGAGATTCTCAAGGATGATGGGATTTAATTTTTATAAGCTTG
CE163:
AATTCAAGCTTATAAAAATTAAATCCCATCATCCTTGAGAATCTCAACTAGTAAAGGAACGATCCTG
FPV挿入ベクターの構築 プラスミドpRW731−15は、ゲノムDNAからクローン化された10kbpのPvuII−PvuII断片をコードしている。3660bpのPvuII−EcoRV断片の両方の鎖に関してヌクレオチド配列が決定され図11に示されている(配列番号67)。ここでF8と名付けられたオープンリーディングフレームの限定が決定された。プラスミドpRW761は2430bpのEcoRV−EcoRV断片を含むpRW731−15のサブクローンである。F8オープンリーディングフレームの全体は、pRW761中のXbaI部位およびSspI部位の間に含まれている。TROVACゲノムDNAとの組換えにおいてF8ORFを取り除く挿入プラスミドを創出するために、以下の工程が行われた。プラスミドpRW761を完全にXbaIで、部分的にSspIで消化した。ゲルから単離された3700bpのXbaI−SspIバンドをアニーリングさせた2本鎖オリゴヌクレオチドJCA017(配列番号37)およびJCA018(配列番号38)と連結した。
JCA017:5’
CTAGACACTTTATGTTTTTTAATATCCGGTCTTAAAAGCTTCCCGGGGATCCTTATACGGGGAATAAT
JCA018:5’
ATTATTCCCCGTATAAGGATCCCCCGGGAAGCTTTTAAGACCGGATATTAAAAAACATAAAGTGT
このライゲーションの結果生成されたプラスミドはpJCA002と命名された。
ALVAC組換体の構築
本実施例では、カナリアポックスウイルスベクター、ALVACおよびALVAC−RG(vCP65)と命名されたカナリアポックス−狂犬病組換体の開発およびその安全性と効率を記述する。
GCTTCCCGGGAATTCTAGCTAGCTAGTTT
により置換された。この置換用配列はHindIII、SmaI、およびEcoRI挿入部位および、それに続くワクシニアウイルスRNAポリメラーゼによって認識される翻訳停止および転写終結シグナル(Yuen et al.,1987)を有している。C5 ORF欠失は以下に記述されるように行った。プラスミドpRW764.5を部分的にRsaIで切断し、線状産物を単離した。RsaI線状断片を再びBglIIで切断し、今、位置156から位置462へのRsaIからBglIIへの欠失のあるpRW764.5断片を単離し、以下の合成ヌクレオチドのためのベクターとして用いた:
(配列番号40および41)
(1)11日経過白レグホン胚から調製された初代ニワトリ胚繊維芽(CEF)細胞;
(2)Vero細胞−アフリカミドリザル腎臓細胞の連続系統(ATCC#CCL81);
(3)MRC−5細胞−ヒト胎児肺組織から派生した二倍体細胞系統(ATCC#CCL171)。
(a)Vero、アフリカミドリザル腎臓細胞、ATCC#CCL81;
(b)MRC−5、ヒト胚肺、ATCC#CCL171;
(c)WISH、ヒト羊膜、ATCC#CCL25;
(d)デトロイト−532、ヒト包皮、ダウン症、ATCC#CCL54;および
(e)初代CEF細胞。
3枚のそれぞれの細胞系統のディッシュを5pfu/細胞のウイルスで試験のもとで接種し、別の1つのそれぞれの細胞系統のディッシュを接種しないでおいた。1つのディッシュは40μg/mlのシトシンアラビノシド(Ara C)の存在下で保温した。37℃、60分間の吸収期間の後、接種物を取り除き、単層を吸収されなかったウイルスを除去するために2回洗浄した。培地(Ara Cが存在もしくは不在)は続いて置換された。1つのディッシュ(Ara C無し)からの細胞を、時間0の試料として回収した。残りのディッシュは、37℃で72時間保温し、そのとき細胞を回収しDNA蓄積分析に用いた。2×106 の細胞それぞれは、0.5mlの40mMEDTAを含むリン酸緩衝食塩水(PBS)に再懸濁し、37℃で5分間保温した。等量の、42℃で予め保温された120mMのEDTAを含む1.5%アガロースを細胞懸濁液に加え、穏やかに混合した。懸濁液はアガロースプラグ型へ移され少なくとも15分固定化された。アガロースプラグを続いて取り除き、プラグを完全にカバーする量の溶解バッファー(1%サルコシル(sarkosyl)、100μg/mlプロテイナーゼK、10mMトリス塩酸pH7.5、200mMEDTA)中で12−16分間50℃で保温した。溶解バッファーを、続いて5.0mlの滅菌0.5xTBE(44.5mMトリス−ホウ酸、44.5mMホウ酸、0.5mMEDTA)で置換し、4℃で6時間にわたり3回TBEバッファーを交換して平衡化した。プラグ内のウイルスDNAをパルスフィールド電気泳動により細胞RNAおよびDNAと分画した。電気泳動は20時間にわたり180Vで50−90秒のランプで、15℃、0.5xTBE中で行った。DNAはラムダDNA分子量スタンダードとともに泳動した。電気泳動後、ウイルスDNAバンドはエチジウムブロミド染色により可視化した。DNAは続いてニトロセルロース膜に移され、精製ALVACゲノムDNAから調製された放射性標識プローブで探索した。
投入多重度を0.1pfu/細胞にしたこと以外は、正確に上記のようにディッシュに接種した。感染後72時間で、3回連続の凍結融解サイクルにより破砕した。ウイルス収量はCEF単層上のプラーク滴定により評価した。
多重度10pfu/細胞で、組換体もしくは親ウイルスをディッシュに接種し、さらに付加的は1つのディッシュをウイルス感染させていない対照とした。1時間の吸収期間の後、培地を取り除きメチオニンフリーの培地で置換した。30分の期間の後に、この培地を25μCi/mlの35S−メチオニンを含むメチオニンフリー培地で置換した。感染させた細胞を一晩標識し、続いてバッファーA溶解バッファーを添加して溶解させた。免疫沈降を、狂犬病G特異的モノクローナル抗体を用いて、以前に記述されたように(Taylor et al.,1990)行った。
カナリアポックスウイルス(ALVAC−RG;vCP65)
を用いたヒトの免疫化
ALVAC−RG(vCP65)は実施例9および図9Aおよび9Bで記述されたように開発された。ワクチン製造をスケールアップするために、ALVAC−RG(vCP65)を特定の病原体のない卵から派生した初代CEF細胞中で増殖させた。細胞を多重度0.01で感染させ、37℃で3日間保温した。
様々なワクシニアウイルス株との比較
マウス 雄の異系交配されたスイスウェブスターマウス(Swiss Webster mice)を、タコニック ファーム(Taconic Farm,Germantown,NY)より購入し、マウス用食事および水 ad libitumで生後3週間で使用するまで育てた(「通常」マウス)。両方の性の新生異系交配スイスウェブスターマウスを、タコニックファームで行われた続いての定期の妊娠により得た。用いられた全ての新生マウスは2日間の期間内に配達された。
NYVAC−RG(vP879)の評価
免疫沈降 事前に形成された鳥類もしくは非鳥類細胞の単層に10pfu/細胞の親NYVAC(vP866)もしくはNYVAC−RG(vP879)ウイルスを接種した。接種をメチオニンフリーで2%の透析牛胎児血清を添加したEMEM中で接種を行った。1時間の保温の後で、接種物を除去し、培地を20μCi/mlの35S−メチオニンを含むEMEM(メチオニンフリー)で置換した。一晩、約16時間の保温の後、細胞をバッファーA(1%Nonidet P−40、10mMトリスpH7.4、150mM NaCl、1mM EDTA、0.01%アジ化ナトリウム、500単位/mlのアプロチニンおよび0.02%フェニルメチルスルフォニルフロリド)の添加により溶解させた。免疫沈降を、C.Trimarchi博士、Griffith研究所、ニューヨーク州保健部、Albany、ニューヨークによって供給された24−3F10と命名された、狂犬病糖タンパク質特異的モノクローナル抗体、およびベーリンガーマンハイム社より得られたラット抗マウス接合体(型番#605−500)を用いて行った。ファルマシアLKBバイオテクノロジー社、Piscataway、ニュージャージーより得られたプロテインAセファロースCL−48を支持マトリクスとして用いた。免疫沈降沈殿物は、Dreyfuss et al.(1984)の方法に従って10%ポリアクリルアミドゲルにより分画した。ゲルを固定化し、間接撮影用に1Mサリチル酸ナトリウムで1時間処理し、免疫沈降されたタンパク質種を視覚化するためにコダックXAR−2フイルムに曝露した。
血球凝集素糖タンパク質を発現する
TROVAC組換体の構築
この実施例はトリインフルエンザウイルスの3つの血清型の血球凝集素遺伝子を発現するニワトリポックスウイルス組換体の開発を記述する。
(+トランスメンブレン)エピトープを発現するALVAC組
換え体の構築
HIV1(IIIB)cDNA配列(Ratner他、1985)を含有するプラスミドpHXB2DをRobert Gallo氏(米国NIH、NCI)から入手した。gag遺伝子の5′末端をコードする配列をワクシニアウイルスtk隣接アームの間にクローニングした。これを行うために、pHXB2Dの1,625bpのBglII断片(gag遺伝子の5′末端を含有する)をpSD542VCVQの4,075bpのBglII断片内にクローニングした。この操作によって得られたプラスミドをpHIVG2と呼ぶ。
特徴 開始点 全長 内容
frag 1 56 C3隣接アーム
frag 162 76 (c) HIV1 (IIIB) env トランスメンフ゛レン 領域
frag 1728 163 (c) HIV1 (MN) gp120遺伝子
frag 1853 1729 (c) ワクシニアH6フ゜ロモーター
frag 1925 1983 ワクシニアI3Lフ゜ロモーター
frag 1984 3746 HIV1 (IIB) gag/フ゜ロ遺伝子
frag 1753 3808 C3 隣接アーム
ALVAC C3隣接アームのDNA配列(配列番号79)は、図15A〜Fに示されている。この図15A〜15Fは、pVQH6CP3LにおけるC3遺伝子座のヌクレオチド配列を示している。
特徴 開始点 全長 内容
frag 1 1460 C3 隣接アーム
frag 1461 1501 クローニンク゛ 部位
frag 1630 1502 H6フ゜ロモーター
frag 1717 4291 C3 隣接アーム
pHIV32を用いてALVACをレスキューウイルスとするインビトロ組換えを行いvCP205を得た。
)(+トランスメンブレン)および2つのnef(BRU)エ
ピトープを発現するALVAC組換体の構築
次に、2つのnef CTLエピトープ、CTL1(アミノ酸66〜147)およびCTL2(アミノ酸182〜206)(Wain-Hobson他、1985;NixonおよびMcMichael、1991)をコードする発現カセットをvCP205に挿入した。nef CTLエピトープを含有する挿入プラスミドp2−60−HIV.3の構築は次のように行った。I13LをプロモーターとするCTLエピトープをpBSH6にクローニングした。これを実施するため、255bpのHindIII−XhoI PCR断片(I3LをプロモーターとするCTL2エピトープを含有する)を、pBSH6の3,100bpのHindIII−XhoI断片にクローニングした。(この255bpのPCRは次にように調製した。すなわち、オリゴヌクレオチドフライマーVQPCRI3(配列番号80)
:5′−ATCATCAAGCTTAATTAATTAGTTATTAGACAAGGTGAAAACGAAACTATTTGTAGCTTAATTATTAGACATCATGCAGTGGTTAAAC−3′)
およびI3PCRCTL(配列番号81)
:5′−CTAGCTACGTGATGAAATGCTAATCTAGAATCAAATCTCCACTCCATGATTAAACCTAAATAATTGTAC −3′)
を用いてプラスミドpMPI3Hから、I3LプロモーターおよびCTL2エピトープの5′末端を有する216bpのPCR断片を調製した。次に、この216bpのPCR断片を鋳型として、オリゴヌクレオチドプライマーVQPCRI3およびCTLPCR(配列番号82)
:5′−GAATTCCTCGAGGATCCTCTAGATTAACAATTTTTAAAATATTCAGGATGTAATTCTCTAGCTACGTGATGAAATGC−3′)
を用いて第2のPCR反応を行い、I3LをプロモーターとするCTL2エピトープを含有するPCR断片を調製し、これをHindIIIXhoIで酵素分解して、pBSH6にクローニングした。)この操作により得られたプラスミドをp2−60−HIV.1と呼ぶ。
:5′−ACTACTAAGCTTCTTTATTCTATACTTAAAAAGTG−3′)
およびNCCPCR1(配列番号84)
:5′−CAGCTGCTTTGTAAGTCATTGGTCTTAAAGGTACTTGAGGTGTTACTGGAAAACCTACCATTACGATACAAACTTAACGGATATCGCG−3′)
を用いて、プラスミドpH6T2から、H6プロモーターおよびCTL1エピトープの5′末端を含有する195bpのPCR断片を得た。次に、この195bpのPCR断片およびオリゴヌクレオチドNCC174A(配列番号85)
:5′−ACTTACAAAGCAGCTGTAGATCTTTCTCACTTTTTAAAAGAAAAAGGAGGTTTAGAAGGGCTAATTCATTCTCAACGAAGACAAGATATTCTTGATTTGTGG−3′)
およびNCC17B(配列番号86)
:5′−CCACAAATCAAGAATATCTTGTCTTCGTTGAGAATGAATTAGCCCTTCTAAACCTCCTTTTTCTTTTAAAAAGTGAGAAAGATCTACAGCTGCTTTGTAAGT−3′)
を鋳型とし、オリゴヌクレオチドプライマーH6PCRおよびNCCPCR2(配列番号87)
:5′−CTGCCAATCAGGAAAATATCCTTGTGTATGATAAATCCACAAATCAAGAATATC−3′
を用いて第2のPCR反応を行った。得られた317bpのPCR断片(H6プロモーターおよび大部分のCTL1エピトープを含有する)ならびにオリゴヌクレオチドNCC291A(配列番号88)
:5′−GGATATTTTCCTGATTGGCAGAATTACACACCAGGACCAGGAGTCAGATACCCATTAACCTTTGGTTGGTGCTACAAGC−3′)
およびNCC291B(配列番号89)
:5′−GCTTGTAGCACCAACCAAAGGTTAATGGGTATCTGACTCCTGGTCCTGGTGTGTAATTCTGCCAATCAGGAAAATATCC−3′)
を鋳型とし、オリゴヌクレオチドプライマーH6PCR1およびNCCPCR3(配列番号90)
:5′−ACTACTGAATTCTCGAGAAAAATTATGGTACTAGCTTGTAGCACCAACC−3′)
を用いて第3のPCR反応を行い、H6をプロモーターとするCTL1エピトープを含有するPCR断片を得て、これをNruIおよびEcoRIで酵素分解し、p2−6−HIV.1にクローニングした。)この操作により得られたプラスミドをp2−60−HIV.2と呼ぶ。
特徴 開始点 全長 内容
frag 1 51 C6左側アーム
pept 175 98 (C)nef CTL2
frag 275 176 (C)I3Lフ゜ロモーター
frag 337 460 H6フ゜ロモーター
pept 461 709 1 nef CTL1
frag 751 801 C6右側アーム
ALVAC C6隣接アームのDNA配列(配列番号92)は、図17A〜Cに示されている。この図17A〜17Cは、pC6LにおけるC6遺伝子座のヌクレオチド配列を示す。
特徴 開始点 全長 内容
frag 1 381 C6隣接アーム
frag 382 467 クローニンク゛部位
frag 448 1615 C6隣接アーム
p2−60HIV.3を用いてvCP205をレスキューウイルスとするインビトロ組換えを行いvCP264を得た。
)(+トランスメンブレン)および2つのnef(BRU)お
よび3つのpol(IIIB)CTLエピトープを含有する領
域を発現するALVAC組換体の構築
次に、このvCP264に、3つのpolCTLエピトープ、pol1(アミノ酸172〜219)、pol2(アミノ酸325〜383)およびpol3(アミノ酸461〜519)(Ratner他、1985;NixonおよびMcMichael、1991)をコードする発現カセットを挿入した。3つのpolCTLエピトープを含有する挿入プラスミドpC5POLT5Aの調製は、948bpのXhoI−BamHI断片(H6をプロモーターとするpol1エピトープ、I3Lをプロモーターとするpol2,42Kをプロモーターとするpol3を含有)を、pBSK+の2,940bpのXhoI−BamHI断片にクローニングすることにより行った。(なお、948bpのXhoI−BamHI断片は次のように調製した。プラスミドpHXB2Dから、オリゴヌクレオチドプライマーPlA(配列番号93)
;5′−TTTGTATCGTAATGATTGAGACTGTACCAGTAAAATTAAAGCC−3′)
およびP1B(配列番号94)
;5′−GGGCTGCAGGAATTCTAATCAATTAAGGCCCAATTTTTGAAATTTTCCCTTCCTTTTCCATCTCTG−3′)
を用いて、183bpのPCR断片(pol1エピトープを含有)を調製した。プラスミドpHXB2Dから、オリゴヌクレオチドプライマーP2A(配列番号95)
;5′−ACAAAGTACAATTATTTAGGTTTAATCATGGCAATATTCCAAAGTAGCATGAC−3′)
およびP2B(配列番号96)
;5′−ATCATCCTCGAGAAAAATTAGGTAAGTCCCCACCTCAACAGATG−3′)
を用いて、224bpのPCR断片(pol2エピトープを含有)を調製した。プラスミドpHXB2Dから、オリゴヌクレオチドプライマーP3A(配列番号97)
;5′−AAAATATATAATTACAATATAAAATGCCACTAACAGAAGAAGCAGAGCTAGAACTGGC−3′)
およびP3B(配列番号98)
;5′−ATCATCTCTAGACTCGAGGATCCATAAAAATTATCCTGTTTTCAGATTTTTAAATGGCTC−3′)
を用いて、236bpのPCR断片(pol3エピトープを含有)を調製した。プラスミドp2−60−HIV.2からオリゴヌクレオチドプライマーP2IVH(配列番号99)
;5′−GTCATGCTACTTTTGAATATTGCCATGATTAAACCTAAATAATTGTACTTTG−3′)
およびIVHP1(配列番号100)
;5′−TTTAATTTTACTGGTACAGTCTCAATCATTACGATACAAACTTAACGGATATCGCG−3′)
を用いて、340bpのPCR断片(I13LプロモーターとH6プロモーターを頭部対頭部の関係で含有)を調製した。プラスミドpVQ42KTh4.1から、オリゴヌクレオチドプライマーEPS42K(配列番号101)
;5′−AATTGATTAGAATTCCTGCAGCCCGGGTCAAAAAAATATAAATG−3′)
および42KP3B(配列番号102)
5′−CCAGTTCTAGCTCTGCTTCTTCTGTTAGTGGCATTTTATATTGTAATTATATATTTTC−3′)
を用いて、168bpのPCR断片(42Kプロモーターを含有)を調製した。224bpのPCR断片(pol2エピトープを含有)、および340bpのPCR断片(I3LプロモーターおよびH6プロモーターを含有)を鋳型とし、オリゴヌクレオチドプライマーP2BおよびIVHP1を用いてPCR反応を行い、H6プロモーターおよびI3Lをプロモーターとするpol2エピトープを含有する511bpのPCR断片を調製した。168bpのPCR断片(42Kプロモーターを含有)および236bpのPCR断片(pol3エピトープを含有)を鋳型とし、オリゴヌクレオチドプライマーIPS42KおよびP3Bを用いて、PCR反応を行い、42Kをプロモーターとするpol3エピトープを含有する347bpのPCR断片を調製した。183bpのPCR断片(pol1エピトープを含有)および347bpのPCR断片(42Kをプロモーターとするpol3エピトープを含有)を鋳型とし、オリゴヌクレオチドプライマーPIAおよびP3Bを用いてPCR反応を行い、pol1エピトープおよび42Kをプロモーターとするpol3エピトープを含有する506bpのPCR断片を調製した。511bpのPCR断片(H6プロモーターおよびI3Lをプロモーターとするpol2エピトープを含有)および506bpのPCR断片(pol1エピトープおよび42Kをプロモーターとするpol3を含有)を鋳型とし、オリゴヌクレオチドプライマーP2BおよびP3Bを用いてPCR反応を行い、H6をプロモーターとするpol1エピトープ、I3Lをプロモーターとするpol2および42Kをプロモーターとするpol3エピトープを含有する977bpのPCR断片を調製した。次いで、この977bpのPCR断片をXhoIとBamHIで酵素分解し、pBSK+の2,940bpのXhoI−BamHI断片にクローニングした。)この操作により得られたプラスミドをpBSPOLT5と称する。
5′−ATCATCGGATCCAAGCTTACATCATGCAGTGG−3′)
およびFIXPOL2(配列番号104)
;5′−ATCATCCTCGAGCTATTCAATTAGGTTGTAAGTCCCCACCTCAAC−3′)
を用いてPCR反応を行った。得られたPCR断片(I3Lをプロモーターとする訂正pol2エピトープを含有)をHindIIIとXhoIで酵素分解し、pBSPOLT5の3,650bpのHindIII−XhoI断片にクローニングした。この操作により得られたプラスミドをpBSPOLT5Aと呼ぶ。
特徴 開始点 全長 内容
frag 1 50 C5左側アーム
pept 272 92 (C)POL 2
frag 372 273 (C)I3Lフ゜ロモーター
frag 377 500 H6フ゜ロモーター
pept 501 647 POL 1
frag 676 782 42kフ゜ロモーター
pept 783 962 POL 3
frag 986 1035 C5右側アーム
ALVAC C5隣接アームのDNA配列(配列番号106)は図19A〜Cに示されている。この図19A〜19CはpNC5L−SP5におけるC5遺伝子座のヌクレオチド配列を示すものである。
特徴 開始点 全長 内容
frag 1 1849 C5隣接アーム
frag 1550 1637 クローニンク゛部位
frag 1638 2049 C5隣接アーム
pC5POLT5Aを用いてvCP264をレスキューウイルスとするインビトロ組換えを行いvCP300を得た。
vCP300内でHIV1配列が正しい遺伝子座に存在していることを確認するために、制限酵素分析を行った。ALVAC、vCP205、vCP264およびvCP300のDNAをHindIII、PstIまたはXhoIで分解し、得られた断片をアガロースゲル上で分画化した。得られた断片の大きさを比較したところ、予期されたように、gag(+プロ)遺伝子およびgp120(+トランスメンブレン)遺伝子はC3遺伝子座に挿入され、nefエピトープはC6遺伝子座に挿入され、また、polエピトープはC5遺伝子座に挿入されていたことが確認された。
実施例18−免疫原性の検討
2匹のウサギまたはモルモットから成る各グループに、108pfuのALVAC、VCP205、または0.1mgのペプチドCLTB−36(GPKEPFRDYVDRFYKNKRKRIHIGPGRAFYTTKN)(配列番号107)(0.05mgのQS−21をアジュバントとする)を次の表(表21)に示すスケジュールに従って筋肉内接種した。
ELDKWAエピトープを含むHIV1 gp120+TMを
発現するALVAC組換体)の構築
gp120 V3ループ領域に挿入されたHIV1のgp41由来の2つのELDKWAエピトープとともにHIV1 gp120+TMを発現するALVAC組換体であるvCP1307を以下のように作成した。ELDKWA要素およびV3ループの一部をコードする配列をpBSK+(米国カリフォルニア州La JollaのStratagene社製)にクローニングした。これを行うために、EcoRI−SacIで酵素分解した225bpのPCR断片(ELDKWA−V3ループ配列の一部を含有)を、pBSK+の2,900bpのEcoRI−SacI断片にクローニングした。(なお、該PCR断片は、プラスミドpBSHIVMN120Tから、プライマーHIVP72(配列番号110)
;5′−TTATTACCATTCCAAGTACTATT−3´
およびHIVP74(配列番号111)
;5′−TCTGTACAAATTAATTGTACAAGACCCAACTACGAGCTCGACAAATGGGCCCATATAGGACCAGGGAGAGAATTGGATAAGTGGGCGAATATAATAGGAACTATAAGAC−3′)
から調製した。)この操作により得られたプラスミドをpHIV55と呼ぶ。
;5′−GGGTTATTAATGATCTGTAG−3′)
およびHIV3B2(配列番号113)
;5′−GAATTACAGTAGAAGAATTCCCCTCCACAATTAAAAC−3′)
を用いて、プラスミドpMN1.8−9から調製した。この操作により得られたプラスミドpBSMIDMNと呼ぶ。
;5′−GTTTTAATTGTGGAGGGGAATTCTTCTACTGTAATTC−3′)
およびHIVMN4(配列番号115)
;5′−ATCATCGAGCTCCTATCGCTGCTC−3′)
を用いて調製した。また、1,050bpのPCR断片は、プラスミドpMN1.8−9から、オリゴヌクレオチドHIVMN5(配列番号116)
;5′−ATCATCGAGCTCTGTTCCTTGGGTTCTTAG−3′)
およびHIVMN3P(配列番号117)
;5′−ATCATCTCTAGAATAAAAATTATAGCAAAGCCCTTTCCAAGCC−3´)
を用いて調製した。このような操作により得られたプラスミドをpBSMID3MNと呼ぶ。
;5′−TTATTCATAATGATAGTAGGAGGCTTGGTAGGTTTAAGAATAGTTTTTGCTGTACTCTCTGTAGTGAATAGAGTTAGGCAGGGATAA−3′)
およびHIVTM2(配列番号119)
;5′−TTATCCCTGCCTAACTCTATTCACTACAGAGAGTACAGCAAAAACTATTCTTAAACCTACCAAGCCTCCTACTATCATTATGAATAA−3′
から、オリゴヌクレオチドHIV3B1(配列番号114)およびHIVTM3(配列番号120)
;5′−ATCATCTCTAGAATAAAAATTATCCCTGCCTAACTCTATTCAC−3′)
を用いて調製した。PCR−MN11は、プラスミドpH6HMNEから、オリゴヌクレオチドHIV3B1(配列番号114)およびHIVMN18(配列番号121)
;5′−GCCTCCTACTATCATTATGAATAATCTTTTTTCTCTCTG−3′)
から調製した。)この操作により得られたプラスミドをpBSHIVMN120Tと呼ぶ。
LDKWAエピトープを含むgp120+TMを発現させるN
YVAC組換体)の構築
gp120 V3ループに挿入したHIV1 gp41からの2つのELDKWAエピトープを含むHIV1 gp120+TMを発現させるNYVAC組換体であるvP1313を次の方法により作成した。ELDKWAエピトープおよびV3ループの一部をコードする配列をpBSK+ にクローニングした。これを行うため、EcoRI−SacIで酵素分解した225bpのPCR断片(ELDKWA−V3ループ配列の一部を含有する)を、pBSK+ の2,900bpのEcoRI−SacI断片にクローニングした。(なお、このPCR断片は、プラスミドpBSHIVMN120Tから、プライマーHIVP72(配列番号130)
;5′−TTATTACCATTCCAAGTACTATT−3′)
およびHIVP74(配列番号131)
;5′−TCTGTACAAATTAATTGTACAAGACCCAACTACGAGCTCGACAAATGGGCCCATATAGGACCAGGGAGAGAATTGGATAAGTGGGCGAATATAATAGGAACTATAAGAC−3′)
を用いて調製した。)この操作により得られたプラスミドをpHIV55と呼ぶ。
;5′−GGGTTATTAATGATCTGTAG−3′)
およびHIV3B2(配列番号113)
;5′−GAATTACAGTAGAAGAATTCCCCTCCACAATTAAAAC−3′)
を用いて調製した。)この操作により得られたプラスミドをpBSMIDMNと呼ぶ。
;5′−GTTTTAATTGTGGAGGGGAATTCTTCTACTGTAATTC−3′)
およびHIVMN4(配列番号115)
;5′−ATCATCGAGCTCCTATCGCTGCTC−3′)
を用いて調製した。また、1,050bpのPCR断片は、プラスミドpMN1.8−9からオリゴヌクレオチドHIVMN5(配列番号116)
;5′−ATCATCGAGCTCTGTTCCTTGGGTTCTTAG−3′)
およびHIVMN3P(配列番号117)
;5′−ATCATCTCTAGAATAAAAATTATAGCAAAGCCCTTTCCAAGCC−3´)
を用いて調製した。)この操作により得られたプラスミドをpBSMID3MNと呼ぶ。
;5′−TTATTCATAATGATAGTAGGAGGCTTGGTAGGTTTAAGAATAGTTTTTGCTGTACTCTCTGTAGTGAATAGAGTTAGGCAGGGATAA−3′)
およびHIVTM2(配列番号114)
;5′−TTATCCCTGCCTAACTCTATTCACTACAGAGAGTACAGCAAAAACTATTCTTAAACCTACCAAGCCTCCTACTATCATTATGAATAA−3′)
から、オリゴヌクレオチドHIV3B1(配列番号120)およびHIVTM3(配列番号114)
;5′−ATCATCTCTAGAATAAAAATTATCCCTGCCTAACTCTATTCAC−3′)
を用いて調製した。PCRMN−11は、プラスミドpH6HMNEから、オリゴヌクレオチドHIV3B1(配列番号114)およびHIVMN18(配列番号121)
;5′−GCCTCCTACTATCATTATGAATAATCTTTTTTCTCTCTG−3′)
を用いて調製した。)このようにして得られたプラスミドをpBSHIVMN120Tと呼ぶ。
;5′−TGATAGTACCAGCTATAGGTTGAT−3′)
およびHIVP75(配列番号123)
;5′−TTTGTCGAGCTCGTAGTTGGGTCTTGTACAATT−3′)
を用いて調製した。)この操作によって得られたプラスミドをpHIV56と呼ぶ。
KWAエピトープを含むHIV1 gp120+TMを発現す
るCOPAK組換体)の構築
gp120 V3ループに挿入されたHIV1のgP41由来の2つのELDKWAを含むHIV1 gp120+TMを発現するCOPAK組換体であるvP1319を次のように作成した。ELDKWAエピトープとV3ループの一部をコードする配列をpBSK+にクローニングした。これは、EcoRI−SacI酵素分解した225bpのPCR断片(ELDKWA−V3ループ配列の一部を含有する)を、pBSK+ の2,900bpのEcoRI−SacI断片にクローニングすることにより行った。(なお、該PCR断片は、プラスミドpBSHIVMN120Tから、プライマーHIVP72(配列番号110)
;5′−TTATTACCATTCCAAGTACTATT−3′)
およびHIVP74(配列番号111)
;5′−TCTGTACAAATTAATTGTACAAGACCCAACTACGAGCTCGACAAATGGGCCCATATAGGACCAGGGAGAGAATTGGATAAGTGGGCGAATATAATAGGAACTATAAGAC −3′)
を用いて調製した。)このような操作により得られたプラスミドをpHIV55と呼ぶ。
;5′−GGGTTATTAATGATCTGTAG−3′)
およびHIV3B2(配列番号113)
;5′−GAATTACAGTAGAAGAATTCCCCTCCACAATTAAAAC −3′)
を用いて調製した。)この操作により得られたプラスミドをpBSMIDMNと呼ぶ。
;5′−GTTTTAATTGTGGAGGGGAATTCTTCTACTGTAATTC −3′)
およびHIVMN4(配列番号115)
;5′−ATCATCGAGCTCCTATCGCTGCTC−3′
を用いて調製した。また、1,050bpのPCR断片は、プラスミドpMN1.8−9から、オリゴヌクレオチドHIVMN5(配列番号116)
;5′−ATCATCGAGCTCTGTTCCTTGGGTTCTTAG−3′)
およびHIVMN3P(配列番号117)
;5′−ATCATCTCTAGAATAAAAATTATAGCAAAGCCCTTTCCAAGCC −3′)
を用いて調製した。)このような操作により得られたプラスミドをpBSMID3MNと呼ぶ。
;5′−TTATTCATAATGATAGTAGGAGGCTTGGTAGGTTTAAGAATAGTTTTTGCTGTACTCTCTGTAGTGAATAGAGTTAGGCAGGGATAA −3′)
およびHIVTM2(配列番号119)
;5′−TTATCCCTGCCTAACTCTATTCACTACAGAGAGTACAGCAAAAACTATTCTTAAACCTACCAAGCCTCCTACTATCATTATGAATAA −3′)
から、オリゴヌクレオチドHIV3B1(配列番号114)およびHIVTM3(配列番号120)
;5′−ATCATCTCTAGAATAAAAATTATCCCTGCCTAACTCTATTCAC −3′)
を用いて調製した。また、PCRMN11は、プラスミドpH6HMNEから、オリゴヌクレオチドHIV3B1(配列番号114)およびHIVMN18(配列番号121)
;5′−GCCTCCTACTATCATTATGAATAATCTTTTTTCTCTCTG −3′)
を用いて調製した。)このような操作によって得られるプラスミドをpBSHIVMN120Tと呼ぶ。]
次にELDKWAおよびV3ループをコードする他方の部分をpBSK+ にクローニングした。これを行うために、HindIII−SacI酵素分解した300bpのPCR断片(ELDKWA−V3ループ配列の一部を含有する)を、pBSK+ の2,900bpのHindIII−SacI断片にクローニングした。(なお、該PCR断片は、プラスミドpBSHIVMN120TからプライマーHIVP69(配列番号122)
;5′−TGATAGTACCAGCTATAGGTTGAT−3′)
およびHIVP75(配列番号123)
;5′−TTTGTCGAGCTCGTAGTTGGGTCTTGTACAATT −3′)
を用いて調製した。)この操作により得られたプラスミドをpHIV56と呼ぶ。
実施例22−vCP205をサル科マカク属のサルに筋肉内投与した場合の接種効果と免疫原性
実験動物:Macaca fascicularis (カニクイザル)(成体、野生捕獲)
数:8匹
性別:オス
捕獲源:モーリシャス島。B型肝炎、フィロウイルスおよび結核症が存在しないと考えられる。
4匹のオスのカニクイザル(Cynomolgus macaque)に、1ヶ月間隔で5回、ALVAC−HIV(vCP205)(1回投与当たり105.6 TCID50を含有)を筋肉内投与した。この注入(注射)によって、何らの症状も外傷も引き起こされなかった。注入によるサルの体重変化はなかった。4匹のコントロール(対照)用サルには、プラセボ(placebo )(DMEM−F12(25%)、ラクトグルタメート(25%)および凍結乾燥用基質(50%)) を注入した。一般的な状態を毎日監視し、各接種後、1、2、3、4および7日目に注射部位の観察を行った。体重を週に1回測定した。
・200ngのV3MNペプチド
・130ngの精製「p24」HIV(大腸菌P25 LAIから単離、バッチ672Cl、Transgene)
インキュベートは、全て、最終体積が100μlになるように行い、その後、リン酸緩衝生理食塩水(pH7.1)/0.1%Tween 20を用いて3回洗浄した。リン酸緩衝生理食塩水(pH7.1)/0.05%Tween 20/5%(w/v)粉状スキムミルク(Gloria)の150μlを用いて37℃で1時間、プレートのブロッキングを行った。
OD値の範囲は0.2から1である。
血液学的および生化学的状態を監視するためのパラメーター(赤血球数、平均血球体積、ヘマトクリット、クレアチニンおよびアラニンアミノトランスフェラーゼなど)の大部分は平常な範囲内で変化した。
実施例23−vCP300をサル科マカク属のサルに投与した場合の接種効果と免疫原性
実験動物:
種:Macaca fascicularis(カニクイザル)
数:8匹
性別:オス
捕獲源:モーリシャス島。B型肝炎、フィロウイルスおよび結核症が存在しないと考えられる。
投与薬剤:プラセボ
投与方法:左または右の三角筋に交互に筋肉内投与。
注射回数:5回(0、4、8、12および16週)
グループ#2
投与薬剤:ALVAC−HIV
投与方法:右または右の三角筋に交互に筋肉内投与。
投与量:106.16TCID50
注射回数:5回(0、4、8、12および16週)
グループ#1および#2
20週目。
投与方法:静脈内投与
量:1ml
投与量:106.16TCID50
注射回数:1回
臨床観察:各接種の後、1、2、3、4および7日目に注射部位の観察を行った。1週間に1度、サルの体重測定を行った。
1)0.129Mクエン酸ナトリウム緩衝チューブ内に1.8ml(プロトロンビン分析用)。
3)0.17MのEDTA K3チューブに2ml(血液学的分析用)。
血液学的分析として、血球数および分化白血球数(differential leucocyte counts )、ヘモグロビン、血小板、プロトロンビン、網状赤血球および沈降率等を測定した。
Maxisorp F96 NUNCプレートに、0.1Mの炭酸塩緩衝液(pH9.6)で稀釈した以下の抗原の1つを37℃で1時間、次いで4℃で一晩被覆した。
・200ngのV3MNペプチド、
・130ngの精製p24 HIV(大腸菌p25 LAI単離物、バッチ672C1、Transgene製)。
力価=log(OD490−650 ×10)/(1/稀釈倍数)
OD値の範囲は0.2〜1.2である。
注入によって何等の症状も外傷も引き起こさなかった。注入によりサルの体重変化はなかった。血液学的パラメーターは有意に変化しておらず、また、生化学的分析によれば、腎臓機能や肝臓機能に変化は認められなかった。
Claims (5)
- gp120 V3ループ上に自然に存在しないエピトープを含有するようにV3ループ中で改変され、前記エピトープが、B-細胞エピトープ、ELDKWAまたはLDKWからなる群より選択されることを特徴とするHIV-1 gp120またはgp160タンパク質。
- 前記エピトープが、B-細胞エピトープであることを特徴とする請求項1記載のgp160またはgp120タンパク質。
- 前記エピトープが、ELDKWAまたはLDKWであることを特徴とする請求項1記載のgp160またはgp120タンパク質。
- ELDKWAまたはLDKWエピトープの少なくとも1つを含有するように改変されたgp120であることを特徴とする請求項1記載のgp160またはgp120タンパク質。
- 前記gp120が、前記エピトープを含有するようにV3ループ中で改変されることを特徴とする請求項4記載のgp120タンパク質。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US22384294A | 1994-04-06 | 1994-04-06 | |
| US08/417,210 US5863542A (en) | 1991-03-07 | 1995-04-05 | Recombinant attenuated ALVAC canaryopox virus containing heterologous HIV or SIV inserts |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP52637895A Division JP4007456B2 (ja) | 1994-04-06 | 1995-04-06 | 免疫不全組換えポックスウイルス |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2007297396A JP2007297396A (ja) | 2007-11-15 |
| JP4138848B2 true JP4138848B2 (ja) | 2008-08-27 |
Family
ID=26918187
Family Applications (4)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP52637895A Expired - Fee Related JP4007456B2 (ja) | 1994-04-06 | 1995-04-06 | 免疫不全組換えポックスウイルス |
| JP2006032543A Expired - Fee Related JP4098811B2 (ja) | 1994-04-06 | 2006-02-09 | 免疫不全組換えポックスウイルス |
| JP2007162393A Expired - Fee Related JP4138848B2 (ja) | 1994-04-06 | 2007-06-20 | 免疫不全組換えポックスウイルス |
| JP2007186820A Pending JP2008000138A (ja) | 1994-04-06 | 2007-07-18 | 免疫不全組換えポックスウイルス |
Family Applications Before (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP52637895A Expired - Fee Related JP4007456B2 (ja) | 1994-04-06 | 1995-04-06 | 免疫不全組換えポックスウイルス |
| JP2006032543A Expired - Fee Related JP4098811B2 (ja) | 1994-04-06 | 2006-02-09 | 免疫不全組換えポックスウイルス |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2007186820A Pending JP2008000138A (ja) | 1994-04-06 | 2007-07-18 | 免疫不全組換えポックスウイルス |
Country Status (7)
| Country | Link |
|---|---|
| US (4) | US5863542A (ja) |
| EP (1) | EP0752887B1 (ja) |
| JP (4) | JP4007456B2 (ja) |
| AT (1) | ATE395932T1 (ja) |
| CA (1) | CA2187031A1 (ja) |
| DE (1) | DE69535757D1 (ja) |
| WO (1) | WO1995027507A1 (ja) |
Families Citing this family (53)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7767449B1 (en) | 1981-12-24 | 2010-08-03 | Health Research Incorporated | Methods using modified vaccinia virus |
| US5505941A (en) * | 1981-12-24 | 1996-04-09 | Health Research, Inc. | Recombinant avipox virus and method to induce an immune response |
| US7374768B1 (en) | 1990-09-25 | 2008-05-20 | Xenova Research Limited | Viral vaccines |
| US5863542A (en) * | 1991-03-07 | 1999-01-26 | Virogenetics Corporation | Recombinant attenuated ALVAC canaryopox virus containing heterologous HIV or SIV inserts |
| DE69229390T2 (de) * | 1991-08-26 | 1999-11-11 | Immuno Ag, Wien | Direkt molekuläre Klonierung eines modifizierten Genoms eines Chordopocken-Virus |
| AU694519B2 (en) * | 1994-04-29 | 1998-07-23 | Immuno Aktiengesellschaft | Recombinant poxviruses with foreign polynucleotides in essential regions |
| JP2001500738A (ja) * | 1996-09-17 | 2001-01-23 | カイロン コーポレイション | 細胞内疾患を処置するための組成物および方法 |
| US7255862B1 (en) * | 1996-11-14 | 2007-08-14 | Connaught Technology Corporation | ALVAC/FIV constructs |
| US5990091A (en) * | 1997-03-12 | 1999-11-23 | Virogenetics Corporation | Vectors having enhanced expression, and methods of making and uses thereof |
| WO2000000216A2 (en) * | 1998-06-26 | 2000-01-06 | Aventis Pasteur | Use of poxviruses as enhancer of specific immunity |
| US20040002472A1 (en) * | 2000-01-21 | 2004-01-01 | Audonnet Jean-Christophe Francis | Vaccination or immunization using a prime-boost regimen |
| KR20070073987A (ko) * | 2000-01-31 | 2007-07-10 | 글락소스미스클라인 바이오로지칼즈 에스.에이. | Hiv에 대한 예방 또는 치료용 면역화를 위한 백신 |
| US8623379B2 (en) * | 2000-03-02 | 2014-01-07 | Emory University | Compositions and methods for generating an immune response |
| US20040105871A1 (en) * | 2000-03-02 | 2004-06-03 | Robinson Harriet L. | Compositions and methods for generating an immune response |
| US20020106798A1 (en) | 2000-03-02 | 2002-08-08 | Robinson Harriet L. | DNA expression vectors and methods of use |
| JP2003531865A (ja) * | 2000-04-28 | 2003-10-28 | アメリカ合衆国 | Dnaおよびワクシニアウイルスベクターワクチンの組み合わせを用いた免疫原性の改善 |
| US6582920B2 (en) * | 2000-09-01 | 2003-06-24 | Gen-Probe Incorporated | Amplification of HIV-1 RT sequences for detection of sequences associated with drug-resistance mutations |
| EP1350100A2 (en) | 2000-09-26 | 2003-10-08 | Health Research, Incorporated | Analysis of hiv-1 coreceptor use in the clinical care of hiv-1-infected patients |
| ES2334335T3 (es) * | 2001-03-08 | 2010-03-09 | The Government Of The Usa, As Represented By The Secretary, Department Of Health And Human Services | Mva que expresa genes de envoltura, gag y pol de vih modificados. |
| EP1281767A3 (en) | 2001-07-31 | 2003-05-28 | Aladar A. Szalay | Light emitting microorganisms and cells for diagnosis and therapy of tumors |
| CN1880457B (zh) | 2001-10-11 | 2010-05-26 | 麦克公司 | Ad6重组核酸 |
| WO2004041997A2 (en) * | 2002-05-01 | 2004-05-21 | National Institutes Of Health | Immunotherapy regimens in hiv-infected patients |
| US7501127B2 (en) | 2002-05-16 | 2009-03-10 | Bavarian Nordic A/S | Intergenic regions as novel sites for insertion of HIV DNA sequences in the genome of Modified Vaccinia virus Ankara |
| US20030228261A1 (en) * | 2002-06-05 | 2003-12-11 | Aladar Szalay | Light emitting microorganisms and cells for diagnosis and therapy of diseases associated with wounded or inflamed tissue |
| EP1369491A1 (en) * | 2002-06-05 | 2003-12-10 | Aladar A. Szalay | Light emitting microorganisms and cells for diagnosis and therapy of diseases associated with wounded or inflamed tissue |
| US7135556B2 (en) * | 2002-07-19 | 2006-11-14 | Schering Corporation | NPC1L1 (NPC3) and methods of use thereof |
| SG179291A1 (en) | 2003-06-18 | 2012-04-27 | Genelux Corp | Modified recombinant vaccinia viruses and other microorganisms, uses thereof |
| WO2005017208A1 (en) * | 2003-07-31 | 2005-02-24 | George Mason Intellectual Properties, Inc. | Compositions and methods for treating or preventing hiv infection |
| JP5475279B2 (ja) | 2005-06-17 | 2014-04-16 | イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・エルレ・エルレ | C型肝炎ウイルス核酸ワクチン |
| CN101313216A (zh) * | 2005-09-22 | 2008-11-26 | 普洛茨股份有限公司 | 酵母突变体中产生的糖基化多肽及其使用方法 |
| EP1989326A4 (en) | 2006-01-17 | 2009-09-30 | Health Research Inc | HETERO DUPLEX TRACKING TEST |
| JP2009535027A (ja) * | 2006-04-27 | 2009-10-01 | シンガポール ヘルス サービシーズ ピーティーイー リミテッド | 抗菌ペプチド |
| WO2007136763A2 (en) * | 2006-05-19 | 2007-11-29 | Sanofi Pasteur, Inc. | Immunological composition |
| TW200819540A (en) | 2006-07-11 | 2008-05-01 | Genelux Corp | Methods and compositions for detection of microorganisms and cells and treatment of diseases and disorders |
| WO2008025015A2 (en) * | 2006-08-25 | 2008-02-28 | The Government Of The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Epitope-protein scaffolds and their use |
| US20090098529A1 (en) * | 2006-10-16 | 2009-04-16 | Nanhai Chen | Methods for attenuating virus strains for diagnostic and therapeutic uses |
| WO2008150496A2 (en) * | 2007-05-31 | 2008-12-11 | Genelux Corporation | Assay for sensitivity to chemotherapeutic agents |
| PL2212345T3 (pl) * | 2007-11-19 | 2016-07-29 | Transgene Sa | Onkolityczne wektory pokswirusowe |
| EP2236520A1 (en) * | 2009-03-31 | 2010-10-06 | Leukocare Ag | Stabilizing composition for immobilized biomolecules |
| DK2414386T3 (en) | 2009-04-03 | 2016-02-22 | Merial Ltd | Birds of vaccines with newcastle disease virus vectors |
| CA2760315C (en) | 2009-04-30 | 2019-05-28 | Centre Hospitalier Universitaire Vaudois Lausanne (Chuv) | Modified immunization vectors |
| MX351643B (es) | 2009-12-28 | 2017-10-23 | Merial Ltd | Antigeno ndv recombinante y usos del mismo. |
| US20130052221A1 (en) | 2010-02-26 | 2013-02-28 | The Govt. of the U.S, as represented by The Sec. of The Dept. of Health and Human Services | Dna-protein vaccination protocols |
| PL2611460T3 (pl) | 2010-08-31 | 2017-02-28 | Merial, Inc. | Szczepionki przeciw herpeswirusom przenoszone z pomocą wirusa choroby newcastle |
| WO2012129295A1 (en) | 2011-03-21 | 2012-09-27 | Vaxin Inc. | Rapid and prolonged immunologic-therapeutic |
| EP2788021B1 (en) | 2011-12-09 | 2017-01-18 | Bavarian Nordic A/S | Poxvirus vector for the expression of bacterial antigens linked to tetanus toxin fragment c |
| ES2959642T3 (es) | 2015-05-13 | 2024-02-27 | The Us Secretary Of The Department Of Health And Human Services | Metodos y composiciones para inducir una respuesta inmunitaria mediante construcciones con elementos conservados |
| WO2016196366A1 (en) | 2015-05-29 | 2016-12-08 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Extension of replicative lifespan in diseases of premature aging using p53 isoforms |
| EA201890110A1 (ru) | 2015-06-23 | 2018-05-31 | Мериал, Инк. | Рекомбинантные вирусные векторы, содержащие минорные белки prrsv, и способы их получения и применения |
| CN113453712A (zh) * | 2018-12-28 | 2021-09-28 | 特兰斯吉恩股份有限公司 | M2缺陷型痘病毒 |
| JPWO2020230785A1 (ja) | 2019-05-14 | 2020-11-19 | ||
| WO2022107705A1 (ja) | 2020-11-17 | 2022-05-27 | 国立大学法人鳥取大学 | 新規遺伝子組換えワクシニアウイルス及びその利用 |
| US20240033347A1 (en) | 2022-07-08 | 2024-02-01 | Viromissile, Inc. | Oncolytic vaccinia viruses and recombinant viruses and methods of use thereof |
Family Cites Families (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DE10399031I1 (de) * | 1987-08-28 | 2004-01-29 | Health Research Inc | Rekombinante Viren. |
| GB8923123D0 (en) * | 1989-10-13 | 1989-11-29 | Connaught Lab | A vaccine for human immunodeficiency virus |
| AU672359B2 (en) * | 1991-03-07 | 1996-10-03 | Virogenetics Corporation | Genetically engineered vaccine strain |
| AU672581B2 (en) * | 1991-03-07 | 1996-10-10 | Virogenetics Corporation | Immunodeficiency virus recombinant poxvirus vaccine |
| US5863542A (en) * | 1991-03-07 | 1999-01-26 | Virogenetics Corporation | Recombinant attenuated ALVAC canaryopox virus containing heterologous HIV or SIV inserts |
| JPH08511007A (ja) * | 1993-06-09 | 1996-11-19 | コノート ラボラトリーズ リミテッド | タンデム合成hiv−1ペプチド |
-
1995
- 1995-04-05 US US08/417,210 patent/US5863542A/en not_active Expired - Lifetime
- 1995-04-06 EP EP95916151A patent/EP0752887B1/en not_active Expired - Lifetime
- 1995-04-06 CA CA002187031A patent/CA2187031A1/en not_active Abandoned
- 1995-04-06 AT AT95916151T patent/ATE395932T1/de not_active IP Right Cessation
- 1995-04-06 WO PCT/US1995/003989 patent/WO1995027507A1/en active Application Filing
- 1995-04-06 DE DE69535757T patent/DE69535757D1/de not_active Expired - Fee Related
- 1995-04-06 JP JP52637895A patent/JP4007456B2/ja not_active Expired - Fee Related
-
1998
- 1998-08-14 US US09/136,159 patent/US6596279B1/en not_active Expired - Lifetime
-
2003
- 2003-05-20 US US10/441,788 patent/US20030223987A1/en not_active Abandoned
-
2006
- 2006-02-09 JP JP2006032543A patent/JP4098811B2/ja not_active Expired - Fee Related
-
2007
- 2007-06-20 JP JP2007162393A patent/JP4138848B2/ja not_active Expired - Fee Related
- 2007-07-18 JP JP2007186820A patent/JP2008000138A/ja active Pending
- 2007-08-08 US US11/891,027 patent/US20080188640A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| CA2187031A1 (en) | 1995-10-19 |
| JPH09511649A (ja) | 1997-11-25 |
| JP2008000138A (ja) | 2008-01-10 |
| JP2006141407A (ja) | 2006-06-08 |
| DE69535757D1 (de) | 2008-07-03 |
| US5863542A (en) | 1999-01-26 |
| JP2007297396A (ja) | 2007-11-15 |
| JP4007456B2 (ja) | 2007-11-14 |
| EP0752887A1 (en) | 1997-01-15 |
| JP4098811B2 (ja) | 2008-06-11 |
| EP0752887A4 (en) | 1997-08-20 |
| ATE395932T1 (de) | 2008-06-15 |
| US20030223987A1 (en) | 2003-12-04 |
| US20080188640A1 (en) | 2008-08-07 |
| WO1995027507A1 (en) | 1995-10-19 |
| EP0752887B1 (en) | 2008-05-21 |
| US6596279B1 (en) | 2003-07-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4138848B2 (ja) | 免疫不全組換えポックスウイルス | |
| JP4108742B2 (ja) | ポックスウイルス−イヌジステンパーウイルス(cdv)組み換え体類および組成物類および前記組み換え体類を用いる方法 | |
| KR100242671B1 (ko) | 유전학적으로 처리한 백신 균주 | |
| JP3940959B2 (ja) | 組換えポックスウイルス−サイトメガロウイルス組成物および使用 | |
| EP0592546B1 (en) | Immunodeficiency virus recombinant poxvirus vaccine | |
| US5766598A (en) | Recombinant attenuated ALVAC canarypoxvirus expression vectors containing heterologous DNA segments encoding lentiviral gene products | |
| JP2007082551A (ja) | 組換えポックスウイルス−カリシウイルス[ウサギ出血疾患ウイルス(rhdv)]組成物および使用 | |
| JP2007254489A (ja) | Htlv抗原を発現する組換え弱毒化ポックスウイルスを含有する免疫原性組成物 | |
| JP3745370B2 (ja) | イヌヘルペスウイルスgB、gCおよびgDの、核酸およびアミノ酸配列とその利用 | |
| CA2105277C (en) | Genetically engineered vaccine strain | |
| AU747139B2 (en) | Immunodeficiency recombinant poxvirus | |
| AU2002300582B2 (en) | Immunodeficiency Recombinant Poxvirus | |
| AU702634B2 (en) | Immunodeficiency recombinant poxvirus | |
| KR100263279B1 (ko) | Trovac 계두 바이러스 재조합체 및 그것의 사용 | |
| AU2006201066A1 (en) | Immunodeficiency recombinant poxvirus | |
| AU4700096A (en) | Immunogenic compositions containing recombinant attenuated poxviruses expressing htlv antigens |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20071023 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20080123 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20080131 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20080225 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20080229 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20080324 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20080327 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20080423 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20080520 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20080605 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110613 Year of fee payment: 3 |
|
| LAPS | Cancellation because of no payment of annual fees |