JP3955352B2 - 破骨細胞形成阻害剤 - Google Patents
破骨細胞形成阻害剤 Download PDFInfo
- Publication number
- JP3955352B2 JP3955352B2 JP05546897A JP5546897A JP3955352B2 JP 3955352 B2 JP3955352 B2 JP 3955352B2 JP 05546897 A JP05546897 A JP 05546897A JP 5546897 A JP5546897 A JP 5546897A JP 3955352 B2 JP3955352 B2 JP 3955352B2
- Authority
- JP
- Japan
- Prior art keywords
- amino acid
- seq
- acid sequence
- osteoclast formation
- cells
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
- 210000002997 osteoclast Anatomy 0.000 title claims description 65
- 230000015572 biosynthetic process Effects 0.000 title claims description 61
- 239000003112 inhibitor Substances 0.000 title claims description 27
- 102000003810 Interleukin-18 Human genes 0.000 claims description 93
- 108090000171 Interleukin-18 Proteins 0.000 claims description 93
- 150000001413 amino acids Chemical class 0.000 claims description 68
- 201000010099 disease Diseases 0.000 claims description 19
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 19
- 239000003814 drug Substances 0.000 claims description 9
- 238000001727 in vivo Methods 0.000 claims description 9
- 229940079593 drug Drugs 0.000 claims description 8
- 230000002401 inhibitory effect Effects 0.000 claims description 8
- 108090000623 proteins and genes Proteins 0.000 claims description 8
- 239000000872 buffer Substances 0.000 claims description 5
- 102000004169 proteins and genes Human genes 0.000 claims description 5
- 239000003381 stabilizer Substances 0.000 claims description 5
- 210000004899 c-terminal region Anatomy 0.000 claims description 4
- 238000012217 deletion Methods 0.000 claims description 2
- 230000037430 deletion Effects 0.000 claims description 2
- 125000003275 alpha amino acid group Chemical group 0.000 claims 7
- 150000001720 carbohydrates Chemical class 0.000 claims 1
- 210000004027 cell Anatomy 0.000 description 57
- 238000000034 method Methods 0.000 description 57
- 101000960954 Homo sapiens Interleukin-18 Proteins 0.000 description 34
- 108020004511 Recombinant DNA Proteins 0.000 description 30
- 102000043959 human IL18 Human genes 0.000 description 30
- 108020004414 DNA Proteins 0.000 description 27
- 108010074328 Interferon-gamma Proteins 0.000 description 23
- 238000004519 manufacturing process Methods 0.000 description 21
- 102100037850 Interferon gamma Human genes 0.000 description 20
- 239000002585 base Substances 0.000 description 20
- 238000007796 conventional method Methods 0.000 description 20
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 17
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 17
- 235000018417 cysteine Nutrition 0.000 description 16
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 16
- 208000035896 Twin-reversed arterial perfusion sequence Diseases 0.000 description 15
- 230000031018 biological processes and functions Effects 0.000 description 13
- 239000013641 positive control Substances 0.000 description 13
- 208000006386 Bone Resorption Diseases 0.000 description 12
- 230000024279 bone resorption Effects 0.000 description 12
- 239000002609 medium Substances 0.000 description 11
- 108090000765 processed proteins & peptides Proteins 0.000 description 11
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 10
- 239000003795 chemical substances by application Substances 0.000 description 10
- 239000002299 complementary DNA Substances 0.000 description 10
- 235000001014 amino acid Nutrition 0.000 description 9
- 229940024606 amino acid Drugs 0.000 description 9
- 210000004698 lymphocyte Anatomy 0.000 description 9
- 239000000203 mixture Substances 0.000 description 9
- 229920001184 polypeptide Polymers 0.000 description 9
- 102000004196 processed proteins & peptides Human genes 0.000 description 9
- 241001465754 Metazoa Species 0.000 description 8
- 230000000694 effects Effects 0.000 description 8
- 208000002966 Giant Cell Tumor of Bone Diseases 0.000 description 7
- 208000037147 Hypercalcaemia Diseases 0.000 description 7
- 101000960949 Mus musculus Interleukin-18 Proteins 0.000 description 7
- 208000001132 Osteoporosis Diseases 0.000 description 7
- 238000012258 culturing Methods 0.000 description 7
- 230000000148 hypercalcaemia Effects 0.000 description 7
- 208000030915 hypercalcemia disease Diseases 0.000 description 7
- 230000001939 inductive effect Effects 0.000 description 7
- 230000005764 inhibitory process Effects 0.000 description 7
- 239000008363 phosphate buffer Substances 0.000 description 7
- 238000002360 preparation method Methods 0.000 description 7
- 239000000047 product Substances 0.000 description 7
- 241000282412 Homo Species 0.000 description 6
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 6
- 235000004279 alanine Nutrition 0.000 description 6
- 238000004113 cell culture Methods 0.000 description 6
- 239000013642 negative control Substances 0.000 description 6
- 239000002773 nucleotide Substances 0.000 description 6
- 125000003729 nucleotide group Chemical group 0.000 description 6
- 230000009471 action Effects 0.000 description 5
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 5
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 5
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 5
- 235000011130 ammonium sulphate Nutrition 0.000 description 5
- 229960000723 ampicillin Drugs 0.000 description 5
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 5
- GMRQFYUYWCNGIN-NKMMMXOESA-N calcitriol Chemical compound C1(/[C@@H]2CC[C@@H]([C@]2(CCC1)C)[C@@H](CCCC(C)(C)O)C)=C\C=C1\C[C@@H](O)C[C@H](O)C1=C GMRQFYUYWCNGIN-NKMMMXOESA-N 0.000 description 5
- 239000003638 chemical reducing agent Substances 0.000 description 5
- 238000004587 chromatography analysis Methods 0.000 description 5
- 239000013611 chromosomal DNA Substances 0.000 description 5
- XEYBRNLFEZDVAW-ARSRFYASSA-N dinoprostone Chemical compound CCCCC[C@H](O)\C=C\[C@H]1[C@H](O)CC(=O)[C@@H]1C\C=C/CCCC(O)=O XEYBRNLFEZDVAW-ARSRFYASSA-N 0.000 description 5
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 5
- 238000000338 in vitro Methods 0.000 description 5
- 210000000963 osteoblast Anatomy 0.000 description 5
- 125000003607 serino group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C(O[H])([H])[H] 0.000 description 5
- 241000588724 Escherichia coli Species 0.000 description 4
- 102000006992 Interferon-alpha Human genes 0.000 description 4
- 108010047761 Interferon-alpha Proteins 0.000 description 4
- 102100039898 Interleukin-18 Human genes 0.000 description 4
- 241000699666 Mus <mouse, genus> Species 0.000 description 4
- 241000699670 Mus sp. Species 0.000 description 4
- 210000004369 blood Anatomy 0.000 description 4
- 239000008280 blood Substances 0.000 description 4
- 238000003501 co-culture Methods 0.000 description 4
- 238000001502 gel electrophoresis Methods 0.000 description 4
- 239000007924 injection Substances 0.000 description 4
- 238000002347 injection Methods 0.000 description 4
- 229920002401 polyacrylamide Polymers 0.000 description 4
- 239000002243 precursor Substances 0.000 description 4
- 235000018102 proteins Nutrition 0.000 description 4
- 239000006228 supernatant Substances 0.000 description 4
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 description 3
- SXAMGRAIZSSWIH-UHFFFAOYSA-N 2-[3-[2-(2,3-dihydro-1H-inden-2-ylamino)pyrimidin-5-yl]-1,2,4-oxadiazol-5-yl]-1-(2,4,6,7-tetrahydrotriazolo[4,5-c]pyridin-5-yl)ethanone Chemical compound C1C(CC2=CC=CC=C12)NC1=NC=C(C=N1)C1=NOC(=N1)CC(=O)N1CC2=C(CC1)NN=N2 SXAMGRAIZSSWIH-UHFFFAOYSA-N 0.000 description 3
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 3
- 108020004705 Codon Proteins 0.000 description 3
- 102000004127 Cytokines Human genes 0.000 description 3
- 108090000695 Cytokines Proteins 0.000 description 3
- 102000004190 Enzymes Human genes 0.000 description 3
- 108090000790 Enzymes Proteins 0.000 description 3
- 102000008070 Interferon-gamma Human genes 0.000 description 3
- 102000000589 Interleukin-1 Human genes 0.000 description 3
- 108010002352 Interleukin-1 Proteins 0.000 description 3
- 108010076504 Protein Sorting Signals Proteins 0.000 description 3
- 239000012980 RPMI-1640 medium Substances 0.000 description 3
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 description 3
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 description 3
- 238000004458 analytical method Methods 0.000 description 3
- 210000004102 animal cell Anatomy 0.000 description 3
- 210000000988 bone and bone Anatomy 0.000 description 3
- 210000001185 bone marrow Anatomy 0.000 description 3
- 210000002798 bone marrow cell Anatomy 0.000 description 3
- 238000003776 cleavage reaction Methods 0.000 description 3
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 3
- 239000002552 dosage form Substances 0.000 description 3
- 229940088598 enzyme Drugs 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- 229960003130 interferon gamma Drugs 0.000 description 3
- 230000007246 mechanism Effects 0.000 description 3
- 239000012533 medium component Substances 0.000 description 3
- 238000001471 micro-filtration Methods 0.000 description 3
- 239000002504 physiological saline solution Substances 0.000 description 3
- 239000002244 precipitate Substances 0.000 description 3
- 230000007017 scission Effects 0.000 description 3
- 239000011780 sodium chloride Substances 0.000 description 3
- 239000000243 solution Substances 0.000 description 3
- 239000000126 substance Substances 0.000 description 3
- OHVLMTFVQDZYHP-UHFFFAOYSA-N 1-(2,4,6,7-tetrahydrotriazolo[4,5-c]pyridin-5-yl)-2-[4-[2-[[3-(trifluoromethoxy)phenyl]methylamino]pyrimidin-5-yl]piperazin-1-yl]ethanone Chemical compound N1N=NC=2CN(CCC=21)C(CN1CCN(CC1)C=1C=NC(=NC=1)NCC1=CC(=CC=C1)OC(F)(F)F)=O OHVLMTFVQDZYHP-UHFFFAOYSA-N 0.000 description 2
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 2
- 101100007328 Cocos nucifera COS-1 gene Proteins 0.000 description 2
- 108010062580 Concanavalin A Proteins 0.000 description 2
- 108010054576 Deoxyribonuclease EcoRI Proteins 0.000 description 2
- 108010010803 Gelatin Proteins 0.000 description 2
- 108090000723 Insulin-Like Growth Factor I Proteins 0.000 description 2
- 102000003815 Interleukin-11 Human genes 0.000 description 2
- 108090000177 Interleukin-11 Proteins 0.000 description 2
- 102000004889 Interleukin-6 Human genes 0.000 description 2
- 108090001005 Interleukin-6 Proteins 0.000 description 2
- 108091092195 Intron Proteins 0.000 description 2
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 2
- TWRXJAOTZQYOKJ-UHFFFAOYSA-L Magnesium chloride Chemical compound [Mg+2].[Cl-].[Cl-] TWRXJAOTZQYOKJ-UHFFFAOYSA-L 0.000 description 2
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 2
- RJKFOVLPORLFTN-LEKSSAKUSA-N Progesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H](C(=O)C)[C@@]1(C)CC2 RJKFOVLPORLFTN-LEKSSAKUSA-N 0.000 description 2
- 102000013275 Somatomedins Human genes 0.000 description 2
- 239000013543 active substance Substances 0.000 description 2
- 238000006243 chemical reaction Methods 0.000 description 2
- 239000012228 culture supernatant Substances 0.000 description 2
- 239000002274 desiccant Substances 0.000 description 2
- 239000012153 distilled water Substances 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 239000003889 eye drop Substances 0.000 description 2
- 239000012894 fetal calf serum Substances 0.000 description 2
- 239000012634 fragment Substances 0.000 description 2
- 239000008273 gelatin Substances 0.000 description 2
- 229920000159 gelatin Polymers 0.000 description 2
- 235000019322 gelatine Nutrition 0.000 description 2
- 235000011852 gelatine desserts Nutrition 0.000 description 2
- 239000001963 growth medium Substances 0.000 description 2
- -1 immune aids Substances 0.000 description 2
- 229940074383 interleukin-11 Drugs 0.000 description 2
- 229940100601 interleukin-6 Drugs 0.000 description 2
- 239000007788 liquid Substances 0.000 description 2
- 239000012092 media component Substances 0.000 description 2
- 229930182817 methionine Natural products 0.000 description 2
- 244000005700 microbiome Species 0.000 description 2
- 239000007923 nasal drop Substances 0.000 description 2
- 239000002674 ointment Substances 0.000 description 2
- 230000011164 ossification Effects 0.000 description 2
- 239000006072 paste Substances 0.000 description 2
- 239000013600 plasmid vector Substances 0.000 description 2
- 239000000843 powder Substances 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 238000001243 protein synthesis Methods 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 238000011218 seed culture Methods 0.000 description 2
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 2
- 210000004989 spleen cell Anatomy 0.000 description 2
- 235000000346 sugar Nutrition 0.000 description 2
- 150000005846 sugar alcohols Chemical class 0.000 description 2
- 208000024891 symptom Diseases 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 230000014616 translation Effects 0.000 description 2
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 2
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 2
- FVVCFHXLWDDRHG-UPLOTWCNSA-N (2s,3r,4s,5r,6r)-2-[(2r,3s,4r,5r,6r)-6-[(2s,3s,4s,5r)-3,4-dihydroxy-2,5-bis(hydroxymethyl)oxolan-2-yl]oxy-4,5-dihydroxy-2-(hydroxymethyl)oxan-3-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O[C@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)O)[C@@H](CO)O1 FVVCFHXLWDDRHG-UPLOTWCNSA-N 0.000 description 1
- MZOFCQQQCNRIBI-VMXHOPILSA-N (3s)-4-[[(2s)-1-[[(2s)-1-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-methyl-1-oxopentan-2-yl]amino]-5-(diaminomethylideneamino)-1-oxopentan-2-yl]amino]-3-[[2-[[(2s)-2,6-diaminohexanoyl]amino]acetyl]amino]-4-oxobutanoic acid Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@@H](N)CCCCN MZOFCQQQCNRIBI-VMXHOPILSA-N 0.000 description 1
- 102100025573 1-alkyl-2-acetylglycerophosphocholine esterase Human genes 0.000 description 1
- DBTMGCOVALSLOR-UHFFFAOYSA-N 32-alpha-galactosyl-3-alpha-galactosyl-galactose Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(OC2C(C(CO)OC(O)C2O)O)OC(CO)C1O DBTMGCOVALSLOR-UHFFFAOYSA-N 0.000 description 1
- MGYWDJWGVICAEL-UHFFFAOYSA-N 5-fluoro-3-(oxolan-2-yl)-1h-pyrimidine-2,4-dione Chemical compound O=C1C(F)=CNC(=O)N1C1OCCC1 MGYWDJWGVICAEL-UHFFFAOYSA-N 0.000 description 1
- 108020005075 5S Ribosomal RNA Proteins 0.000 description 1
- PVXPPJIGRGXGCY-DJHAAKORSA-N 6-O-alpha-D-glucopyranosyl-alpha-D-fructofuranose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OC[C@@H]1[C@@H](O)[C@H](O)[C@](O)(CO)O1 PVXPPJIGRGXGCY-DJHAAKORSA-N 0.000 description 1
- 206010000871 Acute monocytic leukaemia Diseases 0.000 description 1
- 208000031261 Acute myeloid leukaemia Diseases 0.000 description 1
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 1
- 208000006820 Arthralgia Diseases 0.000 description 1
- 208000036487 Arthropathies Diseases 0.000 description 1
- 108010024976 Asparaginase Proteins 0.000 description 1
- COVZYZSDYWQREU-UHFFFAOYSA-N Busulfan Chemical compound CS(=O)(=O)OCCCCOS(C)(=O)=O COVZYZSDYWQREU-UHFFFAOYSA-N 0.000 description 1
- 125000001433 C-terminal amino-acid group Chemical group 0.000 description 1
- 0 C[C@](*C=NC)C=CC Chemical compound C[C@](*C=NC)C=CC 0.000 description 1
- 102000055006 Calcitonin Human genes 0.000 description 1
- 108060001064 Calcitonin Proteins 0.000 description 1
- 108010001789 Calcitonin Receptors Proteins 0.000 description 1
- 102100038520 Calcitonin receptor Human genes 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- OPSXJNAGCGVGOG-DKWTVANSSA-L Calcium L-aspartate Chemical compound [Ca+2].[O-]C(=O)[C@@H](N)CC([O-])=O OPSXJNAGCGVGOG-DKWTVANSSA-L 0.000 description 1
- UXVMQQNJUSDDNG-UHFFFAOYSA-L Calcium chloride Chemical compound [Cl-].[Cl-].[Ca+2] UXVMQQNJUSDDNG-UHFFFAOYSA-L 0.000 description 1
- QMBJSIBWORFWQT-DFXBJWIESA-N Chlormadinone acetate Chemical compound C1=C(Cl)C2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@@](C(C)=O)(OC(=O)C)[C@@]1(C)CC2 QMBJSIBWORFWQT-DFXBJWIESA-N 0.000 description 1
- 102000029816 Collagenase Human genes 0.000 description 1
- 108060005980 Collagenase Proteins 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- UHDGCWIWMRVCDJ-CCXZUQQUSA-N Cytarabine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@@H](O)[C@H](O)[C@@H](CO)O1 UHDGCWIWMRVCDJ-CCXZUQQUSA-N 0.000 description 1
- 241000701022 Cytomegalovirus Species 0.000 description 1
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 1
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 1
- RXVWSYJTUUKTEA-UHFFFAOYSA-N D-maltotriose Natural products OC1C(O)C(OC(C(O)CO)C(O)C(O)C=O)OC(CO)C1OC1C(O)C(O)C(O)C(CO)O1 RXVWSYJTUUKTEA-UHFFFAOYSA-N 0.000 description 1
- ZCLAHGAZPPEVDX-UHFFFAOYSA-N D-panose Natural products OC1C(O)C(O)C(OC(C(O)CO)C(O)C(O)C=O)OC1COC1C(O)C(O)C(O)C(CO)O1 ZCLAHGAZPPEVDX-UHFFFAOYSA-N 0.000 description 1
- 102000012410 DNA Ligases Human genes 0.000 description 1
- 108010061982 DNA Ligases Proteins 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 229920001353 Dextrin Polymers 0.000 description 1
- 239000004375 Dextrin Substances 0.000 description 1
- 108700024394 Exon Proteins 0.000 description 1
- 229920001917 Ficoll Polymers 0.000 description 1
- GHASVSINZRGABV-UHFFFAOYSA-N Fluorouracil Chemical compound FC1=CNC(=O)NC1=O GHASVSINZRGABV-UHFFFAOYSA-N 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 108010024636 Glutathione Proteins 0.000 description 1
- 108091006905 Human Serum Albumin Proteins 0.000 description 1
- 102000008100 Human Serum Albumin Human genes 0.000 description 1
- 206010020850 Hyperthyroidism Diseases 0.000 description 1
- 102000003996 Interferon-beta Human genes 0.000 description 1
- 108090000467 Interferon-beta Proteins 0.000 description 1
- 102000013462 Interleukin-12 Human genes 0.000 description 1
- 108010065805 Interleukin-12 Proteins 0.000 description 1
- 102000000588 Interleukin-2 Human genes 0.000 description 1
- 108010002350 Interleukin-2 Proteins 0.000 description 1
- 102000000646 Interleukin-3 Human genes 0.000 description 1
- 108010002386 Interleukin-3 Proteins 0.000 description 1
- 102000004388 Interleukin-4 Human genes 0.000 description 1
- 108090000978 Interleukin-4 Proteins 0.000 description 1
- SFBODOKJTYAUCM-UHFFFAOYSA-N Ipriflavone Chemical compound C=1C(OC(C)C)=CC=C(C2=O)C=1OC=C2C1=CC=CC=C1 SFBODOKJTYAUCM-UHFFFAOYSA-N 0.000 description 1
- AYRXSINWFIIFAE-SCLMCMATSA-N Isomaltose Natural products OC[C@H]1O[C@H](OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C=O)[C@@H](O)[C@@H](O)[C@@H]1O AYRXSINWFIIFAE-SCLMCMATSA-N 0.000 description 1
- 208000012659 Joint disease Diseases 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 229920001491 Lentinan Polymers 0.000 description 1
- 102000004083 Lymphotoxin-alpha Human genes 0.000 description 1
- 108090000542 Lymphotoxin-alpha Proteins 0.000 description 1
- XJCCHWKNFMUJFE-CGQAXDJHSA-N Maltotriitol Chemical compound O[C@@H]1[C@@H](O)[C@@H](O[C@@H]([C@H](O)[C@@H](O)CO)[C@H](O)CO)O[C@H](CO)[C@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 XJCCHWKNFMUJFE-CGQAXDJHSA-N 0.000 description 1
- 208000029725 Metabolic bone disease Diseases 0.000 description 1
- 208000035489 Monocytic Acute Leukemia Diseases 0.000 description 1
- 101000746372 Mus musculus Granulocyte-macrophage colony-stimulating factor Proteins 0.000 description 1
- 208000033776 Myeloid Acute Leukemia Diseases 0.000 description 1
- 125000001429 N-terminal alpha-amino-acid group Chemical group 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 108091028043 Nucleic acid sequence Proteins 0.000 description 1
- 206010049088 Osteopenia Diseases 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 108090000445 Parathyroid hormone Proteins 0.000 description 1
- 102000003982 Parathyroid hormone Human genes 0.000 description 1
- 102000007079 Peptide Fragments Human genes 0.000 description 1
- 108010033276 Peptide Fragments Proteins 0.000 description 1
- 206010036790 Productive cough Diseases 0.000 description 1
- 239000004373 Pullulan Substances 0.000 description 1
- 229920001218 Pullulan Polymers 0.000 description 1
- MUPFEKGTMRGPLJ-RMMQSMQOSA-N Raffinose Natural products O(C[C@H]1[C@@H](O)[C@H](O)[C@@H](O)[C@@H](O[C@@]2(CO)[C@H](O)[C@@H](O)[C@@H](CO)O2)O1)[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 MUPFEKGTMRGPLJ-RMMQSMQOSA-N 0.000 description 1
- 229920002684 Sepharose Polymers 0.000 description 1
- 108010071390 Serum Albumin Proteins 0.000 description 1
- 102000007562 Serum Albumin Human genes 0.000 description 1
- 229920002125 Sokalan® Polymers 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 239000012505 Superdex™ Substances 0.000 description 1
- 102000007591 Tartrate-Resistant Acid Phosphatase Human genes 0.000 description 1
- 108010032050 Tartrate-Resistant Acid Phosphatase Proteins 0.000 description 1
- 108020005038 Terminator Codon Proteins 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 1
- MUPFEKGTMRGPLJ-UHFFFAOYSA-N UNPD196149 Natural products OC1C(O)C(CO)OC1(CO)OC1C(O)C(O)C(O)C(COC2C(C(O)C(O)C(CO)O2)O)O1 MUPFEKGTMRGPLJ-UHFFFAOYSA-N 0.000 description 1
- LUEWUZLMQUOBSB-UHFFFAOYSA-N UNPD55895 Natural products OC1C(O)C(O)C(CO)OC1OC1C(CO)OC(OC2C(OC(OC3C(OC(O)C(O)C3O)CO)C(O)C2O)CO)C(O)C1O LUEWUZLMQUOBSB-UHFFFAOYSA-N 0.000 description 1
- 229930003316 Vitamin D Natural products 0.000 description 1
- QYSXJUFSXHHAJI-XFEUOLMDSA-N Vitamin D3 Natural products C1(/[C@@H]2CC[C@@H]([C@]2(CCC1)C)[C@H](C)CCCC(C)C)=C/C=C1\C[C@@H](O)CCC1=C QYSXJUFSXHHAJI-XFEUOLMDSA-N 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 231100000215 acute (single dose) toxicity testing Toxicity 0.000 description 1
- 238000011047 acute toxicity test Methods 0.000 description 1
- 239000003513 alkali Substances 0.000 description 1
- GUBGYTABKSRVRQ-ASMJPISFSA-N alpha-maltose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-ASMJPISFSA-N 0.000 description 1
- BBDAGFIXKZCXAH-CCXZUQQUSA-N ancitabine Chemical compound N=C1C=CN2[C@@H]3O[C@H](CO)[C@@H](O)[C@@H]3OC2=N1 BBDAGFIXKZCXAH-CCXZUQQUSA-N 0.000 description 1
- 229950000242 ancitabine Drugs 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QUYVBRFLSA-N beta-maltose Chemical compound OC[C@H]1O[C@H](O[C@H]2[C@H](O)[C@@H](O)[C@H](O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@@H]1O GUBGYTABKSRVRQ-QUYVBRFLSA-N 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 230000037396 body weight Effects 0.000 description 1
- 210000004271 bone marrow stromal cell Anatomy 0.000 description 1
- 229960002092 busulfan Drugs 0.000 description 1
- BBBFJLBPOGFECG-VJVYQDLKSA-N calcitonin Chemical compound N([C@H](C(=O)N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N1[C@@H](CCC1)C(N)=O)C(C)C)C(=O)[C@@H]1CSSC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N1 BBBFJLBPOGFECG-VJVYQDLKSA-N 0.000 description 1
- 229960004015 calcitonin Drugs 0.000 description 1
- 239000011612 calcitriol Substances 0.000 description 1
- 239000011575 calcium Substances 0.000 description 1
- 229960005069 calcium Drugs 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 239000001110 calcium chloride Substances 0.000 description 1
- 229910001628 calcium chloride Inorganic materials 0.000 description 1
- 229960002713 calcium chloride Drugs 0.000 description 1
- 235000011148 calcium chloride Nutrition 0.000 description 1
- FUFJGUQYACFECW-UHFFFAOYSA-L calcium hydrogenphosphate Chemical compound [Ca+2].OP([O-])([O-])=O FUFJGUQYACFECW-UHFFFAOYSA-L 0.000 description 1
- MKJXYGKVIBWPFZ-UHFFFAOYSA-L calcium lactate Chemical compound [Ca+2].CC(O)C([O-])=O.CC(O)C([O-])=O MKJXYGKVIBWPFZ-UHFFFAOYSA-L 0.000 description 1
- 239000001527 calcium lactate Substances 0.000 description 1
- 229960002401 calcium lactate Drugs 0.000 description 1
- 235000011086 calcium lactate Nutrition 0.000 description 1
- 159000000007 calcium salts Chemical class 0.000 description 1
- 239000002775 capsule Substances 0.000 description 1
- 239000000969 carrier Substances 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 1
- 229960001616 chlormadinone acetate Drugs 0.000 description 1
- 238000011098 chromatofocusing Methods 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 108090001092 clostripain Proteins 0.000 description 1
- 229960002424 collagenase Drugs 0.000 description 1
- 239000003246 corticosteroid Substances 0.000 description 1
- 239000013078 crystal Substances 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 229960000684 cytarabine Drugs 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- 235000019425 dextrin Nutrition 0.000 description 1
- 235000019700 dicalcium phosphate Nutrition 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- BNIILDVGGAEEIG-UHFFFAOYSA-L disodium hydrogen phosphate Chemical compound [Na+].[Na+].OP([O-])([O-])=O BNIILDVGGAEEIG-UHFFFAOYSA-L 0.000 description 1
- 108010007093 dispase Proteins 0.000 description 1
- 229940011871 estrogen Drugs 0.000 description 1
- 239000000262 estrogen Substances 0.000 description 1
- 229940012356 eye drops Drugs 0.000 description 1
- 239000012091 fetal bovine serum Substances 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 229960002949 fluorouracil Drugs 0.000 description 1
- 230000037406 food intake Effects 0.000 description 1
- 239000012737 fresh medium Substances 0.000 description 1
- 235000021255 galacto-oligosaccharides Nutrition 0.000 description 1
- 150000003271 galactooligosaccharides Chemical class 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 1
- 239000008187 granular material Substances 0.000 description 1
- 235000013402 health food Nutrition 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 230000013632 homeostatic process Effects 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 210000005260 human cell Anatomy 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 239000000411 inducer Substances 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 229960001388 interferon-beta Drugs 0.000 description 1
- 229940117681 interleukin-12 Drugs 0.000 description 1
- 229940076264 interleukin-3 Drugs 0.000 description 1
- 229940028885 interleukin-4 Drugs 0.000 description 1
- 229960005431 ipriflavone Drugs 0.000 description 1
- DLRVVLDZNNYCBX-RTPHMHGBSA-N isomaltose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OC[C@@H]1[C@@H](O)[C@H](O)[C@@H](O)C(O)O1 DLRVVLDZNNYCBX-RTPHMHGBSA-N 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 239000000832 lactitol Substances 0.000 description 1
- VQHSOMBJVWLPSR-JVCRWLNRSA-N lactitol Chemical compound OC[C@H](O)[C@@H](O)[C@@H]([C@H](O)CO)O[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O VQHSOMBJVWLPSR-JVCRWLNRSA-N 0.000 description 1
- 235000010448 lactitol Nutrition 0.000 description 1
- 229960003451 lactitol Drugs 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 229940115286 lentinan Drugs 0.000 description 1
- 229910001629 magnesium chloride Inorganic materials 0.000 description 1
- 238000012423 maintenance Methods 0.000 description 1
- 239000000845 maltitol Substances 0.000 description 1
- VQHSOMBJVWLPSR-WUJBLJFYSA-N maltitol Chemical compound OC[C@H](O)[C@@H](O)[C@@H]([C@H](O)CO)O[C@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O VQHSOMBJVWLPSR-WUJBLJFYSA-N 0.000 description 1
- 235000010449 maltitol Nutrition 0.000 description 1
- 229940035436 maltitol Drugs 0.000 description 1
- UYQJCPNSAVWAFU-UHFFFAOYSA-N malto-tetraose Natural products OC1C(O)C(OC(C(O)CO)C(O)C(O)C=O)OC(CO)C1OC1C(O)C(O)C(OC2C(C(O)C(O)C(CO)O2)O)C(CO)O1 UYQJCPNSAVWAFU-UHFFFAOYSA-N 0.000 description 1
- LUEWUZLMQUOBSB-OUBHKODOSA-N maltotetraose Chemical compound O[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@H](CO)O[C@@H](O[C@@H]2[C@@H](O[C@@H](O[C@@H]3[C@@H](O[C@@H](O)[C@H](O)[C@H]3O)CO)[C@H](O)[C@H]2O)CO)[C@H](O)[C@H]1O LUEWUZLMQUOBSB-OUBHKODOSA-N 0.000 description 1
- FYGDTMLNYKFZSV-UHFFFAOYSA-N mannotriose Natural products OC1C(O)C(O)C(CO)OC1OC1C(CO)OC(OC2C(OC(O)C(O)C2O)CO)C(O)C1O FYGDTMLNYKFZSV-UHFFFAOYSA-N 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- 229910000403 monosodium phosphate Inorganic materials 0.000 description 1
- 235000019799 monosodium phosphate Nutrition 0.000 description 1
- 229940100662 nasal drops Drugs 0.000 description 1
- 239000006218 nasal suppository Substances 0.000 description 1
- 102000042567 non-coding RNA Human genes 0.000 description 1
- 229940053934 norethindrone Drugs 0.000 description 1
- VIKNJXKGJWUCNN-XGXHKTLJSA-N norethisterone Chemical compound O=C1CC[C@@H]2[C@H]3CC[C@](C)([C@](CC4)(O)C#C)[C@@H]4[C@@H]3CCC2=C1 VIKNJXKGJWUCNN-XGXHKTLJSA-N 0.000 description 1
- 201000008482 osteoarthritis Diseases 0.000 description 1
- 201000008968 osteosarcoma Diseases 0.000 description 1
- ZCLAHGAZPPEVDX-MQHGYYCBSA-N panose Chemical compound O[C@H]1[C@H](O)[C@@H](O)[C@@H](O[C@H]([C@H](O)CO)[C@H](O)[C@@H](O)C=O)O[C@@H]1CO[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 ZCLAHGAZPPEVDX-MQHGYYCBSA-N 0.000 description 1
- 239000000199 parathyroid hormone Substances 0.000 description 1
- 229960001319 parathyroid hormone Drugs 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 108010001062 polysaccharide-K Proteins 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 239000000186 progesterone Substances 0.000 description 1
- 229960003387 progesterone Drugs 0.000 description 1
- 230000035755 proliferation Effects 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 235000019423 pullulan Nutrition 0.000 description 1
- MUPFEKGTMRGPLJ-ZQSKZDJDSA-N raffinose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO[C@@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)O)O1 MUPFEKGTMRGPLJ-ZQSKZDJDSA-N 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 206010039073 rheumatoid arthritis Diseases 0.000 description 1
- 108020004418 ribosomal RNA Proteins 0.000 description 1
- 229920006395 saturated elastomer Polymers 0.000 description 1
- AJPJDKMHJJGVTQ-UHFFFAOYSA-M sodium dihydrogen phosphate Chemical compound [Na+].OP(O)([O-])=O AJPJDKMHJJGVTQ-UHFFFAOYSA-M 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 239000000600 sorbitol Substances 0.000 description 1
- 229960002920 sorbitol Drugs 0.000 description 1
- 235000010356 sorbitol Nutrition 0.000 description 1
- 241000894007 species Species 0.000 description 1
- 208000024794 sputum Diseases 0.000 description 1
- 210000003802 sputum Anatomy 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 239000003826 tablet Substances 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 235000019166 vitamin D Nutrition 0.000 description 1
- 239000011710 vitamin D Substances 0.000 description 1
- 150000003710 vitamin D derivatives Chemical class 0.000 description 1
- MECHNRXZTMCUDQ-RKHKHRCZSA-N vitamin D2 Chemical compound C1(/[C@@H]2CC[C@@H]([C@]2(CCC1)C)[C@H](C)/C=C/[C@H](C)C(C)C)=C\C=C1\C[C@@H](O)CCC1=C MECHNRXZTMCUDQ-RKHKHRCZSA-N 0.000 description 1
- 229940046008 vitamin d Drugs 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 229930010764 α-maltose Natural products 0.000 description 1
- FYGDTMLNYKFZSV-BYLHFPJWSA-N β-1,4-galactotrioside Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@H](CO)O[C@@H](O[C@@H]2[C@@H](O[C@@H](O)[C@H](O)[C@H]2O)CO)[C@H](O)[C@H]1O FYGDTMLNYKFZSV-BYLHFPJWSA-N 0.000 description 1
Images



Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/52—Cytokines; Lymphokines; Interferons
- C07K14/54—Interleukins [IL]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P19/00—Drugs for skeletal disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P19/00—Drugs for skeletal disorders
- A61P19/08—Drugs for skeletal disorders for bone diseases, e.g. rachitism, Paget's disease
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Pharmacology & Pharmacy (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Animal Behavior & Ethology (AREA)
- Zoology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Toxicology (AREA)
- General Chemical & Material Sciences (AREA)
- Gastroenterology & Hepatology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Genetics & Genomics (AREA)
- Molecular Biology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Physical Education & Sports Medicine (AREA)
- Orthopedic Medicine & Surgery (AREA)
- Rheumatology (AREA)
- Immunology (AREA)
- Epidemiology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Description
【発明の属する技術分野】
この発明は、インターロイキン−18(以下、「IL−18」と略記する。)又はその機能性誘導体を含んでなる破骨細胞形成阻害剤に関する。
【0002】
【従来の技術】
健常な生体においては、骨芽細胞による骨形成と破骨細胞による骨吸収はバランスよく保たれ、その結果、骨組織は基本的形状を変えることなく常に新生骨と置換されて健常な状態が保たれている。また、このバランスは、血中のカルシウム濃度を一定に保つなど生体の恒常性維持のためにも重要な役割を果たしている。一方、このバランスが崩れた場合、特に骨吸収量が骨形成量を上回るような場合には、骨関連の疾患のみならずその他の種々の疾患が惹き起こされることとなる。したがって、生体における一連の骨吸収機構の解明、とりわけ、破骨細胞の形成機構の解明は、科学的に意義があるばかりではなく、臨床的にも極めて意義のあることであり、多大な注目を集めている。
【0003】
しかしながら、破骨細胞の形成の機構は、例えば、促進因子としてインターロイキン−1や阻害因子としてインターロイキン−4等が確認されているにも拘わらず、未だ完全に解明されたと言える状況にはない。これは、生体における破骨細胞の形成も、他の生体内での種々の現象と同様に、数多くの促進因子や阻害因子が複雑に且つ密接に関わり合って制御されているためと考えられている。これらのことから、科学的見地のみならず、臨床的見地からも、効果ある破骨細胞形成阻害剤の確立が希求されている。
【0004】
【発明が解決しようとする課題】
斯かる状況に鑑み、この発明の目的は、効果ある、新規な破骨細胞形成阻害剤を提供することにある。
【0005】
【課題を解決するための手段】
【0006】
IL−18は免疫系における情報伝達物質であるサイトカインの一種である。本サイトカインは、その発見当初には、特開平8−27189号公報、特開平8−193098号公報及びハルキ・オカムラら『ネイチャー』、第378巻、第6,552号、88乃至91頁(1995年)に見られるように、インターフェロン−γ誘導因子として記載されていたが、その後、シンペイ・ウシオら『ザ・ジャーナル・オブ・イムノロジー』、第156巻、4,274乃至4,279頁(1996年)における提案にしたがって、IL−18と呼称されるようになった。IL−18は、免疫担当細胞において生理活性物質として有用なインターフェロン−γ(以下、「IFN−γ」と略記する。)や顆粒球マクロファージコロニー刺激因子(以下、「GM−CSF」と略記する。)の産生を誘導する性質、さらには、キラー細胞の細胞障害性を増強したり、キラー細胞の生成を誘導する性質を兼備している。
【0007】
一方、本発明者らは、上記課題を解決すべく研究を進める過程で、イン・ビトロで前駆破骨細胞からの破骨細胞の形成を阻害する能力のある、マウス骨髄由来のある種のストローマ細胞株において、特定の遺伝子が特異的に多量に発現していることを見出した。そしてさらに詳細に解析したところ、当該遺伝子は、配列表における配列番号7に示すアミノ酸配列を有するIL−18をコードするものであることが判明した。これら知見に基づき、さらに鋭意研究したところ、当該アミノ酸配列を有するIL−18及びその機能性誘導体が破骨細胞の形成を顕著に阻害すること及び、この阻害は主として当該IL−18により誘導され産生されたGM−CSFの作用によっていることが判明した。この発明は、以上の独自の知見に基づき完成されたものである。
【0008】
すなわちこの発明は、上記の課題をIL−18又はその機能性誘導体を含んでなる破骨細胞形成阻害剤により解決するものである。
【0009】
【発明の実施の形態】
この発明の破骨細胞形成阻害剤はIL−18又はその機能性誘導体を含んでなるものである。ここでいうIL−18とは、IL−18としての上述の如き性質を有するポリペプチドすべてを包含するものであり、その出所・由来は問わない。この発明で用いるIL−18としては、例えば、中間部の部分アミノ酸配列として配列表における配列番号1、2及び3と、さらに配列番号4及び5に示すアミノ酸配列を有し、全体としては配列番号6又は7に示すアミノ酸配列を有しているIL−18を挙げることができる。また、本明細書でいう機能性誘導体とは、上述したようなIL−18の性質のうち、破骨細胞の形成を阻害する性質を実質的に失わない範囲で、そのアミノ酸配列におけるアミノ酸の1個又は2個以上を他のアミノ酸で置換したもの、これらのアミノ酸配列におけるN末端及び/又はC末端に1個又は2個以上のアミノ酸を付加したもの、これらアミノ酸配列における中間部に1個又は2個以上のアミノ酸を挿入したもの、これらアミノ酸配列におけるN末端及び/又はC末端のアミノ酸が1個又は2個以上欠失したもの、及び、これらアミノ酸配列における中間部のアミノ酸が1個又は2個以上欠失したものを意味する。斯かる機能性誘導体としては、例えば、同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書に、IL−18としての性質を実質的に保持しつつその安定性を向上せしめた機能性誘導体のアミノ酸配列について詳述されている。さらにまた、本明細書でいう機能性誘導体とは、以上のようなIL−18又はその機能性誘導体に糖鎖の付加されてなるポリペプチドをも包含する。以上のようなIL−18又はその機能性誘導体(以下、特に断らない限りIL−18とその機能性誘導体の両者を含めて「当該IL−18」と略記する。)は、細胞培養法等により天然の給源から分離したものであっても、組換えDNA技術やペプチド合成法により人工的に合成したものであっても構わない。
【0010】
経済的見地に立てば、組換えDNA技術による方法が有利であり、斯かる方法においては、通常、微生物又は動植物由来の適宜宿主に当該IL−18をコードするDNAを導入して形質転換体となし、これを常法により培養後、培養物をサイトカインを精製するための斯界における慣用の方法により精製して当該IL−18を得る。ここで用いられるDNAは、当該IL−18をコードするDNAを含んでなるものであればいずれでもよく、破骨細胞形成阻害剤の使用の目的や適用する方法に応じて適宜の配列のDNAを選択することができる。例えば、同じ特許出願人による特開平8−193098号公報、特開平8−231598号公報及び特開平8−27189号公報には、マウス及びヒト由来のIL−18をコードするcDNAを含むDNAを導入した形質転換体微生物を培養して得る当該IL−18の製造方法が、また、同じ特許出願人による特願平8−185305号(特願平9−187418号において優先権主張)の明細書には、ヒト由来のIL−18をコードする、染色体DNAを含むDNAを導入した形質転換体動物細胞を培養して得る当該IL−18の製造方法が詳述されている。さらに、同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書には、ヒトIL−18機能性誘導体をコードするDNAを含むDNAを導入した形質転換体動物細胞を培養して得る当該IL−18の製造方法が詳述されている。
【0011】
以上のような組換えDNA技術は、上述のように経済性において有利である反面、斯かる方法により得られる当該IL−18は、用いる宿主やDNAの配列によっては、本来生体内で産生され機能するIL−18とは理化学的性質に若干の相違の生じる場合もある。しかるに、同じ特許出願人による特願平8−67434号(特願平8−269105号(特開平9−289896号公報)において優先権主張)の明細書には、天然の給源として培養株化されたヒト細胞を用いたIL−18の製造方法が、また、同じ特許出願人による特願平8−213267号(特願平9−213885号において優先権主張)の明細書には、インターロイキン−1β変換酵素を用いたIL−18の製造方法が詳述されており、これらの方法により得られるIL−18は、生体内で産生され機能するIL−18と理化学的性質が実質的に同一か又は、極めて同一に近いと考えられるため、産生量としてはやや低いものの、例えば、ヒトを含む温血動物への投与を前提とする医薬品等として用いる場合には副作用の低さの点で有利である。なお、同じ特許出願人による特開平8−231598号公報に開示された、当該IL−18に対して特異的なモノクローナル抗体を用いた精製方法を適用するときには、高純度の当該IL−18を最小限のコストと労力で得ることができる。
【0012】
この発明の破骨細胞形成阻害剤は、斯くして得られる当該IL−18を含んでなるものであり、イン・ビトロ、イン・ビボを問わず、破骨細胞の形成を阻害するために用いられるすべての形態を包含する。例えば、破骨細胞の形成を阻害して、所望の細胞の維持、増殖及び/又は分化を良好ならしめる、動物細胞等の細胞培養用の培地成分として、骨関連の疾患剤のスクリーニング用キットの構成物として、骨吸収調整剤として、また、破骨細胞関連疾患剤として有利に用いることができる。ここでいう骨吸収調整剤とは、生体内での破骨細胞の形成を阻害することにより骨吸収を正常な域に調整して、比較的軽微な関節痛などの体調不良を改善する薬剤及び健康食品等を包含する。また、ここでいう破骨細胞関連疾患剤とは、生体内での破骨細胞の過剰な形成及び/又は機能に伴うすべての疾患を予防及び/又は治療するための薬剤を包含するものであり、対象疾患としては、例えば、高カルシウム血症、破骨細胞腫、ペーチェット病、骨肉腫、関節症、慢性関節リウマチ、変形性骨炎、原発性甲状腺機能亢進症、骨減少症、骨粗鬆症等を挙げることができる。剤型ならびに使用対象にもよるが、この発明の破骨細胞形成阻害剤は、通常、液状、ペースト状又は固状に調製され、当該IL−18を0.000002乃至100%(w/w)、望ましくは、0.0002乃至0.5%(w/w)含んでなる。
【0013】
この発明の破骨細胞形成阻害剤は当該IL−18単独の形態はもとより、それ以外の、例えば、担体、賦形剤、希釈剤、免疫助成剤、抗生物質、安定剤として血清アルブミンやゼラチンなどの蛋白質のほか、グルコース、マルトース、マルトトリオース、マルトテトラオース、トレハロース、スクロース、イソマルトース、ラクトース、パノース、エルロース、パラチノース、ラクトスクロース、ラフィノース、フラクトオリゴ、ガラクトオリゴ糖、レンチナン、デキストリン及びプルラン等の糖類やソルビトール、マルチトール、ラクチトール及びマルトトリイトール等の糖アルコール類を始めとする糖質、燐酸塩又はクエン酸塩を主体とする緩衝剤、2−メルカプトエタノール、ジチオトレイトール及び還元型グルタチオン等の還元剤、さらには、必要に応じて、インターフェロン−α、インターフェロン−β、インターフェロン−γ、インターロイキン2、インターロイキン3、インターロイキン6、インターロイキン12、TNF−α、TNF−β、GM−CSF、エストロゲン、プロジェステロン、酢酸クロルマジノン、カルシトニン、ソマトカイン、ソマトメジン、インスリン様成長因子、イプリフラボン、パラサイロイドホルモン、ノルエチステロン、ブスルファン、アンシタビン、シタラビン、フルオロウラシル、テトラヒドロフリルフルオロウラシル、メトトレキセート、ビタミンD2 、活性型ビタミンD、クレスチン、L−アスパラギナーゼ及びピシバニールを始めとする他の生理活性物質や、乳酸カルシウム、塩化カルシウム、燐酸水素カルシウム、L−アスパラギン酸カルシウムを始めとするカルシウム塩の1種又は2種以上との組成物としての形態をも包含する。なお、この発明の破骨細胞形成阻害剤を、ヒトを含む温血動物に投与する、例えば、破骨細胞関連疾患剤として用いる場合には、上記のうちから生理的に許容される物質を適宜に選んで組成物とするのが望ましい。
【0014】
さらに、この発明の破骨細胞形成阻害剤は、ヒトを含む温血動物に投与して使用する際の、投薬単位形態の薬剤をも包含し、その投薬単位形態の薬剤とは、当該IL−18を、例えば、1回当りの用量又はその整数倍(4倍まで)若しくはその約数(1/40まで)に相当する量を含んでなり、投薬に適する物理的に分離した一体の剤型にある薬剤を意味する。このような投薬形態の薬剤としては、注射剤、液剤、散剤、顆粒剤、錠剤、カプセル剤、舌下剤、点眼剤、点鼻剤、坐剤などが挙げられる。
【0015】
この発明の、破骨細胞形成阻害剤としての破骨細胞形成阻害剤は、経口的に投与しても非経口的に投与しても、いずれの場合にも、破骨細胞関連疾患の治療・予防に効果を発揮する。疾患の種類や症状にも依るが、具体的には、患者の症状や投与後の経過を観察しながら、成人当たり約0.5μg乃至100mg/回、望ましくは、約2μg乃至10mg/回の当該IL−18を2乃至6回/日又は2乃至10回/週の用量で1日乃至1年間に亙って経口投与するか、皮内、皮下、筋肉内又は静脈内に非経口投与すればよい。
【0016】
次に実験例を示し、当該IL−18の調製及び当該IL−18の理化学的性質並びに生物作用について説明する。
【0017】
【実験例1】
〈ヒトIL−18の調製〉
同じ特許出願人による特開平8−231598号公報に記載された方法にしたがって、ヒトIL−18をコードするcDNAが連結された自律複製可能な組換えDNA『pKGFHH2』を調製した。ジデオキシ法により解析したところ、図1に示すように、この組換えDNAにおいては、配列表における配列番号8に示す塩基配列を含むcDNA『KGFHH2 cDNA』がTacプロモータ『Ptac』の下流に連結されていた。また、この組換えDNA『pKGFHH2』は、配列表における配列番号1、2、3、4及び5に示したアミノ酸配列をコードする塩基配列を全て含むものであった。すなわち、これらアミノ酸配列はそれぞれ、その配列番号8に示した塩基配列における第46乃至63番目、第88乃至105番目、第400乃至420番目、第151乃至165番目及び第214乃至228番目の塩基によりコードされていた。
【0018】
さらに同じく特開平8−231598号公報に記載された方法にしたがって、この組換えDNA『pKGFHH2』を大腸菌Y1090株(ATCC 37197)に導入し、培養し、産生されたポリペプチドをイムノアフィニティークロマトグラフィーにより精製したところ、純度95%以上の精製ヒトIL−18が、培養液1l当たり約25mgの収量で得られた。この精製ヒトIL−18を、同じ特許出願人による特開平8−193098号公報に記載された方法にしたがって、次に示すように分析してその生物作用と理化学的性質を確認した。すなわち、この精製ヒトIL−18の各種濃度の存在下で常法により健常人より採取したヒトリンパ球を培養すると、存在させた精製ヒトIL−18の濃度に依存したIFN−γの産生が認められ、この精製IL−18が、免疫担当細胞であるリンパ球におけるIFN−γの産生を誘導する生物作用を有することが確認された。ユー・ケー・レムリが『ネイチャー』、第227巻、680乃至685頁(1970年)に報告した方法に準じて非還元条件下でSDS−ポリアクリルアミドゲル電気泳動すると、分子量18,500±3,000ダルトンに相当する位置にIFN−γ誘導能ある主たるバンドを示す一方、常法にしたがいクロマトフォーカシングすると、4.9±1.0に等電点を示した。また、そのN末端を常法にしたがいパーキン・エルマー製のプロテイン・シーケンサー『473A型』を用いて分析したところ、配列表の配列番号8に併記したアミノ酸配列におけるN末端にメチオニンが結合した配列番号9に示すアミノ酸配列を有していることが確認された。
【0019】
【実験例2】
〈ヒトIL−18の調製〉
同じ特許出願人による特願平8−67434号(特願平8−269105号(特開平9−289896号公報)において優先権主張)の明細書に記載の方法にしたがって、生後間もないハムスターの新生児の背部皮下にヒト急性単球性白血病由来の骨髄単球系細胞株の一種であるTHP−1細胞(ATCC TIB202)を移植し、3週間飼育した。そして、皮下に生じた腫瘍塊(約15g/匹)を摘出し、分散させた後、細胞を破砕し、産生されたポリペプチドをイムノアフィニティークロマトグラフィーにより精製して、精製ヒトIL−18をハムスター1匹当たり約50ngの収量で得た。
【0020】
この精製ヒトIL−18を、同じく特願平8−67434号の明細書に記載の方法にしたがって、次に示すように分析してその生物作用と理化学的性質を確認した。すなわち、この精製ヒトIL−18の各種濃度の存在下で常法により健常人より採取したヒトリンパ球を培養すると、存在させた精製ヒトIL−18の濃度に依存したIFN−γの産生が認められ、この精製IL−18が、免疫担当細胞であるリンパ球におけるIFN−γの産生を誘導する生物作用を有することが確認された。ユー・ケー・レムリが『ネイチャー』、第272巻、680乃至685頁(1970年)に報告した方法に準じて、還元剤として2%(w/v)ジチオトレイトール存在下でSDS−ポリアクリルアミドゲル電気泳動すると、分子量18,000乃至19,500ダルトンに相当する位置にIFN−γ誘導能ある主たるバンドを示した。さらに同じく特願平8−67434号の明細書にその方法が記載されているペプチドマップにしたがって、この精製ヒトIL−18をシグマ製クロストリパイン剤処理することにより得られるポリペプチド断片を、トーソー製『ODS−120T』のカラムを用いた高速液体クロマトグラフィーに供して分取し、個々のペプチド断片のアミノ酸配列をN末端側から分析したところ、配列表における配列番号10、11、12及び13に示すアミノ酸配列が得られた。これらアミノ酸配列は、それぞれ、配列表における配列番号6に示したアミノ酸配列における第148乃至157番目、第1乃至13番目、第45乃至58番目及び第80乃至96番目の配列と完全に一致した。以上の結果は、この実験例2の方法で得たIL−18が配列表における配列番号6に示すアミノ酸配列を有するものであり、その配列番号1、2、3、4及び5に示した部分アミノ酸配列を全て有していることを示している。
【0021】
【実験例3】
〈機能性誘導体の調製〉
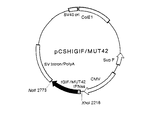
同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書に記載の方法にしたがって、配列表の配列番号6に示すアミノ酸配列における第38番目のシステインをセリンに、第68番目のシステインをセリンに、第76番目のシステインをアラニンに置換したヒトIL−18機能性誘導体をコードするDNAが連結された、自律複製可能な組換えDNA『pCSHIGIF/MUT35』を調製した。ジデオキシ法により解析したところ、図2に示すように、この組換えDNAにおいては、カーステン・ヘンコらが『ジャーナル・オブ・モレキュラー・バイオロジー』、第185巻、227乃至260頁(1985年)に報告しているヒトインターフェロン−αにおけるサブタイプα2bのシグナルペプチドをコードする塩基配列の下流に、同じ読み枠で、配列表における配列番号14に示した塩基配列を有するDNA『IGIF/MUT35』が連結され、さらにその下流に蛋白質合成の終始コドンが存在していた。このDNAがコードするアミノ酸配列は、配列表における配列番号14に併記したごとく、配列表における配列番号6におけるアミノ酸配列の第38番目のシステインをセリンに、第68番目のシステインをセリンに、第76番目のシステインをアラニンに置換してなるものであった。また、このDNAは、配列表における配列番号1、2、3及び4に示したすべてのアミノ酸配列と、配列番号5に示したアミノ酸配列の第5のアミノ酸のシステインがアラニンに置換されたアミノ酸配列をコードする塩基配列を含むものであった。すなわち、これらアミノ酸配列はそれぞれ、その配列番号14に示した塩基配列における第46乃至63番目、第88乃至105番目、第400乃至420番目、第151乃至165番目及び第214乃至228番目の塩基によりコードされていた。
【0022】
さらに同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書に記載の方法にしたがって、この組換えDNA『pCSHIGIF/MUT35』を、アフリカミドリザルの腎臓に由来する株化細胞の一種であるCOS−1細胞(ATCC CRL1650)に導入し、培養し、産生されたポリペプチドをイムノアフィニティークロマトグラフィーにより精製して、精製ヒトIL−18機能性誘導体を培養培地1ml当たり約40ngの収量で得た。そしてさらに、同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書に記載の方法にしたがって、この精製ヒトIL−18機能性誘導体を、次に示すように分析してその生物作用と理化学的性質を確認した。すなわち、この精製ヒトIL−18機能性誘導体の各種濃度の存在下でヒト急性骨髄性白血病に由来する株化細胞の一種であるKG−1細胞(ATCC CCL246)を培養すると、存在させた精製ヒトIL−18機能性誘導体の濃度に依存したIFN−γの産生が認められ、この精製IL−18機能性誘導体が、免疫担当細胞としてのKG−1細胞におけるIFN−γの産生を誘導する生物作用を有することが確認された。ユー・ケー・レムリが『ネイチャー』、第227巻、680乃至685頁(1970年)に報告した方法に準じて、還元剤として2%(w/v)ジチオトレイトール存在下でSDS−ポリアクリルアミドゲル電気泳動すると、分子量18,000乃至19,500ダルトンに相当する位置にIFN−γ誘導能ある主たるバンドを示した。また、そのN末端を常法にしたがいパーキン・エルマー製のプロテイン・シーケンサー『473A型』を用いて分析したところ、配列表の配列番号14に併記したアミノ酸配列におけるN末端部分のアミノ酸配列と完全に一致する、配列表における配列番号15に示すアミノ酸配列を有していることが確認された。
【0023】
【実験例4】
〈機能性誘導体の調製〉
同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書に記載の方法にしたがって、配列表の配列番号6に示すアミノ酸配列における第38番目のシステインをセリンに、第68番目のシステインをセリンに、第76番目のシステインをアラニンに、第127番目のシステインをセリンに置換したヒトIL−18機能性誘導体をコードするDNAが連結された、自律複製可能な組換えDNA『pCSHIGIF/MUT42』を調製した。ジデオキシ法により解析したところ、図3に示すように、この組換えDNAにおいては、カーステン・ヘンコらが『ジャーナル・オブ・モレキュラー・バイオロジー』、第185巻、227乃至260頁(1985年)に報告しているヒトインターフェロン−αにおけるサブタイプα2bのシグナルペプチドをコードする塩基配列の下流に、同じ読み枠で、配列表における配列番号16に示した塩基配列を有するDNA『IGIF/MUT42』が連結され、さらにその下流に蛋白質合成の終始コドンが存在していた。このDNAがコードするアミノ酸配列は、配列表における配列番号16に併記したごとく、その配列番号6におけるアミノ酸配列の第38番目のシステインをセリンに、第68番目のシステインをセリンに、第76番目のシステインをアラニンに、第127番目のシステインをセリンに置換してなるものであった。また、このDNAは、配列表における配列番号1、2、3及び4に示したすべてのアミノ酸配列と、配列表における配列番号5に示したアミノ酸配列の第5のアミノ酸のシステインがアラニンに置換されたアミノ酸配列をコードする塩基配列を含むものであった。すなわち、これらアミノ酸配列はそれぞれ、配列表における配列番号16に示した塩基配列における第46乃至63番目、第88乃至105番目、第400乃至420番目、第151乃至165番目及び第214乃至228番目の塩基によりコードされていた。
【0024】
さらに同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書に記載の方法にしたがって、この組換えDNA『pCSHIGIF/MUT42』を、COS−1細胞に導入し、培養し、産生されたポリペプチドをイムノアフィニティークロマトグラフィーにより精製して、精製ヒトIL−18機能性誘導体を培養培地1ml当たり約20ngの収量で得た。そしてさらに、同じ特許出願人による特願平9−20906号(特願平9−329715号において優先権主張)の明細書に記載の方法にしたがって、この精製ヒトIL−18機能性誘導体を、次に示すように分析してその生物作用と理化学的性質を確認した。すなわち、この精製ヒトIL−18機能性誘導体の各種濃度の存在下でKG−1細胞を培養すると、存在させた精製ヒトIL−18機能性誘導体の濃度に依存したIFN−γの産生が認められ、この精製IL−18機能性誘導体が、免疫担当細胞としてのKG−1細胞におけるIFN−γの産生を誘導する生物作用を有することが確認された。ユー・ケー・レムリが『ネイチャー』、第227巻、680乃至685頁(1970年)に報告した方法に準じて、還元剤として2%(w/v)ジチオトレイトール存在下でSDS−ポリアクリルアミドゲル電気泳動すると、分子量18,000乃至19,500ダルトンに相当する位置にIFN−γ誘導能ある主たるバンドを示した。また、そのN末端を常法にしたがいパーキン・エルマー製のプロテイン・シーケンサー『473A型』を用いて分析したところ、配列表の配列番号16に併記したアミノ酸配列におけるN末端部分のアミノ酸配列と完全に一致する、配列表における配列番号15に示すアミノ酸配列を有していることが確認された。
【0025】
【実験例5】
〈ヒトIL−18の調製〉
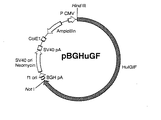
同じ特許出願人による特願平8−185305号(特願平9−187418号において優先権主張)の明細書に記載の方法にしたがって、ヒトIL−18をコードする染色体DNAが連結された、自律複製可能な組換えDNA『pBGHuGF』を調製した。ジデオキシ法により解析したところ、図4に示すようにこの組換えDNAにおいては、ヒトIL−18をコードする染色体DNAである、配列表における配列番号17に示す塩基配列のDNA『HuIGIF』が、制限酵素Hind IIIによる切断部位の下流に連結されていた。配列表における配列番号17に示すように、この染色体DNA『HuIGIF』は11,464bpよりなり、この配列の5′末端より第83乃至1,453番目、第1,466乃至4,848番目、第4,984乃至6,317番目及び第6,452乃至11,224番目に位置する4個のイントロンによりエクソンが分断されている構成であった。これらイントロンを除いた配列のうち、5′末端より第3乃至11443番目の塩基はヒトIL−18前駆体をコードする部分であり、さらにこの内、第4866乃至4983番目の塩基は活性あるヒトIL−18をコードする部分であった。また、このDNAは、配列表における配列番号1、2、3、4及び5に示したアミノ酸配列をコードする塩基配列を全て含むものであった。すなわち、これらアミノ酸配列はそれぞれ、その配列番号17に示す塩基配列における第4,911乃至4,928番目、第4,953乃至4,970番目、第11,372乃至11,392番目、第6,350乃至6,364番目及び第6,413乃至6,427番目の塩基によりコードされていた。
【0026】
さらに同じ特許出願人による特願平8−185305号(特願平9−187418号において優先権主張)の明細書に記載の方法にしたがって、この組換えDNA『pBGHuGF』を、チャイニーズ・ハムスター卵巣細胞由来の株化細胞であるCHO−K1細胞(ATCC CCL61)に導入し、培養し、培養上清にTHP−1細胞を培養して得た細胞破砕物の上清を作用させて、生成したポリペプチドをイムノアフィニティークロマトグラフィーにより精製して、精製ヒトIL−18を培養物1l当たり約15mgの収量で得た。そしてさらに、この精製ヒトIL−18を、同じ特許出願人による特願平8−185305号(特願平9−187418号において優先権主張)の明細書に記載の方法にしたがって、次に示すように分析してその生物作用と理化学的性質を確認した。すなわち、この精製ヒトIL−18の各種濃度の存在下で常法により健常人より採取したヒトリンパ球を培養すると、存在させた精製ヒトIL−18の濃度に依存したIFN−γの産生が認められ、この精製IL−18が、免疫担当細胞であるリンパ球におけるIFN−γの産生を誘導する生物作用を有することが確認された。ユー・ケー・レムリが『ネイチャー』、第227巻、680乃至685頁(1970年)に報告した方法に準じて、還元剤として2%(w/v)ジチオトレイトール存在下でSDS−ポリアクリルアミドゲル電気泳動すると、分子量18,000乃至19,500ダルトンに相当する位置にIFN−γ誘導能ある主たるバンドを示した。また、そのN末端は、配列表の配列番号17に併記したアミノ酸配列のうち、活性あるIL−18のN末端部分のアミノ酸配列と完全に一致する配列表における配列番号15に示すアミノ酸配列を有していた。
【0027】
【実験例6】
〈マウスIL−18の調製〉
0.5ml容反応管に25mM塩化マグネシウムを8μl、10×PCR緩衝液を10μl、25mM dNTPミックスを1μl、2.5単位/μlアンプリタックDNAポリメラーゼを1μl、特開平8−27189号公報に記載された方法にしたがってファージDNAクローンから調製した、配列表における配列番号18に示す塩基配列を有し、配列番号7に示すアミノ酸配列のマウスIL−18をコードするDNAを含む組換えDNAを1ng、配列表の配列番号7におけるN末端及びC末端付近のアミノ酸配列に基づき化学合成した5´−ATAGAATTCAAATGAACTTTGGCCGACTTCACTG−3´及び5´−ATAAAGCTTCTAACTTTGATGTAAGTT−3´で表わされる塩基配列のセンスプライマー及びアンチセンスプライマーの適量を加え、滅菌蒸留水で100μlとした。常法により、この混合物を94℃で1分間、43℃で1分間、72℃で1分間、この順序でインキュベートするサイクルを3回繰返した後、さらに、94℃で1分間、60℃で1分間、72℃で1分間、この順序でインキュベートするサイクルを40回繰返してPCR反応させた。
【0028】
このPCR産物とストラタジーン製プラスミドベクター『pCR−Script SK (+)』を常法にしたがってDNAリガーゼにより連結して組換えDNAとし、これをコンピテントセル法によりストラタジーン製大腸菌株『XL−1 Blue MRF´Kan』に導入して形質転換した。形質転換体を50μg/mlアンピシリンを含むL−ブロス培地(pH7.2)に接種し、37℃で18時間振盪培養した後、培養物を遠心分離して形質転換体を採取し、通常のアルカリ−SDS法を適用して組換えDNAを単離した。この組換えDNAの一部をとり、ジデオキシ法により分析したところ、配列表の配列番号18に示す塩基配列における5´末端及び3´末端にそれぞれEco RI切断部位及びHind III切断部位を含み、さらに、その配列番号18に併記したアミノ酸配列におけるN末端及びC末端のそれぞれ直前及び直後に対応する部位にポリペプチド合成開始のためのメチオニンコドン及びポリペプチド合成終止のためのTAGコドンを有するDNAを含んでいた。また、この組換えDNAは、配列表における配列番号1、2、3、4及び5に示したアミノ酸配列をコードする塩基配列を全て含むものであった。すなわち、これらアミノ酸配列はそれぞれ、その配列番号18に示した塩基配列における第46乃至63番目、第85乃至102番目、第394乃至414番目、第148乃至162番目及び第211乃至225番目の塩基によりコードされていた。
【0029】
そこで、常法にしたがって残りの組換えDNAを制限酵素Eco RI及びHind IIIで切断後、宝酒造製DNAライゲーションキット『DNAライゲーション・キット・バージョン2』を使用して、得られたEco RI−Hind III DNA断片0.1μgと予め同じ制限酵素で切断しておいたファルマシア製プラスミドベクター『pKK223−3』10ngを16℃で30分間反応させて連結して複製可能な組換えDNA『pKGFMH2』を得た。コンピテントセル法により、この組換えDNA pKGFMH2で大腸菌Y1090株(ATCC37197)を形質転換し、得られた形質転換体『KGFMH2』を50μg/mlアンピシリンを含むL−ブロス培地(pH7.2)に接種し、37℃で18時間振盪培養した。培養物を遠心分離して形質転換体を採取し、その一部に通常のSDS−アルカリ法を適用して組換えDNA『pKGFMH2』を抽出した。ジデオキシ法により分析したところ、図5に示すように、組換えDNA『pKGFMH2』においては、配列表における配列番号18に示す塩基配列を含むcDNA『KGFMH2 cDNA』がTacプロモータ『Ptac』の下流に連結されていた。
【0030】
オートクレーブにより滅菌したL−ブロス培地(pH7.2)に、アンピシリンを濃度50μg/mlとなるように添加し、37℃に冷却後、形質転換体KGFMH2を接種し、振盪下、37℃で18時間種培養した。20l容ジャーファーメンタに新鮮な同一培地を18lとり、同様に滅菌し、アンピシリンを添加し、37℃に冷却後、上記で得た種培養物を1%(v/v)接種し、37℃で8時間通気撹拌培養した。培養物を遠心分離して菌体を採取し、150mM塩化ナトリウム、16mM燐酸水素二ナトリウム及び4mM燐酸二水素ナトリウムを含む混液(pH7.3)に浮遊させ、超音波破砕後、遠心分離により菌体破砕物を除去し、上清約2lを採取した。
【0031】
得られた上清約2lに硫酸アンモニウムを含む10mM燐酸緩衝液(pH7.3)を硫酸アンモニウムが40%飽和になるように加え、沈殿物を遠心分離にて除去後、さらに上清に硫酸アンモニウムが85%飽和になるまで加え、4℃で18時間放置後、約8,000rpmで30分間遠心分離して沈澱を採取した。次にこの沈澱を1.5M硫酸アンモニウムを含む10mM燐酸緩衝液(pH6.6)に溶解して約1,300mlとし、濾過後、予め新鮮な同一緩衝液で平衡化させておいたファルマシア製『フェニルセファロース CL−4B』約800mlのカラムに負荷し、カラムを新鮮な同一緩衝液で洗浄後、1.5Mから0Mに下降する硫酸アンモニウムの濃度勾配下、10mM燐酸緩衝液(pH6.6)をSV1.5で通液した。硫酸アンモニウム濃度が1M付近のときに溶出された画分を採取し、膜濃縮し、10mM燐酸緩衝液(pH6.5)に対して4℃で18時間透析後、予め10mM燐酸緩衝液(pH6.5)で平衡化させておいたファルマシア製『DEAE−5PW』約55mlのカラムに負荷した。カラムを新鮮な同一緩衝液で洗浄後、0Mから0.5Mに上昇する塩化ナトリウムの濃度勾配下、カラムに10mM燐酸緩衝液(pH6.5)をSV5.5で通液し、塩化ナトリウム濃度が0.2M付近で溶出された画分を採取した。その後、採取した溶出液を上記と同様に濃縮して約9mlとし、PBSに対して4℃で18時間透析後、予め新鮮なPBSで平衡化させておいたファルマシア製『スーパーデックス 75』のカラムに負荷した。さらにカラムに新鮮なPBSを通液してIFN−γ誘導能ある画分を採取し、膜濃縮して精製マウスIL−18を得た。この精製マウスIL−18の収量は、培養液1l当たり約350μgであった。
【0032】
この精製マウスIL−18を、特開平8−27189号公報に記載の方法にしたがって、次に示すように分析してその生物作用と理化学的性質を確認した。すなわち、この精製マウスIL−18の各種濃度の存在下で常法により採取したマウス脾細胞を培養すると、存在させた精製マウスIL−18の濃度に依存したIFN−γの産生が認められ、この精製IL−18が、免疫担当細胞である牌細胞におけるIFN−γの産生を誘導する生物作用を有することが確認された。ユー・ケー・レムリが『ネイチャー』、第227巻、680乃至685頁(1970年)に報告した方法に準じて、非還元条件下でSDS−ポリアクリルアミドゲル電気泳動すると、分子量19,000±5,000ダルトンに相当する位置にIFN−γ誘導能ある主たるバンドを示した。また、そのN末端は、配列表の配列番号18に併記したアミノ酸配列におけるN末端部に相当する、配列番号19に示すアミノ酸配列を有していた。
【0033】
次に実験例7を示してこの発明で用いるIL−18の生物作用についてさらに詳細に説明し、実験例8を示してその毒性について説明する。
【0034】
【実験例7】
〈生物作用〉
【0035】
【実験例7−1】
〈GM−CSFの産生の誘導〉
ヘパリン加注射器により健常者から血液を採取し、血清無含有のRPMI1640培地(pH7.4)により2倍希釈した。血液をフィコール上に重層し、遠心分離して採取したリンパ球を10%(v/v)ウシ胎児血清を補足したRPMI1640培地(pH7.4)により洗浄した後、新鮮な同一培地に細胞密度1×106 個/mlになるように浮遊させ、12ウェルプレートに2ml/ウェルずつ分注した。
【0036】
別途、実験例1の方法により得たIL−18を10%(v/v)ウシ胎児血清を補足したRPMI1640培地(pH7.4)により濃度1μg/mlに調製して、上記プレートに20乃至200μl/ウェルずつ分注し、500μg/mlのコンカナバリンAを含む新鮮な上記と同一培地を10μl/ウェルずつ加えた後、5%CO2 インキュベータ中、37℃で48時間培養した。培養後、各ウェルから培養上清を0.1mlずつ採取し、通常の酵素免疫測定法によりGM−CSF含量を測定した。同時に、IL−18のみを省略した系を設け、上記と同様に処置して対照とした。結果を表1に示す。
【0037】
【表1】

表1の結果は、補因子としてコンカナバリンAの共存下でIL−18を作用させると、免疫担当細胞としてのリンパ球がIL−18の濃度に依存してGM−CSFを産生したことを示している。なお、実験例2乃至5の方法で得たIL−18乃至その機能性誘導体をそれぞれ別個に同様にこの操作に供した場合にも、いずれも同じくGM−CSFの産生を誘導することが確認された。一方、実験例6の方法で得たIL−18については、実験例7−1で用いたヒトの血液より採取したリンパ球に代えて常法によりマウスより採取した脾細胞を用いたこと以外は実験例7−1に準じて試験したところ、同じくGM−CSFの産生を誘導することが確認された。
【0039】
【実験例7−2】
〈破骨細胞形成の阻害〉
【0040】
【実験例7−2(a)】
ティー・ジェー・マーチンら、『ジャーナル・オブ・セルラー・バイオケミストリー』、第56巻(1994年)、357乃至366頁等にも記載されているように、一般に破骨細胞の前駆細胞が分化し破骨細胞が形成されるためには、骨髄の造血幹細胞に由来する前駆破骨細胞が骨芽細胞や骨髄ストローマ細胞と接触することが必要条件であるとされている。また、ジー・ディー・ルードマン、『エンドクリン・レビュー』、第17巻(1996年)、308乃至332頁等に記載されているように、破骨細胞の特徴は、多核であること、酒石酸耐性酸性ホスファターゼ(以下、「TRAP」と略記する。)活性を有すこと、カルシトニン・レセプターを有すこと等であると一般に認識されている。一方、エヌ・ウダガワら、『ジャーナル・オブ・エキスペリメンタル・メディシン』、第182巻(1995年)、1461乃至1468頁に記載の骨芽細胞と骨髄細胞の共培養系においては、例えば、1α,25−ジヒドロキシビタミンD3、プロスタグランジンE2、副腎皮質ホルモン、インターロイキン1、インターロイキン6又はインターロイキン11のうちのいずれかの因子に応答して破骨細胞様の細胞(以下、「OCL」と略記することもある。)の形成が認められる。ここで形成されるOCLは生体内の破骨細胞の特徴を備えている。したがってこの共培養系は生体内における破骨細胞の形成の過程をイン・ビトロでよく再現するものであり、この系を用いることにより破骨細胞の形成やそれに対する阻害剤についての実験を行うことができる。
【0041】
この共培養系を用いて、当該IL−18の破骨細胞形成の阻害作用を調べた。骨芽細胞は、常法に従い新生マウスの頭蓋冠を0.1%(w/v)コラゲナーゼ(オーストラリア国、ワーシントン・バイオケミカル・カンパニー製)及び0.2%(w/v)ディスパーゼ(合同酒精製)処理して調製した。骨髄細胞は、常法に従い成熟したマウスより調製した。48ウエル・プレートの1ウエル当たり、10%(v/v)ウシ胎児血清を補足したα−MEM培地(以下、この実験例4−2を通して、単に「培地」という。)0.4mlで、2×104 個の初代骨芽細胞と5×105 個の骨髄細胞を、5%CO2 インキュベーター内で37℃で7日間共培養した。これを陰性対照とした。一方、別のウエルでは、1α,25−ジヒドロキシビタミンD3(和光純薬製)及びプロスタグランジンE2(アメリカ国、シグマ製)を、濃度がそれぞれ10-8M及び10-7Mとなるように添加した培地を用いた以外は全て陰性対照と同一の方法で培養した。これを陽性対照とした。また、さらに別のウエルでは、陽性対照と同濃度の1α,25−ジヒドロキシビタミンD3及びプロスタグランジンE2とともに実験例6の方法で調製したIL−18を0.01乃至10ng/mlのいずれかの濃度となるように添加した培地を用いた以外は全て陽性対照と同一の方法で培養した。いずれの場合も培養3日目に、それぞれのウェルでそれまでに用いていたのと同一組成の新鮮な培地と交換した。6日間培養した後の細胞を、エヌ・ウダガワら、『ジャーナル・オブ・エキスペリメンタル・メディシン』、第182巻(1995年)、1461乃至1468頁に記載の方法に従って、固定し、TRAP活性に基づき染色し、1ウエルあたり染色された細胞(以下、「TRAP陽性細胞」という。)の数を数えた。この実験例4−2を通して、全て同一の培養系を4ウエルずつ設け、1ウエル当たりのTRAP陽性細胞数の平均値を求めた。結果を表2に示す。
【0042】
【表2】
表2に示すように、陰性対照ではTRAP陽性細胞の形成はほとんど認められなかったのに対し、陽性対照ではTRAP陽性細胞の形成は顕著であった。一方、陽性対照にさらにIL−18を添加した系では、その濃度に依存してTRAP陽性細胞の形成が阻害され、IL−18濃度が8ng/ml以上のときその阻害は最大で、TRAP陽性細胞数は陰性対照と同等の値となった。以上のことは、当該IL−18には確かにイン・ビトロにおけるOCL形成を阻害する作用のあることを示しており、さらに当該IL−18が破骨細胞形成を阻害することをも強く示唆している。
【0044】
【実験例7−2(b)】
先にも述べたように、この実験例7−2を通して用いられる共培養系において破骨細胞様の細胞の形成を促す因子には種々のものがあると確認されている。そこでこの実験例7−2(b)では、実験例7−2(a)で示された破骨細胞形成に対する当該IL−18の阻害作用が、ある種の因子に特異的なものか否かを調べた。すなわち、1α,25−ジヒドロキシビタミンD3 、プロスタグランジンE2 、副甲状腺ホルモン、インターロイキン1又はインターロイキン11のいずれかを、それぞれ濃度10-8M、10-7M、200ng/ml、100ng/ml又は20ng/mlとなるように添加した培地を用いた以外は、全て実験例7−2(a)の陰性対照と同一の方法で培養した。これらを陽性対照とした。一方、別のウエルでは、この陽性対照と同濃度のいずれかの因子に加え、さらに実験例6の方法で得たIL−18を10ng/mlとなるように添加した培地を用いた以外は全て陽性対照と同一の方法で培養した。培養後実験例7−2(a)と同じくTRAP陽性細胞の数を比較した。結果を表3に示す。
【0045】
【表3】
表3に示すように、いずれも陽性対照では顕著なTRAP陽性細胞の形成を認めた。これに対し、陽性対照にIL−18を添加した場合にはいずれもTRAP陽性細胞の形成はほぼ完全に阻害された。このことは、当該IL−18が破骨細胞形成の要因によらず、広く一般的に破骨細胞の形成を阻害する作用を有することを強く示唆している。
【0047】
【実験例7−2(c)】
次に、以上実験例7−2(a)及び実験例7−2(b)により確認された、当該IL−18による破骨細胞の形成の阻害が、当該IL−18により産生の誘導されたGM−CSFの作用によるものか否かを調べた。陰性対照及び陽性対照は、それぞれ実験例7−2(a)で示したのと同一の系を用いた。別のウエルでは、この陽性対照と同じ濃度の1α,25−ジヒドロキシビタミンD3及びプロスタグランジンE2を添加するとともに、さらに(i)抗マウスGM−CSFポリクローナル抗体(アメリカ国、アール・アンド・ディー・システムズ製)を濃度10μg/mlとなるように添加するか、(ii)実験例6の方法で得たIL−18を濃度10ng/mlとなるよう添加するか、(iii)iiに加えさらに抗マウスGM−CSFポリクローナル抗体を濃度10μg/mlとなるよう添加するか、(iv)マウスGM−CSF(アメリカ国、アール・アンド・ディー・システムズ製)を濃度0.1ng/mlとなるように添加するか、又は(v)ivに加えさらに抗マウスGM−CSFポリクローナル抗体を濃度10μg/mlとなるように添加した培地のいずれかを用いたこと以外は、全て陽性対照と同じ方法で培養した。培養後実験例7−2(a)と同じくTRAP陽性細胞の数を比較した。結果を表4に示す。なお、表4中に示したi乃至vの符号は、ここで説明した対照系以外の培養系の符号と一致している。
【0048】
【表4】
表4に示すように、TRAP陽性細胞の形成はIL−18によりほぼ完全に阻害された(培養系ii)が、この阻害は、抗マウスGM−CSFポリクローナル抗体の添加によりほぼ完全に解除された(培養系iii)。一方マウスGM−CSFにも、IL−18と同様にTRAP陽性細胞の形成を阻害する作用が認められた(培養系iv)が、この阻害は、抗マウスGM−CSFポリクローナル抗体の添加によりほぼ完全に解除された(培養系v)。また、当該抗体単独ではTRAP陽性細胞の形成には何の影響も与えなかった(培養系i)。以上の結果は、当該IL−18の破骨細胞形成に対する阻害は、主として、当該IL−18により誘導され産生したGM−CSFの作用によるものであることを強く示唆している。
【0050】
【実験例8】
〈急性毒性試験〉
常法にしたがって、8週齢のマウスに実験例1乃至6の方法で得た当該IL−18のいずれかをそれぞれ別個に経皮、経口又は腹腔内に注射投与した。その結果、これら当該IL−18のLD50は、いずれの投与経路によっても約1mg/kgマウス体重以上であった。このことは当該IL−18がヒトを始めとする温血動物への投与を前提とする医薬品に配合して安全であることを裏付けている。
【0051】
また、当該IL−18により誘導され産生されるGM−CSFは、日経BP社発行、『日経バイオ年鑑96』(1995年)、498乃至499頁に記載されているように、日本国内ではまだ臨床応用されるに至ってはいないものの、米国や欧州では既に臨床応用されており、その安全性については問題がないといえる。以上のことは、この発明の破骨細胞形成阻害剤が重篤な副作用を惹起することなくヒトを始めとする温血動物に長期連用でき、破骨細胞の形成及び/又は機能が関与する疾患の治療・予防に効果を発揮することを示している。
【0052】
以下に実施例を示し、この発明の破骨細胞形成阻害剤を説明する。
【0053】
【実施例1】
〈液剤〉
安定剤として1%(w/v)ヒト血清アルブミンを含む生理食塩水に実験例1乃至6の方法により得たいずれかの当該IL−18を2mg/mlになるように溶解し、常法にしたがって精密濾過により滅菌して液剤を得た。
【0054】
本品はいずれも安定性に優れ、細胞培養用の培地成分として有用であり、また、骨吸収調整剤や、高カルシウム血症、破骨細胞腫及び骨粗鬆症等を治療・予防するための破骨細胞関連疾患剤としての注射剤、点眼剤、点鼻剤として有用である。
【0055】
【実施例2】
〈乾燥剤〉
安定剤として1%(w/v)精製ゼラチンを含む生理食塩水100mlに実験例1乃至6の方法により得たいずれかの当該IL−18を50mg溶解し、常法にしたがって精密濾過により滅菌し、バイアル瓶に1mlずつ分注し、凍結乾燥後、密栓した。
【0056】
本品はいずれも安定性に優れ、細胞培養用の培地成分として有用であり、また、骨吸収調整剤や、高カルシウム血症、破骨細胞腫及び骨粗鬆症等を治療・予防するための破骨細胞関連疾患剤としての乾燥注射剤として有用である。
【0057】
【実施例3】
〈乾燥剤〉
安定剤として1%(w/v)トレハロースを含む生理食塩水100mlに実験例1乃至6の方法により得たいずれかの当該IL−18を50mg溶解し、常法にしたがって精密濾過により滅菌し、バイアル瓶に1mlずつ分注し、凍結乾燥後、密栓した。
【0058】
本品はいずれも安定性に優れ、細胞培養用の培地成分として有用であり、また、骨吸収調整剤や、高カルシウム血症、破骨細胞腫及び骨粗鬆症等を治療・予防するための破骨細胞関連疾患剤としての乾燥注射剤として有用である。
【0059】
【実施例4】
〈軟膏剤〉
滅菌蒸留水に和光純薬工業製カルボキシビニルポリマー『ハイビスワコー104』及び高純度トレハロースをそれぞれ濃度1.4%(w/w)及び2.0%(w/w)になるように溶解し、実験例1乃至6の方法により得たいずれかの当該IL−18を均一に混合後、pH7.2に調製して、1g当たり当該IL−18を約1mg含むペースト状物を得た。
【0060】
本品はいずれも延展性と安定性に優れ、骨吸収調整剤や、高カルシウム血症、破骨細胞腫及び骨粗鬆症等を治療・予防するための破骨細胞関連疾患剤としての軟膏剤として有用である。
【0061】
【実施例5】
〈錠剤〉
林原製無水結晶α−マルトース粉末『ファイントース』に実験例1乃至6の方法により得たいずれかの当該IL−18と細胞賦活剤としてのルミンを均一に混合し、得られる混合物を常法により打錠して製品1錠(約200mg)当たり当該IL−18及びルミンをそれぞれ約2mg含む錠剤を得た。
【0062】
本品はいずれも摂取性、安定性に優れ、しかも細胞賦活作用をも有し、骨吸収調整剤や、高カルシウム血症、破骨細胞腫及び骨粗鬆症等を治療・予防するための破骨細胞関連疾患剤としての錠剤として有用である。
【0063】
【発明の効果】
以上説明したように、この発明の破骨細胞形成阻害剤は、イン・ビトロ、イン・ビボを問わず、破骨細胞の形成を顕著に阻害するので、細胞培養用の培地成分として有用であり、また、骨吸収調整剤や、高カルシウム血症、破骨細胞腫及び骨粗鬆症を始めとする破骨細胞関連疾患を治療・予防するための疾患剤としても効果を発揮する。
【0064】
この発明は、斯くも顕著な作用効果を発揮するものであり、斯界に貢献すること誠に多大な意義のある発明といえる。
【0065】
【配列表】
【図面の簡単な説明】
【図1】組換えDNA pKGFHH2の構造を示す図である。
【図2】組換えDNA pCSHIGIF/MUT35の構造を示す図である。
【図3】組換えDNA pCSHIGIF/MUT42の構造を示す図である。
【図4】組換えDNA pBGHuGFの構造を示す図である。
【図5】組換えDNA pKGFMH2の構造を示す図である。
【符号の説明】
KGFHH2 cDNA 当該IL−18をコードするcDNA
IGIF/MUT35 当該IL−18をコードするDNA
IGIF/MUT42 当該IL−18をコードするDNA
HuIGIF 当該IL−18をコードする染色体DNA
KGFMH2 cDNA 当該IL−18をコードするcDNA
5S 5SリボゾームRNA遺伝子
Ptac tacプロモータ
rrnBT1T2 リボゾームRNAオペロンの転写終止領域
AmpR アンピシリン耐性遺伝子
pBR322ori 大腸菌における複製開始点
CMV サイトメガロウイルス・プロモーター
IFNss ヒトインターフェロン−αにおけるサブタイプ
α2bのシグナルペプチドをコードする配列
Claims (8)
- インターロイキン−18、又は、破骨細胞の形成を阻害する性質を実質的に失わない範囲で、インターロイキン−18のアミノ酸配列におけるアミノ酸の1個又は2個以上を他のアミノ酸で置換したもの、N末端及び/又はC末端に1個又は2個以上のアミノ酸を付加したもの、中間部に1個又は2個以上のアミノ酸を挿入したもの、N末端及び/又はC末端のアミノ酸が1個又は2個以上欠失したもの、又は、中間部のアミノ酸が1個又は2個以上欠失したものを含んでなる破骨細胞形成阻害剤。
- インターロイキン−18が部分アミノ酸配列として配列表における配列番号1、2及び3に示すアミノ酸配列を有している請求項1に記載の破骨細胞形成阻害剤。
- インターロイキン−18が部分アミノ酸配列として配列表における配列番号4及び5に示すアミノ酸配列を有している請求項1又は2に記載の破骨細胞形成阻害剤。
- インターロイキン−18が配列表における配列番号6に示すアミノ酸配列を有している請求項1、2又は3に記載の破骨細胞形成阻害剤。
- インターロイキン−18がヒト起源である請求項1、2、3又は4に記載の破骨細胞形成阻害剤。
- インターロイキン−18が配列表における配列番号7に示すアミノ酸配列を有している請求項1、2又は3に記載の破骨細胞形成阻害剤。
- 生体内での破骨細胞の過剰な形成及び/又は機能に伴う疾患を治療するための薬剤としての請求項1、2、3、4、5又は6に記載の破骨細胞形成阻害剤。
- 安定剤として、蛋白質、緩衝剤又は糖質をさらに含んでなる請求項1、2、3、4、5、6又は7に記載の破骨細胞形成阻害剤。
Priority Applications (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP05546897A JP3955352B2 (ja) | 1997-02-25 | 1997-02-25 | 破骨細胞形成阻害剤 |
| TW087102640A TW570801B (en) | 1997-02-25 | 1998-02-24 | Osteoclastgenic inhibitory agent |
| DE69816502T DE69816502T2 (de) | 1997-02-25 | 1998-02-24 | Ein Interleukin-18 enthaltender Agent, der Osteoclastzellen inhibiert |
| EP98301352A EP0861663B1 (en) | 1997-02-25 | 1998-02-24 | Osteoclastgenic inhibitory agent comprising interleukin-18 |
| KR1019980005963A KR100520723B1 (ko) | 1997-02-25 | 1998-02-25 | 파골(破骨)세포 형성 억제제 |
| US09/030,061 US20030095946A1 (en) | 1997-02-25 | 1998-02-25 | Il-18 as an osteoclastgenic inhibitor |
| US10/100,057 US6896880B2 (en) | 1997-02-25 | 2002-03-19 | Method for treating a disease associated with excessive osteoclast formation |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP05546897A JP3955352B2 (ja) | 1997-02-25 | 1997-02-25 | 破骨細胞形成阻害剤 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JPH10236974A JPH10236974A (ja) | 1998-09-08 |
| JPH10236974A5 JPH10236974A5 (ja) | 2004-12-24 |
| JP3955352B2 true JP3955352B2 (ja) | 2007-08-08 |
Family
ID=12999446
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP05546897A Expired - Fee Related JP3955352B2 (ja) | 1997-02-25 | 1997-02-25 | 破骨細胞形成阻害剤 |
Country Status (6)
| Country | Link |
|---|---|
| US (2) | US20030095946A1 (ja) |
| EP (1) | EP0861663B1 (ja) |
| JP (1) | JP3955352B2 (ja) |
| KR (1) | KR100520723B1 (ja) |
| DE (1) | DE69816502T2 (ja) |
| TW (1) | TW570801B (ja) |
Families Citing this family (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6790442B1 (en) | 1996-06-27 | 2004-09-14 | Kabushiki Kaisha Hayashibara Seibutsu Kagaku Kenkyujo | Genomic DNA encoding a polypeptide capable of inducing the production of interferon-γ |
| US6060283A (en) * | 1996-06-27 | 2000-05-09 | Kabushiki Kaisha Hayashibara Seibutsu Kagaku Kenkyujo | Genomic DNA encoding human interleukin-18 (IL-18, interferon-γ inducing factor) |
| JP2001103964A (ja) * | 1999-10-05 | 2001-04-17 | Teijin Ltd | 破骨細胞形成抑制方法または骨吸収抑制方法 |
| WO2002066063A1 (en) * | 2001-02-23 | 2002-08-29 | Nippon Organon K.K. | Remedies for metabolic bone diseases |
| MY143582A (en) | 2001-06-26 | 2011-05-31 | Amgent Fremont Inc | Antibodies to opgl |
| KR100454508B1 (ko) * | 2002-07-05 | 2004-11-03 | 허명준 | 소취기능과 다제내성균에 대한 멸균력을 갖는 자연기능수및 그의 제조방법 |
| DE60219495T2 (de) * | 2002-12-27 | 2007-12-13 | Universitätsklinikum Münster | Verwendung von Interleukin-18 zur Behandlung von UV-assoziierten Hautkrankheiten |
| KR101170852B1 (ko) * | 2003-09-30 | 2012-08-02 | 유키지루시 메그밀크 가부시키가이샤 | 골 형성 촉진 및/또는 골 흡수 억제제 |
| WO2005075648A1 (en) * | 2004-01-28 | 2005-08-18 | Gifu University | Interleukin-18 mutant proteins |
| US8168165B2 (en) * | 2008-12-23 | 2012-05-01 | Abbott Laboratories | Alkylated interleukin-18 compositions |
| SG11202001384TA (en) | 2017-09-06 | 2020-03-30 | Univ Yale | Interleukin-18 variants and methods of use |
| US12029778B2 (en) | 2019-05-13 | 2024-07-09 | Yale University | Interleukin-18 mimics and methods of use |
| CA3197145A1 (en) * | 2020-11-02 | 2022-05-05 | Aaron RING | Interleukin-18 variants and methods of use |
| WO2024102587A2 (en) * | 2022-11-11 | 2024-05-16 | Mural Oncology, Inc. | Interleukin 18 variants |
| WO2024238832A2 (en) * | 2023-05-16 | 2024-11-21 | Fred Hutchinson Cancer Center | Pro-inflammatory cytokine enhanced steap1 recombinant receptors |
Family Cites Families (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4588585A (en) | 1982-10-19 | 1986-05-13 | Cetus Corporation | Human recombinant cysteine depleted interferon-β muteins |
| US5714585A (en) | 1987-10-26 | 1998-02-03 | Sterling Winthrop, Inc. | Antibodies that are immunoreactive with interleukin-7 |
| DE69519454T2 (de) * | 1994-07-14 | 2001-05-03 | Kabushiki Kaisha Hayashibara Seibutsu Kagaku Kenkyujo, Okayama | Protein, das Interferon-Gamma Herstellung induziert und monoklonaler Antikörper dagegen |
| JP3109018B2 (ja) | 1994-07-14 | 2000-11-13 | 株式会社林原生物化学研究所 | インターフェロン−γの産生を誘導する蛋白質 |
| JP2952750B2 (ja) | 1995-02-23 | 1999-09-27 | 株式会社林原生物化学研究所 | モノクローナル抗体 |
| US6207641B1 (en) * | 1995-03-10 | 2001-03-27 | Kabushiki Kaisha Hayashibara Seibutsu Kagaku Kenkyujo | Pharmaceutical composition containing IFN-γ inducing polypeptide or factor for treating and/or preventing IFN-γ susceptive diseases |
| JP4004088B2 (ja) | 1995-09-26 | 2007-11-07 | 株式会社林原生物化学研究所 | 免疫担当細胞においてインターフェロン−γの産生を誘導する蛋白質 |
| JP2724987B2 (ja) | 1994-11-15 | 1998-03-09 | 株式会社林原生物化学研究所 | インターフェロン−γの産生を誘導するポリペプチド |
| TW581771B (en) * | 1994-11-15 | 2004-04-01 | Hayashibara Biochem Lab | Recombinant production of a polypeptide for inducing interferon-gamma production, and monoclonal antibody against the polypeptide |
| EP0721780A3 (en) * | 1994-12-16 | 1997-12-17 | Kabushiki Kaisha Hayashibara Seibutsu Kagaku Kenkyujo | Agent for promoting platelet and/or leukocyte production |
| US5776731A (en) | 1996-02-21 | 1998-07-07 | Immunex Corporation | DNA encoding type-I interleukin-I receptor-like protein designated 2F1 |
| EP0914453A1 (en) | 1996-05-20 | 1999-05-12 | Schering Corporation | HUMAN INTERLEUKIN-1j AND ANTAGONISTS THEREOF |
| JP4024366B2 (ja) | 1996-11-29 | 2007-12-19 | 株式会社林原生物化学研究所 | ポリペプチド |
-
1997
- 1997-02-25 JP JP05546897A patent/JP3955352B2/ja not_active Expired - Fee Related
-
1998
- 1998-02-24 TW TW087102640A patent/TW570801B/zh not_active IP Right Cessation
- 1998-02-24 DE DE69816502T patent/DE69816502T2/de not_active Expired - Fee Related
- 1998-02-24 EP EP98301352A patent/EP0861663B1/en not_active Expired - Lifetime
- 1998-02-25 KR KR1019980005963A patent/KR100520723B1/ko not_active Expired - Fee Related
- 1998-02-25 US US09/030,061 patent/US20030095946A1/en not_active Abandoned
-
2002
- 2002-03-19 US US10/100,057 patent/US6896880B2/en not_active Expired - Fee Related
Also Published As
| Publication number | Publication date |
|---|---|
| EP0861663B1 (en) | 2003-07-23 |
| EP0861663A3 (en) | 2000-03-29 |
| DE69816502T2 (de) | 2004-06-09 |
| US20030095946A1 (en) | 2003-05-22 |
| US20020150555A1 (en) | 2002-10-17 |
| EP0861663A2 (en) | 1998-09-02 |
| US6896880B2 (en) | 2005-05-24 |
| KR100520723B1 (ko) | 2007-12-21 |
| TW570801B (en) | 2004-01-11 |
| KR19980071708A (ko) | 1998-10-26 |
| DE69816502D1 (de) | 2003-08-28 |
| JPH10236974A (ja) | 1998-09-08 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP3955352B2 (ja) | 破骨細胞形成阻害剤 | |
| US5892001A (en) | Epithelium-derived T-cell factor antibodies | |
| US7875709B2 (en) | Nucleic acids encoding interleukin-18 mutants | |
| JP4024366B2 (ja) | ポリペプチド | |
| US7705140B2 (en) | DNAs encoding TNF receptor family members | |
| JPH09289896A (ja) | 免疫担当細胞においてインターフェロン−γの産生を誘導する蛋白質 | |
| US6207641B1 (en) | Pharmaceutical composition containing IFN-γ inducing polypeptide or factor for treating and/or preventing IFN-γ susceptive diseases | |
| AU680909B2 (en) | Interleukin-15 | |
| US5728548A (en) | Retinoid receptor-1 (RR1) and DNA encoding RR1 | |
| US7135458B1 (en) | Interferon-γ inducing polypeptide, pharmaceutical composition thereof, monoclonal antibody thereto, and methods of use | |
| US7625553B2 (en) | Method of treating cancer by administering an epithelum-derived T-cell factor | |
| JP2697725B2 (ja) | 悪性腫瘍治療用キット | |
| JP4026923B2 (ja) | ポリペプチド | |
| JP3993652B2 (ja) | 感受性疾患剤 | |
| JP2007143555A (ja) | ポリペプチド | |
| JP3056758B2 (ja) | グリコシル化インターロイキン‐2含有医薬 | |
| JPH06192124A (ja) | 血液細胞増加剤 | |
| JP2000236884A (ja) | ペプチド | |
| JPS63190830A (ja) | 抗悪性腫瘍剤 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20040122 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20040122 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20070123 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070326 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20070424 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20070502 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| LAPS | Cancellation because of no payment of annual fees |