ES2703256T3 - Microorganismo que tiene una mejor capacidad de producción de ornitina y método de producción de ornitina usando el mismo - Google Patents
Microorganismo que tiene una mejor capacidad de producción de ornitina y método de producción de ornitina usando el mismo Download PDFInfo
- Publication number
- ES2703256T3 ES2703256T3 ES11846603T ES11846603T ES2703256T3 ES 2703256 T3 ES2703256 T3 ES 2703256T3 ES 11846603 T ES11846603 T ES 11846603T ES 11846603 T ES11846603 T ES 11846603T ES 2703256 T3 ES2703256 T3 ES 2703256T3
- Authority
- ES
- Spain
- Prior art keywords
- ornithine
- microorganism
- glutamate
- strain
- sequence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/10—Citrulline; Arginine; Ornithine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
- C12N15/77—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora for Corynebacterium; for Brevibacterium
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/34—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Corynebacterium (G)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
- C12N1/205—Bacterial isolates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0008—Oxidoreductases (1.) acting on the aldehyde or oxo group of donors (1.2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1003—Transferases (2.) transferring one-carbon groups (2.1)
- C12N9/1018—Carboxy- and carbamoyl transferases (2.1.3)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1025—Acyltransferases (2.3)
- C12N9/1029—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1096—Transferases (2.) transferring nitrogenous groups (2.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1217—Phosphotransferases with a carboxyl group as acceptor (2.7.2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y102/00—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2)
- C12Y102/01—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2) with NAD+ or NADP+ as acceptor (1.2.1)
- C12Y102/01038—N-Acetyl-gamma-glutamyl-phosphate reductase (1.2.1.38)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y201/00—Transferases transferring one-carbon groups (2.1)
- C12Y201/03—Carboxy- and carbamoyltransferases (2.1.3)
- C12Y201/03003—Ornithine carbamoyltransferase (2.1.3.3)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y203/00—Acyltransferases (2.3)
- C12Y203/01—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
- C12Y203/01035—Glutamate N-acetyltransferase (2.3.1.35)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y206/00—Transferases transferring nitrogenous groups (2.6)
- C12Y206/01—Transaminases (2.6.1)
- C12Y206/01013—Ornithine aminotransferase (2.6.1.13)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y207/00—Transferases transferring phosphorus-containing groups (2.7)
- C12Y207/02—Phosphotransferases with a carboxy group as acceptor (2.7.2)
- C12Y207/02008—Acetylglutamate kinase (2.7.2.8)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/01—Bacteria or Actinomycetales ; using bacteria or Actinomycetales
- C12R2001/15—Corynebacterium
Abstract
Un microorganismo que pertenece a Corynebacterium sp., que tiene una capacidad mejorada para producir ornitina en comparación con su cepa parental, en el que se modifican las actividades de la ornitina carbamoiltransferasa y una proteína que participa en la exportación del glutamato (NCgl1221) para que sean atenuadas mediante una eliminación parcial o completa de los genes que codifican la ornitina carbamoil transferasa y NCg11221, en comparación con las actividades de la ornitina carbamoiltransfereasa y NCg11221 que posee la cepa parental en su estado nativo, en el que la cepa parental no tiene ninguna modificación en las actividades de la ornitina carbamoiltransferasa ni de NCg11221.
Description
DESCRIPCIÓN
Microorganismo que tiene una mejor capacidad de producción de ornitina y método de producción de ornitina usando el mismo
[Campo técnico]
La presente invención se refiere a un microorganismo que tiene una mejor capacidad de producción de ornitina y a un método de producción de ornitina usando el mismo.
[Antecedentes de la técnica]
La ornitina es una sustancia que se encuentra ampliamente en plantas, animales y microorganismos, y que se usa como un precursor en la biosíntesis de arginina, prolina y poliaminas. Como aminoácido no esencial, la ornitina no se encuentra en las proteínas, pero está presente en los antibióticos peptídicos tales como la tirosidina y la gramicidina. La ornitina desempeña un papel importante en la vía de excreción de la urea producida a partir del aminoácido o del amoníaco a través del ciclo de la ornitina en el metabolismo in vivo de animales superiores.
La ornitina ayuda a construir los músculos y a reducir la grasa corporal, y por lo tanto, se usa como suplemento nutricional. El ornitina-alfa quetoglutarato (OKG) que contiene ornitina y el ácido alfa-cetoglutárico a una proporción de 2:1 se usa como un potenciador inmunitario. La ornitina también se usa como fármacos para mejorar la cirrosis hepática y los trastornos de la función hepática, ya que ayuda a eliminar el amoníaco dañino del hígado. Los métodos conocidos para producir ornitina son el tratamiento de la caseína láctea con enzimas digestivas y el uso de E. coli transformada o microorganismos industriales pertenecientes a Corynebacterium sp., que se usan ampliamente en la producción de aminoácidos, ácidos nucleicos, enzimas y sustancias similares a antibióticos.
En los microorganismos que pertenecen a Corynebacterium sp., L-arginina es sintetizada a partir del glutamato por una enzima expresada de un gen en el operón de arginina en forma de argCJBDFRGH. Los genes del operón de arginina, que desempeñan el papel más importante en la biosíntesis de la arginina, sintetizan la arginina usando glutamato intracelular (L-glutamato) como sustrato, y se produce ornitina como producto intermedio durante la síntesis de la arginina. Específicamente, como en la FIG. 2, que ilustra esquemáticamente una vía sintética de arginina a partir de glutamato en un microorganismo perteneciente a Corynebacterium sp., se sabe que argJ codifica una enzima que convierte el glutamato en A/-acetil-glutamato, argB codifica una enzima que convierte el A/-acetil-glutamato en N-acetilglutamil-fosfato, argC codifica una enzima que convierte el N-acetilglutamil-fosfato en semialdehído de N-acetilglutamato, argD codifica una enzima que convierte el semialdehído de N-acetil-glutamato en N-acetil-ornitina, argJ codifica una enzima que convierte la N-acetil-ornitina en ornitina, argF codifica una enzima que convierte la ornitina en citrulina, argG codifica una enzima conversora de la citrulina en argininosuccinato, y argH codifica una enzima conversora del argininosuccinato en arginina en la vía sintética de la arginina, y la vía sintética de ornitina se incluye en la vía sintética de la arginina.
Las cepas productoras de arginina conocidas se han desarrollado mediante la introducción de una mutación en el operón de arginina o mediante una mutación en el promotor para aumentar los niveles de expresión de las enzimas que participan en la biosíntesis de la arginina. Entre ellos, argR que controla y suprime la expresión del operón de arginina y argB inhibida por el nivel de arginina han sido ampliamente estudiadas como dianas para aumentar la producción de arginina (publicación de patente coreana n.° 2010-0060909).
Para mejorar la productividad de la ornitina, se sabe que la producción de ornitina es aumentada por la acción de la ornitina ciclodesaminasa (ocd) mediante el cultivo del microorganismo Corynebacterium en un medio complementado con prolina, o mediante la modificación de los impulsores y las condiciones de cultivo durante el cultivo del microorganismo. Además, cuando se usa una E. coli transformada, la productividad de la ornitina se mejora cultivando las cepas con argF y argR eliminados en un medio complementado con glutamato, o usando una cepa transformada con la eliminación del gen proB que codifica la Y-glutamilquinasa que participa en la primera etapa de la vía sintética de la prolina a partir del glutamato en lugar de la vía sintética de la ornitina a partir del glutamato.
Además, se ha usado sistemáticamente Corynebacterium glutamicum para la producción de alto rendimiento de glutamato como un precursor de la ornitina. Se sabe que la excreción de glutamato a partir de Corynebacterium glutamicum aumenta mediante la limitación de la biotina o el tratamiento con penicilina G o un tensioactivo de éster de ácido graso. Dado que estos tratamientos se correlacionan con el daño en la pared celular, anteriormente se creía que el glutamato se filtra de forma pasiva a través de la pared celular dañada.
La proteína NCgl1221 derivada de Corynebacterium glutamicum de tipo silvestre (Cgl 13032) proporciona un eflujo de betaína y su secuencia de aminoácidos es similar a la de la proteína del canal mecanosensible de E. coli, yggB (publicación de patente coreana n.° 2010-0017581). Hwang Joong-Hee et al. (J. of Microbiology and Biotechnology, 2008, 18, 704-710) desvelan el efecto del aumento de la disponibilidad de glutamato en la producción de L-ornitina en Corynebacterium glutamicum con la eliminación de argF, argR y probB. Se concluye que la concentración intracelular de glutamato no es una etapa limitante de la velocidad en C. glutamicum, pero que es limitante de la velocidad en E.
coli (sumario, Figura 1, página 705, 2° párrafo de la izquierda).
[DIVULGACIÓN]
[Problema técnico]
En este contexto, los presentes inventores han hecho muchos esfuerzos para desarrollar una cepa de Corynebacterium capaz de producir la ornitina útil a un rendimiento superior. Como resultado de ello, encontraron que se puede desarrollar una cepa sobreproductora de ornitina mediante el bloqueo de la vía biosintética de la arginina a partir de la ornitina, mediante el bloqueo de una proteína que participa en la exportación del glutamato para aumentar el nivel de glutamato intracelular y la mejora de la vía biosintética de la ornitina a partir del glutamato, completando de esta forma la presente invención.
[Solución técnica]
Un objeto de la presente invención es proporcionar un microorganismo que tenga una mejor capacidad de producción de ornitina. Otro objeto de la presente invención es proporcionar un método de producción de ornitina usando el microorganismo.
[Efecto ventajoso]
El microorganismo de la presente invención que tiene una mejor capacidad de producción de ornitina se puede usar de manera más eficaz en una amplia variedad de aplicaciones para la producción de ornitina.
[Breve descripción de los dibujos]
La FIG. 1 muestra la vía biosintética de ornitina y los genes relacionados de Corynebacterium glutamicum transformada de la presente invención;
la FIG. 2 muestra la vía biosintética de arginina conocida de Corynebacterium glutamicum; y
la FIG. 3 muestra un vector pDZ que se introducirá en el cromosoma del microorganismo perteneciente a Corynebacterium sp.
[Mejor modo]
En un aspecto para conseguir los objetos anteriores de la presente invención, la presente invención proporciona un microorganismo que pertenece al género Corynebacterium sp. que tiene una mejor capacidad de producción de ornitina, en el que se modifican las actividades de la ornitina carbamoiltransferasa y una proteína que participa en la exportación del glutamato (NCgl1221) para que sean atenuadas mediante una eliminación parcial o completa de los genes que codifican la ornitina carbamoiltransferasa y NCg11221, en comparación con sus actividades endógenas. Como se usa en el presente documento, “ornitina carbamoiltransferasa (OCT)” se refiere a una enzima catalítica que media la reacción entre el fosfato de carbamoílo y la ornitina para sintetizar citrulina y ácido fosfórico. La OCT está presente en el hígado de animales que excretan urea, así como en plantas y en microorganismos; y en los microorganismos, participa en la síntesis de la arginina. La enzima OCT comprende un dominio catalítico y un dominio regulador, y cuando la ornitina se une al dominio regulador, la actividad de la enzima se inhibe.
La cepa de E. coli K12 tiene dos tipos de OCT (ArgF y ArgI), y los microorganismos intestinales que incluyen las cepas E. coli B y W tienen una proteína OCT similar a ArgI. Las OCT codificadas por argFy argI tienen diferentes secuencias de aminoácidos entre sí, pero se consideran isoenzimas que tienen la misma función (EMBO J. (1982) 1:853-857). La cepa de Corynebacterium sp. solo tiene OCT codificada por el gen argF. La OCT solo actúa en la vía sintética de la ornitina a la arginina y, por lo tanto, si la actividad de la OCT se debilita, el nivel de ornitina intracelular puede aumentar. Para la acumulación de ornitina intracelular, la presente invención proporciona un microorganismo de Corynebacterium en el que se bloquea la vía sintética de la arginina a partir de la ornitina. Para conseguir esto, se preparó una cepa transformada con una eliminación del gen que codifica la ornitina carbamoiltransferasa. En este sentido, la ornitina carbamoiltransferasa puede ser, pero sin limitarse en particular a, una proteína que tiene una secuencia de aminoácidos de SEQ ID NO: 18, o una proteína que tiene un 70% o más de homología con la secuencia, más preferentemente, un 80 % o más de homología con la secuencia, y mucho más preferentemente un 90 % o más de homología con la secuencia.
Como se usa en el presente documento, "homología" se refiere a la similitud en las secuencias de nucleótidos o secuencias de aminoácidos del gen que codifica una proteína. Cuando la homología es suficientemente alta, los productos del gen correspondiente pueden ser iguales o tener una actividad similar.
Como se usa en el presente documento, "proteína que participa en la exportación de glutamato" se refiere a un tipo de canales mecanosensibles que funcionan para exportar el glutamato producido intracelularmente al entorno extracelular. La presente invención proporciona un microorganismo de Corynebacterium que tiene mejor productividad de ornitina. Para esta labor, se prepara una cepa transformada capaz de mantener un alto nivel de glutamato intracelular eliminando un gen que codifica la proteína que funciona para excretar el glutamato, que es una materia prima para la síntesis de ornitina.
Al aumentar el nivel intracelular de glutamato, es decir, un precursor de la ornitina, se puede estimular una vía biosintética de ornitina. En la presente invención, la exportación de glutamato puede reducirse o inhibirse disminuyendo la actividad de NCgl1221. La proteína eliminada que participa en la exportación del glutamato puede ser una proteína que tenga una secuencia de aminoácidos de s Eq ID NO: 20 o una secuencia de aminoácidos que tenga una homología del 70 % o más, y más preferentemente, que tenga una homología del 80 % o más, incluso más preferentemente, que tenga una homología del 90 % o más, pero sin limitación.
La actividad de la ornitina carbamoiltransferasa y la proteína que participa en la exportación del glutamato se puede disminuir mediante un método seleccionado del grupo que consiste en (1) una eliminación parcial o completa de un gen que codifica la proteína; (2) la modificación de una secuencia reguladora de la expresión para suprimir la expresión génica; (3) la modificación de la secuencia de nucleótidos en el cromosoma para disminuir la actividad de la proteína; y 4) una combinación de las mismas, pero sin limitación.
Se puede realizar una eliminación parcial o completa de un polinucleótido que codifica la proteína introduciendo un vector para la inserción cromosómica en un microorganismo, reemplazando así el polinucleótido que codifica una proteína diana endógena en el cromosoma con un polinucleótido parcialmente eliminado o un gen marcador. La longitud "parcial" puede variar dependiendo del tipo de polinucleótido, pero específicamente se refiere a una longitud de 1 a 300 nucleótidos, preferentemente de 1 a 100 nucleótidos y, más preferentemente, 1 a 50 nucleótidos.
Además, la modificación de una secuencia reguladora de la expresión para reducir la expresión del polinucleótido puede realizarse induciendo una modificación en la secuencia reguladora de la expresión a través de la eliminación, inserción, sustitución no conservativa o conservativa de la secuencia de nucleótidos, o una combinación de las mismas para disminuir la actividad de la secuencia reguladora de la expresión, o reemplazando la secuencia reguladora de la expresión con una secuencia de nucleótidos que tiene una actividad más débil. La secuencia reguladora de la expresión incluye un promotor, una secuencia de operador, una secuencia que codifica el sitio de unión al ribosoma y una secuencia que regula la terminación de la transcripción y la traducción.
Por otra parte, la modificación de una secuencia de polinucleótidos en el cromosoma, que codifica la enzima de la presente invención, se puede realizar mediante la inducción de una mutación en la secuencia a través de la eliminación, inserción, sustitución no conservativa o conservativa de la secuencia de polinucleótido, o una combinación de las mismas para disminuir la actividad enzimática, o reemplazando la secuencia con una secuencia de polinucleótidos que se modifica para tener una actividad más débil.
Como se usa en el presente documento, "actividad endógena" se refiere a la actividad de enzima que posee un microorganismo en su estado nativo. En la presente invención, la actividad endógena se refiere a la actividad de la ornitina carbamoil transferasa y una proteína que participa en la exportación del glutamato, es decir, NCgl1221, que un microorganismo posee de manera natural. Además, como se usa en el presente documento, "modificadas para que tenga una actividad más débil que una actividad endógena" se refiere al estado en el que una ornitina carbamoil transferasa y una proteína que participa en la exportación del glutamato, es decir, NCgl1221, no funcionan correctamente debido a la eliminación o mutación de genes, y de ese modo, la actividad de la ornitina carbamoil transferasa y una proteína que participa en la exportación del glutamato, es decir, NCgl1221, que un microorganismo posee de manera natural, se debilita.
Como se usa en el presente documento, la expresión "microorganismo que tiene una mejor capacidad de producción de ornitina" se refiere a un microorganismo que tiene una capacidad de producción de ornitina superior a la de la cepa parental, y el microorganismo que tiene una mejor capacidad de producción de ornitina puede ser, pero sin limitarse en particular a, un microorganismo que se transforma adicionalmente para tener una mayor actividad de la acetilglutamato sintasa (ArgJ), que convierte el glutamato en acetil-glutamato (W-acetil-glutamato) o la ornitina acetiltransferasa (ArgJ), que convierte la acetil-ornitina en ornitina, la acetilglutamato quinasa (ArgB), que convierte el acetil-glutamato en acetilglutamil-fosfato (W-acetilglutamil-fosfato), la acetil-gamma-glutamil-fosfato reductasa (ArgC), que convierte el acetil-glutamil-fosfato en semialdehído de acetil-glutamato (semialdehído de W-acetil-glutamato), la acetilornitina aminotransferasa (ArgD), que convierte el semialdehído de acetil-glutamato en acetilornitina (W-acetilornitina) o similares en comparación con la actividad endógena, para mejorar la vía biosintética de la ornitina a partir del glutamato.
El microorganismo transformado se preparó mediante un método diferente al método conocido de desarrollo de una cepa productora de ornitina, es decir, el método conocido se realiza eliminando o atenuando la función de ArgR que actúa como un inhibidor de la transcripción en la vía biosintética de la arginina para aumentar la producción de ornitina, y eliminando además el gen de la ornitina carbamoiltransferasa e introduciendo la W-acetilglutamato sintasa resistente a la retroalimentación para aumentar la producción de ornitina (publicación de patente coreana n.° 2010-0060909).
En este sentido, la acetil gamma-glutamil-fosfato reductasa (ArgC), la acetilglutamato sintasa o la ornitina acetiltransferasa (argJ), la acetilglutamato quinasa (ArgB) y la acetilornitina aminotransferasa (argD) pueden tener preferentemente, pero sin limitarse en particular a, las secuencias de aminoácidos de SEQ ID NO: 23, 25, 27 y 29, o un 70 % o más de homología con la secuencia, más preferentemente, un 80 % o más de homología con la secuencia, y mucho más preferentemente, un 90 % o más de homología con la secuencia, respectivamente. El aumento en su actividad se puede realizar mediante uno o más métodos seleccionados del grupo que consiste en 1) un aumento en el número de copias de un polinucleótido que codifica la proteína; y 2) un aumento en la expresión del polinucleótido mediante la modificación de la secuencia de control de la expresión; 3) la potenciación de la actividad enzimática mediante la modificación de la secuencia polinucleotídica en el cromosoma; y 4) la potenciación mediante una
combinación de las mismas.
Para ser específicos, se pueden usar varios métodos para aumentar la actividad enzimática en un microorganismo en general. Por ejemplo, el nivel de expresión de un polinucleótido puede aumentarse aumentando el número de copias del polinucleótido a través de la transformación que implica la inserción del plásmido, la recombinación homóloga, la conjugación y la translocación; modificando una secuencia reguladora de la expresión del polinucleótido; amplificando un gen que codifica un factor regulador que estimula la expresión del polinucleótido; o eliminando o inhibiendo un gen que codifica un factor regulador que suprime la expresión del polinucleótido. Para ser más específicos, el nivel de expresión de un polinucleótido puede aumentarse uniendo operativamente un fragmento de gen que comprende el polinucleótido a un vector multicopia que puede replicarse en cepas de Corynebacterium sp., introduciendo copias únicas o múltiples del polinucleótido en el cromosoma, o reemplazando la secuencia reguladora de la expresión del polinucleótido por una que tenga una mejor actividad que incluya un promotor potente.
Por ejemplo, el grupo de genes argCJBD puede transformarse en un microorganismo usando el vector pHC139T para preparar un microorganismo con una productividad de ornitina significativamente mejorada. Como alternativa, se puede preparar un microorganismo en el que se mejore la vía biosintética de la ornitina mejorando una región promotora reguladora de la expresión del gen argCJBD en el cromosoma del microorganismo o reemplazando una región promotora por un promotor con una actividad más mejorada. En particular, un método para mejorar la región promotora puede implicar, para reemplazar un promotor dentro del cromosoma, preparar un fragmento génico que comprenda secuencias de nucleótidos de ambos sitios terminales adyacentes al sitio diana en el cromosoma y una secuencia promotora para insertarse de la misma forma que en el cromosoma original y siguiendo el mismo método de eliminación de genes usando un vector pDZ publicado por la publicación de patente coreana n.° 2009-0082702, pero sin limitación. Aquí, el promotor mejorado puede ser preferentemente, pero sin limitación, el promotor pcj7 (o P(CJ7)) que tiene una secuencia de nucleótidos de SEQ ID NO: 30 (patente coreana con n.° de registro 0620092). El vector pDZ puede ser preferentemente, pero sin limitación, un vector representado por un mapa de escisión de la Figura 3.
Como se usa en el presente documento, "vector" se refiere a una construcción de ADN que comprende una secuencia de nucleótidos del gen que está unida operativamente a una secuencia reguladora de la expresión apropiada para expresar un gen diana en una célula hospedadora adecuada. La secuencia reguladora de la expresión comprende un promotor que puede iniciar la transcripción, una secuencia de operador opcional para regular la transcripción, una secuencia que codifica un sitio de unión al ribosoma de ARNm adecuado y una secuencia que regula la terminación de la transcripción y la traducción.
Los ejemplos de vectores convencionales incluyen un plásmido natural o recombinante, un cósmido, un virus y un bacteriófago. Por ejemplo, pWE15, M13, AEMBL3, AEMBL4, AFIXII, ADASHII, AZAPII, Agt10, Agt11, Charon4A y Charon21A pueden usarse como vector de fago o vector de cósmido. Como vector plásmido, se pueden usar el vector pDZ, tipo pBR, tipo pUC, tipo pBluescriptlI, tipo pGEM, tipo pTZ, tipo pCL y tipo pET. El vector que se puede usar no se limita a uno en particular, y puede usarse cualquier vector de expresión conocido, preferentemente, se puede usar el vector pDZ.
Paralelamente, el microorganismo de la presente invención puede ser, pero sin limitarse en particular a, un microorganismo preparado mediante la transformación de un microorganismo que pertenece a Escherichia sp., Shigella sp., Citrobacter sp., Salmonella sp., Enterobacter sp., Yersinia sp., Klebsiella sp., Erwinia sp., Corynebacterium sp., Brevibacterium sp., Lactobacillus sp., Selenomanas sp., o Vibrio sp., que tiene las actividades de la ornitina carbamoiltransferasa y una proteína que participa en la exportación del glutamato (NCgl1221).
Preferentemente, el microorganismo de la presente invención puede ser la cepa de Corynebacterium sp., y más preferentemente, Corynebacterium glutamicum. Para ser más específicos, se puede usar una cepa de tipo silvestre de Corynebacterium glutamicum ATCC 13032 o una cepa sobreproductora de glutamato KCCM-10785P (publicación de patente coreana n.° 2008-0034334), pero sin limitación. La cepa KCCM-10785P es una cepa sobreproductora de glutamato generada mediante la eliminación de los genes cg2624 (NCBI LOCUS ID YP_226636) y cg2115 (NCBI LOCUS ID YP_226173) en una cepa productora de glutamato (KFCC-11074), que se generó usando un mutágeno tal como N-metil-N'-nitro-N-nitrosoguanidina (NTG). Aunque la sobreproducción de glutamato mediante la eliminación de cg2624 y cg2115 no se ha identificado antes de la publicación anterior, cg2624 se identifica como pcaR, que es una proteína reguladora de la familia IclR, y cg2115 se identifica como sugR, que es un regulador de la transcripción del metabolismo del azúcar.
De acuerdo con una realización de la presente invención, se prepararon una cepa de Corynebacterium glutamicum con una eliminación del gen argF (ATCC 13032 AargF y KCCM-10785P AargF) (Ejemplo 1), una cepa de Corynebacterium glutamicum con la eliminación de los genes argF y NCgl1221 (ATCC 13032 AargF ANCgl1221 y KCCM-10785P AargF ANCgl1221) (Ejemplo 2), una cepa de Corynebacterium glutamicum con la eliminación de los genes argF y NCgl1221 y con una introducción del gen argCJBD (ATCC 13032 AargF ANCgl1221/pHC139T-argCJBD(Cgl) y KCCM-10785P AargF ANCgl1221/pHC139T-argCJBD(Cgl)) (Ejemplo 3-1), y una cepa de Corynebacterium glutamicum con la eliminación de los genes argF y NCgl1221 y con el reemplazo del promotor de la agrupación de genes argCJBD en el cromosoma (ATCC 13032 AargF ANCgl1221 P(CJ7)-argCJBD y KCCM-10785P
AargF ANCgl1221 P(CJ7)-argCJBD) (Ejemplo 3-2). El resultado de comparar sus productividades de ornitina mostró que la cepa Corynebacterium glutamicum con las eliminaciones de los genes argF y NCgl1221 y con el reemplazo del promotor de la agrupación de genes argCJBD en el cromosoma (ATCC 13032 AargF ANCgl1221 P(CJ7)-argCJBD y KCCM-10785P AargF ANCgl1221 P(CJ7)-argCJBD) tiene una productividad de ornitina excelente (Tablas 5 y 6). Por lo tanto, la cepa productora de ornitina que tiene una mejor capacidad de producción de ornitina se designó como "CC01-0061 (At Cc 13032 AargF ANCgl122l P(CJ7)-argCJBD)", y se depositó en virtud del Tratado de Budapest en el Centro de Cultivos de Microorganismos de Corea, cuya dirección es Hongje-1-dong, Seodaemun-gu, Seúl, el 24 de noviembre de 2010, con el número de acceso KCCM11137P.
En otro aspecto de la presente invención para conseguirse los objetos anteriores, la presente invención proporciona un método para producir ornitina, incluyendo las etapas de (i) cultivar el microorganismo que tiene una mejor capacidad de producción de ornitina para obtenerse un cultivo; y (ii) recuperar la ornitina del microorganismo cultivado o del cultivo.
En el método, el cultivo del microorganismo puede realizarse preferentemente mediante cultivo discontinuo, cultivo continuo y cultivo discontinuo alimentado, conocidos en la técnica, pero sin limitación. Además, en cuanto a las condiciones de cultivo, se puede mantener un pH óptimo de 5 a 9, preferentemente de 6 a 8, y lo más preferentemente un pH 6,8 usando un agente químico básico (por ejemplo: hidróxido sódico, hidróxido potásico o amoniaco) o un agente químico ácido (por ejemplo: ácido fosfórico o ácido sulfúrico). Además, pueden mantenerse condiciones aeróbicas añadiendo oxígeno o una mezcla gaseosa que contenga oxígeno al cultivo celular. La temperatura de cultivo puede mantenerse a de 20 °C a 45 °C, y preferentemente a de 25 °C a 40 °C. Además, es preferible cultivar durante aproximadamente 10 a 160 horas. La ornitina producida por el cultivo anterior puede excretarse a un medio de cultivo o permanecer dentro de la célula.
Además, el medio para el cultivo puede comprender azúcar e hidratos de carbono (por ejemplo: glucosa, sacarosa, lactosa, fructosa, maltosa, melaza, almidón y celulosa), aceite y grasa (por ejemplo: aceite de soja, aceite de semillas de girasol, aceite de cacahuete y aceite de coco), ácido graso (por ejemplo: ácido palmítico, ácido esteárico y ácido linoleico), alcohol (por ejemplo: glicerol y etanol) y ácido orgánico (por ejemplo: ácido acético) individualmente o en combinación como fuente de carbono; compuesto orgánico que contiene nitrógeno (por ejemplo: peptona, extracto de levadura, jugo de carne, extracto de malta, solución de maíz, harina de soja en polvo y urea) o compuesto inorgánico (por ejemplo: sulfato de amonio, cloruro de amonio, fosfato de amonio, carbonato de amonio y nitrato de amonio) individualmente o en combinación, como fuente de nitrógeno; dihidrogenofosfato de potasio, fosfato de dipotasio o una sal que contenga sodio correspondiente al mismo, individualmente o en combinación, como una fuente de fósforo; otras sustancias esenciales estimulantes del crecimiento, incluyendo las sales metálicas (por ejemplo: sulfato de magnesio o sulfato de hierro), aminoácidos y vitaminas.
[Modo para la invención]
En lo sucesivo, en el presente documento, la presente invención se describirá con más detalle con referencia a los ejemplos. Sin embargo, estos ejemplos son únicamente con fines ilustrativos, y no se pretende que la invención esté limitada por estos ejemplos.
Ejemplo 1: Preparación de la cepa de Corynebacterium glutamicum con argF eliminado
En este ejemplo, se preparó una cepa con argF eliminado a partir de la cepa de Corynebacterium glutamicum ATCC 13032 y una cepa sobreproductora de glutamato KCCM-10785P que se generó mediante la eliminación de los genes cg2624 y cg2115 en una cepa productora de glutamato KFCC-11074 generada mediante el uso de mutágenos tales como n Tg (publicación de patente coreana n.° 2008-0034334) para bloquear una vía sintética de arginina a partir de ornitina. Los genes biosintéticos de arginina de Corynebacterium glutamicum ATCC 13032 están organizados en un operón que tiene una forma de argCJBDFRGH, y está presente un gen argF diana de eliminación (SEQ ID NO: 17) adyacente a los genes que codifican enzimas que participan en la vía sintética de la ornitina en el cromosoma. Por lo tanto, se preparó un plásmido para eliminar el gen argF basándose en la secuencia de nucleótidos de argD y argR que se encuentran adyacentes al gen argF diana de eliminación.
Para ser específicos, basados en la secuencia de nucleótidos de argD y argR de la cepa ATCC 13032, se construyeron un fragmento de recombinación homóloga adyacente a la secuencia N-terminal de argF y un fragmento de recombinación homóloga adyacente a la secuencia C-terminal de argF. Para ello, se obtuvo el fragmento adyacente a la secuencia N-terminal de argF mediante PCR usando el ADN genómico de la cepa ATCC 13032 como molde, y los cebadores (SEQ ID NO: 1 y 2) (28 ciclos de desnaturalización durante 30 segundos a 94 °), hibridación durante 30 segundos a 55 °C y extensión durante 30 segundos a 72 °C). Asimismo, se obtuvo el fragmento adyacente a la secuencia C-terminal de argF mediante PCR usando el ADN genómico de la cepa ATCC 13032 como molde, y los cebadores (SEQ ID NO: 3 y 4) en las mismas condiciones de PCR (Tabla 1).
T l 1

El fragmento de recombinación homóloga preparado anteriormente adyacente a la secuencia N-terminal de argF fue digerido con BamHI y SalI, y el fragmento de recombinación homóloga adyacente a la secuencia C-terminal de argF fue digerido con SalI y XbaI. Luego, se insertó cada uno de los fragmentos escindidos en el vector pDZ que también se digirió con BamHI y XbaI, produciendo así un plásmido pDZ-argF (K/O).
El plásmido PDZ-argF (K/S) preparado anteriormente se transformó en la cepa ATCC 13032 y en la cepa KCCM-10785P. A continuación, se sembraron las cepas transformadas en placas y se cultivaron en una placa BHIS (infusión de cerebro y corazón a 37 g/l, sorbitol a 91 g/l, agar al 2 %) que contiene kanamicina (25 mg/ml) y X-gal (5-bromo-4-cloro-3-indolin-p-D-galactósido), mientras que las colonias crecen en la placa. Entre las colonias formadas en la placa, se recogieron colonias de color azul para seleccionar la cepa insertada con el plásmido pDZ-argF (K/O).
Las cepas seleccionadas anteriores se cultivaron con agitación en medio CM (glucosa 10 g/l, polipeptona a 10 g/l, extracto de levadura 5 g/l, extracto de carne a 5 g/l, NaCl a 2,5 g/l, urea a 2 g/l, pH 6,8) a 30 °C durante 8 horas. Posteriormente, cada cultivo celular se diluyó en serie de 10'4 a 10'10. A continuación, se sembraron las muestras diluidas en placas y se cultivaron en un medio sólido que contenía X-gal, dejando que las colonias crecieran. Entre las colonias formadas en la placa, solo se recogieron las colonias blancas que aparecían con una frecuencia relativamente baja para seleccionar las cepas con argF eliminado.
La inserción exitosa del plásmido pDZargF (K/O) en las cepas seleccionadas anteriormente se confirmó realizando una PCR usando el ADN cromosómico de la cepa seleccionada anteriormente como molde y los cebadores de SEQ ID NO. 1 y 4. A través de esta confirmación por PCR, se confirmó que la cepa seleccionada anteriormente es la cepa con argF eliminado (es decir, ATCC 13032 ÁargF y KCCM-10785P AargF).
Ejemplo 2: Preparación de la cepa de Corynebacterium glutamicum con argF y NCgl1221 eliminados
El gen NCgl1221 codificante de la proteína que participa en la exportación del glutamato se eliminó además en la cepa ATCC 13032 AargF y en la cepa KCCM-10785P AargF obtenidas en el Ejemplo 1 para aumentar el nivel intracelular del glutamato, que es un precursor de ornitina.
Para ser específicos, basado en la secuencia de nucleótidos (SEQ ID NO: 19) de NCgl1221 de la cepa ATCC 13032, se construyeron un fragmento de recombinación homóloga adyacente a la secuencia N-terminal de NCgl1221 y un fragmento de recombinación homóloga adyacente a la secuencia C-terminal de NCgl1221. Para ello, se generó el fragmento adyacente a la secuencia adyacente de N-terminal de NCgl1221 por PCR usando el ADN genómico de la cepa ATCC 13032 como molde y cebadores (SEQ ID NO: 5 y 6), y se generó el fragmento adyacente a la secuencia C-terminal de NCgl1221 por PCR usando el A d N genómico de la cepa ATCC 13032 como molde y cebadores (SEQ ID NO: 7 y 8) en las mismas condiciones de PCR que en el Ejemplo 1 (Tabla 2).
T l 2

El fragmento de recombinación homóloga preparado anteriormente adyacente a la secuencia N-terminal de NCgl1221 fue digerido con BamHI y SalI. Asimismo, el fragmento de recombinación homóloga adyacente a la secuencia C-terminal de NCgl1221 se digirió con SalI y XbaI. Luego, se insertó cada uno de los fragmentos escindidos en el vector pDZ que se escindió con BamHI y XbaI, produciéndose así un plásmido pDZ-NCgl1221 (K/O).
El plásmido PDZ-NCgl1221 (K/S) preparado anteriormente se transformó en la cepa ATCC 13032 AargF y en la cepa KCCM-10785P AargF. A continuación, se sembraron las cepas transformadas en placas y se cultivaron en una placa BHIS (infusión de cerebro y corazón a 37 g/l, sorbitol a 91 g/l, agar al 2 %) que contiene kanamicina (25 mg/ml) y X-gal (5-bromo-4-cloro-3-indolin-p-D-galactósido), mientras que las colonias crecen en la placa. Entre las colonias formadas en la placa, se recogieron colonias de color azul para seleccionar la cepa insertada con el plásmido pDZ-NCgl1221 (K/O).
Las cepas seleccionadas anteriores se cultivaron con agitación en medio CM a 30 °C durante 8 horas. Posteriormente, cada cultivo celular se diluyó en serie de 10-4 a 10-10. A continuación, se sembraron las muestras diluidas en placas y se cultivaron en un medio sólido que contenía X-gal, dejando que las colonias crecieran. Entre las colonias formadas en la placa, solo se recogieron las colonias blancas que aparecieron con una frecuencia relativamente baja para seleccionar las cepas con NCgl1221 eliminado.
La inserción exitosa del plásmido pDZ-NCgl1221(K/O) en las cepas seleccionadas anteriormente se confirmó realizando una PCR usando el ADN cromosómico de la cepa seleccionada anteriormente como molde y los cebadores de SEQ ID NO. 5 y 8. Por consiguiente, las cepas con NCgl1221 eliminado seleccionadas se nombraron como ATCC 13032 AargF ANCgl1221 o KCCM-10785P AargF ANCgl1221.
Ejemplo 3: Preparación de cepa de Corynebacterium glutamicum con argCJBD introducido
Ejemplo 3-1: Clonación del gen argCJBD y preparación del transformante
En este ejemplo, se preparó un vector insertado con los genes argC, argJ, argB, y argD (SEQ ID NO: 22, 24, 26 y 28 respectivamente), y se preparó un transformante mediante la introducción del mismo, para potenciar la vía sintética de la ornitina mediante el aumento del número de copias del operón de argCJBD (SEQ ID NO: 21, que comprende la región promotora) que codifica las enzimas que participan en una vía sintética de la ornitina a partir del glutamato.
En primer lugar, se realizó la PCR, obteniéndose el gen argCJBD mediante el uso del cromosoma de la cepa ATCC 13032 como molde y cebadores (SEQ ID NO: 9 y 10, Tabla 3) (30 ciclos de desnaturalización durante 40 segundos a 95 °C, hibridación durante 40 segundos a 55 °C y extensión durante 150 segundos a 72 °C), obteniéndose así un fragmento génico que tiene un tamaño de 4.900 pb.
T l 1

Se procesó el fragmento del gen preparado anteriormente a través de electroforesis en gel sobre gel de agarosa al 0,8 %, y se cortó una banda del tamaño diana y se aisló una muestra de ADN de la misma. El ADN aislado se digirió con KpnI y XbaI, obteniéndose un fragmento, luego se clonó el fragmento escindido en un vector pHC139T-gfp (publicación de patente coreana n.° 2008-0074286), produciéndose así un vector de expresión pHC139T-argCJBD(Cgl).
Posteriormente, se introdujo el vector de expresión pHC139T-argCJBD(Cgl) preparado para aumentar el nivel de producción de ornitina en la célula en la cepa ATCC 13032 AargF ANCgl1221 y la cepa KCCM-10785P AargF ANCgl1221 a través de electroporación. A continuación, se seleccionó un transformante exitoso sembrando en placas las células transformadas en una placa de BHIS que contenía 25 pg/mt de kanamicina. Por último, cada uno de los transformantes seleccionados se denominó correspondientemente ATCC 13032 AargF ANCgl1221/pHC139T-argCJBD(Cgl) y KCCM-10785P AargF ANCgl1221/pHC139T-argCJBD(Cgl).
Ejemplo 3-2: Sustitución del promotor del gen argCJBD en el cromosoma
En este ejemplo, se sustituyó un promotor de argCJBD con el promotor CJ7 que fue desarrollado recientemente por el presente solicitante en el cromosoma, para aumentar el nivel de expresión mediante la eliminación de la regulación del gen argCJBD que codifica las enzimas que participan en una vía sintética de la ornitina a partir del glutamato.
En primer lugar, se preparó un fragmento de recombinación homóloga que comprendía un promotor CJ7 y una secuencia de nucleótidos de ambos sitios terminales del promotor.
Para ser específicos, se obtuvo la secuencia de nucleótidos del sitio 5'-terminal del promotor CJ7 realizando una PCR con el uso del ADN genómico de la cepa ATCC 13032 como molde y cebadores (Se Q ID NO: 11 y 12) (28 ciclos de desnaturalización durante 30 segundos a 94 °C, hibridación durante 30 segundos a 55 °C y extensión durante 30 segundos a 72 °C). Asimismo, se obtuvo la secuencia de nucleótidos de la región del promotor CJ7 mediante PCR usando cebadores (SEQ ID NO: 13 y 14) en las mismas condiciones de PCR, y se obtuvo la secuencia de nucleótidos del sitio 3'-terminal del promotor CJ7 mediante PCR usando el ADN genómico de las cepas ATCC 13032 como molde y cebadores (SEQ ID NO: 15 y 16) en las mismas condiciones de PCR.
T l 41


El fragmento de sitio 5'-terminal preparado anteriormente de promotor (argC-L) se digirió con BamHI y EcoRI, el fragmento de la región del promotor CJ7 se digirió con EcoRI y XbaI, y el fragmento del sitio 3'-terminal del promotor (argCR) se digirió con XbaI y SalI. Luego, se clonó cada uno de los productos de la PCR escindidos en el vector pDZ que también se digirió con BamHI y SalI, produciéndose así un vector de expresión pDZ-CJ7(arg) en el que el promotor de argCJBD estaba sustituido con el promotor CJ7.
El vector de expresión anteriormente preparado PDZ-CJ7(arg) se transformó en la cepa ATCC 13032 AargF ANCgl1221 y KCCM-10785P AargF ANCgl1221 a través de electroporación. A continuación, se cultivaron los transformantes con agitación en medio CM (30 °C, 8 horas) y se diluyó el cultivo celular en serie de 10-4 a 10-10. A continuación, se sembraron y se cultivaron las muestras diluidas en una placa de BHIS que contenía 25 pg/ml de kanamicina y X-gal, dejando que las colonias crecieran.
Se aislaron las colonias de color blanco que aparecieron a baja frecuencia de la mayoría de las colonias azules, seleccionando de este modo solamente la cepa en la que el promotor arg fue sustituido con éxito con el promotor CJ7 a través de un entrecruzamiento doble.
La sustitución con éxito del promotor argCJBD en el cromosoma mediante el vector de expresión introducido PDZ-CJ7(arg) se confirmó mediante la realización de la PCR usando el ADN genómico de las cepas seleccionadas anteriormente como molde y cebadores (SEQ ID NO: 13 y 16) (28 ciclos de desnaturalización durante 30 segundos a 94 °C, hibridación durante 30 segundos a 55 °C y extensión durante 60 segundos a 72 °C). Por último, las cepas confirmadas se nombraron como ATCC 13032 AargF ANCgl1221 P(CJ7)-argCJBD y KCCM-10785P AargF ANCgl1221 P(CJ7)-argCJBD, correspondientemente.
Ejemplo 4: Mejora de la productividad de ornitina mediante la eliminación de los genes argF y NCgl1221 y mejora del nivel de expresión de argCJBD
Ejemplo 4-1: Productividad de ornitina de la cepa derivada de Corynebacterium glutamicum ATCC 13032
Para examinar si la productividad de la ornitina se ve afectada por la eliminación de los genes argF y NCgl1221 y la mejora del nivel de expresión de argCJBD en las cepas derivadas de Corynebacterium glutamicum ATCC 13032, se comparó la capacidad de producción de ornitina entre las cepas preparadas en los Ejemplos 2 y 3.
En detalle, se dispersó cada una de las cepas preparadas en los Ejemplos 2 y 3 (ATCC 13032 AargF ANCgl1221, ATCC 13032 AargF ANCgl1221/pHC139T-argCJBD(Cgl), ATCC 13032 AargF ANCgl1221 P(CJ7)-argCJBD) en una placa de CMA que contenía arginina 1 mM, y se cultivó a 37 °C durante 24 horas. Se inoculó cada una de las cepas cultivadas en 25 ml de medio de titulación (glucosa al 2 % (p/v), polipeptona al 1 % (p/v), extracto de levadura al 0,5 % (p/v), (NH4)2SO4 al 0,5 % (p/v), urea al 0,15 % (p/v), KH2PO4 al 0,4 % (p/v), K2HPO4 al 0,8 % (p/v), MgSO4 al 0,05 % (p/v), 100 pg/l de biotina y 1 mg/l de tiamina) que contenía arginina 1 mM, y luego se cultivó con agitación a 30 °C y 200 rpm durante 48 horas, y se determinó la concentración de ornitina producida en cada cultivo, y se comparó entre sí (Tabla 5). En este momento, la cepa ATCC 13032 sin modificación genómica se usó como grupo de control.
T l
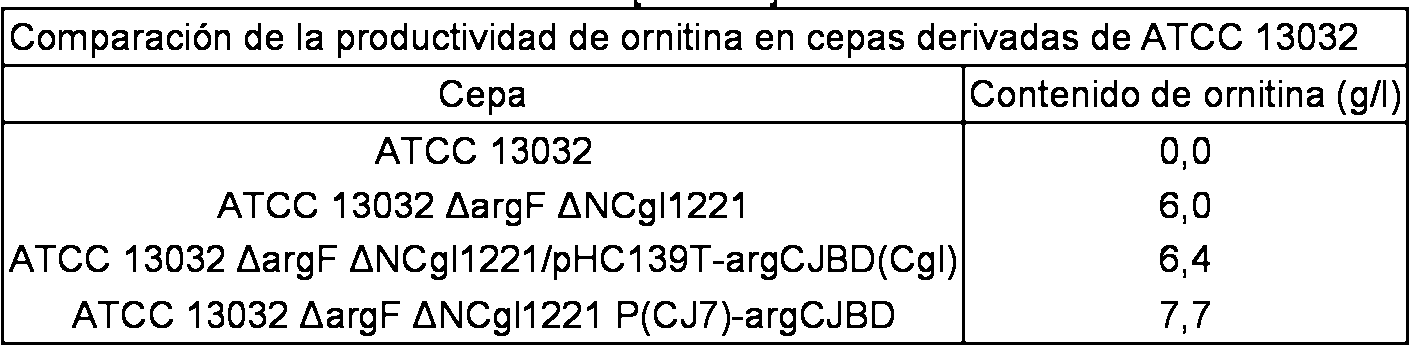
Como se muestra en la Tabla 5, la cepa con argF y NCgl1221 eliminados produjeron 6,0 g/l de ornitina, que no fueron producidos por la cepa de tipo silvestre. Con respecto al aumento del nivel de expresión del gen argCJBD, cuando el gen argCJBD se introdujo en forma de vector, la concentración de la ornitina producida fue de 6,4 g/l, y cuando se reemplazó el promotor argCJBD en el cromosoma por CJ7, la concentración de la ornitina producida aumentó ligeramente hasta 7,7 g/l.
Ejemplo 4-2: Productividad de ornitina de la cepa derivada de Corynebacterium glutamicum KCCM-10785P
Claims (8)
1. Un microorganismo que pertenece a Corynebacterium sp., que tiene una capacidad mejorada para producir ornitina en comparación con su cepa parental, en el que se modifican las actividades de la ornitina carbamoiltransferasa y una proteína que participa en la exportación del glutamato (NCgl1221) para que sean atenuadas mediante una eliminación parcial o completa de los genes que codifican la ornitina carbamoil transferasa y NCg11221, en comparación con las actividades de la ornitina carbamoiltransfereasa y NCg11221 que posee la cepa parental en su estado nativo, en el que la cepa parental no tiene ninguna modificación en las actividades de la ornitina carbamoiltransferasa ni de NCg11221.
2. El microorganismo de acuerdo con la reivindicación 1, en el que la ornitina carbamoiltransferasa tiene una secuencia de aminoácidos de SEQ ID NO: 18, o una secuencia de aminoácidos que tiene un 70 % o más de homología con la secuencia.
3. El microorganismo de acuerdo con la reivindicación 1, en el que la proteína que participa en la exportación del glutamato tiene una secuencia de aminoácidos de SEQ ID NO: 20, o una secuencia de aminoácidos que tiene un 70 % o más de homología con la secuencia.
4. El microorganismo de acuerdo con la reivindicación 1, en el que las actividades de la acetil gamma-glutamil-fosfato reductasa (ArgC), la acetilglutamato sintasa o la ornitina acetiltransferasa (argJ), la acetilglutamato quinasa (ArgB) y la acetilornitina aminotransferasa (ArgD) se potencian aumentando el número de copias de polinucleótidos que codifican las proteínas, en comparación con las actividades de las enzimas que posee la cepa parental en su estado nativo.
5. El microorganismo de acuerdo con la reivindicación 4, en el que ArgC, ArgJ, ArgB y ArgD tienen las secuencias de aminoácidos de SEQ ID NO: 23, 25, 27 y 29, respectivamente, o una secuencia de aminoácidos que tiene un 70 % o más de homología con las respectivas secuencias.
6. El microorganismo de acuerdo con la reivindicación 1, en el que el microorganismo que pertenece a Corynebacterium sp. es Corynebacterium glutamicum.
7. El microorganismo de acuerdo con la reivindicación 1, en el que el microorganismo que pertenece a Corynebacterium sp. es Corynebacterium glutamicum (KCCM11137P).
8. Un método de producción de ornitina, que comprende las etapas de:
(i) cultivar el microorganismo de la reivindicación 1; y
(ii) recuperar la ornitina del microorganismo cultivado o del cultivo.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR20100124866 | 2010-12-08 | ||
| KR1020110130594A KR101372635B1 (ko) | 2010-12-08 | 2011-12-07 | 오르니틴 생산능이 향상된 미생물 및 이를 이용하여 오르니틴을 생산하는 방법 |
| PCT/KR2011/009477 WO2012077994A2 (ko) | 2010-12-08 | 2011-12-08 | 오르니틴 생산능이 향상된 미생물 및 이를 이용하여 오르니틴을 생산하는 방법 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2703256T3 true ES2703256T3 (es) | 2019-03-07 |
Family
ID=46684297
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES11846603T Active ES2703256T3 (es) | 2010-12-08 | 2011-12-08 | Microorganismo que tiene una mejor capacidad de producción de ornitina y método de producción de ornitina usando el mismo |
Country Status (12)
| Country | Link |
|---|---|
| US (1) | US9303277B2 (es) |
| EP (1) | EP2650358B1 (es) |
| JP (1) | JP5860476B2 (es) |
| KR (1) | KR101372635B1 (es) |
| CN (1) | CN103282486B (es) |
| AU (1) | AU2011339171B2 (es) |
| BR (1) | BR112013014441B1 (es) |
| ES (1) | ES2703256T3 (es) |
| MY (1) | MY165897A (es) |
| PL (1) | PL2650358T3 (es) |
| RU (1) | RU2567669C2 (es) |
| WO (1) | WO2012077994A2 (es) |
Families Citing this family (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR101526047B1 (ko) * | 2013-04-26 | 2015-06-04 | 상지대학교산학협력단 | 아미노트랜스퍼라제 활성 강화를 통하여 l-오르니틴 생산능이 향상된 미생물 및 이를 이용한 l-오르니틴의 제조방법 |
| KR101835164B1 (ko) | 2013-07-17 | 2018-03-06 | 씨제이제일제당 (주) | 향상된 퓨트레신 생산능을 가지는 변이된 오르니틴 디카복실레이즈 단백질 및 이의 용도 |
| KR101835935B1 (ko) | 2014-10-13 | 2018-03-12 | 씨제이제일제당 (주) | L-아르기닌을 생산하는 코리네박테리움 속 미생물 및 이를 이용한 l-아르기닌의 제조 방법 |
| US10544435B2 (en) | 2015-03-12 | 2020-01-28 | Chrysea Limited | L-ornithine production in eukaryotic cells |
| KR101813759B1 (ko) * | 2015-06-24 | 2018-01-02 | 씨제이제일제당 (주) | 퓨트레신 또는 오르니틴 생산 미생물 및 이를 이용한 퓨트레신 또는 오르니틴 생산방법 |
| KR101735935B1 (ko) | 2015-07-20 | 2017-05-16 | 씨제이제일제당 (주) | 퓨트레신 또는 오르니틴 생산 미생물 및 이를 이용한 퓨트레신 또는 오르니틴 생산방법 |
| CN105950524A (zh) * | 2016-04-27 | 2016-09-21 | 齐鲁工业大学 | 一种高产l-赖氨酸的谷氨酸棒状杆菌工程菌的构建方法 |
| KR101937569B1 (ko) * | 2017-06-14 | 2019-01-11 | 씨제이제일제당 (주) | 신규 폴리펩타이드 및 이를 이용한 오르니틴계 산물 생산방법 |
| KR101941775B1 (ko) | 2017-06-14 | 2019-01-24 | 씨제이제일제당 (주) | 신규 폴리펩타이드 및 이를 이용한 오르니틴계 산물 생산방법 |
| CN110195088B (zh) * | 2018-02-26 | 2022-09-27 | 中国科学院分子植物科学卓越创新中心 | 一种精氨酸水解酶及其编码基因和应用 |
| KR20230136407A (ko) * | 2022-03-18 | 2023-09-26 | 씨제이제일제당 (주) | 카르바모일 인산 합성효소의 활성이 약화된 l-아미노산을 생산하는 미생물 및 이를 이용한 l-아미노산 생산 방법 |
| KR20230136408A (ko) * | 2022-03-18 | 2023-09-26 | 씨제이제일제당 (주) | 카르바모일 인산 합성효소 라지 서브유닛 변이체 및 이를 이용한 l-오르니틴 생산 방법 |
Family Cites Families (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7344874B2 (en) * | 2004-03-04 | 2008-03-18 | Ajinomoto Co., Inc. | L-glutamic acid-producing microorganism and a method for producing L-glutamic acid |
| BRPI0516176B1 (pt) * | 2004-09-28 | 2020-12-29 | Kyowa Hakko Bio Co., Ltd. | Polipeptídeo, dna, microorganismo, e, processo para produzir l-arginina, l-ornitina ou lcitrulina |
| CN103468758B (zh) * | 2004-12-28 | 2015-10-28 | 味之素株式会社 | 产生l-谷氨酸的微生物和产生l-谷氨酸的方法 |
| EP3170889A1 (en) * | 2006-09-15 | 2017-05-24 | CJ Cheiljedang Corporation | A corynebacteria having enhanced l-lysine productivity and a method of producing l-lysine using the same |
| ES2589378T3 (es) * | 2008-04-10 | 2016-11-14 | Korea Advanced Institute Of Science And Technology | Microorganismo mutante con capacidad elevada para producir putrescina, y preparación de putrescina usando el mismo |
| KR20100060909A (ko) * | 2008-11-28 | 2010-06-07 | 전북대학교산학협력단 | 전사억제인자의 기능 감소에 의한 오르니틴의 생산성을 증가시킨 미생물 및 이를 이용한 오르니틴의 생산 방법 |
-
2011
- 2011-12-07 KR KR1020110130594A patent/KR101372635B1/ko active IP Right Grant
- 2011-12-08 MY MYPI2013002102A patent/MY165897A/en unknown
- 2011-12-08 WO PCT/KR2011/009477 patent/WO2012077994A2/ko active Application Filing
- 2011-12-08 AU AU2011339171A patent/AU2011339171B2/en active Active
- 2011-12-08 US US13/992,241 patent/US9303277B2/en active Active
- 2011-12-08 CN CN201180064409.4A patent/CN103282486B/zh active Active
- 2011-12-08 RU RU2013131039/10A patent/RU2567669C2/ru active
- 2011-12-08 BR BR112013014441-6A patent/BR112013014441B1/pt active IP Right Grant
- 2011-12-08 PL PL11846603T patent/PL2650358T3/pl unknown
- 2011-12-08 ES ES11846603T patent/ES2703256T3/es active Active
- 2011-12-08 EP EP11846603.6A patent/EP2650358B1/en active Active
- 2011-12-08 JP JP2013543102A patent/JP5860476B2/ja active Active
Also Published As
| Publication number | Publication date |
|---|---|
| JP5860476B2 (ja) | 2016-02-16 |
| BR112013014441A2 (pt) | 2016-09-13 |
| EP2650358B1 (en) | 2018-10-17 |
| KR101372635B1 (ko) | 2014-03-13 |
| KR20120064045A (ko) | 2012-06-18 |
| RU2567669C2 (ru) | 2015-11-10 |
| RU2013131039A (ru) | 2015-01-20 |
| WO2012077994A3 (ko) | 2012-09-07 |
| AU2011339171B2 (en) | 2016-02-11 |
| EP2650358A2 (en) | 2013-10-16 |
| US20130344545A1 (en) | 2013-12-26 |
| CN103282486B (zh) | 2016-02-17 |
| CN103282486A (zh) | 2013-09-04 |
| JP2013544532A (ja) | 2013-12-19 |
| MY165897A (en) | 2018-05-18 |
| US9303277B2 (en) | 2016-04-05 |
| EP2650358A4 (en) | 2015-07-01 |
| BR112013014441B1 (pt) | 2021-08-17 |
| AU2011339171A1 (en) | 2013-07-18 |
| PL2650358T3 (pl) | 2019-05-31 |
| WO2012077994A2 (ko) | 2012-06-14 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2703256T3 (es) | Microorganismo que tiene una mejor capacidad de producción de ornitina y método de producción de ornitina usando el mismo | |
| KR101348461B1 (ko) | 퓨트레신을 생산하는 미생물 및 이를 이용하여 퓨트레신을 생산하는 방법 | |
| US11365400B2 (en) | Microorganism for producing ornithine and process for producing ornithine using them | |
| AU2015331160B2 (en) | A Microorganism Of Genus Corynebacterium Having An Ability To Produce L-Arginine And A Method For Producing L-Arginine Using The Same | |
| JP6297134B2 (ja) | プトレシン生産性を有する微生物及びそれを用いたプトレシン生産方法 | |
| ES2739002T3 (es) | Microorganismos para producir putrescina u ornitina y proceso para producir putrescina u ornitina usando los mismos | |
| TW201942360A (zh) | 新穎啟動子及使用該啟動子製造l-胺基酸的方法 | |
| ES2711857T3 (es) | Cepa variante productora de putrescina y método de producción que la utiliza | |
| TWI657140B (zh) | 具有屍胺生產力之微生物及使用該微生物製造屍胺之方法 | |
| ES2717600T3 (es) | Microorganismo productor de diamina y método para producir diamina usando el mismo |