-
TECHNISCHES
GEBIET
-
Die vorliegende Erfindung betrifft
allgemein Verfahren und Zusammensetzungen zum Analysieren von Nukleinsäuremolekülen und,
genauer gesagt, Markierungen, die in einer breiten Vielfalt von
Nukleinsäurereaktionen
eingesetzt werden können,
bei denen die Abtrennung von Nukleinsäuremolekülen auf der Grundlage ihrer
Größe erforderlich
ist.
-
HINTERGRUND
DER ERFINDUNG
-
Nachweis und Analyse von Nukleinsäuremolekülen sind
mit die wichtigsten Techniken in der Biologie. Solche Techniken
sind wesentlich in der Molekularbiologie und spielen eine sich schnell
erweiternde Rolle im Rest der Biologie.
-
Im allgemeinen umfaßt eine
Art von Analyse von Nukleinsäurereaktionen
die Abtrennung von Nukleinsäuremolekülen auf
der Grundlage ihrer Länge.
Zum Beispiel ist eine im breiten Umfang verwendete Technik, die
Polymerasekettenreaktion (PCR) (siehe U.S.-Patente Nrn. 4,683,195,
4,683,202 und 4,800,159), eine im breiten Umfang eingesetzte Technik
geworden, um sowohl in einer Probe vorliegende Sequenzen zu identifizieren
als auch DNA-Moleküle
für weitere
Manipulation zu synthetisieren.
-
Kurz gesagt, werden bei der PCR DNA-Sequenzen
durch eine enzymatische Reaktion amplifiziert, die neue DNA-Stränge in entweder
einer geometrischen oder linearen Art und Weise synthetisiert. Im
Anschluß an
die Amplifikation müssen
die DNA-Sequenzen nachgewiesen und identifiziert werden. Wegen nicht-spezifischer
Amplifikationen, die ansonsten die Analyse durcheinanderbringen
würden,
oder einer Reinheitsanforderung werden die PCR-Reaktionsprodukte
im allgemeinen vor dem Nachweis einer Trennung unterzogen. Die Trennung
auf der Grundlage der Größe (d. h.
Länge)
der Produkte liefert die nützlichste
Information. Das Verfahren, das die höchste Auflösung von Nukleinsäuremolekülen ergibt,
ist die elektrophoretische Trennung. Bei diesem Verfahren wird jede
einzelne PCR-Reaktion auf ein geeignetes Gel aufgebracht und einem
Spannungspotential ausgesetzt. Die Anzahl von Proben, die verarbeitet
werden können,
ist durch die Anzahl von Näpfen
im Gel beschränkt.
Auf den meisten Gelvorrichtungen können ungefähr 10 bis 64 Proben in einem
einzigen Gel getrennt werden. Somit ist die Verarbeitung einer großen Anzahl
von Proben sowohl arbeits- als auch materialintensiv.
-
Elektrophoretische Trennung muß mit irgendeinem
Nachweissystem gekoppelt werden, um Daten zu erhalten. Nachweissysteme
für Nukleinsäuren verwenden
im allgemeinen, und fast ausschließlich, einen Einlagerungsfarbstoff
oder eine radioaktive Markierung und weniger häufig eine nicht-radioaktive
Markierung. Einlagerungsfarbstoffe, wie etwa Ethidiumbromid, sind
einfach zu verwenden. Der Farbstoff wird während der Elektrophorese in
die Gelmatrix einbezogen oder das Gel wird, im Anschluß an die
Elektrophorese, in einer farbstoffhaltigen Lösung getränkt. Der Farbstoff kann in
einigen Fällen
direkt sichtbar gemacht werden, wird aber häufiger, und insbesondere für Ethidiumbromid,
mit Licht (z. B. UV) angeregt, um zu fluoreszieren. Trotz dieser
offensichtlich einfachen Anwendung haben solche Farbstoffe einige
merkbare Nachteile. Erstens sind die Farbstoffe unempfindlich und
es muß eine
große
Menge an Nukleinsäuremolekülen vorliegen,
um die Produkte sichtbar zu machen. Zweitens sind die Farbstoffe
typischerweise mutagen oder karzinogen.
-
Eine empfindlichere Nachweistechnik
als Farbstoffe verwendet eine radioaktive (oder nicht-radioaktive) Markierung.
Typischerweise wird entweder ein radioaktiv markiertes Nukleotid
oder ein radioaktiv markierter Primer in die PCR-Reaktion einbezogen.
Im Anschluß an
die Trennung wird die radioaktive Markierung durch Autoradiographie "sichtbar gemacht". Obgleich empfindlicher
leidet der Nachweis an Filmbeschränkungen, wie etwa Abweichung
vom Reziprozitätsgesetz
und Nicht-Linearität.
Diese Beschränkungen
können überwunden
werden, indem die Markierung durch Phosphorbildanalyse nachgewiesen
wird. Radioaktive Markierungen haben jedoch hohe Anforderungen an
die Sicherheit, was die Ausnutzung von Resourcen erhöht und spezialisierte
Ausrüstung
und Personaltraining erfordert. Aus solchen Gründen hat die Verwendung von nicht-radioaktiven
Markierungen zunehmend an Popularität gewonnen. In solchen Systemen
enthalten Nukleotide eine Markierung, wie etwa ein Fluorphor, Biotin
oder Digoxin, die durch einen Antikörper oder ein anderes Molekül (z. B.
anderes Teil eines Ligandenpaares) nachgewiesen werden kann, der
(das) mit einem Enzym markiert ist, das mit einem chromogenen Substrat
reaktiv ist. Diese Systeme haben nicht die Sicherheitsbedenken,
wie oben beschrieben, verwenden aber Komponenten, die oft labil
sind und nicht-spezifische Reaktionen liefern können, was zu einem hohen Hintergrund
führt (d.
h. niedrigem Signal-Rausch-Verhältnis).
-
US-A-5118605 betrifft Verfahren zum
Testen eines Nukleinsäureanalyten.
WO-A-9504160 betrifft ein Markierungsreagenz und ein Testverfahren.
-
ZUSAMMENFASSUNG
DER ERFINDUNG
-
Kurz gesagt stellt die vorliegende
Erfindung Zusammensetzungen und Verfahren zur Verfügung, die in
einer breiten Vielfalt von Ligandenpaarreaktionen eingesetzt werden
können,
bei denen die Trennung von interessierenden Molekülen, wie
etwa Nukleinsäuremolekülen; auf
der Grundlage ihrer Größe erforderlich
ist. Repräsentative
Beispiele für
Verfahren, die mit dem Hintergrund der hierin zur Verfügung gestellten
Offenbarung verbessert werden können,
schließen
PCR, Differential Display, RNA-Fingerprinting,
PCR-SSCP, Oligo-Ligationsassays, Nukleaseverdauungsverfahren (z.
B. Assays auf der Basis von Exo- und Endonuklease) und Didesoxy-Fingerprinting
ein. Die hierin beschriebenen Verfahren können in einem weiten Bereich
von Feldern eingesetzt werden, einschließlich z. B. bei der Entwicklung
von Diagnostika für
klinische Anwendung oder Forschung, der Bestimmung von Polymorphismen
und der Entwicklung genetischer Karten.
-
In einem Aspekt der vorliegenden
Erfindung wird eine Verbindung der Formel: Tms-L-X'
-
Zur Verfügung gestellt, worin Tms eine organische Gruppe ist, die durch
Massenspektrometrie nachweisbar ist, umfassend Kohlenstoff, mindestens
eines von Wasserstoff und Fluor, und gegebenenfalls Atome, ausgewählt aus
Sauerstoff, Stickstoff, Schwefel, Phosphor und Jod;
-
L ist eine organische Gruppe, die
es einer einmaligen Tms-enthaltenden Gruppe
ermöglicht,
von dem Rest der Verbindung abgespalten zu werden, wobei die Tms-enthaltende Gruppe eine funktionelle Gruppe
umfaßt,
die einen einzelnen ionisierten Ladungszustand unterstützt, wenn
die Verbindung Massenspektrometrie unterzogen wird, und die tertiäres Amin,
quaternäres
Amin oder organische Säure
ist; und
-
X ist eine funktionelle Gruppe, ausgewählt aus
Phosphoramidit und H-Phosphonat.
-
In einem anderen Aspekt stellt die
vorliegende Erfindung ein Verfahren zur Bestimmung der Anwesenheit
eines Single Nukleotide Polymorphismus in einem Nukleinsäureziel
zur Verfügung,
umfassend:
-
- a) Amplifizieren einer Sequenz eines Nukleinsäureziels,
das einen Single Nukleotide Polymorphismus enthält;
- b) Erzeugen einer Einzelstrangform des Ziels;
- c) Kombinieren einer markierten Nukleinsäuresonde, die die Struktur
Tms-L-X' aufweist
mit den amplifizierten Zielnukleinsäuremolekülen unter Bedingungen und für eine Zeitdauer,
die ausreichend ist, um eine Hybridisierung der markierten Nukleinsäuresonde
an komplementäre
amplifizierte ausgewählte
Zielnukleinsäuremoleküle zu ermöglichen,
wobei die Markierung mit einem bestimmten Single Nukleotide Polymorphismus
korrelierbar ist und durch Spektrometrie oder Potentiometrie nachweisbar
ist;
- d) Abtrennen von nicht hybridisierter markierter Probe von hybridisierter
markierter Probe durch eine Größenbestimmungs-Methodologie;
- e) Abspalten des Markers von der Sonde; und
- f) Nachweisen des Markers durch Spektrometrie oder Potentiometrie
und Bestimmen der Anwesenheit des Single Nukleotide Polymorphismus.
-
In einem anderen Aspekt stellt die
vorliegende Erfindung ein Verfahren zur Bestimmung der Anwesenheit
eines Single Nukleotide Polymorphismus in einem Nukleinsäureziel
zur Verfügung,
umfassend:
-
- a) Amplifizieren einer Sequenz eines Nukleinsäureziels,
das einen Single Nukleotide Polymorphismus enthält;
- b) Kombinieren eines markierten Nukleinsäureprimers, der die Struktur
Tms-L-X' aufweist,
mit den amplifizierten Zielnukleinsäuremolekülen unter Bedingungen und für eine Zeitdauer,
die ausreichend ist, um ein Annealing des markierten Nukleinsäureprimers
an komplementäre
amplifizierte ausgewählte
Zielnukleinsäuremoleküle zu ermöglichen,
wobei der Oligonukleotidprimer eine zu der Wildtyp-Sequenz oder
dem Single Nukleotide Polymorphismus komplementäre äußerst 3'-liegende Base aufweist, wobei der Marker
mit einem bestimmten Single Nukleotide Polymorphismus korrelierbar
ist und durch Spektrometrie oder Potentiometrie nachweisbar ist;
- c) Verlängern
des Primers, wobei ein zu dem Ziel komplementärer Strang synthetisiert wird,
wenn die am weitesten 3'-liegende
Base des Primers mit dem Ziel komplementär ist;
- d) Abtrennen von nicht verlängertem
markierten Primer von verlängertem
markierten Primer durch eine Größenbestimmungs-Methodologie;
- e) Abspalten des Markers von den Primern oder verlängerten
Primern; und
- f) Nachweisen des Markers durch Spektrometrie oder Potentiometrie,
und daraus Bestimmen daraus der Anwesenheit des Single Nukleotide
Polymorphismus.
-
In einem anderen Aspekt stellt die
vorliegende Erfindung ein Verfahren zur Bestimmung der Menge eines
spezifischen mRNA-Moleküls
in einer Nukleinsäurepopulation
zur Verfügung,
umfassend:
-
- a) Überführen einer
RNA-Population in eine cDNA-Population;
- b) Hinzufügen
einer einzelsträngigen
Nukleinsäure
(interner Standard), die eine Vielzahl von Single Nukleotide Polymorphismen
enthält,
die ansonsten identisch zu dem cDNA-Ziel ist;
- c) Amplifizieren einer spezifischen Sequenz des cDNA-Ziels;
- d) Coamplifizieren des internen Standards, wobei der interne
Standard dieselbe Länge
aufweist, wie das cDNA-Amplicon;
- e) Erzeugen einer einzelsträngigen
Form des Ziels;
- f) Kombinieren eines Sets von markierten Nukleinsäuresonden,
die die Struktur Tms-L-X' aufweisen,
mit der amplifizierten Ziel-cDNA und dem amplifizierten internen
Standard unter Bedingungen für
eine Zeitdauer, die ausreichend ist, um eine Hybridisierung der
markierten Nukleinsäuresonde
an komplementäre
ausgewählte Ziel-cDNA
und interne Standardsequenzen zu erlauben, wobei der Marker mit
einer bestimmten cDNA-Sequenz korrelierbar ist und ein zweiter Marker
ist mit dem internen Standard korrelierbar, und ist durch Spektrometrie
oder Potentiometrie nachweisbar;
- g) Abtrennen von nicht hybridisierter markierter Sonde von hybridisierter
markierter Sonde durch eine Größenbestimmungs-Methodologie;
- h) Abspalten des Markers von den Sonden;
- i) Nachweisen der Marker durch Spektrometrie oder Potentiometrie;
und
- j) Nehmen des Verhältnisses
von Marker, der zu cDNA korreliert ist, gegenüber Marker, der mit dem internen Standard
korreliert ist und daraus bestimmen der Menge der cDNA, wodurch
die Menge der spezifischen mRNA in einer Nukleinsäurepopulation
bestimmt wird.
-
In den oben genannten Verfahren weist
die markierte Nukleinsäure
die Struktur Tms-L-X' auf, wobei X' die Nukleinsäure ist und Tms und
L wie oben definiert sind.
-
Innerhalb eines Aspekt der vorliegenden
Erfindung werden Verfahren zum Bestimmen der Identität eines
Nukleinsäuremoleküls zur Verfügung gestellt,
die die Schritte umfassen von (a) Erzeugen markierter Nukleinsäuremoleküle aus einem
oder mehreren ausgewählten
Target-Nukleinsäuremolekülen, wobei
eine Markierung mit einem bestimmten Nukleinsäurefragment korrelativ und
durch Nicht-Fluoreszenz-Spektrometrie oder Potentiometrie nachweisbar
ist, (b) Abtrennen der markierten Fragmente nach Größe, (c)
Abspalten der Markierungen von den markierten Fragmenten und (d)
Nachweisen der Markierungen durch Nicht-Fluoreszenz-Spektrometrie
oder Potentiometrie und Bestimmen der Identität der Nukleinsäuremoleküle daraus.
-
In einem verwandten Aspekt der Erfindung
werden Verfahren zum Nachweisen eines ausgewählten Nukleinsäuremoleküls zur Verfügung gestellt,
die die Schritte umfassen: (a) Kombinieren markierter Nukleinsäuresonden
mit Target-Nukleinsäuremolekülen unter
Bedingungen und für
einen Zeitraum, die ausreichend sind, um Hybridisierung einer markierten
Nukleinsäuresonde
an eine komplementäre
ausgewählte
Target-Nukleinsäuresequenz
zu ermöglichen,
wobei eine markierte Nukleinsäuresonde
durch Nicht-Fluoreszenzspektrometrie
oder Potentiometrie nachweisbar ist, (b) Verändern der Größe hybridisierter
markierter Sonden, nicht-hybridisierter Sonden oder Target-Moleküle oder
der Sonden:Target-Hybride, (c) Abtrennen der markierten Sonden nach
Größe, (d)
Abspalten der Markierungen von den markierten Sonden und (e) Nachweisen
der Markierungen durch Nicht-Fluoreszenz-Spektrometrie oder Potentiometrie
und Nachweisen des ausgewählten
Nukleinsäuremoleküls daraus.
-
In weiteren Aspekten werden Verfahren
zum Genotypisieren eines ausgewählten
Organismus zur Verfügung
gestellt, die die Schritte umfassen: (a) Erzeugen markierter Nukleinsäuremoleküle aus einem
ausgewählten
Target-Molekül,
wobei eine Markierung mit einem bestimmten Fragment korrelativ ist
und durch Nicht-Fluoreszenz-Spektrometrie oder Potentiometrie nachgewiesen
werden kann, (b) Abtrennen der markierten Moleküle nach Sequenzlänge, (c)
Abspalten der Markierung vom markierten Molekül und (d) Nachweisen der Markierung
durch Nicht-Fluoreszenz-Spektrometrie oder Potentiometrie und Bestimmen
des Genotyps des Organismus daraus.
-
In einem weiteren Aspekt werden Verfahren
zum Genotypisieren eines ausgewählten
Organismus zur Verfügung
gestellt, die die Schritte umfassen: (a) Kombinieren eines markierten
Nukleinsäuremoleküls mit einem
ausgewählten
Target-Molekül
unter Bedingungen und für
einen Zeitraum, die ausreichend sind, um Hybridisierung des markierten
Moleküls
an das Target-Molekül
zu ermöglichen,
wobei eine Markierung mit einem bestimmten Fragment korrelativ ist
und durch Nicht-Fluoreszenz-Spektrometrie oder Potentiometrie nachgewiesen
werden kann, (b) Abtrennen der markierten Fragmente nach Sequenzlänge, (c)
Abspalten der Markierung vom markierten Fragment und (d) Nachweisen der
Markierung durch Nicht-Fluoreszenz-Spektrometrie oder Potentiometrie
und Bestimmen des Genotyps des Organismus daraus.
-
Im Kontext der vorliegenden Erfindung
sollte "biologische
Proben" so verstanden
werden, daß diese nicht
nur Proben einschließen,
die von lebenden Organismen erhalten werden (z. B. Säugern, Fischen,
Bakterien, Parasiten, Viren, Pilzen und dergleichen) oder aus der
Umwelt (z. B. Luft, Wasser oder Feststoffproben), sondern auch biologische
Materialien, die künstlich
oder synthetisch hergestellt sein können (z. B. Phagenbibliotheken,
Bibliotheken organischer Moleküle,
Pools genomischer Klone, cDNA-Klone, RNA-Klone oder dergleichen).
Repräsentative
Beispiele für
biologische Proben schließen
biologische Flüssigkeiten
(z. B. Blut, Samen, Hirn-Rückenmarks-Flüssigkeit,
Urin), biologische Zellen (z. B. Stammzellen, B- oder T-Zellen, Leberzellen, Fibroblasten
oder dergleichen) und biologische Gewebe ein. Schließlich schließen repräsentative
Beispiele für
Organismen, die genotypisiert werden können, praktisch jeden einzelligen
oder mehrzelligen Organismus ein, wie etwa Warmblüter, Säuger oder
Wirbeltiere (z. B. Menschen, Schimpansen, Makaken, Pferde, Kühe, Schweine,
Schafe, Hunde, Katzen, Ratten und Mäuse sowie Zellen von irgendeinem
dieser), Bakterien, Parasiten, Viren, Pilze und Pflanzen.
-
In verschiedenen Ausführungsformen
der oben beschriebenen Verfahren können die Nukleinsäuresonden
und/oder -moleküle
der vorliegenden Erfindung durch z. B. einen Ligations-, Spaltungs-
oder Extensions(z. B: PCR)-Reaktion erzeugt werden. In weiteren
verwandten Aspekten können
die Nukleinsäuresonden oder
-moleküle
mit nicht-3'-markierten
Oligonukleotid-Primern (z. B. 5'-markierten
Oligonukleotid-Primern) oder Didesoxynukleotidterminatoren markiert
sein.
-
In weiteren Ausführungsformen der Erfindung
können
4, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 200,
250, 300, 350, 400, 450 oder mehr als 500 unterschiedliche und einzigartig
markierte Moleküle
in einer gegebenen Reaktion gleichzeitig eingesetzt werden, wobei
jede Markierung für
ein ausgewähltes Nukleinsäuremolekül oder -fragment,
oder -sonde, einzigartig
ist und separat identifiziert werden kann.
-
Die Markierungen) werden durch Massenspektrometrie
nachgewiesen. Repräsentative
Beispiele für schließen time-of-flight-Massenspektrometrie,
Quadrapol-Massenspektrometrie, Magnetsektor-Massenspektrometrie
und Elektrosektor-Massenspektrometrie ein. Spezifische Ausführungsformen
solcher Techniken schließen
Ionenfallen-Massenspektrometrie, Elektrosprayionisations-Massenspektrometrie,
Ionenspray-Massenspektrometrie, Flüssigionisations-Massenspektrometrie,
Atmosphärendruckioni-sations-Massenspektrometrie,
Elektronenionisations-Massenspektrometrie, Massenspektrometrie mit
Ionisation durch Beschuß mit schnellen
Atomen, MALDI-Massenspektrometrie, Photoionisations-Flugzeit-Massenspektrometrie,
Laser-Tröpfchen-Massenspektrometrie,
MALDI-TOF-Massenspektrometrie, APCI-Massenspektrometrie, Nanospray-Massenspektrometrie,
Massenspektrometrie mit Ionisation zerstäubten Sprays, Massenspektrometrie mit
chemischer Ionisation, Resonanzionisations-Massenspektrometrie,
Sekundärionisations-Massenspektrometrie
und Thermospray-Massenspektrometrie ein.
-
In noch weiteren Ausführungsformen
der Erfindung können
die Target-Moleküle,
hybridisierten markierten Sonden, nicht-hybridisierten Sonden oder
Target-Moleküle,
Sonden:Target-Hybride oder markierten Nukleinsäuresonden oder -moleküle von anderen
Molekülen
unter Verwendung von Verfahren abgetrennt werden, die zwischen der
Größe von Molekülen unterscheiden
(entweder tatsächliche
lineare Größe oder
dreidimensionale Größe). Repräsentative
Beispiele solcher Verfahren schließen Gelelektrophorese, Kapillarelektrophorese,
Mikrokanalelektrophorese, HPLC, Ausschlußchromatographie, Filtration,
Polyacrylamidgelelektrophorese, Flüssigchromatographie, Umkehrausschlußchromatographie,
Ionenaustauschchromatographie, Umkehrphasenflüssigchromatographie, Pulsfeldelektrophorese,
Feldinversionselektrophorese, Dialyse und Fluoreszenz-aktivierte
Flüssigtröpfchensortierung.
Alternativ können
die Target-Moleküle,
hybridisierten markierten Sonden, nicht-hybridisierten Sonden oder
Target-Moleküle,
Sonden:Target-Hybride oder markierten Nukleinsäuresonden oder -moleküle an einen
festen Träger
(z. B. Hohlfasern (Amicon Corporation, Danvers, Mass.), Perlen (Polysciences,
Warrington, Pa.), Magnetperlen (Robbin Scientific, Mountain View,
Calif.), Platten, Schalen und Kolben (Corning Glass Works, Corning,
N.Y.), Maschengitter (Becton Dickinson, Mountain View, Calif.),
Siebe und feste Fasern (siehe Edelman et al., U.S.-Patent Nr. 3,843,324;
siehe auch Kuroda et al., U.S.-Patent
Nr. 4,416,777), Membranen (Millipore Corp., Bedford, Mass.) und
Tauschstäbchen)
gebunden sein. Wenn das erste oder zweite Teil oder freiliegende
Nukleinsäuren
an einen festen Träger
sind, können
die hierin offenbarten Verfahren in bestimmten Ausführungsformen
der Erfindung außerdem
den Schritt umfassen, daß nicht-gebundenes
Material vom Träger
abgewaschen wird.
-
In weiteren Ausführungsformen können die
markierten Nukleinsäuremoleküle oder
-sonden mit solchen Verfahren gespalten werden, wie chemischen,
Oxidations-, Reduktions-, säurelabilen,
basenlabilen, enzymatischen, elektrochemischen, wärme- und
photolabilen Verfahren. In weiteren Ausführungsformen können die
Schritte Abtrennen, Spalten und Nachweisen in einer kontinuierlichen
Art und Weise durchgeführt
werden, z. B. auf einer einzigen Vorrichtung, die automatisiert
sein kann.
-
In bestimmten Ausführungsformen
der Erfindung werden die Größe der hybridisierten
markierten Sonden, nicht-hybridisierten Sonden oder Targetmoleküle oder
Sonden:Target-Hybride
mit einem Verfahren verändert,
das ausgewählt
ist aus der Gruppe, die aus Polymerase-Extension, Ligation, Exonuklease-Verdau,
Endonuklease-Verdau, Restriktionsenzym-Verdau, ortsspezifischem
Rekombinase-Verdau, Ligation, Mismatch-Verdau mit spezifischer Nuklease, Methylierungs-spezifischer
Nuklease-Verdau, kovalente Bindung von Sonde an Target und Hybridisierung
besteht.
-
Die Verfahren und Zusammensetzungen,
die hierin beschrieben sind, können
in einer breiten Vielfalt von Anwendungen eingesetzt werden, einschließlich z.
B. Identifizieren von PCR-Amplikons,
RNA-Fingerprinting, Differentialdisplay, Einzelstrangkonformationspolymorphismusnachweis,
Didesoxy-Fingerprinting, Restriktionskarten- und Restriktionsfragment längenpolymorphismen,
DNA-Fingerprinting, Genotypisierung, Mutationsnachweis, Oligonukleotidligationsassay,
Sequenz-spezifische Amplifikationen, für Diagnose, Forensik, Identifikation,
Entwicklungsbiologie, Biologie, Molekularmedizin, Toxikologie, Tierzüchtung.
-
Diese und weitere Aspekte der vorliegenden
Erfindung werden bei Bezugnahme auf die folgende detaillierte Beschreibung
und die beigefügten
Zeichnungen deutlich werden.
-
KURZE BESCHREIBUNG
DER ZEICHNUNGEN
-
1 zeigt
das Flußdiagramm
für die
Synthese von Pentafluorphenylestern chemisch spaltbarer Massenspektroskopiemarkierungen,
um Markierungen mit Carbonsäureamid-Endgruppen freizusetzen.
-
2 zeigt
das Flußdiagramm
für die
Synthese von Pentafluorphenylestern chemisch spaltbarer Massenspektroskopiemarkierungen,
um Markierungen mit Carbonsäure-Endgruppen freizusetzen.
-
3-6 und 8 zeigen das Flußdiagramm für die Synthese von Tetrafluorphenylestern
eines Satzes von 36 photochemisch spaltbaren Massenspektroskopiemarkierungen.
-
7 zeigt
das Flußdiagramm
für die
Synthese eines Satzes von 36 photochemisch spaltbaren Massenspektroskopiemarkierungen
mit Amin-Endgruppen.
-
9 zeigt
die Synthese von 36 photochemisch spaltbaren Massenspektroskopie-markierten
Oligonukleotiden, hergestellt aus dem entsprechenden Satz von 36
Tetrafluorphenylestern photochemisch spaltbarer Massenspektroskopiemarkierungssäuren.
-
10 zeigt
die Synthese von 36 photochemisch spaltbaren Massenspektroskopie-markierten Oligonukleotiden,
hergestellt aus dem entsprechenden Satz von 36 photochemisch spaltbaren
Massenspektroskopiemarkierungen mit Amin-Endgruppen.
-
11 veranschaulicht
den gleichzeitigen Nachweis von mehreren Markierungen durch Massenspektrometrie.
-
12 zeigt
das Massenspektrogramm der Alpha-Cyano-Matrix allein.
-
13 zeigt
ein modular konstruiertes markiertes Nukleinsäurefragment.
-
14A–14I zeigen die Abtrennung von DNA-Fragmenten
durch HPLC unter der Verwendung einer Vielzahl von verschiedenen
Pufferlösungen.
-
15 zeigt
eine schematische Darstellung von genetischem Fingerprinting- und
Differential-Display-Systemen in Übereinstimmung mit einer beispielhaften
Ausführungsform
der vorliegenden Erfindung.
-
16 zeigt
eine schematische Darstellung von genetischem Fingerprinting- und
Differential-Display-Systemen in Übereinstimmung mit einer beispielhaften
Ausführungsform
der vorliegenden Erfindung.
-
17 zeigt
eine schematische Darstellung von Testsystemen in Übereinstimmung
mit einer exemplarischen Ausführungsform
der vorliegenden Erfindung.
-
18 ist
eine schematische Darstellung von Assay-Systemen in Übereinstimmung
mit einer exemplarischen Ausführungsform
der vorliegenden Erfindung.
-
19A und 19B zeigen die Herstellung
eines abspaltbaren Markers der vorliegenden Erfindung.
-
20A und 20B illustrieren die Herstellung
eines abspaltbaren Markers der vorliegenden Erfindung.
-
21 zeigt
die Herstellung einer intermediären
Verbindung; die bei der Herstellung eines spaltbaren Markers der
Erfindung brauchbar ist.
-
22A, 22B und 22C zeigen die Synthese-Methodologie
für die
Herstellung eines photo-abspaltbaren Massenspektrometrie-nachweisbaren
Markers gemäß der vorliegenden
Erfindung.
-
23 zeigt
die Ergebnisse aus einem Assay, der die Genexpression mit CMST-markierten
ODNs verfolgte.
-
24-28 zeigen Phosphoramidit-Chemie,
die in einem Beispiel hier vollständiger beschrieben wird.
-
DETAILLIERTE
BESCHREIBUNG DER ERFINDUNG
-
Wie oben angegeben, stellt die vorliegende
Erfindung Zusammensetzungen und Verfahren zum Analysieren von Nukleinsäuremolekülen zur
Verfügung,
bei denen die Trennung von Nukleinsäuremolekülen auf der Grundlage ihrer
Größe erforderlich
ist. Die vorliegenden Verfahren ermöglichen den gleichzeitigen
Nachweis von interessierenden Molekülen, die Nukleinsäuren und
Fragmente, Proteine, Peptide, usw. einschließen.
-
Die vorliegende Erfindung stellt
eine neue Klasse von Markern für
genomische Messungen zur Verfügung,
die eine Assay-Plattform zur Verfügung stellen, die mit dem Ausmaß von Messungen
kompatibel ist, die für
die Analyse von komplexen Genomen erforderlich ist. Diese neue Markertechnologie
ist aus Massenspektrometrie, Markern, die mit einem Standard-Quadrapol-Massenspektrometerdetektor
(MSD) unter der Verwendung von atmosphärischem Druck chemischer Ionisierung
(positiver Modus) nachgewiesen werden, zusammengesetzt. Die Technologieplattform
verwendet einen MSD zum Nachweis der bekannten Molekulargewichts-Massenspektrometermarkern,
ganz ähnlich
einem Dioden-Array-Detektor.
Die Macker können
durch kombinatorische Chemie-Ansätze
unter der Verwendung eines primären
Gerüsts
synthetisiert werden, auf das spezifische Masse-Einsteller angefügt sind. Die Marker sind so
aufgebaut, um reversibel an Oligonukleotide angebracht zu sein,
die entweder als Primer im PCR-Ansatz verwendet werden können oder
als Sonden in Hybridisierungsassays verwendet werden. Bei der Vervollständigung
jeder Zahl von Assay-Schritten wird der Marker/die Sonde oder Marker/Primer
einer Abspaltungsreaktion unterzogen, bevorzugterweise Photoabspaltung,
und wenn die Marker-Massenspektrometrie-nachweisbar
sind, werden die Macker durch APCI ionisiert und die Massenidentität des Markers
wird durch Massenspektrometrie nachgewiesen. Die Macker können dazu
verwendet werden, um die Identität
einer Sequenz und die Probenidentifizierung zu kartieren.
-
Kurz gesagt, stellt die vorliegende
Erfindung in einem Aspekt Verbindungen zur Verfügung, bei denen ein interessierendes
Molekül,
oder eine Vorstufe dazu, über
eine labile Bindung (oder labile Bindungen) an eine Markierung gebunden
ist. So können
Verbindungen der Erfindung so betrachtet werden, daß sie die
allgemeine Formel: Tms-L-X
-
Aufweisen, in der T die Markierungskomponente
ist, L die Linkerkomponente ist, die entweder eine labile Bindung
ist oder enthält,
und X entweder die Komponente "Interessierendes
Molekül" (MOI) oder eine Komponente
mit einer funktionellen Gruppe (Lh), durch
die das MOI mit T-L verknüpft
werden kann, ist. Verbindungen der Erfindung können daher durch die spezifischeren
allgemeinen Formeln dargestellt werden: T-L-MOI und T-L-Lh
-
Aus unten genauer beschriebenen Gründen können Sätze von
T-L-MOI-Verbindungen gezielt Bedingungen unterworfen werden, die
bewirken, daß die
labile(n) Bindungen) bricht (brechen), wodurch eine Markierungseinheit
vom Rest der Verbindung freigesetzt wird. Die Markierungseinheit
wird dann durch eine oder mehrere analytische Techniken charakterisiert,
um dadurch direkte Information über
die Struktur der Markierungseinheit zu liefern und (am wichtigsten)
indirekte Information über
die Identität
des entsprechenden MOI.
-
Als ein einfaches veranschaulichendes
Beispiel für
eine repräsentative
Verbindung der Erfindung, bei der L eine direkte Bindung ist, wird
Bezug genommen auf die folgende Struktur (i):
-
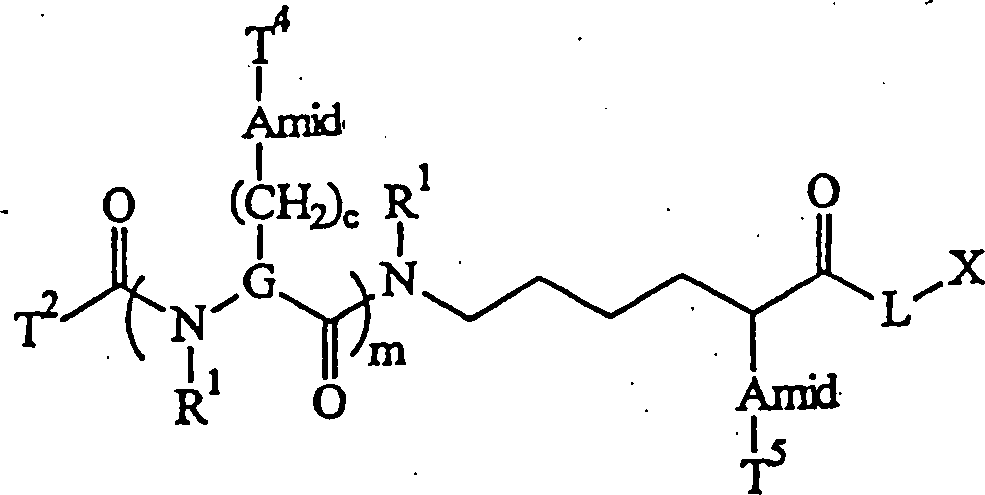
In Struktur (i) ist T eine stickstoffhaltige
polycyclische aromatische Einheit, die an eine Carbonylgruppe gebunden
ist, X ist ein MOI (und spezifisch ein Nukleinsäurefragment, das in einer Amingruppe
endet) und L ist die Bindung, die eine Amidgruppe bildet. Die Amidbindung
ist relativ zu den Bindungen in T labil, weil eine Amidbindung,
wie im Stand der Technik anerkannt, durch saure oder basische Bedingungen
chemisch gespalten (gebrochen) werden kann, die die Bindungen in
der Markierungskomponente unverändert
lassen. So kann eine Markierungseinheit (d. h. das Spaltprodukt,
das T enthält)
freigesetzt werden, wie unten dargestellt:
-
Der Linker L kann jedoch mehr sein
als lediglich eine direkte Bindung, wie in dem folgenden veranschaulichenden
Beispiel gezeigt, in dem Bezug genommen wird auf eine weitere repräsentative
Verbindung der Erfindung der unten dargestellten Struktur (ii):
-
Es ist gut bekannt, daß Verbindungen
mit einer ortho-Nitrobenzylamin-Einheit (siehe eingerahmte Atome
in Struktur (ii)) photolytisch insofern instabil sind, als die Einwirkung
aktinischer Strahlung einer spezifizierten Wellenlänge auf
solche Verbindungen eine selektive Spaltung der Benzylamin-Bindung
(siehe Bindung, die in Struktur (ii) mit einer dicken Linie bezeichnet
ist) bewirkt. So hat Struktur (ii) dieselben T- und MOI-Gruppen wie
Struktur (i) die Linker-Gruppe enthält jedoch mehrere Atome und
Bindungen, innerhalb derer sich eine besonders labile Bindung befindet.
Photoyse von Struktur (ii) setzt somit eine Markierungseinheit (T-enthaltende Einheit)
vom Rest der Verbindung frei, wie unten dargestellt.
-
-
-
Die Erfindung stellt somit Verbindungen
zur Verfügung,
die, bei Einwirkung geeigneter Spaltungsbedingungen, eine Spaltungsreaktion
durchlaufen, so daß eine
Markierungseinheit vom Rest der Verbindung freigesetzt wird. Verbindungen
der Erfindung können
als die Markierungseinheit, das MOI (oder eine Vorstufe dazu, Lh) und die labile(n) Bindung(en), die die
zwei Gruppen miteinander verbindet (verbinden), beschrieben werden.
Alternativ können
die Verbindungen der Erfindung als die Komponenten beschrieben werden,
aus denen sie gebildet sind. So können die Verbindungen wie folgt
als das Reaktionsprodukt eines Markierungs-Reaktanten, eines Linker-Reaktanten
und eines MOI-Reaktanten beschrieben werden.
-
Der Markierungs-Reaktant besteht
aus einem chemischen Griff (Th) und einer
variablen Komponente (Tvc), so daß der Markierungs-Reaktant
so betrachtet wird, daß er
die allgemeine Struktur hat: Tvc-Th
-
Um diese Nomenklatur zu veranschaulichen,
kann Bezug genommen werden auf Struktur (iii), die einen Markieiungs-Reaktanten
zeigt, der verwendet werden kann, um die Verbindung von Struktur
(ii) herzustellen. Der Markierungs-Reaktant mit Struktur (iii) enthält eine
variable Markierungskomponente und einen Markierungsgriff, wie unten
dargestellt:
-
In Struktur (iii) liefert der Markierungsgriff
(-C(=O)-A) nur einen Weg für
die Reaktion des Markierungs-Reaktanten mit dem Linker-Reaktanten,
um eine T-L-Einheit zu bilden. Die Gruppe "A" in
Struktur (iii) zeigt, daß die
Carboxylgruppe sich in einem chemisch aktiven Zustand befindet,
so daß sie
für Kopplung
mit anderen Griffen bereitsteht. "A" kann
z. B. eine Hydroxylgruppe oder Pentafluorphenoxy sein, unter vielen
anderen Möglichkeiten.
Die Erfindung liefert eine große
Anzahl möglicher
Markierungsgriffe, die an eine variable Markierungskomponente gebunden
werden können,
wie im Detail unten diskutiert. Die variable Markierungskomponente
ist somit ein Teil von "T" in der Formel T-L-X
und wird auch Teil der Markierungseinheit sein, die sich aus der
Reaktion bildet, die L spaltet.
-
Wie ebenfalls unten genauer diskutiert,
wird die variable Markierungskomponente so genannt, weil es bei
der Herstellung von Sätzen
von Verbindungen gemäß der Erfindung
erwünscht
ist, daß Mitglieder
eines Satzes einzigartige variable Komponenten aufweisen, so daß die einzelnen
Mitglieder voneinander mit einer analytischen Technik unterschieden
werden können.
Als ein Beispiel kann die variable Markierungskomponente von Struktur
(iii) Mitglied des folgenden Satzes sein, in dem die Mitglieder
des Satzes durch ihre UV- oder Massenspektren unterschieden werden
können:
-
In ähnlicher Weise kann der Linker-Reaktant
anhand seiner chemischen Griffe (es gibt notwendigerweise wenigstens
zwei, von denen jeder als Lh bezeichnet
werden kann), beschrieben werden, die eine labile Linkerkomponente
flankieren, wobei die labile Linkerkomponente aus der erforderlichen
labilen Einheit (L2) und fakultativen labilen
Einheiten (L1 und L3)
besteht, wobei die fakultativen labilen Einheiten effektiv dazu
dienen, L2 von den Griffen Lh zu
trennen, und die erforderliche labile Einheit dazu dient, eine labile
Bindung innerhalb der labilen Linkerkomponente bereitzustellen.
So kann der Linker-Reaktant
so betrachtet werden, daß er
die allgemeine Formel hat: Lh-L1-L2-L3-Lh
-
Die Nomenklatur, die verwendet wird,
um den Linker-Reaktanten zu beschreiben, kann im Hinblick auf Struktur
(iv) veranschaulicht werden, die sich wieder von der Verbindung
von Struktur (ii) ableitet:
-
Wie Struktur (iv) veranschaulicht,
können
Atome in mehr als eine funktionelle Rolle erfüllen. So dient, in Struktur
(iv), der Benzylstickstoff als ein chemischer Griff, um zu ermöglichen,
daß der
Linker-Reaktant sich an den Markierungs-Reaktanten über eine
Amidbildungsreaktion bindet, und dient anschließend auch insofern als ein
notwendiger Teil der Struktur der labilen Einheit L2,
als die Benzyl-Kohlenstoff-Stickstoff-Bindung besonders anfällig ist
für photolytische
Spaltung. Struktur (iv) veranschaulicht auch, daß ein Linker-Reaktant eine L3-Gruppe aufweisen kann (in diesem Fall eine
Methylengruppe), obgleich er keine L1-Gruppe
hat. In ähnlicher Weise
können
Linker-Reaktanten eine L1-Gruppe aufweisen,
aber keine L3-Gruppe, oder können L1- und L3-Gruppen
aufweisen oder können
weder L1- noch L3-Gruppen
aufweisen. In Struktur (iv) zeigt das Vorhandensein der Gruppe "P" neben der Carbonylgruppe, daß die Carbonylgruppe
vor der Reaktion geschützt
ist. Bei Vorgabe dieser Konfiguration kann die aktivierte Carboxylgruppe
des Markierungs-Reaktanten
(iii) sauber mit der Amingruppe des Linker-Reaktanten (iv) reagieren,
um eine Amidbindung zu bilden und eine Verbindung der Formel T-L-Lh, zu ergeben.
-
Der MOI-Reaktant ist eine in geeigneter
Weise reaktive Form eines interssierenden Moleküls. Wenn das interessierende
Molekül
ein Nukleinsäurefragment
ist, ist ein geeigneter MOI-Reaktant
ein Nukleinsäurefragment,
das durch seine 5'-Hydroxylgruppe
an eine Phosphodiestergruppe gebunden ist und anschließend an
eine Alkylen-Kette, die in einer Aminogruppe endet. Diese Aminogruppe
kann dann mit der Carbonylgruppe von Struktur (iv) reagieren (natürlich nach
Entfernen der Schutzgruppe der Carbonylgruppe und vorzugsweise nach
anschließender
Aktivierung der Carbonylgruppe für
die Reaktion mit der Amingruppe), um dadurch das MOI an den Linker
zu binden.
-
Wenn sie in einer chronologischen
Reihenfolge betrachtet wird, kann die Erfindung so gesehen werden,
daß sie
einen Markierungs-Reaktanten (mit einem chemischen Markierungsgriff
und einer variablen Markierungskomponente), einen Linker-Reaktanten
(mit zwei chemischen Linkergriffen, einer erforderlichen labilen
Einheit und 0–2
fakultativen labilen Einheiten) und einen MOI-Reaktanten (mit einer
Komponente "Interessierendes
Molekül" und einem chemischen
Griff des interessierenden Moleküls)
nimmt, um T-L-MOI zu bilden. So werden, um T-L-MOI zu bilden, entweder
der Markierungs-Reaktant und der Linker-Reaktant zunächst miteinander
umgesetzt, um T-L-Lh, zu liefern, und anschließend wird
der MOI-Reaktant mit T-L-Lh umgesetzt, um T-L-MOI
zu liefern, oder (weniger bevorzugt) werden der Linker-Reaktant
und der MOI-Reaktant zunächst
miteinander umgesetzt, um Lh-L-MOI zu liefern, und anschließend wird
Lh-L-MOI mit dem Markierungs-Reaktanten
umgesetzt, um T-L-MOI zu liefern. Aus Gründen der Bequemlichkeit werden
Verbindungen mit der Formel T-L-MOI als der Markierungs-Reaktant,
der Linker-Reaktant und der MOI-Reaktant beschrieben werden, die verwendet
werden können,
um solche Verbindungen zu bilden. Dieselben Verbindungen von Formel
T-L-MOI könnten
natürlich mit
anderen (typischerweise aufwendigeren) Verfahren hergestellt werden
und würden
immer noch unter den Schutzumfang der erfinderischen T-L-MOI-Verbindungen
fallen.
-
In jedem Falle sorgt die Erfindung
dafür,
daß eine
T-L-MOI-Verbindung Spaltungsbedingungen unterworfen wird, so daß eine Markierungseinheit
vom Rest der Verbindung freigesetzt wird. Die Markierungseinheit wird
wenigstens die variable Markierungskomponente umfassen und wird
typischerweise zusätzlich
einige oder alle Atome vom Markierungsgriff, einige oder alle Atome
vom Linkergriff, die verwendet wurden, um das Markierungs-Reaktant
an den Linker-Reaktanten zu binden, die fakultative labile Einheit
L1, wenn diese Gruppe in T-L-MOI vorhanden
war, umfassen und wird vielleicht einen Teil der erforderlichen
labilert Einheit L2 enthalten, in Abhängigkeit
von der genauen Struktur von L2 und der
Natur der Spaltungschemie. Aus Gründen der Bequemlichkeit kann
die Markierungseinheit als die T-enthaltende Einheit bezeichnet
werden, weil T typischerweise den Hauptteil (im Hinblick auf die
Masse) der Markierungseinheit ausmachen wird.
-
Nach dieser Einführung in einen Aspekt der vorliegenden
Erfindung werden die verschiedenen Komponenten T, L und X im Detail
beschrieben werden. Diese Beschreibung beginnt mit den folgenden
Definitionen bestimmter Begriffe, die im weiteren bei der Beschreibung
von T, L und X verwendet werden.
-
Wie hierin verwendet, bedeutet der
Begriff "Nukleinsäurefragment" ein Molekül, das komplementär zu einem
ausgewählten
Target-Nukleinsäuremolekül ist (d.
h. komplementär
zu dem gesamten Molekül
oder einem Teil desselben) und aus der Natur gewonnen oder synthetisch
oder rekombinant hergestellt werden kann, einschließlich nicht
natürlich
vorkommender Moleküle,
und kann je nach Eignung in doppel- oder einzelsträngiger Form
vorliegen; und schließt
ein Oligonukleotid (z. B. DNA oder RNA), einen Primer, eine Sonde,
ein Nukleinsäureanalog
(z. B. PNA), ein Oligonukleotid, das in einer 5'-nach-3'-Richtung mit einer Polymerase verlängert ist,
eine Nukleinsäure,
die chemisch oder enzymatisch gespalten ist, eine Nukleinsäure, die
mit einem Didesoxyterminator endet oder am 3'- oder 5'-Ende mit einer Verbindung abgedeckt
ist, die Polymerisation am 5'-
oder 3'-Ende verhindert,
oder Kombinationen derselben ein. Die Komplimentarität eines
Nukleinsäurefragments
zu einem ausgewählten
Target-Nukleinsäuremolekül bedeutet
im allgemeinen, daß wenigstens
etwa 70% spezifische Basenpaarung über die gesamte Länge des
Fragments gezeigt wird. Vorzugsweise zeigt das Nukleinsäurefragment
wenigstens etwa 80% spezifische Basenpaarung; und am bevorzugtesten
wenigstens etwa 90%. Assays zum Bestimmen des prozentualen Mismatch
(und somit der prozentualen spezifischen Basenpaarung) sind im Stand
der Technik gut bekannt und beruhen auf dem prozentualen Mismatch
als einer Funktion des Tm, wenn bezogen auf eine Kontrolle mit vollständiger Basenpaarung.
-
Wie hierin verwendet, bezieht sich
der Begriff "Alkyl", allein oder in
Kombination, auf einen gesättigten, geradekettigen
oder verzweigtkettigen Kohlenwasserstoffrest, der von 1 bis 10,
vorzugsweise von 1 bis 6 und bevorzugter von 1 bis 4 Kohlenstoffatome
enthält.
Beispiele für
solche Reste schließen
Methyl, Ethyl, n-Propyl, iso-Propyl, n-Butyl, iso-Butyl, sec.-Butyl,
tert.-Butyl, Pentyl, iso-Amyl, Hexyl, Decyl und dergleichen ein,
sind aber nicht hierauf beschränkt.
Der Begriff "Alkylen" bezieht sich auf
einen gesättigten,
geradkettigen oder verzweigtkettigen Kohlenwasserstoffdirest, der
von 1 bis 10, vorzugsweise von 1 bis 6 und bevorzugter von 1 bis 4
Kohlenstoffe enthält.
Beispiele für
solche einen Direst schließen
Methylen, Ethylen (-CH2-CH2-),
Propylen und dergleichen ein, sind aber nicht hierauf beschränkt.
-
Der Begriff "Alkenyl", allein oder in Kombination, bezieht
sich auf einen geradkettigen oder verzweigtkettigen Kohlenwasserstoffrest
mit wenigstens einer Kohlenstoff-Kohlenstoff-Doppelbindung in insgesamt von 2 bis
10, vorzugsweise von 2 bis 6 und bevorzugter von 2 bis 4 Kohlenstoffatomen.
Beispiele für
solche Reste schließen
Ethenyl, E- und Z-Propenyl, Isopropenyl, E- und Z-Butenyl, E- und
Z-Isobutenyl, E- und Z-Pentenyl, Decenyl und dergleichen ein, sind
aber nicht hierauf beschränkt.
Der Begriff "Alkenylen" bezieht sich auf
einen geradkettigen oder verzweigtkettigen Kohlenwasserstoffdirest
mit wenigstens einer Kohlenstoff-Kohlenstoff-Doppelbindung in insgesamt
von 2 bis 10, vorzugsweise von 2 bis 6 und bevorzugter von 2 bis
4 Kohlenstoffatomen. Beispiele für
solche Direste schließen Methyliden
(=CH2), Ethyliden (-CH=CH-), Propyliden (-CH2-CH=CH-) und dergleichen ein, sind aber
nicht hierauf beschränkt.
-
Der Begriff "Alkinyl", allein oder in Kombination, bezieht
sich auf einen geradekettigen oder verzweigtkettigen Kohlenwasserstoffrest
mit wenigstens einer Kohlenstoff-Kohlenstoff-Dreifachbindung in insgesamt von 2 bis
10, vorzugsweise von 2 bis 6 und bevorzugter von 2 bis 4 Kohlenstoffatomen.
Beispiele für
solche Reste schließen
Ethinyl (Acetylenyl), Propinyl (Propargyl), Butinyl, Hexinyl, Decinyl
und dergleichen ein, sind aber nicht hierauf beschränkt. Der
Begriff "Alkinylen", allein oder in
Kombination, bezieht sich auf einen geradkettigen oder verzweigtkettigen
Kohlenwasserstoffdirest mit wenigstens einer Kohlenstoff-Kohlenstoff-Dreifachbindung
in insgesamt von 2 bis 10, vorzugsweise von 2 bis 6 und bevorzugter
von 2 bis 4 Kohlenstoffatomen. Beispiele für solche Reste schließen Ethinylen
(-C≡C-),
Propinylen (-CH2-C≡C-) und dergleichen ein, sind aber
nicht hierauf beschränkt.
-
Der Begriff "Cycloalkyl", allein oder in Kombination, bezieht
sich auf eine gesättigte
cyclische Anordnung von Kohlenstoffatomen, mit einer Anzahl von
3 bis 8 und vorzugsweise von 3 bis 6 Kohlenstoffatomen. Beispiele
für solche
Cycloalkylreste schließen
Cyclopropyl, Cyclobutyl, Cyclopentyl, Cyclohexyl und dergleichen
ein, sind aber nicht hierauf beschränkt. Der Begriff "Cycloalkylen" bezeichnet eine
Direstform eines Cycloalkyls.
-
Der Begriff "Cycloalkenyl", allein oder in Kombination, bezieht
sich auf einen cyclischen Kohlenstoffring, der von 4 bis 8, vorzugsweise
5 oder 6, Kohlenstoffatome und eine oder mehrere Doppelbindungen
enthält.
Beispiele für
solche Cycloalkenylreste schließen
Cyclopentenyl, Cyclohexenyl, Cyclopentadienyl und dergleichen ein,
sind aber nicht hierauf beschränkt.
Der Begriff "Cycloalkenylen" bezieht sich auf
eine Direstform eines Cycloalkenyls.
-
Der Begriff "Aryl" bezieht
sich auf eine carbocyclische (vollständig aus Kohlenstoff und Wasserstoff
bestehende) aromatische Gruppe, die ausgewählt ist aus der Gruppe, die
aus Phenyl, Naphthyl, Indenyl, Indanyl, Azulenyl, Fluorenyl und
Anthracenyl besteht; oder eine heterocyclische aromatische Gruppe,
die ausgewählt ist
aus der Gruppe, die aus Furyl, Thienyl, Pyridyl, Pyrrolyl, Oxazolyl,
Thiazolyl, Imidazolyl, Pyrazolyl, 2-Pyrazolinyl, Pyrazolidinyl,
Isoxazolyl, Isothiazolyl, 1,2,3-Oxadiazolyl, 1,2,3-Triazolyl, 1,3,4-Thiadiazolyl,
Pyridazinyl, Pyrimidinyl, Pyrazinyl, 1,3,5-Triazinyl, 1,3,5-Trithianyl,
Indolizinyl, Indolyl, Isoindolyl, 3H-Indolyl, Indolinyl, Benzo[b]furanyl,
2,3-Dihydrobenzofuranyl, Benzo[b]thiophenyl, 1H-Indazolyl, Benzimidazolyl,
Benzthiazolyl, Purinyl, 4H-Chinolizinyl, Chinolinyl, Isochinolinyl,
Cinnolinyl, Phthalazinyl, Chinazolinyl, Chinoxalinyl, 1,8-Naphthyridinyl, Pteridinyl,
Carbazolyl, Acridinyl, Phenazinyl, Phenothiazinyl und Phenoxazinyl
besteht.
-
"Aryl"-Gruppen, wie in
dieser Anmeldung definiert, können
unabhängig
einen bis vier Substituenten enthalten, die unabhängig ausgewählt sind
aus der Gruppe, die aus Wasserstoff, Halogen, Hydroxyl, Amino, Nitro,
Trifluormethyl, Trifluormethoxy, Alkyl, Alkenyl, Alkinyl, Cyano,
Carboxy, Carboalkoxy, 1,2-Dioxyethylen, Alkoxy, Alkenoxy oder Alkinoxy,
Alkylamino, Alkenylamino, Alkinylamino, aliphatischem oder aromatischem Acyl,
Alkoxycarbonylamino, Alkylsulfonylamino, Morpholinocarbonylamino,
Thiomorpholinocarbonylamino, N-Alkylguanidino, Aralkylaminosulfonyl;
Aralkoxyalkyl; N-Aralkoxyharnstoff;
N-Hydroxylharnstoff; N-Alkenylharnstoff; N,N-(Alkyl, Hydroxyl)harnstoff
Heterocyclyl; Thioaryloxy-substituiertem Aryl; N,N-(Aryl, Alkyl)hydrazino;
Ar'-substituiertem
Sulfonylheterocyclyl; Aralkyl-substituiertem Heterocyclyl; Cycloalkyl-
und Cycloalkenyl-substituiertem Heterocyclyl; Cycloalkyl-kondensiertem Aryl;
Arlyoxy-substituiertem Alkyl; Heterocyclylamino; aliphatischem oder
aromatischem Acylaminocarbonyl; aliphatischem oder aromatischem
Acyl-substituiertem Alkenyl; Ar'-substituiertem
Aminocarbonyloxy; Ar',
Ar'-disubstituiertem
Aryl; aliphatischem oder aromatischem Acyl-substituiertem Acyl;
Cycloalkylcarbonylalkyl; Cycloalkyl-substituiertem Amino; Aryloxycarbonylalkyl;
Phosphorodiamidylsäure
oder -ester besteht.
-
"Ar" ist eine carbocyclische
oder heterocyclische Arylgruppe, wie oben definiert, mit einem bis
drei Substituenten, die ausgewählt
sind aus der Gruppe, die aus Wasserstoff, Halogen, Hydroxyl, Amino,
Nitro, Trifluormethyl, Trifluormethoxy, Alkyl, Alkenyl, Alkinyl,
1,2-Dioxymethylen,
1,2-Dioxyethylen, Alkoxy, Alkenoxy, Alkinoxy, Alkylamino, Alkenylamino
oder Alkinylamino, Alkylcarbonyloxy, aliphatischem oder aromatischem Acyl,
Alkylcarbonylamino, Alkoxycarbonylamino, Alkylsulfonylamino, N-Alkyl-
oder N,N-Dialkylharnstoff
besteht.
-
Der Begriff "Alkoxy", allein oder in Kombination, bezieht
sich auf ein Alkyletherrest, wobei der Begriff "Alkyl" wie oben definiert ist. Beispiele für geeignete
Alkyletherreste schließen
Methoxy, Ethoxy, n-Propoxy, iso-Propoxy, n-Butoxy, iso-Butoxy, sec.-Butoxy,
tert.-Butoxy und dergleichen ein, sind aber nicht hierauf beschränkt.
-
Der Begriff "Alkenoxy", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkenyl-O-, wobei der Begriff "Alkenyl" wie oben definiert
ist, vorausgesetzt, daß der
Rest kein Enolether ist. Beispiele für geeignete Alkenoxyreste schließen Allyloxy,
E- und Z-3-Methyl-2-propenoxy
und dergleichen ein, sind aber nicht hierauf beschränkt.
-
Der Begriff "Alkinyloxy", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkinyl-O-, wobei der Begriff "Alkinyl" wie oben definiert
ist, vorausgesetzt, daß der
Rest kein Inolether ist. Beispiele für geeignete Alkinoxyreste schließen Propargyloxy,
2-Butinyloxy und
dergleichen ein, sind aber nicht hierauf beschränkt.
-
Der Begriff "Thioalkoxy" bezieht sich auf einen Thioetherrest
der Formel Alkyl-S-, wobei Alkyl wie oben definiert ist.
-
Der Begriff "Alkylamino", allein oder in Kombination, bezieht
sich auf einen Mono- oder Dialkyl-substituierten Aminorest (d. h.
einen Rest der Formel Alkyl-NH- oder (Alkyl)2-N-), wobei
der Begriff "Alkyl" wie oben definiert
ist. Beispiele für
geeignete Alkylaminoreste schließen Methylamino, Ethylamino,
Propylamino, Isopropylamino, tert.-Butylamino, N,N-Diethylamino und
dergleichen ein, sind aber nicht hierauf beschränkt.
-
Der Begriff "Alkenylamino", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkenyl-NH- oder (Alkenyl)2N-, wobei der Begriff "Alkenyl" wie oben definiert ist, vorausgesetzt,
daß der
Rest kein Enamin ist. Ein Beispiel für solche Alkenylaminoreste
ist der Allylaminorest.
-
Der Begriff "Alkinylamino", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkinyl-NH- oder (Alkinyl)2N-, in der der Begriff "Alkinyl" wie oben definiert ist, vorausgesetzt,
daß der
Rest kein Inamin ist. Ein Beispiel für solche Alkinylaminoreste
ist der Propargylaminorest.
-
Der Begriff "Amid" bezieht
sich entweder auf -N(R1)-C(=O)- oder -C(=O)-N(R1)-, wobei R1 hierin
so definiert ist, daß es
Wasserstoff sowie andere Gruppen einschließt. Der Begriff "substituiertes Amid" bezieht sich auf
die Situation, in der R1 nicht Wasserstoff
ist, während
der Begriff "unsubstituiertes
Amid" sich auf die
Situation bezieht, in der R1 Wasserstoff
ist.
-
Der Begriff "Aryloxy", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Aryl-O-, wobei Aryl wie oben definiert
ist. Beispiele für
Aryloxyreste schließen
Phenoxy, Naphthoxy, Pyridyloxy und dergleichen ein, sind aber nicht
hierauf beschränkt.
-
Der Begriff "Arylamino", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Aryl-NH-, in der Aryl wie oben definiert
ist. Beispiele für
Arylaminoreste schließen
Phenylamino (Anilido), Naphthylamino, 2-, 3- und 4-Pyridylamino
und dergleichen ein, sind aber nicht hierauf beschränkt.
-
Der Begriff "Aryl-kondensiertes Cycloalkyl", allein oder in
Kombination, bezieht sich auf einen Cycloalkylrest, der zwei benachbarte
Atome mit einem Arylrest teilt, wobei die Begriffe "Cycloalkyl" und "Aryl" wie oben definiert
sind. Ein Beispiel eines Aryl-kondensierten
Cycloalkylrestes ist der Benzo-kondensierte Cyclobutylrest.
-
Der Begriff "Alkylcarbonylamino", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkyl-CONH, wobei der Begriff "Alkyl" wie oben definiert
ist.
-
Der Begriff "Alkoxycarbonylamino", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkyl-OCONH-, wobei der Begriff "Alkyl" wie oben definiert
ist.
-
Der Begriff "Alkylsulfonylamino", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkyl-SO2NH-,
wobei der Begriff "Alkyl" wie oben definiert
ist.
-
Der Begriff "Arylsulfonylamino", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Aryl-SO2NH-,
wobei der Begriff "Aryl" wie oben definiert
ist.
-
Der Begriff "N-Alkylharnstoff", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Alkyl-NH-CO-NH-, wobei der Begriff "Alkyl" wie oben definiert
ist.
-
Der Begriff "N-Arylharnstoff", allein oder in Kombination, bezieht
sich auf einen Rest der Formel Aryl-NH-CO-NH-, wobei der Begriff "Aryl" wie oben definiert
ist.
-
Der Begriff "Halogen" bedeutet Fluor, Chlor, Brom und Iod.
-
Der Begriff "Kohlenwasserstoffrest" bezieht sich auf
eine Anordnung von Kohlenstoff- und Wasserstoffatomen, die nur ein
einziges Wasserstoffatom benötigt,
um ein unabhängiges stabiles
Molekül
zu sein. Somit hat ein Kohlenwasserstoffrest eine offene Valenzstelle
an einem Kohlenstoffatom, durch die der Kohlenwasserstoffrest an
ein anderes Atom (andere Atome) gebunden werden kann. Alkyl, Alkenyl,
Cycloalkyl, etc. sind Beispiele für Kohlenwasserstoffreste.
-
Der Begriff "Kohlenwasserstoffdirest" bezieht sich auf
eine Anordnung von Kohlenstoff- und Wasserstoffatomen, die zwei
Wasserstoffatome benötigt,
um ein unabhängiges
stabiles Molekül
zu sein. Somit hat ein Kohlenwasserstoffrest zwei offene Valenzstellen
an einem oder zwei Kohlenstoffatomen, durch die der Kohlenwasserstoffrest
an ein anderes Atom (andere Atome) gebunden werden kann. Alkylen,
Alkenylen, Alkinylen, Cycloalkylen, etc. sind Beispiele für Kohlenwasserstoffdireste.
-
Der Begriff "Hydrocarbyl" bezieht sich auf jede stabile Anordnung,
die vollständig
aus Kohlenstoff und Wasserstoff besteht, mit einer einzelnen Valenzstelle,
an der sie an eine andere Einheit gebunden ist, und schließt somit
Reste ein, die als Alkyl, Alkenyl, Alkinyl, Cycloalkyl, Cycloalkenyl,
Aryl (ohne Einbeziehung von Heteroatomen in den Arylring), Arylalkyl,
Alkylaryl und dergleichen ein. Kohlenwasserstoffrest ist ein anderer Name
für Hydrocarbyl.
-
Der Begriff "Hydrocarbylen" bezieht sich auf jede stabile Anordnung,
die vollständig
aus Kohlenstoff und Wasserstoff besteht, mit zwei Valenzstellen,
an der sie an andere Einheiten gebunden ist, und schließt somit
Alkylen, Alkenylen, Alkinylen, Cycloalkylen, Cycloalkenylen, Arylen
(ohne Einbeziehung von Heteroatomen in den Arylring), Arylalkylen,
Alkylarylen und dergleichen ein. Kohlenwasserstoffdirest ist ein
anderer Name für Hydrocarbylen.
-
Der Begriff "Hydrocarbyl-O-hydrocarbylen" bezieht sich auf
eine Hydrocarbyl-Gruppe, die an ein Sauerstoffatom gebunden ist,
wobei das Sauerstoffatom in ähnlicher
Weise an eine Hydrocarbylen-Gruppe an einer der zwei Valenzstellen
gebunden ist, an der die Hydrocarbylen-Gruppe an andere Einheiten
gebunden ist. Die Begriffe "Hydrocarbyl-S-hydrocarbylen", "Hydrocarbyl-NH-hydrocarbylen" und "Hydrocarbyl-Amid-hydrocarbylen" haben äquivalente
Bedeutungen, wobei Sauerstoff durch Schwefel, -NH- bzw. eine Amidgruppe
ersetzt worden ist.
-
Der Begriff N-(Hydrocarbyl)hydrocarbylen
bezieht sich auf eine Hydrocarbylen-Gruppe, wobei eine der zwei
Valenzstellen an ein Stickstoffatom gebunden ist und dieses Stickstoffatom
gleichzeitig an einen Wasserstoff und eine Hydrocarbyl-Gruppe gebunden
ist.
-
Der Begriff N,N-Di(hydrocarbyl)hydrocarbylen
bezieht sich auf eine Hydrocarbylengruppe, wobei eine der zwei Valenzstellen
an ein Stickstoffatom gebunden ist und dieses Stickstoffatom gleichzeitig
an zwei Hydrocarbyl-Gruppen gebunden ist.
-
Der Begriff "Hydrocarbylacyl-hydrocarbylen" bezieht sich auf
eine Hydrocarbyl-Gruppe, die durch eine Acyl(-C(≡O)-)-Gruppe an eine der zwei
Valenzstellen einer Hydrocarbylen-Gruppe gebunden ist.
-
Die Begriffe "Heterocyclylhydrocarbyl" und "Heterocyclyl" beziehen sich auf
eine stabile cyclische Anordnung von Atomen, die Kohlenstoffatome
und bis zu vier Atome (als Heteroatome bezeichnet) einschließt, die
ausgewählt
sind aus Sauerstoff, Stickstoff, Phosphor und Schwefel. Die cyclische
Anordnung kann in der Form eines monocyclischen Ringes mit 3–7 Atomen
oder eines bicyclischen Ringes mit 8–11 Atomen vorliegen. Die Ringe
können
gesättigt
oder ungesättigt
sein (einschließlich
aromatischer Ringe) und können
fakultativ Benzo-kondensiert sein. Stickstoff- und Schwefelatomen
im Ring können
in jeder oxidierten Form vorliegen, einschließlich der quaternisierten Form
von Stickstoff. Ein Heterocyclylhydrocarbyl kann an irgendeinem endocyclischen
Kohlenstoff oder Heteroatom gebunden sein, das zur Schaffung einer
stabilen Struktur führt. Bevorzugte
Heterocyclylhydrocarbyle schließen
5- bis 7-gliedrige monocyclische Heterocyclen ein, die ein oder
zwei Stickstoff-Heteroatome enthalten.
-
Ein substituiertes Heterocyclylhydrocarbyl
bezieht sich auf ein Heterocyclyhydrocarbyl, wie oben definiert,
wobei wenigstens ein Ringatom davon an einen angegebenen Substituenten
gebunden ist, der sich vom Ring weg erstreckt.
-
Bei Bezugnahme auf Hydrocarbyl- und
Hydrocarbylen-Gruppen bezieht sich der Begriff "Derivate von irgendeinem der vorstehenden,
wobei ein oder mehr Wasserstoffe durch eine gleiche Anzahl Fluoride
ersetzt ist" auf
Moleküle,
die Kohlenstoff-, Wasserstoff- und Fluoridatome enthalten, aber
keine anderen Atome.
-
Der Begriff "aktivierter Ester" ist ein Ester, der eine "Abgangsgruppe" enthält, die
leicht durch ein Nukleophil verdrängbar ist, wie etwa ein Amin
und Alkohol oder ein Thiol-Nukleophil.
Solche Abgangsgruppen sind gut bekannt und schließen, ohne
Beschränkung,
N-Hydroxysuccinimid,
N-Hydroxybenzotriazol, Halogen (Halogenide), Alkoxy einschließlich Tetrafluorphenolaten,
Thioalkoxy und dergleichen ein. Der Begriff "geschützter Ester" bezieht sich auf eine Estergruppe,
die maskiert oder in anderer Weise unreaktiv ist. Siehe z. B. Greene, "Protecting Groups
In Organic Synthesis".
-
Angesichts der obigen Definitionen
können
weitere chemische Begriffe, die in dieser Anmeldung verwendet werden,
von den Fachleuten leicht verstanden werden. Begriffe können allein
oder in irgendeiner Kombination derselben verwendet werden. Die
bevorzugten und weiter bevorzugten Kettenlängen der Reste gelten für alle solche
Kombinationen.
-
A. ERZEUGUNG MARKIERTER
NUKLEINSÄUREFRAGMENTE
-
Wie oben angegeben, stellt ein Aspekt
der vorliegenden Erfindung ein allgemeines Schema zur DNA-Sequenzierung
bereit, das die Verwendung von mehr als 16 Markierungen in jeder
Spur ermöglicht;
bei kontinuierlichem Nachweis können
die Markierungen nachgewiesen und die Sequenz gelesen werden, wenn die
Trennung nach Größe abläuft, ebenso
wie bei herkömmlicher
Sequenzierung auf Fluoreszenz-Basis. Dieses Schema ist anwendbar
auf alle DNA-Sequenzierungstechniken, die auf Trennung der markierten
Moleküle nach
Größe beruhen.
Geeignete Markierungen und Linker zur Verwendung in der vorliegenden
Erfindung sowie Verfahren zur Sequenzierung von Nukleinsäuren sind
unten detaillierter diskutiert.
-
1. Markierungen
-
"Markierung", wie hierin verwendet,
bezieht sich allgemein auf eine chemische Einheit, die verwendet wird,
um ein "interessierendes
Molekül" in eindeutiger Weise
zu identifizieren, und bezieht sich, genauer gesagt, auf die variable
Markierungskomponente sowie auf alles, was in dem Markierungs-Reaktanten,
der Markierungs-Komponente und/oder der Markierungs-Einheit am dichtesten
daran gebunden ist.
-
Eine Markierung, die in der vorliegenden
Erfindung nützlich
ist, besitzt mehrere Eigenschaften:
-
- 1) Sie kann von allen anderen Markierungen unterschieden
werden. Diese Unterscheidung von anderen chemischen Einheiten basiert
auf ihren spektroskopischen Eigenschaften d. h. in der Massenspektroskopie.
- 2) Die Markierung kann nachgewiesen werden, wenn sie mit 10–22 bis
10–6 Mol
vorliegt.
- 3) Die Markierung besitzt einen chemischen Griff, durch den
sie an das MOI gebunden werden kann, welches die Markierung in eindeutiger
Weise identifizieren soll. Die Bindung kann direkt an das MOI erfolgen
oder indirekt durch eine "Linker"-Gruppe.
- 4) Die Markierung ist chemisch stabil gegenüber allen Manipulationen, denen
sie unterworfen wird, einschließlich
Bindung und Spaltung vom MOI, und allen Manipulationen des MOI,
während
die Markierung daran gebunden ist.
- 5) Die Markierung beeinflußt
die Manipulationen, die am MOI durchgeführt werden, während die
Markierung daran gebunden ist, nicht in signifikanter Weise. Wenn
die Markierung z. B. an ein Oligonukleotid gebunden ist, darf die
Markierung irgendwelche Hybridisierungs- oder enzymatischen Reaktionen
(z. B. PCR-Sequenzierungsreaktionen), die am Nukleotid durchgeführt werden,
nicht signifikant beeinflussen. In ähnlicher Weise darf, wenn die
Markierung an einen Antikörper
gebunden ist, sie die Antigen-Erkennung durch den Antikörper nicht
signifikant beeinflussen.
-
Eine Markierungs-Einheit, die mit
einem bestimmten spektroskopischen oder potentiometrischen Verfahren
nachgewiesen werden soll, sollte Eigenschaften besitzen, die die
Empfindlichkeit und Spezifität
des Nachweises mit diesem Verfahren erhöht. Typischerweise wird die
Markierungs-Einheit diese Eigenschaften haben, weil sie in die variable
Markierungs-Komponente hineinkonstruiert worden sind, die typischerweise
den Hauptteil der Markierungs-Einheit darstellen wird. In der folgenden
Diskussion bezieht sich die Verwendung des Wortes "Markierung" typischerweise auf
die Markierungs-Einheit (d. h. das Spaltprodukt, das die variable Markierungs-Komponente
enthält),
kann jedoch auch so betrachtet werden, daß es die variable Markierungs-Komponente
selbst bezeichnet, weil diese der Teil der Markierungs-Einheit ist,
der typischerweise dafür verantwortlich
ist, die eindeutig nachweisbaren Eigenschaften bereitzustellen.
In Verbindungen der Formel T-L-X wird der "T"-Teil
die variable Markierungs-Komponente enthalten, der mit Massenspektrometrie
charakterisiert wird, der "T"-Teil von T-L-X wird
daher als Tms bezeichnet. In ähnlicher
Weise kann das Spaltprodukt von T-L-X, das T enthält, als
die Tms-enthaltende Einheit bezeichnet werden.
Die folgenden spektroskopischen und potentiometrischen Verfahren
können
verwendet werden, um Tms-enthaltende Einheiten
zu charakterisieren.
-
a. Eigenschaften von MS-Markierungen
-
Der Marker ist durch Massenspektrometrie
analysierbar (d. h. ist eine MS-lesbare Markierung, hierin auch
als eine MS-Markierung oder "Tms-enthaltende Einheit" bezeichnet). Das wesentliche Merkmal
der Markierung ist, daß sie
ionisiert werden kann. Es ist somit ein bevorzugtes Element bei
der Konstruktion von MS-lesbaren Markierungen, darin eine chemische
Funktionalität
einzubauen, die unter Ionisationsbedingungen im MS eine positive
oder negative Ladung tragen kann. Dieses Merkmal verleiht verbesserte
Effizienz der Ionenbildung und höhere
Gesamtempfindlichkeit des Nachweises, insbesondere bei Elektrosprayionisation. Die
chemische Funktionalität,
die eine ionisierte Ladung unterstützt, kann sich von Tms oder L oder beiden ableiten. Faktoren,
die die relative Empfindlichkeit eines durch Massenspektrometrie
nachzuweisenden Analyten erhöhen
kann, sind in z. B. Sunner, J., et al., Anal. Chem. 60: 1300–1307 (1988),
diskutiert.
-
Eine bevorzugte Funktionalität, um das
Tragen einer negativen Ladung zu erleichtern, ist eine organische
Säure,
wie etwa phenolisches Hydroxyl, Carbonsäure, Phosphonat, Phosphat,
Tetrazol, Sulfonylharnstoff Perfluoralkohol und Sulfonsäure.
-
Bevorzugte Funktionalität, um das
Tragen einer positiven Ladung unter Ionisationsbedingungen zu erleichtern,
sind aliphatische oder aromatische Amine. Beispiele für funktionelle
Amingruppen, die erhöhte
Nachweisbarkeit von MS-Markierungen ergeben, schließen quartäre Amine
(d. h. Amine, die vier Bindungen, jede zu Kohlenstoffatomen, aufweisen,
siehe Aebersold, U.S.-Patent Nr. 5,240,859) und tertiäre Amine
ein (d. h. Amine, die drei Bindungen, jede zu Kohlenstoffatomen,
aufweisen, was C=N-C-Gruppen einschließt, wie sie in Pyridin vorhanden
sind, siehe Hess et al., Anal. Biochem. 224: 373, 1995; Bures et
al., Anal. Biochem. 224–364,
1995). Gehinderte tertiäre
Amine sind besonders bevorzugt. Tertiäre und quartäre Amine
können
Alkyl oder Aryl sein. Eine Tms-enthaltende
Einheit muß wenigstens
eine ionisierbare Spezies tragen, kann aber mehr als eine ionisierbare
Spezies besitzen. Der bevorzugte Ladungszustand ist eine einzelne
ionisierte Spezies pro Markierung.
-
Demgemäß ist es bevorzugt, daß jede Tms-enthaltende Einheit (und jede variable
Markierungs-Komponente) nur ein einziges gehindertes Amin oder eine
einzige organische Säuregruppe
enthält.
-
Geeignete Amin-enthaltende Reste,
die Teil der T
ms-enthaltenden Einheit bilden
können,
schließen
die folgenden ein:
-
Die Identifizierung einer Markierung
durch Massenspektrometrie beruht vorzugsweise auf ihrem Verhältnis Molekülmasse zu
Ladung (m/z). Der bevorzugte Molekülmassenbereich für MS-Markierungen
ist von etwa 100 bis 2.000 Daltons, und vorzugsweise hat die Tms-enthaltende
Einheit eine Masse von wenigstens etwa 250 Daltons, bevorzugter
wenigstens etwa 300 Daltons und noch bevorzugter wenigstens etwa
350 Daltons. Es im allgemeinen schwierig für Massenspektrometer, zwischen
Einheiten zu unterscheiden, die Molekülionen unter etwa 200–250 Daltons
haben (in Abhängigkeit
von dem genauen Instrument), und so haben bevorzugte Tms-enthaltende
Einheiten der Erfindung Massen oberhalb dieses Bereiches.
-
Wie oben erklärt, kann die Tms-enthaltende
Einheit andere Atome als diejenigen enthalten, die in der variablen
Markierungs-Komponente vorhanden sind und tatsächlich ändere als in Tms selbst
vorhanden sind. Demgemäß kann die
Masse von Tms selbst niedriger sein als
etwa 250 Daltons, so lange die Tms-enthaltende Einheit
eine Masse von wenigstens etwa 250 Daltons hat. So kann die Masse
von Tms von 15 (d. h. einem Methylrest)
bis etwa 10.000 Daltons reichen und reicht vorzugsweise von etwa
100 bis etwa 5.000 Daltons und reicht bevorzugter von etwa 200 bis
etwa 1.000 Daltons.
-
Es ist relativ schwierig, Markierungen
durch Massenspektrometrie zu unterscheiden, wenn diese Markierungen
Atome enthalten, die mehr als ein Isotop in signifikanter Menge
aufweisen. Demgemäß enthalten T-Gruppen,
die für
massenspektroskopische Identifizierung (Tms-Gruppen)
gedacht sind, Kohlenstoff, wenigstens eines von Wasserstoff und
Fluorid und fakultative Atome, die ausgewählt sind aus Sauerstoff, Stickstoff, Schwefel,
Phosphor und Iod. Obgleich weitere Atome im Tms vorhanden
sein können,
kann ihr Vorhandensein die Analyse der Massenspektrometerdaten etwas
schwieriger machen. Vorzugsweise haben die Tms-Gruppen nur
Kohlenstoff-, Stickstoff- und Sauerstoffatome, zusätzlich zu
Wasserstoff und/oder Fluorid.
-
Fluorid ist ein fakultatives, aber
bevorzugtes Atom für
die Einbeziehung in eine Tms-Gruppe. Im
Vergleich zu Wasserstoff ist Fluorid natürlich viel schwerer. So führt das
Vorhandensein von Fluoridatomen statt Wasserstoffatomen zu Tms-Gruppen mit höherer Masse, wodurch ermöglicht wird,
daß die
Tms-Gruppe eine Masse von mehr als 250 Daltons
erreicht und übersteigt,
was wünschenswert
ist, wie oben erläutert.
Zusätzlich verleiht
der Ersatz von Wasserstoff durch Fluorid der Tms-enthaltenden
Einheit größere Flüchtigkeit,
und größere Flüchtigkeit
des Analyten erhöht
die Empfindlichkeit, wenn Massenspektrometrie als das Nachweisverfahren
verwendet wird.
-
Die Molekülformel von Tms fällt in den
Bereich C1-5
00N0
-100O0-100O0-
10P0
-
10HαFβIδ,
wobei die Summe von α, β und δ ausreichend
ist, um die sonst ungesättigten
Valenzen der C-, N-, O-, S- und P-Atome abzusättigen. Die Bezeichnung C1-500N0-100O0-100O0-
10P
0-
10HαFβIδ bedeutet,
daß Tms wenigstens ein Kohlenstoffatom enthält und jede
Zahl von 1 bis 500 Kohlenstoffatome enthalten kann, zusätzlich dazu,
daß es
so viel wie 100 Stickstoffatome ("N0-" bedeutet, daß Tms keinerlei Stickstoffatome enthalten muß), und
so viel wie 100 Sauerstoffatome und so viel wie 10 Schwefelatome
und so viel wie 10 Phosphoratome enthält. Die Symbole α, β und δ stellen die
Anzahl von Wasserstoff-, Fluorid- und Iodidatomen in Tms dar,
wobei jeweils zwei dieser Zahlen Null sein können und wobei die Summe dieser
Zahlen der Gesamtheit der sonst ungesättigten Valenzen der C-, N-,
O-, S- und P-Atome entspricht. Vorzugsweise hat Tms eine
Molekülformel,
die in den Bereich C1-50N0-10 O
0-10HαFβ fällt, wobei
die Summe von α und β der Anzahl
von Wasserstoff- bzw. Fluoridatomen entspricht, die in der Einheit
vorhanden sind.
-
Zusätzlich zu den obigen Eigenschaften
ist es bevorzugt, daß die
Markierung eine modulare chemische Struktur besitzt. Dies hilft
bei der Konstruktion einer großen
Anzahl von strukturell verwandten Markierungen unter Verwendung
der Techniken der kombinatorischen Chemie. Die Tms-Gruppe
hat z. B. wünschenswerterweise
mehrere Eigenschaften. Sie enthält
wünschenswerterweise
eine funktionelle Gruppe, die einen einzelnen ionisierten Ladungszustand
unterstützt,
wenn die Tms-enthaltende Einheit Massenspektrometrie
unterzogen wird (einfacher als eine "Massenspektrometrieempfindlichkeitsverstärker"-Gruppe oder MSSE
bezeichnet). Auch kann sie wünschenswerterweise
als ein Mitglied in einer Familie von Tms-enthaltenden
Einheiten dienen, in der die Mitglieder der Familie jedes ein unterschiedliches
Masse/Ladungs-Verhältnis
haben, jedoch ungefähr
dieselbe Empfindlichkeit im Massenspektrometer aufweisen. Mitglieder
der Familie haben somit wünschenswerterweise
denselben MSSE. Um die Schaffung von Familien von Verbindungen zu
ermöglichen,
hat es sich als geeignet erwiesen, Markierungs-Reaktanten über ein
modulares Syntheseschema zu erzeugen, so daß die Markierungs-Komponenten
selbst als Module-umfassend angesehen werden können.
-
In einem bevorzugten modularen Ansatz
für die
Struktur der Tms-Gruppe hat Tms die
Formel T2-(J-T3-)n
wobei T2 eine organische Einheit ist, die aus Kohlenstoff
und einem oder mehreren von Wasserstoff, Fluorid, Iodid, Sauerstoff,
Stickstoff, Schwefel und Phosphor gebildet wird, mit einem Massebereich
von 15 bis 500 Daltons; T3 eine organische
Einheit ist, die aus Kohlenstoff und einem oder mehreren von Wasserstoff,
Fluorid, Iodid, Sauerstoff, Stickstoff, Schwefel und Phosphor gebildet
wird, mit einem Massebereich von 50 bis 1.000 Daltons; J eine direkte
Bindung oder eine funktionelle Gruppe ist, wie etwa Amid, Ester,
Amin, Sulfid, Ether, Thioester, Disulfid, Thioether, Harnstoff,
Thioharnstoff, Carbamat, Thiocarbamat, Schiff'sche Base, reduzierte Schiff'sche Base, Imin,
Oxim, Hydrazon, Phosphat, Phosphonat, Phosphoramid, Phosphonamid,
Sulfonat, Sulfonamid oder Kohlenstoff-Kohlenstoff-Bindung; und n
eine ganze Zahl aus dem Bereich von 1 bis 50 ist, so daß, wenn
n größer als
1 ist, jedes T3 und J unabhängig ausgewählt wird.
-
Die modulare Struktur T2-(J-T3)n- liefert einen
geeigneten Zugang zu Familien von T-L-X-Verbindungen, in denen jedes Mitglied
der Familie eine verschiedene T-Gruppe besitzt. Wenn T z. B. Tms ist und jedes Familienmitglied wünschenswerterweise
denselben MSSE besitzt, kann eine der T3-Gruppen
diese MSSE-Struktur liefern. Um für Variabilität zwischen
Mitgliedern einer Familie im Hinblick auf die Masse von Tms zu sorgen, kann die T2-Gruppe
unter den Familienmitgliedern variiert werden. Ein Familienmitglied
kann z. B. T2 = Methyl haben, während ein
anderes T2 = Ethyl hat und ein anderes T2 = Propyl hat, etc.
-
Um "grobe" oder große Sprünge in der Masse bereitzustellen,
kann eine T3-Gruppe konstruiert werden,
die in signifikantem Maße
(z. B. eine oder mehrere hundert) Masseeinheiten zu T-L-X hinzufügt. Solch eine
T3-Gruppe kann als eine Molekulargewichtsbereicheinstellungsgruppe
("WRA") bezeichnet werden.
Eine WRA ist ziemlich nützlich,
wenn man mit einem einzigen Satz von T2-Gruppen
arbeitet, die Massen haben werden, die sich über einen beschränkten Bereich
erstrecken. Ein einzelner Satz von T2-Gruppen
kann verwendet werden, um Tms-Gruppen mit
einem weiten Massebereich zu schaffen, einfach indem eine oder mehrere WRA-T3-Gruppen in die Tms einbezogen
werden. Als ein einfaches Beispiel liefert somit, wenn ein Satz
von T2-Gruppen einen Massebereich von 250–340 Daltons
für die
Tms bereitstellt, das Hinzufügen einer
einzigen WRA mit einer beispielhaften Zahl von 100 Daltons, als
eine T3-Gruppe Zugang zum Massebereich 350–440 Daltons,
während
derselbe Satz von T2-Gruppen verwendet wird.
In ähnlicher
Weise stellt das Hinzufügen
von zwei MWA-Gruppen mit 100 Daltons (jede als eine T3-Gruppe)
Zugang zum Massebereich von 450–540
Daltons bereit, wobei dieses inkrementale Hinzufügen von WRA-Gruppen fortgesetzt werden kann, um
Zugang zu einem sehr großen
Massebereich für
die Tms-Gruppe zu liefern. Bevorzugte Verbindungen
der Formel T2-(J-T3-)n-L-X haben die Formel RVWC-(RWRA)w-RMSSE-L-X,
in der VWC eine "T2"-Gruppe
ist und jede der WRA- und MSSE-Gruppen "T3"-Gruppen sind. Diese
Struktur ist in 12 veranschaulicht
und stellt einen modularen Ansatz für die Herstellung von Tms dar.
-
In der Formel T2-(J-T3-)n- sind T2 und T3 vorzugsweise
ausgewählt
aus Hydrocarbyl, Hydrocarbyl-O-hydrocarbylen, Hydrocarbyl-S-hydrocarbylen,
Hydrocarbyl-NH-hydrocarbylen,
Hydrocarbyl-amid-hydrocarbylen, N-(Hydrocarbyl hydrocarbylen, N,N-Di(hydrocarbyl)hydrocarbylen,
Hydrocarbylacylhydrocarbylen, Heterocyclylhydrocarbyl, wobei das
(die) Heteroatom(e) ausgewählt
ist (sind) aus Sauerstoff, Stickstoff, Schwefel und Phosphor, substituiertem
Heterocyclylhydrocarbyl, wobei das (die) Heteroatom(e) ausgewählt ist
(sind) aus Sauerstoff, Stickstoff, Schwefel und Phosphor und die
Substituenten ausgewählt
sind aus Hydrocarbyl, Hydrocarbyl-O-hydrocarbylen, Hydrocarbyl-NH-hydrocarbylen,
Hydrocarbyl-S-hydrocarbylen, N-(Hydrocarbyl)hydrocarbylen, N,N-Di(hydrocarbyl)hydro'carbylen und Hydrocarbylacyl-hydrocarbylen.
Zusätzlich
kann T2 und/oder T3 ein
Derivat irgendeiner der zuvor aufgelisteten potentiellen T2/T3-Gruppen sein,
wobei ein oder mehrere Wasserstoffe durch Fluoride ersetzt sind.
-
Ebenfalls im Hinblick auf die Formel
T2-(J-T3-)n- hat ein bevorzugtes T3 die
Formel -G(R2)-, in der G eine C1-6-Alkylenkette
mit einem einzelnen R2-Substituenten ist.
So kann, wenn G Ethylen (-CH2-CH2-) ist, einer der zwei Ethylen-Kohlenstoffe
einen R2-Substituenten tragen und R2 ist ausgewählt aus Alkyl, Alkenyl, Alkinyl, Cycloalkyl,
Aryl-kondensiertem Cycloalkyl, Cycloalkenyl, Aryl, Aralkyl, Aryl-substituiertem
Alkenyl oder Alkinyl, Cycloalkyl-substituiertem Alkyl, Cycloalkenyl-substituiertem
Cycloalkyl, Biaryl, Alkoxy, Alkenoxy, Alkinoxy, Aralkoxy, Aryl-substituiertem
Alkenoxy oder Alkinoxy, Alkylamino, Alkenylamino oder Alkinylamino,
Aryl-substituiertem Alkylamino, Aryl-substituiertem Alkenylamino
oder Alkinylamino, Aryloxy, Arylamino, N-Alkylharnstoff-substituiertem
Alkyl, N-Arylharnstoff-substituiertem Alkyl, Alkylcarbylamino-substituiertem
Alkyl, Aminocarbonyl-substituiertem Alkyl, Heterocyclyl, Heterocyclyl-substituiertem
Alkyl, Heterocyclyl-substituiertem Amino, Carboxyalkyl-substituiertem
Aralkyl, Oxocarbocyclyl-kondensiertem
Aryl und Heterocyclylalkyl; Cycloalkenyl, Aryl-substituiertem Alkyl
und Aralkyl, Hydroxy-substituiertem Alkyl, Alkoxy-substituiertem
Alkyl, Aralkoxy-substituiertem
Alkyl, Alkoxy-substituiertem Alkyl, Aralkoxy-substituiertem Alkyl,
Amino-substituiertem
Alkyl, (Aryl-substituiertem Alkyloxycarbonylamino)-substituiertem
Alkyl, Thiol-substituiertem, Alkylsulfonyl-substituiertem Alkyl,
(Hydroxy-substituiertem Alkylthio)-substituiertem Alkyl, Thioalkoxy-substituiertem
Alkyl, Hydrocarbylacylamino-substituiertem
Alkyl, Heterocyclylocylamino-substituiertem Alkyl, Hydrocarbyl-substituiertem-heterocyclylacylamino-substituiertem
Alkyl, Alkylsulfonylamino-substituiertem
Alkyl, Arylsulfonylamino-substituiertem Alkyl, Morpholinoalkyl,
Thiomorpholinoalkyl, Morpholinocarbonyl-substituiertem Alkyl, Thiomorpholinocarbonyl-substituiertem Alkyl,
[N-(Alkyl, Alkenyl oder Alkinyl)- oder N,N-[Dialkyl, Dialkenyl,
Dialkinyl oder (Alkyl, Alkenyl)-amino]carbonyl-substituiertem Alkyl,
Heterocyclylaminocarbonyl, Heterocyclylalkylenaminocarbonyl, Heterocyclylaminocarbonyl-substituiertem Alkyl,
Heterocyclylalkylenaminocarbonyl-substituiertem Alkyl, N,N-[Dialkyl]alkylenaminocarbonyl,
N,N-[Dialkyl]alkylenaminocarbonyl-substituiertem Alkyl, Alkyl-substituiertem
Heterocyclylcarbonyl, Alkyl-substituiertem Heterocyclylcarbonylalkyl,
Carboxyl-substituiertem Alkyl, Dialkylamino-substituiertem Acylaminoalkyl
und Aminosäureseitenketten,
ausgewählt
aus Arginin, Asparagin, Glutamin, S-Methylcystein, Methionin und
entsprechenden Sulfoxid- und Sulfonderivaten davon, Glycin, Leucin,
Isoleucin, allo-Isoleucin, tert.-Leucin, Norleucin, Phenylalanin,
Tyrosin, Tryptophan, Prolin, Alanin, Ornithin, Histidin, Glutamin,
Valin, Threonin, Serin, Asparaginsäure, beta-Cyanoalanin und allo-Threonin; Alinyl
und Heterocyclylcarbonyl, Aminocarbonyl, Amido, Mono- oder Dialkylaminocarbonyl,
Mono- oder Diarylaminocarbonyl, Alkylarylaminocarbonyl, Diarylaminocarbonyl,
Mono- oder Diacylaminocarbonyl, aromatischem oder aliphatischem
Acyl, Alkyl, fakultativ substituiert mit Substituenten, die ausgewählt sind
aus Amino, Carboxy, Hydroxy, Mercapto, Mono- oder Dialkylamino,
Mono- oder Diarylamino,
Alkylarylamino, Diarylamino, Mono- oder Diacylamino, Alkoxy, Alkenoxy,
Aryloxy, Thioalkoxy, Thioalkenoxy, Thioalkinoxy, Thioaryloxy und Heterocyclyl.
-
Eine bevorzugte Verbindung der Formel
T
2-(J-T
3-)
n-L-X hat die Struktur:
in der G(CH
2)
1-6 ist, so daß ein Sauerstoff an einer und
nur einer der CH
2-Gruppen, die durch ein
einziges "G" dargestellt werden,
ersetzt ist durch -(CH
2)
c-Amid-T
4; T
2 und T
4 organische Einheiten der Formel C
2–25N
0
–9O
0–9H
αF
β sind,
so daß die
Summe von α und β ausreichend
ist, um die sonst ungesättigten
Valenzen der C-, N- und O-Atome abzusättigen; Amid
ist; R
1 Wasserstoff
oder C
1-10-Alkyl ist; c eine ganze Zahl
aus dem Bereich von 0 bis 4 ist; und n eine ganze Zahl aus dem Bereich
von 1 bis 50 ist, so daß,
wenn n größer als
1 ist, G, c, Amid, R
1 und T
4 unabhängig ausgewählt werden.
-
In einer weiter bevorzugten Ausführungsform
hat eine Verbindung der Formel T
2-(J-T
3-)
n-L-X die Struktur:
in der T
5 eine
organische Einheit der Formel C
1-25N
0
-9O
0-9H
αF
β ist,
so daß die
Summe von α und β ausreichend ist,
um die sonst ungesätigten
Valenzen der C-, N- und O-Atome abzusättigen; und T
5 ein
tertiäres
oder quartäres
Amin oder eine organische Säure
einschließt;
m eine ganze Zahl aus dem Bereich von 0–49 ist und T
2, T
4, R
1, L und X zuvor
definiert worden sind.
-
Eine weitere bevorzugte Verbindung
mit der Formel T
2-(J-T
3-)
n-L-X hat die besondere Struktur:
in der T
5 eine
organische Einheit der Formel C
1-25N
0
-9O
0-9H
αF
β ist,
so daß die
Summe von α und β ausreichend ist,
um die sonst ungesättigten
Valenzen der C-, N- und O-Atome abzusättigen; und T
5 ein
tertiäres
oder quartäres
Amin oder eine organische Säure
einschließt;
m eine ganze Zahl aus dem Bereich von 0–49 ist und T
2, T
4, c, R
1, "Amid", L und X zuvor definiert
worden sind.
-
In den obigen Strukturen, die eine
T
5-Gruppe besitzen, ist -Amid-T
5 vorzugsweise
eine der folgenden Gruppen, die geeigneterweise hergestellt werden,
indem organische Säuren
mit freien Aminogruppen umgesetzt werden, die an "G" gebunden sind:
Wenn die obigen Verbindungen
eine T
5-Gruppe besitzen und die "G"-Gruppe eine freie Carboxylgruppe besitzt (oder
ein reaktives Äquivalent
davon), dann sind die folgenden Gruppen die bevorzugte -Amid-T
5-Gruppen, die geeigneterweise hergestellt
werden können,
indem das geeignete organische Amin mit einer freien Carboxylgruppe
umgesetzt wird, die an eine "G"-Gruppe gebunden
ist:
-
In drei bevorzugten Ausführungsformen
der Erfindung hat T-L-MOI die Struktur:
in denen
T
2 und T
4 organische
Einheiten der Formel C
1-25N
0-9O
0-9S
0-3P
0
-3H
αF
βI
δ sind,
so daß die
Summe von α, β und δ ausreichend
ist, um die sonst ungesättigten
Valenzen der C-, N-, O-, S- und P-Atome abzusättigen; G(CH
2)
1-6 ist, wobei ein und nur ein Wasserstoff
auf den CH
2-Gruppen, die durch jedes G dargestellt
werden, durch -(CH
2)
c-Amid-T
4 ersetzt ist; Amid
ist; R
1 Wasserstoff
oder C
1-10-Alkyl ist; c eine ganze Zahl
aus dem Bereich von 0 bis 4 ist; "C
2-C
10" eine
Hydrocarbylengruppe mit von 2 bis 10 Kohlenstoffatomen darstellt, "ODN-3'-OH" ein
Nukleinsäurefragment
mit einer endständigen
3'-Hydroxylgruppe
darstellt (d. h. ein Nukleinsäurefragment,
das an (C
1-C
10)
am anderen als dem 3'-Ende
des Nukleinsäurefragments
gebunden ist); und n eine ganze Zahl aus dem Bereich von 1 bis 50
ist, so daß,
wenn n größer als
1 ist, dann G, c, Amid, R
1 und T
4 unabhängig
ausgewählt
werden. Vorzugsweise sind nicht drei Heteroatome an ein einziges
Kohlenstoffatom gebunden.
-
In Strukturen, wie oben angegeben,
die eine T2-C(=O)-N(R1)-Gruppe
enthalten, kann diese Gruppe dadurch gebildet werden, daß ein Amin
der Formel HN(R1)- mit einer organischen
Säure umgesetzt
wird, die aus den folgenden ausgewählt ist, die nur beispielhaft
sind und keine erschöpfende
Liste möglicher
organischer Säuren
darstellen: Ameisensäure,
Essigsäure,
Propiolsäure,
Propionsäure,
Fluoressigsäure,
2-Butinsäure,
Cyclopropancarbonsäure,
Buttersäure,
Methoxyessigsäure,
Difluoressigsäure,
4-Pentinsäure,
Cyclobutancarbonsäure,
3,3-Dimethylacrylsäure,
Valeriansäure,
N,N-Dimethylglycin, N-Formyl-Gly-OH, Ethoxyessigsäure, (Methylthio)essigsäure, Pyrrol-2-carbonsäure, 3-Furancarbonsäure, Isoxazol-5-carbonsäure, trans-3-Hexensäure, Trifluoressigsäure, Hexansäure, Ac-Gly-OH,
2-Hydroxy-2-methylbuttersäure, Benzoesäure, Nicotinsäure, 2-Pyrazincarbonsäure, 1-Methyl-2-pyrrolcarbonsäure, 2-Cyclopenten-1-essigsäure, Cyclopentylessigsäure, (S)-(–)-2-Pyrrolidon-5-carbonsäure, N-Methyl-L-prolin,
Heptansäure,
Ac-b-Ala-OH, 2-Ethyl-2-hydroxybuttersäure, 2-(2-Methoxyethoxy)essigsäure, p-Tolylsäure, 6-Methylnicotinsäure, 5- Methyl-2-pyrazincarbonsäure, 2,5-Dimethylpyrrol-3-carbonsäure, 4-Fluorbenzoesäure, 3,5-Dimethylisoxazol-4-carbonsäure, 3-Cyclopentylpropionsäure, Octansäure, N,N-Dimethylsuccinamidsäure, Phenylpropiolsäure, Zimtsäure, 4-Ethylbenzoesäure, p-Anissäure; 1,2,5-Trimethylpyrrol-3-carbonsäure, 3-Fluor-4-methylbenzoesäure, Ac-DL-Propargylglycin,
3-(Trifluormethyl)buttersäure,
1-Piperidinpropionsäure,
N-Acetylprolin, 3,5-Difluorbenzoesäure, Ac-L-Val-OH,
Indol-2-carbonsäure,
2-Benzofurancarbonsäure,
Benzotriazol-5-carbonsäure,
4-n-Propylbenzoesäure,
3-Dimethylaminobenzoesäure,
4-Ethoxybenzoesäure, 4-(Methylthio)benzoesäure, N-(2-Furoyl)glycin,
2-(Methylthio)nicotinsäure, 3-Fluor-4-methoxybenzoesäure, Tfa-Gly-OH, 2-Naphthoesäure, Chinaldinsäure, Ac-L-Ile-OH,
3-Methylinden-2-carbonsäure,
2-Chinoxalincarbonsäure, 1-Methylindol-2-carbonsäure, 2,3,6-Trifluorbenzoesäure, N-Formyl-L-Met-OH,
2-[2-(2-Methoxyethoxy)ethoxy]essigsäure, 4-n-Butylbenzoesäure, N-Benzoylglycin,
5-Fluorindol-2-carbonsäure, 4-n-Propoxybenzoesäure, 4-Acetyl-3,5-dimethyl-2-pyrrolcarbonsäure, 3,5-Dimethoxybenzoesäure, 2,6-Dimethoxynicotinsäure, Cyclohexanpentansäure, 2-Naphthylessigsäure, 4-(1H-Pyrrol-1-yl)benzoesäure, Indol-3-propionsäure, m-Trifluormethylbenzoesäure, 5-Methoxyindol-2-carbonsäure, 4-Pentylbenzoesäure, Bz-b-Ala-OH,
4-Diethylaminobenzoesäure,
4-n-Butoxybenzoesäure,
3-Methyl-5-CF3-isoxazol-4-carbonsäure, (3,4-Dimethoxyphenyl)essigsäure, 4-Biphenylcarbonsäure, Pivaloyl-Pro-OH,
Octanoyl-Gly-OH, (2-Naphthoxy)essigsäure, Indol-3-buttersäure, 4-(Trifluormethyl)phenylessigsäure, 5-Methoxyindol-3-essigsäure, 4-(Trifluormethoxy)benzoesäure, Ac-L-Phe-OH,
4-Pentyloxybenzoesäure,
Z-Gly-OH14-Carboxy-N-(fur-2-ylmethyl)pyrrolidin-2-on,
3,4-Diethoxybenzoesäure,
2,4-Dimethyl-5-CO2Et-pyrrol-3-carbonsäure, N-(2-Fluorphenyl)succinamidsäure, 3,4,5-Trimethoxybenzoesäure, N-Phenylanthranilsäure, 3-Phenoxybenzoesäure, Nonanoyl-Gly-OH, 2-Phenoxypyridin-3-carbonsäure, 2,5-Dimethyl-l-phenylpyrrol-3-carbonsäure, trans-4-(Trifluormethyl)zimtsäure, (5-Methyl-2-phenyloxazol-4-yl)essigsäure, 4-(2-Cyclohexenyloxy)benzoesäure, 5-Methoxy-2-methylindol-3-essigsäure, trans-4-Cotinincarbonsäure, Bz-5-Aminovaleriansäure, 4-Hexyloxybenzoesäure, N-(3-Methoxyphenyl)succinamidsäure, Z-Sar-OH,
4-(3,4-Dimethoxyphenyl)buttersäure,
Ac-o-Fluor-DL-Phe-OH, N-(4-Fluorphenyl)glutaminsäure, 4'-Ethyl-4-biphenylcarbonsäure, 1,2,3,4-Tetrahydroacridincarbonsäure, 3-Phenoxyphenylessigsäure, N-(2,4-Difluorphenyl)succinamidsäure, N-Decanoyl-Gly-OH,
(+)-6-Methoxy-a-methyl-2-naphthalinessigsäure, 3-(Trifluormethoxy)zimtsäure, N-Formyl- DL-Trp-OH, (R)-(+)-a-Methoxy-a-(trifluormethyl)phenylessigsäure, Bz-DL-Leu-OH,
4-(Trifluormethoxy)phenoxyessigsäure, 4-Heptyloxybenzoesäure, 2,3,4-Trimethoxyzimtsäure, 2,6-Dimethoxybenzoyl-Gly-OH,
3-(3,4,5-Trimethoxyphenyl)propionsäure, 2,3,4,5,6-Pentafluorphenoxyessigsäure, N-(2,4-Difluorphenyl)glutaminsäure, N-Undecanoyl-Gly-OH,
2-(4-Fluorbenzoyl)benzoesäure, 5-Trifluormethoxyindol-2-carbonsäure, N-(2,4-Difluorphenyl)diglycolamidsäure, Ac-L-Trp-OH, Tfa-L-Phenylglycin-OH,
3-Iodbenzoesäure,
3-(4-n-Pentylbenzoyl)propionsäure, 2-Phenyl-4-chinolincarbonsäure, 4-Octyloxybenzoesäure, Bz-L-Met-OH, 3,4,5-Triethoxybenzoesäure, N-Lawoyl-Gly-OH,
3,5-Bis(trifluormethyl)benzoesäure,
Ac-5-Methyl-DL-Trp-OH, 2-Iodphenylessigsäure, 3-Iod-4-methylbenzoesäure, 3-(4-n-Hexylbenzoyl)propionsäure, N-Hexanoyl-L-Phe-OH,
4-Nonyloxybenzoesäure,
4'-(Trifluormethyl)-2-biphenylcarbonsäure, Bz-L-Phe-OH,
N-Tridecanoyl-Gly-OH, 3,5-Bis(trifluormethyl)phenylessigsäure, 3-(4-n-Heptylbenzoyl)propionsäure, N-Heptanoyl-L-Phe-OH,
4-Decyloxybenzoesäure,
N-(α,α,α-Trifluor-m-tolyl)anthranilsäure, Nifluminsäure, 4-(2-Hydroxyhexafluorisopropyl)benzoesäure, N-Myristoyl-Gly-OH,
3-(4-n-Octylbenzoyl)propionsäure,
N-Octanoyl-L-Phe-OH, 4-Undecyloxybenzoesäure, 3-(3,4,5-Trimethoxyphenyl)propionyl-Gly-OH,
8-Iodnaphthoesäure,
N-Pentadecanoyl-Gly-OH, 4-Dodecyloxybenzoesäure, N-Palmitoyl-Gly-OH und
N-Stearoyl-Gly-OH. Diese organischen Säuren sind von einer der folgenden
Firmen erhältlich:
Advanced ChemTech, Louisville, KY; Bachem Bioscience Inc., Tonance,
CA; Calbiochem-Novabiochem Corp., San Diego, CA; Farchan Laboratories
Inc., Gainesville FL; Lancaster Synthesis, Windham, NH; und MayBridge
Chemical Company (c/o Ryan Scientific), Columbia, SC. Die Kataloge
dieser Firmen verwenden die Abkürzungen, die
oben verwendet werden, um die Säuren
zu identifizieren.
-
f. Kombinatorische Chemie
als ein Mittel zur Herstellung von Markierungen
-
Kombinatorische Chemie ist eine Art
von Synthesestrategie, die zur Herstellung großer chemischer Bibliotheken
führt (siehe
z. B. PCT-Anmeldung Veröffentlichungs-Nr.
WO 94/08051). Diese kombinatorischen Bibliotheken können als
Markierungen zur Identifizierung von interessierenden Molekülen (MOIs)
verwendet werden. Kombinatorische Chemie kann definiert werden als
die systematische und wiederholte, kovalente Verknüpfung eines
Satzes unterschiedlicher "Bausteine" variierender Struktur
miteinander, um eine große
Anzahl verschiedener Moleküleinheiten
zu liefern. Bausteine können
viele Formen annehmen, sowohl natürlich vorkommende als auch
synthetische, wie etwa Nukleophile, Elektrophile, Diene, Alkylierungs-
oder Acylierungsmittel, Diamine, Nukleotide, Aminosäuren, Zucker,
Lipide, organische Monomere, Synthons und Kombinationen der obigen.
Chemische Reaktionen, die verwendet werden, um die Bausteine zu
verknüpfen,
können
Alkylierung, Acylierung, Oxidation, Reduktion, Hydrolyse, Substitution,
Elimination, Addition, Zyklisierung, Kondensation und dergleichen
einschließen.
Dieser Prozeß kann
Bibliotheken von Verbindungen erzeugen, die oligomer, nicht-oligomer
oder Kombinationen davon sind. Wenn oligomer, können die Verbindungen verzweigt, unverzweigt
oder zyklisch sein. Beispiele für
oligomere Strukturen, die mit kombinatorischen Verfahren hergestellt
werden können,
schließen
Oligopeptide, Oligonukleotide, Oligosaccharide, Polylipide, Polyester,
Polyamide, Polyurethane, Polyharnstoffe, Polyether, Poly(phosphorderivate),
z. B. Phosphate, Phosphonate, Phosphoramide, Phosphonamide, Phosphite,
Phosphinamide, etc., und Poly(schwefelderivate), z. B. Sulfone,
Sulfonate, Sulfate, Sulfonamide, Sulfenamide, etc., ein.
-
Ein üblicher Typ von oligomerer
kombinatorischer Bibliothek ist die kombinatorische Peptid-Bibliothek. Kürzliche
Innovationen in der Peptidchemie und Molekularbiologie haben Bibliotheken
ermöglicht,
die aus einigen zehn bis einigen hundert von Millionen verschiedenen
Peptidsequenzen bestehen, die hergestellt und verwendet werden.
Solche Bibliotheken können
in drei große
Kategorien unterteilt werden. Eine Kategorie von Bibliotheken umfaßt die chemische
Synthese von löslichen,
nicht an einen Träger
gebundenen Peptidbibliotheken (z. B. Houghten et al., Nature 354:
84, 1991). Eine zweite Kategorie umfaßt die chemische Synthese von
trägergebundenen
Peptidbibliotheken, präsentiert
auf festen Trägern,
wie etwa Kunststoffnadeln, Harzperlen oder Baumwolle (Geysen et
al., Mol. Immunol. 23: 709, 1986; Lam et al., Nature 354: 82, 1991;
Eichler und Houghten, Biochemistry 32: 11035, 1993). In diesen ersten
zwei Kategorien sind die Bausteine typischerweise L-Aminosäuren, D-Aminosäuren, nicht-natürliche Aminosäuren oder
irgendeine Mischung oder Kombination davon. Eine dritte Kategorie
verwendet molekularbiologische Ansätze, um Peptide oder Proteine
auf der Oberfläche
filamentöser
Phagenteilchen oder Plasmide herzustellen (Scott und Craig, Curr.
Opinion Biotech. 5: 40, 1994). Lösliche,
nicht an Träger
gebundene Peptidbibliotheken scheinen für eine Reihe von Anwendungen
geeignet zu sein, einschließlich
Verwendung als Markierungen. Das verfügbare Repertoire von chemischen
Diversitäten
in Peptidbibliotheken kann durch solche Schritte wie Permethylierung
ausgedehnt werden (Ostresh et al., Proc. Natl. Acad. Sci., USA 91:
11138, 1994).
-
Zahlreiche Varianten von kombinatorischen
Peptidbibliotheken sind möglich,
in denen die Peptid-Rückgratkette
modifiziert wird und/oder die Amidbindungen durch mimetische Gruppen
ersetzt worden sind. Amid-mimetische Gruppen, die verwendet werden
können,
schließen
Harnstoffe, Urethane und Carbonylmethylengruppen ein. Die Umstrukturierung
der Rückgratkette,
so daß Seitenketten
von den Amid-Stickstoffen jeder Aminosäure statt von den Alpha-Kohlenstoffatomen
ausgehen, ergeben Bibliotheken von Verbindungen, die als Peptoide
bekannt sind (Simon et al., Proc. Natl. Acad. Sci., USA 89: 9367,
1992).
-
Ein weiterer üblicher Typ von oligomerer
kombinatorischer Bibliothek ist die kombinatorische Oligonukleotidbibliothek,
in der die Bausteine irgendeine Form von natürlich vorkommenden oder nicht-natürlichen
Nukleotid- oder Polysaccharid-Derivaten sind, einschließlich die
Fälle,
in denen verschiedene organische und anorganische Gruppen die Phosphatverknüpfung ersetzen
können
und Stickstoff oder Schwefel Sauerstoff in einer Etherverknüpfung ersetzen
können
(Schneider et al., Biochem. 34: 9599, 1995; Freier et al., J. Med.
Chem. 38: 344, 1995; Frank, J. Biotechnology 41: 259, 1995; Schneider
et al., veröffentlichte
PCT WO 942052; Ecker et al., Nucleic Acids Res. 21: 1853, 1993).
-
Vor kurzem ist die kombinatorische
Herstellung von Sammlungen nicht-oligomerer, kleiner Molekülverbindungen
beschrieben worden (DeWitt et al., Proc. Natl. Acad. Sci. USA 90:
690, 1993; Bunin et al., Proc. Natl. Acad. Sci., USA 91: 4708, 1994).
Strukturen, die für
die Einstellung von Kleinmolekül-Bibliotheken
geeignet sind, umfassen eine breite Vielfalt organischer Moleküle, z. B.
Heterozyklen, Aromaten, Alizyklen, Aliphaten, Steroide, Antibiotika,
Enzyminhibitoren, Liganden, Hormone, pharmazeutische Wirkstoffe,
Alkaloide, Opioide, Terpene, Porphyrine, Toxine, Katalysatoren sowie
Kombinationen davon.
-
g. Spezifische Verfahren
zur kombinatorischen Synthese von Markierungen
-
Zwei Verfahren zur Herstellung und
Verwendung eines unterschiedlichen Satzes Amin-enthaltender MS-Markierungen sind unten
umrissen. In beiden Verfahren wird Festphasensynthese eingesetzt,
um simultane parallele Synthese einer großen Anzahl markierter Linker
zu ermöglichen,
unter Verwendung der Techniken der kombinatorischen Chemie. Beim
ersten Verfahren führt
die letztendliche Abspaltung der Markierung vom Oligonukleotid zur
Freisetzung eines Carbonsäureamids.
Beim zweiten Verfahren erzeugt die Abspaltung der Markierung eine
Carbonsäure.
Die chemischen Komponenten und Verknüpfungselemente, die in diesen
Verfahren verwendet werden, werden wie folgt abgekürzt:
| R | Harz |
| FMOC | Fluorenylmethoxycarbonyl-Schutzgruppe |
| All | Allyl-Schutzgruppe |
| CO2H | Carbonsäuregruppe |
| CONH2 | Carbonsäureamidgruppe |
| NH2 | Aminogruppe |
| OH | Hydroxylgruppe |
| CONH | Amidverknüpfung |
| COO | Esterverknüpfung |
| NH2-Rink-CO2H | 4-[(α-Amino)-2,4-dimethoxybenzyl]-phenoxybuttersäure( Rink-Linker) |
| OH-1MeO-CO2H | (4-Hydroxymethyl)phenoxybuttersäure |
| OH-2MeO-CO2H | (4-Hydroxymethyl-3-methoxy)phenoxyessigsäure |
| NH2-A-COOH | Aminosäure mit
aliphatischer oder aromatischer Aminfunktionalität in der Seitenkette |
| X1
. . . Xn-COOH | Satz
von n verschiedenen Carbonsäuren
mit einzigartigen relativen Molekülmassen |
| Oligo1
. . . Oligo(n) | Satz
von n Oligonukleotiden |
| HBTU | O-Benzotriazol-1-yl-N,N,N',N'-tetramethyluroniumhexafluorophosphat |
-
Die Abfolge der Schritte in Verfahren
1 ist wie folgt:
-
Die Abfolge der Schritte in Verfahren
2 ist wie folgt:
-
h. Phosphoramidit und
verwandte Verfahren zur Marker-Synthese
-
Festphasesynthese von natürlichen
Polymeren wurde ursprünglich
gleichzeitig durch Merrifield (Merrifield, 1963) und für Peptidchemie
entwickelt und anschließend
durch Letsinger (Letsinger und Mahadevan 1965) an die Oligonukleotidsynthese
angepaßt.
Das Konzept weist vier grundsätzliche
Aspekte auf: I. Das Oligonukleotid wird synthetisiert, während es
kovalent an einen festen Träger
gebunden ist; II. Überschüssige lösliche geschützte Nukleotide
und Kopplungsreagenz können
eine Reaktion fast bis zur Vervollständigung antreiben; III. Die
Reaktion wird in einem einzelnen Reagenz durchgeführt, um
mechanische Verluste aufgrund der festen Träger-Manipulation zu verringern,
was eine Synthese mit geringen Mengen an Ausgangsmaterialien ermöglicht;
und IV. Die heterogenen Reaktionen werden standardisiert und diese
Verfahren werden leicht automatisiert.
-
Das am meisten verbreitete Verfahren
zur Synthese von Oligonukleotiden ist der Phosphit-Triester-Ansatz.
Ein weiteres jedoch weniger verbreitetes Verfahren der Synthese
ist der H-Phosphonat-Ansatz.
-
Der Phosphit-Triester-Ansatz
zur Oligonukleotid-Synthese
-
Die Entwicklung dieses Verfahrens
wurde 1975 begonnen, als Letsinger (Letsinger et al. 1975) das symmetrische
Posphit-Reagenz Methoxyphospodichloridit einführten. Obwohl die Kopplungszeiten
dramatisch verringert wurden, weist diese Verbindung den Nachteil
auf, daß sie
zu reaktiv ist, was die Handhabung bei Raumtemperatur sehr schwierig
macht und die Lagerung von Phophitmonomeren sogar bei –10°C unmöglich macht.
Die Reaktion mit geschützten
Nukleotiden führt
zu der Herstellung von großen
Mengen von symmetrischem 3'-3'-Dimer (Letsinger et al., 1982).
-
1981 löste die Einführung des
neuen phosphitylierenden Mittels N,N-Dimethylaminomethoxyphosphin (Beaucage
und Caruthers 1981) nicht nur das Problem der Bildung von 3'-3'-Dimeren während der
Phophitylierung, sondern führte
auch zur Erzeugung von Desoxyribonukleosid-Phosphitderivaten, die
bis zu einem bestimmten Ausmaß stabil
gegenüber
Sauerstoff und atmosphärischer
Luftfeuchtigkeit bei Raumtemperatur sind. Als die am meisten brauchbare
Verbindung erwies sich N,N-Diisopropylamin (Adams et al. 1983, McBride und
Caruthers 1983) das leicht auf Silicagelsäulen gereinigt werden kann
und das als trockene Pulver bei Raumtemperatur stabil ist.
-
Die durch dieses Verfahren verwendete
5'-Schutzgruppe
ist das Dimethoxytrityl (DMT). Diese Gruppe wird vollständig durch
Behandlung mit Trichloressigsäure
(1–3%
w/v) in Dichlormethan in weniger als einer Minute abgespalten. Sobald
die Schutzgruppe entfernt ist, ist das freie 5'-Hydroxyl zur Kopplung an den nächsten Nukleosidbaustein
erhältlich.
-
Anders als die Phosphodichloridite
in Letsingers Ansatz und das Posphomonochloridit/Tetrazol in der Ursprungsarbeit
von Caruthers (Caruthers et al. 1980) können die Phosphoramidite nicht
direkt mit einer freien 5'-Hydroxylfunktion
auf einer wachsenden Kette reagieren. Sie müssen zunächst durch Behandlung mit einer schwachen
Säure,
wie zum Beispiel Tetrazol, aktiviert werden. Tetrazol wurde als
eine zweifache Rolle spielend gezeigt (Berner et al., 1990; Dahl
et al., 1990): sie protoniert die Dialklyaminogruppe der Phosphoramiditfunktion
und wirkt als nächstes
als ein Nukleophil wodurch ein sehr reaktives Tetrazolphosphan-Intermediat erzeugt
wird. Die Kopplungsreaktionen mit diesen Desoxynukleosid-Phosphoramiditreagenzien
sind sehr schnell (weniger als 2 Minuten) und fast quantitativ.
-
Weil die Kopplungsreaktion in einer
begrenzten Zeitdauer nicht quantitativ sein kann, wird bei jedem Kopplungsschritt
ein kleiner Prozentsatz von verkürzten
Sequenzen hergestellt. Diese Reaktionsfehlschläge enthalten 5'-Hydroxyle. Falls
diesen falschen Sequenzen ermöglicht
würde,
weiter zu reagieren, wäre
es schwierig, das Produkt aus dem Sequenzgemisch zu isolieren. Dieses
Problem wird größtenteils
durch maskieren der verbleibenden freien 5'-Hydroxyle durch Acetylierung überwunden.
Dieser Maskierungsschritt wird mit dem starken Acetylierungsreagenz
N-Acetyldimethylaminopyridiniumion erreicht, das sich bei Reaktion von äquimolaren
Mengen von Essigsäureanhydrid
und 4-Dimethylaminopyridin (DMAP) bildet. Die Reaktion ist in 0,5
Minuten fast quantitativ. Unter der Verwendung von N-Methylimidazol
anstelle von DMAP wird das Oligonukleotid verbesserte biologische
Eigenschaften aufweisen (Fenance et al., 1989).
-
Die neu gebildete Phosphit-Internukleotidverbindung
ist instabil und sowohl gegenüber
saurer als auch basischer Spaltung empfindlich. Daher wird der trivalente
Phosphitester nach der Maskierung zu einem stabilen pentavalenten
Phosphattriester oxidiert. Jod wird als ein mildes Oxidationsmittel
in basischer Tetrahydrofwanlösung
mit Wasser als dem Sauerstoff-Donor verwendet. Die Reaktionen sind
extrem schnell, wobei sie in 30 Sekunden quantitativ ist. Die Oxidation
vervollständigt
den Nukleotidadditionszyklus. Die Kettenverlängerung kann nach Entfernen
der Dimethoxytritylgruppe und dem 5'-Ende der wachsenden Kette und dem wiederholen
eines weiteren Zyklus von der Nukleotidaddition fortschreiten. Die
Vervollständigung
der Abspaltung von dem Träger
und die gleichzeitige Phasen- und Phosphatentschützung werden durch Behandlung
mit konzentriertem Ammoniumhydroxyd erreicht.
-
H-Phosphonat-Verfahren
der Oligonukleotid-Synthese
-
Die Verwendung eines Nukleosid-H-Phosphonats
wurde zuerst durch Todd und Mitarbeiter berichtet (Hall et al. 1957).
Danach blieb die H-Phosphonat-Chemie bis in die 80er Jahre unerforscht.
1985 und 1986 wurde das Wiederaufleben dieses Ansatzes durch Garegg
et al. (1985, 1986a, 1986b) und Froehler et al. (1986a und 1986b)
eingeführt.
-
In diesem Verfahren ist das aktivierbare
Monomer ein 5'-DNT-Base-geschütztes-Nukleosid-3'-Hydrogenphosphonat. In diesen Monomeren
macht die Anwesenheit der H-Phosphonatgruppe
den Phosphatschutz überflüssig.
-
Die selben Basenschutzgruppen werden
verwendet, wie im Phosphat-Triester-Ansatz. Daher ist die Schutzstrategie
dieselbe für
die Hydroxyl- und die exocyclischen Amine auf den Heterocyclen der
Entschützung.
-
H-Phosphonat-Synthese-Zyklus. Es
gibt keinen Oxidationsschritt während
der Kettenverlängerung
in der Oliognukleotid-Synthese gemäß den H-Phosphonat-Verfahren.
Die Oxidation wird am Ende der Synthese durchgeführt.
-
Der Kopplungsprozess der H-Phosphonat-Synthese
wird durch ein gehindertes Acylchlorid aktiviert und das gebildete
Anhydrid wird dazu verwendet, um mit einem freien Oligonukleotid
5'-Hydroxylende
zu reagieren, wobei ein H-Phosphonat-Analog der Internukleotid-Brücke gebildet
wird. Die Ausbeuten sind ungefähr 96–99%. Von
Pivaloylchlorid und 1-Adamantancarbonylchlorid (Andrus et al., 1988)
wurde berichtet, daß dies die
besten Aktivatoren sind. Jedoch werden einige Nebenreaktionen zwischen
den kondensierende Reagenzien und den Ausführungsmaterial während der
Kondensation beobachtet und dies führt zu abnehmenden Ausbeuten
der gewünschten
Verbindung. Insbesondere führte
die Präaktivierung
eines nukleosidischen 3'-H-Phosphonats,
gefolgt von der Addition an einer OH-Komponente, die gewöhnlicherweise
bei der Synthese auf Polymerträgern
stattfindet, zu geringeren Ausbeuten der H-Phosphonat-Diester. Die
andere Seite der Reaktion ist eine Modifikation von heterozyklischen
Basen von Nukleotiden (Acylierung oder Phosphitylierung von Guanin
und Thymin), während
der Kondensation). Das Maskierungsreagenz der Phosphoramidit-Chemie (Essigsäureanhydrid/N- Methylimidazol) ist
für den
H-Phosphonat-Ansatz nicht geeignet. Cyanoethyl-H-Phosphonat (Gaffney
und Jones, 1988) oder Iso-Propyl-H-Phosphonat (Andres et al., 1988),
aktiviert durch Acetylchlorid oder PFPC, kann verwendet werden.
-
Nach Vervollständigung der Sequenz werden
alle H-Phosphonat-Bindungen gleichzeitig zu Phosphodiesterverbindungen
oxidiert. Anstelle der Oxidation mit Jod sind H-Phosphonat-Desoxyribonukleotide
in der Lage, in DNA-Analoga zu Phosphorthioaten, Phosphoramidaten
oder Phosphotriestern konvertiert zu werden (Froehler, 1986c, Froehler
et al., 1988). Die Vorteile dieses Verfahrens sind eine erhöhte Monomerstabilität, die Präparation
von 35Smarkierten Oligonukleotiden (Stein
et al., 1990) und die Möglichkeit
der Wiederverwendung des Überschusses
von aktiviertem Nukleosid, das nicht reagiert hat (Seliger und Rösch, 1990).
H-Phosphonate sind käuflich
erhältlich
von Glen Research (Herndon, VA).
-
Die vorliegender Erfindung stellt
Marker zur Verfügung
die leicht in die oben beschriebenen Standardoligonukleotidsynthesen
inkorporiert werden können.
Die vorliegende Erfindung stellt einen CMST-Marker zur Anbringung
an eine Nukleinsäure
während
der Festphase-Synthese
zur Verfügung.
Der CMST-Marker ist ein Phosphoramidit eines CMST.
-
Ein Verfahren gemäß der vorliegenden Erfindung
schließt
die Anbringung eines CMST-Markers
an eine Nukleinsäure
während
Festphase-Synthese ein. Genauer gesagt wird ein Phosphoramidit eines
CMST an ein Träger-gebundenes
Oligodesoxynukleotid kondensiert. Die Anpassung von Festphase-Phosphoramidit-Chemie
an CMST-Technologie vereinfacht die Reinigung und erleichtert die
Automatisierung des Verfahrens, wodurch der Durchsatz erhöht wird.
Dieses ermöglicht
die einfache Herstellung der markierten Moleküle.
-
Wie oben beschrieben wurde das Phosphoramiditverfahren
der Oligonukleotid-Synthese automatisiert und wird weit verwendet.
Ein Phosphoramidit eines CMST-Markers stellt ein bequemes Verfahren
der Markierung von Oligonukleotiden mit CMST-Markern zur Verfügung. Gemäß der vorliegenden
Erfindung wird ein Phosphoramidit eines CMST- Markers an eine Oligonukleotidkette
kondensiert, die an einen festen Träger gebunden wurde. Der Träger kann
von jeder für
die Festphase von Nukleinsäuren
geeigneten Art sein. Das Polynukleosid kann ein Ribonukleosid- oder
ein Desoxyribonuklosid-Phosphodiester sein. Auch können Polynuklosid-Analoga,
einschließlich,
jedoch nicht begrenzt auf, Phosphorthioate, Methylphosphonate, usw.,
verwendet werden.
-
Ein CMST-Phosphoamidid gemäß der vorliegenden
Erfindung kann von der generellen Struktur TMS-L-X
sein, wobei TMS durch Massenspektrometrie
nachgewiesen wird, L ein photochemischer Linker ist und x eine Gruppe
ist, die ein Koppeln an Polynukleosid an einem festen Träger ermöglicht.
-
Die automatisierte Synthese von Oligonukleotiden
verwendete Phosphoramidit-Chemie1 und eine Vielzahl
von Phosphoamidit-Reagenzien (z. B. Biotin2)
wurden entwickelt, die vorteilhafterweise auf dieser Chemie beruhen,
um Oligonukleotide zu markieren (siehe 24)3. Die Anpassung
dieser Chemie, um eine Massenmarkierung von Festphase-getragenen
Oligonukleotiden zu ermöglichen,
vereinfacht die Reinigung und erleichtert die Automatisierung des
Verfahrens, wodurch der Durchsatz erhöht wird.
-
Ein vorgeschlagenes Schema, das von
der bereits hergestellten Markersäure ausgeht, ist in 25 gezeigt.
-
2. Linker
-
Eine "Linker"-Komponente (oder L), wie hierin verwendet,
bedeutet entweder eine direkte kovalente Bindung oder eine organische
chemische Gruppe, die verwendet wird, um eine "Markierung" (oder T) an ein "interessierendes Molekül" (oder MOI) durch
kovalente chemische Bindungen zu binden. Zusätzlich ist die direkte Bindung
selbst oder eine oder mehrere Bindungen innerhalb der Linker-Komponente
unter Bedingungen spaltbar, die ermöglichen, daß T vom Rest der T-L-X-Verbindung
(einschließlich
der MOI-Komponente) freigesetzt (mit anderen Worten abgespalten)
wird. Die variable Markierungskomponente, die innerhalb von T vorhanden
ist, sollte gegenüber
den Spaltungsbedingungen stabil sein. Vorzugsweise kann die Spaltung
schnell durchgeführt
werden; innerhalb weniger Minuten und vorzugsweise innerhalb etwa
15 Sekunden oder weniger.
-
Im allgemeinen wird ein Linker verwendet,
um jede aus einem großen
Satz von Markierungen mit jedem aus einem ähnlich großen Satz von MOIs zu verknüpfen. Typischerweise
wird eine einzige Markierungs-Linker-Kombination mit jedem MOI verknüpft (um
verschiedene T-L-MOI
zu ergeben), aber in einigen Fällen
kann mehr als eine Markierungs-Linker-Kombination mit jedem individuellen
MOI verknüpft
werden (um verschiedene (T-L)n-MOI zu ergeben). In einer weiteren
Ausführungsform
der vorliegenden Erfindung werden zwei oder mehr Markierungen an
einen einzigen Linker durch mehrere, unabhängige Stellen auf dem Linker gebunden,
und diese Mehrfachmarkierungs-Linker-Kombination wird dann an ein
einzelnes MOI gebunden (um verschiedene (T)n-L-MOI zu ergeben).
-
Nach verschiedenen Manipulationen
des Satzes markierter MOIs werden spezielle chemische und/oder physikalische
Bedingungen verwendet, um eine oder mehrere kovalente Bindungen
im Linker zu spalten, was zur Freisetzung der Markierungen von den
MOIs führt.
Die spaltbare(n) Bindungen) kann (können) einige derselben Bindungen
sein, die gebildet wurden, als die Markierung, der Linker und das
MOI miteinander verknüpft
wurden, oder nicht. Die Konstruktion des Linkers wird zum großen Teil
die Bedingungen bestimmen, unter denen die Spaltung durchgeführt werden
kann. Demgemäß können Linker
durch die Spaltungsbedingungen identifiziert werden, gegenüber denen
sie besonders empfindlich sind. Wenn ein Linker photolabil ist (d.
h. zur Spaltung durch Einwirkung von aktinischer Strahlung neigt),
kann dem Linker die Bezeichnung Lhυ gegeben
werden. In ähnlicher
Weise können
die Bezeichnungen LSäure, LBase,
L[O], L[R], Lenz, Lelc, LΔ und
Lss verwendet werden, um Linker zu bezeichnen,
die besonders empfindlich sind gegenüber der Spaltung durch Säure, Base,
chemische Oxidation, chemische Reduktion, die katalytische Aktivität eines
Enzyms (einfacher "Enzym"), elektrochemische
Oxidation oder Reduktion, erhöhte
Temperatur ("thermisch") bzw. Thiolaustausch.
-
Bestimmte Linker-Typen sind gegenüber einem
einzigen Typ von Spaltungsbedingung labil, wohingegen andere gegenüber mehreren
Typen von Spaltungsbedingungen labil sind. Zusätzlich kann jede der Markierungs-Bindungsstellen
in Linkern, die mehrere Markierungen binden können (um (T)n-L-MOI-Strukturen
zu ergeben), gegenüber
unterschiedlichen Spaltungsbedingungen labil sein. Bei einem Linker
mit zwei daran gebundenen Markierungen kann z. B. eine der Markierungen
nur gegenüber
Base labil sein und die andere nur gegenüber Photolyse.
-
Ein Linker, der in der vorliegenden
Erfindung nützlich
ist, besitzt mehrere Eigenschaften:
-
- 1) Der Linker besitzt einen chemischen Griff (Lh),
durch den er mit einem MOI verknüpft
werden kann.
- 2) Der Linker besitzt einen zweiten, separaten chemischen Griff
(Lh), durch den die Markierung an den Linker gebunden
wird. Wenn mehrere Markierungen an einen einzigen Linker gebunden
werden ((T)n-L-MOI-Strukturen), dann existiert für jede Markierung ein separater
Griff.
- 3) Der Linker ist gegenüber
allen Manipulationen stabil, denen er unterworfen wird, mit Ausnahme
der Bedingungen, die eine Spaltung ermöglichen, so daß eine T-enthaltende
Einheit vom Rest der Verbindung, einschließlich MOI, freigesetzt wird.
Somit ist der Linker während
der Bindung der Markierung an den Linker, der Bindung des Linkers
an das MOI und allen Manipulationen des MOIs, während die Markierung und der
Linker (T-L) daran gebunden sind, stabil.
- 4) Der Linker beeinflußt
die Manipulationen, die am MOI durchgeführt werden, während das
T-L daran gebunden ist, nicht signifikant. Wenn das T-L z. B. an
ein Oligonukleotid gebunden ist, darf das T-L irgendwelche Hybridisierungs-
oder enzymatischen Reaktionen (z. B. PCR), die am Oligonukleotid
durchgeführt
werden, nicht signifikant beeinflussen. In ähnlicher Weise darf, wenn das
T-L an einen Antikörper
gebunden ist, es nicht die Antigen-Erkennung durch den Antikörper beeinflussen.
- 5) Die Abspaltung der Markierung vom Rest der Verbindung tritt
in einer in hohem Maße
kontrollierten Art und Weise ein, unter Verwendung physikalischer
oder chemischer Verfahren, die die Nachweisbarkeit der Markierung
nicht nachteilig beeinflussen.
-
Für
jeden gegebenen Linker ist es bevorzugt, daß der Linker mit einer breiten
Vielfalt von MOIs verknüpfbar
ist und daß eine
breite Vielfalt von Markierungen mit dem Linker verknüpfbar ist.
Eine solche Flexibilität
ist vorteilhaft, weil sie erlaubt, daß eine Bibliothek von T-L-Konjugaten,
wenn sie erst einmal hergestellt ist, mit verschiedenen unterschiedlichen
Sätzen
von MOIs verwendet werden kann.
-
Wie oben erläutert, hat ein bevorzugter
Linker die Formel Lh-L1-L2-L3-Lh
-
in der jedes Lh ein
reaktiver Griff ist, der verwendet werden kann, um den Linker mit
einem Markierungs-Reaktanten und einem Reaktanten aus einem interessierenden
Molekül
zu verknüpfen.
L2 ist ein wesentlicher Teil des Linkers,
weil L2 dem Linker Labilität verleiht.
L1 und L3 sind fakultative
Gruppen, die wirkungsvoll dazu dienen, L2 von
den Griffen Lh zu trennen.
-
L1 (das definitionsgemäß näher an T
ist als L3) dient dazu, T von der erforderlichen
labilen Einheit L2 zu trennen. Diese Trennung
kann nützlich
sein, wenn die Spaltungsreaktion besonders reaktive Spezies (z.
B. freie Radikale) erzeugt, die zufällige Veränderungen in der Struktur der
T-enthaltenden Einheit bewirken könnten. Da die Spaltungsstelle
weiter von der T-enthaltenden Einheit getrennt wird, besteht eine
verringerte Wahrscheinlichkeit, daß reaktive Spezies, die an
der Spaltungsstelle gebildet werden, die Struktur der T-enthaltenden
Einheit aufbrechen werden. Auch können, da die Atome in L1 typischerweise in der T-enthaltenden Einheit vorhanden sein
werden, diese L1-Atome der T-enthaltenden
Einheit eine wünschenswerte
Qualität
verleihen. Wenn die T-enthaltende Einheit z. B. eine Tms-enthaltende
ist und ein gehindertes Amin wünschenswerterweise
als Teil der Struktur der Tms-enthaltenden Einheit
vorhanden ist (um z.B. als ein MSSE zu dienen), kann das gehinderte
Amin in der labilen L1-Einheit vorhanden
sein.
-
In anderen Fällen können L1 und/oder
L3 in einer Linkerkomponente vorhanden sein,
weil der kommerzielle Lieferant eines Linkers sich entschieden hat,
den Linker in einer Form mit solch einer L1-
und/oder L3-Gruppe zu vertreiben. In solch
einem Fall besteht kein Nachteil darin, Linker mit L1-
und/oder L3-Gruppen zu verwenden (so lange
diese Gruppen die Spaltungsreaktion nicht hemmen), selbst dann,
wenn sie den Verbindungen, die sie umfassen, keine besonderen Leistungsvorteile
verleihen. Somit erlaubt die vorliegende Erfindung, daß L1- und/oder L3-Gruppen
in der Linkerkomponente vorhanden sind.
-
L1- und/oder
L3-Gruppen können eine direkte Bindung sein
(wobei in diesem Falle die Gruppe effektiv nicht vorhanden ist),
eine Hydrocarbylen-Gruppe (z. B. Alkylen, Arylen, Cycloalkylen,
etc.), -O-Hydrocarbylen (z. B. -O-CH2-,
O-CH2CH(CH3)-, etc.)
oder Hydrocarbylen-(O-hydrocarbylen)w ,
wobei w eine ganze Zahl aus dem Bereich von 1 bis etwa 10 ist (z.
B. -CH2-O-Ar-, -CH2-(O-CH2CH2)4-,
etc.).
-
Mit dem Auftauchen der Festphasensynthese
ist eine große
Menge an Literatur zu Linkem entstanden, die gegenüber spezifischen
Reaktionsbedingungen labil sind. Bei der typischen Festphasensynthese
wird ein fester Träger
durch einen labilen Linker an eine reaktive Stelle gebunden, und
ein zu synthetisierendes Molekül wird
an der reaktiven Stelle erzeugt. Wenn das Molekül vollständig synthetisiert ist, wird
das Konstrukt aus festem Träger,
Linker und Molekül
Spaltungsbedingungen unterworfen, die das Molekül vom festen Träger freisetzen.
Die labilen Linker, die zur Verwendung in diesem Zusammenhang entwickelt
worden sind (oder die in diesem Zusammenhang verwendet werden können), können auch
ohne weiteres als der Linker-Reaktant in der vorliegenden Erfindung
verwendet werden.
-
Lloyd-Williams, P., et al., "Convergent Solid-Phase
Peptide Synthesis",
Tetrahedron Report Nr. 347, 49(48): 11065–11133 (1993) liefert eine
ausführliche
Diskussion von Linkern, die gegenüber aktinischer Strahlung (d.
h. Photolyse) sowie sauren, basischen und anderen Spaltungsbedingungen
labil sind. Zusätzliche
Informationsquellen über
labile Linker sind leicht erhältlich.
-
Wie oben beschrieben, werden unterschiedliche
Linkerkonstruktionen unter unterschiedlichen spezifischen physikalischen
oder chemischen Bedingungen Spaltbarkeit ("Labilität") verleihen. Beispiele für Bedingungen,
die dazu dienen, verschiedene Linkerkonstruktionen zu spalten, schließen saure,
basische, Oxidations-, Reduktions-, Fluorid-, Thiolaustausch-, Photolyse-
und enzymatische Bedingungen ein.
-
Beispiele für spaltbare Linker, die die
allgemeinen Kriterien für
Linker erfüllen,
die oben aufgelistet sind, werden den Fachleuten gut bekannt sein
und schließen
diejenigen ein, die im Katalog zu finden sind, der von Pierce (Rockford,
IL) erhältlich
ist. Beispiele schließen
ein:
-
- – Ethylenglycobis(succinimidylsuccinat)
(EGS), ein aminreaktives Vernetzungsreagens, das mit Hydroxylamin
spaltbar ist (1 M bei 37°C
für 3–6 Stunden);
- – Disuccinimidyltartrat
(DST) und Sulfo-DST, die aminreaktive Vernetzungsreagentien sind,
spaltbar mit 0,015 M Natriumperiodat;
- – Bis[2-(succinimidyloxycarbonyloxy)ethyl]sulfon
(BSOCOES) und Sulfo-BSOCOES,
die aminreaktive Vernetzungsmittel sind, spaltbar durch Base (pH
11,6);
- – 1,4-Di[3'-(2'-pyridyldithio(propionamido))butan
(DPDPB), ein Pyridyldithiol-Vernetzer,
der durch Thiolaustausch oder Reduktion spaltbar ist;
- – N-[4-(p-Azidosalicylamido)-butyl]-3'-(2'-pyridyldithio)propionamid
(APDP), ein Pyridyldithiol-Vernetzer, der durch Thiolaustausch oder
Reduktion spaltbar ist;
- – Bis-[beta-4-(azidosalicylamido)ethyl]-disulfid,
ein photoreaktiver Vernetzer, der durch Thiolaustausch oder Reduktion
spaltbar ist;
- N-Succinimidyl-(4-azidophenyl)-1,3'-dithiopropionat (SADP), ein photoreaktiver
Vernetzer, der durch Thiolaustausch oder Reduktion spaltbar ist;
- – Sulfosuccinimidyl-2-(7-azido-4-methylcumarin-3-acetamid)ethyl-1,3'-dithiopropionat (SAED), ein photoreaktiver
Vernetzer, der durch Thiolaustausch oder Reduktion spaltbar ist;
- – Sulfosuccinimidyl-2-(m-azido-o-nitrobenzamido)-ethyl-1,3'-dithiopropionat
(SAND), ein photoreaktiver Vernetzer, der durch Thiolaustausch oder
Reduktion spaltbar ist.
-
Andere Beispiele für spaltbare
Linker und die Spaltungsbedingungen, die verwendet werden können, um
Markierungen freizusetzen, sind wie folgt. Eine Silyl-Verknüpfungsgruppe
kann durch Fluorid oder unter sauren Bedingungen gespalten werden.
Eine 3-, 4-, 5- oder 6-substituierte
2-Nitrobenzyloxy- oder 2-, 3-, 5- oder 6-substituierte 4-Nitrobenzyloxy-Verknüpfungsgruppe
kann durch eine Photonenquelle (Photolyse) gespalten werden. Eine
3-, 4-, 5- oder 6-substituierte 2-Alkoxyphenoxy- oder 2-, 3-, 5-
oder 6-substituierte 4-Alkoxyphenoxy-Verknüpfungsgruppe
kann durch Ce(NH4)2(NO3)6 (Oxidation) gespalten
werden. Ein NCO2(Urethan)-Linker kann durch
Hydroxid (Base), Säure
oder LiAlH4 (Reduktion) gespalten werden.
Eine 3-Pentenyl-, 2-Butenyl- oder 1-Butenyl-Verknüpfungsgruppe kann durch O3, OsO4/IO4
- oder KMnO4 (Oxidation) gespalten werden. Eine 2-[3-,
4- oder 5-substituierte Furyl]oxy-Verknüpfungsgruppe kann durch O2, Br2, MeOH oder
Säure gespalten
werden.
-
Bedingungen für die Spaltung weiterer labiler
Verknüpfungsgruppen
schließen
ein: tert.-Alkyloxy-Verknüpfungsgruppen
können
durch Säure
gespalten werden; Methyl(dialkyl)methoxy- oder 4-substituierte 2-Alkyl-l,3-dioxan-2-yl-Verknüpfungsgruppen
können
durch H3O+ gespalten
werden; 2-Silylethoxy-Verknüpfungsgruppen
können
durch Fluorid oder durch Säure
gespalten werden; 2-(X)-Ethoxy (wobei X = Keto, Esteramid, Cyano,
NO2, Sulfid, Sulfoxid, Sulfon ist)-Verknüpfungsgruppen
können
unter alkalischen Bedingungen gespalten werden; 2-, 3-, 4-, 5- oder
6-substituierte Benzyloxy-Verknüpfungsgruppen
können
durch Säure
oder unter reduktiven Bedingungen gespalten werden; 2-Butenyloxy-Verknüpfungsgruppen
können
durch (Ph3P)3RhCl(H)
gespalten werden; 3-, 4-, 5- oder 6-substituierte 2-Bromphenoxy-Verknüpfungsgruppen
können
durch Li, Mg oder BuLi gespalten werden; Methylthiomethoxy-Verknüpfungsgruppen
können
durch Hg2+ gespalten werden; 2-(X)-Ethyloxy(wobei
X = ein Halogen)-Verknüpfungsgruppen können durch
Zn oder Mg gespalten werden; 2-Hydroxyethyloxy-Verknüpfungsgruppen
können
durch Oxidation (z. B. mit Pb(OAc)4) gespalten
werden.
-
Bevorzugte Linker sind diejenigen,
die durch Säure
oder Photolyse gespalten werden. Mehrere der säurelabilen Linker, die für Festphasen-Peptidsynthese
entwickelt worden sind, sind zur Verknüpfung von Markierungen mit
MOIs nützlich.
Einige dieser Linker sind bei einem kürzlich erschienenen Überblick
von Lloyd-Williams et al. beschrieben (Tetrahedron 49: 11065–11133,
1993). Ein nützlicher
Linker-Typ beruht auf p-Alkoxybenzylalkoholen, von denen zwei, 4-Hydroxymethylphenoxyessigsäure und
4-(4-Hydroxymethyl-3-methoxyphenoxy)buttersäure, von
Advanced ChemTech (Louisville, KY) kommerziell erhältlich sind. Beide
Linker können über eine
Esterbindung mit dem Benzylalkohol an eine Markierung gebunden werden
und über
eine Amidbindung mit der Carbonsäure
an ein aminhaltiges MOI. Markierungen, die durch diese Moleküle gebunden
sind, werden vom MOI mit variierenden Konzentrationen in Trifluoressigsäure freigesetzt.
Die Spaltung dieser Linker führt
zur Freisetzung einer Carbonsäure
auf der Markierung. Säurespaltung
von Markierungen, die durch verwandte Linker gebunden sind, wie
etwa 2-Dimethoxy-4'-(carboxymethyloxy)-benzhydrylamin (erhältlich von
Advanced ChemTech in FMOCgeschützter
Form), führt
zur Freisetzung eines Carbonsäureamids
auf der freigesetzten Markierung.
-
Die photolabilen Linker, die für diese
Anwendung nützlich
sind, sind ebenfalls größtenteils
für die
Festphasen-Peptidsynthese entwickelt worden (siehe Übersicht
von Lloyd-Williams). Diese Linker beruhen üblicherweise auf 2-Nitrobenzylestern
oder 2-Nitrobenzylamiden. Zwei Beispiele für photolabile Linker, über die vor
kurzem in der Literatur berichtet worden ist, sind 4-(4-(1-FMOC-Amino)ethyl)-2-methoxy-5-nitrophenoxy)butansäure (Holmes
und Jones, J. Org. Chem. 60: 2318–2319, 1995) und 3-(FMOC-Amino)-3-(2-nitrophenyl)propionsäure (Brown
et al., Molecular Diversity 1: 4–12, 1995). Beide Linker können über die
Carbonsäure an
ein Amin auf dem MOI gebunden werden. Die Bindung der Markierung
an den Linker erfolgt durch Bildung eines Amids zwischen einer Carbonsäure auf
der Markierung und dem Amin auf dem Linker. Die Spaltung photolabiler
Linker erfolgt üblicherweise
mit UV-Licht mit 350 nm Wellenlänge
bei Intensitäten
und Zeiträumen,
die den Fachleuten bekannt sind. Die Spaltung der Linker führt zu einer
Freisetzung eines primären
Amids auf der Markierung. Beispiele für photospaltbare Linker schließen Nitrophenylglycinester,
exo- und endo-2-Benzonorborneylchloride
und Methansulfonate und 3-Amino-3(2-nitrophenyl)propionsäure ein.
Beispiele für
enzymatische Spaltung schließen
Esterasen, die Esterbindungen spalten werden, Nuklearen, die Phosphodiester
spalten werden, Proteasen, die Peptidbindungen spalten werden, etc.
ein.
-
Eine bevorzugte Linkerkomponente
besitzt eine ortho-Nitrobenzyl-Struktur, wie unten dargestellt:
in der ein Kohlenstoffatom
an den Positionen a, b, c, d oder e mit -L
3-X
substituiert ist und L
1 (das vorzugsweise eine
direkte Bindung ist) links von N(R
1) in
der obigen Struktur vorhanden ist. Solch eine Linkerkomponente ist empfindlich
gegenüber
selektiver photo-induzierter Spaltung der Bindung zwischen dem Kohlenstoff
mit der Bezeichnung "a" und N(R
l).
Die Identität
von R
1 ist typischerweise für die Spaltungsreaktion
nicht kritisch, R
1 wird jedoch vorzugsweise
ausgewählt
aus Wasserstoff und Hydrocarbyl. Die vorliegende Erfindung sorgt
dafür, daß in der
obigen Struktur -N(R
1)- durch -O- ersetzt
werden könnte.
Auch können
in der obigen Struktur eine oder mehrere der Positionen b, e, d
oder e fakultativ mit Alkyl, Alkoxy, Fluorid, Chlorid, Hydroxyl,
Carboxylat oder Amid substituiert sein, wobei diese Substituenten
bei jedem Auftreten unabhängig
ausgewählt
werden.
-
Eine weitere bevorzugte Linkerkomponente
mit einem chemischen Griff L
h hat die folgende
Struktur:
in der eine oder mehrere
der Positionen b, c, d oder e mit Wasserstoff, Alkyl, Alkoxy, Fluorid,
Chlorid, Hydroxyl, Carboxylat oder Amid substituiert ist, R
1 Wasserstoff oder Hydrocarbyl ist und R
2 -OH oder eine Gruppe ist, die eine Carbonsäure zur
Kopplung mit einer weiteren Einheit entweder schützt oder aktiviert. Fluorkohlenstoff- und
Fluorkohlenwasserstoffgruppen sind bevorzugte Gruppen, die Carbonsäure zu einer
Kopplung mit einer. anderen Einheit aktivieren.
-
3. Interessierendes Molekül (MOI)
-
Beispiele für MOIs schließen Nukleinsäuren oder
Nukleinsäureanaloge
(z. B. PNA), Fragmente von Nukleinsäuren (d. h. Nukleinsäurefragmente),
synthetische Nukleinsäuren
oder Fragmente, Oligonukleotide (z. B. DNA oder RNA), Proteine,
Peptide, Antikörper
oder Antikörperfragmente,
Rezeptoren, Rezeptorliganden, Teile eines Ligandenpaares, Cytokine,
Hormone, Oligosaccharide, synthetische organische Moleküle, pharmazeutische
Wirkstoffe und Kombinationen derselben ein.
-
Bevorzugte MOIs schließen Nukleinsäurefragmente
ein. Bevorzugte Nukleinsäurefragmente
sind Primersequenzen, die zu den Vektoren vorhandenen Sequenzen
komplementär
sind, wobei die Vektoren zur Basensequenzierung verwendet werden.
Vorzugsweise ist ein Nukleinsäurefragment
direkt oder indirekt an eine Markierung an einem anderen als dem
3'-Ende des Fragments
gebunden; am bevorzugtesten am 5'-Ende
des Fragmentes. Nukleinsäurefragmente
können
gekauft oder hergestellt werden auf der Basis genetischer Datenbanken
(z. B. Dib et al., Nature 380: 152–154, 1996 und CEPH Genotype
Database, http://www.cephb.fr) und von kommerziellen Anbietern (z.
B. Promega, Madison, WI).
-
Wie hierin verwendet, schließt MOI Derivate
eines MOIs ein, die eine Funktioalität enthalten, die nützlich ist
bei der Verknüpfung
des MOIs mit einer T-L-Lh-Verbindung. Ein
Nukleinsäurefragment,
das einen Phosphodiester am 5'-Ende
aufweist, wobei der Phosphodiester auch an ein Alkylenamin gebunden
ist, ist z. B. ein MOI. Solch ein MOI ist in z. B. U.S.-Patent 4,762,779
beschrieben, das hierin durch Bezugnahme mit einbezogen wird. Ein
Nukleinsäurefragment
mit einer internen Modifikation ist ebenfalls ein MOI. Eine beispielhafte interne
Modifikation eines Nukleinsäurefragmentes
ist, daß die
Base (z. B. Adenin, Guanin, Cytosin, Thymidin, Uracil) modifiziert
worden ist, um eine reaktive funktionelle Gruppe hinzuzufügen. Solche
intern modifizierten Nukleinsäurefragmente
sind kommerziell erhältlich
von z. B. Glen Research, Herndon, VA. Eine weitere beispielhafte
interne Modifikation eines Nukleinsäurefragments ist, daß ein abasisches
Phosphoramidat verwendet wird, um einen modifizierten Phosphodiester
zu synthetisieren, der zwischen einem Zucker und einer Phosphatgruppe
eines Nukleinsäurefragments
angeordnet ist. Das abasische Phosphoramidat enthält eine
reaktive Gruppe, die ermöglicht,
daß ein
Nukleinsäurefragment,
das diese Phosphoramidat-abgeleitete Einheit enthält, mit
einer weiteren Einheit verknüpft
wird z. B. mit einer T-L-Lh-Verbindung.
Solche abasischen Phosphoramidate sind kommerziell erhältlich von
z. B. Clonetech Laboratories, Inc., Palo Alto, CA.
-
4. Chemische Griffe (Lh)
-
Ein chemischer Griff ist eine stabile,
aber reaktive Atomanordnung, die als ein Teil eines ersten Moleküls vorhanden
ist, wo der Griff eine chemische Reaktion mit einem komplementären chemischen
Griff durchlaufen kann, der als Teil eines zweiten Moleküls vorhanden
ist, um eine kovalente Bindung zwischen den zwei Molekülen zu bilden.
Der chemische Griff kann z. B. eine Hydroxylgruppe sein und der
komplementäre
chemische Griff kann eine Carbonsäuregruppe (oder ein aktiviertes
Derivat, z. B. ein Hydrofluoroarylester) sein, woraufhin eine Reaktion
zwischen diesen zwei Griffen eine kovalente Bindung (spezifisch
eine Estergruppe) bildet, die die zwei Moleküle miteinander verknüpft.
-
Chemische Griffe können in
einer großen
Anzahl von kovalente Bindungen bildenden Reaktionen verwendet werden,
die für
die Bindung von Markierungen an Linker und Linker in MOIs geeignet
sind. Solche Reaktionen schließen
Alkylierung (z. B. um Ether, Thioether zu bilden), Acylierung (z.
B. um Ester, Amide, Carbamate, Harnstoffe, Thioharnstoffe zu bilden),
Phosphorylierung (z. B. um Phosphate, Phosphonate, Phosphoramide,
Phosphonamide zu bilden), Sulfonylierung (z. B. um Sulfonate, Sulfonamide
zu bilden), Kondensation (z. B. um Imine, Oxime, Hydrazone zu bilden),
Silylierung, Disulfidbildung und Erzeugung reaktiver Zwischenstufen,
wie etwa Nitrene oder Carbene, durch Photolyse ein. Im allgemeinen
sind Griffe und bindungsbildende Reaktionen, die zur Bindung von
Markierungen an Linker geeignet sind, auch zur Bindung von Linkem
an MOIs geeignet und umgekehrt. In einigen Fällen kann das MOI eine vorherige
Modifikation oder Derivatisierung durchlaufen, um den für die Bindung
des Linkers benötigten
Griff bereitzustellen.
-
Ein Bindungstyp, der besonders nützlich ist
zur Bindung von Linkem an MOIs, ist die Disulfidbindung. Ihre Bildung
erfordert das Vorhandensein einer Thiolgruppe ("Griff") auf dem Linker und einer weiteren
Thiolgruppe auf dem MOI. Milde oxidierende Bedingungen reichen dann
aus, um die zwei Thiole als ein Disulfid miteinander zu verbinden.
Disulfidbildung kann auch durch Verwendung eines Überschusses
an einem geeigneten Disulfidaustauschreagens, z. B. Pyridyldisulfide,
induziert werden. Weil Disulfidbildung leicht reversibel ist, kann
das Disulfid auch als die spaltbare Bindung zur Freisetzung der
Markierung verwendet werden, falls gewünscht. Dies wird typischerweise
unter ähnlich
milden Bedingungen durchgeführt,
unter Verwendung eines Überschusses
eines geeigneten Thiolaustauschreagens, z. B. Dithiothreitol.
-
Von besonderem Interesse für die Bindung
von Markierungen (oder Markierungen mit Linkem) an Oligonukleotide
ist die Bildung von Amidbindungen. Griffe aus primärem aliphatischen
Amin können
leicht mit Phosphoramiditen, wie etwa 6-Monomethoxytritylhexylcyanoethyl-N,N-düsopropylphosphoramidit
(erhältlich von
Glen Research, Sterling, VA), in synthetische Oligonukleotide eingeführt werden.
Die in natürlichen
Nukleotiden anzutreffenden Amine, wie etwa Adenosin und Guanosin,
sind praktisch nicht-reaktiv, verglichen mit dem eingeführten primären Amin.
Dieser Unterschied in der Reaktivität bildet die Grundlage für die Fähigkeit, selektiv
Amide und verwandte Bindungsgruppen (z. B. Harnstoffe, Thioharnstoffe,
Sulfonamide) mit dem eingeführten
primären
Amin und nicht mit den Nukleotid-Aminen zu bilden.
-
Wie in dem Katalog von Molecular
Probes (Eugene, OR) aufgelistet, schließt eine teilweise Aufzählung von
aminreaktiven funktionellen Gruppen aktivierte Carbonsäureester,
Isocyanate, Isothiocyanate, Sulfonylhalogenide und Dichloitriazene
ein. Aktivester sind hervorragende Reagentien zur Aminmodifizierung,
da die gebildeten Amidprodukte sehr stabil sind. Diese Reagentien
zeigen auch gute Reaktivität
mit aliphatischen Aminen und geringe Reaktivität mit den Nukleotid-Aminen
von Oligonukleotiden. Beispiele für Aktivester schließen N-Hydroxysuccinimidester,
Pentafluorphenylester, Tetrafluorphenylester und p-Nitrophenylester
ein. Aktivester sind nützlich,
weil sie aus praktisch jedem Molekül hergestellt werden können, das
eine Carbonsäure
enthält. Verfahren
zur Herstellung von Aktivestern sind aufgelistet in Bodansky (Principles
of Peptide Chemistry (2. Aufl.), Springer-Verlag, London, 1993).
-
Die "X"-Gruppe
in den durch TMS-L-X bezeichneten Molekülen dient
als ein chemischer Griff, der es ermöglicht, daß das Molekül an ein Biomolekül, z. B.
ein Nukleinsäuremolekül angebracht
wird. Die X-Gruppe ist ein Phosphoramidit oder eine H-Phosphonat-Gruppe.
Wenn X eines dieser zwei Funktionalitäten ist, dann kann das T-L-X-Molekül an das
Ende eines Oligonukleotids angefügt
werden, das durch jedes der gut bekannten Phosphoramidit(auch als
Phosphit-Triester bekannt) oder H-Phosphonat-Synthese-Methodologien
für die Oligonukleotid-Synthese
synthetisiert wurde, hinzugefügt
werden.
-
Zum Beispiel, wenn X eine Phosphoramiditgruppe
ist, kann das T-L-X-Molekül
die Struktur die Formel aufweisen,
wobei ein bevorzugtes T-L-X-Molekül mit X
als einem Phosphoramidit die Struktur
-
-
aufweist.
-
In den oben genannten Phosphoramidit-enthaltenden
T-L-X-Molekülen
ist R typischerweise eine Alkylgruppe, wie zum Beispiel eine C1-C6-Alkylgruppe,
oder eine Alkylgruppe, die einen Substituenten anstelle eines Wasserstoff
der Alkylgruppe aufweist, wobei geeignete einen Substituenten Cyano-(CN)-Gruppe
einschließen.
Daher kann "OR" im Phosphorami lit
OCH2CH2CN sein und
NR2 kann N(isopropyl)2 sein,
die die zwei am meisten bei c er Herstellung von Oligonukleotiden
unter der Verwendung von Phosphoramidit-Chen ie angewendeten Gruppen
sind. NR2 kann alternativ, zum Beispiel,
eine Morpholingruppe sein In einer Ausführungsform ist R eine Alkylgruppe
oder eine substituierte Alkylgruppe, die einen oder mehrere Substituenten ausgewählt aus
Halogen und Cyano aufweist und die zwei R-Gruppen von NR2 können
miteinander verbunden sein, um eine Cycloalkylgruppe zu bilden.
-
Ein zweiter Ansatz für die Synthese
von Oligonukleotiden basiert vorteilhafterweise auf H-Phosphonat-Chemie.
Ein T-L-X-Molekül
der vorliegenden Erfindung kann einen chemisch n Griff/X-Gruppe
aufweisen, die eine H-Phosphonat-Gruppe ist und daher eine Struktur
wie folgt aufweisen:
-
In der oben genannten H-Phosphonatgruppe
stellt R3 drei Alkylgruppen dar, die typischerweise
1-6 Kohlenstoffatome in jeder Alkylgruppe aufweisen. Ethyl ist eine
herkömmliche
R-Gruppe in H-Phosphonat-Reagenzien, die in der Oligonukleotid-Synthese
verwendet werden und ist daher eine bevorzugte R-Gruppe in T-L-X-Molekülen der
Erfindung, worin X eine H-Phosphonat-Gruppe ist.
-
5. Linker-Bindung
-
Typischerweise wird ein einziger
Linker-Typ verwendet, um einen bestimmten Satz oder eine bestimmte
Familie von Markierungen mit einem bestimmten Satz oder einer bestimmten
Familie von MOIs zu verbinden. In einer bevorzugten Ausführungsform
der Erfindung kann ein einziger, gleichförmiger Prozeß befolgt werden,
um alle verschiedenen T-L-MOI-Strukturen
zu schaffen. Dies ist insbesondere dann vorteilhaft, wenn der Satz
von T-L-MOI-Strukturen
groß ist,
weil es ermöglicht,
daß der
Satz unter Verwendung von Verfahren der kombinatorischen Chemie
oder einer anderen Parallelverarbeitungstechnologie hergestellt
werden kann. In einer ähnlichen
Art und Weise ermöglicht
die Verwendung eines einzigen Linker-Typs, das ein einziger, gleichförmiger Prozeß zur Spaltung
aller verschiedenen T-L-MOI-Strukturen
eingesetzt werden kann. Wieder ist dies für einen großen Satz von T-L-MOI-Strukturen vorteilhaft,
weil der Satz auf eine parallele, wiederholte und/oder automatisierte
Art und Weise verarbeitet werden kann.
-
Es gibt jedoch eine weitere Ausführungsform
der vorliegenden Erfindung, in der zwei oder mehr Linker-Typen verwendet
werden, um unterschiedliche Untersätze von Markierungen mit entsprechenden
Untersätzen
von MOIs zu verbinden. In diesem Fall können selektive Spaltungsbedingungen
verwendet werden, um jeden der Linker unabhängig zu spalten, ohne die auf
den anderen Untersätzen
von MOIs vorhandenen Linker zu spalten.
-
Eine große Anzahl von kovalente Bindungen
bildenden Reaktionen sind für
die Bindung von Markierungen an Linker und Linker an MOIs geeignet.
Solche Reaktionen schließen
Alkylierung (z. B. um Ether, Thioether zu bilden), Acylierung (z.
B. um Ester, Amide, Carbamate, Harnstoffe, Thioharnstoffe zu bilden),
Phosphorylierung (z. B. um Phosphate, Phosphonate,l5hosphoramide,
Phosphonamide zu bilden), Sulfonylierung (z. B. um Sulfonate, Sulfonamide
zu bilden), Kondensation (z. B. um Imine, Oxime, Hydrazone zu bilden),
Silylierung, Disulfidbildung und Erzeugung reaktiver Zwischenstufen,
wie etwa Nitrene oder Carbene, durch Photolyse ein. Im allgemeinen
sind Griffe und bindungsbildende Reaktionen, die zur Bindung von
Markierungen an Linker geeignet sind, auch zur Bindung von Linkem
an MOIs geeignet und umgekehrt. In einigen Fällen kann das MOI eine vorherige
Modifikation oder Derivatisierung durchlaufen, um den zur Bindung
des Linkers notwendigen Griff bereitzustellen.
-
Ein Bindungstyp, der besonders nützlich für die Bindung
von Linker an MOIs ist, ist die Disulfidbindung. Ihre Bildung erfordert
das Vorhandensein einer Thiolgruppe ("Griff") auf dem Linker und einer weiteren
Thiolgruppe auf dem MOI. Milde oxidierende Bedingungen reichen dann
aus, um die zwei Thiole miteinander als ein Disulfid zu verbinden.
Disulfidbildung kann auch durch die Verwendung eines Überschusses
eines geeigneten Disulfidaustauschreagens, z. B. Pyridyldisulfide,
induziert werden. Weil Disulfidbildung leicht reversibel ist, kann
das Disulfid auch als die spaltbare Bindung zur Freisetzung der
Markierung verwendet werden, falls gewünscht. Dies wird typischerweise
unter ähnlich
milden Bedingungen durchgeführt,
unter Verwendung eines Überschusses
eines geeigneten Thiolaustauschreagenz, z. B. Dithiothreitol.
-
Von besonderem Interesse für die Bindung
von Markierungen an Oligonukleotide ist die Bildung von Amidbindungen.
Griffe aus primärem
aliphatischen Amin können
leicht mit Phosphoramiditen, wie etwa 6-Monomethoxytritylhexylcyanoethyl-N,N-disopropylphosphoramidit
(erhältlich
von Glen Research, Sterling, VA), in synthetische Oligonukleotide
eingeführt
werden. Die in natürlichen
Nukleotiden anzutreffenden Amine, wie etwa Adenosin und Guanosin,
sind praktisch nicht-reaktiv, verglichen mit dem eingeführten primären Amin. Dieser
Unterschied in der Reaktivität
bildet die Grundlage für
die Fähigkeit,
selektiv Amide und verwandte Bindungsgruppen (z. B. Harnstoffe,
Thioharnstoffe, Sulfonamide) mit dem eingeführten primären Amin und nicht mit den
Nukleotid-Aminen zu bilden.
-
Wie in dem Katalog von Molecular
Probes (Eugene, OR) aufgelistet, schließt eine teilweise Aufzählung von
aminreaktiven funktionellen Gruppen aktivierte Carbonsäureester,
Isocyanate, Isothiocyanate, Sulfonylhalogenide und Dichlortriazene
ein. Aktivester sind hervorragende Reagentien zur Aminmodifizierung,
da die gebildeten Amidprodukte sehr stabil sind. Diese Reagentien
zeigen auch gute Reaktivität
mit aliphatischen Aminen und geringe Reaktivität mit den Nukleotid-Aminen
von Oligonukleotiden. Beispiele für Aktivester schließen N-Hydroxysuccinimidester,
Pentafluorphenylester, Tetrafluorphenylester und p-Nitrophenylester
ein. Aktivester sind nützlich,
weil sie aus praktisch jedem Molekül hergestellt werden können, das
eine Carbonsäure
enthält. Verfahren
zur Herstellung von Aktivestern sind aufgelistet in Bodansky (Principles
of Peptide Chemistry (2. Aufl.), Springer-Verlag, London, 1993).
-
Zahlreiche kommerzielle Vernetzungsreagentien
existieren, die als Linker dienen können (siehe z. B. Pierce Cross-linkers,
Pierce Chemical Co., Rockford, IL). Unter diesen sind homobifunktionelle
-aminreaktive Vernetzungsreagentien, wie beispielhaft homobifunktionelle
Imidoester und N-Hydroxysuccinimidyl(NHS)-Ester. Es gibt auch heterobifunktionelle
Vernetzungsreagentien, die zwei oder mehr unterschiedliche reaktive Gruppen
besitzen, die sequentielle Reaktionen ermöglichen. Imidoester reagieren
schnell mit Aminen bei alkalischem pH. NHS-Ester ergeben stabile
Produkte, wenn sie mit primären
oder sekundären
Aminen umgesetzt werden. Maleimide, Alkyl- und Arylhalogenide, Alpha-Haloacyle und Pyridyldisulfide
sind thiolreaktiv. Maleimide sind spezifisch für Thiol(Sulfhydryl)-Gruppen
im pH-Bereich von 6,5 bis 7,5 und können bei alkalischem pH aminreaktiv
werden. Die Thioether-Bindung ist unter physiologischen Bedingungen
stabil. Alpha-Haloacetyl-Vernetzungsreagentien enthalten die Iodacetylgruppe
und sind gegenüber
Sulfhydrylen reaktiv. Die Imidazole können mit der Iodacetyleinheit
reagieren, aber die Reaktion ist sehr langsam. Pyridyldisulfide
reagieren mit Thiolgruppen, um eine Disulfidbindung zu bilden. Carbodiimide
koppeln Carboxyle an primäre
Amine von Hydraziden, was zur Bildung einer Acyl-Hydrazin-Bindung
führt.
Die Arylazide sind Photoaffinitätsreagentien, die
chemisch inert sind, bis sie UV- oder sichtbarem Licht ausgesetzt
werden. Wenn solche Verbindungen bei 250–460 nm photolysiert werden,
wird ein reaktives Arylnitren gebildet. Das reaktive Arylnitren
ist relativ unspezifisch. Glyoxale sind reaktiv gegenüber dem
Guanidinyl-Teil von Arginin.
-
In einer typischen Ausführungsform
der vorliegenden Erfindung wird eine Markierung zunächst an
einen Linker gebunden, anschließend
die Kombination von Markierung und Linker an ein MOI gebunden, um
die Struktur T-L-MOI zu schaffen. Alternativ wird dieselbe Struktur
gebildet, indem zunächst
ein Linker an ein MOI gebunden wird und anschließend die Kombination von Linker
und MOI an eine Markierung gebunden wird. Ein Beispiel ist, wenn
das MOI ein DNA-Primer oder Oligonukleotid ist. In diesem Falle
wird die Markierung typischerweise zunächst an einen Linker gebunden,
anschließend
das T-L an einen DNA-Primer
oder ein Nukleotid gebunden, was dann z. B. in einer Sequenzierungsreaktion
verwendet wird.
-
Eine nützliche Form, in der eine Markierung
reversibel an ein MOI (z. B. ein Oligonukleotid oder einen DNA-Sequenzierungsprimer)
gebunden werden kann, ist durch einen chemisch labilen Linker. Eine
bevorzugte Konstruktion für
den Linker ermöglicht,
daß der
Linker gespalten werden kann, wenn er einer flüchtigen organischen Säure ausgesetzt
wird, z. B. Trifluoressigsäure
(TFA). TFA ist besonders kompatibel mit den meisten Verfahren zur
MS-Ionisierung,
einschließlich
Elektrospray.
-
Wie im Detail unten beschrieben,
liefert die Erfindung eine Methodik für die Genotypisierung. Eine
Zusammensetzung, die beim Genotypisierungsverfahren nützlich ist,
umfaßt
eine Mehrzahl von Verbindungen der Formel:
Tms-L-MOI in der Tms eine durch Massenspektrometrie nachweisbare
organische Gruppe ist, die Kohlenstoff, wenigstens eines von Wasserstoff
und Fluorid und fakultative Atome, die ausgewählt sind aus Sauerstoff, Stickstoff, Schwefel,
Phosphor und Iod, umfaßt.
In der Formel ist L eine organische Gruppe, die ermöglicht,
daß eine Tms-enthaltende Einheit vom Rest der Verbindung
abgespalten werden kann, wobei die Tms-enthaltende
Einheit eine funktionelle Gruppe umfaßt, die einen einzelnen ionisierten
Ladungszustand unterstützt,
wenn die Verbindung Massenspektrometrie unterworfen wird, und ausgewählt ist
aus tertiärem
Amin, quartärem
Amin und organischer Säure.
In der Formel ist MOI ein Nukleinsäurefragment, wobei L an das
MOI an einer anderen Stelle als dem 3'-Ende des MOI konjugiert ist. In der
Zusammensetzung haben wenigstens zwei Verbindungen dieselbe Tms, aber die MOI-Gruppen dieser Moleküle besitzen
nicht-identische Nukleotidlängen.
-
Eine weitere Zusammensetzung, die
beim Genotypisierungsverfahren nützlich
ist, umfaßt
eine Mehrzahl von Verbindungen der Formel: Tms-L-MOI in der Tms eine durch Massenspektrometrie nachweisbare
organische Gruppe ist, die Kohlenstoff, wenigstens eines von Sauerstoff
und Fluorid und fakultative Atome, die ausgewählt sind aus Sauerstoff, Stickstoff,
Schwefel, Phosphor und Iod, umfaßt. In der Formel ist L eine
organische Gruppe, die es ermöglicht,
daß eine
Tms-enthaltende Einheit vom Rest der Verbindung
abgespalten werden kann, wobei die Tms-enthaltende
Einheit eine funktionelle Gruppe umfaßt, die einen einzelnen ionisierten
Ladungszustand unterstützt,
wenn die Verbindung Massenspektrometrie unterworfen wird, und ausgewählt ist
aus tertiärem
Amin, quartärem
Amin und organischer Säure.
In der Formel ist MOI ein Nukleinsäurefragment, wobei L an das
MOI an einer anderen Stelle als dem 3'-Ende des MOI konjugiert ist. In der
Zusammensetzung haben wenigstens zwei Verbindungen dieselbe Tms, aber diese Verbindungen haben nicht-identische
Elutionszeiten bei Säulenchromatographie.
-
Eine weitere Zusammensetzung, die
beim Genotypisierungsverfahren verwendet werden kann, umfaßt eine
Mehrzahl von Verbindungen der Formel: Tms-L-MOI
in
der Tms eine durch Massenspektrometrie nachweisbare
organische Gruppe ist, die Kohlenstoff, wenigstens eines von Sauerstoff
und Fluorid und fakultative Atome, die ausgewählt sind aus Sauerstoff, Stickstoff,
Schwefel, Phosphor und Iod, umfaßt. In der Formel ist L eine
organische Gruppe, die es ermöglicht,
daß eine
Tms-enthaltende Einheit vom Rest der Verbindung
abgespalten werden kann, wobei die Tms-enthaltende
Einheit eine funktionelle Gruppe umfaßt, die einen einzelnen ionisierten
Ladungszustand unterstützt,
wenn die Verbindung Massenspektrometrie unterworfen wird, und ausgewählt ist
aus tertiärem
Amin, quartärem
Amin und organischer Säure.
In der Formel ist MOI ein Nukleinsäurefragment, wobei L an das
MOI an einer anderen Stelle als dem 3'-Ende des MOI konjugiert ist. In der
Zusammensetzung haben keine zwei Verbindungen, die dieselbe MOI-Nukleotidlänge aufweisen,
auch dasselbe Tms.
-
In der obigen Zusammensetzung ist
die Mehrzahl vorzugsweise größer als
2 und vorzugsweise größer als
4. Auch hat das Nukleinsäurefragment
im MOI eine zu einem Abschnitt eines Vektors komplementäre Sequenz,
wobei das Fragment zum Priming einer Polynukleotidsynthese in der
Lage ist. Vorzugsweise unterscheiden sich die Tms-Gruppen
von Mitgliedern der Mehrzahl um wenigstens 2 amu und können sich
um wenigstens 4 amu unterscheiden.
-
Die Erfindung stellt auch eine Zusammensetzung
zur Verfügung,
die eine Mehrzahl von Sätzen
von Verbindungen umfaßt,
wobei jeder Satz von Verbindungen die Formel besitzt: Tms-L-MOI in der Tms eine durch Massenspektrometrie nachweisbare
organische Gruppe ist, die Kohlenstoff wenigstens eines von Wasserstoff
und Fluorid und fakultative Atome, die ausgewählt sind aus Sauerstoff, Stickstoff, Schwefel,
Phosphor und Iod, umfaßt.
In der Formel ist L eine organische Gruppe, die es ermöglicht,
daß eine Tms-enthaltende Einheit vom Rest der Verbindung
abgespalten werden kann, wobei die Tms-enthaltende
Einheit eine funktionelle Gruppe umfaßt, die einen einzelnen ionisierten
Ladungszustand unterstützt,
wenn die Verbindung Massenspektrometrie unterworfen wird, und ausgewählt ist
aus tertiärem
Amin, quartärem
Amin und organischer Säure.
In der Formel ist MOI auch ein Nukleinsäurefragment, wobei L an das
MOI an einer anderen Stelle als dem 3'-Ende des MOI konjugiert ist. In der
Zusammensetzung haben Mitglieder innerhalb eines ersten Satzes von
Verbindungen identische Tms-Gruppen, haben
jedoch nicht-identische MOI-Gruppen mit unterschiedlichen Anzahlen
von Nukleotiden im MOI, und es gibt wenigstens zehn Mitglieder im
ersten Satz, wobei zwischen den Sätzen die Tms-Gruppen
sich um wenigstens 2 amu unterscheiden. Die Mehrzahl ist vorzugsweise
wenigstens 3 und bevorzugter wenigstens 5.
-
Die Erfindung stellt auch eine Zusammensetzung
zur Verfügung,
die eine Mehrzahl von Sätzen
von Verbindungen umfaßt,
wobei jeder Satz von Verbindungen die Formel hat Tms-L-MOI in der Tms eine durch Massenspektrometrie nachweisbare
organische Gruppe ist, die Kohlenstoff, wenigstens eines von Wasserstoff
und Fluorid und fakultative Atome, die ausgewählt sind aus Sauerstoff, Stickstoff, Schwefel,
Phosphor und Iod, umfaßt.
In der Formel ist L eine organische Gruppe, die es ermöglicht,
daß eine Tms-enthaltende Einheit vom Rest der Verbindung
abgespalten werden kann, wobei die Tms-enthaltende
Einheit eine funktionelle Gruppe umfaßt, die einen einzelnen ionisierten
Ladungszustand unterstützt,
wenn die Verbindung Massenspektrometrie unterworfen wird, und ausgewählt ist
aus tertiärem
Amin, quartärem
Amin und organischer Säure.
In der Formel ist MOI ein Nukleinsäurefragment, wobei L an das
MOI einer anderen Stelle als dem 3'-Ende des MOI konjugiert ist. In der
Zusammensetzung haben die Verbindungen innerhalb eines Satzes dieselbe
Elutionszeit, aber nicht-identische Tms-Gruppen.
-
Zusätzlich stellt die Erfindung
einen Kit zum Genotypisieren zur Verfügung. Der Kit umfaßt eine
Mehrzahl von Amplifikationsprimerpaaren, wobei wenigstens einer
der Primer die Formel hat: Tms-L-MOI in
der Tms eine durch Massenspektrometrie nachweisbare
organische Gruppe ist, die Kohlenstoff, wenigstens eines von Wasserstoff
und Fluorid und fakultative Atome, die ausgewählt sind aus Sauerstoff, Stickstoff, Schwefel,
Phosphor und Iod, umfaßt.
In der Formel ist L eine organische Gruppe, die es ermöglicht,
daß eine Tms-enthaltende Einheit vom Rest der Verbindung
abgespalten werden kann, wobei die Tms-enthaltende
Einheit eine funktionelle Gruppe umfaßt, die einen einzelnen ionisierten
Ladungszustand unterstützt,
wenn die Verbindung Massenspektrometrie unterworfen wird, und ausgewählt ist
aus tertiärem
Amin, quartärem
Amin und organischer Säure.
In der Formel ist MOI ein Nukleinsäurefragment, wobei L an das
MOI an einer anderen Stelle als dem 3'-Ende des MOI konjugiert ist; und jedes
Primerpaar assoziiert mit einem unterschiedlichen Locus. Im Kit
ist die Mehrzahl vorzugsweise wenigstens 3 und bevorzugter wenigstens
5.
-
Wie oben angegeben, stellt die vorliegende
Erfindung Zusammensetzungen und Verfahren zur Bestimmung der Sequenz
von Nukleinsäuremolekülen zur
Verfügung.
Kurz gesagt umfassen solche Verfahren allgemein die Schritte: (a)
Erzeugen markierter Nukleinsäurefragmente,
die komplementär
zu einem ausgewählten
Nukleinsäuremolekül (z. B.
markierte Fragmente) von einem ersten Ende bis zu einem zweiten
Ende eines Nukleinsäuremoleküls) sind,
wobei eine Markierung mit einem bestimmten oder ausgewählten Nukleotid korrelativ
ist und mit irgendeinem aus einer Vielzahl von Verfahren nachgewiesen
werden kann, (b) Abtrennen der markierten Fragmente nach Sequenzlänge, (c)
Abspalten einer Markierung von einem markierten Fragment und (d)
Nachweisen der Markierungen und Bestimmen der Sequenz des Nukleinsäuremoleküls daraus. Jeder
der Aspekte wird unten detaillierter diskutiert werden.
-
B. DIAGNOSEVERFAHREN
-
1. Einführung
-
Wie oben angegeben, stellt die vorliegende
Erfindung auch eine breite Vielfalt von Verfahren zur Verfügung, in
denen die oben beschriebenen Markierungen und/oder Linker anstelle
herkömmlicher
Markierungen (z. B. radioaktiv oder enzymatisch) eingesetzt werden
können,
um die Spezifität,
Empfindlichkeit oder Anzahl von Proben, die gleichzeitig analysiert
werden können,
in einem vorgegebenen Verfahren zu erhöhen. Repräsentative Beispiele für solche
Verfahren, die verbessert werden können, schließen z. B.
RNA-Amplifikation (siehe Lizardi et al., Bio/Technology 6: 1197–1202, 1988;
Kramer et al., Nature 339: 401–402,
1989; Lomeli et al., Clinical Chem. 35(9): 1826–1831, 1989; U.S.-Patent Nr.
4,786,600) und DNA-Amplifikation
unter Verwendung von LCR oder Polymerasekettenreaktion ("PCR") (siehe U.S. Patente
Nrn. 4,683,195, 4,683,202 und 4,800,159) ein.
-
Die CMST-Technologieplattform kann
in einer Zahl von Anwendungen verwendet werden, in denen Nukleinsäure-Messungen
in einem großen
Maßstab
durchgeführt
werden. Diese Technologieplattform kann mit oder ohne einer Auftrennungs-
oder Größen-Methodologie
verwendet werden. Ein Beispiel eines nicht-Größen Assays würde Single
Nucleotide Polymorphism (SNP) Assays einschließen, in denen Oligonukleotide
verwendet werden, um die Anwesenheit oder Abwesenheit einer Basenveränderung
in einer Ziel-Nukleinsäure
nachzuweisen. Alternativ kann ein HPLC- oder Auftrennungssystem
an den Massenspektrometrie-Detektor (MSD) angehängt werden, in dem Nukleinsäurefragmente
nach ihrer Größe sortiert
werden können
und daher wird ein Massenspektrometrie-Marker mit einer Retentionszeit
kombiniert, um eine Sequenz zu identifizieren. Die HPLC von Nukleinsäuren (Huber
et al., 1993, Anal. Biochem 212, 5.351; Huber et al., 1993, Nuc.
Acids Res., 21, 5.1061; Huber et al., 1993, Biotechniques, 16, 5.898)
oder denaturierende HPLC (DHPLC) ist ein bei der Auftrennung von
DNA-Duplices, die sich in ihrer Identität um eine oder mehrere Basenpaare
unterscheiden, allgemein erfolgreiches Verfahren und ist für das Screening
von Mutanten allgemein brauchbar. Ein allgemeines Verfahren wurde
unter der Verwendung von 100 mM Triethylammoniumacetat als dem Ionen-paarenden
Reagenz entwickelt, in dem Oligonukleotide über HPLC erfolgreich auf alkyliertem
nicht-porösen
2,3 μM Poly(styrol-divinylbenzol)
aufgetrennt wurden (Oefner et al., 1994, Anal. Biochem., 223, S.39). Diese
Technik erlaubte auch die Auftrennung von PCR-Produkten, die sich
in Ihrer Länge
in nur 4 bis 8 Basenpaaren innerhalb eines Größenbereichs von 50 bis 200
Nukleotide unterscheiden.
-
DHPLC hat die Suche nach SNPs signifikant
beschleunigt (Oefner und Underhill, Am. J. Hum. Genetics, 57, A266,
1995). Zahlreiche Anwendungen der DHPLC-Technik schließen die
Identifizierung von Polymorphismen auf dem menschlichen Y-Chromosom
ein, um evolutionäre
Untersuchungen zu erleichtern (PNAS USA, 93, 196–200, 1996), und die schnelle
Identifizierung von Erkrankungen die Mutationen auf dem Chromosom
19 verursachen, die Ataxia verursachen (Cell, 87, 543–552, 1996).
-
Während
es schwierig war genetische Tests für multifaktorielle Erkrankungen
zu entwickeln, werden mehr als 200 bekannte menschliche Erkrankungen
durch einen Defekt in einem einzelnen Gen verursacht, oftmals eine
Veränderung
eines einzelnen Aminosäurerests
(Olsen, Biotechnology: An industry comes of age, National Academic
Press, 1986).
-
Sensitive Mutations-Nachweistechniken
bieten außergewöhnliche
Möglichkeiten
für das
Mutationsscreening. Effiziente genetische Tests können auch
ein Screening auf onkogene Mutationen in Zellen ermöglichen,
die aus dem respiratorischen Trakt oder der Blase abgestoßen werden,
in Zusammenhang mit Gesundheitsvorsorgeuntersuchungen (Sidransky
et al., Science 252: 706, 1991). Auch, wenn ein unbekanntes Gen eine
genetische Erkrankung verursacht, sind Verfahren brauchbar, um DNA-Sequenzvarianten
zu überwachen,
um die Vererbung der Erkrankung durch genetische Kopplungsanalyse
zu untersuchen. Mehrere verschiedenen Ansätze wurden verfolgt, jedoch
ist keiner weder effizient noch billig genug um wirklich weit verbreitet
angewendet zu werden (Cotton. RGH, (1997), Mutation Detection, Oxford
University Press, New York). Mutationen, die ein einzelnes Nukleotid
einschließen,
können
in einer Probe durch physikalische, chemische oder enzymatische
Mittel identifiziert werden. Im allgemeinen, können Verfahren zum Mutationsnachweis
in Scann-Techniken
(Fearon, 1997, Science, 278, p 1043–1050) eingeteilt werden, die
geeignet sind, um bisher unbekannte Mutationen zu modifizieren und
Techniken, die dazu aufgebaut sind, um bekannten Sequenzvarianten
nachzuweisen, zu unterscheiden oder zu quantifizieren (Hotzman et
al., 1997, Science, 278, 602–605).
-
Verschiedenen Scan-Techniken zum
Mutationsnachweis wurden für
Heteroduplices entwickelt, wo die Anwesenheit eines Fehlpaares ein
abnormales Verhalten induziert, wenn die Duplex teilweise denaturiert
wird. Dieses Phänomen
wird in den denaturierenden und Temperaturgradientgelelektrophoreseverfahren
(jeweils DGGE und TGGE) ausgenutzt. Duplexe, die in sogar einer
einzelnen Nukleotidposition fehlgepaart sind, können teilweise denaturieren,
was zu verzögerter
Wanderung führt,
wenn in einem ansteigenden denaturierenden Gradientengel elektrophoriert
(Orita, Genomics 5: 874, 1989; Keen, Trends Genet. 7: 5, 1991, Myers
et al., Nature 313: 495, 1985; Abrams et al., Genomics 7: 463, 1990;
henco et al., Nucl. Acids Res. 18: 6733, 1990). Obwohl Mutationen
nachgewiesen werden können,
sind keine Informationen im Hinblick auf den genauen Ort oder die
Sequenz um die Mutation herum erhalten.
-
Fehlgepaarte Basen in einer Duplex
sind ebenfalls gegenüber
chemischer Modifikation empfindlich. Solche Modifikation kann die
Stränge
gegenüber
der Spaltung an der Stelle der Fehlpaarung empfindlich machen oder
verursachen, daß eine
Polymerase in einer anschließenden
Verlängerungsreaktion
anhält.
Die chemische Spaltungstechnik erlaubt die Identifizierung einer
Mutation in vielen Sequenzen von bis zu 2 kb und stellt Informationen über den
ungefähren
Ort der fehlgepaarten Nukleotide zur Verfügung (Cotton et al., PNAS USA
85: 4397, 1988; Ganguly et al., Nucl. Acids Res. 18: 3933, 1991).
-
Eine alternative Strategie zum Nachweis
einer Mutation in einem DNA-Strang ist durch Substituierung (während der
Synthese) eines der normalen Nukleotide in einem modifizierten Nukleotid,
das das Molekulargewicht oder andere physikalische Parameter des
Produkts verändert.
Ein Strang mit einer erhöhten
oder verringerten Zahl diesen modifizierten Nukleotids relativ zu
der Wildtyp-Sequenz zeigt eine veränderte elektrophoretische Mobilität (Naylor
et al., Lancet 337: 635, 1991). Wiederum weist diese Technik die
Anwesenheit einer Mutation nach, stellt jedoch nicht den Ort zur
Verfügung.
-
Alle der oben genannten Techniken
zeigen die Anwesenheit einer Mutation in einem begrenzten Segment
von DNA an, und einige dieser ermöglichen die ungefähre Lokalisierung
innerhalb des Segments. Jedoch ist eine Sequenzanalyse immer noch
erforderlich, um die genaue Position der Basenveränderung
zu lokalisieren.
-
Eine große Anzahl von anderen Techniken
wurden entwickelt, um bekannte Sequenzvarianten oder Single Nucleotide
Polymorphismen zu analysieren. Die Automatisierung und Wirtschaftlichkeit
und sehr wichtige Überlegungen
für diese
Typen von Analysen, die zum Screening von Individuen und der allgemeinen
Population angewendet werden können.
Mutationen können über ihre
destabilisierenden Effekte auf die Hybridisierung von kurzen Oligonukleotidsonden
an eine Zielsequenz identifiziert werden (siehe Wetmur, Crit. Rev. Biochem.
Mol. Biol., 26: 227, 1991). Im allgemeinen, schließt diese
Technik, Allel-spezifische Oligonukleotidhybridisierung, die Amplifikation
von Zielsequenzen ein, und die anschließende Hybridisierung mit kurzen
Oligonukleotidsonden. Der Oligonukleotid-Ligations-Assay ist eine Erweiterung
von PCR-basiertem Screening, der einen ELISAbasierten Assay verwendet
(OLA, Nickerson et al., Proc. Natl. Acad. Sci. USA 87: 8923, 1990), um
die PCR-Produkte nachzuweisen, die Zielsequenz enthalten. Daher
werden sowohl Gelelektrophorese und Koloniehybridisierung beseitigt.
-
Wie oben angegeben stellt die CMST-Technologie
ebenfalls eine breite Vielzahl von Verfahren zur Verfügung, wo
die abspaltbaren Marker und/oder Linker anstelle von herkömmlichen
Markierungen (z. B. radioaktiv, Fluoreszenz oder enzymatisch) verwendet
werden können,
um die Spezifität,
Sensitivität
oder Zahl von Proben, die gleichzeitig analysiert werden können, innerhalb
eines bestimmten Verfahrens zu erhöhen. Repräsentative Beispiele von solchen
Verfahren, die verbessert werden können, schließen zum
Beispiel Standard-Nukleinsäurehybridisierungsreaktionen
(siehe Sambrook et al., supra), diagnostische Reaktionen wie zum
Beispiel die Cycling-Probe-Technology (CPT) (siehe U.S.-Patente
Nrn. 4,876,187 und 5,011,769) oder Oligonukleotid-Ligations-Assay
(OLA) (Burket et al., Science 196: 180, 1987).
-
Die CMST-Technologie, kombiniert
mit Hybridisierung kann für
die Forensik verwendet werden. Die DNA-Analyse ermöglicht leicht
die Ableitung von Verwandtschaft zwischen Individuen, wie es bei
Vaterschaftstests erforderlich ist. Die genetische Analyse hat sich
als sehr brauchbar bei der Knochenmarkstransplantation erwiesen,
wo es erforderlich ist zwischen engen Verwandten Donor- und Empfängerzellen
zu unterscheiden. Zwei Typen von Sonden sind nun für DNA-Fingerprinting
und -Typisierung in Gebrauch. Polymorphe Minisatelliten-DNA-Sonden
identifizieren multiple DNA-Sequenzen, die jeweils in variablen
Formen in verschiedenen Individuen vorhanden sind, wodurch Muster
erzeugt werden, die komplex und zwischen den Individuen hoch variabel
sind. VNTR-Sonden identifizieren einzelne Sequenzen in einem Genom,
jedoch können
diese Sequenzen in bis zu 30 verschiedene Formen in der menschlichen
Population vorhanden sein, wie durch die Größe der identifizierten Fragmente
unterschieden (Bennett und Todd, 1996, Ann. Rev. Genetics, 30, S.
343–70).
-
Tumordiagnose und -Phasenbestimmung
(Goodfellow und Wells, 1995, J. Natl. Cancer Inst., 87, S. 1515–23) ist
eine weitere Anwendung der CMST-Technologie-Plattform. Der Nachweis
von Onkogenen und deren jeweiligen Polymorphismen ist ein wichtiges
Feld der Nukleinsäurediagnose.
Die zellulären
Onkogene können
durch spezifische Modifikationen, wie zum Beispiel Punktmutationen,
aktiviert werden (wie bei dem c-K-ras-Onkogen in Blasenkarzinom
und in colorektalen Tumoren), Promotor-Induktion, Gen-Amplifikation (wie
in dem N-myc-Onkogen im Fall von Neuroblastom) oder der Neuanordnung
von Chromosomen (wie bei der Translokation des c-abl-Onkogens von
Chromosom 9 zu Chromosom 22 im Fall von chronischer Myeloid-Leukämie). Die
CMST-Technologie kann auch für
die Transplantationsanalyse, Genomdiagnose (4% aller Neugeborenen
werden mit genetischen Defekten geboren) angewendet werden. Von
den 3.500 beschriebenen Erbkrankheiten, die durch die Modifikation
von nur einem einzelnen Gen verursacht werden, sind die primären molekularen
Effekte nur für
ungefähr
400 bekannt. Die Verwendung von DNA-Sonden mit abspaltbaren Markern
kann dazu verwendet werden, um die Anwesenheit oder Abwesenheit
von Mikroorganismen in jedem Typ von Probe oder Spezimen nachzuweisen.
-
Die CMST-Technologie-Plattform kann
mit verschiedenen größenabhängigen Techniken
gekoppelt werden. Kapillarelektrophorese (CE) in ihren verschiedenen
Ausführungen
(freie Lösung,
Isotachophorese, isoelektrische Fokussierung, Polyacrylamidgel,
mizellare Elektrokinetik "Chromatographie") entwickelt sich
als ein Verfahren für
schnelle hochauflösende
Abtrennungen von sehr kleinen Probenvolumen von komplexen Gemischen.
In Kombination mit der inhärenten
Aktivität
und Selektivität
von MS, ist CE-MS eine potentiell leistungsfähige Technik für die Bioanalyse.
In der hier beschriebenen neuen Anwendung könnte die Überlappung dieser zwei Verfahren
zu überlegenen
DNA-Sequenzierungsverfahren
führen,
die die augenblickliche Rate-Verfahren der Sequenzierung um mehrere
Größenordnungen übersteigt.
-
Die Korrespondenz zwischen CE und
Elektrosprayionisierung-(ESI)-Flußraten und die Tatsache, daß beide
durch (und primär
verwendet werden für)
ionische Spezies in Lösung unterstützt werden,
stellt die Base für
eine extrem attraktive Kombination zur Verfügung. Die Kombination von sowohl
Kapillarzonenelektrophorese (CZE) und Kapillarisotachophorese mit
Quadrapolmassenspektrometer, basierend auf ESI, wurden beschrieben
(Olivares et al., Anal. Chem. 59: 1230, 1987; Smith et al., Anal.
Chem. 60: 436, 1988; Loo et al., Anal. Chem. 179: 404, 1989; Edmonds
et al., J. Chroma. 474:21, 1989; Loo et al., J. Microcolumn Sep.
1: 223, 1989; Lee et al.; J. Chromatog. 458: 313, 1988; Smith et
al., J. Chromatog. 480: 211, 1989; Grese et al., J. Am. Chem. Soc.
111: 2835, 1989).
-
Das wirksamste Abtrennverfahren für DNA-Fragmente
ist Polyacrylamidgelelektrophorese (PAGE), im allgemeinen in einem
Scheibengelformat. Jedoch ist die hauptsächliche Begrenzung der augenblicklichen Technologie
die relativ lange Zeit, die erforderlich ist, um die Gelelektrophorese
von DNA-Fragmenten durchzuführen,
die in den Sequenzreaktionen produziert wurden. Eine Erhöhung um
eine Größenordnung
(10-fach) kann mit der Verwendung von Kapillarelektrophorese eneicht
werden, die ultradünne
Gele verwendet. In Polyacrylamidgelen werden DNA-Fragmente gesiebt
und wandern als eine Funktion der Länge und dieser Ansatz wurde
nun auf CE angewendet. Eine bemerkenswerte Plattenzahl pro Meter
wurde nun mit kreuz-vernetztem Polyacrylamid (10–7 Platten
pro Meter, Cohen et al., Proc. Natl. Acad. Sci., USA 85: 9660, 1988)
eneicht. Solche CE-Säulen,
wie bisher beschrieben, können
für die
DNA-Sequenzierung verwendet werden. Smith und andere (Smith et al.,
Nuc. Acids. Res. 18: 4417, 1990) haben die parallele Verwendung
von vielen Kapillaren vorgeschlagen, um den Durchsatz zu erhöhen. Ebenfalls
haben Mathies und Huang (Mathies und Hang, Nature 359: 167, 1992)
eine Kapillarelektrophorese eingeführt, in der die Abtrennungen über eine
parallele Anordnung von Kapillaren durchgeführt wird, und eine Hochdurchsatz-Sequenzierung
gezeigt (Huang et al., Anal. Chem. 64: 967, 1992, Huang et al.,
Anal. Chem 64: 2149, 1992). Da es keinen Grund gibt, parallele Spesen
laufen zu lassen, besteht kein Grund ein Scheibengel zu verwenden.
Daher kann man ein Röhrengelformat
für das
elektrophoretische Abtrennverfahren anwenden. Grossuran (Grossuran
et al., Genet. Anal. Tech. Appl. 9: 9, 1992) hat gezeigt, daß ein beträchtlicher
Vorteil erhalten wird, wenn ein Röhrengelformat anstelle eines
Scheibengelformats verwendet wird. Dies liegt an der größeren Fähigkeit,
Joulewärme
in einem Röhrenformat
abzuleiten, verglichen mit einem Scheibengel, was zu schnelleren
Laufzeiten führt
(um 50%) und sehr viel höhere
Auflösung
von hoch molekular Gewichts-DNA-Fragmenten (größer als 1.000 Nukleotide).
Lange Ablesbarkeit ist bei der genomischen Sequenzierung wichtig.
Daher weist die Verwendung von abspaltbaren Mackern bei der Sequenzierung
den weiteren Vorteil auf, daß es
dem Anwender ermöglicht
wird, das am meisten effiziente und sensitive DNA-Abtrennverfahren
zu verwenden, daß auch
die höchste
Auflösung
besitzt.
-
Das hinter der Verwendung von mikro-fabrizierten
Vonichtungen stehende Konzept ist die Fähigkeit, die Informationsdichte
bei der Elektrophorese zu erhöhen,
durch Miniaturisierung der Spurendimensionen auf ungefähr 100 Mikrometer.
Die elektronische Industrie verwendet routinegemäß Mikro-Fabrikation, um die Schaltkreise
mit Merkmalen herzustellen von weniger als einem Mikron Größe. Die
augenblickliche Dichte von Kapillaren-Anays ist auf den äußeren Durchmesser
der Kapillarenröhre
begrenzt. Die Mikro-Fabrikation von Kanälen stellt eine höherer Array-Dichte
zur Verfügung.
Die Mikro-Fabrikation erlaubt auch physikalische Anordnungen, die
mit Glasfasern nicht möglich
nicht und koppelt die Kanäle
direkt zu anderen Vonichtungen auf einem Chip. Ein paar Vonichtungen
wurden auf Microchips für
Abtrenntechnologien konstruiert. Ein Gaschromatograph (Teny e al.,
IEEE Trans. Electron Devic, ED-26: 1880, 1979) und ein Flüssigkeitschromatograph (Manz
et al., Sens. Actuators B1: 249, 1990) wurden auf Siliziumchips
hergestellt, jedoch wurden diese Vonichtungen nicht weiter verwendet.
Verschiedene Gruppen haben die Abtrennung von fluoreszenten Farbstoffen
und Aminosäuren
auf Mikro-hergestellten Vonichtungen berichtet (Manz et al., J.
Chromatography 593: 253, 1992, Effenhauser et al., Anal. Chem. 65:2637,
1993). Vor kurzem haben Woolley und Mathies (Woolley und Mathies,
Proc. Natl. Acad. Scci. 91: 11348, 1994) gezeigt, daß Photolithographie
und chemisches Ätzen dazu
verwendet kann, um eine große
Zahl von Abtrennkanälen
auf Glassubstraten herzustellen. Diese Kanäle werden mit Hedroxyethylzellulose-(HEC)-Abtrennmatrices
gefüllt.
-
Hybotrope werden in Tests und anderen
Verfahren vorteilhaft angewendet, wobei ein markiertes Oligonukleotid
an ein komplementäres
oder semi-komplementäres
(d. h. fast, jedoch nicht genau dieselbe Sequenz, wie das markierte
ODN) Nukleinsäurefragment hybridisiert
wird. Hybotrope sind genauer beschrieben in zum Beispiel U.S.-Patentanmeldungen
Nrn. 60/026,621 (angemeldet 24. September 1996); 08/719,132 (angemeldet
24. September 1996); 08/933,924 (angemeldet 23. September 1997);
09/002,051 (angemeldet 31. Dezember 1997) und PCT Internationale
Publikationsnr. WO 98/13527, die alle in ihrer Gesamtheit hier aufgenommen
werden.
-
Die Beobachtung, daß Δ-Tm sich als eine Funktion der Konzentration
von Hybotropen nicht verändert, weist
wesentliche Brauchbarkeit zur Verwendung in DNA, RNA oder Nukleinsäureamplifikation
auf, die auf der Primerverlängerung
durch Polymerase basieren (z. B. Polymerasekettenreaktion, siehe
U.S.-Patente Nrn. 4,683,195; 4,683,202 und 4,800,159, Cycling Probe
Technology, NASBA), Ligation (LCR, Ligationskettenreaktion) und
RNA-Amplifikation
(siehe Lizardi et al., Bio/Technologe 6: 1197, 1988; Kramer et al.,
Natrure 339: 401, 1989; Lomeli et al., Clin. Chem. 35: 1826, 1989;
U.S.-Patent Nr. 3,786,600). Die Beobachtung, daß WT (Wildtyp) und MT (Mutanten)
30-mer Oligonukleotide (30 verbundene Nukleotide im Oligonukleotid)
auf der Basis von thermischen Schmelzen in 0,5 M LiTCA unterschieden
werden kann, erlaubt die Möglichkeit
einer wesentlichen Verbesserung bei der Priming-Effizienz in der
PCR. In seiner augenblicklichen Anordnung wird der PCR-Puffer eher
für die
Polymerase aktiviert, als für
ein spezifisches Priming gemeint ist, mit der Einführung der
Technik wurden Bedingungen entwickelt, die die Performance der Polymerase über die
Performance der Spezifität
von Priming mit Oligonukleotiden bevorzugen. Daher stellt der PCR-Puffer,
wie augenblicklich käuflich
erhältlich,
keinen hohen Spiegel von Stringenz von Hybridisierung von PCR-Primern
zur Verfügung,
noch unterstützt
er diese.
-
Käuflich
erhältliche
PCR-Puffer werden im Hinblick auf das Schmelzverhalten von 24-mer
Oligonukleotiden in sowohl den Wildtyp-(WT) und Mutanten-(MT)-Formen
untersucht. Alternativ wird das Primen in einer hybotropen Lösung durchgeführt und
die Kettenverlängerung
wird in einem getrennten Puffer durchgeführt, der die Polymerase unterstützt. Zum
Beispiel könnte
eine Festphasen-PCR verwendet werden, wobei die feste Phase durch
zwei Lösungen
hindurch bewegt wird. Ein Primen würde in einiger geeigneten Konzentrationen von
LiTCA oder TMATCA auftreten und dann würde die Polymerasekttenreaktion
in einem verschiedenen PCR-Puffer stattfinden, der die Polymerase
enthält.
Es ist auch möglich,
die ersten paar Runden in der Amplifikation in einer Hybotrop basierten
Hybridisierungslösung
durchzuführen
und die restlichen Runden in einem normalen PCR-Puffer durchzuführen (im
allgemeinen sind nur die ersten paar Runden für die Spezifität wichtig).
-
Die Verwendung von abasisch modifizierten
Oligonukleotiden erhöht
auch die Spezifität
des Primens in der PCR (siehe, z. B. U.S.-Patentanmeldungen Nrn.
60/026,621 (angemeldet 24. September 1996); 08/719,132 (angemeldet
24. September 1996); 08/933,924 (angemeldet 23. September 1997);
09/002,051 (angemeldet 31. Dezember 1997); und PCT Internationale
Publikation Nr. WO 98/13527). Eine in einem Oligonukleotid eingefügte abasische
Substitution verringert den HCT um 2,5°C. Zwei Oligonukleotidsonden,
die drei abasische Stellen pro 24-mer enthalten, weisen eine HCT-Abnahme
um 8°C relativ
zu der nicht substituierten Kontrolle auf. Diese Abnahme in der
HCT erhöht
den Spiegel der Spezifität
von Primen in der PCR-Reaktion dramatisch. Dies liegt an der Reduktion
von falschen oder Miß-Primen
während
der ersten 10 Zyklen der PCR. Gemeint ist, daß die Enthalpie der abasischen
substituierten Oligonukleotide relativ zu dem nicht substituierten
Primer ansteigt, wodurch sich die Spezifität des Primens erhöht. Der
Primer ist bevorzugterweise 6 bis 36 Basen lang und enthält 1 bis
6 abasische Stellen. Die abasischen Stellen sind bevorzugterweise
4, 5, 6, 7 oder 8 Nukleotide von einander getrennt und können bis
zu 12 zu 24 Nukleotide von einander getrennt sein. Die Substitutionen
sind bevorzugterweise am 3'-Ende des Primers
gehäuft,
um die Spezifität
der Primerverlängerung
durch Nukleinsäurepolymerasen
sicherzustellen.
-
Weiterhin verringert die Konzentration
einer abasischen Stelle in einem PCR-Primer und die Verwendung einer
hybotropen Salzlösung,
die einen hohen Enthalpiewert für
die Primerduplex unterstützt,
signifikant die Δ-HTC
der Primer-Duplex. Wie oben diskutiert, wenn die Δ-HTC abnimmt,
dann steigt der Stringenzfaktor an und ein hochdiskriminatorisches
Priming der Polymerasekettenreaktion kann stattfinden. Dies sind Bedingungen,
die für
Multiplex-PCRs erforderlich sind. Der Ausdruck Multiplex betrtfft
die Fähigkeit,
mehr als ein Set von Primern in einer PCR-Reaktion zu verwenden
und multiple Produkte zu erzeugen, oder die Fähigkeit mehr als eine Zielnukleinsäure pro
Set von PCR-Primern
zu verwenden. Die Verwendung des Hybotrops Tetramethylammoniumtrichloracetat
ist von besonderer Brauchbarkeit, da die Abhängigkeit des G + C-Gehalts
auf Tm (Stabilität) neutralisiert wird.
-
Die beschriebenen hybotropen Lösungen werden
dazu verwendet, um die Spezifität
des Primens in der PCR zu erhöhen.
Es gibt verschiedene Optionen im Hinblick auf einen Mechanismus
durch den die Spezifität
des Priming Schritts verbessert werden kann. Die erste ist durch
die Verwendung eines festen Trägers, an
den einer der PCR-Primer (kovalent) angebracht ist. Der feste Träger kann
viele Formen annehmen, wie zum Beispiel Perlen, Membranen, usw.
Der Priming Schritt kann in dem Hybotropen stattfinden und dann
kann der feste Träger
gewaschen werden und in einer Lösung
bewegt werden, die die Polymerasekettenreaktion unterstützt. Der
feste Träger
wird dann zurück
in das Nesstrope für
die Priming Reaktion bewegt und der Zyklus wiederholt. Der Kreislauf
des festen Trägers
zwischen zwei verschiedenen Lösungen
muß nur
bis zu einer begrenzten Zahl von Zeiten (1-15 Zyklen) erfolgen, wonach es dem herkömmlichen
Amplifikationszyklus in einem standardisiertem PCR-Puffer erlaubt
werden kann, fortzuschreiten. Alternativ werden die Zielnukleinsäuren von
Interesse zwischen der Priming Lösung
und der Polymeraseverlängerungslösung unter
der Verwendung elektrischer Felder (d. h. Elektrophorese) hin und
her bewegt.
-
Die Verwendung von Hybotropen und/oder
abasischen oder anukleosidischen Oligonukleotidsonden kann dazu
verwendet werden, die Spezifität
und Effizienz von isothermalen Anwendungen von Polymerasen für die Amplifikation
von Nukleinsäuresequenzen
zu erhöhen.
Die Anwendungen von isothermalen Bedingungen zur Verwendung von
Nukleinsäurepolymerasen
schließen
Nukleinsäuresequenzierung,
Genotypisierung, Mutationsnachweis, Oligonukleotid-Ligationsassays,
Mutationsnachweis und ähnliches
ein.
-
Ein anderes Verfahren, das dazu verwendet
wird, die Spezifität
in Hybridisierungsreaktionen zu verbessern, erzeugt Basenfehlpaarungen
unter der Verwendung von Basenanaloga um jedes der A-, G-, C- oder T-Nukleotide
zu ersetzen. Die Forschung hat gezeigt, daß einige Primer, die eine Basenpaarfehlpaarung
enthalten, eine erhöhte
Spezifität
aufweisen, wenn die Fehlpaarung an präzisen Stellen plaziert wird
(siehe Wenham et al., Clinical Chemistry 37: 241, 1991; Newton et
al., Nucleic Acids Research 17: 2503, 1989; Ishikawa et al., Human
Immunology 42: 315, 1995). Jedoch sind auch Unterschiede von so
wenig wie 0,5°C
in den Schmelztemperaturen gleichfalls zwischen perfekt passenden
Hybriden verbreitet und durch das selbe Hybrid mit einer einzelnen
Basenfehlpaarung eingefügt
(siehe Tibanyenda et al., European Journal of Biochemistry 139:
19, 1984; Werntges et al., Nucleic Acids Research 14: 3773, 1986).
Sogar noch bessere Spezifität
wurde zwischen einem und zwei fehlgepaarten Duplices bemerkt, als
diese zwischen einer perfekt gepaarten Duplex und der selben Duplex
mit einer einzelnen Fehlpaarung beobachtet wurde (siehe Guo et al.,
Nature Biotechnology 15: 31, 1997). Guo et al. fanden eine ΔTm von 4°C zwischen
null und einer Fehlpaarung und eine ΔTm von 13°C zwischen einer und zwei aneinander
grenzenden Fehlpaarungen für
eine 20-mer Duplex. Jedoch, auch mit zwei Fehlpaarungen gibt es
oft immer noch wenig Destabilisierung der Duplex. Diese Unfähigkeit, konsistent
zwischen Fehlpaarungen zu unterscheiden, trägt zur fehlenden Spezifität bei PCR
bei.
-
Die Verwendung von mehr als einer
Basenfehlpaarung pro Hybridisierung, wobei mindestens ein Nukleotidanalogon
verwendet wurde, wurde untersucht (siehe Guo et al., Nature Biotechnology
15: 331, 1997). In diesem Fall besteht die analoge Verbindung aus
einer 3-Nitropyrrole-Ersatz
der Purin- oder Pyrimidin-Basen. 3-Nitropyrrol hat die Fähigkeit
minimale Wasserstoffbrücken
mit allen vier Basen auszubilden (sie Nichols et al., Nature 369:
492, 1994; Bergstorm et al., Journal of the Amercan Cheical Society
117: 1201, 1995). Durch Einfügung
einer künstlichen
Fehlpaarung treten große
Unterschiede in den Duplexschmelztemperaturen auf, die von ungefähr 5°C bis 15°C reichen,
wobei die größte Differenz
auftritt, wenn die Fehlpaarung in der Mitte des 15-mer hybridisierenden
Oligos lokalisiert ist. Signifikante Unterschiede in ΔTm treten
auf, wenn ein künstliches
Nukleotid in eine Duplex eingeführt
wird, die bereits eine Basenfehlpaarung enthält, was eine zwei- Fehlpaarungsduplex
erzeugt. Das Ausmaß der
Destabilisierung hängt
von dem Typ von Basenfehlpaarung ab (z. B. G/T) und der Trennung
zwischen den beiden Fehlpaarungen. Bei experimenteller Untersuchung
reichte das Basenanalogon-Nukleotid von 1 bis 7 Basen von der 3'-Seite der Basenfehlpaarung,
die in der Mitte des 15-mers gehalten wurde. Die Unterschiede in ΔTm für die drei
verschiedenen Basen-fehlgepaarten 15-Mere reichten von einer 2°C Stabilisierung
(nur im Falle der C/T-Fehlpaarung und wenn die Fehlpaarungen aneinander
angrenzen), bis zu einer 7°C
weiteren Destabilisierung, wobei die maximale Destabilisierung konsistenter
weise bei einer 3 oder 4 Basenfehlpaarungstrennung auftrat (siehe
Guo et al., Nature Biotechnology 15: 331, 1997).
-
Wenn zwei künstliche Fehlpaarungen eingefügt werden,
beeinflußt
die Umgebung der künstlichen
Basen das Ausmaß der
Destabilisierung stark. Die zwei künstlichen Fehlpaarungen wurden
auf der Mitte einer 21-Mer-Duplex zentriert, beginnend mit einem
Abstand von 6 Basenpaaren. Die Destabilisierung, oder ΔTm, ist mindestens
12°C verglichen
mit der perfekt gepaarten Duplex. Der größte Unterschied von über 20°C tritt auf,
wenn die zwei künstlichen
Fehlpaarungen 10 Basen von einander beabstandet sind. Dieser Unterschied entspricht
einer Helixwindung und zeigt an, daß eine Art von Interaktion
zwischen den zwei künstlichen
Basen auftritt, die die Stabilität
der Duplex verringert.
-
Experimentell, wenn verwendete PCR-Primer
eine oder zwei künstliche
Fehlpaarungen zwischen dem Primer und der DNA-Probe enthielten,
ergab die PCR Ergebnisse, wie sie für einen perfekt passenden Primer erwartet
würden
(siehe Gua et al. Nature Biotechnology 15: 331, 1997). Jedoch, wenn
der Primer sowohl eine echte und eine künstliche Fehlpaarung enthielt,
versagte die PCR dabei, irgendein meßbares Ergebnis zu produzieren.
Hingegen produzierten die PCR-Reaktionen mit perfekt passenden und
echten Fehlpaarungen alle meßbare
Mengen von PCR-Produkt. Die selbe Untersuchung fand ähnliche
Ergebnisse, wenn Hybridisierungssonden verwendet wurden: diejenigen
mit perfekter Übereinstimmung,
echten Fehlpaarungen und künstlichen
Fehlpaarungen lagerten sich an, während die Sonden, die künstliche
und echte Fehlpaarungen enthielten, dies nicht taten. Diese Untersuchungen
zeigten, daß eine
größere Spezifität erzeugt
wird, wenn künstliche Basenfehlpaarungen
in Hybridisierungsreaktionen eingefügt werden, als wenn natürlich auftretende
Basenfehlpaarungen auftreten. Diese thermodynamisch weniger stabil
als eine perfekt passende Hybridisierungsreaktion und daher ist
weniger wahrscheinlich, daß sie
ein falsch positives Ergebnis in einem Test oder PCR hervorrufen.
Interessanter weise ist jedoch der oben angegebene Unterschied in
der thermodynamischen Stabilität
für Duplexes,
die nur eine künstliche
Fehlpaarung enthalten, nicht in der experimentellen Situation manifestiert.
-
Ein weiteres Mittel zur Hervorrufung
der Hybridisierungsunterscheidung ist durch Unterschiede in der Stabilität zwischen
Hybridisierungsduplices, die Nicks und Lücken enthalten. In diesen Reaktion
werden Duplices aus in Tandem-gestapelten kurzen Oligomeren gebildet,
die an einen längeren
Strang hybridisieren, die sich entweder kontinuierlich oder nicht-kontinuierlich
anlagern, wobei eine Lücke
von ein paar Basen zurückgelassen
wird. Hybridisierungen, die zu einem Nick führen werden "Stapel-Hybridisierung" unterzogen, wobei ein
anderer DNA-Strang über
die Nick-Stelle hinweg hybridisiert. Stapel-Hybridisierung tritt nicht auf, wenn
Lücken
in den nicht-kontinuierlichen Oligomeren vorhanden sind. Das Stapeln
hat den Effekt von erhöhter
Unterscheidung, wie durch abnehmende Dissozüerungsraten und größere thermodynamische
Stabilität
nachgewiesen, anders als bei den nicht-kontinuierlichen Gegenspielern
(siehe Lane et al., Nucleic Acids Res. 25: 611, 1997). Thermodynamische
Messungen zeigen, daß die
Unterschiede zwischen der Hybridisierungs-gestapelte Duplices-Standard-freie
Energieveränderung
(ΔG) und
den lückenhaften
Duplices 1,4 bis 2,4 kcal/Mol sind. Daher kann eine Unterscheidung
bei der Hybridisierung durch die Verwendung von vielen kurzen Sonden
ermöglicht
werden.
-
Die meisten der augenblicklich verwendeten
Basenmimetika sind das Ergebnis des Streben nach einer universellen
Base. Viele verwenden Nitroazol-Basenanaloga und haben eine verringerte
Unterscheidung bei der Basenpaarung gezeigt. Eine Serie von Nitroazol-Nukleobasenanaloga
wurden in einem Versuch untersucht, zusätzliche Einsicht in die Signifikanz
von elektronischer Struktur und heterozyklischer Größe bei der Basenpaarung
für die
Entwicklung von effektiveren universellen Basen zu erhalten (siehe
Bergstrom et al., Nucleic Acids Res. 25: 1935, 1997). In dieser
Arbeit wurden die thermodynamischen Eigenschaften der Desoxyribonukleoside
von 3-Nitropyrrol, 4-Nitropyrazol, 4-Nitoimidazol und 5-Nitroindol
gemessen. Zum Vergleich wurden thermodynamische Messungen auch mit
den Desoxyribonukleosiden von Hypoxanthin und Pyrazol sowie einem
abasischem Spacer, 1,2-Didesoxyribose durchgeführt. Vier Oligonukleotide wurden
für jedes
modifizierte Nukleosid synthetisiert, um Duplices zu erhalten, in
denen jede der vier natürlichen
Basen gegenüber dem
Basenmimetikum plaziert war. Alle der analysierten Basenmimetika
erwiesen sich als viel weniger stabil als die natürlichen
Basenpaarungen (A + T: TM = 65,7°C,
C + G: Tm = 70,5°C),
wobei die Tms von 35–46°C für 5-Nitroindol
bis 18–29°C für die anderen
analysierten Nitroazolbasen reichten. Die einzige Ausnahme war 4-Nitroimidazol,
gepaart mit dGTP, wobei die Tm 40,9°C war. Bei der Analyse der freien
Energie für
die Duplexschmelzung wurde von dem 3-Nitropyrrol-Basenmimetikum
gefunden, welches die geringste Unterscheidung aufweist, wenn mit
jeder der vier natürlich
auftretenden Basen gepaart, mit einer gesamten ΔG 0,4 von kcal/Mol. Die nächste am
wenigsten unterscheidende Base war 5-Nitroindol mit einem ΔG von 0,8
kcal/Mol. Beide dieser Werte sind weniger als die ΔG von 1,1
kcal/Mol, die zwischen den natürlichen
Basenpaarungen von A + T und G + C gefunden wurden. 4-Nitropyrazol
zeigen eine leichte Präferenz
für Paarung
mit A, mit einem ΔG
1 kcal/Mol, die stabiler als die C, G und T freien Energien war.
Zuletzt zeigte 4-Nitroimidazol
eine hohe Selektivität
für Paarung
mit G (wie durch ihren hohen Tm-Wert nachgewiesen wurde), aufgrund
der Fähigkeit
des Imidazol N3 mit dem Desoxyguanosin N1 eine Wasserstoffbrücke auszubilden.
Es sollte jedoch bemerkt werden, daß die oben genannten Werte
von den nächsten
Basennachbarn des Mimetikums abhängig sind.
Weitere Untersuchungen veränderten
die nächsten
Nachbarn und fanden, daß 3-Nitropyrrol
und 5-Nitroindol
relative nicht-diskriminatorische Basenpaarungspartner sind.
-
Von Interesse wurden die Enthalpie-
und Entropie-Veränderungen
als einander folgend gefunden (d. h. eine große Enthalpie-Veränderung
korreliert mit einer großen
Entropie-Veränderung),
unabhängig
vom verwendeten Basenmimetikum, was impliziert, daß die korrelation
zwischen ΔS
und ΔH unabhängig von
der Art der Assoziierung der Basen ist. Was beobachtet wurde war,
daß Enthalpie-Veränderungen
in den nicht-Wasserstoffbindenden Basenmimetika gefunden wurden.
Die geringen Werte für
die Entropie- Veränderung
reflektiert das größer Ausmaß an Bewegungsfreiheit,
das für
Basen möglich
ist, die nicht in die Duplex durch Wasserstoffbrücken-Interaktionen eingeschlossen
sind. Die kleinen Enthalpie-Veränderungen
reflektieren Veränderungen
in den Wasserstoffbrücken-Interaktionen als
eine Ergebnis des Verlust von Wasserstoffbrücken-Interaktionen für die Base
gegenüber
des Basenmimetikums. Falls eine natürliche Base in der Helix gestapelt verblieb,
ohne einen gegenüberliegenden
Wasserstoffbrücken-bindenden
Partner, dann hat sie die Wasserstoffbrücken-Interaktion mit Wasser
verloren, ohne einen neuen Donor-Akzeptor-Partner wiederzuerhalten.
-
Eine ähnliche Untersuchung, schloß die Untersuchung
von azyklischen Nukleosidanaloga mit Carboxamido- oder Nitro-substituierten
heterozyklischen Basen ein (siehe Aerschot et al., Nucleic Acids
Res. 23: 4363, 1995). Die Verwendung von azyklischen Nukleosiden
stattet die Konstrukte mit genug Flexibilität aus, um ein gutes Basenstapeln
zu ermöglichen,
sowie es den Basenmimetika zu ermöglichen, eine Orientierung zu
erreichen, um am besten mit der entsprechenden Base Basenpaarung
auszubilden. Die untersuchten heterozyklischen Basen schlossen ein:
4,5-Immidzoldicarboxamid, 4-Nitroimidazol und 5-Nitroindazol. Diese Komplexe
wurden gegen azyklisches Hypoxanthin, 1-(2(-Desoxy-(-D-ribofuransosyl)-3-nitropyrrol, 5-Nitroindol,
und 2(-Desoxyinosin gemessen. Alle neuen azkylischen Komplexe wiesen
Spuren von 7–20°C weniger als
diejenigen auf, die für
die natürlichen
Basen beobachtet wurden. 5-Nitroindol, und 2(-Desoxyinosin gemessen.
Alle neuen azkylischen Komplexe wiesen Spuren von 7–20°C weniger
als diejenigen auf, die für
die natürlichen
Basen beobachtet wurden. 5-Nitroindazol, wenn gegen jedes der vier
natürlichen
Basen gepaart, wie die kleinste Divergenz mit ΔTm nur 2,2°C auf, während das 4-Nitroimidazol einen
Unterschied von 8,0°C
aufwies, wobei dG signifikant außerhalb der Linie der anderen
drei Basen lag, wie oben ähnlich
beobachtet wurde. Von den Referenzverbindungen wies Desoxyinosin
eine ΔTm
von 5,6°C
auf, 5-Nitroindol ΔTm
war 1,0°C, 1-(2(-Desoxy-(-D-ribofuranosyl)-3-nitropyrrol
wies eine ΔTm
von 5,1°C
auf und die ΔTm
von azyklischen Hypoxanthin war 4,8°C. Jedoch zeigten alle Basenmimetika
ungefähr
die selbe Destabilisierung (ΔTm
45°C), wenn
in einem Oligo plaziert, das fast ausschließlich aus Adenosinen bestand,
mit der Ausnahme von 4-Nitroimidazol und azyklischem Desoxyinosin,
die Tms von jeweils 7,0°C
und 8,9°C
aufwiesen.
-
Aershot und Mitarbeiter untersuchten
auf dem Effekt der Einfigung von mehrfachen Basenmimetika in ein
Oligo (siehe Aershot et al., Nucleic Acids Res. 23: 4363, 1995).
Insgesamt sanken die Schmelztemperaturen insgesamt ab, jedoch am
deutlichsten bei der Einfügung
von drei Basenmimetika. Die Nitroindole zeigten jedoch die geringste
Menge von Temperaturunterschied.
-
Ein anderes Basenmimetikum, 1-(2(-Desoxy-(-D-ribofuranosyl)imidazol-4-carboxiamid
(Nukleosid 1), ahmt bevorzugterweise dA-, sowie dC-Nukleoside (siehe
Johnson et al., Nucleic Acids Res. 25: 559, 1997). Die Fähigkeit
für sowohl
dA und dC zu substituieren ergibt sich aus der Rotation um die Carböxiamid-/Imidazolbindung
sowie der Bindung zwischen dem Imidazol- und Furanose-Ring. Wenn
das Imidazol anti gegenüber der
Furanose ist und die Carboxiamidgruppe anti zu dem Imidazol ist,
ist das einzelne Paar auf dem Sauerstoff und einem der Amid-NH-Wasserstoffe
in einer Position, die das NH2 und N-1 von
Adenosin nachahmt. Die Imidazolrotation um die glycosidische Bindung
zu der Sinnorientierung plaziert die Amidgruppe in einer Position, die
ungefähr
den Positionen des NH2 und N-3 von Cytosin
entspricht.
-
Wenn Nukleosid 1 für jedes
natürlich
auftretende Nukleosid substituiert wird, erhöht sich die Enthalpie, mit
der größten Erhöhung für eine dG-Substitution
für die
1-C-Paarung (von ΔH
= 74,7 (kcal/Mol)/ ΔG
= –16,5 (kcal/Mol)
für die
G/C-Paarung von ΔH
= –45,5
(kcal/Mol)/ΔG –5,8 (kcal/Mol)).
Die kleinste Enthalpie-Veränderung
tritt für
eine dA-Substitution auf (ΔH –72,9 (kcal/Mol)/ ΔG = –15,4 (kcal/Mol)
für A/T-Paarung
von ΔH = –66,7 (kcal/Mol)/ΔG = –11,7 (kcal/Mol)
für die
1-T-Paarung. Entsprechend nimmt die Tm signifikant von 65,7°C und 70,5°C für jeweils
die A-T- und C-G-Paare ab auf 46,6°C für die 1-T-Paarung, 43,4°C für 1-G, 27,6°C für 1-A und
14,6°C für 1-C.
-
Wenn in einer PCR-Reaktion verwendet,
werden Nukleosid 1 und sein N-Propylderivat bevorzugterweise als
dATP-Analoga inkorporiert (siehe Sala et al., Nucleic Acids Res.
24: 3302, 1996). Jedoch, sobald in ein DNA-Templat inkorporiert,
gab ihr mehrdeutiges Wasserstoffbindungspotential eine Miß-Inkorporation
jeder der natürlich
auftretenden Basen bei Sequenzen von 3 × 10–2 pro
Base pro Amplifikation. Die meisten der Substitutionen (die primär aus G
bestanden) waren ein Ergebnis der Rotation um die Carboxyamidgruppe, wenn
Teil des Templats. Zwischen 11 bis 15% der Substitutionen lagen
an der Rotation der Imidazolgruppe um die glycosidische Bindung.
Als Teil eines DNA-Templats wirkte das N-Propylderivat auf die selbe Weise wie
1, trotz seiner Propylgruppe. Diese Untersuchung zeigt, daß während 1
bevorzugterweise als dATP wirkt, es die Fähigkeit in einer PCR-Typ-Umgebung aufweist,
sich auch als alle vier natürlich
auftretenden Nukleotide zu benehmen. Aus dieser und den oben genannten
Untersuchungen ist ersichtlich, daß ein breiter Bereich von Duplex-Stabilität durch
Variationen in Basenmimetika und deren Plazierung innerhalb eines
Oligonukleotids erhalten werden kann.
-
In einem Aspekt der vorliegenden
Erfindung werden Verfahren zur Bestimmung der Identität eines
Nukleinsäuremoleküls oder
-fragments (oder zum Nachweisen des Vorhandenseins eines ausgewählten Nukleinsäuremoleküls oder
-fragments) zur Verfügung
gestellt, die die Schritte umfassen: (a) Erzeugen markierter Nukleinsäuremoleküle aus einem
oder mehreren ausgewählten
Target-Nukleinsäuremolekülen, wobei
eine Markierung mit einem bestimmten Nukleinsäuremolekül korrelativ und mit Nicht-Fluoreszenz-Spektrometrie oder
Potentiometrie nachweisbar ist, (b) Abtrennen der markierten Moleküle nach
Größe, (c)
Abspalten der Markierungen von den markierten Molekülen und
(d) Nachweisen der Markierungen mit Nicht-Fluoreszenz-Spektrometrie
oder Potentiometrie und Bestimmen der Identität der Nukleinsäuremoleküle daraus.
-
In einem verwandten Aspekt der. Erfindung
werden Verfahren zum Nachweisen eines ausgewählten Nukleinsäuremoleküls zur Verfügung gestellt,
die die Schritte umfassen: (a) Kombinieren markierter Nukleinsäuresonden
mit Target-Nukleinsäuremolekülen unter
Bedingungen und für
einen Zeitraum, die ausreichend sind, um Hybridisierung einer markierten
Nukleinsäuresonde
an eine komplementäre
ausgewählte
Target-Nukleinsäuresequenz
zu ermöglichen,
wobei eine markierte Nukleinsäuresonde
mit Nicht-Fluoreszenz-Spektrometrie
oder Potentiometrie nachweisbar ist, (b) Verändern der Größe hybridisierter
markierter Sonden, nicht-hybridisierter Sonden oder Target-Moleküle oder
der Sonden:Target-Hybride, (c) Abtrennen der markierten Sonden nach
Größe, (d)
Abspalten der Markierungen von den markierten Sonden und (e) Nachweisen
der Markierungen mit Nicht-Fluoreszenz-Spektrometrie
oder Potentiometrie und Bestimmen des ausgewählten Nukleinsäuremoleküls daraus.
Diese und andere verwandte Techniken werden detaillierter unten
diskutiert.
-
2. PCR
-
PCR kann eine gewünschte DNA-Sequenz eines beliebigen
Ursprungs (Virus, Bakterium, Pflanze oder Mensch) Hunderte von Millionen
mal innerhalb einiger Stunden amplifizieren. PCR ist insbesondere
wertvoll, weil die Reaktion hoch-spezifisch, leicht zu automatisieren
und zum Amplifizieren kleiner Probenmengen fähig ist. Aus diesen Gründen hat
PCR eine große
Bedeutung für
die klinische Medizin, die Diagnose genetischer Erkrankungen, die
forensische Wissenschaft und die Evolutionsbiologie gehabt.
-
Kurz gesagt ist PCR ein Verfahren,
das auf einer spezialisierten Polymerase beruht, die einen komplementären Strang
zu einem gegebenen DNA-Strang in einer Mischung synthetisieren kann,
die die 4 DNA-Basen und 2 DNA-Fragmente (Primer, jeder etwa 20 Basen
lang), die die Target-Sequenz flankieren, enthält. Die Mischung wird erhitzt,
um die Stränge
aus doppelsträngiger
DNA, die die Target-Sequenz enthalten, voneinander, und anschließend abgekühlt, um
zu ermöglichen,
daß (1)
die Primer ihre komplementären
Sequenzen auf den voneinander getrennten Strängen finden und sich an sie
binden, und (2) daß die
Polymerase die Primer zu neuen komplementären Strängen verlängert. Wiederholte Zyklen von
Erhitzen und Abkühlen multiplizieren
die Target-DNA exponentiell, da jeder neue Doppelstrang sich so
auftrennt, daß er
zwei Matrizen für
eine weitere Synthese wird. In etwa 1 Stunde können 20 PCR-Zyklen das Target
um ein Millionenfaches amplifizieren.
-
In einer Ausführungsform der Erfindung werden
Verfahren zum Bestimmen der Identität eines Nukleinsäuremoleküls oder
zum Nachweisen des ausgewählten
Nukleinsäuremoleküls in z.
B. einer biologischen Probe unter Verwendung der PCR-Technik zw
Verfügung
gestellt. Kurz gesagt umfassen solche Verfahren die Schritte: Erzeugen
einer Reihe markierter Nukleinsäurefragmente
oder -moleküle
während
der PCR und Auftrennen der resultierenden Fragmente nach Größe. Der
Größenauftrennschritt
kann unter Verwendung irgendeiner der hierin beschriebenen Techniken
durchgeführt
werden, einschließlich
z. B. Gelelektrophorese (z. B. Polyacrylamidgelelektrophorese) oder
vorzugsweise HPLC. Die Markierungen werden anschließend von den
aufgetrennten Fragmenten abgespalten und mit der entsprechenden
Nachweistechnologie nachgewiesen Beispiele für solche Technologien sind
hierin beschrieben worden und schließen z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV-Spektrometrie ein.
-
3. RNA-Fingerprinting
und Differential Display
-
Wenn die Matrize RNA ist, ist der
erste Schritt beim Fingerprinting reverse Transkription. Liang und Pardee
(Science 257: 967, 1992) waren die ersten, die ein RNA-Fingerprinting-Protokoll beschrieben
haben, das einen Primer für
reverse Transkription auf der Basis von Oligo(dT) verwendet, aber
mit einem "Anker" aus zwei Basen am
5'-Ende (z. B. Oligo-5'-(dT11)CA-3'). Priming tritt
hauptsächlich
am 5'-Ende des Poly(rA)-Schwanzes
auf und hauptsächlich
in Sequenzen, die 5'-UpG-poly(rA)-3' enden, mit einer
Selektivität,
die sich einer aus 12 polyadenylierten RNAs nähert. Nach reverser Transkription
und Denaturierung wird willkürliches
Priming auf dem resultierenden ersten Strang aus cDNA durchgeführt. PCR
kann jetzt verwendet werden, um einen Fingerprint von Produkten
zu erzeugen, die am besten mit den Primern zusammenpassen und die
vom 3'-Ende der
mRNAs und polyadenylierten heterogenen RNAs abgeleitet werden. Dieses
Protokoll ist als "Differential
Display" bezeichnet
worden.
-
Alternativ kann ein willkürlicher
Primer im ersten Schritt der reversen Transkription verwendet werden, wobei
diejenigen für
die RNA-internen Regionen ausgewählt
werden, die 6-8 Basenmatches mit dem 3'-Ende des Primers haben. Dem folgt willkürliches
Priming des resultierenden ersten Stranges aus cDNA mit demselben
oder einem unterschiedlichen willkürlichen Primer und anschließend PCR.
Dieses besondere Protokoll sampled irgendwo in der RNA, einschließlich offene
Leserahmen (Welsh et al., Nuc. Acids. Res. 20: 4965, 1992). Zusätzlich kann
es auf RNAs verwendet werden, die nicht polyadenyliert sind, wie
etwa viele bakterielle RNAs. Diese Variante von RNA-Fingerprinting
durch PCR mit willkürlichem
Primer ist RAP-PCR genannt worden.
-
Wenn Fingerprinting von RNA durch
PCR mit willkürlichem
Primer auf Proben durchgeführt
wird, die aus Zellen, Geweben oder anderem biologischen Material
gewonnen sind, die verschiedenen experimentellen Behandlungen unterzogen
worden sind oder verschiedene Entwicklungsgeschichten haben, können Unterschiede
in der Genexpression zwischen den Proben nachgewiesen werden. Für jede Reaktion
wird angenommen, daß dieselbe
Anzahl von effektiven PCR-Verdopplungsereignissen auftritt und alle
Unterschiede in den anfänglichen
Konzentrationen von cDNA-Produkten als ein Verhältnis von Intensitäten im letztendlichen
Fingerprint konserviert werden. Es gibt keine sinnvollen Beziehungen
zwischen den Intensitäten
von Banden innerhalb einer einzigen Spur auf einem Gel, die eine
Funktion von Match und Menge sind. Jedoch wird das Verhältnis zwischen
Spuren für
jede gesamplete RNA konserviert, was ermöglicht, daß differentiell exprimierte RNAs
nachgewiesen werden können.
Das Verhältnis
von Ausgangsmaterialien zwischen Proben wird aufrechterhalten, selbst
wenn die Anzahl von Zyklen ausreichend ist, um zu ermöglichen,
daß die
PCR-Reaktion absättigt. Dies
beruht darauf, daß die
Anzahl von Verdopplungen, die benötigt wird, um Sättigung
zu erreichen, beinahe vollständig
durch die invarianten Produkte gesteuert wird, die den Großteil des
Fingerprint ausmachen. In dieser Hinsicht ist PCR-Fingerprinting
verschieden von herkömmlicher
PCR eines einzelnen Produkts, bei der das Verhältnis von Ausgangsmaterialien
zwischen Proben nicht konserviert wird, sofern nicht Produkte in
der exponentiellen Phase der Amplifikation gesampled werden.
-
In einer Ausführungsform der Erfindung werden
Verfahren zum Bestimmen der Identität eines Nukleinsäuremoleküls oder
zum Nachweisen eines ausgewählten
Nukleinsäuremoleküls in z.
B. einer biologischen Probe unter Verwendung der Technik des RNA-Fingerprinting
zur Verfügung
gestellt. Kurz gesagt umfassen solche Verfahren allgemein die Schritte:
Erzeugen einer Reihe markierter Nukleinsäurefragmente. Die Fragmente,
die mit PCR oder ähnlichen
Amplifikationsschemen erzeugt werden, werden dann anschließend nach Größe getrennt.
Der Schritt der Größentrennung
kann z. B. jede der hierin beschriebenen Techniken sein, einschließlich z.
B. Gelelektrophorese (z. B. Polyacrylamidgelelektrophorese) oder
vorzugsweise HPLC. Die Markierungen werden dann von den aufgetrennten
Fragmenten abgespalten, und anschließend werden die Markierungen
mit der entsprechenden Nachweistechnologie nachgewiesen. Repräsentative
Beispiele für
geeignete Technologien schließen
Massenspektrometrie, Infrarotspektrometrie, potentiostatische Amperometrie oder
UV-Spektrometrie ein. Die relativen Mengen aller gegebenen Nukleinsäurefragmente
sind nicht wichtig, aber die Größe der Bande
ist informativ, wenn sie mit einer Kontrollprobe in Beziehung gesetzt
wird.
-
4. PCR-Einzelstrankonformationspolymorphismus
auf Fluoreszenz-Basis (PCR-SSCP)
-
Eine Reihe von Verfahren, zusätzlich zum
RFLP-Ansatz, sind für
die Analyse von Basensubstitutionspolymorphismen verfügbar. Orita
et al. haben einen Weg zur Analyse dieser Polymorphismen auf der
Basis von Konformationsunterschieden denaturierter DNA entwickelt.
Kurz gesagt wird Restriktionsenzymverdau oder PCR verwendet, um
relativ kleine DNA-Fragmente zu erzeugen, die anschließend denaturiert
und durch Elektrophorese auf nicht-denaturierenden Polyacrylamidgelen
aufgetrennt werden. Konformationsunterschiede in den einzelsträngigen DNA-Fragmenten,
die aus Basensubstitutionen resultieren, werden durch Elektrophoresemobilitätsverschiebung nachgewiesen.
Basenpaarung innerhalb des Strangs schafft Einzelstrangkonformationen,
die hoch sequenzspezifisch und in der Elektrophoresemobilität unterschiedlich
sind. Nachweisraten in verschiedenen Studien unter Verwendung von
herkömmlichem
SSCP reichen jedoch von 35% bis nahezu 100%, wobei die höchsten Nachweisraten
am häufigsten
mehrere unterschiedliche Bedingungen erfordern. Im Prinzip könnte das
Verfahren auch verwendet werden, um Polymorphismen auf der Basis
kurzer Insertionen oder Deletionen zu analysieren. Dieses Verfahren
ist eines der leistungsfähigsten
Tools zur Identifizierung von Punktmutationen und -deletionen in
DNA (SSCP-PCR, Dean et al., Cell 61: 863, 1990).
-
In einer Ausführungsform der Erfindung werden
Verfahren zum Bestimmen der Identität eines Nukleinsäuremoleküls oder
zum Nachweisen eines ausgewählten
Nukleinsäuremoleküls in z.
B. einer biologischen Probe unter Verwendung der PCR-SSP-Technik
zur Verfügung
gestellt. Kurz gesagt umfassen solche Verfahren allgemein die Schritte:
Erzeugen einer Reihe markierter Nukleinsäurefragmente. Die mit PCR erzeugten Fragmente
werden dann nach Größe getrennt.
Vorzugsweise ist der Größentrennschritt
nicht-denaturierend, und die Nukleinsäurefragmente werden vor der
Trennmethodik denaturiert. Der Größentrennschritt kann z. B. mit
Gelelektrophorese (z. B. Polyacrylamidgelelektrophorese) oder vorzugsweise
HPLC durchgeführt
werden. Markierungen werden anschließend von den abgetrennten Fragmenten
abgespalten, und anschließend
werden die Markierungen mit der entsprechenden Nachweistechnologie
nachgewiesen (z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV-Spektrometrie).
-
5. Didesoxy-Fingerprinting
(ddF)
-
Ein weiteres Verfahren ist beschrieben
worden (ddF, Sarkar et al., Genomics 13: 441, 1992), das 100% von
Einzelbasenveränderungen
im Gen des Humanfaktors IX nachgewiesen hat, wenn getestet in einer
retrospektiven und prospektiven Art und Weise. Insgesamt wurden
84 von 84 unterschiedlichen Sequenzveränderungen nachgewiesen, wenn
genomische DNA von Patienten mit Haemophilie B analysiert wurde.
-
Kurz gesagt ist, bei den Anwendungen
von Markierungen zum Genotypisieren oder anderen Zwecken, ein Verfahren,
das verwendet werden kann, Didesoxy-Fingerprinting. Dieses Verfahren
setzt einen Didesoxyterminator in einer Sanger-Sequenzierungsreaktion
ein. Das Prinzip des Verfahrens ist wie folgt: Eine Target-Nukleinsäure, die
sequenziert werden soll, wird in eine Reaktion gegeben, die einen
Didesoxy-Terminator aufweist, der komplementär zu der Base ist, von der
bekannt ist, daß sie
in der Target-Nukleinsäure
mutiert ist. Wenn die Mutation z. B. zu einer A → G-Änderung führt, würde die Reaktion in einer C-Didesoxy-Terminator-Reaktion
durchgeführt
werden. PCR-Primer werden verwendet, um die interessierende Targetsequenz
zu lokalisieren und zu amplifizieren. Wenn die hypothetische Targetsequenz
die A → G-Änderung
enthält,
wird die Größe einer
Population von Sequenzen aufgrund des Einbaus eines Didesoxy-Terminators
in die amplifizierten Sequenzen verändert. In dieser besonderen
Anwendung von Markierungen würde
ein Fragment erzeugt werden, das eine vorhersagbare Größe im Fall
einer Mutation besäße. Die
Markierungen wären
an das 5'-Ende der PCR-Primer
gebunden und würden
eine "Karte" für den Probentyp
und den Didesoxy-Terminatortyp liefern. Eine PCR-Amplifikationsreaktion
würde stattfinden,
die resultierenden Fragmente würden
nach Größe mit z. B.
HPLC oder PAGE getrennt werden. Am Ende des Trennverfahrens werden
die DNA-Fragmente in einem temporären Bezugsrahmen gesammelt,
die Markierungen werden abgespalten, und das Vorhandensein oder Nichtvorhandensein
von Mutation wird durch die Kettenlänge aufgrund frühzeitigen
Kettenterminators durch den Einbau eines gegebenen Didesoxy-Terminators
bestimmt.
-
Es ist wichtig anzumerken, daß ddF zu
einem Gewinn oder Verlust eines Didesoxy-Terminationssegments und/oder einer
Verschiebung der Mobilität
wenigstens eines der Terminationssegmente oder -produkte führt. Daher
wird bei diesem Verfahren eine Suche nach der Verschiebung von einer
Fragmentmobilität
vor einem hohen Hintergrund an Fragmenten mit anderer relativer
Molekülmasse
durchgeführt.
Ein Vorteil ist die Vorkenntnis der Länge des Fragmentes, die mit
einer gegebenen Mutation assoziiert ist.
-
In einer Ausführungsform der Erfindung werden
Verfahren zum Bestimmen der Identität eines Nukleinsäuremoleküls oder
zum Nachweisen eines selektierenden Nukleinsäuremoleküls in z. B. einer biologischen Probe
unter Verwendung der ddF-Technik zur Verfügung gestellt. Kurz gesagt
umfassen solche Verfahren allgemein die Schritte: Erzeugung einer
Reihe markierter Nukleinsäurefragmente,
gefolgt von der Trennung auf der Grundlage der Größe. Vorzugsweise
ist der Größentrennschritt
nicht-denaturierend, und die Nukleinsäurefragmente werden vor der
Trennmethodik denaturiert. Der Größentrennschritt kann z. B.
mit Gelelektrophorese (z. B. Polyacrylamidgelelektrophorese) oder
vorzugsweise HPLC durchgeführt
werden. Die Markierungen werden anschließend von den aufgetrennten
Fragmenten abgespalten, und anschließend werden die Markierungen
mit der entsprechenden Nachweistechnologie nachgewiesen (z. B. Massenspektrometrie,
Infrarotspektrometrie, potentiostatische Amperometrie oder UV-Spektrometrie).
-
6. Restriktionskarten
und RFLPs
-
Restriktionsendonukleasen erkennen
kurze DNA-Sequenzen und schneiden DNA-Moleküle an diesen spezifischen Stellen.
Einige Restriktionsenzyme (Rare-Cutters) schneiden DNA sehr selten,
wodurch eine kleine Anzahl sehr großer Fragmente (mehrere tausend
bis eine Million bp) erzeugt werden. Die meisten Enzyme schneiden
DNA häufiger,
wodurch eine große
Anzahl kleiner Fragmente (weniger als einhundert bis mehr als eintausend
bp) erzeugt wird. Im Mittel werden Restriktionsenzyme mit 4-Basen-Erkennungsstellen
Stücke
liefern, die 256 Basen lang sind, 6-Basen-Erkennungsstellen werden
Stücke
liefern, die 4.000 Basen lang sind, und 8-Basen-Erkennungsstellen
werden Stücke
liefern, die 64.000 Basen lang sind. Da hunderte von verschiedenen
Restriktionsenzymen charakterisiert worden sind, kann DNA in viele
verschiedene kleine Fragmente geschnitten werden.
-
Eine breite Vielfalt von Techniken
ist für
die Analyse von DNA-Polymorphismen entwickelt worden. Das am weitesten
verwendete Verfahren, der Restriktionsfragmentlängenpolymorphismus(RFPL)-Ansatz,
kombiniert Restriktionsenzymverdau, Gelelektrophorese, Blotting
auf eine Membran und Hybridisierung an eine klonierte DNA-Sonde.
Polymorphismen werden als Variationen in den Längen der markierten Fragmente
auf den Blots nachgewiesen. Der RFLP-Ansatz kann verwendet werden,
um Basensubstitutionen zu analysieren, wenn die Sequenzveränderung
in eine Restriktionsenzymstelle fällt, oder um Minisatelliten/(VNTRs)
durch Auswahl von Restriktionsenzymen zu analysieren, die außerhalb
der Wiederholungseinheiten schneiden. Die Agarosegele liefern üblicherweise
nicht die notwendige Auflösung,
um Minisatellit/VNTR-Allele zu unterscheiden, die sich um eine einzige
Wiederholungseinheit unterscheiden, aber viele der Minisatelliten/VNTRs
sind so variabel, daß dennoch
hochinformative Marker erhalten werden können.
-
In einer Ausführungsform der Erfindung werden
Verfahren zum Bestimmen der Identität eines Nukleinsäuremoleküls oder
zum Nachweisen eines selektierenden λlukleinsäuremoleküls in z. B. einer biologischen Probe
unter Verwendung der Technik der Restriktionskartierung oder RFLPs
bereitgestellt. Kurz gesagt umfassen solche Verfahren allgemein
die Schritte: Erzeugen einer Reihe markierter Nukleinsäurefragmente,
wobei die erzeugten Fragmente mit Restriktionsenzymenen verdaut
werden. Die markierten Fragmente werden erzeugt, indem ein Hybridisierungsschritt
der markierten Sonden mit der verdauten Target-Nukleinsäure durchgeführt wird.
Der Hybridisierungsschritt kann vor oder nach dem Restriktionsnukleaseverdau
stattfinden. Die resultierenden verdauten Nukleinsäurefragmente
werden dann nach Größe getrennt.
Der Größentrennschritt kann
z. B. mit Gelelektrophorese (z. B. Polyacrylamidgelelektrophorese)
oder vorzugsweise HPLC durchgeführt
werden. Die Markierungen werden anschließend von den getrennten Fragmenten
abgespalten, und anschließend
werden die Markierungen mit der entsprechenden Nachweistechnologie
nachgewiesen (z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV-Spektrometrie).
-
7. DNA-Fingerprinting
-
DNA-Fingerprinting umfaßt das Display
eines Satzes von DNA-Fragmenten von einer spezifischen DNA-Probe.
Eine Vielzahl von DNA-Fingerprintingtechniken sind gegenwärtig verfügbar (Jeffreys
et al., Nature 314: 67, 1985; Zabeua und Vos, 1992); "Selective Restriction Fragment
Amplification: A General Method for DNA Fingerprinting," European Patent
Application 92402629.7; Vos et al., "DNA FINGERPRINTING: A New Technique
for DNA Fingerprinting. "Nucl.
Acids Res. 23: 4407–4414,
1996; Bates, S. R. E., Knorr, D. A., Weller, J. W., und Ziegele,
J. S., "Instrumentation
for Automated Molecular Marker Acquisition and Analysis." Kapitel 14, Seiten
239-255, in The Impact of Plant Molecular Genetics, edited by B.
W. S. Sobral, veröffentlicht
von Birkhauser, 1996.
-
Daher werden in einer Ausführungsform
der Erfindung Verfahren zum Bestimmen der Identität eines Nukleinsäuremoleküls oder
zum Nachweis eines selektierenden Nukleinsäuremoleküls in z. B. einer biologischen
Probe unter Verwendung der Technik des DNA-Fingerprintings bereitgestellt.
Kurz gesagt umfassen solche Verfahren allgemein die Schritte: Erzeugen
einer Reihe markierter Nukleinsäurefragmente,
gefolgt von Trennung der Fragmente nach Größe. Der Größentrennschritt kann z. B.
mit Gelelektrophorese (z. B. Polyacrylamid-gelelektrophorese) oder
vorzugsweise HPLC durchgeführt
werden. Die Markierungen werden dann von den aufgetrennten Fragmenten
abgespalten, und anschließend
werden die Markierungen mit der entsprechenden Nachweistechnologie
nachgewiesen (z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV-Spektrometrie).
-
Kurz gesagt, basiert DNA-Fingerprinting
auf der selektiven PCR-Amplifikation von Restriktionsfragmenten
aus einem Gesamtverdau von genomischer DNA: Die Technik schließt drei
Schritte ein. 1) Restriktion der DNA-Fragmente und anschließende Ligation
von Oligonukleotid-Adaptoren, 2) Die selektive Amplifikation von
Sets von Restriktionsfragmenten, 3) Gelanalyse der amplifizierten
Fragmente. Die PCR-Amplifikation der Restriktionsfragmente wird
durch Verwendung der Adapter- und Restriktionsstellen-Sequenz als Zielstellen
für die
Primer Anhybridisierung erreicht. Die selektive Amplifikation wird
durch die Verwendung von Primern erreicht, die sich in die Restriktionsfragmente
hinein erstrecken und nur die Fragmente amplifizieren, in denen die
Primerverlängerungen
den Nukleotiden entsprechen, die die Restriktionsstellen flankieren.
-
Dieses Verfahren ergibt daher Sets
von Restriktionsfragmenten, die durch eine Vielzahl von Verfahren visualisiert
werden können
(d. h. PAGE, HPLC oder andere Typen von Spektrometrie), ohne vorheriges
Wissen der Nukleotidsequenz. Das Verfahren ermöglicht auch die Co-Amplifikation
von großen
Zahlen von Restriktionsfragmenten. Die Zahl von Fragmenten ist jedoch
abhängig
von der Auflösung
des Nachweissystems. Typischerweise werden 50–100 Restriktionsfragmente
amplifiziert und auf denaturierenden Polyacrylamidgelen nachgewiesen.
In der hier beschriebenen Anmeldung wird die Trennung durch HPLC
durchgeführt.
-
Die DNA-Fingerprinting-Technik basiert
auf der Amplifikation von Untergruppen von genomischen Restriktionsfragmenten
unter der Verwendung von PCR. Die DNA wird mit Restriktionsenzymen
geschnitten und Doppelstrang-Adapter werden an die Enden der DNA-Fragmente ligiert,
um Templat-DNA für
die einplifikationsreaktionen zu erzeugen. Die Sequenz der ligierten
Adapter und die angrenzenden Restriktionsenzyme (-stellen) dienen
als Bindungsstellen für
das DNA-Fingerprinting von Primern für die PCR-basierte Amplifikation.
Selektive Nukleotide werden am 3'-Ende
der PCR-Primer eingeschlossen, die dadurch nur eine DNA-Synthese
von einer Untergruppe der Restriktionsstellen primen können. Nur
Restriktionsfragmente, in denen die Restriktionsstelle flankierende
Nukleotide zu den selektiven Nukleotiden passen, werden amplifiziert
werden. Der DNA-Fingerprintprozess
ergibt "Fingerprint"-Muster von verschiedenen
Fragmentlängen,
die charakteristisch und reproduzierbar für einen individuellen Organismus
sind. Diese Fingerprints können
dazu verwendet werden, sogar sehr eng verwandte Organismen zu unterscheiden,
einschließlich
nahezu isogener Linien. Die Unterschiede in den Fragmentlängen können auf
Basenveränderungen
in der Restriktionsstelle oder der Primerverlängerungsstelle oder auf Insertionen
oder Deletionen im Körper
des DNA-Fragments
zurückgeführt werden.
-
Die Abhängigkeit von dem Wissen um
die Sequenz des Zielgenoms wird durch die Verwendung von Adaptoren
von bekannter Sequenz eleminiert, die an die Restriktionsfragmente
ligiert werden. Die PCR-Primer sind für die bekannten Sequenzen der Adaptoren
und der Restriktionsstellen spezifisch. Die Schritte des genetischen
Fingerprintingverfahrens sind unten beschrieben.
-
- 1) Restriktion und Ligation. Die Restriktionsfragmente von
genomischer DNA werden durch Verwendung zweier verschiedener Restriktionsenzymen
erzeugt: ein selten schneidendes Enzym (das sechs-Base-erkennende Enzym
EcoRI) und ein häufig
schneidendes Enzym (das vier-Basen-erkennende Enzym MseI). Drei
verschiedene Typen von Fragmenten werden produziert: einige mit
EcoRI-Schnitten an beiden Enden, einige MseI-Schnitten an beiden
Enden und einige mit einem EcoRI-Schnitt an einem Ende und mit einem MseI-Schnitt am anderen
Ende. Doppelsträngige
Adaptoren werden dann an die überstehenden
Enden der DNA-Fragmente ligiert, was Templat-DNA für die Amplifikation
erzeugt. Die Adaptoren sind spezifisch für entweder die EcoRI-Stelle
oder die MseI-Stelle. Die Restriktion und Ligation findet in einer
einzelnen Reaktion statt. Die Ligation des Adaptors an die verdaute
DNA verändert
die Restriktionsstelle, um so zu verhindern, daß eine zweite Restriktions
stattfindet, nachdem die Ligation stattgefunden hat.
- 2) Präselektive
Amplifikation. Die Sequenzen der Adaptoren und Restriktionsstellen
dienen als Primerbindungsstellen für die "präselektive
PCR-Amplifikation".
Die präselektiven
Primer weisen jeder ein "selektives" Nukleotid auf, das
das Unterset von Restriktionsfragmenten erkennen wird, die das passenden
Nukleotid stromabwärts
von der Restriktionsstelle aufweisen. Die primären Produkte der präselektiven
PCR sind diejenigen Fragmente, die einen MseI-Schnitt und EcoRI-Schnitt
aufweisen und auch das passende interne Nukleotid aufweisen. Die
präselektive
Amplifikation erreicht eine 16-fache
Verringerung der Komplexität
des Fragmentgemisches.
- 3) Selektive Amplifikation mit CMST-markierten Primern. Die
Komplexität
des PCR-Produkts
wird weiter reduziert (256-fach) und Fragmente werden mit einem
Set von CMSTs durch Durchführen
einer zweiten PCR unter der Verwendung von selektiven Primern, markiert
mit CMSTs, markiert. Es ist möglich,
aus unter 64 verschiedenen Primerpaaren (die sich aus allen möglichen
Kombinationen von acht MseI- und acht EcoRI-Primern ergeben) für die Amplifikation
auszuwählen.
Jeder dieser Primer besitzt drei selektive Nukleotide. Das erste
ist dasselbe, wie das in der präselektiven
Amplifikation verwendete, die anderen können jede von den 16 möglichen
Kombinationen der vier Nukleotide sein. Nur die Untergruppe von
Fragmenten, die passende Nukleotide an allen drei Positionen aufweist,
wird zu dieser Phase in der Amplifikation amplifiziert werden.
-
8. Anwendung spaltbarer
Marker auf die Genotypisierung und den Polymorphismus-Nachweis
-
a. Einführung
-
Obgleich einige bekannte Human-DNA-Polymorphismen
auf Insertionen, Deletionen oder anderen Umordnungen nicht-wiederholter
Sequenzen beruhen, beruht die große Mehrzahl auf entweder Einzelbasensubstitutionen
oder auf Variationen in der Anzahl von Tandemwiederholungen. Basensubstitutionen
sind im menschlichen Genom reichlich vorhanden, wobei sie im Mittel
einmal alle 200–500
bp auftreten. Längenvariationen
in Blöcken
von Tandemwiederholungen sind ebenfalls im Genom üblich, mit
wenigstens einigen 10.000 eingestreuten polymorphen Stellen (Laci
genannt). Wiederholungslängen
für Tandem-Wiederholungs-Polymorphismen
reichen von 1 bp in (dA)n(dT)n-Sequenzen
bis wenigstens 170 bp in α-Satelliten-DNA. Tandem-Wiederholungs-Polymorphismen
können
in zwei große
Gruppen unterteilt werden, die aus Minisatelliten/variable Anzahl
von Tandem-Wiederholungen
(VNTRs), mit typischen Wiederholungslängen von einigen zehn Basenpaaren
und mit einigen 10.000 Gesamtwiederholungseinheiten, und Mikrosatelliten,
mit Wiederholungslängen
von bis zu 6 bp und mit maximalen Gesamtlängen von etwa 70 bp, bestehen.
Die meisten Mikrosatelliten-Polymorphismen, die bisher identifiziert
worden sind, haben auf (dC-dA)n- oder (dG-dT)n-Dinukleotidwiederholungssequenzen beruht.
Die Analyse von Mikrosatelliten-Polymorphismen umfaßt die Amplifikation mit
der Polymerasekettenreaktion (PCR) eines kleinen DNA-Fragments,
das einen Block von Wiederholungen enthält, gefolgt von Elektrophorese
der amplifizierten DNA auf denaturierendem Polyacrylamidgel. Die PCR-Primer
sind komplementär
zu einzigartigen Sequenzen, die die Wiederholungsblöcke flankieren.
Polyacrylamidgele, anstelle von Agarosegelen, werden traditionellerweise
für Mikrosatelliten
verwendet, weil die Allele sich in der Größe oft nur um eine einzige
Wiederholung unterscheiden.
-
So werden in einem Aspekt der vorliegenden
Erfindung Verfahren zum Genotypisieren eines ausgewählten Organismus
zur Verfügung
gestellt, die die Schritte umfassen: (a) Erzeugen markierter Nukleinsäuremoleküle aus einem
ausgewählten
Targetmolekül,
wobei eine Markierung mit einem bestimmten Fragment korrelativ ist
und mit Nicht-Fluoreszenz-Spektrometrie
oder Potentiometrie nachgewiesen werden kann, (b) Abtrennen der
markierten Moleküle
nach Sequenzlänge,
(c) Abspalten der Markierung vom markierten Molekül und (d)
Nachweisen der Markierung mit Nicht-Fluoreszenz-Spektrometrie oder
Potentiometrie und Bestimmen des Genotyps des Organismus daraus.
-
In einem weiteren Aspekt werden Verfahren
zum Genotypisieren eines ausgewählten
Organismus zur Verfügung
gestellt, die die Schritte umfassen: (a) Kombinieren eines markierten
Nukleinsäuremoleküls mit einem
ausgewählten
Targetmolekül
unter Bedingungen und für
einen Zeitraum, die ausreichend sind, um Hybridisierung des markierten
Moleküls
an das Targetmolekül
zu ermöglichen,
wobei eine Markierung mit einem bestimmten Fragment korrelativ ist
und mit Nicht-Fluoreszenz-Spektrometrie oder Potentiometrie nachgewiesen
werden kann, (b) Abtrennen der markierten Fragmente nach Sequenzlänge, (e)
Abspalten der Markierung vom markierte Fragment und (d) Nachweisen
der Markierung mit Nicht-Fluoreszenz-Spektrometrie
oder Potentiometrie und Bestimmen des Genotyps des Organismus daraus.
-
b. Anwendung spaltbarer
Markierungen auf die Genotypisierung
-
Ein PCR-Ansatz, um Restriktionsfragmentlängenpolymorphismus
(RFPL) zu identifizieren, kombiniert Gelelektrophorese und den Nachweis
von Markierungen, die mit spezifischen PCR-Primern assoziiert sind.
Im allgemeinen wird ein PCR-Primer eine spezifische Markierung aufweisen.
Die Markierung wird daher einen Satz von PCR-Primern repräsentieren
und daher eine vorbestimmte DNA-Fragmentlänge. Polymorphismen werden
als Variationen in der Länge
der markierten Fragmente in einem Gel oder aus einem Gel eluierend nachgewiesen.
Polyacrylamidgelelektrophorese wird üblicherweise die notwendige
Auflösung
liefern, um Minisatellit/VNTR-Allele zu unterscheiden, die sich
um eine einzige Wiederholungseinheit unterscheiden. Die Analyse
von Mikrosatellitenpolymorphismen umfaßt die Amplifikation mit der
Polymerasekettenreaktion (PCR) eines kleinen DNA-Fragments, das einen Wiederholungsblock
enthält,
gefolgt von der Elektrophorese der amplifizierten DNA auf denaturierendem
Polyacrylamidgel oder gefolgt von der Trennung von DNA-Fragmenten mit
HPLC. Die amplifizierte DNA wird unter Verwendung von Primern markiert
werden, die spaltbare Markierungen am 5'-Ende des Primers aufweisen. Die Primer
werden in die neu synthetisierten Stränge durch Kettenverlängerung
eingebaut. Die PCR-Primer sind komplementär zu einzigartigen Sequenzen,
die die Wiederholungsblöcke
flankieren. Minisatelliten/VNTR-Polymorphismen können ebenfalls amplifiziert
werden, weitgehend so wie bei den oben beschriebenen Mikrosatelliten.
-
Beschreibungen vieler Arten von DNA-Sequenzpolymorphismen
haben die grundlegende Basis für das
Verständnis
der Struktur des menschlichen Genoms geliefert (Botstein et al.,
Am. J. Human Genetics 32: S. 314, 1980; Donis-Keller, Cell 51: 319,
1987; Weissenbach et al., Nature 359: 794). Die Konstruktion umfangreicher
Kopplungskarten-Systeme ist durch die Verwendung dieser DNA-Polymorphismen
erleichtert worden und hat ein praktisches Mittel zur Lokalisierung
von Krankheitsgenen durch Kopplung geliefert. Mikrosatelliten-Dinukleotidmarker
haben sich als sehr leistungsfähige
Tools bei der Identifizierung menschlicher Gene erwiesen, bei denen
sich gezeigt hat, daß sie
Mutationen enthalten und in einigen Fällen Krankheiten verursachen.
Genomische Dinukleotidwiederholungen sind hochpolymorph (Weber,
1990, Genomic Analysis, Vol. 1, S. 159–181, Cold Spring Laboratory
Press, Cold Spring Harbor, NY; Weber und Wong, 1993, Hum. Mol. Genetics,
2, S. 1123) und können
bis zu 24 Allele besitzen. Mikrosatelliten-Dinukleotidwiederholungen
können
mit PCR unter Verwendung von Primern amplifiziert werden, die zu
den einzigartigen Regionen komplementär sind, die das Dinukleotid
umgeben. Im Anschluß an
die Amplifikation können
mehrere amplifizierte Loci vor einem Größentrennschritt kombiniert
(vervielfältigt)
werden. Das Verfahren der Anwendung der amplifizierten Mikrosatellitenfragmente
auf einen Größentrennschritt
und des anschließenden
Identifizierens der Größe und daher
des Allels ist als Genotypisierung bekannt. Über chromosomenspezifische
Marker, die ein hohes Vervielfältigungsniveau
erlauben, ist zum Durchführen
vollständiger
Genomscans fur Kopplungsanalyse berichtet worden (Davies et al.,
1994, Nature, 371, S. 130).
-
Markierungen können mit großer Wirkung
beim Genotypisieren mit Mikrosatelliten verwendet werden. Kurz gesagt,
werden die PCR-Primer konstruiert, um Markierungen zu tragen, und
in einer sorgfältig
ausgewählten
PCR-Reaktion verwendet, um Di-, Tri- oder Tetranukleotid-Wiederholungen
zu amplifizieren. Die Amplifikationsprodukte werden anschließend nach
Größe mit Verfahren,
wie etwa HPLC oder PAGE getrennt. Die DNA-Fragmente werden anschließend in
einer temporären
Art und Weise gesammelt, die Markierungen von ihren entsprechenden
DNA-Fragmenten abgespalten und die Länge aus einem Vergleich mit
internen Standards im Größentrennschritt
abgeleitet. Allel-Identifizierung
wird aus der Bezugnahme zur Größe der amplifizierten
Produkte durchgeführt.
-
Beim Ansatz mit spaltbaren Markierungen
für die
Genotypisierung ist es möglich,
mehrere Proben in einem einzigen Trennschritt zu kombinieren. Es
gibt zwei allgemeine Wege, auf denen dies durchgeführt werden
kann. Das erste allgemeine Verfahren für ein Screening mit hohem Durchsatz
ist der Nachweis eines einzelnen Polymorphismus in einer großen Gruppe
von Individuen. In diesem Szenario wird ein einzelner oder „nested" Satz von PCR-Primern
verwendet, und jede Amplifikation wird mit einem DNA-Probentyp pro
Reaktion durchgeführt.
Die Anzahl von Proben, die im Trennschritt kombiniert werden können, ist
proportional zur Anzahl spaltbarer Markierungen, die pro Nachweistechnologie
erzeugt werden können
(d. h. 400–600
für Massenspektrometrie-Markierungen).
Es ist daher möglich, 1
bis mehrere Polymorphismen einer großen Gruppe von Individuen gleichzeitig
zu identifizieren. Der zweite Ansatz ist, mehrere Sätze von
PCR-Primern zu verwenden, die zahlreiche Polymorphismen auf einer
einzigen DNA-Probe identifizieren können (ein Individuum z. B.
genotypisieren können).
Bei diesem Ansatz werden PCR-Primer in einer einzigen Amplifikationsreaktion kombiniert,
die PCR-Produkte unterschiedlicher Länge erzeugt. Jedes Primerpaar
oder jeder eingebaute Satz ist durch eine spezifische spaltbare
Markierung codiert, was impliziert, daß jedes PCR-Fragment mit einer
spezifischen Markierung codiert sein wird. Die Reaktion wird in
einem einzigen Trennschritt durchgeführt (siehe unten). Die Anzahl
von Proben, die im Trennschritt kombiniert werden können, ist
proportional zur Anzahl spaltbarer Markierungen, die pro Nachweistechnologie
erzeugt werden können
(d. h. 400–600
für Massenspektrometer-Markierungen).
-
Die Genotypisierung kann auch auf.
landwirtschaftliche Proben angewendet werden. Zum Beispiel ermöglicht die
Amplifizierter Polymorphismus Längen
Polymorphismus (AFLP)-Analyse
eine landwirtschaftlich bedeutsame Gruppierung des Keimplasmas.
Die Zusammenfassung von QTL-Effekten kann verschiedene Regionen
des Genoms ergeben, die konsistent wichtige Determinanten von vielen
quantitativen Eigenschaften sind. Die Assoziierung von AFLP- und
QTL-Polymorphismen sollte von prädiktiver
Brauchbarkeit beim Erstellen von Paarungsgruppen sein. Weiterhin
sollte das Wissen im Hinblick auf die genomische Architektw von
verwandten und nicht verwandten Keimplasmen in Schlüsselregionen
des Genoms eine systematische Trennung auf Basis von Selektionsantworten
in verwandten Keimplasmen ermöglichen.
Es sollte auch die systematische Introgression von exotischen Keimplasmen
erleichtern. Zuletzt sollte eine übergreifende Beschreibung von Keimplasma
auf dem molekularen Niveau, gekoppelt mit extensiver QTL-Information,
es Züchtern
ermöglichen,
eine mittlere Performance beizubehalten und gleichzeitig genetische
Diversität
zu erreichen.
-
Sobald Werkzeuge für die Routinecharakterisierung
zur Verwendung von genetischen Ressourcen erhältlich sind, wird es einen
größeren Impetus
für die
Konservierung von genetischen Ressourcen geben.
-
Lander und Botstein (Lander, E. S.
und D. Botstein, "Mapping
Mendelian factors underlying quantitative traits using RFLP linkage
maps" Genetics 121:
185–189,
1987, beschrieben die Entwicklung von Gedanken, die zum Abgleich
der Mendelschen Theorie der bestimmten Vererbung und der Beobachtung
führten,
daß viele
Marker eine kontinuierliche Variation zeigen. Die Entwicklung von
molekularen Markertechnologien wird die Entwicklung von ausreichend
dichten Karten ermöglichen,
um die Abtrennung von quantitativen Markern zu initiieren. Ein hauptsächlicher
Impetus für
den QTL-Nachweis ist es, die zugrunde liegenden Determinanten in
einem angewandten Züchterkontext
zu manipulieren. Parterson et al. "DNA markers in plant improvement" Adv. in Agron. 46:
39–90,
1991; und Dedley, J. W. "Comparison
of genetic distance estimators using molecular marker data", Proc. Second Plant
Breeding Symposium of the Crop Sci. Soc. Amer. and Amer. Soc. Hort.,
Corvallis, OR, Amer. Soc. Hort Sci. Alexandria, VA, 1994, stellten
alle ausgezeichnete Überblicke über die
potentiellen Anwendungen dieser Techniken zur Verfügung. Zum
Beispiel kann die Assoziierung von quantitativer Markerexpression
mit kartierten Markern als eine Basis für Entscheidungen im Hinblick
auf die Manipulation von Züchtungspopulationen,
für die
molekulare Marker unterstützte
Selektion (MMAS) oder zur Entdeckung von wünschenswerten Genen in exotischen
Keimplasmen (siehe auch Tanksley et al. "Advanced backcross QTL analysis: a method
for the simultaneous discovery and transfer of valuable QTLs form
unadapted germplasm into elite breeding lines", Theor. Appl. Genet. 92: 191–203, 1996)
führen.
QTL für
einen Bereich von Markern, z. B. Ausbeute, Malzqualität, Winterhärte, und
Erkrankungsresistenz, wurden in einer Zahl von Gerste Keimplasmaquellen
lokalisiert (zusammengefaßt
von Hayes et al., "Barley
genome mapping and itsapplicaitons" in P. P. Jauhar (ed.) Methods for Genome
Analysis in Plants, CRC Press, Boca Raton, USA, 1996; und Hayes,
P. M. et al., "Multiple
disease resistance loci and their relationship to agronomic and
quality loci in a spring barley population", JQTL, 1996, see http://probe.nalusda.gov:8000/otherdocs/jqtl/index.htm).
Diese Berichte, wie diejenigen in anderen Getreidsorten, waren größtenteils
beschreibend.
-
Die genetische Basis dieser QTL,
obwohl sie von größerem theoretischem
Interesse und signifikanter praktischer Wichtigkeit sind, ist im
allgemeinen nicht bekannt. Es wurde vorgeschlagen, daß QTL Konsequenzen
von Allelen an Loci sind, wo die Effekte von anderen Allelen auf
eine einfache Mendelsche Weise interpretiert werden können. Falls
das der Fall wäre,
dann sollten sich QTL-Allele in einer additiven Weise verhalten und
sie sollten von einem Set von Keimplasma zu einem anderen transferierbar
sein. Alternativ können
komplexe Typen die Endergebnisse von komplexen Signalwegen sein
(siehe z. B. Dawkins, R. "The
extended phenotype" Oxford
University Press, Oxford. UK, 1982). Falls dem so ist, stellt QTL
die Effekte von Allelen dar, die den komplexen Signalweg stören, der
in quantitativen Phänotypen
gipfelt. In diesem Szenario wäre
es einfach, den quantitativen Phänotyp
zu verändern
(d. h. die Anwesenheit eines nicht wünschenswerten Allels an jedem Punkt
in dem Signalweg wäre
ausreichend, den Signalweg zum Entgleisen zu bringen), jedoch schwierig
den quantitativen Phänotyp
zu erhalten (d. h. ein Wirt mit gewünschten Allelen an multiplen
Loci würde
erforderlich sein, um den quantitativen Phänotyp zu erhalten). Falls dies
der Fall wäre,
dann wäre
die Introgression von QTL-Allelen in nicht verwandtes Keimplasma
ein relativer Erfolg- oder Fehlschlagansatz: Keimplasma, das nicht
mit der kartierenden Referenz-Population
verwandt wäre,
könnte
gut nicht wünschenswerte
Allele an Loci aufweisen, wo vorteilhafte Allele in der kartierenden
Referenz-Population fixiert wären.
-
Die Komplexität der Kontrolle wird möglicherweise
mit der untersuchten Eigenschaft varieren. Zuletzt sollte sogar
der komplexeste Phänotyp
letztendlich auf eine Serie von diskreten, wenn auch hoch interaktiven, Schritten
reduzierbar sein. Relativ wenige QTL sind für komplexe Phänotypen
nachgewiesen, wie zum Beispiel Ausbeute, Komponenten der Malzqualität und die
quantitative Resistenz gegenüber
biotischen und abiotischen Stressfaktoren (zusammengefaßt durch
Hayes et al., "Barley
genome mapping and its applications" in P. P. Jauhar (ed.) Methods of Genome
Analysis in Plants: CRC Press, Boca Raton, USA, 1996). In einigen
Fällen
können
Kandidatengene als QTL-Determinanten angegeben werden: Schüttelresistenz
als eine Determinante der Ausbeute (Hayes et al., 1993, wie oben)
und hydrolytische Enzyme als Determinanten von Malzextrakt (die
Menge von löslichen
Kohlenhydraten, die als ein Substrat für die Fermentation erhältlich sind)
(siehe Hayes, P. M., 1996m, oben). Barley genome mapping: new insights
into the malting quality of the world's oldest crop. (siehe, z. B. MBAA 33:
223-225).
-
In anderen Fällen können bis jetzt keine Kandidatengene
als Determinanten von komplexen Phänotypen angegeben werden. Ein
Beispiel ist die quantitive adulte Planzenresistenz gegenüber Streifenrost,
wobei Resistenz-QTL nicht mit den berichteten Kartenlokationen von
Resistenzgenen zusammenfallen, die Muster von Mendelscher Vererbung
zeigen (siehe Hayes, P. M. et al., "Multiple disease resistance loci and
their relationship to agronomic and quality loci in a spring barley
population", JQTL,
1996 und http://probe.nalusda.gov:8000/otherdocs/jqtl/index.htm).
-
DNA-Niveau-Polymorphismen können auch
dazu verwendet werden, Themen der genetischen Diversität zu untersuchen.
Genetische Diversität
wurde in einer Zahl von Gerste-Keimplasma-Arrays
gemessen, unter der Verwendung von Werkzeugen, die von Abstammungsanalyse
(siehe, z. B. Eslick, R. F. et al., "Genetic engineering as a key to ater
use efficiency",
Agric. Meteor. 14: 13–23,
1974), über
morphologische Marker (siehe, z. B., Tolbert, D. M. et al., "A diversity analysis
of a world collection of barley" Crop.
Sci. 19: 789-794,
1979) bis zu molekularen Markern verschiedener Sorten (Melchinger,
A. E. et al. "Relationships
among European barley germplasm: I. Genetic diversity among winter
and spring cultivars revealed by RFLPs" Crop Sci. 34: 1191–1199, 1994; Saghai-Maroof,
M. A. et al. "RFLPs
in cultivated barley and their application in the evaluation of
malting quality cultivars" Hereditas
121: 2129, 1994) reichten. Molekulare Marker sind aus dieser Hinsicht besonders
attraktiv, da sie reichlich vorhandene adaptiv neutrale Referenzpunkte
zur Verfügung
stellen. Im Fall von molekularen Polymorphismen waren die Zeit,
die Ressourcen die erforderlich waren, die Daten zu erzeugen, eine
Begrenzung.
-
Die Kopplungs-Karten-Konstruktion
ist der erste Schritt in einer systematischen QTL-Analyse. Werkzeuge,
wie zum Beispiel die CMST-Technologie-Plattform wie hier bei der
Kartierung verwendet beschrieben, sollten auch dazu brauchbar sein
es den Züchtern
zu ermöglichen,
die genetische Diversität
auszudehnen ohne den Wert des Arbeits-Keimplasmapools zu verringern. Bis jetzt
war nicht ausreichend Information erhältlich, um eine Charakterisierung
der Diversität
mit den Ergebnissen von QTL-Analysen zu integrieren. Der Schlüssel für solch
eine integrative Strategie ist eine Markertechnologie, die schnell
und Kosten-effektiv ist und ausreichend Polymophismen über das
Genom hinweg zur Verfügung
stellt. Die vorliegende Erfindung stellt diesen Schlüssel zur
Verfügung.
-
c. Enzymatischer Mutationsnachweis
und die Anwendungen von Markierungen.
-
In dieser besonderen Anwendung oder
diesem besonderen Verfahren werden Fehlpaarungen in Heteroduplices
durch enzymatische Spaltung von Fehlpaarungs-Basenpaaren in einer
gegebenen Nukleinsäureduplex
nachgewiesen. DNA-Sequenzen, die auf das Vorhandensein von einer
Mutation getestet werden sollen, werden mit PCR unter Verwendung
eines spezifischen Satzes von Primern amplifiziert, die amplifizierten Produkte
werden denaturiert und mit denaturierten Bezugsfragmenten vermischt
und hybridisiert, was zur Bildung von Heteroduplices führt. Die
Heteroduplices werden anschließend
mit Enzymen behandelt, die die Duplex erkennen und spalten, wenn
eine Fehlpaarung vorhanden ist. Solche Enzyme sind Nuklease S1,
Mungobohnen-Nuklease, "Resolvasen", T4-Endonuklease
IV, etc.. Im wesentlichen kann jedes Enzym verwendet werden, das
Fehlpaarungen in vitro erkennt und die resultierende Fehlpaarung
spaltet. Nach Behandlung mit dem geeigneten Enzym werden die Duplices
nach Größe mit z.
B. HPLC oder PAGE getrennt. Die DNA-Fragmente werden temporär gesammelt.
Markierungen werden abgespalten und nachgewiesen. Das Vorhandensein
einer Mutation wird durch die Verschiebung der Mobilität des Fragments
relativ zu einem Wildtyp-Bezugsfragment nachgewiesen.
-
d. Anwendungen von Markierungen
auf dem Oligonukleotid-Ligationsassay (OLA).
-
Der Oligonukleotid-Ligationsassay,
wie er ursprünglich
von Landegren et al. (Landegren et al., Science 241: 487, 1988)
beschrieben ist, ist eine nützliche
Technik zur Identifizierung von (bekannten) Sequenzen in sehr großen und
komplexen Genomen. Das Prinzip der OLA-Reaktion beruht auf der Fähigkeit
von Ligase, zwei diagnostische Oligonukleotide kovalent zu verknüpfen, wenn
sie benachbart zueinander auf einem gegebenen DNA-Target hybridisieren.
Wenn die Sequenzen an der Sondenverbindungsstelle nicht perfekt basengepaart
sind, werden die Sonden nicht von der Ligase verknüpft werden.
Die Fähigkeit
einer thermostabilen Ligase, potentielle Einzelbasenpaarunterschiede
zu unterscheiden, wenn sie am 3'-Ende
der "stromaufwärtigen" Sonde angeordnet
sind, liefert die Möglichkeit
zur Einzelbasenpaarauflösung
(Barony, PNAS USA 88: 189, 1991). Bei der Anwendung von Markierungen
würden
die Markierungen an die Sonde gebunden werden, die an das amplifizierte
Produkt ligiert wird. Nach Abschluß der OLA werden die Fragmente
auf der Grundlage ihrer Größe getrennt,
die Markierungen abgespalten und mit Massenspektrometrie nachgewiesen.
-
e. Sequenzspezifische
Amplifikation
-
PCR-Primer mit einem 3'-Ende, das entweder
zu einer Mutanten- oder normalen Oligonukleotidsequenz komplementär ist, können verwendet
werden, um das eine oder das andere Allel selektiv zu amplifizieren
(Newton et al., Nuc. Acids Res., 17, S. 2503; et al., 1989, Genomics,
5, S. 535; Okayama et al., 1989, J. Lab. Clin. Med., 114, S. 105;
Sommer et al., 1989, Mayo Clin. Proc., 64, 1361; Wu et al., PNAS
USA, 86, S. 2757). Üblicherweise
werden die PCR-Produkte nach der Amplifikation mit PAGE sichtbar
gemacht, aber das Prinzip sequenzspezifischer Amplifikation kann
auf Festphasenformate angewendet werden.
-
f. Potentielle Anwendung
von Markierungen auf einige Assays auf Amplifikationsbasis.
-
Genotypisierung von Viren: Eine potentielle
Anwendung von Markierungen ist die Genotypisierung oder Identifizierung
von Viren durch Hybridisierung mit markierten Sonden. F+-RNA-Coliphagen
können
z. B. nützliche
Kandidaten als Indikatoren für
Enteroviruskontamination sein. Genotypisierung mit Nukleinsäurehybridisierungsverfahren
ist eine verläßliche,
schnelle, einfache und preiswerte Alternative zur Serotypisierung (Kafatos
et al., Nucleic Acids Res. 7: 1541, 1979). Amplifikationstechniken
und Nukleinsäurehybridisierungstechniken
sind mit Erfolg verwendet worden, um eine Vielzahl von Mikroorganismen
zu klassifizieren, einschließlich
E. coli (Feng, Mol. Cell Probes 7: 151, 1993), Rotavirus (Sethabutr
et al., J. Med. Virol. 37: 192, 1992), Hepatitis-C-Virus (Stuyver
et al., J. Gen Virol. 74: 1093, 1993) und Herpes-simplex-Virus (Matsumoto et
al., J. Virol. Methods 40: 119, 1992).
-
Prognostische Anwendungen von Mutationsanalyse
bei Krebserkrankungen: Genetische Veränderungen sind in einer Vielzahl
experimenteller Säuger-
und Human-Neoplasmen beschrieben worden und stellen die morphologische
Grundlage für
die Abfolge morphologischer Veränderungen
dar, die bei Karzinogenese beobachtet wird (Vogelstein et al., NEJM
319: 525, 1988). In den letzten Jahren, mit dem Aufkommen von molekularbiologischen
Techniken, sind Allelverluste auf bestimmten Chromosomen oder Mutation
von Tumorsuppressorgenen sowie Mutationen in mehreren Onkogenen
(z. B., c-myc, c-jun
und die ras-Familie) die am häufigsten
untersuchten Einheiten gewesen.
-
Frühere Arbeiten (Finkelstein
et al., Arch Surg. 128: 526, 1993) haben eine Korrelation zwischen
spezifischen Typen von Punktmutationen im K-ras-Onkogen und dem
Stadium bei Diagnose bei kolorektalem Karzinom identifiziert. Die
Ergebnisse legen nahe, daß die
Mutationsanalyse wichtige Informationen über Tumoraggressivität liefern
könnte,
einschließlich
dem Muster und der Verteilung von Metastasen. Der prognostische Werte
von TP53- und K-ras-2-Mutationsanalyse bei Stufe III-Karzinomen
des Colons ist vor kurzem nachgewiesen worden (Pricolo et al., Am.
J. Surg. 171: 41, 1996). Es ist daher offensichtlich, daß die Genotypisierung von
Tumoren und präkanzerösen Zellen
und spezifischer Mutationsnachweis bei Behandlung von Krebserkrankungen
bei Menschen zunehmend wichtig werden wird.
-
9. Einzel-Nukleotid
Verlängerungsassay
-
Die Primerverlängerungstechnik für den Nachweis
von Einzelnukleotiden in genomischer DNA wurde erst durch Sokolov
1989 beschrieben (Nucleic Acids Res. 18(12): 3671, 1989). In diesem
Artikel beschrieb Sokolov die Einzelnukleotidverlängerung
von 30-Meren und 20- Meren,
die zu der bekannten Sequenz des zystischen Fibrosegens komplementär waren.
Es wurde gezeigt, daß das
Verfahren die Fähigkeit
aufwies, konekt eine einzelne Nukleotidveränderung innerhalb des Gens
zu identifizieren. Das Verfahren basierte auf der Verwendung von
radiomarkierten Desoxynukleotiden für ein Markierungsverfahren
in dem Einzelnukleotidverlängerungsassay.
-
Spätere Publikationen beschrieben
die Verwendung von Einzelnukleotidverlängerungsassays für genetische
Erkrankungen, wie zum Beispiel Hämopholie
B (Faktor IX) und das zystische Fibrosegen (siehe, z. B., Kuppuswamy
et al., PNAS USA 88: S. 1143–1147,
1991). Kuppuswamy et al. zeigten, daß der Einzelnukleotidverlängerungsassay
dazu verwendet werden könnte,
um genetische Erkrankungen nachzuweisen, wobei die Anwendung der
Nachweis von Hämophilie
B (Faktor IX) und das zystische Fibrosegen waren. Wiederum basiert
dieses Verfahren auf dem einzelnen Nukleotidprimer, der an eine
Sequenz hybridisiert wird, die an einen bekannten Single Nucleotide
Polymorphismus angrenzt. Die geprimte genomische DNA wird dann Bedingungen
unterzogen, in denen Taq-Polymerase ein 32P-markiertes dNTP hinzufügen wird,
falls die Stelle hinter der Stelle von Interesse komplementär zu dem
alpha markierten Reaktionsgemisch ist.
-
Kürzlich
wurden die Parameter des Einzelnukleotidverlängerungsassays im Hinblick
auf den quantitativen Bereich, die Variabilität und Multiplex-Analyse genau
beschrieben. Daher beschrieben 1996 Greenwood und Burke (Greenwood,
A. D. und Burke, D. T. (1996) Genome Research, 6, S. 336–348) genau
die Parameter des Einzelnukleotidverlängerungsassays im Hinblick
auf den quantitativen Bereich, die Variabilität und die Multiplex-Analyse.
RNA diente als ein Templat für
die PCR-Amplifikation von einer Sequenz von Interesse, die einen
einzel-Base Unterschied zwischen zwei Allelen aufweist. Jedes PCR-erzeugte
Templat wird auf die Anwesenheit, Abwesenheit oder relative Menge
jedes einzelnen Allels durch Annealen eines Primers analysiert,
der eine Base 5' zu
dem Polymorphismus liegt und sich um eine markierte Base verlängert (oder
eine markierte Base verwendet). Nur wenn die korrekte Base in der
Reaktion erhältlich
ist, wird ein Einbau am 3'-Ende
des Primers auftreten. Die Verlängerungsprodukte
werden dann analysiert (herkömmlich
durch PAGE).
-
Daher basiert diese Strategie auf
der Genauigkeit der DNA-Polymerase, nur das korrekt gepaarte Nukleotid
an das 3'-Ende des
Templat-hybridisierten Primers anzufügen. Da nur ein Didesoxy-Terminatornukleotid
pro Reaktion hinzugefügt
wird, ist dies eine einfache Weise herauszubekommen, welcher Primer
in allen vier Typen von dNTPs verlängert wurde.
-
Daher werden innerhalb eines Aspekts
der Erfindung Verfahren zum Nachweis einer einzelnen ausgewählten Nukleotids
innerhalb eines Nukleinsäuremoleküls zur Verfügung gestellt,
umfassend die Schritte von (a) Hybridisieren in mindestens zwei
getrennten Reaktionen einen markierten Primer und ein Zielnukleinsäuremolekül unter
Bedingungen und für
eine Zeitdauer, die ausreichend ist, eine Hybridisierung des Primers an
das Zielnukleinsäuremolekül zu ermöglichen,
wobei jede Reaktion ein Enzym enthält, das einen Nukleotidkettenterminator
hinzufügen
wird, und einen Nukleinkettenterminator, komplementär zu Adenosin,
Cytosin, Guanosin, Thymidin oder Uracil, und wobei jede Reaktion
einen verschiedenen Nukleotidkettenterminator enthält, (b)
Auftrennen von markierten Primern mittels Größe, (c) Abspalten des Markers
von dem markierten Primer und (d) Nachweisen des Markers durch nicht-fluoreszente
Spektrometrie oder Potentiometrie und daraus Nachweisen der Anwesenheit
des ausgewählten
Nukleotids innerhalb des Nukleinsäuremoleküls.
-
Wie hier angegeben, kann eine große Vielzahl
von Abtrennverfahren verwendet werden, einschließlich, z. B. Flüssigkeitschromatographie-Mitteln,
wie zum Beispiel HPLC. Zusätzlich
kann eine große
Vielzahl von Nachweisverfahren verwendet werden, einschließlich, zum
Beispiel, Massenspektrometrie, Infrarotspektrometrie, Ultraviolett-Spektrometrie oder
potentiostatische Amperometrie. Auch können mehrere verschiedene Enzyme
verwendet werden (z. B. eine Polymerase) sowie jede der hier zur
Verfügung
gestellten Markierungen. Innerhalb bestimmter bevorzugter Ausführungsformen
weist jeder Primer, der innerhalb einer Reaktion verwendet wird,
einen verschiedenen einmaligen Marker auf. Auf diese Weise können multiple
Proben (oder multiple Stellen) gleichzeitig auf die Anwesenheit
von ausgewählten
Nukleotiden hin getestet werden.
-
Einzelnukleotidtests, so wie diejenigen
hier beschrieben, können
dazu verwendet werden, um polymorphe Varianten nachzuweisen oder
eine biologische Probe auf die Anwesenheit eines spezifischen Nukleotids
innerhalb oder nahe einer bekannten Sequenz hin zu untersuchen.
Die Zielnukleinsäuremoleküle schließen nicht
nur DNA ein (z. B. genomische DNA), sondern auch RNA.
-
Im allgemeinen schießt dieses
Verfahren das Hybridisieren eines Primers an die Ziel-DNA-Sequenz auf solche
Weise ein, daß das
3'-Ende des Primers
unmittelbar an die Mutation, die nachgewiesen und identifiziert
werden soll, angrenzt. Das Verfahren ist ähnlich zu der Sanger Sequenzreaktion,
mit der Ausnahme, daß nur
das Didesoxynukleotid eines bestimmten Nukleotids zum Reaktionsgemisch
hinzugefügt
wird. Jedes Didesoxynukleotid wird mit einem einmaligen Marker markiert.
Von den vier Reaktionsgemischen wird nur eines einen Didesoxyterminator
an die Primersequenz anfügen.
Falls die Mutation vorhanden ist, wird sie durch den einmaligen
Marker auf dem Didesoxynukleotid nachgewiesen werden und seine Identität etabliert
werden. Auch können
multiple Mutationen durch Markieren des DNA-Primers mit einem einmaligen Marker
gleichzeitig ermittelt werden. Innerhalb eines Aspekts der Erfindung
werden Verfahren zur Analyse von Einzelnukleotid-Mutationen aus
einer bestimmten biologischen Probe zur Verfügung gestellt, umfassend die
Schritte von Exponieren von Nukleinsäuren aus einer biologischen
Probe und Kombinieren der ausgesetzten Nukleinsäuren mit einer oder mehreren
ausgewählten
Nukleinsäuren,
die markiert oder nicht markiert sein können, unter Bedingungen und
für eine
Zeitdauer, die ausreichend ist, für die Sonden an die Nukleinsäuren zu
hybridisieren, wobei der Marker, falls verwendet, mit einer bestimmten
Nukleinsäuresonde
korrelativ ist und durch nicht-fluoreszente
Spektrometrie oder Potentiometrie nachweisbar ist. Die DNA-Fragmente
werden in vier getrennten Reaktionen reagiert, die jede einen verschieden
markierten Didesoxyterminator enthalten, wobei der Marker mit einem
bestimmten Didesoxynukleotid korrelativ ist und durch nicht-fluoreszente
Spektrometrie oder Potentiometrie nachweisbar ist. Die DNA-Fragmente
werden gemäß der Größe durch
zum Beispiel Gelelektrophorese (z. B. Polyacrylamidgelelektrophorese)
oder, bevorzugterweise, HPLC aufgetrennt. Die Marker werden von
den aufgetrennten Fragmenten aufgespalten und durch die jeweilige Nachweistechnologie
(z. B. Massenspektrometrie, Infrarotspektrometrie, potentiostatische
Amperometrie oder UV/sichtbare Spektrophotometrie) nachgewiesen.
Die nachgewiesenen Marker können
mit den bestimmten zu analysierenden DNA-Fragmenten sowie der Identität des Mutanten-Nukleotids
korreliert werden.
-
10. Amplifiziertes Längenpolymorphismus
(AFLP)
-
AFLP wurde als ein hoch sensitives
Verfahren für
DNA-Fingerprinting entwickelt, das in einer Vielzahl von Gebieten
verwendet werden soll, einschließlich Pflanzen- und Tierzucht,
medizinische Diagnose, forensische Analyse und mikrobieller Typisierung,
nur um ein paar zu nennen (Vos et al., Nucleic Acids Res. 23: 4407–4414, 1995).
Die Stärke
von AFLP basiert auf den molekulargenetischen Variationen, die zwischen
eng verwandten Spezies, Sorten oder Kulturen bestehen. Diese Variationen
in der DNA-Sequenz werden durch die genetische Fingerprint-Technologie
ausgenutzt, so daß "Fingerprints" von bestimmten Genotypen
routinemäßig erzeugt
werden können.
Diese "Fingerprints" sind einfache RFLPs,
die durch selektive PCR-Amplifikation von DNA-Restriktionsfragmenten
visualisiert werden. Kurz gesagt, besteht die genetische Fingerprinting-Technologie
aus den folgenden Schritten: genomische DNA wird bis zur Vollständigkeit
durch zwei verschiedene Restriktionsenzyme verdaut. Spezifische
Doppelstrang-Oligonukleotid-Adaptoren (~25–30 bp) werden an die verdauten
DNA-Fragmente ligiert. Zu den Adaptoren homologe Oligonukleotidprimer,
die jedoch Verlängerungen
an dem 3'-Ende aufweisen,
werden dazu verwendet, um ein Unterset der DNA-Fragmente zu amplifizieren (ein
Präamplifikationsschritt
kann auch durchgeführt
werden, wenn diese Verlängerung
nur eine Länge
von 1 bp aufweist. Die Amplifikation mit dem Primer, der eine 3-Basenpaar
Verlängerung
aufweist, würde
folgen.) Diese Verlängerungen
können
in ihrer Länge
von einem bis drei Basenpaare variieren, sind jedoch für einen bestimmten
Primer von definierter Länge.
Die Sequenz der Verlängerung
kann auch von einem Primer zu einem anderen variieren, ist jedoch
für einen
bestimmten Primer von definierter Länge. Die Sequenz der Verlängerung
kann auch von einem Primer zum anderen variieren, ist jedoch innerhalb
eines Primers von einer einzelnen, definierten Sequenz. Die selektive
Natur der AFLP-PCR basiert auf den 3'-Verlängerungen an den Oligonukleotid- Primern. Da diese
Verlängerungen
nicht homolog zu der Adaptersequenz sind, werden nur DNA-Fragmente
amplifiziert werden, die komplementär zu den Verlängerungen
sind, aufgrund der Unfähigkeit der
Taq-DNA-Polymerase, anders als einige andere DNA-Polymerasen, DNAs zu verlängern, falls
eine Fehlpaarung am 3'-Ende
eines Moleküls
auftritt, das synthetisiert wird. Daher wird in jeder Reaktion nur
eine Untergruppe des gesamten Genoms amplifiziert. Zum Beispiel,
falls zwei Basenpaar (bp)-Verlängerungen
verwendet werden, wird nur eines in 256 Molekülen amplifiziert. Um die Zahl
von Fragmenten weiter zu begrenzen, die wirklich visualisiert werden
(so daß eine
handhabbare Zahl beobachtet wird), wird nur eine dieser Primer markiert.
Zuletzt werden die amplifizierten DNAs auf einem Polyacrylamidgel
(Sequenziertyp) aufgetrennt und ein Autoradiogramm oder Phosphorimage
wird erzeugt.
-
Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zur Bestimmung der Identität eines Nukleinsäuremoleküls zur Verfügung gestellt
oder zum Nachweis eines ausgewählten
Nukleinsäuremoleküls in, z.
B. einer biologischen Probe, wobei die Technik des genetischen Fingerprinting
verwendet wird. Kurz gesagt, umfassen solche Verfahren im allgemeinen
die Schritte von Verdauen (z. B. genomischer DNA) bis zur Vollständigkeit
durch zwei verschiedene Restriktionsenzyme. Spezifische Doppelstrang-Oligonukleotid-Adapter (~25–30 bp)
werden an die verdauten DNA-Fragmente ligiert. Gegebenenfalls kann
ein Präamplifikation
unter der Verwendung von Primern mit einer 1 bp Verlängerung
durchgeführt
werden. Das PCR-Produkt wird dann verdünnt und markierte Primer, die
homolog zu den Adaptern sind, jedoch Verlängerungen an dem 3'-Ende aufweisen,
werden dazu verwendet, durch PCR eine Untergruppe der DNA-Fragmente
zu amplifizieren. Die sich ergebenden PCR-Produkte werden dann in
ihrer Größe nach
aufgetrennt. Der Größentrennschritt
kann dann durch eine Vielzahl von Verfahren durchgeführt werden,
einschließlich,
z. B. HPLC. Die Marker werden dann von den aufgetrennten Fragmenten
abgespalten und durch die jeweilige Nachweistechnologie (z. B. Massenspektrometrie,
Infrarotspektrometrie, potentiostatische Amperometrie oder UV/sichtbare
Spektrophotometrie) nachgewiesen.
-
11. Genexpressionsanalyse
-
Eine der hier beschrieben Erfindung
ist ein Hochdurchsatzverfahren zur Messung der Expression von zahlreichen
Genen (1–2.000)
in einer einzigen Messung. Das Verfahren hat auch die Fähigkeit,
parallel durchgeführt
zu werden mit mehr als 100 Proben pro Verfahren. Das Verfahren ist
für Wirkstoffscreening,
Entwicklungsbiologie, molekulare medizinische Studien und ähnliches
anwendbar. Innerhalb eines Aspekts der Erfindung werden Verfahren
zur Verfügung
gestellt zur Analyse der Muster von Genexpression aus einer ausgewählten biologischen
Probe, umfassend die Schritte von (a) Exponieren von Nukleinsäuren aus
einer biologischen Probe, (b) Kombinieren der exponierten Nukleinsäuren mit
einer oder mehrerer ausgewählten
markierten Nukleinsonden unter Bedingungen und für eine Zeitdauer,, die ausreichend
ist, für
die Sonden, an den Nukleinsäuren
zu hybridisieren, wobei der Marker mit einer bestimmten Nukleinsäuresonde
korrelativ ist und nachweisbar durch nicht-fluoreszente Spektrometrie
oder Potentiometrie, (c) Abtrennen von hybridisierten Sonden von
nicht-hybridisierten
Sonden, (d) Abspalten des Markers von dem markiertem Fragment, (e)
Nachweisen des Markers durch nicht-fluoreszente Spektrometrie oder
Potentiometrie und Nachweisen des Muster der Genexpression der biologischen
Probe daraus.
-
Innerhalb einer besonders bevorzugten
Ausführungsform
der Erfindung werden Tests oder Verfahren zur Verfügung gestellt,
die wie folgt beschrieben werden: RNA aus einer Zielquelle wird
an einen festen Träger durch
einen spezifischen Hybridisierungsschritt gebunden (d. h. Fangen
von poly(A) mRNA durch eine gebundene Oligo(dT)-Fangsonde). Der
feste Träger
wird dann gewaschen und die cDNA wird auf dem festen Träger unter
der Verwendung von Standardverfahren (d. h. reverser Transkriptase)
synthetisiert. Der RNA-Strang
wird dann über
Hydrolyse entfernt. Das Ergebnis ist die Erzeugung einer DNA-Population, die kovalent
an den festen Träger
immobilisiert ist, die die Diversitäts-Häufigkeit und -komplexität der RNA
widerspiegelt, aus der die cDNA synthetisiert wurde. Der feste Träger wird
dann mit 1 bis mehrere tausend Sonden untersucht (hybridisiert),
die komplementär
zu einer Gensequenz von Interesse sind. Jeder Sondentyp ist mit
einem abspaltbaren Massenspektrometriemarker oder anderem Typ von
abspaltbarem Marker markiert. Nach dem Untersuchungsschritt wird überschüssige oder
nicht hybridisierte Sonde weggewaschen, der feste Träger wird,
zum Beispiel, in die Vertiefung einer Mikrotiterplatte plaziert
und der Massenspektrometriemarker von dem festen Träger abgespalten.
Der feste Träger
wird aus der Vertiefung des Probenbehälters entfernt und die Inhalte
der Vertiefung werden mit einem Massenspektrometer gemessen. Das
Auftreten von spezifschen Massenspektrometermarkern zeigt die Anwesenheit
von RNA in der Probe an und weist nach, daß ein spezifisches Gen in einer
bestimmten biologischen Probe exprimiert wird. Das Verfahren kann
auch quantifizierbar sein.
-
Die Zusammensetzung und Verfahren
zur schnellen Messung von Genexpression und der Verwendung von spaltbaren
Markern können
genau wie folgt beschrieben werden. Kurz gesagt, kann Gewebe (Leber, Muskel,
usw.) primäre
oder transformierte Zellinien, isolierte oder gereinigte Zelltypen
oder jede andere Quelle von biologischem Material, in der die Bestimmung
von genetischer Expression brauchbar ist, als eine Quelle von RNA
verwendet werden. In den bevorzugten Verfahren wird das biologische
Quellenmaterial in der Anwesenheit eines Chaotrops lysiert, um Nukleasen
und Proteasen zu unterdrücken
und die stringente Hybridisierung von Zielnukleinsäure an den
festen Träger
zu unterstützen.
Gewebe, Zellen und biologische Quellen können effektiv in 1 bis 6 molar
chaotropischen Salzen (Guanidinhydrochlorid, Guanidinthocyanat,
Natriumperchlorat, usw.) lysiert werden. Nachdem die quellen-biologische
Probe lysiert ist, wird die Lösung
mit einem festen Träger
gemischt, um ein Fange von Zielnukleinsäure zu bewirken, die in dem
Lysat vorhanden ist. In einer Permutation des Verfahrens wird RNA
unter der Verwendung einer gebundenen Oligo(dT)-Fangprobe gefangen.
Feste Träger
können
Nylonperlen, Polystyrolmikroperlen, Glasperlen und Glasoberflächen oder
jeden anderen Typ von festem Träger
einschließen,
an den Oligonukleotide kovalent angebracht werden können. Die festen
Träger
sind bevorzugterweise mit einem Amin-Polymer, wie zum Beispiel Polyethylen(imin),
Acrylamid, Amin-Dendrimeren, usw. beschichtet. Die Amine der Polymere
werden dazu verwendet, um kovalent Oligonukleotide zu immobilisieren.
Die Oligonukleotide werden bevorzugterweise mit einem 5'-Amin synthetisiert
(im allgemeinen ein Hexylamin, das einen 6-Kohlenstoff-Spacer-Arm und ein
distales Amin einschließt).
Die Oligonukleotide können
15 bis 50 Nukleotide in der Länge
betragen. Die Oligonukleotide werden durch homo-bifunktionelle oder
hetero-bifunktionelle kreuz-vernetzende Reagenzien, wie zum Beispiel
Cyanurchlorid, aktiviert. Die aktivierten Oligonukleotide werden
dann von einem überschüssigen kreuzvernetzendem
Reagenz (z. B. Cyanurchlorid) durch Ausschlußchromatographie gereinigt.
Das aktivierte Oligonukleotid wird dann mit den festen Trägern gemischt,
um eine kovalente Anbindung zu bewirken. Nach der kovalenten Anbringung
der Oligonukleotide werden die nicht reagierten Amine des festen
Trägers
maskiert (d. h. mit Succinianhydrid), um die positive Ladung auf
dem festen Träger
zu eliminieren. Die festen Träger
können
parallel verwendet werden und sind bevorzugterweise in einem 96-Vertiefungen-
oder 384-Vertiefungen-Format
konfiguriert. Die festen Träger
können
an Zapfen, Stämmchen
oder Stäben
in einer 96-Vertiefungen- oder 384-Vertiefungen-Konfiguration angebracht
werden, wobei die festen Träger
entweder ablösbar
oder, alternativ, integral mit der besonderen Konfiguration sind.
Die besondere Konfiguration des festen Träger ist für die Funktion des Assays nicht von
kritischer Wichtigkeit, sondern beeinflußt eher die Fähigkeit
des Tests, an eine Automatisierung angepaßt zu werden. Die festen Träger werden
mit dem Lysat für
15 Minuten bis mehrere Stunden gemischt, um ein Fangen der Zielnukleinsäure auf
dem festen Träger
zu bewirken. Im allgemeinen geschieht das "Fangen" der Zielnukleinsäure durch komplementäre Basenpaarung
von Ziel-RNA und der auf dem festen Träger immobilisierten Fangsonde.
Eine Permutation verwendet den 3'-Poly(A)-Schwanz,
der an den meisten eukaryontischen Messenger-RNAs gefunden wird,
um an ein immobilisiertes Oligo(dT) auf dem festen Träger zu hybridisieren. Eine
andere Permutation ist es, ein spezifisches Oligonukleotid zu verwenden
oder lange Sonden (länger
als 50 Basen), um eine RNA zu fangen, die eine definierte Sequenz
enthält.
Eine andere Möglichkeit
ist es, degenerierte Primer (Oligonukleotide) zu verwenden, was
das Fangen von zahlreichen verwandten Sequenzen in der Ziel-RNA-Population
bewirken würde.
Die Sequenzkomplexität
der RNA-Population
und der Typ von verwendeter Fangsonde bestimmen die Hybridisierungszeiten.
Die Hybridisierungstemperaturen werden durch den Typ von verwendetem
Chaotrop und der finalen Konzentration von Chaotrop diktiert (siehe
Van Ness und Chen, Nuc. Acids Res. 1991, für allgemeine Anweisungen).
Das Lysat wird bevorzugterweise kontinuierlich mit dem festen Träger bewegt,
um eine Diffusion der Ziel-RNA zu bewirken. Sobald der Schritt des
Fangens der Zielnukleinsäure
vervollständigt
ist, wird das Lysat vom festen Träger gewaschen und alle Chaotrope
oder Hybridisierungslösung
wird entfernt. Der feste Träger
wird bevorzugterweise mit Lösungen
gewaschen, die ionische oder nicht-ionische Detergenzien, Puffer
und Salze enthalten. Der nächste
Schritt ist die Synthese von DNA, die komplementär zu der gefangenen RNA ist.
In diesem Schritt dient das befestigte Fangoligonukleotid als der
Verlängerungsprimer
für die
reverse Transkriptase. Die Reaktion wird im allgemeinen bei 25–37°C durchgeführt und
im allgemeinen während
der Polymerisierungsreaktion geschüttelt. Nachdem die cDNA synthetisiert
ist, wird sie kovalent an den festen Träger angebracht, da das Fangoligonukleotid
als der Verlängerungsprimer
dient. Die RNA wird dann aus der cDNA-/RNA-Duplex hydrolysiert.
Der Schritt kann durch die Verwendung von Hitze durchgeführt werden,
der die Duplex denaturiert oder der Verwendung von Base (d. h. 0,1 N
NaOH), um die RNA chemisch zu hydrolysieren. Das Schlüsselergebnis
an diesem Schritt ist es, die cDNA für die anschließende Hybridisierung
mit definierten Sonden erhältlich
zu machen. Der feste Träger
oder das Set von festen Trägern
wird dann weiter gewaschen, um RNA oder RNA-Fragmente zu entfernen.
An diesem Punkt enthält
der feste Träger
eine ungefähre
repräsentative
Population von cDNA-Molekülen,
die die RNA-Population im Hinblick auf Sequenzhäufigkeit, -komplexität und -diversität widerspiegeln.
Der nächste Schritt
ist es, ausgewählte
Sonden an den festen Träger
zu hybridisieren, um die Anwesenheit oder Abwesenheit und die relative
Häufigkeit
von spezifischen cDNA-Sequenzen zu identifizieren. Die Sonden sind
bevorzugterweise Oligonukleotide mit einer Länge von 15–50 Nukleotiden. Die Sequenz
der Sonden wird durch die Endverwender des Assays bestimmt. Zum
Beispiel, falls der Endverwender beabsichtigt, eine Genexpression in
einer entzündlichen
Antwort in einem Gewebe zu untersuchen, würden die Sonden komplementär zu zahlreichen
Cytokin mRNAs, RNAs, die für
Enzyme kodieren, die Lipide modulieren, RNAs, die für Faktoren
kodieren, die Zellen regulieren, die an einer entzündlichen
Antwort beteiligt sind, usw., ausgewählt werden. Sobald ein Set
von definierten Sequenzen für
die Untersuchung definiert ist, wird jede Sequenz in einer Oligonukleotidsonde
hergestellt und jeder Sonde ein spezifischer spaltbarer Marker zugeordnet.
Der Marker/die Marker werden dann an die jeweiligen Oligonukleotid(e)
angebracht. Das Oligonukleotid/die Oligonukleotide werden an die
cDNA auf dem festen Träger
unter geeigneten Hybridisierungsbedingungen hybridisiert. Nach Vervollständigung
des Hybridisierungsschrittes wird der feste Träger gewaschen, um jede nicht-hybridisierte Sonde
zu entfernen. Der feste Träger
oder das Array von Trägern
wird dann erhitzt, um die kovalente Bindung zwischen der cDNA und
dem festen Träger
zu spalten. Die markierten cDNA-Fragmente werden dann gemäß ihrer
Größe durch
Gelelektrophorese getrennt (z. B. Polyacrylamidgelelektrophorese)
oder bevorzugterweise HPLC. Die Marker werden dann von den DNA-Molekülsonden
abgespalten und durch die jeweilige Nachweistechnologie (z. B. Massenspektrometrie,
Infrarotspektrometrie, potentiostatische Amperometrie oder UV/sichtbare
Spektrometrie) nachgewiesen. Jeder vorhandene Marker wird identifiziert
und die Anwesenheit (und Häufigkeit)
oder Anwesenheit einer exprimierten mRNA wird nachgewiesen.
-
Ein alternatives Verfahren würde die
markierten DNA-Sonden direkt an die befestigten mRNA-Zielmoleküle unter
den geeigneten Hybridisierungsbedingungen hybridisieren. Nach Vervollständigung
des Hybridisierungsschrittes wird der feste Träger gewaschen, um jede nicht
hybridisierte Probe zu entfernen. Die RNA wird dann aus der DNA-Sonden/RNA-Duplex hydrolysiert.
Der Schritt kann durch die Verwendung von Hitze erfolgen, die die
Duplex denaturiert oder die Verwendung von Base (d. h. 0,1 N NaOH),
um die RNA chemisch zu hydrolysieren. Dieser Schritt wird freie
mRNA und ihre entsprechenden DNA-Sonden zurücklassen, die dann durch einen
Größenabtrennschritt
isoliert werden können,
der im allgemeinen aus Gelelektrophorese besteht (z. B. Polyacrylamidgelelektrophorese)
oder bevorzugterweise HPLC. Die Marker werden dann von den DNA-Molekülsonden
abgespalten und durch die jeweilige Nachweistechnologie (z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV/sichtbare Spektrophotometrie)
nachgewiesen. Jeder vorhandene Marker wird identifiziert und die
Anwesenheit (und Häufigkeit)
oder Abwesenheit einer exprimierten mRNA wird bestimmt.
-
Ein bevorzugter Genexpressionstests
der vorliegenden Erfindung verwendet markierte Oligonukleotide in
Verbindung mit PCR (oder anderen gleichfalls effektiven Techniken),
Lambda-Exonuklease, Ultrafiltration und einem internen Standard
um quantitative Information über
die Genexpression zu erhalten. Dieser bevorzugte Genexpressionsassay
und Unterverfahren davon werden als nächstes beschrieben.
-
a. BESCHREIBUNG DES INTERNEN
STANDARDVERFAHRENS
-
Die PCR ist ein hoch sensitives Verfahren
zum Nachweis von kleinen Mengen von DNA oder RNA (durch RT-PCR).
Jedoch ist die genaue und präzise
Quantifizierung des Ziels schwierig, da die Menge von amplifiziertem
Produkt nicht immer proportional zu der Menge von Templat ist. Dies
liegt daran, daß PCR
eine "Plateau-Phase" erreicht, in der
fast die gesamte Menge von amplifiziertem Produkt erhalten wird,
unabhängig von
der Menge von Templat, nach einer bestimmten Zahl von Zyklen. Die
Verwendung eines internen Standards während der PCR hilft dabei,
dieses Problem durch Co-Amplifikation von bekannten Mengen eines
internen Standardtemplats zu überwinden,
wobei das selbe Primerset wie das Zieltemplat verwendet wird. Da beide
Template das selbe Set von Primern verwenden, spiegelt das Verhältnis zwischen
den Mengen der zwei amplifizierten Produkte das anfängliche
Verhältnis
zwischen der Menge von Ziel- und dem internen Standardtemplat vor
der PCR-Amplifikation wider. Die Menge von Zieltemplat kann dann
aus der bekannten Menge des internen Standardtemplats berechnet
werden. Das bevorzugte interne Standardtemplat für einen PCR-Test wird sich mit einer identischen
Effizienz im Vergleich zu dem Zieltemplat amplifizieren lassen.
Identische Primerstellen werden in das interne Standardtemplat eingebaut,
um eine Co-Amplifikation sicherzustellen. Die Region zwischen den
Primerstellen und dem internen Standardtemplat sind typischerweise
von dem Zieltemplat verändert
(z. B. Deletion oder Addition von 10–20 Basenpaaren) um die Template
durch Gelelektrophorese oder durch Restriktionsenzyme unterscheidbar
zu machen. Modifikationen dieses Typs verursachen jedoch Unterschiede
in der Amplifikationseffizienz der Template. Typischerweise wird
eine Zahl von internen Standardtemplaten hergestellt und getestet,
bis eine nahezu identische Amplifikation gefunden wird.
-
Ein internes Standardassayformat
wird unter der Verwendung der CMST-Marker und Hybotropen entwickelt.
Hybotrope sind beschrieben in, z. B., U.S.-Anmeldung Nrn. 60/026,621
(angemeldet 24. September 1996); 08/719,132 (angemeldet 24. September
1996); 08/933,924 (angemeldet 23. September 1997); 09/002,051 (angemeldet
31. Dezember 1997); und Internationale Publikation Nr. WO 98/13527,
die alle hier in ihrer Gesamtheit mit aufgenommen sind. Grundsätzlich betrifft
Hybotrope jede chemische Substanz, die die Enthalpie einer Nukleinsäureduplex
um 20% oder mehr erhöhen
kann, wenn mit einer Standardsalzlösung (d. h. 0,165 M NaCl) verglichen.
Eine chemische Substanz zeigt hybotrope Eigenschaften, wenn als
eine Lösung einer
18 bp Oligonukleotid-Duplex, die 50% G + C ist, eine helikale-coil
Transition (HCT) von 15°C
oder weniger aufweist. Die HCT ist der Unterschied zwischen den
Temperaturen, bei denen 80% und 20% der Duplex einzelsträngig sind.
Die Temperatur zur Anlagerung wird dann als diejenige Unterscheidungstemperatur
gewählt,
die eine Temperatur ist, bei der eine Hybridisierungsreaktion durchgeführt wird,
die eine nachweisbare Unterscheidung zwischen einer fehlgepaarten
Duplex und einer perfekt gepaarten Duplex ermöglicht. Ein Bereich von Temperaturen
erfüllt
diese Kriterien einer Unterscheidungstemperatur.
-
Da hoch spezifische Hybridisierung.
unter der Verwendung von Hybotropen durchgeführt werden kann, kann ein bevorzugtes
internes Standardtemplat verwendet werden, das dieselbe Lange aufweist
und nur ein Basenpaar unterschiedlich von dem Zieltemplat ist. Dies
ist ein bevorzugter interner Standard, da er a priori identisch
mit dem Zieltemplat co-amplifizieren wird. Ein bevorzugtes Verfahren
des Nachweises dieses Typs von Standard und Zielamplikongemisch
ist es, hybotrope Puffer zu verwenden, um ein spezifische Hybridisierung
von Amplikon-spezifisch markierten Oligonukleotiden (die bevorzugterweise
CMST-markiert sind, d. h. durch Massenspektrometerie nachweisbare
Marker aufweisen), an deren jeweilige Amplikons, um die Menge von
Amplikon nachzuweisen, wird es Amplikonspezifische Oligonukleotid
mit einem einmaligen Massenmarker markiert. Die Amplikon-Quantifizierung wird
durch Messen des Massenspektrometriesignals abgeleitet, das mit
dem internen Standard und den Zielamplikons assoziiert ist. Eine
bekannte Menge von internem Standard wird in eine Probe gegeben,
und die Menge von Zielamplikon wird unter der Verwendung der folgenden
Gleichung und der Verwendung der Daten bestimmt, die im Anschluß an die
Lambda-Exonuklease und Ultrafiltrationsverfahren erhalten werden,
die als nächstes
beschrieben werden.
-
ZielMassensignal/interner StandardMassensignal) × interner
StandardMenge = ZielMenge
-
b. BESCHREIBUNG DES LAMBDA
EXONUKLEASE VERFAHRENS
-
Nach Inkubation mit einer DNA-Duplex
verdaut Lambda-Exonuklease selektiv einen Strang von einem 5'-phosphoryliertem
Ende her, was ein für
DNA-Sequenzierung geeignetes einzelsträngiges Templat zurückläßt. Die
Lambda-Exonuklease stellt eine einzelsträngige Sequenzierungstemplate
her, ohne die Mühe
von herkömmlichen
biologischen Verfahren oder der Lästigkeit der Optimierung von
asymmetrischer PCR. Durch Amplifikation von DNA in der Anwesenheit
eines Primers, der ein 5'-terminales
Phosphat enthält,
kann ein Fachmann einer DNA-Duplex mit einem 5'-phophoryliertem Ende erzeugen. Danach
wird das PCR-Produkt entweder
durch Fällung
oder Gelfiltration gereinigt, um restliche Primer und andere Reaktionskomponenten
zu entfernen. Der phosphorylierte Strang der DNA-Duplex wird dann
selektiv mit Lambda-Exonuklease verdaut, was ein einzelsträngiges nicht-phosphoryliertes
Templat zurückläßt, das
für die
Sequenzierung geeignet ist. Nach der Hitzeinaktivierung der Lambda-Endonuklease
kann die konzentrierte, einzelsträngige DNA direkt zu einer Hybridierungsreaktion
hinzugefügt
werden, die herkömmliche
Techniken anwendet.
-
c. BESCHREIBUNG DES ULTRAFILTRATIONSVERFAHRENS
-
Genomische DNA, wie aus weißen Blutzellen
geerntet, kann gemäß den Standardverfahren
von RNase-Behandlung, Proteinase-K-Verdau und Phenol : Chloroform-Extraktion,
gefolgt durch Fällung
mit Natriumacetat und Ethanol, gereinigt werden. Die Konzentrationen
werden durch Spektrophotometrie bestimmt und Arbeitsverdünnungen
von 0,01 μg/μl werden
hergestellt. 50 μl
der DNA-Proben werden in 96-Vertiefungs-"Mutter"-Platten ausgebracht. "Tochter"-Platten, die für die Amplifikationsreaktion
verwendet werden, werden durch transferieren von 1,5 μl von DNA
aus der Mutterplatte aus den Vertiefungen von entweder einer 96-Vertiefungsplatte
oder 192-Vertiefungsplatte in einer identischen Konfiguration unter
der Verwendung von 8-Kanal-Pipetten präpariert. Eine 18 μl Lage von
flüssigem
Wachs (MJ Research) wird dann hinzugefügt und dann wird diese Platte
bei 4 Grad gelagert, wobei sich das Wachs verfestigt, was eine Verdampfung
verhindert.
-
Um die PCR anzusetzen, werden die
Tochterplatten entfernt und auf Eis plaziert, um das Wachs festzuhalten.
Dieses bildet eine Barriere zwischen der Templat-DNA und anderen
Komponenten der Reaktion, bis die Platte in dem Thermal-Cycler plaziert
wird und die Reaktion durch Erhitzen während des ersten Zyklus initiiert
wird. Die PCR wird in einer Gesamtzahl von 10–50 μl pro Reaktion durchgeführt. Mastermixlösungen werden
vorher in Vorratslösungen
hergestellt und in Röhrchen
aufgeteilt, die alle der Komponenten der PCR, außer den Marker-spezifischen
Primern enthalten. Die Hauptzahl der Reaktionen werden mit M13-verlängerten Primern
durchgeführt.
M13-verlängerte
Primer sind eine Modifikation des Standard-PCR-Primerpaars. Die
Modifikation ist die Hinzufügung
von 17 Nukleotiden an das 5'-Ende
des Vorwärtsprimers.
Die 17-Nukleotidsequenz ist komplementär zu dem m13-Sequenzierungsprimer
und besitzt die Sequenz: 5'-(NH2)-AGG GTT TTCCCA GTC ACG AC-3'. Die Modifikation
erlaubt die Verwendung eines dritten Oligonukleotidprimers in einer PCR-Reaktion.
Der dritte Primer ist typischerweise gemäß den hier beschriebenen Verfahren
markiert.
-
Ultrafiltration ist das herkömmliche
Verfahren zur Konzentration und Entsalzung von Proteinen. Es ist auch
ein effiziente Alternative zur Ethanolfällung von Nukleinsäuren, insbesondere
für kleine
Mengen und besonders, wenn immer Nukleinsäuren nur gefällt werden,
um Lösungsmittel
auszuwechseln. Für
Proben, die Phosphat oder 10 mM EDTA enthalten, kann die Ultrafiltration
eine bemerkenswert zeitsparende Methodologie sein. Herkömmlich erforderten
solche Proben die vorhergehende Dialyse, um die Co-Fällung von
Salzen mit Nukleinsäuren
während
der Ethanolfällung
zu vermeiden. Zentrifugal- Mikrokonzentratoren
entsalzen und konzentrieren Oligonukleotidsonden oder einzelsträngige Amplikons
(und Nukleinsäuren)
in einem einfachen Schritt.
-
Microcon® Mikrokonzentratoren sind ideal
zur Konzentration von 50–500 μl Proben.
Bei der zentrifugalen Ultrafiltration wird die DNA durch die Membran
zurückgehalten.
Lösungsmittel
und Salze passieren und werden entfernt. Eine zweite umgedrehte
Zentrifugation von Microcon stellt einen maximalen DNA-Rückerhalt der
Sonde oder Nukleinsäure
von Interesse sicher. Bei der Konzentration der Oligomerproben ist
es wichtig, hohe Salzkonzentrationen zu vermeiden, die eine Bindung
von einzelsträngigen
Nukleinsäure
an Zellulose-basierte Ultrafiltrationsmembranen unterstützen.
-
Typische Ultrafiltrationsbedingungen
sind wie folgt: eine DNA-Lösung
(500 μl)
wird einem Microcon-30-Konzentrator für 10 Minuten bei 12.000 × g zentrifugiert;
eine 500 μl
Lösung
von Oligonukleotiden in TE-Puffer wird in einer Microcon-3-Einheit
für 45
Minuten bei 12.000 × g
zentrifugiert. Die Überstände können durch
Umdrehen der Vorrichtungen und Zentrifugieren bei 500–1.000 × g für 2 Minuten
zurückerhalten
werden. Im Gegensatz dazu erfordert Gelelektrophorese Fraktionierung,
Elution und Entsalzung des Fragments aus der Gelscheibe. Sie erfordert
auch genügend
Material, um zu visualisieren, was manchmal schwierig zu erhalten
ist, insbesondere im Fall von cDNA. Beide Verfahren (Gelelektrophorese
und Dialyse) sind zeitaufwendig und schließen viele Probenverarbeitungsschritte
ein.
-
In einer typischen Hybridisierungsreaktion
werden die Sonden in 50–100-fachen
molaren Überschoß von DNA-Fragmentkonzentration
hinzugefügt.
Aus diesem Grund ist es erforderlich, die überschüssige nicht-hybridisierte Sonde
zu entfernen. Herkömmliche
Verfahren zur Sondenentfernung schließen Gelfiltrationchromatographie
oder Gelelektrophorese ein. Die Gelfiltration erfordert, daß viele
Fraktionen gesammelt, analysiert, vereinigt und gefällt werden,
was für
Hochdurchsatzassays nicht anwendbar ist. Jedoch ist die Ultrafiltration
eine effektive Alternative für
ein schnelles Entfernen von überschüssiger Sonde
von PCR-Primern. Bei Amicon's
Centricon wegwerfbarem Konzentrator, würde das Reaktionsgemisch durch
ein Ultrafiltrationsmembran filtriert, was zu der Entfernung von
Puffer und nicht-hybridisierter Sonde oder nicht-verlängerten
Primern führt.
Die konzentrierten Fragmente werden durch die Membran zurückgehalten.
Antreibende Kraft für die
Filtration wird durch die Zentrifugation in einem Rotor mit fixiertem
Winkel bei 1.000-5.000
g zur Verfügung gestellt.
Herkömmliche
Verfahren erfordern typischerweise 24 Stunden für die Probenverarbeitung und
2–3 Stunden
Arbeitszeit. Bei Centricon benötigen
die Proben typischerweise weniger als ein paar Stunden, um diese
zu reinigen. Die Probenhandhabung ist minimal und viele Proben können zur
selben Zeit verarbeitet werden. Als Referenz siehe Krowczynska,
A. M., "Efficient
Purification of PCR Products using Ultrafiltration" BioTechniques 13(2):
286–289,
1992.
-
12. Hybridisierungstechniken
-
Die erfolgreiche Klonierung und Sequenzierung
eines Gens führt
zu der Untersuchung seiner Struktur und der Expression dadurch,
daß es
möglich
gemacht wird, das Gen nachzuweisen oder seine mRNA in einem großen Pool
von nicht-verwandten DNA- oder RNA-Molekülen. Die Menge von mRNA; die
für ein
spezifisches Protein in Gewebe kodiert, ist ein wichtiger Parameter
für die
Aktivität
eines Gens und kann signifikant mit der Aktivität von funktionierenden Systemen
zusammenhängen.
Seine Regulation ist abhängig
von der Interaktion von Sequenzen innerhalb des Gens (cis-wirkende
Elemente) und Sequenz-spezifischen
DNA-bindenden Proteinen (traps-wirkenden Faktoren), die Gewebe-spezifisch
aktiviert werden, oder durch Hormone und Second Messenger Systeme.
-
Verschiedene Techniken zur Analyse
eines bestimmten Gens, seine regulatorischen Sequenzen, seiner spezifischen
mRNA und der Regulation seiner Expression sind erhältlich, diese
schließen
Southern- oder Northern-Blot-Analyse und Ribonuklease-(Rnase)-Protection-Assay ein.
-
Veränderungen in der Nukleotidzusammensetzung
eines bestimmten Gens können
von großer
pathophysiologischer Relevanz sein. Wenn in den nicht-kodierenden
Regionen (5'-, 3'-flankierenden Regionen und Intron) lokalisiert,
können
sie die Regulation der Genexpressioin beeinflussen, was abnormale
Aktivierung oder Inhibierung verursacht. Wenn in den kodierenden
Regionen des Gens (Exone) lokalisiert, können sie zu Veränderung
der Proteinfunktion oder disfunktionellen Proteinen führen. Daher
kann eine bestimmte Sequenz innerhalb eines Gens mit einer spezifischen
Erkrankung korrelieren und kann als ein Marker der Erkrankung brauchbar
sein. Ein primäres
Ziel der Forschung im medizinischen Bereich ist es daher, diejenigen
genetischen Variationen als diagnostische Tools nachzuweisen und
wichtige Informationen für
das Verständnis
von pathophysiologischen Phänomenen
zu erhalten. Das Basisverfahren für die Analyse einer Population
im Hinblick auf die Variationen innerhalb eines bestimmten Gens
ist DNA-Analyse unter der Verwendung der Southern-Blot-Technik.
Kurz gesagt, wird präparierte
genomische DNA mit einem Restriktionsenzym (RE) verdaut, was zu
einer großen
Zahl von DNA-Fragmenten von verschiedenen Längen führt, die durch die Anwesenheit
der spezifischen Erkennungsstelle des RE im Genom bestimmt werden.
Allele eines bestimmten Gens mit Mutationen innerhalb dieser Restriktionsstelle
werden in Fragmente verschiedener Zahl und Länge gespalten werden. Dies
wird Restriktionslängenpolymorphismus
(RFLP) genannt und kann ein wichtiger diagnostischer Marker mit
vielen Anwendungen sein. Das zu analysierende Fragment muß aus dem
Pool von DNA-Fragmenten abgetrennt werden und von anderen DNA-Spezies
unter der Verwendung einer spezifischen Sonde unterschieden werden.
Daher wird die DNA einer elektrophoretischen Fraktionierung unter
Verwendung eines Agarosegels unterzogen, gefolgt von Transfer und
Fixierung an eine Nylon- oder Nitrozellulosemembran. Die fixierte,
Einzelsträngige
DNA wird an eine markierte DNA hybridisiert, die komplementär zu der
nachzuweisenden DNA ist. Nach Entfernen von nicht-spezifischen Hybridisierungen
kann das DNA-Fragment von Interesse gemäß den Sondencharakteristika
(Autoradiagraphie oder Phosphorimageanalyse) visualisiert werden.
-
Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zur Bestimmung der Identität eines Nukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Nukleinsäuremolekül in, zum
Beispiel, einer biologischen Probe zur Verfügung gestellt, wobei die Techniken ähnlich zu
Southern-Blotting verwendet werden. Kurz gesagt umfassen solche
Verfahren im allgemeinen die Schritte von Erzeugen einer Reihe von
markierten Nukleinsäurefragmenten,
in denen die erzeugten Fragmente mit Restriktionsenzymen verdaut
werden. Die markierten Fragmente werden durch Durchführen eines
Hybridisierungsschritts der markierten Sonden mit der verdauten
Zielnukleinsäure
erzeugt. Der Hybridisierungsschritt kann vor oder nach dem Restriktionsnukleaseverdau
erfolgen. Die sich ergebenden verdauten Nukleinsäurefragmente werden dann nach
ihrer Größe getrennt.
Der Größentrennungsschritt
kann dann durch zum Beispiel Gelelektrophorese (z. B. Polyacrylamidgelelektrophorese)
oder, bevorzugterweise HPLC, erfolgen. Die Marker werden dann von
den abgetrennten Fragmenten abgespalten und dann werden die Marker
durch die jeweilige Nachweistechnologie nachgewiesen (z. B. Massenspektrometrie,
Infrarotspektrometrie, potentiostatische Amperometrie oder UV/sichtbare Spektrophotometrie).
-
Die Anwesenheit und Quantifizierung
eines spezifischen Gentranskripts und seine Regulierung durch physiologische
Parameter können
mittels Northern-Blot und RNase-Protection-Assay
analysiert werden. Die prinzipielle Basis dieser Verfahren ist die
Hybridisierung eines Pools von gesamter zellulären DNA an eine spezifische
Sonde. Bei der Northern-Blot-Technik wird Gesamt-RNA eines Gewebes
unter der Verwendung eines HPLC- oder LC-Verfahrens fraktioniert,
an eine markierte Antisense-RNA (cRNA) hybridisiert, die komplementär zu der
nachzuweisenden RNA ist. Durch Anwendung von stringenten Waschbedingungen
werden nicht-spezifisch gebundene Moleküle eliminiert. Spezifisch gebundene
Moleküle
würden
anschließend
gemäß des Typs
von verwendeter Sonde (Massenspektrometrie oder mit einem elektrochemischen
Detektor) nachgewiesen. Zusätzlich
kann die Spezifität
durch Vergleichen der Größe der nachgewiesenen
mRNA und der vorhergesagten Länge
der mRNA von Interesse kontrolliert werden.
-
Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zum Nachweis der Identität eines Ribonukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Ribonukleinsäuremoleküls in, zum
Beispiel, einer biologischen Probe zur Verfügung gestellt, wobei die Techniken ähnlich zu
Northern-Blotting verwendet werden. Kurz gesagt umfassen solche
Verfahren im allgemeinen die Schritte von Erzeugen einer Reihe von
markierten RNA-Molekülen durch
Durchführen
eines Hybridisierungsschrittes der markierten Sonden mit der Ziel-RNA.
Die Ziel-RNA-Moleküle
werden dann nach ihrer Größe aufgetrennt.
Der Größenauftrennungsschritt
kann dann, zum Beispiel, bevorzugterweise, HPLC erreicht werden.
Die Marker werden dann von den getrennten RNA-Molekülen abgespalten
und dann werden die Marker durch die jeweilige Nachweistechnologie
(z. B. Massenspektrometrie, Infrarotspektrometrie, potentiostatische
Amperometrie oder UV/sichtbare Spektrophotometrie) nachgewiesen.
-
Das meisten spezifische Verfahren
zum Nachweis einer mRNA-Spezies ist der RNase-Protection-Assay. Kurz gesagt, wird
Gesamt-RNA aus einem Gewebe oder einer Zellkultur an eine markierte
spezifische mRNA von vollständiger
Homologie hybridisiert. Die Spezifität wird durch anschließenden RNase-Verdau
erreicht. Nicht-hybridisierte Einzel-strängige RNAs und nicht-spezifisch
hybridisierte Fragmente mit sogar kleinen Fehlpaarungen werden erkannt
und gespalten, während
doppelsträngige
RNA von vollständiger
Homologie nicht für
das Enzym zugänglich
ist, und geschützt
werden wird. Nach Entfernen der RNase durch Proteinase-K-Verdau
und Phenol-Extraktion kann das spezifisch geschützte Fragment von den Abbauprodukten
abgetrennt werden, gewöhnlicherweise
durch HPLC.
-
Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zur Bestimmung der Identität eines Ribonukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Ribonukleinsäuremolekül in, zum
Beispiel, einer biologischen Probe zur Verfügung gestellt, wobei die Technik
des RNase-Protection-Assays verwendet wird. Kurz gesagt umfassen
solche Verfahren im allgemeinen die Schritte von Gesamt-RNA aus
einem Gewebe oder einer Zellkultur, die an eine markierte spezifische
cRNA von vollständiger
Homologie hybridisiert wird, einen RNase-Verdau, Behandlung mit
Proteinase K und eine Phenol-Extraktion. Das markierte geschützte RNA-Fragment
wird aus den Abbauprodukten isoliert. Der Größenabtrennschritt kann zum
Beispiel durch LC oder HPLC erreicht werden. Der Marker wird dann
von dem abgetrennten RNA-Molekül
abgespalten und wird dann durch die jeweilige Nachweistechnologie
(z. B. Massenspektrometrie, Infrarotspektrometrie, potentiostatische
Amperometrie oder UV/sichtbare Spektrophotometrie) nachgewiesen.
-
13. Mutationsnachweistechniken
-
Der Nachweis von Erkrankungen ist
bei der Vorbeugung und Behandlungen zunehmend von Wichtigkeit. Während es
für multifaktorielle
Erkrankungen schwierig ist genetische Tests dafür zu entwickeln, werden mehr
als 200 bekannte menschliche Erkrankungen durch einen Defekt in
einem einzelnen Gen, oft eine Veränderung eines einzelnen Aminosäurerests
(Olsen, Biotechnology: An industry comes of age, National Academic
Press, 1996) verursacht. Viele dieser Mutationen führen zu
einer veränderten
Aminosäure,
die einen Erkrankungszustand verursacht.
-
Sensitive Mutationsnachweistechniken
bieten außerordentliche
Möglichkeiten
für Mutationsscreening. Zum
Beispiel, können
Analysen durchgeführt
werden sogar vor der Implantation eines befruchteten Eis (Holding
und Monk, Lacet 3: 532, 19889). Erhöhte effiziente genetische Tests
können
auch ein Screening auf onkogene Mutationen in Zellen ennöglichen,
die vom respiratorischen Trakt oder der Blase abgestoßen wurden, in
Verbindung mit Vorsorgeuntersuchungen (Sidransky et al., Science
252: 706). Auch sind, wenn ein unbekanntes Gen eine genetische Erkrankung
verwsacht, Verfahren zur Überwachung
von DNA-Sequenzvarianten brauchbar, um die Vererbung von Erkrankung
durch genetische Kopplungsanalyse zu untersuchen. Jedoch verursacht
der Nachweis und die Diagnose von Mutationen in einzelnen Genen
technische und ökonomische Herausforderungen.
Mehrere verschiedene Ansätze
wurden verfolgt, jedoch ist keiner sowohl effizient und kostengünstig genug
für eine
wirklich weit verbreitete Anwendung.
-
Mutationen, die ein einzelnes Nukleotid
einschließen,
können
in einer Probe durch physikalische, chemische oder enzymatische
Mittel identifiziert werden. Im allgemeinen können Verfahren zum Mutationsnachweis
in Scan-Techniken eingeteilt werden, die geeignet sind, um vorher
unbekannte Mutationen zu identifizieren und Techniken, die so aufgebaut
sind, um bekannte Sequenzvarianten nachzuweisen, zu unterscheiden oder
zu quantifizieren. Verschiedenen Scan-Techniken für den Mutationsnachweis
wurden in Heteroduplices von fehlgepaarten komplementären DNA-Strängen, die
von Wildtyp- und Mutantensequenzen abgeleitet waren und ein abnormales
Verhalten, insbesondere wenn denaturiert, zeigten, entwickelt. Diese
Phänomen
wird bei der denaturierenden und Temperaturgradient-Gelelektrophorese-(DGGE
und TGGE)-Verfahren ausgenutzt. Duplices, die sogar in einer einzelnen
Nukleotidposition fehlgepaart sind, können teilweise denaturieren, was
zu einer verzögerten
Wanderung führt,
wenn in einem ansteigend denaturierenden Gradienten gelelektrophoriert
(Myers et al., Nature 313: 495, 1985; Abrades et al., Genomics 7:
463, 1990; Henco et al., Nucl. Acids Res. 18: 6733, 1990). Obwohl
Mutationen nachgewiesen werden können,
wird keine Information im Hinblick der präzisen Lokalisierung einer Mutation
erhalten. Die Mutanten-Formen müssen
weiterhin isoliert werden und einer DNA-Sequenzanalyse unterzogen werden. Alternativ
kann RNase A eine Heteroduplex einer RNA-Sonde und eines Zielstranges an einer
Position spalten, wo die zwei Stränge nicht genau gepaart sind. Diese
Spaltstelle kann dann durch Elektrophorese der denaturierten Probe
bestimmt werden. Jedoch können einige
Mutationen dem Nachweis entkommen, da nicht alle Fehlpaarungen durch
RNase A effizient gespalten werden. Fehlgepaarte Basen in einer
Duplex sind ebenfalls gegenüber
chemischer Modifikation empfindlich. Solche Modifikationen können die
Stränge
gegenüber
Spaltung an der Stelle der Fehlpaarung empfindlich machen oder dazu
führen,
daß eine
Polymerase in einer anschließenden
Verlängerungsreaktion
dort stoppt. Die chemische Spalttechnik ermöglicht die Identifizierung
einer Mutation in Zielsequenzen von bis zu 2 kb und sie stellt Informationen über die
ungefähre
Lokalisierung von fehlgepaarten Nukleotiden zur Verfügung (Cotton
et al., PNAS LISA 85: 4397, 1988; Ganguly et al., Nucl. Acids Res.
18: 3933, 1991). Jedoch ist diese Technik arbeitsintensiv und kann
die präzise
Lokalisierung der Mutation vielleicht nicht identifizieren.
-
Eine alternative Strategie zum Nachweis
einer Mutation in einem DNA-Strang ist durch Substituierung (während der
Synthese) eines der normalen Nukleotide mit einem modifizierten
Nukleotid, wobei das Molekulargewicht oder andere physikalische
Parameter des Produkts verändert
werden. Ein Strang mit einer erhöhten oder
verringerten Zahl dieses modifizierten Nukleotids in Bezug auf die
Wildtyp-Sequenz zeigt eine veränderte elektrophoretische
Mobilität
(Naylor et al., Lancet 337: 636, 1991). Diese Technik weist die
Anwesenheit von einer Mutation nach, stellt jedoch nicht die Lokalisierung
zur Verfügung.
-
Zwei andere Strategien visualisieren
Mutationen in einem DNA-Fragment durch veränderte Gelmigration. In der
Einzel-Strang-Polymorphismustechnik (SSCP), verursachen Mutationen
meiner Meinung nach, daß denaturierte
Stränge
verschiedener Sekundärstrukturen
annehmen, wodurch die Mobilität
während
nativer Gelelektrophorese beeinflußt wird. Heteroduplex-DNA-Moleküle, die
interne Fehlpaarungen aufweisen, können auch von korrekt gepaarten
Molekülen
durch Elektrophorese abgetrennt werden (Orita, Genomics 5: 874,
1989; Keen, Trends Genet. 7: 5, 1991). Wie mit den oben diskutierten
Techniken, kann die Anwesenheit einer Mutation bestimmt werden,
jedoch nicht Lokalisierung. Genauso können viele dieser Techniken
nicht zwischen einer einzelnen und multiplen Mutationen unterscheiden.
Alle der oben genannten Techniken zeigen die Anwesenheit einer Mutation
in einem begrenzten Segment von DNA an und einige von ihnen ermöglichen
die ungefähre
Lokalisierung innerhalb des Segments Jedoch ist immer noch Sequenzanalyse
erforderlich, um den Effekt der Mutation auf das Kodierungspotential
des Fragments zu ermitteln. Sequenzanalyse ist sehr wirksam, was
zum Beispiel ein Screening auf die selbe Mutation in anderen Individuen
einer beeinträchtigten
Familie ermöglicht,
wodurch ein Erkrankungsfortschreiten im Falle von maligner Erkrankung
beim Nachweis von restlichen malignen Zellen im Knochenmark vor
autologer Transplantation verfolgt wird. Trotz dieser Vorteile ist
es aufgrund der hohen Aufgaben, die enthalten sind, unwahrscheinlich,
daß das
Verfahren als ein Routine-diagnostisches
Verfahren angenommen werden wird.
-
Eine große Anzahl von anderen Techniken
wurden entwickelt, um bekannte Sequenzvarianten zu analysieren.
Die Automatisierung und Wirtschaftlichkeit sind sehr wichtige Überlegungen
für diese
Typen von Analysen, die für
das Screening von Individuen und der allgemeinen Population angewendet
werden können.
Keine der unten diskutierten Techniken kombiniert Wirtschaftlichkeit
und Automatisierung mit der erforderlichen Spezifität.
-
Mutationen können über ihre destabilisierenden
Effekte auf die Hybridisierung von kurzen Oligonukleotidsonden an
eine Zielsequenz identifiziert werden (siehe Wetmur, Crit. Rev.
Biochem. Mol. Biol. 26: 227, 1991). Im allgemeinen schließt diese
Technik, Allel-spezifische Oligonukleotidhybridisierung, die Amplifikation von
Zielsequenzen und anschließende
Hybridisierung von kurzen Oligonukleotidsonden ein. Ein amplifiziertes Produkt
kann daher auf viele mögliche
Sequenzvarianten durch Bestimmen seines Hybridisierungsmusters an Array
von immobilisierten Oligonukleotidsonden gescannt werden. Jedoch
hängt das
Etablieren von Bedingungen, die zwischen einer Zahl von anderen
Strategien zur Nukleotidsequenz-Unterscheidung unterscheiden, alle
von Enzymen ab, um Sequenzunterschiede zu identifizieren (Saiki,
PNAS USA 86: 6230, 1989; Zhang, Nucl. Acids Res. 19: 3929, 1991).
-
Zum Beispiel erkennen Restriktionsenzyme
Sequenzen von ungefähr
4–8 Nukleotiden.
Basierend auf einem durchschnittlichen G+C-Gehalt können ungefähr die Hälfte der
Nukleotidpositionen in einem DNA-Fragment mit einem Panel mit 100
Restriktionsenzymen überwacht
werden. Als eine Alternative können
künstliche Restriktionsenzyme-Erkennungssequenzen
um eine variable Position herum durch Verwendung von teilweise fehlgepaarten
PCR-Primern erzeugt werden. Mit dieser Technik kann entweder die
Mutanten oder die Wildtyp-Sequenz allein erkannt werden und durch
ein Restriktionsenzym nach Amplifikation gespalten werden (Chen
et al., Anal. Biochem. 195: 51, 1991; Levi et al., Cancer Res. 51:
3497, 1991). Ein anderes Verfahren. nutzt die Fähigkeit aus, daß ein Oligonukleotidprimer,
der an der 3'-vorletzten
Position mit einer Zielsequenz fehlgepaart ist, eine verringerte
Kapazität
aufweist, als ein Primer in der PCR zu dienen. Jedoch sind einige 3'-Fehlpaarungen, bemerkenswerter
weise G-T, weniger inhibitorisch als andere, was dessen Brauchbarkeit begrenzt.
In einem Ansatz um dieses Verfahren zu verbessern, werden zusätzliche
Fehlpaarungen in dem Primer an der dritten Position von dem 3'-Ende eingefügt. Dieses
führt zu
zwei fehlgepaarten Positionen in dem 3'-Nukleotiden des Primers, der mit einer
allelischen Variante hybridisiert und eine Fehlpaarung in der dritten Position
von dem 3'-Ende
wenn der Primer an die andere allelische Variante hybridisiert (Newton
et al., Nucl. Acids Res. 17: 2503, 1989). Es ist erforderlich, Amplifikationsbedingungen
zu definieren, die signifikant die Amplifikation einer 1 bp – Fehlpaarung
bevorzugen.
-
DNA-Polymerasen wurden auch dazu
verwendet, um allelische Sequenzvarianten zu unterscheiden, durch
Bestimmen welches Nukleotid an den Oligonukleotidprimer angefügt wird,
unmittelbar stromaufwärts
einer variablen Position in dem Zielstrang.
-
Ein Ligationsassay wurde entwickelt.
In diesem Verfahren werden zwei Oligonukleotidsonden, die in unmittelbarer
Nebeneinanderstellung auf einem Zielstrang hybridisieren durch eine
DNA-Ligase verbunden. Die Ligation ist inhibiert, falls eine Fehlpaarung
auftritt, wo die zwei Oligonukleotidsonden aneinander grenzen.
-
14. Assays für den Mutationsnachweis
-
Mutationen sind eine Einzel-Basenpaar-Veränderung
in genomischer DNA. Innerhalb des Kontext dieser Erfindung werden
die meisten solcher Veränderungen
leicht durch Hybridisierung mit Oligonukleotiden nachgewiesen, die
zu der fraglichen Sequenz komplementär sind. In dem hier beschriebenen
System werden zwei Oligonukleotide verwendet um eine Mutation nachzuweisen.
Ein Nukleotid besitzt die Wildtyp-Sequenz und das andere Oligonukleotid
besitzt die Mutantensequenz. Wenn die zwei Oligonukleotide als ein
Sonden auf einer Wildtyp-zielgenomischen Sequenz verwendet werden,
wird das Wildtyp-Oligonukleotid eine perfekt Basen-gepaarte Struktur
ausbilden und die Mutanten-Oligonukleotidsequenz
wird eine Duplex mit einer Einzelbasepaarfehlpaarung bilden. Wie
oben diskutiert – erlaubt
eine 6 bis 7°C
Differenz in der Tm einer Wildtyp gegenüber einer fehlgepaarten Duplex
die leichte Identifizierung und Unterscheidung der zwei Typen von
Duplices. Um diese Unterscheidung zu bewirken, wird eine Hybridisierung
bei der Tm der fehlgepaarten Duplex in der jeweiligen hybotropen
Lösung
durchgeführt
(siehe, z. B. U.S.-Anmeldung
Nrn. 60/026,621 (angemeldet 24. September 1996); 08/719,132 (angemeldet
24. September 1996); 08/933,924 (angemeldet 23. September 1997);
09/002,051 (angemeldet 31. Dezember 1997) und Internationale Publikations-Nr.
WO 98/13527, die alle hier in ihrer Gesamtheit für eine Beschreibung von hybotropen
Lösungen
aufgenommen sind). Das Ausmaß der
Hybridisierung wird dann für
das Set von Oligonukleotidsonden gemessen. Wenn das Verhältnis des Ausmaßes der
Hybridisierung der Wildtyp-Sonde zu der fehlgepaarten Sonde gemessen
wird, wird ein Wert von 10/1 bis größer als 20/1 erhalten. Diese
Typen von Ergebnissen ermöglichen
die Entwicklung von robusten Assays für den Mutationsnachweis.
-
Für
beispielhafte Zwecke verwendet ein Testformat für den Mutationsnachweis eine
Zielnukleinsäure (z.
B. genomische DNA) und Oligonukleotidsonden, die den Bereich von
Interesse überspannen.
Die Oligonukleotidsonden sind größer oder
gleich zu 24 Nukleotiden in ihrer Länge (mit einem Maximum von
ungefähr
36 Nukleotiden) und markiert mit einem Fluorochrom an dem 3' oder 5'-Ende der Oligonukleotidsonde.
Die Zielnukleinsäure
wird über
die Lyse von Gewebekulturzellen, Geweben, Organismen, usw., in der
jeweiligen Hybridisierungslösung
erhalten. Die lysierte Lösung
wird dann auf eine Temperatur erhitzt, die die Zielnukleinsäure denaturiert
(15–25°C oberhalb
der Tm der Zielnukleinsäureduplex).
Die Oligonukleotidsonden werden bei der Denaturierungstemperatur
hinzugefügt
und die Hybridisierung wird bei der Tm der fehlgepaarten Duplex
für 0,5
bis 24 Stunden durchgeführt.
Die genomische DNA wird dann gesammelt und durch einen GF/C (GF/B
und ähnliches)
Glasfaserfilter hindurch passiert. Der Filter wird dann mit der
jeweiligen Hybridisierungslösung
gewaschen, um alle nicht-hybridisierten Oligonukleotidsonden zu
entfernen (RNA, kurze Oligos und Nukleinsäure bindet nicht an Glasfaserfilter
und unter diesen Bedingungen). Die Hybridisierungsoligosonde kann
dann von der Ziel-DNA termisch eluiert werden und gemessen werden
(durch Fluoreszenz z. B.). Für
Tests, die sehr hohe Spiegel an Sensitivität erfordern, werden die Sonden
konzentriert und gemessen.
-
Andere hochsensitive Hybridisierungsprotokolle
können
verwendet werden. Die Verfahren der folgenden Erfindung ermöglichen
es einem, leicht auf eine Nukleinsäure zu testen, die eine Mutation
enthält,
von der vermutet wird, daß sie
in Zellen, Proben, usw. vorhanden ist, d. h. einer Zielnukleinsäure. Die
Zielnukleinsäure enthält eine
Nukleotidsequenz von Desoxyribonukleinsäure (DNA) oder Ribonukleinsäure (RNA),
deren Anwesenheit von Interesse ist und deren Anwesenheit oder Abwesenheit
in dem Hybridisierungsassay nachgewiesen werden soll. Die Hybridisierungsverfahren
der folgenden Erfindung können
auch auf ein komplexes biologisches Gemisch von Nukleinsäuren (RNA
und/oder DNA) angewendet werden. Solch ein komplexes biologisches
Gemisch schließt
einen weiten Bereich von eukaryontischen und prokaryontischen Quellen,
einschließlich
Protoplasten ein, und/oder andere biologische Materialien, die Polynukleotidnukleinsäure enthalten.
Das Verfahren ist daher auf Gewebekulturzellen, tierische Zellen,
tierisches Gewebe, Blutzellen (z. B. Retikulozyten, Lymphozyten),
Pflanzenzellen, Bakterien, Hefen, Viren, Mykoplasmen, Protozoen,
Pilze und ähnliches
anwendbar. Durch Nachweisen einer spezifischen Hybridisierung zwischen
Nukleinsäuresonden
einer bekannten Quelle kann die spezifische Anwesenheit einer Zielnukleinsäure etabliert
werden. Ein typisches Hybridisierungsassayprotokoll zum Nachweis
einer Zielnukleinsäure
in einer komplexen Population von Nukleinsäuren wird wie folgt geschrieben:
Zielnukleinsäure
werden in ihrer Größe auf einer
LC oder HPLC aufgetrennt, kloniert und isoliert, unterteilt in Pools
oder als eine komplexe Population belassen. Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zum Nachweis der Identität eines
Nukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Nukleinsäuremoleküls in, z.
B. einer biologischen Probe zur Verfügung gestellt, wobei die allgemeinen
Techniken von Hybridisierungsassay verwendet werden. Kurz gesagt
umfassen solche Verfahren im allgemeinen die Schritte von Klonierung
und Isolierung von Zielnukleinsäuren,
Unterteilung in Pools oder Belassen als eine komplexe Population.
Die Zielnukleinsäuren
werden dann mit markierten Oligonukleotidsonden unter oben beschriebenen
Bedingungen hybridisiert. Die Zielnukleinsäuren werden ihrer Größe nach
durch LC oder HPLC aufgetrennt. Die Marker werden von den aufgetrennten
Fragmenten abgespalten und dann werden die Marker durch die jeweilige
Nachweistechnologie (z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV/sichtbare Spektrophotometrie)
nachgewiesen.
-
15. Sequenzierung
durch Hybridisierung
-
Die DNA-Sequenzanalyse wird herkömmlicherweise
durch Hybridisieren eines Primers an Ziel-DNA und Durchführen von
Kettenverlängerungen
unter der Verwendung einer Polymerase durchgeführt. Spezifische Stops werden
durch den Einbau eines Didesoxydnukleotids kontrolliert. Die Spezifität des Primens
bei diesem Typ von Analyse kann durch Einschließen eines Hybotrops in dem
Annealingpuffer und/oder in Kooperation eines abasischen Rests in
dem Primer und Annealing bei einer unterscheidenden Temperatur erhöht werden.
-
Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zur Bestimmung der Identität eines Nukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Nukleinsäuremoleküls in, z.
B., einer biologischen Probe zur Verfügung gestellt, wobei die allgemeinen
Techniken der Sequenzierung durch Hybridisierung unter der Verwendung
des Sanger-Verfahren verwendet werden. Kurz umfassen solche Verfahren
im allgemeinen die Schritte von Hybridisieren eines markierten Primers
an Ziel-DNA und Durchführung
von Kettenverlängerungen
unter der Verwendung einer Polymerase. Spezifische Stops werden
durch den Einbau eines Didesoxynukleotids kontrolliert, das auch
markiert sein kann. Die Zielnukleinsäuren werden gemäß ihrer
Größe durch
HPLC aufgetrennt. Die Marker werden von den aufgetrennten Fragmenten
abgespalten und werden durch die jeweilige Nachweistechnologie (z.
B. Massenspektrometrie, Infrarotspektrometrie, potentiostatische Amperometrie
oder UV/sichtbare Spektrophotometrie) nachgewiesen. Andere Sequenz-Analyseverfahren schließen die
Hybridisierung des Ziels mit einer Auswahl von zufälligen,
kurzen Oligonukleotiden ein. Die Sequenz wird durch überlappende
Hybridisierungsanalyse konstruiert. Bei dieser Technik ist präzise Hybridisierung
essentiell. Die Verwendung von hybotropen oder abasischen Resten
und ein Annealing bei unterscheidenden Temperatur ist für dieses
Verfahren vorteilhaft, um fehlgepaarte Hybridisierung zu verringern
oder auszuschließen.
Das Ziel ist es, automatisierte Hybridisierungsverfahren zu entwickeln,
um große
Arrays von Oligonukleotidsonden oder große Arrays von Nukleinsäureproben
zu testen. Die Anwendungen von solchen Technologien schließen Genkartierung,
Kloncharakterisierung, medizinische Genetik und Genentdeckung, DNA-Sequenzanalyse
durch Hybridisierung und zuletzt Sequenzverifikation. Viele Parameter
müssen
kontrolliert werden, um Oligonukleotidsonden zu automatisieren oder
einen Multiplex durchzuführen.
Die Stabilität
der jeweiligen Sonden muß ähnlich sein,
das Ausmaß von
Fehlpaarung mit der Zielnukleinsäure,
die Temperatur, Ionenstärke,
der A + T-Gehalt der Sonde (oder des Ziels) sowie andere Parameter
wenn die Sonde kurz ist (d. h. 6–50 Nukleotide) sollten ähnlich sein.
Gewöhnlicherweise
werden die Bedingungen des Experiments und Sequenz der Sonde eingestellt,
bis die Bildung der perfekten Basen-gepaarten Sonde thermodynamisch über jede
Duplex, die eine Fehlpaarung enthält, bevorzugt wird. Die Anwendungen
von Sonden in sehr großem Maßstab, wie
z. B. bei Sequenzierung durch Hybridisierung (SBH) oder dem Testen
von hochpolymorphen Loci, wie z. B. dem zystische Fibrose Trans-Membran
Proteinlocus erfordern ein stringenteres Niveau der Kontrolle von
Multiplex-Sonden. Innerhalb einer Ausführungsform der Erfindung werden
Verfahren zum Nachweis der Identität eines Nukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Nukleinsäuremoleküls in, z. B.
einer biologischen Probe zur Verfügung gestellt, welche die allgemeinen
Techniken von Sequenzierung durch Hybridisierung anwenden. Kurz
gesagt, umfassen solche Verfahren im allgemeinen ein Hybridisieren
einer Reihe von markierten Primern an ein DNA-Ziel oder eine Reihe
von Ziel-DNA-Fragmenten unter sorgfältig kontrollierten Bedingungen.
Die Zielnukleinsäuren
werden nach ihrer Größe durch
HPLC aufgetrennt. Die Marker werden dann von den aufgetrennten Fragmenten
abgespalten und durch die jeweilige Nachweistechnologie (z. B. Massenspektrometrie,
Infrarotspektrometrie, potentiostatische Amperometrie oder UV/sichtbare Spektrophotometrie)
nachgewiesen.
-
16. Oligonukleotid-Ligationsassay
-
Der Oligonukleotid-Ligationsassay
ist eine Erweiterung von PCR-basiertem Screening, die einen ELISA-basierten
Assay verwendet (OLA, Nickerson et al., Proc. Natl. Acad. Sci. USA
87: 8923, 1990), um die PCR-Produkte nachzuweisen, die die Zielsequenz
enthalten. Daher werden sowohl Gelelektrophorese und Koloniehybridisierung
beseitigt. Kurz gesagt verwendet die OLA zwei angrenzende Oligonukleotide:
eine „Reporter"-Sonde (markiert
an 5'-Ende) und
eine 5'-phosphorylierte/3'-biotinylierte „Verankerungs"-Sonde. Die zwei
Oligonukleotide, die zu Sequenzen innerhalb der PCR-Primer komplementär sind,
werden an Ziel-DNA annealed und, falls perfekte Komplementarität vorhanden
ist, werden die zwei Sonden durch T4-DNA-Ligase ligiert. Ein Fangen
der biotinylierten Verankerungssonde auf mobilisierten Streptavidin
und die Analyse auf die kovalent gebundene Reportersonde testet
auf die Anwesenheit oder Abwesenheit der Zielsequenzen unter den
PCR-Produkten. Innerhalb einer Ausführungsform der Erfindung werden
Verfahren zum Nachweis der Identität eines Nukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Nukleinsäuremoleküls, in,
z. B., einer biologischen Probe zur Verfügung gestellt, die die Technik
des Oligonukleotid-Ligationsassays verwenden. Kurz gesagt, umfassen
solche Verfahren im allgemeinen die Schritte von Durchführen einer
PCR auf der Ziel-DNA, gefolgt durch Hybridisierung mit der 5'-markierten „Reporter"-DNA-Sonde und einer
5'-phosphorylierten/nicht-biotinylierten
Sonde. Die Probe wird mit T4-DNA-Ligase inkubiert. Die DNA-Stränge mit
ligierten Proben können
von der DNA mit nicht-ligierten Proben durch, z. B., bevorzugterweise
LC oder HPLC abgetrennt werden. Die Marker werden dann von den abgetrennten
Fragmenten abgespalten und dann werden die Marker durch die jeweilige
Nachweistechnologie (z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV/sichtbare Spektrophotometrie)
nachgewiesen. Kürzliche
Fortschritte im OLA-Assay erlauben die gleichzeitige Analyse von
multiplen Proben und multiplen Mutationen. (Baron et al., Nature
Biotechnology 87: 1279, 1996). Kurz gesagt besteht das Verfahren
aus der Amplifikation des Genfragments, das die Mutation von Interesse
enthält,
mit PCR. Das PCR-Produkt wird dann mit einer gewöhnlichen und zwei Allel-spezifischen
Oligonukleotidsonden (wobei eine die Mutation enthält, während die
andere dies nicht tut) hybridisiert, so daß die 3'-Enden der Allel-spezifischen Sonden
unmittelbar angrenzend zu dem 5'-Ende
der gewöhnlichen
Sonde sind. Dieses erstellt einen kompetetiven Hybridisierungs-Ligationsprozeß zwischen
den zwei allelischen Proben und der herkömmlichen Probe an jedem Locus.
Die thermostabile DNA-Ligase unterscheidet dann zwischen Einzel-Base-Fehlpaarungen
an der Verbindungsstelle, wodurch Allel-spezifische Ligationsprodukte
erzeugt werden. Die herkömmliche
Probe wird mit einem von vier Fluorophoren markiert und die Allel-spezifischen
Sonden werden jede mit einem oder mehreren Pentaethylenoxid-Mobilitäts-modifizierenden
Verlängerungen
markiert, die einen Größenunterschied
zwischen den verschiedenen Allel-spezifischen Sonden zur Verfügung stellen.
Die Proben werden dann durch Gelelektrophorese basierend auf der
Länge der
modifizierenden Enden aufgetrennt und durch den fluoreszenten Marker
an der herkömmlichen
Sonde nachgewiesen. Durch die Verwendung in Größenunterschieden auf den Allel-spezifischen
Sonden und vier für
die herkömmliche
Probe erhältlichen
vier Fluorophore können
viele Proben auf einer Spur des elektrophoretischen Gels analysiert
werden. Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zum Nachweis der Identität eines
Nukleinsäuremoleküls oder
zum Nachweis eines ausgewählten
Nukleinsäuremoleküls in, z.
B. einer biologischen Probe zur Verfügung gestellt, wobei die Technik
des Oligonukleotid-Ligationsassays für gleichzeitigen multiplen
Probennachweis verwendet wird. Kurz gesagt, umfassen solche Verfahren
allgemein die Schritte von Durchführung von PCR auf der markierten
DNA, gefolgt von Hybridisierung mit der herkömmlichen Probe (nicht markiert)
und den zwei Allel-spezifischen Sonden, die gemäß den Beschreibungen der Erfindung
markiert sind. Die Probe wird mit DNA-Ligase inkubiert und die Fragmente,
z. B. bevorzugterweise durch LC oder HPLC aufgetrennt. Die Marker
werden von den aufgetrennten Fragmenten abgespalten und dann werden
die Marker durch die jeweilige Nachweistechnologie (z. B. Massenspektrometrie,
Infrarotspektrometrie, potentiostatische Amperometrie oder UV/sichtbare
Spektrophotometrie) nachgewiesen.
-
17. Differential Display
-
a. Überblick
-
Säugetiere,
wie z. B. der Mensch, weisen ungefähr 100.000 verschiedene Gene
in ihrem Genom auf, von denen nur ein kleiner Teil, möglicherweise
15%, in jeder einzelnen Zelle exprimiert werden. Die Auswahl von
exprimierten Genen bestimmt den biochemischen Charakter jeder bestimmten
Zelle oder Gewebe. Das Verfahren von normalen zellulären Wachstum
und Differenzierung sowie die pathologischen Veränderungen, die in Erkrankungen
wie Krebs entstehen, werden alle durch Veränderungen in der Genexpression
angetrieben. Differential-Display-Verfahren ermöglichen die Identifizierung
von Genen, die spezifisch in einzelnen Zelltypen exprimiert werden.
-
Die Differential-Display-Technik
amplifiziert die 3'-terminalen
Teile von entsprechenden cDNAs durch Verwendung eines Primers, der
so aufgebaut ist, um an die 5'-Grenze
eines Poly(A)-Schwanzes zu binden und Primern mit beliebiger Sequenz,
die stromaufwärts
binden. Amplifizierte Populationen mit jedem Primerpaar werden durch
ein Größentrennverfahren
(PAGE HPLC, usw.) visualisiert, was einen direkten Vergleich der mRNAs
zwischen zwei biologischen Proben von Interesse ermöglicht.
Das Differential-Display-Verfahren weist das Potential auf, um alle
der exprimierten Gene (ungefähr
10.000 bis 15.000 mRNA Spezies) in einer Säugerzelle zu visualisieren
und die Sequenzanalyse zu ermöglichen.
Es ist möglich,
zu vergleichen: (1) die Gesamtzahl von in den Eltern amplifizierten
Peaks, (2) die Zahl von polymorphen Peaks zwischen Eltern und (3)
die Segregationsverhältnisse
von polymorphen Peaks in den Nachkommen von Kreuzungen von Tieren
oder Pflanzen. Differential Display wird auch zur Identifizierung
von Hoch- und Herunter-regulierten bekannten oder unbekannten Genen
nach einer Vielzahl von Stimuli verwendet. Differential-Display-PCR-Fragmente können auch
als Sonden für
die cDNA-Klonierung verwendet werden (wobei ein unbekanntes Gen
aus einer cDNA oder genomischen Bibliothek entdeckt wird).
-
Kurz gesagt sind die Schritte beim
Differential-Display wie folgt: 1) RNA wird aus der biologischen
Probe von Interesse isoliert. Gesamt-RNA, zytoplasmatische RNA oder
mRNA kann verwendet werden. 2) Erster Strang cDNAs werden unter
der Verwendung eines verankerten OligodT (OligodTdN, wobei N A,
C oder G ist) erzeugt. 3) Amplifikation von cDNA unter der Verwendung
von OligoTdN und kurzen Primern mit beliebiger Sequenz. Für eine vollständige Differential-Display-Analyse
von zwei Zellpopulationen oder zwei Proben von Interesse werden
9 verschiedene Primer benötigt.
Die Nachweisgrenze von Differential-Display für eine spezifische mRNA ist
weniger als 0,001% der gesamten mRNA-Population.
-
Aufgrund der Einfachheit, Sensitivität und Reproduzierbarkeit
des hier beschriebenen Verfahrens ist die CMST Differential-Display-Methode
ein signifikanter Fortschritt über
herkömmliche
gelbasierte Systeme. Mit der CMST-basierten Differential-Display-Analyse
von zwei Zelltypen können
einschließlich
64 × 24 PCR-Läufe schnell
vervollständigt
werden, im Gegensatz zu einer arbeitsintensiven lang andauernden
Zeit mit dem herkömmlichen Verfahren.
Darüber
hinaus wurde von Sequenzheterogenität von Banden, die aus Differential-Display-Gelen isoliert
wurden gefunden, daß sie
ein zu der hohen Fehlerrate dieser Technik beitragender Faktor sind.
Dies wird mit der hier beschriebenen CMST-basierten Differential-Display-Methodologie
vollständig
vermieden.
-
b. CMST-basiertes Differential-Display
Beispiel:
-
Das Ausgangsmaterial für Differential-Display
ist RNA, die aus zwei verschiedenen Populationen von Zellen isoliert
wurde. Im allgemeinen sind die Zellen von ähnlichem Ursprung und unterscheiden
sich im Hinblick auf ihre Behandlung mit Wirkstoffen, ihrem „normalen" gegenüber „transformierten" Zustand oder ihrer Expression
von verschiedenen eingefügten
Genen.
-
Pflanzengewebe oder tierisches Material
(2–3 g)
wird geerntet und in sterile Petrischalen zerhackt oder zerschnitten.
Das Material wird dann zu einem feinen Pulver in einem vorgekühlten Mörser- und
Stöße unter
flüssigem
N2 zerrieben. Ein Gramm von gefrorenem Pulver
wird zu einem 12 ml Poly-Propylen-Röhrchen (1 g = ca. 5 ml Pulver),
enthaltend 8 ml eines heißen
(80°C) 1
: 1 Gemisches von RNA-Extraktionspuffer (100 mM Tris-HCl (pH 8,0),
100 mM LiCl, 10 mM EDTA, 1,0% LIDS) und Phenol (Basisvolumen.: 4
ml) transferiert. Die Probe wird bei hoher Geschwindigkeit für mindestens
30 Sekunden gemischt (gevortext). Ein Volumen von Chloroform wird
hinzugefügt
(4 ml) und nochmals gemischt und zentrifugiert für 20 Minuten in einer Zentrifuge bei
5000 bis 10.000 × g.
Die wäßrige Phase
wird in ein neues 12 ml Röhrchen
transferiert und 1/3 Volumen von kaltem 8M LiCl wird hinzugefügt, um die
RNA zu fällen
(3 h bei 0°C).
Die RNA wird bei 0°C
für 20
Minuten zentrifugiert und in 1 ml H2O suspendiert.
Restliche genomische DNA wird durch Behandlung der RNA-Probe mit
DNase I beseitigt. Reverse Transkription wird durchgeführt auf
jeder RNA-Probe unter der Verwendung von 500 ng DNA-freier RNA in
1x reverse Transkriptasepuffer, 10 mM DTT, 20 μM dNTPs, 0,2 μM 5'RS H-711 C (ein basenverankerter
Primer mit 5'-Restriktionsstelle),
200 Einheiten MoMuLV reverser Transkriptase und 1,5 Einheiten RNA
Guard pro 20 μl
Reaktionsvolumen. Obwohl die Verwendung eines stromabwärts-Primers
die Zahl von cDNA-Subfraktionen dreifach vemngert, reduziert sie
nicht die Zahl von PCR-Reaktionen,
die erforderlich sind, um die meisten der cDNA-Spezies, die in dem
Pool vorhanden sind, darzustellen. Im Gegenteil verringert sie die
theoretische Chance, diejenigen cDNA-Spezies zu identifizieren,
die vorhanden sind. Die besten Ergebnisse werden unter der Verwendung
einer Kombination von neun verschiedenen Primern des Typs DMO-VV,
wobei V A, G, C sein kann, jedoch nicht T, erhalten. Mit einem T
in der terminalen 3'-Position führt die
unvollständige
Hybridisierung des Primers zu einem Verschmieren von Banden auf
dem Gel. Die optimale Konzentration von RNA ist 200–300 ng
pro cDNA-Synthese.
-
Das CMST-basierte Differential-Display
wird im wesentlichen wie vorher beschrieben (Liang and Pardee, Science
257: 967–971,
1992) durchgeführt,
mit der Ausnahme für
den Aufbau von für
die reverse Transkription und de Amplifikationsschritte verwendeten
Primer und die Auswahl von radiomarkiertem Nukleotid. Eine vollständige Differential-Display-Analyse
der cDNAs aus zwei biologischen Proben von Interesse unter der Verwendung
von neun Stromabwärts-Primern
und 24 Stromaufwärts-Primern
würde 9 × 24 × 2 CMST-basierte
Differential-Display-Reaktionen erzeugen. Die Amplifikationsprodukte
können
durch HPLC aufgetrennt werden und, falls gewünscht, reamplifiziert werden.
-
Im Anschluß an die Inkubation der RNA
bei 65°C
für 5 Minuten
werden die Proben auf Eis gekühlt,
zu den Reverse-Transkriptionsmix hinzugefügt und für 60 Minuten bei 37°C inkubiert,
gefolgt von 95°C
für 5 Minuten.
Duplikat cDNA-Proben werden dann unter der Verwendung desselben
5'-Primers in Kombination
mit einer Reihe von 13-Meren von beliebiger, jedoch definierter
Sequenz: H-AP: AAGCTTCGACTGT, H-AP: AAGCTTTGGTCAG, H-AP4: AAGCTTCTCAACG,
H-AP5: AAGCTTAGTAGGC amplifiziert.
-
Die Modifikation wird in Raktionsgemischen
durchgeführt,
die 0,1 Volumen von reverser Transkriptionsreaktion enthalten, 1 × PCR-Puffer
(10 × PCR-Puffer
= 100 mM Tris-HCl, 15 mM MgCl2, 10 mM KCl,
pH 8,3) 2 μM
dNTPs, 0,2 μM
RS H-T11C verankertem Primer, 0,2 μM geeignetem beliebigem Primer,
1,5 Einheiten ExpandTM High Fidelity DNA-Polymerase
und Wasser bis zu einem finalen Volumen von 20 μl. Die Amplifikation der cDNA
wird unter den folgenden Bedingungen durchgeführt: 94°C (1 Minute), gefolgt von 40
Zyklen von 94°C
(30 Sekunden), 40°C
(2 Minuten), 72°C
(30 Sekunden) und beendet mit 72°C
(5 Minuten).
-
Die Amplifikation für jedes
Gen wird mit genspezifischen Primern durchgeführt, die eine bekannte Intron/Exon-Grenze überspannen
(siehe unten). Alle Amplifikationen werden in 20 μl Volumen
durchgeführt,
die 10 mM Tris HCl pH 8,3, 1 mM NH4Cl, 1,5 M MgCl2, 100 mM KCl,
0,125 mM NTPs, 10 ng/ml der jeweiligen Oligonukleotidprimer und
0,75 Einheiten von Taq-DNA-Polymesse (Gibco-BRL) enthalten. Die
Zyklusparameter waren 94°C
Vorheizschritt für
5 Minuten, gefolgt von 94°C
Denaturierungsschritt für
1 Minute, 55°C
Annealingschritt für
2 Minuten und einem 72°C
Verlängerungsschritt
für 30
Sekunden bis 1 Minute und einer finalen Verlängerung von 72°C für 10 Minuten.
Die Zahl der Amplifikationszyklen lag im allgemeinen von 30–45.
-
Die Amplifikationsprodukte werden
Gel-gereinigt (Zhen and Swank, BioTechniques 14: 894-898, 1993) auf 1%
Agarosegelen, die in 0,04 M Tris-Acetat, 0,001 M EDTA (1 × TEA)-Puffer
laufen gelassen wurden und mit Ethidiumbromid gefärbt wurden.
Eine Mulde wird gerade vor der Bande von Interesse eingeschnitten
und mit 50–200 μl von 10%
PEG in 1 × TAE-Puffer
gefüllt.
Die Elektrophorese wird fortgeführt,
bis die Bande vollständig
in die Mulde eingetreten ist. Die Inhalte werden dann entfernt und
mit Phenol, Chloroform extrahiert und in 0,1 Volumen von 7,5 M Ammoniumacetat
und 2,5 Volumen 100% EtOH gefällt.
Die Proben werden mit 75% EtOH gewaschen und kurz bei Raumtemperatur
getrocknet. Die Quantifizierung der Ausbeute wird durch Elektrophorese
eines kleinen Aliquots auf 1% Agarosegel in 1 × TBE-Puffer mit Ethidiumbromidfärbung und Vergleich
mit einem bekannten Standard durchgeführt.
-
Die Produkte aus den Amplifikationsreaktionen
werden durch HPLC analysiert. Die HPLC wird unter der Verwendung
von automatisierter HPLC-Instrumentierung (Rainin, Emeryville, CA,
oder Hewlett Packard, Palo Alto, CA) durchgeführt. Ungereinigte DNA-Fingerprinting-Produkte, die für 3 Minuten
bei 95°C
vor der Injektion in eine HPLC denaturiert werden, werden mit linearen
Acetonitril (ACN, J. T. Baker, NJ) Gradienten von 1,8%/Minute bei
einer Flußrate
von 0,9 ml/Minute eluiert. Die Start- und Endpunkte werden gemäß der Größe der amplifizierten
Produkte eingestellt. Die für
die erfolgreiche Auflösung
der Moleküle,
die während
der DNA-Fingerprinting-Technik erzeugt wurden erforderliche Temperatur
ist 50°C.
-
Der Efflux aus der HPLC wird dann
in einen Massenspektrometer (Hewlett Packard, Palo Alto, CA) zum
Nachweis der Marker umgeleitet.
-
Der Vergleich der Chromatogramme
(Massenspektrometrie-basiert) zeigt, daß Banden bei 220 bp und 468
bp in den stimulierten Jurkat-Zellen beobachtet werden und nicht
in den nicht-stimulierten Jurkat-Zellen beobachtet werden.
-
C. TRENNUNG VON NUKLEINSÄUREFRAGMENTEN
-
Eine Probe, die eine Analyse erfordert,
ist oft eine Mischung aus vielen Komponenten in einer komplexen
Matrix. Für
Proben, die unbekannte Verbindungen enthalten, müssen die Komponenten voneinander
getrennt werden, so daß jede
einzelne Komponente mit anderen analytischen Verfahren identifiziert
werden kann. Die Trenneigenschaften der Komponenten in einer Mischung
sind unter konstanten Bedingungen konstant, und wenn sie daher erst
einmal bestimmt sind, können
sie verwendet werden, um jede der Komponenten zu identifizieren
und zu quantifizieren. Solche Verfahren sind typisch bei chromatographischen
oder elektrophoretischen analytischen Trennungen.
-
1. Hochleistungsflüssigkeitschromatrographie
(HPLC)
-
Hochleistungsflüssigkeitschromatographie (HPLC)
ist eine chromatographische Trenntechnik, um Verbindungen zu trennen,
die in Lösung
gelöst
sind. HPLC-Instrumente bestehen aus einem Reservoir aus mobiler
Phase, einer Pumpe, einem Injektor, einer Trennsäule und einem Detektor. Verbindungen
werden durch Injizieren eines aliquoten Teils der Probenmischung
auf die Säule
getrennt. Die unterschiedlichen Verbindungen gehen aufgrund der
Unterschiede in ihrem Verteilungsverhalten zwischen der mobilen
flüssigen Phase
und der stationären
Phase mit unterschiedlichen Geschwindigkeiten durch die Säule hindurch.
-
Vor kurzem ist gezeigt worden, daß IP-RO-HPLC
auf nicht-porösen
PS/DVB-Teilchen mit chemisch gebundenen Alkylketten schnelle Alternativen
zu Kapillarelektrophorese bei der Analyse von sowohl einzel- als auch
doppelsträngigen
Nukleinsäuren
ist, die ähnliche
Auflösungsgrade
liefert (Huber et al., Anal. Biochem. 212: 351, 1993; Huber et al.,
1993, Nuc. Acids Res. 21: 1061; Huber et al, Biotechniques 16: 898,
1993). Im Gegensatz zu Ionenaustauschchromatographie, die nicht
immer doppelsträngige
DNA als eine Funktion der Stranglänge zurückhält (da AT-Basenpaare mit der
positiv geladenen stationären
Phase stärker
als GC-Basenpaare wechselwirken), ermöglicht IP-RP-HPLC eine streng
größenabhängige Trennung.
-
Ein Verfahren unter Verwendung von
100 mM Triethylammoniumacetat als Ionenpaarungsreagens ist entwickelt
worden, Phosphodiester-Oligonukleotide konnten mit Erfolg auf alkylierten
nicht-porösen
2,3 μM Poly(styrol-divinylbenzol)-Teilchen
mittels Hochleistungsflüssigkeitschromatographie
getrennt werden (Oefner et al., Anal. Biochem. 223: 39, 1994). Die
beschriebene Technik ermöglichte
die Trennung von PCR-Produkten, die sich nur 4 bis 8 Basenpaare
in der Länge
unterscheiden, in einem Größenbereich
von 50 bis 200 Nukleotiden.
-
2. Elektrophorese
-
Elektrophorese ist eine Trenntechnik,
die auf der Mobilität
von Ionen (oder DNA, wie dies der hierin beschriebene Fall ist)
in einem elektrischen Feld beruht. Negativ geladene DNA wandert
zu einer positiven Elektrode und positiv geladene Ionen wandern
zu einer negativen Elektrode. Aus Sicherheitsgründen ist üblicherweise eine Elektrode
geerdet und die andere positiv oder negativ vorgespannt. Geladene
Spezies haben unterschiedliche Wanderungsgeschwindigkeiten in Abhängigkeit
von ihrer Gesamtladung, Größe und Gestalt und
können
daher getrennt werden. Ein Elektrodenapparat besteht aus einer Hochspannungs-Stromquelle, Elektroden,
Puffer und einem Träger
für den
Puffer, wie etwa Polyacrylamidgel, oder eine Kapillarröhre. Offene Kapillarröhren werden
für viele
Arten von Proben verwendet und die anderen Gelträger werden üblicherweise für biologische
Proben verwendet, wie etwa Proteinmischungen oder DNA-Fragmente.
-
3. Kapillarelektrophorese
(CE)
-
Kapillarelektrophorese (CE) in ihren
verschiedenen Manifestationen (freie Lösung, Isotachophorese, isoelektrische
Fokussierung, Polyacrylamidgel, mizelläre elektrokinetische "Chromatographie") entwickelt sich als
ein Verfahren für
schnelle Hochauflösungstrennungen
sehr kleiner Probenvolumina komplexer Mischungen. In Kombination
mit der inhärenten
Empfindlichkeit und Selektivität
von MS ist CE-MS eine potentielle leistungsfähige Technik für die Bioanalyse.
In der hierin offenbarten neuartigen Anwendung wird das Zusammenspiel
dieser zwei Verfahren zu überlegenen
DNA-Sequenzierungsverfahren führen,
die die Sequenzierungsverfahren mit der augenblicklichen Geschwindigkeit
um mehrere Größenordnungen übertreffen.
-
Die Beziehung zwischen CE und Durchflüssen bei
Elektrosprayionisierung (ESI) und die Tatsache, daß beide
durch Ionenspezies in Lösung
erleichtert werden (und primär
dafür verwendet
werden), liefert die Grundlage für
eine extrem attraktive Kombination. Die Kombination von sowohl Kapillarzonenelektrophorese (CZE)
und Kapillarisotachophorese mit Quadrapol-Massenspektrometern auf
der Basis von ESI ist beschrieben worden (Olivares et al., Anal.
Chem. 59: 1230, 1987; Smith et al., Anal. Chem. 60: 436, 1988; Loo
et al., Anal. Chem. 179: 404, 1989; Edmonds et al., J. Chroma. 474:
21, 1989; Loo et al., J. Microcolumn Sep. 1: 223, 1989; Lee et al.,
J. Chromatog. 458: 313, 1988; Smith et al., J. Chromatog. 480: 211,
1989; Grese et al., J. Am. Chem. Soc. 111: 2835, 1989). Kleine Peptide
sind einer CZE-Analyse mit guter Empfindlichkeit (im Femtomol-Bereich)
leicht zugänglich.
-
Das leistungsfähigste Trennverfahren für DNA-Fragmente
ist Polyacrylamidgelelektrophorese (PAGE), im allgemeinen in einem
Slabgel-Format. Die wesentliche Beschränkung der gegenwärtigen Technologie ist
jedoch die relativ lange Zeit, die erforderlich ist, um die Gelelektrophorese
von DNA-Fragmenten, die in den Sequenzierungsreaktionen hergestellt
werden, durchzuführen.
Eine Erhöhung
der Größenordnung
(10-fach) kann mit der Verwendung von Kapillarelektrophorese erreicht
werden, die ultradünne
Gele einsetzt. In freier Lösung
wandert, in einer ersten Annäherung,
die gesamte DNA mit derselben Mobilität, da die Zugabe einer Base
zw Kompensation von Masse und Ladung führt. In Polyacrylamidgelen
sieben und wandern DNA-Fragmente als eine Funktion der Länge, und
dieser Ansatz ist nunmehr auf CE angewendet worden. Bemerkenswerte
Bodenzahlen pro Meter sind jetzt mit quervernetztem Polyacrylamid
erreicht worden (10+7 Böden pro Meter, Cohen et al.,
Proc. Natl. Acad. Sci., USA 85: 9660, 1988). Solche CE-Säulen, wie
beschrieben, können
zur DNA-Sequenzierung eingesetzt werden. Das CE-Verfahren ist im
Prinzip 25-mal schneller als Slabgel-Elektrophorese in einem Standard-Sequenzer. Zum Beispiel
können
etwa 300 Basen pro Stunde gelesen werden. Die Trennungsgeschwindigkeit
ist bei Slabgel-Elektrophorese durch die Größenordnung des elektrischen
Feldes beschränkt,
das ohne übermäßige Wärmeproduktion
an das Gel angelegt werden kann. Daher wird die größere Geschwindigkeit
von CE durch die Verwendung höherer
Feldstärken
erreicht (300 V/cm bei CE gegenüber
10 V/cm bei Slabgel-Elektrophorese).
Das Kapillarformat verringert den Stromfluß und damit die Leistung und
die resultierende Wärmeerzeugung.
-
Smith und andere (Smith et al., Nuc.
Acids Res. 18: 4417, 1990) haben den Einsatz mehrerer Kapillaren
in Parallelschaltung vorgeschlagen, um den Durchsatz zu erhöhen. In ähnlicher
Weise haben Mathies und Huang (Mathies und Huang, Nature 359: 167,
1992) Kapillarelektrophorese eingeführt, bei der Trennungen auf einer
Parallelanordnung von Kapillaren durchgeführt werden, und zeigten Sequenzierung
mit hohem Durchsatz (Huang et al., Anal. Chem. 64: 967, 1992, Huang
et al., Anal. Chem. 64: 2149, 1992). Der wichtigste Nachteil von
Kapillarelektrophorese ist die begrenzte Probenmenge, die auf die
Kapillare geladen werden kann. Durch Konzentration einer großen Probenmenge
am Beginn der Kapillare, vor der Trennung, wird die Beladbarkeit
erhöht,
und Nachweisgrenzen können
um mehrere Größenordnungen
gesenkt werden. Das populärste Verfahren
zur Vorkonzentration bei CE ist Probenstapelung. Probenstapelung
ist vor kurzem in einer Übersicht dargestellt worden
(Chien und Burgi, Anal. Chem. 64: 489A, 1992). Probenstapelung hängt ab von
dem Matrixunterschied (pH, Ionenstärke) zwischen dem Probenpuffer
und dem Kapillarpuffer, so daß das
elektrische Feld über
die Probenzone stärker
ist als in der Kapillarregion. Bei Probenstapelung wird ein großes Probenvolumen
in einem Puffer mit geringer Konzentration zur Vorkonzentrierung
am Kopf der Kapillarsäule
eingeführt. Die
Kapillare wird mit einem Puffer derselben Zusammensetzung gefüllt, aber
mit höherer
Konzentration. Wenn die Proben-Ionen den Kapillarpuffer und das
niedrigere elektrische Feld erreichen, stapeln sie sich in eine
konzentrierte Zone hinein. Probenstapelung hat die Nachweisbarkeiten
um 1–3
Größenordnungen
erhöht.
-
Ein weiteres Verfahren zur Vorkonzentration
ist die Anwendung von Isotachophorese (ITP) vor der Freizonen-CE-Trennung
von Analyten. ITP ist eine Elektrophoresetechnik, die es ermöglicht,
daß Mikroliter-Probenvolumina
auf die Kapillare geladen werden können, im Gegensatz zu den Injektionsvolumina
von wenigen nl, die typischerweise mit CE assoziiert sind. Die Technik
beruht auf der Einführung
der Probe zwischen zwei Puffern (Leading- und Trailing-Elektrolyten)
mit höherer
bzw. niedrigerer Mobilität
als der Analyt. Die Technik ist inhärent eine Konzentrationstechnik,
bei der die Analyten sich in reine Zonen hineinkonzentrieren, die
mit derselben Geschwindigkeit wandern. Die Technik ist gegenwärtig weniger
populär
als die Stapelverfahren, die oben beschrieben sind, wegen des Bedarfs
mehrerer Wahlmöglichkeiten
von Leading- und Trailing-Elektrolyten und der Fähigkeit, nur kationische oder
anionische Spezies während
eines Trennverfahrens zu trennen.
-
Das Herz des DNA-Sequenzierungsverfahrens
ist die bemerkenswert selektive elektrophoretische Trennung von
DNA- oder Oligonukleotid-Fragmenten. Sie ist bemerkenswert, weil
jedes Fragment aufgelöst wird
und sich nur um Nukleotide unterscheidet. Trennungen von bis zu
1.000 Fragmenten (1.000 bp) sind erzielt worden. Ein weiterer Vorteil
der Sequenzierung mit spaltbaren Markierungen ist wie folgt. Es
gibt kein Erfordernis, ein Slabgel-Format zu verwenden, wenn DNA-Fragmente
mit Polyacrylamidgelelektrophorese getrennt werden, wenn spaltbare
Markierungen eingesetzt werden. Da zahlreiche Proben kombiniert
werden (4 bis 2.000), besteht keine Notwendigkeit, Proben parallel
laufen zu lassen, wie dies der Fall bei gegenwärtigen Farbstoff-Primer- oder
Farbstoff-Terminator-Verfahren ist (d. h. ABI373-Sequenzer). Da
es keinen Grund gibt, Parallelspuren laufen zu lassen, gibt es keinen
Grund, ein Slabgel zu verwenden. Daher kann man ein Röhrengel-Format
für das
Elektrophoresetrennverfahren einsetzen. Grossman (Grossman et al.,
Genet. Anal. Tech. Appl. 9: 9, 1992) hat gezeigt, daß ein beträchtlicher
Vorteil gewonnen wird, wenn ein Röhrengel-Format anstelle eines
Slabgel-Formats verwendet wird. Dies beruht auf der größeren Fähigkeit,
Joule-Wärme
in einem Röhrenformat
abzuleiten, verglichen mit einem Slabgel, was zu schnelleren Laufzeiten
führt (um
50%) und viel höherer
Auflösung
von DNA-Fragmenten mit hoher relativer Molekülmasse (mehr als 1.000 nt).
Lange Ablesungen sind bei genomischer Sequenzierung entscheidend.
Daher hat die Verwendung spaltbarer Markierungen beim Sequenzieren
den zusätzlichen
Vorteil, daß sie
dem Benutzer erlaubt, das effizienteste und empfindlichste DNA-Trennverfahren
einzusetzen, das auch die höchste
Auflösung
besitzt.
-
4. Mikrosysteme
-
Kapillarelektrophorese (CE) ist ein
leistungsfähiges
Verfahren zur DNA-Sequenzierung, forensischen Analysen, PCR-Produktanalyse
und Restriktionsfragmentgrößenbestimmung.
CE ist weit schneller als traditionelle Slab-PAGE, da bei Kapillargelen
ein weit höheres
Spannungsfeld angelegt werden kann. CE hat jedoch den Nachteil,
daß es
die Verarbeitung von nur einer Probe pro Gel erlaubt. Das Verfahren
kombiniert die schnelleren Trennzeiten von CE mit der Fähigkeit,
mehrere Proben parallel zu analysieren. Das zugrundeliegende Konzept
hinter der Verwendung von Mikrosystemen ist die Fähigkeit,
die Informationsdichte in der Elektrophorese durch Miniaturisierung
der Spwabmessung auf etwa 100 Mikrometer zu erhöhen. Die Elektronikindustrie
verwendet routinemäßig Mikrotechnologie,
um Schaltungen mit Merkmalen von weniger als 1 Mikron in der Größe herzustellen.
Die gegenwärtige
Dichte von Kapillaranordnungen ist beschränkt durch den Außendurchmesser
der Kapillarröhre.
Mikrotechnologisch hergestellte Kanäle erzeugen eine höhere Dichte
von Anordnungen. Mikrotechnologie ermöglicht auch physikalische Baugruppen,
die mit Glasfasern nicht möglich sind,
und verbindet die Kanäle
direkt mit anderen Vorrichtungen auf einem Chip. Mehrere Vonichtungen
sind auf Mikrochips für
Trenntechnologien konstruiert worden. Ein Gaschromatograph (Teny
et al., IEEE Trans. Electron Device, ED-26: 1880, 1979) und ein
Flüssigkeitschromatograph
(Manz et al., Sens. Actuators B 1: 249, 1990) sind auf Siliciumchips
hergestellt worden, aber diese Vorrichtungen sind nicht in breitem
Umfang eingesetzt worden. Mehrere Gruppen haben über die Trennung von Fluoreszenzfarbstoffen
und Aminosäuren auf
mikrotechnologisch hergestellten Vorrichtungen berichtet (Manz et
al., J. Chromatography 593: 253, 1992, Effenhauser et al., Anal.
Chem. 65: 2637, 1993). Vor kurzem haben Woolley und Mathies (Woolley
und Mathies, Proc. Natl. Acad. Sci. 91: 11348, 1994) gezeigt, daß Photolithographie
und chemisches Ätzen
verwendet werden können,
um eine große
Anzahl von Trennkanälen
auf Glassubstraten herzustellen. Die Kanäle werden mit Hydroxyethylcellulose(HEC)trennmatrizes
gefüllt.
Es wurde gezeigt, daß DNA-Restriktionsfragmente
in so wenig wie zwei Minuten getrennt werden konnten.
-
D. ABSPALTUNG VON MARKIERUNGEN
-
Wie oben beschrieben, werden unterschiedliche
Linkerkonstruktionen Spaltbarkeit ("Labilität") unter unterschiedlichen spezifischen
physikalischen oder chemischen Bedingungen verleihen. Beispiele
für Bedingungen,
die dazu dienen, verschiedene Linkerkonstruktionen zu spalten, schließen saure,
basische, Oxidations-, Reduktions-, Fluorid-, Thiolaustausch-, Photolyse-
und enzymatische Bedingungen ein.
-
Beispiele für spaltbare Linker, die die
allgemeinen Kriterien für
Linker, die oben aufgelistet sind, erfüllen, werden den Fachleuten
gut bekannt sein und schließen
diejenigen ein, die im von Pierce (Rockford, IL) erhältlichen
Katalog zu finden sind. Beispiele schließen ein:
-
- – Ethylenglycobis(succinimidylsuccinat)
(EGS), ein aminreaktives Vernetzungsreagens, das mit Hydroxylamin
spaltbar ist (1 M bei 37°C
für 3–6 Stunden);
- – Disuccinimidyltartrat
(DST) und Sulfo-DST, die aminreaktive Vernetzungsreagentien sind,
spaltbar mit 0,015 M Natriumperiodat;
- – Bis[2-(succinimidyloxycarbonyloxy)ethyl]sulfon
(BSOCOES) und Sulfo-BSOCOES,
die aminreaktive Vernetzungsmittel sind, spaltbar durch Base (pH
11,6);
- – 1,4-Di[3'-(2'-pyridyldithio(propionamido))butan
(DPDPB), ein Pyridyldithiol-Vernetzer,
der durch Thiolaustausch oder Reduktion spaltbar ist;
- – N-[4-(p-Azidosalicylamido)-butyl]-3'-(2'-pyridyldithio)propionamid
(APDP), ein Pyridyldithiol-Vernetzer, der durch Thiolaustausch oder
Reduktion spaltbar ist;
- – Bis-[beta-4-(azidosalicylamido)ethyl]-disulfid,
ein photoreaktiver Vernetzer, der durch Thiolaustausch oder Reduktion
spaltbar ist;
- – N-Succinimidyl-(4-azidophenyl)-1,3'-dithiopropionat
(SADP), ein photoreaktiver Vernetzer, der durch Thiolaustausch oder
Reduktion spaltbar ist;
- – Sulfosuccinimidyl-2-(7-azido-4-methylcumarin-3-acetamid)ethyl-1,3'-dithiopropionat
(SAED), ein photoreaktiver Vernetzer, der durch Thiolaustausch oder
Reduktion spaltbar ist;
- – Sulfosuccinimidyl-2-(m-azido-o-nitrobenzamido)-ethyl-1,3'-dithiopropionat
(SAND), ein photoreaktiver Vernetzer, der durch Thiolaustausch oder
Reduktion spaltbar ist.
-
Andere Beispiele für spaltbare
Linker und die Spaltungsbedingungen, die verwendet werden können, um
Markierungen freizusetzen, sind wie folgt. Eine Silyl-Verknüpfungsgruppe
kann durch Fluorid oder unter sauren Bedingungen gespalten werden.
Eine 3-, 4-, 5- oder 6-substituierte
2-Nitrobenzyloxy- oder 2-, 3-, 5- oder 6-substituierte 4-Nitrobenzyloxy-Verknüpfungsgruppe
kann durch eine Photonenquelle (Photolyse) gespalten werden. Eine
3-, 4-, 5- oder 6-substituierte 2-Alkoxyphenoxy- oder 2-, 3-, 5-
oder 6-substituierte 4-Alkoxyphenoxy-Verknüpfungsgruppe
kann durch Ce(NH4)2(NO3)6 (Oxidation) gespalten
werden. Ein NCO2(Urethan)-Linker kann durch
Hydroxid (Base), Säure
oder LiAlH4 (Reduktion) gespalten werden.
Eine 3-Pentenyl-, 2-Butenyl- oder 1-Butenyl-Verknüpfungsgruppe kann durch O3, OsO4/IO4
– oder KmnO4 (Oxidation)
gespalten werden. Eine 2-[3-, 4- oder 5-substituierte Furyl]oxy-Verknüpfungsgruppe
kann durch O2, Br2,
McOH oder Säure
gespalten werden.
-
Bedingungen für die Spaltung weiterer labiler
Verknüpfungsgruppen
schließen
ein: tert.-Alkyloxy-Verknüpfungsgruppen
können
durch Säure
gespalten werden; Methyl(dialkyl)methoxy- oder 4-substituierte 2-Alkyl-1,3-dioxan-2-yl-Verknüpfungsgruppen
können
durch H3O+ gespalten
werden; 2-Silylethoxy-Verknüpfungsgruppen
können
durch Fluorid oder durch Säure
gespalten werden; 2-(X)-Ethoxy (wobei X = Keto, Esteramid, Cyano,
NO2, Sulfid, Sulfoxid, Sulfon ist)-Verknüpfungsgruppen
können
unter alkalischen Bedingungen gespalten werden; 2-, 3-, 4-, 5- oder
6-substituierte Benzyloxy-Verknüpfungsgruppen
können
durch Säure
oder unter reduktiven Bedingungen gespalten werden; 2-Butenyloxy-Verknüpfungsgruppen
können
durch (Ph3P)3RhCl(H)
gespalten werden; 3-, 4-, 5- oder 6-substituierte 2-Bromphenoxy-Verknüpfungsgruppen
können
durch Li, Mg oder BuLi gespalten werden; Methylthiomethoxy-Verknüpfungsgruppen
können
durch Hg2+ gespalten werden; 2-(X)-Ethyloxy(wobei
X = ein Halogen)-Vernüpfungsgruppen
können
durch Zn oder Mg gespalten werden; 2-Hydroxyethyloxy-Verknüpfungsgruppen
können
durch Oxidation (z. B. mit Pb(OAc)4) gespalten
werden.
-
Bevorzugte Linker sind diejenigen,
die durch Säure
oder Photolyse gespalten werden. Mehrere der säurelabilen Linker, die für Festphasen-Peptidsynthese
entwickelt worden sind, sind zur Verknüpfung von Markierungen mit
MOIs nützlich.
Einige dieser Linker sind bei einem kürzlich erschienenen Überblick
von Lloyd-Williams et al. beschrieben (Tetrahedron 49: 11065–11133,
1993). Ein nützlicher
Linker-Typ beruht auf p-Alkoxybenzylalkoholen, von denen zwei, 4-Hydroxymethylphenoxyessigsäure und
4-(4-Hydroxymethyl-3-methoxyphenoxy)buttersäure, von
Advanced ChemTech (Louisville, KY) kommerziell erhältlich sind. Beide
Linker können über eine
Esterbindung mit dem Benzylalkohol an eine Markierung gebunden werden
und über
eine Amidbindung mit der Carbonsäure
an ein aminhaltiges MOI. Markierungen, die durch diese Moleküle gebunden
sind, werden vom MOI mit variierenden Konzentrationen in Trifluoressigsäure freigesetzt.
Die Spaltung dieser Linker führt
zur Freisetzung einer Carbonsäure
auf der Markierung. Säurespaltung
von Markierungen, die durch verwandte Linker gebunden sind, wie
etwa 2-Dimethoxy-4'-(carboxymethyloxy)-benzhydrylamin (erhältlich von
Advanced ChemTech in FMOC-geschützter Form),
führt zur
Freisetzung eines Carbonsäureamids
auf der freigesetzten Markierung.
-
Die photolabilen Linker, die für diese
Anwendung nützlich
sind, sind ebenfalls größtenteils
für die
Festphasen-Peptidsynthese entwickelt worden (siehe Übersicht
von Lloyd-Williams). Diese Linker beruhen üblicherweise auf 2-Nitrobenzylestern
oder 2-Nitrobenzylamiden. Zwei Beispiele für photolabile Linker, über die vor
kurzem in der Literatur berichtet worden ist, sind 4-(4-(1-FMOC-Amino)ethyl)-2-methoxy-5-nitrophenoxy)butansäure (Holmes
und Jones, J. Org. Chem. 60: 2318–2319, 1995) und 3-(FMOC-Amino)-3-(2-nitrophenyl)propionsäure (Brown
et al., Molecular Diversity 1: 4–12, 1995). Beide Linker können über die
Carbonsäure an
ein Amin auf dem MOI gebunden werden. Die Bindung der Markierung
an den Linker erfolgt durch Bildung eines Amids zwischen einer Carbonsäure auf
der Markierung und dem Amin auf dem Linker. Die Spaltung photolabiler
Linker erfolgt üblicherweise
mit UV-Licht mit 350 nm Wellenlänge
bei Intensitäten
und Zeiträumen,
die den Fachleuten bekannt sind. Die Spaltung der Linker führt zu einer
Freisetzung eines primären
Amids auf der Markierung. Beispiele für photospaltbare Linker schließen Nitrophenylglycinester,
exo- und endo-2-benzonorborneylchloride
und Methansulfonate und 3-Amino-3(2-nitrophenyl)propionsäure ein.
Beispiele für
enzymatische Spaltung schließen
Esterasen, die Esterbindungen spalten werden, Nukleasen, die Phosphodiester
spalten werden, Proteasen, die Peptidbindungen spalten werden, etc.
ein.
-
Geeignete Vorrichtungen, die dazu
verwendet werden können,
die Photospaltung von markierten Molekülen durchzuführen, schließen die
Vorrichtung ein, die unter der Abkürzung „PHRED" bekannt ist, was für Photochemischer Reaktor für den verstärkten Nachweis
steht und von Aura Industries, Staten Island, NY erhältlich ist
(erhältlich
mit sowohl einer 254 nm und 366 nm Lampe), PhotoBlaster System –1 mit dem
LuxTube Aufbau, von Agrenetics, 81 Salem Street, Wilmington, MA,
USA 01187 (erhältlich
mit einer 366 nm Lampe, jedoch enthält sie Röhre einen Photokatalysator,
der durch 366 nm Licht aktiviert wird, was zu einer Emission von
einem Bereich von Wällenlängen führt, der
254 nm einschließt).
-
E. NACHWEIS VON MARKIERUNGEN
-
Nachweisverfahren beruhen typischerweise
auf der Absorption und Emission in bestimmten Spektraltypen. Wenn
Atome oder Moleküle
Licht absorbieren, regt die einstrahlende Energie eine quantisierte
Struktur in ein höheres
Energieniveau an. Der Anregungstyp hängt von der Wellenlänge des
Lichtes ab. Elektronen werden durch ultraviolettes oder sichtbares
Licht auf höhere
Orbitale angehoben, Molekülvibrationen
werden durch Infrarotlicht angeregt und Rotationen werden durch
Mikrowellen angeregt. Ein Absorptionsspektrum ist die Lichtabsorption
als eine Funktion der Wellenlänge.
Das Spektrum eines Atoms oder Moleküls hängt von seiner Energieniveaustruktur
ab. Absorptionsspektren sind nützlich
für die
Identifizierung von Verbindungen. Spezifische absorptionsspektroskopische
Verfahren schließen
Atomabsorptionsspektroskopie (AA), Infrarotspektroskopie (IR) und
UV-vis-Spektroskopie
(UV-vis) ein.
-
Atome oder Moleküle, die auf hohe Energieniveaus
angeregt werden, können
durch Abgabe von Strahlung auf niedrigere Niveaus absteigen. Diese
Lichtemission wird Fluoreszenz genannt, wenn der Übergang
zwischen Zuständen
mit demselben Spin erfolgt, und Phosphoreszenz, wenn der Übergang
zwischen Zuständen
mit unterschiedlichem Spin auftritt. Die Emissionsintensität des Analyten
ist linear proportional zur Konzentration (bei niedrigen Konzentrationen)
und ist nützlich
zur Quantifizierung der emittierenden Spezies. Spezifische emissionsspektroskopische
Verfahren schließen
Atomemissionsspektroskopie (AES), Atomfluoreszenzspektroskopie (AFS),
molekulare laserinduzierte Fluoreszenz (LIF) und Röntgenfluoreszenz
(XRF) ein.
-
Wenn elektromagnetische Strahlung
durch die Materie hindurchgeht, setzt der größte Teil der Strahlung seinen
Weg in seiner ursprünglichen
Richtung fort, ein kleinerer Teil wird aber in andere Richtungen
gestreut. Licht, das mit derselben Wellenlänge wie das einstrahlende Licht
gestreut wird, wird Rayleigh-Streuung genannt. Licht, das in durchsichtigen
Feststoffen aufgrund von Vibrationen (Phononen) gestreut wird, wird
Brillouin-Streuung genannt. Brillouin-Streuung ist typischerweise
um 0,1 bis 1 Wellenzahl vom einstrahlenden Licht verschoben. Licht,
das aufgrund von Vibrationen in Molekülen oder optischen Phononen
in undurchsichtigen Feststoffen gestreut wird, wird Raman-Streuung
genannt. Raman-gestreutes Licht wird um so viel wie 4.000 Wellenzahlen
vom einstrahlenden Licht verschoben. Spezifische streuungsspektroskopische
Verfahren schließen
Raman-Spektroskopie ein.
-
IR-Spektroskopie ist die Messung
der Wellenlänge
und Intensität
der Absorption von Licht im mittleren Infrarotbereich durch eine
Probe. Licht im mittleren Infrarotbereich (2,5–50 μm, 4.000–200 cm–1)
ist energetisch ausreichend, um Molekülvibrationen auf höhere Energieniveaus
anzuregen. Die Wellenlänge
der IR-Absorptionsbanden ist charakteristisch für spezifische Typen von chemischen
Bindungen und IR-Spektroskopie ist im allgemeinen am nützlichsten
zur Identifizierung organischer und metallorganischer Moleküle.
-
Absorptionsspektroskopie im nähen Infrarotbereich
(NIR) ist die Messung der Wellenlänge und Intensität der Absorption
von Licht im nahen Infrarotbereich durch eine Probe. Licht im nahen
Infrarotbereich überspannt
den Bereich 800 nm bis 2,5 μm
(12.500 bis 4.000 cm–1) und ist energetisch
ausreichend, um Obertöne und
Kombinationen von Molekülvibrationen
auf höhere
Energieniveaus anzuregen. NIR-Spektroskopie wird typischerweise
zw quantitativen Messung organischer funktioneller Gruppen verwendet,
insbesondere O-H, N-H und C=O. Die Komponenten und die Konstruktion
der NIR-Instrumentierung sind ähnlich
zu UV-vis-Absorptionsspektrometern.
Die Lichtquelle ist üblicherweise
eine Wolframlampe und der Detektor ist üblicherweise ein PbS-Festkörperdetektor.
Probenhalter können
Quarz oder Glas sein und typische Lösungsmittel sind CCl4 und CS2. Die bequeme
Instrumentierung der NIR-Spektroskopie
macht sie für
on-line-Überwachung
und Prozeßkontrolle
geeignet.
-
Absorptionsspektroskopie im ultravioletten
und sichtbaren Bereich ( UV-vis) ist die Messung der Wellenlänge und
Intensität
der Absorption von Licht im nahen ultravioletten und sichtbaren
Bereich durch eine Probe. Die Absorption im Vakuum-UV tritt bei
100–200
nm (105–50.000
cm–1),
Quarz-UV bei 200–350
nm (50.000–28.570
cm–1)
und im Sichtbaren bei 350–800
nm (28.570–12.500
cm–1)
auf und wird von dem Beer-Lambert-Bouguet-Gesetz beschrieben. Ultraviolettes
und sichtbares Licht sind energetisch ausreichend, um äußere Elektronen
in höhere
Energieniveaus zu heben. UV-vis-Spektroskopie kann üblicherweise
auf Moleküle
und anorganische Ionen oder Komplexe in Lösung angewendet werden. Die
UV-vis-Spektren
sind durch die breiten Merkmale der Spektren begrenzt. Die Lichtquelle
ist üblicherweise
eine Wasserstoff- oder Deuteriumlampe für UV-Messungen und eine Wolframlampe
für Messungen
im Sichtbaren. Die Wellenlängen
dieser kontinuierlichen Lichtquellen werden mit einem Wellenlängenseparator
ausgewählt,
wie etwa einem Prisma oder Gittermonochromator. Spektren werden
durch Abtasten des Wellenlängenseparators
erhalten und quantitative Messungen können aus einem Spektrum oder
bei einer einzigen. Wellenlänge
durchgeführt
werden.
-
Massenspektrometer nutzen den Unterschied
im Masse/Ladungs-Verhältnis
(m/z) ionisierter Atome oder Moleküle, um sie voneinander zu trennen.
Massenspektrometrie ist daher nützlich
zur Quantifizierung von Atomen oder Molekülen und auch zur Bestimmung
chemischer und struktureller Informationen über Moleküle. Moleküle haben deutliche Fragmentierungsmuster,
die Strukturinformation liefern, um Verbindungen zu identifizieren.
Die allgemeinen Betriebsweisen eines Massenspektrometers sind wie
folgt. Gasphasen-Ionen werden erzeugt, die Ionen werden in Raum
oder Zeit auf der Grundlage ihres Masse/Ladungs-Verhältnisses
getrennt und die Menge an Ionen jeden Masse/Ladungs-Verhältnisses
wird gemessen. Die Ionentrennkraft eines Massenspektrometers wird
durch die Auflösung
beschrieben, die definiert ist als R = m/delta m, wobei m die Ionenmasse
und delta m der Unterschied in der Masse zwischen zwei auflösbaren Peaks
in einem Massenspektrum ist. Ein Massenspektrometer mit einer Auflösung von
1.000 kann z. B. ein Ion mit einem m/z von 100,0 von einem Ion mit
einem m/z von 100,1 auflösen.
-
Im allgemeinen besteht ein Massenspektrometer
(MS) aus einer Ionenquelle, einem masseselektiven Analysator und
einem Ionendetektor. Die Magnetsektor-, Quadrapol- und Flugzeit-Konstruktionen
erfordern auch Extraktions- und Beschleunigungs-Ionenoptiken, um
Ionen aus dem Quellenbereich in den Masseanalysatorbereich zu überführen. Die
Details mehrerer Masseanalysatorkonstruktionen (fiir Magnetsektor-MS,
Quadrapol-MS oder Flugzeit-MS) sind unten diskutiert. Einzelfokussierungs-Analysatoren
für Magnetsektor-MS nutzen
einen Teilchenstrahlweg von 180, 90 oder 60 Grad. Die verschiedenen
Kräfte,
die das Teilchen beeinflussen, trennen Ionen mit unterschiedlichen
Masse/Ladungs-Verhältnissen.
Bei Doppelfokussierungsanalysatoren wird ein elektrostatischer Analysator
in diesem Instrumententyp hinzugefügt, um Teilchen mit Unterschieden
in den kinetischen Energien zu trennen.
-
Ein Quadrapol-Massenfilter für Quadrapol-MS
besteht aus vier Metallstäben,
die parallel angeordnet sind. Die angelegten Spannungen beeinflussen
die Bahn von Ionen, die sich entlang des Flugweges bewegen, der
zwischen den vier Stäben
zentriert ist. Für
gegebene Gleich- und Wechselspannungen gehen nur Ionen eines bestimmten
Masse/Ladungs-Verhältnisses
durch den Quadrapolfilter hindurch und alle anderen Ionen werden
aus ihrem ursprünglichen
Weg geworfen. Ein Massenspektrum wird durch Überwachen der Ionen erhalten,
die durch den Quadrapolfilter hindurchgehen, wenn die Spannungen
an den Stäben
variiert werden.
-
Ein Flugzeit-Massenspektrometer nutzt
die Unterschiede in der Durchgangszeit durch eine "Driftregion", um Ionen unterschiedlicher
Massen zu trennen. Er arbeitet in einem Pulsmodus, so daß Ionen
in Pulsen erzeugt und/oder in Pulsen extrahiert werden müssen. Ein
gepulstes elektrisches Feld beschleunigt alle Ionen in einer feldfreien
Driftregion mit einer kinetischen Energie qV, wobei q die Ionenladung
und V die angelegte Spannung ist. Da die kinetische Ionenenergie
0,5 mV2 beträgt, haben leichtere Ionen eine
höhere
Geschwindigkeit als schwerere Ionen und eneichen den Detektor am
Ende der Driftregion früher.
Der Ausgang eines Ionendetektors wird auf einem Oszilloskop als
eine Funktion der Zeit dargestellt, um das Massenspektrum zu erzeugen.
-
Der Ionenbildungsprozeß ist der
Startpunkt für
massenspektrometrische Analysen. Chemische Ionisierung ist ein Verfahren,
das ein Reagens-Ion einsetzt, um mit den Analytmolekülen (Markierungen)
zu reagieren, um Ionen durch entweder einen Protonen- oder Hydridtransfer
zu bilden. Die Reagens-Ionen werden erzeugt durch Einbringen eines
großen Überschusses
Methan (relativ zur Markierung) in eine Elektronenstoß(EI)-Ionenquelle.
Elektronenzusammenstöße erzeugen
CH4
+ und CH3
+, die weiter mit
Methan reagieren, um CH5
+ und
C2H5
+ zu
bilden. Ein weiteres Verfahren, um Markierungen zu ionisieren, ist
durch Plasma und Glühentladung.
Plasma ist ein heißes,
teilweise ionisiertes Gas, das Atome wirkungsvoll anregt und ionisiert. Eine
Glühentladung
ist ein Niederdruckplasma, das zwischen zwei Elektroden gehalten
wird. Elektronenstoßionisation
setzt einen Elektronenstrahl ein, üblicherweise von einem Wolframfaden
erzeugt, um Gasphasen-Atome oder -Moleküle zu ionisieren. Ein Elektron
aus dem Strahl schlägt
ein Elektron aus Analyt-Atomen oder
-Molekülen,
um Ionen zu erzeugen. Elektrosprayionisierung verwendet eine sehr
feine Nadel und eine Reihe von Abstreifern. Eine Probenlösung wird
in die Quellenkammer gesprüht,
um Tröpfchen
zu bilden. Die Tröpfchen
tragen Ladung, wenn sie aus der Kapillare austreten, und wenn das
Lösungsmittel
verdampft, verschwinden die Tröpfchen,
wobei sie hochgeladene Analytmoleküle zurücklassen. ESI ist besonders
nützlich für große biologische
Moleküle,
die schwierig zu verdampfen oder zu ionisieren sind. Bombardierung
mit schnellen Atomen (FAB) verwendet einen Hochenergiestrahl neutraler
Atome, typischerweise Xe oder Ar, der auf eine feste Probe trifft,
wodurch Desorption und Ionisierung bewirkt wird. Es wird für große biologische
Moleküle
verwendet, die schwierig in die Gasphase zu bringen sind. FAB bewirkt
geringe Fragmentierung und gibt üblicherweise
einen großen
Molekülionenpeak,
was sie nützlich
zur Bestimmung der relativen Molekülmasse macht. Der Atomstrahl
wird durch Beschleunigung von Ionen aus einer Ionenquelle durch
eine Ladungsaustauschzelle erzeugt. Ionen nehmen in Kollisionen
mit neutralen Atomen ein Elektron auf, um einen Strahl aus Hochenergie-Atomen
zu bilden. Laserionisierung (LIMS) ist ein Verfahren, bei dem ein
Laserpuls Material von der Oberfläche einer Probe ablöst und ein
Mikroplasma schafft, das einen Teil der Probenbestandteile ionisiert. MatriX-unterstützte Laserdesorptionsionisation
(MALDI) ist ein LIMS-Verfahren zum Verdampfen und Ionisieren großer biologischer
Moleküle,
wie etwa Proteine oder DNA-Fragmente. Die biologischen Moleküle werden in
einer festen Matrix, wie etwa Nicotinsäure, dispergiert. Ein UV-Laserpuls
trägt Material
von der Matrix ab, was einen Teil der großen Moleküle in einer ionisierten Form
in die Gasphase bringt, so daß sie
in einen Massenspektrometer extrahiert werden können. Plasmadesorptionsionisation
(PD) verwendet den Zerfall von 252Cf, was
zwei Fissionsfragmente erzeugt, die sich in entgegengesetzte Richtungen
bewegen. Ein Fragment trifft auf die Probe, wodurch 1–10 Analyt-Ionen
herausgeschlagen werden. Das andere Fragment trifft auf einen Detektor
und triggert den Start der Datenaufnahme. Dieses Ionisationsverfahren
ist besonders nützlich
für große biologische
Moleküle.
Resonanzionisation (RIMS) ist ein Verfahren, bei dem ein oder mehrere
Laserstrahlen in Resonanz auf Übergänge eines
Gasphasen-Atoms oder -Moleküls
eingestellt werden, um es schrittweise über sein Ionisationspotential
zu heben, um ein Ion zu erzeugen. Sekundärionisation (SIMS) verwendet
einen Ionenstrahl, wie etwa 3He+, 16O+ oder 40Ar+, wird auf die
Oberfläche
einer Probe fokussiert und zerstäubt Material
in die Gasphase hinein. Funkenquelle ist ein Verfahren, das Analyte
in feste Proben durch Pulsen eines elektrischen Stroms über zwei
Elektroden ionisiert.
-
Eine Markierung kann vor, während oder
nach der Abspaltung vom Molekül,
an die sie gebunden ist, mit Ladung versehen werden. Ionisationsverfahren
auf der Basis von Ionen-"Desorption", der direkten Bildung oder
Emission von Ionen von festen oder flüssigen Oberflächen haben
zunehmende Anwendung auf nicht-flüchtige und thermisch labile Verbindungen
ermöglicht.
Diese Verfahren eliminieren die Notwendigkeit der Verflüchtigung
neutraler Moleküle
vor der Ionisation und minimieren im allgemeinen die thermische
Zersetzung der Molekülspezies.
Diese Verfahren schließen
Felddesorption (Becky, Principles of Field Ionization and Field
Desorption Mass Spectrometry, Pergamon, Oxford, 1977), Plasmadesorption
(Sundgvist und MacFarlane, Mass Spectrom. Rev. 4: 421, 1985), Laserdesorption
(Karas und Hillenkamp, Anal. Chem. 60: 2299, 1988; Karas et al.,
Angew. Chem. 101: 805, 1989), Bombardierung mit schnellen Teilchen
(z. B. Bombardierung mit schnellen Atomen, FAB, und Sekundärionen-Massenspektrometrie,
SIMS, Barher et al., Anal. Chem. 54: 645A, 1982), und Thermospray(TS)-Ionisation
(Vestal, Mass Spectrom. Rev. 2: 447, 1983) ein. Thermospray wird
in breitem Umfang für
die on-line-Kombination mit Flüssigkeitschromatographie
angewendet. Die FAB-Verfahren mit kontinuierlichem Durchfluß (Caprioli
et al., Anal. Chem. 58: 2949, 1986) haben ebenfalls signifikantes
Potential gezeigt. Eine vollständigere
Auflistung von Ionisations/Massenspektrometrie-Kombinationen ist
Ionenfallen-Massenspektrometrie, Elektrosprayionisations-Massenspektrometrie,
Ionenspray-Massenspektrometrie, Flüssigionisations-Massenspektrometrie,
Massenspektrometrie mit Ionisation bei atmosphärischem Druck, Elektronenionisations-Massenspektrometrie,
Massenspektrometrie mit Ionisation durch Bombardierung mit metastabilen
Atomen, Massenspektrometrie mit Ionisation durch Bombardierung mit schnellen
Atomen, MALDI-Massenspektrometrie,
Photoionisations-Flugzeit-Massenspektrometrie, Lasertröpfchen-Massenspektrometrie,
MALDI-TOF-Massenspektrometrie, APCI-Massenspektrometrie, Nanospray-Massenspektrometrie,
Nebelsprayionisations-Massenspektrometrie, Massenspektrometrie mit
chemischer Ionisation, Resonanzionisations-Massenspektrometrie,
Sekundärionisations-Massenspektrometrie, Thermospray-Massenspektrometrie.
-
Die Ionisationsverfahren, die für nicht-flüchtige biologische
Verbindungen geeignet sind, haben überlappende Anwendbarkeitsbereiche.
Ionisationswirkungsgrade sind in hohem Maße abhängig von Matrixzusammensetzung
und Verbindungstyp. Gegenwärtig
verfügbare
Ergebnisse weisen darauf hin, daß die obere Molekülmasse für TS etwa
8.000 Daltons beträgt
(Jones und Krolik, Rapid Comm. Mass Spektrom. 1: 67, 1987). Da TS
hauptsächlich
mit Quadrapol-Massenspektrometern durchgeführt wird, leidet die Empfindlicheit typischerweise disproportional
bei höheren
Masse/Ladungs-Verhältnissen
(m/z). Flugzeit(TOF)-Massenspektrometer
sind kommerziell erhältlich
und besitzen den Vorteil, daß der
m/z-Bereich nur
durch den Detektorwirkungsgrad begrenzt ist. Vor kurzem sind zwei
zusätzliche
Ionisationsverfahren eingeführt
worden. Diese zwei Verfahren werden nun als Matrix-unterstützte Laserdesorption
(MALDI, Karas und Hillenkamp, Anal. Chem. 60: 2299, 1988; Karas
et al., Angew. Chem. 101: 805, 1989) und Elektrosprayionisation
(ESI) bezeichnet. Beide Methodiken haben einen sehr hohen Ionisationswirkungsgrad
(d. h. sehr hohes [erzeugte Molekülionen]/[verbrauchte Moleküle]). Die
Empfindlichkeit, die das letztendliche Potential der Technik definiert,
ist abhängig
von der Probengröße, der
Menge an Ionen, der Durchflußgeschwindigkeit,
dem Nachweisgrad und dem tatsächlichen
Ionisationswirkungsgrad.
-
Elektrospray-MS beruht auf einer
Idee, die als erstes in den 1960igern vorgeschlagen wurde (Dole
et al., J. Chem. Phys. 49: 2240, 1968). Elektrosprayionisation (ESI)
ist ein Mittel, um geladene Moleküle für die Analyse durch Massenspektroskopie
zu erzeugen. Kurz gesagt erzeugt Elektrosprayionisation hochgeladene Tröpfchen durch
Vernebeln von Flüssigkeiten
in einem stark elektrostatischen Feld. Die hochgeladenen Tröpfchen,
die im allgemeinen in einem Trockenbadgas bei atmosphärischem
Druck gebildet werden, schrumpfen durch Verdampfung von neutralem
Lösungsmittel,
bis die Ladungsabstoßung
die kohäsiven
Kräfte überwindet, was
zu einer "Coulomb-Explosion" führt. Der
genaue Mechanismus der Ionisation wird kontrovers diskutiert und
mehrere Gruppen haben Hypothesen vorgelegt (Blades et al., Anal.
Chem. 63: 2109–14,
1991; Kebarle et al., Anal. Chem. 65: A972–86, 1993; Fenn, J. Am. Soc.
Mass. Spektrom. 4: 524–35,
1993). Ungeachtet des letztendlichen Ionenbildungsprozesses erzeugt
ESI geladene Moleküle
aus Lösung
unter milden Bedingungen.
-
Die Fähigkeit, nützliche Massenspektrumdaten
von geringen Mengen eines organischen Moleküls zu erhalten, beruht auf
der effizienten Erzeugung von Ionen. Der Ionisationswirkungsgrad
für ESI
steht im Zusammenhang mit dem Ausmaß der positiven Ladung, die
mit dem Molekül
assoziiert ist. Die Verbesserung der Ionisation auf experimentellem
Wege hat üblicherweise
die Verwendung saurer Bedingungen mit sich gebracht. Ein weiteres
Verfahren, um die Ionisation zu verbessern, ist gewesen, quartäre Amine
zu verwenden, wenn möglich
(siehe Aebersold et al., Protein Science 1: 494–503, 1992; Smith et al., Anal.
Chem. 60: 436–41, 1988).
-
Elektrosprayionisation wird detaillierter
wie folgt beschrieben. Elektrospray-Ionenerzeugung erfordert zwei
Schritte: Dispersion hochgeladener Tröpfchen bei nahezu atmosphärischem
Druck, gefolgt von Zuständen,
um Verdampfung zu induzieren. Eine Lösung von Analyt-Molekülen wird
durch eine Nadel geleitet, die bei hohem elektrischen Potential
gehalten wird. Am Ende der Nadel dispergiert die Lösung in
einen Nebel von kleinen hochgeladenen Tröpfchen hinein, die die Analyt-Moleküle enthalten.
Die kleinen Tröpfchen
verdampfen schnell, und mit einem Verfahren der Felddesorption oder
Restverdampfung werden protonierte Proteinmoleküle in die Gasphase freigesetzt.
Ein Elektrospray wird im allgemeinen durch Anlegen eines hohen elektrischen
Feldes an einen geringen Flüssigkeitsstrom
(im allgemeinen 1–10 μl/min) aus
einer Kapillarröhre
erzeugt. Eine Potentialdifferenz von 3–6 kV wird typischerweise zwischen
der Kapillare und der Gegenelektrode angelegt, die 0,2 bis 2 cm
entfernt angeordnet ist (wo Ionen, geladene Cluster und sogar geladene
Tröpfchen, in
Abhängigkeit
vom Ausmaß der
Desolvatisierung, vom MS durch eine kleine Öffnung gesampelt werden können). Das
elektrische Feld führt
zu einer Ladungsakkumulation auf der Flüssigkeitsoberfläche des
Kapillarendes; somit sind die Flüssigkeitsdurchflußgeschwindigkeit,
der spezifische Widerstand und die Oberflächenspannung wichtige Faktoren
bei der Tröpfchenerzeugung.
Das hohe elektrische Feld führt
zum Aufbrechen der Flüssigkeitsoberfläche und
zw Bildung hochgeladener Flüssigkeitströpfchen.
Positiv oder negativ geladene Tröpfchen
können
in Abhängigkeit
von der Kapillarvorspannung erzeugt werden. Der negative Ionenmodus
erfordert das Vorhandensein eines Elektronenfängers, wie etwa Sauerstoff,
um elektrische Entladung zu hemmen.
-
Ein breiter Bereich von Flüssigkeiten
kann elektrostatisch in ein Vakuum gesprüht werden, oder mit Hilfe eines
Vernebelungsmittels. Die Verwendung von ausschließlich elektrischen Feldern
zur Vernebelung führt
zu einigen praktischen Beschränkungen
des Bereichs der Flüssigkeitsleitfähigkeit
und dielektrischen Konstante. Lösungsleitfähigkeit
von weniger als 10–5 Ohm ist bei Raumtemperatur
für ein
stabiles Elektrospray bei brauchbaren Flüssigkeitsdurchflußgeschwindigkeiten
erforderlich, was einer wäßrigen Elektrolytlösung von < 10–4 M
entspricht. Bei dem Modus, der sich für ESI-MS als am nützlichsten
erwiesen hat, führt
eine geeignete Flüssigkeitsdurchflußgeschwindigkeit
zu Dispersion der Flüssigkeit
als ein feiner Nebel. In kurzer Entfernung von der Kapillare ist
der Tröpfchendurchmesser
oft ziemlich gleichförmig
und in der Größenordnung von
1 μm. Von
besonderem Interesse ist, daß der
Elektrospray-Ionenstrom für
höhere
Flüssigkeitsdurchflußgeschwindigkeiten
nur insgesamt leicht ansteigt. Es gibt Hinweise, daß Erwärmung für die Manipulation
des Elektrosprays nützlich
ist. Leichte Erwärmung
ermöglicht
zum Beispiel, daß wäßrige Lösungen leicht
in Elektrospray überführt werden
können,
vermutlich aufgrund der verringerten Viskosität und Oberflächenspannung. Sowohl
thermisch unterstützte
als auch Gasvernebelungs-unterstützte
Elektrosprays ermöglichen,
daß höhere Flüssigkeitsdurchflußraten verwendet
werden können,
senken aber das Ausmaß der
Tröpfchenladung.
Die Bildung von Molekülionen
erfordert Bedingungen, die eine Verdampfung der anfänglichen
Tröpfchenpopulation bewirken.
Dies kann bei höheren
Drücken
durch einen Trockengasstrom bei mäßigen Temperaturen (< 60°C), durch
Erhitzen während
des Transports durch die Schnittstelle und (insbesondere im Falle
von Ioneneinfangverfahren) durch energetische Kollisionen bei relativ
niedrigem Druck durchgeführt
werden.
-
Obgleich die detaillierten Prozesse,
die ESI zugrundeliegen, unsicher bleiben, scheinen die sehr kleinen
Tröpfchen,
die durch ESI erzeugt werden, zu ermöglichen, daß nahezu jede Spezies, die
in Lösung
eine Nettoladung trägt,
nach Verdampfung von Restlösungsmittel
in die Gasphase überführt werden
kann. Massenspektrometrischer Nachweis erfordert dann, daß Ionen
nach Desolvatisierung einen verfolgbaren m/z-Bereich haben (<4.000 Daltons für Quadrapol-Instrumente)
sowie mit ausreichendem Wirkungsgrad erzeugt und überführt werden
können.
Der breite Bereich von gelösten
Stoffen, für
den bereits festgestellt wurde, daß ESI-MS dafür geeignet
ist, und das Fehlen einer substantiellen Abhängigkeit des Ionisationswirkungsgrades
von der relativen Molekülmasse
weist auf ein in hohem Maße
nicht-diskriminierendes und breit anwendbares Ionisationsverfahren
hin.
-
Die Elektrospray-Ionen-"Quelle" arbeitet bei nahezu
atmosphärischem
Druck. Die Elektrospray-"Quelle" ist typischerweise
eine Metall- oder Glaskapillare, die ein Verfahren zur elektrischen
Vorspannung der flüssigen
Lösung
relativ zu einer Gegenelektrode umfaßt. Lösungen, typischerweise Wasser-Methanol-Mischungen,
die den Analyten und oft weitere Additive, wie etwa Essigsäure, enthalten,
strömen
zum Kapillarende. Eine ESI-Quelle ist beschrieben worden (Smith
et al., Anal. Chem. 62: 885, 1990), die im wesentlichen jedes Lösungsmittelsystem
aufnehmen kann. Typische Durchflußgeschwindigkeiten für ESI sind
1 bis 10 μl/min.
Das prinzipielle Erfordernis einer ESI-MS-Schnittstelle ist, Ionen
aus dem Hochdruckbereich so wirkungsvoll wie möglich in das MS zu sampeln
und zu transportieren.
-
Der Wirkungsgrad von ESI kann sehr
hoch sein, was die Grundlage für
extrem empfindliche Messungen liefert, was für die hierin beschriebene Erfindung
nützlich
ist. Die augenblickliche Instrumentenleistung kann einen Gesamtionenstrom
am Detektor von etwa 2 × 10–12 A
oder etwa 107 Zählimpulse/s für einfach
geladene Spezies liefern. Auf der Basis der Instrumentenleistung
werden so niedrige Konzentrationen wie 10–10 M
oder etwa 10–18 mol/s
einer einfach geladenen Spezies einen nachweisbaren Ionenstrom (etwa
10 Zählimpulse/s)
ergeben, wenn der Analyt vollständig
ionisiert ist. Nachweisgrenzen im unteren Attomol-Bereich sind z. B.
für quartäre Ammonium-Ionen
unter Verwendung einer ESI-Schnittstelle mit Kapillarzonenelektrophorese erhalten
worden (Smith et al., Anal. Chem. 59: 1230, 1988). Für eine Verbindung
mit einer relativen Molekülmasse
von 1.000 ist die mittlere Zahl von Ladungen 1, die mittlere Zahl
von Ladungszuständen
1, die Peakbreite (m/z) 1 und die maximale Intensität (Ionen/s)
1 × 1012.
-
Eine bemerkenswert kleine Probe wird
beim Erstellen eines ESI-Massenspektrums tatsächlich verbraucht (Smith et
al., Anal. Chem. 60: 1948, 1988). Substantielle Gewinne könnten auch
durch die Verwendung von Array-Detektoren mit Sektorinstrumenten
erzielt werden, was gleichzeitigen Nachweis von Teilen des Spektrums
ermöglicht.
Da gegenwärtig
nur etwa 10–5 aller
Ionen, die von ESI gebildet werden, nachgewiesen werden, kann eine
Fokussierung auf die Faktoren, die die Instrumentenleistung begrenzen,
eine Grundlage für verbesserte
Empfindlichkeit liefern. Es wird den Fachleuten deutlich sein, daß die vorliegende
Erfindung Verbesserungen für
Ionisations- und Nachweismethodiken überdenkt und anpaßt.
-
Vorzugsweise wird zwischen die Trennvonichtung
(z. B. Gel) und den Detektor (z. B. Massenspektrometer) eine Schnittstelle
gesetzt. Die Schnittstelle hat vorzugsweise die folgenden Eigenschaften:
(1) die Fähigkeit,
die DNA-Fragmente in diskreten Zeitintervallen zu sammeln, (2) die
DNA-Fragmente zu konzentrieren, (3) die DNA-Fragmente aus den Elektrophoresepuffern
und dem Milieu zu entfernen, (4) die Markierung vom DNA-Fragment
abzuspalten, (5) die Markierung vom DNA-Fragment zu trennen, (6)
das DNA-Fragment zu verwerfen, (7) die Markierung in eine flüchtige Lösung zu
geben, (8) die Markierung zu verflüchtigen und zu ionisieren und
(9) die Markierung in eine Elektrosprayvorrichtung einzubringen
oder dorthin zu transportieren, die die Markierung in das Massenspektrometer
einführt.
-
Die Schnittstelle hat auch die Fähigkeit,
DNA-Fragmente zu "sammeln", wenn sie aus dem
Boden eines Gels eluieren. Das Gel kann aus einem Slabgel, einem
Röhrengel,
einer Kapillare, etc. bestehen. Die DNA-Fragmente können mit
verschiedenen Verfahren gesammelt werden. Das erste Verfahren ist
dasjenige der Verwendung eines elektrischen Feldes, wobei DNA-Fragmente
auf oder nahe einer Elektrode gesammelt werden. Ein zweites Verfahren
ist dasjenige, bei dem die DNA-Fragmente durch Leiten eines Flüssigkeitsstroms
am Boden eines Gels vorbei gesammelt werden. Aspekte beider Verfahren
können
kombiniert werden, wobei DNA in einen fließenden Strom hinein gesammelt
wird, der später
unter Verwendung eines elektrischen Feldes konzentriert werden kann.
Das Endergebnis ist, daß DNA-Fragmente
aus dem Milieu entfernt werden, in dem das Trennverfahren durchgeführt wurde.
Das heißt,
daß DNA-Fragmente
durch Verwendung eines elektrischen Feldes aus einem Lösungstyp
in einen anderen "gezogen" werden können.
-
Wenn die DNA-Fragmente erst einmal
in der geeigneten Lösung
sind (kompatibel mit Elektrospray und Massenspektrometrie), kann
die Markierung vom DNA-Fragment abgespalten werden. Das DNA-Fragment (oder
Reste desselben) kann von der Markierung durch das Anlegen eines
elektrischen Feldes getrennt werden (vorzugsweise hat die Markierung
die entgegengesetzte Ladung zu derjenigen der DNA-Markierung). Die Markierung
wird anschließend
durch die Verwendung eines elektrischen Feldes oder einer strömenden Flüssigkeit
in die Elektrosprayvonichtung eingebracht.
-
Die Nachweis-Vorrichtung ist bevorzugterweise
ein Massenspektrometer. Weil Massenspektrometer den Unterschied
in den Masse-zu-Ladungsverhältnis
(m/e) von ionisierten Spezies verwenden, um spezifisch Moleküle zu identifizieren,
ist diese Nachweistechnik zur Quantifizierung von kleinen Molekülen brauchbar
und auch zur Bestimmung von chemischen und strukturellen Informationen über Moleküle. Einige
Moleküle
weisen distinkte Fragmentierungsmuster auf, die Information zur
Verfügung
stellen können,
um strukturelle Komponenten zu identifizieren. Jedoch, für die hier
beschriebene Verwendung wird das MSD im wesentlichen als ein "Anay-Detektor" für den Nachweis,
die Messung und die Quantifizierung von Markern von bekannten Molekulargewicht
verwendet. Daher kann ein Massenspektrometer so ähnlich verwendet werden, wie
Wissenschaftler im Augenblick einen Dioden-Array-Detektor in einen
UVNIS-Spektrometer verwenden, um kleine Moleküle mit bekannten Extinktionskoeffizienten
zu messen. In der hier beschriebenen Anwendung werden die Marker
dazu verwendet, um die Anwesenheit oder Abwesenheit von spezifischen
Nukleinsäuresequenzen
zu identifizieren und die Probenidentifikation zu kartieren.
-
Ein Quadrapol-Massendetektor besteht
aus vier parallelen Metallstäben.
Zwei gegenüberliegende Stäbe weisen
ein angelegtes Potential von (U-Vcos(wt)) auf und die anderen zwei
Stäbe weisen
ein Potential von –(U-Vcos(wt))
auf, worin U eine DC-Spannung und Vcos(wt) eine AC-Spannung ist.
Die angelegten Spannungen beeinträchtigen die Trajektion von
Ionen, die entlang der Flugstrecke wandern, die zwischen den vier Stäben zentriert
ist. Für
gegebene DC- und AC-Spannungen passieren nur die Ionen eines bestimmten
Masse-zu-Ladungsverhältnisses
durch den Quadrapol-Filter und alle anderen Ionen werden aus ihrer
Ausgangsbahn beworfen. Ein Masenspektrum wird durch Überwachen
der Ionen erhalten, die durch den Quadrapol-Filter passieren, während die
Spannungen an den Stäben
variiert werden. Die Ionentrennfähigkeit
eines Massenspektrometers wird durch die Auflösung beschrieben, die die Differenz
in der Masse zwischen zwei auflösbaren
Peaks in einem Massenspektrum ist. Gemeint ist, ein Massenspektrometer
mit einer Auflösung
von 1000 kann ein Ion mit einer m/e von 100,0 von einem Ion mit
einer m/e von 100,1 auflösen.
Der allgemeine Betrieb eines Massenspektrometers ist es, zuerst
Gas-Phase-Ionen zu erzeugen, zweitens, die Ionen im Raum oder in
der Zeit basierend auf ihren Masse-zu-Ladungsverhältnis zu
trennen und drittens, die Menge von Ionen von jedem Masse-zu-Ladungsverhältnis zu
messen.
-
Das Massenspektrometer ist ideal
als ein Spektrometer für
Anwendungen in Genomics geeignet, da es die gleichzeitige Messung
von hunderten von Markern erlaubt. Die augenblickliche Zahl von
genomischen Anwendungen (Sequenzierung, Kartierung, Genotypisierung)
verwendet an Markern, ist ungefähr
4, was sich aus den überlappenden
Emissionsspektrum von fluoreszenten Markern ergibt, die zwischen
300 nm und 700 nm plaziert werden können. Im Unterschied dazu können mit
augenblicklich verwendeten Quadrapol-Instrumenten (sowie dem Micromass
MS, dem Hewlett Packard LC/MSD 1100, den PE Sciex API 165 LC/MS
oder dem Finnigan Navigator) ungefähr 400 Marker in den Spektren
von 50–3000
amu plaziert werden. Die MS-Instrumente weisen mindestens 0,1 amu
auf Lösung
auf. Das neue Meßsystem,
das wir beschreiben (d. h. markierte Biomoleküle) kann in Verbindung mit
fast allen kommerziellen Massenspektrometern und HPLC-Systemen mit
geringen Modifikationen verwendet werden. Idealer weise wird ein
Software-Paket mit der "Driver"-Software beigefügt werden,
um die molekularbiologischen, genetischen oder genomischen Anwendungen zu
erarbeiten. Geeignete Software-Pakete sind beschrieben in z. B.
U.S. Provisional Patent Application Nr. 60/053,429, angemeldet am
22. Juli 1997.
-
Atmosphärischer Druck chemischer Ionisierung
(APCI) kann auf eine Vielzahl von Markertypen angewendet werden
und wird im allgemeinen dazu verwendet, die Häufigkeit des molekularen Ions
(Moleküle
Formen seltener Addukte während
APCI) zu verstärken.
Der brauchbare Molekulargewichtsbereich von APCI ist 50 bis 3000
amu (im allgemeinen). Die Genauigkeit der Messung bei niedriger
Auflösungsfähigkeit
ist 0,1 amu und in dem hochauflösenden
Modus, 5 ppm. Die APCI verwendet ein Reagenz-Ion, um mit den Analytmolekülen zu reagieren,
um Ionen durch entweder einen Protonen- oder Hydridtransfer zu bilden.
Im Augenblick werden nur ungefähr
10–4 bis
10–5 von
allen durch APCI gebildeten Ionen nachgewiesen. Dieser wichtige
Parameter (der die Instrumenten-Performance begrenzt) kann eine
Basis für
erhöhte
Sensitivität
zur Verfügung stellen.
Diese Ionsierungstechnik ist ein kontinuierliches Verfahren, daß zur Verwendung
als eine Schnittstelle mit HPLC oder Kapillar-Elektrophorese geeignet
ist.
-
Es gibt alternative Formen von Ionisation,
die für
die CMST-Technologie angewendet werden können. Die Elektrospray-Ionisation
(ESI) ermöglicht
die Produktion von Molekularionen, die direkt aus Proben in Lösung. Die
Elektrospray-Ionisation ist ein Verfahren, das mit einzelnen Quadrapol-Instrumenten
kompatibel ist. Sehr wenig Probe wird während des Erhalts eines ESI-Massenspektrums
(Smith et al., Anal. Chem. 60: 1948, 1988) gebraucht, jedoch bleibt
die Gesamteffizienz der Ioneneinführung in das MS-Vakuumsystem
relativ ineffizient. Die ESI leidet auch an den Effekten von denaturierenden
Mitteln und Detergentien, die den Ionisationsschritt nachteilig
beeinflussen. Sie kann auch für
Biopolymere kleinen und großen
Molekulargewichts (Peptide, Proteine, Kohlenhydrate und DNA-Fragmente)
und Lipide verwendet werden. Anders als MALDI, die gepulst ist,
ist ESI ein kontinuierliches Ionisationsverfahren. Mit ESI als der
Ionisationsmethode werden mehrfach geladene Ionen normalerweise
hergestellt (Moleküle
neigen mehr zur Adduktbildung).
-
Ein alternatives Ionisationsverfahren
ist die Matrix-unterstützte
Laser-Desorption (MALDI), die dazu verwendet werden kann, um das
Molekulargewicht von hier beschriebenen Markern zu bestimmen (Peptide, Proteine,
Oligonukleotide und andere Verbindungen von biologischen Ursprung,
sowie von kleinen synthetischen Polymeren können auch gemessen werden).
Die Menge von erforderlicher Probe ist sehr gering (pmol oder weniger).
Die Analyse kann auf eine lineare Weise (hohe Masse, niedriger Auflösung) bis
zu einem Molekulargewicht von m/z 300.000 (in seltenen Fällen) oder
Reflektronmodus (geringere Masse, höhere Auflösung) bis zu einem Molekulargewicht
von 10.000 durchgeführt
werden.
-
APCI und ESI sollten im allgemeinen
als ein Komplement zu MALDI betrachtet werden. MALDI weist den schweren
Nachteil auf, ein nicht-quantifizierbares Ionisationsverfahren zu
sein. Die Elektrospray-Ionisation wwde auf HP-Quadrapol, den Finnigan
LCQ, den Finnigan TSQ 7000 (Medicine), den PE/Sciex-Instrument und
dem Micromass-Instrument installiert. Sowohl APCI und ESI weisen
den Vorteil über
MALDI auf, daß sie in
der Lage sind, Echtzeitmessungen von Markern während jeder Art von Abtrenntechnologie
(d. h. HPLC und Elektrophorese) durchzuführen.
-
Fluoreszenz-Markierungen können am
direktesten durch ihre Absorptions- und Fluoreszenzemissionswellenlängen und
-intensitäten
identifiziert und quantifiziert werden.
-
Während
ein herkömmlicher
Spektrofluorometer extrem flexibel ist, wobei er kontinuierliche
Bereiche für
die Anregungs- und Emissionswellenlängen liefert (IEX,
IS1, IS2), können spezialisiertere
Instrumente, wie etwa durchflußcytometer
und Laserabtastmikroskope, Sonden erfordern, die bei einer einzigen
fixierten Wellenlänge
anregbar sind. In heutigen Instrumenten ist dies üblicherweise
die 488-nm-Linie des Argonlasers.
-
Die Fluoreszenzintensität pro Sondenmolekül ist proportional
zum Produkt von E und QY. Der Bereich dieser Parameter unter Fluorophoren
von gegenwärtiger
praktischer Bedeutung ist ungefähr
10.000 bis 100.000 cm–1M–1 für e und
0,1 bis 1,0 für
QY. Wenn die Absorption durch Hochintensitätsbeleuchtung zur Sättigung
getrieben wird, wird die irreversible Zerstörung des angeregten Fluorophors
(Photobleichung) der Faktor, der die Fluoreszenznachweisbarkeit
begrenzt. Die praktische Auswirkung der Photobleichung hängt von der
jeweiligen Fluoreszenznachweistechnik ab.
-
Es wird einem Fachmann deutlich sein,
daß eine
Vorrichtung (eine Schnittstelle) zwischen die Trenn- und Nachweisschritte
geschaltet werden kann, um den kontinuierlichen Betrieb von Größentrennung
und Markierungsnachweis (in Echtzeit) zu ermöglichen. Dies vereinheitlicht
die Trennmethodik und -instrumentierung mit der Nachweismethodik
und -instrumentierung, unter Bildung einer einzigen Vorrichtung.
Eine Schnittstelle ist z. B. zwischen eine Trenntechnik und den
Nachweis durch Massenspektrometrie oder potentiostatischer Amperometrie
geschaltet.
-
Die Funktion der Schnittstelle ist
primär
die Freisetzung der (z. B. Massenspektrometrie)-Markierung vom Analyten. Es gibt mehrere
repräsentative
Implementierungen der Schnittstelle. Die Konstruktion der Schnittstelle
ist abhängig
von der Auswahl der spaltbaren Linker. Im Falle von licht- oder
photospaltbaren Linkern ist eine Energie- oder Photonenquelle erforderlich.
Im Falle eines säurelabilen
Linkers, eines baselabilen Linkers oder eines Disulfid-Linkers ist
die Zugabe von Reagens in der Schnittstelle erforderlich. Im Falle
wärmelabiler
Linker ist eine Energiewärmequelle
erforderlich. Enzymzugabe ist erforderlich für einen enzymempfindlichen
Linker, wie etwa eine spezifische Protease und einen Peptid-Linker,
einen Nuklease- und einen DNA- oder RNA-Linker, eine Glycosylase,
HRP oder Phosphatase oder einen Linker, der nach Spaltung instabil
ist (z. B. ähnlich
zu Chemilumineszenzsubstraten). Andere Eigenschaften der Schnittstelle
schließen
minimale Bandenverbreiterung, Trennung der DNA von Markierungen
vor Injektion in einen Massenspektrometer -ein. Trenntechniken schließen diejenigen
ein, die auf Elektrophoreseverfahren und -techniken, Affinitätstechniken,
Größenretention
(Dialyse), Filtration und dergleichen beruhen.
-
Es ist auch möglich, die Markierungen (oder
das Nukleinsäure-Linker-Markierungs-Konstrukt) zu konzentrieren,
elektrophoretisch einzufangen und dann in einen alternativen Reagensstrom
freizusetzen, der mit dem bestimmten ausgewählten Ionisationsverfahrenstyp
kompatibel ist. Die Schnittstelle kann auch in der Lage sein, die
Markierungen (oder Nukleinsäure-Linker-Markierungs-Konstrukte)
auf Mikroperlen einzufangen, die Perle(n) in eine Kammer einzuschließen und
anschließend
Laserdesorption/Verdampfung durchzuführen.
-
Es ist auch möglich, im Strom in einen alternativen
Puffer hinein zu extrahieren (z. B. aus Kapillarelektrophoresepuffer
in hydrophoben Puffer durch eine permeable Membran). In einigen
Anwendungen kann es auch wünschenswert
sein, Markierungen intermittierend in den Massenspektrometer einzuführen, was
eine weitere Funktion der Schnittstelle umfassen würde. Eine
weitere Funktion der Schnittstelle ist, Markierungen aus mehreren
Säulen
in einen Massenspektrometer zuzuführen, mit einem rotierenden
Zeitslot für
jede Säule. Es
ist auch möglich,
Markierungen aus einer einzigen Säule in mehrere MS-Detektoren
einzuführen,
getrennt nach Zeit, jeden Satz von Markierungen für ein paar
Millisekunden zu sammeln und dann einem Massenspektrometer zuzuführen.
-
Die folgende ist eine Liste repräsentativer
Anbieter für
Trenn- und Nachweistechnologien, die in der vorliegenden Erfindung
verwendet werden können.
Hoefer Scientific Instruments (San Francisco, CA) stellt Elektrophoreseausrüstung (Two
Step® ,
Poker Face®II)
für Sequenzierungsanwendungen
her. Pharmacia Biotech (Piscataway, NJ) stellt Elektrophoreseausrüstung für DNA-Trennungen
und -sequenzierung her (PhastSystem für PCR-SSCP-Analyse, MacroPhor
System für
DNA-Sequenzierung). Perkin Elmer/Applied Biosystems Division (ABI,
Foster City, CA) stellt halbautomatisierte Sequenzer auf der Basis
von Fluoreszenzfarbstoffen her (ABI373 und ABI377). Analytical Spectral
Devices (Boulder, CO) stellt UV-Spektrometer her. Hitachi Instruments
(Tokio, Japan) stellt Atomabsorptionsspektrometer, Fluoreszenzspektrometer,
LC- und GC-Massenspektrometer, NMR-Spektrometer und UV-vis-Spektrometer
her. PerSeptive Biosystems (Framingham, MA) stellt Massenspektrometer
(Voyager® Elite)
her. Bruker Instrumens, Inc. (Manning Park, MA) stellt FTIR-Spektrometer
(Vector 22), FT-Raman-Spektrometer, Flugzeit-Massenspektrometer (Reflex II®,
Ionenfallen-Massenspektrometer (Esquire®) und
einen Maldi-Massenspektrometer her. Analytical Technology Inc. (ATI,
Boston, MA) stellt Kapillargelelektrophoreseeinheiten, UV-Detektoren
und Diodenarray-Detektoren her. Teledyne Electronic Technologies
(Mountain View, CA) stellt einen Ionenfallen-Massenspektrometer her (3DQ Discovery® und
den 3DQ Apogee®).
Perkin Elmer/Applied Biosystems Division (Foster City, CA) stellt
einen Sciex-Massenspektrometer (Dreifachquadrapol LC/MS/MS, den
API 100/300) her, der mit Elektrospray kompatibel ist.
-
Hewlett-Packard (Santa Clara, CA)
stellt massenselektive Detektoren (HP 5972A), MALDI-TOF-Massenspektrometer
(HP G2025A), Diodenarray-Detektoren, CE-Einheiten, HPLC-Einheiten (HP 1090)
sowie UV-Spektrometer her. Finnigan Corporation (San Jose, CA) stellt
Massenspektrometer (Magnetsektor (MAT 95 S®),
Quadrapol-Spektrometer (MAT 95 SQ®) und
vier weitere verwandte Massenspektrometer) her. Rainin (Ameryville,
CA) stellt HPLC-Instrumente
her.
-
Die Verfahren und Zusammensetzungen,
die hierin beschrieben sind, erlauben die Verwendung von abgespaltenen
Markierungen, um als Karten für
bestimmte Probentypen und Nukleotididentitäten zu dienen. Zu Beginn jedes
Sequenzierungsverfahrens wird ein bestimmter (ausgewählter) Primer
einer bestimmten einzigartigen Markierung zugeordnet. Die Markierungen
kartieren auf entweder einen Probentyp, einen Didesoxyterminatortyp
(im Falle einer Sanger-Sequenzierungsreaktion) oder vorzugsweise
beides. Genauer gesagt kartiert die Markierung auf einen Primertyp,
der seinerseits auf einen Vektortyp kartiert, der seinerseits auf
eine Probenidentität
kartiert. Die Markierung kann auch auf einen Didesoxyterminatortyp
(ddTTP, ddCTP, ddGTP, ddATP) durch Bezugnahme, in welche Didesoxynukleotidreaktion
der markierte Primer gegeben wird, kartieren. Die Sequenzierungsreaktion
wird dann durchgeführt,
und die resultierenden Fragmente werden sequentiell über die
Zeit nach Größe getrennt.
-
Die Markierungen werden von den Fragmenten
in einem Zeitrahmen abgespalten und in einem Zeitrahmen gemessen
und aufgezeichnet. Die Sequenz wird durch Vergleich der Markierungskarte
mit dem Zeitrahmen konstruiert. Das heißt, daß alle Markierungsidentitäten über die
Zeit nach dem Bemessungsschritt aufgezeichnet und miteinander in
einem Zeitrahmen in Beziehung gesetzt werden. Der Bemessungsschritt
trennt die Nukleinsäurefragmente
nach Inkrementen von einem Nukleotid, und die damit zusammenhängenden
Markierungsidentitäten
werden daher nach Inkrementen von einem Nukleotid getrennt. Durch
Vorkenntnis des Didesoxyterminators, oder der Nukleotidkarte und
des Probentyps kann die Sequenz leicht auf lineare Art und Weise
abgeleitet werden.
-
Ein genetisches Fingerprinting-System
der vorliegenden Erfindung besteht im allgemeinen aus einer Probeneinführungsvorrichtung,
einer Vorrichtung, um die markierten Proben von Interesse zu trennen,
eine Aufteilvorrichtung, um eine veränderliche Menge der Probe zu
einem Fraktionssammler umzuleiten, eine Vorrichtung, um die Marker
von den Proben von Interesse abzuspalten, eine Vorrichtung zum Nachweis
des Markers und ein Softwareprogramm, um die gesammelten Daten zu
analysieren und sie in einem Differential-Display-Modus wiederzugeben. Es wird
dem Fachmann ersichtlich sein, wenn er im Besitz der vorliegenden
Beschreibung ist, daß diese
allgemeine Beschreibung viele Varianten jeder der angeführten Komponenten
aufweisen kann. Wie am besten in 15 gesehen,
besteht ein beispielhaftes genetisches Fingerprinting-System 10 der
vorliegenden Erfindung aus einer Probeneinführvorrichtung 12,
einer Trennvorrichtung 14, die die Proben durch High-Performance-Liquid-Chromatographie
(HPLC) trennt, eine Aufteilvorrichtung 13, einen Fraktionssammler 15,
eine Photospaltungsvorrichtung 16, um die Marker von den
Proben von Interesse abzuspalten, eine Nachweisvorrichtung 18,
die die Marker mittels, eines elektrochemischen Detektors nachweist
und eine Daten verarbeitende Vorrichtung 20, mit einem
Datenanalyse-Softwareprogramm, das die Ergebnisse aus der Nachweisvorrichtung
analysiert. Jede Komponente wird im folgenden genauer diskutiert.
-
Die Probeneinführvorrichtung 12 nimmt
automatisch einen gemessenen Aliquot 22 des PCR-Produkts, das in
dem genetischen Fingerprinting-Verfahren erzeugt wurde und liefert
sie über
eine herkömmliche Leitung 24 zu
der Trennvorrichtung 14 (im allgemeinen eine HPLC). Die
Probeneinführvorrichtung 12 der
exemplarischen Ausführungsform
besteht aus einem Temperatur kontrollierten Autosampler 26,
der Mikrotiterplatten aufnehmen kann. Der Autosampler 26 ist
Temperatur kontrolliert, um die Integrität der erzeugten Nukleinsäureproben
aufrechtzuerhalten und ist in der Lage, 25 μl oder weniger an Probe zu injizieren.
Hersteller von diesem Typ von Probeneinführungsvorrichtung 12 sind,
zum Beispiel, Gilson (Middleton, WI).
-
Die Probeneinführvorrichtung 12 ist
operativ in Reihe mit der Auftrennvorrichtung 14 durch eine herkömmliche
Leitung 24 verbunden. Die PCR-Produkte in den gemessenen
Aliquot 22, die in der Auftrennvorrichtung 14 aufgenommen
werden, werden temporal durch High- Performance-Liquid-Chromatographie getrennt,
um getrennte DNA-Fragmente zur Verfügung zu stellen. Die High-Performance-Liquid-Chromatographie
kann eine isokratische, binäre
oder quaternäre
Pumpe(n) 27 aufweisen und kann von vielen Herstellern erworben
werden (z. B. Hewlett Packard (Palo Alto, CA) HP 1100 oder 1090
Serie, Beckman Instruments Inc. (800-142-2345), Bioanalytical Systems,
Inc. (800-845-4246), ESA, Inc. (508) 250-700), Perkin-Eimer Corp. (800-762-4000),
Varian Instruments (800-926-3000), Waters Corp. (800-254-4752)).
-
Die Auftrennvorrichtung 14 schließt eine
analytische HPLC-Säule 28 ein,
die zur Verwendung zur Auftrennung der Oligonukleotide geeignet
ist. Die Säule 28 ist
eine analytische HPLC, zum Beispiel modifizierter nicht-poröses Polystyroldivinylbenzol-(2,2 μm Partikelgröße)-fester
Träger,
der innerhalb eines pH-Bereichs von 2–12, Drücken von bis zu 3000 psi und
einem Temperaturbereich von 10 bis 70°C betrieben werden kann. Eine
Temperatur-Kontrollvorrichtung (z. B. ein Säulenofen) (nicht gezeigt) kann
dazu verwendet werden, um die Temperatur der Säule zu kontrollieren. Solche
Temperatur-Kontrollvorrichtungen
sind im Stand der Technik bekannt und können zum Beispiel von Rainin
Instruments (Tochterunternehmen von Varian Instruments, Palo Alto,
CA) erworben werden. Eine geeignete Säule 28 ist unter dem
kommerziellen Namen DNASep® erhältlich und ist von Serasep
(San Jose, CA) erhältlich.
Andere geeignete analytische HPLC-Säulen sind von anderen Herstellern
erhältlich
(z. B. Hewlett Packard (Palo Alto, CA), Beckman Industries (Brea,
CA), Waters Corp. (Milford, MA) und Supelco (Bellefonte, PA)).
-
Die Auftrennvorrichtung 14 in
der gezeigten Ausführungsform
enthält
den Probenteiler 13 und der Probenteiler ist mit dem Flußstrom der
Proben verbunden. Der Probenteiler 13 ist dazu angepaßt, um auf
eine herkömmliche
Weise variable Mengen von Probe zum Fraktionssammler 15 entweder
für weitere
Analyse oder Lagerung umzuleiten. Der Fraktionssammler 15 muß in der
Lage sein, kleine Volumen aufzunehmen, eine Temperaturkontrolle
zu niedrigen Temperaturen aufzuweisen und einstellbare Probenzeiten
zu haben. Hersteller von In-line-Abteilern schließen Upchurch
(Oak Harbor, WA) ein.
-
Der Fraktionssammler 15 ist
an die HPLC/LC-Vorrichtung über
eine Fluß-Aufteilungsleitung 29 angebracht.
Die Fraktionssammler 15 erlauben die Sammlung von spezifischen
Peaks, DNAs, RNAs und Nukleinsäurefragmenten
oder Molekülen
von Interesse in Röhrchen,
Vertiefungen von Mikrotiterplatten oder Behältern. Zusätzlich können die Fraktionssammler 15 alles
oder einen Teil eines Sets von Nukleinsäurefragmenten sammeln, die
durch HPLC oder LC getrennt wurden. Hersteller von Fraktionssammlern
schließen
Gilson (Middleton; WI) und Isco (Lincoln, NE) ein. Die Verwendung
eines Fraktionssammlers 15 in dieser Technologie stellt
bemerkenswerte wesentliche Vorteile über die Gel-basierten Systeme
zur Verfügung.
Zum Beispiel ist es möglich,
direkt Nukleinsäurefragmente
zu klonieren, die durch die HPLC- oder LC-Verfahren erhalten wurden. Zusätzlich ist
es möglich,
Nukleinsäurefragmente
durch PCR zu amplifizieren, die durch HPLC- oder LC-Verfahren erhalten
wurden. Diese zwei Verfahren ermöglichen
die schnelle Identifizierung von Nukleinsäurefragmenten von Interesse
auf einem Sequenzniveau. Beide Verfahren sind mühsam und ineffektiv, wenn in
Verbindung mit Gel-basierten Systemen verwendet.
-
In der dargestellten Ausführungsform
ist der Fraktionssammler 15 eine einzelne Komponente vom
genetischen Fingerprinting-System 10. In einer alternativen
Ausführungsform
(nicht gezeigt) ist der Fraktionssammler 15 in der Probeneinführungsvorrichtung 12 integriert.
Demzufolge richtet die Fluß-Teilungsleitung 32 die
geteilte Probe von dem Probenteiler 13 zurück zu der
Probeneinführvorrichtung 12.
-
Ein Strom der getrennten DNA-Fragmente
(z. B. Sequenzreaktionsprodukte) fließt durch eine herkömmliche
Leitung 30 von der Trennvorrichtung 14 stromabwärts des
Probenteilers 13 zu der Spaltungsvorrichtung 16.
Jedes der DNA-Fragmente ist mit einem einmaligen abspaltbaren (z.
B. Photo-abspaltbaren) Marker markiert. Der Flußstrom von aufgetrennten DNA-Fragmenten
passiert durch oder entlang der Spaltvorrichtung 16, wo
der Marker für
den Nachweis (z. B. durch Massenspektrometrie oder mit einem elektrochemischen Detektor)
entfernt wird. In der beispielhaften Ausführungsform ist die Spaltvorrichtung 16 eine
photospaltende Einheit, so daß der
Flußstrom
der Probe gegenüber
ausgewählter
Lichtenergie und Wellenlänge
ausgesetzt wird. In einer Ausführungsform
tritt die Probe in die photospaltende Einheit 16 ein und
wird gegenüber
der ausgewählten
Lichtquelle für
eine ausgewählte
Zeitdauer ausgesetzt. In einer alternativen Ausführungsform wird der Flußstrom der
Probe angrenzend an die Lichtquelle entlang eines Flußpfades
getragen, der ein ausreichendes Aussetzen gegenüber seiner Lichtenergie zur
Verfügung
stellt, um die Marker von den getrennten DNA-Fragmenten abzuspalten.
Eine photospaltende Einheit ist von Supelco (Bellefonte, PA) erhältlich.
Photospaltung kann bei vielen Wellenlängen mit einer Quecksilber/Xenon-Lichtbogenlampe
durchgeführt
werden. Die Wellenlängengenauigkeit
ist ungefähr
2 nm mit einer Bandbreite von 10 nm. Der bestrahlte Bereich ist kreisförmig und
typischerweise ein Bereich von 10–100 Quadratzentimetern. In
alternativen Ausführungsformen
können
andere Spaltvorrichtungen, die durch Säure, Base, Oxidation, Reduktion,
Fluorid, Thiolaustausch, Photolyse oder enzymatische Bedingungen
spalten, dazu verwendet werden, um die Marker von den aufgetrennten
DNA-Fragmenten zu entfernen.
-
Nachdem die Spaltvorrichtung 16 die
Marker von den aufgetrennten DNA-Fragmenten abspaltet, fließen die
Marker durch eine herkömmliche
Röhre 32 zur
Nachweisvorrichtung 18 zum Nachweis jedes Markers. Der
Nachweis der Marker kann auf dem Unterschied im elektrochemischen
Potential zwischen jedem der Marker basiert sein, die dazu verwendet
wurden, jede Art von DNA zu markieren, die in dem PCR-Schritt erzeugt wurde.
Der elektrochemische Detektor 18 kann auf entweder coulometrischen
oder amperometrischen Prinzipien beruhen. Der bevorzugte elektromechanische
Detektor 18 ist der coulometrische Detektor, der aus einem Durchfluß- oder
porösen
Carbongraphit amperometrischen Detektor besteht, wobei die Probe
durch die Elektrode hindurch passiert, was zu einer 100%-igen Nachweiseffizienz
führt.
Um jede Komponente vollständig nachzuweisen,
wird ein Array von 16 coulometrischen Detektoren, die jeder
bei einem unterschiedlichen Potential (im allgemeinen bei 60 mV
anstiegen) gehalten wird. Beispiele von Herstellern dieses Typs
von Detektor sind ESA (Bedford, MA) und Bioanalytical Systems Inc.
(800-845-4246).
-
In einer alternativen Ausführungsform,
die schematisch in 16 dargestellt
ist, sind die Probeneinführungsvorrichtung 12,
die Abtrennvorrichtung 14 und die Spaltvorrichtung 16 in
Reihe wie oben diskutiert miteinander verbunden, um den Fluß der Probe
aufrechtzuerhalten. Die Spaltvorrichtung 16 ist mit einer
Nachweisvorrichtung 18 verbunden, die ein Massenspektrometer 40 oder ähnliches
ist, die die Markierungen basierend auf dem Unterschied im Molekulargewicht
zwischen jedem der Marker, die dazu verwendet wurden, um jede Art
von DNA zu markieren, die in dem PCR-Schritt erzeugt wurde, nachzuweisen.
Der Beste auf Unterschieden in der Masse basierende Detektor ist
das Massenspektrometer. Für
diese Verwendung wird das Massenspektrometer 40 typischerweise
eine atmosphärischer
Druckionisations (API)-Schnittstelle mit entweder Elektrospray-
oder chemischer Ionisation, einen Quadrapol-Massenanalyser und einen
Massenbereich von mindestens 50 bis 2600 m/z aufweisen. Beispiele
von Herstellern eines geeigneten Massenspektrometers sind: Hewlett
Packard (Palo Alto, CA), HP 1100 LC/MSD, Hitachi Instruments (San
Jose, CA), M-1200H LC/MS, Perkin Elmer Corporation, Applied Biosystems
Division (Foster City, CA) API 100 LC/MS oder API 300 LC/MS/MS,
Finnigan Corporation (San Jose, CA) LCQ, MAT 95 S, Bruker Analytical
Systems, Inc. (Billerica, MA) APEX, BioAPEX und ESQUIRE und Micromass
(U.K.).
-
Die Nachweisvorrichtung 18 ist elektronisch
mit einem Datenprozessor und Analysen 20 verbunden, der
Daten von der Nachweisvorrichtung empfängt. Der Datenprozessor und
der Analysen r schließen
ein Softwareprogramm ein, das den nachgewiesenen Marken identifiziert.
Der Datenprozessor und Analysen 230 in alternativen Ausführungsformen
ist operativ mit der Injektionsvorrichtung 12, der Trennvorrichtung 14,
den Fraktionssammler 15 und/oder der Spaltvorrichtung 16 verbunden,
um die verschiedenen Komponenten des genetischen Fingerprinting-Systems 10 zu
kontrollieren.
-
Das Software-Paket kartiert die elektrochemische
Signatur eines bestimmten Markers mit einem spezifischen Primer
und einer Retentionszeit. Software erzeugte Nukleinsäureprofile
werden dann verglichen (Länge
zu Länge,
Fragment zu Fragment) und die Ergebnisse werden dem Verwender berichtet.
Die Software wird sowohl Ähnlichkeit
und Unterschiede in den Nukleinsäurefragmentprofilen
hervorheben. Die Software wird auch in der Lage sein, die Sammlung
von spezifischen Nukleinsäurefragmenten
durch den Fraktionssammler 15 zu steuern.
-
Das Software-Paket kartiert die m/z-Signatur
eines bestimmten Markers mit einem bestimmten Primer und einer Retentionszeit.
Software erzeugte Nukleinsäureprofile
werden dann verglichen (Länge
zu Länge, Fragment
zu Fragment) und die Ergebnisse dem Verwender berichtet. Die Software
wird sowohl die Ähnlichkeiten
als auch die Unterschiede in den Nukleinsäurefragmentprofilen hervorheben.
Die Software wird auch in der Lage sein, die Sammlung von spezifischen
Nukleinsäurefragmenten
durch den Fraktionssammler zu steuern.
-
Das System 18 in Übereinstimmung
mit der vorliegenden Erfindung wird durch operatives miteinander Verbinden
der multiplen Komponenten des Systems zur Verfügung gestellt. Demzufolge können eine
oder mehrere Systemkomponenten, wie zum Beispiel die Probeneinführvorrichtung 12 und
die Nachweisvorrichtung 18, die in einem Labor in Betrieb
sind, mit den anderen Komponenten des Systems, kombiniert werden (z.
B. der Abtrennvorrichtung 14, der Spaltvorrichtung 16 und
dem Datenprozessor und Analyser 20, um das Labor mit dem
DNA-Sequenzierungssystem 10 der vorliegenden Erfindung
auszurüsten.
-
Eine weitere Ausführungsform der vorliegenden
Erfindung stellt ein Differential-Display-System zur Verfügung, das im allgemeinen aus
einer Probeneinführvorrichtung,
eine Vorrichtung, um die markierten Proben von Interesse zu trennen,
einer Aufteilvorrichtung, um eine variable Menge der Probe zu einem
Fraktionssammler umzuleiten, eine Einheit, um die Marker von den
Proben von Interesse abzuspalten, eine Vorrichtung zum Nachweis
des Markers und ein Softwareprogramm, um die gesammelten Daten zu
analysieren und sie in einem Differential-Displaymodus wiederzugeben,
besteht. Es wird dem Fachmann ersichtlich sein, der sich in Besitz
der vorliegenden Offenbarung befindet, daß die allgemeine Beschreibung
viele Varianten für
jede der aufgelisteten Komponenten haben kann. Das Differential-Displaysystem
einer beispielhaften Ausführungsform der
vorliegenden Erfindung besteht aus ähnlichen Komponenten, die in 15 dargestellt sind, einschließlich der
Probeneinführvorrichtung 12,
der Abtrennvorrichtung 14 zur Abtrennung der Proben durch
High-Performance-Liquid-Chromatographie (HPLC), der Abteilvorrichtung 13,
dem Fraktionssammler 15, der Photospaltungsvorrichtung 16,
um die Macker von den Proben von Interesse abzuspalten, der Nachweisvorrichtung 18 zum
Nachweis der Macker durch Elektrochemie und den Datenprozessor und
Analyser 20, mit einem Softwareprogramm. Jede Komponente
wird unten genauer diskutiert.
-
In dem Differential-Displaysystem
nimmt die Probeneinführvorrichtung 12 automatisch
einen gemessenen Aliquot 22 des PCR-Produkts, der in dem
Differential-Displayverfahren erzeugt wurde und führt sie über eine
herkömmliche
Leitung 24 zu der Abtrennvorrichtung 14 (im allgemeinen
ein HPLC) zu. Die Probeneinführvorrichtung 12 der
beispielhaften Ausführungsform
besteht aus einem Temperatur kontrollierten Autosampler 26,
der Mikrotiterplatten aufnehmen kann. Der Autosampler 26 muß Temperatur-kontrolliert
sein, um die Integrität
der erzeugten Nukleinsäureproben
aufrechtzuerhalten und in der Lage sein 25 μl oder weniger an Probe zu injizieren.
Hersteller dieses Typs von Produkt sind zum Beispiel durch Gilson
(Middleton, WI repräsentiert.
-
Die Probeneinführvorrichtung ist operativ
in Reihe mit der Abtrennvorrichtung durch die herkömmliche Leitung 24 verbunden.
Die PCR-Produkte in den gemessenen Aliquots 22, die in
der Auftrennvorrichtung 14 aufgenommen werden, werden temporal durch
High-Performance-Liquid-Chromatographie
abgetrennt, um abgetrennte DNA-Fragmente zur Verfügung zu
stellen. Die High-Performance-Liquid-Chromatographie kann eine isokratische,
binäre
oder quaternäre
Pumpe(n) 27 aufweisen und kann von zahlreichen Herstellern
erworben werden (z. B. Hewlett Packard (Palo Alto, CA) HP 1100 oder
1090 Serien, Analytical Technology Inc. (Madison, WI), Perkin Elmer,
Waters, usw.). Die Auftrennvorrichtung 14 schließt eine
analytische HPLC-Säule 28 ein,
die zur Verwendung zur Abtrennung der Oligonukleotide geeignet ist.
Die Säule 28 ist
eine analytische HPLC, zum Beispiel, nicht poröses Polystyroldivinylbenzol
(2,2 μl
Partikelgröße) fester
Träger,
modifiziert, die innerhalb eines pH-Bereichs von 2 bis 12, drücken von
bis zu 3000 psi und einem Temperaturbereich von 10 bis 70°C betrieben
werden kann. Eine Temperatur-Kontrollvorrichtung
(z. B. ein Säulenofen)
(nicht gezeigt) kann dazu verwendet werden, um die Temperatur der
Säule zu
kontrollieren. Solche Temperatur-Kontrollvorrichtungen sind im Stand
der Technik bekannt und können
zum Beispiel von Rainin Instruments (Tochterunternehmen von Varian
Instruments, Palo Alto, CA) erworben werden. Eine geeignete Säule 28 ist
unter dem kommerziellen Namen DNAsep® erhältlich und
ist von Serasep (San Jose, CA) erhältlich. Andere geeignete analytische
HPLC-Säulen
sind von anderen Herstellern erhältlich
(z. B. Hewlett Packard (Palo Alto, CA), Beckman Industries (Brea,
CA), Waters Corp. (Milford, MA) und Supelco (Bellefonte, PA)).
-
In der dargestellten Ausführungsform
ist der Fraktionssammler 15 eine einzelne Komponente des
Differential-Displaystems 10, die an die anderen Komponenten
des Systems gekoppelt ist. In einer alternativen Ausführungsform
ist der Fraktionssammler 15 in die Probeneinführungsvorrichtung 12 integriert.
Demzufolge leitet die Fluß-Aufteilungsleitung 32 die
getrennten Proben von dem Probenteiler 13 zurück zu der
Probeneinführvorrichtung 12.
-
Die Auftrennvorrichtung 14 in
der dargestellten Ausführungsform
enthält
den Probenteiler 13, der mit dem Flußstrom der Probe verbunden
ist. Der Probenteiler 13 ist so angepaßt, um auf eine herkömmliche
Weise veränderliche
Mengen von Probe zu dem Fraktionssammler 15 entweder für die weitere
Analyse oder Lagerung abzuteilen. Der Fraktionssammler 15 muß in der
Lage sein, kleine Volumen aufzunehmen, eine Temperaturkontrolle
zu niedrigen Temperaturen aufzuweisen und einstellbare Probenzeiten
aufzuweisen. Hersteller von In-line-Abteilern schließen Upchurch (Oak Harbor, WA)
ein.
-
Ein Fluß der getrennten DNA-Fragmente
fließt
durch eine herkömmliche
Leitung 30 von der Auftrennvorrichtung 14 stromabwärts des
Probenteilers 113 zu der Spaltvorrichtung 16.
Jedes der PCR-Produkte ist mit einem einmaligen spaltbaren (z. B.
photospaltbaren) Marker markiert. Der Flußstrom von getrennten DNA-Fragmenten
passiert durch oder entlang der Spaltvorrichtung 16, wo
der Marker zum Nachweis durch elektrochemischen Nachweis entfernt
wird. In der beispielhaften Ausführungsform
ist die Spaltvorrichtung 16 eine photospaltende Einheit,
so daß der
Flußstrom
der Probe gegenüber
einer ausgewählten
Lichtenergie ausgesetzt wird. In einer Ausführungsform tritt die Probe
in die photospaltende Einheit 16 ein und wird gegenüber der
ausgewählten
Lichtquelle für
eine ausgewählte
Zeitdauer ausgesetzt. In einer alternativen Ausführungsform wird der Strom an
Probe in einem geeigneten Rohrteil oder ähnlichen in Angrenzung an die
Lichtquelle entlang eines Weges geführt, der ein ausreichendes
Aussetzen gegenüber
der Lichtquelle zur Verfügung
stellt, um die Marker von dem abgetrennten DNA-Fragmenten abzuspalten.
Eine photospaltende Einheit ist erhältlich von Supelco (Bellefonte,
PA). Die Photospaltung kann bei vielen Wellenlängen mit einer Quecksilber/Xenon-Bogenlampe
durchgeführt
werden. Die Wellenlängengenauigkeit
ist ungefähr
2 nm mit einer Bandbreite von 10 nm. Der bestrahlte Bereich ist
kreisförmig
und typischerweise ein Bereich von 10–100 Quadratzentimetern. In
alternativen Ausführungsformen
können
andere Spaltvorrichtungen, die durch Säure, Base, Oxidation, Reduktion,
Fluorid, Thiolaustausch, Photolyse oder enzymatische Bedingungen
spalten, dazu verwendet werden, die Marker von den abgetrennten
DNA-Fragmenten zu entfernen.
-
Nachdem die Spaltvorrichtung 16 die
Marker von den abgetrennten DNA-Fragmenten abspaltet, fließen die
Marker durch ein konventionelles Röhrchen 32 zur der
Nachweisvorrichtung 18 zum Nachweis jedes Markers. Der
Nachweis der Marker basiert auf dem Unterschied in dem elektrochemischen
Potential zwischen jedem der Marker, die dazu verwendet werden,
jede in dem PCR-Schritt erzeugte Art von DNA zu markieren. Der elektrochemische
Detektor 18 kann auf entweder coulometrischen oder amperometrischen
Prinzipien operieren. Der bevorzugte elektromechanische Detektor 18 ist
der coulometrische Detektor, der aus einem Durchfluß oder porösen Carbongraphit
amperometrischen Detektor besteht, wobei das Säuleneluat durch die Elektrode
hindurch passiert, was zu einer 100%-igen Nachweiseffizienz führt. Um
jede Komponente vollständig nachzuweisen,
wird ein Array von 16 coulometrischen Detektoren, wobei jeder bei
einem unterschiedlichen Potential gehalten wird (im allgemeinen
bei 60 mV anstiegen) verwendet. Die Hersteller dieses Typs von Detektor
schließen
ESA (Bedford, MA) und Bioanalytical Systems Inc. (800-845-4246)
ein.
-
In einer alternativen Ausführungsform
des schematisch in 16 dargestellten
Differential-Displaysystems
sind die Probeneinführungsvorrichtung 12,
die Abtrennvorrichtung 14 und die Spaltvorrichtung 16 in Reihe
verbunden, wie oben diskutiert, um den Fluß an Probe aufrechtzuerhalten.
Die Abspaltvorrichtung 16 ist mit einer Nachweisvorrichtung 18 verbunden,
die die Marker basieren auf den Unterschied im Molekulargewicht
zwischen jedem der Marker, die dazu verwendet werden, jede Art von
in PCR-Schritt erzeugte DNA zu markieren, basiert. Der Beste auf
Unterschieden in der Masse basierende Detektor ist das Massenspektrometer 40.
Für diese
Verwendung wird das Massenspektrometer typischerweise eine atmosphärische Druckionisations
(API)-Schnittstelle mit entweder Elektrospray oder chemischer Ionisation,
einen Quadrapol-Massenanalyser und einen Massenbereich von mindestens
50 bis 2600 m/z aufweisen. Beispiele von Herstellern von geeigneten
Massenspektrometern sind: Hewlett Packard (Palo Alto, CA), HP 1100
LC/MSD, Hitachi Instruments (San Jose, CA), M-1200H LC/MS, JEOL
USA, Inc. (Peabody, MA), Perkin Elmer Corporation, Applied Biosystems
Division (Foster City, CA) API 100 LC/MS oder API 300 LC/MS/MS,
Finnigan Corporation (San Jose, CA) LCQ, MAT 95 S, MAT 95 Q, MAT
900 S, MAT 900 SQ und SSQ 7000, Bruker Analytical Systems, Inc. (Billerica,
MA) APEX, BioAPEX und ESQUIRE.
-
Die Nachweisvorrichtung 18 ist
elektronisch mit einem Datenprozessor und Analyser 20 verbunden, der
Daten von der Nachweisvorrichtung empfängt. Der Datenprozessor und
der Analyser 20 schließen
ein Softwareprogramm ein, das den nachgewiesenen Marker und seine
Position in der DNA-Sequenz identifiziert. Der Datenprozessor und
Analyser 20 ist in alternativen Ausführungsformen operativ mit der
Injektionsvorrichtung 12, der Abtrennvorrichtung 14,
den Fraktionssammler 15 und/oder der Spaltvomchtung 16 verbunden,
um die verschiedenen Komponenten des Differential-Displaysystems
zu kontrollieren.
-
Das Softwarepaket kartiert die Signatur
eines gegebenen Markers auf einen spezifischen Primer und eine Retentionszeit.
Software erzeugte Nukleinsäureprofile
werden dann verglichen (Länge
zu Länge,
Fragment zu Fragment) und die Ergebnisse werden dem Verwender berichtet.
Die Software wird sowohl Ähnlichkeiten
als auch Unterschiede in den Nukleinsäurefragmentprofilen hervorheben.
Die Software wird auch in der Lage sein, die Sammlung von spezifischen
Nukleinsäurefragmenten
durch den Fraktionssammler 15 zu steuern.
-
Das Softwarepaket kartiert die m/z-Signatur
eines bestimmten Markers auf einen spezifischen Primer und einer
Retentionszeit. Die Software erzeugten Nukleinsäureprofile werden dann verglichen
(Länge
zu Länge,
Fragment zu Fragment) und die Ergebnisse dem Verwender berichtet.
Die Software hebt sowohl Ähnlichkeiten
als auch Unterschiede in den Nukleinsäurefragmentprofilen hervor.
Die Software ist auch in der Lage, die Sammlung von spezifischen
Nukleinsäurefragmenten
durch den Fraktionssammler zu steuern.
-
Das Differential-Displaysystem wird
durch operatives miteinander Verbinden der zahlreichen Komponenten
des Systems zur Verfügung
gestellt. Demzufolge können
eine oder mehrere Komponenten des Systems, wie zum Beispiel die
Probeneinführvorrichtung 12 und
die Nachweisvorrichtung 18, die in einem Labor in Betrieb
sind, mit den anderen Komponenten des Systems kombiniert werden
(z. B. der Abtrennvorrichtung 14, der Spaltvorrichtung 16 und
dem Datenprozessor und Analyser 20, um das Labor mit einem
System in Übereinstimmung
mit der vorliegenden Erfindung auszurüsten.
-
Einzel-Nukleotid-Verlängerungsassay,
Oligo-Ligationsassay oder Oligonukleotidsonde basierte Assaysysteme
der vorliegenden Erfindung bestehen aus, im allgemeinen, einer Probeneinführvorrichtung,
einer Vorrichtung um die markierten Proben von Interesse aufzutrennen,
einer Vorrichtung um die Marker von den Proben von Interesse abzuspalten,
einer Vorrichtung zum Nachweis des Markers und ein Softwareprogramm, um
die gesammelten Daten zu analysieren. Es wird dem Fachmann ersichtlich
sein, wenn in Besitz der vorliegenden Beschreibung, daß die allgemeine
Beschreibung viele Varianten für
jede der aufgelisteten Komponenten aufweisen kann. Wie am besten
in 17 gesehen wird
(brauche Figur #) besteht ein bevorzugtes Einzel-Nukleotid-Verlängerungsarray,
Oligoligationsassay oder Oligonukleotidsonde basiertes Assaysystem 200 aus
einer Probenzuführvorrichtung 212,
einer Abtrennvorrichtung 214, daß die Proben durch High-Performance-Liquid-Chromatographie
(HPLC) trennt, einer Spaltvorrichtung 216, um die Marker
von den Proben von Interesse abzuspalten, einer Nachweisvorrichtung 218 zum
Nachweis der Marker durch Massenspektrometrie und einen Datenprozessor
und Analyser 220, der ein Softwareprogramm enthält. Jede
Komponente wird unten genauer diskutiert.
-
Die Probenzuführvorrichtung 212 nimmt
automatisch ein gemessenes Aliquot 222 des durch eine Vielzahl
von Verfahren (PCR, Ligationen, Verdau, Nuklearen, usw.) erzeugten
Nukleinsäurefragments
und führt sie über eine
herkömmliche
Leitung 224 zu einer Abtrennvorrichtung 214 (im
allgemeinen ein HPLC) zu. Die Probenzuführvorrichtung 212 der
beispielhaften Ausführungsform
besteht aus einem Temperatur kontrollierten Autosampler 226,
der Mikrotiterplatten aufnehmen kann. Der Autosampler 226 muß Temperatur
kontrolliert sein, um die Integrität der erzeugten Nukleinsäureproben
aufrechtzuerhalten und in der Lage sein, 25 μl oder weniger an Probe zu injizieren.
Hersteller dieses Produkts sind zum Beispiel durch Gilson (Middleton,
WI) repräsentiert.
-
Die Probenzuführvorrichtung ist operativ
in Reihe mit der Abtrennvorrichtung durch die herkömmliche Leitung
224 verbunden. Die Nukleinsäureprodukte
(die durch PCR, Ligationsreaktionen, Verdau, Nukleasen usw. hergestellt
werden können)
in den gemessenen Aliquots 222, die in der Auftrennvorrichtung 214 anfangen
werden, werden temporal durch High-Performance-Liquid-Chromatographie
aufgetrennt. Die High-Performance-Liquid-Chromatographie kann eine isokratische,
binäre
oder quaternäre
Pumpe(n) 227 aufweisen und kann von zahlreichen Herstellern
erworben werden (z. B. Hewlett Packard (Palo Alto, CA) HP 1100 oder
1090 Serie, Beckman Instruments Inc. (800-742-2435), Bioanalytical
Systems, Inc. (800-845-4246), ESA, Inc. (508) 250-700), Perkin Elmer
Corp. (800-762-4000), Varian Instruments (800-926-3000), Waters
Corp. (800-254-4752)).
-
Die Abtrennvorrichtung 214 schließt eine
analytische HPLC-Säule 228 ein,
die zur Verwendung zur Abtrennung der Nukleinsäurefragmente geeignet ist.
Die Säule 228 ist
eine analytische HPLC, zum Beispiel, nicht poröses Polystyroldivinylbenzol
(2,2 μm
Partikelgröße) fester
Träger,
der innerhalb eines pH-Bereichs von 2 bis 12, drücken von bis zu 3000 psi und
einem Temperaturbereich von 10 bis 70°C betrieben werden kann. Eine
Temperatur-Kontrollvorrichtung (z. B. ein Säulenofen) (nicht gezeigt) kann
dazu verwendet werden, die Temperatur der Säule zu kontrollieren. Solche
Temperatur-Kontrollvorrichtungen sind im Stand der Technik bekannt
und können
zum Beispiel von Rainin Instruments (Tochterunternehmen von Varian
Instruments, Palo Alto, CA) erworben werden. Eine geeignete Säule 228 ist
unter dem kommerziellen Namen DNAsep® erhältlich und
ist von Serasep (San Jose, CA) erhältlich. Eine große Vielzahl
von HPLC-Säulen 228 kann
für diese
bestimmte technologische Einheit verwendet werden, da eine Einzel-Basenpaarauflösung nicht
nötiger
weise erforderlich ist. Andere geeignete analytische HPLC-Säulen sind
von anderen Herstellern (z. B. Hewlett Packard (Palo Alto, CA),
Beckman Instruments, Inc. (Brea, CA) und Waters Corp. (Milford,
MA)) erhältlich.
-
Ein Strom der getrennten DNA-Fragmente
(z. B. Sequenzierungs-Reaktionsprodukte) fließt durch eine herkömmliche
Leitung 230 von der Trennvorrichtung 214 zu der
Spaltvorrichtung 216. Jedes der DNA-Fragmente ist mit einem
einmaligen spaltbaren (z. B. Photo-spaltbaren) Marker markiert.
Der Flußstrom
von getrennten DNA-Fragmenten passiert durch oder entlang der Spaltvorrichtung 216,
wobei der Marker zum Nachweis durch Massenspektrometrie oder mit
einem elektrochemischen Detektor entfernt wird. Die Photospaltungseinheit
ist von Supelco (Bellefonte, PA) erhältlich. Die Photospaltung kann
bei vielfachen Wellenlängen mit
einer Quecksilber/Xenon-Bogenlampe durchgeführt werden. Die Wellenlängengenauigkeit
ist ungefähr
2 nm mit einer Bandbreite von 10 nm. Der bestrahlte Bereich ist
kreisförmig
und typischerweise ein Bereich von 10–100 Quadratzentimetern. In
alternativen Ausführungsformen
können
andere Spaltvorrichtungen, die durch Säure, Base, Oxidation, Reduktion,
Fluorid, Thiolaustausch, Photolyse oder enzymatische Bedingungen
spalten, dazu verwendet werden, die Marker von den getrennten DNA-Fragmenten
zu entfernen.
-
Nachdem die Spaltvorrichtung 216 die
Marker von den getrennten DNA-Fragmenten abspaltet, fließen die
Marker durch eine herkömmliche
Leitung 232 zur Nachweisvorrichtung 218 zum Nachweis
jedes Markers. Der Nachweis der Marker kann auf dem Unterschied
im Molekulargewicht zwischen jedem der Marker, die dazu verwendet
wurden, jede Art von den in den verschiedenen Assay-Schritten erzeugten
DNA zu markieren, basiert sein. Der beste Detektor, der auf Unterschieden
in der Masse basiert, ist das Massenspektrometer. Für diese
Verwendung wird das Massenspektrometer typischerweise ein atmosphärische Druck-Ionisations (API)-Schnittstelle
mit entweder Elektrospray oder chemischer Ionisation, ein Quadrapol-Massenanalyser
und einen Massenbereich von mindestens 50 bis 2600 m/z aufweisen.
Beispiele von Herstellern eines geeigneten Massenspektrometers sind:
Hewlett Packard (Palo Alto, CA), HP 1100 LC/MSD, Hitachi Instruments
(San Jose, CA) M-1200H LC/MS, Perkin Elmer Corporation, Applied
Biosystems Division (Foster City, CA) API 100 LC/MS oder API 300
LC/MS/MS, Finnigan Corporation (San Jose, CA) LCQ, Bruker Analytical
Systems, Inc. (Billerica, MA), ESQUIRE und Micromers (U.K.).
-
In einer schematisch in 16 dargestellten alternativen
Ausführungsform
sind die Probeneinführungsvorrichtung 212,
die Abtrennvorrichtung 214 und die Spaltvorrichtung 216 in
Reihe wie oben diskutiert miteinander verbunden, um den Fluß der Probe
aufrechtzuerhalten. Die Spaltvorrichtung 216 ist mit einer Nachweisvorrichtung 218 verbunden,
die ein elektrochemischer Detektor 240 ist, der die Marker
basierend auf dem Unterschied im elektrochemischen Potential zwischen
jedem der Marker, der dazu verwendet wurde, jede Art von DNA, die
in dem Sequenzierungs-Reaktionsschritt erzeugt wurde, zu markieren,
nachweist. Der elektrochemische Detektor 240 der beispielhaften
Ausführungsform
kann auf entweder coulometrischen oder amperometrischen Prinzipien
arbeiten. Der bevorzugte elektrochemische Detektor 240 ist
der coulometrische Detektor, der aus einem Durchfluß- oder
porösen
Carbongraphit amperometrischen Detektor besteht, wobei das Säulen-Eluat
durch die Elektrode hindurch passiert, was zu einer 100% Nachweiseffizienz
führt.
Um jede Komponente voll nachzuweisen, wird ein Array von 16 coulometrischen
Detektoren, die jeder bei einem unterschiedlichen Potential gehalten
werden (im allgemeinen bei 60 mV anstiegen) verwendet. Beispiele
von Herstellern dieses Typs von Detektoren sind ESA (Bedford, MA)
und Bioanalytical Systems Inc. (800-845-4246). Zusätzliche
Hersteller von elektrochemischen Detektoren können in der Liste von anderen
Herstellern gefunden werden, die unten gefunden wird.
-
Der elektrochemische Detektor 240 ist
elektrisch mit dem Datenprozessor und Analyser 220 mit
dem oben diskutierten Softwarepaket verbunden. Das Softwarepaket
kartiert die nachgewiesene Eigenschaft (z. B. die Masse oder elektrochemische
Signatur) eines gegebenen Markers auf einem spezifischen Proben-ID.
Die Software wird in der Lage sein, das Nukleinsäurefragment von Interesse zu
identifizieren und die ID-Information in jeweilige Datenbanken zu
laden.
-
Die hier beschriebenen Analysesysteme
weisen zahlreiche Vorteile gegenüber
den herkömmlichen Gel-basierten
Systemen auf. Einer der prinzipiellen Vorteile ist, daß die Systeme
vollautomatisiert werden können.
Bei Verwendung eines HPLC-basierten Auftrennungssystems können die
Proben automatisch in die HPLC initiiert werden, wohingegen Gel-basierte
Systeme eine manuelle Beladung erfordern. Es gibt auch signifikante
Zeitersparnisse, die bei der Aufbauzeit gefunden werden (keine Gelformen
sind zu reinigen, keine Gele zu gießen) und der Analysezeit (mehr
als 4 Stunden für
ein großes
Gel gegenüber
viel kürzeren
Zeiten (5 Minuten bis 1 Stunde) für eine HPLC-Analyse. Zusätzlich gibt
es einen Probendurchsatzvorteil. Bei Verwendung der in dieser Erfindung
beschriebenen Marker können
viele Proben in einer Charge analysiert werden (potentiell 384 Proben/Spur),
wohingegen die Gel-basierten Analysen oft auf die 4 erhältlichen
Fluorophore oder eine Probe/Spur begrenzt sind. Die verwendeten
Gele sind inhärent
empfindlich und können
leicht brechen oder eine Luftblase oder einen anderen Mangel enthalten,
der das gesamte Gel oder einige Spuren unbrauchbar macht. HPLC-Säulen sind
solide und, wenn prä-gepackt
erworben, frei von Packdefekten, was einen konsistenten, im allgemeinen
gleichmäßigen Auftrennweg
erzeugt. Die HPLC-Systeme verleihen auch eine bessere Qualitätssicherung,
in das interne Standards verwendet werden können, aufgrund der Reproduzierbarkeit
der HPLC-Säulen. Die
Gelqualität
ist sowohl zwischen Gelen als auch innerhalb eines Gels konsistent,
was die Verwendung von Standards nahe zu unmöglich macht. Zuletzt sind sowohl
die Massenspektrometrie und elektrochemischen Detektoren sensitiver
als die Detektoren, die in den Gel-basierten Systemen verwendet
werden, was geringer Nachweisgrenzen und die Analyse von weniger
Probe ermöglicht,
was für nicht-PCR-basierte
Analysen brauchbar wäre.
-
Markierte Proben
in Array-basierten Assays
-
Arrays mit kovalent angebundenen
Oligonukleotiden wurden hergestellt, die dazu verwendet wurden, um
eine DNA-Sequenz-Analyse durch Hybridisierung durchzuführen (Southern
et al., Genenomics 13: 1008, 1992; Drmanac et al., Science 260:
1649, 1993), Expressionsprofile zu bestimmen, auch Mutationen zu
screenen und ähnliches.
Im allgemeinen verwendet der Nachweis für diese Teste fluoreszente
oder radioaktive Markierungen. Fluoreszente Markierungen können am
direktesten durch ihre Absorption und Fluoreszenz-Emissions-Wellenlängen und
-intensitäten
identifiziert und quantifiziert werden. Ein Mikrokoskop/Kameraaufbau,
unter der Verwendung einer Fluoreszenzlichtquelle ist ein bequemes
Mittel zum Nachweis von Fluoreszenzmarker. Radioaktive Marker können durch
Standard-Autoradiographie,
Phosphor-Image-Analyse oder CCD-Detektor visualisiert werden. Für solche
Marker ist die Zahl von verschiedenen Reaktionen, die zu einer einzelnen Zeit
nachgewiesen werden können,
begrenzt. Zum Beispiel begrenzt die Verwendung von 4 fluoreszenten
Molekülen,
wie zum Beispiel herkömmlich
bei der DNA-Sequenz-Analyse verwendet, die Analyse auf zwei Proben zur
gleichen Zeit. Im wesentlichen, aufgrund dieser Begrenzung muß jede Reaktion
einzeln untersucht werden, wenn diese Detektorverfahren verwendet
werden.
-
Ein vorteilhafteres Verfahren zum
Nachweis ermöglicht,
daß Vereinigen
der Probenreaktionen auf mindestens einem Array und den gleichzeitigen
Nachweis der Produkte. Durch Verwendung eines Markers, sowie denjenigen
wie hier beschrieben, der ein unterschiedliches Molekulargewicht
oder andere physikalische Eigenschaft in jeder Reaktion aufweist,
kann das gesamte Set von Reaktionsprodukten zusammen geerntet und analysiert
werden.
-
Wie oben angegeben sind die oben
beschriebenen Verfahren für
eine Vielzahl von Zwecken anwendbar. Zum Beispiel können die
Arrays von Oligonukleotiden dazu verwendet werden, um auf die Qualität von Herstellung
von Anays, zur Quantifizierung oder qualitativen Analyse von Nukleinsäuremolekülen, zum
Nachweis von Mutationen, zum Nachweis von Expressionsmolekülen, für toxikologische
Tests und ähnliches
verwendet werden.
-
1. Sondenquantifizierung
oder -typisierung
-
In dieser Ausführungsform werden die Oligonukleotide
pro Element in einem Array immobilisiert, wobei jedes Oligonukleotid
in dem Element eine verschiedene oder verwandte Sequenz ist. Bevorzugterweise besitzt
jedes Element ein bekanntes oder verwandtes Set von Sequenzen. Die
Hybridisierung einer markierten Sonde an solchen einen Array ermöglicht die
Charakterisierung einer Sonde und die Identifizierung und Quantifizierung
der Sequenzen, die in einer Sondenpopulation enthalten sind.
-
Ein generalisiertes Assay-Format,
das in den bestimmten Anwendungen, die unten diskutiert werden, verwendet
werden kann, ist ein Sandwich-Assay-Format. In diesem Format werden eine
Vielzahl von Oligonukleotiden von bekannter Sequenz auf einem festen
Substrat immobilisiert. Das immobilisierte Oligonukleotid wird dazu
verwendet, um eine Nukleinsäure
(z. B. RNA, rRNA, ein PCR-Produkt, fragmentierte DNA) zu fangen
und dann wird eine Signalsonde an einen verschiedenen Teil der gefangenen
Zielnukleinsäure
hybridisiert.
-
Ein weiteres generalisiertes Assay-Format
ist ein sekundäres
Nachweissystem. In diesem Format werden die Arrays dazu verwendet,
um markierte Nukleinsäuren
zu identifizieren und zu quantifizieren und die in einem primären Bindungsassay
verwendet wurden. Zum Beispiel, falls ein Assay zu einer markierten
Nukleinsäure
führt,
kann die Identität
dieser Nukleinsäure
durch Hybridisierung an einen Array bestimmt werden. Diese Assay-Formate
sind insbesondere brauchbar, wenn mit abspaltbaren Massenspektrometrie-Narkern kombiniert.
-
2. Mutationsnachweis
-
Mutationen, die ein einzelnes Nukleotid
einschließen,
können
in einer Probe durch Scanning-Techniken
identifiziert werden, die dafür
geeignet sind, um vorher unbekannte Mutationen zu identifizieren
oder durch Techniken, die so aufgebaut sind, um bekannte Sequenzvarianten
nachzuweisen, zu unterscheiden oder zu quantifizieren. Verschiedene
Scanning-Techniken für
den Mutationsnachweis wurden entwickelt, basierend auf der Beobachtung,
daß Heteroduplices
von fehlgepaarten komplementären
DNA-Strängen,
die von Wildtyp- und Mutantensequenzen abgeleitet sind, ein abnormales
Wanderungsverhalten aufweisen.
-
Die hier beschriebenen Verfahren
können
zum Mutationsscreening verwendet werden. Eine Strategie zum Nachweis
einer Mutation in einem DNA-Strang ist durch Hybridisierung der
Testsequenz an Zielsequenzen, die Wildtyp- oder Mutantensequenzen
sind. Eine fehlgepaarte Sequenz weist einen destabilisierenden Effekt
auf die Hybridisierung von kurzen Oligonukleotidsonden an eine Zielsequenz
auf (siehe Wetmur, Crit. Rev. Biochem. Mol. Biol. 26: 227, 1991).
Die Test-Nukleinsäurequelle
kann genomische DNA, RNA, cDNA oder die Amplifikation von jeder
dieser Nukleinsäuren
sein. Bevorzugterweise wird die Amplifikation von Testsequenzen zuerst
durchgeführt,
gefolgt von Hybridisierung mit kurzen Oligonukleotidsonden, die
auf einem Array immobilisiert sind. Ein Amplifikationsprodukt kann
auch viele mögliche
Sequenzvarianten durch Bestimmen seines Hybridisierungsmusters an
einen Array von immobilisierten Oligonukleotidsonden gescannt werden.
-
Ein Marker, wie zum Beispiel hier
beschrieben, wird im allgemeinen in das finale Amplifikationsprodukt inkorporiert
durch Verwendung eines markierten Nukleotids oder durch Verwendung
eines markierten Primers. Das Amplifikationsprodukt wird denaturiert
und an den Array hybridisiert. Nicht-gebundenes Produkt wird abgewaschen
und an den Array gebundener Marker wird durch eine der Verfahren
hier nachgewiesen. Zum Beispiel, wenn abspaltbare Massenspektrometriemarker
verwendet werden, können
viele Produkte gleichzeitig nachgewiesen werden.
-
3. Expressionsprofile/Differential
Display
-
Säuger,
wie zum Beispiel der Mensch, weisen etwa 100.000 verschiedene Gene
in ihrem Genom auf, von denen nur ein kleiner Anteil, möglicherweise
15%, in jeder individuellen Zelle exprimiert werden. Differential-Display-Techniken
erlauben die Identifizierung von Genen, die für individuelle Zelltypen spezifisch
sind. Kurz gesagt werden im Differential Display die 3'-terminalen Teile
von mRNAs amplifiziert und auf der Basis ihrer Größe identifiziert.
Unter der Verwendung eines Primers, der so aufgebaut ist, um an
die 5'-Grenze poly(A)-Schwanzes
für die
reverse Transkription zu binden, gefolgt von Amplifikation der cDNA
unter der Verwendung von stromaufwärts liegenden willkürlichen
Sequenzprimern, werden mRNA-Subpopulationen erhalten.
-
Wie hier beschrieben, wird ein Hochdurchsatzverfahren
zur Messung der Expression von zahlreichen Genen, z. B. 1–2.000,
zur Verfügung
gestellt. Innerhalb einer Ausführungsfonn
der Erfindung werden Verfahren zur Analyse des Muster zur Genexpression
von einer ausgewählten
biologischen Probe zur Verfügung
gestellt, umfassend die Schritte von (a) Amplifizieren von cDNA
aus einer biologischen Probe unter der Verwendung von einem oder
mehreren markierten Primern, wobei der Marker korrelativ mit einer
bestimmten Nukleinsäuresonde
ist und nachweisbar durch nicht-fluoreszente Spektrometrie oder
Potentiometrie ist, (b) Hybridisieren von amplifizierten Fragmenten
an ein Array von Oligonukleotiden, wie hier beschrieben, (c) Abwaschen von
nicht-hybridisierten Material und (d) Nachweisen des Markers durch
nicht-fluoreszente Spektrometrie oder Potentiometrie, und daraus
Bestimmen des Musters von Genexpression der biologischen Probe.
Marker-basiertes Differential Display, insbesondere unter der Verwendung
von abspaltbaren Massenspektrometriemarkern auf festen Substraten
ermöglicht
die Charakterisierung von unterschiedlich exprimierten Genen.
-
4. Einzelnukleotidverlängerungsassay
-
Die Primerverlängerungstechnik kann für den Nachweis
von einzelnen Nukleotidaustauschen in einem Nukleinsäure-Templat
verwendet werden (Sokolov, Nucleic Acids Res. 18: 3671, 1989). Die
Technik ist im allgemeinen auf den Nachweis von jeder einzelnen
Basenmutation anwendbar (Kuppuswamy et al., Proc. Natl. Acad. Sci.
USA 88: 1143–1147,
1991). Kurz gesagt hybridisierte dieses Verfahren zuerst einen Primer
an einer Sequenz angrenzend an einen bekannten einzelnen Nukleotidpolymorphismus.
Die geprimte DNA wird dann Bedingungen unterzogen, in denen eine
DNA-Polymerase ein markiertes dNTP, typischerweise ein ddNTP, hinzufügt, falls
die nächste
Base in dem Templat komplementär
zu dem markierten Nukleotid in dem Reaktionsgemisch ist. In einer
Modifikation wird cDNA zuerst auf eine Sequenz von Interesse amplifiziert,
die einen einzelnen Basen-Unterschied zwischen zwei Allelen enthält. Jedes
amplifizierte Produkt wird dann auf die Anwesenheit, Abwesenheit
oder relativen Menge von jedem Allel durch Annealing eines Primers
analysiert, der eine Base 5' zu
dem Polymorphismus ist und Verlängern
durch eine markierte Base (im allgemeinen durch ein Didesoxynukleotid).
Nur wenn die korrekte Base in der Reaktion erhältlich ist, wird eine Base
am 3'-Ende des Primers
inkorporiert werden. Die Verlängerungsprodukte
werden dann durch Hybridisierung an einen Array von Nukleotiden
analysiert, so daß ein
nicht-verlängertes
Produkt nicht hybridisieren wird.
-
Kurz gesagt, wird in der vorliegenden
Erfindung jedes Didesoxynukleotid mit einem einmaligen Marker markiert.
Von den vier Reaktionsgemischen wird nur eine einen Didesoxy-Terminator
an die Primer-Sequenz anfügen.
Falls die Mutation vorhanden ist, wird sie durch den einmaligen
Marker, auf dem Didesoxynukleotid nach der Hybridisierung an den Array
nachgewiesen werden. Multiple Mutationen können gleichzeitig durch Agieren
des DNA-Primers mit einem einmaligen Marker auch bestimmt werden.
Daher werden die DNA-Fragmente
in vier verschiedenen Reaktion reagiert, die jede einen verschieden
markierten Didesoxy-Terminator enthalten, wobei der Marker mit einem
bestimmten Didesoxynukleotid korrelativ ist und die nicht-fluoreszente Spektrometrie
oder Potentiometrie nachweisbar ist. Die DNA-Fragmente werden dann
an einen Array hybridisiert und nicht-hybridisiertes Material wird
weg gewaschen. Die Marker werden von den hybridisierten Fragmenten
abgespalten und durch die jeweilige Nachweistechnologie (z. B. Massenspektrometrie,
Infrarotspektrometrie, potentiostatische Amperometrie oder UV/sichtbare
Spektrophotometrie) nachgewiesen. Die nachgewiesenen Marker können mit
den zu untersuchenden bestimmten DNA-Fragment korreliert werden
sowie der Identität
des Mutanten-Nukleotids.
-
5. Oligonukleotid-Ligationsassay
-
Der Oligonukleotid-Ligationsassay,
(OLA) (Landegren et al., Science 241: 487, 1988) wird für die Identifizierung
von bekannten Sequenzen in sehr großen und komplexen Genomen verwendet.
Das Prinzip der OLA-Reaktion beruht auf der Fähigkeit von Ligase, zwei diagnostische
Oligonukleotide kovalent zu verknüpfen, wenn sie benachbart zueinander
auf einem gegebenen DNA-Target hybridisieren. Wenn die Sequenzen an
der Sondenverbindungsstelle nicht perfekt basengepaart sind, werden
die Sonden nicht von der Ligase verknüpft werden. Bei der Anwendung
von Markierungen sind diese an die Sonde angebracht, die an das
amplifizierte Produkt ligiert wird. Nach Abschluß der OLA werden die Fragmente
an einen Array von komplementären
Sequenzen hybridisiert, die Markierungen abgespalten und mit Massenspektrometrie
nachgewiesen.
-
Innerhalb einer Ausführungsform
der Erfindung werden Verfahren zum Nachweis der Identität eines Nukleinsäuremoleküls oder
zum Nachweis eines selektierenden Nukleinsäuremoleküls in, zum Beispiel, einer biologischen
Probe, zur Verfügung
gestellt, wobei die Technik des Oligonukleotid-Ligationsassays verwendet wird.
Kurz gesagt umfassen solche Verfahren allgemein die Schritte von
Durchführen
einer Amplifikation auf der Ziel-DNAS, gefolgt von der Hybridisierung
mit der 5'-Ende
markierten Reporter-Sonde und einer 5'-phosphorylierten Sonde. Die Probe wird
mit T4-DNA-Ligase inkubiert. Die DNA-Stränge mit ligierten Proben werden auf
dem Array durch Hybridisierung an ein Array gefangen, wobei nicht-ligierte
Produkte nicht hybridisieren. Die Marker werden von den abgetrennten
Fragmenten abgespalten und dann werden die Marker durch die jeweilige
Nachweistechnologie (z. B. Massenspektrometrie, Infrarotspektrometrie,
potentiostatische Amperometrie oder UV/sichtbare Spektrophotometrie)
nachgewiesen.
-
6. Andere Assays
-
Die hier beschriebenen Verfahren
können
auch dazu verwendet werden, um zu genotypisieren oder zur Identifizierung
von Viren oder Mikroben. Zum Beispiel können F + RNA-Coliphagen brauchbare
Kandidaten als Indikatoren für
enterische Virus-Kontamination sein. Die Genotypisierung durch Nukleinsäureamplifikations-
und Hybridisierungsverfahren sind verläßliche, schnelle, einfache
und billige Alternativen zur Serotypisierung (Kafatos et al., Nucleic
Acids Res. 7: 1541, 1979). Die Amplifikationstechniken und Nukleinsäurehybridisierungstechniken
wurden erfolgreich dazu verwendet, um eine Vielzahl von Mikroorganismen
einschließlich
E. coli (Feng, Mol. Cell Probes 7: 151, 1993), Rotavirus (Sethaburt
et al., J. Med Virol. 37: 192, 1992), Hepatitis-C-Virus (Stuyver
et al., J. Gen Virol. 74: 1093, 1993) und Herpex-Simplex-Virus (Matsumoto
et al., J. Virol. Methods 40: 119, 1992) zu klassifizieren.
-
Genetische Änderungen wurden in einer Vielzahl
von experimentellen Säuger-
und menschlichen Neoplasmen beschrieben und stellen die morphologische
Basis für
die Sequenz von morphologischen Veränderungen dar, die bei der
Karzinogenese beobachtet wurden Vogelstein et al., NEJM 319: 525,
1988). In den letzten Jahren mit dem Auftauchen von molekularbiologischen
Techniken, waren allelische Verluste auf bestimmten Chromosomen
oder Mutationen von Tumorsuppressorgenen sowie Mutationen in bestimmten
Onkogenen (z. B. c-myc, c jun. und der ras-Familie) die am meisten
untersuchten Gruppen. Vorherige Arbeit (Finkelstein et al., Arch
Surg. 128: 526, 1993) identifizierte eine Korrelation zwischen verschiedenen
spezifischen Typen von Punktmutationen in den K-ras-Onkogenen und
der Phase bei der Diagnose in den colorectalen Karzinomen. Die Ergebnisse
lassen vermuten, daß die
Mutationsanalyse wichtige Informationen über die Tumoragressivität zur Verfügung stellen
könnte,
einschließlich
des Musters und der Verbreitung von Metastasen. Der prognostische
Wert von TP53- und K-ras-2-Mutationsanalyse in Phase III-Karzinomen
des Colons wurde kürzlich gezeigt
(Pricolo et al., Am. J. Surg. 171: 41, 1996). Es ist daher ersichtlich,
daß die
Genotypisierung von Tumoren und prä-kanzerösen Zellen und der spezifische
Mutationsnachweis bei der Behandlung von Krebsarten in Menschen
zunehmend wichtig sein werden.
-
Markierte Sonden
in Array-basierten Sonden
-
Die hier beschriebenen markierten
Biomoleküle
können
dazu verwendet werden, um (nicht-markierte) Arrays
von Biomolekülen
zu untersuchen. Bevorzugte Arrays von Biomolekülen enthalten ein festes Substrat, das
eine Oberfläche
umfaßt,
wobei die Oberfläche
mindestens teilweise mit einer Lage von Poly(ethylenimin) (PEI)
bedeckt ist. Die PEI-Lage umfaßt
eine Vielzahl von diskreten ersten Regionen, die an eine kontinuierlich zweite
Region angrenzen und umgeben sind. Die ersten Regionen werden durch
die Anwesenheit eines Biomoleküls
und PEI definiert, während
die zweite Region durch die Anwesenheit von PEI und das wesentliche Fehlen
des Biomoleküls
definiert ist. Bevorzugterweise ist das Substrat eine Glasplatte
oder ein Silizium-Wafer. Jedoch kann das Substrat zum Beispiel Quarz,
Gold, Nylon-6,6, Nylon oder Polystyrol sowie Verbundmaterialien
davon sein, wie oben beschrieben.
-
Die PEI-Beschichtung enthält bevorzugterweise
PEI, das ein Molekulargewicht im Bereich von 100 bis 100.000 aufweist.
Die PEI-Beschichtung kann direkt an das Substrat gebunden sein,
unter der Verwendung von zum Beispiel silyliertem PEI. Alternativ
kann ein Reaktionsprodukt eines bifunktionellen Kopplungsmittels zwischen
der Substratoberfläche
und der PEI-Beschichtung zwischengelagert sein, wobei das Reaktionsprodukt
kovalent an sowohl die Oberfläche
und die PEI-Beschichtung gebunden ist und die PEI-Beschichtung die Oberfläche sichert.
Das bifunktionelle Kopplungsmittel enthält eine erste und eine zweite
reaktive funktionelle Gruppe, wobei die erste reaktive funktionelle
Gruppe zum Beispiel ein Tri(O-C1-C5alkyl)silan ist, und die zweite reaktive
funktionelle Gruppe wie zum Beispiel ein Epoxid, Isocyanat, Isothiocyanat
und Anhydridgruppe ist. Bevorzugte bifunktionelle Kopplungsmittel
schließen
2-(3,4-Epoxycyclohexyl)ethyltrimethoxysilan; 3,4-Epoxybutyltrimethoxysilan; 3-Isocyanatopropyltriethoxysilan,
3-(triethoxysilyl)-2-methylpropylsuccinianhydrid
und 3-(2,3-Epoxypropoxy)propyltrimethoxysilan ein.
-
Der Array der Erfindung enthält erste
Biomolekül-enthaltende
Regionen, wobei jede Region einen Bereich innerhalb des Bereichs
von ungefähr
1.000 Quadratmikron bis ungefähr
100.000 Quadratmikron aufweist. In einer bevorzugten Ausführungsform
weisen die ersten Regionen Bereiche auf, die von ungefähr 5.000 Quadratmikron
bis ungefähr
25.000 Quadratmikron reichen.
-
Die ersten Regionen sind bevorzugterweise
im wesentlichen kreisförmig,
wobei die Kreise einen durchschnittlichen Durchmesser von ungefähr 10 Mikron
bis 200 Mikron aufweisen. Ob kreisförmig oder nicht, sind die Grenzen
der ersten Regionen im wesentlichen voneinander (durch die zweite
Region) durch einen durchschnittlichen Abstand von mindestens ungefähr 25 Micron
beabstandet, jedoch durch nicht mehr als ungefähr 1 cm (und bevorzugterweise
durch nicht mehr als ungefähr
1.000 Mikron). In einem bevorzugten Array sind die Grenzen von benachbarten
ersten Regionen durch eine durchschnittliche Distanz von ungefähr 25 Mikron
bis ungefähr
100 Mikron beabstandet, wobei diese Distanz bevorzugterweise durch
das Array hindurch konstant ist und die ersten Regionen sind bevorzugterweise
in sich wiederholenden geometrischen Muster angeordnet, wie in den
hier beigefügten
Figuren gezeigt. In einem bevorzugten parallel wiederholenden geometrischen
Muster sind alle benachbarten ersten Regionen durch ungefähr den selben
Abstand (ungefähr
25 Mikron bis ungefähr
100 Mikron) getrennt.
-
In bevorzugten Ausführungsformen
gibt es von 10 bis 50 erste Regionen auf dem Substrat. In anderen Ausführungsformen
gibt es zwischen 50 und 400 erste Regionen auf dem Substrat. In noch
einer anderen bevorzugten Ausführungsform
gibt es 400–800
erste Regionen auf dem Substrat.
-
Das in den ersten Regionen lokalisierte
Biomolekül
ist bevorzugterweise ein Nukleinsäurepolymer. Ein bevorzugtes
Nukleinsäurepolymer
ist ein Oligonukleotid, das ungefähr 15 bis 50 Nukleotide aufweist.
Das Biomolekül
kann Amplifikationsreaktionsprodukte sein, die von ungefähr 50 bis
1.000 Nukleotide aufweisen. In jeder ersten Region ist das Biomolekül bevorzugterweise
bei einer durchschnittlichen Konzentration im Bereich von 105–109 Biomoleküle pro 2.000 Quadratmikron
einer ersten Region vorhanden. Weiter bevorzugt reicht die durchschnittliche
Konzentration von Biomolekül
von 107 bis 109 Biomolekülen pro
2.000 Quadratmikron. In der zweiten Region ist das Biomolekül bevorzugterweise
bei einer durchschnittlichen Konzentration von weniger als 103 Biomolekülen pro 2.000 Quadratmikron
der zweiten Region vorhanden und weiter bevorzugt bei einer durchschnittlichen
Konzentration von weniger als 102 Biomolekülen pro
2.000 Quadratmikron. Am meisten bevorzugt enthalten die zweiten
Regionen überhaupt
kein Biomolekül.
-
Die zur Anheftung der Lage von PEI
an das Substrat verwendete Chemie hängt zu einem wesentlichen Teil
von der chemischen Identität
des Substrats ab. Der Stand der Technik stellt zahlreiche Beispiele
von geeigneten Chemien zur Verfügung,
die PEI an einen festen Träger
anheften können.
Zum Beispiel, wenn das Substrat Nylon-6,6 ist, kann die PEI-Beschichtung
durch die in Van Ness, J. et al. Nucleic Acids Res. 19: 3345–3350, 1991
und die PCT Internationale Publikation WO 94/00600, die beide durch
Bezugnahme hierdurch aufgenommen sind, beschriebenen Verfahren aufgetragen
werden. Wenn der feste Träger
Glas oder Silizium ist, werden geeignete Verfahren zur Aufbringung
einer Lage von PEI gefunden in, z. B. Wassermann, B.P. Biotechnology
and Bioengineering XXII: 271–287,
1980; und D'Souza,
S. F. Biotechnology Letters 8: 643–648, 1986.
-
Bevorzugterweise wird die PEI-Beschichtung
kovalent an das feste Substrat angebracht. Wenn das feste Substrat
Glas oder Silizium ist, kann die PEI-Beschichtung kovalent an das
Substrat unter der Verwendung von silysierender Chemie angebunden
werden. Zum Beispiel ist PEI, das reaktive Siloxy-Endgruppen aufweist,
käuflich
von Gelest, Inc. (Tullytown, PA) erhältlich. Solches reaktives PEI
kann mit einem Glasträger oder
Silizium-Wafer in Kontakt gebracht werden und nach sanftem Schütteln wird
das PEI an das Substrat adherieren. Alternativ kann ein bifunktionelles
silysierendes Reagenz verwendet werden. Gemäß diesem Verfahren wird das
Glas- oder Silizium-Substrat mit dem bifunktionellen silysierenden
Reagenz behandelt, um das Substrat mit einer reaktiven Oberfläche auszustatten.
Das PEI wird dann mit der reaktiven Oberfläche in Kontakt gebracht und
bindet kovalent an die Oberfläche
durch das bifunktionelle Reagenz.
-
Die in das Array-Format plazierten
Biomoleküle
sind auch ursprünglich
in einer sogenannten "Arraying-Lösung" vorhanden. Um Biomoleküle in diskreten
Regionen auf dem PEI-beschichteten
Substrat zu plazieren, enthält
die Arraying-Lösung
bevorzugterweise ein Verdickungsmittel bei einer Konzentration von
ungefähr
35 Vol% bis 80 Vol%, basierend auf dem Gesamtvolumen der Zusammensetzung,
ein Biomolekül,
das bevorzugterweise ein Oligonukleotid ist, bei einer Konzentration,
die von 0,001 μg/ml
bis 10 μg/ml
reicht und Wasser.
-
Die Konzentration des Verdickungsmittel
ist 35% V/V bis 80% V/V für
flüssige
Verdickungsmittel, wie zum Beispiel Glyzerin. Die bevorzugte Konzentration
von Verdickungsmittel in der Zusammensetzung hängt zu einigem Ausmaß von der
Temperatur ab, bei der das Arraying durchgeführt wird. Je geringer die Arraying-Temperatur
ist, desto geringer die Konzentration von Verdickungsmittel, das
verwendet zu werden braucht. Die Kombination von Temperatur und
flüssiger
Verdickungsmittel-Kombination-Kontrolle ermöglicht es, Arrays auf den meisten
Typen von festen Trägern
(z. B. Glas, Wafern, Nylon 6/6, Nylonmembranen, usw.) herzustellen.
-
Die Anwesenheit eines Verdickungsmittels
weist den zusätzlichen
Vorteil von Ermöglichen
der konkunierenden Anwesenheit von niedrigen Konzentrationen von
verschiedenen anderen Materialien auf, die in Kombination mit dem
Biomolekül
vorhanden sind. Zum Beispiel können
0,001% V/V bis 1% V/V von Detergenzien in der Anaying-Lösung vorhanden
sein. Dies ist brauchbar, da PCR-Puffer eine kleine Menge von Tween-20
oder NP-40 enthält
und es ist häufig
wünschenswert,
Probennukleinsäuren
direkt aus einem PCR-Gefäß ohne vorherige
Reinigung der Amplikons anzuordnen. Die Verwendung eines Verdickungsmittels ermöglicht die
Anwesenheit von Salzen (zum Beispiel NaCl, KCl oder MgCl2), Puffern (zum Beispiel Tris) und/oder
Chelat-bildenden Reagenzien (zum Beispiel EDTA) als auch in der
Anaying-Lösung
vorhanden zu sein. Die Verwendung eines Verdickungsmittels weist
auch den weiteren Vorteil von Ermöglichen der Verwendung von
kreuzvernetzenden Reagenzien und/oder organischen Lösungsmitteln
auf, die in der Anaying-Lösung
vorhanden sein können.
Wie käuflich
erhalten, werden kreuzvernetzende Reagenzien herkömmlich in
organischen Lösungsmitteln
wie zum Beispiel DMSO, DMF, NMP, Methanol, Ethanol und ähnliches
gelöst.
Herkömmlich
verwendete organische Lösungen
können
bei Anaying-Lösungen der
Erfindung in den Spiegeln von 0,05% bis 20% (V/V) verwendet werden,
wenn Verdickungsmittel verwendet werden.
-
Im allgemeinen verleihen die Verdickungsmittel
eine erhöhte
Viskosität
an die Arraying-Lösung. Wenn eine
geeignete Viskosität
in der Arraying-Lösung
erhalten wird, hat der erste Tropfen im wesentlichen die selbe Größe, wie
zum Beispiel der aufgebrachte 100ste Tropfen. Wenn eine ungeeignete
Viskosität
in der Arraying-Lösung
verwendet wird, sind die ersten aufgetragenen Tropfen signifikant
größer als
die späteren
Tropfen, die aufgetragen werden. Die gewünschte Viskosität liegt
zwischen derjenigen von reinem Wasser und reinem Glyzerin.
-
Das Biomolekül in dem Array kann ein Nukleinsäurepolymer
oder ein Analog davon sein, wie zum Beispiel PNA, Phosphorthioate
und Methylphosphonate. Nukleinsäure
betrifft sowohl Ribonukleinsäure
als auch Desoxyribonukleinsäure.
Das Biomolekül
kann unnatürliche
und/oder synthetische Basen umfassen. Das Biomolekül kann ein
Einzel- oder doppelsträngiges
Nukleinsäurepolymer
sein.
-
Ein bevorzugtes Biomolekül ist ein
Nukleinsäurepolymer,
das Oligonukleotide enthält
(bis zu ungefähr 100
Nukleotidbasen) und Polynukleotide (über ungefähr 100 Basen). Ein bevorzugtes
Nukleinsäurepolymer wird
aus 15–50
Nukleotidbasen gebildet. Ein anderes bevorzugtes Nukleinsäurepolymer
weist 50–1.000
Nukleotidbasen auf. Das Nukleinsäurepolymer
kann ein PCR-Produkt, PCR-Primer oder Nukleinsäure-Duplex sein, um nur ein
paar Beispiele aufzulisten. Jedoch kann im wesentlichen jeder Nukleinsäuretyp kovalent
an eine PEI-beschichtete Oberfläche
angebracht werden, wenn die Nukleinsäure ein primäres Amin
enthält,
wie unten diskutiert. Die typische Konzentration von Nukleinsäurepolymer
in der Arrraying-Lösung
ist 0,001–10 μg/ml, bevorzugterweise
0,01–1 μg/ml und
am meisten bevorzugt 0,05–0,5 μg/ml.
-
Bevorzugte Nukleinsäuremoleküle sind "Amin-modifiziert", in das sie modifiziert
wurden um ein primäres
Amin an ein 5'-Ende
des Nukleinsäurepolymers
zu enthalten, bevorzugterweise mit einer oder mehrerer Methylen
(-CH2-) Gruppen, angeordnet zwischen dem
primären
Amin und dem Nukleinsäureteil
des Nukleinsäurepolymers.
Sechs ist eine bevorzugte Zahl von Methylengruppen. Amin-modifizierte
Nukleinsäurepolymere
sind bevorzugt, da sie kovalent an einen festen Träger durch
die 5'-Amingruppe
gekoppelt werden können. PCR-Produkte können unter
der Verwendung von 5'-Hexylamin-modifizierten
PCR-Primern angeordnet werden. Die Nukleinsäure-Duplices können nach
der Einführung
von Aminen durch Nick-Translation unter der Verwendung von Aminoallyl-dUTP
(Sigma, St. Louis, MO) angeordnet werden. Die Amine können in
Nukleinsäuren
durch Polymerasen, wie zum Beispiel terminale Transferase und Aminoallyl-dUTP
oder durch Ligation von kurzen Amin-enthaltenden Nukleinsäurepolymeren
auf Nukleinsäuren
durch Ligaren eingeführt
werden.
-
Bevorzugterweise wird das Nukleinsäurepolymer
aktiviert, bevor es mit der PEI-Beschichtung
in Kontakt gebracht wird. Dies kann bequem durch Kombinieren von
Aminfunktionalisiertem Nukleinsäurepolymer
mit einer multi-funktionellen Amin-reaktiven Chemikalie, wie zum
Beispiel Trichlortriazin eneicht werden. Wenn das Nukleinsäurepolymer
eine 5'-Aminogruppe
enthält,
kann diese 5'-Amin
mit Trichlortriazin reagiert werden, auch bekannt als Cyanurchlorid
(Van Ness et al., Nucleic Acids Res. 19(2): 3345–3350, 1991). Bevorzugterweise
wird ein Überschuß von Cyanurchlorid
zu der Nukleinsäurepolymerlösung hinzugefügt, wobei
ein 10- bis 1.000-facher molarer Überschuß von Cyanurchlorid über die
Zahl von Aminen in dem Nukleinsäurepolymer
in der Anaying-Lösung bevorzugt
ist. Auf diese Weise hat die Hauptzahl von Amin-terminierten Nukleinsäurepolymeren
mit einem Molekül
von Trichlortriazin reagiert, so daß das Nukleinsäurepolymer
mit Trichlortriazin beendet wird.
-
Bevorzugterweise wird die Arraying-Lösung unter
der Verwendung eines herkömmlichen
Puffers, wie zum Beispiel Natriumphosphat, Natriumborat, Natriumcarbonat
und Tris HCl gepuffert. Ein bevorzugter pH-Bereich für die Anaying-Lösung ist
7 bis 9, wobei ein bevorzugter Puffer frisch präpariertes Natriumborat bei
pH 8,3 bis pH 8,5 ist. Um eine typische Arraying-Lösung herzustellen,
wird Hexylamin-modifiziertes Nukleinsäurepolymer in 0,2 M Natriumborat,
pH 8,3, bei 0,1 μg/ml
auf ein Gesamtvolumen von 50 μl
plaziert. 10 μl
einer 15 mg/ml Lösung
von Cyanwchlorid werden dann hinzugefügt und der Reaktion wird ermöglicht,
für eine
Stunde bei 25°C
mit konstantem Schütteln
fortzuschreiten. Glyzerin (Gibco Brl®, Gran Island, NY) wird bis
zu einer finalen Konzentration von 56% hinzugefügt.
-
Die biomolekularen Anaying-Lösungen können auf
die PEI-Beschichtung durch jede einer Zahl von Techniken, die im
Augenblick bei der Mikrofabrikation verwendet werden, aufgetragen
werden. Zum Beispiel können
die Lösungen
in einem Inkjet-Druckkopf plaziert werden und aus solch einem Kopf
auf die Beschichtung ausgestoßen
werden.
-
Ein bevorzugter Ansatz zur Zuführung von
biomolekularer Lösung
auf die PEI-Beschichtung verwendet eine modifizierte Springprobe.
Springproben sind von verschiedenen Verkäufern erhältlich, einschließlich Everett
Charles (Pomona, CA), Interconnect Devices Inc. (Kansas City, Kansas)
und Test Connections Inc., (Upland, CA). Damit die wie oben beschriebenen
erhältlichen
Springsonden zufriedenstellend als flüssige Zubringvorrichtungen
gemäß der vorliegenden
Erfindung funktionieren, müssen
ungefähr
1/1000 bis 5/1000 eines Inchs von Metallmaterial von der Spitze
der Sonde entfernt werden. Das Verfahren muß zu einer flachen Oberfläche führen, die
senkrecht zu der Längsachse
der Springsonde ist. Das Entfernen von ungefähr 1/1000 bis 5/1000 eines
Inchs von Material von dem Boden der Spitze ist bevorzugt und kann
leicht mit einem sehr feingekörnten
Schleifstein durchgeführt
werden. Spezifische Springsonden, die käuflich erhältlich sind und die modifiziert
werden können,
um eine planare Spitze wie oben beschrieben zur Verfügung zu
stellen, schließen die
XP54-Sonde ein, die durch Ostby Barton (eine Abteilung von Everett
Charles (Pomona, CA)); die SPA-Sonde 25P, die durch Everett Charles
hergestellt wird (Pomona, CA) und 43-P-"fluted"-Springsonde von Test Connections Inc.,
(Upland, CA).
-
Die wie oben beschriebenen Anaying-Lösungen können direkt
in einem Arraying-Verfahren verwendet werden. Gemeint ist, die aktivierten
Nukleinsäurepolymere
brauchen nicht vor dem Druckschritt von nicht reagiertem Cyanurchlorid
weg gereinigt zu werden. Typischerweise wird der Reaktion, die die
aktivierte Nukleinsäure
an den festen Träger
anbringt ermöglicht,
für 1 bis
20 Stunden bei 20 bis 50°C
fortzuschreiten. Bevorzugterweise ist die Reaktionszeit 1 Stunde
bei 25°C.
-
Die hier beschriebenen Anays sind
besonders brauchbar bei der Durchführung von Hybridisierungstests,
zum Beispiel unter der Verwendung von CMST-markierten Sonden. Jedoch
müssen
die Amine auf dem festen Träger,
um solche Tests durchzuführen,
vor der Durchführung
des Hybridisierungsschrittes geschützt werden. Dies kann durch
Reagieren des festen Trägers
mit 0,1–2,0
M Succinianhydrid eneicht werden. Die bevorzugten Reaktionsbedingungen
sind 1,0 M Succinianhydrid in 70% m-Pyrol und 0,1 M Natriumborat.
Der Reaktion wird typischerweise erlaubt für 15 Minuten bis 4 Stunden
zu erfolgen, mit einer bevorzugten Reaktionszeit von 30 Minuten
bei 25°C.
Restliches Succinianhydrid wird mit einer 3x-Wasser-Waschung entfernt.
-
Der feste Träger wird dann mit einer Lösung inkubiert,
die 0,1–5
M Glycin in 0,1–10,0
M Natriumborat bei pH 7–9
enthält.
Dieser Schritt "schützt" jedes Dichlortriazin,
das kovalent an die PEI-Oberfläche
gebunden sein kann, durch Überführung in
Monochlortriazin. Die bevorzugten Bedingungen sind 0,2 M Glycin
in 0,1 M Natriumborat bei pH 8,3. Der feste Träger kann dann Detergenz-enthaltenen
Lösungen
gewaschen werden, um nicht-gebundene Materialien zu entfernen, zum
Beispiel Spuren von NMP. Bevorzugterweise wird der feste Träger auf
95°C in
0.01 M NaCl, 0,05 EDTA und 0,1 M Tris, pH 8,0 für 5 Minuten erhitzt. Dieser
Erhitzungsschritt entfernt nicht-kovalent angebrachte Nukleinsäurepolymere,
wie zum Beispiel PCR-Produkte, in dem Fall wo Doppelstrang-Nukleinsäure angeordnet
wird, hat dieser Schritt auch den Effekt von Überführen des Doppelstrangs in die
einzelsträngige
Form (Denaturierung).
-
Die Arrays können dann durch Sonden (z.
B. Oligonukleotide, Nukleinsäurefragmente,
PCR-Produkte, usw.)
untersucht werden, die mit zum Beispiel CMST-Markern, wie hier beschrieben,
Radioisotopen, Fluorophoren oder Biotin markiert sein können. Die
Verfahren zur Biotinylierung von Nukleinsäuren sind im Stand der Technik
gut bekannt und werden geeignet durch Pierce (Avidin-Biotin Chemistry:
A Handbookk, Pierce Chemical Company, 1992, Rockford Illinois) beschrieben.
Die Sonden werden im allgemeinen bei 0,1 ng/ml bis 10 μg/ml in Standard-Hybridisierungslösungen verwendet,
die GuSCN, GuHCl, Formamid, usw. (siehe Van Ness und Chen, Nucleic
Acids Res. 19: 5143–5151,
1991) enthalten.
-
Um das Hybridisierungsereignis (d.
h. die Anwesenheit des Biotins) nachzuweisen wird der feste Träger mit
Streptavidin-/Rettichperoxidase-Konjugat inkubiert. Solche Enzymkonjugate
sind käuflich
erhältlich von,
zum Beispiel, Vector Laboratories (Burlingham, CA). Das Streptavidin
bindet mit hoher Affinität
an das Biotinmolekül,
was die Rettichperoxidase in die Nähe der hybridisierten Sonde
bringt. Nicht-gebundenes Streptavidin-/Rettichperoxidase-Konjugat wird in
einem einzelnen Waschschritt weg gewaschen. Die Anwesenheit von
Rettichperoxidaseenzym wird dann unter der Verwendung eines ausfällenden
Substrats in der Anwesenheit von Peroxid und den geeigneten Puffern
nachgewiesen.
-
Ein blaues Enzymprodukt, das auf
einer reflektierenden Oberfläche,
wie zum Beispiel einem Wafer aufgetragen wird, weist einen vielfach
niedrigeren Spiegel des Nachweises (LLD), verglichen mit demjenigen, der
für ein
colorimetrischen Substrat erwartet wird, auf. Weiterhin ist die
LLD für
verschieden gefärbte
Enzymprodukte sehr verschieden. Zum Beispiel ist die LLD für 4-Methoxynaphthol
(was ein ausgefälltes
blaues Produkt produziert) pro 50 μM Durchmesser-Fleck ungefähr 1.000
Moleküle,
wohingegen ein rot ausfallendes Substrat eine LLD von ungefähr 1.000-fach
höher bei
1.000.000 Moleküle
pro 50 μM
Durchmesser-Fleck ergibt. Die LLD wird durch Untersuchen der Oberfläche mit
einem Mikroskop (wie zum Beispiel des Axiotech-Mikroskop, käuflich erhältlich von
Zeiss), ausgerüstet
mit einer sichtbaren Lichtquelle und einer CCD-Kamera (Princeton
Instruments, Princeton, NJ) nachgewiesen. Ein Bild von ungefähr 10.000 μM × 10.000 μM kann zur selben
Zeit gescannt werden.
-
Um das blaue colorimetrische Nachweisschema
zu verwenden, muß die
Oberfläche
nach der enzymatischen Reaktion wieder rein sein und der Wafer oder
der Glasträger
muß in
einem trockenen Zustand gescannt werden. Zusätzlich muß die enzymatische Reaktion
vor der Sättigung
der Referenzflecken gestoppt werden. Für Rettichperoxidase ist dies
ungefähr
2–5 Minuten.
-
Es ist ebenfalls möglich, chemilumineszente
Substrate für
alkalische Phosphatase oder Rettichperoxidase (HRP) oder Fluoreszenz-Substrate
für HRP
oder alkalische Phosphatase zu verwenden. Beispiele schließen die
Dioxethan-Substrate für
alkalische Phosphatase, erhältlich
von Perkin Elmer oder Attophos HRP-Substrate von JBL Scientific
(San Luis, Obispo, CA) ein.
-
Die folgenden Beispiele werden zur
Veranschaulichung und nicht zur Beschränkung angeboten.
-
Sofern nicht anders angegeben, können Chemikalien,
wie sie in den Beispielen verwendet werden, von Aldrich Chemical
Company, Milwaukee, WI erhalten werden. Die folgenden Abkürzungen,
mit den angegebenen Bedeutungen, werden hierin verwendet.
ANP
= 3-(Fmoc-Amino)-3-(2-nitrophenyl)propionsäure
NBA = 4-(Fmoc-Aminomethyl)-3-nitrobenzoesäure
HATU
= O-7-Azabenzotriazol-1-yl-N,N,N',N'-tetramethyluroniumhexafluorphosphat
DIEA
= Diisopropylethylamin
MCT = Monochlortriazin
NMM = 4-Methylmorpholin
NMP
= N-Methylpyrrolidon
ACT375 = ACT375-Peptidsynthesizer von
Advanced ChemTech, Inc., Louisville, KY
ACT = Advanced ChemTech,
Inc., Louisville, KY
NovaBiochem = CalBiochem-NovaBiochem International,
San Diego, CA
TFA = Trifluoressigsäure
Tfa = Trifluoracetyl
iNIP
= N-Methylisonipecotinsäure
Tfp
= Tetrafluorphenyl
DIAEA = 2-(Diisopropylamino)ethylamin
MCT
= Monochlortriazen
5'-AH-ODN
= Oligodesoxynukleotid mit 5'-Aminohexyl-Schwanz
-
BEISPIELE
-
BEISPIEL 1
-
HERSTELLUNG VON SÄURELABILEN
LINKERN ZUR VERWENDUNG BEI DER SEQUENZIERUNG EINES SPALTBAREN MG-IDENTIFIKATORS
-
A. Synthese von Pentafluorphenylestern
von chemisch spaltbaren Massenspektroskopiemarkierungen, um Markierungen
mit Carbonsäureamid-Enden
freizusetzen
-
1 zeigt
das Reaktionsschema.
-
Schritt A. TentaGel-S-AC-Harz (Verbindung
II; erhältlich
von ACT; 1 Äq.)
wird mit DMF im Sammelgefäß des ACT357-Peptidsynthesizers
(ACT) suspendiert. Verbindung I (3 Äq.), HATU (3 Äq.) und
DIEA (7,5 Äq.) in
DMF werden zugegeben und das Sammelgefäß für 1 h geschüttelt. Das Lösungsmittel
wird entfernt und das Harz mit NMP (2X), McOH (2X) und DMF (2X)
gewaschen. Die Kopplung von I an das Harz und die Waschschritte
werden wiederholt, um Verbindung III zu ergeben.
-
Schritt B. Das Harz (Verbindung III)
wird mit 25%igem Piperidin in DMF vermischt und für 5 min
geschüttelt.
Das Harz wird filtriert, anschließend mit 25%igem Piperidin
in DMF vermischt und für
10 min geschüttelt.
Das Lösungsmittel
wird entfernt, das Harz mit NMP (2X), McOH (2X) und DMF (2X) gewaschen
und direkt in Schritt C verwendet.
-
Schritt C. Das Harz ohne Schutzgruppe
aus Schritt B wird in DMF suspendiert und dazu werden eine Aminosäure mit
FMOC-Schutzgruppe, die Aminofunktionalität in ihrer Seitenkette enthält (Verbindung
IV, z. B. Alpha-N-FMOC-3-(3-Pyridyl)-alanin (erhältlich von Synthetech, Albany,
OR; 3 Äq.),
HATU (3 Äq.)
und DIEA (7,5 Äq.)
in DMF zugegeben. Das Gefäß wird für 1 h geschüttelt. Das
Lösungsmittel
wird entfernt und das Harz mit NMP (2X), MeOH (2X) und DMF (2X)
gewaschen. Die Kopplung von IV an das Harz und die Waschschritte werden
wiederholt, um Verbindung V zu ergeben.
-
Schritt D. Das Harz (Verbindung V)
wird mit Piperidin behandelt, wie in Schritt B beschrieben, um die FMOC-Gruppe
zu entfernen. Das Harz ohne Schutzgruppe wird anschließend gleichmäßig vom
ACT357 aus dem Sammelgefäß in 16
Reaktionsgefäße aufgeteilt.
-
Schritt E. Die 16 Aliquote aus Harz
ohne Schutzgruppe aus Schritt D werden in DMF suspendiert. Zu jedem
Reaktionsgefäß werden
die geeignete Carbonsäure
VI1
-16 (R1-16CO2H; 3 Äq.), HATU
(3 Äq.)
und DIEA (7,5 Äq.)
in DMF zugegeben. Die Gefäße werden
für 1 h
geschüttelt.
Das Lösungsmittel
wird entfernt und die Harz-Aliquote mit NMP (2X), McOH (2X) und
DMF (2X) gewaschen. Die Kopplung von VI1-16 an
die Harz-Aliquote und die Waschschritte werden wiederholt, um die
Verbindungen VII1-16 zu ergeben.
-
Schritt F. Die Harz-Aliquote (Verbindungen
VII1-16) werden mit CH2Cl2 (3X) gewaschen. Zu jedem der Reaktionsgefäße wird
1% TFA in CH2Cl2 zugegeben
und die Gefäße für 30 min
geschüttelt.
Das Lösungsmittel wird
aus den Reaktionsgefäßen in einzelne
Röhren
filtriert. Die Harz-Aliquote werden mit CH2Cl2 (2X) und McOH (2X) gewaschen und die Filtrate
in die einzelnen Röhren
hinein zusammengeführt.
Die einzelnen Röhren
werden im Vakuum eingedampft, was die Verbindungen VIII1-16 liefert.
-
Schritt G. Jede der freien Carbonsäuren VIII1-16 wird in DMF gelöst. Zu jeder Lösung wird
Pyridin (1,05 Äq.)
zugegeben, gefolgt von Pentafluorphenyltrifluoracetat (1,1 Äq.). Die
Mischungen werden für
45 min bei Raumtemperatur gerührt.
Die Lösungen
werden mit EtOAc verdünnt,
mit 1 M wäßriger Zitronensäure (3X)
und 5% wäßrigem NaHCO3 (3X) gewaschen, über Na2SO4 getrocknet, filtriert und in vacuo eingedampft,
was die Verbindungen IX1-16 liefert.
-
B. Synthese von Pentafluorphenylestern
von chemisch spaltbaren Massenspektroskopiemarkierungen, um Markierungen
mit Carbonsäureenden
freizusetzen
-
2 zeigt
das Reaktionsschema.
-
Schrift A. 4-(Hydroxymethyl)phenoxybuttersäure (Verbindung
I; 1 Äq.)
wird zusammengebracht mit DIEA (2,1 Äq.) und Allylbromid (2,1 Äq.) in CHCl3 und für
2 h unter Rückfluß erhitzt.
Die Mischung wird mit EtOAc verdünnt,
mit 1 N HCl (2X), pH 9,5 Carbonatpuffer (2X) und Salzlösung (IX)
gewaschen, über
Na2SO4 getrocknet
und in vacuo eingedampft, um den Allylester von Verbindung I zu
ergeben.
-
Schritt B. Der Allylester von Verbindung
I aus Schritt A (1,75 Äq.)
wird in CH2Cl2 mit
einer Aminosäure mit
FMOC-Schutzgruppe, die Aminofunktionalität in ihrer Seitenkette enthält (Verbindung
II, z. B. Alpha-N-FMOC-3-(3-Pyridyl)-alanin, erhältlich von Synthetech, Albany,
OR; 1 Äq.),
N-Methylmorpholin (2,5 Äq.) und
HATU (1,1 Äq.)
zusammengebracht und bei Raumtemperatur für 4 h gerührt. Die Mischung wird mit CH2Cl2 verdünnt, mit
1 M wäßriger Zitronensäure (2X),
Wasser (1X) und 5%iger wäßriger NaHCO3 (2X) gewaschen, über Na2SO4 getrocknet und in vacuo eingedampft. Verbindung
III wird durch Flashchromatographie (CH2Cl2 → EtOAc)
isoliert.
-
Schritt C. Verbindung III wird in
CH2Cl2 gelöst, Pd(PPh3)4 (0,07 Äq.) und
N-Methylalanin (2 Äq.)
werden hinzugegeben und die Mischung wird bei Raumtemperatur für 4 h gerührt. Die
Mischung wird mit CH2Cl2 verdünnt, mit
1 M wäßriger Zitronensäure (2X)
und Wasser (1X) gewaschen, über
Na2SO4 getrocknet
und in vacuo eingedampft. Verbindung IV wird durch Flashchromatographie
(CH2Cl2 → EtOAc +
HOAc) isoliert.
-
Schritt D. TentaGel-S-AC-Harz (Verbindung
V; 1 Äq.)
wird mit DMF im Sammelgefäß des ACT357-Peptidsynthesizers
(Advanced ChemTech Inc. (ACT), Louisville, KY) suspendiert. Verbindung
IV (3 Äq.),
HATU (3 Äq.)
und DIEA (7,5 Äq.)
in DMF werden hinzugefügt
und das Sammelgefäß für 1 h geschüttelt. Das
Lösungsmittel
wird entfernt und das Harz mit NMP (2X), McOH (2X) und DMF (2X)
gewaschen. Die Kopplung von IV an das Harz und die Waschschritte
werden wiederholt, um Verbindung VI zu ergeben.
-
Schritt E. Das Harz (Verbindung VI)
wird mit 25% Piperidin in DMF vermischt und für 5 min geschüttelt. Das
Harz wird filtriert, dann mit 25% Piperidin in DMF vermischt und
für 10
min geschüttelt.
Das Lösungsmittel wird
entfernt und das Harz mit NMP (2X), McOH (2X) und DMF (2X) gewaschen.
Das Harz ohne Schutzgruppe wird dann gleichmäßig durch den ACT357 aus dem
Sammelgefäß in 16
Reaktionsgefäße aufgeteilt.
-
Schritt F. Die 16 Aliquote aus Harz
ohne Schutzgruppe aus Schritt E werden in DMF suspendiert. Zu jedem
Reaktionsgefäß werden
die geeignete Carbonsäure
VII1
-16 (R1-16CO2H; 3 Äq.), HATU
(3 Äq.)
und DIEA (7,5 Äq.)
in DMF zugegeben. Die Gefäße werden
für 1 h
geschüttelt.
Das Lösungsmittel
wird entfernt und die Harz-Aliquote mit NMP (2X), McOH (2X) und
DMF (2X) gewaschen. Die Kopplung von VII1-16 an
die Harz-Aliquote und die Waschschritte werden wiederholt, um die
Verbindungen VIII1-16 zu ergeben.
-
Schritt G. Die Harz-Aliquote (Verbindungen
VIII1-16) werden mit CH2Cl2 (3X) gewaschen. Zu jedem der Reaktionsgefäße wird
1% TFA in CH2Cl2 zugegeben
und die Gefäße werden
für 30
min geschüttelt.
Das Lösungsmittel
wird aus den Reaktionsgefäßen in einzelne
Röhren
filtriert. Die Harz-Aliquote werden mit CH2Cl2 (2X) und McOH (2X) gewaschen und die Filtrate
in die einzelnen Röhren
hinein zusammengegeben. Die einzelnen Röhren werden im Vakuum verdampft,
was die Verbindungen IX1-16 liefert.
-
Schritt H. Jede der freien Carbonsäuren IX1
-16 wird in DMF
gelöst.
Zu jeder Lösung
wird Pyridin (1,05 Äq.)
hinzugegeben, gefolgt von Pentafluorphenyltrifluoracetat (1,1 Äq.). Die
Mischungen werden für
45 min bei Raumtemperatur gerührt.
Die Lösungen
werden mit EtOAc verdünnt,
mit 1 M wäßriger Zitronensäure (3X)
und 5% wäßriger NaHCO3 (3X) gewaschen, über Na2SO4 getrocknet, filtriert und in vacuo eingedampft,
was die Verbindungen X1-16 liefert.
-
BEISPIEL 2
-
NACHWEIS PHOTOLYTISCHER
SPALTUNG VON T-L-X
-
Eine T-L-X-Verbindung, wie hergestellt
in Beispiel 11, wurde mit Licht im nahen UV-Bereich für 7 min bei Raumtemperatur
bestrahlt. Eine Rayonett-Fluoreszenz-UV-Lampe (Southern New England
Ultraviolet Co., Middletown, CT) mit einem Emissionspeak bei 350
nm wird als eine UV-Lichtquelle verwendet. Die Lampe wird in einem
Abstand von 15 cm von den Petrischalen mit Proben gestellt. SDS-Gelelektrophorese
zeigt, daß >85% des Konjugats unter
diesen Bedingungen gespalten werden.
-
BEISPIEL 3
-
HERSTELLUNG VON FLUORESZENZMARKIERTEN
PRIMERN UND NACHWEIS DER SPALTUNG VON FLUOROPHOR (betrifft nicht
den beanspruchten Gegenstand)
-
Synthese und Reinigung
von Oligonukleotiden
-
Die Oligonukleotide (ODNs) werden
auf automatisierten DNA-Synthesizern unter Verwendung der Standard-Phosphoramidit-Chemie,
geliefert vom Anbieter, oder der H-Phosphonat-Chemie (Glen Research Sterling,
VA) hergestellt. In geeigneter Weise blockierte dA-, dG-, dC- und
T-Phosphoramidite sind in diesen Formen kommerziell erhältlich und
synthetische Nukleoside können
leicht in die geeignete Form überführt werden.
Die Oligonukleotide werden unter Verwendung der Standard-Phosphoramidit-Chemie,
geliefert vom Anbieter, oder der H-Phosphonatchemie hergestellt.
Oligonukleotide werden durch Adaptionen von Standardverfahren gereinigt.
Oligonukleotide mit 5'-Tritylgruppen
werden auf HPLC unter Verwendung einer 12 Mikrometer, 300 # Rainin
(Emeryville, CA) Dynamax C-8 4,2 × 250 mm Umkehrphasensäule unter
Verwendung eines Gradienten von 15% bis 55% MeCN in 0,1 N Et3NH+OAc-,
pH 7,0, über
20 min chromatographiert. Wenn die Detritylierung durchgeführt wird,
werden die Oligonukleotide durch Gelausschlußchromatographie weitergereinigt.
Analytische Überprüfungen der
Qualität
der Oligonukleotide werden mit einer PRP-Säule (Alltech, Deerfield, IL)
bei alkalischem pH und mit PAGE durchgeführt.-Herstellung von 2,4,6-Trichlortriazin-abgeleiteten
Oligonukleotiden: 10 bis 1.000 μg
5'-terminal aminverknüpftes Oligonukleotid
werden mit einem Überschuß umkristallisierter
Cyanurolsäurechloride
in 10% N-Methylpyrrolidon in alkalischem Puffer (vorzugsweise pH
8,3 bis 8,5) bei 19°C
bis 25°C
für 30
bis 120 Minuten umgesetzt. Die endgültigen Reaktionsbedingungen
bestehen aus 0,15 M Natriumborat bei pH 8,3, 2 mg/ml umkristallisiertem
Cyanurolsäurechlorid
und 500 μg/ml
entsprechendem Oligonukleotid. Das nicht-umgesetzte Cyanurolsäurechlorid
wird durch Ausschlußchromatographie auf
einer G-50-Sephadex-Säule (Pharmacia,
Piscataway, NJ) entfernt.
-
Das aktivierte gereinigte Oligonukleotid
wird dann mit einem 100-fachen molaren Überschuß Cystamin in 0,15 M Natriumborat
bei pH 8,3 für
1 Stunde bei Raumtemperatur umgesetzt. Das nicht-umgesetzte Cystamin
wird durch Ausschlußchromatographie
auf einer G-50-Sephadex-Säule entfernt.
Die abgeleiteten ODNs werden dann mit aminreaktiven Fluorochromen
umgesetzt. Die abgeleitete ODN-Zubereitung wird in 3 Portionen aufgeteilt
und jede Portion wird mit entweder (a) einem 20-fachen molaren Überschuß von Texasrot-Sulfonylchlorid (Molecular
Probes, Eugene, OR), (b) einem 20-fachen molaren Überschuß von Lissamin-Sulfonylchlorid
(Molecular Probes, Eugene, OR), (c) einem 20-fachen molaren Überschuß von Fluoresceinisothiocyanat
umgesetzt. Die endgültigen
Reaktionsbedingungen bestehen aus 0,15 M Natriumborat bei pH 8,3
für 1 Stunde
bei Raumtemperatur. Die nicht-umgesetzten
Fluorochrome werden durch Ausschlußchromatographie auf einer
G-50-Sephadex-Säule entfernt.
-
Um das Fluorochrom vom Nukleotid
abzuspalten, werden die ODNs auf 1 × 10–5 molar
eingestellt und dann Verdünnungen
(12,3-fache Verdünnungen)
in TE hergestellt (TE ist 0,01 M Tris, pH 7,0, 5 mM EDTA). Zu 100 μl-Volumina
von ODNs werden 25 μl
0,01 M Dithiothreitol (DTT) zugegeben. Zu einem identischen Satz von
Kontrollzusammensetzungen wird kein DTT zugegeben. Die Mischung
wird für
15 Minuten bei Raumtemperatur inkubiert. Fluoreszenz wird in einer
schwarzen Mikrotiterplatte gemessen. Die Lösung wird aus den Inkubationsröhren (150 μl) entfernt
und in eine schwarze Mikrotiterplatte (Dynatek Laboratories, Chantilly,
VA) gegeben. Die Platten werden dann direkt unter Verwendung eines
Fluoroskan-II-Fluorometers (Flow Laboratories, McLean, VA) unter
Verwendung einer Anregungswellenlänge von 495 nm und Überwachung
der Emission bei 520 nm für
Fluorescein, unter Verwendung einer Anregungswellenlänge von
591 nm und Überwachung
der Emission bei 612 nm für
Texasrot und unter Verwendung einer Anregungswellenlänge von
570 nm und Überwachung
der Emission bei 590 nm für
Lissamin abgelesen, wobei die Ergebnisse in Tabelle I angegeben
sind.
-
-
Die Daten zeigen, daß es einen
etwa 200-fachen Anstieg der relativen Fluoreszenz gibt, wenn das
Fluorochrom vom ODN abgespalten wird.
-
BEISPIEL 4
-
HERSTELLUNG VON MARKIERTEN
M13-SEQUENZPRIMERN UND NACHWEIS DER ABSPALTUNG VON MARKIERUNGEN
-
Herstellung von 2,4,6-Trichlortriazin-abgeleiteten
Oligonukleotiden: 1.000 μg
5'-terminales aminverknüpftes Oligonukleotid
(5'-Hexylamin-TGTAAAACGACGGCCAGT-3") (Seq. ID Nr. 1)
werden mit einem Überschuß umkristallisierten
Cyanursäurechlorids
in 10% N-Methylpyrrolidon
in alkalischem Puffer (vorzugsweise pH 8,3 bis 8,5) bei 19 bis 25°C für 30 bis
120 Minuten umgesetzt. Die endgültigen
Reaktionsbedingungen bestehen aus 0,15 M Natriumborat bei pH 8,3,
2 mg/ml umkristallisiertem Cyanursäurechlorid und 500 μg/ml entsprechendem
Oligonukleotid. Das nicht-umgesetzte Cyanursäurechlorid wird durch Ausschlußchromatographie
auf einer G-50-Sephadex-Säule
entfernt.
-
Das aktivierte gereinigte Oligonukleotid
wird anschließend
mit einem 100-fachen molaren Überschuß Cystamin
in 0,15 M Natriumborat bei pH 8,3 für 1 Stunde bei Raumtemperatur
umgesetzt. Das nicht-umgesetzte Cystamin wird durch Ausschlußchromatographie
auf einer G-50-Sephadex-Säule
entfernt. Die abgeleiteten ODNs werden anschließend mit einer Vielzahl von
Amiden umgesetzt. Die abgeleitete ODN-Zubereitung wird in 12 Portionen
aufgeteilt und je Portion wird (25-molarer Überschuß) mit den Pentafluorphenylestern
von: (1) 4-Methoxybenzoesäure,
(2) 4-Fluorbenzoesäure,
(3) Tolylsäure,
(4) Benzoesäure,
(5) Indol-3-essigsäure, (6)
2,6-Difluorbenzoesäure,
(7) Nicotinsäure-N-oxid,
(8) 2-Nitrobenzoesäure, (9)
5-Acetylsalicylsäure,
(10) 4-Ethoxybenzoesäure,
(11) Zimtsäure
oder (12) 3-Aminorricotinsäure
umgesetzt. Die Reaktion läuft
für 2 Stunden
bei 37°C
in 0,2 M NaBorat pH 8,3. Die abgeleiteten ODNs werden mit Gelausschlußchromatographie
auf G-50-Sephadex
gereinigt.
-
Um die Markierung vom Oligonukleotid
abzuspalten, werden die ODNs auf 1 × 10–5 molar
eingestellt und anschließend
werden Verdünnungen
(12,3-fache Verdünnungen)
in TE hergestellt (TE ist 0,01 M Tris, pH 7,0, 5 mM EDTA) mit 50%
EtOH (v/v). Zu 100 μl-Volumina ODNs werden
25 μl 0,01
M Dithiothreitol (DTT) zugegeben. Zu einem identischen Satz von
Kontrollzusammensetzungen wird kein DTT zugegeben. Die Inkubation
erfolgt für
30 Minuten bei Raumtemperatur. NaCI wird anschließend auf
0,1 M zugegeben und 2 Volumina EtOH werden zugegeben, um die ODNs
auszufällen.
Die ODNs werden aus Lösung
durch Zentrifugation bei 14.000 × G bei 4°C für 15 Minuten entfernt. Die Überstände werden
aufbewahrt, vollständig
getrocknet. Der Pellet wird anschließend in 25 μl McOH gelöst. Der Pellet wird anschließend mit
Massenspektrometrie auf das Vorhandensein von Markierungen getestet.
-
Der Massenspektrometer, der bei diesen
Arbeiten verwendet wurde, ist ein Fouriertransformations-Massenspektrometer
(FTMS) mit externer Ionenquelle. Proben, die für MALDI-Analyse hergestellt
wurden, werden auf der Spitze einer direkten Sonde abgeschieden
und in die Ionenquelle eingeführt.
Wenn die Probe mit einem Laserpuls bestrahlt wird, werden Ionen
aus der Quelle extrahiert und in eine lange Quadrapol- Ionenführung geleitet,
die sie fokussiert und zu einer FTMS-Analysatorzelle transportiert,
die innerhalb der Bohrung eines supraleitfähigen Magneten angeordnet ist.
-
Die Spektren liefern die folgende
Information. Peaks die in der Intensität von 25 bis 100 relativen
Intensitätseinheiten
variieren, bei den folgenden relativen Molekülmassen: (1) 212,1 amu, was
4-Methoxybenzoesäure-Derivat
anzeigt, (2) 200,1, was 4-Fluorbenzoesäure-Derivat anzeigt, (3) 196,1
amu, was Tolylsäure-Derivat
anzeigt, (4) 182,1 amu, was Benzoesäure-Derivat anzeigt, (5) 235,2 amu, was
Indol-3-essigsäure-Derivat
anzeigt, (6) 218,1 amu, was 2,6-Difluorbenzoesäure-Derivat anzeigt, (7) 199,1
amu, was Nikotinsäure-N-oxid-Derivat
anzeigt, (8) 227,1 amu, was 2-Nitrobenzamid anzeigt, (9) 179,18
amu, was 5-Acetylsalylsäure-Derivat
anzeigt, (10) 226,1 amu, was 4-Ethoxybenzoesäure-Derivat. anzeigt, (11)
209,1 amu, was Zimtsäure-Derivat
anzeigt, (12) 198,1 amu, was 3-Aminorricotinsäure-Derivat hinweist.
-
Die Ergebnisse zeigten, daß die MW-Identifikatoren
von den Primern abgespalten werden und mit Massenspektrometrie nachweisbar
sind.
-
BEISPIEL 5
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL R1-36-LYS(εINIP)-ANP-TFP
-
3 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbindungen (X
= Lh), wobei Lh ein
aktivierter Ester ist (genauer Tetrafluorphenylester), L2 eine ortho-Nitrobenzylamin-Gruppe ist, wobei L3 eine Methylengruppe ist, die Lh und
L2 verknüpft,
T eine modulare Struktur hat, wobei die Carbonsäuregruppe von Lysin mit dem
Stickstoffatom der L2-Benzylamin-Gruppe
verknüpft
worden ist, um eine Amidbindung zu bilden, und eine Komponente R1-36 mit variabler Masse (wobei die R-Gruppen
T2 entsprechen, wie hierin definiert, und über jede
der spezifischen Carbonsäuren,
die hierin aufgelistet sind, eingeführt werden können) durch
die α-Aminogruppe
des Lysins gebunden ist, während
eine Massensepktrometrieempfindlichkeitsverstärkergruppe (eingeführt über N-Methylisonipecotinsäure) durch
die ε-Aminogruppe
des Lysins gebunden ist.
-
Bezug nehmend auf 3:
-
Schritt A. NovaSyn-HMP-Harz (erhältlich von
NovaBiochem; 1 Äq.)
wird mit DMF im Sammelgefäß des ACT357
suspendiert. Verbindung I (ANP, erhältlich von ACT; 3 Äq.), HATU
(3 Äq.)
und NMM (7,5 Äq.)
in DMF werden zugegeben und das Sammelgefäß für 1 h geschüttelt. Das Lösungsmittel
wird entfernt und das Harz mit NMP (2X), McOH (2X) und DMF (2X)
gewaschen. Die Kopplung von I an das Harz und die Waschschritte werden
wiederholt, um Verbindung II zu ergeben.
-
Schritt B. Das Harz (Verbindung II)
wird mit 25% Piperidin in DMF vermischt und für 5 min geschüttelt. Das
Harz wird filtriert, anschließend
mit 25% Piperidin in DMF vermischt und für 10 min geschüttelt. Das
Lösungsmittel
wird entfernt, das Harz mit NMP (2X), McOH (2X) und DMF (2X) gewaschen
und direkt in Schritt C verwendet.
-
Schrift C. Das Harz ohne Schutzgruppe
aus Schritt B wird in DMF suspendiert und dazu wird eine Aminosäure aus
FMOC-Schutzgruppe hinzugegeben, die eine geschützte Aminfunktionalität in ihrer
Seitenkette enthält
(Fmoc-Lysin(Aloc)-OH, erhältlich
von PerSeptive Biosystems; 3 Äq.),
HATU (3 Äq.)
und NMM (7,5 Äq.) in
DMF. Das Gefäß wird für 1 h geschüttelt. Das
Lösungsmittel
wird entfernt und das NMP (2X), McOH (2X) und DMF (2X) gewaschen.
Die Kopplung von Fmoc-Lys(Aloc)-OH an das Harz und die Waschschritte
werden wiederholt, um Verbindung IV zu ergeben.
-
Schritt D. Das Harz (Verbindung IV)
wird mit CH2Cl2 (2X)
gewaschen und anschließend
in einer Lösung von
(PPh3)4Pd(0) (0,3 Äq.) und
PhSiH3 (10 Äq.) in CH2Cl2 suspendiert. Die Mischung wird für 1 h geschüttelt. Das
Lösungsmittel
wird entfernt und das Harz mit CH2Cl2 gewaschen. Der Palladiumschritt wird wiederholt. Das
Lösungsmittel
wird entfernt und das Harz wird mit CH2Cl2 (2X), N,N-DÜsopropylethylammoniumdiethyldithiocarbamat
in DMF (2X), DMF (2X) gewaschen, um Verbindung V zu ergeben.
-
Schritt E. Das Harz ohne Schutzgruppe
aus Schritt D wird mit N-Methylisonipecotinsäure gekoppelt, wie in Schritt
C beschrieben, um Verbindung VI zu ergeben.
-
Schritt F. Das Fmoc-geschützte Harz
VI wird gleichmäßig durch
den ACT357 aus dem Sammelgefäß in 36
Reaktionsgefäße aufgeteilt,
um die Verbindungen VI1
-36 zu
ergeben.
-
Schritt G. Das Harz (Verbindungen
VI1-
36) wird mit
Piperidin behandelt, wie beschrieben in Schritt B, um die FMOC-Gruppe
zu entfernen.
-
Schritt H. Die 36 Aliquote des Harzes
ohne Schutzgruppe aus Schritt G werden in DMF suspendiert. Zu jedem
Reaktionsgefäß werden
die entsprechende Carbonsäure
(R1-36CO2H; 3 Äq.), HATU
(3 Äq.)
und NMM (7,5 Äq.)
in DMF zugegeben. Die Gefäße werden
für 1 h
geschüttelt.
Das Lösungsmittel
wird entfernt und die Harz-Aliquote mit NMP (2X), McOH (2X) und
DMF (2X) gewaschen. Die Kopplung von R1-36CO2H an die Harz-Aliquote und die Waschschritte
werden wiederholt, um die Verbindungen VIII1
-36 zu ergeben.
-
Schritt I. Die Harz-Aliquote (Verbindungen
VIII1-
36) werden
mit CH2Cl2 (3X)
gewaschen. Zu jedem der Reaktionsgefäße wird TFA : H2O
: CH2Cl2 im Verhältnis 90
: 5 : 5 zugegeben und die Gefäße für 120 min
geschüttelt.
Das Lösungsmittel
wird aus den Reaktionsgefäßen in einzelne
Röhren
filtriert. Die Harz-Aliquote werden mit CH2Cl2 (2X) und McOH (2X) gewaschen und die Filtrate
in die einzelnen Röhren
hinein zusammengeführt.
Die einzelnen Röhren
werden in vacuo eingedampft, was die Verbindungen IX1-
36 liefert.
-
Schritt J. Jede der freien Carbonsäure IX1
-36 wird in DMF
gelöst.
Zu jeder Lösung
wird Pyridin (1,05 Äq.)
zugegeben, gefolgt von Tetrafluorphenyltrifluoracetat (1,1 Äq.). Die
Mischungen werden für
45 min bei Raumtemperatur gerührt.
Die Lösungen
werden mit EtOAc verdünnt,
mit 5% wäßrigem NaHCO3 (3X) gewaschen, über Na2SO4 getrocknet, filtriert und in vacuo eingedampft,
was die Verbindungen X1-
36 liefert.
-
BEISPIEL 6
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL R1-36-LYS(ε-iNIP)-NBA-TFP
-
4 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbindungen (X
= Lh), wobei Lh ein
aktivierter Ester ist (genauer Tetrafluorphenylester), L2 eine ortho-Nitrobenzylamin-Gruppe ist, wobei L3 eine direkte Bindung zwischen Lh und L2 ist, wobei
Lh direkt mit dem aromatischen Ring der
L2-Gruppe verknüpft ist, T eine modulare Struktur
hat, wobei die Carbonsäuregruppe
von Lysin mit dem Stickstoffatom der L2-Benzylamin-Gruppe
verknüpft
ist, um eine Amidbindung zu bilden, und eine Komponente R1-36 mit variabler Masse (wobei diese R-Gruppen
T2 entsprechen, wie hierin definiert, und über jede
der spezifischen Carbonsäuren,
die hierin aufgelistet ist, eingeführt werden können) durch
die α-Aminogruppe des Lysins
gebunden ist, während
eine Massenspektrometrieverstärkergruppe
(eingeführt über N-Methylisonipecotinsäure) durch die ε-Aminogruppe
des Lysins gebunden ist.
-
Bezug nehmend auf 4
-
Schritt A. NovaSyn-HMP-Harz wird
mit Verbindung I (NBA hergestellt gemäß dem Verfahren von Brown et
al., Molecular Diversity, 1, 4 (1995)) gemäß dem in Schritt A von Beispiel
5 beschriebenen Verfahren gekoppelt, um Verbindung II zu ergeben.
-
Schritte B–J. Das Harz (Verbindung II)
wird behandelt, wie beschrieben in den Schritten B–J von Beispiel
5, um die Verbindungen X1-
36 zu
ergeben.
-
BEISPIEL 7
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL iNIP-LYS(ε-R1-36)-ANP-TFP
-
5 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbindungen (X
= Lh), wobei Lh ein
aktivierter Ester ist (genauer Tetrafluorphenylester), L2 eine ortho-Nitrobenzylamin-Gruppe ist, wobei L3 eine Methylengruppe ist, die Lh und
L2 verknüpft,
T eine modulare Struktur hat, wobei die Carbonsäuregruppe von Lysin mit dem
Stickstoffatom der L2-Benzylamin-Gruppe
verknüpft
worden ist, um eine Amidbindung zu bilden, und eine Komponente R1-36 mit variabler Masse (wobei diese R-Gruppen
T2 entsprechen, wie hierin definiert, und über jede
der spezifischen Carbonsäuren,
die hierin aufgelistet sind, eingeführt werden können) durch
die ε-Aminogruppe
des Lysins gebunden ist, während
eine Massenspektrometrieempfindlichkeitsverstärkergruppe (eingeführt über N-Methylisonipecotinsäure) durch
die α-Aminogruppe
des Lysins gebunden ist.
-
Bezug nehmend auf 5:
-
Schritte A–C. Dieselben wie in Beispiel
5.
-
Schritt D. Das Harz (Verbindung IV)
wird mit Piperidin behandelt, wie in Schritt B von Beispiel 5 beschrieben,
um die FMOC-Gruppe zu entfernen.
-
Schritt E. Das α-Amin ohne Schutzgruppe auf
dem Harz in Schritt D wird mit N-Methylisonipecotinsäure gekoppelt,
wie beschrieben in Schritt C von Beispiel 5, um Verbindung V zu
ergeben.
-
Schritt F. Derselbe wie in Beispiel
5.
-
Schritt G. Das Harz (Verbindungen
VI1
-36) wird behandelt
mit Palladium, wie beschrieben in Schritt D von Beispiel 5, um die
Aloc-Gruppe zu entfernen.
-
Schritte H–J. Die Verbindungen X1
-36 werden auf dieselbe
Art und Weise hergestellt wie in Beispiel 5.
-
BEISPIEL 8
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL R1-36-GLU(γ-DIAEA)-ANP-TFP
-
6 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbidnungen (X
= Lh), wobei Lh ein
aktivierter Ester ist (genauer Tetrafluorphenylester), L2 eine ortho-Nitrobenzylamin-Gruppe ist, wobei L3 eine Methylengruppe ist, die Lh und
L2 verknüpft,
T eine modulare Struktur hat, wobei die α-Carbonsäuregruppe von Glutaminsäure mit
dem Stickstoffatom der L2-Benzylamin-Gruppe
verknüpft
worden ist, um eine Amidbindung zu bilden, und eine Komponente R1-
36 mit variabler
Masse (wobei diese R-Gruppen T2 entsprechen, wie
hierin definiert, und über
eine der spezifischen Carbonsäuren,
die hierin aufgelistet sind, eingeführt werden können) durch
die α-Aminogruppe
der Glutaminsäure
gebunden ist, während
eine Massenspektrometrieempfindlichkeitsverstärkergruppe (eingeführt über 2-(Diisopropylamino)ethylamin)
durch die γ-Carbonsäure der Glutaminsäure gebunden
ist.
-
Bezug nehmend auf 6:
-
Schritte A–B. Dieselben wie in Beispiel
5.
-
Schritt C. Das Harz ohne Schutzgruppe
(Verbindung III) wird an Fmoc-Glu-(OAl)-OH unter Verwendung des
Kopplungsverfahrens, das in Schritt C von Beispiel 5 beschrieben
ist, gekoppelt, um Verbindung IV zu ergeben.
-
Schritt D. Der Allylester auf dem
Harz (Verbindung IV) wird mit CH2Cl2 (2X) gewaschen und mit einer Lösung von
(PPh3)4Pd(0) (0,3 Äq.) und
N-Methylanilin (3 Äq.)
in CH2Cl2 vermischt.
Die Mischung wird für
1 h geschüttelt.
Das Lösungsmittel
wird entfernt und das Harz wird mit CH2Cl2 (2X) gewaschen. Der Palladiumschritt wird
wiederholt. Das Lösungsmittel
wird entfernt und das Harz wird mit CH2Cl2 (2X), N,N-Diisopropylethylammoniumdiethyldithiocarbamat
in DMF (2X), DMF (2X) gewaschen, um Verbindung V zu ergeben.
-
Schritt E. Das Harz ohne Schutzgruppe
aus Schritt D wird in DMF suspendiert und durch Vermischen mit HATU
(3 Äq.)
und NMM (7,5 Äq.)
aktiviert. Die Gefäße werden
für 15
min geschüttelt.
Das Lösungsmittel wird
entfernt und das Harz mit NMP (1X) gewaschen. Das Harz wird mit
2-(Diisopropylamino)ethylamin (3 Äq.) und NMM (7,5 Äq.) vermischt.
Die Gefäße werden
für 1 Stunde
geschüttelt.
Die Kopplung von 2-(Diisopropylamino)ethylamin an das Harz und die
Waschschritte werden wiederholt, um Verbindung VI zu ergeben.
-
Schritte F–J. Dieselben wie in Beispiel
5.
-
BEISPIEL 9
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL R1-36-LYS(ε-iNIP)-ANP-LYS(ε-NH2)-NH2
-
7 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbindungen (X
= Lh), wobei Lh ein
Amin ist (genauer die ε-Aminogruppe
einer von Lysin abgeleiteten Einheit), L2 eine
ortho-Nitrobenzylamin-Gruppe ist, wobei L3 eine
Carboxamido-substituierte Alkylenaminoacylalkylen-Gruppe ist, die
Lh und L2 verknüpft, T eine
modulare Struktur hat, wobei die Carbonsäuregruppe von Lysin mit dem
Stickstoffatom der L2-Benzylamin-Gruppe
verknüpft
worden ist, um eine Amidbindung zu bilden, und eine Komponente R1-36 mit variabler Masse (wobei diese R-Gruppen
T2 entsprechen, wie hierin definiert, und über eine
der spezifischen Carbonsäuren,
die hierin aufgelistet sind, eingeführt werden können) durch
die α-Aminogruppe
des Lysins gebunden ist, während
eine Massenspektrometrieempfindlichkeitsverstärkergruppe (eingeführt über N-Methylisonipecotinsäure) durch
die ε-Aminogruppe
des Lysins gebunden ist.
-
Bezug nehmend auf 7:
-
Schritt A. Fmoc-Lys(Boc)-SRAM-Harz
(erhältlich
von ACT; Verbindung I) wird mit 25% Piperidin in DMF vermischt und
für 5 min
geschüttelt.
Das Harz wird filtriert, dann mit 25% Piperidin in DMF vermischt
und für
10 min geschüttelt.
Das Lösungsmittel
wird entfernt, das Harz mit NMP (2X), McOH (2X) und DMF (2X) gewaschen
und direkt in Schritt B verwendet.
-
Schritt B. Das Harz (Verbindung II),
ANP (erhältlich
von ACT; 3 Äq.),
HATU (3 Äq.)
und NMM (7,5 Äq.) in
DMF werden zugegeben und das Sammelgefäß für 1 h geschüttelt. Das Lösungsmittel
wird entfernt und das Harz mit NMP (2X), McOH (2X) und DMF (2X)
gewaschen. Die Kopplung von I an das Harz und die Waschschritte
werden wiederholt, um Verbindung III zu ergeben.
-
Schritte C–J. Das Harz (Verbindung III)
wird behandelt wie in den Schritten B–I in Beispiel 5, um die Verbindungen
X1-36 zu ergeben.
-
BEISPIEL 10
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL R1-
36-LYS(ε-TFA)-LYS(ε-INP)-ANP-TFP
-
8 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbindungen (X
= Lh), wobei Lh ein
aktivierter Ester ist (genauer Tetrafluorphenylester), L2 eine ortho-Nitrobenzylamin-Gruppe ist, wobei L3 eine Methylengruppe ist, die Lh und
L2 verknüpft,
T eine modulare Struktur hat, wobei die Carbonsäuregruppe eines ersten Lysins
mit dem Stickstoffatom der L2-Benzylamin-Gruppe
verknüpft
worden ist, um eine Amidbindung zu bilden, eine Massenspektrometrieempfindlichkeitsverstärkergruppe
(eingeführt über N-Methylisonipecotinsäure) durch
die ε-Aminogruppe
des ersten Lysins gebunden ist, ein zweites Lysinmolekül mit dem
ersten Lysin über
die a-Aminogruppe des ersten Lysins verknüpft worden ist, eine Gruppe
zur Einstellung der relativen Molekülmasse (mit einer Trifluoracetylstruktur)
durch die ε-Aminogruppe
des zweiten Lysins gebunden ist und eine Komponente R1-36 mit
variabler Masse (wobei diese R-Gruppen T2 entsprechen,
wie hierin definiert, und über
eine der spezifischen Carbonsäuren,
die hierin aufgelistet sind, eingeführt werden können) durch
die α-Aminogruppe
des zweiten Lysins gebunden ist.
-
Bezug nehmend auf 8:
-
Schritte A–E. Diese Schritte sind identisch
mit den Schritten A–E
in Beispiel 5.
-
Schritt F. Das Harz (Verbindung VI)
wird mit Piperidin behandelt wie beschrieben in Schritt B in Beispiel 5,
um die FMOC-Gruppe zu entfernen.
-
Schritt G. Das Harz ohne Schutzgruppe
(Verbindung VII) wird an Fmoc-Lys(Tfa)-OH unter Verwendung des Kopplungsverfahrens,
das in Schritt C von Beispiel 5 beschrieben ist, gekoppelt, um Verbindung
VIII zu ergeben.
-
Schritte H–K. Das Harz (Verbindung VIII)
wird behandelt, wie in den Schritten F–J in Beispiel 5, um die Verbindungen
XI1-
36 zu ergeben.
-
BEISPIEL 11
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL R1-36-LYS(ε-iNIP)-ANP-5'-AH-ODN
-
9 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbindungen (X
= MOI, wobei MOI ein Nukleinsäurefragment
ist, ODN), abgeleitet von den Estern von Beispiel 5 (dasselbe Verfahren könnte bei
anderen T-L-X-Verbindungen verwendet werden, in denen X ein aktivierter
Ester ist). Das MOI ist an T-L durch das 5'-Ende des MOIs konjugiert, über eine
Phosphodiester-Alkylenamin-Gruppe.
-
Bezug nehmend auf 9:
-
Schritt A. Die Verbindungen XII1
-36 werden hergestellt
gemäß einem
modifizierten Biotinylierungsverfahren in Van Ness et al., Nuleic
Acids Res., 19, 3345 (1991). Zu einer Lösung eines der 5'-Aminohexyl-Oligonukleotide
(Verbindungen XI1
-36,
1 mg) in 200 mM Natriumborat (pH 8,3, 250 ml) wird einer der Tetrafluorphenylester
zugegeben (Verbindungen X1
-36 aus
Beispiel A, 100-facher molarer Überschuß in 250
ml NMP). Die Reaktion wird über
Nacht bei Umgebungstemperatur inkubiert. Die nicht-umgesetzten und
hydrolysierten Tetrafluorphenylester werden von den Verbindungen
XII1-36 durch Sephadex-G-50-Chromatographie getrennt.
-
BEISPIEL 12
-
HERSTELLUNG EINES SATZES
VON VERBINDUNGEN DER FORMEL R1-36-LYS(ε-iNIP)-ANP-LYS(ε-(MCT-5'-AH-ODN))-NH2
-
10 veranschaulicht
die parallele Synthese eines Satzes von 36 T-L-X-Verbindungen (X
= MOI, wobei MOI ein Nukleinsäurefragment
ist, ODN), abgeleitet von den Aminen von Beispiel 11 (dasselbe Verfahren könnte bei
anderen T-L-X-Verbindungen verwendet werden, in denen X ein Amin
ist). Das MOI ist an T-L durch das 5'-Ende des MOIs konjugiert, über eine
Phosphodiester-Alkylenamin-Gruppe.
-
Bezug nehmend auf 10:
-
Schritt A. Die 5'-[6-(4,6-Dichlor-1,3,5-triazin-2-ylamino)hexyl]-Oligonukleotide
XII1-
36 werden hergestellt
wie beschrieben in Van Ness et al., Nucleic Acids Res., 19, 3345
(1991).
-
Schritt B. Zu einer Lösung eines
der 5'-[6-(4,6-Dichlor-1,3,5-triazin-2-ylamino)hexyl]-Oligonukleotide (Verbindungen
XII1-
36) in einer
Konzentration von 1 mg/ml in 100 mM Natriumborat (pH 8,3) wurde
ein 100-facher molarer Überschuß eines
primäre
Amins zugegeben, das ausgewählt
ist aus R1-36-Lys(ε-iNIP)-ANP-Lys(ε-NH2)-NH2 (Verbindungen
X1-
36 aus Beispiel
11). Die Lösung
wird über
Nacht bei Umgebungstemperatur gemischt. Das nicht-umgesetzte Amin
wird durch Ultrafiltration durch eine Membran mit einem 3.000 MG-Schnitt (Amicon,
Beverly, MA) unter Verwendung von H2O als
der Waschlösung
(3X) entfernt. Die Verbindungen XIII1
-36 werden durch Verringerung des Volumens
auf 100 ml isoliert.
-
BEISPIEL 13
-
DEMONSTRATION
DES GLEICHZEITIGEN NACHWEISES MEHRERER MARKIERUNGEN DURCH MASSENSPEKTROMETRIE
-
Dieses Beispiel liefert eine Beschreibung
der Fähigkeit,
mehrere Verbindungen (Markierungen) durch Massenspektrometrie gleichzeitig
nachzuweisen. In diesem besonderen Beispiel werden 31 Verbindungen
mit einer Matrix vermischt, auf einem festen Träger abgeschieden und getrocknet
und anschließend
mit einem Laser desorbiert. Die resultierenden Ionen werden anschließend in
einen Massenspektrometer eingeführt.
-
Die folgenden Verbindungen (bezogen
von Aldrich, Milwaukee, WI) werden auf einer äquimolaren Basis auf eine Endkonzentration
von 0,002 M (pro Verbindung) zusammengemischt: Benzamid (121,14),
Nicotinamid (122,13), Pyrazinamid (123,12), 3-Amino-4-pyrazolcarbonsäure (127,10),
2-Thiophencarboxamid (127,17), 4-Aminobenzamid (135,15), Tolumid
(135,17), 6-Methylnicotinamid (136,15), 3-Aminonicotinamid (137,14),
Nicotinamid-N-Oxid (138,12), 3-Hydropicolinamid (138,13), 4-Fluorbenzamid
(139,13), Zimtsäureamid
(147,18), 4-Methoxybenzamid (151,17), 2,6-Difluorbenzamid (157,12),
4-Amino-5-imidazolcarbonsäureamid
(162,58), 3,4-Pyridindicarbonsäureamid
(165,16), 4-Ethoxybenzamid
(165,19), 2,3-Pyrazindicarboxamid (166,14), 2-Nitrobenzamid (166,14),
3-Fluor-4-methoxybenzoesäure (170,4),
Indol-3-acetamid (174,2), 5-Acetylsalicylamid (179,18), 3,5-Dimethoxybenzamid
(181,19), 1-Naphthalinacetamid (185,23), 8-Chlor-3,5-diamino-2-pyrazincarboxyamid
(187,59), 4-Trifluormethylbenzamid (189,00), 5-Amino-5-phenyl-4-pyrazolcarboxamid
(202,22), 1-Methyl-2-benzylmalonamat (207,33), 4-Amino-2,3,5,6-tetrafluorbenzamid
(208,11), 2,3-Naphthalindicarbonsäure (212,22). Die Verbindungen
werden in der oben beschriebenen Konzentration in DMSO gegeben.
Ein μl des
Materials wird dann mit Alpha-Cyano-4-hydroxyzimtsäure-Matrix
(nach einer 1 : 10.000-Verdünnung) vermischt
und auf einem festen Träger
aus rostfreiem Stahl abgeschieden.
-
Das Material wird anschließend mit
einem Laser unter Verwendung des Protein-TOF-Massenspektrometers (Bruker, Manning
Park, MA) desorbiert und die resultierenden Ionen werden in sowohl
dem linearen als auch dem Reflektron-Betriebsmodus gemessen. Die
folgenden m/z-Werte werden beobachtet (
11):
-
Die Daten zeigen, daß 22 von
31 Verbindungen im Spektrum mit der vorhergesagten Masse erschienen,
9 von 31 Verbindungen im Spektrum mit einer n + H-Masse (1 Atommasseneinheit,
amu) über
der vorhergesagten Masse erschienen. Das letztere Phänomen beruht
wahrscheinlich auf der Protonierung eines Amins in den Verbindungen.
Daher werden 31 von 31 Verbindungen mit MALDI-Massenspektroskopie
nachgewiesen. Noch wichtiger ist, daß das Beispiel zeigt, daß mehrere
Markierungen gleichzeitig mit einem spektroskopischen Verfahren
nachgewiesen werden können.
-
Die Alpha-Cyano-Matrix allein (11) ergab Peaks bei 146,2,
164,1, 172,1, 173,1, 189,1, 190,1, 191,1, 192,1, 212,1, 224,1, 228,0,
234,3. Weitere identifizierte Massen im Spektrum beruhen auf Verunreinigungen
in den gekauften Verbindungen, da keine Anstrengungen unternommen
wurden, die Verbindungen weiter zu reinigen.
-
BEISPIEL 14
-
MIKROSATELLITEN-MARKER:PCR-AMPLIFIKATIONEN.
-
Die Mikrosatelliten-Marker werden
amplifiziert unter Verwendung der folgenden Standard-PCR-Bedingungen.
Kwz gesagt, werden PCR-Reaktionen in einem Gesamtvolumen von 50 μl durchgeführt, das
40 ng genomische DNA, 50 pmol von jedem Primer, 0,125 mM dNTPs und
1 Einheit Taq-Polymerase enthält.
1X-Amplifikationspuffer enthält
10 mM Tris-Base, pH 9, 50 mM KCl, 1,5 mM MgCl2,
0,1% Triton X-100 und 0,01% Gelatine. Die Reaktionen werden unter
Verwendung eines "Heißstart"-Verfahrens durchgeführt: Taq-Polymerase wird
erst nach einem ersten Denatwierungsschritt von 5 Minuten bei 96°C zugegeben.
Amplifikation wird über 35
Zyklen durchgeführt:
Denaturierung (94°C
für 40
s) und Annealing (55°C
für 30
s). Ein Verlängerungsschritt (72°C für 2 Minuten)
beendet das Verfahren nach dem letzten Annealing. Da die Amplifikationsprodukte,
die erhalten werden sollen, kurz sind (90 bis 350 Basenpaare lang)
und das Zeitintervall, um die Temperatur von 55°C auf 94°C anzuheben (erhalten mit einer
Anstiegsgeschwindigkeit von 1°C/Sekunde),
lang genug ist, kann die Vervollständigung der DNA-Verlängerung
ohne einen Schritt bei 72°C
erreicht werden.
-
BEISPIEL 15
-
AUFRENNUNG VON DNA-FRAGMENTEN
-
Instrumentierung
-
Die Auftrennung von DNA-Fragmenten
kann unter Verwendung eines HPLC-Systems durchgeführt werden,
das aus verschiedenen Stundardkomponenten zusammengesetzt ist. Diese
Komponenten schließen ein
Minimum von zwei Hochdruckpumpen ein, die Lösungsmittel durch einen Hochdruckgradientenmischer,
einen Injektor, einer HPLC-Säule
und einen Detektor pumpen. Der Injektor ist ein automatisierter
programmierbarer Autosampler, der in der Lage ist, typischerweise
zwischen 80 und 100 Proben bei oder unterhalb von Raumtemperaturen
zu Lagern, um die Stabilität
der Probenkomponenten aufrechtzuerhalten. Der Autoinjektor ist ebenfalls
in der Lage, μl-Größen-Injektionen
auf eine reproduzierbare Weise vollständig ohne Kontrolle durchzuführen. Die
HPLC-Säule
ist in einem geheizten Säulenkompartiment
enthalten, das in der Lage ist, eine definierte Temperatur auf innerhalb
von 0,1°C
zu halten. Die in den Beispielen unten verwendetete Säule wurde
von SeraSep (San Jose, CA) unter dem Namen DNASep erworben. Die
Säule ist
eine 55 × 4,6
mm Säule
mit einem 2,2 μm
nicht-porösen
Polystyroldivinylbenzol-Copolymer-Partikel, der mit C18 alkyliert ist.
Das Packmaterial ist innerhalb eines pH-Bereichs von 2–12 stabil
und toleriert Temperaturen so hoch wie 70°C. Der Nachweis von Analyt wurde
unter der Verwendung eines Einzel- oder multiple Wellenlängen UV-Detektors oder
Dioden-Arraydetektor
durchgeführt.
-
Verfahren
-
Die in diesem Beispiel zur Abtrennung
von DNA-Fragmenten angewendeten Verfahren verwenden Ionen-Paarchromatographie,
eine Form von Chromatographie, in der Ionen in Lösung gepaart oder neutralisiert werden
können
und als ein Ionenpaar auf eine Umkehrphasensäule getrennt werden können. Der
lipophile Charakter und die Konzentration des Gegenions bestimmen
das Ausmaß der
Retention des Analyten. Im Fall eines DNA-Moleküls paart die lipophile kationische
Pufferkomponente mit anionischen Phosphatgruppen auf dem DNA-Rückgrat.
Die Pufferkomponenten interagieren auch mit den Alkylgruppen der
stationären
Phase. Die gepaarte DNA eluiert dann gemäß ihrer Größe, während die mobile Phase fortschreitend
organischer mit ansteigenden Konzentrationen von Acetonitril gemacht
wird. Die Evaluierung der Geeignetheit von verschiedenen Aminsalzen
wurde unter der Verwendung von enzymatischen Verdaus von Plasmiden
oder kommerziell erhältlichen
DNA-Leitern untersucht. Der Bereich von Acetonitril, der erforderlich
ist um die DNA zu eluieren, sowie die Temperatur des Säulenkompartiments
variierte mit jedem untersuchten Puffer.
-
Puffer
-
Die auf ihre Ionen-Paarungsfähigkeit
hin untersuchten Puffer wurden aus Vorratslösungen hergestellt. Um die
Konzentration von Ionen-Paaneagenz während des Gradienten konstant
zu halten, wurde das Ionen-Paaneagenz zu sowohl der Wasser- und
der Acetonitril-mobilen Phase hinzugefügt. Die Säue wurde mit einer neuen mobilen
Phase für
ungefähr
18 Stunden bei einer Flußrate
von 50 μl/Minute
vor dem Versuch jeglicher Trennung äquilibriert. Sobald eine mobile
Phase untersucht wurde, wurde sie entfernt und die Säule mit einer
Spülung
von 800 ml 0,1% Ameisensäure
und 50% Acetonitril, gefolgt von einer Spülung mit 800 ml 0,1% Essigsäure und
50% Acetonitril vor der Äquilibrierung
mit einer neuen mobilen Phase gereinigt.
-
A. nn-Dimethyloctylammoniumtrifluoracetat
-
Eine Vorratslösung von 1 molar von Dimethyloctylammoniumtrifluoracetat
wurde durch Mischen einer Hälfte
eines Äquivalents
von Trifluoressigsäure
in einem geeigneten Volumen von Wasser und langsamem Hinzufügen eines Äquivalents
von nn-Dimethyloctylamin hergestellt. Der pH dieser Vorratslösung ist
7. Die Vorratslösung
wurde mit einem geeigneten Volumen von Wasser oder Acetonitril auf
Arbeitskonzentration verdünnt.
-
B. nn-Dimethylheptylammoniumacetat
-
Eine Vorratslösung von 1 molar Dimethylheptylammoniumacetat
wurde durch Mischen eines Äquivalents
von Eisessig in einem geeigneten Volumen von Wasser und langsamem
Hinzufügen
eines Äquivalents von
nn-Dimethylheptylamin hergestellt. Der pH dieser Vorratslösung ist
6,6. Die Vorratslösung
wurde mit einer geeigneten Menge von Wasser oder Acetonitril auf
Arbeitskonzentration verdünnt.
-
C. nn-Dimethylhexylammoniumacetat
-
Eine Vorratslösung von 1 molar Dimethylhexylammoniumacetat
wurde durch Mischen eines Äquivalents
von Eisessig in einem geeigneten Volumen von Wasser und langsamem Hinzufügen eines Äquivalents von
nn-Dimethylhexylamin hergestellt. Der pH dieser Vorratslösung ist
6,5. Die Vonatslösung
wurde mit einem geeigneten Volumen von Wasser oder Acetonitril auf
Arbeitskonzentration verdünnt.
-
D. nn-Dimethylbutylammoniumacetat
-
Eine Vorratslösung von 1 molar Dimethylbutylammoniumacetat
wurde durch Mischen eines Äquivalents
von Eisessig in einem geeigneten Volumen von Wasser und langsamem
Hinzufügen
eines Äquivalents von
nn-Dimethylbutylamin hergestellt. Der pH der Vorratslösung ist
6,9. Die Vorratslösung
wurde mit einem geeigneten Volumen von Wasser oder Acetonitril auf
Arbeitskonzentration verdünnt.
-
E. nn-Dimethylisopropylammoniumacetat
-
Eine Vorratslösung von 1 molar Dimethylisopropylammoniumacetat
wurde durch Mischen eines Äquivalents
von Eisessig in einem geeigneten Volumen von Wasser und langsamem
Hinzufügen
eines Äquivalents von
nn-Dimethylisopropylamin hergestellt. Der pH der Vorratslösung ist
6,9. Die Vonatslösung
wurde mit einem geeigneten Volumen von Wasser oder Acetonitril auf
Arbeitskonzentration verdünnt.
-
F. nn-Dimethylcyclohexylammoniumacetat
-
Eine Vorratslösung von 1 molar Dimethylcyclohexylammoniumacetat
wurde durch Mischen eines Äquivalents
von Eisessig in einem geeigneten Volumen von Wasser und langsamem
Hinzufügen
eines Äquivalents
von nn-Dimethylcyclohexylamin hergestellt. Der pH der Vorratslösung ist
6,5. Die Vorratslösung
wurde mit einem geeigneten Volumen von Wasser oder Acetonitril auf
Arbeitskonzentration verdünnt.
-
G. Methylpiperidinacetat
-
Eine Vorratslösung von 1 molar Methylpiperidinacetat
wurde durch Mischen eines Äquivalents
von Eisessig in einem geeigneten Volumen von Wasser und langsamen
Hinzufügen
eines Äquivalents
von 1-Methylpiperidin hergestellt. Der pH der Lösung ist 7. Die Vonatslösung wurde
mit einem geeigneten Volumen von Wasser oder Acetonitril auf Arbeitskonzentration
verdünnt.
-
H. Methylpynolidinacetat
-
Eine Vorratslösung von 1 molar Methylpyrrolidinacetat
wurde durch Mischen eines Äquivalents
von Eisessig in einem geeigneten Volumen von Wasser und langsamem
Hinzufügen
eines Äquivalents
1-Methylpyrrolidin hergestellt. Der pH der Vorratslösung ist
7. Die Vonatslösung
wurde in einem geeigneten Volumen von Wasser oder Acetonitril auf
Arbeitskonzentration verdünnt.
-
I. Triethylammoniumacetat
-
Eine Vorratslösung von 2 molar Triethylammoniumacetat
pH 7,0 wurde von Glenn Research Sterling, Virginia, erworben. Die
Vonatslösung
wurde mit einem geeigneten Volumen von Wasser oder Acetonitril auf Arbeitskonzentration
verdünnt.
-
BEISPIEL 16
-
DNA-FINGERPRINT
-
DNA-Fingerprintingadaptoren werden
umfassend das Folgende hergestellt: Eine Kernsequenz und eine Enzym-spezifische
Sequenz. Die Struktur des EcoRI-Adapters ist 5'-CTCGTAGACTGCGTACC
(SEQ ID NO. _), die Struktur des MseI-Adapters ist 5'-GACGATGAGTCCTGAG.
-
Adaptoren für die selten schneidenden Enzyme
waren identisch zu dem EcoRI mit der Ausnahme, daß kohäsive Enden
verwendet wurden. ALPH-Primer bestehen aus drei Teilen: Einer Kernsequenz,
einer Enzym-spezifische Sequenz und einer selektiven Verlängerungssequenz.
Die EcoRI- und MseI-Primer werden wie folgt beschrieben: EcoRI:
5'-gactgcgtaaa-aattc-NNN
(SEQ ID NO. _); MseI: 5'-gatgagtcctgag-taa-NNN (SEQ
ID NO. _).
-
Genomische DNA wurde für 1 Stunde
bei 37°C
mit 5 Einheiten EcoRI und 5 Einheiten MseI und 40 μl Volumen
mit 10 mM Tris-Acetat pH 7,5, 10 mM MgAce, 50 mM KActate, 5 mM DTT,
50 ng/Mikroliter BSA, 5 mM DTT inkubiert. Als nächstes wurden 10 μl einer Lösung, die
5 pMol EcoRI-Adaptoren, 50 pMol MseI-Adaptoren, 1 Einheit T4-Ligase,
1 mM ATP in 10 mM Tris-Acetat pH 7,5, 10 mM MgAce, 50 mM KAcetate,
5 mM DTT, 50 ng/Mikroliter BSA enthielt, hinzugeführt und
die Inkubation wurde für
3 Stunden bei 37°C
fortgeführt. Die
Adaptoren wurden durch Hinzufügen
von äquimolaren
Mengen von beiden Strängen
hergestellt: Die Adaptoren waren nicht phosphoryliert. Nach der
Ligation wurde das Reaktionsgemisch auf 500 μl mit 10 mM Tris-HCl, 0,1 mM
EDTA pH 8,0 verdünnt
und bei – 20°C gelagert.
-
Genetische Fingerprintingreaktionen:
Amplifikationsreaktionen werden beschrieben, die DNA-Template für die Enzymkombination
EcoRI/MseI verwenden. Genomische Fingerprints mit anderen Enzymkombinationen
wurden mit geeigneten Primern durchgeführt. Die Amplifikationsreaktionen
verwendeten im allgemeinen zwei Oligonukleotide, eines entsprechend
den EcoRI-Enden und eines entsprechend den MseI-Enden. Einer der
zwei Primer war mit dem CMST-Marker markiert, bevorzugterweise der
EcoRI-Primer. Die PCRs wurden unter der Verwendung von 5 ng markiertem
EcoRI-Primer, 30 ng MseI-Primer, 5 Mikroliter Templat DNA, 0,4 Einheiten
Taq-Polymerase, 10 mM Tris-HCl, pH 8,3, 1,5 mM MgCl2, 50 mM KCl,
0,2 mM von dATP, dGTP, dCTP, dTTP durchgeführt. Die PCR-Reaktionen unterschieden
sich in Abhängigkeit
von der Art der selektiven Amplifikationsverlängerung der für die Amplifikation
verwendeten DNA-Fingerprintingprimer.
Die DNA-Fingerprintingreaktionen mit Primern, die zwei oder drei
selektive Nukleotide aufweisen, wurden für 36 Zyklen mit dem folgenden
Zyklusprofil durchgeführt:
Ein 30 Sekunden DNA-Denaturierungsschritt bei 94°C, ein 30 Sekunden Annealingschritt
bei 55°C
und dann ein 1 Minute Verlängerungsschritt
bei 72°C
für 1 Minute.
Die Annealingtemperatur im ersten Schritt war 65°C und wurde anschließend für jeden Zyklusschritt
um 0,7°C
in den nächsten
12 Zyklen verringert und wurde bei 56°C für die restlichen 23 Zyklen
fortgeführt.
Alle Amplifikationen wurden in einem MJ-Thermocycler (Watertown,
MA) durchgeführt.
-
Das DNA-Fingerprinting der komplexen
Genome (wie zum Beispiel bei Menschen) schloß zwei Amplifikationsschritte
ein. Die Präamplifikation
wurde mit zwei DNA-Fingerprintings durchgeführt, die ein einzelnes selektives
Nukleotid, wie oben beschrieben, aufwiesen, mit der Ausnahme, daß 30 ng
von beiden DNA-Fingerprintingprimern verwendet wurden und daß diese
Primer nicht mit CMST markiert wurden, nach dem Preamplifikationsschritt
wurden die Reaktionsgemische 10-fach mit 10 mM Trts-HCl, 0,1 mM
EDTA, pH 8,0 verdünnt
und als Template für
die zweite Amplifikationsreaktion verwendet. Die zweite Amplifikationsreaktion
wurde wie oben beschrieben für
die DNA-Fingerprintingreaktionen mit Primern durchgeführt, die
die längeren
selektiven Verlängerungen
aufwiesen.
-
Die Produkte aus den Amplifikationsreaktionen
wurden durch HPLC analysiert. Die HPLC wurde unter der Verwendung
von automatisierter HPLC-Instrumentierung (Rainin, Emeryville, CA
oder Hewlett Packard, Palo Alto, CA) durchgeführt. Nicht-gereinigte DNA-Fingerprintingprodukte,
die für
3 Minuten bei 95°C
vor der Injektion in einer HPLC denaturiert wurden, wurden mit einem
linearen Acetonitril (ACN, J. T. Baker, NJ) Gradienten von 1,8%/Minute
bei einer Flußrate
von 0,9 ml/Minute eluiert. Die Start- und Endpunkte wurden gemäß der Größe der amplifizierten
Produkte eingestellt. Die für
die erfolgreiche Auflösung
der während
der DNA-Fingerprinting-Technik erzeugten Moleküle erforderliche Temperatur
war 50°C.
Der Ausfluß aus
der HPLC wurde dann in ein Massenspektrometer zum Nachweis der Markierungen
geleitet (Hewlett Packard, Palo Alto, CA).
-
Die folgenden Fragmente eluierten
in der präsentierten
Reihenfolge (die zitierte Nummer sind die Positionen innerhalb des
Lambda-Genoms, bei denen eine Spaltstelle auftrat): 47, 78, 91,
733, 1456, 2176, 3275, 3419, 4349, 444, 5268, 5709, 6076, 6184,
6551, 7024, 7949, 8062, 8200, 8461, 9079, 9253, 9692, 9952, 11083,
11116, 11518, 11584, 12619, 12967, 14108, 14892, 15628, 15968, 16034,
16295, 16859, 18869, 19137, 19482, 20800, 21226, 21441, 2635, 21702,
21903, 21948, 22724, 23048, 23084, 23111, 23206, 23297, 23285, 23479, 23498,
23555, 23693, 23887, 23979, 23987, 24073, 24102, 24751, 24987, 25170,
25255, 25353, 25437, 26104, 25578, 25746, 25968, 26133, 26426, 26451,
26483, 26523, 26585, 26651, 26666, 26679, 26693, 26763, 26810, 26984,
26993, 27038, 27092, 27203, 27317, 27683, 28456, 28569, 28922, 28972,
29374, 29981, 30822, 30620, 30639, 30722, 30735, 30756, 31169, 31747,
31808, 32194, 32218, 32641, 32704, 33222, 33351, 33688, 33736, 33748,
33801, 34202, 34366, 34406, 34590, 34618, 34684, 34735, 34753, 34831,
35062, 35269, 35534, 35541, 36275, 36282, 36303, 36430, 36492, 36531,
36543, 36604, 36736, 36757, 36879, 37032, 37442, 37766, 37783, 37882,
37916, 37994, 36164, 38287, 38412, 38834, 39168, 44972, 39607, 39835,
40127, 40506, 40560, 40881, 41017, 41423, 41652, 41715, 42317, 42631,
42651, 42673, 42814, 43410, 43492, 43507, 43528, 43593, 44424, 44538,
44596, 45151, 45788, 46033, 46408, 46556, 46804, 46843, 46853, 46896,
46952, 47256, 47274, 47287, 47430, 47576, 47699, 47799, 48059, 48125,
48227, 48359, 48378. Die durchschnittliche Fragmentlänge war
ungefähr
160 nt. Die beobachteten Stellen der Spaltung waren größtenteils
(>95%) mit denjenigen
aus einer MseI/EcoRI Verdaukarte vorhergesagten kompatibel.
-
BEISPIEL 17
-
EINZELNUKLEOTID-VERLÄNGERUNGSASSAYS
-
RNA-Präparation: Gesamt-RNA wurde
isoliert und wurde aus Jurkat-Zellen (ausgehend von 1 × 109 Zellen in exponentiellen Wachstum) unter
der Verwendung eines RNA-Isolierungskits
von Promega (WI) präpariert.
Die RNA wurde in zwei Aliquots gelagert: 1) Vorratsaliquot in Diethylpyrocarbonat-behandeltem
ddH2O wurde bei –20°C gelagert und 2) Langzeitlagerung
als eine Suspension in 100% H2O.
-
Reverse Transkription: Poly(dT)-geprimte
reverse Transkription von Gesamt-RNA wurde wie beschrieben in Ausubel
et al. (Ausubel et al., in Current Protocols in Molecular Biology,
1991, Greene Publishing Associates/Wiley Interscience, NY, NY.)
durchgeführt
mit der Ausnahme, daß die
Reaktionen) auf die Verwendung von 1 μg von Anfangs-Gesamt-RNA eingestellt
wurden. 20–50
Einheiten von reverser Transkriptase (Promega) wurden 10-fach in
10% Glyzerin, 10 mM KP04, pH 7,4, 0,2% Triton X-100, und 2 mM DTT
verdünnt
und auf Eis für
30 Minuten plaziert, vor der Zugabe zu den Reaktionen. Gen-spezifische
reverse Transkription für GADPH
und andere Kontrollgene, wie unten beschrieben, wurden unter der
Verwendung von 1 μg
Gesamt Jurkat RNA durchgeführt,
die in 10 mM Tris-HCl pH 8,3, 50 mM KCl, M MgCl2, 1 mM dNTPs, 2
U/μl RNAsin
(Gibco-BRL), 0,1 M Oligomer und 0,125 U/μl von M-MLV reverse Transkriptase
(Gibco-BRL) in 20 μl
Reaktionen revers transkribiert wurde. Die Reaktionen wurden bei
42°C für 15 Minuten
inkubiert, bei 95°C
für 5 Minuten Hitze-inaktiviert
und auf 100 μl
mit einem Mastermix von (10 mM Tris HCl pH 8,3, 1 mM NH4Cl, 1,5
M MgCl2, 100 mM KCl), 0,125 mM NTPs, 10 ng/ml der jeweiligen Oligonucleotidprimer
und 0,75 Einheiten von Taq-Polymerase (Gibco-BRL) in Vorbereitung
auf die PCR-Amplifikation verdünnt.
-
PCR: Die PCR für jedes Gen wurde mit genspezifischen
Primern durchgeführt,
die eine bekannte Intron/Exon-Grenze überspannten (siehe unten).
Alle PCRs wurden in 20 μl
Volumen durchgeführt,
die 10 mM Tris HCl pH 8,3, 1 mM NH4Cl, 1,5
M MgCl2, 100 mM KCl), 0,125 mM NTPs, 10
ng/ml der jeweiligen Oligonukleotidprimer und 0,75 Einheiten Taq-Polymerase
(Gibco-BRL) enthielten. Die Zyklusparameter waren 94°C Vor-Erhitzungsschritt
für 5 Minuten,
gefolgt von 94°C
Denaturierungsschritt für
1 Minute, 55°C
Annealingschritt für
2 Minuten und ein 72°C
Verlängerungsschritt
für 30
Sekunden bis 1 Minute und einer finalen Verlängerung bei 72°C für 10 Minuten.
Die Amplifikationen waren im allgemeinen 30–5 an der Zahl.
-
Reinigung von Templaten: Die PCR-Produkte
wurden wie durch Zhen und Swank (Zhen und Swank, BioTechniques,
14(6): 894–898,
1993) beschrieben gelgereinigt. Die PCR-Produkte wurden auf 1% Agarosegelen
getrennt, die in 0,04 M Tris-Acetat, 0,001 M EDTA (1 × TAE) Puffer
laufen gelassen wurden und mit Ethidiumbromid gefärbt wurden,
während
sie mit einer UV Lichtquelle sichtbar gemacht wurden. Eine Vertiefung wurde
gerade vor der Bunde von Interesse eingeschnitten und mit 50–200 μl von 10%
PEG in 1 × TAE
Puffer gefüllt.
Die Elektrophorese wurde fortgeführt,
bis die Bunde vollständig
in die Vertiefung eingetreten war. Die Inhalte wurden dann entfernt
und extrahiert mit Phenol, Chloroform extrahiert, und dann in 0,1
Volumen von 7,5 M Ammoniumacetat und 2,5 Volumen 100% EtOH gefällt. Die
Proben. wurden mit 75% EtOH gewaschen und kurz bei Raumtemperatur
getrocknet. Die Quantifizierung der Ausbeute wurde durch Elektrophorese
eines kleinen Aliquots auf 1% Agarosegel in 1 × TBE Puffer mit Ethidiumbromidfärbung und
Vergleich mit einem bekannten Standard durchgeführt.
-
Jede SNuPE-Reaktion wurde in einem
50 μl Volumen,
das ungefähr
100 ng des amplifizierten DNA Fragments, 1 μM des SNuPE-Primers, 2 Einheiten
Taq-Polymerase und 1 μl
des geeigneten dNTP enthielt, durchgeführt. Alle dNTPs waren in diesem
Typ Assay nicht-markiert. Der verwendete Puffer war 10 mM Tris-HCl
(pH 8,3), mit 50 mM KCl, SmM MgCl2 und 0,001% (Gew/Vol) Gelatine.
Die Proben wurden einem Zyklus unterzogen, der aus einer 2-Minuten
Denaturierungsperiode bei 95°C,
einer 2 Minuten Annealingperiode bei 60°C und einer 2-Minuten Primerverlängerungsperiode
bei 72°C
bestand. Die Sequenz des SNUPE-Primers für jede Familie ist unten beschrieben.
-
Primerverlängerungen: Einzel-Nucleotidprimerverlängerung
wurden wie in Singer-Sam et al., (Singer-Sam et al., PCR Methods
und Applications 1: 160–163,
1992) durchgeführt,
mit der Ausnahme, daß 1
mM Mg++, 0,1 μM
Primer, und 0,05 μM
jedes dNTP-Typs in jedem Reaktionstyp verwendet wurden. Nach jeder oben
beschriebenen Primerverlängerung
wurde ein fünftel
Volumen eines Ladefarbstoffs (80% Formamid, 0,1% Bromophenolblau,
0,1% Xylencyanol, 2 mM EDTA) hinzugefügt und die gesamte Probe in
15% denaturierendem Polyacrylamidgel elektrophoriert. Die Gele wurden
in 10% Glyzerin, 10% Methanol, 10% Eisessig unter konstantem Schütteln fixiert,
gefolgt von Waschschritten mit 10% Glyzerin. Die Gele wurden dann
bei 55°C
für 3–5 Stunden
getrocknet.
-
Die in diesem Experiment beschriebenen
Primer sind durch Rychlik beschrieben (Rychlik, BioTechniques 18:
84–90,
1995). Die Primer können
synthetisiert werden oder als Gel-Filtrationsgradprimer von Midland Certified
Reagent Company (Midland, Texas) erhalten werden. Die Amplifikationen
sind entweder Taq DNA-Polymerase-basiert (10 mM Tris-HCl pH 8,3,
1,5 mM MgCl2, 50 mM KCl) oder Pfu-DNA-Polymerase-basiert (20 mM
Tris-HCl pH 8,3, 2,0 mM MgCl2, 10 mM KCl, 10 mM (NH4)2SO4, 0,1%
Triton X-100, 0,1 mg/ml Rinderserumalbumin). Die Gesamt-Nukleosidtriphosphat
(NTPs) Konzentration in den Reaktionen ist 0,8 mM, die Primerkonzentration
ist 200 nM (falls nicht anders angegeben) und die Templatmenge ist
0,25 ng von Bakteriophagen Lambda-DNA pro 20 μl Reaktion. Die Zyklusparameter
waren 94°C
Vor-Heizschritt für
5 Minuten, gefolgt von 94°C
Denaturierungsschritt für
1 Minute, 55°C
Annealingschritt für
2 Minuten und einem 72°C
Verlängerungsschritt
für 30
Sekunden bis 1 Minute und einer finalen Verlängerung bei 72°C für 10 Minuten.
Die Amplifikationen waren im allgemeinen 30–45 an der Zahl.
-
Zwei Regionen im Bakteriophagen Lambda-Genom
(GenBank Zugangsnummer # J02459) wurden als die Startstellen für die Amplifikation
ausgesucht. Der 5'-Primer
wies ein stabiles GC-reiches 3'-Ende
auf: Der 3'-Primer
is so ausgewählt,
daß sich
ein 381 bp Produkt ergeben wird. Der 5'-Vorwärtsprimer ist H17: 5'-GAACGAAAACCCCCCGC
(SEQ ID NO: _). Der 3'-rückwärtige Primer
ist RP 17: 5'-GATCGCCCCCAAAACACATA
(SEQ ID NO: _).
-
Das amplifizierte Produkt wurde dann
auf die Anwesenheit eines Polymorphismus an Position 31245 getestet.
Der folgende Primer wurde in vier Einzel-Nukleotidverlängerungstests verwendet; SNE17: 5'-GAACGAAAACCCCCCGC
(SEQ ID NO: _). Die vier Einzel-Nukleotidverlängerungsassays wurden dann wie
oben beschrieben durchgeführt.
All die Reaktionen wurden dann vereinigt und 5 μ1 des vereinigten Materials
wurde auf die HPLC-Säule
(SeraSep, San Jose, CA) ohne weitere Reinigung injiziert.
-
Die HPLC wurde unter der Verwendung
von automatisierter HPLC-Instrumentierung (Rainin, Emeryville, CA.,
oder Hewlett Packard, Palo Alto, CA) durchgeführt. Nicht-gereinigte SNEA-Produkte,
die für
3 Minuten bei 95°C
vor der Injektion in eine HPLC denaturiert wurden, wurden mit einem
linearen Acetonitrilgradienten (ACN, J. T. Baker, NJ) von 1,8%/Minute
bei einer Flußrate
von 0,9 ml/Minute eluiert. Die Start und Endpunkte wurden gemäß der Größe des SNEA-Produkts
eingestellt. Die für
die erfolgreiche Auflösung
der SNEA-Moleküle
erforderliche Temperatur war 50°C.
Der Ausfluß aus
der HPLC wurde dann in ein Massenspektrometer geleitet (Hewlett
Packard, Palo Alto, CA) zum Nachweis von Markierungen, wobei die
Ergebnisse in Tabelle 2 angegeben sind.
-
-
Die Ergebnisse zeigen daher an, daß der Massenspektrometrie-Marker
(CMST) bei einer Retentionszeit von 4,6 Minuten nachgewiesen wurde,
was anzeigt, daß der
SNE17-Primer durch eine Base (ddCTP) verlängert wurde und daher der Polymorphismus
an der Position 31245 in diesem Falle ein "G" war.
Die SNE17-487, SNE17-496, und SNE17-555 markierten Primer wurden
nicht verlängert
und ihre Retentionszeit auf der HPLC war jeweils 2,5 Minuten.
-
BEISPIEL 18
-
In diesem Beispiel (18) wurden alle
Reaktionen in Folien-abgedeckten Flaschen durchgeführt. Die
Sequenz der Reaktionen A → F,
beschrieben in diesem Beispiel, ist in 19A und 19B dargestellt.
Die in diesem Beispiel angegebenen Verbindungsnummern beziehen sich
auf Verbindungen derselben Nummern in 19A und 19B.
-
A. Zu einer Lösung eines ANP-Linkers (Verbindung
1, 11,2 mMol) und Diisopropylethylamin (22,4 mMol) in CHCl3 (60 ml) wurde Allylbromid (22,4 mMol) hinzugefügt. Das
Reaktionsgemisch wurde für
3 Stunden refluxiert, für
19 Stunden bei Raumtemperatur gerührt, verdünnt mit CHCl3 (200 ml) und
mit 1,0 M HCl (2 × 150
ml) und H2O (2 × 150 ml) gewaschen. Die organischen
Extrakte wurden getrocknet (MgSO4) und das Lösungsmittel
abgedampft, um Verbindung 2 als gelben Feststoff zu ergeben.
-
Zu einem Gemisch von Verbindung 2
in CH2Cl2 (70 ml),
wurde Tris(2-aminoethyl)amin (50 ml) hinzugefügt und das Reaktionsgemisch
bei Raumtemperatur für
18 Stunden gerührt.
Die Reaktion wurde mit CH2Cl2 (150
ml) verdünnt
und mit pH 6,0 Phosphatpuffer (2 × 150 ml) gewaschen. Die organischen
Extrakte wurden getrocknet (MgSO4) und das
Lösungsmittel abgedampft.
Der Rest wurde Säulenchromatographie
(Hexan/EtOAc) unterzogen, um 1,63 g (58%) von Verbindung 3 zu ergeben: 1H NMR (DMSO-d6): δ 7,85 (dd,
2H), 7,70 (t, 1H), 7,43 (t, 1H), 5,85 (m, 1H); 5,20 (q, 2H), 4,58
(q, 1H), 4,50 (d, 2H), 2,70 (m, 2H), 2,20 (br s, 2H).
-
B. Zu einer Lösung von Boc-5-aminopentanonsäure (1,09
mMol) und NMM (3,27 m/Mol) in trockenem DMF (6 ml) wurde HATU (1,14
mMol) hinzugefügt
und das Reaktionsgemisch bei Raumtemperatur für 0,5 Stunden gerührt. Eine
Lösung
von Verbindung 3 (1,20 mMol) in trockenem DMF (1 ml) wurde hinzugefügt und das
Reaktionsgemisch bei Raumtemperatur für 18 Stunden gerührt. Die
Reaktion wurde mit EtOAc (50 ml) verdünnt und mit 1,0 M HCl (2 × 50 ml)
und D.I. H2O (2 × 50 ml) gewaschen. Die organischen
Extrakte wurden getrocknet (MgSO4) und zur
Trocknung abgedampft. Der Rest wurde Säulenchromatographie unterzogen,
um 420 mg (91%) von Verbindung 4 zu ergeben: 1H
NMR (DMSO-d6): δ 8,65 (d, 1H), 7,88 (d, 1H),
7,65 (m, 2H), 7,48 (t, 1H), 6,73 (br s, 1H), 5,85 (m, 1H), 5,55
(m, 1H), 5,23 (q, 2H), 4,55 (d, 2H), 2,80 (m, 2H), 2,05 (t, 2H), 1,33
(s, 9H), 1,20–1,30
(m, 4H).
-
C. Eine Lösung von Verbindung 4 (0,9
mMol) in HCl·1,4-Dioxan
(20 mMol) wurde bei Raumtemperatur für 2 Stunden gerührt. Das
Reaktionsgemisch wurde konzentriert, in MeOH und Toluol gelöst und nochmals konzentriert
(5 × 5
ml) um 398 mg (quantitativ) der Verbindung 5 zu ergeben: 1H NMR (DMSO-d6): δ 8,75 (d, 1H).
7,88 (d, 1H), 7,65 (m, 2H), 7,51 (t, 1H), 7,22 (m, 2H), 5,85 (m,
1H), 5,57 (m, 1H), 5,23 (q, 2H), 4,55 (d, 2H), 2,80 (m, 2H), 2,71
(m, 2H), 2,07 (s, 2H), 1,40–1,48
(br s, 4H).
-
D. Zu einer Lösung von Verbindung 21 (0,48
mMol, hergestellt gemäß Beispiel
20) und NMM (1,44 mMol) in trockenem DMF (3 ml) wurde HATU (0,50
mMol) hinzugefügt
und das Reaktionsgemisch bei Raumtemperatur für 0,5 Stunden gerührt. Eine
Lösung
von Verbindung 5 (0,51 mMol) in trockenem DMF (3 ml) wurde hinzugefügt und die
Reaktion bei Raumtemperatur für
18 Stunden gerührt.
Das Reaktionsgemisch wurde mit EtOAc (75 ml) verdünnt und
mit 5% Na2CO3 (3 × 50 ml)
gewaschen. Die organischen Extrakte wurden getrocknet (MgSO4) und das Lösungsmittel verdampft, um 281
mg (78%) von Verbindung 6 zu ergeben: 1H
NMR (DMSO-d6): δ 8,65 (d, 1H), 8,17 (d, 1H),
7,82–7,95
(m, 4H), 7,68 (m, 3H), 7,50 (t, 1H), 6,92 (d, 1H), 5,85 (m, 1H),
5,57 (m, 1H), 5,20 (q, 2H), 4,55 (d, 2H), 4,30 (q, 1H), 4,05 (q,
2H), 2,95 (m, 4H), 2,80 (m, 2H), 2,72 (m, 2H), 2,05 (s, 3H), 2,01
(t, 2H), 1,58–1,77
(m, 3H), 1,50 (m, 4H), 1,30 (q, 3H), 1,17–1,40 (m, 9H).
-
E. Zu einem Gemisch von Verbindung
6 (0,36 mMol) in THF (4 ml), wurden 1 M NaOH (1 mMol) hinzugefügt und die
Reaktion bei Raumtemperatur für
2 Stunden gerührt.
Das Reaktionsgemisch wurde auf pH 7,0 mit 1,0 M HCl (1 ml) angesäuert und
das Lösungsmittel
abgedampft, um Verbindung 7 zu ergeben (quantitativ): 1H
NMR (DMSO-d6): δ 8,65 (d, 1H), 8,17 (d, 1H),
7,82–7,95
(m, 4H), 7,68 (m, 3H), 7,50 (t, 1H), 6,92 (d, 1H), 5,52 (m, 1H),
4,30 (q, 1H), 4,05 (q, 2H), 2,95 (m, 4H), 2,80 (m, 2H), 2,72 (m,
2H), 2,05 (s, 3H), 2,01 (t, 2H), 1,58–1,77 (m, 3H), 1,50 (m, 4H),
1,30 (q, 3H), 1,17–1,40
(m, 9H).
-
F. Zu einer Lösung von Verbindung 7 (0,04
mMol) und NMM (0,12 mMol) in trockenem DMF (0,4 ml), wurde HATU
(0,044 mMol) hinzugefügt
und die Reaktion bei Raumtemperatur für 0,5 Stunden gerührt. Allylamin
(0,12 mMol) wurde hinzugefügt
und das Reaktionsgemisch bei Raumtemperatur für 5 Stunden gerührt. Das
Reaktionsgemisch wurde mit EtOAc (15 ml) verdünnt und mit 5% Na2CO3 (3 × 10
ml) gewaschen. Die organischen Extrakte wurden getrocknet (MgSO4) und das Lösungsmittel abgedampft, um
15 mg (49%) von Verbindung 8 zu ergeben: 1H
NMR (DMSO-d6) δ 8,49 (d, 1H), 8,17 (d, 1H),
7,82–7,95
(m, 4H), 7,68 (m, 3H), 7,50 (t, 1H), 6,92 (d, 1H), 5,72 (m, 1H),
5,50 (m, 1H), 5,03 (q, 2H), 4,37 (d, 2H), 4,30 (q, 1H), 4,05 (q,
2H), 2,95 (m, 4H), 2,80 (m, 2H), 2,72 (m, 2H), 2,05 (s, 3H), 2,01
(t, 2H), 1,581,77 (m, 3H), 1,50 (m, 4H), 1,30 (q, 3H), 1,17–1,40 (m,
9H).
-
BEISPIEL 19
-
Die Sequenz von Reaktionen A → G wie in
diesem Beispiel 19 beschrieben, wird in 20A und 20B dargestellt.
Die in diesem Beispiel angegebenen Verbindungsnummern beziehen sich
auf die Verbindungen der selben Nummern, wie in 20A und 20B.
-
A. Zu einer Lösung von Fmoc-Lys(Boc)-OH (Verbindung
9, 33,8 mMol) in CHCl3 (200 ml), wurde Diisopropylethylamin
(67,5 mMol) und Allylbromid (67,5 mMol) hinzugefügt. Das Reaktionsgemisch wurde
für 6 Stunden
refluxiert, bei Raumtemperatur für
16 Stunden gerührt,
verdünnt
mit CHCl3, gewaschen mit 1,0 M HCl (2 × 150 ml),
gesättigtem
NaHCO3 (1 × 150 ml) und D.I. H2O (2 × 150
ml). Die organischen Extrakte wurden getrocknet (MgSO4)
und das Lösungsmittel
abgedampft, um Verbindung 10 zu ergeben.
-
Zu einer Lösung von Verbindung 10 in CHCl3 (90 ml) wurde Pyrrolidin (10 äq.) hinzugefügt und die
Reaktion wurde bei Raumtemperatur für 2,5 Stunden gerührt. Das
Reaktionsgemisch wurde mit CHCl3 (150 ml) verdünnt und
mit gesättigtem
NaHCO3 (3 × 250 ml) gewaschen. Die organischen
Extrakte wurden getrocknet (MgSO4) und das
Lösungsmittel
abgedampft. Der Rest wurde Säulenchromatographie
unterzogen (EtOAc/McOH) um 6,52 g (67%) von Verbindung 11 zu ergeben: 1H NMR (CDCl3): δ 5,90 (m,
1H), 5,27 (m, 2H), 4,60 (d, 2H), 3,48 (t, 1H), 3,10 (d, 2H), 1,40–1,78 (m,
9H), 1,40 (s, 9H).
-
B. Zu einer Lösung von N-Methylisonipekotinsäure (1,60
mMol) und N-Methylmorpholin (4,80 mMol) in trockenem DMF (5 ml)
wurde HATU (1,67 mMol) hinzugefügt.
Nach 0,5 Stunden wurde eine Lösung
der Verbindung 11 (1,75 mMol) in trockenem DMF (2 ml) hinzugefügt und das
Reaktionsgemisch bei Raumtemperatur für 18 Stunden gerührt. Das
Reaktionsgemisch wurde mit CH2CL2 (60 ml) verdünnt und mit gesättigtem Na2CO3 (3 × 40 ml)
gewaschen. Die organischen Extrakte wurden getrocknet (MgSO4) und das Lösungsmittel abgedampft. Der
Rest wurde Säulenchromatographie
unterzogen (CH2Cl2/McOH/
Triethylamin), um 580 mg (88%) von Verbindung 12 zu ergeben: 1H NMR (DMSO): δ 8,12 (d, 1H), 6,77 (t, 1H),
5,90 (m, 1H), 5,27 (m, 2H), 4,53 (d, 2H), 4,18 (m, 1H), 2,62–2,90 (m,
5H), 2,13 (s, 3H), 1,85 (m, 2H), 1,57 (m, 5H), 1,35 (s, 9H), 1,00 (t,
2H).
-
C. Ein Gemisch von Verbindung 12
(1,39 mMol) in HCl·1,4-Dioxan
(20 mMol) wurde bei Raumtemperatur für 4 Stunden gerührt. Das
Reaktionsgemisch wurde konzentriert, in McOH gelöst, mit Toluol co-abgedampft
(5 × 5
ml), um 527 mg (quantitativ) von Verbindung 13 zu ergeben: 1H NMR (DMSO-d6): δ 8,12 (d,
1H), 6,77 (t, 1H), 5,90 (m, 1H), 5,27 (m, 2H), 4,53 (d, 2H), 4,18
(m, 1H), 2,65–3,00
(m, 8H), 2,23 (s, 3H), 1,85 (m, 2H), 1,57 (m, 5H), 1,00 (t, 2H).
-
D. Zu einer Lösung von 4-Ethoxybenzoesäwe (1 äq.) in trockenem
DMF wird NMM (3 äq.)
und HATU (1,05 äq.)
hinzugefügt.
Nach 0,5 Stunden wird eine Lösung
von Verbindung 13 in trockenem DMF hinzugefügt. Nach der Vervollständigung
der Reaktion und der basischen Aufarbeitung wird die Verbindung
14 isoliert und gereinigt.
-
E. Zu einer Lösung von Verbindung 14 in THF
wird 1N NaOH hinzugefügt
und das Reaktionsgemisch bei Raumtemperatur gerührt. Nach der Vervollständigung
der Reaktion und Ansäuerung
wird die Verbindung 15 isoliert.
-
F. Zu einer Lösung von Verbindung 15 (1 äq.) in trockenem
DMF, wird NMM (3 äq.)
und HATU (1,05 äq.)
hinzugefügt.
Nach 0,5 Stunden wird eine Lösung
von Verbindung 21 (ANP-Allylester,
gemäß Beispiel
20 hergestellt) in trockenem DMF hinzugefügt. Nach der Vervollständigung
der Reaktion und der basischen Aufarbeitung wird die Titelverbindung
16 isoliert und gereinigt.
-
G. Zu einer Lösung von Verbindung 16 in THF
wird 1N NaOH hinzugefügt
und das Reaktionsgemisch bei Raumtemperatur gerührt. Nach der Vervollständigung
der Reaktion und der Ansäuerung
wird die Verbindung 17 isoliert.
-
BEISPIEL 20
-
Die Sequenz der Reaktion A bis D,
wie in diesem Beispiel 20 beschrieben, wird in 21 dargestellt. Die wie in diesem Beispiel
angegebenen Verbindungsnummern, sowie in den Beispielen 18 und 19
beziehen sich auf die Verbindungen mit derselben Nummer wie in 21.
-
A. Zu einer Lösung von 4-Ethoxybenzoesäwe (7,82
mMol) und N-Methylmorpholin (20,4 mMol) in CH2Cl2 (10 ml) wurde HATU (7,14 mMol) hinzugefügt. Nach
0,25 Stunden wurde eine Lösung
der Verbindung 11 (6,8 mMol) in CH2Cl2 (6 ml) hinzugefügt und das Reaktionsgemisch
bei Raumtemperatur für
18 Stunden gerührt.
Die Reaktion wurde mit CH2Cl2 (150
ml) verdünnt
und mit 1,0 M HCl (3 × 50
ml) und gesättigtem NaHCO3 (3 × 50 ml)
gewaschen. Die organischen Extrakte wurden getrocknet (MgSO4) und das Lösungsmittel abgedampft. Der
Rest wurde Säulenchromatographie
unterzogen (CH2Cl2/MeOH),
um 2,42 g (82%) von Verbindung 18 zu ergeben: 1H
NMR (CDCl3): δ 7,78 (d, 2H), 6,91 (d, 2H),
6,88 (d, 1 H), 5,83–5,98
(m, 1 H), 5,21–5.38
(m, 2H), 4,80 (q, 1 H), 4,66 (d, 2H), 4,06 (q, 2H), 3,11 (q, 2H),
1,90–2.04
(m, 1H), 1,68–1,87
(m, 1H), 1,39 (t, 3H), 1,34 (s, 9H), 1,32–1,58 (m, 4H).
-
B. Ein Gemisch von Verbindung 18
(5,5 mMol) in HCl·1,4-Dioxan
(14,3 mMol) wurde bei Raumtemperatur für 1 Stunde gerührt. Das
Reaktionsgemisch wurde konzentriert, in MeOH gelöst, azetropiert mit Toluol und
wiederum konzentriert (5 × 5
ml), um eine quantitative Ausbeute von Verbindung 19 zu ergeben.
-
C. Zu einer Lösung von N-Methylisonipekotinsäure (6,21
mMol) in trockenem DMF (15 ml) wurde NMM (21,6 mMol) und HATU (5,67
mMol) hinzugefügt.
Nach 0,5 Stunden wurde eine Lösung
von Verbindung 19 (5,4 mMol) in trockenem DMF (10 ml) hinzugefügt und die
Reaktion bei Raumtemperatur für
18 Stunden gerührt. Das
Reaktionsgemisch wurde auf pH 12 mit 1N NaOH (20 ml) gebracht und
mit CHCl3 (2 × 200 ml) extrahiert. Die organischen
Extrakte wurden getrocknet (MgSO4) und das
Lösungsmittel
abgedampft, um 2,2 g (89%) von Verbindung 20 zu ergeben: 1H NMR (DMSO-d6): δ 8,52 (d,
1H), 7,84 (d, 2H), 7,72 (t, 1H), 6,95 (d, 2H), 5,80–5,95 (m,
1H), 5,18–5,31
(dd, 2H), 4,58 (d, 2H), 4,37 (q, 1H), 4,08 (q, 2H), 3,01 (d, 2H),
2,08 (s, 3H), 1,95 (m, 1H), 1,63–1,82 (m, 4H), 1,51 (m, 4H),
1,32 (t, 3H), 1,22–1,41
(m, 6H).
-
D. Zu einer Lösung von Verbindung 20 (4,4
mMol) in THF (10 ml), wird 1N NaOH (4,4 mMol) hinzugefügt und das
Reaktionsgemisch bei Raumtemperatur für 1 Stunde gerührt. Die
Reaktion wurde konzentriert, in THF/Toluol (2 × 5 ml) gelöst, konzentriert, in CH2Cl2/Toluol (1 × 5 ml)
gelöst
und noch mal konzentriert, um eine quantitative Ausbeute von Verbindung
21 zu ergeben: 1H NMR (DMSO-d6): δ 7,76 (d,
2H), 6,96 (d, 2H), 4,04 (q, 2H), 3,97 (d, 1H), 2,97 (d, 2H), 2,64
(d, 2H), 2,08 (s, 3H), 1,95 (m, 1H), 1,58–1,79 (m, 4H), 1,44 (m, 6H), 1,30
(t, 3H), 1,11–1,35
(m, 4H).
-
BEISPIEL 21
-
Die Synthese der CMSTs (spaltbarer
Massenspektrometrie-nachweisbarer Marker) können, wie in 22 beschrieben,
auf einem kombinatorischen Ansatz basiert sein. Der allgemeine Ansatz
ist so aufgebaut, um mit den Entwicklungen der Massenspektrometer
und den Veränderungen
und Verbesserungen in den Ionisationstechnologien kompatibel zu
sein. Ein zentrales Gerüst
wird zuerst auf seine Kompatibilität mit dem Typ von Ionisation
getestet, der in dem Verfahren verwendet werden wird. Es ist wichtig,
daß das
Gerüst
nicht gegenüber
Fragmentierung, Hitzedegradation, oder Dimerisierung zur Adduktbildung
empfindlich ist. Mit den augenblicklichen APCUQuadrapol-Massenspektrometern
werden ungefähr
400 Markern das Spektrum füllen, wobei
isotopische Kontamination berücksichtigt
wird, die zu einer minimalen Beabstandung von ungefähr 4 AMU
zwingt.
-
Die CMSTs wurden auf eine modulare
Weise aufgebaut, so daß die
Macker mit einem kombinatorischen Chemie-Ansatz aufgebaut werden
können.
Es gibt 5 "Module" (funktionell getrennte
Atomgruppen) in dem vollständigen
Marker-Oligonukleotid. Das erste Modul ist das Oligonukleotid (ODN),
das von jeder Länge und
Sequenz sein kann und das ein 5'-Hexylamin zur Kopplung
an den Linker besitzt, der bevorzugterweise ein photospaltbarer
Linker ist. Das ODN kann entweder als eine Sonde für die Hybridisierung
oder als ein Primer in einem Verlängerungs-, Ligationss- oder
Enzym-basierten Assay sein. Das zweite Modul ist der Linker, bevorzugterweise
ein photolabiler Linker, der den Macker an die Oligonukleotidsonde
oder den Primer bindet. In dem hier genau beschriebenen Ansatz ist
der photolabile Linker ein o-Nitrobenzylaminosäurederivat (siehe Lebl M. et
al. "Dynamic database
of references in molecular diversity", der die Internet-Adresse http://www.5z.com
für eine
Zusammenfassung hat). Das dritte Modul ist der Ionisationsverstärker. Dieses
Modul ist das Gerüst,
auf das der CMST synthetisiert wird und stellt im allgemeinen Funktionalität zur Verfügung, die
für den
Typ von Ionisationsverfahren, das verwendet wird (d. h. APCI (positiv
oder negativer Modus) Elektrospray, MALDI, usw.), spezifisch ist.
Das vierte Modul ist der Gesamtmasseneinsteller, womit die Masse
in großen
Schritten von 200–500
amu verändert
werden kann. Dieses Modul erlaubt die Wiederverwendung der variablen
Gewichtseinsteller. Das fünfte
Modul ist der variable Gewichtseinsteller, auch als variable Masseneinheiten
(VMU) bezeichnet. Die variablen Gewichtseinsteller sind chemische Seitenketten,
die zu dem Markergerüst
hinzugefügt
werden. Diese variablen Gewichtseinsteller stellen das Gewicht der
CMSTs fein ein. Das Gewicht der Marker ist mindestens jede 4 amu
voneinander beabstandet, um überlappende
Spektren aufgrund der isotopischen Kontamination zu vermeiden. Dieselben
VMU-Seitenarme können
wiederholt mit den verschiedenen Typen von Ionisationsgerüsten verwendet
werden. Zusammengefaßt
sind das Ionisationsmodul und die variablen Masseneinsteller so
aufgebaut, um ein vorhersagbares Verhalten in dem MSD zu bewirken, die
Photospaltung des photolabilen Linkers ist schnell (unten beschrieben)
und die CMST/Oligonukleotidkonjugate sind mit PCR und HPLC und anderen
in Assay-Formaten gefundenen Manipulationen kompatibel.
-
Eine genaue synthetische Route der
Marker ist unten und in 22 beschrieben.
Die synthetische Route in Richtung eines CMST beginnt mit der Esterfizierung
des photosensitiven ANP-Linkers (3-Amino-3-(2-nitrophenyl)propionsäure, (1)
um das Ethylesterhydrochlorid (2) mit 84% Ausbeute zu ergeben. Ein wichtiger
Schritt in dem Verfahren ist die enzymatische Transformation (2),
um den Ethylester als ein einzelnes Isomer zur Verfügung zu
stellen. Nachdem das Ethylesterhydrochlorid zum freien Amin basifiziert
wurde und konzentriert wurde, wurde der ölige Rest in pH 7 Phosphatpuffer
aufgenommen und auf neutralen pH mit 2N HCl eingestellt. Das Amano
PS Esteraseenzym wurde als eine Phosphatpufferschlämme hinzugefügt. Nach der
Vervollständigung
der Reaktion entfernte eine basische Aufarbeitung das hydrolysierte
ANP-Nebenprodukt (4) und der Einzel-Isomerethylester (3 > 99% e.e.) wurde zurückerhalten
(92% des erhältlichen
Materials).
-
Das Koppeln von (3) mit einem a-BOC-e-alloc-lysin
(5), unter der Verwendung von EDAC und HOBT, ergab das geschützte ANP-Lysin
(6) in 91% Ausbeute. Das Entfernen von BOC mit TFA stellt den Amino-e-alloc-lysin
ANP-Ester (7) als einen weißen
Feststoff zur Verfügung.
Methyl-isonipecotische Säure
Hydrochloride wurde an 7 gekoppelt, unter der Verwendung von EDAC
und Triethylamin, um ungereinigte Alloc-geschützte Kernstruktur (8) zu ergeben,
die mit Diethylamin, Triphenylphosphin und Palladiumacetat bei 50°C geschützt wurde.
Die sich ergebene Kernstruktur (9) wurde aus dem Reaktionsgemisch
kristallisiert und durch Filtration in 95% Ausbeute als ein gelber
Feststoff zurückgehalten.
-
Eine Vielzahl von Carboxylsäuren, die
als variable Masseneinheiten (VMU) bezeichnet wurden, wurden an
(9) gekoppelt, unter der Verwendung von HATU und N-Methylmorpholin.
Ein Set von VMU's
wurden so aufgebaut, um Gleich-Massenmarker zur Verfügung zu
stellen und 4 amu Beabstandung, um eine isotopische Kontamination
zu vermeiden. Die Folgenden wurden als Ausschlußkriterien bei der Auswahl
bestimmter VMU's
der Zielmassen verwendet: 1) Funktionalitäten, die mit der synthetischen
Sequenz inkompatibel waren (z. B. Ester); 2) Elemente mit multiplen
Isotopen (z. B. Cl, Br, S); 3) Funktionalitäten, die vielleicht zu störenden Photoprozessen
führen
(Jodid, Acyl- und Ary1-phenone);
4) racemische Säuren
und 5) die Erhältlichkeit
von Lieferanten.
-
Nach der Reinigung durch Säulenchromatographie
wurde der CMST-Ethylester (10) in veränderlichen Ausbeuten zurückgehalten.
Basische Hydrolyse von (10) mit NaOH ergab die CMST-Säure (11)
in quantitativer Ausbeute. Der letzte Schritt, die Bildung des aktivierten
Esters, wurde unter Verwendung von Tetrafluorophenoltrifluoroacetat
und Hunnich's Base
erreicht und führte
zu dem CMST-TFP-Ester (12) in verschiedenen Ausbeuten.
-
Es ist bequem, die Marker an das
5'-Ende der Oligonukleotide
anzubringen (im allgemeinen), so daß das 3'-Hydroxyl in der Polymerasekettenreaktion
verlängert
werden kann oder für
andere enzymatische Modifikationen zugänglich ist. Auch, wenn direkt
als Sonden wie in der Anwendung hier beschrieben, verwendet, werden
die Marker bevorzugterweise an das 5'-Ende
der Oligonukleotide angebracht. Die durch Lukhtanov et. al., beschriebenen
Richtlinien "Oligodeoxyribonucleotides
with conjugated dihydropyrroloindole oligopeptides: preparation
und hybridization properties",
Bioconjug Chem 6(4): 418–26,
Jul–Aug,
1995, können
befolgt werden, um die markierten Oligonucleotide aus CMSTs und
Oligonukleotiden herzustellen.
-
BEISPIEL 22
-
Eine PHRED-Photospaltungseinheit
wird zwischen die HPLC und das Massenspektrometer oder einen Autoinjektor
und den MSD plaziert. "PHRED", das für Photochemical
Reactor for Enhanced Detection steht, und erhältlich ist von Aura Industries,
Staten Islund, NY (erhältlich mit
sowohl einer 254 nm und 366 nm Lampe, die 254 nm Lampe wurde verwendet).
Eine in-line-Vorrichtung
wird bevorzugterweise zwischen die Abtrenninstrumentierung (z. B.
HPLC oder Gel) und der Detektor plaziert. Die Schnittstelle weist
bevorzugterweise die folgenden Eigenschaften auf: Die Fähigkeit,
die DNA-Fragmente zu diskreten Zeitintervallen zu sammeln, die DNA-Fragmente
zu konzentrieren, die DNA-Fragmente aus den Elektrophoresepuffern
und dem Milieu zu entfernen, die MW-Identifizierer von dem DNA-Fragment abzuspalten,
die MW-Identifizierer von dem DNA-Fragment abzutrennen, das DNA-Fragment
zu verwerfen, den Marker in eine flüchtige Lösung zu plazieren, den Marker
zu verflüchtigen
und ionisieren, was den Marker in das Massenspektrometer einführt.
-
Eine geeignete Konfiguration der
Photospaltungsvorrichtung ist eine 300 cm lange, 8 Watt, UV-keimabtötende Lampe
(G8T50) (mit einer Emission um 366 nm) unter die eine 801 Spule
von 0,01 Inch ID Tefzel-Röhre
plaziert wird. Flußraten
von 800 l pro Minute sind im allgemeinen geeignet. Die Lösungszusammensetzungen,
die mit APCUMS kompatibel sind und die nur geringe Konzentrationen
von Acetronitril und Puffer, wie zum Beispiel Tris-HCl enthaltend,
sind bevorzugt. Mit diesem Aufbau gibt es kein Erfordernis, die DNA
von dem Marker vor dem Ionisationsschritt abzutrennen. Der photolabile
Linker spaltet sich sehr schnell unter diesen Bedingungen. Die Wärmequelle
in der APCI-Kammer trägt
zu der Spaltung des photolabilen Linkers bei.
-
Durch Variieren der Länge der
Tefzel-Röhrenspule
unter der UV-Quelle und Konstanthalten der Flußrate wurde die Verweildauer
unter der UV-Quelle von den 0,75 bis 6 Sekunden variiert. Der Antwortfaktor
(der integrierte Ionenstrom, der pro Mol Macker produziert wurde,
das in den Flußstrom
injiziert wurde, in der "Flußinjektionsanalyse" (FIA)) der Macker
wurde für
einen Pool von 6 Markierungen bestimmt, die alle an einer einzelnen
Oligonukleotidsequenz angebracht waren (21-mer, M13 Vorwärtssequenzierungsprimer).
Der Antwortfaktor ist das Integral der Effizienz der Ionisation,
Ioneneinführung
in die Vakuumkammer und dann anschließender Nachweis durch den MSD.
Jede Markierung in dem Pool war bei einer Konzentration von 100 fmol
pro μl vorhanden.
Das Verdünnungsmittel
war tRNA (Boehringer Mannheim) bei einer Konzentration von 1 μg/ml in HPLC-Grad
Wasser. In Tabelle 3 sind der Antwortfaktor und die Prozentsätze des beobachteten
Signals relativ zu dem längsten
Aussetzen (6 Sekunden) auf einer Pro-Markierungsbasis aufgelistet. Die Ergebnisse
zeigen an, daß der
Marker schnell wird (weniger als 2 Sekunden) von dem befestigten
Oligonukleotid abgespalten wird. Im Zeitrahmen von 1,9 bis 6 Sekunden
gab es wenig Abnahme im beobachteten Antwortfaktor mit 6 verschiedenen
markierten ODNs. Bei der kürzesten
Aussetzungszeit, die getestet wurde (0,75 Sekunden) gab es bis zu
einer 25%igen Abnahme in dem beobachteten Antwortfaktor.
-
-
Es gab eine ungefähr 40%-ige Abnahme in dem Antwortfaktor
der sechs Marker, wenn die Lampe in der Photospaltungseinheit (Daten
nicht gezeigt). Es wurde nachgewiesen, daß der photolabile Linker thermolabil
ist und der Marker offensichtlich während des APC-Ionisationsschrittes
abgespalten wird, wenn die Verdampfungstemperatur 450°C ist.
-
BEISPIEL 23
-
KOLLEKTIVES MARKERVERHALTEN
UND RELATIVE STABILITÄT
-
Die Marker wurden so aufgebaut, um
ein einzelnes parenterales Ion zur Verfügung zu stellen, das nicht in
Tochter-Ionen zerfiel oder Addukte bildete. Die Antwort der Marker
wurde für
einen Pool von 43 Markern bestimmt, die alle an eine einzelne Oligonukleotidsequenz
befestigt waren (20-mer, M13 Sequenz). Jeder Marker in dem Pool
war bei einer Konzentration von 100 fmol pro μl vorhanden (die Konzentration
wurde bestimmt durch Anwenden eines Verdünnungsfaktors auf eine Oligo/Makervorratslösung, die spektrophotometrisch
bei 260 nm gemessen wurde. Eine Korrektur auf den Markerbeitrag
zur Absorption bei 260 nm wurde nicht verwendet. Das Verdünnungsmittel
war tRNA (Boehringer Mannheim) bei einer Konzentration von 1 μg/ml in HPLC-Grad
Wasser. Der Pool wurde bei 4°C
im Dunkeln gelagert. Die CMST-markierten Oligonukleotide können unter
normalen Labor-Lichtbedingungen gehandhabt werden, ohne das Auftreten
von wahrnehmbarer Degradation.
-
Vor der Analyse wurde ein 55 μl Aliquot
aus der Vorratslösung
entfernt und in ein 200 μl
Polypropylen-Autosamplergefäß plaziert
und durch Falten geschlossen. Fünf
Injektionen von jeweils 5 μl
aus dem Pool und 3 Injektionen von jeweils 5 μl von dem Verdünnungsmittel
wurden durch den HPLC HP1100 ALS durchgeführt. Die APCI-MS-Kammer Parameter
waren wie folgt: 20 PSI für
den Verneblungsdruck, die Verdampfungstemperatur war 450°C, der Trocknungsgasfluß war 3
l/min, die Trockengastemperatur war 350°C, Coronastrom war 4 uA, Fragmentorspannung
125 V, die Verstärkung
wurde auf 1 gesetzt und die Peakweite 0,07 Minuten. Die Flußrate war
0,8 ml/min, das "Todvolumen" der Photospaltungseinheit
war 80 μl
(0,01" ID Tefzel). Die
Lampe der Photospaltungseinheit arbeitete bei 366 nm. Jeder Marker
wurde durch die Extraktion des SIM-Ions aus dem TIC quantifiziert.
Die Peaks wurden unter den folgenden Parametern integriert: Steigungssensitivität bei 2500,
Minimum Peakfläche
bei 800, Minimum Peakhöhe
bei 100, Peakbreite bei 0,15, die Schultereinstellung bei "aus". Die Peakfläche für jeden
Marker wurde für
alle fünf
Injektionen innerhalb eines Experiments aufgezeichnet und die durchschnittliche
Fläche
wurde zusammen mit der Standabweichung und dem Variationskoeffizienten
berechnet. Durchschnittliche Bereiche, Standabweichungen und CVs
wurden auch für
einen einzelnen Tag, über
eine zwei Tage Zeitdauer und für
eine drei Tage-Zeitdauer hinweg berechnet. Innerhalb eines einzelnen
Tages variierte der Koeffizient der Variation zwischen 2,0% und
9,9% zwischen verschiedenen Markern. Über eine drei Tage-Zeitdauer
variierte der Koeffizient der Variation zwischen 4,0 und 9,8% zwischen
verschiedener, Markern. Die Marker sind daher im Hinblick auf Lagerung
und Poolen stabil, wie in Tabellen 4a, 4b und 4c gezeigt.
-
-
-
-
Sensitivität und untere
Nachweisgrenze
-
Spannungsmassen-produzierte Quadrapole
weisen Sensitivitäten
auf, die mit Fluoreszenz-basierter Sequenzierung
vergleichbar sind. Die Sensitivität kann im Hinblick auf die
unter Nachweisgrenze der Masse des CMST angegeben werden. Wir definieren
die untere Nachweisgrenze als 3 Standardabweichungen oberhalb des
Hintergrunds des Assay-Systems. Die untere Nachweisgrenze des Pools
von 43 Konjugaten wurde für
den 30-Ion SIM Modus bestimmt.
-
Ein Set von 10 zweifachen Verdünnungen
wurde in dem tRNA/Wasser-Verdünnungsmittel
präpariert, um
500, 250, 125, 62,5, 31,2, 15,6, 7,8, 3,9, 1,9 und 0,9 Femtomol
von Material pro Injektion (5–10 μl) zu ergeben.
Die Daten wurden in dem 30-Ion SIM Modus unter oben beschriebenen
Bedingungen erhalten: Die LLD für
jeden Marker ist in Tabelle 5 gezeigt.
-
TABELLE
5
UNTERE NACHWEISGRENZE FÜR
43 CMSTs.
-
-
Die LLD für dieses bestimmte Set von
Markern ist ungefähr
4–30 Femtomol
pro Injektion im Anschluß an
den Photospaltungsschritt des Oligonukleotids: (30 × 10–15 Mole
@ 500 MW → 5 × 10–11 Gramm → 50 Picogramm
Marker, 100 × 10–15 Mole
bei 330 × 400
nt × 2
= 3 × 10-8 Gramm = 30 Nanogramm
eines 400 nt doppelsträngigen
PCR-Produkts). Daher, unter der Annahme, daß eine durchschnittliche 25 μl PCR-Reaktion 300
ng eines doppelsträngigen
PCR-Produkts enthält,
wird ungefähr
1/10 dieses Produkts dazu verwendet, um ein Signal zu erzeugen,
das statistisch oberhalb des Hintergrunds der Messung liegt. Zu
Vergleichszwecken wurden ungefähr
100–200
ng DNA pro Spur auf dem ABI 377 Sequenzierer verwendet.
-
Die untere Nachweisgrenze des Markierungssystems
beeinträchtigt
den Grad an Multiplexierung, das in einer einzelnen Injektion eneicht
werden kann. Im Augenblick haben wir kein Problem mit dem Meßprozeß festgestellt,
wenn bis zu 10 μg
an PCR-Produkt pro Injektion verwendet wird. Die Implikation diese
Zahl ist, daß 10.000/30
oder ungefähr
300 Reaktionen unter der Verwendung des Markierungssystems der vorliegenden
Erfindung multiplexiert werden können.
Daher ist für
das HP-MSD 1.100 das Maß von
Multiplexierung ungefähr
dasselbe, wie die Marker, die in das Spektrum des Quadrapol MS plaziert
werden können.
-
Markerinterferenz
-
Es gibt wenig oder keine Interferenz
im Hinblick auf die Antwort (wobei Antwort der meßbare Ionenstrom
ist, der das Ausmaß an
Ionisation, Ausmaß von
Fragmentierung oder Abbau, Adduktbildung, usw. widerspiegelt) bei
der Multiplexierung großer
Zahlen von CMSTs der vorliegenden Erfindung. Die Multiplexierung von
Markern beeinträchtigt
nicht die Antwort von einzelnen Markern und daher beeinträchtigt die
Multiplexierung nicht die Ionisation oder den gesamten Ionenstrom.
-
Antwortfaktor
als eine Funktion des Injektionsvolumens
-
Wie oben beschrieben, wurde ein Pool
von 43 Konjugaten gemessen, um den Antwortfaktor als eine Funktion
des Injektionsvolumens zu bestimmen. 50 fmol pro Injektion und 500
fmol pro Injektion wurden in 5, 10, 20, 50 und 100 μl Volumen
gemessen. Die APCI-MS Parameter waren identisch zu denjenigen, die
in dem Markerstabilitätsexperiment
verwendet wurden. Die in Tabelle 4 gezeigten Werte sind ein Mittelwert
von 5-Replikatinjektionen für
jedes gemessene Volumen. Es gab keinen Anstieg im RF zwischen 5 μl und 10 μl Volumina.
Bei den 20 μl
Injektionsvolumina war der RF 90–97% des 5 μl Volumens. Wenn 50 μl Volumina
gemessen wurden, war der RF 54%–75%
des 5 μl
Volumens. Es gab kein nachweisbares Signal unter der Verwendung der
100 μl Injektionsvolumina.
-
BEISPIEL 24
-
P4502D6 POLYMORPHISMUSNACHWEIS
-
Die hier beschriebene CMST-Technologie-Plattform
kann dazu verwendet werden, den CYP2D6 Polymorphismus zu messen,
der für
den Stoffwechsel von Debrisoquin-4-hydroxylase verantwortlich ist. Dieses bestimmte
P450 Cytochrom ist im Stoffwechsel von mehr als 30 Wirkstoffen und
xenobiotischen Verbindungen wichtig. CYP2D6 (P450-2D6) ist geschätzter weise
für den
Stoffwechsel von ungefähr
25% der im Augenblick verschriebenen Wirkstoffe verantwortlich.
Die CYP2D6 ist auch als Debrisoquin/Spartein-Hydroxylase bekannt.
Sache, Am. J. Human Genetics, 60: 284–295, 1997, hat geschätzt, daß bis zu
10% aller Kaukasier schlechte Metabolisierer sind, aufgrund entweder
inaktivierenden Mutationen in beiden Allelen oder dem vollständigen Fehlen
des Gens. Es gibt auch einen breiten Bereich von CYP2D6-Aktivitäten, zusätzlich zu schlechten
Metabolisieren. Die für
die Untersuchung ausgewählten
Mutationen schließen
C188T, G212A, delT1795, G1846T/A, G1934A, delA2637, C29387, und
G4268C ein. Die RFLPS wurden durch Gelelektrophorese nachgewiesen,
wie vorher beschreiben (siehe Gough et. al., Nature 347: 773–776, 1990).
Die für
die RFLP verwendeten Primer entsprechen denjenigen, die durch Sachse
et al, Am. J. Hum. Genet. 60: 284–295, 1997 verwendet wurden.
Die für
die Sequenzierung verwendeten Primer entsprechen denjenigen, die
von Meyer et al., Pharmacogenetics 5: 373–384, 1995 verwendet wurden.
-
Das Prinzip des CMST-basierten Assays
war es, ein Strang des amplifizierten CYP2D6-Exons auf einer festen Phase (z. B.
magnetischen Partikeln) zu immobilisieren, die Oligonukleotidsonden
zu hybridisieren, nicht hybridisiertes Material abzuwaschen, die
hybridisierte Sonde zu eluieren und dann den Massenspektrometermarker
durch Massenspektrometrie (nach Spalten des Markers von der Probe)
nachzuweisen.
-
Die Amplifikationsbedingungen waren
wie folgt: Primer, die das 2D6-Gen flankierten (Sachse et. al., Am.
J Hum. Genet. 60: 284–295,
1997) wurden dazu verwendet, um ein 4,681 bp genomisches DNA-Fragment zu
amplifizieren, das alles der relevanten Gensequenz enthält. Die
PCR-Reaktion wurde aus 1 × Expand
HF Puffer, 1,5 mM MgCl2, 200 μM
dNTP's, TM 0,5 μM Primern,
P100 und P200, 0,5% Formamid, 100 ng gDNA, und 1,1 U ExpandTM High Fidelity Enzym-Mix (Boehringer Mannheim)
zusammengesetzt. Die Thermozyklusbedingungen waren wie folgt: 94°C für 3 Minuten;
10 Zyklen von 94°C
für 30
Sekunden, 62°C
für 30
Sekunden und 68°C
für 4 Minuten,
20 Zyklen von 94°C
für 30
Sekunden, 62°C
für 30
Sekunden und 68°C
für 4 Minuten +
20 Sekunden/Zyklus; 68°C
für 10
Minuten. Die Produkte wurden auf einem 1% Agarosegel sichtbar gemacht, das
mit Ethidiumbromid gefärbt
wurde.
-
Das Assayformat wird wie folgt beschrieben:
Streptavidin magnetische Partikel (Promega Magnasphere, Bindungskapazität von 80
pmol Biotin/100 μg
Partikeln) wurden mit einem Niedrigsalz und Bindungspuffer gewaschen
(LSWBB, 100 mM NaCl, 1 mM EDTA, 10 mM Tris, pH 7,5) und dann in
einem Hochsalz Wasch- und Bindungspuffer (HSWBB, 2 M NaCl, 1 mM
EDTA, und 10 mM Tris pH 7,5) bei einer Konzentration von 2000 μg/ml resuspendiert.
Die biotinylierten PCR-Produkte wurden mit den Steptavidinpartikeln
für 2 Stunden bei
21°C mit
konstantem Rotormischen inkubiert. Die Partikel wurden zweimal mit
200 μl HSWBB
gewaschen und einmal mit 200 μl
LSWBB. Die gebundenen PCR-Amplikons
wurden dann durch Behandlung mit 50 μl 0,1 N NaOH für 10 Minuten
bei 21°C
denaturiert. Die Partikel wurden dann einmal mit 50 μl 0,1 N NaOH
und dreimal mit 200 μl
LSWBB gewaschen. Die Partikel-gebundenen Amplikons wurden dann mit äquimolaren
Gemischen von Wild-typ (wt) und Mutanten (mt) Sonden hybridisiert,
die verschiedene Massenmarker besaßen. Fünfzig Picomol von jeweiliger
Sonde wurden in 200 μl
von 2 m GuSCN, 5 mM EDTA und 10 mM Tris pH 7,5 plaziert und 50 μl der Hybridisierungslösung wurden
zu den Partikeln plaziert. Die Hybridisierung war für 1 Stunde
bei 21°C
mit konstantem rotierendem Mischen. Die Partikel wurden fünfmal mit
LSWBB gewaschen und die Röhrchen
wurden nach der zweiten Waschung gewechselt. Die hybridisierten
Sonden wurden von den Partikeln durch Behandlung der Partikel mit
50 μl 0,1
N NaOH und einem anschließenden
Waschschritt von 9 μl
von 0,1 N NaOH eluiert. Die Lösung
wurde dann mit 3 μl
1 M Essigsäure
neutralisiert. 5 μl
dieser Lösung wurden
dann in das Massenspektrometer injiziert (HP 1100 Serie LC/MS, ausgerüstet mit
Vakuum-Entgaser, binärer
Pumpe, Autosampler und Diodenarraydetektor). Das Massenspektrometer
wurde mit der APCI Quellenoption verwendet. HP LC/MSD Chemstation-Software
wurde für
die Systemkontrolle, Datenaufnahme und Datenanalyse, installiert
auf einem HP vectra XA mit dem Windows NT Workstation Version 4.0
Betriebssystem) verwendet. Der Flußstrom in das MS besteht aus
50% Acetonitril in ultra-reinem Wasser bei einer Flußrate von
800 μl/Minute.
Die photochemische Spaltvorrichtung bestand aus einer 254 nm Niederdruck
Quecksilberlampe, einer UV-transparenten Reaktorspule und einem
Lampenhalter (Aura Industries).
-
Repräsentative Ergebnisse sind in
Tabelle 6 gezeigt.
-
-
BEISPIEL 25
-
GENEXPRESSIONSVERFOLGUNG
MIT CMST-MARKIERTEN ODNs
-
Gesamt-RNA (1–2 μg) aus der A549 menschlichen
Zelllinie wurde unter der Verwenudung von Superscript II reverser
Transkriptase und OligodT-Primer in einem finalen Volumen von 22 μl gemäß den Anweisungen
des Herstellers (Life Technologies; Gaithersburg, MD) transkribiert.
Eine 75 bp-Region der Apoptose-verwandten menschliches DAD-1 Gen
kodierenden Region, die eine Intron-Exon-Grenze überspannte, wurde durch Taq-Polymerasekettenreaktion
(PCR) amplifiziert; eine anfängliche
Denaturierung bei 95°C
(5 Minuten) wurde von 25–60
Zyklen gefolgt (Annealing bei 45°C
für 15
Sekunden und Denaturierung bei 95°C
für 5 Sekunden).
Die PCR-Reaktion (20–200 μl) enthielt
0,5 μM AD-1
reverser Primer (5'-Biotin-CCA
GGA AAT TCA AAG AGT GA-3'),
0,0125 μm
oder 0,5 μM
DAD-1 phosphorylierter Vorwärtsprimer
(5'-TTG GCT GAA
TCA TTC TCA TT-3'),
7 X 102–1,2
X 107 Moleküle internem Standard (5'-CC AGG AAA TTC AAA
GAG TGA ACA TTC TTT TTG TGT CG-3'),
1 μl A549
cDNA oder 8 X 104- X 1011 Moleküle von WT
Mimetikum (5'-CCA
GGA AAT TCA AAG AGT GAA CAT TCT TTT AGT CTC CTA CTC CTC AAT TAA
GTA AAT GAG AAT GAT TCA GCC AA-3'),
0,8 U Taq-Polymerase, 0,2 mM dATP, 0,2 mM dCTP, 0,2 mM dGTP, 0,2mM
dGTP, 1,5 mM MgCl2, 50 mM KCl, und 10 mM Tris-HCl pH 8,3.
-
Das Amplifikationsprodukt (50 μl) wurde
einzelsträngig
durch entweder asymmetrische Amplifikationsbedingungen (oben) oder
durch Verdau mit 2,5 U Lambda Exonuklease (Boehringer Mannheim;
Indianapolis, 1N) für
15 min (37°C)
in 18 mM Tris-HCl pH 9,5, 1,8 mM MgCl2, 28 mM KCl gemacht. Das verdaute
Amplikon oder die asymmetrische PCR-Reaktion wurden bei 70°C für 5 Min.
plaziert, kurz gekühlt
(Raumtemperatur) und auf 2,1 M Guanidinisothiocyanate, 50 mM Tris-HCl
pH 7,5, 0,1% Sarkosyl, 1 μg/ml
tRNA, 5 μg/ml
WT 394 Sonde (5'-394
MW CMST Marker-TTG AGG AGT AGG AGA CTA AAA-3'), 5 μg/ml
IS 390 probe (5'-390
MW CMST Marker TTGACGACTACGACACAAAAA-3') eingestellt.
-
Die Hybridisierungsreaktion wurde
für 10
Min inkubiert (RT) und auf einen 30K MWCO Spin-Filter (Millipore
Corp.; Bedford, MA) transferiert, der 7 μl von 286 μg/m1 Avidin-DN (Vector; Burlingame,
CA) und 0,7 pg/ml tRNA enthielt. Der Spin-Filter wurde für 5–10 Min.
inkubiert (RT) und zentrifugiert (4,000 × g, 10 min). Der Spin-Filter
wurde zweimal mit 400 μl
kaltem HPLC-Grad dH2O gewaschen und zentrifugiert
(4,000 × g,
10 min). Fünfundzwanzig
Mikroliters von 1 μg/m1
tRNA wurden zu dem Spin-Filter hinzugefügt, um das zurückgehaltene
Hybrid zu eluieren. Die 25 μ1
Retentat wurden in 50% Acetonitril über eine Photolyseeinheit in
ein HP Massenspektrometer (APCI positiver Modus) injiziert. Einzel-Ionenmessung
wurden für
Marker gemacht, die Molekulargewichte von 390 und 394 aufwiesen,
und die Ergebnisse als Peakbereiche dargestellt. Das Verhältnis der
390/394 Markersignale war proportional zu der Menge von Amplikon,
das entweder aus dem internen Standard oder der A549 DAD-1 cDNA
oder dem WT Mimetikum erzeugt wurde. Die Berechnung der unbekannten
Zahl von RNA-Molekülen
in der A549 Gesamt-RNA oder Zahl von WT Mimetika konnte auf einer Standardkurve
gemacht werden, wie auf dem log/log-Plot der Zahl von Eingangs-internen
Standardmolekülen, gegenüber dem
Verhältnis
der 390/394 Markersignale gezeigt. Die Unbekannte ist ein Wert von
x, wenn y = 0. Alternativ ist die Unbekannte = das Verhältnis von
390/394 × der
Zahl von Eingangs-interner Standardmoleküle, wenn das Verhältnis zwischen
0,3 und 3 ist.
-
Die Ergebnisse sind in 23 gezeigt.
-
BEISPIEL 26
-
EINZELNUKLEOTID-VERLÄNGERUNGSASSY
-
RNA-Präparation: Gesamt-RNA wurde
isoliert, die aus Jurkat-Zellen (beginnend mit 1 × 109 Zellen in exponentiellem Wachstum) unter
der Verwendung eines RNA-Isolierungskits von Promega (WI) hergestellt wurde.
Die RNA wurde in zwei Aliquots gelagert: (1) Vorratsaliquots in
Diethylpyrocarbonat-behandeltem ddH2O wurden
bei –20°C gelagert
und (2) Langzeitlagerung als eine Suspension in 100% H2O.
-
Reverse Transkription: Poly(dT) geprimte
reverse Transkription von Gesamt-RNA wurde wie beschrieben in Ausubel
et al. (Ausubel et. al., in Cunent Protocols in Molecular Biology,
1991, Greene Publishing Associates/Wiley Interscience, NY, NY.)
durchgeführt,
mit der Ausnahme, daß die
Reaktionen) auf die Verwendung von 1 μg von Anfangs-Gesamt-RNA eingestellt
wurden. 2–50
Einheiten von reverser Transkriptase (Promega) wurden 10-fach in
10% Glyzerin, 10 mM KPO4, pH 7,4, 0,2% Triton X-100, und 2 mM DTT
verdünnt und
auf Eis für
30 Minuten plaziert, vor der Zugabe zu den Reaktionen. Gen-spezifische
reverse Transkription für
GADPH und andere Kontrollgene, wie beschrieben in den Figuren und
Tabellen, wurden unter Verwendung von 1 μg von Gesamt Jurkat RNA, revers
transkribiert in 10 mM Tris-HCl pH 8,3, 50 mM KCl, M MgCl2, 1 mM dNTPs,
2 U/μl RNAsin
(Gibco-BRL), 0,1 μM Oligomer
und 0,125 U/μl
von M-MLV reverser Transkriptase (Gibco-BRL) in 20 μl Reaktionen
durchgeführt.
Die Reaktionen wurden bei 42°C
für 15
Minuten inkubiert, bei 95°C für 5 Minuten
Hitze-inaktiviert und auf 100 μl
mit einem Mastermix von 10 mM Tris HCl pH 8,3, 1 mM NH4Cl, 1,5 M
MgCl2, 100 mM KCl), 0,125 mM NTPs, 10 ng/ml der jeweiligen Oligonucleotidprimer
und 0,75 Einheiten von Taq-Polymerase (Gibco-BRL) in Vorbereitung
auf die PCR-Amplifikation verdünnt.
-
PCR: Die PCR für jedes Gen wurde mit genspezifischen
Primern durchgeführt,
die eine bekannte Intron/Exon-Grenze überspannten (siehe Tabellen
unten). Alle PCRs wurden in 2Oμ1
Volumina durchgeführt,
die 10 mM Tris HCl pH 8,3, 1 mM NH4Cl, 1,5
M MgCl2, 100 mM KCl), 0,125 mM NTPs, 10
ng/ml der jeweiligen Oligonukleotidprimer und 0,75 Einheiten Taq-Polymerase
(Gibco-BRL) enthielten. Die Zyklusparameter waren 94°C Vor-Erhitzungsschritt
für 5 Minuten,
gefolgt von 94°C
Denaturierungsschritt für
1 Minute, 55°C
Annealingschritt für
2 Minuten und einem 72°C
Verlängerungsschritt
für 30
Sekunden bis 1 Minute und einer finalen Verlängerung bei 72°C für 10 Minuten.
Die Amplifikationen waren im allgemeinen 30–45 an der Zahl.
-
Reinigung von Templaten: Die PCR-Produkte
wurden wie durch Zhen und Swank (Zhen und Swank, BioTechniques,
14(6): 894–898,
1993) beschrieben gelgereinigt. Die PCR-Produkte wurden auf 1% Agarosegel
getrennt, die in 0,04 M Tris-Acetat, 0,001 M EDTA (1 × TAE) Puffer
laufen gelassen wurden und mit Ethidiumbromid gefärbt wurden,
während
sie mit einer UV Lichtquelle sichtbar gemacht wurden. Eine Vertiefung wurde
gerade vor der Bande von Interesse eingeschnitten und mit 50–200 μl von 10%
PEG in 1 × TAE
Puffer gefüllt.
Die Elektrophorese wurde fortgeführt,
bis die Bande vollständig
in die Vertiefung eingetreten war. Die Inhalte wurden dann entfernt
und extrahiert mit Phenol, Chloroform extrahiert und dann in 0,1
Volumen 7,5 M Ammoniumacetat und 2,5 Volumen 100% EtOH gefällt. Die
Proben wurden mit 75% EtOH gewaschen und kwz bei Raumtemperatur
getrocknet. Die Quantifizierung der Ausbeute wurde durch Elektrophorese
eines kleinen Aliquots auf 1% Agarosegel in 1 × TBE Puffer mit Ethidiumbromid-Färbung und
Vergleich mit einem bekannten Standard durchgeführt.
-
Jede SNuPE-Reaktion wurde in einem
50 μl Volumen,
das ungefähr
100 ng des amplifizierten DNA Fragments, 1 μM des SNuPE-Primers, 2 Einheiten
Taq-Polymerase und 1 μl
des geeigneten dNTP enthielt, durchgeführt. Alle dNTPs waren in diesem
Typ Assay nicht-markiert. Der verwendete Puffer war 10 mM Tris-HCl
(pH 8,3), mit 50 mM KCl, 5 mM MgCl2 und 0,001% (Gew/vol) Gelatine.
Die Proben wurden einem Zyklus unterzogen, der aus einer 2-Minuten
Denaturierungsperiode bei 95°C,
einer 2 Minuten Annealingperiode bei 60°C und einer 2-Minuten Primerveilängerungsperiode
bei 72°C
bestand. Die Sequenz des SNuPE-Primers für jede Familie ist unten beschrieben.
-
Primerverlängerungen: Einzel-Nukleotidprimerverlängerung
wurden wie in Singer-Sam et al., (Singer-Sam et al., PCR Methods
und Applications 1: 160–163,
1992) durchgeführt,
mit der Ausnahme, daß 1
mM Mg++, 0,1 μM
Primer, und 0,05 μM
jedes dNTP-Typs in jedem Reaktionstyp verwendet wurden. Nach jeder oben
beschriebenen Primerverlängerung
wurde ein fünftel
Volumen eines Ladefarbstoffs (80% Formamid, 0,1% Bromphenolblau,
0,1% Xylencyanol, 2 mM EDTA) hinzugefügt und die gesamte Probe in
einem 15% denaturierenden Polyacrylamidgel elektrophoriert. Die
Gele wurden in 10% Glyzerin, 10% Methanol, 10% Eisessig unter konstantem
Schütteln
fixiert, gefolgt von Waschschritten mit 10% Glyzerin. Die Gele wurden
dann bei 55°C
für 3–5 Stunden
getrocknet.
-
Die in diesem Experiment beschriebenen
Primer sind durch Rychlik beschrieben (Rychlik, BioTechniques 18:
84–90,
1995). Die Primer können
synthetisiert werden oder als Gel-Filtrationsgrad-Primer von Midland Certified
Reagent Company (Midland, Texas) erhalten werden. Die Amplifikationen
sind entweder Taq DNA-Polymerase-basiert (10 mM Tris-HCl pH 8,3,
1,5 mM MgCl2, 50 mM KCl) oder Pfu-DNA-Polymerase-basiert (20 mM
Tris-HCl pH 8,3, 2,0 mM MgCl2, 10 mM KCl, 10 mM (NH4)2SO4,
0,1% Triton X-100, 0,1 mg/ml Rinderserumalbumin). Die Gesamtnukleosidtriphosphat
(NTPs) Konzentration in den Reaktionen ist 0,8 mM, die Primerkonzentration
ist 200 nM (falls nicht anders angegeben) und die Templatmenge ist
0,25 ng von Bakteriophagen Lambda-DNA pro 20 μl Reaktion. Die Zyklusparameter
waren 94°C
Vorheizschritt für
5 Minuten, gefolgt von 94°C
Denaturierungsschritt für
1 Minute, 55°C
Annealingschritt für
2 Minuten und einem 72°C
Verlängerungsschritt
für 30
Sekunden bis 1 Minute und einer finalen Verlängerung bei 72°C für 10 Minuten.
Die Amplifikationen waren im allgemeinen 30–45 an der Zahl.
-
Zwei Regionen des Bakteriophagen
Lambda-Genoms (GenBank Zugangsnummer # J02459) wurden als die Startstellen
für die
Amplifikation ausgesucht. Der 5'-Primer
weist ein stabiles GC-reiches 3'-Ende
auf: Der 3'-Primer
ist so ausgewählt,
daß sich
ein 381 bp Produkt ergeben wird. Der 5'-Vorwärtsprimer ist H17: 5'-GAACGAAAACCCCCCGC.
Der 3'-rückwärtige Primer
ist RP 17: 5'-GATCGCCCCCAAAACACATA.
-
Das amplifizierte Produkt wurde dann
auf die Anwesenheit eines Polymorphismus an Position 31245 hin getestet.
Der folgende Primer wurde in vier Einzel-Nukleotidverlängerungsassays verwendet; SNE17: 5'-GAACGAAAACCCCCCGC.
Die vier Einzel-Nukleotidverlängerungsassays
wurden dann wie oben beschrieben durchgeführt. All die Reaktionen wurden
dann vereinigt und 5 μl
des vereinigten Materials wurde auf die HPLC-Säule (SeraSep, San Jose, CA)
ohne weitere Reinigung injiziert.
-
Die HPLC wurde unter der Verwendung
von automatisierter HPLC-Instrumentierung (Rainin, Emeryville, CA.
oder Hewlett Packard, Palo Alto, CA) durchgeführt. Nicht-gereinigte SNEA-Produkte, die für 3 Minuten
bei 95°C
vor der Injektion in eine HPLC denaturiert wurden, wurden mit einem
linearen Acetonitrilgradienten (ACN, J. T. Baker, NJ) von 1,8%/Minute
bei einer Flußrate
von 0,9 ml/Minute eluiert. Die Start und Endpunkte wurden gemäß der Größe des SNEA-Produkts
eingestellt. Die für
die erfolgreiche Auflösung
der SNEA-Moleküle
erforderliche Temperatur war 50°C.
Der Ausfluß aus
der HPLC wurde dann in ein Massenspektrometer (Hewlett Packard,
Palo Alto, CA) zum Nachweis von Markierungen geleitet, wobei die
Ergebnisse in Tabelle 7 angegeben sind.
-
-
Die Ergebnisse von Tabelle 7 zeigen,
daß der
Massenspektrometer-Marker (CMST) bei einer Retentionszeit von 4,6
Minuten nachgewiesen wurde, was anzeigte, daß der SNE17-Primer um eine
Base (ddCTP) verlängert
wurde und daher der Polymorphismus an Position 31245 in diesem Falle
ein "G" war. Die SNE17-487,
SNE17-496, und SNE17-555 markierten Primer wurden nicht verlängert und
ihre Retentionszeiten waren auf der HPLC jeweils 2,5 Minuten.
-
BEISPIEL 27
-
PHOSPHORAMIDIT-CHEMIE
FÜR MARKIERTE
MOLEKÜLSYNTHESE
-
Herstellung von Aminohexyl-verlängerten
Markern
-
Wie in 26 gezeigt, wurde die Herstellung von
reinem Aminohexyl-verlängertem
Marker (8) unter der Verwendung der TFP-Methodologie erhalten.
Der TFP-Ester wurde durch die Reaktion des Lithiumsalz von Marker
166 (9) mit Überschuß-TFP-TFA
und Hunigs Base in DMF präpariert.
Nach einer Aufarbeitung wurde ein Dichlormethanlösung-TFP-Ester mit 6-Aminohexanol behandelt.
Ein Präzipitat
von (8) wurde sofort gebildet und wurde durch Filtration
isoliert.
-
Herstellung von Marker-Phosphoramiditen
-
Die Reaktion des Aminohexyl-verlängerten
Markers (5) mit Chlorphosphoramidit ergab das gewünschte Phosphoramidit
(17), wobei durch Massenspektrometeranalyse kein Phosphonat
nachgewiesen wurde.
-
-
Überschüssiges phosphotylierendes
Mittel wurde mit deuteriertem Methanol abgelöscht (H3-Methanol wird normalerweise verwendet,
jedoch erlaubt die Verwendung von deuteriertem Methanol in diesem
Fall die eindeutige Identifizierung des Ursprungs der Löschprodukte),
was (18) hervorrief. Dieses Material überlebte die anschließende wäßrige Aufarbeitung
und ist selber ein Phosphoramidit, das das Potential aufweist, in
den anschließenden
Reaktionen zu interferieren.
-
Reaktion von Phosphoramidit
mit Polymer-getragenem Thymidin
-
Das Thymidin wird an kontrollierte-Poren
Glasperlen gekoppelt, die innerhalb einer Plastikkartusche enthalten
sind. Die verwendeten Kartuschen enthalten 1000 nM (1 × 10–6 Mol)
von getragenem Thymidin. Das 5'-Hydroxyl
der Base wird als der Dimethoxytritylether geschützt, was mit 3% Trichloressigsäure in Dichlormethan
vor der Reaktion mit Phosphoramidit (27)
entfernt wird. Nach der Reaktion mit dem Phosphoramidit wird der
Zwischenprodukt-Phosphidtriester (19) zu dem stabilieren
Phosphotriester (20) oxidiert. Anschließendes Entfernen der Cyanoethyl-Schutzgruppe
und Abspaltung von dem festen Träger
wird mit Ammonium durchgeführt,
um das markierte Thymidin (21) zu ergeben.
-
Das Rohmaterial aus der Phosphoramididherstelltung
wurde in dieser Sequenz verwendet und das von dem Träger abgespaltene
Material durch Massenspektrometrie analysiert. Die hauptsächlichen
beobachteten Ionen im positiven Modus, zusammen mit den Zuordnungen
sind in Tabelle 8 gezeigt.
-
-
Es scheint, daß unter den verwendeten Bedingungen
die Phosphat-Esterbrücke
zur Fragmentierung neigt. Die Analyse im negativen Modus zeigte
einen Peak bei 338 amu, was der Verbindung (22) entspricht, dies
stammt aus der Reaktion des Ablöschprodukts
(18) (28)
die eine hauptsächliche
Kontaminante des ungereinigten Produktes ist.
-
Der Marker-Phosphoramidit (17)
konnte von der nicht-polaren Kontaminante (18) durch Silica-Gelchromatographie
(10% Methanol/89% Dichlormethan/1% Triethylamin) abgetrennt werden
und 115 mg von (17) wurden erhalten. Dieses Material wurde
in dem Reaktionszyklus verwendet, um (21) zu ergeben, das als sein Spaltprodukt
(5) in der Massenspektrometrie (die HPLC-Analyse des Rohprodukts
zeigte, daß (5)
nicht vorhanden war, daher muß der
Peak bei 711 amu in der Massenspektrometrie durch die Spaltung des
gewünschten
Produkts (21) in dem Instrument entstanden sein) nachgewiesen
wurde. Auch vorhanden war nicht-reagiertes Thymidin, was anzeigte,
daß die
Reaktion nicht bis zur Vervollständigung
fortgeschritten war. Das Material wurde auch durch das UV-Spaltungsflußsystem
hindurch passiert und das Photospaltungsprodukt (23) durch Massenspektrometrie
nachgewiesen.
-
-
Daher ist es möglich, ein Massenspektrometrie-Marker-Phosphoramidit
herzustellen und zu reinigen und es mit einem Polymer-gebundenem
Substrat reagieren zu lassen, das anschließen oxidiert, entschützt und
von dem Träger
abgespalten werden kann, um eine markierte Basis zu ergeben.
-
- 1. "Oligonucleotide
Synthesis, a Practical Approach" ed
M. J. Gait, IRL Press 1990.
- 2. R. T Pon, Tetrahedron lett, 1991, 1715.
- 3. Available from Glen Research Cat no 10-1953-xx.
- 4. "Principles
of Peptide Synthesis" M.
Bodansky, Springer-Verlag 1984.
- 5. A. J Cocuzza, Tetrahedron lett, 1989, 6287.
- 6. N. N. Poluskin, B. Chen, L. W. Anderson and J. S Cohen, J.
Org. Chem, 1993, 58, 4606.
-
REFERENZEN
-
Adams, S. P., Kavka, K. S., Wykes,
E. J., Holder, S. B., and Galluppi, G. R. (1983). J. Amer. Chem. Soc.
105, 661–663.
-
Andrus, A., Efcavitch, J. W., McBride,
L. T., und Giusti, B. (1988). *Tetrahedron Lett. 29, 861-864.
-
Beaucage, S. L., und Caruthers, M.
H. (1981). Tetrahedron Lett. 22, 1859 1862.
-
Caruthers, M. H., Beaucage, S. L.,
Efcavitch, J. W., Fisher, E. F., Matteuci, M. D,. und Stabinsky (1980).
Nucleic Acids Res. Symp. Ser. 7, 215–223.
-
Ferrance, I. K., Eadie, J. S., und
Ivarie, R.. (1989). Nucleic Acids Res. 17, 1232–1245.
-
Froehler, B. C., Ng, P. G., und Matteucci,
M. D. (1986a). Nucleic Acids Res. 14, 5399–5407.
-
Froehler, B. C., und Matteucci, M.
D. (1986b). Tetrahedron Lett. 27, 469472.
-
Froehler, B. C. (1986c). Tetrahedron
Lett. 27, 5575-5578.
-
Froehler, B. C., Ng, P. G., und Matteucci,
M. D. (1988). Nucleic Acids Res. 16, 4831–4839.
-
Gaffney, B. L. und Jones, R. A..
(1988). Tetrahedron Lett. 29, 2619–2622.
-
Garegg, P. J., Regberg, T., Stawinski,
J. und Stromberg, R. (1985). Chem. Scripta 25, 280–282.
-
Garegg, P. J., Regberg, T., Stawinski,
J. und Stromberg, R. (1986a). Chem. Scripta 26, 59–62.
-
Garegg, P. J., Lidh, L, Regberg,
T., Stawinski, J. und Stromberg, R. (1986b). Tetrahedron Lett. 27, 4051–4054.
-
Hall, R. H., Todd, A. R., und Webb,
R. F.. (1957). J. Chem. Soc., 3291–3296.
-
Letsinger, R. L., und Mahadevan,
V. J. (1965). J. Am. Chem. Soc. 87, 3526–3227.
-
Letsinger, R. L., Finnan, J. L.,
Heavner, G. A. und Lunsford, W. B. (1975). J. Amer. Chem. Soc. 97, 3278.
-
Letsinger, R. L., Groody, E. P.,
und Tanaka, T. (1982). J. Am. Chem. Soc. 104, 6805–6806.
-
McBride, L. J., und Caruthers, M.
H. (1983). Tetrahedron Lett. 24, 245–248.
-
Merrifield, R. B. (1963. J. Am. Chem.
Soc. 85, 2149–2154.
-
Seliger, H. und Roesch, R. (1990).
DNA und Cell Biol., 9, 691–696.
-
Stein, C. A., Iversen, C. A., Subashinge,
C., Cohen, J. S., Stec, W. J. und Zon, G. (1990). Analytical Biochem.
188, 11–16.
-
Aus dem Voranstehenden wird es ersichtlich
sein, daß,
obwohl spezifische Ausführungsformen
der Erfindung hier zum Zweck der Verdeutlichung beschrieben wurden,
verschiedene Modifikationen gemacht werden können, ohne sich von dem Geist
und Bereich der Erfindung zu entfernen.


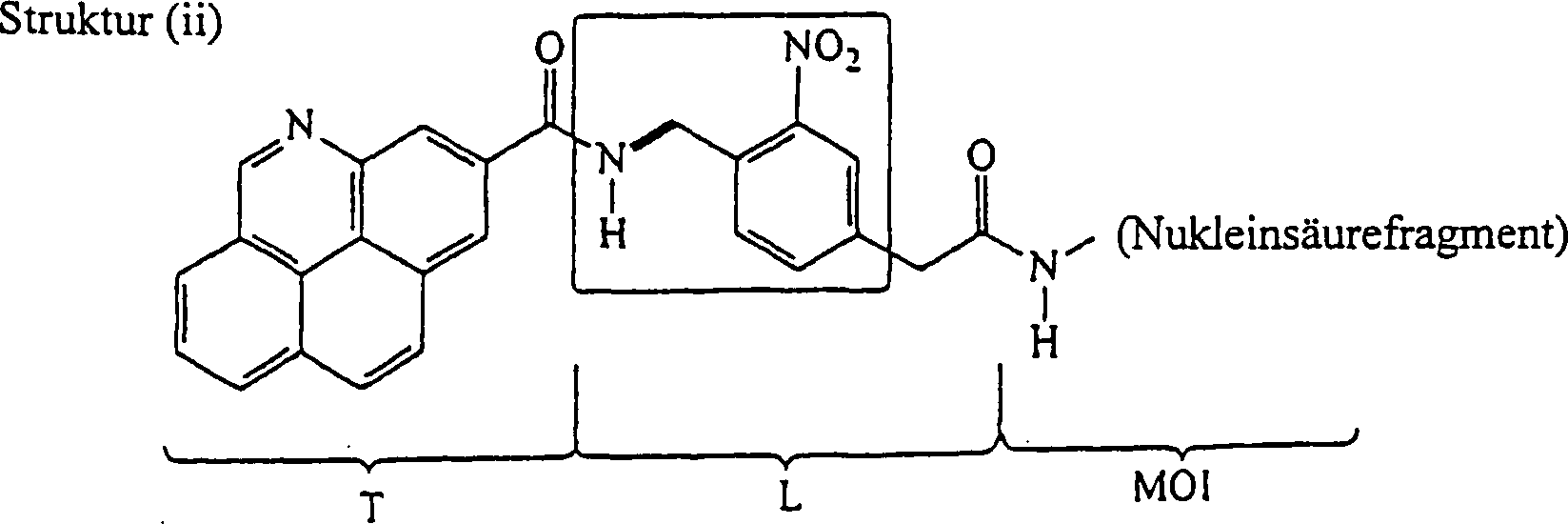







 ist; R1 Wasserstoff oder C1-10-Alkyl ist; c eine ganze Zahl aus dem Bereich von 0 bis 4 ist; und n eine ganze Zahl aus dem Bereich von 1 bis 50 ist, so daß, wenn n größer als 1 ist, G, c, Amid, R1 und T4 unabhängig ausgewählt werden.
ist; R1 Wasserstoff oder C1-10-Alkyl ist; c eine ganze Zahl aus dem Bereich von 0 bis 4 ist; und n eine ganze Zahl aus dem Bereich von 1 bis 50 ist, so daß, wenn n größer als 1 ist, G, c, Amid, R1 und T4 unabhängig ausgewählt werden.





 ist; R1 Wasserstoff oder C1-10-Alkyl ist; c eine ganze Zahl aus dem Bereich von 0 bis 4 ist; "C2-C10" eine Hydrocarbylengruppe mit von 2 bis 10 Kohlenstoffatomen darstellt, "ODN-3'-OH" ein Nukleinsäurefragment mit einer endständigen 3'-Hydroxylgruppe darstellt (d. h. ein Nukleinsäurefragment, das an (C1-C10) am anderen als dem 3'-Ende des Nukleinsäurefragments gebunden ist); und n eine ganze Zahl aus dem Bereich von 1 bis 50 ist, so daß, wenn n größer als 1 ist, dann G, c, Amid, R1 und T4 unabhängig ausgewählt werden. Vorzugsweise sind nicht drei Heteroatome an ein einziges Kohlenstoffatom gebunden.
ist; R1 Wasserstoff oder C1-10-Alkyl ist; c eine ganze Zahl aus dem Bereich von 0 bis 4 ist; "C2-C10" eine Hydrocarbylengruppe mit von 2 bis 10 Kohlenstoffatomen darstellt, "ODN-3'-OH" ein Nukleinsäurefragment mit einer endständigen 3'-Hydroxylgruppe darstellt (d. h. ein Nukleinsäurefragment, das an (C1-C10) am anderen als dem 3'-Ende des Nukleinsäurefragments gebunden ist); und n eine ganze Zahl aus dem Bereich von 1 bis 50 ist, so daß, wenn n größer als 1 ist, dann G, c, Amid, R1 und T4 unabhängig ausgewählt werden. Vorzugsweise sind nicht drei Heteroatome an ein einziges Kohlenstoffatom gebunden.































 R1 ist Wasserstoff oder C1-10 Alkyl; c ist eine ganze Zahl im Bereich von 0–4; X ist gemäß Anspruch 1 definiert; und n ist eine ganze Zahl im Bereich von 1 bis 50, so daß, wenn n größer als 1 ist, G, c, Amid, R1 und T4 unabhängig voneinander ausgewählt sind.
R1 ist Wasserstoff oder C1-10 Alkyl; c ist eine ganze Zahl im Bereich von 0–4; X ist gemäß Anspruch 1 definiert; und n ist eine ganze Zahl im Bereich von 1 bis 50, so daß, wenn n größer als 1 ist, G, c, Amid, R1 und T4 unabhängig voneinander ausgewählt sind.