-
Gebiet der Erfindung
-
Die
Zusammensetzungen der Erfindung betreffen das Gebiet der Proteine,
welche die Serin-Protease-Aktivität inhibieren.
Die Erfindung betrifft auch das Gebiet der Nukleinsäure-Konstrukte, Vektoren
und Wirtszellen für
das Herstellen von Serin-Protease inhibierenden Proteinen, das Protein
enthaltende pharmazeutische Zusammensetzungen, und Verfahren für deren
Verwendung.
-
Stand der Technik
-
Angesprochenes Problem
-
Blutverlust
ist eine ernsthafte Komplikation bei größeren Operationen, wie Operationen
am offenen Herzen und anderen komplizierten Operationen. Auf Kardiochirurgie-Patienten
entfällt
ein signifikanter Anteil des transfusionierten Donatorbluts. Die
Bluttransfusion trägt
die Risiken der Krankheitsübertragung
und unerwünschter
Reaktionen. Zusätzlich
ist Donatorblut teuer und die Nachfrage übersteigt oft das Angebot.
Pharmakologische Verfahren für
das Reduzieren des Blutverlustes und der resultierende Bedarf für die Transfusion sind
beschrieben worden (besprochen von Scott et al., Ann. Thorac. Surg.
50: 843–851,
1990).
-
Protein Serin-Protease-Inhibitoren
-
Aprotinin,
ein boviner Serin-Protease Inhibitor der Kunitz-Familie, ist der
aktive Wirkstoff des Medikaments Trasylol®. Es
ist berichtet worden, dass Aprotinin (Trasylol®) beim
Verringern des perioperativen Blutverlustes wirksam ist (Royston
et al., Lancet ii: 1289–1291,
1987; Dietrich et al., Thorac. Cardiavasc. Surg. 37: 92–98, 1989;
Fraedrich et al., Thorac. Cardiovasc. Surg. 37: 89–91, 1989);
W. van Oeveren et al. (1987), Ann Thorac Surg. 44, S. 640–645; Bistrup
et al., (1988) Lancet I, 366–367),
aber es sind unerwünschte
Wirkungen, einschließlich
Hypotension und Erröten
(Bohrer et al., Anesthesia 45: 853–854, 1990) und allergische
Reaktionen (Dietrich et al., supra) berichtet worden. Die Verwendung
von Aprotinin in Patienten, die früher diesem ausgesetzt waren,
ist nicht empfohlen (Dietrich et al., Supra). Trasylol® ist
für die
Behandlung von hyperfibrinolytischer Hämorrhagie und traumatischem
hämorrhagischen
Schock verwendet worden.
-
Aprotinin
ist bekannt, mehrere Serin-Proteasen, einschließlich Trypsin, Chymotrypsin,
Plasmin und Kallikrein zu inhibieren, und wird bei der Behandlung
von akuter Pankreatitis, verschiedenen Stadien des Schocksyndroms,
hyperfibrinolytischer Hämorrhagie
und Myokardinfarkt therapeutisch verwendet (Trapnell et al., (1974)
Brit J. Surg. 61:177; J. McMichan et al., (1982) Circulatory Shock
9:107; Auer et al., (1979) Acta Neurochir. 49:207; Sher (1977) Am
J. Obstet. Gynecol. 129:164; Schneider (1976), Artzneim.-Firsch.
26: 1606). Es wird im Allgemeinen angenommen, dass Trasylol® den
Blutverlust in vivo durch die Inhibierung von Kallikrein und Plasmin
reduziert. Es ist festgestellt worden, dass Aprotinin (3–58, Arg
15, Ala 17, Ser 42) eine verbesserte Plasma-Kallikrein inhibitorische
Wirksamkeit im Vergleich zu nativen Aprotinin selbst, zeigt (WO 89/10374).
-
Probleme mit Aprotinin
-
Da
Aprotinin bovinen Ursprungs ist, gibt es ein begrenztes Risiko in
menschlichen Patienten eine Anaphylaxie bei Reexposition zu diesem
Arzneimittel auszulösen.
Somit wäre
ein humanes funktionelles Aquivalent zu Aprotinin, aufgrund des
geringeren Risikos der Anaphylaxie, äußerst nützlich und wünschenswert
zu haben.
-
Aprotinin
ist auch in Nagetieren und Hunden nephrotoxisch, wenn es wiederholt
bei hoher Dosis verabreicht wird (Bayer, Trasylol®, Inhibitor
of Proteinase; Glaser et al., in „Verhandlungen der Deutschen
Gesellschaft für
Innere Medizin, 78. Kongress",
Bergmann, München,
1972, S. 1612–1614).
Eine Hypothese schreibt diese Wirkung der Akkumulation von Aprotinin
in den negativ-geladenen proximalen Tubuli der Niere zu, aufgrund
seiner hohen positiven Nettoladung (WO 93/14120).
-
Dementsprechend
ist es ein Ziel der vorliegenden Erfindung menschliche Proteine
mit einer zu Aprotinin ähnlich
funktionellen Aktivität
zu identifizieren. Es war auch ein Ziel der vorliegenden Erfindung
humane Proteine zu identifizieren, die weniger geladen wären, dennoch
die gleiche, eine sehr ähnliche,
oder eine verbesserte Protease-Spezifitäten wie für Aprotinin gefunden wird,
zeigen, insbesondere hinsichtlich der Wirksamkeit der Plasmin und
Kallikrein-Inhibierung.
Solche Inhibitioren könnten
dann wiederholt als Medikamente in menschlichen Patienten mit verringertem
Risiko von unerwünschten
Immunreaktionen und reduzierter Nephrotoxizität verwendet werden.
-
Obwohl
ein Protein mit inhibitorischer Aktiviät auf die Protease-Aktivität von Leberzellen-Wachstumfaktor-Aktivator
in EP-A-0758682 offenbart wurde, ist die darin offenbarte Nukleotidsequenz
mit der in der vorliegenden Erfindung beanspruchten Sequenzen nicht
identisch. Die in EP-A-0758682 offenbarten Sequenzen sind größer als
die beanspruchten Sequenzen der vorliegenden Erfindung.
-
Kurze Zusammenfassung
der Erfindung
-
Die
vorliegende Erfindung sieht einen gereinigten menschlichen Serin-Protease
Inhibitor vor, welcher spezifisch Kallikrein inhibieren kann, welches
aus menschlichem Plazenta-Gewebe mittels Affinitätchromatographie isoliert wurde.
-
Die
vorliegen Erfindung stellt ein neu-identifiziertes menschliches
Protein bereit, hierin menschliches plazentales Bikunin genannt,
welches zwei Serin-Protease Inhibitordomänen der Kunitz-Klasse enthält.
-
Die
vorliegende Erfindung stellt Proteine, pharmazeutische Zusammensetzungen,
Nukleinsäuren, Vektoren,
Verfahren und Verwendungen bereit, wie in den Ansprüchen dargelegt
ist.
-
In
einer bevorzugten Ausführungsform
sieht die vorliegende Erfindung ein natives menschliches plazentales
Bikunin-Protein vor, welches die Aminosäuresequenz aufweist:
oder
-
-
In
einem Aspekt ist die biologische Aktivität des Proteins der vorliegenden
Erfindung, dass es an Trypsin, humane Plasma- und Gewebe Kallikreine,
humanes Plasmin und Faktor XIIa binden, und deren biologische Aktivität inhibieren
kann. In einer bevorzugten Ausführungsform
sieht die vorliegende Erfindung ein natives humanes plazentales
Bikunin-Protein in glycosylierter Form vor. In einer weiteren Ausführungsform
umfasst die vorliegende Erfindung natives humanes Bikunin-Protein,
welches so gebildet wurde, dass es zumindest eine Cystein-Cystein-Disulfidbindung enthält. In einer
bevorzugten Ausführungsform
enthält
das Protein zumindest eine intraketten Cystein-Cystein-Disulfidbindung,
die zwischen einem Paar von Cysteinen gebildet ist, ausgewählt aus
der Gruppe bestehend aus CY11-CYS61, CYS20-CYS44, CY36- CYS57, CYS106-CYS156,
CYS115-CYS139, und CYS131-CYS152, wobei die Cysteine entsprechend
der Aminosäuresequenz
des nativen humanen plazentalen Bikunins nummeriert sind. Der Durchschnittsfachmann
wird erkennen, dass das Protein der vorliegenden Erfindung in die
richtige drei-dimensionale Konformation falten kann, so dass die
biologische Aktivität
des nativen humanen Bikunin aufrechterhalten wird, wo keine, eine
oder mehrere, oder alle der nativen intraketten Cystein-Cystein-Disulfidbindungen
vorhanden sind. In einer am meisten bevorzugten Ausführungsform
ist das Protein der vorliegenden Erfindung richtig gefalten und
ist mit allen der richtigen nativen Cystein-Cystein-Disulfidbindungen
gebildet.
-
Ein
aktives Protein der vorliegenden Erfindung kann durch Reinigung
aus menschlichem Gewebe, wie Plazenta, oder über synthetische Proteinchemie-Techniken,
wie durch die Beispiele unten illustriert ist, erhalten werden.
Es ist auch verständlich,
dass das Protein der vorliegenden Erfindung unter Verwendung molekular-biologischer
Techniken erhalten werden kann, wo selbst-replizierende Vektoren
in der Lage sind, das Protein der vorliegenden Erfindung in transformierten
Zellen zu exprimieren. Ein solches Protein kann als nicht-sezernierende,
oder sezernierende Formen von transformierten Zellen hergestellt
werden. Um die Sekretion aus transformierten Zellen zu fördern, um
die funktionelle Stabilität
des translatierten Proteins zu erhöhen, oder um die Faltung des
Bikunin-Proteins zu unterstützen,
können
bestimmte Signalpeptid-Sequenzen dem NH2-terminal Abschnitt des
nativen humanen Bikunin-Proteins hinzugefügt werden.
-
In
einer Ausführungsform
stellt die vorliegende Erfindung somit das native humane Bikunin-Protein mit zumindest
einem intakten Abschnitt der nativen Signalpeptid-Sequenz bereit.
Somit stellt eine Ausführungsform
der Erfindung ein natives humanes Bikunin bereit, welches zumindest
einen Teil des Signalpeptids aufweist, mit der Aminosäuresequenz:
-
In
einem hierin verwendeten, bevorzugten Nummerierungssystem wird die
mit +1 nummerierte Aminosäure
dem NH2-Terminus der Aminosäuresequenz
des nativen humanen plazentalem Bikunin zugewiesen. Man wird leicht
erkennen, dass funktionelle Proteinfragmente von nativem humanen
plazentalen Bikunin abgeleitet werden können, welche zumindest teilweise
die biologische Aktivität
von nativem menschlichem plazentalen Bikunin beibehalten und als
Serin-Protein-Inhibitoren wirken.
-
In
einer Ausführungsform
weist das Protein der vorliegenden Erfindung ein Fragment des nativen
humanen plazentalen Bikunins auf, welches zumindest eine funktionelle
Kunitz-artige Domäne
enthält,
die die Aminosäuresequenz
der nativen humanen plazentalen Bikunin-Aminosäuren 7–159 aufweist, nachstehend „Bikunin
(7–159)" genannt. Somit enthält die vorliegende
Erfindung ein Protein mit der Aminosäure-Sequenz:
wo die
Aminosäure-Nummerierung
der Aminosäure-Sequenz
des nativen humanen plazentalen Bikunins entspricht. Eine andere
funktionelle Variante dieser Ausführungsform kann das Fragment
des nativen humanen plazentalen Bikunins sein, welches zumindest
eine funktionelle Kunitz-artige Domäne enthält, die die Aminosäure-Sequenz
von nativem humanen plazentalen Bikunin Aminosäuren 11–156 aufweist, Bikunin (11–156):
-
Man
kann erkennen, dass die einzelnen Kunitz-artigen Domänen ebenfalls
Fragmente des nativen plazentalen Bikunins sind.
-
Der
Durchschnittsfachman wird erkennen, dass Fragmente des nativen humanen
Bikunin-Proteins hergestellt
werden können,
die zumindest einen Teil der biologischen Aktivität des nativen
Proteins beibehalten. Solche Fragmente können auch in unterschiedlicher
Orientierung, oder mehrere Kombinationen kombiniert werden, um alternative
Proteine, welche einen Teil der die gleiche, oder eine höhere biologische
Aktivität des
nativen humanen Bikunin-Proteins beibehalten.
-
Man
wird leicht erkennen, dass biologisch aktives Protein der vorliegenden
Erfindung eine oder mehrere der vorliegenden Kunitz-artigen Domänen in Kombination
mit zusätzlichen
Kunitz-artigen Domänen
aus anderen Quellen aufweisen kann. Biologisch aktives Protein der
vorliegenden Erfindung kann eine oder mehrere der vorliegenden Kunitz-artigen
Domänen
in Kombination mit zusätzlichen
Proteindomänen
aus anderen Quellen mit einer Vielzahl von biologischen Aktivitäten aufweisen.
Die biologische Aktivität
des Proteins der vorliegenden Erfindung kann mit der von einem anderen
bekannten Protein oder Proteinen kombiniert werden, um multifunktionelle
Fusionsproteinen mit voraussagbarer biologischer Aktivität bereitzustellen.
-
In
einer Ausführungsform,
stellt die vorliegende Erfindung im Wesentlichen gereinigtes, oder
rekombinant hergestelltes natives humanes Bikunin-Protein mit einem
intakten Abschnitt der Leitsequenz, und zumindest einem intakten
Abschnitt der nativen Transmembranregion bereit. Somit bietet eine
Ausführungsform
der vorliegenden Erfindung natives humanes Bikunin mit einer intakten
Leitsequenz und mit zumindest einem Teil der Transmembran-Domäne (unterstrichen)
bereit, welches eine Aminosäure-Sequenz
aufweist, die ausgewählt
ist aus:
wobei
Sequenz 1) EST abgeleitete Konsensus SEQ ID Nr: 45 ist, 2) PCR Klon
SEQ ID Nr:47 ist, 3) Lambda cDNA Klon SEQ ID Nr: 49 ist. In einer
bevorzugten Ausführungsform
weist ein Protein der vorliegenden Erfindung eine der Aminosäure-Sequenz
der SEQ ID Nr: 45, 47 oder 49 auf, wobei das Protein in der Region
zwischen dem Ende der letzten Kunitz-Domäne und der Transmembranregion
gespalten wurde.
-
Die
Protein-Aminosäure-Sequenzen
der vorliegenden Erfindung lehren einen Fachmann eindeutig die dazugehörigen Nukleinsäuresequenzen,
die in molekular-biologischen Techniken verwendet werden können, um
die Protein der vorliegenden Erfindung herzustellen. In einer Ausführungsform
stellt die vorliegende Erfindung eine Konsensus Nukleinsäure-Sequenz
der 4C (SEQ ID Nr: 51) bereit, welche eine Aminosäure-Sequenz
der 4D (SEQ ID Nr. 45) kodiert.
-
In
einer bevorzugten Ausführungsform
stellt die vorliegende Erfindung eine Nukleinsäure-Sequenz bereit, die natives humanes
plazentales Bikunin mit der DNA Sequenz der 4F (SEQ
ID Nr: 48) kodiert, welche die Protein-Sequenz der SEQ ID Nr: 49
kodiert. In einer anderen Ausführungsform
stellt die vorliegende Erfindung eine Nukleinsäure-Sequenz der 4E (SEQ ID Nr: 46) bereit, welche eine Proteinsequenz
der SEQ ID Nr: 47 kodiert.
-
Man
kann leicht erkennen, dass bestimmte allelische Mutationen, und
in der Nukleinsäure-Sequenz gemachte
konservative Substitutionen durchgeführt werden können, die
nach wie vor zu einer Protein-Aminosäure-Sequenz führen, die
durch die vorliegende Erfindung umfasst ist. Der Fachmann wird erkennen,
dass bestimmte natürliche
allelische Mutationen des Proteins der vorliegenden Erfindung, und
konservative Substitutionen von Aminosäuren im Protein der vorliegenden
Erfindung die biologische Aktivität des Proteins nicht signifikant
verändern
wird, und durch die vorliegende Erfindung umfasst sind.
-
Die
vorliegende Erfindung stellt auch pharmazeutische Zusammensetzungen
bereit, die humanes plazentales Bikunin und Fragmente davon enthalten,
die für
die Reduktion des perioperativen Blutverlustes in einem, sich einer
Operation unterziehenden, Patienten nützlich sind.
-
Die
vorliegende Erfindung stellt auch pharmazeutische Zusammensetzungen
für das
Verringern des perioperativen Blutverlustes in einem, sich einer
Operation unterziehenden, Patienten bereit, wobei eine wirksame
Menge der offenbarten humanen Serin-Protease-Inhibitoren der vorliegenden Erfindung
in einem biologisch-kompatiblen Vehikel dem Patienten verabreicht
wird.
-
Die
vorliegende Erfindung stellt auch Varianten des plazentalen Bikunins
und der oben beschriebenen spezifischen Kunitz-Domänen bereit,
die Aminosäure-Substitutionen
enthalten, die die Protease-Spezifität verändern. Die bevorzugten Substitionsstellen
sind unten als Positionen Xaa1 bis Xaa32 in der Aminosäure-Sequenz des nativen plazentalen
Bikunins angegeben. Die Substitutionen an Xaa1 bis
Xaa16 sind für Varianten von Bikunin (7–64) auch bevorzugt,
während
Substitutionen von Xaa17 bis Xaa32 für
die Varianten von Bikunin (102–159)
bevorzugt sind.
-
Der
Begriff „natürlich vorkommender
Aminosäurerest" ist, in dem vorliegenden
Inhalt beabsichtigt, auf eine beliebige der 20 im Allgemeinen vorkommenden
Aminosäuren
hinzuweisen, d.h. Ala, Arg, Asn, Asp, Cys, Gln, Glu, Gly, His, Ile,
Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr und Val.
-
Durch
Substituieren einer oder mehrerer Aminosäuren an einer oder mehreren
der oben angegebenen Positionen, kann es möglich sein, das Inhibitor-Spezifitätsprofil
des nativen plazentalen Bikunin oder das der individuellen Kunitz-artigen
Domänen,
des Bikunins (7–64)
oder des Bikunins (102–159)
zu verändern,
so dass es vorzugsweise andere Serin-Proteasen inhibiert, wie, ohne
darauf beschränkt
zu sein, die Enzyme der Komplement-Kaskade, TF/FVIIa, FXa, Thrombin,
neutrophile Elastase, Cathepsin G oder Proteinase-3.
-
Zu
Beispielen von bevorzugten plazentalen Bikunin-Varianten zählen jene,
in welchen Xaa1 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus His, Glu, Pro, Ala, Val oder Lys,
insbesondere wobei Xaa1 His oder Pro ist;
oder wobei Xaa2 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Val, Thr, Asp, Pro, Arg, Tyr, Glu,
Ala, Lys, insbesondere wobei Xaa2 Val oder
Thr ist; oder wobei Xaa3 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Arg, Pro, Ile, Leu, Thr, insbesondere
wobei Xaa3 Arg oder Pro ist; oder wobei
Xaa4 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Arg, Lys und Ser, Gln, insbesondere
wobei Xaa4 Arg oder Lys ist; oder wobei
Xaa5 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Ala, Gly, Asp, Thr, insbesondere wobei
Xaa5 Ala ist; oder wobei Xaa6 ein
Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Ser, Ile, Tyr, Asn, Leu, Val, Arg,
Phe, insbesondere wobei Xaa6 Ser oder Arg
ist; oder wobei Xaa7 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Met, Phe, Ile, Glu, Leu, Thr und
Val, insbesondere wobei Xaa7 Met oder Ile
ist; oder wobei Xaa8 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Pro, Lys, Thr, Gln, Asn, Leu, Ser
oder Ile, insbesondere wobei Xaa8 Pro oder
Ile ist; oder wobei Xaa9 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Arg, Lys oder Leu, insbesondere
wobei Xaa9 Arg ist; oder wobei Xaa10 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Val, Ile, Lys, Ala, Pro, Phe, Trp,
Gln, Leu und Thr, insbesondere wobei Xaa10 Val
ist; oder wobei Xaa11 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Gly, Ser und Thr, insbesondere
wobei Xaa11 Gly ist; oder wobei Xaa12 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Asp, Arg, Glu, Leu, Gln, Gly, insbesondere
wobei Xaa12 Asp oder Arg ist; oder wobei
Xaa13 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Gly und Ala; oder wobei Xaa14 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Asn oder Lys; oder wobei Xaa15 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Gly, Asp, Leu, Arg, Glu, Thr, Tyr,
Val und Lys, insbesondere wobei Xaa15 Leu
oder Lys ist; oder wobei Xaa16 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Val, Gln, Asp, Gly, Ile, Ala, Met,
und Val, insbesondere wobei Xaa16 Val oder
Ala ist; oder wobei Xaa17 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus His, Glu, Pro, Ala, Lys und Val,
insbesondere wobei Xaa17 Glu oder Pro ist;
oder wobei Xaa18 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Val, Thr, Asp, Pro, Arg, Tyr, Glu,
Ala oder Lys, insbesondere wobei Xaa18 Thr
ist; oder wobei Xaa19 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Arg, Pro, Ile, Leu oder Thr, insbesondere
wobei Xaa19 Pro ist; oder wobei Xaa20 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Arg, Lys, Gln und Ser, insbesondere
wobei Xaa20 Arg oder Lys ist; oder wobei
Xaa21 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Ala, Asp, Thr oder Gly; insbesondere wobei
Xaa21 Ala ist; oder wobei Xaa22 ein
Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Ser, Ile, Thy, Asn, Leu, Val, Arg,
oder Phe, insbesondere wobei Xaa22 Ser oder
Arg ist; oder wobei Xaa23 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Met, Phe, Ile, Glu, Leu, Thr und
Val, insbesondere wobei Xaa23 Phe oder Ile
ist; oder wobei Xaa24 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Pro Lys, Thr, Asn, Leu, Gln, Ser
oder Ile, insbesondere wobei Xaa24 Pro oder
Ile ist; oder wobei Xaa25 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Arg, Lys oder Leu, insbesondere
wobei Xaa25 Arg ist; oder wobei Xaa26 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Val, Ile, Lys, Leu, Ala, Pro, Phe,
Gln, Trp und Thr, insbesondere wobei Xaa26 Val
oder Ile ist; oder wobei Xaa27 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Gly, Ser und Thr, insbesondere
wobei Xaa27 Gly ist; oder wobei Xaa28 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Asp, Arg, Glu, Leu, Gly oder Gln; insbesondere
wobei Xaa28 Arg ist; oder wobei Xaa29 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Gly und Ala; oder wobei Xaa30 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Asn oder Lys; oder wobei Xaa31 ein Aminosäurerest ist, der ausgewählt ist
aus der Gruppe bestehend aus Gly, Asp, Leu, Arg, Glu, Thr, Tyr,
Val und Lys, insbesondere wobei Xaa31 Arg
oder Lys ist; oder wobei Xaa32 ein Aminosäurerest
ist, der ausgewählt
ist aus der Gruppe bestehend aus Val, Gln, Asp, Gly, Ile, Ala, Met
und Thr, insbesondere wobei Xaa32 Gln oder Ala
ist.
-
Beschreibung der Zeichnungen
-
Die
Erfindung wird durch die Betrachtung der folgenden detaillierten
Beschreibung und Ansprüche,
in Verbindung mit den Zeichnungen besser verstanden werden, wobei
-
1 die
Nukleotidsequenz von EST R35464 (SEQ ID Nr: 12) und die Translation
dieser DNA Sequenz (SEQ ID Nr:13) zeigt, welche einen offenen Leserahmen
mit einer gewissen Sequenzähnlichkeit
zu Aprotinin ergab. Das Translationsprodukt enthält 5 der 6 Cysteine im richtigen
Abstand, der für
Kunitz-artige Inhibitordomänen
(in fett dargestellt) kennzeichnend ist. Die durch das verbleibende
Cystein normalerweise besetze Position (bei Kodon 38) enthält stattdessen
ein Phenylalanin (durch Stern gekennzeichnet).
-
2 die
Nukleotidsequenz von EST R74593 (SEQ ID Nr:14), und die Translation
dieser DNA Sequenz (SEQ ID Nr:15) zeigt, welche einen offenen Leserahmen
mit Homologie zu der Kunitz-Klasse von Serin-Protease-Inhibitordomänen ergab.
Das Translationsprodukt enthielt 6 Cysteine im richtigen Abstand,
was für
Kunitz-artige Inhibitordomänen
(in fett dargestellt) kennzeichnend ist. Dieser offene Leserahmen
enthält
jedoch Stoppkodons bei Kodon 3 und 23.
-
3 zeigt
eine abgeleitete, als „Konsensus" bezeichnete, Nukleinsäure-Sequenz
des humanen plazentalen Bikunins (SEQ ID Nr:9), und abgeglichen
mit der als „translatiert" bezeichneten Protein-Aminosäure-Sequenz
(SEQ ID Nr: 10). Ebenfalls als Vergleich sind die Nukleinsäure-Sequenz
von ESTs H94519 (SEQ ID Nr: 16), N39798 (SEQ ID Nr: 17), R74593
(SEQ ID Nr: 14) und R35464 (SEQ ID Nr: 12) gezeigt. Die unterstrichenen
Nukleotide in der Konsensus-Sequenz entsprechen den in den Beispielen
beschriebenen PCR-Primer Stellen. Die unterstrichenen Aminosäuren in
der translatierten Konsensus-Sequenz sind Reste, deren Identität durch
Aminosäure-Sequenzierung
von gereinigtem nativen humanen plazentalen Bikunin bestätigt wurde.
Der Nukleotid- und Aminosäure-Code
sind der Standart-Einbuchstabencode, „N" in dem Nukleinsäure-Code weist auf eine unbestimmte
Nukleinsäure
hin, und „*" weist auf ein Stoppkodon
in der Aminosäure-Sequenz
hin.
-
4A zeigt
die ursprüngliche Überlappung
einer Reihe von ESTs mit einer gewissen Nukleinsäure-Sequenzhomologie zu ESTs,
die humanes plazentales Bikunin, oder Abschnitte davon kodieren.
Für Vergleichszwecke
sind die relativen Abschnitte von Bikunin (7–64) und Bikunin (102–159), die
als KID1 bzw. KID2 bezeichnet sind, gezeigt.
-
4B zeigt
eine nachfolgende umfassendere EST Überlappung, die zusätzliche
ESTs einbezieht. Die Zahlen auf der oberen X-Achse beziehen sich
auf die Länge
in Basenpaaren, beginnend mit der ersten Base von den meisten 5'EST-Sequenzen. Die
Länge jedes
Balkens ist in Proportion zu der Länge in Basenpaaren der individuellen
ESTs, einschließlich
von Spalten. Die EST Zugangsnummer sind rechts von ihren entsprechenden
EST-Balken angegeben.
-
4C zeigt
die entsprechende Ausrichtung der Oligonukleotid-Sequenzen jeder
der in 4B schematisch gezeigten überlappenden
ESTs. Die obere, als Bikunin bezeichnete, Sequenz (SEQ ID Nr: 51)
stellt die Konsensus Oligonukleotid-Sequenz dar, die von den überlappenden
Nukleotide an jeder Position abgeleitet ist. Die Zahlen beziehen
sich auf die Basenpaar-Position innerhalb der EST-Karte. Die Oligonukleotide
in EST R74593, die fett unterstrichen sind (bei Kartenpositionen
994 und 1005) sind Basen-Insertionen, die in R74593 beobachtet werden,
die in jeder der anderen überlappenden
ESTs konsistent abwesend waren.
-
4D zeigt die Aminosäure-Translation der Konsensus-Oligonukleotid-Sequenz
für Bikunin,
die in 4C gezeigt ist (SEQ ID Nr: 45).
-
4E zeigt die Nukleotid-Sequenz (SEQ ID Nr: 46)
und die entsprechende Aminosäure-Translation (SEQ
ID Nr: 47) einer plazentales Bikunin kodierender Sequenz, die von
einer humanen plazentalen cDNA-Bibliothek durch PCR-basierende Amplifikation
abgeleitet wurde.
-
4F zeigt die Nukleotid-Sequenz (SEQ ID Nr: 48)
und die entsprechende Aminosäure-Translation (SEQ
ID Nr: 49) eines humanen plazentalen Bikunin kodierenden Klons,
der von einer humanen plazentalen-lambda-cDNA-Bibliothek durch Kolonie-Hybridisierung
isoliert wurde.
-
4G vergleicht die Ausrichtung der Aminosäure translatierten
Oligonukleotid-Sequenzen für
plazentales Bikunin, welches durch EST-Überlappung (SEQ ID Nr: 48),
PCR-basierende Klonierung (SEQ ID Nr: 47), und konventionelle Lambda-Kolonie-Hybridisierung
(SEQ ID Nr: 49) erhalten wurde.
-
5 zeigt
ein Diagramm der Reinigung von humanem plazentalem Bikunin aus plazentalem
Gewebe nach Superdex 75 Gel-Filtration. Das Diagramm ist eine Überlappung
des bei OD 280 nm gemessenen Protein-Elutionsprofils (durchgezogene
Linie), der Aktivität
des eluierten Proteins in einer Trypsin-Inhibierungsuntersuchung
(% Inhibierung, gezeigt durch Kreise), und der Aktivität des eluierten
Proteins in einer Kallikrein-Inhibierungsuntersuchung (% Inhibierung,
gezeigt durch Quadrate).
-
6 zeigt ein Digramm, welches die Reinigung
des humanen plazentalen Bikunins aus plazentalmen Gewebe unter Verwendung
der C18-Umkehrphasen-Chromatographie graphisch darstellt. Das Diagramm
ist eine Überlagerung
des bei OD 215 nm gemessenen Protein-Elutionsprofils (durchgezogene Linie),
der Aktivität
des eluierten Proteins in einer Trypsin-Inhibierungsuntersuchung (% Inhibierung,
gezeigt durch Kreise), und der Aktivität des eluierten Proteins in
einer Kallikrein-Inhibierungsuntersuchung (% Inhibierung, gezeigt
durch Quadrate).
-
7 zeigt
ein Silber-gefärbtes
SDS-PAGE Gel von hochgereinigtem plazentalem Bikunin (Bahn 2), und
eine Reihe molekularer Größenmarkerproteine
(Bahn 1) der angegebenen Größen in Kilodaltons.
Die Wanderung erfolgte von oben nach unten.
-
8 zeigt
die Menge an Trypsin-inhibierender Aktivität, die in der zellfreien Fermentationsbrühe von dem
Wachstum der Hefestämme
SC101 (Feld 8A) oder WHL341 (Feld 8B) vorhanden ist, die mit einem
Plasmid (pS604) stabil transformiert wurden, das die Expression
von plazentalem Bikunin (102–159)
leitet.
-
9 zeigt
sowohl eine Silber-gefärbte
SDS-PAGE (linkes Feld) als auch einen Western Blot mit anti-plazentalem
Bikunin (102–159)
pAb (rechtes Feld) einer zellfreien Fermentationsbrühe von dem
Wachstum des Hefestamms SC101 (Rekombinante 2.4 und 2.5), der mit
einem Plasmid, welches die Expression von entweder bovinem Aprotinin,
oder plazentalem Bikunin (102–159)
leitet, stabil transformiert wurde. Die Wanderung erfolgte von oben
nach unten.
-
10 ist eine Photographie, die eine Silber-gefärbte SDS-PAGE
von hochgereinigtem plazentalem Bikunin (102–159) (Bahn 2), und eine Reihe
molekularer Größenmarkerproteine
(Bahn 1) der angegebenen Größen in Kilodaltons
zeigt. Die Wanderung erfolgte von oben nach unten.
-
11 ist eine Photographie, die die Ergebnisse von
Northern Blots von mRNA aus verschiedenen menschlichen Geweben zeigt,
die mit 32P markierten cDNA-Sonden hybridisiert
wurden, die entweder plazentales Bikunin (102–159) (Feld 11A) oder plazentales
Bikunin (1–213)
(Feld 11B) kodieren. Die Wanderung erfolgte von oben nach unten.
Die Nummern rechts von jedem Blot beziehen sich auf die Größe in Kilobasen
des benachbarten RNA Markers. Die Organe aus welchen die mRNA abgeleitet
wurde, ist unter jeder Bahn des Blots beschrieben.
-
12 zeigt ein Immunoblot von plazental-stammendem
plazentalem Bikunin mit Kaninchen Antiserum, welches entweder gegen
synthetisches reduziertes plazentales Bikunin (7–64) (Feld A) oder 120–159 (Feld
B) gezüchtet
wurde. Für
jedes Feld waren die Inhalte: molekulare Größenmarker (Bahnen 1); aus humaner
Plazenta isoliertes natives plazentales Bikunin (Bahnen 2), synthetisches
plazentales Bikunin (7–64)
(Bahnen 3) und synthetisches plazentales Bikunin (102–159) (Bahnen
4). Tricin 10–20%
SDS-PAGE-Gels wurden geblottet und mit Protein A- gereinigtem primären polyklonalem
Antikörper
(8 ug IgG in 20 ml 0,1 % BSA/Tris-gepufferte Salzlösung (pH
7,5), gefolgt von alkalische Phosphatase-konjugierter Ziege Anti-Kaninchen
sekundärem
Antikörper
entwickelt. Die Wanderung erfolgte von oben nach unten.
-
13 zeigt ein Coomassie Blue gefärbtes 10–20% Tricin
SDS-PAGE-Gel von 3 Mikrogramm von hochgereinigtem plazentalem Bikunin
(1–170),
welches von einem Baculovirus/Sf9 Expressionssystem abgeleitet ist
(Bahn 2). Bahn 1 enthält
molekulare Größenmarker.
Die Wanderung erfolgte von oben nach unten.
-
14 zeigt einen Vergleich der Wirkung des Erhöhens der
Konzentrationen von entweder Sf9-abgeleitetem humanem plazentalem
Bikunin (1–170)
(volle Kreise), synthetischem plazentalem Bikunin (102–159 (offene
Kreise), oder Aprotinin (offene Quadrate) auf die aktivierte partielle
Thromoplastinzeit von menschlichem Plasma. Die Gerinnung wurde mit
CaCl2 ausgelöst. Die Konzentration von Proteinen
wurde gegen die vielfache Verlängerung
der Gerinnungszeit graphisch dargestellt. Die nicht-inhibierte Gerinnungszeit
war 30,8 Sekunden.
-
Detaillierte Beschreibung
der Erfindung
-
Die
vorliegende Erfindung umfasst ein neu-identifiziertes humanes Protein,
welches hierin als humanes plazentales Bikunin bezeichnet wird,
welches zwei Serin-Protease Inhibitor-Domänen
der Kunitz-Klasse enthält.
Die vorliegende Erfindung umfasst auch pharmazeutische Zusammensetzungen,
die plazentales Bikunin und Fragmente davon enthalten, die für die Reduktion
des perioperativen Blutverlustes in einem Patienten nützlich sind,
der sich einer Operation unterzieht oder eine schwere Verletzung
aufweist.
-
Eine
bevorzugte Anwendung für
plazentales Bikunin, isolierter Domänen, und anderer Variationen
ist die Reduktion von Blutverlust, der von einer Verletzung oder
Operation herrührt,
die ein Verlustpotential von großen Blutvolumina haben. Diese
Verfahren oder Zusammensetzungen reduzieren oder eliminieren den
Bedarf an Donator-Vollblut oder an Blutprodukten, wodurch das Risiko
von Infektionen und anderer unerwünschter Nebenwirkung, sowie
die Operationskosten verringert werden. Die Proteine sind folglich
für das
Reduzieren des Blutverlustes in normalen Patienten nützlich,
d.h. jene die nicht an angeborenen oder präoperativen Mängeln an
Koagulationsfaktoren leiden. Die Reduktion des Blutverlustes kann
als eine Reduktion des Blutverlustes während der Operation, als verringerte
postoperative Drainage oder beides gesehen werden. Zu bevorzugten
Verwendungen zählen,
ohne darauf beschränkt
zu sein, Verwendungen in Brust- und abdominalen Operationen, gesamte
oder teilweise Hüftersatz-Operationen
und Operationen, um einen Patient mit einer epithelialen Läsion des
Auges zu behandeln. Zu bevorzugten thorakalen Verwendungen zählen, ohne
darauf beschränkt
zu sein, aortokoronarer Venen-Bypass oder kardiales oder aortisches
Aneurysma, Operation von Ösophagusvarizen,
und aortokoronarer Bypass-Operation. Zu bevorzugten abdominale Verwendungen
zählen,
ohne darauf beschränkt
zu sein, Lebertransplantationen, radikale Prostatektomie, Divertikulitis
des Kolons, Tumor-Debulking, an der abdominalen Aorta und Zwölffingerdarmgeschwür, und Wiederherstellung
einer Leber- oder Milzverletzung. Zu bevorzugten Verwendungen für die Behandlung
einer Verletzung zählen,
ohne darauf beschränkt
zu sein, die Verwendung bei der Stabilisierung von schwerverletzten
Patienten an der Unfallstelle, die z.B. an Gliedverlust oder schweren
thorakalen/abdominalen Wunden leiden. Im Falle der Verwendung für die Reduktion
des von der Operation resultierenden Blutverlustes wird es bevorzugt
das plazentale Bikunin, die isolierten Domänen oder andere Varianten vor
und während
der Operation zu verabreichen, wohingegen im Falle der Verwendung
bei Verletzungen die plazentale Bikunin-Variante, die isolierten
Domänen oder
andere Varianten so schnell wie möglich im Anschluss an die Verletzung
verabreicht werden soll, und sollte in Notfallfahrzeugen, welche
zu der Unfallstelle fahren, enthalten sein.
-
Faktor
XII (auch als Hageman Faktor bekannt) ist eine Serin-Protease, die
im Blutkreislauf in einer Zymogen-Form (80 kD) bei etwa 29–40 μg/ml gefunden
wird (siehe Pixley, et al. (1993) Meth in Enz., 222, 51–54) und
wird durch Gewebe und Plasma-Kallikrein aktiviert. Nach der Aktivierung
beteiligt es sich an der intrinsischen Blutgerinnungskaskade, welche
aktiviert wird, wenn Blut oder Plasma mit einer „fremden" oder anionischen Oberfläche in Kontakt
tritt. Nach der Aktivierung, kann Faktor XIIa etliche andere Plasma-Proteasen spalten
und aktivieren, einschließlich
Faktor XI, Präkallikrein
und C1 des Komplementsystems. Folglich kann Faktor XII beim Verursachen
von hypotensiven Reaktionen eingebunden sein, da aktiviertes Kallikrein
Kininogen spalten kann, dadurch wird Bradykinin freigesetzt (siehe
Colman, (1984) J. Clin. Invest., 73, 1249).
-
Sepsis
ist eine Krankheit, die von einer bakteriellen Infektion aufgrund
von bakteriellem Endotoxin oder Lipopolysaccharid (LPS) herrührt. Die
Exposition von Faktor XII zu LPS führt zu der Aktivierung von
Faktor XII. Patienten mit Sepsis zeigen häufig Symptome der intravaskulären Gerinnung,
welche auch aufgrund der Aktivierung von Faktor XII durch LPS sein
kann. Septischer Schock kann von einer bakteriellen Infektion herrühren und
ist mit Fieber, einem niederen systemischen Gefäßwiderstand, und einem niederen
arteriellen Druck assoziiert. Es ist eine häufige Todesursache auf Intensivstationen
in den Vereinigten Staaten, wo fünfundsiebzig
Prozent der Patienten, die an septischem Schock sterben, eine durchgehende
Hypotonie aufweisen (siehe Parillo, et al. (1989 Ann Rev. Med.,
40, 469–485).
-
Atemnotsyndrom
der Erwachsenen ist durch Lungenödem,
Hypoxämie
und verringerter Lungencompliance gekennzeichnet. Die Pathogenese
der Krankheit ist derzeit unbekannt, trotzdem wird vermutet, dass proteolytische
Gerinnungswege und Fibrinolyse eine Rolle spielen (siehe Carvalho
et al., (1988) J. Lab Clin. Med., 112; 270–277).
-
Die
Proteine der vorliegenden Erfindung sind auch ein neuartiger humaner
Kunitz-artiger Inhibitor von Kallikrein, einem Aktivator von Faktor
XII. Somit wird ein auch ein Verfahren für die prophylaktische oder
therapeutische Behandlung von systemischen entzündlichen Reaktionen wie septischer
Schock, Atemnotsyndrom der Erwachsenen (ARDS), Präeklampsie,
mehrfaches Organversagen, und disseminierte intravasale Gerinnung
(DIC) beschrieben. Die therapeutische oder prophylaktische Verabreichung
der Peptide der vorliegenden Erfindung würde zu der Modulation dieser
entzündlichen
Zustände
führen
und würde
für den
Patienten von Vorteil sein.
-
Plasmin
spielt eine wichtige Rolle in der extrazellulären Matrixdegradation und der
Aktivierung von Matix-Metalloprotease (MMP)-Kaskaden. Insgesamt
vermitteln diese Proteasen die Migration und die Gewebsinvasion
von sowohl Endothelzellen während
der Aniogenese/Neovaskularisation, und Krebszellen während der Metastasierung.
Die Neovaskularisation ist essentiell, um das Tumorwachstum zu unterstützen und
die Metastasierung ist ein Vorgang, welcher die Verbreitung der
Tumore vermittelt und welches mit einer extrem-schlechten Patientenprognose
verbunden ist.
-
Mehrere
präklinische
Studien weisen darauf hin, dass Kunitz-artige Serin-Protease-Inhibitoren mit einer
Protease-Spezifität ähnlich zu
Aprotinin als Medikamente für
Krebs nützlich
sind. Zum Beispiel reduzierte Aprotinin das Tumorwachstum und die
Invasion, mit erhöhter
Tumornekrose, wenn es an Hamster, die ein äußerst invasives Fibrosarcom
tragen oder an Mäuse,
die ein ähnlich
malignes Brustkarzinom tragen, verabreicht wird (Latner et al.,
(1974), Br. J. Cancer 30: 60–67;
Latner and Turner, (1976), Br. J. Cancer (33: 535–538). Weiters
reduzierte die Verabreichung von 200.000 KIU Aprotinin i.p. an männliche
C57B1/6 Cr Mäuse
an den Tagen 1 bis 14 nach der Inokulation mit Lewis-Lungenkarzinom-Zellen
die Lungenmetastasierung um 50%, wenngleich es keine Wirkung auf
die primäre
Tumormasse hatte (Giraldi et al., (1977) Eur. J. Cancer, 13: 1321–1323).
Auf ähnliche
Weise inhibierte die Verabreichung von 10.000 KIU i.p. an jedem
der Tage 13–16 nach
der Inokulation von C57BL/6J Mäusen
mit Lewis Tumorzellen die Lungenmetastasierung um 90%, ohne das
primäre
Tumorwachstum zu beeinflussen (Uetsuji et al, (1992), Jpn. J. Surg.
22: 429–442).
In derselben Studie wurde argumentiert, dass die Verabreichung von
Plasmin oder Kallikrein mit dem gleichen Dosierungsplan die Anzahl
der Lungenmetastasen erhöht.
Diese Ergebnisse veranlassten die Autoren darauf hinzuweisen, dass
die perioperative Verabreicherung von Aprotintin an Krebspatienten
die Wahrscheinlichkeit der Metastasierung reduzieren kann. Black
und Steger (1976, Eur. J. Pharmacol., 38: 313–319) haben festgestellt, dass
Aprotinin das Wachstum des eingepflanzten Nagetier Murphy-Strum
Lymphosarkoms in Ratten inhibierte und wiesen darauf hin, dass die
Wirkung, an der Inhibierung des Kinin-bildenden Enzymsystems beteiligt
ist. Die zweimal tägliche
i.p. Injektion von weiblichen ddY Mäusen mit 10.000 KIU Aprotinin
für 7 Wochen
an Mäuse,
von welchen jede ein einzelnes autochtones Plattenzellenkarzinom
trägt,
welches von einer 3-Methylcholanthren-Behandlung resultierte, reduzierte
die Tumorrate des primären
Tumors um 90%. Bei einigen Tieren wurde eine Tumor-Regression beobachtet.
Während
alle Vehikel-behandelten Tiere innerhalb sieben Wochen gestorben
sind, blieben alle der Aprotinin-Behandlungsgruppe am Leben. Das
verringerte Tumorwachstum war mit Hyperkerastose assoziiert (Ohkoshi,
Gann (1980), 71: 246–250).
-
Klinisch
zeigten eine operativ geheilte Gruppe von 26 Patienten, die Aprotinin
i.v. erhielten, eine 70% Überlebensrate
zwei Jahre nach der Operation mit keiner Tumorwiederkehr, wohingegen
eine Placebo-Gruppe von 26 Patienten zur selben Zeit nur eine 38% Überlebensrate
mit einer signifikanten Rate an Tumorwiederkehr zeigten (Freeman
et al. Br. Soc. Gastroenterol (1980) Supplement A: 902). In einer
Fallstudie (Guthrie et al., Br. J. Clin. Pract (1981) 35: 330–332) verursachte
die Verabreichung von Bromocriptin plus Aprotinin an einen Patienten
mit fortgeschrittenem Zervix-Krebs Remission. Aprotinin wurde als
ein 500,000 KIU i.p. Bolus alle acht Stunden gleichzeitig mit einer
kontinuierlichen i.v. Infusion von Aprotinin mit einer Rate von
200.000 KIU pro 6 Stunden für
insgesamt sieben Tage, einmal im Monat verabreicht. Die Behandlung
wurde am Ende des vierten Monats aufgrund der Entwicklung einer
allergischen Reaktion gegen Aprotinin beendet. Jüngere Beweise haben die Rolle
von Plasmin als ein Ziel für
diese Wirkungen von Aprotinin auf die Metastasierung weiter unterstrichen.
-
Der
Mechanismus für
diese Vorgänge
könnte
mit der Tatsache in Verbindung stehen, dass Aprotinin das invasive
Potential von Krebs-Zelllinien blockiert (Liu G., et al., Int. J.
Cancer (1995, 60: 501–506).
Weiters, da die Proteine der vorliegenden Erfindung auch wirksame
Inhibitioren von Plasmin und Kallikrein sind, werden sie für die Verwendung
als Anti-Krebs-Mittel
in Erwägung
gezogen. Zum Beispiel werden sie für die Verwendung beim Blockieren
des primären
Tumorwachstums in Erwägung
gezogen, indem die Neovaskularisierung und die primäre Tumor-Invasion
beschränkt
wird und durch Blockieren der Metastasierung durch Inhibierung der
Gewebeinfiltartion. Die Verbindungen können lokal an Tumore oder systemisch
verabreicht werden. In einer bevorzugten Behandlungsart würde das
Protein perioperativ während
dem Tumor-Debulking verabreicht werden, um das Risiko der Metastasierung
zu minimieren. Bei einem solchen Behandlungsplan würden die blutsparenden
Eigenschaften der Verbindung weiters von Vorteil sein, in dem sie
einen klareren Operationssehbereich bereitstellen. Eine andere bevorzugte
Verabreichungsart würde
als eine Kombinationstherapie mit entweder MMP Inhibitoren oder
Chemotherapie sein. Eine zusätzlich
bevorzugte Verabreichungsart würde
als eine lokal verabreichte Gentherapie sein, die so ausgelegt ist,
um eine selektive Expression von plazentalem Bikunin innerhalb der
Tumorzellen, oder ihrer assoziierten Stroma und den Gefäßbetten
zu erreichen.
-
Bevorzugte
Krebsarten auf die die Therapie gerichtet ist würden Gefäß-abhänigie solide Tumore wie Brust-,
Kolon-, Lungen-, Prostata- und Eierstockkarzinome sein, welche ein
hohes metastatisches Potential zeigen, und für welche eine lokale Abgabe
einer hohen Konzentration des Proteins möglich ist wie Lungenkrebs durch
die pulmonale Abgabe, Kolonkarzinome durch hepatische Abgabe an
Lebermetastasen oder Hautkrebs wie Kopf- und Nackenkarzinome oder
Melanome durch subkutane Abgabe. Da die Proteine der vorliegenden
Verbindung menschlichen Ursprungs sind, würden sie weniger wahrscheinlich
mit allergischen oder anaphylaktischen Reaktion der Art, die von
Guthrie et al, supra, bei Wiederverwendung beobachtet wurden, in Verbindung
gebracht.
-
Zusätzlich werden
die Proteine der vorliegenden Erfindung für die Verwendung in der Reduktion
von thromboembolischen Komplikationen, die mit der Aktivierung der
intrinsischen Gerinnungswege assoziert sind, in Erwägung gezogen.
Diese würden
die Verhinderung von pulmonaler Embolie in Spätstadium-Krebspatienten, einer
häufigen
Todesursache, einschließen
(Donati MB., (1994), Haemostasis 24: 128–131).
-
Ödem des
Hirns und des Rückenmarks
ist eine Komplikation, die von traumatischen Hirn oder Rückenmarkverletzungen,
Schlaganfall, zerebraler Ischämie,
zerebraler und subarachnoidaler Hämhorrhagie, Operation (einschließlich Operation
am offenen Herzen), Infektionskranheiten, wie Encephalitis und Meningitis,
granulomatöse
Krankheiten, wie Sarkoid und fokale oder diffuse Karzinome, und
ist ein Mitwirkender für die
hohen Morbiditäts
und Todesniveaus im Anschluss an diese Vorgänge. Bradykinin ist bekannt
dafür,
die Blut-Hirn-Barriere
experimentell zu unterbrechen (Greenwood J., (1991), Neuroradiology,
33: 95–100;
Whittle et al., (1992) Acta Neurochir., 115: 53–59), und eine Bradykinin-Infusion
in die interne Halsschlagader, löste ein
Hirnödem
in spontan hypertensiven Ratten (SHR), die einem gewöhnlichen
Halsschlagader-Verschluss unterzogen wurde, aus (Kamiya, (1990),
Nippon Ika Daigaku Zasshi. 57: 180–191). Erhöhte Bradykinin-Konzentrationen
werden in extrazellulären
Flüssigkeiten
im Anschluss an Trauma in einem Modell, in welches traumatisiertes
Ratten-Rückenmark
eingebunden war (Xu et al., (1991), J Neurochem, 57: 975–980), und
im Plasma und im Gewebe von Ratten mit einem von einer zerebralen
Ischämie
resultierendem Hirnödem
gefunden (Kamiya et al., (1993), Stroke, 24: 571–575). Bradykinin wird aus
hochmolekularem Kininogen durch Serin-Proteasen, einschließlich Kallikrein
freigesetzt (Coleman (1984) J. Clin. Invest, 73:1249), und es wurde
festgestellt, dass der Serin-Protease-Inhibitor Aprotinin die Größenordung
des Hirnödems
blockiert, welches von einer zerebralen Ischämie in SHR Ratten (Kamiya,
(1990), Nippon Ika Daigaku Zasshi. 57: 180–191; Kamiya et al., (1993),
Stroke, 24: 571–575)
und Kaninchen, die einer kalten Läsion des Hirns unterzogen wurden
(Unterberg et al., (1986), J. Neurosurgery, 64: 269–276) herrührt.
-
Diese
Beobachtungen weisen darauf hin, dass Hirnödem von der proteolytischen
Freisetzung von Kininen wie Bradykinin aus Kininogen mit hohem Molekulargewicht,
gefolgt von der Bradykinin-induzierten Erhöhung der Blut-Hirn-Barriere-Permeabilität resultiert.
Dementsprechend werden plazentales Bikunin und Fragmente davon als
Medikamente für
die Prävention
von Ödemen
in Patienten in Erwägung
gezogen, die für diese
Zustände
gefährdet
sind, insbesondere jene mit hohem Risiko an Mortalität oder Hirnverletzung.
Dies würde
Kopf- und Wirbelsäule-Trauma
Patienten, Polytrauma Patienten, Patienten die einer Operation des
Hirn oder Rückenmark
und ihrer assoziierten Gefäße oder
anderen allgemeinen Operationen unterzogen werden, wie Operation
am offenen Herzen, Patienten, die an einem Schlaganfall, zerebraler
oder sub-arachnoider Hämorrhage,
infektiösen
Erkrankung des Hirns, granulomatösen
Erkrankung des Hirns oder diffusen oder fokalen Karzinomen und Tumoren
des Hirn oder jedem beliebigen Zustand leiden, wie Multiple Sklerose,
einschließlich des
Zusammenbruchs der Blut-Hirn-Barriere
oder Patienten, die an jeglichen anderen entzündlichen Prozessen des Hirns
oder Rückenmarks
leiden. Patienten würden
eine Verabreichung von plazentalem Bikunin entweder als Infusion
oder als Bolus-Injektion, intravenös oder intrakranial, bekommen.
Zusätzliche
Dosen an plazentalem Bikunin könnte
intermittierend über
die nächsten
ein bis drei Wochen verabreicht werden. Die Dosiskonzentrationen
würden
so auslegt sein, um zirkulierende Konzentrationen zu erhalten, die
höher sind,
als jene, die notwendig sind, um die Erhöhungen in Plasmakonzentrationen
oder Bradykinin und anderer vasoaktiver Peptide zu neutralisieren,
die durch die Wirkung der Serinproteasen gebildet wurden, und ausreichend sind,
um das Ödem
zu reduzieren. Da das Protein menschlichen Ursprungs ist, würde die
wiederholte Verabreichung im Verlauf der Therapie nicht zu der Entwicklung
einer Immunreaktion gegen das Protein führen. Plazentales Bikunin und
Fragmente davon, würden
für die
Monotherapie oder Prophylaxe sowie für die Verwendung in Kombinationen
mit anderen Medikamenten, wie Neurotherapeutika und Neuroschutzhilfsmittel
in Erwägung
gezogen werden.
-
Jüngste Indizien
(Dela Cadena R.A. et al., (1995), FASEB J. 9: 446–452) weisen
darauf hin, dass der Kontaktaktivierungsweg zu der Pathogenese von
Arthritis und Anämie
beitragen kann, und dass Kallikrein-Inhibitoren von therapeutischem
Nutzen sein können.
Dementsprechend werden Protease-Inhibitoren der vorliegenden Erfindung,
gemäß ihrer
Fähigkeit
humanes Kallikrein zu inhibieren, als Medikamente für die Behandlung
von Arthritis und Anämie
in Menschen in Erwägung
gezogen.
-
Die
Behandlung von männlichen
nicht-Insulin diabetischen (NIDDM) Patienten mit Aprotinin verbesserte
die Gesamtglucoseaufnahme signifikant und verringerte die metabolische
Klärungsrate
von Insulin (Laurenti et al., (1996), Diabetic Medicine 13: 642–645). Dementsprechend
werden die humanen Proteine der vorliegenden Erfindung für die chronische
Verwendung als Medikamente für
die Behandlung von NIDDM in Erwägung
gezogen.
-
Die
tägliche
Behandlung von Patienten, die in der Gefahr einer Frühgeburt
sind, mit Urinary Trypsin Inhibitor für zwei Wochen reduzierte die
wiederkehrenden Gebärmutterkontraktionen
signifikant (Kanayama et al., (1996), Eur J. Obstet. Gynecol. & Reprod. Biol.
67: 133–138).
Dementsprechend werden die menschlichen Proteine der vorliegenden
Erfindung für
die Verwendung in der Prävention
von Frühgeburten
in Erwägung
gezogen.
-
Es
wurde gezeigt, dass Aprotinin die Differenzierung von Maus-Myoblasten
in Kulturen stimuliert (Wells and Strickland, Development, (1994),
120: 3639–3647)),
einem Prozess der durch TGFb inhibiert wird. TGFb existiert als
ein inaktives Pro-Polypeptid, welches durch limitierte Proteolyse
aktivitert wird. Es ist vorgeschlagen worden, dass der Aprotinin-Wirkungsmechanismus
die Inhibierung der Proteasen einschließt, welche Pro-TGFb in die
reife aktive Form umwandeln. Es ist gezeigt worden, dass TGFb in
verschiedenen fibrotischen Läsionen
hinauf reguliert ist und es wird seit längerem als potentielles Ziel
für anti-fibrotische
Therapien angenommen. In einem Rattenmodell einer Lungenfibrose
zum Beispiel verliefen die TGF-b-Konzentrationen parallel zum Ausmaß der Bleomycin-induzierten
Entzündung.
Weiters stimmten die Plasmin-Konzentrationen in dem alveolaren Makrophagen
mit den reifen TGF-b Konzentrationen überein, und der Zusatz des
Plasmin-Inhibitiors a-2-Antiplasmin hob die posttranslatorische
Aktivierung von Pro-TGFb durch den Makrophagen auf (Khal et al.,
(1996), Am. J. Respir. Cell Mol. Biol. 15: 252–259). Die Daten weisen darauf
hin, dass Plasmin zu der Bildung von aktivem TGFb durch alveolare
Makrophagen beiträgt
und dass dieser Prozess eine pathologische Rolle in der Bleomycin-induzierten
Lungenentzündung
spielt.
-
Angesichts
dieser Beobachtungen werden plazentales Bikunin und Fragmente davon
als Therapeutika für
verschiedene fibrotische Erkrankungen, einschließlich pulmonale, hepatische,
renale und dermale (Sklerodermie) Fibrose in Erwägung gezogen
-
Es
wurde gezeigt, dass aerosolisiertes Aprotinin > 50% der mit letalen Dosen an entweder
Influenza-Viren oder Paramyxovirus infizierten Mäuse schützt (Ovcharenko and Zhirnov,
Antiviral Research, (1994), 23: 107–118). Eine Unterdrückung der
Entwicklung von tödlicher
hämorrhagischer
Bronchopneumonie und eine Normalisierung des Körpergewichtzuwachses wurden
auch bei einer aerosilisierter Aprotinin-Behandlung beobachtet.
Angesichts dieser Beobachtungen werden plazentales Bikunin und Fragmente
davon als Therapeutika für
verschiedene respiratorischbezogene Influenza-artige Krankheiten
in Erwägung
gezogen.
-
Das
humane plazentale Bikunin, isolierte Domänen und andere Varianten der
Erfindung werden für
die Verwendung in den medizinischen/therapeutischen Anwendungen
in Erwägung
gezogen, die für
natives Aprotinin oder Aprotinin-Analoga mit anderen inhibitorischen
Profilen vorgeschlagen wurden, insbesondere jene die die Verwendung
von hohen Dosen erfordern. Diese würden Krankheiten einschließen, für welche
die Verwendung des humanen Proteins aufgrund seiner Fähigkeit
angezeigt ist, humane Serin-Proteasen, wie Trypsin, Plasmin, Kallikrein,
Elastase, Cathepsin G und Proteinase-3 zu inhibieren; zu diesen
zählen,
ohne darauf beschränkt
zu sein: akute Pankreatitis (pankreatische Elastase und Trypsin),
Entzündung,
Thrombozytopenie, Bewahrung der Blutplättchenfunktion, Organkonservierung,
Wundheilung, verschiedene Arten von Schock, einschließlich Schocklunge,
Endotoxinschock und postoperative Komplikationen; Störungen der
Blutgerinnung, wie hyperfibrinolytische Hämorrhagie; akute und chronische
entzündliche
Reaktionen, insbesondere für die
Therapie und Prophylaxe von Organläsionen, wie zum Beispiel Pankreatitis
und Strahlen-induzierte Enteritis, Komplexvermittelte inflammatorische
Reaktionen wie Immunvaskulitis, Glomerulonephritis, und Arten von Arthritis;
Kollagenosen, insbesondere rheumatoide Arthritis; Arten von Arthritis,
die durch stoffwechselbezogene Ablagerungen verursacht werden (zum
Beispiel Gicht), Degeneration von elastischen Bestandteilen von Bindegewebeteilen
von Organen, wie bei Artherosklerose (Serumelastase) oder pulmonales
Emphysem (Neutrophil-Elastase); Atemnotsyndrom der Erwachsenen,
entzündliche
Darmkrankheiten, und Psoriasis.
-
Ein
bedeutendes, unerwartetes Ergebnis war, dass die synthetischen Peptide,
welche Bikunin (7–64) und
Bikunin (102–159)
kodieren, richtig in die korrekte drei-dimensionale Konformation
mit aktiver Protease-Inhibitor-Bioaktivität falten können (Beispiele 2 bzw. 1).
Bei der Faltung war jedes dieser Bikunin-Fragmente einer Reduktion
der Masse um 6 Masseneinheiten ausgesetzt, in Einklang mit der Bildung
in jedem Fall von drei Intraketten-Disulfidbindungen zwischen sechs
Cysteinresten jedes Fragments. Ein anderes überraschendes Ergebnis ist,
dass die synthetischen Peptide, welche Bikunin (7–64), Bikunin
(102–159)
und Bikunin (1–170) kodieren,
stark inhibitorisch für
Plasmin und sowohl Gewebe als auch Plasma-Kallikrein sind (Beispiel
4, 3, bzw. 10). Von der Inhibierung von Plasmin und Kallikrein durch
Trasylol® wird
vermutet, dass sie in den Mechanismus durch welchen Trasylol® den
Blutverlust während
einer Operation am offenen Herzen reduziert, eingebunden ist. Unsere
unerwarteten Ergebnisse der Spezifizität der Kunitz-Domänen der
vorliegenden Erfindungen machen diese zu geeigneten therapeutischen
Mitteln, um Blut während
der Operation oder Trauma, bei welcher es einen signifikanten Blutverlust
gibt zu sparen, oder für
jeden anderen Zustand, bei welchen die Inhibierung von Plasmin und/oder
Kallikrein von Nutzen sein würde.
-
Weiters
haben wir in dieser Offenbarung (Beispiel 10) gezeigt, dass plazentales
Bikunin (1–170)
ein wirksamer Inhibitor von Faktor XIa und ein mäßiger Inhibitor des Faktors
Xa ist. Faktor XIa spielt eine wesentliche Rolle in intrinsischen
Gerinnungswegen, welcher dazu dient, den inaktiven Faktor IX und
den aktiven Faktor IXa ineinander umzuwandeln. Somit inhibiert plazentales
Bikunin zwei Schlüsselenzyme
des intrinsischen Weges, Kallikrein und Faktor XIa. Übereinstimmend
mit diesen Beobachtungen, zeigten wir auch, dass plazentales Bikunin
(1–170)
ein wirksamer Inhibitor der aktivierten partiellen Thromboplastinzeit
ist, welche ein Maß für die Geschwindigkeit
der durch den intrinsischen Weg angetriebenen Gerinnung ist. Andererseits
zeigten wir, dass plazentales Bikunin (1–170) ein sehr schwacher Inhibitor
des Gewebefaktor VIIa-Komplexes ist, was darauf hinweist, dass es
in der Regulierung der extrinsischen Gerinnungskaskade nicht wichtig
ist. Auf diesen unerwarteten Ergebnissen basierend, wird plazentales
Bikunin als ein Medikament für
Krankheiten in Erwägung
gezogen, bei welchen die Aktivierung der intrinsischen Gerinnungskaskade
signifikant zu dem Krankheitsmechansimus beiträgt. Zu Beispielen von solchen
Krankheiten würden
der post-traumatische Schock und die disseminierte intravasale Koagulapathie
zählen.
-
Ein
wesentlicher Vorteil der Kunitz-Domänen der vorliegenden Erfindung
ist, dass sie humane Proteine sind, und auch weniger positiv geladen
sind als Trasylol® (Beispiel 1), wodurch
das Risiko einer Nierenschädigung
bei Verabreicherung hoher Dosen des Proteins reduziert wird. Da
es menschlichen Ursprungs ist, kann das Protein der vorliegenden
Erfindung somit an menschliche Patienten mit einem signifikant verringertem
Risiko von unerwünschten
immunologischen Reaktionen verabreicht werden, im Vergleich zu der
Verabreichung ähnlicher
Dosen an Trasylol®. Es wurde weiters gefunden,
dass Bikunin (102–159),
Bikunin (7–64),
und Bikunin (1–170)
signifikant wirksamere Inhibitoren von Plasma-Kallikrein sind, als
Trasylol® in
vitro (Beispiel 3, 4 und 10). Somit wird erwartet, dass Bikunin
und Fragmente davon in vivo, beim Senken des Blutverlustes bei Patienten
wirksamer sind.
-
Die
Menge des verabreichten Serin-Protease-Inhibitors sollte ausreichend
sind, um eine supra-normale Plasmakonzentration bereitzustellen.
Für die
prophylaktische Reduktion des Blutens während einer und im Anschluss
an eine aortokoronare Venen-Bypass-Operation (CABG) können die
Proteine der vorliegenden Erfindung anstelle von Trasylol® verwendet
werden, solange die Wirksamkeitsunterschiede berücksichtigt werden. Die Verwendung
von Trasylol® wird
im Physicians Desk Reference (1995), Auflistung für Trasylol® Ergänzung A
behandelt. Kurz gesagt, wird einem Patient in Rückenlage, die Loading dose
an plazentalem Bikunin, isolierter Domänen oder anderer Varianten
langsam über
etwa 20 bis 30 Minuten nach der Induktion der Narkose, aber vor
der Sternotomie, verabreicht. Im Allgemeinen wird eine Gesamtdosis
zwischen etwa 2 × 106 KIU (Kallikrein-Inhibierungseinheiten)
und 8 × 106 KIU, in Abhängigkeit von solchen Faktoren
wie das Patientengewicht und die Operationslänge, verwendet. Bevorzugte
Loading doses sind jene, die insgesamt 1 bis 2 Millionen Kallikrein-Inhibierungseinheiten
(KIU) enthalten. Wenn die Loading dose beendet ist, wird sie von
einer konstanten Infusionsdosis gefolgt, die fortgesetzt wird, bis
die Operation abgeschlossen ist und der Patient den Operationssaal
verlässt.
Bevorzugte konstante Infusionsdosen sind im Bereich von etwa 250.000
bis 500.000 KIU pro Stunde. Die Anfangsdosis der Primingflüssigkeit
(pump prime dose) wird der Primingflüssigkeit des kardiopulmonalen
Bypass-Kreislaufs durch Austausch eines Aliquots der Primingflüssigkeit
vor der Einrichtung des kardiopulmonalen Bypasses hinzugefügt. Bevorzugte „Pump prime
doses" sind jene,
die insgesamt ein bis zwei Millionen KIU enthalten.
-
Die
Proteine der vorliegenden Erfindung werden in einer im Fach bekannten
Art und Weise formulierten pharmazeutischen Zusammensetzungen eingesetzt.
Solche Zusammensetzungen enthalten Wirkstoff(e) plus ein oder mehrere
pharmazeutisch akzeptable Träger,
Verdünnungsmittel,
Füllstoffe,
Bindemittel und andere Exzipienten, in Abhängigkeit von der in Erwägung gezogen
Verabreichungsart und Dosierungsform. Zu Beispielen von therapeutisch
inerten anorganischen oder organischen Trägern, die Fachleuten bekannt
sind, zählen,
ohne darauf beschränkt
zu sein, Lactose, Maisstärke
oder Derivate davon, Talkum, Pflanzenöle, Wachse, Fette, Polyole
wie Polyethylenglycol, Wasser, Saccharose, Alkohole, Glycerin und
dergleichen. Verschiedene Konservierungsmittel, Emulgatoren Dispergiermittel,
Aromastoffe, Benetzungsmittel, Antioxidantien, Süßstoffe, Farbstoffe, Stabilisatoren,
Salze, Puffersubstanzen und dergleichen können ebenfalls hinzugefügt werden,
wie erforderlich ist, um bei der Stabilisierung der Formulierung
zu assistieren oder um beim Erhöhen der
Bioverfügbarkeit
des(r) Wirkstoffe(s) zu helfen, oder um eine Formulierung eines
annehmbaren Geschmackes oder Geruches im Falle einer oralen Dosierung
zu ergeben. Der in einer solchen Zusammensetzung eingesetzte Inhibitor
kann in Form der ursprünglichen
Verbindung selbst, oder gegebenenfalls in der Form eines pharmazeutisch
annehmbaren Salzes sein. Die Proteine der vorliegenden Erfindung
können
allein, oder in verschiedenen Kombinationen, und in Kombination
mit anderen therapeutischen Zusammensetzungen verabreicht werden.
Die so formulierten Zusammensetzungen werden, wie benötigt, für die Verabreichung
des Inhibitors durch jede einem Fachmann bekannte, geeignete Art
ausgewählt.
-
Zu
parenteralen Verabreichungsarten zählen intravenöse (i.v.),
subkutane (s.c), intraperitoneale (i.p.) und intramuskuläre (i.m)
Wege. Intravenöse
Verbreichung kann verwendet werden, um eine akute Regulation der
Peak-Plasmakonzentrationen des Arzneimittels zu erhalten, wie es
erforderlich sein kann. Alternativ kann das Arzneimittel mit einer
gewünschten
Rate kontinuierlich durch einen i.v. Katheter verabreicht werden.
Zu geeigneten Vehikeln zählen
sterile, nicht-pyrogene wässrige
Verdünnungsmittel,
wie steriles Wasser für
die Injektion, steril-gepufferte Lösungen oder sterile Salzlösungen.
Die resultierende Zusammensetzung wird dem Patienten vor und/oder
während
der Operation durch intravenöse
Injektion oder Infusion verabreicht.
-
Verbesserte
Halbwertszeit und Abziehlen des Wirkstoffes auf Phagosome, wie Neutrophile
und Makrophagen, die in die Entzündung
eingebunden sind, kann durch den Einbau des Wirkstoffs in Liposome
unterstützt
werden. Es sollte möglich
sein, die Selektivität
des liposomalen „Abziehlens" durch Einbau von
Liganden in die Außenseite
von Liposomen, die an für
Zielorgane/Gewebe, wie GI-Trakt und Lungen, spezifische Makromoleküle binden,
verbessert werden. Alternativ könne
i.m. oder s.c Ablagerungsinjektion mit oder ohne Einkapselung des
Wirkstoffs in abbaubare Mikrokugeln (z.B., Poly-DL-Lactid-co-Glycolid
enthaltend) oder Kollagen enthaltende, schützende Formulierungen verwendet
werden, um eine verlängerte,
verzögerte
Arzneimittelfreisetzung zu erhalten. Für eine verbesserte Annehmlichkeit
der Dosierungsform ist es möglich
ein i.p. implantiertes Reservoir und Septum, wie das Percuseal-System
zu verwenden. Eine verbesserte Annehmlichkeit für und Zustimmung durch den
Patienten kann auch erreicht werden, indem entweder Injektorstifte
(z.B. der Novo Pin oder Q-pen) oder nadelfreie Jet-Injektoren (z.B.,
von Bioject, Mediject oder Becton Dickinson) verwendet werden. Eine
präzis-kontrollierte
Freisetzung kann auch unter Verwendung implantierbarer Pumpen mit
Lieferung an die gewünschte
Stelle über
eine Kanüle
erreicht werden. Zu Beispielen zählen
subkutan implantierte osmotische Pumpen, die von ALZA erhältlich sind,
wie die ALZET osmotische Pumpe.
-
Die
nasale Abgabe kann durch Einbau des Arzneimittels in bioadhäsive partikuläre Träger (< 200 nm) erreicht
werden, wie jene, die Celluose, Polyacrylat oder Polycarbophil in
Verbindung mit geeigneten Absorptionsverstärkern, wie Phospholipide oder
Acylcarnitine aufweisen. Kommerziell erhältliche Systeme beinhalten jede,
die von Dan Biosys und Scios Nova entwickelt wurden.
-
Die
pulmonale Abgabe stellt eine nicht-parenterale Verabreichungsart
des Wirkstoffes an die Zirkulation dar. Die unteren Atemwegs-Epithelien
sind für
eine Vielzahl von Proteinen mit einem Molekulargewicht bis zu etwa
20 kDa höchst
permeabel. Mikron-große
Trockenpulver, die das Medikament in einem geeigneten Träger, wie
Mannitol, Saccharaose oder Lactose enthalten können zu der distalen alveolaren
Oberfläche
unter Verwendung von Trockenpulver-Inhalatoren geliefert werden, wie jenen
von InhaleTM, DuraTM,
Fisons (SpinhalerTM), und Glaxo (RotahalerTM), oder dem Astra (TurbohalerTM)
Treibgas-basierenden Inhalator für
gemessene Dosis. Lösungsformulierung
mit oder ohne Liposome können
unter Verwendung von Ultraschallverneblern abgegeben werden.
-
Die
orale Abgabe kann durch Einarbeiten des Wirkstoffs in Tabletten, überzoge
Tabletten, Dragees, Hart- und Weichgelatinekapseln, Lösungen,
Emulsionen, Suspensionen, oder enterischüberzogene Kapseln, die so ausgelegt
sind, um das Arzneimittel in das Kolon freisetzen, wo digestive
Proteaseaktivität
niedrig ist. Zu Beispielen des letzteren zählen das OROS-CT/OsmetTM-System
von ALZA, und das PULSINCAPTM System von
Scherer Drug Delivery Systems. Andere Systeme verwenden Azo-vernetzte
Polymere, die durch Darm-spezifische bakterielle Azoreduktasen abgebaut
werden, oder pH sensitive Polyacrylate-Polymere, die durch den pH-Anstieg
im Dickdarm aktiviert werden. Die obigen Systeme können in
Verbindung mit einer Vielzahl von erhältlichen Absorptionsverstärkern verwendet
werden. Die rektale Abgabe kann erreicht werden, indem das Arzneimittel
in Suppositorien eingearbeitet wird.
-
In
seiner bevorzugten medizinischen Anwendung, für die Reduktion des perioperativen
Blutverlustes ist die bevorzugte Verabreichungsart von plazentalen
Bikunin-Varianten der vorliegenden Erfindung parenteral, vorzugsweise
durch i.v Wege durch eine zentrale Linie.
-
Die
Menge der einzusetzenden pharmazeutischen Zusammensetzung hängt vom
Empfänger
und dem zu behandelnden Zustand ab. Die erforderliche Menge kann
ohne unnötige
Experimentierung mittels einem Fachmann bekannter Protokolle bestimmt
werden. Alternativ kann die erforderliche Menge basierend auf der
Bestimmung der Menge einer Zielprotease berechnet werden, wie Plasmin
oder Kallikrein, die inhibiert werden muss, um den Zustand zu behandeln.
Da die in dieser Erfindung betrachteten aktiven Materialien (Wirkstoffe)
als nichttoxisch erachtet werden, beinhaltet die Behandlung vorzugsweise
die Verabreicherung eines Überschusses
der optimal-notwendigen Menge an aktiven Wirkstoff.
-
Zusätzlich können plazentales
Bikunin, isolierte Domänen
oder andere Varianten verwendet werden, um natürliche Substanzen wie ihre
verwandten Proteasen aus humanem Material unter Verwendung affinitätsbasierender
Trennverfahren zu isolieren, sowie um Antikörper gegen die Protease herauszulocken,
die weiters verwendet werden können,
um die Gewebsverteilung und nützliche
Funktionen von plazentalem Bikunin zu erforschen.
-
Durchsuchen von humanen
Sequenzdaten
-
Die
Existenz eines eindeutigen humanen Proteins, welches eine homologe
Funktion zu Aprotinin hat, wurde einer einzigartigen Analyse von
Sequenzeinträgen
in der Expressed-Sequenz-Tag-Datenbank
(hiernach dbEST genannt) am NCBI (National Center for Biological
Information, Maryland) folgend abgeleitet. Unter Verwendung des
TBlastN-Algorithmus (BLAST, oder Basic Local Alignment Search Tool
verwendet das Verfahren von Altschul et al., (1990) J. Mol Biol
215: 403–410,
um auf Ähnlichkeiten
zwischen einer Abfrage-Sequenz und allen Sequenzen in einer Datenbank,
Protein oder Nukleinsäure
in einer beliebigen Kombination zu durchsuchen), wurde die Datenbank
auf Nukleotidsequenzen untersucht, die eine Homologie zu der Sequenz des
bovinen Prä-Pro-Aprotinin,
Trasylol® tragen.
Diese Suche von zahlreichen Klonen wurde selektiv auf zwei bestimmte
Klone eingegrenzt, welche möglicherweise
eine abgeleitete Aminosäure-Sequenz
kodieren könnten,
welche einem menschlichen Protein entsprechen, das eine homologe
Funktion zu Aprotinin hat. Die ausgewählten Nukleinsäuresequenzen
waren R35464 (SEQ ID Nr: 12) und R74593 (SEQ ID Nr: 14), die aus
einer humanen plazentalen Nukleinsäure-Bibliothek erzeugt wurden.
Der translatierten Proteinsequenz fehlte im längsten offenen Leseraster für R35464
(SEQ ID Nr: 13) eines der 6 Cysteine, die für die Bildung der kovalenten-Kunitz-Domänstruktur
kritisch ist, was bedeutet, dass die Nukleinsäure-Sequenz von R35464 keinen
funktionellen Inhibitor ergeben konnte. Auf ähnliche Weise enthielt der
längste
translatierte offene Leserahmen des Klons R74596 (SEQ ID Nr: 15)
ein Stoppkodon 5' zu
der Region, die die Kunitz-artige Sequenz kodiert, was bedeutet,
dass diese Sequenz nicht translatiert werden konnte, um eine funktionell-sezernierte
Kunitz-Domäne zu ergeben.
Die Signifikanz dieser Sequenzen alleine war unklar. Es war möglich, dass
sie a) die Produkte von Pseudogenen, b) Regionen nicht translatierter
mRNA, oder c) die Produkte von brauchbarer mRNA darstellen, die
inkorrekt sequenziert wurden.
-
Entdeckung von humanem
Bikunin
-
Um
die aktuelle humane Sequenz spezifisch zu isolieren und zu bestimmen,
wurden cDNA Primer konstruiert, die in der Lage sind an Sequenzen
zu hybridisieren, die 5' und
3' zu den cDNA-Sequenzen
liegen, die unsere vorgeschlagenen Kunitz-artigen Sequenzen kodieren,
die innerhalb R35464 und R74593 gefunden werden. Die Primer, die
verwendet wurden, um ein Fragment zu amplifizieren, das die Kunitz-artige
Sequenz von R74593 kodieren, waren CGAAGCTTCATCTCCGAAGCTCCAGACG
(der 3' Primer mit
einer HindIII Stelle; SEQ ID Nr: 33) und AGGATCTAGACAATAATTACCTGACCAAGGA
(der 5' Primer mit
einer Xba I Stelle; SEQ ID Nr: 34).
-
Diese
Primer wurden verwendet, um ein 500 Basenpaar Produkt aus einer
humanen plazentalen cDNA Bibliothek von Clontech (MATCHMAKER, Kat.Nr.:
HL4003AB, Clontech Laboratories, Palo Alto, CA) mittels PCR (30
Zyklen) zu amplifizieren, welches in Bluescript-SK subkloniert wurde und mit dem T3
Primer mit einem SequenaseTM-Kit Version
2.0 sequenziert wurde. Überraschenderweise
war die unter Verwendung unserer Primer erhaltene Sequenz des Fragments
unterschiedlich zu der in der dbEST Datenbank für den Klon R74593 aufgelisteten
Sequenz. Insbesondere enthielt unsere neue Sequenz eine zusätzliche
Guanosin-Base, welche
3' zum mutmaßlichen
Stoppkodon, aber 5' zu
dem Segement eingefügt
wurde, welches die Kunitz-artige Sequenz kodiert (3).
Die Insertion eines zusätzlichen
Gs verschob das Stoppkodon aus dem Leserahmen der Kunitz-artigen
Domäne
heraus (G bei Basenpaar 114 der korrigierten Sequenz für R74593; 3).
-
Eine
nachfolgende Abfrage der dbEST für
Sequenzen, die homolog zu der Kunitz-artigen Peptidsequenz von R74593
sind, ergab H94519, der aus der humanen Retina-Bibliothek und N39798
abgeleitet ist. Diese Sequenzen enthielten eine Kunitz-artige Sequenz,
die beinahe identisch zu der in R35464 kodierten Kunitz-artigen
Domäne
war, mit Ausnahme dass es alle der sechs charakteristischen Cysteine
enthielt. Die Überlappung
von jeder der Nukleotidsequenzen mit der von R74593 (korrigiert
durch die Insertion von G bei Basenpaar 114) und R35464 wurde verwendet,
um eine Konsensus-Nukleotid-Sequenz für ein partielles humanes plazentales
Bikunin (SEQ ID Nr: 9; 3) zu erhalten. Die translatierte
Konsensus-Sequenz ergab einen offenen Leserahmen, der sich von den
Resten –18
bis +179 erstreckt (3; volle Translation SEQ ID
Nr: 10), welcher zwei vollständige
Kunitz-artige Domänsequenzen,
innerhalb der Region der Aminosäurereste
17–64 bzw.
102–159,
enthielt.
-
Weitere
Bemühungen
wurden unternommen, um zusätzliche
5' Sequenzen durch
Abfragen von dbEST mit der Sequenz von R35464 zu erhalten. Mögliche Übereinstimmungen
von solchen Suchen, die zusätzliche
5' Sequenzen besitzen
wurden dann wiederum verwendet, um die dbEST erneut abzufragen.
Mit einer solchen iterativen Art und Weise wurde eine Reihe von überlappenden
5' Sequenzen identifiziert,
welche die Klone H16866, T66058, R34808, R87894, N40851 und N39876
beinhaltet (4). Die Ausrichtung von einigen
dieser Sequenzen wies auf das Vorhandensein eines 5' ATGs hin, welches
als Startstelle für
die Synthese der Konsensus translatierten Proteinsequenz dienen
könnte.
Von diesen ausgewählten
Informationen war es möglich,
auf Nukleinssäure-
und Polypeptidsequenzen eines menschlichen Proteins mit einer homologen
Funktion zu Aprotinin zu screenen und diese zu bestimmen.
-
Die
erneute Abfrage von dbEST offenbarte etliche neue EST Einträge, die
schematisch in 4B gezeigt sind. Die Überlappung
mit diesen zusätzlichen
ESTs erlaubte uns, eine viel längere
Konsensus-Oligonukleotid-Sequenz zu konstruierten (4C),
die sich sowohl über
das 5' als auch
das 3' Ende der
in 3 dargestellten ursprünglichen Oligonukleotid-Sequenz
erstreckt. Tatsächlich
erstreckt sich die neue Sequenz mit einer Gesamtlänge von
1,6 Kilobasen über
den gesamten Bereich bis zu dem 3' Poly-A-Schwanz. Die erhöhte Anzahl
von überlappenden
ESTs an jeder Basenpaar Position entlang der Sequenz verbesserte
das Vertrauensniveau in bestimmten Regionen, wie die Sequenz, die
mit dem 3' Ende
von EST R74593 überlappt
(3). Mehrere überlappende
ESTs in dieser Region bestätigten
zwei kritische Basen-Deletionen in Bezug auf R74593 (gefunden als
fett-unterstrichen in 4C, Kartenpositionen 994 und
1005). Die Translation der neuen Konsensus-Sequenz (4D) in dem Bikunin-kodierenden Rahmen ergab eine
Form des plazentalen Bikunin, die länger war (248 Aminosäure) als
die reife Sequenz (179 Aminosäuren),
welche von der ursprünglichen Konsensus
(SEQ ID Nr: 1) kodiert wird, und wurde durch ein Stoppkodon im Rahmen
innerhalb des Oligonukleotid-Konsensus beendet. Die Größenzunahme
war infolge einer Rahmenverschiebung in der 3' kodierenden Region, welcher aus der
Entfernung von zwei für
EST R74593 einzigartige Baseninsertionen resultiert. Die Rahmenverschiebung
verschob das Stoppkodon des ursprünglichen Konsensus (3)
aus dem Rahmen hinaus, wodurch ermöglicht wird, dass in einen
neuen Rahmen durchgelesen wird, der die zusätzliche Aminosäuresequenz
kodiert. Das neue Translationsprodukt (4D)
war identisch zu der ursprünglichen
Proteinkonsensus-Sequenz (SEQ ID Nr: 1) zwischen den Resten +1 bis
+175 (kodieren die Kunitz-Domänen),
enthielten jedoch eine neue C-terminale Erweiterung, welche eine
mutmaßliche
24 Reste lange Transmembran-Domäne
(unterstrichen in 4D), gefolgt von einer kurzen
cytoplasmatischen Domäne
mit 31 Resten darstellt. Die genaue Sequenz um das Initiator-Methionin
und Signalpeptid war aufgrund der beträchtlichen Heterogenität unter
den überlappenden
ESTs in dieser Region etwas provisorisch.
-
Die
Analyse der Proteinsequenz durch GeneworksTM hob
Asparagin-Reste an den Postionen 30 und 67 als Konsensusstellen
für eine
mutmaßliche
N-gebundenen Glycosylierung hervor. Asparagin 30 wurde während der
N-terminalen Sequenzierung des aus humaner Plazenta isolierten Proteins
mit voller Länge
nicht beobacht, was damit übereinstimmt,
dass es glycosyliert ist.
-
Klonierung von humanem
Bikunin
-
Die
Existenz einer humanen mRNA, die der mutmaßlichen humanen Bikunin Nukleotidsequenz
entspricht, die aus der Analyse der 3 entnommen
wurde, wurde wie folgt bestätigt.
Der Nukleinsäure-Primer, der
5' zu der Kunitz-kodierenden
cDNA Sequenz von R35454 (b.p. 3–27
der Konsensus-Nukleotid-Sequenz in 3) hybridisiert:
GGTCTAGAGGCCGGGTCGTTTCTCGCCTGGCTGGGA (ein 5' Primer abgeleitet aus der R35464 Sequenz
mit einer XbaI Stelle, SEQ ID Nr: 35) und der Nukleinsäure-Primer,
der 3' zu der Kunitz-kodierenden
Sequenz von R74593 (b.p. 680–700
der Konsensus-Nukleotid-Sequenz in 3) hybridisiert,
wurde verwendet um, aus einer Clontech humanen plazentalen Bibliothek,
ein Fragment der Größe (ca.
670 bp), die aus einer cDNA Konsensus Nukleotid-Sequenz, welche die plazentale Bikunin
Sequenz der 3 kodiert, geschätzt wird,
mittels PCR zu amplifizieren (in 4A schematisch
gezeigt).
-
Unter
Verwendung eines 5' Primers,
der mit einer Sequenz in R87894 hybridisiert, die 126 bp 5' zu der oben diskutierten,
mutmaßlichen
ATG Startstelle ist (in 4A bei
bp 110 schematisch gezeigt), plus des gleichen 3' Primers zu R74593 wie oben verwendet,
war es möglich
ein Fragment der erwarteten Größe (etwa
872 bp) aus der Clontech humanen plazentalen Bibliothek zu amplifizieren,
wie durch eine EST Überlappung
vorhergesagt wurde (in 4 schematisch
gezeigt).
-
Die
Sequenzierung des 872 bp Fragments zeigte, dass es ein Nukleotidsegment
enthält,
welches zu den bp 110 bis 218 des EST R87894 an seinem 5' Ende und den bp
310 bis 542 der Konsensus-Sequenz für plazentales Bikunin, welche
aus der EST Überlappungsanalyse
(der 3) abgeleitet wurde, an seinem 3' Ende entspricht.
Diese 3' Nukleotidsequenz
enthielt alle der Kunitz-artigen Domänen, die durch das plazentale Bikunin
(102–159)
kodiert sind.
-
Um
eine cDNA zu erhalten, die die gesamte extrazelluläre Region
des Proteins kodiert, wurde der folgende 5' PCR Primer:
CACCTGATCGCGAGACCCC
(SEQ ID Nr: 36)
der so konstruiert ist, dass er mit einer Sequenz
innerhalb von EST R34808 hybridisert, mit dem gleichen 3'Primer zu EST 74593
verwendet, um ein etwa 780 Basenpaar cDNA Produkt aus der humanen
plazentalen cDNA Bibliothek zu amplifizieren (30 Zyklen). Dieses
Produkt wurde Gel-gereinigt und in den TA Vektor (Invitrogen) kloniert,
für die
DNA Sequenzierung durch das Dideoxy-Verfahren (Sanger F., et al
(1977) Proc. Natl. Acad. Sci (USA), 74: 5463–5467) mit folgenden Primer:
| Vektor
spezifisch: | GATTTAGGTGACACTATAG
(SP6) (SEQ ID Nr: 37) |
| | TAATACGACTCACTATAGGG
(T7) (SEQ ID Nr: 38) |
| Gen
spezifisch: | TTACCTGACCAAGGAGGAGTGC
(SEQ ID Nr: 39)
AATCCGCTGCATTCCTGCTGGTG (SEQ ID Nr: 40)
CAGTCACTGGGCCTTGCCGT
(SEQ ID Nr: 41) |
-
Die
resultierende cDNA Sequenz ist in 4E zusammen
mit ihrem Translationsprodukt dargestellt. Auf der Nukleotid-Ebene
zeigte die Sequenz nur geringe Unterschiede zu der Konsensus EST-Sequenz (4D). Die Translation der Sequenz ergab eine kodierende
Sequenz, die eine Initiator ATG-Stelle im Rahmen, ein Signalpeptid
und die reife plazentale Bikunin-Sequenz und die Transmembran-Domäne enthält. Der translatierten
Sequenz des PCR Produkts fehlten die letzten 12 Aminosäure-Reste
der cytoplasmatischen Domäne
als Folge der Wahl der Selektion des 3' Primers für die PCR Amplifikation. Diese
Wahl des 3' PCR
Primers (konstruiert, basierend auf der R74593 Sequenz) war auch
für die
Einführung
einer künstlichen
S nach F Mutation an der Aminosäure-Position
211 der translatierten PCR-abgeleiteten Sequenz verantwortlich.
Das von der Translation des PCR Fragments abgeleitete Signalpeptid
war zu jenem des EST Konsensus etwas verschieden.
-
Um
eine plazentale Bikunin cDNA mit voller Länge zu erhalten, wurde das
PCR abgeleitete Produkt (4E)
Gel-gereinigt und verwendet, um ein nicht-PCR basierender Klon mit
voller Länge,
welcher die Bikunin-Sequenz darstellt, zu isolieren. Die PCR abgeleitete
cDNA Sequenz wurde mit 32P-CTP durch High
Prime (Boehringer Mannheim) markiert und verwendet, um eine plazentale
cDNA Bibliothek (Stratagen, UnizapTM λ Bibliothek)
unter Verwendung von Kolonie-Hybridisierungstechniken zu sondieren.
Ungefähr
2 × 106 Phagenplaques waren 3 Screening- und Plaque-Reinigungsrunden
ausgesetzt. Zwei Klone wurden als „mit voller Länge (~ 1,5
Kilobasen) erachtet, wie durch Restriktionenzym-Analyse und basierend
auf Vergleich mit der Größe der EST
Konsensus-Sequenz (siehe oben) bestimmt wurde. Die Sequenzierung
von einem dieser Klone durch das Dideoxy-Verfahren ergab die in 4F abgebildete Oligonukleotid-Sequenz. Das Translationsprodukt
von dieser Sequenz ergab ein Protein mit einem Initiator Methionin
im Rahmen, einem Signalpeptid und einer reifen plazentalen Bikunin-Sequenz. Die reife
plazentale Bikunin-Sequenz war identisch mit der Sequenz des reifen
Proteins, welches durch die Translation der EST Konsensus abgeleitet
ist, obwohl die Signalpeptid-Sequenzlängen und Sequenzen unterschiedlich
waren. In Gegensatz zu dem PCR-abgeleiteten
Produkt enthielt die durch die Koloniehybridiserung-abgeleitete
cDNA die gesamte Ektodomäne,
Transmembran-Domäne,
cytoplasmatische Domäne
und Stoppkodon im Rahmen. Tatsächlich
erstreckte sich der Klon über
den gesamten Bereich bis zu dem Poly-A-Schwanz. Das Initiator-Methionin
wurde von einem hydrophoben Signalpeptid gefolgt, welches identisch
zu dem Signalpeptid war, welches durch den PCR-abgeleiteten Klon
kodiert wird. Im Anschluss daran exprimierten und reinigten wir
ein lösliches
Fragment des plazentalen Bikunins, Bikunin (1–170), aus Sf9 Zellen (Beispiel
9) und stellten fest, dass es ein funktioneller Protease-Inhibitor
(Beispiel 10) ist. Weiters isolierten wir aus der menschlichen Plazenta
ein lösliches
Fragment des plazentalen Bikunin, welches ebenfalls ein aktiver
Protease-Inhibitor (Beispiel 7) war. Sowohl das natürliche Protein
als auch die in Sf9 Zellen exprimierte Proteinform sind wahrscheinlich
am Asparagin-Rest bei Position 30 glycosyliert, basierend auf dem
Wiedergewinnen der PTH-Aminosäuren während der
N-terminalen Sequenzierung (Beispiele 7 und 9).
-
Basierend
auf den obigen Beobachtungen scheint es, dass plazentales Bikunin
mit voller Länge
die Fähigkeit
hat, als ein Transmembran-Protein an der Oberfläche von Zellen sowie als lösliches
Protein zu bestehen. Andere Kunitz-Domänen enthaltende Transmembran-Proteine
sind bekannt, dass sie einer proteolytischen Bearbeitung ausgesetzt
sind, um eine Mischung aus löslichen
und Membran-assoziierter Formen zu ergeben. Diese beinhalten zwei
Formen des amyloiden Vorläufer
Proteins, die APP751 (Esch F., et al., (1990) Science, 248: 1122–1124) und
APP770 (Wang R., et al., (1991), J. Biol Chem, 266: 16960–16964)
bezeichnet werden.
-
Die
Kontaktaktivierung ist ein Prozess, welcher durch Exposition von
geschädigten
vaskulären
Oberflächen
mit Komponenten der Gerinnungskaskade aktiviert wird. Angiogenese
ist ein Prozess, in welchen die lokale Aktivierung von Plasmin an
endothelialen Oberflächen
eingebunden ist. Die Spezifität
des plazentalen Bikunins und seiner mutmaßlichen Fähigkeit an Zelloberflächen zu
binden, weist darauf hin, dass die physiologischen Funktionen des
transmembranen plazentalen Bikunins die Regulation der Kontaktaktivierung
und Angiogenese beinhalten können.
-
Die
Aminosäure-Sequenzen
für plazentales
Bikunin (7–64),
Bikunin (102–159)
und plazentals Bikunin mit voller Länge (4F)
wurden gegen die PIR (Vers. 46.0) und PatchX (Vers. 46.0) Protein-Datenbanken sowie
die GeneSeq (Vers. 20.0) Protein-Datenbank von patentierten Sequenzen
unter Verwendung des Genetics Computer Group Programm FastA durchsucht.
Unter Verwendung des Genetics Computer Group Programms TFastA (Pearson
and Lipman, 1988, Proc. Natl. Acad. Sci. USA 85: 2444–2448),
wurden diese Proteinsequenzen versus die sechs-Rahmen Translationen
der GenBank (Vers 92.0 mit Updates 1/26/96) und EMBL (veränderte Vers
45.0) Nukleotid-Datenbanken sowie die GenSeq (Vers. 20.0) Nukleotid-Datenbanken von
patentierten Sequenzen durchsucht. Die EST und STS Teilmengen der
GenBank und EMBL wurden in diesen Suchsatz nicht eingebunden. Die
beste Übereinstimmung,
die von diesen Suchen resultierten, enthielten Sequenzen, die über die
gesamte Länge
nur etwa zu 50% identisch waren mit der 58-Aminosäure Proteinsequenz,
die aus unserer Analyse der Klone R74593 und R35464 abgeleitet wurde.
-
Isolierung
von menschlichem Bikunin
-
Wie
oben erwähnt
ist, könnten
synthetische Peptide, die Bikunin (7–64) und Bikunin (102–159) entsprechen,
wie aus der translatierten Konsensus-Sequenz für Bikunin (3)
bestimmt wurde, rückgefalten werden
(Beispiele 2 bzw. 1), um aktives Kallikrein Inhibitor Protein (Beispiel
4 bzw. 3) zu ergeben. Wir nutzten diese unerwartete Eigenschaft
aus, um ein Reinigungsschema zu entwickeln, um natives plazentales
Bikunin aus humanem Gewebe zu isolieren.
-
Unter
Verwendung eines Reinigungsschemas, welches Kallikrein-Sepharose-Affinitätschromatograpie
als einen ersten Schritt verwendet, wurde hochgereinigter, nativer,
wirksamer Kallikrein-Inhibitor isoliert. Das isolierte native humane
Bikunin hatte einen identischen N-Terminus (sequenziert für 50 Aminosäurereste) wie
die durch die Translation der Konsensus-Nukleinsäure-Sequenz (3)-Aminosäure-Reste
+1 bis +50 (Beispiel 7) vorhergesage Sequenz. Dies bestätigte zum
ersten Mal die Existenz eines neuartigen, aus humaner Plazenta isolierten,
Kallikrein-Inhibitors.
-
Bekannte
Kunitz-artige Domänen
sind unten aufgelistet. Reste von den angenommen wird, dass sie mit
Zielproteasen in Kontakt treten sind markiert, da sie von speziellem
Interesse sind (fett/unterstrichen). Diese besonderen Reste werden
für spezifische
Referenz mit Positionen Xaa
1-16 bezeichnet,
wie durch die Markierung Xaa unten gezeigt ist:
wobei
Sequenznummer 1) Bikunin (7–64)
ist (SEQ ID Nr: 4); Sequenz 2) ist Bikunin (102–159) (SEQ ID Nr: 6); Sequenz
3) ist Gewebefaktor Weg Inhibitor Vorläufer 1 (SEQ ID Nr: 18); Sequenz
4) ist Gewebefaktor Weg Inhibitor Vorläufer 1 (SEQ ID Nr. 19); Sequenz
5) ist Gewebefaktor Weg Inhibitor Vorläufer (SEQ ID Nr. 20); Sequenz
6) ist Gewebefaktor Weg Inhibitor Vorläufer 2 (SEQ ID Nr. 21); Sequenz
7) ist Gewebefaktor Weg Inhibitor Vorläufer 2 (SEQ ID Nr. 22); Sequenz
8) ist amyloider Vorläuferprotein-Homolog
(SEQ ID Nr. 23); Sequenz 9) ist Aprotinin (SEQ ID Nr. 24); Sequenz
10) ist Inter-α-Trypsin-Inhibitor-Vorläufer (SEQ
ID NR. 25); Sequenz 11) ist Inter-α-Trypsin-Inhibitor-Vorläufer (SEQ
ID Nr. 26); Sequenz 12) ist amyloides Vorläuferprotein (SEQ ID Nr. 27);
Sequenz 13) ist Kollagen α-3(VI)
Vorläufer
(SEQ ID Nr. 28); und Sequenz 14) ist HKI-B9 (SEQ ID Nr. 29).
-
Es
kann gesehen werden, dass jedes der plazentalen Bikunine (7–64) und
(102–159)
die gleiche Anzahl (sechs) und Abstand der Cystein-Reste aufweist,
wie bei den Mitgliedern der Kunitz-Klasse von Serin-Protease-Inhibitoren.
Die genaue Bindung der Cystein-Reste, um die drei intraketten Disulfidbrücken zu
bilden, ist bekannt und für
die vorher-bekannten Kunitz-Familienmitglieder
invariant (Laskowski, M et al., 1980, Ann. Rev. Biochem. 49: 593–626). Basierend
auf diesem bekannten Bindungsmuster und der Tatsache, das die Faltung von
plazentalem Bikunin (7–64)
und (102–159)
in aktive Protease-Inhibitoren, durch eine Massenreduktion begleitet
wird, was mit der Bildung von drei intraketten-Disulfidbindungen (Beispiele
2 und 1) übereinstimmt,
ist es höchst
wahrscheinlich, dass die Disulfidbindungen innerhalb der Kunitz-Domänen von
plazentalem Bikunin zwischen den Cystein-Resten: C11 und C61, C20
und C44; C36 und C57; C106 und C156; C115 und C139; C131 und C152
auftreten. Weiters ist dieses Disulfidbindungsmuster in größeren plazentalen
Bikunin-Formen, die beide Kunitz-Domänen enthält, höchst wahrscheinlich, da solche
Formen des Proteins ebenfalls aktive Serin-Protease-Inhibitoren
sind und weil die N-terminale Sequenzierung (Beispiel 7) des nativen
plazentalen Bikunins für
50 Zyklen eine Sequenz ergab, die an Positionen an welchen die Cystein-Reste
erwartet werden, still war.
-
Das
plazentale Bikunin, isolierte Domänen oder andere Varianten der
vorliegenden Erfindung können durch
Standard Festphasen-Peptidsynthese unter Verwendung von entweder
der t-Boc Chemie, wie von Merrifield R. B. and Barany G., in: The
peptides, Analysis, Synthesis, Biology, 2, Gross E. et al., Hrsg.
Academic Press (1980) Kapitel 1 beschrieben; oder mittels F-moc
Chemie wie von Carpino L.A., and Han G.Y., (1970), J. Amer. Chem
Soc., 92,5748–5749,
beschrieben und in Beispiel 2 illustriert ist, hergestellt werden.
Alternativ kann die Expression einer DNA, welche die plazentale
Bikunin-Variant kodiert, verwendet werden, um rekombinante plazentale
Bikunin-Varianten zu erzeugen.
-
Die
Erfindung betrifft auch DNA Konstrukte, die die plazentalen Bikunin-Proteinvarianten
der vorliegenden Erfindung kodieren. Diese Konstrukte können durch
synthetische Verfahren, wie jene die beschrieben sind in Beaucage
S.L. and Caruthers M.H., (1981) Tetrahedron Lett, 22, S. 1859–1862; Matteucci
M.D. and Caruthers M.H., (1981), J. Am. Chem. Soc. 103, S. 3185;
oder aus genomischer oder cDNA hergestellt werden, welche durch
Screenen von genomischer oder cDNA Bibliotheken mit cDNA-Sonden
erhalten werden können,
die so konstruiert sind, dass sie mit plazentalem Bikunin kodierender
DNA Sequenz hybridisieren. Die genomische oder cDNA Sequenz kann
an einer oder mehrere Stellen modifiziert sein, um eine cDNA zu
erhalten, die jede der in dieser Offenbarung beschriebenen Aminosäure-Substitutionen
oder Deletionen kodiert.
-
Die
vorliegende Erfindung betrifft auch Expressionsvektoren, welche
das DNA Konstrukt enthalten, welches das plazentale Bikunin, isolierte
Domänen
oder andere Varianten der vorliegenden Erfindung kodiert, das für die Herstellung
von rekombinanten plazentalen Bikunin-Varianten verwendet werden kann. Die
cDNA sollte mit einer geeigneten Promotor-Sequenz verbunden sein,
die eine transkriptionale Aktivität in der Wirtszelle der Wahl
zeigen, einen geeignenten Terminator und ein Poly-Adenylierungssignal
besitzen. Die die plazentale Bikunin-Variante kodierende cDNA kann mit einem
5' Signalpeptid
fusioniert werden, dass dazu führt, dass
ein durch diese cDNA kodiertes Protein sezerniert wird. Das Signalpeptid
kann eines sein, das durch den Wirtsorganismus erkannt wird. Im
Falle von einer Säugetierwirtszelle
kann das Signalpeptid das natürliche
Signalpeptid sein, welches in plazentalem Bikunin mit voller Länge vorhanden
ist. Die verwendeten Verfahren, um solche Vektoren für die Expression
von plazentalen Bikunin-Varianten herzustellen, sind im Fach bekannt und
sind zum Beispiel in Sambrook et al., Molecular Cloning: A laboratory
Manual, Cold Spring Harbor, New York, (1989) beschrieben.
-
Die
vorliegende Erfindung betrifft auch transformierte Zellen, die das
DNA Konstrukt enthalten, welches das plazentale Bikunin, isolierte
Domänen
oder andere Varianten der vorliegenden Erfindung kodieren, die für die Herstellung
von rekombinanten plazentalen Bikunin-Varianten verwendet werden
können.
Eine Vielzahl von Kombinationen an Expressionsvektoren und Wirtsorganismus
existieren, die für
die Herstellung von plazentalen Bikunin-Varianten verwendet werden
können.
Zu geeigneten Wirtszellen zählen
Baculovirusinfizierte Sf9 Insektenzellen, Säugetierzellen, wie BHK, CHO,
Hela und C-127, Bakterien wie E. coli, und Hefen wie Saccharomyces
cervisiae. Die für
die Verwendung von Säugetier-,
Insekten und mikrobiellen Expressionssystemen benötigten Verfahren,
um eine Expression von plazentalem Bikunin zu erreichen, sind mit
Fach bekannt und sind zum Beispiel in Ausubel F. M. et al., Current
protocols in Molecular Biology, John Wiley & Sons" (1995), Kapitel 16 beschrieben. Für Fragmente
von plazentalem Bikunin, die eine einzelne Kunitz-Inhibitor-Domäne enthalten,
wie Bikunin (7–64)
und (102–159)
werden Hefe- und E.coli Expressionssysteme bevorzugt, wobei Hefe-Systeme
am meisten bevorzugt werden. Üblicherweise
würde die
Hefeexpression ausgeführt
werden, wie im US Patent 5.164.482 für Aprotinin-Varianten beschrieben
ist und im Beispiel 5 der vorliegenden Spezifikation für plazentales
Bikunin (102–159)
angepasst wurde. Die E. coli Expression könnte unter Verwendung der Verfahren,
die im US Patent 5.032.573 beschrieben sind, ausgeführt werden.
Die Verwendung von Säugetier-
und Hefesystemen sind am meisten für die Expression von größeren plazentalen
Bikunin-Varianten bevorzugt, die beide Inhibitordomänen enthalten,
wie die Variante Bikunin (7–159).
-
DNA,
die plazentale Bikunin-Varianten kodiert, die Aminosäuresubstitutionen
der natürlichen
Aminosäure-Sequenz
besitzen, kann für
die Expression von rekombinantem Protein unter Verwendung der Verfahren von
Kunkel T.A., (1985) Proc. Natl. Acad. Sci USA 82: 488–492 hergestellt
werden. Kurz gesagt wird die zu mutagenisierende DNA in einen einzelsträngigen Bakteriophagenvektor,
wie M13 kloniert. Ein Oligonukleotid, welches den zu verändernden
Bereich umspannt, und die Substitutionen kodiert wird mit der einzelsträngigen DNA
hybridisiert und durch Standard molekularbiologische Techniken doppelsträngig gemacht.
-
Diese
DNA wird dann in einen geeigneten bakteriellen Wirt transformiert
und durch Dideoxynukleotid-Sequenzierung verifiziert. Die korrekte
DNA wird dann in das Expressionsplasmid kloniert. Alternativ kann die
Ziel-DNA durch Standard-PCR-Techniken mutagenisiert werden, sequenziert
und in das entsprechende Expressionsplasmid eingefügt werden.
-
Die
folgenden bestimmten Beispiele sind für Illustrationszwecke, und
nicht als Einschränkung
bestimmter Aspekte und bevorzugter Ausführungsformen der vorliegenden
Erfindung angeführt.
-
Beispiel 1 – Kein Beispiel
der Erfindung
-
Herstellung von synthetischem
plazentalem Bikunin (102–159)
-
Materialien
und Verfahren/Verwendete Reagenzien: Das fluoregene Substrat Tos-Gly-Pro-Lys-AMC wurde von
Bachem BioScience INc (King of Prussia, PA) gekauft. PNGB, Pro-Phe-Arg-AMC, Ala-Ala-Pro-Met-AMC,
bovines Trypsin (Typ III), humanes Plasma Kallikrein, und humanes
Plasmin wurden von Sigma (St. Louis, MO) gekauft.
-
Rekombinantes
Aprotinin (Trasylol®) war von Bayer AG (Wuppertal,
Germany). Vorgeladenes Gln-Wang-Harz war von Novabiochem (La Jolla,
CA). Thioanisole, Ethandithiol und t-Butylmethylether war von Aldrich
(Milwaukee, WI).
-
Quantifizierung
von funktionellem plazentalem Bikunin (7–64) und (102–159) Die
Menge an Trypsin-inhibitorischer Aktivität, die in der rückgefaltenen
Probe bei verschiedenen Reinigungsstufen vorhanden war, wurde unter
Verwendung von GPK-AMC als Substrat gemessen. Bovines Trypsin (200
pmol) wurde für
5 Minuten bei 37°C
mit Bikunin (7–64)
oder (102–159)
von verschiedenen Reinigungsstufen, in Puffer A (50 mM Hepes, pH
7,5, 0,1 M NaCl, 2 mM CaCl2 und 0,01% Triton
X-100) inkubiert. GPK-AMC wurde hinzgefügt (20 μM am Ende) und die Menge an
erzeugtem Cumarin wurde durch Messen der Floureszenz (Ex = 370 nm,
Em = 432 nm) auf einem Perkin-Elmer LS-50B Fluorimeter über eine
Periode von 2 Minuten bestimmt. Für die zu testenden Proben wurde
für jede
Probe die % Inhibierung gemäß Gleichung
1 berechnet; wobei Ro die Rate der Fluoreszenzzunahme
in Gegenwart des Inhibitors ist und R1 ist
die Rate, die in Abwesenheit der hinzugefügten Probe bestimmt wurde.
Eine Aktivitätseinheit
des Inhibitors wird als die Menge definiert, die benötigt wird,
um eine 50% Inhibierung in der Untersuchung unter Verwendung der
beschriebenen Bedingungen, zu erreichen. % Inhibierung = 100 × [1 – Ro/R1] (1)
-
Synthese:
Plazentales Bikunin (102–159)
wurde auf einem Applied Biosystems Modell 420 A Peptidsynthetisierer
unter Verwendung der NMP-HBTU Fmoc-Chemie synthetisiert. Das Peptid
wurde auf einem vorgeladenen Gln-Harz mit einem 8-fachen Überschuss
an Aminosäure
für jede
Kopplung synthetisiert. Spaltung und Schutzeliminierung wurde in
84,6% Trifluoressigsäure
(TFA), 4,4% Thioanisol, 2,2% Ethandithiol, 4,4% verflüssigtes
Phenol und 4,4% Wasser für
2 Stunden bei Raumtemperatur durchgeführt. Das Rohpeptid wurde ausgefällt, zentrifugiert
und zweimal mit t-Butylmethylether gewaschen. Das Peptid wurde an
einer Dynamax 60A C18 Umkehrphasen HPLC Säule mittels eines TFA/Acetonitril-Gradienten
gereinigt. Die letzte Ausfällung (61,0
mg) ergab die korrekte Aminosäurezusammensetzung
und Molekulargewicht mittels Elektronenspray-Massenspektroskopie
(MH+ = 6836,1; berechnet = 6835,5) für die vorhergesagte Sequenz:
YEEYCTANAV
TGPCRASFPR WYFDVERNSC NNFIYGGCRG NKNSYRSEEA CMLRCFRQ (SEQ ID Nr.
6)
-
Reinigung:
Die Rückfaltung
des plazentalem Bikunins (102–159)
wurde gemäß dem Verfahren
von Tam et al., (J. Am. Chem. Soc. 1991, 113: 6657–62) durchgeführt. Ein
Teil des gereinigten Peptids (15,2 mg) wurde in 4,0 ml 0,1 M Tris,
pH 6,0 und 8 M Harnstoff aufgelöst.
Die Oxidation der Disulfide wurde durch die tropfenweise Zugabe
einer 23 % DMSO und 0,1 M Tris, pH 6,0 enthaltenden Lösung erreicht,
um eine Endkonzentration von 0,5 mg/ml Peptid in 20 % DMSO, 0,1
M Tris, pH 6,0 und 1 M Harnstoff zu erreichen. Der Lösung wurde
erlaubt für
24 Stunden bei 25 °C
zu rühren,
nach welchem, die Lösung
1:10 in 50 mM Tris, pH 8,0 und 0,1 M NaCl enthaltendem Puffer verdünnt wurde.
Das Material wurde unter Verwendung einer Kallikrein-Affinitätssäule gereinigt,
welche durch kovalentes Binden von 30 mg bovinem pankreatischem
Kallikrein (Bayer AG) an 3,5 ml CNBr aktivierte Sepharose (Pharmacia),
gemäß den Herstellerangaben
hergestellt wurde. Das rückgefaltete
Material wurde auf die Affinitätssäule mit
einer Flussrate von 1 ml/min geladen und mit 50 mM Tris, pH 8,0
und 0,1 M NaCl gewaschen, bis die Absorption der Waschung bei 280
nm nicht mehr nachgewiesen werden konnte. Die Säule wurde mit 3 Volumina, jeweils
von 0,2 M Essigsäure,
pH 4,0 und 1,7 eluiert. Aktive Fraktionen wurden zusammengefasst
(siehe unten) und der pH der Lösung
wurde auf 2,5 eingestellt. Das Material wurde direkt auf eine Vydac
C18 Umkehrphasen-Säule
(5 Mikron, 0,46 × 25
cm) aufgetragen, die in 22,5% Acetonitril in 0,1 % TFA äquilibriert
wurde. Die Trennung wurde unter Verwendung eines linearen Gradienten
von 22,5 bis 40% Acetonitril in 0,1 % TFA bei 1,0 m/min über 40 Minuten
erreicht. Aktive Fraktionen wurden zusammengefasst, lyophilisiert,
erneut in 0,1 % TFA aufgelöst
und bis zum Bedarf bei –20°C gelagert.
-
Ergebnisse:
Synthetisches plazentales Bikunin (102–159) wurde unter Verwendung
von 20% DMSO als Oxidationsmittel, wie oben beschrieben, rückgefaltet,
und durch ein 2-stufiges Reinigungsverfahren, wie unten gezeigt,
gereinigt, um einen aktiven Trypsin Inhibitor (Tabelle 1, unten)
zu ergeben.
-
Tabelle
1 Reinigungstabelle
für die
Isolierung von synthetischem plazentalem Bikunin (102–159)
-
- a Protein bestimmt
durch AAA.
- b Protein bestimmt durch OD280 nm, unter
Verwendung des Extinktionskoeffizienten, welcher für das gereinigte
Protein bestimmt wurde (1,7 × 104 Lmol–1 cm–1)
- c Eine Einheit ist definiert als die
Menge an Material, die erforderlich ist, um 50% der Trypsin-Aktivität in einer Standarduntersuchung
zu inhibieren.
-
Chromatographie
des rohen rückgefaltenen
Materials über
eine immobilisierte bovine pankreatische Kallikrein-Säule isolierte
selektiv 6,0% des Proteins und 97 % der vorhandenen Trypsin inhibitorischen
Aktivität.
Eine nachfolgende Chromatographie unter Verwendung einer C18 Umkehrphase
ergab eine weitere 2fache Reinigung, mit einer Gesamt-Ausbeute von
74%. Auf einer RPHPLC zeigte das reduzierte und rückgefaltete
plazentale Bikunin (102–159)
Elutionszeiten von 26,3 bzw. 20,1 Minuten. Die Massenspektroskopie-Analyse
des gereinigten Materials offenbarte ein Molekulargewicht von 6829,8;
einem Verlust von 6 Masseneinheiten des Ausgangsmaterials. Dies
demonstriert die vollständige
Bildung der 3 von der Peptidsequenz vorhergesagten Disulfide.
-
Die
isoelektrischen Punkte des gereinigten, rückgefaltenen synthetischen
plazentalen Bikunins (102–159)
wurden unter Verwendung eines Multiphor II Elektrophorese-Systems
(Pharmacia), welches gemäß den Herstellerangaben
betrieben wurde, zusammen mit pI Standards, unter Verwendung eines
vorgefertigten Ampholine® PAGplate (pH 3,5 bis
9,5) und 1,5 stündiger
Fokussierung bestimmt. Nach dem Färben wurde die Migrationsdistanz
von dem kathodischen Rand des Gels zu den unterschiedlichen Proteinbanden
gemessen. Der pI jedes unbekannten Proteins wurde mittels einer
Standardkurve bestimmt, die durch die graphische Darstellung der
Migrationsdistanzen von Standards gegen die entsprechenden pIs erzeugt
wurde. Mit dieser Technik wurde der pI des plazentalem Bikunins
(102–159)
mit 8,3 bestimmt, was in Übereinstimmung
mit dem aus der Aminosäure-Sequenz
vorhergesagten Wert ist. Dieser ist niedriger als der Wert von 10,5,
der für
Aprotinin festgelegte pI (Tenstad et al., 1994, Acta Physiol. Scand.
152: 33–50).
-
Beispiel 2 – kein Beispiel
der Erfindung
-
Herstellung von synthetischem
plazentalem Bikunin (7–64)
-
Plazentales
Bikunin (7–64)
wurde im Wesentlichen wie für
plazentales Bikunin (102–159)
beschrieben synthetisiert, rückgefaltet
und gereinigt, jedoch mit folgenden Modifikationen: während dem
Rückfalten
wurde das synthetische Peptid für
30 Stunden als eine Lösung
in 20% DMSO bei 25°C
gerührt;
Reinigung durch C18 RP-HPLC wurde mit einem linearen Gradient von
25 bis 45 % Acetonitril in 0,1 % TFA über 40 Minuten (1 ml/ml) erreicht.
Die aktiven Fraktionen des ersten C18-Laufes wurden erneut auf die
Säule aufgetragen
und mit einem linearen Gradienten (60 min, 1 ml/min) von 20 bis
40% Acetonitril in 0,1 % TFA fraktioniert.
-
Ergebnisse:
Das endgültige
gereinigte, reduzierte Peptid zeigte eine MH+ = 6563, übereinstimmend mit
der Sequenz:
IHDFCLVSKV VGRCRASMPR WWYNVTDGSC QLFVYGGCDG NSNNYLTKEE
CLKKCATV (SEQ ID Nr: 4)
-
Die
Rückfaltung
und Reinigung ergab eine funktionelle Kunitz-Domäne, die als Trypsin-Inhibitor aktiv war
(Tabelle 2 unter).
-
Tabelle
2A Reinigungstabelle
für die
Isolierung von synthetischem plazentalem Bikunin (7–64)
-
Das
gereinigte rückgefaltete
Protein zeigte einen MH+ = 6558, d.h. 5 ± 1 Masseneinheiten weniger
als für
das reduzierte Peptid. Dies demonstriert, dass die Rückfaltung
die Bildung von zumindest einer entsprechenden Disulfidbindung hervorruft.
-
Der
pI von plazentalem Bikunin (7–64)
wurde unter Verwendung von Verfahren die für das Bestimmen des pIs von
plazentalem Bikunin (102–159)
eingesetzt wurden, bestimmt. Plazentales Bikunin (7–64) zeigte einen
pI, der viel höher
als der vorhergesagte Wert (pI = 7,9) war. Rückgefaltetes plazentales Bikunin
(7–64) wanderte
zu dem kathodischen Rand des Gels (pH 9,5) und ein genauer pI konnte
unter diesen Bedingungen nicht bestimmt werden.
-
Kontinuierliche
Herstellung von synthetischem plazentalem Bikunin (7–64).
-
Da
das synthetische plazentale Bikunin (7–64) nicht einer vollständigen Schutzeliminierung
vor der Reinigung und Rückfaltung
unterzogen werden konnte, wurde die Rückfaltung unter Verwendung
von Protein wiederholt, welches sicherlich vollständig schutzeliminiert
war. Plazentales Bikunin (7–64)
wurde im Wesentlichen wie für
plazentales Bikunin (102–159)
beschrieben, synthetisiert, rückgefaltet
und gereinigt, jedoch mit den folgenden Modifikationen: während der
Rückfaltung
wurde das synthetische Peptid für
30 Stunden als eine Lösung
in 20% DMSO bei 25°C
gerührt;
Reinigung durch C18 RP-HPLC wurde mit einem linearen Gradient von
22,5 bis 50% Acetonitril in 0,1 % TFA über 40 Minuten (1 ml/ml) erreicht.
-
Ergebnisse:
Das endgültig
gereinigte, reduzierte Peptid zeigte einen MH+ = 6567,5, welches übereinstimmt
mit der Sequenz:
IHDFCLVSKV VGRCRASMPRW WYNVTDGSC QLFVYGGCDG
NSNNYLTKEE CLKKCATV (SEQ ID Nr: 4)
-
Das
Rückfalten
und die Reinigung ergab eine funktionale Kunitz-Domäne, die
gleich aktiv, wie ein Trypsin-Inhibitor war (Tabelle 2B, unten).
-
Tabelle
2B Reinigungstabelle
für die
Isolierung von synthetischem plazentalem Bikunin (7–64)
-
Das
gereinigte rückgefaltete
Protein zeigte einen MH+ = 6561,2, d.h. 6,3 Masseneinheiten weniger
als für
das reduzierte Peptid. Dies demonstriert, dass die Rückfaltung
die Bildung der erwarteten 3 Disulfidbindung hervorruft.
-
Der
pI von rückgefaltetem,
plazentalem Bikunin (7–64)
wurde unter Verwendung der Verfahren die für das Bestimmen des pIs von
plazentalem Bikunin (102–159)
eingesetzt wurden, bestimmt. Rückgefaltetes,
plazentales Bikunin (7–64)
zeigte einen pI von 8,85, der etwas höher als der vorhergesagte Wert
(pI = 7,9) war.
-
Beispiel 3 – Kein Beispiel
der Erfindung
-
In vitro Spezifität von funktionellem
plazentalem Bikunin-Fragment (102–159)
-
Proteasen:
Bovines Trypsin, humanes Plasmin und bovine pankreatische Kallikrein-Quantifizierungen wurden
durch Active-Site-Titration unter Verwendung von p-Nitrophenyl p'-guanidinobenzoat HCl, wie früher beschrieben
(Chase, T., und Shaw, E., (1970) Methods Enzmol., 19: 20–27) durchgeführt. Humanes
Kallikrein wurde durch Active-Site-Tritation mittels bovinem Aprotinin
als ein Standard und PFR-AMC als ein Substrat, unter der Annahme
einer 1:1 Komplexbildung, quantifiziert. Der Km für GPK-AMC
mit Trypsin und Plasmin unter den für jedes Enzym verwendeten Bedingungen
war 29 μM
bzw. 726 μM;
der Km für
PFR-AMC mit humanem Plasma Kallikrein und bovinem pankreatischem
Kallikrein war 457 μM
bzw. 81,5 μM;
der Km für
AAPR-AMC mit Elastase war 1600 μM.
Die Quantifizierung des humanen Gewebe-Kallikreins (Bayern, Deutschland)
wurde durch Active-Site-Titration mittels p-Nitrophenyl p'-guanidinobenzoat HCl, wie früher beschrieben (Chase,
T., und Shaw, E., (1970) Methods Enzmol., 19: 20–27) durchgeführt.
-
Inhibierungskinetiken:
Die Inhibierung von Trypsin durch plazentales Bikunin (102–159) oder
Aprotinin wurde durch Inkubieren von 50pM Trypsin mit plazentalem
Bikunin (102–159)
(0–2 nM)
oder Aprotinin (0–3
nM) in Puffer A in einem Gesamtvolumen von 1,0 ml gemessen. Nach
5 Minuten bei 37°C
wurden 15 μl
2 mM GPK-AMC hinzugefügt
und die Änderung
der Fluoreszenz (wie oben) wurde überwacht. Die Inhibierung von humanem
Plasmin durch plazentales Bikunin (102–159) und Aprotinin wurde mit
Plasmin (50 pM) und plazentalem Bikunin (102–159) (0–10nM) oder Aprotinin (0–4 nM) in
50 mM Tris-HCl (pH 7,5), 0,1 M NaCl, und 0,02% Triton X-100 enthaltendem
Puffer bestimmt. Nach einer Inkubation von 5 Minuten bei 37°C wurden
25 μl 20 mM
GPK-AMC hinzugefügt
und die Änderung
der Fluoreszenz wurde überwacht.
Die Inhibierung von humanem Plasma-Kallikrein durch plazentales
Bikunin (102–159)
und Aprotinin wurde unter Verwendung von Kallikrein (2,5 nM) und
plazentalem Bikunin (102–159)
(0–3 nM)
oder Aprotinin (0–45
nM) in 50 mM Tris-HCl (pH 8,0), 50 mM NaCl, und 0,02% Triton X-100
enthaltendem Puffer bestimmt. Nach 5 Minuten bei 37°C wurden 15 μl 20 mM PFR-AMC
hinzugefügt
und die Änderung
der Fluoreszenz wurde überwacht.
Die Inhibierung des bovinen pankreatischen Kallikreins durch plazentales
Bikunin (102–159)
und Aprotinin wurde auf ähnliche
Weise mit Kallikrein (92 pm), plazentalem Bikunin (102–159) (0–1,6 nM)
und Aprotinin (0–14
pM) und einer Substrat-Endkonzentration von 100 μM bestimmt. Die offensichtliche
Inhibierungskonstante Ki* wurde unter Verwendung
einer nicht-linearen Regression-Datenanalysenprogramm-Enzfitter-Software
(Biosoft, Cambridge, UK) bestimmt: die kinetischen Daten von jedem
Experiment wurde in Form der Gleichung für einen festbindenden Inhibitor
analysiert: Vi/Vo = 1 – (Eo + Io + Ki* – [(Eo + Io + Ki*)2 – 4EoIo]1/2)2Eo (2)
-
Wobei
Vi/Vo die fraktionale
Enzymaktivität
ist (inhibierte vs. nicht-inhibierte Rate), und Eo und
Io sind die Gesamtkonzentrationen des Enzyms
bzw. Inhibitors. Ki-Werte wurden durch Korrigieren
für die
Wirkung des Substrats gemäß der Gleichung: Ki =
Ki*/(1 + [So]/Km) (3) erhalten
(Boudier, C., and Bieth, J.G. (1989) Biochim Biophys Acta, 995:
36–41).
-
Für die Inhibierung
der humanen neutrophilen Elastase durch plazentales Bikunin (102–159) und Aprotinin
wurde Elastase (19 nM) mit plazentalem Bikunin (102–159) (150
nM) oder Aprotinin (0–7,5 μM) in 0,1 M
Tris-HCl (pH 8,0) und 0,05% Triton X-100) enthaltendem Puffer inkubiert.
Nach 5 Minuten bei 37°C
wurde AAPM-AMC (500 μM
oder 1000 μM)
hinzugefügt
und die Fluoreszenz über
eine zwei-minütige
Periode gemessen. Ki-Werte wurden von Dixon-Plots der Form 1/V
versus [I], durchgeführt
bei zwei unterschiedlichen Substrat-Konzentrationen, bestimmt (Dixon et
al., 1979).
-
Die
Inhibierung des humanen Gewebe-Kallikreins durch Aprotinin, plazentales
Bikunin Fragment (7–64)
oder plazentales Bikunin Fragment (102–159) wurde durch die Inkubation
von 0,35 nM humanes Gewebe-Kallikrein mit plazentalem Bikunin (7–64) (0–40 nM)
oder plazentalem Bikunin (102–159)
(0–2,5
nM) oder Aprotinin (0–0,5
nM) in einem 1 ml Reaktionsvolumen gemessen, welches 50 mM Tris-HCl-Puffer
pH 9,0, 80 mM NaCl, und 0,1 % Trition X-100 enthält. Nach 5 Minuten bei 37°C wurde 5μl 2 mM PFR-AMC
hinzugefügt, um
einer Endkonzentration von 10 nM zu erreichen, und die Änderung
der Fluoreszenz wurde überwacht.
Der Km für
PFR-AMC mit humanem Gewebe-Kallikrein under den eingesetzten Bedingungen
war 5,7 μM.
Die Inhibierung des humanen Faktors Xa (American Diagnositica, Inc,
Greenwich, CT) durch synthetisches plazentales Bikunin (102–159) rekombinantes
plazentales Bikunin, und Aprotinin wurde durch die Inkubation von
0,87 nM humanem Faktor Xa mit steigenden Mengen an Inhibitor in
20 mm Tris (pH 7,5), 0,1 M NaCl und 0,1 % BSA enthaltendem Puffer
gemessen. Nach 5 Minuten bei 37°C
wurde 30 μl
20 mM LGR-AMC (Sigma) hinzugefügt
und die Änderung
der Fluoreszenz überwacht.
Die Inhibierung der humanen Urokinase (Sigma) durch Kunitz-Inibitoren
wurde durch die Inkubation der Urokinase (2,7 ng) mit Inhibitoren
in einem Gesamtvolumen von 1 ml Puffer, der 50 mM Tris-HCl (PH 8,0)),
50 mM NaCl, und 0,1 % Triton X-100 enthält, gemessen. Nach 5 Minuten
bei 37°C
wurden 35 μl
20 mM GGR-AMC (Sigma) hinzugefügt
und die Änderung
der Fluoreszenz überwacht.
Die Inhibierung von Faktor XIa (von Enzyme Research Labs, Southbend,
IN) wurde durch Inkubieren von FXIa (0,1 nM) mit entweder 0 bis
800 nM plazentalem Bikunin (7–64),
0 bis 140 nM plazentalem Bikunin (102–159) oder 0 bis 40 μM Aprotinin
in 50 mM Hepes pH 7,5, 100 mM NaCl, 2 mM CaCl2,
0,01% Trition X-100, und 1% BSA in einem Gesamtvolumen von 1 ml
gemessen. Nach 5 Minuten bei 37°C
wurde 10 ul 40 mM Boc-Glu(OBzI)-Ala-Arg-AMC (Bachem Biosciences,
King of Prussia, PA) hinzugefügt
und die Änderung der
Fluoreszenz überwacht.
-
Ergebnisse:
Ein direkter Vergleich der Inhibierungsprofile von plazentalem Bikunin
(102–159)
und Aprotinin wurde durch Messen ihrer Inhibierungskonstanten mit
verschiedenen Proteasen unter identischen Bedingungen durchgeführt. Die
Ki-Werte sind in Tabelle 3, unten, aufgelistet.
-
Tabelle
3 K
i-Werte für
die Inhibierung von verschiedenen Proteasen durch Bikunin (102–159)
-
Plazentales
Bikunin (102–159)
und Aprotinin inhibieren bovines Trypsin und humanes Plamin unter den
angewendeten Bedingungen zu einem vergleichbaren Ausmass. Aprotinin
inhibierte Elastase mit einem Ki von 8,5 μM. Plazentales Bikunin (102–159) inhibierte
Elastase mit einem Ki von 323 nM. Der Ki-Wert für die plazentale Bikunin (102–159) Inhibierung
des bovinen pankreatischen Kallikreins war 20fach höher als
der der Aprotinin-Inhibierung. Im Gegensatz dazu ist plazentales
Bikunin (102–159)
ein wirksamerer Inhibitor des humanen Plasma Kallikreins als Aprotinin
und bindet mit einer 56fach höheren
Affinität.
-
Da
plazentales Bikunin (102–159)
mehr als 50-mal wirksamer ist als Trasylol® als
ein Inhibitor von Kallikrein ist, werden geringere Mengen an humanem
plazentalem Bikunin, oder Fragmenten davon (d.h. plazentales Bikunin
(102–159))
als Trasylol® benötigt, um
die wirksame Patientendosis des Inhibitors in KIU aufrechtzuerhalten.
Dies reduziert die Kosten pro Dosis des Arzneimittels und reduziert
die Wahrscheinlichkeit von unerwünschten
nephrotoxischen Wirkungen bei Re-Exposition des Patienten mit dem
Medikament. Weiters ist das Protein von Menschen abgeleitet, und
somit in Menschen viel weniger immunogen als Aprotinin, welches von
Kühen abgeleitet
ist. Dies führt
zu einer signifikanten Abnahme des Risikos des Zuziehens unerwünschter immunologischer
Vorgänge
bei der Re-Exposition des Patienten mit dem Medikament.
-
Beispiel 4 – Kein Beispiel
der Erfindung
-
In vitro Spezifität von funktionellem
humanen plazentalem Bikunin-Fragment (7–64)
-
Die
in vitro Spezifität
von funktionellem plazentalem Bikunin (7–64) wurde unter Verwendung
der in den oberen Beispielen verwendeten Materialien und Verfahren
bestimmt.
-
Ergebnisse:
Die Tabelle unten zeigt die Wirksamkeit von plazentalem Bikunin
(7–64)
als ein Inhibitor von verschiedenen Serin-Proteasen in vitro. Die
Daten werden im Vergleich zu Daten gezeigt, die für das Screenen
der Inhibierung mit entweder plazentalem Bikunin (102–159) oder
Aprotinin (Trasylol®) erhalten wurden.
-
Tabelle
4A Ki-Werte
für die
Inhibierung von verschiedenen Proteasen durch Bikunin (7–64)
-
Die
Ergebnisse zeigen, dass die das plazentale Bikunin (7–64) kodierende
Aminosäure-Sequenz rückgefaltet
werden kann, um einen aktiven Serin-Protease-Inhibitor zu erhalten,
der zumindest gegen vier Trypsin-artige Serin-Proteasen wirksam
ist.
-
Tabelle
4B, unten, zeigt auch die Wirksamkeit des rückgefaltenen plazentalen Bikunins
(7–64)
als ein Inhibitor verschiedener Serin-Proteasen in vitro. Rückgefaltetes
plazentales Bikunin (7–64)
wurde aus Protein hergestellt, das sicherlich vor der Reinigung
und Rückfaltung
vollständig
schutzeliminiert wurde. Die Daten werden im Vergleich zu Daten,
die für
das Screenen der Inhibierung mit entweder plazentalem Bikunin (102–159) oder
Aprotinin (Trasylol®) erhalten wurden, gezeigt.
-
Tabelle
4B Ki-Werte
für die
Inhibierung von verschiedenen Proteasen durch rückgefaltenes Bikunin (7–64)
-
Überraschenderweise,
war plazentales Bikunin (7–64)
wirksamer als Aprotinin bei der Inhibierung des humanen Plasma Kallikreins,
und zumindest ähnlich
wirksam wie ein Plasmin Inhibitor. Diese Daten zeigen, dass plazentales
Bikunin (7–64)
zumindest so wirksam ist wie Aprotinin unter Verwendung von in vitro
Untersuchungen, und man könnte
erwarten, dass die Wirksamkeit in vivo besser oder ähnlich ist.
-
Beispiel 5 – Kein Beispiel
der Erfindung
-
Expression der plazentalem
Bikunin-Variante (102–159)
in Hefe
-
Die
DNA Sequenz, welche das plazentale Bikunin (102–159) kodiert (SEQ ID Nr: 6)
wurde unter Verwendung synthetischer Oligonukleotide hergestellt.
Das endgültige
DNA Produkt bestand (5' zu
3') aus 15 Nukleotiden
der Hefe α-Paarungsfaktor-Propeptid-Sequenz,
welches mit der im-Rahmen cDNA-Sequenz fusioniert ist, die das plazentale
Bikunin (102–159)
kodiert, gefolgt von einem im-Rahmen Stoppkodon. Bei der Klonierung
in einen Hefe-Expressionsvektor pS604 würde die cDNA die Expression
eines Fusionsproteins steuern, welches ein N-terminales Hefe-α-Paarungsfaktor-Propeptid,
das mit der 58 Aminosäure-Sequenz
des plazentalen Bikunins (102–159)
fusioniert ist, aufweist. Die Bearbeitung dieses Fusionsproteins
an der KEX-2- Spaltstelle
and der Verbindungsstelle zwischen dem α-Paarungsfaktor und der Kunitz-Domäne war so ausgelegt,
das die Kunitz-Domäne
an ihrem nativen N-Terminus freigesetzt wird.
-
Ein
5' Sinn-Oligonukleotid,
welches eine HindIII-Stelle für
das Klonieren enthält,
der folgenden Sequenz wurde synthetisiert:
-
Ein
3' Gegensinn-Oligonukleotid,
welches sowohl eine BamHI-Stelle für das Klonieren als auch ein Stoppkodon
enthält,
der folgenden Sequenz wurde synthetisiert:
-
Die
Oligonukleotide wurden in 1 mM EDTA enthaltendem 10 mM Tris-Puffer
pH 8,0 aufgelöst
und 12 ug von jedem Oligo wurden vereinigt hinzugefügt und in
0,25 M NaCl eingebracht. Um zu hybridisieren, wurden die Oligonukleotide
durch Kochen für
5 Minuten denaturiert und es wurde ihnen ermöglicht von 65°C auf Raumtemperatur über 2 Stunden
abzukühlen.
Die Überhänge wurden
unter Verwendung des Klenow-Fragments erweitert und mit HindIII
und BamHI verdaut. Das resultierende, verdaute doppelsträngige Fragment wurde
in pUC19 kloniert und die Sequenz bestätigt. Ein Klon, der das Fragment
der korrekten Sequenz enthält wurde
mit BamHI/HindIII verdaut, um das Bikunin enthaltende Fragment mit
der folgenden +Strang-Sequenz freizusetzen:

welches
dann Gel-gereinigt und in BamHI/HindIII geschnittenen pS604 ligiert
wurde. Die Ligationsmischung wurde in Phenol/Chloroform extrahiert
und über
eine S-200 Minispin-Säule gereinigt.
Das Ligationsprodukt wurde in die Hefestämme SC101 und WHL341 gerichtet
transformiert und auf ura-Selektionsplatten ausplattiert. Zwölf Kolonien
von jedem Stamm wurden auf ura-Dropout-Platten erneut ausgestrichen.
Eine Einzelkolonie wurde in 2 ml ura DO-Medium inokuliert und über Nacht
bei 30°C
kultiviert. Die Zellen wurden für
2 Minuten bei 14000 × g
pelletiert und die Überstände auf
ihren Inhalt an plazentalem Bikunin (102–159) evaluiert.
-
Nachweis der Expression
von plazentalem Bikunin (102–159)
in transformierter Hefe
-
Erstens
wurden die Überstände (50 μl pro Untersuchung)
auf ihre Fähigkeit
evaluiert, die in vitro Aktivität
von Trypsin unter Verwendung der in Beispiel 1 beschriebenen Untersuchungsverfahren
zu inhibieren (1 ml Untersuchungsvolumen). Eine reine unbenutzte
Mediumprobe sowie ein Hefeklon, welcher eine inaktive Aprotinin-Variante
exprimiert, dienten als Negativkontrollen. Ein Hefeklon, welcher
natürliches
Aprotinin exprimiert, diente als Positivkontrolle und wird für Vergleichszwecke
gezeigt.
-
Das
zweite Verfahren, um die plazentale Bikunin (102–159) Expression zu quantifizieren,
nutzte die Verwendung von polyklonalen Antikörpern (pAbs) gegen das synthetische
Peptid aus, um die Akkumulation des rekombinanten Peptids mittels
Western-Blots zu überwachen.
Diese Studien wurden nur mit Rekombinanten durchgeführt, die
von Stamm SC101 abgeleitet sind, da diese eine höhere inhibitorische Aktivität als aus Stamm
WHL341 abgeleitete Rekombinanten erzeugen.
-
Um
die pAb herzustellen, wurde zwei 6–8 Wochen alte weiße weibliche
Neuseeland-Kaninchen
(Hazelton Research Labs, Denver, Pa) am Tag null mit 250 ug gereinigtem,
reduziertem, synthetischem plazentalem Bikunin (102–159) in
vollständigem
Freund's-Adjuvans
immunisiert, gefolgt von Auffrischungen an den Tagen 14, 35, und
56 und 77 mit jeweils 125 ug desselben Antigens in unvollständigem Freund's Adjuvans. Das in
den vorliegenden Studien verwendete Antiserum wurde nach der dritten
Auffrischung durch etablierte Verfahren gesammelt. Die polyklonalen
Antikörper
wurden aus dem Antiserum über
Protein A gereinigt.
-
Die
Kolonien 2.4 und 2.5 von der Hefe SC101-Transformation (8)
sowie eine Aprotinin-Kontrolle wurden über Nacht in 50 ml ura-DO-Medium
bei 30°C
gezüchtet.
Die Zellen wurden pelletiert und der Überstand mittels eines Centriprep
3 (Amicon, Beverly, MA) Konzentrators 100fach konzentriert. Von
jedem wurden Proben (30 μl)
wurde einer SDS-PAGE auf 10–20%
Tricin-gepufferten Gelen (Novex, San Diego, CA) unter Verwendung
der Herstellerverfahren unterzogen. Duplikatgele wurden entweder
mit einem Silberfärbungssatz (Integrated
Separation Systems, Nantick, MA) entwickelt oder auf eine Nitrocellulose überführt und
mit dem gereinigten polyklonalen Antikörper, welcher gegen synthetisches
Bikunin (102–159)
abgeleitet wurde, entwickelt. Alkalische Phosphatase konjugierter
Ziegen Anti-Kaninchen Antikörper
wurde als sekundärer
Antikörper gemäß den Herstellervorschriften
verwendet (Kirkegaard and Perry, Gaithersburg, MD).
-
Reinigung von plazentalem
Bikunin (102–159)
aus einem transformierten SC101-Stamm
-
Die
Fermentationsbrühe
einer 1L-Kultur des SC101 Stammes 2.4 wurde durch Zentrifugation
(4.000g × 30
Minuten) geerntet, dann auf eine 1,0 ml Anhydrochymotrypsin-Sepharose-Säule (Takara
Biochemical, Inc., CA) aufgetragen, die vorher mit 0,1 M NaCl, 2
mM CaCl2 und 0,01 % (Vol.-%/Vol-%) Triton X-100) enthaltendem 50
mM Hepes-Puffer, pH 7,5, äquilibriert
wurde. Die Säule
wurde mit demselben, jedoch 1,0 M NaCl enthaltenden Puffer gewaschen,
bis die A280 nm auf Null abgefallen ist, woraufhin die Säule mit
0,1 M Ameisensäure,
pH 2,5, eluiert wurde. Die eluierten Fraktion wurden zusammengefasst
und auf eine C 18 Säule
(Vydac, 5 um, 4,6 × 250
mm) aufgetragen, die vorher mit 0,1 % TFA äquilibriert wurde, und mit
einem 50 minütigen, linearen
Gradienten von 20 bis 80% Acetonitirl in 0,1 % TFA eluiert. Die
das plazentale Bikunin (102–159)
enthaltenden Fraktionen wurden zusammengefasst und auf einer C18
nochmals chromatographiert, wobei eine Elution mit einem linearen
22,5 bis 50% Acetonitril-Gradienten in 0,1 % TFA verwendet wurde.
-
Ergebnisse. 8 zeigt
die prozentuelle Trypsin-Aktivität,
die durch zwölf
Kolonien inhibiert wurde, die von der Transformation von jedem der
Stämme
SC101 und WHL341 abgeleitet sind. Die Ergebnisse zeigen, dass alle
zwölf Kolonien
des Hefestamms SC101, der mit dem Trypsin-Inhibitor plazentalem
Bikunin (102–159) transformiert
sind, die Fähigkeit
besaßen,
eine beachtliche Menge der Trypsin inhibitorischen Aktivität zu erzeugen,
im Vergleich zu den Negativkontrollen, von denen beide keine Fähigkeit
zeigten, Trypsin zu inhibieren. Die Aktivität ist deshalb auf die Expression
eines spezifischen Inhibitors in den plazentalen Bikunin-Variante (102–159) transformierten
Zellen bezogen. Die Hefe WHL341-Proben enthielten eine minimale
Trypsin inhibitorische Aktivität.
Die kann mit dem für
diesen Stamm, unter den angewendeten Bedingungen, beobachteten langsamen
Wachstum korrelieren.
-
9 zeigt
die SDS-PAGE und Western-Analyse der Hefe SC101-Überstände. Silbergefärbte SDS-PAGE
der Überstände, die
aus rekombinanten, plazentales Bikunin (102–159) exprimierenden Hefen
2.4 und 2.5, sowie aus der Aprotinin-exprimierenden Hefe stammten,
ergaben eine Proteinbande, welche bei ungefähr 6 kDa verläuft, welche
der Größe entspricht,
die für
jede rekombinante Kunitz-Inhibitor-Domäne erwartet wird. Die Western- Analyse zeigte, dass
die durch die Stämme
2.4 und 2.5 exprimierten 6 kDa-Banden mit dem pAb reagierten, der
gegen das plazentale Bikunin (102–159) abgeleitet ist. Dieselbe
6 kDa Bande in der Aprotinin-Kontrolle reagierte nicht mit dem selben
Antikörper,
welches die Spezifität
des Antikörpers
für die
plazentale Bikunin-Variante (102–159) demonstriert.
-
Die
endgültige
Zubereitung der plazentalen Bikunin-C-terminalen Domäne war durch
silbergefärbte SDS-PAGE
sehr rein (10). Die Gesamtausbeute der
aus der Brüheabgeleiteten
Trypsin inhibitorischen Aktivität
in der letzten Zubereitung war 31 %. Die N-terminale Sequenzierung des gereinigten
Inhibitors zeigte, dass 40% des Proteins richtig bearbeitet ist,
um den korrekten N-Terminus für
plazentales Bikunin (102–159) zu
ergeben, während
etwa 60% des Materials einen Teil des Hefe α-Paarungsfaktors enthielt. Das
gereinigte Material wies einen aktiven Serin-Protease-Inhibitor
auf, der einen scheinbaren Ki von 0,35 nM für die in vitro Inhibierung
von Plasma Kallikrein zeigte.
-
Schließlich lieferte
die Akkumulation sowohl einer Protease Inhibitor-Aktivität als auch
eines Proteins, welches immunochemisch mit synthetischem Bikunin
(102–159)
verwandt ist, in der Fermentationsbrühe sowie die Isolierung eines
plazentalen Bikunins (102-159) aus einer der transformierten Linien,
Beweise für
die Expression von plazentalem Bikunin in hierin beschriebenen,
rekombinanten Hefestämmen,
welches für
das erste Mal die Nützlichkeit
von Hefen für
die Herstellung von plazentalen Bikunin-Fragmenten zeigt.
-
Zusätzliche
Konstrukte wurden in einer Anstrengung hergestellt, um das Expressionsniveau
der Kunitz-Domäne,
die innerhalb des plazentalen Bikunins 102–159 enthalten ist, zu erhöhen, sowie
die Ausbeute des Proteins mit dem korrekten N-Terminus zu erhöhen. Wir
nahmen an, dass die N-terminalen Reste des plazentalen Bikunins
102–159
(YEEY-) eine Spaltungstelle darstellen können, die nur schlecht durch
die Hefe KEX-2-Protease erkannt wird, die die Hefe-a-Faktor Proregion
enzymatisch entfernt. Deshalb stellten wir Hefe-Expressionskonstrukte
für die
Herstellung von plazentalem Bikunins 103–159 (N-Terminus von EEY...), 101–159 (N-Terminus von NYEEY...)
und 98–159
(DMFNYEEY...) her, um die P' Substellen,
welche die KEX-2 Spaltstelle umgeben, zu modifizieren. Um die rekombinanten
Proteinexpressionsniveaus zu erhöhen
zu versuchen, verwendeten wir auch beim Herstellen von einigen der
unten beschriebenen Konstrukte die von der Hefe bevorzugten Kodons,
im Gegensatz zu den von Säugetieren
bevorzugten Kodons. Die Konstrukte wurden im Wesentlichen wie oben
für plazentales
Bikunin 102–159
(definiert als Konstrukt Nr. 1) beschrieben hergestellt, jedoch
mit den folgenden Modifikationen:
-
Konstrukt Nr. 2: plazentales
Bikunin 103–159,
Hefe-Kodon-Verwendung
-
Ein
5' Sinn Oligonukleotid
und ein
3' Antisense Oligonukleotid
wurden
wie für
die Herstellung eines Expressionskonstruktes (Konstrukt Nr. 1, oben)
für die
Expression von plazentalem Bikunin 102–159 manipuliert.
-
Konstrukt Nr. 3 plazentales
Bikunin 101–159,
Hefe-Kodon-Verwendung
-
Ein
5' Sinn Oligonukleotid
und das
gleiche 3' Gegensinn-Oligonukleotid
wie für
Konstrukt Nr. 2 verwendet wurde, wurde wie für die Herstellung eines Expressionskonstrukts
(Konstrukt Nr. 1, oben) für
die Expression von plazentalem Bikunin 102–159 manipuliert.
-
Konstrukt Nr. 4 plazentales
Bikunin 98–159,
Hefe-Kodon-Verwendung
-
Ein
5' Sinn Oligonukleotid
und das
gleiche 3' Gegensinn-Oligonukleotid
wie für
Konstrukt Nr. 2 verwendet wurde, wurde wie für die Herstellung eines Expressionskonstrukts
(Konstrukt Nr. 1, oben) manipuliert.
-
Der
Hefestamm SC101 (MATα,
ura 3–52,
suc2) wurde mit den Plasmiden transformiert, die jede der obigen
cDNAs enthält,
und Proteine wurden unter Verwendung der Verfahren exprimiert, die
oben für
die Herstellung von plazentalem Bikunin 102–159 mit humaner Kodon-Verwendung beschrieben
wurden. Etwa 250 ml von jeder Hefe-Kultur wurden geerntet, und der Überstand
aus der Zentrifugation (15 Minuten × 3000 Umdrehungen pro Minute)
wurde einer getrennten Reinigung über 1 ml Kallikrein-Sepharose-Säulen, wie
oben beschrieben, unterzogen. Die relative Menge an Trypsin-inhibitorische
Aktivität
im Aufzutragendem, die Menge an gewonnenem gereinigten Protein und
die N-terminale Sequenz des gereinigten Proteins wurden bestimmt und
sind in Tabelle 7 unten aufgelistet.
-
Tabelle
7 Relative
Herstellungsniveaus von unterschiedlichen Proteinen, die die C-terminale
Kunitz-Domäne des plazentalen
Bikunins enthalten.
-
Die
Ergebnisse zeigen, dass plazentale Bikunin-Fragmente unterschiedlicher
Länge,
die die C-terminale Kunitz-Domäne
enthalten, eine große
Variation in der Kapazität
funktionellsezerniertes Protein zu exprimieren zeigen. Konstrukte,
die die Fragemente 101–159,
und 103–159
exprimieren ergaben kaum eine oder eine geringe enzymatische Aktivität in den Überständen vor
der Reinigung, und die N-terminale Sequenzierung von 0,05 ml Aliquoten
jeder gereinigten Fraktion ergab nicht-nachweisbare Mengen an Inhibitor.
Andererseits ergab die Expression entweder von plazentalem Bikunin
102–159
oder 98–159
signifikante Mengen an Protease Aktivität vor der Reinigung. Die N-terminale
Sequenzierung zeigte jedoch, dass das aus der Expression von 102–159 gewonnene
gereinigte Protein wieder größtenteils
inkorrekt verarbeitet war, und zeigt einen N-Terminus der mit der
Bearbeitung der Mehrheit des Präproteins
an einer Stelle innerhalb der Hefe-α-Paarungsfaktor Pro-Sequenz übereinstimmt.
Das gereinigte Protein, welches aus der Expression von plazentalem Bikunin
98–159
gewonnen wurde, war jedoch gänzlich
an der korrekten Stelle bearbeitet, um den korrekten N-Terminus
zu ergeben. Weiters wurde beinahe zweimal so viel Protein gewonnen,
wie im Vergleich zu der Gewinnung von plazentalem Bikunin 102–159. Somit
stellt plazentales Bikunin 98–159
eine bevorzugte Fragment-Länge
für die
Herstellung der C-terminalen Kunitz-Domäne des plazentalen Bikunins
durch die α-Paarungsfaktor-Prä-Pro-Sequenz/KEX-2
Bearbeitungssystem von S. Cerevisiae dar.
-
Beispiel 6
-
Alternative Verfahren
für die
Hefe-Expression
-
Das
58 Aminosäure
Peptid, welches aus dem R74593 Translationsprodukt abgeleitet ist,
kann ebenfalls von entweder dem R87894-R74593 PCR Produkt, welches
in den TA vectorTM (Invitrogen, San Diego,
CA) kloniert wurde nach der DNA Sequenzierung oder von der humanen
plazentalen cDNA PCR amplifiziert werden. Das amplifizierte DNA
Produkt wird aus 19 Nukleotiden der Hefe α-Paarungsfaktor Leitsequenz,
welche mit der R74593 Sequenz gepaart ist, die für YEEY-CFRQ (58 Reste) kodiert,
bestehen, so dass das Translationsprodukt im Rahmen ist, was ein α-Paarungsfaktor/Kunitz-Domäne Fusionsproteins
erzeugt. Die Protein-Sequenz
enthält
auch eine Kex2-Spaltstelle, welche die Kunitz-Domäne an ihrem
nativen N-Terminus
freisetzt.
-
Das
5' Sinn-Oligonukleotid,
welches eine HindIII-Stelle für
das Klonieren enthält,
wird die folgende Sequenz enthalten:
GCCAAGCTTG GATAAAAGAT
ATGAAGAAT ACTGCACCGC CAACGCA (SEQ ID Nr. 30)
-
Das
3' Gegensinn-Oligonukleotid
enthält
eine BamHI-Stelle für
das Klonieren sowie ein Stoppkodon und besitzt die folgende Sequenz:
GGGGATCCTC
ACTGCTGGCG GAAGCAGCGG AGCAT (SEQ ID Nr. 31)
-
Die
in den Hefe-Expressionsvektor zu klonierende, vollständige 260
Nukleotid-cDNA Sequenz besitzt die folgende Sequenz:
-
Nach
der PCR Amplifikation wird diese DNA mit HindIII, BamHI verdaut
und in den Hefeexpressionsvektor pMT15 (siehe US Patent 5.164.482,
zur Gänze
durch Bezugnahme aufgenommen), ebenfalls mit HindIII und BamHI verdaut,
kloniert. Der resultierende Plasmidvektor wird verwendet, um den
Hefestamm SC 106 unter Verwendung der im US Patent 5.164.482 beschriebenen
Verfahren zu transformieren. Die URA 3+ Hefe-Transformanten werden
isoliert und unter induzierenden Bedingungen kultiviert. Die Ausbeute
an rekombinanten plazentalen Bikunin-Varianten wird gemäß der Menge
an Trypsin inhibitorischer Aktivität, die in den Kulturüberständen über die
Zeit akkumulierten unter Verwendung des oben beschriebenen in vitro
Untersuchungsverfahrens bestimmt. Die Fermentationsbrühen werden
bei 9000 Umdrehungen pro Minute für 30 Minuten zentrifugiert.
Der Überstand
wird dann durch einen 0,4, dann einen 0,2 μm Filter filtriert, auf eine
Leitfähigkeit
von 7,5 ms verdünnt
und mit Zitronensäure
auf pH 3 eingestellt. Die Probe wird dann chargenweise auf eine
200 ml S-Sepharose
fast flow (Pharmacia) in 50 mM Natriumzitrat pH 3 absorbiert und
für 60
Minuten gerührt.
Das Gel wird im Anschluss daran nacheinander mit jeweils 2 L von:
50 mM Natriumzitrat pH 3,0; 50 mM Tris-HCl, pH 9,0; 20 mM HEPES
pH 6,0 gewaschen. Das gewaschene Gel wird in eine geeignete Säule übertragen
und mit einem linearen Gradient von 0 bis 1 M Natriumchlorid in
20 mM HEPES pH 6,0 eluiert. Eluierte, in vitro Trypsin inhibitorische
Aktivität
enthaltende Fraktionen werden dann zusammengefasst und weiters gereinigt
durch entweder a) Chromatographie über eine Säule von immobilisierten Anhydrotrypsin
(im Wesentlichen wie in Beispiel 2 beschrieben); durch Chromatographie über eine
Säule von
immobilisierten bovinen Kallikrein; oder c) eine Kombination von
herkömmlichen
chromatographischen Schritten, einschließlich Gel-Filtration und/oder
Anionenaustauch-Chromatographie.
-
Beispiel 7
-
Isolierung und Charakterisierung
von nativem humanem plazentalem Bikunin aus Plazenta
-
Bikunin-Protein
wurde zur scheinbaren Homogenität
aus ganzer, gefrorenen Plazenta gereinigt (Analytical Biological
Services, Inc, Wilmington, DE). Die Plazenta (740 g) wurde auf Raumtemperatur
auftauen gelassen und in 0,5 bis 1,0 cm Stücke geschnitten, auf Eis platziert
und mit 600 ml PBS Puffer gewaschen. Die Waschung wurde dekantiert
und 240 ml Plazenta-Stücke wurden
in einen Waring-Mixer platziert. Nach dem Hinzufügen von 300 ml Buffer, bestehend
aus 0,1 M Tris (pH 8,0) und 0,1 M NaCl, wurde die Mischung bei hoher
Geschwindigkeit für
2 Minuten gemixt, in ein 750,0 ml Zentrifügenröhrchen dekantiert und auf Eis
gelegt. Dies Verfahren wurde wiederholt, bis das gesamte Material
verarbeitet war. Der kombinierte Schlamm wurde bei 4500 × g für 60 Minuten
bei 4°C
zentrifugiert. Der Überstand
wurde durch ein Käsetuch
filtriert und das plazentale Bikunin mittels einer Kallikrein Affinitätssäule, welche
durch kovalentes Binden von 70 mg bovinen pankreatischen Kallikreins
(Bayer AG) an 5,0 ml CNBr-aktivierte Sepharose (Pharmacia) gemäß den Herstellerinstruktionen
hergestellt wurde, gereinigt. Das Material wurde auf die Affinitätssäule bei
einer Flussrate von 2 ml/min geladen und mit 0,1 M Tris (pH 8,0),
0,1 M NaCl gewaschen bis die Absorption bei 280 nm der Waschung
nicht länger
nachgewiesen werden konnte. Die Säule wurde weiters mit 0,1 M
Tris (pH 8,0), 0,5 M NaCl gewaschen und mit 3 Volumina 0,2 M Essigsäure, pH
4,0 eluiert. Die Fraktionen, die Kallikrein und Trypsin inhibitorische
Aktivität
(siehe unten) enthalten wurden zusammengefasst, gefroren und lyophilisiert.
Plazentales Bikunin wurde weiters durch Gelfiltrations-Chromatographie
unter Verwendung einer Superdex 75 10/30 (Pharmacia)-Säule, welche
an ein Beckman System Gold HPLC-System angeschlossen ist, gereinigt.
Kurz gesagt, die Säule
wurde in 0,1 M Tris, 0,15 M NaCl, und 0,1 % Trition X-100 bei einer
Flussrate von 0,5 ml/min äquilibriert.
Die lyophiliserte Probe wurde in 1,0 ml 0,1 M Tris, pH 8,0 wieder
hergestellt und auf die Gelfiltrationssäule in 200 μl Aliquoten injiziert. Die Fraktionen
wurden gesammelt (0,5 ml) und auf Trypsin und Kallikrein inhibitorische
Aktivität
untersucht. Aktive Fraktionen wurden zusammengefasst und der pH
der Lösung
wurde durch Zugabe von TFA auf 2,5 eingestellt. Das Material wurde
direkt auf eine Vydac C18 Umkehrphasen-Säule (5 Mikron, 0,46 × 25 cm)
aufgetragen, die in 20% Acetonitril in 0,1 % TFA äquilibriert
wurde. Die Trennung wurde unter Verwendung eines linearen Gradienten
von 20 bis 80% Acetonitril in 0,1 % TFA bei 1,0 ml/min über 50 Minuten,
nach einer anfänglichen
20 minütigen
Waschung mit 20 % Acetonitril in 0,1 %TFA erreicht. Fraktionen (1
ml) wurden gesammelt und auf Trypsin und Kallikrein inhibitorsche
Aktivität
untersucht. Inhibitorische Aktivität enthaltende Fraktionen wurde
unter Verwendung eines speed-vac Konzentrators (Savant) konzentriert
und einer N-terminalen Sequenzanalyse unterzogen.
-
Funktionelle Untersuchungen
für plazentales
Bikunin
-
Die
Identifizierung von funktionellem plazentalem Bikunin wurde durch
Messen seiner Fähigkeit
das bovine Trypsin und das humane Plasma Kallikrein zu inhibieren,
erreicht. Die Trypsin inhibitorische Aktivität wurde in Untersuchungpuffer
(50 mM Hepes, pH 7,5, 0,1 M NaCl, 2,0 mM CaCl2,
0,1 % Trition X-100) bei Raumtemperatur in einer 96-Kammer-Mikrotiterplaate
(Perkin Elmer) mittels Gly-Pro-Lys-Aminomethylcumarin als Substrat
durchgeführt.
Die Menge an durch Trypsin produziertem Cumarin wurde durch Messen
der Fluoreszenz (Ex = 370 nm, Em = 432 nm) auf einem Perkin-Elmer
LS-50B Fluorimeter, mit einem Plattenleser ausgestattet, bestimmt.
Trypsin (23 μg
in 100 μl
Puffer) wurde mit 20 μl
der zu testenden Probe vermischt und für 10 Minuten bei 25°C inkubiert.
Die Reaktion wurde durch die Zugabe von 50 μl des GPK-AMC Substrats (33 μM Ende) in
Untersuchungspuffer gestartet. Die Fluoreszenz-Intensität wurde
gemessen und die % Inhibierung für
jede Fraktion wurde bestimmt durch: % Inhibierung
= 100 × [1 – Fo/F1]wobei
Fo die Fluoreszenz des Unbekannten ist und F1 ist die Fluoreszenz
der nur Trypsin-Kontrolle.
Die Kallikrein inhibitorische Aktivität der Fraktionen wurde auf ähnliche
Weise unter Verwendung von 7,0 nM Kallikrein in Untersuchungspuffer
(50 mM Tris, pH 8,0, 50 mM NaCl, 0,1 % Triton X-100) und 66,0 μM Pro-Phe-Arg-AMC als
Substrat gemessen.
-
Bestimmung der in vitro
Spezifität
von plazentalem Bikunin
-
Die
in vitro Spezifität
von nativem humanem plazentalem Bikunin wurde unter Verwendung der
in den vorangehenden Beispielen, oben, beschriebenen Materialien
und Verfahren bestimmt. Plazentales Bikunin wurde durch Active-Site-Titration
gegen eine bekannte Trypsin-Konzentration
unter Verwendung von GPK-AMC als ein Substrat quantifiziert, um
die Fraktion des ungebundenen Trypsins zu überwachen.
-
Protein-Sequenzierung
-
Die
1 ml Fraktion (C18-29 Delaria) wurde auf einer Speed-Vac auf 300
ml Volumen reduziert, um die Menge an organischem Lösungsmittel
zu reduzieren. Die Probe wurde dann auf eine Hewlett-Packard Miniatur biphasische
Reaktionsäule
geladen und mit 1 ml 2% Trifluoressigsäure gewaschen. Die Probe wurde
dann auf einem Hewlett-Packard Modell G1005A Protein-Sequenzierungssystem
mittels Edman-Abbau sequenziert. Version 3.0 Sequenzierungsverfahren
und alle Reagenzien wurde von Hewlett-Packard geliefert. Die Sequenz wurde
für 50
Zyklen bestätigt.
-
Ergebnisse:
Plazentales Bikunin wurde zur scheinbaren Homogenität der Reihe
nach durch Kallikrein-Affinität,
Gelfiltration und Unkehrphasen-Chromatographie gereinigt (siehe
Reinigungstabelle unten)
-
Tabelle
5 Reinigungstabelle
für natives
plazentales Bikunin (1–179)
-
- a Eine Einheit ist definiert als
die Menge, die 50 % der Trypsinaktivität in einer Standarduntersuchung
inhibiert.
-
Die
Mehrheit der Kallikrein und Trypsin inhibitorischen Aktivität eluierte
von der Kallikrein-Affinitätssäule in der
pH 4,0 Elution. Eine nachfolgende Gelfiltration-Chromatographie (5) ergab
einen Peak der Kallikrein und Trypsin inhibitorischen Aktivität mit einem
Molekulargewichtsbereich von 10 bis 40 kDa, wie durch eine Standardkurve
beurteilt wurde, die erzeugt wurde, indem Molekulargewichtstandards
unter identischen Bedingungen laufen gelassen wurden. Umkehrphasen
C18 Chromatographie (6) ergab 4 inhibitorische Aktivitäts-Peaks,
wobei die wirksamste bei etwa 30 % Acetonitril eluierte. Die Aktivität, die mit
dem Peak assoziiert ist, der als erster von der C18 eluierte (Fraktion
29) zeigte eine Aminosäure-Sequenz, beginnend
mit Aminosäure
1 der vorhergesagten Aminosäuresequenz
für plazentales
Bikunin (ADRER...; SEQ ID Nr: 1), und war identisch mit der vorhergesagten
Sequenz für
die 50 Sequenzierungszyklen (unterstrichene Aminosäuren in 3).
Cystein-Reste innerhalb dieses Sequenzabschnittes waren still, wie
für das
Sequenzieren von oxidiertem Protein erwartet wird. Die Cystein-Reste
an den Aminosäure-Positionen
11 und 20 des reifen plazentalen Bikunins wurden später durch
Sequenzieren des S-pyridylethylierten Proteins identifiziert, woraufhin PTH-Pyridylethyl-Cystein
bei Zyklen 11 und 20 gefunden wurde.
-
Interessanterweise
war das Asparagin bei Aminosäure-Restnummer
30 der Sequenz (3) still, was zeigt, dass diese
Stelle wahrscheinlich glykosyliert ist. Fraktion 29 ergab eine Hauptsequenz,
welche der des plazentalen Bikunins entspricht, beginnend bei Rest
Nr. 1 (27 pmol beim 1 Zyklus) und eine Nebensequenz (2 pmol), die
auch von dem plazentalen Bikunin stammt, beginnend bei Rest 6 (SIHD...).
Dies zeigt, das die endgültige
Zubereitung, die in Fraktion 29 sequenziert wurde, hoch rein ist,
und am wahrscheinlichsten für
die Protease inhibitorische Aktivität verantwortlich sein, wird
die mit dieser Fraktion assoziiert ist (6).
-
Dementsprechend
war die endgültige
Zubereitung von plazentalem Bikunin der C18 Chromatographie äußerst rein,
basierend auf einer silbergefärbten
SDS-PAGE-Analyse (7), in welcher das Protein
mit einem scheinbaren Mr von 24 kDa auf einem 10 bis 20% Acrylamin-Tricin-Gel
(Novex, San Diego, CA) wanderte, welches mit den folgenden Molekulargewicht
Marker kalibriert wurde: Insulin (2,9 kDa); boviner Trypsin Inhibitor
(5,8 kDa); Lysozym (14,7 kDa); β-Lactoglobulin
(18,4 kDa), carbonische Anhydrase (29 kDa) und Ovalbumin (43 kDa).
Die obige Größe des plazentalen
Bikunins auf der SDS-PAGE ist übereinstimmend
mit der von der Kodiersequenz mit voller Länge vorhergesagten (4F).
-
Wie
basierend auf den oben beschriebenen N-terminalen Sequenzierungsergebnissen
erwartet wird, reagiert das gereinigte Protein mit einem Antikörper, der
von plazentalem Bikunin (7–64)
stammt, um eine Bande mit demselben Mr (12A)
zu ergeben, wie für
die gereinigte Zubereitung beobachtet wird, wie auf Gelen durch
Silberfärbung
nachgewiesen wurde (7). Wenn jedoch dieselbe Zubereitung
mit einem Antikörper
reagiert wird, der gegen synthetisches plazentales Bikunin (102–159) abgeleitet
ist, wurde keine Bande beobachtet, die dem Protein mit voller Länge entspricht.
Stattdessen wurde ein Fragment von etwa 6 kDa, welches mit synthetischem
Bikunin (102–159)
mitwandert, beobachtet. Die einfachste Interpretation dieser Ergebnisse ist,
dass die gereinigte Zubereitung in Anschluss an die Reinigung einen
Abbau durchgemacht hat, um ein N-terminales Fragment zu ergeben,
welches die N-terminale Domäne
und eine die C-terminales Fragment, welches die C-terminale Domäne enthält, aufweist.
Unter der Annahme, dass das Fragment, welches gegen Antiserum gegen
plazentales Bikunin (7–64)
reaktiv ist, frei von dem C-terminalen Ende des Proteins mit voller Länge ist,
würde die
Größe (24 kDa)
auf einen hohen Glyosylierungszustand hinweisen.
-
Tabelle
6, unten, zeigt die Wirksamkeit der in vitro Inhibierung von verschiedenen
Serin-Proteasen durch
plazentales Bikunin. Die Daten werden mit jenen verglichen, die
mit Aprotinin (Trasylol®) erhalten wurden.
-
Tabelle
6 Ki-Werte
für die
Inhibierung von verschiedenen Proteasen durch plazentales Bikunin
-
Die
Ergebnisse zeigen, dass aus natürlichen
Quellen (humane Plazenta) isoliertes plazentales Bikunin ein wirksamer
Inhibitor der Trypsin-artigen Serin-Proteasen ist.
-
Beispiel 8
-
Expressionsmuster von
plazentalem Bikunin unter verschiedenen humanen Organen und Geweben
-
Ein
Multiple Tissue Northern wurde von Clontech gekauft, welches 2 μg PolyA +
RNA aus menschlichem Herz, Hirn, Plazenta, Lunge, Leber, Skelettmuskel,
Niere, und Pankreas enthält.
Zwei unterschiedliche cDNA Sonden wurden verwendet: 1) eine Gel-gereinigte
cDNA, welche plazentales Bikunin (102–159) kodiert; 2) das 780 Basenpaar
PCR-abgeleitete cDNA (4E), welche aus einem TA Klon
durch Verdau mit EcoRI freigesetzt und Gel-gereinigt wurde. Jede
Sonde wurde mittels 32P-dCTP markiert und
ein Random-Priming-Labeling-Kit von Boehringer Mannheim Biochemicals
(Indiana) wurde dann verwendet, um mit den Multiple Tissue Northern
gemäß den Herstellerangaben
zu hybridisieren. Autoradiogramme wurde unter Verwendung eines Biomax-Films
mit einer 18 stündigen
Belichtungszeit erzeugt, und mittels eines Umax-Scanners entwickelt und mittels Adobe
Photoshop gescannt.
-
Ergebnisse:
Das unter Verwendung einer plazentalen Bikunin (102–159) Sonde
(11A) oder einer größeren Sonde,
welche beide Kunitz-Domänen
des placentalen Bikuin (11B) enthält, beobachtete
Gewebeexpressionsmuster war im Wesentlichen das Gleiche wie man
erwarten könnte.
Die plazentale Bikunin mRNA war am meisten in Pankreas und Plazenta
vorhanden. Signifikante Konzentrationen wurden auch in Lunge, Hirn
und Nieren beobachtet, während
niedere Konzentrationen in Herz und Leber beobachtet wurden, und
die mRNA war im Skelettmuskel nicht nachweisbar. Die Transkriptgröße war in
allen Fällen
1,95 Kilobasen, welches in enger Übereinstimmung mit der vorhergesagten
Größe von plazentalem
Bikunin war, welche sowohl von der EST Überlappung als auch der Klonierung
von cDNA mit voller Länge,
wie in vorangehenden Abschnitten beschrieben, gefolgert wurde.
-
Die
ausgedehnte Gewebe-Verteilung der mRNA zeigt, dass plazentales Bikuin
weitverbreitet exprimiert wird. Da das Protein auch eine Leitsequenz
enthält
würde es
eine ausgiebige Exposition zu dem menschlichen Immunsystem haben,
welches erfordert, dass es als Selbst-Protein erkannt wird. Zusätzliche
Beweise für
eine ausgedehnte Gewebsverteilung der plazentalen Bikunin mRNA-Expression
wurde von der Tatsache abgeleitet, dass einige der EST Einträge mit Homologie
zu plazentalem Bikunin (4B) von
menschlichem Erwachsenen- und
Kinderhirn, und menschlicher Retina, Brust, Eierstock, olfaktorischem
Epithelium und Plazenta abgeleitet wurden. Es wird deshalb daraus
geschlossen, dass es unwahrscheinlich sein würde, dass die Verabreichung
des nativen humanen Proteins an humane Patienten, eine Immunreaktion
auslöst.
-
Interessanterweise
erinnerte das Expressionsmuster von plazentalem Bikunin ein wenig
an jenes für das
bovine Aprotinin, welches in hohen Konzentrationen in boviner Lunge
und Pankreas gefunden wird. Um das Expressionsmuster des plazentalen
Bikunins weiter aufzuklären,
wurde die RT-PCR von der Gesamt-RNA aus den folgenden menschlichen
Zellen bestimmt: nicht-stimulierte humane Nabelvenen Endothelzellen
(HUVECs), HK-2 (Linie, die von proximalen Nierentubuli abstammt),
TF-1 (Erythroleukämie-Line)
und Phorbolester (PMA)-stimulierte
humane periphere Blutleukozyten. Die verwendeten Sonden:
CACCTGATCGCGAGACCCC
(Sinn; SEQ ID Nr: 59);
CTGGCGGAAGCAGCGGAGCATGC (Gegensinn;
SEQ ID Nr: 60);
wurden konstruiert, um ein 600 bp plazentales
Bikunin kodierendes cDNA Fragment zu amplifizieren. Vergleiche wurden
durch Einbeziehung von Actin-Primer normalisiert, um ein 800pb Actinfragment
zu amplifzieren. Während
das 800 bp Fragment, welches auf Agarose-Gelen mit Ethidiumbromid identifziert
wurde, in allen Bahnen die gleiche Intensität hatte, war das 600 bp plazentale
Bikunin-Fragment in den HUVECs abwesend, aber in jeder anderen Zelllinie
in signifikanten Mengen vorhanden. Wir schließen daraus, dass plazentales
Bikunin zumindest in einigen Endothelzellen nicht exprimiert wird,
jedoch in einigen Leukozyt-Populationen
exprimiert wird.
-
Beispiel 9
-
Reinigung und Eigenschaften
von plazentalem Bikunin (1–170),
welches aus einem Baculovirus/Sf9 Expressionsystem hoch gereinigt
wurde.
-
Ein
großes
plazentales Bikunin-Fragment, welches beide Kunitz-Domänen (plazentales
Bikunin 1–170)
enthält,
wurde in Sf9 Zellen wie folgt exprimiert. Plazentale Bikunin-cDNA,
durch PCR erhalten (4E) und innerhalb eines TA-Vektors
(sie voriges Beispiel) enthalten, wurde durch Verdau mit HindIII
und Xba 1 freigestetzt, welcher ein Fragment ergab, welches durch
eine 5' XbaI-Stelle
und eine 3' HindIII-Stelle flankiert
ist. Dieses Fragment wurde Gelgereinigt und dann in den M13mp19
Vektor (New England Biolabs, Beverly, MA) koniert. In vitro Mutagenese
(Kunkel T.A., (1985) Proc. Natl. Acad. Sci. USA, 82: 488–492) wurde verwendet,
um eine Pst1 Stelle 3' zu
der XbaI Stelle am 5' Ende,
aber 5' zu der Sequenz,
welche die ATG Startstelle, das natürliche plazentale Bikunin-Signalpeptid
und die reife plazentale Bikunin-Kodiersequenz kodiert, zu erzeugen.
Das für
die Mutagenese verwendete Oligonukleotid hatte die Sequenz:
5' CGC GTC TCG GCT
GAC CTG GCC CTG CAG ATG GCG CAC GTG TGC GGG 3' (SEQ ID Nr. 61)
-
Ein
Stoppkodon (TAG) und BglII/XmaI Stelle wurde auf ähnliche
Weise am 3' Ende
der cDNA erzeugt, unter Verwendung des Oligonukleotids:
5' CTG CCC CTT GGC
TCA AAG TAG GAA GAT CTT CCC CCC GGG GGG GTG GTT CTG GCG GGG CTG 3' (SEQ ID Nr. 62)
-
Das
Stoppkodon war im Rahmen mit der das plazentale Bikunin kodierenden
Sequenz und verursachte die Termination unmittelbar nach dem Lysin
beim Aminosäure-Rest
170, und kodiert somit ein verkürztes plazentales
Bikunin-Fragment, welchem die mutaßliche Transmembrandomäne fehlt.
Das Produkt der Verdauung mit Pst1 und BglII wurde isoliert und
in den BacPac8 Vektor für
die Expression des plazentalen Bikunin-Fragments (1–170) kloniert,
welches beide Kunitz-Domänen
enthält,
aber welches unmittelbar N-termial zu dem mutmaßlichen Transmembransegment
verkürzt
ist.
-
Die
Expression von Bikunin durch Sf-9 Insektenzellen war bei einer Infektionsdosis
von 1 zu 1 optimal, wenn das Medium 72 Stunden nach der Infektion
geerntet wurde. Nach der Ernte wurde er Baculovirus-Zellkultur-Überstand
(2L) durch die Zugabe von Tris-HCl auf pH 8,0 eingestellt. Bikunin
wurde durch Chromatographie mittels einer 5 ml bovinen pankreatischen
Kallikrein-Affinitätssäule, wie
vorher in Beispiel 7 für
die Reinigung von nativem plazentalem Bikunin aus Plazenta beschrieben
wurde, gereinigt. Eluiertes Material wurde mit TFA auf pH 2,5 eingestellt
und einer Chromatographie auf einer C18 Umkehrphasen-Säule (1,0 × 25 cm), welche
in 10 % Acetonitril in 0,1 % TFA bei einer Flussrate von 1 ml/min äquilibriert
wurde, unterzogen. Das Bikunin wurde mit einem linearen Gradienten
von 10 bis 80 % Acetonitril in 0,1 % TFA über 40 Minuten eluiert. Aktive
Fraktionen wurden zusammengefasst, lyophilisiert, in 50 mM Hepes
(pH 7,5), 0,1 M NaCl, 2 mM CaCl2 und 0,1%
Trition X-100 wieder aufgelöst,
und bei –20°C bis zum
Gebrauch gelagert. Die Konzentration an rekombinanten Bikunin wurde
durch Aminosäure-Analyse
bestimmt.
-
Ergebnisse:
Rekombinantes Bikunin wurde aus Baculovirus-Zellkultur-Überstand
unter Verwendung eines 2-stufigen Reinigungsprotokolls, wie unten
gezeigt, gereinigt, um einen aktiven Trypsin-Inhibitor zu ergeben
(Tabelle 8 unten).
-
Tabelle
8 Reinigung
von rekombinantem Bikunin aus transformierten Kultur-Überstand
-
Chromatographie
des Rohmaterials über
eine immobilisierte bovine pankreatische Kallikrein-Affinitätssäule isolierte
selektiv 0,013% des Proteins und 0,67% der vorhandenen Trypsin inhibitorischen
Aktivität.
Die Mehrheit der in Ausgangsüberstand
vorhandenen Trypsin inhibitorischen Aktivität band nicht an das immobilisierte
Kallikrein und ist nicht mit Bikunin verwandt (Ergebnisse nicht
gezeigt). Die nachfolgende Chromatographie mittels einer C 18 Umkehrphase
ergab eine weitere 5fache Reinigung, mit einer Ausbeute von 0,2%.
Die endgültige
Zubereitung war durch SDS-PAGE (13) äußerst rein,
zeigte ein Mr von 21,3 kDa, und reagierte auf Immunoblots mit Kaninchen-Anti-plazentalem
Bikunin 102–159
(nicht gezeigt). Die N-terminale Sequenzierung (26 Zyklen) ergab
die erwartete Sequenz für
reifes plazentales Bikunin (4F),
beginnend bei Rest +1 (ADRER...), was zeigt, dass das Signalpeptid
in Sf9 Zellen richtig bearbeitet wurde.
-
Gereinigtes
plazentales Bikunin aus Sf9-Zellen (100 pmol) war pyridylethylalkyliert,
CNBr verdaut, und dann ohne Auflösung
der resuliertenden Fragmente sequenziert. Die Sequenzierung für 20 Zyklen
ergab die folgenden N-Terminii:
-
Somit
wurden N-Termini gewonnen, die jedem der vier erwarteten Fragmente
entsprechen. Dies bestätigt,
dass das Sf9 exprimierte Protein, die gesamte Ektodomän-Sequenz
von plazentalem Bikunin (1–170) enthielt.
Die N-terminale Sequenzierung (50 Zyklen) einer zusätzlichen
Probe von unverdautem plazentalem Bikunin (1–170) führte zu einer Aminosäure-Sequenz,
die bei Zyklus 30 frei von jeglicher PTH-Aminosäure (PTH-Asparagin wurde erwartet)
war. Ein ähnliches
Ergebnis wurde bei der Sequenzierung des natürlichen Proteins aus humaner
Plazenta (Beispiel 7) erhalten und ist damit übereinstimmend, dass dieser
Rest glykosyliert ist, wie aus der Aminosäuresequenz vorhergesagt wurde,
die diesen Asparagin-Rest umgibt. Weiters waren die Cysteinreste
innerhalb dieser Region auch still, was übereinstimmend war mit ihrer
Teilnahme an Disulfidbindungen.
-
Beispiel 10
-
Inhibierungsspezifität von aus
Sf9 Zellen abgeleitetem, gereinigtem plazentalem Bikunin
-
Die
in vitro Spezifität
von rekombinantem Bikunin wurde unter Verwendung der in den Beispielen
3, 4 und 7 beschriebenen Materialien und Verfahren bestimmt. Zusätzlich wurde
die Inhibierung von humanem Gewebekallikrein durch Bikunin durch
die Inkubierung von 0,35 nM humanem Gewebekallikein mit rekombinantem
Bikunin in 50 mM Tris (pH 9,0), 50 mM NaCl, und 0,01% Triton X-100
enthaltendem Puffer, gemessen. Nach 5 Minuten bei 37°C wurde 5μl 2 mM PFR-AMC
hinzugefügt
und die Änderung
der Fluoreszenz überwacht.
-
Die
Inhibierung von Gewebe Plasminogen-Aktivator (tPA) wurde auch bestimmt
wie folgt: tPA (Einzelkettenform aus menschicher Melanom-Zellkultur
von Sigma Chemical Co, St. Louis, MO) wurde mit Inhibitor für 2 Stunden
bei Raumtemperatur in 150 mM NaCl, und 0,02% Natriumazid enthaltendem
20 mM Tris-Puffer, pH 7,2 vorinkubiert. Die Reaktionen wurde anschließend durch
den Transfer in ein Reaktionssystem ausgelöst, welches die folgenden anfänglichen
Komponentenkonzentrationen aufweist: tPA (7,5 nM), Inhibitor 0 bis
6,6 mM, DIle-Lpro-Larg-pNitroanilin (1 mM) in 28 mM Tris-Puffer,
pH 8,5, der 0,004 % (Vol-%/Vol-%) Trition X-100 und 0,005 % (Vol-%/Vol-%)
Natriumazid enthält.
Die Bildung von p-Nitroanilin wurde im Anschluß an eine Inkubation für 2 Stunden
bei 37°C
durch Messen der A405nm bestimmt.
-
Die
Tabelle, unten, zeigt die Wirksamkeit des rekombinanten Bikunins
als ein Inhibitor von verschiedenen Serin-Proteasen in vitro. Die
Daten werden im Vergleich zu Daten gezeigt, die für das Screenen
der Inhibierung mit entweder rekombinantem Bikunin oder Aprotinin
erhalten wurden.
-
Tabelle
9 Vergleich
der Ki-Werte für
die Inhibierung von verschiedenen Proteasen durch rekominantes plazentales
Bikunin (1–170)
oder Aprotintin
-
Diese
Ergebnisse zeigen, dass rekombinantes Bikunin in Insektenzellen
exprimiert werden kann, um einen aktiven Protease-Inhibitor zu ergeben,
der wirksam gegen zumindest fünf
unterschiedliche Serin-Protease-Inhibitoren ist. Rekombinantes Bikunin
war wirksamer als Aprotinin gegenüber humanem Plasma-Kallikrein,
Trypsin und Plasmin. Überraschenderweise
war das rekombinante Bikunin wirksamer als die synthetisch hergeleiteten
Bikunin-Fragmente (7–64)
und (102–159)
gegenüber
allen getesteten Enyzmen. Diese Daten zeigen, dass rekombinantes
Bikunin wirksamer ist als Aprotinin, unter Verwendung von in vitro
Untersuchungen, und dass man eine besser in vivo Wirksamkeit erwarten
würde.
-
Außer dem
Messen der Wirksamkeiten gegen spezifische Proteasen, wurde die
Kapazität
von plazentalem Bikunin (1–170),
die aktivierte partielle Thromboplastinzeit (APTT) zu verlängern, evaluiert
und mit der mit Aprotinin-assoziierten Aktivität verglichen. Der Inhibitor
wurde in 150 mM NaCl, und 0,02% Natriumazid enthaltendem 20 mM Tris-Puffer,
pH 7,2 verdünnt
und zu einer Küvette
hinzugefügt
(0,1 ml), welche innerhalb eines MLA ElectraR 800
automatischen Gerinnungsmesser-Koagulometers (Medical Laboratory
Automation, Inc, Pleasantville, N.Y) enthalten ist. Das Instrument
wurde auf den APTT-Modus mit einer Aktivierungszeit von 300s und
den Duplikatmodus gestellt. Im Anschluß an die Zugabe von 0,1 ml
Plasma (speziell untersuchtes Referenzplasma Los 1-6-5185, Helena
Laboratories, Beaumont, TX), wurden das APTT Reagens (Automated APTT-Los
102345, von Organon Teknika Corp. Durhan, NC) und 25 mM CaCl2 automatisch verabreicht, um die Gerinnung
auszulösen,
und die Gerinnungszeit wurde automatisch überwacht. Die Ergebnisse (14) zeigten, dass eine Verdopplung der Gerinnungszeit
etwa 2 μM
Endkonzentration Aprotinin, jedoch nur 0,3 μM Sf9-abgeleitetes plazentales
Bikunin erfordert. Diese Daten zeigen, dass plazentales Bikunin
ein wirksames Antikoagulans ist, und als Medikament für Krankheiten
nützlich
ist, bei denen die pathologische Aktivierung der intrinischen Gerinnungskaskade
eingebunden ist.
-
Obwohl
bestimmte Ausführungsformen
der Erfindung im Detail für
Illustrationszwecke beschrieben wurden, ist es für den Fachman leicht erkennbar,
das die hierin beschriebenen Verfahren und Formulierungen, ohne
vom Wesen und Umfang der Erfindung abzuweichen, modifiziert werden
können.
Dementsprechend ist die Erfindung nicht beschränkt, außer durch die angeschlossenen
Ansprüche.







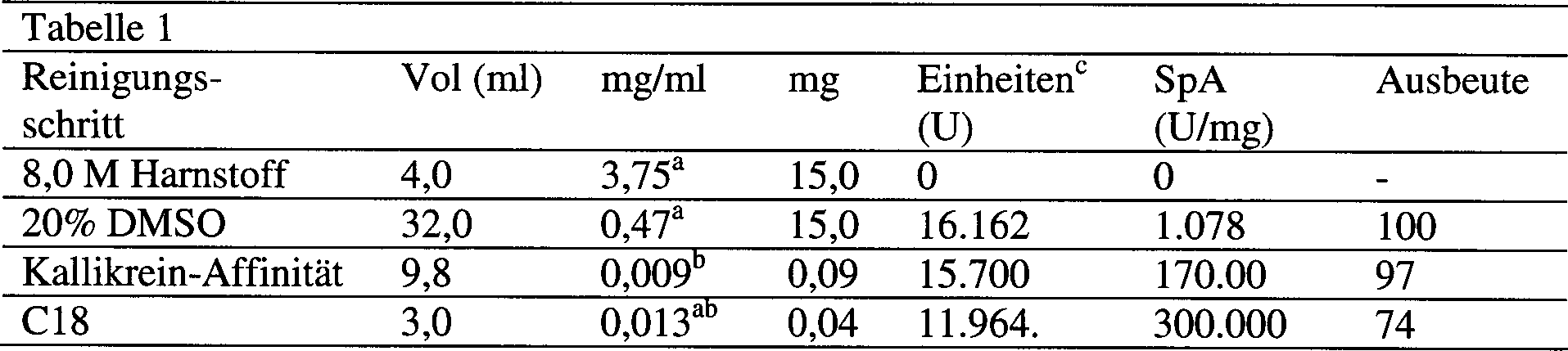







 wurden wie für die Herstellung eines Expressionskonstruktes (Konstrukt Nr. 1, oben) für die Expression von plazentalem Bikunin 102–159 manipuliert.
wurden wie für die Herstellung eines Expressionskonstruktes (Konstrukt Nr. 1, oben) für die Expression von plazentalem Bikunin 102–159 manipuliert.

















 für die Herstellung eines Medikaments für die Behandlung von Hirnödem, Rückenmarködem, Multiple Sklerose, Ischämie, Perioperativer Blutverlust, Sepsis, Septischer Schock, Fibrose, pathologische Blutkoagulation oder Gerinnung, Schlaganfall, zerebrale oder subarachnoidale Hämorrhagie, Entzündung des Gehirns, Entzündung des Rückenmarks, zerebrale Infektion, zerebrale Granulomatose, spinale Infektion, spinale Granulomatose, gastrischer Krebs, zervikaler Krebs oder zur Prävention von Metastasen.
für die Herstellung eines Medikaments für die Behandlung von Hirnödem, Rückenmarködem, Multiple Sklerose, Ischämie, Perioperativer Blutverlust, Sepsis, Septischer Schock, Fibrose, pathologische Blutkoagulation oder Gerinnung, Schlaganfall, zerebrale oder subarachnoidale Hämorrhagie, Entzündung des Gehirns, Entzündung des Rückenmarks, zerebrale Infektion, zerebrale Granulomatose, spinale Infektion, spinale Granulomatose, gastrischer Krebs, zervikaler Krebs oder zur Prävention von Metastasen.