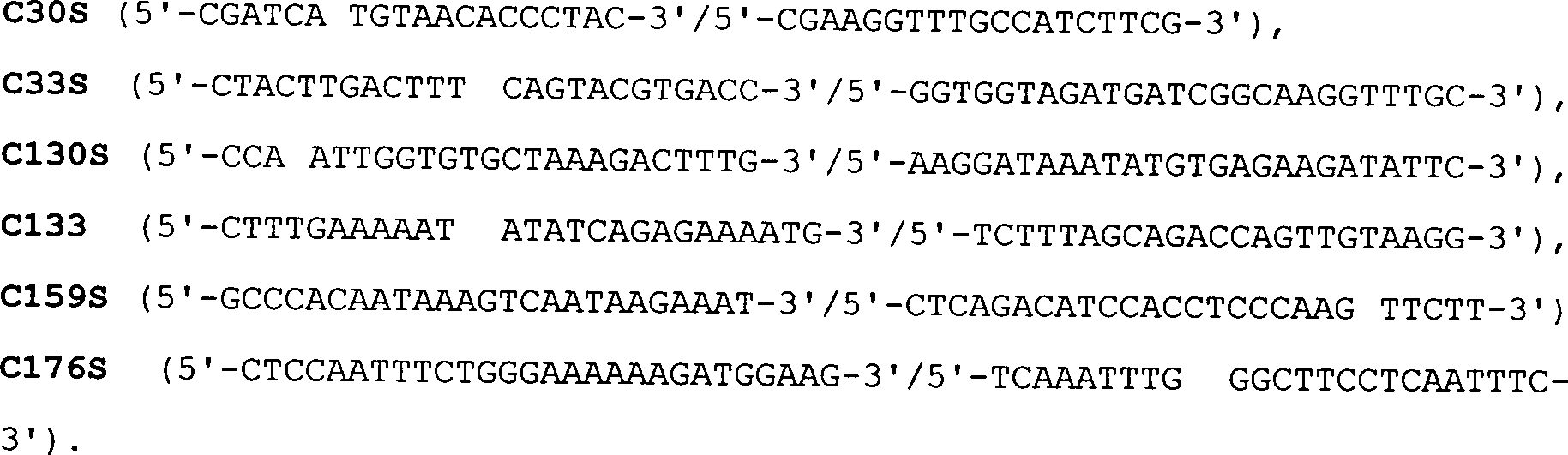-
Die Erfindung betrifft rekombinante,
FAD-abhängige
Sulfhydryloxidasen, in denen mindestens ein Cysteinrest derart mutiert
wurde, dass dies zu einer Veränderung
des Absorptionssmaximums im Vergleich zum Wildtypprotein führt, die
korrespondierenden Nukleinsäuremoleküle sowie
ein Verfahren zu ihrer Herstellung. Ferner betrifft die Erfindung
die Verwendung dieser Proteine als Biokatalysatoren für die Synthese
von Disulfidbrückenbindungen
und deren Übertragung
auf Substrate sowie als regulierbare Indikatoren für Redoxreaktionen.
-
Sulfhydryloxidasen sind Enzyme, welche
die Einführung
von Disulfidbrückenbindungen
in Proteinsubstrate katalysieren (Übersicht in [1]; [2–3]). Dabei
werden Thio-Verbindungen (R-SH)
nach der folgenden Gleichung in die korrespondierenden Disulfid-Verbindungen
umgesetzt werden: 2 R-SH + O2 → R-S-S-R
+ H2O2.
-
Prinzipiell unterscheidet man zwischen
Metalloproteinen (eisen- oder kupferhaltig) und FAD (Flavinadenindinukleotid)-abhängigen Sulfhydryloxidasen,
wobei die Erfindung letztere Gruppe betrifft.
-
FAD-abhängige Sulfhydryloxidasen zeichnen
sich gegenüber
anderen Enzymen, die Dithiol/Disulfid-Transferreaktionen katalysieren,
dadurch aus, dass sie Disulfidbindungen de novo synthetisieren können. Neben
FAD als nicht-kovalent gebundenem Cofaktor ist den Vertretern dieser
Gruppe gemein, dass sie (1) als Homodimere vorliegen, (2) Sauerstoff
als terminalen Elektronenakzeptor verwenden und (3) in ihrer Sequenz ein
konserviertes CXXC-Motiv besitzen (C = Cystein, X = beliebige Aminosäure), das
an der primären
Redoxreaktion beteiligt ist (Übersicht
in [1]; [4]). Die beiden bislang am besten untersuchten Vertreter
dieser Klasse sind die Quiescin-Sulfhydryloxidasen aus Huhn und
Mensch (Übersicht
in [1]).
-
Eine eigene Untergruppe innerhalb
der FAD-abhängigen
Sulfhydryloxidasen stellt die Familie der Erv/Alr-Sulfhydryloxidasen
dar, die nach Erv1p aus Saccharomyces cerevisiae sowie dessen humanem
Homolog Alrp benannt ist (Übersicht
in [1]). Diese beiden Enzyme sind auch die ersten dieser Gruppe,
deren DNA- und Proteinsequenzen bekannt sind [5, 6]. Ihre Aktivität ist essentiell
für das Überleben
der Zellen. Sie spielen eine wichtige Rolle in der Biogenese von
Mitochondrien, insbesondere hinsichtlich einer intakten Membranmorphologie,
sowie in der Versorgung cytoplasmatischer Proteine mit Eisen/Schwefel-Clustern,
die in den Mitochondrien gebildet werden (Übersicht in [1]; [7]). Im endoplasmatischen
Reticulum von Saccharomyces cerevisiae wurde vor kurzem eine zweite
Sulfhydryloxidase gefunden, die als Erv2p bezeichnet wurde [8, 9].
-
Die C-terminalen Domänen von
Erv1p und Erv2p, die das redoxaktive Zentrum und die FAD-Bindedomäne enthalten,
sind einander sehr ähnlich
(30% Identität).
Sie sind insbesondere durch ein konserviertes YPCXXC-Motiv (Y =
Tyrosin, P = Prolin, C = Cystein, X = beliebige Aminosäure) gekennzeichnet,
das für
die enzymatische Aktivität
sowie die Interaktion mit FAD essentiell ist. Bei Erv1p entspricht
dieses Motiv den Aminosäureresten
128–133.
-
Ein weiteres Charakteristikum dieser
Enzyme ist ihre wenig konservierte N-terminale Domäne, die
für die
subzelluläre Lokalisation
der Proteine, die Homodimerbildung sowie möglicherweise auch für die Substratinteraktion
wichtig sind [10]. Erv1p weist in dieser Region ein zweites CXXC-Motiv
auf (Aminosäuren
30–33), das
Erv2p fehlt. Letzteres enthält
allerdings an seinem C-Terminus ein möglicherweise funktionell homologes CGC-Motiv
(G = Glycin). Die genauen strukturellen bzw. funktionellen Aufgaben,
welche die einzelnen Cysteinreste in Erv1p bzw. Erv2p innehaben,
sind bislang noch nicht völlig
verstanden, werden aber derzeit intensiv untersucht.
-
In vielen Anwendungsbereichen, z.B.
bei der Teigherstellung für
Backwaren oder der Entfernung von Fremdaromen aus Milch oder Bier,
ist die Oxidation freier Sulfhydrylgruppen zu Disulfiden erwünscht. Da
enzymkatalysierte Reaktionen im Vergleich zum Einsatz unspezifischer
Oxidationsmittel (etwa Wasserstoffperoxid, Perazide oder Bromide)
keine unerwünschten
Nebenprodukte bilden und die Reaktion auch wesentlich schneller
abläuft,
wurde die Verwendung von Sulfhydryloxidasen für zahlreiche Applikationen
beschrieben.
-
Die Europäische Patentanmeldung
EP 0 705 538 A1 offenbart
eine Enzymzusammensetzung aus einer Sulfhydryloxidase und einer
Hemicellulase zur Verbesserung der Teigeigenschaften von Brot und
anderen Backwaren. Das US Patent 5,547,690 beschreibt eine weitere
Enzymzusammensetzung, bestehend aus einer Sulfhydryloxidase und
einer Glucoseoxidase, welche die rheologischen Eigenschaften, insbesondere
die Stabilität,
von Teigzubereitungen verbessert.
-
Die Deutsche Patentanmeldung
DE 198 40 489 A1 beschreibt
stabile Wirkstoffzubereitungen für
Lebensmittel, Futtermittel oder pharmazeutische Anwendungen, bei
denen der/die Wirkstoffe von einer Proteinschicht umgeben sind.
Zur Herstellung dieser Zubereitungen wird das Hüllprotein enzymatisch quervernetzt, u.a.
durch die Sulfhydryloxidasekatalysierte Synthese von Disulfidbrückenbindungen.
-
Bislang wurden Sulfhydryloxidasen
hauptsächlich
aus Milch bzw. aus verschiedenen Pilzen isoliert. U.S. Patent 4,087,328
beschreibt ein mehrstufiges, konventionelles Reinigungsverfahren
für Sulfhydryloxidase
aus Rohmilch von Kühen.
Ein anderes Verfahren zur Reinigung von Sulfhydryloxidase aus dem
Schimmelpilz Aspergillus niger ist im U.S. Patent 4,894,340 offenbart.
Daneben wurde in der Europäischen
Patentanmeldung
EP
0 565 172 A1 auch ein rekombinantes Verfahren zur Herstellung
von Sulfhydryloxidasen aus filamentösen Pilzen, vorzugsweise wiederum
aus Aspergillus spec., in biotechnologischem Maßstab offenbart.
-
Bislang war es allerdings nicht möglich, Sulfhydryloxidasen
herzustellen, deren katalytische Aktivität reguliert werden kann, obwohl
eine derartige enzymatische Steuerungsmöglichkeit von Redoxreaktionen
für eine
Vielzahl von Anwendungen in der Grundlagenforschung, aber z.B, auch.
in der Lebensmitteltechnologie wünschenswert
wäre.
-
Aufgabe der Erfindung ist es daher,
die katalytische Aktivität
von Sulfhydryloxidasen steuern zu können.
-
Dieses Ziel wird durch die Bereitstellung
neuartiger Sulfhydryloxidasen als molekularen Werkzeuge sowie die
rekombinante Herstellung solcher FAD-abhängiger Sulfhydryloxidasen in
ausreichend großen
Mengen gemäß Anspruch
1 erreicht. Bei diesen erfindungsgmäßen Sulfhydryloxidasen wurde
mindestens ein Cysteinrest der Amino-säuresequenz derart mutiert,
dass das Absorptionssmaximum gegenüber dem Wildtypprotein verändert ist.
-
Die Erfindung beruht dabei auf der überraschenden
Entdeckung, dass der Austausch bestimmter Cysteinreste in der Aminosäuresequenz
einer Sulfhydryloxidase aus Saccharomyces cerevisiae nicht nur die
Enzymaktivität
und die Homodimerbildung beeinflusste, sondern auch eine Farbveränderung
des Proteins zur Folge hatte. FAD-abhängige Sulfhydryloxidasen erscheinen
aufgrund des gebundenen FAD-Cofaktors
gelb. Infolge des Austausches eines einzelnen Cysteinrests ist jedoch
eine Farbänderung
zu beobachten und so wurde u.a. ein schwarzes Protein erhalten,
das weiterhin katalytisch aktiv war.
-
Ferner wurde festgestellt, dass die
Farbgebung reversibel durch Einstellung des Redoxpotentials gesteuert
werden kann. Da sich so Redoxreaktionen sehr leicht anhand der Farbänderung
verfolgen lassen, sind die Enzyme der Erfindung ideale Werkzeuge
zur gezielten Steuerung von Redoxreaktionen.
-
Die Erfindung schließt alle
modifizierten Sulfhydryloxidasen ein, bei denen das Absorptionsmaximum gegenüber dem
Wildtypprotein verändert
ist. Das Absorptionsmaximum bezeichnet die Wellenlänge ("Farbe") des Lichtes, bei
der eine Substanz beim Durchleuchten die Intensität am stärksten reduziert.
Licht anderer Wellenlängen
wird besser durchgelassen. Im sichtbaren Licht erscheint dadurch
die Substanz in der Komplementärfarbe
zu ihrem Absorptionsmaximum. Eine Veränderung des Absorptionsmaximums
gemäß der Erfindung
umfasst dabei alle Veränderungen
des Absorptionsverhaltens, die eine Farbänderung des Sulfhydryloxidase-Proteins
zur Folge haben.
-
Die Sulfhydryloxidasen gemäß der Erfindung
können
mit Hilfe rekombinanten DNA-Technologie hergestellt werden, die
eine vollständige
Kontrolle der Sequenz und damit auch der Eigenschaften eines bestimmten
Enzyms erlaubt. Mutationen können
sehr einfach unter Verwendung etablierter Standardverfahren [20]
in die Aminosäuresequenz
eingeführt
werden.
-
Die Erfindung schließt alle
rekombinanten Sulfhydryloxidasen ein, in denen ein oder mehrere
Cysteinreste ersetzt wurden, wobei Cystein prinzipiell gegen eine
beliebige Aminosäure
ausgetauscht werden kann. In bevorzugten Ausführungsformen wird der Cysteinrest
durch einen Serinrest ersetzt. Bislang konnte jedoch seitens der
Erfinder noch nicht abschließend
geklärt
werden, ob die spezifische Einführung
eines Serinrests in die Aminosäuresequenz
in direktem Zusammenhang mit den beobachteten Farbänderungen
der mutierten Proteine steht. Es sollte entsprechend dem Verständnis der
Erfinder deshalb möglich
sein, auch durch Substitution eines oder mehrerer Cysteinreste durch
andere Aminosäuren
Farbänderungen
der mutierten Proteine hervorzurufen, so z.B. durch den Austausch
von Cystein gegen Alanin oder Threonin, die beide zu Serin ähnliche
Eigenschaften besitzen. Daneben könnte dies aber durchaus auch
durch die Einführung
z.B. eines Arginin- oder
eines Leucinrests anstelle eines Cysteinrests bewirkt werden.
-
Grundsätzlich können alle in einer bestimmten
Sulfhydryloxidase vorkommenden Cysteinreste gegen eine andere Aminosäure ausgetauscht
werden. In bevorzugten Ausführungsformen
der Erfindung werden jedoch ein oder mehrere Cysteinreste ersetzt,
die redoxreaktiv sind. Unter "redoxreaktiven" Cysteinresten versteht
man erfindungsgemäß dabei
diejenigen Reste, die im Verlauf der katalytischen Reaktion einen
Zyklus aus Oxidation und Reduktion durchlaufen und dabei eine Disulfidbrücke ausbilden.
Beispiele für
redoxreaktive Cysteinreste sind etwa C130 und C133 in Erv1p aus
Saccharomyces cerevisiae.
-
Die Erfindung schließt ferner
Sulfhydryloxidasen ein, die sich von der als "Wildtyp" anerkannten Sequenz aufgrund von alternativem
Splicing einer gemeinsamen prä-mRNA
unterscheiden, aber nach wie vor katalytisch aktiv sind.
-
Bevorzugte Ausführungsformen der Erfindung
umfassen rekombinante Sulfhydryloxidasen aus der Familie der Evr/Alr-Sulfhydryloxidasen,
wobei wiederum Erv1p aus Saccharomyces cerevisiae (SEQ ID. Nr. 1) bevorzugt
wird.
-
Bevorzugt sind ferner Sulfhydryloxidasen,
die von Erv1p aus Saccharomyces cerevisiae abgeleitet sind, und
bei denen mindestens ein redoxaktiver Cysteinrest ersetzt wurde,
insbesondere die Enzyme mit einer Cystein → Serin-Mutation an Position
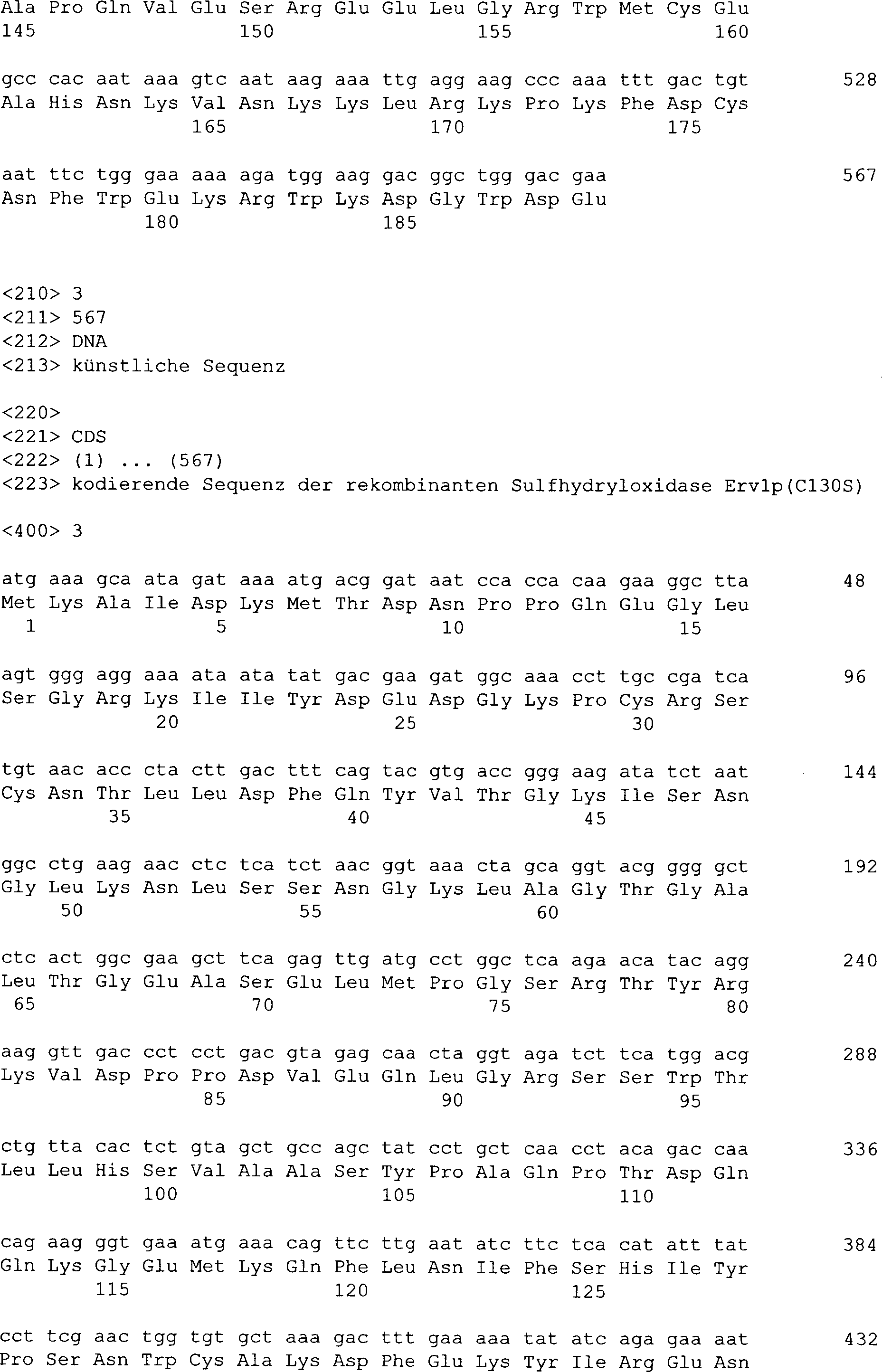
30 (C30S; SEQ ID Nr. 2) bzw. einer Cystein → Serin-Mutation an Position 130
(C130S; SEQ ID Nr. 3).
-
Eine weitere bevorzugte Ausführungsform
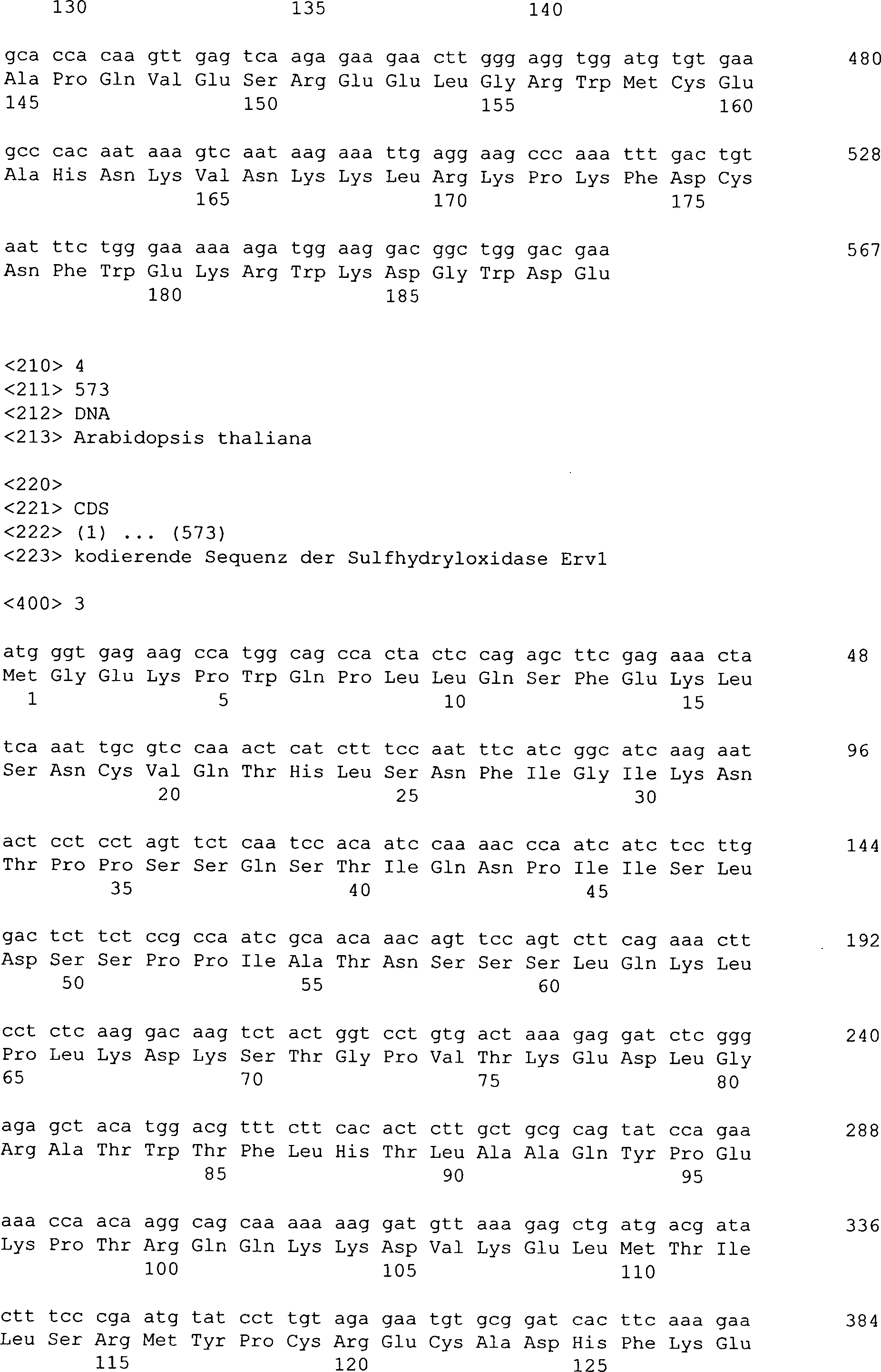
der Erfindung umfasst rekombinante Erv1-Sulfhydryloxidasen aus Arabidopsis
thaliana (SEQ ID Nr. 4).
-
In einer weiteren bevorzugten Ausführungsform
der Erfindung enthält
die rekombinante Sulfhydryloxidase ein Protein- oder Peptidaffinitätsepitop
an ihrem N-Terminus und/oder C-Terminus,
das eine einfache Detektion und/oder Reinigung des Proteins er-möglicht.
Geeignete Epitope sind zum Beispiel das myc-Epitop, das FLAG-Epitop,
das His6-Epitop oder das HA-Epitop.
-
Ferner betrifft die Erfindung Nukleinsäuremoleküle (DNA
und RNA), die Nukleinsäuresequenzen
umfassen, welche die hier offenbarten Sulfhydryloxidasen kodieren.
Da es aufgrund der Degeneriertheit des genetischen Codes möglich ist,
bestimmte Codons durch andere Codons zu ersetzen, welche die gleiche
Aminosäure
kodieren, ist die Erfindung nicht auf ein spezifisches Nukleinsäuremolekül beschränkt, das
eine Sulfhydryloxidase gemäß der Erfindung
kodiert, sondern schließt
alle Nukleinsäuremoleküle ein,
die eine funktionelle Sulfhydryloxidase kodieren.
-
Die Erfindung schließt ferner
Nukleinsäuremoleküle ein,
die sich aufgrund von alternativem Splicing eines gemeinsamen prä-mRNA Moleküls von der
als "Wildtyp" anerkannten Sequenz
unterscheiden. Derartige Splicing-Mechanismen umfassen die alternative
Verwendung von Exons (i.e. Nukleinsäuresequenzen, die eine Aminosäuresequenz
kodieren), die alternative Anordnung von Exons sowie das "Nicht-entfernen" von Introns (i.e,
intervenierende Sequenzen, die normalerweise keine Aminosäuresequenz
kodieren) aus dem mRNA-Molekül.
-
In bevorzugten Ausführungsformen
der Erfindung kodieren die Nukleinsäuremoleküle rekombinante Sulfhydryloxidasen
aus der Familie der Evr/Alr-Sulfhydryloxidasen, wobei insbesondere
Erv1p aus Saccharomyces cerevisiae (SEQ ID. Nr. 1) bevorzugt wird.
-
Andere bevorzugte Ausführungsformen
umfassen Nukleinsäuresequenzen
für Sulfhydryloxidasen,
in denen ein redoxaktiver Cysteinrest ersetzt wurde. Besonders bevorzugt
werden Nukleinsäuresequenzen,
die von Erv1p aus Sacharomyces cerevisiae abgeleitet sind, und hier
insbesondere die Nukleinsäuresequenzen, die
für eine
Cystein → Serin-Mutation
an Position 30 (C30S; SEQ ID Nr. 2) bzw. eine Cystein → Serin-Mutation an
Position 130 (C130S; SEQ ID Nr. 3) kodieren.
-
Eine weitere bevorzugte Ausführungsform
umfasst Nukleinsäuremoleküle, welche
die Erv1-Sulfhydryloxidase aus Arabidopsis thaliana (SEQ ID Nr.
4) kodieren.
-
Ein hier offenbartes Nukleinsäuremolekül kann "operativ" mit einer regulatorischen
Sequenz verknüpft sein,
um die Expression des Nukleinsäuremoleküls zu ermöglichen.
-
Ein Nukleinsäuremolekül wird als "fähig
zur Expression einer Nukleinsäuresequenz" bezeichnet, wenn es
Sequenzelemente umfasst, die Informationen hinsichtlich der Regulation
von Transkription und/oder Translation enthalten, und diese Elemente "operativ" mit der das Polypeptid
kodierenden Nukleinsäuresequenz
verknüpft
sind. Eine operative Verknüpfung
ist eine Verknüpfung,
bei der die regulatorischen Sequenzelemente und die proteinkodierende
Sequenz derart verbunden sind, dass Genexpression möglich ist.
Die genaue Beschaffenheit der zur Genexpression erforderlichen regulatorischen
Bereiche kann zwischen verschiedenen Spezies variieren. In der Regel
umfassen diese Bereiche jedoch einen Promotor, der in Prokaryonten
aus dem Promotor per se besteht, i.e. DNA-Elemente, welche die Transkriptionsinitiation
steuern, sowie aus DNA-Elementen, die nach ihrer Transkription in
mRNA den Beginn der Translation regulieren. Solche Promotoren schließen normalerweise
5' nicht-kodierende
Sequenzen ein, die an der Initiation von Transkription und Translation
beteiligt sind, wie zum Beispiel die –35/–10-Elemente und das Shine-Dalgarno Element
in Prokaryonten oder die TATA-Box, CAAT-Sequenzen und 5'-Capping-Elemente
in Eukaryonten. Diese Regionen können
ferner auch Enhancer- oder Repressorelemente enthalten sowie translatierte Signalsequenzen,
um die native Polypeptidkette in ein spezielles Kompartiment der
Wirtszelle zu dirigieren Zusätzlich
können
auch die 3' nicht-kodierenden
Regionen regulatorische Elemente enthalten, die an der Termination
der Transkription, der Polyadenylierung o.ä. beteiligt sind. Falls diese
Terminationssequenzen in einer speziellen Wirtszelle nicht oder
nur unzureichend funktionell sind, können sie durch Signale ersetzt
werden, die in der betreffenden Zelle ausreichend funktionell sind.
-
Ein Nukleinsäuremolekül gemäß der Erfindung kann demnach
eine regulatorische Sequenz, insbesondere eine Promotorsequenz umfassen.
In einer anderen bevorzugten Ausführungsform umfasst das Nukleinsäuremolekül gemäß der Erfindung
eine Promotorsequenz sowie eine Transkriptionsterminationssequenz. Geeignete
prokaryontische Promotoren sind zum Beispiel der lacUV5-Promotor
oder der T7-Promotor. Beispiele für geeignete eukaryontische
Promotoren sind der SV40-Promotor oder der CMV-Promotor.
-
Die Nukleinsäuremoleküle gemäß der Erfindung können ferner
in einem Vektor oder einem anderen Klonierungsvehikel enthalten
sein, wie zum Beispiel Phagen, Phagemiden, Cosmiden, Baculoviren
oder künstlichen
Chromosomen. In einer bevorzugten Ausführungsform ist das Nukleinsäuremolekül in einem
Vektor enthalten, insbesondere in einem Expressionsvektor. Ein derartiger
Expressionsvektor kann neben den oben bereits beschriebenen regulatorischen
Sequenzen und der Nukleinsäuresequenz,
die eine Sulfhydryloxidase gemäß der Erfindung
kodiert, Replikations- und
Kontrollsequenzen umfassen, die von einem Organismus stammen, der
mit dem zur Expression verwendeten Wirt kompatibel ist, sowie weiterhin
mindestens einen Selektionsmarker, der einen selektierbaren Phänotyp auf
eine transformierte Zelle überträgt. Eine
große
Zahl geeigneter Vektoren, z.B. pBluescript, pUC18, pET oder pcDNA3,
ist detailliert beschrieben und kommerziell erhältlich.
-
DNA-Moleküle, die eine Sulfhydryloxidase
gemäß der Erfindung
kodieren, und insbesondere ein Vektor, der die kodierende Sequenz
einer solchen Sulfhydryloxidase enthält, können in eine entsprechende
Wirtszelle transformiert werden, die zur Expression dieser DNA-Moleküle geeignet
ist. Die Transformation kann dabei mit Hilfe etablierter Standardverfahren
[20] durchgeführt
werden. Die Erfindung betrifft daher auch eine Wirtszelle, die ein
hier offenbartes Nukleinsäuremolekül enthält.
-
Die transformierten Wirtszellen werden
in der Folge unter Bedingungen kultiviert, die zur Expression der
Nukleotidsequenzen, die eine Sulfhydryloxidase gemäß der Erfindung
kodieren, geeignet sind. Die verwendeten Wirtszellen können prokaryontischen
Ursprungs sein, wie z.B. Escherichia coli (E. coli) oder Bacillus subtilis,
oder eukaryontischen Ursprungs, wie etwa Saccharomyces cerevisiae,
Pichia pastoris, SF9 oder High5 Insektenzellen, immortalisierte
Säugetierzelllinien
(z.B. HeLa-Zellen oder CHO-Zellen) oder primäre Säugetierzellen.
-
Die Erfindung betrifft auch ein Verfahren
zur rekombinanten Herstellung einer Sulfhydryloxidase gemäß der Erfindung.
Dieses Verfahren umfasst die folgenden Schritte:
- (a)
Klonieren eines Nukleinsäuremoleküls, das
eine Sulfhydryloxidase kodiert, in einen geeigneten Vektor, und
- (b) Einbringen des in (a) erhaltenen rekombinanten Vektors in
eine geeignete Wirtszelle oder einen geeigneten Zellextrakt.
-
Schritt (a) kann dabei mit einem
Nukleinsäuremolekül durchgeführt werden,
das nur die Sulfhydryloxidase kodiert. Alternativ kann er aber auch
mit einem Nukleinsäuremolekül durchgeführt werden,
in dem die Sulfhydryloxidase operativ mit einer regulatorischen
Sequenz verknüpft
ist. Optional kann das Nukleinsäuremolekül gemäß der Erfindung
auch mit einem Fusionspartner verknüpft sein, wie etwa einem Affinitätsepitop, das
eine einfache Reinigung und/oder Detektion des rekombinanten Proteins
erlaubt. Die Expression der so klonierten Nukleinsäuremoleküle kann
in einer rekombinanten Zelle oder einem Zellextrakt erfolgen, die/der alle
für die
Transkription und Translation erforderlichen Faktoren enthält.
-
Die Erfindung betrifft weiterhin
eine Zusammensetzung, die mindestens eine rekombinante Sulfhydryloxidase
gemäß der Erfindung
enthält.
Die Zusammensetzung kann dabei ausschließlich Sulfhydryloxidasen umfassen,
aber auch weitere enzymatisch aktive Proteine enthalten, wie z.B.
Lipoxygenasen, Proteindisulfidisomerasen, Lysyloxidasen, Tyrosinoxidasen,
Hemicellulasen oder Glucoseoxidasen. Ferner kann die Zusammensetzung
je nach Verwendungszweck auch Trägerstoffe
(z.B. Stärke,
Laktose, Gelatine, Fette, Wachse, Öle oder Polyole), verschiedenen
Zusatzstoffe (etwa Füllstoffe,
Stabilisierungsmittel, Emulgiermittel oder Konservierungsmittel,
Puffersubstanzen, Lösungsmittel
oder Lösungsvermittler)
oder Transportvehikel umfassen. Die Zusammensetzung kann in fester
Form vorliegen, z.B. lyophilisiert oder kristallin, oder aber gelöst, z.B.
als Suspension oder Emulsion.
-
Außerdem wird die Verwendung
der rekombinanten Sulfhydryloxidasen als Biokatalysatoren für die Synthese
von Disulfidbrückenbindungen
sowie deren Übertragung
auf Substrate offenbart, z.B. auf Dithiothreitol, Cystein oder Homocystein.
Im Vergleich zu herkömmlichen
Sulfhydryloxidasen besitzen die gentechnologisch modifizierten Enzymen
der Erfindung den entscheidenden Vorteil, dass sie regulierbar sind,
und diese Regulation anhand von Farbänderungen des Proteins leicht
verfolgt werden kann. Die Steuerung der Farbgebung des Proteins
erfolgt dabei reversibel durch Einstellung des Redoxpotentials über Dithiothreitol, β-Mercaptoethanol bzw. über Luftsauerstoff.
Dadurch wird es möglich,
eine Disulfidbindung gezielt auf ein bestimmtes Substratmolekül zu übertragen
und die Reaktion anhand der Farbänderung
des Proteins zu verfolgen.
-
Die Besonderheit der Erv1/Alr Sulfhydryloxidasen
ist ihre Substratspezifität
für Dithiole
mit einem definierten Abstand, da sie so in der zellulären Umgebung
unabhängig
vom Glutathionspiegel gezielte Disulfidbrücken einfügen können. Jüngste Untersuchungen der Erfinder
zeigen folgende Reihenfolge von künstlichen in vitro Substraten:
Thioredoxin (reduziertes CXXC) > Dithiothreitol > Di-Mercaptoethanol
Lysozym (reduziert) > Cystein.
Demzufolge scheinen Erv/Alr Sulfhydryloxidasen eine starke Präferenz für Dithiole
in Proteinen mit einem genau definierten Abstand zu haben. Thioredoxin
der Haupt-Redoxregulator der Zelle bei den aufgeführten Phänomenen.
Dabei wirkt Erv/Alrp als Thioredoxin-Oxidase und kann so in die
Regulation eingreifen.
-
Die Enzyme der Erfindung können z.B.
klinisch bei der Entwicklung diagnostischer Reagenzien und Testsysteme
von Bedeutung sein. Reduziertes Thioredoxin ist z.B. ein essentieller
Cofaktor der intrazellulären Signalübertragung,
der Steuerung von Apoptose und Nekrosenbildung sowie der Modulation
der Fragilität
roter Blutkörperchen.
Auch als Antioxidationsmittel ist es von großer Bedeutung. Die dabei stattfindende
Oxidation schützt
u.a. das vaskuläre
Endothel vor einer Schädigung
durch freie Radikale. Die gezielte Steuerung bzw. der Nachweis einer
erfolgten Disulfidbrückenbildung
können
daher für
die Diagnose, aber auch Therapie einer Reihe von Krankheiten vorteilhaft
sein Die rekombinanten Sulfhydryloxidasen der Erfindung können ferner
als regulierbare Sensoren/Indikatoren für Redoxreaktionen verwendet
werden. SO kann z. B. die Sauerstoffkonzentration des Mediums über das
Redoxpotential die Farbe des Proteins beeinflussen.
-
Umgekehrt kann aber auch die Farbe
des Proteins durch Änderung
des Redoxpotentials variiert werden. Die Enzyme gemäß der Erfindung
können
demnach auch als redox-regulierte Pigmente verwendet werden. Ein
Protein der Erfindung kann z.B. zwischen zwei Glasscheiben eingebracht
werden, welche mit Elektroden verbunden sind. Durch elektrische
Steuerung des Redoxpotentials kann das Protein die Glasfläche einfärben und
somit als Lichtschutz fungieren.
-
Da die hier offenbarten Enzyme nach
den bislang vorliegenden Ergebnissen für Zellen nicht toxisch zu sein
scheinen, sollten sie auch in vivo als Lichtschutzfaktor einsetzbar
sein, da sich die Lichtdurchlässigkeit von
Zellen wiederum über
das Redoxpotential regulieren lässt.
-
Die rekombinanten Enzyme der Erfindung,
und insbesondere die C30S-Variante von Erv1p aus Saccharomyces cerevisiae
(SEQ ID Nr. 2) können
weiterhin als Akzeptormoleküle
in Resonanzenergietransfer (RET)-Applikationen zur Bestimmung von
Abständen
zwischen Makromolekülen
eingesetzt werden. Beispiele derartiger Verfahren sind Fluoreszenz-RET
(FRET) und Biolumineszenz-RET (BRET) [17, 18]. Das Prinzip beruht
dabei auf dem strahlungslosen Transfer von Lichtenergie von einem
direkt angeregten Donormolekül
auf ein sich in der Nähe
befindliches Akzeptormoleül.
Damit ein Energietransfer stattfinden kann, müssen die Emmissions- bzw. Absorptionsspektren
von Donor bzw. Akzeptor überlappen.
Aufgrund seiner schwarzen Farbe und des breiten Absorptionsspektrums
kann die Q30S-Variante von Erv1p als universelle Akzeptordomäne eingesetzt
werden, die sich mit einem beliebigen Donormolekül kombinieren lässt.
-
Die Erfindung wird ferner durch die
folgenden nicht-einschränkenden
Figuren und Beispiele veranschaulicht.
-
Dabei zeigt 1 die Expression von Wildtyp (WT)-Erv1p
sowie verschiedener mutierter Varianten (C = Cystein, S = Serin).
Die gereinigten Proteine (vgl. auch Beispiel 1) tragen an ihrem
C-Terminus jeweils ein His6-Affinitätsepitop,
das sowohl eine schnelle Reinigung als auch den immunchemischen
Nachweis erlaubt. Pro Spur wurden 200 ng gereinigtes Protein mit
(+) bzw. ohne (–)
20 mM DTT im Probenpuffer auf 4%–12% nicht-reduzierende SDS-Polyacrylamiggele
(Novex/Invitrogen) aufgetragen. Die Proteine wurden über einen primären anti-His5 Antikörper
und einen AP-gekoppelten
sekundären
Antikörper
chemiluminometrisch nachgewiesen.
-
2 zeigt
die Bestimmung der in vitro-Aktivität von Wildtyp (WT)-Erv1p sowie
verschiedener mutierter Varianten. Die Enzymaktivität der gereinigten
Proteine (vgl. auch Beispiel 2) wurde photometrisch mit DTT (entsprechend
50 nmol reduzierte Thiol-Gruppen) als Substrat gemessen. Der Substratumsatz
wurde dabei pro proteingebundenes FAD-Molekül (gemäß der spektroskopischen Bestimmung)
angegeben. Die Ergebnisse repräsentieren
die Mittelwerte ± Standardabweichung
aus drei unabhängigen
Experimenten.
-
3 zeigt
die unterschiedlichen Färbungen
von gereinigtem Wildtyp (WT)-Erv1p sowie verschiedener mutierter
Varianten. Aufgrund von unterschiedlicher Expression in E. coli
variieren die Proteinkonzentrationen der einzelnen Lösungen.
Erv1p (5.9 mg/ml), C30S (36 mg/ml), C33S (10.8 mg/ml), C130S (14.1
mg/ml), C133S (4.1 mg/ml), C159S (9.9 mg/ml), C176S (6.0 mg/ml)
und ΔN-Erv1p
(4.1 mg/ml).
-
4 zeigt
Spektren für
Wildtyp (WT)-Erv1p sowie die beiden Cystein → Serin Varianten C30S und C130S.
Die gereinigten Proteine wurden in einem Zeiss S10 Dioden-Array
Photometer in einem Wellenlängebereich
von 320 nm bis 700 nm analysiert (vgl. auch Beispiel 3). Die verwendeten
Proteinlösungen
hatten die folgenden Konzentrationen: 2.2 mg/ml (Erv1p), 5.1 mg/ml
(C30S) und 4.2 mg/ml (C130S). Da der FAD-Gehalt der Proben unterschiedlich
ist, und auch die molaren Absorptionskoeffizienten für die mutierten
Proteine variieren können,
werden die Spektren nur qualitativ bewertet.
-
Beispiel 1
-
Klonierung und Reinigung
von rekombinanten Erv1p-Varianten aus Saccharomyces cerevisiae
-
Die cDNAs für das vollständige ERV1-Gen
sowie eine 5'-terminal verkürzte Variante
(ΔN-Erv1p;
entspricht Aminosäure
73–189
des Wildtyps) wurden mittels etablierter Standardverfahren [20]
amplifiziert und als NdeI/XhoI- Fragmente
in den Vektor pET24a(+) (Novagen) kloniert [5]. Der rekombinante
Vektor, der die vollständige
cDNA-Sequenz kodierte, wurde im folgenden zur oligonukleotid-gerichteten
Mutagenese ("Excite" PCR-Based Site-Directed
Mutagenesis Kit/Stratagene) verwendet. Die zur Einführung der
einzelnen Mutationen (C = Cystein, S = Serin) verwendeten Primer
(jeweils vorwärts/rückwärts) haben
die folgenden Sequenzen:
-
Die Identität der mutierten Sequenzen wurde
anhand von Restriktions- bzw. Sequenzanalysen verifiziert.
-
Die gereinigten Plasmide (Plasmid
Purification Kit/QIAGEN) wurden in E. coli BL21(+) (Novagen) transformiert
und amplifiziert [20]. Die Reinigung der einzelnen Proteine erfolgte über ein
His6-Affinitätsepitop (in der Sequenz von
pET24a(+) kodiert) an deren C-Termini. Die Reinigung wurde mit Hilfe
von Ni2+ NTA-Agarose unter nativen Bedingungen
in Phosphatpuffer (50mM NaCl, 50mM KH2PO4, 10mM Imidazol, pH 7.5) nach Herstelleranweisung
(QIAGEN) durchgeführt.
Die Homogenität
der Proteinpräparationen
wurde mittels SDS-Polyacrylamidgelelektrophorese
und Western-Analyse verifiziert. Dazu wurden jeweils 200 ng Protein
in 4%–12%
nicht-reduzierenden SDS-Polyacrylamidgelen (Novex/Invitrogen) aufgetrennt.
Die Bildung von Protein-Dimeren (oder -Multimeren) wurde durch Zugabe
bzw. Weglassen von DTT (20mM) im Probenpuffer untersucht (siehe 1).
-
Aus 1 ist
ersichtlich, dass die N-terminal verkürzte Erv1p ausschließlich als
Monomer, das Wildtypprotein jedoch als eine Mischung aus Monomeren
und Dimeren vorliegt. Dies zeigt, dass die N-terminale Domäne für die Bildung
von Dimeren entscheidend ist. Unter nicht-reduzierenden Bedingungen
findet man mit Ausnahme von C133S keine Monomere. Die mutierten
Proteine C30S, C33S und in geringerem Maße C130S liegen vornehmlich
als Dimere vor, d.h. zumindest die beiden Cysteinreste an Position
30 bzw. 33 sind an der Dimerbildung beteiligt. Das Vorliegen hochmolekularer
Proteinmultimere bei den Varianten C133S, C159S und C176S deutet
hingegen eine unspezifische Aggregation dieser Proteine an.
-
Beispiel 2
-
Analyse der in vitro-Enzymaktivität von Erv1p-Varianten
-
Zur Aktivitätsbestimmung wurden die Konzentrationen
der einzelnen Proteine auf 10 pmol proteingebundenes FAD pro 100 μl Reaktionsansatz
eingestellt.
-
Die Enzymreaktion wurde in PBS (68mM
NaCl, 75mM Kaliumphosphatpuffer, pH 7.5, der 3mM EDTA enthält) mit
DTT als Substrat (entsprechend 50 nmol reduzierter Thiolgruppen)
durchgeführt.
Für jede
Messung wurde ein 100 μl
Aliquot dieser Reaktionsmischung entnommen. Zur Bestimmung des Thiolgehalts
wurden die Aliquots mit 800 μl
PBS und 100 μl
DTNB-Lösung
(5,5'-Dithio-bis-2-nitrobenzoesäure, Ellman's Reagens; Endkonzentration
10mM) versetzt. Nach 2 min Inkubation wurde die Extinktion bei 412
nm gemessen und der Thiolgehalt berechnet (molarer Extinktionskoeffizient
14mM–1 cm–1 [21]).
Der Thiolgehalt in der Ausgangsmischung wurde in einer Probe ohne
Enzymzugabe ermittelt. Der Substratumsatz wurde pro proteingebundenes
FAD-Molekül
angegeben. Die in 2 dargestellten
Ergebnisse repräsentieren
die Mittelwerte ± Standardabweichung
aus drei unabhängigen
Experimenten.
-
Die mutierten Proteine zeigen unter
in vitro Bedingungen keine (C130S, C133S) oder nur eine stark verminderte
(C30S, C33S, C159S, C176S) Enzymaktivität, sodass alle sechs getesteten
Cysteinreste für
die volle Aktivität
der Erv1p-Sulfhydryloxidase
wichtig zu sein scheinen. Die Aktivität der N-terminal verkürzten Erv1p
ist erstaunlicherweise mit der des Wildtypproteins vergleichbar.
Eine weitere Studie [19] hat jedoch gezeigt, dass das verkürzte Protein
den Wildtyp in vivo nicht ersetzen kann.
-
Beispiel 3
-
Spektroskopische Analyse
von Erv1p-Varianten
-
Der Austausch von Cystein- gegen
Serinreste hatte in einigen Fällen
auch eine Veränderung
der Proteinfärbung
zur Folge (siehe 3).
Während
das Wildtypprotein und die N-terminal verkürzte Erv1p aufgrund des gebundenen
FAD intensiv gelb erscheinen, führt
eine Mutation an Position 30 zu einer Schwarzfärbung des Proteins, eine Mutation
an Position 130 hingegen zu einer Orangefärbung. Erv1p(C159S) erscheint
farblos, und die anderen drei Varianten (C33S, C133C und C176S)
sind ähnlich
dem Wildtyp gelb gefärbt.
-
Alle Farben verschwinden nach Reduktion
der Proteine mit Dithionit. Nach Reoxidation in der Luft verfärben sich
die Proteinlösungen
gelb. Auch bei längerer
Lagerung (über
mehrere Tage) verändern
sich die Farben der mutierten Proteine hin zum Gelb des Wildtyps,
vermutlich ebenfalls durch Oxidation mit Luftsauerstoff. Im Stand
der Technik gelang es nach einer derartigen Verfärbung nicht, die ursprüngliche
Farbe eines mutierten Proteins wiederherzustellen.
-
Die Farbveränderungen der beiden Erv1p-Varianten
C30S und C130S wurden in einem Zeiss S10 Dioden-Array Photometer
im Detail untersucht. Es wurden Spektren in einem Wellenlängebereich
von 320 nm bis 700 nm aufgenommen (vgl. 4). Die verwendeten Proteinlösungen hatten
die folgenden Konzentrationen: 2.2 mq/ml (WT-Erv1p), 5.1 mg/ml (C30S)
und 4.2 mg/ml (C130S). Da der FAD-Gehalt der Proben unterschiedlich
ist, und auch die molaren Absorptionskoeffizienten für die mutierten
Proteine variieren können,
werden die Spektren nur qualitativ bewertet. Auffällig sind
bei der C30S-Variante ein zusätzliches
Absorptionsmaximum bei 580 nm sowie eine Verbreiterung der Absorption
bei 460 nm (entspricht FAD) bei der C130S-Variante. Die Spektren aller anderen
mutierten Proteine entsprechen dem Wildtypprotein (nicht gezeigt).
-
Referenzen:
-
- [1] Thorpe, C. et al. (2002) Arch. Biochem.
Biophys. 405, 1-12.
- [2] Lash, L.H. and Jones, D.P. (1983) Arch. Biochem. Biophys.
225, 344–352.
- [3] Hoober, K.L. et al. (1999) J. Biol. Chem. 274, 22147-22150.
- [4] Hoober, K.L. et al. (1999) J. Biol. Chem. 274, 31759-31762.
- [5] Lee, J.E. et al. (2000) FEBS Lett. 477, 62–66.
- [6] Giorda, R. (1996) Mol. Med. 2, 97–108.
- [7] Lange, H. et al. (2001) EMBO Rep. 2, 715–720.
- [8] Sevier, C.S. et al. (2001) Nat. Cell Biol. 3, 874–882.
- [9] Gerber, J. et al. (2001) J. Biol. Chem. 276, 23486-23491.
- [10] Hofhaus, G. and Lisowsky, T. (2002) Methods Enzymol. 348,
314–324.
- [11] Europäisches
Patentanmeldung EP
0 705 538 A1
- [12] U.S. Patent 5,547,690
- [13] Deutsches Patentanmeldung DE 198 40 489 A1
- [14] U.S. Patent 4,087,328
- [15] U.S. Patent 4,894,340
- [16] Europäisches
Patent EP 0 565 172
A1
- [17] Boute, N. et al. (2002) Trends Pharmacol. Sci. 23, 351-354.
- [18] Heyduk, T. (2002) Curr. Opin. Biotechnol. 13, 292–296.
- [19] Lisowsky, T. (1996) Yeast 12, 1501–1510.
- [20] Sambrook, J. et al. (1989) Molecular Cloning: A Laboratory
Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, NY.
- [21] Riddles, P.W. et al. (1979) Anal. Biochem. 94, 75–81.
-
-
-
-
-