WO2012104243A1 - Verfahren zur fermentativen herstellung von 2,3-butandiol - Google Patents
Verfahren zur fermentativen herstellung von 2,3-butandiol Download PDFInfo
- Publication number
- WO2012104243A1 WO2012104243A1 PCT/EP2012/051426 EP2012051426W WO2012104243A1 WO 2012104243 A1 WO2012104243 A1 WO 2012104243A1 EP 2012051426 W EP2012051426 W EP 2012051426W WO 2012104243 A1 WO2012104243 A1 WO 2012104243A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- strain
- production
- acetolactate synthase
- butanediol
- klebsiella
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/02—Preparation of oxygen-containing organic compounds containing a hydroxy group
- C12P7/04—Preparation of oxygen-containing organic compounds containing a hydroxy group acyclic
- C12P7/18—Preparation of oxygen-containing organic compounds containing a hydroxy group acyclic polyhydric
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1022—Transferases (2.) transferring aldehyde or ketonic groups (2.2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y202/00—Transferases transferring aldehyde or ketonic groups (2.2)
- C12Y202/01—Transketolases and transaldolases (2.2.1)
- C12Y202/01006—Acetolactate synthase (2.2.1.6)
Definitions
- the invention relates to a process for the fermentative production of 2,3-butanediol (2,3-BDL) by means of an improved microorganism strain having a 2 to 68-fold increased acetolactate synthase activity compared to the non-improved starting strain.
- chemical raw materials (so-called chemical synthesis building blocks) from renewable raw materials are ethanol ⁇ C2 building block), glycerol, 1, 3-propanediol, 1, 2-propanediol (C3 building blocks) or succinic acid, 1-butanol, 2 ⁇ Butanol, 1,4-butanediol or 2, 3-butanediol (C4 building blocks).
- chemical building blocks are the biogenic starting compounds from which further basic chemicals can be chemically produced.
- a C4 building block accessible by fermentation is 2,3-butanediol.
- the state of the art for fermentative 2,3-butanediol production is summarized in Celinska and Grajek (Biotechnol. Advances ⁇ 2009) 27: 715-725).
- 2, 3-butanediol is a possible starting material for petrochemical products with four carbon atoms (C4 building blocks) such as acetoin, diacetyl, 1,3-butadiene, 2-butanone (methyl ethyl ketone, MEK).
- 2,3-butanediol dehydrogenase Reaction of acetoin reductase (2, 3-butanediol dehydrogenase): NADH-dependent reduction of acetoin to 2,3-butanediol.
- 2,3-butanediol Various natural producers of 2,3-butanediol are known, e.g. B, from the genera Klebsiella, Raoultella, Enterobacter, Aerobacter, Aeromonas, Serratia, Bacillus, Paenibacillus, Lactobacillus, Lactococcus etc. But also yeasts are known as producers (eg bakers yeast).
- Known 2, 3 -BDL production strains from the biological safety level Sl are strains of the species Klebsiella terrigena, Klebsiella planticola, strains of the genus Bacillus (or Paenibacillus) such as Bacillus polymyxa or Bacillus licheni ormis.
- Bacillus or Paenibacillus
- the species Klebsiella terrigena and Klebsiella planticola are also synonymous as Raoultella terrigena and Raoultella due to a taxonomic renaming designated planticola. These are strains of the same species.
- 2,3-butanediol yields of not more than 57 g / l (637 mM, production time 60 h) have hitherto been reported for these strains classified in safety level Sl (Nakashimada et al., J. Bioscience and Bioengineering (2000) 90: 661- 664). These yields are far too low for economical production.
- the minimum fermentation yield for economical production is considered to be> 80 g / l 2, 3-butanediol (fermentation duration maximum 72 h), preferably> 100 g / l 2, 3-butanediol.
- biosafety in Microbiological and Biomedical Laboratories p. 9ff. (Table 1), Centers for Disease Control and Prevention (US) (Editor), Public Health Service (US) (Editor), National Institutes of Health (Editor),
- the object of the invention was to provide production strains for the production of 2, 3-butanediol, which allow significantly higher 2,3-Butandiolausbeuten than the output stem.
- the object was achieved by a production strain which can be produced from an initial strain, characterized in that the production strain has an acetolactate synthase activity 2 to 68 times above the starting strain.
- a parent strain In the present invention, a distinction is made between a parent strain and a production strain.
- the starting trunk At the starting trunk it may be a non-optimized, but capable of 2,3-butanediol production wild-type strain or an already further optimized wild-type strain.
- an already optimized wild-type strain eg, through genetic engineering
- the acetolactate synthase activity is not affected by the optimization.
- a production strain is to be understood as meaning a starting strain which has been optimized with respect to the production of 2,3-butanediol and which is distinguished by a
- Starting strain is characterized by increased activity of the enzyme acetolactate synthase (ALS).
- the production strain is made from the parent strain. If an already optimized starting strain is to be further improved by an increase in acetolactate synthase activity, it is of course also possible first to increase the acetolactate synthase activity in an unmodified strain and then to introduce further improvements.
- the increase in acetolactate synthase activity in the production strain can be caused by any mutation in the genome of the parent strain (for example a promoter activity-increasing mutation), an enzyme activity-increasing mutation in the acetolactate synthase gene or by overexpression of a homologous or heterologous acetolactate synthase gene in the parent strain.
- the overexpression of a homologous or a heterologous Acetolactatsynthasegens in the parent strain is preferred.
- the acetolactate synthase activity is preferably increased by a factor of 3 to 64, more preferably by a factor of 3 to 30 and particularly preferably by a factor of 4 to 20, in comparison to the starting starch.
- This increased acetolactate synthase activity is particularly preferred due to an increased expression compared to the parent strain.
- a homologous or heterologous gene coding for an acetolactate synthase enzyme is particularly preferred due to an increased expression compared to the parent strain.
- the parent strain can be any 2,3-butanediol producing strain. It is preferably a strain of the genus Klebsiella, Raoultella, Bacillus or Lactobacillus.
- a strain of the species Klebsiella ⁇ Raoultella ⁇ terrigena, Klebsiella (Raoultella) planticola, Bacillus (Paenibacillus) polymyxa or Bacillus licheniformis with a strain of the species Klebsiella (Raoultella) terrigena or Klebsiella (Raoultella) planticola is again preferred.
- overexpression of the acetolactate synthase by a factor of 3.2 up to a factor of 68.4 is suitable for this purpose 2, 3 ⁇ Butandiolausbeute ⁇ determined as volume yield of 2,3-butanediol in g / 1) in shake flasks by more than 25%, preferably more than 30% and in particular by more than 40% (see Example 4) and in the fermentation to more than 15%, preferably more than 20%, and especially preferably more than 30% (see 5 and 6, Example) to increase.
- Acetolactate synthase is an enzyme from the enzyme class EC 2.2.1.6. It may be any gene-encoded enzyme which, according to formula (I), causes the synthesis of acetolactate from two molecules of pyruvate.
- the gene of the acetolactate synthase is derived from a bacterium of the genus Klebsiella (Raoultella) or Bacillus.
- the gene of acetolactate synthase is derived from a strain of the species Klebsiella terrigena, Klebsiella planticola, Bacillus ⁇ Paenibacillus ⁇ polymyxa or Bacillus licheniformis and in particular from a strain of the species Klebsiella terrigena, Klebsiella planticola or Bacillus licheni ormi. These strains are all commercially available, e.g.
- the strain according to the invention thus makes it possible to increase the fermentative production of acetolactate.
- the invention thus enables not only the production of 2, 3-butanediol but also the production of other metabolites, which can be derived as acetonate, such as 2, 3-butanediol.
- a production strain according to the invention is also characterized in that it is produced from an exit as defined in the application and produces an acetolactate synthase in recombinant form, with the result that its 2,3-BDL production ( Volume production expressed in g / 1 2,3-BDL) compared to the non-genetically optimized starting strain by at least 25%, preferably 50%, more preferably 75% and particularly preferably increased by 100%, wherein the 2,3-butanediol yield of the parent strain at least 90 g / 1.
- the production strain according to the invention is preferably prepared by introducing a gene construct into one of the mentioned parent strains.
- the gene construct in its simplest form is defined as consisting of the acetolactate synthase structural gene operatively linked to a promoter upstream.
- the gene construct may also comprise a terminator downstream of the acetolactate synthase structural gene.
- Preferred is a strong promoter, which leads to a strong transcription.
- Preferred among the strong promoters is the so-called "tac promoter" familiar to the person skilled in the art from the molecular biology of E. coli.
- the gene construct can be present in a manner known per se in the form of an autonomously replicating plasmid, it being possible for the copy number of the plasmid to vary.
- a variety of plasmids are known to those skilled in the art, depending on their genetic
- Structure in a given production strain can autonomously replicate.
- the gene construct can also be integrated in the genome of the production strain, with each gene locus along the genome being suitable as an integration site.
- the gene construct either in plasmid form or with the aim of genomic integration, is introduced into the production strain in a manner known per se by genetic transformation.
- Various methods of genetic transformation are known to the person skilled in the art (Aune and Aachmann, Appl. Microbiol. Biotechol. (2010) 85: 1301-1313), including, for example, the
- Selection markers for the selection of transformants with the desired gene construct are selected from antibiotic resistance markers or from the auxotrophy complementing selection markers.
- antibiotic resistance markers Preference is given to antibiotic resistance markers, more preferably those which confer resistance to antibiotics selected from ampicillin, tetracycline, kanamycin, chloramphenicol or zeocin.
- a production strain according to the invention contains the gene construct, either in plasmid form or integrated into the genome, and produces an acetolactate synthase enzyme in recombinant form.
- the recombinant acetolactate synthase enzyme is capable of producing two molecules of pyruvate acetolactate by cleavage of C0 2 . It has now surprisingly been found that by suitable recombinant overexpression of the acetolactate synthase enzyme in the production strain, the 2,3-butanediol yield can be significantly increased compared to the starting strain.
- the parent strain may be a non-optimized wild type strain.
- the parent strain may have been previously optimized, or further optimized as in the production of acetolactate synthase-producing production strain.
- the optimization of a production strain according to the invention which comprises acetolactate synthase according to the invention can be achieved on the one hand by mutagenesis and selection of Mutants with improved production properties take place.
- the optimization can also be carried out by genetic engineering by additional expression of one or more genes which are suitable for improving the production properties. Examples of such genes are the already mentioned 2,3-butanediol biosynthesis genes acetolactate decarboxylase and acetoin reductase.
- genes can be expressed in a manner known per se as separate gene constructs or in combination as an expression unit (as a so-called operon) in the production strain.
- an expression unit as a so-called operon
- Klebsiella terrigena all three biosynthesis genes of 2, 3-butanediol ⁇ so-called. BUD operon, Blomqvist et al. , J. Bacteriol. (1993) 175: 1392-1404), or in strains of the genus Bacillus the genes of acetoacetate synthase and acetolactate decarboxylase are organized in an operon (enna et al., J. Bacteriol (1993) 175: 3863-3875).
- the production strain can be optimized by inactivating one or more genes whose gene products adversely affect 2,3-butanediol production.
- genes whose gene products are responsible for by-product formation include z. B. the lactate dehydrogenase (lactic acid formation), the acetaldehyde dehydrogenase (ethanol formation) or else the phosphotransacetylase, or the acetate kinase (acetate formation).
- the invention comprises a process for the production of 2,3-butanediol with the aid of a production strain according to the invention.
- the method is characterized in that cells of a production strain according to the invention are cultivated in a growth medium.
- biomass of the production strain and on the other hand the product 2,3-BDL are formed.
- the formation of biomass and 2,3-BDL can be time correlated or temporally decoupled from each other.
- the cultivation takes place in a manner familiar to the person skilled in the art.
- the cultivation in shake flasks (Labormass tab) or by fermentation (production scale ⁇ . Preference is given to a process on a production scale by fermentation, with a fermentation volume greater than 10 l being particularly preferred as the production scale and a fermentation volume greater than 300 l being particularly preferred.
- Cultivation media are familiar to the person skilled in the art of microbial cultivation. They typically consist of a carbon source (C source), a nitrogen source (N source), and additives such as vitamins, salts, and trace elements, which optimize cell growth and 2,3-BDL product formation.
- C sources are those that can be used by the production base for 2,3-BDL product formation. These include all forms of monosaccharides, including C6- ucker such. Glucose, mannose or fructose, and C5 sugar such as glucose. Xylose, arabinose, ibose or galactose.
- the production process according to the invention also comprises all C sources in the form of disaccharides, in particular sucrose, lactose, maltose or cellobiose.
- the production process according to the invention furthermore also comprises all C sources in the form of higher saccharides, glycosides or carbohydrates with more than two sugar units, such as, for example, As maltodextrin, starch, cellulose, hemicellulose, pectin or by hydrolysis ⁇ enzymatically or chemically) released monomers or oligomers.
- the hydrolysis of the higher C sources may precede the production process according to the invention or take place in situ during the production process according to the invention.
- C sources other than sugars or carbohydrates are acetic acid (or derived acetate salts), ethanol, glycerol, citric acid (and salts thereof) or
- C sources which are affected by the production process according to the invention comprise both the isolated pure substances and, for reasons of greater economic efficiency, not further purified mixtures of the individual C sources, such as can be obtained as hydrolyzates by chemical or enzymatic digestion of the vegetable raw materials , These include z. B.
- Hydrolysates of starch ⁇ monosaccharide glucose), sugar beet (monosaccharides glucose, fructose and arabinose), sugar cane (disaccharide sucrose), pectin (monosaccharide galacturonic acid) or lignocellulose (monosaccharide glucose from cellulose, monosaccharides xylose, arabinose, mannose , Galactose from hemicellulose and the non-carbohydrate lignin). Furthermore, can serve as C sources and waste products from the digestion of vegetable raw materials, such. As molasses (sugar beet) or bagasse (sugar cane).
- Preferred C sources for cultivating the production strains are glucose, fructose, sucrose, mannose, xylose, arabinose and plant hydrolysates which can be obtained from starch, lignocellulose, sugar cane or sugar beet.
- C source is glucose, either in isolated form or as part of a vegetable hydrolyzate.
- N sources are those that can be used by the production base for biomass production. These include ammonia, gaseous or in aqueous solution as NH 4 OH or else its salts such. As ammonium sulfate, ammonium chloride, ammonium phosphate, ammonium acetate or ammonium nitrate. Furthermore, suitable N-source are the known nitrate salts such. B. KN0 3 , NaN0 3 , ammonium nitrate, Ca (N0 3 ) 2 , Mg ⁇ N0 3 ) 2 and other N sources such as urea.
- the N sources also include complex amino acid mixtures such as yeast extract, proteose peptone, malt extract, soy peptone, casamino acids, corn steep liquor (corn steep liquor, liquid or dried as so-called CSD) as well as NZ amines and Yeast Nitrogen Base.
- the cultivation can take place in the so-called batch mode, whereby the growth medium is inoculated with a starter culture of the production strain and then the cell growth takes place without further feeding of nutrient sources.
- Cultivation can also take place in the so-called fed-batch mode, with additional nutrient sources being fed in after an initial phase of growth in batch mode (feed) in order to compensate for their consumption.
- the feed can consist of the C source, the N source, one or more important for the production of vitamins, or trace elements or a combination of the aforementioned.
- the feed components can be used together as a mixture or else separately
- Feed lines are added.
- other media components as well as specific 2,3-BDL production enhancing additives may be added to the feed.
- the feed can be fed continuously or in portions (batchwise) or else in a combination of continuous and discontinuous feed. Preference is the ' breeding after the fed-batch mode.
- Preferred C sources in the feed are glucose, sucrose, molasses, or vegetable hydrolysates, which can be obtained from starch, lignocellulose, sugar cane or sugar beet.
- Preferred N sources in the feed are ammonia, gaseous or in aqueous solution as NH 4 0H and its salts ammonium sulfate, ammonium phosphate, ammonium acetate and ammonium chloride, furthermore urea, KN0 3 , NaN0 3 and ammonium nitrate, yeast extract, proteose peptone, malt extract, soya peptone, casamino acids Corn Steep Liquor as well as N -Amine and Yeast Nitrogen Base.
- N sources in the feed are ammonia or ammonium salts, urea, yeast extract, soya peptone, malt extract or corn steep liquor (in liquid or in dried form).
- the cultivation takes place under pH and temperature conditions which favor the growth and the 2,3-BDL production of the production strain.
- the useful pH range is from pH 5 to pH 8.
- Preferred is a pH range of pH 5.5 to pH 7.5.
- Particularly preferred is a pH range of pH 6.0 to pH 7.
- the preferred temperature range for the growth of the production strain is 20 ° C to 40 ° C. Particularly preferred is the temperature range of 25 ° C to 35 ° C.
- the growth of the production strain can optionally take place without oxygen supply (anaerobic cultivation) or else with oxygen supply (aerobic cultivation) .
- oxygen supply is preferred, oxygen supply being ensured by introduction of compressed air or pure oxygen Cultivation by entry of compressed air.
- the cultivation time for 2,3-BDL production is between 10 h and 200 h. Preferred is a cultivation period of 20 h to 120 h. Particularly preferred is a cultivation time of 30 h to 100 h.
- Cultivation batches obtained by the method described above contain the 2,3-BDL product, preferably in the culture supernatant.
- the 2,3-BDL product contained in the cultivation mixtures can either be used further directly without further work-up or else be isolated from the cultivation batch.
- known Verhrens intimide are available, including centrifugation, decantation, filtration, extraction, distillation or crystallization, or precipitation.
- These process steps can be combined in any desired form in order to isolate the 2,3-BDL product in the desired purity.
- the degree of purity to be achieved depends on the further use of the 2,3-BDL product.
- Various analytical methods for the identification, quantification and determination of the purity of the 2,3-BDL product are available, including NMR, gas chromatography, HPLC, mass spectrometry or a combination of these analytical methods.
- FIG. 1 shows the 4.9 kb acetolactate synthase expression vector pBudBkt produced in Example 1.
- FIG. 2 shows the 4.9 kb acetolactate synthase expression vector pALSbl prepared in Example 1.
- FIG. 3 shows the 2.9 kb expression vector p Kj produced in Example 1.
- Fig. 5 shows the 6kb size epitaxial vector pALSbl-tet prepared in Example 1.
- Fig. 6 shows the 6 kb plasmid pBudBkt-tet (rev) used in Example 1.
- FIG. 7 shows the 5.3 kb epic expansion vector pAC-BudBkt produced in Example 1.
- Fig. 8 shows the plasmid pACYC18 used in Example 1.
- the acetolactate synthase genes from K. terrigena and B. licheniformis were used.
- the DNA sequence of the acetolactate synthase gene from K. terrigena is disclosed in the "GenBank” gene database under accession number L04507, bp 969-2648. It was isolated in a PCR reaction (Taq DNA polymerase, Qiagen) from genomic K. terrigena DNA (strain DSM 2687, commercially available from the DSMZ German Collection of Microorganisms and Cell Cultures
- the DNA sequence of the B. licheniformis acetolactate synthase gene is disclosed in the "GenBank" gene database under accession number NC_006270, which discloses the entire genomic sequence of B. licheniformis, in which the acetolactate synthase gene is in complementary form of bp 3675290 - It was found in a PCR reaction ⁇ Taq DNA polymerase, Qiagen) from genomic B lichenotypic DNA ⁇ strain DSM 13, commercially available from DSMZ GmbH) with the primers BLals-lf and BLals-2r as DNA Fragment of 1.7 kb size isolated.
- the genomic DNA used for the PCR reactions was previously in a conventional manner with a DNA isolation kit (Qiagen) from cells of the culture of. terrigena DSM 2687 and B. licheniformis DSM 13 in LB medium ⁇ 10 g / 1 tryptone, 5 g / 1 yeast extract, 5 g / 1 NaCl).
- pKKj is a derivative of the expression vector pKK223-3.
- the DNA sequence of pKK223-3 is disclosed in the "GenBank" gene database accessed by M77749.1, and approximately 1.7 kb were removed from the 4.6 kb plasmid (bp 262-1947 of M77749.1 of DNA sequence) resulting in the 2.9 kb expression vector pKKj ( Figure 3).
- the expression vectors pBudBkt and pALSbl were modified by incorporation of an expression cassette for the tetracycline resistance gene.
- the tetracycline resistance gene was first extracted from the plasmid pACYC184 (The DNA sequence of pACYC184 is accessible in the "Genbank” gene database under the accession number
- Primer tetlf SEQ ID NO: 5
- tet2r SEQ ID NO: 6
- the plasmid pBudBkt-tet (rev) was first cut with Hind III and the 2 kb expression cassette for the acetolactate synthase gene was isolated.
- pBudBkt-tet (rev) had been isolated upon cloning of the vector pBudBkt-tet and contained the tetracycline expression cassette in opposite orientation to the acetolactate synthase expression cassette ( Figure 6).
- the DNA TM fragment was cloned into the Hind III cut vector pACYC-LH.
- pACYC-LH is a 3.2 kb derivative of pACYC184 ( Figure 8) from which the chloramphenicol resistance marker was removed.
- the 4.2 kb vector pACYC184 was cut with Sca I and Bst 11071 and then religated.
- Plasmid DNA of the expression vectors pBudBkt-tet, pALSbl-tet and pAC-BudBkt was transformed into E. coli strain JM105 according to methods known per se.
- the control was E. coli JM105 transformed with the vector pACYC-LH.
- One clone each was selected and cultured in a shake flask culture.
- a preculture was prepared in LBtet medium (10 g / 1 tryptone, 5 g / 1 yeast extract, 5 g / 1 NaCl, 15 g / ml tetracycline) (cultivation at 37 ° C. and 120 ° C.) rpm overnight).
- acetolactate synthase 50 ml of the E. coli cells were centrifuged (10 min 15000 rpm, Sorvall RC5C centrifuge equipped with an SS34 rotor), the cell pellet in 2 ml KPi buffer (0.1 M potassium phosphate, 0, 1 M NaCl, pH 7.0), in a conventional manner with a so-called.
- KPi buffer 0.1 M potassium phosphate, 0, 1 M NaCl, pH 7.0
- acetolactate synthase activity was carried out in a manner known per se (Bauerle et al., Biochim Biophys Acta (1964) 92: 142-149).
- 1 U acetolactate synthase activity is defined as the amount of enzyme which produces 1 ⁇ acetolactate / min under test conditions.
- enzyme extract was adjusted to a volume of 2.5 ml together with assay buffer (100 mM KPi, 10 mM MgCl 2, pH 7.5) containing 2.5 ml of 80 mM Na-pyruvate, 160 ⁇ g / ml of thiamine pyruvate. dissolved in H20, mixed and incubated at 37 ° C.
- assay buffer 100 mM KPi, 10 mM MgCl 2, pH 7.5
- the amount of acetolactate formed was determined from a standard curve previously prepared with acetoin.
- the protein concentration of the cell extracts was determined in a manner known per se using the so-called "BioRad protein assay" from BioRad. 3rd example: Expression of acetolactate synthase in Klebsiella terrigena
- the starting strain was Klebsiella terrigena DSM 2687.
- the transformation with the plasmids pBudBkt-tet, pAC-BudBkt and pALSbl-tet was carried out in a manner known per se analogously to the methods for the transformation of E. coli known to the person skilled in the art.
- the control strain used was the untransformed wild-type strain Klebsiella terrigena DSM 2687.
- Transformants were isolated and tested for acetolactate synthase activity by shake flask culture. For this purpose, in each case 50 ml of FM2tet medium (without tetracycline in the K. terrigena wild-type control strain) were inoculated with a transformant and incubated for 24 h at 30 ° C. and 140 rpm (Infors shaker).
- FM2tet medium contained glucose 60 g / l; 10g / 1; Yeast Extract (Oxoid) 2.5 g / 1; Ammonium sulfate 5 g / 1; NaCl 0.5 g / 1; FeS0 4 x 7 H 2 0 75 mg / 1 ? Na 3 citrate x 2 H 2 0 1 g / 1; CaCl 2 ⁇ 2 H 2 O 14.7 mg / 1; MgS0 4 x 7 H 2 0 0.3 g / 1; KH 2 P0 4 1.5 g / 1; Trace element mix 10 ml / 1 and tetracycline 15 mg / 1.
- the pH of the FM2tet medium was adjusted to 6.0 before starting the culture.
- the trace element mix had the composition H 3 B0 3 2.5 g / l; CoCl 2 x 6 H 2 O 0.7 g / 1; CuSO 4 ⁇ 5 H 2 O 0.25 g / 1; MnCl 2 ⁇ 4 H 2 O 1.6 g / 1; ZnS0 4 ⁇ 7 H 2 0 0.3 g / 1 and Na 2 Mo0 4 ⁇ 2 H 2 0 0.15 g / 1.
- Example 2 The cells were analyzed as described in Example 2 for E. coli. Klebsiella cells were digested with the " French® Press" and the cell extracts analyzed for acetolactate synthase activity The specific acetolactate synthase activity in crude extracts of the various strains (determined as described in Example 2) is shown in Table 1. Table 1: Comparison of acetolactate synthase activity in recombinant K. terrigena strains
- Example 4 2, 3 -BDL production by shake flask cultivation of recombinant K. terrigena strains
- the determination of the 2,3-butanediol content in culture supernatants was carried out in a manner known per se by 1 H-NMR.
- an aliquot of the culture was centrifuged (10 min 5000 rpm, Eppendorf Labofuge) and 0.1 ml of the culture supernatant with 0.6 ml of TSP (3- (trimethylsilyl) ropionsäure- 2, 2, 3-d 4 sodium salt) standard solution defined content (internal standard, typically 5 g / 1) in D 2 0 mixed.
- TSP trimethylsilyl
- Feed medium (74% glucose, w / v) corresponding to the glucose consumption metered in via a peristaltic pump
- an alk Vegetable-based oxylated fatty acid ester commercially available under the name Struktol J673 from Schill & Seilacher (diluted 20-25% v / v in water).
- Klebsiella terrigena wild type strain control strain from Example 3
- the acetolactate synthase overproducing strains Klebsiella terrigena pBudBkt-tet, Klebsiella terrigena pAC-BudBkt and Klebsiella terrigena-pALSbl-tet see Example 4
- Fermentation medium was FM2tet medium (see 3rd example, medium without tetracycline for the K. terrigena wild-type control strain). 1.35 L of the medium was inoculated with 150 ml of preculture. The
- Preculture of the strains to be fermented was produced by 24 h shake flask cultivation in Baten fermentation medium.
- the Fermentation conditions were: temperature 30 ° C, stirrer speed 1000 rpm, aeration with 1 vvm, pH 6.0.
- the fermenter was sampled at regular intervals to analyze the following parameters:
- the cell density OD600 as a measure of the biomass formed was determined photometrically at 600 nm (BioRad photometer SmartSpec TM 3000).
- the glucose content was determined as described in Example 4.
- the 2, 3 -BDL content was determined by NMR as described in Example 4.
- the product was pumped by means of a pump (peristaltic pump 101 U / R of the Fa.
- Watson Marlow added a 74% (w / v) glucose solution.
- the feeding rate was determined by the current glucose consumption rate.
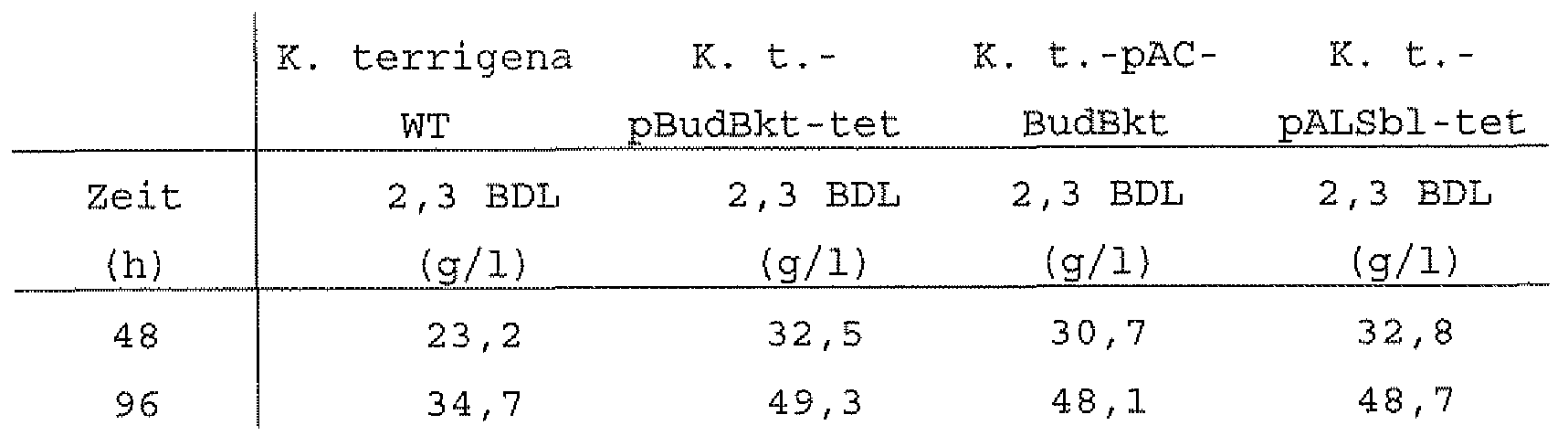
- Table 3 shows the time-dependent 2, 3 -BDL formation in the K. terrigena control strain and in the acetolactate synthase overproducing, recombinant Klebsiella terrigena strains. As already observed in the shake flask experiments (4.
- Example 6 2,3-BDL fermentation in 330 l scale
- the strain Klebsiella terrigena-pALSbl-tet was fermented (see 4th and 5th examples).
- An inoculum of Klebsiella terrigena pALSbl- tet in LBtet medium was prepared by 2 x 100 ml LBtet medium, each in a 1 1 Erlenmeyer flask, each with 0.25 ml of a Glycerol culture (overnight culture of the strain in LBtet medium, mixed with glycerol in a final concentration of 20% v / v and stored at -20 ° C) was seeded for 7 h at 30 ° C and 120 rpm on a Infors orbital shaker (cell density OD SO o / ml of 0.5-2.5) .100 ml of preculture used to inoculate 8 1 fermenter medium. Inoculated were two Vorfermenter with 8 1 fermenter medium.
- Prefermenters The fermentation was performed three fermenters the company Sartorius BBI Systems GmbH in two Biostat ® C-DCU. Fermentation medium was FM2tet (see Example 3) The fermentation was carried out in the so-called batch mode.
- Main fermentor The fermentation was ® in a Biostat D 500 fermenter (working volume 330 1, 500 1 tank volume) of the company Sartorius BBI Systems GmbH performed. Fermentation medium was FM2tet (see 3rd example). The fermentation took place in the so-called fed-batch mode.
- the glucose consumption was reduced by off-line glucose Measurement with a glucose analyzer from the company YSI determined (see 4th example). As soon as the glucose concentration of the fermentation seed was about 20 g / l (8-10 h after inoculation), the metered addition of a 60% w / w glucose feed solution was started. The flow rate of the feed was chosen so that during the production phase a glucose concentration of 10 - 20 g / 1 could be maintained. After completion of the fermentation, the volume in the fermenter 330 was 1. The analysis of the fermentation parameters was carried out as described in the 5th example. The production process is shown in Table 4.
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description






Claims
Priority Applications (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US13/979,878 US20130330794A1 (en) | 2011-01-31 | 2012-01-30 | Method for producing 2,3-butanediol by fermentation |
| BR112013019524A BR112013019524A2 (pt) | 2011-01-31 | 2012-01-30 | método para a produção de 2,3-butanodiol por fermentação |
| EP12701151.8A EP2670836A1 (de) | 2011-01-31 | 2012-01-30 | Verfahren zur fermentativen herstellung von 2,3-butandiol |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| DE102011003394A DE102011003394A1 (de) | 2011-01-31 | 2011-01-31 | Verfahren zur fermentativen Herstellung von 2,3-Butandiol |
| DE102011003394.7 | 2011-01-31 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2012104243A1 true WO2012104243A1 (de) | 2012-08-09 |
Family
ID=45531426
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/EP2012/051426 WO2012104243A1 (de) | 2011-01-31 | 2012-01-30 | Verfahren zur fermentativen herstellung von 2,3-butandiol |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20130330794A1 (de) |
| EP (1) | EP2670836A1 (de) |
| BR (1) | BR112013019524A2 (de) |
| DE (1) | DE102011003394A1 (de) |
| WO (1) | WO2012104243A1 (de) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN103497917A (zh) * | 2013-10-11 | 2014-01-08 | 山东大学 | 一株可利用木质纤维素原料发酵生产2,3-丁二醇的嗜热地衣芽孢杆菌 |
| DE102013224909A1 (de) | 2013-12-04 | 2015-06-11 | Wacker Chemie Ag | Verfahren zur Isolierung von 2,3-Butandiol aus Fermentationsansätzen |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DE102011004915A1 (de) | 2011-03-01 | 2012-09-06 | Wacker Chemie Ag | Verfahren zur Extraktion von 2,3-Butandiol aus einer wässrigen Mischung |
| DE102013204728A1 (de) | 2013-03-18 | 2014-09-18 | Wacker Chemie Ag | Mikroorganismenstamm und Verfahren zur fermentativen Herstellung von C4-Verbindungen aus C5-Zuckern |
| DE102013205986A1 (de) | 2013-04-04 | 2014-10-23 | Wacker Chemie Ag | Mikroorganismenstamm und Verfahren zur fermentativen Herstellung von C4-Verbindungen aus C5-Zuckern |
| DE102013216658A1 (de) | 2013-08-22 | 2015-02-26 | Wacker Chemie Ag | Verfahren zur fermentativen Herstellung von C4-Produkten und dafür geeignete Mikroorganismenstämme |
| DE102013223176A1 (de) | 2013-11-14 | 2015-05-21 | Wacker Chemie Ag | Verfahren zur fermentativen Herstellung von C4-Produkten und dafür geeignete Mikroorganismenstämme |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20070292927A1 (en) * | 2006-05-02 | 2007-12-20 | Donaldson Gail K | Fermentive production of four carbon alcohols |
| US20090305363A1 (en) * | 2008-06-05 | 2009-12-10 | E. I. Du Pont De Nemours And Company | Enhanced pyruvate to acetolactate conversion in yeast |
| WO2010037114A1 (en) * | 2008-09-29 | 2010-04-01 | Butamax™ Advanced Biofuels LLC | Enhanced pyruvate to 2,3-butanediol conversion in lactic acid bacteria |
| DE102010001399A1 (de) | 2010-01-29 | 2011-08-04 | Wacker Chemie AG, 81737 | Verfahren zur Herstellung von Carbonsäuren mit 1-3 Kohlenstoff-atomen aus nachwachsenden Rohstoffen |
-
2011
- 2011-01-31 DE DE102011003394A patent/DE102011003394A1/de not_active Withdrawn
-
2012
- 2012-01-30 BR BR112013019524A patent/BR112013019524A2/pt not_active Application Discontinuation
- 2012-01-30 WO PCT/EP2012/051426 patent/WO2012104243A1/de active Application Filing
- 2012-01-30 EP EP12701151.8A patent/EP2670836A1/de not_active Withdrawn
- 2012-01-30 US US13/979,878 patent/US20130330794A1/en not_active Abandoned
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20070292927A1 (en) * | 2006-05-02 | 2007-12-20 | Donaldson Gail K | Fermentive production of four carbon alcohols |
| US20090305363A1 (en) * | 2008-06-05 | 2009-12-10 | E. I. Du Pont De Nemours And Company | Enhanced pyruvate to acetolactate conversion in yeast |
| WO2010037114A1 (en) * | 2008-09-29 | 2010-04-01 | Butamax™ Advanced Biofuels LLC | Enhanced pyruvate to 2,3-butanediol conversion in lactic acid bacteria |
| DE102010001399A1 (de) | 2010-01-29 | 2011-08-04 | Wacker Chemie AG, 81737 | Verfahren zur Herstellung von Carbonsäuren mit 1-3 Kohlenstoff-atomen aus nachwachsenden Rohstoffen |
Non-Patent Citations (20)
| Title |
|---|
| "Biosafety in Microbiological and Biomedical Laboratories", December 2009, pages: 9FF |
| AUNE; AACHMANN, APPL. MICROBIOL. BIOTECHNOL., vol. 85, 2010, pages 1301 - 1313 |
| BAUERLE ET AL., BIOCHIM. BIOPHYS. ACTA, vol. 92, 1964, pages 142 - 149 |
| BLOMQVIST ET AL., J. BACTERIOL., vol. 175, 1993, pages 1392 - 1404 |
| BLOMQVIST K ET AL: "CHARACTERIZATION OF THE GENES OF THE 2,3-BUTANEDIOL OPERONS FROM KLEBSIELLA TERRIGENA AND ENTEROBACTER AEROGENES", JOURNAL OF BACTERIOLOGY, vol. 175, no. 5, 1 March 1993 (1993-03-01), pages 1392 - 1404, XP000577596, ISSN: 0021-9193 * |
| CELINSKA E ET AL: "Biotechnological production of 2,3-butanediol-Current state and prospects", BIOTECHNOLOGY ADVANCES, vol. 27, no. 6, 1 November 2009 (2009-11-01), pages 715 - 725, XP027205958, ISSN: 0734-9750, [retrieved on 20090512] * |
| CELINSKA; GRAJEK, BIOTECHNOL. ADVANCES, vol. 27, 2009, pages 715 - 725 |
| DAVID R. NIELSEN ET AL: "Metabolic engineering of acetoin and meso-2, 3-butanediol biosynthesis in E. coli", BIOTECHNOLOGY JOURNAL, vol. 5, no. 3, 1 March 2010 (2010-03-01), pages 274 - 284, XP055024039, ISSN: 1860-6768 * |
| DRANCOURT ET AL., INT. J. SYST. EVOL. MICROBIOL., vol. 51, 2001, pages 925 - 932 |
| JI X J ET AL: "Enhanced 2,3-butanediol production by Klebsiella oxytoca using a two-stage agitation speed control strategy", BIORESOURCE TECHNOLOGY, vol. 100, no. 13, 1 July 2009 (2009-07-01), pages 3410 - 3414, XP026063298, ISSN: 0960-8524, [retrieved on 20090317] * |
| NAKASHIMADA ET AL., J. BIOSCIENCE AND BIOENGINEERING, vol. 90, 2000, pages 661 - 664 |
| NAKASHIMADA YUTAKA ET AL: "Enhanced 2,3-butanediol production by addition of acetic acid in Paenibacillus polymyxa", JOURNAL OF BIOSCIENCE AND BIOENGINEERING, vol. 90, no. 6, December 2000 (2000-12-01), pages 661 - 664, XP055023577, ISSN: 1389-1723 * |
| PLATTEEUW C ET AL: "Metabolic engineering of Lactococcus lactis", APPLIED AND ENVIRONMENTAL MICROBIOLOGY, vol. 61, no. 11, 1 November 1995 (1995-11-01), pages 3967 - 3971, XP002078393, ISSN: 0099-2240 * |
| PLATTEEUW ET AL., APPL. ENVIRON. MICROBIOL., vol. 61, 1995, pages 3967 - 3971 |
| QIN J ET AL: "Production of 2,3-Butanediol by Klebsiella Pneumoniae Using Glucose and Ammonium Phosphate", CHINESE JOURNAL OF CHEMICAL ENGINEERING, vol. 14, no. 1, 1 February 2006 (2006-02-01), pages 132 - 136, XP022856384, ISSN: 1004-9541, [retrieved on 20060201] * |
| RENNA ET AL., J. BACTERIOL, vol. 175, 1993, pages 3863 - 3875 |
| RENNA M C ET AL: "REGULATION OF THE BACILLUS SUBTILIS ALSS, ALSD, AND ALSR GENES INVOLVED IN POST-EXPONENTIAL-PHASE PRODUCTION OF ACETOIN", JOURNAL OF BACTERIOLOGY, vol. 175, no. 12, 1 June 1993 (1993-06-01), pages 3863 - 3875, XP009011718, ISSN: 0021-9193 * |
| UI ET AL: "The production of D-acetoin by a transgenic Escherichia coli", LETTERS IN APPLIED MICROBIOLOGY, vol. 26, no. 4, 1 April 1998 (1998-04-01), pages 275 - 278, XP055024040, ISSN: 0266-8254 * |
| UI SADAHARU ET AL: "Molecular generation of an Escherichia coli strain producing only the meso-isomer of 2,3-butanediol", JOURNAL OF FERMENTATION AND BIOENGINEERING, vol. 84, no. 3, 1 January 1997 (1997-01-01), pages 185 - 189, XP008150599, ISSN: 0922-338X, [retrieved on 19980206] * |
| ZHI-KUI NIE ET AL: "An Effective and Simplified Fed-Batch Strategy for Improved 2,3-Butanediol Production by Klebsiella oxytoca", APPLIED BIOCHEMISTRY AND BIOTECHNOLOGY, vol. 163, no. 8, 1 April 2011 (2011-04-01), pages 946 - 953, XP055024045, ISSN: 0273-2289 * |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN103497917A (zh) * | 2013-10-11 | 2014-01-08 | 山东大学 | 一株可利用木质纤维素原料发酵生产2,3-丁二醇的嗜热地衣芽孢杆菌 |
| CN103497917B (zh) * | 2013-10-11 | 2016-02-24 | 山东大学 | 一株可利用木质纤维素原料发酵生产2,3-丁二醇的嗜热地衣芽孢杆菌 |
| DE102013224909A1 (de) | 2013-12-04 | 2015-06-11 | Wacker Chemie Ag | Verfahren zur Isolierung von 2,3-Butandiol aus Fermentationsansätzen |
Also Published As
| Publication number | Publication date |
|---|---|
| EP2670836A1 (de) | 2013-12-11 |
| DE102011003394A1 (de) | 2012-08-02 |
| BR112013019524A2 (pt) | 2018-06-26 |
| US20130330794A1 (en) | 2013-12-12 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP2670837A1 (de) | Verfahren zur fermentativen herstellung von 2,3-butandiol | |
| DE112014000710B4 (de) | Bakterienstamm mit hoher Ausbeute an 5-Aminolävulinsäure, Herstellungsverfahren und Verwendungen dafür | |
| WO2012104243A1 (de) | Verfahren zur fermentativen herstellung von 2,3-butandiol | |
| DE112008000243B4 (de) | Verfahren zur Herstellung eines Fermentationsproduktes aus Kohlenstoffquellen, die Glycerin enthalten, unter Verwendung von Corynebacteria | |
| DE60025976T2 (de) | DNA die für eine mutierte Isopropylmalatsynthase kodiert, Microorganismus, das L-Leucin produziert, und Verfahren zur Herstellung von L-Leucin | |
| EP2670838A1 (de) | Verfahren zur fermentativen herstellung von 2,3-butandiol | |
| US10119152B2 (en) | Fermentation process for producing chemicals | |
| EP1583838B1 (de) | Herstellung von milchsäure aus einem pentose-enthaltenden substrat | |
| DE69931162T2 (de) | Metabolisch veränderte mikrobiellen zelle mit veränderten metabolitherstellung | |
| DE10129711A1 (de) | Verfahren zur fermentativen Herstellung von Pyruvat | |
| DE102013204728A1 (de) | Mikroorganismenstamm und Verfahren zur fermentativen Herstellung von C4-Verbindungen aus C5-Zuckern | |
| DE102013216658A1 (de) | Verfahren zur fermentativen Herstellung von C4-Produkten und dafür geeignete Mikroorganismenstämme | |
| WO2014161723A1 (de) | Mikroorganismenstamm und verfahren zur fermentativen herstellung von c4-verbindungen aus c5-zuckern | |
| US5686277A (en) | Fermentation process for preparing xylitol using mutant cells | |
| US5792630A (en) | Cellulose-producing microorganism transformed with a gene for an enzyme involved in sucrose metabolism | |
| WO2012104233A1 (de) | Verfahren zur fermentativen herstellung von 2, 3-butandiol | |
| KR101576183B1 (ko) | 부산물을 이용한 에리스리톨 제조 방법, 에리스리톨 생산성이 우수한 모닐리엘라 폴리니스 변이 균주 및 이의 이용 | |
| DE102013223176A1 (de) | Verfahren zur fermentativen Herstellung von C4-Produkten und dafür geeignete Mikroorganismenstämme | |
| DE10333144B4 (de) | Verfahren zur biotechnologischen Herstellung von Citronensäure mit einer genetisch veränderten Hefe Yarrowia lipolytica | |
| DE10220234B4 (de) | Verfahren sowie Mikroorganismen zur mikrobiellen Herstellung von Pyruvat aus Kohlenhydraten sowie Alkoholen | |
| WO2023111053A2 (de) | Mischungen von glucose und xylose zur fermentativen herstellung von ortho-aminobenzoesäure | |
| EP1499728A2 (de) | Für eine mannitol-2-dehydrogenase codierende nukleotidsequenz sowie verfahren zur herstellung von d-mannitol | |
| WO2001011052A2 (de) | Ein- oder mehrzellige organismen zur herstellung von riboflavin | |
| DE112008003111B4 (de) | Corynebakterien, die Glyzerin enthaltende Kohlenstoffquellen verwerten, und Verfahren zur Herstellung eines Fermentationsprodukts unter Verwendung derselben | |
| WO2009053489A1 (de) | Fermentative herstellung von alpha-ketoglutarsäure |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 12701151 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 13979878 Country of ref document: US |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2012701151 Country of ref document: EP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| REG | Reference to national code |
Ref country code: BR Ref legal event code: B01A Ref document number: 112013019524 Country of ref document: BR |
|
| ENP | Entry into the national phase |
Ref document number: 112013019524 Country of ref document: BR Kind code of ref document: A2 Effective date: 20130731 |