PLANT GENES INVOLVED IN DEFENSE AGAINST PATHOGENS
Cross-Reference to Related Applications
This application claims the benefit of he filing date of U.S. application Serial No. 60/213,634, filed on June 23, 2000, U.S. application Serial No. 60/214,926, filed on June 23, 2000, U.S. application Serial No. 60/261,320, filed on January 12, 2001, U.S. application Serial No. 60/264,353, filed on January 26, 2001, and U.S. application Serial No. 60/273,879, filed on March 7, 2001 under 35 U.S.C. § 119(e).
Field of the Invention
The present invention generally relates to the field of plant molecular biology, and more specifically to the regulation of gene expression in plants in response to pathogen exposure.
Background ofthe Invention
Plants are capable of activating a large array of defense mechanisms in response to pathogen attack, some of which are preexisting and others are inducible. Pathogens must specialize to circumvent the defense mechanisms ofthe host, especially those biotrophic pathogens that derive their nutrition from an inthnate association with living plant cells. If the pathogen can cause disease, the interaction is said to be compatible, but if the plant is resistant, the interaction is said to be incompatible. A crucial factor determining the success of these mechanisms is the speed of their activation. Consequently, there is considerable interest in understanding how plants recognize pathogen attack and control expression of defense mechanisms. Some potential pathogens trigger a very rapid resistance response called gene-for-gene resistance. This occurs when the pathogen carries an avirulence (avr) gene that triggers specific recognition by a corresponding host resistance (R) gene. R gene specificity is generally quite narrow, in most cases only pathogens carrying a particular avr gene are recognized. Recognition is thought to be mediated by ligand-receptor binding. R genes have been studied extensively in recent years. For a review of R genes, see Ellis et al. (1998); Jones et al. (1997); and Ronald (1998).
One ofthe defense mechanisms triggered by gene-for-gene resistance is programmed cell death at the infection site. This is called the hypersensitive response, or HR. Pathogens that induce the HR, or cause cell death by other means, activate a systemic resistance response
called systemic acquired resistance (SAR). During SAR, levels of salicylic acid (SA) rise throughout the plant, defense genes such as pathogenesis related (PR) genes are expressed, and the plant becomes more resistant to pathogen attack. SA is a crucial component of this response. Plants that cannot accumulate SA due to the presence of a transgene that encodes an SA-degrading enzyme (nahG), develop a HR in response to challenge by avirulent pathogens, but do not exhibit systemic expression of defense genes and do not develop resistance to subsequent pathogen attack (Ryals et al., 1996). The nature ofthe systemic signal that triggers SAR is a subject of debate (Shulaev et al., 1995; Vernooji et al., 1994). SA clearly moves from the site ofthe HR to other parts ofthe plant, but if this is the signal, it must be effective at extremely low concentration (Willitset et al., 1998).
SAR is quite similar to some reactions that occur locally in response to attack by virulent (those that cause disease) or avirulent (those that trigger gene-for-gene resistance) pathogens. In general, activation of defense gene expression occurs more slowly in response to virulent pathogens than in response to avirulent pathogens. Some pathogens trigger expression of defense genes tlirough a different signaling pathway that requires components of the jasmonic acid (JA) and ethylene signaling pathways (Creelman et al., 1997).
One approach to understanding the signal transduction networks that control defense mechanisms is to use genetic methods to identify signaling components and determine their roles within the network. Considerable progress has been made using this approach in Arabidopsis-pathogen model systems. R gene signal transduction
Genes such as NDRI and EDSl, as well as DND1 and the lesion-mimic genes, likely act in signal transduction pathways downstream from R-αvr recognition. NDRI and EDSl are required for gene-for-gene mediated resistance to avirulent strains ofthe bacterial pathogen Pseudomonas syringae and the oomycete pathogen Peronospora parasitica. Curiously, ndrl mutants are susceptible to one set of avirulent pathogens, whereas edsl mutants are susceptible to a non-overlapping set (Aarts et el., 1998). The five cloned R genes that require EDSl all belong to the subset ofthe nucleotide binding site-leucine rich repeat (NBS-LRR) class of R genes that contain sequences similar to the cytoplasmic domains of Drosophila Toll and mammalian interleukin 1 transmembrane receptors (TIR-NBS-LRR). The two genes that require NDRI belong to the leucine-zipper (LZ-NBS-LRR) subclass of NBS-LRR genes. There is another LZ-NBS-LRR gene, RPP8, that does not require EDSl or NDRI, so the correlation between R gene structure and requirement for EDSl or NDRI is not perfect.
Nevertheless, these results show that R genes differ in their requirements for downstream factors and that these differences are correlated with R gene structural type.
NDRI encodes a protein with two predicted transmembrane domains (Century et al. 1997). RPMI, which requires NDRI to mediate resistance, is membrane-associated, despite the fact that its primary sequence does not include any likely membrane-integral stretches
(Boyes et al., 1998). It is possible that part ofthe function of NDRI is to hold R proteins close to the membrane. EDSl encodes a protein with blocks of homology to triacyl glycerol lipases (Falk et al, 1999). The significance of this homology is not known, but it is tempting to speculate that EDSl is involved in synthesis or degradation of a signal molecule. EDSl expression is inducible by SA and pathogen infection, suggesting that EDSl may be involved in signal amplification (Falk et al., 1999).
It has been extremely difficult to isolate mutations in genes other than the R genes that are required for gene-for-gene resistance. A selection procedure was devised (McNellis et al., 1998) on the basis of precisely controlled inducible expression ofthe avr gene avrRpt2 in plants carrying the corresponding resistance gene RPS2. Expression of avrRpt2 in this background is lethal, as it triggers a systemic HR. It is now possible to select for mutants with subtle defects in gene-for-gene signaling by requiring growth on a concentration of inducer slightly higher than the lethal dose.
Putative plant receptor proteins encoded by RPP genes (recognition of P. parasitica) mediate specific recognition of Peronospora isolates and trigger defense reactions. Recently, McDowell et al. (2000) reported that two members of this class, RPP7 and RPP8 (the latter of which encodes a LZ-NBS-LRR type R protein) were not significantly suppressed by mutations in either EDSl or NDRI, and that RPP7 resistance was also not compromised by mutations in EIN2, JAR1 or COI1, which affect ethylene or jasmonic acid signaling, or in coil /nprl or coil/NahG backgrounds. The authors suggested that RPP7 initiates resistance through a novel signaling pathway that is independent of salicylic acid accumulation or jasmonic acid response components. SA-dependent signaling
SA levels increase locally in response to pathogen attack, and systemically in response to the SAR-inducing signal. SA is necessary and sufficient for activation of PR gene expression and enhanced disease resistance. Physiological analyses and characterization of certain lesion-mimic mutants strongly suggest that there is a positive autoregulatory loop affecting SA concentrations (Shirasu et al., 1997; Hunt et al., 1997; Weymann et al., 1995).
Several mutants with defects in SA signaling have been characterized. These include nprl, in which expression of Pi? genes in response to SA is blocked; cprl, cpr5, and cprό, which constitutively express PR genes; the nprl suppressor ssil; pad4, which has a defect in S A accumulation; and eds5, which has a defect in PRl expression. Expression ofthe defense genes PRl, BG2, and PR5 in response to SA treatment requires a gene called NPRl or NIM1. Mutations in nprl abolish SAR, and cause enhanced susceptibility to infection by various pathogens (Cao et al., 1994; Delaney et al., 1995; Glazebrook et al, 1996; Shah et al., 1997). NPRl appears to be a positive regulator of PP gene expression that acts downstream from SA. NPRl encodes a novel protein that contains ankyrin repeats (which are often involved in protein-protein interactions (Cao et al., 1997; Ryals et al., 1997), and that is localized to the nucleus in the presence of SA (Dong et al., 1998). Consequently, it is unlikely that NPRl acts as a transcription factor to directly control PR gene expression, but its nuclear localization suggests that it may interact with such transcription factors. PAD4 appears to act upstream from SA. \npad4 plants infected with a virulent P. syringae strain, SA levels, synthesis ofthe antimicrobial compound camalexin, and Pi expression are all reduced (Zhou et al., 1998). SA is necessary, but not sufficient, for activation of camalexin synthesis (Zhou et al, 1998; Zhao et al., 1996). The camalexin defect iupad4 plants is reversible by exogenous SA (Zhou et al., 1998). Mutations vsxpad4 do not affect SA levels, camalexin synthesis, or PRl when plants are infected with an aviralent P. syringae strain (Zhou et al., 1998). Taken together, these results suggest that PAD4 is required for signal amplification to activate the SA pathway in response to pathogens that do not elicit a strong defense response (Zhou et al., 1998). JA-dependent signaling JA signaling affects diverse processes including fruit ripening, pollen development, root growth, and response to wounding (Creelman et al., 1997). Vas, jar 1 and coil mutants fail to respond to JA (Feys et al., 1994; Staswick et al., 1992). COI1 has been cloned, and found to encode protein containing leucine-rich repeats and a degenerate F-box motif (Xie et al., 1998). These features are characteristic of proteins that function in complexes that ubiquitinate protein targeted for degradation.
In the past few years it has become apparent that JA plays an important role in regulation of pathogen defenses. For example, the induction ofthe defensin gene PDF1.2 after
inoculation of Arabidopsis with the avirulent pathogen Alternaria brassicicola does not require SA or NPRl, but does require ethylene and JA signaling (Penninck et al., 1996). SA signaling and JA signaling pathways are interconnected in complicated ways. Studies in other systems have shown that SA signaling and JA signaling are mutually inhibitory (Creelman et al., 1997; Harms et al., 1998). However, synthesis of camalexin in response to P. syringae infection is blocked in nahG (Zhou et al., 1998; Zhao et al., 1996) and coil (Glazebrook, 1999) plants, strongly suggesting that camalexin synthesis requires both SA and JA signaling. Induced systemic resistance (ISR) Some rhizosphere-associated bacteria promote disease resistance (van Loon et al.,
1998). This phenomenon, called ISR, has been studied using Pseudomonas fluorescens strain WCS417r to colonize Arabidopsis roots (Pieterse et al., 1996). Colonized plants are more resistant to infection by the fungal pathogen Fusarium oxysporum f sp raphani and P. syringae (Pieterse et al., 1996). ISR occurs in nahG plants, indicating that it is not a SA-dependent phenomenon (Pieterse et al., 1996). Rather, ISR appears to be JA- and ethylene-dependent. The observation that ethylene can induce ISR jarl mutants led to the hypothesis that ISR requires a JA signal followed by an ethylene signal (Pieterse et al., 1998). No changes in gene expression associated with ISR have been detected (Pieterse et al., 1998), suggesting that it is different from activation of PDF 1.2 expression by A. brassicicola. Curiously, ISR requires NPRl (Pieterse et al., 1996). This was unexpected in light of the fact that NPRl was previously known to be involved only in SA-dependent processes and ISR is SA-independent. If the SA-dependent signal is received, NPRl mediates a resistance response characterized by PRl expression, whereas if the ISR signal is received, NPRl mediates a different resistance response. It is difficult to imagine how this could occur, unless NPRl is interacting with different 'adapter' molecules to mediate the different signals. The ankyrin repeats found in NPRl could function in protein-protein interactions between NPRl and adapter proteins. Identification of proteins that interact with NPRl, and characterization of plants with loss-of-function mutations affecting those proteins, would be very helpful for understanding how NPRl acts in each pathway. It would also be worthwhile to determine if the ssil or cprό mutations suppress the ISR defect of nprl mutants. Relevance to disease resistance
Characterization ofthe effects of various mutations on resistance to different pathogens has revealed that there is considerable variation in the extent to which pathogens are affected
by defense mechanisms. SAR is known to confer resistance to a wide array of pathogens, including bacteria, fungi, oomycetes, and viruses. JA signaling is important for limiting the growth of certain fungal pathogens. In. Arabidopsis, the SA pathway mutants nprl and pad4 show enhanced susceptibility to P. syringae and P. parasitica (Cao et al., 1994; Delaney et al., 1995; Shah et al., 1997; Zhou et al., 1998; Glazebrook et al., 1997).
Overexpression of rate-limiting defense response regulators may cause the signaling network to respond faster or more strongly to pathogen attack, thereby improving resistance. For example, overexpression of NPRl caused increased resistance to P. syringae and P. parasitica in a dosage dependent manner (Cao et al., 1998). Moreover, NPP -overexpression had no obvious deleterious effects on plant growth, in contrast to mutations that lead to constitutive overexpression of defense responses, which generally cause dwarfism. Promoters for gene expression of plant pathogen defense genes
Promoters (and other regulatory components) from bacteria, viruses, fungi and plants have been used to control gene expression in plant cells. Numerous plant transformation experiments using DNA constructs comprising various promoter sequences fused to various foreign genes (for example, bacterial marker genes) have led to the identification of useful promoter sequences. It has been demonstrated that sequences up to 500-1,000 bases in most instances are sufficient to allow for the regulated expression of foreign genes. However, it has also been shown that sequences much longer than 1 kb may have useful features which permit high levels of gene expression in transgenic plants. The expression of genes encoding proteins that are useful for protecting plants from pathogen attack may have deleterious effects on plant growth if expressed constitutively. Consequently, it is desirable to have promoter sequences that control expression of these gene(s) in such a way that expression is absent or very low in the absence of pathogens, and high in the presence of pathogens. Thus, what is needed is the identification of plant genes useful to confer resistance to a pathogen(s) and plant promoters, the expression of which is altered in response to pathogen attack.
Summary of the Invention The invention generally provides an isolated nucleic acid molecule (polynucleotide) comprising a plant nucleotide sequence obtained or isolatable from a gene, the expression of which is altered, either increased or decreased, in response to pathogen infection. In one embodiment, the plant nucleotide sequence comprises an open reading frame, while in another
embodiment, the plant nucleotide sequence comprises a promoter. A promoter sequence ofthe invention directs transcription of a linked nucleic acid segment, e.g., a linked plant DNA comprising an open reading frame for a stractural or regulatory gene, in a host cell, such as a plant cell, in response to pathogen infection of that cell. As used herein, a "pathogen" includes bacteria, fungi, oomycetes, viruses, nematodes and insects, e.g., aphids (see Hammond-Kosack and Jones, 1997) . Moreover, the expression of a plant nucleotide sequence of the invention comprising a promoter may be altered in response to one or more species of bacteria, nematode, fungi, oomycete, virus, or insect. Likewise, the expression of a plant nucleotide sequence ofthe invention comprising an open reading frame may be useful to confer tolerance or resistance of a plant to one or more species of bacteria, nematode, fungi, oomycete, virus or insect.
The nucleotide sequence preferably is obtained or isolatable from plant DNA. In particular, the nucleotide sequence is obtained or isolatable from a gene encoding a polypeptide which is substantially similar, and preferably has at least 70%, e.g., 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, and even 90% or more, e.g., 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, and 99%, amino acid sequence identity, to a polypeptide encoded by an Arabidopsis gene comprising any one of SEQ ID NOs: 1-953 and 2137-2661 or a fragment (portion) thereof which encodes a partial length polypeptide having substantially the same activity ofthe full-length polypeptide, a rice gene comprising one of SEQ ID NOs:2000-2129 or SEQ ID NOs:2662- 6813, or a Chenopodium gene comprising one of SEQ ID NOs:1954-1966.
The present invention also provides an isolated nucleic acid molecule comprising a plant nucleotide sequence that directs transcription of a linked nucleic acid segment in a host cell, e.g., a plant cell. The nucleotide sequence preferably is obtained or isolatable from plant genomic DNA.In particular the plant DNA is obtained or isolatable from a gene encoding a polypeptide which is substantially similar, and preferably has at least 10%, e.g., 71%), 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, and even 90% or more, e.g., 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, and 99%, amino acid sequence identity, to a polypeptide encoded by an Arabidopsis gene comprising any one of SEQ ID NOs: 1-953, a rice gene comprising one of SEQ ID NOs:2000-2129 or SEQ ID NOs:2662-4737, or a Chenopodium gene having any one of SEQ ID NOs:1954-1966, the expression of which is increased or decreased in response to pathogen infection. Prefened promoters comprise DNA obtained or isolatable from a gene encoding a polypeptide which is
substantially similar, and preferably has at least 70%, e.g., 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, and even 90% or more, e.g., 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, and 99%, amino acid sequence identity, to a polypeptide encoded by an Arabidopsis gene comprising a promoter according to SEQ ID NOs:2137-2661, a rice gene comprising a promoter accordint to SEQ ID NOs:4738- 6813 or a fragment thereof (i.e., promoters isolatable from any one of SEQ ID NOs:2137-2661 or SEQ ID NOs:4738-6813) which increases or decreases transcription of a linked nucleic acid segment in response to pathogen infection.
The invention also provides uses for an isolated nucleic acid molecule, e.g., DNA or RNA, comprising a plant nucleotide sequence comprising an open reading frame encoding a polypeptide which is substantially similar, and preferably has at least 70%, e.g., 71%>, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, and even 90% or more, e.g., 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, and 99%, amino acid sequence identity, to a polypeptide encoded by an Arabidopsis, Chenopodium or rice gene comprising an open reading frame comprising any one of SEQ ID NOs: 1-953, 1954- 1966, 2000-2129, 2662-4737, or the complement thereof. For example, these open reading frames may be useful to prepare plants that over- or under-express the encoded product or to prepare knockout plants.
The promoters and open reading frames ofthe invention can be identified by any method. For example, they can be identified by employing an anay of nucleic acid samples, e.g., each sample having a plurality of oligonucleotides, and each plurality conesponding to a different plant gene, on a solid substrate, e.g., a DNA chip, and probes corresponding to nucleic acid which is up- or down-regulated in response to pathogen infection in one or more ecotypes or species of plant relative to a control (e.g., a water control, nucleic acid from an uninfected plant or nucleic acid from a mutant plant). Thus, genes that are upregulated or downregulated in response to pathogen infection can be systematically identified.
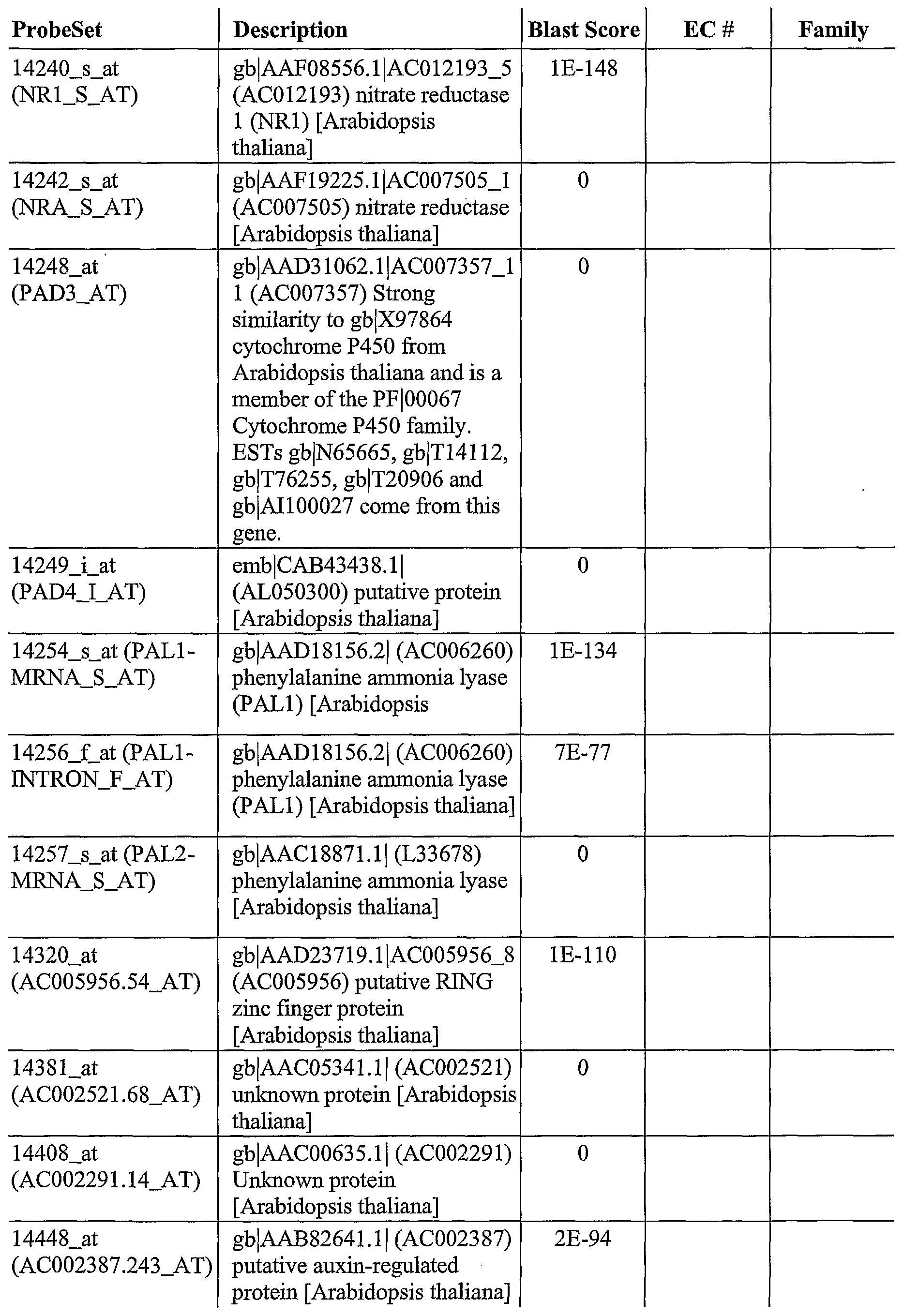
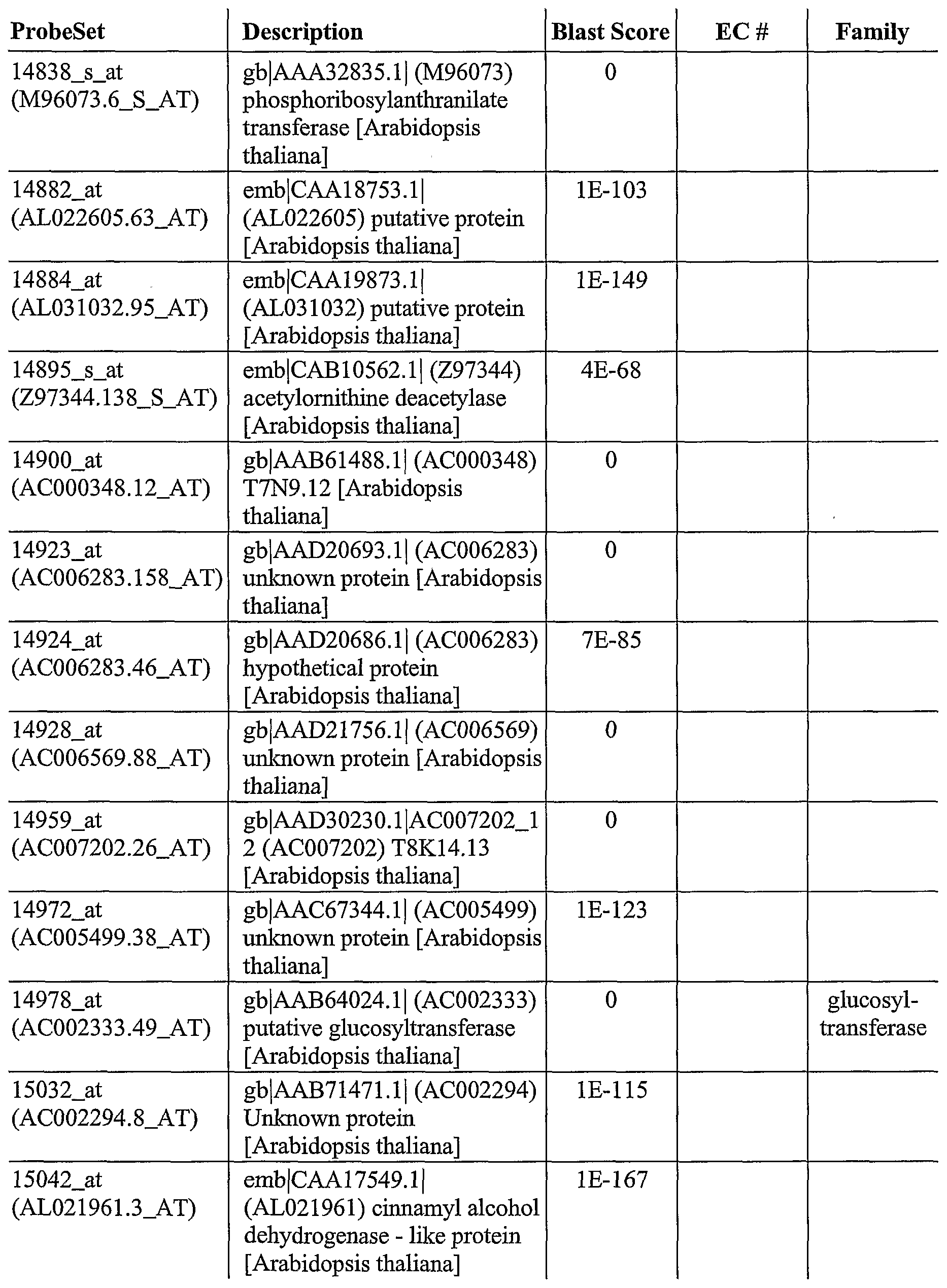
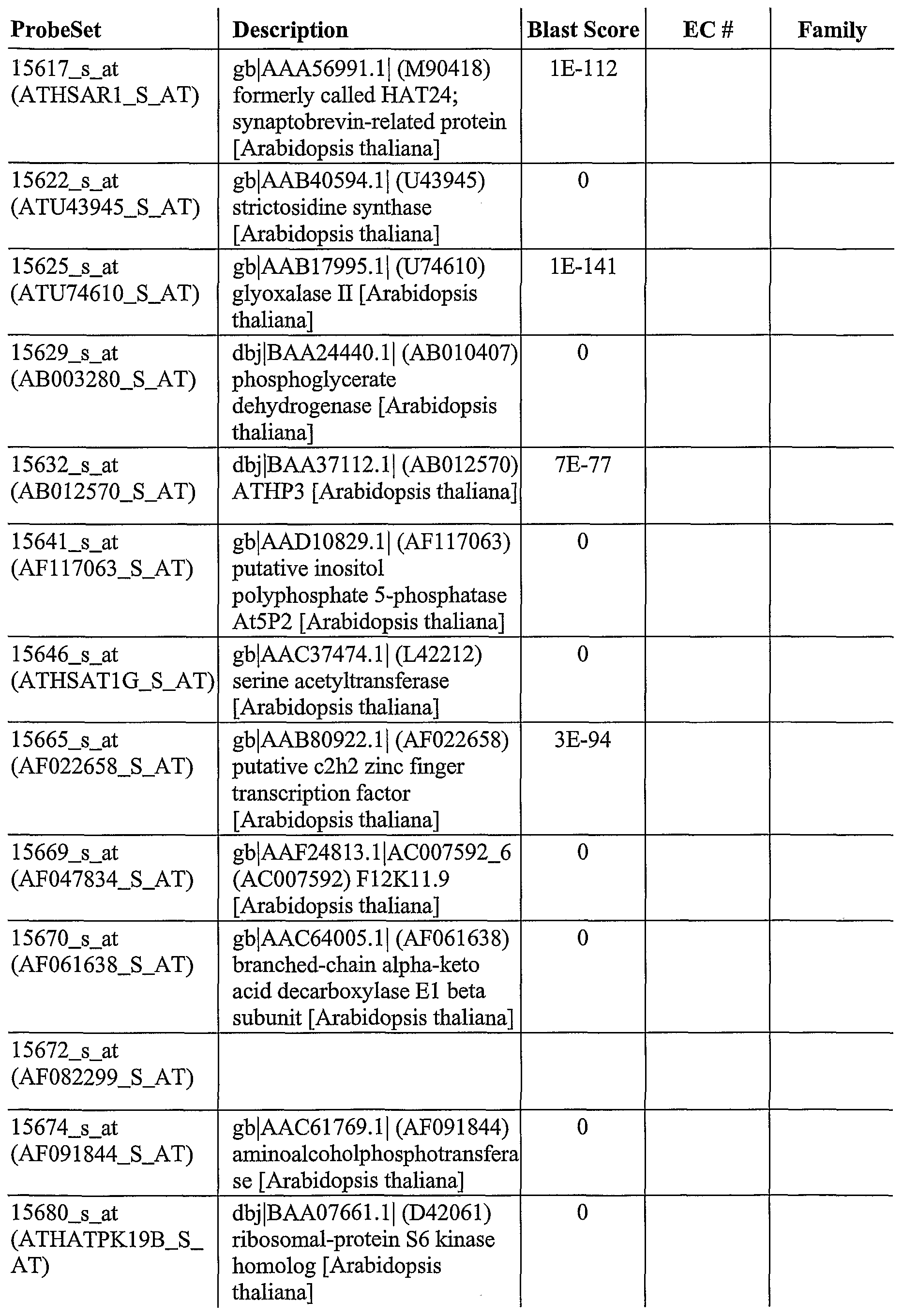
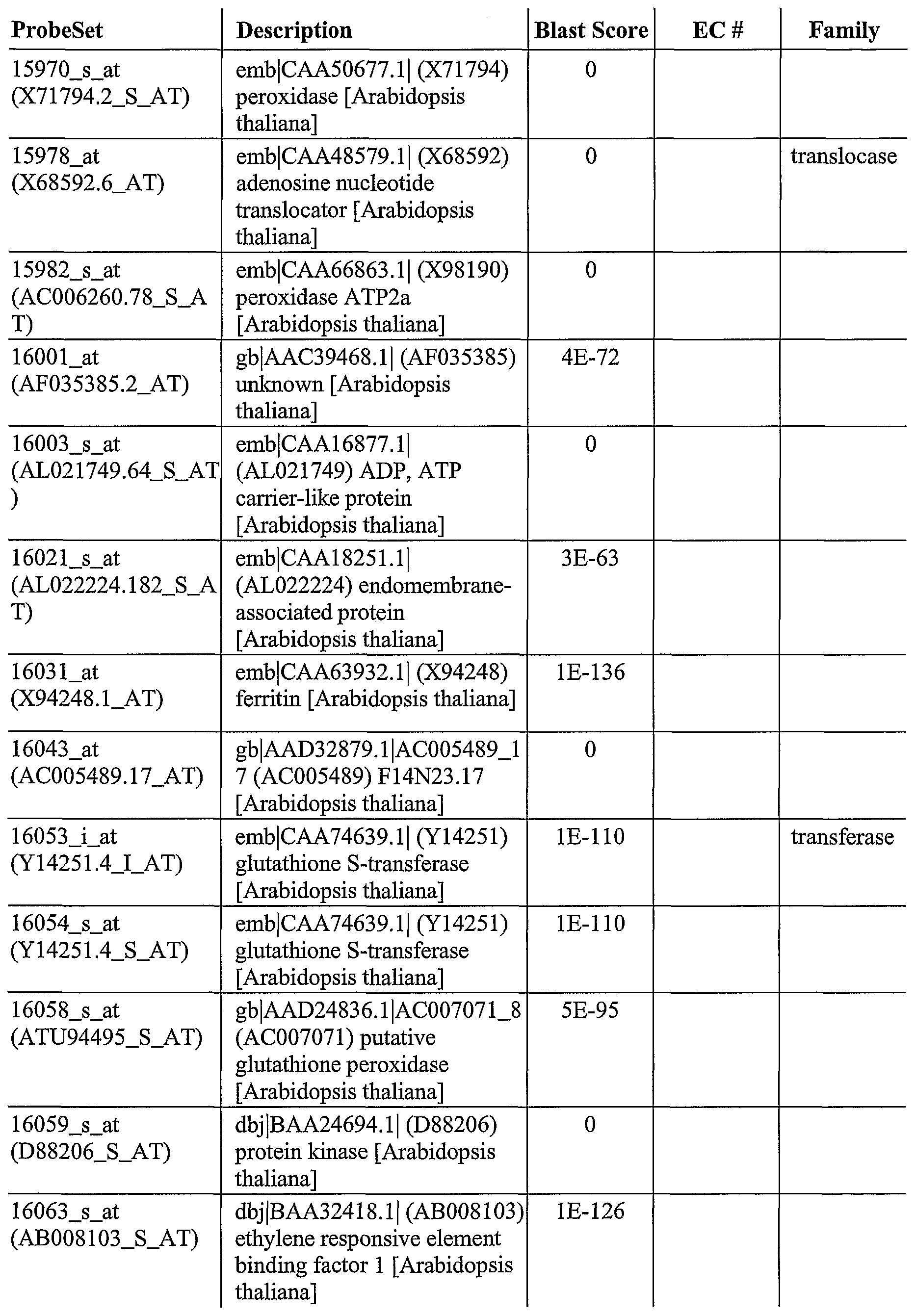
As described herein, GeneChip® technology was utilized to discover a plurality of genes, the expression of which is altered after pathogen infection. The Arabidopsis oligonucleotide probe anay consists of probes from about 8,100 unique Arabidopsis genes, which covers approximately one third ofthe genome. This genome array permits a broader, more complete and less biased analysis of gene expression. Using labeled cRNA probes, expression levels were determined by laser scanning and genes generally selected for expression levels that were > 2 fold over the control.
For example, using this approach, 953 genes were identified, the expression of which was altered after infection of wild-type Arabidopsis plants with a pathogen (SEQ ID NOs: 1- 953). In addition, 745 genes were identified, the expression which was increased after infection of wild-type Arabidopsis with Pseudomonas syringae (SEQ ID NOs: 2-6, 8-13, 16, 18, 22-23, 25, 28-29, 31-32, 35-37, 39-43, 45-47, 49-50, 52, 54-55, 57-58, 60-66, 70-72, 74, 76-77, 79, 81, 83, 85, 87-90, 92, 94, 97, 100-107, 111-115, 117-125, 127-135, 138-140, 142- 153, 156-158, 160, 162-165, 168-170, 173-181, 183-184, 186-188, 190-198, 200-201, 203- 211, 214-215, 218-224, 227-232, 234-249, 251-262, 264, 266-268, 270, 272-275, 277-281, 283, 286-294, 297-298, 302, 304-306, 308-326, 328-339, 341, 344-345, 347, 350-351, 353- 358, 361-371, 373-377, 379-386, 388-390, 392, 394-400, 402-406, 408-410, 412-417, 419- 427, 429-433, 435-443, 445-452, 454-457, 459-460, 462-464, 466-470, 473-475, 478-479, 481-482, 484-187, 489-494, 496-498, 500-501, 503-506, 508, 510, 512-515, 517-523, 526, 528-529, 531-538, 540, 544-548, 550-558, 560, 563-568, 570, 572-577, 579-580, 582-585, 588-594, 596, 598-600, 602-603, 605-606, 608-612, 614-617, 619-624, 626-630, 632-639, 642, 644, 646-651, 653-657, 659-665, 667-671, 673-678, 681-689, 691-693, 695-713, 715- 717, 719, 721-727, 729-733, 736-738, 740, 742, 744, 746, 748-752, 755-756, 758-760, 762- 769, 771, 774, 776-781, 783-788, 790-796, 798-799, 802, 804-808, 810-815, 817-831, 833- 848, 850-855, 857-869, 871-880, 882-900, 903-907, 909, 911-915, 918-920, 922-925, 927, 929, 931-938, 940, 943-945, 947, and 950-953). Ofthe 745 genes, the expression of 530 of those genes was altered in at least one mutant Arabidopsis after infection with Pseudomonas syringae (SEQ ID NOs: 2, 4-6, 11-13, 18, 22-23, 28, 31, 36, 39-43, 45, 47, 49-50, 52, 54-55, 57-58, 60-61, 63-66, 71-72, 74, 77, 81, 83, 85, 87-89, 92, 97, 100-107, 111-112, 114-115, 117- 120, 122, 125, 127-128, 134, 128-140, 143-144, 148-151, 153, 156-157, 160, 165, 168-170, 173-174, 176-180, 183, 187-188, 191, 193-194, 197-198, 200, 203-210, 214, 219-224, 227, 230-232, 235-237, 239-240, 243-246, 248-249, 251-254, 256-258, 261, 264, 266-268, 270, 272-275, 277-278, 280, 283, 286-287, 290-293, 297, 302, 305-306, 308-310, 312-316, 321- 326, 328-331, 333, 336-339, 341, 345, 351, 353, 355-358, 361-363, 365-366, 368-371, 373, 375, 377, 379-381, 384-385, 388-390, 392, 394-400, 402-406, 410, 412, 415-416, 419-420, 422-425, 429-433, 435-439, 441-443, 445-452, 454, 459-460, 463, 466, 468-470, 473, 481- 482, 485-486, 489, 491-494, 497-498, 500-501, 503, 505-506, 508, 510, 513-515, 517, 520- 521, 523, 528-529, 531, 533-538, 540, 545-548, 550-551, 553-554, 556-558, 560, 566-567, 575, 580, 582-584, 588-593, 596, 598-600, 602-603, 605-606, 608-610, 612, 614, 616, 620- 622, 627-629, 633-634, 636-639, 644, 646, 648-651, 654-657, 659, 661-663, 667, 669, 673-
674, 677, 682, 684-687, 689, 691-693, 697, 699, 701, 703-708, 713, 717, 719, 721-727, 730- 733, 736, 740, 744, 746, 749-752, 755-756, 758-760, 762-764, 766-769, 774, 776-778, 780- 781, 786, 788, 791-796, 799, 802, 804-808, 810-812, 815, 818-821, 823-825, 827-829, 831, 833-836, 838-843, 845, 847-848, 852-853, 855, 858, 860-869, 871-874, 876, 878-880, 884- 887, 889, 892-894, 896-900, 904-907, 911-915, 918-920, 922-924, 931, 933, 938, 943-945, 947, and 950-952). Ofthe 530, 81 encode regulatory factors (SEQ ID NOs: 39, 52, 60, 63, 81, 83, 106, 107, 115, 117, 118, 168, 174, 176, 179, 204, 207, 208, 220, 221, 248, 258, 268, 275, 280, 309, 323, 326, 329, 351, 419, 422, 429, 430, 432, 459, 460, 468, 469, 473, 500, 505, 506, 508, 529, 531, 533, 535, 538, 545, 553, 602, 606, 608, 610, 614, 616, 634, 654, 655, 684, 686, 687, 691, 717, 751, 752, 766, 777, 815, 831, 834, 835, 839, 841, 847, 876, 884, 906, 920, and 924).
As also described herein, 333 genes were identified that are useful to confer improved resistance to plants to bacterial infection (SEQ ID NOs: 12-13, 18, 23, 36, 39-40, 43, 45, 50, 52, 57-58, 60-61, 64, 71-72, 81, 87-89, 97, 100, 102-105, 107, 111-112, 115, 119-120, 122, 125, 127-128, 140, 144, 148-150, 153, 165, 168-169, 176-177, 179, 183, 188, 191, 193-194, 197-198, 203-206, 208-209, 214, 219-222, 227, 230, 232, 237, 244-246, 248-249, 251-253, 258, 261, 264, 266, 268, 273-275, 283, 287, 290, 293, 297, 302, 305-306, 308, 312-315, 321- 322, 324, 326, 330, 333, 338, 341, 345, 353, 356-358, 362-363, 366, 369, 371, 375, 377, 380, 384-385, 389, 392, 394-395, 398-399, 402-404, 406, 410, 415, 419, 422, 425, 429-430, 433, 435-439, 443, 445-452, 454, 463, 466, 468-470, 473, 486, 489, 491-492, 4894, 498, 500-501, 503, 508, 513-514, 517, 529, 533-538, 548, 550, 553-554, 4556-558, 566, 575, 580, 582-583, 590-591, 593, 600, 602, 609-610, 612, 614, 620-622, 627-629, 637-638, 644, 649, 654-657, 659, 663, 667, 669, 673-674, 677, 684-685, 689, 691-693, 699, 703-705, 708, 719, 721, 724- 726, 730-732, 744, 746, 749-750, 752, 755-756, 758, 760, 762-764, 767, 769, 774, 780-781, 786, 788, 791-792, 794-796, 799, 804-808, 810-812, 815, 818-819, 823, 828-829, 833,
840841, 843, 847, 852-853, 858, 860, 862-865, 867-868, 872-874, 876, 885-887, 889, 892- 894, 896-900, 904-905, 907, 911-914, 918-920, 922-924, 931, 933, 938, 947, 950, and 952).
Further, 296 genes were identified that are useful to confer improved resistance to plants to fungal infection (SEQ ID NOs: 2, 4, 6, 11-13, 18, 22-23, 31, 41-43, 49-50, 54, 57-58, 61, 64-66, 71-72, 74, 77, 85, 87, 89, 92, 97, 101, 103, 106-107, 112, 114, 117-119, 125, 128, 134, 138, 143, 149, 151, 156-157, 165, 169-170, 174, 176-180, 187-188, 191, 193, 206, 208, 219-220, 222, 224, 231, 236, 239, 243-245, 251-254, 256-257, 267, 272, 287, 290, 292, 297, 302, 312-313, 315-316, 321-322, 324-325, 328, 330, 345, 351, 353, 355-357, 362-363, 366,
368-371, 373, 375, 379, 381, 384, 388-390, 392, 395-400, 405, 410, 415-416, 419, 422, 424, 431-432, 435-436, 438-439, 447, 459-460, 470, 473, 481-482, 489, 491, 493-494, 500-501, 505-506, 513-514, 517, 520-521, 523, 528-529, 531, 535, 537-538, 540, 545-548, 551, 553- 554, 557-558, 566, 575, 580, 582, 584, 589, 591, 593, 596, 598-599, 603, 605, 608-609, 612, 628, 633-634, 636-637, 639, 646, 648, 650-651, 656, 661, 663, 667, 674, 685-687, 689, 691, 693, 697, 699, 701, 705, 707, 713, 723-724, 726, 736, 740, 749, 751-752, 756, 758-759, 764, 766-768, 774, 776, 778, 780, 792-796, 799, 802, 806, 810-812, 818, 820-821, 825, 827-829, 833-836, 838-839, 841-843, 848, 855, 860-861, 866, 868-869, 871, 873-874, 876, 878-880, 889, 892, 898-900, 904-905, 907, 915, 918, 922, 924, 933, 943-945, 947, and 951). In addition, 288 genes were identified that are useful to confer improved resistance to plants to infection with more than one pathogen, e.g., pathogens that include bacteria, oomycetes and viruses (SEQ ID NOs: 12-13, 18, 23, 36, 39-40, 43, 45, 50, 52, 57-58, 60-61, 64, 71-72, 81, 87-88, 100, 102-105, 107, 111-112, 115, 119-120, 122, 125, 127-128, 140, 148- 150, 153, 168-169, 176-177, 188, 191, 193-194, 197-198, 203-206, 209, 219-222, 227, 232, 237, 244-246, 248-249, 251-253, 258, 261, 264, 266, 268, 273-275, 283, 287, 290, 293, 297, 302, 305-306, 308, 312-315, 324, 326, 330, 333, 341, 345, 353, 356, 358, 366, 371, 375, 377, 380, 385, 389, 392, 394, 398, 402-404, 406, 410, 415, 419, 425, 429-430, 433, 435-438, 443, 445.447, 449-452, 454, 463, 466, 468-470, 473, 486, 489, 492, 494, 498, 500-501, 503, 508, 513-514, 517, 533-538, 548, 550, 553-554, 57-558, 566, 575, 582-583, 590-591, 593, 600, 602, 609-610, 612, 620-622, 627-629, 637-638, 644, 649, 654-657, 659, 667, 669, 673, 677, 684, 689, 692-693, 703-705, 719, 721, 724-726, 730-732, 744, 746, 749-750, 752, 755-756, 760, 762-764, 767, 769, 774, 780-781, 786, 788, 791-792, 795-796, 805-808, 810-812, 815, 818-819, 823, 828, 833, 840-841, 843, 852-853, 858, 860, 862-865, 867-868, 872-874, 876, 887, 889, 893-894, 896-898, 900, 905, 907, 911-914, 918-920, 922-923, 931, 933, 938, 947, 950, and 952).
Using the same approach described above, 25 genes were identified (SEQ ID NOs: 1, 15, 19, 20, 24, 26, 27, 34, 38, 51, 56, 59, 67-69, 99, 116, 155, 159, 182, 212, 284, 372, 444, and 789), the expression of which was decreased at 6 hours in an avr2 plant. Also identified were 33 genes (SEQ ID NOs: 17, 70, 76, 81, 84, 109, 123, 144, 160, 230, 265, 268, 269, 271, 323, 333, 385, 427, 428, 430, 457, 505, 569, 597, 602, 606, 616, 708, 730, 741, 812, 862, and 942), the expression of which was elevated in an incompatible or a compatible interaction in four Arabidopsis ecotypes infected with bacteria. Eight ofthe genes were upregulated by 3 hours in an incompatible interaction, 18 ofthe genes were upregulated by 6 hours, but not at 3
hours, in an incompatible interaction, and 6 ofthe genes were upregulated in a compatible interaction.
Further identified were 33 genes, the expression of which was induced early after infection (SEQ ID NOs:17, 21, 80, 81, 156, 174, 176, 221, 227, 296, 302, 303, 306, 333, 340, 360, 500, 505, 524, 575, 601, 602, 614, 628, 687, 733, 782, 811, 835, 862, 900, 905, and 912), 10 genes, the expression of which was decreased early after infection (SEQ ID NOs:30, 73, 282, 541, 640, 679, 761, 870, 917, and 930), and 135 genes, 107 of which were induced at 3 and/or 6 hours after infection, and 28 of which were decreased after infection (SEQ ID NOs:7, 21, 33, 44, 46, 60, 82, 86, 91, 93, 106, 110, 119, 122, 130, 131, 136, 141, 154, 161, 166-168, 171, 176, 185, 189, 199, 200, 202, 203, 213, 225, 227, 248, 261, 262, 266, 274, 285, 300, 301, 302, 320, 326, 341, 345, 348, 349, 360, 366, 378, 406, 409, 422, 425, 434, 441, 443, 446, 449, 454, 461, 471, 475, 476, 483, 485, 499, 500, 511, 512, 516, 527, 530, 533, 543, 545, 549, 550, 552, 567, 575, 578, 586, 590, 608, 611, 615, 618, 625, 631, 643, 656, 658, 659, 666, 668, 671, 680, 690, 694, 704, 706, 711, 714, 718, 721, 728, 734, 738, 757, 770, 772, 791, 807, 811, 813, 816, 827, 857, 864, 868, 875, 881, 893, 901, 905, 908, 912, 916, 939, 941, 951, and 952). In a similar approach, 48 genes that were upregulated in response to infection, e.g., bacterial or fungal infection, as well as 46 ofthe conesponding promoter containing regions, were identified. Thirty-six ofthe genes were upregulated in response to bacterial, e.g., Pseudomonas, infection (the promoters for genes conesponding to SEQ ID NOs: 104-106, 119, 123, 129, 131, 151-152, 183, 191, 198, 200, 227, 249, 274, 302, 358, 415, 481, 547, 566, 582, 628, 633, 639, 656, 673, 793, 818, 827, 864, 874, 880, and 904-905), while 23 ofthe genes were upregulated in response to fungal, e.g., Botrykis, infection (SEQ ID NOs: 18, 71, 119, 123, 129, 151, 191, 244, 245, 302, 545, 547, 562, 566, 637, 653, 747, 756, 774, 793, 842, 864, and 905). Twenty-five ofthe genes were upregulated only in response to bacterial, e.g., Pseudomonas, infection (the promoters for genes conesponding to SEQ ID NOs: 104-106,
131, 152, 183, 198, 200, 227, 249, 274, 358, 415, 481, 582, 628, 633, 639, 656, 673, 818, 827, 874, 880, and 904 are provided in SEQ ID NOs:1001-1025), 10 ofthe genes were upregulated only in response to fungal, e.g., Botrytis, infection (the promoters for genes conesponding to SEQ ID NOs: 18, 71, 244, 245, 545, 562, 637, 653, 747, 756, 774, and 842 are provided in SEQ ID NOs : 1026- 1035), and 11 genes were upregulated in response to both bacterial and fungal infection (the promoters for genes conesponding to SEQ ID NOs: 119, 123, 129, 151, 191, 302, 547, 566, 793, 864, and 905 are provided in SEQ ID NOs:1036-1046).
As also described hereinbelow, 129 Arabidopsis genes (SEQ ID NOs: 3, 51, 54, 60, 61, 66, 75, 76, 78, 88, 95, 96, 101, 106, 108, 123, 126, 128, 129, 131, 137, 145-147, 150, 158, 169, 170, 172, 173, 197, 200, 216, 219, 224, 230, 233, 237, 249, 250, 263, 274, 275, 276, 299; 307, 323, 333, 342, 346, 359, 382, 383, 387, 391, 393, 401, 411, 415, 427, 442, 455, 459, 466 477, 481, 485, 487, 502, 511, 515, 525, 534, 539, 542, 560, 571, 577, 579, 584, 587, 595, 600 627, 638, 645, 654, 659, 668, 681, 688, 695, 696, 706, 708, 730, 742, 753, 775, 785, 786, 791 797, 800, 801, 809, 817, 819, 820, 823, 827, 847, 856, 875, 885, 896, 902, 910, 921, 922, 923 925, 926, 928, 946, and 952) were identified that were upregulated in response to viral infection, and 46 Arabidopsis genes were identified that were downregulated in response to viral infection (SEQ ID NOs: 14, 48, 53, 98, 217, 226, 295, 327, 343, 352, 369, 404, 407, 418, 453, 458, 465, 472, 480, 488, 495, 507, 509, 513, 514, 559, 561, 581, 604, 607, 613, 641, 652, 672, 720, 735, 739, 743, 745, 754, 773, 803, 832, 849, 948, and 949).
Also provided are nucleic acid molecules comprising a nucleotide sequence comprising an open reading frame expressed in response to pathogen infection comprising SEQ ID NOs:209, 216, 262, 267, 317, 386, 425, 440 and 800. These sequences are useful to over- or under-express the encoded product, or prepare knock-out plants which have an altered response to pathogen infection.
The invention therefore provides a method in which the open reading frame of a plant pathogen resistance gene, e.g., a gene that is associated with a response to pathogen infection, which is altered in a plant in response to infection is identified and isolated. A transgene comprising the isolated open reading frame may be introduced to and expressed in a transgenic plant, e.g., prior to infection, e.g., constitutively, or early and/or rapidly after infection, or in regulatable (inducible) fashion, e.g., after exposure to a chemical or using a promoter that is upregulated after infection, so as to confer resistance to that transgenic plant to the pathogen relative to a conesponding plant which does not have the transgene. The expression ofthe transgene is preferably at higher than normal levels, and under the regulation of a promoter that allows very fast and high induction in response to the presence of a pathogen or under cycling promoters (e.g., circadian clock regulated promoters), such that the encoded gene product(s) is maintained at sufficiently high levels to provide enhanced resistance or tolerance. The invention further provides a method in which a gene in a plant which is downregulated in response to infection, is disrupted or the expression of that gene is further downregulated, e.g., using antisense expression, so as to result in a plant that has enhanced resistance to infection,
and which disruption or downregulation preferably has little or no detrimental effect(s) on the host plant.
As also described herein, it was found that the early incompatible response was similar to the late compatible response, suggesting that early expression of plant pathogen-resistance genes is important for resistance. Also, various plant strains were found to respond differently to the same pathogen, but there was also an identifiable global pattern of response. Thus, the comparison ofthe expression patterns in incompatible and compatible interactions in one or more ecotypes can lead to identifying subsets of key responding genes and clusters of genes that are key (early) responders. In addition, the observed global expression pattern indicated that the least resistant strain tested (Ws) had a low basal level of pathogen-upregulated genes and a high level of pathogen-downregulated genes compared to the most resistant strain (Ler). Thus, plant strains that are more resistant to pathogens have a gene expression phenotype in which genes that are upregulated in response to infection are already expressed at a higher than normal basal level, and those genes that are downregulated are expressed at a lower than normal basal level.
The genes and promoters described hereinabove can be used to identify orthologous genes and their promoters which are also likely useful to enhance resistance of plants to pathogens. Moreover, the orthologous promoters are useful to express linked open reading frames. In addition by aligning the promoters of these orthologs, novel cis elements can be identified that are useful to generate synthetic promoters.
Hence, the isolated nucleic acid molecules ofthe invention include the orthologs ofthe Arabidopsis, Chenopodium and rice sequences disclosed herein, i.e., the conesponding nucleic acid molecules in organisms other than Arabidopsis, Chenopodium and rice, including, but not limited to, plants other than Arabidopsis, Chenopodium and rice, preferably cereal plants, e.g., corn, wheat, rye, turfgrass, sorghum, millet, sugarcane, soybean, barley, alfalfa, sunflower, canola, soybean, cotton, peanut, tobacco, sugarbeet, or rice. An ortholog is a gene from a different species that encodes a product having the same function as the product encoded by a gene from a reference organism. Databases such GenBank or one found at http://bioserver.myongjiac.kr/rjce.htinl (for rice) may be employed to identify sequences related to the Arabidopsis or Chenopodium sequences, e.g., orthologs in cereal crops such as rice. Alternatively, recombinant DNA techniques such as hybridization or PCR may be employed to identify sequences related to the Arabidopsis sequences. The encoded ortholog products likely have at least 70% sequence identity to each other. Hence, the invention
includes an isolated nucleic acid molecule comprising a nucleotide sequence encoding a polypeptide having at least 70% identity to a polypeptide encoded by one or more ofthe Arabidopsis, Chenopodium or rice sequences disclosed herein. For example, promoter sequences within the scope ofthe invention are those which direct expression of an open reading frame which encodes a polypeptide that is substantially similar to an Arabidopsis polypeptide encoded by a gene comprising SEQ ID NOs: 1-953.
The genes and promoters described hereinabove can be used to identify orthologous genes and their promoters which are also likely expressed in a particular tissue and/or development manner. Moreover, the orthologous promoters are useful to express linked open reading frames. In addition, by aligning the promoters of these orthologs, novel cis elements can be identified that are useful to generate synthetic promoters. Hence, the isolated nucleic acid molecules ofthe invention include the orthologs ofthe Arabidopsis sequences disclosed herein, i.e., the conesponding nucleotide sequences in organisms other than Arabidopsis, including, but not limited to, plants other than Arabidopsis, preferably cereal plants, e.g., com, wheat, rye, turfgrass, sorghum, millet, sugarcane, soybean, barley, alfalfa, sunflower, canola, soybean, cotton, peanut, tobacco, sugarbeet, or rice. An orthologous gene is a gene from a different species that encodes a product having the same or similar function, e.g., catalyzing the same reaction as a product encoded by a gene from a reference organism. Thus, an ortholog includes polypeptides having less than, e.g., 65% amino acid sequence identity, but which ortholog encodes a polypeptide having the same or similar function. Databases such
GenBank or one found at http://bioserver.myongjiac.kr/rjce.html (for rice) may be employed to identify sequences related to the Arabidopsis sequences, e.g., orthologs in cereal crops such as rice, wheat, sunflower or alfalfa. SEQ ID NOs: 6286 and 4210, for example, are the rice promoter and open reading frame for rice peroxidase, the ortholog ofthe Arabidopsis gene comprising SEQ ID NO: 50. SEQ ID NOs: 3311, 5387, 3791 and 5867 are rice orthologs of the Arabidopsis gene comprising SEQ ID NO:609; SEQ ID NOs: 2699, 4775, 3463, 5539, 3584, 5660, 4451, 6527, 4595 and 6671 are rice orthologs of fhe Arabidopsis gene comprising SEQ ID NO: 139.
Preferably, the promoters ofthe invention include a consecutive stretch of about 25 to 2000, including 50 to 500 or 100 to 250, and up to 1000 or 1500, contiguous nucleotides, e.g., 40 to about 743, 60 to about 743, 125 to about 743, 250 to about 743, 400 to about 743, 600 to about 743, of any one of SEQ ID NOs:2137-2661, SEQ ID NOs:4738-6813 or the promoter orthologs thereof, which include the minimal promoter region. Preferably, the nucleotide
sequence that includes the promoter region includes at least one copy of a TATA box. Thus, the invention provides plant promoters, including orthologs of Arabidopsis promoters conesponding to genes comprising any one of SEQ ID NOs: 1-953. The present invention further provides a composition, an expression cassette or a recombinant vector containing the nucleic acid molecule ofthe invention, and host cells comprising the expression cassette or vector, e.g., comprising a plasmid. In particular, the present invention provides an expression cassette or a recombinant vector comprising a promoter ofthe invention linked to a nucleic acid segment which, when present in a plant, plant cell or plant tissue, results in transcription ofthe linked nucleic acid segment. In its broadest sense, the term "substantially similar" when used herein with respect to a nucleotide sequence means that the nucleotide sequence is part of a gene which encodes a polypeptide having substantially the same stracture and function as a polypeptide encoded by a gene for the reference nucleotide sequence, e.g., the nucleotide sequence comprises a promoter from a gene that is the ortholog ofthe gene conesponding to the reference nucleotide sequence, as well as promoter sequences that are structurally related the promoter sequences particularly exemplified herein, i.e., the substantially similar promoter sequences hybridize to the complement ofthe promoter sequences exemplified herein under high or very high stringency conditions. The term "substantially similar" thus includes nucleotide sequences wherein the sequence has been modified, for example, to optimize expression in particular cells, as well as nucleotide sequences encoding a variant polypeptide comprising one or more amino acid substitutions relative to the (unmodified) polypeptide encoded by the reference sequence, which substitution(s) does not alter the activity ofthe variant polypeptide relative to the unmodified polypeptide. In its broadest sense, the term "substantially similar" when used herein with respect to polypeptide means that the polypeptide has substantially the same stracture and function as the reference polypeptide. The percentage of amino acid sequence identity between the substantially similar and the reference polypeptide is at least 65%, 66%, 67%, 68%, 69%, 70%, e.g., 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, and even 90% or more, e.g., 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, up to at least 99%, wherein the reference polypeptide is a polypeptide encoded by an Arabidopsis gene comprising any one of SEQ ID NOs: 1-953, a
Chenopodium gene comprising any one of SEQ ID NOs:1954-1966, or a rice gene comprising any one of SEQ ID NOs:2000-2129 or 2662-4737. One indication that two polypeptides are substantially similar to each other, besides having substantially the same function, is that an
agent, e.g., an antibody, which specifically binds to one ofthe polypeptides, specifically binds to the other.
Sequence comparisons maybe canied out using a Smith- Waterman sequence alignment algorithm (see e.g., Waterman (1995) or http://www hto.usc.edu/software/seqaln/index.html). The localS program, version 1.16, is preferably used with following parameters: match: 1, mismatch penalty: 0.33, open-gap penalty: 2, extended-gap penalty: 2. Further, a nucleotide sequence that is "substantially similar" to a reference nucleotide sequence hybridizes to the reference nucleotide sequence in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in 2X SSC, 0.1% SDS at 50°C, more desirably in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in IX SSC, 0.1% SDS at 50°C, more desirably still in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in 0.5X SSC, 0.1% SDS at 50°C, preferably in 7% sodium dodecyl sulfate (SDS), 0.5 MNaPO4, 1 mM EDTA at 50°C with washing in 0.1X SSC, 0.1% SDS at 50°C, more preferably in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in 0. IX SSC, 0.1 % SDS at 65°C.
Hence, the present invention further provides an expression cassette or a vector containing the nucleic acid molecule comprising an open reading frame ofthe invention operably linked to a promoter, or comprising a promoter ofthe invention operably linked to an open reading frame or portion thereof, and the vector may be a plasmid. Such cassettes or vectors, when present in a plant, plant cell or plant tissue result in transcription ofthe linked nucleic acid fragment in the plant. The expression cassettes or vectors ofthe invention may optionally include other regulatory sequences, e.g., transcription terminator sequences, operator, repressor binding site, transcription factor binding site, and/or an enhancer and may be contained in a host cell. The expression cassette or vector may augment the genome of a transformed plant or may be maintained extrachromosomally. The expression cassette or vector may further have a Ti plasmid and be contained in an Agrobacterium tumefaciens cell; it may be carried on a microparticle, wherein the microparticle is suitable for ballistic transformation of a plant cell; or it may be contained in a plant cell or protoplast. Further, the expression cassette can be contained in a transformed plant or cells thereof and the plant may be a dicot or a monocot. In particular, the plant may be a cereal plant.
The invention also provides sense and anti-sense nucleic acid molecules conesponding to the open reading frames identified herein as well as their orthologs. Also provided are expression cassettes, e.g., recombinant vectors, and host cells, comprising the nucleic acid
molecule ofthe invention, e.g., one which comprises a nucleotide sequence which encodes a polypeptide the expression of which is altered in response to pathogen infection.
The present invention further provides a method of augmenting a plant genome by contacting plant cells with a nucleic acid molecule ofthe invention, e.g., one isolatable or obtained from a plant gene encoding a polypeptide that is substantially similar to a polypeptide encoded by an Arabidopsis, Chenopodium or rice gene comprising a sequence comprising any one of SEQ ID NOs: 1-953, 1954-1966, 2000-2129 or 2662-4737 so as to yield transformed plant cells; and regenerating the transformed plant cells to provide a differentiated transformed plant, wherein the differentiated transformed plant expresses the nucleic acid molecule in the cells ofthe plant. The nucleic acid molecule may be present in the nucleus, chloroplast, mitochondria and/or plastid ofthe cells ofthe plant. The present invention also provides a transgenic plant prepared by this method, a seed from such a plant and progeny plants from such a plant including hybrids and inbreds. Prefened transgenic plants are transgenic maize, soybean, barley, alfalfa, sunflower, canola, soybean, cotton, peanut, sorghum, tobacco, sugarbeet, rice, wheat, rye, turfgrass, millet, sugarcane, tomato, or potato.
The invention also provides a method of plant breeding, e.g., to prepare a crossed fertile transgenic plant. The method comprises crossing a fertile transgenic plant comprising a particular nucleic acid molecule ofthe invention with itself or with a second plant, e.g., one lacking the particular nucleic acid molecule, to prepare the seed of a crossed fertile transgenic plant comprising the particular nucleic acid molecule. The seed is then planted to obtain a crossed fertile transgenic plant. The plant may be a monocot or a dicot. In a particular embodiment, the plant is a cereal plant.
The crossed fertile transgenic plant may have the particular nucleic acid molecule inherited through a female parent or through a male parent. The second plant may be an inbred plant. The crossed fertile transgenic may be a hybrid. Also included within the present invention are seeds of any of these crossed fertile transgenic plants.
The various breeding steps are characterized by well-defined human intervention such as selecting the lines to be crossed, directing pollination ofthe parental lines, or selecting appropriate progeny plants. Depending on the desired properties different breeding measures are taken. The relevant techniques are well known in the art and include but are not limited to hybridization, inbreeding, backcross breeding, multiline breeding, variety blend, interspecific hybridization, aneuploid techniques, etc. Hybridization techniques also include the sterilization of plants to yield male or female sterile plants by mechanical, chemical or biochemical means.
Cross pollination of a male sterile plant with pollen of a different line assures that the genome ofthe male sterile but female fertile plant will uniformly obtain properties of both parental lines. Thus, the transgenic plants according to the invention can be used for the breeding of improved plant lines that for example increase the effectiveness of conventional methods such as herbicide or pesticide treatment or allow to dispense with said methods due to their modified genetic properties. Alternatively new crops with improved stress tolerance can be obtained that, due to their optimized genetic "equipment", yield harvested product of better quality than products that were not able to tolerate comparable adverse developmental conditions. The nucleic acid molecules ofthe invention, their encoded polypeptides and compositions thereof, are: for open reading frames, useful to provide resistance to pathogens to alter expression of a particular gene conesponding to the open reading frame by decreasing or eliminating expression of that plant gene or by overexpressing a particular gene product, and as a diagnostic for the presence or absence ofthe pathogen by coπelating the expression level or pattern of expression of one or more of the nucleic acid molecules or polypeptides of the invention; and for promoters, useful to alter the expression of a linked open reading frame in response to pathogen infection. As one embodiment ofthe invention includes isolated nucleic acid molecules that have increased expression in response to pathogen infection, the invention further provides compositions and methods for enhancing resistance to pathogen infection. The compositions ofthe invention include plant nucleic acid sequences and the amino acid sequences for the polypeptides or partial-length polypeptides encoded thereby which are described herein, or other plant nucleic acid sequences and the amino acid sequences for the polypeptides or partial-length polypeptides encoded thereby which are operably linked to a promoters are useful to provide tolerance or resistance to a plant to a pathogen, preferably by preventing or inhibiting pathogen infection. Methods ofthe invention involve stably transforming a plant with one or more of at least a portion of these nucleotide sequences which confer tolerance or resistance operably linked to a promoter capable of driving expression of that nucleotide sequence in a plant cell. By "portion" or "fragment", as it relates to a nucleic acid molecule, sequence or segment ofthe invention, when it is linked to other sequences for expression, is meant a sequence comprising at least 80 nucleotides, more preferably at least 150 nucleotides, and still more preferably at least 400 nucleotides. If not employed for expressing, a "portion" or "fragment" means at least 9, preferably 12, more preferably 15, even more preferably at least 20, consecutive nucleotides, e.g., probes and primers
(oligonucleotides), conesponding to the nucleotide sequence ofthe nucleic acid molecules of the invention. By "resistant" is meant a plant which exhibits substantially no phenotypic changes as a consequence of infection with the pathogen. By "tolerant" is meant a plant which, although it may exhibit some phenotypic changes as a consequence of infection, does not have a substantially decreased reproductive capacity or substantially altered metabolism. A method of combating a pathogen in an agricultural crop is also provided. The method comprises introducing to a plant, plant cell, or plant tissue an expression cassette comprising a nucleic acid molecule ofthe invention comprising an open reading frame so as to yield a transformed differentiated plant, transformed cell or transfonned tissue. Transformed cells or tissue can be regenerated to provide a transformed differentiated plant. The transformed differentiated plant preferably expresses the nucleic acid molecule in an amount that confers resistance to the transformed plant to pathogen infection relative to a conesponding nontransformed plant. The present invention also provides a transformed plant prepared by the method, progeny and seed thereof. Examples of plant viruses which may be combated by the present invention include single stranded RNA viruses (with and without envelope), double stranded RNA viruses, and single and double stranded DNA viruses such as (but not limited to) tobacco mosaic virus, cucumber mosaic virus, turnip mosaic virus, turnip vein clearing virus, oilseed rape mosaic viras, tobacco rattle viras, pea enation mosaic virus, barley stripe mosaic viras, potato viruses X and Y, carnation latent virus, beet yellows virus, maize chlorotic viras, tobacco necrosis viras, turnip yellow mosaic viras, tomato bushy stunt virus, southern bean mosaic virus, barley yellow dwarf virus, tomato spotted wilt virus, lettuce necrotic yellows virus, wound tumor virus, maize streak virus, and cauliflower mosaic virus. Other pathogens within the scope ofthe invention include, but are not limited to, fungi such as Cochliobolus carbonum, Phytophthora infestans, Phytophthora sojae, Collesosichum, Melampsora lini, cladosporiumfulvum, Heminthosporium maydia, Peronospora parasitica, Puccinia sorghi, and Puccinia polysora; bacteria such as Phynchosporium secalis, Pseudomonas glycinea, Xanthomonas oryzae and, Fusarium oxyaporium; and nematodes such as Globodera rostochiensis.
For example, the invention provides a nucleic acid molecule comprising a plant nucleotide sequence comprising at least a portion of a key effector gene(s) responsible for host resistance to particular pathogens. To provide resistance or tolerance to a pathogen in a plant, this sequence may be overexpressed individually, in the sense or antisense orientation, or in combination with other sequences to confer improved disease resistance or tolerance to a plant
relative to a plant that does not comprise and/or express the sequence. The overexpression may be constitutive, or it may be preferable to express the effector gene(s) in a tissue-specific manner or from an inducible promoter including a promoter which is responsive to external stimuli, such as chemical application, or to pathogen infection, e.g., so as to avoid possible deleterious effects on plant growth if the effector gene(s) was constitutively expressed. In one embodiment ofthe invention, the promoter employed may be one that is rapidly and transiently and/or highly transcribed after pathogen infection.
A transfoπned (transgenic) plant ofthe invention includes plants, the genome of which is augmented by a nucleic acid molecule ofthe invention, or in which the conesponding gene has been disrupted, e.g., to result in a loss, a decrease or an alteration, in the function ofthe product encoded by the gene, which plant may also have increased yields, e.g., under conditions of pathogen infection, and/or produce a better-quality product than the conesponding wild-type plant. The nucleic acid molecules ofthe invention are thus useful for targeted gene disraption, as well as markers and probes. For example, the invention includes a pathogen, e.g., virus, tolerant or resistant plant and seed thereof having stably integrated and expressed within its genome, a nucleic acid molecule ofthe invention. The normal fertile transformed (transgenic) plant may be selfed to yield a substantially homogenous line with respect to viral resistance or tolerance. Individuals ofthe line, or the progeny thereof, may be crossed with plants which optionally exhibit the trait. In a particular embodiment ofthe method, the selfing and selection steps are repeated at least five times in order to obtain the homogenous (isogenic) line. Thus, the invention also provides transgenic plants and the products ofthe transgenic plants.
The invention further includes a nucleotide sequence which is complementary to one (hereinafter "test" sequence) which hybridizes under low, moderate or stringent conditions with the nucleic acid molecules ofthe invention as well as RNA which is encoded by the nucleic acid molecule. When the hybridization is performed under stringent conditions, either the test or nucleic acid molecule of invention is preferably supported, e.g., on a membrane or DNA chip. Thus, either a denatured test or nucleic acid molecule ofthe invention is preferably first bound to a support and hybridization is effected for a specified period of time at a temperature of, e.g., between 55 and 70°C, in double strength citrate buffered saline (SC) containing 0.1%> SDS followed by rinsing ofthe support at the same temperature but with a buffer having a reduced SC concentration. Depending upon the degree of stringency required
such reduced concentration buffers are typically single strength SC containing 0.1% SDS, half strength SC containing 0.1% SDS and one-tenth strength SC containing 0.1% SDS.
The invention further provides a method to identify an open reading frame in the genome of a plant cell, the expression of which is altered by pathogen infection of that cell. The method comprises contacting a solid substrate comprising a plurality of samples comprising isolated plant nucleic acid of a probe comprising plant nucleic acid, e.g., cRNA, isolated from a pathogen infected plant so as to form a complex. Each individual sample comprises one or more nucleic acid sequences (e.g., oligonucleotides) corresponding to at least a portion of a plant gene. The method may be employed with nucleic acid samples and probes from any organism, e.g., any prokaryotic or eukaryotic organism. Preferably, the nucleic acid sample and probes are from a plant, such as a dicot or monocot. More preferably the nucleic acid samples and probes are from a cereal plant. Even more preferably the nucleic acids and probes are from a crop plant. A second plurality of samples on a solid substrate, i.e., a DNA chip, each comprising a plurality of samples comprising isolated plant nucleic acid is contacted with a probe comprising plant nucleic acid isolated from an uninfected or infected control (mutant) plant so as to form a complex. Then complex formation between the samples and probes comprising nucleic acid from infected or control cells compared. For example, potato virus X, tobacco mosaic virus, tobravirus, cucumber mosaic viras and gemnivirus are known to infect Arabidopsis. Thus, Arabidopsis genes, the expression of which is altered in response to infection by any of these viruses, can be identified. Regions that are 5N to the start codon for the gene can then be identified and/or isolated.
The invention further provides a method for identifying a plant cell infected with a pathogen. The method comprises contacting nucleic acid obtained from a plant cell suspected of being infected with a pathogen with oligonucleotides conesponding to a portion of a plurality of sequences selected from SEQ ID NOs:l-953, 1954-1966, 2000-2129 or 2662-4737 under conditions effective to amplify those sequences. Then the presence ofthe amplified product is detected or detennined. The presence of two or more amplified products, e.g., in an amount that is different than the amount ofthe conesponding amplified products from an uninfected plant, each conesponding to two or more SEQ ID NOs: 1-953, 1954-1966, 2000- 2129, or 2662-4737 is indicative of pathogen infection.
The invention further provides a method for identifying a plant cell infected with a pathogen. The method comprises contacting a protein sample obtained from a plant cell suspected of being infected with a pathogen with an agent that specifically binds a polypeptide
encoded by an open reading frame comprising SEQ ID NOs:l-953, 1954-1966, 2000-2129 or 2662-4737 so as to form a complex. Then the presence or amount of complex formation is detected or determined.
The invention provides an additional method for identifying a plant cell infected with a pathogen. The method comprises hybridizing a probe selected from SEQ ID NOs: 1-953, 1954- 1966, 2000-2129 or 2662-4737 to nucleic acid obtained from a plant cell suspected of being infected with a pathogen. The amount ofthe probe hybridized to nucleic acid obtained from a cell suspected of being infected with a viras is compared to hybridization ofthe probe to nucleic acid isolated from an uninfected cell. A change in the amount of at least two probes that hybridize to nucleic acid isolated from a cell suspected of being infected by a virus relative to hybridization of at least two probes to nucleic acid isolated from an uninfected cell is indicative of viral infection.
A method to shuffle the nucleic acids ofthe invention is provided. This method involves fragmentation of a nucleic acid conesponding to a nucleic acid sequence listed in SEQ ID NOs: 1-953, 1954-1966, 2000-2129 or 2662-4737, the orthologs thereof, and the conesponding genes, followed by religation. This method allows for the production of polypeptides having altered activity relative to the native form ofthe polypeptide. Accordingly, the invention provides cells and transgenic plants containing nucleic acid segments produced through shuffling that encode polypeptides having altered activity relative to the conesponding native polypeptide.
A computer readable medium containing the nucleic acid sequences ofthe invention as well as methods of use for the computer readable medium are provided. This medium allows a nucleic acid segment conesponding to a nucleic acid sequence listed in SEQ ID NOs: 1-953, 2137-2661, 1954-1966, 2000-2129, 2662-4737 or 4738-6813 to be used as a reference sequence to search against databases. This medium also allows for computer-based manipulation of a nucleic acid sequence conesponding to a nucleic acid sequence listed in SEQ ID NOs :l-953, 2137-2661, 1954-1966, 2000-2129, 2662-4737 or 4738-6813, and the conesponding gene and polypeptide encoded by the nucleic acid sequence.
Therefore, another embodiment ofthe present invention provides a method of using known inducers or inhibitors of genes identified as being important in plant-pathogen interactions to induce genes that are important in resistance, or to inhibit genes that are downregulated in resistance.
Thus, some ofthe isolated nucleic acid molecules ofthe invention are useful in a method of combating a pathogen in an agricultural crop. The method comprises introducing to a plant an expression cassette comprising a nucleic acid molecule ofthe invention so as to yield a transformed differentiated plant. The transformed differentiated plant expresses the nucleic acid molecule in an amount that confers resistance to the transformed plant to infection relative to a corresponding nontransformed plant.
Detailed Description of the Invention I. Definitions
The term "gene" is used broadly to refer to any segment of nucleic acid associated with a biological function. Thus, genes include coding sequences and/or the regulatory sequences required for their expression. For example, gene refers to a nucleic acid fragment that expresses mRNA or functional RNA, or encodes a specific protein, and which includes regulatory sequences. Genes also include nonexpressed DNA segments that, for example, form recognition sequences for other proteins. Genes can be obtained from a variety of sources, including cloning from a source of interest or synthesizing from known or predicted sequence information, and may include sequences designed to have desired parameters.
The term "native" or "wild type" gene refers to a gene that is present in the genome of an untransformed cell, i.e., a cell not having a known mutation. A "marker gene" encodes a selectable or screenable trait.
The term "chimeric gene" refers to any gene that contains 1) DNA sequences, including regulatory and coding sequences, that are not found together in nature, or 2) sequences encoding parts of proteins not naturally adjoined, or 3) parts of promoters that are not naturally adjoined. Accordingly, a chimeric gene may comprise regulatory sequences and coding sequences that are derived from different sources, or comprise regulatory sequences and coding sequences derived from the same source, but ananged in a manner different from that found in nature.
A "transgene" refers to a gene that has been introduced into the genome by transformation and is stably maintained. Transgenes may include, for example, genes that are either heterologous or homologous to the genes of a particular plant to be transformed. Additionally, transgenes may comprise native genes inserted into a non-native organism, or chimeric genes. The term "endogenous gene" refers to a native gene in its natural location in
the genome of an organism. A "foreign" gene refers to a gene not normally found in the host organism but that is introduced by gene transfer.
An "oligonucleotide" conesponding to a nucleotide sequence ofthe invention, e.g., for use in probing or amplification reactions, may be about 30 or fewer nucleotides in length (e.g., 9, 12, 15, 18, 20, 21 or 24, or any number between 9 and 30). Generally specific primers are upwards of 14 nucleotides in length. For optimum specificity and cost effectiveness, primers of 16 to 24 nucleotides in length may be prefened. Those skilled in the art are well versed in the design of primers for use processes such as PCR. If required, probing can be done with entire restriction fragments ofthe gene disclosed herein which may be 100's or even lOOO's of nucleotides in length.
The terms "protein," "peptide" and "polypeptide" are used interchangeably herein. The nucleotide sequences ofthe invention can be introduced into any plant. The genes to be introduced can be conveniently used in expression cassettes for introduction and expression in any plant of interest. Such expression cassettes will comprise the transcriptional initiation region ofthe invention linked to a nucleotide sequence of interest. Prefened promoters include constitutive, tissue-specific, developmental-specific, inducible and/or viral promoters. Such an expression cassette is provided with a plurality of restriction sites for insertion ofthe gene of interest to be under the transcriptional regulation ofthe regulatory regions. The expression cassette may additionally contain selectable marker genes. The cassette will include in the 5'-3' direction of transcription, a transcriptional and translational initiation region, a DNA sequence of interest, and a transcriptional and translational termination region functional in plants. The termination region may be native with the transcriptional initiation region, may be native with the DNA sequence of interest, or may be derived from another source. Convenient termination regions are available from the Ti- plasmid of A. tumefaciens, such as the octopine synthase and nopaline synthase termination regions. See also, Guerineau et al., 1991; Proudfoot, 1991; Sanfacon et al., 1991; Mogen et al., 1990; Munroe et al, 1990; Ballas et al., 1989; Joshi et al., 1987.
"Coding sequence" refers to a DNA or RNA sequence that codes for a specific amino acid sequence and excludes the non-coding sequences. It may constitute an "uninterrupted coding sequence", i.e., lacking an intron, such as in a cDNA or it may include one or more introns bounded by appropriate splice junctions. An "intron" is a sequence of RNA which is contained in the primary transcript but which is removed through cleavage and re-ligation of the RNA within the cell to create the mature mRNA that can be translated into a protein.
The teπns "open reading frame" and "ORF" refer to the amino acid sequence encoded between translation initiation and termination codons of a coding sequence. The terms "initiation codon" and "termination codon" refer to a unit of three adjacent nucleotides ('codon') in a coding sequence that specifies initiation and chain termination, respectively, of protein synthesis (mRNA translation).
A "functional RNA" refers to an antisense RNA, ribozyme, or other RNA that is not translated.
The term "RNA transcript" refers to the product resulting from RNA polymerase catalyzed transcription of a DNA sequence. When the RNA transcript is a perfect complementary copy ofthe DNA sequence, it is refened to as the primary transcript or it may be a RNA sequence derived from posttranscriptional processing ofthe primary transcript and is refened to as the mature RNA. "Messenger RNA" (mRNA) refers to the RNA that is without introns and that can be translated into protein by the cell. "cDNA" refers to a single- or a double-stranded DNA that is complementary to and derived from mRNA. "Regulatory sequences" and "suitable regulatory sequences" each refer to nucleotide sequences located upstream (5' non-coding sequences), within, or downstream (3' non-coding sequences) of a coding sequence, and which influence the transcription, RNA processing or stability, or translation ofthe associated coding sequence. Regulatory sequences include enhancers, promoters, translation leader sequences, introns, and polyadenylation signal sequences. They include natural and synthetic sequences as well as sequences which may be a combination of synthetic and natural sequences. As is noted above, the term "suitable regulatory sequences" is not limited to promoters.
"5' non-coding sequence" refers to a nucleotide sequence located 5' (upstream) to the coding sequence. It is present in the fully processed mRNA upstream ofthe initiation codon and may affect processing ofthe primary transcript to mRNA, mRNA stability or translation efficiency (Turner et al., 1995).
"3' non-coding sequence" refers to nucleotide sequences located 3' (downstream) to a coding sequence and include polyadenylation signal sequences and other sequences encoding regulatory signals capable of affecting mRNA processing or gene expression. The polyadenylation signal is usually characterized by affecting the addition of polyadenylic acid tracts to the 3' end ofthe mRNA precursor. The use of different 3' non-coding sequences is exemplified by Ingelbrecht et al., 1989.
The term "translation leader sequence" refers to that DNA sequence portion of a gene
between the promoter and coding sequence that is transcribed into RNA and is present in the fully processed mRNA upstream (5') ofthe translation start codon. The translation leader sequence may affect processing ofthe primary transcript to mRNA, mRNA stability or translation efficiency. The term "mature" protein refers to a post-translationally processed polypeptide without its signal peptide. "Precursor" protein refers to the primary product of translation of an mRNA. "Signal peptide" refers to the amino terminal extension of a polypeptide, which is translated in conjunction with the polypeptide forming a precursor peptide and which is required for its entrance into the secretory pathway. The term "signal sequence" refers to a nucleotide sequence that encodes the signal peptide.
The term "intracellular localization sequence" refers to a nucleotide sequence that encodes an intracellular targeting signal. An "intracellular targeting signal" is an amino acid sequence that is translated in conjunction with a protein and directs it to a particular subcellular compartment. "Endoplasmic reticulum (ER) stop transit signal" refers to a carboxy- terminal extension of a polypeptide, which is translated in conjunction with the polypeptide and causes a protein that enters the secretory pathway to be retained in the ER. "ER stop transit sequence" refers to a nucleotide sequence that encodes the ER targeting signal. Other intracellular targeting sequences encode targeting signals active in seeds and/or leaves and vacuolar targeting signals. "Pathogen" as used herein includes but is not limited to bacteria, fungi, yeast, oomycetes and viras, e.g., American wheat striate mosaic viras mosaic (A WSMV), barley stripe mosaic virus (BSMV), barley yellow dwarf virus (BYDV), Brome mosaic viras (BMV), cereal chlorotic mottle virus (CCMV), com chlorotic vein banding virus (CCVBV), maize chlorotic mottle virus (MCMV), maize dwarf mosaic virus (MDMV), A or B, wheat streak mosaic viras (WSMV), cucumber mosaic viras (CMV), cynodon chlorotic streak viras
(CCSV), Johnsongrass mosaic viras (JGMV), maize bushy stunt or mycoplasma-like organism (NjTLO), maize chlorotic dwarf virus (MCDV), maize chlorotic mottle virus (MCMV), maize dwarf mosaic viras (MDMV) strains A, D, E and F, maize leaf fleck virus (MLFV), maize line virus (NELV), maize mosaic virus (MMV), maize mottle and chlorotic stunt viras, maize pellucid ringspot virus (MPRV), maize raya graesa virus (MRGV), maize rayado fino viras (MRFV), maize red leaf and red stripe virus (MRSV), maize ring mottle virus (MRMV), maize rio cuarto viras (MRCV), maize rough dwarf virus (MRDV), maize sterile stunt virus (strains of barley yellow striate viras), maize streak virus (MSV), maize chlorotic stripe, maize
hoja Maize stripe virus blanca, maize stunting viras, maize tassel abortion virus (MTAV), maize vein enation viras (MVEV), maize wallaby ear virus (MAVEV), maize white leaf virus, maize white line mosaic viras (NTWLMV), millet red leaf viras (NMV), Northern cereal mosaic virus (NCMV), oat pseudorosette viras, oat sterile dwarf virus (OSDV), rice black- streaked dwarf viras (RBSDV), rice stripe viras (RSV), sorghum mosaic viras (SrMV), formerly sugarcane mosaic virus (SCMV) strains H, I and M, sugarcane Fiji disease virus (FDV), sugarcane mosaic viras (SCMV) strains A, B, D, E,SC, BC, Sabi andNM vein enation viras, and wheat spot mosaic virus (WSMV).
Bacterial pathogens include but are not limited to Pseudomonas avenae subsp. avenae, Xanthomonas campestris pv. holcicola, Enterobacter dissolvens, Erwinia dissolvens, Ervinia carotovora subsp. carotovora, Erwinia chrysanthemi pv. zeae, Pseudomonas andropogonis, Pseudomonas syringae pv. coronafaciens, Clavibacter michiganensis subsp., Corynebacterium michiganense pv. nebraskense, Pseudomonas syringae pv. syringae, Hemiparasitic bacteria (see under fungi), Bacillus subtilis, Erwinia stewartii, and Spiroplasma kunkelii. Fungal pathogens include but are not limited to Collelotrichum graminicola,
Glomerella graminicola Politis, Glomerella lucumanensis, Aspergillusflavus, Rhizoctonia solani Kuhn, Thanatephorus cucumeris, Acremonium strictum W. Gams, Cephalosporium acremonium Auct. non Cor da Black Lasiodiplodia theobromae = Bolr odiplodiay theobromae Borde bianco Marasmiellus sp., Physoderma maydis, Cephalosporium Corticium sasakii, Curvularia clavata, C. maculans, Cochhobolus eragrostidis, Curvularia inaequahs, C. intermedia (teleomorph Cochhobolus intermedius), Curvularia lunata (teleomorph: Cochliobolus lunatus), Curvularia pallescens (teleomorph - Cochlioboluspallescens), Curvularia senegalensis, C. luberculata (teleomoφh: Cochliobolus tuberculatus), Didymella exitalis Diplodiaftumenti (teleomorph - Botryosphaeriafestucae), Diplodia maydis = Stenocarpella maydis, Stenocarpella macrospora = Diplodia macrospora, Sclerophthora rayssiae var. zeae, Sclerophthora macrospora = Sclerospora macrospora, Sclerospora graminicola, Peronosclerospora maydis = Sclerospora maydis, Peronosclerospora philippinensis, Sclerospora philippinensis, Peronosclerospora sorghi = Sclerospora sorghi, Peronosclerospora spontanea = Sclerospora spontanea, Peronosclerospora sacchari = Sclerospora sacchari, Nigrospora oryzae (teleomoφh: Khuskia oryzae) A. Iternaria alternala -A. tenuis, Aspergillus glaucus, A. niger, Aspergillus spp., Botrytis cinerea, Cunninghamella sp., Curvulariapallescens, Doratomyces slemonitis = Cephalotrichum slemonitis, Fusarium culmorum, Gonatobotrys simplex, Pithomyces maydicus, Rhizopus microsporus Tiegh., R.
stolonifer = R. nigricans, Scopulariopsis brumptii, Claviceps gigantea (anamoφh: Sphacelia sp.) Aureobasidium zeae = Kabatiella zeae, Fusarium subglutinans = F. moniliforme var. subglutinans, Fusarium moniliforme, Fusarium avenaceum (teleomorph - Gibberella avenacea), Botryosphaeria zeae = Physalospora zeae (anamorph: Allacrophoma zeae), Cercospora sorghi = C. sorghi var. maydis, Helminthosporium pedicellatum (teleomoφh: Selosphaeriapedicellata), Cladosporium cladosporioides - Hormodendrum cladosporioides, C herbarum (teleomorph - Mycosphaerella tassiana), Cephalosporium maydis, A. Iternaria alternata, A. scochyta maydis, A. tritici, A. zeicola, Bipolaris victoriae, Helminthosporium victoriae (teleomoφh Cochhoholus victoriae), C sativus (anamoφh: Bipolaris sorokiniana = H sorokinianum = H. sativum), Epicoccum nigrum, Exserohilum prolatum = Drechslera prolata (teleomoφh: Setosphaeriaprolata), Graphium penicillioides, Leptosphaeria maydis, Leptothyrium zeae, Ophiosphaerella herpotricha (anamoφh - Scolecosporiella sp.), Pataphaeosphaeria michotii, Phoma sp., Septoria zeae, S. zeicola, S. zeina Setosphaeria turcica, Exserohilzim turcicum = Helminthosporiumfurcicum, Cochhoholus carbonum, Bipolaris zeicola = Helminthosporium carhonum, Penicilhum spp., P. chrysogenum, P. expansum, P. oxalicum, Phaeocytostroma ambiguum, Phaeocylosporella zeae, Phaeosphaeria maydis = Sphaerulina maydis, Botryosphaeriafestucae — Physalospora zeicola (anamoφh: Diplodiaftumenfϊ) , Hemiparasitic bacteria and fungi Pyrenochaeta Phoma terrestris = Pyrenochaeta terrestris, Pythium spp., P. arrhenomanes, P. graminicola, Pythium aphanidermatum = P. hutleri L., Rhizoctonia zeae (teleomoφh: Waitea circinata),
Rhizoctonia solani, minor A Iternaria alternala, Cercospora sorghi, Dictochaetaftrtilis, Fusarium acuminatum (teleomoφh Gihherella acuminata), E. equiseti (teleomoφh: G. intricans), E. oxysporum, E. pallidoroseum, E. poae, E. roseum, G cyanogena (anamoφh.- E. sulphureum), Microdochium holleyi, Mucor p., Periconia circinata, Phytophthora cactorum, P. drechsleri, P. nicotianae var. parasitica, Rhizopus arrhizus, Setosphaeria rostrata, Exserohilum rostratum = Helminthosporium rostratum, Puccinia sorghi, Physopella pallescens, P. zeae, Sclerotium rofsii Sacc. (teleomoφh- Athelia rotfsiϊ), Bipolaris sorokiniana, B. zeicola = Helminthosporium carbonum, Diplodia maydis, Exserohilum pedicillatum, Exserohilum furcicum = Helminthosporium turcicum, Fusarium avenaceum, E. culmorum, E. moniliforme, Gibberella zeae (anamoφh - E. graminearum),
Macrophominaphaseolina, Penicillium spp., Phomopsis sp., Pythium spp., Rhizoctonia solani, R. zeae, Sclerotium rolfsfi, Spicaria sp., Selenophoma sp., Gaeumannomyces graminis, Myrothecium gramineum, Monascus purpureus, M. ruber Smut, Ustilago zeae = U. maydis
Smut, Ustilaginoidea virens Smut, Sphacelotheca reiliana = Sporisorium hold, Cochliobolus heterostrophus (anamoφh: Bipolaris maydis = Helminthosporium maydis), Stenocarpella macrospora = Diplodia macrospora, Cercospora sorghi, Fusarium episphaeria, E. merismoides, F. oxysporum Schlechtend, E. poae, E. roseum, E. solani (teleomoφh: Nectria haematococca), F. tricinctum, Mariannaea elegans, Mucor sp., Rhopographus zeae, Spicaria sp., Aspergillus spp., Penicillium spp., Trichoderma viride = Tlignorum teleomorph: Hypocrea sp., Stenocarpella maydis = Diplodia zeae, Ascochyta ischaemi, Phyllosticta maydis (telomoφh: Mycosphaerella zeae-maydis), and Gloeocercospora sorghi.
Parasitic nematodes include but are not limited to Awl Dolichodorus spp., D. heterocephalus Bulb and stem (Europe), Ditylenchus dipsaci Burrowing Radopholus similis Cyst Heterodera avenae, H. zeae, Punctodera chalcoensis Dagger Xiphinema spp., X americanum, X meditenaneum False root-knot Nacobbus dorsalis Lance, Columbia Hoplolaimus columbus Lance Hoplolaimus spp., H. galeatus Lesion Pratylenchus spp., P. brachyuras, P. crenalus, P. hexincisus, P. neglectus, P. penetrans, P. scribneri, P. thornei, P. zeae Needle Longidoras spp., L. breviannulatus Ring Criconemella spp., C ornata Root-knot Meloidogyne spp., M. chitwoodi, M. incognita, M. javanica Spiral Helicotylenchus spp., Belonolaimus spp., B. longicaudatus Stubby-root Paratrichodoras spp., P. christiei, P. minor, Ouinisulcius aculus, and Trichodoras spp.
"Promoter" refers to a nucleotide sequence, usually upstream (5') to its coding sequence, which controls the expression ofthe coding sequence by providing the recognition for RNA polymerase and other factors required for proper transcription. "Promoter" includes a minimal promoter that is a short DNA sequence comprised of a TATA box and other sequences that serve to specify the site of transcription initiation, to which regulatory elements are added for control of expression. "Promoter" also refers to a nucleotide sequence that includes a minimal promoter plus regulatory elements that is capable of controlling the expression of a coding sequence or functional RNA. This type of promoter sequence consists of proximal and more distal upstream elements, the latter elements often refened to as enhancers. Accordingly, an "enhancer" is a DNA sequence which can stimulate promoter activity and may be an innate element ofthe promoter or a heterologous element inserted to enhance the level or tissue specificity of a promoter. It is capable of operating in both orientations (noπnal or flipped), and is capable of functioning even when moved either upstream or downstream from the promoter. Both enhancers and other upstream promoter elements bind sequence-specific DNA-binding proteins that mediate their effects. Promoters
may be derived in their entirety from a native gene, or be composed of different elements derived from different promoters found in nature, or even be comprised of synthetic DNA segments. A promoter may also contain DNA sequences that are involved in the binding of protein factors which control the effectiveness of transcription initiation in response to physiological or developmental conditions.
The "initiation site" is the position suπounding the first nucleotide that is part ofthe transcribed sequence, which is also defined as position +1. With respect to this site all other sequences ofthe gene and its controlling regions are numbered. Downstream sequences (i.e., further protein encoding sequences in the 3' direction) are denominated positive, while upstream sequences (mostly ofthe controlling regions in the 5' direction) are denominated negative.
Promoter elements, particularly a TATA element, that are inactive or that have greatly reduced promoter activity in the absence of upstream activation are refened to as "minimal or core promoters." In the presence of a suitable transcription factor, the minimal promoter functions to permit transcription. A "minimal or core promoter" thus consists only of all basal elements needed for transcription initiation, e.g., a TATA box and/or an initiator.
"Constitutive expression" refers to expression using a constitutive or regulated promoter. "Conditional" and "regulated expression" refer to expression controlled by a regulated promoter. "Constitutive promoter" refers to a promoter that is able to express the open reading frame (ORF) that it controls in all or nearly all ofthe plant tissues during all or nearly all developmental stages ofthe plant. Each ofthe transcription-activating elements do not exhibit an absolute tissue-specificity, but mediate transcriptional activation in most plant parts at a level of >1% ofthe level reached in the part ofthe plant in which transcription is most active. "Regulated promoter" refers to promoters that direct gene expression not constitutively, but in a temporally- and/or spatially-regulated manner, and includes both tissue-specific and inducible promoters. It includes natural and synthetic sequences as well as sequences which may be a combination of synthetic and natural sequences. Different promoters may direct the expression of a gene in different tissues or cell types, or at different stages of development, or in response to different environmental conditions. New promoters of various types useful in plant cells are constantly being discovered, numerous examples may be found in the compilation by Okamuro et al. (1989). Typical regulated promoters useful in plants include but are not limited to safener-inducible promoters, promoters derived from the tetracyclme-
inducible system, promoters derived from salicylate-inducible systems, promoters derived from alcohol-inducible systems, promoters derived from glucocorticoid-inducible system, promoters derived from pathogen-inducible systems, and promoters derived from ecdysome- inducible systems. "Tissue-specific promoter" refers to regulated promoters that are not expressed in all plant cells but only in one or more cell types in specific organs (such as leaves or seeds), specific tissues (such as embryo or cotyledon), or specific cell types (such as leaf parenchyma or seed storage cells). These also include promoters that are temporally regulated, such as in early or late embryogenesis, during fruit ripening in developing seeds or fruit, in fully differentiated leaf, or at the onset of senescence.
"Inducible promoter" refers to those regulated promoters that can be turned on in one or more cell types by an external stimulus, such as a chemical, light, hormone, stress, or a pathogen.
"Operably-linked" refers to the association of nucleic acid sequences on single nucleic acid fragment so that the function of one is affected by the other. For example, a regulatory DNA sequence is said to be "operably linked to" or "associated with" a DNA sequence that codes for an RNA or a polypeptide if the two sequences are situated such that the regulatory DNA sequence affects expression ofthe coding DNA sequence (i.e., that the coding sequence or functional RNA is under the transcriptional control ofthe promoter). Coding sequences can be operably-linked to regulatory sequences in sense or antisense orientation.
"Expression" refers to the transcription and/or translation of an endogenous gene, ORF or portion thereof, or a transgene in plants. For example, in the case of antisense constructs, expression may refer to the transcription ofthe antisense DNA only. In addition, expression refers to the transcription and stable accumulation of sense (mRNA) or functional RNA. Expression may also refer to the production of protein.
"Specific expression" is the expression of gene products which is limited to one or a few plant tissues (spatial limitation) and/or to one or a few plant developmental stages (temporal limitation). It is acknowledged that hardly a trae specificity exists: promoters seem to be preferably switch on in some tissues, while in other tissues there can be no or only little activity. This phenomenon is known as leaky expression. However, with specific expression in this invention is meant preferable expression in one or a few plant tissues.
The "expression pattern" of a promoter (with or without enhancer) is the pattern of expression levels which shows where in the plant and in what developmental stage
transcription is initiated by said promoter. Expression patterns of a set of promoters are said to be complementary when the expression pattern of one promoter shows little overlap with the expression pattern ofthe other promoter. The level of expression of a promoter can be determined by measuring the 'steady state' concentration of a standard transcribed reporter mRNA. This measurement is indirect since the concentration ofthe reporter mRNA is dependent not only on its synthesis rate, but also on the rate with which the mRNA is degraded. Therefore, the steady state level is the product of synthesis rates and degradation rates.
The rate of degradation can however be considered to proceed at a fixed rate when the transcribed sequences are identical, and thus this value can serve as a measure of synthesis rates. When promoters are compared in this way techniques available to those skilled in the art are hybridization SI -RNAse analysis, northern blots and competitive RT-PCR. This list of techniques in no way represents all available techniques, but rather describes commonly used procedures used to analyze transcription activity and expression levels of mRNA. The analysis of transcription start points in practically all promoters has revealed that there is usually no single base at which transcription starts, but rather a more or less clustered set of initiation sites, each of which accounts for some start points ofthe mRNA. Since this distribution varies from promoter to promoter the sequences ofthe reporter mRNA in each of the populations would differ from each other. Since each mRNA species is more or less prone to degradation, no single degradation rate can be expected for different reporter mRNAs. It has been shown for various eukaryotic promoter sequences that the sequence surrounding the initiation site ('initiator') plays an important role in determining the level of RNA expression directed by that specific promoter. This includes also part ofthe transcribed sequences. The direct fusion of promoter to reporter sequences would therefore lead to suboptimal levels of transcription.
A commonly used procedure to analyze expression patterns and levels is through determination ofthe 'steady state' level of protein accumulation in a cell. Commonly used candidates for the reporter gene, known to those skilled in the art are 3-glucuronidase (GUS), chloramphenicol acetyl transferase (CAT) and proteins with fluorescent properties, such as green fluorescent protein (GFP) from Aequora victoria. In principle, however, many more proteins are suitable for this puφose, provided the protein does not interfere with essential plant functions. For quantification and determination of localization a number of tools are suited. Detection systems can readily be created or are available which are based on, e.g.,
immunochemical, enzymatic, fluorescent detection and quantification. Protein levels can be determined in plant tissue extracts or in intact tissue using in situ analysis of protein expression.
Generally, individual transformed lines with one chimeric promoter reporter construct will vary in their levels of expression ofthe reporter gene. Also frequently observed is the phenomenon that such transformants do not express any detectable product (RNA or protein). The variability in expression is commonly ascribed to 'position effects', although the molecular mechanisms underlying this inactivity are usually not clear.
The term "average expression" is used here as the average level of expression found in all lines that do express detectable amounts of reporter gene, so leaving out ofthe analysis plants that do not express any detectable reporter mRNA or protein.
"Root expression level" indicates the expression level found in protein extracts of complete plant roots. Likewise, leaf, and stem expression levels, are determined using whole extracts from leaves and stems. It is acknowledged however, that within each ofthe plant parts just described, cells with variable functions may exist, in which promoter activity may vary.
"Non-specific expression" refers to constitutive expression or low level, basal ('leaky') expression in nondesired cells or tissues from a 'regulated promoter'.
"Altered levels" refers to the level of expression in transgenic organisms that differs from that of normal or untransformed organisms.
"Overexpression" refers to the level of expression in transgenic cells or organisms that exceeds levels of expression in normal or untransformed (nontransgenic) cells or organisms.
"Antisense inhibition" refers to the production of antisense RNA transcripts capable of suppressing the expression of protein from an endogenous gene or a transgene. "Co-suppression" and "transwitch" each refer to the production of sense RNA transcripts capable of suppressing the expression of identical or substantially similar transgene or endogenous genes (U.S. Patent No. 5,231,020).
"Gene silencing" refers to homology-dependent suppression of viral genes, transgenes, or endogenous nuclear genes. Gene silencing may be transcriptional, when the suppression is due to decreased transcription ofthe affected genes, or post-transcriptional, when the suppression is due to increased turnover (degradation) of RNA species homologous to the affected genes (English et al., 1996). Gene silencing includes virus-induced gene silencing (Ruiz et al. 1998).
"Silencing suppressor" gene refers to a gene whose expression leads to counteracting gene silencing and enhanced expression of silenced genes. Silencing suppressor genes may be of plant, non-plant, or viral origin. Examples include, but are not limited to HC-Pro, Pl-HC- Pro, and 2b proteins. Other examples include one or more genes in TGMV-B genome. The terms "heterologous DNA sequence," "exogenous DNA segment" or "heterologous nucleic acid," as used herein, each refer to a sequence that originates from a source foreign to the particular host cell or, if from the same source, is modified from its original foπn. Thus, a heterologous gene in a host cell includes a gene that is endogenous to the particular host cell but has been modified through, for example, the use of DNA shuffling. The terms also include non-naturally occuning multiple copies of a naturally occurring DNA sequence. Thus, the terms refer to a DNA segment that is foreign or heterologous to the cell, or homologous to the cell but in a position within the host cell nucleic acid in which the element is not ordinarily found. Exogenous DNA segments are expressed to yield exogenous polypeptides. A "homologous" DNA sequence is a DNA sequence that is naturally associated with a host cell into which it is introduced.
"Homologous to" in the context of nucleotide sequence identity refers to the similarity between the nucleotide sequence of two nucleic acid molecules or between the amino acid sequences of two protein molecules. Estimates of such homology are provided by either DNA-DNA or DNA-RNA hybridization under conditions of stringency as is well understood by those skilled in the art (as described in Haines and Higgins (eds.), Nucleic Acid
Hybridization, IRL Press, Oxford, U.K.), or by the comparison of sequence similarity between two nucleic acids or proteins.
The term "substantially similar" refers to nucleotide and amino acid sequences that represent functional and/or stractural equivalents of Arabidopsis sequences disclosed herein. For example, altered nucleotide sequences which simply reflect the degeneracy ofthe genetic code but nonetheless encode amino acid sequences that are identical to a particular amino acid sequence are substantially similar to the particular sequences. In addition, amino acid sequences that are substantially similar to a particular sequence are those wherein overall amino acid identity is at least 65% or greater to the instant sequences. Modifications that result in equivalent nucleotide or amino acid sequences are well within the routine skill in the art. Moreover, the skilled artisan recognizes that equivalent nucleotide sequences encompassed by this invention can also be defined by their ability to hybridize, under low, moderate and/or stringent conditions (e.g., 0.1X SSC, 0.1% SDS, 65°C), with the nucleotide
sequences that are within the literal scope ofthe instant claims.
"Target gene" refers to a gene on the replicon that expresses the desired target coding sequence, functional RNA, or protein. The target gene is not essential for replicon replication. Additionally, target genes may comprise native non- viral genes inserted into a non-native organism, or chimeric genes, and will be under the control of suitable regulatory sequences. Thus, the regulatory sequences in the target gene may come from any source, including the virus. Target genes may include coding sequences that are either heterologous or homologous to the genes of a particular plant to be transformed. However, target genes do not include native viral genes. Typical target genes include, but are not limited to genes encoding a structural protein, a seed storage protein, a protein that conveys herbicide resistance, and a protein that conveys insect resistance. Proteins encoded by target genes are known as "foreign proteins". The expression of a target gene in a plant will typically produce an altered plant trait. The term "altered plant trait" means any phenotypic or genotypic change in a transgenic plant relative to the wild-type or non-transgenic plant host.
"Transcription Stop Fragment" refers to nucleotide sequences that contain one or more regulatory signals, such as polyadenylation signal sequences, capable of terminating transcription. Examples include the 3' non-regulatory regions of genes encoding nopaline synthase and the small subunit of ribulose bisphosphate carboxylase.
"Replication gene" refers to a gene encoding a viral replication protein. In addition to the ORF ofthe replication protein, the replication gene may also contain other overlapping or non-overlapping ORF(s), as are found in viral sequences in nature. While not essential for replication, these additional ORFs may enhance replication and/or viral DNA accumulation. Examples of such additional ORFs are AC3 and AL3 in ACMV and TGMV geminivirases, respectively.
"Chimeric trans-acύng replication gene" refers either to a replication gene in which the coding sequence of a replication protein is under the control of a regulated plant promoter other than that in the native viral replication gene, or a modified native viral replication gene, for example, in which a site specific sequence(s) is inserted in the 5' transcribed but untranslated region. Such chimeric genes also include insertion ofthe known sites of
replication protein binding between the promoter and the transcription start site that attenuate transcription of viral replication protein gene.
"Chromosomally-integrated" refers to the integration of a foreign gene or DNA construct into the host DNA by covalent bonds. Where genes are not "chromosomally integrated" they may be "transiently expressed." Transient expression of a gene refers to the expression of a gene that is not integrated into the host chromosome but functions independently, either as part of an autonomously replicating plasmid or expression cassette, for example, or as part of another biological system such as a virus.
"Production tissue" refers to mature, harvestable tissue consisting of non-dividing, terminally-differentiated cells. It excludes young, growing tissue consisting of germline, meristematic, and not-fully-differentiated cells.
"Geπnline cells" refer to cells that are destined to be gametes and whose genetic material is heritable.
"Trans-activation" refers to switching on of gene expression or replicon replication by the expression of another (regulatory) gene in trans.
The term "transformation" refers to the transfer of a nucleic acid fragment into the genome of a host cell, resulting in genetically stable inheritance. Host cells containing the transformed nucleic acid fragments are refened to as "transgenic" cells, and organisms comprising transgenic cells are refened to as "transgenic organisms". Examples of methods of transformation of plants and plant cells include Agrobacterium-mediated transformation (De Blaere et al., 1987) and particle bombardment technology (Klein et al. 1987; U.S. Patent No. 4,945,050). Whole plants may be regenerated from transgenic cells by methods well known to the skilled artisan (see, for example, Fromm et al., 1990).
"Transformed," "transgenic," and "recombinant" refer to a host organism such as a bacterium or a plant into which a heterologous nucleic acid molecule has been introduced.
The nucleic acid molecule can be stably integrated into the genome generally known in the art and are disclosed in Sambrook et al, 1989. See also Innis et al., 1995 and Gelfand, 1995; and Innis and Gelfand, 1999. Known methods of PCR include, but are not limited to, methods using paired primers, nested primers, single specific primers, degenerate primers, gene-specific primers, vector-specific primers, partially mismatched primers, and the like. For example, "transformed," "transformant," and "transgenic" plants or calli have been tlirough the transformation process and contain a foreign gene integrated into their chromosome. The term "untransformed" refers to normal plants that have not been through the transformation process.
"Transiently transformed" refers to cells in which transgenes and foreign DNA have been introduced (for example, by such methods as Agrobacterium-mediated transformation or biolistic bombardment), but not selected for stable maintenance.
"Stably transformed" refers to cells that have been selected and regenerated on a selection media following transformation.
"Transient expression" refers to expression in cells in which a virus or a transgene is introduced by viral infection or by such methods as Agrobacterium-mediated transformation, electroporation, or biolistic bombardment, but not selected for its stable maintenance.
"Genetically stable" and "heritable" refer to chromosomally-integrated genetic elements that are stably maintained in the plant and stably inherited by progeny tlirough successive generations.
"Primary transformant" and "TO generation" refer to transgenic plants that are ofthe same genetic generation as the tissue which was initially transformed (i.e., not having gone tlirough meiosis and fertilization since transformation). "Secondary transformants" and the "TI, T2, T3, etc. generations" refer to transgenic plants derived from primary transformants through one or more meiotic and fertilization cycles. They may be derived by self-fertilization of primary or secondary transformants or crosses of primary or secondary transformants with other transformed or untransformed plants.
"Wild-type" refers to a viras or organism found in nature without any known mutation. "Genome" refers to the complete genetic material of an organism.
The term "nucleic acid" refers to deoxyribonucleotides or ribonucleotides and polymers thereof in either single- or double-stranded form, composed of monomers (nucleotides) containing a sugar, phosphate and a base which is either a purine or pyrimidine. Unless specifically limited, the term encompasses nucleic acids containing known analogs of natural nucleotides which have similar binding properties as the reference nucleic acid and are metabolized in a manner similar to naturally occuning nucleotides. Unless otherwise indicated, a particular nucleic acid sequence also implicitly encompasses conservatively modified variants thereof (e.g., degenerate codon substitutions) and complementary sequences as well as the sequence explicitly indicated. Specifically, degenerate codon substitutions may be achieved by generating sequences in which the third position of one or more selected (or all) codons is substituted with mixed-base and/or deoxyinosine residues (Batzer et al., 1991; Ohtsuka et al., 1985; Rossolini et al. 1994). A "nucleic acid fragment" is a fraction of a given nucleic acid molecule. In higher plants, deoxyribonucleic acid (DNA) is the genetic material
while ribonucleic acid (RNA) is involved in the transfer of information contained within DNA into proteins. The term "nucleotide sequence" refers to a polymer of DNA or RNA which can be single- or double-stranded, optionally containing synthetic, non-natural or altered nucleotide bases capable of incoφoration into DNA or RNA polymers. The terms "nucleic acid" or "nucleic acid sequence" may also be used interchangeably with gene, cDNA, DNA and RNA encoded by a gene.
The invention encompasses isolated or substantially purified nucleic acid or protein compositions. In the context ofthe present invention, an "isolated" or "purified" DNA molecule or an "isolated" or "purified" polypeptide is a DNA molecule or polypeptide that, by the hand of man, exists apart from its native environment and is therefore not a product of nature. An isolated DNA molecule or polypeptide may exist in a purified form or may exist in a non-native environment such as, for example, a transgenic host cell. For example, an "isolated" or "purified" nucleic acid molecule or protein, or biologically active portion thereof, is substantially free of other cellular material, or culture medium when produced by recombinant techniques, or substantially free of chemical precursors or other chemicals when chemically synthesized. Preferably, an "isolated" nucleic acid is free of sequences (preferably protein encoding sequences) that naturally flank the nucleic acid (i.e., sequences located at the 5' and 3' ends ofthe nucleic acid) in the genomic DNA ofthe organism from which the nucleic acid is derived. For example, in various embodiments, the isolated nucleic acid molecule can contain less than about 5 kb, 4 kb, 3 kb, 2 kb, 1 kb, 0.5 kb, or 0.1 kb of nucleotide sequences that naturally flank the nucleic acid molecule in genomic DNA ofthe cell from which the nucleic acid is derived. A protein that is substantially free of cellular material includes preparations of protein or polypeptide having less than about 30%, 20%, 10%, 5%, (by dry weight) of contaminating protein. When the protein ofthe invention, or biologically active portion thereof, is recombinantly produced, preferably culture medium represents less than about 30%), 20%), 10%), or 5% (by dry weight) of chemical precursors or non-protein of interest chemicals.
The nucleotide sequences ofthe invention include both the naturally occurring sequences as well as mutant (variant) forms. Such variants will continue to possess the desired activity, i.e., either promoter activity or the activity ofthe product encoded by the open reading frame ofthe non- variant nucleotide sequence.
Thus, by "variants" is intended substantially similar sequences. For nucleotide sequences comprising an open reading frame, variants include those sequences that, because of
the degeneracy ofthe genetic code, encode the identical amino acid sequence ofthe native protein. Naturally occuning allelic variants such as these can be identified with the use of well-known molecular biology techniques, as, for example, with polymerase chain reaction (PCR) and hybridization techniques. Variant nucleotide sequences also include synthetically derived nucleotide sequences, such as those generated, for example, by using site-directed mutagenesis and for open reading frames, encode the native protein, as well as those that encode a polypeptide having amino acid substitutions relative to the native protein. Generally, nucleotide sequence variants ofthe invention will have at least 40, 50, 60, to 70%, e.g., preferably 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, to 79%, generally at least 80%, e.g., 81%-84%, at least 85%, e.g., 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%o, to 98%> and 99%) nucleotide sequence identity to the native (wild type or endogenous) nucleotide sequence.
"Conservatively modified variations" of a particular nucleic acid sequence refers to those nucleic acid sequences that encode identical or essentially identical amino acid sequences, or where the nucleic acid sequence does not encode an amino acid sequence, to essentially identical sequences. Because ofthe degeneracy ofthe genetic code, a large number of functionally identical nucleic acids encode any given polypeptide. For instance the codons CGT, CGC, CGA, CGG, AGA, and AGG all encode the amino acid arginine. Thus, at every position where an arginine is specified by a codon, the codon can be altered to any ofthe conesponding codons described without altering the encoded protein. Such nucleic acid variations are "silent variations" which are one species of "conservatively modified variations." Every nucleic acid sequence described herein which encodes a polypeptide also describes every possible silent variation, except where otherwise noted. One of skill will recognize that each codon in a nucleic acid (except ATG, which is ordinarily the only codon for methionine) can be modified to yield a functionally identical molecule by standard techniques. Accordingly, each "silent variation" of a nucleic acid which encodes a polypeptide is implicit in each described sequence.
The nucleic acid molecules ofthe invention can be "optimized" for enhanced expression in plants of interest. See, for example, EPA 035472; WO 91/16432; Perlak et al., 1991; and Murray et al., 1989. In this manner, the open reading frames in genes or gene fragments can be synthesized utilizing plant-prefened codons. See, for example, Campbell and Gowri, 1990 for a discussion of host-prefened codon usage. Thus, the nucleotide sequences can be optimized for expression in any plant. It is recognized that all or any part of
the gene sequence may be optimized or synthetic. That is, synthetic or partially optimized sequences may also be used. Variant nucleotide sequences and proteins also encompass sequences and protein derived from a mutagenic and recombinogenic procedure such as DNA shuffling. With such a procedure, one or more different coding sequences can be manipulated to create a new polypeptide possessing the desired properties. In this manner, libraries of recombinant polynucleotides are generated from a population of related sequence polynucleotides comprising sequence regions that have substantial sequence identity and can be homologously recombined in vitro or in vivo. Strategies for such DNA shuffling are known in the art. See, for example, Stemmer, 1994; Stemmer, 1994; Crameri et al., 1997; Moore et al., 1997; Zhang et al., 1997; Crameri et al., 1998; and U.S. Patent Nos. 5,605,793 and 5,837,458.
By "variant" polypeptide is intended a polypeptide derived from the native protein by deletion (so-called truncation) or addition of one or more amino acids to the N-terminal and/or C-terminal end ofthe native protein; deletion or addition of one or more amino acids at one or more sites in the native protem; or substitution of one or more amino acids at one or more sites in the native protein. Such variants may result from, for example, genetic polymoφhism or from human manipulation. Methods for such manipulations are generally known in the art.
Thus, the polypeptides may be altered in various ways including amino acid substitutions, deletions, truncations, and insertions. Methods for such manipulations are generally known in the art. For example, amino acid sequence variants ofthe polypeptides can be prepared by mutations in the DNA. Methods for mutagenesis and nucleotide sequence alterations are well known in the art. See, for example, Kunkel, 1985; Kunkel et al., 1987; U. S. Patent No. 4,873,192; Walker and Gaastra, 1983 and the references cited therein. Guidance as to appropriate amino acid substitutions that do not affect biological activity ofthe protein of interest may be found in the model of Dayhoff et al. (1978). Conservative substitutions, such as exchanging one amino acid with another having similar properties, are prefened.
Individual substitutions deletions or additions that alter, add or delete a single amino acid or a small percentage of amino acids (typically less than 5%>, more typically less than 1%) in an encoded sequence are "conservatively modified variations," where the alterations result in the substitution of an amino acid with a chemically similar amino acid. Conservative substitution tables providing functionally similar arnino acids are well known in the art. The following five groups each contain amino acids that are conservative substitutions for one another: Aliphatic: Glycine (G), Alanine (A), Valine (V), Leucine (L), Isoleucine (I);
Aromatic: Phenylalanine (F), Tyrosine (Y), Tryptophan (W); Sulfur-containing: Methionine (M), Cysteine (C); Basic: Arginine I, Lysine (K), Histidine (H); Acidic: Aspartic acid (D), Glutamic acid (E), Asparagine (N), Glutamine (Q). See also, Creighton, 1984. In addition, individual substitutions, deletions or additions which alter, add or delete a single amino acid or a small percentage of amino acids in an encoded sequence are also "conservatively modified variations."
"Expression cassette" as used herein means a DNA sequence capable of directing expression of a particular nucleotide sequence in an appropriate host cell, comprising a promoter operably linked to the nucleotide sequence of interest which is operably linked to termination signals. It also typically comprises sequences required for proper translation ofthe nucleotide sequence. The coding region usually codes for a protein of interest but may also code for a functional RNA of interest, for example antisense RNA or a nontranslated RNA, in the sense or antisense direction. The expression cassette comprising the nucleotide sequence of interest may be chimeric, meaning that at least one of its components is heterologous with respect to at least one of its other components. The expression cassette may also be one which is naturally occurring but has been obtained in a recombinant form useful for heterologous expression. The expression ofthe nucleotide sequence in the expression cassette may be under the control of a constitutive promoter or of an inducible promoter which initiates transcription only when the host cell is exposed to some particular external stimulus. In the case of a multicellular organism, the promoter can also be specific to a particular tissue or organ or stage of development.
"Vector" is defined to include, inter alia, any plasmid, cosmid, phage or Agrobacterium binary vector in double or single stranded linear or circular form which may or may not be self transmissible or mobilizable, and which can transform prokaryotic or eukaryotic host either by integration into the cellular genome or exist extrachromosomally (e.g. autonomous replicating plasmid with an origin of replication).
Specifically included are shuttle vectors by which is meant a DNA vehicle capable, naturally or by design, of replication in two different host organisms, which may be selected from actinomycetes and related species, bacteria and eukaryotic (e.g. higher plant, mammalian, yeast or fungal cells).
Preferably the nucleic acid in the vector is under the control of, and operably linked to, an appropriate promoter or other regulatory elements for transcription in a host cell such as a microbial, e.g. bacterial, or plant cell. The vector may be a bi-functional expression vector
which functions in multiple hosts. In the case of genomic DNA, this may contain its own promoter or other regulatory elements and in the case of cDNA this may be under the control of an appropriate promoter or other regulatory elements for expression in the host cell.
"Cloning vectors" typically contain one or a small number of restriction endonuclease recognition sites at which foreign DNA sequences can be inserted in a determinable fashion without loss of essential biological function ofthe vector, as well as a marker gene that is suitable for use in the identification and selection of cells transformed with the cloning vector. Marker genes typically include genes that provide tetracycline resistance, hygromycin resistance or ampicillin resistance. A "transgenic plant" is a plant having one or more plant cells that contain an expression vector.
"Plant tissue" includes differentiated and undifferentiated tissues or plants, including but not limited to roots, stems, shoots, leaves, pollen, seeds, tumor tissue and various forms of cells and culture such as single cells, protoplast, embryos, and callus tissue. The plant tissue may be in plants or in organ, tissue or cell culture.
The following terms are used to describe the sequence relationships between two or more nucleic acids or polynucleotides: (a) "reference sequence", (b) "comparison window", (c) "sequence identity", (d) "percentage of sequence identity", and (e) "substantial identity".
(a) As used herein, "reference sequence" is a defined sequence used as a basis for sequence comparison. A reference sequence may be a subset or the entirety of a specified sequence; for example, as a segment of a full length cDNA or gene sequence, or the complete cDNA or gene sequence.
(b) As used herein, "comparison window" makes reference to a contiguous and specified segment of a polynucleotide sequence, wherein the polynucleotide sequence in the comparison window may comprise additions or deletions (i.e., gaps) compared to the reference sequence (which does not comprise additions or deletions) for optimal alignment ofthe two sequences. Generally, the comparison window is at least 20 contiguous nucleotides in length, and optionally can be 30, 40, 50, 100, or longer. Those of skill in the art understand that to avoid a high similarity to a reference sequence due to inclusion of gaps in the polynucleotide sequence a gap penalty is typically introduced and is subtracted from the number of matches. Methods of alignment of sequences for comparison are well known in the art. Thus, the determination of percent identity between any two sequences can be accomplished using a mathematical algorithm. Prefened, non-lhriiting examples of such mathematical algorithms
are the algorithm of Myers and Miller, 1988; the local homology algorithm of Smith et al. 1981; the homology alignment algorithm of Needleman and Wunsch 1970; the search-for- similarity-method of Pearson and Lipman 1988; the algorithm of Karlin and Altschul, 1990, modified as in Karlin and Altschul, 1993. Computer implementations of these mathematical algorithms can be utilized for comparison of sequences to determine sequence identity. Such implementations include, but are not limited to: CLUSTAL in the PC/Gene program (available from Intelligenetics, Mountain View, California); the ALIGN program (Version 2.0) and GAP, BESTFIT, BLAST, FASTA, and TFASTA in the Wisconsin Genetics Software Package, Version 8 (available from Genetics Computer Group (GCG), 575 Science Drive, Madison, Wisconsin, USA). Alignments using these programs can be performed using the default parameters. The CLUSTAL program is well described by Higgins et al. 1988; Higgins et al. 1989; Coφet et al. 1988; Huang et al. 1992; and Pearson et al. 1994. The ALIGN program is based on the algorithm of Myers and Miller, supra. The BLAST programs of Altschul et al., 1990, are based on the algorithm of Karlin and Altschul supra .
Software for performing BLAST analyses is publicly available through the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/). This algorithm involves first identifying high scoring sequence pairs (HSPs) by identifying short words of length W in the query sequence, which either match or satisfy some positive- valued threshold score T when aligned with a word ofthe same length in a database sequence. T is refened to as the neighborhood word score threshold (Altschul et al., 1990). These initial neighborhood word hits act as seeds for initiating searches to find longer HSPs containing them. The word hits are then extended in both directions along each sequence for as far as the cumulative alignment score can be increased. Cumulative scores are calculated using, for nucleotide sequences, the parameters M (reward score for a pair of matching residues; always > 0) and N (penalty score for mismatching residues; always < 0). For amino acid sequences, a scoring matrix is used to calculate the cumulative score. Extension ofthe word hits in each direction are halted when the cumulative alignment score falls off by the quantity X from its maximum achieved value, the cumulative score goes to zero or below due to the accumulation of one or more negative-scoring residue alignments, or the end of either sequence is reached.
In addition to calculating percent sequence identity, the BLAST algorithm also performs a statistical analysis ofthe similarity between two sequences (see, e.g., Karlin & Altschul (1993). One measure of similarity provided by the BLAST algorithm is the smallest
sum probability (P(N)), which provides an indication ofthe probability by which a match between two nucleotide or amino acid sequences would occur by chance. For example, a test nucleic acid sequence is considered similar to a reference sequence if the smallest sum probability in a comparison ofthe test nucleic acid sequence to the reference nucleic acid sequence is less than about 0.1, more preferably less than about 0.01, and most preferably less than about 0.001.
To obtain gapped alignments for comparison puφoses, Gapped BLAST (in BLAST 2.0) can be utilized as described in Altschul et al. 1997. Alternatively, PSI-BLAST (in BLAST 2.0) can be used to perform an iterated search that detects distant relationships between molecules. See Altschul et al., supra. When utilizing BLAST, Gapped BLAST, PSI- BLAST, the default parameters ofthe respective programs (e.g. BLASTN for nucleotide sequences, BLASTX for proteins) can be used. The BLASTN program (for nucleotide sequences) uses as defaults a wordlength (W) of 11, an expectation (E) of 10, a cutoff of 100, M=5, N=-4, and a comparison of both strands. For amino acid sequences, the BLASTP program uses as defaults a wordlength (W) of 3, an expectation (E) of 10, and the BLOSUM62 scoring matrix (see Henikoff & Henikoff, 1989). See http://www.ncbi.nlm.nih.gov. Alignment may also be performed manually by inspection.
For piuposes ofthe present invention, comparison of nucleotide sequences for determination of percent sequence identity to the promoter sequences disclosed herein is preferably made using the BlastN program (version 1.4.7 or later) with its default parameters or any equivalent program. By "equivalent program" is intended any sequence comparison program that, for any two sequences in question, generates an alignment having identical nucleotide or amino acid residue matches and an identical percent sequence identity when compared to the conesponding alignment generated by the prefened program. (c) As used herein, "sequence identity" or "identity" in the context of two nucleic acid or polypeptide sequences makes reference to the residues in the two sequences that are the same when aligned for maximum conespondence over a specified comparison window. When percentage of sequence identity is used in reference to proteins it is recognized that residue positions which are not identical often differ by conservative amino acid substitutions, where amino acid residues are substituted for other amino acid residues with similar chemical properties (e.g., charge or hydrophobicity) and therefore do not change the functional properties ofthe molecule. When sequences differ in conservative substitutions, the percent sequence identity may be adjusted upwards to conect for the conservative nature ofthe
substitution. Sequences that differ by such conservative substitutions are said to have "sequence similarity" or "similarity." Means for making this adjustment are well known to those of skill in the art. Typically this involves scoring a conservative substitution as a partial rather than a full mismatch, thereby increasing the percentage sequence identity. Thus, for example, where an identical amino acid is given a score of 1 and a non-conservative substitution is given a score of zero, a conservative substitution is given a score between zero and 1. The scoring of conservative substitutions is calculated, e.g., as implemented in the program PC/GENE (Intelligenetics, Mountain View, California).
(d) As used herein, "percentage of sequence identity" means the value determined by comparing two optimally aligned sequences over a comparison window, wherein the portion of the polynucleotide sequence in the comparison window may comprise additions or deletions (i.e., gaps) as compared to the reference sequence (which does not comprise additions or deletions) for optimal alignment ofthe two sequences. The percentage is calculated by determining the number of positions at which the identical nucleic acid base or arnino acid residue occurs in both sequences to yield the number of matched positions, dividing the number of matched positions by the total number of positions in the window of comparison, and multiplying the result by 100 to yield the percentage of sequence identity.
(e)(i) The term "substantial identity" of polynucleotide sequences means that a polynucleotide comprises a sequence that has at least 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, or 79%, preferably at least 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, or 89%, more preferably at least 90%, 91%, 92%, 93%, or 94%, and most preferably at least 95%, 96%, 97%, 98%, or 99% sequence identity, compared to a reference sequence using one ofthe aligmnent programs described using standard parameters. One of skill in the art will recognize that these values can be appropriately adjusted to determine conesponding identity of proteins encoded by two nucleotide sequences by taking into account codon degeneracy, amino acid similarity, reading frame positioning, and the like. Substantial identity of amino acid sequences for these puφoses normally means sequence identity of at least 10%, more preferably at least 80%, 90%, and most preferably at least 95%.
Another indication that nucleotide sequences are substantially identical is if two molecules hybridize to each other under stringent conditions (see below). Generally, stringent conditions are selected to be about 5°C lower than the thermal melting point (Tm) for the specific sequence at a defined ionic strength and pH. However, stringent conditions encompass temperatures in the range of about 1°C to about 20°C, depending upon the desired
degree of stringency as otherwise qualified herein. Nucleic acids that do not hybridize to each other under stringent conditions are still substantially identical if the polypeptides they encode are substantially identical. This may occur, e.g., when a copy of a nucleic acid is created using the maximum codon degeneracy permitted by the genetic code. One indication that two nucleic acid sequences are substantially identical is when the polypeptide encoded by the first nucleic acid is immunologically cross reactive with the polypeptide encoded by the second nucleic acid.
(e)(ii) The term "substantial identity" in the context of a peptide indicates that a peptide comprises a sequence with at least 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, or 79%, preferably 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, or 89%, more preferably at least 90%, 91%, 92%, 93%, or 94%, or even more preferably, 95%, 96%, 97%, 98% or 99%, sequence identity to the reference sequence over a specified comparison window. Preferably, optimal alignment is conducted using the homology alignment algorithm of Needleman and Wunsch (1970). An indication that two peptide sequences are substantially identical is that one peptide is immunologically reactive with antibodies raised against the second peptide. Thus, a peptide is substantially identical to a second peptide, for example, where the two peptides differ only by a conservative substitution.
For sequence comparison, typically one sequence acts as a reference sequence to which test sequences are compared. When using a sequence comparison algorithm, test and reference sequences are input into a computer, subsequence coordinates are designated if necessary, and sequence algorithm program parameters are designated. The sequence comparison algorithm then calculates the percent sequence identity for the test sequence(s) relative to the reference sequence, based on the designated program parameters.
As noted above, another indication that two nucleic acid sequences are substantially identical is that the two molecules hybridize to each other under stringent conditions. The phrase "hybridizing specifically to" refers to the binding, duplexing, or hybridizing of a molecule only to a particular nucleotide sequence under stringent conditions when that sequence is present in a complex mixture (e.g., total cellular) DNA or RNA. "Bind(s) substantially" refers to complementary hybridization between a probe nucleic acid and a target nucleic acid and embraces minor mismatches that can be accommodated by reducing the stringency ofthe hybridization media to achieve the desired detection ofthe target nucleic acid sequence.
"Stringent hybridization conditions" and "stringent hybridization wash conditions" in the context of nucleic acid hybridization experiments such as Southern and Northern hybridization are sequence dependent, and are different under different environmental parameters. The Tm is the temperature (under defined ionic strength and pH) at which 50% of the target sequence hybridizes to a perfectly matched probe. Specificity is typically the function of post-hybridization washes, the critical factors being the ionic strength and temperature ofthe final wash solution. For DNA-DNA hybrids, the Tm can be approximated from the equation of Meinkoth and Wahl, 1984; Tm 81.5°C + 16.6 (log M) +0.41 (%GC) - 0.61 (% form) - 500/L; where M is the molarity of monovalent cations, %GC is the percentage of guanosine and cytosine nucleotides in the DNA, % form is the percentage of formamide in the hybridization solution, and L is the length ofthe hybrid in base pairs. Tm is reduced by about 1°C for each 1% of mismatching; thus, Tm, hybridization, and/or wash conditions can be adjusted to hybridize to sequences ofthe desired identity. For example, if sequences with >90% identity are sought, the Tm can be decreased 10°C. Generally, stringent conditions are selected to be about 5°C lower than the thermal melting point I for the specific sequence and its complement at a defined ionic strength and pH. However, severely stringent conditions can utilize a hybridization and/or wash at 1, 2, 3, or 4°C lower than the thermal melting point I; moderately stringent conditions can utilize a hybridization and/or wash at 6, 7, 8, 9, or 10°C lower than the thermal melting point I; low stringency conditions can utilize a hybridization and/or wash at 11, 12, 13, 14, 15, or 20°C lower than the thermal melting point I. Using the equation, hybridization and wash compositions, and desired T, those of ordinary skill will understand that variations in the stringency of hybridization and/or wash solutions are inherently described. If the desired degree of mismatching results in a T of less than 45°C (aqueous solution) or 32°C (formamide solution), it is prefened to increase the SSC concentration so that a higher temperature can be used. An extensive guide to the hybridization of nucleic acids is found in Tijssen, 1993. Generally, highly stringent hybridization and wash conditions are selected to be about 5°C lower than the thermal melting point Tm for the specific sequence at a defined ionic strength and pH.
An example of highly stringent wash conditions is 0.15 M NaCl at 72°C for about 15 minutes. An example of stringent wash conditions is a 0.2X SSC wash at 65°C for 15 minutes (see, Sambrook, infra, for a description of SSC buffer). Often, a high stringency wash is preceded by a low stringency wash to remove background probe signal. An example medium
stringency wash for a duplex of, e.g., more than 100 nucleotides, is IX SSC at 45°C for 15 minutes. An example low stringency wash for a duplex of, e.g., more than 100 nucleotides, is 4-6X SSC at 40°C for 15 minutes. For short probes (e.g., about 10 to 50 nucleotides), stringent conditions typically involve salt concentrations of less than about 1.5 M, more preferably about 0.01 to 1.0 M, Na ion concentration (or other salts) at pH 7.0 to 8.3, and the temperature is typically at least about 30°C and at least about 60°C for long robes (e.g., >50 nucleotides). Stringent conditions may also be achieved with the addition of destabilizing agents such as formamide. In general, a signal to noise ratio of 2X (or higher) than that observed for an unrelated probe in the particular hybridization assay indicates detection of a specific hybridization. Nucleic acids that do not hybridize to each other under stringent conditions are still substantially identical if the proteins that they encode are substantially identical. This occurs, e.g., when a copy of a nucleic acid is created using the maximum codon degeneracy permitted by the genetic code.
Very stringent conditions are selected to be equal to the Tm for a particular probe. An example of stringent conditions for hybridization of complementary nucleic acids which have more than 100 complementary residues on a filter in a Southern or Northern blot is 50% formamide, e.g., hybridization in 50% formamide, 1 M NaCl, 1%> SDS at 37°C, and a wash in 0. IX SSC at 60 to 65°C. Exemplary low stringency conditions include hybridization with a buffer solution of 30 to 35% formamide, 1 M NaCl, 1% SDS (sodium dodecyl sulphate) at 37°C, and a wash in IX to 2X SSC (20X SSC = 3.0 M NaCl/0.3 M trisodium citrate) at 50 to 55°C. Exemplary moderate stringency conditions include hybridization in 40 to 45% formamide, 1.0 M NaCl, 1% SDS at 37°C, and a wash in 0.5X to IX SSC at 55 to 60°C.
The following are examples of sets of hybridization wash conditions that may be used to clone orthologous nucleotide sequences that are substantially identical to reference nucleotide sequences ofthe present invention: a reference nucleotide sequence preferably hybridizes to the reference nucleotide sequence in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in 2X SSC, 0.1% SDS at 50°C, more desirably in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in IX SSC, 0.1% SDS at 50°C, more desirably still in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in 0.5X SSC, 0.1 % SDS at 50°C, preferably in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in 0.1X
SSC, 0.1% SDS at 50°C, more preferably in 7% sodium dodecyl sulfate (SDS), 0.5 M NaPO4, 1 mM EDTA at 50°C with washing in 0.1X SSC, 0.1% SDS at 65°C.
"DNA shuffling" is a method to introduce mutations or reanangements, preferably randomly, in a DNA molecule or to generate exchanges of DNA sequences between two or more DNA molecules, preferably randomly. The DNA molecule resulting from DNA shuffling is a shuffled DNA molecule that is a non-naturally occuning DNA molecule derived from at least one template DNA molecule. The shuffled DNA preferably encodes a variant polypeptide modified with respect to the polypeptide encoded by the template DNA, and may have an altered biological activity with respect to the polypeptide encoded by the template DNA.
"Recombinant DNA molecule' is a combination of DNA sequences that are joined together using recombinant DNA technology and procedures used to join together DNA sequences as described, for example, in Sambrook et al., 1989.
The word "plant" refers to any plant, particularly to seed plant, and "plant cell" is a stractural and physiological unit ofthe plant, which comprises a cell wall but may also refer to a protoplast. The plant cell may be in form of an isolated single cell or a cultured cell, or as a part of higher organized unit such as, for example, a plant tissue, or a plant organ.
"Significant increase" is an increase that is larger than the margin of enor inherent in the measurement technique, preferably an increase by about 2-fold or greater. "Significantly less" means that the decrease is larger than the margin of enor inherent in the measurement technique, preferably a decrease by about 2-fold or greater. II. DNA Sequences for Transformation
Virtually any DNA composition may be used for delivery to recipient plant cells, e.g., monocotyledonous cells, to ultimately produce fertile transgenic plants in accordance with the present invention. For example, DNA segments in the form of vectors and plasmids, or linear DNA fragments, in some instances containing only the DNA element to be expressed in the plant, and the like, may be employed. The construction of vectors which may be employed in conjunction with the present invention will be known to those of skill ofthe art in light ofthe present disclosure (see, e.g., Sambrook et al, 1989; Gelvin et al., 1990). Vectors, plasmids, cosmids, YACs (yeast artificial chromosomes), BACs (bacterial artificial chromosomes) and DNA segments for use in fransforming such cells will, of course, generally comprise the cDNA, gene or genes which one desires to introduce into the cells. These DNA constructs can further include structures such as promoters, enhancers,
polylinkers, or even regulatory genes as desired. The DNA segment or gene chosen for cellular introduction will often encode a protein which will be expressed in the resultant recombinant cells, such as will result in a screenable or selectable trait and/or which will impart an improved phenotype to the regenerated plant. However, this may not always be the case, and the present invention also encompasses transgenic plants incoφorating non- expressed transgenes.
In certain embodiments, it is contemplated that one may wish to employ replication- competent viral vectors in monocot transformation. Such vectors include, for example, wheat dwarf viras (WDV) "shuttle" vectors, such as pWl-11 and PW1-GUS (Ugaki et al., 1991). These vectors are capable of autonomous replication in maize cells as well as E. coli, and as such may provide increased sensitivity for detecting DNA delivered to transgenic cells. A replicating vector may also be useful for delivery of genes flanked by DNA sequences from transposable elements such as Ac, Ds, or Mu. It has been proposed (Laufs et al., 1990) that transposition of these elements within the maize genome requires DNA replication. It is also contemplated that transposable elements would be useful for introducing DNA fragments lacking elements necessary for selection and maintenance ofthe plasmid vector in bacteria, e.g., antibiotic resistance genes and origins of DNA replication. It is also proposed that use of a transposable element such as Ac, Ds, or Mu would actively promote integration ofthe desired DNA and hence increase the frequency of stably transformed cells. The use of a transposable element such as Ac, Ds, or Mu may actively promote integration ofthe DNA of interest and hence increase the frequency of stably transformed cells. Transposable elements may be useful to allow separation of genes of mterest from elements necessary for selection and maintenance of a plasmid vector in bacteria or selection of a transformant. By use of a transposable element, desirable and undesirable DNA sequences may be transposed apart from each other in the genome, such that through genetic segregation in progeny, one may identify plants with either the desirable undesirable DNA sequences.
DNA useful for introduction into plant cells includes that which has been derived or isolated from any source, that may be subsequently characterized as to stracture, size and/or function, chemically altered, and later introduced into plants. An example of DNA "derived" from a source, would be a DNA sequence that is identified as a useful fragment within a given organism, and which is then chemically synthesized in essentially pure form. An example of such DNA "isolated" from a source would be a useful DNA sequence that is excised or removed from said source by chemical means, e.g., by the use of restriction endonucleases, so
that it can be further manipulated, e.g., amplified, for use in the invention, by the methodology of genetic engineering. Such DNA is commonly refened to as "recombinant DNA."
Therefore useful DNA includes completely synthetic DNA, semi-synthetic DNA, DNA isolated from biological sources, and DNA derived from introduced RNA. Generally, the introduced DNA is not originally resident in the plant genotype which is the recipient ofthe DNA, but it is within the scope ofthe invention to isolate a gene from a given plant genotype, and to subsequently introduce multiple copies ofthe gene into the same genotype, e.g., to enhance production of a given gene product such as a storage protein or a protem that confers tolerance or resistance to water deficit. The introduced DNA includes but is not limited to, DNA from plant genes, and non- plant genes such as those from bacteria, yeasts, animals or viruses. The introduced DNA can include modified genes, portions of genes, or chimeric genes, including genes from the same or different maize genotype. The term "chimeric gene" or "chimeric DNA" is defined as a gene or DNA sequence or segment comprising at least two DNA sequences or segments from species which do not combine DNA under natural conditions, or which DNA sequences or segments are positioned or linked in a manner which does not normally occur in the native genome of untransformed plant.
The introduced DNA used for transformation herein may be circular or linear, double- stranded or single-stranded. Generally, the DNA is in the form of chimeric DNA, such as plasmid DNA, that can also contain coding regions flanked by regulatory sequences which promote the expression ofthe recombinant DNA present in the resultant plant. For example, the DNA may itself compri.se or consist of a promoter that is active in a plant which is derived from a source other than that plant, or may utilize a promoter already present in a plant genotype that is the transformation target. Generally, the introduced DNA will be relatively small, i.e., less than about 30 kb to niimmize any susceptibility to physical, chemical, or enzymatic degradation which is known to increase as the size ofthe DNA increases. As noted above, the number of proteins, RNA transcripts or mixtures thereof which is introduced into the plant genome is preferably preselected and defined, e.g., from one to about 5-10 such products ofthe introduced DNA may be formed.
Two principal methods for the control of expression are known, viz.: overexpression and underexpression. Overexpression can be achieved by insertion of one or more than one extra copy ofthe selected gene. It is, however, not unknown for plants or their progeny,
originally transformed with one or more than one extra copy of a nucleotide sequence, to exhibit the effects of underexpression as well as overexpression. For underexpression there are two principle methods which are commonly refened to in the art as "antisense downregulation" and "sense downregulation" (sense downregulation is also refened to as "cosuppression"). Generically these processes are refened to as "gene silencing". Both of these methods lead to an inhibition of expression ofthe target gene.
Obtaining sufficient levels of transgene expression in the appropriate plant tissues is an important aspect in the production of genetically engineered crops. Expression of heterologous DNA sequences in a plant host is dependent upon the presence of an operably linked promoter that is functional within the plant host. Choice ofthe promoter sequence will determine when and where within the organism the heterologous DNA sequence is expressed.
Furthermore, it is contemplated that promoters combining elements from more than one promoter may be useful. For example, U.S. Patent No. 5,491,288 discloses combining a Cauliflower Mosaic Virus promoter with a histone promoter. Thus, the elements from the promoters disclosed herein may be combined with elements from other promoters.
Promoters which are useful for plant transgene expression include those that are inducible, viral, synthetic, constitutive (Odell et al., 1985), temporally regulated, spatially regulated, tissue-specific, and spatio-temporally regulated.
Where expression in specific tissues or organs is desired, tissue-specific promoters may be used. In contrast, where gene expression in response to a stimulus is desired, inducible promoters are the regulatory elements of choice. Where continuous expression is desired throughout the cells of a plant, constitutive promoters are utilized. Additional regulatory sequences upstream and/or downstream from the core promoter sequence may be included in expression constructs of transformation vectors to bring about varying levels of expression of heterologous nucleotide sequences in a transgenic plant. A. Transcription Regulatory Sequences 1. Promoters
The choice of promoter will vary depending on the temporal and spatial requirements for expression, and also depending on the target species. In some cases, expression in multiple tissues is desirable. While in others, tissue-specific, e.g., leaf-specific, seed-specific, petal-specific, anther-specific, or pith-specific, expression is desirable. Although many promoters from dicotyledons have been shown to be operational in monocotyledons and vice versa, ideally dicotyledonous promoters are selected for expression
in dicotyledons, and monocotyledonous promoters for expression in monocotyledons. However, there is no restriction to the provenance of selected promoters; it is sufficient that they are operational in driving the expression ofthe nucleotide sequences in the desired cell. These promoters include, but are not limited to, constitutive, inducible, temporally regulated, developmentally regulated, spatially-regulated, chemically regulated, stress- responsive, tissue-specific, viral and synthetic promoters. Promoter sequences are known to be strong or weak. A strong promoter provides for a high level of gene expression, whereas a weak promoter provides for a very low level of gene expression. An inducible promoter is a promoter that provides for the turning on and off of gene expression in response to an exogenously added agent, or to an environmental or developmental stimulus. A bacterial promoter such as the Ptac promoter can be induced to varying levels of gene expression depending on the level of isothiopropylgalactoside added to the transformed bacterial cells. An isolated promoter sequence that is a strong promoter for heterologous nucleic acid is advantageous because it provides for a sufficient level of gene expression to allow for easy detection and selection of transformed cells and provides for a high level of gene expression when desired.
Within a plant promoter region there are several domains that are necessary for full function ofthe promoter. The first of these domains lies immediately upstream ofthe stractural gene and forms the "core promoter region" containing consensus sequences, normally 70 base pairs immediately upstream ofthe gene. The core promoter region contains the characteristic CAAT and TATA boxes plus sunounding sequences, and represents a transcription initiation sequence that defines the transcription start point for the stractural gene. The presence ofthe core promoter region defines a sequence as being a promoter: if the region is absent, the promoter is non-functional. Furthermore, the core promoter region is insufficient to provide full promoter activity. A series of regulatory sequences upstream ofthe core constitute the remainder ofthe promoter. The regulatory sequences determine expression level, the spatial and temporal pattern of expression and, for an important subset of promoters, expression under inductive conditions (regulation by external factors such as hght, temperature, chemicals, hormones). A range of naturally-occurring promoters are known to be operative in plants and have been used to drive the expression of heterologous (both foreign and endogenous) genes in plants: for example, the constitutive 35S cauliflower mosaic virus (CaMV) promoter, the ripening-enhanced tomato polygalacturonase promoter (Bird et al., 1988), the E8 promoter
(Diekman & Fischer, 1988) and the fruit specific 2A1 promoter (Pear et al, 1989) and many others, e.g., U2 and U5 snRNA promoters from maize, the promoter from alcohol dehydrogenase, the Z4 promoter from a gene encoding the Z4 22 kD zein protein, the Z10 promoter from a gene encoding a 10 kD zein protein, a Z27 promoter from a gene encoding a 27 kD zein protein, the A20 promoter from the gene encoding a 19 kD -zein protein, inducible promoters, such as the light inducible promoter derived from the pea rbcS gene and the actin promoter from rice, e.g., the actin 2 promoter (WO 00/70067); seed specific promoters, such as the phaseolin promoter from beans, may also be used. The nucleotide sequences of this invention can also be expressed under the regulation of promoters that are chemically regulated. This enables the nucleic acid sequence or encoded polypeptide to be synthesized only when the crop plants are treated with the inducing chemicals. Chemical induction of gene expression is detailed in EP 0 332 104 (to Ciba-Geigy) and U.S. Patent 5,614,395. A prefened promoter for chemical induction is the tobacco PR- la promoter.
Examples of some constitutive promoters which have been described include the rice actin 1 (Wang et al, 1992; U.S. Patent No. 5,641,876), CaMV 35S (Odell et al., 1985), CaMV 19S (Lawton et al., 1987), nos, Adh, sucrose synthase; and the ubiquitin promoters.
Examples of tissue specific promoters which have been described include the lectin (Vodkin, 1983; Lindstrom et al., 1990) com alcohol dehydrogenase 1 (Vogel et al., 1989; Dennis et al., 1984), com light harvesting complex (Simpson, 1986; Bansal et al., 1992), corn heat shock protein (Odell et al., 1985), pea small subunit RuBP carboxylase (Poulsen et al., 1986), Ti plasmid mannopine synthase (Langridge et al., 1989), Ti plasmid nopaline synthase (Langridge et al., 1989), petunia chalcone isomerase (vanTunen et al., 1988), bean glycine rich protein 1 (Keller et al., 1989), truncated CaMV 35s (Odell et al., 1985), potato patatin (Wenzler et al., 1989), root cell (Yamamoto et al., 1990), maize zein (Reina et al., 1990; Kriz et al., 1987; Wandelt et al., 1989; Langridge et al., 1983 ; Reina et al, 1990), globulin- 1 (Belanger et al., 1991), α-tubulin, cab (Sullivan et al., 1989), PEPCase (Hudspeth & Grula, 1989), R gene complex-associated promoters (Chandler et al., 1989), histone, and chalcone synthase promoters (Franken et al., 1991). Tissue specific enhancers are described in Fromm et al. (1989). Inducible promoters that have been described include the ABA- and turgor-inducible promoters, the promoter ofthe auxin-binding protein gene (Schwob et al., 1993), the UDP glucose flavonoid glycosyl-transferase gene promoter (Ralston et al., 1988), the MPI
proteinase inhibitor promoter (Cordero et al., 1994), and the gly ceraldehyde-3 -phosphate dehydrogenase gene promoter (Kohler et al., 1995; Quigley et al, 1989; Martinez et al., 1989).
Several other tissue-specific regulated genes and/or promoters have been reported in plants. These include genes encoding the seed storage proteins (such as napin, cruciferin, beta- conglycinin, and phaseolin) zein or oil body proteins (such as oleosin), or genes involved in fatty acid biosynthesis (including acyl carrier protein, stearoyl-ACP desaturase. And fatty acid desaturases (fad 2-1)), and other genes expressed during embryo development (such as Bce4, see, for example, EP 255378 and Kridl et al., 1991). Particularly useful for seed-specific expression is the pea vicilin promoter (Czako et al., 1992). (See also U.S. Pat. No. 5,625,136, herein incoφorated by reference.) Other useful promoters for expression in mature leaves are those that are switched on at the onset of senescence, such as the SAG promoter from Arabidopsis (Gan et al., 1995).
A class of fruit-specific promoters expressed at or during antithesis through fruit development, at least until the beginning of ripening, is discussed in U.S. 4,943,674. cDNA clones that are preferentially expressed in cotton fiber have been isolated (John et al., 1992). cDNA clones from tomato displaying differential expression during fruit development have been isolated and characterized (Mansson et al., 1985, Slater et al., 1985). The promoter for polygalacturonase gene is active in fruit ripening. The polygalacturonase gene is described in U.S. Patent No. 4,535,060, U.S. Patent No. 4,769,061, U.S. Patent No. 4,801,590, and U.S. Patent No. 5,107,065, which disclosures are incoφorated herein by reference.
Other examples of tissue-specific promoters include those that direct expression in leaf cells following damage to the leaf (for example, from chewing insects), in tubers (for example, patatin gene promoter), and in fiber cells (an example of a developmentally-regulated fiber cell protein is E6 (John et al., 1992). The E6 gene is most active in fiber, although low levels of transcripts are found in leaf, ovule and flower.
The tissue-specificity of some "tissue-specific" promoters may not be absolute and may be tested by one skilled in the art using the diphtheria toxin sequence. One can also achieve tissue-specific expression with "leaky" expression by a combination of different tissue-specific promoters (Beals et al., 1997). Other tissue-specific promoters can be isolated by one skilled in the art (see U.S. 5,589,379). Several inducible promoters ("gene switches") have been reported. Many are described in the review by Gatz (1996) and Gatz (1997). These include tetracycline repressor system, Lac repressor system, copper-inducible systems, salicylate- inducible systems (such as the PRla system), glucocorticoid- (Aoyama et al., 1997) and
ecdysome-inducible systems. Also included are the benzene sulphonamide- (U.S. Patent No. 5,364,780) and alcohol-(WO 97/06269 and WO 97/06268) inducible systems and glutathione S-transferase promoters. Other studies have focused on genes inducibly regulated in response to environmental stress or stimuli such as increased salinity. Drought, pathogen and wounding. (Graham et al., 1985; Graham et al., 1985, Smith et al., 1986). Accumulation of metallocarboxypeptidase-inhibitor protein has been reported in leaves of wounded potato plants (Graham et al., 1981). Other plant genes have been reported to be induced methyl jasmonate, elicitors, heat-shock, anaerobic stress, or herbicide safeners.
Regulated expression ofthe chimeric transacting viral replication protein can be further regulated by other genetic strategies. For example, Cre-mediated gene activation as described by Odell et al. 1990. Thus, a DNA fragment containing 3 ' regulatory sequence bound by lox sites between the promoter and the replication protein coding sequence that blocks the expression of a chimeric replication gene from the promoter can be removed by C/'e-mediated excision and result in the expression ofthe trαns-acting replication gene. In this case, the chimeric Cre gene, the chimeric trαw^-acting replication gene, or both can be under the control of tissue- and developmental- specific or inducible promoters. An alternate genetic strategy is the use of tRNA suppressor gene. For example, the regulated expression of a tRNA suppressor gene can conditionally control expression of a transacting replication protein coding sequence containing an appropriate termination codon as described by Ulmasov et al. 1997. Again, either the chimeric tRNA suppressor gene, the chimeric transacting replication gene, or both can be under the control of tissue- and developmental-specific or inducible promoters.
Frequently it is desirable to have continuous or inducible expression of a DNA sequence throughout the cells of an organism in a tissue-independent manner. For example, increased resistance of a plant to infection by soil- and airborne-pathogens might be accomplished by genetic manipulation ofthe plant's genome to comprise a continuous promoter operably linked to a heterologous pathogen-resistance gene such that pathogen- resistance proteins are continuously expressed throughout the plant's tissues.
Alternatively, it might be desirable to inhibit expression of a native DNA sequence within a plant's tissues to achieve a desired phenotype. In this case, such inhibition might be accomplished with transformation ofthe plant to comprise a constitutive, tissue-independent promoter operably linked to an antisense nucleotide sequence, such that constitutive
expression ofthe antisense sequence produces an RNA transcript that interferes with translation ofthe mRNA ofthe native DNA sequence.
To define a minimal promoter region, a DNA segment representing the promoter region is removed from the 5' region ofthe gene of interest and operably linked to the coding sequence of a marker (reporter) gene by recombinant DNA techniques well known to the art. The reporter gene is operably linked downstream ofthe promoter, so that transcripts initiating at the promoter proceed through the reporter gene. Reporter genes generally encode proteins which are easily measured, including, but not limited to, chloramphenicol acetyl transferase (CAT), beta-glucuronidase (GUS), green fluorescent protein (GFP), beta-galactosidase (beta- GAL), and luciferase.
The construct containing the reporter gene under the control ofthe promoter is then introduced into an appropriate cell type by transfection techniques well known to the art. To assay for the reporter protein, cell lysates are prepared and appropriate assays, which are well known in the art, for the reporter protein are performed. For example, if CAT were the reporter gene of choice, the lysates from cells transfected with constructs containing CAT under the control of a promoter under study are mixed with isotopically labeled chloramphenicol and acetyl-coenzyme A (acetyl-CoA). The CAT enzyme transfers the acetyl group from acetyl-CoA to the 2- or 3-ρosition of chloramphenicol. The reaction is monitored by thin-layer chromatography, which separates acetylated chloramphenicol from unreacted material. The reaction products are then visualized by autoradiography.
The level of enzyme activity conesponds to the amount of enzyme that was made, which in turn reveals the level of expression from the promoter of interest. This level of expression can be compared to other promoters to determine the relative strength ofthe promoter under study. In order to be sure that the level of expression is determined by the promoter, rather than by the stability ofthe mRNA, the level ofthe reporter mRNA can be measured directly, such as by Northern blot analysis.
Once activity is detected, mutational and or deletional analyses may be employed to determine the minimal region and/or sequences required to initiate transcription. Thus, sequences can be deleted at the 5' end ofthe promoter region and/or at the 3' end ofthe promoter region, and nucleotide substitutions introduced. These constracts are then introduced to cells and their activity determined.
In one embodiment, the promoter may be a gamma zein promoter, an oleosin olelό promoter, a globulinl promoter, an actin I promoter, an actin cl promoter, a sucrose synthetase
promoter, an INOPS promoter, an EXM5 promoter, a globulin2 promoter, a b-32, ADPG- pyrophosphorylase promoter, an Ltpl promoter, an Ltp2 promoter, an oleosin olel7 promoter, an oleosin olel8 promoter, an actin 2 promoter, a pollen-specific protein promoter, a pollen- specific pectate lyase promoter, an anther-specific protein promoter (Huffman), an anther- specific gene RTS2 promoter, a pollen- specific gene promoter, a tapeturn-specific gene promoter, tapetum- specific gene RAB24 promoter, a anthranilate synthase alpha subunit promoter, an alpha zein promoter, an anthranilate synthase beta subunit promoter, a dihydrodipicolinate synthase promoter, a Thil promoter, an alcohol dehydrogenase promoter, a cab binding protein promoter, an H3C4 promoter, a RUBISCO SS starch branching enzyme promoter, an ACCase promoter, an actin3 promoter, an actin7 promoter, a regulatory protein GF14-12 promoter, a ribosomal protein L9 promoter, a cellulose biosynthetic enzyme promoter, an S-adenosyl-L-homocysteine hydrolase promoter, a superoxide dismutase promoter, a C-kinase receptor promoter, a phosphoglycerate mutase promoter, a root-specific RCc3 mRNA promoter, a glucose-6 phosphate isomerase promoter, a pyrophosphate-fructose 6-phosphatelρhosphotransferase promoter, an ubiquitin promoter, a beta-ketoacyl-ACP synthase promoter, a 33 kDa photosystem 11 promoter, an oxygen evolving protein promoter, a 69 kDa vacuolar ATPase subunit promoter, a metallothionein-like protein promoter, a glyceraldehyde-3 -phosphate dehydrogenase promoter, an ABA- and ripening- inducible-like protein promoter, a phenylalanine ammonia lyase promoter, an adenosine triphosphatase S- adenosyl-L-homocysteine hydrolase promoter, an a- tubulin promoter, a cab promoter, a PEPCase promoter, an R gene promoter, a lectin promoter, a light harvesting complex promoter, a heat shock protein promoter, a chalcone synthase promoter, a zein promoter, a globulin- 1 promoter, an ABA promoter, an auxin-binding protein promoter, a UDP glucose flavonoid glycosyl-transferase gene promoter, an NTI promoter, an actin promoter, an opaque 2 promoter, a b70 promoter, an oleosin promoter, a CaMV 35S promoter, a CaMV 19S promoter, a histone promoter, a turgor-inducible promoter, a pea small subunit RuBP carboxylase promoter, a Ti plasmid mannopine synthase promoter, Ti plasmid nopaline synthase promoter, a petunia chalcone isomerase promoter, a bean glycine rich protein I promoter, a CaMV 35S transcript promoter, a potato patatin promoter, or a S-E9 small subunit RuBP carboxylase promoter.
2. Other Regulatory Elements
In addition to promoters, a variety of 5N and 3N transcriptional regulatory sequences are also available for use in the present invention. Transcriptional terminators are responsible
for the termination of transcription and conect mRNA polyadenylation. The 3N nontranslated regulatory DNA sequence preferably includes from about 50 to about 1,000, more preferably about 100 to about 1,000, nucleotide base pairs and contains plant transcriptional and translational termination sequences. Appropriate transcriptional terminators and those which are known to function in plants include the CaMV 35S terminator, the tml terminator, the nopaline synthase terminator, the pea rbcS E9 terminator, the tenninator for the T7 transcript from the octopine synthase gene of Agrobacterium tumefaciens, and the 3N end ofthe protease inhibitor I or II genes from potato or tomato, although other 3N elements known to those of skill in the art can also be employed. Alternatively, one also could use a gamma coixin, oleosin 3 or other terminator from the genus Coix.
Preferred 3' elements include those from the nopaline synthase gene of Agrobacterium tumefaciens (Bevan et al., 1983), the terminator for the T7 transcript from the octopine synthase gene of Agrobacterium tumefaciens, and the 3' end ofthe protease inhibitor I or II genes from potato or tomato. As the DNA sequence between the transcription initiation site and the start ofthe coding sequence, i.e., the untranslated leader sequence, can influence gene expression, one may also wish to employ a particular leader sequence. Prefened leader sequences are contemplated to include those which include sequences predicted to direct optimum expression ofthe attached gene, i.e., to include a prefened consensus leader sequence which may increase or maintain mRNA stability and prevent inappropriate initiation of translation. The choice of such sequences will be known to those of skill in the art in light ofthe present disclosure. Sequences that are derived from genes that are highly expressed in plants will be most prefened.
Other sequences that have been found to enhance gene expression in transgenic plants include intron sequences (e.g., from Adhl, bronzel, actinl, actin 2 (WO 00/760067), or the sucrose synthase intron) and viral leader sequences (e.g., from TMV, MCMV and AMV). For example, a number of non-translated leader sequences derived from viruses are known to enhance expression. Specifically, leader sequences from Tobacco Mosaic Virus (TMV), Maize Chlorotic Mottle Virus (MCMV), and Alfalfa Mosaic Virus (AMV) have been shown to be effective in enhancing expression (e.g., Gallie et al., 1987; Skuzeski et al., 1990). Other leaders known in the art include but are not limited to: Picornavirus leaders, for example, EMCV leader (Encephalomyocarditis 5 noncoding region) (Elroy-Stein et al., 1989); Potyvirus leaders, for example, TEV leader (Tobacco Etch Virus); MDMV leader (Maize Dwarf Mosaic
Virus); Human immunoglobulin heavy-chain binding protein (BiP) leader, (Macejak et al., 1991); Untranslated leader from the coat protein mRNA of alfalfa mosaic virus (AMV RNA 4), (Jobling et al., 1987; Tobacco mosaic viras leader (TMV), (Gallie et al., 1989; and Maize Chlorotic Mottle Virus leader (MCMV) (Lo mel et al., 1991. See also, Della-Cioppa et al., 1987.
Regulatory elements such as Adh intron 1 (Callis et al., 1987), sucrose synthase intron (Vasil et al., 1989) or TMV omega element (Gallie, et al., 1989), may further be included where desired.
Examples of enhancers include elements from the CaMV 35S promoter, octopine synthase genes (Ellis el al., 1987), the rice actin I gene, the maize alcohol dehydrogenase gene (Callis et al., 1987), the maize shrunken I gene (Vasil et al., 1989), TMV Omega element (Gallie et al., 1989) and promoters from non-plant eukaryotes (e.g. yeast; Ma et al., 1988).
Vectors for use in accordance with the present invention may be constracted to include the ocs enhancer element. This element was first identified as a 16 bp palindromic enhancer from the octopine synthase (ocs) gene of ultilane (Ellis et al., 1987), and is present in at least 10 other promoters (Bouchez et al., 1989). The use of an enhancer element, such as the ocs element and particularly multiple copies ofthe element, will act to increase the level of transcription from adjacent promoters when applied in the context of monocot transformation. Ultimately, the most desirable DNA segments for introduction into for example a monocot genome may be homologous genes or gene families which encode a desired trait (e.g., increased yield per acre) and which are introduced under the control of novel promoters or enhancers, etc., or perhaps even homologous or tissue specific (e.g., root-, collar/sheath-, whorl-, stalk-, earshank-, kernel- or leaf-specific) promoters or control elements. Indeed, it is envisioned that a particular use ofthe present invention will be the targeting of a gene in a constitutive manner or a root-specific manner. For example, insect resistant genes may be expressed specifically in the whorl and collar/sheath tissues which are targets for the first and second broods, respectively, of ECB. Likewise, genes encoding proteins with particular activity against rootworm may be targeted directly to root tissues.
Vectors for use in tissue-specific targeting of genes in transgenic plants will typically include tissue-specific promoters and may also include other tissue-specific control elements such as enhancer sequences. Promoters which direct specific or enhanced expression in certain plant tissues will be known to those of skill in the art in light ofthe present disclosure. These include, for example, the rbcS promoter, specific for green tissue; the ocs, nos and mas
promoters which have higher activity in roots or wounded leaf tissue; a truncated (-90 to +8) 35S promoter which directs enhanced expression in roots, an alpha-tubulin gene that directs expression in roots and promoters derived from zein storage protein genes which direct expression in endosperm. It is particularly contemplated that one may advantageously use the 16 bp ocs enhancer element from the octopine synthase (ocs) gene (Ellis et al., 1987; Bouchez et al., 1989), especially when present in multiple copies, to achieve enhanced expression in roots.
Tissue specific expression may be functionally accomplished by introducing a constitutively expressed gene (all tissues) in combination with an antisense gene that is expressed only in those tissues where the gene product is not desired. For example, a gene coding for the crystal toxin protein from B. thuringiensis (Bt) may be introduced such that it is expressed in all tissues using the 35S promoter from Cauliflower Mosaic Viras. Expression of an antisense transcript ofthe Bt gene in a maize kernel, using for example a zein promoter, would prevent accumulation ofthe Bt protein in seed. Hence the protein encoded by the introduced gene would be present in all tissues except the kernel.
Expression of some genes in transgenic plants will be desired only under specified conditions. For example, it is proposed that expression of certain genes that confer resistance to environmental stress factors such as drought will be desired only under actual stress conditions. It is contemplated that expression of such genes throughout a plants development may have detrimental effects. It is known that a large number of genes exist that respond to the environment. For example, expression of some genes such as rbcS, encoding the small subunit of ribulose bisphosphate carboxylase, is regulated by light as mediated through phytochrome. Other genes are induced by secondary stimuli. For example, synthesis of abscisic acid (ABA) is induced by certain environmental factors, including but not limited to water stress. A number of genes have been shown to be induced by ABA (Skriver and Mundy, 1990). It is also anticipated that expression of genes confening resistance to insect predation would be desired only under conditions of actual insect infestation. Therefore, for some desired traits inducible expression of genes in transgenic plants will be desired.
Expression of a gene in a transgenic plant will be desired only in a certain time period during the development ofthe plant. Developmental timing is frequently conelated with tissue specific gene expression. For example, expression of zein storage proteins is initiated in the endosperm about 15 days after pollination.
Additionally, vectors may be constructed and employed in the intracellular targeting of a specific gene product within the cells of a transgenic plant or in directing a protein to the extracellular environment. This will generally be achieved by joining a DNA sequence encoding a transit or signal peptide sequence to the coding sequence of a particular gene. The resultant transit, or signal, peptide will transport the protein to a particular intracellular, or extracellular destination, respectively, and will then be post-translationally removed. Transit or signal peptides act by facilitating the transport of proteins through intracellular membranes, e.g., vacuole, vesicle, plastid and mitochondrial membranes, whereas signal peptides direct proteins through the extracellular membrane. A particular example of such a use concerns the direction of a herbicide resistance gene, such as the EPSPS gene, to a particular organelle such as the chloroplast rather than to the cytoplasm. This is exemplified by the use ofthe rbcs transit peptide which confers plastid- specific targeting of proteins. In addition, it is proposed that it may be desirable to target certain genes responsible for male sterility to the mitochondria, or to target certain genes for resistance to phytopathogenic organisms to the extracellular spaces, or to target proteins to the vacuole.
By facilitating the transport ofthe protein into compartments inside and outside the cell, these sequences may increase the accumulation of gene product protecting them from proteolytic degradation. These sequences also allow for additional mRNA sequences from highly expressed genes to be attached to the coding sequence ofthe genes. Since mRNA being translated by ribosomes is more stable than naked mRNA, the presence of translatable mRNA in front ofthe gene may increase the overall stability ofthe mRNA transcript from the gene and thereby increase synthesis ofthe gene product. Since transit and signal sequences are usually post- translationally removed from the initial translation product, the use of these sequences allows for the addition of extra translated sequences that may not appear on the final polypeptide. Targeting of certain proteins may be desirable in order to enhance the stability of the protein (U.S. Patent No. 5,545,818).
It may be useful to target DNA itself within a cell. For example, it may be useful to target introduced DNA to the nucleus as this may increase the frequency of transformation. Within the nucleus itself it would be useful to target a gene in order to achieve site specific integration. For example, it would be useful to have an gene introduced through transformation replace an existing gene in the cell.
Other elements include those that can be regulated by endogenous or exogenous agents, e.g., by zinc finger proteins, including naturally occurring zinc finger proteins or chimeric zinc finger proteins (see, e.g., U.S. Patent No. 5,789,538, WO 99/48909; WO 99/45132; WO 98/53060; WO 98/53057; WO 98/53058; WO 00/23464; WO 95/19431; and WO 98/54311) or myb-like transcription factors. For example, a chimeric zinc finger protein may include amino acid sequences which bind to a specific DNA sequence (the zinc finger) and amino acid sequences that activate (e.g., GAL 4 sequences) or repress the transcription ofthe sequences linked to the specific DNA sequence.
3. Prefened Nucleic Acid Molecules ofthe Invention The invention relates to an isolated plant, e.g., Arabidopsis, Chenopodium and rice, nucleic acid molecule comprising a gene having an open reading frame, the expression of which is altered in response to pathogen infection, as well as the endogenous plant promoters for those genes. However, the expression of these genes may also be altered in response to non-pathogens, e.g., in response to environmental stiumuli. The nucleic acid molecules can be used in pathogen control strategies, e.g., by overexpressing nucleic acid molecules which can confer tolerance to a cell, or by altering the expression of host genes which are required for pathogen infection, e.g., by "knocking out" the expression of at least one genomic copy ofthe gene. Plants having genetic disruptions in host genes may be less susceptible to infection, e.g., due to a decrease or absence of a host protein needed for infection, or, alternatively, hypersusceptible to infection. Plants that are hypersusceptible to infection may be useful to prepare transgenic plants as the expression ofthe gene(s) which was disrupted may be related to gene silencing.
Preferred sources from which the nucleic acid molecules ofthe invention can be obtained or isolated include, but are not limited to, corn (Zea mays), Brassica sp. (e.g., B. napus, B. rapa, B. juncea), particularly those Brassica species useful as sources of seed oil, alfalfa (Medicago sativa), rice (Oryza sativa), rye (Secale cereale), sorghum (Sorghum bicolor, Sorghum vulgare), millet (e.g., pearl millet (Pennisetum glaucum), proso millet (Panicum miliaceum), foxtail millet (Setaria italica), finger millet (Eleusine coracana)), sunflower (Helianthus annuus), safflower (Carthamus tinctorius), wheat (Triticum aestivum), soybean (Glycine max), tobacco (Nicotiana tabacum), potato (Solanum tuberosum), peanuts (Arachis hypogaea), cotton (Gossypium barbadense, Gossypium hirsutum), sweet potato (Ipomoea batatus), cassava (Manihot esculenta), coffee (Cofea spp.), coconut (Cocos nucifera), pineapple (Ananas comosus), citrus trees (Citrus spp.), cocoa (Theobroma cacao),
tea (Camellia sinensis), banana (Musa spp.), avocado (Per sea ultilane), fig (Ficus casica), guava (Psidium guajava), mango (Mangifera indica), olive (Olea europaea), papaya (Carica papaya), cashew (Anacardium occidentale), macadamia (Macadamia integrifolia), almond (Prunus amygdalus), sugar beets (Beta vulgaris), sugarcane (Saccharum spp.), oats, duckweed (Lemna), barley, vegetables, ornamentals, and conifers.
Duckweed (Lemna, see WO 00/07210) includes members ofthe family Lemnaceae. There are known four genera and 34 species of duckweed as follows: genus Lemna (L. aequinoctialis, L. disperma, L. ecuadoriensis, L. gibba, L. japonica, L. minor, L. miniscula, L. obscura, L. perpusilla, L. tenera, L. trisulca, L. turionifera, L. valdiviana); genus Spirodela (S. intermedia, S. polyrrhiza, S. punctata); genus Woffia (Wa. Angusta, Wa. Arrhiza, Wa.
Australina, Wa. Borealis, Wa. Brasiliensis, Wa. Columbiana, Wa. Elongata, Wa. Globosa, Wa. Microscopica, Wa. Neglecta) and genus Wofiella (WL ultila, WL ultilane n, WI. gladiata, WL ultila, WI. lingulata, WL repunda, WL rotunda, and WL neotropica). Any other genera or species of Lemnaceae, if they exist, are also aspects ofthe present invention. Lemna gibba, Lemna minor, and Lemna miniscula axe prefened, with Lemnα minor and Lemnα miniscula being most prefened. Lemna species can be classified using the taxonomic scheme described by Landolt, Biosystematic Investigation on the Family of Duckweeds: The family of Lemnaceae - A Monograph Study. Geobatanischen Institut ETH, Stiftung Rubel, Zurich (1986)). Vegetables from which to obtain or isolate the nucleic acid molecules ofthe invention include, but are not limited to, tomatoes (Lycopersicon esculentum), lettuce (e.g., Lactuca sativa), green beans (Phaseolus vulgaris), lima beans (Phaseolus limensis), peas (Lathyrus spp.), and members ofthe genus Cucumis such as cucumber (C. sativus), cantaloupe (C. cantalupensis), and musk melon (C melo). Ornamentals from which to obtain or isolate the nucleic acid molecules ofthe invention include, but are not limited to, azalea (Rhododendron spp.), hydrangea (Macrophylla hydrangea), hibiscus (Hibiscus rosasanensis), roses (Rosa spp.), tulips (Tulipa spp.), daffodils (Narcissus spp.), petunias (Petunia hybrida), carnation (Dianthus caryophyllus), poinsettia (Euphorbia pulcherrima), and chrysanthemum. Conifers that may be employed in practicing the present invention include, for example, pines such as loblolly pine (Pinus taeda), slash pine (Pinus elliotii), ponderosa pine (Pinus ponderosa), lodgepole pine (Pinus contorta), and Monterey pine (Pinus radiata), Douglas-fir (Pseudotsuga menziesii); Western hemlock (Tsuga ultilane); Sitka spruce (Picea glauca); redwood (Sequoia sempervirens); trae firs such as silver fir (Abies amabilis) and balsam fir (Abies balsamea);
and cedars such as Western red cedar (Thuja plicata) and Alaska yellow-cedar (Chamaecyparis nootkatensis). Leguminous plants from which the nucleic acid molecules of the invention can be isolated or obtained include, but are not limited to, beans and peas. Beans include guar, locust bean, fenugreek, soybean, garden beans, cowpea, mungbean, lima bean, fava bean, lentils, chickpea, and the like. Legumes include, but are not limited to, Arachis, e.g., peanuts, Vicia, e.g., crown vetch, hairy vetch, adzuki bean, mung bean, and chickpea, Lupinus, e.g., lupine, trifolium, Phaseolus, e.g., common bean and lima bean, Pisum, e.g., field bean, Melilotus, e.g., clover, Medicago, e.g., alfalfa, Lotus, e.g., trefoil, lens, e.g., lentil, and false indigo. Papaya, garlic, pea, peach, pepper, petunia, strawberry, sorghum, sweet potato, turnip, safflower, corn, pea, endive, gourd, grape, snap bean, chicory, cotton, tobacco, aubergine, beet, buckwheat, broad bean, nectarine, avocado, mango, banana, groundnut, ρotato,peanut, lettuce, pineapple, spinach, squash, sugarbeet, sugarcane, sweet com, chrysanthemum.
Other prefened sources ofthe nucleic acid molecules ofthe invention include Acacia, aneth, artichoke, arugula, blackberry, canola, cilantro, Clementines, escarole, eucalyptus, fennel, grapefruit, honey dew, jicama, kiwifruit, lemon, lime, mushroom, nut, okra, orange, parsley, persimmon, plantain, pomegranate, poplar, radiata pine, radicchio, Southern pine, sweetgum, tangerine, triticale, vine, yams, apple, pear, quince, cherry, apricot, melon, hemp, buckwheat, grape, raspberry, chenopodium, blueberry, nectarine, peach, plum, strawberry, watermelon, eggplant, pepper, cauliflower, Brassica, e.g., broccoli, cabbage, ultilan sprouts, onion, carrot, leek, beet, broad bean, celery, radish, pumpkin, endive, gourd, garlic, snapbean, spinach, squash, turnip, ultilane, and zucchini.
Yet other sources of nucleic acid molecules are ornamental plants including, but not limited to, impatiens, Begonia, Pelargonium, Viola, Cyclamen, Verbena, Vinca, Tagetes, Primula, Saint Paulia, Agertum, Amaranthus, Antihinhinum, Aquilegia, Cineraria, Clover, Cosmo, Cowpea, Dahlia, Datura, Delphinium, Gerbera, Gladiolus, Gloxinia, Hippeastrum, Mesembryanthemum, Salpiglossos, and Zinnia, and plants such as those shown in Table 1.
Table 1
Prefened forage and turfgrass nucleic acid sources for the nucleic acid molecules of the invention include, but are not limited to, alfalfa, orchard grass, tall fescue, perennial ryegrass, creeping bent grass, and redtop. Yet other prefened sources include, but are not limited to, crop plants and in particular cereals (for example, corn, alfalfa, sunflower, rice, Brassica, canola, soybean, barley, soybean, sugarbeet, cotton, safflower, peanut, sorghum, oat, rye, rape, wheat, millet, tobacco, and the like), and even more preferably corn, rice and soybean.
According to one embodiment, the present invention is directed to a nucleic acid molecule comprising a nucleotide sequence isolated or obtained from any plant which encodes a polypeptide having at least 70% amino acid sequence identity to a polypeptide encoded by a gene comprising any one of SEQ ID NOs:l-953, 1954-1966, 2000-2129 or 2662-4737, or a gene comprising SEQ ID NOs:2137-2661 or 4738-6813. Based on the Arabidopsis, Chenopdoium and rice nucleic acid sequences ofthe present invention, orthologs may be identified or isolated from the genome of any desired organism, preferably from another plant, according to well known techniques based on their sequence similarity to the Arabidopsis, Chenopodium and rice nucleic acid sequences, e.g., hybridization, PCR or computer generated sequence comparisons. For example, all or a portion of a particular Arabidopsis, Chenopodium and rice nucleic acid sequence is used as a probe that selectively hybridizes to other gene sequences present in a population of cloned genomic DNA fragments or cDNA fragments (i.e., genomic or cDNA libraries) from a chosen source organism. Further, suitable genomic and cDNA libraries may be prepared from any cell or tissue of an organism. Such techniques include hybridization screening of plated DNA libraries (either plaques or colonies; see, e.g., Sambrook et al., 1989) and amplification by PCR using oligonucleotide primers
preferably conesponding to sequence domains conserved among related polypeptide or subsequences ofthe nucleotide sequences provided herein (see, e.g., Innis et al., 1990). These methods are particularly well suited to the isolation of gene sequences from organisms closely related to the organism from which the probe sequence is derived. The application of these methods using the Arabidopsis sequences as probes is well suited for the isolation of gene sequences from any source organism, preferably other plant species. In a PCR approach, oligonucleotide primers can be designed for use in PCR reactions to amplify conesponding DNA sequences from cDNA or genomic DNA extracted from any plant of interest. Methods for designing PCR primers and PCR cloning are generally known in the art. In hybridization techniques, all or part of a known nucleotide sequence is used as a probe that selectively hybridizes to other conesponding nucleotide sequences present in a population of cloned genomic DNA fragments or cDNA fragments (i.e., genomic or cDNA libraries) from a chosen organism. The hybridization probes may be genomic DNA fragments, cDNA fragments, RNA fragments, or other oligonucleotides, and may be labeled with a detectable group such as 32P, or any other detectable marker. Thus, for example, probes for hybridization can be made by labeling synthetic oligonucleotides based on the sequence ofthe invention. Methods for preparation of probes for hybridization and for construction of cDNA and genomic libraries are generally known in the art and are disclosed in Sambrook et al. (1989). In general, sequences that hybridize to the sequences disclosed herein will have at least 40% to 50%, about 60% to 70% and even about 80% 85%, 90%, 95% to 98% or more identity with the disclosed sequences. That is, the sequence similarity of sequences may range, sharing at least about 40% to 50%, about 60% to 70%, and even about 80%, 85%, 90%, 95% to 98% sequence similarity.
The nucleic acid molecules ofthe invention can also be identified by, for example, a search of known databases for genes encoding polypeptides having a specified amino acid sequence identity or DNA having a specified nucleotide sequence identity. Methods of alignment of sequences for comparison are well known in the art and are described hereinabove.
4. Methods for Mutagenizing DNA It is specifically contemplated by the inventors that one could mutagenize DNA having a promoter or open reading frame to, for example, potentially improve the utility ofthe DNA for expression of transgenes in plants. The mutagenesis can be canied out at random and the mutagenized sequences screened for activity in a trial-by-eπor procedure. Alternatively,
particular sequences which provide the promoter with desirable expression characteristics, or a promoter with expression enhancement activity, could be identified and these or similar sequences introduced into the sequences via mutation. It is further contemplated that one could mutagenize these sequences in order to enhance their expression of transgenes in a particular species.
The means for mutagenizing a DNA segment ofthe cunent invention are well-known to those of skill in the art. As indicated, modifications may be made by random or site-specific mutagenesis procedures. The DNA may be modified by altering its stracture through the addition or deletion of one or more nucleotides from the sequence which encodes the conesponding un-modified sequences.
Mutagenesis may be performed in accordance with any ofthe techniques known in the art, such as, and not limited to, synthesizing an oligonucleotide having one or more mutations within the sequence of a particular regulatory region. In particular, site-specific mutagenesis is a technique useful in the preparation of promoter mutants, through specific mutagenesis ofthe underlying DNA. The technique further provides a ready ability to prepare and test sequence variants, for example, incoφorating one or more ofthe foregoing considerations, by introducing one or more nucleotide sequence changes into the DNA. Site-specific mutagenesis allows the production of mutants through the use of specific oligonucleotide sequences which encode the DNA sequence ofthe desired mutation, as well as a sufficient number of adjacent nucleotides, to provide a primer sequence of sufficient size and sequence complexity to form a stable duplex on both sides ofthe deletion junction being traversed. Typically, a primer of about 17 to about 75 nucleotides or more in length is prefened, with about 10 to about 25 or more residues on both sides of the junction ofthe sequence being altered.
In general, the technique of site-specific mutagenesis is well known in the art, as exemplified by various publications. As will be appreciated, the technique typically employs a phage vector which exists in both a single stranded and double stranded form. Typical vectors useful in site-directed mutagenesis include vectors such as the Ml 3 phage. These phage are readily commercially available and their use is generally well known to those skilled in the art. Double stranded plasmids also are routinely employed in site directed mutagenesis which eliminates the step of transfening the gene of interest from a plasmid to a phage. In general, site-directed mutagenesis in accordance herewith is performed by first obtaining a single-stranded vector or melting apart of two strands of a double stranded vector which includes within its sequence a DNA sequence which encodes the promoter. An
oligonucleotide primer bearing the desired mutated sequence is prepared, generally synthetically. This primer is then annealed with the single-stranded vector, and subjected to DNA polymerizing enzymes such as E. coli polymerase I Klenow fragment, in order to complete the synthesis ofthe mutation-bearing strand. Thus, a heteroduplex is formed wherein one strand encodes the original non-mutated sequence and the second strand bears the desired mutation.
This heteroduplex vector is then used to transfonn or transfect appropriate cells, such as E. coli cells, and cells are selected which include recombinant vectors bearing the mutated sequence anangement. Vector DNA can then be isolated from these cells and used for plant transformation. A genetic selection scheme was devised by Kunkel et al. (1987) to enrich for clones incorporating mutagenic oligonucleotides. Alternatively, the use of PCR with commercially available thermostable enzymes such as Taq polymerase may be used to incorporate a mutagenic oligonucleotide primer into an amplified DNA fragment that can then be cloned into an appropriate cloning or expression vector. The PCR-mediated mutagenesis procedures of Tomic et al. (1990) and Upender et al. (1995) provide two examples of such protocols. A PCR employing a thennostable ligase in addition to a thermostable polymerase also may be used to incorporate a phosphorylated mutagenic oligonucleotide into an amplified DNA fragment that may then be cloned into an appropriate cloning or expression vector. The mutagenesis procedure described by Michael (1994) provides an example of one such protocol.
The preparation of sequence variants of DNA segments using site-directed mutagenesis is provided as a means of producing potentially useful species and is not meant to be limiting as there are other ways in which sequence variants of DNA sequences may be obtained. For example, recombinant vectors encoding the desired promoter sequence may be treated with mutagenic agents, such as hydroxylamine, to obtain sequence variants.
In addition, an unmodified or modified nucleotide sequence of the present invention can be varied by shuffling the sequence ofthe invention. To test for a function of variant DNA sequences according to the invention, the sequence of interest is operably linked to a selectable or screenable marker gene and expression of the marker gene is tested in transient expression assays with protoplasts or in stably transformed plants. It is known to the skilled artisan that DNA sequences capable of driving expression of an associated nucleotide sequence are build in a modular way. Accordingly, expression levels from shorter DNA fragments may be different than the one from the longest fragment and may be different from_each other. For
example, deletion of a down-regulating upstream element will lead to an increase in the expression levels of the associated nucleotide sequence while deletion of an up-regulating element will decrease the expression levels of the associated nucleotide sequence. It is also known to the skilled artisan that deletion of development-specific or a tissue-specific element will lead to a temporally or spatially altered expression profile of the associated nucleotide sequence.
As used herein, the term "oligonucleotide directed mutagenesis procedure" refers to template-dependent processes and vector-mediated propagation which result in an increase in the concentration of a specific nucleic acid molecule relative to its initial concentration, or in an increase in the concentration of a detectable signal, such as amplification. As used herein, the term "oligonucleotide directed mutagenesis procedure" also is intended to refer to a process that involves the template-dependent extension of a primer molecule. The term template- dependent process refers to nucleic acid synthesis of an RNA or a DNA molecule wherein the sequence ofthe newly synthesized strand of nucleic acid is dictated by the well- known rules of complementary base pairing (see, for example, Watson and Rarnstad, 1987). Typically, vector mediated methodologies involve the introduction ofthe nucleic acid fragment into a DNA or RNA vector, the clonal amplification ofthe vector, and the recovery ofthe amplified nucleic acid fragment. Examples of such methodologies are provided by U.S. Patent No. 4,237,224. A number of template dependent processes are available to amplify the target sequences of interest present in a sample, such methods being well known in the art and specifically disclosed herein below.
Where a clone comprising a promoter has been isolated in accordance with the instant invention, one may wish to delimit the essential promoter regions within the clone. One efficient, targeted means for preparing mutagenizing promoters relies upon the identification of putative regulatory elements within the promoter sequence. This can be initiated by comparison with promoter sequences known to be expressed in similar tissue-specific or developmentally unique manner. Sequences which are shared among promoters with similar expression patterns are likely candidates for the binding of transcription factors and are thus likely elements which confer expression patterns. Confirmation of these putative regulatory elements can be achieved by deletion analysis of each putative regulatory region followed by functional analysis of each deletion constract by assay of a reporter gene which is functionally
attached to each constract. As such, once a starting promoter sequence is provided, any of a number of different deletion mutants ofthe starting promoter could be readily prepared.
As indicated above, deletion mutants, deletion mutants ofthe promoter ofthe invention also could be randomly prepared and then assayed. With this strategy, a series of constructs are prepared, each containing a different portion ofthe clone (a subclone), and these constructs are then screened for activity. A suitable means for screening for activity is to attach a deleted promoter or intron construct which contains a deleted segment to a selectable or screenable marker, and to isolate only those cells expressing the marker gene. In this way, a number of different, deleted promoter constructs are identified which still retain the desired, or even enhanced, activity. The smallest segment which is required for activity is thereby identified through comparison ofthe selected constructs. This segment may then be used for the construction of vectors for the expression of exogenous genes. B. Marker Genes In order to improve the ability to identify transformants, one may desire to employ a selectable or screenable marker gene as, or in addition to, the expressible gene of interest.
"Marker genes" are genes that impart a distinct phenotype to cells expressing the marker gene and thus allow such transformed cells to be distinguished from cells that do not have the marker. Such genes may encode either a selectable or screenable marker, depending on whether the marker confers a trait which one can "select" for by chemical means, i.e., through the use of a selective agent (e.g., a herbicide, antibiotic, or the like), or whether it is simply a trait that one can identify through observation or testing, i.e., by 'screening' (e.g., the R-locus trait, the green fluorescent protein (GFP)). Of course, many examples of suitable marker genes are known to the art and can be employed in the practice ofthe invention.
Included within the terms selectable or screenable marker genes are also genes which encode a "secretable marker" whose secretion can be detected as a means of identifying or selecting for transformed cells. Examples include markers which encode a secretable antigen that can be identified by antibody interaction, or even secretable enzymes which can be detected by their catalytic activity. Secretable proteins fall into a number of classes, including small, diffusible proteins detectable, e.g., by ELISA; small active enzymes detectable in extracellular solution (e.g., alpha-amylase, beta-lactamase, phosphinothricin acetyltransferase); and proteins that are inserted or trapped in the cell wall (e.g., proteins that include a leader sequence such as that found in the expression unit of extensin or tobacco PR-S).
With regard to selectable secretable markers, the use of a gene that encodes a protein that becomes sequestered in the cell wall, and which protein includes a unique epitope is considered to be particularly advantageous. Such a secreted antigen marker would ideally employ an epitope sequence that would provide low background in plant tissue, a promoter- leader sequence that would impart efficient expression and targeting across the plasma membrane, and would produce protein that is bound in the cell wall and yet accessible to antibodies. A normally secreted wall protein modified to include a unique epitope would satisfy all such requirements.
One example of a protein suitable for modification in this manner is extensin, or hydroxyproline rich glycoprotein (HPRG). For example, the maize HPRG (Steifel et al., 1990) molecule is well characterized in terms of molecular biology, expression and protein structure. However, any one of a variety of ultilane and/or glycine-rich wall proteins (Keller et al., 1989) could be modified by the addition of an antigenic site to create a screenable marker.
One exemplary embodiment of a secretable screenable marker concerns the use of a maize sequence encoding the wall protein HPRG, modified to include a 15 residue epitope from the pro-region of murine interleukin, however, virtually any detectable epitope may be employed in such embodiments, as selected from the extremely wide variety of antigen- antibody combinations known to those of skill in the art. The unique extracellular epitope can then be straightforwardly detected using antibody labeling in conjunction with chromogenic or fluorescent adjuncts.
Elements ofthe present disclosure may be exemplified in detail through the use ofthe bar and or GUS genes, and also through the use of various other markers. Of course, in light of this disclosure, numerous other possible selectable and/or screenable marker genes will be apparent to those of skill in the art in addition to the one set forth hereinbelow. Therefore, it will be understood that the following discussion is exemplary rather than exhaustive. In light ofthe techniques disclosed herein and the general recombinant techniques which are known in the art, the present invention renders possible the introduction of any gene, including marker genes, into a recipient cell to generate a transformed plant. 1. Selectable Markers Possible selectable markers for use in connection with the present invention include, but are not limited to, a neo gene which codes for kanamycin resistance and can be selected for using kanamycin, G418, paromomycin, and the like; a bar gene which codes for bialaphos or phosphinothricin resistance; a gene which encodes an altered EPSP synthase protein (Hinchee
et al., 1988) thus confening glyphosate resistance; a nitrilase gene such as bxn from Klebsiella ozaenae which confers resistance to bromoxynil (Stalker et al., 1988); a mutant acetolactate synthase gene (ALS) which confers resistance to imidazolinone, sulfonylurea or other ALS- inhibiting chemicals (European Patent Application 154,204, 1985); a methotrexate-resistant DHFR gene (ThiUet et al., 1988); a dalapon dehalogenase gene that confers resistance to the herbicide dalapon; a mutated anthranilate synthase gene that confers resistance to 5-methyl tryptophan. Prefened selectable marker genes encode phosphinothricin acetyltransferase; glyphosate resistant EPSPS, aminoglycoside phosphotransferase; hygromycin phosphotransferase, or neomycin phosphotransferase. Where a mutant EPSP synthase gene is employed, additional benefit may be realized through the incorporation of a suitable chloroplast transit peptide, CTP (European Patent Application 0,218,571, 1987).
An illustrative embodiment of a selectable marker gene capable of being used in systems to select transformants is the genes that encode the enzyme phosphinothricin acetyltransferase, such as the bar gene from Streptomyces hygroscopicus or the pat gene from Streptomyces viridochromogenes. The enzyme phosphinothricin acetyl transferase (PAT) inactivates the active ingredient in the herbicide bialaphos, phosphinothricin (PPT). PPT inhibits glutamine synthetase, (Murakami et al., 1986; Twell et al., 1989) causing rapid accumulation of ammonia and cell death. The success in using this selective system in conjunction with monocots was particularly surprising because ofthe major difficulties which have been reported in transformation of cereals.
Where one desires to employ a bialaphos resistance gene in the practice ofthe invention, a particularly useful gene for this purpose is the bar ox pat genes obtainable from species of Streptomyces (e.g., ATCC No. 21,705). The cloning ofthe bar gene has been described (Murakami et al., 1986; Thompson et al., 1987) as has the use ofthe bar gene in the context of plants other than monocots (De Block et al., 1987; De Block et al.,_1989).
Selection markers resulting in positive selection, such as a phosphomannose isomerase gene, as described in patent application WO 93/05163, may also be used. Alternative genes to be used for positive selection are described in WO 94/20627 and encode xyloisomerases and phosphomanno-isomerases such as mannose-6-phosphate isomerase and mannose- 1 -phosphate isomerase; phosphomanno mutase; mannose epimerases such as those which convert carbohydrates to mannose or mannose to carbohydrates such as glucose or galactose; phosphatases such as mannose or xylose phosphatase, mannose-6-phosρhatase and mannose- 1 -phosphatase, and permeases which are involved in the transport of mannose, or a
derivative, or a precursor thereof into the cell. Transformed cells are identified without damaging or killing the non-transformed cells in the population and without co-introduction of antibiotic or herbicide resistance genes. As described in WO 93/05163, in addition to the fact that the need for antibiotic or herbicide resistance genes is eliminated, it has been shown that the positive selection method is often far more efficient than traditional negative selection. 2. Screenable Markers
Screenable markers that may be employed include, but are not limited to, a beta- glucuronidase (GUS) or uidA gene which encodes an enzyme for which various chromogenic substrates are known; an R-locus gene, which encodes a product that regulates the production of anthocyanin pigments (red color) in plant tissues (Dellaporta et al., 1988); a beta-lactamase gene (Sutcliffe, 1978), which encodes an enzyme for which various chromogenic substrates are known (e.g., PAD AC, a chromogenic cephalosporin); axylE gene (Zukowsky et al., 1983) which encodes a catechol dioxygenase that can convert chromogenic catechols; an V-amylase gene (Ikuta et al., 1990); a tyrosinase gene (Katz et al., 1983) which encodes an enzyme capable of oxidizing tyrosine to DOPA and dopaquinone which in turn condenses to form the easily detectable compound melanin; a 5-galactosidase gene, which encodes an enzyme for which there are chromogenic substrates; a luciferase (lux) gene (Ow et al., 1986), which allows for bioluminescence detection; or even an aequorin gene (Prasher et al., 1985), which may be employed in calcium-sensitive bioluminescence detection, or a green fluorescent protein gene (Niedz et al., 1995).
Genes from the maize R gene complex are contemplated to be particularly useful as screenable markers. The R gene complex in maize encodes a protein that acts to regulate the production of anthocyanin pigments in most seed and plant tissue. A gene from the R gene complex was applied to maize transformation, because the expression of this gene in transformed cells does not harm the cells. Thus, an R gene introduced into such cells will cause the expression of a red pigment and, if stably incorporated, can be visually scored as a red sector. If a maize line is canies dominant ultila for genes encoding the enzymatic intemiediates in the anthocyanin biosynthetic pathway (C2, Al, A2, Bzl and Bz2) (Roth et al., 1990), but carries a recessive allele at the R locus, transformation of any cell from that line with R will result in red pigment formation. Exemplary lines include Wisconsin 22 which contains the rg-Stadler allele and TRI 12, a K55 derivative which is r-g, b, PI. Alternatively any genotype of maize can be utilized if the Cl and R alleles are introduced together.
It is further proposed that R gene regulatory regions may be employed in chimeric constructs in order to provide mechanisms for controlling the expression of chimeric genes. More diversity of phenotypic expression is known at the R locus than at any other locus (Coe et al., 1988). It is contemplated that regulatory regions obtained from regions 5' to the stractural R gene would be valuable in directing the expression of genes, e.g., insect resistance, drought resistance, herbicide tolerance or other protein coding regions. For the purposes ofthe present invention, it is believed that any ofthe various R gene family members may be successfully employed (e.g., P, S, Lc, etc.). However, the most preferred will generally be Sn (particularly Sn:bol3). Sn is a dominant member ofthe R gene complex and is functionally similar to the R and B loci in that Sn controls the tissue specific deposition of anthocyanin pigments in certain seedling and plant cells, therefore, its phenotype is similar to R.
A further screenable marker contemplated for use in the present invention is firefly luciferase, encoded by the lux gene. The presence ofthe lux gene in transformed cells may be detected using, for example, X-ray film, scintillation counting, fluorescent spectrophotometry, low-light video cameras, photon counting cameras or multiwell luminometry. It is also envisioned that this system may be developed for populational screening for bioluminescence, such as on tissue culture plates, or even for whole plant screening. Where use of a screenable marker gene such as lux or GFP is desired, benefit may be realized by creating a gene fusion between the screenable marker gene and a selectable marker gene, for example, a GFP-NPTII gene fusion. This could allow, for example, selection of transformed cells followed by screening of transgenic plants or seeds.
C. Exogenous Genes for Modification of Plant Phenotypes Genes of interest are reflective ofthe commercial markets and interests of those involved in the development ofthe crop. Crops and markets of interest changes, and as developing nations open up world markets, new crops and technologies will also emerge. In addition, as the understanding of agronomic traits and characteristics such as yield and heterosis increase, the choice of genes for transformation will change accordingly. General categories of genes of interest include, for example, those genes involved in information, such as zinc fingers, those involved in communication, such as kinases, and those involved in housekeeping, such as heat shock proteins. More specific categories of transgenes, for example, include genes encoding important traits for agronomics, insect resistance, disease resistance, herbicide resistance, sterility, grain characteristics, and commercial products. Genes of interest include, generally, those involved in starch, oil, carbohydrate, or nutrient
metabolism, as well as those affecting kernel size, sucrose loading, zinc finger proteins, see, e.g., US. Patent No. 5,789,538, WO 99/48909; WO 99/45132; WO 98/53060; WO 98/53057; WO 98/53058; WO 00/23464; WO 95/19431; and WO 98/54311, and the like.
One skilled in the art recognizes that the expression level and regulation of a transgene in a plant can vary significantly from line to line. Thus, one has to test several lines to find one with the desired expression level and regulation. Once a line is identified with the desired regulation specificity of a chimeric Cre transgene, it can be crossed with lines carrying different inactive replicons or inactive transgene for activation.
Other sequences which may be linked to the gene of interest which encodes a polypeptide are those which can target to a specific organelle, e.g., to the lnitochondria, nucleus, or plastid, within the plant cell. Targeting can be achieved by providing the polypeptide with an appropriate targeting peptide sequence, such as a secretory signal peptide (for secretion or cell wall or membrane targeting, a plastid transit peptide, a chloroplast transit peptide, e.g., the chlorophyll ab binding protein, a mitochondrial target peptide, a vacuole targeting peptide, or a nuclear targeting peptide, and the like. For example, the small subunit of ribulose bisphosphate carboxylase transit peptide, the EPSPS transit peptide or the dihydrodipicolinic acid synthase transit peptide may be used. For examples of plastid organelle targeting sequences (see WO 00/12732). Plastids are a class of plant organelles derived from proplastids and include chloroplasts, leucoplasts, aravloplasts, and chromoplasts. The plastids are major sites of biosynthesis in plants. In addition to photosynthesis in the chloroplast, plastids are also sites of lipid biosynthesis, nitrate reduction to ammonium, and starch storage. And while plastids contain their own circular genome, most ofthe proteins localized to the plastids are encoded by the nuclear genome and are imported into the organelle from the cytoplasm. Transgenes used with the present invention will often be genes that direct the expression of a particular protein or polypeptide product, but they may also be non-expressible DNA segments, e.g., transposons such as Ds that do no direct their own transposition. As used herein, an "expressible gene" is any gene that is capable of being transcribed into RNA (e.g., mRNA, antisense RNA, etc.) or translated into a protein, expressed as a trait of interest, or the like, etc., and is not limited to selectable, screenable or non-selectable marker genes. The invention also contemplates that, where both an expressible gene that is not necessarily a marker gene is employed in combination with a marker gene, one may employ the separate
genes on either the same or different DNA segments for transformation. In the latter case, the different vectors are delivered concurrently to recipient cells to maximize cotransformation. The choice ofthe particular DNA segments to be delivered to the recipient cells will often depend on the purpose ofthe transformation. One ofthe major purposes of transformation of crop plants is to add some commercially desirable, agronomically important traits to the plant. Such traits include, but are not limited to, herbicide resistance or tolerance; insect resistance or tolerance; disease resistance or tolerance (viral, bacterial, fungal, nematode); stress tolerance and/or resistance, as exemplified by resistance or tolerance to drought, heat, chilling, freezing, excessive moisture, salt stress; oxidative stress; increased yields; food content and makeup; physical appearance; male sterility; drydown; standability; prolificacy; starch properties; oil quantity and quality; and the like. One may desire to incorporate one or more genes conferring any such desirable trait or traits, such as, for example, a gene or genes encoding pathogen resistance.
In certain embodiments, the present invention contemplates the transformation of a recipient cell with more than one advantageous transgene. Two or more transgenes can be supplied in a single transformation event using either distinct transgene-encoding vectors, or using a single vector incorporating two or more gene coding sequences. For example, plasmids bearing the bar and aroA expression units in either convergent, divergent, or colinear orientation, are considered to be particularly useful. Further prefened combinations are those of an insect resistance gene, such as a Bt gene, along with a protease inhibitor gene such as pinll, or the use of bar in combination with either ofthe above genes. Of course, any two or more transgenes of any description, such as those conferring herbicide, insect, disease (viral, bacterial, fungal, nematode) or drought resistance, male sterility, drydown, standability, prolificacy, starch properties, oil quantity and quality, or those increasing yield or nutritional quality may be employed as desired. 1. Herbicide Resistance
The genes encoding phosphinothricin acetyltransferase (bar andpat), glyphosate tolerant EPSP synthase genes, the glyphosate degradative enzyme gene gox encoding glyphosate oxidoreductase, deh (encoding a dehalogenase enzyme that inactivates dalapon), herbicide resistant (e.g., sulfonylurea and imidazolinone) acetolactate synthase, and bxn genes (encoding a nitrilase enzyme that degrades bromoxynil) are good examples of herbicide resistant genes for use in transformation. The bar and pat genes code for an enzyme, phosphinothricin acetyltransferase (PAT), which inactivates the herbicide phosphinothricin
and prevents this compound from inhibiting glutamine synthetase enzymes. The enzyme 5- enolpyravylshikimate 3-phosphate synthase (EPSP Synthase), is normally inhibited by the herbicide N-(phosphonomethyl)glycine (glyphosate). However, genes are known that encode glyphosate-resistant EPSP Synthase enzymes. These genes are particularly contemplated for use in monocot transformation. The deh gene encodes the enzyme dalapon dehalogenase and confers resistance to the herbicide dalapon. The bxn gene codes for a specific nitrilase enzyme that converts bromoxynil to a non-herbicidal degradation product. 2. Insect Resistance An important aspect ofthe present invention concerns the introduction of insect resistance-conferring genes into plants. Potential insect resistance genes which can be introduced include Bacillus thuringiensis crystal toxin genes or Bt genes (Watrud et al., 1985). Bt genes may provide resistance to lepidopteran or coleopteran pests such as European Corn Borer (ECB) and corn rootworm (CRW). Prefened Bt toxin genes for use in such embodhnents include the CryIA(b) and CryΙA(c) genes. Endotoxin genes from other species of P. thuringiensis which affect insect growth or development may also be employed in this regard.
The poor expression of Pt toxin genes in plants is a well-documented phenomenon, and the use of different promoters, fusion proteins, and leader sequences has not led to significant increases in Bt protein expression (Vaeck et al., 1989; Barton et al., 1987). It is therefore contemplated that the most advantageous Bt genes for use in the transformation protocols disclosed herein will be those in which the coding sequence has been modified to effect increased expression in plants, and more particularly, those in which maize prefened codons have been used. Examples of such modified Bt toxin genes include the variant Bt CryIA(b) gene tenned labό (Perlak et al., 1991) and the synthetic CryΙA(c) genes termed 1800a and 1800b.
Protease inhibitors may also provide insect resistance (Johnson et al., 1989), and will thus have utility in plant transformation. The use of a protease inhibitor II gene, pinll, from tomato or potato is envisioned to be particularly useful. Even more advantageous is the use of a pinll gene in combination with a Bt toxin gene, the combined effect of which has been discovered by the present inventors to produce synergistic insecticidal activity. Other genes which encode inhibitors ofthe insects' digestive system, or those that encode enzymes or co-
factors that facilitate the production of inhibitors, may also be useful. This group may be exemplified by oryzacystatin and amylase inhibitors, such as those from wheat and barley.
Also, genes encoding lectins may confer additional or alternative insecticide properties. Lectins (originally termed phytohemagglutinins) are multivalent carbohydrate-binding proteins which have the ability to agglutinate red blood ceUs from a range of species. Lectins have been identified recently as insecticidal agents with activity against weevils, ECB and rootworm (Murdock et al., 1990; Czapla and Lang, 1990). Lectin genes contemplated to be useful include, for example, barley and wheat germ agglutinin (WGA) and rice lectins (Gatehouse et al., 1984), with WGA being prefened. Genes controlling the production of large or small polypeptides active against insects when introduced into the insect pests, such as, e.g., lytic peptides, peptide hormones and toxins and venoms, form another aspect ofthe mvention. For example, it is contemplated that the expression of juvenile hormone esterase, directed towards specific insect pests, may also result in insecticidal activity, or perhaps cause cessation of metamorphosis (Hammock et al., 1990). Transgenic plants expressing genes which encode enzymes that affect the integrity of the insect cuticle form yet another aspect ofthe invention. Such genes include those encodmg, e.g., chitinase, proteases, Upases and also genes for the production of nikkomycin, a compound that inhibits chitin synthesis, the introduction of any of which is contemplated to produce insect resistant maize plants. Genes that code for activities that affect insect molting, such those affecting the production of ecdysteroid UDP-glucosyl transferase, also fall within the scope ofthe useful transgenes ofthe present invention.
Genes that code for enzymes that facilitate the production of compounds that reduce the nutritional quality ofthe host plant to insect pests are also encompassed by the present invention. It may be possible, for instance, to confer insecticidal activity on a plant by altering its sterol composition. Sterols are obtained by insects from their diet and are used for hormone synthesis and membrane stability. Therefore alterations in plant sterol composition by expression of novel genes, e.g., those that directly promote the production of undesirable sterols or those that convert desirable sterols into undesirable forms, could have a negative effect on insect growth and/or development and hence endow the plant with insecticidal activity. Lipoxygenases are naturally occurring plant enzymes that have been shown to exhibit anti-nutritional effects on insects and to reduce the nutritional quality of their diet. Therefore, further embodiments ofthe invention concern transgenic plants with enhanced lipoxygenase activity which may be resistant to insect feeding.
The present invention also provides methods and compositions by which to achieve qualitative or quantitative changes in plant secondary metabolites. One example concerns transforming plants to produce DIMBOA which, it is contemplated, wdl confer resistance to European corn borer, rootworm and several other maize insect pests. Candidate genes that are particularly considered for use in this regard include those genes at the bx locus known to be involved in the synthetic DIMBOA pathway (Dunn et al., 1981). The introduction of genes that can regulate the production of maysin, and genes involved in the production of dhurrin in sorghum, is also contemplated to be of use in facilitating resistance to earworm and rootworm, respectively. Tripsacum dactyloides is a species of grass that is resistant to certain insects, including corn root worm. It is anticipated that genes encoding proteins that are toxic to insects or are involved in the biosynthesis of compounds toxic to insects will be isolated from Tripsacum and that these novel genes will be useful in confening resistance to insects. It is known that the basis of insect resistance in Tripsacum is genetic, because said resistance has been transfened to Zea mays via sexual crosses (Branson and Guss, 1972).
Further genes encoding proteins characterized as having potential insecticidal activity may also be used as transgenes in accordance herewith. Such genes include, for example, the cowpea trypsin inhibitor (CpTI; Hilder et al., 1987) which may be used as a rootworm detenent; genes encoding avermectin (Campbell, 1989; Ikeda et al., 1987) which may prove particularly useful as a corn rootworm detenent; ribosome inactivating protein genes; and even genes that regulate plant stractures. Transgenic maize including anti-insect antibody genes and genes that code for enzymes that can covert a non-toxic insecticide (pro-insecticide) applied to the outside ofthe plant into an insecticide inside the plant are also contemplated. 3. Environment or Stress Resistance Improvement of a plant's ability to tolerate various environmental stresses such as, but not limited to, drought, excess moisture, chilling, freezing, high temperature, salt, and oxidative stress, can also be effected through expression of heterologous, or overexpression of homologous genes. Benefits may be realized in terms of increased resistance to freezing temperatures through the introduction of an "antifreeze" protein such as that ofthe Winter Flounder (Cutler et al., 1989) or synthetic gene derivatives thereof. Improved chilling tolerance may also be confened through increased expression of glycerol-3-phosphate acetyltransferase in chloroplasts (Murata et al., 1992; Wolter et al., 1992). Resistance to oxidative stress (often exacerbated by conditions such as chilling temperatures in combination
with high light intensities) can be confened by expression of superoxide dismutase (Gupta et al., 1993), and may be improved by glutathione reductase (Bowler et al., 1992). Such strategies may allow for tolerance to freezing in newly emerged fields as well as extending later maturity higher yielding varieties to earlier relative maturity zones. Expression of novel genes that favorably effect plant water content, total water potential, osmotic potential, and turgor can enhance the ability ofthe plant to tolerate drought. As used herein, the terms "drought resistance" and "drought tolerance" are used to refer to a plants increased resistance or tolerance to stress induced by a reduction in water availability, as compared to normal circumstances, and the ability ofthe plant to function and survive in lower-water environments, and perform in a relatively superior manner. In this aspect ofthe invention it is proposed, for example, that the expression of a gene encoding the biosynthesis of osmotically-active solutes can impart protection against drought. Within this class of genes are DNAs encoding mannitol dehydrogenase (Lee and Saier, 1982) and trehalose-6-phosphate synthase (Kaasen et al., 1992). Through the subsequent action of native phosphatases in the cell or by the introduction and coexpression of a specific phosphatase, these introduced genes will result in the accumulation of either mannitol or trehalose, respectively, both of which have been well documented as protective compounds able to mitigate the effects of stress. Mannitol accumulation in transgenic tobacco has been verified and preliminary results indicate that plants expressing high levels of this metabolite are able to tolerate an applied osmotic stress (Tarczynski et al., cited supra (1992), 1993).
Similarly, the efficacy of other metabolites in protecting either enzyme function (e.g. alanopine or propionic acid) or membrane integrity (e.g., alanopine) has been documented (Loomis et al., 1989), and therefore expression of gene encoding the biosynthesis of these compounds can confer drought resistance in a manner similar to or complimentary to mannitol. Other examples of naturally occurring metabolites that are osmotically active and/or provide some direct protective effect during drought and/or desiccation include sugars and sugar derivatives such as fructose, erythritol (Coxson et al., 1992), sorbitol, dulcitol (Karsten et al., 1992), glucosylglycerol (Reed et al., 1984; Erdmann et al., 1992), sucrose, stachyose (Koster and Leopold, 1988; Blackman et al., 1992), ononitol and pinitol (Vernon and Bohnert, 1992), and raffinose (Bernal-Lugo and Leopold, 1992). Other osmotically active solutes which are not sugars include, but are not limited to, proline and glycine-betaine (Wyn- Jones and Storey, 1981). Continued canopy growth and increased reproductive fitness during times of stress can be augmented by introduction and expression of genes such as those controlling the
osmotically active compounds discussed above and other such compounds, as represented in one exemplary embodiment by the enzyme myoinositol 0-methyltransferase.
It is contemplated that the expression of specific proteins may also increase drought tolerance. Three classes of Late Embryogenic Proteins have been assigned based on structural similarities (see Dure et al., 1989). AU three classes of these proteins have been demonstrated in maturing (i.e., desiccating) seeds. Within these 3 types of proteins, the Type-II (dehydrin- type) have generally been implicated in drought and/or desiccation tolerance in vegetative plant parts (i.e. Mundy and Chua, 1988; Piatkowski et al., 1990; Yamaguchi-Shinozaki et al., 1992). Recently, expression of a Type-Ill LEA (HVA-1) in tobacco was found to influence plant height, maturity and drought tolerance (Fitzpatrick, 1993). Expression of stractural genes from all three groups may therefore confer drought tolerance. Other types of proteins induced during water stress include thiol proteases, aldolases and transmembrane transporters (Guenero et al., 1990), which may confer various protective and/or repair-type functions during drought stress. The expression of a gene that effects lipid biosynthesis and hence membrane composition can also be useful in conferring drought resistance on the plant.
Many genes that improve drought resistance have complementary modes of action. Thus, combinations of these genes might have additive and/or synergistic effects in improving drought resistance in maize. Many of these genes also improve freezing tolerance (or resistance); the physical stresses incuned during freezing and drought are similar in nature and may be mitigated in similar fashion. Benefit may be confened via constitutive expression of these genes, but the prefened means of expressing these novel genes may be through the use of a turgor-induced promoter (such as the promoters for the turgor-induced genes described in Guenero et al. 1990 and Shagan et al., 1993). Spatial and temporal expression patterns of these genes may enable maize to better withstand stress. Expression of genes that are involved with specific morphological traits that allow for increased water extractions from drying soil would be of benefit. For example, introduction and expression of genes that alter root characteristics may enhance water uptake. Expression of genes that enhance reproductive fitness during times of stress would be of significant value. For example, expression of DNAs that improve the synchrony of pollen shed and receptiveness ofthe female flower parts, i.e., silks, would be of benefit. In addition, expression of genes that minimize kernel abortion during times of stress would increase the amount of grain to be harvested and hence be of value. Regulation of cytokinin levels in monocots, such as maize, by introduction and expression of an isopentenyl transferase gene with appropriate regulatory
sequences can improve monocot stress resistance and yield (Gan et al., Science. 270: 1986 (1995)).
Given the overall role of water in determining yield, it is contemplated that enabling plants to utilize water more efficiently, through the introduction and expression of novel genes, will improve overall performance even when soil water availability is not limiting. By introducing genes that improve the ability of plants to maximize water usage across a full range of stresses relating to water availability, yield stability or consistency of yield performance may be realized. 4. Disease Resistance It is proposed that increased resistance to diseases may be realized through introduction of genes into plants period. It is possible to produce resistance to diseases caused by viruses, bacteria, fungi, root pathogens, insects and nematodes. It is also contemplated that control of mycotoxin producing organisms may be realized through expression of introduced genes. Resistance to viruses may be produced through expression of novel genes. For example, it has been demonstrated that expression of a viral coat protein in a transgenic plant can impart resistance to infection ofthe plant by that virus and perhaps other closely related viruses (Cuozzo et al., 1988, Hemenway et al., 1988, Abel et al., 1986). It is contemplated that expression of antisense genes targeted at essential viral functions may impart resistance to said viras. For example, an antisense gene targeted at the gene responsible for replication of viral nucleic acid may inhibit said replication and lead to resistance to the virus. It is believed that interference with other viral functions through the use of antisense genes may also increase resistance to viruses. Further it is proposed that it may be possible to achieve resistance to viruses through other approaches, including, but not limited to the use of satellite viruses.
It is proposed that increased resistance to diseases caused by bacteria and fungi may be realized through introduction of novel genes. It is contemplated that genes encoding so-called "peptide antibiotics," pathogenesis related (PR) proteins, toxin resistance, and proteins affecting host-pathogen interactions such as morphological characteristics will be useful. Peptide antibiotics are polypeptide sequences which are inhibitory to growth of bacteria and other microorganisms. For example, the classes of peptides refened to as cecropins and magainins inhibit growth of many species of bacteria and fungi. It is proposed that expression of PR proteins in plants may be useful in confening resistance to bacterial disease. These genes are induced following pathogen attack on a host plant and have been divided into at least five classes of proteins (Bol et al., 1990). Included amongst the PR proteins are beta-1,3-
glucanases, chitinases, and osmotin and other proteins that are believed to function in plant resistance to disease organisms. Other genes have been identified that have antifungal properties, e.g., UDA (stinging nettle lectin) and hevein (Broakgert et al., 1989; Barkai-Golan et al., 1978). It is known that certain plant diseases are caused by the production of phytotoxins. Resistance to these diseases could be achieved through expression of a novel gene that encodes an enzyme capable of degrading or otherwise inactivating the phytotoxin. Expression novel genes that alter the interactions between the host plant and pathogen may be useful in reducing the ability the disease organism to invade the tissues ofthe host plant, e.g., an increase in the waxiness ofthe leaf cuticle or other morphological characteristics. Plant parasitic nematodes are a cause of disease in many plants. It is proposed that it would be possible to make the plant resistant to these organisms through the expression of novel genes. It is anticipated that control of nematode infestations would be accomplished by altering the ability ofthe nematode to recognize or attach to a host plant and/or enabling the plant to produce nematicidal compounds, including but not limited to proteins. 5. Mycotoxin Reduction Elimination
Production of mycotoxins, including aflatoxin and fumonisin, by fungi associated with plants is a significant factor in rendering the grain not useful. These fungal organisms do not cause disease symptoms and/or interfere with the growth ofthe plant, but they produce chemicals (mycotoxins) that are toxic to animals. Inhibition ofthe growth of these fungi would reduce the synthesis of these toxic substances and, therefore, reduce grain losses due to mycotoxin contamination. Novel genes may be introduced into plants that would inhibit synthesis ofthe mycotoxin without interfering with fungal growth. Expression of a novel gene which encodes an enzyme capable of rendering the mycotoxin nontoxic would be useful in order to achieve reduced mycotoxin contamination of grain. The result of any ofthe above mechanisms would be a reduced presence of mycotoxins on grain. 6. Grain Composition or Quality
Genes may be introduced into plants, particularly commercially important cereals such as maize, wheat or rice, to improve the grain for which the cereal is primarily grown. A wide range of novel transgenic plants produced in this manner may be envisioned depending on the particular end use ofthe grain.
For example, the largest use of maize grain is for feed or food. Introduction of genes that alter the composition ofthe grain may greatly enhance the feed or food value. The primary components of maize grain are starch, protein, and oil. Each of these primary
components of maize grain may be improved by altering its level or composition. Several examples may be mentioned for illustrative purposes but in no way provide an exhaustive list ofpossibilities.
The protein of many cereal grains is suboptimal for feed and food purposes especially when fed to pigs, poultry, and humans. The protein is deficient in several amino acids that are essential in the diet of these species, requiring the addition of supplements to the grain. Limiting essential amino acids may include lysine, methionine, tryptophan, threonine, valine, arginine, and histidine. Some amino acids become limiting only after the grain, is supplemented with other inputs for feed formulations. For example, when the grain is supplemented with soybean meal to meet lysine requirements, methionine becomes limiting. The levels of these essential amino acids in seeds and grain may be elevated by mechanisms which include, but are not limited to, the introduction of genes to increase the biosynthesis of the amino acids, decrease the degradation ofthe amino acids, increase the storage ofthe amino acids in proteins, or increase transport ofthe amino acids to the seeds or grain. One mechanism for increasing the biosynthesis ofthe amino acids is to introduce genes that deregulate the amino acid biosynthetic pathways such that the plant can no longer adequately control the levels that are produced. This may be done by deregulating or bypassing steps in the amino acid biosynthetic pathway which are normally regulated by levels ofthe amino acid end product ofthe pathway. Examples include the introduction of genes that encode deregulated versions ofthe enzymes aspartokinase or dihydrodipicolinic acid (DHDP)- synthase for increasing lysine and threonine production, and anthranilate synthase for increasing tryptophan production. Reduction ofthe catabolism ofthe amino acids may be accomplished by introduction of DNA sequences that reduce or eliminate the expression of genes encoding enzymes that catalyse steps in the catabolic pathways such as the enzyme lysine-ketoglutarate reductase.
The protein composition ofthe grain may be altered to improve the balance of amino acids in a variety of ways including elevating expression of native proteins, decreasing expression of those with poor composition, changing the composition of native proteins, or introducing genes encoding entirely new proteins possessing superior composition. DNA may be introduced that decreases the expression of members ofthe zein family of storage proteins. This DNA may encode ribozymes or antisense sequences directed to impairing expression of zein proteins or expression of regulators of zein expression such as the opaque-2 gene product. The protein composition ofthe grain may be modified through the phenomenon of
cosuppression, i.e., inhibition of expression of an endogenous gene tlirough the expression of an identical stractural gene or gene fragment introduced through transformation (Goring et al., 1991). Additionally, the introduced DNA may encode enzymes which degrade seines. The decreases in zein expression that are achieved may be accompanied by increases in proteins with more desirable amino acid composition or increases in other major seed constituents such as starch. Alternatively, a chimeric gene may be introduced that comprises a coding sequence for a native protein of adequate amino acid composition such as for one ofthe globulin proteins or 10 kD zein of maize and a promoter or other regulatory sequence designed to elevate expression of said protein. The coding sequence of said gene may include additional or replacement codons for essential amino acids. Further, a coding sequence obtained from another species, or, a partially or completely synthetic sequence encoding a completely unique peptide sequence designed to enhance the amino acid composition ofthe seed may be employed.
The introduction of genes that alter the oil content ofthe grain may be of value. Increases in oil content may result in increases in metabohzable energy content and density of the seeds for uses in feed and food. The introduced genes may encode enzymes that remove or reduce rate-limitations or regulated steps in fatty acid or lipid biosynthesis. Such genes may include, but are not limited to, those that encode acetyl-CoA carboxylase, ACP-acyltransferase, beta-ketoacyl-ACP synthase, plus other well known fatty acid biosynthetic activities. Other possibilities are genes that encode proteins that do not possess enzymatic activity such as acyl canier protein. Additional examples include 2-acetyltransferase, oleosin pyruvate dehydrogenase complex, acetyl CoA synthetase, ATP citrate lyase, ADP-glucose pyrophosphorylase and genes ofthe carnitine-CoA- acetyl-CoA shuttles. It is anticipated that expression of genes related to oil biosynthesis will be targeted to the plastid, using a plastid transit peptide sequence and preferably expressed in the seed embryo. Genes may be introduced that alter the balance of fatty acids present in the oil providing a more healthful or nutritive feedstuff. The introduced DNA may also encode sequences that block expression of enzymes involved in fatty acid biosynthesis, altering the proportions of fatty acids present in the grain such as described below. Genes may be introduced that enhance the nutritive value ofthe starch component of the grain, for example by increasing the degree of branching, resulting in improved utilization ofthe starch in cows by delaying its metabolism.
Besides affecting the major constituents ofthe grain, genes may be introduced that affect a variety of other nutritive, processing, or other quality aspects ofthe grain as used for feed or food. For example, pigmentation ofthe grain may be increased or decreased. Enhancement and stability of yellow pigmentation is desirable in some animal feeds and may be achieved by introduction of genes that result in enhanced production of xanthophylls and carotenes by eliminating rate-limiting steps in their production. Such genes may encode altered forms ofthe enzymes phytoene synthase, phytoene desaturase, or lycopene synthase. Alternatively, unpigmented white corn is desirable for production of many food products and may be produced by the introduction of DNA which blocks or eliminates steps in pigment production pathways.
Feed or food comprising some cereal grains possesses insufficient quantities of vitamins and must be supplemented to provide adequate nutritive value. Introduction of genes that enhance vitamin biosynthesis in seeds may be envisioned including, for example, vitamins A, E, B12, choline, and the like. For example, maize grain also does not possess sufficient mineral content for optimal nutritive value. Genes that affect the accumulation or availability of compounds containing phosphoras, sulfur, calcium, manganese, zinc, and iron among others would be valuable. An example may be the introduction of a gene that reduced phytic acid production or encoded the enzyme phytase which enhances phytic acid breakdown. These genes would increase levels of available phosphate in the diet, reducing the need for supplementation with mineral phosphate.
Numerous other examples of improvement of cereals for feed and food purposes might be described. The improvements may not even necessarily involve the grain, but may, for example, improve the value ofthe grain for silage. Introduction of DNA to accomplish this might include sequences that alter lignin production such as those that result in the "brown midrib" phenotype associated with superior feed value for cattle.
In addition to direct improvements in feed or food value, genes may also be introduced which improve the processing of grain and improve the value ofthe products resulting from the processing. The primary method of processing certain grains such as maize is via wetmilling. Maize may be improved though the expression of novel genes that increase the efficiency and reduce the cost of processing such as by decreasing steeping time.
Improving the value of wetmilling products may include altering the quantity or quality of starch, oil, corn gluten meal, or the components of corn gluten feed. Elevation of starch may be achieved through the identification and elimination of rate limiting steps in starch
biosynthesis or by decreasing levels ofthe other components ofthe grain resulting in proportional increases in starch. An example ofthe former may be the introduction of genes encoding ADP-glucose pyrophosphorylase enzymes with altered regulatory activity or which are expressed at higher level. Examples ofthe latter may include selective inhibitors of, for example, protein or oil biosynthesis expressed during later stages of kernel development.
The properties of starch may be beneficially altered by changing the ratio of amylose to amylopectin, the size ofthe starch molecules, or their branching pattern. Through these changes a broad range of properties may be modified which include, but are not limited to, changes in gelatinization temperature, heat of gelatinization, clarity of films and pastes, Theological properties, and the like. To accomplish these changes in properties, genes that encode granule-bound or soluble starch synthase activity or branching enzyme activity may be introduced alone or combination. DNA such as antisense constructs may also be used to decrease levels of endogenous activity of these enzymes. The introduced genes or constracts may possess regulatory sequences that time their expression to specific intervals in starch biosynthesis and starch granule development. Furthermore, it may be advisable to introduce and express genes that result in the in vivo derivatization, or other modification, ofthe glucose moieties ofthe starch molecule. The covalent attachment of any molecule may be envisioned, limited only by the existence of enzymes that catalyze the derivatizations and the accessibility of appropriate substrates in the starch granule. Examples of important derivations may include the addition of functional groups such as amines, carboxyls, or phosphate groups which provide sites for subsequent in vitro derivatizations or affect starch properties through the introduction of ionic charges. Examples of other modifications may include direct changes of the glucose units such as loss of hydroxyl groups or their oxidation to aldehyde or carboxyl groups. Oil is another product of wetmilling of corn and other grains, the value of which may be improved by introduction and expression of genes. The quantity of oil that can be extracted by wetmilling may be elevated by approaches as described for feed and food above. Oil properties may also be altered to improve its performance in the production and use of cooking oil, shortenings, lubricants or other oil-derived products or improvement of its health attributes when used in the food-related applications. Novel fatty acids may also be synthesized which upon extraction can serve as starting materials for chemical syntheses. The changes in oil properties may be achieved by altering the type, level, or lipid arrangement ofthe fatty acids present in the oil. This in rum may be accomplished by the addition of genes that encode
enzymes that catalyze the synthesis of novel fatty acids and the lipids possessing them or by increasing levels of native fatty acids while possibly reducing levels of precursors. Alternatively DNA sequences may be introduced which slow or block steps in fatty acid biosynthesis resulting in the increase in precursor fatty acid intermediates. Genes that might be added include desaturases, epoxidases, hydratases, dehydratases, and other enzymes that catalyze reactions involving fatty acid intermediates. Representative examples of catalytic steps that might be blocked include the desaturations from stearic to oleic acid and oleic to linolenic acid resulting in the respective accumulations of stearic and oleic acids.
Improvements in the other major cereal wetmilling products, gluten meal and gluten feed, may also be achieved by the introduction of genes to obtain novel plants. Representative possibilities include but are not limited to those described above for improvement of food and feed value.
In addition it may further be considered that the plant be used for the production or manufacturing of useful biological compounds that were either not produced at all, or not produced at the same level, in the plant previously. The novel plants producing these compounds are made possible by the introduction and expression of genes by transformation methods. The possibilities include, but are not limited to, any biological compound which is presently produced by any organism such as proteins, nucleic acids, primary and intermediary metabolites, carbohydrate polymers, etc. The compounds may be produced by the plant, extracted upon harvest and/or processing, and used for any presently recognized useful purpose such as pharmaceuticals, fragrances, industrial enzymes to name a few.
Further possibilities to exemplify the range of grain traits or properties potentially encoded by introduced genes in transgenic plants include grain with less breakage susceptibility for export purposes or larger grit size when processed by dry milling through introduction of genes that enhance gamma-zein synthesis, popcorn with improved popping quality and expansion volume through genes that increase pericarp thickness, corn with whiter grain for food uses though introduction of genes that effectively block expression of enzymes involved in pigment production pathways, and improved quality of alcoholic beverages or sweet corn through introduction of genes which affect flavor such as the shrunken gene (encoding sucrose synthase) for sweet com. 7. Plant Agronomic Characteristics
Two ofthe factors determining where plants can be grown are the average daily temperature during the growing season and the length of thne between frosts. Within the areas
where it is possible to grow a particular plant, there are varying limitations on the maximal time it is allowed to grow to maturity and be harvested. The plant to be grown in a particular area is selected for its ability to mature and dry down to harvestable moisture content within the required period of time with maximum possible yield. Therefore, plant of varying maturities are developed for different growing locations. Apart from the need to dry down sufficiently to permit harvest is the desirability of having maximal drying take place in the field to miriimize the amount of energy required for additional drying post-harvest. Also the more readily the grain can dry down, the more time there is available for growth and kernel fill. Genes that influence maturity and/or dry down can be identified and introduced into plant lines using transformation techniques to create new varieties adapted to different growing locations or the same growing location but having improved yield to moisture ratio at harvest. Expression of genes that are involved in regulation of plant development may be especially useful, e.g., the liguleless and rough sheath genes that have been identified in plants.
Genes may be introduced into plants that would improve standability and other plant growth characteristics. For example, expression of novel genes which confer stronger stalks, improved root systems, or prevent or reduce ear droppage would be of great value to the com farmer. Introduction and expression of genes that increase the total amount of photoassimilate available by, for example, increasing light distribution and/or interception would be advantageous. In addition the expression of genes that increase the efficiency of photosynthesis and/or the leaf canopy would further increase gains in productivity. Such approaches would allow for increased plant populations in the field.
Delay of late season vegetative senescence would increase the flow of assimilate into the grain and thus increase yield. Overexpression of genes within plants that are associated with "stay green" or the expression of any gene that delays senescence would achieve be advantageous. For example, a non-yellowing mutant has been identified in Festuca pratensis (Davies et al., 1990). Expression of this gene as well as others may prevent premature breakdown of chlorophyll and thus maintain canopy function. 8. Nutrient Utilization The ability to utilize available nutrients and minerals may be a limiting factor in growth of many plants. It is proposed that it would be possible to alter nutrient uptake, tolerate pH extremes, mobilization through the plant, storage pools, and availability for metabolic activities by the introduction of novel genes. These modifications would allow a plant to more efficiently utilize available nutrients. It is contemplated that an increase in the activity of, for
example, an enzyme that is normally present in the plant and involved in nutrient utilization would increase the availability of a nutrient. An example of such an enzyme would be phytase. It is also contemplated that expression of a novel gene may make a nutrient source available that was previously not accessible, e.g., an enzyme that releases a component of nutrient value from a more complex molecule, perhaps a macromolecule.
9. Male Sterility
Male sterility is useful in the production of hybrid seed. It is proposed that male sterility may be produced through expression of novel genes. For example, it has been shown that expression of genes that encode proteins that interfere with development ofthe male inflorescence and/or gametophyte result in male sterility. Chimeric ribonuclease genes that express in the anthers of transgenic tobacco and oilseed rape have been demonstrated to lead to male sterility (Mariani et al, 1990).
For example, a number of mutations were discovered in maize that confer cytoplasmic male sterility. One mutation in particular, refened to as T cytoplasm, also conelates with sensitivity to Southern corn leaf blight. A DNA sequence, designated TURF- 13 (Levings, 1990), was identified that correlates with T cytoplasm. It would be possible through the introduction of TURF- 13 via transformation to separate male sterility from disease sensitivity. As it is necessary to be able to restore male fertility for breeding purposes and for grain production, it is proposed that genes encoding restoration of male fertility may also be introduced.
10. Negative Selectable Markers
Introduction of genes encoding traits that can be selected against may be useful for eliminating undesirable linked genes. When two or more genes are introduced together by cotransformation, the genes will be linked together on the host chromosome. For example, a gene encoding a Bt gene that confers insect resistance on the plant may be introduced into a plant together with a bar gene that is useful as a selectable marker and confers resistance to the herbicide Ignite® on the plant. However, it may not be desirable to have an insect resistant plant that is also resistant to the herbicide Ignite®. It is proposed that one could also introduce an antisense bar gene that is expressed in those tissues where one does not want expression of the bar gene, e.g., in whole plant parts. Hence, although the bar gene is expressed and is useful as a selectable marker, it is not useful to confer herbicide resistance on the whole plant. The bar antisense gene is a negative selectable marker.
Negative selection is necessary in order to screen a population of transformants for rare homologous recombinants generated through gene targeting. For example, a homologous recombinant may be identified through the inactivation of a gene that was previously expressed in that cell. The antisense gene to neomycin phosphotransferase II (nptll) has been investigated as a negative selectable marker in tobacco (Nicotiana tabacum) and Arabidopsis thaliana (Xiang and Guena, 1993). In this example both sense and antisense nptll genes are introduced into a plant through transformation and the resultant plants are sensitive to the antibiotic kanamycin. An introduced gene that integrates into the host cell chromosome at the site ofthe antisense nptll gene, and inactivates the antisense gene, will make the plant resistant to kanamycin and other aminoglycoside antibiotics. Therefore, rare site specific recombinants may be identified by screening for antibiotic resistance. Similarly, any gene, native to the plant or introduced through transformation, that when inactivated confers resistance to a compound, may be useful as a negative selectable marker.
It is contemplated that negative selectable markers may also be useful in other ways. One application is to construct transgenic lines in which one could select for transposition to unlinked sites. In the process of tagging it is most common for the transposable element to move to a genetically linked site on the same chromosome. A selectable marker for recovery of rare plants in which transposition has occurred to an unlinked locus would be useful. For example, the enzyme cytosine deaminase may be useful for this purpose (Stouggard, 1993). In the presence of this enzyme the compound 5-fluorocytosine is converted to 5-fluoraracil which is toxic to plant and animal cells. If a transposable element is linked to the gene for the enzyme cytosine deaminase, one may select for transposition to unlinked sites by selecting for transposition events in which the resultant plant is now resistant to 5-fluorocytosine. The parental plants and plants containing transpositions to linked sites will remain sensitive to 5- fluorocytosine. Resistance to 5-fluorocytosine is due to loss ofthe cytosine deaminase gene through genetic segregation ofthe transposable element and the cytosine deaminase gene. Other genes that encode proteins that render the plant sensitive to a certain compound will also be useful in this context. For example, T-DNA gene 2 from Agrobacterium tumefaciens encodes a protein that catalyzes the conversion of alpha-naphthalene acetamide (NAM) to alpha-napthalene acetic acid (NAA) renders plant cells sensitive to high concentrations of NAM (Depicker et al., 1988).
It is also contemplated that negative selectable markers may be useful in the construction of transposon tagging lines. For example, by marking an autonomous
transposable element such as Ac, Master Mu, or En/Spn with a negative selectable marker, one could select for transformants in which the autonomous element is not stably integrated into the genome. This would be desirable, for example, when transient expression ofthe autonomous element is desired to activate in trans the transposition of a defective transposable element, such as Ds, but stable integration ofthe autonomous element is not desired. The presence ofthe autonomous element may not be desired in order to stabilize the defective element, i.e., prevent it from further transposing. However, it is proposed that if stable integration of an autonomous transposable element is desired in a plant the presence of a negative selectable marker may make it possible to eliminate the autonomous element during the breeding process .
11. Non-Protein-Expressing Sequences a. RNA-Expressing DNA may be introduced into plants for the purpose of expressing RNA transcripts that function to affect plant phenotype yet are not translated into protein. Two examples are antisense RNA and RNA with ribozyme activity. Both may serve possible functions in reducing or eliminating expression of native or introduced plant genes.
Genes may be constructed or isolated, which when transcribed, produce antisense RNA that is complementary to all or part(s) of a targeted messenger RNA(s). The antisense RNA reduces production ofthe polypeptide product ofthe messenger RNA. The polypeptide product may be any protein encoded by the plant genome. The aforementioned genes will be refened to as antisense genes. An antisense gene may thus be introduced into a plant by transformation methods to produce a novel transgenic plant with reduced expression of a selected protein of interest. For example, the protein may be an enzyme that catalyzes a reaction in the plant. Reduction ofthe enzyme activity may reduce or eliminate products ofthe reaction which include any enzymatically synthesized compound in the plant such as fatty acids, amino acids, carbohydrates, nucleic acids and the like. Alternatively, the protein may be a storage protein, such as a zein, or a stractural protein, the decreased expression of which may lead to changes in seed amino acid composition or plant morphological changes respectively. The possibilities cited above are provided only by way of example and do not represent the full range of applications .
Genes may also be constracted or isolated, which when transcribed produce RNA enzymes, or ribozymes, which can act as endoribonucleases and catalyze the cleavage of RNA molecules with selected sequences. The cleavage of selected messenger RNA's can result in
the reduced production of their encoded polypeptide products. These genes may be used to prepare novel transgenic plants which possess them. The transgenic plants may possess reduced levels of polypeptides including but not limited to the polypeptides cited above that may be affected by antisense RNA. It is also possible that genes may be introduced to produce novel transgenic plants which have reduced expression of a native gene product by a mechanism of cosuppression. It has been demonstrated in tobacco, tomato, and petunia (Goring et al, 1991; Smith et al., 1990; Napoli et al., 1990; van der Krol et al., 1990) that expression ofthe sense transcript of a native gene will reduce or eliminate expression ofthe native gene in a manner similar to that observed for antisense genes. The introduced gene may encode all or part ofthe targeted native protein but its translation may not be required for reduction of levels of that native protein. b. Non-RNA-Expressing For example, DNA elements including those of transposable elements such as Ds, Ac, or Mu, may be inserted into a gene and cause mutations. These DNA elements may be inserted in order to inactivate (or activate) a gene and thereby "tag" a particular trait. In this instance the transposable element does not cause instability ofthe tagged mutation, because the utility ofthe element does not depend on its ability to move in the genome. Once a desired trait is tagged, the introduced DNA sequence may be used to clone the correspondmg gene, e.g., using the introduced DNA sequence as a PCR primer together with PCR gene cloning techniques (Shapiro, 1983; Dellaporta et al., 1988). Once identified, the entire gene(s) for the particular trait, including control or regulatory regions where desired may be isolated, cloned and manipulated as desired. The utility of DNA elements introduced into an organism for purposed of gene tagging is independent ofthe DNA sequence and does not depend on any biological activity ofthe DNA sequence, i.e., transcription into RNA or translation into protein. The sole function ofthe DNA element is to disrapt the DNA sequence of a gene.
It is contemplated that unexpressed DNA sequences, including novel synthetic sequences could be introduced into cells as proprietary "labels" of those cells and plants and seeds thereof. It would not be necessary for a label DNA element to disrapt the function of a gene endogenous to the host organism, as the sole function of this DNA would be to identify the origin ofthe organism. For example, one could introduce a unique DNA sequence into a plant and this DNA element would identify all cells, plants, and progeny of these cells as having arisen from that labeled source. It is proposed that inclusion of label DNAs would
enable one to distinguish proprietary germplasm or germplasm derived from such, from unlabelled germplasm.
Another possible element which may be introduced is a matrix attachment region element (MAR), such as the chicken lysozyme A element (Stief et al., 1989), which can be positioned around an expressible gene of interest to effect an increase in overall expression of the gene and diminish position dependant effects upon incorporation into the plant genome (Stief et al., 1989; Phi- Van et al., 1990).
III. Transformed (Transgenic) Plants of the Invention and Methods of Preparation Plant species may be transformed with the DNA constract ofthe present invention by the DNA-mediated transformation of plant cell protoplasts and subsequent regeneration ofthe plant from the transformed protoplasts in accordance with procedures well known in the art. Any plant tissue capable of subsequent clonal propagation, whether by organogenesis or embryogenesis, may be transformed with a vector ofthe present invention. The term "organogenesis," as used herein, means a process by which shoots and roots are developed sequentially from meristematic centers; the term "embryogenesis," as used herein, means a process by which shoots and roots develop together in a concerted fashion (not sequentially), whether from somatic cells or gametes. The particular tissue chosen will vary depending on the clonal propagation systems available for, and best suited to, the particular species being transformed. Exemplary tissue targets include leaf disks, pollen, embryos, cotyledons, hypocotyls, megagametophytes, callus tissue, existing meristematic tissue (e.g., apical meristems, axillary buds, and root meristems), and induced meristem tissue (e.g., cotyledon meristem and ultilane meristem).
Plants ofthe present invention may take a variety of fonns. The plants may be chimeras of transformed cells and non-transformed cells; the plants may be clonal transformants (e.g., all cells transformed to contain the expression cassette); the plants may comprise grafts of transformed and untransformed tissues (e.g., a transformed root stock grafted to an untransformed scion in citrus species). The transformed plants may be propagated by a variety of means, such as by clonal propagation or classical breeding techniques. For example, first generation (or TI) transformed plants may be selfed to give homozygous second generation (or T2) transformed plants, and the T2 plants further propagated through classical breeding techniques. A dominant selectable marker (such as npt II) can be associated with the expression cassette to assist in breeding.
Thus, the present invention provides a transformed (transgenic) plant cell, in planta or ex planta, including a transformed plastid or other organelle, e.g., nucleus, mitochondria or chloroplast. The present invention may be used for transformation of any plant species, including, but not limited to, cells from corn (Zea mays), Brassica sp. (e.g., B. napus, B. rapa, B.juncea), particularly those Brassica species useful as sources of seed oil, alfalfa (Medicago sativa), rice (Oryza sativa), rye (Secale cereale), sorghum (Sorghum bicolor, Sorghum vulgare), millet (e.g., pearl millet (Pennisetum glaucum), proso millet (Panicum miliaceum), foxtail millet (Setaria italica), finger millet (Eleusine coracana)), sunflower (Helianthus annuus), safflower (Carthamus tinctorius), wheat (Triticum aestivum), soybean (Glycine max), tobacco (Nicotiana tabacum), potato (Solanum tuberosum), peanuts (Arachis hypogaea), cotton (Gossypium barbadense, Gossypium hirsutum), sweet potato (Ipomoea batatus), cassava (Manihot esculenta), coffee (Cofea spp.), coconut (Cocos nucifera), pineapple (Ananas comosus), citrus trees (Citrus spp.), cocoa (Theobroma cacao), tea (Camellia sinensis), banana (Musa spp.), avocado (Persea ultilane), fig (Ficus casica), guava (Psidium guajava), mango (Mangifera indica), olive (Olea europaea), papaya (Carica papaya), cashew (Anacardium occidentale), macadamia (Macadamia integrifolia), almond (Prunus amygdalus), sugar beets (Beta vulgaris), sugarcane (Saccharum spp.), oats, duckweed (Lemna), barley, vegetables, ornamentals, and conifers.
Duckweed (Lemna, see WO 00/07210) includes members ofthe family Lemnaceae. There are known four genera and 34 species of duckweed as follows: genus Lemna (L. aequinoctialis, L. disperma, L. ecuadoriensis, L. gibba, L. japonica, L. minor, L. miniscula, L. obscura, L. perpusilla, L. tenera, L. trisulca, L.turionifera, L. valdiviana); genus Spirodela (S. intermedia, S. polyrrhiza, S. punctata); genus Woffia (Wa. Angusta, Wa. Arrhiza, Wa. Australina, Wa. Borealis, Wa. Brasiliensis, Wa. Columbiana, Wa. Elongata, Wa. Globosa, Wa. Microscopica, Wa. Neglecta) and genus Wofiella (WL ultila, WL ultilanen, WL gladiata, WL ultila, WL lingulata, WL repunda, WL rotunda, and WL neotropica). Any other genera or species of Lemnaceae, if they exist, are also aspects ofthe present invention. Lemna gibba, Lemna minor, and Lemna miniscula are prefened, with Lemna minor and Lemna miniscula being most prefened. Lemna species can be classified using the taxonomic scheme described by Landolt, Biosystematic Investigation on the Family of Duckweeds: The family of
Lemnaceae - A Monograph Study. Geobatanischen Institut ETH, Stiftung Rubel, Zurich (1986)).
Vegetables within the scope ofthe invention include tomatoes (Lycopersicon esculentum), lettuce (e.g., Lactuca sativa), green beans (Phaseolus vulgaris), lima beans (Phaseolus limensis), peas (Lathyrus spp.), and members ofthe genus Cucumis such as cucumber (C. sativus), cantaloupe (C cantalupensis), and musk melon (C. melo). Ornamentals include azalea (Rhododendron spp.), hydrangea (Macrophylla hydrangea), hibiscus (Hibiscus rosasanensis), roses (Rosa spp.), tulips (Tulipa spp.), daffodils (Narcissus spp.), petunias (Petunia hybrida), carnation (Dianthus caryophyllus), poinsettia (Euphorbia pulcherrima), and chrysanthemum. Conifers that may be employed in practicing the present invention include, for example, pines such as loblolly pine (Pinus taeda), slash pine (Pinus elliotii), ponderosa pine (Pinus ponderosa), lodgepole pine (Pinus contorta), and Monterey pine (Pinus radiata), Douglas-fir (Pseudotsuga menziesii); Western hemlock (Tsuga ultilane); Sitka spruce (Picea glauca); redwood (Sequoia sempervirens); trae firs such as silver fir (Abies amabilis) and balsam fir (Abies balsamea); and cedars such as Western red cedar (Thuja plicata) and Alaska yellow-cedar (Chamaecyparis nootkatensis). Leguminous plants include beans and peas. Beans include guar, locust bean, fenugreek, soybean, garden beans, cowpea, mungbean, lima bean, fava bean, lentils, chickpea, etc. Legumes include, but are not limited to, Arachis, e.g., peanuts, Vicia, e.g., crown vetch, hairy vetch, adzuki bean, mung bean, and chickpea, Lupinus, e.g., lupine, trifolium, Phaseolus, e.g., common bean and lima bean, Pisum, e.g., field bean, Melilotus, e.g., clover, Medicago, e.g., alfalfa, Lotus, e.g., trefoil, lens, e.g., lentil, and false indigo. Prefened forage and turfgrass for use in the methods ofthe invention include alfalfa, orchard grass, tall fescue, perennial ryegrass, creeping bent grass, and redtop.
Papaya, garlic, pea, peach, pepper, petunia, strawberry, sorghum, sweet potato, ttirnip, safflower, corn, pea, endive, gourd, grape, snap bean, chicory, cotton, tobacco, aubergine, beet, buckwheat, broad bean, nectarine, avocado, mango, banana, groundnut, potato,peanut, lettuce, pineapple, spinach, squash, sugarbeet, sugarcane, sweet corn, chrysanthemum.
Other plants within the scope ofthe invention include Acacia, aneth, artichoke, aragula, blackberry, canola, cilantro, Clementines, escarole, eucalyptus, fennel, grapefruit, honey dew, jicama, kiwifiuit, lemon, lime, mushroom, nut, okra, orange, parsley, pers rimon, plantain, pomegranate, poplar, radiata pine, radicchio, Southern pine, sweetgum, tangerine, triticale, vine, yams, apple, pear, quince, cherry, apricot, melon, hemp, buckwheat, grape, raspberry, chenopodium, blueberry, nectarine, peach, plum, strawberry, watermelon, eggplant, pepper, cauliflower, Brassica, e.g., broccoli, cabbage, ultilan sprouts, onion, canot, leek, beet,
broad bean, celery, radish, pumpkin, endive, gourd, garlic, snapbean, spinach, squash, turnip, ultilane, and zucchini.
Ornamental plants within the scope ofthe invention include impatiens, Begonia, Pelargonium, Viola, Cyclamen, Verbena, Vinca, Tagetes, Primula, Saint Paulia, Agertum, Amaranthus, Ajitihirrhinum, Aquilegia, Cineraria, Clover, Cosmo, Cowpea, Dahlia, Datura, Delphinium, Gerbera, Gladiolus, Gloxinia, Hippeastrum, Mesembryanthemum, Salpiglossos, and Zinnia. Other plants within the scope ofthe invention are shown in Table 1 (above).
Preferably, transgenic plants ofthe present invention are crop plants and in particular cereals (for example, corn, alfalfa, sunflower, rice, Brassica, canola, soybean, barley, soybean, sugarbeet, cotton, safflower, peanut, sorghum, wheat, millet, tobacco, etc.), and even more preferably corn, rice and soybean.
Transformation of plants can be undertaken with a single DNA molecule or multiple DNA molecules (i.e., co-transformation), and both these techniques are suitable for use with the expression cassettes ofthe present invention. Numerous transformation vectors are available for plant transformation, and the expression cassettes of this invention can be used in conjunction with any such vectors. The selection of vector will depend upon the prefened transformation technique and the target species for transformation.
A variety of techniques are available and known to those skilled in the art for introduction of constracts into a plant cell host. These techniques generally include transfonnation with DNA employing A. tumefaciens or A. rhizogenes as the transforming agent, liposomes, PEG precipitation, electroporation, DNA injection, direct DNA uptake, microprojectile bombardment, particle acceleration, and the like (See, for example, EP 295959 and EP 138341) (see below). However, cells other than plant cells may be transformed with the expression cassettes ofthe invention. The general descriptions of plant expression vectors and reporter genes, and Agrobacterium and Agrobacterium-mediated gene transfer, can be found in Gruber et al. (1993).
Expression vectors containing genomic or synthetic fragments can be introduced into protoplasts or into intact tissues or isolated cells. Preferably expression vectors are introduced into intact tissue. General methods of culturing plant tissues are provided for example by Maki et al., (1993); and by Phillips et al. (1988). Preferably, expression vectors are introduced into maize or other plant tissues using a direct gene transfer method such as microprojectile- mediated delivery, DNA injection, electroporation and the like. More preferably expression vectors are introduced into plant tissues using the microprojectile media delivery with the
biolistic device. See, for example, Tomes et al. (1995). The vectors ofthe invention can not only be used for expression of stractural genes but may also be used in exon-trap cloning, or promoter trap procedures to detect differential gene expression in varieties of tissues, (Lindsey et al., 1993; Auch & Reth et al.). It is particularly prefened to use the binary type vectors of Ti and Ri plasmids of
Agrobacterium spp. Ti-derived vectors transform a wide variety of higher plants, including monocotyledonous and dicotyledonous plants, such as soybean, cotton, rape, tobacco, and rice (Pacciotti et al., 1985: Byrne et al., 1987; Sukhapinda et al., 1987; Park et al., 1985: Hiei et al., 1994). The use of T-DNA to transform plant cells has received extensive study and is amply described (EP 120516; Hoekema, 1985; Knauf, et al., 1983; and An et al., 1985). For introduction into plants, the chimeric genes ofthe invention can be inserted into binary vectors as described in the examples.
Other transformation methods are available to those skilled in the art, such as direct uptake of foreign DNA constracts (see EP 295959), techniques of electroporation (Fromm et al., 1986) or high velocity ballistic bombardment with metal particles coated with the nucleic acid constracts (Kline et al., 1987, and U.S. Patent No. 4,945,050). Once transformed, the cells can be regenerated by those skilled in the art. Of particular relevance are the recently described methods to transform foreign genes into commercially important crops, such as rapeseed (De Block et al., 1989), sunflower (Everett et al., 1987), soybean (McCabe et al., 1988; Hinchee et al, 1988; Chee et al., 1989; Christou et al., 1989; EP 301749), rice (Hiei et al., 1994), and corn (Gordon Kamm et al., 1990; Fromm et al., 1990).
Those skilled in the art will appreciate that the choice of method might depend on the type of plant, i.e., monocotyledonous or dicotyledonous, targeted for transformation. Suitable methods of transforming plant cells include, but are not limited to, microinjection (Crossway et al., 1986), electroporation (Riggs et al., 1986), Agrobacterium-mediated transformation (Hinchee et al., 1988), direct gene transfer (Paszkowski et al., 1984), and ballistic particle acceleration using devices available from Agracetus, Inc., Madison, Wis. And BioRad, Hercules, Calif, (see, for example, Sanford et al., U.S. Pat. No. 4,945,050; and McCabe et al., 1988). Also see, Weissinger et al., 1988; Sanford et al., 1987 (onion); Christou et al., 1988 (soybean); McCabe et al., 1988 (soybean); Datta et al., 1990 (rice); Klein et al., 1988 (maize); Klein et al., 1988 (maize); Klein et al., 1988 (maize); Fromm et al., 1990 (maize); and Gordon- Kamm et al., 1990 (maize); Svab et al., 1990 (tobacco chloroplast); Koziel et al., 1993 (maize); Shimamoto et al, 1989 (rice); Christou et al., 1991 (rice); European Patent
Application EP 0 332 581 (orchardgrass and other Pooideae); Vasil et al., 1993 (wheat); Weeks et al., 1993 (wheat). In one embodiment, the protoplast transformation method for maize is employed (European Patent Application EP 0 292435, U. S. Pat. No. 5,350,689). In another embodiment, a nucleotide sequence ofthe present invention is directly transformed into the plastid genome. Plastid transformation technology is extensively described in U.S. Patent Nos. 5,451,513, 5,545,817, and 5,545,818, in PCT application no. WO 95/16783, and in McBride et al., 1994. The basic technique for chloroplast transformation involves introducing regions of cloned plastid DNA flariking a selectable marker together with the gene of interest into a suitable target tissue, e.g., using biolistics or protoplast transformation (e.g., calcium chloride or PEG mediated transformation). The 1 to 1.5 kb flanking regions, termed targeting sequences, facilitate orthologous recombination with the plastid genome and thus allow the replacement or modification of specific regions ofthe plastome. Initially, point mutations in the chloroplast 16S rRNA and rpsl2 genes conferring resistance to spectinomycin and/or streptomycin are utilized as selectable markers for transformation (Svab et al., 1990; Staub et al., 1992). This resulted in stable homoplasmic transformants at a frequency of approximately one per 100 bombardments of target leaves. The presence of cloning sites between these markers allowed creation of a plastid targeting vector for introduction of foreign genes (Staub et al., 1993). Substantial increases in transformation frequency are obtained by replacement ofthe recessive rRNA or r-protein antibiotic resistance genes with a dominant selectable marker, the bacterial aadA gene encoding the spectinomycin-detoxifying enzyme aminoglycoside-3N-adenyltransferase (Svab et al., 1993). Other selectable markers useful for plastid transformation are known in the art and encompassed within the scope ofthe invention. Typically, approximately 15-20 cell division cycles following transformation are required to reach a homoplastidic state. Plastid expression, in which genes are inserted by orthologous recombination into all ofthe several thousand copies ofthe circular plastid genome present in each plant cell, takes advantage of the enoπnous copy number advantage over nuclear-expressed genes to penriit expression levels that can readily exceed 10% ofthe total soluble plant protein. In a prefened embodiment, a nucleotide sequence ofthe present invention is inserted into a plastid targeting vector and transformed into the plastid genome of a desired plant host. Plants homoplastic for plastid genomes containing a nucleotide sequence ofthe present invention are obtained, and are preferentially capable of high expression ofthe nucleotide sequence.
Agrobacterium tumefaciens cells containing a vector comprising an expression cassette ofthe present invention, wherein the vector comprises a Ti plasmid, are useful in methods of making transformed plants. Plant cells are infected with an Agrobacterium tumefaciens as described above to produce a transformed plant cell, and then a plant is regenerated from the transformed plant cell. Numerous Agrobacterium vector systems useful in carrying out the present invention are known.
For example, vectors are available for transformation using Agrobacterium tumefaciens. These typically carry at least one T-DNA border sequence and include vectors such as pBIN19 (Bevan, 1984). In one prefened embodiment, the expression cassettes ofthe present invention may be inserted into either ofthe binary vectors pCIB200 and pCIB2001 for use with Agrobacterium. These vector cassettes for Agrobacterium-mediated transformation wear constracted in the following manner. PTJS75kan was created by Narl digestion of pTJS75 (Schmidhauser & Helinski, 1985) allowing excision ofthe tetracycline-resistance gene, followed by insertion of an Accl fragment from pUC4K carrying an NPTII (Messing & Vierra, 1982; Bevan et al., 1983; McBride et al., 1990). Xhol linkers were ligated to the
EcoRV fragment of ρCIB7 which contains the left and right T-DNA borders, a plant selectable nos/nptll chimeric gene and the pUC polylinker (Rothstein et al., 1987), and the Xhol- digested fragment was cloned into Sail-digested pTJS75kan to create pCIB200 (see also EP 0 332 104, example 19). PCIB200 contains the following unique polylinker restriction sites: EcoRI, Sstl, Kpnl, Bglll, Xbal, and Sail. The plasmid pCIB2001 is a derivative of pCIB200 which was created by the insertion into the polylinker of additional restriction sites. Unique restriction sites in the polylinker of pCIB2001 are EcoRI, Sstl, Kpnl, Bglll, Xbal, Sail, M , Bell, Avrll, Apal, Hpal, and Stul. PCIB2001, in addition to containing these unique restriction sites also has plant and bacterial kanamycin selection, left and right T-DNA borders for Agrobacterium-mediated transformation, the RK2-derived trfA function for mobilization between E. coli and other hosts, and the OriT and OriV functions also from RK2. The pCIB2001 polylinker is suitable for the cloning of plant expression cassettes containing their own regulatory signals.
An additional vector useful fox Agrobacterium-mediated transformation is the binary vector pCIB 10, which contains a gene encoding kanamycin resistance for selection in plants, T-DNA right and left border sequences and incorporates sequences from the wide host- range plasmid pRK252 allowing it to replicate in both E. coli and Agrobacterium. Its construction is described by Rothstein et al., 1987. Various derivatives of pCIBlO have been constructed
which incorporate the gene for hygromycin B phosphotransferase described by Gritz et al., 1983. These derivatives enable selection of transgenic plant cells on hygromycin only (pCIB743), or hygromycin and kanamycin (pCIB715, pCIB717).
Methods using either a form of direct gene transfer ox Agrobacterium-mediated transfer usually, but not necessarily, are undertaken with a selectable marker which may provide resistance to an antibiotic (e.g., kanamycin, hygromycin or methotrexate) or a herbicide (e.g., phosphinothricin). The choice of selectable marker for plant transformation is not, however, critical to the invention.
For certain plant species, different antibiotic or herbicide selection markers may be preferred. Selection markers used routinely in transformation include the nptll gene which confers resistance to kanamycin and related antibiotics (Messing & Vierra, 1982; Bevan et al., 1983), the bar gene which confers resistance to the herbicide phosphinothricin (White et al., 1990, Spencer et al., 1990), the hph gene which confers resistance to the antibiotic hygromycin (Blochinger & Diggelmann), and the dhfr gene, which confers resistance to methotrexate (Bourouis et al., 1983).
One such vector useful for direct gene transfer techniques in combination with selection by the herbicide Basta (or phosphinothricin) is pCIB3064. This vector is based on the plasmid pCIB246, which comprises the CaMV 35S promoter in operational fusion to the E. coli GUS gene and the CaMV 35S transcriptional terminator and is described in the PCT published application WO 93/07278, herein incorporated by reference. One gene useful for confening resistance to phosphinothricin is the bar gene from Streptomyces viridochromogenes (Thompson et al., 1987). This vector is suitable for the cloning of plant expression cassettes containing their own regulatory signals.
An additional transformation vector is pSOG35 which utilizes the E. coli gene dihydrofolate reductase (DHFR) as a selectable marker confening resistance to methotrexate. PCR was used to amplify the 35S promoter (about 800 bp), intron 6 from the maize Adhl gene (about 550 bp) and 18 bp ofthe GUS untranslated leader sequence from pSOGlO. A 250 bp fragment encoding the E. coli dihydrofolate reductase type II gene was also amplified by PCR and these two PCR fragments were assembled with a Sacl-Pstl fragment from pBI221 (Clontech) which comprised the pUC 19 vector backbone and the nopaline synthase tenninator. Assembly of these fragments generated pSOG19 which contains the 35S promoter in fusion with the intron 6 sequence, the GUS leader, the DHFR gene and the nopaline synthase terminator. Replacement ofthe GUS leader in pSOG19 with the leader sequence from Maize
Chlorotic Mottle Viras check (MCMV) generated the vector pSOG35. pSOG19 and pSOG35 carry the pUC-derived gene for ampicillin resistance and have Hindlll, Sphl, Pstl and EcoRI sites available for the cloning of foreign sequences. IV. Production and Characterization of Stably Transformed Plants Transgenic plant cells are then placed in an appropriate selective medium for selection of transgenic cells which are then grown to callus. Shoots are grown from callus and plantlets generated from the shoot by growing in rooting medium. The various constracts normally will be joined to a marker for selection in plant cells. Conveniently, the marker may be resistance to a biocide (particularly an antibiotic, such as kanamycin, G418, bleomycin, hygromycin, chloramphemcol, herbicide, or the like). The particular marker used will allow for selection of transformed cells as compared to cells lacking the DNA which has been introduced. Components of DNA constracts including transcription cassettes of this invention may be prepared from sequences which are native (endogenous) or foreign (exogenous) to the host. By "foreign" it is meant that the sequence is not found in the wild-type host into which the constract is introduced. Heterologous constracts will contain at least one region which is not native to the gene from which the transcription-initiation-region is derived.
To confirm the presence ofthe transgenes in transgenic cells and plants, a variety of assays may be performed. Such assays include, for example, "molecular biological" assays well known to those of skill in the art, such as Southern and Northern blotting, in situ hybridization and nucleic acid-based amplification methods such as PCR or RT-PCR; "biochemical" assays, such as detecting the presence of a protein product, e.g., by immunological means (ELISAs and Western blots) or by enzymatic function; plant part assays, such as leaf or root assays; and also, by analyzing the phenotype ofthe whole regenerated plant, e.g., for disease or pest resistance. DNA may be isolated from cell lines or any plant parts to determine the presence ofthe preselected nucleic acid segment through the use of techniques well known to those skilled in the art. Note that intact sequences will not always be present, presumably due to rearrangement or deletion of sequences in the cell.
The presence of nucleic acid elements introduced through the methods of this invention may be determined by polymerase chain reaction (PCR). Using this technique discreet fragments of nucleic acid are amplified and detected by gel electrophoresis. This type of analysis permits one to determine whether a preselected nucleic acid segment is present in a stable transformant, but does not prove integration ofthe introduced preselected nucleic acid
segment into the host cell genome. In addition, it is not possible using PCR techniques to determine whether transfoπnants have exogenous genes introduced into different sites in the genome, i.e., whether transformants are of independent origin. It is contemplated that using PCR techniques it would be possible to clone fragments ofthe host genomic DNA adjacent to an introduced preselected DNA segment.
Positive proof of DNA integration into the host genome and the independent identities of transformants may be determined using the technique of Southern hybridization. Using this technique specific DNA sequences that were introduced into the host genome and flanking host DNA sequences can be identified. Hence the Southern hybridization pattern of a given transformant serves as an identifying characteristic of that transformant. In addition it is possible through Southern hybridization to demonstrate the presence of introduced preselected DNA segments in high molecular weight DNA, i.e., confirm that the introduced preselected DNA segment has been integrated into the host cell genome. The technique of Southern hybridization provides information that is obtained using PCR, e.g., the presence of a preselected DNA segment, but also demonstrates integration into the genome and characterizes each individual transformant.
It is contemplated that using the techniques of dot or slot blot hybridization which are modifications of Southern hybridization techniques one could obtain the same information that is derived from PCR, e.g., the presence of a preselected DNA segment. Both PCR and Southern hybridization tecliniques can be used to demonstrate transmission of a preselected DNA segment to progeny. In most instances the characteristic Southern hybridization pattern for a given transformant will segregate in progeny as one or more Mendelian genes (Spencer et al., 1992); Laursen et al., 1994) indicating stable inheritance ofthe gene. The nonchimeric nature ofthe callus and the parental transformants (Ro) was suggested by germline transmission and the identical Southern blot hybridization patterns and intensities ofthe fransforming DNA in callus, R0 plants and Rt progeny that segregated for the transformed gene.
Whereas DNA analysis techniques may be conducted using DNA isolated from any part of a plant, RNA may only be expressed in particular cells or tissue types and hence it will be necessary to prepare RNA for analysis from these tissues. PCR techniques may also be used for detection and quantitation of RNA produced from introduced preselected DNA segments. In this application of PCR it is first necessary to reverse transcribe RNA into DNA, using enzymes such as reverse transcriptase, and then through the use of conventional PCR
techniques amplify the DNA. In most instances PCR techniques, while useful, will not demonstrate integrity ofthe RNA product. Further information about the nature ofthe RNA product may be obtained by Northern blotting. This technique will demonstrate the presence of an RNA species and give information about the integrity of that RNA. The presence or absence of an RNA species can also be determined using dot or slot blot Northern hybridizations. These techniques are modifications of Northern blotting and will only demonstrate the presence or absence of an RNA species.
While Southern blotting and PCR may be used to detect the preselected DNA segment in question, they do not provide information as to whether the preselected DNA segment is being expressed. Expression may be evaluated by specifically identifying the protein products ofthe introduced preselected DNA segments or evaluating the phenotypic changes brought about by their expression.
Assays for the production and identification of specific proteins may make use of physical-chemical, stractural, functional, or other properties ofthe proteins. Unique physical- chemical or structural properties allow the proteins to be separated and identified by electrophoretic procedures, such as native or denaturing gel electrophoresis or isoelectric focusing, or by chromatographic techniques such as ion exchange or gel exclusion chromatography. The unique stractures of individual proteins offer opportunities for use of specific antibodies to detect their presence in formats such as an ELISA assay. Combinations of approaches may be employed with even greater specificity such as Western blotting in which antibodies are used to locate individual gene products that have been separated by electrophoretic techniques. Additional techniques may be employed to absolutely confirm the identity ofthe product of interest such as evaluation by amino acid sequencing following purification. Although these are among the most commonly employed, other procedures may be additionally used.
Assay procedures may also be used to identify the expression of proteins by their functionality, especially the ability of enzymes to catalyze specific chemical reactions involving specific substrates and products. These reactions may be followed by providing and quantifying the loss of substrates or the generation of products ofthe reactions by physical or chemical procedures. Examples are as varied as the enzyme to be analyzed.
Very frequently the expression of a gene product is determined by evaluating the phenotypic results of its expression. These assays also may take many forms including but not limited to analyzing changes in the chemical composition, moφhology, or physiological
properties ofthe plant. Moφhological changes may include greater stature or thicker stalks. Most often changes in response of plants or plant parts to imposed treatments are evaluated under carefully controlled conditions teπned bioassays. V. Uses of Transgenic Plants Once an expression cassette ofthe invention has been transformed into a particular plant species, it may be propagated in that species or moved into other varieties ofthe same species, particularly including commercial varieties, using traditional breeding techniques. Particularly prefened plants ofthe invention include the agronomically important crops listed above. The genetic properties engineered into the transgenic seeds and plants described above are passed on by sexual reproduction and can thus be maintained and propagated in progeny plants. The present invention also relates to a transgenic plant cell, tissue, organ, seed or plant part obtained from the transgenic plant. Also included within the invention are transgenic descendants ofthe plant as well as transgenic plant cells, tissues, organs, seeds and plant parts obtained from the descendants. Preferably, the expression cassette in the transgenic plant is sexually transmitted. In one prefened embodiment, the coding sequence is sexually transmitted through a complete normal sexual cycle ofthe R0 plant to the Rl generation. Additionally prefened, the expression cassette is expressed in the cells, tissues, seeds or plant of a transgenic plant in an amount that is different than the amount in the cells, tissues, seeds or plant of a plant which only differs in that the expression cassette is absent.
The transgenic plants produced herein are thus expected to be useful for a variety of commercial and research puφoses. Transgenic plants can be created for use in traditional agriculture to possess traits beneficial to the grower (e.g., agronomic traits such as resistance to water deficit, pest resistance, herbicide resistance or increased yield), beneficial to the consumer ofthe grain harvested from the plant (e.g., improved nutritive content in human food or animal feed; increased vitamin, amino acid, and antioxidant content; the production of antibodies (passive immunization) and nutriceuticals), or beneficial to the food processor (e.g., improved processing traits). In such uses, the plants are generally grown for the use of their grain in human or animal foods. Additionally, the use of root-specific promoters in transgenic plants can provide beneficial traits that are localized in the consumable (by animals and humans) roots of plants such as canots, parsnips, and beets. However, other parts ofthe plants, including stalks, husks, vegetative parts, and the like, may also have utility, including use as part of animal silage or for ornamental puφoses. Often, chemical constituents (e.g., oils
or starches) of maize and other crops are extracted for foods or industrial use and transgenic plants may be created which have enhanced or modified levels of such components.
Transgenic plants may also find use in the commercial manufacture of proteins or other molecules, where the molecule of interest is extracted or purified from plant parts, seeds, and the like. Cells or tissue from the plants may also be cultured, grown in vitro, or fermented to manufacture such molecules.
The transgenic plants may also be used in commercial breeding programs, or may be crossed or bred to plants of related crop species. Improvements encoded by the expression cassette may be transfened, e.g., from maize cells to cells of other species, e.g., by protoplast fusion.
The transgenic plants may have many uses in research or breeding, including creation of new mutant plants through insertional mutagenesis, in order to identify beneficial mutants that might later be created by traditional mutation and selection. An example would be the introduction of a recombinant DNA sequence encoding a transposable element that may be used for generating genetic variation. The methods ofthe invention may also be used to create plants having unique "signature sequences" or other marker sequences which can be used to identify proprietary lines or varieties.
Thus, the transgenic plants and seeds according to the invention can be used in plant breeding which aims at the development of plants with improved properties conferred by the expression cassette, such as tolerance of drought, disease, or other stresses. The various breeding steps are characterized by well-defined human intervention such as selecting the lines to be crossed, directing pollination ofthe parental lines, or selecting appropriate descendant plants. Depending on the desired properties different breeding measures are taken. The relevant techniques are well known in the art and include but are not limited to hybridization, inbreeding, backcross breeding, ultilane breeding, variety blend, interspecific hybridization, aneuploid techniques, etc. Hybridization techniques also include the sterilization of plants to yield male or female sterile plants by mechanical, chemical or biochemical means. Cross pollination of a male sterile plant with pollen of a different line assures that the genome ofthe male sterile but female fertile plant will uniformly obtain properties of both parental lines. Thus, the transgenic seeds and plants according to the invention can be used for the breeding of improved plant lines which for example increase the effectiveness of conventional methods such as herbicide or pesticide treatment or allow to dispense with said methods due to their modified genetic properties. Alternatively new crops with improved stress tolerance can be
obtained which, due to their optimized genetic "equipment", yield harvested product of better quality than products which were not able to tolerate comparable adverse developmental conditions.
VI. A Computer Readable Medium The invention also provides a computer readable medium having stored thereon a data stracture containing nucleic acid sequences having at least 70% sequence identity to a nucleic acid sequence selected from those listed in SEQ ID Nos: 1-953, 2137-2661, 1954-1966, 2000- 2129, 2662-4737 and 4738-6813, as well as complementary, ortholog, and variant sequences thereof. Storage and use of nucleic acid sequences on a computer readable medium is well known in the art. (See for example U.S. Patent Nos. 6,023,659; 5,867,402; 5,795,716)
Examples of such medium include, but are not limited to, magnetic tape, optical disk, CD- ROM, random access memory, volatile memory, non- volatile memory and bubble memory. Accordingly, the nucleic acid sequences contained on the computer readable medium may be compared through use of a module that receives the sequence infonnation and compares it to other sequence information. Examples of other sequences to which the nucleic acid sequences ofthe invention may be compared include those maintained by the National Center for Biotechnology Information (NCBI)(http://www.ncbi.nlm.nih.gov/) and the Swiss Protein Data Bank. A computer is an example of such a module that can read and compare nucleic acid sequence information. Accordingly, the invention also provides the method of comparing a nucleic acid sequence ofthe invention to another sequence. For example, a sequence ofthe invention may be submitted to the NCBI for a Blast search as described herein where the sequence is compared to sequence information contained within the NCBI database and a comparison is returned. The invention also provides nucleic acid sequence information in a computer readable medium that allows the encoded polypeptide to be optimized for a desired property. Examples of such properties include, but are not limited to, increased or decreased: thermal stability, chemical stability, hydrophylicity, hydrophobicity, and the like. Methods for the use of computers to model polypeptides and polynucleotides having altered activities are well known in the art and have been reviewed. (Lesyng et al., 1993; Surles et al., 1994; Koehl et al., 1996; Rossi et al., 2001). The invention will be further described by the following non-limiting examples.
Example 1
GeneChip Standard Protocol Quantitation of total RNA
Total RNA from plant tissue is extracted and quantified. 1. Quantify total RNA using GeneQuant
1OD260=40 mg RNA/ml; A260/A280=1.9 to about 2.1 2. Run gel to check the integrity and purity ofthe extracted RNA
Synthesis of double-stranded cDNA Gibco/BRL Superscript Choice System for cDNA Synthesis (Cat# 1 B090-019) was employed to prepare cDNAs. T7-(dT)24 oligonucleotides were prepared and purified by HPLC. (5'-GGCCAGTGAATTGTAATACGACTCACTATAGGGAGGCGG-(dT)24-3' SEQ ID NO:2136).
Step 1. Primer hybridization: Incubate at 70 °C for 10 minutes
Quick spin and put on ice briefly Step 2. Temperature adjustment:
Incubate at 42°C for 2 minutes Step 3. First strand synthesis: DEPC-water- 1 μl
RNA (10 μg final)-10 μl T7=(dT)24 Primer (100 pmol fmal)-l μl pmol 5X 1st strand cDNA buffer-4 μl 0.1M DTT (10 mM final)- 2 μl 10 mM dNTP mix (500 μM final) -1 μl
Superscript II RT 200 U/μl- 1 μl Total of 20 μl Mix well
Incubate at 42°C for 1 hour Step 4. Second strand synthesis:
Place reactions on ice, quick spin
DEPC-water- 91 μl
5X 2nd strand cDNA buffer- 30 μl
mM dNTP mix (250 mM final) - 3 μl E. coli DNA ligase (10 U/μl)-l μl E. coli DNA polymerase 1-10 U/μl- 4 μl RnaseH 2U/μl -l μl T4 DNA polymerase 5 U/μl-2 μl
0.5 M ΕDTA (0.5 M final)-10 μl Total 162 μl
Mix/spin down/incubate 16°C for 2 hours Step 5. Completing the reaction: Incubate at 16°C for 5 minutes
Purification of double stranded cDNA
1. Centrifuge PLG (Phase Lock Gel, Eppendorf 5 Prime, Inc., PI-188233) at 14,000X, transfer 162 μl of cDNA to PLG 2. Add 162 μl of Phenol:Chloroform:Isoamyl alcohol (pH 8.0), centrifuge 2 minutes
3. Transfer the supernatant to a fresh 1.5 ml tube, add Glycogen (5 mg/ml) 2
0.5 M NH4OAC (0.75xVol) 120
ETOH (2.5xVol, -20 C) 400 4. Mix well and centrifuge at 14,000X for 20 minutes
5. Remove supernatant, add 0.5 ml 80% EtOH (-20°C)
6. Centrifuge for 5 minutes, air dry or by speed vac for 5-10 minutes 7. Add 44 μl DEPC H2O
Analyze of quantity and size distribution of cDNA Run a gel using 1 μl ofthe double -stranded synthesis product
Synthesis of biotinylated cRNA
(use Enzo BioArray High Yield RNA Transcript Labeling Kit Cat#900182)
Purified cDNA 22 μl 1 OX Hy buffer 4 μl
1 OX biotin ribonucleotides 4 μl
1 OX DTT 4 μl
1 OX Rnase inhibitor mix 4 μl
20X T7 RNA polymerase 2_μl
Total 40 μl
Centrifuge 5 seconds, and incubate for 4 hours at 37EC Gently mix every 30-45 minutes
Purification and quantification of cRNA
(use Qiagen Rneasy Mini kit Cat# 74103)
Determine concentration and dilute to 1 μg/μl concentration
Fragmentation of cRNA cRNA (l μg/μl) 15 μl
5X Fragmentation Buffer* 6 μl
DEPC H2O 9_μi
30 μl
*5x Fragmentation Buffer
MgOAc 0.64 g
KOAC 0.98 g DEPC H2O
Total 20 ml
Filter Sterilize
Array wash and staining Stringent Wash Buffer* *
Non-Stringent Wash Buffer*** SAPE Stain**** Antibody Stain*****
Wash on fluidics station using the appropriate antibody amplification protocol
**Stringent Buffer: 12X MES 83.3 ml, 5 MNaCl 5.2 ml, 10% Tween 1.0 ml, H2O 910 ml, Filter Sterilize
***Non-Stringent Buffer: 20X SSPE 300 ml, 10% Tween 1.0 ml, H2O 698 ml, Filter
Sterilize, Antifoam 1.0. ****SAPE stain: 2X Stain Buffer 600 μl, BSA 48 μl, SAPE 12μl, H 2O 540 μl. *****Antibody Stain: 2X Stain Buffer 300 μl, H2O 266.4 μl, BSA 24 ul, Goat IgG 6 μl, Biotinylated Ab 3.6 μl
Image analysis and data mining 1. Two text files are included in the analysis: a. One with Absolute analysis: giving the status of each gene, either absent or present in the samples b. The other with Comparison analysis: comparing gene expression levels between two samples
Example 2
Analysis ofthe RPS2 Mediated Interaction in Arabidopsis The identification and cloning of resistance genes is extremely important for the treatment of crops. For example, bacterial blight disease caused by Xanthomonas spp. infects virtually all crop plants and leads to extensive crop losses worldwide. Therefore, it is of mterest to identify diverse and abundant plant resistance genes for use as future crop treatments for pathogen resistance, e.g., to identify particular pathogen resistance (R) genes in a plant.
Differential gene expression analysis was used to identify pathogen resistance (R) genes in a plant. This method takes advantage ofthe HR-associated disease resistance. One model plant-pathogen interaction is that of Arabidopsis thaliana and Pseudomonas syringae pv tomato. There are four possible genetic interactions of a P. syringae infection of Arabidopsis when analyzing HR-associated disease resistance (Table 2). However, there are only two possible outcomes: a compatible outcome occurs when there is disease, and an incompatible outcome occurs when there is no disease. An incompatible outcome, or disease resistance, occurs only when the plant possesses the resistance gene, e.g., RPS2, and the pathogen posesses the conesponding avr gene, e.g., avrRpt2. RPS2 belongs to the NBS-LRR class of R genes, which can confer resistance to a wide variety of phytopathogens. It has been suggested that AvrRpt2 is delivered to the plant via the bacteria's type III secretion system and recognized by a surveillance system involving RPS2 inside the plant cell. The plant response during an incompatible interaction includes a change in ion flux across the plasma membrane,
generation of reactive oxygen species, induction of defense genes, induction of HR, fortification ofthe cell wall, accumulation of salicylic acid, and anti-microbial compounds.
Methods
Differential Expression
Analysis of differential gene expression is a classic and very powerful tool in experimental biology not only to study large trends in gene regulation but also small differences among similar responses. Historically, methods for analysis only allowed the comparison of a very few genes in each experiment. However, with new methods to identify and quantitate differential mRNA profiles, such as long distance differential display PCR, cDNA microarrays, and gene chips, one can much more quickly and comprehensively identify and analyze differentially expressed genes. By analyzing and comparing the expression profile of genes in the above 4-way matrix, a number of types of genes can be identified that are involved in the resistance pathway. Resistance genes would be highly expressed or strongly downregulated in outcome number 2 in the four way matrix and less oppositely expressed in outcome numbers 1, 3, and 4. Genes that are highly expressed or strongly downregulated in outcome numbers 1 and 2 and oppositely expressed or not expressed above baseline in outcome numbers 3 and 4 are of interest as being associated with the reaction of a plant having resistance genes to a bacterial infection, regardless ofthe avr genot pe ofthe bacterium. Such a comparison is very useful in identifying strong candidates for different roles in plant/pathogen interactions, as are numerous other kinds of outcomes in the four- way plant/pathogen interaction analysis of gene expression. Such genes include those involved in recognition of pathogen (unrelated to virulence status); genes involved in recognition of pathogen having a virulence or avirulence gene (regardless ofthe status ofthe conesponding plant); genes related to the status ofthe
plant, regardless ofthe status ofthe pathogen; and genes that do not change expression during plant-pathogen interaction.
Use of a Gene Chip to Study Gene Regulation n. Arabidopsis in Response to Exposure to Pathogen Initially isogenic strains of Arabidopsis thaliana ecotype Col-0 were used, one having the wild type RPS2 gene that confers resistance, and one having the rps2 mutant that confers susceptibility to attack by Pseudomonas syringae pathovar tomato (Pst). Subsequently, comparisons between ecotypes, mutant Arabidopsis, and infection with different pathogens were made. After infection, the RNA was isolated and a probe produced using the Affymetrix GeneChip™ protocol. A gene anay representing approximately 8,100 Arabidopsis thaliana genes was used to carry out global gene expression profiling in response to exposure to a particular pathogen.
Initially, the analysis involved comparing all four ofthe interactions to a water control
(plants "infected" with water). In the initial analysis, the mRNA levels of approximately 1,600 genes were significantly affected (> 2.5-fold change in expression) by exposure to the bacterial pathogen. This suggested a dramatic change in the molecular biology ofthe cell and a more detailed analysis was performed.
Results
A. Comparison Of Compatible To Incompatible Infections Two different types of interactions between Arabidopsis and Pseudomonas syringae were analyzed. In one type of experiment, a gene for gene interaction conditioned by the plant resistance (R) gene RPS2 and the bacterial avirulence gene avrRpt2 at a relatively early stage was analyzed. When the pathogen has an avr gene and the plant has the corresponding R gene, the plant is resistant to the pathogen and the interaction is called incompatible. When the plant-pathogen system lacks either or both genes, the plant is susceptible to the pathogen and the interaction is called compatible. A hypersensitive response (HR, localized rapid cell death ofthe plant) is one aspect of resistance.
Isogenic strains of Arabidopsis thaliana ecotype Col-0 were used, one having the wild type RPS2 gene that confers resistance, and one having the mutant rps2 mutant that confers susceptibility to attack by Pseudomonas syringae pathovar tomato (Pst) carrying avrRpt2.
Two strains of Pseudomonas syringae were used, one having the avr gene avrRpt2 and the other having no avr. The avr gene is canied on a plasmid.
A gene array having 8,700 probe sets representing approximately 8,100 Arabidopsis thaliana genes was used to carry out global gene expression profiling of each ofthe infection outcomes. The pairings were as follows:
1. RPS2 WT plant; P. syringae (no avr) 2. RPS2 WT plant; P. syringae! avrRpt2
3. φs2- 101 C mutant plant; P. syringae (no avr)
4. φs2-101C mutant plant; P. syringae/avrRpt2 Additionally, two controls were used:
5. RPS2 WT plant; water control 6. Rps2- 101 C mutant plant; water control
Data were processed such that genes having a difference in mRNA levels that was greater than 2.5-fold increased or reduced, compared with controls were selected. The fold change for each gene was log-scaled and normalized.
1. Data analysis: identification of expression clusters Data analysis was carried out by comparing expression of each gene in interactions 1-4
(Table 2), plotting that expression level, and identifying the genes of interest, i.e., those that show more than a 2.5X change in expression (about 1,600 genes). Classification of patterns, or expression clusters were as follows: a) Genes strongly induced (> 2.5X change in expression level) only in the resistant (incompatible) response; b) Genes responding weakly only in the resistance response, but strongly induced in the compatible response; c) Genes that show a high level of expression in all outcomes; d) Genes that show a high level of repression in all outcomes; e) Genes that show a very high level of repression only when the bacterial avr is expressed; and f) Genes that show a very different level of expression in the presence ofthe plant resistance compared to the level in the absence ofthe plant resistance (the mutant rps2).
Genes that fall within groups la and lb, i.e., those that are differentially expressed only when an incompatible interaction occurs, include genes directly involved in resistance to pathogens. These genes show a peak (either up or down) only during plant-pathogen interaction 2. The differential expression can be of two types: upregulated (increased expression of this gene is potentially important in the incompatible interaction) or
downregulated (decreased expression of this gene is potentially important in the incompatible interaction).
2. Heat shock proteins and transcription factors
AU major heat shock proteins (HSPs) were identified to be upregulated only during the incompatible interaction. Heat shock factors (HSFs) are transcription factors which control the transcription ofthe HSP genes. Eight HSF genes are known in Arabidopsis. HSF4 and HSF21 were identified as being upregulated when the plant was infected with P. syringae. HSF4 showed strong induction that was restricted to resistance, and HSF4 was the only HSF specifically upregulated during the incompatible interaction. The data suggests that the upregulation of HSPs is downstream of upregulation of HSF4.
To analyze whether the response was a more general one, or specific to a given ecotype, expression of HSF4 was analysed in two different Arabidopsis ecotypes, A. thaliana, ecotypes Col-0 and Ws. HSF4 was also upregulated in the response of Ws ecotype to infection and, specifically, was upregulated during an incompatible response. HSF21 is thus a prefened protein for resistance applications, and HSF4, a protein which is expressed in all plants, is especially preferred for engineering resistance.
A transgene containing the ACT2 promoter and the HSF4 open reading frame was introduced to Arabidopsis and transgenic HSF4 Arabidopsis lines generated to overexpress and underexpress HSF4. The expression of HSF4 during pathogen infection may cause lower general resistance to P. syringae.
Conditional overexpression lines were also generated using the estradiol-inducible promoter system. Infiltration of 20 :M estradiol into the intercellular space ofthe leaves of transgenic plants induced expression of HSF4 mRNA for a short time (down by 4 hours). Addition of 20 estradiol to the hydroponic medium yielded sustained HSF4 mRNA accumulation.
B. Genes Involved in Arabidopsis Responses to Pathogens
A number of mutations in Arabidopsis thaliana that disrapt expression of pathogen- induced genes and cause enhanced disease susceptibility have been identified. Pathogen- induced genes whose expression is altered in these enhanced disease susceptibility mutants are likely to play important roles in conferring disease resistance.
To identify such genes, wild type and various mutant plants were infected with strain Psm ES4326 at a dose of 10,000 colony forming units per square centimeter of leaf tissue. Control plants were mock-infected. After thirty hours, tissue samples were collected and used
to prepare RNA. Three sets of experiments were carried out. Each set of experiments included three independent replicate experiments. RNA from replicate experiments was pooled to reduce enors arising from the effects of variations in environmental conditions. Each RNA sample was used to prepare a fluorescently-labelled probe which was applied to an Affymetrix GeneChip™, allowing the expression level of each gene represented on the GeneChip™ to be determined for each sample. The plant genotypes included in each experiment were as follows:
Experiment #1 Wild-type (ecotype Columbia) nahG pad4-l eds5-l eds4 pad2-l nprl-1 nprl -3
Experiment #2 Wild-type (ecotype Columbia) coil ein2 padl
FN1-3 eds3 eds8
Experiment #3
Wild-type (ecotype Columbia) pad4-l nahG sid2 eds5-3
FN1-9 FN3-2
1. Data analysis
Expression values that were less than 5 were set to five. This ensures that no values are 0 or negative. Such values interfere with subsequent analysis steps. To obtain a list of pathogen-induced genes, the ratios of infected wild-type to mock infected wild type were calculated for each experiment. Then genes were selected in which expression levels were infected wild-type/mock wild-type > 2.5, and infected wild-type > 50 for at least 2 of 3
experiments. The ratio of 2.5 was chosen because the false positive rate for the GeneChip™ is essentially 0 at this level of stringency, and the absolute value of 50 was chosen to eliminate expression values below the detection limit ofthe GeneChip™. The result of this analysis was a list of 745 probe sets representing genes that are induced by infection in wild-type plants (note that some genes are represented by more than one probe set, so the number of different genes is somewhat fewer) (see Table 3 below). Hence, the expression of genes comprising SEQ ID NOs:2-6, 8-13, 16, 18, 22-23, 25, 28-29, 31-32, 35-37, 39-43, 45-47, 49-50, 52, 54- 55, 57-58, 60-66, 70-72, 74, 76-77, 79, 81, 83, 85, 87-90, 92, 94, 97, 100-107, 111-115, 117- 125, 127-135, 138-140, 142-153, 156-158, 160, 162-165, 168-170, 173-181, 183-184, 186- 188, 190-198, 200-201, 203-211, 214-215, 218-224, 227-232, 234-249, 251-262, 264, 266- 268, 270, 272-275, 277-281, 283, 286-294, 297-298, 302, 304-306, 308-326, 328-339, 341, 344-345, 347, 350-351, 353-358, 361-371, 373-377, 379-386, 388-390, 392, 394-400, 402- 406, 408-410, 412-417, 419-427, 429-433, 435-443, 445-452, 454-457, 459-460, 462-464, 466-470, 473-475, 478-479, 481-482, 484-187, 489-494, 496-498, 500-501, 503-506, 508, 510, 512-515, 517-523, 526, 528-529, 531-538, 540, 544-548, 550-558, 560, 563-568, 570, 572-577, 579-580, 582-585, 588-594, 596, 598-600, 602-603, 605-606, 608-612, 614-617, 619-624, 626-630, 632-639, 642, 644, 646-651, 653-657, 659-665, 667-671, 673-678, 681- 689, 691-693, 695-713, 715-717, 719, 721-727, 729-733, 736-738, 740, 742, 744, 746, 748- 752, 755-756, 758-760, 762-769, 771, 774, 776-781, 783-788, 790-796, 798-799, 802, 804- 808, 810-815, 817-831, 833-848, 850-855, 857-869, 871-880, 882-900, 903-907, 909, 911- 915, 918-920, 922-925, 927, 929, 931-938, 940, 943-945, 947, and 950-953 is increased after infection of wild-type Arabidopsis with Pseudomonas syringae.
To identify pathogen-induced genes whose expression is affected by the mutations, genes for which the ratio of infected mutant/infected wild-type was < 0.5 or > 2 for at least one mutant were selected from the list of 745 pathogen-inducible probe sets. The limits of 5 and 2 were chosen because changes of at least 2-fold are likely to be significant for impact on disease resistance, and because the false positive rate for the GeneChip™ at 2-fold is 0.2%. This selection yielded a list of 530 probe sets conesponding to genes, the expression of which is induced by Pseudomonas infection in wild-type plants and perturbed in at least one mutant plant (see Tables 4a and 4b below). Thus, the expression of genes comprising SEQ ID NOs:2, 4-6, 11-13, 18, 22-23, 28, 31, 36, 39-43, 45, 47, 49-50, 52, 54-55, 57-58, 60-61, 63-66, 71-72, 74, 77, 81, 83, 85, 87-89, 92, 97, 100-107, 111-112, 114-115, 117-120, 122, 125, 127-128, 134, 138-140, 143-144, 148-151, 153, 156-157, 160, 165, 168-170, 173-174, 176-180, 183,
187-188, 191, 193-194, 197-198, 200, 203-210, 214, 219-224, 227, 230-232, 235-23?, 239- 240, 243-246, 248-249, 251-254, 256-258, 261, 264, 266-268, 270, 272-275, 277-278, 280, 283, 286-287, 290-293, 297, 302, 305-306, 308-310, 312-316, 321-326, 328-331, 333, 336- 339, 341, 345, 351, 353, 355-358, 361-363, 365-366, 368-371, 373, 375, 377, 379-381, 384- 385, 388-390, 392, 394-400, 402-406, 410, 412, 415-416, 419-420, 422-425, 429-433, 435- 439, 441-443, 445-452, 454, 459-460, 463, 466, 468-470, 473, 481-482, 485-486, 489, 491- 494, 497-498, 500-501, 503, 505-506, 508, 510, 513-515, 517, 520-521, 523, 528-529, 531, 533-538, 540, 545-548, 550-551, 553-554, 556-558, 560, 566-567, 575, 580, 582-584, 588- 593, 596, 598-600, 602-603, 605-606, 608-610, 612, 614, 616, 620-622, 627-629, 633-634, 636-639, 644, 646, 648-651, 654-657, 659, 661-663, 667, 669, 673-674, 677, 682, 684-687, 689, 691-693, 697, 699, 701, 703-708, 713, 717, 719, 721-727, 730-733, 736, 740, 744, 746, 749-752, 755-756, 758-760, 762-764, 766-769, 774, 776-778, 780-781, 786, 788, 791-796, 799, 802, 804-808, 810-812, 815, 818-821, 823-825, 827-829, 831, 833-836, 838-843, 845, 847-848, 852-853, 855, 858, 860-869, 871-874, 876, 878-880, 884-887, 889, 892-894, 896- 900, 904-907, 911-915, 918-920, 922-924, 931, 933, 938, 943-945, 947, and 950-952 is increased after infection of wild-type Arabidopsis, and altered after infection of at least one mutant Arabidopsis, with Pseudomonas syringae. 2. Data inteφretation
Genes that encode regulatory proteins such as transcription factors, protein kinases, calcium binding proteins and the like, are likely to play important roles in disease resistance, as they are likely to affect the expression of multiple defense effector genes. The list of 530 probe sets include 81 that conespond to genes encoding regulatory factors. These are likely to be useful for engineering plants to respond more quickly to pathogen attack by activating expression of defense responses (see Table 5 below). Thus, the expression of genes comprising SEQ ID NOs:39, 52, 60, 63, 81, 83, 106, 107, 115, 117, 118, 168, 174, 176, 179, 204, 207, 208, 220, 221, 248, 258, 268, 275, 280, 309, 323, 326, 329, 351, 419, 422, 429, 430, 432, 459, 460, 468, 469, 473, 500, 505, 506, 508, 529, 531, 533, 535, 538, 545, 553, 602, 606, 608, 610, 614, 616, 634, 654, 655, 684, 686, 687, 691, 717, 751, 752, 766, 777, 815, 831, 834, 835, 839, 841, 847, 876, 884, 906, 920, and 924 is increased after infection of wild-type Arabidopsis, and altered after infection of at least one mutant Arabidopsis, with Pseudomonas syringae.
The mutations nahG, pad4-l, eds5-l, eds4, pad2-l, nprl-1, nprl-3, padl, FN1-3, eds3, eds8, sid2, eds5-3, FN1-3 and FN3-2 cause enhanced susceptibility to Pseudomonas syringae. Consequently, pathogen-inducible genes whose expression is reduced by one of these
mutations are likely to be important for resistance to Pseudomonas syringae and possibly other bacterial pathogens. These 333 probe sets are shown in Table 6 (below). Therefore, the expression of genes comprising SEQ ID NOs:12-13, 18, 23, 36, 39-40, 43, 45, 50, 52, 57-58, 60-61, 64, 71-72, 81, 87-89, 97, 100, 102-105, 107, 111-112, 115, 119-120, 122, 125, 127- 128, 140, 144, 148-150, 153, 165, 168-169, 176-177, 179, 183, 188, 191, 193-194, 197-198, 203-206, 208-209, 214, 219-222, 227, 230, 232, 237, 244-246, 248-249, 251-253, 258, 261, 264, 266, 268, 273-275, 283, 287, 290, 293, 297, 302, 305-306, 308, 312-315, 321-322, 324, 326, 330, 333, 338, 341, 345, 353, 356-358, 362-363, 366, 369, 371, 375, 377, 380, 384-385, 389, 392, 394-395, 398-399, 402-404, 406, 410, 415, 419, 422, 425, 429-430, 433, 435-439, 443, 445-452, 454, 463, 466, 468-470, 473, 486, 489, 491-492, 494, 498, 500-501, 503, 508, 513-514, 517, 529, 533-538, 548, 550, 553-554, 4556-558, 566, 575, 580, 582-583, 590-591, 593, 600, 602, 609-610, 612, 614, 620-622, 627-629, 637-638, 644, 649, 654-657, 659, 663, 667, 669, 673-674, 677, 684-685, 689, 691-693, 699, 703-705, 708, 719, 721, 724-726, 730- 732, 744, 746, 749-750, 752, 755-756, 758, 760, 762-764, 767, 769, 774, 780-781, 786, 788, 791-792, 794-796, 799, 804-808, 810-812, 815, 818-819, 823, 828-829, 833, 840-841, 843, 847, 852-853, 858, 860, 862-865, 867-868, 872-874, 876, 885-887, 889, 892-894, 896-900, 904-905, 907, 911-914, 918-920, 922-924, 931, 933, 938, 947, 950, and 952 is increased after infection of wild-type Arabidopsis, and altered after infection of at least one mutant Arabidopsis having a mutation that results in enhanced susceptibility to Pseudomonas (nahG, pad 4-1, eds 5-1, eds4, pad2-l, np41-1, npr 1-3, padl, FNl-3, eds3, eds8, sid2, eds5-3, NFl-3 and FN3-2).
The mutations coil and ein2 block jasmonate and ethylene signaling, respectively. Jasmonate and ethylene-dependent disease resistance responses are known to be important for resistance to the fungal pathogens Alternaria brassicicola and Botrytis cinerea, and may also be important for resistance to other necrotrophic fungal pathogens. Alternaria and Botrytis are distantly related, yet plant resistance to these fungi is controlled similarly, suggesting that jasmonate- and ethylene-dependent responses function to limit growth of a wide range of fungal pathogens. Consequently, pathogen-induced genes whose expression is reduced in coil and ein2 mutants are likely to be important for resistance to these necrotrophic fungal pathogens. These 296 probe sets are shown in Table 7 (see below). Hence, the expression of genes comprising SEQ ID NOs:2, 4, 6, 11-13, 18, 22-23, 31, 41-43, 49-50, 54, 57-58, 61, 64- 66, 71-72, 74, 77, 85, 87, 89, 92, 97, 101, 103, 106-107, 112, 114, 117-119, 125, 128, 134, 138, 143, 149, 151, 156-157, 165, 169-170, 174, 176-180, 187-188, 191, 193, 206, 208, 219-
220, 222, 224, 231, 236, 239, 243-245, 251-254, 256-257, 267, 272, 287, 290, 292, 297, 302, 312-313, 315-316, 321-322, 324-325, 328, 330, 345, 351, 353, 355-357, 362-363, 366, 368- 371, 373, 375, 379, 381, 384, 388-390, 392, 395-400, 405, 410, 415-416, 419, 422, 424, 431- 432, 435-436, 438-439, 447, 459-460, 470, 473, 481-482, 489, 491, 493-494, 500-501, 505- 506, 513-514, 517, 520-521, 523, 528-529, 531, 535, 537-538, 540, 545-548, 551, 553-554, 557-558, 566, 575, 580, 582, 584, 589, 591, 593, 596, 598-599, 603, 605, 608-609, 612, 628, 633-634, 636-637, 639, 646, 648, 650-651, 656, 661, 663, 667, 674, 685-687, 689, 691, 693, 697, 699, 701, 705, 707, 713, 723-724, 726, 736, 740, 749, 751-752, 756, 758-759, 764, 766- 768, 774, 776, 778, 780, 792-796, 799, 802, 806, 810-812, 818, 820-821, 825, 827-829, 833- 836, 838-839, 841-843, 848, 855, 860-861, 866, 868-869, 871, 873-874, 876, 878-880, 889, 892, 898-900, 904-905, 907, 915, 918, 922, 924, 933, 943-945, 947, and 951 is increased after infection of wild-type Arabidopsis, and altered after infection of at least one mutant Arabidopsis having a mutation in a gene whose expression is important for resistance to necrotrophic fungi (a mutation that blocks or interferes with jasmonate and ethylene signaling such as co/1 and ein2). Accordingly, these genes are useful to improve the resistance of plants to fungal infection.
The mutations nahG,pad4-l, sid2, eds5-l, eds5-3, and eds4 axe known to interfere with salicylic acid dependent signaling. Such signaling is known to be important for resistance to the bacterial pathogen Pseudomonas syringae, the oomycete pathogen Peronospora parasitica, the viral pathogen tobacco mosaic viras, as well as various other plant pathogens. Consequently, pathogen-induced genes whose expression is reduced by one ofthe mutations that block salicylate signaling are likely to be important for disease resistance, and useful for engineering improved disease resistance. These 288 probesets are shown in Table 8 (see below). Therefore, the expression of genes comprising SEQ ID NOs: 12-13, 18, 23, 36, 39-40, 43, 45, 50, 52, 57-58, 60-61, 64, 71-72, 81, 87-88, 100, 102-105, 107, 111-112, 115, 119-120, 122, 125, 127-128, 140, 148-150, 153, 168-169, 176-177, 188, 191, 193-194, 197-198, 203- 206, 209, 219-222, 227, 232, 237, 244-246, 248-249, 251-253, 258, 261, 264, 266, 268, 273- 275, 283, 287, 290, 293, 297, 302, 305-306, 308, 312-315, 324, 326, 330, 333, 341, 345, 353, 356, 358, 366, 371, 375, 377, 380, 385, 389, 392, 394, 398, 402-404, 406, 410, 415, 419, 425, 429-430, 433, 435-438, 443, 445-447, 449-452, 454, 463, 466, 468-470, 473, 486, 489, 492, 494, 498, 500-501, 503, 508, 513-514, 517, 533-538, 548, 550, 553-554, 57-558, 566, 575, 582-583, 590-591, 593, 600, 602, 609-610, 612, 620-622, 627-629, 637-638, 644, 649, 654- 657, 659, 667, 669, 673, 677, 684, 689, 692-693, 703-705, 719, 721, 724-726, 730-732, 744,
746, 749-750, 752, 755-756, 760, 762-764, 767, 769, 774, 780-781, 786, 788, 791-792, 795- 796, 805-808, 810-812, 815, 818-819, 823, 828, 833, 840-841, 843, 852-853, 858, 860, 862- 865, 867-868, 872-874, 876, 887, 889, 893-894, 896-898, 900, 905, 907, 911-914, 918-920, 922-923, 931, 933, 938, 947, 950, and 952 which is increased after infection of wild-type Arabidopsis and altered after infection of at least one mutant Arabidopsis having a mutation in a gene that interferes with salicylic acid dependent signaling (nahG,pad4-l, sid2, eds5-l, eds 5-3 and eds4). Thus, these genes are particularly useful to improve the resistance of plants to infection by more than one pathogen including bacteria, oomycetes and viruses, such as TMV.
Example 3
Further Analysis ofthe Pathogen Response and Comparison ofthe Response in Different
Ecotypes Materials and Methods Arabidopsis ecotypes (or accessions) (the wild-types of all the Arabidopsis ecotypes used here have wild-type alleles of RPS2 and RPMI). Col, Columbia-0 Ler, Landsberg erecta Ws, Wassilewskija Arabidopsis mutants and transgenics
Col φs2-101C, a loss-of-function mutant ofthe resistance gene RPS2 in
Col background. NahG, transgene for salicylic acid hydroxylase (inactivating salicylic acid). Col background. ndrl-1, null mutant allele of NDRI (non-race specific disease resistance).
The mutation strongly affects RPS2-mediated resistance and partially affects RPMI -mediated resistance. Col background. Bacterial strains
Pst, Pseudomonas syringae pv. tomato DC3000 (virulent strain of Arabidopsis)
Psm, P. syringae pv. maculicola ES4326 (another virulent strain of
Arabidopsis) Psp, P. syringae pv. phaseolicola NPS3121 (very weak pathogen of
Arabidopsis) Avirulence (ayr) genes of P. syrinsae avrRpt2: conesponding to the Arabidopsis resistance (R) gene RPS2 avrB: corresponding to the Arabidopsis resistance (R) gene RPMI Experimental Protocols
A. Gene for gene resistance (6 hours after treatment) plant treatment
Col WT H2O
Col WT Pst Col WT Pst/avrRpt2
Col φs2-101C H2O
Col φs2-101C Pst
Col φs2- 101 C Pst/avrRpt2
Ws WT H2O Ws WT Pst
Ws WT Pst/avrRpt2
B. Differences in the response to bacterial pathogens among ecotypes (3, 6, and 9 hours after treatment)
Plant treatment Col H2O
Col Pst
Col Pst/avrRpt2
Ler H2O
Ler Pst Ler Pst/avrRpt2
Ws H2O
Ws Pst
Ws Pst/avrRpt2 Note that overall results for Cvi were very similar to Ler. C. Genetic factors that affect the plant response to incompatible interactions (3, 6, and 9 hours after treatment) plant treatment
Col H2O
Col Pst
Col Pst/avrRpt2
Col Pst/avrB
Col Psm
Col Psm/avrRpt2
Col Psp (not 9 hours)
Col Psp/avrRpt2 (not 9 hours)
Col NahG Pst
Col NahG Pst/avrRpt2
Col NahG Pst/avrB
Col ndrl -1 Pst
Col ndrl-1 Pst/avrRpt2
Col ndrl-1 Pst/avrB
Results
Four hundred sixty-five genes were specifically/preferentially induced in the incompatible interaction (WT and Pst/avrRpt2), and 616 genes were specifically/preferentially repressed in the incompatible interaction. Examples of these genes are provided in Tables 10 and 13. Gene expression patterns in the incompatible interaction in Col and Ws were significantly different, indicating that the genetic diversity among ecotypes can affect gene regulation during the incompatible interaction significantly. In comparison, a relatively small number of genes (314 genes for induction, 167 genes for repression) were affected at this thne point during the compatible interactions (but not preferential to the incompatible interactions). A comparison ofthe results in three genetically different compatible interactions (WT and Pst, φs2 and Pst, φs2 and avrRpt2) revealed that 25 genes were repressed in an avrRpt2-dependent manner (see Table 9). Thus, the expression of genes comprising SEQ ID NOs:l, 15, 19, 20, 24, 26, 27, 34, 38, 51, 56, 59, 67-69, 99, 116, 155, 159, 182, 212, 284, 372, 444, and 789 is downregulated (repressed) in an avrRpt-2-dependent manner in Arabidopsis. These genes are good candidates to be involved in avrRpt2 virulence functions (in φs2 plants).
Genes that were induced in rps2 plants after infection irrespective of avrRpt2 indicate a function of RPS2 other than an interaction with avrRpt2. Thus, global gene expression profiling can identify large and minor trends in gene regulation and is useful in gene discovery.
One general phenomenon when plants are resistant to a pathogen is the early response of pathogen-responsive (induced or repressed) genes compared to plants that are susceptible to
infection. This has been proposed based on observing expression of a very limited number of genes, but it has not been proven as a global trend. To examine the results from early incompatible interactions and late compatible interactions, 4 week old Col-0 plants with well expanded leaves were infected with a high dose (OD6oo = 0.02) or low dose (OD600 = 0.002) of P. syringae and samples collected at 6 or 30 hours, respectively. The two expression patterns were similar. The conelation values between the late compatible and incompatible interaction at either 6 hours, 9 hours or the average of 3-9 hour time points was 0.71, 0.72 and 0.75, respectively.
The majority of genes that did not respond within 9 hours after infection of a virulent strain but that responded in 30 hours (Pst or Psm, for Pseudomonas syringae pv. tomato DC3000 and Pseudomonas syringae pv. maculicola ES4326, respectively; the plant is susceptible to these strains) responded within 6 hours after infection of an avirulent strain (Pst/avrRpt2; Pst carrying the avirulence gene avrRpt2; the plant is resistant to this strain). This strongly suggests that early response ofthe pathogen-responsive genes is crucial for the plant to be resistant.
A comparison ofthe differences in the expression patterns ofthe 2 primary ecotypes of Arabidopsis'' response to infection provides a further way to identify which genes have a more universal role (unchanged expression pattern) and which may be very specific to a particular plant ecotype involved in a very specific gene-for-gene interaction. For example, responses that are common between two ecotypes may be important for resistance. Genes that show the same pattern in both ecotypes may be part of more universal, or commonly-used, mechanisms involved in plant-pathogen interactions. Responses that are different may indicate that the two ecotypes use different combinations of responses to achieve resistance. This implies that a variety of genes can participate in plant-pathogen interactions. Nevertheless, ecotype-specific responses are expected to have counteφarts in other plant species.
The differences in resistance response between ecotypes can be used for improving resistance in plants. In responses that are different between ecotypes, using the methods and compounds ofthe invention, such a response can be added to (induced or repressed) the response seen in the ecotype which does not normally use that response. This will likely give the plant a more robust or a wider range of resistance.
Table shows a comparison of gene expression in 4 ecotypes, i.e., Col-0, Ws-2, Cvi and Ler in response to infection. Table 10A shows the expression data for 9 probe sets conesponding to genes that are specifically induced at 3 hours after incompatible infection of
four different ecotypes of Arabidopsis with P. syringae pv. tomato DC3000. Table 10B shows expression data for 18 probe sets conesponding to genes that are induced by 6 hours but not at 3 hours after incompatible infection of four different ecotypes of Arabidopsis with three different bacterial strains, i.e., P. syringae pv. tomato DC3000. Table IOC illustrates the expression data for 6 probe sets conesponding to genes that are activated by P. syringae at 6 hours post-infection. Most ofthe genes are compatible interaction-specific or -preferential.
Four week old plants with fully expanded leaves were infected and samples collected at 3 or 6 hours post-infection (OD600 = 0.02). Some common patterns were observed. At 3 hours after infection of an avirulent strain, Pst/avrRpt2, the overall qualitative gene expression patterns were very similar for all the ecotypes tested. Common responses to Pst/avrRpt2 could be important for gene-for-gene resistance and so may be useful to identify targets for reverse genetics. Quantitative and qualitative differences in the response were noted, indicating that there are qualitative and/or quantitative differences in the signal transduction mechanisms that regulate the response among the ecotypes. Such signal transduction mechanism differences are attributed to genetic differences among the ecotypes.
In particular, early inducible genes (3 hours) in the incompatible interaction were identified (70 genes are common in all the ecotypes, and 360 genes if selected for induced in at least one ecotype). One group ofthe early genes (38 genes in Col) were repressed to the control level by 6 hours. These genes did not respond in the compatible interaction at 3 hours and were repressed below the control level in the compatible interaction by 6 hours. This suggests that shutting down these genes in the incompatible interaction by 6 hours could be caused by defense response inhibiting factor(s) delivered by bacteria. Another group ofthe early genes were expressed even higher at 6 hours in the incompatible interaction. One hundred eighty-eight genes showed significant induction or repression at 3 hours in the compatible interaction in at least one ofthe ecotypes. Of these, 3 induced genes and 3 repressed genes were induced or repressed in all three ecotypes.
At 3 hours in the incompatible interaction, a major difference among the ecotypes was quantitative; overall expression patterns were very similar, but overall fold change amplitudes were clearly in the order of Ws>Col>Ler. Thus, in this type of analysis it is not appropriate to analyze datasets by comparing the genes from different datasets that are selected by a certain cut-off value (e.g., 2.5-fold difference). This fold change difference was mainly caused by differences in the basal expression of these genes. In fact, a strong negative conelation in each gene was found between the relative basal expression level in Ws (relative to the other
ecotypes; Pearson correlation -0.78) and response in the incompatible interaction (especially at 3 hours) and a moderate positive conelation between the relative basal expression level in Ler and response in the incompatible interaction (Person conelation 0.38) (almost no conelation for the relative basal expression level in Col; Person conelation 0.10). These observations indicate that Ws has the tightest regulation of these incompatible interaction-responsive genes, and Ler has the loosest. Another interesting observation is that the relative susceptibility to a virulent strain (Pst) was in the order of Ws>Col>Ler. Although it is unknown whether these two phenomena are controlled by same gene(s), it is conceivable that leaky expression of early response genes (in Ler) confers relative resistance to a virulent strain. At 6 hours in the incompatible interaction, the gene expression pattern for Col was significantly different from the other ecotypes.
Moreover, different ecotypes may use a different but overlapping set of responses to achieve resistance against the same pathogen. Gene expression profiling can thus reveal ecotype differences. Therefore, it is possible to isolate the genes responsible for these differences in regulatory mechanisms using ecotype differences in gene expression as a phenotype, by a map-based cloning approach.
For example, a majority ofthe incompatible response-inducible genes have lower basal levels in ecotype Ws and higher basal levels in ecotype Ler. Among the numerous genes, a few genes that display large differences in the basal level in two ecotypes are chosen. The large differences in expression level constitute easy-to-score phenotypic markers. Ws and Ler are crossed to obtain F2 populations. The larger the F2 population is, the better resolution in the map position can be obtained. For each ofthe F2 plants, expression levels ofthe chosen phenotypic marker genes are measured and physical markers that distinguish these ecotype genomes are scored. The map position ofthe responsible gene is determined by analyzing the linkage between the phenotype and the physical markers. If more than a single gene is responsible for the ecotype difference and each ofthe genes has a quantitative effect on the phenotype, quantitative trait locus (QTL) analysis can be used for mapping. Instead of using F2 populations, the use of recombinant inbred lines (RILs) between the ecotypes of interest may facilitate the analysis, especially using RILs that are already mapped for recombination points. Once the gene(s) responsible for the phenotype is mapped, a combination of increasing the map resolution, sequencing the chromosomal region identified by mapping in both ecotypes, and gene transfer from one ecotype to the other leads to isolation ofthe gene.
If the phenotype of interest in gene expression depends on bacterial infection, such as expression of ecotype Col-specific inducible genes at 6 hours after infection of Pst/avrRpt2, expression ofthe conesponding phenotypic marker genes (e.g., genes that show good difference in induction between Col and Ler) can be measured at an appropriate time after bacterial infection.
Differences in gene expression patterns between two virulent strain backgrounds (Pst and Psm) are relatively small. Gene expression patterns for Pst/avrRpt2 and Pst/avrB were quite similar at 3 hours, but the difference increased at 6 hours. Psp (no avr) shows similar expression pattern to incompatible bacteria although the amplitude of fold difference was smaller in general. This suggests that Psp, which does not induce the HR in the plant, is still recognized by the plant and induce major part ofthe defense response seen during the incompatible interaction. It also suggests that plants monitor the effect ofthe defense response and that if it seems effective (bacteria do not grow like Psp), the plant does not go for a fullblown defense response. Prefened Genes
Preferred early inducible genes were selected as induced > 2.5 fold (except for 2 fold for Psp at 6 hours) in all ofthe following datasets: Pst/avrRρt2 at 3 hours in Col, Ws, and Ler; Pst/avrRpt2 at 3 hours, Psm/avrRpt2 at 3 hours, Psp at 6 hours, and Pst avrB at 3 hours, relative to the water control, as well as estradiol-inducible (avrRpml at 0, 45, and 120 minutes and avrRpt2 at 0, 45, and 120 minutes, where the fold change was relative to the appropriate resistance gene mutant carrying the same transgenes. Among these genes, the genes were ranked according to genes that are not induced by S A or BTH and not induced in late time points with Psm. Regulatory genes were given higher rankings (see Table 11). Hence, the expression of genes comprising SEQ ID NOs:17, 70, 76, 81, 84, 109, 123, 144, 160, 230, 265, 268, 269, 271, 323, 333, 385, 427, 428, 430, 457, 505, 569, 597, 602, 606, 616, 708, 730, 741, 812, 862, and 942 is induced early after infection of different Arabidopsis ecotypes with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121 (at 3 or 6 hours) or is estradiol inducible (at 45 or 120 minutes).
Prefened early repressible genes were selected as repressed > 2.5 fold (except for > 2 fold for Psp at 6 hours) in all ofthe following datasets: Pst/avrRpt2 at 3 hours, Psm/avrRpt2 at 3 hours, Psp at 6 hours, and Pst/avrB at 3hours) and Pst/avrRpt2 at 3 hours in Col (the fold change was relative to the appropriate water controls). Among them, the genes were ranked in order of expression (highest to lower levels of expression) (see Table 12). Thus, the
expression for genes comprising SEQ ID NOs:30, 73, 282, 541, 640, 679, 761, 870, 917, and 930 is repressed early after infection of Arabidopsis with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121.
Other genes are induced/repressed during incompatible interactions at 3 and/or 6 hours after inoculation of bacteria. Prefened genes in this group were selected as induced/repressed > 2.5 fold in the incompatible interaction compared to water inoculated control and 2 > fold compared to the conesponding compatible interaction at 3 and/or 6 hours after inoculation with Pst/avrRpt2 and Pst/avrB, and Psm/avrRpt2 and Pst/avrRpt2, in all four ecotypes (see Tables 13a and 13b). Hence, the expression of genes comprising SEQ ID NOs:21, 44, 46, 60, 86, 91, 93, 106, 110, 119, 122, 130, 131, 161, 166, 167, 168, 171, 176, 200, 203, 213, 225, 227, 248, 261, 262, 266, 274, 285, 300, 301, 302, 320, 326, 341, 345, 348, 349, 360, 366, 378, 615, 618, 406, 409, 422, 425, 441, 443, 446, 449, 454, 461, 475, 476, 485, 500, 511, 512, 527, 533, 543, 545, 549, 550, 552, 567, 575, 590, 608, 611, 625, 643, 656, 659, 666, 668, 671, 680, 690, 704, 706, 711, 721, 728, 738, 757, 791, 807, 811, 813, 827, 857, 864, 868, 875, 881, 893, 901, 905, 908, 912, 939, 941, 951, and 952 is induced in an incompatible interaction at 3 and/or 6 hours after infection of fou Arabidopsis ecotypes with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121, while the expression of genes comprising SEQ ID NOs:7, 33, 82, 136, 141, 154, 185, 189, 199, 202, 434, 471, 483, 499, 516, 530, 578, 586, 631, 658, 694, 714, 718, 734, 770, 772, 816, and 916 is decreased in an incompatible interaction at 3 and/or 6 hours after infection of four Arabidopsis ecotypes with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121.
Garlic T-DNA insertion lines conesponding to these genes are searched by BLAST. Global expression profiling after infection with one of two different pathogens (P. syringae and Alternaria brassicicola) may be employed as a phenotyping method. Transgenic plants for overexpression, underexpression, and conditional overexpression of selected genes are also prepared.
Example 4 Promoters of Genes Responsive to Pathogen Infection
In many cases the major outcomes of plant-pathogen interactions are largely determined by how plants react in an early stage. Therefore, it is useful to isolate promoters
that rapidly react to pathogen attack for use in expressing proteins that provide tolerance or resistance to pathogen attack.
Genes were selected according to the conditions described below based on the results of a GeneChip™ analysis. These genes were particularly selected for a high level of induction in the avrRpt2-RPS2 interaction and for a very low mRNA level in the absence of pathogen attack among four Arabidopsis ecotypes tested (Col, Ws, Ler, and Cvi). The genes were also analyzed to determine if their expression was similar in other combinations of incompatible interactions (three different bacterial strain backgrounds: P. syringae pv. tomato DC3000, P. syringae pv. maculicola ES4326, and P. syringae p '. phaseolicola NP3121; three different avirulence genes: avrRpt2, avrB, and avrRpml; and direct expression of avirulence genes in plants using an estradiol-inducible system). For each gene, the 1.2-kb sequence upstream of the initiation codon is provided in SEQ ID NOs: 1047-1095. Prefened Highly Inducible Promoters
Promoters were selected that had low basal expression level (i.e., uninduced level) in all the ecotypes (Col, Ler, Ws, and Cvi) and high inducibility in Col. Five such promoters of genes represented by the probe sets in Table 14 were identified: the promoters of germin precursor-like oxalate oxidase gene, extra-large G protein gene, PR-1, EREBP5 gene, and a C2H2-type zinc finger protein gene were chosen. The promoters for the germin-precursor like oxalate oxidase gene and PR-1 gene are relatively slow response promoters (no induction 3 hours after infection), but have high induction by 6 hours. The extralarge G protein gene is an intermediate in terms of response time, but maintains high expression over time. The other two are useful as early transient response promoters (good induction by 3 hours, but shut down by 6 hours) in the incompatible interaction (wild type plant infected with Pst/avrRρt2). Promoter sequences comprising SEQ ID NOs: 1046-1095 and 1047-1055 correspond to genes comprising one of SEQ ID Nos: 17, 21, 80, 81, 109, 156, 174, 176, 221, 227, 296, 302, 303, 306, 333, 340, 360, 500, 505, 524, 575, 601, 602, 614, 628, 687, 733, 782, 811, 835, 862, 900, 905, 912, and 109, 306, 524, 600, 875, 912, 913, 941 and 942, respectively. Promoter-LUC reporter fusions are prepared and tested in a transient expression system using biolistic co- bombardment of avrRρt2 gene. Promoters Responsive to Particular Pathogens
Proteins that are useful for protecting plants from pathogen attack may have deleterious effects on plant growth if expressed constitutively. Consequently, it is desirable to have
promoter sequences that control gene expression in such a way that expression is absent or very low in the absence of pathogens, and high in the presence of pathogens.
Wild-type Arabidopsis plants (ecotype Columbia) were either mock-infected or infected with the bacterial pathogen Pseudomonas syringae pv. maculiola strain ES4326 (2 x 10 cfu per square centimeter of leaf). After 30 hours, samples were collected, and RNA was purified. This procedure was repeated three times independently, and the RNAs from conesponding samples were pooled, in order to reduce the impact of variation due to uncontrolled variables. The two pools of RNA representing mock-infected and infected plants were then used for gene expression profiling using an Arabidopsis GeneChip®. This entire procedure was repeated three times, yielding three sets of GeneChip data representing a total of nine independent experiments.
To identify promoter sequences that are likely to be useful for driving expression of transgenes in plants in response to pathogen attack, genes were selected whose expression level was less than 40 in all ofthe mock-infected samples and whose expression level was greater than 400 in all ofthe infected samples. The value of 40 was chosen arbitrarily as a low expression level and the value of 400 was chosen arbitrarily as a reasonably high expression level. Thirty-seven genes met these criteria and promoter sequences could be identified for 36 of them. Table 15 indicates the identifying probe set number for these 36 genes, the conesponding Arabidopsis gene, the mean expression level of each gene in mock-infected plants, the mean expression level of each gene in infected plants, and the fold induction in expression of each gene after infection. For 11 genes, expression in mock-infected plants was undetectable, so it was not possible to calculate fold induction. Therefore, the expression of genes comprising SEQ ID NOs: 104-106, 119, 123, 129, 131, 151-152, 183, 191, 198, 200, 227, 249, 274, 302, 358, 415, 481, 547, 566, 582, 628, 633, 639, 656, 673, 793, 818, 827, 864, 874, 880, and 904-905 is induced in Pseudomonas syringae pv. αew/.o/α-infected Arabidopsis.
It is possible that promoters that strongly activate gene expression in response to infection by a bacterial pathogen might be different from promoters that strongly activate gene expression in response to infection by a fungal pathogen. To test this possibility, a second GeneChip® experiment was conducted, in which wild-type Arabidopsis plants (ecotype
Columbia) were mock-infected or infected with the fungus Botrytis cinerea. Samples were collected at 0, 12, 36, 60, and 84 hours after infection, RNA was purified and used for expression profiling using an Arabidopsis GeneChip®. To identify useful promoters, genes
were selected whose expression level was less than 40 in mock-infected samples from all time points and whose expression level was greater than 400 in infected plants at 84 hours after infection. Twenty-three genes met these criteria, and promoter sequences could be identified for 21 of them. These genes are described in Table 16, with their identifying probe set number, the conesponding Arabidopsis gene, the mean expression level of each gene in mock- infected plants, and the expression level of each gene in infected plants at various times after infection. Among these 23 genes, 11 genes were previously identified in the search for genes whose expression was strongly induced by Pseudomonas syringae infection. These 11 genes conespond to identifying codes 12989, 13015, 13100, 13215, 13565, 14609, 16649, 16914, 19284, 19991, and 20356. Hence, the expression of genes comprising SEQ ID NOs:18, 71, 119, 123, 129, 151, 191, 244, 245, 302, 545, 547, 562, 566, 637, 653, 747, 756, 774, 793, 842, 864, and 905 is induced in Botrytis cinerea-mfected Arabidopsis.
The promoter sequences for the 25 genes that were only identified in the P. syringae data set are shown in SEQ ID NOs:1001-1025. The promoter sequences for the 10 genes that were only identified in the P. cinerea data set are listed in SEQ ID NOs: 1026-1035) The promoter sequences ofthe 11 genes that were identified in both data sets are listed in SEQ ID NOs: 1036-1046. The 11 promoter sequences that were identified in both data sets are most likely to be useful for driving expression of transgenes in response to attacks by various pathogens, as these promoters are activated in response to attack by either Pseudomonas syringae or Botrytis cinerea, two very different pathogens. The other promoters may also be useful for driving expression of transgenes that are efficiently expressed in response to infection by certain types of pathogens.
Further, orthologs of the Arabidopsis promoters are also useful to drive expression of transgenes. To identify the orthologous promoter, a BLAST search for orthologous genes was conducted. To identify the ortholog, the alignments from the BLAST search are used to determine the range of nucleotides showing homology to the Arabidopsis gene. The coding sequences shown at the beginning of each search result that contain regions conesponding to the nucleotides showing homology are likely orthologous genes. Orthologous promoter sequences may be isolated by any method known to the art, e.g., cloning of genomic DNA 5' to the ATG in orthologous genes identified in a computer assisted database search or hybridization of a probe comprising any one of SEQ ID NOs: 1001-1046 to genomic plant DNA.
Example 6
Genes the Expression of Which Are Altered by Viral Infection To identify host genes that are commonly up or down regulated during local RNA or DNA viras infection, gene expression profiling was employed. The host genes may include host factors that are induced by viral infection, e.g., activated host defense genes, suppressed by viral infection, e.g., suppressed host defense genes, genes involved in symptom development, as well as genes regulated by virus inducible promoters. Once the genes are identified, the function of each is then determined. Reverse genetics is then employed to examine the effect of mutations on these genes during virus infection. Experimental Procedure
Arabidopsis thaliana (Colurnbia-0 (Col-0) were grown in a Conviron growth chamber to 4 weeks of age. The growth conditions were 22°C, 12 hour day length and 75% relative humidity. At least four rosette leaves of twenty plants were inoculated with one of five viruses or a mock control (120 plants total). The viruses were turnip vein clearing viras (TVCV), a tobamoviras, an oil seed rape mosaic virus (ORMV), a tobamoviras, tobacco rattle tobraviras (TRV), a tobraviras, cucumber mosaic viras strain Y (CMV-Y), a cucumoviras, and turnip mosaic viras (TuMV), a potyvirus. Each viras was diluted to approximately 0.5 to 1.0 μg/ml in 10 mM potassium phosphate buffer pH 7.2 (or 20 mM Tris-HCI pH 8.0 for the TuMV). The phosphate buffer was used as the mock infection control for the experiments. Inoculated Col-0 leaves were first dusted with carborundum then 10 μl of virus solution or phosphate buffer were pipetted onto the leaf surface. The viras solution or phosphate buffer alone were then rubbed into the leaf surface using a gloved finger, and the leaf surfaces were washed with distilled water at about 10 minutes post inoculation.
Inoculated leaf tissue was removed from each plant at 1, 2, 4 and 5 days post inoculation (dpi), weighed, snap frozen in liquid nitrogen and stored at -80°C. Total RNA was extracted from leaf tissue by the RNAwiz method (Ambion, Inc.) and further purified using the RNeasy method (Qiagen, Inc.). RNA was diluted to 1 μg/ml and labeled as a probe for Affymetrix GeneChip hybridization according to Affymetrix protocol for synthesizing labeled copy RNA (cRNA) (see Example 1). Labeled cRNA for each viras or mock treatment was hybridized to an Affymetrix GeneChip containing sequences conesponding to 8775
Arabidopsis genes. The hybridization data was then analyzed using Affymetrix GeneChip software.
Arabidopsis genes that were induced by at least 2-fold in all virus treatments were identified by importing the data into Microsoft Excel and then subjecting the data to selection criteria. Within each time point, the expression level of a gene exceeded 25 and the fold change was greater than 2 by comparison with the mock-infected treatment. Thus, for genes that were induced by all five viruses, the expression level exceeded 25 and the fold change was greater than 2 for all five virases. For genes that were repressed by at least 2-fold, the expression level ofthe gene must exceed 25 in the mock-infected treatment and the fold change must be less than 2 in all ofthe five viras treatments. Results A gene chip from Affymetrix having oligonucleotides conesponding to approximately
8,100 Arabidopsis genes was used with labeled cRNA obtained from plant cells infected with a selected viruses at different days post-infection (dpi). For example, for Arabidopsis, the RNA may be obtained from Arabidopsis infected with potyvirus, tobamoviras, tobraviras, cucumoviras or geminivirus. After hybridization, laser scanning is employed to detect expression levels and the data obtained is then analyzed. For genes that are induced in response to viral infection, genes that are expressed at levels greater than, for example, 2 fold over control, are selected. Alternatively, for genes that are suppressed in response to viral infection, genes that are expressed at levels lower than control are selected. The advantages of a gene chip in such an analysis include a global gene expression analysis, quantitative results, a highly reproducible system, and a higher sensitivity than Northern blot analyses. Moreover, a gene chip with Arabidopsis DNA has a further advantage in that the Arabidopsis genome is well characterized.
Data obtained from probe sets which conespond to genes upregulated or downregulated in response to infection by all 5 virases reveiled forty-six genes that were downregulated and 126 that were upregulated in response to viral infection (Tables 17 and 18). Once the induced and/or suppressed genes are identified, the functions ofthe genes are then characterized by standard methodology.
Therefore, the expression of genes comprising SEQ ID NOs: 14, 48, 53, 98, 217, 226, 295, 327, 343, 352, 369, 404, 407, 418, 453, 458, 465, 472, 480, 488, 495, 507, 509, 513, 514, 559, 561, 581, 604, 607, 613, 641, 652, 672, 720, 735, 739, 743, 745, 754, 773, 803, 832, 849, 948, and 949 is downregulated after viral infection, and the expression of genes comprising SEQ ID NOs:3, 51, 54, 60, 61, 66, 75, 76, 78, 88, 95, 96, 101, 106, 108, 123, 126, 128, 129, 131, 137, 145-147, 150, 158, 169, 170, 172, 173, 197, 200, 216, 219, 224, 230, 233, 237, 249,
250, 263, 274, 275, 276, 299, 307, 323, 333, 342, 346, 359, 382, 383, 387, 391, 393, 401, 411, 415, 427, 442, 455, 459, 466, 477, 481, 485, 487, 502, 511, 515, 525, 534, 539, 542, 560, 571, 577, 579, 584, 587, 595, 600, 627, 638, 645, 654, 659, 668, 681, 688, 695, 696, 706, 708, 730, 742, 753, 775, 785, 786, 792, 797, 800, 801, 809, 817, 819, 820, 823, 827, 847, 856, 875, 885, 896, 902, 910, 921, 922, 923, 925, 926, 928, 946, and 952 is upregulated after viral infection. Rice and other plant orthologs of these Arabidopsis sequences were identified as described in Example 14 below. The results are summarized in Tables 22 and 23.
A conelation ofthe SEQ ID NOs: 1-953 and the probe set designations corresponding to genes, the expression of which is altered after infection of Arabidopsis with a pathogen is shown in Table 19.
Example 7
Identification of Gene Products that are Modulated upon Infection of a Chenopodium Cell with a Virus
Ofthe many disease resistance mechanisms that can be studied, the HR (hypersensitive resistance) system of Chenopodium spp. is attractive because ofthe broad-spectrum virus resistance it confers. This is shown by the ability of members ofthe bromo-, como-, cucumo-, ilar-, alfamo-, nepo-, sobemo-, tombus-, tymo-, carla-, clostero-, hordei-, potex-, poty-, tobra- and tobamoviras groups to elicit local lesion HR on Chenopodium spp. (CMI/AAB Description of Plant Virases, 1984; Cooper et al., (1995)). In many instances, the HR completely blocks viral spread. However, certain virases can break through the hypersensitive response and move from one species of Chenopodium to another. The ability of some viruses to infect more than one species of Chenopodium provides an opportunity to isolate genes that provide a cell with resistance to viral infection.
The genetic mechanisms of Chenopodium spp. HR involve a number of factors. These factors can be studied to further understand the hypersensitive response and the mechanism through which the response acts. There are some similarities between the products of Chenopodium spp. genes and gene products involved in common defense signaling pathways in other plants. These similarities allow comparisons to be made between Chenopodium and these other plants. One example includes genes that are induced upon viral infection during HR in C.foetidum (Visedo et al., (1990).
Additionally, some circumstantial experimental evidence suggests that Chenopodium HR may be somewhat similar to tobacco N gene HR (Whitham et al., 1994). Movement defective tobacco mosaic tobamoviras (TMV) replicates within an inoculated cell of a tobacco plant with an N gene, but fails to move from cell to cell (Cooper et al., 1996). Hypersensitivity is not induced, thus replication alone is not sufficient to induce HR despite the N gene elicitor being mapped to the replicase gene of TMV (Padgett and Beachy, 1993). Therefore, the process of virus movement may trigger hypersensitivity, which implicates intercellular signaling in this type of HR. Support for this position comes from experiments in which cell- to-cell contacts were disrupted in N gene tobacco which resulted in the prevention of necrotic lesion fonnation in infected leaves (Gulyas and Farkas, 1978). Likewise, TMV will not induce HR cell death in ΝΝ tobacco protoplasts where plasmodesmata are not intact (Otsuki et al., 1972), although HR does occur in callus cultures where plasmodesmata are intact (Beachy and Murakishi, 1971). By comparison in C. quinoa, movement defective brome mosaic bromovirus (BMV) replicates but fails to move from cell to cell. Initial infection is not sufficient to induce HR since local lesions do not form (Schmitz and Rao, 1996). Similarly, in C. amaranticolor, cucumber mosaic cucumoviras (CMV) lacking a movement protein replicates within inoculated cells, fails to move and does not elicit cell death (Canto and Palukaitis, 1999). Therefore, like TMV on N gene tobacco, the process of viral spread of BMV and CMV in C. quinoa and C. amaranticolor may induce HR. Methods and Materials Inoculation of Plants
Leaves of 10-week old C. amaranticolor or C. quinoa were inoculated with in vitro transcripts of TMV-MGfus (Heinlein et al, 1995), TMV virions, tobacco rattle tobraviras (TRV), or they were mock-inoculated. TMV-MGfus encodes GFP (green fluorescent protein) fused to the viral movement protein. Infectious spread can be monitored through the detection of GFP. Using an Olympus stereomicroscope fitted with a U-ULH Olympus lamp, infected C. amaranticolor tissue accumulating GFP was excised at 4, 7 and 11 days after inoculation (dai). Leaves inoculated with TRV or TMV were collected at 4 dai, at which point local lesions were forming. Mock-inoculated tissue was collected at the same time. Tissue was frozen in liquid nitrogen and total RΝA was purified from it. Three separate sets of plants were inoculated with TMV-MGfus and yielded three independent preparations of RΝA. cDΝA-AFLP (complementary DΝA-amplified fragment length polymoφhism)
Poly-A+ RNA was isolated from TMV-MGfus infected C. amaranticolor using Qiagen' s Oligotex mRNA purification system (Qiagen, Valencia, CA) and cDNA was generated using cDNA synthesis reagents from Life Technologies (RockviUe, MD). cDNA was used to generate AFLP fragments with the AFLP reagents from Life Technologies and reactions were performed according to the manufacturer's instructions. cDNA made from one microgram of poly- A+ RNA was digested with EcoRI and Msel and the supplied compatible linkers were ligated to the ends ofthe digested molecules. A few modifications were introduced. EcoRI-NN primers (GACTGCGTACCAATTCNN; SEQ ID NO:2134), rather than EcoRI-NNN, were used with the 5' fluorescent label NED (Applied Biosystems, Foster City, CA) and Msel-N and Msel-NN [GATGAGTCCTGAGTAAN(N); SEQ ID NO:2135), rather than Msel-NNN, primers were used (Genosys, The Woodlands, TX), to reduce the complexity ofthe primer sets evaluated. AU possible primer combinations (256 + 64) were used for PCR amplification and products were separated on polyacrylamide gels and visualized using a Genomyx SC fluorescent scanner (Beckman Coulter, Fullerton, CA). Gene fragments that appeared to be upregulated in infected tissues compared to mock-inoculated tissues were tested to see if they were also upregulated by the same primers from a second preparation of cDNA from RNA from a second set of infected plants. Gene fragments that were upregulated in both RNA preparations were excised from the gel, eluted from the gel in water and reamplified by PCR using the appropriate Msel and EcoRI primers and sequenced with 377 ABI sequencers (Applied Biosystems) using dideoxysequencing methods. Quantitative RT-PCR
DNase treated total RNA (2 ng per reaction) from the third independent preparation of TMV-MGfus infected C amaranticolor, the first preparation of TRV infected C. amaranticolor, or the first preparation of TMV C. quinoa, was used with TaqMan One-Step RT-PCR reagents for quantitative analysis in an ABI 7700 (Applied Biosystems). Reactions were performed according to the manufacturer's instructions. Primers and 6-FAM 5' end- labeled probes (6-carboxyfluorescein, Applied Biosystems or Genosys) were designed from the sequences from the C. amaranticolor upregulated gene fragments using Primer Express software (Applied Biosystems) and are listed in SEQ ID Nos:954-1000 and 2130-2135. Expression levels were inteφolated from standard curves with a conelation coefficient of 0.99 or greater and the quantities were normalized to the expression level of actin in each sample.
Results
The interaction ofthe elicitor and the R gene product establishes a cascade of reactions and signaling events that is then manifested in a phenotypic HR. In essence, HR is the end result of disease activated signaling events. In order to detect the early expression of genes induced by viral infection, it was necessary to isolate infected tissue before the onset of local lesion formation. Therefore, C. amaranticolor was infected with RNA transcripts of TMV- MGfus that express GFP (green fluorescent protein) in infected cells. This allowed the spread of viral infection to be monitored over time. Infection foci comprising over 100 cells could be detected at 4 dai and foci of more than 500 cells could be detected at 7 dai. There was no visible appearances of cell death or chlorotic local lesion formation at the infection foci at 4 and 7 dai. By 11 dai, the infection foci were associated with chlorotic local lesions. Virus infected tissue was excised from leaves at each time point and RNA was purified from the tissue and used for cDNA-AFLP as previously described. cDNA-AFLP fragments were separated on polyacrylamide sequencing gels and imaged with a fluorescent scanner. Samples derived from mock-inoculated tissue at 7 dai were run next to samples derived from TMV-MGfus infected tissue at 7 dai for comparison. Ninety- eight bands having intensity in the TMV-MGfus lanes that was greater than that of analogous bands in the mock lanes were easily detected. Thirty out ofthe 98 bands were also upregulated in an independent set of experiments designed to reduce biological variation between experiments. These bands were excised from the gel, reamplified, and sequenced.
The hypothetical protein sequences derived from the reamplified fragments (SEQ ID NOs: 1954-1966) translated from all six reading frames were compared to sequences in the GenBank protein sequence database. The results ofthe BLASTX search (Altschul et al., 1997) are summarized in Table 20a. The results of a BLAST search similar to the one described for Arabidopsis ORFs in Example 6 above is summarized in Table 24. To confirm that the expression levels of DESCA genes were upregulated in infected tissue compared to mock inoculated tissue, the relative amount of DESCA and actin transcript in a third independent set of samples at 4 dai, 7dai, and 11 dai was quantitatively measured (Table 20b).
The expression level of DESCA1 increased the most in the TMV-MGfus infected plants. The expression level of DESCA1 increased 200 times by 4 dai but tapered off drastically by 11 dai. DESCA1 is unrelated to any protein known at this time.
Two sequences, DESCA4 and DESCA10, are both related to pumps found in Arabidopsis and yeast (Sanchez-Fernandez et al., 1998; Smart and Fleming, 1996). DESCA4
is expressed highly at 4 dai but the expression drops off over time whereas DESCAl 0 is only moderately induced and its expression returns to normal by the time ofthe visible appearance of local lesions in C. amaranticolor.
DESCA7 is similar to a salicylate-induced glucosyltransferase gene in tobacco (Horvath and Chua, 1996). DESCA9 is similar to cytochrome P450-like proteins which can produce cytotoxic compounds including phytoalexins that are deployed by a plant to defend against invading microbes. DESCA12 is related to a proanthranilate benzoyltransferase from carnation that plays a direct role in the phytoalexin biosynthesis in carnation (Yang et al., 1998). DESCAl 1 is similar to the tryptophan biosynthetic enzyme phosphoribosylanthranilate transferase whose gene expression is induced in the presence of ozone in Arabidopsis (Conklin and Last, 1995).
DES C A3 is similar to endo-l,4-betaglucanases that have a role in fruit ripening, abscission, and cell elongation (Lashbrook et al., 1994). DESCA3 is highly expressed in the infected C. amaranticolor and remains highly expressed during the appearance of local lesions and necrosis.
Many disease responses are mediated by positive regulators such as transcription factors or kinases that initiate signaling cascades for the activation of defense responses. One gene, DESCA5, is loosely similar to a yeast potential transcriptional regulator. DESCA5 expression is twice as high at the early stages of infection compared to the late stages of infection illustrating an important role played by gene regulation at the early stages of infection. DESCA6 is related to kinases of Arabidopsis. Kinases have essential roles in programmed cell death during viral infection (Dunigan and Madlener, 1995). DESCA2 is the most highly expressed ofthe group suggesting that it is an important regulator at the onset of infection. It is similar to a receptor-like protein kinase in bean that responds to Fusarium solani attack (Lange et al., 1999).
Some R genes have kinase-like regions that may function in initiating a signal cascade during the onset of HR (Song et al.; 1995, Zhou et al., 1997). Global amino acid sequence alignment (Henikoff and Henikoff, 1992) of DESCA2 with Pto or Xa21, R genes with ser/thr kinase domains, reveals a 37% similarity. DESCA8 has a nucleotide binding site and a leucine-rich repeat that is common for many R genes. (Meyers et al., 1999; Leister et al., 1998).
To link DESCA genes to a multivirus resistance pathway, C. amaranticolor was inoculated with TRV (tobacco rattle viras), a viras that is taxonomically distinct from TMV.
Local lesions appeared by 4 dai and RNA was purified from the infected leaves. DESCA gene expression levels in infected tissue were compared to mock inoculated tissue by quantitative RT-PCR and revealed that the same DESCA genes upregulated during a TMV infection are also upregulated during a TRV infection (Table 20b). The gene expression levels in TMV infected C. quinoa were measured using the same
C. amaranticolor-deήved primers in quantitative PCR to deteπnine if DESCA genes were upregulated during HR in another Chenopodium species. Most ofthe DESCA genes were upregulated in C. quinoa and were expressed at levels many times higher than in C. amaranticolor (Table 3). This may be a result ofthe infection of C. quinoa with the aggressive wild-type viras rather than slower moving TMV-MGfus.
The experimental procedure presented here can detect any similar gene involved in the aforementioned signaling pathways such as SA signaling. Except for DESCAl, whose expression is increased the most at 200+ fold, many ofthe fragments have homology to other genes that have been placed in disease resistance pathways in other plants. DESCAl 2 and DESCA9 are respectively similar to hypersensitivity related gene 201, possibly a proanthranilate benzoyltranferase, and p450 monooxygenases, both which are expressed during the hypersensitive response in tobacco upon infection with Pseudomonas solanacearum but are not regulated by SA (Czernic et al., 1996). DESCA7 is similar to a salicylate-induced glucosyltransferase gene in tobacco (Horvath and Chua, 1996). Thus, the disease resistance response in C. amaranticolor involves pathways both dependent and independent of SA signaling.
The suφrising discovery of DESCA4, DESCA7, DESCA9, DESCA 10, and DESCA12, reveal the undeφinnings of an endogenous detoxification system. Briefly, the activation phase involves cytochrome P450 monooxygenases introducing functional groups (e.g. aromatic rings) to potential toxins. The conjugation phase in plants involves the linking of glutathione or glucose to the toxin at which point the conjugated molecule can be recognized by an ATP-binding cassette transporter and pumped into the vacuole, or possibly the neighboring cells, during the elimination phase. The final phase includes either storage or breakdown of such molecules. DESCA9, similar to cytochrome P450, and DESCA12, similar to a gene associated with the production of phytoalexin, may produce potential toxins. In fact, C. amaranticolor produces many such compounds that are antiviral to TMV. DESCA7, similar to a glucosyltransferase, may conjugate such toxins to be transported by the ABC- transporters encoded by DESCA4 or DESCA10. In this particular case, the transported
compound could then be deployed by the infected plant cell as an antiviral agent or cytotoxic compound, stored by noninfected cells in anticipation of infection, or eliminated by noninfected cells neighboring infected cells. Since all of these genes are induced by TMV and TRV in C. amaranticolor, their induced expressions are a result of a specific or general multivirus or disease resistance pathway.
Possessing the R genes that allow C. amaranticolor to initially recognize multiple viruses provides an opportunity to use these genes, and the regulatory elements associated with these genes, to transfer viral resistance to other plants. In addition, possession of genes that produce and transport antiviral and cytotoxic products allows for the transfer of viral resistance through a mechanism involving induced cell death upon viral infection.
Two genes that may be used for early recognition of viral infection are DESCA8 and DESCA2, as these genes may act as signaling components to initiate the resistance cascade. DESCA8 has a nucleotide binding site and a leucine-rich repeat that is common for many R genes and that can be found in other plants (Meyers et al., 1999; Leister et al., 1998). DESCA2 is induced in both Chenopodium species and is similar to other R genes, Xa21 and Pto, which have similar ser/thr kinase domains.
Resistance to viral spread may be transfened between Chenopodium spp. For example, BMV (brome mosaic virus) induces local lesions in the green variety of C. hybridum, however lesion formation does not limit the systemic spread ofthe viras (Verduin, 1978). The systematic spread ofthe BMV virus may be restricted in the green variety of C. hybridum by transformation with a gene from the puφle variety that does limit spread (Komari, 1990). Thus, genes that confer viral resistance may be used for complementation, reverse genetics, overexpression, and gene silencing. Furthermore, as indicated by the functionality ofthe R genes N and Pto after being transfened into heterologous species, (Whitham et al., 1996; Rommens et al., 1995), the Chenopodium genes may function to initiate hjφersensitivity in crops, Arabidopsis or other useful plants.
Example 8
Other Plant-Pathogen Interactions The methods set out hereinabove can be used for any type of comparable resistance interaction. For example any ofthe following plant/pathogen interactions will be produced as compatible and incompatible interactions. The RNA from such an interaction is isolated and subject to a protocol such as one outlined in Example 1, e.g., using a Genechip with a specific plant's genes or microanay, differential display PCR or cDNA-ALFP (Example 7). A four- way analysis is performed and genes which are expressed differently are identified. The plant/pathogen interactions in Table 21 are well known in the art. However, any type of plant/pathogen interaction that involves this type of resistance can be used.
Genes that are upregulated and cause resistance in a wide variety of plants are particularly useful in methods which upregulate or overexpress the gene. One method is to add an exogenous copy, thus providing more ofthe gene product or allowing for a different induction from that used by the plant. Alternatively, the endogenous gene can be upregulated using a known inducer or using artificial methods such as using an artificial induction signal in the endogenous promoter. Examples ofthe two methods are provided in Examples 9 and 10.
Accordingly, embodiments ofthe invention provide the sequences disclosed herein, which sequences can be used in genetic engineering of crops, as probes and markers to study the dynamics of plant/pathogen interactions, and as markers in marker-assisted breeding protocols to identify plants carrying particularly useful combinations of genes associated with pathogen resistance, as well as in plant defense.
Example 9
Transfonnation of Resistance Genes into Plants To produce resistant plants, resistance genes such as those identified herein can be introduced into plant cells to generate transgenic plants having enhanced resistance. While HSF4 is any prefened gene for this embodiment ofthe invention, the invention can be employed with other genes, alone or in combination, whose regulation is strongly responsive to plant/pathogen interactions, such as the genes identified herein. Since some genes are strongly induced and others are strongly repressed in plant/pathogen interactions, and since some genes that are strongly induced in one ecotype can be strongly repressed in another, the invention contemplates use of any ofthe genes and sequences, or fragments thereof, disclosed
herein, in a construct adapted to cause overexpression, repression, or knock out, ofthe genes in a transgenic plant.
Transgenic downregulation of genes associated with pathogen resistance can have several useful applications. In one embodiment, transgenic downregulation of genes that are strongly repressed in resistance interactions can enhance resistance. Such transgenic downregulation can employ the genes disclosed herein, or fragments thereof, in an antisense orientation to interfere with accumulation ofthe products of those genes. Likewise, any other methodology capable of lowering expression of such genes is also included within these embodiments ofthe invention. Plant transformation can be canied out by conventional means, and can include
^-groέαcterwm-mediated transformation, electroporation, particle acceleration, abrasion, and any other useful means leading to expression of a transgene in a plant of interest. Transfonned plant cell are then used to regenerate one or more plants in tissue culture. Subsequent generations of transgenic plants can be used directly or bred with other lines to generate plants having enhanced pathogen resistance.
Example 10
Upregulation of Resistance Genes in Crops Because many or most Arabidopsis genes have orthologs in other plants, the genes and sequences disclosed herein are generally useful in constracts to be up-regulated and cause resistance in a wide variety of plants. As examples, the heat shock proteins, and particularly HSF4, are found throughout the plant kingdom. For many such regulatory and responsive genes it is well known that there exist substances that can induce expression. Chemicals such as dexamethasone have been found to induce mammalian HSF proteins. Likewise, a chemical induction of key plant defense genes can be chemically induced. High throughput screening for chemical inducers ofthe plant HSF4 or other resistance gene is performed. Potentially useful substances are then tested on crop plants and eventually used as a soil additive or sprayed onto plants when needed to induce resistance. Accordingly, embodiments ofthe invention usefully employ the genes disclosed herein, or fragments thereof, for screening to identify useful chemical inducers and/or repressors of gene responsive to pathogenic infections.
Example 11
Identification of Inducers and Repressors of Resistance Genes The yeast two-hybrid method and many methods which use its basic idea, provide a technique to identify proteins which interact with a protein of interest. The method relies on the fact that a protein contains domains which can be separated. Thus the protein of interest is fused to the GAL4 DNA binding region of a known protein. The GAL4 (or another) activation signal is fused in a library to produce a library of fused proteins. If one ofthe proteins from the library interacts with the protein of interest the protein binds and a signal protein is produced, such as luciferase. There are a number of such systems presently, some of which can be used in mammalian cells, allowing for correct processing and folding of certain proteins and others which allow the interaction to occur in the cytoplasm allowing for the identification of other types of proteins. cDNA from HSF4 and any other protein of interest is cloned in fusion to the yeast GAL4 DNA binding domain on a vector. A library containing cDNA from Arabidopsis is fused to the GAL4 or an activation domain of choice. Expression of luciferase conelates with identification of an interacting protein. This protein is then analyzed as to its action as an inducer or repressor.
Example 12 Determination ofthe Minimal Promoter Fragment
A promoter sequence as given in SEQ ID Nos: 2137-2661 and 4738-6813. preferably a promoter sequence of one ofthe genes defined by SEQ ID NOs 536-579 such as SEQ ID Nos: 536; 537; 539-542; 548; 550-553; 555-558; 560; 565-568; 571-576, 578 and 579, or a promoter ortholog thereof is fused to the β-glucuronidase (GUS) gene at the native ATG to obtain a chimeric gene cloned into plasmid DNA. The plasmid DNA is then digested with restriction enzymes to release a fragment comprising the full-length promoter sequence and the GUS gene, which is then used to constract the binary vector. This binary vector is transformed into Agrobacterium tumefaciens, which is in turn used to transform Arabidopsis plants. The above plasmid can also be used to form a series of 5' end deletion mutants having increasingly shorter promoter fragments fused to the GUS gene at the native ATG. Various restriction enzymes are used to digest the plasmid DNA to obtain the binary vectors with different lengths of promoter fragments. In particular, a binary vector 1 is
constructed with a 1,900-bp long promoter fragment; a binary vector 2 is constructed with a 1,300-bp long promoter fragment; a binary vector 3 is constracted with a 1000-bp long promoter fragment; a binary vector 4 is constructed with a 800-bp long promoter fragment; a binary vector 5 is constructed with a 700-bp long promoter fragment; a binary vector 6 is constracted with a 600-bp long promoter fragment; a binary vector 6 is constracted with a
500-bp long promoter fragment; and a binary vector 7 is constracted with a 100-bp long promoter fragment. Like the binary vector comprising the full-length promoter fragment, these 5' end deletion mutants are also transformed into Agrobacterium tumefaciens and, in turn, Arabidopsis plants (for further details of Arbabidopsis transformation and promoter assay procedures see example 5 above) .
The presence ofthe conect hybrid constract in the transgenic lines is confirmed by PCR amplification.
By using the above protocol it can be determined, which portion ofthe promoter or the promoter orthologs thereof is required for gene expression. Minimal promoter fragments having lengths substantially less than the full-length promoter can therefore be operatively linked to coding sequences to form smaller constructs than can be formed using the full-length promoter. As noted earlier, shorter DNA fragments are often more amenable to manipulation than longer fragments. The chimeric gene constructs thus formed can then be transformed into hosts such as crop plants to enable at- will regulation of coding sequences in the hosts.
Example 13
Determination of Promoter Motifs
While a deletion analysis characterizes regions in a promoter that are required overall for its regulation, linker-scanning mutagenesis allows for the identification of short defined motifs whose mutation alters the promoter activity. Accordingly, a set of linker- scanning mutant promoters fused to the coding sequence ofthe GUS reporter gene are constructed. Each of them contains a 8-10-bp mutation located between defined positions and included in a promoter fragment as given in SEQ ID Nos: 2137-2661 and 4738-6813, preferably a promoter sequence of one ofthe genes defined by SEQ ID NOs 536-579 such as SEQ ID Nos: 536; 537; 539-542; 548; 550-553; 555-558; 560; 565-568; 571-576, 578 and 579 or the promoter orthologs thereof.
Each constract is transformed into Arabidopsis and GUS activity is assayed for 19 to 30 independent transgenic lines. The presenceof the conect hybrid consstract in transgenic lines is confirmed by PCR amplification of all lines containing the mutant constracts and by random sampling of lines containing the other constructs. Amplified fragments are digested with restriction enzyme (e.g.Xbal) and separated on high resolution agarose gels to distinguish between the different mutant constracts. constracts. The effect of each mutation on promoter activity is compared to an equivalent number of transgenic lines containing the unmutated constract. Two repetitions resulting from independent plating of seeds are canied out in every case. The sequences mutated in the linker-scanning constracts, in particular those that showed marked differences from the control constract, are then examined more closely.
Example 14
Identifying Orthologs Orthologs were identified through use of BLAST and SCAN software with some additional filters. For the Arabidopsis search, a BLAST database was created that was a subset of GenBank ver 123.0 (released April 15, 2001) that contained all ofthe plant translated regions excluding Arabidopsis thaliana sequences. The subset was created with PERL script. A BLAST search with all ofthe peptide sequences was performed against the GenBank subset. Each query was executed using the "blastall" command with the parameters" "-p blastp", "-v 50", "-b 50", "-F F". The BLAST search results were then processed with SCAN (Sequence Comparison Analysis program, version 1.0k, Los Alamos National Laboratories) using default settings and the orthologs were identified following implementation of an E-value cutoff of <=le-4. The candidate orthologs were further filtered by comparing words in the description to the text ofthe annotation fields: product, function and note. The sequence was considered to have the same or similar function if any ofthe words matched. Words excluded from the filter included: the, like, protein, related, unknown, subunit, hypothetical, and, putative, precursor, clone, homolog, small, beta, class, dna, rna, alpha, gamma, has, not, been, from, to, by, long, type and induced. For the rice search, amino acid sequences were used that resulted from FGENESH
(version l.C) gene prediction results. The peptide sequences were obtained from gene predictions and formatted into a BLAST database. A BLASTP comparison was then performed against the Arabidopsis sequences. The BLASTP results were then filtered through
use of SCAN with the following parameters: "-a 60 60" with an E-value cutoff of le-4. This produced orthologs having 60 or more identities and where 60% ofthe alignments were made up of identities.
The following pages compile Tables 3 to 24 refened to in the examples above.
Table 3 Probe Sets conesponding to genes, the expression of which is increased after infection of wild-type Arabidopsis with Pseudomonas syringae
ProbeSet Description Blast Score EC # Family
16161_s_at gb|AAA91165.1| (U39072) 8E-57 (ATU39072_S_AT) AtGRP2b [Arabidopsis thaliana]
16173_s__at dbj]BAA28539.1| (D78607) (D78607_S__AT) cytochrome P450 monooxygenase [Arabidopsis thaliana]
16203_at gb|AAD46036.1 |AC007519_2 (AC007519.53_AT) 1 (AC007519) Contains similarity to gb|M74161 inositol polyphosphate 5- phosphatase from Homo sapiens and contains a PF|00783 inositol polyphosphate phosphatase catalytic domain. [Arabidopsis thaliana]
16230_at emb|CAB41089.1| (AL049655.78__AT) (AL049655) putative protein [Arabidopsis thaliana]
16232_s_at emb|CAB45796.1| 1E-119 (AL080252.77_S__AT (AL080252) putative protein
) [Arabidopsis thaliana]
16233__at emb|CAB45846.1| (AL080254.83_AT) (AL080254) putative protein [Arabidopsis thaliana]
16236__g_at emb|CAA63346.1| (X92657) 1E-37 (X92657.3_G_AT) cationic amino acid transporter [Arabidopsis thaliana]
16272_at gb|AAD20108.1| (AC006304) (AC006304.136_AT) hypothetical protein [Arabidopsis thaliana]
16288__at gb|AAB80790.2| (AF024504) 1E-143 hydroxylase (AF024504.17_AT) similar to prolyl 4-hydroxylase alpha subunit [Arabidopsis thaliana]
16298_at emb|CAA17152.1| 1E-68 (AL021890.71_AT) (AL021890) putative protein [Arabidopsis thaliana]
16299_at emb|CAA19705.1] 1E-170 (AL024486.185_AT) (AL024486) putative protein [Arabidopsis thaliana]
ProbeSet Description Blast Score EC # Family
18587_s_at gb|AAFl 8667.1 |AC007166_9 0 (AC007166.53_S__A (AC007166) unknown protein T) [Arabidopsis thaliana]
18590_at emb|CAA10955.1| 1E-151 (AJ222713.4__AT) (AJ222713) unnamed protein product [Arabidopsis thaliana]
18591__at emb|CAA52772.1| (X74756) 2E-99 (X74756.2_AT) ATAF2 [Arabidopsis thaliana]
18596_at gb|AAD43614.1|AC005698_l (AC005698.13_AT) 3 (AC005698) T3P18.13 [Arabidopsis thaliana]
18597_at emb|CAB45881.1| (AL080282.74_AT) (AL080282) berberine bridge enzyme-like protein [Arabidopsis thaliana]
18601_s_at gb|AAFl 8602.1 |AC002387_1 8E-45
(AC002387.279_S__A (AC002387) putative
T) microtubule-associated protein [Arabidopsis thaliana]
18604_at gb|AAC19273.1| (AF069298) 3E-82 (AF069298.31_AT) similar to several small proteins (~100 aa) that are induced by heat, auxin, ethylene and wounding such as Phaseolus aureus indole-3- acetic acid induced protein ARG (SW:32292) [Arabidopsis thaliana]
18622__g__at emb|CAA06759.1| 3E-47 (AJ005902.2_G_AT) (AJ005902) vag2 [Arabidopsis thaliana]
18625__at gb|AAC72125.1| (AC005278) (AC005278.22_AT) ESTs gb|H36966, gb|R65511, gb|T42324 and gb|T20569 come from this gene. [Arabidopsis thaliana]
18631_at gb]AAB84346.1] (AC002510) (AC002510.112_AT) unknovra protein [Arabidopsis thaliana]
ProbeSet Description Blast Score EC # Family
19462_s_at emb|CAB75467.1| 0 (AF001168.2_S__AT) (AL138659) serine/threonine- specific kinase lecRKl precursor, lectin
19464_at gb|AAC67338.1| (AC005560) EC 2.7.1.37 kinase (AC005560.51_AT) putative MAP kinase [Arabidopsis thaliana]
19465_at emb|CAAl 6929.11 (AL021768.96_AT) (AL021768) resistance protein RPP5-like [Arabidopsis thaliana]
1953 l_at emb|CAA17531.1|
(AL021960.91_AT) (AL021960) amino acid transport protein AAT1 [Arabidopsis thaliana]
19546_at gb|AAC69380.1| (AC005398) transferase (AC005398.172_AT) putative endoxyloglucan glycosyltransferase [Arabidopsis thaliana]
19555_at gb|AAC13630.1| (AF058919) (AF058919.48_AT) F6N23.8 gene product [Arabidopsis thaliana]
19591_at emb|CAA09330.1| EC 3.5.-, a ido- (AJ010735.4_AT) (AJ010735) grl -protein hydrolase [Arabidopsis thaliana]
19614_at gb|AAC33210.1| (AC003970) dehydro(AC003970.32__AT) Highly similar to cinnamyl genase alcohol dehydrogenase, gi| 1143445 [Arabidopsis thaliana]
19623__at gb|AAB72175.1| (AF000657) 2E-63 (AF000657.40__AT) cytochrome C [Arabidopsis thaliana]
19624_at emb|CAB39628.1| 4E-63 (AL049481.196__AT) (AL049481) cytochrome c [Arabidopsis thaliana]
19625_s_at gb|AAC00610.1| (AC002311)
(AC002311.26__S_A Putative sulphate transporter
T) protein#protein [Arabidopsis thaliana]
19635_at emb|CAB41856.1| (AL049746.38_AT) (AL049746) ABC-type transport-like protein [Arabidopsis thaliana]
Table 4a Probe Sets in addition to those in Table 4b corresponding to genes, the expression of which is induced by Pseudomonas infection in wild-type Arabidopsis plants and perturbed in at least one mutant plant
ProbeSet Description Blast Score
13716__at emb|CAA62665.1| (X91259) lectin like 6E-70
(NOVARTIS 103_RC__AT) protein [Arabidopsis thaliana]
13751_at gb|AAF16751.1|AC010155__4 (AC010155) 2E-27 (NOVARTIS 127_AT) F3M18.8 [Arabidopsis thaliana]
13755 at emb|CAAl 6797.1] (AL021713) receptor 1E-114 (NOVARTIS 15__AT) serine/threonine kinase-like protein [Arabidopsis thaliana]
13763 at gb|AAF24849.1|AC012679_20 (AC012679) 1E-105 (NOVARTIS21_AT) putative calmodulin-binding protein [Arabidopsis thaliana]
13764_at gb|AAD39641.1|AC007591_6 (AC007591) 2E-30 (NOVARTIS22_AT) F9L1.6 [Arabidopsis thaliana]
14139_at gb|AAD09343.1| (AF026538) ABA- 7E-36 (NOVARTIS30_AT) responsive protein [Hordeum vulgare]
14148 at gb| AAF34713.1 ] AF224762_1 (AF224762) 4E-53 (NOVARTIS38__AT) SigA binding protein [Arabidopsis thaliana]
14170 at gb|AAF29406.1 |AC022354_5 (AC022354) 9E-26 (NOVARTIS51__AT) unknown protein [Arabidopsis thaliana]
14197_at No hits found. (NOVARTIS71_AT)
14214_at gb|AAF24849.1|AC012679_20 (AC012679) 2E-92 (NOVARTIS83_AT) putative calmodulin-binding protein [Arabidopsis thaliana]
14223_at (NOVARTIS9_AT) emb|CAAl 9683.1| (AL024486) putative 0 protein [Arabidopsis thaliana]
Table 4b Probe Sets as referred to in Table 3 corresponding to genes, the expression of which is induced by Pseudomonas infection in wild-type Arabidopsis plants and perturbed in at least one mutant plant
11997_at (AC005967.4_AT) 12004_at (AL022023.132_AT) 12007_at (Z99708.249_AT) 12037_at (AC004005.174__AT) 12072_at (AL035396.4_AT) 12079_s_at (A71597.1_S__AT) 12081__at (AC001645.140__AT) 12115__at (AL033545.26_AT) 12136_at (AC007591.60_AT) 12150_at (AC004005.151__AT) 12198_at (AC006954.90__AT) 12216_at (AC007119.56__AT) 12233_at (AJ001807.1_AT) 12307_at (AC002392.162__AT)
12314__at (AC001229.28_AT) 12317 _at (AC004138.27_AT)
12323_at (AC002333.18_AT) 12332. _s_at (AB023448.2_S_AT)
12341__s_at (AL021637.176_S_AT) 12349 _s_at (X84728.6_S_AT)
12369_at (AC002535.59_AT) 12400. _at (X98453.1_AT)
12438_at (AL021710.83_AT) 12449. _s_at (AC002343.179_S_AT)
12454__at (AC006232.164_AT) 12475. _at (Y11794.1__AT)
12487_at (AC004411.126__AT) 12497. _at (AC006533.51_AT)
12500__s_at (AF081067.3_S_AT) 12525. _at (AC006587.85_AT)
12530_at (Z99707.184_AT) 12535. _at (AL035538.156_AT)
12538_at (AF033205.2__AT) 12574. .at (X82624.2_AT)
12584_at (AC004521.233_AT) 12626. _at (AC006234.95_AT)
12645_at (AL021712.56_AT) 12712 _f_at (Z95774_F_AT)
12736_f_at (Z97048_F_AT) 12744. _at (AC001645.15_AT)
12760_g_at (AC005278.32_G_AT) 12764 _f_at (AC004138.69_F__AT)
12772_at (AC005278.34_AT) 12797. _s_at (AC007138.25_S_AT)
12851_s_at (ACCSYN1_S_AT) 12879 _s_at (AIG1_S_AT)
12880_s_at (AIG2_S_AT) 12883. _s_at (APX_S_AT)
12889_s_at (ASA1_S_AT) 12891. .at (ATACS6_AT)
12892_g_at (ATACS6_G__AT) 12904 _s_at (ATERF1_S_AT)
12905_s_at (ATERF2_S_AT) 12911. _s_at (ATG6PDHE5_S__AT)
12921_s_at (ATHHMGCOAR_S_AT) 12951. .at (AC005489.5_AT)
12958_at (AC002332.249_AT) 12965. _at (AL021711.118__AT)
12966_s_at (AL023094.197_S_AT) 12989. _s_at (AC004077.149_S_AT)
13003__s_at (AB021936.1_S_AT) 13014. _at (U93215.87_AT)
13040_at (AC002392.134_AT) 13070. _at (AC006919.171__AT)
13094_at (AL035523.163_AT) 13134 _s_at (AC002337.9_S_AT)
13152_s_at (AC005322.24_S_AT) 13154 _s_at (AC002333.210_S_AT)
13157_at (AC002409.35_AT) 13176. .at (AL031394.56_AT)
13177_at (AL049640.42_AT) 13190. _s_at (ATTHIREDA_S_AT)
1321 l_s_at (BCHI_S_AT) 13212. .s_at (BGL2_S_AT) 13215__s_at (cafferoylcoamethyltrans_S_AT) 13219 _s_at (CHI4_S_AT)
13243_r_at (ELI32_R__AT) 13244. _s_at (ELI32_S_AT) 13255_i_at (gammaglutamyltranspepti_I_AT) 13266 _s_at (GST4_S_AT)
13273_s__at (HSF4_S__AT) 13275. _f_at (HSP174_F_AT)
13277_i_at (HSP 176A__I_AT) 13285. _s_at (HSP83__S_AT)
13312__at (AC006223.75_AT) 13370. .at (AC005322.4_AT)
13381_at (AC006580.8_AT) 13395. .at (AL035528.202_AT)
13435__at (AF003102.3_AT) 13437 _at (AF096371.8_AT)
13467_at (AL096860.198_AT) 13536. _at (AL021636.47_AT)
13538_at (AL080254.75_AT) 13565 _at (AL035601.21_AT)
13588_at (AL021961.24__AT) 13589 _at (AC000132.24__AT)
13617_at (AC006592.64_AT) 13627. _at (AL035394.196_AT)
13645_at (AC000098.8_AT) 13656 _at (AC007138.31_AT)
13659_at (AL022347.46_AT) 13666 _s__at (INDOLE3GPS__S_AT)
13680_s__at (LOXl_S__AT) 13685. _s_at (MLOLIKE2_S_AT)
13688_s_at (MONOPTEROS_S_AT) 13697. _at (NI16_AT)
13705_s_at (AC003671X_S__AT) 13789. _at (AJ132436.2_AT)
13803_at (Z97341.376_AT) 13818 _s_at (AC006218.175_S_AT)
13842_at (AC002396.12_AT) 13848. .at (AC003981.31__AT)
13880__s__at (AL049480.183_S_AT) 13908. _s_at (A71590.1_S_AT)
13918__at (AC005388.29_AT) 13920. _at (AC005990.53_AT)
13949_s__at (Z97343.352_S_AT) 13963. _at (AL021711.26_AT)
13999_at (AF071527.56_AT) 14015 _s_at (A71588.1__S_AT)
14016__s_at (A71596.1_S__AT) 14025. .s__at (AC007293.3_S_AT)
14030_at (AC005970.225_AT) 14032 _at (AL035601.11_AT)
14041_at (AC003970.28__AT) 14052 .at (AC004122.24__AT)
14068_s__at (AC006922.197__S__AT) 14070. .at (AL049658.217__AT)
14089_at (AC006223.65_AT) 14100. _at (AF002109.108_AT)
14110_i_at (AL035528.279_I__AT) 14240. .s_at (NR1_S_AT)
14242_s_at (NRA_S_AT) 14248. .at (PAD3__AT)
14249__Lat (PAD4__I__AT) 14254 _s_at (PAL1-MRNA_S_AT)
14256_f_at (PALl-INTRON__F_AT) 14320. .at (AC005956.54__AT)
14408_at (AC002291.14_AT) 14448. _at (AC002387.243_AT)
14450_at (AC002986.49__AT) 14461. .at (AC006202.73__AT)
14468__at (AC007576.62_AT) 14475 .at (AL021811.121_AT)
14487__at (Z97341.343_AT) 14584. .at (AC007658.25_AT)
14609_at (AC002340.147_AT) 14620. _s__at (PAT1_S__AT)
14635_s_at (PR.1_S_AT) 14638 _s__at (PRXCB_S_AT)
14640_s_at (putativemloHI_S_AT) 14643. ,s_at (RAR047_S__AT)
14663__s_at (trehalaseprecusor_RC_S_AT) 14667 _s_at (TRPB_S_AT)
14672_s_at (TSA1_S_AT) 14673. _s_at (TSB2_S_AT)
14675_s__at (VSP_S__AT) 14705. _i__at (WT77_RC__I__AT)
14706_r_at (WT77_RC_R__AT) 14711. _s__at (ZFPL_S_AT)
14735_s_at (AF008124_S_AT) 14750. _s_at (AF096370.12_S_AT)
14763_at (X86958.1_AT) 14779. _at (AC004680.71__AT)
14780_at (AC004683.103_AT) 14786 .at (AC005397.115_AT)
14793_at (AC006202.10_AT) 14882. _at (AL022605.63_AT)
14900_at (AC000348.12._AT) 14923 _at (AC006283.158_AT)
14924_at (AC006283.46_AT) 14928. _at (AC006569.88_AT)
14959_at (AC007202.26_AT) 14978. _at (AC002333.49_AT)
15052_at (AC002332.103_AT) 15073. _at (AC007069.93_AT)
15088_s_at (AC002311.37_S_AT) 15091. _at (AC004683.97_AT)
15098__s_at (ATU26945_S_AT) 15116. _f_at (AF121356_F_AT)
15123_s_at (ATU40857_S_AT) 15124 _s_at (ATU59508_S__AT)
15125__f_at (D85190_F_AT) 15132 _s_at (AF121878_S_AT)
15137_s_at (ATU57320_S__AT) 15141. _s_at (D85191__S__AT)
15154_s_at (ATHMTGDAS_S_AT) 15161. _s_at (ATU90522_S_AT)
15162_s_at (U01880_S_AT) 15188. _s„at (AF081202_S_AT)
15196_s_at (ATU43412_S_AT) 15199. _s_at (AB005804_S__AT)
1521 l__s_at (ATH243813__S__AT) 15216 _s_at (ATU75191_S_AT)
15342_at (AC006593.101_AT) 15406. _at (AC006931.179_AT)
15431_at (AL030978.64_AT) 15479 _at (AL049483.205_AT)
15483__s_at (AC005819.20_S_AT) 15485 _at (AC006233.109_AT)
15496_at (AC006282.167__AT) 15518 _at (AC005322.28_AT)
15522_ at (AL078637.213_I_AT) 15523 _s_at (AL078637.213_S_AT)
15524_at (AC005508.25_AT) 15526. _at (AC004122.16_AT)
1553 lJLat (AL078637.191_I__AT) 15532 _r_at (AL078637.191_R_AT)
15540_at (AC006585.205_AT) 15543. _at (AF096371.10_AT)
15544_at (AL021633.110_AT) 15547 _at (AC005970.122_AT)
15551_at (AL035440.289_AT) 15594. _s_at (ATU56635_S_AT)
15613_s_at (ATHHOMEOA__S_AT) 15622 _s_at (ATU43945__S_AT)
15625_s_at (ATU74610_S_AT) 15632 _s_at (AB012570_S_AT)
15641_s_at (AFl 17063_S_AT) 15665. _s_at (AF022658_S__AT)
15669_s_at (AF047834_S_AT) 15670. _s_at (AF061638_S__AT)
15672__s_at (AF082299_S_AT) 15778 _at (X98676.2_AT)
15779_g_at (X98676.2__G__AT) 15792. _at (AC002341.106_AT)
15798_at (AC002521.173_AT) 15815 _s_at (Z97342.366_S_AT)
15839_at (AC005662.203_AT) 15859 _at (AC006587.164_AT)
15866_s_at (AC007133.59__S_AT) 15874 _at (AL022223.106_AT)
15886_at (AL078637.204__AT) 15919 _at (AC007060.42_AT)
15921_s_at (AC007067.1__S__AT) 15924. _at (AC007138.61_AT)
15970_s_at (X71794.2__S_AT) 15978 _at (X68592.6__AT)
15982_s_at (AC006260.78_S__AT) 16001. _at (AF035385.2__AT)
16003_s_at (AL021749.64_S_AT) 16021. _s__at (AL022224.182_S_AT)
16031_at (X94248.1_AT) 16043 _at (AC005489.17_AT)
16053_i_at (Y14251.4_I_AT) 16063 _s_at (AB008103_S_AT)
16073_f_at (AF062908_F_AT) 16083 _s_at (AF153283_S_AT)
16091_s_at (ATHHSP83_S_AT) 16103. _s_at (ATU60445_S_AT)
16105_s_at (ATU68017_S__AT) 16108. _s_at (D78604_S__AT)
16130_s_at (AF078683_S_AT) 16173. _s_at (D78607_S__AT)
16203_at (AC007519.53_AT) 16232. _s_at (AL080252.77_S_AT)
16233_at (AL080254.83_AT) 16272. _at (AC006304.136_AT)
16298_at (AL021890.71_AT) 16299. _at (AL024486.185_AT)
16301_s_at (AL031018.105_S_AT) 16306. _at (AL049751.112_AT)
16335_at (AL079347.105__AT) 16340. _at (AC004255.15_AT)
16357_at (AF149413.38_AT) 16363. _at (AC004255.14__AT)
16383_at (AC006300.64_AT) 16398. _s_at (AL022603.3_S_AT)
16405_at (AC005850.9_AT) 16409. _at (AC004393.2_AT)
16440__s_at (AF002109.137_S_AT) 16461. _ at (AC004683.79_I_AT)
16462_s__at (AC004683.79_S_AT) 16465. _at (Y08892.1_AT)
16470_s_at (AF068299.4_S_AT) 16510. _at (AL034567.198_AT)
16522_at (X77500.2__AT) 16526. _at (Z49227.1_AT)
16541_s_at (AB023423_S_AT) 16545. _s_at (AF037229_S_AT)
16553_f_at (AF078821_F_AT) 16570. _s_at (ATHCDPKA_S_AT)
16578_s_at (ATHRPRP1B_S__AT) 16589 _s__at (ATU26937_S_AT)
16594_s_at (ATU39783_S__AT) 16603. _s_at (ATU81293_S_AT)
16609_s_at (AB008104_S_AT) 16611. _s_at (AB008782_S_AT)
16638_s__at (AF139098_S_AT) 16646 _s_at (ATHDHS1_S_AT)
16649_s_at (ATHORF_S_AT) 16701. _at (AC005312.61__AT)
16721_at (AC006533.58_AT) 16747 _at (AL021713.3_AT)
1678 l_at (AC002392.100_AT) 16810. _at (AC002339.46_AT)
16859_at (AL035523.135_AT) 16864. _i_at (AF037367.4_I_AT)
16865_s__at (AF037367.4_S_AT) 16888 _s_at (AC004684.174_S_AT)
16914_s_at (AL049500.57_S_AT) 16916. _s_at (X77199.8_S_AT)
16968_at (AL021961.93__AT) 16989..at (AL030978.46_AT)
16995_at (AC002391.188_AT) 17007 .at (AC005896.26_AT)
17008_at (AC006585.212_AT) 17039. _s_at (D78602_S__AT)
17041_s_at (D89631_S_AT) 17051. _s_at (AF098947_S_AT)
17066_s_at (ATHLIPOXY_S_AT) 17073. _s_at (ATTS4391__S_AT)
17075_s_at (ATU09961_S__AT) 17104 .s_at (D88541__S_AT)
1711 l_s__at (ATHACSC_S_AT) 17119. _s_at (AF132212_S_AT)
17128_s_at (ATHRPRP1A_S_AT) 17180. _at (AF007270.30_AT)
17187_at (AF128396.2_AT) 17300_at (X66017.2_AT) 17303_s_at (AC004683.25_S_AT) 17323__at (U95973.69_AT) 17338_at (AC002535.97_AT) 17341_at (AL021713.89_AT) 17356_s_at (Z97338.190__S_.AT) 17371_at (AF076243.44_AT) 17379_at (AF085279.9_AT) 17413_s_at (AJ006961.4_S_AT) 17451_at (AC002343.47_AT) 17452__g_at (AC002343.47__G_AT) 17484_at (X79052.2__AT) 17485__s_at (Z97340.345_S_AT) 17487__s_at (U18993.2_S_AT) 17511_s_at (AF067605_S_AT) 17514_s_at (AF076277_S_AT) 17522_s_at (D78606_S_AT) 17533_s_at (ATU43488_S_AT) 17544_s_at (ATU40856_S_AT) 17548_s_at (AFl 18823_S__AT) 17585_s_at (AF134487_S__AT) 17595_s_at (AF166352_S__AT) 17648_at (AL021684.43__AT) 17653_at (AL035679.144_AT) 17702_at (AC005700.212_AT) 17719_at (AC006592.17_AT) 17752_at (AC003974.37_AT) 17758_at (AF076243.41_AT) 17775_at (AC004392.2_AT) 17781_at (AL049746.177__AT) 17840_s_at (AC002333.223_.S_AT) 17860_at (AL078467.4_AT) 17876_at (AJ007587.2__AT) 17877_g_at (AJ007587.2_G_AT) 17893_at (AC004401.135_AT) 17899_at (Z97339.197_AT) 17930_s_at (AJ006960.4_S__AT) 17945_at (Z97341.411_AT) 17963_at (AL049730.88_AT) 18012__s__at (AJ002295__S_AT) 18054_at (AJ238846_AT) 18109_s_at (AC002391.206JS_.AT) 18121_s_at (AC002337.21_S_AT) 18122_at (AC002338.110__AT) 18148_at (AC004669.25_AT) 18176_at (AL035540.31_AT) 18194 _at (AL096859.227JLAT) 18213_at (AL022140.126_AT) 18224_s_at (AL021890.57__S_AT) 18228_at (X91259.1_AT) 18236_s_at (AC004683.69__S_AT) 18242__g__at (AC006580.71_G_AT) 18255_at (AC005770.25_AT) 18258__s_at (AC006439.222_S_AT) 18263_at (AC005724.36_AT) 18266_at (AC004684.33_AT) 18267_at (AC006223.23_AT) 18299_s_at (M23872.2_S_AT) 18456_s_at (AC004697.159_S_AT) 18508_s_at (AC006532.89_S_AT) 18544_at (AC007060.14_AT) 18582_s_at (AC003671.36_S__AT) 18587_s_at (AC007166.53_S_AT) 18590_at (AJ222713.4_AT) 18591_at (X74756.2_AT) 18596_at (AC005698.13_AT) 18597_at (AL080282.74_AT) 18604_at (AF069298.31_AT) 18622__g__at (AJ005902.2_G__AT) 18625_at (AC005278.22_AT) 18631_at (AC002510.112__AT) 18636_at (AC006577.22_AT) 18668_at (AJ249794_AT) 18686__s__at (U18126_S_AT) 18698_s_at (X17528_S_AT)
18735__s_at (Z29490__S_AT) 18753_s_at (AF 118222.28_S_AT) 18782__at (AC003040.90__AT) 18803__at (AC005315.94__AT) 18885_at (AC006921.147_AT) 18888_at (AC007591.68__AT) 18899_s_at (X13434.1_S_AT) 18908_i_at (AF055848.2_I__AT) 18909_s_at (AF055848.2__S_AT) 18928_at (AC002333.181_AT) 18930_at (AC005990.57_AT) 18933_at (AC007020.48_AT) 18949_at (Z54136.1_AT) 18953_at (AF077955.1_AT) 18963_at (AC004561.99_AT) 18966_at (AC004561.106__AT) 19019_ at (X82623.2__I_AT) 19060_at (AC003671.34_AT) 19092_at (AL078606.188_AT) 19110_s_at (X86947.2__S_AT) 19137_at (X74755.2__AT) 19140_at (AC005170.24__AT) 19181__s_at (AF053065.2_S_AT) 19207_at (AC006069.117_AT) 19247_at (AF071527.44_AT) 19257__s_at (AC000104.57_S_AT) 19284_at (AC003028.196_AT) 19288_at (AC005824.130_AT) 19325_at (AL022604.42__AT) 19364_at (AL022023.142_AT) 19383_at (AC006200.203_AT) 19395_at (AF007270.32_AT) 19405__at (AJ223803.1__AT) 19407_at (AC004697.81_AT) 19409_at (AC007357.56_AT) 1941 l_at (AC007661.104__AT) 19421_at (X70990.4_AT) 19432_s_at (AL035680.11_S_AT) 19451_at (AC004392.6_AT) 19460_s_at (AC000132.66_S__AT) 19465_at (AL021768.96_AT) 19546_at (AC005398.172_AT) 19555_at (AF058919.48_AT) 19591_at (AJ010735.4_AT) 19614_at (AC003970.32_AT) 19624_at (AL049481.196_AT) 19625_s_at (AC002311.26_S_AT) 19635_at (AL049746.38__AT) 19640_at (AC004561.78_AT) 1964 l_at (AC004561.66_AT) 19645_at (AC004561.70_AT) 19655_at (Y14199.1_AT) 19667_at (AL021710.5_AT) 19672_at (AC005687.19__AT) 19673__g_at (AC005687.19_G__AT) 19700_s_at (AL031326.154_S_AT) 19704__i_at (AJ005927.2_I_AT) 19707_s_at (Z95768.3__S_AT) 19741_at (AL049171.72__AT) 19755_at (AC006593.64_AT) 19762_at (AL035527.204_AT) 19818J_at (AL021749.33__I_AT) 19844_at (AJ007588.2_AT) 19848_s__at (AC004261.94_S_AT) 19851_at (U23794.3__AT) 19881_at (AC004077.49_AT) 19892_at (AC005770.30_AT) 19895_s_at (U77347.4_S__AT) 19944_at (AC002130.4__AT) 19956_at (AC006282.11_AT) 19960_at (AL035527.360_AT) 19970__s__at (AC003674.10_S__AT) 19982_at (AC002986.28_AT) 19991__at (AC007017.124_AT) 20017_at (AC004521.66__AT) 20023_at (AC006577.46_AT)
20030_at (AL078637.51_AT) 20051_at (AC000106.38_AT) 20053__at (AC002292.27_AT) 20096__at (AC004238.31_AT) 20098__at (AC004697.123_AT) 20133 _i_at (AC007178.71_I_AT) 20134__s_at (AC007178.71__S_AT) 20144_at (AL079350.68_AT) 20179_at (AL035538.229_AT) 20189_at (AC005489.2_AT) 20194_at (AC007584.48_AT) 20223_at (AL022347.145_AT) 20238_at (X74514.2_AT) 20239_g_at (X74514.2_G_AT) 20245_s_at (AC005309.97_S__AT) 20247_at (AC004392.4_AT) 20263_at (AB004798.1_AT) 20269_at (AC002387.237_AT) 2027 l_at (Z99707.27__AT) 20287_at (Y14590.5_AT) 20288__g_at (Y14590.5_G_AT) 20291_s_at (M92353.4_S_AT) 20297_at (AC007153.27_AT) 20323_at (AC004561.62__AT) 20348_at (AC005967.35_AT) 20356_at (AC004561.74_AT) 20365__s_at (AC005850.19_S__AT) 20370_at (AC004561.263_AT) 20420_at (AL024486.131_AT) 20421_at (U81294.2__AT) 20422_g_at (U81294.2_G_AT) 20432_at (U43486.2__AT) 20433_at (AC006232.147_AT) 20450_at (AJ005930.2_AT) 2046 l_at (AL049480.157_AT) 20462_at (U82399.2_AT) 20479_ at (AF069495.2JL.AT) 20480_s_at (AF069495.2_S_AT) 20485_at (AC007660.131__AT) 20529__at (Z97341.125_AT) 20572_s_at (AC005560.229_S_AT) 20589_at (AF081066.3_AT) 20646_at (AC002291.20_AT) 20656_at (AL035396.46_AT) 20658_s__at (AL050400.70_S_AT) 20669__s_at (AC002388.6_S_AT) 20685_at (AL049751.46_AT) 20686_at (Y14424.2_AT) 20689_s_at (AC002335.19_S__AT)
Table 5 Probe Sets as referred to in Tables 4a and 4b corresponding to genes encoding regulatory proteins, the expression of which is increased after infection of wild- type Arabidopsis, and altered after infection of at least one mutant Arabidopsis, with Pseudomonas syringae
ProbeSet Regulatory factor
12497_at (AC006533.51_AT) putative receptor-like protein kinase
16409_at (AC004393.2_AT) putative receptor-like kinase
12307_at (AC002392.162_AT) putative receptor-like protein kinase
13659_at (AL022347.46_AT) serine/threonine kinase-like protein
17341_at (AL021713.89_AT) receptor serine/threonine kinase-like protein
12958_at (AC002332.249_AT) putative protein kinase
17758__at (AF076243.41_AT) putative receptor-like protein kinase
16570_s_at (ATHCDPKA_S__AT) CDPK1 calcium dependent protein kinase 1
19092_at (AL078606.188__AT) protein kinase-like protein
17371_at (AF076243.44_AT) putative receptor-like protein kinase
20223_at (AL022347.145_AT) putative ser/thr protein kinase
19655_at (Y14199.1_AT) MAP3K delta- 1 protein kinase
14030_at (AC005970.225_AT) putative protein kinase
20462_at (U82399.2_AT) putative protein kinase PK1
16781_at (AC002392.100_AT) putative receptor-like protein kinase
14763__at (X86958.1_AT) protein kinase catalytic domain (fragment)
17752_at (AC003974.37_AT) putative protein kinase
13755_at (NOVARTIS 15_AT) putative ser/thr kinase
13370_at (AC005322.4_AT) similar to Dsorl protein kinase
20144_at (AL079350.68_AT) receptor kinase-like protein
16357_at (AF149413.38_AT) putative protein kinase
18176_at (AL035540.31_AT) putative protein kinase
15798_at (AC002521.173_AT) putative receptor-like protein kinase
12965_at (AL021711.118_AT) protein kinase-like protein
16398_s_at (AL022603.3__S_AT) putative ser/thr protein kinase
18122_at (AC002338.110_AT) putative protein kinase
17323_at (U95973.69_AT) putative ser/thr protein kinase
18782_at (AC003040.90_AT) putative protein kinase
16405_at (AC005850.9__AT) putative serine/threonine protein kinase
ProbeSet Regulatory factor
141 lOJLat (AL035528.279JLAT) putative R protein. Like Hcr9-9A, Lycopersicon pimpinellifolium F18A5_290 chr.4
14214_at (NOVARTIS83_AT) putative calmodulin-binding protein (duplicate)
13763_at (NOVARTIS21_AT) putative calmodulin-binding protein (duplicate)
12438_at (AL021710.83_AT) membrane-bound small GTP-binding - like protein
19848_s_at (AC004261.94J_._AT) calmodulin-related protein
16103_s_at (ATU60445_S_AT) GRF7 general regulatory factor encoding 14-3-3 protein
14249_ at (PAD4_I_AT) PAD4 phytoalexin deficient 4
19465_at (AL021768.96__AT) RPP5-like NBS-LRR resistance protein
14640_s_at (putativemloHI_S_AT) Mlo-like (duplicate)
18456_s__at (AC004697.159_S_AT) Mlo-like (duplicate)
14320_at (AC005956.54_AT) putative RING zinc finger protein
18054_at (AJ238846_AT) SGPl monomeric G-protein
16130_s_at (AF078683_S_AT) RHAla RING-H2 finger protein
20485_at (AC007660.131_AT) putative two-component response regulator protein
15052_at (AC002332.103_AT) putative calcium-binding EF-hand protein
15632_s_at (AB012570_S_AT) ATHP3 two-compoent phosphorelay mediator with a single HPt domain
16553__f_at (AF078821_F__AT) RHAlb RING-H2 finger protein
13685_s_at (MLOLIKE2_S_AT) Mlo-like 2 (duplicate)
20365_s_at (AC005850.19_S__AT) Mlo-like 2 (duplicate)
13312_at (AC006223.75_AT) putative disease resistance protein
17180_at (AF007270.30_AT) similar to GATA-type zinc fingers
15779_g_at (X98676.2_G__AT) zinc finger protein (duplicate)
15778_at (X98676.2__AT) zinc finger protein (duplicate)
1471 l_s__at (ZFPL_S__AT) hypothetical Cys-3-His zinc finger protein
17379_at (AF085279.9_AT) putative CCCH-type zinc finger protein
12525_at (AC006587.85_AT) putative DOF zinc finger protein
16589_s_at (ATU26937_S__AT) AtMYB7 transcription factor
12712_f_at (Z95774_F_AT) AtMYB51 R2R3 myb transcription factor
13273__s_at (HSF4__S_AT) HSF4 heat shock transcription factor 4 (duplicate)
16105_s_at (ATU68017_S_AT) HSF4 heat shock transcription factor 4 (duplicate)
18803_at (AC005315.94_AT) putative SCARECROW homeobox gene regulator
ProbeSet Regulatory factor
12905_s_at (ATERF2_S_AT) EREBP2 ethylene responsive element binding factor 2 (duplicate)
16609_s_at (AB008104_S_AT) EREBP2 ethylene responsive element binding factor 2 (duplicate)
19755_at (AC006593.64_AT) ethylene reponse factor-like AP2 domain transcription factor
17514_s_at (AF076277_S_AT) ERFl ethylene response factor 1 transcription factor
18121_s_at (AC002337.21_S_AT) AtMYB2 MYB transcription factor
13688_s_at (monopteros_S_AT) transcription factor
12904_s_at (ATERF1_S_AT) EREBPl ethylene responsive element binding factor 1 (duplicate)
16063_s_at (AB008103_S_AT) EREBPl ethylene responsive element binding factor 1 (duplicate)
16638_s_at (AF139098_S_AT) putative zinc finger protein
16545_s_at (AF037229_S_AT) transcription factor
13435_at (AF003102.3_AT) RAP2.9 AP2 domain containing putative transcription factor
15665_s__at (AF022658_S_AT) putative c2h2 zinc finger transcription factor
19673_g_at (AC005687.19_G_AT) RAP2.6 AP2 domain containing putative transcription factor (duplicate)
19672_at (AC005687.19_AT) RAP2.6 AP2 domain containing putative transcription factor (duplicate)
14780_at (AC004683.103_AT) DREB-like AP2 domain transcription factor
17303_s_at (AC004683.25_S_AT) WRKY33 transcription factor
18949_at (Z54136.1_AT) MYB-related protein
19707_s_at (Z95768.3_S_AT) AtMYB44 transcription factor
16073_f_at (AF062908_F_AT) putative transcription factor
12966_s_at (AL023094.197_S_AT) ATB2 bZIP transcription factor
12736_f_at (Z97048_F_AT) AtMYB13 transcription factor
Table 6 Probe Sets as referred to in Tables 4a and 4b corresponding to genes, the expression of which is increased after infection of wild-type Arabidopsis, and altered after infection of at least one mutant Arabidopsis having a mutation that results in enhanced susceptibility to Pseudomonas
12079_s_at (A71597.1_S_AT) 12081_at (AC001645.140_AT) 12115_at (AL033545.26_AT) 12150_at (AC004005.151_AT) 12233_at (AJ001807.1_AT) 12307_at (AC002392.162_AT) 12314_at (AC001229.28_AT) 12332_s_at (AB023448.2_S_AT) 12341_s_at (AL021637.176_S_AT) 12400_at (X98453.1_AT) 12438_at (AL021710.83_AT) 12475_at (Y11794.1_AT) 12487_at (AC004411.126_AT) 12497_at (AC006533.51_AT) 12500_s_at (AF081067.3_S_AT) 12530_at (Z99707.184_AT) 12574__at (X82624.2_AT) 12584_at (AC004521.233_AT) 12712_f_at (Z95774_F_AT) 12760_g_at (AC005278.32_G_AT) 12764_f_at (AC004138.69_F_AT) 12772_at (AC005278.34_AT) 12851_s_at (ACCSYN1_S_AT) 12879_s_at (AIG1__S_AT) 12883_s_at (APX_S_AT) 12889_s_at (ASA1_S_AT) 12891_at (ATACS6_AT) 12892_g_at (ATACS6_G_AT) 12905_s_at (ATERF2_S_AT) 1291 l_s_at (ATG6PDHE5_S_AT) 12921_s_at (ATHHMGCOAR_S_AT) 12958_at (AC002332.249_AT) 12989_s_at (AC004077.149_S_AT) 13003_s_at (AB021936.1_S_AT) 13014_at (U93215.87_AT) 13040_at (AC002392.134_AT) 13070_at (AC006919.171_AT) 13094_at (AL035523.163_AT) 13157_at (AC002409.35_AT) 13177_at (AL049640.42_AT) 13190_s_at (ATTHIREDA_S_AT) 13211_s_at (BCHI_S_AT) 13212_s_at (BGL2_S_AT) 13219_s_at (CHI4__S_AT) 13266_s_at (GST4_S_AT) 13273_s_at (HSF4_S_AT) 13275_f_at (HSP174_F_AT) 13370_at (AC005322.4_AT) 13381_at (AC006580.8_AT) 13435_at (AF003102.3_AT) 13467_at (AL096860.198_AT) 13538_at (AL080254.75_AT) 13565_at (AL035601.21_AT) 13588_at (AL021961.24_AT) 13589_at (AC000132.24_AT) 13617_at (AC006592.64_AT) 13627_at (AL035394.196_AT) 13656_at (AC007138.31_AT) 13659_at (AL022347.46_AT) 13666_s_at (INDOLE3GPS_S_AT) 13680_s_at (LOXl_S_AT) 13688_s_at (MONOPTEROS_S_AT) 13697_at (NI16_AT) 13716_at (NOVARTIS 103_RC_AT) 13751_at (NOVARTIS127_AT) 13755_at (NOVARTIS 15_AT) 13763_at (NOVARTIS21_AT) 13764_at (NOVARTIS22_AT)
13818_s_at (AC006218.175_S_AT) 13842_at (AC002396.12_AT) 13880_s_at (AL049480.183_S_AT) 13920_at (AC005990.53_AT) 14015_s_at (A71588.1_S_AT) 14016_s_at (A71596.1_S_AT) 14025_s_at (AC007293.3_S_AT) 14030_at (AC005970.225_AT) 14032_at (AL035601.11_AT) 14041_at (AC003970.28_AT) 14052_at (AC004122.24_AT) 14068_s_at (AC006922.197_S_AT) 14110_i_at (AL035528.279_I_AT) 14139_at (NOVARTIS30_AT) 14148_at (NOVARTIS38_AT) 14170_at (NOVARTIS51_AT) 14214_at (NOVARTIS83_AT) 14242_s_at (NRA_S_AT) 14248_at (PAD3_AT) 14249_i_at (PAD4_I_AT) 14408_at (AC002291.14_AT) 14450_at (AC002986.49_AT) 1446 l_at (AC006202.73_AT) 14487_at (Z97341.343_AT) 14584_at (AC007658.25_AT) 14609_at (AC002340.147_AT) 14620_s_at (PAT1_S_AT) 14635_s_at (PR.1_S_AT) 14638_s_at (PRXCB_S_AT) 14663_s_at (trehalaseprecusor_RC_S_AT) 14667_s_at (TRPB_S_AT) 14672_s_at (TSA1_S_AT) 14673_s_at (TSB2_S_AT) 14705_i_at (WT77_RC_I_AT) 14706_r_at (WT77_RC_R_AT) 14735_s_at (AF008124_S_AT) 14763_at (X86958.1_AT) 14786_at (AC005397.115_AT) 14882_at (AL022605.63_AT) 14924_at (AC006283.46_AT) 14959_at (AC007202.26_AT) 14978_at (AC002333.49_AT) 15073_at (AC007069.93_AT) 1509 l_at (AC004683.97_AT) 15098_s_at (ATU26945_S_AT) 15116_f_at (AF121356_F_AT) 15124_s_at (ATU59508_S_AT) 15125_f_at (D85190_F_AT) 15137_s_at (ATU57320_S_AT) 15154_s_at (ATHMTGDAS_S_AT) 15162_s_at (UO 1880_S_AT) 15196_s_at (ATU43412_S_AT) 15199_s_at (AB005804_S_AT) 15216_s_at (ATU75191_S_AT) 15406_at (AC006931.179_AT) 15431_at (AL030978.64_AT) 15483_s_at (AC005819.20_S_AT) 15496_at (AC006282.167_AT) 15518_at (AC005322.28_AT) 15522_i_at (AL078637.213_I_AT) 15526_at (AC004122.16_AT) 15531_ at (AL078637.191_I_AT) 15540_at (AC006585.205_AT) 15543_at (AF096371.10_AT) 15544_at (AL021633.110_AT) 15551_at (AL035440.289_AT) 15594_s_at (ATU56635_S_AT) 15622_s_at (ATU43945_S_AT) 15632_s_at (AB012570_S_AT) 15665_s_at (AF022658_S__AT) 15672_s_at (AF082299_S_AT) 15778_at (X98676.2_AT) 15779_g_at (X98676.2_G_AT) 15815_s_at (Z97342.366_S_AT) 15839_at (AC005662.203_AT) 15859_at (AC006587.164_AT)
15866_s_at (AC007133.59_S_AT) 15874_at (AL022223.106_AT) 15886_at (AL078637.204_AT) 15924_at (AC007138.61_AT) 15970_s_at (X71794.2_S_AT) 15978_at (X68592.6_AT) 15982_s_at (AC006260.78_S_AT) 16001_at (AF035385.2_AT) 16003_s_at (AL021749.64_S_AT) 16021_s_at (AL022224.182_S_AT) 16031_at (X94248.1_AT) 16043_at (AC005489.17_AT) 16053_i_at (Y14251.4_I_AT) 16083_s_at (AF153283_S_AT) 16091_s_at (ATHHSP83_S_AT) 16103_s_at (ATU60445_S_AT) 16105_s_at (ATU68017_S_AT) 16108_s_at (D78604_S_AT) 16130_s_at (AF078683_S_AT) 16233_at (AL080254.83_AT) 16272_at (AC006304.136_AT) 16298_at (AL021890.71_AT) 16299_at (AL024486.185_AT) 16306_at (AL049751.112_AT) 16340_at (AC004255.15_AT) 16357_at (AF149413.38_AT) 16363_at (AC004255.14_AT) 16383_at (AC006300.64_AT) 16409_at (AC004393.2_AT) 16461_i_at (AC004683.79_I_AT) 16462_s_at (AC004683.79_S_AT) 16470_s_at (AF068299.4_S_AT) 16545_s_at (AF037229_S_AT) 16570_s_at (ATHCDPKA_S_AT) 16578_s_at (ATHRPRP1B_S_AT) 16589_s_at (ATU26937_S_AT) 16594_s_at (ATU39783_S_AT) 16603_s_at (ATU81293_S_AT) 16609_s_at (AB008104_S_AT) 16701_at (AC005312.61_AT) 16721_at (AC006533.58_AT) 16781_at (AC002392.100_AT) 16810_at (AC002339.46_AT) 16859_at (AL035523.135_AT) 16864_i_at (AF037367.4_I_AT) 16865_s_at (AF037367.4_S_AT) 16914_s_at (AL049500.57_S_AT) 16968_at (AL021961.93_AT) 16989_at (AL030978.46_AT) 16995_at (AC002391.188_AT) 17007_at (AC005896.26_AT) 17051_s_at (AF098947_S__AT) 17066_s_at (ATHLIPOXY_S_AT) 17075_s_at (ATU09961_S_AT) 17128_s_at (ATHRPRP1A_S_AT) 17180_at (AF007270.30_AT) 17338_at (AC002535.97_AT) 17341_at (AL021713.89_AT) 17356_s_at (Z97338.190_S_AT) 17371_at (AF076243.44_AT) 17413_s_at (AJ006961.4_S_AT) 17451_at (AC002343.47_AT) 17452_g_at (AC002343.47_G_AT) 17484_at (X79052.2_AT) 17485_s_at (Z97340.345_S_AT) 17487_s_at (U18993.2_S__AT) 17533_s_at (ATU43488_S_AT) 17544_s_at (ATU40856_S_AT) 17585_s_at (AF134487_S_AT) 17653_at (AL035679.144_AT) 17752_at (AC003974.37_AT) 17758_at (AF076243.41_AT) 17775_at (AC004392.2_AT) 17781_at (AL049746.177_AT) 17840_s_at (AC002333.223_S_AT) 17877__g_at (AJ007587.2_G_AT)
17893_at (AC004401.135_AT) 17899_at (Z97339.197_AT) 17930_s_at (AJ006960.4_S_AT) 17945_at (Z97341.411_AT) 17963_at (AL049730.88_AT) 18054_at (AJ238846_AT) 18109_s_at (AC002391.206_S_AT) 18148_at (AC004669.25_AT) 18176_at (AL035540.31_AT) 18194_i_at (AL096859.227_I_AT) 18213_at (AL022140.126_AT) 18228_at (X91259.1_AT) 18242_g_at (AC006580.71_G_AT) 18255_at (AC005770.25_AT) 18258_s_at (AC006439.222_S_AT) 18267_at (AC006223.23_ AT) 18508_s_at (AC006532.89_S_AT) 18544_at (AC007060.14_AT) 18590_at (AJ222713.4_AT) 18591_at (X74756.2_AT) 18596_at (AC005698.13_AT) 18604_at (AF069298.31_AT) 18622__g_at (AJ005902.2_G_AT) 18625_at (AC005278.22_AT) 18686_s_at (U18126_S_AT) 18698_s_at (X17528_S_AT) 18735_s_at (Z29490_S_AT) 18753_s_at (AFl 18222.28_S_AT) 18803_at (AC005315.94_AT) 18885_at (AC006921.147_AT) 18888_at (AC007591.68_AT) 18899_s_at (X13434.1_S_AT) 18909_s_at (AF055848.2_S_AT) 18928_at (AC002333.181_AT) 18930_at (AC005990.57_AT) 18933_at (AC007020.48_AT) 18953_at (AF077955.1_AT) 18966_at (AC004561.106_AT) 19019_ at (X82623.2_I_AT) 19137_at (X74755.2_AT) 19140_at (AC005170.24_AT) 19181_s_at (AF053065.2_S_AT) 19207_at (AC006069.117_AT) 19247_at (AF071527.44_AT) 19257_s_at (AC000104.57_S_AT) 19288_at (AC005824.130_AT) 19325_at (AL022604.42_AT) 19364_at (AL022023.142_AT) 19383_at (AC006200.203_AT) 19405_at (AJ223803.1_AT) 19407_at (AC004697.81_AT) 19409_at (AC007357.56_AT) 1941 l_at (AC007661.104_AT) 19421_at (X70990.4_AT) 19432_s_at (AL035680.11_S_AT) 19451_at (AC004392.6_AT) 19460_s_at (AC000132.66_S_AT) 19465_at (AL021768.96_AT) 19546_at (AC005398.172_AT) 19555_at (AF058919.48_AT) \9624_at (AL049481.196_AT) 19641_at (AC004561.66_AT) 19645_at (AC004561.70_AT) 19667_at (AL021710.5_AT) 1974 l_at (AL049171.72_AT) 19755_at (AC006593.64_AT) 19818_i_at (AL021749.33_I_AT) 19848_s_at (AC004261.94_S_AT) 19881_at (AC004077.49_AT) 19892_at (AC005770.30_AT) 19944_at (AC002130.4_AT) 19956_at (AC006282.11_AT) 19970_s_at (AC003674.10_S_AT) 19982_at (AC002986.28_AT) 19991_at (AC007017.124_AT) 20017_at (AC004521.66_AT)
20030_at (AL078637.51_AT) 20051_at (AC000106.38_AT) 20098_at (AC004697.123_AT) 20133_i_at (AC007178.71 JLAT) 20134_s_at (AC007178.71_S_AT) 20144_at (AL079350.68__AT) 20238_at (X74514.2_AT) 20239_g_at (X74514.2_G_AT) 20245_s_at (AC005309.97_S_AT) 20247_at (AC004392.4_AT) 20263__at (AB004798.1_AT) 20269_at (AC002387.237_AT) 2027 l_at (Z99707.27_AT) 20287_at (Y14590.5_AT) 20288_g_at (Y14590.5_G_AT) 20291_s_at (M92353.4_S_AT) 20297_at (AC007153.27_AT) 20323_at (AC004561.62_AT) 20348_at (AC005967.35_AT) 20356_at (AC004561.74_AT) 20370_at (AC004561.263_AT) 20420_at (AL024486.131_AT) 2042 l_at (U81294.2_AT) 20422_g_at (U81294.2_G__AT) 20432_at (U43486.2_AT) 20450_at (AJ005930.2_AT) 2046 l_at (AL049480.157_AT) 20462_at (U82399.2_AT) 20479_i_at (AF069495.2_I_AT) 20480_s_at (AF069495.2_S_AT) 20485_at (AC007660.131_AT) 20529_at (Z97341.125_AT) 20572_s_at (AC005560.229_S_AT) 20589_at (AF081066.3_AT) 20669_s_at (AC002388.6_S_AT) 20685_at (AL049751.46_AT) 20689_s_at (AC002335.19_S_AT)
Table 7 Probe Sets as referred to in Tables 4a and 4b corresponding to genes, the expression of which is increased after infection of wild-type Arabidopsis, and altered after infection of at least one mutant Arabidopsis having a mutation in a gene whose expression is important for resistance to necrotrophic fungi
11997_at (AC005967.4_AT) 12004 _at (AL022023.132_AT)
12037_at (AC004005.174_AT) 12072. _at (AL035396.4_AT)
12079_s_at (A71597.1_S_AT) 12081. _at (AC001645.140_AT)
12115_at (AL033545.26_AT) 12136 _at (AC007591.60_AT)
12150_at (AC004005.151_AT) 12216 _at (AC007119.56_AT)
12317_at (AC004138.27_AT) 12323. _at (AC002333.18_AT)
12332_s_at (AB023448.2_S_AT) 12369 _at (AC002535.59_AT)
12400_at (X98453.1_AT) 12449. _s_at (AC002343.179_S_AT)
12475_at (Yl 1794.1_AT) 12487 _at (AC004411.126_AT)
12500_s_at (AF081067.3_S_AT) 12530. _at (Z99707.184_AT)
12535_at (AL035538.156_AT) 12538. _at (AF033205.2_AT)
12574_at (X82624.2_AT) 12584 _at (AC004521.233_AT)
12626_at (AC006234.95_AT) 12645. _at (AL021712.56_AT)
12744_at (AC001645.15_AT) 12760. _g_at (AC005278.32_G_AT)
12772_at (AC005278.34_AT) 12797 _s_at (AC007138.25_S_AT)
12851_s_at (ACCSYNl_S_AT) 12880. _s_at (AIG2_S_AT)
12889_s_at (ASA1_S_AT) 12904 _s_at (ATERF1_S_AT)
12905_s_at (ATERF2_S_AT) 12921. _s_at (ATHHMGCOAR_S_AT)
12951__at (AC005489.5_AT) 12965. _at (AL021711.118_AT)
12966_s_at (AL023094.197_S_AT) 12989. _s_at (AC004077.149_S_AT)
13040_at (AC002392.134_AT) 13094. _at (AL035523.163_AT)
13134_s_at (AC002337.9_S_AT) 13152. _s_at (AC005322.24_S_AT)
13176_at (AL031394.56_AT) 13211. _s_at (BCHI_S_AT) 13215_s_at (cafferoylcoAmethyltrans_S_AT) 13243. _r_at (ELI32_R_AT)
13244_s__at (ELI32_S_AT) 13266 _s_at (GST4_S_AT)
13275_f_at (HSP174_F_AT) 13277 _i_at (HSP176A_I_AT)
13312_at (AC006223.75_AT) 13370. _at (AC005322.4__AT)
13381_at (AC006580.8_AT) 13395. _at (AL035528.202_AT)
13435_at (AF003102.3_AT) 13437. _at (AF096371.8_AT)
13536_at (AL021636.47_AT) 13538. _at (AL080254.75_AT)
13565_at (AL035601.21_AT) 13588. _at (AL021961.24_AT)
13680_s_at (LOXl_S_AT) 13688. _s_at (monoρteros_S_AT)
13751_at (NOVARTIS127_AT) 13755. _at (NOVARTIS 15_AT)
13764_at (NOVARTIS22_AT) 13803. _at (Z97341.376_AT)
13848__at (AC003981.31_AT) 13918_at (AC005388.29_AT) 13949_s_at (Z97343.352_S_AT) 13999_at (AF071527.56_AT) 14015_s_at (A71588.1_S_AT) 14016_s_at (A71596.1_S_AT) 14041_at (AC003970.28_AT) 14052_at (AC004122.24_AT) 14068_s_at (AC006922.197_S_AT) 14070_at (AL049658.217_AT) 14089_at (AC006223.65_AT) 14100_at (AF002109.108_AT) 14197_at (NOVARTIS71_AT) 14240_s_at (NR1_S_AT) 14450_at (AC002986.49_AT) 14461_at (AC006202.73_AT) 14475_at (AL021811.121_AT) 14584_at (AC007658.25_AT) 14609_at (AC002340.147_AT) 14663_s_at (trehalaseprecusor_RC_S_AT) 14667_s_at (TRPB_S_AT) 14673_s_at (TSB2_S_AT) 14675_s_at (VSP_S_AT) 14705_ at (WT77_RC_I_AT) 14706_r_at (WT77_RC_R_AT) 14735_s_at (AF008124_S_AT) 14750_s_at (AF096370.12_S_AT) 14779_at (AC004680.71_AT) 14786_at (AC005397.115_AT) 14978_at (AC002333.49_AT) 15052_at (AC002332.103_AT) 15073_at (AC007069.93_AT) 15088_s_at (AC002311.37_S_AT) 15091_at (AC004683.97_AT) 15098_s_at (ATU26945_S_AT) 15124_s_at (ATU59508_S_AT) 15125_f_at (D85190_F_AT) 15137_s_at (ATU57320_S_AT) 15141_s_at (D85191_S_AT) 15154_s_at (ATHMTGDAS_S_AT) 15161_s_at (ATU90522_S_AT) 15162_s_at (UO 1880_S_AT) 15188_s_at (AF081202_S_AT) 15196_s_at (ATU43412_S_AT) 1521 l_s_at (ATH243813_S_AT) 15342_at (AC006593.101__AT) 15406_at (AC006931.179_AT) 15479_at (AL049483.205_AT) 15483_s_at (AC005819.20_S_AT) 15485_at (AC006233.109_AT) 15496_at (AC006282.167_AT) 15522_i_at (AL078637.213_I_AT) 15523_s_at (AL078637.213_S_AT) 15524_at (AC005508.25_AT) 15526_at (AC004122.16_AT) 15531_ at (AL078637.191_I_AT) 15532_r_at (AL078637.191_R__AT) 15547_at (AC005970.122_AT) 15594_s_at (ATU56635_S_AT) 15622_s_at (ATU43945_S_AT) 15625_s_at (ATU74610_S_AT) 15632_s_at (AB012570_S_AT) 15665_s_at (AF022658_S_AT) 15670_s_at (AF061638_S_AT) 15792_at (AC002341.106_AT) 15798_at (AC002521.173_AT) 15839_at (AC005662.203_AT) 15859_at (AC006587.164_AT) 15874_at (AL022223.106_AT) 15886_at (AL078637.204_AT) 15982_s_at (AC006260.78_S_AT) 16063_s_at (AB008103_S_AT) 16073_f_at (AF062908_F_AT) 16108_s_at (D78604_S_AT) 16130_s_at (AF078683_S_AT) 16173_s_at (D78607_S_AT)
16203_at (AC007519.53_AT) 16272_at (AC006304.136_AT) 16298_at (AL021890.71_AT) 16301_s_at (AL031018.105_S_AT) 16306_at (AL049751.112_AT) 16357_at (AF149413.38_AT) 16363_at (AC004255.14_AT) 16398_s_at (AL022603.3_S_AT) 16405_at (AC005850.9_AT) 16461_ at (AC004683.79_I_AT) 16462_s_at (AC004683.79_S_AT) 16470_s_at (AF068299.4_S_AT) 16510_at (AL034567.198_AT) 16522_at (X77500.2_AT) 16526_at (Z49227.1_AT) 16541_s_at (AB023423_S_AT) 16545_s_at (AF037229_S_AT) 16553 J at (AF078821_F_AT) 16589_s_at (ATU26937_S_AT) 16603_s_at (ATU81293_S_AT) 16609_s_at (AB008104_S_AT) 1661 l_s_at (AB008782_S_AT) 16638_s_at (AF139098_S_AT) 16646_s_at (ATHDHS1_S_AT) 16649_s_at (ATHORF_S_AT) 16701_at (AC005312.61_AT) 16747_at (AL021713.3_AT) 16781_at (AC002392.100_AT) 16810_at (AC002339.46_AT) 16864_i_at (AF037367.4_I_AT) 16865_s_at (AF037367.4_S_AT) 16914_s_at (AL049500.57_S_AT) 16968_at (AL021961.93_AT) 16989_at (AL030978.46_AT) 16995_at (AC002391.188_AT) 17008_at (AC006585.212_AT) 17041_s_at (D89631_S__AT) 17066_s_at (ATHLIPOXY_S_AT) 17075_s_at (ATU09961_S_AT) 17104_s_at (D88541_S_AT) 1711 l_s_at (ATHACSC_S_AT) 17119_s_at (AF132212_S_AT) 17187_at (AF128396.2_AT) 17300_at (X66017.2_AT) 17323_at (U95973.69_AT) 17338_at (AC002535.97_AT) 17356_s_at (Z97338.190_S_AT) 17485_s_at (Z97340.345_S_AT) 1751 l_s_at (AF067605_S_AT) 17514_s_at (AF076277_S_AT) 17522_s_at (D78606_S_AT) 17533_s_at (ATU43488_S_AT) 17548_s_at (AFl 18823_S_AT) 17595_s_at (AF166352_S_AT) 17648_at (AL021684.43_AT) 17702_at (AC005700.212_AT) 17719_at (AC006592.17_AT) 17775_at (AC004392.2_AT) 17860_at (AL078467.4_AT) 17877__g_at (AJ007587.2_G_AT) 17893_at (AC004401.135_AT) 17945_at (Z97341.411_AT) 18109_s_at (AC002391.206_S_AT) 18121_s_at (AC002337.21_S_AT) 18122_at (AC002338.110_AT) 18148_at (AC004669.25_AT) 18176_at (AL035540.31_AT) 18213_at (AL022140.126_AT) 18224_s_at (AL021890.57_S_AT) 18228_at (X91259.1_AT) 18236_s_at (AC004683.69_S_AT) 18258_s_at (AC006439.222_S_AT) 18266_at (AC004684.33_AT) 18299_s_at (M23872.2_S_AT) 18587_s_at (AC007166.53_S_AT) 18590_at (AJ222713.4_AT)
18596_at (AC005698.13_AT) 18636_at (AC006577.22_AT) 18668_at (AJ249794_AT) 18735_s_at (Z29490_S_AT) 18782_at (AC003040.90_AT) 18803_at (AC005315.94_AT) 18888_at (AC007591.68_AT) 18899_s_at (X13434.1_S_AT) 18908 _at (AF055848.2_I_AT) 18933_at (AC007020.48__AT) 18949_at (Z54136.1_AT) 18953_at (AF077955.1_AT) 18963_at (AC0 4561.99_AT) 19019_i_at (X82623.2_I_AT) 19060_at (AC003671.34_AT) 19110_s_at (X86947.2__S_AT) 19137_at (X74755.2_AT) 19257_s_at (AC000104.57_S_AT) 19284_at (AC003028.196_AT) 19288_at (AC005824.130_AT) 19325_at (AL022604.42_AT) 19364_at (AL022023.142_AT) 19383_at (AC006200.203_AT) 19395_at (AF007270.32_AT) 19409_at (AC007357.56_AT) 19432_s_at (AL035680.11_S_AT) 19451_at (AC004392.6_AT) 19460_s_at (AC000132.66_S_AT) 19546_at (AC005398.172_AT) 19591_at (AJ010735.4_AT) 19614_at (AC003970.32_AT) 19635_at (AL049746.38_AT) 19640_at (AC004561.78_AT) 19641_at (AC004561.66_AT) 19645_at (AC004561.70_AT) 19667_at (AL021710.5_AT) 19672_at (AC005687.19_AT) 19673_g__at (AC005687.19_G_AT) 19700_s_at (AL031326.154_S_AT) 19704_i_at (AJ005927.2_I_AT) 19707_s_at (Z95768.3_S_AT) 19755_at (AC006593.64_AT) 19762_at (AL035527.204_AT) 19818_i_at (AL021749.33_I_AT) 19851_at (U23794.3_AT) 19895_s_at (U77347.4_S_AT) 19956_at (AC006282.11_AT) 19960_at (AL035527.360_AT) 20023_at (AC006577.46_AT) 20051_at (AC000106.38__AT) 20053_at (AC002292.27_AT) 20096_at (AC004238.31_AT) 20133_ at (AC007178.71_I_AT) 20134_s_at (AC007178.71_S_AT) 20144_at (AL079350.68_AT) 20179_at (AL035538.229_AT) 20189_at (AC005489.2_AT) 20194_at (AC007584.48_AT) 20247_at (AC004392.4_AT) 20263_at (AB004798.1_AT) 20291_s_at (M92353.4_S_AT) 20297_at (AC007153.27_AT) 20323_at (AC004561.62_AT) 20348_at (AC005967.35_AT) 20356_at (AC004561.74_AT) 20370_at (AC004561.263_AT) 20433_at (AC006232.147_AT) 20450_at (AJ005930.2_AT) 20479_i_at (AF069495.2_I_AT) 20485_at (AC007660.131_AT) 20572_s_at (AC005560.229_S_AT) 20646_at (AC002291.20_AT) 20656_at (AL035396.46_AT) 20658_s_at (AL050400.70_S_AT) 20669_s_at (AC002388.6_S_AT) 20686_at (Y14424.2_AT)
235
Table 8 Probe Sets as referred to in Table 6 corresponding to genes, the expression of which is is increased after infection of wild-type Arabidopsis and altered after infection of at least one mutant Arabidopsis having a mutation in a gene that interferes with salicylic acid dependent signaling
12079_s_at (A71597.1_S_AT) 12081_at (AC001645.140_AT) 12115_at (AL033545.26_AT) 12150_at (AC004005.151_AT) 12233_at (AJ001807.1_AT) 12307_at (AC002392.162_AT) 12314_at (AC001229.28_AT) 12332_s_at (AB023448.2_S_AT) 12341_s_at (AL021637.176_S_AT) 12400_at (X98453.1_AT) 12438_at (AL021710.83_AT) 12475_at (Y11794.1_AT) 12487_at (AC004411.126_AT) 12497_at (AC006533.51_AT) 12500_s_at (AF081067.3__S_AT) 12530_at (Z99707.184_AT) 12574_at (X82624.2_AT) 12584_at (AC004521.233_AT) 12712_f_at (Z95774_F_AT) 12760_g_at (AC005278.32_G_AT) 12764_f_at (AC004138.69_F_AT) 12879_s_at (AIG1_S_AT) 12883_s_at (APX_S_AT) 12889_s_at (ASA1_S_AT) 12891_at (ATACS6_AT) 12892__g_at (ATACS6_G_AT) 12905_s_at (ATERF2_S_AT) 1291 l_s_at (ATG6PDHE5_S_AT) 12921_s_at (ATHHMGCOAR_S_AT) 12958_at (AC002332.249_AT) 12989_s_at (AC004077.149_S_AT) 13003_s_at (AB021936.1_S_AT) 13014_at (U93215.87_AT) 13040_at (AC002392.134_AT) 13070_at (AC006919.171_AT) 13094_at (AL035523.163_AT) 13157_at (AC002409.35_AT) 13190_s_at (ATTHIREDA_S_AT) 13211_s_at (BCHI_S_AT) 13212_s_at (BGL2_S_AT) 13219_s_at (CHI4_S_AT) 13273_s_at (HSF4_S_AT) 13275_f_at (HSP174_F__AT) 13370_at (AC005322.4_AT) 13381_at (AC006580.8_AT) 13538_at (AL080254.75_AT) 13565_at (AL035601.21_AT) 13588_at (AL021961.24_AT) 13589_at (AC000132.24_AT) 13617_at (AC006592.64_AT) 13627_at (AL035394.196_AT) 13656_at (AC007138.31_AT) 13659_at (AL022347.46_AT) 13666_s_at (INDOLE3GPS_S_AT) 13680_s_at (LOXl_S_AT) 13697_at (NI16_AT) 13751_at (NOVARTIS127_AT) 13755_at (NOVARTIS 15_AT) 13763__at (NOVARTIS21_AT) 13764_at (NOVARTIS22_AT) 13818_s_at (AC006218.175_S_AT) 13880_s_at (AL049480.183_S_AT) 13920_at (AC005990.53_AT) 14015_s_at (A71588.1_S_AT) 14016_s_at (A71596.1_S_AT) 14025_s_at (AC007293.3_S_AT) 14030_at (AC005970.225_AT) 14032_at (AL035601.11_AT)
14041_at (AC003970.28_AT) 14052 _at (AC004122.24_AT)
14068_s_at (AC006922.197_S_AT) 14110. _i_at (AL035528.279_I_AT)
14139_at (NOVARTIS30_AT) 14148. _at (NOVARTIS38_AT)
14170_at (NOVARTIS51_AT) 14214..at (NOVARTIS83_AT)
14242_s_at (NRA_S_AT) 14248. _at (PAD3_AT)
14249_ at (PAD4_I_AT) 14408. _at (AC002291.14_AT)
14450_at (AC002986.49_AT) 14461. _at (AC006202.73_AT)
14487_at (Z97341.343_AT) 14584. _at (AC007658.25_AT)
14609_at (AC002340.147_AT) 14620. _s_at (PAT1_S_AT)
14635_s_at (PR.l_S_AT) 14638. _s_at (PRXCB_S_AT) 14663_s_at (trehalaseprecusor_RC_S_AT) 14667. _s_at (TRPB_S_AT)
14672_s_at (TSA1_S_AT) 14673. _s_at (TSB2_S_AT)
14735_s_at (AF008124__S_AT) 14763. _at (X86958.1_AT)
14786_at (AC005397.115_AT) 14882. _at (AL022605.63_AT)
14959_at (AC007202.26_AT) 14978. _at (AC002333.49_AT)
15073_at (AC007069.93_AT) 15091. _at (AC004683.97_AT)
15116_f_at (AF121356_F_AT) 15137. _s_at (ATU57320_S_AT)
15162_s_at (U01880_S_AT) 15196. _s_at (ATU43412_S_AT)
15199_s_at (AB005804_S_AT) 15216. _s_at (ATU75191_S_AT)
1543 l_at (AL030978.64_AT) 15483. _s_at (AC005819.20_S_AT)
15496_at (AC006282.167_AT) 15518. _at (AC005322.28_AT)
15526_at (AC004122.16_AT) 15540. _at (AC006585.205_AT)
15543_at (AF096371.10_AT) 15544..at (AL021633.110_AT)
15551_at (AL035440.289_AT) 15594. _s_at (ATU56635_S_AT)
15622_s_at (ATU43945_S_AT) 15632. _s_at (AB012570_S_AT)
15672_s_at (AF082299_S_AT) 15778.at (X98676.2_AT)
15779_g_at (X98676.2_G_AT) 15815. _s_at (Z97342.366_S_AT)
15839_at (AC005662.203_AT) 15859. _at (AC006587.164_AT)
15866_s_at (AC007133.59_S_AT) 15874. _at (AL022223.106_AT)
15924_at (AC007138.61_AT) 15970. _s_at (X71794.2_S__AT)
15978_at (X68592.6_AT) 15982 _s_at (AC006260.78_S_AT)
16003_s_at (AL021749.64_S_AT) 16021. _s_at (AL022224.182_S_AT)
16031_at (X94248.1_AT) 16043. _at (AC005489.17_AT)
16053 J_at (Y14251.4_I_AT) 16083. _s__at (AF153283_S_AT)
1609 l_s_at (ATHHSP83_S_AT) 16103. _s__at (ATU60445_S_AT)
16105_s_at (ATU68017_S_AT) 16108. _s_at (D78604_S_AT)
16130_s_at (AF078683_S_AT) 16233..at (AL080254.83_AT)
16272_at (AC006304.136_AT) 16299. _at (AL024486.185_AT)
16306_at (AL049751.112_AT) 16340_at (AC004255.15_AT) 16357_at (AF149413.38_AT) 16363_at (AC004255.14_AT) 16383_at (AC006300.64_AT) 16409_at (AC004393.2_AT) 16461_i_at (AC004683.79_I_AT) 16462_s_at (AC004683.79_S_AT) 16470_s_at (AF068299.4_S_AT) 16570_s_at (ATHCDPKA_S_AT) 16578_s_at (ATHRPRP1B_S_AT) 16589_s_at (ATU26937_S_AT) 16594_s_at (ATU39783_S_AT) 16603_s_at (ATU81293_S_AT) 16609_s_at (AB008104_S_AT) 16701_at (AC005312.61_AT) 16721_at (AC006533.58_AT) 16781_at (AC002392.100_AT) 16810_at (AC002339.46_AT) 16864_ at (AF037367.4_I_AT) 16865_s_at (AF037367.4_S_AT) 16914_s_at (AL049500.57_S_AT) 16968_at (AL021961.93_AT) 16995_at (AC002391.188_AT) 17007_at (AC005896.26_AT) 17051_s_at (AF098947_S_AT) 17066_s_at (ATHLIPOXY_S_AT) 17075_s_at (ATU09961_S_AT) 17128_s_at (ATHRPRP1A_S_AT) 17180_at (AF007270.30_AT) 17338_at (AC002535.97_AT) 17341_at (AL021713.89_AT) 17356_s_at (Z97338.190_S_AT) 17413_s_at (AJ006961.4_S_AT) 17451_at (AC002343.47_AT) 17452_g_at (AC002343.47_G_AT) 17484_at (X79052.2_AT) 17485_s_at (Z97340.345_S_AT) 17487_s_at (U18993.2_S_AT) 17533_s_at (ATU43488_S_AT) 17544_s_at (ATU40856_S_AT) 17585_s_at (AF134487_S_AT) 17653_at (AL035679.144_AT) 17752_at (AC003974.37_AT) 17758_at (AF076243.41_AT) 17775_at (AC004392.2_AT) 17781_at (AL049746.177_AT) 17840_s_at (AC002333.223_S_AT) 17893_at (AC004401.135_AT) 17899_at (Z97339.197_AT) 17930_s_at (AJ006960.4_S_AT) 17963_at (AL049730.88_AT) 18054_at (AJ238846_AT) 18148_at (AC004669.25_AT) 18194_i_at (AL096859.227_I_AT) 18213_at (AL022140.126_AT) 18242_g_at (AC006580.71_G_AT) 18255_at (AC005770.25_AT) 18258_s_at (AC006439.222_S_AT) 18508_s_at (AC006532.89_S_AT) 18544_at (AC007060.14_AT) 18590_at (AJ222713.4_AT) 18591_at (X74756.2_AT) 18596_at (AC005698.13_AT) 18604_at (AF069298.31_AT) 18622_g_at (AJ005902.2_G_AT) 18625_at (AC005278.22_AT) 18686_s_at (U18126_S_AT) 18698_s_at (X17528_S_AT) 18735_s_at (Z29490_S_AT) 18753_s_at (AFl 18222.28_S_AT) 18803_at (AC005315.94_AT) 18885_at (AC006921.147_AT) 18888_at (AC007591.68_AT) 18909_s_at (AF055848.2_S_AT) 18928_at (AC002333.181__AT)
18930_at (AC005990.57_AT) 18933_at (AC007020.48_AT) 18953_at (AF077955.1_AT) 18966_at (AC004561.106_AT) 19019 _at (X82623.2_I_AT) 19137__at (X74755.2_AT) 19140_at (AC005170.24_AT) 19181__s_at (AF053065.2_S_AT) 19207_at (AC006069.117_AT) 19247__at (AF071527.44_AT) 19257_s_at (AC000104.57_S_AT) 19325_at (AL022604.42_AT) 19364_at (AL022023.142_AT) 19407_at (AC004697.81_AT) 19409_at (AC007357.56_AT) 19411_at (AC007661.104_AT) 19421_at (X70990.4_AT) 19432_s_at (AL035680.11_S_AT) 19451_at (AC004392.6_AT) 19460_s_at (AC000132.66_S_AT) 19465_at (AL021768.96_AT) 19546_at (AC005398.172_AT) 19555_at (AF058919.48_AT) 19624_at (AL049481.196_AT) 19641_at (AC004561.66_AT) 19667_at (AL021710.5_AT) 19741_at (AL049171.72_AT) 19755_at (AC006593.64_AT) 19818_ at (AL021749.33_I_AT) 19881_at (AC004077.49_AT) 19892_at (AC005770.30_AT) 19944_at (AC002130.4_AT) 19956_at (AC006282.11_AT) 19970_s_at (AC003674.10_S_AT) 19982_at (AC002986.28_AT) 19991_at (AC007017.124_AT) 20017_at (AC004521.66_AT) 20030_at (AL078637.51_AT) 2005 l_at (AC000106.38_AT) 20098_at (AC004697.123_AT) 20133_i_at (AC007178.71_I_AT) 20134_s_at (AC007178.71_S_AT) 20144_at (AL079350.68_AT) 20245_s_at (AC005309.97_S_AT) 20247_at (AC004392.4_AT) 20269_at (AC002387.237_AT) 20271_at (Z99707.27_AT) 20287_at (Y14590.5_AT) 20288_g_at (Y14590.5_G_AT) 20291_s_at (M92353.4_S_AT) 20323_at (AC004561.62_AT) 20356_at (AC004561.74_AT) 20370_at (AC004561.263_AT) 20420_at (AL024486.131_AT) 20421_at (U81294.2_AT) 20422_g_at (U81294.2_G_AT) 20432_at (U43486.2_AT) 20450_at (AJ005930.2_AT) 20461_at (AL049480.157_AT) 20462_at (U82399.2_AT) 20479 _at (AF069495.2_I_AT) 20480_s_at (AF069495.2_S_AT) 20529_at (Z97341.125_AT) 20572_s_at (AC005560.229_S_AT) 20589_at (AF081066.3_AT) 20669_s_at (AC002388.6_S_AT) 20685__at (AL049751.46_AT) 20689_s_at (AC002335.19_S_AT)
Table 9 Probe Sets corresponding to genes, the expression of which is downregulated (repressed) in an avrRpt-2-dependent manner in Arabidopsis
ProbeSet Description
H991_g_at (AC002387.210_G_AT) 11 91_g_at
(AC002387.210_G_AT)gb|AAB82645.11 (AC002387) unknown protein [Arabidopsis thaliana]
1209 l_at (AC004450.116_AT) 12091_at (AC004450.116_AT)gb|AAC64313.1| (AC004450) unknown protein [Arabidopsis thaliana]
12124_s_at (Z97337.149_S_AT) 12124_s_at (Z97337.149_S_AT)emb|CAB10270.1| (Z97337) imidazoleglycerol-phosphate dehydratase [Arabidopsis thaliana]
12125_at (Z97341.99_AT) 12125_at (Z97341.99_AT)emb|CAB10421.11 (Z97341) hypothetical protein [Arabidopsis thaliana]
12160_at (AC006284.117_AT) 12160_at (AC006284.117_AT)gb|AAD17436.1] (AC006284) unknown protein [Arabidopsis thaliana]
12191_at (AC006068.35_AT) 12191_at (AC006068.35_AT)gb|AAD15440.1| (AC006068) unknown protein [Arabidopsis thaliana]
12193_at (AC006072.132_AT) 12193_at (AC006072.132_AT)gb|AAD13708.1| (AC006072) unknown protein [Arabidopsis thaliana]
12223_s_at (AC007168.178_S_AT) 12223_s_at
(AC007168.178_S_AT)gb|AAF18668.1]AC007168_ 1 (AC007168) unknown protein [Arabidopsis thaliana]
12290_at (Y09418.2_AT) 12290_at (Y09418.2_AT)emb|CAA70572.1 ] (Y09418) CDPK-related protein kinase [Arabidopsis thaliana]
12421_at (AJ002414.1_AT) 12421_at (AJ002414. l_AT)emb|CAA05398.11 (AJ002414) hnRNP-like protein [Arabidopsis thaliana]
12460_s_at (AC006920.129_S_AT) 12460_s_at
(AC006920.129_S_AT)gb|AAD22284.1 |AC006920 _8 (AC006920) DNA-directed RNA polymerase II, third largest subunit [Arabidopsis thaliana]
12493_g_at (Y09095.1_G_AT) 12493_g_at (Y09095. l_G_AT)emb|CAA70310.11 (Y09095) chloride channel [Arabidopsis thaliana]
ProbeSet Description
12559_at (AC005727.83_AT) 12559_at (AC005727.83_AT)gb|AAC79586.1| (AC005727) putative DOF zinc finger protein [Arabidopsis thaliana]
12560_at (AC005825.57_AT) 12560_at
(AC005825.57_AT)gb|AAD24598.1 |AC005825_5 (AC005825) putative chloroplast outer membrane protein [Arabidopsis thaliana]
12561_at (AL021687.107_AT) 1256 l__at (AL021687.107_AT)emb|CAAl 6701.11 (AL021687) putative protein [Arabidopsis thaliana]
12855_f_at (ADH_F_AT) 12855_f_at (ADH_F_AT)gb|AAC00625.11 (AC002291) Alcohol Dehydrogenase [Arabidopsis thaliana]
12962_at (AC004697.165_AT) 12962_at (AC004697.165_AT)gb|AAC28988.11 (AC004697) putative ABC transporter [Arabidopsis thaliana]
13221_at (CHS-WHOLE-SEQ_AT) 13221_at (CHS-WHOLE-
SEQ_AT)gb|AAF23561.1|AF112086_l (AF112086) chalcome synthase [Arabidopsis thaliana]
13253_f_at (FPS1_F_AT) 13253_f_at (FPSl_F_AT)gb|AAB49290.1| (U80605) farnesyl diphosphate synthase precursor [Arabidopsis thaliana]
13459_at (AF013294.21_AT) 13459_at (AF013294.21_AT)gb|AAB62852.11 (AF013294) similar to auxin-induced protein [Arabidopsis thaliana]
13708_s_at (AC007259X_S_AT) 13708_s_at
(AC007259X_S_AT)gb|AAD50000.1|AC007259_l 3 (AC007259) Similar to protein kinases [Arabidopsis thaliana]
14428_s_at (AL023094.342_S_AT) 14428_s_at
(AL023094.342_S_AT)emb|CAAl 8852.11 (AL023094) putative protein [Arabidopsis thaliana]
15175_s_at (ATU28215_S_AT) 15175_s_at (ATU28215_S_AT)gb|AAC62130.11 (AC005169) hexokinase (ATHXK2) [Arabidopsis thaliana]
15943_at (AC007202.16_AT) 15943_at
(AC007202.16_AT)gb]AAD30227.1|AC007202_9 (AC007202) Strong similarity to gb|Y14272 3- deoxy-D-manno-2-octulosonate-8-phosphate synthase from Pisum sativum. ESTs gb|AA067485 and gb]AI100551 come from this gene. [Arabidopsis thaliana]
ProbeSet Description
19219_at (AC007019.185_AT) 19219_at (AC007019.185_AT)gb|AAD20413.1| (AC007019) unknovra protein [Arabidopsis thaliana]
Table 10 A: Expression data for 9 probe sets corresponding to genes that are specifically induced during incompatible interaction within 3 hours
Table 10B: Expression data for 18 probe sets corresponding to genes that are specifically induced during incompatible interaction in 6 hours (but not within 3 hours)
Table IOC: Expression data for 6 probe sets corresponding to genes that are activated by P. syringae in 6 hours. Most of them are compatible interaction-specific/preferential
Pst/aR2 ... represents the incompatible interaction Ps ... P. syringae pv. tomato aR2 ... avrRpt2 ud ... undetectable
Table 11 Probe Sets corresponding to genes induced early after infection of different Arabidopsis ecotypes with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121 (at 3 or 6 hours) or by estradiol inducible
Table 12 Probe Sets corresponding to genes repressed early after infection of Arabidopsis with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121
Table 13a Probe Sets corresponding to genes, the expression of which is induced in an incompatible interaction at 3 and/or 6 hours after infection of Tour Arabidopsis ecotypes with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121
Table 13b Probe Sets corresponding to genes, the expression of which is decreased in an incompatible interaction at 3 and/or 6 hours after infection of Tour Arabidopsis ecotypes with Pseudomonas syringae pv tomato DC3000, P. maculicola ES4326 and P. phaseolica NPS3121
Table 14 Probe Sets as referred to in Table 3 corresponding to genes with promoters conferring low basal expression in all ecotypes (Col, Ler, Ws, and Cvi) and high inducibility in Col
Table 15 Probe Sets as referred to in Table 3 corresponding to genes with promoters inducing expression in Pseudomonas syringae pv. αcw.z'o/α-infected Arabidopsis
12891_at (ATACS6_AT) 13015_s_at (X98673.2_S_AT) 13100_at (AC003680.50_AT) 13115_at (AC000375.44_AT) 13217_s_at (CALMODULINLIKE_S_AT) 13467_at (AL096860.198_AT) 13645_at (AC000098.8_AT) 13818_s_at (AC006218.175_S_AT) 14032_at (AL035601.11_AT) 14248_at (PAD3_AT) 14609_at (AC002340.147_AT) 15116_f_at (AF121356_F_AT) 15622_s_at (ATU43945_S_AT) 16173_s_at (D78607_S_AT) 17485_s_at (Z97340.345_S_AT) 17511_s_at (AF067605_S_AT) 17548_s_at (AFl 18823_S_AT) 17775_at (AC004392.2__AT) 17930_s_at (AJ006960.4_S_AT) 19284_at (AC003028.196_AT) 19546_at (AC005398.172_AT) 19640_at (AC004561.78_AT) 20134_s_at (AC007178.71_S_AT) 20194_at (AC007584.48_AT) 20348_at (AC005967.35_AT) 12892_g_at (ATACS6_G_AT) 12904_s_at (ATERF1_S_AT) 12989_s_at (AC004077.149_S_AT) 13215_s_at (cafferoylco Amethyltrans_S_AT) 13565__at (AL035601.21_AT) 13627__at (AL035394.196_AT) 16649_s_at (ATHORF_S_AT) 16914_s_at (AL049500.57_S_AT) 16995_at (AC002391.188_AT) 19991_at (AC007017.124_AT) 20356_at (AC004561.74_AT)
Table 16 Probe Sets corresponding to genes with promoters inducing expression in Botrytis cinere -infected Arabidopsis
Table 17 Probe sets which correspond to genes downregulated in response to infection by 5 different viruses
Table 18 Probe sets which correspond to genes upregulated in response to infection by 5 different viruses
Probeset Description Time Function Blast
19555_at gb|AAC13630.1| (AF058919) 4dpi metabolic probable (AF058919.48_AT) F6N23.8 gene product protein methylene- [Arabidopsis thaliana] tetrahudrofol ate dehydragenas e (NADP+)
15515_r_at gb|AAC 13627.11 (AF058919) 5dpi metabolic putative (AF058919.32_R_AT) F6N23.26 gene product protein phosphoribos [Arabidopsis thaliana] ylanthranilate transferase
[Arabidopsis thaliana]
15606_s_at gb|AACl 5807.11 (AF061517) 5dpi stress (AF061517_S_AT) putative copper/zinc superoxide response dismutase copper chaperone [Arabidopsis thaliana]
18604_at gb|AAC19273.1| (AF069298) 5dpi stress (AF069298.31_AT) similar to several small proteins response (~100 aa) that are induced by heat, auxin, ethylene and wounding such as Phaseolus aureus indole-3 -acetic acid induced protein ARG (SW:32292) [Arabidopsis thaliana]
12002_at gb|AAC79112.1| (AF069442) 4-5dpi protein ubiquitin-like (AF069442.47_AT) hypothetical protein processing [Arabidopsis thaliana]
20480_s_at emb|CAB38908.1| (AL035708) 4-5dpi metabolic (AF069495.2_S_AT) cytochrome P450-like protein protein [Arabidopsis thaliana]
20479_ at emb|CAB38908.1| (AL035708) 5dpi metabolic (AF069495.2JLAT) cytochrome P450-like protein protein [Arabidopsis thaliana]
19247_at gb|AAD11587.1|AAD11587 4-5dpi unknown Ankyrin (AF071527.44_AT) (AF071527) hypothetical protein repeats [Arabidopsis thaliana]
12500_s_at gb|AAC32192.1| (AF081067) 4-5dpi hormone (AF081067.3_S_AT) IAA-Ala hydrolase; IAA-amino response acid hydrolase [Arabidopsis thaliana]
Probeset Description Time Function Blast
20516_at emb|CAB36736.1| (AL035523) 4dpi unknown unknown (AL035523.64_AT) ubiquitin activating enzyme-like protein [Arabidopsis thaliana]
13094_at emb|CAB36745.1| (AL035523) 5dpi protein (AL035523.163_AT) putative protein [Arabidopsis processing thaliana]
14032__at emb|CAB38204.1| (AL035601) 2dpi metabolic (AL035601.11_AT) cytochrome P450-like protein 5dpi protein [Arabidopsis thaliana]
13147_at emb|CAB38794.1| (AL035678) 5dpi metabolic HpnA (AL035678.99_AT) putative protein [Arabidopsis protein protein, thaliana] oxidoreducta se/cinnamyl- alcohol dehydrogenas e
19624_at emb|CAB39628.1| (AL049481) 4dpi metabolic (AL049481.196_AT) cytochrome c [Arabidopsis protein thaliana]
14036_at emb|CAB41088.11 (AL049655) 4dpi protein (AL049655.54_AT) protein disulfide-isomerase-like processing protein [Arabidopsis thaliana]
14249_i_at emb|CAB43438.1| (AL050300) 5dpi stress stress (PAD4_I_AT) putative protein [Arabidopsis response thaliana]
14250_r_at emb|CAB43438.1| (AL050300) 5dpi stress stress (PAD4_R_AT) putative protein [Arabidopsis response thaliana]
16232_s_at emb|CAB45796.1| (AL080252) 5dpi signaling not protein (AL080252.77_S_AT) putative protein [Arabidopsis kinase phosphatse thaliana] 2C, putative
Ser/The phosphatase
2C
13212_s_at emb|CAB68132.1| (AL137080) 2-5dpi stress (BGL2_S_AT) beta-1, 3-glucanase 2 (BG2) response [Arabidopsis thaliana]
16578_s_at emb|CAB68132.1| (AL137080) 2-5dpi stress (ATHRPRP1B_S_AT) beta-1, 3-glucanase 2 (BG2) response [Arabidopsis thaliana]
15622_s_at gb|AAB40594.1| (U43945) 4dpi stress (ATU43945_S_AT) strictosidine synthase response [Arabidopsis thaliana]
Table 19 Correlation of SEQ ID NOs: 1-953 (#) representing genes, the expression of which is altered after pathogen infection of Arabidopsis with ProbeSet designations
# ProbeSet # ProbeSet
11 91_g_at 33 12218_at (AJ242588.2_AT)
1 (AC002387.210_G_AT) 12223_s_at
2 11997_at (AC005967.4_AT) 34 (AC007168.178_S_AT)
3 12002_at (AF069442.47_AT) 35 12227_at (AC007576.18_AT)
4 12004_at (AL022023.132_AT) 36 12233_at (AJ001807.1_AT)
5 12007_at (Z99708.249_AT) 37 12278_at (AJ011674.2_AT)
6 12037_at (AC004005.174_AT) 38 12290_at (Y09418.2_AT)
7 12048_at (AF001308.46_AT) 39 12307_at (AC002392.162_AT)
8 1205 l_at (AL021889.94_AT) 40 12314_at (AC001229.28_AT)
9 12062_at (AC006069.147_AT) 41 12317_at (AC004138.27_AT)
10 12068_at (AF118223.24_AT) 42 12323_at (AC002333.18_AT)
11 12072_at (AL035396.4_AT) 43 12332_s_at (AB023448.2_S_AT)
12 12079_s_at (A71597.1_S_AT) 44 12335_at (AC004411.73_AT)
13 12081_at (AC001645.140_AT) 12341_s_at
14 12086_s_at (AC002409.88_S_AT) 45 (AL021637.176_S_AT)
15 12091_at (AC004450.116_AT) 46 12347_at (AC007258.28_AT)
16 12092_at (AC004793.13_AT) 47 12349_s_at (X84728.6_S_AT)
17 12094_at (AC006223.143_AT) 48 12356_at (X99952.1_AT)
18 12115_at (AL033545.26_AT) 49 12369_at (AC002535.59_AT)
19 12124_s_at (Z97337.149_S_AT) 50 12400_at (X98453.1_AT)
20 12125_at (Z97341.99_AT) 51 12421_at (AJ002414.1_AT)
21 12128_at (AC004261.157_AT) 52 12438_at (AL021710.83_AT)
22 12136_at (AC007591.60_AT) 53 12448_at (AC002337.58_AT)
23 12150_at (AC004005.151_AT) 12449_s__at
54 (AC002343.179_S_AT)
24 12160_at (AC006284.117_AT)
55 12454_at (AC006232.164_AT)
25 12187_at (AC005489.31_AT)
12460_s_at
26 12191_at (AC006068.35_AT) 56 (AC006920.129_S_AT)
27 12193_at (AC006072.132_AT) 57 12475_at (Y11794.1_AT)
28 12198_at (AC006954.90_AT) 58 12487_at (AC004411.126_AT)
29 12203_at (AL021710.268_AT) 59 12493_g_at (Y09095.1_G_AT)
30 12212_at (AL049711.168_AT) 60 12497_at (AC006533.51_AT)
31 12216_at (AC007119.56_AT) 61 12500_s_at (AF081067.3_S__AT)
32 12217_at (AJ223804.1_AT) 62 12521_at (AF049236.28_AT)
ProbeSet # ProbeSet 12525_at (AC006587.85_AT) 12832_f_at (U33014.2_M_F_AT) 12530_at (Z99707.184_AT) 12851_s_at (ACCSYN1_S_AT) 12535_at (AL035538.156_AT) 12854_s_at (ACS1_S_AT) 12538_at (AF033205.2__AT) 12855_f_at (ADH_F_AT) 12559_at (AC005727.83_AT) 12879_s_at (AIG1_S_AT) 12560_at (AC005825.57_AT) 12880_s_at (AIG2_S_AT) 12561_at (AL021687.107_AT) 12883_s_at (APX_S_AT)
12571_s_at 12889_s_at (ASA1_S_AT) (AF149413.18_S_AT) 12891_at (ATACS6_AT) 12574_at (X82624.2_AT) 12892_g_at (ATACS6_G_AT) 12584_at (AC004521.233_AT) 12904_s_at (ATERF1_S_AT) 12609__at (X92975.2_AT) 12905_s_at (ATERF2_S_AT) 12626_at (AC006234.95_AT) 12906_s_at (ATERF3_S_AT) 12627_at (AC006533.99_AT) 12908_s_at (ATERF5_S_AT) 12642_at (AC006920.138_AT) 12909_s_at (ATERF6_S_AT) 12645_at (AL021712.56_AT) 12911_s_at 12656_at (AC006569.43_AT) (ATG6PDHE5_S_AT) 12698_at (AC000106.42_AT) 12921_s_at 12711_f_at (Z95773_F_AT) (ATHHMGCOAR_S_AT) 12712_f_at (Z95774_F_AT) 12930_s_at (ATLLS1_S_AT) 12727_f_at (Z95799_F_AT) 12951_at (AC005489.5_AT) 12736_f_at (Z97048_F_AT) 12958_at (AC002332.249_AT) 12737_f_at (Z97049_F_AT) 12962_at (AC004697.165_AT) 12744_at (AC001645.15_AT) 12965_at (AL021711.118_AT) 12759_at (AC005278.32_AT) 12966_s_at (AL023094.197_S_AT)
12760_g_at (AC005278.32_G_AT) 12989_s_at (AC004077.149_S_AT) 12764_f_at (AC004138.69_F_AT)
13003_s_at (AB021936.1_S_AT) 12772_at (AC005278.34_AT)
13005_at (AC004683.61_AT) 12776_at (AL021811.156_AT)
13014_at (U93215.87_AT) 12790_s_at (AL021635.58_S_AT)
13015_s_at (X98673.2_S_AT)
12797_s_at (AC007138.25_S_AT) 13025_at (AL050400.20_AT) 12801_at (AC005223.34_AT) 13040_at (AC002392.134_AT) 12802_at (AL022373.153_AT) 13048_s_at
126 (AC004138.22_S_AT) 12831_f_at (U33014.2_5_F_AT)
127 13070_at (AC006919.171_AT)
# ProbeSet # ProbeSet
128 13094_at (AL035523.163_AT) 156 13243_r_at (ELI32_R_AT)
129 13100_at (AC003680.50_AT) 157 13244_s_at (ELI32_S_AT)
130 13110_at (AF074021.34_AT) 158 13246_at (ERECTAL_AT)
131 13115_at (AC000375.44_AT) 159 13253_f_at (FPS1_F_AT)
132 13119_at (AC007260.23_AT) 13255_i_at
133 13128_at (AL049607.47_AT) 160 (gammaglutamyltranspepti_I_AT)
134 13134_s_at (AC002337.9_S_AT) 13256_s_at
161 (gammaglutamyltranspepti_S_AT)
135 13137_at (AC007169.86_AT)
13259_s_at
136 13144_at (AC007017.246_AT) 162 (glutathioneperoxidase 1 _S_AT)
137 13147_at (AL035678.99_AT) 13261_s_at
13152_s_at 163 (glutathionereductase 1 _S_AT)
138 (AC005322.24_S_AT) 164 13263_s_at (GST1_RC_S__AT)
13154_s_at 165 13266_s_at (GST4_S_AT)
139 (AC002333.210_S_AT)
166 13270_at (HSF21_AT)
140 13157_at (AC002409.35_AT)
167 13271_g_at (HSF21_G_AT)
141 13161_at (AF002109.89_AT)
168 13273__s_at (HSF4_S_AT)
13163_s_at
142 (AC005560.223_S_AT) 169 13275_f_at (HSP174_F_AT)
143 13176_at (AL031394.56_AT) 170 13277_i_at (HSP176A_I_AT)
144 13177_at (AL049640.42_AT) 171 13279_s_at (HSP176II_S_AT)
145 13187_i_at (ATTHIRED4_I_AT) 172 13284_s_at (HSP70_S_AT)
13188_r_at 173 13285_s_at (HSP83_S_AT)
146 (ATTHIRED4_R_AT) 174 13312_at (AC006223.75_AT)
13189_s_at 175 13367_at (AC004680.97_AT)
147 (ATTHIRED4_S_AT) 176 13370_at (AC005322.4_AT)
148 13190_s_at (ATTHIREDA_S_AT) 177 13381_at (AC006580.8_AT)
149 13211_s_at (BCHI_S_AT) 178 13395_at (AL035528.202_AT)
150 13212_s_at (BGL2_S_AT) 179 13435_at (AF003102.3_AT)
13215_s_at 180 13437_at (AF096371.8_AT) (CAFFEROYLCOAMETHYLTR
151 ANS_S_AT) 181 13450_at (AL049657.33_AT)
13217_s_at 182 13459_at (AF013294.21_AT)
152 (CALMODULINLIKE_S_AT) 183 13467_at (AL096860.198_AT)
153 13219_s_at (CHI4_S_AT) 184 13480_at (AC005223.15_AT)
13220_s_at (CHS- 185 13482_at (AC005896.195_AT)
154 EXONl_S_AT) 186 13534_at (AF149413.36_AT)
13221_at (CHS-WHOLE- 187 13536_at (AL021636.47_AT)
155 SEQ_AT)
188 13538_at (AL080254.75_AT)
# ProbeSet # ProbeSet
189 13547_s_at (AC004450.15_S_AT) 220 13755_at (NOVARTIS 15_AT)
190 13564_at (AC005312.113_AT) 221 13763_at (NOVARTIS21_AT)
191 13565_at (AL035601.21_AT) 222 13764_at (NOVARTIS22_AT)
192 13584_at (AC007127.23_AT) 223 13789_at (AJ132436.2_AT)
193 13588_at (AL021961.24_AT) 224 13803_at (Z97341.376_AT)
194 13589_at (AC000132.24_AT) 225 13806_at (AC002354.15_AT)
195 13604_at (AC000104.20_AT) 13812_s_at
196 13605_at (AL078470.75_AT) 226 (AC005275.104_S_AT)
197 13617_at (AC006592.64_AT) 13818_s_at
227 (AC006218.175_S_AT)
198 13627_at (AL035394.196_AT)
228 13825_s_at (AF104919.22_S_AT)
199 13637_at (AL049482.9_AT)
229 13834_at (AL080237.29_AT)
200 13645_at (AC000098.8__AT)
230 13842_at (AC002396.12_AT)
201 13647_at (AF000657.22_AT)
231 13848_at (AC003981.31_AT)
202 13651_at (AL035538.320_AT)
13880_s_at
203 13656_at (AC007138.31_AT) 232 (AL049480.183_S_AT)
204 13659_at (AL022347.46_AT) 233 13895_at (AC004218.63_AT)
13666_s_at 234 13896_at (AC004473.8_AT)
205 (INDOLE3GPS_S_AT)
235 13908_s_at (A71590.1_S_AT)
206 13680_s_at (LOXl_S_AT)
236 13918_at (AC005388.29_AT)
207 13685_s_at (MLOLIKE2_S_AT)
237 13920_at (AC005990.53_AT)
13688_s_at
208 (MONOPTEROS_S_AT) 238 13944_at (U89959.24_AT)
209 13697_at (NI16_AT) 239 13949_s_at (Z97343.352_S_AT)
210 13705_s_at (AC003671X_S_AT) 240 13963_at (AL021711.26_AT)
211 13706_s_at (AC005724X_S_AT) 241 13964_at (AL021889.3_AT)
212 13708_s_at (AC007259X_S_AT) 242 13966_at (AL022023.172_AT)
13714_at 243 13999_at (AF071527.56_AT)
213 (NOVARTIS101_RC_AT) 244 14015_s_at (A71588.1_S_AT)
13716_at 245 14016_s_at (A71596.1_S_AT)
214 (NOVARTIS 103_RC_AT) 246 14025_s_at (AC007293.3_S_AT)
13718_at 247 14026_at (AC000106.5_AT)
215 (NOVARTIS 105_RC_AT)
248 14030_at (AC005970.225_AT)
216 13722_at (NOVARTIS108_AT)
249 14032_at (AL035601.11_AT)
217 13728_at (NOVARTIS111_AT)
250 14036_at (AL049655.54_AT)
13746_at
218 (NOVARTIS 121_RC_AT) 251 14041_at (AC003970.28_AT)
219 13751_at (NOVARTIS 127_AT) 252 14052_at (AC004122.24_AT)
# ProbeSet # ProbeSet
14068_s_at 286 14448_at (AC002387.243_AT)
253 (AC006922.197_S_AT) 287 14450_at (AC002986.49_AT)
254 14070_at (AL049658.217_AT) 288 14459_at (AC006200.69_AT)
255 14083_at (AC005662.56_AT) 289 14460_at (AC006201.21_AT)
256 14089_at (AC006223.65_AT) 290 14461_at (AC006202.73._AT)
257 14100_at (AF002109.108_AT) 291 14468_at (AC007576.62_AT)
258 14110_i_at (AL035528.279_I_AT) 292 14475_at (AL021811.121_AT)
259 14116_at (AF077407.30_AT) 293 14487_at (Z97341.343_AT)
260 14122_at (AF058826.23_AT) 294 14498_at (AC004261.51_AT)
261 14139_at (NOVARTIS30_AT) 295 14530_at (AL021889.231_AT)
262 14141_at (NOVARTIS31_AT) 296 14573_at (AF069298.35_AT)
263 14145_at (NOVARTIS35_AT) 297 14584_at (AC007658.25_AT)
264 14148_at (NOVARTIS38_AT) 298 14591_at (AL035440.107_AT)
265 14162_at (NOVARTIS46_AT) 299 14595_at (AL022580.163_AT)
266 14170_at (NOVARTIS51_AT) 300 14605_at (AC006193.6_AT)
267 14197_at (NOVARTIS71_AT) 301 14608_at (AC007357.49_AT)
268 14214_at (NOVARTIS83_AT) 302 14609_at (AC002340.147_AT)
14217_at 303 14613_at (AC004669.20_AT)
269 (NOVARTIS85_RC_AT)
304 14614_at (AC004165.66_AT)
270 14223_at (NOVARTIS9_AT)
305 14620_s_at (PAT1_S_AT)
271 14235_at (NOVARTIS97_AT)
306 14635_s_at (PR.1_S_AT)
272 14240_s_at (NR1_S_AT)
307 14636_s_at (PR5_S_AT)
273 14242_s_at (NRA_S_AT)
308 14638_s_at (PRXCB_S_AT)
274 14248_at (PAD3_AT) 14640_s_at
275 14249_i_at (PAD4_I_AT) 309 (PUTATΓVΈMLOHI_S_AT)
276 14250_r_at (PAD4_R_AT) 310 14643_s_at (RAR047_S_AT)
277 14254_s_at (PALl-MRNA_S_AT) 14660_s_at
14256_f_at (PALl- 311 (THIOREDOXL_S_AT)
278 LNTRON_F_AT) 14663_s_at
279 14257_s_at (PAL2-MRNA_S_AT) (TREHALASEPRECUSOR_RC_
280 14320_at (AC005956.54_AT) 312 S_AT)
281 14381_at (AC002521.68_AT) 313 14667_s_at (TRPB_S_AT)
282 14386_at (AC005309.177_AT) 314 14672_s_at (TSA1_S_AT)
283 14408_at (AC002291.14_AT) 315 14673_s_at (TSB2_S_AT)
14428_s_at 316 14675_s_at (VSP_S_AT)
284 (AL023094.342_S_AT) 14682_i_at
285 1443 l_at (AL035394.279_AT) 317 (WT1012A_RC_I_AT)
# ProbeSet # ProbeSet
318 14686_s_at (WT1073_S_AT) 352 15067_at (AC004683.36_AT)
319 14696_at (WT740_RC_AT) 353 15073_at (AC007069.93_AT)
320 14697__g_at (WT740_RC_G_AT) 15085_s_at
321 14705_i_at (WT77_RC_I_AT) 354 (AL031018.274_S_AT)
322 14706_r_at (WT77_RC_R_AT) 355 15088_s_at (AC002311.37_S_AT)
323 1471 l_s_at (ZFPL_S_AT) 356 15091_at (AC004683.97_AT)
324 14735_s_at (AF008124_S_AT) 357 15098_s_at (ATU26945_S_AT)
325 14750_s_at (AF096370.12_S_AT) 358 15116_f_at (AF121356_F_AT)
326 14763_at (X86958.1_AT) 15118_s_at
359 (ATHGLUGRFS_S_AT)
14770_s_at
327 (AC002338.167_S_AT) 360 15120_s_at (ATU10034_S_AT)
328 14779_at (AC004680.71_AT) 361 15123__s_at (ATU40857_S_AT)
329 14780_at (AC004683.103_AT) 362 15124_s_at (ATU59508_S_AT)
330 14786_at (AC005397.115_AT) 363 15125_f_at (D85190_F_AT)
331 14793_at (AC006202.10_AT) 364 15129_s_at (AF030386_S_AT)
332 14838_s_at (M96073.6_S_AT) 365 15132_s_at (AF121878_S_AT)
333 14882_at (AL022605.63_AT) 366 15137_s_at (ATU57320_S_AT)
334 14884_at (AL031032.95_AT) 367 15140_s_at (ATU93845_S_AT)
335 14895_s_at (Z97344.138_S_AT) 368 15141_s_at (D85191_S_AT)
336 14900_at (AC000348.12_AT) 15154__s_at
369 (ATHMTGDAS_S_AT)
337 14923_at (AC006283.158_AT)
370 15161_s_at (ATU90522_S_AT)
338 14924_at (AC006283.46_AT)
371 15162_s_at (U01880_S_AT)
339 14928_at (AC006569.88_AT)
372 15175_s_at (ATU28215_S_AT)
340 1493 l_at (AC006951.173_AT)
373 15188_s_at (AF081202_S_AT)
341 14959_at (AC007202.26_AT)
374 15192_s_at (ATHERD1_S_AT)
342 14964_at (AC001229.8_AT)
375 15196_s_at (ATU43412_S_AT)
343 14965_at (AC002329.22_AT)
376 15197_s_at (ATU52851_S_AT)
344 14972__at (AC005499.38_AT)
377 15199_s_at (AB005804_S_AT)
345 14978_at (AC002333.49_AT)
378 15203_s_at (AB013887_S_AT)
346 14998_at (U93215.42_AT)
379 15211_s_at (ATH243813_S_AT)
347 15032_at (AC002294.8_AT)
380 15216_s_at (ATU75191_S_AT)
348 15039_at (AF001308.67_AT)
381 15342_at (AC006593.101_AT)
15040_g_at
349 (AF001308.67_G_AT) 382 15379_at (AC002335.182_AT)
350 15042_at (AL021961.3_AT) 383 15389_at (AC004786.100_AT)
351 15052_at (AC002332.103_AT) 384 15406_at (AC006931.179_AT)
# ProbeSet # ProbeSet
385 15431_at (AL030978.64_AT) 415 15622_s_at (ATU43945_S_AT)
386 15463_at (AL031326.226_AT) 416 15625_s_at (ATU74610_S_AT)
387 15473_at (AC006836.125_AT) 417 15629_s_at (AB003280_S_AT)
388 15479_at (AL049483.205_AT) 418 1563 l_s_at (AB005805_S_AT)
15483_s_at 419 15632_s_at (AB012570_S_AT)
389 (AC005819.20_S_AT) 420 15641_s_at (AF117063_S_AT)
390 15485_at (AC006233.109_AT) 421 15646_s__at (ATHSATlG_S_AT)
391 15487_at (AC007661.87_AT) 422 15665_s_at (AF022658_S_AT)
392 15496_at (AC006282.167_AT) 423 15669_s_at (AF047834_S_AT)
15515_r_at 424 15670_s_at (AF061638_S_AT)
393 (AF058919.32_R_AT)
425 15672_s_at (AF082299_S_AT)
394 15518_at (AC005322.28_AT)
426 15674_s_at (AF091844_S_AT)
395 15522_i_at (AL078637.213_I_AT)
15680_s_at
15523_s_at 427 (ATHATPK19B_S_AT)
396 (AL078637.213_S_AT)
428 15775_at (AL079344.196_AT)
397 15524_at (AC005508.25_AT)
429 15778_at (X98676.2_AT)
398 15526_at (AC004122.16_AT)
430 15779_g_at (X98676.2_G_AT)
15531_i_at
399 (AL078637.191_I_.AT) 431 15792_at (AC002341.106_AT)
15532_r_at 432 15798_at (AC002521.173_AT)
400 (AL078637.191_R_AT) 433 15815_s_at (Z97342.366_S_AT)
401 15539_at (AC005770.21_AT) 434 15832_at (AC004561.255_AT)
402 15540_at (AC006585.205_AT) 435 15839_at (AC005662.203_AT)
403 15543_at (AF096371.10_AT) 436 15859_at (AC006587.164_AT)
404 15544_at (AL021633.110_AT) 437 15866_s_at (AC007133.59_S_AT)
405 15547_at (AC005970.122_AT) 438 15874_at (AL022223.106_AT)
406 15551_at (AL035440.289_AT) 439 15886_at (AL078637.204_AT)
407 15578_s_at (AF004213_S_AT) 440 15900_at (AC005311.74_AT)
408 15580_s_at (AF057043_S_AT) 441 15919_at (AC007060.42_AT)
409 15582_s_at (ATH131392_S_AT) 442 1592 l_s_at (AC007067.1_S_AT)
410 15594_s_at (ATU56635_S_AT) 443 15924_at (AC007138.61_AT)
411 15606_s_at (AF061517_S_AT) 444 15943_at (AC007202.16_AT)
15613_s_at 445 15970_s_at (X71794.2_S_AT)
412 (ATHHOMEOA_S_AT) 446 15978_at (X68592.6_AT)
15614_s_at 15982_s_at
413 (ATHMERI5B_S_AT) 447 (AC006260.78_S_AT)
414 15617_s_at (ATHSAR1_S_AT) 448 16001_at (AF035385.2_AT)
# ProbeSet # ProbeSet
16003_s_at 481 16173_s_at (D78607_S_AT)
449 (AL021749.64_S_AT) 482 16203_at (AC007519.53_AT)
16021_s_at 483 16229_at (AL049638.148_AT)
450 (AL022224.182_S_AT)
484 16230_at (AL049655.78_AT)
451 1603 l_at (X94248.1_AT)
16232_s_at
452 16043_at (AC005489.17_AT) 485 (AL080252.77_S_AT)
453 16048_at (X78586.2_AT) 486 16233_at (AL080254.83_AT)
454 16053_i_at (Y14251.4_I_AT) 487 16236_g_at (X92657.3_G_AT)
455 16054_s_at (Y14251.4_S_AT) 488 16241_at (AL022604.84_AT)
456 16058_s_at (ATU94495_S_AT) 489 16272_at (AC006304.136_AT)
457 16059_s_at (D88206_S_AT) 490 16288_at (AF024504.17_AT)
458 16062_s_at (AB007789_S_AT) 491 16298_at (AL021890.71_AT)
459 16063_s_at (AB008103_S_AT) 492 16299_at (AL024486.185_AT)
460 16073_f_at (AF062908_F_AT) 16301_s_at
461 16077_s_at (AF085230_S_AT) 493 (AL031018.105_S_AT)
462 16080_f_at (AF 118822_F_AT) 494 16306_at (AL049751.112_AT)
463 16083_s_at (AF153283_S_AT) 495 16327_at (AC002334.12_AT)
16087_s_at 16329_s_at
464 (ATHATPK6A_S_AT) 496 (AF013294.17_S_AT)
465 16090_s_at (ATHFAD8A_S_AT) 497 16335_at (AL079347.105_AT)
466 16091_s_at (ATHHSP83_S_AT) 498 16340_at (AC004255.15_AT)
467 16092_s_at (ATHKAT1_S_AT) 499 16351_at (AL021684.194_AT)
468 16103_s_at (ATU60445_S_AT) 500 16357_at (AF149413.38_AT)
469 16105_s_at (ATU68017_S_AT) 501 16363_at (AC004255.14_AT)
470 16108_s_at (D78604_S_AT) 502 16365_at (AC003974.136_AT)
471 16110_s_at (AB004822_S_AT) 503 16383_at (AC006300.64_AT)
472 16111_f_at (AB007788_F_AT) 504 16391_at (AL050351.194_AT)
473 16130_s_at (AF078683_S__AT) 505 16398_s_at (AL022603.3_S_AT)
474 16133_s_at (AF089810_S_AT) 506 16405_at (AC005850.9_AT)
475 16134_s_at (AF132016_S_AT) 507 16406_at (AC006921.33_AT)
16151_s_at 508 16409_at (AC004393.2_AT)
476 (ATHPRKINA_S_AT) 509 16434_at (AL021711.157_AT)
16153_s_at 16440_s_at
477 (ATHRPRP1C_S_AT) 510 (AF002109.137_S_AT)
478 16159_s_at (ATU37697_S_AT) 511 16442_s_at (AJ002551.2_S_AT)
479 16161_s_at (ATU39072_S_AT) 16457_s_at
480 16164_s_at (ATU47029_S_AT) 512 (AC005397.17_S_AT)
# ProbeSet # ProbeSet
513 16461_i_at (AC004683.79_I_AT) 547 16649_s_at (ATHORF_S_AT)
16462_s_at 548 16701_at (AC005312.61_AT)
514 (AC004683.79_S_AT) 549 16712_at (AC006068.67_AT)
515 16465_at (Y08892.1_AT) 550 16721_at (AC006533.58_AT)
516 16468_at (AF049870.5_AT) 551 16747_at (AL021713.3_AT)
517 16470_s_at (AF068299.4_S_AT) 552 16753_at (AL031032.110_AT)
518 16483_at (X68053_AT) 553 16781_at (AC002392.100_AT)
519 16496_s__at (AF030386.1_S_AT) 554 16810_at (AC002339.46_AT)
520 16510_at (AL034567.198_AT) 555 16817_s_at (AL096882.91_S_AT)
521 16522_at (X77500.2_AT) 556 16859_at (AL035523.135_AT)
522 16524_at (AC006577.38_AT) 557 16864_ at (AF037367.4_I_AT)
523 16526_at (Z49227.1_AT) 558 16865_s_at (AF037367.4_S_AT)
524 16536_s_at (AB008107_S_AT) 559 16868_at (AL035679.123_AT)
525 16537_s_at (AB008111_S_AT) 16888_s_at
526 16538_s_at (AB010259_S_AT) 560 (AC004684.174_S_AT)
527 16539_s_at (AB013301_S_AT) 16891_s_at
528 1654 l_s_at (AB023423_S_AT) 561 (AF080120.33_S_AT)
529 16545_s_at (AF037229_S_AT) 562 16892_at (U37336.3_AT)
530 16548_s_at (AF054617_S_AT) 563 16902_at (AC007119.67_AT)
531 16553_f_at (AF078821_F_AT) 16903_g_at
564 (AC007119.67_G_AT)
16568_s_at
532 (ATHATCDPK_S_AT) 565 16908_at (AC002396.22_AT)
533 16570_s_at (ATHCDPKA_S__AT) 566 16914_s_at (AL049500.57_S_AT)
534 16578_s_at (ATHRPRPlB_S_AT) 567 16916_s__at (X77199.8_S_AT)
535 16589_s_at (ATU26937_S_AT) 568 16927_s_at (AF035384.2_S_AT)
536 16594_s_at (ATU39783_S_AT) 569 16939_at (AC002334.110_AT)
537 16603_s_at (ATU81293_S_AT) 16940_g_at
570 (AC002334.110_G_AT)
538 16609_s_at (AB008104_S_AT)
571 16943_s_at (Z97339.466_S_AT)
539 16610_s_at (AB008490_S_AT)
572 16951_i_at (AC005662.30_I_AT)
540 1661 l_s_at (AB008782_S_AT)
573 16952_s_at (AC005662.30_S_AT)
541 16613_s_at (AF012657_S_AT)
574 16955_at (AL031326.215_AT)
542 16617_s_at (AF029980_S_AT)
575 16968_at (AL021961.93_AT)
543 16620_s_at (AF051338_S_AT)
576 16970_s_at (Y18291.5_S_AT)
544 16635_s_at (AF126057_S_AT)
577 16972_at (AC004261.89_AT)
545 16638_s_at (AF139098_S_AT)
578 16978_g_at (AF076641.2_G__AT)
546 16646_s_at (ATHDHS1_S_AT)
579 16981_s_at (U35829.2_S_AT)
# ProbeSet # ProbeSet
580 16989_at (AL030978.46_AT) 613 17362_s_at (Z97338.181_S_AT)
581 16990_at (AC004684.91_AT) 614 17371_at (AF076243.44_AT)
582 16995_at (AC002391.188_AT) 615 17376_at (AL021890.218_AT)
583 17007_at (AC005896.26_AT) 616 17379_at (AF085279.9_AT)
584 17008_at (AC006585.212_AT) 617 17380_at (AL021961.39_AT)
585 17009_at (AL021633.163_AT) 618 17381_at (Z99708.402_AT)
586 17018_s_at (ATU18929_S_AT) 619 17398_at (AC002535.143_AT)
587 17027_s_at (ATU72958_S_AT) 620 17413_s_at (AJ006961.4_S_AT)
588 17039_s_at (D78602_S_AT) 621 17451_at (AC002343.47_AT)
589 17041_s_at (D89631_S_AT) 17452__g_at
590 1705 l_s__at (AF098947_S_AT) 622 (AC002343.47_G_AT)
17066_s_at 623 17458_at (AC006260.91_AT)
591 (ATHLIPOXY_S_AT) 624 17464_at (AC000132.72_AT)
592 17073_s_at (ATTS4391_S_AT) 625 17477_s_at (X63443.2_S_AT)
593 17075_s_at (ATU09961_S_AT) 626 17482_s_at (Z97343.441_S_AT)
594 17083_s_at (ATU18770_S_AT) 627 17484_at (X79052.2_AT)
595 17097_s_at (ATU66345_S_AT) 628 17485_s_at (Z97340.345_S_AT)
596 17104_s__at (D88541_S_AT) 629 17487_s_at (U18993.2_S_AT)
597 17105_s_at (AF055357_S_AT) 630 17490_s_at (M90416.2_S_AT)
598 1711 l_s_at (ATHACSC_S_AT) 631 17494_s_at (ATU30478_S_AT)
599 17119_s_at (AF132212_S_AT) 17500_s_at
17128_s_at 632 (ATHCALLGA_S_AT)
600 (ATHRPRP1A_S_AT) 633 17511_s__at (AF067605_S_AT)
601 17134_at (AC000106.53_AT) 634 17514_s_at (AF076277_S_AT)
602 17180_at (AF007270.30_AT) 635 17516_s_at (AF072536_S_AT)
603 17187_at (AF128396.2_AT) 636 17522_s_at (D78606_S_AT)
17191_i_at 637 17533_s_at (ATU43488_S_AT)
604 (AL021961.178JLAT) 638 17544_s_at (ATU40856_S_AT)
605 17300_at (X66017.2_AT) 639 17548_s_at (AF118823_S_AT)
17303_s_at 640 17555_s_at (ATU89296_S_AT)
606 (AC004683.25_S_AT)
641 17577_g_at (AF087820_G_AT)
607 17305_at (U25649.3_AT)
642 17578_at (AF093604_AT)
608 17323_at (U95973.69_AT)
643 17579_s_at (AF093753_S_AT)
609 17338_at (AC002535.97_AT)
644 17585_s_at (AF134487_S_AT)
610 17341_at (AL021713.89_AT)
645 17589_at (AF156783_AT)
611 17352_at (AC007127.33_AT)
646 17595_s_at (AF166352_S_AT)
612 17356_s_at (Z97338.190_S_AT)
# ProbeSet # ProbeSet
647 17636_at (AF077409.7_AT) 679 17975_at (AF175998__AT)
648 17648_at (AL021684.43_AT) 680 17990_at (AF178075_AT)
649 17653_at (AL035679.144_AT) 681 18010_s_at (AJ001264_S__AT)
650 17702_at (AC005700.212_AT) 682 18012_s_at (AJ002295_S_AT)
651 17719_at (AC006592.17_AT) 683 18045_at (AJ011976_AT)
652 17743_at ( AC002341.99_AT) 684 18054_at (AJ238846_AT)
17744_s_at 18109_s_at
653 (AC004684.168_S_AT) 685 (AC002391.206_S_AT)
654 17752_at (AC003974.37_AT) 686 18121_s_at (AC002337.21_S_AT)
655 17758_at (AF076243.41_AT) 687 18122_at (AC002338.110_AT)
656 17775_at (AC004392.2_AT) 688 18140_at (Z97341.319_AT)
657 17781_at (AL049746.177_AT) 689 18148_at (AC004669.25_AT)
17823_s_at 18167_s_at
658 (AC006555.10_S_AT) 690 (AL021711.23_S_AT)
17840_s_at 691 18176_at (AL035540.31_AT)
659 (AC002333.223_S_AT) 18194_i_at
660 17854_at (Z99707.366_AT) 692 (AL096859.227_I_AT)
661 17860_at (AL078467.4_AT) 693 18213_at (AL022140.126_AT)
662 17876_at (AJ007587.2_AT) 694 18215_at (97335.114_AT)
663 17877_g_at (J007587.2_G_AT) 695 18216_at (X95573.2_AT)
664 17881_at (AC002391.54_AT) 696 18217_g_at (X95573.2_G_AT)
665 17882_at (AL035523.49_AT) 18224_s_at
666 17886_at (AC004484.151_AT) 697 (AL021890.57_S_AT)
667 17893_at (AC004401.135_AT) 18226_s_at
698 (AC002343.142_S_AT)
668 17894_at (AC005724.44_AT)
699 18228_at (X91259.1_AT)
669 17899_at (Z97339.197_AT)
700 18234_at (AC000348.3_AT)
17900_s_at
670 (AC000106.13_S_AT) 18236_s_at
701 (AC004683.69_S_AT)
17907_s_at
671 (AC004684.165_S_AT) 702 18241_at (AC006580.71_AT)
672 17916_at (U22428.2_AT) 18242_g_at
703 (AC006580.71_G_AT)
673 17930_s_at (AJ006960.4_S_AT)
704 18255_at (AC005770.25_AT)
674 17945_at (Z97341.411_AT)
18258_s_at
675 17955_at (AL021768.242_AT) 705 (AC006439.222__S_AT)
676 17956_i_at (AC005967.32_I_AT) 706 18263_at (18263_AT)
677 17963_at (AL049730.88_AT) 707 18266_at (AC004684.33_AT)
678 17967_at (AL096859.32_AT) 708 18267_at (AC006223.23_AT)
# ProbeSet # ProbeSet
709 18268_s_at (AC006418.38_S_AT) 18681_at (L23573_AT)
710 18280_at (AC007369.2_AT) 18683_s_at (L27158_S_AT)
711 18284_at (AL021961.67_AT) 18686_s_at (U18126_S_AT)
712 18287_at (AC007661.142_AT) 18694_s_at (U89272_S_AT)
713 18299_s_at (M23872.2_S_AT) 18698_s_at (XI 7528_S_AT)
714 18301_s_at (AL022223.48_S_AT) 18716_at (X91916_AT)
715 18314_ at (AL078579.83_I_AT) 18720_s_at (X92419_S_AT)
716 18348_at (AL022603.104_AT) 18735_s_at (Z29490_S_AT)
18456_s_at 18753_s_at
717 (AC004697.159_S_AT) (AF118222.28_S_AT)
718 18471_at (AC006533.103_AT) 18782_at (AC003040.90_AT)
719 18508_s_at (AC006532.89_S__AT) 18803_at (AC005315.94_AT)
720 18515_at (AC007063.215_AT) 18844_at (AC005315.131_AT)
721 18544_at (AC007060.14_AT) 18866_at (AC005917.178__AT)
722 18582_s_at (AC003671.36_S_AT) 18885_at (AC006921.147_AT)
723 18587_s_at (AC007166.53_S_AT) 18888_at (AC007591.68_AT)
724 18590_at (AJ222713.4_AT) 18896_at (AC002329.51_AT)
725 18591_at (X74756.2_AT) 18899_s_at (X13434.1_S_AT)
726 18596_at (AC005698.13_AT) 18908_i_at (AF055848.2_I_AT)
727 18597_at (AL080282.74_AT) 18909_s_at (AF055848.2_S_AT)
728 18600_at (L76926.1_AT) 18916__s_at (X92393.1_S_AT)
18601_s_at 18928_at (AC002333.181_AT)
729 (AC002387.279_S_AT) 18930_at (AC005990.57_AT)
730 18604_at (AF069298.31_AT) 18933_at (AC007020.48_AT)
731 18622_g_at (AJ005902.2_G_AT) 18936_at (AJ003119.4_AT)
732 18625_at (AC005278.22_AT) 18949_at (Z54136.1_AT)
733 18631_at (AC002510.112_AT) 18953_at (AF077955.1_AT)
734 18634_s_at (Z97343.468_S_AT) 18963_at (AC004561.99_AT)
735 18635_at (AC004005.44_AT) 18966__at (AC004561.106_AT)
736 18636_at (AC006577.22_AT) 18976_at (AC000106.31_AT)
18650_s_at 18980_at (U78721.20_AT)
737 (AF013294.25_S_AT)
18984__at (AC003096.100_AT)
18662_s_at
738 (AC002343.20_S_AT) 19017_at (AL035709.69_AT)
739 18667_at (AJ249442_AT) 19019_i_at (X82623.2_I_AT)
740 18668_at (AJ249794_AT) 19044_at (AC004392.38_AT)
741 18672_s_at (D13983_S_AT) 19060_at (AC003671.34_AT)
# ProbeSet # ProbeSet
777 19092_at (AL078606.188_AT) 811 19451_at (AC004392.6_AT)
778 19110_s_at (X86947.2_S_AT) 19460_s_at
19132_s_at 812 (AC000132.66_S_AT)
779 (AL022603.298_S_AT) 813 19462_s_at (AFOO 1168.2_S_AT)
780 19137_at (X74755.2_AT) 814 19464_at (AC005560.51__AT)
781 19140_at (AC005170.24_AT) 815 19465_at (AL021768.96_AT)
782 19150_at (AC006577.20_AT) 816 19494_at (AC007296.26_AT)
783 19161_at (AL078579.9_AT) 817 19531_at (AL021960.91_AT)
784 19171_at (AC002335.160_AT) 818 19546_at (AC005398.172_AT)
785 19178_at (Y18227.2_AT) 819 19555_at (AF058919.48_AT)
786 19181_s_at (AF053065.2_S_AT) 820 19591_at (AJ010735.4_AT)
787 19182_at (AL031804.245_AT) 821 19614_at (AC003970.32_AT)
788 19207_at (AC006069.117_AT) 822 19623_at (AF000657.40_AT)
789 19219_at (AC007019.185_AT) 823 19624_at (AL049481.196_AT)
790 19230_at (AC003113.15_AT) 19625_s_at
791 19247_at (AF071527.44_AT) 824 (AC002311.26_S_AT)
792 19257_s_at (AC000104.57_S_AT) 825 19635_at (AL049746.38_AT)
793 19284_at (AC003028.196_AT) 826 19639_at (AL080252.22_AT)
794 19288_at (AC005824.130_AT) 827 19640_at (AC004561.78_AT)
795 19325_at (AL022604.42_AT) 828 19641_at (AC004561.66_AT)
796 19364_at (AL022023.142_AT) 829 19645_at (AC004561.70_AT)
797 19368_at (AC000348.22_AT) 19646_s_at
830 (AC005819.55_S_AT)
798 19376_at (AF024504.11_AT)
831 19655_at (Y14199.1_AT)
799 19383_at (AC006200.203_AT)
832 19660_at (AC002336.29_AT)
800 19386_at (AC006592.51_AT)
833 19667_at (AL021710.5_AT)
801 19388_at (AC000104.61_AT)
834 19672_at (AC005687.19_AT)
802 19395_at (AF007270.32_AT)
19673_g_at
803 19396_at (AJ001855.2_AT) 835 (AC005687.19_G_AT)
804 19405_at (AJ223803.1__AT) 19700_s_at
805 19407_at (AC004697.81__AT) 836 (AL031326.154_S_AT)
806 19409_at (AC007357.56_AT) 19701_s_at
807 1941 l_at (AC007661.104_AT) 837 (AC005724.67_S_AT)
808 19421_at (X70990.4_AT) 838 19704 _at (AJ005927.2_I_AT)
809 19424_at (AC002396.44_AT) 839 19707_s_at (Z95768.3_S_AT)
19432_s_at 840 19741_at (AL049171.72_AT)
810 (AL035680.11_S_AT) 841 19755_at (AC006593.64_AT)
ProbeSet # ProbeSet
19762_at (AL035527.204_AT) 875 20142_at (AL035521.155__AT)
19818_i_at (AL021749.33_I_AT) 876 20144_at (AL079350.68_AT)
19819_s_at 877 20165_at (AC002311.16_AT) (AL021749.33_S_AT) 878 20179_at (AL035538.229_AT)
19844_at (AJ007588.2_AT) 879 20189_at (AC005489.2_AT)
19845__g_at (AJ007588.2_G_AT) 880 20194_at (AC007584.48_AT)
19848_s_at 881 20199_at (AL050300.89_AT) (AC004261.94_S_AT)
882 20200_at (AL050400.67_AT)
1985 l_at (U23794.3_AT)
883 20215_s_at (AFl 17125.2_S_AT)
19870_s_at (AL021811.48_S_AT) 884 20223_at (AL022347.145_AT)
19878_at (AL080252.102_AT) 885 20238_at (X74514.2_AT)
19879_s_at (Z97338.342_S_AT) 886 20239_g_at (X74514.2_G_AT)
19881_at (AC004077.49_AT) 20245_s_at
887 (AC005309.97_S_AT)
19892_at (AC005770.30_AT)
888 20246_s_at (AF084037.3_S_AT)
19894_at (AJ001809.1_AT)
889 20247_at (AC004392.4_AT)
19895_s_at (U77347.4_S_AT)
890 20258_at (AFl 30252.1_AT)
19903_at (AC007660.40_AT)
891 20262_at (AC002294.26_AT)
19916_at (AC006577.34_AT)
892 20263_at (AB004798.1_AT)
19944_at (AC002130.4_AT)
893 20269_at (AC002387.237_AT)
\9946_at (AC004482.14_AT)
894 20271_at (Z99707.27_AT)
19956_at (AC006282.11_AT)
20285_s_at
19960_at (AL035527.360_AT) 895 (AC003674.18_S_AT)
19970_s_at 896 20287_at (Y14590.5_AT) (AC003674.10_S_AT)
897 20288__g_at (Y14590.5_G_AT)
19982_at (AC002986.28_AT)
898 20291_s_at (M92353.4_S_AT)
19991_at (AC007017.124_AT)
899 20297_at (AC007153.27_AT)
20017_at (AC004521.66_AT)
900 20323_at (AC004561.62_AT)
20023_at (AC006577.46_AT)
901 20335_s_at (Y14208.2_S_AT)
20030_at (AL078637.51_AT)
902 20345_at (AF104919.16_AT)
20051_at (AC000106.38_AT)
903 20346__at (L031135.156_AT)
20053_at (AC002292.27_AT)
904 20348__at (AC005967.35_AT)
2006 l_at (AC005508.23_AT)
905 20356_at (AC004561.74_AT)
20096_at (AC004238.31_AT)
906 20365_s_at (AC005850.19_S_AT)
20098_at (AC004697.123_AT)
907 20370_at (AC004561.263_AT)
20133_i_at (AC007178.71__I_AT)
908 20372_at (AL021713.24_AT)
20134_s_at (AC007178.71_S_AT)
# ProbeSet # ProbeSet
20382_s_at 20572_s_at
909 (AC002338.35_S_AT) 933 (AC005560.229_S_AT)
20409_g_at 934 20577_at (AL078464.72_AT)
910 (AC004077.132_G_AT) 935 20584_at (AC004450.75_AT) 911 20420_at (AL024486.131_AT) 20586_i_at
912 20421_at (U81294.2_AT) 936 (AC005824.195_I_AT)
913 20422_g_at (U81294.2_G_AT) 20587_s_at 914 20432_at (U43486.2_AT) 937 (AC005824.195_S_AT)
915 20433_at (AC006232.147_AT) 938 20589_at (AF081066.3_AT)
916 20442_i_at (AC006341.42_I_AT) 939 20590_at (AL035540.159_AT)
917 20443_s_at (AC006341.42_S_AT) 940 2059 l_at (AL080252.115_AT)
918 20450_at (AJ005930.2_AT) 941 20619_at (AC005896.161__AT)
919 20461_at (20461_AT) 20620_g_at
942 (AC005896.161_G_AT) 920 20462_at (U82399.2_AT) 943 20646_at (AC002291.20_AT) 921 20477_at (AC004238.154_AT) 944 20656_at (AL035396.46_AT) 922 20479_i_at (AF069495.2_I_AT)
20658_s_at
923 20480_s_at (AF069495.2_S_AT) 945 (2AL050400.70__S_AT)
924 20485_at (AC007660.131_AT) 946 20660_s_at (X97488.2_S_AT)
925 20491_at (AC004561.146_AT) 947 20669_s_at (AC002388.6_S_AT)
926 20507_at (AL021635.67_AT) 948 20675_at (AC006234.204_AT)
927 20511_at (AC007290.24_AT) 949 20678_at (AC007296.30_AT)
928 20516_at (AL035523.64_AT) 950 20685_at (AL049751.46_AT)
929 20517_at (Y17722.7_AT) 951 20686_at (Y14424.2_AT)
930 20524_at (AC005698.12_AT) 20689_s_at
931 20529_at (Z97341.125_AT) 952 (AC002335.19_S_AT)
932 2055 l_at (AC006081.211_AT) 953 20715_at (AF079183.1_AT)
Table 20a: cDNA-AFLP gene fragments with similarity to other known proteins
Related accession Blast cDNA*** Known protein Organism number** score*
DESCAl no significant similarity - - -
DESCA2 ser/thr kinase bean AF078082 2e-12
DESCA3 endo- 1 ,4-betaglucanase rape AJ242807 4e-19
DESCA4 pdr 5-abc transporter duckweed Z70524 le-27
DESCA5 transcriptional regulator yeast NP 014933 9e-l
DESCA6 kinase Arabidopsis T00502 7e-l
DESCA7 salicylate-induced glucosyltransferase IS5a tobacco T03747 2e-21
DESCA8 nbs-LRR rice AAF82158 5e-2
DESCA9 cytochrome p450 monooxygenase tobacco X96784 7e-31
DESCA10 MRP -like ABC transporter Arabidopsis U96399 8e-7
DESCAl 1 phosphoribosylanthranilate transferase Arabidopsis AAF18518 8e-17
DESCA12 hypersensitivity-related 201 tobacco X95343 3e-23
DESCAl 3 integral membrane glycoprotein puffer fish AF013613 9e-l
7a tgaa no significant similarity - - - lOd tcg hypothetical protein F3F9.18 Arabidopsis AC013430 7e-15 l la tgca no significant similarity - - - cr. actin actin C. rubrum X92353 7e-45
* NCBI BLASTX translated search (Altschul et al., 1997) ** Accession numbers are listed in SEQ ID Nos 1967-1980 *** DESCA sequences are listed in SEQ ID Nos 1954-1966
Table 20b: Relative gene expression fold changes of cDNA-AFLP gene fragments with similarity to other known proteins at different time points during viral infections as determined by quantitative RT-PCR
Gene expression fold changes
C. amaranticolor C. quinoa cDNA TMV1 TRV2 TMV3
4dai 7dai lldai 4dai 4dai
DESCAl 200 180 6.1 278 not detectable
DESCA2 36 25 27 10 700
DESCA3 23 15 2.7 19 46
DESCA4 21 9.2 4.5 6.8 52
DESCA5 19 8.0 9.1 15 1100
DESCA6 8.4 9.6 12 2.1 not detectable
DESCA7 8.9 2.8 5.6 53 150
DESCA8 5.9 3.2 2.0 5.8 120
DESCA9 5.1 5.9 4.2 3.1 not detectable
DESCAl 0 5.6 1.9 0.95 4.1 not detectable
DESCAl 1 5.5 3.6 3.0 3.9 230
DESCA12 5.0 2.4 1.7 5.6 7.9
DESCAl 3 2.9 1.8 1.9 3.3 34
7a tgaa 1.5 1.4 1.7 not tested not tested lOd tcg 2.2 0.75 0.25 not tested not tested llatgca 1.6 0.54 0.18 not tested not tested cr. actin 1.0 1.0 1.0 1.0 1.0
1 Values are the fold increases in gene expression of TMV-MGfus infected compared to mock- inoculated plants.
2 Values are the fold increases in gene expression of TRV infected compared to mock- inoculated plants.
Values are the fold increases in gene expression of TMV infected compared to mock- inoculated plants.
Table 21 Known plant/pathogen interactions
Plant Pathogen
Tomato Cladosporium fulvum
Maize Rust fungus
Antirrhinium Rust fungus
Flax Melampsora lini
Lettuce Downy mildew
Arabidopsis Peronospora parasitica
Tomato Nematode
Corn Cochliobolus carbonum
Tomato Pseudomonas syringae
Rice Xanfhomonas oryzae pv. Oryzae
Rice Pyricularia oryzae
Tobacco Tobacco Mosaic Virus
Table 22 Rice Open Reading Frames (ORFs) orthologous to specific Arabidopsis ORFs and corresponding rice promoter regions
AORF SEQ ID NO of a specific Arabidopsis ORF
RH SEQ ID NO of homologous rice ORF(s)
RP SEQ ID NO ofthe rice promoter linked to a homologous rice ORF
AORF RH RP AORF RH RP AORF RH RP
3 2808 4884 3475 5551 4307 6383
3066 5142 3567 5643 4458 6534
3820 5896 3632 5708 52 3035 5111
4655 6731 3769 5845 3157 5233
6 2757 4833 3916 5992 3379 5455
3013 5089 4298 6374 3626 5702
3393 5469 19 3184 5260 3990 6066
3899 5975 20 2678 4754 55 2687 4763
7 3227 5303 2926 5002 2869 4945
3579 5655 3736 5812 2984 5060
3851 5927 3864 5940 3054 5130
3890 5966 4124 6200 3180 5256
4516 6592 4414 6490 3561 5637
8 3086 5162 22 2805 4881 4306 6382
3341 5417 3997 6073 4326 6402
3762 5838 4126 6202 4460 6536
3767 5843 4153 6229 4476 6552
4285 6361 4311 6387 4521 6597
4487 6563 4639 6715 4603 6619
4488 6564 32 3718 5794 57 4005 6081
4489 6565- 4345 6421 59 2922 4998
10 3629 5705 37 4493 6569 3833 5909
3914 5990 42 3479 5555 3967 6043
4291 6367 4208 6284 4202 6278
4424 6500 4236 6312 60 2706 4782
11 3996 6072 46 3891 5967 4125 6201
4555 6631 49 4645 6721 4720 6796
14 2809 4885 50 4210 6286 65 2815 4891
3160 5236 51 2864 4940 4112 6188
3201 5277 2964 5040 4288 6364
3455 5531 4218 6294 67 2716 4792
AORF RH RP AORF RH RP AORF RH RP
2928 5004 75 2920 4996 3198 5274
2940 5016 3417 5493 3319 5395
3182 5258 3742 5818 3797 5873
3333 5409 3927 6003 4048 6124
3351 5427 4061 6137 4184 6260
3386 5462 78 2705 4781 4259 6335
3387 5463 2737 4813 102 3563 5639
3445 5521 2738 4814 3683 5759
3530 5606 4654 6730 4108 6184
3581 5657 4681 6757 4168 6244
3727 5803 85 3163 5239 4232 6308
3825 5901 3525 5601 110 2876 4952
3915 5991 3965 6041 3099 5175
4120 6196 4114 6190 3367 5443
4170 6246 91 3138 5214 3610 5686
4267 6343 3745 5821 4363 6439
4280 6356 3803 5879 4618 6694
4350 6426 4233 6309 4656 6732
4479 6555 4275 6351 111 2747 4823
4713 6789 93 4325 6401 2766 4842
68 2764 4840 95 2903 4979 2955 5031
3072 5148 3230 5306 3450 5526
3192 5268 3676 5752 3988 6064
3290 5366 4480 6556 112 2779 4855
3546 5622 4631 6707 3345 5421
3553 5629 97 2959 5035 3799 5875
3658 5734 3105 5181 3801 5877
3747 5823 3222 5298 4216 6292
3972 6048 3838 5914 113 3371 5447
4543 6619 4026 6102 3372 5448
70 3173 5249 4224 6300 3373 5449
72 3245 5321 4523 6599 4022 6098
4329 6405 4646 6722 4319 6395
4412 6488 100 2667 4743 115 2707 4783
73 2862 4938 2751 4827 2871 4947
3012 5088 2948 5024 3261 5337
3506 5582 3167 5243 3452 5528
AORF RH RP AORF RH RP AORF RH RP
4513 6589 4496 6572 3761 5837
116 2833 4909 4593 6669 3869 5945
2939 5015 4629 6705 4609 6685
3200 5276 127 2981 5057 137 2723 4799
3571 5647 3199 5275 3220 5296
4031 6107 3467 5543 4173 6249
118 2944 5020 3963 6039 138 3024 5100
3186 5262 4053 6129 3487 5563
3631 5707 128 3267 5343 3583 5659
3633 5709 4182 6258 4503 6579
3697 5773 4530 6606 4591 6661
3726 5802 130 2890 4966 139 2699 4775
4009 6085 3293 5369 3463 5539
4597 6673 3312 5388 3584 5660
119 3169 5245 3326 5402 4451 6527
3637 5713 3812 5888 4595 6671
3638 5714 3889 5965 140 3042 5118
3639 5715 4134 6210 3175 5251
3656 5732 4254 6330 4337 6413
121 4692 6768 133 2799 4875 148 3558 5634
122 3041 5117 3008 5084 3655 5731
3179 5255 3208 5284 4084 6160
3291 5367 3706 5782 150 3107 5183
4269 6345 3765 5841 3560 5636
4633 6709 134 3005 5081 151 4146 6222
124 3388 5464 3145 5221 4398 6474
3410 5486 3146 5222 4399 6475
3469 5545 3328 5404 152 4368 6444
4316 6392 3511 5587 4483 6559
4449 6525 4572 6648 4550 6627
126 3215 5291 4584 6660 4551 6628
3557 5633 135 2832 4908 4616 6692
3575 5651 3073 5149 158 3023 5099
3847 5923 3257 5333 3880 5956
4044 6120 3352 5428 4237 6313
4400 6476 3378 5454 4492 6568
4444 6520 3555 5631 159 2720 4796
AORF RH RP AORF RH RP AORF RH RP
3115 5191 172 3389 5465 189 2778 4854
3383 5459 3509 5585 3514 5590
3844 5920 3603 5679 4662 6738
3989 6065 3664 5740 190 3020 5096
4157 6233 3665 5741 3088 5164
4627 6703 173 4608 6684 3605 5681
162 2813 4889 175 3754 5830 3606 5682
2927 5003 3835 5911 3992 6068
3346 5422 4015 6091 199 2873 4949
3423 5499 4198 6274 2942 5018
3682 5758 4420 6496 3492 5568
3781 5857 179 2684 4760 3671 5747
3830 5906 4394 6470 3856 5932
4225 6301 180 3158 5234 4030 6106
4475 6551 3635 5711 4100 6176
4622 6698 4557 6633 4253 6329
164 3232 5308 4558 6634 4680 6756
3491 5567 181 2696 4772 202 2711 4787
4590 6666 3850 5926 3015 5091
4625 6701 4549 6625 3774 5850
165 3271 5347 185 3110 5186 3906 5982
3322 5398 3304 5380 3943 6019
3406 5482 3518 5594 203 3135 5211
3681 5757 3594 5670 3318 5394
3973 6049 3853 5929 3443 5519
4231 6307 187 2730 4806 3523 5599
4324 6400 2893 4969 3911 5987
166 2767 4843 3087 5163 3924 6000
2781 4857 3176 5252 4596 6672
3347 5423 4234 6310 204 2851 4927
4093 6169 188 3068 5144 2852 4928
4130 6206 3082 5158 3151 5227
171 2733 4809 3644 5720 3152 5228
2761 4837 4344 6420 4186 6262
3049 5125 4427 6503 205 3078 5154
3402 5478 4468 6544 3984 6060
3945 6021 4636 6712 4085 6161
AORF RH RP AORF RH RP AORF RH RP
206 3051 5127 219 2789 4865 247 3971 6047
3075 5151 2801 4877 250 2971 5047
3117 5193 2802 4878 3696 5772
3527 5603 2803 4879 3719 5795
3756 5832 3703 5779 4035 6111
209 3091 5167 4222 6298 4273 6349
3505 5581 220 2885 4961 4675 6751
3602 5678 3394 5470 252 2909 4985
3921 5997 3395 5471 3738 5814
4679 6755 3396 5472 3831 5907
210 3628 5704 3499 5575 3904 5980
4129 6205 221 2918 4994 4039 6115
212 2736 4812 3225 5301 4219 6295
2966 5042 4453 6529 4246 6322
3888 5964 4534 6610 4467 6543
4240 6316 4553 6630 4495 6571
4484 6560 225 3634 5710 254 2771 4847
213 2822 4898 3673 5749 2874 4950
2916 4992 3920 5996 3413 5489
2961 5037 4440 6516 3510 5586
3102 5178 4668 6744 4143 6219
3255 5331 229 2731 4807 255 2664 4740
215 2709 4785 3466 5542 2740 4816
2804 4880 3897 5973 2792 4868
2993 5069 4395 6471 3171 5247
3174 5250 4664 6740 3368 5444
3852 5928 237 2847 4923 3425 5501
216 2662 4738 3126 5202 3788 5864
2695 4771 3237 5313 4183 6259
3457 5533 3508 5584 4206 6282
3504 5580 3704 5780 4439 6515
3842 5918 3930 6006 256 2798 4874
4580 6656 4006 6082 3109 5185
4583 6659 239 2924 5000 3566 5642
218 3338 5414 4707 6783 3901 5977
3951 6027 240 3211 5287 4190 6266
4051 6127 3751 5827 4501 6577
AORF RH RP AORF RH RP AORF RH RP
4579 6655 4333 6409 3953 6029
4601 6677 4377 6453 3966 6042
258 2952 5028 271 3979 6055 4025 6101
3283 5359 4570 6646 4161 6237
4001 6077 4571 6647 4544 6620
4040 6116 274 3426 5502 289 2668 4744
4397 6473 3534 5610 2746 4822
260 3438 5514 275 2951 5027 3147 5223
3886 5962 3268 5344 3284 5360
4070 6146 3909 5985 3285 5361
4215 6291 4277 6353 3878 5954
4703 6119 4490 6566 4373 6449
263 2752 4828 285 3336 5412 4457 6533
3098 5174 3987 6063 290 2780 4856
3276 5352 4431 6507 2896 4972
3625 5701 286 3430 5506 3569 5645
4309 6385 287 2703 4779 4486 6562
265 2701 4777 3028 5104 4731 6807
2861 4937 3329 5405 295 2785 4861
3164 5240 3595 5671 2807 4883
4187 6263 3667 5743 2921 4997
4270 6346 3895 5971 2957 5033
267 2866 4942 4598 6674 3281 5357
2941 5017 4602 6678 3303 5379
3002 5078 4688 6764 3477 5553
4330 6406 288 2820 4896 3792 5868
270 2875 4951 2977 5053 3925 6001
3001 5077 2991 5067 3946 6022
3061 5137 3007 5083 4175 6251
3282 5358 3057 5133 296 2689 4765
3590 5666 3097 5173 2995 5071
3608 5684 3155 5231 2996 5072
3618 5694 3170 5246 3133 5209
3817 5893 3464 5540 4058 6134
3969 6045 3528 5604 4185 6261
4211 6287 3532 5608 4695 6771
4265 6341 3872 5948 298 2685 4761
AORF RH RP AORF RH RP AORF RH RP
2769 4845 3256 5332 4381 6457
2777 4853 3385 5461 4430 6506
3122 5198 3539 5615 4533 6609
3153 5229 3541 5617 331 2760 4836
3187 5263 4145 6221 2848 4924
3323 5399 314 2839 4915 3103 5179
4542 6618 3240 5316 3419 5495
299 2915 4991 3485 5561 3740 5816
3705 5781 3643 5719 3999 6075
4313 6389 3848 5924 4023 6099
4353 6429 3957 6033 4352 6428
4415 6491 4406 6482 4735 6811
4600 6616 4514 6590 333 2796 4872
4721 6797 316 3119 5195 2958 5034
300 3380 5456 3142 5218 3191 5267
3381 5457 3251 5327 3365 5441
3456 5532 3274 5350 3687 5763
3755 5831 3619 5695 3932 6008
4027 6103 318 2972 5048 4342 6418
301 2679 4755 3875 5951 4360 6436
2865 4941 4152 6228 339 3011 5087
3565 5641 4375 6451 3296 5372
4029 6105 4461 6537 3624 5700
4272 6348 319 4197 6273 3948 6024
302 2836 4912 4334 6410 4620 6696
3236 5312 324 3033 5109 4733 6809
304 2960 5036 3771 5847 341 3248 5324
3111 5187 3910 5986 3942 6018
3252 5328 326 2934 5010 346 4299 6375
3884 5960 3114 5190 348 3000 5076
3885 5961 3806 5882 3137 5213
305 2899 4975 4359 6435 3866 5942
3074 5150 4520 6596 4038 6114
3431 5507 330 3401 5477 4327 6403
3713 5789 3804 5880 4425 6501
4008 6084 4098 6174 360 2989 5065
312 2721 4797 4331 6407 2990 5066
AORF RH RP AORF RH RP AORF RH RP
3090 5166 4546 6622 391 2671 4747
3100 5176 4587 6663 3862 5938
3436 5512 370 4056 6132 3863 5939
3515 5591 372 3414 5490 3955 6031
3522 5598 3471 5547 3956 6032
3636 5712 4247 6323 4133 6209
3883 5959 4482 6558 393 3010 5086
3977 6053 4650 6726 3802 5878
4028 6104 374 2776 4852 3839 5915
4199 6275 3497 5573 3923 5999
4201 6277 3645 5721 4421 6497
4223 6299 3728 5804 4556 6632
4317 6393 3837 5913 4614 6690
4354 6430 378 3808 5884 394 3069 5145
4408 6484 4045 6121 3159 5235
4450 6526 4308 6384 3870 5946
4509 6585 4525 6601 4147 6223
365 3678 5754 380 3207 5283 4512 6588
3941 6017 3773 5849 398 2976 5052
4547 6623 4701 6111 3680 5756
4577 6653 4715 6791 404 4217 6293
4684 6760 4716 6792 405 4116 6192
4714 6790 381 4034 6110 410 2754 4830
366 2691 4767 382 4405 6481 3260 5336
3614 5690 4537 6613 411 3824 5900
3711 5787 385 3490 5566 4004 6080
3763 5839 4065 6141 412 2967 5043
3975 6051 4104 6180 2988 5064
369 2708 4784 4454 6530 3887 5963
3125 5201- 4456 6532 3935 6011
3144 5220 4730 6806 4462 6538
3190 5266 389 2750 4826 418 3223 5299
3370 5446 3529 5605 4376 6452
3432 5508 3620 5696 4504 6580
4067 6143 4575 6651 4505 6581
4355 6431 4694 6770 4663 6739
4410 6486 4723 6799 419 2854 4930
AORF RH RP AORF RH RP AORF RH RP
3085 5161 4640 6716 4049 6125
3442 5518 438 2818 4894 4158 6234
3537 5613 2863 4939 4164 6240
3552 5628 3344 5420 4466 6542
4200 6276 3454 5530 457 4382 6458
4378 6454 3613 5689 460 3166 5242
420 3321 5397 4683 6759 3592 5668
3517 5593 439 2819 4895 4107 6183
3544 5620 2905 4981 4128 6204
3689 5765 2935 5011 463 2850 4926
4243 6319 3949 6025 3488 5564
422 3228 5304 3958 6034 3489 5565
3249 5325 4413 6489 3578 5654
3250 5326 4417 6493 3849 5925
3709 5785 4465 6541 4517 6593
4589 6665 441 3330 5406 4624 6700
426 2919 4995 3708 5784 464 3657 5733
2943 5019 3836 5912 3752 5828
3022 5098 3898 5974 4251 6327
3273 5349 4419 6495 4372 6448
3739 5815 448 2715 4791 4726 6802
4409 6485 2773 4849 465 3071 5147
428 2753 4829 3053 5129 3270 5346
2897 4973 3408 5484 3533 5609
3096 5172 3415 5491 3796 5872
3500 5576 3538 5614 3905 5981
4438 6514 3900 5976 4418 6494
431 3193 5269 4358 6434 4605 6681
432 3029 5105 450 3398 5474 4674 6750
3219 5295 3795 5871 467 3640 5716
3375 5451 451 2992 5068 3734 5810
3376 5452 3183 5259 4447 6523
3623 5699 3269 5345 4464 6540
3968 6044 3300 5376 4515 6591
437 2855 4931 3478 5554 472 3462 5538
2937 5013 4000 6076 3591 5667
4341 6417 4018 6094 3823 5899
AORF RH RP AORF RH RP AORF RH RP
4212 6288 3503 5579 3929 6005
4349 6425 3776 5852 4052 6128
475 2739 4815 4685 6761 4081 6157
2772 4848 497 3669 5745 4089 6165
3168 5244 500 3444 5520 4314 6390
3382 5458 4226 6302 4628 6704
3451 5527 4370 6446 525 4162 6238
3865 5941 4621 6697 526 2840 4916
3974 6050 4651 6727 2994 5070
3982 6058 501 3481 5557 3003 5079
4364 6440 4239 6315 4020 6096
476 2829 4905 502 4610 6686 4653 6729
3343 5419 503 3857 5933 528 3004 5080
3593 5669 4611 6687 3433 5509
3741 5817 505 3165 5241 3674 5750
3829 5905 3242 5318 3789 5865
482 2853 4929 3749 5825 4638 6714
4062 6138 4383 6459 533 3044 5120
4106 6182 4718 6794 3241 5317
4127 6203 507 2763 4839 3646 5722
484 2702 4778 3026 5102 3961 6037
3302 5378 3409 5485 4518 6594
3327 5403 3468 5544 535 3859 5935
3701 5777 3983 6059 4328 6404
4502 6578 509 2759 4835 536 3286 5362
487 3143 5219 3418 5494 3545 5621
3677 5753 3912 5988 3659 5735
4075 6151 4079 6155 4117 6193
4443 6519 4470 6546 4149 6225
4710 6786 517 2672 4748 539 3641 5717
489 2841 4917 2690 4766 4248 6324
3188 5264 2727 4803 4286 6362
3686 5762 2846 4922 540 3093 5169
3867 5943 3278 5354 3127 5203
4673 6749 3337 5413 3325 5401
493 3032 5108 3526 5602 541 2888 4964
3480 5556 3730 5806 3037 5113
AORF RH RP AORF RH RP AORF RH RP
543 3116 5192 4136 6212 3822 5898
3434 5510 553 2666 4742 4010 6086
3435 5511 2945 5021 581 2765 4841
4087 6163 3101 5177 3065 5141
4088 6164 3224 5300 3513 5589
546 2791 4867 4459 6535 4043 6119
2933 5009 556 2929 5005 4416 6492
3735 5811 3810 5886 582 3212 5288
3970 6046 3811 5887 3411 5487
4154 6230 4046 6122 3841 5917
4213 6289 4548 6624 4169 6245
4292 6368 559 2734 4810 4293 6369
4401 6477 3611 5687 583 2712 4788
549 3369 5445 3737 5813 2713 4789
3420 5496 4179 6255 2725 4801
3421 5497 4647 6723 2812 4888
3441 5517 560 3598 5674 3031 5107
3559 5635 3599 5675 3794 5870
4069 6145 3600 5676 4274 6350
4300 6376 3712 5788 4699 6775
4301 6377 4388 6464 586 3746 5822
4561 6637 566 2953 5029 4672 6748
550 3141 5217 2954 5030 595 2821 4897
3361 5437 3615 5691 3275 5351
3377 5453 4386 6462 3473 5549
3939 6015 4387 6463 3877 5953
3960 6036 569 2786 4862 4097 6173
4091 6167 3019 5095 4312 6388
551 3340 5416 3233 5309 4362 6438
3363 5439 3307 5383 4554 6626
4142 6218 4469 6545 599 3055 5131
4159 6235 572 2674 4750 3213 5289
4436 6512 4099 6175 4166 6242
552 3805 5881 4366 6442 4167 6243
3993 6069 577 2825 4901 4562 6638
4016 6092 3006 5082 601 3079 5155
4077 6153 3136 5212 3424 5500
AORF RH RP AORF RH RP AORF RH RP
4241 6317 2999 5075 642 3039 5115
4321 6397 3353 5429 4082 6158
602 4083 6159 3362 5438 4238 6314
603 2775 4851 3472 5548 4437 6513
3685 5761 3723 5799 4563 6639
3934 6010 628 2898 4974 647 2758 4834
4235 6311 3106 5182 2879 4955
4351 6427 3108 5184 3214 5290
4657 6733 4539 6615 3476 5552
605 2724 4800 4540 6616 3564 5640
2784 4860 631 3247 5323 3661 5737
3052 5128 633 3294 5370 3772 5848
3288 5364 3295 5371 651 2867 4943
3422 5498 3391 5467 3244 5320
3731 5807 3453 5529 4017 6093
3944 6020 3621 5697 4191 6267
4007 6083 634 2710 4786 4242 6318
4346 6422 2756 4832 653 3077 5153
4389 6465 3148 5224 654 2663 4739
609 3311 5387 3470 5546 2835 4911
3791 5867 3572 5648 3459 5535
610 4011 6087 4078 6154 3908 5984
4522 6598 4221 6291 4582 6658
612 2795 4871 4365 6441 656 3660 5736
3130 5206 4404 6480 3893 5969
3574 5650 635 3040 5116 4012 6088
4315 6391 3258 5334 4021 6097
4529 6605 3339 5415 4508 6584
613 3162 5238 640 3568 5644 657 4139 6215
619 2974 5050 3790 5866 658 2878 4954
3717 5793 3821 5897 3437 5513
620 3650 5726 4511 6587 3486 5562
3653 5729 641 2719 4795 4002 6078
3675 5751 2793 4869 4160 6236
4526 6602 3556 5632 660 3132 5208
623 2849 4925 3714 5790 3292 5368
2936 5012 3991 6067 4426 6502
AORF RH RP AORF RH RP AORF RH RP
4632 6708 3622 5698 686 2907 4983
4687 6763 3940 6016 2917 4993
662 2969 5045 4367 6443 3540 5616
3535 5611 676 2963 5039 3892 5968
4260 6336 3084 5160 3937 6013
4261 6337 3112 5188 687 3779 5855
4262 6338 3221 5297 3902 5978
664 2704 4780 3332 5408 4569 6645
3766 5842 3507 5583 4615 6691
4094 6170 3570 5646 4722 6798
4689 6765 3699 5775 688 3297 5373
4732 6808 4138 6214 4095 6171
665 2741 4817 4390 6466 4282 6358
2827 4903 611 3439 5515 4433 6509
3056 5132 3440 5516 4446 6522
3181 5257 3952 6028 691 2837 4913
3448 5524 4072 6148 3238 5314
3520 5596 4073 6149 3364 5440
3882 5958 678 2810 4886 3542 5618
4080 6156 2883 4959 3768 5844
4101 6177 3195 5271 692 3585 5661
4171 6247 3266 5342 3586 5662
4176 6252 3427 5503 4658 6734
4205 6281 680 2797 4873 4659 6735
4264 6340 3750 5826 693 2806 4882
4278 6354 4588 6664 2923 4999
4507 6583 683 2735 4811 3043 5119
4676 6752 2970 5046 3229 5305
4691 6161 3760 5836 3320 5396
4734 6810 3778 5854 695 4258 6334
672 3104 5180 4560 6636 698 2842 4918
3907 5983 684 4320 6396 2843 4919
4141 6217 685 3140 5216 2844 4920
4612 6688 3587 5663 2908 4984
4669 6745 3876 5952 2949 5025
675 2717 4793 4119 6195 3554 5630
3149 5225 4727 6803 3670 5746
AORF RH RP AORF RH RP AORF RH RP
3926 6002 4538 6614 3725 5801
4338 6414 4607 6683 4189 6265
700 2947 5023 4704 6780 4402 6478
3246 5322 4705 6781 731 2688 4764
4180 6256 4706 6782 2900 4976
4441 6517 715 3757 5833 2962 5038
4666 6742 3936 6012 2979 5055
4667 6743 4229 6305 3016 5092
702 3216 5292 4230 6306 3048 5124
3549 5625 4528 6604 3070 5146
3654 5730 718 3047 5123 3092 5168
3733 5809 719 2817 4893 3589 5665
4132 6208 2894 4970 3617 5693
706 2691 4773 3076 5152 3720 5796
2770 4846 3314 5390 3782 5858
2788 4864 3759 5835 3962 6038
2845 4921 4092 6168 4155 6231
3156 5232 4271 6347 4174 6250
3234 5310 4302 6378 4474 6550
3744 5820 4303 6379 4510 6586
4013 6089 4304 6380 4545 6621
707 2983 5059 4305 6381 4729 6805
3582 5658 4455 6531 732 3027 5103
3642 5718 4473 6549 3064 5140
4059 6135 720 2930 5006 3710 5786
4076 6152 3062 5138 4524 6600
4194 6270 3172 5248 4566 6642
4686 6762 3289 5365 740 3861 5937
712 3038 5114 4384 6460 741 3196 5272
3392 5468 725 4228 6304 3871 5947
3400 5476 729 2787 4863 744 2732 4808
3512 5588 2975 5051 3210 5286
3758 5834 3095 5171 4255 6331
4110 6186 3494 5570 4256 6332
4118 6194 3495 5571 4347 6423
4177 6253 3496 5572 4374 6450
4422 6498 3702 5778 4500 6576
AORF RH RP AORF RH RP AORF RH RP
4626 6702 4144 6220 782 3783 5859
745 2892 4968 4196 6272 3834 5910
4428 6504 4295 6371 3854 5930
752 2686 4762 4396 6472 4165 6241
2830 4906 4724 6800 4586 6662
2831 4907 765 3231 5307 788 2677 4753
2932 5008 3627 5703 3573 5649
4623 6699 4268 6344 4192 6268
756 3094 5170 4361 6437 4434 6510
3484 5560 4661 6737 790 3349 5425
3597 5673 161 3777 5853 3793 5869
3652 5728 768 3813 5889 4263 6339
3860 5936 3814 5890 792 2877 4953
3879 5955 770 2913 4989 2901 4977
3964 6040 3354 5430 3516 5592
757 2826 4902 3355 5431 3918 5994
2910 4986 3903 5979 3947 6023
3017 5093 4527 6603 4019 6095
3030 5106 771 2906 4982 4279 6355
3067 5143 2911 4987 4296 6372
3080 5156 3131 5207 4356 6432
3113 5189 3458 5534 4643 6719
3178 5254 3874 5950 793 2698 4774
3272 5348 772 2856 4932 2794 4870
3482 5558 2973 5049 2811 4887
4585 6661 3083 5159 3350 5426
4671 6747 3197 5273 794 3919 5995
4711 6787 3217 5293 3954 6030
763 2868 4944 3502 5578 3986 6062
3124 5200 4003 6079 4448 6524
3679 5755 4565 6641 4665 6741
4203 6279 4604 6680 795 2676 4752
4432 6508 116 2670 4746 3058 5134
4498 6574 3209 5285 3139 5215
4712 6788 3780 5856 3690 5766
764 3301 5377 4209 6285 3691 5767
3715 5791 4485 6561 3692 5768
AORF RH RP AORF RH RP AORF RH RP
3693 5769 4559 6635 4369 6445
3928 6004 816 2768 4844 832 2790 4866
4214 6290 3150 5226 833 2824 4900
797 2700 4776 3846 5922 2889 4965
2828 4904 3894 5970 2956 5032
2882 4958 4057 6133 3360 5436
3306 5382 820 3648 5724 3397 5473
3695 5771 3855 5931 3521 5597
4135 6211 4257 6333 3647 5723
798 3358 5434 823 3800 5876 3698 5774
3405 5481 825 2870 4946 4357 6433
3407 5483 4096 6172 4423 6499
3460 5536 4283 6359 4637 6713
3596 5672 4284 6360 836 3243 5319
803 3194 5270 4564 6640 3262 5338
3239 5315 826 2931 5007 3263 5339
3299 5375 3331 5407 3404 5480
3588 5664 3429 5505 4131 6207
3612 5688 3601 5677 4322 6398
4736 6812 4103 6179 4348 6424
804 2692 4768 4494 6570 4592 6668
3059 5135 4541 6617 838 3045 5121
3399 5475 827 2814 4890 3154 5230
3721 5797 3707 5783 3357 5433
3729 5805 3784 5860 840 2800 4876
808 2904 4980 3785 5861 3843 5919
3280 5356 4188 6264 3933 6009
3827 5903 829 3786 5862 3978 6054
4429 6505 830 2673 4749 3998 6074
4660 6736 2982 5058 841 3089 5165
809 2872 4948 3390 5466 3279 5355
4717 6793 4435 6511 3335 5411
811 4033 6109 4649 6725 4641 6717
814 2880 4956 4690 6766 848 2749 4825
2987 5063 831 3334 5410 2912 4988
3416 5492 3543 5619 3254 5330
4055 6131 3743 5819 3976 6052
AORF RH RP AORF RH RP AORF RH RP
4708 6784 4452 6528 3985 6061
852 2680 4756 4599 6675 4391 6467
2859 4935 868 2681 4757 4617 6693
2860 4936 2895 4971 901 3265 5341
3118 5194 2980 5056 3672 5748
3313 5389 4193 6269 3775 5851
4047 6123 4477 6553 3873 5949
4379 6455 4478 6554 4635 6711
4532 6608 4696 6772 904 4266 6342
856 3366 5442 871 3308 5384 908 2891 4967
4481 6557 3309 5385 3009 5085
857 3501 5577 3310 5386 3536 5612
3609 5685 4567 6643 3950 6026
3931 6007 4576 6652 4071 6147
4336 6412 876 3021 5097 909 3348 5424
4702 6778 3161 5237 3651 5727
859 3412 5488 3206 5282 4102 6178
3694 5770 3607 5683 4578 6654
4156 6232 4024 6100 4613 6689
4499 6575 883 3384 5460 911 2823 4899
4568 6644 3819 5895 2857 4933
861 2665 4741 3832 5908 3226 5302
3120 5196 3868 5944 3938 6014
3121 5197 4318 6394 4294 6370
4109 6185 887 3018 5094 912 2718 4794
4140 6216 3913 5989 2816 4892
4163 6239 4066 6142 3185 5261
4574 6650 4407 6483 3662 5738
863 2729 4805 4725 6801 4393 6469
2925 5001 889 4204 6280 914 2728 4804
3576 5652 891 2834 4910 3177 5253
4148 6224 3298 5374 3483 5559
4195 6271 3815 5891 3809 5885
867 2726 4802 4151 6227 4339 6415
4310 6386 4252 6328 918 2968 5044
4332 6408 892 3129 5205 3577 5653
4442 6518 3342 5418 3787 5863
AORF RH RP AORF RH RP
4297 6373 3981 6057
4445 6521 944 2997 5073
919 2985 5061 3046 5122
3356 5432 950 3519 5595
3881 5957 3980 6056
4090 6166 4250 6326
4652 6728 4471 6547
921 2887 4963 4535 6611
926 2946 5022 952 2714 4790
3128 5204 2774 4850
3649 5725 3287 5363
3858 5934 3716 5792
928 4121 6197 4122 6198
4392 6468 4123 6199
4552 6629 4531 6607
929 2743 4819 4536 6612
2745 4821 4573 6649
3666 5742 4634 6710
4276 6352 953 2683 4759
4343 6419 3531 5607
4630 6706 3668 5744
930 4249 6325 3826 5902
932 2965 5041 4150 6226
3014 5090 SEQ
3840 5916 ID
NO:
4041 6117
4137 6213
4506 6582
934 3616 5692
3816 5892
4105 6181
4111 6187
4172 6248
943 2881 4957
2998 5074
3203 5279
3818 5894
Table 23 Plant Open Reading Frames (ORFs) orthologous to specific Arabidopsis ORFs
SEQ ID NO: 2
AAD17487.1 AF049347 Berberis stolonifera
Catalyzes the oxidative cyclization ofthe N-methyl group of (S)-reticuline into the berberine bridge carbon (C-8) of (S)-scoulerine. berberine bridge enzyme, bbel. Covalently flavinylated oxidase of isoquinoline alkaloid biosynthesis in plants.
AAB20352.1 S65550 Eschscholzia californica
(S)-reticuline:oxygen oxidoreductase (methy lene-bridge-foπning). /gene- '(S)- reticuline: oxygen oxidoreductase. This sequence comes from Fig 2; berberine bridge enzyme.
AAC39358.1 AF005655 Eschscholzia californica oxidizes the N-methyl group of (S)-reticuline to the berberine bridge carbon C-8 of (S)- scoulerine in berberine and benzophenanthridine alkaloid biosynthesis, berberine bridge enzyme, bbel. covalently-bound FAD-dependent oxidase; elicitor-inducible.
AAC61839.1 AF025430 Papaver somniferum berberine bridge enzyme, bbel. (S)-reticuline:oxygen oxidoreductase (methylene bridge fonning).
SEQ ID NO: 19 AAC83458.1 AF023140 Thlaspi goesingense imidazoleglycerol phosphate dehydratase. THB1.
AAB67738.1 U49978 Pisum sativum imidazoleglycerol-phosphate dehydratase. PSHIS3.
AAA93197.1 U02690 Triticum aestivum imidazoleglycerolphosphate dehydratase. partial interval represents the confirmed coding region based on homology to sequence U02689, maybe a chimeric molecule.
CAB55393.1 ALl 17264 Oryza sativa zwh0009.1. similar to wheat imidazoleglycerol-phosphate dehydratase (P34048); Method: conceptual translation with partial peptide sequencing.
SEQ ID NO: 32 CAA45066.2 X63464 Pisum sativum dihydrolipoamide dehydrogenase. lpd.
CAA44729.1 X62995 Pisum sativum lipoamide dehydrogenase.
AAG17888.1 AF295339 Solanum tuberosum dihydrolipoamide dehydrogenase precursor. Ipd2. NADH; E3 isoform.
BAB39219.1 AP002869 Oryza sativa putative dihydrolipoamide dehydrogenase precursor. P0554D10.3. contains ESTs
C98433(E0148),AU101058(E0148). SEQ ID NO: 33
CAC03581.1 AJ297566 Zea mays putative 1-deoxy-D-xylulose 5-phosphate reductoisomerase. dxr.
AAD24768.1 AFl 16825 Mentha x piperita l-deoxy-D-xylulose-5-phosphate reductoisomerase. DXR.
AAF65154.1 AF250235 Catharanthus roseus catalyzes the formation of 2-C-methyl-D-erythritol-4-phosphate from l-deoxy-D-xylulose-5- phosphate. l-deoxy-D-xylulose-5-phosphate reductoisomerase. dxr.
AAD56391.2 AF182287 Artemisia annua catalyzes the formation of 2-C-methyl-D-erythritol 4-phosphate from l-deoxy-D-xylulose-5- phosphate. l-deoxy-D-xylulose-5-phosphate reductoisomerase. DXR1.
BAB 16915.1 AP002863 Oryza sativa l-deoxy-d-xylulose-5-phosphate reductoisomerase. P0005A05.19. contains ESTs
AU108198(S11168),D46469(S11168). SEQ ID NO: 37
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein kinase.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAB41878.1 Y18259 Brassica oleracea
SRK5 protein. SRK5. receptor-like kinase.
CAB41879.1 Y18260 Brassica oleracea
SRK15 protein. SRK15. receptor-like kinase.
CAA74661.1 Y14285 Brassica oleracea
SFR1. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl .
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an itnmunoglobulin-like domain.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRK1. Chrkl .
BAB18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) corresponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
CAA79324.1 Z18884 Brassica oleracea S-receptor kinase related protein.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
SEQ ID NO: 38
CAA58750.1 X83869 Daucus carota CDPK-related protein kinase. CRK (or PK421).
BAA12692.1 D84508 Zea mays
CDPK-related protein kinase. Does not require calcium for its activity.
AAG01179.1 AF289237 Zea mays calcium calmodulin dependent protein kinase MCK2. MCK2.
AAB47181.1 S82324 Zea mays
/gene=''calcium/cahnodulin-dependent protein kinase. This sequence comes from Fig. 1.
BAA12691.1 D84507 Zea mays
CDPK-related protein kinase. Does not require calcium for its activity (by similarity).
BAA22410.1 D38452 Zea mays calcium-dependent protein kinase-related kinase.
AAC24961.1 AF009337 Tradescantia virginiana CDPK-related protein kinase. CRK1.
AAF23901.2 AF194414 Oryza sativa calcium-dependent protein kinase. CDPK5. OsCDPK5.
AAF23900.1 AF194413 Oryza sativa calcium-dependent protein kinase. CDPK1. OsCDPKl .
AAC78558.1 AF030879 Solanum tuberosum protein kinase CPK1.
CAA57157.1 X81394 Oryza sativa calcium-dependent protein kinase. OSCPK2.
BAB21081.1 AP002819 Oryza sativa putative calcium-dependent protein kinase. P0501G01.10.
AAC25423.1 AF072908 Nicotiana tabacum calcium-dependent protein kinase. CDPK1.
BAA12715.1 D85039 Zea mays calcium-dependent protein kinase.
AAB49984.1 U90262 Cucurbita pepo calcium-dependent cahnodulin-independent protein kinase CDPK. cpCPKl . serine/threonine protein kinase that is activated by direct binding of calcium.
AAA69507.1 U28376 Zea mays calcium-dependent protein kinase. MZECDPK2.
AAD 17800.1 AF090835 Mesembryanthemum crystallinum Ca2+-dependent protein kinase. CPK1. serine/threonine protein kinase.
AAB80693.1 U69174 Glycine max calmodulin-like domain protein kinase isoenzyme gamma. CDPK gamma. BAA81751.1 ABO 17517 Marchantia polymorpha calcium-dependent protein kinase. CDPK1. CDPK-B; alternative splicing. BAA81749.1 ABO 17515 Marchantia polymorpha calcium-dependent protein kinase. CDPK1. CDPK-B; alternative splicing. CAA39936.1 X56599 Daucus carota calcium- dependent protein kinase. DcPK431.
BAA81748.1 ABO 17515 Marchantia polymorpha calcium-dependent protein kinase. CDPK1. CDPK-A; alternative splicing. BAA81750.1 ABO 17516 Marchantia polymorpha calcium-dependent protein kinase. CDPK1. CDPK-A; alternative splicing. AAA61682.1 L27484 Zea mays calcium-dependent protein kinase. CDPK.
BAB 16888.1 AB042550 Oryza sativa
OsCDPK7. oscdpk7.
AAB88537.1 AF035944 Fragaria x ananassa calcium-dependent protein kinase. MAX17.
AAD28192.2 AFl 15406 Solanum tuberosum calcium-dependent protein kinase. CDPK; catalytic domain.
CAA07481.1 AJ007366 Zea mays calcium-dependent protein kinase. BAA12338.1 D84408 Zea mays calcium dependent protein kinase. ZmCDPKl .
BAA13440.1 D87707 Ipomoea batatas calcium dependent protein kinase. CDPK.
BAA13232.1 D87042 Zea mays Calcium-dependent protein kinase.
CAA65500.1 X96723 Medicago sativa protein kinase. CDPK.
AAB70706.1 U82087 Tortula ruralis calmodulin-like domain protein kinase. TrCPKl .
AAC49405.1 U08140 Vigna radiata calcium dependent protein kinase. CDPK.
BAA85396.1 AP000615 Oryza sativa
ESTs C22369(C12239),C22370(C12239), AU057852(S21844),AU057853(S21844) correspond to a region ofthe predicted gene.; similar to calcium dependent protein kinase. (AF048691).
CAA57156.1 X81393 Oryza sativa calcium-dependent protein kinase. OSCPKII.
AAC05270.1 AF048691 Oryza sativa calcium dependent protein kinase. CDPK12.
AAF21062.1 AF216527 Dunaliella tertiolecta calcium-dependent protein kinase. CPK1; CDPK.
AAB80692.1 U69173 Glycine max calmodulin-like domain protein kinase isoenzyme beta. CDPK beta.
AAK26164.1 AY027885 Cucumis sativus calcium-dependent cahnodulin-independent protein kinase 5. CDPK5. CsCDPK5.
AAA33443.1 L15390 Zea mays calcium-dependent protein kinase. CDPK.
CAA89202.1 Z49233 Chlamydomonas eugametos calcium-stimulated protein kinase.
AAG46110.1 AC073166 Oryza sativa calcium-dependent protein kinase. OSJNBb0064P21.2.
BAA02698.1 D13436 Oryza sativa calcium-dependent protein kinase. spk.
BAA90814.1 AP001168 Oryza sativa
ESTs AU030197(E50746),AU030196(E50746) correspond to a region ofthe predicted gene.;
Similar to calcium-dependent cahnodulin-independent protein kinase CDPK (U90262).
CAB46228.1 Y18055 Arachis hypogaea calcium dependent protein kinase. CDPK.
AAC49008.1 U24188 Lilium longiflorum calcium/calmodulin-dependent phosphorylation activity, calcium/calmodulin-dependent protein kinase. CCaMK. serine/threonine kinase; binds to calcium and calmodulin.
AAF19401.1 AF203479 Glycine max phosphoenolpyruvate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAF06970.1 AF162662 Kalanchoe fedtschenkoi phosphoenolpyruvate carboxylase kinase. calcium-independent protein kinase.
SEQ ID NO: 39 AAG00510.1 AF285172 Phaseolus vulgaris leaf senescence-associated receptor-like protein kinase. SARK.
BAB21175.1 AP002909 Oryza sativa putative serine/threonine-specific receptor protein kinase. P0044F08.3.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLK1. accumulates in mesophyll cells.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1 A.
AAF59906.1 AFl 97947 Glycine max receptor protein kinase-like protein. CLV1B.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml .
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl . Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl.7.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLK1. GmRLKl.
AAK11569.1 AF318493 Lycopersicon hirsutum Pto-like protein kinase D. LhirPtoD.
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAC48932.1 U13923 Lycopersicon pimpinellifolium
Fen. putative serine/threonine protein kinase; similar to product encoded by Lycopersicon pimpinellifolium Pto gene, GenBank Accession Number U02271; Fen is a member ofthe Pto gene family.
AAF76307.1 AF220602 Lycopersicon pimpinellifolium Fen kinase.
AAB47424.1 U59317 Lycopersicon pimpinellifolium serine/threonine protein kinase Fen. Fen, fenthion sensitivity gene from tomato.
AAK11566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhirPtoE. confers resistance to bacterial speck disease.
AAB47423.1 U59315 Lycopersicon pimpinellifolium serine/threonine protein kinase Pto. Pto. bacterial speck disease resistance in tomato; disease resistance gene.
AAC48914.1 U02271 Lycopersicon pimpinellifolium protein kinase.
AAF76306.1 AF220602 Lycopersicon pimpinellifolium
Pto kinase.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) corresponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AFOO 1308).
AAF76313.1 AF220603 Lycopersicon esculentum
Pto kinase. LescPth5.
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
AAF76314.1 AF220603 Lycopersicon esculentum
Fen kinase. Lescfen.
AAK11568.1 AF318492 Lycopersicon hirsutum
Pto-like protein kinase B. LhirPtoB.
AAB47422.1 U59318 Lycopersicon esculentum serine/threonine protein kinase Fen, fen, allele of Fen, fenthion sensitivity gene from tomato.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAA06538.1 D31737 Nicotiana tabacum protein-serine/threonine kinase.
AAK11567.1 AF318491 Lycopersicon hirsutum
Pto-like protein kinase F. LhirPtoF.
AAF34426.1 AF172282 Oryza sativa leucine rich repeat containing protein kinase. DUPR11.16.
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRK1. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461). SEQ ID NO: 41
AAG12987.1 AF166498 Lycopersicon esculentum sucrose transporter-like protein. SUT2. sucrose sensor.
AAF08330.1 AF021809 Vitis vinifera putative sucrose transporter. VvSUC12.
BAA24071.1 D87819 Oryza sativa sucrose transporter. OsSUTl.
AAF90181.1 AF280050 Oryza sativa subsp. indica sucrose proton symporter. sucrose transporter. SUTl.
BAA83501.1 AB008464 Zea mays Sucrose Transporter. ZmSUTl.
CAB75882.1 AJ272309 Hordeum vulgare
Transport sucrose through membrane, sucrose transporter 1. sutl .
AAD55269.1 AFl 82445 Vitis vinifera sucrose transporter.
CAA83436.1 Z31561 Ricinus communis sucrose transport, sucrose carrier. Scrl.
CAA76368.1 Y16767 Daucus carota sucrose/H+ symporter. SUTlb.
CAA76367.1 Y16766 Daucus carota sucrose/H+ symporter. SUTl a.
CAC19688.1 AJ303198 Daucus carota essential for sucrose transport, sucrose/proton symporter. sutl a.
AAG25923.1 AF237780 Solanum tuberosum sucrose transporter SUT4.
AAF08329.1 AF021808 Vitis vinifera putative sucrose transporter. VvSUCl 1.
AAG09270.1 AFl 76950 Lycopersicon esculentum sucrose transporter. SUT4.
AAF65765.1 AF242307 Euphorbia esula sucrose transport protein.
AAF04295.1 AFl 91025 Alonsoa meridionalis sucrose transporter 1. SUTl.
CAA57727.1 X82276 Nicotiana tabacum sucrose transporter. NtSUTla. Var. SNN bp 1-1346, Var. SRI bp 1172-1614.
BAA89458.1 AB036758 Daucus carota sucrose transporter protein. cSUT.
CAB75881.1 AJ272308 Hordeum vulgare transport sucrose through membrane, sucrose transporter 2. sut2.
CAA76369.1 Y16768 Daucus carota sucrose/H+ symporter. SUT2.
CAC19689.1 AJ303199 Daucus carota essential for sucrose transport, sucrose/proton symporter. sut2.
CAB07811.1 Z93774 Vicia faba sugar transport, sucrose transport protein, sut.
CAA47604.1 X67125 Spinacia oleracea sucrose permease. S21.
CAA48915.1 X69165 Solanum tuberosum sucrose transport protein. SUTl .
AAD41024.1 AFl 09922 Pisum sativum sucrose transport protein SUTl .
AAD53000.1 U64967 Beta vulgaris sucrose-proton symporter. Bv8-6.
AAF08331.1 AF021810 Vitis vinifera putative sucrose transporter. VvSUC27.
CAA58730.1 X83850 Beta vulgaris sucrose/proton-symporter. sutl. AAC99332.1 AF063400 Apium graveolens sucrose transporter. SUTl.
AAD45391.1 AF167416 Apium graveolens sucrose transporter SUT2B. SUT2B.
AAD45390.1 AF167415 Apium graveolens sucrose transporter SUT2A. SUT2A.
AAF04294.1 AF191024 Asarina barclaiana sucrose transporter 1. SUTl.
CAA59113.1 X84379 Plantago major SUC 1 -sucrose proton symporter. SUC1.
AAD34610.1 AF149981 Nicotiana tabacum sucrose transporter-like protein. SUT3.
CAA57726.1 X82275 Lycopersicon esculentum sucrose transporter. LeSUTl.
CAA12256.1 AJ224961 Ricinus communis Sucrose transport. Sucrose carrier. SUTl.
CAC33492.1 AJ310643 Ricinus communis sucrose carrier, sucrose carrier, scrl.
AAD45932.1 AF168771 Betula pendula sucrose transport protein. SUC1.
CAA53390.1 X75764 Plantago major sucrose transporter, ptp 1.
AAF22281.1 AFl 67417 Apium graveolens putative sucrose transporter SUTl. SUTl.
BAA76434.1 AB025006 Cicer arietinum sucrose transport protein.
SEQ ID NO: 42
AAC64184.1 AF095577 Prunus persica endopolygalacturonase.
CAA54150.1 X76735 Prunus persica endopolygalacturonase.
AAC28906.1 AFOO 1003 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 5.
TAPG5. expressed in abscission.
AAC28947.1 AF029230 Lycopersicon esculentum polygalacturonase. TPG6.
AAC70951.1 AF072732 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella, polygalacturonase 7. specifically and abundantly expressed in pistils.
AAC28905.1 AFOO 1002 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 4.
TAPG4. expressed in abscission.
AAA80489.1 U23053 Lycopersicon esculentum polygalacturonase precursor.
AAC28903.1 AFOO 1000 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 1.
TAPG1. expressed in abscission.
AAB09575.1 U70480 Lycopersicon esculentum abscission polygalacturonase. TAPG2.
AAC28904.1 AF001001 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 2.
TAPG2. expressed in abscission.
AAB09576.1 U70481 Lycopersicon esculentum abscission polygalacturonase. TAPG4.
AAC28902.2 AF000999 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 3.
TAPG3. expressed in abscission.
AAC26510.1 AF062465 Cucumis melo polygalacturonase precursor. MPG1.
AAC26511.1 AF062466 Cucumis melo polygalacturonase precursor. MPG2.
BAB21092.1 AP002819 Oryza sativa putative endopolygalacturonase. P0501G01.21.
AAA62286.1 U20431 Medicago sativa polygalacturonase.
AAA82167.1 U09717 Gossypium hirsutum polygalacturonase. __^^
AAA58322.1 U09805 Gossypium barbadense polygalacturonase.
CAA50336.1 X71018 Nicotiana tabacum polygalacturonase. NPG. Protein sequence is in conflict with the conceptual translation;
G27Y.
CAA50334.1 X71016 Nicotiana tabacum polygalacturonase. NPG. Protein sequence is in conflict with the conceptual translation;
G27.X.
CAA50337.1 X71019 Nicotiana tabacum polygalacturonase. NPG. Protein sequence is in conflict with the conceptual translation;
G27W.
CAA65072.1 X95800 Brassica napus polygalacturonase.
BAA89478.1 AB029459 Salix gilgiana polygalacturonase. SgPG3.
CAA90272.1 Z49971 Brassica napus Hydrolytic enzyme. Polygalacturonase. pga.
CAA50335.1 X71017 Nicotiana tabacum polygalacturonase. Npgl. Protein sequence is in conflict with the conceptual translation.
CAA50338.1 X71020 Nicotiana tabacum polygalacturonase. Npgl.
BAA89479.1 AB029460 Salix gilgiana polygalacturonase. SgPG4.
AAC14453.1 L12019 Actinidia deliciosa polygalacturonase.
BAA89476.1 AB029457 Salix gilgiana polygalacturonase . SgPGl.
AAC26512.1 AF062467 Cucumis melo polygalacturonase precursor. MPG3.
CAA72003.1 Y11118 Medicago sativa polygalacturonase. PG3.
CAB42886.1 AJ238848 Phleum pratense polygalacturonase. pg.
BAA89477.1 AB029458 Salix gilgiana polygalacturonase. SgPG2.
CAC05658.1 AJ250919 Brassica napus endopolygalacturonase. pegaz.
CAC05657.1 AJ250918 Brassica napus endopolygalacturonase. pgaz.
AAA32914.1 L06094 Persea americana cell wall degradation, polygalacturonase.
AAF71160.1 AF152758 Actinidia chinensis polygalacturonase A. PGA.
AAF61444.1 AF138858 Lycopersicon esculentum hydrolyses polygalacturonic acid, polygalacturonase. XOPG 1.
AAA34178.1 M37304 Lycopersicon esculentum polygalacturonase.
CAA32235.1 X14074 Lycopersicon esculentum polygalacturonase.
CAA29148.1 X05656 Lycopersicon esculentum polygalacturonase (AA 1-457).
AAD46483.1 AF 128266 Glycine max polygalacturonase PG1.
BAA88472.1 AB035890 Cucumis sativus polygalacturonase. CUPGl.
AAD46484.1 AF 128267 Glycine max polygalacturonase PG2.
CAA54448.1 X77231 Prunus persica polygalacturonase. PG.
CAA47055.1 X66426 Persea americana polygalacturonase.
CAA47052.1 X66422 Zea mays polygalacturonase. PG.
SEQ ID NO: 44
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil. Ptil kinase.
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S13927).
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
AAF91336.1 AF249317 Glycine max Ptil kinase-like protein. Ptila. protein kinase.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) corresponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAC27894.1 AF023164 Zea mays leucine-rich repeat transmembrane protein kinase 1. ltkl .
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) corresponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAC27895.1 AF023165 Zea mays leucine-rich repeat transmembrane protein kinase 2. Itk2.
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) correspond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APK1 gene for protein tyrosine-serine-threonine kinase.(D12522).
AAG25966.1 AF302082 Nicotiana tabacum cyto-άnin-regulated kinase 1. CRK1. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
AAF43496.1 AF131222 Lophopyrum elongatum protein serme/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAB09771.1 U67422 Zea mays
CRINKLY4 precursor. cr4. receptor kinase homolog.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
AAF76313.1 AF220603 Lycopersicon esculentum Pto kinase. LescPth5.
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
AAB47423.1 U59315 Lycopersicon pimpinellifolium serine/tlireonine protein kinase Pto. Pto. bacterial speck disease resistance in tomato; disease resistance gene.
AAF76306.1 AF220602 Lycopersicon pimpinellifolium
Pto kinase.
AAC48914.1 U02271 Lycopersicon pimpinellifolium protein kinase.
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
AAF66615.1 AF142596 Nicotiana tabacum
LRR. receptor-like protein kinase.
AAK11566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhirPtoE. confers resistance to bacterial speck disease.
AAK11567.1 AF318491 Lycopersicon hirsutum Pto-like protein kinase F. LhirPtoF. CAB51836.1 AJ243961 Oryza sativa
Putitive Ser/Thr protein kinase. 11332.7.
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSXNBa0004G10.30.
BAB18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
BAA92836.1 AB032473 Brassica oleracea
S18 S-locus receptor kinase. SRK18. SEQ ID NO: 45
AAB72110.1 U79958 Pisum sativum
BP-80 vacuolar sorting receptor.
AAF80450.1 AF161719 Triticum aestivum vacuolar targeting receptor bp-80.
AAG60258.1 AY017377 Physcomitrella patens
EGF receptor-like protein. ELP. PPELP; similar to putative vacuolar sorting receptor.
AAF22842.1 AF209910 Prunus dulcis vacuolar sorting receptor protein. BP-80 type protein.
AAK31596.1 AY029172 Helianthus annuus
EGF receptor-like protein. ELP.
AAB72113.1 U79961 Zea mays vacuolar sorting receptor homolog. similar to Pisum sativum BP-80 vacuolar sorting receptor,
GenBank Accession Number U79958.
BAA92985.1 AP001550 Oryza sativa
EST D15617(C0960A) corresponds to a region ofthe predicted gene.; Similar to Arabidopsis thaliana chromosome 4, BAC clone F18F4; vacuolar sorting receptor-like protein
(AL021637). SEQ ID NO: 47
BAA00885.1 D10001 Pisum sativum phenylalanine ammonia-lyase.
AAK15640.1 AF326116 Agastache rugosa phenylalanine ammonia-lyase. PAL.
CAB42794.1 AJ238754 Citrus Clementina x Citrus reticulata phenylalanine-ammonia lyase. pa!2.
BAA00886.1 D10002 Pisum sativum phenylalanine ammonia-lyase. PALI.
BAA00887.1 D10003 Pisum sativum phenylalanine ammonia-lyase. PAL2.
AAB67733.1 U43338 Citrus limon phenylalanine ammonia-lyase. pal6.
AAF40224.1 AF237955 Rubus idaeus phenylalanine ammonia-lyase 2. PAL2. PAL; phenylpropanoid; multigene; flavonoid.
AAAI 7993.1 M91192 Trifolium subterraneum phenylalanine ammonia-lyase. PALI.
CAA57057.1 X81159 Petroselinum crispum phenylalanine ammonia-lyase 3. PAL3. tetramere subunit.
AAF40223.1 AF237954 Rubus idaeus phenylalanine ammonia-lyase 1. PALI. PAL; phenylpropanoid; multigene; flavonoid.
CAA68938.1 Y07654 Petroselinum crispum
PALI protein, pall gene.
CAA57056.1 X81158 Petroselinum crispum phenylalanine ammonia-lyase 2. PAL2. deaminase subunit.
CAB60719.1 AJ250836 Cicer arietinum phenylpropanoid pathway, phenylalanine ammonia-lyase. pal.
CAA05251.1 AJ002221 Digitalis lanata phenylalanine ammonia lyase.
AAC78457.1 AF036948 Prunus avium phenylalanine ammonia-lyase. PALI.
BAA23367.1 D85850 Daucus carota phenylalanine ammonia-lyase. gDcPALl.
CAA68256.1 X99997 Bromheadia fmlaysoniana phenylalanine ammonia-lyase. pal.
AAA33805.1 LI 1747 Populus x generosa phenylalanine ammonia lyase. PAL.
BAA24928.1 D83075 Lithospermum erythrorhizon phenylalanine ammonia-lyase.
CAA34226.1 XI 6099 Oiyza sativa subsp. japonica phenylalanine ammonia-lyase.
CAA41169.1 X58180 Medicago sativa phenylalanine ammonia-lyase. PAL.
AAA34176.1 M90692 Lycopersicon esculentum phenylalanine ammonia-lyase. PAL5.
BAA07860.1 D43802 Populus kitakamiensis phenylalanine ammonia-lyase.
AAA84889.1 U39792 Pinus taeda phenylalanine ammonia-lyase. lpPAL.
CAA73065.1 Y12461 Helianthus annuus phenylalanine ammonia lyase. PAL.
BAA95629.1 AB042520 Catharanthus roseus phenylalanine ammonia lyase.
BAA05643.1 D26596 Camellia sinensis phenylalanine ammonia-lyase.
BAA24929.1 D83076 Lithospermum erythrorhizon phenylalanine ammonia-lyase.
BAA21643.1 D30656 Populus kitakamiensis phenylalanine ammonia-lyase.
CAA37129.1 X52953 Glycine max phenylalanine ammonia-lyase. PALI.
AAA34122.1 M84466 Nicotiana tabacum phenylalanine ammonia lyase. tpal.
BAA22948.1 AB008200 Nicotiana tabacum phenylalanine ammonia-lyase. palB.
AAA99500.1 L36822 Stylosanthes humilis phenylalanine ammonia lyase. PALI 7.1.
CAB42793.1 AJ238753 Citrus Clementina x Citrus reticulata phenylalanine-ammonia lyase. pall.
AAG49585.1 AF325496 Ipomoeanil phenylalanine ammonia-lyase.
CAA55075.1 X78269 Nicotiana tabacum phenylalanine ammonia-lyase.
BAA22963.1 D 17467 Nicotiana tabacum phenylalanine ammonia-lyase. TOBPALl.
BAA22947.1 AB008199 Nicotiana tabacum phenylalanine ammonia-lyase. palA.
AAA34179.2 M83314 Lycopersicon esculentum deamination of phenylalanine to coumarate. phenylalanine ammonia lyase. pal.
BAA11459.1 D78640 Ipomoea batatas
Phenylalanine Ammonia-Lyase.
AAA33389.1 M29232 Ipomoea batatas phenylalanine ammonia-lyase.
CAA68036.1 X99705 Triticum aestivum phenylalanine ammonia-lyase. PAL.
CAA61198.1 X87946 Oryza sativa phenylalanine ammonia-lyase. ZB8.
BAA06337.1 D30657 Populus kitakamiensis phenylalanine ammonia-lyase.
AAD45384.1 AF165998 Vigna unguiculata phenylalanine ammonia-lyase.
CAA53733.1 X76130 Cucumis melo phenylanaline ammonia-lyase. pal.
AAA51873.1 Ul 6130 Persea americana phenylalanine ammonia lyase. PAL.
BAB19128.1 AB041361 Dianthus caryophyllus phenylalanine ammonia-lyase. Dcpall.
CAA34715.1 X16772 Petroselinum crispum phenylalanine ammonia-lyase (AA 137 - 716) (1 is 3rd base in codon).
BAA07861.1 D43803 Populus kitakamiensis phenylalanine ammonia-lyase.
SEQ ID NO: 48
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
CAA66037.1 X97351 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
BAA06335.1 D30653 Populus kitakamiensis peroxidase.
AAA34108.1 J02979 Nicotiana tabacum lignin-forming peroxidase precursor (EC 1.11.1.7).
BAA01992.1 D11396 Nicotiana tabacum 'peroxidase'.
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI-1.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
BAA11853.1 D83225 Populus nigra peroxidase.
CAA66034.1 X97348 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
BAA01877.1 D11102 Populus kitakamiensis peroxidase. prxAl.
BAA11852.1 D83224 Populus nigra peroxidase.
CAA66035.1 X97349 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
AAB47602.1 L07554 Linum usitatissimum peroxidase. FLXPER1.
AAD37427.1 AF149277 Phaseolus vulgaris peroxidase 1 precursor. FBP1. secretory peroxidase.
CAA66036.1 X97350 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
AAB97734.1 AF014502 Glycine max seed coat peroxidase precursor. Ep. H2O2 oxidoreductase; class III plant peroxidase.
BAA06334.1 D30652 Populus kitakamiensis peroxidase.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
CAA62225.1 X90692 Medicago sativa peroxidaselA. prxlA.
CAA62227.1 X90694 Medicago sativa peroxidase IC. prxlC.
AAB41811.1 L36157 Medicago sativa peroxidase. pxdC. amino acid feature: conserved domains, aa 123 .. 129, 191 .. 198; amino acid feature: heme-binding domain, aa 68 .. 73.
AAB41810.1 L36156 Medicago sativa peroxidase. pxdA. amino ; icid feature: conserved motifs, aa 181 .. 188; amino acid feature: heme-binding domain, aa 60 .. 65.
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
BAA02840.1 D13683 Populus kitakamiensis peroxidase. HPOX14.
BAA14144.1 D90116 Armoracia rusticana peroxidase isozyme.
AAA33129.1 M91372 Cucumis sativus peroxidase. pre-peroxidase.
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
BAA08499.1 D49551 Oryza sativa peroxidase. poxN.
BAA03373.1 D14482 Oryza sativa putative peroxidase.
AAA34101.1 L02124 Nicotiana tabacum peroxidase.
BAA82306.1 AB027752 Nicotiana tabacum peroxidase.
CAA40796.1 X57564 Armoracia rusticana peroxidase. peroxidase precursor.
AAA33121.1 M32742 Cucumis sativus peroxidase (CuPer2).
CAA76680.1 Y17192 Cucurbita pepo peroxidase. aprx. type III peroxidase.
BAA92500.1 AP001383 Oryza sativa
ESTs D39300(R3292),AU030751(E60187) correspond to a region ofthe predicted gene.
Similar to peroxidase ATP6a. (X98774).
BAA77388.1 AB024438 Scutellaria baicalensis peroxidase 2.
AAA33127.1 M91373 Cucumis sativus peroxidase. pre-peroxidase :. putative.
AAD43561.1 AFl 55124 Gossypium hirsutum bacterial-induced peroxidase precursor. Perx_Goshiko.
AAB06183.1 M37636 Arachis hypogaea cationic peroxidase. PNC1
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl5 precursor, type III peroxidase.
CAA71492.1 Y10466 Spinacia oleracea peroxidase. prxr5.
AAB02554.1 L37790 Stylosanthes humilis cationic peroxidase.
BAA94962.1 AB042103 Asparagus officinalis peroxidase. AspPOXl.
BAA77389.1 AB024439 Scutellaria baicalensis peroxidase 3.
SEQ ID NO: 50
CAA40796.1 X57564 Armoracia rusticana peroxidase. peroxidase precursor.
BAA03373.1 D14482 Oryza sativa putative peroxidase.
BAA08499.1 D49551 Oryza sativa peroxidase. poxN.
CAA66037.1 X97351 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
BAA84764.1 D84400 Oryza sativa peroxidase. poxA.
BAA03372.1 D14481 Oryza sativa putative peroxidase.
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
BAA06335.1 D30653 Populus kitakamiensis peroxidase.
BAA11853.1 D83225 Populus nigra peroxidase.
AAD37427.1 AF149277 Phaseolus vulgaris peroxidase 1 precursor. FBP1. secretory peroxidase.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
BAA11852.1 D83224 Populus nigra peroxidase.
CAA66035.1 X97349 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
CAA66034.1 X97348 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
BAA92500.1 AP001383 Oryza sativa
ESTs D39300(R3292),AU030751(E60187) correspond to a region ofthe predicted gene.
Similar to peroxidase ATP6a. (X98774).
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI-1.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
CAA66036.1 X97350 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
AAB47602.1 L07554 Linum usitatissimum peroxidase. FLXPER1.
CAA62226.1 X90693 Medicago sativa peroxidase IB. prxlB.
AAA34108.1 J02979 Nicotiana tabacum lignin-forming peroxidase precursor (EC 1.11.1.7).
CAA62227.1 X90694 Medicago sativa peroxidase IC, . prxlC.
BAA06334.1 D30652 Populus kitakamiensis peroxidase.
BAA01992.1 D11396 Nicotiana tabacum 'peroxidase'.
AAD43561.1 AFl 55124 Gossypium hirsutum bacterial-induced peroxidase precursor. Perx_Goshiko.
BAA82306.1 AB027752 Nicotiana tabacum peroxidase.
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
AAB97734.1 AF014502 Glycine max seed coat peroxidase precursor. Ep. H2O2 oxidoreductase; class III plant peroxidase.
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl5 precursor, type III peroxidase.
CAA62225.1 X90692 Medicago sativa peroxidaselA. prxlA.
BAA01877.1 D11102 Populus kitakamiensis peroxidase. prxAl.
AAF63026.1 AF244923 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl4 precursor, type III peroxidase.
BAA14144.1 D90116 Armoracia rusticana peroxidase isozyme.
AAB41810.1 L36156 Medicago sativa peroxidase. pxdA. amino acid feature: conserved motifs, aa 181 .. 188; amino acid feature: heme-binding domain, aa 60 .. 65.
AAA34050.1 M74103 Nicotiana sylvestris anionic peroxidase.
CAA62597.1 X91172 Raphanus sativus korean-radish isoperoxidase. prxkl .
AAB02554.1 L37790 Stylosanthes humilis cationic peroxidase.
CAA76680.1 Y17192 Cucurbita pepo peroxidase. aprx. type III peroxidase.
CAA71492.1 Y10466 Spinacia oleracea peroxidase. prxr5.
BAA94962.1 AB042103 Asparagus officinalis peroxidase. AspPOXl.
BAA77389.1 AB024439 Scutellaria baicalensis peroxidase 3.
AAD37428.1 AF149278 Phaseolus vulgaris peroxidase 3 precursor. FBP3. secretory peroxidase.
AAB41811.1 L36157 Medicago sativa peroxidase. pxdC. amino acid feature: conserved domains, aa 123 .. 129, 191 .. 198; amino acid feature: heme-binding domain, aa 68 .. 73.
AAB06183.1 M37636 Arachis hypogaea cationic peroxidase. PNC1.
SEQ ID NO: 52
BAA02112.1 D12544 Pisum sativum GTP-binding protein.
CAA98184.1 Z73956 Lotus japonicus GTP-binding protein. RABl IH. rabl IH.
AAK15703.1 AF327517 Oryza sativa GTP-binding protein.
BAA02904.1 D13758 Oryza sativa ras-related GTP binding protein. ss230.
BAA02111.1 D 12543 Pisum sativum GTP-binding protein.
BAA02113.1 D 12545 Pisum sativum GTP-binding protein.
CAA98180.1 Z73952 Lotus japonicus GTP-binding protein. RABl ID. rabl ID.
CAB65172.1 AJ245570 Lycopersicon esculentum putative role in secretion of cell wall modifying enzymes. Rabl 1 GTPase. Rabl la.
BAA02114.1 D12546 Pisum sativum GTP-binding protein.
CAA98181.1 Z73953 Lotus japonicus GTP-binding protein. RABl IE. rabl IE.
CAA95859.1 Z71276 Mangifera indica small GTPase. rabX. homologous to Rabl 1.
CAA55865.1 X79278 Medicago sativa GTP binding protein. Rab.
CAA89049.1 Z49190 Beta vulgaris GTP-binding. small G protein.
CAA98179.1 Z73951 Lotus japonicus GTP-binding protein. RABl IC. rabl IC.
BAA02437.1 D13152 Oryza sativa GTP binding protein. rgp2.
BAA06701.1 D31905 Zea mays mgpl GTP-binding protein, mgpl.
BAA06702.1 D31906 Zea mays mgp2 GTP-binding protein. mgp2.
BAA02110.1 D12542 Pisum sativum GTP-binding protein.
CAA98177.1 Z73949 Lotus japonicus GTP-binding protein. RABl 1 A. rabl 1 A.
CAA41966.1 X59276 Oryza sativa GTP-binding protein, rgpl.
AAB97114.1 U58853 Glycine max small GTP-binding protein, sral.
CAA98185.1 Z73957 Lotus japonicus GTP-binding protein. RABl II. rabl II.
CAA67153.1 X98540 Fagus sylvatica FSGTP1.
CAA98183.1 Z73955 Lotus japonicus GTP-binding protein. RAB11G. rabllG.
CAA98182.1 Z73954 Lotus japonicus GTP-binding protein. RABl IF. rabl IF.
CAA54506.1 X77301 Glycine max GTPase. gmr2.
BAA02108.1 D12540 Pisum sativum GTP-binding protein.
AAA68983.1 L12395 Brassica napus signal transduction, membrane vehicle traffic, small GTP-binding protein, bra. putative.
AAD48018.1 AF165095 Gossypium hirsutum
Rab GTP-binding protein Rabl la.
BAA02109.1 D12541 Pisum sativum GTP-binding protein.
CAA98186.1 Z73958 Lotus japonicus GTP-binding protein. RABl 1 J. rabl 1 J.
BAA84640.1 AB007911 Pisum sativum PRA2. pra2. light-repressible GTP binding protein.
AAD48019.1 AF165096 Gossypium hirsutum Rab GTP-binding protein Rabl lb.
CAA98178.1 Z73950 Lotus japonicus GTP-binding protein. RABl IB. rabl IB.
AAA63901.1 U22432 Zea mays GTP binding protein. rab2.
CAA98165.1 Z73937 Lotus japonicus GTP-binding protein. RAB2A. rab2A.
AAA34253.1 L08130 Volvox carteri GTP-binding protein. yptV4.
AAA90955.1 U32185 Glycine max vesicular transport, guanine nucleotide regulatory protein. rab2. GTP-binding protein; soyrab.
AAA63902.1 U22433 Zea mays GTP binding protein. rab2.
SEQ ID NO: 53
CAA64327.1 X94624 Brassica napus acyl-CoA synthetase.
CAA96523.1 Z72153 Brassica napus acyl CoA synthetase.
CAC19877.1 AJ401089 Brassica napus activation of free fatty acids, long chain acyl-CoA synthetase. acs6. activity confirmed by expression in E. coli.
CAA06820.1 AJ006025 Cicer arietinum acyl-coA synthetase.
BAA08365.1 D49366 Lithospermum erythrorhizon 4-coumarate:CoA ligase.
CAA36850.1 X52623 Oryza sativa 4-coumarate-CoA ligase.
AAF37734.1 AF052223 Lolium perenne 4-coumarate~CoA ligase 4CL3.
AAF91309.1 AF239686 Rubus idaeus 4-coumarate:coA ligase 2. adeny late-forming enzyme; 4CL2.
AAC24503.1 AF041049 Populus tremuloides 4-coumarate:CoA ligase.
BAA08366.2 D49367 Lithospermum erythrorhizon 4-coumarate:CoA ligase.
AAF91308.1 AF239685 Rubus idaeus 4-coumarate:coA ligase 3. adenylate-forming enzyme; 4CL3.
AAC39366.1 AF008184 Populus x generosa 4-coumarate:CoA ligase 1. 4CL1.
CAA31696.1 XI 3324 Petroselinum crispum 4-coumarate:CoA ligase Pc4Cl-l (AA 1-544).
AAC39365.1 AF008183 Populus x generosa 4-coumarate: Co A ligase 2. 4CL2.
AAA33842.1 M62755 Solanum tuberosum 4-coumarate— CoA ligase. St4Cl-L
CAA31697.1 XI 3325 Petroselinum crispum 4-coumarate: CoA ligase Pc4Cl-2 (AA 1-544).
AAB42383.1 U39405 Pinus taeda 4-coumarate: CoA ligase. lp4CL-l.
AAB42382.1 U39404 Pinus taeda 4-coumarate:CoA ligase. lp4CL-2.
AAA92669.1 U12013 Pinus taeda 4-coumarate-CoA ligase enzyme.
AAF91310.1 AF239687 Rubus idaeus 4-coumarate:coA ligase 1. adenylate-forming enzyme; 4CL1.
AAA92668.1 U12012 Pinus taeda 4-coumarate-CoA ligase enzyme.
AAF37732.1 AF052221 Lolium perenne 4-coumarate— CoA ligase 4CL1.
CAA49575.1 X69954 Glycine max 4-coumarate— CoA ligase.
AAC24504.1 AF041050 Populus tremuloides 4-coumarate:CoA ligase.
AAF37733.1 AF052222 Lolium perenne 4-coumarate— CoA ligase 4CL2.
CAB97359.1 AJ278455 Juglans nigra 4-coumarate-CoA ligase. 4CL.
AAA69580.1 L43362 Oryza sativa 4-coumarate: CoA ligase isoform 2. 4cl.2. putative.
AAG46175.1 ACO 18727 Oryza sativa putative 4-coumarate CoA ligase. OSJNBa0056G17.30.
AAF73997.2 AF144504 Picea smithiana 4-coumarate: CoA ligase. 4CL.
AAF73995.2 AF144502 Pinus armandii 4-coumarate:CoA ligase. 4CL.
AAF73998.2 AF144505 Cathaya argyrophylla 4-coumarate: CoA ligase. 4CL.
AAF73994.2 AF144501 Pinus armandii 4-coumarate: CoA ligase. 4CL.
AAF73996.2 AF144503 Pinus armandii 4-coumarate: CoA ligase. 4CL.
SEQ ID NO: 55
CAA87068.1 Z46944 Citrus sinensis non-photosynthetic ferredoxin.
AAK15005.1 AF233452 Impatiens balsamina ferredoxin. plastidal protein.
AAA33461.1 M73831 Zea mays ferredoxin.
CAB65696.1 AJ270962 Lycopersicon esculentum electron transfer, putative ferredoxin. ferredoxin.
BAA90760.1 AB038037 Ipomoea nil non-photosynthetic ferredoxin.
BAA06456.1 D30794 Oryza sativa ferredoxin.
AAB61593.1 AF003125 Mesembryanthemum crystallinum ferredoxin I precursor.
AAA33665.1 M31713 Pisum sativum ferredoxin I precursor.
AAD02175.1 AF039662 Capsicum annuum delays the haφin-mediated hypersensitive response, ferredoxin-like protein, apl.
CAA73265.1 Y12734 Physcomitrella patens ferredoxin.
CAA52980.1 X75089 Triticum aestivum ferredoxin. petF.
BAA06436.1 D30763 Oryza sativa ferredoxin.
CAA99756.1 Z75520 Lycopersicon esculentum transfer of electrons in a wide variety of metabolic reactions. ferredoxin-I. precursor.
AAA33462.1 M73828 Zea mays ferredoxin.
AAA34028.1 M35660 Spinacia oleracea ferredoxin I precursor.
CAA26281.1 X02432 Silene latifolia subsp. alba ferredoxin precursor.
AAA33459.1 M73829 Zea mays ferredoxin.
AAA33460.1 M73830 Zea mays ferredoxin.
BAA32348.1 AB016810 Zea mays ferredoxin. pFD2.
BAA19865.1 D83660 Oryza sativa root ferredoxin.
AAA33085.1 L10349 Chlamydomonas reinhardtii ferredoxin. precursor.
AAC49171.1 U29516 Chlamydomonas reinhardtii ferredoxin precursor. Method: conceptual translation supplied by author.
AAB65699.1 AF010320 Oryza sativa ferredoxin.
AAK14422.1 AC087851 Oryza sativa putative ferredoxin. OS JNBb0072E24.16.
SEQ ID NO: 57
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
CAA66037.1 X97351 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
BAA06335.1 D30653 Populus kitakamiensis peroxidase.
AAA34108.1 J02979 Nicotiana tabacum lignin-forming peroxidase precursor (EC 1.11.1.7).
BAA01992.1 D11396 Nicotiana tabacum 'peroxidase'.
CAA66034.1 X97348 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
AAB47602.1 L07554 Linum usitatissimum peroxidase. FLXPER1.
BAA11853.1 D83225 Populus nigra peroxidase.
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI-1.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
BAA11852.1 D83224 Populus nigra peroxidase.
CAA66035.1 X97349 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
AAD37427.1 AF149277 Phaseolus vulgaris peroxidase 1 precursor. FBP1. secretory peroxidase.
BAAO 1877.1 D11102 Populus kitakamiensis peroxidase. prxAl.
CAA66036.1 X97350 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
CAA62225.1 X90692 Medicago sativa peroxidaselA. prxlA.
AAB97734.1 AF014502 Glycine max seed coat peroxidase precursor. Ep. H2O2 oxidoreductase; class III plant peroxidase.
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
CAA62227.1 X90694 Medicago sativa peroxidaselC. prxl C .
BAA06334.1 D30652 Populus kitakamiensis peroxidase.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
AAB41810.1 L36156 Medicago sativa peroxidase. pxdA. amino acid feature: conserved motifs, aa 181 .. 188; arnino acid feature: heme-binding domain, aa 60 .. 65.
AAB41811.1 L36157 Medicago sativa peroxidase. pxdC. amino acid feature: conserved domains, aa 123 .. 129, 191 .. 198; amino acid feature: heme-binding domain, aa 68 .. 73.
BAA14144.1 D90116 Armoracia rusticana peroxidase isozyme.
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
BAA02840.1 D13683 Populus kitakamiensis peroxidase. HPOX14.
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
AAA33129.1 M91372 Cucumis sativus peroxidase. pre-peroxidase.
BAA08499.1 D49551 Oryza sativa peroxidase. poxN.
BAA03373.1 D14482 Oryza sativa putative peroxidase.
AAA34101.1 L02124 Nicotiana tabacum peroxidase.
CAA76680.1 Y17192 Cucurbita pepo peroxidase. aprx. type III peroxidase.
CAA40796.1 X57564 Armoracia rusticana peroxidase. peroxidase precursor.
AAA33121.1 M32742 Cucumis sativus peroxidase (CuPer2).
BAA82306.1 AB027752 Nicotiana tabacum peroxidase.
AAD43561.1 AF155124 Gossypium hirsutum bacterial-induced peroxidase precursor. Perx_Goshiko.
AAB06183.1 M37636 Arachis hypogaea cationic peroxidase. PNC1.
AAA33127.1 M91373 Cucumis sativus peroxidase. pre-peroxidase. putative.
CAA71492.1 Y10466 Spinacia oleracea peroxidase. prxr5.
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 5 precursor, type III peroxidase.
AAA33128.1 M91374 Cucumis sativus peroxidase. putative.
BAA77389.1 AB024439 Scutellaria baicalensis peroxidase 3.
BAA77388.1 AB024438 Scutellaria baicalensis peroxidase 2.
AAB02554.1 L37790 Sτylosanthes humilis cationic peroxidase.
AAF63026.1 AF244923 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prx!4 precursor, type III peroxidase. SEQ ID NO: 58
AAG49002.1 AYO 13246 Hordeum vulgare putative ABC transporter. 635P2.4a; GC splice donor confirmed by cDNA and comparative sequencing.
AAG45492.1 AY013245 Oryza sativa
3615.4. putative ABC transporter; GC splice donor confirmed by cDNA alignment and comparative sequence.
AAG49003.1 AYO 13246 Hordeum vulgare putative ABC transporter. 635P2.4b; GC splice donor confirmed by cDNA alignment and comparative sequence.
BAA83352.1 AP000391 Oryza sativa
ESTs AU067992(C11433),AU077424(C11433) correspond to a region ofthe predicted gene.;
Similar to ABC transporter-7 (U43892).
BAB17113.1 AP002866 Oryza sativa putative white protein; ATP-binding cassette transporter. P0410E01.34.
BAA90508.1 AP001111 Oryza sativa similar to ABC transporter of Arabidopsis thaliana (AC004697).
BAA90507.1 AP001111 Oryza sativa similar to ABC transporter of Arabidopsis thaliana (AC004697).
BAB40032.1 AP003046 Oryza sativa putative ABC transporter. P0445D12.3.
BAB16495.1 AP002861 Oryza sativa putative ABC transporter ATP-binding protein. P0665D10.21.
BAA94511.1 AB041505 Populus nigra
ABC transporter homolog. PnATH.
BAB21275.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.6.
BAB21276.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.7. contains EST D22472(C1173).
BAB21279.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.10. contains ESTs
AU065360(R3463),AU101680(R3463).
CAA94437.1 Z70524 Spirodela polyrrhiza multidrug resistance protein. PDR5-like ABC transporter.
BAB21273.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.4. SEQ ID NO: 59
CAA71369.1 Y10338 Solanum tuberosum chloride channel Stele 1. putative.
AAD29679.1 AF133209 Nicotiana tabacum CLC-Nt2 protein, putative chloride channel.
SEQ ID NO: 60
BAB40094.1 AP003210 Oryza sativa putative receptor protem kinase. OSJNBaOOlOKOl .7.
AAG52992.1 U77888 Ipomoea nil receptor-like protein kinase INRPKla. inφkl.
AAF59906.1 AF197947 Glycine max receptor protein kinase-like protein. CLV1B.
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAB36558.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl . inφkl . leucine-rich repeat.
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) correspond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) correspond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl . GmRLKl .
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1 A. AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl. accumulates in mesophyll cells.
CAC20842.1 AJ250467 Pinus sylvestris receptor protein kinase. upk.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat receptor protein kinase. Irk2.
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
AAG52994.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl c. impkl.
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSJNBa0004G10.30.
BAB18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
BAB07903.1 AP002835 Oryza sativa putative receptor kinase. P0417G05.10. contains ESTs
AU032341(R3918),AU071016(R10613).
AAF34426.1 AF172282 Oryza sativa leucine rich repeat containing protein kinase. DUPR11.16.
BAA94519.1 AP001800 Oryza sativa
ESTs AU032341(R3918),AU071016(R10613) correspond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome 4, BAC F9D16; putative receptor kinase
(AL035394).
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAA82393.1 AP000367 Oryza sativa
EST C96716(C 10608) corresponds to a region ofthe predicted gene.; Similar to putative receptor protein kinase. (AC002334).
BAB03631.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.32.
AAB82755.1 U72725 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member Al; downstream of microsatellite region; disease resistance gene family member.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
AAK27817.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.16.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) corresponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB03621.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.21.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S 11214) corresponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
BAB39421.1 AP002901 Oryza sativa putative receptor kinase. P0456F08.21.
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
CAB51836.1 AJ243961 Oryza sativa Putitive Ser/Thr protein kinase. 11332.7.
BAB17139.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.31.
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
SEQ ID NO: 62
BAA25753.1 AB012932 Vigna radiata Ca2+/H+ exchanger. VCAX1.
AAF91350.1 AF256229 Zea mays calcium/proton exchanger CAXl-like protein.
BAA75232.1 AB018526 Ipomoea nil H+/Ca2+ exchanger 2. CAX2.
SEQ ID NO: 63
BAA85655.1 AB026297 Pisum sativum elicitor-responsive Dof protein ERDP.
AAB70119.1 U82230 Zea mays transcription factor, prolamin box binding factor. PBF. endosperm-specific DOF protein;
DNA binding.
CAB89831.1 AJ242853 Solanum tuberosum DNA binding protein. Dof zinc finger protein, dofl.
CAA08755.1 AJ009594 Nicotiana tabacum Dof zinc finger protein. BBF1.
CAA66601.1 X97942 Nicotiana tabacum Zn finger protein. NtBBF 1.1.
CAA66604.1 X97945 Nicotiana tabacum Zn finger protein. NtBBF2a.
BAA78575.1 AB028132 Oryza sativa Dof zinc finger protein.
BAA78572.1 AB028129 Oryza sativa Dof zinc finger protein.
BAA78573.1 AB028130 Oryza sativa
Dof zinc finger protein.
CAA66606.1 X97947 Nicotiana tabacum
Zn finger protein. BBF3
CAA66605.1 X97946 Nicotiana tabacum
Zn finger protein. NtBBF2b.
BAA78574.1 AB028131 Oryza sativa
Dof zinc finger protein. Rice cDNA for protein that binds to AT-rich sequence of rice carboxypeptidase-3 promoter.
CAA56287.1 X79934 Zea mays
Dof2. No start codon.
CAA56288.1 X79935 Zea mays
Dof3 gene, no start codon.
BAA78576.1 AB028133 Oryza sativa
Dof zinc finger protein.
SEQ ID NO: 64
CAA11219.1 AJ223281 Manihot esculenta alpha-hydroxynitrile lyase. HNL4.
BAB19413.1 AP002870 Oryza sativa putative acetone-cyanohydrin lyase. P0458A05.22.
AAC49184.1 U40402 Hevea brasiliensis hydroxynitrile lyase. hnl.
CAA82334.1 Z29091 Manihot esculenta alpha-hydroxynitrile lyase.
CAA11428.1 AJ223506 Manihot esculenta alpha-hydroxynitrile lyase. HNL24.
SEQ ID NO: 67
CAB89831.1 AJ242853 Solanum tuberosum
DNA binding protein. Dof zinc finger protein, dofl.
CAA56288.1 X79935 Zea mays
Dof3 gene, no start codon.
BAA85655.1 AB026297 Pisum sativum elicitor-responsive Dof protein ERDP.
BAA78575.1 AB028132 Oryza sativa
Dof zinc finger protein.
AAB70119.1 U82230 Zea mays transcription factor, prolamin box binding factor. PBF. endosperm-specific DOF protein; DNA binding.
CAA08755.1 AJ009594 Nicotiana tabacum
Dof zinc finger protein. BBF1.
CAA66601.1 X97942 Nicotiana tabacum
Zn finger protein. NtBBFl .1.
CAA66606.1 X97947 Nicotiana tabacum Zn finger protein. NtBBF3.
BAA78573.1 AB028130 Oryza sativa Dof zinc finger protein.
BAA78572.1 AB028129 Oryza sativa Dof zinc finger protein.
CAA56287.1 X79934 Zea mays Dof2. No start codon.
CAA66604.1 X97945 Nicotiana tabacum Zn finger protein. NtBBF2a.
CAA66605.1 X97946 Nicotiana tabacum Zn finger protein. NtBBF2b.
BAA78574.1 AB028131 Oryza sativa
Dof zinc finger protein. Rice cDNA for protein that binds to AT-rich sequence of rice carboxypeptidase-3 promoter.
BAA78576.1 AB028133 Oryza sativa Dof zinc finger protein.
SEQ ID NO: 68
CAA83453.1 Z31581 Pisum sativum chloroplast outer envelope protein 86.
AAA53276.1 L36857 Pisum sativum component of chloroplast outer membrane protein import apparatus. GTP-binding protein.
IAP86.
AAF75761.1 AF262939 Pisum sativum chloroplast protein import component Tocl59. major receptor ofthe chloroplast outer envelope membrane protein import apparatus.
AAG48839.1 AC084218 Oryza sativa similar to Arabidopsis thaliana putative chloroplast outer envelope 86-like protein
(AC002330).
AAC25785.1 L36856 Pisum sativum component of chloroplast outer membrane protein import apparatus. GTP-binding protein.
IAP34.
CAB77551.1 AJ271049 Zea mays a component ofthe protein translocon at the outer envelope of chloroplast. Toc34-2 protein. toc34B.
CAB65537.1 AJ245968 Zea mays a component ofthe protein translocon at the outer envelope of chloroplast. Toc34-l protein. toc34A. SEQ ID NO: 70
AAB71887.1 AF020791 Hordeum vulgare inserts Fe2+ into protopoφhyrin IX. ferrochelatase. hemH. protoheme IX ferro-lyase.
BAA05101.1 D26105 Hordeum vulgare ferrochelatase. hemH.
BAA05102.1 D26106 Cucumis sativus ferrochelatase. hemH.
BAB20760.1 AB037113 Cucumis sativus ferrochelatase. hemH.
BAA22284.1 AB007120 Oryza sativa ferrochelatase. hemH.
CAA06705.1 AJ005802 Solanum tuberosum tetrapyrrole biosynthesis, ferrochelatase.
AAK16728.1 AF332962 Chlamydomonas reinhardtii ferrochelatase.
AAK16729.1 AF332963 Polytomella sp. 'Pringsheim 198.80' ferrochelatase.
AAC84139.1 AF101426 Cichorium intybus ferrochelatase.
SEQ ID NO: 71
CAA64442.1 X94986 Manihot esculenta beta glucosidase. bglA.
AAB22162.1 S35175 Manihot esculenta linamarase. linamarase. beta-glucosidase; Method: conceptual translation with partial peptide sequencing; This sequence comes from Fig. 4.
AAA93032.1 U50201 Prunus serotina hydrolysis ofthe cyanogenic glucoside (R)-prunasin. prunasin hydrolase precursor, located in protein bodies of Prunus seeds; encodes 8 putative N-glycosylation sites (N-X-S/T); encodes
NEP and ITENG motifs characteristic ofthe BGA family of beta-glucosidases.
AAA91166.1 U39228 Prunus avium beta-glucosidase.
AAF34650.1 AF221526 Prunus serotina hydrolysis ofthe cyanogenic glucoside (R)-prunasin. prunasin hydrolase isoform PHA precursor, beta-glucosidase; contains 6 potential N-glycosylation sites (N-X-S/T); glycosyl hydrolase family 1 member.
AAF03675.1 AF149311 Rauvolfia seφentina hydrolyses the glucoalkaloid raucaffricine. raucaffricine-O-beta-D-glucosidase. beta glucosidase; RG; part ofthe Ajmaline biosynthesis pathway; belongs to family 1 ofthe glucosyl hydrolases.
CAA57913.1 X82577 Brassica napus beta-glucosidase. bgl.
BAA78708.1 AB003089 Polygonum tinctorium beta-glucosidase.
BAA11831.1 D83177 Costus speciosus saponin metabolite, fiirostanol glycoside 26-O-beta-glucosidase (F26G). functional expression in E. coli; one ofthe F26G isozymes.
AAG25897.1 AFl 70087 Cucurbita pepo silverleaf whitefly-induced protein 3. SLW3. similar to beta-glucosidase.
AAF04007.1 AF163097 Dalbergia cochmchinensis beta-fucosidase beta-glucosidase. dalcoclώiin 8'-O-beta-glucoside beta-glucosidase precursor.
BGLU1. rotenoid beta-glucosidase.
AAC69619.1 AF072736 Pinus contorta beta-glucosidase.
AAB38784.1 U72154 Brassica nigra beta-glucosidase. psr3.1. PSR3.1; phosphate-starvation responsive enzyme.
AAD02839.1 AF082991 Avena sativa beta-D-glucosidase beta subunit precursor. P60b. avenacosidase.
AAA87339.1 L41869 Hordeum vulgare beta-glucosidase. BGQ60. expression specific to starchy endosperm of seed.
AAB71381.1 U95298 Manihot esculenta linamarase. pLIN-GEN. beta-glucosidase.
CAA55196.1 X78433 Avena sativa beta-D-glucosidase.
AAC49177.1 U33817 Sorghum bicolor beta-glucosidase, catalyzes the hydrolysis ofthe cyanogenic beta-glucoside dhurrin.
(murrinase.
AAD09850.1 U44087 Zea mays beta-D-glucosidase precursor. glu2. product subunit structure: autodimer of 58.4 kDa monomers Allele: glu2-B73.
AAG00614.1 AF293849 Secale cereale beta-glucosidase.
AAF28800.1 AFl 12888 Catharanthus roseus plays a role in secondary metabolism by hydrolyzing strictosidine to cathenamine during indole alkaloid biosynthesis, strictosidine beta-glucosidase. localized in the endoplasmic reticulum.
CAA40058.1 X56734 Trifolium repens beta-glucosidase. non-cyanogenic. CAA40057.1 X56733 Trifolium repens beta-glucosidase. Li.
AAD10503.1 U33816 Zea mays functions in defense of young plant parts against pests via the production of hydroxamic acids from hydroxamic acid glucosides. beta-D-glucosidase.
AAA65946.1 U25157 Zea mays functions in defense of young plant parts against pests via the production of hydroxamic acids from hydroxamic acid glucosides. beta-D-glucosidase.
CAA52293.1 X74217 Zea mays beta-glucosidase. p60.1.
AAB03266.1 U44773 Zea mays beta-D-glucosidase. glul. autodimer of 58.4 kD monomers.
AAK07429.1 AF321287 Musa acuminata beta-glucosidase.
CAA79989.2 Z21977 Brassica napus beta thioglucosidase. myrosinase, thioglucoside glucohydrolase. Myrl.Bnl.
AAF34651.1 AF221527 Prunus serotina putative prunasin hydrolase precursor, beta-glucosidase; glycosyl hydrolase family 1 member.
AAA84906.1 U28047 Oryza sativa catalyzes the release of either giberellin or cyanogenic substances from their glucoconjugates. beta glucosidase. beta-D-glucoside glucohydrolase; dimer of 60 kDa monomers; localized in the plastid.
CAC08209.1 AJ005950 Cicer arietinum beta-glucosidase.
SEQ ID NO: 72 CAA56570.1 X80301 Nicotiana tabacum axi 1.
AAB72114.1 U81288 Pisum sativum
PsRT17-l . similar to the Nicotiana tabacum axi 1 gene product encoded by the sequence presented in GenBank Accession Number X80301. SEQ ID NO: 74
AAG23130.1 AF198260 Lycopersicon esculentum diacylglycerol kinase variant A. Dgkl. DGK-1; alternatively spliced.
AAG23129.1 AF198259 Lycopersicon esculentum phosphorylates diacylglycerol to yield phosphatidic acid, diacylglycerol kinase. DGK1.
LeDGKl; lipid kinase; non-calmodulin-binding isoform; alternatively spliced product.
AAG23131.1 AF 198260 Lycopersicon esculentum diacylglycerol kinase variant B. Dgkl. CBDGK; calmodulin-binding; alternatively spliced.
AAG23128.1 AF198258 Lycopersicon esculentum phosphorylates diacylglycerol to yield phosphatidic acid, calmodulin-binding diacylglycerol kinase. DGKL LeCBDGK; lipid kinase; alternatively spliced product.
SEQ ID NO: 75 CAC09580.1 AJ298992 Fagus sylvatica
Abscisic acid (ABA) and calcium induced protein kinase. protein kinase (PK). pkl.
AAA34002.1 M67449 Glycine max protein kinase. PK6.
AAG31141.1 AF305911 Oryza sativa
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAG31142.1 AF305912 Hordeum vulgare
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
CAA73722.1 Y13273 Lycopersicon esculentum putative protein kinase.
AAD46406.1 AF096250 Lycopersicon esculentum ethylene-responsive protein kinase TCTR1. ER50. serine/threonine kinase; similar to Arabidopsis thaliana negative regulator ofthe ethylene response pathway encoded by GenBank Accession Number L08789.
AAD10056.1 AFl 10518 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTR1.
AAD 10057.1 AFl 10519 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTRlv.
CAA06334.1 AJ005077 Lycopersicon esculentum protein kinase. TCTR2 protein. TCTR2.
AAK30005.1 AY029067 Rosa hybrid cultivar
CTR2 protein kinase.
AAK11734.1 AY027437 Arachis hypogaea serine/threonine/tyrosine kinase.
CAA73067.1 Y12464 Sorghum bicolor serine/threonine kinase. SNFL1.
CAA73068.1 Y12465 Sorghum bicolor serine/threonine kinase. SNFL2.
AAK31267.1 AC079890 Oryza sativa putative protein kinase. OSJNBb0089A17.2.
AAF66615.1 AFl 42596 Nicotiana tabacum LRR receptor-like protein kinase.
BAB39451.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.24.
BAB 17126.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.16.
BAB17321.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.1.
AAF68398.1 AF237568 Oryza sativa receptor-like protein kinase. RLG2.
BAB17129.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.20.
BAB 17348.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.32.
CAA57898.1 X82548 Hordeum vulgare SNF1 -related protein kinase. BKIN2.
BAB17345.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.29.
BAB17116.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.3.
BAB39441.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.11.
CAC20842.1 AJ250467 Pinus sylvestris receptor protein kinase. upk.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRK1. Chrkl.
BAB 16918.1 AP002863 Oryza sativa putative protein kinase. P0005A05.22.
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl. GmRLKl.
BAB 17342.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.26.
AAF78044.1 AF248493 Oryza sativa receptor-like kinase. RLG18. protein kinase.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1 A.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAD46917.1 AF164021 Oryza sativa receptor kinase.
AAF59906.1 AF197947 Glycine max receptor protein kinase-like protein. CLV1B.
AAF78018.1 AF238474 Oryza sativa receptor-like kinase. RLG16. protein kinase.
BAB39438.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.7.
BAA05649.1 D26602 Nicotiana tabacum protein kinase.
AACO 1746.1 AF044489 Oryza sativa receptor-like protein kinase. drpkL
CAA71142.1 Y10036 Cucumis sativus SNF1 -related protein kinase.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
SEQ ID NO: 77
AAD03693.1 AF084554 Brassica napus fibrillin.
CAAl 0372.1 AJ131455 Plastid Solanum demissum carotenoid-associated. fibrillin. c40.4.
CAA50750.1 X71952 Capsicum annuum fibrillin.
SEQ ID NO: 79
AAB53155.1 U43629 Beta vulgaris putative sugar transporter, integral membrane protein, member of major facilitator superfamily.
AAF74566.1 AF215852 Nicotiana tabacum transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAG00995.1 AF286906 Mesembryanthemum crystallfnum putative glucose translocator. metabolite transporter; targeted to plastid inner envelope membrane.
AAF74565.1 AF215851 Spinacia oleracea transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAF74567.1 AF215853 Solanum tuberosum transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAF74568.1 AF215854 Zea mays transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAG46179.1 ACO 18727 Oryza sativa putative sugar transporter protein. OSJNBa0056G17.3.
AAB88879.1 AF000952 Prunus armeniaca putative sugar transporter.
AAG43998.1 AF215837 Apium graveolens var. dulce mannitol transporter. Matl .
CAA68813.1 Y07520 Chlorella kessleri
H(+)/hexose cotransporter (AA 1-533).
CAA53192.1 X75440 Chlorella kessleri hexose transporter like protein. HUP3.
CAB52689.1 AJ132224 Lycopersicon esculentum hexose transporter. ht2.
CAA39036.1 X55349 Chlorella kessleri
H(+)/hexose-cotransporter. HUP1.
CAA47324.1 X66856 Nicotiana tabacum monosaccharid transporter. MST1.
AAB68028.1 U64902 Beta vulgaris
BvcDNA-205. putative sugar transporter; member of major facilitative superfamily; integral membrane protein.
AAB68029.1 U64903 Beta vulgaris
BvcDNA-397. putative sugar transporter; member of major facilitative superfamily; integral membrane protein.
CAA09419.1 AJ010942 Lycopersicon esculentum hexose transporter protein.
CAB07812.1 Z93775 Vicia faba sugar transport, monosaccharid transport protein, hext.
BAB19864.1 AB052885 Oryza sativa monosaccharide transporter 3. OsMST3.
AAA79761.1 L08196 Ricinus communis hexose transport, sugar carrier protein. RCSTC.
AAK13147.1 AC083945 Oryza sativa Putative sugar transporter. OSJNBa0058E19.22.
AAB06594.1 U38651 Medicago truncatula sugar transporter.
CAA04511.1 AJ001061 Vitis vinifera hexose uptake, hexose transporter.
AAC61852.1 AF061106 Petunia x hybrida putative monosaccharide transporter 1. pmtl . similar to hexose transporter protein; PMTl .
CAA70777.1 Y09590 Vitis vinifera hexose transporter.
AAA79769.1 L08197 Ricinus communis sugar transport, sugar carrier protein. RCSTA. putative.
CAB06079.1 Z83829 Picea abies monosaccharide transporter. PaMst-1. PaMst-1.
AAAI 8534.1 L21753 Saccharum hybrid cultivar H65-7052 glucose transporter, putative.
CAC00697.1 AJ278765 Lycopersicon esculentum putative sugar transporter. st3.
AAA79857.1 L08188 Ricinus communis hexose transport, hexose carrier protein. HEX6.
BAB19863.1 AB052884 Oryza sativa monosaccharide transporter 2. OsMST2.
BAB19862.1 AB052883 Oryza sativa monosaccharide transporter 1. OsMSTl.
AAF91432.1 AF280432 Mesembryanthemum crystallinum putative Na+/myo-inositol symporter. Itr2. membrane transport protein.
CAB52688.1 AJ132223 Lycopersicon esculentum hexose transporter, htl .
BAA85398.1 AP000615 Oryza sativa similar to sugar transporter protein. (AL022604).
AAG46115.1 AC073166 Oryza sativa putative sugar transporter. OSJNBb0064P21.3.
CAB52690.1 AJ132225 Lycopersicon esculentum hexose transporter. ht3.
AAA18533.1 L21752 Saccharum hybrid cultivar H65-7052 glucose transporter, putative.
AAD55054.1 AF173655 Beta vulgaris glucose transporter. Gt.
BAB39246.1 AP002869 Oryza sativa putative transport protein homolog. P0554D10.30.
AAA79764.1 L08191 Ricinus communis sugar transport, sugar carrier protein. RCSTC. Nucleotides 1 to 23 and 226 to 246 are originated fromdegenerated primers; putative.
AAA79762.1 L08189 Ricinus communis sugar transport, sugar carrier protein. RCSTA. Nucleotides 1 to 23 and 226 to 246 are originated from degenerated primers; putative.
AAA79767.1 L08194 Ricinus communis sugar transport, sugar carrier protein. RCSTG. Nucleotides 1 to 23 and 226 to 246 are originated from degenerated primers; putative.
AAA79766.1 L08193 Ricinus communis sugar transport, sugar carrier protein. RCSTF. Nucleotides 1 to 23 and 226 to 246 are originated from degenerated primers; putative.
SEQ ID NO: 82 CAA78386.1 Z13996 Petunia x hybrida
DNA binding protein; transcriptional activator, protein 1. myb.Ph3. Product related to animal myb proto-oncoproteins. Sequence from nucleotide 992 is not included in clone cPFl and has been obtained by PCR amplification of cDNA.
CAB43399.1 AJ006292 Antirrhinum majus Myb-related transcription factor mixta-like 1. mybmll. CAA67600.1 X99210 Lycopersicon esculentum myb-related transcription factor. THM16.
BAA93038.1 AP001552 Oryza sativa
EST AU082058(C12976) corresponds to a region ofthe predicted gene. Similar to
Arabidopsis thaliana putative transcription factor (AF062916).
BAA23337.1 D88617 Oryza sativa transfactor. OSMYBl. Osmybl.
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related. CAA72186.1 Y11351 Oryza sativa myb factor, myb.
AAF22256.1 AF161711 Pimpinella brachyca a myb-related transcription factor.
AAC04720.1 AF034134 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-O. similar to MYB A encoded by GenBank Accession Number L04497.
AAA82943.1 U39448 Picea mariana
MYB-like transcriptional factor MBFl. putative DNA binding region highly similar to the maize Cl.
BAB39987.1 AP003020 Oryza sativa putative transcription factor (myb). P0498A12.16. contains ESTs
AU097474(S5087),D40175(S1959).
BAB39972.1 AP003018 Oryza sativa putative transcription factor (myb). OSJNBa0004B13.27. contains ESTs
AU097474(S5087),D40175(S1959). ____
BAA23338.1 D88618 Oryza sativa transfactor. OSMYB2. Osmyb2.
CAA67575.1 X99134 Lycopersicon esculentum transcription factor. THM6. myb-related.
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, transcriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins.
AAA19821.1 L19495 Zea mays transcriptional activator for anthocyanin synthesis, transcriptional activator.
AAC49394.1 U57002 Zea mays
P protein. P. allele P-wr; Myb-like transcriptional regulator with a putative zinc-finger at the
C-terminal.
AAA33500.1 M73028 Zea mays myb-like transcription factor. P.
AAG36774.1 AF210616 Zea mays
P2 protein. P2. myb-like transcriptional factor; similar to Zea mays P gene.
BAA88222.1 AB028650 Nicotiana tabacum myb-related transcription factor LBM2. Ibm2.
CAA72185.1 Y11350 Oryza sativa myb factor, myb.
AAG13574.1 AC037425 Oryza sativa myb factor. OSJNBa0055P24.4.
AAB41101.1 U72762 Nicotiana tabacum transcription factor Mybl. mybl. TMV-inducible Myb homolog; contains helix-turn-helix motif; contains redox-sensitive cysteine.
BAA88223.1 AB028651 Nicotiana tabacum myb-related transcription factor LBM3. Ibm3.
AAA19819.1 L19496 Zea mays transcriptional activator for anthocyanin synthesis, transcriptional activator.
BAA88224.1 AB028652 Nicotiana tabacum myb-related transcription factor LBM4. Ibm4.
AAB67720.1 AF015268 Zea mays activator of anthocyanin structural genes. PL transcription factor. PI.
AAA33492.1 L13454 Zea mays transcriptional activator for anthocyanin biosynthesis. Pl-Bh (Blotchedl).
SEQ ID NO: 83
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, transcriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins. ^^
BAA81736.1 AB029165 Glycine max GmMYB29B2.
BAA81732.1 AB029161 Glycine max GmMYB29A2.
BAA81731.1 AB029160 Glycine max GmMYB29Al.
BAA81730.1 AB029159 Glycine max GmMYB29Al.
BAA88221.1 AB028649 Nicotiana tabacum myb-related transcription factor LBM1. lbml.
BAA88224.1 AB028652 Nicotiana tabacum myb-related transcription factor LBM4. lbm.4.
CAA66952.1 X98308 Lycopersicon esculentum THM18. myb-related transcription factor.
BAA81733.2 AB029162 Glycine max GmMYB29A2.
CAA72217.1 Y11414 Oryza sativa myb.
BAA88222.1 AB028650 Nicotiana tabacum myb-related transcription factor LBM2. Ibm2.
AAB41101.1 U72762 Nicotiana tabacum transcription factor Mybl. mybl. TMV-inducible Myb homolog; contains helix-turn-helix motif; contains redox-sensitive cysteine.
BAA88223.1 AB028651 Nicotiana tabacum myb-related transcription factor LBM3. Ibm3.
CAA72185.1 Y11350 Oryza sativa myb factor, myb.
AAG13574.1 AC037425 Oryza sativa myb factor. OSJNBa0055P24.4.
AAK19616.1 AF336283 Gossypium hirsutum
GHMYB25. ghmyb25. similar to myb; contains an unspliced intron.
CAA78386.1 Z13996 Petunia x hybrida
DNA binding protein; transcriptional activator, protein 1. myb.Ph3. Product related to animal myb proto-oncoproteins. Sequence from nucleotide 992 is not included in clone cPFl and has been obtained by PCR amplification of cDNA.
CAA72218.1 Y11415 Oryza sativa myb.
AAA33067.1 L04497 Gossypium hirsutum
MYB A; putative.
CAA68235.1 X99973 Hordeum vulgare myb4 transcription factor. myb4. _________^
AAC49394.1 U57002 Zea mays
P protein. P. allele P-wr; Myb-like transcriptional regulator with a putative zinc-finger at the
C-terminal.
AAA33500.1 M73028 Zea mays myb-like transcription factor. P.
CAA67600.1 X99210 Lycopersicon esculentum myb-related transcription factor. THM16.
AAG36774.1 AF210616 Zea mays
P2 protein. P2. myb-like transcriptional factor; similar to Zea mays P gene.
AAF22256.1 AF161711 Pimpfnella brachycaφa myb-related transcription factor.
CAA72187.1 Y11352 Oryza sativa myb factor, myb.
AAC04718.1 AF034132 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-J. similar to MYB A encoded by GenBank Accession Number L04497.
CAB43399.1 AJ006292 Antirrhinum majus
Myb-related transcription factor mixta-like 1. mybmll .
AAK19615.1 AF336282 Gossypium hirsutum GHMYBIO. ghmyblO. similar to myb.
AAK19618.1 AF336285 Gossypium hirsutum GHMYB38. ghmyb38. similar to myb.
CAA72186.1 Y11351 Oryza sativa myb factor, myb.
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related.
CAA50221.1 X70876 Hordeum vulgare MybHv5. myb2.
BAA23338.1 D88618 Oryza sativa transfactor. OSMYB2. Osmyb2.
AAK19619.1 AF336286 Gossypium hirsutum GHMYB9. ghmyb9. similar to myb.
AAA82943.1 U39448 Piceamariana
MYB-like transcriptional factor MBFl. putative DNA binding region highly similar to the maize Cl.
AAC04720.1 AF034134 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-O. similar to MYB A encoded by GenBank Accession Number L04497.
AAK19611.1 AF336278 Gossypium hirsutum BNLGH.233. bnlghi6233. similar to myb.
BAA23337.1 D88617 Oryza sativa transfactor. OSMYB1. Osmybl.
BAB39987.1 AP003020 Oryza sativa putative transcription factor (myb). P0498A12.16. contains ESTs
AU097474(S5087),D40175(S 1959).
BAB39972.1 AP003018 Oryza sativa putative transcription factor (myb). OSJNBa0004B13.27. contains ESTs
AU097474(S5087),D40175(S1959).
CAA65525.1 X96749 Oryza sativa myb7.
AAK19617.1 AF336284 Gossypium hirsutum GHMYB36. ghmyb36. similar to myb.
CAA67575.1 X99134 Lycopersicon esculentum transcription factor. THM6. myb-related.
CAA50224.1 X70879 Hordeum vulgare MybHvl. mybl.
SEQ ID NO: 84
BAA88222.1 AB028650 Nicotiana tabacum myb-related transcription factor LBM2. Ibm2.
BAA88221.1 AB028649 Nicotiana tabacum myb-related transcription factor LBM1. lbml.
BAA88224.1 AB028652 Nicotiana tabacum myb-related transcription factor LBM4. Ibm4.
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, transcriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins.
CAA66952.1 X98308 Lycopersicon esculentum THM18. myb-related transcription factor.
AAB41101.1 U72762 Nicotiana tabacum transcription factor Mybl. mybl. TMV-inducible Myb homolog; contains helix-turn-helix motif; contains redox-sensitive cysteine.
BAA88223.1 AB028651 Nicotiana tabacum myb-related transcription factor LBM3. Ibm3.
BAA81733.2 AB029162 Glycine max GmMYB29A2.
BAA81731.1 AB029160 Glycine max GmMYB29Al.
BAA81730.1 AB029159 Glycine max GmMYB29Al.
BAA81736.1 AB029165 Glycine max GmMYB29B2.
CAA72217.1 Y11414 Oryza sativa myb.
BAA81732.1 AB029161 Glycine max GmMYB29A2.
CAA72185.1 Y11350 Oryza sativa myb factor, myb.
AAG13574.1 AC037425 Oryza sativa myb factor. OSJNBa0055P24.4.
CAA72218.1 Y11415 Oryza sativa myb.
CAA78386.1 Z13996 Petunia x hybrida
DNA binding protein; transcriptional activator, protein 1. myb.Ph3. Product related to animal myb proto-oncoproteins. Sequence from nucleotide 992 is not included in clone cPFl and has been obtained by PCR amplification of cDNA.
AAC49394.1 U57002 Zea mays
P protein. P. allele P-wr; Myb-like transcriptional regulator with a putative zinc-finger at the
C-terminal.
CAB43399.1 AJ006292 Antirrhinum majus
Myb-related transcription factor mixta-like 1. mybmll .
AAK19616.1 AF336283 Gossypium hirsutum
_ G_H_M__Y_B_2_5_. g_h_m_y_b2_5_._ simila _r_ to_ m_yb_; contains an unspliced intron.
P2 protein. P2. myb-like transcriptional factor; similar to Zea mays P gene.
AAA33500.1 M73028 Zea mays myb-like transcription factor. P.
CAA68235.1 X99973 Hordeum vulgare myb4 transcription factor. myb4.
CAA72187.1 Y11352 Oryza sativa myb factor, myb.
AAA33067.1 L04497 Gossypium hirsutum MYB A; putative.
CAA72186.1 Y11351 Oryza sativa myb factor, myb.
CAA67600.1 X99210 Lycopersicon esculentum myb-related transcription factor. THM16.
AAK19618.1 AF336285 Gossypium hirsutum GHMYB38. ghmyb38. similar to myb.
AAK19611.1 AF336278 Gossypium hirsutum BNLGHJ233. bnlgh.6233. similar to myb.
AAC04718.1 AF034132 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-J. similar to MYB A encoded by GenBank Accession Number L04497.
AAK19619.1 AF336286 Gossypium hirsutum GHMYB9. ghmyb9. similar to myb.
AAK19615.1 AF336282 Gossypium hirsutum GHMYB10. ghmyblO. similar to myb.
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related.
BAA23338.1 D88618 Oryza sativa transfactor. OSMYB2. Osmyb2.
BAA23337.1 D88617 Oryza sativa transfactor. OSMYB1. Osmybl.
CAA67575.1 X99134 Lycopersicon esculentum transcription factor. THM6. myb-related.
CAA65525.1 X96749 Oryza sativa myb7.
BAB39987.1 AP003020 Oryza sativa putative transcription factor (myb). P0498A12.16. contains ESTs
AU097474(S5087),D40175(S 1959).
BAB39972.1 AP003018 Oryza sativa putative transcription factor (myb). OSJNBa0004B13.27. contains ESTs
AU097474(S5087),D40175(S 1959).
CAA50221.1 X70876 Hordeum vulgare MybHv5. myb2.
AAA82943.1 U39448 Piceamariana
MYB-like transcriptional factor MBFl . putative DNA binding region highly similar to the maize Cl.
AAF22256.1 AF161711 Pimpinella brachycaφa myb-related transcription factor.
AAK19617.1 AF336284 Gossypium hirsutum GHMYB36. ghmyb36. similar to myb.
CAA50222.1 X70877 Hordeum vulgare MybHyl. mybl.
CAA50224.1 X70879 Hordeum vulgare MybHvl. mybl.
SEQ ID NO: 85
AAD 11575.1 AF064029 Helianthus tuberosus lectin 1. LECHeltubal; agglutinin.
AAD11578.1 AF064030 Helianthus tuberosus lectin 2. LECHeltuba2; agglutinin.
AAD 11577.1 AF064032 Helianthus tuberosus lectin HE 17.
AAG10403.1 AF233284 Convolvulus arvensis mannose-binding lectin. cr8. Conarva.
AAD 11576.1 AF064031 Helianthus tuberosus lectin 3.
AAB82776.2 AF001527 Musa acuminata ripening-associated protein, similar to lectin.
AAC49564.1 U56820 Calystegia sepium lectin.
CAB40792.1 AJ237754 Hordeum vulgare putative lectin. hl#2.
SEQ ID NO: 88
AAB65163.1 AF002692 Solanum commersonii glutathione S-transferase, class-phi. GSTl. low temperature induced.
CAA55039.1 X78203 Hyoscyamus muticus glutathione transferase.
AAA33930.1 M84968 Silene vulgaris glutathione-S-transferase.
AAA33931.1 M84969 Silene vulgaris glutathione-S-transferase.
AAF65767.1 AF242309 Euphorbia esula glutathione S-transferase. putative auxin-binding GST.
BAAO 1394.1 D 10524 Nicotiana tabacum glutathione S-transferase. parB.
CAA96431.1 Z71749 Nicotiana plumbaginifolia glutathione S-transferase.
AAF61392.1 AF133894 Persea americana glutathione S-transferase. GTH.
CAB38119.1 AJ010296 Zea mays Glutathione transferase 111(b). gst3b.
CAB38118.1 AJ010295 Zea mays Glutathione transferase IΙI(a). gst3a.
BAB39935.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.17.
CAA09190.1 AJ010451 Alopecurus myosuroides glutathione transferase. GST2a.
CAA09192.1 AJ010453 Alopecurus myosuroides glutathione transferase. GST2c.
CAA09193.1 AJ010454 Alopecurus myosuroides glutathione transferase. GST2d.
CAA09191.1 AJ010452 Alopecurus myosuroides glutathione transferase. GST2b.
BAB39941.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.23.
AAG34811.1 AF243376 Glycine max glutathione S-transferase GST 21.
BAB39939.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.21.
AAG32476.1 AF309383 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF4.
AAG34814.1 AF243379 Glycine max glutathione S-transferase GST 24.
AAG34812.1 AF243377 Glycine max glutathione S-transferase GST 22.
BAB39929.1 AP002914 Oryza sativa putative glutathione transferase. P0493G01.7.
CAA39487.1 X56012 Triticum aestivum glutathione transferase. gstAl.
AAD56395.1 AFl 84059 Triticum aestivum glutathione S-transferase. GSTl.
BAB39927.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.1. contains ESTs
AU031696(R0596),C97559(C60386),C28218(C60386), D28287(R0596).
AAA20585.1 U12679 Zea mays glutathione S-transferase IV. GSTIV.
CAA56047.1 X79515 Zea mays glutathione transferase. GST27.
CAA39480.1 X56004 Triticum aestivum glutathione transferase. gstA2.
AAA33469.1 M16902 Zea mays glutathione S-transferase I.
AAG32475.1 AF309382 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF5.
AAA33470.1 M16901 Zea mays glutathione S-transferase I.
AAG32477.1 AF309384 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF3.
CAA68993.1 Y07721 Petunia x hybrida conjugates glutathione to anthocyanin to facilitate transport to the vacuole. glutathione S- transferase. an9 locus.
BAB39940.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.22.
AAC64007.1 AF062403 Oryza sativa glutathione S-transferase II.
AAG34823.1 AF244680 Zea mays glutathione S-transferase GST 15.
AAG34817.1 AF244674 Zea mays glutathione S-transferase GST 9.
CAB66333.1 AJ279691 Betula pendula glutathione-S-transferase. gst.
AAG34820.1 AF244677 Zea mays glutathione S-transferase GST 11.
AAG34821.1 AF244678 Zea mays glutathione S-transferase GST 13.
CAA05354.1 AJ002380 Oryza sativa glutathione S-transferase. Rgst I.
AAG34816.1 AF244673 Zea mays glutathione S-transferase GST 8.
AAG34818.1 AF244675 Zea mays glutathione S-transferase GST 10.
CAA05355.1 AJ002381 Oryza sativa glutathione S-transferase. Rgst II.
AAG34824.1 AF244681 Zea mays glutathione S-transferase GST 16.
SEQ ID NO: 91
AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
CAA71516.1 Y10492 Glycine max putative cytochrome P450.
BAA12159.1 D83968. Glycine max Cytochrome P-450 (CYP93A1).
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl. P450 enzyme.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3'-hydroxylase. Htl. cytochrome P450; CYP75B2.
BAB12433.1 AB025030 Coptis japonica p450.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
CAA50155.1 X70824 Solanum melongena flavonoid hydroxylase (P450). CYP75.
AAB 17562.1 U72654 Eustoma grandiflorum flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75.
AAF05621.1 AF191772 Papaver somniferum hydroxylase involved in benzylisoquinoline alkaloid biosynthesis. (S)-N-methylcoclaurine 3'' hydroxylase. CYP80B1. cytochrome P450-dependent monooxygenase.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
BAA74466.1 AB022733 Glycyrrhiza echinata cytochrome P450. CYP Ge-51.
CAA50648.1 X71657 Solanum melongena P450 hydroxylase.
BAA22423.1 AB001380 Glycyrrhiza echinata cytochrome P450. CYP93B1.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
AAB94588.1 AF022459 Glycine max
CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
AAG14961.1 AF214007 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
AAC39452.1 AFO 14800 Eschscholzia californica hydroxylase involved in the biosynthesis of tetrahydrobenzylisoquinoline alkaloids in plants. (S)-N-methylcoclaurine 3'-hydroxylase. CYP80B1. cytochrome P-450-dependent monooxygenase; CYP80Blvl .
AAG14962.1 AF214008 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2.
BAA84072.1 AB028152 Torenia hybrida flavone synthase II. cytochrome P450. TFNS5.
AAC39453.1 AFO 14801 Eschscholzia californica hydroxylase involved in the biosynthesis of tetrahydrobenzylisoquinoline alkaloids in plants. (S)-N-methylcoclaurine 3'-hydroxylase. CYP80B1. cytochrome P-450-deρendent monooxygenase; CYP80Blv2.
AAD47832.1 AF166332 Nicotiana tabacum cytochrome P450.
SEQ ID NO: 92
AAG42490.1 AF321001 Suaeda maritima subsp. salsa S-adenosylmethionine sythetase 2.
AAG17666.1 AF271220 Brassica juncea S-adenosylmethionine synthetase. MSAMS2.
BAA96637.1 AP002482 Oryza sativa
Similar to Oryza sativa S-adenosylmethionine synthetase 1 (P46611).
AAG17036.1 AF187821 Pinus contorta catalyzes the reaction between methionine and ATP to S-adenosylmethionine. S- adenosylmethionine synthetase. sams2.
BAA94605.1 AB041534 Camellia sinensis s-adenosylmethionine synthetase. SAM.
AAA81377.1 U17239 Actinidia chinensis S-adenosylmethionine synthetase.
AAB38500.1 U79767 Mesembryanthemum crystallinum S-adenosylmethionine synthetase. methionine adenosyltransferase.
AAA81378.1 U17240 Actinidia chinensis S-adenosylmethionine synthetase.
BAA09895.1 D63835 Hordeum vulgare S-adenosylmethionine synthetase.
AAA33274.1 M61882 Dianthus caryophyllus S-adenosyhnetMonine synthetase. CARSAM2.
AAA58773.1 L36681 Pisum sativum
S-adenosylmethionine synthase. precursor for ethylene and polyamine biosynthesis.
AAA58772.1 L36680 Pisum sativum precursor for ethylene and polyamine biosynthesis. S-adenosylmethionine synthase.
AAA81379.1 U17241 Actinidia chinensis S-adenosylmethionine synthetase.
AAA33857.1 M62758 Petroselinum crispum S-adenosylmethionine synthetase. SMS-1.
AAG17035.1 AF187820 Pinus contorta catalyzes the reaction between methionine and ATP to S-adenosylmethionine. S- adenosyhnethionine synthetase. samsl.
AAB71833.1 AF008568 Chlamydomonas reinhardtii S-adenosylmethionine synthetase. CHRSAMS .
AAA33858.1 M62757 Petroselinum crispum S-adenosylmethionine synthetase. SMS-2.
BAA21726.1 AB006187 Nicotiana tabacum S-adenosylmethionine synthase. BYJ90.
AAF78525.1 AF195233 Pyrus pyrifolia S-adenosylmethionine synthase. SAMS.
SEQ ID NO: 94
AAB36543.1 U77935 Phaseolus vulgaris
DnaJ-like protein, synthesis and expression are regulated by heavy metal stress, viras infection and wounding treatment, suggesting that DnaJ-like protein plays a role in plant defense.
SEQ ID NO: 95
AAB36546.1 U77940 Phaseolus vulgaris polyubiquitin. expression is regulated by heavy metal stress, UV, virus infection, heat shock and wounding treatment, this suggests that ubiquitin plays a role in plant defense.
SEQ ID NO: 98 AAF22109.1 AFl 19411 Lupinus albus ethylene biosynthesis enzyme. 1-aminocyclopropane-l -carboxylate synthase 1. ACSl. ACC synthase; S-adenosyl-L-methionine methylthioadenosine-lyase.
AAC83146.1 AF057562 Nicotiana glutinosa
1 -aminocy clopropane- 1 -carboxylate synthase. ACSl. ACC synthase.
AAB06724.1 U64804 Petunia x hybrida ethylene biosynthesis enzyme. 1-aminocyclopropane-l -carboxylate synthase. PH-ACS1.
CAA79478.1 Z18953 Petunia x hybrida 1 -aminocyclopropane- 1 -carboxylate synthase.
AAB17279.1 U72390 Lycopersicon esculentum converts S-adenosyhnethionine to 1-aminocyclopropane-l -carboxylate hormone biosynthesis.
1-aminocyclopropane-l -carboxylate synthase. LE-ACS1B.
BAA94600.1 AB033503 Populus euramericana 1 -aminocyclopropane- 1 -carboxylate synthase. peacs-2.
BAA96743.1 AB044662 Prunus persica 1-aminocycloρropane-l -carboxylate synthase. PP-ACS1.
CAB60831.1 AJ012696 Citrus sinensis ACC synthase. acs2.
CAA09477.1 AJO 11095 Citrus sinensis ACC synthase. acs-1.
CAA41855.1 X59139 Lycopersicon esculentum 1 -aminocyclopropane 1 -carboxylate synthase. LE-Acc2.
BAA90549.1 AB031026 Prunus mume
ACC synthase. PM-ACS1. 1-aminocyclopropane-l -carboxylic acid synthase.
AAA81580.1 M34289 Lycopersicon esculentum 1 -aminocyclopropane- 1 -carboxylate synthase.
CAA44397.1 X62536 Lycopersicon esculentum 1 -aminocyclopropane 1 -carboxylate synthase.
CAA46797.1 X65982 Nicotiana tabacum 1 -aminocyclopropane 1 -carboxylate synthase.
BAA78333.1 AB015625 Pyrus pyrifolia 1-aminocyclopropane-l -carboxylic acid synthase. pPPACS3.
CAA41856.1 X59145 Lycopersicon esculentum 1 -aminocyclopropane 1-carboxylate synthase. LE-ACC2.
AAB17278.1 U72389 Lycopersicon esculentum converts S-adenosylmethionine to 1-aminocyclopropane-l -carboxylate hormone biosynthesis.
1-aminocyclopropane-l -carboxylate synthase. LE-ACS1A.
AAF97614.1 U18056 Lycopersicon esculentum converts SAM to ACC; plant hormone biosynthesis. 1-amino-cyclopropane-l -carboxylate synthase. LE-ACS1A. ACC synthase; direct precursor of ethylene; one member of a multigene family.
BAB 16433.1 AB041521 Solanum tuberosum
ACC synthase. ppACSl.
BAA34923.1 AB013100 Lycopersicon esculentum
1 -aminocyclopropane- 1-carboxylate synthase. LE-ACS6. ACC synthase.
BAA25916.1 AB013346 Lycopersicon esculentum
1 -aminocyclopropane- 1-carboxylate synthase.
AAF97615.1 U 18057 Lycopersicon esculentum converts SAM to ACC; plant hormone biosynthesis. 1-amino-cyclopropane-l -carboxylate synthase. LE-ACSIB. ACC synthase; direct precursor of ethylene; one member of a multigene family.
BAA92350.1 AB034992 Malus x domestica
1-aminocyclopropane-l -carboxylate synthase. MdACS-5A.
BAA92351.1 AB034993 Malus x domestica
1-aminocyclopropane-l -carboxylate synthase. MdACS-5B.
BAA93712.1 AB032935 Cucumis melo
1-aminocyclopropane-l -carboxylate synthase. CMe-ACSl.
BAA94599.1 AB033502 Populus euphratica
1-aminocyclopropane-l -carboxylate synthase. peacs-1.
BAA33374.1 AB006803 Cucumis sativus
ACC synthase. CS-ACS1.
BAA93714.1 AB032937 Cucumis sativus
1-aminocyclopropane-l -carboxylate synthase. CS-ACS1.
AAC49153.1 U17229 Pelargonium x hortorum
1 -aminocyclopropane 1-carboxylate synthase. ACC synthase.
CAA06288.1 AJ005002 Nicotiana tabacum 1 -cyclopropane- 1-carboxylate synthase. acs2.
CAB65314.1 AJ131836 Nicotiana tabacum 1 -aminocyclopropane- 1-carboxylate synthase. acs2.
AAC15777.1 AF061605 Nicotiana glutinosa ACC synthase. ACS3.
AAC83147.1 AF057563 Nicotiana glutinosa 1-aminocyclopropane-l -carboxylate synthase. ACS2. ACC synthase.
BAA06464.1 D30805 Cucumis melo 1 -aminocyclopropane- 1 -carboxylate synthase.
BAA83618.1 AB025906 Cucumis melo
1-arninocyclopropane- 1-carboxylate synthase. CMe-ACSl (ME-ACS1).
AAB70885.1 U88971 Pelargonium x hortorum
1-aminocyclopropane-l -carboxylate synthase. GACS2. similar to Pelargonium hortorum clone pGAC-2, Genbank Accession Number U17231.
AAA33113.1 M58323 Cucurbita pepo 1 -aminocyclopropane- 1 -carboxylate synthase.
AAA33111.1 M61195 Cucurbita pepo
1 -aminocyclopropane- 1-carboxylate synthase. CP-ACC1A.
CAA57724.1 X82273 Brassica oleracea 1 -aminocyclopropane- 1 -carboxylate synthase.
CAA47474.1 X67100 Glycine max
1 -aminocyclopropane 1-carboxylate synthase.
AAA33112.1 M61195 Cucurbita pepo 1-aminocyclopropane-l -carboxylate synthase. CP-ACC1B.
AAC98809.1 U68216 Carica papaya ACC synthase. fruit specific; ripening related.
CAA51227.1 X72676 Brassica juncea 1-aminocyclopropane-l -carboxylate synthase. MACC.
BAA00838.1 D01032 Cucurbita maxima 1-aminocyclopropane-l -carboxylate synthase. accW.
AAA34131.1 M63490 Lycopersicon esculentum enzyme. 1-aminocyclopropane-l -carboxylate synthase.
AAA03164.1 M88487 Lycopersicon esculentum 1-aminocyclopropane-l -carboxylate synthase. ACC4.
CAA41857.1 X59146 Lycopersicon esculentum 1 -aminocyclopropane 1-carboxylate synthase. LE-ACC4.
CAA77688.1 Z11613 Vignaradiata 1 -aminocyclopropane 1-carboxylate synthase.
CAA67118.1 X98492 Nicotiana tabacum ACC synthase. ACCS2.
SEQ ID NO: 99
BAA22976.1 D63457 Arabis gemmifera alcohol dehydrogenase. Adh. ADH.
BAA22973.1 D63454 Arabis gemmifera alcohol dehydrogenase. Adh. ADH.
AAF23537.1 AFl 10439 Arabis glabra alcohol dehydrogenase.
BAA22978.1 D63459 Arabis gemmifera alcohol dehydrogenase. Adh. ADH.
BAA22974.1 D63455 Arabis gemmifera alcohol dehydrogenase. Adh. ADH.
AAF23540.1 AFl 10442 Arabidopsis halleri alcohol dehydrogenase.
AAF23551.1 AFl 10453 Arabidopsis lyrata subsp. petraea alcohol dehydrogenase.
AAF23539.1 AFl 10441 Halimolobos peφlexa var. lemhiensis alcohol dehydrogenase.
BAA22975.1 D63456 Arabis gemmifera alcohol dehydrogenase. Adh. ADH.
AAF23546.1 AFl 10448 Arabis lyallii alcohol dehydrogenase.
AAF23548.1 AFl 10450 Arabis parishii alcohol dehydrogenase.
AAF23550.1 AFl 10452 Arabidopsis lyrata subsp. petraea alcohol dehydrogenase.
BAA22972.1 D63453 Arabis gemmifera alcohol dehydrogenase. Adh. ADH.
BAA22971.1 D63452 Arabis gemmifera alchohol dehydrogenase. Adh. ADH.
BAA22977.1 D63458 Arabis gemmifera alcohol dehydrogenase. Adh. ADH.
AAF23549.1 AFl 10451 Arabis pauciflora alcohol dehydrogenase.
AAF23545.1 AFl 10447 Arabis lignifera alcohol dehydrogenase.
AAF23536.1 AF110438 Arabis fendleri alcohol dehydrogenase.
AAF23541.1 AFl 10443 Arabis hirsuta alcohol dehydrogenase.
AAF23531.1 AF110433 Arabis blepharophylla alcohol dehydrogenase.
AAF23530.1 AFl 10432 Arabis blepharophylla alcohol dehydrogenase.
AAF23523.1 AFl 10425 Aubrieta deltoidea alcohol dehydrogenase.
AAF23529.1 AFl 10431 Arabis blepharophylla alcohol dehydrogenase.
AAF23547.1 AFl 10449 Arabidopsis lyrata subsp. lyrata alcohol dehydrogenase.
AAF23533.1 AFl 10435 Capsella rubella alcohol dehydrogenase.
AAF23534.1 AFl 10436 Arabis drummondii alcohol dehydrogenase.
AAF23532.1 AF110434 Brassica oleracea alcohol dehydrogenase.
AAF23556.1 AFl 10458 Barbarea vulgaris alcohol dehydrogenase.
AAF23555.1 AFl 10457 Arabis turrita alcohol dehydrogenase.
AAF23538.1 AFl 10440 Arabidopsis griffithiana alcohol dehydrogenase.
AAF23524.1 AFl 10426 Arabis alpina alcohol dehydrogenase.
AAF23543.1 AFl 10445 Arabis hirsuta alcohol dehydrogenase.
AAF23525.1 AFl 10427 Arabis alpina alcohol dehydrogenase.
AAF23527.1 AFl 10429 Arabis alpina alcohol dehydrogenase.
AAF23535.1 AFl 10437 Arabis drummondii alcohol dehydrogenase.
AAF23553.1 AFl 10455 Arabis procurrens alcohol dehydrogenase.
AAF23544.1 AFl 10446 Arabis jacquinii alcohol dehydrogenase.
AAF23526.1 AFl 10428 Arabis alpina alcohol dehydrogenase.
AAF23528.1 AFl 10430 Cardamine amara alcohol dehydrogenase.
AAF23552.1 AFl 10454 Arabis procurrens alcohol dehydrogenase.
AAF23542.1 AFl 10444 Arabis hirsuta alcohol dehydrogenase.
AAC79418.1 AF037560 Leavenworthia stylosa alcohol dehydrogenase 3. Adh3.
BAA34682.1 ABO 15504 Arabidopsis griffithiana alcohol dehydrogenase. Adh.
BAA34685.1 AB015507 Arabidopsis suecica alcohol dehydrogenase. Adh.
BAA34683.1 AB015505 Arabidopsis korshinskyi alcohol dehydrogenase. Adh.
CAB72921.1 AJ251281 Arabidopsis lyrata subsp. petraea alcohol dehydrogenase. adh.
CAB72920.1 AJ251280 Arabidopsis lyrata subsp. petraea alcohol dehydrogenase. adh.
CAB72919.1 AJ251279 Arabidopsis lyrata subsp. petraea alcohol dehydrogenase. adh.
CAB72918.1 AJ251278 Arabidopsis lyrata subsp. petraea alcohol dehydrogenase. adh.
CAB72917.1 AJ251277 Arabidopsis lyrata subsp. petraea alcohol dehydrogenase. adh.
SEQ ID NO: 100
AAD09518.1 U64925 Nicotiana tabacum NTGP4. similar to AIG1; geranylgeranylated protein.
SEQ ID NO: 102
BAA12039.1 D83669 Spinacia oleracea stromal ascorbate peroxidase.
BAA19611.1 D77997 Spinacia oleracea thylakoid-bound ascorbate peroxidase.
BAA24610.1 AB002467 Spinacia oleracea stromal ascorbate peroxidase. APX2.
BAA24609.1 AB002467 Spinacia oleracea thylakoid-bound ascorbate peroxidase. APX2.
AAC19394.1 AF069316 Mesembryanthemum crystallinum stromal L-ascorbate peroxidase precursor.
AAC 19393.1 AF069315 Mesembryanthemum crystallinum thylakoid-bound L-ascorbate peroxidase precursor.
BAA78553.1 AB022274 Chloroplast Nicotiana tabacum stromal ascorbate peroxidase.
BAA78552.1 AB022273 Chloroplast Nicotiana tabacum thylakoid-bound ascorbate peroxidase.
BAA22196.1 D88420 Cucurbita sp. a hydrogen peroxide-scavenging enzyme, stromal ascorbate peroxidase.
BAA12029.1 D83656 Cucurbita sp.
2AsA+H202 — > 2MDA+2H2O. thylakoid-bound ascorbate peroxidase.
CAAl 1265.1 AJ223325 Chlamydomonas reinhardtii ascorbate peroxidase. apxl.
BAA83595.1 AB009084 Chlamydomonas sp. W80 chloroplast ascorbate peroxidase.
AAD30294.1 AF139190 Mesembryanthemum crystallinum cytosolic ascorbate peroxidase. similar to LO-328, R6-3, L72-2.
AAB52954.1 U37060 Gossypium hirsutum ascorbate peroxidase. glyoxysomal membrane-bound protein.
AAD43334.1 AFl 59254 Zantedeschia aethiopica ascorbate peroxidase. apx2. putative glyoxysomal membrane-bound protein.
AAC08576.1 AF053474 Zantedeschia aethiopica cytosolic ascorbate peroxidase. apx.
AAD43338.1 AF158654 Fragaria x ananassa cytosolic ascorbate peroxidase APX26.
AAD43336.1 AFl 58652 Fragaria x ananassa cytosolic ascorbate peroxidase. ApxSC. corresponds to mRNA sequence deposited in
GenBank Accession Number AF039953.
AAB95222.1 AF039953 Fragaria x ananassa cytosolic ascorbate peroxidase. ApxSC.
AAD41408.1 AF159633 Fragaria x ananassa cytosolic ascorbate peroxidase. APX.
AAD41407.1 AF159632 Fragaria x ananassa cytosolic ascorbate peroxidase. APX.
AAD41403.1 AF159628 Fragaria x ananassa cytosolic ascorbate peroxidase. APX.
AAD41402.1 AF159627 Fragaria x ananassa cytosolic ascorbate peroxidase. APX.
AAD43337.1 AF158653 Fragaria x ananassa cytosolic ascorbate peroxidase APX19.
AAD41406.1 AF159631 Fragaria x ananassa cytosolic ascorbate peroxidase. APX.
AAD41404.1 AF159629 Fragaria x ananassa cytosolic ascorbate peroxidase. APX.
AAB94574.1 AF022213 Fragaria x ananassa cytosolic ascorbate peroxidase. APX-c.
AAD41405.1 AF159630 Fragaria x ananassa cytosolic ascorbate peroxidase. APX.
BAA08264.1 D45423 Oryza sativa ascorbate peroxidase.
BAA13671.1 D88649 Cucumis sativus cytosolic ascorbate peroxidase.
CAA55209.1 X78452 Raphanus sativus L-ascorbate peroxidase. APX.
BAA12890.1 D85864 Spinacia oleracea cytosolic ascorbate peroxidase.
AAA99518.1 L20864 Spinacia oleracea ascorbate peroxidase.
BAA12918.1 D85912 Nicotiana tabacum cytosolic ascorbate peroxidase.
AAB03844.1 U61379 Vigna unguiculata ascorbate peroxidase.
CAB58361.1 Y16773 Lycopersicon esculentum ascorbate peroxidase. Apx. H2O2 scavenger, cytosolic protein.
CAA57140.1 X81376 Capsicum annuum L-ascorbate peroxidase.
CAA06996.1 AJ006358 Hordeum vulgare ascorbate peroxidase. apx.
AAF22246.1 AF159380 Pimpinella brachycaφa ascorbate peroxidase. APX.
AAA86689.1 U15933 Nicotiana tabacum reduction of hydrogen peroxide, ascorbate peroxidase. APX.
CAA84406.1 Z34934 Zea mays cytosolic ascorbate peroxidase. apx.
CAA43992.1 X62077 Pisum sativum L-ascorbate peroxidase. Appxl.
AAA33645.1 M93051 Pisum sativum hydrogen peroxide removal; oxidative stress, ascorbate peroxidase. Apxl.
AAB01221.1 U56634 Glycine max ascorbate peroxidase 2. APx2.
AAA61779.1 L10292 Glycine max ascorbate peroxidase. Automated Edman sequencing ofthe N-terminal amino acids confirmed that the protein was authentic ascorbate peroxidase; putative.
BAB20889.1 AB053297 Oryza sativa L-ascorbate peroxidase. APXb.
CAA72247.1 Y11461 Brassica napus L-ascorbate peroxidase. APX.
BAB17666.1 AB050724 Oryza sativa subsp. japonica ascorbate peroxidase. APXb.
AAD20022.1 AF 127804 Glycine max ascorbate peroxidase. apxl. AsPOX; cytosolic.
AAB94927.1 AF038839 Brassica juncea ascorbate peroxidase.
SEQ ID NO: 103
AAA74900.1 L34343 Ruta graveolens anthranilate synthase alpha subunit.
AAA74901.1 L34344 Ruta graveolens anthranilate synthase alpha subunit.
AAC27795.1 AF079168 Nicotiana tabacum feedback-insensitive anthranilate synthase alpha-2 chain. ASA2.
BAA82095.1 AB022603 Oryza sativa anthranilate synthase alpha 2 subunit. OsASA2.
BAA82094.1 AB022602 Oryza sativa anthranilate synthase alpha 1 subunit. OsASAl.
CAC29060.1 AJ250008 Catharanthus roseus enzyme in tryptophan biosynthesis, anthranilate synthase alpha subunit. asa.
CAA06837.1 AJ006065 Catharanthus roseus isochorismate synthase.
SEQ ID NO: 104
CAA57724.1 X82273 Brassica oleracea 1 -aminocyclopropane- 1 -carboxylate synthase.
CAA51227.1 X72676 Brassica juncea
1 -aminocyclopropane- 1 -carboxylate synthase. MACC.
AAC83147.1 AF057563 Nicotiana glutinosa 1-aminocyclopropane-l -carboxylate synthase. ACS2. ACC synthase.
CAB60722.1 AJ012551 Citrus sinensis ACC synthase. acsl.
CAB60721.1 AJ012550 Citrus sinensis ACC synthase. acsl.
CAA06288.1 AJ005002 Nicotiana tabacum 1 -cyclopropane- 1-carboxylate synthase. acs2.
CAB65314.1 AJ131836 Nicotiana tabacum 1-aminocyclopropane-l -carboxylate synthase. acs2.
BAA78333.1 AB015625 Pyrus pyrifolia
1 -aminocyclopropane- 1 -carboxylic acid synthase. pPPACS3.
AAB17279.1 U72390 Lycopersicon esculentum converts S-adenosylmetMonine to 1-aminocyclopropane-l -carboxylate hormone biosynthesis.
1-aminocyclopropane-l -carboxylate synthase. LE-ACSIB.
AAF97614.1 U18056 Lycopersicon esculentum converts SAM to ACC; plant hormone biosynthesis. 1-amino-cyclopropane-l -carboxylate synthase. LE-ACS1A. ACC synthase; direct precursor of ethylene; one member of a multigene family.
AAF97615.1 U18057 Lycopersicon esculentum converts SAM to ACC; plant hormone biosynthesis. 1-amino-cyclopropane-l -carboxylate synthase. LE-ACSIB. ACC synthase; direct precursor of ethylene; one member of a multigene family.
AAB 17278.1 U72389 Lycopersicon esculentum converts S-adenosylmethionine to 1-aminocyclopropane-l -carboxylate hormone biosynthesis.
1-aminocyclopropane-l -carboxylate synthase. LE-ACS1A.
CAA46797.1 X65982 Nicotiana tabacum
1 -aminocyclopropane 1-carboxylate synthase.
BAA06464.1 D30805 Cucumis melo
1 -aminocyclopropane- 1 -carboxylate synthase.
AAB06724.1 U64804 Petunia x hybrida ethylene biosynthesis enzyme. 1-aminocyclopropane-l -carboxylate synthase. PH-ACS1.
CAA79478.1 Z18953 Petunia x hybrida
1 -aminocyclopropane- 1 -carboxylate synthase.
BAA94599.1 AB033502 Populus euphratica
1-aminocyclopropane-l -carboxylate synthase. peacs-1.
CAA67118.1 X98492 Nicotiana tabacum
ACC synthase. ACCS2.
CAA79477.1 Z18952 Dianthus caryophyllus
1 -aminocyclopropane 1-carboxylate synthase.
AAA33275.1 M66619 Dianthus caryophyllus
1-aminocyclopropane-l -carboxylate synthase. CARACC.
BAA83618.1 AB025906 Cucumis melo
1-aminocyclopropane-l -carboxylate synthase. CMe-ACSl (ME-ACS1).
BAA34923.1 AB013100 Lycopersicon esculentum
1-aminocyclopropane-l -carboxylate synthase. LE-ACS6. ACC synthase.
BAA25916.1 ABO 13346 Lycopersicon esculentum
1 -aminocyclopropane- 1 -carboxylate synthase.
CAB60831.1 AJ012696 Citrus sinensis
ACC synthase. acs2.
CAA41855.1 X59139 Lycopersicon esculentum
1 -aminocyclopropane 1-carboxylate synthase. LE-Acc2.
CAA44397.1 X62536 Lycopersicon esculentum
1 -aminocyclopropane 1-carboxylate synthase.
CAA41856.1 X59145 Lycopersicon esculentum 1 -aminocyclopropane 1-carboxylate synthase. LE-ACC2.
AAA81580.1 M34289 Lycopersicon esculentum 1 -aminocyclopropane- 1 -carboxylate synthase.
AAC98809.1 U68216 Carica papaya ACC synthase. fruit specific; ripening related.
BAA92350.1 AB034992 Malus x domestica
1 -aminocyclopropane- 1 -carboxylate synthase. MdACS-5A.
BAA92351.1 AB034993 Malus x domestica 1-aminocyclopropane-l -carboxylate synthase. MdACS-5B.
BAA94600.1 AB033503 Populus euramericana 1-aminocyclopropane-l -carboxylate synthase. peacs-2.
CAA77688.1 Z11613 Vigna radiata 1 -aminocyclopropane 1-carboxylate synthase.
CAA47474.1 X67100 Glycine max
1 -aminocyclopropane 1-carboxylate synthase.
AAB70885.1 U88971 Pelargonium x hortorum
1-aminocyclopropane-l -carboxylate synthase. GACS2. similar to Pelargonium hortorum clone pGAC-2, Genbank Accession Number U17231.
AAD04199.1 AF016459 Pisum sativum
1-aminocyclopropane-l -carboxylate synthase. ACS2. ACC synthase.
BAB 18464.1 D86242 Cucumis melo
1-aminocyclopropane-l -carboxylate synthase. CMe-ACS2 (ME-ACS2).
BAA93713.1 AB032936 Cucumis melo
1-aminocyclopropane-l -carboxylate synthase. CMe-ACS2.
BAA93715.1 AB032938 Cucumis sativus
1-aminocyclopropane-l -carboxylate synthase. CS-ACS2.
BAA33375.1 AB006804 Cucumis sativus
ACC synthase. CS-ACS2.
BAB16433.1 AB041521 Solanum tuberosum
ACC synthase. ppACSl.
AAC15777.1 AF061605 Nicotiana glutinosa ACC synthase. ACS3.
CAA72191.1 Y11357 Caricapapaya 1 -aminocyclopropane- 1 -carboxylate synthase.
BAA96743.1 AB044662 Prunus persica 1-aminocyclopropane-l -carboxylate synthase. PP-ACS1.
BAA90549.1 AB031026 Prunus mume
ACC synthase. PM-ACSl. 1-aminocyclopropane-l -carboxylic acid synthase.
BAA93712.1 AB032935 Cucumis melo 1-aminocyclopropane-l -carboxylate synthase. CMe-ACSl.
AAF22109.1 AFl 19411 Lupinus albus ethylene biosynthesis enzyme. 1-aminocyclopropane-l-carboxylate synthase 1. ACSl. ACC synthase; S-adenosyl-L-methionine methylthioadenosine-lyase.
AAC83146.1 AF057562 Nicotiana glutinosa
1-aminocyclopropane-l -carboxylate synthase. ACSl. ACC synthase.
CAA09477.1 AJ011095 Citrus sinensis
ACC synthase. acs-1.
BAA93714.1 AB032937 Cucumis sativus
1-aminocyclopropane-l -carboxylate synthase. CS-ACS1.
SEQ ID NO: 106 AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
BAA97122.1 AB016264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserfZ.
AAC50047.1 U89255 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti4.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive transcription factor.
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBPl.
BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding protein 1 homolog. McEREBPl.
AAC49740.1 U89256 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti5.
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACRE1. similar to EREBP transcription factors.
BAA97124.1 AB016266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
AAB38748.1 U81157 Nicotiana tabacum
S25-XP1 DNA binding protein.
AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. OSJNBa0027P10.12.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
AAC49741.1 U89257 Lycopersicon esculentum
DNA-binding protein binds the GCC box pathogenesis-related promoter element. Pti6.
CAC12822.1 AJ299252 Nicotiana tabacum
AP2 domain-containing transcription factor. ap2.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
AAC14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nser_3.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. similar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
AAF05606.1 AF190770 Oryza sativa
EREBP-like protein, tshl. TSH1; induced by ethylene.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor. AAC29516.1 U77655 Solanum tuberosum
DNA binding protein homolog. ST WAAEIRD.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like transcription factor.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal .
AAD45623.1 AF084185 Brassica napus dehydration responsive element binding protein. DNA binding protein; DRE binding protein.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP transcription factors.
BAB21218.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.24.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
BAB21211.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.17.
AAKO 1088.1 AF298230 Hordeum vulgare
CBFl-like protein BCBF1. BCBF1. AP2 domain protein; putative DRE binding factor.
AAK01089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor. SEQ ID NO: 107
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
AAC50047.1 U89255 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti4. BAA97122.1 AB016264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive transcription factor.
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBPl.
BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding proteinl homolog. McEREBPl.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2 -domain DNA-binding protein. orca3.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
AAB38748.1 U81157 Nicotiana tabacum
S25-XP1 DNA binding protein.
AAC49740.1 U89256 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti5.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREl. similar to EREBP transcription factors.
BAA97124.1 AB016266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. OSJNBa0027P10.12.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf3.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. similar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
AAC14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
CAC12822.1 AJ299252 Nicotiana tabacum AP2 domam-containing transcription factor. ap2.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
AAF05606.1 AF190770 Oryza sativa
ERE BP-like protein, tshl. TSH1; induced by ethylene.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor. AAC49741.1 U89257 Lycopersicon esculentum
DNA-binding protein binds the GCC box pathogenesis-related promoter element. Pti6.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like transcription factor.
AAC29516.1 U77655 Solanum tuberosum
DNA binding protein homolog. STWAAEIRD.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal.
BAA78738.1 AB023482 Oryza sativa
EST AU055776(S20048) conesponds to a region ofthe predicted gene.; Similar to
Arabidopsis thaliana AP2 domain containing protein RAP2.10 mRNA, partial cds.(AF003103).
AAD45623.1 AF084185 Brassica napus dehydration responsive element binding protein. DNA binding protein; DRE binding protein.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP transcription factors.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
BAA99376.1 AP002526 Oryza sativa
ESTs AU093391(E60370),AU091593(C60458), AU093392(E60370) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana BAC F21 J9; AP2 domain protein.
(AC000103).
BAB21218.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.24.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A 17.22.
BAB21211.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.17.
AAK01088.1 AF298230 Hordeum vulgare
CBFl-like protein BCBF1. BCBF1. AP2 domain protein; putative DRE binding factor.
AAG59618.1 AF239616 Hordeum vulgare
CRT/DRE-bindfng factor. CBF.
AAK01089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor.
SEQ ID NO: 109
BAA97124.1 ABO 16266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREl. similar to EREBP transcription factors.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. QSJNBa0027P10.12.
BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding proteinl homolog. McEREBPl.
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
AAC50047.1 U89255 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti4.
BAA97122.1 AB016264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive transcription factor.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
AAC49740.1 U89256 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti5.
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBPl.
AAB38748.1 U81157 Nicotiana tabacum
S25-XP1 DNA binding protein.
CAC12822.1 AJ299252 Nicotiana tabacum
AP2 domain-containing transcription factor. ap2.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserB.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. similar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
AAC14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
AAC49741.1 U89257 Lycopersicon esculentum
DNA-binding protein binds the GCC box pathogenesis-related promoter element. Pti6.
AAC29516.1 U77655 Solanum tuberosum
DNA binding protein homolog. STWAAEIRD.
AAF05606.1 AFl 90770 Oryza sativa
EREBP-like protein, tshl. TSH1; induced by ethylene.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like transcription factor.
BAA78738.1 AB023482 Oryza sativa
EST AU055776(S20048) conesponds to a region ofthe predicted gene.; Similar to
Arabidopsis thaliana AP2 domain containing protein RAP2.10 mRNA, partial cds.(AF003103).
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal .
AAD45623.1 AF084185 Brassica napus dehydration responsive element binding protein. DNA binding protein; DRE binding protein.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IA. ACREl 11 A. similar to EREBP transcription factors.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
AAK01088.1 AF298230 Hordeum vulgare
CBFl-like protein BCBF1. BCBF1. AP2 domain protein; putative DRE binding factor.
BAA99376.1 AP002526 Oryza sativa
ESTs AU093391(E60370),AU091593(C60458), AU093392(E60370) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana BAC F21 J9; AP2 domain protein.
(AC000103).
AAG59619.1 AF243384 Oryza sativa
CRT/PRE binding factor. CBF. DREB.
BAB21218.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.24.
AAG32659.1 AF253971 Picea abies
APETALA2-related transcription factor 2. AP2L2. PaAP2L2.
AAC49567.1 U41466 Zea mays
Glossy 15. Glossy 15. AP2 DNA-binding domain protein; similar to the Arabidopsis floral homeotic gene APETALA2; maize AP2-domain regulator of leaf epidermal traits; homeotic regulator of leaf epidermal cell identity; allele: W64A; putative transcription factor.
SEQ ID NO: 110
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREl. similar to EREBP transcription factors.
BAA97124.1 ABO 16266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. OSJNBa0027P10.12.
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
BAA97122.1 ABO 16264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2. BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding proteinl homolog. McEREBPl.
AAC50047.1 U89255 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti4.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive transcription factor.
AAC49740.1 U89256 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti5.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBPl.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
AAB38748.1 U81157 Nicotiana tabacum
S25-XP1 DNA binding protein.
CAC12822.1 AJ299252 Nicotiana tabacum
AP2 domain-containing transcription factor. ap2.
AAF05606.1 AF190770 Oryza sativa
EREBP-like protein, tshl. TSH1; induced by ethylene.
AAC29516.1 U77655 Solanum tuberosum
DNA binding protein homolog. STWAAEIRD.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like transcription factor.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf3. BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. similar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
AAC49741.1 U89257 Lycopersicon esculentum
DNA-binding protein binds the GCC box pathogenesis-related promoter element. Pti6.
AAC14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal.
AAD45623.1 AF084185 Brassica napus dehydration responsive element binding protein. DNA binding protein; DRE binding protein.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP transcription factors.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
BAB21218.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.24.
AAK01088.1 AF298230 Hordeum vulgare
CBFl-like protein BCBF1. BCBF1. AP2 domain protein; putative DRE binding factor.
AAG59619.1 AF243384 Oryza sativa
CRT/PRE binding factor. CBF. DREB.
BAB21211.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.17.
AAK01089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor. SEQ ID NO: 111
AAF87216.1 AF231351 Nicotiana tabacum plastidic glucose 6-phosphate dehydrogenase. G6PDHP2.
CAA67782.1 X99405 Nicotiana tabacum glucose-6-phosphate dehydrogenase. G6PD.
AAB69317.1 AF012861 Petroselinum crispum plastidic glucose-6-phosphate dehydrogenase. pG6PDH.
CAB52708.1 AJ010712 Solanum tuberosum glucose-6-ρhosphate 1 -dehydrogenase. g6pd.
CAB52685.1 AJ132346 Dunaliella bioculata plastidic glucose-6-phosphate dehydrogenase. g6PD.
CAA04994.1 AJ001772 Nicotiana tabacum glucose-6-phosphate dehydrogenase. TCG18.
CAA58775.1 X83923 Solanum tuberosum glucose-6-phosphate dehydrogenase.
CAA03941.1 AJ000184 Spinacia oleracea Glucose-6-phosphate dehydrogenase. G6PD.
CAA03939.1 AJ000182 Spinacia oleracea Glucose-6-phosphate dehydrogenase. G6PD.
CAA03940.1 AJ000183 Spinacia oleracea Glucose-6-phosphate dehydrogenase. G6PD.
AAB41552.1 U18238 Medicago sativa subsp. sativa glucose-6-phosphate dehydrogenase.
AADl 1426.1 AF097663 Mesembryanthemum crystallinum cytoplasmic glucose-6-phosphate 1 -dehydrogenase. G6PD.
AAB69319.1 AF012863 Petroselinum crispum cytosolic glucose-6-phosphate dehydrogenase 2. cG6PDH2.
CAA52442.1 X74421 Solanum tuberosum glucose-6-phosphate 1 -dehydrogenase. g6pdh. homotetramer.
AAB69318.1 AFO 12862 Petroselinum crispum cytosolic glucose-6-phosphate dehydrogenase 1. cG6PDHl.
BAA97662.1 AB029454 Triticum aestivum glucose-6-phosphate dehydrogenase. gόpdh.
BAA97663.1 AB029455 Triticum aestivum glucose-6-phosphate dehydrogenase. gόpdh.
CAA04993.1 AJ001770 Nicotiana tabacum glucose-6-phosphate dehydrogenase. TCG9.
CAA04992.1 AJ001769 Nicotiana tabacum glucose-6-phosphate dehydrogenase. TCG6.
BAA97664.1 AB029456 Triticum aestivum glucose-6-phosphate dehydrogenase. g6pdh.
AAG23802.1 AF260736 Cucurbita pepo plastidic glucose-6-phosphate dehydrogenase.
CAB66330.1 AJ279688 Betula pendula glucose-6-phosphate dehydrogenase. g6pd.
BAA82155.1 AB011441 Triticum aestivum glucose-6-phosphate dehydrogenase. WESR5. salt-stress responding gene.
CAA06200.1 AJ004900 Glycine max pentose phosphate pathway oxidoreductase generating NADPH. glucose-6-phosphate- dehydrogenase.
SEQ ID NO: 112
CAA48611.1 X68652 Raphanus sativus hydroxymethylglutaryl-CoA reductase (NADPH). HMG2.
CAA48610.1 X68651 Raphanus sativus hydroxymethylglutaryl-CoA reductase (NADPH). HMG1.
AAC05089.1 AF038046 Gossypium hirsutum catalyzes the synthesis of mevalonate from hmg-coA. 3-hydroxy-3-methylglutaryl-coenzyme
A reductase 2. hmg2. hmg-coA reductase 2; HMGR2.
AAA33108.1 M96068 Catharanthus roseus hydroxymethylglutaryl-CoA reductase. HMGR.
BAA36291.1 AB021862 Cucumis melo HMG-CoA reductase. Cm-HMGR. putative.
AAB52551.1 U51985 Solanum tuberosum HMG-CoA reductase.
AAA93498.1 L01400 Solanum tuberosum convert HMG-CoA into mevalonate. hydroxymethylglutaryl coenzyme A reductase. hmgr. putative.
AAB87727.1 U60452 Nicotiana tabacum hydroxy-methylglutaryl-coenzyme A reductase. HMGR1.
CAA70440.1 Y09238 Zea mays 3-hydroxy-3-methylglutaryl coenzyme A reductase.
CAA45181.1 X63649 Nicotiana sylvestris catalyses synthesis of mevalonate. 3-hydroxy-3-methylglutaryl-coenzyme A reductase.
HMGR. endoplasmic reticulum location.
AAD28179.1 AFl 10383 Capsicum annuum
3-hydroxy-3-methylglutaryl-coenzyme A reductase. HMGR2M. HMGR.
AAB52552.1 U51986 Solanum tuberosum
HMG-CoA reductase.
BAA93631.1 AB022690 Solanum tuberosum
3-hydroxy-3-methylglutaryl coenzyme A reductase. HMG3.
AAB62581.1 U68072 Lycopersicon esculentum 3-hydroxy-3-methylglutaryl CoA reductase 2. HMG2.
AAB69726.1 U72145 Camptotheca acuminata converts HMGCoA to mevalonate. 3-hydroxy-3-methylglutaryl coenzyme a reductase. hmg3.
HMGR.
AAB53748.1 U95816 Oryza sativa 3-hydroxy-3-methylglutaryl-CoA reductase. HMGR.
BAB20771.1 AB041031 Solanum tuberosum 3-hydroxy-3-methylglutaryl coenzyme A reductase. HMG2.
AAB69727.1 U72146 Camptotheca acuminata
3-hydroxy-3-methylglutaryl coenzyme A reductase. converts HMGCoA in mevalonate precursor for isoprenoid compounds; HMGR; hmg2; similar to protein encoded by GenBank Accession Number LI 0390.
AAD47596.1 AF142473 Artemisia annua HMG-CoA reductase. HMGR1.
AAA34169.1 M63642 Lycopersicon esculentum 3-hydroxy-3-methylglutaryl coenzyme A reductase. hmg2.
AAA68966.1 U14625 Artemisia annua 3-hydroxy-3-methylglutaryl coenzyme A reductase. AHM4.
AAA33358.1 M74798 Hevea brasiliensis 3-hydroxy-3-methylglutaryl-coenzyme A reductase. hmgr3.
AAA68965.1 U14624 Artemisia annua 3-hydroxy-3-methylglutaryl coenzyme A reductase. AHM1.
AAD08820.1 U43961 Oryza sativa
3 -hydroxy-3 -methylglutaryl=CoA reductase. HMGR.
AAA33360.1 M74800 Hevea brasiliensis
3 -hydroxy-3 -methy lglutaryl-coenzyme A reductase. hmgr3.
CAA92821.1 Z68504 Oryza sativa 3 -hydroxy-3 -methy lglutaryl-Co A reductase.
AAC05088.1 AF038045 Gossypium hirsutum catalyzes the synthesis of mevalonate fromhmg-coA. 3 -hydroxy-3 -methy lglutaryl-coenzyme
A reductase 1. hmgl. hmg-coA reductase 1; HMGR1.
AAC15475.1 AF034760 Tagetes erecta
3 -hydroxy-3 -methylglutaryl coenzyme A reductase.
AAC 15476.1 AF034761 Tagetes erecta
3 -hydroxy-3 -methylglutaryl coenzyme A reductase.
CAA38469.1 X54659 Hevea brasiliensis hydroxymethylglutaryl-CoA reductase. HMGR1.
CAA38467.1 X54657 Hevea brasiliensis hydroxymethylglutaryl-CoA reductase. HMGR1.
AAD38873.1 AFl 10382 Oryza sativa
3 -hydroxy-3 -methy lglutaryl-coenzyme A reductase. HMGR3G. HMG-CoA reductase.
AAA33040.1 L10390 Camptotheca acuminata 3 -hydroxy-3 -methylglutaryl coA reductase.
AAD03789.1 U43711 Moras alba catalyzes the final step in mevalonate pathway. 3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. HMGR.
AAA21720.1 L28995 Oryza sativa conversion of hydroxymethylglutaryl coenzyme A to mevalonate. 3 -hydroxy-3 -methylglutaryl coenzyme A reductase. putative.
AAC72378.1 AF096838 Solanum tuberosum 3-hydroxy-3-methylglutaryl coenzyme A reductase.
AAB04043.1 L40938 Lycopersicon esculentum HMGR CoA reductase. HMGR1.
CAA38468.1 X54658 Hevea brasiliensis hydroxymethylglutaryl-CoA reductase. HMGR2.
CAA52787.1 X74783 Lithospermum erythrorhizon
3-hydroxy-3-methyl glutaryl coenzyme A reductase. Lehmgrl.
AAD09278.1 U97683 Glycine max catalyzes the synthesis of mevalonate, the specific precursor of all isoprenoid compounds present in plants. 3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. HMG-CoA reductase.
AAG43469.1 AF196964 Bixa orellana catalyzes mevalonate synthesis from hmg-CoA. 3 -hydroxy-3 -methylglutaryl-coenzyme A reductase.
BAA09705.1 D63389 Cucumis sativus
3-hydroxy-3-methylglutaryl CoA reductase.
AAB47161.1 S82272 Gossypium barbadense
3 -hydroxy-3 -methylglutaryl coenzyme A reductase. /gene="3-hydroxy-3-methylglutaryl coenzyme A reductase,. This sequence comes from Fig. 2; 3 -hydroxy-3 -methylglutaryl CoA reductase; HMGR.
AAA33359.1 M74799 Hevea brasiliensis
3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. hmgr3.
AAC37432.1 L34825 Solanum tuberosum
HMG-CoA reductase. hmgl gene family.
AAC37434.1 L34827 Solanum tuberosum
HMG-CoA reductase. hmgl gene family.
AAC37431.1 L34823 Solanum tuberosum
HMG-CoA reductase. hmgl gene family.
AAC37433.1 L34826 Solanum tuberosum HMG-CoA reductase. hmgl gene family. AAC37435.1 L34828 Solanum tuberosum
HMG-CoA reductase. hmgl gene family.
AAC37436.1 L34829 Solanum tuberosum
HMG-CoA reductase. hmgl gene family.
SEQ ID NO: 113 AAC49676.1 U77345 Zea mays lethal leaf-spot 1. 11s 1. Allele: wild-type; LLS1; similar to bacterial ring-hydroxylating dioxygenase.
AAG03051.1 AF284781 Oryza sativa
LLS1 protein. Llsl. SEQ ID NO: 115
BAA82379.1 AP000367 Oryza sativa
Similar to putative receptor kinase. (AC002332).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
AAG25966.1 AF302082 Nicotiana tabacum cytokimn-regulated kinase 1. CRKl. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
AAC27894.1 AF023164 Zea mays leucine-rich repeat transmembrane protein kinase 1. ltkl .
AAC27895.1 AF023165 Zea mays leucine-rich repeat transmembrane protein kinase 2. Itk2.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
AAF66615.1 AF142596 Nicotiana tabacum
LRR receptor-like protein kinase.
CAB41878.1 Y18259 Brassica oleracea
SRK5 protein. SRK5. receptor-like kinase.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
CAB41879.1 Y18260 Brassica oleracea
SRK15 protein. SRK15. receptor-like kinase.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
CAA74661.1 Y14285 Brassica oleracea
SFR1. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK 1.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
CAA47962.1 X67733 Zea mays receptor-like protein kinase. PK1.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
AAB09771.1 U67422 Zea mays
CRINKLY4 precursor. cr4. receptor kinase homolog.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
BAA06538.1 D31737 Nicotiana tabacum protem-serine/threonine kinase.
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
AAKl 1674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl . accumulates in mesophyll cells.
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APK1 gene for protein tyrosine-serine-threonine kinase.(D12522).
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
AAB47422.1 U59318 Lycopersicon esculentum serine/threonine protein kinase Fen, fen, allele of Fen, fenthion sensitivity gene from tomato.
BAA21132.1 D88193 Brassica rapa
S-receptor kinase. SRK9 (B.c).
SEQ ID NO: 117
AAA34002.1 M67449 Glycine max protein kinase. PK6.
AAG31141.1 AF305911 Oryza sativa
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAG31142.1 AF305912 Hordeum vulgare
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
CAC09580.1 AJ298992 Fagus sylvatica
Abscisic acid (ABA) and calcium induced protein kinase. protein kinase (PK). pkl.
CAA06334.1 AJ005077 Lycopersicon esculentum protein kinase. TCTR2 protein. TCTR2.
AAD10057.1 AFl 10519 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTRlv.
AAD10056.1 AFl 10518 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTR1.
AAD46406.1 AF096250 Lycopersicon esculentum ethylene-responsive protein kinase TCTR1. ER50. serine/threonine kinase; similar to
Arabidopsis thaliana negative regulator ofthe ethylene response pathway encoded by
GenBank Accession Number L08789.
CAA73722.1 Y13273 Lycopersicon esculentum putative protein kinase.
AAK11734.1 AY027437 Arachis hypogaea serine/threonine/tyrosine kinase.
AAK30005.1 AY029067 Rosa hybrid cultivar CTR2 protein kinase.
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
CAA73068.1 Y12465 Sorghum bicolor serine/threonine kinase. SNFL2.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
BAA83688.1 AB011967 Oryza sativa OsPK4. OsPK4. protein kinase.
AAF34436.1 AF172282 Oryza sativa similar to mitogen-activated protein kinases. DUPR11.32.
AAF22219.1 AF141378 Zea mays protein kinase PK4. ZmPK4.
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl .7.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
BAA34675.1 ABO 11670 Triticum aestivum wpk4 protein kinase. wpk4.
BAA92970.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F28A21 ; putative protein kinase.
(AL035526).
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl. GmRLKl.
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
CAA73067.1 Y12464 Sorghum bicolor serine/threonine kinase. SNFL1.
AAF59906.1 AF197947 Glycine max receptor protein kinase-like protein. CLV1B.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1 A.
BAB16918.1 AP002863 Oryza sativa putative protein kinase. P0005A05.22.
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml .
BAA83689.1 AB011968 Oryza sativa OsPK7. OsPK7. protein kinase.
CAA74646.1 Y14274 Sorghum bicolor putative serine/threonine protein kinase. SNFL3.
BAA95893.1 AP002071 Oryza sativa
Similar to Arabidopsis thaliana wak4 gene; wall-associated kinase 4. (AJ009695).
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPK10) mRNA. (L27821).
AAC83393.1 U83625 Zea mays protein kinase ZmMEKl . mitogen-activated; ERK-activating protein kinase (MEK) homolog.
BAA92972.1 AP001551 Oryza sativa
ESTs AU056183(S20356),AU056881(S20950) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome 4 BAC clone F6I18 ; putative protein kinase.
(AL022198).
BAB18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAB40015.1 AP003021 Oryza sativa putative wall-associated kinase 1. P0503E05.18.
BAA05648.1 D26601 Nicotiana tabacum protein kinase.
AAC27489.1 AF077130 Oryza sativa receptor-like protein kinase.
AAC02535.1 AF044260 Oryza sativa receptor serine/threonine kinase. protein kinase.
AAG40578.1 AF216314 Oryza sativa MAP kinase kinase 1. protein kinase; MEK1.
CAA08997.1 AJ010093 Brassica napus MAP3K beta 1 protein kinase. MAP3K beta 1.
SEQ ID NO: 118
AAD21199.1 AF127797 Capsicum chinense putative bZIP DNA-binding protein.
CAC00658.1 AJ292745 Petroselinum crispum bZIP type transcription factor, common plant regulatory factor 7. cprf7.
CAC00657.1 AJ292744 Petroselinum crispum bZIP type transcription factor, common plant regulatory factor 6. cprfδ.
CAA74023.1 Y13676 Antinhinum majus bZIP DNA-binding protein.
CAA74022.1 Y13675 Antinhinum majus bZIP DNA-binding protein.
BAA22204.1 D63951 Nicotiana tabacum TBZ17. tbzl7. bZIP protein.
AAD55394.1 AF176641 Lycopersicon esculentum bZIP DNA-binding protein.
CAA44607.1 X62745 Zea mays ocs-binding factor 1. OBF1.
AAK25822.1 AF350505 Phaseolus vulgaris bZip transcription factor.
AAK01953.1 AY026054 Phaseolus acutifolius bZIP. transcription factor.
CAA71687.1 Y10685 Glycine max bZIP DNA-binding protein. G/HBF-1. G/HBF-1.
AAC37418.1 L34551 Oryza sativa transcriptional activator protein. RITA-1.
BAA36492.1 AB021736 Oryza sativa bZIP protein.
BAA11431.1 D78609 Oryza sativa bZIP protein.
CAA41453.1 X58577 Petroselinum crispum DNA-binding protein; bZIP type. CPRF2.
CAA71768.1 Y10809 Petroselinum crispum bZIP DNA-binding protein. CPRF4a.
AAD42938.1 AF084972 Catharanthus roseus
G-Box binding protein 2. GBF2. basic leucine zipper; trans-regulatory factor.
AAC49556.1 U04295 Oryza sativa DNA-binding factor of bZIP class. osZIP-la.
BAA07289.1 D38111 Triticum aestivum transcription factor HBP-la(17).
CAA71795.1 Y10834 Hordeum vulgare bZIP transcription factor 2. Blz2.
CAA70216.1 Y09013 Triticum aestivum transcriptional activator. SPA.
CAA40101.1 X56781 Triticum aestivum transcription factor. HBP- la. la- 17.
AAA80169.1 U10270 Zea mays G-box binding factor 1. GBF1.
CAAl 1499.1 AJ223624 Spinacia oleracea basic leucine zipper protein. bZIP.
AAC49474.1 U41817 Phaseolus vulgaris regulator of MAT2. ROM2. Repressor of seed-specific lectin (phytohemagglutinin) gene; bZIP transcriptional repressor.
AAB40291.1 U42208 Oryza sativa
OSBZ8. G-box binding protein; GBF type bZIP protein.
BAA02304.1 D 12920 Triticum aestivum transcription factor HBP-la(cl4).
SEQ ID NO: 119
BAA96200.1 AP002093 Oryza sativa
EST D40368(S2303) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome II putative cytochrome P450 (AC004077).
BAA96158.1 AP002092 Oryza sativa
EST D40368(S2303) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome II BAC T31E10; putative cytochrome P450 (AC004077).
BAA96196.1 AP002093 Oryza sativa
ESTs AU086027(S2303),D40339(S2251) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome II BAC T31E10; putative cytochrome P450 (AC004077).
BAA96154.1 AP002092 Oryza sativa
ESTs AU086027(S2303),D40339(S2251) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome II BAC T31E10; putative cytochrome P450
(AC004077).
BAA96193.1 AP002093 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC T31E10; putative cytochrome P450
(AC004077).
BAA96151.1 AP002092 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC T31E10; putative cytochrome P450
(AC004077).
BAA96194.1 AP002093 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC T31E10; putative cytochrome P450
(AC004077).
BAA96152.1 AP002092 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC T31E10; putative cytochrome P450
(AC004077).
AAG41777.1 AF212991 Cucurbita maxima ent-kaurene oxidase. CYP88A2. cytochrome P450; similar to maize Dwarf3 protein.
AAK11616.1 AF326277 Hordeum vulgare ent-kaurenoic acid oxidase. KAO1. cytochrome P450; CYP88A; HvKAOl.
AAK00946.1 AF318211 Taxus cuspidata
5-alpha-taxadienol-lO-beta-hydroxylase. cytochrome P450-like protein.
AAC49659.1 U74319 Sorghum bicolor obtusifoliol 14-alpha demethylase CYP51. CYP51. cytochrome P450 catalyzing the 14-alpha demethylation of obtusifoliol in plants.
AAA17746.1 L19075 Catharanthus roseus cytochrome P450. CYP72C. putative.
AAA33106.1 L10081 Catharanthus roseus cytochrome P-450 protein. CYP72. putative; CYP72 protein.
AAA17732.1 L19074 Catharanthus roseus cytochrome P450. CYP72B.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
AAB 17070.1 U54770 Lycopersicon esculentum cytochrome P450 homolog. dwarf.
CAB41490.1 AJ238439 Cicer arietinum cytochrome P450 monooxygenase. cyp81E3v2.
BAB19089.1 AP002744 Oryza sativa putative cytochrome P450. P0006C01.31. contains ESTs D24685(R2374),AU031882(R2374).
AAD44150.1 AF124815 Mentha spicata cytochrome p450.
BAB19110.1 AP002839 Oryza sativa putative cytochrome P450. P0688A04.16. contains ESTs
D24685(R2374),AU031882(R2374).
BAA74465.1 AB022732 Glycyrrhiza echinata cytochrome P450. CYP Ge-31.
BAA22422.1 AB001379 Glycynhiza echinata cytochrome P450. CYP81E1.
CAA10067.1 AJ012581 Cicer arietinum cytochrome P450. cyp81E3.
BAB40322.1 AB036772 Triticum aestivum cytochrome P450. N-l.
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
CAB56742.1 AJ249800 Cicer arietinum cytochrome P450 monooxygenase. cyp81E5.
AAF89209.1 AF279252 Vigna radiata cytochrome P450. CipCYP.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
BAB19107.1 AP002839 Oryza sativa putative cytochrome P450. P0688A04.13. contains ESTs
AU100635(C10787),D22354(C10787).
BAB 19086.1 AP002744 Oryza sativa putative cytochrome P450. P0006C01.28. contains ESTs
AU100635(C10787),D22354(C10787).
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81Blc.
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'- race).
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
BAB21156.1 AP002899 Oryza sativa putative cytochrome P450. P0456A01.12.
AAG13498.1 AC068924 Oryza sativa putative cytochrome P450. OSJNBa0026L12.20.
AAF34534.1 AF195813 Lupinus albus isoflavone synthase 1. ifsl. cytochrome P450.
CAA83941.1 Z33875 Mentha x piperita cytochrome P-450 oxidase.
AAF34533.1 AF195812 Pisum sativum isoflavone synthase 1. ifsl. cytochrome P450.
AAF45142.1 AFl 95818 Glycine max isoflavone synthase 1. ifsl. cytochrome P450.
SEQ ID NO: 120
CAB42052.1 AJ242045 Lycopersicon esculentum mcotianamine synthase. chin.
BAA74581.1 AB011264 Hordeum vulgare nicochianainine synthase 3. hvnas3.
AAD32651.1 AF136942 Hordeum vulgare nicotianamine synthase 2. nashor2.
BAA74582.1 AB011265 Hordeum vulgare mcochianamine synthase 2. hvnas2.
BAB17824.1 AB023819 Oryza sativa nicotianamine synthase 3. osnas3.
AAD32650.1 AF136941 Hordeum vulgare mcotianamine synthase 1. nashorl .
BAA74583.1 ABO 11266 Hordeum vulgare nicotianamine Synthase 4. hvnas4.
BAA74586.1 AB011269 Hordeum vulgare mcotianamine Synthase 6. hvnas6.
BAA74587.1 ABO 19525 Hordeum vulgare nicotianamine synthase 7. hvnas7.
BAB17826.1 AB046401 Oryza sativa nicotianamine synthase 2. OsNAS2.
BAB17823.1 AB023818 Oryza sativa nicotianamine synthase 2. osnas2.
BAB17825.1 AB046401 Oryza sativa nicotianamine synthase 1. OsNASl.
BAA74588.2 AB021746 Oryza sativa nicotianamine synthase 1. osnasl.
BAA74580.1 AB010086 Hordeum vulgare nicotianamine synthase 1. hvnasl.
BAA74585.1 AB011268 Hordeum vulgare mcotianamine Synthase 5-2. hvnas5-2.
BAA74584.1 ABO 11267 Hordeum vulgare nicotianamine synthase 5-1. hvnas5-l.
SEQ ID NO: 122
AADO 1804.1 AF026480 Dianthus caryophyllus lipase. lipid-protein-particle associated.
BAB39417.1 AP002901 Oryza sativa putative lipase. P0456F08.17. contains ESTs
C99390(E11001),AU101109(E0858),AU101332(E11001).
AAB07724.1 U55867 Ipomoea nil Pn47p. lipase-like protein.
AAK31273.1 AC079890 Oryza sativa putative lipase. OSJNBb0089A17.13.
SEQ ID NO: 123
BAA21923.1 AB006601 Petunia x hybrida ZPT2-14. C2H2 zinc finger protein, 2 finger.
BAA21922.1 AB006600 Petunia x hybrida ZPT2-13. C2H2 zinc finger protein, 2finger.
BAA21921.1 AB006599 Petunia x hybrida ZPT2-12. C2H2 zinc finger protein, 2 finger.
BAA19110.1 AB000451 Petunia x hybrida PEThy;ZPT2-5. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA21926.1 AB006604 Petunia x hybrida ZPT2-9. C2H2 zinc finger protein, 2 finger.
BAA21925.1 AB006603 Petunia x hybrida ZPT2-8. C2H2 zinc finger protein, 2 finger.
BAA21924.1 AB006602 Petunia x hybrida ZPT2-7. C2H2 zinc finger protein, 2finger.
BAA21920.1 AB006598 Petunia x hybrida ZPT2-11. C2H2 zinc finger protein, 2finger.
CAA60828.1 X87374 Pisum sativum putative zinc finger protein.
BAA19111.1 AB000452 Petunia x hybrida PEThy;ZPT2-6. Cys(2) His(2) zinc finger protein, 2 fingers.
CAB77055.1 Y18788 Medicago sativa putative TFIIIA (or kruppel)-like zinc finger protein.
BAA96071.1 AB035133 Petunia x hybrida C2H2 zinc-finger protein ZPT3-3. ZPT3-3.
BAA21927.1 AB006605 Petunia x hybrida ZPT3-3. C2H2 zinc finger protein, 3 finger.
BAA96070.1 AB035132 Petunia x hybrida C2H2 zinc-finger protein ZPT2- 10. PEThy;ZPT2-10.
BAA21919.1 AB006597 Petunia x hybrida ZPT2-10. C2H2 zinc finger protein, 2 finger.
AAK01713.1 AF332876 Oryza sativa zinc finger transcription factor ZF1.
AAC06243.1 AF053077 Nicotiana tabacum transcription factor, osmotic stress-induced zinc-finger protein, zfp.
BAA05079.1 D26086 Petunia x hybrida zinc-finger protein.
BAA05078.1 D26085 Petunia x hybrida zinc-finger DNA binding protein.
AAB39638.1 U68763 Glycine max putative transcription factor. SCOF-1. scof-1. zinc-finger protein.
BAA05076.1 D26083 Petunia x hybrida zinc-finger DNA binding protein.
BAA05077.1 D26084 Petunia x hybrida zinc-finger DNA binding protein.
BAA21928.1 AB006606 Petunia x hybrida ZPT4-4. C2H2 zinc finger protein, 4 finger.
BAA19114.1 AB000455 Petunia x hybrida PEThy;ZPT4-l. Cys(2) His(2) zinc finger protein, 4 fingers.
AAD26942.1 AFl 19050 Datisca glo erata zinc-finger protein 1. zfpl. DgZFPl.
BAA19112.1 AB000453 Petunia x hybrida PEThy;ZPT3-l. Cys(2) His(2) zinc finger protein, 3 fingers.
AAB53260.1 U76554 Brassica rapa transcription factor, zinc-finger protein- 1. BR42.
AAB53261.1 U76555 Brassica rapa zinc-finger protein BcZFPl. BcZFPl(3-2z).
BAA19926.1 AB000456 Petunia x hybrida PEThy; ZPT4-2. C2H2 zinc finger protein, 4 finger.
SEQ ID NO: 126
CAA10134.1 AJ012693 Cicer arietinum basic blue copper protein.
CAB65280.1 AJ248323 Medicago sativa subsp. x varia basic blue protein, babl.
AAC32448.1 U76296 Spinacia oleracea plantacyanin. member of a strictly plant-specific family of mononuclear blue copper proteins known as phytocyanins, which are further classified into three distinct subfamilies: Uclacyanins, Stellacyanins, and Plantacyamns; member ofthe subfamily of Plantacyamns; non-glycosylated.
AAF66243.1 AF243181 Lycopersicon esculentum plantacyanin. naturally occurring cupredoxin with a Val residue in the position ofthe axial ligand Met; member ofthe plantacyanin subfamily of a strictly plant-specific family of mononuclear blue copper proteins known as phytocyanins.
AAD10251.1 AF031195 Triticum aestivum blue copper-binding protein homolog. S85.
AAF66242.1 AF243180 Lycopersicon esculentum dicyanin. binuclear blue copper protein; contains two stellacyanins linked together.
CAA80963.1 Z25471 Pisum sativum blue copper protein.
AAC64163.1 AF093537 Zea mays blue copper protein, similar to pea blue copper protein in GenBank Accession Number
Z25471.
AAC32421.1 U65511 Cucumis sativus putative oxygen activation and/or lignin formation, stellacyanin. member of a strictly plant- specific family of mononuclear blue copper proteins known as phytocyanins, which are further classified into three distinct subfamilies: Uclacyanins, Stellacyanins, and Plantacyanins; member ofthe subfamily of Stellacyanins; similar to Rhus vernicifera stellacyanin: SwissProt Accession Number P00302; similar to umecyanin: SwissProt Accession Number P42849; similar to mavicyanin; SwissProt Accession Number P80728; similar to BCB encoded by GenBank Accession Number Z15058; similar to F18A8.9, encoded by GenBank Accession Number AC003105; similar to F7F1.27 encoded by GenBank Accession Number AC004669; similar to F9D12.16 encoded by GenBank Accession Number AF077407; glycoprotein.
SEQ ID NO: 127
AAA17000.1 L08632 Glycine max pyruvate kinase.
CAA37727.1 X53688 Solanum tuberosum pyruvate kinase.
CAA82628.1 Z29492 Nicotiana tabacum glycolytic enzyme, pyruvate kinase.
AAF44707.1 AF242871 Lilium longiflorum cytosolic pyruvate kinase.
BAA88185.1 AP000836 Oryza sativa
ESTs AU081247(C50004),AU068940(C51113) conespond to a region ofthe predicted gene.
Similar to pyruvate kinase (Q42954).
BAA76433.1 AB025005 Cicer arietinum pyruvate kinase.
CAA49996.1 X70653 Nicotiana tabacum pyruvate kinase. PKTL7. monomer.
CAA82223.1 Z28374 Nicotiana tabacum glycolytic enzyme. Pyruvate kinase; plastid isozyme.
CAA82222.1 Z28373 Nicotiana tabacum Glycolytic enzyme, pyruvate kinase; plastid isozyme.
AAA33871.1 M64737 Ricinus communis ATP:pyruvate phosphotransferase.
AAA33870.1 M64736 Ricinus communis ATP.-pyruvate phosphotransferase.
SEQ ID NO: 129
BAB12433.1 AB025030 Coptis japonica p450.
AAF05621.1 AF191772 Papaver somniferum hydroxylase involved in benzylisoquinoline alkaloid biosynthesis. (S)-N-methylcoclaurine 3'- hydroxylase. CYP80B1. cytochrome P450-dependent monooxygenase.
AAC39452.1 AF014800 Eschscholzia californica hydroxylase involved in the biosynthesis of tetrahydrobenzylisoquinoline alkaloids in plants.
(S)-N-methylcoclaurine 3 '-hydroxylase. CYP80B1. cytochrome P-450-dependent monooxygenase; CYP80Blyl.
AAC39453.1 AF014801 Eschscholzia californica hydroxylase involved in the biosynthesis of tetrahydrobenzylisoquinoline alkaloids in plants.
(S)-N-methylcoclaurine 3 '-hydroxylase. CYP80B1. cytochrome P-450-dependent monooxygenase; CYP80Blv2.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
AAAI 9701.1 L24438 Thlaspi arvense cytochrome P450.
CAA50648.1 X71657 Solanum melongena P450 hydroxylase.
AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
AAC48987.1 U09610 Berberis stolonifera cytochrome P-450 CYP80.
AAD56282.1 AFl 55332 Petunia x hybrida flavonoid 3'-hydroxylase. Htl. cytochrome P450; CYP75B2.
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl. P450 enzyme.
AAB 17562.1 U72654 Eustoma grandiflorum flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75.
BAA84916.1 AB032833 Cicer arietinum cytochrome P450. CYP76D1.
AAB94588.1 AF022459 Glycine max
CYP71D10ρ. CYP71D10. cytochrome P450 monooxygenase.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
CAA50155.1 X70824 Solanum melongena flavonoid hydroxylase (P450). CYP75.
AAC39318.1 AF029858 Sorghum bicolor second multifunctional cytochrome P450 in the biosynthetic pathway ofthe cyanogenic glucoside dhunin. Catalyzes the conversion of p-hydroxyphenylacetaldoxime to p- hydroxymandelonitrile. cytochrome P450 CYP71E1. CYP71E1. No EST#s identified.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
CAA71514.1 Y10490 Glycine max putative cytochrome P450.
BAA84071.1 AB028151 Antinhinum majus flavone synthase II. cytochrome P450. AFNS2.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71A6.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
BAA84072.1 AB028152 Torenia hybrida flavone synthase II. cytochrome P450. TFNS5.
CAA71516.1 Y10492 Glycine max putative cytochrome P450.
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
SEQ ID NO: 132
CAB55396.1 ALl 17264 Oryza sativa zwhl2.1. similar to Arabidopsis putative UDP-galactase-4-epimerase (AC007060); Method: conceptual translation with partial peptide sequencing.
SEQ ID NO: 133
AAG43835.1 AF213455 Zea mays protein phosphatase tyρe-2C. ρp2c-l. PP2C-1.
AAG13599.1 AC051633 Oryza sativa putative protein phosphatase-2C. OSJNBb0015I11.26.
AAG46118.1 AC073166 Oryza sativa putative protein phosphatase-2C. OSJNBb0064P21.12. tRNA-Lys.
BAB12036.1 AP002820 Oryza sativa putative protein phosphatase. P0702D12.18.
AAC36698.1 AF075580 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
CAC10359.1 AJ277087 Nicotiana tabacum protein phosphatase 2C. PP2C2.
CAB90633.1 AJ277743 Fagus sylvatica protein phpsphatase 2C (PP2C). pp2Cl. ABA-induced protein.
CAC10358.1 AJ277086 Nicotiana tabacum protein phosphatase 2C. PP2C1.
CAA72341.1 Y11607 Medicago sativa protein phosphatase 2C. MP2C.
AAC36697.1 AF075579 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
CAB61839.1 AJ242803 Sporobolus stapfianus putative serine/threonine phosphatase type 2c.
AAC36700.1 AF075582 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
AAD 17804.1 AF092431 Lotus japonicus nodule-enhanced protein phosphatase type 2C. NPP2C1.
AAD17805.1 AF092432 Lotus japonicus protein phosphatase type 2C. PP2C2.
CAC09575.1 AJ298987 Fagus sylvatica protein phosphatase 2C (PP2C). pp2Cfl.
AAC36699.1 AF075581 Mesembryanthemum crystallinum protein ρhosphatase-2C. PP2C.
AAD 11430.1 AF097667 Mesembryanthemum crystallinum protein phosphatase 2C homolog. PP2C.
CAB90634.1 AJ277744 Fagus sylvatica protein phosphatase 2C (PP2C). pρ2C2. ABA and calcium induced protein.
AAC26828.1 AF075603 Oryza sativa kinase interaction domain mediates phosphorylation-dependent binding to a subset of active receptor-like protein kinases. kinase associated protein phosphatase. kapp. type 2C protein phosphatase.
AAC35951.1 AF079355 Mesembryanthemum crystallinum protein phosphatase-2c. PP2C.
AAK20060.1 AC025783 Oryza sativa putative protein phosphatase 2C. OSJNBa0001O14.1.
AAB93832.1 U81960 Zea mays kinase interaction domain mediates phosphorylation-dependent binding to a subset of active receptor-like protein kinases; type 2C protein phosphatase. kinase associated protein phosphatase. KAPP.
CAC09576.1 AJ298988 Fagus sylvatica protein phosphatase 2C (PP2C). pp2Cf2. SEQ ID NO: 134
AAD26116.1 AFl 06954 Brassica napus galactinol synthase.-GS. UDP-D-galactose:myo-inositol-D- galactosyltransferase.
CAB51130.1 AJ243815 Pisum sativum role in alpha galactoside synthesis, putative galactinol synthase.
CAB51533.1 AJ237693 Ajugareptans galactosyl transfer from UDP-galactose to myo-inositol to form galactinol. galactinol synthase, isoform GolS-1. GolS.
CAB51534.1 AJ237694 Ajugareptans galactosyl transfer from UDP-galactose to myo-inositol. galactinol synthase, isoform GolS-2.
GolS.
AAD55726.1 AF178569 Vitis riparia galactinol synthase. WSI76. water stress induced protein.
SEQ ID NO: 135
AAB57734.1 U64818 Lycopersicon esculentum fructokinase. Frk2.
AAB51108.1 U62329 Lycopersicon esculentum fructokinase. FK.
AAA80675.1 U37838 Beta vulgaris fructokinase.
CAA78283.1 Z12823 Solanum tuberosum fructokinase.
AAB57733.1 U64817 Lycopersicon esculentum fructokinase. Frkl .
SEQ ID NO: 138
BAA94601.1 AB033504 Populus euramericana 1-aminocyclopropane-l -carboxylate oxidase. peaco-1.
AAA33697.1 L21978 Petunia x hybrida
1 -aminocyclopropane- 1-carboxylate oxidase. ACO3.
AAC48977.1 U07953 Pelargonium x hortorum 1 -aminocyclopropane- 1 -carboxylate oxidase.
CAA54449.1 X77232 Prunus persica 1-aminocyclopropane-l -carboxylate oxidase. PAO1.
AAF36483.1 AF129073 Prunus persica 1-aminocyclopropane-l -carboxylate oxidase. ACOl.
AAC33524.1 AF026793 Prunus armeniaca 1-aminocyclopropane-l -carboxylate oxidase. ACC oxidase.
AAB70884.1 U67861 Pelargonium x hortorum 1-aminocyclopropane-l -carboxylate oxidase. GACO3.
AAG49361.1 AF321533 Citrus sinensis ACC oxidase.
BAA90550.1 AB031027 Prunus mume
ACC oxidase. PM-ACOl. 1 -aminocyclopropane- 1-carboxylic acid oxidase.
AAA99792.1 U54565 Nicotiana glutinosa oxidation of 1-aminocyclopropane-l -carboxylic acid. 1-aminocyclopropane-l -carboxylic acid oxidase. NGACOl. ACC oxidase.
AAB05171.1 U62764 Nicotiana glutinosa oxidation of 1-aminocyclopropane-l -carboxylic acid. ACC oxidase. NGACO3.
AAC37381.1 L21976 Petunia x hybrida
1-aminocyclopropane-l -carboxylate oxidase. ACOl.
CAA71738.1 Y10749 Betula pendula
1-aminocyclopropane-l -carboxylate oxidase. ACO.
CAA86468.1 Z46349 Nicotiana tabacum
1 -aminocyclopropane- 1 -carboxylate deaminase.
BAA83466.1 AB012857 Nicotiana tabacum ACC oxidase.
AAC98808.1 U68215 Carica papaya ACC oxidase. fruit specific; ripening related.
BAA06526.1 D31727 Cucumis melo 1 -aminocyclopropane- 1 -carboxylate oxidase.
CAA64797.1 X95551 Cucumis melo ACC oxidase.
CAA58232.1 X83229 Nicotiana tabacum ethylene forming enzyme. 1-amniocyclopropane- 1-carboxylate oxidase.
BAA34924.1 AB013101 Lycopersicon esculentum
1-aminocyclopropane-l -carboxylate oxidase. LE-ACO4. ACC oxidase.
AAF64528.1 AF254125 Carica papaya
1-aminocyclopropane-l -carboxylate oxidase. ACC oxidase.
AAA33698.1 L21979 Petunia x hybrida
1-aminocyclopropane-l -carboxylate oxidase. ACO4.
CAA41212.1 X58273 Lycopersicon esculentum conversion of ACC to ethylene. 1 -Aminocyclopropane- 1 -carboxylic acid oxidase. LEACOl.
BAA21541.1 AB003514 Actinidia deliciosa l-arninocyclopropane-l -carboxylic acid oxidase.
AAB71421.1 L29405 Helianthus annuus
1-aminocyclopropapne-l -carboxylic acid oxidase. ACC oxidase.
AAA99793.1 U54566 Nicotiana glutinosa oxidation of 1-aminocyclopropane-l -carboxylic acid. 1-aminocyclopropane-l -carboxylic acid oxidase. NGACO2. ACC oxidase.
AAF36484.1 AF129074 Prunus persica 1-aminocyclopropane-l -carboxylate oxidase. ACO2. CAA64799.1 X95553 Cucumis melo
ACC oxidase.
CAA90904.1 Z54199 Lycopersicon esculentum catalyses the final step in ethylene biosynthesis. 1-aminocyclopropane-l -carboxylic acid oxidase. ACO3.
CAA68538.1 Y00478 Lycopersicon esculentum conversion of ACC to ethylene. 1-aminocyclopropane-l -carboxylate oxidase. LEACO2.
CAB97173.1 AJ297435 Mangifera indica ethene biosynthesis, putative 1-aminocyclopropane-l -carboxylic acid oxidase. acol.
CAA82646.1 Z29529 Nicotiana tabacum oxidation of 1-aminocyclopropane-l -carboxylic acid, ethylene forming enzyme (EFE).
AAC12934.1 AF053354 Phaseolus vulgaris
1-aminocyclopropane-l -carboxylic acid oxidase. ACOl. ACC oxidase.
AAB70883.1 U19856 Pelargonium x hortorum
1-aminocyclopropane-l -carboxylate oxidase.
AAC67233.1 AF033582 Cucumis sativus ACC oxidase 2. Cs-ACO2.
AAB02051.1 L76283 Carica papaya formation of ethylene. 1-aminocyclopropane-l -carboxylate oxidase. putative.
BAA33377.1 AB006806 Cucumis sativus
ACC oxidase. CS-ACO1.
BAA33378.1 AB006807 Cucumis sativus ACC oxidase. CS-ACO2.
CAA71140.1 Y10034 Rumex palustris 1-aminocyclopropane-l -carboxylic acid oxidase.
AAA33644.1 M98357 Pisum sativum convert ACC to ethylene. 1-aminocyclopropane-l -carboxylate oxidase.
AAC48921.1 U06046 Vigna radiata
1-arninocylopropane- 1-carboxylate oxidase homolog.
AAK07883.1 AF315316 Vigna radiata ACC oxidase. ACO.
AAA33273.1 L35152 Dianthus caryophyllus amino-cyclopropane carboxylic acid oxidase.
CAA74328.1 Y14005 Malus x domestica Converts ACC into ethylene in apple fruit. ACC oxidase.
SEQ ID NO: 139
AAB65777.1 U97522 Vitis vinifera class IV endochitinase. VvChi4B.
AAB65776.1 U97521 Vitis vinifera class IV endochitinase. VvChi4A.
CAC 17793.1 AJ301671 Nicotiana sylvestris hydrolysis of chitin. endochitinase. chnb. class I chitinase. AAA34070.1 Ml 5173 Nicotiana tabacum endochitinase precursor (EC 3.2.1.14).
CAA30142.1 X07130 Solanum tuberosum endochitinase.
CAA53626.1 X76041 Triticum aestivum endochitinase. CHI.
SEQ ID NO: 140
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
BAA92894.1 AB006790 Petunia x hybrida cytochrome P450. IMT-2.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3 '-hydroxylase. Htl. cytochrome P450; CYP75B2.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
CAA50312.1 X70981 Solanum melongena P450 hydroxylase. CYPEG2.
CAA50155.1 X70824 Solanum melongena flavonoid hydroxylase (P450). CYP75.
AAB 17562.1 U72654 Eustoma grandiflorum flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75.
BAA84071.1 AB028151 Antinhinum majus flavone synthase II. cytochrome P450. AFNS2.
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl. P450 enzyme.
AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
AAB94588.1 AF022459 Glycine max
CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
AAD37433.1 AFl 50881 Lycopersicon esculentum x Lycopersicon peravianum catalyzes the hydroxylation of ferulic acid to 5-hydroxyferulic acid, ferulate-5-hydroxylase. CYP84. cytochrome P450-dependent monooxygenase; F5H; FAH1.
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
BAA13414.1 D87520 Glycynhiza echinata putative tians-cinnamic acid 4-hydroxylase. cytochrome P4S0 (CYP73A14). CYP Ge-1.
AAA19701.1 L24438 Thlaspi arvense cytochrome P450.
CAA50645.1 X71654 Solanum melongena P450 hydroxylase.
BAA03635.1 D14990 Solanum melongena Cytochrome P-450EG4.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
AAC05148.1 AF049067 Pinus radiata cytochrome P450. PRE74.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71A6.
AAG10196.1 AF286647 Gossypium arboreum cinnamate-4-hydroxylase. LP89. P450.
AAB94584.1 AF022157 Glycine max capable of catalyzing the metabolism of phenylurea herbicides. CYP71A10. CYP71A10. cytochrome P450 monooxygenase.
CAA50648.1 X71657 Solanum melongena P450 hydroxylase.
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
SEQ ID NO: 141
AAB97167.1 AF030882 Zea mays
SU1 isoamylase. sugaryl. starch debranching enzyme.
AAA91298.1 U18908 Zea mays
Sulp. Sugaryl. similar to Pseudomonas sp. isoamylase, Swiss-Prot Accession Number
P26501.
AAD33889.1 AF142589 Hordeum vulgare isoamylase 1.
BAA29041.1 ABO 15615 Oryza sativa isoamylase.
AAD33891.1 AF142591 Solanum tuberosum isoamylase 1.
AAD33890.1 AF142590 Triticum aestivum isoamylase 1.
AAD53260.1 AF142588 Hordeum vulgare isoamylase 1.
SEQ ID NO: 145
AAG35777.1 AF273844 Brassica oleracea var. alboglabra thioredoxin-h-like protein 1. THL1.
AAB53694.1 U59379 Brassica napus thioredoxin-h-like- 1. THL-1. thioredoxin-h homolog.
CAA61908.1 X89759 Brassica oleracea pollen coat protein. bopcl7.
BAA25681.1 AB010434 Brassica rapa Thioredoxin. PEC-2.
BAB20886.1 AB053294 Oryza sativa thioredoxin h. RTRXH2.
AAB53695.1 U59380 Brassica napus thioredoxin-h-like-2. THL-2. thioredoxin-h homolog.
AAF88067.1 AF286593 Triticum aestivum thioredoxin H. similar to wheat thioredoxin H.
CAA94534.1 Z70677 Ricinus communis thioredoxin.
CAA05081.1 AJ001903 Triticum turgidum subsp. durum thioredoxin H.
CAA49540.1 X69915 Triticum aestivum unnamed protein product.
BAA13524.1 D87984 Fagopyrum esculentum thioredoxin.
CAA41415.1 X58527 Nicotiana tabacum thioredoxin.
CAA77847.1 Z11803 Nicotiana tabacum THIOREDOXIN.
AAC32111.1 AF051206 Piceamariana probable thioredoxin H. Sb09. similar to Nicotiana tabacum thioredoxin HI encoded by
GenBank Accession Number X58527.
BAA05546.1 D26547 Oryza sativa rice thioredoxin h.
BAA04864.1 D21836 Oryza sativa thioredoxin h. encoding rice phloem sap 13kD protein- 1.
AAB51522.1 U92541 Oryza sativa thioredoxin h.
AAD49232.1 AF159387 Lolium perenne thioredoxin-like protein. Trx.
AAD56954.1 AF186240 Secale cereale thioredoxin-like protein. Trx.
AAD49231.1 AF159386 Secale cereale thioredoxin-like protein. Trx.
AAD49230.1 AF159385 Hordeum bulbosum thioredoxin-like protein. Trx.
AAD49234.1 AFl 59389 Phalaris coerulescens thioredoxin-like protein. Trx.
AAD49233.1 AFl 59388 Phalaris coerulescens thioredoxin-like protein. Trx.
BAB39913.1 AP002912 Oryza sativa thioredoxin-like protein. P0028E10.17. contains EST C72705(E2091).
CAA55399.1 X78822 Chlamydomonas reinhardtii thioredoxin h. Trx.
CAA56850.1 X80887 Chlamydomonas reinhardtii thioredoxin h. Trx h.
AAD33596.1 AF133127 Hevea brasiliensis thioredoxin h.
CAA35826.1 X51462 Spinacia oleracea thioredoxin M precursor (AA -67 to 114).
CAA35827.1 X51463 Spinacia oleracea thioredoxin M precursor (AA -67 to 114).
CAA45098.1 X63537 Pisum sativum thioredoxin F. isoform.
AAC49357.1 U35830 Pisum sativum thioredoxin f.
AAC19392.1 AF069314 Mesembryanthemum crystallinum thioredoxin F precursor.
AAC04671.1 AF018174 Brassica napus thioredoxin-f. TRXF.
AAB47556.1 U87141 Mesembryanthemum crystallinum thioredoxin h.
CAA53900.1 X76269 Pisum sativum thioredoxin m.
AAC49358.1 U35831 Pisum sativum thioredoxin m. chloroplastic.
CAA33082.1 X14959 Spinacia oleracea pre-thioredoxin f (AA -77 to 113).
CAA06736.1 AJ005841 Oryza sativa thioredoxin M.
CAA55398.1 X78821 Chlamydomonas reinhardtii thioredoxin m. Trx.
CAA56851.1 X80888 Chlamydomonas reinhardtii thioredoxin m. Trx m.
CAA44209.1 X62335 Chlamydomonas reinhardtii thioredoxin Ch2. Trx.
AAA92464.1 L40957 Zea mays regulation of activities of photosynthetic enzymes, thioredoxin M. putative.
CAA06735.1 AJ005840 Triticum aestivum thioredoxin M.
AAB52409.1 U76831 Brassica napus thioredoxin-m.
AAD45358.1 AF160870 Brassica napus thioredoxin-m precursor.
CAA71103.1 Y09987 Solanum tuberosum
CDSP32 protein (Chloroplast Drought-induced Stress Protein of 32kDa).
AAA32662.1 M82973 Medicago sativa putative endomembrane protein; putative.
CAA77575.1 Z11499 Medicago sativa protein disulfide isomerase.
SEQ ID NO: 148
AAA33376.1 L36129 Helianthus annuus NADPH thioredoxin reductase.
SEQ ID NO: 150
AAA92013.1 U49454 Prunus persica beta-l,3-glucanase. Gnsl.
CAA54952.1 X77990 Brassica rapa beta-l,3-glucanase. bgl.
AAF33405.1 AF230109 Populus x canescens beta-1,3 glucanase. BGLUC.
AAA33946.1 M37753 Glycine max beta-1, 3 -endoglucanase (EC 3.2.1.39).
CAA03908.1 AJ000081 Citrus sinensis glucan hydrolase. beta-1,3- -glucanase. gnsl.
CAB91554.1 AJ277900 Vitis vinifera beta 1-3 glucanase. gl.
AAB03501.1 U41323 Glycine max beta-l,3-glucanase. SGN1.
AAA34078.1 M63634 Nicotiana plumbaginifolia regulator of beta(l,3)-glucanase. beta(l,3)-glucanase regulator.
CAA30261.1 X07280 Nicotiana plumbaginifolia beta-glucanase.
AAA51643.1 M23120 Nicotiana plumbaginifolia beta-glucanase precursor.
AAA87456.1 U22147 Hevea brasiliensis beta-l,3-glucanase. HGN1. hydrolytic enzyme.
CAB38443.1 AJ133470 Hevea brasiliensis beta-l,3-glucanase. hgnl.
AAA03618.1 M80608 Lycopersicon esculentum beta- 1 ,3-glucanase.
AAAI 8928.1 U01901 Solanum tuberosum catalyzes the hydrolysis of 1,3 -beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). glub2. plant defense gene; induced expression in response to infection, elicitor, ethylene, wounding.
AAC19114.1 AF067863 Solanum tuberosum
1,3-beta-glucan glucanohydrolase. glucanase.
CAA92278.1 Z68154 Gossypium hirsutum 1 ,3-beta-glucanase.
AAG24921.1 AF311749 Hevea brasiliensis beta-l,3-glucanase.
AAA63539.1 M60402 Nicotiana tabacum glucan beta-l,3-glucanase. glucanase GLA.
AAA63540.1 M60403 Nicotiana tabacum glucan-l,3-beta-glucosidase. glucanase GLB.
AAA88794.1 U01900 Solanum tuberosum catalyzes the hydrolysis of 1,3-beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). gluBl. plant defense gene; induced expression i response to infection, elicitor, ethylene, wounding.
AAA63541.1 M59442 Nicotiana tabacum basic beta-1, 3-glucanase. glucanase.
AAB82772.2 AFOO 1523 Musa acuminata beta-1, 3-glucananse. similar to beta-1, 3-glucanase.
CAA37289.1 X53129 Phaseolus vulgaris
1 ,3 ,-beta-D-glucanase.
AAF08679.1 AF004838 Musa acuminata beta-l,3-glucanase.
AAD33881.1 AF141654 Nicotiana tabacum beta-l,3-glucanase. GGL4.
AAD33880.1 AF141653 Nicotiana tabacum beta-l,3-glucanase. GGL1.
AAA33648.1 L02212 Pisum sativum beta-1, 3-glucan hydrolysis, beta-1, 3-glucanase. putative.
AAA34082.1 M20620 Nicotiana tabacum prepro-beta- 1 ,3-glucanase precursor.
AAA19111.1 U01902 Solanum tuberosum catalyzes the hydrolysis of 1,3-beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). gluB3. plant defense gene; induced expression in response to infection, elicitor, ethylene, wounding.
CAA57255.1 X81560 Nicotiana tabacum
(l-)-beta-glucanase. Sp41a.
AAA34053.1 M60464 Nicotiana tabacum beta- 1 ,3-glucanase.
AAA63542.1 M59443 Nicotiana tabacum acidic beta- 1,3 -glucanase. glucanase. AAB24398.1 S51479 Pisum sativum beta-1, 3-glucanase. beta-1, 3-glucanase. This sequence comes from Fig. IB.
AAB41551.1 U27179 Medicago sativa subsp. sativa acidic glucanase.
AAD10384.1 U72253 Oryza sativa beta-1, 3-glucanase precursor. Gns7.
AAA03617.1 M80604 Lycopersicon esculentum beta-l,3-glucanase.
~~ ~~__~ SEQ ID NO: 151
BAA19102.1 AB000408 Populus kitakamiensis o-methyltransferase. caffeoyl-CoA 3-O-methyltransferase.
AAC28973.1 U20736 Medicago sativa subsp. sativa synthesis of feruloyl-CoA from caffeoyl-CoA and S-adenosyl-L-methionine. S-adenosyl-L- methionine:trans-cafFeoyl-CoA 3-O-methyltransferase. CCOMT.
CAA12198.1 AJ224894 Populus balsamifera subsp. trichocaφa methylates lignin precursors. caffeoyl-CoA 3-O-methyltransferase.
CAAl 1496.1 AJ223621 Populus balsamifera subsp. trichocaφa caffeoyl CoA 3-O-methyltransferase. CCoAOMTl.
AAB80931.1 AF022775 Nicotiana tabacum caffeoyl-CoA 3-O-methyltransferase 5. CCoAOMT-5. implicated in lignification and defense reaction against pathogens.
CAA83943.1 Z33878 Petroselinum crispum caffeoyl-CoA 3-O-methyltransferase.
AAA33851.1 M69184 Petroselinum crispum caffeoyl-CoA 3-O-methyltransferase. CCoAMT.
CAA90894.1 Z54183 Petroselinum crispum trans-caffeoyl-CoA 3-O-methyltransferase. CCoAOMT.
CAA90969.1 Z54233 Vitis vinifera plant defense and lignification. caffeoyl-CoA O-methyltransferase.
AAA59389.1 U13151 Zinnia elegans
S-adenosyl-L-methionine:trans-caffeoyl-CoA 3-O-methyltransferase. CCoAOMT. CAB05369.1 Z82982 Nicotiana tabacum methylation of caffeoyl-CoA in lignin biosynthesis. caffeoyl-CoA O-methyltransferase 5.
CCoAOMT-5.
AAA80651.1 U27116 Populus tremuloides caffeoyl-CoA 3-O-methyltransferase. S-adenyosyl-methionine caffeoyl-CoA 3-O- methyltransferase; similar to Swiss-Prot Accession Number P28034; similar to proteins encoded by GenBank accession Numbers U20736, U13151, and L22203; Mr = 27.9 kDa and pi = 5.16.
AAC08395.1 AF053553 Mesembryanthemum crystallinum caffeoyl-CoA O-methyltransferase.
CAA12200.1 AJ224896 Populus balsamifera subsp. trichocaφa methylates lignin precursors. caffeoyl-CoA 3-O-methyltransferase.
CAA12199.1 AJ224895 Populus balsamifera subsp. trichocaφa methylates lignin precursors. caffeoyl-CoA 3-O-methyltransferase.
CAAl 1495.1 AJ223620 Populus balsamifera subsp. trichocaφa caffeoyl CoA 3-O-methyltransferase. CCoAQMT2.
AAD50443.1 AFl 68780 Eucalyptus globulus caffeoyl-CoA O-methyltransferase. CCoAOMT2.
AAF44689.1 AF240466 Populus tomentosa caffeoyl-CoA O-methyltransferase. CCoAOMT.
AAC49913.1 U38612 Nicotiana tabacum methylation of caffeoyl-CoA in feruloyl-coA in phenylpropanoid pathway, caffeoyl - coenzyme A O-methyltransferase. CCoAOMT- 1.
CAA72911.1 Y12228 Eucalyptus gunnii caffeoyl-CoA O-methyltransferase. COO AMT.
AAC49916.1 U62736 Nicotiana tabacum methylation of caffeoyl-CoA in feruloyl-CoA in phenylpropanoid pathway (lignin biosynthesis). caffeoyl-CoA O-methyltransferase 4. CCoAOMT-4.
BAA78733.1 AB023482 Oryza sativa
ESTs AU058067(E20733), AAU058070(E20873) conespond to a region ofthe predicted gene.; Similar to Populus tremuloides caffeoyl-CoA 3-O-methyltransferase rnRNA, complete cds.(U27116).
CAA91228.1 Z56282 Nicotiana tabacum plant defense and lignification. caffeoyl-CoA O-methyltransferase. NTCCOAOMT.
AAC49914.1 U62734 Nicotiana tabacum methylation of caffeoyl-CoA in feruloyl-CoA in phenylpropanoid pathway (lignin biosynthesis). caffeoyl-CoA O-methyltransferase 2. CCoAOMT-2.
AAC26191.1 AF046122 Eucalyptus globulus catalyses the methylation of caffeoyl CoA in lignin biosynthesis. caffeoyl-CoA 3-O- methyltransferase. CCOMT. S-adenosyl-L-methionine:caffeoyl-CoA 3-O-methyltransferase.
AAC49915.1 U62735 Nicotiana tabacum methylation of caffeoyl-CoA in feruloyl-CoA in phenylpropanoid pathway (lignin biosynthesis). caffeoyl-CoA O-methyltransferase 3. CCoAOMT-3.
AAD02050.1 AF036095 Pinus taeda caffeoyl-CoA O-methyltransferase. CCoAOMT. lignin pathway O-methyltransferase.
AAK16714.1 AF327458 Populus alba x Populus glandulosa caffeoyl-CoA 3-O-methyltransferase. CCoAOMT.
CAB45150.1 AJ242981 Zea mays lignin synthesis. Caffeoyl CoA O-methyltransferase. ccoAOMT.
CAB45149.1 AJ242980 Zea mays lignin synthesis. Caffeoyl CoA O-methyltransferase. ccoAOMT.
AAB61680.1 L22203 Stellaria longipes
S-adenosyl-L-methionine:trans-caffeoyl-CoA 3-O-methyltransferase. 26.7-kDa; pl=5.3.
BAA88234.1 AB035144 Citrus natsudaidai
Methylation of caffeoyl-coA in feruloyl-coA in phenylpropanoid pathway. caffeoyl-CoA 3-O- methyltransferase. CCoAMT.
BAA81776.1 AP000364 Oryza sativa
ESTs C98431(E0144),C71728(E0144) conespond to a region ofthe predicted gene.; Similar to Medicago sativa S-adenosyl-L-methionine. (U20736).
BAA81774.1 AP000364 Oryza sativa
ESTs AU030740(E60171),AU030739(E60171) conespond to a region ofthe predicted gene.;
Similar to Populus tremuloides caffeoyl-CoA 3-O-methyltransferase. (U27116).
BAA81777.1 AP000364 Oryza sativa
Similar to Petroselinum crispum caffeoyl-CoA 3-O-methyltransferase. (S49342).
CAA10217.1 AJ130841 Populus balsamifera subsp. trichocaφa methylates lignin precursors. caffeoyl-CoA 3-O-methyltransferase.
CAA04769.1 AJ001447 Fragaria vesca caffeoyl-CoA 3-O-methyltransferase. putative.
AAD50441.1 AF168778 Eucalyptus globulus caffeoyl-CoA O-methyltransferase. CCoAOMT 1.
AAD50442.1 AFl 68779 Eucalyptus globulus caffeoyl-CoA O-methyltransferase. CCoAOMT2.
AAC15067.1 AF060180 Nicotiana tabacum plant lignification and defense, caffeoyl-coenzyme A trunc2. truncated caffeoyl-coenzyme A.
SEQ ID NO: 152
AAK11255.1 AF329729 Nicotiana tabacum regulator of gene silencing. rgs-CaM; calmodulin-related protein.
AAD10245.1 AF030033 Phaseolus vulgaris calmodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
CAA62150.1 X90560 Physcomitrella patens
Calmodulin. CaM.
BAA94696.1 AB041711 Chara corallina calmodulin. cccaml.
BAA96536.1 AB044286 Chara corallina calmodulin. ccam.
BAA94697.1 AB041712 Chara corallina calmodulin. cccam2.
BAA87825.1 AP000815 Oryza sativa
ESTs AU030013(E50493),AU081341(E50493) conespond to a region ofthe predicted gene.
Similar to O.sativa gene encoding calmodulin. (Z12828).
CAA61980.1 X89890 Bidens pilosa
Calmodulin.
AAA19571.1 U10150 Brassica napus calcium binding, calmodulin. bcml .
AAA87347.1 M88307 Brassica juncea calmodulin.
CAA74111.1 Y13784 Mougeotia scalaris
Calmodulin.
AAA92677.1 U13736 Pisum sativum binds calcium, calmodulin-like protein.
AAA34015.1 L01433 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-4. putative.
AAA33083.1 M20729 Chlamydomonas reinhardtii calmodulin.
AAK25753.1 AF334833 Castanea sativa calmodulin. CAM2.
AAF73157.1 AF150059 Brassica napus calmodulin. CaMl. involved in seed germination.
CAA74307.1 Y13974 Zea mays calmodulin.
AAA34238.1 L20507 Vigna radiata calmodulin.
AAA34237.1 L20691 Vigna radiata calmodulin.
AAC49587.1 U49105 Triticum aestivum calmodulin TaCaM4- 1. calcium-binding protein.
AAC49586.1 U49104 Triticum aestivum calmodulin TaCaM3-3. calcium-binding protein.
AAC49585.1 U49103 Triticum aestivum cahnodulin TaCaM3-2. calcium-binding protein.
AAC49584.1 U48693 Triticum aestivum cahnodulin TaCaM3-l. calcium-binding protein.
AAC49580.1 U48689 Triticum aestivum cahnodulin TaCaMl-3. calcium-binding protein.
AAC49579.1 U48688 Triticum aestivum cahnodulin TaCaMl-2. calcium binding protein.
AAC49578.1 U48242 Triticum aestivum calmodulin TaCaM 1 - 1. calcium-binding.
AAA34014.1 L01432 Glycine max calcium-binding regulatory protein, cahnodulin. SCaM-3. putative.
AAA03580.1 L01431 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-2. putative.
AAA34013.1 L01430 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-1. putative.
AAB36130.1 S81594 Vigna radiata auxin-regulated calmodulin. auxin-regulated calmodulin, arCaM. This sequence comes from
Fig. 1; arCaM.
AAA33901.1 L18913 Oryza sativa calcium binding protein, signal transduction. calmodulin. putative.
AAA92681.1 U13882 Pisum sativum calcium-binding protein, calmodulin.
AAA33706.1 M80836 Petunia x hybrida calmodulin. CAM81.
AAA33705.1 M80831 Petunia x hybrida calmodulin-related protein. CAM53.
CAA78287.1 Z12827 Oryza sativa calcium binding protein, signal transduction. calmodulin.
CAA46150.1 X65016 Oryza sativa calmodulin. cam.
CAA36644.1 X52398 Medicago sativa calmodulin (AA 1-149).
CAA43143.1 X60738 Malus x domestica Calmodulin. CaM.
CAA78301.1 Z12839 Lilium longiflorum calcium binding protein, signal transduction. calmodulin.
AAA33397.1 LI 8912 Lilium longiflorum calcium binding protein, signal transduction. calmodulin. putative.
AAB68399.1 U79736 Helianthus annuus calmodulin. HaCaM.
CAA42423.1 X59751 Daucus carota calmodulin. Ccam-1.
AAA32938.1 M27303 Hordeum vulgare calmodulin.
BAA88540.1 AP000969 Oryza sativa
ESTs AU081349(E61253), D41425(S3918) conespond to a region ofthe predicted gene.
Similar to calmodulin. (AF042840).
AAG27432.1 AF295637 Elaeis guineensis cahnodulin.
AAG11418.1 AF292108 Prunus avium calmodulin.
AAC36059.1 AF042840 Oryza sativa calmodulin. CaMl.
SEQ ID NO: 153
CAA40474.1 X57187 Phaseolus vulgaris chitinase. Chi4.
AAB65776.1 U97521 Vitis vinifera class IV endochitinase. VvChi4A.
CAA61281.1 X88803 Vigna unguiculata chitinase class 4. CHI4.
AAB65777.1 U97522 Vitis vinifera class IV endochitinase. VvChi4B.
BAA22966.1 D45182 Chenopodium amaranticolor chitinase.
BAA22968.1 D45184 Chenopodium amaranticolor chitinase.
BAA22965.1 D45181 Chenopodium amaranticolor chitinase.
CAA43708.1 X61488 Brassica napus chitinase.
BAA22967.1 D45183 Chenopodium amaranticolor chitinase.
CAA53544.1 X75945 Beta vulgaris chitinase. Ch4.
AAC49435.1 U52845 Daucus carota class IV chitinase EP3-1/H5. EP3.
AAB08468.1 U52846 Daucus carota class IV chitinase EP3-2/H1. EP3.
AAB08470.1 U52848 Daucus carota class IV chitinase EP3B/E6. EP3.
AAB08469.1 U52847 Daucus carota class IV chitinase EP3-3/E7. EP3.
AAA33445.1 M84165 Zea mays chitinase B. seed chitinase.
AAA33444.1 M84164 Zea mays chitinase A. seed chitinase.
AAA32916.1 L25826 Beta vulgaris chitinase. SP2.
AAD28733.1 AFl 12966 Triticum aestivum chitinase IV precursor. Cht4.
BAB21377.1 AB054811 Oryza sativa PR-3 class IV chitinase. Cht4. Catalytic domain.
BAB21374.1 AB054687 Oryza sativa PR-3 class IV chitinase. Cht4. catalytic domain.
BAA19793.1 AB003194 Oryza sativa chitinase lib.
AAA85364.1 L42467 Picea glauca chitinase. chi.
AABO 1665.1 U21848 Brassica napus chitinase class IV. LSC222.
AAC35981.1 AF090336 Citrus sinensis chitin hydrolase. chitinase CHU. chil.
AAD28730.1 AFl 12963 Triticum aestivum chitinase II precursor. Cht2.
AAF04454.1 AF000966 Poa pratensis chitinase. Chi2.
CAC17793.1 AJ301671 Nicotiana sylvestris hydrolysis of chitin. endochitinase. chnb. class I chitinase.
AAF04453.1 AF000964 Poa pratensis chitinase. Chil.
CAA34812.1 XI 6938 Nicotiana tabacum chitinase precursor.
CAA34813.1 X16939 Nicotiana tabacum chitinase precursor (AA -23 to 306).
CAA45822.1 X64519 Nicotiana tabacum chitinase B class I. CHN200.
CAA35945.1 X51599 Nicotiana tabacum chitinase. CHN50.
AAB23374.1 S44869 Nicotiana tabacum basic chitinase. basic chitinase. This sequence comes from Fig. 1.
AAA34070.1 Ml 5173 Nicotiana tabacum endochitinase precursor (EC 3.2.1.14).
CAA30142.1 X07130 Solanum tuberosum endochitinase.
CAA33517.1 X15494 Solanum tuberosum pre-chitinase (AA -26 to 302).
AAG53609.1 AF280437 Secale cereale
31.7 kDa class I endochitinase-antifreeze protein precursor. cht9.
CAB01591.1 Z78202 Persea americana hydrolysis ofthe 1,4-beta-linkages of chitin. endochitinase. chil.
CAA53626.1 X76041 Triticum aestivum endochitinase. CHI.
CAA78845.1 Z15140 Lycopersicon esculentum chitinase. Encodes 30 kD basic intracellular chitinase.
AAC16010.1 AF061805 Elaeagnus umbellata acidic chitinase.
AAA32640.1 M94106 Allium sativum chitinase. chitinase.
AAA32641.1 M94105 Allium sativum chitinase. chitinase.
AAA56787.1 L34211 Hordeum vulgare hydrolysis of chitin. chitinase. CHI33.
AAAI 7409.1 U02607 Solanum tuberosum catalyzes the random hydrolysis of l,4-beta-(2-acetamido-2-deoxy-D-glucoside) linkages in chitin; plant defense gene, chitinase. chtB3. induced expression in response to infection, elicitor, ethylene, wounding; preproprotern; gene product subunit is monomer.
AAAI 8332.1 U02605 Solanum tuberosum catalyzes the random hydrolysis of l,4-beta-(2-acetamido-2-deoxy-D-glucoside) linkages in chitin; plant defense gene, chitinase. chtBl. induced expression in response to infection, elicitor, ethylene, wounding; preproprotein; gene product subunit is a monomer.
CAA45821.1 X64518 Nicotiana tabacum chitinase C class I. CHN14. SEQ ID NO: 154
AAB35812.1 S80554 Arabidopsis chalcone synthase. chalcone synthase, CHS. This sequence comes from Fig. 5; CHS.
AAF23570.1 AFl 12095 Arabidopsis halleri chalcone synthase. CHS.
AAF23568.1 AFl 12093 Arabidopsis griffithiana chalcone synthase. CHS.
AAG43351.1 AF144533 Arabidopsis korshinskyi chalcone synthase. chs.
AAF23581.1 AF112106 Capsella rubella chalcone synthase. CHS.
AAF23569.1 AFl 12094 Halimolobos peφlexa var. peφlexa chalcone synthase. CHS.
AAG43349.1 AF144531 Arabidopsis himalaica chalcone synthase. chs.
AAF23575.1 AFl 12100 Arabidopsis lyrata subsp. lyrata chalcone synthase. CHS.
AAF23567.1 AFl 12092 Arabidopsis griffithiana chalcone synthase. CHS.
AAF23578.1 AFl 12103 Arabidopsis lyrata subsp. petraea chalcone synthase. CHS.
AAF23576.1 AF112101 Arabis parishii chalcone synthase. CHS.
AAF23574.1 AFl 12099 Arabis lyallii chalcone synthase. CHS.
AAF23566.1 AFl 12091 Arabis glabra chalcone synthase. CHS.
AAF23565.1 AFl 12090 Arabis fendleri chalcone synthase. CHS.
AAF23563.1 AFl 12088 Arabis drummondii chalcone synthase. CHS.
AAF23564.1 AFl 12089 Arabis drummondii chalcone synthase. CHS.
AAF23579.1 AFl 12104 Arabidopsis lyrata subsp. petraea chalcone synthase. CHS.
AAF23573.1 AFl 12098 Arabis lignifera chalcone synthase. CHS.
AAF23560.1 AFl 12085 Cardamine amara chalcone synthase. CHS.
AAG43348.1 AFl 44530 Rorippa amphibia chalcone synthase. chs.
AAG43356.1 AF144538 Cardamine penzesii chalcone synthase. chs.
AAG43359.1 AF144541 Sisymbrium irio chalcone synthase. chs.
AAG43352.1 AF144534 Lepidium campestre chalcone synthase. chs.
CAA32495.1 X14314 Sinapis alba chalcone synthase (AA 1-395).
AAG43357.1 AF144539 Cardamine rivularis chalcone synthase. chs.
AAF23583.1 AF112108 Barbarea vulgaris chalcone synthase. CHS.
AAC31914.1 AF076336 Brassica napus chalcone synthase B2. CHSB2.
AAC31912.1 AF076334 Brassica napus chalcone synthase A2. CHSA2.
AAF23577.1 AFl 12102 Arabis pauciflora chalcone synthase. CHS.
AAG43350.1 AF144532 Cochlearia danica chalcone synthase. chs.
CAA34460.1 XI 6437 Sinapis alba chalcone synthase.
CAA35600.1 X17577 Matthiola incana chalcone synthase (AA 1-394).
AAG43358.1 AF144540 Cardamine pratensis chalcone synthase. chs.
AAG43353.1 AF144535 Thlaspi arvense chalcone synthase. chs.
AAC31913.1 AF076335 Brassica napus chalcone synthase Bl. CHSB1.
AAF23571.1 AFl 12096 Arabis hirsuta chalcone synthase. CHS.
AAF23582.1 AFl 12107 Arabis turrita chalcone synthase. CHS.
AAG43406.1 AF174529 Aubrieta deltoidea chalcone synthase. chs.
AAG43355.1 AF144537 Alliaria petiolata chalcone synthase. chs.
AAF23580.1 AFl 12105 Arabis procunens chalcone synthase. CHS.
AAF23572.1 AFl 12097 Arabis jacquinii chalcone synthase. CHS.
AAF23562.1 AFl 12087 Arabis blepharophylla chalcone synthase. CHS.
AAF23584.1 AFl 12109 Aubrieta deltoidea chalcone synthase. CHS.
AAG43354.1 AF144536 Microthlaspi perfoliatiim chalcone synthase. chs.
AAF23557.1 AFl 12082 Aethionema grandiflora chalcone synthase. CHS.
AAF23558.1 AFl 12083 Arabis alpina chalcone synthase. CHS.
AAF23559.1 AFl 12084 Arabis alpina chalcone synthase. CHS.
AAB87072.1 AF031922 Raphanus sativus chalcone synthase. CHS.
AAG43360.1 AF144542 Ionopsidium abulense chalcone synthase. chs.
AAC31911.1 AF076333 Brassica napus chalcone synthase Al . CHSA1.
SEQ ID NO: 156
AAD10327.1 U63534 Fragaria x ananassa catalyzes the reduction of ciimamylaldehydes leading to monolignols. cinnamyl alcohol dehydrogenase. CAD. involved with lignin biosynthesis.
AAK28509.1 AF320110 Fragaria x ananassa cinnamyl alcohol dehydrogenase.
AAB38503.1 U79770 Mesembryanthemum crystallinum cinnamyl-alcohol dehydrogenase Eli3.
CAA48028.1 X67817 Petroselinum crispum Eli3.
AAC 15467.1 U24561 Apium graveolens converts mannitol to mannose. mannitol dehydrogenase. Mtd. 1 -oxidoreductase; induced with sodium salicylate; similar to the plant defense gene ELB in Arabidopsis thaliana, PIR Accession Number S28044; EC number unassigned; MTD.
AAC35846.1 AF083333 Medicago sativa cinnamyl-alcohol dehydrogenase. MsaCadl .
AAC61854.1 AF067082 Apium graveolens oxidizes mannitol to mannose. mannitol dehydrogenase. Mtd. mannitol 1 -oxidoreductase.
AAA74882.1 L36823 Stylosanthes humilis cinnamyl-alcohol dehydrogenase. CADI.
AAA74883.1 L36456 Stylosanthes humilis cinnamyl-alcohol dehydrogenase. CAD3.
AAF72100.1 AF146691 Lycopersicon esculentum ELB. Eli3. similar] to cinnamyl alcohol dehydrogenase.
CAA86072.1 Z37991 Pinus taeda cinnamyl alcohol dehydrogenase.
CAA05095.1 AJ001924 Picea abies cinnamyl alcohol dehydrogenase. cad2.
AAB38774.1 U62394 Pinus radiata cinnamyl alcohol dehydrogenase. CAD.
CAA51226.1 X72675 Picea abies cinnamyl-alcohol dehydrogenase.
CAA05097.1 AJ001926 Picea abies cinnamyl alcohol dehydrogenase. cad8.
CAA05096.1 AJ001925 Picea abies cinnamyl alcohol dehydrogenase. cad7.
AAC31166.1 AF060491 Pinus radiata cinnamyl alcohol dehydrogenase. CAD.
CAA86073.1 Z37992 Pinus taeda cinnamyl alcohol dehydrogenase.
CAA44216.1 X62343 Nicotiana tabacum cinnamyl-alcohol dehydrogenase. CAD 14.
CAA44217.1 X62344 Nicotiana tabacum cinnamyl-alcohol dehydrogenase. CAD19.
BAA03099.1 D13991 Aralia cordata cinnamyl alcohol dehydrogenase. cadacl.
CAA79625.1 Z19573 Medicago sativa lignin biosynthesis, cinnamyl alcohol dehydrogenase.
AAF43140.1 AF217957 Populus tremuloides cinnamyl alcohol dehydrogenase. CAD.
AAC35845.1 AF083332 Medicago sativa cinnamyl-alcohol dehydrogenase. MsaCad2.
CAC07423.1 AJ295837 Populus balsamifera subsp. trichocaφa lignin monomer biosynthesis, cinnamyl alcohol dehydrogenase. cad.
CAA79622.1 Z 19568 Populus deltoides lignin biosynthesis, cinnamyl alcohol dehydrogenase.
AAC07987.1 AF038561 Eucalyptus globulus catalyses the reduction of cinnamaldehydes to the conesponding cinnamyl alcohols as the last step in the production of lignin monomers, cinnamyl alcohol dehydrogenase. CAD.
AAG15553.1 AF294793 Eucalyptus saligna cinnamyl alcohol dehydrogenase. cad. CAD.
AAK00679.1 AF229407 Brassica napus
Eli3 product. ELB-BN-2. protein identity based on similarity to Arabidopsis sequence; intron/exon junctions deduced from alignments to DNA or mRNA sequences already present in the GenBank database.
CAA46585.1 X65631 Eucalyptus gunnii cinnamyl-alcohol dehydrogenase. cad.
CAA53211.1 X75480 Eucalyptus gunnii cinnamyl-alcohol dehydrogenase. CAD.
AAK00681.1 AF229409 Brassica napus
Eli3 product. ELB-BN-4. protein identity based on similarity to Arabidopsis sequence; intron/exon junctions deduced from alignments to DNA or mRNA sequences already present in the GenBank database.
AAB70908.1 AFO 10290 Lolium perenne cinnamyl alcohol dehydrogenase. CAD.
AAK00682.1 AF229410 Brassica oleracea
Eli3 product. EL -BO- 1. protein identity based on similarity to Arabidopsis sequence; intron/exon junctions deduced from alignments to DNA or mRNA sequences already present in the GenBank database.
CAA74070.1 Y13733 Zea mays cinnamyl alcohol dehydrogenase. cad.
CAA06687.1 AJ005702 Zea mays cinnamyl alcohol dehydrogenase. cad.
AAK00678.1 AF229406 Brassica napus
Eli3 product. ELI3-BN-1. protein identity based on similarity to Arabidopsis sequence; intron exon junctions deduced from alignments to DNA or mRNA sequences already present in the GenBank database.
CAA13177.1 AJ231135 Saccharum officinarum lignin biosynthesis, cinnamyl alcohol dehydrogenase. cad.
AAK00684.1 AF229412 Brassica rapa
Eli3 product. ELB-BR-2. protein identity based on similarity to Arabidopsis sequence; intron/exon junctions deduced from alignments to DNA or mRNA sequences already present in the GenBank database.
AAK00680.1 AF229408 Brassica napus
Eli3 product. ELB-BN-3. protein identity based on similarity to Arabidopsis sequence; intron exon junctions deduced from alignments to DNA or mRNA sequences already present in the GenBank database.
AAK00683.1 AF229411 Brassica rapa
Eli3 product. ELB-BR-1. protein identity based on similarity to Arabidopsis sequence; intron/exon junctions deduced from alignments to DNA or mRNA sequences already present in the GenBank database.
BAA19487.1 D86590 Zinnia elegans cinnamyl alcohol dehydrogenase. ZCAD1.
BAA04046.1 D16624 Eucalyptus botryoides cinnamyl alcohol dehydrogenase. Cadi :Eb: 1.
AAD18000.1 AF109157 Eucalyptus globulus cinnamyl alcohol dehydrogenase. CAD.
AAF23409.1 AF207552 Brassica napus cinnamyl alcohol dehydrogenase. CADa-1.
AAF23412.1 AF207555 Brassica rapa cinnamyl alcohol dehydrogenase. CADa.
AAF23411.1 AF207554 Brassica oleracea cinnamyl alcohol dehydrogenase. CADa.
AAF23410.1 AF207553 Brassica napus cinnamyl alcohol dehydrogenase. CADa-2.
AAF23416.1 AF207559 Brassica rapa cinnamyl alcohol dehydrogenase. CADb.
AAF23415.1 AF207558 Brassica oleracea cinnamyl alcohol dehydrogenase. CADb.
SEQ ID NO: 158
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AFOO 1308).
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APKl gene for protein tyrosine-serine-threonine kinase.(D12522).
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
AAC27894.1 AF023164 Zea mays leucine-rich repeat transmembrane protein kinase 1. ltkl.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. PQ456F08.9. contains EST C23560(R0290).
AAC27895.1 AF023165 Zea mays leucine-rich repeat transmembrane protein kinase 2. Itk2.
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S 13927).
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
AAF91336.1 AF249317 Glycine max Ptil kinase-like protein. Ptila. protein kinase.
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
AAB09771.1 U67422 Zea mays
CRINKLY4 precursor. cr4. receptor kinase homolog.
BAB18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSJNBa0004G10.30.
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil . Ptil kinase.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAF34428.1 AF172282 Oryza sativa receptor-like protein kinase. DUPR11.18.
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
AAF66615.1 AF 142596 Nicotiana tabacum LRR receptor-like protein kinase.
CAB51836.1 AJ243961 Oryza sativa Putitive Ser/Thr protein kinase. 11332.7.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAF76313.1 AF220603 Lycopersicon esculentum
Pto kinase. LescPth5.
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
CAA79355.1 Z18921 Brassica oleracea
S-receptor kinase-like protein.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
BAA95893.1 AP002071 Oryza sativa
Similar to Arabidopsis thaliana wak4 gene; wall-associated kinase 4. (AJ009695).
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml . SEQ ID NO: 159
AAA86687.1 U15777 Lupinus albus farnesyl pyrophosphate synthase. fpsl.
AAA87729.1 U20771 Lupinus albus farnesyl pyrophosphate synthase. fpsl.
CAA72793.1 Y12072 Gossypium arboreum farnesyl pyrophosphate synthase. fpsl.
BAB40665.1 AB053486 Humulus lupulus farnesyl pyrophosphate synthase. fpps.
BAB40666.1 AB053487 Humulus lupulus farnesyl pyrophophate synthase. fpps.
CAA57893.1 X82543 Parthenium argentatum farnesyl diphosphate synthase. fps2.
AAC78557.1 AF019892 Helianthus annuus farnesyl pyrophosphate synthase. FPS.
CAA57892.1 X82542 Parthenium argentatum farnesyl diphosphate synthase. fpsl.
AAC49452.1 U36376 Artemisia annua farnesyl diphosphate synthase. fpsl.
CAA59170.1 X84695 Capsicum annuum dimethylallyltransferase. Fps, farnesyl pyrophosphate synthase gene.
AAC73051.1 AF048747 Lycopersicon esculentum synthesis of farnesyl pyrophosphate. farnesyl pyrophosphate synthase. FPSl. prenyl transferase; farnesyl diphosphate synthetase.
AAD17204.1 AFl 12881 Artemisia annua farnesyl diphosphate synthase. BAA19856.1 D85317 Oryza sativa farnesyl pyrophosphate synthase. dimethylallyltransferase; geranyltranstransferase.
BAA36276.1 AB021747 Oryza sativa famesyl diphosphate synthase. FPPS1.
AAD32648.1 AF136602 Artemisia annua farnesyl diphosphate synthase. fbs2.
AAB39276.1 L39789 Zea mays farnesyl pyrophosphate synthetase. fps. putative.
AAD27558.1 AFl 11710 Oryza sativa subsp. indica putative farnesyl pyrophosphate synthase. similar to Oryza sativa EST clones E10230_l A,
C52647__1A, 232.
BAA36347.1 AB021979 Oryza sativa farnesyl diphosphate synthase. FPPS2.
AAD37789.1 AF149257 Artemisia annua farnesyl diphosphate synthase. FPP synthase.
BAB20822.1 AB045713 Taraxacum japonicum putative FPP synthase. TJFPPS.
BAB16688.1 AB041627 Eucommia ulmoides
FPP synthase 2. EUFPPS2. putative.
BAB21061.1 AB046212 Sonchus oleraceus putative FPP synthase. SoFPPS.
AAD45122.1 AFl 64026 Xanthoceras sorbifolium synthesis of farnesyl pyrophosphate. farnesyl pyrophosphate synthase. FPS. prenyl transferase; farnesyl diphosphate synthetase.
AAB93951.1 U97330 Nicotiana tabacum farnesylpyrophosphate synthase. FPPS.
BAB16687.1 AB041626 Eucommia ulmoides
FPP synthase 1. EUFPPS1. putative.
BAB39479.1 AB049086 Youngia japonica putative FPP synthase 1. YjFPPSl.
AAB93984.1 AF005201 Parthenium argentatum farnesyl pyrophosphate synthase. FPS3.
SEQ ID NO: 160 BAB03615.1 AP002522 Oryza sativa putative gamma-glutamyltiansferase. P0009G03.15. contains ESTs AU056150(S20332),AU056151(S20332).
BAB03616.1 AP002522 Oryza sativa putative gamma-glutamyltransferase. P0009G03.16. contains EST AU056150(S20332).
SEQ ID NO: 162
CAB96145.1 AJ250951 Mesembryanthemum crystallinum phospholipid hydroperoxide glutathione peroxidase-like protein, gpxmcl.
CAA42780.1 X60219 Nicotiana sylvestris homologous to animal glutathione peroxidases.
BAB16430.1 AB041518 Nicotiana tabacum glutathione peroxidase Nt-SubC08. Nt-SubC08.
CAA75054.1 Y14762 Lycopersicon esculentum glutathione peroxidase. GPXle-1.
AAB94892.1 AF037051 Gossypium hirsutum glutathione peroxidase.
CAB59893.1 AJ238697 Hordeum vulgare GPX12Hv, glutathione peroxidase-like protein.
BAA22194.1 D63425 Spinacia oleracea phopholipid hydroperoxide glutathione peroxidase-like protein, similar to mammalian phospholipid hydroperoxide glutathione peroxidases.
CAB59895.1 AJ238745 Hordeum vulgare glutathione peroxidase-like protein GPX54Hv.
AAC78466.1 AF053311 Zantedeschia aethiopica glutathione peroxidase. gpx.
CAA04142.1 AJ000508 Pisum sativum phospholipid glutathione peroxidase. plastid-localised.
CAA75009.1 Y14707 Helianthus annuus glutathione peroxidase. GPxha-2.
CAB59894.1 AJ238744 Hordeum vulgare glutathione peroxidase-like protein GPX15Hv.
CAA74775.1 Y14429 Helianthus annuus glutathione peroxidase. GPxha-1.
CAC17628.1 AJ270955 Oryza sativa putative role in antioxidative systems, putative phospholipid hydroperoxide glutathione peroxidase. riPHGPX.
BAA83594.1 AB009083 Chlamydomonas sp. W80 glutathione peroxidase.
AAB66330.1 AFO 14927 Chlamydomonas reinhardtii glutathione peroxidase homolog. gpxh.
CAA75055.1 Y14763 Lycopersicon esculentum glutathione peroxidase. GPXle-2.
CAA09194.1 AJO 10455 Triticum aestivum glutathione peroxidase. PHGPX6.
CAB66331.1 AJ279689 Berula pendula glutahione peroxidase. gpx.
SEQ ID NO: 163
AAF67753.1 AF255651 Brassica rapa subsp. pekinensis conversion of oxidized glutathione to reduced glutathione. cytosolic glutathione reductase.
GR1.
AAC49980.2 AF008441 Brassica rapa glutathione reductase. BcGRl. cytosolic.
BAA11214.1 D78136 Oryza sativa Glutathione Reductase. putative.
CAA66924.1 X98274 Pisum sativum glutathione reductase. cytosolic.
BAA36283.1 D85751 Oryza sativa glutathione reductase.
BAA37092.1 AB009592 Oryza sativa conversion of oxidized glutathione to reduced glutathione. cytosolic glutathione reductase.
RGRC2. Amino Acids 1-496.
BAA07108.1 D37870 Spinacia oleracea
Glutathione Reductase precursor. Chloroplastic glutathione reductase.
CAC13956.1 AJ400816 Mesembryanthemum crystallinum reduction of glutathione. glutathione reductase. grl.
CAB66332.1 AJ279690 Betula pendula glutathione reductase. gr.
CAA53925.1 X76293 Nicotiana tabacum glutathione reductase (NADPH). gor.
CAA42921.1 X60373 Pisum sativum glutathione reductase (NADPH). Protein sequence is in conflict with the conceptual translation.
AAK27157.1 AF349449 Brassica juncea glutathione reductase. GR2.
AAD28177.1 AF109694 Brassica juncea glutathione reductase. GR1.
CAA62482.1 X90996 Pisum sativum glutathione reductase (NADPH). gr. alpha II subunit.
AAF26175.1 AF105199 Glycine max glutathione reductase. GR-5.
AAB70837.1 AF019907 Vitis vinifera glutathione reductase (NADPH). GOR. VvGRl.
AAA33962.1 LI 1632 Glycine max glutathione reductase. GR.
CAA54043.1 X76533 Nicotiana tabacum glutathione reductase (NADPH). gor.
CAA06835.1 AJ006055 Zea mays
NADPH-dependent reduction of glutathione disulphide. glutathione reductase. gorl.
CAA53993.1 X76455 Nicotiana tabacum glutathione reductase. gor.
AAB30526.1 S70187 Glycine max ferric leghemoglobin reductase. ferric leghemoglobin reductase, FLbR. Method: conceptual translation with partial peptide sequencing; This sequence comes from Fig. 3; FLbR.
AAC26053.1 AF074940 Glycine max fenic leghemoglobin reductase-2 precursor. FLbR homolog;FLbR-2.
AAD53185.1 AFl 81096 Vigna unguiculata fe ic leghemoglobin reductase. flbr.
AAA60979.1 U06461 Pisum sativum catalyzes the conversion of monodehydroascorbate to ascorbate, oxidizing NADH in the process, binds to flavin as a single subunit. monodehydroascorbate reductase.
BAA05408.1 D26392 Cucumis sativus monodehydroascorbate reductase.
AAC41654.1 L41345 Lycopersicon esculentum ascorbate free radical reductase. AFRR.
AAD53522.1 AFl 58602 Zantedeschia aethiopica monodehydroascorbate reductase. MDAR. putative.
AAD28178.1 AFl 09695 Brassica juncea monodehydroascorbate reductase. MDAR1.
BAA77214.1 D85764 Oryza sativa cytosolic monodehydroascorbate reductase.
SEQ ID NO: 164
CAA04391.1 AJ000923 Carica papaya glutathione transferase. PGST 1.
AAC18566.1 AF048978 Glycine max 2,4-D inducible glutathione S-transferase. GSTa.
AAG34800.1 AF243365 Glycine max glutathione S-transferase GST 10.
AAF22647.1 AFl 93439 Lycopersicon esculentum glutathione S-transferase/peroxidase. BI-GST/GPX.
CAA71784.1 Y10820 Glycine max glutathione transferase.
AAG34799.1 AF243364 Glycine max glutathione S-transferase GST 9.
AAG16760.1 AY007562 Lycopersicon esculentum putative glutathione S-transferase T5.
CAA48717.1 X68819 Glycine max lactoylglutathione lyase. glyoxalase I.
CAC24549.1 AJ296343 Cichorium intybus x Cichorium endivia glutathione S-transferase. chi-GSTl. auxin-induced GST.
AAC28101.1 AF079511 Mesembryanthemum crystallinum glutathione S-transferase.
AAG34806.1 AF243371 Glycine max glutathione S-transferase GST 16.
AAF23357.1 AF109194 Hordeum vulgare glutathione-S-transferase.
CAA73369.1 Y12862 Zea mays glutathione transferase. GST5.
AAG32470.1 AF309377 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTU5.
CAA74197.1 Y13898 Brassica juncea glutathione-S-transferase. gst.
AAG34827.1 AF244684 Zea mays glutathione S-transferase GST 19.
AAC05216.1 AF050102 Oryza sativa glutathione s-transferase. GSTl.
AAC32118.1 AF051214 Piceamariana probable glutathione S-transferase. Sbl8. similar to Nicotiana tabacum probable glutathione
S-transferase encoded by GenBank Accession Number X56266.
AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-transferase T3.
AAG34798.1 AF243363 Glycine max glutathione S-transferase GST 8.
AAG34842.1 AF244699 Zea mays glutathione S-transferase GST 34.
AAG34807.1 AF243372 Glycine max glutathione S-transferase GST 17.
AAG34809.1 AF243374 Glycine max glutathione S-transferase GST 19.
AAG34839.1 AF244696 Zea mays glutathione S-transferase GST 31.
AAG34804.1 AF243369 Glycine max glutathione S-transferase GST 14.
AAG34844.1 AF244701 Zea mays glutathione S-transferase GST 36.
AAG34831.1 AF244688 Zea mays glutathione S-transferase GST 23.
AAG34797.1 AF243362 Glycine max glutathione S-transferase GST 7.
AAG34832.1 AF244689 Zea mays glutathione S-transferase GST 24.
AAG32471.1 AF309378 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTU4.
AAG34796.1 AF243361 Glycine max glutathione S-transferase GST 6.
AAG34810.1 AF243375 Glycine max glutathione S-transferase GST 20.
AAG34835.1 AF244692 Zea mays glutathione S-transferase GST 27.
AAC32139.1 AF051238 Piceamariana probable glutathione S-transferase. Sb52. similar to Nicotiana tabacum probable glutathione
S-transferase encoded by GenBank Accession Number X56266.
SEQ ID NO: 165
CAA71878.1 Y10984 Brassica juncea
ATP-dependent addition of glycine to gamma-glutamylcysteine. glutathione synthetase. gshll.
AAB71231.1 AFO 17984 Lycopersicon esculentum glutathione synthetase. GSH2.
AAF98157.1 AF258320 Phaseolus vulgaris homoglutathione synthetase. hgshs.
AAF98156.1 AF258319 Pisum sativum putative homoglutathione synthetase. hgshs.
CAB91078.1 AJ272035 Glycine max homoglutathione synthetase. hGS. putatively predicted to be targetted to the chloroplast.
AAF98121.1 AF231137 Pisum sativum glutathione synthetase precursor, gshs. putative mitochondrial protein.
AAD29848.1 AF075699 Medicago truncatula putative glutathione synthetase. GSHS1.
AAD29849.1 AF075700 Medicago truncatula putative glutathione synthetase. GSHS2.
SEQ ID NO: 166
BAA83711.1 AB014484 Nicotiana tabacum heat shock factor. NtHSF2.
AAF37579.1 AF235958 Medicago sativa heat shock transcription factor. HSFA4-6. MsHSFA4-6.
CAA58117.1 X82943 Zea mays heat shock factor, hsfb.
CAA47868.1 X67599 Lycopersicon esculentum heat stress transcription factor 8. hsf8.
CAA47869.1 X67600 Lycopersicon peravianum heat shock transcription factor 8. hsf8.
CAA47870.1 X67601 Lycopersicon peravianum heat stress transcription factor HSF30. hsBO.
AAF74563.1 AF208544 Lycopersicon peravianum heat stress transcription factor A3. HSF A3.
CAA87080.1 Z46956 Glycine max heat shock transcription factor 5. HSF.
CAA87076.1 Z46952 Glycine max heat shock transcription factor 21. HSF.
CAA39034.1 X55347 Lycopersicon peravianum heat stress transcription factor. Lp-HSF24.
BAA83710.1 AB014483 Nicotiana tabacum heat shock factor. NtHSFl.
CAA87077.1 Z46953 Glycine max heat shock transcription factor 34. HSF. conesponds to longest open reading frame; preceeded by four short open reading frames in the 5' leader sequence.
BAB 19067.1 AP002744 Oryza sativa putative heat shock factor protein 1 (HSF 1). P0006C01.9.
CAA09301.1 AJ010644 Pisum sativum heat shock transcription factor (HSFA). hsfA.
CAA87079.1 Z46955 Glycine max heat shock transcription factor 31. HSF.
CAA87075.1 Z46951 Glycine max heat shock transcription factor 29. HSF.
CAA09300.1 AJ010643 Pisum sativum heat shock transcription factor (HSFA). hsfA.
SEQ ID NO: 168
BAA83710.1 AB014483 Nicotiana tabacum heat shock factor. NtHSFl .
CAA39034.1 X55347 Lycopersicon peravianum heat stress transcription factor. Lp-HSF24.
CAA87077.1 Z46953 Glycine max heat shock transcription factor 34. HSF. conesponds to longest open reading frame; preceeded by four short open reading frames in the 5' leader sequence.
CAA87080.1 Z46956 Glycine max heat shock transcription factor 5. HSF.
CAA47869.1 X67600 Lycopersicon peravianum heat shock transcription factor 8. hsf8.
CAA47868.1 X67599 Lycopersicon esculentum heat stress transcription factor 8. hsf .
BAA83711.1 AB014484 Nicotiana tabacum heat shock factor. NtHSF2.
CAA58117.1 X82943 Zea mays heat shock factor, hsfb.
CAA87075.1 Z46951 Glycine max heat shock transcription factor 29. HSF.
CAA87076.1 Z46952 Glycine max heat shock transcription factor 21. HSF.
CAA47870.1 X67601 Lycopersicon peravianum heat stress transcription factor HSF30. hsf30.
AAF74563.1 AF208544 Lycopersicon peravianum heat stress transcription factor A3. HSFA3.
AAF37579.1 AF235958 Medicago sativa heat shock transcription factor. HSFA4-6. MsHSFA4-6.
CAA87079.1 Z46955 Glycine max heat shock transcription factor 31. HSF.
BAB 19067.1 AP002744 Oryza sativa putative heat shock factor protein 1 (HSF 1). P0006C01.9.
CAA09301.1 AJ010644 Pisum sativum heat shock transcription factor (HSF A). hsfA.
CAA09300.1 AJ010643 Pisum sativum heat shock transcription factor (HSFA). hsfA.
SEQ ID NO: 169
AAB72109.1 AF022217 Brassica rapa low molecular weight heat-shock protein. BcHSP17.6. 17.6 kDa; cytosolic class I.
CAB93512.1 AJ243565 Brassica oleracea putative class I small heat shock protein. HSP17.7-a protein. hsp!7.7-a.
CAA37847.1 X53851 Daucus carota heat shock protein.
AAD49336.1 AF166277 Nicotiana tabacum low molecular weight heat-shock protein. LHS-1. TLHS-1.
BAA33062.1 AB017273 Cuscuta japonica low-molecular- weight heat shock protein. CJHSP17.
CAB36910.1 AJ000691 Quercus suber stress protein chaperone. heat shock protein 17.4. hsp!7.
CAA08908.1 AJ009880 Castanea sativa molecular chaperone. cytosolic class I small heat-shock protein HSP17.5. hsp!7.5.
AAA33975.1 Ml 1395 Glycine max small heat shock protein.
CAA25578.1 X01104 Glycine max heat shock protein 6871 (aa 1-153).
AAB03893.1 Ml 1318 Glycine max 17.5 kd heat shock protein Gmhspl7.6L.
CAA41547.1 X58711 Medicago sativa heat shock protein.
AAB63310.1 U46544 Helianthus annuus
18.6 kDa heat-shock protein. Class I low-molecular- weight heat-shock protein.
CAB08441.1 Z95153 Helianthus annuus
17.6 kD class I small heat-shock protein HSP17.6. Ha hspl7.6.
CAA42222.1 X59701 Helianthus annuus 17.6 kDa heat shock protein.
CAA37848.1 X53852 Daucus carota heat shock protein.
AAC39360.1 U63631 Fragaria x ananassa LMW heat shock protein.
AAA33672.1 M33899 Pisum sativum 18.1 kDa heat shock protein (hsp 18.1).
AAB63311.1 U46545 Helianthus annuus
17.7 kDa heat shock protein. Class I low-molecular-weight heat-shock protein.
AAA33974.1 M11317 Glycine max 17.6 kd heat shock protein Gmhspl7.6L.
CAA63903.1 X94193 Pennisetum glaucum heat shock protein 17.9. hsp!7.9.
AAA61632.1 U08601 Papaver somniferum low molecular weight heat-shock protein.
CAB55634.2 AJ237596 Helianthus annuus 17.9 kDa heat-shock protein. hspl7.9.
AAC78392.1 U83669 Oryza sativa low molecular mass heat shock protein Oshspl7.3. OSHSP17.3. class I LMMHSP.
AAA33910.1 M80939 Oryza sativa
16.9 kDa heat shock protein.
BAA02160.1 D12635 Oryza sativa 'low molecular weight heat shock protein'.
CAA43210.1 X60820 Oryza sativa
16.9 KD low molecular weight heat shock protein.
CAA37864.1 X53870 Chenopodium rabram heat-shock protein.
AAA33909.1 M80938 Oryza sativa 16.9 kDa heat shock protein.
AAC78393.1 U83670 Oryza sativa low molecular mass heat shock protein Oshsplδ.O. OSHSP18.0. class I LMMHSP.
AAB39856.1 U81385 Oryza sativa heat shock protein. Oshspl6.9C. class I, low molecular mass.
AAD30454.1 AF123257 Lycopersicon esculentum 17.6 kD class I small heat shock protein. HSP 17.6.
AAA33671.1 M33900 Pisum sativum 17.9 kDa heat shock protein (hspl7.9).
AAC78394.1 U83671 Oryza sativa low molecular mass heat shock protein Oshspl7.7. OSHSP17.7. class I LMMHSP.
CAB93514.1 AJ243567 Brassica oleracea
Putative class I small heat shock protein. HSP 17.x protein. hspl7.x.
CAA63901.1 X94191 Pennisetum glaucum heat shock protein 17.0. hspl7.0.
CAA63902.1 X94192 Pennisetum glaucum heat shock protein 16.9. hspl6.9.
AAD30452.1 AF123255 Lycopersicon esculentum 17.7 kD class I small heat shock protein. HSP17.7.
CAA46641.1 X65725 Zea mays heat shock protein 17.2. Zmhsp 17.2. Class I low molecular weight heat shock protein.
CAA39603.1 X56138 Lycopersicon esculentum small heat shock protein (class I).
CAA63570.1 X92983 Pseudotsuga menziesii low molecular weight heat-shock protein.
AAD30453.1 AF123256 Lycopersicon esculentum 17.8 kD class I small heat shock protein. HSP17.8.
CAA63571.1 X92984 Pseudotsuga menziesii low molecular weight heat-shock protein.
CAA31785.1 XI 3431 Triticum aestivum put, heat shock protein (AA 1 -151).
CAA53286.1 X75616 Oryza sativa heat shock protein 17.8.
SEQ ID NO: 170
AAC14577.1 U72396 Lycopersicon esculentum class II small heat shock protein Le-HSP17.6. heat treatment/chilling tolerance related protein from tomato fruit.
AAA33670.1 M33901 Pisum sativum 17.7 kDa heat shock protein (hspl7.7).
CAA82653.1 Z29554 Helianthus annuus 17.9 kDa heat-shock protein.
AAD41409.1 AF159562 Prunus dulcis cytosolic class II low molecular weight heat shock protein, hsp 17.5.
CAA65020.1 X95716 Petroselinum crispum small heat shock protein, cytoplasmic class II HSP.
AAC36312.1 AF090115 Lycopersicon esculentum cytosolic class II small heat shock protein HCT2. HSP17.4.
AAB01561.1 L47717 Picea glauca heat shock protein 17.0. EMB27.
AAB39336.1 M99430 Ipomoea nil small heat shock protein.
AAB01562.1 L47740 Picea glauca class II cytoplasmic small molecular weight heat shock protein 17.1. EMB29, SMW HSP 17.1.
CAA67206.1 X98617 Medicago sativa
17kD heat shock protein.
BAA99529.1 AP002484 Oryza sativa putative heat shock protein, 18K - maize. P0489A01.20. contains ESTs
C99035(E4351),AU093460(E3974).
CAA41218.1 X58279 Triticum aestivum heat shock protein 17.3. Tahspl7.3.
CAA67726.1 X99346 Picea abies small heat shock protein.
CAA38012.1 X54075 Zea mays 18kDa heat shock protein.
CAA38013.1 X54076 Zea mays 18kDa heat shock protein.
AAB26481.1 S59777 Zea mays
HSP18. HSP18. 18 kda heat shock protein; This sequence comes from Fig. 2B.
AAB39335.1 M99429 Ipomoea nil small heat shock protein
AAD09184.1 AF089845 Funaria hygrometrica cytosolic II small heat shock protein HSP16.4II. HSP16.4II.
BAA04841.1 D21817 Lilium longiflorum small heat shock protein. LIM11.
AAD09185.1 AF089846 Funaria hygrometrica cytosolic II small heat shock protein HSP18.3II. HSP18.3II.
BAA04842.1 D21818 Lilium longiflorum small heat shock protein. LIM12.
CAA63570.1 X92983 Pseudotsuga menziesii low molecular weight heat-shock protein.
CAA63571.1 X92984 Pseudotsuga menziesii low molecular weight heat-shock protein.
AAC39360.1 U63631 Fragaria x ananassa LMW heat shock protein.
AAD09178.1 AF087640 Funaria hygrometrica cytosolic I small heat shock protein HSP17.2IA. HSP17.2IA.
AAD09182.1 AF089843 Funaria hygrometrica cytosolic I small heat shock protein HSP17.2IC. HSP17.2IC.
BAA04840.1 D21816 Lilium longiflorum small heat shock protein. LIM10.
CAB93514.1 AJ243567 Brassica oleracea
Putative class I small heat shock protein. HSP17.X protein. hsp!7.x.
AAD30452.1 AF123255 Lycopersicon esculentum 17.7 kD class I small heat shock protein. HSP17.7.
AAB63311.1 U46545 Helianthus annuus
17.7 kDa heat shock protein. Class I low-molecular- weight heat-shock protein.
CAA31785.1 X13431 Triticum aestivum put, heat shock protein (AA 1 -151).
CAA63901.1 X94191 Pennisetum glaucum heat shock protein 17.0. hspl7.0.
AAA33672.1 M33899 Pisum sativum 18.1 kPa heat shock protein (hsp 18.1).
CAA41547.1 X58711 Medicago sativa heat shock protein.
CAA46641.1 X65725 Zea mays heat shock protein 17.2. Zmhsp 17.2. Class I low molecular weight heat shock protein.
AAB39856.1 U81385 Oryza sativa heat shock protein. Oshspl6.9C. class I, low molecular mass.
AAC78394.1 U83671 Oryza sativa low molecular mass heat shock protein Oshspl7.7. OSHSP17.7. class I LMMHSP.
AAC78392.1 U83669 Oryza sativa low molecular mass heat shock protein Oshspl7.3. OSHSP17.3. class I LMMHSP.
CAA39603.1 X56138 Lycopersicon esculentum small heat shock protein (class I).
AAA33910.1 M80939 Oryza sativa
16.9 kDa heat shock protein.
AAA33909.1 M80938 Oryza sativa 16.9 kDa heat shock protein.
CAA43210.1 X60820 Oryza sativa
16.9 KD low molecular weight heat shock protein.
AAD30453.1 AF123256 Lycopersicon esculentum 17.8 kD class I small heat shock protein. HSP17.8.
AAA34294.1 L14444 Triticum aestivum heat shock protein 16.9C. hspl6.9C.
CAA69172.1 Y07844 Hordeum vulgare
17 kDa class I small heat shock protein, hsp 17. putative.
AAD30454.1 AF123257 Lycopersicon esculentum 17.6 kD class I small heat shock protein. HSP17.6.
SEQ ID NO: 171
CAA82653.1 Z29554 Helianthus annuus 17.9 kDa heat-shock protein.
AAC14577.1 U72396 Lycopersicon esculentum class II small heat shock protein Le-HSP17.6. heat treatment/chilling tolerance related protein from tomato fruit.
CAA65020.1 X95716 Petroselinum crispum small heat shock protein, cytoplasmic class II HSP.
AAB39336.1 M99430 Ipomoea nil small heat shock protein.
AAA33670.1 M33901 Pisum sativum 17.7 kDa heat shock protein (hsp 17.7).
AAD41409.1 AF159562 Prunus dulcis cytosolic class II low molecular weight heat shock protein, hsp 17.5.
CAA67206.1 X98617 Medicago sativa 17kD heat shock protein.
BAA99529.1 AP002484 Oryza sativa putative heat shock protein, 18K - maize. P0489A01.20. contains ESTs
C99035(E4351),AU093460(E3974).
AAC36312.1 AF090115 Lycopersicon esculentum cytosolic class II small heat shock protein HCT2. HSP17.4.
CAA38012.1 X54075 Zea mays 18kDa heat shock protein.
CAA38013.1 X54076 Zea mays 18kDa heat shock protein.
CAA41218.1 X58279 Triticum aestivum heat shock protein 17.3. Tahspl7.3.
AAB26481.1 S59777 Zea mays
HSP18. HSP18. 18 kda heat shock protein; This sequence comes from Fig. 2B.
AAB01561.1 L47717 Picea glauca heat shock protein 17.0. EMB27.
AAB39335.1 M99429 Ipomoea nil small heat shock protein.
AAB01562.1 L47740 Picea glauca class II cytoplasmic small molecular weight heat shock protein 17.1. EMB29, SMW HSP17.1.
CAA67726.1 X99346 Picea abies small heat shock protein.
AAD09184.1 AF089845 Funaria hygrometrica cytosolic II small heat shock protein HSP16.4II. HSP16.4II.
BAA04841.1 D21817 Lilium longiflorum small heat shock protein. LIM 11.
BAA04842.1 D21818 Lilium longiflorum small heat shock protein. LIM12.
AAD09185.1 AF089846 Funaria hygrometrica cytosolic II small heat shock protein HSP18.3II. HSP18.3II.
BAA04840.1 D21816 Lilium longiflorum small heat shock protein. LIM10.
CAA63570.1 X92983 Pseudotsuga menziesii low molecular weight heat-shock protein.
CAA63571.1 X92984 Pseudotsuga menziesii low molecular weight heat-shock protein.
AAD30452.1 AF123255 Lycopersicon esculentum 17.7 kD class I small heat shock protein. HSP17.7.
AAD09178.1 AF087640 Funaria hygrometrica cytosolic I small heat shock protein HSP17.2IA. HSP17.2IA.
AAB63311.1 U46545 Helianthus annuus
17.7 kDa heat shock protein. Class I low-molecular-weight heat-shock protein.
CAA39603.1 X56138 Lycopersicon esculentum small heat shock protein (class I).
AAC39360.1 U63631 Fragaria x ananassa LMW heat shock protein.
AAA33672.1 M33899 Pisum sativum
18.1 kDa heat shock protein (hsp 18.1).
AAD30453.1 AF123256 Lycopersicon esculentum 17.8 kD class I small heat shock protein. HSP17.8.
AAD30454.1 AF123257 Lycopersicon esculentum 17.6 kD class I small heat shock protein. HSP 17.6.
CAA41547.1 X58711 Medicago sativa heat shock protein.
CAA31785.1 X13431 Triticum aestivum put. heat shock protein (AA 1 -151).
AAB39856.1 U81385 Oryza sativa heat shock protein. Oshspl6.9C. class I, low molecular mass.
CAA41546.1 X58710 Medicago sativa heat shock protein.
CAB93514.1 AJ243567 Brassica oleracea
Putative class I small heat shock protein. HSP17.X protem. hsp 17.x.
CAA63901.1 X94191 Pennisetum glaucum heat shock protein 17.0. hspl7.0.
AAA33910.1 M80939 Oryza sativa 16.9 kDa heat shock protein.
AAA33909.1 M80938 Oryza sativa 16.9 kDa heat shock protein.
CAA43210.1 X60820 Oryza sativa
16.9 KD low molecular weight heat shock protein.
AAB03097.1 U21723 Glycine max
Hsp22.3. Gmhsp22.3. low molecular weight heat shock protein.
AAC78394.1 U83671 Oryza sativa low molecular mass heat shock protein Oshspl7.7. OSHSP17.7. class I LMMHSP.
AAC78392.1 U83669 Oryza sativa low molecular mass heat shock protein Oshspl7.3. OSHSP17.3. class I LMMHSP.
AAD09182.1 AF089843 Funaria hygrometrica cytosolic I small heat shock protein HSP17.2IC. HSP17.2IC.
SEQ ID NO: 173
AAD30456.1 AF123259 Lycopersicon esculentum heat shock protein 90. HSP90.
AAF31705.1 AF221856 Euphorbia esula heat-shock protein 80.
AAC32131.1 AF051230 Piceamariana heat shock protein. Sb40. similar to Oryza sativa heat shock protein 82 encoded by GenBank
Accession Number Zl 1920.
AAF64453.1 AF239931 Euphorbia esula putative heat-shock protein 90. GRP94; similar to endoplasmin homolog precursor; contains the endoplasmic reticulum targeting sequence KDEL at the 3 '-tail.
CAA78738.1 Z15018 Oryza sativa heat shock protein, heat shock protein hsp 82. hsp 82.
SEQ ID NO: 174
AAG43546.1 AF211528 Nicotiana tabacum
Avr9/Cf-9 rapidly elicited protein 4. ACRE4. similar to Nicotiana glutinosa Nfr truncated N resistance protein encoded by GenBank Accession Number Ul 5605.
AAA50763.1 U15605 Nicotiana glutinosa virus resistance. N.
CAA08797.1 AJ009719 Solanum tuberosum disease resistance. NL25. nl25.
CAA08798.1 AJ009720 Solanum tuberosum disease resistance. NL27. nl27.
AAG09951.1 AF175388 Glycine max resistance protein LM6.
AAG09954.1 AF175399 Glycine max resistance protein MG13.
AAD25974.1 AF093647 Linum usitatissimum flax rast resistance protein. L.
AAD25966.1 AF093639 Linum usitatissimum flax rast resistance protein. L.
AAD25969.1 AF093642 Linum usitatissimum flax rast resistance protein. L.
AAD25965.1 AF093638 Linum usitatissimum flax rast resistance protein. L.
AAD25968.1 AF093641 Linum usitatissimum flax rast resistance protein. L.
AAA91021.1 U27081 Linum usitatissimum rust resistance. L6tr. L6. The shorter of two alternate protein products of The L6 gene that results from retention of intron 3 in the mRNA; truncated L6 gene product.
AAA91022.1 U27081 Linum usitatissimum rust resistance. L6. L6. The longer of two alternate proteins encoded by the L6 gene.
AAD25967.1 AF093640 Linum usitatissimum flax rast resistance protein. L.
AAK28803.1 AF310958 Linum usitatissimum resistance-like protein PI -A. i -A.
AAK28808.1 AF310961 Linum usitatissimum resistance-like protein P3-A. p3-A.
AAK28805.1 AF310960 Linum usitatissimum resistance-like protein P2-A. p2-A.
AAD25976.1 AF093649 Linum usitatissimum flax rust resistance protein. L.
AAD25973.1 AF093646 Linum usitatissimum flax rust resistance protein. L.
AAD25972.1 AF093645 Linum usitatissimum flax rast resistance protein. L.
AAD25971.1 AF093644 Linum usitatissimum flax rast resistance protein. L.
AAD25970.1 AF093643 Linum usitatissimum flax rast resistance protein. L.
AAD25975.1 AF093648 Linum usitatissimum flax rast resistance protein. L.
AAG48132.1 AF322632 Glycine max putative resistance protein. L20a.
AAG01052.1 AF175395 Glycine max resistance protein MG23.
CAC35330.1 AJ310155 Linum usitatissimum
Nl-D protein. Nl-D. N locus resistance gene homolog: TIR-NBS-LRR protein.
CAC35333.1 AJ310158 Linum usitatissimum
N2-C protein. N2-C. N locus rast resistance gene homolog: TIR-NBS-LRR protein.
AAG48133.1 AF322633 Glycine max putative resistance protein. L33.
AAF61452.1 AF139523 Tagetes erecta disease-resistance protein NRSA1.
CAC35334.1 AJ310159 Linum usitatissimum
N2-D protein. N2-D. N locus rust resistance gene homolog: TIR-NBS-LRR protein.
CAC35337.1 AJ310162 Linum usitatissimum
Nbi-C protein. Nbi-C. N locus rast resistance gene homolog: TIR-NBS-LRR protein.
CAC35339.1 AJ310164 Linum usitatissimum
Nho-C protein. Nho-C. N locus rust resistance gene homolog: TIR-NBS-LRR protein.
CAC35326.1 AJ310151 Linum usitatissimum
Ngc-C protein. Ngc-C. N rast resistance gene homolog: TIR-NBS-LRR protein.
AAG01051.1 AF175394 Glycine max resistance protein LM12.
CAC35338.1 AJ310163 Linum usitatissimum
Nbi-D protein. Nbi-D. N locus rast resistance gene homolog: TIR-NBS-LRR protein.
CAC35329.1 AJ310154 Linum usitatissimum
Nl-C protein. Nl-C. N locus resistance gene homolog: TIR-NBS-LRR protein.
AAK28804.1 AF310959 Linum usitatissimum resistance-like protein Pl-B. pl-B.
AAK28809.1 AF310962 Linum usitatissimum resistance-like protein P3-B. p3-B.
CAC35332.1 AJ310157 Linum usitatissimum
N2-B protein. N2-B. N locus rast resistance gene homolog: TIR-NBS-LRR protein.
CAC35328.1 AJ310153 Linum usitatissimum
Nl-B protein. Nl-B. N locus resistance gene homolog: TIR-NBS-LRR protein.
CAC35325.1 AJ310150 Linum usitatissimum
Ngc-B protein. Ngc-B. N rast resistance gene homolog: TIR-NBS-LRR protein.
C AC35336.1 A J310161 Linum usitatissimum
Nbi-B protein. Nbi-B. N locus rust resistance gene homolog: TIR-NBS-LRR protein.
CAC35321.1 A J310150 Linum usitatissimum
Ngc-D protein. Ngc-D. N rast resistance gene: TIR-NBS-LRR protein.
C AC35331.1 A J310156 Linum usitatissimum
N2-A protein. N2-A. N locus rast resistance gene homolog: TIR-NBS-LRR protein.
CAC35323.1 A J310150 Linum usitatissimum
Ngc-A protein. Ngc-A. N rast resistance gene homolog: TIR-NBS-LRR protein.
AAB47618.1 U73916 Linum usitatissimum rust resistance protein M. nucleotide-binding site, leucine-rich repeat class of plant disease resistance protein.
AAG01053.1 AF175396 Glycine max resistance protein MG55.
CAC35327.1 AJ310152 Linum usitatissimum
Nl-A protein. Nl-A. N locus resistance gene homolog: TIR-NBS-LRR protein.
AAK28811.1 AF310966 Linum usitatissimum resistance-like protein P-B. p-B.
SEQ ID NO: 175
CAB52796.1 AJ245861 Solanum tuberosum respiratory chain, putative internal rotenone-insensitive NADH dehydrogenase. ndal. CAB52797.1 AJ245862 Solanum tuberosum respiratory chain, putative external rotenone-insensitive NADH dehydrogenase. ndbl. SEQ ID NO: 176
BAA05648.1 D26601 Nicotiana tabacum protein kinase.
CAA08997.1 AJ010093 Brassica napus MAP3K beta 1 protein kinase. MAP3K beta 1.
CAA08995.1 AJ010091 Brassica napus MAP3K alpha 1 protein kinase. MAP3K alpha 1.
AAF34436.1 AF172282 Oryza sativa similar to mitogen-activated protein kinases. DUPR11.32.
CAB54520.1 AJ238845 Brassica napus putative role in cell cycle control. MAP3K epsilon 1 protein kinase. MAP3Kel.
CAA08758.1 AJ009609 Brassica napus
BnMAP4K alpha2.
CAA08757.1 AJ009608 Brassica napus BnMAP4K a_phal.
AAC83393.1 U83625 Zea mays protein kinase ZmMEKl. mitogen-activated; ERK-activating protein kinase (MEK) homolog.
AAG53979.1 AF325168 Nicotiana tabacum mitogen-activated protein kinase 2. MEK2. upstream kinase for SIPK and WIPK, two tobacco
MAP kinases.
AAG40578.1 AF216314 Oryza sativa
MAP kinase kinase 1. protein kinase; MEK1.
CAA04261.2 AJ000728 Lycopersicon esculentum
MAP kinase kinase. mekl.
AAF67262.1 AF165186 Nicotiana tabacum
MAP kinase kinase.
AAG45491.1 AY013245 Oryza sativa
3615.3. putative serine/threonine kinase.
BAB32405.1 AB055514 Nicotiana tabacum
NQK1 MAPKK. nqkl .
AAG49001.1 AY013246 Hordeum vulgare putative serine/threonine kinase. 635P2.3.
AAF19403.1 AF203481 Lycopersicon esculentum phosphoenolpyruvate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAF 19402.1 AF203480 Lycopersicon esculentum phosphoenolpyruvate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
CAC24705.1 AJ302651 Nicotiana tabacum protein kinase. MAP kinase. mekl.
BAA06731.1 D31964 Nicotiana tabacum
NPK2. protein kinase.
AAG31141.1 AF305911 Oryza sativa
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAF19401.1 AF203479 Glycine max phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAD46406.1 AF096250 Lycopersicon esculentum ethylene-responsive protein kinase TCTR1. ER50. serine/threonine kinase; similar to
Arabidopsis thaliana negative regulator ofthe ethylene response pathway encoded by
GenBank Accession Number L08789.
CAA73722.1 Y13273 Lycopersicon esculentum putative protein kinase.
BAA05649.1 D26602 Nicotiana tabacum protein kinase.
AAD23582.1 AF128443 Glycine max probably involved in plant stress responses possibly regulates gene expression. SNF-1-like serine/threonine protein kinase. expressed in nodules, roots and leaves.
CAA06334.1 AJ005077 Lycopersicon esculentum protein kinase. TCTR2 protein. TCTR2.
AAD 10057.1 AFl 10519 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTRlv.
CAB89082.1 AJ277534 Asparagus officinalis S6 ribosomal protein kinase. pkl . putative.
AAC78558.1 AF030879 Solanum tuberosum protein kinase CPK1.
AADl 0056.1 AFl 10518 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTR1.
AAK18832.1 AC082645 Oryza sativa putative protein kinase. OSJNBb0033N16.9.
AAG31142.1 AF305912 Hordeum vulgare
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAC24961.1 AF009337 Tradescantia virginiana CDPK-related protein kinase. CRKl .
BAB21278.1 AP002844 Oryza sativa putative MAP kinase. P0410E03.9.
AAA61682.1 L27484 Zea mays calcium-dependent protein kinase. CDPK.
CAA56313.1 X79992 Avena sativa putative pp70 ribosomal protein S6 kinase. Aspkl 1.
AAD31900.1 AFl 45482 Mesembryanthemum crystallinum putative serme/threonine protein kinase.
AAC25423.1 AF072908 Nicotiana tabacum calcium-dependent protein kinase. CDPK1.
CAA71142.1 Y10036 Cucumis sativus SNF1 -related protein kinase.
BAA83689.1 AB011968 Oryza sativa OsPK7. OsPK7. protein kinase.
CAA46554.1 X65604 Hordeum vulgare protein kinase. BKIN12.
CAA46556.1 X65606 Hordeum vulgare protein kinase. BKIN12.
CAA65500.1 X96723 Medicago sativa protein kinase. CDPK.
AAF05112.1 AF158091 Mesembryanthemum crystallinum day/night regulation of carbon fixation by crassulacean acid metabolism pathway, phosphoenolpyravate carboxylase-kinase. SNIK. protein kinase; salt/night induced kinase; phosphorylase; member of Ca2+/Cam protein kinase family; lacks both CAM domain and autoinhibitory domain; Ser/Thr kinase.
AAD17800.1 AF090835 Mesembryanthemum crystallinum Ca2+-dependent protein kinase. CPK1. serine/threonine protein kinase.
CAA07813.1 AJ007990 Hordeum vulgare SnRKl-type protein kinase. kin 12a.
SEQ ID NO: 177
BAB16335.1 AP002818 Oryza sativa putative NAM protein. P0436E04.18. contains ESTs E10793(C19698),E10793(C99379).
BAA84803.1 AP000559 Oryza sativa
Similar to NAM like protein (AC005310).
BAB19365.1 AP002542 Oryza sativa putative NAM (no apical meristem) protein. P0679C08.4.
BAB 16328.1 AP002818 Oryza sativa putative NAM protein. P0436E04.il. contains ESTs R4069(AU032425),R4069(AU082730).
CAA63102.2 X92205 Petunia x hybrida apical meristem formation. NAM.
CAA63101.1 X92204 Petunia x hybrida apical meristem formation. NAM.
AAK13151.1 AC078829 Oryza sativa putative NAM (no apical meristem) protein. OSJNBa0026O12.6.
BAB03447.1 AP002817 Oryza sativa
ESTs C96615(C10106),C26336(C12127),D21959(C10106) conespond to a region of the predicted gene. Similar to Arabidopsis thaliana DNA chromosome 4, BAC clone T6K21;
NAM (no apical meristem) - like protein (AL021889).
BAA92400.1 AP001366 Oryza sativa
ESTs C96615(C10106),C26336(C12127),D21959(C10106) conespond to a region ofthe predicted gene. Similar to NAM (AL021889). SEQ ID NO: 178
BAA25434.1 AB000708 Raphanus sativus
SAUR.
AAG14454.1 AF283706 Tulipa gesneriana auxin-induced protein TGSAUR12. SAUR12. small auxin upregulated RNA.
AAG14455.1 AF283707 Tulipa gesneriana auxin-induced protein TGSAUR21. SAUR21. small auxin upregulated RNA.
AAG14456.1 AF283708 Tulipa gesneriana auxin-induced protein TGSAUR22. SAUR22. small auxin upregulated RNA.
SEQ ID NO: 179 BAA78738.1 AB023482 Oryza sativa
EST AU055776(S20048) conesponds to a region ofthe predicted gene.; Similar to Arabidopsis thaliana AP2 domain containing protein RAP2.10 mRNA, partial cds.(AF003103).
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
AAF76898.1 AF274033 Atriplex hortensis apetala2 domam-containing protein.
CAC12822.1 AJ299252 Nicotiana tabacum AP2 domam-containing transcription factor. ap2.
AAC 14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserB.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
AAC24587.1 AF071893 Prunus armeniaca
AP2 domain containing protein. AP2DCP.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
BAB16083.1 AB036883 Oryza sativa transcriptional repressor. osERF3. osERF3. ERF protein family ERF3 associated repression domain.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal .
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREl. similar to EREBP transcription factors.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP transcription factors.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like transcription factor.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive transcription factor.
BAA97122.1 AB016264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
BAA97124.1 ABO 16266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
BAA94514.2 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4, BAC clone F9D16; putative Ap2 domain protein (AL035394).
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBPl.
AAK01088.1 AF298230 Hordeum vulgare
CBFl-like protein BCBF1. BCBF1. AP2 domain protein; putative DRE binding factor.
BAA99376.1 AP002526 Oryza sativa
ESTs AU093391(E60370),AU091593(C60458), AU093392(E60370) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana BAC F21 J9; AP2 domain protein. (AC000103).
AAK01089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor.
AAG59618.1 AF239616 Hordeum vulgare
CRT/DRE-binding factor. CBF. SEQ ID NO: 181
CAB71134.1 AJ271667 Cicer arietinum putative proteasome regulatory subunit. SEQ ID NO: 185
BAA85440.1 AP000616 Oryza sativa
ESTs AU055729(S20023),AU055730(S20023) conespond to a region ofthe predicted gene.; similar to Medicago nodulin N21-like protein (AC004218).
CAB53493.1 AJ245900 Oryza sativa
CAA303720.1 protein. q3037.20. Similar to Medicago nodulin N21 (MtN21).
BAB17350.1 AP002747 Oryza sativa putative nodulin. P0698G03.34. contains ESTs
D39891(S1543),D41717(S4395),AU033037(S1543). SEQ ID NO: 186
BAA02724.1 D13506 Glycine max early nodulin.
BAA33816.1 AB018378 Glycine max early nodulin. GmENOD93.
BAA83560.1 AP000399 Oryza sativa
EST AU077941(C12908) conesponds to a region ofthe predicted gene. Similar to
OsENOD93a gene for early nodulin (AB018375).
BAA33815.1 AB018377 Oryza sativa early nodulin. OsENOD93b.
BAA83566.1 AP000399 Oryza sativa
ESTs C98280(C1391),D15843(C1391) conespond to a region ofthe predicted gene. Similar to OsENOD93a gene for early nodulin (AB018375).
BAA83568.1 AP000399 Oryza sativa
EST AU077972(C53511) conesponds to a region ofthe predicted gene. Similar to
OsENOD93a gene for early nodulin (AB018375).
BAA83565.1 AP000399 Oryza sativa
ESTs C98096(C0688),C98097(C0688) conespond to a region ofthe predicted gene. Shnilar to OsENOD93a gene for early nodulin (ABO 18375).
AAD30134.1 AF140229 Oryza sativa early nodulin.
BAA33814.1 AB018376 Oryza sativa early nodulin. OsENOD93a.
BAA33813.1 AB018375 Oryza sativa early nodulin. OsENOD93a.
BAA83567.1 AP000399 Oryza sativa
EST C97982(C0324) conesponds to a region ofthe predicted gene. Similar to OsENOD93a gene for early nodulin (AB018375).
BAA83559.1 AP000399 Oryza sativa
EST C97982(C0324) conesponds to a region ofthe predicted gene. Similar to OsENOD93a gene for early nodulin (AB018375).
BAA83557.1 AP000399 Oryza sativa
Similar to OsENOD93a gene for early nodulin (ABO 18375).
SEQ ID NO: 188
AAA33811.1 L02830 Solanum tuberosum calcium-binding protein.
AAG43547.1 AF211529 Nicotiana tabacum
Avr9/Cf-9 rapidly elicited protein 31. ACRE31. similar to Solanum tuberosum CAST calcium binding protein encoded by GenBank Accession Number L02830.
CAB63264.1 AJ251808 Lotus japonicus calcium-binding protein, cbpl.
AAA34015.1 L01433 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-4. putative.
AAA92677.1 U13736 Pisum sativum binds calcium, calmodulin-like protein.
AAF31152.1 AF078680 Olea europaea calcium-binding protein. PCA23. Pca23.
AAA33948.1 L19359 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-5. putative.
AAD10245.1 AF030033 Phaseolus vulgaris calmodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
AAF31151.1 AF078679 Olea europaea calcium-binding protein. PCA18. Peal 8.
AAA19571.1 U10150 Brassica napus calcium binding, calmodulin. bcml.
AAC49587.1 U49105 Triticum aestivum cahnodulin TaCaM4-l. calcium-binding protein.
AAC49586.1 U49104 Triticum aestivum calmodulin TaCaM3-3. calcium-binding protein.
AAC49585.1 U49103 Triticum aestivum calmodulin TaCaM3-2. calcium-binding protein.
AAC49584.1 U48693 Triticum aestivum calmodulin TaCaM3-l . calcium-binding protein.
AAC49580.1 U48689 Triticum aestivum calmodulin TaCaMl-3. calcium-binding protein.
AAC49579.1 U48688 Triticum aestivum calmodulin TaCaMl-2. calcium binding protein.
AAC49578.1 U48242 Triticum aestivum cahnodulin TaCaMl-1. calcium-binding.
AAA85157.1 U20297 Solanum tuberosum calcium-binding protein, calmodulin.
AAA85156.1 U20296 Solanum tuberosum calcium-binding protein, calmodulin.
AAA62351.1 U20295 Solanum tuberosum calcium-binding protein, cahnodulin.
AAA85155.1 U20294 Solanum tuberosum calcium-binding protein, cahnodulin.
AAA34014.1 L01432 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-3. putative.
AAA03580.1 L01431 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-2. putative.
AAA34013.1 L01430 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-1. putative.
AAA33901.1 L18913 Oryza sativa calcium binding protein, signal transduction. calmodulin. putative.
AAA33900.1 L18914 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAA92681.1 U13882 Pisum sativum calcium-binding protein, cahnodulin.
CAA78288.1 Z12828 Oryza sativa calcium binding protein, signal transduction. calmodulin. SEQ ID NO: 190
AAF72197.1 AF263737 Euphorbia esula glutathione S-transferase. theta class GST.
AAG34815.1 AF243380 Glycine max glutathione S-transferase GST 25.
AAG34825.1 AF244682 Zea mays glutathione S-transferase GST 17.
AAG32474.1 AF309381 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTZl.
AAG34826.1 AF244683 Zea mays glutathione S-transferase GST 18.
AAA33277.1 M64268 Dianthus caryophyllus glutathione transferase. CARSR8.
CAA41279.1 X58390 Dianthus caryophyllus glutathione s-tiansferase. CARSR8.
AAD09190.1 AF109714 Triticum aestivum glutathione S-transferase. GST.
AAB60886.1 AF002211 Triticum aestivum glutathione-S-transferase.
AAA51450.1 L05916 Dianthus caryophyllus glutathione s-transferase. GST2.
AAC50036.1 U42463 Coccomyxa sp. PA glutathione S-transferase. GST.
AAG34801.1 AF243366 Glycine max glutathione S-transferase GST 11.
AAG32469.1 AF309376 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTUό.
AAG34850.1 AF244707 Zea mays glutathione S-transferase GST 42.
AAG34807.1 AF243372 Glycine max glutathione S-transferase GST 17.
CAB38119.1 AJ010296 Zea mays Glutathione transferase 111(b). gst3b.
AAF23357.1 AF109194 Hordeum vulgare glutathione-S-transferase.
AAG34849.1 AF244706 Zea mays glutathione S-transferase GST 41.
CAB38118.1 AJ010295 Zea mays Glutathione transferase 111(a). gst3a.
AAG32475.1 AF309382 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF5.
AAB65163.1 AF002692 Solanum commersonii glutathione S-transferase, class-phi. GSTl . low temperature induced.
AAF29773.1 AFl 59229 Gossypium hirsutum glutathione S-transferase. GST.
BAB39927.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.1. contains ESTs
AU031696(R0596),C97559(C60386),C28218(C60386), D28287(R0596).
AAG32473.1 AF309380 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTU2.
CAA09188.1 AJ010449 Alopecurus myosuroides glutathione transferase. GSTlb.
AAG34846.1 AF244703 Zea mays glutathione S-transferase GST 38.
AAG34812.1 AF243377 Glycine max glutathione S-transferase GST 22.
CAA55039.1 X78203 Hyoscyamus muticus glutathione transferase.
AAG34803.1 AF243368 Glycine max glutathione S-transferase GST 13.
CAA09189.1 AJ010450 Alopecurus myosuroides glutathione transferase. GSTlc.
AAG34808.1 AF243373 Glycine max glutathione S-transferase GST 18.
AAG34813.1 AF243378 Glycine max glutathione S-transferase GST 23.
AAD10129.1 AF004358 Aegilops tauschii chloroacetamide herbicide metabolism, glutathione S-transferase TSI-1. GST isozyme.
AAG41204.1 AF321437 Suaeda maritima glutathione transferase.
AAG34810.1 AF243375 Glycine max glutathione S-tiansferase GST 20.
AAG34804.1 AF243369 Glycine max glutathione S-fransferase GST 14.
AAG34814.1 AF243379 Glycine max glutathione S-transferase GST 24.
AAG32472.1 AF309379 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU3.
AAG32471.1 AF309378 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTU4.
CAA09187.1 AJ010448 Alopecurus myosuroides glutathione transferase. GSTla.
AAG34835.1 AF244692 Zea mays glutathione S-transferase GST 27.
AAF64449.1 AF239927 Euphorbia esula glutathione S-transferase. theta class GST.
AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-tiansferase T3.
AAG34848.1 AF244705 Zea mays glutathione S-tiansferase GST 40.
SEQ ID NO: 191
AAD32141.1 AF123503 Nicotiana tabacum Nt-gh3 deduced protein.
CAA42636.1 X60033 Glycine max auxin-responsive GH3 product. GH3.
BAA96221.1 AP002094 Oryza sativa
ESTs C19814(E10971),AU090481(E10971) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, sequence from clones T3F17, Fl 1C10, F13A10; auxin-responsive GH3-like protein (AC006526).
SEQ ID NO: 192
AAG13589.1 AC051633 Oryza sativa putative ubiquitin protein. OSJNBb0015I11.23.
SEQ ID NO: 193
CAC09348.1 AL442007 Oryza sativa putative phosphoglycerate dehydrogenase. H0212B02.4.
CAA79702.2 Z21493 Solanum tuberosum mitochondrial formate dehydrogenase precursor.
BAA77337.1 AB019533 Oryza sativa Nad-dependent formate dehydrogenase.
BAA36181.1 D88272 Hordeum vulgare formate dehydrogenase.
SEQ ID NO: 194
AAD46412.1 AF096262 Lycopersicon esculentum
ER6 protein, ethylene-inducible; similar to sequence F21M12 from Arabidopsis thaliana encoded by GenBank Accession Number AC000132.
SEQ ID NO: 195
AAK13154.1 AC078829 Oryza sativa putative casein kinase. OSJNBa0026O12.5.
BAA92986.1 AP001550 Oryza sativa
ESTs D41826(S4655),C22685(S4655) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana chromosome 4, BAC clone F16A16; protein kinase-like protein
(AL035353).
AAF19807.1 AF180356 Brassica oleracea casein kinase I-like protein. CKlb. strong similarity to Arabidopsis thaliana casein kinase 1.
AAD20819.1 AFl 07592 Dendrobium grex Madame Thong-In putative casein kinase I. otg!6.
AAF 19403.1 AF203481 Lycopersicon esculentum phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAF19402.1 AF203480 Lycopersicon esculentum phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
BAA05648.1 D26601 Nicotiana tabacum protein kinase.
AAF23901.2 AF194414 Oryza sativa calcium-dependent protein kinase. CDPK5. OsCDPK5.
AAC04324.1 U73937 Nicotiana tabacum ethylene signal transduction. PK12 protein kinase. PK12. component ofthe LAMMER family of protein kinases; dual-specificity protein kinase.
AAF23900.1 AF194413 Oryza sativa calcium-dependent protein kinase. CDPK1. OsCDPKl .
BAA34675.1 ABO 11670 Triticum aestivum wpk4 protein kinase. wpk4.
BAA13440.1 D87707 Ipomoea batatas calcium dependent protein kinase. CDPK.
AAD17800.1 AF090835 Mesembryanthemum crystallinum Ca2+-dependent protein kinase. CPK1. serine/threonine protein kinase.
CAA57157.1 X81394 Oryza sativa calcium-dependent protein kinase. OSCPK2.
AAD23582.1 AF128443 Glycine max probably involved in plant stress responses possibly regulates gene expression. SNF-1-like serine/tlneonine protein kinase. expressed in nodules, roots and leaves.
CAA39936.1 X56599 Daucus carota calcium- dependent protein kinase. DcPK431.
BAA12715.1 D85039 Zea mays calcium-dependent protein kinase.
CAA58750.1 X83869 Daucus carota CDPK-related protein kinase. CRK (or PK421).
AAB80693.1 U69174 Glycine max calmodulin-like domain protein kinase isoenzyme gamma. CDPK gamma.
AAF21062.1 AF216527 Dunaliella tertiolecta calcium-dependent protein kinase. CPK1; CDPK.
CAA74646.1 Y14274 Sorghum bicolor putative serine/threonine protein kinase. SNFL3.
AAD28192.2 AFl 15406 Solanum tuberosum calcium-dependent protein kinase. CDPK; catalytic domain.
BAA05649.1 D26602 Nicotiana tabacum protein kinase.
AAA69507.1 U28376 Zea mays calcium-dependent protein kinase. MZECDPK2.
AAB05457.1 U55768 Oryza sativa
SNF1 -related protein kinase. RSkl . Ser/Thr protein kinase homolog.
AAG36872.1 AF239819 Zea mays protein kinase CK2 catalytic subunit CK2 alpha-3.
CAA89202.1 Z49233 Chlamydomonas eugametos calcium-stimulated protein kinase.
CAA72362.1 Y11649 Zea mays protein kinase CK2, alpha subunit.
CAA72290.1 Y11526 Zea mays casein kinase II alpha subunit. CK2.
CAA43659.1 X61387 Zea mays casein kinase II alpha subunit. ZMACK2.
CAA65244.1 X95997 Solanum tuberosum SNFl-related protein kinase. PKIN1.
AAF76187.1 AF271237 Zea mays casein kinase II alpha subunit.
BAB21591.1 AB036788 Oryza sativa casein kinase II alpha subunit. OSCKA2.
BAB21589.1 AB036786 Oryza sativa casein kinase II alpha subunit. QSCKA2.
CAB89082.1 AJ277534 Asparagus officinalis S6 ribosomal protein kinase. pkl. putative.
AAF06970.1 AFl 62662 Kalanchoe fedtschenkoi phosphoenolpyruvate carboxylase kinase. calcium-independent protein kinase.
AAF06969.1 AFl 62661 Kalanchoe fedtschenkoi phosphoenolpyruvate carboxylase kinase. calcium-independent protein kinase.
CAA41172.1 X58194 Oryza sativa cdc2+/CDC28-related protein kinase.
CAA65500.1 X96723 Medicago sativa protein kinase. CDPK.
BAA12691.1 D84507 Zea mays
CDPK-related protein kinase. Does not require calcium for its activity (by similarity).
AAB47181.1 S82324 Zea mays
/gene- 'calcium/calmodulin-dependent protein kinase. This sequence comes from Fig. 1.
CAA07481.1 AJ007366 Zea mays calcium-dependent protein kinase.
BAA99439.1 AP002743 Oryza sativa putative protein kinase. P0710E05.26. contains ESTs C22394(C30013),C22393(C30013).
BAB12687.1 AP002746 Oryza sativa putative protein kinase. P0671B11.2. contains ESTs C22394(C30013),C22393(C30013).
AAF40430.1 AF234652 Mesembryanthemum crystallinum protein kinase MK5.
BAB21081.1 AP002819 Oryza sativa putative calcium-dependent protein kinase. P0501 GO 1.10.
SEQ ID NO: 196
AAF37267.1 AF220406 Vitis riparia
26S proteasome regulatory ATPase subunit SlOb. Revl36-3.
SEQ ID NO: 197
BAA08104.1 D45074 Panicum miliaceum
2-oxoglutarate/malate translocator. mitochondrial 2-oxoghιtarate/malate translocator.
BAA08103.1 D45073 Panicum miliaceum
2-oxoglutarate/malate translocator. mitochondrial 2-oxoglutarate/malate translocator.
BAA08105.1 D45075 Panicum miliaceum
2-oxoglutarate/malate translocator. mitochondrial 2-oxoglutarate/malate translocator.
CAA72107.1 Y11220 Solanum tuberosum mitochondrial uncoupling protein.
CAC 12820.1 AJ299250 Nicotiana tabacum mitochondrial 2-oxoglutarate/malate canier protein, momcl.
AAB71744.1 U75346 Chlamydomonas reinhardtii envelope protein. LIP-36G2. low CO2 inducible carrier protein LIP-36 with a molecular weight of 36 kDa.
AAB71743.1 U75345 Chlamydomonas reinhardtii envelope protein. LIP-36G1. low CO2 inducible carrier protein LIP-36 with a molecular weight of 36 kDa.
CAA07568.1 AJ007580 Ribes nigrum
Mitochondrial carrier protein. prib7.
BAB40117.1 AP003311 Oryza sativa putative peroxisomal Ca-dependent solute canier protein. P0024G09.9.
BAB 16462.1 AP002483 Oryza sativa putative peroxisomal Ca-dependent solute canier protein. P0019D06.21.
CAA56325.1 X80023 Triticum turgidum
ATP/ADP canier protein.
CAA46311.1 X65194 Chlamydomonas reinhardtii mitochondrial APP/ ATP translocator protein. CRANT.
CAA67107.1 X98474 Solanum tuberosum mitochondrial energy transfer protein, brittle 1.
CAC27140.1 AJ132535 Picea abies
ADP, ATP canier protein precursor.
CAA69726.1 Y08499 Betula pendula mitochondrial phosphate translocator.
BAA31583.1 ABO 16064 Zea mays mitochondrial phosphate transporter.
CAB61741.1 AJ275306 Cicer arietinum mitochondrial phosphate transporter.
AAG45489.1 AYO 13245 Oryza sativa
3615.1. putative mitochondrial carrier protein.
BAA31584.1 AB016065 Oryza sativa mitochondrial phosphate transporter.
BAA92520.1 AP001383 Oryza sativa
ESTs AU068633(C30614),AU068634(C30614) conespond to a region ofthe predicted gene.
Similar to Bos taurus mitochondrial solute canier protein. (AF049236).
SEQ ID NO: 199
BAA03455.1 D14605 Daucus carota AXI 1 OP. AXI 10.
SEQ ID NO: 204
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intiacellular kinase domain: from 1413.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl . receptor-like protein kinase; serine/threonine protein kinase.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
AAK02023.1 AC074283 Oryza sativa Putative protein kinase-like. OSJNBa0087H07.5.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRKl . Chrkl .
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
CAB51836.1 AJ243961 Oryza sativa Putitive Ser/Thr protein kinase. 11332.7.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAA94529.2 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPK10) mRNA. (L27821).
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
SEQ ID NO: 211
AAG13478.1 AC026758 Oryza sativa putative trehalose-6-phosphate phosphatase. OSJNBa0015J15.3.
SEQ ID NO: 212
AAC27894.1 AF023164 Zea mays leucine-rich repeat transmembrane protein kinase 1. ltkl.
AAC27895.1 AF023165 Zea mays leucine-rich repeat transmembrane protein kinase 2. Itk2.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil. Ptil kinase.
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S13927).
AAF91336.1 AF249317 Glycine max Ptil kinase-like protein. Ptil a. protein kinase.
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
BAA82394.1 AP000367 Oryza sativa
ESTs D23521(C2939),C22481(C2939) conespond to a region ofthe predicted gene.; Similar to serine/threonine protein kinase like protein. (AL022140).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAF43496.1 AF131222 Lophopyrum elongatum protein serme/tlireonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APK1 gene for protein tyrosine-serine-threonine kinase.(D12522).
AAB09771.1 U67422 Zea mays
CRINKLY4 precursor. cr4. receptor kinase homolog.
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAF66615.1 AF142596 Nicotiana tabacum
LRR receptor-like protein kinase.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSJNBa0004G10.30.
BAB 18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl. accumulates in mesophyll cells.
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
BAB 16326.1 AP002818 Oryza sativa putative receptor ser/thr protein kinase. P0436E04.9. contains ESTs
S10111(AU070304),S10111(AU083519).
BAA82556.1 AB030083 Populus nigra lectin-like protein kinase. PnLPK.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
AAF59906.1 AFl 97947 Glycine max receptor protein kinase-like protein. CLV1B.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1 A.
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12.
SEQ ID NO: 214
AAB51442.1 U63012 Sophora japonica lectin precursor.
CAA93829.1 Z69999 Phaseolus lunatus lectin 3.
BAA36415.1 ABO 12634 Robinia pseudoacacia lectin.
CAA93828.1 Z69998 Phaseolus lunatus lectin 2.
AAC49137.1 U21959 Cladrastis kentukea lectin precursor.
AAC49150.1 U21940 Cladrastis kentukea storage protein precursor, lectin.
CAB96391.1 AJ271873 Phaseolus lunatus carbohydrate-binding, lectin. Ibl5.
CAB96392.1 AJ271874 Phaseolus lunatus carbohydrate-binding, lectin. Ibl6.
CAA76366.1 Y16754 Medicago sativa lectin. Iec2.
BAA82556.1 AB030083 Populus nigra lectin-like protein kinase. PnLPK.
CAA93830.1 Z70000 Phaseolus lunatus lectin 4.
AAG16779.1 AF190633 Ulex europaeus lectin II.
AAC49136.1 U21958 Cladrastis kentukea lectin precursor.
AAB39933.1 U65009 Maackia amurensis lectin precursor.
AAB39934.1 U65010 Maackia amurensis lectin precursor.
AAA33766.1 L26237 Phaseolus lunatus lectin II.
AAA33143.1 M34270 Dolichos bifloras seed lectin.
CAA57697.1 X82216 Medicago truncatula lectin. Iec3.
BAA36413.1 AB012632 Robinia pseudoacacia lectin.
AAA80182.1 U12783 Robinia pseudoacacia lectin.
BAA04604.1 D 17757 Robinia pseudoacacia lectin precursor.
CAA68497.1 Y00440 Pisum sativum lectin-precursor (AA -30 to 245).
AAC49271.1 U24249 Robinia pseudoacacia lectin precursor.
AAA80181.1 U12782 Robinia pseudoacacia lectin.
BAA36416.1 AB012635 Robinia pseudoacacia lectin-related polypeptide.
AAA33676.1 M18160 Pisum sativum lectin.
CAA47011.1 X66368 Pisum sativum Psi lectin. psi.
AAA33141.1 J02721 Dolichos bifloras lectin subunit I precursor.
BAA36414.1 AB012633 Robinia pseudoacacia lectin.
BAA02049.1 D12481 Bauhinia puφurea lectin.
AAA80183.1 U12784 Robinia pseudoacacia lectin.
AAC49272.1 U24250 Robinia pseudoacacia lectin precursor.
AAA82737.1 U18296 Medicago sativa lectin. Mslecl.
AAA74571.1 U22468 Arachis hypogaea agglutinin. galactose-binding lectin precursor, lee, lectin.
AAB51441.1 U63011 Sophora japonica lectin precursor.
AAA74574.1 U22471 Arachis hypogaea agglutinin. galactose-binding lectin precursor, lee. lectin.
AAG00508.1 AF285121 Sophora flavescens lectin.
AAB39932.1 U65008 Maackia amurensis lectin precursor.
SEQ ID NO: 215
CAA64327.1 X94624 Brassica napus acyl-CoA synthetase.
CAA96523.1 Z72153 Brassica napus acyl CoA synthetase.
CAC19877.1 AJ401089 Brassica napus activation of free fatty acids, long chain acyl-CoA synthetase. acs6. activity confirmed by expression in E. coli.
CAA06820.1 AJ006025 Cicer arietinum acyl-coA synthetase.
AAC39365.1 AF008183 Populus x generosa 4-coumarate:CoA ligase 2. 4CL2.
CAA31697.1 X13325 Petroselinum crispum 4-coumarate: CoA ligase Pc4Cl-2 (AA 1-544).
CAA31696.1 XI 3324 Petroselinum crispum 4-coumarate:CoA ligase Pc4Cl-l (AA 1-544).
AAC39366.1 AF008184 Populus x generosa 4-coumarate:CoA ligase 1. 4CL1.
AAF37734.1 AF052223 Lolium perenne 4-coumarate— CoA ligase 4CL3.
AAF91309.1 AF239686 Rubus idaeus
4-coumarate: co A ligase 2. adenylate-forming enzyme; 4CL2.
AAF37733.1 AF052222 Lolium perenne 4-coumarate— CoA ligase 4CL2.
CAA36850.1 X52623 Oryza sativa 4-coumarate-CoA ligase.
BAA07828.1 D43773 Nicotiana tabacum 4-coumarate: coenzyme A ligase.
BAA08365.1 D49366 Lithospermum erythrorhizon
4-coumarate:CoA ligase.
AAB 18637.1 U50845 Nicotiana tabacum
4-coumarate: coenzyme A ligase. 4CL1. Nt4CL-l.
AAD40664.1 AFl 50686 Solanum tuberosum
4-coumarate:coenzyme A ligase. 4CL-2a.
AAA33842.1 M62755 Solanum tuberosum 4-coumarate— CoA ligase. St4Cl-l.
AAB18638.1 U50846 Nicotiana tabacum 4-coumarate: coenzyme A ligase. 4CL2. Nt4CL-19.
AAF91310.1 AF239687 Rubus idaeus
4-coumarate: co A ligase 1. adenylate-forming enzyme; 4CL1.
AAC24503.1 AF041049 Populus tremuloides 4-coumarate: CoA ligase.
AAF37732.1 AF052221 Lolium perenne 4-coumarate~CoA ligase 4CL1.
AAA92669.1 U12013 Pinus taeda 4-coumarate-CoA ligase enzyme.
AAB42382.1 U39404 Pinus taeda 4-coumarate:CoA ligase. lp4CL-2.
AAB42383.1 U39405 Pinus taeda 4-coumarate:CoA ligase. lp4CL-l.
AAF91308.1 AF239685 Rubus idaeus
4-coumarate: co A ligase 3. adenylate-forming enzyme; 4CL3.
AAC24504.1 AF041050 Populus tremuloides 4-coumarate: CoA ligase.
AAA92668.1 U12012 Pinus taeda 4-coumarate-CoA ligase enzyme.
CAC36095.1 X69955 Glycine max 4-coumarate:Coenzyme A ligase isoenzyme 4. 4CL4.
AAG43823.1 AF212317 Capsicum annuum 4-coumarate:coenzyme A ligase. 4CL.
AAA69580.1 L43362 Oryza sativa 4-coumarate:CoA ligase isoform 2. 4cl.2. putative.
BAA08366.2 D49367 Lithospermum erythrorhizon 4-coumarate:CoA ligase.
CAA49575.1 X69954 Glycine max 4-coumarate~CoA ligase.
CAB97359.1 AJ278455 Juglans nigra 4-coumarate-CoA ligase. 4CL.
AAF73995.2 AF144502 Pinus armandii 4-coumarate: CoA ligase. 4CL.
AAF73994.2 AF144501 Pinus armandii 4-coumarate:CoA ligase. 4CL.
AAF74018.2 AF144525 Tsuga canadensis 4-coumarate:CoA ligase. 4CL.
AAF74010.2 AF144517 Abies holophylla 4-coumarate: CoA ligase. 4CL.
AAF74020.2 AF144527 Pseudolarix amabilis 4-coumarate : CoA ligase . 4CL.
AAF74016.2 AFl 44523 Nothotsuga longibracteata 4-coumarate:CoA ligase. 4CL.
AAF73997.2 AF144504 Picea smithiana 4-coumarate: CoA ligase. 4CL.
AAF74013.2 AFl 44520 Abies beshanzuensis 4-coumarate: CoA ligase. 4CL.
AAF74008.2 AF144515 Abies firm 4-coumarate: CoA ligase. 4CL.
AAF74022.2 AFl 44529 Cedras atlantica 4-coumarate: CoA ligase. 4CL.
AAF74005.2 AF144512 Larix gmelini 4-coumarate: CoA ligase. 4CL.
AAF74021.2 AF144528 Pseudolarix amabilis 4-coumarate: CoA ligase. 4CL.
AAF74003.2 AFl 44510 Pseudotsuga sinensis 4-coumarate:CoA ligase. 4CL.
AAF74019.2 AF144526 Tsuga canadensis 4-coumarate: CoA ligase. 4CL.
SEQ ID NO: 221
AAB37246.1 U58971 Nicotiana tabacum calmodulin-binding protein. TCB60.
SEQ ID NO: 224
BAB19413.1 AP002870 Oryza sativa putative acetone-cyanohydrrn lyase. P0458A05.22.
AAC49184.1 U40402 Hevea brasiliensis hydroxynitrile lyase. hnl.
CAAl 1219.1 AJ223281 Manihot esculenta alpha-hydroxyriitrile lyase. H A
CAA82334.1 Z29091 Manihot esculenta alpha-hydroxynitrile lyase.
SEQ ID NO: 225
BAB 16335.1 AP002818 Oryza sativa putative NAM protein. P0436E04.18. contains ESTs E10793(C19698),E10793(C99379).
AAK13151.1 AC078829 Oryza sativa putative NAM (no apical meristem) protein. OSJNBa0026O12.6.
CAA63102.2 X92205 Petunia x hybrida apical meristem formation. NAM.
CAA63101.1 X92204 Petunia x hybrida apical meristem formation. NAM.
BAB16328.1 AP002818 Oryza sativa putative NAM protein. P0436E04.i l. contains ESTs R4069(AU032425),R4069(AU082730).
BAB19365.1 AP002542 Oryza sativa putative NAM (no apical meristem) protein. P0679C08.4.
BAA84803.1 AP000559 Oryza sativa Similar to NAM like protein (AC005310).
BAB03447.1 AP002817 Oryza sativa
ESTs C96615(C10106),C26336(C12127),D21959(C10106) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana DNA chromosome 4, BAC clone T6K21;
NAM (no apical meristem) - like protein (AL021889).
BAA92400.1 AP001366 Oryza sativa
ESTs C96615(C10106),C26336(C12127),D21959(C10106) conespond to a region ofthe predicted gene. Similar to NAM (AL021889).
SEQ ID NO: 226
BAA96221.1 AP002094 Oryza sativa
ESTs C19814(E10971),AU090481(E10971) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, sequence from clones T3F17, Fl 1C10, F13A10; auxin-responsive GH3 -like protein (AC006526).
AAD32141.1 AFl 23503 Nicotiana tabacum Nt-gh3 deduced protein.
CAA42636.1 X60033 Glycine max auxin-responsive GH3 product. GH3.
SEQ ID NO: 227
CAA70403.1 Y09204 Nicotiana tabacum histidinol-phosphate aminotransferase. hpa.
CAC20728.1 AJ278767 Nicotiana plumbaginifolia essential for histidine biosynthesis, histidinol phosphate aminotransferase. hpa. ~~~~~~ SEQ ID NO: 229
AAF33670.1 AF079872 Nicotiana tabacum cyclic nucleotide-gated calmodulin-binding ion channel. CBP4.
AAF33669.1 AF079871 Nicotiana tabacum cyclic nucleotide-gated calmodulin-binding ion channel. CBP7.
AAK16188.1 AC079887 Oryza sativa putative cyclic nucleotide and calmodulin-regulated ion channel protein. OSJNBa0040E01.13.
AAB53255.1 U65390 Nicotiana tabacum cyclic nucleotide gated channel protein. CaMB-channel protein, channel protein homolog.
CAB54856.1 AJ132686 Zea mays potassium channel protein ZMK2. ZMK2.
AAD16278.1 AF099095 Samanea saman pulvinus inward-rectifying channel for potassium SPICKl . similar to Arabidopsis potassium channel AKT3.
CAA71598.1 Y10579 Vicia faba potassium channel.
CAA56175.1 X79779 Solanum tuberosum K+ channel inward rectifying. KST1.
CAC05489.1 AJ271447 Populus tremula x Populus tremuloides potassium channel, potassium channel 2. ptk2.
AAD39492.1 AF145272 Samanea saman pulvinus inward-rectifying channel SPICK2. potassium channel; similar to AKT2/3.
CAA70870.1 Y09699 Solanum tuberosum putative inward rectifying potassium channel. SKT2.
CAB62555.1 AJ249962 Daucus carota potassium channel, kdcl.
BAA96192.1 AP002093 Oryza sativa
Similar to Arabidopsis thaliana potassium channel protein (M86990).
BAA96150.1 AP002092 Oryza sativa
Similar to Arabidopsis thaliana potassium channel protein (M86990).
BAA84085.1 AB032074 Nicotiana paniculata potassium channel. NpKTl.
AAF81251.1 AF267755 Mesembryanthemum crystallinum potassium channel protein Mkt2p.
CAA68912.1 Y07632 Zea mays potassium channel. ZMK1.
CAA60016.1 X86021 Solanum tuberosum potassium channel. SKT1 gene, putative start codon.
CAA65254.1 X96390 Lycopersicon esculentum potassium channel. LKT1.
CAA12645.1 AJ225805 Egeria densa inward potassium channel alpha subunit. homologous to the sequences ofthe family of inwardly rectifying potassium channels in plants which is stracturally related to the shaker family of outwardly rectifying channels in Drosophila.
AAF36832.1 AF207745 Triticum aestivum AKTl-like potassium channel. TaAKTl.
CAC10514.1 AJ299019 Samanea saman potassium release, outwardly rectifying potassium channel, sporkl.
AAF81249.1 AF267753 Mesembryanthemum crystallinum putative potassium channel protein Mktlp.
CAC05488.1 AJ271446 Populus tremula x Populus tremuloides potassium channel, outward rectifying potassium channel, ptork.
SEQ ID NO: 232
AAA80575.1 U13148 Pennisetum ciliare possible apospory-associated protein.
AAF34174.1 AFl 95243 Chlamydomonas reinhardtii apospory-associated protein C. APOC.
SEQ ID NO: 233
AAB97366.1 AF039531 Oryza sativa lysophospholipase homolog. LPL1.
SEQ ID NO: 235
AAC61839.1 AF025430 Papaver somniferam berberine bridge enzyme, bbel. (S)-reticuline: oxygen oxidoreductase (methylene bridge foirning).
AAC39358.1 AF005655 Eschscholzia californica oxidizes the N-methyl group of (S)-reticuline to the berberine bridge carbon C-8 of (S)- scoulerine in berberine and benzophenanthridine alkaloid biosynthesis, berberine bridge enzyme, bbel. covalently-bound FAD-dependent oxidase; elicitor-inducible.
AAB20352.1 S65550 Eschscholzia californica
(S)-reticuline: oxygen oxidoreductase (methylene-bridge-foirning). /gene="(S)~ reticuline:oxygen oxidoreductase. This sequence comes from Fig 2; berberine bridge enzyme.
AAD 17487.1 AF049347 Berberis stolonifera
Catalyzes the oxidative cyclization ofthe N-methyl group of (S)-reticulrne into the berberine bridge carbon (C-8) of (S)-scoulerine. berberine bridge enzyme, bbel. Covalently flavinylated oxidase of isoquinoline alkaloid biosynthesis in plants.
SEQ ID NO: 244 AAD17487.1 AF049347 Berberis stolonifera
Catalyzes the oxidative cyclization ofthe N-methyl group of (S)-reticulrne into the berberine bridge carbon (C-8) of (S)-scoulerine. berberine bridge enzyme, bbel. Covalently flavinylated oxidase of isoquinoline alkaloid biosynthesis in plants.
AAB20352.1 S65550 Eschscholzia californica
(S)-reticuline: oxygen oxidoreductase (methy lene-bridge-forming). /gene="(S)- reticuline: oxygen oxidoreductase. This sequence comes from Fig 2; berberine bridge enzyme.
AAC39358.1 AF005655 Eschscholzia californica oxidizes the N-methyl group of (S)-reticuline to the berberine bridge carbon C-8 of (S)- scoulerine in berberine and benzophenanthridine alkaloid biosynthesis, berberine bridge enzyme, bbel. covalently-bound FAD-dependent oxidase; elicitor-inducible.
AAC61839.1 AF025430 Papaver somniferam berberine bridge enzyme, bbel. (S)-reticuline:oxygen oxidoreductase (methylene bridge forming).
SEQ ID NO: 247 AAF37267.1 AF220406 Vitis riparia
26S proteasome regulatory ATPase subunit SI Ob. Reyl36-3. SEQ ID NO: 248
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APK1 gene for protein tyrosine-serine-threonine kinase.(D12522).
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJN_a0071K19.il.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAF91336.1 AF249317 Glycine max
Ptil kinase-like protein. Ptil a. protein kinase. ^^__^
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S13927).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil. Ptil kinase.
AAG33377.1 AF290411 Oryza meyeriana serine/threonine protein kinase. Rl .
AAC27894.1 AF023164 Zea mays leucine-rich repeat transmembrane protein kinase 1. ltkl.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAC27895.1 AF023165 Zea mays leucine-rich repeat transmembrane protein kinase 2. Itk2.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
AAK11566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhirPtoE. confers resistance to bacterial speck disease.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl. accumulates in mesophyll cells.
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
AAG00510.1 AF285172 Phaseolus vulgaris leaf senescence-associated receptor-like protein kinase. SARK.
AAC48932.1 U13923 Lycopersicon pimpinellifolium
Fen. putative serine/threonine protein kinase; similar to product encoded by Lycopersicon pimpinellifolium Pto gene, GenBank Accession Number U02271 ; Fen is a member of the Pto gene family.
AAB47424.1 U59317 Lycopersicon pimpinellifolium serine/threonine protein kinase Fen. Fen, fenthion sensitivity gene from tomato.
AAF76307.1 AF220602 Lycopersicon pimpinellifolium
Fen kinase.
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
AAK11567.1 AF318491 Lycopersicon hirsutum
Pto-like protein kinase F. LhirPtoF.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
BAB 19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
SEQ ID NO: 249
BAA22422.1 AB001379 Glycynhiza echinata cytochrome P450. CYP81E1.
BAA74465.1 AB022732 Glycynhiza echinata cytochrome P450. CYP Ge-31.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
CAB41490.1 AJ238439 Cicer arietinum cytochrome P450 monooxygenase. cyp81E3v2.
CAA10067.1 AJ012581 Cicer arietinum cytochrome P450. cyp81E3.
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'- race).
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81Blc.
AAK38079.1 AF321855 Lolium rigidum putative cytochrome P450.
AAK38080.1 AF321856 Lolium rigidum putative cytochrome P450.
AAK38081.1 AF321857 Lolium rigidum putative cytochrome P450.
AAC34853.1 AF082028 Hemerocallis hybrid cultivar putative cyt P450-containing fatty acid hydroxylase. senescence-associated protein 3. SA3. mRNA accumulates in senescing petals.
AAB94590.1 AF022461 Glycine max
CYP82Clp. CYP82C1. cytochrome P450 monooxygenase.
CAB56742.1 AJ249800 Cicer arietinum cytochrome P450 monooxygenase. cyp81E5.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3'-hydroxylase. Htl. cytochrome P450; CYP75B2.
AAG09208.1 AF175278 Pisum sativum wound-inducible P450 hydroxylase. CYP82A1.
CAA71515.1 Y10491 Glycine max putative cytochrome P450.
CAA71516.1 Y10492 Glycine max putative cytochrome P450.
AAC49188.2 U29333 Pisum sativum cytochrome P450 monooxygenase. CYP82. new cytochrome P450 family.
CAA71876.1 Y10982 Glycine max putative cytochrome P450.
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
AAC39454.1 AF014802 Eschscholzia californica
(S)-N-methylcoclaurine 3 '-hydroxylase. CYP82B1. cytochrome P-450-dependent monooxygenase; methyl jasmonate-inducible cytochrome P-450-dependent, homologous to wound-inducible CYP82A1 of Pisum sativum GenBank Accession Number U29333.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
CAA71877.1 Y10983 Glycine max putative cytochrome P450.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
AAD38930.1 AF135485 Glycine max cytochjome P450 monooxygenaseCYP93Dl . CYP93E1.
BAA84072.1 AB028152 Torenia hybrida flavone synthase II. cytochrome P450. TFNS5.
BAA74466.1 AB022733 Glycynhiza echinata cytochrome P450. CYP Ge-51.
BAA22423.1 AB001380 Glycynhiza echinata cytochrome P450. CYP93B1.
BAA35080.1 ABO 15762 Nicotiana tabacum putative cytochrome P450. CYP82E1.
BAA92894.1 AB006790 Petunia x hybrida cytochrome P450. IMT-2.
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl. P450 enzyme.
CAB56743.1 AJ249801 Cicer arietinum cytochrome P450 monooxygenase. cyp81E4.
BAA84071.1 AB028151 Antinhinum majus flavone synthase II. cytochrome P450. AFNS2.
SEQ ID NO: 250
AAD55566.1 AFl 10784 Volvox carteri f. nagariensis protein disulfide isomerase precursor, pdf
AAD02069.1 AF036939 Chlamydomonas reinhardtii redox-regulator of 5'UTR psbA mRNA binding complex and translation, protein disulfide isomerase. localized to ER and chloroplast.
AAC49896.1 AF027727 Chlamydomonas reinhardtii involved in the redox-regulated binding of chloroplast poly(A)-binding protein to the 5'-UTR of psbA mRNA; regulates chloroplast translational activation, protein disulfide isomerase RB60. PDI.
AAD28260.1 AF131223 Datisca glomerata protein disulfide isomerase homolog. PDI.
AAB08519.1 L39014 Zea mays protein disulfide isomerase. pdi. putative.
AAA19660.1 U11496 Triticum aestivum protein disulfide isomerase. PDI.
CAC21230.1 AJ277379 Triticum turgidum subsp. durum catalyzes formation of disulfide bonds, protein disulfide isomerase. Pdi.
CAC21228.1 AJ277377 Triticum turgidum subsp. durum catalyzes the formation of disulfide bonds, protein disulfide isomerase. Pdi.
AAA70345.1 L33251 Hordeum vulgare catalyze the formation of disulfide bonds, disulfide isomerase. PDI. putative.
AAA70344.1 L33250 Hordeum vulgare catalyze the formation of disulfide bonds, disulfide isomerase. PDI. putative.
AAB05641.1 U41385 Ricinus communis protein disulphide isomerase PDI. molecular chaperone.
CAA77575.1 Z11499 Medicago sativa protein disulfide isomerase.
CAC21231.1 AJ277380 Triticum turgidum subsp. durum catalyzes formation of disulfide bonds, protein disulfide isomerase. Pdi. CAC21229.1 AJ277378 Triticum turgidum subsp. durum catalyzes formation of disulfide bonds, protein disulfide isomerase. Pdi. BAB 18780.1 AB047268 Cucumis sativus disulfide isomerase.
BAA92322.1 AB039278 Oryza sativa protein disulfide isomerase. Pdi.
AAA70346.1 L33252 Hordeum vulgare catalyze the formation of disulfide bonds, disulfide isomerase. PDI. putative.
CAA72092.1 Y11209 Nicotiana tabacum protein disulfide-isomerase precursor. PDI.
AAG13988.1 AF298829 Primus avium putative protein disulfide-isomerase. PDI.
SEQ ID NO: 251
CAA61275.1 X88797 Eucalyptus gunnii cinnamyl alcohol dehydrogenase. CADI.
AAC06319.1 AF053084 Malus x domestica putative cinnamyl alcohol dehydrogenase. CAD.
SEQ ID NO: 253
CAB61745.1 AJ275311 Cicer arietinum farnesylated protein.
AAD09515.1 U64917 Glycine max putative metal-binding protein. GMFP7. farnesylated protein.
SEQ ID NO: 254
BAB19757.1 AB052785 Glycine max nitrate transporter NRT1-2. NRT1-2.
BAB19756.1 AB052784 Glycine max nitrate tiansporter NRT1-1. NRT1-1.
BAB19760.1 AB052788 Glycine max nitrate tiansporter NRT 1-5. NRT1-5.
AAC32034.1 AF023472 Hordeum vulgare peptide transporter, ptil. PTR1; integral membrane protein.
AAD01600.1 AF016713 Lycopersicon esculentum LeOPTl. LeOPTl. oligopeptide transporter.
BAB40113.1 AP003311 Oryza sativa putative peptide transport protein. P0024G09.4. contains ESTs
D40448(S2437),C71800(E0368),AU102190(E2393),
AU055921(S20154),AU102191(E2393),AU055922(S20154),
C98524(E0368),AU097146(S2437).
BAB16458.1 AP002483 Oryza sativa putative peptide tiansport protein. P0019D06.16. contains ESTs
D40448(S2437),C71800(E0368),AU102190(E2393),
AU055921(S20154),AU102191(E2393),AU055922(S20154),
C98524(E0368),AU097146(S2437).
CAA93316.1 Z69370 Cucumis sativus nitrite transporter. NiTRl .
AAK15441.1 AC037426 Oryza sativa putative nitrate transporter. OSJNBb0014I11.9.
AAG21898.1 AC026815 Oryza sativa putative peptide transport protein. OSJNBa0079L16.13.
CAC00544.1 AJ277084 Nicotiana plumbaginifolia ion transport, putative low-affinity nitrate tiansporter. nrtl.l.
AAG46153.1 AC018727 Oryza sativa putative peptide transporter. OSJNBa0056G17.8.
CAC00545.1 AJ277085 Nicotiana plumbaginifolia ion transport, putative low-affinity nitrate transporter, nrtl .2.
AAG21906.1 AC026815 Oryza sativa putative peptide transport protein. OSJNBa0079L16.9.
AAF20002.1 AF213936 Prunus dulcis amino acid/peptide transporter. PTR2. similar to transporters of nitrogenous compounds.
AAF07875.1 AF140606 Oryza sativa nitrate transporter. NRTL
BAB 16322.1 AP002818 Oryza sativa putative peptide transporter-like protein. P0436E04.4.
BAB19758.1 AB052786 Glycine max putative nitrate transporter NRT 1-3. NRT 1-3.
AAA80582.1 U17987 Brassica napus putative nitrate transporter. RCH2 protein.
CAC07206.1 AJ278966 Brassica napus
Low-affinity nitrate transporter, nitrate transporter, nrtl.
AAG46154.1 AC018727 Oryza sativa putative peptide transporter. OSJNBa0056G17.27.
AAB69642.1 AF000392 Lotus japonicus peptide transporter. LJNOD65.
BAB19759.1 AB052787 Glycine max putative nitrate transporter NRT1-4. NRT1-4.
AAD16016.1 AF080545 Nepenthes alata peptide transporter. PTR1.
AAD42860.1 AFl 54930 Prunus dulcis transporter-like protein. TLP1.
AAG13513.1 AC068924 Oryza sativa putative peptide transporter. OSJNBa0026L12.7. SEQ ID NO: 255
AAB01567.1 L47672 Picea glauca
EMB34. embryo-abundant protein.
SEQ ID NO: 257 AAB71743.1 U75345 Chlamydomonas reinhardtii envelope protein. LIP-36G1. low CO2 inducible carrier protein LIP-36 with a molecular weight of 36 kDa.
AAB71744.1 U75346 Chlamydomonas reinhardtii envelope protein. LIP-36G2. low CO2 inducible carrier protein LIP-36 with a molecular weight of 36 kDa.
BAA92520.1 AP001383 Oryza sativa
ESTs AU068633(C30614),AU068634(C30614) conespond to a region ofthe predicted gene.
Similar to Bos taurus mitochondrial solute carrier protein. (AF049236).
BAB 16462.1 AP002483 Oryza sativa putative peroxisomal Ca-dependent solute carrier protein. P0019D06.21.
BAB40117.1 AP003311 Oryza sativa putative peroxisomal Ca-dependent solute canier protein. P0024G09.9.
CAA07568.1 AJ007580 Ribes nigrum
Mitochondrial carrier protein. prib7.
CAC27140.1 AJ132535 Picea abies
ADP, ATP carrier protein precursor.
CAA56325.1 X80023 Triticum turgidum
ATP/ADP canier protein.
CAC12820.1 AJ299250 Nicotiana tabacum mitochondrial 2-oxoglutarate/malate canier protein, momcl.
AAG48999.1 AYO 13246 Hordeum vulgare putative mitochondrial canier protein. 635P2.1. SEQ ID NO: 258
CAA05276.1 AJ002236 Lycopersicon pimpinellifolium resistance gene. Hcr9-9E. Hcr9-9E.
AAC78591.1 AF053993 Lycopersicon esculentum disease resistance protein. Cf-5.
AAC78596.1 AF053998 Lycopersicon esculentum
Hcr2-5D. Hcr2-5D. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
CAA05279.1 AJ002237 Lycopersicon esculentum
Hcr9-0. Hcr9-0. homologue of Cladosporium fiilvum disease resistance gene Cf-9.
AAC78593.1 AF053995 Lycopersicon esculentum
Hcr2-0B. Hcr2-0B. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAA65235.1 U15936 Lycopersicon pimpinellifolium
Cf-9 precursor. Cf-9. this is the ninth resistance gene to disease caused by Cladosporium fulvum to be isolated.
CAA05274.1 AJ002236 Lycopersicon pimpinellifolium resistance gene. Cf-9. Cf-9.
AAC78592.1 AF053994 Lycopersicon esculentum
Hcr2-0A. Hcr2-0A. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAC78595.1 AF053997 Lycopersicon esculentum
Hcr2-5B. Hcr2-5B. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAC78594.1 AF053996 Lycopersicon pimpinellifolium
Hcr2-2A. Hcr2-2A. similar to Lycopersicon pimpinelhfolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
BAA96776.1 AP002521 Oryza sativa
Similar to Lycopersicon esculentum disease resistance protein (AF053993).
BAB08215.1 AP002539 Oryza sativa
Similar to Lycopersicon esculentum disease resistance protein (AF053993).
CAA05268.1 AJ002235 Lycopersicon hirsutum
Resistance gene. Cf-4. Cf-4.
AAG21897.1 AC026815 Oryza sativa putative disease resistance protein (3' partial). OSJNBa0079L 16.21.
AAD50430.1 AF166121 Hordeum vulgare
Cf2/Cf5 disease resistance protein homolog. Bigl. leucine rich repeat protein.
AAG21917.1 AC026815 Oryza sativa putative disease resistance protein. OSJNBa0079L16.5.
AAG21909.1 AC026815 Oryza sativa putative disease resistance protein. OSJNBa0079L16.3.
CAB55409.1 ALl 17265 Oryza sativa zhbOOOl.l. Incomplete at 5'end,Similar to disease resistance protein; Method: conceptual translation with partial peptide sequencing.
AAC49123.1 U37133 Oryza sativa receptor kinase-like protein. Xa21. Xa21 disease resistance gene.
AAC80225.1 U72723 Oryza longistaminata receptor kinase-like protein. Xa21. disease resistance gene. SEQ ID NO: 259
CAB52689.1 AJ132224 Lycopersicon esculentum hexose transporter. ht2.
CAA09419.1 AJ010942 Lycopersicon esculentum hexose transporter protein.
BAB19864.1 AB052885 Oryza sativa monosaccharide transporter 3. OsMST3.
AAA18534.1 L21753 Saccharum hybrid cultivar H65-7052 glucose transporter, putative.
CAA47324.1 X66856 Nicotiana tabacum monosaccharid transporter. MST1.
AAB06594.1 U38651 Medicago truncatula sugar transporter.
AAA79761.1 L08196 Ricinus communis hexose transport, sugar carrier protein. RCSTC.
CAA04511.1 AJ001061 Vitis vinifera hexose uptake, hexose transporter.
CAA70777.1 Y09590 Vitis vinifera hexose transporter.
A A Λ ΛAAΛΛCCoo6611iioo88oo5522....ii11 A A __--__FFrr00uu66oo11ιι11ιι00uu66oo P P rreeeettiiuuuunnnniiiiaaaa xx xx h h nnyyyybboorinππdduuaaaa ppuuttaattiivvee mmoonnoossaacccchhaarriiddee ttrraannssppoortrteerr 11.. ppmmttll.. g similar to hexose tiansporter protein; PMTl . L T 0 fl8R118 β8» R Pilcfiinmuiso r c-nommmniTuinniisc hexose transport, hexose carrier protein. HEX6.
CAB06079.1 Z83829 Picea abies monosaccharide transporter. PaMst-1. PaMst-1.
BAB 19863.1 AB052884 Oryza sativa monosaccharide transporter 2. OsMST2.
BAA83554.1 AP000399 Oryza sativa
Similar to hexose carrier protein HEX6 &RCCHCP_1 (Q07423).
AAK31286.1 AC079890 Oryza sativa putative hexose carrier protein. OSJNBb0089A17.11. CAA53192.1 X75440 Chlorella kessleri hexose transporter like protein. HUP3.
CAA68813.1 Y07520 Chlorella kessleri H(+)/hexose cotransporter (AA 1-533).
CAA39036.1 X55349 Chlorella kessleri H(+)/hexose-cotransporter. HUPl.
BAB19862.1 AB052883 Oryza sativa monosaccharide transporter 1. OsMSTl.
AAA18533.1 L21752 Saccharum hybrid cultivar H65-7052 glucose transporter, putative.
CAB52688.1 AJ132223 Lycopersicon esculentum hexose transporter, htl.
AAD55054.1 AF173655 Beta vulgaris glucose transporter. Gt.
CAB52690.1 AJ132225 Lycopersicon esculentum hexose transporter. ht3.
AAA33875.1 L31352 Ricinus communis hexose transport, hexose canier. Hex9. putative.
AAK13147.1 AC083945 Oryza sativa Putative sugar transporter. OSJNBa0058E19.22.
AAB68029.1 U64903 Beta vulgaris
BvcDNA-397. putative sugar transporter; member of major facilitative superfamily; integral membrane protein.
AAB68028.1 U64902 Beta vulgaris
BvcDNA-205. putative sugar transporter; member of major facilitative superfamily; integral membrane protein.
AAG43998.1 AF215837 Apium graveolens var. dulce mannitol transporter. Matl .
AAF74568.1 AF215854 Zea mays transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAF74567.1 AF215853 Solanum tuberosum transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAF74566.1 AF215852 Nicotiana tabacum transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAF74565.1 AF215851 Spinacia oleracea transport of hexoses across the plastid inner envelope membrane, hexose transporter. pGlcT.
AAG46179.1 AC018727 Oryza sativa putative sugar transporter protein. OSJNBa0056G17.3.
AAG00995.1 AF286906 Mesembryanthemum crystallinum putative glucose translocator. metabolite tiansporter; targeted to plastid inner envelope membrane.
AAB53155.1 U43629 Beta vulgaris putative sugar transporter, integral membrane protein, member of major facilitator superfamily.
AAB88879.1 AF000952 Prunus armeniaca putative sugar transporter.
AAA33874.1 L31353 Ricinus communis hexose transport, hexose carrier. HexlO. putative.
AAD37424.1 AF149282 Phaseolus vulgaris hexose canier protein 1. HCP1.
AAD45934.1 AF168773 Betula pendula hexose transport protein. HEX2.
SEQ ID NO: 260 AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
AAF34428.1 AFl 72282 Oryza sativa receptor-like protein kinase. DUPR11.18.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPK10) mRNA. (L27821).
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAA94528.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 BAC T20K24; putative receptor-like protein kinase (AC002392).
BAA94517.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
BAA94529.2 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12.
BAA94518.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 section 111 of 255; putative receptor-like protein kinase (AC002392).
CAA47962.1 X67733 Zea mays receptor-like protein kinase. PK1.
BAA83573.1 AP000399 Oryza sativa
Similar to serine/threonine-specific protein kinase PK10 precursor (AL021811).
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
BAB 17345.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.29.
BAB 17348.1 AP002747 Oryza sativa putative receptor kinase . P0698G03.32.
BAB 17342.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.26.
BAB39451.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.24.
BAB 17126.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.16.
BAB 19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl. accumulates in mesophyll cells.
BAB 17339.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.23.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
AAF78016.1 AF238472 Oryza sativa receptor-like kinase. RLG15. protein kinase.
AAD46420.1 AF100771 Hordeum vulgare receptor-like kinase. Hv3ARK. similar to wheat ARK1 AS.
AAC49629.1 U51330 Triticum aestivum rast resistance kinase LrlO. LRK10.
BAB17139.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.31.
BAB 17331.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.12.
AACO 1746.1 AF044489 Oryza sativa receptor-like protein kinase. dφkl.
AAC27489.1 AF077130 Oryza sativa receptor-like protein kinase.
AAC02535.1 AF044260 Oryza sativa receptor serine/threonine kinase. protein kinase.
BAB39434.1 AP003338 Oryza sativa receptor-like kinase. OJ1212_B09.1.
AAF78020.1 AF238476 Oryza sativa receptor-like kinase. RLG11. protein kinase.
BAA92953.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F10M6 ; S-receptor kinase -like protein. (AL021811).
AAF78018.1 AF238474 Oryza sativa receptor-like kinase. RLG16. protein kinase.
AAD46917.1 AF164021 Oryza sativa receptor kinase.
BAB39438.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.7.
BAB39435.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.2.
BAB17129.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.20.
BAB17321.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.1.
AAF68398.1 AF237568 Oryza sativa receptor-like protein kinase. RLG2.
BAB39437.1 AP003338 Oryza sativa receptor-like kinase. OJ1212_B09.6.
AAF78019.1 AF238475 Oryza sativa receptor-like kinase. RLG17. protein kinase.
SEQ ID NO: 261
AAD09343.1 AF026538 Hordeum vulgare AB A-responsive protein.
SEQ ID NO: 263
BAA22813.1 D26015 Nicotiana tabacum aspartic protease activity. CND41, chloroplast nucleoid DNA binding protein. cnd41.
BAB21205.1 AP002913 Oryza sativa nucleoid DNA-binding protein cnd41-like protein. P0480E02.11. contains ESTs
AU166073(E31027),AU029516(E31027).
SEQ ID NO: 270
BAB12719.1 AP002746 Oryza sativa putative regulatory protein NPRl . P0671B11.35.
BAB16860.1 AP002537 Oryza sativa
Arabidopsis thaliana regulatory protein NPRl like protein. P0001B06.13.
SEQ ID NO: 271
AAG35658.1 AF204925 Petroselinum crispum transcription factor WRKY4. WRKY4. binds to W box (TTGACC) elements.
BAA87058.1 AB028022 Nicotiana tabacum WIZZ. wizz. wound-induced transcription factor.
BAA86031.1 AB026890 Nicotiana tabacum transcription factor NtWRKY4.
AAD16139.1 AF096299 Nicotiana tabacum DNA-binding protein 2. WRKY2. transcription factor.
BAA77383.1 AB020590 Nicotiana tabacum transcription factor NtWRKY2.
AAF23898.1 AF193802 Oryza sativa zinc finger transcription factor WRKY1.
AAD55974.1 AF121353 Petroselinum crispum zinc-finger type transcription factor WRKY1. WRKY1.
CAB97004.1 AJ278507 Solanum tuberosum putative transcription factor. WRKY DNA binding protein. WRKY1.
BAA82107.1 AB022693 Nicotiana tabacum transcription factor. NtWRKYl .
AAD32677.1 AF140554 Avena sativa
DNA-binding protein WRKY1. wrkyl. putative transcription factor.
BAB 16432.1 AB041520 Nicotiana tabacum WRKY transcription factor Nt-SubD48. Nt-SubD48.
AAD 16138.1 AF096298 Nicotiana tabacum DNA-binding protein 1. WRKYl. transcription factor.
AAD32676.1 AF 140553 Avena sativa
DNA-binding protein WRKY3. wrky3. putative transcription factor.
AAD27591.1 AF121354 Petroselinum crispum binds sequence specifically to W Boxes (TTGACC). transcription factor. WRKY3. sequence specific DNA-binding protein.
AAF61864.1 AF193771 Nicotiana tabacum
DNA-binding protein 4. WRKY4. transcription factor.
AAG35659.1 AF204926 Petroselinum crispum transcription factor WRKY5. WRKY5. binds to W box (TTGACC) elements.
AAF61863.1 AFl 93770 Nicotiana tabacum DNA-binding protein 3. WRKY3. transcription factor.
SEQ ID NO: 272
BAA07395.1 D38220 Brassica napus nitrate reductase.
BAA07394.1 D38219 Brassica napus nitrate reductase.
AAG30576.1 AF314093 Ricinus communis nitrate reductase. NLA.
CAA32217.1 X14059 Nicotiana tabacum nitrate reductase.
AAA33713.1 L13691 Petunia x hybrida nitrate reductase. putative.
CAA32218.1 X14060 Lycopersicon esculentum nitrate reductase.
AAA33712.1 LI 1563 Petunia x hybrida nitrate reductase apoenzyme. nia.
CAA32216.1 X14058 Nicotiana tabacum nitrate reductase.
CAA56696.1 X80670 Lotus japonicus nitrate reductase (NADH). NIA.
AAB52786.1 U95317 Solanum tuberosum NADH nitrate reductase. StNR3.
AAB 18985.1 U76701 Solanum tuberosum NADH nitrate reductase. StNR2.
AAA95940.1 U01029 Phaseolus vulgaris nitrate reductase. PVNR2.
AAA34033.1 M32600 Spinacia oleracea NADH nitrate reductase.
CAA38031.1 X54097 Betula pendula nitrate reductase (NADH). nial.
BAA13047.1 D86226 Spinacia oleracea nitrate reductase.
AAA33114.1 M33154 Cucurbita maxima nitrate reductase.
AAD19790.1 AF055369 Glycine max nitrate reductase. nr2.
CAA58909.1 X84103 Cichorium intybus nitrate reductase (NADH). nia.
AAA96813.1 U13987 Glycine max inducible nitrate reductase 2. INR2.
CAA37672.1 X53603 Phaseolus vulgaris nitrate reductase.
AAA96727.1 L23854 Glycine max nitrate reductase. INR1.
AAA62316.1 U20450 Zea mays nitrate reductase.
AAD38068.1 AF153448 Zea mays nitrate reductase. NR1.
CAA40975.1 X57844 Hordeum vulgare nitrate reductase. cDNA is 9bp short of atg.
CAA40976.1 X57845 Hordeum vulgare nitrate reductase.
CAA42739.1 X60173 Hordeum vulgare nitrate reductase (NAD(P)H). nar7.
AAB93560.1 AF022780 Glycine max nitrate reductase. BCNR-A.
AAF17595.1 AF203033 Chlamydomonas reinhardtii nitrate reductase. NIT1.
CAA45497.1 X64136 Volvox carteri nitrate reductase (NADH). nitA.
AAC49460.1 U39931 Chlorella vulgaris nitrate reductase.
AAC49459.1 U39930 Chlorella vulgaris nitrate reductase.
CAA29497.1 X06134 Nicotiana tabacum nitrate reductase.
AAAI 8377.1 U08029 Spinacia oleracea reduces nitrate to nitrite with NADH. NADH:nitrate reductase.
AAB39553.1 U64308 Agrostemma githago nitrate reductase. agnrl. NADH; similar to agnr2 product encoded by GenBank Accession
Number U64309 and to agnr3 product encoded by GenBank Accession Number U64310.
AAA03202.1 M27821 Zea mays
NADHmitrate reductase; (EC 1.6.6.1).
AAA33483.1 M77792 Zea mays enzyme, nitiate reductase. NAR1S.
AAB39555.1 U64310 Agrostemma githago nitrate reductase. agnr3. NADH; similar to agnrl product encoded by GenBank Accession
Number U64308 and agnr2 product encoded by GenBank Accession Number U64309.
AAB39554.1 U64309 Agrostemma githago nitrate reductase. agnr2. NADH; similar to agnrl product encoded by GenBank Accession
Number U64308 and agnr3 product encoded by GenBank Accession Number U64310.
CAA33819.1 X15820 Oryza sativa nitrate reductase apoenzyme (AA 472-916); Protein sequence is in conflict with the conceptual translation.
CAA33817.1 X15819 Oryza sativa nitrate reductase apoenzyme.
AAA33998.1 L23853 Glycine max nitiate reductase. mutant.
CAA58908.1 X84102 Cichorium intybus nitiate reductase (NADH). nia.
CAA40090.1 X56771 Chlorella vulgaris nitrate reductase (NADH).
CAA45776.1 X64446 Zea mays nitrate reductase (NAD(P)H). nar.
AAD17694.1 AF077372 Zea mays possible reduction of Fe3+-chelates. cytochrome b5 reductase. NFR.
AAA96242.1 L40147 Avena strigosa nitrate reductase.
AAB20155.1 S61885 Nicotiana plumbaginifolia nitrate reductase heme domain, nitrate reductase heme domain, NR. This sequence comes from fig3; NR.
AAA96245.1 L40151 Hordeum pusillum nitrate reductase.
AAA96247.1 L40153 Hordeum stenostachys nitrate reductase.
SEQ ID NO: 273
BAA07395.1 D38220 Brassica napus nitrate reductase.
BAA07394.1 D38219 Brassica napus nitrate reductase.
AAA33713.1 L13691 Petunia x hybrida nitrate reductase. putative.
CAA32218.1 X14060 Lycopersicon esculentum nitrate reductase.
AAA33712.1 L11563 Petunia x hybrida nitrate reductase apoenzyme. nia.
AAG30576.1 AF314093 Ricinus communis nitiate reductase. NIA.
CAA32217.1 X14059 Nicotiana tabacum nitrate reductase.
CAA32216.1 X14058 Nicotiana tabacum nitrate reductase.
AAA33114.1 M33154 Cucurbita maxima nitrate reductase.
AAB52786.1 U95317 Solanum tuberosum
NADH nitrate reductase. StNR3.
AAB18985.1 U76701 Solanum tuberosum
NADH nitrate reductase. StNR2.
AAA34033.1 M32600 Spinacia oleracea
NADH nitrate reductase.
BAA13047.1 D86226 Spinacia oleracea nitrate reductase.
CAA38031.1 X54097 Betula pendula nitrate reductase (NADH). nial.
CAA56696.1 X80670 Lotus japonicus nitrate reductase (NADH). NIA.
AAA95940.1 U01029 Phaseolus vulgaris nitiate reductase. PVNR2.
CAA58909.1 X84103 Cichorium intybus nitrate reductase (NADH). nia.
AAD19790.1 AF055369 Glycine max nitiate reductase. nr2.
AAA96813.1 U13987 Glycine max inducible nitrate reductase 2. INR2.
CAA40976.1 X57845 Hordeum vulgare nitrate reductase.
AAA96727.1 L23854 Glycine max nitrate reductase. INRl.
CAA37672.1 X53603 Phaseolus vulgaris nitiate reductase.
CAA33819.1 X15820 Oryza sativa nitrate reductase apoenzyme (AA 472-916); Protein sequence is in conflict with the conceptual translation.
AAD38068.1 AFl 53448 Zea mays nitrate reductase. NR1.
CAA40975.1 X57844 Hordeum vulgare nitrate reductase. cDNA is 9bp short of atg.
AAB93560.1 AF022780 Glycine max nitrate reductase. BCNR-A.
AAA62316.1 U20450 Zea mays nitrate reductase.
CAA42739.1 X60173 Hordeum vulgare nitrate reductase (NAD(P)H). nar7.
AAF17595.1 AF203033 Chlamydomonas reinhardtii nitrate reductase. NIT1.
CAA45497.1 X64136 Volvox carteri nitrate reductase (NADH). nitA.
AAC49460.1 U39931 Chlorella vulgaris nitrate reductase.
AAC49459.1 U39930 Chlorella vulgaris nitrate reductase.
CAA29497.1 X06134 Nicotiana tabacum nitiate reductase.
AAA18377.1 U08029 Spinacia oleracea reduces nitrate to nitrite with NADH. NADH:nitrate reductase.
AAB39553.1 U64308 Agrostemma githago nitrate reductase. agnrl . NADH; similar to agnr2 product encoded by GenBank Accession
Number U64309 and to agnr3 product encoded by GenBank Accession Number U64310.
AAA03202.1 M27821 Zea mays NADH.-nitrate reductase; (EC 1.6.6.1).
AAA33483.1 M77792 Zea mays enzyme, nitrate reductase. NAR1 S.
AAB39555.1 U64310 Agrostemma githago nitrate reductase. agnr3. NADH; similar to agnrl product encoded by GenBank Accession
Number U64308 and agnr2 product encoded by GenBank Accession Number U64309.
AAB39554.1 U64309 Agrostemma githago nitiate reductase. agnr2. NADH; similar to agnrl product encoded by GenBank Accession
Number U64308 and agnr3 product encoded by GenBank Accession Number U64310.
CAA33817.1 X15819 Oryza sativa nitrate reductase apoenzyme.
CAA58908.1 X84102 Cichorium intybus nitrate reductase (NADH). nia.
AAA33998.1 L23853 Glycine max nitiate reductase. mutant.
CAA40090.1 X56771 Chlorella vulgaris nitrate reductase (NADH).
CAA45776.1 X64446 Zea mays nitrate reductase (NAD(P)H). nar.
AAD17694.1 AF077372 Zea mays possible reduction of Fe3+-chelates. cytochrome b5 reductase. NFR.
AAA96242.1 L40147 Avena strigosa nitrate reductase.
AAA96250.1 L40149 Hordeum chilense nitiate reductase.
AAA96245.1 L40151 Hordeum pusillum nitrate reductase.
AAA96247.1 L40153 Hordeum stenostachys nitrate reductase.
SEQ ID NO: 274
AAC39318.1 AF029858 Sorghum bicolor second multifunctional cytochrome P450 in the biosynthetic pathway ofthe cyanogenic glucoside dhurrin. Catalyzes the conversion of p-hydroxyphenylacetaldoxime to p- hydroxymandelonitrile. cytochrome P450 CYP71E1. CYP71E1. No EST#s identified.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
AAA19701.1 L24438 Thlaspi arvense cytochrome P450.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
AAB94589.1 AF022460 Glycine max
CYP83Dlρ. CYP83D1. cytochrome P450 monooxygenase.
AAB94588.1 AF022459 Glycine max CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
AAD47832.1 AF166332 Nicotiana tabacum cytochrome P450.
AAK38084.1 AF321860 Lolium rigidum putative cytochrome P450.
AAB94584.1 AF022157 Glycine max capable of catalyzing the metabolism of phenylurea herbicides. CYP71A10. CYP71A10. cytochrome P450 monooxygenase.
AAF27282.1 AF122821 Capsicum annuum cytochrome P450. PepCYP
AAK38083.1 AF321859 Lolium rigidum putative cytochrome P450.
CAA50312.1 X70981 Solanum melongena P450 hydroxylase. CYPEG2.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
AAB61964.1 U48434 Solanum chacoense putative cytochrome P450.
AAK38087.1 AF321863 Lolium rigidum putative cytochrome P450.
CAA71514.1 Y10490 Glycine max putative cytochrome P450.
CAA50645.1 X71654 Solanum melongena P450 hydroxylase.
BAA03635.1 D14990 Solanum melongena Cytochrome P-450EG4.
AAD44151.1 AFl 24816 Mentha x piperita cytochrome p450 isoform PMI 7.
AAD44152.1 AF124817 Mentha x piperita cytochrome p450 isoform PM2.
CAA83941.1 Z33875 Mentha x piperita cytochrome P-450 oxidase.
AAD44150.1 AF124815 Mentha spicata cytochrome p450.
AAB69644.1 AF000403 Lotus japonicus putative cytochrome P450. LJNP450.
BAB40322.1 AB036772 Triticum aestivum cytochrome P450. N-l.
CAC27827.1 AJ295719 Catharanthus roseus geraniol hydroxylase. cytochrome P450. cyp71.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3 '-hydroxylase. Htl. cytochrome P450; CYP75B2.
AAG14963.1 AF214009 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H3.
AAD37433.1 AF150881 Lycopersicon esculentum x Lycopersicon peravianum catalyzes the hydroxylation of feralic acid to 5 -hydroxy feralic acid, ferulate-5-hydroxylase. CYP84. cytochrome P450-dependent monooxygenase; F5H; FAHl.
BAB39252.1 AP002968 Oryza sativa putative cytochrome P450. P0416G11.1.
AAK38088.1 AF321864 Lolium rigidum putative cytochrome P450.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
AAG14961.1 AF214007 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
AAG14962.1 AF214008 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2.
AAB 17562.1 U72654 Eustoma grandifloram flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75.
SEQ ID NO: 279
AAA34122.1 M84466 Nicotiana tabacum phenylalanine ammonia lyase. tpal .
BAA22948.1 AB008200 Nicotiana tabacum phenylalanine ammonia-lyase. palB.
AAA34176.1 M90692 Lycopersicon esculentum phenylalanine ammonia-lyase. PAL5.
AAF40224.1 AF237955 Rubus idaeus phenylalanine ammonia-lyase 2. PAL2. PAL; phenylpropanoid; multigene; flavonoid.
CAA37129.1 X52953 Glycine max phenylalanine ammonia-lyase. PALI.
CAA68036.1 X99705 Triticum aestivum phenylalanine ammonia-lyase. PAL.
AAA33389.1 M29232 Ipomoea batatas phenylalanine ammonia-lyase.
AAA34179.2 M83314 Lycopersicon esculentum deamination of phenylalanine to coumarate. phenylalanine ammonia lyase. pal.
BAA21643.1 D30656 Populus kitakamiensis phenylalanine ammonia-lyase.
AAB67733.1 U43338 Citrus limon phenylalanine ammonia-lyase. pal6.
BAA95629.1 AB042520 Catharanthus roseus phenylalanine ammonia lyase.
BAA05643.1 D26596 Camellia sinensis phenylalanine ammonia-lyase.
CAA73065.1 Y12461 Helianthus annuus phenylalanine ammonia lyase. PAL.
BAA24929.1 D83076 Lithospermum erythrorhizon phenylalanine ammonia-lyase.
BAA24928.1 D83075 Lithospermum erythrorhizon phenylalanine ammonia-lyase.
BAA00885.1 D10001 Pisum sativum phenylalanine ammonia-lyase.
AAA84889.1 U39792 Pinus taeda phenylalanine ammonia-lyase. lpPAL.
CAA61198.1 X87946 Oryza sativa phenylalanine ammonia-lyase. ZB8.
CAA41169.1 X58180 Medicago sativa phenylalanine ammonia-lyase. PAL.
BAA00887.1 D10003 Pisum sativum phenylalanine ammonia-lyase. PAL2.
BAA00886.1 D 10002 Pisum sativum phenylalanine ammonia-lyase. PALI.
AAAI 7993.1 M91192 Trifolium subtenaneum phenylalanine ammonia-lyase. PALI .
AAA33805.1 LI 1747 Populus x generosa phenylalanine ammonia lyase. PAL.
AAC78457.1 AF036948 Prunus avium phenylalanine ammonia-lyase. PALI.
BAA23367.1 D85850 Daucus carota phenylalanine ammonia-lyase. gDcPALl.
CAB42793.1 AJ238753 Citrus Clementina x Citras reticulata phenylalanine-animonia lyase. pall.
AAA99500.1 L36822 Stylosanthes humilis phenylalanine ammonia lyase. PAL 17.1.
CAA55075.1 X78269 Nicotiana tabacum phenylalanine ammonia-lyase.
BAA22963.1 D 17467 Nicotiana tabacum phenylalanine ammonia-lyase. TOBPAL1.
BAA22947.1 AB008199 Nicotiana tabacum phenylalanine ammonia-lyase. palA.
CAA57057.1 X81159 Petroselinum crispum phenylalanine ammonia-lyase 3. PAL3. tetramere subunit.
AAG49585.1 AF325496 Ipomoea nil phenylalanine ammonia-lyase.
CAA57056.1 X81158 Petroselinum crispum phenylalanine ammonia-lyase 2. PAL2. deaminase subunit.
CAB42794.1 AJ238754 Citras Clementina x Citrus reticulata phenylalanine-ammonia lyase. pa!2.
CAA05251.1 AJ002221 Digitalis lanata phenylalanine ammonia lyase.
BAA07860.1 D43802 Populus kitakamiensis phenylalanine ammonia-lyase.
CAB60719.1 AJ250836 Cicer arietinum phenylpropanoid pathway, phenylalanine ammonia-lyase. pal.
CAA68256.1 X99997 Bromheadia finlaysoniana phenylalanine ammonia-lyase. pal.
AAK15640.1 AF326116 Agastache ragosa phenylalanine ammonia-lyase. PAL.
CAA34226.1 XI 6099 Oryza sativa subsp. japonica phenylalanine ammonia-lyase.
AAF40223.1 AF237954 Rubus idaeus phenylalanine ammonia-lyase 1. PALI. PAL; phenylpropanoid; multigene; flavonoid.
BAA11459.1 D78640 Ipomoea batatas
Phenylalanine Ammonia-Lyase.
BAA06337.1 D30657 Populus kitakamiensis phenylalanine ammonia-lyase.
AAD45384.1 AFl 65998 Vigna unguiculata phenylalanine ammonia-lyase.
CAA53733.1 X76130 Cucumis melo phenylanaline ammonia-lyase. pal.
AAA51873.1 U16130 Persea americana phenylalanine ammonia lyase. PAL.
BAB19128.1 AB041361 Dianthus caryophyllus phenylalanine ammonia-lyase. Dcpall.
CAA34715.1 XI 6772 Petroselinum crispum phenylalanine ammonia-lyase (AA 137 - 716) (1 is 3rd base in codon).
BAA07861.1 D43803 Populus kitakamiensis phenylalanine ammonia-lyase.
SEQ ID NO: 280
AAG43550.1 AF211532 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 132. ACREl 32. similar to RING finger proteins.
AAK00436.1 AC060755 Oryza sativa putative zinc finger protein. OSJNBaOQ03O19.23.
BAA78746.1 AB023482 Oryza sativa
Similar to Arabidopsis thaliana RING-H2 finger protein RHXla mRNA, partial cds.(AF079184).
CAA74911.1 Y14573 Hordeum vulgare ring finger protein, putative.
AAG46117.1 AC073166 Oryza sativa putative ring finger protein. OSJNBb0064P21.7.
BAA96875.1 AB045121 Oryza sativa RING finger 1. RRF1.
BAA90357.1 AP001080 Oryza sativa
EST AU070319(S 10788) conesponds to a region ofthe predicted gene. Similar to RING-H2 finger protein RHA2b (AC006200).
BAA90806.1 AP001168 Oryza sativa
ESTs C26000(C11448),AU082130(C11448) conespond to a region ofthe predicted gene.;
Similar to mRNA for zinc-finger protein (Z36749).
SEQ ID NO: 286
AAG14454.1 AF283706 Tulipa gesneriana auxin-induced protein TGSAUR12. SAUR12. small auxin upregulated RNA.
AAG14456.1 AF283708 Tulipa gesneriana auxin-induced protein TGSAUR22. SAUR22. small auxin upregulated RNA.
AAG14455.1 AF283707 Tulipa gesneriana auxin-induced protein TGSAUR21. SAUR21. small auxin upregulated RNA.
AAC08401.1 AF053564 Mesembryanthemum crystallinum auxin-induced protein, similar to auxin-induced proteins from soybean.
SEQ ID NO: 290
AAB65498.1 U73856 Chlamydomonas reinhardtii carbonic anhydrase, alpha type. CAH3.
AAC49983.1 U40871 Chlamydomonas reinhardtii intracellular carbonic anhydrase, alpha type. CAH3.
AAF04292.2 AF190735 Dunaliella salina carbonic anhydrase. CA.
AAC49378.1 U53811 Dunaliella salina carbonic anhydrase. dca.
AAF22644.1 AFl 83939 Dunaliella salina duplicated carbonic anhydrase. DCAl . DCA; carbonic anhydrase gene family member; salt- inducible; intra-duplicated.
AAD51633.1 AF170173 Acetabularia acetabulum putative carbonic anhydrase 2. CA2. AaCA2.
AAD51634.1 AFl 70174 Acetabularia acetabulum putative carbonic anhydrase 1. CAL AaCAl.
AAD51635.1 AFl 70175 Acetabularia acetabulum putative carbonic anhydrase 1. CAL AaCAl.
BAA14232.1 D90206 Chlamydomonas reinhardtii carbonic anhydrase.
BAA28217.1 AB013804 Chlorella sorokiniana soluble carbonic anhydrase precursor. CAHl.
SEQ ID NO: 301
AAG03089.2 AC073405 Oryza sativa similar to an Arabidopsis putative P-type transporting ATPase (ACO 10926).
BAA89544.1 AP001072 Oryza sativa
Similar to chromaffm granule ATPase II homolog. (U75321).
BAA88191.1 AP000836 Oryza sativa
Similar to chromaffm granule ATPase II homolog (U75321).
BAA90510.2 AP001111 Oryza sativa rice EST AU030811, similar to rice Ca+2- ATPase (U82966).
AADl 1618.1 AF050496 Lycopersicon esculentum Ca2+- ATPase. LCA1B; alternative transcript.
AAA34138.1 M96324 Lycopersicon esculentum
The calcium ATPase is a calcium ion pump. Ca2+-ATPase. LCAl .
AADl 1617.1 AF050495 Lycopersicon esculentum Ca2+-ATPase. LCAl A; alternative transcript.
AAF73985.1 AF096871 Zea mays calcium pump, calcium ATPase. cap 1.
AAD31896.1 AF145478 Mesembryanthemum crystallinum calcium ATPase.
AAG28436.1 AF195029 Glycine max plasma membrane Ca2+-ATPase. SCA2.
CAA63790.1 X93592 Dunaliella bioculata P-type ATPase. cal. calcium pumping; CAL
AAG28435.1 AF195028 Glycine max plasma membrane Ca2+- ATPase. SCA1.
CAA68234.1 X99972 Brassica oleracea calmodulin-stimulated calcium-ATPase.
AAB58910.1 U82966 Oryza sativa Ca2+-ATPase.
CAB69824.1 AJ271439 Prunus persica plasma membrane H+ ATPase. PPA1.
AAB60276.1 U09989 Zea mays H(+)-transporting ATPase. Mhal .
BAA01058.1 D10207 Oryza sativa H-ATPase. OSAl.
CAC29435.1 AJ310523 Vicia faba
P-type H+-ATPase. vha4. predominantly expressed in flowers.
AAD20330.1 AFl 10268 Oryza sativa plasma membrane proton- ATPase gene OSA3.
AAA34098.1 M80490 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pma3.
AAB84203.1 AF029257 Kosteletzkya virginica plasma membrane H+-ATPase.
CAC28224.1 AJ286749 Sesbania rostrata p-type H+- ATPase. ha5.
AAD46188.1 AF156691 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma9.
AAA34173.1 M60166 Lycopersicon esculentum H+-ATPase. LHAl.
AAA34094.1 M80489 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pmal .
AAA34052.1 M27888 Nicotiana plumbaginifolia H+-translocating ATPase.
CAC28221.1 AJ286746 Sesbania rostrata p-type H+-ATPase. ha2.
CAA54045.1 X76535 Solanum tuberosum H(+)-transporting ATPase. PHA2.
BAA06629.1 D31843 Oryza sativa plasma membrane H+- ATPase. OSA2.
CAA64406.1 X94936 Phaseolus vulgaris H(+)-transporting ATPase. BHA-2.
AAF98344.1 AF275745 Lycopersicon esculentum plasma membrane H+- ATPase. LHA2. P-type ion pump.
AAD55399.1 AFl 79442 Lycopersicon esculentum plasma membrane H+-ATPase isoform LHA2. LHA2.
CAA54046.1 X76536 Solanum tuberosum H(+)-transρorting ATPase. PHAL
SEQ ID NO: 302
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71A6.
CAA50312.1 X70981 Solanum melongena P450 hydroxylase. CYPEG2.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
BAA03635.1 D14990 Solanum melongena Cytochrome P-450EG4.
CAA50645.1 X71654 Solanum melongena P450 hydroxylase.
CAA83941.1 Z33875 Mentha x piperita cytochrome P-450 oxidase.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
AAB94584.1 AF022157 Glycine max capable of catalyzing the metabolism of phenylurea herbicides. CYP71A10. CYP71A10. cytochrome P450 monooxygenase.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
AAB69644.1 AF000403 Lotus japonicus putative cytochrome P450. LJNP450.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
AAD47832.1 AF166332 Nicotiana tabacum cytochrome P450.
AAB61964.1 U48434 Solanum chacoense putative cytochrome P450.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
CAA71514.1 Y10490 Glycine max putative cytochrome P450.
AAB94588.1 AF022459 Glycine max
CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
AAF27282.1 AF122821 Capsicum annuum cytochrome P450. PepCYP.
CAA50313.1 X70982 Solanum melongena P450 hydroxylase. CYPEG3.
AAA19701.1 L24438 Thlaspi arvense cytochrome P450.
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
AAK38082.1 AF321858 Lolium rigidum putative cytochrome P450.
BAB40322.1 AB036772 Triticum aestivum cytochrome P450. N-l.
CAA71516.1 Y10492 Glycine max putative cytochrome P450.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
AAK38083.1 AF321859 Lolium rigidum putative cytochrome P450.
AAK38084.1 AF321860 Lolium rigidum putative cytochrome P450.
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
CAA72196.1 Y11368 Zea mays cytochrome p450. cyp71c4
CAA57425.1 X81831 Zea mays cytochrome P450. CYP71C4. family CYP71, subfamily CYP71C.
AAC39318.1 AF029858 Sorghum bicolor second multifunctional cytochrome P450 in the biosynthetic pathway ofthe cyanogenic glucoside dhunin. Catalyzes the conversion of p-hydroxyphenylacetaldoxime to p- hydroxymandelonitrile. cytochrome P450 CYP71E1. CYP71E1. No EST#s identified. AAD56282.1 AFl 55332 Petunia x hybrida flavonoid 3 '-hydroxylase. Htl. cytochrome P450; CYP75B2.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
AAK38087.1 AF321863 Lolium rigidum putative cytochrome P450.
CAA57421.1 X81827 Zea mays cytochrome P450. CYP71C1. family CYP71, subfamily CYP71C.
CAA57422.1 X81828 Zea mays cytochrome P450. CYP71C1. family CYP71, subfamily CYP71C.
BAB39252.1 AP002968 Oryza sativa putative cytochrome P450. P0416G11.1.
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
AAB94589.1 AF022460 Glycine max
CYP83Dlp. CYP83D1. cytochrome P450 monooxygenase.
AAD38930.1 AF135485 Glycine max cytochrome P450 monooxygenaseCYP93Dl. CYP93E1.
CAA57423.1 X81829 Zea mays cytochrome P450. CYP71C2. family CYP71, subfamily CYP71C.
CAA72208.1 Y11404 Zea mays cytochrome p450. cyp71c2.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
AAK38088.1 AF321864 Lolium rigidum putative cytochrome P450.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
SEQ ID NO: 303
AAC49826.1 U71604 Catharanthus roseus involved in the second to last step in vindoline biosynthesis, desacetoxyvindoline 4- hydroxylase. CRSD4H. 2-oxoglutarate dependent dioxygenase.
AAB97311.1 AF008597 Catharanthus roseus desacetoxyvindoline-4-hydroxylase. CRSD4H. 2-oxoglutarate dependent dioxygenase; involved in the second to last step in vindoline biosynthesis.
AAC49827.1 U71605 Catharanthus roseus involved in the second to last step in vindoline biosynthesis, desacetoxyvindoline 4- hydroxylase. CRSD4H. 2-oxoglutarate dependent dioxygenase.
BAA95828.1 AP002069 Oryza sativa
ESTs D47168(S12332),D46350(S10967) conespond to a region of he predicted gene. Similar to Prunus armeniaca ethylene-forrning-enzyme-like dioxygenase. (U97530). BAA37127.1 AB012203 Lactuca sativa
2-oxoglutarate-dependent dioxygenase. gibberelin 20-oxidase. Ls20oxl.
CAA54557.1 X77368 Solanum melongena dioxygenase. DIOX.
BAA81862.1 AB026295 Oryza sativa
Similar to leucoanthocyanidin dioxygenase.(AI440611). SEQ ID NO: 304
AAF61647.1 AF190634 Nicotiana tabacum
UDP-glucose: salicylic acid glucosyltransferase. SA-GTase.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltransferase. PHI.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitrile-o- glucosyltiansferase.
AAF98390.1 AF287143 Brassica napus catalyzes the transfer of glucose from UDP-glucose to sinapate and some other hydroxy cinnamates (4-coumarate, caffeate, feralate). UDP-glucose: sinapate glucosyltransferase. SGT1. SGT.
BAA36423.1 AB013598 Verbena x hybrida
UDP-glucose:anthocyanin 5-O-glucosyltransferase. HGT8.
AAK16181.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.16.
BAA36421.1 AB013596 Perilla frutescens
UDP-glucose:anthocysnin 5-O-glucosyltransferase. PF3R4.
BAB07962.1 AP002524 Oryza sativa putative anthocyanin 5-O-glucosyltransferase. P0406H10.16. contains ESTs
AU067881(C10481),AU067882(C10481).
AAK16178.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.5.
BAA93039.1 AB033758 Citras unshiu limonoid UDP -glucosyltransferase. LGTase.
AAK16172.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
BAA36422.1 AB013597 Perilla frutescens
UDP-glucose: anthocyanin 5-O-glucosyltransferase homologue. PF3R6.
BAB17182.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.19.
BAA12737.1 D85186 Gentiana triflora
UDP-glucose:flavonoid-3-glucosyltransferase.
AAG25643.1 AF303396 Phaseolus vulgaris
UDP-glucosyltransferase HRA25. putative; defense associated.
AAB48444.1 U82367 Solanum tuberosum UDP-glucose glucosyltransferase. BAA19659.1 AB002818 Perilla frutescens flavonoid 3-O-glucosyltransferase. UDP glucose.
CAA54612.1 X77462 Manihot esculenta
UTP-glucose glucosyltransferase. CGT5.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltransferase. PGT8.
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
BAA83484.1 AB031274 Scutellaria baicalensis
UDP-glucose: flavonoid 7-O-glucosyltransferase. ufgt.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS 10a.
AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid: glucosyltransferase 2. togtl. glucosyltransferase.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS5a.
AAD04166.1 AF101972 Phaseolus lunatus catalyzes formation of O-glucosylzeatin from zeatin and UDPG; catalyzes formation of O- xylosylzeatin from zeatin and UDPX. zeatin O-glucosyltiansferase. ZOG1. cytolάnin O- glucosyltransferase.
AAD21086.1 AF127218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
CAA54611.1 X77461 Manihot esculenta UTP-glucose glucosyltransferase. CGT2.
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid:glucosyltransferase 1. togtl. glucosyltransferase.
CAA54609.1 X77459 Manihot esculenta UTP-glucose glucosyltransferase. CGT1.
CAA54613.1 X77463 Manihot esculenta UTP-glucose glucosyltransferase. CGT6.
BAB41026.1 AB047099 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. F1UFGT2.
BAB41024.1 AB047097 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT2.
BAB41022.1 AB047095 Vitis vinifera UDP-glucose.-flavonoid 3-O-glucosyltransferase. RuUFGT2.
BAB41017.1 AB047090 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. KyUFGTl. The gene was derived from one ofthe parents V. labruscana cv. Ishiharawase.
BAB41020.1 AB047093 Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. I.UFGT2.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltransferases.
BAB41025.1 AB047098 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. F1UFGT1.
BAB41023.1 AB047096 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. AlUFGTl.
B AB41018.1 AB047091 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. KyUFGT2. The gene originated in one ofthe parents V. vinifera cv. Centennial.
SEQ ID NO: 306
AAB06458.1 U64806 Brassica napus pathogenesis-related protein PRl . Yprl .
AAB01666.1 U21849 Brassica napus PR-la. LSC94.
AAB09587.1 U70666 Brassica napus pathogenesis-related protein PRl. Yprl.
CAA47374.1 X66942 Nicotiana tabacum prb-lb. PRB-1B.
AAK30143.1 AF348141 Capsicum annuum pathogenesis-related protein PR-1 precursor.
CAA36790.1 X52555 Nicotiana tabacum PR-1 protein (AA 1-184).
CAA35666.1 X17681 Nicotiana tabacum pathogenesis-related protein lb (AA 1-168).
CAA31010.1 X12487 Nicotiana tabacum PRlc preprotein.
CAA29023.1 X05454 Nicotiana tabacum PR- 1 c protein.
CAA32228.1 X14065 Nicotiana tabacum PRP 1 precursor (AA -23 to 154).
BAA14220.1 D90196 Nicotiana tabacum PR la protein precursor.
CAA31233.1 X12737 Nicotiana tabacum PR-la protein (AA 1 - 168).
CAA29392.1 X05959 Nicotiana tabacum PR- la precursor (AA -30 to 138).
CAA29660.1 X06361 Nicotiana tabacum PRl a precursor (AA -30 to -1).
CAA09671.1 A JO 11520 Lycopersicon esculentum pathogenesis-related protein PRla (P4). prla (P4).
AAA03615.1 M69247 Lycopersicon esculentum pathogenesis-related protein P4. P4.
CAA30017.1 X06930 Nicotiana tabacum PR-la protein (AA 1 - 168).
CAA31008.1 X12485 Nicotiana tabacum PRla preprotein.
CAA52893.1 X74939 Hordeum vulgare PR- la pathogenesis related protein (Hv-la).
CAB58263.1 AJ250136 Solanum tuberosum pathogenesis related protein PR-1. ρrl-1.
AAB49685.1 U89895 Oryza sativa pathogenesis-related protein class 1. PR-1. induced by pathogen attack in plants.
CAA27183.1 X03465 Nicotiana tabacum
PR- lb precursor; (aa -30-138).
CAA35665.1 X17680 Nicotiana tabacum pathogenesis-related protein lb (AA 1-168).
BAA14221.1 D90197 Nicotiana tabacum PRlb protein precursor.
CAA48672.1 X68738 Lycopersicon esculentum Pl(pl4) protein. pTE28.1.
CAA81229.1 Z26320 Hordeum vulgare pathogenesis-related protein, pathogenesis-related protein.
AAA03616.1 M69248 Lycopersicon esculentum pathogenesis-related protein P6. P6.
CAA70042.1 Y08804 Lycopersicon esculentum PR protein. PRlbl.
AAB05225.1 U49241 Nicotiana glutinosa pathogenesis-related protein- 1.
CAA31009.1 X12486 Nicotiana tabacum PRlb preprotein.
AAC25629.1 U82200 Zea mays pathogenesis related protein- 1. PR-1.
AAF78528.1 AF195237 Pyras pyrifolia pathogenesis-related protein. PR- lb.
AAD33696.1 AFl 36636 Glycine max PRla precursor. PRla.
CAA79703.1 Z21494 Hordeum vulgare Pathogenesis-related protein 1.
CAA52894.1 X74940 Hordeum vulgare PR-lb pathogenesis related protein (Hv-8).
CAA81234.1 Z26333 Hordeum vulgare pathogenesis-related protein, pathogenesis-related protein.
CAA81230.1 Z26321 Hordeum vulgare pathogenesis-related protein, pathogenesis-related protein.
CAA04881.1 AJ001627 Lycopersicon esculentum pathogenesis-related protein. PRld.
CAA07473.1 AJ007348 Triticum aestivum pathogenisis-related protein 1.1. PR-1.1.
CAA07474.1 AJ007349 Triticum aestivum pathogenisis-related protein 1.2. PR-1.2.
CAA50596.1 X71592 Lycopersicon esculentum PR-lal.
CAA70070.1 Y08844 Lycopersicon esculentum PR protein. PRl a2.
CAA38223.1 X54325 Zea mays pathogenesis-related protein. PRms
AAF78527.1 AF195236 Pyras pyrifolia pathogenesis-related proteins. PR- la.
CAC03571.1 AJ278436 Oryza sativa defence response. PRla protein. Prla.
AAG44566.1 AF251277 Oryza sativa subsp. japonica acidic PR-1 type pathogenesis-related protein PR- la. PR- la. induced by pathogen attack.
AAC06244.1 AF053343 Capsicum annuum
PR-1 protein precursor, pathogen-induced PRl protein.
CAA56174.1 X79778 Medicago truncatula PR-1.
SEQ ID NO: 307
AAF06347.1 AF195654 Vitis vinifera SCUTL2. thaumatin-like protein.
BAA28872.1 AB006009 Pyras pyrifolia thaumatin-like protein precursor. PsTLl.
AAB38064.1 U32440 Prunus avium thaumatin-like protein precursor.
BAA95017.1 AB031870 Cestram elegans thaumatin-like protein. CETLP.
BAA74546.2 AB000834 Nicotiana tabacum thaumatin-like protein SE39b.
AAC36740.1 AF090143 Malus x domestica thaumatin-like protein precursor Mdtll . MDTLl . pathogenesis-related.
CAC10270.1 AJ243427 Malus x domestica thaumatin-like protein, tl. allergen, pathogenesis-related.
AAB95118.1 U71244 Brassica rapa pathogenesis-related group 5 protein. BFTP. thaumatin-like protein; PR-5.
CAC09477.1 AL442113 Oryza sativa thaumatin-like protein. H0806H05.10.
CAB62167.1 AJ242828 Castanea sativa antifungal. thaumatin-like protein, til.
CAA06927.1 AJ006233 Nicotiana tabacum putative thaumatin-like protein precursor.
AAF06346.1 AF195653 Vitis vinifera SCUTL1. thaumatin-like protein.
AAB02259.1 U57787 Avena sativa permatin precursor, thaumatin-like protein.
AAD55090.1 AF178653 Vitis riparia thaumatin. osmotin; pathogenesis-related protein.
CAA10492.1 AJ131731 Pseudotsuga menziesii Thaumatin-like protein. 5A1A.16.
CAA09228.1 AJ010501 Cicer arietinum thaumatin-like protein PR-5b.
BAA95165.1 AB029918 Nicotiana tabacum pistil transmitting tissue specific thaumatin (SE39b)-like protein. SE39b.
AAF82264.1 AF227324 Vitis vinifera thaumatin-like protein.
CAB85636.1 AJ237998 Vitis vinifera putative thaumatin-like protein. T12.
AAB53368.1 U77657 Oryza sativa pathogenesis-related thaumatin-like protein.
AAB61590.1 AF003007 Vitis vinifera
WTLl. osmotin-like protein, PR-5 protein; thaumatin-like protein.
AAB53367.1 U77656 Oryza sativa pathogenesis-related thaumatin-like protein.
CAB85637.1 AJ237999 Vitis vinifera putative thaumatin-like protein. Til. alternative name grip 51.
SEQ ID NO: 308
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
BAA14144.1 D90116 Armoracia rusticana peroxidase isozyme.
BAA11853.1 D83225 Populus nigra peroxidase.
CAA66035.1 X97349 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
CAA66036.1 X97350 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
BAA11852.1 D83224 Populus nigra peroxidase.
CAA66034.1 X97348 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
CAA66037.1 X97351 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
AAB47602.1 L07554 Linum usitatissimum peroxidase. FLXPER1.
BAA06334.1 D30652 Populus kitakamiensis peroxidase.
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
AAB97734.1 AF014502 Glycine max seed coat peroxidase precursor. Ep. H2O2 oxidoreductase; class III plant peroxidase.
BAA06335.1 D30653 Populus kitakamiensis peroxidase.
AAD37427.1 AF149277 Phaseolus vulgaris peroxidase 1 precursor. FBP1. secretory peroxidase.
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
CAA62227.1 X90694 Medicago sativa peroxidase IC. prxlC.
AAB41811.1 L36157 Medicago sativa peroxidase. pxdC. amino acid feature: conserved domains, aa 123 .. 129, 191 .. 198; amino acid feature: heme-binding domain, aa 68 .. 73.
CAA62225.1 X90692 Medicago sativa peroxidaselA. prxlA.
AAB41810.1 L36156 Medicago sativa peroxidase. pxdA. amino acid feature: conserved motifs, aa 181 .. 188; amino acid feature: heme-binding domain, aa 60 .. 65.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
AAA34108.1 J02979 Nicotiana tabacum lignin-forming peroxidase precursor (EC 1.11.1.7).
CAA40796.1 X57564 Armoracia rusticana peroxidase. peroxidase precursor.
BAA01877.1 D11102 Populus kitakamiensis peroxidase. prxAl.
BAA01992.1 D11396 Nicotiana tabacum 'peroxidase'.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI-1.
AAD43561.1 AF155124 Gossypium hirsutum bacterial-induced peroxidase precursor. Perx_Goshiko.
BAA92500.1 AP001383 Oryza sativa
ESTs D39300(R3292),AU030751(E60187) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP6a. (X98774).
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 5 precursor, type III peroxidase.
AAB02554.1 L37790 Stylosanthes humilis cationic peroxidase.
AAB06183.1 M37636 Arachis hypogaea cationic peroxidase. PNC
CAA59487.1 X85230 Triticum aestivum peroxidase. pox4.
BAA94962.1 AB042103 Asparagus officinalis peroxidase. AspPOXl.
AAF63026.1 AF244923 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl4 precursor, type III peroxidase.
CAB99487.1 AJ276227 Hordeum vulgare defence against plant pathogens, peroxidase. prx8.
CAA71492.1 Y10466 Spinacia oleracea peroxidase. prxr5.
CAB65334.1 AJ250121 Picea abies peroxidase. SPI2 protein. spi2.
AAA33121.1 M32742 Cucumis sativus peroxidase (CuPer2).
CAA39486.1 X56011 Triticum aestivum peroxidase.
BAA92422.1 AP001366 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to A.thaliana mRNA for peroxidase ATP18a. (X98804).
BAA92497.1 AP001383 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP18a. (X98804).
CAA59485.1 X85228 Triticum aestivum peroxidase. POX2.
CAA76680.1 Y17192 Cucurbita pepo peroxidase. aprx. type III peroxidase.
SEQ ID NO: 309
BAA85400.1 AP000615 Oryza sativa similar to OsMlo-hl. (Z95353).
CAB06083.1 Z83834 Hordeum vulgare Mlo. Mlo.
CAA74909.1 Y14573 Hordeum vulgare Mlo protein. Mlo.
AAG46114.1 AC073166 Oryza sativa putative Mlo (pathogen resistance) protein. OSJNBb0064P21.5.
CAA06487.1 AJ005341 Linum usitatissimum MLO. homolog.
SEQ ID NO: 310
AAC63113.1 AF000307 Brassica napus steroid sulfotransferase 3. BnST3.
AAC63112.1 AF000306 Brassica napus steroid sulfotransferase 2. BnST2.
AAC63111.1 AF000305 Brassica napus steroid sulfotransferase 1. BnSTl.
AAA61638.1 U10275 Flaveria bidentis
O-sulfation of position 3 of flavonols. flavonol 3-sulfotransferase.
AAA33342.2 M84135 Flaveria chloraefolia flavonol 3-sulfotransferase.
AAA87399.1 U10277 Flaveria bidentis transfers sulfate group into flavonol. sulfotiansferase-like flavonol.
AAA33343.1 M84136 Flaveria chloraefolia
O-sulfation of position 4' of flavonol. flavonol 4'-sulfotransferase. SEQ ID NO: 312
AAD22970.1 AF124148 Glycine max trehalase 1 GMTREl. expressed constitutively in many tissues of soybean at a low level; similar to the Arabidopsis thaliana trehalase precursor encoded by GenBank Accession Number AC002343.
AAG13442.1 AC051634 Oryza sativa putative trehalase. OSJNBb0018B10.19.
CAB50901.1 AJ238651 Medicago truncatula trehalase. TRE1 protein, trel.
SEQ ID NO: 313
BAA19928.1 AB003491 Oryza sativa tryptophan synthase B. tipB.
AAA33491.1 M76685 Zea mays tryptophan synthase beta-subunit. TSB2.
AAB97526.1 AF042321 Camptotheca acuminata tryptophan synthase beta. TSB.
AAB97087.1 AF042320 Camptotheca acuminata tryptophan synthase beta subunit
AAA33490.1 M76684 Zea mays tryptophan synthase beta-subunit. TSB 1.
AAC25986.1 AF047024 Chlamydomonas reinhardtii tryptophan synthase beta. MAA7.
SEQ ID NO: 314
AAG42689.1 AF271384 Zea mays putative tryptophan synthase alpha. TSAlike.
CAA54131.1 X76713 Zeamays tryptophan synthase, alpha subunit. tφA.
AAG42688.1 AF271383 Zeamays putative tryptophan synthase alpha. TSAlike.
SEQ ID NO: 315
AAB97526.1 AF042321 Camptotheca acuminata tryptophan synthase beta. TSB.
AAB97087.1 AF042320 Camptotheca acuminata tryptophan synthase beta subunit.
BAA19928.1 AB003491 Oryza sativa tryptophan synthase B. tφB.
AAA33491.1 M76685 Zeamays tryptophan synthase beta-subunit. TSB2.
AAA33490.1 M76684 Zeamays tryptophan synthase beta-subunit. TSB1.
AAC25986.1 AF047024 Chlamydomonas reinhardtii tryptophan synthase beta. MAA7.
SEQ ID NO: 316
AAA33967.1 M76981 Glycine max vegetative storage protein. vspA.
BAA23563.1 D50094 Phaseolus vulgaris pod storage protein.
BAA19152.1 AB000585 Phaseolus vulgaris pod storage protein. PSP.
AAA34020.1 M20037 Glycine max vegetative storage protein.
AAA34022.1 M76980 Glycine max vegetative storage protein. vspB.
AAA34021.1 M20038 Glycine max vegetative storage protein.
SEQ ID NO: 321
BAA87043.1 AB035183 Ipomoea batatas N-hydroxy cinnamoyl/benzoyltransferase . hcbt.
CAB06427.1 Z84383 Dianthus caryophyllus phytoalexin biosynthesis, anthranilate N-hydroxycinnamoyl/benzoyltransferase.
CAB06429.1 Z84385 Dianthus caryophyllus phytoalexin biosynthesis, anthranilate N-hydroxycinnamoyl/benzoyltiansferase.
CAB06430.1 Z84386 Dianthus caryophyllus phytoalexin biosynthesis, anthranilate N-hydroxy cinnamoyl/benzoyltransferase.
CAB11466.1 Z98758 Dianthus caryophyllus carnation phytoalexin biosynthesis, anthranilate N-hydroxycinnamoyl/benzoyltransferase.
CAB06538.1 Z84571 Dianthus caryophyllus phytoalexin biosynthesis, anthranilate N-hy(koxycinnamoyl/benzoyltransferase.
CAB06428.1 Z84384 Dianthus caryophyllus phytoalexin biosynthesis, anthranilate N-hydroxycinnamoyl/benzoyltransferase. SEQ ID NO: 323
BAB12694.1 AP002746 Oryza sativa putative zinc finger transcription factor. P0671B11.10. contains ESTs
AU098331(E31537),C91783(E31537).
SEQ ID NO: 325
BAB03447.1 AP002817 Oryza sativa
ESTs C96615(C10106),C26336(C12127),D21959(C10106) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana DNA chromosome 4, BAC clone T6K21;
NAM (no apical meristem) - like protein (AL021889).
BAA92400.1 AP001366 Oryza sativa
ESTs C96615(C10106),C26336(C12127),D21959(C10106) conespond to a region ofthe predicted gene. Similar to NAM (AL021889).
BAB16335.1 AP002818 Oryza sativa putative NAM protein. P0436E04.18. contains ESTs E10793(C19698),E10793(C99379).
BAB 16328.1 AP002818 Oryza sativa putative NAM protein. P0436E04.il. contains ESTs R4069(AU032425),R4069(AU082730).
CAA63102.2 X92205 Petunia x hybrida apical meristem formation. NAM.
CAA63101.1 X92204 Petunia x hybrida apical meristem formation. NAM.
AAK13151.1 AC078829 Oryza sativa putative NAM (no apical meristem) protein. OSJNBa0026O12.6.
BAB19365.1 AP002542 Oryza sativa putative NAM (no apical meristem) protein. P0679C08.4.
BAA84803.1 AP000559 Oryza sativa
Similar to NAM like protein (AC005310). SEQ ID NO: 326
CAB51836.1 AJ243961 Oryza sativa
Putitive Ser/Thr protein kinase. 11332.7.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
BAA92836.1 AB032473 Brassica oleracea
S18 S-locus receptor kinase. SRK18.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl .
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase. AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
BAA21132.1 D88193 Brassica rapa
S-receptor kinase. SRK9 (B.c).
BAA06285.1 D30049 Brassica rapa
S-receptor kinase SRK9.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
BAB 16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
CAB41879.1 Y18260 Brassica oleracea
SRK15 protein. SRK15. receptor-like kinase.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein kinase.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214). SEQ ID NO: 327
AAC04717.1 AF034131 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-G. similar to MYB A encoded by GenBank Accession Number L04497.
BAA23340.1 D88620 Oryza sativa transfactor. OSMYB4. Osmyb4.
AAK19611.1 AF336278 Gossypium hirsutum BNLGHi233. bnlghi6233. similar to myb.
AAA33482.1 M37153 Zeamays cl locus myb homologue; putative.
AAK09326.1 AF320613 Zea mays activates anthocyanin transcription, anthocyanin regulatory Cl . cl . transcription factor.
AAK09327.1 AF320614 Zeamays activates anthocyanin transcription, anthocyanin regulatory Cl. cl. transcription factor.
AAA82943.1 U39448 Piceamariana
MYB-like transcriptional factor MBFl . putative DNA binding region highly similar to the maize Cl.
AAK19618.1 AF336285 Gossypium hirsutum GHMYB38. ghmyb38. similar to myb.
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related.
AAK19619.1 AF336286 Gossypium hirsutum GHMYB9. ghmyb9. similar to myb.
AAC04720.1 AF034134 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-O. similar to MYB A encoded by GenBank Accession Number L04497.
AAC04718.1 AF034132 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-J. similar to MYB A encoded by GenBank Accession Number L04497. SEQ ID NO: 329
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal.
AAF76898.1 AF274033 Atriplex hortensis apetala2 domain-containing protein.
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREL similar to EREBP transcription factors.
CAC 12822.1 AJ299252 Nicotiana tabacum
AP2 domam-containing transcription factor. ap2.
BAA78738.1 AB023482 Oryza sativa
EST AU055776(S20048) conesponds to a region ofthe predicted gene.; Similar to
Arabidopsis thaliana AP2 domain containing protein RAP2.10 mRNA, partial cds.(AF0031O3).
AAC14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
BAA97122.1 ABO 16264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor. CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2 -related transcription factor. CDBP. stress induced transcription factor.
AAC24587.1 AF071893 Prunus armeniaca
AP2 domain containing protein. AP2DCP.
BAA94514.2 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4, BAC clone F9D16; putative Ap2 domain protein (AL035394).
BAA97124.1 AB016266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
BAA07321.1 D38123 Nicotiana tabacum ERFl. ethylene-responsive transcription factor.
BAB16083.1 AB036883 Oryza sativa transcriptional repressor. osERF3. osERF3. ERF protein family ERF3 associated repression domain.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3. BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf3. AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like transcription factor.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBPl.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP transcription factors.
BAA90812.1 AP001168 Oryza sativa
Similar to mRNA for DREB1 A (AB007787).
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
BAA99376.1 AP002526 Oryza sativa
ESTs AU093391(E60370),AU091593(C60458), AU093392(E60370) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana BAC F21 J9; AP2 domain protein.
(AC000103).
AAK01089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor.
AAG59618.1 AF239616 Hordeum vulgare
CRT/DRE-bindfng factor. CBF.
AAG59619.1 AF243384 Oryza sativa
CRT/PRE binding factor. CBF. DREB.
AAK01088.1 AF298230 Hordeum vulgare
CBF 1 -like protein BCBF1. BCBF1. AP2 domain protein; putative DRE binding factor.
AAC49567.1 U41466 Zea mays
Glossyl5. Glossyl5. AP2 DNA-binding domain protein; shnilar to the Arabidopsis floral homeotic gene APETALA2; maize AP2-domain regulator of leaf epidermal traits; homeotic regulator of leaf epidermal cell identity; allele: W64A; putative transcription factor. SEQ ID NO: 330
AAC67571.1 AF060884 Brassica napus desiccation protein. Cdes. induced by dehydration.
AAA61564.1 U08108 Glycine max putative desiccation protectant protein, homolog of Leal 4, GenBank Accession Number
M88321.
AAA18543.1 M88322 Gossypium hirsutum probable desiccation protectant. Group 4 late embryogenesis-abundant protein. Leal4-A. This CDS is colinear and 66% identical with that in cDNA clone pcC27-45 from Craterostigma plantagineum desiccated leaves (Piatkowski et al, 1990, Plant Physiol. 94: 1682-1688).; putative.
AAA18542.1 M88321 Gossypium liirsutum probable desiccation protectant. Group 4 late embryogenesis-abundant protein. Leal4-A. putative.
AAD25354.1 AF115314 Glycine max possible desiccation protectant. seed maturation protein PM22. PM22. similar to desiccation protectant protein encoded by GenBank Accession Number U08108; late embryogenesis abundant protein; LEA protein.
AAF64451.1 AF239929 Euphorbia esula late-embryogenesis abundant protein, similar to desiccation protectant protein and late- embryogenesis abundant protein LEA14.
AAB96796.1 U77719 Lycopersicon esculentum ethylene-responsive late embryogenesis-like protein. ER5. LEA-like protein; drought- inducible; ABA-inducible; putative desiccation protectant protein; similar to cotton Leal4A product encoded by GenBank Accession Number M88321.
SEQ ID NO: 345 AAF61647.1 AFl 90634 Nicotiana tabacum UDP-glucose:salicylic acid glucosyltransferase. SA-GTase. BAA36423.1 AB013598 Verbena x hybrida
UDP-glucose: anthocyanin 5-O-glucosyltransferase. HGT8.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltransferase. PHI.
BAA36421.1 AB013596 Perilla frutescens
UDP-glucose:anthocysnin 5-O-glucosyltransferase. PF3R4.
BAA93039.1 AB033758 Citrus unshiu limonoid UDP-glucosyltransferase. LGTase.
BAA36422.1 AB013597 Perilla frutescens
UDP-glucose:anthocyanin 5-O-glucosyltransferase homologue. PF3R6.
AAF98390.1 AF287143 Brassica napus catalyzes the transfer of glucose from UDP-glucose to sinapate and some other hydroxycinnamates (4-coumarate, caffeate, feralate). UDP-glucose:sϊnapate glucosyltransferase. SGT1. SGT.
BAB07962.1 AP002524 Oryza sativa putative anthocyanin 5-O-glucosyltransferase. P0406H10.16. contains ESTs
AU067881(C10481),AU067882(C10481).
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose :p-hydroxymandelonitrile-o- glucosyltiansferase.
AAK16172.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS5a.
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid.-glucosyltransferase 1. togtl. glucosyltransferase.
BAA83484.1 AB031274 Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltransferase. ufgt.
AAK16181.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.16.
AAD21086.1 AFl 27218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltiansferase. UFGT.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS 10a.
AAK28304.1 AF346432 Nicotiana tabacum phenylproρanoid:glucosyltransferase 2. togt2. glucosyltransferase.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltransferases.
AAK16178.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.5.
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
AAK16180.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.21.
CAB56231.1 Y18871 Dorotheanthus bellidiformis betanidin-5-O-glucosyltransferase.
BAA12737.1 D85186 Gentiana triflora UDP-glucose:flavonoid-3-glucosyltransferase.
AAG25643.1 AF303396 Phaseolus vulgaris UDP-glucosyltransferase HRA25. putative; defense associated.
B AB41017.1 AB047090 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltiansferase. KyUFGTl. The gene was derived from one ofthe parents V. labruscana cv. Ishiharawase.
AAB81683.1 AF000372 Vitis vinifera UDP ghιcose:flavonoid 3-o-glucosyltiansferase.
BAB41020.1 AB047093 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. ItUFGT2.
BAB41022.1 AB047095 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. RuUFGT2.
BAB41019.1 AB047092 Vitis vinifera U P-glucose:flavonoid 3-O-glucosyltransferase. ItUFGTl.
AAB81682.1 AF000371 Vitis vinifera UDP glucose: flavonoid 3-o-glucosyltransferase.
BAB41025.1 AB047098 Vitis vinifera
UDP-glucose : flavonoid 3-O-glucosyltransferase. F1UFGT1.
BAB41023.1 AB047096 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT1.
BAB41021.1 AB047094 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. RuUFGTl.
BAB41026.1 AB047099 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. F1UFGT2.
BAB41024.1 AB047097 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT2.
B AB41018.1 AB047091 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltiansferase. KyUFGT2. The gene originated in one ofthe parents V. vinifera cv. Centennial.
BAB17182.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.19.
BAA19659.1 AB002818 Perilla frutescens flavonoid 3 -O-glucosyltransferase. UDP glucose.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltransferase. PGT8.
BAB17176.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.13.
AAB86473.1 AF028237 Ipomoea pmpurea
UDP glucose: flavonoid 3-O-glucosyltransferase. UF3GT-FL1.
SEQ ID NO: 350
CAA44216.1 X62343 Nicotiana tabacum cinnamyl-alcohol dehydrogenase. CAD14.
BAA03099.1 D13991 Aralia cordata cinnamyl alcohol dehydrogenase. cadacl.
CAA79622.1 Z19568 Populus deltoides lignin biosynthesis, cinnamyl alcohol dehydrogenase.
CAC07423.1 AJ295837 Populus balsamifera subsp. trichocaφa lignin monomer biosynthesis, cinnamyl alcohol dehydrogenase. cad.
AAF43140.1 AF217957 Populus tremuloides cinnamyl alcohol dehydrogenase. CAD.
CAA44217.1 X62344 Nicotiana tabacum cinnamyl-alcohol dehydrogenase. CAD19.
CAA79625.1 Z19573 Medicago sativa lignin biosynthesis, cinnamyl alcohol dehydrogenase.
AAC35845.1 AF083332 Medicago sativa cinnamyl-alcohol dehydrogenase. MsaCad2.
AAC07987.1 AF038561 Eucalyptus globulus catalyses the reduction of cinnamaldehydes to the conesponding cinnamyl alcohols as the last step in the production of lignin monomers, cinnamyl alcohol dehydrogenase. CAD.
CAA46585.1 X65631 Eucalyptus gunnii cinnamyl-alcohol dehydrogenase. cad.
AAG15553.1 AF294793 Eucalyptus saligna cinnamyl alcohol dehydrogenase. cad. CAD.
CAA53211.1 X75480 Eucalyptus gunnii cinnamyl-alcohol dehydrogenase. CAD.
AAB70908.1 AF010290 Lolium perenne cinnamyl alcohol dehydrogenase. CAD.
CAA74070.1 Y13733 Zeamays cinnamyl alcohol dehydrogenase. cad.
CAA06687.1 AJ005702 Zea mays cinnamyl alcohol dehydrogenase. cad.
CAA13177.1 AJ231135 Saccharum officfnarum lignin biosynthesis, cinnamyl alcohol dehydrogenase. cad.
BAA19487.1 D86590 Zinnia elegans cinnamyl alcohol dehydrogenase. ZCAD1.
CAA51226.1 X72675 Picea abies cinnamyl-alcohol dehydrogenase.
CAA05097.1 AJ001926 Picea abies cinnamyl alcohol dehydrogenase. cad8.
CAA05096.1 AJ001925 Picea abies cinnamyl alcohol dehydrogenase. cad7.
CAA05095.1 AJ001924 Picea abies cinnamyl alcohol dehydrogenase. cad2.
AAB38774.1 U62394 Pinus radiata cinnamyl alcohol dehydrogenase. CAD.
AAC31166.1 AF060491 Pinus radiata cinnamyl alcohol dehydrogenase. CAD.
CAA86073.1 Z37992 Pinus taeda cinnamyl alcohol dehydrogenase.
CAA86072.1 Z37991 Pinus taeda cinnamyl alcohol dehydrogenase.
BAA04046.1 D16624 Eucalyptus botryoides cinnamyl alcohol dehydrogenase. Cadl:Eb:l.
AAD10327.1 U63534 Fragaria x ananassa catalyzes the reduction of cinnamylaldehydes leading to monolignols. cinnamyl alcohol dehydrogenase. CAD, involved with lignin biosynthesis.
AAK28509.1 AF320110 Fragaria x ananassa cinnamyl alcohol dehydrogenase.
AAB38503.1 U79770 Mesembryanthemum crystallinum cinnamyl-alcohol dehydrogenase Eli3.
AAC35846.1 AF083333 Medicago sativa cinnamyl-alcohol dehydrogenase. MsaCadl .
AAA74882.1 L36823 Stylosanthes humilis cinnamyl-alcohol dehydrogenase. CADI.
AAF23409.1 AF207552 Brassica napus cinnamyl alcohol dehydrogenase. CADa-1.
AAC 15467.1 U24561 Apium graveolens converts mannitol to mannose. mannitol dehydrogenase. Mtd. 1 -oxidoreductase; induced with sodium salicylate; similar to the plant defense gene ELB in Arabidopsis thaliana, PIR Accession Number S28044; EC number unassigned; MTD.
AAF23411.1 AF207554 Brassica oleracea cinnamyl alcohol dehydrogenase. CADa.
AAF23412.1 AF207555 Brassica rapa cinnamyl alcohol dehydrogenase. CADa.
AAC61854.1 AF067082 Apium graveolens oxidizes mannitol to mannose. mannitol dehydrogenase. Mtd. mannitol 1 -oxidoreductase.
AAF23410.1 AF207553 Brassica napus cinnamyl alcohol dehydrogenase. CADa-2.
AAA74883.1 L36456 Stylosanthes humilis cinnamyl-alcohol dehydrogenase. CAD3.
AAD18000.1 AF109157 Eucalyptus globulus cinnamyl alcohol dehydrogenase. CAD.
AAF72100.1 AF146691 Lycopersicon esculentum ELB. Eli3. similar] to cinnamyl alcohol dehydrogenase.
CAA63410.1 X92754 Hordeum vulgare cinnamyl alcohol dehydrogenase. CAD.
AAF23416.1 AF207559 Brassica rapa cinnamyl alcohol dehydrogenase. CADb.
SEQ ID NO: 351
AAB71227.1 AF004809 Glycine max
Ca+2-binding EF hand protein. GmPM13. encodes EF-hand motifs.
AAF13743.1 AF109921 Sesamum indicum caleosin. 27 kDa calcium-binding protein.
CAA61981.1 X89891 Oryza sativa EFA27 for EF hand, abscisic acid, 27kD. efa27.
CAB71337.1 AJ250283 Hordeum vulgare putative calcium binding EF-hand protein, bci-4.
CAB42585.1 AJ238627 Chlorella protothecoides putative Ca++ binding protein. dee!12.
SEQ ID NO: 353
AAF60316.1 AF236108 Glycine max putative puφle acid phosphatase precursor. PAP. type 5 acid phosphatase.
AAF60315.1 AF236107 Ipomoea batatas putative puφle acid phosphatase precursor. PAP, type 5 acid phosphatase.
AAF60317.1 AF236109 Phaseolus vulgaris putative puφle acid phosphatase precursor. PAP, type 5 acid phosphatase. SEQ ID NO: 359
CAA55039.1 X78203 Hyoscyamus muticus glutathione transferase.
AAB65163.1 AF002692 Solanum commersonii glutathione S-transferase, class-phi. GSTl. low temperature induced.
CAA96431.1 Z71749 Nicotiana plumbaginifolia glutathione S-tiansferase.
BAA01394.1 D10524 Nicotiana tabacum glutathione S-tiansferase. parB.
AAA33930.1 M84968 Silene vulgaris glutathione-S-transferase.
AAA33931.1 M84969 Silene vulgaris glutathione-S-transferase.
AAF65767.1 AF242309 Euphorbia esula glutathione S-transferase. putative auxin-binding GST.
AAF61392.1 AFl 33894 Persea americana glutathione S-transferase. GTH.
CAB38119.1 AJ010296 Zea mays Glutathione transferase 111(b). gst3b. CAB38118.1 AJ010295 Zeamays
Glutathione transferase IΙI(a). gst3a.
BAB39935.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.17.
AAG34811.1 AF243376 Glycine max glutathione S-transferase GST 21.
BAB39941.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.23.
CAA09190.1 AJ010451 Alopecurus myosuroides glutathione transferase. GST2a.
CAA09192.1 AJ010453 Alopecurus myosuroides glutathione transferase. GST2c.
CAA09193.1 AJO 10454 Alopecurus myosuroides glutathione transferase. GST2d.
AAG34814.1 AF243379 Glycine max glutathione S-transferase GST 24.
CAA09191.1 AJO 10452 Alopecurus myosuroides glutathione transferase. GST2b.
BAB39939.1 AP002914 Oryza sativa putative glutathione S-tiansferase. P0493G01.21.
AAG32476.1 AF309383 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF4.
AAG34812.1 AF243377 Glycine max glutathione S-tiansferase GST 22.
BAB39929.1 AP002914 Oryza sativa putative glutathione transferase. P0493G01.7.
CAA39487.1 X56012 Triticum aestivum glutathione transferase. gstAl.
AAD56395.1 AFl 84059 Triticum aestivum glutathione S-transferase. GSTl.
BAB39940.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.22.
CAA68993.1 Y07721 Petunia x hybrida conjugates glutathione to anthocyanin to facilitate transport to the vacuole. glutathione S- transferase. an9 locus.
AAA33469.1 M16902 Zea mays glutathione S-tiansferase I.
AAG32477.1 AF309384 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF3.
AAA33470.1 M16901 Zea mays glutathione S-transferase I.
AAA20585.1 U12679 Zea mays glutathione S-transferase IV. GSTIV.
CAA56047.1 X79515 Zeamays glutathione transferase. GST27.
CAA39480.1 X56004 Triticum aestivum glutathione transferase. gstA2.
AAC64007.1 AF062403 Oryza sativa glutathione S-transferase II.
BAB39927.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.1. contains ESTs
AU031696(R0596),C97559(C60386),C28218(C60386), D28287(R0596).
AAG32475.1 AF309382 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF5.
AAG34823.1 AF244680 Zeamays glutathione S-transferase GST 15.
AAG34817.1 AF244674 Zea mays glutathione S-transferase GST 9.
CAA05354.1 AJ002380 Oryza sativa glutathione S-transferase. Rgst I.
AAG34820.1 AF244677 Zeamays glutathione S-transferase GST 11.
AAG34821.1 AF244678 Zea mays glutathione S-transferase GST 13.
CAB66333.1 AJ279691 Betula pendula glutathione-S-transferase. gsti
AAG34818.1 AF244675 Zea mays glutathione S-transferase GST 10.
AAG34816.1 AF244673 Zea mays glutathione S-transferase GST 8.
AAG34822.1 AF244679 Zea mays glutathione S-transferase GST 14.
CAA05355.1 AJ002381 Oryza sativa glutathione S-tiansferase. Rgst II.
SEQ ID NO: 360
AAA33710.1 L16977 Petunia x hybrida glutamate decarboxylase. gad.
AAA33709.1 L16797 Petunia x hybrida glutamate decarboxylase. gad.
AAC24195.1 AF020425 Nicotiana tabacum calmodulin binding protein, glutamate decarboxylase isozyme 1. NtGADl. calcium- calmodulin-dependent enzyme.
AAB40608.1 U54774 Nicotiana tabacum glutamate decarboxylase. NtGADl. calmodulin regulated enzyme; calmodulin-binding protein.
AAK18620.1 AF352732 Nicotiana tabacum converts glutamate to gamma-aminobutyric acid, glutamate decarboxylase isozyme 3. GAD;
GAD3; NtGAD3; calcium/calmodulin-dependent enzyme.
AAC39483.1 AF020424 Nicotiana tabacum glutamate decarboxylase isozyme 2. NtGAD2. calcium-cahnodulin-dependent enzyme.
BAB32870.1 AB056062 Oryza sativa glutamate decarboxylase. GAD.
BAB32868.1 AB056060 Oryza sativa glutamate decarboxylase. GAD. BAB32869.1 AB056061 Oryza sativa glutamate decarboxylase. GAD.
BAB32871.1 AB056063 Oryza sativa glutamate decarboxylase. GAD.
CAA56812.1 X80840 Lycopersicon esculentum homology to pyroxidal-5'-phosphate-dependant glutamate decarboxylases; putative start codon.
CAA50719.1 X71900 Lycopersicon esculentum histidine decarboxylase. hdc. pyridoxal 5'-phosρhate dependant.
SEQ ID NO: 362
AAG13467.1 AC026758 Oryza sativa putative proline oxidase. OSJNBa0015J15.31.
AAD48490.1 AF171226 Brassica napus proline dehydrogenase. pdh.
SEQ ID NO: 363
AAA33967.1 M76981 Glycine max vegetative storage protein. vspA.
AAA34022.1 M76980 Glycine max vegetative storage protein. vspB.
AAA34021.1 M20038 Glycine max vegetative storage protein.
BAA23563.1 D50094 Phaseolus vulgaris pod storage protein.
BAA19152.1 AB000585 Phaseolus vulgaris pod storage protein. PSP.
AAA34020.1 M20037 Glycine max vegetative storage protein.
SEQ ID NO: 364
AAB86939.1 AF030387 Oryza sativa NOI protein.
AAC03022.1 AF045033 Zeamays nitrate-induced NOI protein.
AAB86937.1 AF030385 Zea mays nitrate-induced NOI protein.
SEQ ID NO: 366
AAF75824.1 AF101788 Pinus taeda phytocyanin homolog.
AAC32448.1 U76296 Spinacia oleracea plantacyanin. member of a strictly plant-specific family of mononuclear blue copper proteins known as phytocyanins, which are further classified into three distinct subfamilies: Uclacyanins, Stellacyanins, and Plantacyanins; member ofthe subfamily of Plantacyanins; non-glycosylated.
AAC32421.1 U65511 Cucumis sativus putative oxygen activation and/or lignin formation, stellacyanin. member of a strictly plant- specific family of mononuclear blue copper proteins known as phytocyanins, which are further classified into three distinct subfamilies: Uclacyanins, Stellacyanins, and Plantacyanins; member ofthe subfamily of Stellacyanins; shnilar to Rhus vernicifera stellacyanin: SwissProt Accession Number P00302; similar to umecyanin: SwissProt Accession Number P42849; similar to mavicyanin; SwissProt Accession Number P80728; shnilar to BCB encoded by GenBank Accession Number Z 15058; similar to F18A8.9, encoded by GenBank Accession Number AC003105; shnilar to F7F1.27 encoded by GenBank Accession Number AC004669; similar to F9D12.16 encoded by GenBank
Accession Number AF077407; glycoprotein.
AAF66243.1 AF243181 Lycopersicon esculentum plantacyanin. naturally occuning cupredoxin with a Val residue in the position ofthe axial ligand Met; member ofthe plantacyanin subfamily of a strictly plant-specific family of mononuclear blue copper proteins known as phytocyanins.
BAA90481.1 AB035146 Ipomoea nil phytocyanin-related protein. SEQ ID NO: 367
AADl 1617.1 AF050495 Lycopersicon esculentum
Ca2+- ATPase. LCAl A; alternative transcript.
AAA34138.1 M96324 Lycopersicon esculentum
The calcium ATPase is a calcium ion pump. Ca2+-ATPase. LCAL
AAD 11618.1 AF050496 Lycopersicon esculentum Ca2+-ATPase. LCA1B; alternative transcript. CAA63790.1 X93592 Dunaliella bioculata
P-type ATPase. cal. calcium pumping; CAL
AAF73985.1 AF096871 Zea mays calcium pump, calcium ATPase. capl.
AAB58910.1 U82966 Oryza sativa
Ca2+-ATPase.
AAG28435.1 AF 195028 Glycine max plasma membrane Ca2+- ATPase. SCA1.
AAG28436.1 AF195029 Glycine max plasma membrane Ca2+-ATPase. SCA2.
AAB49042.1 U54690 Dunaliella acidophila plasma membrane proton ATPase. dhal . DaDHAl; proton pump.
AAB35314.2 S79323 Vicia faba plasma membrane H(+)-ATPase precursor, plasma membrane H(+)- ATPase. This sequence comes from Fig. 1; conceptual translation presented here differs from translation in publication.
CAB85495.1 AJ132892 Medicago truncatula proton pump. H+- ATPase. hal.
CAB85494.1 AJ132891 Medicago truncatula proton pump. H+- ATPase. hal.
AAB17186.1 U72148 Lycopersicon esculentum plasma membrane H+- ATPase. LHA4. plasma membrane proton pumping ATPase.
CAC29436.1 AJ310524 Vicia faba
P-type H+-ATPase. ha5. predominantly expressed in guard cells and flowers.
AAB41898.1 U84891 Mesembryanthemum crystallinum plasma membrane proton pump. H+-transporting ATPase. PMA.
CAC29435.1 AJ310523 Vicia faba
P-type H+- ATPase. vha4. predominantly expressed in flowers.
AAF98344.1 AF275745 Lycopersicon esculentum plasma membrane H+-ATPase. LHA2. P-type ion pump.
SEQ ID NO: 369
BAB 17726.1 AB050900 Raphanus sativus asparagine synthetase. Asnl.
CAA59138.1 X84448 Brassica oleracea asparagine synthase (glutamine-hydrolysing).
AAC16325.1 AF061740 Elaeagnus umbellata asparagine synthetase. AS.
CAA08913.1 AJ009952 Phaseolus vulgaris asparagine synthesis, asparagine synthetase type II. as2.
AAF02775.1 AF190728 Helianthus annuus asparagine synthetase. HAS1.
AAC49613.1 U77678 Glycine max catalyzes the ATP-dependent transfer ofthe amide group of glutamine to aspartate producing asparagine and glutamate. asparagine synthetase 2. AS2.
AAB81011.1 U89923 Medicago sativa asparagine synthetase.
AAC09952.1 U55874 Glycine max asparagine synthetase.
AAB48058.1 L40327 Medicago sativa asparagine synthetase.
CAA61589.1 X89409 Lotus japonicus asparagine synthase (glutamine-hydrolysing) . AS.
CAA67889.1 X99552 Asparagus officinalis asparagine synthetase.
AAF74755.1 AF263432 Helianthus annuus asparagine synthetase. HAS1.1.
AAD05035.1 AFO 14057 Triphysaria versicolor asparagine synthetase. AS. glutamine-hydrolyzing.
AAD05034.1 AFO 14056 Triphysaria versicolor asparagine synthetase. AS. glutamine-hydrolyzing.
AAD05033.1 AFO 14055 Triphysaria versicolor asparagine synthetase. AS. glutamine-hydrolyzing.
CAA96526.1 Z72354 Vicia faba synthesis of asparagine from aspartate and glutamine. asparagine synthetase. VfASl.
CAA48141.1 X67958 Asparagus officinalis asparagine synthase (glutamine-hydrolysing).
CAA61590.1 X89410 Lotus japonicus asparagine synthase (glutamine-hydrolysing). AS.
CAA36429.1 X52179 Pisum sativum asparagine synthase (glutamine-hydrolysing).
BAA96252.1 AB035248 Astragalus sinicus asparagine synthetase. AsAS2.
CAA36430.1 X52180 Pisum sativum asparagine synthase (glutarnine-hydrolysing).
BAA96251.1 AB035247 Astragalus sinicus asparagine synthetase. AsASl.
CAB57292.1 AJ133522 Phaseolus vulgaris asparagine synthetase (type-I). asl.
AAC49614.1 U77679 Glycine max catalyzes the ATP-dependent transfer ofthe amide group of glutamine to aspartate producing asparagine and glutamate. asparagine synthetase 1. ASL
BAA18951.1 D83378 Oryza sativa asparagine synthetase.
AAB03991.1 U55873 Oryza sativa asparagine synthetase.
AAF02776.1 AFl 90729 Helianthus annuus asparagine synthetase. HAS2.
AAB71532.1 AF005724 Sandersonia aurantiaca role in flower senescence, asparagine synthetase. SAND1.
CAA58052.1 X82849 Zea mays aspanagine synthetase. AS.
AAB91481.1 AF037363 Helianthus annuus asparagine synthetase.
CAA73762.1 Y13321 Pisum sativum asparagine synthetase 1. ASL
CAA73763.1 Y13322 Pisum sativum asparagine synthetase 2. AS2.
BAA96452.1 AB021793 Pyras pyrifolia asparagine synthetase. PPFRU32.
AAA73943.1 L23833 Glycine max production of phosphoribosylamine using glutamine and phosphoribosylpyrophosphate as substrates, glutamine phosphoribosylpyrophosphate amidotransferase.
SEQ ID NO: 370
AAG21985.1 AF271636 Zeamays lysine ketoglutarate reductase/saccharopine dehydrogenase. LKRSDH. bifunctional enzyme;
LKR/SDH; lysine 2-oxoglutarate reductase/saccharopine dehydrogenase.
AAC 18622.2 AF003551 Zea mays lysine-ketoglutarate reductase/saccharopine dehydrogenase bifunctional enzyme.
AAG28387.1 AF191667 Brassica oleracea lysine-ketoglutarate reductase/saccharopine dehydrogenase.
AAG28386.1 AF191666 Brassica napus lysine-ketoglutarate reductase/saccharopine dehydrogenase.
AAB97685.1 AF042184 Brassica napus lysine-ketoglutarate reductase/saccharopine dehydrogenase.
AAG14462.1 AF293461 Brassica napus lysine-ketoglutarate reductase. LKR.
SEQ ID NO: 371 CAB62537.1 AJ012583 Hevea brasiliensis pseudo-hevein.
AAA33357.1 M36986 Hevea brasiliensis hevein (HEV1) precursor.
CAA05978.1 AJ003196 Hevea brasiliensis
N-ace1yl-D-glucosaιmne/N-acetyl-D-neuranrinic acid binding lectin. prohevein.
AAF61435.1 AF137352 Pisum sativum pre-hevein-like protein. PHLP. stress-induced; the coding region is putative in the 5' end.
SEQ ID NO: 372 AAG28503.1 AFl 96966 Citrus sinensis hexokinase.
AAF18584.1 AF118132 Spinacia oleracea chloroplast outer envelope hexokinase 1. Hxkl .
AAF18585.1 AF118133 Nicotiana tabacum chloroplast outer envelope hexokinase 1. Hxkl.
AAF14186.1 AF106068 Solanum tuberosum hexokinase 2.
AAG35735.1 AF208543 Lycopersicon esculentum hexokinase. Hxk2.
CAA63966.1 X94302 Solanum tuberosum hexokinase. hxk.
BAA99425.1 AP002743 Oryza sativa putative chloroplast outer envelope hexokinase 1. P0710E05.10.
SEQ ID NO: 374
AAC83688.2 AF083343 Nicotiana tabacum 101 kDa heat shock protein. HSP101.
AAF01280.1 AF174433 Triticum aestivum heat shock protein 101. HSP101. ClpB family member.
AAD33606.1 AF133840 Zeamays heat shock protein HSP101. HSPIOI. lOl kDa protein.
AAD25223.1 AF077337 Zeamays heat shock protein 101. HSPIOI. ClpB/HsplOO protein homolog; 101 kDa heat shock protein.
AAF91178.1 AF203700 Phaseolus lunatus
ClpB. clpB. heat shock protein HSP100.
AAD22629.1 AF097363 Triticum aestivum heat shock protein 101. HsplOla.
AAC83689.2 AF083344 Triticum aestivum 101 kDa heat shock protein. HSP 101.
AAD26530.1 AF083327 Zeamays
101 kDa heat shock protein. HSPIOI. similar to HSPlOO/ClpB; HSP104.
SEQ ID NO: 375
CAA04611.1 AJ001208 Brassica juncea
APS reductase. apsrδ. putative mitochondrial or plastidic transit peptide.
CAA04610.1 AJ001207 Brassica juncea
APS reductase. apsr2. putative mitochondrial or plastidic transit peptide.
AAB05871.2 U63784 Catharanthus roseus reduction of adenylyl sulfate (APS). PAPS-reductase-like protein. par2neu.
CAB65911.1 AJ249831 Lemna minor
APR reducing enzyme, sulphur assimilation, adenosine 5'-phosphosulphate reductase. lapr.
AAF18999.1 AF212155 Allium cepa
APS-reductase.
AAC26855.1 AF069951 Enteromoφha intestinalis catalyzes the formation of sulfite and 5 -AMP from APS and reduced glutathione. 5'- adenylylsulfate reductase. EAPRl; sulfate assimilation enzyme; similar to Escherichia coli 3'- phosphoadenosine, 5'-phosphosulfate (PAPS) reductase encoded by cysH.
AAD02069.1 AF036939 Chlamydomonas reinhardtii redox-regulator of 5'UTR psbA mRNA binding complex and translation, protein disulfide isomerase. localized to ER and chloroplast.
AAC49896.1 AF027727 Chlamydomonas reinhardtii involved in the redox-regulated binding of chloroplast poly(A)-binding protein to the 5'-UTR of psbA mRNA; regulates chloroplast translational activation, protein disulfide isomerase RB60. PDI.
SEQ ID NO: 376
AAC62017.1 AF077547 Brassica juncea arginine decarboxylase.
AAF26434.1 AF220097 Brassica juncea arginine decarboxylase.
AAF26435.1 AF220098 Brassica juncea arginine decarboxylase.
AAB60880.1 AF002017 Dianthus caryophyllus arginine decarboxylase. gCARADC8.
AAF42972.1 AFl 27241 Nicotiana tabacum arginine decarboxylase 2. ADC2.
BAA25685.1 ABO 12873 Nicotiana sylvestris arginine decarboxylase. NsADC-1.
CAA85773.1 Z37540 Pisum sativum arginine decarboxylase.
CAB64599.1 AJ251898 Datura stramonium polyamine biosynthesis, arginine decarboxylase 1. adcl.
AAD09204.1 U35367 Glycine max arginine decarboxylase.
BAA84799.1 AP000559 Oryza sativa ESTs C99670(E21043),C99671(E21043),
AU078262(R10938),AU078261(R10938),D15282(C0402) conespond to a region ofthe predicted gene.; Similar to arginine decarboxylase (U52851).
AAB67887.1 U63832 Dianthus caryophyllus arginine decarboxylase. ADC.
AAF42971.1 AF127240 Nicotiana tabacum arginine decarboxylase 1. ADCL
AAF42970.1 AFl 27239 Nicotiana tabacum arginine decarboxylase 1. ADCl.
AAC68511.1 AF045666 Theobro a cacao arginine decarboxylase. spe2.
CAA65585.1 X96791 Vitis vinifera arginine decarboxylase. ADC.
AAA61347.1 L16582 Lycopersicon esculentum decarboxylation of L-arginine. arginine decarboxylase.
AAC68530.1 AF045685 Arabidopsis arenosa arginine decarboxylase. spe2.
AAC68529.1 AF045684 Capsella bursa-pastoris arginine decarboxylase. spe2.
AAC68525.1 AF045680 Arabis drummondii arginine decarboxylase. spe2.
AAC68526.1 AF045681 Barbarea vulgaris arginine decarboxylase. spe2.
AAC68535.1 AF045690 Nasturtium officinale arginine decarboxylase. spe2.
AAC68534.1 AF045689 Thellungiella salsuginea arginine decarboxylase. spe2.
AAC68533.1 AF045688 Thlaspi arvense arginine decarboxylase. spe2.
AAC68532.1 AF045687 Stanleya pinnata arginine decarboxylase. spe2.
AAC68531.1 AF045686 Sisymbrium altissimum arginine decarboxylase. spe2.
AAC68510.1 AF045665 Aethionema grandiflora arginine decarboxylase. spe2.
AAC68528.1 AF045683 Brassica oleracea arginine decarboxylase. spe2.
AAC68519.1 AF045674 Arabidopsis arenosa arginine decarboxylase. sρe2.
AAC68527.1 AF045682 Brassica nigra arginine decarboxylase. spe2.
AAC68523.1 AF045678 Thellungiella salsuginea arginine decarboxylase. spe2.
AAC68514.1 AF045669 Arabis drummondii arginine decarboxylase. spe2.
AAC68524.1 AF045679 Nasturtium officinale arginine decarboxylase. spe2.
AAC68522.1 AF045677 Thlaspi arvense arginine decarboxylase. spe2.
AAC68513.1 AF045668 Polanisia dodecandra arginine decarboxylase. spe2.
AAC68518.1 AF045673 Capsella bursa-pastoris arginine decarboxylase. spe2.
AAC68515.1 AF045670 Barbarea vulgaris arginine decarboxylase. spe2.
AAC68521.1 AF045676 Stanleya pinnata arginine decarboxylase. spe2.
AAC68520.1 AF045675 Sisymbrium altissimum arginine decarboxylase. spe2.
AAC68517.1 AF045672 Brassica oleracea arginine decarboxylase. spe2.
AAC68516.1 AF045671 Brassica nigra arginine decarboxylase. spe2.
AAC68512.1 AF045667 Carica papaya arginine decarboxylase. spe2.
CAA40137.1 X56802 Avena sativa arginine decarboxylase. spel.
AAD24801.1 AF132498 Brassica napus arginine decarboxylase. ADC.
BAA21617.1 AB005880 Nicotiana tabacum arginine decarboxylase.
AAB82607.1 AF026809 Ipomoea nil arginine decarboxylase. adc.
SEQ ID NO: 377
AAG22606.1 AF258809 Lycopersicon esculentum aldehyde oxidase. AO2. molybdenum cofactor-binding enzyme.
BAA23226.1 D88451 Zeamays aldehyde oxidase. zmAO-1.
AAG22607.1 AF258810 Lycopersicon esculentum aldehyde oxidase. AO3. molybdenum cofactor-binding enzyme.
AAG22605.1 AF258808 Lycopersicon esculentum aldehyde oxidase. AOl. molybdenum cofactor-binding enzyme.
BAA23227.1 D88452 Zea mays aldehyde oxidase-2. zmAO-2. putative.
AAB41742.1 U82559 Lycopersicon esculentum aldehyde oxidase 1 homolog. TAOl. has sequence properties ofthe molybdenum cofactor containing the hydroxylase genes aldehyde oxidase and xanthine dehydrogenase; belongs to a multigene family.
AAG22608.1 AF259793 Lycopersicon esculentum aldehyde oxidase. AOl. molybdenum cofactor-binding enzyme.
AAB41741.1 U82558 Lycopersicon esculentum aldehyde oxidase 1 homolog. TAOl. has sequence properties ofthe molybdenum cofactor containing the hydroxylase genes aldehyde oxidase and xanthine dehydrogenase; belongs to a multigene family.
SEQ ID NO: 378
BAB21211.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.17.
BAB21218.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.24.
BAA90643.1 AP001129 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC Fl 1F19 genomic sequence, putative
DNA-binding protein RAV2. (AC007017).
BAA85426.1 AP000616 Oryza sativa similar to putative DNA-binding protein RAV2 (AC007017).
SEQ ID NO: 380
CAB65369.1 AJ250832 Pisum sativum germin-like protein, gerl.
AAF03355.1 AF132671 Nicotiana plumbaginifolia nectarin I precursor. NEC1. germin-like protein.
AAD38298.1 AC007789 Oryza sativa putative oxalate oxidase (germin protein). OSJNBa0049B20.25.
BAB18339.1 AP002865 Oryza sativa putative ger in protein. P0034C11.30. contains EST C97263(C53484).
BAA25197.1 AB012138 Lycopersicon esculentum adaptation to Mn-deficiency. germin-like protein. Mdipl.
AAC78470.1 AF067731 Solanum tuberosum germin-like protein. OXAOXA. similar to oxalate oxidase.
AAC04835.1 AF032974 Oryza sativa germin-like protein 4. GER4. similar to wheat and barley oxalate oxidase.
BAB39980.1 AP003020 Oryza sativa probable germin protein 4. P0498A12.8. contains ESTs
AU101991(S4037);AU070167(R0031).
BAB39965.1 AP003018 Oryza sativa probable gennin protein 4. OSJNBa0004B13.19. contains ESTs
AU101991(S4037),AU070167(R0031).
AAC04833.1 AF032972 Oryza sativa germin-like protein 2. GER2. similar to wheat and barley oxalate oxidase.
AAG00425.1 AF250933 Hordeum vulgare germin A. GerA. apoplastic protein; contains prepeptide for targeting into the cell wall.
AAD43972.1 AF141879 Oryza sativa germin-like protein 2 precursor. RGLP2. similar to barley epidermis/papilla-specific oxalate oxidase-like protein.
AAD43973.1 AF141880 Oryza sativa germin-like protein 1 precursor. RGLPl. similar to barley epidermis/papilla-specific oxalate oxidase-like protein.
AAD43971.1 AF141878 Oryza sativa germin-like protein 1 precursor. RGLPl. similar to barley epidermis/papilla-specific oxalate oxidase-like protein.
CAB55559.1 AJ237943 Triticum aestivum germin-like protein. glp2b.
CAB55558.1 AJ237942 Triticum aestivum germin-like protein. glp2a.
AAC04837.1 AF032976 Oryza sativa germin-like protein 6. GER6. similar to wheat and barley oxalate oxidase.
AAC04832.1 AF032971 Oryza sativa germin-like protein 1. GER1. similar to wheat and barley oxalate oxidase.
CAA63659.1 X93171 Hordeum vulgare oxalate oxidase-like protein or germin-like protein.
CAB55394.1 ALl 17264 Oryza sativa zwhθθlθ.1. similar to Arabidopsis germin-like protein 6 (AF032976); Method: conceptual translation with partial peptide sequencing.
AAB97470.1 AF042489 Oryza sativa germin-like protein 16. glplό.
AAC25777.1 AF072694 Oryza sativa gerrnin-like protein 7. GER7. similar to wheat and barley oxalate oxidase.
AAG00427.1 AF250935 Hordeum vulgare germin F. GerF. apoplastic protein; contains prepeptide for targeting into the cell wall.
AAG00426.1 AF250934 Hordeum vulgare germin B. GerB. apoplastic protein; contains prepeptide for targeting into the cell wall.
BAA78563.1 AB024338 Atriplex lentiformis gerrnin-like protein.
AAA20245.1 U01963 Hordeum vulgare germin subunit.
AAG00428.1 AF250936 Hordeum vulgare germin D. GerD. apoplastic protein; contains prepeptide for targeting into the cell wall.
AAC99473.1 AF039201 Pinus caribaea gerrnin-like protein. PcGERl.
AAC04834.1 AF032973 Oryza sativa germin-like protein 3. GER3. similar to wheat and barley oxalate oxidase.
AAA34271.1 M63224 Triticum aestivum germin. germin 9f-3.8.
AAC05146.1 AF049065 Pinus radiata germin-like protein. PRGerl .
AAA34268.1 M21962 Triticum aestivum germin protein precursor.
AAA34270.1 M63223 Triticum aestivum germin. germin 9f-2.8.
CAA71052.1 Y09917 Triticum aestivum germin homolog. ρSBGer3.
AAG00429.1 AF250937 Hordeum vulgare germin E. GerE. apoplastic protein.
BAA86880.1 AB028454 Barbula unguiculata germin-like protein.
CAB65370.1 AJ250833 Pisum sativum germin-like protein. ger2a. 1st variant of this clone.
AAA33030.1 M93041 Mesembryanthemum crystallinum germin-like protein, germin-like protein.
CAB65371.1 AJ250834 Pisum sativum gennin-like protein. ger2b. 2nd variant ofthe clone PsGER2.
CAA71050.1 Y09915 Triticum aestivum germin homolog. pSBGerl .
CAA71051.1 Y09916 Triticum aestivum germin homolog. pSBGer2.
AAA86365.1 U21743 Brassica napus germin-like protein, similar to product encoded by GenBank Accession Number X84786.
CAC34417.1 AJ311624 Pisum sativum
Gerrnin-like protein. glp3.
SEQ ID NO: 389
CAA04703.1 AJ001370 Olea europaea cytochome b5. cytb5-2.
AAA32990.1 M87514 Brassica oleracea cytochrome b-5. cytochrome b-5.
CAA50575.1 X71441 Nicotiana tabacum cytochrome b5.
CAA53366.1 X75670 Oryza sativa cytochrome b5.
CAA04702.1 AJ001369 Olea europaea cytochrome b5. cytb5-l.
AAA62621.1 L22209 Cuscuta reflexa associated with cytokinin-induced haustoria formation in Cuscuta reflexa. cytochrome b5.
AAC49701.1 U79011 Borago officinalis haem-binding protein, cytochrome b5.
CAA56318.1 X80008 Nicotiana tabacum cytochrome b5.
CAA48240.1 X68140 Nicotiana tabacum cytochrome b5.
AAD10774.1 AF098510 Petunia x hybrida involved in anthocyqanin biosynthesis, cytochrome b5 DIF-F. difF. required for full activity of flavonoid 3 ',5' hydroxylase.
AAF60299.1 AF233640 Petunia x hybrida involved in anthocyqanin biosynthesis, cytochrome b5 DIF-F. difF.
CAA11033.1 AJ222981 Physcomitrella patens delta6-acyl-lipid desaturase. des6. des6 represents a fusion between a C-terminal desaturase with a cytochrome b5-related part and a N-terminal extension. SEQ ID NO: 392
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid: glucosyltransferase 1. togtl. glucosyltransferase.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS5a.
BAB17061.1 AP002523 Oryza sativa putative glucosyl transferase. P0013F10.7. contains EST C73149(E2992).
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS 10a. AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid:glucosyltiansferase 2. togt2. glucosyltransferase.
BAB17059.1 AP002523 Oryza sativa putative glucosyl transferase. P0013F10.5.
BAB 17060.1 AP002523 Oryza sativa putative glucosyl transferase. P0013F10.6.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltransferases.
CAB56231.1 Y18871 Dorotheanthus bellidifonnis betanidin-5-O-glucosyltransferase.
BAA83484.1 AB031274 Scutellaria baicalensis
UDP-glucose: flavonoid 7-O-glucosyltransferase. ufgt.
AAB48444.1 U82367 Solanum tuberosum UDP-glucose glucosyltransferase.
BAA36410.1 AB012114 Vigna mungo UDP-gIycose:flavonoid glycosyltransferase. UFGlyT.
CAA54610.1 X77460 Manihot esculenta UTP-glucose glucosyltransferase. CGT4.
CAB88666.1 AJ400861 Cicer arietinum flavonoid glycosyltransferase. putative UDP-glycose.
AAK16180.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.21.
AAD51778.1 AFl 16858 Phaseolus vulgaris utilizes UDPX as the sugar donor and catalyzes the formation of O-xylosylzeatin from zeatin. zeatin O-xylosyltransferase. ZOX1.
AAD04166.1 AF101972 Phaseolus lunatus catalyzes formation of O-glucosylzeatin from zeatin and UDPG; catalyzes formation of O- xylosylzeatin from zeatin and UDPX. zeatin O-glucosyltransferase. ZOGl. cytokinin O- glucosyltransferase.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltransferase. PHI.
CAC09351.1 AL442007 Oryza sativa putative glucosyltransferase. H0212B02.7.
AAB62270.1 AF006081 Solanum berthaultii UDPG glucosyltransferase. PLGT.
AAK16172.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
AAF61647.1 AFl 90634 Nicotiana tabacum
UDP -glucose : salicy lie acid glucosyltransferase. SA-GTase.
CAA54612.1 X77462 Manihot esculenta UTP-glucose glucosyltransferase. CGT5.
AAK16181.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.16.
AAK16178.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.5.
BAA36412.1 AB012116 Vigna mungo UDP-glycose: flavonoid glycosyltransferase. UFGlyT.
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
AAF98390.1 AF287143 Brassica napus catalyzes the transfer of glucose from UDP-glucose to sinapate and some other hydroxycinnamates (4-coumarate, caffeate, feralate). UDP-glucose:sinapate glucosyltransferase. SGT1. SGT.
BAA36423.1 ABO 13598 Verbena x hybrida
UDP-glucose:anthocyanin 5-O-glucosyltransferase. HGT8.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitrile-o- glucosyltransferase.
BAA93039.1 AB033758 Citrus unshiu limonoid UDP -glucosyltransferase. LGTase.
BAB17176.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.13.
BAB 17182.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.19.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltransferase. PGT8.
CAA54558.1 X77369 Solanum melongena glycosyl transferase. GT.
CAA54611.1 X77461 Manihot esculenta
UTP-glucose glucosyltransferase. CGT2.
CAA54609.1 X77459 Manihot esculenta
UTP-glucose glucosyltransferase. CGT1.
BAA12737.1 D85186 Gentiana triflora
UDP-glucose:flavonoid-3-glucosyltransferase.
AAD21086.1 AF127218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
AAG25643.1 AF303396 Phaseolus vulgaris
UDP-glucosyltransferase HRA25. putative; defense associated.
BAA36411.1 AB012115 Vigna mungo
UDP-glycose:flavonoid glycosyltransferase. UFGlyT.
BAA19155.1 AB000623 Nicotiana tabacum glucosyl transferase. JIGT.
AAD55985.1 AF165148 Petunia x hybrida catalyzes the penultimate step of flavonol glucosylgalactoside biosynthesis from UDP- galactose and flavonol aglycones in petunia pollen. UDP-galactose:flavonol 3-O- galactosyltransferase. F3galtase.
CAA54613.1 X77463 Manihot esculenta
UTP-glucose glucosyltransferase. CGT6.
BAA3642L1 AB013596 Perilla frutescens UDP-glucose:anthocysnin 5-O-glucosyltransferase. PF3R4.
BAA19659.1 AB002818 Perilla frutescens flavonoid 3-O-glucosyltransferase. UDP glucose.
AAB81683.1 AF000372 Vitis vinifera UDP glucose.-flavonoid 3-o-glucosyltransferase.
BAB41025.1 AB047098 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. F1UFGT1.
BAB41023.1 AB047096 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. AlUFGTl.
SEQ ID NO: 393
CAB56756.1 AJ011589 Pisum sativum
5, 10-methylenetetrahydrofolate dehydrogenase: 5, 10-methenyltetrahydrofolate cyclohydrolase. bifunctional enzyme.
AAD01907.1 AF030516 Pisum sativum
5, 10-methylenetetrahydrofolate dehydrogenase-5, 10-methenyltetrahydrofolate cyclohydrolase. bifunctional protein; 31.3 kDa protein.
AAG48834.1 AC084218 Oryza sativa similar to Pisum sativum methylenetetrahydrofolate dehydrogenase (NADP+) (EC 1.5.1.5)
(AF030516). SEQ ID NO: 406
AAD46491.1 AF135014 Zea mays dihydrolipoamide S-acetyltransferase. pyruvate dehydrogenase complex E2 subunit.
BAA90623.1 AP001129 Oryza sativa
ESTs AU033004(S0924),C74754(E50863) conespond to a region ofthe predicted gene.;
Similar to Rat mRNA for dihydrolipoamide acetyltransferase. (D 10655).
BAA77024.1 AB026124 Lithospermum erythrorhizon dihydrolipoamide acetyltransferase. SEQ ID NO: 407
AAF69017.1 AF261654 Dianthus caryophyllus ethylene-insensitive 3 -like protein 1. EIL1. EIN3-like protein.
CAC09582.1 AJ298994 Fagus sylvatica gibberellic acid (GA3)-induced. ethylene insensitive (EIN3/EIL)-like transcription regulator. einll.
AAG00419.1 AF247568 Nicotiana tabacum
EIN3. component in ethylene signal transduction pathway.
SEQ ID NO: 408 AAC15870.1 AF002016 Cucurbita sp. acyl CoA oxidase homolog. AAF14635.1 AF202987 Petroselinum crispum acyl-CoA oxidase. ACO. peroxisomal acyl-CoA oxidase.
AAB67883.1 U66299 Phalaenopsis sp. 'True Lady' acyl-CoA oxidase homolog.
AAC32108.1 AF051203 Piceamariana acyl-CoA oxidase homolog. Sb06. shnilar to Phalaenopsis sp. acyl-CoA oxidase homolog encoded by GenBank Accession Number U66299.
CAA04688.1 AJ001341 Hordeum vulgare putative acyl-CoA oxidase.
BAB08201.1 AP002539 Oryza sativa
ESTs AU056822(S20908),C26441(C12328), C28477(C61243) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana putative acyl-coA dehydrogenase (AF049236).
BAA96762.1 AP002521 Oryza sativa
ESTs AU056822(S20908),C26441(C12328), C28477(C61243) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana putative acyl-coA dehydrogenase (AF049236).
CAB55555.1 AJ010946 Pisum sativum auxin binding protein (ABP44). abp44/ivdh. has both auxin binding capability and enzymatic activity; isovaleryl-CoA Dehydrogenase.
CAB55554.1 AJ010945 Pisum sativum auxin binding protein (ABP44). abp44/ivdh. has both auxin binding capability and enzymatic activity; Isovaleryl-CoA Dehydrogenase.
CAC08233.1 AJ278987 Solanum tuberosum leucine catabolism. isovaleryl-CoA dehydrogenase. ivdl . CAC08234.1 AJ278988 Solanum tuberosum leucine catabolism. isovaleryl-CoA dehydrogenase. ivd2.
AAF02449.1 AF127432 Picea abies acyl-CoA oxidase homolog. Sb06.
AAF02451.1 AF127434 Picea abies acyl-CoA oxidase homolog. Sb06. AAF02450.1 AF127433 Picea abies acyl-CoA oxidase homolog. Sb06.
AAC32152.1 AF051733 Picea mariana acyl-CoA oxidase homolog. Sb06.
AAC32153.1 AF051734 Piceamariana acyl-CoA oxidase homolog. Sb06.
CAA58874.1 X84055 Hordeum vulgare mammalian acyl CoA oxidase homologous. cdr29. putative.
SEQ ID NO: 409 CAB64356.1 AJ251511 Populus tremula x Populus tremuloides mitochondrial oxidase. alternative oxidase. aoxl.
AAC60576.1 S71335 Mitochondrion Nicotiana tabacum alternative oxidase. Aoxl. This sequence comes from Fig. 1; AOX.
CAA56163.1 X79768 Nicotiana tabacum alternative oxidase. aox.
CAA48653.1 X68702 Glycine max alternative oxidase. aoxl . putative.
AAC35354.1 AF083880 Glycine max alternative oxidase precursor. Aoxl. AOXl; terminal oxidase of mitochondrial electron transport chain.
BAB21500.1 AB055060 Catharanthus roseus alternative oxidase.
BAA23803.1 AB009395 Catharanthus roseus alternative oxidase.
BAA86963.1 AB007452 Oryza sativa alternative oxidase. Aoxl(Aol-l).
BAA28773.1 AB004864 Oryza sativa alternative oxidase. AOXl a.
BAA28772.1 AB004813 Oryza sativa alternative oxidase. AOXl a.
CAA78823.1 Z15117 Sauromatum guttatum salicylic acid-inducible alternative oxidase. aoxl.
AAA34048.1 M60330 Sauromatum guttatum alternative oxidase protein, aoxl .
AAD51707.1 AFl 74004 Triticum aestivum alternative oxidase.
CAA55892.1 X79329 Mangifera indica alternative oxidase. AOMI 1.
CAB72441.1 AJ271889 Populus tremula x Populus tremuloides mitochondrial oxidase. alternative oxidase. aoxlb.
AAB97285.1 U87906 Glycine max alternative oxidase. Aox2.
BAA28771.1 AB004813 Oryza sativa alternative oxidase. AOXlb.
BAA28774.1 AB004865 Oryza sativa alternative oxidase. AOXlb.
AAB97286.1 U87907 Glycine max alternative oxidase. Aox3.
AAB97839.1 AF040566 Zea mays alternative oxidase. Aox.
AAG33634.1 AF314255 Chlamydomonas reinhardtii alternative oxidase 2. AOX2.
AAG02081.1 AF285187 Chlamydomonas reinhardtii alternative oxidase. AOX2.
AAG33633.1 AF314254 Chlamydomonas reinhardtii alternative oxidase 1. AOXl.
AAC05743.2 AF047832 Chlamydomonas reinhardtii alternative oxidase. AOXl.
BAA23725.1 AB009087 Chlamydomonas sp. W80 alternative oxidase.
AAC34192.1 S81466 Glycine max alternative oxidase Aoxl precursor. Aoxl . This sequence comes from Fig. 1.
AAG18450.1 AF302932 Lycopersicon esculentum plastid quinol oxidase.
AAG18449.1 AF302931 Lycopersicon esculentum plastid quinol oxidase.
AAG02287.1 AFl 77980 Lycopersicon esculentum plastid terminal oxidase. PTOX.
AAG02286.1 AF177979 Lycopersicon esculentum plastid terminal oxidase. PTOX.
AAG02288.1 AF177981 Capsicum annuum plastid terminal oxidase. PTOX.
AAB36072.1 S81470 Glycine max
Aox2. Aox2. alternative oxidase 2; This sequence comes from Fig. 1.
AAC35554.2 AF085174 Oryza sativa oxidase. IM1.
AAG00450.1 AF274001 Triticum aestivum oxidase. IML
SEQ ID NO: 410
CAA69600.1 Y08292 Nicotiana plumbaginifolia NADH glutamate dehydrogenase. GDH A.
CAB94836.1 AJ277949 Nicotiana plumbaginifolia NADH glutamate dehydrogenase. gdhA.
CAC18730.1 AJ303070 Vitis vinifera NADH glutamate dehydrogenase. gdhA.
CAA60507.1 X86924 Vitis vinifera glutamate dehydrogenase. GDH.
BAA08445.1 D49475 Zeamays glutamate dehydrogenase.
AAB51596.1 U93561 Zea mays glutamate dehydrogenase. GDHl. wild type.
AAB51595.1 U93560 Zeamays glutamate dehydrogenase mutant. GDHl. mutant allele.
CAB94837.1 AJ277950 Nicotiana plumbaginifolia NADH-glutamate dehydrogenase. gdhB.
CAA09478.1 AJ011096 Asparagus officinalis glutamate dehydrogenase. gdhb.
CAA69601.2 Y08293 Nicotiana plumbaginifolia NADH glutamate dehydrogenase. GDH B.
AAB39508.1 U48695 Lycopersicon esculentum glutamate dehydrogenase. legdhl.
CAA09456.1 AJO 11006 Asparagus officinalis NADH glutamate dehydrogenase. gdhA.
CAA41635.1 X58831 Chlorella sorokiniana glutamate dehydrogenase (NADP+). gdhANC. There are other coding exons before the first exon given for this gene.
CAA41636.1 X58832 Chlorella sorokiniana glutamate dehydrogenase (NADP+). gdhANC.
SEQ ID NO: 412
CAA65456.2 X96681 Oryza sativa transcription factor. DNA-binding protein. Oshoxl. homeodomain leucine zipper gene.
AAK00416.1 AC069324 Oryza sativa
Putative DNA-binding protein. OSJNBa0071K19.2.
BAA05622.1 D26573 Daucus carota tianscriptional regulator. DNA-binding protein, homeodomain at nt 585-764; leucine zipper at nt 765-851.
BAA05625.1 D26576 Daucus carota transcriptiional regulator. DNA-binding protein, homeodomain at nt 300-479; leucine zipper at nt 480-587.
BAA21017.1 D26578 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 761-940; leucine zipper at nt 941-1048.
BAA05624.1 D26575 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 520-699; leucine zipper at nt 700-805.
BAA05623.1 D26574 Daucus carota trancriptional regulator. DNA-binding protein, homeodomain at nt 498-677; leucine zipper at nt 678-785.
SEQ ID NO: 414
BAA95814.1 AP002069 Oryza sativa
ESTs AU082579(S2069),D40238(S2069) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana vesicle-associated membrane protein 7C; synaptobrevin 7C.
(AF025332).
SEQ ID NO: 415
CAA45025.1 X63431 Rauvolfia mannii strictosidine synthase. strl
CAA68725.1 Y00756 Rauvolfia seφentina strictosidine s; ynthase.
CAA44208.1 X62334 Rauvolfia seφentina strictosidine synthase. strl
CAA37671.1 X53602 Catharanthus roseus strictosidine synthase precursor.
CAA71255.1 Y10182 Catharanthus roseus strictosidine synthase. strl .
CAA43936.1 X61932 Catharanthus roseus strictosidine synthase. sss.
AAF75751.1 AF261141 Lycopersicon esculentum putative strictosidine synthase.
CAB53484.1 AJ245900 Oryza sativa
CAA303711.1 protein. q3037.11. Similar to strictosidine synthase 3 precursor.
SEQ ID NO: 418 AAG22607.1 AF258810 Lycopersicon esculentum aldehyde oxidase. AO3. molybdenum cofactor-binding enzyme.
AAG22606.1 AF258809 Lycopersicon esculentum aldehyde oxidase. AO2. molybdenum cofactor-binding enzyme.
AAG22605.1 AF258808 Lycopersicon esculentum aldehyde oxidase. AOl. molybdenum cofactor-binding enzyme.
BAA23227.1 D88452 Zea mays aldehyde oxidase-2. zmAO-2. putative.
BAA23226.1 D88451 Zea mays aldehyde oxidase. zmAO-1.
AAB41742.1 U82559 Lycopersicon esculentum aldehyde oxidase 1 homolog. TAOl. has sequence properties ofthe molybdenum cofactor containing the hydroxylase genes aldehyde oxidase and xanthine dehydrogenase; belongs to a multigene family.
AAG22608.1 AF259793 Lycopersicon esculentum aldehyde oxidase. AO molybdenum cofactor-binding enzyme.
AAB41741.1 U82558 Lycopersicon esculentum aldehyde oxidase 1 homolog. TAOl. has sequence properties ofthe molybdenum cofactor containing the hydroxylase genes aldehyde oxidase and xanthine dehydrogenase; belongs to a multigene family.
SEQ ID NO: 421
BAA13635.1 D88530 Spinacia oleracea serine acetyltransferase.
BAA13634.1 D88529 Spinacia oleracea serine acetyltransferase.
BAA93050.1 AB040502 Allium tuberosum serine acetyltransferase. ASAT5.
BAA08479.1 D49535 Citrullus lanatus serine acetyltransferase.
BAA12843.1 D85624 Citrullus lanatus serine acetyltransferase. Sat.
BAA21827.1 AB006530 Citrullus lanatus serine acetyltransferase. Sat.
AAF19000.1 AF212156 Allium cepa serine acetyltransferase.
SEQ ID NO: 422
BAA05079.1 D26086 Petunia x hybrida zinc-finger protein.
CAB77055.1 Y18788 Medicago sativa putative TFIIIA (or krappel)-like zinc finger protein.
AAD26942.1 AFl 19050 Datisca glomerata zinc-finger protein 1. zfpl. DgZFPl.
AAB39638.1 U68763 Glycine max putative transcription factor. SCOF-1. scof-1. zinc-finger protein.
AAC06243.1 AF053077 Nicotiana tabacum transcription factor, osmotic stress-induced zinc-finger protein, zfp.
BAA05077.1 D26084 Petunia x hybrida zinc-finger DNA binding protein.
BAA05076.1 D26083 Petunia x hybrida zinc-finger DNA binding protein.
BAA05078.1 D26085 Petunia x hybrida zinc-finger DNA binding protein.
AAK01713.1 AF332876 Oryza sativa zinc finger transcription factor ZF1.
AAB53260.1 U76554 Brassica rapa transcription factor, zinc-finger protein- 1. BR42.
AAB53261.1 U76555 Brassica rapa zinc-finger protein BcZFPl. BcZFPl(3-2z).
BAA96070.1 AB035132 Petunia x hybrida C2H2 zinc-finger protein ZPT2- 10. PEThy;ZPT2-10.
BAA21919.1 AB006597 Petunia x hybrida ZPT2-10. C2H2 zinc finger protein, 2 finger.
BAA96071.1 AB035133 Petunia x hybrida C2H2 zinc-finger protein ZPT3-3. ZPT3-3.
BAA21927.1 AB006605 Petunia x hybrida ZPT3-3. C2H2 zinc finger protein, 3 finger.
BAA19112.1 AB000453 Petunia x hybrida PEThy;ZPT3-l. Cys(2) His(2) zinc finger protein, 3 fingers.
BAA21928.1 AB006606 Petunia x hybrida ZPT4-4. C2H2 zinc finger protein, 4 finger.
BAA21922.1 AB006600 Petunia x hybrida ZPT2-13. C2H2 zinc finger protein, 2finger.
BAA19114.1 AB000455 Petunia x hybrida PEThy;ZPT4-l. Cys(2) His(2) zinc finger protein, 4 fingers.
BAA21920.1 AB006598 Petunia x hybrida ZPT2-11. C2H2 zinc finger protein, 2finger.
BAA21921.1 AB006599 Petunia x hybrida ZPT2-12. C2H2 zinc finger protein, 2 finger.
BAA19110.1 AB000451 Petunia x hybrida PEThy;ZPT2-5. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA21926.1 AB006604 Petunia x hybrida ZPT2-9. C2H2 zinc finger protein, 2 finger.
BAA21925.1 AB006603 Petunia x hybrida ZPT2-8. C2H2 zinc finger protein, 2 finger.
CAA60828.1 X87374 Pisum sativum putative zinc finger protein.
BAA19111.1 AB000452 Petunia x hybrida PEThy;ZPT2-6. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA21923.1 AB006601 Petunia x hybrida ZPT2-14. C2H2 zinc finger protein, 2 finger.
BAA21924.1 AB006602 Petunia x hybrida ZPT2-7. C2H2 zinc finger protein, 2f_nger.
BAA19113.1 AB000454 Petunia x hybrida PEThy;ZPT3-2. Cys(2) His(2) zinc finger protein, 3 fingers.
BAA19926.1 AB000456 Petunia x hybrida PEThy; ZPT4-2. C2H2 zinc finger protein, 4 finger.
SEQ ID NO: 423
AAC49815.1 U87257 Daucus carota 4-hydroxyphenylρyravate dioxygenase.
CAA04245.1 AJ000693 Hordeum vulgare 4-hydroxyphenylpyravate dioxygenase.
SEQ ID NO: 424
AAC72193.1 AF069909 Zeamays pyravate dehydrogenase El beta subunit isoform 2.
AAC72192.1 AF069908 Zea mays pyruvate dehydrogenase El beta subunit isoform 1.
AAC72194.1 AF069910 Zea mays pyruvate dehydrogenase El beta subunit isoform 3.
AAB01223.1 U56697 Pisum sativum pyravate dehydrogenase Elbeta.
AAC32149.1 AF051249 Piceamariana pyravate dehydrogenase El beta subunit. Sb68.
AAF43837.1 AF166114 Chloroplast Mesostigma viride beta subunit of pyravate dehydrogenase El component. odpB.
AAD22077.1 AF124755 Pinus banksiana pyruvate dehydrogenase El beta subunit. Sb68.
CAA75778.1 Y15782 Capsicum annuum transketolase 2.
AAB88295.1 AF024512 Oryza sativa
CLAl tiansketolase-like protein. CLAl. similar to Arabidopsis CLAl product; required for chloroplast development. ^^
SEQ ID NO: 426
AAB53764.1 U96713 Brassica rapa aminoalcoholphosphotiansferase. AAPT1.
AAD56040.1 AF183933 Brassica rapa aminoalcoholphosphotransferase. AAPT3.
AAC79507.1 U96439 Pimpinella brachycaφa aminoalcoholphosphotransferase. AAPTase.
AAA67719.1 U12735 Glycine max
CDP-choline:diacylglycerol cholinephosphotransferase activity and possibly CDP- ethanolamine: diacylglycerol ethanolaminephosphotiansferase activity, aminoalcoholphosphotransferase. AAPT1.
SEQ ID NO: 427
CAA56313.1 X79992 Avena sativa putative pp70 ribosomal protein S6 kinase. Aspkl 1.
CAB89082.1 AJ277534 Asparagus officinalis S6 ribosomal protein kinase. pkl . putative.
AAK18843.1 AC082645 Oryza sativa putative protein kinase. OSJNBb0033N16.3.
AAC05084.1 AF033097 Avena sativa
NPH1-2. NPH1-2. putative serine/threonine protein kinase.
AAC05083.1 AF033096 Avena sativa
NPH1-1. NPH1-1. putative serine/threonine protein kinase.
AAK13156.1 AC078829 Oryza sativa putative protein kinase. OSJNBa0026O12.14.
AAB88817.1 AF033263 Zeamays signal transduction for phototiopism. nonphototiopic hypocotyl 1. nphl. NPHl; putative serine/threonine kinase; similar to oat NPHl proteins.
CAB82852.1 Z30329 Mesembryanthemum crystallinum protein kinase MK6.
BAB18104.1 AB042714 Chlamydomonas reinhardtii cyclic nucleotide dependent protein kinase. CL-PK1.
BAB18105.1 AB042715 Chlamydomonas reinhardtii cyclic nucreotide dependent protein kinase II. CL-PK2.
BAA83689.1 AB011968 Oryza sativa OsPK7. OsPK7. protein kinase.
AAD31900.1 AF 145482 Mesembryanthemum crystallinum putative serine/threonine protein kinase. ,
BAA92970.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F28A21 ; putative protein kinase.
(AL035526).
BAA92972.1 AP001551 Oryza sativa
ESTs AU056183(S20356),AU056881(S20950) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome 4 BAC clone F6I18 ; putative protein kinase.
(AL022198).
BAA83688.1 AB011967 Oryza sativa OsPK4. OsPK4. protein kinase.
AAF22219.1 AF141378 Zea mays protein kinase PK4. ZmPK4.
CAA73067.1 Y12464 Sorghum bicolor serine/threonine kinase. SNFL1.
BAA96628.1 AP002482 Oryza sativa
ESTs D41739(S4522),AU055999(S20214), AU057588(S21592 conespond to a region ofthe predicted gene. Similar to Sorghum bicolor serine/threonine kinase (Y12465).
CAA89202.1 Z49233 Chlamydomonas eugametos calcium-stimulated protein kinase.
CAA73068.1 Y12465 Sorghum bicolor serine/threonine kinase. SNFL2.
BAA34675.1 ABO 11670 Triticum aestivum wpk4 protein kinase. wpk4.
CAA74646.1 Y14274 Sorghum bicolor putative serine/threonine protein kinase. SNFL3.
AAF06970.1 AF162662 Kalanchoe fedtschenkoi phosphoenolpyruvate carboxylase kinase. calcium-independent protein kinase.
AAF06969.1 AFl 62661 Kalanchoe fedtschenkoi phosphoenolpyravate carboxylase kinase. calcium-independent protein kinase.
CAA39936.1 X56599 Daucus carota calcium- dependent protein kinase. DcPK431.
BAA99439.1 AP002743 Oryza sativa putative protein kinase. P0710E05.26. contains ESTs C22394(C30013),C22393(C30013).
BAA90814.1 AP001168 Oryza sativa
ESTs AU030197(E50746),AU030196(E50746) conespond to a region ofthe predicted gene.;
Similar to calcium-dependent calmodulin-independent protein kinase CDPK (U90262).
BAB 12687.1 AP002746 Oryza sativa putative protein kinase. P0671B11.2. contains ESTs C22394(C30013),C22393(C30013).
AAB62693.1 AF004947 Oryza sativa protein kinase.
CAA71142.1 Y10036 Cucumis sativus
SNF1 -related protein kinase.
BAA05649.1 D26602 Nicotiana tabacum protein kinase.
AAC25423.1 AF072908 Nicotiana tabacum calcium-dependent protein kinase. CDPK1.
AAF19403.1 AF203481 Lycopersicon esculentum phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAF21062.1 AF216527 Dunaliella tertiolecta calcium-dependent protein kinase. CPK1; CDPK.
AAF 19402.1 AF203480 Lycopersicon esculentum phosphoenolpyruvate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
CAA65244.1 X95997 Solanum tuberosum
SNF1 -related protein kinase. PKIN1.
CAA57898.1 X82548 Hordeum vulgare
SNFl-related protein kinase. BKI N2.
BAA13608.1 D88399 Oryza sativa serine-threonine kinase. endosperm kinase. REK.
CAA08995.1 AJ010091 Brassica napus
MAP3K alpha 1 protein kinase. MAP3K alpha 1.
AAF19401.1 AF203479 Glycine max phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
BAA05648.1 D26601 Nicotiana tabacum protein kinase.
AAC69450.1 AF032465 Nicotiana tabacum putative serine/threonine protein kinase. WAPK.
AAD23582.1 AF128443 Glycine max probably involved in plant stress responses possibly regulates gene expression. SNF-1-like serine/threonine protein kinase. expressed in nodules, roots and leaves.
AAD00239.1 U73938 Nicotiana tabacum protein kinase. PKl 1-Cl. PKl 1-Cl. induced at the transcriptional level by the abscicic acid plant hormone; similar to serine/threonine protein kinase.
AAG60195.1 AC084763 Oryza sativa protein kinase REK. OSJNBa0027P10.6. SEQ ID NO: 428
CAC17753.1 AJ294543 Dendrobium 'Sonia' metabolic enzyme of cytokinins. cytokinin oxidase. ckol. CAC17752.1 AJ294542 Dendrobium 'Sonia' metabolic enzyme of cytokinins. cytokinin oxidase. ckol.
CAA77151.1 Y18377 Zea mays cytokinin oxidase. cko.
AAC27500.1 AF044603 Zea mays cytokinin oxidase. ckxl .
BAB07927.1 AP002836 Oryza sativa putative cytokinin oxidase. P0512G09.9.
BAB03420.1 AP002816 Oryza sativa
Similar to Zea mays mRNA for cytokinin oxidase. (Y18377).
SEQ ID NO: 429
BAA21922.1 AB006600 Petunia x hybrida ZPT2-13. C2H2 zinc fmger protein, 2finger.
BAA21923.1 AB006601 Petunia x hybrida ZPT2-14. C2H2 zinc finger protein, 2 fmger.
BAA21921.1 AB006599 Petunia x hybrida ZPT2-12. C2H2 zinc finger protein, 2 finger.
BAA19110.1 AB000451 Petunia x hybrida PEThy;ZPT2-5. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA21925.1 AB006603 Petunia x hybrida ZPT2-8. C2H2 zinc finger protein, 2 finger.
BAA21926.1 AB006604 Petunia x hybrida ZPT2-9. C2H2 zinc finger protein, 2 finger.
BAA21924.1 AB006602 Petunia x hybrida ZPT2-7. C2H2 zinc finger protein, 2fmger.
BAA19111.1 AB000452 Petunia x hybrida PEThy;ZPT2-6. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA05077.1 D26084 Petunia x hybrida zinc-finger DNA binding protein.
AAD26942.1 AFl 19050 Datisca glomerata zinc-finger protein 1. zfpl . DgZFPl .
BAA05076.1 D26083 Petunia x hybrida zinc-finger DNA binding protein.
CAB77055.1 Y18788 Medicago sativa putative TFIIIA (or krappel)-like zinc finger protein.
AAC06243.1 AF053077 Nicotiana tabacum transcription factor, osmotic stress-induced zinc-finger protein, zfp.
BAA21927.1 AB006605 Petunia x hybrida ZPT3-3. C2H2 zinc finger protein, 3 fmger.
BAA96071.1 AB035133 Petunia x hybrida C2H2 zinc-finger protein ZPT3-3. ZPT3-3.
BAA21920.1 AB006598 Petunia x hybrida ZPT2-11. C2H2 zinc finger protein, 2fmger.
AAB53261.1 U76555 Brassica rapa zinc-finger protein BcZFPl. BcZFP 1 (3-2z).
AAB53260.1 U76554 Brassica rapa transcription factor, zinc-finger protein- 1. BR42.
CAA60828.1 X87374 Pisum sativum putative zinc finger protein.
AAB39638.1 U68763 Glycine max putative transcription factor. SCOF-1. scof-1. zinc-finger protein.
AAK01713.1 AF332876 Oryza sativa zinc finger transcription factor ZF1.
BAA05079.1 D26086 Petunia x hybrida zinc-finger protein.
BAA96070.1 AB035132 Petunia x hybrida C2H2 zinc-finger protein ZPT2-10. PEThy;ZPT2-10.
BAA21919.1 AB006597 Petunia x hybrida ZPT2-10. C2H2 zinc finger protein, 2 finger.
BAA05078.1 D26085 Petunia x hybrida zinc-finger DNA binding protein.
BAA19112.1 AB000453 Petunia x hybrida PEThy;ZPT3-l. Cys(2) His(2) zinc finger protein, 3 fingers.
BAA19114.1 AB000455 Petunia x hybrida PEThy;ZPT4-l. Cys(2) His(2) zinc finger protein, 4 fingers.
BAA21928.1 AB006606 Petunia x hybrida ZPT4-4. C2H2 zinc finger protein, 4 finger.
BAA19926.1 AB000456 Petunia x hybrida PEThy; ZPT4-2. C2H2 zinc finger protein, 4 finger.
SEQ ID NO: 432
AAC05084.1 AF033097 Avena sativa
NPHl -2. NPHl -2. putative serine/threonine protein kinase.
AAC05083.1 AF033096 Avena sativa
NPH1-1. NPH1-1. putative serine/threonine protein kinase.
CAA82993.1 Z30332 Spinacia oleracea protein kinase.
AAB88817.1 AF033263 Zea mays signal transduction for phototiopism. nonphototiopic hypocotyl 1. nphl. NPHl; putative serine/threonine kinase; similar to oat NPHl proteins.
CAB65325.1 AJ252142 Oryza sativa putative blue light receptor phototropin. non-phototropic hypocotyl NPHl . nphl .
CAA82994.1 Z30333 Mesembryanthemum crystallinum protein kinase.
BAA36192.1 AB012082 Adiantum capillus-veneris
PHY3. PHY3. Ser/Thr protein kinase; chimeric stracture of red/far-red light photoreceptive region (phytochrome)and putative blue light photoreceptor in phototropism of hypocotyl
(NPHl). SEQ ID NO: 446
AAB72047.1 AF006489 Gossypium hirsutum adenine nucleotide translocator 1. CANT1.
CAA05979.1 AJ003197 Lupinus albus transfer of ATP from mitochondria to cytosol. adenine nucleotide translocator. anti.
CAA44054.1 X62123 Solanum tuberosum
ADP /ATP translocator. ant. product has dimeric subunit structure.
AAB49700.1 U89839 Lycopersicon esculentum ADP/ATP translocator.
CAA40782.1 X57557 Solanum tuberosum adenine nucleotide translocator. AAC.
BAA02161.1 D12637 Oryza sativa ATP/ADP translocator.
CAA41812.1 X59086 Zeamays adenine nucleotide translocator. MANT2.
CAA40781.1 X57556 Zea mays adenine nucleotide translocator. MANTl.
CAA33743.1 X15712 Zea mays adenine nucleotide translocator.
CAA33742.1 X15711 Zea mays adenine nucleotide translocator.
CAA65119.1 X95863 Triticum turgidum adenine nucleotide translocator.
CAA26600.1 X02842 Zea mays put. ATP/ADP translocator.
CAA65120.1 X95864 Triticum turgidum adenine nucleotide translocator.
CAA46311.1 X65194 Chlamydomonas reinhardtii mitochondrial ADP/ATP translocator protein. GRANT.
AAA33027.1 M76669 Chlorella kessleri ATP/ADP translocator. AAT.
AAB72048.1 AF006490 Gossypium hirsutum adenine nucleotide translocator 2. CANT2.
BAA08104.1 D45074 Panicum miliaceum
2-oxoglutarate/malate translocator. mitochondrial 2-oxoglutarate/malate translocator.
BAA08103.1 D45073 Panicum miliaceum
2-oxoglutarate/malate translocator. mitochondrial 2-oxoglutarate/malate translocator.
BAA08105.1 D45075 Panicum miliaceum
2-oxoglutarate/malate translocator. mitochondrial 2-oxoglutarate/malate translocator.
SEQ ID NO: 447
BAB16317.1 AB049589 Avicennia marina secretory peroxidase. PER.
AAC83463.1 AF039027 Glycine max
H2O2 oxidoreductase. cationic peroxidase 2. Prx2. class III plant peroxidase.
AAD37374.1 AF145348 Glycine max peroxidase. Prx2b.
CAB71128.2 AJ271660 Cicer arietinum cationic peroxidase.
AAD33072.1 AF149251 Nicotiana tabacum secretory peroxidase. PER.
AAA99868.1 L08199 Gossypium hirsutum peroxidase. putative.
BAA94962.1 AB042103 Asparagus officinalis peroxidase. AspPOXl.
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 5 precursor, type III peroxidase.
AAF63026.1 AF244923 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 4 precursor, type III peroxidase.
BAA92500.1 AP001383 Oryza sativa
ESTs D39300(R3292),AU030751(E60187) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP6a. (X98774).
AAF63025.1 AF244922 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 3 precursor, type III peroxidase.
AAG46133.1 AC082644 Oryza sativa putative peroxidase. OSJNBa0013M12.15.
AAA32973.1 M73234 Hordeum vulgare peroxidase BP 1. Prx5.
CAA71493.1 Y10467 Spinacia oleracea peroxidase. prxr6.
AAG46122.1 AC082644 Oryza sativa putative peroxidase. OSJNBa0013M12.2.
AAG46141.1 AC082644 Oryza sativa putative peroxidase. OSJNBa0013M12.11.
BAA77387.1 AB024437 Scutellaria baicalensis peroxidase 1.
AAG46142.1 AC082644 Oryza sativa putative peroxidase. OSJNBa0013M12.9.
AAB02926.1 U59284 Linum usitatissimum peroxidase. FLXPER3.
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI- 1.
BAA82306.1 AB027752 Nicotiana tabacum peroxidase.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
AAG46125.1 AC082644 Oryza sativa putative peroxidase. OSJNBa0013M12.17.
BAA92422.1 AP001366 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to A.thaliana mRNA for peroxidase ATP18a. (X98804).
BAA92497.1 AP001383 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP18a. (X98804).
CAA59487.1 X85230 Triticum aestivum peroxidase. ρox4.
AAA98491.1 L36981 Petroselinum crispum anionic peroxidase.
CAA71492.1 Y10466 Spinacia oleracea peroxidase. prxr5.
BAA96643.1 AP002482 Oryza sativa
Similar to Arabidopsis thaliana peroxidase ATP19a (X98805).
BAB12025.1 AP002820 Oryza sativa putative peroxidase. P0702D12.1.
AAB02554.1 L37790 Stylosanthes humilis cationic peroxidase.
AAF63024.1 AF244921 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 2 precursor, type III peroxidase.
AAC84140.1 AF101427 Cichorium intybus peroxidase.
AAA65637.1 L13654 Lycopersicon esculentum peroxidase. TPX1.
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
CAA62615.1 X91232 Mercurialis annua PRX. peroxidase.
AAG46130.1 AC082644 Oryza sativa putative peroxidase. OSJNBa0013M12.18.
CAC21392.1 AJ401275 Zeamays peroxidase. ρox2.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
BAB39277.1 AP002971 Oryza sativa putative peroxidase. P0537A05.6. contains ESTs C23550(C52903),C97179(C52903).
CAB65334.1 AJ250121 Picea abies peroxidase. SPI2 protein. spi2.
CAA62597.1 X91172 Raphanus sativus korean-radish isoperoxidase. prxkl .
AAC49820.1 AFO 14469 Oryza sativa peroxidase. POX5.1. wound inducible.
CAA71496.1 Y10470 Spinacia oleracea peroxidase. prxr9.
BAA01950.1 D11337 Vigna angularis peroxidase.
CAA71491.1 Y10465 Spinacia oleracea peroxidase. prxr4.
BAA03911.1 D 16442 Oryza sativa peroxidase.
SEQ ID NO: 449
CAA56325.1 X80023 Triticum turgidum ATP/ADP carrier protein.
CAC27140.1 AJ132535 Picea abies ADP, ATP carrier protein precursor.
BAA92520.1 AP001383 Oryza sativa
ESTs AU068633(C30614),AU068634(C30614) conespond to a region ofthe predicted gene.
Similar to Bos taurus mitochondrial solute canier protein. (AF049236).
BAB 16462.1 AP002483 Oryza sativa putative peroxisomal Ca-dependent solute carrier protein. P0019D06.21.
BAB40117.1 AP003311 Oryza sativa putative peroxisomal Ca-dependent solute canier protein. P0024G09.9.
AAB71744.1 U75346 Chlamydomonas reinhardtii envelope protein. LIP-36G2. low CO2 inducible carrier protein LIP-36 with a molecular weight of 36 kDa.
AAB71743.1 U75345 Chlamydomonas reinhardtii envelope protein. LIP-36G1. low CO2 inducible carrier protein LIP-36 with a molecular weight of 36 kDa.
AAG48999.1 AYO 13246 Hordeum vulgare putative mitochondrial carrier protein. 635P2.1.
CAC12820.1 AJ299250 Nicotiana tabacum mitochondrial 2-oxoglutarate/malate canier protein, momcl. CAA07568.1 AJ007580 Ribes nigrum
Mitochondrial carrier protein. prib7.
AAG45489.1 AY013245 Oryza sativa
3615.1. putative mitochondrial carrier protein. SEQ ID NO: 451
AAB53099.1 U68217 Brassica napus iron binding protein, ferritin. LSC30.
AAA33959.1 M64337 Glycine max ferritin light chain, ferritin.
AAA34016.1 M72894 Glycine max ferritin light chain. SOF-H2.
CAA65771.1 X97059 Medicago sativa iron storage, ferritin. FER. abcissic acid regulated.
AAB18928.1 U31648 Glycine max iron storage protein, fenitin.
AAC06026.1 AF052057 Vigna unguiculata iron storage and mobilization in plants, fenitin subunit cowpea3 precursor.
AAC06027.1 AF052058 Vigna unguiculata iron storage and mobilization in plants, fenitin subunit cowpea2 precursor.
CAA41213.1 X58274 Phaseolus vulgaris ferritin. pfe.
AAD50644.1 AF133814 Solanum tuberosum fenitin 1. Fl.
CAA51786.1 X73369 Pisum sativum fenitin.
CAA45763.1 X64417 Pisum sativum fenitin-precursor.
CAA43663.1 X61391 Zea mays fenitin.
CAA58146.1 X83076 Zea mays fenitin. Ferl .
CAA58147.1 X83077 Zea mays fenitin. Fer2.
CAA43664.1 X61392 Zea mays fenitin.
AAA33958.1 M58336 Glycine max fenitin light chain. SOF-5L.
CAB42587.1 AJ238628 Chlorella protothecoides putative fenitin. dee 188.
BAB17852.1 AB042612 Nicotiana tabacum ferritin 1. tob-fer-1. putative.
AAC15241.1 AF028072 Pinus taeda fenitin.
CAA47982.1 X67754 Vigna unguiculata ferritin 1. pfel .
CAA47984.1 X67756 Vigna unguiculata ferritin 5. ρfe5.
CAA47983.1 X67755 Vigna unguiculata ferritin 2. pfe2.
AAC12282.1 AF052511 Glycine max iron storage and mobilization, ferritin 2. soybean 2; cowpea 2 homolog.
AAC12281.1 AF052513 Glycine max iron storage and mobilization, fenitin 1. soybean 1; cowpea 1 homolog.
SEQ ID NO: 452 CAB56619.1 AJ244024 Nicotiana tabacum putative role in cell plate formation, phragmoplastin. phragmoplastin. dynamin-like protein. SEQ ID NO: 454
CAA55039.1 X78203 Hyoscyamus muticus glutathione transferase.
AAB65163.1 AF002692 Solanum commersonii glutathione S-transferase, class-phi. GST low temperature induced.
BAA01394.1 D 10524 Nicotiana tabacum glutathione S-transferase. parB.
CAA96431.1 Z71749 Nicotiana plumbaginifolia glutathione S-tiansferase.
AAA33931.1 M84969 Silene vulgaris glutathione-S-transferase.
AAA33930.1 M84968 Silene vulgaris glutathione-S-transferase.
AAF65767.1 AF242309 Euphorbia esula glutathione S-transferase. putative auxin-binding GST.
AAF61392.1 AF133894 Persea americana glutathione S-transferase. GTH.
CAB38119.1 AJ010296 Zea mays
Glutathione transferase 111(b). gst3b.
CAB38118.1 AJ010295 Zea mays
Glutathione transferase IΙI(a). gst3a.
BAB39935.1 AP002914 Oryza sativa putative glutathione S-tiansferase. P0493G01.17.
AAG34811.1 AF243376 Glycine max glutathione S-transferase GST 21.
BAB39939.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.21.
AAG34812.1 AF243377 Glycine max glutathione S-tiansferase GST 22.
AAG34814.1 AF243379 Glycine max glutathione S-tiansferase GST 24.
CAA09190.1 AJ010451 Alopecurus myosuroides glutathione transferase. GST2a.
AAG32476.1 AF309383 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTF4.
BAB39929.1 AP002914 Oryza sativa putative glutathione transferase. P0493G01.7.
CAA09193.1 AJO 10454 Alopecurus myosuroides glutathione transferase. GST2d.
BAB39941.1 AP002914 Oryza sativa putative glutathione S-transferase. P0493G01.23.
CAA09191.1 AJO 10452 Alopecurus myosuroides glutathione transferase. GST2b.
CAA09192.1 AJ010453 Alopecurus myosuroides glutathione transferase. GST2c.
AAD56395.1 AFl 84059 Triticum aestivum glutathione S-transferase. GSTl.
CAA39487.1 X56012 Triticum aestivum glutathione transferase. gstAl.
AAG32477.1 AF309384 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTF3.
CAA68993.1 Y07721 Petunia x hybrida conjugates glutathione to anthocyanin to facilitate transport to the vacuole. glutathione S- tiansferase. an9 locus.
AAA33470.1 M16901 Zea mays glutathione S-transferase I.
AAA33469.1 M16902 Zea mays glutathione S-transferase I.
AAA20585.1 U12679 Zea mays glutathione S-transferase IV. GSTIV.
CAA56047.1 X79515 Zea mays glutathione transferase. GST27.
CAB66333.1 AJ279691 Betula pendula glutathione-S-transferase. gst
BAB39927.1 AP002914 Oryza sativa putative glutathione S-tiansferase. P0493G01.1. contains ESTs
AU031696(R0596),C97559(C60386),C28218(C60386), D28287(R0596).
AAC64007.1 AF062403 Oryza sativa glutathione S-transferase II.
CAA39480.1 X56004 Triticum aestivum glutathione transferase. gstA2.
AAG32475.1 AF309382 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTF5.
AAG34823.1 AF244680 Zea mays glutathione S-tiansferase GST 15.
BAB39940.1 AP002914 Oryza sativa putative glutathione S-tiansferase. P0493G01.22.
AAG34818.1 AF244675 Zea mays glutathione S-transferase GST 10.
AAG34820.1 AF244677 Zeamays glutathione S-tiansferase GST 11.
AAG34817.1 AF244674 Zea mays glutathione S-tiansferase GST 9.
AAG34821.1 AF244678 Zeamays glutathione S-tiansferase GST 13.
CAA05354.1 AJ002380 Oryza sativa glutathione S-transferase. Rgst I.
AAG34816.1 AF244673 Zea mays glutathione S-transferase GST 8.
AAG34822.1 AF244679 Zea mays glutathione S-tiansferase GST 14.
CAA05355.1 AJ002381 Oryza sativa glutathione S-transferase. Rgst II.
SEQ ID NO: 458
AAD45623.1 AF084185 Brassica napus dehydration responsive element binding protein. DNA binding protein; DRE binding protein.
BAA90812.1 AP001168 Oryza sativa
Similar to mRNA for DREB1 A (AB007787).
AAK01089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor.
AAG59618.1 AF239616 Hordeum vulgare CRT/DRE-binding factor. CBF.
AAK01088.1 AF298230 Hordeum vulgare
CBF 1 -like protein BCBFl. BCBFl. AP2 domain protein; putative DRE binding factor.
AAG59619.1 AF243384 Oryza sativa CRT/DRE binding factor. CBF. DREB.
AAC29516.1 U77655 Solanum tuberosum DNA binding protein homolog. STWAAEIRD.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserB.
BAA97124.1 AB016266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
AAC50047.1 U89255 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti4.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. similar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
AAC14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. OSJNBa0027P10.12.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
AAC49741.1 U89257 Lycopersicon esculentum
DNA-binding protein binds the GCC box pathogenesis-related promoter element. Ptiβ.
BAA97122.1 AB016264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
BAB21218.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.24.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding proteinl homolog. McEREBPl.
AAC49740.1 U89256 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti5.
AAB38748.1 U81157 Nicotiana tabacum
S25-XP1 DNA binding protein.
BAB21211.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.17.
BAA78732.1 AB023482 Oryza sativa
Similar to Arabidopsis thaliana mRNA for DREBIB, complete cds.(AB007788). SEQ ID NO: 460
AAB67721.1 AF015269 Zeamays
PL transcription factor. PI. nonfunctional due to retrotransposon insertion.
SEQ ID NO: 461 AAD50592.1 AF093752 Triticum aestivum phytochelatin synthase. PCS1.
AAG22095.1 AF308658 Typha latifolia phytochelatin synthase.
SEQ ID NO: 463
AAG13629.1 AC078840 Oryza sativa putative steroid membrane binding protein. OSJNBb0073N24.4.
AAG13623.1 AC078840 Oryza sativa putative steroid membrane binding protein. OSJNBb0073N24.2.
SEQ ID NO: 464
CAA56313.1 X79992 Avena sativa putative pρ70 ribosomal protein S6 kinase. Aspkl 1.
CAB89082.1 AJ277534 Asparagus officinalis S6 ribosomal protein kinase. pkl . putative.
AAK18843.1 AC082645 Oryza sativa putative protein kinase. OSJNBb0033N16.3.
AAC05083.1 AF033096 Avena sativa
NPH1-1. NPHl-1. putative serine/threonine protein kinase.
AAC05084.1 AF033097 Avena sativa
NPHl -2. NPHl -2. putative serine/threonine protein kinase.
AAK13156.1 AC078829 Oryza sativa putative protein kinase. OSJNBa0026O12.14.
CAB82852.1 Z30329 Mesembryanthemum crystallinum protein kinase MK6.
AAB88817.1 AF033263 Zeamays signal transduction for phototropism. nonphototiopic hypocotyl 1. nphl. NPHl; putative serine/threonine kinase; similar to oat NPHl proteins.
BAB18105.1 AB042715 Chlamydomonas reinhardtii cyclic nucreotide dependent protein kinase II. CL-PK2.
BAB18104.1 AB042714 Chlamydomonas reinhardtii cyclic nucleotide dependent protein kinase. CL-PK1.
BAA83689.1 AB011968 Oryza sativa OsPK7. OsPK7. protein kinase.
BAA92970.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F28A21 ; putative protein kinase.
(AL035526).
AAD31900.1 AF145482 Mesembryanthemum crystallinum putative serine/threonine protein kinase.
BAA92972.1 AP001551 Oryza sativa
ESTs AU056183(S20356),AU056881(S20950) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 4 BAC clone F6I18 ; putative protein kinase. (AL022198).
BAA83688.1 AB011967 Oryza sativa
OsPK4. OsPK4. protein kinase.
CAA73067.1 Y12464 Sorghum bicolor serine/threonine kinase. SNFL1.
AAF22219.1 AF141378 Zeamays protein kinase PK4. ZmPK4.
BAA96628.1 AP002482 Oryza sativa
ESTs D41739(S4522),AU055999(S20214), AU057588(S21592 conespond to a region ofthe predicted gene. Similar to Sorghum bicolor serme/threonine kinase (Y12465).
CAA89202.1 Z49233 Chlamydomonas eugametos calcium-stimulated protein kinase.
CAA73068.1 Y12465 Sorghum bicolor serine/threonine kinase. SNFL2.
BAA34675.1 ABO 11670 Triticum aestivum wpk4 protein kinase. wpk4.
BAA99439.1 AP002743 Oryza sativa putative protein kinase. P0710E05.26. contains ESTs C22394(C30013),C22393(C30013).
CAA74646.1 Y14274 Sorghum bicolor putative serine/threonine protein kinase. SNFL3.
BAB12687.1 AP002746 Oryza sativa putative protein kinase. P0671B11.2. contains ESTs C22394(C30013),C22393(C30013).
AAF06969.1 AF162661 Kalanchoe fedtschenkoi phosphoenolpyravate carboxylase kinase. calcium-independent protein kinase.
AAF06970.1 AF162662 Kalanchoe fedtschenkoi phosphoenolpyravate carboxylase kinase. calcium-independent protein kinase.
BAA90814.1 AP001168 Oryza sativa
ESTs AU030197(E50746),AU030196(E50746) conespond to a region ofthe predicted gene.;
Similar to calcium-dependent cahnodulin-independent protein kinase CDPK (U90262).
AAB62693.1 AF004947 Oryza sativa protein kinase.
AAF21062.1 AF216527 Dunaliella tertiolecta calcium-dependent protein kinase. CPK1; CDPK.
CAA71142.1 Y10036 Cucumis sativus
SNF1 -related protein kinase.
CAA39936.1 X56599 Daucus carota calcium- dependent protein kinase. DcPK431.
CAA08995.1 AJ010091 Brassica napus MAP3K alpha 1 protein kinase. MAP3K alpha 1.
BAA05649.1 D26602 Nicotiana tabacum protein kinase.
AAF 19403.1 AF203481 Lycopersicon esculentum phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAC25423.1 AF072908 Nicotiana tabacum calcium-dependent protein kinase. CDPK1.
AAF 19402.1 AF203480 Lycopersicon esculentum phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
CAA65244.1 X95997 Solanum tuberosum
SNFl-related protein kinase. PKINl.
CAA57898.1 X82548 Hordeum vulgare
SNFl-related protein kinase. BKIN2.
AAF19401.1 AF203479 Glycine max phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAD23582.1 AF128443 Glycine max probably involved in plant stress responses possibly regulates gene expression. SNF-1-like serine/threonine protein kinase. expressed in nodules, roots and leaves.
AAF34436.1 AF172282 Oryza sativa similar to mitogen-activated protein kinases. DUPR11.32.
BAA05648.1 D26601 Nicotiana tabacum protein kinase.
SEQ ID NO: 465
CAB85467.1 AJ250316 Brassica juncea chloroplast omega-3 fatty acid desaturase. fad7.
BAA22441.1 D63954 Zea mays fatty acid desaturase. FAD7.
BAA11475.1 D79979 Nicotiana tabacum omega-3 fatty acid desaturase. NtFAD7.
AAA70334.1 U25817 Sesamum indicum omega-3 fatty acid desaturase. linoleic acid and hexadecadienoic acids desaturase.
AAB39387.1 U59477 Perilla frutescens omega-3 fatty acid desaturase.
CAA07638.1 AJ007739 Solanum tuberosum w-3 desaturase.
AAF27933.1 AF222989 Capsicum annuum omega-3 fatty acid desaturase. FAD.
AAB72241.1 U75745 Petroselinum crispum omega-3 fatty acid desaturase. complements the Arabidopsis fad7/8 fatty acid double mutant.
AAA61776.1 L22965 Chloroplast Glycine soj a omega-3 fatty acid desaturase. Fadd.
AAF12821.1 AF200717 Vernicia fordii TnDES2. omega-3 fatty acid desaturase.
AAA86690.1 U17063 Limnanthes douglasii linoleic acid desaturation. delta- 15 lineoyl desaturase. omega-3 -fatty acid desaturase.
AAA73511.1 L25897 Ricinus communis desaturates linoleate to alpha-linolenate. linoleoyl desaturase. fad7. putative.
CAC18722.1 AJ302017 Picea abies putative plastidial w-3 fatty acid desaturase. fad3.
AAD13527.1 AF061027 Vernicia fordii omega-3 fatty acid desaturase precursor.
BAA22442.1 D84409 Zeamays fatty acid desaturase. FAD8.
BAA22440.1 D63953 Zea mays fatty acid desaturase. FAD8.
BAA07785.2 D43688 Triticum aestivum plastid omega-3 fatty acid desaturase. TaFAD7.
AAA61774.1 L22963 Chloroplast Brassica napus omega-3 fatty acid desaturase. Fadd.
AAC98967.1 AF047172 Vernicia fordii omega-3 fatty acid desaturase. Fad3.
CAB45155.1 AJ011004 Vernicia fordii omega-3 desaturase.
AAC 16443.1 AF020204 Pelargonium x hortorum omega-3 desaturase. pxh-15.
AAA61775.1 L22962 Brassica napus omega-3 fatty acid desaturase. Fad3.
AAA61777.1 L22964 Chloroplast Glycine soja omega-3 fatty acid desaturase. Fad3.
AAA32994.1 L01418 Brassica napus desaturation of linoleic acid to linolenic acid, linoleic acid desaturase. fad3.
AAD15744.1 AF047039 Perilla frutescens omega-3 fatty acid desaturase. FAD3.
BAA28358.1 D84678 Triticum aestivum omega-3 fatty acid desaturase. TaFAD3.
BAA05515.1 D26509 Nicotiana tabacum desaturation of dienoic fatty acid, microsomal omega-3 acid desaturase. NtFAD3.
BAA11397.1 D78506 Oryza sativa w-3 fatty acid desaturase.
BAB18135.1 AB051215 Glycine max microsomal omega-3 fatty acid desaturase.
BAA22439.1 D63952 Zea mays fatty acid desaturase. FAD7.
BAA11396.1 D78505 Oryza sativa w-3 fatty acid desaturase.
CAB71341.1 AJ250664 Hordeum vulgare omega-3 fatty acid desaturase. bci-8.
AAD48897.1 AF083613 Dunaliella salina omega-3 fatty acid desaturase. des3-l.
AAF80560.1 AFl 92486 Sesamum indicum omega-6 fatty acid desaturase. FAD2.
CAB64256.1 AJ245938 Calendula officinalis production of calendic acid. (8,ll)-linoleoyl desaturase. des8.11.
AAB80696.1 U86072 Petroselinum crispum omega-6 fatty acid desaturase. functional expression in yeast results in the formation of dienoic fatty acids (18:2 and 16:2).
SEQ ID NO: 467
CAA60016.1 X86021 Solanum tuberosum potassium channel. SKT1 gene, putative start codon.
CAA65254.1 X96390 Lycopersicon esculentum potassium channel. LKT1.
AAF36832.1 AF207745 Triticum aestivum AKTl-like potassium channel. TaAKTl.
CAA68912.1 Y07632 Zea mays potassium channel. ZMK1.
AAF81249.1 AF267753 Mesembryanthemum crystallinum putative potassium channel protein Mktlp.
CAC05489.1 AJ271447 Populus tremula x Populus tremuloides potassium channel, potassium channel 2. ptk2.
AAD 16278.1 AF099095 Samanea saman pulvinus inward-rectifying channel for potassium SPICKl. similar to Arabidopsis potassium channel AKT3.
CAB54856.1 AJ132686 Zea mays potassium channel protein ZMK2. ZMK2.
CAA70870.1 Y09699 Solanum tuberosum putative inward rectifying potassium channel. SKT2.
CAA71598.1 Y10579 Vicia faba potassium channel.
BAA84085.1 AB032074 Nicotiana paniculata potassium channel. NpKTl.
CAA12645.1 AJ225805 Egeria densa inward potassium channel alpha subunit. homologous to the sequences ofthe family of inwardly rectifying potassium channels in plants which is structurally related to the shaker family of outwardly rectifying channels in Drosophila.
AAD39492.1 AF145272 Samanea saman pulvinus inward-rectifying channel SPICK2. potassium channel; similar to AKT2/3.
CAC10514.1 AJ299019 Samanea saman potassium release, outwardly rectifying potassium channel, sporkl .
CAC05488.1 AJ271446 Populus tremula x Populus tremuloides potassium channel, outward rectifying potassium channel, ptork.
AAF81251.1 AF267755 Mesembryanthemum crystallinum potassium channel protein Mkt2p.
CAA70900.1 Y09753 Secale cereale potassium channel, encodes membrane spanning domains H5-S6.
CAA70899.1 Y09752 Secale cereale potassium channel, encodes membrane spanning domains H5-S6.
CAA70947.1 Y09818 Solanum tuberosum putative inward rectifying potassium channel. SKT3.
CAA70895.1 Y09748 Hordeum vulgare potassium channel, encodes membrane spanning domains H5-S6.
CAA70894.1 Y09747 Zea mays potassium channel, encodes membrane spanning domains H5-S6.
CAA70896.1 Y09749 Vicia faba potassium channel, encodes membrane spanning domains H5-S6.
CAA70897.1 Y09750 Plantago major potassium channel, encodes membrane spanning domains H5-S6.
AAF33670.1 AF079872 Nicotiana tabacum cyclic nucleotide-gated calmodulin-binding ion channel. CBP4.
AAF33669.1 AF079871 Nicotiana tabacum cyclic nucleotide-gated calmodulin-binding ion channel. CBP7.
CAA05637.1 AJ002610 Hordeum vulgare putative calmodulin binding transporter protein. CBT1.
CAA70898.1 Y09751 Plantago major potassium channel, encodes membrane spanning domains H5-S6.
AAK16188.1 AC079887 Oryza sativa putative cyclic nucleotide and cahnodulin-regulated ion channel protein. OSJNBa0040E01.13.
SEQ ID NO: 468 AAF76226.1 AF272572 Populus x canescens
14-3-3 protein. 14-3-3P20-1.
AAC04811.1 AF037460 Fritillaria agrestis
GF14 protein. GRF.
AAB40395.1 U80070 Mesembryanthemum crystallinum
G-box binding factor. 14-3 -3 -like protein. GBF.
AAF05737.1 AF191746 Lilium longiflorum
14-3 -3 -like protein.
AAB07457.1 U65957 Oryza sativa
GF14-C protein, rice 14-3-3 protein homolog; osGF14c.
AAB09580.1 U70533 Glycine max
SGF14A. 14-3-3 related protein.
AAA99431.1 L29150 Lycopersicon esculentum
14-3-3 protein homologue.
CAA74592.1 Y14200 Hordeum vulgare
14-3-3 protein.
AAB33304.1 S77133 Zeamays
GF14-6. GRFl. 14-3-3 protein homolog; This sequence comes from Fig. 5.
CAA66309.1 X97724 Solanum tuberosum 14-3-3 protein, leaf specific.
AAB07456.1 U65956 Oryza sativa
GF14-b protein, rice 14-3-3 protein homolog; osGF14b.
AAD27823.2 AF121194 Populus x canescens 14-3-3 protein. 14-3-3P20-2.
AAC49894.1 U91726 Nicotiana tabacum 14-3-3 isoform e. T14-3e.
CAA44259.1 X62388 Hordeum vulgare 14-3-3 protein homologue.
AAD27827.2 AF121198 Picea glauca 14-3-3 protein. 14-3-3EB9D.
CAA63658.1 X93170 Hordeum vulgare Hvl4-3-3b.
CAB42546.2 AJ238681 Pisum sativum 14-3-3-like protein. 14-3-3.
AAA33505.1 M96856 Zea mays regulatory protein. GF14-12.
AAA85817.1 U15036 Pisum sativum 14-3-3-like protein.
AAK26634.1 AF342780 Brassica napus GF14 omega. 14-3-3 protein.
CAA72094.1 Y11211 Nicotiana tabacum 14-3-3-like protein B.
CAA72383.1 Y11687 Solanum tuberosum 14-3-3 protein. 34G.
AAC49892.1 U91724 Nicotiana tabacum 14-3-3 isoform c. T14-3c.
CAA53700.1 X76086 Cucurbita pepo
14-3-3 protein 32kDa endonuclease. A215. single polypeptide.
CAA72382.1 Y11686 Solanum tuberosum 14-3-3 protein. 30G.
CAB42547.1 AJ238682 Pisum sativum 14-3-3-like protein. 14-3-3.
CAA65147.1 X95902 Lycopersicon esculentum 14-3-3 protein. tft3 gene.
AAC49891.1 U91723 Nicotiana tabacum 14-3-3 isoform b. T14-3b.
AAB07458.1 U65958 Oryza sativa
GF14-d protein, rice 14-3-3 protein homolog; osGF14d.
CAA72381.1 Y11685 Solanum tuberosum 14-3-3 protein. 16R.
AAC49895.1 U91727 Nicotiana tabacum 14-3-3 isoform f. T14-3f.
CAA72095.1 Y11212 Nicotiana tabacum 14-3-3-like protein A.
BAB11739.1 AB042193 Triticum aestivum
TaWINl. TaWINl. TaWINl is a member of 14-3-3 protein family.
CAA65146.1 X95901 Lycopersicon esculentum 14-3-3 protein. tft2 gene.
CAA65148.1 X95903 Lycopersicon esculentum 14-3-3 protein. tft5 gene.
AAC 17447.1 AF066076 Helianthus annuus 14-3-3-like protein.
CAA60800.1 X87370 Solanum tuberosum 14-3-3 protein. RA215. root specific.
CAA55964.1 X79445 Chlamydomonas reinhardtii 14-3-3 protein.
CAC03467.1 Y19105 Chlamydomonas reinhardtii 14-3-3 protein.
CAA65149.1 X95904 Lycopersicon esculentum 14-3-3 protein. tft6 gene.
CAB65693.1 AJ270959 Lycopersicon esculentum tft3 14-3-3 protein. tft3.
BAB11740.1 AB042194 Triticum aestivum
TaWIN2. TaWIN2. TaWIN2 is a member of 14-3-3 protein family.
AAA99430.1 L29151 Lycopersicon esculentum 14-3-3 protein homologue.
CAA72384.1 Y11688 Solanum tuberosum 14-3-3 protein. 35G.
CAA65145.1 X95900 Lycopersicon esculentum 14-3-3 protein, tftl gene.
AAC49893.1 U91725 Nicotiana tabacum 14-3-3 isoform d. T14-3d.
SEQ ID NO: 470
AAA19701.1 L24438 Thlaspi arvense cytochrome P450.
AAA32913.1 M32885 Persea americana cytochrome P-45QLXXIA1 (cyp71Al).
AAC39318.1 AF029858 Sorghum bicolor second multifunctional cytochrome P450 in the biosynthetic pathway ofthe cyanogenic glucoside dhunin. Catalyzes the conversion of p-hydroxyphenylacetaldoxime to p- hydroxymandelonitrile. cytochrome P450 CYP71E1. CYP71El. No EST#s identified.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
AAB94589.1 AF022460 Glycine max
CYP83Dlp. CYP83D1. cytochrome P450 monooxygenase.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71A6.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
AAB94588.1 AF022459 Glycine max
CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
CAA50312.1 X70981 Solanum melongena P450 hydroxylase. CYPEG2.
CAA71514.1 Y10490 Glycine max putative cytochrome P450.
AAB94584.1 AF022157 Glycine max capable of catalyzing the metabolism of phenylurea herbicides. CYP71A10. CYP71 AIO. cytochrome P450 monooxygenase.
AAD47832.1 AF166332 Nicotiana tabacum cytochrome P450.
AAF27282.1 AF122821 Capsicum annuum cytochrome P450. PepCYP.
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
CAA83941.1 Z33875 Mentha x piperita cytochrome P-450 oxidase.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
AAB61964.1 U48434 Solanum chacoense putative cytochrome P450.
AAB69644.1 AF000403 Lotus japonicus putative cytochrome P450. LJNP450.
CAA50645.1 X71654 Solanum melongena P450 hydroxylase.
BAA03635.1 D14990 Solanum melongena Cytochrome P-450EG4.
AAD44151.1 AFl 24816 Mentha x piperita cytochrome p450 isoform PM17.
AAD44150.1 AF124815 Mentha spicata cytochrome p450.
AAD44152.1 AF124817 Mentha x piperita cytochrome p450 isoform PM2.
BAB39252.1 AP002968 Oryza sativa putative cytochrome P450. P0416G11.1.
AAK38084.1 AF321860 Lolium rigidum putative cytochrome P450.
AAK38083.1 AF321859 Lolium rigidum putative cytochrome P450.
AAK38087.1 AF321863 Lolium rigidum putative cytochrome P450.
AAK38082.1 AF321858 Lolium rigidum putative cytochrome P450.
CAC27827.1 AJ295719 Catharanthus roseus geraniol hydroxylase. cytochrome P450. cyp71.
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
AAK38088.1 AF321864 Lolium rigidum putative cytochrome P450.
AAD37433.1 AF 150881 Lycopersicon esculentum x Lycopersicon peravianum catalyzes the hydroxylation of feralic acid to 5-hydroxyferalic acid, feralate-5-hydroxylase. CYP84. cytochrome P450-dependent monooxygenase; F5H; FAHL
AAG14963.1 AF214009 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H3.
CAA57425.1 X81831 Zeamays cytochrome P450. CYP71C4. family CYP71, subfamily CYP71C.
CAA72196.1 Y11368 Zeamays cytochrome p450. cyp71c4.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
AAG14962.1 AF214008 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2.
AAG14961.1 AF214007 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3'-hydroxylase. Htl. cytochrome P450; CYP75B2.
SEQ ID NO: 471
AAD17855.1 AF099111 Zea mays sigma factor. sig2. putative sigma subunit of chloroplast Escherichia coli-like RNA polymerase; Sig2. SEQ ID NO: 472
AAD45623.1 AF084185 Brassica napus dehydration responsive element binding protein. DNA binding protein; DRE binding protein.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP transcription factors.
AAKO 1089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor.
AAG59618.1 AF239616 Hordeum vulgare
CRT/DRE-binding factor. CBF.
AAK01088.1 AF298230 Hordeum vulgare
CBF 1 -like protein BCBF BCBF AP2 domain protein; putative DRE binding factor.
AAG59619.1 AF243384 Oryza sativa
CRT/PRE binding factor. CBF. DREB.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
CAC12822.1 AJ299252 Nicotiana tabacum AP2 domain-containing transcription factor. ap2.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor.
AAC29516.1 U77655 Solanum tuberosum DNA binding protein homolog. STWAAEIRD.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBP EREBP/AP2-like transcription factor.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal .
BAB 16083.1 AB036883 Oryza sativa transcriptional repressor. osERF3. osERF3. ERF protein family ERF3 associated repression domain.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
BAA97124.1 ABO 16266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf3.
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREL similar to EREBP transcription factors.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. shnilar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
AAC49741.1 U89257 Lycopersicon esculentum
DNA-binding protein binds the GCC box pathogenesis-related promoter element. Pti6.
AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. OSJNBa0027P10.12.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
AAC14323.1 AF058827 Nicotiana tabacum
TSI1. Tsil. contains putative AP2 DNA-binding domain; similar to Ptiό.
AAC50047.1 U89255 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti4.
BAA97122.1 ABO 16264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding proteinl homolog. McEREBPl.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
BAB21218.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.24.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive transcription factor.
AAC49740.1 U89256 Lycopersicon esculentum binds the GCC box pathogenesis-related promoter element DNA-binding protein. Pti5.
AAB38748.1 U81157 Nicotiana tabacum
S25-XP1 DNA binding protein.
BAB21211.1 AP002913 Oryza sativa putative DNA binding protein RAV2. P0480E02.17. SEQ ID NO: 473
BAA85438.1 AP000616 Oryza sativa similar to RING-H2 finger protein RHAla (AF078683).
AAK00436.1 AC060755 Oryza sativa putative zinc finger protein. OSJNBa0003O19.23.
AAG43550.1 AF211532 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 132. ACRE132. similar to RING finger proteins. SEQ ID NO: 475
AAG43550.1 AF211532 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 132. ACREl 32. similar to RING finger proteins.
BAA78746.1 AB023482 Oryza sativa
Similar to Arabidopsis thaliana RING-H2 finger protein RHXla mRNA, partial cds.(AF079184).
BAA96875.1 AB045121 Oryza sativa RING finger 1. RRF1.
CAA74911.1 Y14573 Hordeum vulgare ring finger protein, putative.
AAK00436.1 AC060755 Oryza sativa putative zinc finger protein. OSJNBa0003O19.23.
AAG46117.1 AC073166 Oryza sativa putative ring finger protein. OSJNBb0064P21.7.
BAA90357.1 AP001080 Oryza sativa
EST AU070319(S 10788) conesponds to a region of the predicted gene. Similar to RING-H2 finger protein RHA2b (AC006200).
BAA77204.1 AB026262 Cicer arietinum ring finger protein.
BAA90806.1 AP001168 Oryza sativa
ESTs C26000(C11448),AU082130(C11448) conespond to a region ofthe predicted gene.
Similar to mRNA for zinc-finger protein (Z36749).
SEQ ID NO: 476
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APK1 gene for protein tyrosine-serine-threonine kinase.(D12522). AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S 13927).
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKL accumulates in mesophyll cells.
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil. Ptil kinase.
BAA87852.1 AP000816 Oryza sativa
Shnilar to putative Ser/Thr protein kinase. (AC004218).
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
AAB09771.1 U67422 Zea mays CRJNKLY4 precursor. cr4. receptor kinase homolog.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAC27894.1 AF023164 Zea mays leucine-rich repeat transmembrane protein kinase 1. ltkl.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
AAF91336.1 AF249317 Glycine max Ptil kinase-like protein. Ptil a. protein kinase.
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
AAG33377.1 AF290411 Oryza meyeriana serine/threonine protein kinase. Rl .
AAC27895.1 AF023165 Zea mays leucine-rich repeat transmembrane protein kinase 2. Itk2.
AAK11566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhirPtoE. confers resistance to bacterial speck disease.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
AAF34428.1 AF172282 Oryza sativa receptor-like protein kinase. DUPR11.18.
AAF76306.1 AF220602 Lycopersicon pimpinellifolium Pto kinase.
AAB47423.1 U59315 Lycopersicon pimpinellifolium serine/threonine protein kinase Pto. Pto. bacterial speck disease resistance in tomato; disease resistance gene.
AAC48914.1 U02271 Lycopersicon pimpinellifolium protein kinase.
AAK11567.1 AF318491 Lycopersicon hirsutum Pto-like protein kinase F. LhirPtoF.
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
AAF76313.1 AF220603 Lycopersicon esculentum Pto kinase. LescPth5.
AAF66615.1 AFl 42596 Nicotiana tabacum LRR receptor-like protein kinase.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPK10) mRNA. (L27821).
BAA92953.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F10M6 ; S-receptor kinase -like protein. (AL021811).
SEQ ID NO: 479
CAA42622.1 X60007 Nicotiana sylvestris nsGRP-2. putative glycine-rich protein of 19.7kDa.
SEQ ID NO: 480
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region of the predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl. GmRLKl.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAB36558.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl. impkl. leucine-rich repeat.
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml .
AAF59906.1 AF197947 Glycine max receptor protein kinase-like protein. CLV1B.
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBa0010K01.7.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1 A.
CAC20842.1 AJ250467 Pinus sylvestris receptor protein kinase. upk.
BAB03629.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.30.
BAB03621.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.21.
BAB03631.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.32.
AAB82755.1 U72725 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member Al; downstream of microsatellite region; disease resistance gene family member.
AAF34426.1 AF172282 Oryza sativa leucine rich repeat containing protein kinase. DUPR11.16.
AAK27806.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.21.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
AAC49123.1 U37133 Oryza sativa receptor kinase-like protein. Xa21. Xa21 disease resistance gene.
AAC80225.1 U72723 Oryza longistaminata receptor kinase-like protein. Xa21. disease resistance gene.
AAK27817.1 AC022457 Oryza sativa putative protein kinase. QSJNBa0006L06.16.
BAA82393.1 AP000367 Oryza sativa
EST C96716(C10608) conesponds to a region ofthe predicted gene.; Similar to putative receptor protein kinase. (AC002334).
BAB03627.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.27.
AAB82756.1 U72724 Oryza sativa receptor kinase-like protein. Xa21 gene family member E.
AAG52992.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl a. impkl.
BAA88636.1 AB029327 Nicotiana tabacum elicitor-inducible LRR receptor-like protein EILP. EILP.
AAB82753.1 U72726 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member D.
AAG52994.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl c. impkl .
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl . accumulates in mesophyll cells.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
SEQ ID NO: 481
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
BAA22422.1 AB001379 Glycynhiza echinata cytochrome P450. CYP81E1.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
BAA74465.1 AB022732 Glycynhiza echinata cytochrome P450. CYP Ge-31.
CAB41490.1 AJ238439 Cicer arietinum cytochrome P450 monooxygenase. cyp81E3v2.
CAA10067.1 AJ012581 Cicer arietinum cytochrome P450. cyp81E3.
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'- race).
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81Blc.
AAK38080.1 AF321856 Lolium rigidum putative cytochrome P450.
AAK38079.1 AF321855 Lolium rigidum putative cytochrome P450.
AAK38081.1 AF321857 Lolium rigidum putative cytochrome P450.
AAG09208.1 AF175278 Pisum sativum wound-inducible P450 hydroxylase. CYP82A1.
CAA71876.1 Y10982 Glycine max putative cytochrome P450.
AAC49188.2 U29333 Pisum sativum cytochrome P450 monooxygenase. CYP82. new cytochrome P450 family.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
CAA71515.1 Y10491 Glycine max putative cytochrome P450.
BAA92894.1 AB006790 Petunia x hybrida cytochrome P450. IMT-2.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
AAD56282.1 AFl 55332 Petunia x hybrida flavonoid 3 '-hydroxylase. Htl. cytochrome P450; CYP75B2.
AAC34853.1 AF082028 Hemerocallis hybrid cultivar putative cyt P450-containing fatty acid hydroxylase. senescence-associated protein 3. SA3. mRNA accumulates in senescing petals.
BAA12159.1 D83968 Glycine max
Cytochrome P-450 (CYP93A1).
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
CAA71877.1 Y10983 Glycine max putative cytochrome P450.
CAA71516.1 Y10492 Glycine max putative cytochrome P450.
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
AAC39454.1 AF014802 Eschscholzia californica
(S)-N-methylcoclaurine 3'-hydroxylase. CYP82B1. cytochrome P-450-dependent monooxygenase; methyl jasmonate-inducible cytochrome P-450-dependent, homologous to wound-inducible CYP82A1 of Pisum sativum GenBank Accession Number U29333.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71A6.
AAB94590.1 AF022461 Glycine max
CYP82Clp. CYP82C1. cytochrome P450 monooxygenase.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
AAA19701.1 L24438 Thlaspi arvense cytochrome P450.
BAA84071.1 AB028151 Antinhinum majus flavone synthase II. cytochrome P450. AFNS2.
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
AAD38930.1 AF135485 Glycine max cytochrome P450 monooxygenaseCYP93Dl. CYP93E1.
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5' -hydroxylase !. Hfl. P450 enzyme.
SEQ ID NO: 487
BAA96181.1 AP002093 Oryza sativa
EST AU056651(S20760) conesponds to a region ofthe predicted gene. Similar to
Arabidopsis thaliana chromosome II BAC F19I3; putative amino acid transporter
(AC004238).
BAA96139.1 AP002092 Oryza sativa
EST AU056651(S20760) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome II BAC F19I3; putative amino acid transporter (AC004238). SEQ ID NO: 495
AAB71528.1 U94784 Helianthus annuus ATPase. unconventional myosin. hamy4. Hamyo4.
AAB71529.1 U94785 Helianthus annuus ATPase. unconventional myosin. hamy5. Hamyo5.
AAB71527.1 U94782 Helianthus annuus ATPase. unconventional myosin. hamy2. Hamyo2.
AAD17931.2 AF104924 Zea mays unconventional myosin heavy chain. MYO1. ATPase; similar to myosin class XI.
AAC27525.1 AF077352 Chlamydomonas reinhardtii myosin heavy chain. MYOl. class XI.
AAK21311.1 AF338254 Petroselinum crispum myosin subfamily XI heavy chain. PCM3.
BAA87057.1 AB034154 Chara corallina unconventional myosin heavy chain, cem.
BAB03273.1 AB007459 Chara corallina cytoplasmic streerning. myosin. ccml. CCM1.
AAF43440.1 AF233886 Vallisneria gigantea unconventional myosin XI. VMYO1.
AAB53062.1 U94398 Acetabularia cliftonii myosin. myo2.
AAD34597.1 AF147739 Zeamays myosin XI. M2. unconventional myosin.
AAB53061.1 U94397 Acetabularia cliftonii myosin. myol.
AAD31926.1 AF147738 Zeamays myosin VIII ZMM3. zmm3. unconventional myosin.
AAB71526.1 U94781 Helianthus annuus unconventional myosin. hamyl. homologous to ATM1.
AAG49341.1 AF319457 Petroselinum crispum myosin subfamily VIII heavy chain.
AAB93521.1 U94783 Helianthus annuus unconventional myosin. hamy3. Hamyo3; similar to class VIII myosin.
CAA47476.1 X67102 Anemia phyllitidis myosin heavy chain, myol 5.
CAA47477.1 X67103 Anemia phyllitidis myosin heavy chain. myo22.
AAF43441.1 AF233887 Vallisneria gigantea unconventional myosin VIII. VMYO2.
AAG13633.1 AC078840 Oryza sativa putative myosin. OSJNBb0073N24.16.
AAA92120.1 U48785 Vigna mungo plant myosin MBM1. mbml . head coding region.
SEQ ID NO: 496
AAA34138.1 M96324 Lycopersicon esculentum
The calcium ATPase is a calcium ion pump. Ca2+-ATPase. LCAL
AAD 11618.1 AF050496 Lycopersicon esculentum Ca2+-ATPase. LCA1B; alternative transcript.
AADl 1617.1 AF050495 Lycopersicon esculentum Ca2+-ATPase. LCAl A; alternative transcript.
BAA90510.2 AP001111 Oryza sativa rice EST AU030811, similar to rice Ca+2-ATPase (U82966).
CAA63790.1 X93592 Dunaliella bioculata P-type ATPase. cal . calcium pumping; CA
AAB58910.1 U82966 Oryza sativa Ca2+-ATPase.
AAF73985.1 AF096871 Zeamays calcium pump, calcium ATPase. capl.
AAG28436.1 AFl 95029 Glycine max plasma membrane Ca2+-ATPase. SCA2.
AAG28435.1 AF195028 Glycine max plasma membrane Ca2+- ATPase. SCA1.
CAA68234.1 X99972 Brassica oleracea calmodulin-stimulated calcium-ATPase.
AAD31896.1 AF145478 Mesembryanthemum crystallinum calcium ATPase.
CAB85494.1 AJ132891 Medicago truncatula proton pump. H+-ATPase. hal.
CAB85495.1 AJ132892 Medicago truncatula proton pump. H+- ATPase. hal.
CAB69824.1 AJ271439 Prunus persica plasma membrane H+ ATPase. PPA1.
CAA52107.1 X73901 Dunaliella bioculata plasma membrane ATPase. pmal.
AAD46188.1 AF156691 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma9.
BAA01058.1 D10207 Oryza sativa H-ATPase. OSA1.
AAA34094.1 M80489 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pmal.
AAA34098.1 M80490 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pma3.
AAA34173.1 M60166 Lycopersicon esculentum H+-ATPase. LHAl.
CAA54046.1 X76536 Solanum tuberosum H(+)-transporting ATPase. PHA1.
AAD55399.1 AF179442 Lycopersicon esculentum plasma membrane H+- ATPase isoform LHA2. LHA2.
AAB49042.1 U54690 Dunaliella acidophila plasma membrane proton ATPase. dhal . DaDHAl ; proton pump.
AAF98344.1 AF275745 Lycopersicon esculentum plasma membrane H+- ATPase. LHA2. P-type ion pump.
BAA08134.1 D45189 Zostera marina plasma membrane H+- ATPase. zhal.
AAA34052.1 M27888 Nicotiana plumbaginifolia H+-translocatrng ATPase.
CAA59800.1 X85805 Zeamays H(+)-transporting ATPase. MHA-2.
AAB17186.1 U72148 Lycopersicon esculentum plasma membrane H+- ATPase. LHA4. plasma membrane proton pumping ATPase.
CAA54045.1 X76535 Solanum tuberosum H(+)-transporting ATPase. PHA2.
CAB69823.1 AJ271438 Prunus persica plasma membrane H+ ATPase. PPA2.
BAA06629.1 D31843 Oryza sativa plasma membrane H+-ATPase. OSA2.
CAC29435.1 AJ310523 Vicia faba
P-type H+-ATPase. vha4. predominantly expressed in flowers.
CAC29436.1 AJ310524 Vicia faba
P-type H+- ATPase. ha5. predominantly expressed in guard cells and flowers.
CAA59799.1 X85804 Phaseolus vulgaris H(+)-transporting ATPase. BHA-1.
AAD46186.1 AFl 56679 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma6.
AAB84202.2 AF029256 Kosteletzkya virginica plasma membrane proton ATPase. ATP1.
AAD46187.1 AFl 56683 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma8.
AAB60276.1 U09989 Zea mays H(+)-transporting ATPase. Mhal.
AAB41898.1 U84891 Mesembryanthemum crystallinum plasma membrane proton pump. H+-transporting ATPase. PMA.
AAB35314.2 S79323 Vicia faba plasma membrane H(+)- ATPase precursor, plasma membrane H(+)- ATPase. This sequence comes from Fig. 1; conceptual translation presented here differs from translation in publication.
AAK31799.1 AY029190 Lilium longiflorum plasma membrane H+ ATPase. LILHA1.
CAA47275.1 X66737 Nicotiana plumbaginifolia plasma membrane H+-ATPase. pma4.
BAA37150.1 AB022442 Vicia faba p-type H+-ATPase. VHA2.
AAG01028.1 AF289025 Cucumis sativus plasma membrane H+-ATPase.
AAA81348.1 U38965 Vicia faba p-type H+-ATPase. VHA2.
AAK32118.1 AF308816 Hordeum vulgare plasmalemma H+- ATPase 1.
AAF97591.1 AF263917 Lycopersicon esculentum plasma membrane proton ATPase. LHA8.
AAK32119.1 AF308817 Hordeum vulgare plasmalemma H+- ATPase 2.
AAA20601.1 U08985 Zeamays plasma-membrane H+ ATPase. Zmpmal.
SEQ ID NO: 497
AAB41742.1 U82559 Lycopersicon esculentum aldehyde oxidase 1 homolog. TAOl. has sequence properties ofthe molybdenum cofactor containing the hydroxylase genes aldehyde oxidase and xanthine dehydrogenase; belongs to a multigene family.
AAB41741.1 U82558 Lycopersicon esculentum aldehyde oxidase 1 homolog. TAOl. has sequence properties ofthe molybdenum cofactor containing the hydroxylase genes aldehyde oxidase and xanthine dehydrogenase; belongs to a multigene family. SEQ ID NO: 498
CAA73134.1 Y12531 Brassica oleracea serine/tlireonine kinase. BRLK.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl . receptor-like protein kinase; serine/threonine protein kinase.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/tlireonine kinase receptor.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
AAC23542.1 U20948 Ipomoea tiifida receptor protein kinase. IRK1.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B . c) .
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
AAF34428.1 AFl 72282 Oryza sativa receptor-like protein kinase. DUPRl 1.18.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRKl. Chrkl.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
BAA94517.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
AAA33915.1 L27821 Oryza sativa receptor type serine/tlireonine kinase. protein kinase.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAA94529.2 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
SEQ ID NO: 499
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml .
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
AAF59906.1 AFl 97947 Glycine max receptor protein kinase-like protein. CLV1B.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1A.
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl. GmRLKl.
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl.7.
CAC20842.1 AJ250467 Pinus sylvestris receptor protein kinase. upk.
AAB36558.1 U77888 Ipomoea nil receptor-like protein kinase INRPKL impkl. leucine-rich repeat.
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAK27806.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.21.
AAF34426.1 AFl 72282 Oryza sativa leucine rich repeat containing protein kinase. DUPRl 1.16.
BAB03627.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.27.
BAB03631.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.32.
AAK27817.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.16.
BAB03621.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.21.
BAB03629.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.30.
AAC80225.1 U72723 Oryza longistaminata receptor kinase-like protein. Xa21. disease resistance gene.
AAC49123.1 U37133 Oryza sativa receptor kinase-like protein. Xa21. Xa21 disease resistance gene.
BAA82393.1 AP000367 Oryza sativa
EST C96716(C10608) conesponds to a region ofthe predicted gene.; Similar to putative receptor protein kinase. (AC002334).
AAB82755.1 U72725 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member Al; downstieam of microsatellite region; disease resistance gene family member.
AAG52992.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl a. impkl.
AAB82756.1 U72724 Oryza sativa receptor kinase-like protein. Xa21 gene family member E.
AAB82753.1 U72726 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member D.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl. accumulates in mesophyll cells.
BAA88636.1 AB029327 Nicotiana tabacum elicitor-inducible LRR receptor-like protein EILP. EILP.
AAG52994.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl c. inφkl.
BAA94519.1 AP001800 Oryza sativa
ESTs AU032341(R3918),AU071016(R10613) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome 4, BAC F9D16; putative receptor kinase
(AL035394).
BAB07903.1 AP002835 Oryza sativa putative receptor kinase. P0417G05.10. contains ESTs
AU032341(R3918),AU071016(R10613).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSJNBa0004G10.30.
BAB18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
SEQ ID NO: 500
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl.7.
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml.
AAF59906.1 AF197947 Glycine max receptor protein kinase-like protein. CLV1B.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1A.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. Gιr_RLK2.
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLK1. GmRLKl .
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
CAC20842.1 AJ250467 Pinus sylvestris receptor protein kinase. upk.
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC0Q4484).
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAK27806.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.21.
AAB36558.1 U77888 Ipomoea nil receptor-like protein kinase INRPKL inφkl. leucine-rich repeat.
BAB03627.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.27.
BAB03629.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.30.
BAB03631.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.32.
BAB03621.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.21.
AAK27817.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.16.
AAF34426.1 AFl 72282 Oryza sativa leucine rich repeat containing protein kinase. DUPRl 1.16.
AAB82755.1 U72725 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member Al; downstream of microsatellite region; disease resistance gene family member.
AAC80225.1 U72723 Oryza longistaminata receptor kinase-like protein. Xa21. disease resistance gene.
AAC49123.1 U37133 Oryza sativa receptor kinase-like protein. Xa21. Xa21 disease resistance gene.
BAA82393.1 AP000367 Oryza sativa
EST C96716(C10608) conesponds to a region ofthe predicted gene.; Similar to putative receptor protein kinase. (AC002334).
AAB82756.1 U72724 Oryza sativa receptor kinase-like protein. Xa21 gene family member E.
BAA88636.1 AB029327 Nicotiana tabacum elicitor-inducible LRR receptor-like protein EILP. EILP.
AAG52992.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl a. impkl.
AAB82753.1 U72726 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member D.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /tlireonine kinase. RLKl. accumulates in mesophyll cells.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
SEQ ID NO: 501
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
AABttss9yy3jj8883__ii4...1ιι U UU8882ZZ4888111 Z __eeeaaa mmmaaayyysss
KI domain interactmg kinase 1. KIKL receptor-like protein kinase; serine/threonine protein kinase.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an nmunoglobulin-like domain.
BAA92836.1 AB032473 Brassica oleracea
S18 S-locus receptor kinase. SRK18.
CAB41878.1 Y18259 Brassica oleracea
SRK5 protein. SRK5. receptor-like kinase.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRKl.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl .
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
CAA74662.1 Y14286 Brassica oleracea
SFR3, extracellular S domain: 123-1346; tiansmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAF34428.1 AF172282 Oryza sativa receptor-like protein kinase. DUPRl 1.18.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRK1. Chrkl.
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJO 10166).
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12.
BAA94518.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 section 111 of 255; putative receptor-like protein kinase (AC002392).
SEQ ID NO: 502
AAC78596.1 AF053998 Lycopersicon esculentum
Hcr2-5D. Hcr2-5D. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAC78591.1 AF053993 Lycopersicon esculentum disease resistance protein. Cf-5.
AAC78593.1 AF053995 Lycopersicon esculentum
Hcr2-0B. Hcr2-0B. similar to Lycopersicon pimpinelhfolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAC78592.1 AF053994 Lycopersicon esculentum
Hcr2-0A. Hcr2-0A. similar to Lycopersicon phnpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAC78594.1 AF053996 Lycopersicon pimpinellifolium
Hcr2-2A. Hcr2-2A. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
CAA05274.1 AJ002236 Lycopersicon pimpinellifolium resistance gene. Cf-9. Cf-9.
AAA65235.1 U15936 Lycopersicon pimpinellifolium
Cf-9 precursor. Cf-9. this is the ninth resistance gene to disease caused by Cladosporium fulvum to be isolated.
CAA05276.1 AJ002236 Lycopersicon pimpinellifolium resistance gene. Hcr9-9E. Hcr9-9E.
AAC78595.1 AF053997 Lycopersicon esculentum
Hcr2-5B. Hcr2-5B. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
CAA05268.1 AJ002235 Lycopersicon hirsutum
Resistance gene. Cf-4. Cf-4.
CAA05279.1 AJ002237 Lycopersicon esculentum
Hcr9-0. Hcr9-0. homologue of Cladosporium fulvum disease resistance gene Cf-9.
BAB08215.1 AP002539 Oryza sativa
Similar to Lycopersicon esculentum disease resistance protein (AF053993).
BAA96776.1 AP002521 Oryza sativa
Similar to Lycopersicon esculentum disease resistance protein (AF053993).
AAG21897.1 AC026815 Oryza sativa putative disease resistance protein (3' partial). OSJNBa0079L16.21.
AAD50430.1 AF166121 Hordeum vulgare
Cf2/Cf5 disease resistance protein homolog. Bigl. leucine rich repeat protein.
AAG21917.1 AC026815 Oryza sativa putative disease resistance protein. OSJNBa0079L16.5.
CAB55409.1 ALl 17265 Oryza sativa zhbOOOl.l. Incomplete at 5'end,Similar to disease resistance protein; Method: conceptual translation with partial peptide sequencing.
AAG21909.1 AC026815 Oryza sativa putative disease resistance protein. OSJNBa0079L16.3.
AAC80225.1 U72723 Oryza longistaminata receptor kinase-like protein. Xa21. disease resistance gene.
AAC49123.1 U37133 Oryza sativa receptor kinase-like protein. Xa21. Xa21 disease resistance gene. SEQ ID NO: 503
CAA05276.1 AJ002236 Lycopersicon pimpinellifolium resistance gene. Hcr9-9E. Hcr9-9E.
AAC78592.1 AF053994 Lycopersicon esculentum
Hcr2-0A. Hcr2-0A. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
CAA05274.1 AJ002236 Lycopersicon pimpinellifolium resistance gene. Cf-9. Cf-9.
AAA65235.1 U15936 Lycopersicon pimpinellifolium
Cf-9 precursor. Cf-9. this is the ninth resistance gene to disease caused by Cladosporium fulvum to be isolated.
AAC78594.1 AF053996 Lycopersicon pimpinellifolium
Hcr2-2A. Hcr2-2A. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
CAA05279.1 AJ002237 Lycopersicon esculentum
Hcr9-0. Hcr9-0. homologue of Cladosporium fulvum disease resistance gene Cf-9.
AAC78595.1 AF053997 Lycopersicon esculentum
Hcr2-5B. Hcr2-5B. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAC78593.1 AF053995 Lycopersicon esculentum
Hcr2-0B. Hcr2-0B. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
CAA05268.1 AJ002235 Lycopersicon hirsutum
Resistance gene. Cf-4. Cf-4.
AAC78596.1 AF053998 Lycopersicon esculentum
Hcr2-5D. Hcr2-5D. similar to Lycopersicon pimpinellifolium disease resistance protein Cf-2.2 encoded by the sequence presented in GenBank Accession Number U42445.
AAC78591.1 AF053993 Lycopersicon esculentum disease resistance protein. Cf-5. BAA96776.1 AP002521 Oryza sativa
Similar to Lycopersicon esculentum disease resistance protein (AF053993).
BAB08215.1 AP002539 Oryza sativa
Similar to Lycopersicon esculentum disease resistance protein (AF053993).
CAB55409.1 ALl 17265 Oryza sativa zhbOOOl.l. Incomplete at 5'end,Similar to disease resistance protein; Method: conceptual translation with partial peptide sequencing.
AAD50430.1 AF166121 Hordeum vulgare
Cf2/Cf5 disease resistance protein homolog. Bigl. leucine rich repeat protein.
AAG21897.1 AC026815 Oryza sativa putative disease resistance protein (3' partial). OSJNBa0079L16.21.
AAB82755.1 U72725 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member Al; downstream of microsatellite region; disease resistance gene family member. SEQ ID NO: 504
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl. GmRLKl.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
CAC20842.1 AJ250467 Pinus sylvestris receptor protein kinase. upk.
AAK27806.1 AC022457 Oryza sativa putative protein kinase. OS JNBa0006L06.21.
AAB36558.1 U77888 Ipomoea nil receptor-like protein kinase INRPKL impkl. leucine-rich repeat.
BAA82393.1 AP000367 Oryza sativa
EST C96716(C 10608) conesponds to a region ofthe predicted gene.; Similar to putative receptor protein kinase. (AC002334).
AAF59906.1 AFl 97947 Glycine max receptor protein kinase-like protein. CLV1B.
BAB03629.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.30.
BAB03627.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.27.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1A.
AAK27817.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.16.
BAB03631.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.32.
BAB03621.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.21.
AAF34426.1 AF172282 Oryza sativa leucine rich repeat containing protein kinase. DUPRl 1.16.
AAF66615.1 AF142596 Nicotiana tabacum
LRR receptor-like protein kinase.
AAB82755.1 U72725 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member Al; downstream of microsatellite region; disease resistance gene family member.
AAC49123.1 U37133 Oryza sativa receptor kinase-like protein. Xa21. Xa21 disease resistance gene.
AAC80225.1 U72723 Oryza longistaminata receptor kinase-like protein. Xa21. disease resistance gene.
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
BAA88636.1 AB029327 Nicotiana tabacum elicitor-inducible LRR receptor-like protein EILP. EILP.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl . accumulates in mesophyll cells.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAB07903.1 AP002835 Oryza sativa putative receptor kinase. P0417G05.10. contains ESTs
AU032341(R3918),AU071016(R10613).
BAA94519.1 AP001800 Oryza sativa
ESTs AU032341(R3918),AU071016(R10613) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome 4, BAC F9D16; putative receptor kinase
(AL035394).
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAB82756.1 U72724 Oryza sativa receptor kinase-like protein. Xa21 gene family member E.
AAB82753.1 U72726 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member D.
SEQ ID NO: 505 CAA73134.1 Y12531 Brassica oleracea serine/tlireonine kinase. BRLK.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
AAB93834.1 U82481 Zeamays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein kinase.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; tiansmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
BAA94517.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRK1. Clrrkl .
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
SEQ ID NO: 507
AAA33509.1 M62985 Zeamays protein kinase. putative; putative.
CAA62476.1 X90990 Solanum tuberosum stpkl protein kinase.
AAK31277.1 AC079890 Oryza sativa putative protein kinase. OSJNBb0089A17.15.
CAA66616.1 X97980 Solanum berthaultii protein kinase.
AAF66637.1 AF143505 Lycopersicon esculentum viroid symptom modulation protein. PKv. protein kinase; induced by viroid infection.
BAA96593.1 AP002481 Oryza sativa
Similar to Solanum berthaultii protein kinase. (X97980).
AAB54117.1 U93559 Brassica rapa putative serine/threonine protein kinase. Bcpkl .
AAB88817.1 AF033263 Zeamays signal transduction for phototiopism. nonphototiopic hypocotyl 1. nphl. NPHl; putative serine/threonine kinase; similar to oat NPHl proteins.
AAC05084.1 AF033097 Avena sativa
NPH1-2. NPH1-2. putative serine/tlireonine protein kinase.
CAA82994.1 Z30333 Mesembryanthemum crystallinum protein kinase.
AAC05083.1 AF033096 Avena sativa NPH1-1. NPH1-1. putative serine/threonine protein kinase.
CAA82992.1 Z30331 Mesembryanthemum crystallinum Protein Kinase.
CAA82993.1 Z30332 Spinacia oleracea protein kinase.
AAA50304.1 M92989 Pisum sativum protein kinase. PK5. homologue.
AAB71418.1 U11553 Pisum sativum PsPK3. putative protein kinase.
BAA93704.1 AB032564 Cucumis sativus cucumber protein kinase CsPK3. CsPK3. putative.
BAA36192.1 AB012082 Adiantum capillus-veneris
PHY3. PHY3. Ser/Thr protein kinase; chimeric stracture of red/far-red light photoreceptive region (phytochrome)and putative blue light photoreceptor in phototropism of hypocotyl
(NPHl).
AAK18843.1 AC082645 Oryza sativa putative protein kinase. OSJNBb0033N16.3.
CAB82852.1 Z30329 Mesembryanthemum crystallinum protein kinase MK6.
CAA50374.1 X71057 Nicotiana tabacum protein kinase. PKTL7.
AAK13156.1 AC078829 Oryza sativa putative protein kinase. OSJNBa0026O12.14.
BAB03409.1 AP002816 Oryza sativa Shnilar to Spii tiacia oleracea protein kinase (S42867).
CAA82991.1 Z30330 Spinacia oleracea protein kinase
AAD50584.1 AF089097 Salvia columbariae protein kinase 1. PKl.
AAB93860.1 U89679 Lycopersicon esculentum protein kinase. LePK2. contains catalytic domain.
AAD50585.1 AF089099 Salvia columbariae protein kinase 3. PK3.
AAD50586.1 AF089100 Salvia columbariae protein kinase 4. PK4.
AAB93859.1 U89678 Lycopersicon esculentum protein kinase. LePKl. contains catalytic domain.
AAA50772.1 M69030 Pisum sativum protein serine/threonine kinase. PsPKl. putative; putative.
AAB93861.1 U89680 Lycopersicon esculentum protein kinase. LePK3. contains catalytic domain.
AAD50587.1 AF089101 Salvia columbariae protein kinase 5. PK5.
AAD50588.1 AF089102 Salvia columbariae protein kinase 6. PK6.
CAA56313.1 X79992 Avena sativa putative pp70 ribosomal protein S6 kinase. Aspkl 1.
AAD50589.1 AF089103 Salvia columbariae protein kinase 7. PK7.
CAB89082.1 AJ277534 Asparagus officinalis S6 ribosomal protein kinase. pkl. putative.
AAB93862.1 U89681 Lycopersicon esculentum protein kinase. LePK4. contains catalytic domain.
BAA92972.1 AP001551 Oryza sativa
ESTs AU056183(S20356),AU056881(S20950) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome 4 BAC clone F6I18 ; putative protein kinase.
(AL022198).
BAB12687.1 AP002746 Oryza sativa putative protein kinase. P0671B11.2. contains ESTs C22394(C30013),C22393(C30013). SEQ ID NO: 508
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; tiansmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
CAA73134.1 Y12531 Brassica oleracea serine/tlireonine kinase. BRLK.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRKl.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRKl. Chrkl.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAK02023.1 AC074283 Oryza sativa Putative protein kinase-like. OSJNBa0087H07.5.
CAA79324.1 Z18884 Brassica oleracea S-receptor kinase related protein.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
CAB51836.1 AJ243961 Oryza sativa Putitive Ser/Thr protein kinase. 11332.7.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK.1.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAA92953.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F10M6 ; S-receptor kinase -like protein. (AL021811).
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPK10) mRNA. (L27821).
AAG16628.1 AY007545 Brassica napus protein serine/thxeonine kinase BNK1.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
SEQ ID NO: 510
AAG30254.1 AF307333 Hordeum vulgare putative nematode-resistance protein. Hsl. similar to Beta procumbens Hslpro protein.
AAB48305.1 U79733 Beta procumbens nematode resistance. Hslpro- 1.
SEQ ID NO: 513
BAA14144.1 D90116 Armoracia rusticana peroxidase isozyme.
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
BAA11853.1 D83225 Populus nigra peroxidase.
CAA66037.1 X97351 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
CAA66034.1 X97348 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
CAA66036.1 X97350 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
CAA66035.1 X97349 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
BAA11852.1 D83224 Populus nigra peroxidase.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
BAA06335.1 D30653 Populus kitakamiensis peroxidase.
AAB47602.1 L07554 Linum usitatissimum peroxidase. FLXPER1.
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
AAD37427.1 AF149277 Phaseolus vulgaris peroxidase 1 precursor. FBP1. secretory peroxidase.
BAA06334.1 D30652 Populus kitakamiensis peroxidase.
AAB97734.1 AF014502 Glycine max seed coat peroxidase precursor. Ep. H2O2 oxidoreductase; class III plant peroxidase.
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
CAA62227.1 X90694 Medicago sativa peroxidase IC. prxlC.
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
CAA62225.1 X90692 Medicago sativa peroxidaselA. prxlA.
AAB41811.1 L36157 Medicago sativa peroxidase. pxdC. amino acid feature: conserved domains, aa 123 .. 129, 191 .. 198; amino acid feature: heme-binding domain, aa 68 .. 73.
AAB41810.1 L36156 Medicago sativa peroxidase. pxdA. amino acid feature: conserved motifs, aa 181 .. 188; amino acid feature: heme-binding domain, aa 60 .. 65.
BAA01877.1 D11102 Populus kitakamiensis peroxidase. prxAl.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI-1.
BAA01992.1 D11396 Nicotiana tabacum 'peroxidase'.
AAA34108.1 J02979 Nicotiana tabacum lignin-forming peroxidase precursor (EC 1.11.1.7).
AAA33127.1 M91373 Cucumis sativus peroxidase. pre-peroxidase. putative.
BAA92500.1 AP001383 Oryza sativa
ESTs D39300(R3292),AU030751(E60187) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP6a. (X98774).
CAB65334.1 AJ250121 Picea abies peroxidase. SPI2 protein. spi2.
CAA40796.1 X57564 Armoracia rusticana peroxidase. peroxidase precursor.
AAB06183.1 M37636 Arachis hypogaea cationic peroxidase. PNC1.
CAA76680.1 Y17192 Cucurbita pepo peroxidase. aprx. type III peroxidase.
AAA33129.1 M91372 Cucumis sativus peroxidase. pre-peroxidase.
CAA71492.1 Y10466 Spinacia oleracea peroxidase. prxr5.
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 5 precursor, type III peroxidase.
AAD43561.1 AF155124 Gossypium hirsutum bacterial-induced peroxidase precursor. Perx_Goshiko.
AAF63026.1 AF244923 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prx!4 precursor, type III peroxidase.
AAA33121.1 M32742 Cucumis sativus peroxidase (CuPer2).
BAA92422.1 AP001366 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to A.thaliana mRNA for peroxidase ATP 18a. (X98804).
BAA92497.1 AP001383 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP 18a. (X98804).
BAA77389.1 AB024439 Scutellaria baicalensis peroxidase 3.
BAA08499.1 D49551 Oryza sativa peroxidase. poxN.
AAB19129.1 U41657 Glycine max seed coat peroxidase isozyme. SPOD4.1. H2O2 oxidoreductase.
BAA03373.1 D14482 Oryza sativa putative peroxidase.
AAB02554.1 L37790 Stylosanthes humilis cationic peroxidase.
SEQ ID NO: 515 AAB88134.1 AF034618 Spinacia oleracea cytosolic heat shock 70 protein. HSC70-1.
AAF34134.1 AF161180 Malus x domestica high molecular weight heat shock protein. Hsp2.
AAB99745.1 AF005993 Triticum aestivum
HSP70. TaHSP70d. 70 kDa heat shock protein, molecular chaperone.
AAA62325.1 L32165 Hordeum vulgare
Molecular chaperone. HSP70. Heat-shock protein HSP70; The predicted amino acid sequence is highly homologous (more than 80% identity) to other plant heat-shock proteins (HSP70s) in the database; however the C terminus is quite unique.; putative.
AAA21808.1 L23551 Spinacia oleracea molecular chaperone. ER-lumenal protein. HSC70.
AAA34139.1 L08830 Lycopersicon esculentum molecular chaperon (precursor), glucose-regulated protein 78. BiP/gφ78. an endoplasmic reticulum residing heat shock protein 70 family member; precursor peptide.
AAB86942.1 AF031241 Glycine max endoplasmic reticulum transport protein; molecular chaperone; roles in protein folding, assembly, and transport, endoplasmic reticulum HSC70-cognate binding protein precursor.
BIP. BiP; similar to HSC70 and GRP78.
AAK21920.1 AF338252 Glycine max molecular chaperone. BiP-isoform D. BiPD. ER-lumenal HSP70; binding protein GRP78.
AAB91473.1 AF035458 Spinacia oleracea heat shock 70 protein. HSC70-11. mitochondrial protein.
AAB96660.1 AF039084 Spinacia oleracea heat shock 70 protein. HSC70-11. molecular chaperone.
AAB91472.1 AF035457 Spinacia oleracea heat shock 70 protein. HSC70-10. mitochondrial protein.
SEQ ID NO: 516 CAA06927.1 AJ006233 Nicotiana tabacum putative thaumatin-like protein precursor.
AAF06346.1 AFl 95653 Vitis vinifera SCUTL1. thaumatin-like protein.
BAA28872.1 AB006009 Pyras pyrifolia thaumatin-like protein precursor. PsTLl.
CAC 10270.1 AJ243427 Malus x domestica thaumatin-like protein, tl. allergen, pathogenesis-related.
AAC36740.1 AF090143 Malus x domestica thaumatin-like protein precursor Mdtll. MDTL1. pathogenesis-related.
CAB62167.1 AJ242828 Castanea sativa antifungal. thaumatin-like protein, til.
AAB38064.1 U32440 Primus avium thaumatin-like protein precursor.
BAA74546.2 AB000834 Nicotiana tabacum thaumatin-like protein SE39b.
AAF06347.1 AF195654 Vitis vinifera SCUTL2. thaumatin-like protein.
CAC09477.1 AL442113 Oryza sativa thaumatin-like protein. HQ806H05.10.
AAB95118.1 U71244 Brassica rapa pathogenesis-related group 5 protein. BFTP. thaumatin-like protein; PR-5.
CAA10492.1 AJ131731 Pseudotsuga menziesii Thaumatin-like protein. 5A1A.16.
BAA95017.1 AB031870 Cestram elegans thaumatin-like protein. CETLP.
BAA95165.1 AB029918 Nicotiana tabacum pistil transmitting tissue specific thaumatin (SE39b)-like protein. SE39b.
AAB61590.1 AF003007 Vitis vinifera
WTLl. osmotin-like protein, PR-5 protein; thaumatin-like protein.
AAD55090.1 AF178653 Vitis riparia thaumatin. osmotin; pathogenesis-related protein.
CAB85637.1 AJ237999 Vitis vinifera putative thaumatin-like protein. Til, alternative name grip 51.
AAF82264.1 AF227324 Vitis vinifera thaumatin-like protein.
AAB02259.1 U57787 Avena sativa permatin precursor, thaumatin-like protein.
AAB53368.1 U77657 Oryza sativa pathogenesis-related thaumatin-like protein.
CAA09228.1 AJ010501 Cicer arietinum thaumatin-like protein PR-5b.
CAA33293.1 X15224 Nicotiana tabacum thaumatin-like protein. E22.
CAA33292.1 XI 5223 Nicotiana tabacum thaumatin-like protein. E2.
SEQ ID NO: 517
CAA71801.1 Y10848 Brassica juncea gamma-glutamylcysteine synthetase. gshl.
AAB71230.1 AF017983 Lycopersicon esculentum gamma-glutamylcysteine synthetase. GSHL
AAC82334.1 AF041340 Medicago truncatula gamma-glutamylcysteine synthetase. putative plastid protein.
AAF22137.1 AF128455 Pisum sativum gamma-glutamylcysteine synthetase precursor, gshl . putative plastid protein.
AAF22136.1 AF128454 Phaseolus vulgaris gamma-glutamylcysteine synthetase precursor, gshl. putative plastid protein.
CAA06613.1 AJ005587 Brassica juncea gamma-glutamylcysteine synthetase.
AAG13459.1 AF128453 Glycine max gamma-glutamylcysteine synthetase precursor, gshl.
CAA64808.1 X95563 Brassica juncea gamma-glutamylcysteine synthetase. gshl.
SEQ ID NO: 518
AAA75414.1 L28005 Glycine max TGACG-motif-binding protein. STGA1.
AAB31250.2 S73827 Solanum tuberosum mas-binding factor MBF3. transcription factor TGAla homolog; This sequence comes from
Fig. 4.
AAB31249.1 S73826 Solanum tuberosum mas-binding factor MBF2. mas-binding factor MBF2. transcription factor TGAla homolog;
This sequence comes from Fig. 4.
CAA34468.1 XI 6449 Nicotiana sp. TGAla protein (AA 1-359).
AAA34091.1 M62855 Nicotiana tabacum ASF-1/G13. leucine-zipper DNA-binding protein.
AAB31251.2 S73828 Solanum tuberosum mas-binding factor MBFl. transcription factor TGAla homolog; This sequence comes from
Fig. 4.
CAA48904.1 X69152 Zea mays ocs-element binding factor 3.2. OBF3.2.
CAA48905.1 X69153 Zea mays ocs-element binding factor 3.1. OBF3.1.
AAC24123.1 AF067187 Cichorium intybus
AAC24122.1 AF067186 Cichorium intybus cAMP responsive element binding protein. CREB1.
AAC49760.1 AF001454 Helianthus annuus Dc3 promoter-binding factor-2. DPBF-2.
SEQ ID NO: 521
CAA10608.1 AJ132228 Ricinus communis amino acid carrier. aap3.
CAA07563.1 AJ007574 Ricinus communis amino acid carrier, aapl.
CAA70778.1 Y09591 Vicia faba amino acid transporter.
AAD16014.1 AF080543 Nepenthes alata amino acid transporter. AAP2.
CAA70969.1 Y09826 Solanum tuberosum amino acid transporter. AAP2. transmembrane protein.
AAD16015.1 AF080544 Nepenthes alata amino acid transporter. AAP3.
CAA70968.1 Y09825 Solanum tuberosum amino acid transporter. AAPL transmembrane protein.
CAA92992.1 Z68759 Ricinus communis amino acid carrier.
AAD 16013.1 AF080542 Nepenthes alata amino acid transporter. AAP 1.
AAF15945.1 AF061435 Vicia faba amino acid transporter b. AAPB.
CAA72006.1 Y11121 Ricinus communis amino acid carrier.
AAF15944.1 AF061434 Vicia faba amino acid transporter a. AAPA.
AAF15946.1 AF061436 Vicia faba amino acid transporter c. AAPC.
AAB48944.1 U31932 Nicotiana sylvestris amino acid permease 1. NSAAPl. amino acid tiansporter; Method: conceptual translation supplied by author.
AAB96830.1 U64823 Nicotiana sylvestris amino acid transporter, amino acid permease. nsaapl.
BAA93437.1 AB022783 Oryza sativa amino acid permease. OsproT.
AAD25162.1 AFO 14810 Lycopersicon esculentum proline transporter 3. LeProT3.
AAD25161.1 AF014809 Lycopersicon esculentum proline transporter 2. LeProT2.
AAD25160.1 AFO 14808 Lycopersicon esculentum proline transporter 1. LeProTl.
AAF76897.1 AF274032 Atriplex hortensis prolrne/glycine betaine transporter.
CAB42599.1 AJ238635 Chlorella protothecoides arnino acid carrier. dee4.
SEQ ID NO: 526
BAA03763.1 D16247 Nicotiana sylvestris RNA helicase like protein DB10.
AAD46404.1 AF096248 Lycopersicon esculentum ethylene-responsive RNA helicase. ER68. putative DEAD box/RNA helicase.
AAF75791.1 AF271892 Pisum sativum DEAD box protein P68. P68. RNA helicase.
AAF40306.1 AF156667 Vigna radiata RNA helicase. VRH1.
CAA68193.1 X99937 Spinacia oleracea
RNA helicase. prh75. DEAD-box protein; homologous to X99938.
AAG13612.1 AC078840 Oryza sativa putative RNA helicase. OSJNBb0073N24.12.
AAG34876.1 AF261021 Nicotiana tabacum putative chloroplast RNA helicase VDL isoform 1. VDL. alternatively spliced.
AAG34873.1 AF261020 Nicotiana tabacum putative chloroplast RNA helicase VDL isoform 1. VDL. essential for chloroplast development; may be involved in post-tianscriptional regulation.
AAG34879.1 AF261024 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 1. VDL'. alternatively spliced.
AAD20980.1 AF079782 Zeamays
ATPase and RNA helicase. translation initiation factor 4A2. tif4A2.
AAG34882.1 AF261027 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 4. VDL'. alternatively spliced.
BAA95705.1 AB042644 Oryza sativa DEAD box RNA helicase OsPLlOb. OsPLlOb.
BAA95704.1 AB042643 Oryza sativa DEAD box RNA helicase OsPLlOa. OsPLlOa.
AAG34883.1 AF261028 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 5. VDL'. alternatively spliced.
AAG38493.1 AF261032 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 1. VDL'. alternatively spliced.
AAG34884.1 AF261029 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 6. VDL'. alternatively spliced.
AAG38496.1 AF261032 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 4. VDL'. alternatively spliced.
AAG38497.1 AF261032 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 5. VDL'. alternatively spliced.
AAG38498.1 AF261032 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 6. VDL'. alternatively spliced.
AAG38500.1 AF261032 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 8. VDL'. alternatively spliced.
AAG34886.1 AF261031 Nicotiana tabacum putative chloroplast RNA helicase VDL' isoform 8. VDL'. alternatively spliced.
AAG48833.1 AC084218 Oryza sativa similar to Arabidopsis thaliana DNA helicase (AJ404475).
SEQ ID NO: 528
CAA65536.1 X96761 Sporobolus stapfianus sulphate tiansporter protein.
AAK27688.1 AF347614 Lycopersicon esculentum sulfate tiansporter 2. ST2.
AAK27687.1 AF347613 Lycopersicon esculentum sulfate transporter 1. STl.
CAA57711.1 X82256 Stylosanthes hamata high affinity sulphate transporter. SHST2.
AAG41419.1 AF309643 Solanum tuberosum high affinity sulfate transporter type 1. STl.
CAA57710.1 X82255 Stylosanthes hamata high affinity sulphate transporter. SHST1.
AAK35215.1 AF355602 Zeamays sulfate transporter STl.
CAA65291.1 X96431 Hordeum vulgare high affinity sulphate transporter. HVST1.
AAA97952.1 U52867 Hordeum vulgare high affinity sulfate transporter HVST1.
CAB42985.1 AJ238244 Aegilops tauschii putative plasma membrane sulfate tiansport. putative high affinity sulfate transporter, stl.
CAB42986.1 AJ238245 Aegilops tauschii putative plasma membrane sulfate tiansport. putative high affinity sulfate tiansporter. st2.
CAA57831.1 X82454 Stylosanthes hamata low affinity sulphate tiansporter. SHST3.
SEQ ID NO: 531
AAK00436.1 AC060755 Oryza sativa putative zinc finger protein. OSJNBa0003O19.23.
BAA85438.1 AP000616 Oryza sativa similar to RING-H2 finger protein RHAla (AF078683).
AAG43550.1 AF211532 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 132. ACRE132. similar to RING finger proteins.
BAA90357.1 AP001080 Oryza sativa
EST AU070319(S 10788) conesponds to a region ofthe predicted gene. Similar to RING-H2 finger protein RHA2b (AC006200).
SEQ ID NO: 532
BAA81751.1 ABO 17517 Marchantia polymoφha calcium-dependent protein kinase. CDPK1. CDPK-B; alternative splicing.
BAA13232.1 D87042 Zea mays Calcium-dependent protein kinase.
BAA81749.1 ABO 17515 Marchantia polymoφha calcium-dependent protein kinase. CDPK1. CDPK-B; alternative splicing.
BAA81750.1 ABO 17516 Marchantia polymorpha calcium-dependent protein kinase. CDPK1. CDPK-A; alternative splicing.
BAA85396.1 AP000615 Oryza sativa
ESTs C22369(C12239),C22370(C12239), AU057852(S21844),AU057853(S21844) conespond to a region ofthe predicted gene.; similar to calcium dependent protein kinase.
(AF048691).
CAA57156.1 X81393 Oryza sativa calcium-dependent protein kinase. OSCPKII.
AAC05270.1 AF048691 Oryza sativa calcium dependent protein kinase. CDPK12.
AAB49984.1 U90262 Cucurbita pepo calcium-dependent cahnodulin-independent protein kinase CDPK. cpCPKl . serine/threonine protein kinase that is activated by direct binding of calcium.
AAB70706.1 U82087 Tortula ruralis calmodulin-like domain protein kinase. TrCPKl.
AAG46110.1 AC073166 Oryza sativa calcium-dependent protein kinase. OSJNBb0064P21.2.
BAA02698.1 D13436 Oryza sativa calcium-dependent protein kinase. spk.
BAA81748.1 AB017515 Marchantia polymoφha calcium-dependent protein kinase. CDPKl. CDPK-A; alternative splicing.
CAA07481.1 AJ007366 Zeamays calcium-dependent protein kinase.
AAB80692.1 U69173 Glycine max calmodulin-like domain protein kinase isoenzyme beta. CDPK beta.
AAC49405.1 U08140 Vigna radiata calcium dependent protein kinase. CDPK.
BAA12338.1 D84408 Zea mays calcium dependent protein kinase. ZmCDPKl.
AAA33443.1 L15390 Zea mays calcium-dependent protein kinase. CDPK.
CAA57157.1 X81394 Oryza sativa calcium-dependent protein kinase. OSCPK2.
BAB21081.1 AP002819 Oryza sativa putative calcium-dependent protein kinase. P0501G01.10.
BAA12715.1 D85039 Zea mays calcium-dependent protein kinase.
CAA65500.1 X96723 Medicago sativa protein kinase. CDPK.
AAC25423.1 AF072908 Nicotiana tabacum calcium-dependent protein kinase. CDPKl .
AAA69507.1 U28376 Zea mays calcium-dependent protein kinase. MZECDPK2.
BAA13440.1 D87707 Ipomoea batatas calcium dependent protein kinase. CDPK.
AAD28192.2 AFl 15406 Solanum tuberosum calcium-dependent protein kinase. CDPK; catalytic domain.
AAB80693.1 U69174 Glycine max calmodulin-like domain protein kinase isoenzyme gamma. CDPK gamma.
AAA61682.1 L27484 Zea mays calcium-dependent protein kinase. CDPK.
AAD17800.1 AF090835 Mesembryanthemum crystallinum Ca2+-dependent protein kinase. CPK1. serine/threonine protein kinase.
CAA39936.1 X56599 Daucus carota calcium- dependent protein kinase. DcPK431.
AAK26164.1 AY027885 Cucumis sativus calcium-dependent cahnodulin-independent protein kinase 5. CDPK5. CsCDPK5.
AAB88537.1 AF035944 Fragaria x ananassa calcium-dependent protein kinase. MAX17.
AAF21062.1 AF216527 Dunaliella tertiolecta calcium-dependent protein kinase. CPK1; CDPK.
CAA89202.1 Z49233 Chlamydomonas eugametos calcium-sthnulated protein kinase.
AAC32116.1 AF051211 Piceamariana probable calcium dependent protein kinase. Sbl5. similar to Vigna radiata calcium dependent protein kinase encoded by U08140.
AAF23900.1 AF194413 Oryza sativa calcium-dependent protein kinase. CDPKL OsCDPKl .
AAF23901.2 AF194414 Oryza sativa calcium-dependent protem kinase. CDPK5. OsCDPK5.
CAB46228.1 Y18055 Arachis hypogaea calcium dependent protein kinase. CDPK.
AAC78558.1 AF030879 Solanum tuberosum protein kinase CPK1.
CAA58750.1 X83869 Daucus carota CDPK-related protein kinase. CRK (or PK421).
AAB47181.1 S82324 Zea mays
/gene="calcium/calmodulin-dependent protein kinase. This sequence comes from Fig. 1.
BAA12691.1 D84507 Zeamays
CDPK-related protein kinase. Does not require calcium for its activity (by similarity).
BAA22410.1 D38452 Zea mays calcium-dependent protein kinase-related kinase. BAA12692.1 D84508 Zea mays
CDPK-related protein kinase. Does not require calcium for its activity.
AAG01179.1 AF289237 Zea mays calcium/calmodulin dependent protein kinase MCK2. MCK2.
AAC24961.1 AF009337 Tradescantia virgfniana
CDPK-related protein kinase. CRKL
BAA90814.1 AP001168 Oryza sativa
ESTs AU030197(E50746),AU030196(E50746) conespond to a region ofthe predicted gene.;
Similar to calcium-dependent cahnodulin-independent protein kinase CDPK (U90262).
AAC49008.1 U24188 Lilium longiflorum calcium/calmodulin-dependent phosphorylation activity, calcium/cahnodulin-dependent protein kinase. CCaMK. serine/threonine kinase; binds to calcium and calmodulin.
AAF19402.1 AF203480 Lycopersicon esculentum phosphoenolpyruvate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands. SEQ ID NO: 535
AAK19619.1 AF336286 Gossypium rsutum
GHMYB9. ghmyb9. similar to myb.
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related.
CAA50224.1 X70879 Hordeum vulgare MybHvl. mybl.
CAA50222.1 X70877 Hordeum vulgare MybHvl. mybl.
AAA33067.1 L04497 Gossypium liirsutum MYB A; putative.
CAA50221.1 X70876 Hordeum vulgare MybHv5. myb2.
BAA23337.1 D88617 Oryza sativa transfactor. OSMYB1. Osmybl.
BAA23338.1 D88618 Oryza sativa transfactor. OSMYB2. Osmyb2.
AAC04720.1 AF034134 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-O. similar to MYB A encoded by GenBank Accession Number L04497.
CAA72218.1 Y11415 Oryza sativa myb.
CAA50225.1 X70880 Hordeum vulgare MybHv5. myb2.
AAA82943.1 U39448 Picea mariana
MYB-like transcriptional factor MBFl. putative DNA binding region highly similar to the maize Cl.
CAA78386.1 Z13996 Petunia x hybrida
DNA binding protein; tianscriptional activator, protein 1. myb.Ph3. Product related to animal myb proto-oncoproteins. Sequence from nucleotide 992 is not included in clone cPFl and has been obtained by PCR amplification of cDNA.
BAA81732.1 AB029161 Glycine max GmMYB29A2.
AAK19616.1 AF336283 Gossypium hirsutum
GHMYB25. ghmyb25. similar to myb; contains an unspliced intron.
AAK19611.1 AF336278 Gossypium hirsutum BNLGHi233. bnlghi6233. similar to myb.
AAK19617.1 AF336284 Gossypium hirsutum GHMYB36. ghmyb36. similar to myb.
BAA93038.1 AP001552 Oryza sativa
EST AU082058(C12976) conesponds to a region ofthe predicted gene. Similar to
Arabidopsis thaliana putative transcription factor (AF062916).
AAK19615.1 AF336282 Gossypium hirsutum GHMYB10. ghmyb 10. similar to myb.
BAB39987.1 AP003020 Oryza sativa putative transcription factor (myb). P0498A12.16. contains ESTs
AU097474(S5087),D40175(S1959).
BAB39972.1 AP003018 Oryza sativa putative transcription factor (myb). OSJNBa0004B 13.27. contains ESTs
AU097474(S5087),D40175(S 1959).
CAB43399.1 AJ006292 Antinhinum majus Myb-related transcription factor mixta-like 1. mybmll.
CAA72185.1 Y11350 Oryza sativa myb factor, myb.
AAF22256.1 AF161711 Pimpinella brachycaφa myb-related transcription factor.
AAG13574.1 AC037425 Oryza sativa myb factor. OSJNBa0055P24.4.
BAA81731.1 AB029160 Glycine max GmMYB29Al.
BAA81730.1 AB029159 Glycine max GmMYB29Al.
CAA72186.1 Y11351 Oryza sativa myb factor, myb.
CAA67600.1 X99210 Lycopersicon esculentum myb-related transcription factor. THM16.
CAA75509.1 Y15219 Oryza sativa subsp. indica transcriptional activator. Cl.
AAC04718.1 AF034132 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-J. similar to MYB A encoded by GenBank Accession Number L04497.
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, transcriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins.
BAA81736.1 AB029165 Glycine max GmMYB29B2.
AAK19618.1 AF336285 Gossypium hirsutum GHMYB38. ghmyb38. similar to myb.
CAA72217.1 Y11414 Oryza sativa myb.
AAC49394.1 U57002 Zea mays
P protein. P. allele P-wr; Myb-like tianscriptional regulator with a putative zinc-finger at the
C-terminal.
AAA33500.1 M73028 Zea mays myb-like transcription factor. P.
BAB20661.1 AP002871 Oryza sativa putative myb-related protein P. P0475H04.31.
AAG36774.1 AF210616 Zeamays
P2 protein. P2. myb-like tianscriptional factor; similar to Zea mays P gene.
BAA88222.1 AB028650 Nicotiana tabacum myb-related transcription factor LBM2. Ibm2.
BAA81733.2 AB029162 Glycine max GmMYB29A2.
BAA23339.1 D88619 Oryza sativa transfactor. OSMYB3. Osmyb3.
BAA88224.1 AB028652 Nicotiana tabacum myb-related transcription factor LBM4. lbm.4.
BAA88221.1 AB028649 Nicotiana tabacum myb-related transcription factor LBM1. lbml.
AAB41101.1 U72762 Nicotiana tabacum transcription factor Mybl. mybl. TMV-inducible Myb homolog; contains helix-turn-helix motif; contains redox-sensitive cysteine.
SEQ ID NO: 536
CAA70968.1 Y09825 Solanum tuberosum amino acid transporter. AAP1. tiansmembrane protein.
AAF15946.1 AF061436 Vicia faba arnino acid transporter c. AAPC.
CAA70969.1 Y09826 Solanum tuberosum amino acid transporter. AAP2. transmembrane protein.
AAB96830.1 U64823 Nicotiana sylvestris amino acid tiansporter. amino acid permease. nsaapl.
CAA07563.1 AJ007574 Ricinus communis amino acid carrier, aapl.
AAB48944.1 U31932 Nicotiana sylvestris amino acid permease 1. NSAAPl. amino acid transporter; Method: conceptual translation supplied by author.
AAD16015.1 AF080544 Nepenthes alata amino acid transporter. AAP3.
CAA70778.1 Y09591 Vicia faba arnino acid transporter.
AAF15944.1 AF061434 Vicia faba amino acid transporter a. AAPA.
AAF15945.1 AF061435 Vicia faba amino acid transporter b. AAPB.
AAF76897.1 AF274032 Atriplex hortensis proline/glycine betaine transporter.
AAD16014.1 AF080543 Nepenthes alata amino acid transporter. AAP2.
AAD25161.1 AF014809 Lycopersicon esculentum proline transporter 2. LeProT2.
CAA10608.1 AJ132228 Ricinus communis amino acid carrier. aap3.
SEQ ID NO: 537
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltransferase. PHI.
BAA36423.1 AB013598 Verbena x hybrida
UDP-glucose:anthocyanin 5-O-glucosyltransferase. HGT8.
BAA36421.1 AB013596 Perilla frutescens
UDP-glucose:anthocysnin 5-O-glucosyltransferase. PF3R4.
BAB07962.1 AP002524 Oryza sativa putative anthocyanin 5-O-glucosyltiansferase. P0406H10.16. contains ESTs
AU067881(C10481),AU067882(C10481).
BAA36422.1 AB013597 Perilla frutescens
UDP-glucose: anthocyanin 5-O-glucosyltransferase homologue. PF3R6.
BAA93039.1 AB033758 Citrus unshiu limonoid UDP-glucosyltransferase. LGTase.
AAF61647.1 AF190634 Nicotiana tabacum
UDP-glucose: salicylic acid glucosyltransferase. SA-GTase.
AAF98390.1 AF287143 Brassica napus catalyzes the transfer of glucose from UDP-glucose to sinapate and some other hydroxycinnamates (4-coumarate, caffeate, feralate). UDP-glucose: sinapate glucosyltransferase. SGT1. SGT.
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitrile-o- glucosyltransferase.
AAK16172.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
BAA83484.1 AB031274 Scutellaria baicalensis
UDP-glucose: flavonoid 7-O-glucosyltransferase. ufgt.
AAK16181.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.16.
AAG25643.1 AF303396 Phaseolus vulgaris
UDP-glucosyltransferase HRA25. putative; defense associated.
AAK16178.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.5.
AAD21086.1 AF127218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
BAA12737.1 D85186 Gentiana triflora
UDP-glucose:flavonoid-3-glucosyltiansferase.
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid:glucosyltiansferase 1. togtl. glucosyltransferase.
CAB56231.1 Y18871 Dorotheanthus bellidiformis betanidin-5-O-glucosyltransferase.
CAA54612.1 X77462 Manihot esculenta
UTP-glucose glucosyltransferase. CGT5.
BAB41019.1 AB047092 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. ItUFGTl .
BAB41020.1 AB047093 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. ItUFGT2.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase . IS5a.
BAB41025.1 AB047098 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. F1UFGT1.
BAB41023.1 AB047096 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. A1UFGT1.
BAB41022.1 AB047095 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. RuUFGT2.
BAB41021.1 AB047094 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltiansferase. RuUFGTl.
BAA19659.1 AB002818 Perilla frutescens flavonoid 3-O-glucosyltransferase. UDP glucose.
BAB41026.1 AB047099 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. F1UFGT2.
BAB41024.1 AB047097 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT2.
AAK16180.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.21.
AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid:glucosyltransferase 2. togt2. glucosyltiansferase.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS 10a.
BAB41017.1 AB047090 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. KyUFGTl. The gene was derived from one ofthe parents V. labrascana cv. Ishiharawase.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltransferase. PGT8.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltransferases. CAA54614.1 X77464 Manihot esculenta
UTP-glucose glucosyltransferase. CGT7.
CAA54611.1 X77461 Manihot esculenta
UTP-glucose glucosyltiansferase. CGT2.
CAA54613.1 X77463 Manihot esculenta
UTP-glucose glucosyltransferase. CGT6. SEQ ID NO: 540
AAK27688.1 AF347614 Lycopersicon esculentum sulfate tiansporter 2. ST2.
AAG41419.1 AF309643 Solanum tuberosum high affinity sulfate transporter type 1. STL
AAK27687.1 AF347613 Lycopersicon esculentum sulfate transporter 1. STl.
AAA97952.1 U52867 Hordeum vulgare high affinity sulfate transporter HVST1.
CAA57711.1 X82256 Stylosanthes hamata high affinity sulphate transporter. SHST2.
CAA65291.1 X96431 Hordeum vulgare high affinity sulphate transporter. HVSTl.
CAA57710.1 X82255 Stylosanthes hamata high affinity sulphate transporter. SHST1.
CAB42985.1 AJ238244 Aegilops tauschii putative plasma membrane sulfate transport, putative high affinity sulfate transporter, stl.
AAK35215.1 AF355602 Zeamays sulfate transporter STl.
CAB42986.1 AJ238245 Aegilops tauschii putative plasma membrane sulfate transport, putative high affinity sulfate transporter. st2.
CAA65536.1 X96761 Sporobolus stapfianus sulphate transporter protein.
CAA57831.1 X82454 Stylosanthes hamata low affinity sulphate tiansporter. SHST3.
CAAl 1413.1 AJ223495 Brassica juncea sulfate permease. spl.
AAB94543.1 AF016306 Zeamays sulfate permease.
SEQ ID NO: 541
AAF36491.1 AF129479 Hordeum vulgare
HAK2. HAK2. similar to Hordeum vulgare K+ transporter HAKl .
BAB32443.1 AB055630 Phragmites austialis high-affinity potassium transporter. PcnHAKl.
BAB32444.1 AB055631 Phragmites austialis high-affinity potassium transporter. PceHAKlA.
BAB32445.1 AB055632 Phragmites austialis high-affinity potassium transporter. PceHAKlB.
BAB32442.1 AB055629 Phragmites australis high-affinity potassium transporter. PcuHAKl.
AAC39315.1 AF025292 Hordeum vulgare putative rtigh-affinity potassium transporter. HvHAKl .
AAF36497.1 AF129485 Oryza sativa
HAK4. HAK4. OsHAK4; similar to Hordeum vulgare K+ tiansporter HAKl .
AAF36496.1 AF129484 Hordeum vulgare
HAK4. HAK4. HvHAK4; similar to Hordeum vulgare K+ tiansporter HAKl .
CAC14883.1 AJ297888 Hordeum vulgare putative potassium transporter, hakl .
CAC14787.1 AJ297886 Hordeum vulgare putative potassium transporter, hakl .
AAF36492.1 AF129480 Hordeum vulgare
HAK1B. HAK1B. HvHAKlB; similar to Hordeum vulgare K+ transporter HAKl.
CAC15061.1 AJ300161 Hordeum vulgare potassium transporter. hak4.
SEQ ID NO: 542
AAA91063.1 M88254 Hevea brasiliensis ethylene-inducible protein. ER1.
SEQ ID NO: 546
CAA75386.1 Y15113 Morinda citrifolia
3-deoxy-D-arabino-heptulosonate 7-phosphate synthase. DS3. 2-dehydro-3- deoxyphosphoheptonate aldolase.
CAA79855.1 Z21792 Lycopersicon esculentum phospho-2-dehydro-3-deoxyheptonate aldolase.
CAA79856.1 Z21793 Lycopersicon esculentum phospho-2-dehydro-3 -deoxyheptonate aldolase.
SEQ ID NO: 548
BAA96751.1 AP002521 Oryza sativa
Similar to Arabidopsis thaliana chromosome4, BAC clone T16H5; lectin like protein
(AL024486).
SEQ ID NO: 551
BAB19096.1 AP002839 Oryza sativa putative DNA-binding protein homolog. P0688A04.2.
BAB19075.1 AP002744 Oryza sativa putative DNA-binding protein homolog. P0006C01.17.
AAK16170.1 AC079887 Oryza sativa putative DNA binding protein. OSJNBa0040E01.4.
AAD32677.1 AF140554 Avena sativa
DNA-binding protein WRKYl . wrkyl. putative transcription factor.
AAD16139.1 AF096299 Nicotiana tabacum DNA-binding protein 2. WRKY2. transcription factor.
CAB97004.1 AJ278507 Solanum tuberosum putative transcription factor. WRKY DNA binding protein. WRKYL
CAA88326.1 Z48429 Avena fatua binds conserved cis-element from cereal alpha- Amy2 promoters. DNA-binding protein.
AAC49527.1 U48831 Petroselinum crispum
WRKYl. contains two WRKY domains; WRKY-type DNA-binding protein; sequence- specific DNA-binding protein.
AAC49529.1 U58540 Petroselinum crispum
WRKY2. Contains two WRKY domains; WRKY-type DNA-binding protein.
AAK16171.1 AC079887 Oryza sativa putative DNA-binding protein. OSJNBa0040E01.10.
AAC37515.1 L44134 Cucumis sativus SPFl-like DNA-binding protein.
AAD16138.1 AF096298 Nicotiana tabacum DNA-binding protein 1. WRKYl. transcription factor.
AAD38283.1 AC007789 Oryza sativa putative WRKY DNA binding protein. OSJNBa0049B20.9.
BAB18313.1 AP002865 Oryza sativa putative WRKY DNA binding protein. P0034C11.1. contains EST C26525(C12525).
BAB40073.1 AP003074 Oryza sativa putative WRKY DNA binding protein. OSJNBa0004G10.20. contains EST C26525(C12525).
AAC49528.1 U56834 Petroselinum crispum
DNA-binding. WRKY3. WRKY-type DNA-binding protein.
BAA77358.1 AB020023 Nicotiana tabacum
WRKY domain Zn-finger type DNA-binding protein. DNA-binding protein NtWRKY3.
AAG46150.1 ACO 18727 Oryza sativa putative DNA-binding protein. OSJNBa0056G17.18.
CAA88331.1 Z48431 Avena fatua binds conserved cis-element from cereal alpha-Amy2 promoters. DNA-binding protein.
AAD32676.1 AF140553 Avena sativa
DNA-binding protein WRKY3. wrky3. putative transcription factor.
CAB66338.1 AJ279697 Betula pendula wrky-type DNA binding protein, wrky.
AAF61863.1 AF193770 Nicotiana tabacum DNA-binding protein 3. WRKY3. transcription factor.
AAF61864.1 AFl 93771 Nicotiana tabacum DNA-binding protein 4. WRKY4. transcription factor.
AAD27591.1 AF121354 Petroselinum crispum binds sequence specifically to W Boxes (TTGACC). transcription factor. WRKY3. sequence specific DNA-binding protein.
BAA87069.1 AB035271 Matricaria chamomilla elicitor-induced DNA-binding protem homolog. McWRKYl .
SEQ ID NO: 553
AAF34428.1 AFl 72282 Oryza sativa receptor-like protein kinase. DUPRl 1.18.
BAA94518.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 section 111 of 255; putative receptor-like protein kinase (AC002392).
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12.
BAA94516.1 AP001800 Oryza sativa
Shnilar to Zea mays S-domain receptor-like protein kinase (AJOl 0166).
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
BAA94517.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
BAA94528.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 BAC T20K24; putative receptor-like protein kinase (AC002392).
BAA94529.2 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2. CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRKl.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl .
BAA92836.1 AB032473 Brassica oleracea SI 8 S-locus receptor kinase. SRK18.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extiacellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
BAA92954.1 AP001551 Oryza sativa Similar to Ory -za sativa protein kinase (OSPKIO) mRNA. (L27821).
SEQ ID NO: 554
CAC05658.1 AJ250919 Brassica napus endopolygalacturonase. pegaz.
CAC05657.1 AJ250918 Brassica napus endopolygalacturonase. pg az.
CAA65072.1 X95800 Brassica napus polygalacturonase.
CAA90272.1 Z49971 Brassica napus Hydrolytic enzyme. Polygalacturonase. pga.
CAA54448.1 X77231 Prunus persica polygalacturonase. PG.
AAC14453.1 L12019 Actinidia deliciosa polygalacturonase.
AAF71160.1 AF152758 Actinidia chinensis polygalacturonase A. PGA
AAA34178.1 M37304 Lycopersicon esculentum polygalacturonase .
CAA32235.1 X14074 Lycopersicon esculentum polygalacturonase.
CAA29148.1 X05656 Lycopersicon esculentum polygalacturonase (AA 1-457).
AAA32914.1 L06094 Persea americana cell wall degradation, polyj galacturonase.
CAA47055.1 X66426 Persea americana polygalacturonase.
AAC26512.1 AF062467 Cucumis melo polygalacturonase precursor. MPG3.
CAAl 1846.1 AJ224147 Rubus idaeus polygalacturonase. RAS3.
AAF61444.1 AF138858 Lycopersicon esculentum hydrolyses polygalacturonic acid, polygalacturonase. XOPGl.
BAA88472.1 AB035890 Cucumis sativus polygalacturonase. CUPGl
AAD46483.1 AF 128266 Glycine max polygalacturonase PG1.
AAD46484.1 AF 128267 Glycine max polygalacturonase PG2.
AAC28905.1 AFOO 1002 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 4.
TAPG4. expressed in abscission.
AAC28906.1 AFOO 1003 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 5.
TAPG5. expressed in abscission.
AAC28903.1 AFOO 1000 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 1.
TAPG1. expressed in abscission.
AAA80489.1 U23053 Lycopersicon esculentum polygalacturonase precursor.
AAC64184.1 AF095577 Prunus persica endopolygalacturonase.
AAC28902.2 AF000999 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 3.
TAPG3. expressed in abscission.
AAB09575.1 U70480 Lycopersicon esculentum abscission polygalacturonase. TAPG2.
AAC28904.1 AFOO 1001 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 2.
TAPG2. expressed in abscission.
CAA54150.1 X76735 Prunus persica endopolygalacturonase.
AAC70951.1 AF072732 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella, polygalacturonase 7. specifically and abundantly expressed in pistils.
AAC26511.1 AF062466 Cucumis melo polygalacturonase precursor. MPG2.
AAB09576.1 U70481 Lycopersicon esculentum abscission polygalacturonase. TAPG4.
AAC28947.1 AF029230 Lycopersicon esculentum polygalacturonase. TPG6.
AAA82167.1 U09717 Gossypium hirsutum polygalacturonase.
AAA58322.1 U09805 Gossypium barbadense polygalacturonase.
CAA47052.1 X66422 Zea mays polygalacturonase. PG.
AAG14416.1 AF248538 Nicotiana tabacum NTS1 protein, similar to polygalacturonase.
CAB42886.1 AJ238848 Phleum pratense polygalacturonase. pg.
CAA40850.1 X57627 Zea mays polygalacturonase.
CAA46679.1 X65844 Zea mays polygalacturonase. PGg6.
CAA40910.1 X57743 Zea mays polygalacturonase.
CAA44249.1 X62385 Zea mays polygalacturonase .
CAA45751.1 X64408 Zea mays polygalacturonase. PG. pollen-preferentially expressed.
CAA40851.1 X57628 Zea mays polygalacturonase.
CAA46680.1 X65845 Zea mays polygalacturonase. PGgl4.
AAC26510.1 AF062465 Cucumis melo polygalacturonase precursor. MPG1.
SEQ ID NO: 556
CAA51734.1 X73301 Vigna mungo alpha-amylase . amyVml .
CAA37217.1 X53049 Vigna mungo alpha-amylase (AA 1-421)
BAA33879.1 AB015131 Phaseolus vulgaris alpha-amylase
AAA16513.1 U06754 Cuscuta reflexa starch hydrolysis, alpha amylase precursor. CUS AMY2.
AAA98615.1 J04202 Hordeum vulgare alpha-amylase . Amy46. precursor.
AAA98790.1 K02637 Hordeum vulgare alpha-amylase type B. Amy6-4. precursor.
CAA33298.1 XI 5226 Hordeum vulgare alpha-amylase
AAA33885.1 M24286 Oryza sativa alpha-amylase (EC 3.2.1.1).
CAA34516.1 X16509 Oryza sativa alpha-amylase
CAA72144.1 Y11277 Hordeum vulgare alpha-amylase . amy.
CAA33299.1 XI 5227 Hordeum vulgare alpha amylase.
AAA32925.1 M17126 Hordeum vulgare alpha-amylase 1.
AAA32927.1 M17128 Hordeum vulgare alpha-amylase 2.
AAA32929.1 J01236 Hordeum vulgare alpha-amylase type A, EC 3.2.1.1.
CAA39777.1 X56337 Oryza sativa alpha-amylase. RAmy3B.
CAA09323.1 AJO 10728 Avena fatua alpha amylase. alpha- Amy2A.
AAA33897.1 M24941 Oryza sativa alpha-amylase precursor (EC 3.2.1.1).
CAA09324.1 AJO 10729 Avena fatua alpha-amylase. alpha-Amy2D.
CAA28803.1 X05166 Hordeum vulgare alpha-amylase type A. Amy32b.
AAA32926.1 M17125 Hordeum vulgare alpha-amylase 1.
CAA39778.1 X56338 Oryza sativa alpha-amylase. RAmy3C.
AAA32935.1 Ml 5208 Hordeum vulgare alpha-amylase.
AAA33895.1 M59351 Oryza sativa alpha-amylase. RAmy3D.
AAA33896.1 M59352 Oryza sativa alpha-amylase. RAmy3E.
AAA50161.1 L25805 Zea mays alpha-amylase.
AAA33894.1 M74177 Oryza sativa alpha-amylase. amy2A.
CAA72143.1 Y11276 Hordeum vulgare alpha-amylase. amy.
CAA39776.1 X56336 Oryza sativa alpha-amylase. RAmy3A.
CAA45903.1 X64619 Oryza sativa alpha-amylase. Amyc2. starts hydrolysis during seed germination.
AAA33886.1 M24287 Oryza sativa alpha-amylase (EC 3.2.1.1).
AAA32928.1 M17127 Hordeum vulgare alpha-amylase 2.
AAA32933.1 K02638 Hordeum vulgare pre-alpha-amylase type B, EC 3.2.1.1.
AAA91883.1 M81682 Solanum tuberosum alpha-amylase. Amy23.
AAA34259.1 M16991 Triticum aestivum alpha-amylase
CAA29252.1 X05809 Triticum aestivum alpha-amylase
AAA91884.1 M79328 Solanum tuberosum alpha-amylase
AAF63239.1 AF153828 Malus x domestica degrades starch, alpha-amylase. alpha-amylase by similarity.
AAA32930.1 M10056 Hordeum vulgare alpha-amylase type B, EC 3.2.1.1.
AAA32931.1 K02635 Hordeum vulgare alpha-amylase type B, EC 3.2.1.1.
CAA36485.1 X52240 Oryza sativa alpha-amylase . OSamy-c.
AAA32932.1 K02636 Hordeum vulgare alpha-amylase type B, EC 3.2.1.1.
SEQ ID NO: 557
CAA90272.1 Z49971 Brassica napus Hydrolytic enzyme. Polygalacturonase. pga.
CAA65072.1 X95800 Brassica napus polygalacturonase.
CAA67020.1 X98373 Brassica napus endo-polygalacturonidase.
CAC05658.1 AJ250919 Brassica napus endopolygalacturonase. pegaz.
CAC05657.1 AJ250918 Brassica napus endopolygalacturonase. pg ;az.
AAC14453.1 L12019 Actinidia deliciosa polygalacturonase .
AAF71160.1 AF152758 Actinidia chinensis polygalacturonase A. PGA
CAA54448.1 X77231 Prunus persica polygalacturonase. PG.
AAA34178.1 M37304 Lycopersicon esculentum polygalacturonase.
CAA29148.1 X05656 Lycopersicon esculentum polygalacturonase (AA 1-457).
CAA32235.1 X14074 Lycopersicon esculentum polygalacturonase.
AAA32914.1 L06094 Persea americana cell wall degradation, poly galacturonase.
CAA47055.1 X66426 Persea americana polygalacturonase.
AAC26512.1 AF062467 Cucumis melo polygalacturonase precursor. MPG3.
CAAl 1846.1 AJ224147 Rubus idaeus polygalacturonase. RAS3.
BAA88472.1 AB035890 Cucumis sativus polygalacturonase. CUPGl.
AAF61444.1 AF138858 Lycopersicon esculentum hydrolyses polygalacturonic acid, polygalacturonase. XOPGl.
AAA80489.1 U23053 Lycopersicon esculentum polygalacturonase precursor.
AAC28903.1 AFOO 1000 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 1.
TAPG1. expressed in abscission.
AAB09575.1 U70480 Lycopersicon esculentum abscission polygalacturonase. TAPG2.
AAC28904.1 AF001001 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 2.
TAPG2. expressed in abscission.
AAC28905.1 AF001002 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 4.
TAPG4. expressed in abscission.
AAC64184.1 AF095577 Prunus persica endopolygalacturonase.
AAD46483.1 AF128266 Glycine max polygalacturonase PG1.
AAC28906.1 AF001003 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 5.
TAPG5. expressed in abscission.
AAD46484.1 AF128267 Glycine max polygalacturonase PG2.
CAA54150.1 X76735 Prunus persica endopolygalacturonase.
AAC26511.1 AF062466 Cucumis melo polygalacturonase precursor. MPG2.
AAC28902.2 AF000999 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella of plant cells, polygalacturonase 3.
TAPG3. expressed in abscission.
AAC70951.1 AF072732 Lycopersicon esculentum hydrolyzes pectin in the cell wall and middle lamella, polygalacturonase 7. specifically and abundantly expressed in pistils.
CAA47052.1 X66422 Zea mays polygalacturonase. PG.
AAC28947.1 AF029230 Lycopersicon esculentum polygalacturonase. TPG6.
AAA82167.1 U09717 Gossypium hirsutum polygalacturonase.
AAB09576.1 U70481 Lycopersicon esculentum abscission polygalacturonase. TAPG4.
CAA40910.1 X57743 Zea mays polygalacturonase.
CAA44249.1 X62385 Zea mays polygalacturonase.
CAA40850.1 X57627 Zea mays polygalacturonase.
AAG14416.1 AF248538 Nicotiana tabacum NTS1 protein. similar to polygalacturonase.
CAA46679.1 X65844 Zea mays polygalacturonase. PGg6.
AAC26510.1 AF062465 Cucumis melo polygalacturonase precursor. MPG1.
AAA58322.1 U09805 Gossypium barbadense polygalacturonase.
CAA46680.1 X65845 Zea mays polygalacturonase. PGgl4,
CAA40851.1 X57628 Zea mays polygalacturonase.
CAA45751.1 X64408 Zeamays polygalacturonase. PG. pollen-preferentially expressed.
CAB42886.1 AJ238848 Phleum pratense polygalacturonase. pg.
SEQ ID NO: 559
CAB43937.1 AJ006348 Fragaria x ananassa cell wall hydrolysis. endo-beta-l,4-glucanase. egl.
AAC95009.1 AF074923 Fragaria x ananassa endo-l,4-beta- -glucanase precursor. Cell. 1,4-beta-glucanohydrolase.
AAC12684.1 U76725 Pinus radiata endo-beta-1,4- -glucanase. PrCell. cellulase; PRCEL1.
AAA69909.1 U13055 Lycopersicon esculentum cell wall hydrolase. endo- 1,4-beta-glucanase precursor. Cel2. cellulase.
CAA65600.1 X96856 Prunus persica endo-beta-1,4- -glucanase. ppEGl.
CAA65597.1 X96853 Prunus persica endo-beta-l,4-glucanase. pcelL
CAA65827.1 X97189 Capsicum annuum endo-beta-l,4-glucanase. cceB. cellulase.
AAC62241.1 AF077339 Lycopersicon esculentum endo-l,4-beta-glucanase. cel5. cellulase; also expressed in fruit and pistils.
BAA85150.1 AB032830 Pisum sativum endo- 1,4-beta-glucanase. EGL2.
AAA80495.1 U20590 Lycopersicon esculentum endo- 1,4-beta-glucanase precursor, cellulase.
BAB32662.1 AB055886 Atriplex lentiformis beta-l,4-glucanase. Al-cell. cellulase.
CAA65828.1 X97190 Capsicum annuum endo-beta-l,4-glucanase. ccel2.
AAC12685.1 U76756 Pinus radiata endo-beta-l,4-glucanase. PrCel2. cellulase; PRCEL2.
BAA77239.1 AB025796 Populus alba endo-l,4-beta glucanase. POPCEL2. cellulase.
CAB59900.1 AJ010950 Capsicum annuum cell wall degradation. endo-beta-l,4-glucanase. eg2.
BAB39483.1 AB049200 Populus alba endo- 1,4-beta-glucanase. PopCel2.
BAB39482.1 AB049199 Populus alba endo- 1 ,4-beta glucanase. PopCell.
CAA72133.1 Y11268 Lycopersicon esculentum endo-l,4-beta-D-glucanase. ce!7.
AAA96135.1 L41046 Pisum sativum endo-1 ,4-beta-glucanase. EGL1.
AAC78504.1 U34754 Phaseolus vulgaris endo- 1 ,4-beta-D-glucanase. cellulase.
AAA02563.1 M57400 Phaseolus vulgaris cellulase precursor. endo-l,4-beta-D-glucanase.
CAA60737.1 X87323 Capsicum annuum catalyzes hydrolysis of cell wall polysaccharides. cellulase. cell. Beta-1, 4- endoglycanohydrolase.
CAA65826.1 X97188 Capsicum annuum endo-beta-l,4-glucanase. ccell. cellulase.
AAA69908.1 U13054 Lycopersicon esculentum cell wall hydrolase. endo- 1,4-beta-glucanase precursor. Cell, cellulase.
CAB43938.1 AJ006349 Fragaria x ananassa cell wall hydrolysis. endo-beta-l,4-glucanase. eg3.
BAA96209.1 AP002094 Oryza sativa
EST C72268(E1328) conesponds to a region ofthe predicted gene. Similar to Fragaria x ananassa endo-beta-l,4-glucanase (AJ006349).
BAA96207.1 AP002094 Oryza sativa
EST C72268(E1328) conesponds to a region ofthe predicted gene. Similar to Fragaria x ananassa endo-beta-l,4-glucanase (AJ006349).
AAD08699.1 AF098292 Lycopersicon esculentum endo-beta-l,4-D-glucanase. Cel8.
BAA94257.1 AB040769 Hordeum vulgare endo- 1,4-beta-glucanase Cell. Cell.
AAC49704.1 U78526 Lycopersicon esculentum endo- 1,4-beta-glucanase. Cel3.
CAB51903.1 AJ242807 Brassica napus endo-l,4-beta-D-glucanase. Cell6. cellulase.
AAA20082.1 U00730 Glycine max CMCase; cellulase; endo-l,4-beta-D-glucanase.
CAA11301.1 AJ223386 Fragaria x ananassa endo-beta-l,4-glucanase. faEGl. partial cDNA obtained by RT-PCR amplification.
CAAl 1302.1 AJ223387 Fragaria x ananassa endo-beta-l,4-glucanase. partial cDNA obtained by RT-PCR amplification.
BAA21111.1 D88417 Gossypium hirsutum endo- 1 ,4-beta-glucanase.
AAA20083.1 U00731 Glycine max CMCase; cellulase; endo-l,4-beta-D-glucanase.
SEQ ID NO: 560
AAB97617.1 U83687 Apium graveolens
NADPH-dependent mannose 6-phosphate reductase. m6pr. aldo-keto reductase; similar to aldose 6-phosphate reductase also known as NADP-sorbitol-6-phosphate dehydrogenase encoded by GenBank Accession Number Dl 1080.
AAC97607.1 AF057134 Malus x domestica synthesizes sorbitol, a major photosynthetic product in many members ofthe Rosaceae family.
NADP -dependent sorbitol 6-phosphate dehydrogenase. S6PDH.
BAA01853.1 D11080 Malus x domestica
NADP -dependent D-sorbitol-6-phosphate dehydrogenase. S6PDH.
SEQ ID NO: 561
CAB43938.1 AJ006349 Fragaria x ananassa cell wall hydrolysis. endo-beta-l,4-glucanase. eg3.
BAA96209.1 AP002094 Oryza sativa
EST C72268(E1328) conesponds to a region ofthe predicted gene. Similar to Fragaria x ananassa endo-beta-l,4-glucanase (AJ006349).
BAA96207.1 AP002094 Oryza sativa
EST C72268(E1328) conesponds to a region ofthe predicted gene. Similar to Fragaria x ananassa endo-beta-l,4-glucanase (AJ006349).
AAD08699.1 AF098292 Lycopersicon esculentum endo-beta-1 ,4-D-glucanase. Cel8.
BAA21111.1 D88417 Gossypium hirsutum endo- 1 ,4-beta-glucanase.
CAA65828.1 X97190 Capsicum annuum endo-beta-1 ,4-glucanase. ccel2.
CAB59900.1 AJOl 0950 Capsicum annuum cell wall degradation, endo-beta-1, 4-glucanase. eg2.
AAC95009.1 AF074923 Fragaria x ananassa endo- 1,4-beta-glucanase precursor. Cell. 1,4-beta-glucanohydrolase.
CAB43937.1 AJ006348 Fragaria x ananassa cell wall hydrolysis, endo-beta-1, 4-glucanase. egl.
AAA80495.1 U20590 Lycopersicon esculentum endo- 1,4-beta-glucanase precursor, cellulase.
BAA85150.1 AB032830 Pisum sativum endo- 1,4-beta-glucanase. EGL2.
AAC12684.1 U76725 Pinus radiata endo-beta-1, 4-glucanase. PrCell. cellulase; PRCEL1.
BAB32662.1 AB055886 Atriplex lentiformis beta-1 ,4-glucanase. Al-cell . cellulase.
AAC62241.1 AF077339 Lycopersicon esculentum endo- 1,4-beta-glucanase. ce!5. cellulase; also expressed in fruit and pistils .
CAA65826.1 X97188 Capsicum annuum endo-beta-1, 4-glucanase. ccell. cellulase.
AAA69909.1 U13055 Lycopersicon esculentum cell wall hydrolase. endo- 1,4-beta-glucanase precursor. Cel2. cellulase.
CAA65597.1 X96853 Prunus persica endo-beta-1, 4-glucanase. pcell.
CAA65600.1 X96856 Prunus persica endo-beta-1, 4-glucanase. ppEGl.
AAC12685.1 U76756 Pinus radiata endo-beta-1, 4-glucanase. PrCel2. cellulase; PRCEL2.
BAB39482.1 AB049199 Populus alba endo-l,4-beta glucanase. PopCell.
CAA65827.1 X97189 Capsicum annuum endo-beta-1, 4-glucanase. ccel3. cellulase.
BAB39483.1 AB049200 Populus alba endo- 1,4-beta-glucanase. PopCel2.
BAA77239.1 AB025796 Populus alba endo-l,4-beta glucanase. POPCEL2. cellulase.
AAA69908.1 U13054 Lycopersicon esculentum cell wall hydrolase. endo- 1,4-beta-glucanase precursor. Cell, cellulase.
AAC78504.1 U34754 Phaseolus vulgaris endo-l,4-beta-D-glucanase. cellulase.
AAA02563.1 M57400 Phaseolus vulgaris cellulase precursor. endo-l,4-beta-D-glucanase.
CAA72133.1 Y11268 Lycopersicon esculentum endo-l,4-beta-D-glucanase. cel7.
AAA96135.1 L41046 Pisum sativum endo- 1,4-beta-glucanase. EGL1.
CAB51903.1 AJ242807 Brassica napus endo-l,4-beta-D-glucanase. Cell6. cellulase.
AAC49704.1 U78526 Lycopersicon esculentum endo- 1,4-beta-glucanase. CeB.
BAA94257.1 AB040769 Hordeum vulgare endo- 1,4-beta-glucanase Cell. Cell.
CAAl 1302.1 AJ223387 Fragaria x ananassa endo-beta-1, 4-glucanase. partial cDNA obtained by RT-PCR amplification.
AAA20082.1 U00730 Glycine max CMCase; cellulase; endo-l,4-beta-D-glucanase.
CAAl 1301.1 AJ223386 Fragaria x ananassa endo-beta-1, 4-glucanase. faEGl. partial cDNA obtained by RT-PCR amplification.
AAC64045.1 AF077340 Lycopersicon esculentum endo- 1,4-beta-glucanase. cel5. cellulase.
SEQ ID NO: 562
AAD53011.1 AF089848 Brassica napus senescence-specific cysteine protease. SAG12-1. BnSAG12-l.
AAD53012.1 AF089849 Brassica napus senescence-specific cysteine protease. SAG12-2. BnSAG12-2.
AAK27968.1 AF242372 Ipomoea batatas cysteine protease. SPCP1.
AAA50755.1 U13940 Alnus glutinosa cysteine proteinase. putative preproprotein.
BAB13759.1 AB040454 Astragalus sinicus cysteine proteinase. AsNODB2. preproprotein putative.
AAC62396.1 AF050756 Ricinus communis cysteine endopeptidase precursor. CysEP.
CAB09698.1 Z97022 Hordeum vulgare cysteine proteinase. putative.
CAA52425.1 X74406 Hemerocallis sp. thiol-protease. SEN102.
BAA83472.1 AB004648 Oryza sativa cysteine endopeptidase. RepA.
CAA56844.1 X80876 Oryza sativa cysteine protease.
BAA88898.1 AB020961 Zeamays cysteine protease component of protease-inhibitor complex. CPPIC. AAC35211.1 U12637 Hemerocallis hybrid cultivar cysteine proteinase. SEN11.
CAB09697.1 Z97021 Hordeum vulgare cysteine endopeptidase EP-A. precursor.
AAB88263.1 AF019147 Zeamays cysteine proteinase Mir3. mir3.
AAD28477.1 AF133839 Sandersonia aurantiaca papain-like cysteine protease. PRT5. senescence-related.
CAB 16317.1 Z99173 Nicotiana tabacum storage protein hydrolysis, cysteine proteinase precursor.
CAB09699.1 Z97023 Hordeum vulgare cysteine endopeptidase EP-A.
AAD10337.1 U94591 Hordeum vulgare cysteine proteinase precursor. EPA.
CAA06243.1 AJ004958 Pisum sativum thiol-protease. pre-pro-TPE4A protein. tpE4A.
CAB53515.1 AJ245924 Solanum tuberosum proteolysis. cysteine protease, cyp.
AAD48496.1 AFl 72856 Lycopersicon esculentum cysteine protease TDI-65. tdi-65. induced by drought; localized in the nuclei and chloroplast
(Tabaeizadeh, Z. et al., 1995. Protoplasma, 186:208-219).
CAA05894.1 AJ003137 Lycopersicon esculentum cysteine protease. CYPl. C14.
AAA79915.1 U17135 Dianthus caryophyllus cysteine proteinase. DCCP1. expressed in senescing flower petals.
AAB37233.1 U34747 Phalaenopsis sp. SM9108 cysteine proteinase.
CAB 17076.1 Z99954 Phaseolus vulgaris protein hydrolysis, cysteine proteinase precursor.
CAA84378.1 Z34895 Vicia sativa storage protein degradation, cysteine proteinase.
CAA53377.1 X75749 Vicia sativa storage protein hydrolysis, cysteine protease.
CAB 17074.1 Z99952 Phaseolus vulgaris degradation of storage proteins, cysteine proteinase precursor.
AAB68374.1 U52970 Phaseolus vulgaris suggested to initiate phaseolin degradation during gennination. cysteine endopeptidase 1.
PvCEP-1.
CAA12118.1 AJ224766 Phaseolus vulgaris phaseolin degradation, cysteine protease.
AAC49455.1 U41902 Pseudotsuga menziesii cysteine protease. Pseudotzain. PM33cysP.
CAA46863.1 X66061 Pisum sativum thiolprotease. tpp. start codon ttg.
AAB41816.1 U44947 Pisum sativum NTH1. PsCypl. cysteine protease homolog.
BAA83473.1 AB004819 Oryza sativa cysteine endopeptidase. Repl.
BAA22544.1 D38532 Ananas comosus precursor of cysteine proteinase. FBSB precursor, stem bromelain precursor in fruit.
BAA11170.1 D76415 Oryza sativa cysteine proteinase.
AAD20453.1 AF099203 Oryza sativa cysteine endopeptidase precursor. EP3 A.
CAA08860.1 AJ009829 Ananas comosus cysteine proteinase precursor, AN8. an8.
AAA85036.1 U19384 Hordeum vulgare cysteine proteinase EPB2 precursor.
AAA85035.1 U19359 Hordeum vulgare cysteine proteinase EPB1 precursor.
SEQ ID NO: 565
AAA97411.1 U51918 Pisum sativum pyravate dehydrogenase El alpha subunit.
CAA81558.1 Z26949 Solanum tuberosum subunit ofthe mitochondrial pyravate dehydrogenase complex. El alpha subunit of pyravate dehydrogenase precursor.
AAC72195.1 AF069911 Zea mays pyravate dehydrogenase El alpha subunit.
AAG43499.1 AF209924 Lycopersicon esculentum pyravate dehydrogenase.
CAB08111.1 Z94180 Lycopersicon esculentum branched chain alpha-keto acid dehydrogenase El -alpha subunit.
SEQ ID NO: 566
AAD55090.1 AF178653 Vitis riparia thaumatin. osmotin; pathogenesis-related protein.
CAA51432.1 X72928 Solanum commersonii osmotin-like protein.
CAA47601.1 X67121 Solanum commersonii osmotin-like protein.
CAC34055.1 AJ297410 Capsicum annuum osmotin-like protein. pr5 p23.
CAA47047.1 X66416 Lycopersicon esculentum tpm 1. Induced during viroid infection. Osmotin-like, antifungal protein homologue.
AAB23375.1 S44889 Nicotiana tabacum osmotin. osmotin. pathogenesis-related protein homolog; This sequence comes from Fig. 3.
AAB22459.2 S40046 Nicotiana tabacum osmotin. abscisic acid-activated, basic PR-like protein; This sequence comes from Fig. 1 A; conceptual translation presented here differs from translation in publication; mismatch(181[R-
>G]).
AAG16625.1 AY007309 Solanum dulcamara cryoprotective osmotin-like protein.
CAA46623.1 X65701 Nicotiana tabacum osmotin. AP24.
CAA46622.1 X65700 Nicotiana tabacum osmotin. AP24.
CAA51431.1 X72927 Solanum commersonii osmotin-like protein.
CAA64620.1 X95308 Nicotiana tabacum PR protein, osmotin.
CAA51430.1 X72926 Solanum commersonii osmotin-like protein.
AAC64171.1 AF093743 Lycopersicon esculentum pathogenesis-related protein osmotin precursor. NP24.
AAB61590.1 AF003007 Vitis vinifera
WTLl. osmotin-like protein, PR-5 protein; thaumatin-like protein.
BAA11180.1 D76437 Nicotiana sylvestris antifungal activity, neutral PR-5 (osmotin-like protein, PR-5d). neutral isoform of pathogenesis-related protein graop 5.
AAA34087.1 M64081 Nicotiana tabacum osmotin-like protein. OLP1.
CAA47669.1 X67244 Solanum commersonii osmotin-like protein.
CAA71883.1 Y10992 Vitis vinifera osmotin-like protein. OSM1.
AAF13707.1 AF199508 Fragaria x ananassa osmotin-like protein, olp.
AAA34089.1 M29279 Nicotiana tabacum osmotin.
CAA43854.1 X61679 Nicotiana tabacum osmotin.
CAA04642.1 AJ001268 Hordeum vulgare antifungal. basic pathogenesis-related protein PR5. osmotin/permatin-like.
CAC22330.1 AJ298304 Fagus sylvatica stress protein, osmotin-like protein. olp2.
CAC22329.1 AJ298303 Fagus sylvatica stress protein, osmotin-like protein, olp.
AAB67852.1 L76377 Oryza sativa osmotin. 14b.
CAB36911.1 AJ000692 Quercus suber stress protein, osmotin-like protein, olp.
SEQ ID NO: 569
BAA95814.1 AP002069 Oryza sativa
ESTs AU082579(S2069),D40238(S2069) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana vesicle-associated membrane protein 7C; synaptobrevin 7C.
(AF025332).
SEQ ID NO: 572
AAA92677.1 U13736 Pisum sativum binds calcium, calmodulin-like protein.
AAA34015.1 L01433 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-4. putative.
AAA33948.1 L19359 Glycine max calcium-binding regulatory protein, cahnodulin. SCaM-5. putative.
CAA66159.1 X97558 Capsicum annuum calmodulin- 1.
CAA09302.1 AJO 10645 Capsicum annuum calcium binding protein, calmodulin 3 protein, calmodulin 3.
AAA34144.1 M67472 Lycopersicon esculentum calmodulin. CALM1LE.
CAA62150.1 X90560 Physcomitrella patens Calmodulin. CaM.
AAB46588.1 U83402 Capsicum annuum cahnodulin.
BAA87825.1 AP000815 Oryza sativa
ESTs AU030013(E50493),AU081341(E50493) conespond to a region ofthe predicted gene.
Similar to O.sativa gene encoding cahnodulin. (Z12828).
AAF65511.1 AF108889 Capsicum annuum calmodulin.
AAA85157.1 U20297 Solanum tuberosum calcium-binding protein, calmodulin.
AAA85156.1 U20296 Solanum tuberosum calcium-binding protein, calmodulin.
AAA62351.1 U20295 Solanum tuberosum calcium-binding protein, calmodulin.
AAA85155.1 U20294 Solanum tuberosum calcium-binding protein, calmodulin.
AAA33900.1 L18914 Oryza sativa calcium binding protein, signal transduction. calmodulin.
CAA78288.1 Z12828 Oryza sativa calcium binding protein, signal transduction. cahnodulin.
AAC49583.1 U48692 Triticum aestivum cahnodulin TaCaM2-3. calcium-binding protein.
AAC49582.1 U48691 Triticum aestivum calmodulin TaCaM2-2. calcium-binding protein.
CAA61980.1 X89890 Bidens pilosa Cahnodulin.
CAA67054.1 X98404 Capsicum annuum cahnodulin-2.
AAA33083.1 M20729 Chlamydomonas reinhardtii cahnodulin.
AAG11418.1 AF292108 Prunus avium calmodulin.
AAF33852.1 AF231026 Oryza sativa cahnodulin-like protein.
AAA92681.1 U13882 Pisum sativum calcium-binding protein, calmodulin.
AAA33706.1 M80836 Petunia x hybrida cahnodulin. CAM81.
AAA33705.1 M80831 Petunia x hybrida calmodulin-related protein. CAM53.
AAA98933.1 U37936 Oryza sativa novel calmodulin-like protein. C-terminal of this sequence contains a CAAX box which is common in signal transport protein, but not in calmodulin.
CAA43143.1 X60738 Malus x domestica Cahnodulin. CaM.
CAA78301.1 Z12839 Lilium longiflorum calcium binding protein, signal transduction. calmodulin.
AAA33397.1 L18912 Lilium longiflorum calcium binding protein, signal transduction. calmodulin. putative.
CAA42423.1 X59751 Daucus carota calmodulin. Ccam-1.
AAF73157.1 AF150059 Brassica napus cahnodulin. CaMl. involved in seed germination.
AAA19571.1 U10150 Brassica napus calcium binding, calmodulin. bcml .
AAA87347.1 M88307 Brassica juncea calmodulin.
AAG27432.1 AF295637 Elaeis guineensis cahnodulin.
BAA94697.1 AB041712 Chara corallina calmodulin. cccam2.
BAA94696.1 AB041711 Chara corallina calmodulin. cccaml.
BAA96536.1 AB044286 Chara corallina calmodulin. ccam.
AAC18355.1 AF064456 Oryza sativa subsp. indica calmodulin-like protein. CAM-like.
AAA34237.1 L20691 Vigna radiata calmodulin.
CAA52602.1 X74490 Zeamays Calmodulin. ZMCALM1.
CAA54583.1 X77397 Zeamays calmodulin. CaM2.
AAC49585.1 U49103 Triticum aestivum calmodulin TaCaM3-2. calcium-binding protein.
AAC49586.1 U49104 Triticum aestivum calmodulin TaCaM3-3. calcium-binding protein.
AAC49587.1 U49105 Triticum aestivum calmodulin TaCaM4-l. calcium-binding protein.
AAC49580.1 U48689 Triticum aestivum calmodulin TaCaMl-3. calcium-binding protein.
AAC49584.1 U48693 Triticum aestivum calmodulin TaCaM3-l. calcium-binding protein.
SEQ ID NO: 575
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid: glucosyltiansferase 1. togtl. glucosyltiansferase.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS5a.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS 10a.
AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid:glucosyltiansferase 2. togt2. glucosyltransferase.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltransferases.
CAB56231.1 Y18871 Dorotheanthus bellidifonnis betanidin-5-O-glucosyltransferase.
BAA83484.1 AB031274 Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltiansferase. ufgt.
AAB48444.1 U82367 Solanum tuberosum UDP-glucose glucosyltransferase.
CAA54610.1 X77460 Manihot esculenta UTP-glucose glucosyltiansferase. CGT4.
AAD04166.1 AF101972 Phaseolus lunatus catalyzes formation of O-glucosylzeatin from zeatin and UDPG; catalyzes formation of O- xylosylzeatin from zeatin and UDPX. zeatin O-glucosyltiansferase. ZOGl. cytokinin O- glucosyltransferase.
BAB17182.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.19.
BAA93039.1 AB033758 Citrus unshiu limonoid UDP-glucosyltransferase. LGTase. AAF61647.1 AFl 90634 Nicotiana tabacum
UDP-glucose: salicylic acid glucosyltiansferase. SA-GTase.
BAB17176.1 AP002843 Oryza sativa putative UTP-glucose glucosyltiansferase. P0407B12.13.
CAC09351.1 AL442007 Oryza sativa putative glucosyltransferase. H0212B02.7.
CAA54611.1 X77461 Manihot esculenta UTP-glucose glucosyltiansferase. CGT2.
AAF98390.1 AF287143 Brassica napus catalyzes the transfer of glucose from UDP-glucose to sinapate and some other hydroxy cinnamates (4-coumarate, caffeate, feralate). UDP-glucose: sinapate glucosyltransferase. SGT 1. SGT.
AAK16181.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.16.
AAK16178.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.5.
CAA54612.1 X77462 Manihot esculenta UTP-glucose glucosyltiansferase. CGT5.
AAK16180.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.21.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitrile-o- glucosyltransferase.
CAA54609.1 X77459 Manihot esculenta UTP-glucose glucosyltiansferase. CGT1.
BAB41025.1 AB047098 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. F1UFGT1.
AAK16172.1 AC079887 Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.14.
BAB41019.1 AB047092 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. ItUFGTl.
BAB41018.1 AB047091 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. KyUFGT2. The gene originated in one ofthe parents V. vinifera cv. Centennial. ^
BAB41023.1 AB047096 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT1. BAB41021.1 AB047094 Vitis vinifera UDP-glucose : flavonoid 3-O-glucosyltransferase. RuUFGTl.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltransferase. PHI.
AAB81682.1 AF000371 Vitis vinifera UDP glucose : flavonoid 3 -o-glucosyltiansferase.
BAB41017.1 AB047090 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. KyUFGTl. The gene was derived from one ofthe parents V. labrascana cv. Ishiharawase.
BAB41022.1 AB047095 Vitis vinifera
UDP-glucose : flavonoid 3-O-glucosyltransferase. RuUFGT2.
BAB41020.1 AB047093 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. ItUFGT2.
BAB41024.1 AB047097 Vitis vinifera
UDP-glucose : flavonoid 3-O-glucosyltiansferase. A1UFGT2.
AAB81683.1 AF000372 Vitis vinifera UDP glucose:flavonoid 3 -o-glucosyltiansferase.
BAB41026.1 AB047099 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. F1UFGT2.
CAA54613.1 X77463 Manihot esculenta UTP-glucose glucosyltransferase. CGT6.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltransferase. PGT8.
BAA12737.1 D85186 Gentiana triflora UDP-glucose : flavonoid-3 -glucosyltiansferase.
SEQ ID NO: 576
CAB60277.1 AJ002586 Solanum tuberosum UCP.
CAA72107.1 Y11220 Solanum tuberosum mitochondrial uncoupling protein.
BAA92172.1 AB024733 Symplocaφus renifolius SfUCPa. SfUCPa.
BAB40658.1 AB049998 Oryza sativa uncoupling protein. OsUCP2.
BAA92173.1 AB024734 Symplocaφus renifolius SfUCPb. SfUCPb.
BAB 16385.1 AB042429 Triticum aestivum uncoupling protein. WhUCPlb.
BAB16384.1 AB042428 Triticum aestivum uncoupling protein. WhUCPla.
BAB40657.1 AB049997 Oryza sativa uncoupling protein. OsUCPl.
SEQ ID NO: 577
AAA19571.1 U10150 Brassica napus calcium binding, calmodulin. bcml.
AAA87347.1 M88307 Brassica juncea calmodulin.
AAA92681.1 U13882 Pisum sativum calcium-binding protein, calmodulin.
AAA33706.1 M80836 Petunia x hybrida calmodulin. CAM81.
CAA43143.1 X60738 Malus x domestica Calmodulin. CaM.
CAA78301.1 Z12839 Lilium longiflorum calcium binding protein, signal transduction. calmodulin.
AAA33397.1 L18912 Lilium longiflorum calcium binding protein, signal transduction. calmodulin. putative.
CAA42423.1 X59751 Daucus carota calmodulin. Ccam-1.
AAG27432.1 AF295637 Elaeis guineensis calmodulin.
AAG11418.1 AF292108 Prunus avium calmodulin.
AAA34237.1 L20691 Vigna radiata calmodulin.
AAC49587.1 U49105 Triticum aestivum calmodulin TaCaM4-l. calcium-binding protein.
AAC49586.1 U49104 Triticum aestivum calmodulin TaCaM3-3. calcium-binding protein.
AAC49585.1 U49103 Triticum aestivum calmodulin TaCaM3-2. calcium-binding protein.
AAC49584.1 U48693 Triticum aestivum calmodulin TaCaM3-l. calcium-binding protein.
AAC49580.1 U48689 Triticum aestivum calmodulin TaCaMl-3. calcium-binding protein.
AAC49579.1 U48688 Triticum aestivum calmodulin TaCaMl-2. calcium binding protein.
AAC49578.1 U48242 Triticum aestivum calmodulin TaCaMl-1. calcium-binding.
AAA03580.1 L01431 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-2. putative.
AAB36130.1 S81594 Vigna radiata auxin-regulated cahnodulin. auxin-regulated calmodulin, arCaM. This sequence comes from
Fig. 1; arCaM.
AAA33901.1 L18913 Oryza sativa calcium binding protein, signal transduction. calmodulin. putative.
CAA78287.1 Z12827 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAB46588.1 U83402 Capsicum annuum calmodulin.
AAA32938.1 M27303 Hordeum vulgare calmodulin.
BAA88540.1 AP000969 Oryza sativa
ESTs AU081349(E61253), D41425(S3918) conespond to a region ofthe predicted gene.
Similar to calmodulin. (AF042840).
AAF65511.1 AF108889 Capsicum annuum cahnodulin.
AAC36059.1 AF042840 Oryza sativa calmodulin. CaMl.
AAA34238.1 L20507 Vigna radiata calmodulin.
AAA34014.1 L01432 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-3. putative.
AAA34013.1 L01430 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-1. putative.
CAA36644.1 X52398 Medicago sativa calmodulin (AA 1-149).
AAB68399.1 U79736 Helianthus annuus calmodulin. HaCaM.
AADl 0244.1 AF030032 Phaseolus vulgaris calmodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
AAA33705.1 M80831 Petunia x hybrida calmodulin-related protein. CAM53.
AAA16320.1 L14071 Bryonia dioica calmodulin. Bc329. calcium-binding sites (amino acid #): 1. (21..33); 2. (57..68); 3. (94..106);
4. (130..141).
CAA61980.1 X89890 Bidens pilosa
Calmodulin.
AAA33900.1 L18914 Oryza sativa calcium binding protein, signal transduction. cahnodulin.
CAA78288.1 Z12828 Oryza sativa calcium binding protein, signal transduction. calmodulin.
CAA74307.1 Y13974 Zeamays cahnodulin.
AAC49583.1 U48692 Triticum aestivum calmodulin TaCaM2-3. calcium-binding protein.
AAC49582.1 U48691 Triticum aestivum calmodulin TaCaM2-2. calcium-binding protein.
AAA85157.1 U20297 Solanum tuberosum calcium-binding protein, calmodulin.
AAA85156.1 U20296 Solanum tuberosum calcium-binding protein, calmodulin.
AAA62351.1 U20295 Solanum tuberosum calcium-binding protein, cahnodulin.
AAA85155.1 U20294 Solanum tuberosum calcium-binding protein, calmodulin.
CAA46150.1 X65016 Oryza sativa cahnodulin. cam.
AAD10246.1 AF030034 Phaseolus vulgaris cahnodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
AAF73157.1 AF150059 Brassica napus calmodulin. CaMl. involved in seed gennination.
CAA54583.1 X77397 Zeamays calmodulin. CaM2.
SEQ ID NO: 578
AAF01764.2 AFl 84277 Glycine max homeodomain-leucine zipper protein 56. Hdl56. transcription factor.
BAA21017.1 D26578 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 761-940; leucine zipper at nt 941-1048.
AAD37697.1 AF145728 Oryza sativa homeodomain leucine zipper protein. Oshox4. transcription factor.
CAB67118.1 Y17306 Lycopersicon esculentum homeodomain protein. h52.
AAF01765.1 AF184278 Glycine max homeodomain-leucine zipper protein 57. Hdl57. transcription factor.
BAA05624.1 D26575 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 520-699; leucine zipper at nt 700-805.
BAA93466.1 AB028078 Physcomitrella patens homeobox protein PpHB7. PpHB7. homeodomain-leucine zipper gene.
BAA93465.1 AB028077 Physcomitrella patens homeobox protein PpHB6. PpHB6. homeodomain-leucine zipper gene.
BAA93464.1 AB028076 Physcomitrella patens homeobox protein PpHB5. PpHB5. homeodomain-leucine zipper gene.
BAA05622.1 D26573 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 585-764; leucine zipper at nt 765-851.
BAA05625.1 D26576 Daucus carota transcriptϋonal regulator. DNA-binding protein, homeodomain at nt 300-479; leucine zipper at nt 480-587.
BAA05623.1 D26574 Daucus carota trancriptional regulator. DNA-binding protein, homeodomain at nt 498-677; leucine zipper at nt 678-785.
AAD37698.1 AFl 45729 Oryza sativa homeodomain leucine zipper protein. Oshox5. transcription factor.
BAA93461.1 AB028073 Physcomitrella patens homeobox protein PpHB2. PpHB2. homeodomain-leucine zipper gene.
BAA93467.1 AB028079 Physcomitrella patens homeobox protein PpHB8. PpHB8. homeodomain-leucine zipper gene.
BAA93468.1 AB028080 Physcomitrella patens homeobox protein PpHB9. PpHB9. homeodomain-leucine zipper gene.
BAA93460.1 AB028072 Physcomitrella patens homeobox protein PpHBl. PpHBl. homeodomain-leucine zipper gene.
AAD37699.1 AF145730 Oryza sativa homeodomain leucine zipper protein. Oshox6. transcription factor.
CAA06717.1 AJ005820 Craterostigma plantagineum transcription factor, homeodomain leucine zipper protein, hb-1.
BAA93463.1 AB028075 Physcomitrella patens homeobox protein PpHB4. PpHB4. homeodomain-leucine zipper gene.
CAA65456.2 X96681 Oryza sativa transcription factor. DNA-binding protein. Oshoxl . homeodomain leucine zipper gene.
AAF19980.1 AF211193 Oryza sativa homeodomain-leucine zipper transcription factor. Hoxl. hoxl.
AAK31270.1 AC079890 Oryza sativa homeodomain leucine zipper protein hoxl. OSJNBb0089A17.12.
CAA06728.1 AJ005833 Craterostigma plantagineum transcription factor, homeodomain leucine zipper protein, hb-2.
AAD37696.1 AF145727 Oryza sativa homeodomain leucine zipper protein. Oshox3. transcription factor. SEQ ID NO: 580
AAD32141.1 AF123503 Nicotiana tabacum
Nt-gh3 deduced protein.
CAA42636.1 X60033 Glycine max auxin-responsive GH3 product. GH3.
BAA96221.1 AP002094 Oryza sativa
ESTs C19814(E10971),AU090481(E10971) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, sequence from clones T3F17, Fl 1C10, F13A10; auxin-responsive GH3-like protein (AC006526).
SEQ ID NO: 581
AAD13632.1 AF059488 Lycopersicon esculentum expansin precursor. Exp4.
AAF32410.1 AF230277 Triphysaria versicolor alpha-expansin 2.
CAA04385.1 AJ000885 Brassica napus
Cell wall extension in plants. Expansin.
CAB46492.1 AJ243340 Lycopersicon esculentum expansin9. exp9.
AAC63088.1 U82123 Lycopersicon esculentum expansin. LeEXPl. fruit ripening regulated expansin.
CAA06271.2 AJ004997 Lycopersicon esculentum expansinlδ. expl8.
AAF62182.1 AF247164 Oryza sativa alpha-expansin OsEXP7. cell wall loosening factor; expressed in internodes and leaves.
AAF35900.1 AF230331 Zinnia elegans expansin. Expl.
AAG13982.1 AF297521 Prunus avium expansin 1. Expl. PraavExpl.
AAC33529.1 U93167 Prunus armeniaca expansin. PA-Expl.
AAF21101.1 AF159563 Fragaria x ananassa expansin. Exp2. ripening regulated.
AAF35901.1 AF230332 Zinnia elegans expansin 2.
AAG13983.1 AF297522 Prunus avium expansin 2. Exp2. PraavExp2.
AAC33530.1 AF038815 Prunus armeniaca expansin. Exp2.
BAB 19676.1 AB029083 Prunus persica expansin. PchExpl.
AAB40637.1 U64893 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession
Numbers U30477 and U30479.
AAB37746.1 U30382 Cucumis sativus expansin S 1 precursor. Cs-EXPl . similar to pollen allergen Lol pl, Lolium perenne, SwissProt Accession Number P14946; former gene name CuExSl; expansin-29 (Ex29) protein.
CAC19184.1 AJ291817 Cicer arietinum expansin.
AAD47901.1 AF085330 Pinus taeda expansin.
AAB40634.1 U64890 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession Numbers U30477 and U30479.
AAF32409.1 AF230276 Triphysaria versicolor alpha-expansin 3.
AAC39512.1 AF043284 Gossypium hirsutum expansin. GhEXl . contains N-terminal signal peptide.
AAB40635.1 U64891 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession
Numbers U30477 and U30479.
AAC96081.1 AF049354 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP5. cell wall protein.
AAB38074.1 U30477 Oryza sativa induces extension (creep) in plant cell walls, expansin Os-EXP2. Os-EXP2. former gene name
RiExB.
AAG32921.1 AFl 84233 Lycopersicon esculentum expansin. Exp 10.
AAB81662.1 U85246 Oryza sativa expansin. Os-EXP4.
AAD49956.1 AFl 67360 Rumex palustris expansin. EXPL
AAB40636.1 U64892 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession Numbers U30477 and U30479.
AAF32411.1 AF230278 Triphysaria versicolor alpha-expansin 1.
AAC96080.1 AF049353 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP4. cell wall protein.
AAF35902.1 AF230333 Zinnia elegans expansin 3.
CAB43197.1 AJ239068 Lycopersicon esculentum cell wall loosening enzyme. expansin2. exp2.
AAC64201.1 AF096776 Lycopersicon esculentum expansin. LeEXP2.
AAF 17570.1 AF202119 Marsilea quadrifolia alpha-expansin. EXPL Mq-EXPl.
AAD13633.1 AF059489 Lycopersicon esculentum expansin precursor. Exp5.
AAF62181.1 AF247163 Oryza sativa alpha-expansin OsEXP6. cell wall loosening factor; expressed in internodes and leaves.
AAF17571.1 AF202120 Regnellidium diphyllum alpha-expansin. EXPL Rd-EXPl.
AAF62180.1 AF247162 Oryza sativa alpha-expansin OsEXP5. cell wall loosening factor; expressed in internodes, leaves, coleoptiles, and roots.
CAC19183.1 AJ291816 Cicer arietinum expansin.
CAC06433.1 AJ276007 Festuca pratensis expansin. exp2.
BAB32732.1 AB049406 Eustoma grandiflorum expansin. Eg Expansin.
AAG01875.1 AF291659 Striga asiatica alpha-expansin 3. Exp3.
AAB37749.1 U30460 Cucumis sativus expansin S2 precursor. Cs-EXP2. similar to pollen allergen Lol pl, Lolium perenne, Swiss-
Prot Accession Number P 14946; former gene name CuExS2; expansin-30 (Ex30) protein.
AAC96077.1 AF049350 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXPl. cell wall protein.
AAC96079.1 AF049352 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP3. cell wall protein.
AAG32920.1 AFl 84232 Lycopersicon esculentum expansin. Exp8.
CAA69105.1 Y07782 Oryza sativa expansin. RiExA.
AAC96078.1 AF049351 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP2. cell wall protein.
SEQ ID NO: 583
BAA85412.1 AP000615 Oryza sativa
ESTs AU065232(E60855),C23624(S1554), AU078241(E60855) conespond to a region ofthe predicted gene.; similar to putative adenylate kinase. (AC005896).
BAA01181.1 D10335 Oryza sativa adenylate kinase-b. Adk-b.
BAA01180.1 D10334 Oryza sativa adenylate kinase-a. Adk-a.
BAA94761.1 AB041773 Oryza sativa adenylate kinase. Adk-a.
AAB68604.1 U82330 Prunus armeniaca adenylate kinase homolog.
AAF23372.1 AFl 87063 Oryza sativa catalyzes the tiansfer of phosphate from ATP to UMP or CMP to form ADP and UDP or
CDP. UMP/CMP kinase b. ura6.
AAF23371.1 AFl 87062 Oryza sativa catalyzes the tiansfer of phosphate from ATP to UMP or CMP to form ADP and UDP or
CDP. UMP/CMP kinase a. ura6.
AAD41679.1 AF086603 Ceratopteris richardii adenylate kinase. ADKL
BAA85443.1 AP000616 Oryza sativa similar to UMP/CMP kinase (AF000147).
SEQ ID NO: 584
BAA87052.2 D88273 Hordeum vulgare nicotianamine aminotransferase A. naat-A.
BAA87055.1 AB024006 Hordeum vulgare nicotianamine aminotransferase. naat-A.
BAA87053.1 AB005788 Hordeum vulgare mcotianamine aminotransferase B. naat-b. NAAT-B.
BAA87054.1 AB024006 Hordeum vulgare nicotianamine aminotransferase. naat-B.
BAA77261.1 AB007405 Oryza sativa alanine aminotransferase. AlaAT.
BAA77260.1 AB007404 Oryza sativa alanine aminotransferase. AlaAT.
CAA49199.1 X69421 Panicum miliaceum alanine aminotransferase. pAlaAT-2.
AAB01685.1 U31975 Chlamydomonas reinhardtii catalyzes the transfer of -NH2 from ala to 2-oxoglutarate. alanine aminotransferase. The translation start site has not been experimentally tested, but a 55 kDa product can be detected in Western blot.
AAC62456.1 AF055898 Zeamays alanine aminotransferase. alt. AlaAT.
CAA81231.1 Z26322 Hordeum vulgare alanine aminotransferase.
SEQ ID NO: 586
AAB94589.1 AF022460 Glycine max
CYP83Dlp. CYP83D1. cytochrome P450 monooxygenase.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
AAA19701.1 L24438 Thlaspi arvense cytochrome P450.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
AAC39318.1 AF029858 Sorghum bicolor second multifunctional cytochrome P450 in the biosynthetic pathway ofthe cyanogenic glucoside dhurrin. Catalyzes the conversion of p-hydroxyphenylacetaldoxime to p- hydroxymandelonitrile. cytochrome P450 CYP71E1. CYP71E1. No EST#s identified.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
AAB61964.1 U48434 Solanum chacoense putative cytochrome P450.
AAD47832.1 AFl 66332 Nicotiana tabacum cytochrome P450.
AAB94588.1 AF022459 Glycine max CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
AAF27282.1 AF122821 Capsicum annuum cytochrome P450. PepCYI >
CAA50312.1 X70981 Solanum melongena P450 hydroxylase. CYPEG2.
CAA83941.1 Z33875 Mentha x piperita cytochrome P-450 oxidase.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71A6.
AAD44151.1 AFl 24816 Mentha x piperita cytochrome p450 isoform PMI 7.
BAA03635.1 D14990 Solanum melongena Cytochrome P-450EG4.
CAA50645.1 X71654 Solanum melongena P450 hydroxylase.
AAB69644.1 AF000403 Lotus japonicus putative cytochrome P450. LJNP450.
AAD37433.1 AF150881 Lycopersicon esculentum x Lycopersicon peravianum catalyzes the hydroxylation of feralic acid to 5-hydroxyferalic acid, ferulate-5-hydroxylase. CYP84. cytochrome P450-dependent monooxygenase; F5H; FAHL
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
AAD44152.1 AF124817 Mentha x piperita cytochrome ρ450 isoform PM2.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
AAG14962.1 AF214008 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2.
AAG14961.1 AF214007 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
AAG14963.1 AF214009 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H3.
AAK38088.1 AF321864 Lolium rigidum putative cytochrome P450.
AAD44150.1 AF124815 Mentha spicata cytochrome p450.
AAK38084.1 AF321860 Lolium rigidum putative cytochrome P450.
AAD56282.1 AFl 55332 Petunia x hybrida flavonoid 3 '-hydroxylase. Htl. cytochrome P450; CYP75B2.
AAK38087.1 AF321863 Lolium rigidum putative cytochrome P450.
AAK38083.1 AF321859 Lolium rigidum putative cytochrome P450.
AAB94584.1 AF022157 Glycine max capable of catalyzing the metabolism of phenylurea herbicides. CYP71 AIO. CYP71 A10. cytochrome P450 monooxygenase.
CAA71514.1 Y10490 Glycine max putative cytochrome P450.
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
BAB39252.1 AP002968 Oryza sativa putative cytochrome P450. P0416G11.1.
CAA50155.1 X70824 Solanum melongena flavonoid hydroxylase (P450). CYP75.
SEQ ID NO: 587
CAA60120.1 X86222 Pisum sativum heat shock protein. hsp22.
AAF37726.1 AF237957 Euphorbia esula
LMW heat shock protein, putative 22 kDa mitochondrial heat shock protein.
BAA32547.1 AB017134 Lycopersicon esculentum mitochondrial small heat shock protein. LEMTSHP.
CAA33388.1 X15333 Chenopodium rabram heat shock protein (AA 1-204).
AAB03096.1 U21722 Glycine max
Hsp23.9. Gmhsp23.9. low molecular weight heat shock protein.
AAC12279.1 AF035460 Zea mays low molecular weight heat shock protein precursor. hsp22.
AAD03604.1 AF104107 Triticum aestivum small heat shock protein Hsp23.5. mRNA not present, or only at very low levels, prior to heat stress; mRNA abundant after two hours at 40C.
AAB01557.1 L47741 Picea glauca mitochondria-localized low molecular weight heat shock protein 23.5. EMB22, SMW
HSP23.5.
AAD03605.1 AFl 04108 Triticum aestivum small heat shock protein Hsp23.6.
CAA38037.1 X54103 Plastid Petunia x hybrida heat shock protein. hsp21.
AAF 19022.1 AFl 97942 Funaria hygrometrica chloroplast-localized small heat shock protein 22. CPsHSP22.
AAB49626.1 U59917 Lycopersicon esculentum chromoplast-associated hsp20. pTOMl 11. small molecular weight heat shock protein.
AAB07023.1 U66300 Chloroplast Lycopersicon esculentum heat shock protein. HSP21.
BAA29064.1 D88584 Nicotiana tabacum heat shock protein 26 (Type I). Nthsp26a. plastid-localized small heat shock protein.
AAF 19021.1 AFl 97941 Funaria hygrometrica chloroplast-localized small heat shock protein. CPsHSP21. CPsHS 21.
BAA78385.1 AB020973 Oryza sativa heat shock protein 26. Oshsp26. chloroplast-localized small heat shock protein.
CAA41219.1 X58280 Triticum aestivum heat shock protein 26.6. Tahsp26.6.
AAC96315.1 AF097657 Triticum aestivum heat shock protein HSP26. hsp26.6. similar to the Triticum aestivum hsp26.6G2 protein encoded by the sequence presented in GenBank Accession Number L41503; 26 kDa protein. AAC96316.1 AF097658 Triticum aestivum heat shock protein HSP26. hsp26.6. 26 kDa protein.
AAC96314.1 AF097656 Triticum aestivum heat shock protein HSP26. hsp26.6. 26 kDa protein.
CAA47745.1 X67328 Triticum aestivum heat shock protein 26.6B. hsp 26.6B.
AAC96317.1 AF097659 Triticum aestivum heat shock protein HSP26. hsp26.6. 26 kDa protein.
AAA33477.1 L28712 Zeamays heat shock protein 26. HSP26. plastid-localized low molecular weight hsp.
BAA29066.1 AB006043 Nicotiana sylvestris heat shock protein 26. hsp26. plastid-localized small heat shock protein.
BAA29067.1 AB006044 Nicotiana tomentosiformis heat shock protein 26. hsp26. plastid-localized small heat shock protein.
BAA29065.1 AB006041 Nicotiana tabacum heat shock protein 26 (Type I). Nthsp26a. plastid-localized small heat shock protein 26.
AAC01570.1 AF019144 Agrostis stolonifera var. palustris low molecular weight heat shock protein.
AAD30452.1 AF123255 Lycopersicon esculentum
17.7 kD class I small heat shock protein. HSP17.7.
CAA39603.1 X56138 Lycopersicon esculentum small heat shock protein (class I).
AAD30453.1 AF123256 Lycopersicon esculentum
17.8 kD class I small heat shock protein. HSP17.8.
AAD30454.1 AF123257 Lycopersicon esculentum
17.6 kD class I small heat shock protein. HSP 17.6.
AAD49336.1 AFl 66277 Nicotiana tabacum low molecular weight heat-shock protein. LHS-1. TLHS-1.
AAF34133.1 AF161179 Malus x domestica low molecular weight heat shock protein. Hspl.
AAC39360.1 U63631 Fragaria x ananassa LMW heat shock protein.
AAA61632.1 U08601 Papaver somniferam low molecular weight heat-shock protein.
CAA08908.1 AJ009880 Castanea sativa molecular chaperone. cytosolic class I small heat-shock protein HSP17.5. hsp!7.5.
AAA33975.1 M11395 Glycine max small heat shock protein.
CAA63570.1 X92983 Pseudotsuga menziesii low molecular weight heat-shock protein.
AAB03893.1 M11318 Glycine max 17.5 kdheat shock protein Gmhspl7.6L.
CAA25578.1 X01104 Glycine max heat shock protein 6871 (aa 1-153).
AAB72109.1 AF022217 Brassica rapa low molecular weight heat-shock protein. BcHSP17.6. 17.6 kDa; cytosolic class I.
CAA46641.1 X65725 Zea mays heat shock protein 17.2. Zmhsp 17.2. Class I low molecular weight heat shock protein.
CAB08441.1 Z95153 Helianthus annuus
17.6 kD class I small heat-shock protein HSP 17.6. Ha hspl 7.6. ^^^
AAA33974.1 M11317 Glycine max 17.6 kd heat shock protein Gmhspl7.6L.
SEQ ID NO: 588
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
AAAI 9701.1 L24438 Thlaspi arvense cytochrome P450.
AAC39318.1 AF029858 Sorghum bicolor second multifunctional cytochrome P450 in the biosynthetic pathway ofthe cyanogenic glucoside dhurrin. Catalyzes the conversion of p-hydroxyphenylacetaldoxime to p- hydroxymandelonitrile. cytochrome P450 CYP71E1. CYP71E1. No EST#s identified.
BAB40323.1 AB037244 Asparagus officinalis cytochrome P450. ASPI-1.
BAB40324.1 AB037245 Asparagus officinalis cytochrome P450. ASPI-2.
AAB94589.1 AF022460 Glycine max
CYP83Dlp. CYP83D1. cytochrome P450 monooxygenase.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
AAB94584.1 AF022157 Glycine max capable of catalyzing the metabolism of phenylurea herbicides. CYP71A10. CYP71A10. cytochrome P450 monooxygenase.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
AAF27282.1 AF122821 Capsicum annuum cytochrome P450. PepCYP.
CAB56503.1 AJ238612 Catharanthus roseus cytochrome P450.
AAD47832.1 AF166332 Nicotiana tabacum cytochrome P450.
AAB94588.1 AF022459 Glycine max
CYP71D10ρ. CYP71D10. cytochrome P450 monooxygenase.
AAB61964.1 U48434 Solanum chacoense putative cytochrome P450.
CAA71517.1 Y10493 Glycine max putative cytochrome P450.
CAA50645.1 X71654 Solanum melongena P450 hydroxylase.
BAA03635.1 D 14990 Solanum melongena Cytochrome P-450EG4.
CAA50312.1 X70981 Solanum melongena P450 hydroxylase. CYPEG2.
CAA71514.1 Y10490 Glycine max putative cytochrome P450.
AAB69644.1 AF000403 Lotus japonicus putative cytochrome P450. LJNP450.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71A6.
AAD44151.1 AFl 24816 Mentha x piperita cytochrome p450 isofonn PMI 7.
AAK38084.1 AF321860 Lolium rigidum putative cytochrome P450.
CAA83941.1 Z33875 Mentha x piperita cytochrome P-450 oxidase.
AAK38083.1 AF321859 Lolium rigidum putative cytochrome P450.
AAD44152.1 AF124817 Mentha x piperita cytochrome p450 isoform PM2.
BAB39252.1 AP002968 Oryza sativa putative cytochrome P450. P0416G11.1.
AAD44150.1 AF124815 Mentha spicata cytochrome p450.
AAK38087.1 AF321863 Lolium rigidum putative cytochrome P450.
AAK38082.1 AF321858 Lolium rigidum putative cytochrome P450.
CAA57423.1 X81829 Zea mays cytochrome P450. CYP71C2. family CYP71, subfamily CYP71C.
CAA72208.1 Y11404 Zea mays cytochrome p450. cyp71c2.
CAC27827.1 AJ295719 Catharanthus roseus geraniol hydroxylase. cytochrome P450. cyp71.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
AAK38088.1 AF321864 Lolium rigidum putative cytochrome P450.
AAG14963.1 AF214009 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H3.
CAA57424.2 X81830 Zea mays cytochrome P450. CYP71C3. family CYP71, subfamily CYP71C.
CAA72207.1 Y11403 Zea mays cytochrome p450. cyp71c3.
AAG14962.1 AF214008 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2. AAG14961.1 AF214007 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
BAB40322.1 AB036772 Triticum aestivum cytochrome P450. N-l.
SEQ ID NO: 593
BAA09645.1 D63331 Nicotiana tabacum
Indole-3 -acetonitrile hydrolysis, nitrilase. TNIT4. Indole-3 -acetonitrile hydolysis,indole-3- acetic acid biosynthesis.
BAA11770.1 D83078 Nicotiana tabacum indole-3 -acetonitrile hydolysis. nitrilase. TNIT4B. indole-3 -acetonitrile hydlysis, indole-3- aceticacid biosynthesis.
BAA77679.1 AB027054 Oryza sativa nitrilase-like protein. ONIT4.
SEQ ID NO: 595
AAG01147.1 AF283816 Pinus taeda calreticulin. calcium-binding protein.
CAA05161.1 AJ002057 Beta vulgaris calreticulin.
AAB71419.1 U74630 Ricinus communis calreticulin.
AAB71420.1 U74631 Ricinus communis endoplasmic reticulum calcium binding protein, calreticulin.
CAA95999.1 Z71395 Nicotiana plumbaginifolia ER Ca2+ binding protein, calreticulin. call .
AAD 17490.1 AF052040 Berberis stolonifera possible molecular chaperon, caketiculin. calcium binding protein ofthe endoplasmic reticulum.
AAD32207.1 AFl 34733 Prunus armeniaca calcium-binding protein calreticulin.
CAA61939.1 X89813 Zea mays Calreticulin precursor.
AAA32948.1 L27348 Hordeum vulgare calcium binding protein, calreticulin. CRHl .
AAA32949.1 L27349 Hordeum vulgare calcium binding protein, calreticulin. CRH2.
AAF01470.1 AF190454 Zeamays calreticulin. CRT, calcium binding protien.
CAB54526.1 AJ000765 Chlamydomonas reinhardtii calreticulin.
AAB70919.1 AF019376 Brassica napus calreticulin. Crtl. calcium binding protein; similar to A. thaliana calreticulin encoded by
GenBank Accession Number U66343.
BAA85118.1 ABO 18243 Solanum melongena calreticulin-like protein. EEF22.
CAA54975.1 X78057 Zea mays calreticulin. CRH.
CAA57914.1 X82578 Parthenium argentatum calreticulin. 111R.
AAK15502.1 AF325720 Pennisetum ciliare calreticulin-like protein. Pcp4.
BAA77025.1 AB026251 Lithospermum erythrorhizon calreticulin.
SEQ ID NO: 596
BAA12206.1 D84061 Spinacia oleracea phosphoserrne aminotransferase.
SEQ ID NO: 597
AAF73124.1 AF148534 Lycopersicon esculentum whitefly-induced gp91-phox. Wfil. mammalian gp91-phox homolog; respiratory burst oxidase homolog; putative NADPH oxidase; similar to neutrophil NADPH oxidase gp91- phox subunit; similar to Arabidopsis thaliana Rboh and Oryza sativa Rboh; similar to
Caenorhabditis elegans PhoX-like proteins.
AAF73104.1 AF 147783 Lycopersicon esculentum whitefly-induced gp91-phox. Wfil. mammalian gp91-phox homolog; respiratory burst oxidase homolog; putative NADPH oxidase; similar to neutrophil NADPH oxidase gpl-phox subunit; similar to Arabidopsis thaliana Rboh and Oryza sativa Rboh.
AAD25300.1 AF088276 Lycopersicon esculentum NADPH oxidase. RBOH1. gp91; phox homolog.
AAB87790.1 AFO 15302 Oryza sativa
RbohAOsp. putative NADPH oxidase; intrinsic plasma membrane protein; similar to the neutrophil NADPH oxidase gp91phox subunit; contains RanGAPl domain; similar to O. sativa NAD(P)H oxidase sequence with GenBank Accession Number X93301.
CAA63704.1 X93301 Oryza sativa NAD(P)H oxidase. rbohA.
AAD24966.1 AFl 09150 Lycopersicon esculentum NADPH oxidase. gp91; phox homolog.
AAD25225.1 AF088279 Potamogeton crispus NADPH oxidase. RBOH1. gp91; phox homolog.
SEQ ID NO: 599
CAC21424.1 AJ278332 Lycopersicon esculentum involved in octadecanoid biosynthesis. 12-oxophytodienoate reductase 3. oρr3.
BAB40340.1 AB044940 Pisum sativum
12-oxophytodienoic acid 10, 11 -reductase. OPDAR.
CAB43506.1 AJ242551 Lycopersicon esculentum involved in jasmonic acid biosynthesis. 12-oxophytodienoate reductase. opr.
CAC21423.1 AJ278331 Lycopersicon esculentum involved in octadecanoid biosynthesis, putative 12-oxophytodienoate reductase 2. opr2.
CAB97122.1 AJ297421 Daucus carota oxidoreductase. putative 12-oxophytodienoate reductase. opr2. SEQ ID NO: 602
AAB37246.1 U58971 Nicotiana tabacum calmodulin-binding protein. TCB60.
SEQ ID NO: 603
AAK15006.1 AF233433 Brassica napus arginase. similar to Arabidopsis thaliana and Glycine max arginase.
AAK07744.1 AF130440 Pinus taeda converts L-arginrne to L-ornithrne and urea, arginase. ARS20. L-arginine amidinohydrolase.
AAC04613.1 AF035671 Glycine max hydrolysis of L- Arg to L-Orn and urea, arginase. pAGl . L-Arg amidinohydrolase.
SEQ ID NO: 605 CAA81210.1 Z26251 Helianthus tuberosus Catalyzes the reduction of cytochrome P450 in microsomes. NADPH-fenihemoprotein reductase.
AAB02721.1 U58629 Helianthus tuberosus
NADPH-fenihemoprotein oxidoreductase. HTR2. NADPH-cytochrome P450 reductase isoform.
CAC27143.1 AJ132538 Picea abies
NADPH-cytochrome P450 reductase.
AAA79131.1 U10545 Chlamydomonas reinhardtii fe_τedoxin-NADP+ reductase. f r.
CAA55406.1 X78851 Chlamydomonas reinhardtii fenedoxin NADP reductase.
AAB40978.1 U22328 Volvox carteri fenedoxm-NADP+ reductase. fnr. FNR.
AAA33029.1 M25528 Mesembryanthemum crystallinum fenedoxin-NADP+ reductase precursor (f rA; EC 1.6.7.1).
CAA30978.1 X12446 Pisum sativum fenedoxin-NADH+ reductase preprotein (AA -52 to 308).
BAA13417.1 D87547 Oryza sativa precursor fenedoxin-NADP+ oxidoreductase.
AAA21758.1 U14956 Vicia faba photosynthetic election transport, fenedoxin NADP+ reductase precursor, fnr.
AAA34029.1 M86349 Spinacia oleracea fenedoxin-NADP oxidoreductase. precursor protein.
CAB71293.1 AJ250378 Capsicum annuum chloroplast fenedoxin-NADP+ oxidoreductase precursor, fhr.
CAA74359.1 Y14032 Nicotiana tabacum fenedoxin~NADP(+) reductase. fhr.
BAA88236.1 AB035644 Zeamays NADP+ oxidoreductase. fenedoxin. L-FNRI.
BAA88237.1 AB035645 Zea mays NADP+ oxidoreductase. fenedoxin. L-FNRU.
BAA20365.1 AB004307 Nicotiana tabacum fenedoxin-NADP oxidoreductase.
BAA07479.1 D38445 Oryza sativa root fenedoxin-NADP+ reductase. root FNR. nitrate assimilation pathway.
BAA02248.1 D12815 Oryza sativa fenedoxin-NADP+ reductase enzyme.
BAA04232.1 D17410 Oryza sativa fenedoxin-NADP+ reductase.
BAA90642.1 AP001129 Oryza sativa
ESTs AU078647(E1557),C72400(E1557) conespond to a region ofthe predicted gene.;
Similar to Oryza sativa, fenedoxin-NADP+ reductase. (D 17790).
BAA85425.1 AP000616 Oryza sativa
ESTs AU078647(E1557),C72400(E1557) conespond to a region ofthe predicted gene.; similar to fenedoxin-NADP+ reductase (D17790).
BAA04616.1 D 17790 Oryza sativa fenedoxin-NADP+ reductase.
AAB40034.1 U10418 Zea mays fenedoxin-NADP reductase precursor.
CAA67796.1 X99419 Pisum sativum fenodoxin NADP oxidoreductase.
AAK09367.1 AF321525 Pisum sativum fenedoxin-NADP+ reductase. FNR.
AAK09370.1 AF321528 Pisum sativum fenedoxin-NADP+ reductase. FNR.
AAK09369.1 AF321527 Pisum sativum fenedoxin-NADP+ reductase. FNR.
AAK09368.1 AF321526 Pisum sativum fenedoxin-NADP+ reductase. FNR.
SEQ ID NO: 606
BAA82107.1 AB022693 Nicotiana tabacum transcription factor. NtWRKYl.
AAC31956.1 AF080595 Pimpinella brachycaφa zinc finger protein. ZFP1. WRKY
AAC49527.1 U48831 Petroselinum crispum
WRKYl. contains two WRKY domains; WRKY-type DNA-binding protein; sequence- specific DNA-binding protein.
AAD55974.1 AF121353 Petroselinum crispum zinc-finger type transcription factor WRKYl . WRKYl .
BAA77383.1 AB020590 Nicotiana tabacum transcription factor NtWRKY2.
AAD32677.1 AF140554 Avena sativa
DNA-binding protein WRKYl. wrkyl. putative transcription factor.
CAA88326.1 Z48429 Avena fatua binds conserved cis-element from cereal alpha-Amy2 promoters. DNA-binding protein.
BAA86031.1 AB026890 Nicotiana tabacum transcription factor NtWRKY4.
AADl 6139.1 AF096299 Nicotiana tabacum DNA-binding protein 2. WRKY2. transcription factor.
AAC37515.1 L44134 Cucumis sativus SPFl-like DNA-binding protein.
AAF23898.1 AF193802 Oryza sativa zinc finger transcription factor WRKYl .
AAK16171.1 AC079887 Oryza sativa putative DNA-binding protein. OSJNBa0040E01.10.
AAC49529.1 U58540 Petroselinum crispum
WRKY2. Contains two WRKY domains; WRKY-type DNA-binding protein.
AADl 6138.1 AF096298 Nicotiana tabacum DNA-binding protein 1. WRKYl. transcription factor.
BAB 19096.1 AP002839 Oryza sativa putative DNA-binding protein homolog. P0688A04.2.
BAB 19075.1 AP002744 Oryza sativa putative DNA-binding protein homolog. P0006C01.17.
AAK16170.1 AC079887 Oryza sativa putative DNA binding protein. OSJNBa0040E01.4.
CAB97004.1 AJ278507 Solanum tuberosum putative transcription factor. WRKY DNA binding protein. WRKYl.
BAB18313.1 AP002865 Oryza sativa putative WRKY DNA binding protein. P0034C11.1. contains EST C26525(C12525).
AAD38283.1 AC007789 Oryza sativa putative WRKY DNA binding protein. OSJNBa0049B20.9.
BAB40073.1 AP003074 Oryza sativa putative WRKY DNA binding protein. OSJNBa0004G10.20. contains EST C26525(C12525).
AAC49528.1 U56834 Petroselinum crispum
DNA-binding. WRKY3. WRKY-type DNA-binding protein.
BAB 16432.1 AB041520 Nicotiana tabacum
WRKY transcription factor Nt-SubD48. Nt-SubD48.
BAA77358.1 AB020023 Nicotiana tabacum
WRKY domain Zn-finger type DNA-binding protein. DNA-binding protein NtWRKY3.
AAD32676.1 AF140553 Avena sativa
DNA-binding protein WRKY3. wrky3. putative transcription factor.
CAA88331.1 Z48431 Avena fatua binds conserved cis-element from cereal alpha- Amy2 promoters. DNA-binding protein.
AAG35658.1 AF204925 Petroselinum crispum transcription factor WRKY4. WRKY4. binds to W box (TTGACC) elements.
AAG46150.1 AC018727 Oryza sativa putative DNA-binding protein. OSJNBa0056G17.18.
AAD27591.1 AF121354 Petroselinum crispum binds sequence specifically to W Boxes (TTGACC). transcription factor. WRKY3. sequence specific DNA-binding protein.
AAG35659.1 AF204926 Petroselinum crispum transcription factor WRKY5. WRKY5. binds to W box (TTGACC) elements.
CAB66338.1 AJ279697 Betula pendula wrky-type DNA binding protein, wrky.
AAF61864.1 AFl 93771 Nicotiana tabacum
DNA-binding protein 4. WRKY4. transcription factor.
AAF61863.1 AF193770 Nicotiana tabacum
DNA-binding protein 3. WRKY3. transcription factor. SEQ ID NO: 608
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
BAA92836.1 AB032473 Brassica oleracea
S18 S-locus receptor kinase. SRK18.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
CAB41878.1 Y18259 Brassica oleracea
SRK5 protein. SRK5. receptor-like kinase.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
AAF76314.1 AF220603 Lycopersicon esculentum Fen kinase. Lescfen.
AAK11568.1 AF318492 Lycopersicon hirsutum Pto-like protein kinase B. LhirPtoB.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; tiansmembrane domain: 1347-1412; intiacellular kinase domain: from 1413.
AAB47424.1 U59317 Lycopersicon pimpinellifolium serine/threonine protein kinase Fen. Fen, fenthion sensitivity gene from tomato.
AAB47422.1 U59318 Lycopersicon esculentum serme/threonine protein kinase Fen. fen. allele of Fen, fenthion sensitivity gene from tomato.
AAC48932.1 U13923 Lycopersicon pimpinellifolium
Fen. putative serine/tlireonine protein kinase; similar to product encoded by Lycopersicon pimpinellifolium Pto gene, GenBank Accession Number U02271; Fen is a member ofthe Pto gene family.
AAF76307.1 AF220602 Lycopersicon pimpinellifolium Fen kinase.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein kinase.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
SEQ ID NO: 610
BAB 17348.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.32.
BAB 17345.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.29.
BAB17126.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.16.
BAB39451.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.24.
BAB 17342.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.26.
AAC27489.1 AF077130 Oryza sativa receptor-like protein kinase.
AAF78018.1 AF238474 Oryza sativa receptor-like kinase. RLG16. protein kinase.
AAC02535.1 AF044260 Oryza sativa receptor serine/tlireonine kinase. protein kinase.
BAB39435.1 AP003338 Oryza sativa putative receptor kinase. OJ1212__B09.2.
AAD46917.1 AF164021 Oryza sativa receptor kinase.
BAB39438.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.7.
BAB17339.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.23.
AAC49629.1 U51330 Triticum aestivum rust resistance kinase LrlO. LRK10.
BAB 17321.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.1.
BAB17129.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.20.
AAF68398.1 AF237568 Oryza sativa receptor-like protein kinase. RLG2.
BAB 17331.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.12.
BAB17139.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.31.
AACO 1746.1 AF044489 Oryza sativa receptor-like protein kinase. chpkl .
AAF78016.1 AF238472 Oryza sativa receptor-like kinase. RLG15. protein kinase.
BAB39441.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.11.
BAB17116.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.3.
AAF78044.1 AF248493 Oryza sativa receptor-like kinase. RLG18. protein kinase.
AAD44029.1 AF085164 Hordeum vulgare receptor-like kinase LRK10.
BAB 17332.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.13. contains ESTs D47575(S13157),AU032665(S13157).
BAB 17347.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.31. contains ESTs D47575(S13157),AU032665(S13157).
BAB 17344.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.28. contains ESTs D47575(S13157),AU032665(S13157).
BAB17337.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.21. contains ESTs D47575(S13157),AU032665(S13157).
BAB 17335.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.17. contains ESTs D47575(S13157),AU032665(S13157).
BAB 17115.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.2.
BAB39440.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.10.
BAB39437.1 AP003338 Oryza sativa receptor-like kinase. OJ1212_B09.6.
AAF78019.1 AF238475 Oryza sativa receptor-like kinase. RLG17. protein kinase.
BAB17127.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.17.
AAD46916.1 AF164020 Oryza sativa receptor kinase.
AAF78021.1 AF238477 Oryza sativa receptor-like kinase. RLG5. protein kinase.
AAD46417.1 AF100767 Oryza sativa receptor-like kinase. 8ARK3. shnilar to wheat ARK1 AS.
AAD46415.1 AF100765 Oryza sativa receptor-like kinase. 8ARK1. similar to wheat ARK1AS.
BAB17128.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.18.
AAF68400.1 AF237570 Oryza sativa receptor-like protein kinase. RLG3.
BAB 17323.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.3.
BAB 17131.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.22.
AAF68397.1 AF237567 Oryza sativa receptor-like protein kinase. RLG1.
AAF78015.1 AF238471 Oryza sativa receptor-like kinase. RLG10. protein kinase.
AAD43962.1 U78762 Triticum aestivum receptor-like kinase ARK1AS. ARK1AS.
SEQ ID NO: 611
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APKl gene for protein tyrosine-serine-threonine kinase.(D12522).
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAF43496.1 AFl 31222 Lophopyrum elongatum protein serine/tlireonine kinase. ESI47. induced in roots by salt stress, osmotic stiess, and
ABA treatment.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AFOO 1308).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
AAK11567.1 AF318491 Lycopersicon hirsutum
Pto-like protein kinase F. LhrrPtoF.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAC48932.1 U13923 Lycopersicon pimpinellifolium
Fen. putative serine/threonine protein kinase; similar to product encoded by Lycopersicon pimpinellifolium Pto gene, GenBank Accession Number U02271; Fen is a member ofthe Pto gene family.
AAB47424.1 U59317 Lycopersicon pimpinellifolium serme/threonine protein kinase Fen. Fen, fenthion sensitivity gene from tomato.
AAF76307.1 AF220602 Lycopersicon pimpinellifolium
Fen kinase.
AAK11566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhirPtoE. confers resistance to bacterial speck disease.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAF76314.1 AF220603 Lycopersicon esculentum
Fen kinase. Lescfen.
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
AAF76313.1 AF220603 Lycopersicon esculentum Pto kinase. LescPth5.
AAB47422.1 U59318 Lycopersicon esculentum serine/threonine protein kinase Fen, fen. allele of Fen, fenthion sensitivity gene from tomato.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAF76306.1 AF220602 Lycopersicon pimpinellifolium Pto kinase.
AAB47423.1 U59315 Lycopersicon pimpinellifolium serine/threonine protein kinase Pto. Pto. bacterial speck disease resistance in tomato; disease resistance gene.
AAC48914.1 U02271 Lycopersicon pimpinellifolium protein kinase.
AAK11568.1 AF318492 Lycopersicon hirsutum Pto-like protein kinase B . LhirPtoB .
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S13927).
AAC27894.1 AF023164 Zeamays leucine-rich repeat transmembrane protein kinase 1. ltkl .
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil. Ptil kinase.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl . Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
AAF91336.1 AF249317 Glycine max Ptil kinase-like protein. Ptil a. protein kinase.
AAF66615.1 AF 142596 Nicotiana tabacum LRR receptor-like protein kinase.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
AAK11569.1 AF318493 Lycopersicon hirsutum Pto-like protein kinase D. LhirPtoD.
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218). ^^
AAC27895.1 AF023165 Zea mays leucine-rich repeat transmembrane protein kinase 2. Itk2.
SEQ ID NO: 612
CAA54613.1 X77463 Manihot esculenta UTP-glucose glucosyltiansferase. CGT6.
CAA54609.1 X77459 Manihot esculenta UTP-glucose glucosyltransferase. CGT1.
CAA54611.1 X77461 Manihot esculenta UTP-glucose glucosyltiansferase. CGT2.
CAA54612.1 X77462 Manihot esculenta UTP-glucose glucosyltransferase. CGT5.
BAB17182.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.19.
BAB17176.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.13.
CAB56231.1 Y18871 Dorotheanthus bellidiformis betanidrn-5-O-glucosyltiansferase.
BAA83484.1 AB031274 Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltransferase. ufgt.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS5a.
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid:glucosyltransferase 1. togtl. glucosyltransferase.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltiansferases.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS 10a.
AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid:glucosyltiansferase 2. togt2. glucosyltransferase.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltiansferase. PHI.
AAG25643.1 AF303396 Phaseolus vulgaris UDP-glucosyltransferase HRA25. putative; defense associated.
AAF61647.1 AF190634 Nicotiana tabacum UDP-glucose:salicylic acid glucosyltransferase. SA-GTase.
BAA93039.1 AB033758 Citras unshiu limonoid UDP-glucosyltransferase. LGTase.
AAB48444.1 U82367 Solanum tuberosum UDP-glucose glucosyltransferase.
AAD21086.1 AF127218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
CAC09351.1 AL442007 Oryza sativa putative glucosyltransferase. H0212B02.7.
AAD04166.1 AF101972 Phaseolus lunatus catalyzes formation of O-glucosylzeatin from zeatin and UDPG; catalyzes formation of O- xylosylzeatin from zeatin and UDPX. zeatin O-glucosyltransferase. ZOGl. cyto_cinin O- glucosyltransferase .
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitrile-o- glucosyltransferase.
BAA12737.1 D85186 Gentiana triflora UDP-glucose : flavonoid-3 -glucosyltransferase.
BAB07962.1 AP002524 Oryza sativa putative anthocyanin 5-O-glucosyltiansferase. P0406H10.16. contains ESTs
AU067881(C10481),AU067882(C10481).
BAA36421.1 AB013596 Perilla frutescens UDP-glucose:anthocysnin 5-O-glucosyltiansferase. PF3R4.
CAA54610.1 X77460 Manihot esculenta UTP-glucose glucosyltiansferase. CGT4.
AAF98390.1 AF287143 Brassica napus catalyzes the tiansfer of glucose from UDP-glucose to sinapate and some other hydroxy cinnamates (4-coumarate, caffeate, feralate). UDP-glucose: sinapate glucosyltransferase. SGT1. SGT.
BAA36423.1 AB013598 Verbena x hybrida UDP-glucose: anthocyanin 5-O-glucosyltiansferase. HGT8.
CAA30760.1 X07937 Zea mays
UDPglucose flavonoid glycosyl transferase. Bz-W22.
CAA30761.1 X07940 Zea mays
UDPglucose flavonoid glycosyl-transferase. Bz-McC.
CAA31855.1 X13500 Zea mays UDPglucose.-flavonol 3-0-glucosyltransferase.
BAA19659.1 AB002818 Perilla frutescens flavonoid 3-O-glucosyltransferase. UDP glucose.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidrn 3-O-glucosyltiansferase. PGT8.
AAK16172.1 AC079887 Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.14.
SEQ ID NO: 613
CAA54609.1 X77459 Manihot esculenta UTP-glucose glucosyltiansferase. CGT1.
CAA54613.1 X77463 Manihot esculenta UTP-glucose glucosyltiansferase. CGT6.
CAA54611.1 Xli 461 Manihot esculenta UTP-glucose glucosyltiansferase. CGT2.
CAA54612.1 X77462 Manihot esculenta UTP-glucose glucosyltiansferase. CGT5.
BAB17182.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.19.
BAB 17176.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.13.
CAB56231.1 Y18871 Dorotheanthus bellidiformis betanidin-5-O-glucosyltransferase.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS5a.
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid:glucosyltransferase 1. togtl. glucosyltiansferase.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS 10a.
AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid:glucosyltransferase 2. togt2. glucosyltiansferase.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltransferases.
BAA83484.1 AB031274 Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltiansferase. ufgt.
AAF61647.1 AFl 90634 Nicotiana tabacum UDP-glucose: salicylic acid glucosyltransferase. SA-GTase.
BAA93039.1 AB033758 Citras unshiu limonoid UDP-glucosyltransferase. LGTase.
AAB48444.1 U82367 Solanum tuberosum UDP-glucose glucosyltransferase.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltransferase. PHI.
BAA36421.1 AB013596 Perilla frutescens UDP-glucose :anthocysnin 5-O-glucosyltransferase. PF3R4.
CAC09351.1 AL442007 Oryza sativa putative glucosyltransferase. H0212B02.7.
BAA12737.1 D85186 Gentiana triflora UDP-glucose:flavonoid-3-glucosyltransferase.
AAG25643.1 AF303396 Phaseolus vulgaris UDP-glucosyltransferase HRA25. putative; defense associated.
AAD04166.1 AFl 01972 Phaseolus lunatus catalyzes formation of O-glucosylzeatin from zeatin and UDPG; catalyzes formation of O- xylosylzeatin from zeatin and UDPX. zeatin O-glucosyltransferase. ZOGl. cytokinin O- glucosyltransferase.
CAA54610.1 X77460 Manihot esculenta UTP-glucose glucosyltiansferase. CGT4.
AAF98390.1 AF287143 Brassica napus catalyzes the transfer of glucose from UDP-glucose to sinapate and some other hydroxycinnamates (4-coumarate, caffeate, feralate). UDP-glucose:sinapate glucosyltransferase. SGT1. SGT.
AAK16172.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
BAA36423.1 AB013598 Verbena x hybrida UDP-glucose: anthocyanin 5-O-glucosyltransferase. HGT8.
AAD21086.1 AFl 27218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitiile-o- glucosyltransferase.
AAK16175.1 AC079887 Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.15.
BAB07962.1 AP002524 Oryza sativa putative anthocyanin 5-O-glucosyltransferase. P0406H10.16. contains ESTs
AU067881(C10481),AU067882(C10481).
CAA31855.1 X13500 Zea mays UDPglucose:flavonol 3-0-glucosyltransferase.
BAA36422.1 AB013597 Perilla frutescens
UDP-glucose: anthocyanin 5-O-glucosyltransferase homologue. PF3R6.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltransferase. PGT8.
SEQ ID NO: 614
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein kinase.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intiacellular kinase domain: 1412-2554.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
AAC23542.1 U20948 Ipomoea tiifida receptor protein kinase. IRKl.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
BAB21001.1 AB054061 Brassicarapa S locus receptor kinase. SRK22.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRKl. Chrkl.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
BAA94529.2 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AFOO 1308).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
AAG03090.1 AC073405 Oryza sativa
Shnilar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
BAA94518.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 section 111 of 255; putative receptor-like protein kinase (AC002392).
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12. SEQ ID NO: 619
CAA72092.1 Y11209 Nicotiana tabacum protein disulfide-isomerase precursor. PDI.
AAG13988.1 AF298829 Prunus avium putative protein disulfide-isomerase. PDI.
AAD02069.1 AF036939 Chlamydomonas reinhardtii redox-regulator of 5'UTR psbA mRNA binding complex and translation, protein disulfide isomerase. localized to ER and chloroplast.
AAC49896.1 AF027727 Chlamydomonas reinhardtii involved in the redox-regulated binding of chloroplast poly(A)-binding protein to the 5'-UTR of psbA mRNA; regulates chloroplast translational activation, protein disulfide isomerase
RB60. PDI.
AAD55566.1 AFl 10784 Volvox carteri f. nagariensis protein disulfide isomerase precursor, pdi.
AAB08519.1 L39014 Zea mays protein disulfide isomerase. pdi. putative.
AAA70344.1 L33250 Hordeum vulgare catalyze the formation of disulfide bonds, disulfide isomerase. PDI. putative.
AAA70345.1 L33251 Hordeum vulgare catalyze the formation of disulfide bonds, disulfide isomerase. PDI. putative.
CAC21230.1 AJ277379 Triticum turgidum subsp. durum catalyzes formation of disulfide bonds, protein disulfide isomerase. Pdi.
AAAI 9660.1 U11496 Triticum aestivum protein disulfide isomerase. PDI.
CAC21231.1 AJ277380 Triticum turgidum subsp. durum catalyzes formation of disulfide bonds, protein disulfide isomerase. Pdi.
CAC21229.1 AJ277378 Triticum turgidum subsp. durum catalyzes formation of disulfide bonds, protein disulfide isomerase. Pdi.
CAC21228.1 AJ277377 Triticum turgidum subsp. durum catalyzes the formation of disulfide bonds, protein disulfide isomerase. Pdi.
AAB05641.1 U41385 Ricinus communis protein disulphide isomerase PDI. molecular chaperone.
CAA77575.1 Z11499 Medicago sativa protein disulfide isomerase.
AAD28260.1 AFl 31223 Datisca glomerata protein disulfide isomerase homolog. PDI.
AAA70346.1 L33252 Hordeum vulgare catalyze the formation of disulfide bonds, disulfide isomerase. PDI. putative.
BAA92322.1 AB039278 Oryza sativa protein disulfide isomerase. Pdi.
BAA77026.1 AB026252 Lithospermum erythrorhizon disulfide-isomerase precursor.
AAC79709.1 AF093614 Acetabularia acetabulum putative protein disulfide isomerase.
SEQ ID NO: 620
CAA64413.1 X94943 Lycopersicon esculentum peroxidase. cevilό.
AAA32676.1 M37637 Arachis hypogaea cationic peroxidase. PNC2.
BAA82307.1 AB027753 Nicotiana tabacum peroxidase.
AAB67737.1 L77080 Stylosanthes humilis cationic peroxidase.
AAD37429.2 AF149279 Phaseolus vulgaris peroxidase 4 precursor. FBP4. secretory peroxidase. CAA71494.1 Y10468 Spinacia oleracea peroxidase. prxr7.
AAD37375.1 AF145349 Glycine max peroxidase. Prx3.
AAF63024.1 AF244921 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 2 precursor, type III peroxidase.
CAA66037.1 X97351 Populus balsamifera subsp. trichocarpa signal for ER. peroxidase.
BAB39274.1 AP002971 Oryza sativa putative peroxidase. P0537A05.2.
AAA65637.1 LI 3654 Lycopersicon esculentum peroxidase. TPX1.
CAA40796.1 X57564 Armoracia rusticana peroxidase. peroxidase precursor.
AADl 1482.1 U51192 Glycine max peroxidase precursor. sEPa2.
CAA80502.1 Z22920 Spirodela polynhiza peroxidase.
BAA77387.1 AB024437 Scutellaria baicalensis peroxidase 1.
CAA59485.1 X85228 Triticum aestivum peroxidase. POX2.
BAA07663.1 D42064 Nicotiana tabacum cationic peroxidase isozyme 38K precursor.
BAA11853.1 D83225 Populus nigra peroxidase.
BAA07664.1 D42065 Nicotiana tabacum cationic peroxidase isozyme 40K precursor.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
AADl 1481.1 U51191 Glycine max peroxidase precursor. sEPal .
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
BAA03644.1 D14997 Oryza sativa peroxidase.
AAD43561.1 AF155124 Gossypium hirsutum bacterial-induced peroxidase precursor. Perx_Goshiko.
BAA06334.1 D30652 Populus kitakamiensis peroxidase.
BAA92500.1 AP001383 Oryza sativa
ESTs D39300(R3292),AU030751(E60187) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP6a. (X98774).
BAA90365.1 AP001081 Oryza sativa
ESTs D24550(R2151),D24265(R1609),AU031848(R2151) conespond to a region ofthe predicted gene. Similar to cationic peroxidase isozyme 40K precursor (D42065).
BAA89584.1 AP001073 Oryza sativa
ESTs D24550(R2151),D24265(R1609),AU031848(R2151) conespond to a region ofthe predicted gene. Similar to cationic peroxidase isozyme 40K precursor (D42065).
AAF34416.1 AF172282 Oryza sativa putative peroxidase. DUPRl 1.5.
AAC49820.1 AFO 14469 Oryza sativa peroxidase. POX5.1. wound inducible.
CAA66034.1 X97348 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
BAB39281.1 AP002971 Oryza sativa putative peroxidase. P0537A05.10. contains ESTs D24657(R2329),AU082066(R2329).
BAA14144.1 D90116 Armoracia rusticana peroxidase isozyme.
AAB97734.1 AF014502 Glycine max seed coat peroxidase precursor. Ep. H2O2 oxidoreductase; class III plant peroxidase.
CAA37713.1 X53675 Triticum aestivum peroxidase.
AAC05277.1 AF049881 Linum usitatissimum peroxidase FLXPER4. PER4.
CAA66035.1 X97349 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
BAA06335.1 D30653 Populus kitakamiensis peroxidase.
CAA39486.1 X56011 Triticum aestivum peroxidase.
BAA03911.1 D16442 Oryza sativa peroxidase.
BAA94962.1 AB042103 Asparagus officinalis peroxidase. AspPOXl.
AAC49821.1 AF014470 Oryza sativa peroxidase. POXgX9. expressed in roots.
AAD37427.1 AF149277 Phaseolus vulgaris peroxidase 1 precursor. FBPL secretory peroxidase.
CAA76374.2 Y16776 Spinacia oleracea peroxidase. prxlO.
BAA08499.1 D49551 Oryza sativa peroxidase. poxN.
CAA66036.1 X97350 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
AAA34108.1 J02979 Nicotiana tabacum lignin-forming peroxidase precursor (EC 1.11.1.7).
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
CAA59487.1 X85230 Triticum aestivum peroxidase. pox4.
SEQ ID NO: 626
CAA98160.1 Z73932 Lotus japonicus GTP-binding protein. RABIC. rablC.
BAA76422.1 AB024994 Cicer arietinum rab-type small GTP-binding protein.
BAA02116.1 D12548 Pisum sativum GTP-binding protein.
CAA69701.1 Y08425 Nicotiana plumbaginifolia small GTP-binding protein. Rabl subfamily.
AAA80678.1 U38464 Lycopersicon esculentum small GTP-binding protein. LeRablA. ; YPTl/Rabl A homolog LeRablA.
BAA02118.1 D12550 Pisum sativum GTP-binding protein.
AAB97115.1 U58854 Glycine max small GTP-binding protein. sra2.
CAA51011.1 X72212 Nicotiana tabacum ras-related GTP-binding protein. ypt2 homologue.
AAF65510.1 AF108883 Capsicum annuum small GTP-binding protein.
AAA80680.1 U38466 Lycopersicon esculentum small GTP-binding protein. LeRablC. ; YPTl/RablA homolog LeRablC.
CAA98161.1 Z73933 Lotus japonicus GTP-binding protein. RABID. rablD.
BAA02117.1 D12549 Pisum sativum GTP-binding protein.
CAA98162.1 Z73934 Lotus japonicus GTP-binding protein. RAB IE. rablE.
AAA50159.1 L27417 Glycine max GTP binding protein.
AAB28535.1 S66160 Oryza sativa ricl. ras-related GTP binding protein possessing GTPase activity; This sequence comes from
Fig. 1.
CAA98159.1 Z73931 Lotus japonicus GTP-binding protein. RAB1B. rablB.
BAA02115.1 D 12547 Pisum sativum GTP-binding protein.
CAA66447.1 X97853 Lotus japonicus GTP-binding protein. RAB1A. rablA.
AAD10389.1 U35026 Petunia x hybrida Rabl -like small GTP-binding protein.
AAA80679.1 U38465 Lycopersicon esculentum small GTP-binding protein. LeRablB. ; Yptl/Rabl A homolog LeRablB.
BAB07961.1 AP002524 Oryza sativa putative GTP-binding protein. P0406H10.17. contains ESTs
D23874(R0480),AU031678(R0480).
CAA98176.1 Z73948 Lotus japonicus GTP-binding protein. RAB8E. rab8E.
CAA89021.1 Z49152 Beta vulgaris GTP-binding. small G protein.
CAA98172.1 Z73944 Lotus japonicus GTP-binding protein. RAB 8 A. rab8A.
CAA04701.1 AJ001367 Daucus carota small GTP-binding protein. Dc-Rab8.
CAA90080.1 Z49900 Pisum sativum small GTP-binding protein.
AAD46405.1 AF096249 Lycopersicon esculentum ethylene-responsive small GTP-binding protein. ER43.
CAA98174.1 Z73946 Lotus japonicus GTP-binding protein. RAB8C. rab8C.
CAA90082.1 Z49902 Pisum sativum small GTP-binding protein.
CAA49600.1 X69980 Lycopersicon esculentum GTP-binding protein. ypt2.
CAA98175.1 Z73947 Lotus japonicus GTP-binding protein. RAB8D. rab8D.
CAA90081.1 Z49901 Pisum sativum small GTP-binding protein.
AAB17726.1 U38471 Brassica rapa small GTP-binding protein rab. BRAB-1. small GTP-binding protein rab family.
AAA34251.1 L08128 Volvox carteri
GTP-binding protein. yptV2.
CAA98173.1 Z73945 Lotus japonicus GTP-binding protein. RAB8B. rab8B.
CAA89049.1 Z49190 Beta vulgaris GTP-binding. small G protein.
CAA98179.1 Z73951 Lotus japonicus GTP-binding protein. RABllC. rabllC.
AAA34253.1 L08130 Volvox carteri GTP-binding protein. yptV4.
CAA98165.1 Z73937 Lotus japonicus GTP-binding protein. RAB2A. rab2A.
AAA63902.1 U22433 Zea mays GTP binding protein. rab2.
AAA90955.1 U32185 Glycine max vesicular transport, guanine nucleotide regulatory protein. rab2. GTP-binding protein; soyrab.
SEQ ID NO: 628
AAA87456.1 U22147 Hevea brasiliensis beta-1, 3-glucanase. HGN1. hydrolytic enzyme.
CAB38443.1 AJ133470 Hevea brasiliensis beta-1 ,3-glucanase. hgnL
AAG24921.1 AF311749 Hevea brasiliensis beta-l,3-glucanase.
AAF44667.1 AF239617 Vitis vinifera hydrolysis of 1,3-beta-D-glucosidic linkages in 1,3-beta-D-glucans. beta- 1,3 -glucanase. fungal pathogen defense-related protein.
AAA33648.1 L02212 Pisum sativum beta- 1,3 -glucan hydrolysis. beta-l,3-glucanase. putative.
AAB41551.1 U27179 Medicago sativa subsp. sativa acidic glucanase.
AAB24398.1 S51479 Pisum sativum beta-l,3-glucanase. beta-1, 3-glucanase. This sequence comes from Fig. IB.
CAA37289.1 X53129 Phaseolus vulgaris 1 ,3 ,-beta-D-glucanase.
AAA34078.1 M63634 Nicotiana plumbaginifolia regulator ofbeta(l,3)-glucanase. beta(l,3)-glucanase regulator.
AAA51643.1 M23120 Nicotiana plumbaginifolia beta-glucanase precursor.
CAA30261.1 X07280 Nicotiana plumbaginifolia beta-glucanase.
AAA03618.1 M80608 Lycopersicon esculentum beta-1 ,3-glucanase.
AAC19114.1 AF067863 Solanum tuberosum 1,3-beta-glucan glucanohydrolase. glucanase.
AAA18928.1 U01901 Solanum tuberosum catalyzes the hydrolysis of 1,3-beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). glub2. plant defense gene; induced expression in response to infection, elicitor, ethylene, wounding.
AAA63539.1 M60402 Nicotiana tabacum glucan beta-1, 3-glucanase. glucanase GLA.
AAA63540.1 M60403 Nicotiana tabacum glucan-l,3-beta-glucosidase. glucanase GLB.
AAA88794.1 U01900 Solanum tuberosum catalyzes the hydrolysis of 1,3-beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). gluBl. plant defense gene; induced expression in response to infection, elicitor, ethylene, wounding.
AAA63541.1 M59442 Nicotiana tabacum basic beta-l,3-glucanase. glucanase.
AAB82772.2 AF001523 Musa acuminata beta-1, 3-glucananse. similar to beta-1, 3-glucanase.
AAF08679.1 AF004838 Musa acuminata beta-l,3-glucanase.
AAA19111.1 U01902 Solanum tuberosum catalyzes the hydrolysis of 1,3-beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). gluB3. plant defense gene; induced expression in response to infection, elicitor, ethylene, wounding.
AAC04710.1 AF034106 Glycine max beta-l,3-glucanase 1. SGlul.
AAC04714.1 AF034113 Glycine max beta- 1,3 -glucanase 8. SGlu8.
CAB91554.1 AJ277900 Vitis vinifera beta 1-3 glucanase. gl.
AAA34082.1 M20620 Nicotiana tabacum prepro-beta-l,3-glucanase precursor.
CAA03908.1 AJ000081 Citras sinensis glucan hydrolase. beta-1, 3-glucanase. gnsl.
AAB03501.1 U41323 Glycine max 'beta- 1, 3-glucanase. SGN1.
AAA92013.1 U49454 Prunus persica beta-1, 3-glucanase. Gnsl.
AAA33946.1 M37753 Glycine max beta- 1 ,3 -endoglucanase (EC 3.2.1.39).
AAA63542.1 M59443 Nicotiana tabacum acidic beta-1, 3-glucanase. glucanase.
AAF34761.1 AF227953 Capsicum annuum basic beta-1, 3-glucanase. BGLU.
AAD33881.1 AF141654 Nicotiana tabacum beta-l,3-glucanase. GGL4.
AAG34080.1 AF294849 Capsicum annuum beta-l,3-glucanase-like protein.
AAF33405.1 AF230109 Populus x canescens beta-1,3 glucanase. BGLUC.
AAD33880.1 AF141653 Nicotiana tabacum beta-1 ,3-glucanase. GGL1.
CAA57255.1 X81560 Nicotiana tabacum (l-)-beta-glucanase. Sp41a.
AAA34053.1 M60464 Nicotiana tabacum beta-1 ,3-glucanase.
SEQ ID NO: 630
AAD37698.1 AF145729 Oryza sativa homeodomain leucine zipper protein. Oshox5. transcription factor.
BAA05624.1 D26575 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 520-699; leucine zipper at nt 700-805.
AAF01765.1 AF184278 Glycine max homeodomain-leucine zipper protein 57. Hdl57. transcription factor.
CAA64417.1 X94947 Lycopersicon esculentum homeobox. VAHOX1.
BAA93465.1 AB028077 Physcomitrella patens homeobox protein PpHB6. PpHB6. homeodomain-leucine zipper gene.
BAB18171.1 AB042769 Zinnia elegans homeobox-leucine zipper protein. ZeHB3. full length.
BAA93460.1 AB028072 Physcomitrella patens homeobox protein PpHBl. PpHBl. homeodomain-leucine zipper gene.
BAA93466.1 AB028078 Physcomitrella patens homeobox protein PpHB7. PpHB7. homeodomain-leucine zipper gene.
BAA93461.1 AB028073 Physcomitrella patens homeobox protein PpHB2. PpHB2. homeodomain-leucine zipper gene.
BAA05625.1 D26576 Daucus carota transcriptiional regulator. DNA-binding protein, homeodomain at nt 300-479; leucine zipper at nt 480-587.
BAA05622.1 D26573 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 585-764; leucine zipper at nt 765-851.
BAA93467.1 AB028079 Physcomitrella patens homeobox protein PpHB8. PpHB8. homeodomain-leucine zipper gene.
BAA93464.1 AB028076 Physcomitrella patens homeobox protein PpHB5. PpHB5. homeodomain-leucine zipper gene.
AAD37697.1 AF145728 Oryza sativa homeodomain leucine zipper protein. Oshox4. transcription factor.
AAF01764.2 AFl 84277 Glycine max homeodomain-leucine zipper protein 56. Hdl56. transcription factor.
BAA21017.1 D26578 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 761-940; leucine zipper at nt 941-1048.
BAB18168.1 AB042766 Zinnia elegans homeobox-leucine zipper protein. ZeHB7. 3'RACE product.
BAA93468.1 AB028080 Physcomitrella patens homeobox protein PpHB9. PpHB9. homeodomain-leucine zipper gene.
BAA05623.1 D26574 Daucus carota trancriptional regulator. DNA-binding protein, homeodomain at nt 498-677; leucine zipper at nt 678-785.
AAD37699.1 AF145730 Oryza sativa homeodomain leucine zipper protein. Oshox6. transcription factor.
AAD38144.1 AF139497 Prunus armeniaca
DNA-binding protein, homeobox leucine zipper protein. HBLZP.
AAA63768.2 AF339748 Helianthus annuus homeobox-leucine zipper protein HAHB-4. Hahb-4.
BAA93463.1 AB028075 Physcomitrella patens homeobox protein PpHB4. PpHB4. homeodomain-leucine zipper gene.
CAA64491.1 X95193 Pimpinella brachycaφa transcription activator, homeobox-leucine zipper protein.
CAA64221.1 X94449 Pimpinella brachycaφa transcription activator, homeobox-leucine zipper protein. PHZ4.
CAA64152.1 X94375 Pimpinella brachycaφa transcription activator, homeobox-leucine zipper protein. AAD37700.1 AF145731 Oryza sativa homeodomain leucine zipper protein. Oshox7. transcription factor.
AAD37695.1 AF145726 Oryza sativa homeodomain leucine zipper protein. Oshox2. transcription factor.
CAA06728.1 AJ005833 Craterostigma plantagineum transcription factor, homeodomain leucine zipper protein, hb-2.
CAA62608.1 X91212 Lycopersicon esculentum
HD-ZIP protein. THOM1.
CAA63222.1 X92489 Glycine max transcription activator, homeobox-leucine zipper protein.
CAA65456.2 X96681 Oryza sativa transcription factor. DNA-binding protein. Oshoxl. homeodomain leucine zipper gene.
AAF19980.1 AF211193 Oryza sativa homeodomain-leucine zipper transcription factor. Hoxl. hoxl. SEQ ID NO: 631
CAC19183.1 AJ291816 Cicer arietinum expansin.
AAD13633.1 AF059489 Lycopersicon esculentum expansin precursor. Exp 5.
AAG13983.1 AF297522 Prunus avium expansin 2. Exp2. PraavExp2.
AAF35902.1 AF230333 Zinnia elegans expansin 3.
AAF32409.1 AF230276 Triphysaria versicolor alpha-expansin 3.
AAC96080.1 AF049353 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP4. cell wall protein.
AAG13982.1 AF297521 Prunus avium expansin 1. Expl. PruavExpl.
AAC33529.1 U93167 Prunus armeniaca expansin. PA-Expl.
AAF32411.1 AF230278 Triphysaria versicolor alpha-expansin 1.
AAF35901.1 AF230332 Zinnia elegans expansin 2.
AAF21101.1 AF159563 Fragaria x ananassa expansin. Exp2. ripening regulated.
BAB 19676.1 AB029083 Prunus persica expansin. PchExpl.
AAD47901.1 AF085330 Pinus taeda expansin.
AAC33530.1 AF038815 Prunus armeniaca expansin. Exp2.
CAC19184.1 AJ291817 Cicer arietinum expansin.
AAB40635.1 U64891 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession Numbers U30477 and U30479.
AAG32921.1 AF184233 Lycopersicon esculentum expansin. Exp 10.
AAD49956.1 AF167360 Rumex palustris expansin. EXPL
AAB40637.1 U64893 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession
Numbers U30477 and U30479.
AAB40634.1 U64890 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession Numbers U30477 and U30479.
AAB37746.1 U30382 Cucumis sativus expansin SI precursor. Cs-EXPl. similar to pollen allergen Lol pi, Lolium perenne, Swiss-
Prot Accession Number P 14946; former gene name CuExSl; expansin-29 (Ex29) protein.
AAB40636.1 U64892 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476,
U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession
Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession
Numbers U30477 and U30479.
AAC39512.1 AF043284 Gossypium hirsutum expansin. GhEXl. contains N-terminal signal peptide.
CAB43197.1 AJ239068 Lycopersicon esculentum cell wall loosening enzyme. expansin2. exp2.
AAC64201.1 AF096776 Lycopersicon esculentum expansin. LeEXP2.
AAC96081.1 AF049354 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP5. cell wall protein.
CAB46492.1 AJ243340 Lycopersicon esculentum expansin9. exp9.
AAF17570.1 AF202119 Marsilea quadrifolia alpha-expansin. EXPL Mq-EXPl.
AAB81662.1 U85246 Oryza sativa expansin. Os-EXP4.
AAF62180.1 AF247162 Oryza sativa alpha-expansin OsEXP5. cell wall loosening factor; expressed in internodes, leaves, coleoptiles, and roots.
AAB38074.1 U30477 Oryza sativa induces extension (creep) in plant cell walls, expansin Os-EXP2. Os-EXP2. former gene name
RiExB.
AAD13632.1 AF059488 Lycopersicon esculentum expansin precursor. Exp4.
AAF32410.1 AF230277 Triphysaria versicolor alpha-expansin 2.
AAG01875.1 AF291659 Striga asiatica alpha-expansin 3. Exp3.
CAA04385.1 AJ000885 Brassica napus Cell wall extension in plants. Expansin.
CAA06271.2 AJ004997 Lycopersicon esculentum expansinl8. expl 8.
BAB32732.1 AB049406 Eustoma grandiflorum expansin. Eg Expansin.
AAC63088.1 U82123 Lycopersicon esculentum expansin. LeEXPl. fruit ripening regulated expansin.
AAF62182.1 AF247164 Oryza sativa alpha-expansin OsEXP7. cell wall loosening factor; expressed in internodes and leaves.
AAB37749.1 U30460 Cucumis sativus expansin S2 precursor. Cs-EXP2. similar to pollen allergen Lol pi, Lolium perenne, Swiss-
Prot Accession Number P14946; former gene name CuExS2; expansin-30 (Ex30) protein.
AAG32920.1 AF184232 Lycopersicon esculentum expansin. Exp 8.
CAC06433.1 AJ276007 Festuca pratensis expansin. exp2.
AAF62181.1 AF247163 Oryza sativa alpha-expansin OsEXP6. cell wall loosening factor; expressed in internodes and leaves.
AAC96079.1 AF049352 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP3. cell wall protein.
AAC96077.1 AF049350 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXPl. cell wall protein.
BAA88200.1 AP000837 Oryza sativa
EST AU078708(E60526) conesponds to a region ofthe predicted gene. Shnilar to expansin
(U85246).
AAF17571.1 AF202120 Regnellidium diphyllum alpha-expansin. EXP Rd-EXPl.
AAC96078.1 AF049351 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. N.-EXP2. cell wall protein.
AAG01874.1 AF291658 Striga asiatica alpha-expansin 2. Exp2.
SEQ ID NO: 634
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
BAA97122.1 AB016264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive transcription factor.
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREl . similar to EREBP transcription factors.
BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding proteinl homolog. McEREBPl.
BAA97124.1 AB016266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBP
CAB93940.1 AJ238740 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein. orca2.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. OSJNBa0027P10.12.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
CAC12822.1 AJ299252 Nicotiana tabacum
AP2 domam-containing transcription factor. ap2.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like transcription factor.
AAF05606.1 AF190770 Oryza sativa
EREBP-like protein, tshl. TSH1; induced by ethylene.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserB.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. similar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
AAG43548.1 AF211530 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP transcription factors.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
AAK01088.1 AF298230 Hordeum vulgare
CBF 1 -like protein BCBFl. BCBFl. AP2 domain protein; putative DRE binding factor.
AAC49567.1 U41466 Zea mays
Glossyl5. Glossyl5. AP2 DNA-binding domain protein; similar to the Arabidopsis floral homeotic gene APETALA2; maize AP2-domain regulator of leaf epidermal tiaits; homeotic regulator of leaf epidermal cell identity; allele: W64A; putative transcription factor. SEQ ID NO: 635
CAC19789.1 AJ251686 Catharanthus roseus putative transcription factor. MYB-like DNA-binding protein, bpf-1.
CAA55693.1 X79086 Zea mays initiator-binding protein. IBP2.
CAA55691.1 X79085 Zea mays initiator binding protein. IBP1.
AAF97508.1 AF242298 Oryza sativa telomere binding protein- 1. TBP1.
SEQ ID NO: 636
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
BAA22422.1 AB001379 Glycynhiza echinata cytochrome P450. CYP81E1.
BAA74465.1 AB022732 Glycynhiza echinata cytochrome P450. CYP Ge-31.
CAA10067.1 AJ012581 Cicer arietinum cytochrome P450. cyp81E3.
CAB41490.1 AJ238439 Cicer arietinum cytochrome P450 monooxygenase. cyp81E3v2.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'- race).
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81Blc.
AAK38080.1 AF321856 Lolium rigidum putative cytochrome P450.
AAK38079.1 AF321855 Lolium rigidum putative cytochrome P450.
AAK38081.1 AF321857 Lolium rigidum putative cytochrome P450.
AAC34853.1 AF082028 Hemerocallis hybrid cultivar putative cyt P450-containing fatty acid hydroxylase. senescence-associated protein 3. SA3. mRNA accumulates in senescing petals.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
CAB56742.1 AJ249800 Cicer arietinum cytochrome P450 monooxygenase. cyp81E5.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3'-hydroxylase. Htl. cytochrome P450; CYP75B2.
CAA71515.1 Y10491 Glycine max putative cytochrome P450.
AAB94590.1 AF022461 Glycine max
CYP82Clρ. CYP82C1. cytochrome P450 monooxygenase.
CAA71516.1 Y10492 Glycine max putative cytochrome P450.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
AAG09208.1 AF175278 Pisum sativum wound-inducible P450 hydroxylase. CYP82A1.
AAC49188.2 U29333 Pisum sativum cytochrome P450 monooxygenase. CYP82. new cytochrome P450 family.
AAD38930.1 AF135485 Glycine max cytochrome P450 monooxygenaseCYP93Dl. CYP93E1.
CAA71876.1 Y10982 Glycine max putative cytochrome P450.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
CAA70576.1 Y09424 Nepeta racemosa cytochrome P450. CYP71 A6.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
CAA71877.1 Y10983 Glycine max putative cytochrome P450.
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
AAC39454.1 AF014802 Eschscholzia californica
(S)-N-methylcoclaurrne 3'-hydroxylase. CYP82B1. cytochrome P-450-dependent monooxygenase; methyl jasmonate-inducible cytochrome P-450-dependent, homologous to wound-inducible CYP82A1 of Pisum sativum GenBank Accession Number U29333.
BAA92894.1 AB006790 Petunia x hybrida cytochrome P450. IMT -2.
BAA84072.1 AB028152 Torenia hybrida flavone synthase II. cytochrome P450. TFNS5.
BAA35080.1 AB015762 Nicotiana tabacum putative cytochrome P450. CYP82E1.
CAB56743.1 AJ249801 Cicer arietinum cytochrome P450 monooxygenase. cyp81E4.
AAB 17562.1 U72654 Eustoma grandiflorum flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75. AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
BAA74466.1 AB022733 Glycynhiza echinata cytochrome P450. CYP Ge-51.
BAA22423.1 AB001380 Glycynhiza echinata cytochrome P450. CYP93B1.
SEQ ID NO: 639
AAC06319.1 AF053084 Malus x domestica putative cinnamyl alcohol dehydrogenase. CAD.
CAA61275.1 X88797 Eucalyptus gunnii cinnamyl alcohol dehydrogenase. CADI.
SEQ ID NO: 640
BAA92916.1 AP001539 Oryza sativa
EST C26826(C50159) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome II BAC F13A10; putative ARFl family auxin responsive transcription factor. (AC006418).
AAK21342.1 AC024594 Oryza sativa putative transcription factor. OSJNBa0093Bl 1.2.
AAG43286.1 AF140228 Oryza sativa auxin response factor 1.
SEQ ID NO: 641
CAC24691.1 AJ132363 Brassica juncea efflux canier of polar auxir i transport, pina.
AAG17172.1 AF190881 Populus tremula x Populus tremuloides PINl-like auxin transport protein, ppll.
AAC39514.1 AF056027 Oryza sativa auxin transport protein REH1. REH1. potential membrane protein.
SEQ ID NO: 642
AAG22044.1 AF305783 Pisum sativum apyrase 2. apy2. phosphatase.
AAF00610.1 AF156781 Dolichos bifloras apyrase. apyrase-2.
AAG32959.1 AF207687 Glycine soja apyrase GS50.
AAG32960.1 AF207688 Glycine soja apyrase GS52.
AAF00609.1 AFl 56780 Lotus japonicus apyrase. nod factor binding lectin-nucleotide phosphohydrolase. LNP.
AAD31285.1 AF139807 Dolichos bifloras apyrase. nod factor binding lectin-nucleotide phosphohydrolase. LNP.
AAF00611.1 AF156782 Medicago sativa apyrase. nod factor binding lectin-nucleotide phosphohydrolase. LNP.
AAK15160.1 AF288132 Medicago truncatula putative apyrase. apyl. nucleotide phosphohydrolase; Mtapyl.
BAB18896.1 AB038669 Pisum sativum apyrase.
BAB18895.1 AB038668 Pisum sativum apyrase.
BAB18894.1 AB038555 Pisum sativum apyrase H-type.
BAB 18893.1 AB038554 Pisum sativum apyrase S-type.
BAB18900.1 AB027614 Pisum sativum apyrase.
BAB40230.1 AB027613 Pisum sativum
S-type apyras. ATP diphosphohydrolase (apyrase) S-type.
BAB18890.1 AB023621 Pisum sativum apyrase S-type. ATP diphosphohydrolase (apyrase) S-type.
BAA75506.1 AB022319 Pisum sativum apyrase. cytoskeleton associated.
BAA89275.1 AB027616 Pisum sativum apyrase.
BAB40231.1 AB027615 Pisum sativum
S-type apyrase. ATP diphosphohydrolase (apyrase) S-type.
AAB02720.1 U58597 Solanum tuberosum catalyzes the hydrolysis of phosphoanhydride bonds of nucleoside tri- and di- phosphates in the presence of divalent cations. ATP-diphosphohydrolase. RROPl. apyrase, Ecto- ATPase, E- type ATPase; NTP-diphosphohydrolase.
AAK15161.1 AF288133 Medicago truncatula putative apyrase. apy4. nucleotide phosphohydrolase; Mtapy4.
BAB18891.1 AB030444 Pisum sativum apyrase. ATP diphosphohydrolase, (EC 3.6.1.5) this sequence is reported in Acc#:AB022319.
BAB18892.1 AB030445 Pisum sativum apyrase. ATP diphosphohydrolase, EC 3.6.1.5 this sequence is reported in Acc#:AB022319,
Acc#:AB027613.
SEQ ID NO: 645
AAG22044.1 AF305783 Pisum sativum apyrase 2. apy2. phosphatase.
AAF00610.1 AF156781 Dolichos bifloras apyrase. apyrase-2.
AAG32959.1 AF207687 Glycine soja apyrase GS50.
AAG32960.1 AF207688 Glycine soja apyrase GS52.
AAD31285.1 AF139807 Dolichos bifloras apyrase. nod factor binding lectin-nucleotide phosphohydrolase. LNP.
AAK15160.1 AF288132 Medicago truncatula putative apyrase. apyl. nucleotide phosphohydrolase; Mtapyl.
AAF00609.1 AFl 56780 Lotus japonicus apyrase. nod factor binding lectin-nucleotide phosphohydrolase. LNP.
BAB18896.1 AB038669 Pisum sativum apyrase.
BAB18895.1 AB038668 Pisum sativum apyrase.
BAB18894.1 AB038555 Pisum sativum apyrase H-type.
BAB 18893.1 AB038554 Pisum sativum apyrase S-type.
BAB 18900.1 AB027614 Pisum sativum apyrase.
BAB40230.1 AB027613 Pisum sativum
S-type apyras. ATP diphosphohydrolase (apyrase) S-type.
BAB18890.1 AB023621 Pisum sativum apyrase S-type. ATP diphosphohydrolase (apyrase) S-type.
BAA75506.1 AB022319 Pisum sativum apyrase. cytoskeleton associated.
AAF00611.1 AF156782 Medicago sativa apyrase. nod factor binding lectin-nucleotide phosphohydrolase. LNP.
BAA89275.1 AB027616 Pisum sativum apyrase.
BAB40231.1 AB027615 Pisum sativum
S-type apyrase. ATP diphosphohydrolase (apyrase) S-type.
AAB02720.1 U58597 Solanum tuberosum catalyzes the hydrolysis of phosphoanhydride bonds of nucleoside tii- and di- phosphates in the presence of divalent cations. ATP-diphosphohydrolase. RROPl. apyrase, Ecto- ATPase, E- type ATPase; NTP-diphosphohydrolase.
AAK15161.1 AF288133 Medicago truncatula putative apyrase. apy4. nucleotide phosphohydrolase; Mtapy4.
BAB18891.1 AB030444 Pisum sativum apyrase. ATP diphosphohydrolase, (EC 3.6.1.5) this sequence is reported in Acc#:AB022319.
BAB18892.1 AB030445 Pisum sativum apyrase. ATP diphosphohydrolase, EC 3.6.1.5 this sequence is reported in Acc#:AB022319,
Acc#:AB027613. SEQ ID NO: 646
AAB80947.1 AF022915 Triticum aestivum ormthine/acetylornithine aminotransferase.
CAA69936.1 Y08680 Alnus glutinosa acetylornithine aminotransferase. agl 18.
AAK11219.1 AF324485 Oryza sativa aminotransferase-like protein.
AAG09278.1 AFl 77590 Vitis vinifera omithine aminotransferase.
AAC78480.1 AF085149 Capsicum chinense putative aminotransferase. pyridoxal phosphate dependent.
AAA02916.1 L08400 Vigna aconitifolia production of pynoline-5-carboxylate by deamination of omithine. omithine aamimnoottrraannssfieerraassee..
AAB59330.1 M31545 Hordeum vulgare glutamate 1-semialdehyde aminotransferase. GSA. precursor.
AAAI 8861.1 U03632 C Chhllaammyyddoommoonnaass rreeiinnhhaarrddttiiii glutamate- 1 -semialdehyde aminotransferase. gsa.
AAA33968.1 L12453 Glycine max catalyzes 5-anιinolevulinic acid formation from GSA. glutamate 1-semialdehyde aminotransferase. Gsa. putative.
AAC48996.1 U20260 Glycine max converts GSA to 5-aminolevulinic acid, glutamate 1-semialdehyde aminotransferase. Gsal.
SEQ ID NO: 650
AAF66982.1 AF247646 Zeamays transposase. similar to Mutator family tiansposases.
SEQ ID NO: 652
AAB41812.1 L36158 Medicago sativa peroxidase. pxdD. amino acid feature: conserved domains, aa 120 .. 126, 188 .. 195; amino acid feature: heme-binding domain, aa 63 .. 68.
CAA71495.1 Y10469 Spinacia oleracea peroxidase. prxrδ.
CAA09881.1 AJOl 1939 Trifolium repens peroxidase. prx2.
CAA62228.1 X90695 Medicago sativa peroxidase2. prx2.
AAA98491.1 L36981 Petroselinum crispum anionic peroxidase.
BAB39281.1 AP002971 Oryza sativa putative peroxidase. P0537A05.10. contains ESTs D24657(R2329),AU082066(R2329).
AAB02926.1 U59284 Linum usitatissimum peroxidase. FLXPER3.
BAA77387.1 AB024437 Scutellaria baicalensis peroxidase 1.
CAA66037.1 X97351 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
CAA71488.1 Y10462 Spinacia oleracea peroxidase. prxrl.
BAA01950.1 D11337 Vigna angularis peroxidase.
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
CAA71490.1 Y10464 Spinacia oleracea peroxidase. prxr3.
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
BAA92497.1 AP001383 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to peroxidase ATP18a. (X98804).
AAC36707.1 AF078691 Manihot esculenta peroxidase.
BAA92422.1 AP001366 Oryza sativa
ESTs AU081576(R0541),AU032412(R4029) conespond to a region ofthe predicted gene.
Similar to A.thaliana mRNA for peroxidase ATP18a. (X98804).
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
AAF63024.1 AF244921 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 2 precursor, type III peroxidase.
AAA32676.1 M37637 Arachis hypogaea cationic peroxidase. PNC2
BAA11853.1 D83225 Populus nigra peroxidase.
CAC21393.1 AJ401276 Zea mays peroxidase. pox3.
AAA65636.1 L13653 Lycopersicon esculentum peroxidase. TPX2.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
CAA62227.1 X90694 Medicago sativa peroxidaselC. prxlC.
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI-1.
AAD37376.1 AF145350 Glycine max peroxidase. Prx4.
AAB67737.1 L77080 Stylosanthes humilis cationic peroxidase.
CAA71489.1 Y10463 Spinacia oleracea peroxidase. prxr2.
CAA71496.1 Y10470 Spinacia oleracea peroxidase. prxr9.
CAA71494.1 Y10468 Spinacia oleracea peroxidase. prxr7.
BAA06334.1 D30652 Populus kitakamiensis peroxidase.
CAA66034.1 X97348 Populus balsamifera subsp. tiichocaφa signal for ER. peroxidase.
BAA94962.1 AB042103 Asparagus officinalis peroxidase. AspPOXl.
CAA80502.1 Z22920 Spirodela polynhiza peroxidase.
CAA66035.1 X97349 Populus balsamifera subsp. trichocaφa signal for ER. peroxidase.
BAA11852.1 D83224 Populus nigra peroxidase.
CAA66036.1 X97350 Populus balsamifera subsp. tiichocaφa signal for ER. peroxidase.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
AADl 1481.1 U51191 Glycine max peroxidase precursor. sEPal .
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
AAA32973.1 M73234 Hordeum vulgare peroxidase BP 1. Prx5.
AAB47602.1 L07554 Linum usitatissimum peroxidase. FLXPER1.
AAB41810.1 L36156 Medicago sativa peroxidase. pxdA. amino acid feature: conserved motifs, aa 181 .. 188; amino acid feature: heme-binding domain, aa 60 .. 65.
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 5 precursor, type III peroxidase.
AAA34108.1 J02979 Nicotiana tabacum lignin-forming peroxidase precursor (EC 1.11.1.7).
AAD43561.1 AF155124 Gossypium hirsutum bacterial-induced peroxids ise precursor. Perx_Goshiko.
SEQ ID NO: 653
AAB97617.1 U83687 Apium graveolens
NADPH-dependent mannose 6-phosphate reductase. m6pr. aldo-keto reductase; similar to aldose 6-phosphate reductase also known as NADP-sorbitol-6-phosphate dehydrogenase encoded by GenBank Accession Number Dl 1080.
BAA01853.1 D11080 Malus x domestica
NADP-dependent D-sorbitol-6-phosphate dehydrogenase. S6PDH.
AAC97607.1 AF057134 Malus x domestica synthesizes sorbitol, a major photosynthetic product in many members ofthe Rosaceae family.
NADP-dependent sorbitol 6-phosphate dehydrogenase. S6PDH. SEQ ID NO: 654
BAA82556.1 AB030083 Populus nigra lectin-like protein kinase. PnLPK.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIK receptor-like protein kinase; serine/threonine protein kinase.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKL accumulates in mesophyll cells.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
AAG16628.1 AY007545 Brassica napus protein serine/tfireonine kinase BNK1.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLK GmRLKl.
BAA94517.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
CAB51836.1 AJ243961 Oryza sativa
Putitive Ser/Thr protein kinase. 11332.7.
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APKl gene for protein tyrosine-serine-threonine kinase.(D12522). AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
BAA92953.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F10M6 ; S-receptor kinase -like protein. (AL021811).
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl.7.
AAF43496.1 AFl 31222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stiess, osmotic stress, and
ABA treatment.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
SEQ ID NO: 655
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKL receptor-like protein kinase; serine/threonine protein kinase.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extiacellular S domain: 123-1346; tiansmembrane domain: 1347-1412; intiacellular kinase domain: from 1413.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRK1. Chrkl .
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
BAA94529.2 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAF34428.1 AFl 72282 Oryza sativa receptor-like protein kinase. DUPRl 1.18.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJOl 0166).
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
SEQ ID NO: 657
AAF43869.1 AF166114 Chloroplast Mesostigma viride probable transport protein. cysA.
BAB17U3.1 AP002866 Oryza sativa putative white protein; ATP-binding cassette tiansporter. P0410E01.34.
BAA57907.1 AB001684 Chlorella vulgaris sulfate tiansport system permease protein. cysA.
AAD54843.1 AF137379 Chloroplast Nephroselmis olivacea probable transport protein. cysA.
BAA90508.1 AP001111 Oryza sativa similar to ABC transporter of Arabidopsis thaliana (AC004697).
BAA90507.1 AP001111 Oryza sativa similar to ABC transporter of Arabidopsis thaliana (AC004697).
BAA83352.1 AP000391 Oryza sativa
ESTs AU067992(C11433),AU077424(C11433) conespond to a region ofthe predicted gene.;
Similar to ABC transporter-7 (U43892).
AAG49003.1 AY013246 Hordeum vulgare putative ABC transporter. 635P2.4b; GC splice donor confirmed by cDNA alignment and comparative sequence.
AAD10836.1 U52079 Solanum tuberosum
P-glycoprotern. pmdrl. binds ATP; ATPase; transporter; tiansmembrane protein.
BAA96612.1 AP002482 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2, BAC F14M4 ; putative ABC transporter
(AC004411).
BAB40032.1 AP003046 Oryza sativa putative ABC transporter. P0445D12.3.
AAG45492.1 AYO 13245 Oryza sativa
3615.4. putative ABC transporter; GC splice donor confirmed by cDNA alignment and comparative sequence.
AAG49002.1 AY013246 Hordeum vulgare putative ABC transporter. 635P2.4a; GC splice donor confirmed by cDNA and comparative sequencing.
BAB21275.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.6.
BAB21273.1 AP002844 Oryza sativa putative ABC tiansporter protein. P0410E03.4.
CAA94437.1 Z70524 Sprrodela polynhiza multidrug resistance protein. PDR5-like ABC transporter.
BAB21276.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.7. contains EST D22472(C1173).
BAB21279.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.10. contains ESTs
AU065360(R3463),AU101680(R3463).
SEQ ID NO: 658
AAD10386.1 U72255 Oryza sativa beta-1, 3-glucanase precursor. Gns9.
BAA89481.1 AB029462 Salix gilgiana beta-l,3-glucanase. SgGNl.
CAB85903.1 AJ251646 Pisum sativum hydrolysis of beta-1,3 conjugated glucans. beta-1,3 glucanase. gns2.
CAA49513.1 X69887 Brassica napus beta- 1,3 -glucanase homologue.
AAA90953.1 U30323 Triticum aestivum beta 1, 3-glucanase. Glcl.
BAB 19363.1 AP002542 Oryza sativa putative beta-1, 3-glucanase. P0679C08.2.
CAA82271.1 Z28697 Nicotiana tabacum beta-1 ,3-glucanase.
CAA30261.1 X07280 Nicotiana plumbaginifolia beta-glucanase.
AAA51643.1 M23120 Nicotiana plumbaginifolia beta-glucanase precursor.
AAA34078.1 M63634 Nicotiana plumbaginifolia regulator of beta(l,3)-glucanase. beta(l,3)-glucanase regulator.
AAB82772.2 AF001523 Musa acuminata beta-1, 3-glucananse. similar to beta-1, 3-glucanase.
AAF08679.1 AF004838 Musa acuminata beta-l,3-glucanase.
CAB71021.1 AJ271598 Hieracium piloselloides putative role in callose degradation, putative beta- 1 ,3-glucanase. glue.
AADl 0383.1 U72252 Oryza sativa beta- 1,3 -glucanase precursor. Gns6.
AAD28732.1 AFl 12965 Triticum aestivum beta-1, 3-glucanase precursor. Glb3.
BAB40807.1 AB052291 Pyras pyrifolia catalyzing the hydrolysis of 1,3-beta-glucosyl linkages. endo-l,3-beta-glucanase-like protein. bgn-1. Ajmino acid alignment ofthe protein(BGN-l) encoded by pear pollen bgn-1 with barley endo-l,3-beta-glucanase(GII. accession number:pdb/lGHS-B/2.3/2/306/N/)(40% identity) and their hydrophobic cluster analysis(HCA)(a overall HCA homology score of 87.1%) showed that it was most likely that the bgn-1 encoded a endo-l,3-beta-glucanase. A higher identity(59.3%) was found between BGN-1 and a putative pea endo-l,3-beta-glucanase (accession number:dad/AJ251646-l).
AAA63539.1 M60402 Nicotiana tabacum glucan beta-1, 3-glucanase. glucanase GLA.
AAA63541.1 M59442 Nicotiana tabacum basic beta- 1,3 -glucanase. glucanase.
AAA63540.1 M60403 Nicotiana tabacum glucan-l,3-beta-glucosidase. glucanase GLB.
AAA32939.1 M62907 Hordeum vulgare hydrolysis of beta-(l-3)-glucan. (1 -3)-beta-glucanase. cBGL32.
AAC14399.1 AF030771 Hordeum vulgare beta-1, 3-glucanase 2. BGL32.
AAA87456.1 U22147 Hevea brasiliensis beta-l,3-glucanase. HGN1. hydrolytic enzyme.
BAA77784.1 AB027429 Oryza sativa beta- 1 ,3 -glucanase.
BAA77785.1 AB027430 Oryza sativa beta-1 ,3-glucanase.
CAB91554.1 AJ277900 Vitis vinifera beta 1-3 glucanase. gl.
AAD 10381.1 U72250 Oryza sativa beta-1 ,3-glucanase precursor. Gns4.
AAA33946.1 M37753 Glycine max beta-1, 3 -endoglucanase (EC 3.2.1.39).
AAD33881.1 AF141654 Nicotiana tabacum beta-l,3-glucanase. GGL4.
AAB86541.1 AF030166 Oryza sativa glucanase. glul .
AAD 10384.1 U72253 Oryza sativa beta-1, 3-glucanase precursor. Gns7.
CAB38443.1 AJ133470 Hevea brasiliensis beta-l,3-glucanase. hgnl.
AAB03501.1 U41323 Glycine max beta-l,3-glucanase. SGN1.
AAAI 8928.1 U01901 Solanum tuberosum catalyzes the hydrolysis of 1,3-beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). glub2. plant defense gene; induced expression in response to infection, elicitor, ethylene, wounding.
AAA88794.1 U01900 Solanum tuberosum catalyzes the hydrolysis of 1,3-beta-D-glucoside linkages in 1,3,-beta-D-glucans (such as laminarin). 1,3-beta-D-glucan glucanohydrolase; endo-l,3-beta-D-glucanase; 1,3-beta- glucanase (basic, class I). gluBl. plant defense gene; induced expression in response to infection, elicitor, ethylene, wounding.
AAC19114.1 AF067863 Solanum tuberosum 1,3-beta-glucan glucanohydrolase. glucanase.
AAG24921.1 AF311749 Hevea brasiliensis beta-l,3-glucanase.
CAA03908.1 AJ000081 Citrus sinensis glucan hydrok ise. beta-1,3- -glucanase. gnsl.
CAA37289.1 X53129 Phaseolus vulgaris 1 ,3,-beta-D-glucanase.
CAA57255.1 X81560 Nicotiana tabacum (l-)-beta-glucanase. Sp41a L.
SEQ ID NO: 659
AAB65776.1 U97521 Vitis vinifera class IV endochitinase. VvChi4A.
AAB65777.1 U97522 Vitis vinifera class IV endochitinase. VvChi4B.
BAA03751.1 D 16223 Oryza sativa endochitinase. Cht-3.
CAA30142.1 X07130 Solanum tuberosum endochitinase.
BAA03749.1 D16221 Oryza sativa endochitinase. Cht-1.
SEQ ID NO: 660
AAF07221.1 AF072519 Nicotiana tabacum centrin. CEN1. caltractin; EF-hand domain calcium-binding protein.
AAF07222.1 AF072520 Nicotiana tabacum centrin. CEN2. caltiactin; EF-hand domain calcium-binding protein.
CAA49153.1 X69220 Scherffelia dubia caltiactin.
AAC04626.1 U92973 Marsilea vestita calcium-binding protein, centrin. MvCenl . caltiactin.
AAB67855.1 U53812 Dunaliella salina caltiactin-like protein.
CAA41039.1 X57973 Chlamydomonas reinhardtii caltiactin.
CAA31163.1 X12634 Chlamydomonas reinhardtii caltractin (AA 1 - 169).
SEQ ID NO: 664
BAB 16432.1 AB041520 Nicotiana tabacum WRKY transcription factor Nt-SubD48. Nt-SubD48.
AAC49528.1 U56834 Petroselinum crispum DNA-binding. WRKY3. WRKY-type DNA-binding protein.
AAD27591.1 AF121354 Petroselinum crispum binds sequence specifically to W Boxes (TTGACC). transcription factor. WRKY3. sequence specific DNA-binding protein.
AAD32676.1 AF140553 Avena sativa
DNA-binding protein WRKY3. wrky3. putative transcription factor.
BAA77358.1 AB020023 Nicotiana tabacum
WRKY domain Zn-finger type DNA-binding protein. DNA-binding protein NtWRKY3.
AAG46150.1 ACO 18727 Oryza sativa putative DNA-binding protein. OSJNBa0056G17.18.
BAB40073.1 AP003074 Oryza sativa putative WRKY DNA binding protein. OSJNBa0004G10.20. contains EST C26525(C12525).
BAB 18313.1 AP002865 Oryza sativa putative WRKY DNA binding protein. P0034C11.1. contains EST C26525(C12525).
AAD38283.1 AC007789 Oryza sativa putative WRKY DNA binding protein. OSJNBa0049B20.9.
BAA77383.1 AB020590 Nicotiana tabacum transcription factor NtWRKY2.
BAA82107.1 AB022693 Nicotiana tabacum transcription factor. NtWRKYl.
CAA88326.1 Z48429 Avena fatua binds conserved cis-element from cereal alpha-Amy2 promoters. DNA-binding protein.
AAD32677.1 AF140554 Avena sativa
DNA-binding protein WRKY wrkyl. putative transcription factor.
AAD55974.1 AF121353 Petroselinum crispum zinc-finger type transcription factor WRKYL WRKYl.
AAC49529.1 U58540 Petroselinum crispum
WRKY2. Contains two WRKY domains; WRKY-type DNA-binding protein.
AAC49527.1 U48831 Petroselinum crispum
WRKYl. contains two WRKY domains; WRKY-type DNA-binding protein; sequence- specific DNA-binding protein. AAC31956.1 AF080595 Pimpinella brachycaφa zinc finger protein. ZFP1. WRKYl.
BAA86031.1 AB026890 Nicotiana tabacum transcription factor NtWRKY4.
AAD16139.1 AF096299 Nicotiana tabacum DNA-binding protein 2. WRKY2. transcription factor.
AAF23898.1 AF193802 Oryza sativa zinc finger transcription factor WRKYl.
AAD16138.1 AF096298 Nicotiana tabacum DNA-binding protein 1. WRKYl. transcription factor.
AAC37515.1 L44134 Cucumis sativus SPFl-like DNA-binding protein.
AAG35658.1 AF204925 Petroselinum crispum transcription factor WRKY4. WRKY4. binds to W box (TTGACC) elements.
CAA88331.1 Z48431 Avena fatua binds conserved cis-element from cereal alpha-Amy2 promoters. DNA-binding protein.
BAB 19075.1 AP002744 Oryza sativa putative DNA-binding protein homolog. P0006C01.17.
BAB19096.1 AP002839 Oryza sativa putative DNA-binding protein homolog. P0688A04.2.
AAK16170.1 AC079887 Oryza sativa putative DNA binding protein. OSJNBa0040E01.4.
AAK16171.1 AC079887 Oryza sativa putative DNA-binding protein. OSJNBa0040E01.10.
AAG35659.1 AF204926 Petroselinum crispum transcription factor WRKY5. WRKY5. binds to W box (TTGACC) elements.
CAB97004.1 AJ278507 Solanum tuberosum putative transcription factor. WRKY DNA binding protein. WRKY
AAF61864.1 AFl 93771 Nicotiana tabacum DNA-binding protein 4. WRKY4. transcription factor.
BAA87069.1 AB035271 Matricaria chamomilla elicitor-rnduced DNA-binding protein homolog. McWRKYl.
AAF61863.1 AFl 93770 Nicotiana tabacum DNA-binding protein 3. WRKY3. transcription factor.
SEQ ID NO: 665
AAD02558.1 AF049933 Petunia x hybrida
PGPS/NH17. PGPS/NH17. protein sec61 gamma subunit homolog; protein tiansport to ER.
SEQ ID NO: 667
AAD51623.1 AF169020 Glycine max seed maturation protein PM35. PM35. similar to Phaseolus vulgaris putative osmoprotector
PvLEA-18.
AAC49859.1 U72764 Phaseolus vulgaris putative osmoprotector. PvLEA-18. Pylea-18. atypical late embryogenesis abundant protein.
AAF81194.1 AF240774 Phaseolus vulgaris
LEA- 18. SEQ ID NO: 672
CAB40743.1 AJOl 1885 Solanum tuberosum starch branching enzyme II. sbe IL
CAB40746.1 AJOl 1888 Solanum tuberosum starch branching enzyme II. SBE II.
CAB40748.1 AJOl 1890 Solanum tuberosum starch branching enzyme II. SBE II.
AAD30186.1 AF076679 Triticum aestivum starch branching enzyme-I. SBE-I.
AAD30187.1 AF076680 Aegilops tauschii starch branching enzyme-I. SBE-I.
BAA82348.1 AB029548 Phaseolus vulgaris branching enzyme 1. kbel.
CAA56319.1 X80009 Pisum sativum starch branching enzyme I. SBEI.
CAB40747.1 AJOl 1889 Solanum tuberosum starch branching enzyme II. SBE II.
CAA03846.1 AJ000004 Solanum tuberosum branches 1,4-alpha glucans. starch branching enzyme II, SBE-II. Sbe-H
BAA03738.1 D16201 Oryza sativa branching enzyme-3 precursor.
AAG27623.1 AF286319 Triticum aestivum starch branching enzyme 2. Sbe2. glucosyltiansferase.
CAA72154.1 Y11282 Triticum aestivum 1,4-alpha-glucan branching enzyme II. sbe2.
AAK26821.1 AF338431 Aegilops tauschii starch branching enzyme Ila. SBEIIa.
AAK26822.1 AF338432 Triticum aestivum starch branching enzyme Ila variant. SBEIIa variant.
AAC33764.1 AF072725 Zeamays starch branching enzyme lib. ae. SBEIIb.
AAAI 8571.1 L08065 Zeamays starch branching enzyme II.
BAA82828.1 AB023498 Oryza sativa starch branching enzyme rbe4. RBE4.
AAC69753.1 AF064560 Hordeum vulgare starch branching enzyme Ila. sbella.
CAA56320.1 X80010 Pisum sativum starch branching enzyme II. SBEII.
AAC69754.1 AF064561 Hordeum vulgare starch branching enzyme lib. sbellb.
AAC36471.1 AF072724 Zea mays starch branching enzyme I. sbel. confirmed by partial peptide sequencing.
AAA82735.1 U17897 Zea mays starch branching enzyme I. sbel.
AAD50279.2 AF169833 Sorghum bicolor seed starch branching enzyme. SBE.
BAA01854.1 D11081 Zea mays branching enzyme-I precursor.
CAA49463.1 X69805 Solanum tuberosum 1,4-alpha-glucan branching enzyme. SBE.
CAA70038.1 Y08786 Solanum tuberosum 1,4-alpha-glucan branching enzyme, sbel.
AAB17086.1 U66376 Triticum aestivum
1,4-alpha-D-glucan 6-alpha-D-(l,4-alρha-D-glucanotransferase. branching enzyme.
AAB67316.1 U65948 Zea mays formation of alpha- 1-6 glucosidic linkage in starch biosynthesis, starch branching enzyme Ila.
Sbe2a. starch branching enzyme isozyme SBEIIa.
BAB40334.1 AB042937 Ipomoea batatas starch branching enzyme. IBE.
BAA01584.1 D10752 Oryza sativa branching enzyme.
AAD28284.1 AF136268 Oryza sativa subsp. japonica starch-branching enzyme I. Rbel.
BAA01616.1 D 10838 Oryza sativa 1,4-alpha-glucan branching enzyme, sbel.
BAA01855.1 D11082 Oryza sativa branching enzyme-I precursor.
CAB40981.1 AJ237897 Triticum aestivum starch branching enzyme I. sbel. alternative.
CAB40979.1 AJ237897 Triticum aestivum starch branching enzyme I. sbel.
CAB40980.1 AJ237897 Triticum aestivum starch branching enzyme I. sbel. alternative.
AAG27622.1 AF286318 Triticum aestivum starch branching enyzyme 1. Sbel A. glucosyltiansferase.
CAA54308.1 X77012 Manihot esculenta 1 ,4-alpha-glucan branching enzyme. SBE.
CAA72987.1 Y12320 Triticum aestivum starch branching enzyme I. Sbel.
AAG27621.1 AF286317 Triticum aestivum starch branching enyzyme 1. SbelD. glucosyltiansferase.
BAA82349.1 AB029549 Phaseolus vulgaris branching enzyme 3. kbe3.
AAB61925.1 AF002820 Triticum aestivum starch branching enzyme I. wSBE I-D2.
CAB40749.1 AJO 11891 Solanum tuberosum starch branching enzyme II. SBE II.
CAB40745.1 AJOl 1887 Solanum tuberosum starch branching enzyme II. SBE II.
CAB40744.1 AJOl 1886 Solanum tuberosum starch branching enzyme II. SBE II.
BAA85762.1 AB028067 Nicotiana tabacum starch branching enzyme. SBE.
CAA49371.1 X69713 Manihot esculenta branching enzyme, r-2.
BAB40335.1 AB042940 Ipomoea batatas starch branching enzyme. IBE.
CAA49370.1 X69712 Manihot esculenta branching enzyme, r-1.
AAC72336.1 AF064563 Hordeum vulgare starch branching enzyme lib. sbellb.
SEQ ID NO: 673
CAA09881.1 AJOl 1939 Trifolium repens peroxidase. prx2.
CAA62228.1 X90695 Medicago sativa peroxidase2. prx2.
CAA71495.1 Y10469 Spinacia oleracea peroxidase. prxrδ.
AAB41812.1 L36158 Medicago sativa peroxidase. pxdD. amino acid feature: conserved domains, aa 120 .. 126, 188 .. 195; amino acid feature: heme-binding domain, aa 63 .. 68.
BAA77387.1 AB024437 Scutellaria baicalensis peroxidase 1.
AAF63024.1 AF244921 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 2 precursor, type III peroxidase.
AADl 1483.1 U51193 Glycine max peroxidase. sEPbl.
AAB67737.1 L77080 Stylosanthes humilis cationic peroxidase.
BAA07663.1 D42064 Nicotiana tabacum cationic peroxidase isozyme 38K precursor.
BAA07664.1 D42065 Nicotiana tabacum cationic peroxidase isozyme 40K precursor.
CAB94692.1 AJ242742 Ipomoea batatas
Removal of H2O2, oxidation of toxic reductants, defence response toward wounding. peroxidase. pod.
CAB67121.1 Y19023 Lycopersicon esculentum peroxidase. cevi-1.
CAA62226.1 X90693 Medicago sativa peroxidaselB. prxlB.
CAA50597.1 X71593 Lycopersicon esculentum peroxidase. CEVI-1.
AADl 1481.1 U51191 Glycine max peroxidase precursor. sEPal.
AADl 1484.1 U51194 Glycine max peroxidase. sEPb2.
BAA82306.1 AB027752 Nicotiana tabacum peroxidase.
AAA65637.1 L13654 Lycopersicon esculentum peroxidase. TPX1.
AADl 1482.1 U51192 Glycine max peroxidase precursor. sEPa2.
CAA62225.1 X90692 Medicago sativa peroxidaselA. prxlA.
AAC98519.1 AF007211 Glycine max peroxidase precursor. GMIPER1. pathogen-induced.
AAD37427.1 AF 149277 Phaseolus vulgaris peroxidase 1 precursor. FBP1. secretory peroxidase.
BAA14144.1 D90116 Armoracia rusticana peroxidase isozyme.
AAA98491.1 L36981 Petroselinum crispum anionic peroxidase.
CAA71488.1 Y10462 Spinacia oleracea peroxidase. prxrl.
AAD43561.1 AFl 55124 Gossypium hirsutum bacterial-induced peroxidase precursor. Perx_Goshiko.
BAA14143.1 D90115 Armoracia rusticana peroxidase isozyme.
CAA71490.1 Y10464 Spinacia oleracea peroxidase. prxr3.
AAB02554.1 L37790 Stylosanthes humilis cationic peroxidase.
CAA66037.1 X97351 Populus balsamifera subsp. tiichocaφa signal for ER. peroxidase.
BAA01877.1 D11102 Populus kitakamiensis peroxidase. prxAl.
CAA62227.1 X90694 Medicago sativa peroxidaselC. prxlC.
BAA07241.1 D38051 Populus kitakamiensis peroxidase. prxA4a.
BAA01950.1 D11337 Vigna angularis peroxidase.
AAF65464.2 AF247700 Oryza sativa peroxidase POC1.
AAF63027.1 AF244924 Spinacia oleracea hydrogen peroxide catabolism. peroxidase prxl 5 precursor, type III peroxidase.
CAC21393.1 AJ401276 Zea mays peroxidase. pox3.
CAA59487.1 X85230 Triticum aestivum peroxidase. pox4.
AAD37430.1 AF149280 Phaseolus vulgaris peroxidase 5 precursor. FBP5. secretory peroxidase.
CAA71491.1 Y10465 Spinacia oleracea peroxidase. prxr4.
AAB41811.1 L36157 Medicago sativa peroxidase. pxdC. amino acid feature: conserved domains, aa 123 .. 129, 191 .. 198; amino acid feature: heme-binding domain, aa 68 .. 73.
BAA03644.1 D14997 Oryza sativa peroxidase.
AAA32676.1 M37637 Arachis hypogaea cationic peroxidase. PNC2
CAA71494.1 Y10468 Spinacia oleracea peroxidase. prxr7.
AAA34050.1 M74103 Nicotiana sylvestris anionic peroxidase.
CAA40796.1 X57564 Armoracia rusticana peroxidase. peroxidase precursor.
SEQ ID NO: 674
AAG31438.1 AF241793 Perilla frutescens lrmonene synthase.
BAA08367.1 D49368 Perilla frutescens limonene cyclase.
BAA21629.1 AB005744 Perilla frutescens catalyzing the cyclization of geranyl pyrophosphate to 1-limonene. 1-limonene synthase. gPFLC. similar to perilla PFLC-1:DDBJ Acc#D49368.
AAG31437.1 AF241792 Perilla frutescens limonene synthase.
AAK06663.1 AF317695 Perilla frutescens var. frutescens limonene synthase.
AAG31435.1 AF241790 Perilla citriodora limonene synthase.
AAF65545.1 AF233894 Perilla citriodora limonene synthase.
AAD50304.1 AF175323 Mentha longifolia limonene synthase. monoteφene synthase.
AAC37366.1 L13459 Mentha spicata 4S-limonene synthase.
AAGO 1140.1 AF282875 Schizonepeta tenuifolia (+)-4R-limonene synthase.
AAC61260.1 AF061285 Capsicum annuum sesquiteφene cyclase. UV induced.
AAG09949.1 AF171216 Lycopersicon esculentum vetispiradiene synthase. LEVS2. sesquiteφene cyclase.
AAF74977.1 AF270425 Gossypium hirsutum
(E,E)-farnesyl diphosphate cyclase. (+)-delta-cadinene synthase. cdn2. sesquiteφene cyclase.
BAA82141.1 AB023816 Solanum tuberosum vetispiradiene synthase. PVS4. potato sesquiteφene cyclase.
BAA82092.1 AB022598 Solanum tuberosum vetispiradiene synthase. PVSL potato sesquiteφene cyclase.
AAF21053.1 AF212433 Capsicum annuum UV-induced sesquiteφene cyclase. SC2.
BAA82109.1 AB022720 Solanum tuberosum vetispiradiene synthase. PVS3. potato sesquiteφene cyclase; PVS3.
BAA82108.1 AB022719 Solanum tuberosum vetispiradiene synthase. PVS2. potato sesquiteφene cyclase; PVS2.
AAC12784.1 U88318 Gossypium hirsutum
(E,E)-farnesyl diphosphate cyclizing. (+)-delta-cadinene synthase. cdnl . sesquiteφene cyclase; delta-cadrnene synthase.
AAG24640.2 AF304444 Artemisia annua sesquiteφene cyclase.
AAK15641.1 AF326117 Capsicum annuum sesquiteφene cyclase. PSC2.
CAC 12731.1 AJ271792 Artemisia annua putative sesquiteφene cyclase. cASC125.
SEQ ID NO: 675
CAA50609.1 X71609 Nicotiana tabacum ras-related GTP-binding protein.
CAA98166.1 Z73938 Lotus japonicus GTP-binding protein. RAB5A. rab5A.
CAC24477.1 AJ296336 Cichorium intybus x Cichorium endivia
GTP binding protein. chi3154.
CAC24476.1 AJ296335 Cichorium intybus x Cichorium endivia
GTP binding protein. chi3152.
CAB57220.1 AJ249866 Cichorium intybus x Cichorium endivia
GTP binding protein. gtp2.
CAB57219.1 AJ249865 Cichorium intybus x Cichorium endivia
GTP binding protein, gtpl.
CAC24475.1 AJ296334 Cichorium intybus x Cichorium endivia
GTP binding protein. chi3153.
CAA46112.1 X64941 Nicotiana plumbaginifolia small GTP binding protein.
CAC24474.1 AJ296333 Cichorium intybus x Cichorium endivia
GTP binding protein. chi3151.
CAC19792.1 AJ292320 Oryza sativa small GTP-binding protein, RAB family. RAB5A protein. rab5A.
AAD28731.1 AFl 12964 Triticum aestivum small GTP-binding protein. Sgp.
CAA98167.1 Z73939 Lotus japonicus
GTP-binding protein. RAB5B. rab5B.
CAA06922.1 AJ006225 Mesembryanthemum crystallinum small GTP-binding protein. rab5B.
AAG42497.1 AF323991 Oryza sativa small GTP-binding protein RAB5B. rab5B.
BAA84717.1 AB032761 Oryza sativa
GTP-binding protein. rab5B.
AAG24438.1 AF304518 Oryza sativa small GTP-binding protein RAB5B. rab5B.
CAB57221.1 AJ249867 Cichorium intybus x Cichorium endivia
GTP binding protein, bl.5.
CAA98180.1 Z73952 Lotus japonicus
GTP-binding protein. RABl ID. rabl ID.
CAA98181.1 Z73953 Lotus japonicus
GTP-binding protein. RABl IE. rabl IE.
BAA02114.1 D12546 Pisum sativum
GTP-binding protein.
BAA02113.1 D 12545 Pisum sativum
GTP-binding protein.
BAA02116.1 D12548 Pisum sativum
GTP-binding protein.
BAA02112.1 D12544 Pisum sativum GTP-binding protein.
BAA02118.1 D12550 Pisum sativum GTP-binding protein. ^^
CAA98159.1 Z73931 Lotus japonicus GTP-binding protein. RABIB. rablB.
CAB65172.1 AJ245570 Lycopersicon esculentum putative role in secretion of cell wall modifying enzymes. Rabl 1 GTPase. Rabl la.
CAA89049.1 Z49190 Beta vulgaris
GTP-binding. small G protein.
BAA02110.1 D12542 Pisum sativum GTP-binding protein.
AAB97114.1 U58853 Glycine max small GTP-binding protein, sral .
BAA02904.1 D13758 Oryza sativa ras-related GTP binding protein. ss230.
CAA98178.1 Z73950 Lotus japonicus GTP-binding protein. RABl IB. rabl IB.
CAA98177.1 Z73949 Lotus japonicus GTP-binding protein. RABl IA. rabl 1 A.
AAK15703.1 AF327517 Oryza sativa GTP-binding protein.
AAF65510.1 AF108883 Capsicum annuum small GTP-binding protein.
BAA76422.1 AB024994 Cicer arietinum rab-type small GTP-binding protein.
CAA98160.1 Z73932 Lotus japonicus GTP-binding protein. RABIC. rablC.
AAA80680.1 U38466 Lycopersicon esculentum small GTP-binding protein. LeRablC. ; YPTl/RablA homolog LeRablC.
AAA80678.1 U38464 Lycopersicon esculentum small GTP-binding protein. LeRablA. ; YPTl/RablA homolog LeRablA.
SEQ ID NO: 676
AAA80499.1 U20594 Lycopersicon esculentum leucine aminopeptidase.
CAA54314.1 X77015 Solanum tuberosum leucine aminopeptidase. LAP.
CAA48038.1 X67845 Solanum tuberosum leucine aminopeptidase. LAP.
AAC49457.1 U50152 Lycopersicon esculentum peptidase. leucine aminopeptidase. Iap2.
AAC49456.1 U50151 Lycopersicon esculentum exoprotease in the defense response, leucine aminopeptidase. lap.
AAA80498.1 U20593 Lycopersicon esculentum leucine aminopeptidase.
CAA68143.1 X99825 Petroselinum crispum cytosol aminopeptidase. leucine aminopeptidase.
BAA90521.1 AB037678 Phaseolus vulgaris leucine aminopeptidase.
SEQ ID NO: 678
CAC09580.1 AJ298992 Fagus sylvatica
Abscisic acid (ABA) and calcium induced protein kinase. protein kinase (PK). pkl.
AAA34002.1 M67449 Glycine max protein kinase. PK6.
AAK11734.1 AY027437 Arachis hypogaea serme/threonine/tyrosine kinase.
BAB16918.1 AP002863 Oryza sativa putative protein kinase. P0005A05.22.
CAA06334.1 AJ005077 Lycopersicon esculentum protein kinase. TCTR2 protein. TCTR2.
AAG31141.1 AF305911 Oryza sativa
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
CAA73722.1 Y13273 Lycopersicon esculentum putative protein kinase.
AAD46406.1 AF096250 Lycopersicon esculentum ethylene-responsive protein kinase TCTR1. ER50. serine/threonine kinase; similar to Arabidopsis thaliana negative regulator ofthe ethylene response pathway encoded by GenBank Accession Number L08789.
AAG31142.1 AF305912 Hordeum vulgare
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAD 10056.1 AF 110518 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTR1.
AAD 10057.1 AF110519 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTRlv.
AAK30005.1 AY029067 Rosa hybrid cultivar
CTR2 protein kinase.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAA06538.1 D31737 Nicotiana tabacum protein-serine/threonine kinase.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAA94528.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 BAC T20K24; putative receptor-like protein kinase (AC002392).
BAB17126.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.16.
BAB39451.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.24.
BAB17348.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.32.
BAB 17321.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.1.
BAB17129.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.20.
AAF68398.1 AF237568 Oryza sativa receptor-like protein kinase. RLG2.
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
BAA94517.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAB17335.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.17. contains ESTs D47575(S13157),AU032665(S13157).
BAB39441.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.11.
BAB 17116.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.3.
AAF78044.1 AF248493 Oryza sativa receptor-like kinase. RLG18. protein kinase.
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAB17127.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.17.
AAF78021.1 AF238477 Oryza sativa receptor-like kinase. RLG5. protein kinase.
AAD46420.1 AF100771 Hordeum vulgare receptor-like kinase. Hv3ARK. similar to wheat ARK1 AS.
AAD46917.1 AF164021 Oryza sativa receptor kinase.
BAB17345.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.29.
BAB39438.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.7.
AAF78018.1 AF238474 Oryza sativa receptor-like kinase. RLG16. protein kinase.
AAC01746.1 AF044489 Oryza sativa receptor-like protein kinase. drpkl .
BAB 17337.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.21. contains ESTs D47575(S13157),AU032665(S13157).
BAB 17347.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.31. contains ESTs D47575(S13157),AU032665(S13157).
BAB 17344.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.28. contains ESTs D47575(S13157),AU032665(S13157).
BAB39437.1 AP003338 Oryza sativa receptor-like kinase. OJ1212_B09.6.
AAF78019.1 AF238475 Oryza sativa receptor-like kinase. RLG17. protein kinase.
BAA05648.1 D26601 Nicotiana tabacum protein kinase.
BAB 17332.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.13. contains ESTs D47575(S13157),AU032665(S13157).
BAB39435.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.2.
SEQ ID NO: 679
CAA78386.1 Z13996 Petunia x hybrida
DNA binding protein; tianscriptional activator, protein 1. myb.Ph3. Product related to animal myb proto-oncoproteins. Sequence from nucleotide 992 is not included in clone cPFl and has been obtained by PCR amplification of cDNA.
CAB43399.1 AJ006292 Antirrhinum majus Myb-related transcription factor mixta-like 1. mybmll.
CAA67600.1 X99210 Lycopersicon esculentum myb-related transcription factor. THM16.
BAA23337.1 D88617 Oryza sativa transfactor. OSMYB1. Osmybl.
AAA82943.1 U39448 Picea mariana
MYB-like transcriptional factor MBFl. putative DNA binding region highly similar to the maize Cl.
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related.
BAA93038.1 AP001552 Oryza sativa
EST AU082058(C12976) conesponds to a region ofthe predicted gene. Similar to
Arabidopsis thaliana putative transcription factor (AF062916).
AAC04720.1 AF034134 Gossypium rsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-O. similar to MYB A encoded by GenBank Accession Number L04497.
CAA72186.1 Y11351 Oryza sativa myb factor, myb.
AAF22256.1 AF161711 Pimpinella brachycaφa myb-related transcription factor.
BAB39987.1 AP003020 Oryza sativa putative transcription factor (myb). P0498A12.16. contains ESTs
AU097474(S5087),D40175(S1959).
BAB39972.1 AP003018 Oryza sativa putative transcription factor (myb). OSJNBa0004B13.27. contains ESTs
AU097474(S5087),D40175(S1959).
BAA23338.1 D88618 Oryza sativa transfactor. OSMYB2. Osmyb2.
BAA88222.1 AB028650 Nicotiana tabacum myb-related transcription factor LBM2. Ibm2.
CAA67575.1 X99134 Lycopersicon esculentum transcription factor. THM6. myb-related.
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, tianscriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins.
CAA66952.1 X98308 Lycopersicon esculentum THM18. myb-related transcription factor.
BAA23339.1 D88619 Oryza sativa transfactor. OSMYB3. Osmyb3.
AAC49394.1 U57002 Zeamays
P protein. P. allele P-wr; Myb-like tianscriptional regulator with a putative zinc-finger at the
C-tenninal.
AAB41101.1 U72762 Nicotiana tabacum transcription factor Mybl. mybl. TMV-inducible Myb homolog; contains helix-turn-helix motif; contains redox-sensitive cysteine.
AAA33500.1 M73028 Zea mays myb-like transcription factor. P.
AAG36774.1 AF210616 Zeamays
P2 protein. P2. myb-like transcriptional factor; similar to Zea mays P gene.
BAA88223.1 AB028651 Nicotiana tabacum myb-related transcription factor LBM3. Ibm3.
BAA88224.1 AB028652 Nicotiana tabacum myb-related transcription factor LBM4. Ibm4.
CAA72187.1 Y11352 Oryza sativa myb factor, myb.
BAA88221.1 AB028649 Nicotiana tabacum myb-related transcription factor LBM1. lbml.
CAA72185.1 Y11350 Oryza sativa myb factor, myb.
AAG13574.1 AC037425 Oryza sativa myb factor. OSJNBa0055P24.4.
SEQ ID NO: 680
AAA34238.1 L20507 Vigna radiata calmodulin.
AAA34014.1 L01432 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-3. putative.
AAA34013.1 L01430 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-1. putative.
CAA36644.1 X52398 Medicago sativa calmodulin (AA 1-149).
AAD10245.1 AF030033 Phaseolus vulgaris calmodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
AAD10244.1 AF030032 Phaseolus vulgaris calmodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
CAA74307.1 Y13974 Zea mays calmodulin.
CAA46150.1 X65016 Oryza sativa calmodulin. cam.
AAC36058.1 AF042839 Oryza sativa cahnodulin. CaM2.
AAD10246.1 AF030034 Phaseolus vulgaris calmodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
CAA54583.1 X77397 Zea mays calmodulin. CaM2.
BAA87825.1 AP000815 Oryza sativa
ESTs AU030013(E50493),AU081341(E50493) conespond to a region ofthe predicted gene. Similar to O.sativa gene encoding calmodulin. (Z12828).
AAA92681.1 U13882 Pisum sativum calcium-binding protein, calmodulin.
AAA33706.1 M80836 Petunia x hybrida calmodulin. CAM81.
AAA33705.1 M80831 Petunia x hybrida calmodulin-related protein . CAM53.
CAA43143.1 X60738 Malus x domestica Calmodulin. CaM.
CAA78301.1 Z12839 Lilium longiflorum calcium binding protein, signal transduction. cahnodulin.
AAA33397.1 L18912 Lilium longiflorum calcium binding protein, signal transduction. calmodulin. putative.
CAA42423.1 X59751 Daucus carota calmodulin. Ccam-1.
CAA67054.1 X98404 Capsicum annuum cahnodulin-2.
AAG27432.1 AF295637 Elaeis guineensis calmodulin.
AAG11418.1 AF292108 Prunus avium calmodulin.
AAA34237.1 L20691 Vigna radiata calmodulin.
AAC49587.1 U49105 Triticum aestivum calmodulin TaCaM4-l. calcium-binding protein.
AAC49586.1 U49104 Triticum aestivum cahnodulin TaCaM3-3. calcium-binding protein.
AAC49585.1 U49103 Triticum aestivum calmodulin TaCaM3-2. calcium-binding protein.
AAC49584.1 U48693 Triticum aestivum cahnodulin TaCaM3-l. calcium-binding protein.
AAC49583.1 U48692 Triticum aestivum calmodulin TaCaM2-3. calcium-binding protein.
AAC49582.1 U48691 Triticum aestivum calmodulin TaCaM2-2. calcium-binding protein.
AAC49580.1 U48689 Triticum aestivum calmodulin TaCaMl-3. calcium-binding protein.
AAC49579.1 U48688 Triticum aestivum calmodulin TaCaMl-2. calcium binding protein.
AAC49578.1 U48242 Triticum aestivum calmodulin TaCaMl-1. calcium-binding.
AAA03580.1 L01431 Glycine max calcium-binding regulatory protein, cahnodulin. SCaM-2. putative.
AAB36130.1 S81594 Vigna radiata auxin-regulated cahnodulin. auxin-regulated cahnodulin, arCaM. This sequence comes from
Fig. 1; arCaM.
AAA33901.1 L18913 Oryza sativa calcium binding protein, signal transduction. calmodulin. putative.
CAA78287.1 Z12827 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAB46588.1 U83402 Capsicum annuum cahnodulin.
CAA61980.1 X89890 Bidens pilosa Calmodulin.
AAA32938.1 M27303 Hordeum vulgare calmodulin.
BAA88540.1 AP000969 Oryza sativa
ESTs AU081349(E61253), D41425(S3918) conespond to a region ofthe predicted gene.
Similar to cahnodulin. (AF042840).
AAF65511.1 AF108889 Capsicum annuum calmodulin.
AAC36059.1 AF042840 Oryza sativa calmodulin. CaMl.
AAA33900.1 L18914 Oryza sativa calcium binding protein, signal transduction. calmodulin.
CAA78288.1 Z12828 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAA34015.1 L01433 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-4. putative.
AAA16320.1 L14071 Bryonia dioica calmodulin. Bc329. calcium-binding sites (amino acid #): 1. (21..33); 2. (57..68); 3. (94..106);
4. (130..141).
AAA19571.1 U10150 Brassica napus calcium binding, calmodulin. bcml.
AAA87347.1 M88307 Brassica juncea cahnodulin.
CAA52602.1 X74490 Zeamays Calmodulin. ZMCALM1.
SEQ ID NO: 683
CAA71800.1 Y10847 Brassica juncea O-acetylserine(thiol) lyase.
CAA71798.1 Y10845 Brassica juncea O-acetylserine(thiol) lyase.
AAC25635.1 AF044172 Solanum tuberosum cysteine synthase. CS-A; O-acetylserrne (thiol) lyase; cytosolic isoform.
BAA01279.1 D10476 Spinacia oleracea O-acetylserine(thiol) lyase.
BAA02438.1 D13153 Triticum aestivum O-acetylserine (thiol) lyase. cysl.
CAA59798.1 X85803 Zeamays
O-acetylserine (thiol) lyase. Mcysp. cysteine synthase.
AAD23907.1 AF073695 Oryza sativa cysteine synthase. rcsl. O-acetylserine(thiol)-lyase.
AAD23909.1 AF073697 Oryza sativa cysteine synthase. rcs3. O-acetylserine(thiol)-lyase.
AAC25636.1 AF044173 Solanum tuberosum cysteine synthase. CS-B; O-acetylserine (thiol) lyase; plastidic isoform.
AAC27794.1 AF078693 Chlamydomonas reinhardtii cysteine biosynthesis, putative O-acetylserine(thiol)lyase precursor. Crcys-IA.
CAA06819.1 AJ006024 Cicer arietinum cysteine synthase, O-acetyl-L-serine (thiol)-lyase.
CAA46086.1 X64874 Capsicum annuum O-acetylserine (thiol)-lyase.
CAA71799.1 Y10846 Brassica juncea O-acetylserine(thiol) lyase.
AAAI 6973.1 L05184 Chloroplast Spinacia oleracea O-acetylserrne-(thiol)-lyase.
AAD23908.1 AF073696 Oryza sativa cysteine synthase. rcs2. O-acetylserine(thiol)-lyase.
AAD23910.1 AF073698 Oryza sativa cysteine synthase. rcs4. O-acetylserine(thiol)-lyase.
SEQ ID NO: 684
AAA68983.1 L12395 Brassica napus signal transduction, membrane vehicle traffic, small GTP-binding protein, bra. putative.
SEQ ID NO: 685
AAB61961.1 L81152 Oryza sativa integral membrane protein. OsNramp2.
AAB36424.1 S81897 Oryza sativa
OsNrampl. OsNrampl. Nrampl homolog/Bcg product homolog; This sequence comes from
Fig. 2.
AAB62273.1 L41217 Oryza sativa integral membrane protein. OsNrampl. putative.
AAC49720.1 U60767 Oryza sativa integral membrane protein OsNramp3. OsNramp3.
SEQ ID NO: 686
AAC04719.1 AF034133 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-N. similar to MYB A encoded by GenBank Accession Number L04497.
AAB58314.1 U33917 Craterostigma plantagineum
Cpm7. cpm7. putative DNA-binding protein; myb-like gene; myb-related transcription factor.
AAB58313.1 U33916 Craterostigma plantagineum
Cpm5. cpm5. putative DNA-binding protein; myb-related gene; myb-related transcription factor.
AAC 13876.1 U33915 Craterostigma plantagineum myb-related transcription factor CpmlO. cpmlO. putative DNA-binding protein.
AAK08983.1 AY026332 Oryza sativa
Myb tianscription factor JAMyb. related to host cell death and defense responses; induced by jasmonic acid, wounding, or infection of rice blast fungus, but not by sahcylic acid or abscisic acid.
BAB 18296.1 AP002860 Oryza sativa putative myb-related transcription factor. P0409B08.23.
CAA71992.1 Y11105 Pisum sativum Myb26.
BAB40790.1 AB058642 Lilium hybrid division I LhMyb.
AAK19618.1 AF336285 Gossypium hirsutum GHMYB38. ghmyb38. shnilar to myb.
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, tianscriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins.
BAA88221.1 AB028649 Nicotiana tabacum myb-related transcription factor LBM1. lbml.
BAA88224.1 AB028652 Nicotiana tabacum myb-related transcription factor LBM4. Ibm4.
AAK19617.1 AF336284 Gossypium hirsutum GHMYB36. ghmyb36. similar to myb.
AAA33067.1 L04497 Gossypium hirsutum MYB A; putative.
AAK19615.1 AF336282 Gossypium hirsutum GHMYB10. ghmyblO. similar to myb.
BAA81733.2 AB029162 Glycine max GmMYB29A2.
BAA81732.1 AB029161 Glycine max GmMYB29A2.
BAA81730.1 AB029159 Glycine max GmMYB29Al.
BAA88223.1 AB028651 Nicotiana tabacum myb-related transcription factor LBM3. Ibm3.
AAB41101.1 U72762 Nicotiana tabacum tianscription factor Mybl. mybl. TMV-inducible Myb homolog; contains helix-turn-helix motif; contains redox-sensitive cysteine. ^^
BAA81736.1 AB029165 Glycine max GmMYB29B2.
BAA81731.1 AB029160 Glycine max GmMYB29Al.
CAA72218.1 Y11415 Oryza sativa myb.
CAA67000.1 X98355 Oryza sativa activator of alpha-amylase gene promoter, tianscription factor GAMyb. Gaml. Myb-like; expression is regulated by gibberellin.
AAK19611.1 AF336278 Gossypium hirsutum BNLGHi233. bnlghi6233. similar to myb.
CAA67575.1 X99134 Lycopersicon esculentum transcription factor. THM6. myb-related.
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related.
AAD31395.1 AFl 14162 Lolium temulentum gibberellin MYB tianscription factor. GAMyb. R2/R3-MYB.
BAA93038.1 AP001552 Oryza sativa
EST AU082058(C12976) conesponds to a region ofthe predicted gene. Similar to
Arabidopsis thaliana putative transcription factor (AF062916).
AAG22863.1 AY008692 Hordeum vulgare transcription factor GAMyb. Gamyb.
BAA96421.1 AB044084 Triticum aestivum transcription activator for gibberellin response. GAMyb protein. Ta-GAMyb.
CAA61021.1 X87690 Hordeum vulgare transcriptional activator of alpha-amylase gene promoter. GAMyb protein. Gaml.
CAA66952.1 X98308 Lycopersicon esculentum THM18. myb-related transcription factor.
AAC49394.1 U57002 Zea mays
P protein. P. allele P-wr; Myb-like transcriptional regulator with a putative zinc-finger at the
C-terminal.
BAA88222.1 AB028650 Nicotiana tabacum myb-related tianscription factor LBM2. Ibm2.
AAG36774.1 AF210616 Zea mays
P2 protein. P2. myb-like tianscriptional factor; similar to Zea mays P gene.
AAA33500.1 M73028 Zea mays myb-like transcription factor. P.
CAC19439.1 AJ237661 Oryza sativa Myb factor protein, myb.
CAB40189.1 AJ133638 Avena sativa transcriptional activator, myb protein, gamyb.
AAK19619.1 AF336286 Gossypium hirsutum GHMYB9. ghmyb9. similar to myb.
AAC04720.1 AF034134 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-O. similar to MYB A encoded by GenBank Accession Number L04497.
AAC04716.1 AF034130 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-D. similar to MYB A encoded by GenBank Accession Number L04497.
AAG28526.1 AF198499 Nicotiana tabacum anther-specific myb-related protein 1. mybASl. NtMYBASl; contains N-terminal R2, R3 myb domain repeats similar to c-myb.
CAA78388.1 Z13998 Petunia x hybrida
DNA-binding protein; transcriptional activator, protein 3. myb.Ph3. related to animal myb proto-oncoproteins .
AAA33482.1 M37153 Zea mays cl locus myb homologue; putative.
AAK09327.1 AF320614 Zea mays activates anthocyanin transcription, anthocyanin regulatory C cl. tianscription factor.
AAK09326.1 AF320613 Zeamays activates anthocyanin transcription, anthocyanin regulatory Cl. cl. transcription factor. SEQ ID NO: 687
BAA92972.1 AP001551 Oryza sativa
ESTs AU056183(S20356),AU056881(S20950) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 4 BAC clone F6I18 ; putative protein kinase. (AL022198).
BAA83689.1 AB011968 Oryza sativa
OsPK7. OsPK7. protein kinase.
BAA83688.1 AB011967 Oryza sativa
OsPK4. OsPK4. protein kinase.
AAF22219.1 AF141378 Zea mays protein kinase PK4. ZmPK4.
BAA34675.1 ABO 11670 Triticum aestivum wpk4 protein kinase. wpk4.
CAA73068.1 Y12465 Sorghum bicolor serine/threonine kinase. SNFL2.
CAA73067.1 Y12464 Sorghum bicolor serine/threonine kinase. SNFL1.
BAA92970.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F28A21 ; putative protein kinase.
(AL035526).
CAA74646.1 Y14274 Sorghum bicolor putative serine/threonine protein kinase. SNFL3.
AAD31900.1 AF 145482 Mesembryanthemum crystallinum putative serine/threonine protein kinase.
AAB62693.1 AF004947 Oryza sativa protein kinase.
BAA96628.1 AP002482 Oryza sativa
ESTs D41739(S4522),AU055999(S20214), AU057588(S21592 conespond to a region ofthe predicted gene. Similar to Sorghum bicolor serine/tlireonine kinase (Y12465).
AAD23582.1 AF128443 Glycine max probably involved in plant stress responses possibly regulates gene expression. SNF-1-like serine/threonine protein kinase. expressed in nodules, roots and leaves.
BAA05649.1 D26602 Nicotiana tabacum protein kinase.
CAA71142.1 Y10036 Cucumis sativus
SNFl-related protein kinase.
AAC99329.1 AF062479 Oryza sativa protein kinase SNF1. Snfl. similar to yeast SNF1.
CAA65244.1 X95997 Solanum tuberosum SNFl-related protein kinase. PKIN1.
CAA57898.1 X82548 Hordeum vulgare SNFl-related protein kinase. BKIN2.
CAA07813.1 AJ007990 Hordeum vulgare SnRKl-type protein kinase. kin!2a.
CAA46556.1 X65606 Hordeum vulgare protein kinase. BKIN12.
AAB05457.1 U55768 Oryza sativa
SNFl-related protein kinase. RSkl. Ser/Thr protein kinase homolog.
CAA46554.1 X65604 Hordeum vulgare protein kinase. BKIN12.
AAD00239.1 U73938 Nicotiana tabacum protein kinase. PKl 1-Cl. PKl 1-Cl. induced at the transcriptional level by the abscicic acid plant hormone; similar to serine/threonine protein kinase.
AAC69450.1 AF032465 Nicotiana tabacum putative serine/threonine protein kinase. WAPK.
AAD00240.1 U73939 Nicotiana tabacum protein kinase. PKl 1-C5. PKl 1-C5. induced at the transcriptional level by the abscicic acid plant hormone; similar to serine/threonine protein kinase.
BAA13608.1 D88399 Oryza sativa serrne-threonine kinase. endosperm kinase. REK.
AAA34017.1 L19360 Glycine max protein kinase 2. SPK-2. putative.
AAG60195.1 AC084763 Oryza sativa protein kinase REK. OSJNBa0027P10.6.
AAA33004.1 L12394 Brassica napus serine/threonine protein kination. serine/threonine protein kinase. BSK2. putative.
BAA19573.1 AB002109 Oryza sativa protein kinase. a novel protein kinase.
AAA33979.1 L01453 Glycine max protein phosphorylation, regulatory protein, protein kinase. SPK-1. putative.
AAA33003.1 L12393 Brassica napus protein kination. serine/threonine protein kinase. BSK1. putative.
AAB68962.1 L38855 Glycine max protein kinase. SPK-4.
AAB68961.1 L19361 Glycine max protein kinase 3. SPK-3. putative.
AAB58348.1 U29095 Triticum aestivum serme-threonine protein kinase. TaPK3.
AAF27340.1 AFl 86020 Vicia faba abscisic acid-activated protein kinase. AAPK.
AAA96325.1 M94726 Triticum aestivum protein kinase. abscisic acid inducible.
CAA81443.1 Z26846 Mesembryanthemum crystallinum protein kinase. MCPK9.
AAG31326.1 AF178575 Vitis vinifera putative serine/threonine kinase GDBrPK. GDBrPK. similar to SNF protein kinase.
CAA06503.1 AJ005373 Craterostigma plantagineum protein kinase. cppkl.
AAF21062.1 AF216527 Dunaliella tertiolecta calcium-dependent protein kinase. CPK1; CDPK.
CAA89202.1 Z49233 Chlamydomonas eugametos calcium-stimulated protein kinase.
SEQ ID NO: 689
AAC49826.1 U71604 Catharanthus roseus involved in the second to last step in vindoline biosynthesis, desacetoxyvindoline 4- hydroxylase. CRSD4H. 2-oxoglutarate dependent dioxygenase.
AAB97311.1 AF008597 Catharanthus roseus desacetoxyvindoline-4-hydroxylase. CRSD4H. 2-oxoglutarate dependent dioxygenase; involved in the second to last step in vindoline biosynthesis.
AAC49827.1 U71605 Catharanthus roseus involved in the second to last step in vindoline biosynthesis, desacetoxyvindoline 4- hydroxylase. CRSD4H. 2-oxoglutarate dependent dioxygenase.
BAA37127.1 ABO 12203 Lactuca sativa
2-oxoglutarate-dependent dioxygenase. gibberelin 20-oxidase. Ls20oxl.
BAA95828.1 AP002069 Oryza sativa
ESTs D47168(S12332),D46350(S10967) conespond to a region ofthe predicted gene.
Similar to Prunus armeniaca ethylene-fonning-enzyme-like dioxygenase. (U97530).
AAD56580.1 AFl 84273 Daucus carota leucoanthocyanidin dioxygenase 1. LDOXl . 2-oxoglutarate dependent dioxygenase.
AAB39995.1 U82432 Dianthus caryophyllus anthocyanidin synthase. allele: S; 2-oxoglutarat-dependent dioxygenase.
AAD56581.1 AFl 84274 Daucus carota leucoanthocyanidin dioxygenase 2. LDOX. 2-oxoglutarate dependent dioxygenase. SEQ ID NO: 691
AAA34002.1 M67449 Glycine max protein kinase. PK6.
CAC09580.1 AJ298992 Fagus sylvatica
Abscisic acid (ABA) and calcium induced protein kinase. protein kinase (PK). pkl.
AAK11734.1 AY027437 Arachis hypogaea serine/threonine/tyrosine kinase.
BAB 16918.1 AP002863 Oryza sativa putative protein kinase. P0005A05.22.
CAA06334.1 AJ005077 Lycopersicon esculentum protein kinase. TCTR2 protein. TCTR2.
AADl 0057.1 AFl 10519 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTRlv.
AAD 10056.1 AF 110518 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTR1.
CAA73722.1 Y13273 Lycopersicon esculentum putative protein kinase.
AAD46406.1 AF096250 Lycopersicon esculentum ethylene-responsive protein kinase TCTR1. ER50. serine/threonine kinase; similar to
Arabidopsis thaliana negative regulator ofthe ethylene response pathway encoded by
GenBank Accession Number L08789.
AAG31141.1 AF305911 Oryza sativa
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAG31142.1 AF305912 Hordeum vulgare
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAK30005.1 AY029067 Rosa hybrid cultivar
CTR2 protein kinase.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLK GmRLKl. ^_____^^
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAK16409.1 AF320086 Zea mays serine threonine kinase 1. stkl. expressed in mature tassel.
BAA06538.1 D31737 Nicotiana tabacum protein-serine/threonine kinase.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
CAB54520.1 AJ238845 Brassica napus putative role in cell cycle control. MAP3K epsilon 1 protein kinase. MAP3Kel.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
CAA08995.1 AJO 10091 Brassica napus MAP3K alpha 1 protein kinase. MAP3K alpha 1.
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl .7.
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAB40021.1 AP003021 Oryza sativa putative wall-associated kinase 2. P0503E05.25. contains EST C24950(S 16264).
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/tlireonine kinase. ESI47. induced in roots by salt stiess, osmotic stiess, and
ABA treatment.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
BAB40010.1 AP003021 Oryza sativa putative wall-associated kinase 2. P0503E05.12.
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml .
AAC23542.1 U20948 Ipomoea tiifida receptor protein kinase. IRKl .
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
BAB 18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
BAA92836.1 AB032473 Brassica oleracea SI 8 S-locus receptor kinase. SRK18.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSJNBa0004Gl 0.30.
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
SEQ ID NO: 692
AAK16172.1 AC079887 Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.14.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitiile-o- glucosyltransferase.
AAF61647.1 AFl 90634 Nicotiana tabacum UDP-glucose: salicylic acid glucosyltransferase. SA-GTase.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltransferase. PHI.
AAF98390.1 AF287143 Brassica napus catalyzes the transfer of glucose from UDP-glucose to sinapate and some other hydroxycinnamates (4-coumarate, caffeate, feralate). UDP-glucose:sinapate glucosyltransferase. SGT1. SGT.
AAD21086.1 AF127218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
BAA93039.1 AB033758 Citras unshiu limonoid UDP-glucosyltransferase. LGTase.
BAA83484.1 AB031274 Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltiansferase. ufgt.
BAA12737.1 D85186 Gentiana triflora UDP-glucose : flavonoid-3 -glucosyltransferase.
CAA54612.1 X77462 Manihot esculenta UTP-glucose glucosyltransferase. CGT5.
BAA19155.1 AB000623 Nicotiana tabacum glucosyl transferase. JIGT.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltiansferase. PGT8.
BAB 17060.1 AP002523 Oryza sativa putative glucosyl transferase. P0013F10.6.
BAA90787.1 AB038248 Ipomoea batatas
UDP glucose: flavonoid 3-O-glucosyltransferase. uf3gt.
AAK16178.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.5.
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS5a.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS 10a.
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid: glucosyltiansferase 1. togtl. glucosyltiansferase.
AAK28304.1 AF346432 Nicotiana tabacum phenylρropanoid:glucosyltransferase 2. togt2. glucosyltransferase.
BAB17061.1 AP002523 Oryza sativa putative glucosyl transferase. P0013F 10.7. contains EST C73149(E2992).
CAA30761.1 X07940 Zea mays
UDPglucose flavonoid glycosyl-tiansferase. Bz-McC.
AAK16410.1 AF320086 Zea mays UDPG-flavonoid 3-O-glucosyl transferase. bz.
CAA31855.1 X13500 Zea mays UDPglucose:flavonol 3-O-glucosyltiansferase.
BAB17059.1 AP002523 Oryza sativa putative glucosyl transferase. P0013F10.5.
BAA36421.1 AB013596 Perilla frutescens UDP-glucose:anthocysnin 5-O-glucosyltransferase. PF3R4.
AAB86473.1 AF028237 Ipomoea puφurea
UDP glucose: flavonoid 3-O-glucosyltransferase. UF3GT-FL1.
CAA30760.1 X07937 Zea mays
UDPglucose flavonoid glycosyl transferase. Bz-W22.
BAB41021.1 AB047094 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. RuUFGTl .
BAB41019.1 AB047092 Vitis vinifera UDP-glucose.-flavonoid 3-O-glucosyltransferase. ItUFGTl.
BAB41025.1 AB047098 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. F1UFGT1.
AAD55985.1 AF165148 Petunia x hybrida catalyzes the penultimate step of flavonol glucosylgalactoside biosynthesis from UDP- galactose and flavonol aglycones in petunia pollen. UDP-galactose:flavonol 3-O- galactosyltransferase. F3galtase.
BAB41018.1 AB047091 Vitis labrusca x Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. KyUFGT2. The gene originated in one ofthe parents V. vinifera cv. Centennial.
BAB41017.1 AB047090 Vitis labrusca x Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. KyUFGTl. The gene was derived from one ofthe parents V. labrascana cv. Ishiharawase.
AAK16181.1 AC079887 Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.16.
BAB41023.1 AB047096 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT1.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltiansferases.
CAA54614.1 X77464 Manihot esculenta UTP-glucose glucosyltransferase. CGT7.
AAB81682.1 AF000371 Vitis vinifera UDP glucose: flavonoid 3 -o-glucosyltransferase.
AAB81683.1 AF000372 Vitis vinifera UDP glucose.-flavonoid 3 -o-glucosyltransferase.
BAB41024.1 AB047097 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltiansferase. A1UFGT2.
CAA54558.1 X77369 Solanum melongena glycosyl transferase. GT.
BAB41026.1 AB047099 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. F1UFGT2.
BAB41020.1 AB047093 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. ItUFGT2.
BAB41022.1 AB047095 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. RuUFGT2.
BAA19659.1 AB002818 Perilla frutescens flavonoid 3-O-glucosyltransferase. UDP glucose.
AAG25643.1 AF303396 Phaseolus vulgaris UDP-glucosyltransferase HRA25. putative; defense associated.
SEQ ID NO: 694
BAB40923.1 AB059401 Oryza sativa putative selenium binding protein. Os SBP.
SEQ ID NO: 695
AAD26942.1 AFl 19050 Datisca glomerata zinc-finger protein 1. zfpl. DgZFPl.
BAA05079.1 D26086 Petunia x hybrida zinc-finger protein.
AAB39638.1 U68763 Glycine max putative transcription factor. SCOF-1. scof-1. zinc-finger protein.
CAB77055.1 Y18788 Medicago sativa putative TFIIIA (or krappel)-like zinc finger protein.
AAC06243.1 AF053077 Nicotiana tabacum transcription factor, osmotic stress-induced zinc-finger protein, zfp.
BAA05077.1 D26084 Petunia x hybrida zinc-finger DNA binding protein.
BAA05076.1 D26083 Petunia x hybrida zinc-finger DNA binding protein.
AAB53260.1 U76554 Brassica rapa transcription factor, zinc-finger protein- 1. BR42.
BAA05078.1 D26085 Petunia x hybrida zinc-finger DNA binding protein.
AAB53261.1 U76555 Brassica rapa zinc-finger protein BcZFPl. BcZFPl(3-2z).
AAK01713.1 AF332876 Oryza sativa zinc finger transcription factor ZF1.
BAA21920.1 AB006598 Petunia x hybrida ZPT2-11. C2H2 zinc finger protein, 2finger.
BAA21922.1 AB006600 Petunia x hybrida ZPT2-13. C2H2 zinc finger protein, 2finger.
BAA19112.1 AB000453 Petunia x hybrida PEThy;ZPT3-l. Cys(2) His(2) zinc finger protein, 3 fingers.
BAA21927.1 AB006605 Petunia x hybrida ZPT3-3. C2H2 zinc finger protein, 3 finger.
BAA96071.1 AB035133 Petunia x hybrida C2H2 zinc-finger protein ZPT3-3. ZPT3-3.
BAA96070.1 AB035132 Petunia x hybrida C2H2 zinc-finger protein ZPT2-10. PEThy;ZPT2-10.
BAA21919.1 AB006597 Petunia x hybrida ZPT2-10. C2H2 zinc finger protein, 2 finger.
BAA19114.1 AB000455 Petunia x hybrida PEThy;ZPT4-l. Cys(2) His(2) zinc finger protein, 4 fingers.
BAA21921.1 AB006599 Petunia x hybrida ZPT2-12. C2H2 zinc fmger protein, 2 fmger.
BAA21928.1 AB006606 Petunia x hybrida ZPT4-4. C2H2 zinc finger protein, 4 finger.
CAA60828.1 X87374 Pisum sativum putative zinc finger protein.
BAA21925.1 AB006603 Petunia x hybrida ZPT2-8. C2H2 zinc finger protein, 2 finger.
BAA19111.1 AB000452 Petunia x hybrida PEThy;ZPT2-6. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA19926.1 AB000456 Petunia x hybrida PEThy; ZPT4-2. C2H2 zinc finger protein, 4 finger.
BAA21924.1 AB006602 Petunia x hybrida ZPT2-7. C2H2 zinc finger protein, 2fmger.
BAA21923.1 AB006601 Petunia x hybrida ZPT2-14. C2H2 zinc finger protein, 2 finger.
BAA21926.1 AB006604 Petunia x hybrida ZPT2-9. C2H2 zinc finger protein, 2 finger.
BAA19110.1 AB000451 Petunia x hybrida PEThy;ZPT2-5. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA19113.1 AB000454 Petunia x hybrida PEThy;ZPT3-2. Cys(2) His(2) zinc finger protein, 3 fingers.
SEQ ID NO: 702
AAK29419.1 AF348319 Zea mays
TERMINAL EARL tel. RNA-binding protein; 3 putative RRM motifs; similar to Schizosaccharomyces pombe Mei2 and Arabidopsis AMLl; tel cDNA sequence is presented in GenBank Accession Number AF047852.
CAA57551.1 X82030 Phaseolus vulgaris chloroplast RNA binding protein. RNP1.
CAA66479.1 X97905 Vicia faba RNA- or ssDNA-binding protein.
CAC01237.1 AJ292767 Nicotiana plumbaginifolia nuclear RNA binding protein. RNA Binding Protein 45. rbp45.
AAF66823.1 AF190655 Nicotiana tabacum poly(A)-binding protein. PABP.
AAK30205.1 AF349964 Daucus carota poly(A)-binding protein. TranslinlP.
AAG59664.1 AC084319 Oryza sativa putative RNA binding protein. OSJNBa0004B24.1.
AAB38974.1 U81318 Triticum aestivum poly(A)-binding protein, wheatpab.
CAA81127.1 Z26042 Anemia phyllitidis
Binding to the poly(A)-tail of eukaryotic mRNAs. poly(A)-mRNA binding protein.
AAA79045.1 U34742 Spinacia oleracea
24 kDa RNA binding protein.
AAC39368.1 AF043297 Chlamydomonas reinhardtii poly(A) binding protein RB47. PABP.
AAF63202.1 AF240679 Cucumis sativus poly(A)-binding protein.
CAAl 1894.1 AJ224325 Hordeum vulgare nucleic acid-binding protein. cp33Hv.
AAF66825.1 AFl 90657 Nicotiana tabacum poly(A)-binding protein. PABP.
CAA06469.1 AJ005286 Hordeum vulgare nucleic acid-binding protein. cp31AHv protein.
SEQ ID NO: 709
AAB36546.1 U77940 Phaseolus vulgaris polyubiquitin. expression is regulated by heavy metal stress, UV, virus infection, heat shock and wounding treatment, this suggests that ubiquitin plays a role in plant defense.
SEQ ID NO: 714
CAA61946.1 X89828 Pisum sativum fructose- 1,6-bisphosphate aldolase.
AAB61592.1 AF003124 Mesembryanthemum crystallinum fructose-biphosphate aldolase.
CAB77243.2 AJ133146 Persea americana glycolytic enzyme, sixth step in glycolysis. fiuctose-bisphosphate aldolase. alf.
BAA08845.1 D50307 Oryza sativa aldolase C-1.
BAA08830.1 D50301 Oryza sativa aldolase C-1.
CAA46649.1 X65742 Spinacia oleracea fiuctose-bisphosphate aldolase.
BAA02729.1 D13512 Oryza sativa cytoplasmic aldolase.
AAG21429.1 AF308587 Fragaria x ananassa cytosolic aldolase. SCA1.
CAA37290.1 X53130 Oryza sativa fructose-diphosphate aldolase (AA 1-358).
CAA31366.1 X12872 Zea mays fructose bisphosphate aldolase.
AAA33435.1 M16220 Zea mays aldolase.
CAA61947.1 X89829 Pisum sativum fructose- 1 ,6-bisphosphate aldolase.
CAA06308.1 AJ005041 Cicer arietinum cytosolic fructose- 1,6-bisphosphate aldolase.
AAK19324.1 AF329673 Dunaliella salina fructose-bisphosphate aldolase isoenzyme 1. salt-induced.
CAA71408.1 Y10380 Solanum tuberosum homologous to plastidic aldolases.
BAA77603.1 AB027002 Nicotiana paniculata plastidic aldolase.
BAA77604.1 AB027001 Nicotiana paniculata plastidic aldolase NPALDP1. NpAldPl.
AAA33642.1 M97476 Pisum sativum aldolase.
AAA33643.1 M97477 Pisum sativum aldolase.
BAA02730.1 D13513 Oryza sativa chloroplastic aldolase.
AAF74220.1 AF216582 Avena sativa fructose 1,6-bisphosphate aldolase precursor.
CAA47293.1 X66814 Spinacia oleracea fructose-bisphosphate aldolase.
AAK19325.1 AF329674 Dunaliella salina fructose-bisphosphate aldolase isoenzyme 2. salt-induced.
CAB46520.1 AJ243524 Phleum pratense sixth step in glycolysis. putative fructose-bisphosphate aldolase.
CAA09669.1 AJOl 1516 Scherffelia dubia fructose-bisphosphate aldolase.
CAA49590.1 X69969 Chlamydomonas reinhardtii fructose-bisphosphate aldolase. ALDCHL.
AAC60574.1 S72951 Chloroplast Chlamydomonas reinhardtii fructosediphophate aldolase. fructosediphophate aldolase. This sequence comes from Fig. 4.
AAB70542.1 AFO 17362 Oryza sativa aldolase.
BAA76430.1 AB025002 Cicer arietinum fructose-bisphosphate aldolase.
AAD20818.1 AF107590 Dendrobium grex Madame Thong-In putative fructose-bisphosphate aldolase. otgll.
BAA11395.1 D78500 Brassica rapa putative aldolase. Sequence homologous to cytoplasmic aldolase of rice (D13512).
BAA78593.1 AU066535 Chlamydomonas sp. HS-5 fructose-bisphosphate aldolase precursor. NaCl inducible.
CAC34412.1 Y18576 Flaveria trinervia fructose-bisphosphate aldolase. alf.
SEQ ID NO: 715
AAK07429.1 AF321287 Musa acuminata beta-glucosidase.
AAC69619.1 AF072736 Pinus contorta beta-glucosidase.
AAF04007.1 AFl 63097 Dalbergia cochinchinensis beta-fucosidase beta-glucosidase. dalcochinin 8'-O-beta-glucoside beta-glucosidase precursor.
BGLU1. rotenoid beta-glucosidase. ___^
AAA93032.1 U50201 Prunus serotina hydrolysis ofthe cyanogenic glucoside (R)-pranasin. prunasin hydrolase precursor, located in protein bodies of Prunus seeds; encodes 8 putative N-glycosylation sites (N-X-S/T); encodes
NEP and ITENG motifs characteristic ofthe BGA family of beta-glucosidases.
BAA78708.1 AB003089 Polygonum tinctorium beta-glucosidase.
AAA91166.1 U39228 Primus avium beta-glucosidase.
BAA11831.1 D83177 Costus speciosus saponin metabolite, furostanol glycoside 26-O-beta-glucosidase (F26G). functional expression in E. coli; one ofthe F26G isozymes.
AAF34650.1 AF221526 Prunus serotina hydrolysis ofthe cyanogenic glucoside (R)-prunasrn. prunasin hydrolase isoform PHA precursor, beta-glucosidase; contains 6 potential N-glycosylation sites (N-X-S/T); glycosyl hydrolase family 1 member.
AAG25897.1 AF170087 Cucurbita pepo silverleaf whitefly-induced protein 3. SLW3. similar to beta-glucosidase.
AAG00614.1 AF293849 Secale cereale beta-glucosidase.
AAB22162.1 S35175 Manihot esculenta linamarase. linamarase. beta-glucosidase; Method: conceptual translation with partial peptide sequencing; This sequence comes from Fig. 4.
AAF03675.1 AF149311 Rauvolfia seφentina hydrolyses the glucoalkaloid raucaffricine. raucaffricine-O-beta-D-glucosidase. beta glucosidase; RG; part ofthe Ajmaline biosynthesis pathway; belongs to family 1 ofthe glucosyl hydrolases.
AAA87339.1 L41869 Hordeum vulgare beta-glucosidase. BGQ60. expression specific to starchy endosperm of seed.
AAC49177.1 U33817 Sorghum bicolor beta-glucosidase, catalyzes the hydrolysis ofthe cyanogenic beta-glucoside dhunin. dhuninase.
CAA64442.1 X94986 Manihot esculenta beta glucosidase. bglA.
AAD02839.1 AF082991 Avena sativa beta-D-glucosidase beta subunit precursor. P60b. avenacosidase.
AAD09850.1 U44087 Zea mays beta-D-glucosidase precursor. glu2. product subunit stracture: autodimer of 58.4 kDa monomers Allele: glu2-B73.
AAD10503.1 U33816 Zeamays functions in defense of young plant parts against pests via the production of hydroxamic acids from hydroxamic acid glucosides. beta-D-glucosidase.
AAB03266.1 U44773 Zea mays beta-D-glucosidase. glul. autodimer of 58.4 kD monomers.
AAA65946.1 U25157 Zeamays functions in defense of young plant parts against pests via the production of hydroxamic acids from hydroxamic acid glucosides. beta-D-glucosidase.
CAA52293.1 X74217 Zea mays beta-glucosidase. p60.1. ^
AAF28800.1 AFl 12888 Catharanthus roseus plays a role in secondary metabolism by hydrolyzing strictosidine to cathenamine during indole alkaloid biosynthesis, strictosidine beta-glucosidase. localized in the endoplasmic reticulum.
CAA40057.1 X56733 Trifolium repens beta-glucosidase. Li.
CAA40058.1 X56734 Trifolium repens beta-glucosidase. non-cyanogenic.
CAA55196.1 X78433 Avena sativa beta-D-glucosidase.
CAA79989.2 Z21977 Brassica napus beta thioglucosidase. myrosinase, thioglucoside glucohydrolase. Myrl.Bnl.
AAB71381.1 U95298 Manihot esculenta linamarase. pLIN-GEN. beta-glucosidase.
CAA57913.1 X82577 Brassica napus beta-glucosidase. bgl.
AAB38784.1 U72154 Brassica nigra beta-glucosidase. psr3.1. PSR3.1; phosphate-starvation responsive enzyme.
AAF34651.1 AF221527 Prunus serotina putative prunasin hydrolase precursor, beta-glucosidase; glycosyl hydrolase family 1 member.
AAA84906.1 U28047 Oryza sativa catalyzes the release of either giberellin or cyanogenic substances from their glucoconjugates. beta glucosidase. beta-D-glucoside glucohydrolase; dimer of 60 kDa monomers; localized in the plastid.
CAC08209.1 AJ005950 Cicer arietinum beta-glucosidase.
SEQ ID NO: 718 AAF61647.1 AFl 90634 Nicotiana tabacum
UDP-glucose: salicylic acid glucosyltransferase. SA-GTase.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltiansferase. PHI.
BAA93039.1 AB033758 Citras unshiu limonoid UDP-glucosyltransferase. LGTase.
BAA36423.1 AB013598 Verbena x hybrida
UDP-glucose: anthocyanin 5-O-glucosyltiansferase. HGT8.
BAA36421.1 AB013596 Perilla frutescens
UDP-glucose :anthocysnin 5-O-glucosyltransferase. PF3R4.
AAF98390.1 AF287143 Brassica napus catalyzes the tiansfer of glucose from UDP-glucose to sinapate and some other hydroxycinnamates (4-coumarate, caffeate, feralate). UDP-glucose:sinapate glucosyltransferase. SGT1. SGT.
BAA36422.1 AB013597 Perilla frutescens
UDP-glucose:anthocyanin 5-O-glucosyltiansferase homologue. PF3R6.
BAB07962.1 AP002524 Oryza sativa putative anthocyanin 5-O-glucosyltiansferase. P0406H10.16. contains ESTs
AU067881(C10481),AU067882(C10481).
AAD21086.1 AF127218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
AAK16178.1 AC079887 Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.5.
BAA12737.1 D85186 Gentiana tiiflora UDP-glucose:flavonoid-3-glucosyltransferase.
AAK16181.1 AC079887 Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.16.
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
AAK16172.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltiansferases.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltiansferase. UDP-glucose:p-hydroxymandelonitiile-o- glucosyltransferase.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltransferase. PGT8.
AAB81683.1 AF000372 Vitis vinifera UDP glucose: flavonoid 3-o-glucosyltiansferase.
BAB41017.1 AB047090 Vitis labrusca x Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. KyUFGTl. The gene was derived from one ofthe parents V. labrascana cv. Ishiharawase.
AAB81682.1 AF000371 Vitis vinifera UDP glucose .-flavonoid 3-o-glucosyltiansferase.
BAB41022.1 AB047095 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. RuUFGT2.
BAB41020.1 AB047093 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltiansferase. ItUFGT2.
BAB41021.1 AB047094 Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase. RuUFGTl.
BAB41019.1 AB047092 Vitis vinifera UDP-glucose:flavonoid 3 -O-glucosyltransferase. ItUFGTl.
BAA83484.1 AB031274 Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltiansferase. ufgt.
BAB41025.1 AB047098 Vitis vinifera UDP-glucose:flavonoid 3 -O-glucosyltiansferase. FlUFGTl.
BAB41023.1 AB047096 Vitis vinifera UDP-glucose.-flavonoid 3-O-glucosyltiansferase. A1UFGT1.
BAA90787.1 AB038248 Ipomoea batatas
UDP glucose: flavonoid 3-O-glucosyltransferase. uf3gt.
BAA19659.1 AB002818 Perilla frutescens flavonoid 3-Q-glucosyltiansferase. UDP glucose.
BAB41018.1 AB047091 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. KyUFGT2. The gene originated in one ofthe parents V. vinifera cv. Centennial.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS 10a.
AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid: glucosyltransferase 2. togt2. glucosyltransferase.
BAB41024.1 AB047097 Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT2.
BAB41026.1 AB047099 Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltiansferase. F1UFGT2.
CAA31855.1 X13500 Zea mays UDPglucose.-flavonol 3-0-glucosyltiansferase.
AAB86473.1 AF028237 Ipomoea puφurea
UDP glucose: flavonoid 3-O-glucosyltiansferase. UF3GT-FL1.
CAA54614.1 X77464 Manihot esculenta UTP-glucose glucosyltransferase. CGT7.
SEQ ID NO: 719
AAC24195.1 AF020425 Nicotiana tabacum calmodulin binding protein, glutamate decarboxylase isozyme 1. NtGADl. calcium- calmodulin-dependent enzyme.
AAK18620.1 AF352732 Nicotiana tabacum converts glutamate to gamma-aminobutyric acid, glutamate decarboxylase isozyme 3. GAD; GAD3; NtGAD3; calcium/calmodulin-dependent enzyme.
AAB40608.1 U54774 Nicotiana tabacum glutamate decarboxylase. NtGADl. cahnodulin regulated enzyme; calmodulin-binding protein.
AAA33710.1 L16977 Petunia x hybrida glutamate decarboxylase. gad.
AAA33709.1 LI 6797 Petunia x hybrida glutamate decarboxylase. gad.
AAC39483.1 AF020424 Nicotiana tabacum glutamate decarboxylase isozyme 2. NtGAD2. calcium-calmodulin-dependent enzyme.
BAB32870.1 AB056062 Oryza sativa glutamate decarboxylase. GAD. ^^
BAB32868.1 AB056060 Oryza sativa glutamate decarboxylase. GAD.
CAA56812.1 X80840 Lycopersicon esculentum homology to pyroxidal-5'-phosphate-dependant glutamate decarboxylases; putative start codon.
BAB32871.1 AB056063 Oryza sativa glutamate decarboxylase. GAD.
BAB32869.1 AB056061 Oryza sativa glutamate decarboxylase. GAD.
CAA50719.1 X71900 Lycopersicon esculentum histidine decarboxylase. hdc. pyridoxal 5'-phosphate dependant.
SEQ ID NO: 720
BAA92713.1 AP001389 Oryza sativa
ESTs AU033035(S1515),D39871(S1515) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome II BAC F22D22 genomic sequence; putative glucan synthase (AC006223).
AAD25952.1 AF085717 Gossypium hirsutum putative callose synthase catalytic subunit. CFL1. cotton FKSl-like protein; similar to
Saccharomyces cerevisiae beta-l,3-glucan synthase subunit FKS1.
SEQ ID NO: 721
CAB55396.1 ALl 17264 Oryza sativa zwhl2.1. similar to Arabidopsis putative UDP-galactase-4-epimerase (AC007060); Method: conceptual translation with partial peptide sequencing.
CAA06339.1 AJ005082 Cyamopsis tetragonoloba UDP-galactose 4-epimerase.
AAA86532.1 U31544 Pisum sativum catabolism of galactose to glucose in Leloir pathway, and in galactose synthesis from glucose.
UDP-galactose-4-epimerase. galE. galactowaldenase.
CAA06338.1 AJ005081 Cyamopsis tetragonoloba UDP-galactose 4-epimerase.
BAB40967.1 AB059568 Pisum sativum biosynthesis of UDP-D-xylose. UDP-D-glucuronate carboxy-lyase. uxsl.
SEQ ID NO: 726
BAA88198.1 AP000837 Oryza sativa
Similar to human dimethylaniline monooxygenase (AC002376).
BAA35120.1 AB008845 Oryza sativa NADH dependent Glutamate Synthase.
AAB41904.1 L37606 Medicago sativa NADH-dependent glutamate synthase.
BAA88195.1 AP000837 Oryza sativa
Similar to human dimethylaniline monooxygenase (AC002376).
AAB46617.1 L01660 Medicago sativa the 3 cysteine residues in this region (amino acid residues 1246-1257) may serve to coordinate the [3Fe-4S] cluster that is probably found in this protein. See Knaff et al. (1991) J. Biol. Chem.266:150. NADH-glutamate synthase. NADH-GOGAT; activity increases dramatically during nodule development; the 3 cysteine residues in this region may serve to coordinate the [3Fe-4S] cluster that is probably found in this protein. See Knaff et al. (1991) J. Biol. Chem.266: 15080- 15084.
SEQ ID NO: 728
AAG43550.1 AF211532 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 132. ACRE132. similar to RING finger proteins.
BAA96875.1 AB045121 Oryza sativa
RING finger l. RRFl.
AAK00436.1 AC060755 Oryza sativa putative zinc finger protein. OSJNBa0003019.23.
BAA78746.1 AB023482 Oryza sativa
Similar to Arabidopsis thaliana RING-H2 fmger protein RHXla mRNA, partial cds.(AF079184).
CAA74911.1 Y14573 Hordeum vulgare ring finger protein, putative.
AAG46117.1 AC073166 Oryza sativa putative ring finger protein. OSJNBb0064P21.7.
BAA85438.1 AP000616 Oryza sativa similar to RING-H2 finger protein RHAla (AF078683).
BAA77204.1 AB026262 Cicer arietinum ring finger protein.
SEQ ID NO: 730
AAA86424.1 U44386 Lycopersicon esculentum heat shock protein. TFHS 1. similar to protein encoded by the arg2 gene in mung bean, encoded by Genbank Accession Number D14411.
SEQ ID NO: 731
CAA06756.1 AJ005899 Nicotiana tabacum
G subunit. G subunit of Vacuolar-type H+- ATPase. vagi.
CAA06757.1 AJ005900 Nicotiana tabacum Subunit G of vacuolar-type H+- ATPase. vag2. vag2.
AAD56039.1 AFl 84068 Citrus limon vacuolar membrane ATPase subunit G. LVMA10. V-ATPase VMA10.
SEQ ID NO: 737
AAF69008.1 AF257779 Oryza sativa stiess-inducible protein. OsSIl.
SEQ ID NO: 739
CAC12996.1 AJ299398 Medicago truncatula putative auxin import, putative AUXl-like permease. Iax2.
AAF21982.1 AFl 15543 Populus tremula x Populus tremuloides
AUXl-like protein. PAX1.
AAG17171.1 AF190880 Populus tremula x Populus tiemuloides putative AUXl-like permease. pax5.
CAC 12995.1 AJ299397 Medicago truncatula putative auxin import, putative AUXl-like permease. laxl.
CAC12997.1 AJ299399 Medicago truncatula putative auxin import, putative AUXl-like permease. Iax3.
CAB65535.1 AJOl 1794 Zeamays AUXl protein, auxl.
SEQ ID NO: 740 CAA65269.1 X96406 Solanum tuberosum 13-lipoxygenase.
AAB65767.1 U37840 Lycopersicon esculentum hpoxygenase. loxD. wound, systemin and methy 1-jasmonate induced. BAA03102.1 D14000 Oryza sativa hpoxygenase. lox2osPil.
AAC12951.1 U56406 Hordeum vulgare methy ljasmonate-inducible hpoxygenase 2.
CAB94852.1 AJ404331 Prunus dulcis hydroperoxydation of polyunsaturated fatty acids, hpoxygenase. lox.
AAG21691.1 AY008278 Lycopersicon esculentum hpoxygenase. oxido-reductase.
AAA79186.1 U36339 Cucumis sativus hpoxygenase.
AAB67858.1 U60200 Solanum tuberosum hpoxygenase. POTLX-1. expressed during early tuberization.
CAA64765.1 X95512 Solanum tuberosum hpoxygenase.
CAB83038.1 AJ271161 Cucumis sativus oxygenase. lipoxygenase-9. lox9. CAA58859.1 X84040 Nicotiana tabacum hpoxygenase. Loxl.
AAB67865.1 U60202 Solanum tuberosum hpoxygenase. POTLX-3. expressed in ABA-treated leaves.
CAA55724.1 X79107 Solanum tuberosum hpoxygenase. Loxl:St:l.
AAB67860.1 U60201 Solanum tuberosum hpoxygenase. POTLX-2. expressed during early tuberization.
CAB65460.1 Y18548 Solanum tuberosum hpoxygenase. loxl-St-2.
AAD04258.1 AF039651 Solanum tuberosum 5-lipoxygenase.
AAB81594.1 AF019613 Solanum tuberosum hpoxygenase. ploxl .
AAA33986.1 J02795 Glycine max lipoxygenase-1.
AAB67732.1 U50075 Glycine max hpoxygenase L-5. vlxB.
CAA47717.1 X67304 Glycine max hpoxygenase.
AAB81595.1 AF019614 Solanum tuberosum hpoxygenase. plox2.
CAA64766.1 X95513 Solanum tuberosum hpoxygenase.
AAB31252.1 S73865 Solanum tuberosum lrnoleate:oxygen oxidoreductase. linoleate:oxygen oxidoreductase, hpoxygenase, LOX. This sequence comes from Fig. 1; hpoxygenase; LOX.
AAA53184.1 U09026 Lycopersicon esculentum hpoxygenase. loxA.
AAB65766.1 U37839 Lycopersicon esculentum hpoxygenase. loxC. expressed during ripening fruit.
CAA65268.1 X96405 Solanum tuberosum 13-lipoxygenase.
AAF15296.2 AF204210 Phaseolus vulgaris hpoxygenase. LOX4.
CAA55319.1 X78581 Pisum sativum hpoxygenase. Loxl:Ps:3.
AAA03728.1 U04526 Glycine max hpoxygenase.
CAA34906.1 X17061 Pisum sativum hpoxygenase (AA 1-864).
CAA55318.1 X78580 Pisum sativum hpoxygenase. Loxl:Ps:2.
BAA03042.1 D13949 Glycine max lacking, lipoxygenase-2. lox2.
AAA33987.1 J03211 Glycine max hpoxygenase (EC 1.13.11.12).
AAB71759.1 U84198 Pisum sativum hpoxygenase. Loxl:Ps:l. < expressed in root nodules.
AAB41272.1 U50081 Glycine max liρoxygenase-3.
CAA39604.1 X56139 Glycine max hpoxygenase. sc514.
AAA96817.1 U26457 Glycine max Hpoxygenase. vlxC.
AAG42354.1 AF234983 Phaseolus vulgaris Hpoxygenase.
AAG18376.1 AF283894 Zantedeschia aethiopica Hpoxygenase. lox2.
AAD39093.1 AF095895 Oryza sativa catalyzes the addition of molecular oxygen to fatty acid, hpoxygenase. CM-LOXl.
CAC04380.1 AJ293015 Pisum sativum Hpoxygenase. loxl:Ps:7.
CAA45088.1 X63525 Phaseolus vulgaris Hpoxygenase. loxA.
AAB 18970.2 U76687 Phaseolus vulgaris Hpoxygenase. PvLOX2.
AAC49159.1 U36191 Glycine max linoleate:oxyg ;en oxidoreductase. Hpoxygenase. lox7.
AAA03726.1 U04785 Glycine max Hpoxygenase.
CAA45086.1 X63521 Phaseolus vulgaris Hpoxygenase.
SEQ ID NO: 741
AAG28436.1 AF195029 Glycine max plasma membrane Ca2+-ATPase. SCA2.
AAG28435.1 AF195028 Glycine max plasma membrane Ca2+-ATPase. SCA1.
CAA68234.1 X99972 Brassica oleracea cahnodulin-stimulated calcium-ATPase.
AAD31896.1 AF145478 Mesembryanthemum crystallinum calcium ATPase.
BAA90510.2 AP001111 Oryza sativa rice EST AU030811, similar to rice Ca+2-ATPase (U82966).
AADl 1618.1 AF050496 Lycopersicon esculentum Ca2+- ATPase. LCA1B; alternative transcript.
AAD 11617.1 AF050495 Lycopersicon esculentum Ca2+-ATPase. LCA1A; alternative transcript.
AAA34138.1 M96324 Lycopersicon esculentum
The calcium ATPase is a calcium ion pump. Ca2+- ATPase. LCAl .
CAA63790.1 X93592 Dunaliella bioculata P-type ATPase. cal. calcium pumping; CAL
AAF73985.1 AF096871 Zea mays calcium pump . calcium ATPase. capl.
AAB58910.1 U82966 Oryza sativa Ca2+-ATPase.
AAD46188.1 AF156691 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma9.
AAB17186.1 U72148 Lycopersicon esculentum plasma membrane H+-ATPase. LHA4. plasma membrane proton pumping ATPase.
CAA47275.1 X66737 Nicotiana plumbaginifolia plasma membrane H+-ATPase. pma4.
CAA54045.1 X76535 Solanum tuberosum H(+)-transportfng ATPase. PHA2.
AAD46186.1 AF156679 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma6.
CAA59800.1 X85805 Zea mays H(+)-transporting ATPase. MHA-2.
BAA06629.1 D31843 Oryza sativa plasma membrane H+- ATPase. OSA2.
AAB35314.2 S79323 Vicia faba plasma membrane H(+)-ATPase precursor, plasma membrane H(+)-ATPase. This sequence comes from Fig. 1; conceptual translation presented here differs from translation in publication.
CAA59799.1 X85804 Phaseolus vulgaris H(+)-tiansporting ATPase. BHA-1.
AAB41898.1 U84891 Mesembryanthemum crystallinum plasma membrane proton pump. H+-transporting ATPase. PMA. AAA34094.1 M80489 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pmal.
AAB84202.2 AF029256 Kosteletzkya virginica plasma membrane proton ATPase. ATP1.
AAA34052.1 M27888 Nicotiana plumbaginifolia H+-tianslocating ATPase.
AAA34173.1 M60166 Lycopersicon esculentum H+-ATPase. LHAl.
AAA34098.1 M80490 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pma3.
AAF98344.1 AF275745 Lycopersicon esculentum plasma membrane H+- ATPase. LHA2. P-type ion pump.
AAD55399.1 AF179442 Lycopersicon esculentum plasma membrane H+-ATPase isoform LHA2. LHA2.
CAC29436.1 AJ310524 Vicia faba
P-type H+-ATPase. ha5. predominantly expressed in guard cells and flowers.
BAA08134.1 D45189 Zostera marina plasma membrane H+- ATPase. zhal.
BAA01058.1 D10207 Oryza sativa H-ATPase. OSAl.
CAB69824.1 AJ271439 Prunus persica plasma membrane H+ ATPase. PPA1.
CAA54046.1 X76536 Solanum tuberosum H(+)-transporting ATPase. PHA1.
AAB60276.1 U09989 Zea mays H(+)-transporting ATPase. Mhal.
CAB69823.1 AJ271438 Prunus persica plasma membrane H+ ATPase. PPA2.
AAK31799.1 AY029190 Lilium longiflorum plasma membrane H+ ATPase. LILHA1.
BAA37150.1 AB022442 Vicia faba p-type H+-ATPase. VHA2.
CAC29435.1 AJ310523 Vicia faba
P-type H+-ATPase. vha4. predominantly expressed in flowers.
CAB85495.1 AJ132892 Medicago truncatula proton pump. H+-ATPase. hal.
CAB85494.1 AJ132891 Medicago truncatula proton pump. H+- ATPase. hal.
AAD46187.1 AFl 56683 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma8.
AAD29712.1 AF140499 Oryza sativa chloroplast envelope calcium ATPase precursor.
AAK32118.1 AF308816 Hordeum vulgare plasmalemma H+- ATPase 1.
AAG01028.1 AF289025 Cucumis sativus plasma membrane H+- ATPase.
AAF97591.1 AF263917 Lycopersicon esculentum plasma membrane proton ATPase. LHA8.
AAA81348.1 U38965 Vicia faba p-type H+-ATPase. VHA2.
AAA20600.1 U08984 Zea mays plasma-membrane H+ ATPase. Zmpmal.
AAA20601.1 U08985 Zea mays plasma-membrane H+ ATPase. Zmpmal.
AAK32119.1 AF308817 Hordeum vulgare plasmalemma H+- ATPase 2.
SEQ ID NO: 742
AAD02548.1 AF049922 Petunia x hybrida
PGPS/D6. PGPS/D6. ER lumen protein retaining receptor homolog; putative HDEL receptor. SEQ IDNO: 744
AAG13424.1 AC051634 Oryza sativa putaive mitochondrial inner membrane protein. OSJNBb0018B10.5.
AAG46068.1 AC079830 Oryza sativa putative inner mitochondrial membrane protein. OSJNBb0009F04.14. SEQ ID NO: 746
BAA32557.1 AB017159 Daucus carota citrate synthase. DcCS.
AAA82743.1 U19481 Citras maxima synthesis of citrate from oxaloacetate and aceτylCoA. citrate synthase precursor, cit.
AAG28777.1 AF302906 Oryza sativa citrate synthase. similar to putative Oryza sativa citrate synthase in GenBank Accession
Number AC004521.
CAA59008.1 X84226 Nicotiana tabacum citrate synthase. citl.
CAA52976.1 X75082 Solanum tuberosum mitochondrial citrate-synthase. ethanolamine ammonia-lyase.
BAA82390.1 AP000367 Oryza sativa
ESTs C96653(C10531),C96654(C10531),C28571(C61641) conespond to a region ofthe predicted gene.; Similar to citrate synthetase. (AC004521).
CAA59010.1 X84228 Beta vulgaris citrate (si)-synthase. citl . CAA59009.1 X84227 Populus x generosa citrate (si)-synthase. citl. BAA07328.1 D38132 Cucurbita sp. conversion of oxaloacetate to citrate in the glyoxylate cycle, glyoxysomal citrate synthase.
SEQ ID NO: 750
AAA85365.1 L42466 Picea glauca ethylene-fonning enzyme. EFE.
AAC95363.1 AFl 04925 Solanum chacoense
2-oxoglutarate-dependent dioxygenase. SPP2. pollination and fertilization induced gene.
BAA75309.1 AB023790 Ipomoea batatas flavanone 3-hydroxyrase. f3h III.
BAA75308.1 AB023789 Ipomoea batatas flavanone 3-hydroxyrase. f3h II.
BAA75306.1 AB023787 Ipomoea batatas anthocyanidin synthase. ans II.
AAC48922.1 U06047 Vigna radiata 1-aminocyclopropane-l -carboxylate oxidase homolog.
SEQ ID NO: 751
BAB40010.1 AP003021 Oryza sativa putative wall-associated kinase 2. P0503E05.12.
BAA95893.1 AP002071 Oryza sativa
Similar to Arabidopsis thaliana wak4 gene; waU-associated kinase 4. (AJ009695).
BAB40015.1 AP003021 Oryza sativa putative wall-associated kinase 1. P0503E05.18.
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
BAB40021.1 AP003021 Oryza sativa putative wall-associated kinase 2. P0503E05.25. contains EST C24950(S 16264).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
AAB09771.1 U67422 Zea mays
CRINKLY4 precursor. cr4. receptor kinase homolog.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAK11566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhirPtoE. confers resistance to bacterial speck disease.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAB40022.1 AP003021 Oryza sativa putative wall-associated kinase 1. P0503E05.26.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl . Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
BAB 16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAF43496.1 AFl 31222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stiess, osmotic stress, and
ABA treatment.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAF76313.1 AF220603 Lycopersicon esculentum
Pto kinase. LescPth5.
AAB47421.1 U59316 Lycopersicon esculentum serine/tlireonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
AAB47423.1 U59315 Lycopersicon pimpinellifolium serine/threonine protein kinase Pto. Pto. bacterial speck disease resistance in tomato; disease resistance gene.
AAC48914.1 U02271 Lycopersicon pimpinellifolium protein kinase.
AAF76306.1 AF220602 Lycopersicon pimpinellifolium Pto kinase.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAK11567.1 AF318491 Lycopersicon hirsutum Pto-like protein kinase F. LhirPtoF.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl. protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
AAK11569.1 AF318493 Lycopersicon hirsutum Pto-like protein kinase D. LhirPtoD.
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; tiansmembrane domain: 1347-1412; intiacellular kinase domain: from 1413.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
AAK11568.1 AF318492 Lycopersicon hirsutum
Pto-like protein kinase B. LhirPtoB.
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APKl gene for protein tyrosine-serine-threonine kinase.(D12522).
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
AAK31267.1 AC079890 Oryza sativa putative protein kinase. OSJNBb0089A17.2.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl.
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
SEQ ID NO: 752
BAB39155.1 AB048713 Pisum sativum SCARECROW. PsSCR.
AAG13663.1 AF263457 Zea mays transcription factor. SCARECROW. SCR. ZmSCR.
BAA90816.1 AP001168 Oryza sativa Similar to SCARECROW (U62798).
AAC98090.1 AF067400 Zeamays
Sell protein. Sell. Scarecrow-like; similar to Zea mays sequence presented in GenBank
Accession Number T 18310.
BAB39156.1 AB048714 Pisum sativum SCARECROW. PsSCR.
SEQ ID NO: 753
AAF21901.1 AF109392 Brassica napus ligand gated channel-like protein, glutamate receptor homolog.
SEQ ID NO: 757
CAA92821.1 Z68504 Oryza sativa 3 -hydroxy-3 -methylglutaryl-CoA reductase.
AAA33360.1 M74800 Hevea brasiliensis
3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. hmgr3.
CAA70440.1 Y09238 Zeamays
3 -hydroxy-3 -methylglutaryl coenzyme A reductase.
AAB69727.1 U72146 Camptotheca acuminata
3 -hydroxy-3 -methylglutaryl coenzyme A reductase. converts HMGCoA in mevalonate precursor for isoprenoid compounds; HMGR; hmg2; similar to protein encoded by GenBank Accession Number L10390.
AAD08820.1 U43961 Oryza sativa
3 -hydroxy-3 -methy lglutaryl=CoA reductase. HMGR.
AAA68965.1 U14624 Artemisia annua
3 -hydroxy-3 -methylglutaryl coenzyme A reductase. AHM1.
AAD47596.1 AF142473 Artemisia annua HMG-CoA reductase. HMGR1.
AAD03789.1 U43711 Moras alba catalyzes the final step in mevalonate pathway. 3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. HMGR.
AAA33108.1 M96068 Catharanthus roseus hydroxymethylglutaryl-CoA reductase. HMGR.
CAA48610.1 X68651 Raphanus sativus hydroxymethylglutaryl-CoA reductase (NADPH). HMGL
AAA93498.1 L01400 Solanum tuberosum convert HMG-CoA into mevalonate. hydroxymethylglutaryl coenzyme A reductase. hmgr. putative.
AAA68966.1 U14625 Artemisia annua
3 -hydroxy-3 -methylglutaryl coenzyme A reductase. AHM4.
AAB52552.1 U51986 Solanum tuberosum HMG-CoA reductase.
CAA48611.1 X68652 Raphanus sativus hydroxymethylglutaryl-CoA reductase (NADPH). HMG2.
BAA93631.1 AB022690 Solanum tuberosum
3 -hydroxy-3 -methylglutaryl coenzyme A reductase. HMG3.
AAB62581.1 U68072 Lycopersicon esculentum 3 -hydroxy-3 -methylglutaryl CoA reductase 2. HMG2.
AAA34169.1 M63642 Lycopersicon esculentum 3 -hydroxy-3 -methylglutaryl coenzyme A reductase. hmg2.
AAB69726.1 U72145 Camptotheca acuminata converts HMGCoA to mevalonate. 3 -hydroxy-3 -methylglutaryl coenzyme a reductase. hmg3.
HMGR.
AAB52551.1 U51985 Solanum tuberosum HMG-CoA reductase.
AAA33040.1 LI 0390 Camptotheca acuminata 3 -hydroxy-3 -methylglutaryl coA reductase.
AAD28179.1 AFl 10383 Capsicum annuum
3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. HMGR2M. HMGR.
BAB20771.1 AB041031 Solanum tuberosum
3 -hydroxy-3 -methylglutaryl coenzyme A reductase. HMG2.
AAB53748.1 U95816 Oryza sativa
3 -hydroxy-3 -methy lglutaryl-Co A reductase. HMGR.
AAC05088.1 AF038045 Gossypium hirsutum catalyzes the synthesis of mevalonate from hmg-coA. 3-hydroxy-3-methylglutaryl-coenzyme
A reductase 1. hmgl. hmg-coA reductase 1; HMGR1.
CAA45181.1 X63649 Nicotiana sylvestris catalyses synthesis of mevalonate. 3 -hydroxy-3 -methylglutaryl-coenzyme A reductase.
HMGR. endoplasmic reticulum location.
CAA38469.1 X54659 Hevea brasiliensis hydroxymethylglutaryl-CoA reductase. HMGR1.
CAA38467.1 X54657 Hevea brasiliensis hydroxymethylglutaryl-CoA reductase. HMGR1.
AAD38873.1 AFl 10382 Oryza sativa
3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. HMGR3G. HMG-CoA reductase.
BAA36291.1 AB021862 Cucumis melo
HMG-CoA reductase. Cm-HMGR. putative.
AAB87727.1 U60452 Nicotiana tabacum hydroxy-methylglutaryl-coe zyme A reductase. HMGR1.
AAC05089.1 AF038046 Gossypium hirsutum catalyzes the synthesis of mevalonate from hmg-coA. 3-hydroxy-3-methylglutaryl-coer_zyme
A reductase 2. hmg2. hmg-coA reductase 2; HMGR2.
AAC15475.1 AF034760 Tagetes erecta
3 -hydroxy-3 -methylglutaryl coenzyme A reductase.
AAC15476.1 AF034761 Tagetes erecta
3 -hydroxy-3 -methylglutaryl coenzyme A reductase.
AAA21720.1 L28995 Oryza sativa conversion of hydroxymethylglutaryl coenzyme A to mevalonate. 3 -hydroxy-3 -methylglutaryl coenzyme A reductase. putative.
AAA33358.1 M74798 Hevea brasiliensis
3 -hydroxy-3 -methylglutaryl-coenzyme A reductase. hmgr3.
AAC72378.1 AF096838 Solanum tuberosum 3 -hydroxy-3 -methylglutaryl coenzyme A reductase.
AAB04043.1 L40938 Lycopersicon esculentum HMGR CoA reductase. HMGR1.
CAA38468.1 X54658 Hevea brasiliensis hydroxymethylglutaryl-CoA reductase. HMGR2.
CAA52787.1 X74783 Lithospennum erythrorhizon
3 -hydroxy-3 -methyl glutaryl coenzyme A reductase. Lehmgrl.
AAD09278.1 U97683 Glycine max catalyzes the synthesis of mevalonate, the specific precursor of all isoprenoid compounds present in plants. 3-hydroxy-3-methylglutaryl-coenzyme A reductase. HMG-CoA reductase.
AAG43469.1 AF196964 Bixa orellana catalyzes mevalonate synthesis from hmg-CoA. 3 -hydroxy-3 -methylglutaryl-coenzyme A reductase.
BAA09705.1 D63389 Cucumis sativus 3 -hydroxy-3 -methylglutaryl CoA reductase.
AAB47161.1 S82272 Gossypium barbadense
3 -hydroxy-3 -methylglutaryl coenzyme A reductase. /gene="3-hydroxy-3-methylglutaryl coenzyme A reductase,. This sequence comes from Fig. 2; 3 -hydroxy-3 -methylglutaryl CoA reductase; HMGR.
AAA33359.1 M74799 Hevea brasiliensis
3-hydroxy-3-methylglutaryl-coenzyme A reductase. hmgr3.
AAC37434.1 L34827 Solanum tuberosum HMG-CoA reductase. hmgl gene family.
AAC37432.1 L34825 Solanum tuberosum HMG-CoA reductase. hmgl gene family.
AAC37435.1 L34828 Solanum tuberosum HMG-CoA reductase. hmgl gene family.
AAC37433.1 L34826 Solanum tuberosum HMG-CoA reductase. hmgl gene family^
AAC37431.1 L34823 Solanum tuberosum HMG-CoA reductase. hmgl gene family.
SEQ ID NO: 761
CAA96512.1 Z71980 Malus x domestica knottedl-like homeobox protein.
BAA25921.1 AB004797 Nicotiana tabacum NTH23. homeobox gene.
AAD09582.1 U76409 Lycopersicon esculentum homeobox 1 protein. THoxl . homeodomain protein.
AAC49918.1 AF000142 Lycopersicon esculentum class II knotted-like homeodomain protein. LeT12.
BAA08552.1 D49704 Oryza sativa OSH45. OSH44 transcript homeobox gene.
BAB 18585.1 AB043957 Ceratopteris richardii CRKNOX3. crknox3. knottedl-like homeodomain protein.
AAD00253.1 U76410 Lycopersicon esculentum homeobox 2 protein. THox2. homeodomain protein.
CAA82314.1 Z29073 Brassica napus homeodomain-containing protein. Bnhdl.
BAA77822.1 AB007628 Oryza sativa HOS59. homeobox gene.
BAA77823.1 AB007629 Oryza sativa HOS66. homeobox gene.
AAB81079.1 AF022390 Hordeum vulgare knotted class 1 homeodomain protein, k. similar to the hooded gene product encoded by GenBank Accession Number X83518; similar to the maize knotted-1 gene product encoded by GenBank Accession Number X61308.
AAF32399.1 AF224499 Triticum aestivum KNOTTED-1-like homeobox protein b. knoxlb. KNOXlb.
BAA76750.1 AB025573 Nicotiana tabacum KNl-type homeobox protein. NTH1.
AAF32400.1 AF224500 Triticum aestivum KNOTTED-1-like homeobox protein d. knoxld. KNOXld.
AAC49917.1 AF000141 Lycopersicon esculentum class I knotted-like homeodomain protein. LeT6.
BAA25546.1 AB004785 Nicotiana tabacum NTH15. homeobox gene.
AAF32398.1 AF224498 Triticum aestivum KNOTTED-1 -like homeobox protein a. knoxla. KNOX1.
AAD13611.1 AF100455 Zeamays knotted class 1 homeodomain protein Hguleless3. Ig3.
AAC84001.1 AF063248 Picea abies homeobox protein.
AAC33008.1 AF080104 Pisum sativum knottedl-like class I homeodomain protein. PsKnl.
AAD00692.1 U90092 Picea mariana homeobox transcription factor SKN2. knottedl-like homeobox gene.
AAC32262.1 AF063307 Pisum sativum Knox class 1 protein. Hopl.
BAA03959.1 D16507 Oryza sativa homeobox protein. OSH1.
AAG27464.1 AF308454 Medicago truncatula knotted class I homeodomain KNOX.
AAD00691.1 U90091 Pice mariana homeobox transcription factor SKN knottedl-like homeobox gene.
AAC32817.1 AF050180 Oryza sativa transcription factor. KNOX class homeodomain protein. Oskn2.
BAA79226.1 AB028885 Oryza sativa knottedl-type homeobox protein OSH71. OSH71.
BAA79224.1 AB028883 Oryza sativa knottedl-type homeobox protein OSH6. OSH6.
BAA77818.1 AB007624 Oryza sativa HOS9. homeobox gene.
BAB19772.1 AP002881 Oryza sativa putative knottedl-type homeobox protein. P0035H10.13.
AAF23753.2 AF193813 Brassica oleracea shoot meristemless. Stm. homeodomain protein.
AAD00251.1 U76407 Lycopersicon esculentum knotted 2 protein. TKn2. homeodomain protein.
CAA96510.1 Z71978 Malus x domestica knl-like protein.
BAA31688.1 AB016071 Oryza sativa OSH15. homeobox gene.
BAA77817.1 AB007623 Oryza sativa HOS3. homeobox gene.
BAB 18582.1 AB043954 Ceratopteris richardii CRKNOX 1. crknox 1. knottedl-like homeodomain protein.
BAA76903.1 AB025713 Nicotiana tabacum homeobox 9. NTH9.
AAD00252.1 U76408 Lycopersicon esculentum knotted 3 protein. TKn3. homeodomain protein.
BAB 18584.1 AB043956 Ceratopteris richardii
CRKNOX2. crknox2. knottedl-like homeodomain protein.
CAA96511.1 Z71979 Malus x domestica knl-like protein.
AAA20882.1 L13663 Glycine max
SBH1. Sbhl. soybean homeobox-containing gene.
CAB88029.1 AJ276389 Dendrobium grex Madame Thong-In tianscription factor, knottedl-like homeobox protein.
SEQ ID NO: 762 AAB65776.1 U97521 Vitis vinifera class IV endochitinase. VvChi4A.
AAB65777.1 U97522 Vitis vinifera class IV endochitinase. VvChi4B. CAA53626.1 X76041 Triticum aestivum endochitinase. CHI.
AAG53609.1 AF280437 Secale cereale
31.7 kDa class I endochitinase-antifreeze protein precursor. cht9.
BAA03750.1 D16222 Oryza sativa endochitinase. Cht-2.
AAA32986.1 M95835 Brassica napus endochitinase. Ch25.
BAA03749.1 D16221 Oryza sativa endochitinase. Cht-1.
AAB01895.1 U48687 Castanea sativa endochitinase.
SEQ ID NO: 763 AAA34065.1 M94135 Nicotiana tabacum chloroplast carbonic anhydrase.
AAA34057.1 LI 9255 Nicotiana tabacum carbonic anhydrase.
AAB65822.1 U55838 Populus tremula x Populus tremuloides carbonic anhydrase. CAlb. EC 4.2.1.1.
AAC49785.1 U55837 Populus tremula x Populus tiemuloides carbonic anhydrase. CAla. EC 4.2.1.1.
AAA34026.1 M27295 Spinacia oleracea carbonic anhydrase precursor. AAA34027.1 J05403 Spinacia oleracea carbonic anhydrase (EC 4.2.1.1).
AAA86993.1 U19738 Flaveria linearis reversible hydration of carbon dioxide, carbonic anhydrase 1.
AAA86942.1 U08402 Flaveria brownii carbonic anhydrase.
AAA86992.1 U19737 Flaveria pringlei reversible hydration of carbon dioxide, carbonic anhydrase.
AAA86939.1 U08398 Flaveria bidentis carbonic anhydrase.
AAD27876.2 AF139464 Vigna radiata carbonic anhydrase. CipCal.
AAA33652.1 M63627 Pisum sativum carbonic anhydrase.
AAD29050.1 AFl 32855 Gossypium hirsutum interconversion of CO2 and HCO3-. carbonic anhydrase isoform 2. CA2. zinc metalloenzyme; carbonate dehydratase.
AAD29049.1 AF132854 Gossypium hirsutum interconversion of CO2 and HCO3-. carbonic anhydrase isoform 1. CAL zinc metalloenzyme; carbonate dehydratase.
AAA86994.1 U19740 Flaveria linearis reversible hydration of carbon dioxide, carbonic anhydrase 2.
CAB43571.1 AJ239132 Glycine max hydration of carbon dioxide, carbonic anhydrase. cal.
CAA63712.1 X93312 Medicago sativa Carbonic anhydrase. cal .
AAA86945.1 U08403 Zea mays carbonic anhydrase.
AAA86944.1 U08401 Zea mays carbonic anhydrase.
AAC41656.1 L36959 Hordeum vulgare carbonic anhydrase. putative.
AAA86943.1 U08404 Oryza sativa carbonic anhydrase. nuclear encoded, localized to chloroplast.
AAD56038.1 AF182806 Oryza sativa carbonic anhydrase 3. ca3.
BAA31953.1 AB016283 Oryza sativa carbonic anhydrase.
AAA69027.1 U19739 Urochloa panicoides reversible hydration of carbon dioxide, carbonic anhydrase 2.
AAA69028.1 U19741 Urochloa panicoides reversible hydration of carbon dioxide, carbonic anhydrase 1.
BAA95793.1 AB009887 Nicotiana tabacum carbonic anhydrase. carbonic anhydrase.
AAF78507.1 AF195204 Pyras pyrifolia carbonic anhydrase isoform 1. CAL
AAC33484.1 U49976 Coccomyxa sp. PA beta-type carbonic anhydrase beta-CAl.
AAAI 8560.1 M95073 Zeamays putative, silimar to carbonic anhydrases.
AAB19184.1 U41190 Chlamydomonas reinhardtii carbonic anhydrase precursor. beta-CA2.
AAB 19183.1 U41189 Chlamydomonas reinhardtii carbonic anhydrase precursor. beta-CAl.
AAC49887.1 U80804 Chlamydomonas reinhardtii beta-carbonic anhydrase. cal. beta-CAl.
AAC49888.1 U80805 Chlamydomonas reinhardtii beta-carbonic anhydrase. ca2. beta-CA2.
SEQ ID NO: 764
AAC06027.1 AF052058 Vigna unguiculata iron storage and mobilization in plants, ferritin subunit cowpea2 precursor.
AAD50644.1 AF133814 Solanum tuberosum fenitin 1. Fl.
AAB53099.1 U68217 Brassica napus iron binding protein, ferritin. LSC30.
AAA33959.1 M64337 Glycine max ferritin light chain, fenitin.
AAA34016.1 M72894 Glycine max fenitin light chain. SOF-H2.
CAA58146.1 X83076 Zea mays fenitin. Ferl .
AAB 18928.1 U31648 Glycine max iron storage protein, ferritin.
CAA65771.1 X97059 Medicago sativa iron storage, ferritin. FER. abcissic acid regulated.
CAA43663.1 X61391 Zea mays fenitin.
CAA58147.1 X83077 Zeamays fenitin. Fer2.
AAC06026.1 AF052057 Vigna unguiculata iron storage and mobilization in plants, fenitin subunit cowpea3 precursor.
CAA43664.1 X61392 Zea mays ferritin.
CAA51786.1 X73369 Pisum sativum fenitin.
CAA45763.1 X64417 Pisum sativum fenitin-precursor.
CAA41213.1 X58274 Phaseolus vulgaris ferritin. pfe.
AAA33958.1 M58336 Glycine max ferritin light chain. SOF-5L.
CAB42587.1 AJ238628 Chlorella protothecoides putative ferritin. dee 188.
BAB17852.1 AB042612 Nicotiana tabacum fenitin 1. tob-fer-1. putative.
AAC15241.1 AF028072 Pinus taeda fenitin.
CAA47983.1 X67755 Vigna unguiculata fenitin 2. pfe2.
CAA47982.1 X67754 Vigna unguiculata ferritin 1. pfe 1.
CAA47984.1 X67756 Vigna unguiculata ferritin 5. pfe5.
AAC12282.1 AF052511 Glycine max iron storage and mobilization, fenitin 2. soybean 2; cowpea 2 homolog.
AAC12281.1 AF052513 Glycine max iron storage and mobilization, ferritin 1. soybean 1; cowpea 1 homolog.
SEQ ID NO: 765 ~
AAC36697.1 AF075579 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
CAC10358.1 AJ277086 Nicotiana tabacum protein phosphatase 2C. PP2C1.
CAB90633.1 AJ277743 Fagus sylvatica protein phpsphatase 2C (PP2C). pp2Cl. ABA-induced protein.
AAD17804.1 AF092431 Lotus japonicus nodule-enhanced protein phosphatase type 2C. NPP2C1.
CAC10359.1 AJ277087 Nicotiana tabacum protein phosphatase 2C. PP2C2.
CAC09575.1 AJ298987 Fagus sylvatica protein phosphatase 2C (PP2C). pp2Cfl.
CAA72341.1 Y11607 Medicago sativa protein phosphatase 2C. MP2C.
CAB61839.1 AJ242803 Sporobolus stapfianus putative serine/threonine phosphatase type 2c.
AAD17805.1 AF092432 Lotus japonicus protein phosphatase type 2C. PP2C2.
AAG43835.1 AF213455 Zeamays protein phosphatase type-2C. pp2c-l . PP2C-1.
AAG46118.1 AC073166 Oryza sativa putative protein phosρhatase-2C. OSJNBb0064P21.12. tRNA-Lys.
AAG13599.1 AC051633 Oryza sativa putative protein phosphatase-2C. OSJNBbOOl 5111.26.
AAC36698.1 AF075580 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
BAB12036.1 AP002820 Oryza sativa putative protein phosphatase. P0702D12.18.
AAC36700.1 AF075582 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
AAC36699.1 AF075581 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
AAK20060.1 AC025783 Oryza sativa putative protein phosphatase 2C. OSJNBa0001O14.1.
AADl 1430.1 AF097667 Mesembryanthemum crystallinum protein phosphatase 2C homolog. PP2C.
CAB90634.1 AJ277744 Fagus sylvatica protein phosphatase 2C (PP2C). pp2C2. ABA and calcium induced protein.
AAC35951.1 AF079355 Mesembryanthemum crystallinum protein phosphatase-2c. PP2C.
AAF19804.1 AFl 80355 Brassica oleracea
ABU protein. ABU, similar to Arabidopsis thaliana ABIL
AAB93832.1 U81960 Zea mays kinase interaction domain mediates phosphorylation-dependent binding to a subset of active receptor-like protein kinases; type 2C protein phosphatase. kinase associated protein phosphatase. KAPP.
AAC26828.1 AF075603 Oryza sativa kinase interaction domain mediates phosphorylation-dependent binding to a subset of active receptor-like protein kinases. kinase associated protein phosphatase. kapp. type 2C protein phosphatase.
CAC09576.1 AJ298988 Fagus sylvatica protein phosphatase 2C (PP2C). pp2Cf2. SEQ ID NO: 766
AAG08959.1 AF122051 Solanum tuberosum tuber-specific and sucrose-responsive element binding factor. TSF. R2R3 MYB class transcription factor.
AAG08960.1 AFl 22052 Solanum tuberosum tuber-specific and sucrose-responsive element binding factor. TSF. R2R3 MYB class transcription factor.
AAG08961.1 AF122053 Solanum tuberosum tuber-specific and sucrose-responsive element binding factor. TSF. R2R3 MYB class transcription factor.
BAA88169.1 AP000836 Oryza sativa
Similar to putative transcription factor (AF062890).
BAA88205.1 AP000837 Oryza sativa
Similar to putative transcription factor (AF062890).
AAF34434.1 AFl 72282 Oryza sativa myb-like protein. DUPRl 1.29.
AAF78890.1 AFl 89788 Hordeum vulgare putative c-myb-like tianscription factor. MYB3R-1. HvMYB3R-l.
AAF78889.1 AF189787 Hordeum vulgare putative c-myb-like tianscription factor. MYB3R-1. HvMYB3R-l.
AAF67053.1 AF190304 Adiantum raddianum c-myb-like tianscription factor. MYB3R-1. contains three MYB repeats.
AAF67052.1 AF190303 Adiantum raddianum c-myb-like tianscription factor. MYB3R-1. contains three MYB repeats.
AAF78888.1 AFl 89786 Physcomitrella patens putative c-myb-like tianscription factor. MYB3R-1. PpMYB3R-l.
AAF78887.1 AFl 89785 Physcomitrella patens putative c-myb-like transcription factor. MYB3R-1. PpMYB3R-l.
AAF43043.1 AF236059 Papaver rhoeas putative Myb-related domain, prnr.
BAA94769.1 AP001859 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4, BAC clone F4D11; putative myb-protein.
(AL022537).
AAF67050.1 AF190301 Secale cereale c-myb-like transcription factor. MYB3R-1. contains three MYB repeats.
AAF67051.1 AF190302 Secale cereale c-myb-like tianscription factor. MYB3R-1. contains three MYB repeats.
CAA78388.1 Z13998 Petunia x hybrida
DNA-binding protein; transcriptional activator, protein 3. myb.Ph3. related to animal myb proto-oncoproteins.
BAB39987.1 AP003020 Oryza sativa putative tianscription factor (myb). P0498A12.16. contains ESTs
AU097474(S5087),D40175(S1959).
BAB39972.1 AP003018 Oryza sativa putative transcription factor (myb). OSJNBa0004B13.27. contains ESTs
AU097474(S5087),D40175(S 1959).
CAA72218.1 Y11415 Oryza sativa myb.
BAA81731.1 AB029160 Glycine max
GmMYB29Al.
BAA81730.1 AB029159 Glycine max GmMYB29Al.
CAA72217.1 Y11414 Oryza sativa myb.
BAA81736.1 AB029165 Glycine max GmMYB29B2.
BAB12688.1 AP002746 Oryza sativa putative MYB family transcription factor. P0671B11.3. contains ESTs AU082307(E0784),C72014(E0784).
BAA99440.1 AP002743 Oryza sativa putative MYB family transcription factor. P0710E05.27. contains ESTs
AU082307(E0784),C72014(E0784). ■
AAB41101.1 U72762 Nicotiana tabacum transcription factor Mybl. mybl. TMV-inducible Myb homolog; contains helix-turn-helix motif; contains redox-sensitive cysteine.
BAA88223.1 AB028651 Nicotiana tabacum myb-related transcription factor LBM3. Ibm3.
BAA23340.1 D88620 Oryza sativa transfactor. OSMYB4. Osmyb4.
BAA93038.1 AP001552 Oryza sativa
EST AU082058(C12976) conesponds to a region ofthe predicted gene. Similar to
Arabidopsis thaliana putative transcription factor (AF062916).
AAA33067.1 L04497 Gossypium hirsutum
MYB A; putative.
BAA81733.2 AB029162 Glycine max
GmMYB29A2.
BAA88222.1 AB028650 Nicotiana tabacum myb-related transcription factor LBM2. Ibm2.
CAB43399.1 AJ006292 Antinhinum majus
Myb-related transcription factor mixta-like 1. mybmll .
BAA81732.1 AB029161 Glycine max
GmMYB29A2.
AAG28525.1 AF198498 Nicotiana tabacum anther-specific myb-related protem 2. mybAS2. N.MYBAS2; contains tandem R2, R3 myb domains similar to c-myb family.
CAA50226.1 X70881 Hordeum vulgare
MybHv33. myb3.
CAA50223.1 X70878 Hordeum vulgare
MybHv33. myb3.
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, tianscriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins.
CAA67000.1 X98355 Oryza sativa activator of alpha-amylase gene promoter, tianscription factor GAMyb. Gaml. Myb-like; expression is regulated by gibberellin.
AAK19616.1 AF336283 Gossypium hirsutum
GHMYB25. ghmyb25. similar to myb; contains an unspliced intron.
AAG28526.1 AFl 98499 Nicotiana tabacum anther-specific myb-related protein 1. ybASl. NtMYBASl; contains N-teiminal R2, R3 myb domain repeats similar to c-myb.
CAA78386.1 Z13996 Petunia x hybrida
DNA binding protein; tianscriptional activator, protein 1. myb.Pb.3. Product related to animal myb proto-oncoproteins. Sequence from nucleotide 992 is not included in clone cPFl and has been obtained by PCR amplification of cDNA.
CAA66952.1 X98308 Lycopersicon esculentum
THM18. myb-related tianscription factor.
BAA23341.1 D88621 Oryza sativa transfactor. OSMYB5. Osmyb5.
CAA50221.1 X70876 Hordeum vulgare
MybHv5. myb2.
AAK19611.1 AF336278 Gossypium hirsutum
BNLGHJ233. bnlghi6233. similar to myb.
AAC04716.1 AF034130 Gossypium hirsutum putative MYB-like transcription factor. MYB-like DNA-binding domain protein. Cmy-D. similar to MYB A encoded by GenBank Accession Number L04497.
CAA61021.1 X87690 Hordeum vulgare transcriptional activator of alpha-amylase gene promoter. GAMyb protein. Gaml.
AAG22863.1 AY008692 Hordeum vulgare transcription factor GAMyb. Gamyb. SEQ ID NO: 767
CAB08111.1 Z94180 Lycopersicon esculentum branched chain alpha-keto acid dehydrogenase El -alpha subunit.
CAA81558.1 Z26949 Solanum tuberosum subunit ofthe mitochondrial pyravate dehydrogenase complex. El alpha subunit of pyruvate dehydrogenase precursor.
AAC72195.1 AF069911 Zeamays pyravate dehydrogenase El alpha subunit.
AAG43499.1 AF209924 Lycopersicon esculentum pyravate dehydrogenase.
AAA97411.1 U51918 Pisum sativum pyravate dehydrogenase El alpha subunit.
CAAl 0992.1 AJ222787 Hordeum vulgare alpha-keto acid dehydrogenase-like protein, homology to branched chain alpha-keto acid dehydrogenase El -alpha subunit. SEQ ID NO: 768
AAF64450.1 AF239928 Euphorbia esula glutathione S-transferase. similar to auxin-inducible GST.
AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-tiansferase T3.
AAG34803.1 AF243368 Glycine max glutathione S-tiansferase GST 13.
AAG16756.1 AY007558 Lycopersicon esculentum putative glutathione S-tiansferase TI.
AAG34796.1 AF243361 Glycine max glutathione S-tiansferase GST 6.
AAG34809.1 AF243374 Glycine max glutathione S-transferase GST 19.
AAG34797.1 AF243362 Glycine max glutathione S-transferase GST 7.
AAG34807.1 AF243372 Glycine max glutathione S-transferase GST 17.
AAG34798.1 AF243363 Glycine max glutathione S-tiansferase GST 8.
AAG34804.1 AF243369 Glycine max glutathione S-tiansferase GST 14.
AAG16759.1 AY007561 Lycopersicon esculentum putative glutathione S-tiansferase T4.
AAG34801.1 AF243366 Glycine max glutathione S-transferase GST 11.
AAG34810.1 AF243375 Glycine max glutathione S-tiansferase GST 20.
AAG16757.1 AY007559 Lycopersicon esculentum putative glutathione S-transferase T2.
AAC18566.1 AF048978 Glycine max 2,4-D inducible glutathione S-transferase. GSTa.
AAG34808.1 AF243373 Glycine max glutathione S-transferase GST 18.
AAG34800.1 AF243365 Glycine max glutathione S-transferase GST 10.
CAA71784.1 Y10820 Glycine max glutathione transferase.
AAG34844.1 AF244701 Zeamays glutathione S-tiansferase GST 36.
AAG32472.1 AF309379 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU3.
AAA68430.1 J03679 Solanum tuberosum glutathione S-tiansferase. gstl. previously called pathogenesis-related protein; prpl-1.
CAA04391.1 AJ000923 Carica papaya glutathione transferase. PGST1.
AAG34831.1 AF244688 Zea mays glutathione S-tiansferase GST 23.
CAA09187.1 AJ010448 Alopecurus myosuroides glutathione transferase. GSTl a.
CAA09188.1 AJO 10449 Alopecurus myosuroides glutathione transferase. GST lb.
AAG34802.1 AF243367 Glycine max glutathione S-tiansferase GST 12.
AAG34805.1 AF243370 Glycine max glutathione S-tiansferase GST 15.
AAG34832.1 AF244689 Zeamays glutathione S-tiansferase GST 24.
AAG34837.1 AF244694 Zea mays glutathione S-transferase GST 29.
AAG34836.1 AF244693 Zea mays glutathione S-transferase GST 28.
AAG32471.1 AF309378 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTU4.
AAG34849.1 AF244706 Zea mays glutathione S-transferase GST 41.
CAC24549.1 AJ296343 Cichorium intybus x Cichorium endivia glutathione S-transferase. chi-GSTl. auxin-induced GST.
AAC32118.1 AF051214 Picea mariana probable glutathione S-tiansferase. Sbl8. similar to Nicotiana tabacum probable glutathione
S-tiansferase encoded by GenBank Accession Number X56266.
AAG34795.1 AF243360 Glycine max glutathione S-transferase GST 5.
AAG34841.1 AF244698 Zeamays glutathione S-tiansferase GST 33.
AAF29773.1 AFl 59229 Gossypium hirsutum glutathione S-tiansferase. GST.
SEQ ID NO: 769
AAG34797.1 AF243362 Glycine max glutathione S-tiansferase GST 7.
AAG34798.1 AF243363 Glycine max glutathione S-tiansferase GST 8.
AAG34803.1 AF243368 Glycine max glutathione S-tiansferase GST 13.
AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-tiansferase T3.
AAF64450.1 AF239928 Euphorbia esula glutathione S-transferase. similar to auxin-inducible GST.
AAG34801.1 AF243366 Glycine max glutathione S-transferase GST 11.
AAG34804.1 AF243369 Glycine max glutathione S-transferase GST 14.
AAG34796.1 AF243361 Glycine max glutathione S-transferase GST 6.
AAG34809.1 AF243374 Glycine max glutathione S-tiansferase GST 19.
AAG16759.1 AY007561 Lycopersicon esculentum putative glutathione S-tiansferase T4.
AAG16757.1 AY007559 Lycopersicon esculentum putative glutathione S-tiansferase T2.
AAG16756.1 AY007558 Lycopersicon esculentum putative glutathione S-transferase TI.
AAG34807.1 AF243372 Glycine max glutathione S-tiansferase GST 17.
AAG34810.1 AF243375 Glycine max glutathione S-tiansferase GST 20.
AAG34844.1 AF244701 Zeamays glutathione S-tiansferase GST 36.
AAG34831.1 AF244688 Zeamays glutathione S-tiansferase GST 23.
AAC18566.1 AF048978 Glycine max 2,4-D inducible glutathione S-transferase. GSTa.
AAG34832.1 AF244689 Zea mays glutathione S-transferase GST 24.
AAG34808.1 AF243373 Glycine max glutathione S-tiansferase GST 18.
AAG34837.1 AF244694 Zeamays glutathione S-tiansferase GST 29.
AAG34800.1 AF243365 Glycine max glutathione S-tiansferase GST 10.
AAG34836.1 AF244693 Zeamays glutathione S-tiansferase GST 28.
CAA04391.1 AJ000923 Carica papaya glutathione transferase. PGST1.
CAA71784.1 Y10820 Glycine max glutathione transferase.
AAG34849.1 AF244706 Zeamays glutathione S-tiansferase GST 41.
AAA68430.1 J03679 Solanum tuberosum glutathione S-tiansferase. gstl. previously called pathogenesis-related protein; prpl-1.
AAG34802.1 AF243367 Glycine max glutathione S-tiansferase GST 12.
CAC24549.1 AJ296343 Cichorium intybus x Cichorium endivia glutathione S-transferase. chi-GSTl. auxin-induced GST.
CAA09187.1 AJO 10448 Alopecurus myosuroides glutathione transferase. GST la.
AAF22518.1 AFl 18925 Papaver somniferam glutathione S-tiansferase 2. GST2.
AAG32471.1 AF309378 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTU4.
CAA09188.1 AJ010449 Alopecurus myosuroides glutathione transferase. GSTlb.
AAF22517.1 AFl 18924 Papaver somniferam glutathione S-tiansferase 1. GST
AAF22647.1 AF193439 Lycopersicon esculentum glutathione S-tiansferase/peroxidase. BI-GST/GPX.
AAG32473.1 AF309380 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU2.
AAG32472.1 AF309379 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU3. SEQ ID NO: 771
AAG46118.1 AC073166 Oryza sativa putative protein phosphatase-2C. OSJNBb0064P21.12. tRNA-Lys.
AAG13599.1 AC051633 Oryza sativa putative protein phosphatase-2C. OSJNBb0015Il 1.26.
BAB 12036.1 AP002820 Oryza sativa putative protein phosphatase. P0702D12.18.
AAC36698.1 AF075580 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
CAA72341.1 Y11607 Medicago sativa protein phosphatase 2C. MP2C.
AAG43835.1 AF213455 Zea mays protein phosphatase type-2C. pp2c-l. PP2C-1.
CAB61839.1 AJ242803 Sporobolus stapfianus putative serine/threonine phosphatase type 2c.
AAD17804.1 AF092431 Lotus japonicus nodule-enhanced protein phosphatase type 2C. NPP2C1.
AAD17805.1 AF092432 Lotus japonicus protein phosphatase type 2C. PP2C2.
AAC36697.1 AF075579 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
CAB90633.1 AJ277743 Fagus sylvatica protein phpsphatase 2C (PP2C). pp2Cl. AB A-induced protein.
CAC 10358.1 AJ277086 Nicotiana tabacum protein phosphatase 2C. PP2C1.
CAC10359.1 AJ277087 Nicotiana tabacum protein phosphatase 2C. PP2C2.
AAC36700.1 AF075582 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
CAC09575.1 AJ298987 Fagus sylvatica protein phosphatase 2C (PP2C). pp2Cfl.
AAK20060.1 AC025783 Oryza sativa putative protein phosphatase 2C. OSJNBa0001O14.1.
CAB90634.1 AJ277744 Fagus sylvatica protein phosphatase 2C (PP2C). pp2C2. ABA and calcium induced protein.
AAC35951.1 AF079355 Mesembryanthemum crystallinum protein phosphatase-2c. PP2C.
AAD 11430.1 AF097667 Mesembryanthemum crystallinum protein phosphatase 2C homolog. PP2C.
AAB93832.1 U81960 Zea mays kinase interaction domain mediates phosphorylation-dependent binding to a subset of active receptor-like protein kinases; type 2C protein phosphatase. kinase associated protein phosphatase. KAPP.
AAC26828.1 AF075603 Oryza sativa kinase interaction domain mediates phosphorylation-dependent binding to a subset of active receptor-like protein kinases. kinase associated protein phosphatase. kapp. type 2C protein phosphatase.
AAC36699.1 AF075581 Mesembryanthemum crystallinum protein phosphatase-2C. PP2C.
CAC09576.1 AJ298988 Fagus sylvatica protein phosphatase 2C (PP2C). pp2Cf2. SEQ ID NO: 777
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RI 20-L
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIK receptor-like protein kinase; serine/threonine protein kinase.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extracellular S domain: 8-1342; tiansmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRKl. Chrkl.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRKl .
CAA74662.1 Y14286 Brassica oleracea
SFR3. extracellular S domain: 123-1346; transmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl .
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OS JNBa0071 K19.l l.
BAB39435.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.2.
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12.
BAA94518.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 section 111 of 255; putative receptor-like protein kinase (AC002392).
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAB 16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
BAA82556.1 AB030083 Populus nigra lectin-like protein kinase. PnLPK.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLK G RLKl.
SEQ ID NO: 779
AAK31284.1 AC079890 Oryza sativa putative quinone oxidoreductase. OSJNBb0089A17.10.
BAA78050.1 AB027757 Cicer arietinum NADPH oxidoreductase homolog.
BAA83082.1 AB030704 Lithospermum erythrorhizon
LEDI-4 protein. LEDI-4. preferentially expressed in darkness; putative NADPH quinone oxidoreductase; similar to zeta-crystallin.
AAG53944.1 AF304461 Triphysaria versicolor quinone-oxidoreductase QR1. TvQRl.
SEQ ID NO: 780
AAK17067.1 AF254558 Oryza sativa NAC6. NAC6.
BAA89800.1 AB028185 Oryza sativa OsNAC6 protein. OsNAC6
BAA89799.1 AB028184 Oryza sativa OsNAC5 protein. OsNAC5
BAA89798.1 AB028183 Oryza sativa OsNAC4 protein. OsNAC4
BAA89797.1 AB028182 Oryza sativa OsNAC3 protein. OsNAC3
BAA89801.1 AB028186 Oryza sativa OsNAC7 protein. OsNAC7
AAF68626.1 AF254124 Medicago truncatula NACL NACL NAC domain containing protein.
BAA89802.1 AB028187 Oryza sativa OsNAC8 protein. OsNAC8
BAA78417.1 AB021178 Nicotiana tabacum NAC-domain protein. TERN, elicitor-responsive gene.
SEQ ID NO: 783
CAA54390.1 X77134 Brassica napus acyl-CoA binding protein.
CAA70200.1 Y08996 Ricinus communis acyl-CoA-binding protein.
CAB56693.1 AJ249833 Digitalis lanata binds medium- and long-chain acyl-coa esters. Acyl-CoA binding protein (ACBP). acbp3.
CAB56694.1 AJ249834 Digitalis lanata binds medium- and long-chain acyl-coa esters. Acyl-CoA binding protein (ACBP). acbp4.
AAB67736.1 U35015 Gossypium hirsutum acyl-CoA-binding protein.
AAB86851.1 AF031541 Fritillaria agrestis acyl-CoA-binding protein, acabp.
SEQ ID NO: 784
CAA58994.1 X84208 Sinapis alba trypsin inhibitor 2. mti-2.
CAA76116.1 Y16190 Sinapis alba trypsin inhibitor 2. mti-2.
SEQ ID NO: 785
AAF66242.1 AF243180 Lycopersicon esculentum dicyanin. binuclear blue copper protein; contains two stellacyanins linked together.
AAC32421.1 U65511 Cucumis sativus putative oxygen activation and/or lignin formation, stellacyanin. member of a strictly plant- specific family of mononuclear blue copper proteins known as phytocyanins, which are further classified into three distinct subfamilies: Uclacyanins, Stellacyanins, and Plantacyanins; member ofthe subfamily of Stellacyanins; similar to Rhus vernicifera stellacyanin: SwissProt Accession Number P00302; similar to umecyanin: SwissProt Accession Number P42849; similar to mavicyanin; SwissProt Accession Number P80728; similar to BCB encoded by GenBank Accession Number Z15058; similar to F18A8.9, encoded by GenBank Accession Number AC003105; similar to F7F1.27 encoded by GenBank Accession Number AC004669; similar to F9D12.16 encoded by GenBank Accession Number AF077407; glycoprotein.
CAA80963.1 Z25471 Pisum sativum blue copper protein.
AAD 10251.1 AF031195 Triticum aestivum blue copper-binding protein homolog. S85.
AAC64163.1 AF093537 Zeamays blue copper protein, similar to pea blue copper protein in GenBank Accession Number
Z25471.
CAA10134.1 AJ012693 Cicer arietinum basic blue copper protein.
AAF66243.1 AF243181 Lycopersicon esculentum plantacyanin. naturally occuning cupredoxin with a Val residue in the position ofthe axial ligand Met; member ofthe plantacyanin subfamily of a strictly plant-specific family of mononuclear blue copper proteins known as phytocyanins.
CAB65280.1 AJ248323 Medicago sativa subsp. x varia basic blue protein, babl.
AAC32448.1 U76296 Spinacia oleracea plantacyanin. member of a stiictly plant-specific family of mononuclear blue copper proteins known as phytocyanins, which are further classified into three distinct subfamilies: Uclacyanins, Stellacyanins, and Plantacyanins; member ofthe subfamily of Plantacyanins; non-glycosylated.
SEQ ID NO: 793
BAA81862.1 AB026295 Oryza sativa
Similar to leucoanthocyanidin dioxygenase.(AI440611).
AAB39995.1 U82432 Dianthus caryophyllus anthocyanidin synthase. allele: S; 2-oxoglutarat-dependent dioxygenase.
BAA36554.1 AB011796 Citrus unshiu flavonol synthase. CitFLS.
AAD56580.1 AF184273 Daucus carota leucoanthocyanidin dioxygenase 1. LDOXl. 2-oxoglutarate dependent dioxygenase.
AAD56581.1 AFl 84274 Daucus carota leucoanthocyanidin dioxygenase 2. LDOX. 2-oxoglutarate dependent dioxygenase.
CAA50498.1 X71360 Malus sp. anthocyanidin hydroxylase. apple equivalent to 'Candi' from Antinhinum majus.
AAD26205.1 API 17269 Malus x domestica anthocyanidin synthase. ANS.
AAB82287.1 AF026058 Matthiola incana anthocyanidin synthase.
CAA80264.1 Z22543 Petunia x hybrida flavonol synthase.
AAF64168.1 AF240764 Eustoma grandiflorum flavonol synthase. fls.
BAA20143.1 AB003779 Perilla frutescens leucoanthocyanidin dioxygenase.
AAB66560.1 AF015885 Callistephus chinensis anthocyanidin synthase.
BAB21477.1 AB044091 Torenia fournieri anthocyanidin synthase. 2-oxoglutarate dependent oxygenase.
CAA63092.1 X92178 Solanum tuberosum flavonol synthase.
AAD26261.1 AFl 19095 Malus x domestica flavonol synthase. FLS.
CAA53580.1 X75966 Vitis vinifera leucoanthocyanidin dioxygenase. LDOX.
BAA75305.1 AB023786 Ipomoea batatas anthocyanidin synthase. ans I.
AAB84049.1 AF028602 Ipomoea puφurea anthocyanidin synthase. ANS-FL1.
BAA75306.1 AB023787 Ipomoea batatas anthocyanidin synthase. ans II.
CAA73094.1 Y12489 Forsythia x intermedia anthocyanidin synthase.
CAA69252.1 Y07955 Oryza sativa anthocyanidin synthase. ANS.
SEQ ID NO: 794
AAD10204.1 AF030260 Vicia sativa
CYP94A1. vaghl 11. cytochrome P450 fatty acid hydroxylase; Method: conceptual translation with partial peptide sequencing.
AAG33645.1 AF092917 Vicia sativa cytochrome P450-dependent fatty acid hydroxylase. CYP94A2.
AAG17470.1 AF123609 Triticum aestivum cytochrome P450.
BAA99523.1 AP002484 Oryza sativa putative cytochrome P450. P0489A01.14.
AAK31592.1 AY029178 Brassica rapa subsp. pekinensis cytochrome P450. mf-CYP450. possible relevance to male-sterility.
BAA99522.1 AP002484 Oryza sativa putative cytochrome P450. P0489A01.13.
BAA83370.1 AP000391 Oryza sativa
ESTs AU056036(S20239),C72753(E2173), AU056035(S20239) conespond to a region ofthe predicted gene.; Similar to putative cytochrome P-450 (AC003680).
CAB41474.1 AJ238402 Catharanthus roseus cytochrome P450. CYP96C1.
AAB94586.1 AF022457 Glycine max
CYP97B2p. CYP97B2. cytochrome P450 monooxygenase.
AAK20054.1 AC025783 Oryza sativa putative cytochrome P450 monooxygenase. OSJNBa0001O14.16.
AAK38086.1 AF321862 Lolium rigidum putative cytochrome P450.
AAK38085.1 AF321861 Lolium rigidum putative cytochrome P450.
AAB94588.1 AF022459 Glycine max
CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
CAA89260.1 Z49263 Pisum sativum cytochrome P450.
AAG09208.1 AFl 75278 Pisum sativum wound-inducible P450 hydroxylase. CYP82A1.
AAC49188.2 U29333 Pisum sativum cytochrome P450 monooxygenase. CYP 82. new cytochrome P450 family.
BAB 19083.1 AP002744 Oryza sativa putative cytochrome P450. P0006C01.25. contains ESTs
AU081507(C12518),C26520(C12518).
BAB19104.1 AP002839 Oryza sativa putative cytochrome P450. P0688A04.10. contains ESTs
AU081507(C12518),C26520(C12518).
AAK38092.1 AF321868 Lolium rigidum putative cytochrome P450.
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'- race).
AAK38091.1 AF321867 Lolium rigidum putative cytochrome P450.
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81Blc.
BAA22423.1 AB001380 Glycynhiza echinata cytochrome P450. CYP93B1.
CAA71876.1 Y10982 Glycine max putative cytochrome P450.
BAB39252.1 AP002968 Oryza sativa putative cytochrome P450. P0416G11.1.
AAA33106.1 LI 0081 Catharanthus roseus cytochrome P-450 protein. CYP72. putative; CYP72 protein.
AAA17746.1 L19075 Catharanthus roseus cytochrome P450. CYP72C. putative.
BAA74466.1 AB022733 Glycynhiza echinata cytochrome P450. CYP Ge-51.
BAB19121.1 AP002839 Oryza sativa putative cytochrome P450. P0688A04.28.
AAA17732.1 L19074 Catharanthus roseus cytochrome P450. CYP72B.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
AAB05376.3 U35226 Nicotiana plumbaginifolia putative cytochrome P-450.
AAB61965.1 U48435 Solanum chacoense putative cytochrome P450.
AAF27282.1 AF122821 Capsicum annuum cytochrome P450. PepCYP.
CAB50768.1 AJ243804 Cicer arietinum putative isoflavone synthase. cytochrome P450. cyp93C3.
AAC34853.1 AF082028 Hemerocallis hybrid cultivar putative cyt P450-containing fatty acid hydroxylase. senescence-associated protein 3. SA3. mRNA accumulates in senescing petals.
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
BAB21156.1 AP002899 Oryza sativa putative cytochrome P450. P0456A01.12.
CAA50648.1 X71657 Solanum melongena P450 hydroxylase.
BAB19112.1 AP002839 Oryza sativa putative cytochrome P450. P0688A04.18. contains ESTs AU067870(C10320),AU067869(C10320).
BAB 19091.1 AP002744 Oryza sativa putative cytochrome P450. P0006C01.33. contains ESTs
AU067870(C10320),AU067869(C10320).
BAA84072.1 AB028152 Torenia hybrida flavone synthase II. cytochrome P450. TFNS5.
BAA76380.1 AB023636 Glycynhiza echinata cytochrome P450. CYP Ge-8.
CAA72208.1 Y11404 Zea mays cytochrome p450. cyp71c2.
CAA57423.1 X81829 Zea mays cytochrome P450. CYP71C2. family CYP71, subfamily CYP71C.
BAB12433.1 AB025030 Coptis japonica p450.
SEQ ID NO: 798
BAB21205.1 AP002913 Oryza sativa nucleoid DNA-binding protein cnd41 -like protein. P0480E02.il. contains ESTs
AU166073 (E31027), AU029516(E31027).
BAA22813.1 D26015 Nicotiana tabacum aspartic protease activity. CND41, chloroplast nucleoid DNA binding protein. cnd41.
SEQ ID NO: 804 AAD46491.1 AF135014 Zea mays dihydrolipoamide S-acetyltransferase. pyravate dehydrogenase complex E2 subunit.
SEQ ID NO: 805
BAA85412.1 AP000615 Oryza sativa
ESTs AU065232(E60855),C23624(S1554), AU078241(E60855) conespond to a region ofthe predicted gene.; similar to putative adenylate kinase. (AC005896).
BAA01181.1 D10335 Oryza sativa adenylate kinase-b. Adk-b.
BAA01180.1 D10334 Oryza sativa adenylate kinase-a. Adk-a.
BAA94761.1 AB041773 Oryza sativa adenylate kinase. Adk-a.
AAB68604.1 U82330 Prunus armeniaca adenylate kinase homolog.
AAF23372.1 AFl 87063 Oryza sativa catalyzes the transfer of phosphate from ATP to UMP or CMP to form ADP and UDP or
CDP. UMP/CMP kinase b. ura6.
AAF23371.1 AF187062 Oryza sativa catalyzes the transfer of phosphate from ATP to UMP or CMP to form ADP and UDP or
CDP. UMP/CMP kinase a. ura6.
AAD41679.1 AF086603 Ceratopteris richardii adenylate kinase. ADK1.
BAA85443.1 AP000616 Oryza sativa similar to UMP/CMP kinase (AF000147).
SEQ ID NO: 808
CAA41774.1 X59046 Oryza sativa sucrose-UDP glucosyltiansferase (isoenzyme 2). RSs2.
BAA89049.1 AB029401 Citras unshiu sucrose synthase. CitSUSl-2.
AAA34196.1 L19762 Lycopersicon esculentum sucrose synthase.
BAA88905.1 AB022092 Citras unshiu sucrose synthase. CitSUSl
AAD28641.1 U73588 Gossypium hirsutum sucrose synthase.
CAA49428.1 X69773 Vicia faba sucrose synthase. VfSucs.
AAC37346.1 M97551 Vicia faba cleavage of sucrose. UDP- glucose:D-fructose-2-glucosyltransferase. putative.
CAA09681.1 AJOl 1535 Lycopersicon esculentum sucrose synthase. sus2.
AAA97572.1 U24088 Solanum tuberosum sucrose synthase.
CAA09593.1 AJ011319 Lycopersicon esculentum sucrose synthase. sus3.
CAB40794.1 AJ131943 Medicago truncatula sucrose synthase. sucSl.
AAC17867.1 AF049487 Medicago sativa sucrose hydrolysis, sucrose synthase.
CAB40795.1 AJ131964 Medicago truncatula sucrose synthase. sucSl.
CAA65640.1 X96939 Tulipa gesneriana sucrose-synthase 21.
AAA97571.1 U24087 Solanum tuberosum sucrose synthase.
CAA63122.1 X92378 Alnus glutinosa sucrose synthase. susl.
AAA33514.1 L22296 Zea mays UDP-glucose:D-fructose 2 -glucosyl-transferase. Susl.
CAA65639.1 X96938 Tulipa gesneriana sucrose-synthase 1.
AAC41682.1 L03366 Oryza sativa sucrose synthase 3. RSs3.
CAA75793.1 Y15802 Hordeum vulgare sucrose synthase 2. Ss2.
CAA49551.1 X69931 Hordeum vulgare sucrose synthase. Ss2.
CAA76056.1 Y16090 Daucus carota sucrose synthase isoform I . Susy*Dcl.
CAA53081.1 X75332 Daucus carota sucrose synthase.
AAA33515.1 L33244 Zea mays sucrose synthase 2. Susl.
BAB20799.1 AB045710 Pyras pyrifolia sucrose synthase 1. PypSUSl.
CAA03935.1 AJ000153 Triticum aestivum sucrose synthase type 2.
AAC39323.1 AF030231 Glycine max sucrose synthase. SS. nodulin-100.
BAA01108.1 D10266 Vigna radiata sucrose synthase. vssl.
CAA09910.1 AJ012080 Pisum sativum sucrose synthase.
AAC28107.1 AF079851 Pisum sativum nodule-enhanced sucrose synthase. ness.
CAC32462.1 AJ311496 Pisum sativum sucrose metabolism, sucrose synthase isoform 3. sus3.
CAA57881.1 X82504 Chenopodium rabrum sucrose synthase. CSS1.
CAA26229.1 X02382 Zea mays sucrose synthase.
CAA26247.1 X02400 Zea mays sucrose synthase.
CAA46017.1 X64770 Oryza sativa sucrose synthase. RSsl.
CAB38022.1 AJl 32000 Craterostigma plantagineum sucrose metabolism, sucrose synthase. Ss2.
CAA78747.1 Z15028 Oryza sativa sucrose synthase.
AAF85966.1 AF263384 Saccharum officinarum sucrose synthase- 1.
CAA46701.1 X65871 Hordeum vulgare sucrose synthase.
CAA04543.1 AJ001117 Triticum aestivum sucrose synthase type I. Ssl.
BAA88904.1 AB022091 Citrus unshiu sucrose synthase. CitSUSA.
BAA88981.1 AB025778 Citrus unshiu sucrose synthase. CitSUSA-2.
CAA04512.1 AJ001071 Pisum sativum second sucrose synthase.
CAA76057.1 Y16091 Daucus carota sucrose synthase isoform I] t. Susy*Dc2.
CAB38021.1 AJl 31999 Craterostigma plantagineum sucrose metabolism, sucrose synthase. Ssl.
CAA57499.1 X81974 Beta vulgaris sucrose synthase. SBSS1.
CAA47264.1 X66728 Hordeum vulgare sucrose synthase.
SEQ ID NO: 809
AAB69317.1 AF012861 Petroselinum crispum plastidic glucose-6-phosphate dehydrogenase. pG6PDH.
AAF87216.1 AF231351 Nicotiana tabacum plastidic glucose 6-phosphate dehydrogenase. G6PDHP2.
CAA67782.1 X99405 Nicotiana tabacum glucose-6-phosphate dehydrogenase. G6PD.
CAB52708.1 AJ010712 Solanum tuberosum ghιcose-6-phosphate 1 -dehydrogenase. g6pd.
CAB52685.1 AJ132346 Dunaliella bioculata plastidic glucose-6-phosphate dehydrogenase. g6PD.
CAA58775.1 X83923 Solanum tuberosum glucose-6-phosphate dehydrogenase.
CAA03941.1 AJ000184 Spinacia oleracea Glucose-6-phosphate dehydrogenase . G6PD.
CAA03939.1 AJ000182 Spinacia oleracea Glucose-6-phosphate dehydrogenase. G6PD.
CAA04994.1 AJ001772 Nicotiana tabacum glucose-6-phosphate dehydrogenase. TCG18.
CAA03940.1 AJ000183 Spinacia oleracea Glucose-6-phosphate dehydrogenase. G6PD.
AADl 1426.1 AF097663 Mesembryanthemum crystallinum cytoplasmic glucose-6-phosphate 1 -dehydrogenase. G6PD.
AAB41552.1 Ul 8238 Medicago sativa subsp. sativa glucose-6-phosphate dehydrogenase.
CAA52442.1 X74421 Solanum tuberosum glucose-6-phosρhate 1 -dehydrogenase. g6pdh. homotetramer.
AAB69318.1 AF012862 Petroselinum crispum cytosolic glucose-6-phosphate dehydrogenase 1. cG6PDHl.
AAB69319.1 AF012863 Petroselinum crispum cytosolic glucose-6-phosphate dehydrogenase 2. cG6PDH2.
CAA04992.1 AJ001769 Nicotiana tabacum glucose-6-phosphate dehydrogenase. TCG6.
CAA04993.1 AJ001770 Nicotiana tabacum glucose-6-phosphate dehydrogenase. TCG9.
BAA97662.1 AB029454 Triticum aestivum glucose-6-phosphate dehydrogenase. g6ρdh.
BAA97663.1 AB029455 Triticum aestivum glucose-6-phosphate dehydrogenase. gόpdh.
BAA97664.1 AB029456 Triticum aestivum glucose-6-phosphate dehydrogenase. g6pdh.
AAG23802.1 AF260736 Cucurbita pepo plastidic glucose-6-phosphate dehydrogenase.
CAB66330.1 AJ279688 Betula pendula glucose-6-phosphate dehydrogenase. g6pd.
BAA82155.1 AB011441 Triticum aestivum glucose-6-phosphate dehydrogenase. WESR5. salt-stress responding gene.
CAA06200.1 AJ004900 Glycine max pentose phosphate pathway oxidoreductase generating NADPH. glucose-6-ρhosphate- dehydrogenase.
SEQ ID NO: 812
BAΛ08910.1 D50407 Cucumis sativus glutamyl-tRNA reductase. hemA.
AAD16897.1 AF105221 Glycine max converts glutamyl-tRNA to glutamate 1-semialdehyde. glutamyl-tRNA reductase precursor. gtrl.
BAA11091.1 D67088 Cucumis sativus glutamyl-tRNA reductase. hemA2.
BAA25003.1 ABO 11416 Oryza sativa glutamyl-tRNA reductase.
AAG13620.1 AC078840 Oryza sativa putative glutamyl-tRNA reductase. OSJNBb0073N24.1.
CAA60054.1 X86101 Hordeum vulgare aminolevulinate synthesis (chlorophyll synthesis), glutamyl tRNA reductase. hemA 1. 1st isoform.
CAA63140.1 X92403 Hordeum vulgare glutamyl-tRNA reductase. hemAl. isoform I.
BAA25167.1 D88382 Hordeum vulgare glutamyl-tRNA reductase. hemAl. isoform 1.
CAA60055.1 X86102 Hordeum vulgare aminolevulinate synthesis (chlorophyll synthesis), glutamyl tRNA reductase. hemA 2. 2nd isoform.
BAA25168.1 D88383 Hordeum vulgare glutamyl-tRNA reductase. hemA3. isoform 3.
AAG41962.1 AF305613 Chlamydomonas reinhardtii glutamyl-tRNA reductase precursor. HemA. pGtr.
AAG02480.1 AF294753 Hordeum vulgare converts glutamyl-tRNA to glutamate 1-semialdehyde in the chlorophyll biosynthetic pathway. glutamyl-tRNA reductase. hemA2.
AAG02479.1 AF294752 Hordeum vulgare converts glutamyl-tRNA to glutamate 1-semialdehyde in the chlorophyll biosynthetic pathway. glutamyl-tRNA reductase. hemAl.
SEQ ID NO: 813
BAA82556.1 AB030083 Populus nigra lectin-like protein kinase. PnLPK.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAF43408.1 AF230515 Oryza sativa subsp. japonica serine/threonine protein kinase. YK35.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl. accumulates in mesophyll cells.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein kinase.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAA92953.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F10M6 ; S-receptor kinase -like protein. (AL021811).
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJO 10166).
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRKl. Chrkl.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
AAD46420.1 AF100771 Hordeum vulgare receptor-like kinase. Hv3ARK. similar to wheat ARK1 AS.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSJNBa0004G10.30.
BAB18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
BAA94517.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
BAA94529.2 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
BAA94528.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 BAC T20K24; putative receptor-like protein kinase (AC002392).
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
SEQ ID NO: 814
AAF23903.1 AFl 94416 Oryza sativa MAP kinase homolog. MAPK2. RMAPK2.
AAD52659.1 AFl 77392 Oryza sativa blast and wounding induced mitogen-activated protein kinase. BWMKl . BWMKl MAP kinase.
AAF23902.1 AF194415 Oryza sativa MAP kinase homolog. MAPK1. RMAPK1.
AAD28617.1 AF129087 Medicago sativa mitogen-activated protein kinase homologue. TDY1.
CAB61750.1 AJ275316 Cicer arietinum MAP kinase protein.
AAB57843.1 U96716 Selaginella lepidophylla MAP kinase-like protein. sdhn-6r.
AAF65766.1 AF242308 Euphorbia esula mitogen-activated protein kinase. regulated by tyrosine and threonine phosphorylation.
BAB18271.1 AB035141 Chlamydomonas reinhardtii mitogen-activated protein kinase. CrMPK2.
CAA58761.1 X83880 Nicotiana tabacum p45Ntf4 serine/threonine protein kinase. ntf4.
CAA47099.1 X66469 Medicago sativa MAP Kinase. MSK7.
AAB41548.1 L07042 Medicago sativa autophosphorylating serine/threonine protein kinase. MAP kinase. MsERKl.
AAB58396.1 U94192 Nicotiana tabacum salicylic acid-activated MAP kinase. NtSIPK.
CAA50036.1 X70703 Pisum sativum MAP kinase homologue. PSMAPKIN.
AAF73236.1 AF153061 Pisum sativum MAP kinase 3. Mapk3. PsMAPK3.
BAB32406.1 AB055515 Nicotiana tabacum
NRKl MAPK. nrkl. A tobacco MAPK that is phosphorylated and activated by NQKl.
CAA58760.1 X83879 Nicotiana tabacum p43Nft6 serine/threonine protein kinase. ntf6.
CAA57721.1 X82270 Medicago sativa protein kinase. MMK4.
AAF81420.1 AF247136 Capsicum annuum
MAP kinase 2. MK2. CAMK2; wound, UV-C, and cold-inducible expression.
AAD37790.1 AFl 49424 Ipomoea batatas MAP kinase.
AAG40580.1 AF216316 Oryza sativa MAP kinase 2. protein kinase; MAP2.
CAB37188.1 AJ224336 Medicago sativa MAP kinase. MMK3.
AAF61238.1 AF241166 Oryza sativa MAP kinase MAPK2.
AAG40581.1 AF216317 Oryza sativa MAP kinase 3. protein kinase; MAP3.
CAB61889.1 AJ251330 Oryza sativa protein kinase. MAPK4 protein. mapk4.
CAA73323.1 Y12785 Petroselinum crispum MAP kinase I.
CAC13967.1 AJ250311 Oryza sativa protein kinase. MAPK2 protein. mapk2.
CAA56314.1 X79993 Avena sativa MAP KINASE. Asmapl.
CAA49592.1 X69971 Nicotiana tabacum serine/threonine protein kinase. NTF3.
CAA58466.1 X83440 Petunia x hybrida MAP/ERK kinase 1. MEKL
AAK01710.1 AF332873 Oryza sativa MAP kinase BIMK1.
AAG40579.1 AF216315 Oryza sativa MAP kinase 1. protein kinase; MAPI.
CAA57719.1 X82268 Medicago sativa protein kinase. MMK2.
AAC28850.1 AF079318 Triticum aestivum protein kinase. MAP kinase homolog. WCK-1.
AAD32204.1 AF134730 Prunus armeniaca putative mitogen-activated protein kinase MAPK. MAP kinase.
BAA74734.1 AB016802 Zeamays MAP kinase 5. ZmMPK5.
AAF73257.1 AF154329 Pisum sativum MAP kinase PsMAPK2. Mapk2.
BAA09600.1 D61377 Nicotiana tabacum WIPK. MAP (mitogen-activated protein) kinase.
AAF81419.1 AF247135 Capsicum annuum
MAP kinase 1. MK1. wound and UV-C inducible expression.
BAA74733.1 AB016801 Zea mays MAP kinase 4. ZmMPK4.
CAA05328.1 AJ002314 Nicotiana tabacum serine/threonine protein kinase. shaggy-like kinase 111. NSK 111.
CAA05329.1 AJ002315 Nicotiana tabacum shaggy-like kinase 59. NSK 59.
CAAl 1861.1 AJ224164 Petunia x hybrida shaggy kinase 6. PSK6.
CAA58595.1 X83620 Petunia x hybrida Petunia Shaggy kinase 6. PSK6.
CAAl 1862.1 AJ224165 Petunia x hybrida shaggy kinase 7. PSK7.
CAA58594.1 X83619 Petunia x hybrida Petunia Shaggy kinase 4. PSK4.
AAA92823.1 U18365 Brassica napus cyclin dependent protein kinase homolog; similar to moth bean p34cdc2 protein, PIR
Accession Number JQ2243.
BAA92214.1 AP001278 Oryza sativa
ESTs C22403(C50132),C22404(C50132) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana shaggy related protein kinase ASK-gamma. (P43289).
CAA67554.1 X99100 Trifolium repens protein kinase. tiK.
SEQ ID NO: 816 CAA65065.1 X95759 Solanum tuberosum glycogen (starch) synthase. CAA64173.1 X94400 Solanum tuberosum soluble-starch-synthase. SSSIII.
AAC14014.1 AF023159 Zeamays starch synthase DULLl . duUl . similar to potato starch synthase SSIII; likely to be the maize starch synthase defined biochernically as SSII.
CAB40374.1 AJ225088 Vigna unguiculata
ADP-glucose-starch glucosyltiansferase. Starch synthase isoform SS III.
AAF88000.1 AF258609 Aegilops tauschii starch synthase III.
AAF87999.1 AF258608 Triticum aestivum starch synthase III. wSSIII.
CAB40375.1 AJ006752 Vigna unguiculata
ADP-glucose starch glucosyltransferase. starch synthase, isoform V.
AAC14015.1 AF023160 Zeamays starch synthase DULLl. dulll. similar to potato starch synthase SSIII; like to be the maize starch synthase defined biochemically as SSII.
AAC 17971.2 AF026422 Chlamydomonas reinhardtii soluble starch synthase. ADP-glucose: alpha- 1, 4-D-glucan-4-alpha-D-glucosyltransferase.
AAD13342.1 AF019297 Zea mays starch synthase isoform zSTSII-2. zSSIIb.
CAB86618.1 AJ269502 Triticum aestivum transfers the glucosyl unit of ADP glucose to the non-reducing end of an alpha 1,4. starch synthase Ila- 1. wSs2a-l.
CAA61269.1 X88790 Pisum sativum glycogen (starch) synthase.
CAB96626.1 AJ269503 Triticum aestivum transfers the glucosyl unit of ADP glucose to the non-reducing end of an alpha 1,4. starch synthase IIa-2. wSs2a-2.
CAB96627.1 AJ269504 Triticum aestivum transfers the glucosyl unit of ADP glucose to the non-reducing end of an alpha 1,4. starch synthase IIa-3. wSs2a-3.
CAA71442.1 Y10416 Solanum tuberosum soluble starch (bacterial glycogen) synthase. SS I.
AAD53263.1 AFl 55217 Triticum aestivum starch synthase IIA.
AAF37876.1 AF234163 Hordeum vulgare starch synthase I. SSI.
CAB99209.1 AJ292521 Triticum aestivum essential for starch synthesis, starch synthase I-L wSsI-1.
AAD54661.1 AF091803 Triticum aestivum starch synthase I.
AAB 17085.1 U66377 Triticum aestivum starch synthase. TaSS. EC 2.4.1.11.
CAB99210.1 AJ292522 Triticum aestivum essential for starch synthesis, starch synthase 1-2. wSsI-2.
AAF03557.1 AF091802 Aegilops tauschii starch synthase I.
AAD13341.1 AF019296 Zeamays starch synthase isoform zSTSII-1. zSSIIa.
AAC17969.2 AF026420 Chlamydomonas reinhardtii
ADP-Glucose: alpha- 1, 4-D-glucan-4-alpha-D-glucosyltiansferase. granule-bound starch synthase I precursor. STA2. GBSSI.
BAA82346.1 AB029546 Phaseolus vulgaris granule-bound starch synthase I. GBSSI.
CAA37732.1 X53694 Oryza sativa starch synthase.
CAA52273.1 X74160 Manihot esculenta starch (bacterial glycogen) synthase. GBSS.
AAF72561.1 AF141954 Oryza sativa granule-bound starch synthase. Waxy.
CAA46294.1 X65183 Oryza sativa glycogen (starch) synthase. waxy gene, starch granule enzyme.
AAF72562.1 AF141955 Oryza sativa granule-bound starch synthase. Waxy.
CAA44065.1 X62134 Oryza sativa starch biosynthesis, starch (bacterial glycogen) synthase. Wx.
AAB02197.1 U48227 Triticum aestivum soluble starch synthase.
AAF13168.1 AF173900 Manihot esculenta granule bound starch synthase II precursor. GBSSII. MEGBSSII.
CAA45472.1 X64108 Oryza sativa starch granule-bound starch synthase. waxy.
AAC61675.2 AF031162 Oryza sativa granule-bound starch synthase. Waxy.
AAC70779.1 AF097922 Astragalus membranaceus granule-bound glycogen (starch) synthase. GBSS.
CAA06958.1 AJ006293 Antinhinum majus granule-bound starch synthase. waxy.
AAC19119.1 AF068834 Ipomoea batatas starch synthase.
AAD49850.1 AF165890 Oryza sativa subsp. japonica soluble starch synthase.
BAA81848.1 AB026295 Oryza sativa
ESTs AU075322(C11109),D22430(C11109) conespond to a region ofthe predicted gene.;
Rice gene for soluble starch synthase (SSSl), complete cds (exonl-15).(D38221).
BAA03739.1 D16202 Oryza sativa soluble starch synthase precursor.
CAA61268.1 X88789 Pisum sativum glycogen (starch) synthase.
AAA86423.1 U44126 Ipomoea batatas starch synthase. SPSS67.
AAF14233.1 AF109395 Triticum aestivum granule-bound starch synthase GBSSII.
CAA41359.1 X58453 Solanum tuberosum glycogen (starch) synthase.. amf. waxy protein,granule-bound starch synthase.
AAG43519.1 AF210699 Perilla frutescens granule-bound starch synthase. GBSSI. waxy protein.
SEQ ID NO: 819
BAA13032.1 D86180 Pisum sativum phosphoribosylanthranilat. : transferase. PAT1.
SEQ ID NO: 822
AAB86850.1 AF031540 Fritillaria agrestis cytochrome C, . cytC.
AAC84135.1 AF101422 Cichorium intybus cytochrome.
BAA02159.1 D12634 Oryza sativa 'cytochrome C.
AAA63515.1 M63704 Oryza sativa cytochrome c. Cc-1.
AAA92712.1 L77113 Helianthus annuus cytochrome c. cytcl. putative.
AAB70265.1 AF017367 Oryza sativa cytochrome C,
AAA33084.1 M35173 Chlamydomonas reinhardtii apocytochrome c (eye).
CAB 16954.1 Z99829 Chlamydomonas reinhardtii cytochrome c. CYCl.
CAA79708.1 Z21499 Stellaria longipes mitochondrial cytochrome c.
SEQ ID NO: 823
BAA02159.1 D12634 Oryza sativa 'cytochrome C
AAA63515.1 M63704 Oryza sativa cytochrome c. Cc-1.
AAB86850.1 AF031540 Fritillaria agrestis cytochrome C.. cytC.
AAC84135.1 AF101422 Cichorium intybus cytochrome.
AAA92712.1 L77113 Helianthus annuus cytochrome c. cytcl. putative.
AAB70265.1 AF017367 Oryza sativa cytochrome C.
AAA33084.1 M35173 Chlamydomonas reinhardtii apocytochrome c (eye).
CAB 16954.1 Z99829 Chlamydomonas reinhardtii cytochrome c. CYCl.
CAA79708.1 Z21499 Stellaria longipes mitochondrial cytochrome c.
SEQ ID NO: 825
BAB17113.1 AP002866 Oryza sativa putative white protein; ATP-binding cassette transporter. P0410E01.34.
AAF43869.1 AF166114 Chloroplast Mesostigma viride probable transport protein. cysA.
BAA90508.1 AP001111 Oryza sativa similar to ABC transporter of Arabidopsis thaliana (AC004697).
AAD54843.1 AF137379 Chloroplast Nephroselmis olivacea probable transport protein. cysA.
BAA57907.1 AB001684 Chlorella vulgaris sulfate tiansport system permease protein. cysA.
BAA90507.1 AP001111 Oryza sativa similar to ABC transporter of Arabidopsis thaliana (AC004697).
BAB40032.1 AP003046 Oryza sativa putative ABC transporter. P0445D12.3.
AAG49003.1 AYO 13246 Hordeum vulgare putative ABC transporter. 635P2.4b; GC splice donor confirmed by cDNA alignment and comparative sequence.
BAB21275.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.6.
AAG45492.1 AY013245 Oryza sativa
3615.4. putative ABC transporter; GC splice donor confirmed by cDNA alignment and comparative sequence.
AAG49002.1 AY013246 Hordeum vulgare putative ABC tiansporter. 635P2.4a; GC splice donor confirmed by cDNA and comparative sequencing.
BAB21276.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.7. contains EST D22472(C1173).
AAD10836.1 U52079 Solanum tuberosum
P-glycoprotein. pmdrl . binds ATP; ATPase; transporter; transmembrane protein.
BAB21279.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.10. contains ESTs
AU065360(R3463),AU101680(R3463).
BAB21273.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.4.
BAA83352.1 AP000391 Oryza sativa
ESTs AU067992(C11433),AU077424(C11433) correspond to a region ofthe predicted gene.;
Similar to ABC tiansporter-7 (U43892).
BAA96612.1 AP002482 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2, BAC F14M4 ; putative ABC tiansporter
(AC004411).
SEQ ID NO: 827
AAG34803.1 AF243368 Glycine max glutathione S-transferase GST 13.
AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-transferase T3.
AAG34798.1 AF243363 Glycine max glutathione S-transferase GST 8.
AAF64450.1 AF239928 Euphorbia esula glutathione S-tiansferase. similar to auxin-inducible GST.
AAG34807.1 AF243372 Glycine max glutathione S-transferase GST 17.
AAG34796.1 AF243361 Glycine max glutathione S-tiansferase GST 6.
AAG16759.1 AY007561 Lycopersicon esculentum putative glutathione S-tiansferase T4.
AAG34797.1 AF243362 Glycine max glutathione S-tiansferase GST 7.
AAG34801.1 AF243366 Glycine max glutathione S-tiansferase GST 11.
AAG34804.1 AF243369 Glycine max glutathione S-tiansferase GST 14.
AAG34809.1 AF243374 Glycine max glutathione S-transferase GST 19.
AAG34808.1 AF243373 Glycine max glutathione S-transferase GST 18.
AAG34810.1 AF243375 Glycine max glutathione S-tiansferase GST 20.
AAG16757.1 AY007559 Lycopersicon esculentum putative glutathione S-transferase T2.
AAG16756.1 AY007558 Lycopersicon esculentum putative glutathione S-transferase TI.
AAG34844.1 AF244701 Zea mays glutathione S-tiansferase GST 36.
AAG34805.1 AF243370 Glycine max glutathione S-transferase GST 15.
AAG34831.1 AF244688 Zeamays glutathione S-transferase GST 23.
AAG34832.1 AF244689 Zeamays glutathione S-transferase GST 24.
AAG34849.1 AF244706 Zeamays glutathione S-transferase GST 41.
AAG34802.1 AF243367 Glycine max glutathione S-tiansferase GST 12.
CAA09187.1 AJ010448 Alopecurus myosuroides glutathione transferase. GSTl a.
AAG34829.1 AF244686 Zea mays glutathione S-tiansferase GST 21.
CAA09188.1 AJO 10449 Alopecurus myosuroides glutathione transferase. GSTlb.
AAA68430.1 J03679 Solanum tuberosum glutathione S-tiansferase. gstl. previously called pathogenesis-related protein; pφl-1.
AAG34836.1 AF244693 Zeamays glutathione S-tiansferase GST 28.
AAG34837.1 AF244694 Zeamays glutathione S-transferase GST 29.
AAG34800.1 AF243365 Glycine max glutathione S-transferase GST 10.
AAC18566.1 AF048978 Glycine max 2,4-D inducible glutathione S-transferase. GSTa.
AAC32118.1 AF051214 Picea mariana probable glutathione S-tiansferase. Sbl8. similar to Nicotiana tabacum probable glutathione
S-transferase encoded by GenBank Accession Number X56266.
AAF22517.1 AFl 18924 Papaver somniferam glutathione S-tiansferase 1. GSTL
AAF22518.1 AFl 18925 Papaver somniferam glutathione S-transferase 2. GST2.
AAG32471.1 AF309378 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU4.
AAG34806.1 AF243371 Glycine max glutathione S-tiansferase GST 16.
CAA04391.1 AJ000923 Carica papaya glutathione transferase. PGST1.
AAG32472.1 AF309379 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU3.
AAG34833.1 AF244690 Zeamays glutathione S-tiansferase GST 25.
CAA71784.1 Y10820 Glycine max glutathione transferase.
AAG34847.1 AF244704 Zea mays glutathione S-transferase GST 39.
AAF22519.1 AFl 18926 Papaver somniferam glutathione S-tiansferase 3. GST3.
SEQ ID NO: 828
AAG34803.1 AF243368 Glycine max glutathione S-transferase GST 13.
AAG34797.1 AF243362 Glycine max glutathione S-transferase GST 7.
AAG34798.1 AF243363 Glycine max glutathione S-tiansferase GST 8.
AAF64450.1 AF239928 Euphorbia esula glutathione S-transferase. similar to auxin-inducible GST.
AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-transferase T3.
AAG34796.1 AF243361 Glycine max glutathione S-tiansferase GST 6.
AAG16759.1 AY007561 Lycopersicon esculentum putative glutathione S-transferase T4.
AAG34801.1 AF243366 Glycine max glutathione S-tiansferase GST 11.
AAG34804.1 AF243369 Glycine max glutathione S-tiansferase GST 14.
AAG34807.1 AF243372 Glycine max glutathione S-transferase GST 17.
AAG34809.1 AF243374 Glycine max glutathione S-transferase GST 19.
AAG34810.1 AF243375 Glycine max glutathione S-transferase GST 20.
AAG16757.1 AY007559 Lycopersicon esculentum putative glutathione S-transferase T2.
AAGl 6756.1 AY007558 Lycopersicon esculentum putative glutathione S-tiansferase TI.
AAG34802.1 AF243367 Glycine max glutathione S-transferase GST 12.
AAG34808.1 AF243373 Glycine max glutathione S-transferase GST 18.
AAG34844.1 AF244701 Zeamays glutathione S-tiansferase GST 36.
CAA09188.1 AJ010449 Alopecurus myosuroides glutathione transferase. GSTlb.
CAA09187.1 AJ010448 Alopecurus myosuroides glutathione transferase. GSTla.
AAG32472.1 AF309379 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU3.
AAA68430.1 J03679 Solanum tuberosum glutathione S-transferase. gstl. previously called pathogenesis-related protein; prpl-1.
AAG34837.1 AF244694 Zeamays glutathione S-tiansferase GST 29.
AAG34800.1 AF243365 Glycine max glutathione S-transferase GST 10.
AAG34831.1 AF244688 Zea mays glutathione S-transferase GST 23.
AAC32118.1 AF051214 Piceamariana probable glutathione S-tiansferase. Sbl8. similar to Nicotiana tabacum probable glutathione
S-transferase encoded by GenBank Accession Number X56266.
AAG34805.1 AF243370 Glycine max glutathione S-transferase GST 15.
AAC18566.1 AF048978 Glycine max 2,4-D inducible glutathione S-transferase. GSTa.
AAG34829.1 AF244686 Zea mays glutathione S-tiansferase GST 2L
CAA04391.1 AJ000923 Carica papaya glutathione transferase. PGST1.
CAA71784.1 Y10820 Glycine max glutathione transferase.
AAG34795.1 AF243360 Glycine max glutathione S-transferase GST 5.
AAG34836.1 AF244693 Zea mays glutathione S-tiansferase GST 28.
AAG34832.1 AF244689 Zeamays glutathione S-transferase GST 24.
AAG34833.1 AF244690 Zeamays glutathione S-transferase GST 25.
AAG34849.1 AF244706 Zeamays glutathione S-tiansferase GST 41.
AAG34806.1 AF243371 Glycine max glutathione S-transferase GST 16.
CAA09189.1 AJ010450 Alopecurus myosuroides glutathione transferase. GSTlc.
SEQ ID NO: 829
AAG34803.1 AF243368 Glycine max glutathione S-transferase GST 13.
AAF64450.1 AF239928 Euphorbia esula glutathione S-transferase. similar to auxin-inducible GST. AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-transferase T3.
AAG34798.1 AF243363 Glycine max glutathione S-tiansferase GST 8.
AAG34801.1 AF243366 Glycine max glutathione S-tiansferase GST 11.
AAG34797.1 AF243362 Glycine max glutathione S-transferase GST 7.
AAG34796.1 AF243361 Glycine max glutathione S-tiansferase GST 6.
AAG34807.1 AF243372 Glycine max glutathione S-tiansferase GST 17.
AAG16759.1 AY007561 Lycopersicon esculentum putative glutathione S-transferase T4.
AAG34804.1 AF243369 Glycine max glutathione S-tiansferase GST 14.
AAG34810.1 AF243375 Glycine max glutathione S-tiansferase GST 20.
AAG34809.1 AF243374 Glycine max glutathione S-transferase GST 19.
AAG16757.1 AY007559 Lycopersicon esculentum putative glutathione S-transferase T2.
AAG16756.1 AY007558 Lycopersicon esculentum putative glutathione S-tiansferase TI.
AAG34805.1 AF243370 Glycine max glutathione S-transferase GST 15.
AAC18566.1 AF048978 Glycine max 2,4-D inducible glutathione S-transferase. GSTa.
AAG34808.1 AF243373 Glycine max glutathione S-tiansferase GST 18.
AAG34800.1 AF243365 Glycine max glutathione S-tiansferase GST 10.
AAG34829.1 AF244686 Zea mays glutathione S-transferase GST 21.
AAG34802.1 AF243367 Glycine max glutathione S-transferase GST 12.
AAG34837.1 AF244694 Zeamays glutathione S-tiansferase GST 29.
CAA09187.1 AJ010448 Alopecurus myosuroides glutathione transferase. GSTl a.
CAA09188.1 AJO 10449 Alopecurus myosuroides glutathione transferase. GSTlb.
AAG34849.1 AF244706 Zea mays glutathione S-transferase GST 41.
AAG34844.1 AF244701 Zeamays glutathione S-tiansferase GST 36.
AAG34806.1 AF243371 Glycine max glutathione S-transferase GST 16.
CAA71784.1 Y10820 Glycine max glutathione transferase.
AAA68430.1 J03679 Solanum tuberosum glutathione S-transferase. gstl. previously called pathogenesis-related protein; pφl-1.
CAA04391.1 AJ000923 Carica papaya glutathione transferase. PGST1.
AAG34836.1 AF244693 Zeamays glutathione S-transferase GST 28.
AAG34831.1 AF244688 Zea mays glutathione S-tiansferase GST 23.
AAG34847.1 AF244704 Zea mays glutathione S-transferase GST 39.
AAC32118.1 AF051214 Piceamariana probable glutathione S-transferase. Sbl8. similar to Nicotiana tabacum probable glutathione
S-tiansferase encoded by GenBank Accession Number X56266.
AAF29773.1 AFl 59229 Gossypium hirsutum glutathione S-transferase. GST.
AAG32472.1 AF309379 Oryza sativa subsp. japonica putative glutathione S-transferase OsGSTU3.
AAG41204.1 AF321437 Suaeda maritima glutathione transferase.
CAC24549.1 AJ296343 Cichorium intybus x Cichorium endivia glutathione S-transferase. chi-GSTl. auxin-induced GST.
SEQ ID NO: 830
AAD37699.1 AF145730 Oryza sativa homeodomain leucine zipper protein. Oshoxδ. tianscription factor.
BAA93461.1 AB028073 Physcomitrella patens homeobox protein PpHB2. PpHB2. homeodomain-leucine zipper gene.
AAF01765.1 AF184278 Glycine max homeodomain-leucine zipper protein 57. Hdl57. tianscription factor.
AAF01764.2 AF184277 Glycine max homeodomain-leucine zipper protein 56. Hdl56. transcription factor.
BAA93466.1 AB028078 Physcomitrella patens homeobox protein PpHB7. PpHB7. homeodomain-leucine zipper gene.
CAB67118.1 Y17306 Lycopersicon esculentum homeodomain protein. h52.
AAF73482.1 AF268422 Brassica rapa subsp. pekrnensis hb-6-like protein, transcription factor; similar to Arabidopsis thaliana hb-6 protein.
AAD37697.1 AF145728 Oryza sativa homeodomain leucine zipper protein. Oshox4. transcription factor.
BAA21017.1 D26578 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 761-940; leucine zipper at nt 941-1048.
BAA05624.1 D26575 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 520-699; leucine zipper at nt 700-805.
BAA93460.1 AB028072 Physcomitrella patens homeobox protein PpHBl. PpHBl. homeodomain-leucine zipper gene.
BAA93464.1 AB028076 Physcomitrella patens homeobox protein PpHB5. PpHB5. homeodomain-leucine zipper gene.
BAA93467.1 AB028079 Physcomitrella patens homeobox protein PpHB8. PpHB8. homeodomain-leucine zipper gene.
BAA93465.1 AB028077 Physcomitrella patens homeobox protein PpHB6. PpHB6. homeodomain-leucine zipper gene.
BAA05625.1 D26576 Daucus carota transcriptiional regulator. DNA-binding protein, homeodomain at nt 300-479; leucine zipper at nt 480-587.
BAA93468.1 AB028080 Physcomitrella patens homeobox protein PpHB9. PpHB9. homeodomain-leucine zipper gene.
BAA05623.1 D26574 Daucus carota trancriptional regulator. DNA-binding protein, homeodomain at nt 498-677; leucine zipper at nt 678-785.
BAA05622.1 D26573 Daucus carota transcriptional regulator. DNA-binding protein, homeodomain at nt 585-764; leucine zipper at nt 765-851.
AAD37698.1 AF145729 Oryza sativa homeodomain leucine zipper protein. Oshox5. transcription factor.
CAA64221.1 X94449 Pimpinella brachycaφa transcription activator, homeobox-leucine zipper protein. PHZ4.
CAA64152.1 X94375 Pimpinella brachycaφa transcription activator, homeobox-leucine zipper protein.
CAA64491.1 X95193 Pimpinella brachycaφa tianscription activator, homeobox-leucine zipper protein.
BAA93463.1 AB028075 Physcomitrella patens homeobox protein PpHB4. PpHB4. homeodomain-leucine zipper gene.
CAA06728.1 AJ005833 Craterostigma plantagineum transcription factor, homeodomain leucine zipper protein, hb-2.
AAD37695.1 AF145726 Oryza sativa homeodomain leucine zipper protein. Oshox2. transcription factor.
CAA65456.2 X96681 Oryza sativa transcription factor. DNA-binding protein. Oshoxl. homeodomain leucine zipper gene.
AAF19980.1 AF211193 Oryza sativa homeodomain-leucine zipper tianscription factor. Hoxl. hoxl.
AAK31270.1 AC079890 Oryza sativa homeodomain leucine zipper protein hoxl. OSJNBb0089A17.12.
CAA63222.1 X92489 Glycine max transcription activator, homeobox-leucine zipper protein.
CAA06717.1 AJ005820 Craterostigma plantagineum transcription factor, homeodomain leucine zipper protein, hb-1.
AAA79778.1 L48485 Helianthus annuus homeodomain protein, putative. SEQ ID NO: 831
CAA06334.1 AJ005077 Lycopersicon esculentum protein kinase. TCTR2 protein. TCTR2.
AAG31141.1 AF305911 Oryza sativa
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAG31142.1 AF305912 Hordeum vulgare
EDR1. EDR1. MAP kinase kinase kinase; similar to Arabidopsis thaliana EDR1.
AAK30005.1 AY029067 Rosa hybrid cultivar
CTR2 protein kinase.
AAD46406.1 AF096250 Lycopersicon esculentum ethylene-responsive protein kinase TCTR1. ER50. serine/threonine kinase; similar to
Arabidopsis thaliana negative regulator ofthe ethylene response pathway encoded by
GenBank Accession Number L08789.
CAA73722.1 Y13273 Lycopersicon esculentum putative protein kinase.
AAD10057.1 AFl 10519 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTRlv.
AAD 10056.1 AF 110518 Lycopersicon esculentum ethylene-inducible CTRl-like protein kinase. protein kinase homolog; ethylene and fruit ripening inducible CTRl-like protein kinase; TCTR1.
AAA34002.1 M67449 Glycine max protein kinase. PK6.
AAK11734.1 AY027437 Arachis hypogaea serine/threonine/tyrosine kinase.
BAB16918.1 AP002863 Oryza sativa putative protein kinase. P0005A05.22.
CAC09580.1 AJ298992 Fagus sylvatica
Abscisic acid (ABA) and calcium induced protein kinase. protein kinase (PK). pkl.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl . Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
AAF59906.1 AF197947 Glycine max receptor protein kinase-like protein. CLV1B.
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1A.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl .7.
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
CAA08995.1 AJ010091 Brassica napus MAP3K alpha 1 protein kinase. MAP3K alpha 1.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
CAA08997.1 AJO 10093 Brassica napus MAP3Kbeta 1 protein kinase. MAP3Kbeta 1.
CAB54520.1 AJ238845 Brassica napus putative role in cell cycle control. MAP3K epsilon 1 protein kinase. MAP3Kel.
AAF34436.1 AF172282 Oryza sativa similar to mitogen-activated protein kinases. DUPRl 1.32.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl . protein kinase; transcript abundance decreases rapidly after cytokinin treatment.
BAA06538.1 D31737 Nicotiana tabacum protem-serine/threonine kinase.
AAF76189.1 AF271206 Rosa hybrid cultivar CTRl-like protein kinase. Raf-like protein kinase.
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484)
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl. GrnRLKl.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
AAF43394.1 AF230501 Oryza sativa subsp. japonica serine/tlireonine protein kinase. YK1.
AAK16409.1 AF320086 Zeamays serine threonine kinase 1. stkl . expressed in mature tassel.
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAB39437.1 AP003338 Oryza sativa receptor-like kinase. OJ1212_B09.6.
AAK11568.1 AF318492 Lycopersicon hirsutum Pto-like protein kinase B. LhirPtoB.
SEQ ID NO: 832
AAF35901.1 AF230332 Zinnia elegans expansin 2.
CAC19184.1 AJ291817 Cicer arietinum expansin.
AAGl 3982.1 AF297521 Prunus avium expansin 1. Exp 1. PruavExp 1.
BAB 19676.1 AB029083 Prunus persica expansin. PchExpl.
AAC33529.1 U93167 Prunus armeniaca expansin. PA-Expl.
AAC33530.1 AF038815 Prunus armeniaca expansin. Exp2.
AAD47901.1 AF085330 Pinus taeda expansin.
AAB37746.1 U30382 Cucumis sativus expansin SI precursor. Cs-EXPl. similar to pollen allergen Lol pi, Lolium perenne, Swiss- Prot Accession Number P 14946; former gene name CuExSl; expansin-29 (Ex29) protein. AAF21101.1 AF159563 Fragaria x ananassa expansin. Exρ2. ripening regulated.
AAB40634.1 U64890 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession
Numbers U30477 and U30479.
AAB40637.1 U64893 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession
Numbers U30477 and U30479.
AAB40635.1 U64891 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession
Numbers U30477 and U30479.
CAB43197.1 AJ239068 Lycopersicon esculentum cell wall loosening enzyme. expansin2. exp2.
AAB40636.1 U64892 Pinus taeda expansin. similar to Arabidopsis expansin encoded by GenBank Accession Numbers U30476, U30478, U30480 and U30481, to Cucumis sativus expansin encoded by GenBank Accession Numbers U30382 and U30460, and to rice expansin encoded by GenBank Accession Numbers U30477 and U30479.
AAC64201.1 AF096776 Lycopersicon esculentum expansin. LeEXP2.
AAD49956.1 AFl 67360 Rumex palustris expansin. EXPL
AAC96081.1 AF049354 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP5. cell wall protein.
AAC39512.1 AF043284 Gossypium hirsutum expansin. GhEXl. contains N-terminal signal peptide.
AAB81662.1 U85246 Oryza sativa expansin. Os-EXP4.
AAG13983.1 AF297522 Prunus avium expansin 2. Exp2. PraavExρ2.
AAF32409.1 AF230276 Triphysaria versicolor alpha-expansin 3.
AAG32921.1 AF184233 Lycopersicon esculentum expansin. Exp 10.
BAB32732.1 AB049406 Eustoma grandiflorum expansin. Eg Expansin.
AAF32411.1 AF230278 Triphysaria versicolor alpha-expansin 1.
AAF35902.1 AF230333 Zinnia elegans expansin 3.
AAB38074.1 U30477 Oryza sativa induces extension (creep) in plant cell walls, expansin Os-EXP2. Os-EXP2. former gene name
RiExB.
AAC96080.1 AF049353 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP4. cell wall protein.
AAF17570.1 AF202119 Marsilea quadrifolia alpha-expansin. EXPL Mq-EXPl.
CAC06433.1 AJ276007 Festuca pratensis expansin. exp2.
AAD13633.1 AF059489 Lycopersicon esculentum expansin precursor. Exp5.
CAC19183.1 AJ291816 Cicer arietinum expansin.
AAF62181.1 AF247163 Oryza sativa alpha-expansin OsEXP6. cell wall loosening factor; expressed in internodes and leaves.
AAF62180.1 AF247162 Oryza sativa alpha-expansin OsEXP5. cell wall loosening factor; expressed in internodes, leaves, coleoptiles, and roots.
CAB46492.1 AJ243340 Lycopersicon esculentum expansin9. exp9.
BAA88200.1 AP000837 Oryza sativa
EST AU078708(E60526) conesponds to a region ofthe predicted gene. Similar to expansin
(U85246).
AAF32410.1 AF230277 Triphysaria versicolor alpha-expansin 2.
AAB37749.1 U30460 Cucumis sativus expansin S2 precursor. Cs-EXP2. similar to pollen allergen Lol pi, Lolium perenne, Swiss-
Prot Accession Number P14946; former gene name CuExS2; expansin-30 (Ex30) protein.
CAA04385.1 AJ000885 Brassica napus Cell wall extension in plants. Expansin.
AAF17571.1 AF202120 Regnellidium diphyllum alpha-expansin. EXP 1. Rd-EXP 1.
AAD13632.1 AF059488 Lycopersicon esculentum expansin precursor. Exp4.
CAA06271.2 AJ004997 Lycopersicon esculentum expansinlδ. exp 18.
AAC63088.1 U82123 Lycopersicon esculentum expansin. LeEXPl. fruit ripening regulated expansin.
A ΔAΔPCQ9ή6m0777.11 A ΛFPn0_49n35<;n0
involved in acid-growth response, alpha-expansin precursor. Nt-EXPl. cell wall protein.
AAF62182.1 AF247164 Oryza sativa alpha-expansin OsEXP7. cell wall loosening factor; expressed in internodes and leaves.
CAC18802.1 AJ289154 Glycine max expansion of cell walls, expansin. dd2/63.
AAC96078.1 AF049351 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP2. cell wall protein.
AAG01875.1 AF291659 Striga asiatica alpha-expansin 3. Exp3.
CAA69105.1 Y07782 Oryza sativa expansin. RiExA.
AAC96079.1 AF049352 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP3. cell wall protein.
SEQ ID NO: 833 ~~~~
AAA34030.1 J03492 Spinacia oleracea glycolate oxidase (EC 1.1.3.15).
AAB40396.1 U80071 Mesembryanthemum crystallinum glycolate oxidase. GOX.
BAA03131.1 D 14044 Cucurbita sp. glycolate oxidase.
AAB82143.1 AF022740 Oryza sativa glycolate oxidase. GOX.
CAA63482.1 X92888 Lycopersicon esculentum conversion of glycolate to glyoxylate + H2O2. glycolate oxidase.
AAC32392.1 AF082874 Medicago sativa glycolate oxidase.
AAC33509.1 U62485 Nicotiana tabacum photorespiration. glycolate oxidase. GLO.
AAF03097.1 AF162196 Lactuca sativa glycolate oxidase.
SEQ ID NO: 838
CAA06770.1 AJ005928 Brassica napus squalene epoxidase homologue. Sqpl;2.
CAA06773.1 AJ005931 Brassica napus squalene epoxidase homologue. Sqpl;l.
BAA24448.1 AB003516 Panax ginseng squalene epoxidase.
CAA06223.1 AJ004923 Lycopersicon esculentum Squalene epoxidase. ERG.
SEQ ID NO: 840
BAB12686.1 AP002746 Oryza sativa putative pyrophosphate-dependent phosphofracto-1-kinase. P0671B11.1. contains ESTs
AU068014(C11507),C28532(C61484),AU090544(C61415).
BAA99438.1 AP002743 Oryza sativa putative pyrophosphate-dependent phosphofructo-1 -kinase. P0710E05.25. contains ESTs
AU068014(C11507),C28532(C61484),AU090544(C61415).
AAB88875.1 U93272 Prunus armeniaca pyrophosphate-dependent phosphofructo- 1 -kinase.
CAA83683.1 Z32850 Ricinus communis pyrophosphate-dependent phosphofructokinase beta subunit. AAC67587.1 AF095521 Citras x paradisi pyrophosphate-dependent phosphofructokinase alpha subunit. PPi-PFKa.
AAA63452.1 M55191 Solanum tuberosum pyrophosphate-fructose 6-phosphate 1 -phosphotransferase beta-subunit. AAC67586.1 AF095520 Citrus x paradisi pyrophosphate-dependent phosphofructokinase beta subunit. PPi-PFKb. PFP.
AAA63451.1 M55190 Solanum tuberosum pyrophosphate-fructose 6-phosphate 1 -phosphotransferase alpha-subunit.
CAA83682.1 Z32849 Ricinus communis pyrophosphate-dependent phosphofructokinase alpha subunit.
SEQ ID NO: 841 AAG60182.1 AC084763 Oryza sativa putative ethylene-responsive element binding protein. OSJNBa0027P10.12.
AAK31279.1 AC079890 Oryza sativa putative ethylene-responsive element binding protein. OSJNBb0089A17.16.
AAG43545.1 AF211527 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 1. ACREl . similar to EREBP transcription factors.
AAF63205.1 AF245119 Mesembryanthemum crystallinum
AP2-related transcription factor. CDBP. stress induced transcription factor.
BAA07321.1 D38123 Nicotiana tabacum
ERFl. ethylene-responsive tianscription factor.
BAA97122.1 AB016264 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf2.
CAB93940.1 AJ238740 Catharanthus roseus putative tianscription factor. AP2-domain DNA-binding protein. orca2.
BAA87068.1 AB035270 Matricaria chamomilla ethylene-responsive element binding proteinl homolog. McEREBPl.
BAA97124.1 AB016266 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf4.
CAB96900.1 AJ251250 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3. CAB96899.1 AJ251249 Catharanthus roseus transcription factor. AP2-domain DNA-binding protein. orca3.
AAC62619.1 AF057373 Nicotiana tabacum transcription factor, ethylene response element binding protein 1. EREBPl.
BAA97123.1 ABO 16265 Nicotiana sylvestris
ERF (EREBP); ethylene-responsive element binding factor for basic PR (Pathogenesis- related) gene of higher plant, ethylene-responsive element binding factor. nserf3.
AAC24587.1 AF071893 Prunus armeniaca
AP2 domain containing protein. AP2DCP.
CAC 12822.1 AJ299252 Nicotiana tabacum
AP2 domain-containing transcription factor. ap2.
AAF76898.1 AF274033 Atiiplex hortensis apetala2 domain-containing protein.
BAA94514.2 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4, BAC clone F9D16; putative Ap2 domain protein (AL035394).
AAC14323.1 AF058827 Nicotiana tabacum
TSH. Tsil. contains putative AP2 DNA-binding domain; similar to Pti6.
AAD00708.1 U91857 Stylosanthes hamata ethylene-responsive element binding protein homolog. similar to EREBPl, -2, -3 and -4 proteins encoded by GenBank Accession Numbers D38123, D38126, D38124, and D38125 respectively.
BAA76734.1 AB024575 Nicotiana tabacum ethylene responsive element binding factor.
BAB03248.1 AB037183 Oryza sativa
ERF protein transcriptional repressor. ethylene responsive element binding factor3. osERF3.
BAB16083.1 AB036883 Oryza sativa transcriptional repressor. osERF3. osERF3. ERF protein family ERF3 associated repression domain.
AAF23899.1 AF193803 Oryza sativa transcription factor EREBPl. EREBP/AP2-like tianscription factor.
AAF05606.1 AFl 90770 Oryza sativa
EREBP-like protein, tshl. TSH1; induced by ethylene.
CAB93939.1 AJ238739 Catharanthus roseus putative transcription factor. AP2-domain DNA-binding protein, orcal.
BAA78738.1 AB023482 Oryza sativa
EST AU055776(S20048) conesponds to a region ofthe predicted gene.; Similar to
Arabidopsis thaliana AP2 domain containing protein RAP2.10 mRNA, partial cds.(AF003103).
AAG43548.1 AF211530 Nicotiana tabacum
Avr9/Cf-9 rapidly elicited protein 111 A. ACREl 11 A. similar to EREBP tianscription factors.
AAG43549.1 AF211531 Nicotiana tabacum
Ayr9/Cf-9 rapidly elicited protein 11 IB. ACREl 1 IB. similar to EREBP transcription factors.
BAA99376.1 AP002526 Oryza sativa
ESTs AU093391(E60370),AU091593(C60458), AU093392(E60370) conespond to a region ofthe predicted gene. Similar to Arabidopsis thaliana BAC F21 J9; AP2 domain protein.
(AC000103).
AAK31271.1 AC079890 Oryza sativa putative transcriptional factor. OSJNBb0089A17.22.
AAKO 1089.1 AF298231 Hordeum vulgare
CBF3-like protein BCBF3. BCBF3. AP2 domain protein; DRE binding factor.
AAG59618.1 AF239616 Hordeum vulgare CRT/DRE-binding factor. CBF.
AAC49567.1 U41466 Zea mays
Glossyl5. Glossyl5. AP2 DNA-binding domain protein; similar to the Arabidopsis floral homeotic gene APETALA2; maize AP2-domain regulator of leaf epidermal tiaits; homeotic regulator of leaf epidermal cell identity; allele: W64A; putative transcription factor.
~^ SEQ ID NO: 842 _______ ~~~^~~~
AAC32034.1 AF023472 Hordeum vulgare peptide transporter, ptrl. PTR1; integral membrane protein.
BAB40113.1 AP003311 Oryza sativa putative peptide transport protein. P0024G09.4. contains ESTs
D40448(S2437),C71800(E0368),AU102190(E2393),
AU055921(S20154),AU102191(E2393),AU055922(S20154),
C98524(E0368),AU097146(S2437).
BAB16458.1 AP002483 Oryza sativa putative peptide transport protein. P0019D06.16. contains ESTs
D40448(S2437),C71800(E0368),AU102190(E2393),
AU055921(S20154),AU102191(E2393),AU055922(S20154),
C98524(E0368),AU097146(S2437).
AAD01600.1 AF016713 Lycopersicon esculentum LeOPTl. LeOPTl. oligopeptide transporter.
AAF07875.1 AF140606 Oryza sativa nitrate transporter. NRTL
AAF20002.1 AF213936 Prunus dulcis amino acid/peptide transporter. PTR2. similar to transporters of nitrogenous compounds.
AAG46153.1 ACO 18727 Oryza sativa putative peptide transporter. OSJNBa0056G17.8.
CAC00544.1 AJ277084 Nicotiana plumbaginifolia ion transport, putative low-affinity nitrate transporter, nrtl.l.
CAC00545.1 AJ277085 Nicotiana plumbaginifolia ion transport, putative low-affinity nitrate transporter, nrtl .2.
AAA80582.1 U17987 Brassica napus putative nitiate transporter. RCH2 protein.
CAC07206.1 AJ278966 Brassica napus Low-affinity nitiate transporter, nitiate transporter, nrtl. AAG21898.1 AC026815 Oryza sativa putative peptide transport protein. OSJNBa0079L16.13.
BAB 19758.1 AB052786 Glycine max putative nitrate tiansporter NRT1-3. NRT1-3.
AAK15441.1 AC037426 Oryza sativa putative nitiate transporter. OSJNBb0014Il 1.9.
AAG21906.1 AC026815 Oryza sativa putative peptide tiansport protein. OSJNBa0079L16.9.
BAB19760.1 AB052788 Glycine max nitiate transporter NRT 1-5. NRT1-5.
AAG46154.1 AC018727 Oryza sativa putative peptide tiansporter. OSJNBa0056G17.27.
BAB 19757.1 AB052785 Glycine max nitiate transporter NRT1-2. NRT1-2.
BAB 19756.1 AB052784 Glycine max nitrate tiansporter NRT1-1. NRT1-1.
BAB 16322.1 AP002818 Oryza sativa putative peptide transporter-like protein. P0436E04.4.
AAB69642.1 AF000392 Lotus japonicus peptide transporter. LJNOD65.
CAA93316.1 Z69370 Cucumis sativus nitrite transporter. NiTRl.
AAD 16016.1 AF080545 Nepenthes alata peptide transporter. PTR1.
BAB 19759.1 AB052787 Glycine max putative nitiate tiansporter NRT 1-4. NRT1-4.
AAD42860.1 AF154930 Prunus dulcis transporter-like protein. TLP 1.
SEQ ID NO: 847
CAA61980.1 X89890 Bidens pilosa Calmodulin.
AAF73157.1 AF150059 Brassica napus calmodulin. CaMl . involved in seed germination.
BAA87825.1 AP000815 Oryza sativa
ESTs AU030013(E50493),AU081341(E50493) conespond to a region ofthe predicted gene.
Similar to O. sativa gene encoding calmodulin. (Z12828).
CAA67054.1 X98404 Capsicum annuum cahnodulin-2.
AAA87347.1 M88307 Brassica juncea calmodulin.
AAA33397.1 L18912 Lilium longiflorum calcium binding protein, signal transduction. calmodulin. putative.
AAG27432.1 AF295637 Elaeis guineensis calmodulin.
CAA42423.1 X59751 Daucus carota calmodulin. Ccam-1.
AAGl 1418.1 AF292108 Prunus avium calmodulin.
AAA92681.1 U13882 Pisum sativum calcium-binding protein, calmodulin.
AAB46588.1 U83402 Capsicum annuum cahnodulin.
AAA33706.1 M80836 Petunia x hybrida cahnodulin. CAM81.
AAF65511.1 AF108889 Capsicum annuum cahnodulin.
CAA43143.1 X60738 Malus x domestica
Calmodulin. CaM.
AAAI 9571.1 UIO 150 Brassica napus calcium binding, calmodulin. bcml .
CAA78301.1 Z12839 Lilium longiflorum calcium binding protein, signal transduction. calmodulin.
BAA88540.1 AP000969 Oryza sativa
ESTs AU081349(E61253), D41425(S3918) conespond to a region ofthe predicted gene.
Similar to calmodulin. (AF042840).
AAB36130.1 S81594 Vigna radiata auxin-regulated calmodulin. auxin-regulated calmodulin, arCaM. This sequence comes from
Fig. 1; arCaM.
AAC36059.1 AF042840 Oryza sativa calmodulin. CaMl.
AAA33901.1 L18913 Oryza sativa calcium binding protein, signal transduction. calmodulin. putative.
AAA33900.1 L18914 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAA34237.1 L20691 Vigna radiata cahnodulin.
CAA78288.1 Z12828 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAA32938.1 M27303 Hordeum vulgare calmodulin.
CAA78287.1 Z12827 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAC49587.1 U49105 Triticum aestivum calmodulin TaCaM4-l. calcium-binding protein.
AAC49586.1 U49104 Triticum aestivum calmodulin TaCaM3-3. calcium-binding protein.
AAC49583.1 U48692 Triticum aestivum calmodulin TaCaM2-3. calcium-binding protein.
AAC49585.1 U49103 Triticum aestivum cahnodulin TaCaM3-2. calcium-binding protein.
AAC49584.1 U48693 Triticum aestivum calmodulin TaCaM3-l. calcium-binding protein.
AAC49582.1 U48691 Triticum aestivum calmodulin TaCaM2-2. calcium-binding protein.
AAC49580.1 U48689 Triticum aestivum calmodulin TaCaMl-3. calcium-binding protein.
AAC49579.1 U48688 Triticum aestivum calmodulin TaCaMl-2. calcium binding protein.
AAC49578.1 U48242 Triticum aestivum calmodulin TaCaM 1 - 1. calcium-binding.
AAA03580.1 L01431 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-2. putative.
AAC36058.1 AF042839 Oryza sativa calmodulin. CaM2.
AAA85156.1 U20296 Solanum tuberosum calcium-binding protein, calmodulin.
AAA34014.1 L01432 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-3. putative.
AAD10244.1 AF030032 Phaseolus vulgaris calmodulin. CaM. EF-hand protein; calcium-dependent protein; functions in calcium signal transduction pathways.
CAA36644.1 X52398 Medicago sativa calmodulin (AA 1-149).
AAA85155.1 U20294 Solanum tuberosum calcium-binding protein, calmodulin. AAB68399.1 U79736 Helianthus annuus cahnodulin. HaCaM.
AAA62351.1 U20295 Solanum tuberosum calcium-binding protein, calmodulin.
AAA34238.1 L20507 Vigna radiata calmodulin.
AAA85157.1 U20297 Solanum tuberosum calcium-binding protein, calmodulin.
AAA34013.1 L01430 Glycine max calcium-binding regulatory protein, cahnodulin. SCaM-1. putative.
AAA33705.1 M80831 Petunia x hybrida calmodulin-related protein. CAM53.
CAA74307.1 Y13974 Zea mays calmodulin.
CAA54583.1 X77397 Zeamays calmodulin. CaM2.
SEQ ID NO: 848
CAA06486.1 AJ005340 Linum usitatissimum IAA amidohydrolase. homolog.
SEQ ID NO: 850
BAB17350.1 AP002747 Oryza sativa putative nodulin. P0698G03.34. contains ESTs D39891(S1543),D41717(S4395),AU033037(S1543).
BAA85440.1 AP000616 Oryza sativa
ESTs AU055729(S20023),AU055730(S20023) conespond to a region ofthe predicted gene.; similar to Medicago nodulin N21 -like protein (AC004218).
CAB53493.1 AJ245900 Oryza sativa
CAA303720.1 protein. q3037.20. Similar to Medicago nodulin N21 (MtN21).
SEQ ID NO: 852
AAD16018.1 AF081514 Taxus canadensis prenyltiansferase. geranylgeranyl diphosphate synthase. geranylgeranyl pyrophosphate synthase.
SEQ ID NO: 853
BAB32588.1 AB055807 Momordica charantia inhibitor against trypsin. bgit.
AAA34180.1 J05094 Lycopersicon peravianum proteinase inhibitor I precursor.
AAA34198.1 M59427 Lycopersicon peravianum proteinase inhibitor I. proteinase inhibitor I.
CAB61327.1 AJl 32473 Amaranthus hypochondriacus Proteinase inhibition, trypsin inhibitor.
AAA60745.1 J04099 Lycopersicon esculentum proteinase inhibitor I. ER1.
CAA78269.1 Z12623 Nicotiana tabacum
Putative precursor of serine proteinase inhibitor type I. Pre-pro-proteinase inhibitor I.
CAA47461.1 X67076 Nicotiana tabacum inhibitor of microbial serine proteinases (major isoform). TIMPa.
CAA78265.1 Z12619 Nicotiana tabacum precursor for serine proteinase inhibitor I. Pre-pro-proteinase inhibitor I.
CAA47460.1 X67075 Nicotiana tabacum inhibitor of microbial serine proteinases (minor isoform). TIMPb. AAA34067.1 M74102 Nicotiana sylvestris pre-pro-proteinase inhibitor I.
AAC49603.1 U30861 Solanum tuberosum serine proteinase inhibitor, wound-inducible proteinase inhibitor I. BAA02823.1 D13662 Nicotiana glauca X Nicotiana langsdorffii genetic tumor-related proteinase inhibitor I precursor. GTI.
AAA34199.1 K03290 Lycopersicon esculentum wound-induced proteinase inhibitor I prepropeptide.
AAA34200.1 M13938 Lycopersicon esculentum proteinase inhibitor I. PIIF.
AAA69780.1 L06137 Solanum tuberosum proteinase inhibitor I. pinl. putative.
AAA72133.1 L06985 Solanum tuberosum proteinase inhibitor I. pinl. The 'a' ofthe first atg is missing.
CAA78259.1 Z12611 Solanum tuberosum proteinase inhibitor I.
AAA69781.1 L06606 Solanum tuberosum proteinase inhibitor I. precursor.
CAA48136.1 X67950 Solanum tuberosum protease inhibitor I. pinl.
CAA47907.1 X67675 Solanum tuberosum proteinase inhibitor I. pinl.
CAB71340.1 AJ250663 Hordeum vulgare putative proteinase inhibitor, bci-7. similarity to subtilisin/chymotrypsin inhibitor.
CAA57677.1 X82187 Zeamays serine proteinase inhibitor, substilin /chymotiypsin-like inhibitor. pis7.
CAA55588.1 X78988 Zea mays proteinase inhibitor. MPI.
CAA49593.1 X69972 Zea mays proteinase inhibitor. MPI.
AAA33816.1 Ml 7108 Solanum tuberosum proteinase inhibitor I. precursor.
CAA57307.1 X81647 Cucurbita maxima Pumpkin fruit trypsin inhibitor. pfiAF4.
CAA57203.1 X81447 Cucurbita maxima Pumpkin Frait Chymotrypsin Inhibitor. pfiBM7.
SEQ ID NO: 859
AAC34855.1 AF082030 Hemerocallis hybrid cultivar senescence-associated protein 5. SA5. mRNA accumulates in senescing petals and accumulation is induced by exogenous ABA.
AAG13616.1 AC078840 Oryza sativa putative senescence-associated protein. OSJNBb0073N24.21.
SEQ ID NO: 864
AAF62403.1 AF212183 Nicotiana tabacum haφin inducing protein, hinl . similar to hinl protein.
CAA68848.1 Y07563 Nicotiana tabacum activated during hypersensitive response, hinl.
AAB97367.1 AF039532 Oryza sativa haφin induced gene 1 homolog. Hinl.
SEQ ID NO: 871
AAC61839.1 AF025430 Papaver somniferam berberine bridge enzyme, bbel. (S)-reticuline:oxygen oxidoreductase (methylene bridge forming).
AAB20352.1 S65550 Eschscholzia californica
(S)-reticuline: oxygen oxidoreductase (methylene-bridge-forming). /gene="(S)- reticulrne:oxygen oxidoreductase. This sequence comes from Fig 2; berberine bridge enzyme.
AAC39358.1 AF005655 Eschscholzia californica oxidizes the N-methyl group of (S)-reticuline to the berberine bridge carbon C-8 of (S)- scoulerrne in berberine and benzophenanthridine alkaloid biosynthesis, berberine bridge enzyme, bbel. covalently-bound FAD-dependent oxidase; elicitor-inducible.
AAD17487.1 AF049347 Berberis stolonifera
Catalyzes the oxidative cyclization of the N-methyl group of (S)-reticuline into the berberine bridge carbon (C-8) of (S)-scoulerine. berberine bridge enzyme, bbel. Covalently flavinylated oxidase of isoquinoline alkaloid biosynthesis in plants.
SEQ ID NO: 872 AAF98369.1 AF158253 Nicotiana tabacum patatin-like protein 3. PAT3. NtPat3.
CAA73328.1 Y12793 Cucumis sativus mobilization of fat during seed germination, patatin-like protein.
CAAl 1042.1 AJ223039 Hevea brasiliensis latex allergen, sequence similarity to patatins.
AAF25553.1 AFl 13546 Hevea brasiliensis latex protein allergen Hev b 7. putative PLA2; similar to Solanum tubulin patatin encoded by
GenBank Accession Number X03932.
CAAl 1041.1 AJ223038 Hevea brasiliensis latex allergen, with sequence similarity to patatins.
AAC27724.1 U80598 Hevea brasiliensis latex patatin homolog. putative PLA2; latex protein allergen; similar to Solanum tubulin patatin encoded by GenBank Accession Number X03932.
AAK27797.1 AF318315 Vigna unguiculata patatin-like protein.
AAK18751.1 AF193067 Vigna unguiculata patatin-like protein.
AAB08428.1 U68484 Nicotiana tabacum patatin homolog.
AAD22170.1 AF061282 Sorghum bicolor patatin-like protein.
AAF98368.1 AF158027 Nicotiana tabacum patatin-like protein 1. PAT1. NtPatl.
AAD22169.1 AF061282 Sorghum bicolor patatin-like protein.
CAA81735.1 Z27221 Solanum tuberosum patatin.
CAA31575.1 X13178 Solanum tuberosum patatin B2 (AA 1 - 386).
AAA33819.1 M18880 Solanum tuberosum patatin.
CAA31576.1 XI 3179 Solanum tuberosum patatin Bl (377 AA) (1 is 3rd base in codon).
CAA27588.1 X03956 Solanum tuberosum patatin.
AAA33828.1 M21879 Solanum tuberosum patatin.
CAA27571.1 X03932 Solanum tuberosum patatin.
CAA25592.1 X01125 Solanum tuberosum patatin.
AAA66198.1 U09331 Solanum brevidens patatin precursor.
AAB08427.1 U68483 Nicotiana tabacum patatin homolog.
AAF98370.1 AF158254 Nicotiana tabacum patatin-like protein 2. PAT2.
AAD22149.1 AF061282 Sorghum bicolor patatin-like protein, similar to the EST sequences E0496(panicle at flowering stage), R2382
(root), R2382 (root), S4036 (shoot), S3728 (shoot), SI 3457 (green shoot).
SEQ ID NO: 875
BAA93022.1 AP001552 Oryza sativa
ESTs C74776(E51022),C26123(C116681) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana cultivar Landsberg extra-large G-protein (AF060942).
SEQ ID NO: 876
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.il.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAA33915.1 L27821 Oryza sativa receptor type serme/threonine kinase. protein kinase.
BAA82556.1 AB030083 Populus nigra lectin-like protein kinase. PnLPK.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and
ABA treatment.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
AAB93834.1 U82481 Zeamays
KI domain interacting kinase 1. KIKl . receptor-like protein kinase; serine/threonine protein kinase.
AAF66615.1 AF 142596 Nicotiana tabacum LRR receptor-like protein kinase.
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl. protein kinase; tianscript abundance decreases rapidly after cytokinin treatment.
AAB09771.1 U67422 Zea mays
CRINKLY4 precursor. cr4. receptor kinase homolog.
AAF34428.1 AF172282 Oryza sativa receptor-like protein kinase. DUPRl 1.18.
BAB07906.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.14.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
BAA94516.1 AP001800 Oryza sativa
Similar to Zea mays S-domain receptor-like protein kinase (AJ010166).
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
CAA73134.1 Y12531 Brassica oleracea serme/threonine kinase. BRLK.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAA06538.1 D31737 Nicotiana tabacum protein-serine/threonine kinase.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extiacellular S domain: 8-1342; transmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl .
BAB07904.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.12.
BAA94518.1 AP001800 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2 section 111 of 255; putative receptor-like protein kinase (AC002392).
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extiacellular S domain: 123-1346; transmembrane domain: 1347-1412; intiacellular kinase domain: from 1413.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB07905.1 AP002835 Oryza sativa putative S-receptor kinase. P0417G05.13.
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
SEQ ID NO: 883 BAA90510.2 AP001111 Oryza sativa rice EST AU030811, similar to rice Ca+2-ATPase (U82966).
AAF73985.1 AF096871 Zea mays calcium pump, calcium ATPase. capl.
CAA63790.1 X93592 Dunaliella bioculata
P-type ATPase. cal. calcium pumping; CAL
AADl 1618.1 AF050496 Lycopersicon esculentum
Ca2+- ATPase. LCA1B; alternative transcript.
AAA34138.1 M96324 Lycopersicon esculentum
The calcium ATPase is a calcium ion pump. Ca2+-ATPase. LCA
AAB58910.1 U82966 Oryza sativa Ca2+-ATPase.
AADl 1617.1 AF050495 Lycopersicon esculentum Ca2+- ATPase. LCAl A; alternative transcript.
AAG28435.1 AF195028 Glycine max plasma membrane Ca2+-ATPase. SCA1.
AAG28436.1 AF195029 Glycine max plasma membrane Ca2+-ATPase. SCA2.
CAA68234.1 X99972 Brassica oleracea calmodulin-stimulated calcium-ATPase.
AAD31896.1 AF 145478 Mesembryanthemum crystallinum calcium ATPase.
AAB60276.1 U09989 Zeamays H(+)-transporting ATPase. Mhal .
CAB69824.1 AJ271439 Prunus persica plasma membrane H+ ATPase. PPA1.
AAD46187.1 AFl 56683 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma8.
BAA01058.1 D10207 Oryza sativa H-ATPase. OSA1.
AAB49042.1 U54690 Dunaliella acidophila plasma membrane proton ATPase. dhal. DaDHAl; proton pump.
AAA34173.1 M60166 Lycopersicon esculentum H+- ATPase. LHA1.
CAA52107.1 X73901 Dunaliella bioculata plasma membrane ATPase. pmal.
AAB35314.2 S79323 Vicia faba plasma membrane H(+)-ATPase precursor, plasma membrane H(+)-ATPase. This sequence comes from Fig. 1; conceptual translation presented here differs from translation in publication.
BAA06629.1 D31843 Oryza sativa plasma membrane H+-ATPase. OSA2.
AAA34094.1 M80489 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pmal .
BAA08134.1 D45189 Zostera marina plasma membrane H+- ATPase. zhal .
CAB85494.1 AJ132891 Medicago truncatula proton pump. H+- ATPase. hal.
CAB85495.1 AJ132892 Medicago truncatula proton pump. H+- ATPase. hal .
AAB84202.2 AF029256 Kosteletzkya virginica plasma membrane proton ATPase. ATP1.
CAA47275.1 X66737 Nicotiana plumbaginifolia plasma membrane H+-ATPase. pma4.
CAA54045.1 X76535 Solanum tuberosum H(+)-transporting ATPase. PHA2.
AAD46186.1 AF 156679 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma6.
AAB17186.1 U72148 Lycopersicon esculentum plasma membrane H+-ATPase. LHA4. plasma membrane proton pumping ATPase.
CAB69823.1 AJ271438 Prunus persica plasma membrane H+ ATPase. PPA2.
AAB41898.1 U84891 Mesembryanthemum crystallinum plasma membrane proton pump. H+-transporting ATPase. PMA.
CAC29436.1 AJ310524 Vicia faba
P-type H+- ATPase. ha5. predominantly expressed in guard cells and flowers.
BAA37150.1 AB022442 Vicia faba p-type H+- ATPase. VHA2.
CAA59800.1 X85805 Zeamays
H(+)-transporting ATPase. MHA-2.
CAA59799.1 X85804 Phaseolus vulgaris
H(+)-transporting ATPase. BHA-1.
CAC29435.1 AJ310523 Vicia faba
P-type H+-ATPase. vha4. predominantly expressed in flowers.
AAD46188.1 AF156691 Nicotiana plumbaginifolia plasma membrane proton ATPase. pma9.
AAA34099.1 M80491 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pma3.
AAA34052.1 M27888 Nicotiana plumbaginifolia H+-tianslocating ATPase.
CAA54046.1 X76536 Solanum tuberosum H(+)-tiansporting ATPase. PHA1.
AAA34098.1 M80490 Nicotiana plumbaginifolia plasma membrane H+ ATPase. pma3.
AAD55399.1 AFl 79442 Lycopersicon esculentum plasma membrane H+- ATPase isoform LHA2. LHA2.
AAF98344.1 AF275745 Lycopersicon esculentum plasma membrane H+-ATPase. LHA2. P-type ion pump.
AAGO 1028.1 AF289025 Cucumis sativus plasma membrane H+- ATPase.
AAK31799.1 AY029190 Lilium longiflorum plasma membrane H+ ATPase. LILHA1.
AAA81348.1 U38965 Vicia faba p-type H+-ATPase. VHA2.
AAK32119.1 AF308817 Hordeum vulgare plasmalemma H+- ATPase 2.
AAK32118.1 AF308816 Hordeum vulgare plasmalemma H+- ATPase 1.
AAA20600.1 U08984 Zeamays plasma-membrane H+ ATPase. Zmpmal.
SEQ ID NO: 884
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl . receptor-like protein kinase; serine/tlireonine protein kinase.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extiacellular S domain: 123-1346; tiansmembrane domain: 1347-1412; intracellular kinase domain: from 1413.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extiacellular S domain: 8-1342; transmembrane domain: 1343-1411; intiacellular kinase domain: 1412-2554.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
BAA92836.1 AB032473 Brassica oleracea S18 S-locus receptor kinase. SRK18.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulin-like domain.
AAA33008.1 M97667 Brassica napus serme/threonine kinase receptor.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRK1. Chrkl.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
AAK02023.1 AC074283 Oryza sativa Putative protein kinase-like. OSJNBa0087H07.5.
CAA79324.1 Z 18884 Brassica oleracea S-receptor kinase related protein.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
CAB51836.1 AJ243961 Oryza sativa Putitive Ser/Thr protein kinase. 11332.7.
AAA33915.1 L27821 Oryza sativa receptor type serine/threonine kinase. protein kinase.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
AAF66615.1 AF142596 Nicotiana tabacum
LRR receptor-like protein kinase.
BAB17139.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.31.
SEQ ID NO: 885
CAA80358.1 Z22645 Solanum tuberosum cleavage of sucrose to glucose and fructose, beta-fructofuranosidase.
CAA79676.1 Z21486 Solanum tuberosum cleavage of sucrose to glucose and fructose, beta-fructofuranosidase.
CAA57428.1 X81834 Nicotiana tabacum beta-fructofuranosidase. Ntbfracl. beta-fructosidase.
CAA49162.1 X69321 Daucus carota beta-fructofuranosidase. Inval*DCl.
CAA57389.1 X81792 Chenopodium rabrum beta-fructofuranosidase. CINl.
AAC17166.1 AF063246 Pisum sativum hydrolyzes sucrose to glucose and fructose, cell wall invertase. bfructl. isoform PcI-2; the
Pcl-l isoform is in the file with GenBank Accession Number X85327; beta- fructofuranosidase.
CAA59677.1 X85327 Pisum sativum hydφlyze sucrose into fructose and glucose, invertase. bfructl. beta-fructofuranosidase.
AAD02263.1 AF043346 Zea mays sucrose hydrolysis, cell wall invertase. incw3. Incw3; beta-fructofuranosidase.
CAA84526.1 Z35162 Vicia faba hydrolyze sucrose, beta-fructofuranosidase; cell wall invertase I; fructosidase. VFCWINV
AAC96065.1 AF030420 Triticum aestivum hydrolyzes sucrose, cell wall invertase. IVR1. hydrolase; beta-fructofuranosidase; fructosidase.
CAA53099.1 X75353 Daucus carota beta-fructofuranosidase.
AAB68679.1 U92438 Phaseolus vulgaris soluble acid invertase. PVSAI; potential vacuolar targeted enzyme; beta-fructofuranosidase.
CAA89992.1 Z49831 Vicia faba hydrolyze sucrose, vacuolar invertase; beta-fructofuranosidase. VFVCINV.
CAA53097.1 X75351 Daucus carota beta-fructofuranosidase .
CAA77267.1 Y 18707 Daucus carota beta-fructofuranosidase, isoform I. Inv*Dc4. soluble acid invertase.
CAA53098.1 X75352 Daucus carota beta-fructofuranosidase.
CAA77266.1 Y18706 Daucus carota beta-fructofuranosidase, isoform II. Inv*Dc5. soluble acid invertase.
AAC96066.1 AF030421 Triticum aestivum hydrolyzes sucrose, cell wall invertase. IVR3. hydrolase; beta-fructofuranosidase; fructosidase.
AAG36943.1 AF274299 Brassica oleracea cleaves sucrose into glucose and fructose at acid pH optima, acid invertase AI7-3. sucrose hydrolysing enzyme; beta-fructofuranosidase. SEQ ID NO: 887
AAD10836.1 U52079 Solanum tuberosum
P-glycoprotein. pmdrl. binds ATP; ATPase; transporter; tiansmembrane protein.
BAA96612.1 AP002482 Oryza sativa
Similar to Arabidopsis thaliana chromosome 2, BAC F14M4 ; putative ABC transporter
(AC004411).
AAG49002.1 AY013246 Hordeum vulgare putative ABC transporter. 635P2.4a; GC splice donor confirmed by cDNA and comparative sequencing.
AAG45492.1 AYO 13245 Oryza sativa
3615.4. putative ABC transporter; GC splice donor confirmed by cDNA alignment and comparative sequence.
BAA83352.1 AP000391 Oryza sativa
ESTs AU067992(C11433),AU077424(C11433) conespond to a region ofthe predicted gene.;
Similar to ABC transporter-7 (U43892).
AAG49003.1 AYO 13246 Hordeum vulgare putative ABC transporter. 635P2.4b; GC splice donor confirmed by cDNA alignment and comparative sequence.
BAB 17113.1 AP002866 Oryza sativa putative white protein; ATP-binding cassette tiansporter. P0410E01.34.
BAA90508.1 AP001111 Oryza sativa similar to ABC transporter of Arabidopsis thaliana (AC004697).
BAA90507.1 AP001111 Oryza sativa similar to ABC tiansporter of Arabidopsis thaliana (AC004697).
BAB16495.1 AP002861 Oryza sativa putative ABC tiansporter ATP-binding protein. P0665D10.21.
BAB21276.1 AP002844 Oryza sativa putative ABC tiansporter protein. P0410E03.7. contains EST D22472(C1173).
BAB21275.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.6.
BAB21273.1 AP002844 Oryza sativa putative ABC transporter protein. P0410E03.4.
BAB40032.1 AP003046 Oryza sativa putative ABC transporter. P0445D12.3. SEQ ID NO: 888
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIKl. receptor-like protein kinase; serine/threonine protein kinase.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRKl.
AAA33000.1 M76647 Brassica oleracea receptor protein kinase. SKR6.
CAB89179.1 AJ245479 Brassica napus subsp. napus ser /thr kinase. S-locus receptor kinase. srk. AAA33008.1 M97667 Brassica napus serine/threonine kinase receptor.
CAA74661.1 Y14285 Brassica oleracea
SFRl. extiacellular S domain: 8-1342; tiansmembrane domain: 1343-1411; intracellular kinase domain: 1412-2554.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2.
AAA62232.1 U00443 Brassica napus
S-receptor kinase. protein contains an immunoglobulm-like domain.
CAA73133.1 Y12530 Brassica oleracea serine /threonine kinase. ARLK.
BAA23676.1 AB000970 Brassica rapa receptor kinase 1. BcRKl.
BAA92836.1 AB032473 Brassica oleracea
S18 S-locus receptor kinase. SRK18.
CAA74662.1 Y14286 Brassica oleracea
SFR3. extiacellular S domain: 123-1346; transmembrane domain: 1347-1412; intiacellular kinase domain: from 1413.
CAA79355.1 Z18921 Brassica oleracea S-receptor kinase-like protein.
CAB41879.1 Y18260 Brassica oleracea SRK15 protein. SRK15. receptor-like kinase.
BAA06285.1 D30049 Brassica rapa S-receptor kinase SRK9.
BAA21132.1 D88193 Brassica rapa S-receptor kinase. SRK9 (B.c).
CAB41878.1 Y18259 Brassica oleracea SRK5 protein. SRK5. receptor-like kinase.
BAA92837.1 AB032474 Brassica oleracea S60 S-locus receptor kinase. SRK60.
BAA07577.2 D38564 Brassica rapa receptor protein kinase SRK12.
BAB21001.1 AB054061 Brassica rapa S locus receptor kinase. SRK22.
BAA07576.1 D38563 Brassica rapa receptor protein kinase SRK8.
AAD52097.1 AF088885 Nicotiana tabacum receptor-like kinase CHRK1. Chrkl.
AAK02023.1 AC074283 Oryza sativa Putative protein kinase-like. OSJNBa0087H07.5.
BAB 18292.1 AP002860 Oryza sativa putative receptor-like protein kinase. P0409B08.19.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AFOO 1308).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
BAB 17331.1 AP002747 Oryza sativa putative receptor kinase. P0698G03.12.
BAB17139.1 AP002867 Oryza sativa putative receptor kinase. P0463F06.31.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
CAB51836.1 AJ243961 Oryza sativa
Putitive Ser/Thr protein kinase. 11332.7.
AAF78016.1 AF238472 Oryza sativa receptor-like kinase. RLG15. protein kinase.
BAB39451.1 AP003338 Oryza sativa putative receptor kinase. OJ1212_B09.24.
SEQ ID NO: 890
AAB47181.1 S82324 Zea mays
/gene- 'calciuni/calmodulin-dependent protein kinase. This sequence comes from Fig. 1.
BAA22410.1 D38452 Zea mays calcium-dependent protein kinase-related kinase.
BAA12692.1 D84508 Zea mays
CDPK-related protein kinase. Does not require calcium for its activity.
AAG01179.1 AF289237 Zea mays calcium/calmodulin dependent protein kinase MCK2. MCK2.
CAA58750.1 X83869 Daucus carota CDPK-related protein kinase. CRK (or PK421).
BAA12691.1 D84507 Zea mays
CDPK-related protein kinase. Does not require calcium for its activity (by similarity).
AAC24961.1 AF009337 Tradescantia virginiana
CDPK-related protein kinase. CRKl.
AAF23901.2 AF194414 Oryza sativa calcium-dependent protein kinase. CDPK5. OsCDPK5.
AAF23900.1 AF194413 Oryza sativa calcium-dependent protein kinase. CDPKl . OsCDPKl.
AAC78558.1 AF030879 Solanum tuberosum protein kinase CPK1.
AAD17800.1 AF090835 Mesembryanthemum crystallinum
Ca2+-dependent protein kinase. CPK1. serine/tlireonine protein kinase.
AAB49984.1 U90262 Cucurbita pepo calcium-dependent cahnodulin-independent protein kinase CDPK. cpCPKl . serine/threoiiine protein kinase that is activated by direct binding of calcium.
CAA07481.1 AJ007366 Zea mays calcium-dependent protein kinase.
BAB21081.1 AP002819 Oryza sativa putative calcium-dependent protein kinase. P0501G01.10.
BAA12715.1 D85039 Zea mays calcium-dependent protein kinase.
AAC25423.1 AF072908 Nicotiana tabacum calcium-dependent protein kinase. CDPKL
CAA39936.1 X56599 Daucus carota calcium- dependent protein kinase. DcPK431.
CAA57157.1 X81394 Oryza sativa calcium-dependent protein kinase. OSCPK2.
AAF21062.1 AF216527 Dunaliella tertiolecta calcium-dependent protein kinase. CPK1; CDPK.
BAA81751.1 ABO 17517 Marchantia polymoφha calcium-dependent protein kinase. CDPK CDPK-B; alternative splicing. BAA81749.1 AB017515 Marchantia polymoφha calcium-dependent protein kinase. CDPKL CDPK-B; alternative splicing.
AAB88537.1 AF035944 Fragaria x ananassa calcium-dependent protein kinase. MAX17.
BAA81750.1 ABO 17516 Marchantia polymoφha calcium-dependent protein kinase. CDPK CDPK-A; alternative splicing. BAA81748.1 AB017515 Marchantia polymoφha calcium-dependent protein kinase. CDPKl . CDPK-A; alternative splicing. AAA69507.1 U28376 Zea mays calcium-dependent protein kinase. MZECDPK2.
AAA61682.1 L27484 Zea mays calcium-dependent protein kinase. CDPK.
AAB80693.1 U69174 Glycine max calmodulin-like domain protein kinase isoenzyme gamma. CDPK gamma.
BAA13232.1 D87042 Zea mays
Calcium-dependent protein kinase.
BAA13440.1 D87707 Ipomoea batatas calcium dependent protein kinase. CDPK. CAA89202.1 Z49233 Chlamydomonas eugametos calcium-stimulated protein kinase.
AAB70706.1 U82087 Tortula ruralis calmodulin-like domain protein kinase. TrCPKl.
AAD28192.2 AFl 15406 Solanum tuberosum calcium-dependent protein kinase. CDPK; catalytic domain.
AAC49405.1 U08140 Vigna radiata calcium dependent protein kinase. CDPK. BAA12338.1 D84408 Zea mays calcium dependent protein kinase. ZmCDPKl .
CAA65500.1 X96723 Medicago sativa protein kinase. CDPK.
BAA85396.1 AP000615 Oryza sativa
ESTs C22369(C12239),C22370(C12239), AU057852(S21844),AU057853(S21844) conespond to a region ofthe predicted gene.; similar to calcium dependent protein kinase. (AF048691).
AAA33443.1 LI 5390 Zea mays calcium-dependent protein kinase. CDPK.
AAB80692.1 U69173 Glycine max cahnodulin-like domain protein kinase isoenzyme beta. CDPK beta.
CAA57156.1 X81393 Oryza sativa calcium-dependent protein kinase. OSCPKII.
AAC05270.1 AF048691 Oryza sativa calcium dependent protein kinase. CDPK12.
AAK26164.1 AY027885 Cucumis sativus calcium-dependent calmodulin-independent protein kinase 5. CDPK5. CsCDPK5.
AAG46110.1 AC073166 Oryza sativa calcium-dependent protein kinase. OSJNBb0064P2L2.
BAA02698.1 D 13436 Oryza sativa calcium-dependent protein kinase. spk.
BAA90814.1 AP001168 Oryza sativa
ESTs AU030197(E50746),AU030196(E50746) conespond to a region ofthe predicted gene.;
Similar to calcium-dependent calmodulin-independent protein kinase CDPK (U90262).
AAF19401.1 AF203479 Glycine max phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
CAB46228.1 Y18055 Arachis hypogaea calcium dependent protein kinase. CDPK.
AAF19403.1 AF203481 Lycopersicon esculentum phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
AAF 19402.1 AF203480 Lycopersicon esculentum phosphoenolpyravate carboxylase kinase. protein kinase; member of Ca2+/CaM kinase family; lacks the autoinhibitory region and EF hands.
SEQ ID NO: 892
AAF20931.1 AF206721 Brassica juncea ascorbate oxidase.
BAA07734.1 D43624 Nicotiana tabacum ascorbate oxidase precursor.
AAA33119.1 J04494 Cucumis sativus ascorbate oxidase precursor (EC 1.10.3.3).
CAA75577.1 Y15295 Medicago truncatula L-ascorbate oxidase. MtN23.
AAF35911.2 AF233594 Cucumis melo ascorbate oxidase AO4. multicopper oxidase.
CAA39300.1 X55779 Cucurbita sp. ascorbate oxidase.
BAA09528.1 D55677 Cucurbita maxima ascorbate oxidase. AAO.
AAF35910.1 AF233593 Cucumis melo ascorbate oxidase AOl. multicopper oxidase.
CAA71275.1 Y10226 Cucumis melo L-ascorbate oxidase. ao3.
AAF20932.1 AF206722 Brassica juncea ascorbate oxidase.
AAF20933.1 AF206723 Brassica juncea ascorbate oxidase.
CAA71273.1 Y10224 Cucumis melo L-ascorbate oxidase. aol.
CAA71274.1 Y10225 Cucumis melo L-ascorbate oxidase. aol.
BAA20520.1 AB004799 Oryza sativa ascorbate oxidase.
AAF33751.1 AF202460 Capsicum annuum ascorbic acid oxidase.
AAB 17193.1 U73105 Lkiodendron tulipifera monolignol polymerization; lignin biosynthesis, laccase. LAC2-3. diphenol oxidase; blue copper oxidase.
AAB17191.1 U73103 Liriodendron tulipifera monolignol polymerization; lignin biosynthesis, laccase. LAC2-1. diphenol oxidase; blue copper oxidase.
AAC49536.1 U43542 Nicotiana tabacum diphenol oxidase. laccase.
AAB17194.1 U73106 Liriodendron tulipifera monolignol polymerization; lignin biosynthesis, laccase. LAC2-4. diphenol oxidase; blue copper oxidase.
AAB17192.1 U73104 Liriodendron tulipifera monolignol polymerization; lignin biosynthesis, laccase. LAC2-2. diphenol oxidase; blue copper oxidase.
AAC04576.1 AF047697 Oryza sativa p-diphenol oxidase. putative high-pi laccase.
AAB09228.1 U12757 Acer pseudoplatanus monolignol polymerization; lignin biosynthesis, laccase. diphenol oxidase.
CAA45554.1 X64257 Brassica napus
BplO. protein homologous to ascorbate oxidase.
AAC49538.1 U45243 Nicotiana tabacum diphenol oxidase. laccase.
AAC49537.1 U43543 Nicotiana tabacum diphenol oxidase. laccase.
AAD02557.1 AF049931 Petunia x hybrida PGPS/NH15. PGPS/NH15. ascorbate oxidase homolog.
SEQ ID NO: 894
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
BAA74465.1 AB022732 Glycynhiza echinata cytochrome P450. CYP Ge-31.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
BAA22422.1 AB001379 Glycynhiza echinata cytochrome P450. CYP81E1.
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'- race).
CAB41490.1 AJ238439 Cicer arietinum cytochrome P450 monooxygenase. cyp81E3v2.
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYPδlBlc.
CAA10067.1 AJ012581 Cicer arietinum cytochrome P450. cyp81E3.
AAK38079.1 AF321855 Lolium rigidum putative cytochrome P450.
AAK38080.1 AF321856 Lolium rigidum putative cytochrome P450.
AAK38081.1 AF321857 Lolium rigidum putative cytochrome P450.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3 '-hydroxylase. Htl. cytochrome P450; CYP75B2.
AAC34853.1 AF082028 Hemerocallis hybrid cultivar putative cyt P450-containing fatty acid hydroxylase. senescence-associated protein 3. SA3. mRNA accumulates in senescing petals.
AAG09208.1 AF175278 Pisum sativum wound-inducible P450 hydroxylase. CYP82A1.
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
CAA71515.1 Y10491 Glycine max putative cytochrome P450.
CAA71876.1 Y10982 Glycine max putative cytochrome P450.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
AAC49188.2 U29333 Pisum sativum cytochrome P450 monooxygenase. CYP82. new cytochrome P450 family.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
CAA71516.1 Y10492 Glycine max putative cytochrome P450.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
AAC39454.1 AF014802 Eschscholzia californica
(S)-N-methylcoclaurine 3'-hydroxylase. CYP82B1. cytochrome P-450-dependent monooxygenase; methyl jasmonate-inducible cytochrome P-450-dependent, homologous to wound-inducible CYP82A1 of Pisum sativum GenBank Accession Number U29333.
AAB94590.1 AF022461 Glycine max
CYP82Clp. CYP82C1. cytochrome P450 monooxygenase.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
CAA71877.1 Y10983 Glycine max putative cytochrome P450.
BAA84072.1 AB028152 Torenia hybrida flavone synthase II. cytochrome P450. TFNS5.
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
BAA84071.1 AB028151 Antinhinum majus flavone synthase II. cytochrome P450. AFNS2.
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl. P450 enzyme.
BAA35080.1 AB015762 Nicotiana tabacum putative cytochrome P450. CYP82E1.
CAA50155.1 X70824 Solanum melongena flavonoid hydroxylase (P450). CYP75.
CAA70575.1 Y09423 Nepeta racemosa cytochrome P450. CYP71A5.
BAA92894.1 AB006790 Petunia x hybrida cytochrome P450. IMT-2.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
SEQ ID NO: 895
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APKl gene for protein tyrosine-serine-threonine kinase.(D12522).
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stiess, osmotic stress, and
ABA treatment.
AAK11674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAGl 6628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619.S11214).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
AAK00425.1 AC069324 Oryza sativa
Putative protein kinase. OSJNBa0071K19.11.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil. Ptil kinase.
AAF91336.1 AF249317 Glycine max Ptil kinase-like protein. Ptil a. protein kinase.
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S13927).
AAB09771.1 U67422 Zea mays
CRXNKLY4 precursor. cr4. receptor kinase homolog.
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
AAG25966.1 AF302082 Nicotiana tabacum cytokinin-regulated kinase 1. CRKl. protein kinase; tianscript abundance decreases rapidly after cytokinin treatment.
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAF76313.1 AF220603 Lycopersicon esculentum Pto kinase. LescPth5.
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
AAC27894.1 AF023164 Zeamays leucine-rich repeat transmembrane protein kinase 1. ltkl .
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKl. accumulates in mesophyll cells.
BAA92954.1 AP001551 Oryza sativa
Similar to Oryza sativa protein kinase (OSPKIO) mRNA. (L27821).
BAA92953.1 AP001551 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4 BAC clone F10M6 ; S-receptor kinase -like protein. (AL021811).
AAKl 1566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhirPtoE. confers resistance to bacterial speck disease.
AAKl 1567.1 AF318491 Lycopersicon hirsutum
Pto-like protein kinase F. LhirPtoF.
BAA82556.1 AB030083 Populus nigra lectin-like protein kinase. PnLPK.
CAA67145.1 X98520 Brassica oleracea receptor-like kinase. SFR2. SEQ ID NO: 899
AAF61647.1 AFl 90634 Nicotiana tabacum
UDP-glucose: salicylic acid glucosyltransferase. SA-GTase.
BAA89009.1 AB027455 Petunia x hybrida anthocyanin 5-O-glucosyltiansferase. PHI.
AAA59054.1 L34847 Zea ays conjugation ofthe growth hormone indole-3 -acetic acid (IAA). LAA-glu synthetase. iaglu.
BAA93039.1 AB033758 Citras unshiu limonoid UDP-glucosyltransferase. LGTase.
BAA36423.1 AB013598 Verbena x hybrida
UDP-glucose: anthocyanin 5-O-glucosyltiansferase. HGT8.
BAA36421.1 ABO 13596 Perilla frutescens
UDP-glucose:anthocysnin 5-O-glucosyltiansferase. PF3R4.
AAF98390.1 AF287143 Brassica napus catalyzes the tiansfer of glucose from UDP-glucose to sinapate and some other hydroxycinnamates (4-coumarate, caffeate, feralate). UDP-glucose: sinapate glucosyltransferase. SGT1. SGT.
BAA36422.1 AB013597 Perilla frutescens
UDP-glucose: anthocyanin 5-O-glucosyltiansferase homologue. PF3R6.
BAB07962.1 AP002524 Oryza sativa putative anthocyanin 5-O-glucosyltiansferase. P0406H10.16. contains ESTs
AU067881(C10481),AU067882(C10481).
AAK16175.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.15.
AAK16181.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.16.
AAK16178.1 AC079887. Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.5.
AAG25643.1 AF303396 Phaseolus vulgaris
UDP-glucosyltransferase HRA25. putative; defense associated.
AAK16172.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
BAA83484.1 AB031274 Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltiansferase. ufgt.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS 10a.
AAK28304.1 AF346432 Nicotiana tabacum phenylpropanoid:glucosyltiansferase 2. togt2. glucosyltransferase.
AAD21086.1 AF 127218 Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds, flavonoid 3-O- glucosyltransferase. UFGT.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltiansferase. IS5a.
BAA12737.1 D85186 Gentiana triflora
UDP-glucose:flavonoid-3-glucosyltiansferase.
AAK28303.1 AF346431 Nicotiana tabacum phenylpropanoid: glucosyltiansferase 1. togtl. glucosyltransferase.
AAF17077.1 AF199453 Sorghum bicolor
UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitrile-o- glucosyltiansferase.
AAK16180.1 AC079887 Oryza sativa putative glucosyltransferase. OSJNBa0040E01.21.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltiansferases.
BAB41017.1 AB047090 Vitis labrusca x Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. KyUFGTl. The gene was derived from one ofthe parents V. labrascana cv. Ishiharawase.
BAB41026.1 AB047099 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. F1UFGT2.
BAB41024.1 AB047097 Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltransferase. A1UFGT2.
BAB41022.1 AB047095 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. RuUFGT2.
BAB41020.1 AB047093 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. ItUFGT2.
BAA89008.1 AB027454 Petunia x hybrida anthocyanidin 3-O-glucosyltiansferase. PGT8.
BAB41018.1 AB047091 Vitis labrusca x Vitis vinifera
UDP-glucose:flavonoid 3-O-glucosyltiansferase. KyUFGT2. The gene originated in one ofthe parents V. vinifera cv. Centennial.
CAB56231.1 Y18871 Dorotheanthus bellidiformis betanidin-5-O-glucosyltiansferase. ^^__^
AAB81683.1 AF000372 Vitis vinifera UDP glucose:flavonoid 3-O-glucosyltransferase.
BAB41019.1 AB047092 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. ItUFGTl.
BAB41025.1 AB047098 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltransferase. FlUFGTl.
BAB41023.1 AB047096 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. AlUFGTl.
BAB41021.1 AB047094 Vitis vinifera
UDP-glucose: flavonoid 3-O-glucosyltiansferase. RuUFGTl.
AAB81682.1 AF000371 Vitis vinifera UDP glucose:flavonoid 3-o-glucosyltiansferase.
BAA19659.1 AB002818 Perilla frutescens flavonoid 3-O-glucosyltiansferase. UDP glucose.
BAA90787.1 AB038248 Ipomoea batatas
UDP glucose: flavonoid 3-O-glucosyltiansferase. uf3gt.
AAD04166.1 AF101972 Phaseolus lunatus catalyzes formation of O-glucosylzeatin from zeatin and UDPG; catalyzes formation of O- xylosylzeatin from zeatin and UDPX. zeatin O-glucosyltiansferase. ZOGl. cytokinin O- glucosyltransferase.
AAB86473.1 AF028237 Ipomoea puφurea
UDP glucose: flavonoid 3-O-glucosyltiansferase. UF3GT-FL1.
SEQ ID NO: 900
AAA33975.1 Ml 1395 Glycine max small heat shock protein.
CAA41547.1 X58711 Medicago sativa heat shock protein.
AAB03893.1 M11318 Glycine max 17.5 kd heat shock protein Gmhspl7.6L.
AAA33672.1 M33899 Pisum sativum 18.1 kDa heat shock protein (hsp 18.1).
CAA25578.1 X01104 Glycine max heat shock protein 6871 (aa 1-153).
CAB55634.2 AJ237596 Helianthus annuus 17.9 kDa heat-shock protein, hsp 17.9.
BAA33062.1 AB017273 Cuscuta japonica low-molecular- weight heat shock protein. CJHSP17.
AAA33974.1 M11317 Glycine max 17.6 kd heat shock protein Gmhspl7.6L.
AAD30454.1 AF123257 Lycopersicon esculentum 17.6 kD class I small heat shock protein. HSP17.6.
AAD30452.1 AF123255 Lycopersicon esculentum 17.7 kD class I small heat shock protein. HSP17.7.
CAA39603.1 X56138 Lycopersicon esculentum small heat shock protein (class I).
AAD30453.1 AF123256 Lycopersicon esculentum 17.8 kD class I small heat shock protein. HSP17.8.
CAA37847.1 X53851 Daucus carota heat shock protein.
AAA33671.1 M33900 Pisum sativum 17.9 kDa heat shock protein (hsp!7.9).
AAF34133.1 AF161179 Malus x domestica low molecular weight heat shock protein. Hspl.
CAA41546.1 X58710 Medicago sativa heat shock protein.
AAB63310.1 U46544 Helianthus annuus
18.6 kDa heat-shock protein. Class I low-molecular-weight heat-shock protein. AAB63311.1 U46545 Helianthus annuus
17.7 kDa heat shock protein. Class I low-molecular-weight heat-shock protein. CAB08441.1 Z95153 Helianthus annuus
17.6 kD class I small heat-shock protein HSP17.6. Ha hsρ!7.6.
CAA42222.1 X59701 Helianthus annuus 17.6 kDa heat shock protein.
AAC39360.1 U63631 Fragaria x ananassa LMW heat shock protein.
AAA33910.1 M80939 Oryza sativa 16.9 kDa heat shock protein.
CAA37848.1 X53852 Daucus carota heat shock protein.
AAA33909.1 M80938 Oryza sativa 16.9 kDa heat shock protein.
CAA43210.1 X60820 Oryza sativa
16.9 KD low molecular weight heat shock protein.
CAA37864.1 X53870 Chenopodium rabrum heat-shock protein.
AAD49336.1 AFl 66277 Nicotiana tabacum low molecular weight heat-shock protein. LHS-1. TLHS-1.
AAA61632.1 U08601 Papaver somniferam low molecular weight heat-shock protein.
AAB72109.1 AF022217 Brassica rapa low molecular weight heat-shock protein. BcHSP17.6. 17.6 kDa; cytosolic class I.
CAA63902.1 X94192 Pennisetum glaucum heat shock protein 16.9. hsp 16.9.
AAB39856.1 U81385 Oryza sativa heat shock protein. Oshspl6.9C. class I, low molecular mass.
CAA46641.1 X65725 Zea mays heat shock protein 17.2. Zmhsp 17.2. Class I low molecular weight heat shock protein.
CAA08908.1 AJ009880 Castanea sativa molecular chaperone. cytosolic class I small heat-shock protein HSP17.5. hsp!7.5.
CAB93512.1 AJ243565 Brassica oleracea putative class I small heat shock protein. HSP17.7-a protein, hsp 17 7-a.
CAA63903.1 X94193 Pennisetum glaucum heat shock protein 17.9. hsp!7.9.
CAB36910.1 AJ000691 Quercus suber stiess protein chaperone. heat shock protein 17.4. hspl7.
CAA63901.1 X94191 Pennisetum glaucum heat shock protein 17.0. hsp 17.0.
AAC78392.1 U83669 Oryza sativa low molecular mass heat shock protein Oshspl7.3. OSHSP17.3. class I LMMHSP.
BAA02160.1 D12635 Oryza sativa
'low molecular weight heat shock protein'.
AAC78393.1 U83670 Oryza sativa low molecular mass heat shock protein Oshspl8.0. OSHSP18.0. class I LMMHSP.
CAA31785.1 X13431 Triticum aestivum put. heat shock protein (AA 1 -151).
CAA63570.1 X92983 Pseudotsuga menziesii low molecular weight heat-shock protein.
AAC78394.1 U83671 Oryza sativa low molecular mass heat shock protein Oshspl7.7. OSHSP17.7. class I LMMHSP.
CAA63571.1 X92984 Pseudotsuga menziesii low molecular weight heat-shock protein.
SEQ ID NO: 901
AAG08959.1 AF122051 Solanum tuberosum tuber-specific and sucrose-responsive element binding factor. TSF. R2R3 MYB class transcription factor.
AAG08960.1 AF122052 Solanum tuberosum tuber-specific and sucrose-responsive element binding factor. TSF. R2R3 MYB class tianscription factor.
AAG08961.1 AF122053 Solanum tuberosum tuber-specific and sucrose-responsive element binding factor. TSF. R2R3 MYB class transcription factor.
AAF67053.1 AF190304 Adiantum raddianum c-myb-like transcription factor. MYB3R-1. contains three MYB repeats.
AAF67052.1 AFl 90303 Adiantum raddianum c-myb-like transcription factor. MYB3R-1. contains three MYB repeats.
AAF34434.1 AF172282 Oryza sativa myb-like protein. DUPRl 1.29.
AAF78888.1 AFl 89786 Physcomitrella patens putative c-myb-like tianscription factor. MYB3R-1. PpMYB3R-l.
AAF78887.1 AFl 89785 Physcomitrella patens putative c-myb-like transcription factor. MYB3R-1. PpMYB3R-l.
AAF43043.1 AF236059 Papaver rhoeas putative Myb-related domain, pmr.
BAA94769.1 AP001859 Oryza sativa
Similar to Arabidopsis thaliana chromosome 4, BAC clone F4D11; putative myb-protein.
(AL022537).
AAF78890.1 AFl 89788 Hordeum vulgare putative c-myb-like tianscription factor. MYB3R-1. HvMYB3R-l.
AAF78889.1 AF189787 Hordeum vulgare putative c-myb-like tianscription factor. MYB3R-1. HvMYB3R-l.
BAA88169.1 AP000836 Oryza sativa
Similar to putative transcription factor (AF062890).
BAA88205.1 AP000837 Oryza sativa
Similar to putative transcription factor (AF062890).
CAA78388.1 Z13998 Petunia x hybrida
DNA-binding protein; transcriptional activator, protein 3. myb.Ph.3. related to animal myb proto-oncoproteins .
AAF67051.1 AFl 90302 Secale cereale c-myb-like tianscription factor. MYB3R-1. contains three MYB repeats.
AAF67050.1 AF190301 Secale cereale c-myb-like tianscription factor. MYB3R-1. contains three MYB repeats.
AAG28525.1 AF198498 Nicotiana tabacum anther-specific myb-related protein 2. mybAS2. MMYBAS2; contains tandem R2, R3 myb domains similar to c-myb family.
BAB 12688.1 AP002746 Oryza sativa putative MYB family transcription factor. P0671B11.3. contains ESTs AU082307(E0784),C72014(E0784).
BAA99440.1 AP002743 Oryza sativa putative MYB family transcription factor. P0710E05.27. contains ESTs
AU082307(E0784),C72014(E0784).
AAK19619.1 AF336286 Gossypium hirsutum
GHMYB9. ghmyb9. similar to myb.
CAA64615.1 X95297 Lycopersicon esculentum transcription factor. THM1. myb-related.
BAA81730.1 AB029159 Glycine max
GmMYB29Al.
CAA50224.1 X70879 Hordeum vulgare
MybHyl. mybl.
CAA50222.1 X70877 Hordeum vulgare
MybHvl. mybl.
CAB40189.1 AJ133638 Avena sativa transcriptional activator, myb protein, gamyb.
AAG28526.1 AFl 98499 Nicotiana tabacum anther-specific myb-related protein 1. mybASl. NtMYBASl; contains N-terminal R2, R3 myb domain repeats similar to c-myb.
AAD31395.1 AFl 14162 Lolium temulentum gibberellin MYB transcription factor. GAMyb. R2/R3-MYB.
BAA81732.1 AB029161 Glycine max GmMYB29A2.
BAA81731.1 AB029160 Glycine max C_MYB29A1.
CAA61021.1 X87690 Hordeum vulgare transcriptional activator of alpha-amylase gene promoter. GAMyb protein. Gaml.
AAG22863.1 AY008692 Hordeum vulgare transcription factor GAMyb. Gamyb. BAA96421.1 AB044084 Triticum aestivum transcription activator for gibberellin response. GAMyb protein. Ta-GAMyb.
BAA81733.2 AB029162 Glycine max
GmMYB29A2.
BAA81736.1 AB029165 Glycine max
GmMYB29B2.
AAA33067.1 L04497 Gossypium hirsutum
MYB A; putative.
BAB39972.1 AP003018 Oryza sativa putative transcription factor (myb). OSJNBa0004B13.27. contains ESTs
AU097474(S5087),D40175(S 1959).
CAA72218.1 Y11415 Oryza sativa myb.
CAA67000.1 X98355 Oryza sativa activator of alpha-amylase gene promoter, tianscription factor GAMyb. Gaml. Myb-like; expression is regulated by gibberellin.
BAB39987.1 AP003020 Oryza sativa putative transcription factor (myb). P0498A12.16. contains ESTs
AU097474(S5087),D40175(S1959).
CAA78387.1 Z13997 Petunia x hybrida
DNA-binding protein, tianscriptional activator, protein 2. myb.Ph2. related to animal myb proto-oncoproteins .
CAA72217.1 Y11414 Oryza sativa myb.
BAA23341.1 D88621 Oryza sativa transfactor. OSMYB5. Osmyb5.
BAA93038.1 AP001552 Oryza sativa
EST AU082058(C12976) conesponds to a region ofthe predicted gene. Similar to
Arabidopsis thaliana putative tianscription factor (AF062916).
CAA64614.1 X95296 Lycopersicon esculentum transcription factor. THM27. myb-related.
AAK19616.1 AF336283 Gossypium hirsutum
GHMYB25. ghmyb25. similar to myb; contains an unspliced intron.
CAA78386.1 Z13996 Petunia x hybrida
DNA binding protein; tianscriptional activator, protein 1. myb.Ph3. Product related to animal myb proto-oncoproteins. Sequence from nucleotide 992 is not included in clone cPFl and has been obtained by PCR amplification of cDNA.
CAA67575.1 X99134 Lycopersicon esculentum transcription factor. THM6. myb-related.
BAA88222.1 AB028650 Nicotiana tabacum myb-related transcription factor LBM2. Ibm2.
SEQ ID NO: 902 CAA57773.1 X82329 Arachis hypogaea chitinase (class II). chi2;l.
AAD54935.1 AF141373 Petroselinum crispum random hydrolysis of l,4-beta-acetamido-2-deoxy-D-glucoside linkages in chitin. chitinase precursor. Chi2-1. class II.
AAD54936.1 AF141374 Petroselinum crispum random hydrolysis of l,4-beta-acetamido-2-deoxy-D-glucoside linkages in chitin. chitinase precursor. Chi2-2. class II.
AAF00131.1 AF147091 Fragaria x ananassa chitin degradation, class II chitinase. Chi2-1.
AAC95376.1 AFl 05426 Cynodon dactylon chitinase. Cht2.
BAA95846.1 AP002070 Oryza sativa
Similar to Oryza sativa clone RGCH7 chitinase. (AF013581).
AAA32986.1 M95835 Brassica napus endochitinase. Ch25.
AAF69783.1 AF135143 Arabis lemmonii class I chitinase.
AAF69775.1 AF135135 Arabis drummondii class I chitinase.
AAF69792.1 AF135152 Arabis parishii class I chitinase.
AAC95375.1 AFl 05425 Cynodon dactylon chitinase. Chtl .
AAF69785.1 AF135145 Arabis lignifera class I chitinase.
AAF69770.1 AF135130 Arabis holboellii class I chitinase.
AAF69781.1 AF135141 Arabis gunnisoniana class I chitinase.
AAF69777.1 AF135137 Arabis fecunda class I chitinase.
AAF69790.1 AF135150 Arabis microphylla class I chitinase.
AAF69787.1 AF135147 Arabis lignifera class I chitinase.
AAF69772.1 AFl 35132 Arabis gunnisoniana class I chitinase.
AAF69782.1 AF135142 Halimolobos peφlexa var. peφlexa class I chitinase.
AAF69784.1 AF135144 Arabis lemmonii class I chitinase.
AAF69788.1 AF135148 Arabis lyallii class I chitinase.
BAA03750.1 D16222 Oryza sativa endochitinase. Cht-2.
AAF69776.1 AF135136 Arabis fecunda class I chitinase.
CAA40107.1 X56787 Oryza sativa chitinase.
BAB 13369.1 AB048531 Psophocaφus tetragonolobus class I chitinase.
AAF69778.1 AF135138 Arabis glabra class I chitinase.
AAF69786.1 AF135146 Arabis lignifera class I chitinase.
BAA82826.1 AB023464 Arabis gemmifera basic endochitinase. ChiB.
AAF69773.1 AF135133 Arabis blepharophylla class I chitinase.
AAF69791.1 AF135151 Arabis microphylla class I chitinase.
AAF69793.1 AF135153 Arabis parishii class I chitinase.
CAA39535.1 X56063 Oryza sativa chitinase.
BAA03749.1 D16221 Oryza sativa endochitinase. Cht-1.
AAF69789.1 AF135149 Arabis microphylla class I chitinase.
CAA71402.1 Y10373 Medicago truncatula chitinase.
AAC16010.1 AF061805 Elaeagnus umbellata acidic chitinase.
CAA53626.1 X76041 Triticum aestivum endochitinase. CHI.
BAA33971.1 AB008892 Nicotiana tabacum chitinase 134. Chnl34.
CAA47921.1 X67693 Solanum tuberosum chitinase. SK2. endochitinase.
AAF69780.1 AF135140 Arabis glabra class I chitinase.
BAB18519.1 AB051578 Secale cereale seed chitinase-a. rsca.
AAA51377.1 L37289 Oryza sativa chitinolytic activity, antifungal activity, chitinase.
AAB41324.1 U83591 Medicago sativa class I chitinase.
CAC 17793.1 AJ301671 Nicotiana sylvestris hydrolysis of chitin. endochitinase. chnb. class I chitinase.
AAB41325.1 U83592 Medicago sativa class I chitinase.
AAB23263.1 S43926 Phaseolus vulgaris chitinase. CH5B. This sequence comes from Fig. 1.
AAA33756.1 M13968 Phaseolus vulgaris chitinase (EC 3.2.1.14).
CAA35945.1 X51599 Nicotiana tabacum chitinase. CHN50.
AAA34070.1 M15173 Nicotiana tabacum endochitinase precursor (EC 3.2.1.14).
CAA45822.1 X64519 Nicotiana tabacum chitinase B class I. CHN200.
SEQ ID NO: 903
BAA78764.1 AB023482 Oryza sativa
ESTs C98382(C2985),D22444(C11129) conespond to a region ofthe predicted gene.; Similar to Arabidopsis thaliana APKl gene for protein tyrosine-serine-threonine kinase.(D12522).
AAF43496.1 AF131222 Lophopyrum elongatum protein serine/threonine kinase. ESI47. induced in roots by salt stress, osmotic stress, and ABA treatment.
AAKl 1674.1 AF339747 Lophopyrum elongatum protein kinase. ESI47.
AAG16628.1 AY007545 Brassica napus protein serine/threonine kinase BNK1.
BAA94509.1 AB041503 Populus nigra protein kinase 1. PnPKl .
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAA94510.1 AB041504 Populus nigra protein kinase 2. PnPK2.
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
BAB03429.1 AP002817 Oryza sativa
EST C22619(S11214) conesponds to a region ofthe predicted gene. Similar to Arabidopsis thaliana chromosome 2, BAC clone T17D12; putative protein kinase (AC006587).
BAB07999.1 AP002525 Oryza sativa putative protein kinase. P0462H08.22. contains EST C22619(S11214).
BAB39409.1 AP002901 Oryza sativa putative protein kinase. P0456F08.9. contains EST C23560(R0290).
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
CAB51834.1 00069 Oryza sativa
11332.5. contains eukaryotic protein kinase domain PF.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs AU056701(S20808),AU056702(S20808).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
AAG03090.1 AC073405 Oryza sativa
Similar to an Arabidopsis somatic embryogenesis receptor-like kinase (AC007504).
BAB19337.1 AP003044 Oryza sativa putative protein kinase. P0038C05.10. contains ESTs
AU056335(S20481),AU056336(S20481).
BAA90808.1 AP001168 Oryza sativa
Similar to putative receptor-like protein kinase (AL035679).
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
AAB47424.1 U59317 Lycopersicon pimpinellifolium serme/tlπeonine protein kinase Fen. Fen, fenthion sensitivity gene from tomato.
AAC48932.1 U13923 Lycopersicon pimpinellifolium
Fen. putative serine/threonine protein kinase; similar to product encoded by Lycopersicon pimpinellifolium Pto gene, GenBank Accession Number U02271; Fen is a member ofthe Pto gene family.
AAF76307.1 AF220602 Lycopersicon pimpinellifolium
Fen kinase.
BAA92221.1 AP001278 Oryza sativa
Similar to Arabidopsis thaliana chromosome II BAC F12L6 genomic sequence, putative protein kinase. (AC004218).
BAA87852.1 AP000816 Oryza sativa
Similar to putative Ser/Thr protein kinase. (AC004218).
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
AAKl 1566.1 AF318490 Lycopersicon hirsutum
Pto-like protein kinase E. LhrrPtoE. confers resistance to bacterial speck disease.
AAKl 1568.1 AF318492 Lycopersicon hirsutum
Pto-like protein kinase B. LhirPtoB.
AAD21872.1 AF078082 Phaseolus vulgaris receptor-like protein kinase homolog RK20-1.
AAF66615.1 AF142596 Nicotiana tabacum LRR. receptor-like protein kinase.
AAC23542.1 U20948 Ipomoea trifida receptor protein kinase. IRK1.
AAF91337.1 AF249318 Glycine max Ptil kinase-like protein. Ptilb. protein kinase.
AAC27894.1 AF023164 Zeamays leucine-rich repeat transmembrane protein kinase 1. ltkl .
AAF76314.1 AF220603 Lycopersicon esculentum Fen kinase. Lescfen.
AAB47422.1 U59318 Lycopersicon esculentum serine/threonine protein kinase Fen, fen, allele of Fen, fenthion sensitivity gene from tomato.
AAF76313.1 AF220603 Lycopersicon esculentum
Pto kinase. LescPth5.
AAB47421.1 U59316 Lycopersicon esculentum serine/threonine protein kinase Pto. pto. allele of Pto bacterial speck disease resistance gene in tomato.
AAF91336.1 AF249317 Glycine max Ptil kinase-like protein. Ptil a. protein kinase.
AAKl 1567.1 AF318491 Lycopersicon hirsutum Pto-like protein kinase F. LhirPtoF.
AAC61805.1 U28007 Lycopersicon esculentum serine/threonine protein kinase. Pto kinase interactor 1. Ptil. Ptil kinase.
AAB93834.1 U82481 Zea mays
KI domain interacting kinase 1. KIK receptor-like protein kinase; serine/threonine protein k kiinnaassee
CAA97692.1 Z73295 Catharanthus roseus receptor-like protein kinase. CRPKl. Autophosphorylation predominantly on Thr, less on Ser.
Mechanism: autophosphorylation in cis.
CAA73134.1 Y12531 Brassica oleracea serine/threonine kinase. BRLK.
AAF76306.1 AF220602 Lycopersicon pimpinellifolium Pto kinase.
AAB47423.1 U59315 Lycopersicon pimpinellifolium serine/threonine protein kinase Pto. Pto. bacterial speck disease resistance in tomato; disease resistance gene.
BAB21241.1 AP002953 Oryza sativa
Putative Pto kinase interactor 1. P0426D06.21. contains ESTs
AU108280(E0721),D48017(S13927).
AAC48914.1 U02271 Lycopersicon pimpinellifolium protein kinase.
SEQ ID NO: 904
AAF65545.1 AF233894 Perilla citriodora limonene synthase.
AAG31437.1 AF241792 Perilla frutescens limonene synthase.
AAK06663.1 AF317695 Perilla frutescens var. frutescens limonene synthase.
AAG31435.1 AF241790 Perilla citriodora limonene synthase.
BAA21629.1 AB005744 Perilla frutescens catalyzing the cyclization of geranyl pyrophosphate to 1-limonene. 1-limonene synthase. gPFLC. similar to perilla PFLC-LDDBJ Acc#D49368.
AAG31438.1 AF241793 Perilla frutescens limonene synthase.
BAA08367.1 D49368 Perilla frutescens limonene cyclase.
AAC37366.1 L13459 Mentha spicata 4S-limonene synthase.
AAD50304.1 AF175323 Mentha longifolia limonene synthase. monoteφene synthase.
AAG01140.1 AF282875 Schizonepeta tenuifolia (+)-4R-limonene synthase.
AAF21053.1 AF212433 Capsicum annuum UV-induced sesquiteφene cyclase. SC2.
BAA82141.1 AB023816 Solanum tuberosum vetispiradiene synthase. PVS4. potato sesquiteφene cyclase.
AAF74977.1 AF270425 Gossypium hirsutum
(E,E)-farnesyl diphosphate cyclase. (+)-delta-cadinene synthase. cdn2. sesquiteφene cyclase.
BAA82092.1 AB022598 Solanum tuberosum vetispiradiene synthase. PVSl. potato sesquiteφene cyclase.
BAA82109.1 AB022720 Solanum tuberosum vetispiradiene synthase. PVS3. potato sesquiteφene cyclase; PVS3.
AAG09949.1 AF171216 Lycopersicon esculentum vetispiradiene synthase. LEVS2. sesquiteφene cyclase.
BAA82108.1 AB022719 Solanum tuberosum vetispiradiene synthase. PVS2. potato sesquiteφene cyclase; PVS2.
AAC12784.1 U88318 Gossypium hirsutum
(E,E)-farnesyl diphosphate cyclizing. (+)-delta-cadinene synthase. cdnl . sesquiteφene cyclase; delta-cadinene synthase.
AAKl 5641.1 AF326117 Capsicum annuum sesquiteφene cyclase. PSC2.
AAC61260.1 AF061285 Capsicum annuum sesquiteφene cyclase. UV induced.
AAG24640.2 AF304444 Artemisia annua sesquiteφene cyclase.
CAC 12731.1 AJ271792 Artemisia annua putative sesquiteφene cyclase. cASC125.
SEQ ID NO: 905
AAF64450.1 AF239928 Euphorbia esula glutathione S-tiansferase. similar to auxin-inducible GST.
AAG34803.1 AF243368 Glycine max glutathione S-tiansferase GST 13.
AAG16759.1 AY007561 Lycopersicon esculentum putative glutathione S-tiansferase T4.
AAG34798.1 AF243363 Glycine max glutathione S-transferase GST 8.
AAG34797.1 AF243362 Glycine max glutathione S-transferase GST 7.
AAG16758.1 AY007560 Lycopersicon esculentum putative glutathione S-transferase T3.
AAG34809.1 AF243374 Glycine max glutathione S-tiansferase GST 19.
AAG34801.1 AF243366 Glycine max glutathione S-tiansferase GST 11.
AAG34796.1 AF243361 Glycine max glutathione S-tiansferase GST 6.
AAG34807.1 AF243372 Glycine max glutathione S-tiansferase GST 17.
AAG16757.1 AY007559 Lycopersicon esculentum putative glutathione S-tiansferase T2.
AAGl 6756.1 AY007558 Lycopersicon esculentum putative glutathione S-tiansferase TI.
AAG34804.1 AF243369 Glycine max glutathione S-tiansferase GST 14.
AAG34810.1 AF243375 Glycine max glutathione S-transferase GST 20.
AAG34808.1 AF243373 Glycine max glutathione S-tiansferase GST 18.
AAG34805.1 AF243370 Glycine max glutathione S-tiansferase GST 15.
AAG34844.1 AF244701 Zea mays glutathione S-transferase GST 36.
AAG34829.1 AF244686 Zea mays glutathione S-transferase GST 21.
AAG34831.1 AF244688 Zeamays glutathione S-tiansferase GST 23.
AAA68430.1 J03679 Solanum tuberosum glutathione S-transferase. gstl. previously called pathogenesis-related protein; prpl-1.
AAC18566.1 AF048978 Glycine max
2,4-D inducible glutathione S-transferase. GSTa.
AAG34832.1 AF244689 Zea mays glutathione S-tiansferase GST 24.
AAG34800.1 AF243365 Glycine max glutathione S-transferase GST 10.
AAG34849.1 AF244706 Zeamays glutathione S-tiansferase GST 41.
AAG34837.1 AF244694 Zea mays glutathione S-tiansferase GST 29.
AAC32118.1 AF051214 Picea mariana probable glutathione S-tiansferase. Sbl8. similar to Nicotiana tabacum probable glutathione
S-tiansferase encoded by GenBank Accession Number X56266.
AAG41204.1 AF321437 Suaeda maritima glutathione transferase.
AAG34802.1 AF243367 Glycine max glutathione S-tiansferase GST 12.
AAF29773.1 AF159229 Gossypium hirsutum glutathione S-transferase. GST.
AAG32472.1 AF309379 Oryza sativa subsp. japonica putative glutathione S-tiansferase OsGSTU3.
AAG34836.1 AF244693 Zea mays glutathione S-tiansferase GST 28.
CAA04391.1 AJ000923 Carica papaya glutathione transferase. PGST1.
CAA71784.1 Y10820 Glycine max glutathione transferase.
CAA09187.1 AJOl 0448 Alopecurus myosuroides glutathione transferase. GSTla.
AAG34795.1 AF243360 Glycine max glutathione S-tiansferase GST 5.
CAA09188.1 AJ010449 Alopecurus myosuroides glutathione transferase. GSTlb.
AAG34806.1 AF243371 Glycine max glutathione S-transferase GST 16.
CAC24549.1 AJ296343 Cichorium intybus x Cichorium endivia glutathione S-transferase. chi-GSTl. auxin-induced GST.
AAC28101.1 AF079511 Mesembryanthemum crystallinum glutathione S-tiansferase.
SEQ ID NO: 907
AAA33280.1 L20475 Datura stramonium
29kDa protein; high homology to aa sequence of tropinone reductases.
AAA33281.1 L20473 Datura stramonium catalyses a stereospecific reduction of tropinone to tropine. tiopinone reductase-I.
CAC34420.1 AJ307584 Solanum tuberosum tropine forming tiopinone reductase. tropinone reductase I. TRI.
BAA85844.1 AB026544 Hyoscyamus niger tropinone reductase-I. til.
BAA13547.1 D88156 Hyoscyamus niger tropinone reductase-I.
AAA33282.1 L20474 Datura stramonium catalyses a stereospecific reduction of tropinone to pseudotropine. tropinone reductase-II.
AAB09776.1 L20485 Hyoscyamus niger tiopinone reductase-II.
BAA85845.1 AB026545 Hyoscyamus niger tropinone reductase-II. tr2.
CAB52307.1 AJ245634 Solanum tuberosum pseudotropine fonning tiopinone reductase. tiopinone reductase II. TRII.
CAC19810.1 AJ292343 Solanum tuberosum pseudotropine forming tropinone reductase. tiopinone reductase II. till.
CAA45866.1 X64566 Cuphea lanceolata beta-ketoacyl-ACP reductase. Clkr27. 3-oxoacyl-[acyl-canier protein] reductase.
CAA45793.1 X64463 Brassica napus 3-oxoacyl-[acyl-carrier protein] reductase.
AAB82767.1 U89509 Zea mays cuticular wax biosynthesis, b-keto acyl reductase. glossy8.
AAB82766.1 U89510 Hordeum vulgare cuticular wax biosynthesis, b-keto acyl reductase. glossy 8.
CAA74176.1 Y13861 Nicotiana tabacum enoyl-ACP reductase. enr-T2.
AAB20114.2 S60064 Brassica napus enoyl-acyl canier protein reductase. This sequence comes from fig3; conceptual translation differs from published sequence.
CAA74177.1 Y13862 Nicotiana tabacum enoyl-ACP reductase. enr-Tl.
AAC78100.1 AF093628 Oryza sativa protochlorophyll.de reductase homolog.
CAA05879.1 AJ003124 Petunia x hybrida enoyl-ACP reductase. pte.
CAA64729.1 X95462 Brassica napus enoyl reductase.
CAA05816.1 AJ003025 Oryza sativa enoyl-ACP reductase.
BAA99570.1 AB036823 Chlorella vulgaris oxidoreductase. oxi.
SEQ ID NO: 909
BAA82107.1 AB022693 Nicotiana tabacum transcription factor. NtWRKYl.
AAC31956.1 AF080595 Pimpinella brachycaφa zinc finger protein. ZFP 1. WRKYL
AAD55974.1 AF121353 Petroselinum crispum zinc-finger type tianscription factor WRKYL WRKYl.
BAA77383.1 AB020590 Nicotiana tabacum transcription factor NtWRKY2.
AAC49527.1 U48831 Petroselinum crispum
WRKYl. contains two WRKY domains; WRKY-type DNA-binding protein; sequence- specific DNA-binding protein.
AAD32677.1 AF140554 Avena sativa
DNA-binding protein WRKYl . wrkyl . putative transcription factor.
CAA88326.1 Z48429 Avena fatua binds conserved cis-element from cereal alpha- Amy2 promoters. DNA-binding protein.
AAD16139.1 AF096299 Nicotiana tabacum
DNA-binding protein 2. WRKY2. tianscription factor.
BAA86031.1 AB026890 Nicotiana tabacum transcription factor NtWRKY4.
AAC37515.1 L44134 Cucumis sativus SPFl-like DNA-binding protein.
AAK16171.1 AC079887 Oryza sativa putative DNA-binding protein. OSJNBa0040E01.10.
AAF23898.1 AF193802 Oryza sativa zinc finger tianscription factor WRKYL
AAD16138.1 AF096298 Nicotiana tabacum DNA-binding protein 1. WRKYl. tianscription factor.
AAC49529.1 U58540 Petroselinum crispum
WRKY2. Contains two WRKY domains; WRKY-type DNA-binding protein.
CAB97004.1 AJ278507 Solanum tuberosum putative transcription factor. WRKY DNA binding protein. WRKYl .
BAB 19075.1 AP002744 Oryza sativa putative DNA-binding protein homolog. P0006C01.17.
BAB19096.1 AP002839 Oryza sativa putative DNA-binding protein homolog. P0688A04.2.
AAK16170.1 AC079887 Oryza sativa putative DNA binding protein. OS JNBa0040E01A
BAB40073.1 AP003074 Oryza sativa putative WRKY DNA binding protein. OSJNBa0004G10.20. contains EST C26525(C12525).
AAD38283.1 AC007789 Oryza sativa putative WRKY DNA binding protein. OSJNBa0049B20.9.
BAB 18313.1 AP002865 Oryza sativa putative WRKY DNA binding protein. P0034C11.1. contains EST C26525(C12525).
CAA88331.1 Z48431 Avena fatua binds conserved cis-element from cereal alpha- Amy2 promoters. DNA-binding protein.
AAC49528.1 U56834 Petroselinum crispum DNA-binding. WRKY3. WRKY-type DNA-binding protein.
AAG35658.1 AF204925 Petroselinum crispum transcription factor WRKY4. WRKY4. binds to W box (TTGACC) elements.
BAB 16432.1 AB041520 Nicotiana tabacum WRKY transcription factor Nt-SubD48. Nt-SubD48.
AAD32676.1 AF140553 Avena sativa
DNA-binding protein WRKY3. wrky3. putative tianscription factor.
AAG46150.1 AC018727 Oryza sativa putative DNA-binding protein. OSJNBa0056G17.18.
BAA77358.1 AB020023 Nicotiana tabacum
WRKY domain Zn-finger type DNA-binding protein. DNA-binding protein NtWRKY3.
AAG35659.1 AF204926 Petroselinum crispum tianscription factor WRKY5. WRKY5. binds to W box (TTGACC) elements.
AAD27591.1 AF 121354 Petroselinum crispum binds sequence specifically to W Boxes (TTGACC). tianscription factor. WRKY3. sequence specific DNA-binding protein.
CAB66338.1 AJ279697 Betula pendula wrky-type DNA binding protein, wrky.
AAF61864.1 AF193771 Nicotiana tabacum DNA-binding protein 4. WRKY4. transcription factor.
BAA87069.1 AB035271 Matricaria chamomilla elicitor-induced DNA-binding protein homolog. McWRKYl.
AAF61863.1 AFl 93770 Nicotiana tabacum DNA-binding protein 3. WRKY3. tianscription factor.
SEQ ID NO: 910
AAC09420.1 M68929 Mitochondrion Marchantia polymoφha φsl4.
CAA33994.1 XI 5901 Plastid Oryza sativa ribosomal protein S14. φs!4.
SEQ ID NO: 911
CAA55128.1 X78325 Nicotiana tabacum chitinase/lysozyme. Pz.
CAA54373.1 X77110 Nicotiana tabacum chitinase, class V. chi-V.
CAA54374.1 X77111 Nicotiana tabacum chitinase, class V. chi-V.
SEQ ID NO: 912
CAB55394.1 ALl 17264 Oryza sativa zwhOOlO.l. similar to Arabidopsis germin-like protein 6 (AF032976); Method: conceptual translation with partial peptide sequencing.
BAA78563.1 AB024338 Atriplex lentiformis germin-like protein.
AAB97470.1 AF042489 Oryza sativa germin-like protein 16. glpl6.
AAA33030.1 M93041 Mesembryanthemum crystallinum germin-like protein, germin-like protein.
CAB65371.1 AJ250834 Pisum sativum germin-like protein. ger2b. 2nd variant ofthe clone PsGER2.
CAB55559.1 AJ237943 Triticum aestivum germin-like protein. glρ2b.
CAB65370.1 AJ250833 Pisum sativum germin-like protein. ger2a. 1st variant of this clone.
CAB55558.1 AJ237942 Triticum aestivum gerrnin-like protein. glp2a.
AAD43971.1 AF 141878 Oryza sativa germin-like protein 1 precursor. RGLPl. similar to barley epidermis/papilla-specific oxalate oxidase-like protein.
AAD43973.1 AF141880 Oryza sativa gerrnin-like protein 1 precursor. RGLPl. similar to barley epidermis/papilla-specific oxalate oxidase-like protein.
AAC04837.1 AF032976 Oryza sativa germin-like protein 6. GER6. similar to wheat and barley oxalate oxidase.
AAG00425.1 AF250933 Hordeum vulgare germin A. GerA. apoplastic protein; contains prepeptide for targeting into the cell wall.
AAD43972.1 AF141879 Oryza sativa germin-like protein 2 precursor. RGLP2. similar to barley epidermis/papilla-specific oxalate oxidase-like protein.
CAA63659.1 X93171 Hordeum vulgare oxalate oxidase-like protein or germin-like protein.
AAC04833.1 AF032972 Oryza sativa germin-like protein 2. GER2. similar to wheat and barley oxalate oxidase.
AAC04834.1 AF032973 Oryza sativa germin-like protein 3. GER3. similar to wheat and barley oxalate oxidase.
AAC04832.1 AF032971 Oryza sativa germin-like protein 1. GER1. similar to wheat and barley oxalate oxidase.
AAG00426.1 AF250934 Hordeum vulgare gennin B. GerB. apoplastic protein; contains prepeptide for targeting into the cell wall.
AAG00427.1 AF250935 Hordeum vulgare germin F. GerF. apoplastic protein; contains prepeptide for targeting into the cell wall.
AAF34811.1 AF005084 Triticum aestivum oxalate oxidase. up-regulated by aluminum.
AAC04835.1 AF032974 Oryza sativa germin-like protein 4. GER4. similar to wheat and barley oxalate oxidase.
BAB39965.1 AP003018 Oryza sativa probable gennin protein 4. OSJNBa0004B13.19. contains ESTs
AU101991(S4037),AU070167(R0031).
BAB39980.1 AP003020 Oryza sativa probable germin protein 4. P0498A12.8. contains ESTs
AU101991(S4037),AU070167(R0031).
AAG00428.1 AF250936 Hordeum vulgare germin P. GerD. apoplastic protein; contains prepeptide for targeting into the cell wall.
AAA20245.1 UOI 963 Hordeum vulgare germin subunit.
CAC 19429.1 AJ291825 Lolium perenne oxalate oxidase. oxOl.
AAA34270.1 M63223 Triticum aestivum germin. gennin 9f-2.8.
AAA34268.1 M21962 Triticum aestivum germin protein precursor.
CAA74595.1 Y14203 Hordeum vulgare oxalate oxidase.
AAG00429.1 AF250937 Hordeum vulgare germin E. GerE. apoplastic protein.
CAB65369.1 AJ250832 Pisum sativum germin-like protein, gerl.
AAA32959.1 LI 5737 Hordeum vulgare oxalate oxidase. This is the sequence ofthe complete mature peptide.
CAA71052.1 Y09917 Triticum aestivum germin homolog. pSBGer3.
AAA34271.1 M63224 Triticum aestivum germin. germin 9f-3.8.
CAA71050.1 Y09915 Triticum aestivum germin homolog. pSBGerl .
BAB18339.1 AP002865 Oryza sativa putative germin protein. P0034C11.30. contains EST C97263(C53484).
AAD38298.1 AC007789 Oryza sativa putative oxalate oxidase (gennin protein). OSJNBa0049B20.25.
BAA86880.1 AB028454 Barbula unguiculata germin-like protein.
AAF03355.1 AF132671 Nicotiana plumbaginifolia nectarin I precursor. NEC 1. germin-like protein.
BAA25197.1 AB012138 Lycopersicon esculentum adaptation to Mn-deficiency. germin-like protein. Mdipl.
AAC78470.1 AF067731 Solanum tuberosum germin-like protein. OXAOXA. similar to oxalate oxidase.
AAC25777.1 AF072694 Oryza sativa germin-like protein 7. GER7. similar to wheat and barley oxalate oxidase.
AAC99473.1 AF039201 Pinus caribaea germin-like protein. PcGERl.
CAA71051.1 Y09916 Triticum aestivum germin homolog. pSBGer2.
AAC05146.1 AF049065 Pinus radiata germin-like protein. PRGerl .
CAC34417.1 AJ311624 Pisum sativum Germin-like protein. glp3.
SEQ ID NO: 915
BAB 19363.1 AP002542 Oryza sativa putative beta-1, 3-glucanase. P0679C08.2.
AAD10386.1 U72255 Oryza sativa beta- 1 ,3-glucanase precursor. Gns9.
CAB85903.1 AJ251646 Pisum sativum hydrolysis of beta-1,3 conjugated glucans. beta-1,3 glucanase. gns2.
AAA90953.1 U30323 Triticum aestivum beta 1,3 -glucanase. Glcl.
BAA89481.1 AB029462 Salix gilgiana beta-1, 3-glucanase. SgGNl.
BAB40807.1 AB052291 Pyras pyrifolia catalyzing the hydrolysis of 1,3-beta-glucosyl linkages. endo-l,3-beta-glucanase-like protein. bgn-1. Amino acid alignment ofthe protein(BGN-l) encoded by pear pollen bgn-1 with barley endo-l,3-beta-glucanase(GII. accession number:pdb/lGHS-B/2.3/2/306/N/)(40% identity) and their hydrophobic cluster analysis(HCA)(a overall HCA homology score of 87.1%) showed that it was most likely that the bgn-1 encoded a endo-l,3-beta-glucanase. A higher identity(59.3%) was found between BGN-1 and a putative pea endo- 1,3 -beta-glucanase
(accession number:dad/AJ251646-l).
AAB82772.2 AFOO 1523 Musa acuminata beta-1, 3-glucananse. similar to beta-1, 3-glucanase. CAA49513.1 X69887 Brassica napus beta-l,3-glucanase homologue.
AAF08679.1 AF004838 Musa acuminata beta- 1 ,3-glucanase.
CAA82271.1 Z28697 Nicotiana tabacum beta-1 ,3-glucanase.
AAD10384.1 U72253 Oryza sativa beta-l,3-glucanase precursor. Gns7.
CAB71021.1 AJ271598 Hieracium piloselloides putative role in callose degradation, putative beta-l,3-glucanase. glue.
CAA30261.1 X07280 Nicotiana plumbaginifolia beta-glucanase.
AAA51643.1 M23120 Nicotiana plumbaginifolia beta-glucanase precursor.
AAA87456.1 U22147 Hevea brasiliensis beta-1, 3-glucanase. HGN1. hydrolytic enzyme.
AAA34078.1 M63634 Nicotiana plumbaginifolia regulator of beta(l,3)-glucanase. beta(l,3)-glucanase regulator.
AAD10385.1 U72254 Oryza sativa beta- 1,3 -glucanase precursor. Gns8.
CAB38443.1 AJ133470 Hevea brasiliensis beta-1 ,3-glucanase. hgnl.
AAB03501.1 U41323 Glycine max beta-1, 3-glucanase. SGN1.
AAA32939.1 M62907 Hordeum vulgare hydrolysis of beta-(l-3)-glucan. (l-3)-beta-glucanase. cBGL32.
AAD33881.1 AF141654 Nicotiana tabacum beta-l,3-glucanase. GGL4.
AAA03617.1 M80604 Lycopersicon esculentum beta-l,3-glucanase.
BAA77786.1 AB027431 Oryza sativa beta-l,3-glucanase.
BAA77787.1 AB027432 Oryza sativa beta-l,3-glucanase.
CAB91554.1 AJ277900 Vitis vinifera beta 1-3 glucanase. gl.
AAC14399.1 AF030771 Hordeum vulgare beta-l,3-glucanase 2. BGL32.
CAA03908.1 AJ000081 Citras sinensis glucan hydrolase. beta-l,3-glucanase. gnsl.
AAG24921.1 AF311749 Hevea brasiliensis beta-1 ,3-glucanase.
AAA33946.1 M37753 Glycine max beta-1, 3-endoglucanase (EC 3.2.1.39).
AAA63542.1 M59443 Nicotiana tabacum acidic beta-1, 3-glucanase. glucanase.
AAD28732.1 AFl 12965 Triticum aestivum beta- 1,3 -glucanase precursor. Glb3.
AAD 10381.1 U72250 Oryza sativa beta-l,3-glucanase precursor. Gns4.
CAA57255.1 X81560 Nicotiana tabacum (l-)-beta-glucanase. Sp41a.
AAD33880.1 AF141653 Nicotiana tabacum beta-l,3-glucanase. GGL1.
AAB86541.1 AF030166 Oryza sativa glucanase. glul .
AAA63539.1 M60402 Nicotiana tabacum glucan beta- 1,3 -glucanase. glucanase GLA.
AAA34053.1 M60464 Nicotiana tabacum beta-l,3-glucanase.
AAD 10380.1 U72249 Oryza sativa beta-1, 3-glucanase precursor. Gns3.
AAA63540.1 M60403 Nicotiana tabacum glucan-l,3-beta-glucosidase. glucanase GLB.
SEQ ID NO: 916
AAA85440.1 U32624 Sorghum bicolor cytochrome P-450. CYP79. P450TYR; N-hydroxylase.
AAF27289.1 AF140613 Manihot esculenta N-hydroxylating cytochrome P450. CYP79D1.
AAF27290.1 AF140614 Manihot esculenta N-hydroxylating cytochrome P450. CYP79D2.
AAD03415.1 AF069494 Sinapis alba converts tyrosine to para-hydrophenylacetaldoxime in para-hydroxybenzylglucosinolate biosynthesis, cytochrome P450. CYP79B1.
AAG59648.1 AC084319 Oryza sativa putative cytochrome p450tyr. OSJNBa0004B24.15.
AAF66543.1 AF140609 Triglochin maritimum cytochrome P450 CYP79E1. CYP79E1.
AAF66544.1 AFl 40610 Triglochin maritimum cytochrome P450 CYP79E2. CYP79E2.
BAA92894.1 AB006790 Petunia x hybrida cytochrome P450. IMT-2.
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl . P450 enzyme.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3'-hydroxylase. Htl. cytochrome P450; CYP75B2.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
CAA50155.1 X70824 Solanum melongena flavonoid hydroxylase (P450). CYP75.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
BAA84071.1 AB028151 Antinhinum majus flavone synthase II. cytochrome P450. AFNS2.
AAD38930.1 AF135485 Glycine max cytochrome P450 monooxygenaseCYP93Dl. CYP93E1.
AAB17562.1 U72654 Eustoma grandiflorum flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75.
AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
BAA93634.1 AB025016 Lotus japonicus cytochrome P450.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
AAB94590.1 AF022461 Glycine max
CYP82Clp. CYP82C1. cytochrome P450 monooxygenase.
CAA71876.1 Y10982 Glycine max putative cytochrome P450.
BAA12159.1 D83968 Glycine max Cytochrome P-450 (CYP93A1).
AAG14962.1 AF214008 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2.
BAA74465.1 AB022732 Glycynhiza echinata cytochrome P450. CYP Ge-31.
BAA22422.1 AB001379 Glycynhiza echinata cytochrome P450. CYP81E1.
AAG14961.1 AF214007 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'' race).
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81Blc.
AAB94593.1 AF022464 Glycine max
CYP77A3p. CYP77A3. cytochrome P450 monooxygenase.
BAA13076.1 D86351 Glycine max cytochrome P-450 (CYP93A2).
CAA50647.1 X71656 Solanum melongena P450 hydroxylase.
SEQ ID NO: 918
CAA06223.1 AJ004923 Lycopersicon esculentum Squalene epoxidase. ERG.
SEQ ID NO: 919
AAF71823.1 AF 153276 Populus tremula x Populus tremuloides pumilio domain-containing protein PPDL PPD similar to Drosophila pumilio protein RNA- binding domain.
SEQ ID NO: 920
BAA82393.1 AP000367 Oryza sativa
EST C96716(C 10608) conesponds to a region ofthe predicted gene.; Similar to putative receptor protein kinase. (AC002334).
CAC20842.1 AJ250467 Pinus sylvestris receptor protein kinase. upk.
AAB36558.1 U77888 Ipomoea nil receptor-like protein kinase INRPKl. inφkl. leucine-rich repeat.
AAF91324.1 AF244890 Glycine max receptor-like protein kinase 3. RLK3. GmRLK3.
BAB40094.1 AP003210 Oryza sativa putative receptor protein kinase. OSJNBaOOlOKOl.7.
AAF91322.1 AF244888 Glycine max receptor-like protein kinase 1. RLKl. GmRLKl.
AAC36318.1 AF053127 Malus x domestica leucine-rich receptor-like protein kinase. LRPKml .
BAA84787.1 AP000559 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
BAA83373.1 AP000391 Oryza sativa
ESTs C22657(S0014),C22656(S0014) conespond to a region ofthe predicted gene.; Similar to receptor protein kinase, ERECTA (AC004484).
AAF59905.1 AF197946 Glycine max receptor protein kinase-like protein. CLV1 A.
AAK27806.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.21.
AAF91323.1 AF244889 Glycine max receptor-like protein kinase 2. RLK2. GmRLK2.
AAF34426.1 AF172282 Oryza sativa leucine rich repeat containing protein kinase. DUPRl 1.16.
AAK27817.1 AC022457 Oryza sativa putative protein kinase. OSJNBa0006L06.16.
CAA61510.1 X89226 Oryza sativa leucine-rich repeat/receptor protein kinase. Irk2.
BAB03621.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.21.
AAB82755.1 U72725 Oryza longistaminata receptor kinase-like protein. Xa21 gene family member Al; downstream of microsatellite region; disease resistance gene family member.
BAB03629.1 AP002522 Oryza sativa putative protein kinase Xa21. P0009G03.30.
AAB61708.1 U93048 Daucus carota somatic embryogenesis receptor-like kinase. SERK.
AAF66615.1 AF142596 Nicotiana tabacum LRR receptor-like protein kinase.
AAB82756.1 U72724 Oryza sativa receptor kinase-like protein. Xa21 gene family member E.
CAB51480.1 Y14600 Sorghum bicolor putative protein serine /threonine kinase. RLKL accumulates in mesophyll cells.
BAB39873.1 AP002882 Oryza sativa putative protein kinase. P0439B06.8. contains ESTs
AU056701(S20808),AU056702(S20808).
BAB18321.1 AP002865 Oryza sativa putative receptor protein kinase. P0034C11.11.
BAB40081.1 AP003074 Oryza sativa putative receptor protein kinase. OSJNBa0004G10.30.
AAD38286.1 AC007789 Oryza sativa putative protein kinase. OSJNBa0049B20.13.
BAB21240.1 AP002953 Oryza sativa
Putative protein kinase. P0426D06.20. contains ESTs C22359(C11461),C22360(C11461).
AAK21965.1 AY028699 Brassica napus receptor protein kinase PERK1.
BAA87853.1 AP000816 Oryza sativa
EST AU030604(E51294) conesponds to a region ofthe predicted gene. Similar to putative
NAK-like Ser/Thr protein kinase. (AF001308).
BAB16871.1 AP002537 Oryza sativa putative protein kinase APKlAArabidopsis thaliana. P0001B06.24. contains ESTs
C22608(R3192),D25110(R3192).
AAC27894.1 AF023164 Zeamays leucine-rich repeat transmembrane protein kinase 1. ltkl.
AAK00425.1 AC069324 Oryza sativa Putative protein kinase. OSJNBa0071K19.11.
AAG59657.1 AC084319 Oryza sativa putative protein kinase. OSJNBa0004B24.20.
BAA94519.1 AP001800 Oryza sativa
ESTs AU032341(R3918),AU071016(R10613) conespond to a region ofthe predicted gene.
Similar to Arabidopsis thaliana chromosome 4, BAC F9D16; putative receptor kinase
(AL035394).
BAB07903.1 AP002835 Oryza sativa putative receptor kinase. P0417G05.10. contains ESTs
AU032341(R3918),AU071016(R10613).
SEQ ID NO: 921
BAA96250.1 AB027617 Pyras pyrifolia UDP-glucose pyrophosphorylase.
BAA25917.1 AB013353 Pyras pyrifolia UDP-glucose pyrophosphorylase.
CAA62689.1 X91347 Hordeum vulgare UDP-glucose pyrophosphorylase.
AAF62555.1 AF249880 Oryza sativa subsp. indica UDP-glucose pyrophosphorylase. UDPGase.
SEQ ID NO: 922
AAD03415.1 AF069494 Sinapis alba converts tyrosine to para-hydrophenylacetaldoxime in para-hydroxybenzylglucosinolate biosynthesis, cytochrome P450. CYP79B1.
AAA85440.1 U32624 Sorghum bicolor cytochrome P-450. CYP79. P450TYR; N-hydroxylase.
AAF27289.1 AF140613 Manihot esculenta N-hydroxylating cytochrome P450. CYP79D1.
AAF27290.1 AF140614 Manihot esculenta N-hydroxylating cytochrome P450. CYP79D2.
AAG59648.1 AC084319 Oryza sativa putative cytochrome p450tyr. OSJNBa0004B24.15.
AAF66543.1 AFl 40609 Triglochin maritimum cytochrome P450 CYP79E1. CYP79E1.
AAF66544.1 AFl 40610 Triglochin maritimum cytochrome P450 CYP79E2. CYP79E2.
BAA92894.1 AB006790 Petunia x hybrida cytochrome P450. IMT-2.
AAD56282.1 AF155332 Petunia x hybrida flavonoid 3'-hydroxylase. Htl. cytochrome P450; CYP75B2.
CAA50155.1 X70824 Solanum melongena flavonoid hydroxylase (P450). CYP75.
AAC32274.1 AF081575 Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl. P450 enzyme.
AAG34695.1 AF313492 Matthiola incana putative cytochrome P450.
AAA32913.1 M32885 Persea americana cytochrome P-450LXXIA1 (cyp71Al).
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
AAB94587.1 AF022458 Glycine max
CYP98A2p. CYP98A2. cytochrome P450 monooxygenase.
AAB 17562.1 U72654 Eustoma grandifloram flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75.
AAG09208.1 AF175278 Pisum sativum wound-inducible P450 hydroxylase. CYP82A1.
AAD37433.1 AF150881 Lycopersicon esculentum x Lycopersicon peravianum catalyzes the hydroxylation of feralic acid to 5-hydroxyferalic acid, feralate-5-hydroxylase.
CYP84. cytochrome P450-dependent monooxygenase; F5H; FAHL
CAA04117.1 AJ000478 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81B11. chimeric sequence (from 5'- race).
CAA04116.1 AJ000477 Helianthus tuberosus fatty acid in-chain hydroxylase. cytochrome P450. CYP81Blc.
CAB43505.1 AJ239051 Cicer arietinum cytochrome P450. cyp81E2.
AAC49188.2 U29333 Pisum sativum cytochrome P450 monooxygenase. CYP82. new cytochrome P450 family.
AAG14963.1 AF214009 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H3.
AAD38930.1 AF135485 Glycine max cytochrome P450 monooxygenaseCYP93Dl. CYP93E1.
AAG14962.1 AF214008 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2.
AAG44132.1 AF218296 Pisum sativum cytochrome P450. P450 isolog.
AAB94588.1 AF022459 Glycine max
CYP71D10p. CYP71D10. cytochrome P450 monooxygenase.
AAG14961.1 AF214007 Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
AAB94590.1 AF022461 Glycine max CYP82Clp. CYP82Cl. cytochrome P450 monooxygenase.
CAA71513.1 Y10489 Glycine max putative cytochrome P450.
BAA84071.1 AB028151 Antinhinum majus flavone synthase II. cytochrome P450. AFNS2.
CAA50442.1 X71130 Petunia x hybrida P450 hydroxylase. PET 1.
SEQ ID NO: 924
BAB20581.1 AB042268 Zea mays response regulator 6. ZmRR6.
AAK14395.1 AF339732 Dianthus caryophyllus response regulator protein. RR.
BAB20580.1 AB042267 Zea mays response regulator 5. ZmRR5.
BAB20579.1 AB042261 Zea mays response regulator 4. ZmRR4.
BAA85113.1 AB031012 Zea mays response regulator 2. ZmRR2. cytokinin-inducible.
BAA82873.1 AB024291 Zea mays response regulator. ZmRR2.
BAA75253.1 AB004882 Zea mays response regulator. ZmCipl, cytokinin-inducible protein.
BAA85112.1 AB031011 Zeamays response regulator 1. ZmRRl. cytokinin-inducible.
BAB17300.1 AB042260 Zeamays response regulator. ZmRR3.
BAB20582.1 AB042269 Zeamays response regulator 7. ZmRR7.
BAB41137.1 AB060130 Zea mays response regulator 8. ZmRR8.
SEQ ID NO: 925
AAA33280.1 L20475 Datura stramonium
29kDa protein; high homology to aa sequence of tropinone reductases.
AAA33281.1 L20473 Datura stramonium catalyses a stereospecific reduction of tiopinone to tropine. tropinone reductase-I.
BAA85844.1 AB026544 Hyoscyamus niger tropinone reductase-I. til.
CAC34420.1 AJ307584 Solanum tuberosum tropine forming tiopinone reductase. tropinone reductase I. TRI.
BAA13547.1 D88156 Hyoscyamus niger tropinone reductase-I.
AAA33282.1 L20474 Datura stramonium catalyses a stereospecific reduction of tiopinone to pseudotropine. tropinone reductase-II.
BAA85845.1 AB026545 Hyoscyamus niger tropinone reductase-II. tr2.
AAB09776.1 L20485 Hyoscyamus niger tiopinone reductase-II.
CAB52307.1 AJ245634 Solanum tuberosum pseudotropine forming tiopinone reductase. tropinone reductase II. TRII.
CAC 19810.1 AJ292343 Solanum tuberosum pseudotropine forming tiopinone reductase. tiopinone reductase II. till.
CAA45866.1 X64566 Cuphea lanceolata beta-ketoacyl-ACP reductase. Clkr27. 3-oxoacyl-[acyl-carrier protein] reductase.
CAA45793.1 X64463 Brassica napus
3-oxoacyl-[acyl-carrier protein] reductase.
AAB82767.1 U89509 Zea mays cuticular wax biosynthesis, b-keto acyl reductase. glossy8.
AAB82766.1 U89510 Hordeum vulgare cuticular wax biosynthesis, b-keto acyl reductase. glossy8.
AAB20114.2 S60064 Brassica napus enoyl-acyl canier protein reductase. This sequence comes from fig3; conceptual translation differs from published sequence.
CAA74176.1 Y13861 Nicotiana tabacum enoyl-ACP reductase. enr-T2.
AAB82764.1 U89511 Allium porrum cuticular wax biosynthesis, b-keto acyl reductase. glossyδ.
AAC78100.1 AF093628 Oryza sativa protochlorophyll.de reductase homolog.
CAA74177.1 Y13862 Nicotiana tabacum enoyl-ACP reductase. enr-Tl.
SEQ ID NO: 927
AAD43046.1 AF124045 Sorghum bicolor
GTP-binding protein-like. 110K5.8. terminal exon, defined by homology to maize ESTs.
SEQ ID NO: 928
CAA09619.1 AJOl 1418 Lycopersicon esculentum ubiquitin activating enzyme.
AAC32140.1 AF051239 Picea mariana probable ubiquitin activating enzyme 2. Sb53. similar to Arabidopsis thaliana ubiquitin activating enzyme 2 encoded by GenBank Accession Number U40566.
AAA34308.1 M55604 Triticum aestivum ubiquitin-activating enzyme El. UBA1.
AAA34265.1 M90663 Triticum aestivum ubiquitin activating enyme. El.
AAA34266.1 M90664 Triticum aestivum ubiquitin activating enzyme. El.
CAA71762.1 Y10804 Nicotiana tabacum Ubiquitin activating enzyme El. UBA1.
SEQ ID NO: 929
CAA55693.1 X79086 Zea mays initiator-binding protein. IBP2.
CAC 19789.1 AJ251686 Catharanthus roseus putative tianscription factor. MYB-like DNA-binding protein, bpf-1.
CAA55691.1 X79085 Zea mays initiator binding protein. IBP1.
AAF97508.1 AF242298 Oryza sativa telomere binding protein- 1. TBPL
SEQ ID NO: 930
AAKl 5447.1 AC037426 Oryza sativa putative flavin-containing monooxygenase. OSJNBb0014I11.12.
AAKl 5439.1 AC037426 Oryza sativa putative flavin-containing monooxygenase. OSJNBb0014I11.8.
BAA88198.1 AP000837 Oryza sativa
Similar to human dimethylaniline monooxygenase (AC002376).
BAA88195.1 AP000837 Oryza sativa
Similar to human dimethylaniline monooxygenase (AC002376).
BAB32703.1 AP002902 Oryza sativa dimethylaniline monooxygenase-like protein. P0492F05.5. contains EST C72123(E1029).
BAB07916.2 AP002835 Oryza sativa dimethylaniline monooxygenase-like protein. P0417G05.24. contains EST C72123(E1029).
SEQ ID NO: 931
AAG38521.1 AF283536 Citras x paradisi cystatin-like protein, cystein proteinase inhibitor.
AAA97905.1 U51853 Glycine max cysteine proteinase inhibitor.
CAA79954.1 Z21954 Vigna unguiculata cysteine proteinase inhibitor.
AAB66355.1 U54702 Oryza sativa thiol proteinase inhibitor; cysteine proteinase inhibitor, oryzacystatin. has potential for insect resistance in rice and for cancer therapeutics.
AAB24010.1 S49967 Oryza oryzacystatin. oryzacystatin. cysteine protease inhibitor; This sequence comes from Fig. 1.
CAA60610.1 X87126 Zeamays cysteine proteinase inhibitor. zmc-IX
BAA09666.1 D63342 Zea mays cysteine proteinase inhibitor. gCC.
AAA32672.1 L16624 Ambrosia artemisiifolia cystatin proteinase inhibitor.
BAA07327.1 D38130 Zea mays inhibition against cysteine proteinases. cystatin II.
AAA97907.1 U51855 Glycine max cysteine proteinase inhibitor.
BAB18766.1 AB038392 Triticum aestivum cysteine proteinase inhibitor. WC61. cystatin.
BAB18767.1 AB038393 Triticum aestivum cysteine proteinase inhibitor. WC92. cystatin.
CAA11899.1 AJ224331 Castanea sativa cysteine proteinase inhibitor, cystatin.
AAB71505.1 U82220 Pyras communis cysteine protease inhibitor.
AAA79239.1 L48182 Brassica rapa cysteine proteinase inhibitor. N-terminal deletion clone; putative.
CAA60634.1 X87168 Sorghum bicolor cysteine proteinase inhibitor. CPU.
AAA68150.1 L42819 Brassica rapa cysteine protenase inhibitor. N-terminal deletion clone; putative.
BAB18765.1 AB038391 Triticum aestivum cysteine proteinase inhibitor. gWC2. cystatin.
BAB18768.1 AB038394 Triticum aestivum cysteine proteinase inhibitor. WC83. cystatin.
AAC37479.1 L41355 Brassica rapa cysteine proteinase inhibitor.
BAA28867.1 AB014760 Cucumis sativus root-specific cystein protease inhibitor, cystein proteinase inhibitor.
AAD33907.1 AF143677 Artemisia vulgaris cysteine proteinase inhibitor. CPI. pollen cystatin.
BAA89582.1 AP001073 Oryza sativa
ESTs AU067919(C10906),AU067918(C 10906) conespond to a region ofthe predicted gene.
Similar to cysteine proteinase inhibitor (D31700).
CAA72790.1 Y12068 Hordeum vulgare cysteine proteinase inhibitor. CPI.
AAA96316.1 U51119 Brassica rapa cysteine proteinase inhibitor. BCPI-2.
AAKl 5090.1 AF240007 Sesamu indicum cystatin. cysteine proteinase inhibitor.
AAF23127.1 AF198389 Lycopersicon esculentum cysteine proteinase inhibitor, cystatin. STC. expression induced by arachidonic acid and gamma linolenic acid.
BAA19610.1 D64115 Glycine max cysteine proteinase inhibitor, cystatin.
BAA19608.1 D31700 Glycine max cysteine proteinase inhibitor, cystatin.
CAA89697.1 Z49697 Ricinus communis cysteine proteinase inhibitor.
AAF72202.1 AF265551 Manihot esculenta cysteine protease inhibitor.
AAA97906.1 U51854 Glycine max cysteine proteinase inhibitor.
CAA50437.1 X71124 Carica papaya cysteine proteinase inhibitor (cystatin).
AAF23126.1 AF198388 Lycopersicon esculentum cysteine proteinase inhibitor, cystatin. LTC. expression induced by arachidonic acid and gamma linolenic acid.
AAD13812.1 AFl 17334 Ipomoea batatas cysteine proteinase inhibitor, cystatin.
AAF64480.1 AF241536 Ipomoea batatas cysteine protease inhibitor. CYSPI.
AAK30004.1 AY028994 Dianthus caryophyllus cysteine proteinase inhibitor. DC-CPIn.
BAB 18769.1 AB038395 Triticum aestivum cysteine proteinase inhibitor. WC81. cystatin.
AAC32853.1 AF083253 Lycopersicon esculentum cysteine protease inhibitor.
SEQ ID NO: 938
CAA06486.1 AJ005340 Linum usitatissimum IAA amidohydrolase. homolog.
SEQ ID NO: 940
BAB17350.1 AP002747 Oryza sativa putative nodulin. P0698G03.34. contains ESTs D39891(S1543),D41717(S4395),AU033037(S1543).
BAA85440.1 AP000616 Oryza sativa
ESTs AU055729(S20023),AU055730(S20023) conespond to a region ofthe predicted gene.; similar to Medicago nodulin N21-like protein (AC004218).
CAB53493.1 AJ245900 Oryza sativa
CAA303720.1 protein. q3037.20. Similar to Medicago nodulin N21 (MtN21).
SEQ ID NO: 941
BAA21921.1 AB006599 Petunia x hybrida ZPT2-12. C2H2 zinc finger protein, 2 finger.
BAA21922.1 AB006600 Petunia x hybrida ZPT2-13. C2H2 zinc finger protein, 2fmger.
BAA19110.1 AB000451 Petunia x hybrida PEThy;ZPT2-5. Cys(2) His(2) zinc finger protein, 2 fingers.
BAA21923.1 AB006601 Petunia x hybrida ZPT2-14. C2H2 zinc finger protein, 2 finger.
BAA21925.1 AB006603 Petunia x hybrida ZPT2-8. C2H2 zinc finger protein, 2 finger.
BAA21924.1 AB006602 Petunia x hybrida ZPT2-7. C2H2 zinc finger protein, 2finger.
BAA21920.1 AB006598 Petunia x hybrida ZPT2-11. C2H2 zinc finger protein, 2fmger.
CAA60828.1 X87374 Pisum sativum putative zinc fmger protein.
BAA19111.1 AB000452 Petunia x hybrida PEThy;ZPT2-6. Cys(2) His(2) zinc fmger protein, 2 fingers.
BAA21926.1 AB006604 Petunia x hybrida ZPT2-9. C2H2 zinc finger protein, 2 fmger.
BAA21927.1 AB006605 Petunia x hybrida ZPT3-3. C2H2 zinc finger protein, 3 finger.
BAA96071.1 AB035133 Petunia x hybrida C2H2 zinc-finger protein ZPT3-3. ZPT3-3.
BAA21919.1 AB006597 Petunia x hybrida ZPT2-10. C2H2 zinc finger protein, 2 fmger.
BAA96070.1 AB035132 Petunia x hybrida C2H2 zinc-finger protein ZPT2-10. PEThy;ZPT2-10.
CAB77055.1 Y18788 Medicago sativa putative TFIIIA (or krappel)-like zinc finger protein.
AAB53260.1 U76554 Brassica rapa transcription factor, zinc-finger protein- 1. BR42.
AAB53261.1 U76555 Brassica rapa zinc-finger protein BcZFPl. BcZFPl(3-2z).
BAA19114.1 AB000455 Petunia x hybrida PEThy;ZPT4-l. Cys(2) His(2) zinc finger protein, 4 fingers.
BAA05078.1 D26085 Petunia x hybrida zinc-finger DNA binding protein.
AAD26942.1 AFl 19050 Datisca glomerata zinc-finger protein 1. zfpl. DgZFPL
BAA05077.1 D26084 Petunia x hybrida zinc-finger DNA binding protein.
AAB39638.1 U68763 Glycine max putative transcription factor. SCOF-1. scof-1. zinc-finger protein.
AAC06243.1 AF053077 Nicotiana tabacum transcription factor, osmotic stress-induced zinc-finger protein, zfp.
BAA19112.1 AB000453 Petunia x hybrida
PEThy;ZPT3-l. Cys(2) His(2) zinc finger protein, 3 fingers.
AAK01713.1 AF332876 Oryza sativa zinc finger transcription factor ZF1.
BAA05076.1 D26083 Petunia x hybrida zinc-finger DNA binding protein.
BAA05079.1 D26086 Petunia x hybrida zinc-finger protein.
BAA21928.1 AB006606 Petunia x hybrida ZPT4-4. C2H2 zinc finger protein, 4 fmger.
BAA19926.1 AB000456 Petunia x hybrida PEThy; ZPT4-2. C2H2 zinc finger protein, 4 finger.
BAA19113.1 AB000454 Petunia x hybrida PEThy;ZPT3-2. Cys(2) His(2) zinc finger protein, 3 fingers.
SEQ ID NO: 945
AAG10793.1 AF296158 Citras unshiu beta-carotene hydroxylase. CHXl .
AAG33636.1 AF315289 Citras unshiu beta-carotene hydroxylase. CHX2. similar to beta-carotene hydroxylase of Citras unshiu encoded by GenBank Accession Number AF296158.
AAGl 0430.1 AF251018 Tagetes erecta beta hydroxylase.
CAB55625.1 Y14809 Lycopersicon esculentum beta-carotene hydroxylase. CrtR-bl.
CAA70888.1 Y09722 Capsicum annuum beta-carotene hydroxylase 2.
CAB55626.1 Y14810 Lycopersicon esculentum beta-carotene hydroxylase. CrtR-b2.
CAA70427.1 Y09225 Capsicum annuum beta-carotene hydrolase.
CAC06712.1 AJ278882 Narcissus pseudonarcissus synthesis of zeaxanthin. beta-carotene hydroxylase.
AAD54243.1 AFl 62276 Haematococcus pluvialis carotenoid hydroxylase.
SEQ ID NO: 946
AAC 18914.1 U94748 Petunia x hybrida
AN11. Anl 1. No functional information available. Protein is involved in transcriptional regulation of anthocyanin biosynthesis in petunia. Protein contains five WD 40 repeats; WD 40 repeat protein.
SEQ ID NO: 948
AAG52887.1 AF333386 Nicotiana tabacum beta-expansin-like protein. PPAL. pollen allergen-like protein.
AAF72986.1 AF261273 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB5.
AAF72990.1 AF261277 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB9. putative group-1 pollen allergen.
BAB20817.1 AB051899 Atriplex lentiformis beta-expansin. A1-EXP1.
AAF72984.1 AF261271 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB3.
AAKl 5453.1 AC037426 Oryza sativa beta-expansin EXPB3. OSJNBbOOl 4111.1.
AAF72991.1 AF261278 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB10. putative group-1 pollen allergen.
AAKl 5442.1 AC037426 Oryza sativa beta-expansin EXPB6. OSJNBb0014IlL3.
AAF72987.1 AF261274 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB6.
AAF72983.1 AF261270 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPBl. putative group-1 pollen allergen
Ory si.
AAF72988.1 AF261275 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB7.
AAF72989.2 AF261276 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB8.
AAF72985.1 AF261272 Oryza sativa putative cell wall loosening activity, beta-expansin. EXPB4.
AAKl 5440.1 AC037426 Oryza sativa beta-expansin EXPB2. OSJNBbOOHIl 1.2.
AAB61710.1 U95968 Oryza sativa beta-expansin. EXPB2. cell wall loosening protein.
AAB37749.1 U30460 Cucumis sativus expansin S2 precursor. Cs-EXP2. similar to pollen allergen Lol pi, Lolium perenne, Swiss-
Prot Accession Number P14946; former gene name CuExS2; expansin-30 (Ex30) protein.
AAD38296.1 AC007789 Oryza sativa putative expansin. OSJNBa0049B20.23.
BAB 18336.1 AP002865 Oryza sativa putative expansin. P0034C 11.27.
CAC 19183.1 AJ291816 Cicer arietinum expansin.
AAG13983.1 AF297522 Prunus avium expansin 2. Exp2. PruavExp2.
AAG01875.1 AF291659 Striga asiatica alpha-expansin 3. Exp3.
AAG32920.1 AFl 84232 Lycopersicon esculentum expansin. Exp 8.
AAC96077.1 AF049350 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXPl. cell wall protein.
AAC96078.1 AF049351 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP2. cell wall protein.
AAF35902.1 AF230333 Zinnia elegans expansin 3.
BAA88200.1 AP000837 Oryza sativa
EST AU078708(E60526) conesponds to a region ofthe predicted gene. Similar to expansin
(U85246).
AAC39512.1 AF043284 Gossypium hirsutum expansin. GhEXl. contains N-terminal signal peptide.
AAD38297.1 AC007789 Oryza sativa putative expansin. OSJNBa0049B20.24.
AAF32411.1 AF230278 Triphysaria versicolor alpha-expansin 1.
BAB18338.1 AP002865 Oryza sativa putative expansin. P0034C 11.29.
AAG32921.1 AFl 84233 Lycopersicon esculentum expansin. Exp 10.
CAC19184.1 AJ291817 Cicer arietinum expansin.
AAC96079.1 AF049352 Nicotiana tabacum involved in acid-growth response, alpha-expansin precursor. Nt-EXP3. cell wall protein.
SEQ ID NO: 950
CAA52213.1 X74115 Picea abies short-chain alcohol dehydrogenase.
AAC35342.1 AF072449 Ipomoea trifida short-chain alcohol dehydrogenase. SSP. self-incompatibility-locus specific stigma protein.
AAC35340.1 AF072447 Ipomoea trifida short-chain alcohol dehydrogenase. SSP. self-incompatibility-locus specific stigma protein.
CAAl 1153.1 AJ223177 Nicotiana tabacum short chain alcohol dehydrogenase.
CAAl 1154.1 AJ223178 Nicotiana tabacum short chain alcohol dehydrogenase. SCANT.
AAK29646.1 AF349916 Solanum tuberosum putative short-chain type alcohol dehydrogenase. GAN; similar to tomato LeertlO and maize
Ts2.
AAC37345.1 L20621 Zeamays alcohol dehydrogenase. short chain.
AAC35341.1 AF072448 Ipomoea trifida short-chain alcohol dehydrogenase. SSP. self-incompatibility-locus specific stigma protein.
AAC35343.1 AF072450 Ipomoea trifida short-chain alcohol dehydrogenase. SSP. self-incompatibility-locus specific stigma protein.
AAB57737.1 U89270 Tripsacum dactyloides short-chain alcohol dehydrogenase. gynomonoecious sex form 1. similar to Zea mays tasselseed 2: SwissProt Accession Number P50160.
AAF89645.1 AF169018 Glycine max seed maturation protein PM34. PM34. similar to bacterial glucose and ribitol dehydrogenase.
AAF04253.1 AF097651 Pisum sativum short-chain alcohol dehydrogenase SAD-C. sadC. contains the entire nucleotide binding motif of 3(alpha), 20(beta)-hydroxysteroid dehydrogenases, GXXXXXXGXXG(A)XGXXXA
(Ghosh et al, 1991, Proc. Natl. Acad. Sci. USA 88, 10064-10068).
AAF04193.1 AF053638 Pisum sativum short-chain alcohol dehydrogenase. sadA. contains the entire nucleotide binding motif of the 3(alfa),20(beta)-hydroxysteroid dehydrogenase: GXXXXXXGXXG(A)XGXXXA (Ghosh et al. (1991) Proc. Natl. Acad. Sci. USA 88, 10064-10068); similar to Lycopersicon esculentum product encoded by GenBank Accession Number U21801 and Streptomyces hydrogenans steroid alcohol dehydrogenase.
AAB57738.1 U89271 Tripsacum dactyloides short-chain alcohol dehydrogenase. gynomonoecious sex form 1. shnilar to Zea mays tasselseed 2: SwissProt Accession Number P50160.
CAB91875.1 AJ277945 Lycopersicon esculentum putative alcohol dehydrogenase. yfe37.
AAF04194.1 AF053639 Pisum sativum short-chain alcohol dehydrogenase. sadB. contains the Prosite pattern no. PS00061 for short- chain alcohol dehydrogenases; contains a deletion in the nucleotide binding motif of the 3(alfa),20(beta)-hydroxysteroid dehydrogenase: GXXXXXXGXXG(A)XGXXXA (Ghosh et al. (1991) Proc. Natl. Acad. Sci. USA 88, 10064-10068).
AAB00109.1 U21801 Lycopersicon esculentum alcohol dehydrogenase homolog. GAD3. mRNA is supressed in the presence of gibberellin; similar to nonmetallo-short-chain alcohol dehydrogenases, PIR Accession Number A47542. SEQ ID NO: 952
CAB63264.1 AJ251808 Lotus japonicus calcium-binding protein, cbpl.
AAG43547.1 AF211529 Nicotiana tabacum
Avr9/Cf-9 rapidly elicited protein 31. ACRE31. similar to Solanum tuberosum CAST calcium bindmg protein encoded by GenBank Accession Number L02830.
AAA34014.1 L01432 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-3. putative.
AAA34013.1 L01430 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-1. putative.
AAA92681.1 U13882 Pisum sativum calcium-binding protein, calmodulin.
CAA78301.1 Z12839 Lilium longiflorum calcium binding protein, signal transduction. calmodulin.
AAA33397.1 L18912 Lilium longiflorum calcium binding protein, signal transduction. calmodulin. putative.
AAAI 9571.1 U10150 Brassica napus calcium binding, calmodulin. bcml .
AAA85157.1 U20297 Solanum tuberosum calcium-binding protein, calmodulin.
AAA85156.1 U20296 Solanum tuberosum calcium-binding protein, calmodulin.
AAA62351.1 U20295 Solanum tuberosum calcium-binding protein, calmodulin.
AAA85155.1 U20294 Solanum tuberosum calcium-binding protein, calmodulin.
AAC49587.1 U49105 Triticum aestivum calmodulin TaCaM4-l. calcium-binding protein.
AAC49586.1 U49104 Triticum aestivum calmodulin TaCaM3-3. calcium-binding protein.
AAC49585.1 U49103 Triticum aestivum calmodulin TaCaM3-2. calcium-binding protein. AAC49584.1 U48693 Triticum aestivum calmodulin TaCaM3-l. calcium-binding protein.
AAC49580.1 U48689 Triticum aestivum calmodulin TaCaMl-3. calcium-binding protein.
AAC49579.1 U48688 Triticum aestivum calmodulin TaCaMl-2. calcium binding protein.
AAC49578.1 U48242 Triticum aestivum cahnodulin TaCaMl-1. calcium-binding.
CAA78287.1 Z12827 Oryza sativa calcium binding protein, signal transduction. calmodulin.
AAA03580.1 L01431 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-2. putative.
AAA33901.1 L18913 Oryza sativa calcium binding protein, signal transduction. cahnodulin. putative.
AAA34015.1 L01433 Glycine max calcium-binding regulatory protein, calmodulin. SCaM-4. putative.
Table 24 Plant Open Reading Frames (ORFs) orthologous to specific Chenopodium ORFs
SEQ ID NO:1956
CAB51903.1 AJ242807Brassica napus endo-l,4-beta-D-glucanase. Cell 6. cellulase.
BAA94257.1 AB040769Hordeum vulgare endo- 1,4-beta-glucanase Cell. Cell.
AAC49704.1 U78526 Lycopersicon esculentum endo- 1,4-beta-glucanase. Cel3.
SEQ ID NO:1957
BAB21273.1 AP002844Oryza sativa putative ABC transporter protein. P0410E03.4.
BAB21275.1 AP002844Oryza sativa putative ABC transporter protein. P0410E03.6.
BAB21276.1 AP002844Oryza sativa putative ABC tiansporter protein. P0410E03.7. contains EST
D22472(C1173).
CAA94437.1 Z70524 Spirodela polynhiza multidrag resistance protein. PDR5-like ABC tiansporter.
BAB21279.1 AP002844Oryza sativa putative ABC transporter protein. P0410E03.10. contains ESTs
AU065360(R3463),AU101680(R3463).
CAA03960.1 AJ000234Hordeum vulgare partial sequence, homology to PDR5-like ABC transporter.
SEQ ID NO:1960
AAD51778.1 AFl 16858Phaseolus vulgaris utilizes UDPX as the sugar donor and catalyzes the formation of O-xylosylzeatin from zeatin. zeatin O-xylosyltransferase. ZOXl.
AAD04166.1 AFl 01972Phaseolus lunatus catalyzes formation of O-glucosylzeatin from zeatin and
UDPG; catalyzes formation of O-xylosylzeatin from zeatin and UDPX. zeatin
O-glucosyltiansferase. ZOGl. cytokinin O-glucosyltiansferase.
BAA36410.1 AB012114Vigna mungo UDP-glycose: flavonoid glycosyltransferase. UFGlyT.
BAB17061.1 AP002523Oryza sativa putative glucosyl transferase. P0013F10.7. contains EST
C73149(E2992).
BAA36412.1 ABO 12116Vigna mungo UDP-glycose:flavonoid glycosyltransferase. UFGlyT.
AAK28303.1 AF34643 lNicotiana tabacum phenylpropanoid:glucosyltiansferase 1. togtl. glucosyltransferase.
BAB17059.1 AP002523Oryza sativa putative glucosyl transferase. P0013F10.5.
AAF17551.1 AF198453Glycine max UDP-glycose:flavonoid glycosyltransferase. UFGlyT.
AAB36653.1 U32644 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS5a.
CAB88666.1 AJ400861Cicer arietinum flavonoid glycosyltransferase. putative UDP-glycose.
AAB36652.1 U32643 Nicotiana tabacum immediate-early salicylate-induced glucosyltransferase. IS 10a.
CAA54612.1 X77462 Manihot esculenta UTP-glucose glucosyltransferase. CGT5.
BAB17060.1 AP002523Oryza sativa putative glucosyl transferase. P0013F10.6.
AAK28304.1 AF346432Nicotiana tabacum phenylpropanoid: glucosyltransferase 2. togt2. glucosyltransferase.
CAA59450.1 X85138 Lycopersicon esculentum twil. homologous to glucosyltiansferases.
BAA83484.1 AB031274Scutellaria baicalensis UDP-glucose: flavonoid 7-O-glucosyltiansferase. ufgt.
CAA54611.1 X77461 Manihot esculenta UTP-glucose glucosyltiansferase. CGT2.
BAA36411.1 AB012115Vigna mungo UDP-glycose:flavonoid glycosyltransferase. UFGlyT.
CAC35167.1 AJ310148Rauvolfia seφentina arbutin synthase. as.
BAB17182.1 AP002843Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.19.
CAB56231.1 Y18871 Dorotheanthus bellidifonnis betanidrn-5-O-glucosyltiansferase.
BAB17176.1 AP002843 Oryza sativa putative UTP-glucose glucosyltransferase. P0407B12.13.
AAK16172.1 AC079887Oryza sativa putative glucosyltransferase. OSJNBa0040E01.14.
CAA54613.1 X77463 Manihot esculenta UTP-glucose glucosyltiansferase. CGT6.
AAF61647.1 AF 190634Nicotiana tabacum UDP-glucose: salicylic acid glucosyltiansferase. SA-GTase.
BAA93039.1 AB033758Citras unshiu limonoid UDP-glucosyltransferase. LGTase.
CAA54610.1 X77460 Manihot esculenta UTP-glucose glucosyltransferase. CGT4.
AAA59054.1 L34847 Zea mays conjugation ofthe growth hormone indole-3 -acetic acid
(IAA). IAA-glu synthetase. iaglu.
BAA36423.1 ABO 13598 Verbena x hybrida UDP-glucose: anthocyanin 5-O-glucosyltiansferase. HGT8.
AAF17077.1 AFl 99453 Sorghum bicolor UDP-glucose glucosyltransferase. UDP-glucose:p-hydroxymandelonitrile-o- glucosyltransferase.
CAA54558.1 X77369 Solanum melongena glycosyl transferase. GT.
AAK16175.1 AC079887Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.15.
CAA81057.1 Z25802 Petunia x hybrida UDP rhamnose: anthocyanidin-3-glucoside rhamnosyltransferase.
CAA50377.1 X71060 Petunia x hybrida anthocyanin: rhamnosyltransferase. rt.
CAA50376.1 X71059 Petunia x hybrida anthocyanin 3 glucoside: rhamnosyltransferase. rt.
AAK16181.1 AC079887Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.16.
AAK16178.1 AC079887Oryza sativa putative glucosyltiansferase. OSJNBa0040E01.5.
AAD21086.1 AF127218Forsythia x intermedia adds glucose residue to position 3 of flavonoid compounds. flavonoid 3-O-glucosyltransferase. UFGT.
BAA89008.1 AB027454Petunia x hybrida anthocyanidin 3-O-glucosyltiansferase. PGT8.
SEQ ID NO: 1962
CAA65580.1 X96784 Nicotiana tabacum cytochrome P450. hsr515.
CAA64635.1 X95342 Nicotiana tabacum cytochrome P450. hsr515. hypersensitivity-related gene.
AAG44132.1 AF218296Pisum sativum cytochrome P450. P450 isolog.
AAG49299.1 AF313489Callistephus chinensis flavonoid 3',5'-hydroxylase.
BAA03438.1 D14588 Petunia x hybrida flavonoid-3',5'-hydroxylase. Hfl.
AAC32274.1 AF081575Petunia x hybrida flavonoid 3',5'-hydroxylase. Hfl. P450 enzyme.
CAA50442.1 X71130 Petunia x hybrida P450 hydroxylase. PET 1.
CAA80266.1 Z22545 Petunia x hybrida flavonoid 3',5'-hydroxylase.
AAG49315.1 AF315465Pelargonium x hortorum flavonoid 3'-hydroxylase.
AAG49298.1 AF313488Callistephus chinensis putative flavonoid 3 '-hydroxylase.
AAB 17562.1 U72654 Eustoma grandiflorum flavonoid 3'5'-hydroxylase. F3'5'H. cytochrome P450; CYP75.
BAB20076.1 ABO 12925Torenia hybrida flavonoid 3',5'-hydroxylase. F3'5'H.
AAG14961.1 AF214007Brassica napus cytochrome p450-dependent monooxygenase. BNF5H1.
CAC26920.1 AJ295586Arabidopsis lyrata subsp. petraea feralate-5-hydroxylase. fahl.
AAG14962.1 AF214008Brassica napus cytochrome p450-dependent monooxygenase. BNF5H2.
References
Aarts et al, Proc Natl Acad Sci 95:10306 (1998).
Alonso et al., Science. 284:2148 (1999)
Altschul et al., J. Mol. Biol. 215:403 (1990). Altschul et al.. Nucleic Acids Res. 25:3389 (1991).
An et al., EMBO J. 4:277 (1985).
Aoyama et al., N-H Plant Journal. 11 :605 (1997).
Bachem et al., Plant X. 9:745 (1996).
Bailey and Elkan In: Proceedings ofthe second international Conference on Intelligent Systems for Molecular Biology (Altman, R., ed), pp 28-36, AAAI press (1994).
Ballas et al., Nucleic Acids Res. 17:7891 (1989).
Balzi et al., J. Biol. Chem.. 269:2206 (1994).
Bansal et al., Proc. Natl. Acad. Sci. USA. 89:3654 (1992). Batzer et al., Nucleic Acid Res.. 19:5081 (1991).
Beachy and Murakishi, Phytopathology. 61:877 (1971).
Beals et al., Plant Cell 9:1527 (1997).
Belanger et al., Genetics. 129:863 (1991).
Bevan et al, Nature. 304:184 (1983). Blochinger & Diggelmann, Mol Cell Biol. 4:2929.
Bourouis et al., EMBO X. 2:1099 (1983).
Boyes et al., Proc Natl Acad Sci USA. 95:15849 (1998).
Byrne et al. Plant Cell Tissue and Organ Culture. 8:3 (1987).
Campbell and Gowri, Plant Physiol.. 92:1 (1990). Canto and Palukaitis, Virology. 265:74 (1999).
Cao et al., Plant Cell. 6:1583 (1994).
Cao et al, Cell, 88:57 (1997).
Century et al., Science. 278:1963 (1997).
Chandler et al., Plant Cell. 1:1175 (1989). Christou et al., Biotechnology. 9:957 (1991).
Christou et al., Plant Physiol.. 87:671 (1988).
Conklin and Last, Plant Physiol.. 109:203 (1995).
Cooper et al., Virology. 206:307 (1995)
Cooper et al., Virology. 216:208 (1996).
Cordero et al., Plant X. 6:141 (1994).
Coφet et al, Nucleic Acids Res. 16:10881 (1988).
Crameri et al., Nature Biotech.. 15:436 (1997). Crameri et al., Nature. 391:288 (1998).
Creelman et al. Annu Rev Plant Phvsiol Plant Mol Biol 48:355 (1997).
Crossway et al., BioTechniques. 4:320 (1986).
Czako et al., Mol. Gen. Genet. 23 5 (1), 33-40 (1992).
Czernic et al., Plant Mol. Biol.. 31:255 (1996). Datta et al., Bio/Technology. 8, 736 (1990).
Dayhoff et al., Atlas of Protein Sequence and Stracture. Natl. Biomed. Res. Found., Washington, CD. (1978).
De Blaere et al, Meth. Enzymol. 143:277 (1987). de Framond, FEBS. 290:103 (1991). Delaney et al.. Proc. Natl. Acad. Sci. USA 92:6602 (1995).
Della-Cioppa et al., Plant Physiology. 84:965 (1987).
De Oliveira et al., Microbios.. 76:213 (1993).
Dennis et al., Nucleic Acids Res.. 12:3983 (1984).
Dong et al., Curr Onin Plant Biol 1:316 (1998). Dunigan and Madlener, Virology. 207:460 (1995).
Durrant et al., The Plant Cell. 12:963 (2000).
Dzelkalns et al., Plant Cell. 5:855 (1993).
Eisen et al., Trends in Plant Sci. 95:14863 (2000).
Ellis and Jones, Cun Onin Plant Bio 1:288 (1998). Elroy-Stein et al, PNAS USA. 86:6126 (1989).
English, et al, Plant Cell 8:179 (1996).
Eulgem et al., Trends in Plant Sci. 5:199 (2000).
Falk et al., Proc Natl Acad Sci USA 96:3292 (1999).
Feys et al., Plant Cell 6:751 (1994). Franken et al., EMBO X. 10:2605 (1991).
Fromm et al., Bio/Technology 8:833 (1990).
Gallie et al, Molecular Biology of RNA. 237 (1989).
Gallie et al, Nucl. Acids Res.. 15:8693 (1987).
Gan et al., Science (1995), 270 (5244), 1986-8).
Gatz Cunent Opinion in Biotechnology. 7:168 (1996).
Gatz, C, Annu. Rev. Plant Physiol. Plant Mol. Biol.. 48:89 (1997).
Gelfand, eds., PCR Strategies (Academic Press, New York (1995)). Glazebrook. X. Cun. Opin. Plat Biology. 2:280 (1999V
Glazebrook et al., Genetics 143:973 (1996).
Glazebrook et al., Proc. Natl. Acad. Sci. USA 91:8955 (1994).
Gordon-Kamm et al., Plant Cell. 2, 603 (1990).
Graham et al., Biochem. Biophys. Res. Comm.. 101:1164 (1981). Graham et al., J. Biol. Chem.. 260:6555 (1985).
Graham et al., J. Biol. Chem.. 260:6561 (1985).
Guerineau et al., Mol. Gen. Genet. 262:141 (1991).
Gulyas and Farkas, Phvtopath. Z.. 91:182 (1978)
Hammand-Kosack and Jones, Ann. Rev. Plant Physiol. Plant Mol. Biol.. 48:575 \ 1997).
Heinlein et al, Science. 270: 1983 (1995).
Henikoff & Henikoff, Proc. Natl. Acad. Sci. USA. 89:10915 (1989).
Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA. 89:10915 (1992).
Hiei et al., Plant J. 6:271 (1994). Higgins et al., Gene 73:237 (1988).
Higgins et al.. CABIOS 5:151 (1989).
Hinchee et al., Biotechnology. 6:915 (1988).
Hoekema, In: The Binary Plant Vector System. Offset-drukkerij Kanters BN.
Horvath and Chua, Plant Mol. Biol.. 31:1061 (1996). Huang et al., CABIOS 8:155 (1992).
Hudspeth & Grala, Plant Molec. Biol.. 12:579 (1989).
Huffman et al., J. Cell. Biochem.. 17B: Abstract.
Hunt et al., Mol. Plant-Microbe Int. 9:261 (1997).
Ingelbrecht et al., Plant Cell. 1:671 (1989). Innis et al., eds., PCR Protocols: A Guide to Methods and Applications (Academic
Press, New York (1995).
Innis and Gelfand, eds., PCR Methods Manual (Academic Press, New York) (1999).
Jobling et al, Nature. 325:622 (1987).
John et al., Proc. Natl. Acad. Sci. USA 89(13): 5769 (1992).
Jones et al., Adv Bot Res 24:89 (1991).
Joshi et al., Nucleic Acid Res. 15:9627 (1987). Joshi, Nucl. Acid Res.. 15, 6643 (197).
Karlin and Altschul, Proc. Natl. Acad Sci. USA 87:2264 (1990).
Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90:5873 (1993).
Keller et al., Genes Dev.. 3:1639 (1989).
Klein et al., Bio/Technology. 6:559 (1988). Klein et al., Nature (London . 327:70 (1987).
Klein et al., Plant Physiol.. 91 :440 (1988).
Klein et al, Proc. Natl. Acad. Sci. USA. 85:4305 (1988).
Kohler et al., Plant Mol. Biol.. 29:1293 (1995).
Knauf, et al., Genetic Analysis of Host Range Expression by Agrobacterium In: Molecular Genetics ofthe Bacteria-Plant Interaction, Puhler, A. ed., Springer-
Verlag, New York, 245 (1983).
Komari, Plant Cell Reports. 9:303 (1990).
Koziel et al., Biotechnology. 11:194 (1993).
Kridl et al., Seed Science Research. 1:209 (1991). Kriz et al., Mol. Gen. Genet.. 207:90 (1987).
Kunkel, Proc. Natl. Acad. Sci. USA. 82:488 (1985).
Kunkel et al., Methods in Enzvmo 154:367 (1987).
Lange et al., Plant Sci.. 142:133 (1999).
Langridge et al., Cell, 34:1015 (1983). Lashbrook et al., Plant Cell. 6:1485 (1994).
Leister et al., P.N.A.S.. (USA), 95:370 (1998).
Leister et al., Proc. Natl. Acad. Sci. USA. 95:370 (1998).
Liang et al., Science. 257:967 (1992).
Lindstiom et al., Der. Genet.. 11 : 160 (1990). Lommel et al.. Virology. 81:382 . 19911
Ly et al., Science. 287:2486 (2000).
Macejak et al., Nature. 353:90 (1991).
Mansson et al., Gen. Genet.. 200:356 (1985).
Martin and Paz-Ares, Trends in Genetics 13:67 (1997).
Martinez et al, J. Mol. Biol.. 208:551 (1989).
McBride et al. Proc. Natl. Acad. Sci. USA. 91:7301 (1994).
McCabe et al, Bio/Technology. 6:923 (1988). McDowell et al., Plant J. 22:523 (2000).
McDowell et al., Plant Cell 10:1861 (1998).
McNellis et al., Plant J. 1998, 14:247-257.
Meinkoth and Wahl, Anal. Biochem.. 138:267 (1984).
Messing & Viena, Gene. 19:259 (1982). Meyers et al., Plant J. 20:317 (1999).
Mogen et al.. Plant Cell 2:1261 (1990).
Moore et al., X Mol. Biol. 272:336 (1997).
Munroe et al., Gene 91:151 (1990).
Munay et al., Nucleic Acids Res.. 17:477 (1989). Myers and Miller, CABIOS 4:11 (1988).
Needleman and Wunsch, X Mol. Biol. 48:443 (1970).
Odell et al., Nature. 313:810 (1985).
Odell et al., Nature. 313:810 (1985).
Ohtsuka et al., J. Biol. Chem.. 260:2605 (1985). Okamuro et al., Biochemistry of Plants. 15:1 (1989).
Otsuki et al., Virology. 50:45 (1972)
Paszkowski et al., EMBO J., 3:2717 (1984).
Pacciotti et al. Bio/Technology 3:241 (1985).
Padgett and Beachy, Plant Cell. 5:577 (1993). Park et al., J. Plant Biol. 38(4):365 (1985).
Pearson et al, Meth. Mol. Biol. 24:307 (1994).
Pearson and Lipman, Proc. Natl. Acad. Sci. 85:2444 (1988).
Penninck et al, Plant Cell 8:2309 (1996).
Perlak et al., Proc. Natl. Acad. Sci. USA. 88:3324 (1991). Pieterse et al., Plant Cell. 10:1571 (1998).
Pieterse et al., Plant Cell. 8:1225 (1996).
Proudfoot, Cell 64:671 (1991).
Quigley et al., J. Mol. Evol.. 29:412 (1989).
Ralston et al., Genetics, 119:185 (1988).
Reina et al., Nucleic Acids Res.. 18:6425 (1990).
Reina et al., Nucleic Acids Res.. 18:7449 (1990).
Riggs et al., Proc. Natl. Acad. Sci. USA. 83:5602 (1986). Rochester et al., (1986).
Rommens et al., Plant Cell, 7:1537 (1995).
Ronald, Cun Opin Plant Biol 1:294 (1998).
Rossolini et al., Mol. Cell. Probes. 8:91 (1994).
Roth et al., Nature Biotechnology. 16:939 (1998). Ruiz et al., Plant Cell 10:937 (1998).
Ryals et al., Plant Cell 8:1809 (1996).
Ryals et al., Plant Cell 9:425 (1997)
Sambrook et al., Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Plainview, New York) (1989). Sanchez-Fernandez et al., Mol. Gen. Genet.. 258:655 (1998).
Sanfacon et al., Genes Dev. 5:141 (1991).
Sanford et al., Particulate Science and Technology, 5:27 (1987).
Schernthaner et al., EMBO J.. 7:1249 (1988).
Schindler et al., Plant Cell 4:1309 (1992). Schmitz and Rao, Virology, 248:323 (1996)
Schwob et al., Plant X. 4:423 (1993).
Shah et al., Mol. Plant-Microbe Interact. 10:69 (1997).
Shimamoto et al., Nature. 338:274 (1989).
Shirasu et al., Plant Cell 9:261 (1997). Shulaev et al., Plant Cell 7:1691 (1995).
Simpson, Plant Mol. Biol.. 19:699 (1985).
Skuzeski et al., Plant Molec. Biol.. 15:65 (1990).
Slater et al., Plant Mol. Biol. 5:137 (1985).
Smart and Fleming, X Biol. Chem.. 271:19351 (1996). Smith et al.. Adv. Appl. Math. 2:482 (1981).
Smith et al., Planta 168:94 (1986).
Song et al., Science. 270: 1804 (1995).
Spencer et al., Theor Appl Genet. 79:625 (1990).
Staswick et al., Proc. Natl. Acad. Sci. USA 89:6837 (1992).
Staub et al., EMBO X. 12:601 (1993).
Staub et al., Plant Cell. 4:39 (1992).
Stemmer, Nature. 370:389 (1994). Stemmer. Proc. Natl. Acad. Sci. USA. 91:10747 (1994).
Sukhapinda et al. Plant Mol. Biol. 8:209 (1987).
Sullivan et al., Mol. Gen. Genet.. 215:431 (1989).
Svab et al, Proc. Natl. Acad. Sci. USA. 87:8526 (1990).
Svab et al, Proc. Natl. Acad. Sci. USA. 90:913 (1993). Thomma et al, Plant Physiol.. 121:1093 (1999).
Thomma et al., Proc. Natl. Acad. sci. U.S.A., 85:15107 (1998).
Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology- Hybridization with Nucleic Acid Probes, part I chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays" Elsevier, New York (1993).
Turner et al., Molecular Biotechnology. 3:225 (1995).
VanTunen et al, EMBO J.. 7:1257 (1988).
Vasil et al.. Biotechnology. 11: 1553 (1993).
Verduin, J. Gen. Virol.. 38:571 (1978). Vernooij et al., Plant Cell 6:959 (1994).
Visedo et al., Phvsiologia Plantarum. 78:218 (1990).
Vodkin, Prog. Clin. Biol. Res.. 138:87 (1983).
Vogel et al. EMBO X. 11:157 (1992).
Walker and Gaastia, eds., Techniques in Molecular Biology. MacMillan Publishing Company, New York (1983).
Wandelt et al., Nucleic Acids Res.. 17:2354 (1989).
Waterman, M.S. Intioduction to Computational Biology: Maps, sequences and genomes. Chapman & Hall. London (1995).
Weeks et al., Plant Physiol.. 102:1077 (1993). Weissinger et al, Annual Rev. Genet.. 22:421 (1988).
Wenzler et al., Plant Mol. Biol. 13:347 (1989).
Weymann et al, Plant Cell 7:2013 (1995).
White et al., Nucl Acids Res, 18:1062 (1990).
Whitham et al., P.N.A.S.. (USA), 93:8776 (1996).
Whitham et al., Cell, 78:1101 (1994).
Wilhtset al., Mol Plant-Microbe Interact 11:795 (1998).
Xie et al.. Science 280:1091 (1998). Yamamoto et al, Nucleic Acids Res.. 18:7449 (1990).
Yang et al., Plant Mol. Biol.. 38:1201 (1998).
Yang & Klessig, Proc. Natl. Acad. Sci. USA 93:14972 (2000).
Yu et al., Proc. Natl. Acad. Sci. USA 95:7819 (1998).
Zhang et al., Proc. Natl. Acad. Sci. USA. 94:4504 (1997). Zhou et al.. EMBO X. 16:3207 .1997V
Zhou et al., Plant Cell 8:2235 (1996).
Zhou et al., Plant Cell 10:1021 (1998).
All publications, patents and patent applications are incoφorated herein by reference.
While in the foregoing specification this invention has been described in relation to certain prefened embodiments thereof, and many details have been set forth for puφoses of illustration, it will be apparent to those skilled in the art that the invention is susceptible to additional embodiments and that certain ofthe details described herein may be varied considerably without departing from the basic principles ofthe invention.