JP7207721B2 - ニューロン細胞培養のための培地組成物 - Google Patents
ニューロン細胞培養のための培地組成物 Download PDFInfo
- Publication number
- JP7207721B2 JP7207721B2 JP2019071296A JP2019071296A JP7207721B2 JP 7207721 B2 JP7207721 B2 JP 7207721B2 JP 2019071296 A JP2019071296 A JP 2019071296A JP 2019071296 A JP2019071296 A JP 2019071296A JP 7207721 B2 JP7207721 B2 JP 7207721B2
- Authority
- JP
- Japan
- Prior art keywords
- concentration
- brainphys
- neurons
- medium
- cells
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images










































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0618—Cells of the nervous system
- C12N5/0619—Neurons
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/05—Inorganic components
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/05—Inorganic components
- C12N2500/10—Metals; Metal chelators
- C12N2500/12—Light metals, i.e. alkali, alkaline earth, Be, Al, Mg
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/05—Inorganic components
- C12N2500/10—Metals; Metal chelators
- C12N2500/12—Light metals, i.e. alkali, alkaline earth, Be, Al, Mg
- C12N2500/16—Magnesium; Mg chelators
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/05—Inorganic components
- C12N2500/10—Metals; Metal chelators
- C12N2500/20—Transition metals
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/05—Inorganic components
- C12N2500/10—Metals; Metal chelators
- C12N2500/20—Transition metals
- C12N2500/22—Zinc; Zn chelators
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/05—Inorganic components
- C12N2500/10—Metals; Metal chelators
- C12N2500/20—Transition metals
- C12N2500/24—Iron; Fe chelators; Transferrin
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/30—Organic components
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/30—Organic components
- C12N2500/32—Amino acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/30—Organic components
- C12N2500/34—Sugars
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/30—Organic components
- C12N2500/38—Vitamins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/10—Growth factors
- C12N2501/13—Nerve growth factor [NGF]; Brain-derived neurotrophic factor [BDNF]; Cilliary neurotrophic factor [CNTF]; Glial-derived neurotrophic factor [GDNF]; Neurotrophins [NT]; Neuregulins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/30—Hormones
- C12N2501/38—Hormones with nuclear receptors
- C12N2501/39—Steroid hormones
Landscapes
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Life Sciences & Earth Sciences (AREA)
- Zoology (AREA)
- Neurology (AREA)
- Organic Chemistry (AREA)
- Biotechnology (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- Wood Science & Technology (AREA)
- Microbiology (AREA)
- Cell Biology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Neurosurgery (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Description
本願は、2013年4月17日に出願された米国仮特許出願第61/813,034号への優先権および利益を主張し、この米国仮特許出願の全体の内容は、本明細書中に参考として援用される。
ヒト疾患を模擬し、緊急的に必要とされる治療を見出すための幹細胞および体細胞の再プログラミングに対して相当量の希望と努力が注がれている。ヒト体細胞を多能性幹細胞に再プログラミングする革命的方法を公表した僅か5年後に山中博士に最近のノーベル生理学医学賞が授与されたことによって、いかに多くの期待が人工多能性幹細胞(IPSC)技術に注がれているかが裏付けられる(Takahashiら、2007年)。その直後に、患者の皮膚細胞に由来するIPSCはin vitroでニューロンに変えることができることが示された(Dimosら、2008年)。神経科学者は神経障害および精神障害を有する患者からin vitroで生きたニューロンの培養物を得るというこの前例のない機会へと一斉に向かい、特定の障害と関連する主要な表現型をディッシュで再現することができることを実証した(Marchettoら、2010年;Brennandら、2011年;Marchettoら、2011年;Nguyenら、2011年;Seiblerら、2011年;Bellinら、2012年;Egawaら、2012年;Israelら、2012年;Shiら、2012年)。
本出願は、培地中で培養した脳細胞の成長および/または維持および/または機能活性を促進する細胞培地を提供する。ある特定の実施形態において、細胞培地は、1種もしくは複数の神経活性無機塩、および/または1種もしくは複数の神経活性アミノ酸、および/または1種もしくは複数のビタミン、および/または1種もしくは複数のアミノ酸、および/または1種もしくは複数のエネルギー基質、および/または1種もしくは複数のpH調節剤を含む。
本発明の実施形態において、例えば以下の項目が提供される。
(項目1)
(a)約70~約150mMの濃度で塩化ナトリウム;
(b)約0.000001~約10mMの濃度で神経活性無機塩;
(c)約0.0001~約0.05mMの濃度でグリシン;
(d)約00001~約0.05mMの濃度でL-アラニン;および
(e)約0.001~約0.03mMの濃度でL-セリン
を含む、細胞培地。
(項目2)
前記神経活性無機塩が、塩化カリウム、塩化カルシウム、硫酸マグネシウム、塩化マグネシウム、硝酸第二鉄、硫酸亜鉛、硫酸第二銅、硫酸第二鉄、およびこれらの組合せからなる群から選択される、項目1に記載の細胞培地。
(項目3)
約0.00001~約0.003mMの濃度でL-アスパラギン酸をさらに含む、項目1に記載の細胞培地。
(項目4)
約0.00001~約0.02mMの濃度でL-グルタミン酸をさらに含む、項目1に記載の細胞培地。
(項目5)
pH調節剤をさらに含み、該調節剤が、無機塩である、項目1に記載の細胞培地。
(項目6)
前記無機塩が、約0.001~約1mMの濃度であり、該無機塩が、リン酸水素二ナトリウム、リン酸二水素ナトリウム、およびこれらの組合せからなる群から選択される、項目5に記載の細胞培地。
(項目7)
前記無機塩が、約1~約35mMの濃度であり、該無機塩が、炭酸水素ナトリウムである、項目5に記載の細胞培地。
(項目8)
1種または複数のアミノ酸をさらに含み、各アミノ酸が、約0.001~約1mMの濃度である、項目1に記載の細胞培地。
(項目9)
前記1種または複数のアミノ酸が、L-アラニル-L-グルタミン、L-アルギニン塩酸塩、L-アスパラギン-H2O、L-システイン塩酸塩-H2O、L-シスチン2HCl、L-ヒスチジン塩酸塩-H2O、L-イソロイシン、L-ロイシン、L-リシン塩酸塩、L-メチオニン、L-フェニルアラニン、L-プロリン、L-トレオニン、L-トリプトファン、L-チロシン二ナトリウム塩二水和物、L-バリン、およびこれらの組合せからなる群から選択される、項目8に記載の細胞培地。
(項目10)
1種または複数のビタミンをさらに含み、各ビタミンは、約0.00001~約1mMの濃度で存在する、項目1に記載の細胞培地。
(項目11)
前記1種または複数のビタミンが、塩化コリン、D-パントテン酸カルシウム(B5)、葉酸(B9)、i-イノシトール、ナイアシンアミド(B3)、ピリドキシン塩酸塩、チアミン塩酸塩、ビタミンB12(シアノコバラミン)、リボフラビン(B2)、およびこれらの組合せからなる群から選択される、項目10に記載の細胞培地。
(項目12)
タンパク質、神経栄養因子、ステロイド、ホルモン、脂肪酸、脂質、ビタミン、硫酸塩鉱物、有機化学化合物、単糖、ヌクレオチド、およびこれらの組合せからなる群から選択される補充剤をさらに含む、項目1に記載の細胞培地。
(項目13)
0.1~約5mMの濃度でエネルギー基質をさらに含む、項目1に記載の細胞培地。
(項目14)
前記エネルギー基質が、糖、ピルビン酸ナトリウム、およびこれらの組合せからなる群から選択される、項目13に記載の細胞培地。
(項目15)
光増感剤をさらに含む、項目1に記載の細胞培地。
(項目16)
前記光増感剤が、約0.0001~約0.0006mMの濃度のリボフラビン(B2)である、項目15に記載の細胞培地。
(項目17)
前記光増感剤が、約1~約10mMの濃度のHEPESである、項目15に記載の細胞培地。
(項目18)
血清を含まない、項目1に記載の細胞培地。
(項目19)
前記培地の容量オスモル濃度が、約280~約330Osm/Lである、項目1に記載の細胞培地。
(項目20)
ニューロン細胞と、項目1から19のいずれか一項に記載の培地とを接触させることを含む、ニューロン細胞を培養する方法。
(項目21)
前記ニューロン細胞が、初代ニューロン細胞である、項目20に記載の方法。
(項目22)
前記ニューロン細胞が、iPSCに由来するニューロン細胞である、項目20に記載の方法。
(項目23)
項目1から19のいずれか一項に記載の培地中で培養した哺乳動物細胞。
(項目24)
ニューロン細胞である、項目23に記載の哺乳動物細胞。
本開示は、培地中で培養された神経細胞の成長および維持、ならびに機能活性を促進する培地の発見に少なくとも部分的に基づいている。特に、本出願は、このような培地が、神経活性無機塩、神経活性アミノ酸、pH調節剤、エネルギー基質、アミノ酸、およびビタミンの組合せを含むことができることを開示する。ある特定の実施形態において、培地の成分は、培地中で培養した神経細胞の成長、維持および神経機能性を促進する量で存在する。
他の規定がない限り、本明細書において使用される技術用語および科学用語は、当業者が一般に理解するのと同じ意味を有する。例えば、Singletonら、DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY、第2版、J. Wiley & Sons(New York、NY、1994年);Sambrookら、MOLECULAR CLONING, A LABORATORY MANUAL、Cold Springs Harbor Press(Cold Springs Harbor、NY、1989年)を参照されたい。本明細書に記載されているものと同様または等しいあらゆる方法、デバイスおよび材料を本発明の実施に使用することができる。下記の定義は、本明細書において頻繁に使用され、本開示の範囲を限定することを意味しない特定の用語の理解を容易にするために提供する。
一態様において、本出願は、細胞の成長および/または維持および/または機能活性を促進する培地を提供する。ある特定の実施形態において、培地は、1種または複数の神経活性無機塩および1種または複数の神経活性アミノ酸を含む。ある特定の実施形態において、培地は、1種もしくは複数のpH調節剤、1種もしくは複数のエネルギー基質、1種もしくは複数のアミノ酸、および/または1種もしくは複数のビタミン、およびこれらの組合せをさらに含む。
ある特定の実施形態において、培地は、塩化ナトリウム(NaCl)、塩化カリウム(KCl)、塩化カルシウム(CaCl2)(例えば、無水CaCl2)、硫酸マグネシウム(MgSO4)(例えば、無水MgSO4)、塩化マグネシウム(MgCl2)(例えば、無水MgCl2)、硝酸第二鉄(Fe(NO3)3-9H2O)、硫酸亜鉛(ZnSO4-7H2O)、硫酸第二銅(CuSO4-5H2O)、硫酸第二鉄(FeSO4-7H2O)、およびこれらの組合せからなる群から選択される神経活性無機塩を含む。
ある特定の実施形態において、培地は、グリシン、L-アラニン、L-アスパラギン酸、L-グルタミン酸、L-セリン、およびこれらの組合せからなる群から選択される神経活性アミノ酸を含む。
ある特定の実施形態において、培地は、無機塩、pH緩衝剤、pH指示薬、またはこれらの組合せからなる群から選択されるpH調節剤を含む。
ある特定の実施形態において、培地は、糖(例えば、D-グルコース(デキストロース))、ピルビン酸ナトリウム、およびこれらの組合せからなる群から選択されるエネルギー基質を含む。ある特定の実施形態において、エネルギー基質は、約0.001~約10mM、または約0.005~約9mM、または約0.01~約8mM、または約0.05~約7mM、または約0.1~約6mM、または約1~約5mM、または約2~約4.5mMの濃度で存在する。ある特定の実施形態において、エネルギー基質は、約5mM未満、または約1mM未満の濃度で存在する。ある特定の実施形態において、エネルギー基質は、0.1~約5mMの濃度で存在する。ある特定の実施形態において、エネルギー基質は、約2.5mMまたは約0.5mMの濃度で存在する。
ある特定の実施形態において、培地は、1種または複数のアミノ酸を含む。ある特定の実施形態において、1種または複数のアミノ酸は、L-アラニル-L-グルタミン、L-アルギニン塩酸塩、L-アスパラギン-H2O、L-システイン塩酸塩-H2O、L-シスチン2HCl、L-ヒスチジン塩酸塩-H2O、L-イソロイシン、L-ロイシン、L-リシン塩酸塩、L-メチオニン、L-フェニルアラニン、L-プロリン、L-トレオニン、L-トリプトファン、L-チロシン二ナトリウム塩二水和物、L-バリン、およびこれらの組合せからなる群から選択される。ある特定の実施形態において、各アミノ酸は、約0.001~約5mM、または約0.005~約3mM、または約0.01~約1mM、または約0.05~約0.8mM、または約0.1~約0.5mM、または約0.001~約1mMの濃度で存在する。
ある特定の実施形態において、培地は、1種または複数のビタミンを含む。ある特定の実施形態において、1種または複数のビタミンは、塩化コリン、D-パントテン酸カルシウム(B5)、葉酸(B9)、i-イノシトール、ナイアシンアミド(B3)、ピリドキシン塩酸塩、チアミン塩酸塩、ビタミンB12(シアノコバラミン)、リボフラビン(B2)、およびこれらの組合せからなる群から選択される。ある特定の実施形態において、各ビタミンは、約0.00001~約1mM、または約0.00005~約0.5mM、または約0.0001~約0.9mM、または約0.0005~約0.8mM、または約0.001~約0.7mM、または約0.005~約0.6mM、または約0.01~約0.5mM、または約0.05~約0.1mMの濃度で存在する。
ある特定の実施形態において、培地は、タンパク質(例えば、ラミニン;BSA、脂肪酸を含まない画分V;カタラーゼ;インスリン;ヒト組換えインスリン;インスリン組換え完全鎖;トランスフェリン;ヒトトランスフェリン;ヒトトランスフェリン(Holo);スーパーオキシドジスムターゼ)、神経栄養因子(例えば、ヒト脳由来神経栄養因子(BDNF);グリア神経栄養因子(GDNF))、ホルモン、ステロイド(例えば、コルチコステロン、プロゲステロン)、甲状腺ホルモン(例えば、T3(トリヨード-I-チロニン))、脂肪酸、必須脂肪酸(例えば、リノール酸、リノレン酸)、脂質(例えば、コレステロール)、ビタミン(例えば、アスコルビン酸(Vit C)、ビオチン(B7)、DLアルファ酢酸トコフェロール、DLアルファ-トコフェロール、ビタミンA(レチノイン酸))、硫酸塩鉱物(例えば、セレナイト)、有機化学化合物(例えば、プトレシン2HCl)、単糖(Monosacharide)(例えば、D-ガラクトース)、ヌクレオチド(例えば、ジブチリル(Dibutyril)cAMPナトリウム塩)、およびこれらの組合せからなる群から選択される1種(on)または複数の補充剤をさらに含む。
ある特定の実施形態において、培地は、約0.05mM未満の濃度でL-アラニン、約0.25mM未満の濃度でグリシン、約0.25mM未満の濃度でL-セリン、約0.15mM未満の濃度でL-プロリン、約0.60mM未満の濃度でL-アルギニン塩酸塩、約2.5mM未満の濃度でL-アラニル-L-グルタミン、約0.41mM超の濃度で硫酸マグネシウム、約1.05mM超の濃度で塩化カルシウム、約4.16mM超の濃度で塩化カリウム、約120.7mM超の濃度で塩化ナトリウム、約17.5mM未満の濃度でD-グルコース、または約5mMの濃度でHEPESを含む。いくつかの実施形態において、培地は、L-アスパラギン酸、L-グルタミン酸、L-シスチン2HCl、CuSO4-5H2O、FeSO4-7H2O、無水塩化マグネシウム、リノール酸、プトレシン2HCl、リポ酸、チミジン、ヒポキサンチンNa、またはビオチンを含まない。
いくつかの実施形態において、培地は、約315mOsmol/L未満の容量オスモル濃度を有する。他の実施形態において、容量オスモル濃度は、約200~約400mOsmol/L、または約220~約380mOsmol/L、または約240~約360mOsmol/L、または約260~約340mOsmol/L、または約280~約330mOsmol/L、または約300~約320mOsmol/Lである。ある特定の実施形態において、容量オスモル濃度は、約300mOsmol/L~約310mOsmol/Lである。いくつかの実施形態において、容量オスモル濃度は、約310mOsmol/Lである。
別の態様において、ニューロン細胞を培養する方法を提供する。この方法は、ニューロン細胞と、その実施形態を含め本明細書で示されるような培地とを接触させ、ニューロン細胞が成長することを可能とし、それによってニューロン細胞を培養することを含む。いくつかの実施形態において、ニューロン細胞は、初代ニューロン細胞である。他の実施形態において、ニューロン細胞は、iPSCに由来するニューロン細胞である。
in vitroでのヒト脳細胞のシナプス活性および機能を支持する神経生理学的培地
要約
人工多能性幹細胞(iPSC)技術は、患者に由来する生きたヒトニューロンへのアクセス、ならびに神経障害および精神障害をin vitroで模擬する新規な代替物を提供する。ニューロン回路の有効なモデルは、ニューロン機能を持続させる生理学的条件を必要とする。したがって、出願人は、ニューロンを培養するために現在使用されている異なる培地中でニューロン活性を特に検査した。げっ歯類およびヒトからのニューロンは、種々の補充物を有する基礎培地中でin vitroにて常習的に成長する。標準的基礎培地(例えば、DMEM、Neurobasal)および血清は、活動電位発射およびシナプスのコミュニケーションを含めた基本的なニューロン機能を強く損なうことが見出された。これらの制限を克服し、in vitroでのヒト脳の生理学的条件をより良好に再現するために、新規な基礎培地(BrainPhys)を設計し、これを化学的に定義された補充物と合わせ、種々のヒトニューロン培養物でこれを完全に試験した。この新規な培地は、典型的にはin vivoで観察される電気生理学的活性のタイプを最適に支持するが、加えて、これはまたin vitroでの長期間の生存を持続させる。BrainPhysは、患者または健康な被験体から得た機能的および成熟ヒトニューロンを培養するように本質的に設計されたが、またげっ歯類ニューロンに対して使用することができる。全体的に、これはヒト神経系疾患を模擬するより現実的および機能的なin vitroの条件を提供する。
・古典的な基礎培地(DMEMおよびNeurobasal)および血清は、in vitroでのヒトニューロンの基本的機能(活動電位および興奮性/抑制性シナプス活性)を強く損なう。
・血清を含まないBrainPhysをベースとする培地は、in vitroでのニューロンの成熟および脳細胞の生存をまた持続させる一方で、最適なニューロン機能(活動電位/シナプスのコミュニケーション)を支持する。
・BrainPhysをベースとする培地は、活性および機能的シナプスの結合性を有する成熟ニューロンの割合を増加させる。
・無血清補充物を有するBrainPhysは、安定的および好ましいニューロン活性を数週間にわたり持続させ、DMEMをベースとする培地において観察される供給サイクルとリンクする変動を低減させることができる。
・BrainPhys組成物は、CSF神経生理学的条件をより良好に模倣し、より現実的な条件をヒト神経系疾患および精神疾患のin vitroのモデルに提供し、トランスレーショナルの成功の機会を増加させる。
培養物中のニューロンの神経ネットワーク活性を評価するために、iPSCまたは胚性幹細胞(ESC)に由来したヒト神経前駆細胞(NPC)を使用した。ヒトNPCは、N2、B27、BDNF、GDNF、アスコルビン酸、cAMPおよびラミニンを補充した様々な基礎培地を有するガラス製カバースリップ上で2~8週間にわたってニューロンに分化した。カバースリップを灌流チャンバーにおいてニューロンと共に移し、様々な細胞外溶液中の細胞の自発性カルシウム活性または電気生理学的特性を短期間に検査した。
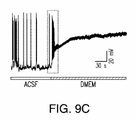
ヒトニューロンのカルシウム活性を、DMEMをベースとする培地中で最初に画像化し、該培地中で該ヒトニューロンを数週間培養した。ごく少数の活性ヒトニューロンが見出されたが、同じ視野をACSF中で画像化したとき、より多くの細胞が自発的活性となった。ACSF溶液の組成は、試験施設間で僅かに異なっている可能性がある。例えばカルシウム濃度は、シナプス放出を人工的に増加させるために生理学的レベルより高いことが多く、これは、潜在的にネットワーク活性を増加させ得る。この問題を明らかにするために、本発明者らのACSFの無機塩濃度、pH、および容量オスモル濃度をDMEM中のそれらと整合させて、これらのカルシウムイメージング実験を繰り返した。これらの注意に関わらず、活性細胞の数は、ACSF中よりDMEM中で有意に少なかったことが確認された(図1a)。
DMEMを事前に改変して、培養中のラット初代ニューロンの生存を最適化した(Brewerら、1993年)。Neurobasal培地と称されるこの改変バージョンにおいて、開発者は、いくつかの興奮性アミノ酸および硫酸第一鉄を本質的に除去し、容量オスモル濃度を低減させた。開発者は、DMEMと比較して、Neurobasalがin vitroでのラットニューロンの生存を改善したことを見出した(Brewerら、1993年)。細胞生存は細胞培養の決定的なパラメーターであるが、基本的電気生理学的特性はNeurobasalで試験されなかった。したがって、ヒトiPSCおよびESCに由来するニューロン(神経成熟培地中で2~8週間)を使用して、ニューロン機能に対するNeurobasalの効果を検査した。DMEMとは異なり、Neurobasalは静止膜電位を脱分極させず、場合により少なくともいくつかの興奮性シナプス事象を観察することができた。それにも関わらず、Neurobasalは、ACSF中で観察される自発性興奮性シナプス活性を強く低減または消失させた(n=5/5成熟ニューロン、ACSF対照:21.3±12.5Hz、Neurobasal-A:0.44±0.17Hz、ACSF回復:15.1±10.5Hz;図1c)。Neurobasalの無機塩の濃度および容量オスモル濃度は、ニューロンが脳内で天然に曝露しているものと非常に異なる(DavsonおよびSegal、1996年)。可能性がある結果として、Neurobasalが、電圧依存性ナトリウム電流および急速に不活性化するカリウム電流を有意に低減させ、誘発活動電位および自発性活動電位の振幅および周波数を損なったことが見出された(n=8/8個の細胞;図1c)。Neurobasal中のNaClの濃度を増加させる試みを最初に試みたが、電圧依存性ナトリウム電流の振幅における僅かな改善にも関わらず、この改変はヒトニューロンの電気的活性およびシナプス活性を最適化するのに十分ではなかった。したがって、DMEM中と同様に、Neurobasal中のいくつかの神経活性構成要素は、神経生理学的活性を防止したことが疑われた。DMEMもNeurobasalも、不可欠な電気生理学的なニューロンの特性、例えば、活動電位発射およびシナプス機能を持続させることができなかったため、ニューロン機能により良好に適合される新規な基礎培地を設計した。
ニューロン機能は、活動電位の発生および伝播に基本的に基づいている。電圧依存性ナトリウム(Nav)およびカリウム(Kv)チャネルは、高い発射率の活動電位を達成するのに決定的である。活動電位は、イオン勾配に負うところが大きいナトリウム電流の大きな流出およびカリウム電流の流入をトリガーする逐次活性化NavおよびKvチャネルを反映する。したがって、BrainPhysの最初の改善は、無機塩の濃度を神経生理学的レベルに近く調整することであった(例えば、Na+、Cl-、K+、Ca2+)。電圧クランプにおける試験は、ACSFおよびBrainPhys中のNavおよびKv電流の振幅が異ならなかったことを示した(図2a)。結果的に、BrainPhys中で試験した単一のニューロンにおける自発性活動電位および誘発活動電位はまた、ACSF中で測定した同様のレベルまでかなり改善した(n=3個の細胞;図2b)。同様に、ACSFおよびBrainPhys中のカルシウムイメージングによるネットワークレベルで測定した自発活性は匹敵するものであった。
統合神経ネットワーク活性および最終的に脳機能は、ニューロン間のシナプスのコミュニケーションによって達成される。したがって、本発明者らは、BrainPhysが興奮性および抑制性シナプス機能の両方を持続させるようにデザインした。興奮性および抑制性シナプス神経伝達物質の合計は、ニューロンが活動電位を発射する可能性を決定する。このように、単純に最適な活動電位発射を支持することによって、BrainPhysは間接的に作用して、シナプス活性を促進させる。さらに、特定のイオン、例えば、カルシウムは、シナプスにおいて速い小胞の放出をトリガーする決定的な役割を果たす。できるだけ現実的なin vitroでの条件を設定することを目的として、本発明者らは、BrainPhys中のカルシウムレベルをin vivoでのヒト脳脊髄液中で報告されるものに近くなるよう(Ca2+約1.1mM)に調整した。しかし、DMEMまたはNeurobasalの灌流によって強いNa+および/またはCl-電流を示す電圧クランプ実験に基づいて、シナプス機能はまた、神経活性構成要素の存在によって損なわれたと本発明者らは強く疑った。
inotropic)チャネルを飽和する神経活性構成要素の存在によるものであったことが疑われた。
主要な目的は、in vitroでのニューロン機能および活性をより良好に支持する培地を設計することであった。しかし、ヒト脳における基礎条件をより良好に模倣する培地を設計することもまた目的であった。これらの改善は、ニューロン活性に直接的に焦点をあてる研究のために重要なだけでなく、より現実的な実験モデルは患者のためのトランスレーショナルの成功を増加させるため、疾患モデリング研究のためにもまた潜在的に必要不可欠である。
異なるタイプのニューロンを使用して、ヒトニューロンをin vitroで研究することができ(例えば、皮質性、ドパミン作動性)、これらのニューロンを得るための方法論的バリエーションは無数である。しかし、特定のタイプのニューロン前駆体が得られた後、通常、細胞は、補充物を有する基礎培地中で成熟する。典型的には、DMEM、Neurobasalまたは両方の混合物を、基礎培地として選ぶ。基礎培地に加える補充物は、実験者の必要性または好みによって調整することができる。一般に使用される補充物の完全なセット(N2、B27、レチノイン酸、BDNF、GDNF、アスコルビン酸、cAMP、ラミニン、およびコレステロール)をBrainPhys基礎培地に短期間に加えることは、発射率にも興奮性/抑制性シナプス活性にも影響を与えないことについて本発明者らは最初に試験した(n=6個の細胞、図12)。しかし、ニューロン培地に場合により加えられる血清(例えば、ウシ胎仔血清、FBS、または化学的に定義された血清)は、ニューロン活性を強く損なったことを本発明者らは見出した(図6a)。したがって、全ての実験は、無血清培地中で行った。さらに、血清は、バッチ毎に可変性をもたらすことができる。同様に、抗生物質は使用せず、実験は無菌状態で行い、上清をマイコプラズマについて毎週試験した。
DMEMおよびNeurobasal培地を特に最適化して、in vitroでの細胞の生存を促進した。異なる基礎培地、DMEM、NeurobasalおよびBrainPhys(全て同じセットの無血清補充物を有する)中での細胞の生存率を試験するために、同様の密度で同時に蒔いたが、数週間異なる基礎培地で支持した同じ細胞系を使用して整合実験(matched experiment)を行った。いずれかの培地中で1カ月後、アポトーシス細胞の割合(活性カスパーゼ3)、全体的な細胞密度(DAPI)、または瀕死の細胞によって上清中に放出されるLDHの濃度は、3群の間で有意に変化しなかったことが見出された(図4a~d)。GABAニューロン、ドパミン作動性ニューロン(TH)またはNeuN陽性ニューロンの割合において有意差は見出されなかった(図4a、e)。同様に、いずれかをベースとする培地中の4週間の成熟の後に、近位樹状突起上のシナプスの点の密度における明らかな差異は観察されなかった(図4f~g)。
BrainPhysが、数週間後に培養物中の全体的な細胞密度を変化させ得るかを評価するために、平均して35日間(21~54日の範囲)、いずれかの培地中で並列させて成長させたニューロンを含有する36のカバースリップを、固定および染色した。同じ密度のDAPI核が、両方の群において見出された(DMEM=1657±239/mm2、BrainPhys=1745±252/mm2、マンホイットニーP値=0.75)。BrainPhysがドパミン作動性ニューロンの割合を変化させ得るかを問うために、12のカバースリップ上で、チロシンヒドロキシラーゼ(TH)について染色で陽性である細胞の数を定量化したが、有意差は見出されなかった(TH+/DAPIについて、DMEM=8.7±2.7%、BrainPhys 12±2.5%、P=0.26)。
BrainPhysはヒトニューロンを培養するために開発されたが、急性マウス脳スライスおよびラット初代ニューロン培養物における電気生理学的記録のために利用することができたことがまた実証された。BrainPhys中でパッチしたマウス海馬顆粒細胞は、完全に機能的であり(図13)、BrainPhysにおいて記録したこれらの特性は、ACSFにおけるそれらと実質的に同一であった(n=4個の細胞)。ラット初代ニューロンはBrainPhys(sm1を補充)中で健全に成長することができ、2週間以内にin vitroで活性および機能的神経ネットワークを形成したことがまた示された(図8)。
この実施例において、基礎培地を、基本的なニューロン機能に対するこれらの影響について完全に試験した。多くの重大な神経生理学的特性は、汎用のDMEMおよびNeurobasal培地中で有意に変化したことが見出された(表1)。したがって、新規な基礎培地を設計し、これをヒトおよびげっ歯類ニューロンで試験した。BrainPhysは、DMEMおよびNeurobasalにおいて同定された問題を克服した。適当な補充物を伴って、BrainPhysを使用して、NPCをニューロンに分化させることができ、in vitroで活発に作動する成熟ニューロンのための条件が提供される。BrainPhysはまた補充物を伴って、または伴わずに使用して、in vitroの培養物において、またはex vivoの脳スライスにおいて、ヒトおよびげっ歯類ニューロンの機能特性を電気生理学的に評価することができる。
BrainPhys中の構成要素の大部分は、基本的な細胞機能のために必須である。BrainPhysは、9種の無機塩、18種のアミノ酸、9種のビタミン、ならびにデキストロース、ピルビン酸ナトリウム、Hepes、コレステロールおよびフェノールレッドを含む5種のその他の構成要素の混合物である(表2)。ニューロンが生きた脳において機能するように、in vitroで機能することを可能とするために、DMEMまたはNeurobasal中に存在する多くの構成要素の濃度を適合させた。例えば、DMEM中の20種の構成要素およびNeurobasal中の39種の構成要素を、BrainPhys中の10%超だけ除去し、加え、または変更した。BrainPhys中の無機塩の濃度は、脳脊髄液において報告される濃度と似ている。Neurobasalの容量オスモル濃度は、神経生理学的レベルより約30%低い(Neurobasal-Aについて約220または250mOsmol/L)。対照的に、BrainPhysの容量オスモル濃度を、生理学的脳脊髄液の容量オスモル濃度(約305mOsmol/L)に整合するように設定した。ニューロンに対して潜在的に毒性であると報告された、DMEMまたはNeurobasal中に存在するいくつかのその他の構成要素を低減させ、または完全に除去した(例えば、酸化ストレスを誘導し得るリポ酸、Na+K+ATPアーゼを低減させ、酸化ストレスを誘導し得るヒポキサンチン(hypoxantine)、HEPES、光毒性であり得るリボフラビン、神経毒性であり、NMDA受容体と相互作用し得るL-システイン塩酸塩、ニューロンにおけるDNA修復を損ない、ニューロンをアミロイドに対して感受性にし得る葉酸およびホモシステインの欠損、その他のシナプスのGABAa受容体を遮断することができる銅、ならびに硫酸第二鉄)。いくつかのアミノ酸およびビタミンの濃度を変化させ、神経生理学的機能を改善させた。BrainPhysのpHを、重要な無機塩濃度を乱すことなく生理学的pH(約7.3~7.5)に調整した。CO2レベルに対して感受性のpH緩衝剤、および低濃度のHepesでpHを着実に維持する。
BrainPhysを、異なるプロトコルを使用することによって得たiPSCおよびESCに由来するニューロンを含めて異なる系のヒトニューロンで広範におよび首尾よく試験した。BrainPhysは、マウス海馬スライスの電気生理学的記録のため、およびラット初代海馬ニューロンを培養するために適合されていることがまた示された。BrainPhysはまた、多様な脳領域からの、および他の種からのニューロンを支持するはずである。
本実施例は、ヒトニューロンを分化させるために使用される利用可能な基礎培地が、生きている脳における条件と非常に異なり、したがって、ニューロンの培養物の電気的活性のために最適以下であることを記載する。ニューロンのin vitroでの成長および持続可能性の両方を最適化する1つの重要な態様は、BrainPhysと同様に、成熟および機能的ニューロンを見出す可能性が、現在の条件において相対的に低いことである。さらに、大部分の神経障害は慢性および進行性であり、ニューロン活性およびシナプス機能と非常に密接に関連する。このように、ヒト神経障害および精神障害をin vitroで模擬するとき、生理学的条件および活性の欠如によって、人工産物をもたらし、または病理の現実の機序をマスクする可能性がある。現在の培地中で分化した患者に由来するニューロン培養物および対照のiPSCに由来するニューロン培養物の間で関連性のある表現型が見出された(Marchettoら、2010年;Brennandら、2011年;Nguyenら、2011年;Seiblerら、2011年;Bellinら、2012年;Egawaら、2012年;Israelら、2012年;Shiら、2012年)が、新規な表現型はこれらの電気生理学的活性を促進する条件においてニューロンを研究することから明らかにし得る。生きている脳を密接に模倣する神経モデルは、患者の脳において起こっている機能障害を再現する可能性が高く、ひいては、神経障害および精神障害に対するより有効な処置の発見をもたらす。BrainPhysは、生理学的神経活性をin vitroで持続させることによってこのような使用のために有利である。
ヒトニューロン:ヒト皮膚線維芽細胞を、レトロウイルスベクターまたは非組み込み型センダイウイルスベクターのいずれかにおいて4種の山中因子(Oct3/4、Sox2、Klf4およびcMyc)で多能性細胞へと再プログラミングした。ヒトiPSCおよびESCは、従前に記載したように神経前駆体(NPC)に分化した(Brennandら、2011年;Boyerら、2012年)。神経成熟のために、NPCは、ポリ-オルニチン(Sigma P3655)およびラミニン(Invitrogen23017-015)でコーティングしたガラス製カバースリップ(Fisher Scientific12-545-80)上に蒔き、24ウェルプレートにおいてニューロン成熟培地(NM、下記の「培地および細胞外溶液」を参照されたい)中で培養した。培地の半分を、1週間に2~3回穏やかに置き換えた。プレートを、インキュベーター中で37℃、5%CO2で保持した。
vivoの脳上で直接試験するために、本発明者らは、マウス海馬(C57Bl6)の300μm厚さのスライスを調製した。















Claims (9)
- (a)121mMの濃度の塩化ナトリウム;
(b)4.2mMの濃度の塩化カリウム;
(c)1.1mMの濃度の塩化カルシウム;
(d)1mMの濃度の硫酸マグネシウム;
(e)0.0015mMの濃度の硫酸亜鉛;
(f)0.000124mMの濃度の硝酸第二鉄;
(g)29mMの濃度の炭酸水素ナトリウム;
(h)0.5mMの濃度のリン酸水素二ナトリウム無水;
(i)0.45mMの濃度のリン酸二水素ナトリウム;
(j)0.416mMの濃度のL-イソロイシン;
(k)0.449mMの濃度のL-トレオニン;
(l)0.451mMの濃度のL-ロイシン;
(m)0.452mMの濃度のL-バリン;
(n)0.499mMの濃度のL-リシン塩酸塩;
(o)0.5mMの濃度のL-アラニル-L-グルタミン;
(p)0.3mMの濃度のL-アルギニン塩酸塩;
(q)0.214mMの濃度のL-チロシン二ナトリウム塩二水和物;
(r)0.215mMの濃度のL-フェニルアラニン;
(s)0.15mMの濃度のL-ヒスチジン塩酸塩-H2O;
(t)0.116mMの濃度のL-メチオニン;
(u)0.06mMの濃度のL-プロリン;
(v)0.1mMの濃度のL-システイン塩酸塩-H2O;
(w)0.0411mMの濃度のL-トリプトファン;
(x)0.05mMの濃度のL-アスパラギン-H2O;
(y)0.002mMの濃度のL-アラニン;
(z)0.002mMの濃度のグリシン;
(aa)0.002mMの濃度のL-セリン;
(ab)0.0641mMの濃度の塩化コリン;
(ac)0.07mMの濃度のi-イノシトール;
(ad)0.0166mMの濃度のナイアシンアミド;
(ae)0.00986mMの濃度のピリドキシン塩酸塩;
(af)0.00644mMの濃度のチアミン塩酸塩;
(ag)0.0047mMの濃度のD-パントテン酸カルシウム;
(ah)0.00601mMの濃度の葉酸;
(ai)0.000502mMの濃度のシアノコバラミン;
(aj)0.000582mMの濃度のリボフラビン;
(ak)2.5mMの濃度のデキストロース;
(al)0.5mMの濃度のピルビン酸ナトリウム;
(am)10μg/mlの濃度のコレステロール;
(an)5mMの濃度のHEPES;および
(ao)0.0215mMの濃度のフェノールレッド
を含む細胞培地であって、(a)から(ao)がそれぞれ、該培地中で培養した神経細胞の生存および神経機能性を維持する濃度で存在する、細胞培地。 - 0.00001mM~0.003mMの濃度でL-アスパラギン酸をさらに含む請求項1に記載の細胞培地。
- 0.00001mM~0.02mMの濃度でL-グルタミン酸をさらに含む請求項1に記載の細胞培地。
- タンパク質、神経栄養因子、ステロイド、ホルモン、脂肪酸、脂質、ビタミン、硫酸塩鉱物、有機化学化合物、単糖、ヌクレオチド、およびこれらの組合せからなる群から選択される補充剤をさらに含む、請求項1に記載の細胞培地。
- 血清を含まない、請求項1に記載の細胞培地。
- 前記培地の容量オスモル濃度が、280~330mOsm/Lである、請求項1に記載の細胞培地。
- ニューロン細胞と、請求項1から6のいずれか一項に記載の培地とを接触させることを含む、ニューロン細胞を培養する方法。
- 前記ニューロン細胞が、初代ニューロン細胞である、請求項7に記載の方法。
- 前記ニューロン細胞が、iPSCに由来するニューロン細胞である、請求項8に記載の方法。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2021092829A JP2021129599A (ja) | 2013-04-17 | 2021-06-02 | ニューロン細胞培養のための培地組成物 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201361813034P | 2013-04-17 | 2013-04-17 | |
| US61/813,034 | 2013-04-17 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2016509110A Division JP2016515403A (ja) | 2013-04-17 | 2014-04-17 | ニューロン細胞培養のための培地組成物 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021092829A Division JP2021129599A (ja) | 2013-04-17 | 2021-06-02 | ニューロン細胞培養のための培地組成物 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2019122408A JP2019122408A (ja) | 2019-07-25 |
| JP7207721B2 true JP7207721B2 (ja) | 2023-01-18 |
Family
ID=51731860
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2016509110A Withdrawn JP2016515403A (ja) | 2013-04-17 | 2014-04-17 | ニューロン細胞培養のための培地組成物 |
| JP2019071296A Active JP7207721B2 (ja) | 2013-04-17 | 2019-04-03 | ニューロン細胞培養のための培地組成物 |
| JP2021092829A Pending JP2021129599A (ja) | 2013-04-17 | 2021-06-02 | ニューロン細胞培養のための培地組成物 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2016509110A Withdrawn JP2016515403A (ja) | 2013-04-17 | 2014-04-17 | ニューロン細胞培養のための培地組成物 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021092829A Pending JP2021129599A (ja) | 2013-04-17 | 2021-06-02 | ニューロン細胞培養のための培地組成物 |
Country Status (9)
| Country | Link |
|---|---|
| US (3) | US20160108361A1 (ja) |
| EP (1) | EP2986715B1 (ja) |
| JP (3) | JP2016515403A (ja) |
| CN (1) | CN105431525B (ja) |
| AU (3) | AU2014253855B2 (ja) |
| CA (1) | CA2909637A1 (ja) |
| HK (1) | HK1216761A1 (ja) |
| IL (2) | IL281665B (ja) |
| WO (1) | WO2014172580A1 (ja) |
Families Citing this family (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2957801C (en) | 2014-08-19 | 2022-08-30 | Cellular Dynamics International, Inc. | Neural networks formed from cells derived from pluripotent stem cells |
| ITUB20152804A1 (it) * | 2015-08-03 | 2017-02-03 | Blast Res S R L | Mezzo di coltura cellulare privo di siero. |
| CN105567635A (zh) * | 2016-02-21 | 2016-05-11 | 中国医科大学附属第一医院 | 一种改良的Neurobasal B27培养基、制备方法及应用 |
| JP7142284B2 (ja) * | 2017-06-06 | 2022-09-27 | 株式会社らいむ | 神経伸長促進剤 |
| JPWO2018235899A1 (ja) * | 2017-06-22 | 2020-04-23 | 国立大学法人 東京医科歯科大学 | 筋サテライト細胞の培養方法 |
| CN111793607B (zh) * | 2017-07-28 | 2022-05-17 | 杨涛 | 定向诱导hiPSC分化为神经细胞体系中培养神经元细胞的基础培养基 |
| FR3098223A1 (fr) | 2019-07-05 | 2021-01-08 | Universite de Bordeaux | Procede de production in vitro de neurones de mammiferes |
| CN111117968B (zh) * | 2019-07-30 | 2020-09-08 | 武汉赛尔朗灵科技有限公司 | 一种基于人脑胶质瘤原代细胞建立荧光裸鼠肿瘤模型的方法 |
| CN110443362B (zh) * | 2019-08-27 | 2024-02-20 | 北京理工大学 | 一种基于机器人化的体外神经网络构建系统 |
| CN112779220B (zh) * | 2021-04-01 | 2023-10-10 | 新疆赛尔托马斯生物科技有限公司 | 一种用于神经干细胞扩增的培养基 |
| WO2023278783A1 (en) * | 2021-06-30 | 2023-01-05 | The Regents Of The University Of California | Methods to generate glial restricted neural progenitor cells and uses thereof |
| EP4286513A1 (en) | 2022-06-03 | 2023-12-06 | IMBA-Institut für Molekulare Biotechnologie GmbH | Triple tissue culture fusion |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20080160006A1 (en) | 2001-10-02 | 2008-07-03 | Brewer Gregory J | Nutrient Medium for Maintaining Neural Cells in Injured Nervous System |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2662697A (en) * | 1996-04-09 | 1997-10-29 | Board Of Trustees Of Southern Illinois University, The | A cultural medium for maintaining neural cells in ambient atmosphere |
| US20120052577A1 (en) * | 2010-08-31 | 2012-03-01 | The Regents Of The University Of California | Culture system for stem cell propagation and neural and oligodendrocyte specification |
| CA2821065C (en) * | 2011-01-12 | 2021-05-18 | Tsuneo KIDO | Culture method to obtain and maintain a pure or enriched population of mammalian neural stem cells and/or neural progenitor cells that are prone to differentiate into oligodendrocyte-lineage cells in vitro |
| WO2012174225A2 (en) * | 2011-06-14 | 2012-12-20 | The University Of North Carolina At Chapel Hill | Isolation, expansion and use of autologous pluripotent stem cells |
-
2014
- 2014-04-17 EP EP14785372.5A patent/EP2986715B1/en active Active
- 2014-04-17 US US14/783,633 patent/US20160108361A1/en not_active Abandoned
- 2014-04-17 CA CA2909637A patent/CA2909637A1/en active Pending
- 2014-04-17 CN CN201480034512.8A patent/CN105431525B/zh active Active
- 2014-04-17 WO PCT/US2014/034565 patent/WO2014172580A1/en active Application Filing
- 2014-04-17 AU AU2014253855A patent/AU2014253855B2/en active Active
- 2014-04-17 JP JP2016509110A patent/JP2016515403A/ja not_active Withdrawn
- 2014-04-17 IL IL281665A patent/IL281665B/en unknown
-
2015
- 2015-10-08 IL IL241990A patent/IL241990B/en active IP Right Grant
-
2016
- 2016-04-27 HK HK16104814.8A patent/HK1216761A1/zh unknown
-
2019
- 2019-04-03 JP JP2019071296A patent/JP7207721B2/ja active Active
-
2020
- 2020-05-14 US US16/874,275 patent/US20200339949A1/en not_active Abandoned
- 2020-08-28 AU AU2020223761A patent/AU2020223761B2/en active Active
-
2021
- 2021-06-02 JP JP2021092829A patent/JP2021129599A/ja active Pending
-
2023
- 2023-04-11 US US18/133,237 patent/US20240076612A1/en active Pending
- 2023-08-28 AU AU2023222834A patent/AU2023222834A1/en active Pending
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20080160006A1 (en) | 2001-10-02 | 2008-07-03 | Brewer Gregory J | Nutrient Medium for Maintaining Neural Cells in Injured Nervous System |
Also Published As
| Publication number | Publication date |
|---|---|
| JP2016515403A (ja) | 2016-05-30 |
| AU2023222834A1 (en) | 2023-09-14 |
| JP2019122408A (ja) | 2019-07-25 |
| AU2014253855A1 (en) | 2015-10-29 |
| EP2986715A4 (en) | 2016-11-02 |
| IL241990B (en) | 2021-04-29 |
| EP2986715B1 (en) | 2018-08-22 |
| US20240076612A1 (en) | 2024-03-07 |
| AU2020223761A1 (en) | 2020-09-17 |
| IL281665B (en) | 2022-07-01 |
| EP2986715A1 (en) | 2016-02-24 |
| US20200339949A1 (en) | 2020-10-29 |
| HK1216761A1 (zh) | 2016-12-02 |
| CN105431525A (zh) | 2016-03-23 |
| CN105431525B (zh) | 2020-04-10 |
| CA2909637A1 (en) | 2014-10-23 |
| AU2014253855B2 (en) | 2020-05-28 |
| IL281665A (en) | 2021-05-31 |
| WO2014172580A1 (en) | 2014-10-23 |
| US20160108361A1 (en) | 2016-04-21 |
| JP2021129599A (ja) | 2021-09-09 |
| AU2020223761B2 (en) | 2023-06-01 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7207721B2 (ja) | ニューロン細胞培養のための培地組成物 | |
| Constantinescu et al. | Neuronal differentiation and long-term culture of the human neuroblastoma line SH-SY5Y | |
| US10017734B2 (en) | Method for producing dopaminergic neurons | |
| US12090252B2 (en) | Method for producing retinal tissues | |
| EP4001403A1 (en) | Methods for differentiating pluripotent cells | |
| Yavin et al. | Survival and maturation of cerebral neurons on poly (L-lysine) surfaces in the absence of serum | |
| Li et al. | Myocyte enhancer factor 2C as a neurogenic and antiapoptotic transcription factor in murine embryonic stem cells | |
| EP3656852A1 (en) | Method for maturation of retinal tissue containing continuous epithelium | |
| US20080227137A1 (en) | Method of in vitro differentiation of neural stem cells, motor neurons and dopamine neurons from primate embryonic stem cells | |
| IL173832A (en) | Method of in vitro differentiation of neural stem cells, motor neurons and dopamine neurons from primate embryonic stem cells | |
| US20080206865A1 (en) | Method of in vitro differentiation of neural stem cells, motor neurons and dopamine neurons from primate embryonic stem cells | |
| KR101696874B1 (ko) | 직접 리프로그래밍을 통한 유도 도파민성 전구세포 제조방법 | |
| EP3683304A1 (en) | Method for amplifying cone photoreceptors or rod photoreceptors using dorsalization signal transmitter or ventralization signal transmitter | |
| EP3105320B1 (en) | Synthetic retina | |
| KR20170051497A (ko) | 소뇌 선구체 조직의 제조 방법 | |
| WO2017126551A1 (ja) | ヒト多能性幹細胞から視床下部ニューロンへの分化誘導 | |
| US9428730B2 (en) | Coatings and culture media for promoting neurogenesis in adipose tissue derived stem cells | |
| Tu | Role of reactive oxygen species in pluripotent stem cells cardiac differentiation and survival | |
| CN117441011A (zh) | 多巴胺能前体细胞及使用方法 | |
| Hu | Inner Ear Stem Cell Niche | |
| Annis | Morphological and electrophysiological development of cerebral cortical neurons, in vitro | |
| 고지윤 | Efficient derivation of neural precursors and dopamine neurons from human embryonic stem cells and their application to a Parkinsonian rat model |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20190403 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20200303 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20200601 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20200731 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20200903 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20210202 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210602 |
|
| C60 | Trial request (containing other claim documents, opposition documents) |
Free format text: JAPANESE INTERMEDIATE CODE: C60 Effective date: 20210602 |
|
| C11 | Written invitation by the commissioner to file amendments |
Free format text: JAPANESE INTERMEDIATE CODE: C11 Effective date: 20210610 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20210712 |
|
| C21 | Notice of transfer of a case for reconsideration by examiners before appeal proceedings |
Free format text: JAPANESE INTERMEDIATE CODE: C21 Effective date: 20210713 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20210910 |
|
| C211 | Notice of termination of reconsideration by examiners before appeal proceedings |
Free format text: JAPANESE INTERMEDIATE CODE: C211 Effective date: 20210914 |
|
| C22 | Notice of designation (change) of administrative judge |
Free format text: JAPANESE INTERMEDIATE CODE: C22 Effective date: 20220405 |
|
| C13 | Notice of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: C13 Effective date: 20220427 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20220726 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20220826 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20220926 |
|
| C22 | Notice of designation (change) of administrative judge |
Free format text: JAPANESE INTERMEDIATE CODE: C22 Effective date: 20221004 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20221027 |
|
| C23 | Notice of termination of proceedings |
Free format text: JAPANESE INTERMEDIATE CODE: C23 Effective date: 20221104 |
|
| C03 | Trial/appeal decision taken |
Free format text: JAPANESE INTERMEDIATE CODE: C03 Effective date: 20221128 |
|
| C30A | Notification sent |
Free format text: JAPANESE INTERMEDIATE CODE: C3012 Effective date: 20221128 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20221223 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7207721 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |