JP7204192B2 - 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 - Google Patents
認知症の予防、軽減、及び/または治療のためのシステムならびに方法 Download PDFInfo
- Publication number
- JP7204192B2 JP7204192B2 JP2018525754A JP2018525754A JP7204192B2 JP 7204192 B2 JP7204192 B2 JP 7204192B2 JP 2018525754 A JP2018525754 A JP 2018525754A JP 2018525754 A JP2018525754 A JP 2018525754A JP 7204192 B2 JP7204192 B2 JP 7204192B2
- Authority
- JP
- Japan
- Prior art keywords
- subject
- mice
- stimulation
- stimulus
- visual
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images


















































































































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61M—DEVICES FOR INTRODUCING MEDIA INTO, OR ONTO, THE BODY; DEVICES FOR TRANSDUCING BODY MEDIA OR FOR TAKING MEDIA FROM THE BODY; DEVICES FOR PRODUCING OR ENDING SLEEP OR STUPOR
- A61M21/00—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61H—PHYSICAL THERAPY APPARATUS, e.g. DEVICES FOR LOCATING OR STIMULATING REFLEX POINTS IN THE BODY; ARTIFICIAL RESPIRATION; MASSAGE; BATHING DEVICES FOR SPECIAL THERAPEUTIC OR HYGIENIC PURPOSES OR SPECIFIC PARTS OF THE BODY
- A61H23/00—Percussion or vibration massage, e.g. using supersonic vibration; Suction-vibration massage; Massage with moving diaphragms
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61H—PHYSICAL THERAPY APPARATUS, e.g. DEVICES FOR LOCATING OR STIMULATING REFLEX POINTS IN THE BODY; ARTIFICIAL RESPIRATION; MASSAGE; BATHING DEVICES FOR SPECIAL THERAPEUTIC OR HYGIENIC PURPOSES OR SPECIFIC PARTS OF THE BODY
- A61H23/00—Percussion or vibration massage, e.g. using supersonic vibration; Suction-vibration massage; Massage with moving diaphragms
- A61H23/02—Percussion or vibration massage, e.g. using supersonic vibration; Suction-vibration massage; Massage with moving diaphragms with electric or magnetic drive
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61N—ELECTROTHERAPY; MAGNETOTHERAPY; RADIATION THERAPY; ULTRASOUND THERAPY
- A61N5/00—Radiation therapy
- A61N5/06—Radiation therapy using light
- A61N5/0613—Apparatus adapted for a specific treatment
- A61N5/062—Photodynamic therapy, i.e. excitation of an agent
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61N—ELECTROTHERAPY; MAGNETOTHERAPY; RADIATION THERAPY; ULTRASOUND THERAPY
- A61N5/00—Radiation therapy
- A61N5/06—Radiation therapy using light
- A61N5/0613—Apparatus adapted for a specific treatment
- A61N5/0622—Optical stimulation for exciting neural tissue
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61M—DEVICES FOR INTRODUCING MEDIA INTO, OR ONTO, THE BODY; DEVICES FOR TRANSDUCING BODY MEDIA OR FOR TAKING MEDIA FROM THE BODY; DEVICES FOR PRODUCING OR ENDING SLEEP OR STUPOR
- A61M21/00—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis
- A61M2021/0005—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis by the use of a particular sense, or stimulus
- A61M2021/0022—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis by the use of a particular sense, or stimulus by the tactile sense, e.g. vibrations
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61M—DEVICES FOR INTRODUCING MEDIA INTO, OR ONTO, THE BODY; DEVICES FOR TRANSDUCING BODY MEDIA OR FOR TAKING MEDIA FROM THE BODY; DEVICES FOR PRODUCING OR ENDING SLEEP OR STUPOR
- A61M21/00—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis
- A61M2021/0005—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis by the use of a particular sense, or stimulus
- A61M2021/0027—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis by the use of a particular sense, or stimulus by the hearing sense
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61M—DEVICES FOR INTRODUCING MEDIA INTO, OR ONTO, THE BODY; DEVICES FOR TRANSDUCING BODY MEDIA OR FOR TAKING MEDIA FROM THE BODY; DEVICES FOR PRODUCING OR ENDING SLEEP OR STUPOR
- A61M21/00—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis
- A61M2021/0005—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis by the use of a particular sense, or stimulus
- A61M2021/0044—Other devices or methods to cause a change in the state of consciousness; Devices for producing or ending sleep by mechanical, optical, or acoustical means, e.g. for hypnosis by the use of a particular sense, or stimulus by the sight sense
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61M—DEVICES FOR INTRODUCING MEDIA INTO, OR ONTO, THE BODY; DEVICES FOR TRANSDUCING BODY MEDIA OR FOR TAKING MEDIA FROM THE BODY; DEVICES FOR PRODUCING OR ENDING SLEEP OR STUPOR
- A61M2210/00—Anatomical parts of the body
- A61M2210/06—Head
- A61M2210/0693—Brain, cerebrum
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61N—ELECTROTHERAPY; MAGNETOTHERAPY; RADIATION THERAPY; ULTRASOUND THERAPY
- A61N5/00—Radiation therapy
- A61N5/06—Radiation therapy using light
- A61N2005/063—Radiation therapy using light comprising light transmitting means, e.g. optical fibres
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61N—ELECTROTHERAPY; MAGNETOTHERAPY; RADIATION THERAPY; ULTRASOUND THERAPY
- A61N5/00—Radiation therapy
- A61N5/06—Radiation therapy using light
- A61N2005/065—Light sources therefor
- A61N2005/0651—Diodes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61N—ELECTROTHERAPY; MAGNETOTHERAPY; RADIATION THERAPY; ULTRASOUND THERAPY
- A61N5/00—Radiation therapy
- A61N5/06—Radiation therapy using light
- A61N2005/0658—Radiation therapy using light characterised by the wavelength of light used
- A61N2005/0662—Visible light
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61N—ELECTROTHERAPY; MAGNETOTHERAPY; RADIATION THERAPY; ULTRASOUND THERAPY
- A61N5/00—Radiation therapy
- A61N5/06—Radiation therapy using light
- A61N5/0613—Apparatus adapted for a specific treatment
- A61N5/0618—Psychological treatment
Landscapes
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Life Sciences & Earth Sciences (AREA)
- Animal Behavior & Ethology (AREA)
- General Health & Medical Sciences (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Pathology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Radiology & Medical Imaging (AREA)
- Biophysics (AREA)
- Psychology (AREA)
- Anesthesiology (AREA)
- Physics & Mathematics (AREA)
- Neurosurgery (AREA)
- Heart & Thoracic Surgery (AREA)
- Hematology (AREA)
- Acoustics & Sound (AREA)
- Epidemiology (AREA)
- Rehabilitation Therapy (AREA)
- Physical Education & Sports Medicine (AREA)
- Pain & Pain Management (AREA)
- Child & Adolescent Psychology (AREA)
- Social Psychology (AREA)
- Psychiatry (AREA)
- Hospice & Palliative Care (AREA)
- Developmental Disabilities (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Radiation-Therapy Devices (AREA)
- Peptides Or Proteins (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Saccharide Compounds (AREA)
- Electrotherapy Devices (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
Description
本発明は、国立衛生研究所によって与えられた認可番号RF1 AG047661の下、政府支援を受けてなされた。政府は本発明に所定の権利を有する。
本出願は、米国特許法第119条(e)の定めにより、2015年11月24日に出願された「System and Methods for Preventing,Mitigating,and/or Treating Dementia」と題する米国出願第62/259,187号の優先権の利益を主張する。当該出願の開示は、参照することによりその全体が本明細書に組み込まれる。
一態様において、本開示は、対象における脳障害または認知機能障害/認知障害を予防、軽減、及び/または治療するための方法、装置、ならびにシステムを提供する。いくつかの実施形態では、該脳障害は認知症である。
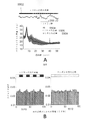
ガンマ振動は、5XFADマウスにおいて疾患早期の海馬SWR中に低下する。
ガンマの障害は、いくつかの神経及び精神障害における複数の脳領域で、ADのヒト患者における自発的ガンマ同期の低下を含め、観察されている。興味深いことに、自発的ガンマの低下はまた、2つのADのマウスモデル(ヒトアミロイド前駆体タンパク質(hAPP)Tgマウス及びアポリポタンパク質E4対立遺伝子(APOE4)ノックインマウス)においてインビボで、ならびにインビトロスライス試験では別のマウスモデル(Tg CRND8マウス)で見出されている。しかしながら、他のADのマウスモデルでガンマ振動が変化するかどうか、それが疾患の進行の早期に生じるかどうか、及びガンマの乱れが疾患の進行に影響を与えるかどうかは不明である。
ADの本マウスモデルにおける当該疾患の進行の初期のSWR中のガンマ障害の所見で、ガンマ振動がADの分子及び細胞病態生理に影響を与え得るかどうかという疑問が生じる。それを試験するため、2.5か月齢の5XFAD/PV-Cre二重トランスジェニックマウスの海馬CA1のFS-PV介在ニューロンにおいて、二重floxed逆方向オープン・リーディング・フレーム(DIO)ChR2-EYFPアデノ随伴ウイルス(AAV)を用い、Cre依存的にChR2を発現させることによって、ガンマ振動を光遺伝学的に駆動した。試験を実施し、マウスの海馬ガンマ振動の遺伝子誘導がADのマウスモデルの分子病理に影響を与えるかどうかを確認した。海馬ガンマ振動は、覚醒行動WT及び5XFADマウスにおいて遺伝子学的に誘導された。
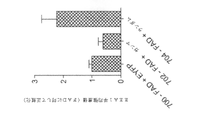
Aβの蓄積は、ADの病変に典型的な複数の神経毒性事象を引き起こすことがある。従って、いくつかの実施形態では、5XFADマウスの全般的なAβペプチドのレベルにおけるガンマ刺激の影響を調べた。3か月齢のマウスには、この段階で海馬に斑が存在しないためにこれらを用い、斑負荷と無関係の可溶性Aβの動態を調べられるようにした。いくつかの実施形態では、FS-PV介在ニューロンの1時間の刺激が、Aβ ELISA分析で測定した、海馬のCA1領域におけるEYFP対照群と比較して、40Hz群において、Aβ1-40を53.22%及びAβ1-42を44.62%低減したことが見出された。
表1



いくつかの実施形態では、さらに40Hzの刺激の細胞及び分子効果を不偏的に検討するため、5XFAD/PV-Cre二重トランスジェニックマウスの40HzのFS-PV介在ニューロン刺激、または無刺激(EYFP)1時間後の海馬CA1組織のゲノムワイドRNA-seqを実施した。RNA-seq実験では、平均26,518,345のシーケンシングリードが3匹の刺激及び3匹の無刺激マウスから得られた。データのQC分析で、エクソン/イントロン比の平均値183、エクソン/遺伝子間比の平均値272、及びリボソームRNAリードのパーセンテージの平均値3.6%が明らかになった。この分析は、523の差次的に発現される遺伝子(DEG)を特定し、このうち130が40Hzの刺激に応答して上方制御され、393が下方制御された。
表2


表3




表4

表5

40Hzでの光遺伝学的刺激によってAβレベルが強く低下したことで、脳において40Hzの振動を誘導する他の方法を探索し、この効果が光遺伝学的操作または侵襲的方法に何らかの形で特異的ではないことを確かめるに至った。光の点滅が視覚野において40Hzの振動を誘導する非侵襲的方法として使用できるかどうかを調べるため、いくつかの実施形態では、動物を暗周期と交互配置された40Hzまたはランダムな点滅、及び連続光の周期に曝露した。
光遺伝学的方法の有効性を考慮して、翻訳的で非侵襲的なアミロイド低減処理を設計した。図46Aは、いくつかの実施形態に従う実験的パラダイムを示す概略図である。図46Aに示すように、ADモデルマウスの第一のサブセットを、40Hzの閃光灯を備えた第一のチャンバー4600に入れ、ADモデルマウスの第二のサブセットを、暗くしておいた第二のチャンバー4602に入れた。第一のチャンバー4600内の動物を約1時間、40Hzの閃光灯に曝露した。
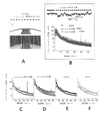
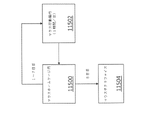
光の点滅による視覚刺激がADに関与する脳回路に影響を与えるかどうかを判定するため、いくつかの実施形態では、海馬、すなわち、ヒトにおいてADの経過の初期に影響を受ける脳領域の1つに対する光の点滅の影響を調べた。図55Aは、いくつかの実施形態に従って、40Hzの光の点滅5502の前及び最中での海馬における局所電場電位の電気トレースならびにパワースペクトル密度のプロットを含む。CA1における暗5504、40Hzの光の点滅5506、及びランダムな光の点滅5508の間のパワースペクトル密度の平均(実線)及び標準偏差(網掛け領域)(5XFADマウスn=2及びWTマウス3)。
40Hzの振動が光遺伝学的に、または光の点滅を介した視覚刺激によってのいずれかで駆動された場合に、前斑5XFADマウスにおいて影響を受けたアミロイド存在量を調べ、本明細書に開示してきた。次に、目的は、この処理がすでに斑負荷を示している動物に有効であるかどうかを判定することとなった。この目的で、いくつかの実施形態では、6か月齢の5XFADマウスを用いたが、これは、それらが視覚野を含む多くの脳領域において広範囲のアミロイド斑病変を発症するためである。非侵襲的ガンマ刺激後、進行したAβ関連の病変に何が起こるかを調べるための試験を実施した。40Hzの点滅1時間に反応したAβ減少の期間を調べるため、いくつかの実施形態では、Aβレベルを視覚野において、40Hzの点滅または暗条件1時間の4、12、及び24時間後に測定した。
いくつかの実施形態に従うガンマ曝露及び/または投与が対象に何らかのストレスを引き起こすかどうかを調べるための試験を実施した。図68Aは、本試験を示す流れ図である。図68Aの6800に示すように、いくつかの実施形態に従って、WTマウスを連続7日間、すなわち1~7日目に、1日1時間、普通室内光(N=8)または40Hzの光の点滅(N=8)のいずれかに曝露した。6802に示す8日目、マウスから採血し、血漿を分離してコルチコステロンレベルを調べた。マウスでは、コルチコステロンは、ストレス反応に関与する主要な糖質コルチコイドである。
T(14)=0.827、p=0.422 (1)
と計算された。
T(18)=-1.652、p=0.11 (2)
と計算された。
T(18)=-2.136、p=0.047 (3)
と計算された。
F(1,14)=4.860、p=0.045 (4)
と計算された。
χ2(4,n=16)=16.088、p=0.003 (5)
と計算された。
U=58.00、Z=2.731、p=0.005 (6)
と計算された。
T(24)=0.577、p=0.569 (7)
と計算された。
F(1,24)=3.106、p=0.091 (8)
と計算された。
T(24)=-2.155、p=0.041 (9)
と計算された。
T(24)=-1.433、p=0.164 (10)
と計算された。
F(1,24)=4.559、p=0.043 (11)
と計算された。
T(24)=1.874、p=0.073 (12)
と計算された。
T(24)=2.223、p=0.036 (13)
と計算された。
F(1,24)=6.646、p=0.017 (14)
と計算された。
いくつかの実施形態に従って、対象の視覚野におけるDNA損傷ならびにニューロンの欠損に対するガンマ曝露及び/または投与の影響を調べるために試験を実施した。本試験のため、p25蓄積の誘導性マウスモデル(すなわち、クレアチンキナーゼカルボキシル末端断片p25Tgマウス(CK-p25Tgマウス))を用いた。CK-p25Tgマウスモデルは、前脳内の重大なニューロン損失、Aβペプチド産生増加、タウ病理、DNA損傷、及び重度の認知機能障害を含むADの重要な病理学的特徴を示す。本モデルでは、ニューロンの損失に先立ってAβペプチドレベルの上昇が認められ、さらに、Aβペプチド産生の低減はCK-p25Tgマウスモデルにおいて記憶障害を軽減することから、この事象がカルボキシル末端断片p25と相乗的に作用し、神経変性及び記憶障害の発現につながることを示している。
いくつかの実施形態では、ガンマ曝露及び/または投与は、聴覚刺激を含む。聴覚刺激は音波パルスまたはクリックを含み得る。音刺激は、1秒当たり約35回の音波パルスまたはクリック(クリック/秒)から約45クリック/秒のクリック・トレインを含み得る。図114は、いくつかの実施形態に従って、クリック・トレイン刺激を示す刺激図である。図114の刺激は、各クリック間が25ミリ秒でクリック頻度40クリック/秒を有し、各クリックが持続時間1ミリ秒を有する。
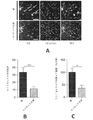
聴覚ガンマ刺激は、いくつかの実施形態によれば、対象のAβのレベルを減少させることが示された。図115の試験を、いくつかの実施形態に従って、6か月齢の5XFAD Tgマウスで繰り返した。8日目、この聴覚野及び海馬を切断した。ELISAを用いて、アイソフォームAβ1-40ペプチド及びアイソフォームAβ1-42ペプチドを含む可溶性及び不溶性のAβアイソフォームのレベルを測定した。不溶性のAβは、5MのグアニジンHClで3時間処理し、斑を可溶化した。
聴覚ガンマ刺激は、いくつかの実施形態によれば、対象の認識を改善することが示された。図125Aは、いくつかの実施形態に従ってクリック・トレイン刺激に曝露された5XFADマウス及び静寂に曝露された5XFADマウスを用いて実施された新奇物体認識試験を示す流れ図である。この試験は、げっ歯類が慣れ親しんだ物体よりも新奇物体を探索するのにより多くの時間を費やす傾向に基づいて、対象が慣れ親しんだ物体から新奇物体を認識する能力(すなわち、認識記憶)を評価する。認識指数RIを用いて対象を比較した:


動物
すべての動物実験は、比較医学部門の実験動物委員会(the Committee for Animal Care of the Division of Comparative Medicine)(マサチューセッツ工科大学、マサチューセッツ州ケンブリッジ)によって承認された。成体(3か月齢)雄二重Tg 5XFAD Creマウスは、5XFAD TgマウスをTg PVまたはCW2プロモーター駆動Cre系統とかけ合わせることによって産生した。成体(5か月齢)雄及び雌APP/PS1マウスは、Tonegawa Laboratory(マサチューセッツ工科大学、マサチューセッツ州ケンブリッジ)から寄贈された。成体(4か月齢)雄TauP301Sマウスは、Jackson Laboratoryより入手した。老齢WTマウス(8か月齢、C57Bl/6)は、Jackson Laboratory(メイン州バーハーバー)より入手した。マウスは、3~5の群で、標準的な12時間の明/12時間の暗サイクルにて飼育し、すべての実験は、明サイクルの間に実施した。餌と水は、他に記載のない限り不断に与えた。同腹仔は、実験者によって各条件にランダムに割り当てられた。実験者には、組織の処理ならびに電気生理学的記録及び分析中、動物の遺伝子型は分からなかった。分析から除外した動物はなかった。
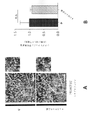
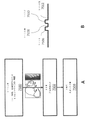
血清型5のアデノ随伴ウイルス粒子は、Vector Core Facility(ノースカロライナ大学、ノースカロライナ州チャペルヒル)から入手した。このAAV5ウイルスは、EF1αプロモーターによって駆動される高感度黄色蛍光タンパク質(EYFP)に融合された二重floxed、逆方向オープン・リーディング・フレーム(DIO)内のChR2を含んでいた(例えば、図9参照)。AAV DIO EYFP構築物は対照として用いた。
3か月齢の5XFAD/PV-CreまたはCW2マウスは、ケタミン(1.1mg kg-1)及びキシラジン(0.16mg kg-1)の混合物の腹腔内注射で麻酔した。2.0mm後葉から泉門点まで及び1.8mm側葉から正中まで左側に小開頭を行った。ウイルスは、硬膜の小切開を介して、Quintessential Stereotaxic Injector(商標)(Stoelting Co.、イリノイ州ウッドデールより入手可能)に取り付けられたガラス製マイクロピペットで供給した。このガラス製マイクロピペットを脳表面の下1.2mmまで下ろした。ウイルス(AAV DIO ChR2-EYFPまたはAAV DIO EYFP、1ml当たり2X1012ウイルス分子)1μlのボーラスを、海馬のCA1領域に0.075μl分-1で注射した。ピペットは、注射後5分間その場に残し、その後脳から後退させた。片側光ファイバーインプラント(コア径300μm、Thorlabs Inc.、ニュージャージー州ニュートンより入手可能)をほぼ注射部位の脳表面の下0.9mmまで下ろした。手術部位の前縁及び後縁に固定した2つの小ネジを歯科用の接着剤で固着し、このインプラントを定位置に固定した。電気生理学的記録については、成体(3か月齢)雄5XFAD/PV-Cre二重トランスジェニックマウス及び5XFAD陰性同腹仔(CA1記録用)、または5XFAD及びそのWT同腹仔(視覚野記録用)マウスをイソフルランで麻酔し、定位フレームに入れた。頭皮を剃毛し、眼軟膏(例えば、Puralube(登録商標)Vet軟膏(Dechra Pharmaceuticals PLC、英国ノースウィッチ))を目に塗り、Betadine(登録商標)消毒剤(Purdue Products L.P.、コネチカット州スタンフォードより入手可能)及び70%エタノールを用いて手術領域を消毒した。CA1の記録については、開頭(mm単位で、泉門点から:-2A/P、1.8M/L)を行い、ウイルス1μLをCA1に供給した(上述の通り)。LFP記録のための標的開頭部位は、頭蓋骨上にマークし(mm単位で、泉門点から:CA1については-3.23A/P、0.98M/L及び視覚野については2.8A/P、2.5M/L)、3つのセルフタッピングネジ(例えば、F000CE094、Morris Precision Screws and Parts、マサチューセッツ州サウスブリッジより入手可能)をこの頭蓋骨に取り付け、カスタムステンレス鋼ヘッドプレートを歯科用セメント(例えば、C&B Metabond(登録商標)、Parkell Inc.、ニューヨーク州エッジウッドより入手可能)を用いて取り付けた。第一の記録期間の日に、歯科用ドリルを用い、第一に頭蓋骨を厚さ約100μmまで薄くし、その後30ゲージの針を用いて小開口を作ることによってLFP開頭術(例えば、300~400μm径)を行った。この開頭は、その後、滅菌シリコーンエラストマー(例えば、Kwik-Sil(商標)接着剤、World Precision Instruments,Inc.、フロリダ州サラソータより入手可能)で、その日の記録まで、及び記録期間の間、密封した。
マウスが回復し、電気生理学用の動物用の行動訓練を受けるための、及びウイルスがニューロンで発現するための時間を与える、ウイルス注射及びインプラント設置の2~4週間後に、海馬CA1ニューロンを光遺伝学的に操作した。200mWの4793nm DPSSレーザを、各端にファイバー・チャンネル/身体的接触コネクタを備えたパッチ・コードに接続した。実験中、1mW(ファイバーの端部から測定)の光刺激を1時間送った。分子及び生化学的分析用に、各動物は、3つの刺激プロトコル、すなわち、8Hz、40Hz、もしくはランダム刺激(光パルスは、平均周波数40Hzで、ポアソン過程により決定されたランダムな間隔で送った)のうちの1つを受けたか、または、電気生理学的記録については、各動物は、記録中、交互配置されたすべての刺激条件を受けた。
実験の15分前、5XFADマウスを生理食塩水(対照)またはピクロトキシン(0.18mg/kg)で処理した。分子及び生化学的分析用に、その後マウスを、LED電球によって照らされる暗室に入れ、5つの刺激条件、すなわち、暗、明、20Hzの点滅、40Hzの点滅、または80Hzの点滅(12.5ミリ秒の光オン、12.5ミリ秒の光オフ)1時間(例えば、図43A参照)のうちの1つに曝露した。電気生理学的記録については、各動物は、記録中、10秒のブロックで交互配置された、暗、明、40Hzの点滅、またはランダム(光パルスは、平均間隔40Hzで、ポアソン過程により決定されたランダムな間隔で送った)刺激条件を受けた。
CA1の記録については、Harveyらに記載の通り、頭部を固定した動物を、エアクッションで支えられた8インチの球状トレッドミル上を仮想現実環境で走らせた。球状トレッドミルの動きは、光学式マウスで測定し、MATLAB(登録商標)コンピュータ環境(ソフトウェアバージョン2013b、MathWorks、マサチューセッツ州ナティックより入手可能)で実行中の仮想現実ソフトウェアに送った。この仮想環境は、直線トラックと、動物が向きを変えることができる端部の2つの小さな囲いからなる。動物は、トラックの各端への交互の訪問に対して、トラックの各端で、加糖練乳(1:2に水で希釈)の報酬を得た。動物は、約1週間以上、この仮想直線トラック上を走行することを学習した。これらの動物を1週間おいて手術から回復させ、1~2日間ハンドリングに馴らし、その後行動訓練を開始した。トレッドミルを操ること、及び試験環境に慣れることを学習するように、訓練の最初の2日間、動物は、仮想現実システムをオフにして球状トレッドミル上に置かれ、希釈されていない加糖練乳の報酬を得た。球状トレッドミル上での訓練の2日目に、動物の餌を制限し、走るための動機づけをした。動物を、その基準体重のせいぜい85%に制限し、通常はその基準体重の88%を超えた。3日目から訓練の終わりまで(通常5~7日)、動物を、時間を延長しつつ(30分~2時間)トレッドミル上に置き、VR直線トラックを走行させた。動物は、トラックの長さを横断した後、直線トラックの端で希釈(1:2)加糖練乳の報酬を得た。記録期間の間、動物は、行動能力を維持するために再訓練期間を与えられた。視覚野の記録については、動物に球状トレッドミル上を走行させ、その間に暗、明、または光の点滅条件(以下のデータ収集に記載)に曝露した。記録に先立って、動物は、球状トレッドミル上に置かれ(仮想現実システムをオフにして)、希釈されていない加糖練乳の報酬を得ることで、トレッドミルを操ること及び試験環境に慣れることを学習した。
記録中のCA1の光遺伝学的刺激については、300μmコアの光ファイバーを、CA1へウイルスを供給するために用いた開頭を介して脳内に900μmの深さまで進めた。1ミリ秒及び1mW(ファイバーの端部から測定)の光パルスを473nmのDPSS(ダイオード励起固体)レーザを介して供給した(上述の通り)。光電アーチファクトを避けるため、神経活動はガラス電極で記録した。LFP電極は、ホウケイ酸ガラス製ピペット(例えば、Warner Instruments、コネチカット州ハムデンより入手可能)から、フィラメント系のマイクロピペットプラー(例えば、P-97 Flaming/Brown(商標)マイクロピペットプラー、Sutter Instrument Co.、カリフォルニア州ナバトより入手可能)にて引き延ばして微細な先端にし、これを次に手で折って直径約10~20μmに戻し、その後滅菌生理食塩水で満たした。CA1の記録については、このLFP電極を、LFP記録用開頭を介して、前頭面に対して後方に60度の角度及び水平面に対して下側45度で、海馬層の錐体層の明確な電気生理学的サイン(動物の走行中、約600~1000μVのシータ波、不動状態では明らかに区別できるSWR、150μVを超える複数のスパイク、例えば、図2A~2B参照)が認められるまで進めた。視覚野の記録については、LFP電極は、LFP記録用開頭を介して垂直に600~900μmの深さまで進め、150μVを超える複数のスパイクが認められた。データは、サンプリングレート20kHz及びバンドパスフィルター1Hz~1kHzで収集した。動物は、長期にわたって球状トレッドミル上を走行させたか、または休息させた。光遺伝学的刺激期間については、いずれかの刺激の開始前30分間データを記録した。その後、刺激をガンマ(40Hz)、ランダム(光遺伝学的刺激で記載)、またはシータ(8Hz)の周波数で10秒間の周期を10秒間のベースライン周期(無刺激)と交互配置して供給した。2匹の動物において、各タイプの刺激またはベースラインを10秒周期の代わりに5分周期で供給した。各30分間の刺激の記録の後、5~30分間の無刺激の記録を行った。視覚の光の点滅刺激期間については、10秒間の光オフの周期を交互配置して、動物ライト周囲のLEDストリップライトをガンマ(40Hz)、ランダム(視覚刺激プロトコルに上述)、シータ(8Hz)、または20Hzの周波数で10秒間の周期で点滅させたか、または、10秒間の連続オンの周期であった。光の点滅の間、脳表面の上でいくつかの記録を行い、記録中にこれらのライトが電気的または光電ノイズを生成しないことを保証した。記録期間は、約3~5時間後に終了させた。動物は、記録時、3~4か月齢であった。電気生理学的記録の分析
スパイクは、300~6000Hzのバンドパスシグナルを閾値化することによって検出した。閾値は、スパイクによる標準偏差測定のコンタミネーションをさけるため、フィルターにかけられたシグナルの中央値プラスフィルターにかけられたシグナルの標準偏差のロバスト推定量の5倍(中央値/0.675)であった(例えば、Rossant et al.,“Spike Sorting for Large,Dense Electrode Arrays,”bioRxiv doi:dx_doi_org_10.1101_015198(Feb.16,2015)参照)。
記録されたトレースを、2kHzに低解像度処理し、1~300Hz間でバンドパスフィルターにかけた。
動物が走行や静止をすると、海馬のネットワークを通じた活動は顕著に変化し、これらの変化はしばしば異なるネットワーク状態と呼ばれる。これらのネットワーク状態は、異なる周波数帯域におけるLFP振動の有無によって明らかに区別できる。動物が走行した際、他が示したように、CA1において大きなシータ(4~12Hz)振動が認められた(例えば、図2A参照)。動物が静止した際、シータ振動はもはや見られず、SWR、すなわち、約50~100ミリ秒続き、集団的活動のバーストと関連する150~250Hzの高周波振動(例えば、図2B参照)が記録された。SWRは、フィルターにかけられたトレースのエンベロープ振幅が、少なくとも15ミリ秒に対する平均の上側4標準偏差を超えた時に検出された(例えば、図4A、4B、5A、5B、6A、6B、7B、及び8参照)。エンベロープ振幅は、フィルターにかけられたLFPのヒルベルト変換の絶対値を取って計算した。本明細書に開示の結果は、SWR検出のため、より高閾値、すなわち、より大きなSWRを検出する、平均の上側6標準偏差を用いた場合に有効であることが確認されている(例えば、図6C及び7C参照)。シータを検出するため(例えば、図3A及び3C参照)、FIR等リップルフィルターを用いて、LFPをシータ(4~12Hz)、デルタ(1~4Hz)、及びベータ(12~30Hz)についてバンドパスフィルターにかけた。デルタ及びベータに対するシータの比(「シータ比」)を、デルタ及びベータエンベロープ振幅の合計で除したシータエンベロープ振幅として計算した。シータ周期は、シータ比が少なくとも2秒間平均の上側1標準偏差を超え、この比が平均の上側少なくとも2標準偏差のピークに達した場合にそのように分類した。非シータ周期は、このシータ比が少なくとも2秒間1未満の場合にそのように分類した。SWR、シータ周期、及び非シータ周期を視覚的に検査し、これらの基準がそれぞれ、SWR、シータ周期、及び非シータ周期を正確に検出することを確認した。
スペクトル解析は、マルチテーパ法を用いて実施した(例えば、Mitra Lab in Cold Spring Harbor Laboratory、ニューヨーク州コールドスプリングハーバーより入手可能のChronuxのオープンソースソフトウェア、時間帯域幅積=3、テーパ数=5)。刺激なしのパワースペクトルを調べるため(例えば、図3A及び3C参照)、シータ周期のみを含めた。5秒の長さを超えるシータ周期を5秒の試験に分割し、平均パワースペクトル密度をこれらの試験にわたって各動物について計算した。光遺伝学的刺激(例えば、図13A及び6C参照)及び視覚刺激(例えば、図43B及び43C参照)中にパワースペクトルを調べるため、データは、各刺激条件またはベースライン周期の10秒試験に分割し、平均パワースペクトル密度をこれら試験にわたって各動物について計算した。
スペクトログラムは、マルチテーパ法を用いて計算した(例えば、Mitra Lab in Cold Spring Harbor Laboratory、ニューヨーク州コールドスプリングハーバーより入手可能のChronuxのオープンソースソフトウェア)。スペクトログラムは、SWRのピークの前後400ミリ秒の期間を含む各SWRについて計算した。その後、zスコアのスペクトログラムは、全記録期間にわたって計算したスペクトログラムの平均及び標準偏差を用いて各周波数帯域で計算し、標準偏差の単位でパワーの正規化された尺度を生成した(例えば、図4A、4B、5A、及び5B参照)。SWR中のガンマ振動の瞬時周波数は、LFPを10~50Hzに対してバンドパスフィルターにかけ、ヒルベルト変換をし、その後変換されたシグナルのピークの差の逆数をとって計算した(例えば、図4A、5A、及び6B参照)。SWRの前、最中、及び後のガンマパワーは、LFPを低ガンマ(20~50Hz)に対してフィルターにかけ、ヒルベルト変換のエンベロープ振幅を取ることで計算し、SWRピークを中心とした100ミリ秒のビンにおける平均ガンマパワーを得た。これは、全記録期間についてのエンベロープ振幅の平均及び標準偏差によって正規化し、各SWRの周囲の各ビンについてのzスコアのガンマパワーを得た(例えば、図6A及び7B参照)。SWR中のガンマによる位相変調は、LFPをガンマ(20~50Hz)に対してバンドパスフィルターにかけ、ヒルベルト変換をし、SWR中に生じた各スパイクに対して得られたシグナルの位相を決定することによって計算した(例えば、図7E参照)。5XFAD及びWT動物間で位相変調の差を測定するため、復元再抽出を用いた。すなわち、各記録から100スパイクのサブセットをランダムに選択して位相変調分布を作成し、これを各記録について500回繰り返した(例えば、図6C及び7A参照)。変調の深さは、その後、各分布に対するピークとトラフの和で除したピークとトラフの差を計算することによって、スパイク-ガンマ位相分布を測定した(例えば、図6C及び7A参照)。刺激中の発火の差:刺激によって誘発されたマルチユニット発火ヒストグラムをプロットするため、パルスの各光の開始後100ミリ秒について、スパイクを2.5ミリ秒のビンにし、各ビンにおけるスパイクの割合を計算した。平均及びSEMをその後周期のすべての光にわたって計算した。条件間のマルチユニット発火率の差を計算するため、発火率を各10秒間の刺激またはベースライン周期について計算した(周期の期間で除した合計スパイク数)。発火率の差は、関連のあるタイプの刺激の隣接する周期間で取った(光遺伝学的刺激についてはガンマ刺激周期マイナスベースラインまたはランダム周期の発火率、光の点滅刺激については、ガンマ刺激周期マイナスベースライン、連続オン、またはランダム周期の発火率)。すべての動物からの差をヒストグラムにプロットし(例えば、図14A及び44A参照)、動物当たりの差の中央値及び四分位数を箱ひげ図にプロットした(例えば、図13B及び44A参照)。
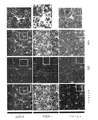
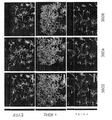
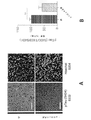
マウスは、深麻酔下で4%のパラホルムアルデヒドでかん流し、脳を4%のパラホルムアルデヒドで一夜後固定した。脳をビブラトーム(例えば、Leica VT100S、Leica Biosystems、イリノイ州バッファローグローブより入手可能)を用いて40μmで切片にした。切片を、室温で1時間、0.2%のTriton X-100及び10%の正常ロバ血清を含むPBSで透過処理及びブロックした。切片を一次抗体のPBS溶液中、4℃で0.2%Triton X-100及び10%正常ロバ血清とともに一夜インキュベートした。一次抗体は、抗EEA1(BD Transduction Laboratories(商標)EEA1(641057)、BD Biosciences、カリフォルニア州サンノゼより入手可能)、抗βアミロイド(例えば、βアミロイド(D54D2)XP(登録商標)、Cell Signaling Technology、マサチューセッツ州ダンバーズより入手可能)、抗Iba1(例えば、019-19741、Wako Chemicals、バージニア州リッチモンドより入手可能)、抗パルブアルブミン(例えば、ab32895、Abcam、マサチューセッツ州ケンブリッジより入手可能)、抗Rab5(ADI-KAp-GP006-E、Enzo Life Sciences Inc.、ニューヨーク州ファーミンデールより入手可能)であった。ELISA実験を確認するため、EEA1との同時標識を可能にすることから抗Aβ抗体D54D2を用い、APPと反応せず、この標識がAβに特異的かどうかについての判断を可能にすることから、抗Aβ抗体12F4を用いた。同時標識実験については、抗Aβ抗体12F4(805501、BioLegend、カリフォルニア州サンディエゴより入手可能)を用いた。一次抗体は、Alexa-Fluor 488及びAlex-Fluor 647二次抗体(Molecular Probes)、Hoechst 33342(94403、Sigma-Aldrich、ミズーリ州セントルイスより入手可能)でのニューロン核で可視化した。画像は、共焦点顕微鏡(LSM 710、Zeiss(商標))を用い、すべての条件に対して同一の設定で取得した。画像は、ImageJ 1.42qを用い、処理群が分からない実験者が定量化した。各実験条件に対して、少なくとも3匹の動物から少なくとも2つの冠状断面を定量化に用いた。海馬CA1画像化については、十分な細胞数を画像化するのに全視野が必要なIba1+細胞分析の場合を除いて、この分析を錐体細胞層に限定した。ImageJを用いて、Iba1+細胞体の直径を測定し、長さ測定の過程をトレースした。さらに、Coloc2プラグインを用いて、Iba1及びAβの共局在化を測定した。3-Dレンダリング用にImaris x64 8.1.2(Bitplane、英国ベルファストより入手可能)を用いた。「斑数」の計数に関しては、10μm以上の沈着物を含めた。
固定した脳を1XPBS中、ビブラトーム(例えば、Leica VT100S、Leica Biosystems、イリノイ州バッファローグローブより入手可能)で100uMの冠状断面にスライスした。Allen Mouse Brain Atlasを参照して視覚野を含む切片を選択し、清澄用緩衝液(pH8.5~9.0、ドデシル硫酸ナトリウム200mM、水酸化リチウム一水和物20mM、ホウ酸4mMのddH2O溶液)中、55℃で振盪しながら2時間インキュベートした。清澄切片を1XPBST(0.1%Triton X-100/1XPBS)で3×10分間洗浄し、ブロッキング用溶液(2%ウシ血清アルブミン/1XPBST)に入れ、室温で一夜振盪した。次に、1XPBST中で、室温にて振盪しながら1時間の洗浄を3回実施した。切片をその後、1:100に1XPBSTで希釈した抗βアミロイド(805501、BioLegend、カリフォルニア州サンディエゴより入手可能)及び抗Iba1(Wako Chemicals、バージニア州リッチモンド、019-19741)一次抗体とともに振盪しながら4℃で2時間インキュベートした。さらに3×1時間の洗浄を1XPBST中で実施した後、切片を、1:100に1XPBSで希釈した二次抗体混合物中、室温にて振盪しながら9時間インキュベートした。断片化ロバ抗ウサギAlexa Fluor(登録商標)488(ab175694)及び抗マウス568(ab150101)二次抗体(いずれもAbcam、マサチューセッツ州ケンブリッジより入手可能)を用いて一次抗体標識を可視化した。このインキュベート期間の途中で、Hoechst33258(Sigma-Aldrich、94403)を各試料中に最終希釈1:250でスパイクした。切片をその後室温で振盪しながら1xPBS中で一夜洗浄した。画像化用にマウントする前に、スライスを、室温で振盪しながら、1時間、RIMS(屈折率整合溶液:Histodenz75g、0.1Mリン酸緩衝液20mL、ddH2O 60mL)中でインキュベートした。Fluoromount G Mounting Medium(Electron Microscopy Sciences、米国ペンシルベニア州ハットフィールド)を用い、カバースリップ(例えば、VistaVision(商標)、VWR International,LLC、ペンシルベニア州ラドナーより入手可能)とともに組織切片を顕微鏡のスライドにマウントした。画像を、Zen Black 2.1ソフトウェア(Carl Zeiss Microscopy、ドイツ、イエナ)付随のZeiss(商標)LSM 880顕微鏡で取得した。3-D復元に用いる断面の概観及び細胞レベルの画像をPlan-Apochromat63x/1.4 Oil DIC対物レンズを用いて撮影した。Imarisx64 8.1.2(Bitplane(商標)(スイス、チューリヒ))を3-Dレンダリング及び分析に用いた。
海馬CA1全細胞溶解物は、3か月齢の雄5XFAD/PV-Creマウスの組織を用いて調製した。組織を手動式ホモジナイザー(Sigma-Aldrich(ミズーリ州セントルイス))を用いて1mlのRIPA(Tris HCl pH8.0 50mM、NaCl 150mM、Np-40 1%、デオキシコール酸ナトリウム0.5%、SDS0.1%)緩衝液中でホモジナイズし、氷上で15分間インキュベートし、4℃で30分間回転させた。14,000rpmで10分間の遠心分離により細胞残屑を単離し廃棄した。溶解物を、ナノドロップを用いて定量化し、25μgのタンパク質を10%アクリルアミドゲルにロードした。100Vで120分間、タンパク質をアクリルアミドゲルからPVDF膜(例えば、Invitrogen(商標)、Thermo Fisher Scientific、マサチューセッツ州ウォルサムより入手可能)に移した。TBS:Tweenで希釈したウシ血清アルブミン(5%w/v)を用いて膜をブロックした。膜を一次抗体中、4℃で一夜、及び二次抗体中、室温で90分間インキュベートした。一次抗体は、抗APP(Invitrogen(商標)PAD CT695、Thermo Fisher Scientific、マサチューセッツ州ウォルサムより入手可能)、抗APP(A8967、Sigma-Aldrich、ミズーリ州セントルイスより入手可能)、抗βアクチン(ab9485、Abcam、マサチューセッツ州ケンブリッジより入手可能)であった。二次抗体は、西洋ワサビペルオキシダーゼ結合(例えば、GE Healthcare、マサチューセッツ州モールバラより入手可能)であった。シグナル強度は、ImageJ 1.46aを用いて定量化し、βアクチンの値に対して正規化した。タウタンパク質の溶解性は、順次タンパク質抽出を用いて調べた。界面活性剤不溶性タウ画分は、Tau5に対する抗体(例えば、AHB0042、Thermo Fisher Scientific、マサチューセッツ州ウォルサムより入手可能)を用いてプローブした。
海馬CA1またはVCを雄のマウスから単離し、PBSまたは5MのグアニジンHClで溶解し、マウス/ヒトAβ1-40またはAβ1-42ELISAキット(例えば、Invitrogen(商標)、Thermo Fisher Scientific、マサチューセッツ州ウォルサムより入手可能)を用い、製造業者の取扱説明書に従ってAβ測定に供した。組織はリン酸緩衝生理食塩水(PBS)中で溶解し、PBS可溶性Aβ画分を抽出した。この可溶性Aβ画分は、恐らくモノマー及びオリゴマーのAβを含んでいた。組織をさらにグアニジン塩酸(HCl)で処理し、不溶性Aβ画分を抽出した。
RNeasy(登録商標)キット(Qiagen、ドイツ、ヒルデンより入手可能)を用いて、海馬CA1単離物から全RNAを抽出した。BIOO NEXTflex(商標)キット(BIOO番号5138-08)を製造業者の取扱説明書の通りに用いるRNA-seqライブラリの調製用に、精製mRNAを用いた。簡潔には、全mRNA1μgを、ポリA精製、断片化、第一のフレックスストランド及び第二のストランド合成、DNA末端アデニル化、ならびにアダプターライゲーションの連続ワークフローに供した。これらライブラリは、15サイクルのPCR反応により濃縮し、Agencourt(登録商標)AMPure XP磁気ビーズ(Beckman Coulter Genomics、マサチューセッツ州ダンバーズより入手可能)で洗浄した。ライブラリの品質は、Advanced Analytical-fragment Analyzerを用いて評価した。バーコード化されたライブラリは、MIT BioMicro Center(マサチューセッツ工科大学、マサチューセッツ州ケンブリッジ)にて、Illumina HiSeq 2000プラットフォームの単一のレーンにおけるシークエンシングのため、均等に混合した。50塩基対シングルエンドシークエンシングリードの生fastqデータを、TopHat2.0ソフトウェア(ジョンズ/ホプキンス大学、メリーランド州ボルチモアの計算生物学センターより入手可能、RNA-seqリードを哺乳類サイズのゲノムに対して、超高スループットショートリードアライナBowtieを用いて整列させ、その後マッピングの結果を分析してエクソン間のスプライス部位を特定するため)を用いてマウスmm9参照ゲノムに対して整列させた。マッピングされたリードをCufflinks2.2ソフトウェア(ワシントン大学Trapnell Lab、ワシントン州シアトルより入手可能)により、UCSCmm9参照遺伝子注釈で処理し、転写物の存在量を推定し、差次的発現を調べた。転写物の相対的存在量は、マッピングされた100万断片当たりのエクソンのキロベース当たりの断片(FPKM)で測定した。処理及び未処理群間の遺伝子の差次的発現試験は、Cuffdiffモジュール(転写物の発現、スプライシング、及びプロモーターの使用における有意な変化を見つけるため。Cufflinks2.2ソフトウェア(ワシントン大学Trapnell Lab、ワシントン州シアトルより入手可能)の一部として含まれる)を用い、統計的有意性に対する調整p値<0.05で実施した(GEO登録:GSE77471)。
CA1は、3か月齢の雄5XFAD/PV-Creマウスの海馬から単離した。液体窒素を用いて組織を急速に凍結し、-80℃で保存し、製造業者のプロトコル(Qiagen(ドイツ、ヒルデン))に従ってRNeasyキットを用いてRNAを抽出した。RNA(3μg)をDNase Iで処理し(4U、Worthington Biochemical Corporation(ニュージャージー州レークウッド))、RNA Clean及びConcentrator-5キット(Zymo Research(カリフォルニア州アーバイン))を用い、製造業者の取扱説明書に従って精製し、14μlのDEPC処理水で溶出した。各試料について、ランダムヘキサマー混合物及びSuperscript III逆転写酵素(50U、Invitrogen(商標)、Thermo Fisher Scientific、マサチューセッツ州ウォルサムより入手可能)を含む20μlの反応体積中、50℃で1時間、1μgのRNAを逆転写した。第一ストランドcDNAを1:10に希釈し、1μlを、プライマー(0.2μM)を含む20μlの反応物(SsoFast(商標)EvaGreen(登録商標)Supermix、Bio-Rad)中でのRT-qPCR増幅に用いた。遺伝子発現の相対的変化を2-ΔΔCt法を用いて評価した。
正規分布しなかった電気生理学的データについては、別段の記載がない限り、結果は中央値及び四分位数として示した。等しい中央値に対する両側ウィルコクソン順位和検定を実施し、分布が有意に異なるかどうかを判定したか、または、ウィルコクソン符号順位検定を実施し、これらが、データが正規分布していると仮定しない場合に分布がゼロから有意差があるかどうかを判定した。ばらつきは、統計的に比較された群間で同様であった。ボンフェローニの方法を用いて多重比較を補正した。分子及び生化学的結果を平均及びSEMで示す。本開示に記載のパーセンテージは群平均である。すべての統計分析は、Prism GraphPadソフトウェア(GraphPad software Inc.、カリフォルニア州ラ・ホーヤ)を用いて実施した。正規性はD’Agostino&Pearsonオムニバス正規性検定を用いて判定した。ばらつきは、統計的に比較された群間で同様であった。2群からなる正規分布したデータに対する比較データは、両側独立t検定によって分析した。3群以上からなる正規分布したデータに対する比較データは、一元配置分散分析に続いてテューキーの多重比較検定によって分析した。非正規分布データに関する比較データは、マン・ホイットニー検定を用いて行った。各実験についての統計検定、正確なP値、及びサンプルサイズ(n)を図の凡例に明記する。分子及び生化学的分析を、条件当たり最低3回の生物学的反復を用いて実施した。
MATLAB(登録商標)プログラミング言語(MathWorks、マサチューセッツ州ナティックより入手可能)に構成される以下のスクリプトは、いくつかの実施形態に従って聴覚クリック・トレイン刺激を生成する1つの方法を示す:

様々な発明に関する実施形態を本明細書に記載及び説明してきたが、当業者には、本明細書に記載の機能を実行し、及び/またはその結果及び/または1つ以上の利点を獲得するための様々な他の方法及び/または構造が容易に想起されるとともに、かかる変化及び/または修正の各々は、本明細書に記載の本発明の実施形態の範囲内であると見なされる。より一般的には、当業者には、本明細書に記載のすべてのパラメータ、寸法、材料、及び構成が例示的であることを意図し、実際のパラメータ、寸法、材料、及び/または構成は、本発明の開示が用いられる特定の用途または複数の用途に依存するということが容易に理解されよう。当業者は、従来の実験だけを用いて、本明細書に記載の特定の発明に関する実施形態の多くの等価物を認識し、またはこれらを確認することができるであろう。従って、前述の実施形態が単なる例示として提示され、添付の特許請求の範囲及びその等価物の範囲内で、発明に関する実施形態が、具体的に記載され、請求される以外の方法で実施され得ることを理解されたい。本開示の発明に関する実施形態は、本明細書に記載の個々の特徴、システム、物品、材料、キット、及び/または方法に関する。さらに、かかる特徴、システム、物品、材料、キット、及び/または方法の2つ以上の組合せは、かかる特徴、システム、物品、材料、キット、及び/または方法が互いに矛盾しない限り、本開示の発明の範囲内に含まれる。
Claims (72)
- その必要がある対象において認知症を治療するための装置であって、
前記装置は、
約35Hz~約45Hzの間の周波数で非侵襲的刺激を放出することができ、及び、
前記対象に前記非侵襲的刺激を投与して、前記対象の少なくとも1つの脳領域において、周波数約35Hz~約45Hzを有する同期ガンマ振動を誘導することにより、前記対象における認知症を治療することができる、
前記装置。 - 前記同期ガンマ振動が、周波数約40Hzを有する、請求項1に記載の装置。
- 前記対象に前記非侵襲的刺激を投与して認知症を治療することが、前記同期ガンマ振動を細胞型特異的に誘導することを含む、請求項1に記載の装置。
- 前記対象に前記非侵襲的刺激を投与して認知症を治療することが、ファスト・スパイキング・パルブアルブミン(FS-PV)介在ニューロンの同期的活動化を誘導することを含む、請求項3に記載の装置。
- 前記対象に前記非侵襲的刺激を投与して認知症を治療することが、前記同期ガンマ振動を脳領域特異的に誘導することを含む、請求項1に記載の装置。
- 前記対象に前記非侵襲的刺激を投与して認知症を治療することが、海馬領域及び感覚野領域の少なくとも一方において同期的活動化を誘導することを含む、請求項5に記載の装置。
- 前記認知症が、アルツハイマー病、血管性認知症、前頭側頭認知症、レビー小体認知症、及び加齢関連認知低下のうちの少なくとも1つと関連する、請求項1に記載の装置。
- 前記対象がヒトである、請求項1に記載の装置。
- 前記装置が、触覚装置、発光装置、及び放音装置のうちの少なくとも1つである、請求項1に記載の装置。
- その必要がある対象において認知症を治療するための装置であって、
前記装置は、
約35Hz~約45Hzの間の周波数で刺激を放出することができ、及び、
前記対象に、前記対象の少なくとも1つの脳領域において周波数約35Hz~約45Hzを有する同期ガンマ振動を誘導するために少なくとも1つの刺激を投与して、前記対象の少なくとも1つの脳領域におけるアミロイドβ(Aβ)ペプチド量を維持又は低減し、それによって前記対象における認知症を治療することができる、
前記装置。 - 前記少なくとも1つの刺激を投与して認知症を治療することが、前記対象の前記少なくとも1つの脳領域においてAβペプチドの生産を低減することを含む、請求項10に記載の装置。
- 前記少なくとも1つの刺激を投与して認知症を治療することが、前記対象の前記少なくとも1つの脳領域においてアミロイド前駆体タンパク質(APP)のC末端断片(CTF)及びN末端断片(NTF)の少なくとも一方の量を低減することを含む、請求項11に記載の装置。
- 前記少なくとも1つの刺激を投与して認知症を治療することが、前記対象の前記少なくとも1つの脳領域においてβセクレターゼ1(BACE1)及びγセクレターゼの少なくとも一方によるAPPのCTF及びNTFへの切断を低減することを含む、請求項12に記載の装置。
- 前記少なくとも1つの刺激を投与して認知症を治療することが、前記対象の前記少なくとも1つの脳領域においてエンドソームの数を低減することを含む、請求項11に記載の装置。
- 前記少なくとも1つの刺激を投与して認知症を治療することが、前記対象の前記少なくとも1つの脳領域においてAβペプチドのクリアランスを促進することを含む、請求項10に記載の装置。
- 前記少なくとも1つの刺激を投与して認知症を治療することが、前記対象の前記少なくとも1つの脳領域においてミクログリアによるAβペプチドの取り込みを増大することを含む、請求項15に記載の装置。
- その必要がある対象においてアルツハイマー病を治療するための装置であって、
前記装置は、
前記対象に周波数約35Hz~約45Hzを有する視覚又は聴覚信号を非侵襲的に送出して、前記対象の少なくとも1つの脳領域において同期ガンマ振動を誘導することにより、前記対象におけるアルツハイマー病を治療することができる、
前記装置。 - 前記同期ガンマ振動が、周波数約40Hzを有する、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記同期ガンマ振動を細胞型特異的に誘導することを含む、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、ファスト・スパイキング・パルブアルブミン(FS-PV)介在ニューロンの同期的活動化を誘導することを含む、請求項19に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記同期ガンマ振動を脳領域特異的に誘導することを含む、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、海馬領域及び感覚野領域の少なくとも一方における同期的活動化を誘導することを含む、請求項21に記載の装置。
- 前記感覚野領域が、視覚野又は聴覚野を含む、請求項22に記載の装置。
- 前記対象が、ヒトである、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の少なくとも1つの脳領域におけるアミロイドβ(Aβ)ペプチド量を維持及び低減することを含み、かつ前記Aβペプチドが、アイソフォームAβ1-40ペプチド及びアイソフォームAβ1-42ペプチドの少なくとも一方を含む、請求項17に記載の装置。
- 前記Aβペプチドが、可溶性Aβペプチド及び不溶性Aβペプチドの少なくとも一方を含む、請求項25に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の前記少なくとも1つの脳領域においてAβペプチドの産生を低減することを含む、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の前記少なくとも1つの脳領域においてアミロイド前駆体タンパク質(APP)のC末端断片(CTF)及びN末端断片(NTF)の少なくとも一方の量を低減することを含む、請求項27に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の前記少なくとも1つの脳領域において、βセクレターゼ1(BACE1)及びγセクレターゼの少なくとも一方によるAPPのCTF及びNTFへの切断を低減することを含む、請求項28に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の前記少なくとも1つの脳領域におけるエンドソームの数を低減することを含む、請求項27に記載の装置。
- 前記エンドソームが、初期エンドソーム抗原1(EEA1)及びRAB5A遺伝子(Rab5)がコードするRas関連タンパク質の少なくとも一方に対して陽性である、請求項30に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の前記少なくとも1つの脳領域におけるAβペプチドのクリアランスを促進することを含む、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の前記少なくとも1つの脳領域において、ミクログリアによるAβペプチドの取り込みを増大することを含む、請求項32に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の前記少なくとも1つの脳領域においてミクログリア細胞の活性に関与する少なくとも1つの差次的に発現される遺伝子を上方制御することを含み、前記差次的に発現される遺伝子が、マウスに対する刺激の違いによって差次的に発現される遺伝子である、請求項17に記載の装置。
- 前記少なくとも1つの差次的に発現される遺伝子が、Nr4a1、Arc、Npas4、Cd68、B2m、Bsr2、Icam1、Lyz2、Irf7、Spp1、Csf1r、及びCsf2raのうちの少なくとも1つを含む、請求項34に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の少なくとも1つの脳領域における、ミクログリア細胞数を増大すること、神経保護状態と一致する前記ミクログリア細胞における形態学的変化を誘導すること、又は前記ミクログリア細胞の活性を促進することを含み、前記神経保護状態と一致する前記ミクログリア細胞における前記形態学的変化が、細胞体のサイズの増加及び過程の長さの減少の少なくとも一方を含む、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出することが、
前記対象を複数の光パルスで刺激し、それにより前記対象の視覚野に同期ガンマ振動を誘導すること;又は前記対象を複数の音パルスで刺激し、それにより前記対象の海馬及び/又は聴覚野に前記同期ガンマ振動を誘導することを含む、請求項17に記載の装置。 - 前記対象に前記視覚又は聴覚信号を送出することが、前記視覚又は聴覚信号を約1時間送出することを含む、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出することが、前記視覚又は聴覚信号をある期間にわたって少なくとも1日1回繰り返し送出することを含み、前記期間が、2日、3日、4日、5日、6日、1週間、2週間、3週間、及び1か月のうちの少なくとも1つである、請求項17に記載の装置。
- 前記装置が、
信号を生成する信号生成器;及び
前記信号生成器に結合され、前記信号生成器により生成された前記信号に基づき、対象に刺激を非侵襲的に投与して、前記対象の少なくとも1つの脳領域において同期ガンマ振動を誘導する、放出体
を含む、請求項17~39のいずれかに記載の装置。 - 前記放出体が、触覚装置、発光装置、及び放音装置のうちの少なくとも1つを含む、請求項40に記載の装置。
- 前記放出体が、発光装置、及び放音装置のうちの少なくとも1つを含み、
前記発光装置が、前記対象の少なくとも一方の目に対する周囲光を低減するための光閉塞装置を含み、前記光閉塞装置が、前記対象の前記少なくとも一方の目に前記視覚信号を放出して、前記対象の視覚野及び海馬の少なくとも1つに同期ガンマ振動を誘導するための発光ユニットを含み、かつ
前記放音装置が、前記対象の少なくとも一方の耳に対する周囲騒音を低減するための雑音消去装置を含み、前記雑音消去装置が、前記対象の前記少なくとも一方の耳に前記聴覚信号を放出し、前記対象の聴覚野及び海馬の少なくとも1つに同期ガンマ振動を誘導するためのスピーカーユニットを含む、
請求項41に記載の装置。 - 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、前記対象の少なくとも1つの脳領域におけるタウリン酸化を低減することを含む、請求項17に記載の装置。
- 前記対象に前記視覚又は聴覚信号を送出してアルツハイマー病を治療することが、
a)前記対象の少なくとも1つの脳領域におけるアミロイドβ(Aβ)ペプチド量を維持又は低減すること;
b)前記対象の少なくとも1つの脳領域における、ミクログリア細胞数を増大すること;神経保護状態と一致する前記ミクログリア細胞の形態学的変化を誘導すること;及び前記ミクログリア細胞の活性を促進すること;及び
c)前記対象の少なくとも1つの脳領域におけるタウリン酸化を低減すること
の1又は2以上を含む、請求項40に記載の装置。 - 対象に周波数約35Hz~約45Hzを有する非侵襲的刺激を非侵襲的に投与して、前記対象の少なくとも1つの脳領域において、同期ガンマ振動を誘導することができる、アルツハイマー病及び/又は認知症の治療のための装置。
- 前記同期ガンマ振動が、周波数約40Hzを有する、請求項45に記載の装置。
- 前記対象が、ヒトである、請求項45に記載の装置。
- 前記対象に前記非侵襲的刺激を投与して、前記対象の少なくとも1つの脳領域において、同期ガンマ振動を誘導することを介して、前記対象における記憶の連合を維持又は強化することができる、請求項45に記載の装置。
- 前記対象に前記非侵襲的刺激を投与して、前記対象の少なくとも1つの脳領域において、同期ガンマ振動を誘導することを介して、前記対象における認識の柔軟性を維持又は強化することができる、請求項45に記載の装置。
- 前記対象に前記非侵襲的刺激を投与して、前記対象の少なくとも1つの脳領域において、同期ガンマ振動を誘導することを介して、前記対象における不安を低減することができる、請求項45に記載の装置。
- 周波数約35Hz~約45Hzを有する信号を生成する、信号生成器;及び
前記信号生成器に結合され、前記信号生成器により生成された前記信号に基づき、対象に刺激を非侵襲的に投与して、前記刺激に応答して前記対象の少なくとも1つの脳領域において同期ガンマ振動を誘導する、放出体を含む、アルツハイマー病及び/又は認知症の治療のための装置。 - 前記放出体が、発光体である、請求項51に記載の装置。
- 前記発光体が、光ファイバー系放出体及び固体光源からなる群から選択される、請求項52に記載の装置。
- 前記発光体が、固体光源であり、少なくとも1つの発光ダイオード(LED)を含む、請求項53に記載の装置。
- 前記放出体が、表示画面を含む、請求項52に記載の装置。
- 前記放出体が、放音体である、請求項51に記載の装置。
- 前記刺激が、所定の頻度の一連のクリックにより構成される音刺激を含む聴覚クリック・トレインである、請求項56に記載の装置。
- 前記放出体が、触覚装置であり、前記刺激が、触覚刺激である、請求項51に記載の装置。
- 前記装置が、
信号を生成する信号生成器;及び
前記信号生成器に結合され、前記信号生成器により生成された前記信号に基づき、対象に前記非侵襲的刺激を非侵襲的に投与して、前記対象の少なくとも1つの脳領域において同期ガンマ振動を誘導する、放出体;
を含む、請求項45~50のいずれかに記載の装置。 - 前記放出体が、発光体である、請求項59に記載の装置。
- 前記発光体が、光ファイバー系放出体及び固体光源からなる群から選択される、請求項60に記載の装置。
- 前記発光体が、固体光源であり、少なくとも1つの発光ダイオード(LED)
を含む、請求項61に記載の装置。 - 前記放出体が、表示画面を含む、請求項59に記載の装置。
- 前記放出体が、放音体である、請求項59に記載の装置。
- 前記刺激が、所定の頻度の一連のクリックにより構成される音刺激を含む聴覚クリック・トレインである、請求項64に記載の装置。
- 前記刺激が、触覚刺激である、請求項45~50のいずれかに記載の装置。
- 対象の少なくとも1つの脳領域において同期ガンマ振動が誘導されるように、放出体を制御して、対象を、前記放出体から生成され、周波数約35Hz~約45Hzを有する非侵襲的刺激に曝露することができる、アルツハイマー病及び/又は認知症の治療のための装置。
- 前記放出体が、発光体であり、
前記発光体を制御して、前記対象を前記非侵襲的刺激としての光刺激に曝露することができる、請求項67に記載の装置。 - 前記放出体が、放音体であり、
前記放音体を制御して、前記対象を前記非侵襲的刺激としての音刺激に曝露することができる、請求項67に記載の装置。 - 前記放出体が、触覚装置であり、
前記触覚装置を制御して、前記対象を前記非侵襲的刺激としての触覚刺激に曝露することができる、請求項67に記載の装置。 - 1又は2以上のコンピュータ実行可能な指示を実行して、前記放出体を制御することができる、請求項67に記載の装置。
- 前記装置が、
信号を生成する信号生成器;及び
前記信号生成器に結合され、前記信号生成器により生成された前記信号に基づき、対象に刺激を非侵襲的に投与して、前記対象の少なくとも1つの脳領域において同期ガンマ振動を誘導する、放出体
を含む、請求項1~16のいずれかに記載の装置。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2021188541A JP2022020829A (ja) | 2015-11-24 | 2021-11-19 | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 |
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201562259187P | 2015-11-24 | 2015-11-24 | |
| US62/259,187 | 2015-11-24 | ||
| PCT/US2016/063536 WO2017091698A1 (en) | 2015-11-24 | 2016-11-23 | Systems and methods for preventing, mitigating, and/or treating dementia |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021188541A Division JP2022020829A (ja) | 2015-11-24 | 2021-11-19 | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2019502429A JP2019502429A (ja) | 2019-01-31 |
| JP2019502429A5 JP2019502429A5 (ja) | 2020-09-10 |
| JP7204192B2 true JP7204192B2 (ja) | 2023-01-16 |
Family
ID=58720012
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2018525754A Active JP7204192B2 (ja) | 2015-11-24 | 2016-11-23 | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 |
| JP2021188541A Pending JP2022020829A (ja) | 2015-11-24 | 2021-11-19 | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 |
| JP2023083776A Pending JP2023101026A (ja) | 2015-11-24 | 2023-05-22 | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 |
Family Applications After (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021188541A Pending JP2022020829A (ja) | 2015-11-24 | 2021-11-19 | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 |
| JP2023083776A Pending JP2023101026A (ja) | 2015-11-24 | 2023-05-22 | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 |
Country Status (11)
| Country | Link |
|---|---|
| US (6) | US10265497B2 (ja) |
| EP (2) | EP4218916B1 (ja) |
| JP (3) | JP7204192B2 (ja) |
| KR (3) | KR102603963B1 (ja) |
| CN (2) | CN115569282A (ja) |
| AU (3) | AU2016361503B2 (ja) |
| CA (1) | CA3003183A1 (ja) |
| DK (1) | DK3383482T3 (ja) |
| ES (1) | ES2943587T3 (ja) |
| IL (3) | IL295071A (ja) |
| WO (1) | WO2017091698A1 (ja) |
Families Citing this family (79)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US12478833B2 (en) | 2012-08-31 | 2025-11-25 | Blue Goji Llc | System and method for targeted neurological therapy using brainwave entrainment with passive treatment |
| US11465013B2 (en) | 2012-08-31 | 2022-10-11 | Blue Goji Llc | System and method for targeted neurological therapy using brainwave entrainment |
| US11612786B2 (en) | 2012-08-31 | 2023-03-28 | Blue Goji Llc | System and method for targeted neurological therapy using brainwave entrainment with passive treatment |
| US10974078B2 (en) * | 2012-12-27 | 2021-04-13 | Brainsonix Corporation | Treating degenerative dementia with low intensity focused ultrasound pulsation (LIFUP) device |
| US10512794B2 (en) | 2016-12-16 | 2019-12-24 | Brainsonix Corporation | Stereotactic frame |
| US9061133B2 (en) | 2012-12-27 | 2015-06-23 | Brainsonix Corporation | Focused ultrasonic transducer navigation system |
| ES2943587T3 (es) | 2015-11-24 | 2023-06-14 | Massachusetts Inst Technology | Sistemas para prevenir, atenuar y/o tratar demencia |
| US10274712B2 (en) * | 2016-01-08 | 2019-04-30 | Optomak, Inc. | Microscope for fluorescence imaging with variable focus |
| WO2018039602A1 (en) | 2016-08-26 | 2018-03-01 | Wisconsin Alumni Research Foundation | Neuromodulation to modulate glymphatic clearance |
| US10905897B2 (en) * | 2016-10-20 | 2021-02-02 | Neuro-Laser Foundation, Inc. | Synergistic treatment methodologies using light therapy |
| US9950189B1 (en) * | 2016-10-20 | 2018-04-24 | Neuro-Laser Foundation Inc. | Treatment methodologies using light therapy |
| KR102612531B1 (ko) | 2016-11-17 | 2023-12-12 | 코그니토 쎄라퓨틱스, 인코포레이티드 | 인지 기능 개선을 위한 시스템 |
| US20220008746A1 (en) * | 2016-11-17 | 2022-01-13 | Cognito Therapeutics, Inc. | Methods and systems for neural stimulation via visual stimulation |
| US20190359661A1 (en) * | 2016-11-25 | 2019-11-28 | Nanoscope Technologies, LLC | Method and device for pain modulation by optical activation of neurons and other cells |
| US11433253B2 (en) | 2017-02-15 | 2022-09-06 | The Regents Of The University Of California | Light therapy system and methods of use |
| JP7458786B2 (ja) | 2017-02-15 | 2024-04-01 | ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア | 改善された光療法システムおよび使用方法 |
| US20190076670A1 (en) * | 2017-09-11 | 2019-03-14 | Alzheimer's Light Llc | Flashing Light Therapy with Image Presentation and Interaction for Treatment of Alzheimer's Disease and Dementia |
| CN111655319A (zh) * | 2017-10-10 | 2020-09-11 | 麻省理工学院 | 用视觉刺激同步脑中γ振荡治疗痴呆 |
| US10960225B2 (en) * | 2017-10-10 | 2021-03-30 | Massachusetts Institute Of Technology | Systems and methods for preventing, mitigating, and/or treating dementia via visual stimulation that binds higher order brain regions, reduces neurodegeneration and neuroinflammation, and improves cognitive function |
| ES3058718T3 (en) * | 2017-10-10 | 2026-03-12 | Massachusetts Inst Technology | Systems for preventing, mitigating, and/or treating dementia |
| US10310959B2 (en) | 2017-11-07 | 2019-06-04 | Bank Of America Corporation | Pre-deployment validation system using intelligent databases |
| WO2019173847A1 (en) * | 2018-03-09 | 2019-09-12 | Georgia Tech Research Corporation | Systems and methods for driving neural activity to control brain signaling and gene expression |
| US20190388020A1 (en) * | 2018-06-20 | 2019-12-26 | NeuroPlus Inc. | System and Method for Treating and Preventing Cognitive Disorders |
| US12029579B2 (en) * | 2018-07-13 | 2024-07-09 | Pst Inc. | Apparatus for estimating mental/neurological disease |
| CN112533672A (zh) * | 2018-08-01 | 2021-03-19 | 坪田实验室股份有限公司 | 基于光刺激的脑波及细胞活性控制装置和方法以及改善、预防或增强脑功能的装置 |
| USD889677S1 (en) | 2018-10-24 | 2020-07-07 | Verilux, Inc. | Lamp with stand |
| US20220040366A1 (en) * | 2018-11-05 | 2022-02-10 | Aleddra Inc. | Multi-Mode Lighting Device |
| US10518063B1 (en) | 2018-11-07 | 2019-12-31 | Symbolics Inc. | Method for stimulating neural activity |
| KR102244367B1 (ko) * | 2018-11-13 | 2021-04-27 | 고려대학교 산학협력단 | 광유전학 기법을 이용한 알츠하이머성 치매에서의 시냅스 가소성 회복 방법 |
| CN109621152A (zh) * | 2019-01-28 | 2019-04-16 | 邹珍友 | 一种用强场脉冲激光无介入性地干预动物认知意识活动与行为的方法 |
| KR102272858B1 (ko) | 2019-05-22 | 2021-07-05 | 경희대학교 산학협력단 | 뇌 이상 단백질 치료를 위한 방사선 시스템 및 이의 사용 방법 |
| JP7355334B2 (ja) * | 2019-08-10 | 2023-10-03 | 智夫 松田 | 電子閃光サングラスからなる認知症治療器具 |
| US20220322993A1 (en) * | 2019-08-16 | 2022-10-13 | The Regents Of The University Of California | Neurofeedback Training to Promote Sharp Wave Ripples |
| CN110681029A (zh) * | 2019-09-29 | 2020-01-14 | 广东省医疗器械研究所 | 一种阿尔茨海默病预防与治疗装置及其实现方法 |
| CN114555166A (zh) | 2019-10-18 | 2022-05-27 | 奥地利科学技术研究所 | 神经元可塑性的刺激 |
| CN110639131A (zh) * | 2019-11-07 | 2020-01-03 | 杭州崇正医疗科技有限公司 | 眼镜型阿尔茨海默症声光治疗设备 |
| CN112791078B (zh) | 2019-11-13 | 2022-12-06 | 润佳(苏州)医药科技有限公司 | 同位素富集的3-氨基-1-丙磺酸及其衍生物的用途 |
| KR102467533B1 (ko) | 2019-11-18 | 2022-11-15 | 중앙대학교 산학협력단 | 알츠하이머병 환자를 위한 광조사 헤드셋 |
| US20210275827A1 (en) * | 2020-03-07 | 2021-09-09 | Reversal Solutions, Inc. | Systems and methodologies for treating or preventing psychiatric disorders, brain trauma, and addiction or dependence by light therapy with modulated frequency |
| WO2021216957A1 (en) * | 2020-04-23 | 2021-10-28 | Massachusetts Institute Of Technology | Systems, devices, and methods for gamma entrainment using sensory stimuli to alleviate cognitive deficits and/or neuroinflammation induced by chemotherapy agents |
| US11759661B2 (en) | 2020-05-20 | 2023-09-19 | Brainsonix Corporation | Ultrasonic transducer treatment device |
| WO2021257548A1 (en) | 2020-06-18 | 2021-12-23 | Diagnostyx, Inc. | Treatment of neurological abnormalities using dynamic electroencephalography |
| KR20240051882A (ko) * | 2020-07-27 | 2024-04-22 | 코그니토 쎄라퓨틱스, 인코포레이티드 | 알츠하이머병 환자의 기능적 능력을 유지하고 수면 품질을 개선하는 감각 감마 자극 치료법 |
| WO2022046764A1 (en) | 2020-08-24 | 2022-03-03 | Brainsonix Corporation | Systems and methods for neuromodulation of neuronal circuits using transcranial focused microwave pulses |
| US11730917B2 (en) | 2020-10-29 | 2023-08-22 | Optoceutics ApS | Gamma stimulation pulsing light source system with dosage adjustment for gaze angle |
| CN112569445A (zh) * | 2020-12-09 | 2021-03-30 | 郑州布恩科技有限公司 | 一种脑功能康复设备 |
| JP6994652B1 (ja) | 2020-12-14 | 2022-01-14 | エクボ株式会社 | ガンマ波誘発装置 |
| IT202100000776A1 (it) | 2021-01-18 | 2022-07-18 | Univ Degli Studi Di Brescia | METODO NON TERAPEUTICO DI STIMOLAZIONE CON γ-TACS |
| CN115335102B (zh) * | 2021-02-08 | 2024-06-04 | 张鸿勋 | 在大脑中产生反馈的方法、系统及大脑键盘 |
| EP4304479A4 (en) * | 2021-03-09 | 2025-04-23 | Cognito Therapeutics, Inc. | METHODS AND SYSTEMS FOR SLOWING DOWN CEREBRAL ATROPHY |
| WO2022212891A1 (en) * | 2021-04-02 | 2022-10-06 | The Board Of Trustees Of The Leland Stanford Junior University | Novel neural control signals for therapeutic behavioral modulation in eating-related disorders |
| CN113082193B (zh) * | 2021-04-08 | 2022-04-01 | 上海南方模式生物科技股份有限公司 | 受调节的内分泌特异性蛋白18及其基因的用途 |
| US12249307B2 (en) * | 2021-04-18 | 2025-03-11 | Tony Jonathan Graber | Methods and systems of sound-stimulation practice |
| CN113368362A (zh) * | 2021-04-20 | 2021-09-10 | 刘书朋 | 用于谐振脑部γ节律震荡波的光声振动波装置及用途 |
| US11948531B2 (en) | 2021-06-21 | 2024-04-02 | Industrial Technology Research Institute | Light source device and display device alternately emitting light beams |
| WO2022272093A1 (en) * | 2021-06-24 | 2022-12-29 | The Regents Of The University Of California | Virtual and augmented reality devices to diagnose and treat cognitive and neuroplasticity disorders |
| EP4399311A4 (en) * | 2021-09-10 | 2025-11-26 | Alnylam Pharmaceuticals Inc | APP ARNI COMPOSITIONS AND THEIR METHODS OF USE FOR TREATMENT OR PREVENTION OF DISEASES CHARACTERIZED BY ENLARGED ENDOSOMES |
| WO2023049467A1 (en) * | 2021-09-27 | 2023-03-30 | Cognito Therapeutics, Inc. | Entertainment device for promoting gamma oscillations |
| CN118354712A (zh) * | 2021-09-27 | 2024-07-16 | 科格尼托治疗公司 | 活动期间增强神经刺激的方法 |
| WO2023108170A2 (en) * | 2021-12-10 | 2023-06-15 | Massachusetts Institute Of Technology | Systems, devices, and methods for enhancing the neuroprotective effects of non-invasive gamma stimulation with pharmacological agents |
| US20250041553A1 (en) * | 2021-12-23 | 2025-02-06 | Georgia Tech Research Corporation | Brain stimulation systems and methods |
| KR102400319B1 (ko) * | 2021-12-31 | 2022-05-24 | (주)리솔 | 객체의 복수의 뇌 영역에서 동기화된 진동을 동조시킴으로써, 뇌를 자극하는 장치 및 방법 |
| US20250099779A1 (en) * | 2022-01-10 | 2025-03-27 | Wenzhou Ventura Technology Co., Ltd. | Device and method for regulating release of adenosine in organism, and use |
| CN114209957B (zh) * | 2022-01-21 | 2024-01-26 | 中国科学院电工研究所 | 一种用于改善老年认知障碍的可穿戴式脉冲调制磁声刺激装置 |
| US12451249B2 (en) * | 2022-01-31 | 2025-10-21 | At&T Intellectual Property I, L.P. | Dynamic home themes for assisted early detection and treatment in healthcare |
| WO2023196610A1 (en) * | 2022-04-08 | 2023-10-12 | Cognito Therapeutics, Inc. | Methods for detecting and monitoring neurological disorders or conditions |
| US20260000910A1 (en) | 2022-06-10 | 2026-01-01 | Tsubota Laboratory, Inc. | Method for treating parkinson's disease through photostimulation, and device used in said method |
| CN115211813A (zh) * | 2022-07-19 | 2022-10-21 | 军事科学院军事医学研究院环境医学与作业医学研究所 | 一种基于闪光诱发并监测γ神经振荡同步化的系统 |
| WO2024071452A1 (ko) * | 2022-09-26 | 2024-04-04 | (주)뉴로니어 | 치매 예방 및 개선을 위한 헤드 마운팅 장치 |
| WO2024081719A1 (en) * | 2022-10-11 | 2024-04-18 | Katherine Quittner | System and method for producing sound and light for therapeutic effect |
| CN116059531B (zh) * | 2022-12-21 | 2024-11-05 | 重庆大学 | 脑深部刺激改善阿尔茨海默症小鼠的记忆功能的神经评估方法及系统 |
| EP4680331A1 (en) * | 2023-03-13 | 2026-01-21 | Nuuron GmbH | Device for inducing high-frequency oscillations in a brain |
| US12168140B1 (en) | 2023-06-20 | 2024-12-17 | Photigen, Inc. | Apparatus for neural stimulation and method of use |
| US12544666B2 (en) * | 2023-06-22 | 2026-02-10 | King.Com Limited | Method and apparatus providing a computer implemented game |
| CN119653539A (zh) | 2023-09-15 | 2025-03-18 | 台达电子工业股份有限公司 | Led驱动器及led照明系统 |
| US20250111800A1 (en) * | 2023-10-02 | 2025-04-03 | Mark Freeman | Methods for conducting memory therapy using facial recognition and vibrotactile stimulation with synchronized music |
| US12458817B2 (en) | 2023-11-21 | 2025-11-04 | Sanmai Technologies, PBC | Wearable closed loop TUS system |
| CN118059391A (zh) * | 2024-01-17 | 2024-05-24 | 中国人民解放军军事科学院军事医学研究院 | 微波调节gamma节律振荡改善认知功能的方法 |
| WO2026003580A1 (en) * | 2024-06-25 | 2026-01-02 | Syntropic Medical Gmbh | Systems and methods for treating mental disorders |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2015519096A (ja) | 2012-04-06 | 2015-07-09 | ニューポート ブレイン リサーチ ラボラトリー インコーポレイテッド | 周波数特異的感覚刺激 |
Family Cites Families (216)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4315502A (en) | 1979-10-11 | 1982-02-16 | Gorges Denis E | Learning-relaxation device |
| US4449047A (en) | 1981-10-26 | 1984-05-15 | Sunoco Energy Development Co., A Wholly-Owned Subsidiary Of Sun Company, Inc. | Airborne mineral exploration system |
| US4456910A (en) | 1982-08-12 | 1984-06-26 | General Electric Company | Non-complementary metal oxide semiconductor (MOS) driver for liquid crystal displays |
| JPS60194422A (ja) | 1984-03-16 | 1985-10-02 | Tanaka Sangyo:Kk | めがねフレ−ムと智の結合構造 |
| US5151687A (en) | 1990-07-16 | 1992-09-29 | Younger George G | Apparatus for logging electronic media-taped material |
| ATE386115T1 (de) | 1991-12-06 | 2008-03-15 | Max Planck Gesellschaft | Verwendung von proteinkinasen zur diagnose und behandlung der alzheimer krankheit |
| US6071229A (en) * | 1992-05-05 | 2000-06-06 | Rubins; Tye | Video or compact disk encoder/decoder for brain wave synchronizer |
| US5534953A (en) | 1994-07-01 | 1996-07-09 | Schmielau; Fritz | Training device for the therapy of patients having perception defects |
| JP3648802B2 (ja) | 1994-09-27 | 2005-05-18 | 松下電工株式会社 | 光治療用高照度光照射装置 |
| US5659287A (en) | 1995-03-21 | 1997-08-19 | General Signal Corporation | Strobe synchronization for averting convulsive reactions to strobe light |
| US5923398A (en) | 1995-06-15 | 1999-07-13 | Enlightened Technologies, Inc. | Interactive light field for non-visual stimulation |
| US6077545A (en) | 1995-10-30 | 2000-06-20 | Matrix Pharmaceuticals, Inc. | Process and composition for therapeutic cisplatin (CDDP) |
| US6206537B1 (en) | 1995-11-15 | 2001-03-27 | Lane T. Hauck | Electrically illuminated attention-attracting devices and method of using same |
| US6463328B1 (en) | 1996-02-02 | 2002-10-08 | Michael Sasha John | Adaptive brain stimulation method and system |
| US6066163A (en) | 1996-02-02 | 2000-05-23 | John; Michael Sasha | Adaptive brain stimulation method and system |
| US6113537A (en) | 1996-04-19 | 2000-09-05 | Castano; Jaime A. | Optical method and device for determining blood glucose levels |
| JP4076232B2 (ja) | 1996-06-06 | 2008-04-16 | フラレックス・テラピューティクス・インコーポレーテッド | 低周波数磁気パルスを使用する電気治療装置 |
| US5934967A (en) | 1997-07-31 | 1999-08-10 | Blue Leaf Design, Inc. | Self-actuating spreading apparatus |
| WO1999034865A1 (en) | 1998-01-08 | 1999-07-15 | Levin Richard B | Eeg based consciousness-alert monitoring system |
| US6299632B1 (en) | 1998-11-30 | 2001-10-09 | Peter Jaillet | Method for changing critical brain activity using light and sound |
| US6231528B1 (en) | 1999-01-15 | 2001-05-15 | Jonathan J. Kaufman | Ultrasonic and growth factor bone-therapy: apparatus and method |
| UA81216C2 (en) * | 1999-06-01 | 2007-12-25 | Prevention and treatment of amyloid disease | |
| US6539263B1 (en) | 1999-06-11 | 2003-03-25 | Cornell Research Foundation, Inc. | Feedback mechanism for deep brain stimulation |
| US20010039012A1 (en) | 1999-06-14 | 2001-11-08 | Lapidus Stanley N. | Methods for diagnostic screening |
| US7446785B1 (en) | 1999-08-11 | 2008-11-04 | Texas Instruments Incorporated | High bit depth display with low flicker |
| GB0010565D0 (en) | 2000-05-02 | 2000-06-21 | Medical Res Council | Assay for acetylcholinesterase inhibitors |
| RU2202323C2 (ru) | 2001-01-24 | 2003-04-20 | Савельев Борис Сергеевич | Способ волновой биомеханотерапии б.с. савельева |
| US20130211238A1 (en) | 2001-01-30 | 2013-08-15 | R. Christopher deCharms | Methods for physiological monitoring, training, exercise and regulation |
| US6733490B1 (en) | 2001-04-12 | 2004-05-11 | Iridex Corporation | Method and apparatus for controlling sub-clinical laser procedures with intra-operative monitoring of electrophysiological changes |
| US7010356B2 (en) * | 2001-10-31 | 2006-03-07 | London Health Sciences Centre Research Inc. | Multichannel electrode and methods of using same |
| CA2477488A1 (en) | 2002-02-04 | 2003-08-14 | Great Lake Biosciences, Llc | Treatment of neurological disorders using electrical stimulation |
| US8328420B2 (en) | 2003-04-22 | 2012-12-11 | Marcio Marc Abreu | Apparatus and method for measuring biologic parameters |
| KR100553516B1 (ko) * | 2002-06-24 | 2006-02-20 | 정종필 | 알파파유도 전기자극기 |
| DE10233960B4 (de) | 2002-07-29 | 2006-11-02 | Forschungszentrum Jülich GmbH | Vorrichtung zur bedarfsgesteuerten Modulation physiologischer und pathologischer neuronaler rhythmischer Aktivität im Gehirn mittels sensorischer Stimulation |
| US8579786B2 (en) | 2002-10-15 | 2013-11-12 | Medtronic, Inc. | Screening techniques for management of a nervous system disorder |
| US11082664B2 (en) | 2004-07-06 | 2021-08-03 | Tseng-Lu Chien | Multiple functions LED night light |
| US7798982B2 (en) | 2002-11-08 | 2010-09-21 | Engineering Acoustics, Inc. | Method and apparatus for generating a vibrational stimulus |
| US7998460B2 (en) * | 2003-04-17 | 2011-08-16 | The Board Of Trustees Of The Leland Stanford Junior University | Prevention of deficits in neurogenesis with anti-inflammatory agents |
| WO2004096343A2 (en) | 2003-04-28 | 2004-11-11 | Azna Health And Wellness, Inc. | Light and magnetic emitting mask |
| EP1624932A4 (en) | 2003-05-01 | 2010-02-10 | Flinders Technologies Pty Ltd | APPARATUS FOR ADMINISTERING LIGHT STIMULATION |
| US8190248B2 (en) * | 2003-10-16 | 2012-05-29 | Louisiana Tech University Foundation, Inc. | Medical devices for the detection, prevention and/or treatment of neurological disorders, and methods related thereto |
| KR101049605B1 (ko) | 2004-04-01 | 2011-07-14 | 윌리엄 씨. 토치 | 눈 움직임을 모니터하는 바이오센서, 커뮤니케이터, 및컨트롤러 및 그들의 사용 방법 |
| US8177702B2 (en) | 2004-04-15 | 2012-05-15 | Neuronetics, Inc. | Method and apparatus for determining the proximity of a TMS coil to a subject's head |
| AU2005265033B2 (en) | 2004-06-18 | 2011-08-04 | Neuronetrix Solutions, Llc | Evoked response testing system for neurological disorders |
| DE102004048982A1 (de) * | 2004-10-01 | 2006-04-06 | Markus Klose | Augenvorsatz für ein Stimulationsgerät zum Stimulieren des Zentralnervensystems und der Gehirnströme eines Menschen |
| US9764135B2 (en) | 2004-10-21 | 2017-09-19 | Advanced Neuromodulation Systems, Inc. | Stimulation design for neuromodulation |
| US7645226B2 (en) | 2004-11-12 | 2010-01-12 | Biogenics Ii L.L.C. | Relaxation device and method |
| JP2008520280A (ja) | 2004-11-15 | 2008-06-19 | デチャームス,クリストファー | 光を使用した神経組織の刺激の適用 |
| US7769439B2 (en) | 2004-11-30 | 2010-08-03 | Infinite Z, Inc. | Brain balancing by binaural beat |
| ATE550912T1 (de) | 2004-12-07 | 2012-04-15 | Elumen Lighting Networks Inc | System und verfahren zur steuerung einer matrix von leuchtdioden und dadurch hergestelltem licht |
| US7361074B1 (en) | 2005-02-18 | 2008-04-22 | Rapid Pro Manufacturing, Martin And Periman Partnership | Rotating light toy |
| AU2006230433A1 (en) | 2005-03-30 | 2006-10-05 | Novavision, Inc. | Method and device for delivering visual stimuli with head mounted display during vision training |
| US7569545B2 (en) * | 2005-05-20 | 2009-08-04 | Academia Sinica | Methods of increasing neurotrophic factor expression |
| CN101262863B (zh) * | 2005-07-08 | 2016-08-31 | Dsmip资产公司 | 用于治疗痴呆和前痴呆相关病症的多不饱和脂肪酸 |
| US20140336514A1 (en) | 2005-08-05 | 2014-11-13 | Gholam A. Peyman | Methods to regulate polarization and enhance function of cells |
| US20190030190A1 (en) | 2005-08-05 | 2019-01-31 | Gholam A. Peyman | Methods to regulate polarization and enhance function of cells |
| US9962558B2 (en) * | 2005-08-05 | 2018-05-08 | Gholam A. Peyman | Methods to regulate polarization and enhance function of cells |
| US20070225773A1 (en) | 2005-09-28 | 2007-09-27 | Yang Shen | Implantable transcranial pulse generator having a collapsible portion |
| US8868177B2 (en) * | 2009-03-20 | 2014-10-21 | ElectroCore, LLC | Non-invasive treatment of neurodegenerative diseases |
| WO2007062367A2 (en) * | 2005-11-21 | 2007-05-31 | The Curators Of The University Of Missouri | Light sensitivity meter and uses thereof |
| CN1991364A (zh) * | 2005-12-26 | 2007-07-04 | 中国科学院上海生命科学研究院 | 预防和治疗阿尔兹海默症的g蛋白偶联受体拮抗剂 |
| US7660636B2 (en) | 2006-01-04 | 2010-02-09 | Accelerated Care Plus Corp. | Electrical stimulation device and method for the treatment of dysphagia |
| US7801601B2 (en) | 2006-01-27 | 2010-09-21 | Cyberonics, Inc. | Controlling neuromodulation using stimulus modalities |
| US10357662B2 (en) * | 2009-02-19 | 2019-07-23 | Pthera LLC | Apparatus and method for irradiating a surface with light |
| JP5089060B2 (ja) | 2006-03-14 | 2012-12-05 | 株式会社ソニー・コンピュータエンタテインメント | エンタテインメントシステムおよびゲームコントローラ |
| US20070253561A1 (en) | 2006-04-27 | 2007-11-01 | Tsp Systems, Inc. | Systems and methods for audio enhancement |
| JP2008029804A (ja) | 2006-06-27 | 2008-02-14 | Family Co Ltd | マッサージ機 |
| US20080004660A1 (en) * | 2006-06-28 | 2008-01-03 | Medtrode Inc. | Systems and methods for improving a cognitive function |
| CA2657762C (en) | 2006-07-25 | 2015-07-21 | Novavision, Inc. | Dynamic stimuli for visual field testing and therapy |
| CA2658802A1 (en) | 2006-08-08 | 2008-02-14 | Mony Paz | Combination dental hand tool |
| EP2050073A4 (en) | 2006-08-11 | 2009-12-30 | Thomson Licensing | ACCURATE REPRESENTATION OF THE MOVEMENT FOR DIGITAL AND OPTICAL PROJECTORS |
| US7828434B2 (en) | 2006-08-31 | 2010-11-09 | Nike, Inc. | Zone switched sports training eyewear |
| US10786669B2 (en) | 2006-10-02 | 2020-09-29 | Emkinetics, Inc. | Method and apparatus for transdermal stimulation over the palmar and plantar surfaces |
| US20080181882A1 (en) | 2006-11-15 | 2008-07-31 | The University Of Pennsylvania | Neurgulin 1 (NRG1) - ErbB4 signaling as a target for the treatment of schizophrenia |
| US8700167B2 (en) | 2006-12-22 | 2014-04-15 | Ebs Technologies Gmbh | Apparatus and method for stimulating a brain of a person |
| US8894696B2 (en) | 2007-01-16 | 2014-11-25 | Katherine L Hurst | Suprachiasmatic nucleus inducing, melatonin suppressing light emitting device to enhance wake cycle |
| US8401609B2 (en) | 2007-02-14 | 2013-03-19 | The Board Of Trustees Of The Leland Stanford Junior University | System, method and applications involving identification of biological circuits such as neurological characteristics |
| WO2008106694A2 (en) | 2007-03-01 | 2008-09-04 | The Board Of Trustees Of The Leland Stanford Junior University | Systems, methods and compositions for optical stimulation of target cells |
| EP2129352B1 (en) | 2007-03-13 | 2016-03-09 | The Feinstein Institute for Medical Research | Treatment of inflammation by non-invasive stimulation |
| US20080255949A1 (en) | 2007-04-13 | 2008-10-16 | Lucid Systems, Inc. | Method and System for Measuring Non-Verbal and Pre-Conscious Responses to External Stimuli |
| US20140324138A1 (en) | 2007-05-09 | 2014-10-30 | Massachusetts Institute Of Technology | Wirelessly-powered illumination of biological tissue |
| CA2686929A1 (en) | 2007-05-11 | 2008-11-20 | Clarimedix Inc. | Visible light modulation of mitochondrial function in hypoxia and disease |
| KR20100039287A (ko) | 2007-05-25 | 2010-04-15 | 토마스 제퍼슨 유니버시티 | 집중력 및/또는 기억력의 향상이 필요한 개체에 집중력 및/또는 기억력을 향상시키는 치료방법 |
| US20090005837A1 (en) * | 2007-06-29 | 2009-01-01 | Ruth Olmstead | Method and apparatus for stimulating the neurochemistry of the brain resulting in increased overall brain function, cognitive performance, and intelligence quota |
| AU2008283857A1 (en) | 2007-08-06 | 2009-02-12 | Great Lakes Biosciences, Llc | Methods and apparatus for electrical stimulation of tissues using signals that minimize the effects of tissue impedance |
| US20100217358A1 (en) | 2007-08-20 | 2010-08-26 | Universite Laval | Artificial light apparatus and its use for influencing a condition in a subject |
| US8380314B2 (en) | 2007-09-26 | 2013-02-19 | Medtronic, Inc. | Patient directed therapy control |
| US8121694B2 (en) | 2007-10-16 | 2012-02-21 | Medtronic, Inc. | Therapy control based on a patient movement state |
| US8577470B2 (en) | 2007-12-18 | 2013-11-05 | Souhile Assaf | Systems and methods for improving memory in Alzheimer's patients |
| GB0725124D0 (en) * | 2007-12-24 | 2008-01-30 | Carr Peter | Photic stimulation for eyes |
| US8396545B2 (en) * | 2008-02-25 | 2013-03-12 | Wisconsin Alumni Research Foundation | Electrophysiological screens for cognitive modulators |
| DE102008012669B8 (de) | 2008-03-05 | 2011-03-03 | Anm Adaptive Neuromodulation Gmbh | Vorrichtung und Verfahren zur visuellen Stimulation |
| DE102008015259B4 (de) | 2008-03-20 | 2010-07-22 | Anm Adaptive Neuromodulation Gmbh | Vorrichtung und Verfahren zur auditorischen Stimulation |
| EP2265328B1 (en) | 2008-03-20 | 2018-05-09 | EBS Technologies GmbH | An apparatus for automatic treatment adjustment after nervous system dysfunction |
| US8636640B2 (en) | 2008-04-11 | 2014-01-28 | Brain Symphony LLC | Method and system for brain entertainment |
| BRPI0911448A2 (pt) * | 2008-04-23 | 2019-02-26 | The Board Of Trustees For The Leland Stanford Junior University | sistemas, métodos e composições para simulação óticas de células alvo |
| US20090270776A1 (en) | 2008-04-28 | 2009-10-29 | Hung-Chu Chang | Massage Chair With Function of Stress Relaxation and Sleep Inducing |
| US8070669B2 (en) | 2008-08-04 | 2011-12-06 | Gestion Ultra International Inc. | Wall integrated multisensory therapy device |
| JP4500369B2 (ja) | 2008-09-19 | 2010-07-14 | パナソニック株式会社 | 注意散漫検出装置、注意散漫検出方法およびコンピュータプログラム |
| US20120065709A1 (en) * | 2008-09-30 | 2012-03-15 | The Regents Of The University Of Colorado | Methods and devices for visible light modulation of mitochondrial function in hypoxia and disease |
| US8022631B2 (en) | 2008-11-03 | 2011-09-20 | General Electric Company | Color control of light sources employing phosphors |
| US8355793B2 (en) | 2009-01-02 | 2013-01-15 | Cochlear Limited | Optical neural stimulating device having a short stimulating assembly |
| ITRM20090027A1 (it) | 2009-01-26 | 2010-07-27 | Alain Rocco | Apparato di stimolazione per il trattamento del dolore e relativo metodo di funzionamento. |
| US8932218B1 (en) | 2009-02-19 | 2015-01-13 | Jeffrey D. Thompson | Methodology, use and benefits of neuroacoustic frequencies for assessing and improving the health and well-being of living organisms |
| US10232178B2 (en) | 2009-03-20 | 2019-03-19 | Electrocore, Inc. | Non-invasive magnetic or electrical nerve stimulation to treat or prevent dementia |
| US10220207B2 (en) | 2009-03-20 | 2019-03-05 | Electrocore, Inc. | Nerve stimulation methods for averting imminent onset or episode of a disease |
| WO2010113164A1 (en) | 2009-04-03 | 2010-10-07 | Neuronix Ltd. | Method and system for neurological treatment |
| US20100274329A1 (en) | 2009-04-24 | 2010-10-28 | Chris Bradley | System and method for skin care using light and microcurrents |
| US20120159656A1 (en) * | 2009-04-24 | 2012-06-21 | Galenea Corp. | Compositions and methods for evaluating cognitive defects |
| CA2765500C (en) | 2009-06-15 | 2019-07-30 | Brain Computer Interface Llc | A brain-computer interface test battery for the physiological assessment of nervous system health. |
| US8267851B1 (en) | 2009-06-16 | 2012-09-18 | James M Kroll | Method and apparatus for electrically generating signal for inducing lucid dreaming |
| US20120220905A1 (en) * | 2009-10-11 | 2012-08-30 | Ads & B Investment Fund L.P | Methods and protocols for providing artificially induced vibrations (aivs) to mammalian patients for therapeutic purposes |
| CN102791332A (zh) | 2009-11-04 | 2012-11-21 | 代理并代表亚利桑那州立大学的亚利桑那董事会 | 调节大脑活动的设备和方法 |
| US8541741B2 (en) | 2009-11-20 | 2013-09-24 | Perkinelmer Health Sciences, Inc. | Photonic measurement instrument using fiber optics |
| US8239030B1 (en) | 2010-01-06 | 2012-08-07 | DJ Technologies | Transcranial stimulation device and method based on electrophysiological testing |
| US8579793B1 (en) | 2010-01-27 | 2013-11-12 | James David Honeycutt | Apparatus to affect brainwave entrainment over premises power-line wiring |
| US9308254B2 (en) | 2010-02-17 | 2016-04-12 | Imperial Innovations Limited | Method for treatment of post operative cognitive dysfunction |
| US9770414B2 (en) | 2010-05-13 | 2017-09-26 | Pacira Pharmaceuticals, Inc. | Sustained release formulation of methotrexate as a disease-modifying antirheumatic drug (DMARD) and an anti-cancer agent |
| WO2011146777A2 (en) | 2010-05-19 | 2011-11-24 | Photothera, Inc. | Device and method for providing a synergistic combination of phototherapy and a non-light energy modality to the brain |
| WO2012024243A1 (en) | 2010-08-16 | 2012-02-23 | Photothera, Inc. | Minimally invasive low level light therapy for neurological disorders |
| US9622911B2 (en) | 2010-09-30 | 2017-04-18 | Cxl Ophthalmics, Llc | Ophthalmic treatment device, system, and method of use |
| JP6355335B2 (ja) | 2010-11-05 | 2018-07-11 | ザ ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティー | 報酬関連行動の光遺伝学的制御 |
| EP2635295B1 (en) * | 2010-11-05 | 2017-12-20 | The Board of Trustees of the Leland Stanford Junior University | Control and characterization of memory function |
| CN106947741A (zh) * | 2010-11-05 | 2017-07-14 | 斯坦福大学托管董事会 | 光活化嵌合视蛋白及其使用方法 |
| GB201103200D0 (en) | 2011-02-24 | 2011-04-13 | Isis Innovation | An optical device for the visually impaired |
| CN103392196A (zh) | 2011-03-02 | 2013-11-13 | 皇家飞利浦有限公司 | 用于用户的认知增强的装置和方法 |
| US20120253236A1 (en) | 2011-04-04 | 2012-10-04 | Snow Buddy L | Methods and apparatuses for delivering external therapeutic stimulation to animals and humans |
| US8892207B2 (en) | 2011-04-20 | 2014-11-18 | Medtronic, Inc. | Electrical therapy for facilitating inter-area brain synchronization |
| CN103501855B (zh) | 2011-04-20 | 2015-12-23 | 美敦力公司 | 基于生物电共振响应来确定电治疗的参数 |
| PL2709522T3 (pl) | 2011-05-20 | 2017-03-31 | Nanyang Technological University | Układ do synergicznej neurofizjologicznej rehabilitacji i/lub rozwoju funkcjonalnego |
| US20130021138A1 (en) | 2011-07-20 | 2013-01-24 | GM Global Technology Operations LLC | Method of evaluating structural integrity of a vehicle component with radio frequency identification tags and system for same |
| US9566426B2 (en) * | 2011-08-31 | 2017-02-14 | ElectroCore, LLC | Systems and methods for vagal nerve stimulation |
| US9268406B2 (en) | 2011-09-30 | 2016-02-23 | Microsoft Technology Licensing, Llc | Virtual spectator experience with a personal audio/visual apparatus |
| CN103889495A (zh) | 2011-10-25 | 2014-06-25 | 学校法人金泽医科大学 | 用于改善痴呆症状和抑郁症状的光照射装置、设置有该光照射装置的房间和用于改善痴呆症状和抑郁症状的照明器具 |
| EP2785377A4 (en) * | 2011-11-29 | 2015-07-08 | Baylor College Medicine | PROCESS FOR IMPROVING THE COGNITION |
| US20130211277A1 (en) | 2012-02-10 | 2013-08-15 | Oyvind Berg | System And Method For Reducing Stress Levels Through A Biofeedback Mediated System Using Color, Sound, And Aroma Therapy |
| CA2864213A1 (en) | 2012-02-17 | 2013-08-22 | Frank M. WANCA | Method, system and apparatus for integrated dynamic neural stimulation |
| US9272118B1 (en) | 2012-03-02 | 2016-03-01 | George Acton | Methods for treating brain malfunctions |
| US9155473B2 (en) | 2012-03-21 | 2015-10-13 | Korea Electrotechnology Research Institute | Reflection detection type measurement apparatus for skin autofluorescence |
| US8918176B2 (en) * | 2012-04-23 | 2014-12-23 | Medtronic, Inc. | Assessing cognitive disorders based on non-motor epileptiform bioelectrical brain activity |
| RU2488388C1 (ru) | 2012-05-24 | 2013-07-27 | Ооо "Валента Интеллект" | Фармацевтическая композиция для профилактики и лечения психических, поведенческих, когнитивных расстройств |
| US20150337030A1 (en) | 2012-05-31 | 2015-11-26 | The Trustees Of Columbia University In The City Of New York | Methods to treat alzheimer's disease using apoe inhibitors |
| US9095014B2 (en) | 2012-06-07 | 2015-07-28 | Wellsino Electric Co., Ltd | LED light string and control method thereof |
| EP2895970B1 (en) | 2012-09-14 | 2018-11-07 | InteraXon Inc. | Systems and methods for collecting, analyzing, and sharing bio-signal and non-bio-signal data |
| JP6046439B2 (ja) * | 2012-10-01 | 2016-12-14 | トヨタ自動車株式会社 | ブレイン・マシン・インターフェース |
| US10537703B2 (en) | 2012-11-26 | 2020-01-21 | Thync Global, Inc. | Systems and methods for transdermal electrical stimulation to improve sleep |
| US9629976B1 (en) | 2012-12-21 | 2017-04-25 | George Acton | Methods for independent entrainment of visual field zones |
| US20150351655A1 (en) | 2013-01-08 | 2015-12-10 | Interaxon Inc. | Adaptive brain training computer system and method |
| US10293160B2 (en) | 2013-01-15 | 2019-05-21 | Electrocore, Inc. | Mobile phone for treating a patient with dementia |
| PL2948040T3 (pl) | 2013-01-28 | 2023-09-25 | Lkc Technologies, Inc. | Urządzenie do elektrofizjologicznego badania wzroku |
| KR20160008162A (ko) | 2013-02-22 | 2016-01-21 | 틴크, 인크. | 개인 그룹의 신경조절을 네트워킹하기 위한 방법 및 장치 |
| US20140303025A1 (en) | 2013-03-15 | 2014-10-09 | The Translational Genomics Research Institute | Methods for the diagnosis and prognosis of neurodegenerative diseases |
| CA3187490A1 (en) | 2013-03-15 | 2014-09-18 | Interaxon Inc. | Wearable computing apparatus and method |
| US20140303424A1 (en) | 2013-03-15 | 2014-10-09 | Iain Glass | Methods and systems for diagnosis and treatment of neural diseases and disorders |
| JP2016515432A (ja) | 2013-04-01 | 2016-05-30 | モーション セラピューティックス インコーポレイテッド | 脳機能または感覚神経運動系機能と組み合わさって働く固有受容皮膚および身体センサー、視覚、前庭および身体感覚を仲介する方法および装置 |
| WO2014162271A2 (en) | 2013-04-03 | 2014-10-09 | Branislav Nikolic | Light therapy device and method |
| US9872968B2 (en) | 2013-04-17 | 2018-01-23 | Sri International | Biofeedback virtual reality sleep assistant |
| WO2014179331A2 (en) * | 2013-04-29 | 2014-11-06 | The Board Of Trustees Of The Leland Stanford Junior University | Devices, systems and methods for optogenetic modulation of action potentials in target cells |
| US20150002025A1 (en) | 2013-06-28 | 2015-01-01 | General Electric Company | Lighting assembly, apparatus and associated method for maintaining light intensities |
| EP3041413A4 (en) | 2013-09-04 | 2017-04-19 | Question Mark Computing Limited | System and method for data anomaly detection process in assessments |
| MX384984B (es) | 2013-10-10 | 2025-03-14 | Beth Israel Deaconess Medical Ct Inc | Proteinas de union del miembro 1 de la familia de transmembrana 4 l-seis (tm4sf1) y metodos de uso de las mismas. |
| EP3057654B1 (en) | 2013-10-18 | 2020-08-19 | Functional Neuromodulation Inc. | Brain stimulation system including diagnostic tool |
| US10625042B2 (en) * | 2013-11-04 | 2020-04-21 | Phoenix Neurostim Therapeutics, Llc | Treatment of central nervous system conditions using sensory stimulus |
| US10188342B2 (en) * | 2013-11-05 | 2019-01-29 | The University Of North Carolina At Chapel Hill | Methods, apparatuses and systems for transcranial stimulation |
| US11203783B2 (en) | 2013-11-21 | 2021-12-21 | Repertoire Genesis Incorporation | T cell receptor and B cell receptor repertoire analysis system, and use of same in treatment and diagnosis |
| US9511228B2 (en) | 2014-01-14 | 2016-12-06 | Cyberonics, Inc. | Implantable neurostimulator-implemented method for managing hypertension through renal denervation and vagus nerve stimulation |
| CN103854613B (zh) | 2014-02-14 | 2016-04-13 | 北京京东方显示技术有限公司 | 光源控制模块、背光模组及其驱动方法、显示装置 |
| WO2015149170A1 (en) | 2014-03-31 | 2015-10-08 | Functional Neuromodulation, Inc. | Systems and methods for determining a trajectory for a brain stimulation lead |
| CN103932701B (zh) * | 2014-04-13 | 2015-09-23 | 北京师范大学 | 一种基于皮层脑电高频Gamma神经振荡的个体化脑功能映射方法 |
| US9955907B2 (en) | 2014-04-23 | 2018-05-01 | Case Western Reserve University | Low frequency non-invasive sensorial stimulation for seizure control |
| WO2016023126A1 (en) | 2014-08-14 | 2016-02-18 | Functional Neuromodulation, Inc. | Brain stimulation system including multiple stimulation modes |
| US20160067087A1 (en) | 2014-09-09 | 2016-03-10 | LumiThera, Inc. | Wearable devices and methods for multi-wavelength photobiomodulation for ocular treatments |
| WO2016044227A1 (en) | 2014-09-15 | 2016-03-24 | Abvitro, Inc. | High-throughput nucleotide library sequencing |
| JP2016072521A (ja) | 2014-09-30 | 2016-05-09 | 富士フイルム株式会社 | バックライトユニット、液晶表示装置、および色度制御方法 |
| US11534608B2 (en) | 2015-01-04 | 2022-12-27 | Ist, Llc | Methods and apparatuses for transdermal stimulation of the outer ear |
| WO2016127183A1 (en) | 2015-02-06 | 2016-08-11 | Noothera Technologies, Llc | Systems and methods for directed energy therapeutics |
| KR102335562B1 (ko) | 2015-03-16 | 2021-12-03 | 매직 립, 인코포레이티드 | 건강 질환 진단과 치료를 위한 방법 및 시스템 |
| KR102460393B1 (ko) | 2015-04-30 | 2022-11-01 | 삼성전자주식회사 | 사운드 출력 기기, 전자 장치 및 그 제어 방법 |
| CN104783788A (zh) * | 2015-05-05 | 2015-07-22 | 刘冬冬 | 脑波诱导控制方法和脑波诱导设备 |
| ITUB20153295A1 (it) | 2015-08-31 | 2017-03-03 | Marposs Spa | Sistema e metodo di elaborazione e trasmissione dati |
| US10076632B2 (en) | 2015-09-11 | 2018-09-18 | International Business Machines Corporation | Sensory feedback system with active learning |
| US10197235B2 (en) | 2015-09-23 | 2019-02-05 | Jenesis International Inc. | Multiple light source artificial moving flame |
| CN105278387B (zh) | 2015-11-17 | 2021-05-18 | 冼光 | 具有改善睡眠及缓解痛症的手环装置 |
| KR102349466B1 (ko) | 2015-11-23 | 2022-01-10 | 사나 헬스, 인크. | 뇌에 자극을 제공하는 방법들 및 시스템들 |
| ES2943587T3 (es) | 2015-11-24 | 2023-06-14 | Massachusetts Inst Technology | Sistemas para prevenir, atenuar y/o tratar demencia |
| US20190143119A1 (en) | 2016-03-28 | 2019-05-16 | Duke University | Systems and Methods for Brain Stimulation |
| RO131383A0 (ro) | 2016-04-11 | 2016-09-30 | Bogdan Constantin Vlădilă | Echipament electromagnetic pentru creşterea elasticităţii ţesuturilor, folosit în cosmetică |
| TWI759301B (zh) | 2016-05-24 | 2022-04-01 | 美商安美基公司 | 聚乙二醇化卡非佐米化合物 |
| US10905897B2 (en) | 2016-10-20 | 2021-02-02 | Neuro-Laser Foundation, Inc. | Synergistic treatment methodologies using light therapy |
| KR102612531B1 (ko) | 2016-11-17 | 2023-12-12 | 코그니토 쎄라퓨틱스, 인코포레이티드 | 인지 기능 개선을 위한 시스템 |
| US20220008746A1 (en) | 2016-11-17 | 2022-01-13 | Cognito Therapeutics, Inc. | Methods and systems for neural stimulation via visual stimulation |
| WO2018134823A1 (en) | 2017-01-20 | 2018-07-26 | Oridion Medical 1987 Ltd. | Dual sensor |
| JP7458786B2 (ja) | 2017-02-15 | 2024-04-01 | ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア | 改善された光療法システムおよび使用方法 |
| US11433253B2 (en) | 2017-02-15 | 2022-09-06 | The Regents Of The University Of California | Light therapy system and methods of use |
| JP6837886B2 (ja) | 2017-03-21 | 2021-03-03 | 株式会社日立ハイテク | プラズマ処理装置およびプラズマ処理方法 |
| US10181241B2 (en) | 2017-04-04 | 2019-01-15 | Qualcomm Incorporated | Modulated warning lights for vehicles |
| US20200171267A1 (en) | 2017-05-17 | 2020-06-04 | Matthew D. Millard | Head worn device for treating neurodegenerative diseases |
| EP3675906A4 (en) | 2017-08-28 | 2021-05-26 | Angiex, Inc. | ANTI-TM4SF1 ANTIBODIES AND METHOD OF USING THEREOF |
| US20190076670A1 (en) | 2017-09-11 | 2019-03-14 | Alzheimer's Light Llc | Flashing Light Therapy with Image Presentation and Interaction for Treatment of Alzheimer's Disease and Dementia |
| ES3058718T3 (en) | 2017-10-10 | 2026-03-12 | Massachusetts Inst Technology | Systems for preventing, mitigating, and/or treating dementia |
| CN111655319A (zh) | 2017-10-10 | 2020-09-11 | 麻省理工学院 | 用视觉刺激同步脑中γ振荡治疗痴呆 |
| US10960225B2 (en) | 2017-10-10 | 2021-03-30 | Massachusetts Institute Of Technology | Systems and methods for preventing, mitigating, and/or treating dementia via visual stimulation that binds higher order brain regions, reduces neurodegeneration and neuroinflammation, and improves cognitive function |
| CA3086907A1 (en) | 2018-01-08 | 2019-07-11 | Casper Sleep Inc. | Interactive portable lighting system |
| CA3011873A1 (en) | 2018-02-19 | 2019-08-19 | Millennium Healthcare Technologies, Inc. | Dental lasing device system and method |
| CN108725462B (zh) | 2018-05-22 | 2024-04-30 | 中交四航局江门航通船业有限公司 | 一种同步移动巨型构件的滚轮台车集成系统及应用方法 |
| CA3103265A1 (en) | 2018-06-12 | 2019-12-19 | Angiex, Inc. | Antibody-oligonucleotide conjugates |
| US20190388020A1 (en) | 2018-06-20 | 2019-12-26 | NeuroPlus Inc. | System and Method for Treating and Preventing Cognitive Disorders |
| WO2020041502A1 (en) | 2018-08-21 | 2020-02-27 | The Regents Of The University Of California | Transcutaneous electrical and/or magnetic spinal stimulation for bladder or bowel control in subjects without cns injury |
| US10518063B1 (en) | 2018-11-07 | 2019-12-31 | Symbolics Inc. | Method for stimulating neural activity |
| US20210030998A1 (en) | 2019-08-04 | 2021-02-04 | Well Being Digital Limited | Apparatus for improving mental well being, and a method thereof |
| WO2021216957A1 (en) | 2020-04-23 | 2021-10-28 | Massachusetts Institute Of Technology | Systems, devices, and methods for gamma entrainment using sensory stimuli to alleviate cognitive deficits and/or neuroinflammation induced by chemotherapy agents |
| WO2021221879A1 (en) | 2020-05-01 | 2021-11-04 | Albert Einstein College Of Medicine | Compositions and methods for using transplanted microglia as a vehicle for widespread delivery of cells and other biologic agents to the brain |
| KR20240051882A (ko) | 2020-07-27 | 2024-04-22 | 코그니토 쎄라퓨틱스, 인코포레이티드 | 알츠하이머병 환자의 기능적 능력을 유지하고 수면 품질을 개선하는 감각 감마 자극 치료법 |
| US20220151864A1 (en) | 2020-11-19 | 2022-05-19 | Massachusetts Institute Of Technology | Systems, Devices, and Methods for Gamma Entrainment using Tactile Stimuli |
| EP4304479A4 (en) | 2021-03-09 | 2025-04-23 | Cognito Therapeutics, Inc. | METHODS AND SYSTEMS FOR SLOWING DOWN CEREBRAL ATROPHY |
| WO2023108170A2 (en) | 2021-12-10 | 2023-06-15 | Massachusetts Institute Of Technology | Systems, devices, and methods for enhancing the neuroprotective effects of non-invasive gamma stimulation with pharmacological agents |
-
2016
- 2016-11-23 ES ES16869248T patent/ES2943587T3/es active Active
- 2016-11-23 JP JP2018525754A patent/JP7204192B2/ja active Active
- 2016-11-23 KR KR1020227037169A patent/KR102603963B1/ko active Active
- 2016-11-23 KR KR1020237039330A patent/KR102929434B1/ko active Active
- 2016-11-23 KR KR1020187017689A patent/KR102460518B1/ko active Active
- 2016-11-23 US US15/360,637 patent/US10265497B2/en active Active
- 2016-11-23 EP EP23150106.5A patent/EP4218916B1/en active Active
- 2016-11-23 CN CN202210905503.8A patent/CN115569282A/zh active Pending
- 2016-11-23 DK DK16869248.1T patent/DK3383482T3/da active
- 2016-11-23 WO PCT/US2016/063536 patent/WO2017091698A1/en not_active Ceased
- 2016-11-23 CA CA3003183A patent/CA3003183A1/en active Pending
- 2016-11-23 IL IL295071A patent/IL295071A/en unknown
- 2016-11-23 EP EP16869248.1A patent/EP3383482B1/en active Active
- 2016-11-23 AU AU2016361503A patent/AU2016361503B2/en active Active
- 2016-11-23 CN CN201680075447.2A patent/CN108697889B/zh active Active
- 2016-11-23 IL IL283779A patent/IL283779B/en unknown
-
2017
- 2017-07-11 US US15/647,157 patent/US10159816B2/en active Active
-
2018
- 2018-05-22 IL IL259533A patent/IL259533B/en active IP Right Grant
-
2019
- 2019-04-04 US US16/375,393 patent/US10682490B2/en active Active
-
2020
- 2020-06-15 US US16/901,592 patent/US12296106B2/en active Active
- 2020-06-15 US US16/901,628 patent/US12318549B2/en active Active
-
2021
- 2021-08-10 AU AU2021215128A patent/AU2021215128B2/en active Active
- 2021-11-19 JP JP2021188541A patent/JP2022020829A/ja active Pending
-
2023
- 2023-05-22 JP JP2023083776A patent/JP2023101026A/ja active Pending
-
2024
- 2024-02-01 AU AU2024200621A patent/AU2024200621A1/en active Pending
-
2025
- 2025-04-07 US US19/172,282 patent/US20250288771A1/en active Pending
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2015519096A (ja) | 2012-04-06 | 2015-07-09 | ニューポート ブレイン リサーチ ラボラトリー インコーポレイテッド | 周波数特異的感覚刺激 |
Non-Patent Citations (1)
| Title |
|---|
| 田中慶太,ほか2名,事象関連コヒーレンス解析を用いた脳磁界聴性40Hz応答の検討,電気学会論文誌C(電子・情報・システム部門誌),2005年,第125巻,第6号,pp.898-903 |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7204192B2 (ja) | 認知症の予防、軽減、及び/または治療のためのシステムならびに方法 | |
| EP3694464B1 (en) | Systems for preventing, mitigating, and/or treating dementia | |
| Zhao et al. | Sleep restoration by optogenetic targeting of GABAergic neurons reprograms microglia and ameliorates pathological phenotypes in an Alzheimer’s disease model | |
| Tinston et al. | Forty‐hertz sensory entrainment impedes kindling epileptogenesis and reduces amyloid pathology in an Alzheimer disease mouse model | |
| KR20260035288A (ko) | 치매를 예방, 경감 및/또는 치료하기 위한 시스템 및 방법 | |
| HK40086450A (zh) | 用於预防、减轻和/或治疗痴呆的系统和方法 | |
| HK1261893B (en) | Systems and methods for preventing, mitigating, and/or treating dementia | |
| HK1261893A1 (en) | Systems and methods for preventing, mitigating, and/or treating dementia | |
| Hackett | The cholinergic system and pharmacological inhibition of MMP-9 in the McGill THY-1APP mouse model of Alzheimer's disease |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20191125 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20200730 |
|
| A871 | Explanation of circumstances concerning accelerated examination |
Free format text: JAPANESE INTERMEDIATE CODE: A871 Effective date: 20200730 |
|
| A975 | Report on accelerated examination |
Free format text: JAPANESE INTERMEDIATE CODE: A971005 Effective date: 20201021 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20201102 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20210120 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20210330 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210422 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20210719 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20211119 |
|
| C60 | Trial request (containing other claim documents, opposition documents) |
Free format text: JAPANESE INTERMEDIATE CODE: C60 Effective date: 20211119 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20211119 |
|
| C11 | Written invitation by the commissioner to file amendments |
Free format text: JAPANESE INTERMEDIATE CODE: C11 Effective date: 20211220 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20220121 |
|
| C21 | Notice of transfer of a case for reconsideration by examiners before appeal proceedings |
Free format text: JAPANESE INTERMEDIATE CODE: C21 Effective date: 20220124 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20220204 |
|
| C211 | Notice of termination of reconsideration by examiners before appeal proceedings |
Free format text: JAPANESE INTERMEDIATE CODE: C211 Effective date: 20220214 |
|
| C22 | Notice of designation (change) of administrative judge |
Free format text: JAPANESE INTERMEDIATE CODE: C22 Effective date: 20220411 |
|
| C13 | Notice of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: C13 Effective date: 20220530 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20220824 |
|
| C23 | Notice of termination of proceedings |
Free format text: JAPANESE INTERMEDIATE CODE: C23 Effective date: 20221027 |
|
| C03 | Trial/appeal decision taken |
Free format text: JAPANESE INTERMEDIATE CODE: C03 Effective date: 20221121 |
|
| C30A | Notification sent |
Free format text: JAPANESE INTERMEDIATE CODE: C3012 Effective date: 20221121 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20221221 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7204192 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| RD02 | Notification of acceptance of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: R3D02 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: R3D04 |