JP5599745B2 - ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 - Google Patents
ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 Download PDFInfo
- Publication number
- JP5599745B2 JP5599745B2 JP2011044525A JP2011044525A JP5599745B2 JP 5599745 B2 JP5599745 B2 JP 5599745B2 JP 2011044525 A JP2011044525 A JP 2011044525A JP 2011044525 A JP2011044525 A JP 2011044525A JP 5599745 B2 JP5599745 B2 JP 5599745B2
- Authority
- JP
- Japan
- Prior art keywords
- nucleic acid
- dna
- stranded
- sequence
- oligonucleotide
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
Images





Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1093—General methods of preparing gene libraries, not provided for in other subgroups
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1037—Screening libraries presented on the surface of microorganisms, e.g. phage display, E. coli display
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/66—General methods for inserting a gene into a vector to form a recombinant vector using cleavage and ligation; Use of non-functional linkers or adaptors, e.g. linkers containing the sequence for a restriction endonuclease
-
- C—CHEMISTRY; METALLURGY
- C40—COMBINATORIAL TECHNOLOGY
- C40B—COMBINATORIAL CHEMISTRY; LIBRARIES, e.g. CHEMICAL LIBRARIES
- C40B40/00—Libraries per se, e.g. arrays, mixtures
- C40B40/02—Libraries contained in or displayed by microorganisms, e.g. bacteria or animal cells; Libraries contained in or displayed by vectors, e.g. plasmids; Libraries containing only microorganisms or vectors
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Biotechnology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- General Health & Medical Sciences (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Crystallography & Structural Chemistry (AREA)
- Virology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Bioinformatics & Computational Biology (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Enzymes And Modification Thereof (AREA)
Description
ンバーを提示し、そしてこのファミリーの多様性の少なくとも一部を集合的に提
示する、遺伝的パッケージのライブラリーを構築することに関する。好ましい実
施形態では、提示されるポリペプチドはヒトFabである。
、この切断された核酸は、本発明のライブラリーの遺伝的パッケージ上に提示さ
れたペプチド、ポリペプチドまたはタンパク質を少なくとも部分的にコードする
。好ましい実施形態では、この遺伝的パッケージは、糸状ファージまたはファー
ジミドである。
遺伝的パッケージのライブラリーをスクリーニングする方法、ならびにこのよう
なスクリーニングによって同定されたペプチド、ポリペプチドおよびタンパク質
に関する。
ペプチド、ポリペプチドまたはタンパク質の多様なファミリーのメンバーを提
示し、そしてこのファミリーの多様性の少なくとも一部を集合的に提示する、遺
伝的パッケージのライブラリーを調製することは現在、当該分野において一般的
な方法である。多くの通常のライブラリーでは、提示されるペプチド、ポリペプ
チドまたはタンパク質は、抗体に関連する。しばしば、これらはFabまたは単
鎖抗体である。
ニングされて遺伝的パッケージの表面に所望のメンバーを提示するように使用さ
れる前に増幅されなければならない。このような増幅は代表的に、正方向プライ
マーおよび逆方向プライマーを利用する。
対して相補的であり得るか、またはそのDNAの5’末端もしくは3’末端に結
合したオリゴヌクレオチドに対して相補的であり得る。増幅されるべきDNAに
対してネイティブである配列に相補的であるプライマーは、提示されるべきファ
ミリーのメンバーを偏らせるという点で不利である。ネイティブなDNA中にこ
のプライマーに対して実質的に相補的である配列を含むメンバーのみが増幅され
る。そうでないメンバーはこのファミリーから存在しなくなる。増幅されるメン
バーについては、プライマー領域内の何らかの多様性が抑制される。
て体細胞変異を含むプロセスを通して生じることが一般に受け入れられている。
大部分の多様性は相補性決定領域(CDR)に生じるが、多様性はまた、より保
存されたフレームワーク領域(FR)において生じ、そしてこの多様性の少なく
ともいくつかは、抗原(Ag)に対する特異的結合を付与または増強する。その
結果、ライブラリーは、可能な限り多くのCDRおよびFR多様性を含むべきで
ある。
の提示のために、増幅されたDNAをクローニングするために、DNAは、ベク
ターへの連結のために適切な末端を生じるように切断されなければならない。こ
のような切断は一般に、プライマーに保有された制限エンドヌクレアーゼ認識部
位を用いてもたらされる。RNAの逆転写から生成されるDNAの5’末端にプ
ライマーが存在する場合、このような制限処理は、増幅されたDNA中に有害な
5’非翻訳領域を残す。これらの領域は、クローニングされた遺伝子の発現を妨
害し、従って、これらによってコードされるペプチド、ポリペプチドおよびタン
パク質の提示を妨害する。
本発明の目的は、ペプチド、ポリペプチドまたはタンパク質の多様なファミリ
ーのメンバーを提示し、そしてこのファミリーの多様性の少なくとも一部を集合
的に提示する、遺伝的パッケージのライブラリーを構築するための新規方法を提
供することである。これらの方法は、増幅のために用いられるプライマーに相補
的であるネイティブな配列を含むDNAには偏っていない。これらの方法はまた
、発現に有害であり得る任意の配列が、クローニングおよび提示の前に、増幅さ
れたDNAから除去されることを可能にする。
供することであり、この方法は、以下の工程を包含する:
(i)この核酸を一本鎖オリゴヌクレオチドと接触させる工程であって、この
オリゴヌクレオチドは、切断が所望される領域においてこの核酸に機能的に相補
的であり、そしてその核酸中の相補体とともに、制限処理の際に所望の位置での
核酸の切断をもたらす制限エンドヌクレアーゼ認識部位を形成する配列を含む、
工程;および
(ii)この核酸のみを、この核酸とこのオリゴヌクレオチドとの相補によっ
て形成された認識部位で切断する工程;
接触工程および切断工程は、実質的に一本鎖形態の核酸を維持するに充分な温度
で行われ、このオリゴヌクレオチドは、二本の鎖が会合し、その結果、選択され
た温度および所望の位置で切断が生じ得るのを可能にするに充分に大きな領域に
わたってこの核酸に対して機能的に相補的であり、そして切断は、選択された温
度で活性である制限エンドヌクレアーゼを用いて行われる。
的な方法を提供することであり、この方法は以下の工程を包含する:
(i)この核酸を部分的に二本鎖のオリゴヌクレオチドと接触させる工程であ
って、このオリゴヌクレオチドの一本鎖領域は、切断が所望される領域において
この核酸に機能的に相補的であり、そしてこのオリゴヌクレオチドの二本鎖領域
は、II−S型制限エンドヌクレアーゼ認識部位を有し、その切断部位は、この
認識部位から既知の距離に位置する、工程;および
(ii)この核酸のみを、この核酸とこのオリゴヌクレオチドの一本鎖領域と
の相補によって形成された切断部位で切断する工程;
接触工程および切断工程は、実質的に一本鎖形態の核酸を維持するに充分な温度
で行われ、このオリゴヌクレオチドは、二本の鎖が会合し、その結果、選択され
た温度および所望の位置で切断が生じ得るのを可能にするに充分に大きな領域に
わたってこの核酸に対して機能的に相補的であり、そして切断は、選択された温
度で活性である制限エンドヌクレアーゼを用いて行われる。
のファミリーの多様性の少なくとも一部を集合的に含む、DNA分子を捕獲する
方法を提供することである。一本鎖形態のこれらのDNA分子は、本発明の方法
のうちの1つによって切断されている。この方法は、このファミリーの個々の一
本鎖DNAメンバーを、部分的に二重鎖のDNA複合体に連結することを含む。
この方法は以下の工程を包含する:
(i)制限エンドヌクレアーゼで切断された一本鎖核酸配列を、部分的に二本
鎖のオリゴヌクレオチドと接触させる工程であって、このオリゴヌクレオチドの
一本鎖領域は、切断後に残る領域においてこの核酸に対して機能的に相補的であ
り、このオリゴヌクレオチドの二本鎖領域は、切断後に残っている配列を、発現
に適切なリーディングフレームに戻すために必要な任意の配列を含み、そしてこ
れらの配列の5’側に制限エンドヌクレアーゼ認識部位を含む、工程;および
(ii)この部分的に二本鎖のオリゴヌクレオチド配列のみを、この部分的に
二本鎖のオリゴヌクレオチドの二本鎖領域内に含まれる制限エンドヌクレアーゼ
認識部位で切断する工程。
ミリーを提示し、そしてこのファミリーの多様性の少なくとも一部を集合的に提
示する、ライブラリーを、上記の方法およびDNAを用いて調製することである
。
ド、ポリペプチドおよびタンパク質を同定し、そしてこれらの物質をヒトの治療
において使用することである。たとえば、本発明は以下を提供する:
(項目1) 一本鎖核酸配列を所望の位置で切断するための方法であって、
該方法は、以下の工程:
(i)該核酸を、一本鎖オリゴヌクレオチドと接触させる工程であって、該オ
リゴヌクレオチドは、切断が所望される領域において該核酸に対して機能的に相
補的であり、そして制限の際に該核酸の該所望の位置での切断を生じる制限エン
ドヌクレアーゼ認識部位を、その補体を用いて該核酸中に形成する配列を含む、
工程;および
(ii)該核酸および該オリゴヌクレオチドの相補性によって形成される該認
識部位で、該核酸を単独で切断する工程;
を包含し、該接触工程および該切断工程は、該核酸を実質的に一本鎖形態で維持
するために十分な温度で行われ、該オリゴヌクレオチドは、切断が選択された該
温度および該所望の位置で起こり得るように、該二本の鎖の会合を可能にするた
めに十分大きな領域にわたって該核酸と機能的に相補的であり、そして該選択さ
れた温度で活性である制限エンドヌクレアーゼを使用して、該切断が行われる、
方法。
(項目2) 一本鎖核酸配列を所望の位置で切断するための方法であって、
該方法は、以下の工程:
(i)該核酸を、部分的に二本鎖のオリゴヌクレオチドと接触させる工程であ
って、該オリゴヌクレオチドの一本鎖領域は、切断が所望される該領域において
該核酸に対して機能的に相補的であり、そして該オリゴヌクレオチドの二本鎖領
域は、II型−S制限エンドヌクレアーゼ認識部位を有し、該切断部位は、該認
識部位から既知の距離に位置する、工程;および
(ii)該核酸および該オリゴヌクレオチドの該一本鎖領域の相補性によって
形成されるII型−S切断部位で、該核酸を単独で切断する工程;
を包含し、該接触工程および該切断工程は、該核酸を実質的に一本鎖形態で維持
するために十分な温度で行われ、該オリゴヌクレオチドは、切断が選択された該
温度および該所望の位置で起こり得るように、該二本の鎖の会合を可能にするた
めに十分大きな領域にわたって該核酸と機能的に相補的であり、そして該選択さ
れた温度で活性である制限エンドヌクレアーゼを使用して、該切断が行われる、
方法。
(項目3) 遺伝的パッケージの表面上のペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示し、そして集合的に、該ファミリーの多様性の少なくとも一部を提示する方法であって、改善が、提示されるペプチド、ポリペプチド、またはタンパク質の少なくとも一部が、
以下の工程:
(i)核酸を、一本鎖オリゴヌクレオチドと接触させる工程であって、該オリ
ゴヌクレオチドは、切断が所望される領域において該核酸に対して機能的に相補
的であり、そして制限の際に該核酸の所望の位置での切断を生じる制限エンドヌ
クレアーゼ認識部位を、その補体を用いて該核酸中に形成する配列を含む、工程
;および
(ii)該核酸および該オリゴヌクレオチドの相補性によって形成される該認
識部位で、該核酸を単独で切断する工程;
を包含する方法によって所望の位置で切断される該核酸によって、少なくとも一
部がコードされることを特徴とし、該接触工程および該切断工程は、該核酸を実
質的に一本鎖形態で維持するために十分な温度で行われ、該オリゴヌクレオチド
は、切断が選択された該温度および該所望の位置で起こり得るように、該二本の
鎖の会合を可能にするために十分大きな領域にわたって該核酸と機能的に相補的
であり、そして該選択された温度で活性である制限エンドヌクレアーゼを使用し
て、該切断が行われる、方法。
(項目4) 遺伝的パッケージの表面上のペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示し、そして集合的に、該ファミリーの多様性の少なくとも一部を提示する方法であって、改善が、提示されるペプチド、ポリペプチド、またはタンパク質が、以下の工程:
(i)該核酸を、部分的に二本鎖のオリゴヌクレオチドと接触させる工程であ
って、該オリゴヌクレオチドの一本鎖領域は、切断が所望される領域において該
核酸に対して機能的に相補的であり、該オリゴヌクレオチドの該二本鎖領域は、
II型−S制限エンドヌクレアーゼ認識部位を有し、該切断部位は、該認識部位
から既知の距離に位置する、工程;および
(ii)該核酸および該オリゴヌクレオチドの該一本鎖領域の相補性によって
形成されるII型−S切断部位で、該核酸を単独で切断する工程;
によって所望の位置で切断される核酸を含むDNA配列によってコードされるこ
とを特徴とし、該接触工程および該切断工程は、該核酸を実質的に一本鎖形態で
維持するために十分な温度で行われ、該オリゴヌクレオチドは、切断が選択され
た該温度および該所望の位置で起こり得るように、該二本の鎖の会合を可能にす
るために十分大きな領域にわたって該核酸と機能的に相補的であり、そして該選
択された温度で活性である制限エンドヌクレアーゼを使用して、該切断が行われ
る、方法。
(項目5) 遺伝的パッケージの表面上のペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示し、そして集合的に、該ファミリーの多様性の少なくとも一部を提示する方法であって、
該方法は、以下の工程:
(i)該多種多様なファミリーのメンバーの少なくとも一部をコードする核酸
の収集物を調製する工程;
(ii)該核酸を一本鎖にする工程;
(iii)以下の工程:
(a)該核酸を、一本鎖オリゴヌクレオチドと接触させる工程であって、該
オリゴヌクレオチドは、切断が所望される領域において該核酸に対して機能的に
相補的であり、そして制限の際に該核酸の所望の位置での切断を生じる制限エン
ドヌクレアーゼ認識部位を、その補体を用いて該核酸中に形成する配列を含む、
工程;および
(b)該核酸および該オリゴヌクレオチドの相補性によって形成される該認
識部位で、該核酸を単独で切断する工程;
を包含する方法によって、所望の位置で該一本鎖核酸を切断する工程であって
、該接触工程および該切断工程は、該核酸を実質的に一本鎖形態で維持するため
に十分な温度で行われ、該オリゴヌクレオチドは、切断が選択された該温度およ
び該所望の位置で起こり得るように、該二本の鎖の会合を可能にするために十分
大きな領域にわたって該核酸と機能的に相補的であり、そして該選択された温度
で活性である制限エンドヌクレアーゼを使用して、該切断が行われる、工程;な
らびに
(iv)該遺伝的パッケージの該表面上の該切断された核酸によって少なくと
も一部がコードされるペプチド、ポリペプチドまたはタンパク質の該ファミリー
のメンバーを提示し、そして集合的に、該ファミリーの該多様性の少なくとも一
部を提示する工程、
を包含する、方法。
(項目6) 遺伝的パッケージの表面上のペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示し、そして集合的に、該ファミリーの多様性の少なくとも一部を提示する方法であって、
該方法は、以下の工程:
(i)該多種多様なファミリーのメンバーの少なくとも一部をコードする核酸
の収集物を調製する工程;
(ii)該核酸を一本鎖にする工程;
(iii)以下の工程:
(a)該核酸を、部分的に二本鎖のオリゴヌクレオチドと接触させる工程で
あって、該オリゴヌクレオチドの一本鎖領域は、切断が所望される領域において
該核酸に対して機能的に相補的であり、該オリゴヌクレオチドの該二本鎖領域は
、II型−S制限エンドヌクレアーゼ認識部位を有し、該切断部位は、該認識部
位から既知の距離に位置する、工程;および
(b)該核酸および該オリゴヌクレオチドの該一本鎖領域の相補性によって
形成されるII型−S切断部位で、該核酸を単独で切断する工程;
を包含する方法によって、所望の位置で該一本鎖核酸を切断する工程であって
、該接触工程および該切断工程は、該核酸を実質的に一本鎖形態で維持するため
に十分な温度で行われ、該オリゴヌクレオチドは、切断が選択された該温度およ
び該所望の位置で起こり得るように、該二本の鎖の会合を可能にするために十分
大きな領域にわたって該核酸と機能的に相補的であり、そして該選択された温度
で活性である切断エンドヌクレアーゼを使用して、該制限が行われる、工程;な
らびに;
(iv)該遺伝的パッケージの該表面上の該切断された核酸によって少なくと
も一部がコードされるペプチド、ポリペプチドまたはタンパク質の該ファミリー
のメンバーを提示し、そして集合的に、該ファミリーの多様性の少なくとも一部
を提示する工程、
を包含する、方法。
(項目7) ペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示する遺伝的パッケージの収集物を含に、そして集合的に、該ファミリーの多様性の少なくとも一部を提示するライブラリーであって、該ライブラリーは、項目3、4、5または6に記載の方法を使用して作製される、ライブラリー。
(項目8) ペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示し、そして集合的に、該ファミリーの少なくとも一部を提示する遺伝的パッケージの収集物を含むライブラリーであって、該提示されるペプチド、ポリペプチドまたはタンパク質は、以下の工程:
(i)該核酸を、一本鎖オリゴヌクレオチドと接触させる工程であって、該オ
リゴヌクレオチドは、切断が所望される領域において該核酸に対して機能的に相
補的であり、そして制限の際に該核酸の所望の位置での切断を生じる制限エンド
ヌクレアーゼ認識部位を、その補体を用いて該核酸中に形成する配列を含む、工
程;および
(ii)該核酸および該オリゴヌクレオチドの相補性によって形成される該認
識部位で、該核酸を単独で切断する工程;
を包含する方法によって、所望の位置で一本鎖核酸配列を切断することによって
生成される配列の少なくとも一部を含むDNA配列によってコードされ、該接触
工程および該切断工程は、該核酸を実質的に一本鎖形態で維持するために十分な
温度で行われ、該オリゴヌクレオチドは、切断が選択された該温度および該所望
の位置で起こり得るように、該二本の鎖の会合を可能にするために十分大きな領
域にわたって該核酸と機能的に相補的であり、そして該選択された温度で活性で
ある制限エンドヌクレアーゼを使用して、該切断が行われる、ライブラリー。
(項目9) ペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示し、そして集合的に、該提示されるペプチド、ポリペプチドまたはタンパク質の該ファミリーの多様性の少なくとも一部を提示する遺伝的パッケージの収集物を含むライブラリーであって、該提示されるペプチド、ポリペプチドまたはタンパク質は、以下の工程:
(i)該核酸を、部分的に二本鎖のオリゴヌクレオチドと接触させる工程であ
って、該オリゴヌクレオチドの一本鎖領域は、切断が所望される領域において該
核酸に対して機能的に相補的であり、該オリゴヌクレオチドの該二本鎖領域は、
II型−S制限エンドヌクレアーゼ認識部位を有し、該切断部位は、該核酸の該
切断が所望される該認識部位から既知の距離に位置する、工程;および
(ii)該核酸および該オリゴヌクレオチドの該一本鎖領域の相補性によって
形成されるII型−S切断部位で、該核酸を単独で切断する工程;
を包含する方法によって、所望の位置で一本鎖核酸配列を切断することによって
生成される配列の少なくとも一部を含むDNA配列によってコードされ、該接触
工程および該切断工程は、該核酸を実質的に一本鎖形態で維持するために十分な
温度で行われ、該オリゴヌクレオチドは、切断が選択された該温度および該所望
の位置で起こり得るように、該二本の鎖の会合を可能にするために十分大きな領
域にわたって該核酸と機能的に相補的であり、そして該選択された温度で活性で
ある制限エンドヌクレアーゼを使用して、該切断が行われる、ライブラリー。
(項目10) 前記核酸が、免疫グロブリンの少なくとも一部をコードする、項目1〜9のいずれか1項に記載の方法。
(項目11) 前記免疫グロブリンが、Fabまたは単鎖Fvを含む、請求項10に記載の方法。
(項目12) 前記免疫グロブリンが、重鎖の少なくとも部分を含む、請求項10または11に記載の方法。
(項目13) 前記重鎖の少なくとも一部がヒトである、項目12に記載の方法。
(項目14) 前記免疫グロブリンが、FR1の少なくとも一部を含む、
項目10または11に記載の方法。
(項目15) 前記FR1の少なくとも一部がヒトである、項目14に記載の方法。
(項目16) 前記免疫グロブリンが、軽鎖の少なくとも一部を含む、請求項10または11に記載の方法。
(項目17) 前記軽鎖の少なくとも一部がヒトである、項目16に記載の方法。
(項目18) 前記核酸配列の少なくとも一部が、少なくとも1つの自己免疫疾患および/または癌に罹患した患者由来である、項目1〜9のいずれか1項に記載の方法。
(項目19) 前記自己免疫疾患が、狼瘡、エリテマトーデス、全身性硬化症、慢性関節リウマチ、抗リン脂質症候群、または脈管炎からなる群から選択される、項目18に記載の方法。
(項目20) 前記核酸の少なくとも一部が、末梢血球、骨髄細胞、脾臓細胞またはリンパ節細胞からなる群から単離される、項目18に記載の方法。
(項目21) さらに、核酸の増幅工程を、工程(i)と工程(ii)との間、工程(ii)と工程(iii)との間、または工程(iii)と工程(iv)との間に包含する、項目5または6に記載の方法。
(項目22) 前記増幅工程が、geneRACETMを使用する、請求項21に記載の方法。
(項目23) 前記温度が、45℃と75℃との間である、項目1〜9のいずれか1項に記載の方法。
(項目24) 前記温度が、50℃と60℃との間である、項目23に記載の方法。
(項目25) 前記温度が、55℃と60℃との間である、項目24に記載の方法。
(項目26) 前記一本鎖オリゴヌクレオチドの長さが、17塩基と30塩基との間である、項目1、3、5または8に記載の方法。
(項目27) 前記一本鎖オリゴヌクレオチドの長さが、18塩基と24塩基との間である、項目26に記載の方法。
(項目28) 前記制限エンドヌクレアーゼが、MaeIII、Tsp451、HphI、BsaJI、AluI、BlpI、DdeI、BglII、MslI、BsiEI、EaeI、EagI、HaeIII、Bst4CI、HpyCH4III、HinfI、MlyI、PleI、MnlI、HpyCH4V、BsmAI、BpmI、XmnI、またはSacIからなる群から選択される、
項目1、3、5または8に記載の方法。
(項目29) 前記制限エンドヌクレアーゼが、Bst4CI、TaaI、HpyCH4III、BlpI、HpyCH4VまたはMslIを含む群から選択される、項目28に記載の方法。
(項目30) 前記部分的に二本鎖のオリゴヌクレオチドの前記一本鎖領域の長さが、14塩基と22塩基との間である、項目2、4、6または9に記載の方法。
(項目31) 前記部分的に二本鎖のオリゴヌクレオチドの前記一本鎖領域の長さが、14塩基と17塩基との間である、項目30に記載の方法。
(項目32) 前記オリゴヌクレオチドの前記一本鎖領域の長さが、18塩基と20塩基との間である、項目31に記載の方法。
(項目33) 前記部分的に二本鎖のオリゴヌクレオチドの前記二本鎖領域の長さが、幹およびそのパリンドロームによって形成される10塩基対と14塩基対との間である、項目2、4、6または9に記載の方法。
(項目34) 前記部分的に二本鎖のオリゴヌクレオチドが、前記幹と前記パリンドロームとの間に、3〜8塩基のループを含む、項目33に記載の方法。
(項目35) 前記II型−S制限エンドヌクレアーゼが、AarICAC、AceIII、Bbr7I、BbvI、BbvII、Bce831、BceAI、BcefI、BciVI、BfiI、BinI、BscAI、BseRI、BsmFI、BspMI、EciI、Eco57I、FauI、FokI、GsuI、HgaI、HphI、MboII、MlyI、MmeI、MnlI、PleI、RleAI、SfaNI、SspD5I、Sthl32I、StsI、TaqII、Tth111II、またはUbaPIを含む群から選択される、請求項2、4、6または9に記載の方法。
(項目36) 前記II型−S制限エンドヌクレアーゼが、FokIである、項目35に記載の方法。
(項目37) ベクターにクローニングするための一本鎖核酸を調製するための方法であって、該方法は、以下の工程:
(i)制限エンドヌクレアーゼで切断された一本鎖核酸配列を、部分的に二本
鎖のオリゴヌクレオチドと接触させる工程であって、該オリゴヌクレオチドの一
本鎖領域は、切断の後に残る領域において該核酸と機能的に相補的であり、該オ
リゴヌクレオチドの該二本鎖領域は、発現のために切断後に残る配列を適切かつ
本来のリーディングフレームに戻すのに必要とされる任意の配列を含み、そして
これらの配列の制限エンドヌクレアーゼ認識部位5’を含む、工程;および
(ii)該部分的に二本鎖のオリゴヌクレオチドの該二本鎖領域内に含まれる
該制限エンドヌクレアーゼ認識部位で、単独で該部分的に二本鎖のオリゴヌクレ
オチド配列を切断する工程、
を包含する、方法。
(項目38) 前記部分的に二本鎖のオリゴヌクレオチドの前記一本鎖部分の長さが、2塩基と15塩基との間である、項目37に記載の方法。
(項目39) 前記部分的に二本鎖のオリゴヌクレオチドの前記一本鎖部分の長さが、7塩基と10塩基との間である、項目38に記載の方法。
(項目40) 前記部分的に二本鎖のオリゴヌクレオチドの前記二本鎖部分の長さが、12塩基対と100塩基対との間である、項目37に記載の方法。
(項目41) 前記部分的に二本鎖のオリゴヌクレオチドの前記二本鎖部分の長さが、20塩基対と100塩基対との間である、項目40に記載の方法。
この出願では、以下の用語および略号を用いる:
センス鎖:通常記載されるように、ds DNAの上の鎖(upper st
rand)。センス鎖では、5’−ATG−3’はMetをコードする。
strand)。アンチセンス鎖では、3’−TAC−5’は、センス鎖にお
けるMetコドンに対応する。
一部に対して相補的であり、そして新たなアンチセンス鎖分子の合成を開始する
。「正方向プライマー」および「下の鎖のプライマー」は等価である。
ンス鎖の一部に対して相補的であり、そして新たなセンス鎖分子の合成を開始す
る。「逆方向プライマー」および「上の鎖のプライマー」は等価である。
ターまたは遺伝子のいずれかにおけるそれらの位置によって特定される。例えば
、「89.1」は、コドン89の最初の塩基であり、89.2は、コドン89の
二番目の塩基である。
に相補的である。
に相同である結合ドメインを有する任意のタンパク質をカバーする。この定義内
の抗体のいくつかの例は、特に、免疫グロブリンンアイソタイプならびにFab
、F(ab1)2、scfv、Fv、dAbおよびFdフラグメント。
を含む単鎖Ab。
本発明の方法において有用である核酸配列(すなわち、本発明の遺伝子パッケ
ージに関して示される、少なくとも一部の個々のペプチド、ポリペプチドおよび
タンパク質)は、天然に存在する配列、合成配列またはその組み合わせであり得
る。これらは、mRNA、DNAまたはcDNAであり得る。好ましい実施形態
において、核酸は抗体をコードする。最も好ましくは、これらはFabをコード
する。
天然に多様性のメンバーに導入され得るか、またはこの多様性は、全体的に合成
であり得る。例えば、合成多様性は、抗体遺伝子の1つ以上のCDRに導入され
得る。
使用を介して作製され得る。TRIM技術は、変化される位置で、どのアミノ酸
型が可能で、そしてどんな割合でかを正確に制御することが可能である。TRI
M技術において、多様化されるコドンは、三ヌクレオチドの混合物を使用して合
成される。これは、任意の組のアミノ酸型が、任意の位置に含まれるのを可能に
する。
ヌクレオチド合成である。TRIM技術を用いて、AlaおよびTrpが可能で
あり得る。混合オリゴヌクレオチド合成を用いて、AlaおよびTrpを含む混
合物はまた、SerおよびGlyもまた必然的に含む。変性された位置において
可能なアミノ酸型は、抗体の構造、または他のペプチド、ファミリーのポリペプ
チドまたはタンパク質、生殖系列遺伝子において観察された多様性、頻繁に観察
される観察された体細胞変異、および変性の所望の領域および型を参照して選択
される。
たはタンパク質の少なくとも1つのCDRまたは他の領域の核酸配列は、mRN
Aからの逆転写によって生成されたcDNAである。より好ましくは、mRNA
は、関連遺伝子の天然に多様性の組のメンバーを発現する末梢血液細胞、骨髄細
胞、脾臓細胞またはリンパ節細胞(例えば、Bリンパ球細胞またはプラズマ細胞
)から得られ得る。より好ましくは、mRNAは、抗体の多様なファミリーをコ
ードする。最も好ましくは、mRNAは、少なくとも1つの自己免疫障害または
癌に罹患する患者から得られる。好ましくは、自己免疫疾患(例えば、全身性エ
リテマトーデス、全身性硬化症、慢性関節リウマチ、抗リン脂質症候群および脈
管炎)の高い多様性を含むmRNAが使用される。
から生成される。この好ましい実施形態において、mRNAは、細胞から分離さ
れ、標準的な方法を使用して分解され、その結果、全長(すなわち、キャップさ
れた)mRNAのみが残る。次いで、キャップは除去され、そしてcDNAを生
成するために逆転写が使用される。
様式で行なわれ得る。例えば、HJ de Haardら、Journal o
f Biological Chemistry,274(26):18218
−30(1999)を参照のこと。mRNAが抗体をコードする本発明の好まし
い実施形態において、抗体遺伝子の定常領域に相補的なプライマーが使用され得
る。これらのプライマーは有用である。なぜなら、これらは、抗体のサブクラス
に対して偏りを生じないためである。別の実施例において、ポリdTプライマー
が使用され得る(そして重鎖遺伝子に対して好ましくあり得る)。あるいは、プ
ライマーに相補的な配列は、アンチセンス鎖の末端に付加され得る。
プライマーであり得、従って、cDNA産物がストレプトアビジン(Sv)ビー
ズに固定されるのを可能にする。固定化はまた、a)遊離アミノ基、b)チオー
ル、c)カルボン酸、またはd)不溶性媒体の既知のパートナーに対して強力な
結合を形成するように反応し得るDNAにおいて見出されない別の基、のうちの
1つを用いて5’末端において標識されたプライマーを使用して行なわれ得る。
例えば、遊離アミン(好ましくは、主にアミン)が、DNAプライマーの5’末
端において提供される場合、このアミンは、標準的なアミド形成化学を使用して
、ポリマービーズ上のカルボン酸と反応され得る。このような好ましい固定が、
逆転写の間に使用される場合、上方鎖RNAは、固定の前または後に、周知の酵
素(例えば、RNAseHおよびRNAseAの組み合わせ)を使用して分解さ
れる。
リペプチド、またはタンパク質を表示するために使用される前に増幅される。増
幅の前に、一本鎖DNAは、前に記載された方法のいずれかを使用して切断され
得る。あるいは、一本鎖DNAは増幅され得、次いで、これらの方法のうちの1
つを使用して切断される。
使用され得る。多様性を最大化し、偏らせない方法が好ましい。核酸配列が、抗
体遺伝子に由来する本発明の好ましい実施形態において、本発明は、好ましくは
、重鎖および軽鎖遺伝子の定常領域中のプライマーならびにセンス鎖の5’末端
に付加された合成配列に対するプライマーを利用する。このような合成配列にお
けるプライミングは、抗体遺伝子の可変領域内の配列の使用を回避する。これら
の可変領域プライミング部位は、まれなサブクラスのいずれかであるか、または
プライミング部位において変異されたV遺伝子に対する偏りを生じる。この偏り
は、プライマー領域内の多様性の抑制に部分的に起因し、そして多数の変異がプ
ライマーに相補的な領域中に存在する場合のプライミングの欠如に部分的に起因
する。本発明において開示された方法は、特定のV遺伝子型についての増幅され
た抗体遺伝子の集団を偏らせない利点を有する。
NA鎖の5’末端に付加され得る。RTキャップ伸長CapExtention
は、1つの好ましい方法である。
重複(上方鎖プライマー(USP−GGG)における5’−...GGG−3’
は、下方鎖における3’−CCC....5’を補完する)および逆転写酵素を
使用して、上方鎖プライマーの逆相補体が、下方鎖に付加される。
a)遊離アミノ基、b)チオール、c)カルボン酸、またはd)不溶性媒体の既
知のパートナーに対して強力な結合を形成するように反応し得るDNAにおいて
見出されない別の基、のうちの1つを用いて、5’末端においてビオチン化また
は標識され得る。次いで、これらは、増幅後の標識された鎖を固定するために使
用され得る。固定されたDNAは、一本鎖または二本鎖のいずれかであり得る。
の3’UTRプライミング合成のポリdT領域に特異的なプライマーを示す。定
常領域において結合するプライマーもまた、適切である。パネルBは、mRNA
に相補的でない3つのCによって、その3’末端において伸長した下方鎖を示す
。パネルCは、3’末端のCCCにハイブリダイズする3つのGGGにおける合
成上方鎖プライマーのアニーリング、および合成プライマー配列の逆相補体によ
る下方鎖を伸長する逆転写の伸長の結果を示す。パネルDは、パネルCの合成プ
ライマーの5’末端を複製する5’ビオチン化合成上方鎖プライマーおよび定常
ドメインの部分に相補的な下方鎖プライマーを使用するPCR増幅の結果を示す
。パネルEは、5’ビオチン化上方鎖プライマーを使用することによって得られ
た固定二本鎖(ds)cDNAを示す。
第1の下方鎖の3’末端プライミング合成の領域またはその近くの定常領域に特
異的なプライマーを示す。ポリdT領域において結合するプライマーはまた、適
切である。パネルBは、mRNAに相補的でない3つのCによって、その3’末
端において伸長した下方鎖を示す。パネルCは、3’末端のCCCにハイブリダ
イズする3つのGGGにおける合成上方鎖プライマーのアニーリング、および合
成プライマー配列の逆相補体による下方鎖を伸長する逆転写の伸長の結果を示す
。パネルDは、パネルCの合成プライマーの5’末端を複製する5’ビオチン化
合成上方鎖プライマーおよび定常ドメインの部分に相補的な下方鎖プライマーを
使用するPCR増幅の結果を示す。下方鎖プライマーはまた、有用な制限エンド
ヌクレアーゼ部位(例えば、AscI)を含む。パネルEは、5’ビオチン化上
方鎖プライマーを使用することによって得られた固定ds cDNAを示す。
シグナル(これには可変領域が続き、これには定常領域が続き、これには3’非
翻訳領域(これは、代表的にはポリAで終わる)が続く)からなる。逆転写のた
めの最初のプライマーは、定常領域または3’UTRのポリAセグメントに対し
て相補的であり得る。ヒト重鎖遺伝子については、15Tプライマーが好ましい
。逆転写酵素は、新しく合成されたDNAの3’末端にいくつかのC残基を付加
する。RTキャップ伸長は、この特徴を利用する。逆転写反応は、第1に、下方
鎖プライマーのみと作動される。約1時間後、プライマーは、GGG(USP−
GGG)で終了し、そして多くのRTaseが添加される。これは、下方鎖cD
NAが、USP−GGGの逆相補体によって最終GGGまで伸長されることを引
き起こす。付加された合成配列の部分に同一な1つのプライマーおよびセンス鎖
の3’末端における既知の配列の領域に相補的な第2のプライマーを使用して、
全てのV遺伝子は、それらのV遺伝子サブクラスに関わらず、増幅される。
プライマーを使用すること、ストレプトアビジン樹脂上のビオチン化産物を捕獲
することによって、DNAを変成することによって、および相補的な鎖を洗い流
すことによって、分離され得る。捕獲されたDNAのどの末端が望まれているか
に依存して、上方(センス)鎖または下方(アンチセンス)鎖のいずれかを固定
することを選択する。
ポリペプチドまたはタンパク質の表示を行なうために、遺伝子パッケージへのク
ローニングのための一本鎖増幅DNAを調製し、クローニングおよび発現のため
に適切な末端を提供するように操作されなければならない。詳細には、任意の5
’非翻訳領域および最小のシグナル配列が除去され、そしてインフレームで、表
示宿主において機能する適切なシグナル配列によって置換され得る。さらに、可
変ドメイン(抗体遺伝子における)の部分は、除去され、そして合成多様性を含
む合成セグメントによって置換される。他の遺伝子ファミリーの多様性は、同様
に、合成多様性を用いて拡大され得る。
めの2つの方法が存在する。第1の方法は、以下の工程:
(i)核酸と一本鎖オリゴヌクレオチドとを接触させる工程であって、オリゴ
ヌクレオチドが、切断が望ましい領域における核酸に機能的に相補的であり、そ
して制限が核酸の切断を所望の位置で生じる制限エンドヌクレアーゼ認識部位を
形成する核酸中にその相補体を有する配列を含む、工程;および
(ii)核酸およびオリゴヌクレオチドの補完によって形成された認識部位の
みで核酸を切断する工程;
を包含し、接触および切断工程は、実質的に一本鎖形態で核酸を維持するのに十
分な温度で行なわれ、オリゴヌクレオチドは、切断が選択された温度および所望
の位置で生じ得るように、2つの鎖が会合するのを可能にするのに十分に大きい
領域にわたって核酸と機能的に相補的であり、そして切断は、選択された温度で
活性である制限エンドヌクレアーゼを使用して行なわれる。
ルさせ、現在の局部的なDNAの2本鎖領域内に形成される制限エンドヌクレア
ーゼ認識部位を切断し得る。特に、1本鎖DNAの実質的な画分の同じ位置に生
じる認識部位は同一である。
を用いてなされ得る。例えば、「http://www.mrc−cpe.ca
m.ac.uk/imt−doc/restricted/ok.html.」
を参照のこと。アップデータは、表題「Amino acid and nuc
leotide sequence alignments」の下でこのサイト
から獲得され得る。他のファミリーに関して、類似の比較が存在し、そして切断
のための適切な領域を選択するために、および多様性を維持するために用いられ
得る。
NA配列を示す。この領域において、この遺伝子は、表200に示される制限エ
ンドヌクレアーゼ認識部位を含む。同じ部位で生殖系列遺伝子の大画分を切断す
る制限エンドヌクレアーゼは、種々の部位で切断するエンドヌクレアーゼ以上に
好ましい。さらに、1本鎖DNA上で短いオリゴヌクレオチドが結合する領域(
例えば、制限エンドヌクレアーゼ認識部位のいずれかの側面の約10塩基)内に
制限エンドヌクレアーゼのための1つのみの部位が存在することが好ましい。
は、フレームワークにおける少数の変異を捕捉するからである。このことは、い
くつかの場合、有利であり得る。しかし、フレームワークの変異が存在し、そし
て抗体結合を付与しそして増強することが周知である。本発明は、適切な制限部
位の選択により、FR3の多様性の全てまたは一部を捕捉することを可能とする
。従って、この方法はまた、広範囲の多様性を捕捉することを可能とする。
ドヌクレアーゼが用いられる。好ましくは、50℃以上で活性な酵素、より好ま
しくは、約55℃で活性な酵素が用いられる。このような温度は、核酸配列が、
実質的に1本鎖形態で切断されることを維持する。
る酵素として、以下が挙げられる:MaeIII(24@4)、Tsp45I(
21@4)、HphI(44@5)、BsaJI(23@65)、AluI(2
3@47)、BlpI(21@48)、DdeI(29@58)、BglII(
10@61)、MslI(44@72)、BsiEI(23@74)、EaeI
(23@74)、EagI(23@74)、HaeIII(25@75)、Bs
t4CI(51@86)、HpyCH4III(51@86)、HinfI(3
8@2)、MlyI(18@2)、PleI(18@2)、MnlI(31@6
7)、HpyCH4V(21@44)、BsmAI(16@11)、BpmI(
19@12)、XmnI(12@30)、およびSacI(11@51)(使用
される表記は、例えば、BsmAIが、FR3の塩基11で開始する制限エンド
ヌクレアーゼ認識部位で16のFR3生殖系列遺伝子を切断することを意味する
)。
、以下のようなものである:Bst4CI(またはTaaIもしくはHpyCH
4III)、BlpI、HpyCH4V、およびMslI。ACNGT(Bst
4CI、TaaI、およびHpyCH4IIIのための制限エンドヌクレアーゼ
認識部位)は、全てのヒトFR3生殖系列遺伝子において一致する部位に見出さ
れるため、これらの酵素の1つは、重鎖CDR3の多様性の捕捉のために最も好
ましい。BlpIおよびHpyCH4Vは相補的である。BlpIは、VH1お
よびVH4ファミリーのほとんどのメンバーを切断し、HpyCH4Vは、VH
3、VH5、VH6、およびVH7のファミリーのほとんどのメンバーを切断す
る。両方の酵素ともVH2を切断しないが、これは非常に小さいファミリーであ
り、3つのメンバーを含むのみである。従って、これらの酵素はまた、本発明の
方法の好ましい実施形態において用いられ得る。
は全て、5’―ACnGT―3’を認識し、そしてnの後の上方のストランドD
NAおよびnに相補的な塩基の前の下方ストランドDNAを切断する。これは、
ヒト重鎖上でのこの方法のために最も好ましい制限エンドヌクレアーゼ認識部位
である。なぜならば、それは、全ての生殖系列遺伝子において見出されるからで
ある。さらに、制限エンドヌクレアーゼ認識領域(ACnGT)は、表206に
示されるような、チロシンコドン(tay)およびそれに続くシステインコドン
(tgy)の第2および第3の塩基に適合する。これらのコドン(特に、成熟抗
体遺伝子におけるシステイン)は、高度に保存されている。
ヌクレオチドを示す。表255Cは、1617の実際の重鎖抗体遺伝子の分析を
示す。これらの中で、1511つはこの部位を有し、そして4つの不適合内で候
補オリゴヌクレオチドの1つに適合する。8つのオリゴヌクレオチドが、ほとん
どの適合の原因であり、表250F.1.に示す。8つのオリゴヌクレオチドは
非常に類似しており、その結果、満足のいく切断が、温度、pH、塩分などを調
整することにより、1つのみのオリゴヌクレオチド(例えば、H43.77.9
7.1−02#1)を用いて達成されるようである。1つまたは2つのオリゴヌ
クレオチドは、生殖系列遺伝子配列がごくわずかしか異ならない場合であっても
、そして特に切断される制限エンドヌクレアーゼ認識領域付近でごくわずかしか
異ならない場合、同様に満足し得る。表255Dは、8つの選択されたオリゴヌ
クレオチドのみを用いた、1617つの実際の重鎖抗体遺伝子の反復分析を示す
。これは、1463つの配列が、4つの不適合内で少なくとも1つのオリゴヌク
レオチドで適合すること、および期待するような部位を有することを示す。7つ
の配列のみが、この領域内に第2のHpyCH4III制限エンドヌクレアーゼ
認識領域を有する。
R1における切断に関する。FR1における切断は、重鎖のCDRの多様性全体
の捕捉を可能にする。
ト生殖系列遺伝子FR1において見出される制限エンドヌクレアーゼ認識部位を
示す。好まし部位は、BsgI(GTGCAG;39@4)、BsoFI(GC
ngc;43@6、11@9、2@3、1@12)、TseI(Gcwgc;4
3@6、11@9、2@3、1@12)、MspAlI(CMGckg;46@
7、2@1)、PvuII(CAGctg;46@7、2@1)、AluI(A
Gct;48@82@2)、DdeI(Ctnag;22@52、9@48)、
HphI(tcacc;22@80)、BssKI(Nccngg;35@39
、2@40)、BsaJI(Ccnngg;32@40、2@41)、BstN
I(CCwgg;33@40)、ScrFI(CCngg;35@40、2@4
1)、Eco0109I(RGgnccy;22@46、11@43)、Sau
96I(Ggncc;23@47、11@44)、AvaII(Ggwcc;2
3@47、4@44)、PpuMI(RGgwccy;22@46、4@43)
、BsmFI(gtccc;20@48)、HinfI(Gantc;34@1
6、21@56、21@77)、TfiI(21@77)、MlyI(GAGT
C;34@16)、MlyI(gactc;21@56)、およびAlwNI(
CAGnnnctg;22@68)である。より好ましい部位は、MspAIお
よびPvuIIである。MspAIおよびPvuIIは、7〜12での46つの
部位および1〜6での2つの部位を有する。両方の部位での切断を回避するため
に、1〜6での部位を完全に覆わないオリゴヌクレオチドが用いられる。従って
、DNAは、その部位で切断されない。本発明者らは、PvuII部位を超えて
3、4、または5塩基延びるDNAが効果的に切断され得ることを示した。
FR1における切断に関する。表300は、ヒトκFR1生殖系列を示し、そし
て表302は、一致する部位で実質的に多くのヒトκFR1生殖系列遺伝子にお
いて見出される制限エンドヌクレアーゼ認識部位を示す。列挙される制限エンド
ヌクレアーゼ認識部位の中で、BsmAIおよびPflFIが最も好ましい酵素
である。BsmAI部位は、40の生殖系列遺伝子の中の35において塩基18
にて見出される。PflFI部位は、40の生殖系列遺伝子の中の35において
塩基12にて見出される。
R1における切断に関する。表400は、31の公知のヒトλFR1生殖系列遺
伝子配列を示す。表405は、ヒトλFR1生殖系列遺伝子において見出される
制限エンドヌクレアーゼ認識部位を示す。HinfIおよびDdeIが、FR1
におけるヒトλ鎖を切断するための最も好ましい制限エンドヌクレアーゼである
。
いオリゴヌクレオチドが、1つかまたは組み合わせて、選択された認識部位を機
能的に補うために調製される。このオリゴヌクレオチドはまた、増幅された遺伝
子の大部分において認識部位に隣接する配列を含む。この隣接配列は、この配列
が、選択された部位に特異的な制限エンドヌクレアーゼにより切断されるのに十
分に、1本鎖DNAにアニールすることを可能にする。
よび切断のために用いられる状態に依存する。この長さは、オリゴヌクレオチド
が、2つのストランドが結合し、その結果、選択された温度および所望の位置の
みでの切断が生じるのに十分大きな領域にわたって、1本鎖DNAに機能的に相
補性であるために十分でなければならない。
クレオチド長である。約17塩基未満では、アニーリングが弱すぎ、そして30
塩基より上では、特異性の喪失が存在し得る。好ましい長さは、18〜24塩基
である。
はない。むしろ、生じる数個の不適合が寛容され得る。しかし、好ましくは、1
〜3の不適合のみが許容される。このような不適合は、1本鎖DNAに対するこ
のオリゴヌクレオチドのアニーリングに逆に影響しない。従って、この2つのD
NAは、機能的に相補性であると呼ばれる。
方法は、以下の工程を包含する:
(i)核酸と部分的に2本鎖のオリゴヌクレオチドを接触される工程であっ
て、このオリゴヌクレオチドの1本鎖領域が、切断が所望される領域においてこ
の核酸に機能的に相補的であり、そしてこのオリゴヌクレオチドの2本鎖領域が
、II−S型制限エンドヌクレアーゼ認識部位を有し、それらの切断部位が、認
識部位から既知の距離で配置される、工程;および
(ii)核酸とオリゴヌクレオチドの1本鎖領域との補完により形成される
切断部位のみでこの核酸を切断する工程。
接触工程および切断工程は、この核酸を実質的に1本鎖形態で維持するために十
分な温度にて実施され、オリゴヌクレオチドは、2つのストランドが結合し、そ
の結果、選択された温度および所望の位置で切断が生じ得るのに十分に大きな領
域にわたってこの核酸に対して機能的に相補性であり、そして切断は、選択され
た温度で活性な制限エンドヌクレアーゼを用いて実施される。
いる。UREは、部分的に2本鎖オリゴヌクレオチドである。1本鎖部分または
UREの重複は、1本鎖DNAにおいて切断されるためにこの配列に対して機能
的に相補性であるDNAアダプターからなる。2本鎖部分は、II−S型制限エ
ンドヌクレアーゼ認識部位でからなる。
つか(例えば、1〜3)の不適合を寛容し得る(すなわち、それはその領域に対
して機能的に相補性である)。さらに、UREが用いられる条件が調整され、そ
の結果、増幅されるほとんどの遺伝子が切断され、それらの遺伝子から生成され
るライブラリー中の偏りを減少させ得る。
〜22塩基からなる。しかし、より長いかまたはより短いアダプターが用いられ
得る。大きさは、アダプターが1本鎖DNAの機能的な相補体と結合する能力、
ならびにII−S型酵素を用いてこのDNAを切断するために用いられる温度に
てUREと1本鎖DNAを接触させるために用いられる温度に依存する。アダプ
ターは、2つのストランドが結合し、その結果、選択された温度および所望の位
置で切断が生じ得るのに十分に大きな領域にわたって1本鎖DNAに対して機能
的に相補性でなければならない。本発明者らは、14〜17塩基長、より好まし
くは18〜20塩基長の1本鎖部分または重複部分を好む。
Aのファミリーにおいて実質的に保存される部位である。本発明の第1の切断方
法と比較する場合、これらの部位は、エンドヌクレアーゼ認識部位である必要が
ない。しかし、第1の方法のように、選択される部位は、ネイティブのDNAに
存在するというよりも合成性であり得る。このような部位は、公知の抗体または
遺伝子の他のファミリーの配列に関する参照により選択され得る。例えば、多く
の生殖系列遺伝子の配列は、http://www.mrc−cpe.cam.
ac.uk/imt−doc/restricted/ok.htmlにて報告
されている。例えば、1つの好ましい部位は、FR3の末端付近(コドン89〜
コドン93の第2の塩基)で生じる。CDR3は、コドン95で開始する。
m.nih.gov/entre2/nucleotide.htmlにて利用
可能である。このサイトは、本発明の方法に従うURE切断のための適切な配列
を同定するために用いられ得る。例えば、表8Bを参照のこと。
列情報を用いて同定される。これらの配列は、共に、増幅されたDNAの実質的
な画分に存在する。例えば、複数の配列が、生殖系列遺伝子における公知の多様
性または頻度の高い体細胞変異を可能にするために用いられ得る。合成性の変性
配列もまた、用いられ得る。好ましくは、2〜3の不適合のみを有する試験され
た遺伝子の少なくとも65%において生じる配列が、選択される。
るように作製される。UREを使用する条件は、経験的に決定される。これらの
条件は、2〜3の不適合のみを有する機能的に相補性の配列を含むDNAの切断
を可能にするが、このような配列を欠くDNAの切断を可能にしない。
識部位が挙げられる。一本鎖DNAを実質的にその形態で保持し、そして一本鎖
DNAが所望される部位で切断されるのを可能にするために、UREの一本鎖D
NAアダプター部分が十分な長さでアニーリングすることを可能にするのに必要
な温度で活性な任意のII−S型酵素が使用され得る。
Aの非対称的な切断を提供する。とりわけ、表800にこれらの酵素を列挙する
。もっとも好ましいII−S型酵素が、FokIである。
かの条件を使用することが好ましい:
1)酵素を活性化するための標的DNA全体のUREの発現が存在するべきで
ある。標的DNAに対して等モル量しか存在しないUREは、ssDNAの少な
い切断を生じる。なぜなら、利用可能な活性な酵素の量は限られているからであ
る。
ことなく二量体化させ得る。適切なアクチベーターの例を、表510に示す。
度、そして最も好ましくは55℃より高い温度で実施される。
塩基のステム(FokI部位を含む)、それに続く10塩基のステムのパリンド
ロームを含んだ。このようなUREを本発明の本方法で使用し得るが、本発明の
好ましいUREはまた、セグメントを含むFokI制限エンドヌクレアーゼ認識
部位の間の3〜8塩基(1つのループ)のセグメントも含む。好ましい実施形態
において、ステム(FokI部位を含む)およびそのパリンドロームはまた、1
0塩基よりも長い。好ましくは、これらは、10〜14塩基長である。これらの
「ロリポップ(lollipop)」UREアダプターの例を表5に示す。
R3領域が挙げられる。表508は、示されるURE認識配列を有する840の
全長成熟ヒト重鎖の分析を示す。大部分(718/840=0.85)は、5つ
のURE(VHS881−1.1、VHS881−1.2、VHS881−2.
1、VHS881−4.1、およびVHS881−9.1)を使用して、2以下
のミスマッチを認識される。これらのそれぞれは、生殖系列の遺伝子に相補的な
20塩基のアダプター配列、FokI部位を含む10塩基のステムセグメント、
5つの塩基ループ、および第一のステムセグメントの逆の相補体を有する。これ
らのアダプターを、単独でまたは組合せて一本鎖アンチセンス重鎖DNAにアニ
ーリングし、そして例えば、アクチベーターFOKIactの存在下でFokI
を用いて処理することによって、指示された位置でアンチセンス鎖の切断を生じ
る。
1領域が挙げられる。表512は、4つの示された19塩基のプローブ配列によ
る一致についての182の全長ヒトκ鎖の分析を示す。96%の配列は、2以下
のミスマッチを有するプローブの1つと一致する。表512に示されるUREア
ダプターは、κ鎖のセンス鎖の切断のためである。従って、アダプター配列は、
生殖系列の遺伝子配列の逆の相補体である。UREは、10塩基のステム、5塩
基のループ、ステムの逆の相補体およびその相補配列から構成される。ループは
、ここではTTGTTを示すが、他の配列が使用され得る。その機能は、ロリポ
ップ単量体の形成が二量体化よりも好まれるように、ステムのパリンドロームを
妨げることである。表512はまた、センス鎖が切断される場所を示している。
られる。表515は、4つの示された19塩基のプローブを一致させるための1
28のヒトλ軽鎖の分析を示している。3以下のミスマッチを有し、128のう
ちの88(69%)の鎖がプローブの1つと一致する。表515はまた、これら
のプローブに対応するUREアダプターを示している。これらのアダプターを、
λ鎖の上側の鎖ssDNAにアニーリングし、そしてFOKIactの存在下、
45℃またはそれより高い温度で、FokIで処理することによって、これらの
鎖の特異的かつ正確な切断が生じる。
鎖DNAと接触する条件下で、経験的に決定され得る。この条件は、一本鎖DN
Aが、実質的に一本鎖の形態で保持されるようでなければならない。より詳細に
は、これらの条件は、一本鎖DNAが、ループを形成しないような条件でなけれ
ばならず、このループは、それが関係するオリゴヌクレオチド配列もしくはUR
Eを妨害し得るか、または選択的制限エンドヌクレアーゼによってループ自身が
切断の部位を提供し得る。
よび特異性は、UREアダプター/オリゴヌクレオチドならびに基質DNAの濃
度、温度、pH、金属イオン濃度、イオン強度、カオトロープ(chaotro
pe)(例えば、尿素およびホルムアミド)の濃度、制限エンドヌクレアーゼ(
例えば、FokI)の濃度、および消化の時間を制御することによって調整され
得る。これらの条件は、以下を有する合成オリゴヌクレオチドを用いて最適化さ
れ得る:1)標的生殖系列遺伝子配列、2)成熟標的遺伝子配列、または3)幾
分関連する非標的配列。目標は、標的配列のほとんどを切断しかつ非標的の最小
量を切断することである。
温度を使用して、実質的に一本鎖DNAの形態で維持される。より好ましくは、
50℃〜60℃の間であり、最も好ましくは55℃〜60℃の間の温度が使用さ
れる。これらの温度は、DNAがオリゴヌクレオチドまたはUREと接触する場
合、および本発明の方法を使用してDNAを切断する場合の両方で使用される。
DNAのファミリーの個々のメンバーを、1つの実質的に保存されたエンドヌク
レアーゼ認識部位で単独に切断することを可能にする。この方法はまた、逆転写
または増幅プライマーに構築されるエンドヌクレアーゼ認識部位を必要としない
。ファミリー中の任意のネイティブな部位または合成部位が使用され得る。
鎖DNAが、天然に存在するか、または合成で構築されたエンドヌクレアーゼ認
識部位を有さない位置で切断されることを可能にする。
3’プライマーの使用を可能にし、次いでクローニングおよびディスプレイの前
に所望でないか、または有害な配列を除去するような切断を可能にする。
はクローニングのために調製される。これは、部分的に二重鎖の合成DNAアダ
プター(この末端配列は、特定の切断部位に基づいており、増幅されたDNAは
、この部位で切断される)を使用することによってなされる。
ペプチド、ポリペプチドまたは遺伝子パッケージの表面タンパク質をディスプレ
イするように正確なリーディングフレームに発現されることを可能にするように
設計される。好ましくは、アダプターの二本鎖部分は、ペプチド、ポリペプチド
または切断部位までのタンパク質のファミリーのアミノ酸配列の特徴をコードす
るいくつかのコドン配列を含む。ヒト重鎖について、好ましくは、3〜23のフ
レームワークのアミノ酸を使用して、切断されたDNAの発現に必要な配列を提
供する。
好ましくは、約20〜100塩基が使用される。アダプターの二本鎖領域はまた
、好ましくは、DNAを適切なディスプレイベクター(または多様性を保存する
ために使用されるレシピエントベクター)にクローニングするために有用な少な
くとも1つのエンドヌクレアーゼ認識部位を含む。このエンドヌクレアーゼ制限
部位は、DNA配列を伸長するために使用される生殖系列遺伝子配列に対してネ
イティブであり得る。これはまた、ネイティブな生殖系列遺伝子配列に対して縮
重した配列を使用して構築され得る。または、これは全合成であり得る。
このオーバーラップは、約2塩基〜約15塩基までであり得る。オーバーラップ
がより長くなるにつれて、連結がより効率的になるようである。オーバーラップ
について好ましい長さは、7〜10である。これは、領域における多様性が捕捉
され得るようにこの領域のいくつかのミスマッチを可能にする。
なぜなら、これは、DNAが選択された部位で切断されることを可能にするが、
捕捉される他のフラグメントは切断されないからである。このようなフラグメン
トは、適切な抗体に折り畳まれず、そして非特異的粘着性であるらしい遺伝子コ
ード配列を有するライブラリーを汚染する。
、このようなアダプターを上記のように、5’−ACnGT−3’において、H
pyCH4III、Bst4CIまたはTaaIを使用して切断されるヒトFR
3領域に連結する工程を包含する。
ターの二本鎖部分の下側の鎖を示している。HpyCH4III部位は、右(表
206に示されるような)に対して非常に遠いので、AflII部位およびXb
aI部位を含む配列が、付加され得る。部分的に二重鎖のアダプターのこの下側
の鎖部分(H43.XAExt)は、XbaIおよびAflII部位の両方を組
込む。アダプターの二重鎖部分の上側の鎖は、いずれの部位も有さない(H43
.XAExtのXbaIおよびAflII部位に逆のセグメントにおける計画さ
れたミスマッチのため)が、H43.XAExtに非常に強固アニーリングする
。H43AExtは、AflII部位のみを含み、そして上側鎖H43.ABr
1およびH43.ABr2(これは、AflII部位を破壊するために意図的な
変更を有する)とともに使用される。
場合、好ましい実施形態において、抗体遺伝子の下流の定常領域に対するプライ
マー、およびアダプターの二本鎖領域の部分に対するプライマーを使用する。こ
れらのプライマーはまた、増幅されたDNAのクローニングに使用するための制
限エンドヌクレアーゼ部位を保有する。
の後、複合体DNAは、選択された5’側および3’側エンドヌクレアーゼ認識
部位で切断される。
ージミドおよび抗体遺伝子におけるその利用可能な部位に依存する。表1は、7
5のヒト軽鎖についての制限エンドヌクレアーゼのデータを提供する。表2は、
79のヒト重鎖についての対応するデータを示す。各表において、エンドヌクレ
アーゼは、切断の頻度が増す順番で並べられている。これらの表において、Nc
hは、酵素によって切断される鎖の数であり、Nsは、部位の数(いくつかの鎖
は1より多い部位を有する)である。
が非常に適切である。SfiIおよびNotIは、好ましくは、重鎖ディスプレ
イセグメントを挿入するためのpCES1において使用される。ApaLIおよ
びAscIは、好ましくは、軽鎖ディスプレイセグメントを挿入するためにpC
ES1において使用される。
遺伝子において、重鎖遺伝子の54/79(68%)のみが、BstEII部位
を含み、そしてこれらの7/61が、2つの部位を含む。従って、47/79(
59%)が、1つのBstEII部位を含む。BstEIIを使用することの代
替策は、JH末端でのUREを介する切断、およびCH1の部分をコードする合
成オリゴヌクレオチドへの連結である。
ヒトCDR3多様性を捕捉することが挙げられる。上記のように、種々の自己免
疫の患者からのmRNAは、cDNAへ逆転写される。mRNAが分解された後
、cDNAを固定化し、そして短いオリゴヌクレオチドを使用してcDNA上流
のCDR3を切断する。次いで、部分的に二重鎖の合成DNAアダプターは、D
NAにアニーリングされ、そしてこのDNAは、アダプターに対するプライマー
および定常領域(FR4の後)に対するプライマーを使用して増幅される。次い
で、DNAはBstEII(FR4中)および部分的二本鎖アダプター(例えば
、XbaIおよびAflII(FR3中))に適切な制限エンドヌクレアーゼを
使用して切断される。次いで、DNAは合成VH骨格(例えば、3〜23)に連
結される。
トκ鎖が挙げられる。この鎖のセンス鎖の切断部位は、表512に示される。オ
リゴヌクレオチドkapextUREは、オリゴヌクレオチド(kaBR01U
R、kaBR02UR、kaBR03UR、およびkaBR04UR)にアニー
リングされて部分的二重鎖DNAを形成する。次いで、このDNAは、切断され
た可溶性のκ鎖に連結される。次いで、連結産物は、プライマーkapextU
REPCRおよびCKForeAsc(これは、Cκ末端後のAscI部位に挿
入する)を使用して増幅される。次いで、この産物は、ApaLIおよびAsc
Iを用いて切断され、そして同様にレシピエントベクターを切断するために連結
される。
ダー(extender)(ON_LamEx133)オリゴヌクレオチドおよ
び4つのブリッジ(bridge)オリゴヌクレオチド(ON_LamB1−1
33、ON_LamB2−133、ON_LamB3−133およびON_La
mB4−133)は、アニーリングされて部分的二重鎖DNAを形成する。この
DNAは、切断されたλ鎖センス鎖に連結される。連結後、DNAは、ON_L
am133PCRおよびλ定常ドメインに特異的な前進プライマー(例えば、C
L2ForeAscまたはCL7ForeAsc(表130))を用いて増幅さ
れる。
常に大きな画分中の定常位置で生じるBstEII部位で切断され得る(下流、
すなわち、CDR3のセンス鎖の3’末端へ向かって)。次いで、CDR3の多
様性のみが捕捉される場合、FR3中の部位が、CDR2およびCDR3の多様
性が所望される場合、FR2中の部位が、またはCDRの多様性全てが所望され
る場合、FR1中の部位が必要である。これらの部位は、好ましくは、部分的二
重鎖のアダプターの一部として挿入される。
能にする部位を有するレシピエントベクターを提供することである。このような
ベクターは、周知であり、当該分野で広く使用される。本発明に従った好ましい
ファージ提示ベクターは、ファージMALIA3である。これは、遺伝子III
において提示する。ファージMALIA3の配列を、表120A(注釈付)およ
び表120B(要約)に示す。
ずれかを非常に稀にしか切断しないエンドヌクレアーゼを使用してベクターに移
入し得る。例えば、軽鎖は、ApaLIおよびAscIによって捕捉される。重
鎖遺伝子は、好ましくは、SfiI、NcoI、XbaI、AflII、Bst
EII、ApaIおよびNotI部位を有するレシピエントベクターにクローニ
ングされる。軽鎖は、好ましくは、ApaLI−AscIフラグメントとしてラ
イブラリー中に移される。重鎖は、好ましくは、SfiI−NotIフラグメン
トとしてライブラリーに移される。
最も好ましいベクターは、M13の全ての遺伝子、抗生物質耐性遺伝子、および
提示カセットを含む。好ましいベクターは、遺伝子の多様なファミリーのメンバ
ーの導入および切除を可能にする制限部位をカセットとして提供される。好まし
いベクターは、ファージを増幅させるために使用される増殖条件下での再編成に
対して安定である。
IIIタンパク質上にペプチド、ポリペプチドまたはタンパク質を提示するファ
スミドベクター(例えば、pCES1)において提示され得る。このようなベク
ターはまた、他のベクターまたはファージを使用する引き続く提示のための多様
性を保存するために使用され得る。
に対する短いリンカーを介し得る。M13 III(「IIIスタンプ」)の最
終部分である1つのアンカードメイン、全長III成熟タンパク質である第二の
アンカー、およびM13 VIII成熟タンパク質である第三のアンカー。
十分に含むが、感染性の媒介に関するドメインを含まない。野生型のIIIおよ
びVIIIタンパク質が存在することから、このファージは、抗体遺伝子を除去
しないようであるが、非常に低い増殖の利点のみを受けるこれらのセグメントを
欠失する。各々のアンカードメインのために、このDNAは野生型のAA配列を
コードするが、非常に高い程度で野生型のDNA配列とは異なる。これは、提示
アンカーと野生型遺伝子(これもまた存在する)との間の相同組換えの可能性を
非常に減少させる。
遺伝子)および提示カセットを保有する完全なファージを使用する。野生型ii
iおよびviii遺伝子が存在することから、野生型タンパク質もまた存在する
。この提示カセットは、調節可能なプロモーター(例えば、PLacZ)から転
写される。調節可能なプロモーターの使用は、対応する野生型コートタンパク質
に対する融合提示遺伝子の比を制御することを可能にする。この比は、ファージ
(またはファスミド)粒子当たりの提示融合の平均コピー数を決定する。
ク質(および特にFab)を提示する方法である。最も好ましい実施形態におい
て、この方法は、FABを提示し、以下を包含する:
a)以下のエレメントをコードするDNAの断片の送達を捕捉するカセットを得
る工程:
Preg::RBS1::SS1::VL::CL::停止::RBS2::
SS2::VH::CH1::リンカー::アンカー::停止::、
ここで、Pregは、調節可能なプロモーターであり、PBS1は、第一のリボ
ソーム結合部位であり、SS1は、宿主株において作動可能なシグナル配列であ
り、VLは、軽鎖可変領域の種々のセットのメンバーであり、CLは、軽鎖定常
領域であり、停止は、1以上の停止コドンであり、PBS2は、第二のリボソー
ム結合部位であり、SS2は、宿主株において作動可能な第二のシグナル配列で
あり、VHは、重鎖可変領域の種々のセットのメンバーであり、CH1は、抗体
重鎖の第一の定常領域であり、リンカーは、1〜約50残基のアミノ酸の配列で
あり、アンカーは、糸状ファージ粒子に構築されるタンパク質であり、そして停
止は、1以上の停止コドンの第二の例である;ならびに
b)ファージの生存性を最大化し、そしてこのカセットまたはその部分の欠失の
可能性を最小化するためにファージゲノム内にカセットを配置する工程。
(または密接に関連する)アミノ酸配列(ファージのコートタンパク質の1つに
見出されるような)をコードするが、別個のDNA配列を有するように設計され
る。これは、野生型遺伝子との望まない相同組換えを防止する。さらに、このカ
セットは、遺伝子間領域に配置されるべきである。提示カセットの配置および方
向は、ファージの挙動に影響し得る。
トの第二の停止(例えば、Trp)の後に配置され得る。これは、提示カセット
とファージ抗体提示ベクター(PADV)中の他の遺伝子との間の相互作用を減
少させる。
Fab以外のタンパク質を、他のタンパク質遺伝子で置換することによって、上
記のFab部分を提示し得る。
し得る。このような宿主は、当該分野で周知である。好ましい実施形態において
、Fabが提示される場合、好ましい宿主は30℃で増殖し、そしてRecA−
(望まない遺伝子組み換えを減少させるため)およびEndA−(RF DNA
の回収を容易化するため)であり得る。エレクトロポレーションによって容易に
形質転換される宿主株もまた好ましい。
、30℃ではあまり増殖しない。XL1−Blue MRF’は、38℃でゆっ
くりと増殖し、従って、許容可能な宿主である。TG−1はまた、中間宿主とし
てより好ましいが、RecA+およびEndA+である。XL1−Blue M
RF’は、ライブラリーの最終構築の前の多様性を蓄積するために使用される中
間宿主としてより好ましい。
てスクリーニングし得る。次いで、選択されたペプチド、ポリペプチドまたはタ
ンパク質を、疾患を処置するために使用し得る。一般に、治療または薬学的組成
物における使用のためのペプチド、ポリペプチドまたはタンパク質を、選択され
るライブラリーのメンバーから所望のペプチド、ポリペプチドまたはタンパク質
をコードするDNAを単離することによって生成する。次いで、このDNAを、
慣用的な方法において使用して、適切な宿主細胞、好ましくは、哺乳動物細胞(
例えば、CHO細胞)においてコードされるペプチド、ポリペプチドまたはタン
パク質を生成する。単離後、ペプチド、ポリペプチドまたはタンパク質を、単独
でか、または疾患を処置するための治療において薬学的に受容可能な組成物と共
に使用する。
(実施例1:BsmAIを用いるκ鎖の捕捉)
ヒトκ鎖mRNAのレパートリーを、種々の自己免疫疾患を有する患者の集団
から単離した総RNAまたはポリ(A+)RNAを仔ウシ腸ホスファターゼで処
理して、例えば、リボソームRNA、フラグメント化mRNA、tRNAおよび
ゲノムDNAを有する全ての分子から5’−リン酸を除去することによって調製
した。全長のmRNA(保護的7−メチルキャップ構造を含む)は、影響されな
い。次いで、RNAを、タバコ酸ピロホスファターゼで処理して、5’−一リン
酸基を残して全長のmRNAからキャップ構造を除去する。
、そしてGenerRACETM方法およびキット(Invitrogen)を
使用して増幅した。アダプターに相補的な5’ビオチン化プライマー、および構
築物の領域の一部に相補的な3’プライマーを使用した。
κ鎖(Igκ)遺伝子RACE材料を、200マイクロリットル(μL)のSe
radyn磁気ビーズ上に固定した。下方の鎖を、2アリコートの200μLの
0.1M NaOH(pH13)で、第一のアリコートについて3分間、次いで
第二のアリコートについて30秒間、DNAを洗浄することによって除去した。
これらのビーズを、200μLの10mM Tris(pH7.5)100mM
NaClで中和した。表525に示した短いオリゴヌクレオチドを、100μ
LのNEB緩衝液2(50mM NaCl、10mM Tris−HCl、10
mM MgCl2、1mM ジチオトレイトール pH7.9)中に40倍モル
過剰に、乾燥ビーズに添加した。この混合物を、95℃で5分間インキュベート
し、次いで、30分かけて55℃まで冷却した。過剰なオリゴヌクレオチドを、
NEB緩衝液3(100mM NaCl、50mM Tris−HCl、10m
M MgCl2、1mM ジチオトレイトール pH7.9)で2回洗浄して除
去した。10単位のBsmAI(NEB)を、NEB緩衝液3に添加し、そして
55℃で1時間インキュベートした。切断された下流のDNAを、収集し、そし
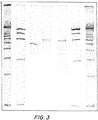
てQiagen PCR精製カラムで精製した(図3および図4)。
して調製した。このアダプターを、1000単位のT4 DNAリガーゼ(NE
B)と共に100倍モル過剰に一本鎖DNAに添加し、そして16℃で一晩イン
キュベートした。過剰なオリゴヌクレオチドを、Qiagen PCR精製カラ
ムで除去した。この連結材料を、表525に示したプライマーkapPCRt1
およびkapforを使用するPCRによって、表530に示したプログラムで
10サイクル増幅した。
ンドを示した(図5および図6)。このDNAを、酵素ApaLIおよびAsc
Iで切断し、ゲル精製し、そして同様に切断したベクターpCES1に連結した
。正確なサイズの挿入物の存在を、図15に示したように、いくつかのクローン
でPCRによってチェックした。
1は、この手順によって捕捉された第二の配列を示す。最も近い架橋配列は、配
列5’−agccacc−3’に相補的であったが、捕捉された配列は5’−T
gccacc−3’と読め、これは、重複領域中にいくつかのミスマッチを寛容
したことを示す。
CDR2の多様性の構築)
合成相補性決定基領域(CDR)1および2の多様性を、二段階のプロセスで
3−23 VHフレームワークにおいて構築した:第一に、3−23 VHフレ
ームワークを含むベクターを構築し、次いで、合成CDR1および2を、構築し
てこのベクター中にクローニングした。
のPCRプライマー(表600に示した長いオリゴヌクレオチド:TOPFR1
A、BOTFR1B、BOTFR2、BOTFR3、F06、BOTFR4、O
N−vgC1、およびON−vgC2ならびにプライマー:SFPREMTおよ
びBOTPCRPRIM)を、V323 VHのGenbankの配列に基づい
て設計した。この設計は、表600に示されるような、各々のフレームワークに
おける少なくとも1つの有用な制限部位を取り込んだ。表600において、合成
された断片を太字として示し、重複する領域に下線を付し、そして各末端でのP
CRプライマー領域に下線を付した。これら8つのオリゴの混合物を、20μl
のポリメラーゼ連鎖反応(PCR)反応中の2.5μMの最終濃度で組み合わせ
た。このPCR混合物は、200μM dNTP、2.5mM MgCl2、0
.02U Pfu TurboTMDNAポリメラーゼ、1U Qiagen
HotStart Taq DNA ポリメラーゼ、および1×Qiagen
PCR緩衝液を含んだ。このPCRプログラムは、94℃で30秒、55℃で3
0秒、および72℃で30秒の10サイクルからなった。次いで、100μl
PCR反応における最初のPCRからの10倍希釈の2.5μlを使用して、構
築されたV3−23 DNA配列を増幅した。このPCR反応は、200μM
dNTP、2.5mM MgCl2、0.02U Pfu TurboTMDN
Aポリメラーゼ、1U Qiagen HotStart Taq DNA ポ
リメラーゼ、1×Qiagen PCR緩衝液、および1μMの濃度での2つの
外側のプライマー(SFPRMETおよびBOTPCRPRIM)を含んだ。こ
のPCRプログラムは、94℃で30秒、55℃で30秒、および72℃で60
秒の23サイクルからなった。このV3−23 VH DNA配列を、設計し、
そしてSfiIおよびBstEII制限エンドヌクレアーゼ部位を使用してpC
ES1(ファスミドベクター)にクローニングした(本明細書中で記載した全て
の制限酵素は、New England BioLabs、Beverly、M
Aによって供給され、そして製造業者の指示通りに使用した)。
r)配列(表610および表620に示す)を、pCES1に導入してCDR1
/CDR2配列(BspEI制限酵素部位とXbaI制限酵素部位との間の90
0塩基)およびCDR3配列(AflIIとBstEIIとの間の358塩基)
を置換した。新規のベクターは、pCES5であり、そしてその配列を、表62
0に与える。CDRの代わりにスタッファーを有することは、親配列がライブラ
リーにおいて過剰に提示される危険性を回避する。このCDR1−2スタッファ
ーは、BglII、Bsu36I、BclI、XcmI、MluI、PvuII
、HpaIおよびHincIIについての制限部位、ベクターpCES5内に独
自である、強調された領域を含む。CDR3を置換するこのスタッファーは、独
自の制限エンドヌクレアーゼ部位、RsrIIを含む。このスタッファー配列は
、E.coliのペニシリン遺伝子由来のフラグメントである。
をコードする4つの重複オリゴヌクレオチド(表600および表630に示した
ON−vgC1、ON_Br12、ON_CD2Xba、およびON−vgC2
)を設計した。これら4つのオリゴの混合物を、40μl PCR反応物中2.
5μMの最終濃度で組み合わせた。4つのオリゴのうち2つは、CDR1および
CDR2に位置する、変化させた配列を含んだ。このPCR混合物は、200μ
M dNTP、2.5U Pwo DNAポリメラーゼ(Roche)、および
2mM MgSO4を含む1×Pwo PCR緩衝液を含んだ。このPCRプロ
グラムは、94℃で30秒、60℃で30秒、および72℃で60秒の10サイ
クルからなった。これは、100μl PCR反応物中の2.5μlの混合物を
使用して、増幅されたCDR1/2 DNA配列を構築した。このPCR反応物
は、200μM dNTP、2.5U Pwo DNAポリメラーゼ、2mM
MgSO4を含む1×Pwo PCR緩衝液、および1μMの濃度で2つの外側
のプライマーを含んだ。このPCRプログラムは、94℃で30秒、60℃で3
0秒、および72℃で60秒の10サイクルからなった。これらの変化された配
列を、消化し、そしてCDR1/2スタッファーの代わりにV3−23フレーム
ワーク中にクローニングした。
者らは、ドナー集団または合成DNAのいずれかからCDR3多様性をクローニ
ングし得る。
から逸脱することなく当業者によって種々の改変がなされ得ることが理解される。

Claims (16)
- ペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーを提示し、そして該ファミリーの少なくとも一部を集合的に提示する、遺伝的パッケージの収集物を含むライブラリーであって、該提示されるペプチド、ポリペプチドまたはタンパク質は、以下:
(a)式S−X1−Y−X2−M−X3−のアミノ酸配列を有する重鎖CDR1であって、ここで、X1、X2およびX3は、A、D、E、F、G、H、I、K、L、M、N、P、Q、R、S、T、V、WおよびYからなる群より独立して選択される、重鎖CDR1、ならびに
(b)式X4−I−X5−X6−S−G−G−X7−T−X8−Y−A−D−S−V−K−G−のアミノ酸配列を有する重鎖CDR2であって、ここで、X4およびX5は、Y、R、W、V、GおよびSからなる群より独立して選択され、X6は、PおよびSからなる群より選択され、そしてX7およびX8は、A、D、E、F、G、H、I、K、L、M、N、P、Q、R、S、T、V、WおよびYからなる群より独立して選択される、重鎖CDR2、
をコードする配列、ならびに
(c)重鎖CDR3の多様性のコードする配列
を含む核酸配列によってコードされる、ライブラリー。 - 前記核酸配列が、VH 3−23のフレームワーク領域をコードする配列をさらに含む、請求項1に記載のライブラリー。
- 前記遺伝的パッケージがM13ファージである、請求項1または2に記載のライブラリー。
- 前記M13ファージが、抗生物質耐性遺伝子を含む、請求項3に記載のライブラリー。
- 前記M13ファージが、前記ペプチド、ポリペプチドまたはタンパク質の多種多様なファミリーのメンバーの導入および切除を可能にする制限部位を含む、請求項3に記載のライブラリー。
- 前記核酸配列がファージベクター中にある、請求項1〜5のいずれか一項に記載のライブラリー。
- 前記ファージベクターが、抗生物質耐性遺伝子を含む、請求項6に記載のライブラリー。
- 前記抗生物質耐性遺伝子がアンピシリン耐性遺伝子である、請求項7に記載のライブラリー。
- 前記ファージベクターが、調節可能なプロモーターを含む、請求項6に記載のライブラリー。
- 前記DNA配列が、ファージミドベクター中にある、請求項1〜5のいずれか一項に記載のライブラリー。
- 前記提示されるペプチド、ポリペプチドまたはタンパク質が、M13 IIIの最終部分に対して短いリンカーを介して提示される、請求項1〜10のいずれか一項に記載のライブラリー。
- 前記提示されるペプチド、ポリペプチドまたはタンパク質が、全長成熟IIIタンパク質に対して短いリンカーを介して提示される、請求項1〜10のいずれか一項に記載のライブラリー。
- 前記提示されるペプチド、ポリペプチドまたはタンパク質が、全長成熟VIIIタンパク質に対して短いリンカーを介して提示される、請求項1〜10のいずれか一項に記載のライブラリー。
- 前記重鎖CDR3の多様性が、天然のヒトCDR3の多様性を含む、請求項1〜13のいずれか一項に記載のライブラリー。
- 前記天然の多様性が、自己免疫患者から捕捉される、請求項14に記載のライブラリー。
- 前記重鎖CDR3多様性が、合成多様性を含む、請求項1〜13のいずれか一項に記載のライブラリー。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US19806900P | 2000-04-17 | 2000-04-17 | |
| US60/198,069 | 2000-04-17 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2001577464A Division JP5149476B2 (ja) | 2000-04-17 | 2001-04-17 | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013255876A Division JP5956413B2 (ja) | 2000-04-17 | 2013-12-11 | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2011135893A JP2011135893A (ja) | 2011-07-14 |
| JP5599745B2 true JP5599745B2 (ja) | 2014-10-01 |
Family
ID=22731873
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2001577464A Expired - Lifetime JP5149476B2 (ja) | 2000-04-17 | 2001-04-17 | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 |
| JP2011044525A Expired - Lifetime JP5599745B2 (ja) | 2000-04-17 | 2011-03-01 | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 |
| JP2013255876A Expired - Lifetime JP5956413B2 (ja) | 2000-04-17 | 2013-12-11 | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2001577464A Expired - Lifetime JP5149476B2 (ja) | 2000-04-17 | 2001-04-17 | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013255876A Expired - Lifetime JP5956413B2 (ja) | 2000-04-17 | 2013-12-11 | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 |
Country Status (10)
| Country | Link |
|---|---|
| US (2) | US20040029113A1 (ja) |
| EP (4) | EP1276855B1 (ja) |
| JP (3) | JP5149476B2 (ja) |
| AU (3) | AU5358901A (ja) |
| CA (2) | CA2406236C (ja) |
| DK (3) | DK2308982T3 (ja) |
| ES (3) | ES2400302T3 (ja) |
| HK (2) | HK1145515A1 (ja) |
| PT (1) | PT2308982E (ja) |
| WO (1) | WO2001079481A2 (ja) |
Families Citing this family (21)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6696251B1 (en) * | 1996-05-31 | 2004-02-24 | Board Of Trustees Of The University Of Illinois | Yeast cell surface display of proteins and uses thereof |
| US6759243B2 (en) * | 1998-01-20 | 2004-07-06 | Board Of Trustees Of The University Of Illinois | High affinity TCR proteins and methods |
| EP1276855B1 (en) * | 2000-04-17 | 2012-10-24 | Dyax Corp. | Methods of constructing display libraries of genetic packages for members of a diverse family of peptides |
| US8288322B2 (en) | 2000-04-17 | 2012-10-16 | Dyax Corp. | Methods of constructing libraries comprising displayed and/or expressed members of a diverse family of peptides, polypeptides or proteins and the novel libraries |
| EP1326882A2 (en) * | 2000-06-19 | 2003-07-16 | Dyax Corp. | Enterokinase cleavage sequences and their use |
| ES2430857T3 (es) * | 2000-12-18 | 2013-11-22 | Dyax Corp. | Bibliotecas focalizadas de paquetes genéticos |
| AU2013205033B2 (en) * | 2001-04-17 | 2016-06-09 | Takeda Pharmaceutical Company Limited | Novel Methods of Constructing Libraries Comprising Displayed and/or Expressed Members of a Diverse Family of Peptides, Polypeptides or Proteins and the Novel Libraries |
| AU2016225923B2 (en) * | 2001-04-17 | 2018-07-05 | Takeda Pharmaceutical Company Limited | Novel Methods of Constructing Libraries Comprising Displayed and/or Expressed Members of a Diverse Family of Peptides, Polypeptides or Proteins and the Novel Libraries |
| US20040005709A1 (en) * | 2001-10-24 | 2004-01-08 | Hoogenboom Henricus Renerus Jacobus Mattheus | Hybridization control of sequence variation |
| US7135310B2 (en) | 2002-04-24 | 2006-11-14 | The Regents Of The University Of California | Method to amplify variable sequences without imposing primer sequences |
| US20040067532A1 (en) | 2002-08-12 | 2004-04-08 | Genetastix Corporation | High throughput generation and affinity maturation of humanized antibody |
| EP1726657A4 (en) * | 2004-02-10 | 2008-08-20 | Japan Science & Tech Agency | METHOD OF CONVERTING A BASE IN A DNA SEQUENCE |
| CN101855242B (zh) | 2007-09-14 | 2014-07-30 | 阿迪马布有限责任公司 | 合理设计的合成抗体文库及其用途 |
| US8877688B2 (en) | 2007-09-14 | 2014-11-04 | Adimab, Llc | Rationally designed, synthetic antibody libraries and uses therefor |
| FR2924440B1 (fr) * | 2007-12-04 | 2015-01-09 | Pf Medicament | Nouveau procede de generation et de criblage d'une banque d'anticorps |
| US9873957B2 (en) | 2008-03-13 | 2018-01-23 | Dyax Corp. | Libraries of genetic packages comprising novel HC CDR3 designs |
| ES2528963T3 (es) | 2008-04-24 | 2015-02-13 | Dyax Corp. | Bibliotecas de paquetes genéticos que comprenden nuevos diseños de CDR1, CDR2, y CDR3 de HC y nuevos diseños de CDR1, CDR2, y CDR3 de LC |
| WO2010003316A1 (en) * | 2008-07-10 | 2010-01-14 | Si Lok | Methods for nucleic acid mapping and identification of fine-structural-variations in nucleic acids |
| WO2011032181A2 (en) * | 2009-09-14 | 2011-03-17 | Dyax Corp. | Libraries of genetic packages comprising novel hc cdr3 designs |
| KR102024922B1 (ko) | 2010-07-16 | 2019-09-25 | 아디맵 엘엘씨 | 항체 라이브러리 |
| SG11201609055RA (en) * | 2014-05-08 | 2016-11-29 | Fluidigm Corp | Integrated single cell sequencing |
Family Cites Families (70)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US596271A (en) * | 1897-12-28 | l-anohestee | ||
| US5118605A (en) * | 1984-10-16 | 1992-06-02 | Chiron Corporation | Polynucleotide determination with selectable cleavage sites |
| WO1986005803A1 (fr) * | 1985-03-30 | 1986-10-09 | Marc Ballivet | Procede d'obtention d'adn, arn, peptides, polypeptides ou proteines, par une technique de recombinaison d'adn |
| US6492107B1 (en) * | 1986-11-20 | 2002-12-10 | Stuart Kauffman | Process for obtaining DNA, RNA, peptides, polypeptides, or protein, by recombinant DNA technique |
| US5618920A (en) * | 1985-11-01 | 1997-04-08 | Xoma Corporation | Modular assembly of antibody genes, antibodies prepared thereby and use |
| US5223409A (en) * | 1988-09-02 | 1993-06-29 | Protein Engineering Corp. | Directed evolution of novel binding proteins |
| US5688666A (en) * | 1988-10-28 | 1997-11-18 | Genentech, Inc. | Growth hormone variants with altered binding properties |
| EP0368684B2 (en) * | 1988-11-11 | 2004-09-29 | Medical Research Council | Cloning immunoglobulin variable domain sequences. |
| US6969586B1 (en) * | 1989-05-16 | 2005-11-29 | Scripps Research Institute | Method for tapping the immunological repertoire |
| US6291158B1 (en) * | 1989-05-16 | 2001-09-18 | Scripps Research Institute | Method for tapping the immunological repertoire |
| US6291159B1 (en) * | 1989-05-16 | 2001-09-18 | Scripps Research Institute | Method for producing polymers having a preselected activity |
| US6291161B1 (en) * | 1989-05-16 | 2001-09-18 | Scripps Research Institute | Method for tapping the immunological repertiore |
| US6680192B1 (en) * | 1989-05-16 | 2004-01-20 | Scripps Research Institute | Method for producing polymers having a preselected activity |
| US6291160B1 (en) * | 1989-05-16 | 2001-09-18 | Scripps Research Institute | Method for producing polymers having a preselected activity |
| EP0440147B2 (de) * | 1990-02-01 | 2014-05-14 | Siemens Healthcare Diagnostics Products GmbH | Herstellung und Verwendung von Genbanken menschlicher Antikörper ("Human-Antikörper-Bibliotheken") |
| DE4002897A1 (de) * | 1990-02-01 | 1991-08-08 | Behringwerke Ag | Herstellung und verwendung von genbanken synthetischer menschlicher antikoerper ("synthetische human-antikoerper-bibliotheken") |
| CA2079802C (en) * | 1990-04-05 | 2001-09-18 | Roberto Crea | Walk-through mutagenesis |
| IE911295A1 (en) * | 1990-04-18 | 1991-10-23 | Gist Brocades Nv | Mutated beta-lactam acylase genes |
| US6172197B1 (en) * | 1991-07-10 | 2001-01-09 | Medical Research Council | Methods for producing members of specific binding pairs |
| US6916605B1 (en) * | 1990-07-10 | 2005-07-12 | Medical Research Council | Methods for producing members of specific binding pairs |
| GB9015198D0 (en) * | 1990-07-10 | 1990-08-29 | Brien Caroline J O | Binding substance |
| US7063943B1 (en) * | 1990-07-10 | 2006-06-20 | Cambridge Antibody Technology | Methods for producing members of specific binding pairs |
| GB9206318D0 (en) * | 1992-03-24 | 1992-05-06 | Cambridge Antibody Tech | Binding substances |
| US5780279A (en) * | 1990-12-03 | 1998-07-14 | Genentech, Inc. | Method of selection of proteolytic cleavage sites by directed evolution and phagemid display |
| DE69129154T2 (de) * | 1990-12-03 | 1998-08-20 | Genentech, Inc., South San Francisco, Calif. | Verfahren zur anreicherung von proteinvarianten mit geänderten bindungseigenschaften |
| ATE269401T1 (de) * | 1991-04-10 | 2004-07-15 | Scripps Research Inst | Bibliotheken heterodimerer rezeptoren mittels phagemiden |
| US6492160B1 (en) * | 1991-05-15 | 2002-12-10 | Cambridge Antibody Technology Limited | Methods for producing members of specific binding pairs |
| US5871907A (en) * | 1991-05-15 | 1999-02-16 | Medical Research Council | Methods for producing members of specific binding pairs |
| US5858657A (en) * | 1992-05-15 | 1999-01-12 | Medical Research Council | Methods for producing members of specific binding pairs |
| US6225447B1 (en) * | 1991-05-15 | 2001-05-01 | Cambridge Antibody Technology Ltd. | Methods for producing members of specific binding pairs |
| US5962255A (en) * | 1992-03-24 | 1999-10-05 | Cambridge Antibody Technology Limited | Methods for producing recombinant vectors |
| WO1993003151A1 (en) * | 1991-08-10 | 1993-02-18 | Medical Research Council | Treatment of cell populations |
| ES2136092T3 (es) * | 1991-09-23 | 1999-11-16 | Medical Res Council | Procedimientos para la produccion de anticuerpos humanizados. |
| US5872215A (en) * | 1991-12-02 | 1999-02-16 | Medical Research Council | Specific binding members, materials and methods |
| DE69233782D1 (de) * | 1991-12-02 | 2010-05-20 | Medical Res Council | Herstellung von Autoantikörpern auf Phagenoberflächen ausgehend von Antikörpersegmentbibliotheken |
| US5733743A (en) * | 1992-03-24 | 1998-03-31 | Cambridge Antibody Technology Limited | Methods for producing members of specific binding pairs |
| US5869644A (en) | 1992-04-15 | 1999-02-09 | The Johns Hopkins University | Synthesis of diverse and useful collections of oligonucleotidies |
| ATE233814T1 (de) | 1992-09-30 | 2003-03-15 | Scripps Research Inst | Humaner neutralisierender monoklonaler antikörper gegen den die immunschwäche beim menschen hervorringenden virus |
| US5837242A (en) * | 1992-12-04 | 1998-11-17 | Medical Research Council | Multivalent and multispecific binding proteins, their manufacture and use |
| GB9225453D0 (en) * | 1992-12-04 | 1993-01-27 | Medical Res Council | Binding proteins |
| US5422253A (en) * | 1992-12-07 | 1995-06-06 | Wisconsin Alumni Research Foundation | Method of site specific nucleic acid cleavage |
| ES2199959T3 (es) * | 1993-02-04 | 2004-03-01 | Borean Pharma A/S | Procedimiento mejorado para el replegamiento de proteinas. |
| US5714320A (en) * | 1993-04-15 | 1998-02-03 | University Of Rochester | Rolling circle synthesis of oligonucleotides and amplification of select randomized circular oligonucleotides |
| GB9313509D0 (en) * | 1993-06-30 | 1993-08-11 | Medical Res Council | Chemisynthetic libraries |
| JPH09503759A (ja) * | 1993-09-22 | 1997-04-15 | メディカル リサーチ カウンシル | 抗体のリターゲッティング |
| US5702892A (en) | 1995-05-09 | 1997-12-30 | The United States Of America As Represented By The Department Of Health And Human Services | Phage-display of immunoglobulin heavy chain libraries |
| US6696248B1 (en) * | 1995-08-18 | 2004-02-24 | Morphosys Ag | Protein/(poly)peptide libraries |
| WO1997008320A1 (en) * | 1995-08-18 | 1997-03-06 | Morphosys Gesellschaft Für Proteinoptimierung Mbh | Protein/(poly)peptide libraries |
| US5972693A (en) | 1995-10-24 | 1999-10-26 | Curagen Corporation | Apparatus for identifying, classifying, or quantifying DNA sequences in a sample without sequencing |
| US5854033A (en) * | 1995-11-21 | 1998-12-29 | Yale University | Rolling circle replication reporter systems |
| FR2741892B1 (fr) | 1995-12-04 | 1998-02-13 | Pasteur Merieux Serums Vacc | Procede de preparation d'une banque multicombinatoire de vecteurs d'expression de genes d'anticorps, banque et systemes d'expression d'anticorps "coliclonaux" obtenus |
| US5962271A (en) * | 1996-01-03 | 1999-10-05 | Cloutech Laboratories, Inc. | Methods and compositions for generating full-length cDNA having arbitrary nucleotide sequence at the 3'-end |
| DE19624562A1 (de) | 1996-06-20 | 1998-01-02 | Thomas Dr Koehler | Verfahren zur Bestimmung von Konzentrationsverhältnissen zweier sich um mindestens ein Nucleotid unterscheidender Nucleinsäuren und Testkit zur Durchführung des Verfahrens |
| JP3841360B2 (ja) * | 1996-06-24 | 2006-11-01 | ツェットエルベー・ベリング・アクチェンゲゼルシャフト | リーサスd抗原に対して特異性を有する抗原結合性構造を形成し得るポリペプチド、それらをコードするdna及びその調製方法並びに使用 |
| GB9712818D0 (en) * | 1996-07-08 | 1997-08-20 | Cambridge Antibody Tech | Labelling and selection of specific binding molecules |
| US5858671A (en) * | 1996-11-01 | 1999-01-12 | The University Of Iowa Research Foundation | Iterative and regenerative DNA sequencing method |
| WO1998047343A2 (en) * | 1997-04-04 | 1998-10-29 | Biosite Diagnostics, Inc. | Antibodies or binding protein libraries displayed on phage, cells, or other replicatable genetic packages |
| US6057098A (en) * | 1997-04-04 | 2000-05-02 | Biosite Diagnostics, Inc. | Polyvalent display libraries |
| WO1999006834A2 (en) | 1997-08-04 | 1999-02-11 | Ixsys, Incorporated | Methods for identifying ligand specific binding molecules |
| GB9722131D0 (en) * | 1997-10-20 | 1997-12-17 | Medical Res Council | Method |
| US7244826B1 (en) | 1998-04-24 | 2007-07-17 | The Regents Of The University Of California | Internalizing ERB2 antibodies |
| WO2000018905A1 (en) * | 1998-09-25 | 2000-04-06 | G.D. Searle & Co. | Method of producing permuteins by scanning permutagenesis |
| US6531580B1 (en) * | 1999-06-24 | 2003-03-11 | Ixsys, Inc. | Anti-αvβ3 recombinant human antibodies and nucleic acids encoding same |
| EP1990409A3 (en) * | 1999-07-20 | 2011-05-18 | MorphoSys AG | Bacteriophage |
| US6238904B1 (en) * | 1999-09-23 | 2001-05-29 | New England Biolabs, Inc. | Type II restriction endonuclease, HpyCH4III, obtainable from Helicobacter pylori CH4 and a process for producing the same |
| EP1276855B1 (en) | 2000-04-17 | 2012-10-24 | Dyax Corp. | Methods of constructing display libraries of genetic packages for members of a diverse family of peptides |
| US8288322B2 (en) * | 2000-04-17 | 2012-10-16 | Dyax Corp. | Methods of constructing libraries comprising displayed and/or expressed members of a diverse family of peptides, polypeptides or proteins and the novel libraries |
| US20050158838A1 (en) * | 2000-06-19 | 2005-07-21 | Dyax Corp., A Delaware Corporation | Novel enterokinase cleavage sequences |
| ES2430857T3 (es) * | 2000-12-18 | 2013-11-22 | Dyax Corp. | Bibliotecas focalizadas de paquetes genéticos |
| DE10230997A1 (de) * | 2001-10-26 | 2003-07-17 | Ribopharma Ag | Medikament zur Erhöhung der Wirksamkeit eines Rezeptor-vermittelt Apoptose in Tumorzellen auslösenden Arzneimittels |
-
2001
- 2001-04-17 EP EP01927108A patent/EP1276855B1/en not_active Expired - Lifetime
- 2001-04-17 WO PCT/US2001/012454 patent/WO2001079481A2/en active Application Filing
- 2001-04-17 CA CA2406236A patent/CA2406236C/en not_active Expired - Lifetime
- 2001-04-17 DK DK10179786T patent/DK2308982T3/da active
- 2001-04-17 AU AU5358901A patent/AU5358901A/xx active Pending
- 2001-04-17 CA CA2798625A patent/CA2798625C/en not_active Expired - Lifetime
- 2001-04-17 ES ES01927108T patent/ES2400302T3/es not_active Expired - Lifetime
- 2001-04-17 US US09/837,306 patent/US20040029113A1/en not_active Abandoned
- 2001-04-17 ES ES10156326T patent/ES2393535T3/es not_active Expired - Lifetime
- 2001-04-17 PT PT101797868T patent/PT2308982E/pt unknown
- 2001-04-17 ES ES10179786.8T patent/ES2531551T3/es not_active Expired - Lifetime
- 2001-04-17 EP EP10179786.8A patent/EP2308982B1/en not_active Expired - Lifetime
- 2001-04-17 AU AU2001253589A patent/AU2001253589B2/en not_active Expired
- 2001-04-17 EP EP10156326A patent/EP2199393B1/en not_active Expired - Lifetime
- 2001-04-17 DK DK10156326.0T patent/DK2199393T3/da active
- 2001-04-17 JP JP2001577464A patent/JP5149476B2/ja not_active Expired - Lifetime
- 2001-04-17 DK DK01927108.9T patent/DK1276855T3/da active
- 2001-04-17 EP EP10179777A patent/EP2295580A1/en not_active Withdrawn
-
2006
- 2006-02-28 US US11/365,556 patent/US9382535B2/en not_active Expired - Fee Related
-
2007
- 2007-08-21 AU AU2007211861A patent/AU2007211861B2/en not_active Expired
-
2010
- 2010-12-22 HK HK10111983.4A patent/HK1145515A1/en not_active IP Right Cessation
-
2011
- 2011-03-01 JP JP2011044525A patent/JP5599745B2/ja not_active Expired - Lifetime
- 2011-10-11 HK HK11110739A patent/HK1156656A1/xx not_active IP Right Cessation
-
2013
- 2013-12-11 JP JP2013255876A patent/JP5956413B2/ja not_active Expired - Lifetime
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5599745B2 (ja) | ペプチドの多種多様なファミリーのメンバーとしての、遺伝的パッケージの提示ライブラリーを構築する方法 | |
| EP1578903B2 (en) | Novel methods of constructing libraries comprising displayed and/or expressed members of a diverse family of peptides, polypeptides or proteins and the novel libraries | |
| AU2001253589A1 (en) | Methods of constructing display libraries of genetic packages for members of a diverse family of peptides | |
| JP2022550497A (ja) | バーコードに基づいた核酸配列アセンブリ | |
| US20140045728A1 (en) | Orthogonal Amplification and Assembly of Nucleic Acid Sequences | |
| US20040009507A1 (en) | Concatenated nucleic acid sequence | |
| US9523092B2 (en) | Compatible display vector systems | |
| US20100113304A1 (en) | Compatible display vector systems | |
| CA2737351A1 (en) | Single chain antibody library design | |
| Class et al. | Patent application title: Orthogonal Amplification and Assembly of Nucleic Acid Sequences Inventors: George M. Church (Brookline, MA, US) Sriram Kosuri (Cambridge, MA, US) Sriram Kosuri (Cambridge, MA, US) Nikolai Eroshenko (Boston, MA, US) Assignees: President and Fellows of Harvard College | |
| JP4581079B2 (ja) | 連結dnaの調製方法及びトランスフォーマントの調製方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20110301 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20120220 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20130117 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20130410 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20130415 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20130517 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20130522 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20130522 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20130909 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20130912 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20131211 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20140106 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20140507 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20140624 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20140717 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20140813 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 5599745 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |