FR2848064A1 - Plantes legumineuses transplastomiques fertiles - Google Patents
Plantes legumineuses transplastomiques fertiles Download PDFInfo
- Publication number
- FR2848064A1 FR2848064A1 FR0215490A FR0215490A FR2848064A1 FR 2848064 A1 FR2848064 A1 FR 2848064A1 FR 0215490 A FR0215490 A FR 0215490A FR 0215490 A FR0215490 A FR 0215490A FR 2848064 A1 FR2848064 A1 FR 2848064A1
- Authority
- FR
- France
- Prior art keywords
- expression cassette
- transformation
- legume
- plastoma
- transplastomic
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8201—Methods for introducing genetic material into plant cells, e.g. DNA, RNA, stable or transient incorporation, tissue culture methods adapted for transformation
- C12N15/8214—Plastid transformation
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- Chemical & Material Sciences (AREA)
- General Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Cell Biology (AREA)
- Microbiology (AREA)
- Plant Pathology (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Physics & Mathematics (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Pretreatment Of Seeds And Plants (AREA)
Abstract
L'invention concerne la transformation des plastes de plantes, et plus précisément l'obtention de plantes légumineuses transplastomiques fertiles, en particulier de soja transplastomique fertile.
Description
<Desc/Clms Page number 1>
Plantes légumineuses transplastomiques fertiles [0001] L'invention concerne la transformation des plastes de plantes, et plus précisément l'obtention de plantes légumineuses transplastomiques fertiles, en particulier de soja transplastomique fertile.
Etat de la technique [0002] L'information génétique des plantes est distribuée dans trois compartiments cellulaires : noyau, les mitochondries et les plastes. Chacun de ces compartiments porte son propre génome.
Depuis quelques années, les plastes des végétaux supérieurs sont une cible attractive pour les manipulations génétiques. Les plastes de plantes (les chloroplastes, siège de la photosynthèse, les amyloplastes accumulant de l'amidon, élaioplastes, étioplastes, chromoplastes accumulant des caroténoïdes, etc...) sont des centres majeurs de biosynthèse qui, outre la photosynthèse, sont responsables de la production de composés industriellement importants tels que les acides aminés, les carbohydrates, les acides gras et les pigments. Les plastes sont dérivés d'un précurseur indifférencié commun, le proplaste, et possèdent donc, dans une espèce de plante donnée, le même contenu génétique.
] Le génome plastidial, ou plastome, des végétaux supérieurs est constitué d'une molécule d'ADN circulaire double brin de 120-160 kilobases, portant une large séquence répétée et inversée (environ 25 kb). Une caractéristique remarquable du génome plastidial réside dans la présence de nombreuses copies identiques de ce génome dans toutes les cellules et tous les types de plastes. Selon le stade de développement, une cellule de feuille de tabac peut contenir jusqu'à
10 000 copies de plastomes. Il est donc possible de manipuler des cellules de plantes contenant jusqu'à 20 000 copies d'un gène d'intérêt, lequel peut potentiellement résulter dans un fort niveau d'expression de gène hétérologue.
10 000 copies de plastomes. Il est donc possible de manipuler des cellules de plantes contenant jusqu'à 20 000 copies d'un gène d'intérêt, lequel peut potentiellement résulter dans un fort niveau d'expression de gène hétérologue.
] La transformation des génomes plastidiaux de plantes offre un potentiel énorme pour les biotechnologies végétales et de nombreux avantages très attractifs par rapport à la transformation conventionnelle du génome nucléaire. Le premier avantage réside dans le mécanisme même de la transformation plastidiale. En effet, l'intégration d'un transgène dans le plastome procède d'un phénomène de double recombinaison homologue. Ce processus permet de cibler précisément la région du plastome où l'on souhaite intégrer le gène d'intérêt, notamment grâce à l'utilisation de séquences plastidiales positionnées de part et d'autre du
<Desc/Clms Page number 2>
transgène sur le vecteur de transformation. Ce ciblage précis évite l'effet dit de position, fréquemment observé dans les événements de transgénèse nucléaire.
Le deuxième avantage réside dans le nombre élevé de copies de transgènes par plaste.
Les cellules de plantes peuvent être manipulées de manière à contenir jusqu'à 20 000 copies d'un gène d'intérêt. Cette caractéristique permet de forts niveaux d'expression de transgènes pouvant résulter dans une accumulation de protéines transgènes allant jusqu'à 40% des protéines cellulaires totales solubles (De Cosa et al., 2001, Nat. Biotechnol. 19,71-74).
La nature procaryotique du plaste constitue un autre atout, notamment en permettant l'expression de gènes organisés en opérons et la traduction efficace d'ARNm polycistroniques.
Cette particularité facilite l'intégration et le fonctionnement coordonné de plusieurs transgènes, tout en limitant le nombre d'étapes de transformation et le recours à de multiples marqueurs de sélection (Daniell, 1998, Nat. Biotechnol. 16, 345-8; De Cosa et al., 2001, Nat. Biotechnol. 19, 71-74).
Un autre avantage de la transformation plastidiale par rapport à la transformation nucléaire réside dans le contrôle de la dispersion des transgènes dans l'environnement. Chez de nombreux Angiospermes, les plastes ont une hérédité maternelle stricte, et l'ADN plastidial n'est pas transmis via le pollen. Cette particularité limite donc fortement le risque de dispersion du transgène dans l'environnement, et sa propagation potentielle à des végétaux avoisinants.
De nombreuses applications de la transformation plastidiale ont permis de confirmer les avantages de cette technologie sur la transformation nucléaire. Ainsi, la surexpression à partir du plastome de tabac de gènes de tolérance à des herbicides tels que le glyphosate (Daniell, 1998, Nat. Biotechnol. 16, 345-8; W099/10513; Ye et al., 2000 ; WO 01/04331, WO 01/04327), ou la phosphinothricine (Basta) (Lutz et al., 2001, Physiol. Plant 125,1585-1590), confère une excellente tolérance à ces herbicides. D'autres applications ont conduit à l'obtention de plantes transplastomiques tolérantes aux insectes ou surproduisant des protéines thérapeutiques (McBride et al., 1995; US Patent 5,451,513; Staub et al., 2000, Nat. Biotech. 18, 333-338).
] Pour obtenir une transformation plastidiale, l'ADN transformant doit traverser la paroi cellulaire, la membrane plasmique et la double membrane de l'organite avant d'atteindre le stroma. A ce titre, la technique la plus couramment utilisée pour transformer le génome
<Desc/Clms Page number 3>
plastidial est celle du bombardement de particules (Svab et Maliga, 1993, Proc. Natl. Acad. Sci.
USA, Feb 1, 90(3): 913-7).
Actuellement, chez les végétaux supérieurs, la transformation stable des plastes est pratiquée de manière courante seulement chez le tabac, Nicotiana tabacum (Svab et Maliga, 1990 Proc. Natl. Acad. Sci. USA 87, 8526-8530; Svab et Maliga, 1993, Proc. Natl. Acad. Sci.
USA, Feb 1, 90(3): 913-7). Bien que cette technique ait démontré son efficacité chez le tabac, sa transposition à des espèces végétales de grandes cultures semble confrontée à des obstacles techniques. Un de ces obstacles pourrait être, non pas une difficulté de transformation, mais probablement une limitation dans les systèmes de culture in vitro de tissus actuellement disponibles et dans les protocoles de transformation et de régénération de plantes transplastomiques. Quelques progrès récents ont néanmoins été réalisés avec la transformation des plastes de riz (Khan M. S. et Maliga, 1999, Nat. Biotechnol. 17,910-915), d'Arabidopsis thaliana (Sikdar et al, 1998, Plant Cell Reports 18 :20-24), de pomme de terre (Sidorov et al, 1999, Plant J. 19(2): 209-216), de Brassica napus (Chaudhuri et al, 1999) et de tomates (Ruf et al., 2001, Nat. Biotechnol. 19,870-875).
Récemment, Zhang et al. (2001, J. Plant Biotechnol. 3,39-44) ont décrit une technique de transformation de plastes d'une suspension cellulaire de soja à très faible fréquence. Toutefois, cette technique donne des tissus incapables de régénérer des plantes. A la connaissance des inventeurs, aucune légumineuse transplastomique fertile, et plus particulièrement aucun soja transplastomique fertile n'a été obtenu à ce jour.
] De très nombreuses espèces cultivées appartiennent à la famille des Légumineuses, notamment des protéagineux comme le pois, la féverole, le haricot, le pois chiche, les lentilles, des oléo-protéagineux comme le soja et l'arachide, et des fourrages comme la luzerne ou le trèfle. Une propriété fondamentale des Légumineuses, qui est largement responsable de leur intérêt agronomique, est leur forte teneur en protéines. Cette propriété en fait des plantes de choix pour sur-exprimer des protéines d'intérêt.
] Le soja, essentiellement cultivé en Amérique du Nord et Latine, ainsi qu'en Chine, est exporté en majorité vers l'Europe. Ces dernières années, des caractères de résistance à un herbicide ou aux insectes ravageurs, ont été introduits au niveau du génome nucléaire du soja.
Ces manipulations génétiques au niveau du génome nucléaire de soja ont été accomplies grâce à
Ces manipulations génétiques au niveau du génome nucléaire de soja ont été accomplies grâce à
<Desc/Clms Page number 4>
la technique de bombardement de particules. De nombreux génotypes ont ainsi été produits présentant une augmentation de la tolérance à des herbicides (Roundup Ready Soybean, Pagette et al., 1995, Crop Sci. 35,1451-1461) ou à des insectes ravageurs (Stewart et al., 1996, Plant Physiol. 112 : 121-129), ou une amélioration des traits de qualité tels que les acides gras, le phytate, les acides aminés (Soy 2000, 8th biennial Conference of the cellular and Molecular biology of the soybean, Lexington, Kentucky).
Dans ce contexte, et au vu des avantages techniques de la transformation plastidiale mentionnés plus haut, il devient crucial de mettre au point une technique fiable de transformation et de régénération de Légumineuses transplastomiques fertiles, en particulier de sojas. Ainsi, les inventeurs ont développé une méthode de transformation à haute fréquence des plastomes de soja conduisant à des plantes fertiles. Cette méthode peut aisément être adaptée à la transformation d'autres Légumineuses d'intérêt agronomique.
<Desc/Clms Page number 5>
Description [0015] La présente invention concerne une plante Légumineuse transplastomique fertile.
Selon la présente invention, on entend par Légumineuse une plante de la famille des Fabaceae. Des Légumineuses préférées selon l'invention sont les Légumineuses d'intérêt agronomique comme le pois (Pisum sativum), la fève (Viciafaba major), la féverole (Viciafaba minor), les lentilles (Lens culinaris), le haricot (Phaseolus vulgaris), le pois chiche (Cicer arietinum), le soja (Glycine max), l'arachide (Arachis hypogea), la luzerne (Medicago sativa), ou le trèfle (Trifolium sp.) [0017] Selon un mode de réalisation préféré de l'invention, la Légumineuse transplastomique fertile est le soja, Glycine max.
Par "transplastomique", on entend selon l'invention des plantes ayant intégré de manière stable dans leur plastome au moins une cassette d'expression fonctionnelle dans les plastes. Le plastome est constitué par le génome des organites cellulaires autres que le noyau et les mitochondries. Une cassette d'expression selon l'invention comprend, entre autres éléments, au moins un promoteur fonctionnel dans les plastes de cellules végétales, une séquence codant pour une protéine d'intérêt et un terminateur fonctionnel dans les plastes de cellules végétales. Ladite cassette d'expression peut contenir des éléments génétiques originaires de la plante transformée ou de tout autre organisme.
[0019] De préférence, les plantes Légumineuses transplastomiques selon l'invention sont à l'état d'homoplasmie. L'état d'homoplasmie correspond à un état selon lequel toutes les cellules contiennent une population de plastomes identiques. Selon l'invention, des plantes transplastomiques sont à l'état d'homoplasmie lorsque toutes leurs cellules ne contiennent que des copies de plastomes transformés, et plus aucune copies de plastomes non-transformés. Cet état est généralement obtenu par sélection des copies de plastomes ayant intégré la cassette d'expression, notamment grâce à l'association de ladite cassette d'expression avec un gène codant pour un marqueur de sélection. Les plastomes n'ayant pas intégré le marqueur de sélection sont alors éliminés lorsque les tissus transformés sont mis en contact avec l'agent de sélection correspondant.
<Desc/Clms Page number 6>
] Selon l'invention, les Légumineuses transplastomiques sont fertiles. Une plante fertile est une plante capable de produire une descendance viable par le biais d'un cycle de reproduction sexuée. En particulier, une plante fertile selon l'invention est une plante transplastomique capable de transmettre la cassette d'expression intégrée dans son plastome à sa descendance.
L'invention comprend également des vecteurs de transformation adaptés à la transformation des plastes. On entend par "vecteur adapté à la transformation des plastes", un vecteur capable d'intégrer de manière stable la (ou les) cassette(s) d'expression(s) qu'il contient dans le plastome des cellules végétales. De manière avantageuse, un vecteur adapté à la transformation des plastes selon l'invention est un vecteur comprenant au moins deux séquences homologues d'une zone du plastome de la légumineuse à transformer, lesdites séquences homologues encadrant au moins une cassette d'expression. Selon un mode de réalisation préféré, lesdites séquences homologues encadrent, en plus d'une cassette d'expression codant pour une protéine d'intérêt, au moins une autre cassette d'expression codant pour un marqueur de sélection. Avec de tels vecteurs, l'intégration de la (ou des) cassette(s) d'expression(s) dans le plastome est réalisée par double recombinaison homologue des deux séquences homologues d'une zone du plastome de la légumineuse à transformer présentes sur le vecteur avec celles correspondantes dans le plastome de la légumineuse à transformer. De manière avantageuse, les deux séquences homologues d'une zone du plastome de la légumineuse à transformer permettent l'intégration de la (ou des) cassette (s) d'expression (s) dans une zone intergénique du génome plastidial sans interrompre l'intégrité ni le fonctionnement des gènes plastidiaux. De manière préférée, cette zone correspond à la région de l'opéron des ARN ribosomiques du plastome.
Selon un mode particulier de réalisation de l'invention, les séquences homologues d'une zone du plastome de la légumineuse à transformer correspondent à des séquences présentant 80% d'identité avec les séquences correspondantes dans le plastome de la légumineuse à transformer, de préférence 90% d'identité, de préférence 95%, et de préférence 99% d'identité.
Selon un mode préféré de réalisation de l'invention, les séquences homologues d'une zone du plastome de la légumineuse à transformer correspondent à des séquences présentant 100% d'identité avec les séquences correspondantes dans le plastome de la légumineuse à transformer.
L'invention concerne donc un vecteur adapté à la transformation des plastes, caractérisé en ce que les deux séquences homologues d'une zone du plastome de la légumineuse à transformer correspondent à des séquences permettant l'intégration de la cassette d'expression
<Desc/Clms Page number 7>
dans une région intergénique de plastome. Selon un mode de réalisation préféré, ladite zone correspond à la région de l'opéron des ARN ribosomiques du plastome.
L'invention comprend également une Légumineuse transplastomique fertile caractérisée en ce qu'elle comprend au moins une cassette d'expression insérée dans une région intergénique de plastome. Selon un mode de réalisation préféré, ladite région intergénique est sélectionnée dans la région de l'opéron des ARN ribosomiques du plastome.
Selon un mode de réalisation particulier de l'invention, l'une des deux séquences homologues comprend les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV), et l'autre séquence homologue comprend la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7. L'invention concerne donc un vecteur adapté à la transformation des plastes, caractérisé en ce que l'une des deux séquences homologues comprend les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV). et que l'autre séquence homologue comprend la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7.
L'invention comprend donc également une Légumineuse transplastomique fertile caractérisée en ce qu'elle comprend au moins une cassette d'expression insérée dans une région intergénique de plastome, ladite région intergénique de plastome est située entre le gène ARNtV et l'opéron rpsl2/7.
Selon un mode préféré de réalisation de l'invention, la Légumineuse à transformer est le soja. Selon ce mode de réalisation, la séquence comprenant les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV) correspond à la séquence représentée par l'identificateur SEQ ID NO:1, et la séquence comprenant la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7 correspond à la séquence représentée par l'identificateur SEQ ID NO:2. L'invention concerne donc un vecteur adapté à la transformation des plastes, caractérisé en ce que la séquence homologue comprenant les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV) est représentée par l'identificateur de séquences SEQ ID NO: 1, et que la séquence homologue comprenant la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7 est représentée par l'identificateur de séquences SEQ ID NO:2.
<Desc/Clms Page number 8>
] Selon un mode de réalisation particulier, l'invention comprend donc un soja transplastomique fertile, caractérisé en ce qu'il comprend au moins une cassette d'expression insérée dans une région intergénique de plastome, ladite cassette d'expression étant insérée entre les séquences du plastome de soja correspondant aux identificateurs SEQ ID NO:1et SEQID NO:2.
Selon un mode préféré de réalisation de l'invention, la séquence homologue comprenant les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV) est positionnée en 5' de la cassette d'expression, et la séquence homologue comprenant la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7 est positionnée en 3' de la cassette d'expression.
Les vecteurs de transformation adaptés à la transformation des plastes selon l'invention comprennent au moins une cassette d'expression. Une cassette d'expression selon l'invention comprend au moins, liés entre eux de manière opérationnelle, un promoteur fonctionnel dans les plastes de cellules végétales, une séquence codant pour une protéine d'intérêt et un terminateur fonctionnel dans les plastes de cellules végétales. L'expression "liés entre eux de manière opérationnelle" signifie que lesdits éléments de la cassette d'expression sont liés entre eux de manière à ce que leur fonctionnement soit coordonné et permette l'expression de la séquence codante. A titre d'exemple, un promoteur est lié de manière opérationnelle à une séquence codante lorsqu'il est capable d'assurer l'expression de ladite séquence codante. La construction d'une cassette d'expression selon l'invention et l'assemblage de ses différents éléments est réalisable par l'emploi de techniques bien connues de l'homme du métier, notamment celles décrites dans Sambrook et al. (1989, Molecular Cloning : Laboratory Manual, Nolan C. ed., New York : Spring Harbor Laboratory Press). Le choix des éléments régulateurs constituant la cassette d'expression est essentiellement fonction de la plante et du type de plaste dans lesquelles ils doivent fonctionner, et l'homme du métier est capable de sélectionner des éléments régulateurs fonctionnels dans une plante donnée.
Parmi les promoteurs fonctionnels dans les plastes de cellules végétales, on peut noter à titre d'exemple le promoteur du gène psbA, codant pour la protéine Dl du PSII (Staub et al., 1993, EMBO Journal 12 (2): ou le promoteur constitutif de l'opéron des ARN ribosomaux, Prrn (Staub et al., 1992, Plant Cell 4: 39-45). D'une manière générale, tout promoteur issu d'un gène de plastome de plantes conviendra, et l'homme du métier saura faire le
<Desc/Clms Page number 9>
choix adéquat parmi les différents promoteurs disponibles de manière à obtenir un mode d'expression désiré (constitutif ou inductible). Un promoteur préféré selon l'invention comprend le promoteur Prrn de Tabac associé à une portion 5' de la région 5' non-traduite du gène rbcL de tabac (Svab et Maliga, 1993, Proc. Natl. Acad. Sci. 90 : 913-917).[0032] Parmi les terminateurs fonctionnels dans les plastes de cellules végétales, on peut noter à titre d'exemple le terminateur du gène psbA de tabac (Shinozaki et al., 1986; Staub et al., 1993).
D'une manière générale, tout terminateur issu d'un gène de plastome de plantes conviendra, et l'homme du métier saura faire le choix adéquat parmi les différents terminateurs disponibles.
De manière avantageuse, le vecteur utilisé dans la présente invention peut contenir, en plus d'une cassette d'expression comprenant une séquence codant pour une protéine d'intérêt, au moins une autre cassette d'expression comprenant une séquence codant pour un marqueur de sélection. Le marqueur de sélection permet de sélectionner les plastes et les cellules effectivement transformées, c'est-à-dire celles ayant incorporé la (ou les) cassette (s) dans leur plastome. Il permet également d'obtenir des plastes transplastomiques fertiles à l'état d'homoplasmie. Parmi les séquences utilisables codant pour des marqueurs de sélection, on peut citer celles des gènes de résistance aux antibiotiques tel que, par exemple, celle du gène aadA codant pour une amino glycoside 3"-adényltransférase, qui confère une résistance à la spectinomycine et à la streptomycine (Svab et al., 1993; Staub et al., 1993), ou celle du gène de l'hygromycine phosphotransférase (Gritz et al., 1983, Gene 25 :179-188), mais également celles des gènes de tolérance aux herbicides tel que le gène bar (White et al., 1990, Nucleic Acid Res.
18(4):1062) pour la tolérance au bialaphos, le gène EPSPS (US 5,188,642) pour la tolérance au glyphosate ou encore le gène HPPD (WO 96/38567) pour la tolérance aux isoxazoles. On peut également utiliser des séquences de gènes rapporteurs codant pour des enzymes facilement identifiables comme l'enzyme GUS, des séquences de gènes codant pour des pigments ou des enzymes régulant la production de pigments dans les cellules transformées. De tels gènes sont notamment décrits dans les demandes de brevet WO 91/02071, WO 95/06128, WO 96/38567, et WO 97/04103.
] Selon un mode de réalisation préféré de l'invention, le gène codant pour un marqueur de sélection est le gène aadA codant pour une amino glycoside 3"-adényltransférase, qui confère aux cellules et aux plastes transformés une résistance à la spectinomycine et à la streptomycine (Svab et al., 1993; Staub et al., 1993).
<Desc/Clms Page number 10>
L'invention concerne également un procédé d'obtention de légumineuses transplastomiques fertiles. Ce procédé comprend les étapes de : (a) Transformation de tissus embryogènes obtenus à partir d'embryons immatures de légumineuses avec un vecteur adapté à la transformation des plastes (b) Sélection des tissus transformés (c) Régénération de plantes transplastomiques fertiles à partir des tissus transformés [0036] Pour mettre en #uvre le procédé selon l'invention, l'étape (a) de transformation doit s'effectuer sur des tissus embryogènes obtenus à partir d'embryons immatures de légumineuses.
De préférence, les tissus embryogènes sont des cals ou tout autre tissu comportant des cellules ayant conservé un état de totipotence.
La transformation des tissus embryogènes peut s'effectuer par toute méthode de transformation directe (ADN nu) ou indirecte de cellules végétales. Parmi les méthodes de transformation utilisables pour obtenir des plantes transplastomiques selon l'invention, une de celles-ci consiste à mettre les cellules ou tissus des plantes à transformer en présence de polyéthylène glycol (PEG) et du vecteur de transformation (Chang and Cohen, 1979, Mol. Gen.
Genet. 168(1), 111-115 ; Mercenier and Chassy, 1988, Biochimie 70(4), 503-517). L'électroporation est une autre méthode qui consiste à soumettre les cellules ou tissus à transformer et les vecteurs à un champ électrique (Andreason and Evans, 1988, Biotechniques 6 (7), 650-660; Shigekawa and Dower, 1989, Aust. J. Biotechnol. 3(1), 56-62). Une autre méthode consiste à injecter directement les vecteurs dans les cellules ou les tissus par microinjection (Gordon and Ruddle, 1985, Gene 33(2), 121-136). La transformation de plastomes peut également se faire à l'aide de bactéries du genre Agrobacterium, de préférence par infection des cellules ou tissus desdites plantes par A. tumefaciens (Knopf, 1979, Subcell. Biochem. 6, 143- 173; Shaw et al., 1983, Gene 23(3):315-330) ou A. rhizogenes (Bevan et Chilton, 1982, Annu.
Rev. Genet. 16:357-384; Tepfer and Casse-Delbart, 1987, Microbiol. Sci. 4(1). 24-28). De manière préférentielle, la transformation de cellules ou tissus végétaux par Agrobacterium tumefaciens est réalisée selon le protocole décrit par Ishida et al. (1996, Nat. Biotechnol. 14(6), 745-750). Pour la transformation des plastomes, la souche d'Agrobacterium utilisée doit être préalablement transformée de manière à diriger spécifiquement son ADN-T dans les plastes.
Selon un mode de réalisation préféré du procédé selon l'invention, la méthode dite de
<Desc/Clms Page number 11>
bombardement de particules sera utilisée. Elle consiste à bombarder les tissus embryogènes avec des particules, de préférence d'or ou de tungstène, sur lesquelles sont adsorbés les vecteurs selon l'invention (Bruce et al., 1989, Proc. Natl. Acad. Sci. USA 86 (24), Finer et al., 1992, Plant Cell Rep. 11, 232-238; Klein et al., 1992, Biotechnology 10(3), 286-291; US Patent No.
4,945,050).
Selon le présent procédé d'obtention de légumineuses transplastomiques fertiles, les tissus embryogènes sont transformés avec un vecteur adapté à la transformation des plastes, tel que décrit dans la présente invention.
Lors de l'étape (a) de transformation des tissus embryogènes, tous les tissus soumis à la technique de transformation n'intègrent pas le vecteur. L'étape (b) de sélection des tissus transformés transplastomiques s'effectue par mise en contact des tissus soumis à l'étape (a) de transformation avec l'agent de sélection correspondant au gène marqueur de sélection employé. Durant cette phase, seules les cellules ayant intégré le gène marqueur de sélection vont survivre au contact de l'agent de sélection et former des cals verts. La durée de mise en contact des tissus avec l'agent de sélection dépend du marqueur et de l'agent de sélection employés, et peut aisément être déterminée par un homme du métier. De préférence, cette durée correspond à une période allant jusqu'à la formation desdits cals verts à partir des tissus transformés.
L'étape (c) de régénération de plantes transplastomiques fertiles à partir des tissus transformés s'effectue en induisant la formations d'embryons à partir des tissus transplastomiques sélectionnés à l'étape (b). L'induction de formation d'embryon s'effectue généralement par mise en contact desdits tissus avec un milieu d'embryogenèse approprié. De tels milieux sont connus de l'homme du métier. Un milieu préféré selon l'invention est le milieu décrit dans Finer et McMullen (1991).
Une fois induits, les embryons formés sont mis à germer dans un milieu approprié. De préférence, le milieu approprié à la germination est un milieu gélosé comprenant les éléments nutritifs nécessaires à la germination. Les jeunes plantules formées sont ensuite plantées dans un substrat adapté à la croissance des plantes. Un substrat préféré est la terre, ou un mélange à base de terre.
L'invention comprend également des parties des légumineuses transplastomiques fertiles,
<Desc/Clms Page number 12>
et la descendance de ces plantes. On entend par "parties", tout organe de ces plantes, qu'il soit aérien ou souterrain. Les organes aériens sont les tiges, les feuilles, et les fleurs comprenant les organes reproducteurs mâles et femelles. Les organes souterrains sont principalement les racines, mais ils peuvent également être des tubercules. Par "descendance", on entend principalement les graines contenant les embryons issus de la reproduction de ces plantes entreelles. Par extension, le terme "descendance" s'applique à toutes les graines formées à chaque nouvelle génération issue de croisements dont au moins un des parents est une plante transformée selon l'invention. Une descendance peut également être obtenue par multiplication végétative desdites plantes transformées. Les graines selon l'invention peuvent être enrobées d'une composition agrochimique comprenant au moins un produit actif possédant une activité sélectionnée parmi les activités fongicide, herbicide, insecticide, nématicide, bactéricide ou virucide.
Parmi les séquences codant pour une protéine d'intérêt qui peuvent être intégrées dans les légumineuses transplastomiques selon l'invention, on peut citer les séquences codantes de gènes codant une enzyme de résistance à un herbicide, comme par exemple le gène bar codant pour l'enzyme PAT (White et al., NAR 18:1062, 1990) conférant une tolérance au bialaphos, le gène codant pour une enzyme EPSPS (WO 97/04103) conférant une tolérance au glyphosate, ou le gène codant pour une enzyme HPPD (WO 96/38567) conférant une tolérance aux isoxazoles. On peut également citer un gène codant pour une toxine insecticide, par exemple un gène codant pour une 8-endotoxine de la bactérie Bacillus thuringiensis (WO 98/40490). Peuvent également être introduits dans ces plantes des gènes de résistance aux maladies. par exemple un gène codant pour l'enzyme oxalate oxydase tel que décrit dans la demande de brevet EP 0 531 498 ou le brevet US 5,866,778, ou un gène codant pour un autre peptide antibactérien et/ou antifongique tels que ceux décrits dans les demandes de brevets WO 97/30082, WO 99/24594, WO 99/02717, WO 99/53053, et W099/91089. On peut également introduire des gènes codant pour des caractères agronomiques de la plante, en particulier un gène codant pour une enzyme delta-6 désaturase tel que décrit dans les brevets US 5,552,306, US 5,614,313, et demandes de brevets WO 98/46763 et WO 98/46764, ou un gène codant pour une enzyme sérine acétyltransférase (SAT) tel que décrit dans les demandes de brevets WO 00/01833 et WO 00/36127.
Selon un mode particulier de réalisation de l'invention, les légumineuses transplastomiques selon l'invention peuvent être transformées avec une cassette d'expression codant pour une protéine d'intérêt pharmaceutique ou vétérinaire. A titre d'exemple, une telle
<Desc/Clms Page number 13>
protéine peut être un anticoagulant (sérum protéase, hirudine), un interféron, l'albumine de sérum humaine. Les protéines produites par les plantes selon l'invention peuvent également être des anticorps, ou des protéines servant de base à des vaccins.
<Desc/Clms Page number 14>
Les exemples ci-après permettent d'illustrer la présente invention, sans toutefois en limiter la portée.
Exemples Exemple 1 : Construction d'un vecteur adapté à la transformation des plastes de soja
Le plasmide pCLT312 contient une cassette d'expression hétérologue AADA-312, encadrée par deux fragments d'ADN plastidial de soja, RRHD (Région de Recombinaison Homologue Droite) et RRHG (Région de Recombinaison Homologue Gauche) permettant l'intégration ciblée dans la région de l'opéron des ARN ribosomiques du plastome de soja. Cette région d'insertion est différente de celle utilisée par Zhang et al. (2001). La région RRHD contient les gènes codant pour l'ARNrl6S (sous le contrôle du promoteur de l'opéron des ARN ribosomaux noté Prrn) et l'ARNtV (SEQ ID NO:1). La région RRHG contient la région intergénique comprise entre le gène ARNtV et l'opéron rpsl2/7 (SEQ ID NO:2). Aucun gène plastidial n'est interrompu après recombinaison homologue avec ces séquences.
Le plasmide pCLT312 contient une cassette d'expression hétérologue AADA-312, encadrée par deux fragments d'ADN plastidial de soja, RRHD (Région de Recombinaison Homologue Droite) et RRHG (Région de Recombinaison Homologue Gauche) permettant l'intégration ciblée dans la région de l'opéron des ARN ribosomiques du plastome de soja. Cette région d'insertion est différente de celle utilisée par Zhang et al. (2001). La région RRHD contient les gènes codant pour l'ARNrl6S (sous le contrôle du promoteur de l'opéron des ARN ribosomaux noté Prrn) et l'ARNtV (SEQ ID NO:1). La région RRHG contient la région intergénique comprise entre le gène ARNtV et l'opéron rpsl2/7 (SEQ ID NO:2). Aucun gène plastidial n'est interrompu après recombinaison homologue avec ces séquences.
La cassette d'expression du vecteur pCLT312 (AADA-312) contient un gène chimérique composé, de 5' en 3', du promoteur "court" de l'opéron des ARN ribosomiques de tabac (PrrnC, nucléotides 102 561 à 102 677 du plastome de Nicotiana tabacum; Shinozaki et al., 1986), d'une portion 5'rbcL de la région 5' non traduite du gène rbcL de tabac (nucléotides 57 569 à 57 584 du plastome de Nicotiana tabacum; Shinozaki et al., 1986), de la séquence codante du gène aadA et du terminateur 3'psbA de tabac (nucléotides 533à 146 du plastome de N. tabacum; Shinozaki et al., 1986). Le produit du gène aadA, une amino glycoside 3"-adényltransférase, confère la résistance à la spectinomycine et à la streptomycine aux plantes transformées au niveau de leur génome plastidial (Svab et al., 1993; Staub et al., 1993).
Le vecteur pCLT312 a été obtenu comme décrit ci-dessous.
Les deux fragments d'ADN plastidial de soja (constituant les régions de recombinaison homologues RRHD et RRHG) ont été amplifiés par PCR à partir d'ADN total de Glycine max (cv. Jack) (PWO DNA polymérase, Stratagene). La région RRHD a été obtenue en utilisant les oligonucléotides OSSD5 (SEQ ID NO:4) et OSSD3 (SEQ ID NO:3). L'appariement (à une température de 60 C) de ce couple d'amorce a conduit à l'amplification d'un fragment de 1800 pb. De plus, la séquence de ces amorces génère des sites de restrictions en 5' et en 3' permettant les clonages ultérieurs. La région RRHG a été amplifiée en utilisant les amorces OSSG5 (SEQ
<Desc/Clms Page number 15>
ID N0:6) et OSSG3 (SEQ ID NO:5) conçues de manière à insérer des sites de restriction en 5' et en 3'. Au cours de cycles de réaction PCR, la température d'appariement appliquées est de 60 C.
Le produit PCR obtenu, de environ 1400 pb, a une taille supérieure à celle attendue (de 1180 pb) déterminée d'après la séquence du plastome de soja publiée dans GeneBank (X07675). Le séquençage des fragments PCR de ces deux régions montre la présence d'une insertion de 217 pb dans la région RRHG. Cette région insérée, d'après la séquence analysée, ne contient aucune ORF et s'avère être une région intergénique.
Après purification sur gel d'agarose, ces deux fragments PCR, RRHG et RRHD, ont été clonés dans le vecteur pPCRscript (Stratagene) pour donner les vecteurs pCLT309 et pCLT308, respectivement. La région RRHG excisé du vecteur pCLT309 par digestion KpnI a été cloné dans pCLT308 digéré au préalable par cette enzyme. Dans le vecteur pCLT300 obtenu, une cassette d'expression hétérologue plastidiale de tabac a ensuite été clonée à l'aide des enzymes XhoI et HindIII pour donner le vecteur pCLT311. Cette cassette contient un gène chimérique composé, de 5' en 3', du promoteur "court" PrrnC de l'opéron des ARN ribosomiques de tabac d'une portion 5'rbcL de la région 5' non traduite du gène rbcL de tabac, de la séquence codante d'un gène d'intérêt et du terminateur 3'psbA de tabac. Le gène d'intérêt présent dans pCLT311a été excisé par digestion avec les enzymes NcoI et XbaI, puis remplacé par le gène aadA libéré par ces mêmes enzymes du plasmide pCLT115. Le vecteur de transformation plastidial obtenu est nommé pCLT312.
Exemple 2 : Transformation par bombardement des génomes plastidiaux de soja
La technique de transformation du soja utilisée est le bombardement de particules. Elle est appliquée sur des tissus embryogènes de soja. Des tissus embryogènes de Glycine max (cv. Jack) ont été obtenus (préparés en condition stériles) en deux phases: une phase d'induction, et une phase de multiplication.
La technique de transformation du soja utilisée est le bombardement de particules. Elle est appliquée sur des tissus embryogènes de soja. Des tissus embryogènes de Glycine max (cv. Jack) ont été obtenus (préparés en condition stériles) en deux phases: une phase d'induction, et une phase de multiplication.
Des gousses de soja sont récoltées en serre lorsque les embryons sont encore immatures (3 mm de long maximum). Elles sont décontaminées à l'eau de javel diluée et rincées à l'eau stérile. Les gousses sont ouvertes sous hotte, en conditions stériles et les embryons sont récupérés. Les deux cotylédons sont séparés et déposés face externe sur un milieu d'induction gélosé D40. Le milieu D40 est un milieu Murashige and Skoog décrit dans Murashige and Skoog (1962, A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol.
Plant. 15 : 473-497). Il comprend (en mg/1): NH4NO3: 1650, H3BO3: 6,2; CaCl2, 2H2O: 332,2: CoCl2, 6H2O: 0,025; CUS04, 5H2O: 0,025 ; Na2EDTA: 37,26; FeS04, 7H2O: 27,8 ; MnS04, 7H2O:

16,9; Na2MoO4, 2H,O: 0,25; KI: 0,83; KNO;: 1900; KH2po4: 170; ZnSO4, 7H0: 8,6; de la
16,9; Na2MoO4, 2H,O: 0,25; KI: 0,83; KNO;: 1900; KH2po4: 170; ZnSO4, 7H0: 8,6; de la
<Desc/Clms Page number 16>
vitamine Gamborg B5 (Gamborg, Miller and Ojima, 1968, Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 50 : composée de (en mg/1) : myo- inositol : 100; acide nicotinique: 1; Pyridoxine-Hcl: 1; Thiamine-Hcl: 10), ainsi que de 40 mg/1 de 2,4-D; 6% saccharose; et 0,3% gelrite, pH 7,0.
Ce milieu est riche en sucre et en 2,4-D, des substances nécessaires à l'induction des embryons somatiques. Les embryons sont laissés sur ce milieu durant 3 semaines à 28 C, avec une luminosité et photopériode déterminées (24 C, avec 16 heures de jour et 8 heures de nuit).
Les embryons somatiques qui se sont développés à la surface des cotylédons sont récupérés puis étalés sur du milieu D20 qui comprend essentiellement les mêmes éléments que le milieu D40, à l'exception de la concentration en 2,4D qui est de 20 mg/1 et de la concentration en saccharose qui est abaissée de 60 g/1 à 30 g/1 à pH 5,7. Cette phase d'amplification dure 2 semaines sur le milieu D20 à 28 C.
Les embryons sont ensuite régulièrement repiqués sur un milieu FNL dérivé de celui décrit par Samoylov et al. (1998). Le milieu FNL modifié comprend (en mg/1): Na2EDTA: 37,24 ; FeS04, 7H2O: 27,84; MgS04, 7H20: 370; MnS04, H2O: 16,9; ZnSO4, H2O: 8,6; CuS04,
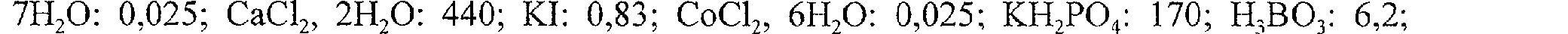
7H20: 0,025; CaCl2, 2H20: 440; KI: 0,83; COC'2, 6II20: 0,025; KH2po4: 170; H;B03: 6,2; NaMoO4, 2H20: 0,25; myo-inositol : 100 : acide nicotinique: 1; pyridoxine-HCL : 1; thiamineHCL : 10 ; (NH4)2SO4: 460; KN03: 2820 ; asparagine : 670 ; 1% sucrose; 2,4-D : 10 ; 0,3% gelrite; pH 5,7. Ce milieu, moins riche en sucre et 2,4-D, permet d'obtenir des cals compétents pour une transformation à très haute fréquence en 3 ou 4 repiquages réalisés tous les 15 jours environ.
7H20: 0,025; CaCl2, 2H20: 440; KI: 0,83; COC'2, 6II20: 0,025; KH2po4: 170; H;B03: 6,2; NaMoO4, 2H20: 0,25; myo-inositol : 100 : acide nicotinique: 1; pyridoxine-HCL : 1; thiamineHCL : 10 ; (NH4)2SO4: 460; KN03: 2820 ; asparagine : 670 ; 1% sucrose; 2,4-D : 10 ; 0,3% gelrite; pH 5,7. Ce milieu, moins riche en sucre et 2,4-D, permet d'obtenir des cals compétents pour une transformation à très haute fréquence en 3 ou 4 repiquages réalisés tous les 15 jours environ.
Pour la transformation des plastes de soja par bombardement, les tissus embryogènes "FNL" de soja sont mis à 4 C pendant 16 à 20h. Ces cals sont ensuite déposés dans une capsule métallique grillagée, puis bombardés sur leurs deux faces (recto/verso) en utilisant un canon de type "PIG" (Particule Inflow Gun) tel que décrit dans Finer et al. (1992, Plant Cell Rep. 11,232- 238). Des micro-particules d'or (particules de 0,6 m de diamètre) sont complexées à l'ADN (vecteur pCLT312, 5 g/ tir) en présence de CaCl2 (0,8 à 1 M) et de spermidine (14 à 16 mM) selon les procédés décrit dans la littérature (Russel et al, 1992). Les cals embryogènes de soja bombardés sont ensuite découpés en petits morceaux de 1,5 à 2 mm et transférés sur un milieu FNL gélosé contenant l'agent de sélection.
<Desc/Clms Page number 17>
Exemple 3 : des lignées transplastomiques de soja
3.1. Evaluation de la sensibilité du soja à la spectinomycine
Actuellement, seul le gène aadA conférant la résistance à la spectinomycine a été utilisé avec succès comme marqueur de sélection d'évènements transplastomiques. Nous avons dans un premier temps vérifié la sensibilité du soja à la spectinomycine. En effet, certaines espèces végétales comme le riz sont naturellement résistantes à la spectinomycine car elles possèdent un ARNrl6S muté. Dans cette optique, des cals embryogènes de soja ont été placés sur du milieu FNL suppléé en spectinomycine à une concentration de 100mg/l, 300 mg/1 (dose employée dans l'art antérieur pour la sélection de la pomme de terre-Sidorov et al., 1999-), 500 mg/1 (dose employée dans l'art antérieur pour la sélection du tabac -Svab et Maliga, 1990 ; Svabet al., 1993), 600 mg/1 et 700 mg/1. Ces cals ont été repiqués sur le même milieu au bout de trois semaines. Pour toutes ces concentrations, les tissus commencent à blanchir au bout de deux semaines environ, ce qui montre la sensibilité naturelle du soja pour la spectinomycine.
3.1. Evaluation de la sensibilité du soja à la spectinomycine
Actuellement, seul le gène aadA conférant la résistance à la spectinomycine a été utilisé avec succès comme marqueur de sélection d'évènements transplastomiques. Nous avons dans un premier temps vérifié la sensibilité du soja à la spectinomycine. En effet, certaines espèces végétales comme le riz sont naturellement résistantes à la spectinomycine car elles possèdent un ARNrl6S muté. Dans cette optique, des cals embryogènes de soja ont été placés sur du milieu FNL suppléé en spectinomycine à une concentration de 100mg/l, 300 mg/1 (dose employée dans l'art antérieur pour la sélection de la pomme de terre-Sidorov et al., 1999-), 500 mg/1 (dose employée dans l'art antérieur pour la sélection du tabac -Svab et Maliga, 1990 ; Svabet al., 1993), 600 mg/1 et 700 mg/1. Ces cals ont été repiqués sur le même milieu au bout de trois semaines. Pour toutes ces concentrations, les tissus commencent à blanchir au bout de deux semaines environ, ce qui montre la sensibilité naturelle du soja pour la spectinomycine.
3. 2. Sélection de lignées tranplastomiques
Après 2 jours sur milieu FNL, les cals embryogènes de soja bombardés avec pCLT312 (comme décrit ci-dessus) sont recoupés, puis transférés sur une gaze stérile de criblage de manière à être en contact direct avec un milieu de sélection FNL gélosé contenant 200 mg/1 de spectinomycine. Les tissus sont repiqués sur ce même milieu au bout de 15 jours, puis, au bout de 15 jours supplémentaires, sur un milieu FNL gélosé contenant 300 mg/1 de spectinomycine.
Après 2 jours sur milieu FNL, les cals embryogènes de soja bombardés avec pCLT312 (comme décrit ci-dessus) sont recoupés, puis transférés sur une gaze stérile de criblage de manière à être en contact direct avec un milieu de sélection FNL gélosé contenant 200 mg/1 de spectinomycine. Les tissus sont repiqués sur ce même milieu au bout de 15 jours, puis, au bout de 15 jours supplémentaires, sur un milieu FNL gélosé contenant 300 mg/1 de spectinomycine.
Après 20 jours, ils sont à nouveau repiqués sur ce dernier milieu. Selon ce procédé de sélection, seuls les tissus transformés restent verts. Les premiers cals verts, résistants à la spectinomycine, apparaissent au bout de 1,5 à 2 mois. Ces transformants plastidiaux putatifs sont ensuite entretenus sur un milieu FNL suppléé de 150 mg/1 de spectinomycine.
A partir de 4 bombardements (15 cals en moyenne par bombardement), onze événements résistants à la spectinomycine (200 mg/1) ont été obtenus. Les premiers transformants putatifs sont apparus au bout de 63 jours (2 mois). Ces cals ont ensuite été amplifiés en milieu SBP6 liquide (contenant de la spectinomycine 150 mg/1) pour permettre la régénération de plantes et les analyses moléculaires. Le milieu SBP6 est décrit dans Finer et Nagasawa (1988,
Development of an embryogénie suspension culture of soybean (Glycine max Merill. ) Plant Cell.
Development of an embryogénie suspension culture of soybean (Glycine max Merill. ) Plant Cell.
Tissue and Organ Culture 15 : 125-136). Il contient les ingrédients suivants (en mg/1) :
Na2EDTA: 37,24; FeSO4, 7H20: 27,84; MgSO4, 7H2O: 370; MnSO4, H2O: 16,9; ZnS04, H2O:
8,6 ; CUS04, 7H2O: 0,025 ; CaCl2, 2H20: 440; KI: 0,83; CoCl2, 6H20: 0,025; KH2PO4: 170 ;
Na2EDTA: 37,24; FeSO4, 7H20: 27,84; MgSO4, 7H2O: 370; MnSO4, H2O: 16,9; ZnS04, H2O:
8,6 ; CUS04, 7H2O: 0,025 ; CaCl2, 2H20: 440; KI: 0,83; CoCl2, 6H20: 0,025; KH2PO4: 170 ;
<Desc/Clms Page number 18>
H3BO3: 6,2; Na2MoO4, 2H2O: 0,25; myo-inositol : 100 ; acide nicotinique: 1: pyridoxine-HCL : 1 ; thiamine-HCL : 10; NH4N03: 800; KNO3: 3000; asparagine : 670 ; 6% sucrose; 2,4-D : 5; pH 5,7.
Exemple 4: Identification des lignées transplastomiques de soja et étude de l'état d'homoplasmie de ces différentes lignées par Southern Blot.
L'identification des lignées transplastomiques a été réalisée par Southern Blot (Sambrook et al., 1989, Molecular Cloning : A Laboratory Manual, Nolan C. ed., New York : Spring Harbor Laboratory Press) sur des cals puis sur les plantes issues de ces cals.
L'ADN total de 10 cals des 11cals résistants à la spectinomycine a été extrait avec un kit commercial (Qiagen: "Dneasy Plant Mini Kit"). Toutefois, toutes techniques d'extraction d'ADN connue de l'homme du métier peut être valablement employée (Sambrook et al., 1989, Molecular Cloning : A Laboratory Manual, Nolan C. ed., New York : Spring Harbor Laboratory Press) Un g d'ADN extrait de chacun de ces 10 cals a ensuite été digéré par l'enzyme de restriction EcoRI (Biolabs). Cette digestion permet de générer des fragments d'intérêt de taille exploitable en Southern Blot, en particulier un fragment de 4042 pb pour les plastomes transformés, un fragment de 2667 pb pour les plastomes sauvages, et un fragment de 2452 pb pour les plastomes transformés ayant subis une recombinaison entre les deux PrrnC (tabac et soja). En effet, les mécanismes de recombinaison au sein du plaste étant très actifs, l'éventualité d'une recombinaison entre ces deux éléments de séquence très homologue, orientés dans le même sens, est possible.
Les fragments d'ADN sont séparés par électrophorèse avec une migration lente pendant la nuit à 55V dans un gel d'agarose 0,8% (QA Agarose TM Multipurpose , QBIOGENE). Le transfert a ensuite été réalisé de manière classique (Maniatis et al, 1989). Ces fragments d'ADN sont révélés par hybridation avec des sondes radioactives (marquées au 32P) qui sont de deux sortes : une sonde hybridant sur le transgène aadA (sonde ne révélant que les transplastomes), et une sonde hybridant sur une portion de la région intergénique de l'ADN plastidial (sonde permettant de visualiser les 3 formes de plastomes, correspondant aux nucléotides 2293 à 3068 du plastome de Glycine max; Genebank X07675). Ces deux sondes ont été amplifiées par PCR (avec le couple OSSG5 -SEQ ID NO:6- et OSSG310 -SEQ ID NO:7- pour la sonde hybridant sur la région intergénique de l'ADN plastidial, et le couple OAAX3-SEQ ID NO:8- et OAAN5 SEQ ID NO:9- pour la sonde aadA), puis marquées au 32P (kit Megaprime, AMERSHAM). Les deux membranes ont été lavées avec des solutions de stringence croissante (6xSSC, puis 2xSSC- 0,1% SDS, et O,IxSSC-0,1% SDS à 65 C). Après deux heures d'exposition à -80 C, avec un écran intensificateur, l'autoradiogramme a révélé la présence d'une bande attendue de 2607 pb
<Desc/Clms Page number 19>
(correspondant au plastome transformé par aadA) dans chacun des 10 cals résistants à la spectinomycine testés. Tous les événements de soja tolérants à la spectinomycine testés sont donc transplastomiques. Contrairement à la transformation plastidiale de toutes les espèces obtenues à ce jour (Svab et al., 1993 ; Staub et al., 1993 ; Sidorov et al., 1999 ; Sikdar S.R. et al., 1998), aucun mutant spontané résistant à cet antibiotique, dû à des mutations spécifiques dans le gène plastidial de l'ARNr16S n'a été observé lors de nos expériences de transformation du soja.
De plus, neuf des dix événements sont à l'état d'homoplasmie (ou au moins très proche) puisque seul le cal numéro 1 possède encore des copies de plastomes sauvages visibles en Southern Blot. Aucun événement de recombinaison entre les deux Prrn consécutifs (PrrnC de tabac et Prrn natif de soja), orientés en sens, n'a été détecté par cette analyse.
Exemple 5 : Régénération des plantes transplastomiques de soja
Les plantes transplastomiques de soja ont été régénérées de la manière suivante. Lorsque suffisamment de tissus sont produits en milieu FNL, ils sont ensuite convertis en embryons en employant un milieu décrit par Finer et McMullen dans : Transformation of soybean via particle bombardment of embryogénie suspension culture tissue. In Vitro Cell. Dev. Biol. 27P : 1991. Après 3 à 4 transferts sur ce milieu contenant 150 mg/1 de spectinomycine, les embryons sont séchés à l'air dans une boîte de Petri pendant 2 jours avant germination sur un milieu Murashige et Skoog (vitamines B5) à demi force ionique (50% des quantités du milieu MS) avec 15 g/1 de saccharose, 150 mg/1 de spectinimycine, 7 g/1 de phytagar, pH 5,7. Lorsque les jeunes plantes sont bien développées (stade 3 folioles) et enracinées, elles sont alors transférées dans un substrat à base de tourbe "jiffy pot" durant une période de 10-15 jours pour une phase d'acclimatation avant d'être transférées en serre. Les plantes sont ensuite élevées en serre avec des conditions de culture identiques à celles de soja non transplastomiques. Au cours de la floraison, le pollen est prélevé pour réaliser une pollinisation artificielle des plantes non transgéniques afin de vérifier la non-transmission du caractère de résistance à la spectinomycine par ces organes reproducteurs.
Les plantes transplastomiques de soja ont été régénérées de la manière suivante. Lorsque suffisamment de tissus sont produits en milieu FNL, ils sont ensuite convertis en embryons en employant un milieu décrit par Finer et McMullen dans : Transformation of soybean via particle bombardment of embryogénie suspension culture tissue. In Vitro Cell. Dev. Biol. 27P : 1991. Après 3 à 4 transferts sur ce milieu contenant 150 mg/1 de spectinomycine, les embryons sont séchés à l'air dans une boîte de Petri pendant 2 jours avant germination sur un milieu Murashige et Skoog (vitamines B5) à demi force ionique (50% des quantités du milieu MS) avec 15 g/1 de saccharose, 150 mg/1 de spectinimycine, 7 g/1 de phytagar, pH 5,7. Lorsque les jeunes plantes sont bien développées (stade 3 folioles) et enracinées, elles sont alors transférées dans un substrat à base de tourbe "jiffy pot" durant une période de 10-15 jours pour une phase d'acclimatation avant d'être transférées en serre. Les plantes sont ensuite élevées en serre avec des conditions de culture identiques à celles de soja non transplastomiques. Au cours de la floraison, le pollen est prélevé pour réaliser une pollinisation artificielle des plantes non transgéniques afin de vérifier la non-transmission du caractère de résistance à la spectinomycine par ces organes reproducteurs.
D'autre part, un contrôle de la bonne transmission de la cassette d'expression et de l'état d'homoplasmie de la descendance est effectué par PCR et Southern blot. Les graines issues des différentes lignées transplastomiques ont été semées sur un milieu de type Murashige et Skoog à demi force ionique contenant 15 g/1 de saccharose et 500 mg/1 de spectinomycine. Toutes les graines ont germé et produit des plantes tolérantes à la spectinomycine au contraire de graines sauvages. Cette expérience démontre ainsi la stabilité et la transmission de la cassette d'expression à la descendance. De plus, toutes les plantes transplastomiques de soja obtenues
<Desc/Clms Page number 20>
sont fertiles. Il s'agit donc du premier rapport décrivant la production d'une plante transplastomique fertile autre que le tabac et la tomate (Ruf et al., 2001). En effet, d'une part tous les événements transplastomiques d'A. thaliana et de riz produits à ce jour étaient tous stériles (Sikdar et al., 1998 ; Khan and Maliga, 1999), et d'autre part, les cellules de soja transformées n'avaient jamais pu être régénérées en plantes fertiles (Zhang et al., 2001).
Claims (16)
1- Légumineuse transplastomique fertile
2- Légumineuse transplastomique fertile selon la revendication 1, caractérisée en ce qu'il s'agit du soja
3- Légumineuse transplastomique fertile selon l'une des revendications 1 ou 2, caractérisée en ce qu'elle comprend au moins une cassette d'expression insérée dans une région intergénique de plastome
4- Légumineuse transplastomique fertile selon la revendication 3, caractérisée en ce que ladite région intergénique de plastome est située entre le gène ARNtV et l'opéron rpsl2/7.
5- Légumineuse transplastomique fertile selon l'une des revendications 2 à 4, caractérisée en ce que ladite cassette d'expression est insérée entre les séquences du plastome de soja correspondant aux identificateurs SEQ ID NO:1et SEQID NO:2.
6- Légumineuse transplastomique fertile selon l'une des revendications 1 à 5, caractérisée en ce que ladite cassette d'expression comprend au moins, liés entre eux de manière opérationnelle, un promoteur fonctionnel dans les plastes de cellules végétales, une séquence codant pour une protéine et un terminateur fonctionnel dans les plastes de cellules végétales
7- Vecteur de transformation adapté à la transformation des plastes de légumineuses, caractérisé en ce qu'il comprend au moins deux séquences homologues d'une zone du plastome de la légumineuse à transformer, lesdites séquences homologues encadrant au moins une cassette d'expression
8- Vecteur selon la revendication 7, caractérisé en ce que les deux séquences homologues d'une zone du plastome de la légumineuse à transformer correspondent à des séquences permettant l'intégration de la cassette d'expression dans une région intergénique de plastome
<Desc/Clms Page number 22>
9- Vecteur selon l'une des revendications 7 ou 8, caractérisé en ce que ladite zone correspond à la région de l'opéron des ARN ribosomiques du plastome
10- Vecteur selon la revendication 9, caractérisé en ce que l'une des deux séquences homologues comprend les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV), et que l'autre séquence homologue comprend la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7
11- Vecteur selon la revendication 10, caractérisé en ce que la séquence homologue comprenant les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV) est représentée par l'identificateur de séquences SEQ ID NO:1 et que la séquence homologue comprenant la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7 est représentée par l'identificateur de séquences SEQ ID NO:2.
12- Vecteur selon l'une des revendications 10 ou 11, caractérisé en ce que la séquence homologue comprenant les gènes codant pour l'ARN ribosomique 16S (ARNrl6S) et l'ARN de transfert V (ARNtV) est positionnée en 5' de la cassette d'expression, et que la séquence homologue comprend la région intergénique située entre le gène ARNtV et l'opéron rpsl2/7 est positionnée en 3' de la cassette d'expression
13- Vecteur selon l'une des revendications 7 à 12, caractérisé en ce que lesdites séquences homologues encadrent, en plus d'une cassette d'expression comprenant une séquence codant pour une protéine d'intérêt, au moins une autre cassette d'expression comprenant une séquence codant pour un marqueur de sélection
14- Procédé d'obtention de légumineuses transplastomiques fertiles, caractérisé en ce qu'il comprend les étapes de : (a) Transformation de tissus embryogènes obtenus à partir d'embryons immatures de légumineuses avec un vecteur adapté à la transformation des plastes (b) Sélection des tissus transformés (c) Régénération de plantes transplastomiques fertiles à partir des tissus transformés
15- Procédé selon la revendication 14, caractérisé en ce que la méthode de transformation employée est la méthode dite de bombardement de particules
<Desc/Clms Page number 23>
16- Procédé selon l'une des revendications 14 ou 15, caractérisé en ce que le vecteur adapté à la transformation des plastes est un vecteur selon l'une des revendications 7 à 13
Priority Applications (9)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0215490A FR2848064B1 (fr) | 2002-12-06 | 2002-12-06 | Plantes legumineuses transplastomiques fertiles |
| ARP030104480A AR042302A1 (es) | 2002-12-06 | 2003-12-04 | Plantas leguminosas transplastomicas fertiles |
| PCT/EP2003/015007 WO2004053133A1 (fr) | 2002-12-06 | 2003-12-08 | Plantes legumineuses transplastomiques fertiles |
| BRPI0314915A BRPI0314915B1 (pt) | 2002-12-06 | 2003-12-08 | método para a obtenção de plantas de soja e vetor de transformação |
| AU2003298257A AU2003298257A1 (en) | 2002-12-06 | 2003-12-08 | Fertile transplastomic leguminous plants |
| US10/538,130 US20070039075A1 (en) | 2002-12-06 | 2003-12-08 | Fertile transplastomic leguminous plants |
| CA2502424A CA2502424C (fr) | 2002-12-06 | 2003-12-08 | Plantes de soja transplastomiques fertiles |
| BR0314915-3A BR0314915A (pt) | 2002-12-06 | 2003-12-08 | Planta leguminosa, vetor de transformação e método para a obtenção de plantas leguminosas |
| US16/374,557 US20190390209A1 (en) | 2002-12-06 | 2019-04-03 | Fertile transplastomic leguminous plants |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0215490A FR2848064B1 (fr) | 2002-12-06 | 2002-12-06 | Plantes legumineuses transplastomiques fertiles |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| FR2848064A1 true FR2848064A1 (fr) | 2004-06-11 |
| FR2848064B1 FR2848064B1 (fr) | 2006-01-13 |
Family
ID=32320082
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| FR0215490A Expired - Fee Related FR2848064B1 (fr) | 2002-12-06 | 2002-12-06 | Plantes legumineuses transplastomiques fertiles |
Country Status (7)
| Country | Link |
|---|---|
| US (2) | US20070039075A1 (fr) |
| AR (1) | AR042302A1 (fr) |
| AU (1) | AU2003298257A1 (fr) |
| BR (2) | BRPI0314915B1 (fr) |
| CA (1) | CA2502424C (fr) |
| FR (1) | FR2848064B1 (fr) |
| WO (1) | WO2004053133A1 (fr) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2005005643A2 (fr) * | 2003-07-01 | 2005-01-20 | Biolex, Inc. | Transformation de chloroplastes de pourpier |
Families Citing this family (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7150993B2 (en) | 2003-08-05 | 2006-12-19 | Monsanto Technology Llc | Method for excision of plant embryos for transformation |
| US7560611B2 (en) | 2003-08-05 | 2009-07-14 | Monsanto Technology Llc | Method and apparatus for substantially isolating plant tissues |
| EP1679374A1 (fr) | 2005-01-10 | 2006-07-12 | Bayer CropScience GmbH | Plantes transformées exprimant un mutane sucrase et synthétisant un amidon modifie |
| WO2007031547A1 (fr) | 2005-09-16 | 2007-03-22 | Bayer Cropscience Sa | Plantes transplastomiques exprimant une protéine à destination du lumen |
| US8044260B2 (en) | 2007-03-09 | 2011-10-25 | Monsanto Technology Llc | Method of meristem excision and transformation |
| AR074941A1 (es) | 2009-01-07 | 2011-02-23 | Bayer Cropscience Sa | Plantas transplastomicas exentas de marcador de seleccion |
| ES2727575T3 (es) | 2009-01-22 | 2019-10-17 | Syngenta Participations Ag | Polipéptidos de Hidroxifenilpiruvato Dioxigenasa mutantes, y Métodos de uso |
| US9347046B2 (en) | 2009-01-22 | 2016-05-24 | Syngenta Participations Ag | Hydroxyphenylpyruvate dioxygenase polypeptides and methods of use |
| US9187762B2 (en) | 2010-08-13 | 2015-11-17 | Pioneer Hi-Bred International, Inc. | Compositions and methods comprising sequences having hydroxyphenylpyruvate dioxygenase (HPPD) activity |
| JP6815322B2 (ja) * | 2015-02-04 | 2021-01-20 | モンサント テクノロジー エルエルシー | 色素体形質転換方法 |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1998011235A2 (fr) * | 1996-09-12 | 1998-03-19 | Novartis Ag | Plantes transgeniques exprimant des enzymes cellulolytiques |
| WO1999010513A1 (fr) * | 1997-08-07 | 1999-03-04 | Auburn University | Vecteurs universels d'integration et d'expression de chloroplastes, plantes transformees et produits obtenus |
Family Cites Families (14)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4945050A (en) * | 1984-11-13 | 1990-07-31 | Cornell Research Foundation, Inc. | Method for transporting substances into living cells and tissues and apparatus therefor |
| EP0218571B1 (fr) * | 1985-08-07 | 1993-02-03 | Monsanto Company | Plantes résistant au glyphosate |
| US7129391B1 (en) * | 1988-09-26 | 2006-10-31 | Auburn University | Universal chloroplast integration and expression vectors, transformed plants and products thereof |
| US5451513A (en) * | 1990-05-01 | 1995-09-19 | The State University of New Jersey Rutgers | Method for stably transforming plastids of multicellular plants |
| US5877402A (en) * | 1990-05-01 | 1999-03-02 | Rutgers, The State University Of New Jersey | DNA constructs and methods for stably transforming plastids of multicellular plants and expressing recombinant proteins therein |
| AU656761B2 (en) * | 1991-02-25 | 1995-02-16 | Syngenta Limited | Newly characterised oxalate oxidase and uses therefor |
| US6229065B1 (en) * | 1991-03-05 | 2001-05-08 | Rhone-Poulenc Agrochimie | Production of plants resistant to attacks by Sclerotinia sclerotiorum by the introduction of a gene encoding an oxalate oxidase |
| PH31293A (en) * | 1991-10-10 | 1998-07-06 | Rhone Poulenc Agrochimie | Production of y-linolenic acid by a delta6-desaturage. |
| US5614313A (en) * | 1994-07-07 | 1997-03-25 | Imperial Chemical Industries Plc | Polymeric film having a layer comprising calcined silicone particles and china clay particles |
| FR2734842B1 (fr) * | 1995-06-02 | 1998-02-27 | Rhone Poulenc Agrochimie | Sequence adn d'un gene de l'hydroxy-phenyl pyruvate dioxygenase et obtention de plantes contenant un gene de l'hydroxy-phenyl pyruvate dioxygenase, tolerantes a certains herbicides |
| FR2736926B1 (fr) * | 1995-07-19 | 1997-08-22 | Rhone Poulenc Agrochimie | 5-enol pyruvylshikimate-3-phosphate synthase mutee, gene codant pour cette proteine et plantes transformees contenant ce gene |
| FR2745004B1 (fr) * | 1996-02-16 | 1998-03-27 | Rhone Poulenc Agrochimie | Peptide antibacterien et antifongique |
| FR2766207B1 (fr) * | 1997-07-11 | 2000-12-08 | Rhone Poulenc Agrochimie | Gene chimere codant pour la drosomycine, vecteur le contenant pour la transformation des cellules vegetales et plantes transformees obtenues resistantes aux maladies |
| FR2770853B1 (fr) * | 1997-11-07 | 1999-12-31 | Rhone Poulenc Agrochimie | Gene codant pour la thanatine, vecteur le contenant et plantes transformees obtenues resistantes aux maladies |
-
2002
- 2002-12-06 FR FR0215490A patent/FR2848064B1/fr not_active Expired - Fee Related
-
2003
- 2003-12-04 AR ARP030104480A patent/AR042302A1/es active IP Right Grant
- 2003-12-08 WO PCT/EP2003/015007 patent/WO2004053133A1/fr not_active Application Discontinuation
- 2003-12-08 AU AU2003298257A patent/AU2003298257A1/en not_active Abandoned
- 2003-12-08 BR BRPI0314915A patent/BRPI0314915B1/pt unknown
- 2003-12-08 BR BR0314915-3A patent/BR0314915A/pt active IP Right Grant
- 2003-12-08 CA CA2502424A patent/CA2502424C/fr not_active Expired - Lifetime
- 2003-12-08 US US10/538,130 patent/US20070039075A1/en not_active Abandoned
-
2019
- 2019-04-03 US US16/374,557 patent/US20190390209A1/en not_active Abandoned
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1998011235A2 (fr) * | 1996-09-12 | 1998-03-19 | Novartis Ag | Plantes transgeniques exprimant des enzymes cellulolytiques |
| WO1999010513A1 (fr) * | 1997-08-07 | 1999-03-04 | Auburn University | Vecteurs universels d'integration et d'expression de chloroplastes, plantes transformees et produits obtenus |
Non-Patent Citations (2)
| Title |
|---|
| BOCK R: "TRANSGENIC PLASTIDS IN BASIC RESEARCH AND PLANT BIOTECHNOLOGY", JOURNAL OF MOLECULAR BIOLOGY, LONDON, GB, vol. 3, no. 312, 21 September 2001 (2001-09-21), pages 425 - 438, XP001084937, ISSN: 0022-2836 * |
| ZHANG, X.-H. ET AL.: "Plastid transformation of soybean suspension cultures", JOURNAL OF PLANT BIOTECHNOLOGY, vol. 3, no. 1, January 2001 (2001-01-01), pages 39 - 44, XP002253093 * |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2005005643A2 (fr) * | 2003-07-01 | 2005-01-20 | Biolex, Inc. | Transformation de chloroplastes de pourpier |
| WO2005005643A3 (fr) * | 2003-07-01 | 2005-09-15 | Biolex Inc | Transformation de chloroplastes de pourpier |
Also Published As
| Publication number | Publication date |
|---|---|
| US20070039075A1 (en) | 2007-02-15 |
| AU2003298257A1 (en) | 2004-06-30 |
| CA2502424C (fr) | 2018-02-27 |
| CA2502424A1 (fr) | 2004-06-24 |
| BRPI0314915B1 (pt) | 2019-08-27 |
| BR0314915A (pt) | 2005-08-02 |
| US20190390209A1 (en) | 2019-12-26 |
| FR2848064B1 (fr) | 2006-01-13 |
| AR042302A1 (es) | 2005-06-15 |
| WO2004053133A1 (fr) | 2004-06-24 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP1186666B1 (fr) | Utilisation d'inhibiteurs d' hydroxylphenyl pyruvate dioxygenase (HPPD) comme agents de sélection dans la transformation de plantes | |
| US20190390209A1 (en) | Fertile transplastomic leguminous plants | |
| EP2935592B1 (fr) | Transformation de soja à graine fendue améliorée pour la production d'événement transgénique efficace et à rendement élevé | |
| EP1537216B1 (fr) | Plantes transformees a biosynthese de prenylquinones amelioree | |
| FR2848568A1 (fr) | Gene chimere permettant l'expression d'une hydroxy-phenyl pyruvate dioxygenase dans les plastes et plantes transplastomiques contenant un tel gene tolerantes aux herbicides | |
| CZ20013860A3 (cs) | Herbicidně rezistentní rostliny | |
| EP2016180B1 (fr) | Plateformes végétales de chromosomes artificiels au moyen d'un troncage du télomère | |
| CZ20013856A3 (cs) | Herbicidně rezistentní rostliny | |
| CA2548484C (fr) | Methode pour modifier l'expression genique d'un champignon phytopathogene | |
| JP2013537412A (ja) | 植物形質転換率を高めるように改変されたアグロバクテリウム(Agrobacterium)株 | |
| Mehrotra et al. | Agrobacterium-mediated gene transfer in plants and biosafety considerations | |
| MX2014007161A (es) | Metodo para la transformacion mejorada usando agrobacterium. | |
| CN102741415A (zh) | 使用hppd抑制剂作为选择剂的大豆转化 | |
| EP1042491B1 (fr) | Promoteur h3c4 de mais associe au premier intron de l'actine de riz, gene chimere le comprenant et plante transformee | |
| AU776046B2 (en) | Transgenic plants and methods for production thereof | |
| MXPA01012750A (es) | Transformacion de algodon de alta eficiencia mediada por agrobacterium utilizando explantes de peciolo. | |
| Meng et al. | Effects of hygromycin on cotton cultures and its application in Agrobacterium-mediated cotton transformation | |
| FR2880356A1 (fr) | Plantes transplastomiques exemptes du gene marqueur de selection | |
| JP3964701B2 (ja) | 病害抵抗性イネ科植物 | |
| JP2003511049A (ja) | プロトポルフィリノーゲンオキシダーゼ遺伝子を利用した作物の収量またはバイオマスの増大方法 | |
| DE60028751T2 (de) | Herbizidresistente pflanzen | |
| CN117425401A (zh) | 用于递送基因组编辑试剂的嫁接的植物 | |
| FR2784393A1 (fr) | Procede d'obtention de plantes transgeniques exprimant une proteine a activite productrice de peroxyde d'hydrogene par transformation par agrobacterium rhizogenes |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| TP | Transmission of property |
Owner name: BAYER SAS, FR Effective date: 20110906 |
|
| ST | Notification of lapse |
Effective date: 20130830 |