ES2931540T3 - Mutante de transaminasa y aplicación del mismo - Google Patents
Mutante de transaminasa y aplicación del mismo Download PDFInfo
- Publication number
- ES2931540T3 ES2931540T3 ES18903278T ES18903278T ES2931540T3 ES 2931540 T3 ES2931540 T3 ES 2931540T3 ES 18903278 T ES18903278 T ES 18903278T ES 18903278 T ES18903278 T ES 18903278T ES 2931540 T3 ES2931540 T3 ES 2931540T3
- Authority
- ES
- Spain
- Prior art keywords
- transaminase
- immobilized
- enzyme
- mutant
- carrier
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1096—Transferases (2.) transferring nitrogenous groups (2.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N11/00—Carrier-bound or immobilised enzymes; Carrier-bound or immobilised microbial cells; Preparation thereof
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N11/00—Carrier-bound or immobilised enzymes; Carrier-bound or immobilised microbial cells; Preparation thereof
- C12N11/02—Enzymes or microbial cells immobilised on or in an organic carrier
- C12N11/08—Enzymes or microbial cells immobilised on or in an organic carrier the carrier being a synthetic polymer
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N11/00—Carrier-bound or immobilised enzymes; Carrier-bound or immobilised microbial cells; Preparation thereof
- C12N11/02—Enzymes or microbial cells immobilised on or in an organic carrier
- C12N11/08—Enzymes or microbial cells immobilised on or in an organic carrier the carrier being a synthetic polymer
- C12N11/082—Enzymes or microbial cells immobilised on or in an organic carrier the carrier being a synthetic polymer obtained by reactions only involving carbon-to-carbon unsaturated bonds
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N11/00—Carrier-bound or immobilised enzymes; Carrier-bound or immobilised microbial cells; Preparation thereof
- C12N11/02—Enzymes or microbial cells immobilised on or in an organic carrier
- C12N11/08—Enzymes or microbial cells immobilised on or in an organic carrier the carrier being a synthetic polymer
- C12N11/082—Enzymes or microbial cells immobilised on or in an organic carrier the carrier being a synthetic polymer obtained by reactions only involving carbon-to-carbon unsaturated bonds
- C12N11/087—Acrylic polymers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N11/00—Carrier-bound or immobilised enzymes; Carrier-bound or immobilised microbial cells; Preparation thereof
- C12N11/02—Enzymes or microbial cells immobilised on or in an organic carrier
- C12N11/10—Enzymes or microbial cells immobilised on or in an organic carrier the carrier being a carbohydrate
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P17/00—Preparation of heterocyclic carbon compounds with only O, N, S, Se or Te as ring hetero atoms
- C12P17/10—Nitrogen as only ring hetero atom
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P17/00—Preparation of heterocyclic carbon compounds with only O, N, S, Se or Te as ring hetero atoms
- C12P17/10—Nitrogen as only ring hetero atom
- C12P17/12—Nitrogen as only ring hetero atom containing a six-membered hetero ring
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02P—CLIMATE CHANGE MITIGATION TECHNOLOGIES IN THE PRODUCTION OR PROCESSING OF GOODS
- Y02P20/00—Technologies relating to chemical industry
- Y02P20/50—Improvements relating to the production of bulk chemicals
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Molecular Biology (AREA)
- General Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- Enzymes And Modification Thereof (AREA)
- Immobilizing And Processing Of Enzymes And Microorganisms (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Pyrrole Compounds (AREA)
Abstract
Se proporciona un mutante de transaminasa y una aplicación del mismo. La secuencia de aminoácidos del mutante de transaminasa se forma después de la mutación de la secuencia de aminoácidos como se muestra en SEQ ID NO: 1, y los sitios de aminoácidos mutados comprenden un sitio T7C+S47C. El mutante de transaminasa que tiene los sitios mutados se puede preparar adicionalmente en una enzima inmovilizada por medio de la tecnología de inmovilización, la actividad y la estabilidad de la enzima inmovilizada son relativamente altas, y la enzima inmovilizada se puede reciclar varias veces y es aplicable a la reacción de flujo continuo en un lecho empacado. (Traducción automática con Google Translate, sin valor legal)
Description
DESCRIPCIÓN
Mutante de transaminasa y aplicación del mismo
Campo técnico
La invención se refiere al campo de la ingeniería enzimática, en particular a un mutante de transaminasa y aplicación del mismo.
Antecedentes
La w-transaminasa (w-TA) pertenece a las transferasas y cataliza el intercambio de un grupo amino con un grupo ceto similar a otras transaminasas. En la mayoría de los casos, la w-transaminasa se refiere a una clase de enzima, siempre que el sustrato o producto de la reacción no contenga a-aminoácido en una reacción de transaminación catalizada por enzimas, la enzima puede llamarse w-transaminasa. Las w-transaminasas pueden producir eficazmente aminas quirales mediante transaminación estereoselectiva utilizando cetonas como materia prima. Las aminas quirales enantioméricas son productos intermedios clave para muchos compuestos farmacéuticos con amplia actividad biológica (ChemBioChem. 9, 2008, 363-365, Chem. Commun. 46, 2010, 5569-5571, Biotechnol. Bioeng. 108, 2011, 1479-1493). Debido al sustrato relativamente barato y la alta pureza del producto, ha llamado cada vez más la atención de los investigadores (Green Chemistry, 2017, 19, 2: 333-360). Y las transaminasas se han mostrado prometedoras para la producción de aminas quirales (Organic Process Research & Development, 2010, 14, 234-237).
Aunque se ha prestado mucha atención al progreso en la producción de aminas quirales con transaminasas, existen muchos problemas en la aplicación de métodos enzimáticos en la producción a escala. Por ejemplo, la baja actividad enzimática y la gran cantidad de enzima conducen a un aumento en el coste de la fermentación y son fáciles de desnaturalizar e inactivar mediante la influencia de disolventes orgánicos en el sistema de reacción.
Además, en el proceso de separación de la amina del producto, la enzima solo puede desnaturalizarse e inactivarse para formar una precipitación y luego se retira, se desecha y no puede reutilizarse, o bien el producto se extrae de la solución acuosa con disolvente orgánico y la enzima continúa existiendo en la solución acuosa, pero en ese momento, debido a la influencia de muchas condiciones adversas como el pH y el disolvente, la enzima se inactiva y no puede volver a utilizarse.
En la técnica anterior, se ha informado que la enzima se inmoviliza mediante tecnología de inmovilización para mejorar la recuperación y reutilización de la enzima. Sin embargo, en la actualidad, existen muchas investigaciones sobre la tecnología de inmovilización en los campos de la lipasa, la penicilina acilasa, la amilasa y otras, ya que estas enzimas poseen una mejor estabilidad que otras enzimas y la pérdida de actividad enzimática después de la inmovilización es menor. Para la mayor parte de las transaminasas, la estabilidad es deficiente, en especial cuando existe una fase orgánica en el sistema, la pérdida de actividad enzimática se puede provocar fácilmente en un proceso de operación de inmovilización, por lo que hay menos investigación sobre la inmovilización de la transaminasa y la investigación sobre la transaminasa inmovilizada adecuada para la reacción continua es aún menor.
En cuanto a la transaminasa capaz de catalizar la reacción de conversión de amino del siguiente sustrato 1 y sustrato 2, si la reacción es catalizada por la transaminasa libre, la enzima libre no puede recuperarse y solo puede utilizarse una vez. Además, debido a la existencia de proteína enzimática en el sistema de reacción, el fenómeno de emulsificación posterior al tratamiento es extremadamente grave y la separación del producto es difícil.

Si la transaminasa utilizada se inmoviliza, la recuperación y utilización de la enzima puede realizarse teóricamente, pero la recuperación de la actividad enzimática sigue siendo baja después de inmovilizar la transaminasa existente utilizada para la catalización del sustrato 1 o del sustrato 2. Además, como la mayor parte de las cetonas sustrato existentes (amino receptores) son poco solubles en agua, la reacción continua no puede llevarse a cabo en una fase acuosa pura. Para llevar a cabo la reacción continua, se debe añadir suficiente codisolvente orgánico para disolver el sustrato, pero la transaminasa existente tiene una tolerancia deficiente a la temperatura, el pH y el disolvente orgánico, y el disolvente orgánico puede inactivar fácilmente la transaminasa. Por lo tanto, es difícil llevar a cabo el tratamiento de inmovilización.
Por lo tanto, es necesario mejorar las transaminasas existentes que pueden catalizar los sustratos anteriores a fin de mejorar su estabilidad deficiente y aplicación limitada en entornos extremos, incluidos los disolventes orgánicos. Sumario
La invención tiene como objetivo principal proporcionar un mutante de transaminasa y la aplicación del mismo, a fin de resolver el problema de que la aplicación está limitada debido a la tolerancia deficiente de la actividad de la transaminasa en un entorno extremo en la técnica anterior.
Para lograr el objetivo anterior, de acuerdo con un aspecto de la presente invención, se proporciona un mutante de transaminasa, el mutante de transaminasa tiene una secuencia en la que se produce una mutación de aminoácidos en la secuencia que se muestra en la SEQ ID NO: 1, y los sitios de mutación de aminoácidos comprenden T7C+S47C.
Además, los sitios de mutación de aminoácidos comprenden además cualquiera o más de los siguientes: M356L, F364L, C404L, M430L, R405E/A, K90G, K219T, K304D, K51R, A95P, E368P, Q346E, H333K, D371G, E246A, C328A, N412G, T402P, T107F/A, G110P, K69N, G201C, Q380L, K193I, I297L, R305H, F111Y, K190E y A286T, en donde "/" representa "o".
Además, los sitios de mutación de aminoácidos incluyen además cualquiera de los siguientes sitios de mutación combinados: K51R+W187Y, R405E+A95P, R405E+A95P+K304D, R405E+A95P+K304D+Q380L, R405E+K90G+A95P+K304D+Q380L, R405E+K90G+A95P+K304D+Q380L+E368P, R405E+K90G+A95P+K304D+Q380L+Q346E, R405E+K90G+A95P+K304D+Q380L+H333K, R405E+K90G+A95P+K304D+Q380L+D371G, R405E+K90G+A95P+K304D+Q380L+E246A, R405E+K90G+A95P+K304D+Q380L+C328A, R405E+K90G+A95P+K304D+Q380L+N412G, R405E+K90G+A95P+K304D+Q380L+T402P, R405E+K90G+A95P+K304D+Q380L+T107F, R405E+K90G+A95P+K304D+Q380L+T107A, R405E+K90G+A95P+K304D+Q380L+G110P, R405E+K90G+A95P+K304D+Q380L+I297L, R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A, R405E+K90G+A95P+K304D+Q380L+I297L+A286T R405E+K90G+A95P+K304D+Q380L+I297L+E368P R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+K69N, R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+G201C y R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+A286T.
Para lograr el objetivo anterior, de acuerdo con el segundo aspecto de la presente invención, se proporciona una molécula de ADN que codifica cualquiera de los mutantes de transaminasa descritos con anterioridad.
De acuerdo con el tercer aspecto de la presente invención, se proporciona un plásmido recombinante que se ha unido a cualquiera de las moléculas de a Dn descritas con anterioridad.
De acuerdo con el cuarto aspecto de la presente invención, se proporciona una transaminasa inmovilizada que comprende cualquiera de los mutantes de transaminasa descritos con anterioridad.
Además, la transaminasa inmovilizada es un agregado enzimático reticulado de transaminasa del mutante de transaminasa; preferiblemente, el mutante de transaminasa se precipita para obtener un agregado de transaminasa, y un grupo amino libre, un grupo fenólico, un grupo imidazolilo o un grupo sulfhidrilo en el agregado de transaminasa se reticula además con un agente de reticulación para obtener el agregado enzimático reticulado de transaminasa, en donde el agente de reticulación se selecciona entre cualquiera de glutaraldehído, N,N-metilenbisacrilamida, bismaleimida y dextrano; preferiblemente, el agregado enzimático reticulado de transaminasa es un agregado enzimático reticulado de un mutante de transaminasa que contiene los siguientes sitios de mutación de aminoácidos en base a la secuencia de aminoácidos que se muestra en la SEQ ID NO:1: T7C+S47C, T7C+S47C+A95P, T7C+S47C+R405E+K90G+A95P+K304D+Q380L, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+A286T, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+K69N, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+H333K, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+E368P, T7C+S47C+K51R+W187Y, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+Q346E, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+C328A y T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L; preferiblemente, el dextrano tiene un peso molecular de 6 KDa - 200 KDa; preferiblemente, el agregado de transaminasas se obtiene por precipitación con etanol del mutante de transaminasa; preferiblemente, un grupo amino libre en el agregado de transaminasas se reticula con glutaraldehído para obtener agregados enzimáticos reticulados de transaminasas.
Además, la transaminasa inmovilizada es una enzima inmovilizada incrustada-reticulada de transaminasa; preferiblemente, la enzima incrustada-reticulada de transaminasa es una enzima inmovilizada incrustada-reticulada del mutante de transaminasa que contiene los siguientes sitios de mutación de aminoácidos en base a la secuencia de aminoácidos que se muestra en la SEQ ID NO: 1: T7C+S47C, T7C+S47C+R405E+K90G+A95P+K304D+Q380L, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T 107A, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+Q346E y T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L; preferiblemente, un grupo amino libre en el mutante de transaminasa forma una base de Schiff mediante la reticulación con glutaraldehído para obtener una enzima reticulada de transaminasa, y la enzima reticulada de transaminasa se incrusta en una red de gel de poliacrilamida para obtener la enzima inmovilizada incrustada-reticulada de transaminasa.
Además, la transaminasa inmovilizada es una enzima inmovilizada covalente en la que el mutante de transaminasa está conectado covalentemente con el portador; preferiblemente, la enzima inmovilizada covalentemente es una enzima inmovilizada covalentemente de un mutante de transaminasa que contiene los siguientes sitios de mutación de aminoácidos en base a la secuencia de aminoácidos que se muestra en la SEQ ID NO:1: T7C+S47C, T7C+S47C+A95P, T7C+S47C+Q380L, T7C+S47C+R405E, T7C+S47C+K51R+W187Y, T7C+S47C+R405E+A95P+K304D,
T7C+S47C+R405E+K90G+A95P+K304D+Q380L+E368P, T7C+S47C+R405E+K90G+A95P+K304D+Q380L, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+H333K, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A y T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+A286T; preferiblemente, el portador es un portador de quitosano y un portador de resina; más preferiblemente, el portador de quitosano está unido covalentemente al mutante de transaminasa mediante un grupo hidroxilo y/o amino para formar la enzima inmovilizada covalente; más preferiblemente, el portador de resina comprende una matriz y un grupo funcional enlazado con la matriz, en donde la matriz se selecciona de cualquiera de los copolímeros de estireno y metacrilato, resinas de poliestireno y resinas de polimetacrilato, y el grupo funcional enlazado con la matriz se selecciona de un grupo amino de cadena corta C2, un grupo amino de cadena media C4, un grupo amino de cadena larga C6 o un grupo epoxi; más preferiblemente, el portador de resina se selecciona de ECR8309, ECR8315, EC-HFA, LX-1000HA, LX-1000EA, ECR8409, ECR8415, EC-EP, ECEP403, EXE119, LX-1000EP, Immobead-150A, Immobead-150P, Immobead350A, ECR8206, ECR8209, ECR8215 o ECR8285.
Además, la transaminasa inmovilizada es una enzima inmovilizada quelante formada mediante la quelación de un mutante de transaminasa y un portador a través de iones metálicos; preferiblemente, la enzima inmovilizada quelante es una enzima inmovilizada quelante de un mutante de transaminasa que contiene los siguientes sitios de mutación de aminoácidos en base a la secuencia de aminoácidos que se muestra en la SEQ ID NO:1: T7C+S47C, T7C+S47C+A95P, T7C+S47C+R405E+K90G+A95P+K304D+Q380L, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+G201C y T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+A286T; preferiblemente, el portador es un portador de vidrio poroso; más preferiblemente, el portador de vidrio poroso es EziG-101, EziG-102 o EziG-103.
Además, la transaminasa inmovilizada es una enzima inmovilizada por adsorción formada por el mutante de transaminasa y el portador mediante adsorción física; preferiblemente, la enzima inmovilizada por adsorción es una enzima inmovilizada covalente de un mutante de transaminasa que contiene los siguientes sitios de mutación de aminoácidos en base a la secuencia de aminoácidos que se muestra en la SEQ ID NO:1: T7C+S47C, T7C+S47C+A95P, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+H333K, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+Q346E, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+N412G, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A, T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+A286T y T7C+S47C+R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+G201C; preferiblemente, el portador es un portador de resina; más preferiblemente, el portador de resina comprende una matriz y un grupo funcional conectado con la matriz, la matriz se selecciona de cualquiera de un copolímero de estireno y metacrilato, una resina de poliestireno y una resina de polimetacrilato, y el grupo funcional conectado con la matriz es un octadecilo; más preferiblemente, el portador de resina se selecciona de ECR8806, ECR1030, ECR1090, ECR1061, ECR1091, ECR8804, Immobead-EC1, Immobead-S60S, Immobead-S861, X17S0409, EXE120 o Diaion HP2MG.
De acuerdo con el quinto aspecto de la presente invención, se proporciona un método para la producción de una amina quiral, que comprende la etapa de reacción de transaminación entre un compuesto de cetona y un donante de amino catalizada por una transaminasa, en donde la transaminasa es cualquiera de los mutantes de transaminasa anteriores o cualquiera de las transaminasas inmovilizadas anteriores.
Además, la transaminasa es cualquiera de los mutantes de transaminasa, y el método es una reacción por lotes; preferiblemente, el sistema de reacción de la reacción por lotes es un sistema de reacción en fase acuosa.
Además, la transaminasa es cualquier transaminasa inmovilizada y el método es una reacción continua; preferiblemente, el sistema de reacción de la reacción continua es un sistema de reacción en fase orgánica.
Además, el compuesto de cetona es

en donde R1 y R2 son cada uno independientemente alquilo C1-C8, cicloalquilo C5-C10, arilo C6-C10 o heteroarilo C5-C10, o R1 y R2 junto con el carbono en el carbonilo forman un heterociclilo C5-C10, carbociclilo C5-C10 o un heteroarilo C5-C10, los heteroátomos en el heterociclilo C5-C10 y el heteroarilo C5-C10 se seleccionan cada uno independientemente de al menos uno de nitrógeno, oxígeno y azufre, el arilo en el arilo C6-C10, el heteroarilo en el heteroarilo C5-C10, el carbociclilo en el carbociclilo C5-C10 o el heterociclilo en el heterociclilo C5-C10 están cada uno independientemente sustituidos o no sustituidos con al menos uno de halógeno, alcoxi o alquilo, preferiblemente, el compuesto de cetona es

y el producto de la reacción de transaminación es

De acuerdo con el esquema técnico proporcionado por la invención, una serie de mutantes de transaminasas con actividad enzimática y/o estabilidad muy mejoradas se seleccionan realizando una evolución direccional en la transaminasa de la secuencia de aminoácidos que se muestra en la SEQ ID NO: 1, en donde la secuencia de aminoácidos de los mutantes es una secuencia de aminoácidos que está mutada en base a la secuencia de aminoácidos que se muestra en la SEQ ID NO: 1, y los sitios de aminoácidos mutados comprenden los sitios T7C S47C. El mutante de transaminasa que contiene los sitios de mutación anteriores se puede aplicar en un entorno relativamente extremo.
Descripción detallada de la invención
Cabe señalar que los ejemplos y las características de los ejemplos de la presente pueden combinarse entre sí sin conflicto. La presente invención se describirá en detalle a continuación en combinación con los ejemplos.
Explicación de términos:
Mutagénesis dirigida al sitio: se refiere a la introducción de cambios deseados (que generalmente caracterizan cambios en direcciones favorables) en los fragmentos de ADN objetivo (ya sean genomas o plásmidos) mediante la reacción en cadena de la polimerasa (PCR), incluida la adición, eliminación, mutación puntual, etc. de bases La mutagénesis dirigida al sitio puede mejorar rápida y eficazmente las propiedades y la caracterización de las proteínas diana expresadas por el ADN, y es un medio muy útil en la investigación genética.
La introducción de mutación dirigida al sitio mediante la PCR del plásmido completo es sencilla y eficaz, y es un método ampliamente utilizado en la actualidad. El principio es el siguiente: un par de cebadores que contienen sitios de mutación (directos e inversos), y el plásmido molde se hibridan, luego se "extienden en ciclo" mediante la polimerasa, la denominada extensión cíclica significa que la polimerasa extiende los cebadores de acuerdo con el molde, y luego regresa al extremo 5' de los cebadores después de un círculo, después de ciclos de calentamiento e hibridación repetidos, esta reacción es diferente de la amplificación del círculo rodante, no formará múltiples copias en tándem. Los productos de extensión del cebador directo y el cebador inverso se hibridan y emparejan para formar un plásmido circular abierto fragmentado. Dpn I digiere el producto de extensión, dado que el plásmido molde original deriva de escherichia coli (E. Coli) convencional, sometido a modificación de metilación dam y sensible a Dpn I, se corta y el plásmido con la secuencia mutante sintetizado in vitro no se corta porque no hay metilación, de modo que el plásmido se transforma con éxito en la transformación posterior y se pueden obtener clones del plásmido mutante. El plásmido mutante se transforma en una célula huésped para inducir la expresión de la proteína diana.
PCR propensa a errores: se refiere a la PCR en condiciones propensas a errores; es una técnica de PCR que causa errores fácilmente en las secuencias de ADN copiadas, también conocida como PCR incompatible o PCR propensa a errores. En particular, se refiere a un método para inducir la variación de la secuencia de ADN in vitro con ADN polimerasa Taq de baja exactitud y cambiar las condiciones de reacción de la PCR, lo que reduce la exactitud de la replicación del ADN y aumenta la incompatibilidad de bases en el proceso de síntesis de la nueva cadena de ADN, lo que resulta en más mutaciones puntuales en los productos amplificados.
La PCR propensa a errores es la técnica más sencilla y eficaz para la mutagénesis aleatoria de genes in vitro. Su principio es que la isomerización de la base proporciona la posibilidad de una incompatibilidad. Los tautómeros aparecen en las cuatro bases que componen el ADN, en donde tres bases que contienen oxígeno, guanina (G), citosina (C) y timina (T), tienen dos tautómeros de forma ceto y forma enol. Las dos bases que contienen nitrógeno, adenina (A) y timina, tienen una forma amina y una forma imina. G, C y T existen principalmente en una estructura en forma ceto, la relación de la estructura en forma enol es extremadamente baja, y los átomos de nitrógeno en las bases A y T que contienen nitrógeno existen principalmente en un estado amino (NH2), y la relación de los átomos de nitrógeno en el estado de imina (NH) es extremadamente bajo. Las diferentes posiciones de los átomos de hidrógeno entre diferentes isómeros y las diferentes direcciones de desviación de las nubes de electrones en la misma posición pueden cambiar las formas de emparejamiento de las bases, de modo que pueden producirse
incompatibilidades en las subcadenas replicadas. Por ejemplo, cuando la timina está presente en una estructura en forma ceto, emparejada con adenina, y cuando la timina está en una estructura en forma enol, emparejada con guanina, dando así lugar a un par de bases inestable en donde A puede emparejarse con C y T puede emparejarse con G, dando lugar a una incompatibilidad.
Entre varias polimerasas de ADN con termotolerancia conocidas, la ADN polimerasa Taq tiene la tasa de incompatibilidad más alta. La ADN polimerasa Taq tiene la actividad más alta entre las polimerasas de ADN de termotolerancia halladas. Tiene actividad exonucleasa 5'-3', pero no actividad exonucleasa 3'-5', por lo tanto, no tiene función de corrección para algunas incompatibilidades de mononucleótidos durante la síntesis, por lo que tiene una mayor probabilidad de incompatibilidad que las ADN polimerasas con actividad de corrección 3'-5'. La exactitud de las ADN polimerasas se puede reducir mediante una variedad de métodos, incluido el uso de cuatro dNTP de diferentes concentraciones, la adición de Mn2+, el aumento de la concentración de Mg2+, etc. Varios métodos de mutagénesis conducen a diferentes mecanismos de variación de bases en la cadena de ADN amplificada. MnCh es un factor mutagénico de las ADN polimerasas, añadir Mn2+ puede reducir la especificidad de la polimerasa al molde y mejorar la tasa de incompatibilidad; el desequilibrio de las concentraciones de los cuatro dNTP puede mejorar la probabilidad de una incorporación errónea de bases y concretar la incompatibilidad; Mg2+ tiene la función de activar la enzima Taq, el aumento de la concentración de Mg2+ para exceder la dosis normal puede estabilizar los pares de bases no complementarios; el aumento de la dosis de ADN polimerasa Taq y el tiempo de extensión de cada ciclo pueden aumentar la probabilidad de una extensión terminal incompatible; la disminución de la concentración de molde inicial aumentará la proporción de moldes variantes en el siguiente ciclo de PCR.
Enzima inmovilizada: se refiere a una enzima cuya catálisis puede usarse repetida y continuamente dentro de un cierto rango espacial. Por lo general, la reacción catalizada por enzimas se lleva a cabo en soluciones acuosas, mientras que las enzimas inmovilizadas se tratan física o químicamente para hacer que las enzimas solubles en agua sean insolubles, pero sigan siendo enzimáticamente activas. Después de la inmovilización, la estabilidad general de la enzima aumenta, es fácil de separar del sistema de reacción, fácil de controlar, puede usarse muchas veces, es fácil de transportar y almacenar y es favorable para la producción automática, pero la actividad y el rango de uso esta reducido.
Matriz portadora de enzimas inmovilizadas: se refiere a un material que forma la estructura principal de un portador de enzimas inmovilizado.
Como se usa en la presente, 1 en peso se refiere a 1 g de células húmedas recombinantes mutantes de transaminasa necesarias para transformar 1 g de sustrato.
En la presente solicitud, el 1V al que se hace referencia es igual al volumen del sistema de reacción/masa del sustrato.
Para resolver el problema de que la solicitud está limitada debido a la escasa tolerancia de la actividad de la transaminasa en un entorno extremo en el estado de la técnica, una realización típica de la aplicación lleva a cabo la evolución dirigida de la transaminasa que está mutada en el sitio R416T de la chromobacterium violaceum para obtener el mutante de la transaminasa, en donde el mutante de transaminasa tiene una secuencia en la que se produce una mutación de aminoácidos en la secuencia mostrada en la SEQ ID NO: 1; el sitio en el que se produce la mutación de aminoácidos incluye el sitio T7C+S47C. En el entorno relativamente extremo, la actividad transaminasa mutacional de la transaminasa con mutación que se produce en el sitio R416T+T7C+S47C fue significativamente mayor que la del mutante R416T.
A continuación, las soluciones técnicas y los efectos anteriores se describirán con referencia a experimentos.
I. Selección de mutantes con tolerancia mejorada a entornos extremos
La transaminasa derivada de Chromobacterium violaceum se modificó en la presente invención para obtener el mutante R416T con actividad enzimática mejorada, y la secuencia de aminoácidos del mutante R416T se muestra en la SEQ ID NO: 1. La actividad de la enzima es buena, pero su estabilidad no es ideal. Para mejorar la estabilidad de la enzima, se diseñaron cinco grupos de mutaciones de doble punto Q78C+A330C, V137C+G313C, A217C+Y252C, T7C+S47C y L295C+328C con el mutante R416T como molde y se diseñó una secuencia de cebadores con la página web QuikChange Primer Design. El sitio mutante se introdujo en el mutante R416T mediante PCR de plásmido completo, y el plásmido mutante con el nuevo sitio de mutación se obtuvo con pET-22b(+) como vector de expresión.
El plásmido mutante se transformó en células de E. coli y se indujo durante la noche en condiciones óptimas de expresión inducida por transaminasas a 25 °C y 0,1 mM de IPTG, y luego se obtuvo la enzima sin procesar mediante ultrasonidos de las células. Después de tratar la solución enzimática expresada por la cepa mutante durante 1 hora en un entorno extremo de 45-50 °C, pH 9,5 y DMSO al 20 %, se añadió el sustrato 1 o el sustrato 2, y se utilizó la cantidad de enzima de 1 en peso para continuar reaccionando bajo esta condición durante 16 horas, y luego se
detectó la tasa de transformación. El mutante con estabilidad mejorada se seleccionó mediante este método, en el que la actividad del mutante (R416T+T7C+S47C) en los sitios T7C+S47C fue significativamente mayor que la del mutante R416T. Bajo esta condición, la tasa de transformación catalizada por R416T fue del 15 %, mientras que la catalizada por el mutante R416T+T7C+S47C fue del 72 %.
Además, mediante el uso del mutante R416T+T7C+S47C como progenitor femenino, 33 pares de mutagénesis dirigida al sitio (cebadores específicos diseñados por la página web QuikChange Primer Design) (M356L, w360L, F364L, C404L, M430L, M438L, C445A, F449V, R405E, R405A, K90G, K190R, K219T, K304D, K51R, W187Y, K193E, K143R, N151M, S8P, A33P, A95P, E368P, Q346E, H333K, D371G, E246A, C328A, N412G, T402P, T107F, T107A, G110P), las secuencias de los cebadores se diseñaron mediante la página web QuikChange Primer Design y el plásmido mutante con el gen diana se obtuvo mediante mutagénesis dirigida al sitio con pET-22b (+) como vector de expresión. El plásmido mutante se transformó en células de E. coli y se indujo durante la noche en condiciones óptimas de expresión inducida por transaminasas a 25°C y 0,1 mM de IPTG. Luego se obtuvo la enzima sin procesar mediante ultrasonidos de las células.
Después de tratar la solución enzimática expresada por la cepa mutante durante 1 hora en un entorno extremo de 30-45°C, pH 9,5-10 y DMSO al 50 %, se añadió el sustrato 1 o el sustrato 2, y se continuó con la reacción en estas condiciones durante 16 horas, y luego se detectó la tasa de transformación. Los mutantes con mayor tolerancia a la temperatura, el pH y el disolvente orgánico se seleccionaron de esta manera. Los resultados de la selección son: la tolerancia del mutante con sitio de mutación en M356L, F364L, C404L, M430L, R405E, R405A, K90G, K219T, K304D, K51R,A95P, E368P, Q346E, H333K, D371G, E246A, C328A, N412G, T402P, T107F, T107A, G110P en un entorno de 30°C, pH 9,5 y DMSO al 50 % fue entre un 8 % y un 40 % superior al del mutante R416T+T7C+S47C. La tolerancia de algunos mutantes hasta un entorno de 45°C, pH 9,5, DMSO al 50 % fue 1,7-2,1 veces mayor que la del mutante R416T+T7C+S47C, y la tolerancia de algunos mutantes hasta un entorno de 40°C, pH 10, el 50 % de DMSO fue 3,7-3,9 veces mayor que la del mutante R416T+T7C+S47C.
Con el mutante R416T+T7C+S47C como progenitor femenino, se seleccionaron mutantes específicos con tolerancia mejorada en diferentes entornos extremos mediante mutagénesis dirigida al sitio, como se muestra en la Tabla 1 y la Tabla 2 a continuación.
Tabla 1: mutantes de punto único con tolerancia mejorada hasta el entorno de 30 °C, pH 9,5, 50 % de DMSO ni m i n m n i iri i l ii .

Tabla 2: mutantes de punto único con tolerancia mejorada hasta el entorno de 40°C, pH 10, 50 % de DMSO ni m i n m n i iri i l ii .

Con el fin de seleccionar de forma sencilla y eficaz más mutantes ideales, el mutante R416T+T7C+S47C se muta aleatoriamente adoptando una tecnología de PCR propensa a errores en la presente invención.
En esta solicitud, el fragmento del gen diana se unió al vector pET-22b mediante un método de PCR propenso a errores y se obtuvo un plásmido mutante con el gen diana. El plásmido mutante se transformó en células de E. coli y se indujo durante la noche en condiciones óptimas de expresión inducida por transaminasas a 25°C y 0,1 mM de IPTG. Finalmente, se obtuvo la enzima sin procesar mediante ultrasonidos de las células.
Después de tratar la solución enzimática expresada por la cepa mutante durante 1 hora en un entorno extremo de 30-45 °C, pH 9-10 y una concentración de disolvente orgánico de DMSO al 50 % o MeOH al 35 %, se añadió el sustrato 1 o el sustrato 2, y continuó con la reacción en esta condición durante 16 horas, luego se detectó la tasa de transformación. Los mutantes con mayor tolerancia a la temperatura, el pH y los disolventes orgánicos se seleccionaron de esta manera. Los resultados de la selección mostraron que la tolerancia del mutante con sitio de mutación en K69N, G201C, Q380L, K193I, I297L, R305H, F111Y, K190E, A286T hasta el entorno de 30 °C, pH 9,5 y 50 % de DMSO fue 16 % - 45 % superior a la del mutante R416T+T7C+S47C. La tolerancia de algunos mutantes al entorno de 40°C, pH 10, DMSO al 50% fue 117 % - 537 % mayor que la del mutante R416T+T7C+S47C, y la tolerancia de algunos mutantes un entorno de 30°C, pH 8, MeOH al 35 % fue un 233 % - 649 % mayor que la del progenitor femenino.
Con el mutante R416T+T7C+S47C como progenitor femenino, se seleccionaron mutantes específicos con tolerancia mejorada en diferentes entornos extremos mediante PCR propensa a errores, como se muestra en la Tabla 3 a la Tabla 5 a continuación.
Tabla 3: mutantes de punto único con tolerancia mejorada hasta el entorno de 30 °C, pH 9,5, 50 % de DMSO ni m i n P R r n rr r .

Tabla 4: mutantes de punto único con tolerancia mejorada hasta el entorno de 40°C, pH 10, 50 % de DMSO ni m i n P R r n rr r .

Tabla 5: mutantes de punto único con tolerancia mejorada hasta el entorno de 30 °C, pH 8, 35 % de MeOH ni m i n P R r n rr r .

Para seguir con la evolución de las transaminasas con mejor estabilidad y tolerancia, en la presente invención, los sitios con estabilidad y tolerancia mejoradas de las transaminasas se someten a mutación combinada multipunto, y luego los mutantes multipuntos con estabilidad y tolerancia mejoradas se obtienen mediante un método de selección dirigida.
Los sitios de mutación para mutaciones combinadas fueron de K51 R, W187Y, R405E, K90G, A95P, K304D, Q380L, E368P, Q346E, H333K, D371G, E246A, C328A, N412G, T402P, T107F, T107A, G110P, I297L, K69N, G201C y A286T.
La mutación combinada es cualquier combinación de estos sitios. En particular, la mutación combinada incluye, entre otros, lo siguiente: K51R+W187Y, R405E+A95P, R405E+A95P+K304D, R405E+A95P+K304D+Q380L, R405E+K90G+A95P+K304D+Q380L, R405E+K90G+A95P+K304D+Q380L+E368P, R405E+K90G+A95P+K304D+Q380L+Q346E, R405E+K90G+A95P+K304D+Q380L+H333K, R405E+K90G+A95P+K304D+Q380L+D371G, R405E+K90G+A95P+K304D+Q380L+E246A, R405E+K90G+A95P+K304D+Q380L+C328A, R405E+K90G+A95P+K304D+Q380L+N412G, R405E+K90G+A95P+K304D+Q380L+T402P, R405E+K90G+A95P+K304D+Q380L+T107F, R405E+K90G+A95P+K304D+Q380L+T107A, R405E+K90G+A95P+K304D+Q380L+G110P, R405E+K90G+A95P+K304D+Q380L+1297L, R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A, R405E+K90G+A95P+K304D+Q380L+I297L+A286T, R405E+K90G+A95P+K304D+Q380L+I297L+E368P,
R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+K69N, R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+G201C y R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+A286T.
El plásmido mutante se transformó en células de E. coli y se indujo durante la noche en condiciones óptimas de expresión inducida por transaminasas a 25 °C y 0,1 mM de IPTG. Luego se obtuvo la enzima sin procesar mediante ultrasonidos de las células.
Después de tratar la solución enzimática durante 1 hora en un entorno más extremo de 45 °C, pH 9,5-10 y que contenía 50 % de DMSO o 35 % de MeOH, se añadió el sustrato 1 y se continuó con la reacción en estas condiciones durante 16 horas, luego se detectó la tasa de transformación. La tolerancia de algunos mutantes combinados hasta el entorno de 40 °C, pH 10, 50 % de DMSO fue 231 % - 610 % mayor que la del progenitor femenino, la tolerancia de algunos mutantes combinados hasta el entorno de 30 °C, pH 8, 35 % de MeOH fue 213 % - 990 % mayor que la del progenitor femenino, y la tolerancia de algunos mutantes combinados hasta el entorno de 45 °C, pH 8, 40 % de MeOH fue 3000 % mayor que la del progenitor femenino. Los mutantes combinados específicos con tolerancia mejorada se muestran en las Tablas 6 a 8 a continuación.
T l : m n m in n l r ni m r h l n rn 4 ° H 1 DM

T l 7: m n min n l r ni m r h l n rn ° H M H.

°

continuación

II. Inmovilización del mutante de transaminasa de la solicitud
2.1 Agregados enzimáticos reticulados de transaminasas (CLEA)
En la solicitud, las transaminasas del mutante R416T+T7C+S47C progenitor femenino, los mutantes de punto único y los mutantes combinados seleccionados en base al mutante progenitor femenino se inmovilizaron mediante el método de reticulación para preparar agregados enzimáticos reticulados de transaminasas.
En general, la preparación de los agregados enzimáticos reticulados se lleva a cabo principalmente en dos etapas: (1) formación de un precipitado de agregación de proteínas enzimáticas; (2) reticulación entre precipitados.
La proteína enzimática puede coagularse y precipitarse mediante un fraccionamiento de sales, un método de precipitación isoeléctrica, un método de precipitación de sales de metales pesados o un método de precipitación de disolventes orgánicos para obtener agregados de proteínas enzimáticas. Normalmente, el precipitado de proteína enzimática es un precipitado reversible que se puede volver a disolver en solución acuosa.
Y después de la precipitación de proteínas, agregar un agente de reticulación para conectar aún más los precipitados de proteínas a través de enlaces covalentes para formar agregados de enzimas reticuladas precipitadas insolubles en agua. El agente de reticulación utilizado es un reactivo bifuncional o multifuncional; el reactivo bifuncional comprende glutaraldehído, N,N-metileno bisacrilamida (MBA), bismaleimida y similares, y el reactivo multifuncional dextrano (el peso molecular es 6 KDa-200 KDa). El grupo amino libre, el grupo fenol, el grupo imidazol y el grupo sulfhidrilo de la proteína enzimática pueden participar en la reacción de reticulación.
El tampón para preparar la solución de enzimas inmovilizadas contiene PLP de 0,4 a 1 mg/ml y el valor de pH de la solución enzimática es de 7,0 a 8,0. El precipitante utilizado para la preparación del precipitado de proteína enzimática es etanol, isopropanol y/o sulfato de amonio, y la concentración final del precipitante es del 90 %. El agente de reticulación utilizado para la preparación de los agregados reticulados de las proteínas enzimáticas fue una solución de glutaraldehído al 25 % con una concentración final de glutaraldehído de 200 mM - 500 mM.
El agregado enzimático reticulado preparado puede someterse directamente a una reacción catalítica en una fase acuosa con la enzima reticulada acuosa obtenida por filtración, o la enzima reticulada acuosa puede liofilizarse para obtener un polvo seco y luego aplicarse. El polvo liofilizado también se puede aplicar a la reacción en la fase de disolvente orgánico. Después de que el agregado enzimático reticulado se utiliza una vez, el agregado enzimático reticulado puede recuperarse mediante centrifugación o filtración y similares y luego utilizarse nuevamente; los tiempos de uso repetido se cuentan en el rango en el que la pérdida de actividad es inferior al 5 % en comparación con el primer uso, el agregado enzimático reticulado se utiliza en una fase acuosa. En comparación con la enzima libre, la recuperación de la actividad es superior al 80 %, y el progenitor femenino R416T+T7C+S47C se puede reutilizar 3 veces, los tiempos de reutilización de mutantes de punto único y/o mutantes combinados sobre la base del progenitor femenino fueron significativamente superiores a los de los progenitores femeninos, y los tiempos de reutilización de los mejores mutantes fueron de hasta 13 veces.
Las enzimas reticuladas de algunos mutantes se pueden reutilizar al menos 6 veces en un sistema que contenga 35 % de metanol. Se mejoró la estabilidad de la actividad enzimática libre del mutante, y también se incrementaron los tiempos de recuperación y reutilización de la actividad enzimática después de la inmovilización.
El fenómeno de emulsificación obviamente se redujo con enzima reticulada para catalizar la reacción y extraer el producto de la fase acuosa con disolvente orgánico después de la reacción. La preparación de la enzima reticulada tiene las ventajas de no tener portador, bajo coste, reacción catalizadora con la enzima reticulada, más veces de uso repetido, tiempos de uso amplios, cantidad reducida de enzima y menor coste que la enzima libre. El polvo liofilizado de enzimas reticuladas se hace reaccionar en un disolvente de fase 100 % orgánica, y cuando el progenitor femenino se utiliza repetidamente por segunda vez, la actividad se pierde en más del 10 % en comparación con la primera vez. Sin embargo, algunos mutantes podrían reutilizarse hasta 5 veces con menos del 5 % de pérdida de actividad en comparación con la primera vez.
2.2 Método de inmovilización de la transaminasa inmovilizada reticulada
Los CLEA no tienen soporte portador, las partículas de enzima inmovilizadas son pequeñas (<10 |jm) y la resistencia mecánica es deficiente, la enzima se endurece fácilmente en el proceso de filtración y recuperación de la enzima, y la enzima no se puede dispersar bien en el sistema de reacción en el siguiente uso. A fin de resolver el problema, se puede utilizar glutaraldehído como agente de reticulación en combinación con dos tecnologías de reticulación e incrustación, el glutaraldehído se añade por goteo a una solución mixta que contiene enzima líquida, acrilamida y metileno diacrilamida para permitir que el glutaraldehído y la enzima libre formen la base de Schiff para preparar un agregado enzimático reticulado, se añade un persulfato de amonio iniciador para formar un gel de poliacrilamida, y el agregado enzimático reticulado se incrusta en una matriz de gel de poliacrilamida para obtener una enzima inmovilizada estable.
En la solicitud, las transaminasas del mutante R416T+T7C+S47C progenitor femenino, los mutantes de punto único y los mutantes combinados seleccionados en base al mutante progenitor femenino se inmovilizaron mediante el método de reticulación incrustada para preparar la enzima incrustada-reticulada.
La enzima incrustada-reticulada preparada, cuya recuperación de actividad enzimática es superior al 80 %, después de que la reacción en fase acuosa se utiliza una vez, se puede recuperar fácilmente mediante filtración y similares y luego se utiliza nuevamente, los tiempos de uso repetidos se cuentan en el rango que la pérdida de actividad es inferior al 5 % en comparación con el primer uso, el progenitor femenino R416T+T7C+S47C se puede reutilizar 8 veces, los tiempos de reutilización de mutantes de punto único y/o mutantes combinados sobre en base a progenitores femeninos fueron significativamente superiores a los progenitores femeninos, y los tiempos de reutilización de los mejores mutantes fueron de hasta 18 veces.
Las enzimas reticuladas incrustadas de algunos mutantes se pueden reutilizar al menos 12 veces en un sistema que contenga 35 % de metanol. Al extraer el producto de la fase acuosa con disolvente orgánico después de la reacción, el fenómeno de emulsificación se redujo de manera evidente.
2.3 Método de inmovilización de transaminasa mediante adsorción
La enzima inmovilizada adsorbida se puede preparar mediante la adsorción y combinación de la molécula enzimática y el portador insoluble en agua mediante interacción electrostática, enlace de hidrógeno, interacción hidrofóbica, etc. El método tiene condiciones suaves y no es fácil provocar la desnaturalización de la enzima, pero la enzima se separa fácilmente del portador en solución acuosa y no se puede reciclar, por lo que la enzima inmovilizada preparada por el método de adsorción se utiliza principalmente en la reacción en disolventes orgánicos. Los portadores que se pueden utilizar para adsorber enzimas inmovilizadas se pueden dividir en dos tipos: portadores inorgánicos y portadores macromoleculares. Los portadores inorgánicos incluyen carbón activado, vidrio poroso, caolín , arcilla blanqueadora, caolinita, alúmina, gel de sílice, bentonita, hidroxiapatita, fosfato de calcio, óxidos metálicos y similares; los portadores macromoleculares incluyen almidón, glutelina, resinas sintéticas macroporosas, cerámica, etc.
El portador utilizado en la solicitud es un portador de resina sintética macroporosa, que comprende una matriz y un grupo funcional opcional que modifica la matriz, en donde la matriz incluye, entre otras, resina de poliestireno, resina de polimetacrilato o copolímero de estireno-metacrilato. Además, se pueden modificar varios tipos de portadores con el grupo funcional octadecilo. La siguiente Tabla 9 se enumera como el portador adecuado para la adsorción e inmovilización de la transaminasa en la solicitud.
Tabla 9. Portador de ti o de adsorción

En esta solicitud, la transaminasa y el portador de resina macroporosa se combinan directamente en modos físicos tales como enlaces hidrofóbicos, enlaces de hidrógeno y similares.
Las transaminasas del mutante R416T+T7C+S47C progenitor femenino, los mutantes de punto único y los mutantes combinados seleccionados en base al mutante progenitor femenino se inmovilizaron mediante el método de unión por adsorción física.
El tampón utilizado para la preparación de la solución enzimática contiene 0,4-1 mg/ml de PLP, el pH del tampón es 7,0-8,0 y la sal tampón es Na2HPO4-NaH2PO4, Tris-CI o ácido bórico-hidróxido de sodio.
La enzima inmovilizada por adsorción preparada puede ser secada por soplado de nitrógeno, secado al vacío, liofilización, etc.
En esta solicitud, la transaminasa se une al portador por adsorción y la recuperación de la actividad es superior al 80 %; al reaccionar en disolvente orgánico, la enzima inmovilizada es recuperada por filtración o aspiración de líquido mediante jeringa, que puede reutilizarse. En comparación con el primer uso, el número de usos repetidos se contó en el rango de pérdida de actividad inferior al 5 %. Para algunas enzimas inmovilizadas mutantes, la enzima inmovilizada podría reutilizarse 6 veces y la pérdida de actividad fue inferior al 5 %. 2.4 Método de inmovilización covalente de transaminasas
La inmovilización covalente de una enzima consiste en que un grupo no esencial de una proteína enzimática se une irreversiblemente con un portador insoluble en agua a través de un enlace covalente, y los grupos de proteínas capaces de acoplarse en condiciones suaves comprenden: amino, carboxilo, sulfhidrilo de cisteína, imidazolilo de histidina, grupo fenólico de tirosina, hidroxilo de serina y treonina. El grupo unido covalentemente con el transportador no suele ser el grupo necesario para que la enzima exprese su actividad.
2.4.1 Portador de enzima inmovilizada covalente
El portador de enzima inmovilizada puede ser materiales inorgánicos como sílice, vidrio, minerales, celite y similares; materiales orgánicos naturales tales como carboximetilcelulosa, dextrano, agarosa, pectina, quitosano y similares; y polímeros sintéticos orgánicos no naturales tales como resinas de poliestireno, resinas de polimetacrilato o copolímeros de estireno y metacrilato. Estos portadores pueden funcionalizarse adicionalmente para facilitar la unión a la molécula de proteína, por ejemplo mediante la incorporación de grupos amino, hidroxilo, epoxi, octadecilo y otros grupos funcionales en el portador. El portador con funcionalidad amino y el portador con funcionalidad hidroxilo pueden unirse a la molécula de proteína enzimática a través de enlaces iónicos, el portador de tipo amino también puede unirse covalentemente a la proteína enzimática, el portador con funcionalidad epoxi se une principalmente a la proteína enzimática a través de enlaces covalentes, y el portador con funcionalidad octadecilo se une a la molécula de enzima a través de una interacción hidrofóbica. El portador puede adoptar cualquier aspecto o forma, como una película, un tubo, una lámina, una perla, una partícula, un chip, una fibra óptica, etc.
El portador utilizado en la presente solicitud es quitosano, resina y vidrio poroso.
El quitosano se puede utilizar como portador para la inmovilización de enzimas debido a su buena biocompatibilidad, alta plasticidad de forma (que se puede convertir en gel, película, fibra y otras formas), no toxicidad, fácil modificación química y otras características (ProcessBiochem, 2005; 40: 2833-40). El propio quitosano es soluble en agua y debe prepararse en partículas portadoras insolubles en agua mediante evaporación de disolventes, emulsificación, coacervación y otros métodos (MacromolBiosci, 2003; 3: 511-20). Las partículas portadoras producidas por emulsificación son pequeñas y uniformes y generalmente son las preferidas. Las moléculas de quitosano tienen grupos hidroxilo activos, grupos amino y similares, la enzima se puede adsorber y unir a través de enlaces iónicos, enlaces de hidrógeno, fuerza de Van der Waals y otros, pero el efecto de adsorción es débil, la enzima cae con facilidad, y los agentes de reticulación comunes tales como formaldehído, glutaraldehído y similares se activan y luego se unen covalentemente a la enzima.
El portador de resina utilizado en la solicitud comprende una matriz y grupos funcionales que modifican la matriz, en donde la matriz incluye, entre otras, resinas de poliestireno, resinas de polimetacrilato y copolímeros de estirenometacrilato. Los grupos funcionales adecuados portados por tales matrices incluyen, entre otros, grupos amino de cadena corta, grupos amino de cadena larga y grupos epoxi. La siguiente Tabla 10 se enumera como el portador adecuado para la inmovilización de la transaminasa en la solicitud.
Tabla 10:

En la solicitud, la transaminasa se une directamente a la resina con el grupo funcional epoxi mediante enlace covalente, y a la resina con grupo funcional amino activado por glutaraldehído mediante enlace covalente.
2.4.2 Método de inmovilización covalente
En la solicitud, las transaminasas del mutante R416T+T7C+S47C progenitor femenino, los mutantes de punto único y los mutantes combinados seleccionados en base al mutante progenitor femenino se inmovilizaron mediante el método de unión covalente.
La solución tampón utilizada para la preparación de la solución enzimática contiene 0,4-1 mg/ml de PLP, el pH de la solución tampón es 7,0-8,0 y la sal tampón es Na2HPO4-NaH2PO4, Tris-CI o ácido bórico-hidróxido de sodio. El peso molecular del quitosano utilizado en la presente solicitud incluye, entre otros, 300-500 KDa, preparar un portador mediante un método de emulsificación, después activar el portador con glutaraldehído, añadir solución enzimática, incubar durante 6 horas a 20 °C, recolectar el precipitado por filtración o centrifugación y enjuagar el precipitado con tampón.
Inmovilizado covalentemente el portador de tipo amino, primero activar el portador con glutaraldehído, luego añadir solución enzimática, incubar durante la noche a 20 °C, recolectar el precipitado por filtración y enjuagar el precipitado con tampón. Inmovilizado covalentemente en un portador basado en epoxi, mezclar directamente la solución enzimática con el portador, incubar durante la noche a 20 °C, luego dejar reposar durante 20 horas, recolectar el precipitado por filtración y enjuagar el precipitado con tampón.
La enzima inmovilizada preparada puede ser secada por soplado de nitrógeno, secado al vacío, liofilización, etc.
En la solicitud, la transaminasa se inmoviliza en los portadores anteriores mediante unión covalente y se inmoviliza en un portador de quitosano y la recuperación de la actividad es del 50 % - 60 %; inmovilizado en un portador de amino de cadena corta, la recuperación de la actividad es del 50% - 70 %; inmovilizado en un portador de tipo amino de cadena larga, la recuperación de la actividad es del 70 % - 80 %; inmovilizado en un portador basado en epoxi, la recuperación de la actividad es del 40 % - 60 %. La actividad de la enzima inmovilizada del mutante obviamente mejora en comparación con la enzima inmovilizada del progenitor femenino. La enzima inmovilizada se recupera por filtración o aspiración de líquido mediante jeringa, que puede reutilizarse. En comparación con el primer uso, el número de usos repetidos se contó en el rango de pérdida de actividad inferior al 5 %, para la enzima inmovilizada en el portador de quitosano, la enzima inmovilizada podría reutilizarse 3 veces y la pérdida de actividad fue inferior a 5 %. Inmovilizadas en un portador de tipo amino de cadena corta, algunas enzimas se pueden reutilizar 5 veces; inmovilizada en un portador de tipo amino de cadena larga, la enzima de algunos mutantes se podría reutilizar 11 veces; inmovilizadas en un portador a base de epoxi, algunas enzimas se pueden reutilizar 6 veces. Los tiempos de reutilización de la enzima mutante inmovilizada fueron significativamente más altos que los de la enzima inmovilizada progenitora femenina. Algunas transaminasas después de la inmovilización covalente pueden desempeñar una función catalítica en un disolvente orgánico al 100 %, algunos mutantes pueden reaccionar en un disolvente orgánico después de la inmovilización y la pérdida de actividad es inferior al 5 % después de un uso repetido 3 veces. Desempeñando una función catalítica en una solución de metanol al 35 %, algunos mutantes se reutilizaron 5 veces después de la inmovilización y la pérdida de actividad fue inferior al 5 %.
2.5 Método de inmovilización quelante de iones metálicos
El vidrio poroso es muy adecuado para la inmovilización de enzimas debido a su material inerte y buena permeabilidad al agua. Los grupos silanol en el vidrio y la superficie del canal del mismo se utilizan como sitios de unión para unirse a la enzima y lograr la inmovilización, pero el grupo silanol en la superficie del vidrio tradicional tiene una densidad limitada y una distribución desigual, alto impedimento estérico de unión a la enzima, baja carga de proteína (Science, 2010, 329, 305-309; JChromatogr, 1976, 125, 115-127) y facilidad para inactivar la enzima. Cubrir la superficie interior y exterior del vidrio poroso con una capa de película de polímero orgánico puede formar un entorno más favorable para la inmovilización de enzimas. (Langmuir, 2004, 20, 10639-10647). La película de polímero se puede modificar adicionalmente según se desee para añadir varios grupos funcionales adecuados para la inmovilización en la superficie.
La proteína etiquetada con histidina se puede purificar mediante cromatografía de afinidad con metales en fase sólida, los residuos de histidina en la proteína se pueden quelar con iones metálicos (Ni2+, Co2+, Fe3+) quelados en una matriz insoluble en agua y luego se puede eluir la proteína diana con un tampón que contiene imidazol (Nature, 1975, 258, 598-599). Según esta tecnología, la enzima marcada con histidina se puede quelar específicamente con el portador con iones metálicos quelados al final, y se puede lograr el objetivo de la inmovilización. Al mismo tiempo, la especificidad del método es alta y difícilmente se pueden inmovilizar proteínas diversas.
El portador de vidrio utilizado en la solicitud está recubierto en la superficie interna y externa del vidrio poroso mediante una película de polímero, y la película de polímero puede ser un polímero hidrofílico, como un polímero acrílico, un polímero semihidrofílico de estireno y acrilonitrilo y un polímero hidrofóbico tal como un polímero de clorometilestireno. La superficie de la película se modifica con amino y luego se acila mediante 2,4-dihidroxiacetofenona, y el hidroxilo de la 2,4-dihidroxiacetofenona se quela con iones metálicos, de modo que un extremo de la 2,4-dihidroxiacetofenona se une al portador y el otro extremo de la 2,4-dihidroxiacetofenona se quela con los iones metálicos a través de la acción del grupo de la 2,4-dihidroxiacetofenona, de modo que el extremo del portador de vidrio se proporciona con los iones metálicos y puede unirse por afinidad a la proteína marcada con
histidina para lograr una inmovilización específica. (ChemicalCommunications, 2014, 50 (65): 9134-7). Los iones metálicos quelados incluyen, entre otros, Ni2+, Co2+ y Fe3+. La siguiente Tabla 11 se enumera como el portador adecuado para la inmovilización de la transaminasa en la solicitud.
Tabla 11

En la solicitud, las transaminasas del mutante R416T+T7C+S47C progenitor femenino, los mutantes de punto único y los mutantes combinados seleccionados en base al mutante progenitor femenino se inmovilizaron mediante el método de unión covalente.
La solución tampón utilizada para la preparación de la solución enzimática contiene 0,4-1 mg/ml de PLP, el pH de la solución tampón es 7,0-8,0 y la sal tampón es Na2HPO4-NaH2PO4, Tris-CI o ácido bórico-hidróxido de sodio.
Inmovilizado covalentemente el portador de vidrio poroso, mezclar directamente la solución de enzima con el portador, incubar durante 40-60 minutos a 20 °C, recolectar el precipitado por filtración y enjuagar el precipitado con el tampón.
La enzima inmovilizada preparada puede ser secada por soplado de nitrógeno, secado al vacío, liofilización, etc. En esta solicitud, la transaminasa se une al portador por quelación y la recuperación de la actividad es del 70 % - 80 %. La enzima inmovilizada se recupera por filtración o aspiración de líquido mediante jeringa, que puede reutilizarse. Reaccionando en un disolvente acuoso, en comparación con el primer uso, el número de veces de reutilización se contó en el rango de pérdida de actividad inferior al 5 %. Para algunas enzimas inmovilizadas mutantes, la enzima inmovilizada podría reutilizarse 12 veces y la pérdida de actividad fue inferior al 5%; al reaccionar en disolventes orgánicos, en comparación con el primer uso, el número de veces de reutilización se contó en el rango de pérdida de actividad inferior al 5 %. Para algunas enzimas inmovilizadas mutantes, la enzima inmovilizada podría reutilizarse 8 veces.
III. Método de aplicación de la transaminasa inmovilizada
La transaminasa inmovilizada de la presente solicitud puede transformar los aceptores de amino que se muestran en el sustrato 1 y el sustrato 2 en las aminas primarias correspondientes, y el donante amino utilizado es isopropilamina.
La transaminasa inmovilizada de la solicitud se puede aplicar en los siguientes disolventes: solución acuosa al 100 %, disolvente que contenga 20% - 50% de DMSO, disolvente que contenga 35% de metanol, o disolvente orgánico saturado de agua al 100 % (por ejemplo, puede ser éter terc-butílico de metilo saturado de agua al 100 % o acetato de isopropilo saturado de agua al 100 %).
La enzima inmovilizada divulgada en la invención se puede aplicar a reacciones por lotes en un modo de agitación y también se puede aplicar a reacciones de flujo continuo llenas en un reactor de tubería.
El modo de funcionamiento de reacción de agitación por lotes es el siguiente: añadir las materias primas, es decir, el aceptor amino, el donante amino, la enzima inmovilizada, la coenzima PLP y el disolvente en el recipiente de reacción al mismo tiempo y hacer reaccionar durante más de 16 horas por medios de agitación mecánica. Después de la reacción, la enzima inmovilizada se recuperó por filtración y se aplicó a la siguiente ronda de reacción.
El modo de funcionamiento de la reacción continua es el siguiente: llenar la enzima inmovilizada en un reactor tubular, disolver por completo las materias primas, es decir, un aceptor amino, un donante amino y la coenzima PLP, en una solución de reacción con un solvente adecuado, inyectar la solución de reacción en el reactor tubular lleno con la enzima inmovilizada a un caudal adecuado con una bomba de émbolo, y recibir una solución de producto con el disolvente en una salida.
Durante la operación de reacción continua, el disolvente puede ser una solución de metanol al 35 % o éter terciario de metilo saturado con agua al 100 %.
Los beneficios de la presente solicitud se ilustran adicionalmente mediante los siguientes ejemplos específicos. Ejemplo 1: Prueba de tolerancia de los mutantes a 30 °C, pH 9,5 y 50 % de DMSO
La enzima sin procesar se trató durante 1 hora a 30 °C, pH 9,5, concentración de DMSO del 50%, luego se añadieron 0,1 g de sustrato 1, 4 eq. de hidrocloruro de isopropilamina y 0,6-1 mg de PLP (piridoxal-5'-fosfato) a un matraz de reacción de 10 ml, luego se añadieron 5 mg de la enzima tratada y se agitó termostáticamente durante 16 horas en el entorno de 30 °C, pH 9,5, concentración de DMSO del 50 %. La tasa de transformación se midió por HPLC y los datos de la reacción del mutante se muestran en la siguiente Tabla 12.
Tabla 12

Ejemplo 2: Prueba de tolerancia de los mutantes a 45°C, pH 10 y 50 % de DMSO
La enzima sin procesar se trató durante 1 hora a 45 °C, pH 10, concentración de DMSO del 50 %, luego se añadieron 0,1 g de sustrato 1, 4 eq. de hidrocloruro de isopropilamina y 0,6-1 mg de PLP (piridoxal-5'-fosfato) a un matraz de reacción de 10 ml, luego se añadieron 5 mg de la enzima tratada y se agitó termostáticamente durante 16 horas en el entorno de 45°C, pH 10, concentración de DMSO del 50 %. La tasa de transformación se midió por HPLC y los datos de la reacción del mutante se muestran en la siguiente Tabla 13.
Tabla 13

Ejemplo 3: Prueba de tolerancia de los mutantes a 30 °C, pH 8 y 35 % de metanol
La enzima sin procesar se trató durante 1 hora a 30 °C, pH 8, concentración de MeOH del 35%, luego se añadieron 0,1 g de sustrato 1, 4 eq. de hidrocloruro de isopropilamina y 0,6-1 mg de PLP (piridoxal-5'-fosfato) en un matraz de reacción de 10 ml, luego se añadieron 5 mg de la enzima tratada y se continuó agitando termostáticamente durante
16 horas en el entorno de 30 °C, pH 8, concentración de MeOH del 35 %. La tasa de transformación se midió por HPLC y los datos de la reacción del mutante se muestran en la siguiente Tabla 14.
Tabla 14

Ejemplo 4: Prueba de tolerancia de los mutantes a 45°C, pH 8 y 40% de metanol
La enzima sin procesar se trató durante 1 hora a 45°C, pH 8, concentración de MeOH del 40%, luego se añadieron 0,1 g de sustrato 1, 4 eq. de hidrocloruro de isopropilamina y 0,6-1 mg de PLP (piridoxal-5'-fosfato) en un matraz de reacción de 10 ml, luego se añadieron 5 mg de la enzima tratada y se continuó agitando termostáticamente durante 16 horas en el entorno de 45°C, pH 8, concentración de MeOH del 40%. La tasa de transformación se midió por HPLC y los datos de la reacción del mutante se muestran en la siguiente Tabla 15.
Tabla 15

Ejemplo 5: Inmovilización por reticulación
Se disolvieron 0,1 g de enzima en polvo en 2 ml de tampón de fosfato (0,1 M de PB, pH 7,0-8,0, que contenía 0,4-1 mg/ml de PLP (piridoxal-5'-fosfato)), se añadieron 18 ml de etanol o 18 ml de isopropanol o sulfato de amonio (saturación final del 90 %) como precipitante con agitación en un baño de hielo y agua, y después de 10 minutos de agitación, se añadieron 1,1-2,7 ml de solución de glutaraldehído al 25 % (concentración final de 200-500 mM), se centrifugó o filtró después de agitar en el baño de hielo-agua durante 30-40 minutos, el precipitado se lavó tres
veces con un tampón de fosfato, se almacenó a 4 °C y se aplicó directamente a una reacción en fase acuosa. O el agregado enzimático reticulado se liofiliza, y el polvo liofilizado del agregado enzimático reticulado después de la liofilización se puede aplicar en reacciones de fase acuosa y fase orgánica.
Ejemplo 6: Reacción de agregados enzimáticos reticulados en fase acuosa para evaluar la actividad
A un matraz de reacción de 10 ml, se añadieron 0,3 ml de DMSO, se disolvieron 0,1 g de sustrato 1, se añadieron 4 eq. de clorhidrato de isopropilamina y 1,0 mg de PLP (piridoxal-5'-fosfato), se añadió 0,1 M de PB 7,0 hasta que el volumen final de la solución de reacción fue de 1 ml, y se añadieron 5 mg de enzima o agregado enzimático reticulado en húmedo preparado a partir de 5 mg de enzima o agregado enzimático reticulado en polvo liofilizado y se agitó a 45 °C durante 16 horas. La tasa de transformación se midió por HPLC y los datos de la reacción se muestran en la Tabla 16 a continuación.
Tabla 16

continuación

Ejemplo 7: Reacción de enzima reticulada en fase orgánica para evaluar la actividad
A un matraz de reacción de 10 ml, se añadió 1 ml de éter terc-butílico de metilo saturado de agua, seguido de 10 mg de sustrato 1 y 4 eq. De isopropilamina, luego se añadieron agregados enzimáticos reticulados liofilizados preparados a partir de 10 mg de enzima en polvo, y se agitó durante 16 horas a 30 °C. La tasa de transformación se midió por HPLC y los datos de la reacción se muestran en la Tabla 17 a continuación.
Tabla 17

Ejemplo 8: Reacción de la enzima inmovilizada incrustada-reticulada de transaminasa en fase acuosa para evaluar la actividad
A un matraz de reacción de 10 ml, se añadieron 0,4 ml de DMSO (la concentración final de DMSO era del 40 %), se disolvieron 0,1 g de sustrato 1, se añadieron 4 eq. De clorhidrato de isopropilamina y 1,0 mg de PLP (piridoxal-5'-fosfato), se añadió 0,1 M de PB 7,0 hasta que el volumen final de la solución de reacción fue de 1 ml, y se añadieron 5 mg de enzima o de enzima inmovilizada incrustada-reticulada preparada a partir de 5 mg de enzima y se agitó a 45 °C durante 16 horas. La tasa de transformación se midió por HPLc y los datos de la reacción se muestran en la Tabla 18 a continuación.
Tabla 18

continuación

Ejemplo 9: Reacción de la enzima inmovilizada incrustada-reticulada de transaminasa en una solución de agua con metanol al 35 % para evaluar la actividad
En un matraz de reacción de 10 ml, se añadieron 0,35 ml de metanol (la concentración final de metanol era del 40 %), se disolvieron 0,1 g de sustrato 1, se añadieron 4 eq. de clorhidrato de isopropilamina y 1,0 mg de PLP (piridoxal-5'-fosfato), se añadió 0,1 M de PB 7,0 hasta que el volumen final de la solución de reacción fue de 1 ml, y se añadieron 10 mg de enzima o de enzima inmovilizada incrustada-reticulada preparada a partir de 10 mg de enzima y se agitó a 30 °C durante 16 horas. La tasa de transformación se midió por HPLC y los datos de reacción se muestran en la Tabla 19 a continuación.
Tabla 19

Ejemplo 10: Método de inmovilización de transaminasa mediante adsorción
Se añadieron al portador 4 ml de tampón PB que contenía 0,4 mg/ml de PLP (100 mM, pH 7,0) y al mismo tiempo se añadió 0,1 g de enzima, se agitó a una velocidad baja de 80 rpm durante la noche a 20 °C. El sobrenadante se eliminó, el precipitado se lavó de 3 a 4 veces con tampón, el sobrenadante se eliminó, el precipitado se secó con nitrógeno o se secó mediante liofilización y se almacenó a 4 °C.
Ejemplo 11: Reacción de la enzima inmovilizada por adsorción de transaminasa en fase orgánica para evaluar la actividad
En un matraz de reacción de 10 ml, se añadió 1 ml de éter terc-butílico de metilo saturado de agua, seguido de 10 mg de sustrato 1 y 4 eq. de isopropilamina, luego se añadió transaminasa inmovilizada preparada a partir de 20 mg de enzima y se agitó durante 16 horas a 30 °C. La tasa de transformación se midió por HPLC y los datos de reacción se muestran en la Tabla 20 a continuación.
Tabla 20

continuación

Ejemplo 12: Inmovilización de transaminasas con quitosano como portador
Preparación del portador de quitosano: Se añadieron 5 g de quitosano (peso molecular 300 KDa-500 KDa) a 250 ml de solución de ácido acético al 1 % y se disolvieron calentando en un horno de microondas para preparar la fase acuosa. La fase oleosa se preparó mezclando uniformemente 300 ml de tolueno con 2,2 g de Span 80, 1,2 ml de nhexanol y agitación a temperatura ambiente durante 2 horas. Se añadió por goteo lentamente una fase acuosa a una fase oleosa con agitación para preparar una emulsión, la emulsión se vertió en 1,5 L de una solución de NaOH al 12 %, se agitó durante 3 horas, se añadió 1 L de etanol, se filtró y el remanente del filtro se limpió en profundidad con agua purificada para obtener aproximadamente 40 g de portador húmedo, el portador húmedo se sumerge en 140 ml de agua pura y se almacena a 4 °C.
Activación de un portador: Se añadieron 1,1 ml de glutaraldehído al 25 % (concentración final de glutaraldehído del 2,5 %) por mililitro de portador húmedo.
Inmovilización: Se añadieron 0,2 g de enzima al portador activado, se agitó durante 6 horas a 20-25 °C, se lavó el portador, se centrifugó para eliminar el sobrenadante y el precipitado es la enzima inmovilizada y se almacenó a 4 °C.
Ejemplo 13: Inmovilización de la transaminasa en el transportador amino C6
Activación de un portador: Se lavó 1 g de ECR8409 de 1 a 2 veces con 20 mM de tampón de baja fuerza iónica, se eliminó el sobrenadante, se añadieron 4 ml de glutaraldehído al 2 % (preparado a partir de reactivo de dilución de tampón de baja fuerza iónica de 20 mM y glutaraldehído al 25 %), a 20 °C, se activó a 80 rpm durante 1 hora, se lavó de 1 a 2 veces con 20 mM de tampón y el sobrenadante se eliminó.
Inmovilización: Se añadieron al portador activado 4 ml de tampón PB que contenía 0,4 mg/ml de PLP (20 mM, pH 7,0) y al mismo tiempo se añadió 0,1-0,2 g de enzima, se agitó a una velocidad baja de 80 rpm durante la noche a 20 °C. El sobrenadante se eliminó, el precipitado se lavó de 3 a 4 veces con tampón, el sobrenadante se eliminó y el precipitado se secó con nitrógeno o se secó mediante liofilización y se almacenó a 4 °C.
Ejemplo 14: Inmovilización de transaminasas sobre portador epoxi
Se lavó 1 g de portador ECR8285 de 1 a 2 veces con tampón PB 100 mM, el líquido transparente se eliminó, se añadieron 4 ml de tampón (PB 100 mM, pH 7,0, que contenía NaCl 1 M), se añadieron de 0,1 g a 0,2 g de enzima a al mismo tiempo, a 20 °C, se agitó a baja velocidad de 80 rpm durante la noche (18 a 20 horas), y luego se dejó reposar a 4 °C durante 20 horas. El sobrenadante se eliminó y el precipitado se lavó de 3 a 4 veces con tampón, se secó con nitrógeno y se almacenó a 4°C.
Ejemplo 15: Reacción de transaminasa inmovilizada covalente en fase acuosa para evaluar la actividad
En un matraz de reacción de 10 ml, se añadieron 04 ml de DMSO (la concentración final de metanol era del 40 %), se disolvieron 0,1 g de sustrato 1, se añadieron 4 eq. de clorhidrato de isopropilamina y 1,0 mg de PLP (piridoxal-5'-fosfato), se añadió 0,1 M de PB 7,0 hasta que el volumen final de la solución de reacción fue de 1 ml, y se añadieron 5 mg de enzima o transaminasa inmovilizada preparada a partir de 5 mg de enzima y se agitó a 45 °C durante 16 horas. La tasa de transformación se midió por HPLC y los datos de reacción se muestran en la Tabla 21 a continuación.
Tabla 21

continuación

Ejemplo 16: Reacción de transaminasa inmovilizada covalente en solución de agua con metanol al 35 % para evaluar la actividad
En un matraz de reacción de 10 ml, se añadieron 0,35 ml de metanol (la concentración final de metanol era del 40 %), se disolvieron 0,1 g de sustrato 1, se añadieron 4 eq. de clorhidrato de isopropilamina y 1,0 mg de PLP (piridoxal-5'-fosfato), se añadió 0,1 M de PB 7,0 hasta que el volumen final de la solución de reacción fue de 1 ml, y se añadieron 10 mg de enzima o de transaminasa inmovilizada preparada a partir de 10 mg de enzima y se agitó a 30 °C durante 16 horas. La tasa de transformación se midió por HPLC y los datos de reacción se muestran en la Tabla 22 a continuación.
Tabla 22

continuación

Ejemplo 17: Reacción de transaminasa inmovilizada covalente en fase orgánica para evaluar la actividad
En un matraz de reacción de 10 ml, se añadió 1 ml de acetato de isopropilo saturado de agua, seguido de 10 mg de sustrato 1 y 4 eq. de isopropilamina, luego se añadieron 20 mg de enzima o transaminasa inmovilizada preparada a partir de 20 mg de enzima y se agitó durante 16 horas a 30 °C La tasa de transformación se midió por HPLC y los datos de reacción se muestran en la Tabla 23 a continuación.
Tabla 23

Ejemplo 18: Reacción continua de enzima inmovilizada covalente en lecho empaquetado de transaminasas (metanol como codisolvente para sustrato)
Se introdujeron en el reactor 75 g de enzima inmovilizada preparada por el mutante R405E+K90G+A95P+K304D+Q380L+1297L inmovilizado en el portador ECR8409, el volumen de la columna (CV) fue de 150 ml, y se inyectó en el lecho empaquetado el tampón 2CV (0,1 M de PB 7,0 con 5 mg/ml de PLP, clorhidrato de isopropilamina 2 M) con una bomba de émbolo. La solución de reacción formulada (sustrato 10,5 M, clorhidrato de isopropilamina 2 M, PLP 5 mg/ml, MeOH al 35 %) se inyectó en un lecho empaquetado con una bomba de émbolo, baño de agua a 40 °C, un caudal de 0,25 ml/minutos, tiempo de retención de aproximadamente 600 min, tasa de transformación > 98 % y una operación continua durante 240 horas sin reducción en la tasa de transformación.
Ejemplo 19: Transaminasa inmovilizada quelante del portador de vidrio
Se lavaron 1 g de portadores de vidrio poroso EziG-101, EziG-102 o EziG-103 de 1 a 2 veces con tampón (20 mM Tris-CI 8,5), el sobrenadante se eliminó, se añadieron 20 ml de tampón a los portadores mientras se añadía la enzima en polvo o la solución de enzima, se agitó a 80 rpm durante 1 hora a 20 °C, el sobrenadante se eliminó, el precipitado se lavó de 3 a 4 veces con tampón, se filtró y se secó al vacío, y se almacenó a 4 °C.
Ejemplo 20: Reacción de la transaminasa inmovilizada quelada sobre el portador de vidrio poroso EziG-101 en fase acuosa para evaluar la actividad
En un matraz de reacción de 10 ml, se añadieron 0,2 ml de DMSO, se disolvieron 0,1 g de sustrato 1, se añadieron 4 eq. de clorhidrato de isopropilamina y 0,5 mg de PLP (piridoxal-5'-fosfato), se añadió 0,1 M de PB 7,0 hasta que el volumen final de la solución de reacción fue de 1 ml, y se añadieron 0,1 g de enzima en polvo o transaminasa inmovilizada preparada a partir de 0,1 g de enzima en polvo mediante el método de quelación del portador EziG-101 y se agitó a 45 °C durante 16 horas. La tasa de transformación se midió por HPLC y los datos de reacción se muestran en la Tabla 24 a continuación.
Tabla 24

Ejemplo 21: Reacción de la transaminasa inmovilizada quelada sobre un portador de vidrio poroso en fase orgánica para evaluar la actividad
En un matraz de reacción de 10 ml, se añadió 1 ml de éter terc-butílico de metilo saturado de agua, seguido de 10 mg de sustrato 1 y 4 eq. de isopropilamina, luego se añadieron 20 mg de enzima o transaminasa inmovilizada preparada a partir de 20 mg de enzima y se agitó durante 16 horas a 30 °C. La tasa de transformación se midió por HPLC y los datos de reacción se muestran en la Tabla 25 a continuación.
Tabla 25:

Claims (18)
1. Un mutante de transaminasa, en donde el mutante de transaminasa que tiene una actividad transaminasa tiene una secuencia de mutación de aminoácidos de la SEQ ID NO:1, y los sitios de la mutación de aminoácidos comprenden T7C+S47C.
2. El mutante de transaminasa según la reivindicación 1, en donde los sitios de mutación de aminoácidos comprenden además uno o más de M356L, F364L, C404L, M430L, R405E/A, K90G, K219T, K304D, K51R, A95P, E368P, Q346E, H333K, D371G, E246A, C328A, N412G, T402P, T107F/A, G110P, K69N, G201C, Q380L, K193I, I297L, R305H, F111Y, K190E y A286T, en donde "/" representa "o".
3. El mutante de transaminasa según la reivindicación 1, en donde los sitios de mutación de aminoácidos comprenden además cualquiera de los siguientes sitios mutantes de combinación: K51R+W187Y, R405E+A95P, R405E+A95P+K304D, R405E+A95P+K304D+Q380L, R405E+K90G+A95P+K304D+Q380L, R405E+K90G+A95P+K304D+Q380L+E368P, R405E+K90G+A95P+K304D+Q380L+Q346E, R405E+K90G+A95P+K304D+Q380L+H333K, R405E+K90G+A95P+K304D+Q380L+D371G, R405E+K90G+A95P+K304D+Q380L+E246A, R405E+K90G+A95P+K304D+Q380L+C328A, R405E+K90G+A95P+K304D+Q380L+N412G, R405E+K90G+A95P+K304D+Q380L+T402P, R405E+K90G+A95P+K304D+Q380L+T107F, R405E+K90G+A95P+K304D+Q380L+T107A, R405E+K90G+A95P+K304D+Q380L+G110P, R405E+K90G+A95P+K304D+Q380L+1297L, R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A, R405E+K90G+A95P+K304D+Q380L+I297L+A286T, R405E+K90G+A95P+K304D+Q380L+I297L+E368P, R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+K69N, R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A+G201C y R405E+K90G+A95P+K304D+Q380L+I297L+E368P+T107A A286T.
4. Una molécula de ADN, en donde la molécula de ADN codifica el mutante de transaminasa según cualquiera de las reivindicaciones 1 a 3.
5. Un plásmido recombinante, en donde el plásmido recombinante está unido a la molécula de ADN según la reivindicación 4.
6. Una transaminasa inmovilizada, en donde la transaminasa inmovilizada comprende el mutante de transaminasa según cualquiera de las reivindicaciones 1 a 3.
7. La transaminasa inmovilizada según la reivindicación 6, en donde la transaminasa inmovilizada es un agregado enzimático reticulado de transaminasa del mutante de transaminasa.
8. La transaminasa inmovilizada según la reivindicación 6, en donde la transaminasa inmovilizada es una enzima inmovilizada incrustada-reticulada de transaminasa;
un grupo amino libre en el mutante de transaminasa y el glutaraldehído forman una base de Schiff a través de la reticulación para obtener una enzima reticulada de transaminasa, la enzima reticulada de transaminasa se incrusta en una red de gel de poliacrilamida para obtener la enzima inmovilizada incrustada-reticulada de transaminasa.
9. La transaminasa inmovilizada según la reivindicación 6, en donde la transaminasa inmovilizada es una enzima inmovilizada covalente en la que el mutante de transaminasa está unido covalentemente con un portador;
el portador es un portador de quitosano o un portador de resina.
10. La transaminasa inmovilizada según la reivindicación 9, en donde el portador de resina comprende una matriz y un grupo funcional conectado con la matriz, en donde la matriz se selecciona de cualquiera de un copolímero de estireno y metacrilato, una resina de poliestireno y una resina de polimetacrilato, y el grupo funcional conectado con la matriz se selecciona de un grupo amino de cadena corta C2, un grupo amino de cadena media C4 y un grupo amino de cadena larga C6 o un grupo epoxi.
11. La transaminasa inmovilizada según la reivindicación 10, en donde el portador de resina se selecciona de ECR8309, ECR8315, EC-HFA, LX-1000HA, LX-1000EA, ECR8409, ECR8415, EC-EP, EC EP403, EXE119, LX-1000EP, Immobead- 150A, Immobead-150P, Immobead350A, ECR8206, ECR8209, ECR8215 o ECR8285.
12. La transaminasa inmovilizada según la reivindicación 6, en donde la transaminasa inmovilizada es una enzima inmovilizada quelante formada mediante la quelación de un mutante de transaminasa y un portador a través de iones metálicos;
el portador es un portador de vidrio poroso.
13. La transaminasa inmovilizada según la reivindicación 6, en donde la transaminasa inmovilizada es una enzima inmovilizada adsorbente formada por adsorción física entre el mutante de transaminasa y el portador;
el portador de resina comprende una matriz y un grupo funcional conectado con la matriz, en donde la matriz se selecciona de cualquiera de un copolímero de estireno y metacrilato, una resina de poliestireno y una resina de polimetacrilato, y el grupo funcional conectado con la matriz es un octadecilo.
14. La transaminasa inmovilizada según la reivindicación 13, en donde
el portador de resina se selecciona de ECR8806, ECR1030, ECR1090, ECR1061, ECR1091, ECR8804, Immobead-EC1, Immobead-S60S, Immobead-S861, X17S0409, EXE120 o Diaion HP2MG.
15. Un método para la producción de una amina quiral, que comprende una etapa de reacción de transaminación entre un compuesto de cetona y un donante amino catalizada por una transaminasa, en donde la transaminasa es el mutante de transaminasa según cualquiera de las reivindicaciones 1 a 3 o la transaminasa inmovilizada según cualquiera una de las reivindicaciones 6 a 14.
16. El método según la reivindicación 15, en donde la transaminasa es el mutante de transaminasa según cualquiera de las reivindicaciones 1 a 3, y el método es una reacción por lotes; un sistema de reacción de la reacción por lotes es un sistema de reacción en fase acuosa o la transaminasa es la transaminasa inmovilizada según cualquiera de las reivindicaciones 6 a 14, y el método es una reacción continua; un sistema de reacción de la reacción continua es un sistema de reacción en fase orgánica.
17. El método según cualquiera de las reivindicaciones 15 a 16, en donde el compuesto de cetona es
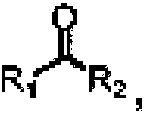
en donde R1 y R2 son cada uno de manera independiente un alquilo C1~C8, un cicloalquilo C5~C10, un arilo C6~C10 o un heteroarilo C5~C10,
o R1 y R2 junto con el carbono en un carbonilo forman un heterociclilo C5~C10, un carbociclilo C5 ~ C10 o un heteroarilo C5 ~ C10,
uno o más heteroátomos en el heterociclilo C5-C10 y el heteroarilo C5-C10 se seleccionan respectivamente de al menos uno de nitrógeno, oxígeno y azufre, y el arilo en arilo C6-C10, el heteroarilo en el heteroarilo C5~C10, el carbociclilo en el carbociclilo C5~C10 o el heterociclilo en el heterociclilo C5~C10 está independientemente sustituido o sin sustituir por al menos un radical de halógeno, alcoxi o alquilo.
18. El método según la reivindicación 17, en donde el compuesto de cetona es

en donde un producto de la reacción de transaminación es

; el donante amino es isopropil amina.
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PCT/CN2018/075272 WO2019148494A1 (zh) | 2018-02-05 | 2018-02-05 | 转氨酶突变体及其应用 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2931540T3 true ES2931540T3 (es) | 2022-12-30 |
Family
ID=67479474
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES18903278T Active ES2931540T3 (es) | 2018-02-05 | 2018-02-05 | Mutante de transaminasa y aplicación del mismo |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US11359219B2 (es) |
| EP (1) | EP3733689B1 (es) |
| JP (1) | JP7105307B2 (es) |
| ES (1) | ES2931540T3 (es) |
| WO (1) | WO2019148494A1 (es) |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20230227797A1 (en) * | 2019-10-28 | 2023-07-20 | Asymchem Laboratories (Tianjin) Co., Ltd. | Transaminase mutant and use thereof |
| CN110950788B (zh) * | 2020-02-26 | 2020-06-16 | 凯莱英医药集团(天津)股份有限公司 | 含氟手性胺类化合物的合成方法 |
| CN110982856B (zh) * | 2020-02-26 | 2020-06-23 | 凯莱英医药集团(天津)股份有限公司 | 手性二胺化合物的合成方法 |
| KR102534312B1 (ko) * | 2020-11-30 | 2023-05-19 | (주)네오크레마 | 고정화된 이눌리나제를 이용한 돼지감자로부터 이눌린의 제조 방법 |
| CN113789310B (zh) * | 2021-01-25 | 2023-11-10 | 上海予君生物科技发展有限公司 | 一种转氨酶及其在制备莫西沙星或其中间体中的应用 |
| WO2023087270A1 (zh) * | 2021-11-19 | 2023-05-25 | 辽宁凯莱英医药化学有限公司 | 酶固定化载体及其制备方法、固定化酶及其制备方法 |
| WO2023175002A1 (en) * | 2022-03-15 | 2023-09-21 | Univerza V Ljubljani | Microfluidics-based preparation of cross-linked enzyme aggregates |
| WO2023215742A2 (en) * | 2022-05-02 | 2023-11-09 | Aditya Kunjapur | Enzymatic reduction or reductive amination of aryl and heterocyclic carboxylic acids and aldehydes |
Family Cites Families (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20130305398A1 (en) | 2012-02-16 | 2013-11-14 | Marie Coffin | Genes and uses for plant enhacement |
| US20090265841A1 (en) | 2008-04-28 | 2009-10-29 | Ferrara Vincent R | Chinstrap assembly |
| EP2271760A2 (en) * | 2008-04-29 | 2011-01-12 | Monsanto Technology LLC | Genes and uses for plant enhancement |
| DK3354727T3 (da) * | 2009-01-08 | 2020-11-16 | Codexis Inc | Transaminasepolypeptider |
| EP2739744A1 (de) | 2011-08-05 | 2014-06-11 | Evonik Degussa GmbH | Oxidation und aminierung von sekundären alkoholen |
| US20160201097A1 (en) | 2014-12-22 | 2016-07-14 | Invista North America S.á.r.l. | Materials and Methods of Producing 7-Carbon Monomers |
| KR101869432B1 (ko) * | 2015-04-08 | 2018-06-20 | 연세대학교 산학협력단 | 케톤에 대한 활성이 증대된 오메가 트랜스아미네이즈의 변이체 및 이것을 이용한 광학활성 아민의 제조방법 |
| CN104894148A (zh) | 2015-04-13 | 2015-09-09 | 浙江科技学院 | 一种ω-转氨酶突变体基因及其应用 |
| CN106676142B (zh) * | 2016-11-01 | 2021-01-22 | 凯莱英医药集团(天津)股份有限公司 | 一种手性氨基杂环化合物及其衍生物的制备方法 |
-
2018
- 2018-02-05 JP JP2020531690A patent/JP7105307B2/ja active Active
- 2018-02-05 WO PCT/CN2018/075272 patent/WO2019148494A1/zh unknown
- 2018-02-05 EP EP18903278.2A patent/EP3733689B1/en active Active
- 2018-02-05 ES ES18903278T patent/ES2931540T3/es active Active
- 2018-02-05 US US16/966,528 patent/US11359219B2/en active Active
Also Published As
| Publication number | Publication date |
|---|---|
| US11359219B2 (en) | 2022-06-14 |
| WO2019148494A1 (zh) | 2019-08-08 |
| JP7105307B2 (ja) | 2022-07-22 |
| EP3733689B1 (en) | 2022-10-26 |
| EP3733689A1 (en) | 2020-11-04 |
| EP3733689A4 (en) | 2021-12-15 |
| JP2021505184A (ja) | 2021-02-18 |
| US20210054428A1 (en) | 2021-02-25 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2931540T3 (es) | Mutante de transaminasa y aplicación del mismo | |
| CN108384767B (zh) | 转氨酶突变体及其应用 | |
| Wang et al. | Immobilization of cellulase on polyamidoamine dendrimer-grafted silica | |
| CN107828751A (zh) | 转氨酶突变体及其应用 | |
| WO2019153183A1 (zh) | 一种单加氧酶突变体及其制备方法和应用 | |
| Yao et al. | Heterologous expression and purification of Arabidopsis thaliana VIM1 protein: in vitro evidence for its inability to recognize hydroxymethylcytosine, a rare base in Arabidopsis DNA | |
| CN105219745A (zh) | 一种固定化转氨酶及其在合成西他列汀中间体中的应用 | |
| CN102559648B (zh) | 一种以改性的环氧基树脂为载体的固定化酶及其制备方法和应用 | |
| JP7022819B2 (ja) | トランスアミナーゼ突然変異体及びその使用 | |
| He et al. | Immobilization and stabilization of cephalosporin C acylase on aminated support by crosslinking with glutaraldehyde and further modifying with aminated macromolecules | |
| WO2021077425A1 (zh) | 转氨酶突变体及其应用 | |
| WO2023004969A1 (zh) | 酯酶突变体及其应用 | |
| CA1306965C (en) | Stabilized enzymes | |
| WO2023186186A1 (zh) | 一种高稳定性固定化酶的制备方法 | |
| CN117070514B (zh) | 非天然rna的制备方法及产品 | |
| JP7263557B2 (ja) | アミノ基転移酵素突然変異体及びその応用 | |
| Ma et al. | Intracellular Delivery of mRNA for Cell‐Selective CRISPR/Cas9 Genome Editing using Lipid Nanoparticles | |
| JPH02109979A (ja) | エポキシ活性化ビーズに固定化されたポリヌクレオチドホスホリラーゼ | |
| CN106032543B (zh) | 一种制备s-氰醇的方法 | |
| CN113817699B (zh) | 转氨酶突变体及其应用 | |
| WO2016153053A1 (ja) | 改変ポリメラーゼ | |
| WO2023035357A1 (zh) | 脂肪酶突变体及其应用 | |
| Chang et al. | Immobilization of leuconostoe mesenteroides dextransucrase to porous phenoxyacetyl cellulose beads | |
| Gaggero et al. | Cascade Biocatalytic Processes through Combined Cross-Linked Enzyme Aggregates | |
| WO2020093191A1 (zh) | 单加氧酶突变体及其应用 |