WO2016056566A1 - 形質転換体およびその製造方法、ならびに乳酸の製造方法 - Google Patents
形質転換体およびその製造方法、ならびに乳酸の製造方法 Download PDFInfo
- Publication number
- WO2016056566A1 WO2016056566A1 PCT/JP2015/078394 JP2015078394W WO2016056566A1 WO 2016056566 A1 WO2016056566 A1 WO 2016056566A1 JP 2015078394 W JP2015078394 W JP 2015078394W WO 2016056566 A1 WO2016056566 A1 WO 2016056566A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- gene
- transformant
- lactic acid
- host
- pombe
- Prior art date
Links
Images





Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/40—Preparation of oxygen-containing organic compounds containing a carboxyl group including Peroxycarboxylic acids
- C12P7/56—Lactic acid
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/14—Fungi; Culture media therefor
- C12N1/145—Fungal isolates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/88—Lyases (4.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01027—L-Lactate dehydrogenase (1.1.1.27)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y401/00—Carbon-carbon lyases (4.1)
- C12Y401/01—Carboxy-lyases (4.1.1)
- C12Y401/01001—Pyruvate decarboxylase (4.1.1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/645—Fungi ; Processes using fungi
Definitions
- the present invention relates to a transformant, a method for producing the transformant, and a method for producing lactic acid.
- Schizosaccharomyces pombe (hereinafter also referred to as S. pombe) is a gene encoding a lactate dehydrogenase gene (lactate dehydrogenase (LDH), hereinafter also referred to as “LDH gene”. ) And a part of the gene group encoding pyruvate decarboxylase is deleted or inactivated, a method for producing the transformant, and the transformant in a culture solution And lactic acid is obtained from the culture solution.
- LDH lactate dehydrogenase
- Lactic acid is widely used for chemical raw materials such as food use, medical treatment, and cosmetics.
- Polylactic acid obtained by using lactic acid is attracting attention as a biodegradable plastic that is finally decomposed into carbon dioxide and water by microorganisms and the like. Therefore, it is necessary to produce lactic acid at low cost with high productivity.
- lactic acid As a method for producing lactic acid, a biological method is known in which sugar is fermented by lactic acid bacteria and produced.
- lactic acid bacteria have low acid resistance, it is necessary to neutralize lactic acid produced by fermentation with an alkali to obtain lactate in order to obtain high productivity by this method.
- Such neutralization with alkali requires a step of returning from lactate to lactic acid, which complicates the manufacturing process and increases the manufacturing cost.
- Patent Document 1 discloses S.I. In Pombe, an LDH gene derived from a mammal such as a human is incorporated, and By culturing transformants in which a part of the gene group encoding the pyruvate decarboxylase encoding Pombe host is deleted or inactivated, lactic acid can be produced with high productivity without performing a neutralization step with alkali. It is disclosed that it can be manufactured.
- Patent Document 2 the L-lactate dehydrogenase gene of Lactobacillus plantarum was introduced into Saccharomyces cerevisiae, which essentially does not produce ethanol when cultured in a culture medium. It is disclosed that L-lactic acid can be obtained by culturing a transformant.
- Patent Document 3 includes S.I. Pombe as a host, the LDH gene (LpLDH gene) of Lactobacillus pentosus is incorporated, and A transformant in which a part of the gene group encoding the pyruvate decarboxylase encoding the pombe host is deleted or inactivated is also produced by the same or more lactic acid as the transformant described in Patent Document 1.
- a LpLDH gene and a human-derived LDH gene HsLDH gene
- HsLDH gene human-derived LDH gene
- a transformant excellent in both lactic acid production ability and proliferation ability is preferable.
- a mutant strain into which an exogenous LDH gene has been introduced tends to have a lower growth ability as the strain has a higher lactic acid-producing ability.
- lactic acid can be produced with high productivity without the need for neutralization with an alkali, and S. coli having both excellent lactic acid production ability and proliferation ability.
- the present invention is directed to a pombe transformant and a method for producing the transformant.
- Another object of the present invention is to produce lactic acid with high productivity using the transformant without performing a neutralization step with an alkali.
- lactic acid refers to L-lactic acid obtained by a biological method.
- the first transformant according to the present invention uses Schizosaccharomyces pombe as a host, incorporates 3 to 5 copies of a human-derived LDH gene, and encodes the pyruvate decarboxylase of the Schizosaccharomyces pombe host. It is characterized in that a part of the gene group to be deleted is inactivated or inactivated.
- the second transformant according to the present invention has an initial bacterial cell concentration of 0.1% in 100 mL of a liquid medium containing 1% yeast extract, 2% peptone, and 6% glucose in a 500 mL Sakaguchi flask.
- the bacterial cell concentration of the bacterial culture was inoculated at a temperature of 32 ° C., 110 rpm, and a shaking width of 7 cm for 20 hours after inoculating the bacterial cells to 04 g (converted into dry cells) / L, and the bacterial cell concentration was 4.0 g (dry bacteria
- the initial bacterial cell concentration is 0.04 g (dried cells) in 100 mL of a liquid medium consisting of 1% yeast extract, 2% peptone, and 6% glucose in a 500 mL Sakaguchi flask.
- the bacterial cells obtained by inoculating the bacterial cells so as to have a conversion / L and culturing for 20 hours under shaking conditions of a temperature of 32 ° C., 110 rpm, and a swinging width of 7 cm were 18 mm in diameter and 15 in length. Inoculate 4.5 mL of 11.1% glucose aqueous solution in a mm test tube so that the initial cell concentration is 36 g (converted to dry cell) / L, temperature 32 ° C., elevation angle 43.5 °, 110 rpm And the lactic acid density
- variety is characterized by being 80 g / L or more.
- Shizo Saccharomyces pombe is a host, at least one lactate dehydrogenase gene is integrated, and a part of a gene group encoding pyruvate decarboxylase of the Schizosaccharomyces pombe host Those in which the gene is deleted or inactivated are preferred.
- the deleted or inactivated gene encoding pyruvate decarboxylase is preferably a PDC2 gene.
- the LDH gene is preferably integrated into the chromosome of Schizosaccharomyces pombe.
- the method for producing a transformant according to the present invention comprises using an expression cassette containing a promoter, terminator, and human-derived lactate dehydrogenase gene functioning in Schizosaccharomyces pombe as a host.
- a human-derived lactate dehydrogenase gene is used in which 3 or more genes of a gene group encoding pyruvate decarboxylase of the transformant obtained above are deleted or inactivated.
- the expression cassette is composed of a region from 10000 bp upstream to 10000 bp downstream from the eno101 locus in the chromosome of Schizosaccharomyces pombe, a region from 10,000 bp downstream from the leu1 locus upstream from 10000 bp, and It is preferably introduced into a region selected from the region of 10000 bp upstream to 10000 bp downstream of the gpm1 locus.
- the deleted or inactivated gene encoding pyruvate decarboxylase is preferably a PDC2 gene.
- the transformant is cultured in a culture solution, and lactic acid is obtained from the culture solution.
- the culture is preferably performed using a culture solution containing glucose having a concentration of 1 to 50% by mass.
- cultivation after the pH of the said culture solution will be 3.5 or less with the lactic acid produced by the said transformant.
- Pombe transformants are excellent in growth ability and can produce lactic acid with high productivity without the need for neutralization with alkali. It is also suitable for lactic acid production in the presence of a high concentration of sugar, particularly glucose, fructose, sucrose, or maltose, and also suitable for high density culture. Furthermore, the ability to produce lactic acid in a long-term continuous culture under high oxygen conditions is also high.
- the transformant can be easily obtained by the method for producing a transformant according to the present invention.
- the manufacturing method of lactic acid which concerns on this invention can manufacture lactic acid with high productivity, without performing the neutralization process by an alkali.
- FIG. 1 is a schematic diagram of the structure of a recombinant vector pSM-HsLDH vector.
- FIG. 1 is a schematic diagram of the structure of a recombinant vector pSN-HsLDH vector.
- FIG. Lactic acid concentration (g / L) 3 hours after fermentation of each transformant in Example 1 (vertical axis) and bacterial cell concentration after 20 hours of culture (g (converted to dry cells (dcw)) / L) It is the dispersion
- Example is a view showing a measurement result of the concentration of lactic acid fermentation liquid after OD 660 and 3 hours fermentation broth after 60 hours of culture of each transformant (g / L) in 2. It is the figure which showed the time-dependent change of the lactic acid density
- the first transformant according to the present invention is S. cerevisiae.
- Pombe is a host, 3-5 copies of human-derived lactate dehydrogenase gene is integrated, and a part of the genes encoding pyruvate decarboxylase of the Schizo Saccharomyces pombe host is deleted or lost. It is characterized by being alive.
- the first transformant according to the present invention has a sufficiently high growth ability even though it has a high ability to produce lactic acid. For this reason, it is very suitable as a lactic acid-producing bacterium for mass-producing industrial lactic acid.
- the gene encoding the deleted or inactivated pyruvate decarboxylase is preferably a PDC2 gene.
- the LDH gene is preferably integrated into the chromosome of Schizosaccharomyces pombe.
- the second transformant according to the present invention has an initial bacterial cell concentration of 0.04 g in 100 mL of a liquid medium composed of 1% yeast extract, 2% peptone and 6% glucose in a 500 mL Sakaguchi flask.
- the bacterial cell concentration was 4.0 g (converted to dry cells) after inoculating the cells to a dry cell equivalent) / L and culturing for 20 hours under shaking at a temperature of 32 ° C., 110 rpm, and a swing width of 7 cm.
- the initial bacterial cell concentration is 0.04 g (in terms of dry cells) in 100 mL of a liquid medium consisting of yeast extract 1%, peptone 2%, and glucose 6% in a 500 mL Sakaguchi flask. / L was inoculated and cultured for 20 hours under shaking at a temperature of 32 ° C., 110 rpm, and a swinging width of 7 cm.
- the second transformant according to the present invention has a sufficiently high proliferation ability even though the ability to produce lactic acid is high. For this reason, it is very suitable as a lactic acid-producing bacterium for mass-producing industrial lactic acid.
- S. Pombe is the host, the LDH gene is integrated, and A transformant in which a part of genes of the gene group encoding Pombe host pyruvate decarboxylase is deleted or inactivated is preferred.
- the first transformant is particularly preferable.
- S. host Pombe is a yeast belonging to the genus Schizosaccharomyces (fission yeast), and is a microorganism particularly excellent in acid resistance compared to other yeasts.
- S. Pombe was found to be superior in lactic acid productivity under high concentrations of glucose compared to other yeasts such as Saccharomyces cerevisiae and suitable for high-density culture (culture using a large amount of yeast). Therefore, S. By using the Pombe transformant, lactic acid can be produced with extremely high productivity.
- the entire base sequence of the pombe chromosome is recorded and published in the database "Pombase (http://www.pombase.org/)". As described in S.A.
- the sequence data of the pombe gene can be obtained by searching from the database by gene name or strain name.
- a gene encoding pyruvate decarboxylase in Pombe includes a gene encoding pyruvate decarboxylase 1 (hereinafter referred to as “PDC1 gene”).
- a gene encoding pyruvate decarboxylase 2 (hereinafter referred to as “PDC2 gene”), a gene encoding pyruvate decarboxylase 3 (hereinafter referred to as “PDC3 gene”), pyruvate decarboxylase
- PDC4 gene A gene encoding pyruvate decarboxylase 2 (hereinafter referred to as “PDC2 gene”), a gene encoding pyruvate decarboxylase 3 (hereinafter referred to as “PDC3 gene”), pyruvate decarboxylase
- PDC4 gene There are four types of genes encoding 4 (hereinafter referred to as “PDC4 gene”). In particular, S.M. In Pombe, the PDC2 gene and the PDC4 gene are PDC genes having major functions. The system name of each PDC gene is as follows.
- PDC1 gene (Pdc1); SPAC13A11.06 PDC2 gene (Pdc2); SPAC1F8.07c PDC3 gene (Pdc3); SPAC186.09 PDC4 gene (Pdc4); SPAC3G9.11c
- the sequence data of the PDC gene is the S.P. It can be obtained by searching by gene name or strain name from the Pombe gene database.
- wild type S. cerevisiae Since Pombe does not have a functioning LDH gene, there is no route for producing lactic acid from pyruvic acid. On the other hand, LDH expressed from the incorporated LDH gene produces lactic acid by reducing pyruvate to lactic acid. Therefore, wild type S. cerevisiae. Even if the LDH gene is incorporated into the pombe so that lactic acid can be produced, both ethanol fermentation and lactic acid fermentation are performed as it is, so that the lactic acid productivity is not sufficiently increased.
- the transformant according to the present invention has a chromosome in which a part of the gene group encoding the pyruvate decarboxylase is deleted or inactivated. Since a part of the PDC gene group of the transformant is deleted or inactivated, the efficiency of ethanol fermentation of the transformant is reduced and the amount of pyruvate converted to ethanol is reduced. Productivity is improved. However, if all genes in the PDC gene group are deleted or inactivated, ethanol fermentation cannot be performed at all and growth is inhibited. Therefore, deletion or inactivation is limited to a part of the genes in the PDC gene group. .
- the PDC gene to be deleted or inactivated is particularly preferably a PDC2 gene.
- the PDC2 gene is a PDC gene having a particularly major function. As described above, if the PDC gene is completely deleted or inactivated, the transformant cannot perform ethanol fermentation, and thus growth is inhibited. Therefore, deletion or inactivation of the PDC gene can reduce the ethanol fermentation ability and improve the fermentation efficiency of lactic acid while leaving a sufficient amount of transformant, leaving the ethanol fermentation ability necessary for growth. Must be done as follows. As a result of studies by the present inventors on the problem, when the PDC2 gene is deleted or inactivated, the PDC4 gene is activated to some extent and has a high ethanol fermentation ability to obtain a sufficient amount of transformant. It was found that the production of lactic acid with fermentation efficiency is compatible.
- Deletion or inactivation of the PDC gene can be performed by a known method.
- the PDC gene can be deleted by using the Latour method (described in Nucleic Acids Res, 2006, 34, e11, WO 2007/063919 pamphlet, etc.).
- the PDC gene can be inactivated by causing deletion, insertion, substitution, or addition in a part of the base sequence of the PDC gene. Mutation due to deletion, insertion, substitution, or addition may cause only one of them, or may cause two or more.
- a known method can be used as a method for introducing the mutation into a part of the PDC gene.
- mutation isolation method using mutagen yeast Molecular Genetics Experimental Method, 1996, Society Press Center
- random mutation method using PCR polymerase chain reaction
- PCR Methods Appl PCR Methods Appl
- the PDC gene into which a mutation is partially introduced may express a temperature-sensitive mutant pyruvate decarboxylase.
- a temperature-sensitive mutant pyruvate decarboxylase is an enzyme that exhibits the same activity as wild-type pyruvate decarboxylase at a certain culture temperature, but loses or decreases its activity at a specific culture temperature or higher. It is.
- the mutant strain expressing the mutant pyruvate decarboxylase exhibits a growth rate equivalent to that of wild-type yeast under conditions where the activity is not limited by temperature, and has a growth rate under specific temperature conditions where the activity is limited. It can be obtained by selecting one that significantly decreases.
- the transformant according to the present invention has at least one LDH gene.
- S.M. Pombe does not have an LDH gene that inherently exhibits strong enzyme activity. Therefore, S.
- the LDH gene of organisms other than Pombe can be obtained by genetic engineering.
- a transformant is obtained by introducing into pombe.
- the species of the LDH gene incorporated into the second transformant according to the present invention is not particularly limited. Further, the LDH gene incorporated into the second transformant according to the present invention may be one, or two or more. When a plurality of LDH genes are introduced into a host, a plurality of LDH genes derived from the same species may be introduced, or LDH genes derived from different species may be introduced in appropriate combination. S. host. By appropriately adjusting the species and copy number of the LDH gene to be introduced into the pombe, a transformant having a high lactic acid production ability can be obtained without excessively degrading the growth ability.
- HsLDH gene As the transformant according to the present invention, S. Using Pombe as a host, 3 to 5 copies of the HsLDH gene (GenBank accession number: X02152.1) are incorporated, and A gene in which a part of the gene group encoding the pyruvate decarboxylase of the pombe host is deleted or inactivated (that is, the first transformant) is preferable.
- the HsLDH gene contained in the transformant according to the present invention is more preferably 3 to 4 copies, and most preferably 3 copies. As shown in Example 1 below, a part of the gene group encoding pyruvate decarboxylase is deleted or inactivated.
- the transformant according to the present invention is an S. cerevisiae in which a part of genes of the PDC gene group is deleted or inactivated.
- the LDH gene was transferred to S. cerevisiae by a genetic engineering method. It is obtained by introducing it into Pombe.
- the PDC gene cluster is not deleted or inactivated.
- Pombe as a host An LDH gene is introduced into pombe by a genetic engineering method to obtain a transformant, and then a part of the PDC gene group of the obtained transformant is deleted or inactivated, and the trait according to the present invention is obtained.
- a conversion body can also be obtained.
- the target transformant was produced by the former method, but almost the same transformant can be produced by the latter method.
- the pombe may be a wild type or a mutant type in which a specific gene is deleted or inactivated depending on the use.
- a method for deleting or inactivating a specific gene a known method can be used. Specifically, the gene can be deleted by using the Latour method (Nucleic Acids Research, Vol. 34, e11 page, 2006; described in International Publication No. 2007/063919 pamphlet, etc.).
- the gene can be inactivated by introducing a mutation into a part of the gene.
- yeasts belonging to the genus Schizosaccharomyces from which a specific gene has been deleted or inactivated are described in, for example, WO 2002/101038 and WO 2007/015470.
- the part where a specific gene is deleted or inactivated may be an ORF (open reading frame) part or an expression regulatory sequence part.
- a particularly preferred method is a method of deletion or inactivation by PCR-mediated homologous recombination method (Yeast, Vol. 14, pages 943-951, 1998) in which the ORF portion of the structural gene is replaced with a marker gene.
- a mutant in which the PDC gene is deleted or inactivated can be preferably used as a host for producing the transformant according to the present invention. Furthermore, in addition to the PDC gene, a specific gene other than the PDC gene was deleted or inactivated. Pombe can be the host. Deletion or inactivation of a protease gene or the like can increase the expression efficiency of a heterologous protein, and application to the host in the present invention can be expected to improve the production efficiency of lactic acid.
- S. cerevisiae used as a host. It is preferable to use a pombe having a marker for selecting a transformant. For example, it is preferable to use a host in which a specific nutritional component is essential for growth because a certain gene is missing.
- the transformant is produced by transforming with a vector containing the target gene sequence, the transformant is equipped with the missing gene (auxotrophic complementary marker), so that the transformant has a host nutritional requirement. Sex disappears. Due to the difference in auxotrophy between the host and the transformant, the transformant can be obtained by distinguishing both. For example, an orotidine phosphate decarboxylase gene (ura4 gene) has been deleted or inactivated and has become uracil-requiring.
- ura4 gene an orotidine phosphate decarboxylase gene
- a transformant incorporating the vector can be obtained by selecting those that have lost uracil requirement.
- the gene that becomes auxotrophic due to deletion in the host is not limited to the ura4 gene as long as it is used for selection of transformants, and may be an isopropylmalate dehydrogenase gene (leu1 gene) or the like.
- the PDC gene cluster is not deleted or inactivated.
- Pombe can also be used as a host for the production of transformants.
- a host in this case a host in which the above-described genes (such as an auxotrophic marker and a protease gene) other than the PDC gene are deleted or inactivated can be used.
- a transformant according to the present invention can be obtained by deleting or deactivating a part of the PDC gene group of the obtained transformant.
- HsLDH gene introduction method As a method for introducing the HsLDH gene into a host by a genetic engineering method, a known method can be used. S. Examples of methods for introducing a heterologous protein structural gene into pombe as a host include, for example, JP-A-5-15380, WO95 / 09914, JP-A-10-234375, JP-A-2000-262284. The methods described in Japanese Patent Laid-Open No. 2005-198612 and International Publication No. 2011/021629 can be used.
- An expression cassette is a combination of DNAs necessary for expressing a target protein, and includes a structural gene encoding the target protein, a promoter functioning in the host, and a terminator.
- the HsLDH gene expression cassette is composed of HsLDH gene and S. cerevisiae.
- a promoter that functions in Pombe Includes a terminator that functions within the pombe.
- the expression cassette may contain any one or more of 5′-untranslated region and 3′-untranslated region.
- the auxotrophic complementary marker may be included.
- a preferred expression cassette is an expression cassette containing an HsLDH gene, a promoter, a terminator, a 5′-untranslated region, a 3′-untranslated region, and an auxotrophic complementary marker.
- a plurality of HsLDH genes may be present in one expression cassette. For example, 2 to 5 HsLDH genes may be contained in one expression cassette.
- the gene encoded by the wild type may be used as it is.
- the wild type gene sequence was transformed into S. cerevisiae. It may be modified to a codon frequently used in pombe.
- S. Promoters and terminators that function in pombe can maintain the expression of LDH by functioning in the transformant even when lactic acid is accumulated by the transformant according to the present invention and becomes acidic (pH 6 or lower). Anything is acceptable.
- S. Examples of promoters that function in pombe include S. cerevisiae. Pombe's inherent promoter (preferably having high transcription initiation activity) or S. A promoter (eg, a virus-derived promoter) that Pombe does not originally have can be used. Two or more promoters may be present in the vector.
- promoters inherent to Pombe include alcohol dehydrogenase gene promoter, nmt1 gene promoter involved in thiamine metabolism, fructose-1, 6-bisphosphatase gene promoter involved in glucose metabolism, and invertase gene involved in catabolite repression. Examples include promoters (see International Publication No. 99/23223 pamphlet), heat shock protein gene promoters (see International Publication No. 2007/26617 pamphlet), and the like.
- promoters that Pombe does not originally include are promoters derived from animal cell viruses described in JP-A-5-15380, JP-A-7-163373, and JP-A-10-234375, and hCMV A promoter, SV40 promoter is preferred.
- S. Pombe's inherent terminator and S. A terminator that Pombe does not have can be used. Two or more terminators may be present in the vector. Examples of the terminator include human-derived terminators described in JP-A-5-15380, JP-A-7-163373, and JP-A-10-234375, and the terminator of human-derived lipocortin I is preferable. .
- the transformant according to the present invention has an expression cassette containing the HsLDH gene in the chromosome or as an extrachromosomal gene.
- Having an expression cassette in the chromosome means that the expression cassette is incorporated at one or more locations in the host cell chromosome, and having as an extrachromosomal gene means having a plasmid containing the expression cassette in the cell. It is.
- a transformant containing an expression cassette containing the HsLDH gene is used as a host using a vector containing the expression cassette of the HsLDH gene. It is obtained by transforming pombe.
- the vector can be produced by incorporating the expression cassette into a vector having a circular DNA structure or a linear DNA structure.
- the vector may contain a sequence for replication in the host cell, that is, an autonomously replicating sequence (AutonomouslynomReplicating). Sequence: ARS) is preferred.
- ARS autonomously replicating sequence
- the vector is assumed to have a linear DNA structure and no ARS. It is preferable to be introduced into.
- the vector may be a vector composed of linear DNA, or a vector having a circular DNA structure provided with a restriction enzyme recognition sequence for cleavage into linear DNA upon introduction into a host.
- the vector is a plasmid having ARS, it can be introduced into the host after forming a linear DNA structure by deleting the ARS part or a linear DNA structure in which the function of ARS is inactivated by cleaving the ARS part.
- the vector preferably has a marker for selecting a transformant.
- the marker include ura4 gene (auxotrophic complementary marker) and isopropylmalate dehydrogenase gene (leu1 gene).
- HsLDH gene is S. It is preferably introduced into the pombe chromosome. By introducing the HsLDH gene into the chromosome, a transformant excellent in passage maintenance stability can be obtained. Among the transformants according to the present invention, those having 3 to 5 copies of the HsLDH gene may have 3 to 5 HsLDH genes introduced into one place in the chromosome. One HsLDH gene may be introduced in each.
- a known method can be used as a method for introducing the HsLDH gene into the chromosome.
- a plurality of HsLDH genes can be introduced into the chromosome by the method described in JP-A-2000-262284.
- one HsLDH gene can be introduced into the chromosome by this method.
- one or a plurality of LDH genes can be introduced into a plurality of locations on the chromosome.
- the HsLDH gene was transformed into S. cerevisiae.
- a method for introduction into the pombe chromosome a method of introduction by homologous recombination using a vector having an expression cassette having an HsLDH gene and a recombination site is preferred.
- the recombination site of the vector is S. cerevisiae. This is a site having a base sequence that allows homologous recombination to be performed on a target site for homologous recombination in the pombe chromosome.
- the target site is S. pneumoniae. This is a target site for integrating the expression cassette in the pombe chromosome.
- the target site can be freely set by setting the recombination site of the vector to a base sequence that allows homologous recombination to be performed on the target site.
- the homology between the base sequence of the recombination site and the base sequence of the target site needs to be 70% or more.
- the homology between the base sequence of the recombination site and the base sequence of the target site is preferably 90% or more, and more preferably 95% or more from the viewpoint that homologous recombination is likely to occur.
- the expression cassette is incorporated into the target site by homologous recombination.
- the length (number of bases) of the recombination site is preferably 20 to 2000 bp. If the length of the recombination site is 20 bp or more, homologous recombination is likely to occur.
- the length of the recombination site is 2000 bp or less, it is easy to prevent the vector from becoming too long and causing homologous recombination to hardly occur.
- the length of the recombination site is more preferably 100 bp or more, and further preferably 200 bp or more. Further, the length of the recombination site is more preferably 800 bp or less, and further preferably 400 bp or less.
- the vector may have other DNA regions in addition to the expression cassette and the recombination site.
- a replication initiation region called “ori” necessary for replication in E. coli, an antibiotic resistance gene (neomycin resistance gene, etc.) can be mentioned. These are genes usually required when constructing a vector using Escherichia coli.
- the replication initiation region is preferably removed when the vector is integrated into the host chromosome as described later.
- the vector When the LDH gene is integrated into the chromosome, the vector is S. cerevisiae. When introducing into a pombe cell, it is preferable to introduce it in a linear DNA structure. That is, in the case of a vector having a circular DNA structure such as a commonly used plasmid DNA, S. It is preferable to introduce into pombe cells. In this case, the position for opening the vector having a circular DNA structure is within the recombination site. As a result, the recombination sites partially exist at both ends of the opened vector, and the entire vector is integrated into the target site of the chromosome by homologous recombination.
- the vector may be constructed by a method other than the method of cutting a vector having a circular DNA structure, as long as it can have a linear DNA structure in which a part of the recombination site exists at each end.
- the vector for example, plasmids derived from E. coli such as pBR322, pBR325, pUC118, pUC119, pUC18, and pUC19 can be preferably used.
- the plasmid vector used for homologous recombination preferably has a replication initiation region called “ori” that is necessary for replication in E. coli.
- ori replication initiation region
- the method for constructing the vector from which the replication initiation region has been removed is not particularly limited, but the method described in JP-A-2000-262284 is preferably used.
- a method is preferred in which a precursor vector in which a replication initiation region is inserted at the cleavage site in the recombination site is constructed so that the replication initiation region is excised at the same time as the linear DNA structure as described above. Thereby, a vector from which the replication initiation region has been easily removed can be obtained. Further, by applying the expression vector and the construction method thereof described in JP-A-5-15380, JP-A-7-163373, WO96 / 23890, JP-A-10-234375, etc.
- a method may be used in which a precursor vector having an expression cassette and a recombination site is constructed, and a vector used for homologous recombination is obtained by removing the replication initiation region from the precursor vector by a normal genetic engineering technique.
- Target sites that incorporate the vector are S. cerevisiae. It may be present only at one location in the pombe chromosome, or may be present at two or more locations. When two or more target sites exist, S.P.
- the vector can be integrated at two or more positions on the pombe chromosome.
- a plurality of LDH genes are included in one expression cassette, a plurality of LDH genes can be incorporated into one target site.
- an expression cassette can be incorporated into two or more target sites using two or more vectors having recombination sites corresponding to the respective target sites. In this manner, S. A plurality of LDH genes can be incorporated into the pombe chromosome, thereby increasing the expression level of LDH and improving lactic acid productivity.
- the target site described in the method described in JP-A No. 2000-262284 can be used.
- Two or more types of vectors having different integration sites can be used to integrate the vectors at different target sites.
- this method is complicated when the vector is integrated at two or more sites on the chromosome.
- Base sequences that are substantially identical to each other means that the homology of the base sequences is 90% or more.
- the homology between the target sites is preferably 95% or more.
- the length of the base sequences that are substantially identical to each other is a length that includes the recombination site of the vector, and is preferably 1000 bp or more.
- target sites into which the expression cassette is incorporated include, for example, near the ura4 locus, near the leu1 locus, near the adh1 locus, There are three positions selected from the vicinity of the gpd1 locus, the vicinity of the eno101 locus, the vicinity of the leu1 locus, and the vicinity of the gpm1 locus.
- near the X locus refers to a range from the upstream end of the ORF of the X gene in the chromosome to 10 kbp upstream (10000 bp) to the downstream end of the ORF to 10 kbp downstream of the ORF of another gene. Means an area that does not contain
- the HsLDH gene is S. cerevisiae. Those introduced into a target site selected from the vicinity of the eno101 locus, the vicinity of the leu1 locus, and the vicinity of the gpm1 locus in the pombe chromosome are preferred. That is, as a transformant according to the present invention, the HsLDH gene is expressed as S. cerevisiae.
- Tf2 is the S.T.
- Tf2 is the S.T.
- the length (number of bases) is about 4900 bp, and the base sequence homology between these genes is 99.7%. It is known (see the following literature). Nathan J. Bowen et al, “Retrotransposons and Their Recognition of pol II Promoters: A Comprehensive Survey of the Transposable Elements From the Complete Genome Sequence of Schizosaccharomyces pombe”, Genome Res. 2003 13: 1984-1997
- a vector can be incorporated into only one location of Tf2 existing in 13 locations on a chromosome.
- a transformant having two or more LDH genes can be obtained by incorporating a vector having two or more LDH genes.
- a transformant having two or more LDH genes can be obtained by incorporating a vector into two or more locations of Tf2.
- a transformant having more LDH genes can be obtained by incorporating a vector having two or more LDH genes.
- Transformation method Any known transformation method may be used as the transformation method.
- the transformation method include conventionally known methods such as lithium acetate method, electroporation method, spheroplast method, glass bead method, and the method described in JP-A-2005-198612.
- a commercially available yeast transformation kit may also be used.
- S. As a method for transforming the pombe host by the homologous recombination method, a known homologous recombination method can be used.
- a transformation method in producing the transformant according to the present invention S. cerevisiae in which a part of genes of the above-mentioned PDC gene group has been deleted or inactivated.
- a method in which the expression cassette is integrated by homologous recombination using the above-mentioned vector as a host and pombe as a host is preferable. According to this method, the transformant according to the present invention can be easily produced.
- the obtained transformant is usually selected after homologous recombination.
- the selection method include the following methods. Screening is performed with a medium capable of selecting transformants using the auxotrophic marker, and a plurality of colonies obtained are selected. Next, after separately culturing them in liquid, the expression level of the heterologous protein (HsLDH in the present invention) in each culture solution is examined, and a transformant with a higher expression level of the heterologous protein is selected.
- HsLDH in the present invention the expression level of the heterologous protein in each culture solution is examined, and a transformant with a higher expression level of the heterologous protein is selected.
- the number of vectors integrated into the chromosome and the number of expression cassettes can be examined. The number of vectors integrated into the chromosome can be adjusted to some extent by adjusting the integration conditions. Depending on the size (number of bases) and structure of the vector, the integration efficiency and the number of integrations will also vary.
- the present inventors have a higher lactic acid production efficiency even when a relatively small number of expression cassettes of an appropriate size are incorporated into the chromosome.
- Pombe transformants It was considered necessary to select a foreign LDH gene with high expression efficiency in pombe and high activity of expressed LDH and to introduce an appropriate copy number.
- 3-5 copies of the HsLDH gene have been deleted or inactivated from a part of the PDC gene group.
- the method for producing lactic acid according to the present invention is a method for producing lactic acid by culturing the transformant according to the present invention in a culture solution and obtaining lactic acid from the culture solution.
- a culture solution containing sugar By culturing the transformant according to the present invention in a culture solution containing sugar, pyruvic acid obtained from the sugar is reduced by LDH by a glycolysis system to produce lactic acid, and the lactic acid produced in the culture solution Lactic acid can be produced by obtaining from the culture solution.
- a known yeast culture medium containing sugar can be used. Contains a nitrogen source, inorganic salts, etc. that can be utilized by Pombe; Any material that can efficiently culture pombe is acceptable.
- a natural medium or a synthetic medium may be used.
- the sugar that is a carbon source include sugars such as glucose, fructose, sucrose, and maltose.
- the nitrogen source include inorganic acids such as ammonia, ammonium chloride, and ammonium acetate, or ammonium salts of inorganic acids, peptone, casamino acid, yeast extract, and the like.
- inorganic salts include magnesium phosphate, magnesium sulfate, sodium chloride and the like.
- a fermentation promoting factor such as proteolipid can be included.
- a culture solution containing glucose particularly as sugar it is preferable to use a culture solution containing glucose particularly as sugar.
- the glucose concentration in the culture medium (100% by mass) at the initial stage of culture is preferably 1% by mass or more, more preferably 1 to 50% by mass, and further preferably 2 to 16% by mass. It is preferable to continue the culture by adding glucose as necessary, because the glucose concentration is lowered by the culture.
- the glucose concentration at the end of the culture may be 1% by mass or less.
- the productivity of lactic acid is further improved.
- the production efficiency of lactic acid improves more by making glucose in a culture solution into 16 mass% or less.
- the initial cell concentration of the transformant in the culture solution is preferably 0.1 to 5 g / L in terms of dry cell weight. More preferably, the initial bacterial cell concentration of the transformant in the culture solution is 0.2 to 2 g / L in terms of dry cell weight.
- High productivity can be achieved in a short time by increasing the initial cell concentration.
- the initial bacterial cell concentration is too high, problems such as bacterial cell aggregation and a reduction in purification efficiency may occur. Incidentally, cell concentration indicated in the Examples etc.
- OD 660 1 at 660 nm corresponds to 0.2 g of fission yeast dry weight and 0.8 g of wet weight.
- a known yeast culture method can be used, for example, shake culture, stirring culture, or the like.
- the culture temperature is preferably 23 to 37 ° C.
- the culture time can be determined as appropriate.
- the culture may be batch culture or continuous culture.
- the cells can be separated from the culture solution to obtain a culture solution containing lactic acid.
- the continuous culture method for example, a part of the culture solution is extracted from the culture tank being cultured, lactic acid is separated from the extracted culture solution, and the culture supernatant is recovered, and glucose and new A method of continuously culturing by repeatedly adding a culture solution or the like and returning it to the culture tank can be mentioned. By performing continuous culture, the productivity of lactic acid is further improved.
- lactic acid can be produced without neutralization even when the pH is lowered due to the accumulation of lactic acid (about pH 2 to 4). Therefore, even after the pH of the culture solution becomes 3.5 or less, lactic acid can be produced by continuous culture that continues the culture.
- the pH at the end of culture and the pH in continuous culture are preferably 3.5 or less, and particularly preferably 2.3 to 3.5. In order to increase the productivity of lactic acid, it is preferable to continue the culture after the pH of the culture solution becomes 3.5 or lower. Since the transformant according to the present invention has excellent acid resistance, the culture can be continued without neutralizing lactic acid in the culture solution produced by the transformant.
- a known method can be used to obtain lactic acid from the culture solution.
- the lactic acid in the culture medium is neutralized, and then the culture liquid and lactate are separated to obtain lactic acid.
- lactic acid can also be obtained by converting lactic acid in a culture solution into a calcium salt or a lithium salt and crystallizing the neutralized salt.
- the above-described method for producing lactic acid according to the present invention is an S.A. Since a transformant using pombe as a host is used, lactic acid can be easily produced with high productivity without neutralization with alkali. In addition, since the efficiency of ethanol fermentation is reduced due to deletion or inactivation of some genes in the PDC gene group, the yield of lactic acid versus sugar (the amount of lactic acid produced relative to the amount of consumed sugar) Ratio) is improved. In the present invention, the yield of lactic acid with respect to sugar can be easily set to 50% or more. In some cases, the sugar yield of lactic acid reaches 70% or more.
- the method for producing lactic acid according to the present invention is also suitable for high-density culture in the presence of a high concentration of glucose and a high concentration of transformants.
- Example 1 S. Using Pombe as a host, a transformant in which the PDC2 gene was deleted and 3 copies of the HsLDH gene were introduced was prepared.
- UP regions with UF and UR, OL regions with OF and OR, and DN regions with DF and DR were prepared by PCR using KOD-Dash (manufactured by Toyobo Co., Ltd.).
- Full length deletion fragments were prepared by the same PCR method using FF and FR as templates.
- two synthetic oligo DNAs manufactured by Operon
- the ura4 region fragment was also used as a template.
- the Pombe PDC2 gene deleted strain (IGF543 strain, h - leu1-32ura4-D18 pdc2-D23) had a slow growth rate. Therefore, in order to recover the growth rate, the IGF543 strain was streaked on a YES plate (yeast extract 0.5% / glucose 3% / SP supplement) and cultured at 25 ° C., and the obtained colonies were cultured in YPD medium (yeast 1% extract / 2% peptone / 2% glucose) and cultured at 25 ° C., a glycerol stock was prepared using a well-grown culture, and stored at ⁇ 80 ° C. The above operation was repeated until an appropriate growth rate was obtained, and a strain with a restored growth rate was produced (name inherited from IGF543).
- the expression cassette is introduced in the vicinity of the leu1 locus on the genome, one copy of the HsLDH gene controlled by the hCMV promoter in the vicinity of the gpm1 locus, and the HsLDH gene controlled by the ihc promoter in the vicinity of the Leu1 locus
- a transformant in which a total of 2 copies of HsLDH gene was introduced was prepared.
- the transformed strain (HsLDH gene 2-copy introduced strain) was designated as ASP4121 strain.
- an HsLDH expression cassette was introduced near the eno101 locus on the genome, one copy of the HsLDH gene controlled by the hCMV promoter near the gpm1 locus, and an HsLDH gene controlled by the ihc promoter near the Leu1 locus
- a transformant was prepared in which a total of 3 copies of the HsLDH gene were introduced, one copy of the HsLDH gene controlled by the hCMV promoter in the vicinity of the eno101 locus.
- the transformed strain (HsLDH gene 3-copy introduced strain) was designated as ASP4956 strain.
- the growth ability and lactic acid production ability of the ASP5019 strain, the ASP3509 strain, the ASP2914 strain, the ASP3619 strain, the ASP3621 strain, the ASP3631 strain, the ASP3622 strain, and the ASP3623 strain described in Patent Document 3 were compared.
- the ASP3509 strain is a strain in which one copy of the HsLDH gene is introduced into the IGF543 strain
- the ASP2914 strain is a strain in which two copies of the HsLDH gene are introduced into the IGF543 strain
- the ASP3619 strain is one copy of the HsLDH gene and the LbLDH gene (Lactobacillus gene).
- ASP3621 strain has one copy of HsLDH gene and one copy of SaLDH gene (LDH gene of Staphylococcus aureus) in IGF543 strain
- the ASP3631 strain is a strain obtained by introducing one copy of the HsLDH gene and one copy of the LpLDH gene into the IGF543 strain.
- Two strains are strains in which one copy of the HsLDH gene and one copy of the LplLDH gene (LDH gene of Lactobacillus plantarum) are introduced into the IGF543 strain, and the ASP3623 strain is one copy of the HsLDH gene and the PaLDH gene (in the IGF543 strain). This is a strain into which 1 copy of Pediococcus acidilactici (LDH gene) has been introduced.
- LplLDH gene LplLDH gene of Lactobacillus plantarum
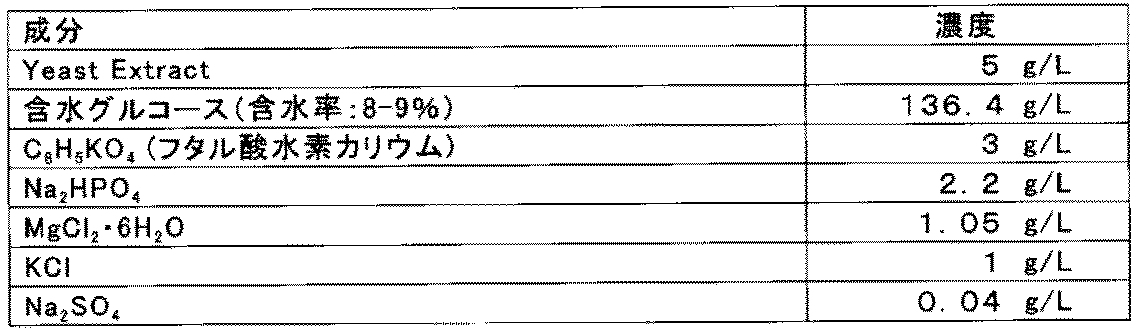
- Each transformed strain was added to a 100 mL YPD6 liquid medium (1% yeast extract, 2% peptone, 6% glucose) in a 500 mL Sakaguchi flask (manufactured by AGC Techno Glass) at an initial cell concentration of 0.04 g (The cells were inoculated so as to have a dry cell equivalent) / L, and cultured for 20 hours under shaking conditions of a temperature of 32 ° C., a shaking speed of 110 rpm, and a swinging width of 7 cm. After measuring the OD 660 of the culture solution, the doubling was terminated and the cells were collected.
- YPD6 liquid medium 1% yeast extract, 2% peptone, 6% glucose
- a 4.5 mL 11.1% glucose aqueous solution in a test tube having a diameter of 18 mm and a length of 150 mm is inoculated so that the initial bacterial cell concentration is 36 g (converted to dry cells) / L, and the temperature is 32 ° C. and the elevation angle is 43 Fermentation was performed for 3 hours under shaking conditions of 5 °, 110 rpm, and a swing width of 7 cm, and after completion, the concentration of lactic acid (g / L) in the fermentation broth was measured.
- Table 3 shows the cell concentration (g (converted to dry cell (dcw)) / L) and the lactic acid concentration (g / L) after 3 hours fermentation of each transformant after culturing for 20 hours.
- the transformant having a higher lactic acid production ability tends to have a lower growth ability.
- the straight line is a straight line connecting plots of the ASP3509 strain and the ASP3631 strain. From this tendency, the transformed strain introduced with 3 copies of the HsLDH gene has higher lactate production ability and lower growth ability than the transformed strain introduced with 2 copies of the HsLDH gene (ASP2914 strain). If the transformed strain introduced with 3 copies of the HsLDH gene has a higher lactic acid-producing ability than the ASP3631 strain, it is expected that the growth ability of the strain will be lower than that of the ASP3631 strain. Nevertheless, the ASP5019 strain into which 3 copies of the HsLDH gene had been introduced had a high lactic acid production ability almost the same as the ASP3631 strain and a sufficient growth ability almost the same as the ASP3509 strain.
- Example 2 The ASP5019 strain and ASP3631 strain used in Example 1 were cultured in flasks, and the growth ability and lactic acid production ability were compared. Specifically, each transformant is treated with a YES liquid medium (yeast extract 5 g / L, glucose 30 g / L, adenine 1 g / L, histidine 1 g / L, leucine 1 g / L, uracil 1 g / L, lysine 1 g / L). And the cells were cultured for 30 hours under conditions of a temperature of 32 ° C. and a shaking speed of 110 rpm.
- a YES liquid medium yeast extract 5 g / L, glucose 30 g / L, adenine 1 g / L, histidine 1 g / L, leucine 1 g / L, uracil 1 g / L, lysine 1 g / L.
- FIG. 4 shows the measurement results of OD 660 of the culture solution after 60-hour culture of each transformant and the lactic acid concentration (g / L) of the fermentation solution after 7-hour fermentation.
- the ASP5019 strain was confirmed to have a high lactic acid-producing ability as well as the ASP3631 strain, and the proliferation ability was significantly superior to the ASP3631 strain.
- the dilution rate was 0.066 (1 / h).
- the microbial cells were refluxed into the tank and recycled during the continuous fermentation over 200 hours.
- the pH was not neutralized with alkali.
- FIG. 5 shows the change over time of the lactic acid concentration (g / L) in the fermentation broth during continuous fermentation.
- the lactic acid concentration of the ASP5019 strain and the ASP3631 strain is almost the same until 120 hours have passed since the start of fermentation, and remained almost constant, and after 150 hours have passed since the start of fermentation. Tended to decline.
- the tendency of the lactic acid concentration to decrease was more gentle in the ASP5019 strain than in the ASP3631 strain. From the results, it was found that the ASP5019 strain has a significantly high ability to produce lactic acid under long-term operation and high oxygen conditions.
- the transformant according to the present invention and the method for producing lactic acid using the same can be suitably used as an industrial method for producing lactic acid because lactic acid can be produced with high productivity without neutralization with alkali even at low pH. be able to.
- the entire content of the specification, claims, abstract, and drawings of Japanese Patent Application No. 2014-209048 filed on October 10, 2014 is cited here as disclosure of the specification of the present invention. Incorporated.
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- Microbiology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Biophysics (AREA)
- General Chemical & Material Sciences (AREA)
- Physics & Mathematics (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Plant Pathology (AREA)
- Botany (AREA)
- Mycology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Virology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
また、前記形質転換体を用いて、アルカリによる中和工程を行わずに高い生産性で乳酸を製造する方法を目的とする。
なお、本発明において乳酸とは生物学的方法で得られるL-乳酸をいう。
該形質転換体としては、シゾサッカロミセス・ポンベを宿主とし、少なくとも1の乳酸脱水素酵素遺伝子が組み込まれ、かつ前記シゾサッカロミセス・ポンベ宿主のピルビン酸脱炭酸酵素をコードする遺伝子群の一部の遺伝子が欠失または失活しているものが好ましい。
本発明に係る形質転換体の製造方法においては、前記発現カセットを、シゾサッカロミセス・ポンベの染色体中のeno101遺伝子座上流10000bpから下流10000bpの領域、leu1遺伝子座上流10000bpから下流10000bpの領域、およびgpm1遺伝子座上流10000bpから下流10000bpの領域から選ばれる領域に導入することが好ましい。
また、本発明に係る形質転換体の製造方法においては、前記欠失または失活しているピルビン酸脱炭酸酵素をコードする遺伝子がPDC2遺伝子であることが好ましい。
本発明に係る乳酸の製造方法において、濃度1~50質量%のグルコースを含む培養液を使用して培養を行うことが好ましい。また、前記形質転換体により産生される乳酸により、前記培養液のpHが3.5以下になった後にさらに培養を継続することが好ましい。
さらに、前記形質転換体により産生された培養液中の乳酸を中和することなく培養を継続することが好ましく、また、培養液中の乳酸を中和することなく培養液から乳酸を分離することが好ましい。
前記形質転換体は、本発明に係る形質転換体の製造方法により簡便に得ることができる。
また、本発明に係る乳酸の製造方法は、アルカリによる中和工程を行わずに高い生産性で乳酸を製造できる。
本発明に係る第1の形質転換体は、S.ポンベを宿主とし、ヒト由来の乳酸脱水素酵素遺伝子が3~5コピー組み込まれ、かつ前記シゾサッカロミセス・ポンベ宿主のピルビン酸脱炭酸酵素をコードする遺伝子群の一部の遺伝子が欠失または失活していることを特徴とする。本発明に係る第1の形質転換体は、乳酸産生能が高いにもかかわらず、増殖能も充分に高い。このため、工業上の乳酸を量産するための乳酸産生菌として非常に好適である。
上記欠失または失活しているピルビン酸脱炭酸酵素をコードする遺伝子はPDC2遺伝子であることが好ましい。さらに、上記LDH遺伝子はシゾサッカロミセス・ポンベの染色体に組み込まれていることが好ましい。
宿主であるS.ポンベは、シゾサッカロミセス属に属する酵母(分裂酵母)であり、他の酵母に比べて特に耐酸性に優れる微生物である。また、S.ポンベは、サッカロミセス・セレビシエ等の他の酵母に比べ、高濃度のグルコース下における乳酸の生産性に優れ、高密度培養(大量の酵母を用いた培養)にも適していることがわかった。そのため、S.ポンベの形質転換体を用いることにより、極めて高い生産性で乳酸を製造できる。
なお、S.ポンベの染色体の全塩基配列は、データベース「Pombase (http://www.pombase.org/)」に、収録され、公開されている。本明細書記載のS.ポンベの遺伝子の配列データは前記データベースから遺伝子名または前記系統名で検索して、入手できる。
S.ポンベにおけるピルビン酸脱炭酸酵素をコードする遺伝子(ピルビン酸脱炭酸酵素遺伝子、以下「PDC遺伝子」ともいう。)群には、ピルビン酸脱炭酸酵素1をコードする遺伝子(以下、「PDC1遺伝子」という。)、ピルビン酸脱炭酸酵素2をコードする遺伝子(以下、「PDC2遺伝子」という。)、ピルビン酸脱炭酸酵素3をコードする遺伝子(以下、「PDC3遺伝子」という。)、ピルビン酸脱炭酸酵素4をコードする遺伝子(以下、「PDC4遺伝子」という。)の4種類がある。なかでも、S.ポンベにおいては、PDC2遺伝子とPDC4遺伝子が主要な機能を持つPDC遺伝子である。各PDC遺伝子の系統名は以下の通りである。
PDC1遺伝子(Pdc1);SPAC13A11.06
PDC2遺伝子(Pdc2);SPAC1F8.07c
PDC3遺伝子(Pdc3);SPAC186.09
PDC4遺伝子(Pdc4);SPAC3G9.11c
前記PDC遺伝子の配列データは、前記S.ポンベ遺伝子データベースから遺伝子名または系統名で検索して入手できる。
一方、組み込まれたLDH遺伝子から発現されるLDHは、ピルビン酸を乳酸に還元することにより乳酸を産生させる。そのため、野生型S.ポンベにLDH遺伝子を組み込んで乳酸を産生できるようにしても、そのままではエタノール発酵と乳酸発酵の両方を行うことになるため、乳酸の生産性が充分に高くならない。
前述のように、PDC遺伝子を全て欠失または失活させてしまうと、その形質転換体はエタノール発酵が行えなくなるために生育が阻害される。そのため、PDC遺伝子の欠失または失活は、生育に必要なエタノール発酵能を残して充分な形質転換体量が得られるようにしつつ、エタノール発酵能を低下させて乳酸の発酵効率を向上させられるように行わなければならない。該課題に対して本発明者等が検討を行った結果、PDC2遺伝子を欠失または失活させるとPDC4遺伝子がある程度活性化し、充分な形質転換体量が得られる程度のエタノール発酵能と、高い発酵効率での乳酸の生産が両立できることを見い出した。
また、PDC遺伝子の塩基配列の一部に欠失、挿入、置換、付加を起こすことにより、該PDC遺伝子を失活させることもできる。欠失、挿入、置換、付加による変異は、それらのいずれか1つのみを起こしてもよく、2つ以上を起こしてもよい。
該変異型ピルビン酸脱炭酸酵素を発現する突然変異株は、温度により活性が制限されない条件下では野生型酵母と同等の生育速度を示し、活性が制限される特定の温度条件下において生育速度が著しく低下するものを選択することで得ることができる。
本発明に係る形質転換体は、少なくとも1のLDH遺伝子を有する。前記のように、S.ポンベは本来強い酵素活性を示すLDH遺伝子を有していない。したがって、S.ポンベ以外の生物のLDH遺伝子を遺伝子工学的方法でS.ポンベに導入して形質転換体を得る。
本発明に係る形質転換体は、PDC遺伝子群の一部の遺伝子が欠失または失活したS.ポンベを宿主とし、これにLDH遺伝子を遺伝子工学的方法でS.ポンベに導入して得られる。また、PDC遺伝子群が欠失も失活もしていないS.ポンベを宿主とし、該S.ポンベにLDH遺伝子を遺伝子工学的方法で導入して形質転換体を得て、その後得られた形質転換体のPDC遺伝子群の一部の遺伝子を欠失または失活させて、本発明に係る形質転換体を得ることもできる。後述の実施例では、前者の方法で目的の形質転換体を製造したが、後者の方法でもほぼ同等の形質転換体を製造できる。
宿主とするS.ポンベは、野生型であってもよく、用途に応じて特定の遺伝子を欠失または失活させた変異型であってもよい。特定の遺伝子を欠失または失活させる方法としては、公知の方法を用いられる。具体的には、Latour法(Nucleic Acids Research誌、第34巻、e11ページ、2006年;国際公開第2007/063919号パンフレット等に記載)を用いることにより遺伝子を欠失させられる。また、変異剤を用いた突然変異分離法(酵母分子遺伝学実験法、1996年、学会出版センター)、PCR(ポリメラーゼ連鎖反応)を利用したランダム変異法(PCR Methods Application誌、第2巻、28-33ページ、1992年)等により遺伝子の一部に変異を導入することにより該遺伝子を失活させられる。特定遺伝子を欠失または失活させたシゾサッカロミセス属酵母宿主としては、たとえば、国際公開第2002/101038号、国際公開第2007/015470号等に記載されている。
また、特定の遺伝子の削除または不活性化を行う部分はORF(オープンリーディングフレーム)部分であってもよく、発現調節配列部分であってもよい。特に好ましい方法は、構造遺伝子のORF部分をマーカー遺伝子に置換するPCR媒介相同組換え法(Yeast誌、第14巻、943-951ページ、1998年)による削除または不活性化の方法である。
たとえば、オロチジンリン酸デカルボキシラーゼ遺伝子(ura4遺伝子)が欠失または失活してウラシル要求性となっているS.ポンベを宿主とし、ura4遺伝子(栄養要求性相補マーカー)を有するベクターにより形質転換した後、ウラシル要求性が消失したものを選択することにより、ベクターが組み込まれた形質転換体を得ることができる。宿主において欠落により栄養要求性となる遺伝子は、形質転換体の選択に用いられるものであればura4遺伝子には限定されず、イソプロピルリンゴ酸デヒドロゲナーゼ遺伝子(leu1遺伝子)等であってもよい。
遺伝子工学的方法で宿主にHsLDH遺伝子を導入する方法としては、公知の方法を使用できる。S.ポンベを宿主としてこれに異種蛋白質の構造遺伝子を導入する方法としては、たとえば、特開平5-15380号公報、国際公開第95/09914号、特開平10-234375号公報、特開2000-262284号公報、特開2005-198612号公報、国際公開第2011/021629号などに記載の方法を使用できる。
発現カセットとは、目的の蛋白質を発現するために必要なDNAの組み合わせであり、目的の蛋白質をコードする構造遺伝子と宿主内で機能するプロモーターとターミネーターを含む。本発明に係る形質転換体の製造において、HsLDH遺伝子の発現カセットは、HsLDH遺伝子とS.ポンベ内で機能するプロモーターとS.ポンベ内で機能するターミネーターを含む。該発現カセットは、5’-非翻訳領域、3’-非翻訳領域のいずれか1つ以上が含まれていてもよい。さらに、前記栄養要求性相補マーカーが含まれていてもよい。好ましい発現カセットは、HsLDH遺伝子、プロモーター、ターミネーター、5’-非翻訳領域、3’-非翻訳領域、栄養要求性相補マーカーを含む発現カセットである。1の発現カセットには複数のHsLDH遺伝子が存在していてもよい。たとえば、1の発現カセット中に2~5個のHsLDH遺伝子を含んでいてもよい。
S.ポンベが本来有しないプロモーターとしては、たとえば、特開平5-15380号公報、特開平7-163373号公報、特開平10-234375号公報に記載されている動物細胞ウイルス由来のプロモーターが挙げられ、hCMVプロモーター、SV40プロモーターが好ましい。
ターミネーターとしては、たとえば、特開平5-15380号公報、特開平7-163373号公報、特開平10-234375号公報に記載されているヒト由来のターミネーターが挙げられ、ヒト由来リポコルチンIのターミネーターが好ましい。
本発明に係る形質転換体は、HsLDH遺伝子を含む発現カセットを、染色体中に有するか、または染色体外遺伝子として有する。発現カセットを染色体中に有するとは、宿主細胞の染色体中の1箇所以上に発現カセットが組み込まれていることであり、染色体外遺伝子として有するとは、発現カセットを含むプラスミドを細胞内に有することである。HsLDH遺伝子を含む発現カセットを含む形質転換体は、HsLDH遺伝子の発現カセットを含むベクターを用いて宿主であるS.ポンベを形質転換することにより得られる。
HsLDH遺伝子をS.ポンベの染色体に導入する方法としては、HsLDH遺伝子を有する発現カセットと組換え部位とを有するベクターを用い、相同組換え法により導入する方法が好ましい。
前記組換え部位の塩基配列と標的部位の塩基配列との相同性は70%以上とすることが必要である。また、組換え部位の塩基配列と標的部位の塩基配列との相同性は、相同組換えが起きやすくなる点から、90%以上とすることが好ましく、95%以上であることがより好ましい。該組換え部位を有するベクターを用いることにより、発現カセットが相同組換えにより標的部位に組み込まれる。
組換え部位の長さ(塩基数)は、20~2000bpであることが好ましい。組換え部位の長さが20bp以上であれば、相同組換えが起きやすくなる。また、組換え部位の長さが2000bp以下であれば、ベクターが長くなりすぎて相同組換えが起き難くなることを防ぎやすい。組換え部位の長さは100bp以上であることがより好ましく、200bp以上であることがさらに好ましい。また、組換え部位の長さは800bp以下であることがより好ましく、400bp以下であることがさらに好ましい。
この場合、環状DNA構造を有するベクターを切り開く位置は、組換え部位内とする。これにより、切り開かれたベクターの両端にそれぞれ組換え部位が部分的に存在することとなり、相同組換えによりベクター全体が染色体の標的部位に組み込まれる。
ベクターは、両端それぞれに組換え部位の一部が存在するような線状DNA構造とすることができれば、環状DNA構造を有するベクターを切り開く方法以外の方法で構築してもよい。
この場合、相同組換えに用いる際のプラスミドベクターは、大腸菌内での複製のために必要な「ori」と呼ばれる複製開始領域が除去されていることが好ましい。これにより、上述したベクターを染色体に組み込む際に、その組み込み効率を向上させることができる。
複製開始領域が除去されたベクターの構築方法は特に限定されないが、特開2000-262284号公報に記載されている方法を用いることが好ましい。すなわち、組換え部位内の切断箇所に複製開始領域が挿入された前駆体ベクターを構築しておき、前述のように線状DNA構造とすると同時に複製開始領域が切り出されるようにする方法が好ましい。これにより、簡便に複製開始領域が除去されたベクターを得ることができる。
また、特開平5-15380号公報、特開平7-163373号公報、国際公開第96/23890号パンフレット、特開平10-234375号公報等に記載された発現ベクターおよびその構築方法を適用して、発現カセットおよび組換え部位を有する前駆体ベクターを構築し、さらに通常の遺伝子工学的手法で該前駆体ベクターから複製開始領域を除去して相同組換えに用いるベクターを得る方法であってもよい。
ベクターを組み込む標的部位は、S.ポンベの染色体中の1箇所のみに存在していてもよく、2箇所以上に存在していてもよい。標的部位が2箇所以上存在している場合、S.ポンベの染色体の2箇所以上に該ベクターを組み込める。また、1の発現カセット中のLDH遺伝子を複数とした場合には、標的部位の1箇所に複数のLDH遺伝子を組み込むことができる。さらに、2種以上の標的部位に、それぞれの標的部位に対応する組換え部位を有する2種以上のベクターを用いて、発現カセットを組み込むこともできる。該方法で、S.ポンベの染色体に複数のLDH遺伝子を組み込むことができ、これによりLDHの発現量を増大させ、乳酸の生産性を向上させることができる。
Nathan J. Bowen et al, “Retrotransposons and Their Recognition of pol II Promoters: A Comprehensive Survey of the Transposable Elements From the Complete Genome Sequence of Schizosaccharomyces pombe”, Genome Res. 2003 13: 1984-1997
形質転換方法は、公知の形質転換方法がいずれも用いられる。該形質転換方法としては、たとえば、酢酸リチウム法、エレクトロポレーション法、スフェロプラスト法、ガラスビーズ法など従来周知の方法、特開2005-198612号公報記載の方法が挙げられる。また、市販の酵母形質転換用キットを用いてもよい。
染色体に組み込まれるベクターの数は組み込み条件などを調整することによりある程度は調整可能である。ベクターの大きさ(塩基数)および構造により、組み込み効率および組み込み数も変化すると考えられる。
本発明に係る乳酸の製造方法は、前記本発明に係る形質転換体を培養液中で培養し、該培養液から乳酸を取得する乳酸の製造方法である。
本発明に係る形質転換体を、糖を含む培養液中で培養することにより、解糖系により該糖から得られるピルビン酸がLDHにより還元されて乳酸が産生され、培養液に産出された乳酸を培養液から取得することで乳酸を製造できる。
炭素源である糖としては、たとえば、グルコース、フルクトース、スクロース、マルトース等の糖が挙げられる。窒素源としては、たとえば、アンモニア、塩化アンモニウム、酢酸アンモニウム等の無機酸または無機酸のアンモニウム塩、ペプトン、カザミノ酸、イーストエキス等が挙げられる。無機塩類としては、たとえば、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム等が挙げられる。さらには、プロテオリピド等の発酵促進因子などを含ませることができる。
なお、後述の実施例等で示す菌体濃度は、日本分光社製可視紫外分光器V550によって測定した波長660nmの光の吸光度(OD660)から換算した値である。660nmにおけるOD660=1は、分裂酵母乾燥重量の0.2g、湿重量の0.8gに相当する。
また、培養温度は、23~37℃であることが好ましい。また、培養時間は適宜決定できる。
また、培養は、回分培養であってもよく、連続培養であってもよい。たとえば、回分培養で培養を行った後、菌体を培養液から分離して、乳酸を含む培養液を取得できる。また、連続培養法では、たとえば、培養中の培養槽から培養液の一部を抜き出し、抜き出した培養液から乳酸を分離するとともに、培養上清を回収し、該培養上清にグルコースおよび新たな培養液等を加えて培養槽に戻すことを繰り返して、連続的に培養する方法が挙げられる。連続培養を行うことにより、乳酸の生産性がより向上する。
S.ポンベを宿主とし、PDC2遺伝子が削除され、かつHsLDH遺伝子が3コピー導入された形質転換体を作製した。
S.ポンベのウラシル要求性株ARC010株(遺伝子型:h- 、leu1-32、ura4-D18)(国際公開第2007/015470号パンフレット参照。)を、pdc2削除断片で、東田らの方法(米国特許第6,235,499号明細書参照。)に従って形質転換し、該断片がS.ポンベのゲノム上のpdc2遺伝子座近傍に導入された形質転換体を得た。該形質転換体を、さらにLatour法(Nucleic Acids Res.誌、2006年、34巻、e11頁、国際公開第2007/063919号パンフレットに記載)に従ってFOA処理し、PDC2遺伝子(系統名:SPAC1F8.07c)を削除した削除株(IGF543株)を作製した。
pdc2削除断片(2811bp、配列番号11)の作製には、S.ポンベのARC032株(遺伝子型:h-)(国際公開第2007/015470号パンフレット参照。)からDNeasy(キアゲン社製)によって調製した全ゲノムDNAを鋳型とし、表1に示す塩基配列を有する8種の合成オリゴDNA(オペロン社製)を使用した。


まず、HsLDH遺伝子発現カセットを保持する単座組込型組換えベクターpSM-HsLDHベクター(4562bp、図1)を作製した。pSM-HsLDHベクターは、DNA合成により、配列番号12に示す塩基配列からなるDNA断片として作製した。
次いで、IGF543株を、pSM-HsLDHベクターで形質転換した。該操作により、HsLDH発現カセットがゲノム上のgpm1遺伝子座近傍に導入された。該形質転換株(HsLDH遺伝子1コピー導入株)をASP3494株とした。
ASP3494株を、HsLDH遺伝子発現カセットを保持し、ihc1プロモーターを有する単座組込型組換えベクターpSL17-HsLDH(特許文献3参照。)の制限酵素BsiWI消化物で、Bahlerらの方法(Yeast誌、1998年、14巻、943-951頁)に従い形質転換した。該操作により、該発現カセットがゲノム上のleu1遺伝子座近傍に導入され、gpm1遺伝子座近傍にhCMVプロモーターによって制御されたHsLDH遺伝子を1コピーと、Leu1遺伝子座近傍にihcプロモーターによって制御されたHsLDH遺伝子を1コピーとの合計2コピーのHsLDH遺伝子が導入された形質転換株を作製した。該形質転換株(HsLDH遺伝子2コピー導入株)をASP4121株とした。
pSM-HsLDHベクターによりS.ポンベのゲノムに導入されたura4遺伝子は、2つのhCMVプロモーター配列に挟まれている為、ホモロガスリコンビネーションによりゲノム上から脱落しやすい。
そこで、ASP4121株をFOA処理し、ウラシルの要求性を復帰させた。
まず、HsLDH遺伝子発現カセットを保持する単座組込型組換えベクターpSN-HsLDHベクター(4535bp、図2)を作製した。pSN-HsLDHベクターは、DNA合成により、配列番号13に示す塩基配列からなるDNA断片として作製した。
次いで、ASP4121株をFOA処理してウラシルの要求性を復帰させた形質転換株を、pSN-HsLDHベクターで形質転換した。該操作により、HsLDH発現カセットがゲノム上のeno101遺伝子座近傍に導入され、gpm1遺伝子座近傍にhCMVプロモーターによって制御されたHsLDH遺伝子を1コピーと、Leu1遺伝子座近傍にihcプロモーターによって制御されたHsLDH遺伝子を1コピーと、eno101遺伝子座近傍にhCMVプロモーターによって制御されたHsLDH遺伝子を1コピーとの合計3コピーのHsLDH遺伝子が導入された形質転換株を作製した。該形質転換株(HsLDH遺伝子3コピー導入株)をASP4956株とした。
pSM-HsLDHベクターによる場合と同様に、pSN-HsLDHベクターによりS.ポンベのゲノムに導入されたura4遺伝子は、2つのhCMVプロモーター配列に挟まれている為、ホモロガスリコンビネーションによりゲノム上から脱落しやすい。
そこで、ASP4956株をFOA処理し、ウラシルの要求性を復帰させた。
ASP4956株をFOA処理してウラシルの要求性を復帰させた形質転換株を、ura4遺伝子を含むDNA断片(3277bp、配列番号14)で形質転換し、ウラシル要求性が補完された形質転換株を作製した。該形質転換株(HsLDH遺伝子3コピー導入栄養要求補完株)をASP5019株とした。
ASP5019株と、特許文献3に記載のASP3509株、ASP2914株、ASP3619株、ASP3621株、ASP3631株、ASP3622株、およびASP3623株の増殖能と乳酸産生能を比較した。ASP3509株はIGF543株にHsLDH遺伝子1コピーを導入した株であり、ASP2914株はIGF543株にHsLDH遺伝子2コピーを導入した株であり、ASP3619株はIGF543株にHsLDH遺伝子1コピーとLbLDH遺伝子(ラクトバチルス・ブルガリクス(Lactobacillus bulgaricus)のLDH遺伝子)1コピーを導入した株であり、ASP3621株はIGF543株にHsLDH遺伝子1コピーとSaLDH遺伝子(スタヒロコッカス・アウレウス(Staphylococcus aureus)のLDH遺伝子)1コピーを導入した株であり、ASP3631株はIGF543株にHsLDH遺伝子1コピーとLpLDH遺伝子1コピーを導入した株であり、ASP3622株はIGF543株にHsLDH遺伝子1コピーとLplLDH遺伝子(ラクトバチルス・プランタラム(Lactobacillus plantarum)のLDH遺伝子)1コピーを導入した株であり、ASP3623株はIGF543株にHsLDH遺伝子1コピーとPaLDH遺伝子(ペディオコッカス・アシディラクティシ(Pediococcus acidilactici)のLDH遺伝子)1コピーを導入した株である。

例1で用いたASP5019株とASP3631株について、フラスコ培養し、増殖能と乳酸産生能を比較した。
具体的には、各形質転換株をYES液体培地(イーストエキス5g/L、グルコース30g/L、アデニン1g/L、ヒスチジン1g/L、ロイシン1g/L、ウラシル1g/L、リシン1g/L)に植菌して、温度32℃、振盪速度110rpmの条件下で30時間培養を行った。培養終了後、菌体を回収し、表4に示す組成に微量元素類およびビタミン類を適当量加えた培養培地を張り込んだ、3L容ジャーファーメンターに接種し、温度32℃、ODカスケード制御下にて60時間流加培養を行い、終了後、培養液のOD660を測定した。
次いで、培養終了後の培養液から遠心分離処理により菌体を分離し、該菌体を11.1%グルコース水溶液に初発菌体濃度が36g(乾燥菌体換算)/L(OD660=180)になるように接種したものを発酵液として、7時間試験管にて発酵させた。発酵終了後の発酵液の乳酸濃度(g/L)を測定した。

例1で用いたASP5019株とASP3631株について、連続培養し、乳酸産生能を比較した。
具体的には、例2と同様の方法を用いて得られた60時間流加培養菌体を回収し、該菌体を表5に示す組成に微量元素類およびビタミン類を適当量加えた発酵培地に、初発菌体濃度が36g(乾燥菌体換算)/L(OD660=180)になるように接種したものを発酵液とした。該発酵液0.5Lを、1L容ジャーファーメンター中に移し、クロスフロー方式の精密濾過膜を通して循環させた。そして、一定流量で発酵培地の供給および膜濾過液の引抜きを行う連続発酵を、28℃で200時間超行った。その際、希釈率は0.066(1/h)とした。該連続発酵では、菌体のサイズよりも小さい細孔径の精密濾過膜を用いているため、菌体は槽内へ還流され、200時間超の連続発酵中においてリサイクルされた。アルカリを用いたpHの中和は行わなかった。
なお、2014年10月10日に出願された日本特許出願2014-209048号の明細書、特許請求の範囲、要約書および図面の全内容をここに引用し、本発明の明細書の開示として、取り入れるものである。
Claims (15)
- シゾサッカロミセス・ポンベを宿主とし、ヒト由来の乳酸脱水素酵素遺伝子が3~5コピー組み込まれ、かつ前記シゾサッカロミセス・ポンベ宿主のピルビン酸脱炭酸酵素をコードする遺伝子群の一部の遺伝子が欠失または失活していることを特徴とする形質転換体。
- 前記欠失または失活しているピルビン酸脱炭酸酵素をコードする遺伝子がPDC2遺伝子である、請求項1に記載の形質転換体。
- 前記乳酸脱水素酵素遺伝子がシゾサッカロミセス・ポンベの染色体に組み込まれている、請求項1または2に記載の形質転換体。
- 500mL容の坂口フラスコに入った、イーストエクストラクト1%、ペプトン2%、およびグルコース6%からなる液体培地100mLに初発菌体濃度を0.04g(乾燥菌体換算)/Lになるように菌体を接種し、温度32℃、110rpmかつ振り幅7cmの振盪条件下で20時間培養した培養液の菌体濃度が4.0g(乾燥菌体換算)/L以上であり、
500mL容の坂口フラスコに入った、イーストエクストラクト1%、ペプトン2%、およびグルコース6%からなる液体培地100mLに初発菌体濃度を0.04g(乾燥菌体換算)/Lになるように菌体を接種し、温度32℃、110rpmかつ振り幅7cmの振盪条件下で20時間培養し得られた菌体を、直径18mmかつ長さ150mmの試験管に入った4.5mLの11.1%グルコース水溶液に初発菌体濃度が36g(乾燥菌体換算)/Lになるように接種し、温度32℃、仰角43.5°、110rpm、およびふり幅7cmの振盪条件下で3時間発酵させた発酵液の乳酸濃度が80g/L以上であることを特徴とする形質転換体。 - シゾサッカロミセス・ポンベを宿主とし、少なくとも1の乳酸脱水素酵素遺伝子が組み込まれ、かつ前記シゾサッカロミセス・ポンベ宿主のピルビン酸脱炭酸酵素をコードする遺伝子群の一部の遺伝子が欠失または失活している、請求項4に記載の形質転換体。
- 前記欠失または失活しているピルビン酸脱炭酸酵素をコードする遺伝子がPDC2遺伝子である、請求項5に記載の形質転換体。
- 前記乳酸脱水素酵素遺伝子がシゾサッカロミセス・ポンベの染色体に組み込まれている、請求項5または6に記載の形質転換体。
- シゾサッカロミセス・ポンベを宿主とし、シゾサッカロミセス・ポンベ内で機能するプロモーターとターミネーターとヒト由来の乳酸脱水素酵素遺伝子とを含む発現カセットを、前記宿主の染色体中の3~5箇所に導入した形質転換体を得ること、および、
前記宿主としてピルビン酸脱炭酸酵素をコードする遺伝子群の一部の遺伝子が欠失または失活した宿主を用いるか、または前記得られた形質転換体のピルビン酸脱炭酸酵素をコードする遺伝子群の一部の遺伝子を欠失または失活させること
を特徴とする、ヒト由来の乳酸脱水素酵素遺伝子が3~5コピー組み込まれかつピルビン酸脱炭酸酵素をコードする遺伝子群の一部の遺伝子が欠失または失活している形質転換体の製造方法。 - 前記発現カセットを、シゾサッカロミセス・ポンベの染色体中のeno101遺伝子座上流10000bpから下流10000bpの領域、leu1遺伝子座上流10000bpから下流10000bpの領域、およびgpm1遺伝子座上流10000bpから下流10000bpの領域から選ばれる領域に導入する、請求項8に記載の形質転換体の製造方法。
- 前記欠失または失活しているピルビン酸脱炭酸酵素をコードする遺伝子がPDC2遺伝子である、請求項8または9に記載の形質転換体の製造方法。
- 請求項1~7のいずれかに記載の形質転換体を培養液中で培養し、該培養液から乳酸を取得する乳酸の製造方法。
- 濃度1~50質量%のグルコースを含む培養液を使用して培養を行う、請求項11に記載の乳酸の製造方法。
- 前記形質転換体により産生される乳酸により、前記培養液のpHが3.5以下になった後にさらに培養を継続する、請求項11または12に記載の乳酸の製造方法。
- 前記形質転換体により産生された培養液中の乳酸を中和することなく培養を継続する、請求項11~13のいずれか一項に記載の乳酸の製造方法。
- 前記形質転換体により産生された培養液中の乳酸を中和することなく、培養液から乳酸を分離する、請求項11~14のいずれか一項に記載の乳酸の製造方法。
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US15/506,960 US10214754B2 (en) | 2014-10-10 | 2015-10-06 | Transformant and its production process, and method for producing lactic acid |
| EP15849633.1A EP3205721B1 (en) | 2014-10-10 | 2015-10-06 | Transformant and method for producing same, and method for producing lactic acid |
| JP2016553124A JP6620375B2 (ja) | 2014-10-10 | 2015-10-06 | 形質転換体およびその製造方法、ならびに乳酸の製造方法 |
| ES15849633T ES2759269T3 (es) | 2014-10-10 | 2015-10-06 | Transformante y procedimiento para la producción del mismo, y procedimiento para la producción de ácido láctico |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2014-209048 | 2014-10-10 | ||
| JP2014209048 | 2014-10-10 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2016056566A1 true WO2016056566A1 (ja) | 2016-04-14 |
Family
ID=55653177
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2015/078394 WO2016056566A1 (ja) | 2014-10-10 | 2015-10-06 | 形質転換体およびその製造方法、ならびに乳酸の製造方法 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US10214754B2 (ja) |
| EP (1) | EP3205721B1 (ja) |
| JP (1) | JP6620375B2 (ja) |
| AR (1) | AR102238A1 (ja) |
| ES (1) | ES2759269T3 (ja) |
| WO (1) | WO2016056566A1 (ja) |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2019536445A (ja) * | 2016-11-01 | 2019-12-19 | ピーティーティー グローバル ケミカル パブリック カンパニー リミテッド | D−乳酸またはその塩を生産するための発酵プロセス |
| JPWO2019240243A1 (ja) * | 2018-06-15 | 2021-06-24 | キッコーマン株式会社 | セレノネインの製造方法 |
| JP2022504813A (ja) * | 2018-10-08 | 2022-01-13 | エスケー イノベーション カンパニー リミテッド | アルコール生成が抑制された組換え耐酸性酵母及びこれを用いた乳酸の製造方法 |
| US11898173B2 (en) | 2020-06-24 | 2024-02-13 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast having improved lactic-acid-producing ability |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2011021629A1 (ja) * | 2009-08-21 | 2011-02-24 | 旭硝子株式会社 | 形質転換体およびその製造方法、ならびに乳酸の製造方法 |
| WO2014030655A1 (ja) * | 2012-08-24 | 2014-02-27 | 旭硝子株式会社 | 形質転換体およびその製造方法、ならびに乳酸の製造方法 |
Family Cites Families (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001007572A2 (en) | 1999-07-23 | 2001-02-01 | The Regents Of The University Of California | Dna recombination in eukaryotic cells by the bacteriophage phic31 recombination system |
| WO2001092513A1 (en) | 2000-05-30 | 2001-12-06 | Johnson & Johnson Research Pty Limited | METHODS FOR MEDIATING GENE SUPPRESION BY USING FACTORS THAT ENHANCE RNAi |
| CN1955299A (zh) * | 2000-11-22 | 2007-05-02 | 内特尔沃克公司 | 合成有机产物的方法和材料 |
| JP4049364B2 (ja) | 2002-03-22 | 2008-02-20 | 独立行政法人科学技術振興機構 | 多コピー型・ゲノム挿入型の選択両用ベクター |
| BR0311452A (pt) * | 2002-05-30 | 2005-03-29 | Cargill Dow Llc | Processos e materiais para a produção de ácido lático em levedura |
| US7514253B2 (en) | 2003-05-16 | 2009-04-07 | Glycofi, Inc. | URA5 gene and methods for stable genetic integration in yeast |
| US20050112737A1 (en) | 2003-11-20 | 2005-05-26 | A. E. Staley Manufacturing Co. | Lactic acid producing yeast |
| EP2147976B1 (en) | 2005-10-14 | 2014-07-23 | Toray Industries, Inc. | Yeast and method of producing L-lactic acid |
| JP5424871B2 (ja) | 2006-05-19 | 2014-02-26 | グライコフィ, インコーポレイテッド | 組換えベクター |
| JP2008092862A (ja) | 2006-10-12 | 2008-04-24 | Institute Of Physical & Chemical Research | 組換え開始酵素認識配列の低侵襲染色体導入による減数分裂期組換え分布の制御法 |
| CA2818499A1 (en) | 2010-11-22 | 2012-06-07 | Novozymes, Inc. | Compositions and methods for 3-hydroxypropionic acid production |
| KR102208959B1 (ko) | 2013-08-09 | 2021-01-28 | 삼성전자주식회사 | 락테이트 데히드로게나제가 활성화된 효모 세포 및 그를 이용한 락테이트를 생산하는 방법 |
-
2015
- 2015-10-06 US US15/506,960 patent/US10214754B2/en not_active Expired - Fee Related
- 2015-10-06 EP EP15849633.1A patent/EP3205721B1/en not_active Not-in-force
- 2015-10-06 JP JP2016553124A patent/JP6620375B2/ja not_active Expired - Fee Related
- 2015-10-06 ES ES15849633T patent/ES2759269T3/es active Active
- 2015-10-06 WO PCT/JP2015/078394 patent/WO2016056566A1/ja active Application Filing
- 2015-10-09 AR ARP150103283A patent/AR102238A1/es unknown
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2011021629A1 (ja) * | 2009-08-21 | 2011-02-24 | 旭硝子株式会社 | 形質転換体およびその製造方法、ならびに乳酸の製造方法 |
| WO2014030655A1 (ja) * | 2012-08-24 | 2014-02-27 | 旭硝子株式会社 | 形質転換体およびその製造方法、ならびに乳酸の製造方法 |
Non-Patent Citations (1)
| Title |
|---|
| FUTOSHI HARA ET AL.: "Lactate production using Schizosaccharomyces pombe", THE 61ST ABSTRACTS OF THE ANNUAL MEETING OF THE SOCIETY FOR BIOTECHNOLOGY, 2009, Japan, pages 190, XP008151368 * |
Cited By (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2019536445A (ja) * | 2016-11-01 | 2019-12-19 | ピーティーティー グローバル ケミカル パブリック カンパニー リミテッド | D−乳酸またはその塩を生産するための発酵プロセス |
| JPWO2019240243A1 (ja) * | 2018-06-15 | 2021-06-24 | キッコーマン株式会社 | セレノネインの製造方法 |
| JP7210577B2 (ja) | 2018-06-15 | 2023-01-23 | キッコーマン株式会社 | セレノネインの製造方法 |
| JP2022504813A (ja) * | 2018-10-08 | 2022-01-13 | エスケー イノベーション カンパニー リミテッド | アルコール生成が抑制された組換え耐酸性酵母及びこれを用いた乳酸の製造方法 |
| JP7530891B2 (ja) | 2018-10-08 | 2024-08-08 | エスケー イノベーション カンパニー リミテッド | アルコール生成が抑制された組換え耐酸性酵母及びこれを用いた乳酸の製造方法 |
| US12084665B2 (en) | 2018-10-08 | 2024-09-10 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast in which alcohol production is inhibited and method for producing lactic acid by using same |
| US11898173B2 (en) | 2020-06-24 | 2024-02-13 | Sk Innovation Co., Ltd. | Recombinant acid-resistant yeast having improved lactic-acid-producing ability |
Also Published As
| Publication number | Publication date |
|---|---|
| US20170356017A1 (en) | 2017-12-14 |
| EP3205721B1 (en) | 2019-08-28 |
| JP6620375B2 (ja) | 2019-12-18 |
| EP3205721A1 (en) | 2017-08-16 |
| US10214754B2 (en) | 2019-02-26 |
| EP3205721A4 (en) | 2018-04-11 |
| JPWO2016056566A1 (ja) | 2017-07-27 |
| ES2759269T3 (es) | 2020-05-08 |
| AR102238A1 (es) | 2017-02-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5772594B2 (ja) | 形質転換体およびその製造方法、ならびに乳酸の製造方法 | |
| DE60315028T2 (de) | Verfahren und materialien zur produktion von d-milchsäure in hefe | |
| JP6176251B2 (ja) | 形質転換体およびその製造方法、ならびに乳酸の製造方法 | |
| JP6620375B2 (ja) | 形質転換体およびその製造方法、ならびに乳酸の製造方法 | |
| JP2018157814A (ja) | シュードザイマ・アンタクティカの新規菌株 | |
| US20120034654A1 (en) | Method for transforming schizosaccharomyces pombe, transformant of schizosaccharomyces pombe and method for producing heterologous protein | |
| JP6499587B2 (ja) | 形質転換体およびその製造方法、ならびに乳酸の製造方法 | |
| WO2014030644A1 (ja) | シゾサッカロミセス・ポンベ変異体の形質転換体、およびクローニングベクター | |
| JP6620373B2 (ja) | 形質転換体およびその製造方法、ならびに炭素数4のジカルボン酸の製造方法 | |
| JP6488666B2 (ja) | クローニングベクター、発現ベクターおよび形質転換体の製造方法 | |
| WO2016084857A1 (ja) | 3-ヒドロキシプロピオン酸の製造方法、および形質転換体 | |
| JP7452900B2 (ja) | 乳酸耐性の向上を有する酵母およびその使用 | |
| JP2006280268A (ja) | 細胞増殖の促進によりアルコール生産効率を向上させる方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 15849633 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2016553124 Country of ref document: JP Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 15506960 Country of ref document: US |
|
| REEP | Request for entry into the european phase |
Ref document number: 2015849633 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2015849633 Country of ref document: EP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |