WO2014171570A2 - 화농연쇄구균 유래 신규 nadh 산화효소 및 l-아라비니톨 산화효소와의 커플링에 의한 l-자일룰로스의 생산 - Google Patents
화농연쇄구균 유래 신규 nadh 산화효소 및 l-아라비니톨 산화효소와의 커플링에 의한 l-자일룰로스의 생산 Download PDFInfo
- Publication number
- WO2014171570A2 WO2014171570A2 PCT/KR2013/003316 KR2013003316W WO2014171570A2 WO 2014171570 A2 WO2014171570 A2 WO 2014171570A2 KR 2013003316 W KR2013003316 W KR 2013003316W WO 2014171570 A2 WO2014171570 A2 WO 2014171570A2
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- nadh oxidase
- water
- producing
- gene
- xylulose
- Prior art date
Links
Images







Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0012—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7)
- C12N9/0036—Oxidoreductases (1.) acting on nitrogen containing compounds as donors (1.4, 1.5, 1.6, 1.7) acting on NADH or NADPH (1.6)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/315—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Streptococcus (G), e.g. Enterococci
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
Definitions
- the present invention relates to a method for producing L-xylulose by coupling a reaction of NADH oxidase and L-arabinitol oxidase to produce novel water derived from P. aureus, more specifically P. aureus-derived.
- NADH oxidase producing a novel water, nucleic acid molecule encoding the same, a vector comprising the nucleic acid molecule, a transformant comprising the vector and a combination of NADH oxidase and L-arabinitol oxidase using the same
- It relates to a method for preparing L-xylulose through a ring reaction.
- redox reactions are not only important for asymmetric reactions, but are also areas of interest in chemicals and pharmaceuticals with optical purity.
- such redox reactions require expensive coenzymes such as NAD (P) + or NAD (P) H, which makes industrial applications difficult. Therefore, it is necessary to economically secure such expensive coenzymes and use them continuously.
- Regeneration of coenzymes is usually accomplished by adding chemical, electrical or photocatalytic substances or adding enzymes during the enzyme reaction. There are two ways to regenerate coenzymes using enzymes. The first method involves regenerating enzymes and chemical additives. The second method involves regenerating coenzymes by adding and coupling two enzymes.
- NAD + and NAD (P) H are well known methods using formate dehydrogenase and glucose dehydrogenase.
- byproducts are generated during the reaction to reduce the purity of the product and increase the purification cost.
- NADH oxidase that produces water can ensure high economy because only water is produced as a by-product.
- the present invention proposes a novel water-producing NADH oxidase, which is coupled with L-arabinitol dehydrogenase to regenerate coenzyme to produce L-xylulose from L-arabinitol in high yield and continuously.
- L-arabinitol dehydrogenase By presenting the optimum reaction conditions, it is intended to provide a method for mass production of L-xylulose inexpensively and in high yield.
- a second object of the present invention is to provide a NADH oxidase that produces water expressed from the gene.
- a fifth object of the present invention is to provide a NADH oxidase that produces a recombinant using transformed recombinant E. coli.
- a sixth object of the present invention is to propose the optimum conditions for producing L-xylulose by regenerating the coenzyme by coupling the enzyme with L-arabinitol dehydrogenase.
- the present invention in the production method of L-xylulose, purulent streptococci NADH oxidase and Hippocrea zecorina, which produce water of the strain Hypocrea jecorina L-arabinitol dehydrogenase derived from the) is characterized in that to regenerate the coenzyme.
- the present invention is characterized by cloning the NADH oxidase gene that produces water from Streptococcus through Southern hybridization and colony hybridization.
- the present invention provides a NADH oxidase that produces water having an amino acid sequence of SEQ ID NO: 4 or a functional fragment thereof.
- the water-producing NADH oxidase is preferably derived from P. aureus, but is not limited thereto.
- the molecular weight of the enzyme of the present invention is characterized in that 50 kDa.
- the present invention also provides a NADH oxidase gene for producing water encoding the enzyme of the present invention.
- the gene of the present invention preferably has a nucleotide sequence of SEQ ID NO: 3.
- the present invention provides a method for producing NADH oxidase to produce water by culturing the strain transformed with a recombinant expression vector containing the water-producing NADH oxidase gene.
- the present invention also relates to the L-arabinitol L-arabinitol by regenerating the NADH oxidase of the present invention through the coupling reaction with Hypocrea jecorina L-arabinitol dehydrogenase.
- Provided is a method for preparing xylulose at low cost and high yield.
- the present invention provides a composition for producing L-xylulose comprising the enzyme of the present invention as an active ingredient.
- NADH oxidase producing water of the present invention is characterized by having the amino acid sequence represented by SEQ ID NO: 4.
- at least one amino acid sequence of SEQ ID NO: 4 may be deleted, substituted, or added to one or more amino acids within a range in which the NADH oxidase activity that generates the water indicated by the protein having these amino acid sequences is not impaired.
- Variant introduced NADH oxidase is also included in the NADH oxidase producing water according to the present invention.
- the present invention includes an NADH oxidase gene encoding a NADH oxidase that generates water having an amino acid sequence of SEQ ID NO: 4, and the gene sequence thereof is represented by SEQ ID NO: 3.
- generates the above-mentioned variant water obtained by changing these nucleotide sequences of SEQ ID NO: 3 is also included in the NADH oxidase gene which produces the water which concerns on this invention.
- the present invention includes a recombinant vector containing the NADH oxidase gene to produce the water, transformants transformed by the recombinant vector.
- the present invention also includes a method for producing NADH oxidase for producing water, which is characterized by separating NADH oxidase for producing water from the culture obtained by culturing the transformant.
- the NADH oxidase gene which produces the water of this invention is isolate
- chromosomal DNA is obtained from a strain having a NADH oxidase gene that produces water.
- PCR polymerase chain reaction
- PCR amplification fragments are labeled with appropriate reagents, and colony hybridization is performed on the chromosomal DNA library to select NADH oxidase genes that produce water (Current Protocols in Molecular Biology, Vol. 1, p. 603). , 1994).
- the transformed microorganism of the present invention is obtained by introducing the recombinant vector of the present invention into a host suitable for the expression vector used in the preparation of the recombinant vector.
- a host suitable for the expression vector used in the preparation of the recombinant vector for example, when a bacterium such as E. coli is used as a host, the recombinant vector according to the present invention is capable of autonomous replication in the host, and at the same time, contains a DNA and a transcription sequence containing a NADH oxidase gene that generates a promoter and water. It is preferable to have what is necessary for expression of these.
- pET28a was used as the expression vector used in the present invention, any expression vector satisfying the above requirements can be used.
- Production of the NADH oxidase producing water according to the present invention comprises culturing a transformant obtained by transforming a host with a recombinant vector having a gene encoding the same, and generating a gene product in a culture (culture medium or culture supernatant). It is performed by generating and accumulating NADH oxidase which produces phosphorus, and obtaining an enzyme from the culture.
- Acquisition and purification of NADH oxidase producing water can be carried out by recovering the cells of the obtained culture by centrifugation, and by using cell disruption, affinity chromatography, cation or anion exchange chromatography alone or in combination. .
- the present inventors cloned the gene of NADH oxidase which produces water from Streptococcus to develop NADH oxidase which produces water.
- the coupling reaction between the recombinant NADH oxidase and the hypocreatic zecorina L-arabinitol dehydrogenase incorporating the above-described genes can be used to prepare L-xylulose in high yield from L-arabinitol.
- it was confirmed that the production of by-products can be greatly reduced, and the present invention was completed.
- the present invention clones a gene encoding NADH oxidase from a gene of P. aureus to produce NADH oxidase producing industrially useful water, and analyzes the nucleotide sequence of the gene and the amino acid sequence inferred therefrom.
- NADH can be oxidized by the recombinant strain inserted with the above gene, and can be used for the purpose of not generating by-products except water in regenerating the coenzymes NADH and NAD + .
- the optimum conditions for producing L-xylulose through the coupling reaction between water-producing NADH oxidase and hypocreas zecorina L-arabinitol dehydrogenase were found to be 4.9 U / ml, 8.2 mM NAD + , The reaction is characterized in that the pH 8.0, the reaction temperature 30.9 °C.
- the NADH oxidase which produces P. aureus derivative isolated from the present invention can be used to regenerate the coenzyme NAD + , and in particular, the dehydration of L-arabinitol derived from NADH oxidase and Hypocrea zecolina which produce water.
- L-xylulose can be produced efficiently by the coupling reaction of the digestive enzymes.
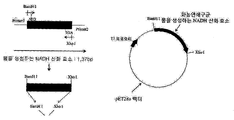
- Figure 1 is a vector map of the vector pUC-LOX to find a fragment containing a NADH oxidase gene that produces water in the chromosome of Streptococcus, and cloned into a vector used in E. coli.
- Figure 2 is a view showing a method for producing an expression vector containing the NADH oxidase gene to produce water derived from P. aureus strains.
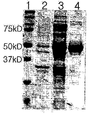
- FIG. 3 is a SDS-PAGE gel photograph of NADH oxidase producing water derived from P. aureus strains. One; Size markers, 2; Insoluble protein of NADH oxidase, which produces water, 3; Water soluble protein of NADH oxidase to produce water, 4; NADH oxidase to produce purified water.
- FIG. 4 shows the conversion of L-xylulose produced from L-aravinitol with the concentration of L-arabinitol.
- FIG. 5 to 7 are analysis of the conversion rate of L-xylulose produced from L-arabinitol through reaction surface analysis.
- FIG. 5 Correlation between L-arabinitol dehydrogenase concentration (U / ml) and NAD concentration (mM)
- FIG. 6 L-arabinitol dehydrogenase concentration (U / ml) and pH Correlation between
- FIG. 7 Correlation between L-aravinitol dehydrogenase concentration (U / ml) and production temperature (° C.).
- FIG. 8 is a diagram showing the results of analysis by high pressure liquid chromatography (HPLC) that L-xylulose is produced from L-arabinitol during L-xylulose production using a coupling reaction.
- HPLC high pressure liquid chromatography
- FIG. 9 is a diagram showing the production of L-xylulose over time under optimized conditions through a coupling reaction.
- the bacterium Pseudomonas aeruginosa (KCTC 3208) was used to obtain the nucleotide sequence of the NADH oxidase gene that produces L-arabinitol lyase.
- genes with similar functions are known to be somewhat similar in size to each nucleotide sequence. Therefore, it is assumed that the gene of NADH oxidase that produces water of Streptococcus pneumoniae (KCTC 3208) is about 1.4 kb in size, and is based on the NADH oxidase nucleotide sequence that generates water known from other bacteria.
- the entire gene of NADH oxidase producing (KCTC 3208) water was cloned.
- E. coli XL1-Blue and pUC18 vectors were used for cloning.
- LB medium having a general composition was used, and the peptone agar medium (Malt extract peptone agar) was used for culturing P. aureus (KCTC 3208).
- P. aureus KCTC 3208.
- plate medium of E. coli plates containing LB agar, 3-5% sugar, 0.3-0.5% beef extract, 0.9-1.1% bactopeptone, and 1.3-1.7% agar were used. 50 ⁇ g / ml ampicillin was added as needed.
- the culture method was inoculated into 250 ml Erlenmeyer flask containing 50 ml of P. aeruginosa (KCTC 3208) and incubated at 37 ° C and 200 rpm for 1 day, and E. coli at 37 ° C and 200 rpm. Incubated for 16 hours.
- KCTC 3208 P. aeruginosa
- RNA extraction of P. aeruginosa was performed using a Qiagen plant total RNA kit (QIAGEN), and a reverse transcriptase for cDNA synthesis was used with Oligo-dT RT-mix (intron).
- KCTC 3208 chromosome was isolated to clone the NADH oxidase gene producing water.
- Degenerated primer, DhLOX F-5'-TCH TTY based on the NADH oxidase sequencing, which produces water known from other bacteria to amplify a portion of the NADH oxidase gene in P. aeruginosa (KCTC 3208) YTH TCW TGT GGD AT -3 '(SEQ ID NO: 1) and DhLOX R-5'-CGH AYV GCR TTV GTD GCY A -3' (SEQ ID NO: 2) were prepared.
- the genomic DNA of Streptococcus pneumoniae (KCTC 3208) was completely cleaved using restriction enzymes BamHI, EcoRI, HindIII, SalI, and XbaI which do not have a cleavage site in the amplified partial sequences. And a radiolabeled probe was made using a DNA fragment obtained through the polymerase chain reaction. Using this, we searched for DNA fragments containing genes to be searched by Southern hybridization. When the chromosome was cut with BamHI, HindIII, and XbaI, the DNA having the NADH oxidase gene, which produced water as a result of Southern hybridization, was about 20 to 23 kb.
- the desired gene was searched for using the fragment cut with EcoRI of about 2.5 kb and the fragment with SalI of about 5.8 kb.
- the 2.5 kb DNA fragment and the 5.8 kb DNA fragment separated by SalI were cloned into pUC18 after chromosome cleavage with EcoRI and named as pUC-LOX (FIG. 1).
- Colonies hybridization was performed using the 1 kb probe made in the pUC-LOX library to determine clones with the gene of NADH oxidase that produced the desired water.
- the nucleotide sequence of the determined clone was analyzed to find 1,371 bp of the entire gene sequence of NADH oxidase generating water (SEQ ID NO: 3). As expected, it was similar in size to the NADH oxidase gene that produced water found in many other bacteria.
- NADH oxidase that produces P. aureus (KCTC 3208) water has a nucleotide sequence common to other water-producing NADH oxidase.
- the enzyme gene was inserted into the BamHI and XhoI sites of the expression vector pET28a (Novagen, USA) to express NADH oxidase in large amounts in E. coli.
- E. coli BL21 (DE3) (NEB, UK) was then transformed (FIG. 2).
- the recombinant strain prepared in Example 3 was inoculated in LB medium and incubated at 37 ° C. for 24 hours, and then the protein expressed in the SDS-PAGE gel was confirmed (FIG. 3).
- the recombinant strain culture solution was centrifuged (8000 ⁇ g, 10 minutes) to collect only the cells, and then subjected to sonication to disrupt the cell wall of Escherichia coli. The precipitate was removed by centrifugation at 20,000 x g for 20 minutes to obtain a supernatant. After obtaining, Ni-NTA His-tag binding chromatography (Qiagen, Germany) was finally performed to purely isolate the NADH oxidase producing the recombinant product.
- the concentration of NADH oxidase producing new water was 5.0 U / ml
- the concentration of L-arabinitol dehydrogenase was 4.9 U / ml
- the concentration of NAD + was 8.2 mM
- the pH was 8
- the temperature was 30 ° C and L.
- Enzyme activity was compared by varying the production temperature of xylulose from 50 to 300 mM.
- the conversion rate to L-xylulose at an L-aribinitol concentration of 50 mM to 150 mM showed an optimum conversion rate of 90% or more.
- the concentration of NADH oxidase producing water was 6.25 U / ml
- the concentration of L-arabinitol was 250 mM
- the concentration of L-arabinitol dehydrogenase was 1.37-66.8 U / ml
- the concentration of NAD + was 2.5-12.5.
- the pH was 2.5 to 12.5 and the production temperature was varied from 20 to 40 ° C. to compare the conversion of L-Xylulose to L-Xylulose (Table 1).
- Table 1 is a table of experimental conditions for the optimization of enzyme activity.
- Example 4 Based on the conditions of Example 4 (2), NAD concentration, pH, reaction temperature, L-arabinitol dehydrogenation in order to optimize the conversion efficiency of L-xylulose from L-arabinitol through the coupling reaction. Different concentrations of enzymes were tested. To optimize the production of L-xylulose, 30 experiments were designed through the Reaction Surface Methodology. The L-xylulose conversion rate through each experiment was as shown in Table 2, and the correlation of each experimental variable was as shown in Figs.
- Table 2 is a table of optimization of L-xylulose production through RSM.
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- General Engineering & Computer Science (AREA)
- Gastroenterology & Hepatology (AREA)
- Biophysics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Enzymes And Modification Thereof (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
본 발명은 신규 NADH 산화효소를 생산하는 화농연쇄구균 (Streptococcus pyogenes), 그 균주로부터 유래한 물을 생성하는 NADH 산화효소와 유전자 및 그 효소의 생산방법에 관한 것으로, 보다 상세하게는 물을 생성하는 NADH 산화효소, 이를 코딩하는 핵산 분자, 상기 핵산 분자를 포함하는 벡터, 상기 벡터를 포함하는 형질전환체, 상기 물을 생성하는 NADH 산화효소를 이용한 조효소의 재생에 관한 것이다. 또한 L-희소당을 생성하는 L-아라비니톨 탈수소화효소와 신규 물을 생성하는 NADH 산화효소와의 커플링 반응을 통하여 고가의 조효소를 재생시켜 반복 사용함으로써 경제적으로 L-희소당을 생산할 수 있음을 입증하였으며, 물을 생성하는 NADH 산화효소를 이용함으로써 물 이외에 부산물이 생성되지 않아 정제 공정을 간소화할 수 있다.
Description
본 발명은 화농연쇄구균 유래의 신규 물을 생성하는 NADH 산화효소와 L-아라비니톨 산화효소를 커플링 반응시켜 L-자일룰로스를 제조하는 방법에 관한 것으로서, 보다 상세하게는 화농연쇄구균 유래의 신규 물을 생성하는 NADH 산화효소, 이를 코딩하는 핵산 분자, 상기 핵산 분자를 포함하는 벡터, 상기 벡터를 포함하는 형질전환체 및 이를 이용하여 NADH 산화효소와 L-아라비니톨 산화효소와의 커플링 반응을 통한 L-자일룰로스의 제조방법에 관한 것이다.
생화학적으로 산화환원 반응은 비대칭 반응에서 중요할 뿐만아니라 광학적 순도를 가지는 화학물질이나 의약학에서 주목하고 있는 분야이다. 그러나 이러한 산화환원 반응에는 NAD(P)+ 또는 NAD(P)H와 같은 고가의 조효소가 필요로 하기 때문에 산업적인 응용이 어려운 실정이다. 따라서 이러한 고가의 조효소를 재생하여 연속적으로 사용하는 것이 경제적 확보를 위해 필요하다. 보통 조효소의 재생은 효소 반응 시에 화학적, 전기적, 광촉매적 물질을 첨가하거나 효소를 첨가하여 이루어진다. 효소를 사용한 조효소 재생에는 두 가지 방법이 있다. 첫 번째 방법은 효소와 화학첨가물을 넣어 재생하는 것이고, 두 번째 방법은 두 개의 효소를 넣고 커플링시켜 조효소를 재생하는 방법이다. NAD(P)+ 및 NAD(P)H를 재생하기 위한 대표적 방법으로는 formate dehydrogenase와 glucose dehydrogenase를 이용한 방법이 잘 알려져 있으나, 반응 중 부산물이 생성되어 산물의 순도를 낮추고 정제비용을 높이는 문제점이 있다. 하지만 물을 생성하는 NADH 산화효소를 사용하면 부산물로 물만 생성되기 때문에 높은 경제성을 확보할 수 있다.
본 발명에서는 신규 물을 생성하는 NADH 산화효소를 제시하고, 이를 L-아라비니톨 탈수소화효소와 커플링하여 조효소를 재생시킴으로써 L-아라비니톨로부터 L-자일룰로스를 고수율 및 연속적으로 생산할 수 있는 최적 반응 조건을 제시함으로써, L-자일룰로스를 높은 수율로 저렴하게 대량 생산할 수 있는 방법을 제공하고자 한다.
본 발명의 첫 번째 목적은 화농연쇄구균으로부터 유래된 물을 생성하는 NADH 산화효소의 유전자를 제공하는 것이다.
본 발명의 두 번째 목적은 상기 유전자로부터 발현된 물을 생성하는 NADH 산화효소를 제공하는 것이다.
본 발명의 세 번째 목적은 상기 물을 생성하는 NADH 산화효소의 유전자를 포함한 재조합 발현벡터를 제공하는 것이다.
본 발명의 네 번째 목적은 형질전환된 재조합 대장균을 포함하는 모든 형질전환 균주를 제공하는 것이다.
본 발명의 다섯 번째 목적은 형질전환된 재조합 대장균을 이용한 재조합 물을 생성하는 NADH 산화효소를 제공하는 것이다.
본 발명의 여섯 번째 목적은 상기 효소와 L-아라비니톨 탈수소화효소를 커플링 시켜 조효소를 재생시킴으로써 L-자일룰로스를 생산하는 최적 조건을 제시하는 것이다.
본 발명의 다른 목적 및 이점은 하기의 발명의 상세한 설명, 청구범위 및 도면에 의해 보다 명확하게 된다.
본 발명은 L-자일룰로스의 생산방법에 있어서, 화농연쇄구균 균주의 물을 생성하는 NADH 산화효소와 히포크레아 제코리나(Hypocrea jecorina) 유래의 L-아라비니톨 탈수소화효소를 커플링 시켜 조효소를 재생시키는 것을 특징으로 한다. 또한 본 발명은 서어던 하이브리다이제이션과 콜로니 혼성화를 통하여 화농연쇄구균으로부터 물을 생성하는 NADH 산화효소 유전자를 클로닝하는 것을 특징으로 한다.
상기의 목적을 달성하기 위하여 본 발명은 서열번호 4의 아미노산 서열 또는 그 기능적 단편을 가지는 물을 생성하는 NADH 산화효소를 제공한다.
본 발명의 일 구체예에 있어서 상기 물을 생성하는 NADH 산화효소는 화농연쇄구균에서 유래한 것이 바람직하나 이에 한정되지 아니한다.
본 발명의 일 구체예에 있어서, 본 발명의 상기 효소의 분자량은 50 kDa인 것을 특징으로 한다.
또한 본 발명은 본 발명의 상기 효소를 코딩하는 물을 생성하는 NADH 산화효소 유전자를 제공한다.
본 발명의 상기 유전자는 서열번호 3의 염기서열을 가지는 것이 바람직하다.
또한 본 발명은 본 발명의 상기 물을 생성하는 NADH 산화효소 유전자를 포함하는 재조합 발현벡터로 형질전환된 균주를 배양하여 물을 생성하는 NADH 산화효소를 제조하는 방법을 제공한다.
또한 본 발명은 본 발명의 상기 물을 생성하는 NADH 산화효소를 히포크레아 제코리나(Hypocrea jecorina) L-아라비니톨 탈수소화효소와 커플링 반응을 통하여 조효소를 재생시킴으로써 L-아라비니톨로부터 L-자일룰로스를 저비용 및 고수율로 제조하는 방법을 제공한다.
또한 본 발명은 상기 본 발명의 효소를 유효성분으로 포함하는 L-자일룰로스 생산용 조성물을 제공한다.
이하, 본 발명을 설명한다.
본 발명의 물을 생성하는 NADH 산화효소는 서열번호 4로 표시되는 아미노산서열을 가진 것을 특징으로 한다. 또, 서열번호 4의 아미노산 서열에 대해서, 이들 아미노산서열을 가진 단백질이 표시하는 물을 생성하는 NADH 산화효소 활성이 손상되지 않는 범위 내에서, 1이상의 아미노산의 결실, 치환 및 부가의 적어도 1종의 변이가 도입된 변이체 NADH 산화효소도 본 발명에 관한 물을 생성하는 NADH 산화효소에 포함된다.
또, 본 발명에는 서열번호 4의 아미노산서열을 가진 물을 생성하는 NADH 산화효소를 코딩하는 NADH 산화효소 유전자가 포함되고, 그 유전자 서열로서는 서열번호 3으로 표시되는 것을 들 수 있다. 또, 이들 서열번호 3의 염기서열을 변이시켜서 얻게 되는 상기한 변이체 물을 생성하는 NADH 산화효소를 코딩하는 변이 유전자도 본 발명에 관한 물을 생성하는 NADH 산화효소 유전자에 포함된다.
또, 본 발명에는 상기 물을 생성하는 NADH 산화효소 유전자를 함유하는 재조합벡터, 상기 재조합벡터에 의해서 형질전환된 형질전환체가 포함된다. 또한, 본 발명에는 이 형질전환체를 배양하여 얻게 되는 배양물로부터 물을 생성하는 NADH 산화효소를 분리하는 것을 특징으로 하는 물을 생성하는 NADH 산화효소의 제조방법이 포함된다.
이하, 본 발명을 더욱 상세히 설명한다.
본 발명의 물을 생성하는 NADH 산화효소 유전자는 화농연쇄구균의 균체로부터 분리된 것이다. 먼저 물을 생성하는 NADH 산화효소 유전자를 보유한 균주로부터 염색체 DNA를 취득한다.
다음에, 설계한 올리고뉴클레오타이드를 프라이머로 하고 화농연쇄구균 균주의 염색체 DNA를 주형으로해서 폴리머라제 연쇄반응(PCR)을 행하여, 물을 생성하는 NADH 산화효소 유전자를 부분적으로 증폭한다. 이와같이 얻은 PCR 증폭 단편은 화농연쇄구균 균주의 물을 생성하는 NADH 산화효소 유전자에 100% 상동성을 가진 단편으로서, 콜로니 하이브리디제이션을 행할 때의 프로브로서 높은 S/N비를 기대할 수 있는 동시에, 하이브리다이제이션의 스트린전시(stringency) 제어를 용이하게 한다. 상기의 PCR 증폭 단편을 적당한 시약을 사용해서 표지하고, 상기 염색체 DNA라이브러리에 대해서 콜로니 하이브리다이제이션을 행하여, 물을 생성하는 NADH 산화효소 유전자를 선발한다 (Current Protocols in Molecular Biology, 1권, 603페이지, 1994년).
상기의 방법에 의해 선발된 대장균으로부터 알칼리법 (Current Protocols in Molecular Biology, 1권, 161페이지, 1994년)을 사용해서 플라스미드를 회수함으로써, 물을 생성하는 NADH 산화효소 유전자를 함유하는 DNA단편을 얻을 수 있다. 또한, 상기 방법에 의해 염기서열을 결정한 후에는, 상기 염기서열을 가진 DNA 단편의 제한효소 분해에 의해 조제한 DNA단편을 프로브로 해서 하이브리다이즈함으로써 본 발명의 전체 유전자를 얻는 것이 가능하다. 서열번호 3에는 본 발명의 물을 생성하는 NADH 산화효소 유전자의 염기서열을, 서열번호 4에는 상기 유전자가 코딩하는 아미노산 서열을 표시한다.
본 발명의 형질전환된 미생물은, 본 발명의 재조합벡터를 상기 재조합벡터의 제작 시 사용한 발현벡터에 적합한 숙주 속에 도입함으로써 얻게 된다. 예를 들면 대장균 등의 세균을 숙주로서 사용하는 경우, 본 발명에 관한 재조합벡터는 그 자신이 숙주 속에서 자율복제 가능한 동시에, 프로모터와 물을 생성하는 NADH 산화효소 유전자를 함유하는 DNA 및 전사종결서열 등의 발현에 필요한 구성을 가진 것임이 바람직하다. 본 발명에 사용된 발현벡터로서는 pET28a를 사용하였으나 상기의 요건을 만족하는 발현벡터이면 어느 것이나 사용가능하다.
본 발명에 관한 물을 생성하는 NADH 산화효소의 제조는, 이를 코딩하는 유전자를 가진 재조합벡터에 의해 숙주를 형질전환해서 얻은 형질전환체를 배양하고, 배양물(배양균체 또는 배양상청액) 속에 유전자 산물인 물을 생성하는 NADH 산화효소를 생성 축적시켜, 배양물로부터 효소를 취득함으로써 행하여진다.
물을 생성하는 NADH 산화효소의 취득 및 정제는, 얻게 되는 배양물의 균체 를 원심분리를 통해 회수하고, 균체파쇄, 친화성크로마토그래피, 양이온 또는 음이온교환 크로마토그래피 등을 단독으로 또는 조합함으로써 행할 수 있다.
본 발명자는 물을 생성하는 NADH 산화효소를 개발하고자 화농연쇄구균으로부터 물을 생성하는 NADH 산화효소의 유전자를 클로닝하였다. 전기 유전자를 삽입한 재조합 NADH 산화효소와 하이포크레아 제코리나 L-아라비니톨 탈수소화효소와의 커플링 반응을 통해 L-아라비니톨로부터 높은 수율로 L-자일룰로스를 제조할 수 있을 뿐만 아니라, 부산물의 생성을 크게 감소시킬 수 있음을 확인하고 본 발명을 완성하였다.
본 발명은 산업적으로 유용한 물을 생성하는 NADH 산화효소를 제조하기 위하여 화농연쇄구균의 유전자로부터 NADH 산화효소를 암호화하는 유전자를 클로닝하고, 전기 유전자의 염기서열 및 그로부터 유추되는 아미노산 서열을 분석한다.
전기 유전자를 삽입한 재조합 균주에 의해 NADH를 산화할 수 있으며, 조효소인 NADH와 NAD+를 재생하는데 있어서 물을 제외한 부산물을 생성하지 않는 목적으로 사용할 수 있다.
또한 물을 생성하는 NADH 산화효소와 하이포크레아 제코리나 L-아라비니톨 탈수소화효소의 커플링 반응을 통해 L-자일룰로스를 생산하는 최적 조건은 효소 농도 4.9 U/ml, 8.2 mM NAD+, 반응 pH 8.0, 반응온도 30.9℃인 것을 특징으로 한다.
본 발명에서 분리한 화농연쇄구균 유래 물을 생성하는 NADH 산화효소는 조효소인 NAD+를 재생하는데 이용될 수 있고, 특히 물을 생성하는 NADH 산화효소와 하이포크레아 제코리나 유래의 L-아라비니톨 탈수소화효소의 커플링 반응에 의해 L-자일룰로스를 효율적으로 생산할 수 있다.
도 1은 벡터 pUC-LOX의 벡터맵으로 화농연쇄구균의 염색체에서 물을 생성하는 NADH 산화효소 유전자를 지니고 있는 단편을 찾아 대장균에서 이용되는 벡터에 클로닝한 것이다.
도 2는 화농연쇄구균 균주로부터 유래된 물을 생성하는 NADH 산화효소 유전자를 포함하는 발현벡터의 제조방법을 나타내는 도면이다.
도 3은 화농연쇄구균 균주로부터 유래된 물을 생성하는 NADH 산화효소의 SDS-PAGE 젤 사진이다. 1; 사이즈 마커, 2; 물을 생성하는 NADH 산화효소의 불용성 단백질, 3; 물을 생성하는 NADH 산화효소의 수용성 단백질, 4; 정제된 물을 생성하는 NADH 산화효소.
도 4는 L-아라비니톨의 농도에 따른 L-아라비니톨로부터 생산되는 L-자일룰로스의 전환율.
도 5 내지 7은 반응표면분석을 통하여 L-아라비니톨로부터 생산되는 L-자일룰로스의 전환율의 분석. (도 5) L-아라비니톨 탈수소화효소 농도 (U/ml)와 NAD의 농도 (mM) 사이의 상관관계, (도 6) L-아라비니톨 탈수소화효소 농도 (U/ml)와 pH 사이의 상관관계, (도 7) L-아라비니톨 탈수소화효소 농도 (U/ml)와 생산 온도 (℃) 상이의 상관관계.
도 8은 커플링 반응을 통하여 사용하여 L-자일룰로스 생산 시 L-아라비니톨로부터 L-자일룰로스가 생산되는 것을 고압 액체 크로마토그래피 (HPLC)로 분석한 결과를 나타낸 도이다. A. 커플링 반응전 B. 커플링 반응 후
도 9는 커플링 반응을 통해 최적화된 조건에서 시간대별 L-자일룰로스의 생산량을 타나낸 도이다.
이하, 본 발명을 다음의 실시예에 의하여 더욱 상세히 설명하나, 본 발명이 실시예에 한정되는 것은 아니다.
실시예 1: 물을 생성하는 NADH 산화효소 생산균의 선별
물을 생성하는 NADH 산화효소를 생산하는 고활성 콜로니를 분리하기 위하여 화농연쇄구균의 배양액 10 ul를 생리식염수 10 ml에 현탁하고, 현탁액의 10 ul (1× 104 cfu ml-1)를 취하여 3% malt extract가 첨가된 펩톤 한천배지 (Malt extract peptone agar)에 도말한 후, 37 ℃에서 2일간 배양하였다. 고체 펩톤배지에서 콜로니가 형성된 후 콜로니를 취하여 물을 생성하는 NADH 산화효소 활성을 갖는 콜로니를 선별하는 방법으로 물을 생성하는 NADH 산화효소를 생산하는 고활성 화농연쇄구균을 탐색하였다.
실시예 2: 화농연쇄구균 (KCTC 3208)으로부터 신규 물을 생성하는 NADH 산화효소 유전자의 클로닝
L-아라비니톨 분해 효소인 물을 생성하는 NADH 산화효소 유전자의 염기서열을 얻기 위해 세균인 화농연쇄구균 (KCTC 3208)을 사용하였다. 일반적으로 유사한 기능을 지니는 유전자의 경우에는 각 염기서열과 크기가 어느 정도 유사하다고 알려져 있다. 따라서 화농연쇄구균 (KCTC 3208)의 물을 생성하는 NADH 산화효소의 유전자도 약 1.4 kb 정도의 크기를 지녔을 것으로 추정하고 다른 세균의 이미 알려진 물을 생성하는 NADH 산화효소 염기서열을 바탕으로 화농연쇄구균 (KCTC 3208)의 물을 생성하는 NADH 산화효소 전체 유전자를 클로닝 하였다.
클로닝에는 대장균 XL1-Blue와 pUC18 벡터를 사용하였다. 대장균의 배양 배지로는 일반적 조성의 LB 배지를 사용하였고, 화농연쇄구균 (KCTC 3208)의 배양에는 상기 펩톤한천배지(Malt extract peptone agar)를 사용하였다. 대장균의 평판(plate) 배지로는 각각 LB 아가(agar)와 3~5% 설탕, 0.3~0.5% 쇠고기 추출물, 0.9~1.1% 박토 펩톤, 1.3~1.7% 아가 조성의 플레이트를 사용하였다. 필요에 따라 50 ㎍/ml 엠피실린(amipicillin)을 첨가하였다.
배양 방법은 화농연쇄구균 (KCTC 3208)의 경우, 배지 50 ml이 들어 있는 250 ml의 삼각 플라스크에 접종하여 37℃, 200 rpm 조건에서 1일간 배양하였고, 대장균의 경우에는 37℃, 200 rpm 조건에서 16 시간 배양하였다.
대부분의 DNA는 아가로스겔(TAE buffer, 0.5%) 전기영동법으로 확인하였고, 겔 상에서 DNA 밴드의 정제는 QiaXII 겔 추출장지(QIAGEN, USA)를 이용하였으며, DNA간의 연결(ligation) 반응은 T4 DNA 연결효소(NEB)를 이용하였다. 또한 화농연쇄구균 (KCTC 3208)의 RNA 추출은 Qiagen plant total RNA kit(QIAGEN)를 이용하였으며, cDNA 합성을 위한 역전사 효소는 Oligo-dT RT-mix(intron)를 이용하였다.
물을 생성하는 NADH 산화효소 유전자를 클로닝하기 위하여 화농연쇄구균 (KCTC 3208) 염색체를 분리하였다. 화농연쇄구균 (KCTC 3208) 내의 NADH 산화효소 유전자의 일부분을 증폭하기위해 다른 세균에서 이미 알려진 물을 생성하는 NADH 산화효소 염기서열을 바탕으로 비특이적 프라이머(degenerated primer), DhLOX F-5'- TCH TTY YTH TCW TGT GGD AT -3' (서열번호 1)와 DhLOX R-5'- CGH AYV GCR TTV GTD GCY A -3' (서열번호 2)를 제작하였다. 이를 이용하여 연쇄중합반응에 의해 784 bp 크기에 해당하는 NADH 산화효소 유전자 일부를 화농연쇄구균 (KCTC 3208) 염색체에서 증폭하였다.
그리고 증폭된 상기의 부분 염기서열 중 그 절단 부위가 존재하지 않는 제한 효소인 BamHI, EcoRI, HindIII, SalI, XbaI을 이용하여 화농연쇄구균 (KCTC 3208)의 genomic DNA를 완전히 절단하였다. 그리고 앞서 중합효소 연쇄반응을 통하여 얻은 DNA 단편을 이용하여 방사능 표지된 탐침자(probe)를 만들었다. 이를 이용하여 서어던 하이브리다이제이션으로 찾고자 하는 유전자를 지닌 DNA 단편을 탐색하였다. BamHI, HindIII, XbaI으로 염색체를 자른 경우에 있어서는 서어던 하이브리다이제이션의 결과 나타난 물을 생성하는 NADH 산화효소의 유전자를 지닌 DNA의 크기가 약 20~23 kb정도 되어 너무 큰 관계로 이용하지 않았고, 2.5kb 정도의 EcoRI으로 잘린 조각과 약 5.8 kb정도의 SalI으로 잘린 조각을 이용하여 원하는 유전자를 탐색하였다. 염색체를 EcoRI으로 절단한 후 분리한 2.5kb 정도 크기의 DNA 조각과 SalI으로 절단한 5.8kb 정도의 DNA 단편들을 pUC18에 클로닝하고 이를 pUC-LOX라고 명명하였다(도 1).
pUC-LOX 라이브러리에서 앞서 만든 1 kb 크기의 탐침자를 이용하여 콜로니 혼성화를 수행하여 원하는 물을 생성하는 NADH 산화효소의 유전자를 지닌 클론을 결정하였다. 결정한 클론의 염기서열을 분석하여 물을 생성하는 NADH 산화효소 전체 유전자 염기서열 1,371 bp를 밝혔다 (서열번호 3). 이는 앞서 예상한 바와 같이 다른 여러 세균에서 밝혀진 물을 생성하는 NADH 산화효소 유전자와 크기가 비슷하였다. 또한 화농연쇄구균 (KCTC 3208) 물을 생성하는 NADH 산화효소는 다른 물을 생성하는 NADH 산화효소에서 공통적으로 나타나는 염기서열을 지니고 있음을 확인하였다.
실시예 3: 재조합 발현 벡터 및 재조합 균주 제조
실시예 2에 따른 물을 생성하는 NADH 산화효소를 암호화하는 유전자를 이용하여, NADH 산화효소를 대장균에서 대량으로 발현시키기 위하여 발현 벡터 pET28a(Novagen, 미국)의 BamHⅠ과 XhoI 부위에 상기 효소 유전자를 삽입한 후 대장균 BL21(DE3)(NEB, 영국)에 형질 전환시켰다 (도 2).
실시예 4: 재조합 물을 생성하는 NADH 산화효소의 발현 및 순수 분리
상기 실시예 3에서 제조된 재조합 균주를 LB 배지에 접종하고 37℃에서 24시간 동안 배양한 다음 SDS-PAGE 젤에서 발현된 단백질을 확인하였다 (도 3).
상기 실시예 4의 방법으로 발현시킨 재조합 물을 생성하는 NADH 산화효소를 정제하기 위하여, 재조합 균주 배양액을 원심분리 (8000×g, 10분)하여 균체만을 모은 후, 초음파 처리하여 대장균의 세포벽을 파쇄하고, 20,000×g에서 20분간 원심분리하여 침전물(균체)을 제거하고 상등액을 수득하였다. 수득 후, 최종적으로 Ni-NTA His-tag 결합 크로마토그래피(Qiagen, 독일)를 수행하여, 재조합 물을 생성하는 NADH 산화효소를 순수 분리하였다.
실시예 4: NADH 산화효소와 L-아라비니톨 탈수소화효소의 커플링 반응을 통한 L-자일룰로스의 최적 생산 조건 실험
(1) 커플링 반응을 통한 L-자일룰로스의 최적생산에 L-아라비니톨의 농도가 미치는 영향
신규 물을 생성하는 NADH 산화효소의 농도는 5.0 U/ml, L-아라비니톨 탈수소화효소효소의 농도는 4.9 U/ml, NAD+ 농도는 8.2 mM, pH는 8, 온도는 30℃ 및 L-자일룰로스의 생산온도를 50 ~ 300 mM로 달리하여 효소 활성을 비교하였다. L-아라비니톨의 농도를 달리하여 실험한 결과 도 4와 같았고, 50 mM ~ 150 mM의 L-아리비니톨 농도에서 L-자일룰로스로 전환되는 수율이 90%이상으로 최적 전환율을 보였다.
(2) 커플링 반응을 통한 L-자일룰로스의 최적생산 실험 조건
물을 생성하는 NADH 산화효소의 농도는 6.25 U/ml, L-아라비니톨 농도는 250 mM, L-아라비니톨 탈수소화효소의 농도는 1.37~6.86 U/ml, NAD+ 농도는 2.5~12.5, pH는 2.5~12.5 및 생산온도를 20 ~ 40℃로 달리하여 L-아리비니톨로부터 L-자일룰로스의 전환율을 비교하였다 (표 1).
표 1
| 암호화된 수치 | |||||
| -α | -1 | 0 | +1 | + α | |
| 효소 농도 (U/ml) | 1.37 | 2.74 | 4.12 | 5.49 | 6.86 |
| NAD+농도 (mM) | 2.5 | 5.0 | 7.5 | 10.0 | 12.5 |
| pH | 6 | 7 | 8 | 9 | 10 |
| 온도 (℃) | 20 | 25 | 30 | 35 | 40 |
표 1은 효소활성 최적화를 위한 조건 실험 표이다.
(3) 커플링 반응에 의한 L-자일룰로스의 전환효율에 최적화
상기 실시예 4(2)의 조건을 바탕으로 커플링 반응을 통하여 L-아라비니톨로부터 L-자일룰로스의 전환 효율을 최적화하기 위하여 NAD 농도, pH, 반응 온도, L-아라비니톨 탈수소화효소의 농도를 달리하여 실험하였다. L-자일룰로스의 생산을 최적화하기 위하여 Reaction Surface Methodology를 통하여 30개의 실험을 디자인 하였다. 각각의 실험을 통한 L-자일룰로스 전환율은 표 2와 같았으며, 각 실험 변수들의 상관관계는 도 5 내지 7과 같았다.
표 2
| 번호 | HjLAD (U/mL) | NAD+(mM) | pH | 온도 (oC) | 전환율 (%) | 예상전환율 (%) |
| 1 | 4.12 | 7.5 | 8 | 30 | 76.94 | 76.94 |
| 2 | 5.49 | 5 | 9 | 25 | 63.71 | 65.87 |
| 3 | 4.12 | 12.5 | 8 | 30 | 70.74 | 73.07 |
| 4 | 2.75 | 5 | 7 | 25 | 61.39 | 61.01 |
| 5 | 2.75 | 10 | 7 | 25 | 70.36 | 71.04 |
| 6 | 2.75 | 5 | 9 | 35 | 50.00 | 50.52 |
| 7 | 5.49 | 10 | 9 | 25 | 72.04 | 69.33 |
| 8 | 5.49 | 10 | 7 | 35 | 68.87 | 68.53 |
| 9 | 4.12 | 7.5 | 8 | 30 | 76.94 | 76.94 |
| 10 | 2.75 | 5 | 7 | 35 | 63.33 | 65.03 |
| 11 | 5.49 | 10 | 9 | 35 | 69.07 | 70.02 |
| 12 | 5.49 | 10 | 7 | 25 | 62.16 | 62.22 |
| 13 | 2.75 | 10 | 9 | 25 | 64.29 | 64.07 |
| 14 | 4.12 | 7.5 | 8 | 30 | 76.94 | 76.94 |
| 15 | 4.12 | 7.5 | 8 | 30 | 76.94 | 76.94 |
| 16 | 4.12 | 7.5 | 6 | 30 | 67.52 | 66.37 |
| 17 | 2.75 | 10 | 7 | 35 | 73.62 | 72.04 |
| 18 | 2.75 | 5 | 9 | 25 | 52.80 | 52.12 |
| 19 | 4.12 | 7.5 | 8 | 30 | 76.94 | 76.94 |
| 20 | 5.49 | 5 | 9 | 35 | 71.26 | 69.57 |
| 21 | 2.75 | 10 | 9 | 35 | 61.39 | 59.46 |
| 22 | 4.12 | 7.5 | 8 | 30 | 76.94 | 76.94 |
| 23 | 4.12 | 2.5 | 8 | 30 | 64.48 | 62.59 |
| 24 | 4.12 | 7.5 | 8 | 40 | 60.00 | 60.57 |
| 25 | 5.49 | 5 | 7 | 35 | 69.19 | 70.01 |
| 26 | 4.12 | 7.5 | 8 | 20 | 56.00 | 55.87 |
| 27 | 1.37 | 7.5 | 8 | 30 | 63.32 | 64.04 |
| 28 | 6.86 | 7.5 | 8 | 30 | 74.56 | 74.27 |
| 29 | 5.49 | 5 | 7 | 25 | 59.77 | 60.68 |
| 30 | 4.12 | 7.5 | 10 | 30 | 57.38 | 58.97 |
표 2는 RSM을 통한 L-자일룰로스 생산 최적화 표이다.
(4) 최적 조건에서 커플링을 통한 L-아라비니톨로부터 L-자일룰로스의 생산실험
본 발명 균주 화농연쇄구균 (KCTC 3208) 유래의 물을 생성하는 NADH 산화효소와 히포크레아 제코리나 유래의 L-아라비니톨 탈수소화효소를 이용한 커플링 실험을 수행하였다. 커플링 반응액 내의 물을 생성하는 NADH 산화효소의 농도는 6.25 U/ml, L-아라비니톨 농도는 250 mM, L-아라비니톨 탈수소화효소의 농도는 4.9 U/ml, NAD+ 농도는 8.2 mM, pH는 8.0, 온도는 30.9℃로 조절하였다. 상기 최적화된 커플링 반응에서 생성된 L-자일룰로스를 HPLC에서 확인하였다 (도 8). 최적화 된 조건에서 실험한 결과 반응 시간 별 전환수율을 도 7에 나타내었으며, 8시간 반응시켰을 때 78.4%의 최대 전환율을 나타내었다 (도 9).
Claims (10)
- 서열번호 4의 아미노산 서열을 가지는 물을 생성하는 신규한 NADH 산화효소.
- 제 1항에 있어서, 상기 물을 생성하는 NADH 산화효소는 NADH 산화 시에 물을 생성하는 하는 것을 특징으로 하는 신규한 NADH 산화효소.
- 제 1항 내지 제 2항 중 어느 한 항의 효소를 코딩하는 유전자.
- 제 3항에 있어서, 상기 유전자는 서열번호 3의 염기서열을 가지는 유전자.
- 제 3항 내지 제 4항 중 어느 한 항의 물을 생성하는 NADH 산화효소 유전자를 포함하는 재조합 발현벡터.
- 제 5항의 재조합 발현벡터로 형질전환된 균주를 배양하여 물을 생성하는 NADH 산화효소를 제조하는 방법.
- 기질에 제 1항 내지 제 2항 중 어느 한 항의 효소를 처리하여 기질을 산화하는 방법.
- 제 1항 내지 제 2항 중 어느 한 항의 효소를 이용하여 NADH로부터 NAD+를 재생하는 방법.
- 물을 생성하는 제 1항 내지 제 2항 중 어느 한 항의 NADH 산화효소와 L-아라비니톨 탈수소효소를 커플링시켜 L-자일룰로스를 생산하는 방법.
- 제 1항 내지 제 2항 중 어느 한 항의 효소를 유효성분으로 포함하는 L-자일룰로스 생산용 조성물.
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PCT/KR2013/003316 WO2014171570A2 (ko) | 2013-04-18 | 2013-04-18 | 화농연쇄구균 유래 신규 nadh 산화효소 및 l-아라비니톨 산화효소와의 커플링에 의한 l-자일룰로스의 생산 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PCT/KR2013/003316 WO2014171570A2 (ko) | 2013-04-18 | 2013-04-18 | 화농연쇄구균 유래 신규 nadh 산화효소 및 l-아라비니톨 산화효소와의 커플링에 의한 l-자일룰로스의 생산 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| WO2014171570A2 true WO2014171570A2 (ko) | 2014-10-23 |
| WO2014171570A3 WO2014171570A3 (ko) | 2015-05-07 |
Family
ID=51731936
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/KR2013/003316 WO2014171570A2 (ko) | 2013-04-18 | 2013-04-18 | 화농연쇄구균 유래 신규 nadh 산화효소 및 l-아라비니톨 산화효소와의 커플링에 의한 l-자일룰로스의 생산 |
Country Status (1)
| Country | Link |
|---|---|
| WO (1) | WO2014171570A2 (ko) |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20050064570A1 (en) * | 2001-08-16 | 2005-03-24 | Degussa Ag | NADH oxidase from lactobacillus |
| US20130030164A1 (en) * | 2010-01-20 | 2013-01-31 | Kaneka Corporation | Nadh oxidase mutant having improved stability and use thereof |
-
2013
- 2013-04-18 WO PCT/KR2013/003316 patent/WO2014171570A2/ko active Application Filing
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20050064570A1 (en) * | 2001-08-16 | 2005-03-24 | Degussa Ag | NADH oxidase from lactobacillus |
| US20130030164A1 (en) * | 2010-01-20 | 2013-01-31 | Kaneka Corporation | Nadh oxidase mutant having improved stability and use thereof |
Non-Patent Citations (3)
| Title |
|---|
| DATABASE NCBI 21 December 2012 Database accession no. YP-060186.1 * |
| GAO, HUI ET AL.: 'Characterization of H2O-forming NADH Oxidase from Streptococcus Pyogenes and its Application in 1-rare Sugar Production' BIOORG. MED. CHEM. LETT. vol. 22, no. 5, 26 January 2012, pages 1931 - 1935 * |
| KAWASAKI, SHINJI ET AL.: 'Purification and Characterization of an H20-forming NADH Oxidase from Clostridium Aminovalericum: Existence of an Oxygen-detoxifying Enzyme in an Obligate Anaerobic Bacteria' ARCH. MICROBIOL. vol. 181, no. 4, 11 March 2004, pages 324 - 330 * |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2014171570A3 (ko) | 2015-05-07 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4561010B2 (ja) | D−ヒダントインハイドロラーゼをコードするdna、n−カルバミル−d−アミノ酸ハイドロラーゼをコードするdna、該遺伝子を含む組み換えdna、該組み換えdnaにより形質転換された細胞、該形質転換細胞を用いるタンパク質の製造方法、および、d−アミノ酸の製造方法 | |
| WO2018093153A1 (ko) | 신규한 d-사이코스 3-에피머화 효소 및 이를 이용한 d-사이코스의 제조 방법 | |
| CN107858340B (zh) | 高催化活性的d-果糖-6-磷酸醛缩酶a突变体、重组表达载体、基因工程菌及其应用 | |
| CN112301013A (zh) | 复合酶及其在制备麦角硫因中的应用 | |
| WO2019098723A1 (ko) | 신규한 d-사이코스 3-에피머화 효소 및 이를 이용한 d-사이코스의 제조 방법 | |
| EP0877084B1 (en) | Thermostable diaphorase gene | |
| WO2015137565A1 (ko) | 신규 포름알데하이드 탈수소효소 및 이를 이용한 포름알데하이드의 생산방법 | |
| WO2014171570A2 (ko) | 화농연쇄구균 유래 신규 nadh 산화효소 및 l-아라비니톨 산화효소와의 커플링에 의한 l-자일룰로스의 생산 | |
| JP4307609B2 (ja) | コエンザイムq10の製造法 | |
| CN115747183A (zh) | 一种酮还原酶突变体及其应用 | |
| WO2014171635A1 (ko) | 신규 d-솔비톨 탈수소화효소 및 이를 이용한 l-소르보스의 생산방법 | |
| KR101228974B1 (ko) | 락토바실러스 람노수스 유래 내열성물 생성 nadh 산화효소 및 그 생산 방법 | |
| CN115975964A (zh) | 一种高活性酮基泛解酸内酯还原酶突变体及其编码基因和应用 | |
| WO2014171636A1 (ko) | 신규 리비톨 탈수소화효소 및 이를 이용한 l-리불로스의 생산방법 | |
| WO2015190633A1 (ko) | 활성이 개선된 대장균 유래의 돌연변이 당 이성질화 효소 및 그를 이용한 l-굴로스의 생산 | |
| WO2010090359A1 (ko) | 신규 l-아라비니톨 탈수소화효소 및 이를 이용한 l-리불로스의 생산방법 | |
| KR101479134B1 (ko) | 화농연쇄구균 유래 신규 nadh 산화효소 및 l-아라비니톨 산화효소와의 커플링에 의한 l-자일룰로스의 생산 | |
| KR101325057B1 (ko) | 안정성 및 활성이 개선된 돌연변이 nadh 산화효소 | |
| WO2005123921A1 (ja) | 新規グリセロール脱水素酵素、その遺伝子、及びその利用法 | |
| KR101479135B1 (ko) | 아스페르기루스 플라버스 유래 솔비톨 탈수소화효소와 nadh 산화효소와의 커플링에 의한 l-자일룰로스의 생산 | |
| WO2017065457A1 (ko) | L-쓰레오닌 생산능을 가지는 미생물 및 그를 이용하여 l-쓰레오닌을 생산하는 방법 | |
| AU2021100409A4 (en) | Recombinant low-temperature catalase, recombinant vector and engineered strain thereof | |
| CN113249348B (zh) | 羰基还原酶、其基因、含有该基因的重组表达转化体及其应用 | |
| CN111575258B (zh) | 一种羰基还原酶EbSDR8突变体及其构建方法和应用 | |
| WO2011099657A1 (ko) | 신규 자일리톨 탈수소화효소 및 이를 이용한 l-자일룰로즈의 생산방법 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 13882338 Country of ref document: EP Kind code of ref document: A2 |
|
| 122 | Ep: pct application non-entry in european phase |
Ref document number: 13882338 Country of ref document: EP Kind code of ref document: A2 |