WO2011016102A1 - 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 - Google Patents
変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 Download PDFInfo
- Publication number
- WO2011016102A1 WO2011016102A1 PCT/JP2009/063743 JP2009063743W WO2011016102A1 WO 2011016102 A1 WO2011016102 A1 WO 2011016102A1 JP 2009063743 W JP2009063743 W JP 2009063743W WO 2011016102 A1 WO2011016102 A1 WO 2011016102A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- amino acid
- formate dehydrogenase
- substitution
- mutant
- isoleucine
- Prior art date
Links
Images














Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/26—Preparation of nitrogen-containing carbohydrates
- C12P19/28—N-glycosides
- C12P19/30—Nucleotides
- C12P19/36—Dinucleotides, e.g. nicotineamide-adenine dinucleotide phosphate
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0008—Oxidoreductases (1.) acting on the aldehyde or oxo group of donors (1.2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y102/00—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2)
- C12Y102/01—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2) with NAD+ or NADP+ as acceptor (1.2.1)
- C12Y102/01002—Formate dehydrogenase (1.2.1.2)
Definitions
- the present invention relates to a mutant formate dehydrogenase having a substitution mutation at a specific site in wild type formate dehydrogenase, a gene encoding the mutant formate dehydrogenase, and a method for producing NADH.
- Formate dehydrogenase reduces NAD + to reduced nicotinamide adenine dinucleotide (NADH) in the presence of oxidized nicotinamide adenine dinucleotide (NAD + ), formic acid and water. Oxidizes formic acid to carbon dioxide. Based on this enzymatic reaction, formate dehydrogenase is used in a system for regenerating NADH from NAD + .
- formate dehydrogenase formate dehydrogenase derived from Candida boidinii (ATCC32195) as disclosed in, for example, JP-A-2003-180383, disclosed in JP-A-2002-233395 NAD + -dependent formate dehydrogenase derived from the genus Bacillus, formate dehydrogenase derived from Mycobacterium vaccae as disclosed in Japanese Patent Application No. 10-023896 has been known .
- BIOCHEMISTRYow Moscow
- Vol. 69, No. 11, 2004, pp. 1252-1267 English translation of Biokhimiya, Vol. 69, No. 11, 2004, pp. 1537-1554.
- various microorganisms and plant-derived formate dehydrogenases are disclosed.
- formate dehydrogenase has a low specific activity among various enzymes.
- it can be said that the NADH production method using the reduction reaction to NADH by formate dehydrogenase has poor productivity due to the low specific activity of formate dehydrogenase.
- formate dehydrogenase various research results have been accumulated so far and reports on functional modification by site-directed mutagenesis have been made (Biomolecular Engineering, 23, (2006) 98-110).
- all of the conventionally known formate dehydrogenases have low specific activity and durability, and must be evaluated as insufficient for the production of NADH using formate dehydrogenase.
- the present inventors previously filed a patent application relating to a formate dehydrogenase exhibiting a high specific activity unpredictable from conventional knowledge, and a method for producing NADH using the formate dehydrogenase.
- Japanese Patent Application No. 2008-1000044 The present invention searches for a useful substitution mutation for the formate dehydrogenase according to the patent application and the conventionally known formate dehydrogenase, and is more durable than the conventionally known mutant formate dehydrogenase. It is an object of the present invention to provide a mutant formate dehydrogenase excellent in the above, a gene encoding the mutant formate dehydrogenase, and a method for producing NADH using the mutant formate dehydrogenase.
- amino acid substitution mutation at a specific site in the formate dehydrogenase and the conventionally known formate dehydrogenase according to the above-mentioned patent application is the durability of formate dehydrogenase. As a result, the present invention has been completed.
- mutant formate dehydrogenase according to the present invention comprises any one of the following amino acid sequences A) to C) and has improved durability.
- Amino acid sequence, and C) amino acid residues corresponding to the 73rd glycine, the 99th valine, the 153rd valine and the 159th glutamine in the formate dehydrogenase consisting of the amino acid sequence shown in SEQ ID NO: 2 are the other amino acids.
- the mutant formate dehydrogenase according to the present invention includes the amino acid sequence substituted with With respect to the amino acid sequence of A), the 31st isoleucine, the 50th arginine, the 60th glutamic acid, the 64th isoleucine, the 73rd glycine, the 82nd lysine, the 136th isoleucine, the 159th glutamine, 239 Amino acid residues corresponding to 1 to 3 amino acids selected from the group consisting of the 1st methionine, the 286th aspartic acid, the 287th histidine, the 293th glutamic acid and the 343th asparagine are further substituted with other amino acids. It preferably contains a sequence.
- the mutant formate dehydrogenase according to the present invention has a 31st isoleucine, 50th arginine, 60th glutamic acid, 64th isoleucine, 82th lysine, 136 with respect to the amino acid sequence of B) above.
- 1st to 3 amino acids selected from the group consisting of: th isoleucine, 159th glutamine, 239th methionine, 286th aspartic acid, 287th histidine, 293th glutamic acid, 298th asparagine and 343th asparagine
- the amino acid residue corresponding to is preferably an amino acid sequence further substituted with another amino acid.
- the mutant formate dehydrogenase according to the present invention comprises a 31st isoleucine, a 50th arginine, a 60th glutamic acid, a 64th isoleucine, an 82th lysine with respect to the amino acid sequence of C) above. 1 to 3 selected from the group consisting of 136th isoleucine, 155th histidine, 239th methionine, 286th aspartic acid, 287th histidine, 293th glutamic acid, 298th asparagine and 343th asparagine It is preferable that the amino acid residue corresponding to an amino acid includes an amino acid sequence further substituted with another amino acid.
- the amino acid after substitution mutation is not particularly limited.
- the amino acid after substitution of the amino acid corresponding to the 31st isoleucine is leucine
- the amino acid after substitution of the amino acid corresponding to the 50th arginine is The amino acid after substitution of the amino acid corresponding to the 60th glutamic acid is valine
- the amino acid after substitution of the amino acid corresponding to the 64th isoleucine is valine, and corresponds to the 73rd glycine.
- the amino acid after the amino acid substitution is alanine
- the amino acid after the amino acid substitution corresponding to the 82nd lysine is arginine
- the amino acid after the amino acid substitution corresponding to the 99th valine is isoleucine
- the amino acid after substitution of the amino acid corresponding to the 153rd valine is isoleucine
- the amino acid after substitution of the amino acid corresponding to the 155th histidine is glutamine or lysine
- the amino acid after substitution of the amino acid is leucine or asparagine
- the amino acid after substitution of the amino acid corresponding to the 239th methionine is leucine
- the amino acid after substitution of the amino acid corresponding to the 286th aspartic acid is phenylalanine
- the amino acid after substitution of the amino acid corresponding to the 287th histidine is glutamine or pro
- the NADH production method according to the present invention is a method in which the above-described mutant formate dehydrogenase according to the present invention is allowed to act on a reaction system containing formic acid and NAD + .
- the mutant formate dehydrogenase according to the present invention has a characteristic that durability is significantly improved as compared with that before mutation for having a novel substitution mutation.
- NADH known as a very expensive substance can be produced with excellent productivity.
- industrial production of NADH becomes possible.
- FIG. 3 is a schematic diagram showing a process for constructing GzFDH / pET23b (+).
- Formate dehydrogenase The mutant formate dehydrogenase according to the present invention has an amino acid sequence in which a specific amino acid residue in the formate dehydrogenase is substituted.
- the formate dehydrogenase is not particularly limited as long as it has a substitution target amino acid residue, which will be described in detail later, and has formate dehydrogenase activity. That is, formate dehydrogenase is an enzyme classified as EC 1.2.1.2 in the enzyme classification, and has an activity of catalyzing the reaction of oxidizing formate ions to carbon dioxide and reducing NAD + ions to NADH.
- the formate dehydrogenase may be a plant-derived enzyme, an animal-derived enzyme, or a microorganism-derived enzyme.
- Examples of the formate dehydrogenase derived from microorganisms include various formate dehydrogenases disclosed in a review on formate dehydrogenase (Biomolecular Engineering 23 (2006) 98-110). More specifically, formate dehydrogenase derived from Staphylococcus aureus (Baba, T. et al. Lancet 359, 1819-1827, 2002), Mycobacterium avium subsp. Paratuberculosis str.k10 (Li et al. Proc) Natl. Acad. Sci.
- the present invention can also be applied to a formate dehydrogenase derived from Paracoccus sp. 12-A.
- the present invention to the formate dehydrogenase derived from Gibberella zeae filed earlier (Japanese Patent Application No. 2008-1000044).
- the formate dehydrogenase derived from Gibberella zeae exhibits a remarkably superior specific activity as compared with the above-mentioned conventionally known formate dehydrogenase.
- the base sequence of the gene encoding formate dehydrogenase derived from Gibberella zeae is shown in SEQ ID NO: 1
- the amino acid sequence of the formate dehydrogenase derived from Gibberella zeae is shown in SEQ ID NO: 2.
- red mold stored in the American Type Culture Collection (ATCC) as ATCC numbers 10910, 20271, 20272, 20274, 24689, 28106 or 48063 can be used.
- ATCC American Type Culture Collection
- the registered name may be stored as Fusarium graminearum, but when Gibberella zeae is registered as an alias, these can also be used.
- Gibberella zeae indicates the complete generation (Teleomorph) of Fusarium graminearum.
- NBRC Biological Genetic Resources Department
- formate dehydrogenase may be obtained by using a red mold originally isolated from the natural world, not a strain stored in an organization such as ATCC or NBRC.
- the amino acid sequence of formate dehydrogenase derived from Pseudomonas sp. 101 is shown in SEQ ID NO: 3.
- the amino acid sequence of formate dehydrogenase derived from Mycobacterium vaccae N10. Is shown in SEQ ID NO: 4.
- the base sequence of the gene encoding formate dehydrogenase derived from Candida boidini is shown in SEQ ID NO: 5, and the amino acid sequence of formate dehydrogenase derived from Candida boidini is shown in SEQ ID NO: 6.
- the base sequence of the gene encoding formate dehydrogenase derived from Candida methylica is shown in SEQ ID NO: 7, and the amino acid sequence of formate dehydrogenase derived from Candida methylica is shown in SEQ ID NO: 8.
- the base sequence of the gene encoding formate dehydrogenase derived from Saccharomyces cerevisiae is shown in SEQ ID NO: 9, and the amino acid sequence of the formate dehydrogenase derived from Saccharomyces cerevisiae is shown in SEQ ID NO: 10.
- the base sequence of the gene encoding formate dehydrogenase derived from Paracoccus sp. 12-A is shown in SEQ ID NO: 11, and the amino acid sequence of formate dehydrogenase derived from Paracoccus sp. 12-A is shown in SEQ ID NO: 12.
- the formate dehydrogenase that can be used in the present invention is not limited to the amino acid sequence shown in SEQ ID NO: 2, 3, 4, 6, 8, 10 or 12, and for example, SEQ ID NO: 2, 3, 4, 6, 8, 10, or 1 or more amino acids are deleted except replacement amino acid residues described in detail below in the amino acid sequence shown in 12, substituted, added or inserted in the amino acid sequence, formic acid and NAD + It may have catalytic activity in a reaction using carbon dioxide and NADH as a substrate as a product.
- the plurality of amino acids for example, 1 to 30, preferably 1 to 20, more preferably 1 to 10, still more preferably 1 to 5, particularly preferably 1 to 3 means.
- amino acid deletion, substitution, or addition can be performed by modifying the base sequence encoding the gene by a technique known in the art.
- Mutation can be introduced into a base sequence by a known method such as Kunkel method or Gapped duplex method or a method according thereto, for example, a mutation introduction kit (for example, Mutant-) using site-directed mutagenesis. Mutations are introduced using K, Mutant-G (both trade names, manufactured by TAKARA) or the like, or LA PCR in vitro Mutagenesis series kit (trade name, manufactured by TAKARA).
- the homology value means a value obtained by default setting using a computer program in which the blast algorithm is implemented and a database storing gene sequence information.
- a polynucleotide that hybridizes under stringent conditions to a polynucleotide complementary to part or all of the base sequence shown in SEQ ID NO: 1, 5, 7, 9 or 11 is used.
- An encoded protein that has catalytic activity in a reaction using formic acid and NAD + as substrates and carbon dioxide and NADH as products can be used as formate dehydrogenase.
- hybridizing under stringent conditions means maintaining binding at 60 ° C. under 2 ⁇ SSC washing conditions.
- Hybridization can be performed by a conventionally known method such as the method described in J. Sambrook et al. Molecular Cloning, A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory (1989).
- the mutant formate dehydrogenase according to the present invention has a plurality of predetermined amino acid residues in the formate dehydrogenase described above, and the durability is significantly improved compared to the formate dehydrogenase before amino acid substitution.
- the amino acid residue to be substituted can be specified as a numerical value counted from the N-terminus with reference to formate dehydrogenase derived from Gibberella zeae having the amino acid sequence shown in SEQ ID NO: 2.
- the substitution target amino acid residue specified by specific numerical values based on the amino acid sequence shown in SEQ ID NO: 2 will be expressed by different numerical values depending on the type of formate dehydrogenase.
- the amino acid residue corresponding to the predetermined amino acid residue in the amino acid sequence shown in SEQ ID NO: 2 is a multiple alignment for a plurality of amino acid sequences including the amino acid sequence shown in SEQ ID NO: 2. It can be specified by performing analysis. Multiple alignment analysis is not particularly limited, but CLUSTAL W (1.83) multiple sequence alignment program (available at DDBJ of the National Institute of Genetics (http://clustalw.ddbj.nig.ac.jp/top-j.html) ) Can be easily implemented by those skilled in the art.
- amino acid sequence shown in SEQ ID NO: 2 is aligned with another different amino acid sequence, and the amino acid residue corresponding to the predetermined amino acid residue in the amino acid sequence shown in SEQ ID NO: 2 It can also be specified in other different amino acid sequences.
- SEQ ID NO: 4 derived from Candida boidini Formate dehydrogenase (SEQ ID NO: 6), formate dehydrogenase derived from Candida methylica (SEQ ID NO: 8), formate dehydrogenase derived from Saccharomyces cerevisiae (SEQ ID NO: 10), and formate dehydrogenase derived from Paracoccus sp.12-A
- SEQ ID NO: 12 The result of the multiple alignment analysis for (SEQ ID NO: 12) is shown in FIG.
- these specific formate dehydrogenases other than formate dehydrogenase can be similarly subjected to multiple alignment analysis, and can be compared with a formate dehydrogenase derived from Gibberella zeae (SEQ ID NO: 2) and the like. .
- the amino acid to be substituted is expressed based on the amino acid sequence shown in SEQ ID NO: 2, ie, formate dehydrogenase derived from Gibberella zeze.
- the numerical value representing the amino acid position varies depending on the formate dehydrogenase.
- the mutant formate dehydrogenase according to the present invention is one in four mutant formate dehydrogenases having substitution mutations at four amino acid residues described below, and one for the four mutant formate dehydrogenase. Mutant formate dehydrogenase further having up to 3 substitution mutations is included.
- Quadruple mutant formate dehydrogenase The quadruple mutant formate dehydrogenase according to the present invention has any of the amino acid sequences shown in A) to C) below.
- other amino acids are not particularly limited as long as the durability of formate dehydrogenase after mutation is significantly improved compared to the durability of formate dehydrogenase before mutation. Any amino acid may be used.
- the durability can be evaluated by, for example, the residual activity after being left at 37 ° C. for 500 hours.
- the durability may be evaluated under the accelerated test of the test that is allowed to stand at 37 ° C. for 500 hours, for example, the condition that the test is left at 52 to 58 ° C. for 30 to 50 minutes.
- the improvement in durability means that the residual activity after mutation introduction is statistically significantly higher than the residual activity of wild-type formate dehydrogenase before mutation introduction.
- a conventionally known method can be appropriately used as the enzyme activity of the mutant formate dehydrogenase and the formate dehydrogenase before substitution mutation. For example, by measuring directly or indirectly the amount of NADH produced according to the following reaction formula and the amount of components consumed according to the following reaction formula, the enzyme activity of formate dehydrogenase contributing to the following reaction formula is measured. be able to.
- the 73rd glycine is substituted with alanine
- the 99th valine is substituted with isoleucine
- the 153rd valine is substituted with isoleucine
- the 155th histidine is substituted.
- the 159th glutamine is substituted with leucine
- the 298th asparagine is preferably substituted with tyrosine.
- reference (2) Henikoff S., Henikoff JG, Amino-acid substitution matrices from protein blocks, Proc. Natl. Acad.Sci. USA, USA89, 10915-10919 (1992), Fig. A score matrix (BLOSUM) for substitution mutations of amino acid residues was proposed and widely used in .2.
- amino acid substitution between similar side chain chemical properties is based on the knowledge that structural and functional changes given to the whole protein are reduced.
- the side chain groups of amino acids considered in multiple alignment can be considered based on indicators such as chemical properties and physical size. This is shown in the score matrix (BLOSUM) disclosed in reference (2) as an amino acid group with a score of 0 or more, preferably a group of amino acids with a value of 1 or more.
- amino acids with similar properties can be classified into the following 8 groups. Therefore, the amino acid after substitution is preferably an amino acid classified into the group including the amino acids exemplified above.
- the 99th valine in the formate dehydrogenase derived from Gibberella zeae is preferably substituted with isoleucine.
- isoleucine is classified.
- Leucine and methionine classified into the aliphatic hydrophobic amino acid group Alternatively, valine may be substituted.
- the 155th histidine in the formate dehydrogenase derived from Gibberella zeae is preferably substituted with glutamine or lysine. However, glutamine and lysine are classified later.
- Methylene group polar group It may be substituted with glutamic acid or arginine.
- glutamine at position 159 in formate dehydrogenase derived from Gibberella zeae is preferably substituted with leucine, leucine is classified as described below. 1) Isoleucine and methionine classified into aliphatic hydrophobic amino acid groups Alternatively, valine may be substituted.
- the 298th asparagine in the formate dehydrogenase derived from Gibberella zeae is preferably substituted with tyrosine, it will be described later 7) Phenylalanine or tryptophan classified into the aromatic group, or 8) It may be substituted with histidine classified into a cyclic & polar group.
- Aliphatic hydrophobic amino acid group This group is a group of amino acids having a hydrophobic side chain of a fat attribute among the neutral nonpolar amino acids shown in the above reference (1), V (Val, valine), L (Leu, leucine) , I (Ile, isoleucine) and M (Met, methionine).
- V Val, valine
- L Leu, leucine
- I Ile, isoleucine
- M Metal, methionine
- FGACWP is not included in this “aliphatic hydrophobic amino acid group” for the following reasons. This is because G (Gly, glycine) and A (Ala, alanine) are less than a methyl group and have a weak nonpolar effect.
- C Cys, cysteine
- F Phenylalanine
- W Trp, tryptophan
- P Pro, proline
- ST group Group with hydroxymethylene group
- S Ser, serine
- T Thr, threonine
- Acidic amino acids This group is a group of amino acids having an acidic carboxyl group in the side chain, and is composed of D (Asp, aspartic acid) and E (Glu, glutamic acid).
- KR group This group is a group of basic amino acids and is composed of K (Lys, lysine) and R (Arg, arginine). These K and R are positively charged and have basic properties over a wide pH range. On the other hand, H (His, histidine) classified as a basic amino acid is not classified into this group because it is hardly ionized at pH 7.
- Methylene group polar group (DHN group) This group has a feature that a methylene group is bonded as a side chain to a carbon element at the ⁇ -position and has a polar group at the tip.
- Dimethylene group polar group (EKQR group) This group has a feature that a linear hydrocarbon having a dimethylene group or higher as a side chain is bonded to the ⁇ -position carbon element and has a polar group at the end.
- E Glu, glutamic acid, polar group is carboxyl group
- K Lis, lysine, polar group is amino group
- Q Gln, glutamine, polar group is amide group
- R Arg, arginine, polar group is imino group
- Aromatic (FYW Group) This group is an aromatic amino acid with a benzene nucleus in the side chain and is characterized by aromatic chemical properties. It consists of F (Phe, phenylalanine), Y (Tyr, tyrosine), W (Trp, tryptophan).
- Circular & polar (HY group) This group is an amino acid that has a cyclic structure in the side chain and also a polarity, H (H, histidine, both cyclic structure and polar group are imidazole groups), Y (Tyr, tyrosine, cyclic structure is polar with benzene nucleus The group consists of hydroxyl).
- Arginine 60th glutamic acid, 64th isoleucine, 73rd glycine, 82th lysine, 136th isoleucine, 155th histidine, 159th glutamine, 239th methionine, 286th aspartic acid, 287th Histidine, 293th glutamic acid, 298th asparagine and 343th asparagine amino acid.
- amino acid residues listed here are not particularly limited as long as the durability of the formate dehydrogenase after mutation is significantly improved as compared with the durability of wild-type formate dehydrogenase.
- An amino acid may be substituted.
- the 31st isoleucine is mutated to leucine
- the 50th arginine is mutated to glycine
- the 60th glutamic acid is mutated to valine
- the 64th isoleucine is mutated to valine
- 82nd lysine is substituted with arginine
- 136th isoleucine is substituted with valine
- 155th histidine is substituted with glutamine or lysine
- 159th glutamine is leucine or asparagine.
- the 239th methionine is replaced with leucine
- the 286th aspartic acid is replaced with phenylalanine
- the 287th histidine is replaced with glutamine or proline
- the 293rd glutamic acid is replaced with aspartic acid.
- the 298th a Aspartic replaces mutated to tyrosine
- 343 asparagine is preferably substituted mutated to arginine.
- the kind of preferable amino acid after substitution mutation was disclosed about the amino acid residue enumerated here, similarly to the above, the kind of amino acid after substitution mutation is classified into the eight groups classified into amino acids having similar properties. Based on this, it can be selected as appropriate.
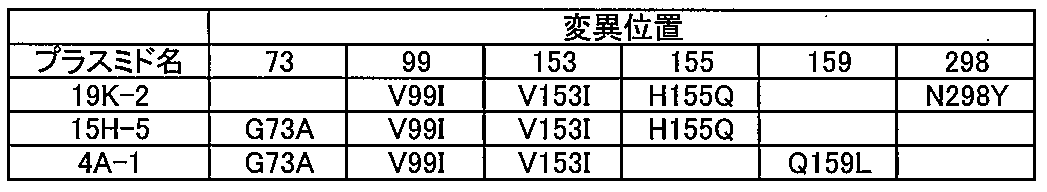
- the substitution mutation of glycine at position 73 to alanine (abbreviated as G73A using the single letter code of amino acid, the same shall apply hereinafter), V99I, V153I, H155Q and N298Y Mutated formate dehydrogenase having These five mutant formate dehydrogenases with G73A, V99I, V153I, H155Q, and N298Y have the same specific activity as wild-type formate dehydrogenase, but have excellent durability as defined above. Can show gender.
- examples of the quintuple mutant formate dehydrogenase include mutant formate dehydrogenases having G73A, V99I, V153I, H155Q, and H287Q. These five mutant formate dehydrogenases having G73A, V99I, V153I, H155Q and H287Q can also exhibit excellent durability.
- examples of the quintuple mutant formate dehydrogenase include mutant formate dehydrogenases having G73A, V99I, V153I, H155K and N298Y. These five mutant formate dehydrogenases having G73A, V99I, V153I, H155K and N298Y can also exhibit excellent durability.
- the quintuple mutant formate dehydrogenase having G73A, V99I, V153I, H155K and N298Y and the quintuple mutant formate dehydrogenase having G73A, V99I, V153I, H155Q and N298Y shows superior durability. Therefore, it can be understood that the amino acid after substitution mutation of the 155th histidine is preferably lysine rather than glutamine.
- mutant formate dehydrogenase having G73A, V99I, V153I, Q159L and N298Y mutant formate dehydrogenase having V99I, V153I, H155Q, H287Q and N298Y are used.
- These quintuple mutant formate dehydrogenases also have superior durability compared to the wild-type formate dehydrogenase equivalent to the quintuple mutant formate dehydrogenases having G73A, V99I, V153I, H155Q and N298Y. Can show.
- mutant formate dehydrogenase having E60V, G73A, V99I, V153I and H155Q mutant formate dehydrogenase having E60V, V99I, V153I, H155Q and N298Y are used. Can be mentioned. These quintuple mutant formate dehydrogenases can also exhibit superior durability compared to wild-type formate dehydrogenase.
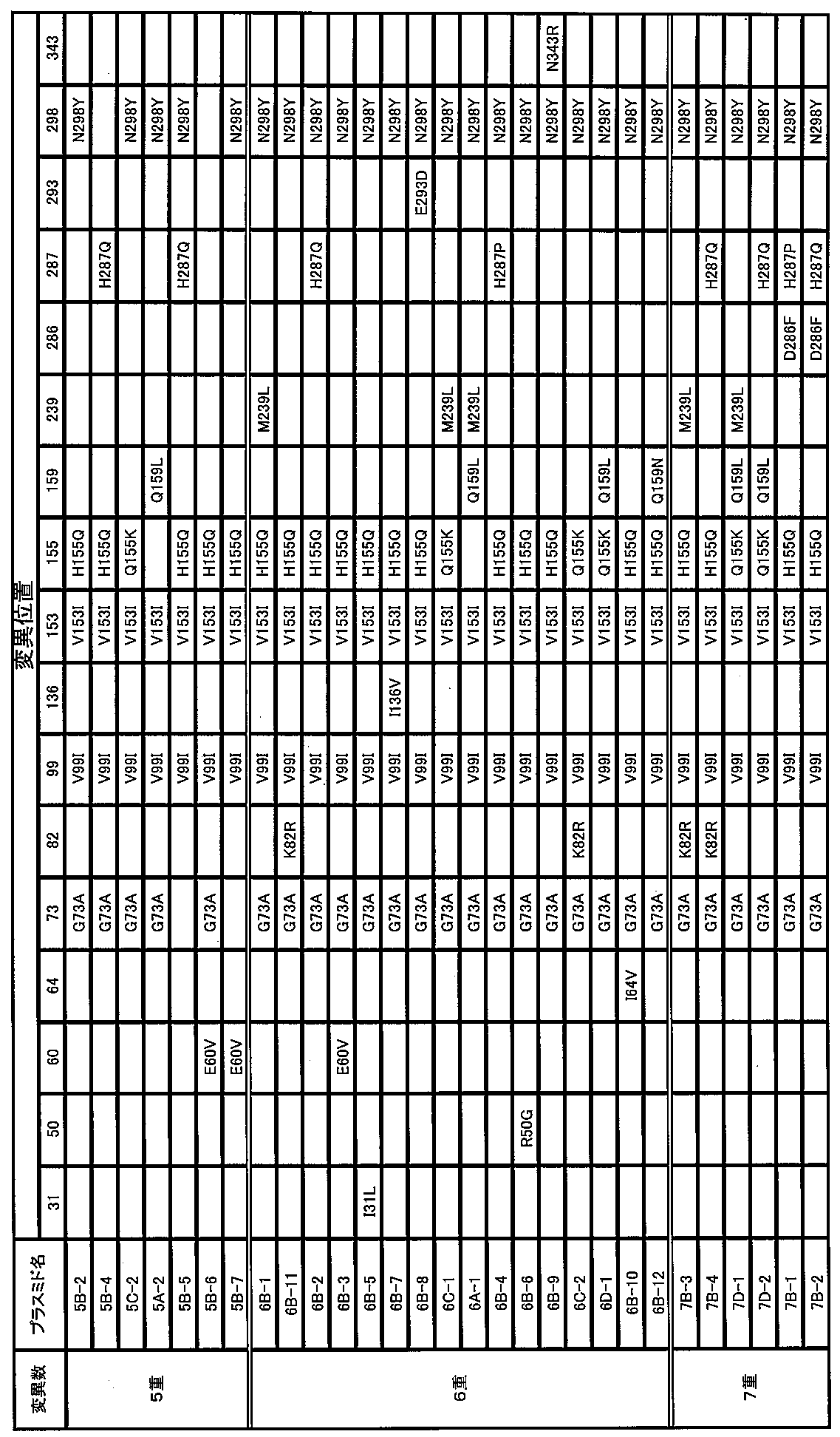
- 6-fold mutant formate dehydrogenase M239L, K82R, H287Q, E60V, I31L, I136V and E293D are also used for the 5-fold mutant formate dehydrogenase having G73A, V99I, V153I, H155Q and N298Y. It is preferable that one substitution mutation selected from is introduced.
- These 6-fold mutant formate dehydrogenases can exhibit superior durability compared to the 5-fold mutant formate dehydrogenases having G73A, V99I, V153I, H155Q and N298Y.
- a sixfold mutant formate dehydrogenase having G73A, V99I, V153I, H155Q, M239L and N298Y a sixfold mutant formate dehydrogenase having E60V, G73A, V99I, V153I, H155Q and N298Y, and G73A, V99I
- the hexamutated formate dehydrogenase having V153I, H155K, M239L, and N298Y not only has excellent durability as described above, but also exhibits excellent specific activity as compared with wild-type formate dehydrogenase. it can.
- examples of the 6-fold mutant formate dehydrogenase include those obtained by introducing M239L into the above-described 5-fold mutant formate dehydrogenase having G73A, V99I, V153I, Q159L, and N298Y.
- M239L the specific activity can be improved while maintaining the same durability as that of the quintuple mutant formate dehydrogenase having G73A, V99I, V153I, Q159L and N298Y described above.
- examples of the 6-fold mutant formate dehydrogenase include those in which H287P is introduced into a modified quinary formate dehydrogenase having G73A, V99I, V153I, H155Q and N298Y.
- the specific activity decreased compared to the 6-fold mutant formate dehydrogenase in which H287Q was introduced to the 5 mutant formate dehydrogenase having G73A, V99I, V153I, H155Q and N298Y. It is recognized that Therefore, it can be understood that glutamine is preferable to proline as the amino acid after substitution mutation of 287th histidine.
- examples of the 6-fold mutant formate dehydrogenase include those obtained by introducing R50G or N343R into the 5-fold mutant formate dehydrogenase having G73A, V99I, V153I, H155Q and N298Y. These six-fold mutant formate dehydrogenase can also exhibit the same durability as the five-fold mutant formate dehydrogenase having G73A, V99I, V153I, H155Q, and N298Y.
- examples of the 6-fold mutant formate dehydrogenase include those obtained by introducing K82R or Q159L into the 5-fold mutant formate dehydrogenase having G73A, V99I, V153I, H155K, and N298Y. These six-fold mutant formate dehydrogenase can exhibit superior durability compared to the five-fold mutant formate dehydrogenase having G73A, V99I, V153I, H155K, and N298Y.
- mutant formate dehydrogenase having I64V, G73A, V99I, V153I, H155Q and N298Y, and mutant formate having G73A, V99I, V153I, H155Q, Q159N and N298Y Mention may be made of dehydrogenases. These quintuple mutant formate dehydrogenases can also exhibit superior durability compared to wild-type formate dehydrogenase.
- examples of the 7-fold mutant formate dehydrogenase include those obtained by introducing K82R into the 6-fold mutant formate dehydrogenase having G73A, V99I, V153I, H155Q, M239L and N298Y described above. .
- This 7-fold mutant formate dehydrogenase exhibits superior durability and superior ratio compared to the 6-fold mutant formate dehydrogenase having G73A, V99I, V153I, H155Q, M239L and N298Y described above. Activity can be demonstrated.
- the seven-fold mutant formate dehydrogenase includes G73A, K82R, V99I, V153I, H155Q, H287Q and N298Y, G73A, V99I, V153I, H155Q, Q159L, M239L and N298Y, G73A, V99I , V153I, H155Q, Q159L, H287Q and N298Y.
- These seven-mutated formate dehydrogenases can also exhibit superior durability compared to wild-type formate dehydrogenase.
- examples of the seven-fold mutant formate dehydrogenase include those having G73A, V99I, V153I, H155Q, D286F, H287P and N298Y, and those having G73A, V99I, V153I, H155Q, D286F, H287P and N298Y. it can. These seven-fold formate dehydrogenases can also exhibit superior durability compared to wild-type formate dehydrogenase.
- mutant formate dehydrogenase derived from red mold formate dehydrogenase is very Show high specific activity. Therefore, mutant formate dehydrogenase derived from red mold formate dehydrogenase has a higher durability and / or specific activity than mutant formate dehydrogenase derived from formate dehydrogenase derived from other organisms. Because it is expensive, it is preferable.
- red mold-derived formate dehydrogenase cannot be obtained by a conventionally known protein production method. Therefore, it is difficult to obtain the mutant mold formate dehydrogenase derived from red mold by a conventionally known protein production method, and can be produced according to the method described below. That is, the method for producing red mold formate dehydrogenase prepares a host into which a vector in which a red mold derived formate dehydrogenase gene is placed under the control of an inducible promoter is introduced. Then, the host is cultured, and the expression of formate dehydrogenase gene is induced when the logarithmic growth phase has passed. Next, formate dehydrogenase is expressed in the host by culturing at a temperature lower than the optimum growth temperature of the host and at which the host can survive.
- the inducible promoter is not particularly limited, and a conventionally known one can be used.
- an inducible promoter exhibiting transcriptional activity in the presence of isopropyl- ⁇ -thiogalactopyranoside (IPTG) can be used.
- IPTG isopropyl- ⁇ -thiogalactopyranoside
- promoters include Trp promoter, Lac promoter, Trc promoter, and Tac promoter.
- other promoters that exhibit transcriptional activity in the presence of inducers other than IPTG and other promoters that exhibit transcriptional activity according to culture conditions such as medium components and temperature can also be used as inducible promoters.
- the vector in the method for producing formic acid dehydrogenase derived from red mold, is not particularly limited as long as it can replicate in the above-mentioned host, and any vector can be used.
- the vector when E. coli is used as the host, the vector may be a plasmid vector or a phage vector. Specific examples of the vector include pCDF series, pRSF series, and pET series.
- the host is not particularly limited as long as it can be transcribed from a promoter incorporated in an expression vector.
- E. coli BL21 (DE3) may be used. it can.
- transformation methods As a technique for introducing the above-described vector into a host, various techniques generally known as transformation methods can be applied. As a specific method, for example, a calcium phosphate method, an electroporation method, a lipofection method, or the like can be applied.
- the vector-introduced host is cultured, and the expression of the formate dehydrogenase gene is induced when the logarithmic growth phase has passed.
- the culture conditions of the host are not limited in any way, and may be appropriately set in consideration of, for example, the optimum growth temperature and optimum growth pH of the host.
- the growth of the host is observed while continuing the culture, and the culture conditions are changed to satisfy the following conditions when the so-called logarithmic growth phase has passed. That is, as the condition 1, the expression of the formate dehydrogenase gene is induced, and as the condition 2, the culture is performed at a temperature lower than the optimum growth temperature of the host and at which the host can survive.
- the culture curve can be prepared by measuring OD600 nm in the medium.
- the stationary phase is entered.
- the stationary period is a period in which the slope of the tangent of the above-described growth curve is substantially zero.
- the optimum growth temperature of the host is known as a temperature range different for each host.
- the optimum growth temperature is 37 ° C.
- the temperature at which growth is possible is 15 to 37 ° C. Therefore, when Escherichia coli B strain is used as a host, it means 15 to 37 ° C. as a temperature lower than the optimum growth temperature of the host and at which the host can survive.
- Escherichia coli B strain is used as a host, it is preferable to continue the culture at about 20 ° C. after the logarithmic growth phase.
- the formate dehydrogenase gene is expressed by setting the temperature within the above range, and a formate dehydrogenase having a very high specific activity is produced in the host.
- the cells or cells are disrupted to prepare a crude enzyme suspension.
- This crude enzyme suspension contains formate dehydrogenase having a very high specific activity. Therefore, the obtained crude enzyme suspension can be used as it is. Formate dehydrogenase can also be isolated and purified from the resulting crude enzyme suspension.
- the isolated and purified formate dehydrogenase can be used in a state suspended in a buffer solution having a predetermined pH.
- NADH regeneration systems can be used as a form of utilization of mutant formate dehydrogenase.
- NADH is used in various enzyme reactions and converted to NAD + .
- NADH is used, for example, as a coenzyme when biologically synthesizing optical isomers in the chemical and pharmaceutical industries.
- the NADH regeneration system means that NAD + remaining in the reaction system is reduced to NADH, and NADH is recovered and used again for the enzyme reaction.
- NADH can be efficiently produced from NAD + contained in the reaction system.
- mutated formate dehydrogenase exhibits extremely superior durability compared to conventionally known formate dehydrogenase, and therefore has a higher activity over a longer period of time than formate dehydrogenase before mutation introduction. Can be maintained.
- the productivity of NADH can be significantly improved as compared with the case where a conventionally known formate dehydrogenase is used.
- Example 1 In this example, a red mold-derived formate dehydrogenase gene was cloned, a random mutation was introduced into the gene, and durability and specific activity of the formate dehydrogenase derived from the obtained mutant gene were evaluated.
- KPB pH 7.5 -Potassium phosphate buffer (KPB) pH 7.5 ⁇ Solution A> 0.5M KH 2 PO 4 13.6g / 200ml ⁇ Solution B> 0.5M K 2 HPO 4 26.13g / 300ml 0.5M KPB pH7.5 was prepared by mixing SolutionA and SolutionB in the following ratio.
- LB medium LB Broth 20g / L. Used by autoclaving. Before use, ampicillin (manufactured by SIGMA) was added to 50 ⁇ g / ml.
- a cDNA synthesis reaction was performed in a reaction cycle of 50 ° C. for 2 hours, then 99 ° C. for 5 minutes, and then 4 ° C.
- PCR was performed using the synthesized cDNA as a template and Pyrobest® DNA polymerase.
- the component composition in 50 ⁇ l of the reaction solution is shown in Table 2.
- Sterile water was added to adjust the volume to 50 ⁇ l.
- Sterile water was added to make a total volume of 20 ⁇ l.
- WT described in the column of the template in Table 4 means wild type FDH, for example, V99I means a mutant FDH in which 99th valine in FDH is substituted with isoleucine.
- a single mutant formate dehydrogenase gene in which a V99I mutation was introduced into wild type FDH was first prepared, and a V153I / H155Q mutation or a V153I / Q159L mutation was introduced into this to form a triple mutant formate.
- a dehydrogenase gene was prepared, and a quadruple mutant formate dehydrogenase was prepared by introducing N298Y mutation, G73A mutation, H287Q mutation or E60V mutation into the triple mutant formate dehydrogenase gene.
- reaction solution having the above composition 94 cycles at 94 ° C. for 2 minutes, followed by 30 cycles of 94 ° C. for 15 seconds and 68 ° C. for 1 minute 30 seconds, then 68 ° C. for 2 minutes, then 4 ° C. PCR was performed in a reaction cycle.
- a fragment of about 1.4 kbp amplified by PCR was purified by MinElute PCR-Purification Kit and used for translation reaction.
- the formic acid decomposition reaction by the FDH catalyst is represented by the following formula.
- the yellow formazan can be quantified by measuring the absorbance at 430 nm with a plate reader (Spectrafluor Plus: manufactured by TECAN), and the activity of FDH having a plurality of amino acid mutations adjusted in 4-1-2 can be evaluated.
- the culture solution was once put on ice and cooled, and then collected.
- the culture was divided into two 500 ml tubes and centrifuged at 5200 rpm for 10 minutes at 4 ° C. Discard the supernatant, add 60 ml of 10 mM KPB (pH 7.5) to each cell and suspend them, and suspend 120 ml of the suspension into three 50 ml multi-bead shocker dedicated crushing tubes (manufactured by Yasui Instruments Co., Ltd.). Separately, centrifugation was performed again at 4 ° C. for 5 minutes at 5600 rpm to obtain bacterial cells.
- (4-3-2) Disruption of expression-inducing cells
- the cells obtained in (4-3-1) were disrupted while cooling to prepare a crude extract.
- Wet cell weight after collection is measured, 10 mM KPB (pH 7.5) 10 times the amount (a ratio of 10 ml to 1 g of wet cell weight), 0.1 mm glass beads for multi beads shocker (manufactured by Yasui Instruments Co., Ltd.) ) was added in the same amount as KPB (a ratio of 10 g to 10 ml of KPB).
- a multi-bead shocker manufactured by Yasui Instruments Co., Ltd. was used for crushing.
- the crude extract was adjusted according to the procedure. First, the process of shaking at 6000 rpm for 120 seconds, and after shaking for 60 seconds at the same condition was repeated three times. Thereafter, the mixture was centrifuged at 6000 rpm for 20 minutes at 4 ° C., and the supernatant was collected. Thereafter, the mixture was centrifuged at 6000 rpm for 10 minutes at 4 ° C., and the supernatant was filtered through a 0.45 ⁇ m filter bottle (Millipore) to prepare a crude extract.
- the fraction in which the peak of FDH activity was observed was used as a sample for secondary purification using a hydroxyapatite column.
- equilibration was performed by supplying 30 ml of 1 mM KPB (pH 7.2) to the column at a flow rate of 1 ml / min.
- the desalted sample was supplied to the column at a flow rate of 0.5 ml / min (circulated twice) to bind protein components.
- 10 ml of 1 mM KPB (pH 7.2) was supplied to the column at a flow rate of 1 ml / min for washing.
- 10 ml each of 1 mM to 10 mM KPB (pH 6.8) was supplied to the column at a flow rate of 0.5 ml / min for elution.
- fractionation total 49 fractions was performed so that one fraction was 0.5 ml.
- 5 ml of 500 mM KPB (pH 6.8) was supplied to the column at a flow rate of 1 ml / min for washing.
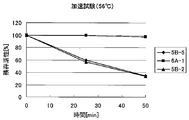
- FIG. 3 shows the results of the durability test performed in (4-4).
- the five-fold formate dehydrogenase (G73A, V99I, V153I, H155Q and N298Y) shown in plasmid name 5B-2 is almost completely inactivated by wild-type formate dehydrogenase. It was revealed that 80% or more of the activity can be maintained even after a long period of time.
- the 5-mutated formate dehydrogenase shown in the plasmid name 5B-2 can be said to be an enzyme having extremely excellent durability.
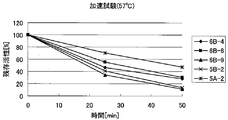
- FIG. 4 shows the results of the accelerated durability test conducted in (4-2) for the three types of quadruple mutant formate dehydrogenase obtained in (3-1). Moreover, the result of having measured the specific activity about these three types of 4 type
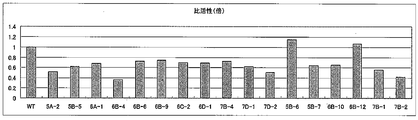
- FIGS. 15 and 16 show relative values when the specific activity of wild-type formate dehydrogenase is 1.
- FIG. 15 collectively shows mutant formate dehydrogenases that exhibit specific activity equivalent to that of wild-type formate dehydrogenase.
- FIG. 16 shows a mutant formate dehydrogenase that exhibits specific activity that is slightly inferior to that of wild-type formate dehydrogenase.
- mutant formate dehydrogenases shown in 6C-1 and 7B-3 were found to exhibit excellent durability while exhibiting the same specific activity as the wild-type formate dehydrogenase.
- mutant formate dehydrogenases shown in the plasmid names 5B-6, 5B-7, 6B-10, 6B-12, 7B-1 and 7B-2 have the mutations described above. Although it was inferior in specific activity and / or durability as compared with the type formate dehydrogenase, it was revealed that it showed superior durability compared with the wild type formate dehydrogenase.
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Medicinal Chemistry (AREA)
- Biomedical Technology (AREA)
- Enzymes And Modification Thereof (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
従来の知見からは予測できないほどに高い比活性を示すギ酸脱水素酵素を利用し、当該ギ酸脱水素酵素の耐久性を向上させる。アカカビ由来のギ酸脱水素酵素に対して特定のアミノ酸置換を導入する。変異型ギ酸脱水素酵素は、野生型のギ酸脱水素酵素と比較して極めて優れた耐久性を示すため、この変異型ギ酸脱水素酵素を利用して製造するNADHの生産性を向上させることができる。
Description
本発明は、野性型ギ酸脱水素酵素における特定の部位に置換型変異を有する変異型ギ酸脱水素酵素、当該変異型ギ酸脱水素酵素をコードする遺伝子及びNADHの製造方法に関する。
ギ酸脱水素酵素(EC.1.2.1.2)は、酸化型ニコチンアミドアデニンジヌクレオチド(NAD+)、ギ酸及び水の存在下で、NAD+を還元型ニコチンアミドアデニンジヌクレオチド(NADH)に還元するとともにギ酸を二酸化炭素に酸化する。この酵素反応に基づいてギ酸脱水素酵素は、NAD+からNADHを再生する系に利用されている。従来、ギ酸脱水素酵素としては、例えば特開2003-180383号公報に記載されるようなカンジダ ボイジニイ(Candida boidinii)(ATCC32195)由来のギ酸脱水素酵素、特開2002-233395号公報に開示されるようなバチルス属細菌由来のNAD+依存性ギ酸脱水素酵素、特願平10-023896号公報に開示されるようなマイコバクテリウム・バッカエ(Mycobacterium vaccae)由来のギ酸脱水素酵素が知られていた。
また、BIOCHEMISTRY (Moscow), Vol. 69, No. 11, 2004, pp. 1252-1267.(Biokhimiya, Vol. 69, No. 11, 2004, pp. 1537-1554.の英訳)には、これら以外にも種々の微生物や植物由来のギ酸脱水素酵素が開示されている。しかしながら、本文献に記述されているように、ギ酸脱水素酵素は、様々な酵素のなかで、比活性はそれほど高くない。換言すると、ギ酸脱水素酵素によるNADHへの還元反応を利用したNADHの生産方法は、ギ酸脱水素酵素の比活性の低さに起因して生産性が悪いといえる。
ところで、ギ酸脱水素酵素に関しては、現在までに種々の研究結果が蓄積され、部位特異的変異導入による機能改変に関する報告がなされている(Biomolecular Engineering, 23, (2006) 98-110)。しかしながら、従来公知のギ酸脱水素酵素は、いずれも比活性や耐久性が低く、ギ酸脱水素酵素を利用したNADHの製造には不十分であると評価せざるを得ない。
上述した実情に鑑み、本発明者らは先に、従来の知見からは予測できないほどに高い比活性を示すギ酸脱水素酵素、当該ギ酸脱水素酵素を利用したNADHの製造方法に関する特許出願を行った(特願2008-100448号)。そして、本発明は、当該特許出願に係るギ酸脱水素酵素及び従来公知のギ酸脱水素酵素について有用な置換型変異を探索し、従来公知の変異型ギ酸脱水素酵素と比較して、より耐久性に優れた変異型ギ酸脱水素酵素、当該変異型ギ酸脱水素酵素をコードする遺伝子及び当該変異型ギ酸脱水素酵素を利用したNADHの製造方法を提供することを目的とする。
上述した目的を達成するため本発明者らが鋭意検討した結果、上記特許出願に係るギ酸脱水素酵素及び従来公知のギ酸脱水素酵素における特定の部位におけるアミノ酸置換変異が、ギ酸脱水素酵素の耐久性を大幅に向上させることを見いだし本発明を完成するに至った。
すなわち、本発明に係る変異型ギ酸脱水素酵素は、以下のA)~C)のうちいずれか1つのアミノ酸配列を含み耐久性が向上したものである。
A)配列番号2に示すアミノ酸配列からなるギ酸脱水素酵素における、99番目のバリン、153番目のバリン、155番目のヒスチジン及び298番目のアスパラギンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列
B)配列番号2に示すアミノ酸配列からなるギ酸脱水素酵素における、73番目のグリシン、99番目のバリン、153番目のバリン及び155番目のヒスチジンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列、及び
C)配列番号2に示すアミノ酸配列からなるギ酸脱水素酵素における、73番目のグリシン、99番目のバリン、153番目のバリン及び159番目のグルタミンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列
また、本発明に係る変異型ギ酸脱水素酵素は、上記A)のアミノ酸配列に対して、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、73番目のグリシン、82番目のリシン、136番目のイソロイシン、159番目のグルタミン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸及び343番目のアスパラギンからなる群から選ばれる1~3つのアミノ酸に相当するアミノ酸残基が他のアミノ酸に更に置換したアミノ酸配列を含むものであることが好ましい。
B)配列番号2に示すアミノ酸配列からなるギ酸脱水素酵素における、73番目のグリシン、99番目のバリン、153番目のバリン及び155番目のヒスチジンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列、及び
C)配列番号2に示すアミノ酸配列からなるギ酸脱水素酵素における、73番目のグリシン、99番目のバリン、153番目のバリン及び159番目のグルタミンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列
また、本発明に係る変異型ギ酸脱水素酵素は、上記A)のアミノ酸配列に対して、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、73番目のグリシン、82番目のリシン、136番目のイソロイシン、159番目のグルタミン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸及び343番目のアスパラギンからなる群から選ばれる1~3つのアミノ酸に相当するアミノ酸残基が他のアミノ酸に更に置換したアミノ酸配列を含むものであることが好ましい。
さらに、本発明に係る変異型ギ酸脱水素酵素は、上記B)のアミノ酸配列に対して、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、82番目のリシン、136番目のイソロイシン、159番目のグルタミン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸、298番目のアスパラギン及び343番目のアスパラギンからなる群から選ばれる1~3つのアミノ酸に相当するアミノ酸残基が他のアミノ酸に更に置換したアミノ酸配列を含むものであることが好ましい。
さらにまた、本発明に係る変異型ギ酸脱水素酵素は、上記C)のアミノ酸配列に対して、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、82番目のリシン、136番目のイソロイシン、155番目のヒスチジン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸、298番目のアスパラギン及び343番目のアスパラギンからなる群から選ばれる1~3つのアミノ酸に相当するアミノ酸残基が他のアミノ酸に更に置換したアミノ酸配列を含むものであることが好ましい。
ここで、置換変異後のアミノ酸は特に限定されないが、例えば、上記31番目のイソロイシンに相当するアミノ酸の置換後のアミノ酸がロイシンであり、上記50番目のアルギニンに相当するアミノ酸の置換後のアミノ酸がグリシンであり、上記60番目のグルタミン酸に相当するアミノ酸の置換後のアミノ酸がバリンであり、上記64番目のイソロイシンに相当するアミノ酸の置換後のアミノ酸がバリンであり、上記73番目のグリシンに相当するアミノ酸の置換後のアミノ酸がアラニンであり、上記82番目のリシンに相当するアミノ酸の置換後のアミノ酸がアルギニンであり、上記99番目のバリンに相当するアミノ酸の置換後のアミノ酸がイソロイシンであり、上記136番目のイソロイシンに相当するアミノ酸の置換後のアミノ酸がバリン、上記153番目のバリンに相当するアミノ酸の置換後のアミノ酸がイソロイシンであり、上記155番目のヒスチジンに相当するアミノ酸の置換後のアミノ酸がグルタミン又はリシンであり、上記159番目のグルタミンに相当するアミノ酸の置換後のアミノ酸がロイシン又はアスパラギンであり、上記239番目のメチオニンに相当するアミノ酸の置換後のアミノ酸がロイシンであり、上記286番目のアスパラギン酸に相当するアミノ酸の置換後のアミノ酸がフェニルアラニンであり、上記287番目のヒスチジンに相当するアミノ酸の置換後のアミノ酸がグルタミン又はプロリンであり、上記293番目のグルタミン酸に相当するアミノ酸の置換後のアミノ酸がアスパラギン酸であり、上記298番目のアスパラギンに相当するアミノ酸の置換後のアミノ酸がチロシンであり、上記343番目のアスパラギンに相当するアミノ酸の置換後のアミノ酸がアルギニンとすることができる。
また、本発明に係るNADHの製造方法は、ギ酸及びNAD+を含む反応系に上述した本発明に係る変異型ギ酸脱水素酵素を作用させるものである。
本発明に係る変異型ギ酸脱水素酵素は、新規な置換型変異を有するための変異前と比較して大幅に耐久性が向上するといった特徴を有している。本発明に係る変異型ギ酸脱水素酵素を利用することによって、非常に高価な物質として知られるNADHを優れた生産性で製造することができる。本発明に係る変異型ギ酸脱水素酵素を利用してNADHを製造することによって、NADHの工業的製造が可能となる。
以下、本発明を図面を参照して詳細に説明する。
ギ酸脱水素酵素
本発明に係る変異型ギ酸脱水素酵素は、ギ酸脱水素酵素における特定のアミノ酸残基を置換したアミノ酸配列を有する。ここで、ギ酸脱水素酵素としては、詳細を後述する置換対象アミノ酸残基を有し、ギ酸脱水素酵素活性を有するものであれば特に限定されない。すなわち、ギ酸脱水素酵素とは、酵素分類においてEC1.2.1.2に分類され、ギ酸イオンを二酸化炭素に酸化するとともに、NAD+イオンをNADHに還元する反応を触媒する活性を有する酵素である。
本発明に係る変異型ギ酸脱水素酵素は、ギ酸脱水素酵素における特定のアミノ酸残基を置換したアミノ酸配列を有する。ここで、ギ酸脱水素酵素としては、詳細を後述する置換対象アミノ酸残基を有し、ギ酸脱水素酵素活性を有するものであれば特に限定されない。すなわち、ギ酸脱水素酵素とは、酵素分類においてEC1.2.1.2に分類され、ギ酸イオンを二酸化炭素に酸化するとともに、NAD+イオンをNADHに還元する反応を触媒する活性を有する酵素である。
ギ酸脱水素酵素としては、植物由来の酵素であっても良いし、動物由来の酵素であっても良いし、微生物由来の酵素であっても良い。微生物由来のギ酸脱水素酵素としては、例えば、ギ酸脱水素酵素に関する総説(Biomolecular Engineering 23 (2006) 98-110)に開示されている種々のギ酸脱水素酵素を挙げることができる。より具体的に、近年のゲノム解析から明らかとなったStaphylococcus aureus由来のギ酸脱水素酵素(Baba, T.らLancet 359、1819-1827、2002)、Mycobacterium avium subsp. paratuberculosis str.k10 (LiらProc. Natl. Acad. Sci. U.S.A. 102、12344-12349、2005)、Bordetella由来のギ酸脱水素酵素(ParkhillらNat. Genet. 35, 32-40.、2003)、Legionella由来のギ酸脱水素酵素(ChienらScience 305, 1966v1968、2004及びCazaletらNat. Genet. 36, 1165-1173、2004)、Francisella tularensis subsp. tularensis SCHU S4由来のギ酸脱水素酵素(LarssonらNat. Genet. 37, 153-159、2005)、Histoplasma capsulatum由来のギ酸脱水素酵素(HwangらMol. Biol. Cell 14, 2314-2326、2003)及びCryptococcus neoformans var. neoformans JEC21 (LoftusらScience 307, 1321-1324、2005)等に対して本発明を適用することができる。
また、上記総説に開示されているPseudomonas sp.101由来のギ酸脱水素酵素、Mycobacterium vaccae N10.由来のギ酸脱水素酵素、Candida boidini由来のギ酸脱水素酵素、Candida methylica由来のギ酸脱水素酵素Saccharomyces cerevisiae由来のギ酸脱水素酵素及びParacoccus sp.12-A由来のギ酸脱水素酵素に対しても本発明を適用することができる。
特に、先に出願した(特願2008-100448号)Gibberella zeae由来のギ酸脱水素酵素に本発明を適用することが好ましい。Gibberella zeae由来のギ酸脱水素酵素は、上述した従来公知のギ酸脱水素酵素と比較して著しく優れた比活性を示すものである。Gibberella zeae由来のギ酸脱水素酵素をコードする遺伝子の塩基配列を配列番号1に示し、Gibberella zeae由来のギ酸脱水素酵素のアミノ酸配列を配列番号2に示す。ここで、ギ酸脱水素酵素を取得するためには、アカカビ(学名:Gibberella zeae)として保存されている、従来公知の種々の菌株を使用することができる。例えば、American Type Culture Collection (ATCC)にATCC番号10910、20271、20272、20274、24689、28106又は48063として保存されているアカカビを使用することができる。また、ATCCには、登録名がFusarium graminearumとして保存されている場合もあるが、別名としてGibberella zeaeが登録されている場合にはこれらも使用することができる。なお、Gibberella zeaeは、Fusarium graminearumの完全世代(テレオモルフ)のことを示している。また、独立行政法人製品評価技術基盤機構の生物遺伝資源部門(NBRC)には、NBRC番号4474、5269、6608、7160、7520、7772、8850又は9462として保存されているアカカビを使用することができる。さらに、ATCCやNBRCといった機関に保存された菌株ではなく、自然界から独自に単離したアカカビを使用してギ酸脱水素酵素を取得しても良い。
また、Pseudomonas sp.101由来のギ酸脱水素酵素のアミノ酸配列を配列番号3に示す。Mycobacterium vaccae N10.由来のギ酸脱水素酵素のアミノ酸配列を配列番号4に示す。Candida boidini由来のギ酸脱水素酵素をコードする遺伝子の塩基配列を配列番号5に示し、Candida boidini由来のギ酸脱水素酵素のアミノ酸配列を配列番号6に示す。Candida methylica由来のギ酸脱水素酵素をコードする遺伝子の塩基配列を配列番号7に示し、Candida methylica由来のギ酸脱水素酵素のアミノ酸配列を配列番号8に示す。Saccharomyces cerevisiae由来のギ酸脱水素酵素をコードする遺伝子の塩基配列を配列番号9に示し、Saccharomyces cerevisiae由来のギ酸脱水素酵素のアミノ酸配列を配列番号10に示す。Paracoccus sp.12-A由来のギ酸脱水素酵素をコードする遺伝子の塩基配列を配列番号11に示し、Paracoccus sp.12-A由来のギ酸脱水素酵素のアミノ酸配列を配列番号12に示す。
本発明において使用可能なギ酸脱水素酵素は、配列番号2、3、4、6、8、10又は12に示したアミノ酸配列からなるものに限定されず、例えば、配列番号2、3、4、6、8、10又は12に示すアミノ酸配列において詳細を後述する置換対象アミノ酸残基を除く1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するものであっても良い。ここで、複数個のアミノ酸としては、例えば、1から30個、好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1個から5個、特に好ましくは1個から3個を意味する。なお、アミノ酸の欠失、置換若しくは付加は、上記遺伝子をコードする塩基配列を、当該技術分野で公知の手法によって改変することによって行うことができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA社製)を用いて変異が導入される。
また、本発明においては、配列番号2、3、4、6、8、10又は12に示すアミノ酸配列に対して、例えば85%以上、好ましくは90%以上、より好ましくは95%以上、最も好ましくは98%以上の相同性を有するアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するギ酸脱水素酵素を使用することもできる。ここで、相同性の値は、blastアルゴリズムを実装したコンピュータプログラム及び遺伝子配列情報を格納したデータベースを用いてデフォルトの設定で求められる値を意味する。
さらに、本発明においては、配列番号1、5、7、9又は11に示す塩基配列の一部又は全部に対して相補的なポリヌクレオチドに対してストリンジェントな条件下でハイブリダイズするポリヌクレオチドにコードされたタンパク質であってギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質をギ酸脱水素酵素として使用することができる。ここで、ストリンジェントな条件下でハイブリダイズするとは、60℃で2×SSC洗浄条件下で結合を維持することを意味する。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
置換変異
本発明に係る変異型ギ酸脱水素酵素は、上述したギ酸脱水素酵素における所定のアミノ酸残基を複数置換し、アミノ酸置換前のギ酸脱水素酵素と比較して耐久性が有意に向上したものである。ここで、置換対象のアミノ酸残基は、配列番号2に示すアミノ酸配列からなるGibberella zeae由来のギ酸脱水素酵素を基準として、N末端から数えた数値として特定することができる。しかしながら、配列番号2に示すアミノ酸配列を基準として具体的数値で特定された置換対象のアミノ酸残基は、ギ酸脱水素酵素の種類によっては異なる数値で表されることとなる。したがって、『配列番号2に示すアミノ酸配列におけるX番目のアミノ酸残基』と表記した場合、配列番号2に示すアミノ酸配列とは異なるアミノ酸配列を有するギ酸脱水素酵素についてはX番目とはならず、異なる数値として表現されることとなる。
本発明に係る変異型ギ酸脱水素酵素は、上述したギ酸脱水素酵素における所定のアミノ酸残基を複数置換し、アミノ酸置換前のギ酸脱水素酵素と比較して耐久性が有意に向上したものである。ここで、置換対象のアミノ酸残基は、配列番号2に示すアミノ酸配列からなるGibberella zeae由来のギ酸脱水素酵素を基準として、N末端から数えた数値として特定することができる。しかしながら、配列番号2に示すアミノ酸配列を基準として具体的数値で特定された置換対象のアミノ酸残基は、ギ酸脱水素酵素の種類によっては異なる数値で表されることとなる。したがって、『配列番号2に示すアミノ酸配列におけるX番目のアミノ酸残基』と表記した場合、配列番号2に示すアミノ酸配列とは異なるアミノ酸配列を有するギ酸脱水素酵素についてはX番目とはならず、異なる数値として表現されることとなる。
配列番号2に示すアミノ酸配列と異なるアミノ酸配列において、配列番号2に示すアミノ酸配列における所定のアミノ酸残基に相当するアミノ酸残基は、配列番号2に示すアミノ酸配列を含む複数のアミノ酸配列についてマルチプルアラインメント解析を行うことで特定することができる。マルチプルアラインメント解析は、特に限定されないがCLUSTAL W (1.83) multiple sequence alignmentプログラム(国立遺伝学研究所のDDBJで使用できる(http://clustalw.ddbj.nig.ac.jp/top-j.html))を用いて当業者が容易に実施することができる。なお、ペアワイズアライメント解析法を用いて、配列番号2に示すアミノ酸配列に対して他の異なるアミノ酸配列をアラインメントし、配列番号2に示すアミノ酸配列における所定のアミノ酸残基に相当するアミノ酸残基を当該他の異なるアミノ酸配列において特定することもできる。
Gibberella zeae由来のギ酸脱水素酵素(配列番号2)、Pseudomonas sp.101由来のギ酸脱水素酵素(配列番号3)、Mycobacterium vaccae N10.由来のギ酸脱水素酵素(配列番号4)、Candida boidini由来のギ酸脱水素酵素(配列番号6)、Candida methylica由来のギ酸脱水素酵素(配列番号8)、Saccharomyces cerevisiae由来のギ酸脱水素酵素(配列番号10)及びParacoccus sp.12-A由来のギ酸脱水素酵素(配列番号12)についてマルチプルアラインメント解析した結果を図1に示す。なお、これら具体的なギ酸脱水素酵素以外のギ酸脱水素酵素についても、同様にマルチプルアラインメント解析に供することができ、Gibberella zeae由来のギ酸脱水素酵素(配列番号2)等と比較することができる。
以下の説明において、置換対象アミノ酸は、配列番号2に示すアミノ酸配列、すなわちGibberella zeae由来のギ酸脱水素酵素を基準として表記する。しかし、上述したように、アミノ酸の位置を表す数値は、ギ酸脱水素酵素に応じて異なる数値となる点に留意する。本発明に係る変異型ギ酸脱水素酵素は、以下に説明する4箇所のアミノ酸残基に置換変異を有する4重変異型ギ酸脱水素酵素、及び当該4重変異型ギ酸脱水素酵素に対して1~3箇所の置換変異を更に有する変異型ギ酸脱水素酵素が含まれる。
4重変異型ギ酸脱水素酵素
本発明に係る4重変異型ギ酸脱水素酵素は以下のA)~C)に示すいずれかのアミノ酸配列を有している。
本発明に係る4重変異型ギ酸脱水素酵素は以下のA)~C)に示すいずれかのアミノ酸配列を有している。
A)99番目のバリン、153番目のバリン、155番目のヒスチジン及び298番目のアスパラギンが他のアミノ酸に置換したアミノ酸配列
B)73番目のグリシン、99番目のバリン、153番目のバリン及び155番目のヒスチジンが他のアミノ酸に置換したアミノ酸配列、及び
C)73番目のグリシン、99番目のバリン、153番目のバリン及び159番目のグルタミンが他のアミノ酸に置換したアミノ酸配列
ここで、上記A)~C)のアミノ酸配列において、他のアミノ酸としては、変異後のギ酸脱水素酵素の耐久性が変異前のギ酸脱水素酵素における耐久性と比較して有意に向上するものである限り、特に限定されず、如何なるアミノ酸であっても良い。耐久性とは、例えば37℃で500時間放置した後の残存活性により評価することができる。なお、耐久性は、37℃で500時間放置する試験の加速試験、例えば52~58℃で30~50分放置する条件で評価しても良い。また、耐久性が向上するとは、変異導入前の野生型のギ酸脱水素酵素の残存活性と比較して変異導入後の残存活性が統計的に有意に高いことを意味する。ここで、変異型ギ酸脱水素酵素及び置換変異前のギ酸脱水素酵素の酵素活性は、従来公知の手法を適宜使用することができる。例えば、下記反応式に従って生成するNADH量や、下記反応式に従って消費される成分の量を直接的又は間接的に測定することで、下記反応式に寄与するギ酸脱水素酵素の酵素活性を測定することができる。
B)73番目のグリシン、99番目のバリン、153番目のバリン及び155番目のヒスチジンが他のアミノ酸に置換したアミノ酸配列、及び
C)73番目のグリシン、99番目のバリン、153番目のバリン及び159番目のグルタミンが他のアミノ酸に置換したアミノ酸配列
ここで、上記A)~C)のアミノ酸配列において、他のアミノ酸としては、変異後のギ酸脱水素酵素の耐久性が変異前のギ酸脱水素酵素における耐久性と比較して有意に向上するものである限り、特に限定されず、如何なるアミノ酸であっても良い。耐久性とは、例えば37℃で500時間放置した後の残存活性により評価することができる。なお、耐久性は、37℃で500時間放置する試験の加速試験、例えば52~58℃で30~50分放置する条件で評価しても良い。また、耐久性が向上するとは、変異導入前の野生型のギ酸脱水素酵素の残存活性と比較して変異導入後の残存活性が統計的に有意に高いことを意味する。ここで、変異型ギ酸脱水素酵素及び置換変異前のギ酸脱水素酵素の酵素活性は、従来公知の手法を適宜使用することができる。例えば、下記反応式に従って生成するNADH量や、下記反応式に従って消費される成分の量を直接的又は間接的に測定することで、下記反応式に寄与するギ酸脱水素酵素の酵素活性を測定することができる。
HCOO- + NAD+ → CO2 + NADH
特に、上記A)~C)のアミノ酸配列において、73番目のグリシンはアラニンに置換変異し、99番目のバリンはイソロイシンに置換変異し、153番目のバリンはイソロイシンに置換変異し、155番目のヒスチジンはグルタミン又はリシンに置換変異し、159番目のグルタミンはロイシンに置換変異し、298番目のアスパラギンがチロシンに置換変異していることが好ましい。
特に、上記A)~C)のアミノ酸配列において、73番目のグリシンはアラニンに置換変異し、99番目のバリンはイソロイシンに置換変異し、153番目のバリンはイソロイシンに置換変異し、155番目のヒスチジンはグルタミン又はリシンに置換変異し、159番目のグルタミンはロイシンに置換変異し、298番目のアスパラギンがチロシンに置換変異していることが好ましい。
なお、上述したように、置換後のアミノ酸として具体的に好ましいアミノ酸を例示したが、置換後のアミノ酸は上記例示に限定されるものではない。参考文献(1)(「マッキー生化学」第3版 5章アミノ酸・ペプチド・タンパク質 5.1アミノ酸、監修:市川厚、監訳:福岡伸一、発行者:曽根良介、発行所:(株)化学同人、ISBN4-7598-0944 -9)でも記載されているように、アミノ酸は同様の性質(化学的性質や物理的大きさ)を持つ側鎖に従って分類される事がよく知られる。また、タンパク質の活性を保持したまま、所定のグループに分類されるアミノ酸残基間における分子進化上の置換が頻度高く起こることがよく知られる。この考えを基に、参考文献(2): Henikoff S., Henikoff J.G., Amino-acid substitution matrices from protein blocks, Proc. Natl. Acad. Sci. USA, 89, 10915-10919 (1992)中の、Fig.2でアミノ酸残基の置換変異のスコアマトリックス(BLOSUM)が提唱され、広く使用されている。参考文献(2)では、側鎖の化学的性質が似たもの同士のアミノ酸置換は、タンパク質全体に与える構造や機能変化が少なくなると言う知見に基づくものである。上記参考文献(1)及び(2)によれば、マルチプルアラインメントで考慮するアミノ酸の側鎖のグループは、化学的性質や物理的大きさなどの指標を基にして考えることができる。これは、参考文献(2)に開示されたスコアマトリックス(BLOSUM)において、スコアの0以上の値を持つアミノ酸、好ましくは1以上の値を持つアミノ酸のグループとして示される。
以上のような知見に基づいて、性質の類似したアミノ酸を下記の8つのグループに分類することができる。したがって、置換後のアミノ酸としては、上記例示したアミノ酸が含まれるグループに分類されるアミノ酸とすることが好ましい。例えば、Gibberella zeae由来のギ酸脱水素酵素における99番目のバリンは、イソロイシンに置換することが好ましいとしたが、イソロイシンが分類されている後記1)脂肪族疎水性アミノ酸グループに分類されるロイシン、メチオニン又はバリンに置換されても良い。同様に、Gibberella zeae由来のギ酸脱水素酵素における155番目のヒスチジンは、グルタミン又はリシン置換することが好ましいとしたが、グルタミン及びリシンが分類されている後記6)メチレン基=極性基グループに分類されるグルタミン酸又はアルギニンに置換されても良い。また、Gibberella zeae由来のギ酸脱水素酵素における159番目のグルタミンは、ロイシンに置換することが好ましいとしたが、ロイシンが分類されている後記1)脂肪族疎水性アミノ酸グループに分類されるイソロイシン、メチオニン又はバリンに置換されても良い。さらに、Gibberella zeae由来のギ酸脱水素酵素における298番目のアスパラギンは、チロシンに置換することが好ましいとしたが、チロシンが分類されている後記7)芳香族グループに分類されるフェニルアラニン又はトリプトファン、若しくは後記8)環状&極性グループに分類されるヒスチジンに置換されても良い。
1)脂肪族疎水性アミノ酸グループ(ILMVグループ)
このグループは、上記参考文献(1)で示された中性非極性アミノ酸のうち、脂肪属性の疎水性側鎖をもつアミノ酸のグループであり、V(Val、バリン)、L(Leu、ロイシン)、I(Ile、イソロイシン)及びM(Met、メチオニン)から構成される。参考文献(1)による中性非極性アミノ酸と分類されるもののうちFGACWPは以下理由で、この「脂肪族疎水性アミノ酸グループ」には含めない。G(Gly、グリシン)やA(Ala、アラニン)はメチル基以下の大きさで非極性の効果が弱いからである。C(Cys、システイン)はS-S結合に重要な役目を担う場合があり、また、酸素原子や窒素原子と水素結合を形成する特性があるからである。F(Phe、フェニルアラニン)やW(Trp、トリプトファン)は側鎖がとりわけ大きな分子量をもち、かつ、芳香族の効果が強いからである。P(Pro、プロリン)はイミノ酸効果が強く、ポリペプチドの主鎖の角度を固定してしまうからである。
このグループは、上記参考文献(1)で示された中性非極性アミノ酸のうち、脂肪属性の疎水性側鎖をもつアミノ酸のグループであり、V(Val、バリン)、L(Leu、ロイシン)、I(Ile、イソロイシン)及びM(Met、メチオニン)から構成される。参考文献(1)による中性非極性アミノ酸と分類されるもののうちFGACWPは以下理由で、この「脂肪族疎水性アミノ酸グループ」には含めない。G(Gly、グリシン)やA(Ala、アラニン)はメチル基以下の大きさで非極性の効果が弱いからである。C(Cys、システイン)はS-S結合に重要な役目を担う場合があり、また、酸素原子や窒素原子と水素結合を形成する特性があるからである。F(Phe、フェニルアラニン)やW(Trp、トリプトファン)は側鎖がとりわけ大きな分子量をもち、かつ、芳香族の効果が強いからである。P(Pro、プロリン)はイミノ酸効果が強く、ポリペプチドの主鎖の角度を固定してしまうからである。
2)ヒドロキシメチレン基をもつグループ(STグループ)
このグループは、中性極性アミノ酸のうちヒドロキシメチレン基を側鎖に持つアミノ酸のグループであり、S(Ser、セリン)とT(Thr、スレオニン)から構成される。SとTの側鎖に存在する水酸基は、糖の結合部位であるため、あるポリペプチド(タンパク質)が特定の活性を持つために重要な部位である場合が多い。
このグループは、中性極性アミノ酸のうちヒドロキシメチレン基を側鎖に持つアミノ酸のグループであり、S(Ser、セリン)とT(Thr、スレオニン)から構成される。SとTの側鎖に存在する水酸基は、糖の結合部位であるため、あるポリペプチド(タンパク質)が特定の活性を持つために重要な部位である場合が多い。
3)酸性アミノ酸(DEグループ)
このグループは、酸性であるカルボキシル基を側鎖に持つアミノ酸のグループであり、D(Asp、アスパラギン酸)とE(Glu、グルタミン酸)から構成される。
このグループは、酸性であるカルボキシル基を側鎖に持つアミノ酸のグループであり、D(Asp、アスパラギン酸)とE(Glu、グルタミン酸)から構成される。
4)塩基性アミノ酸(KRグループ)
このグループは、塩基性アミノ酸のグループであり、K(Lys、リジン)とR(Arg、アルギニン)から構成される。これらKとRは、pHの広い範囲で正に帯電し塩基性の性質をもつ。一方、塩基性アミノ酸に分類されるH(His、ヒスチジン)はpH7においてほとんどイオン化されないので、このグループには分類されない。
このグループは、塩基性アミノ酸のグループであり、K(Lys、リジン)とR(Arg、アルギニン)から構成される。これらKとRは、pHの広い範囲で正に帯電し塩基性の性質をもつ。一方、塩基性アミノ酸に分類されるH(His、ヒスチジン)はpH7においてほとんどイオン化されないので、このグループには分類されない。
5)メチレン基=極性基(DHNグループ)
このグループは、全てα位の炭素元素に側鎖としてメチレン基が結合しその先に極性基を有すると言う特徴を持つ。非極性基であるメチレン基の物理的大きさが酷似している特徴を持ち、N(Asn、アスパラギン、極性基はアミド基)、D(Asp、アスパラギン酸、極性基はカルボキシル基)とH(His、ヒスチジン、極性基はイミダゾール基)から成る。
このグループは、全てα位の炭素元素に側鎖としてメチレン基が結合しその先に極性基を有すると言う特徴を持つ。非極性基であるメチレン基の物理的大きさが酷似している特徴を持ち、N(Asn、アスパラギン、極性基はアミド基)、D(Asp、アスパラギン酸、極性基はカルボキシル基)とH(His、ヒスチジン、極性基はイミダゾール基)から成る。
6)ジメチレン基=極性基(EKQRグループ)
このグループは、全てα位の炭素元素に側鎖としてジメチレン基以上の直鎖炭化水素が結合しその先に極性基を有すると言う特徴を持つ。非極性基であるジメチレン基の物理的大きさが酷似している特徴を持つ。E(Glu、グルタミン酸、極性基はカルボキシル基)、K(Lys、リジン、極性基はアミノ基)、Q(Gln、グルタミン、極性基はアミド基)、R(Arg、アルギニン、極性基はイミノ基とアミノ基)から成る。
このグループは、全てα位の炭素元素に側鎖としてジメチレン基以上の直鎖炭化水素が結合しその先に極性基を有すると言う特徴を持つ。非極性基であるジメチレン基の物理的大きさが酷似している特徴を持つ。E(Glu、グルタミン酸、極性基はカルボキシル基)、K(Lys、リジン、極性基はアミノ基)、Q(Gln、グルタミン、極性基はアミド基)、R(Arg、アルギニン、極性基はイミノ基とアミノ基)から成る。
7)芳香族(FYWグループ)
このグループには、側鎖にベンゼン核を持つ芳香族アミノ酸であり、芳香族特有の化学的性質を特徴とする。F(Phe、フェニルアラニン)、Y(Tyr、チロシン)、W(Trp、トリプトファン)から成る。
このグループには、側鎖にベンゼン核を持つ芳香族アミノ酸であり、芳香族特有の化学的性質を特徴とする。F(Phe、フェニルアラニン)、Y(Tyr、チロシン)、W(Trp、トリプトファン)から成る。
8)環状&極性(HYグループ)
このグループには、側鎖に環状構造を持つと同時に極性も持つアミノ酸で、H(H、ヒスチジン、環状構造と極性基は共にイミダゾール基)、Y(Tyr、チロシン、環状構造はベンゼン核で極性基は水酸基)から成る。
このグループには、側鎖に環状構造を持つと同時に極性も持つアミノ酸で、H(H、ヒスチジン、環状構造と極性基は共にイミダゾール基)、Y(Tyr、チロシン、環状構造はベンゼン核で極性基は水酸基)から成る。
5重、6重及び7重変異型ギ酸脱水素酵素
上述した4重変異型ギ酸脱水素酵素に対して、更に1~3つの置換変異を導入することにより5重、6重及び7重変異型ギ酸脱水素酵素を取得することができる。ここで、更なる1~3つの置換変異としては、上記A)~C)に示したアミノ酸配列における変異箇所以外であって、配列番号2に示すアミノ酸配列における、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、73番目のグリシン、82番目のリシン、136番目のイソロイシン、155番目のヒスチジン、159番目のグルタミン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸、298番目のアスパラギン及び343番目のアスパラギンのアミノ酸を挙げることができる。
上述した4重変異型ギ酸脱水素酵素に対して、更に1~3つの置換変異を導入することにより5重、6重及び7重変異型ギ酸脱水素酵素を取得することができる。ここで、更なる1~3つの置換変異としては、上記A)~C)に示したアミノ酸配列における変異箇所以外であって、配列番号2に示すアミノ酸配列における、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、73番目のグリシン、82番目のリシン、136番目のイソロイシン、155番目のヒスチジン、159番目のグルタミン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸、298番目のアスパラギン及び343番目のアスパラギンのアミノ酸を挙げることができる。
ここで列挙したアミノ酸残基については、変異後のギ酸脱水素酵素の耐久性が野生型のギ酸脱水素酵素における耐久性と比較して有意に向上するものである限り、特に限定されず、如何なるアミノ酸に置換しても良い。特に、31番目のイソロイシンはロイシンに置換変異し、50番目のアルギニンはグリシンに置換変異し、60番目のグルタミン酸はバリンに置換変異し、64番目のイソロイシンはバリンに置換変異し、73番目のグリシンはアラニンに置換変異し、82番目のリシンはアルギニンに置換変異し、136番目のイソロイシンはバリンに置換変異し、155番目のヒスチジンはグルタミン又はリシンに置換変異し、159番目のグルタミンはロイシン又はアスパラギンに置換変異し、239番目のメチオニンはロイシンに置換変異し、286番目のアスパラギン酸はフェニルアラニンに置換変異し、287番目のヒスチジンはグルタミン又はプロリンに置換変異し、293番目のグルタミン酸はアスパラギン酸に置換変異し、298番目のアスパラギンはチロシンに置換変異し、343番目のアスパラギンはアルギニンに置換変異することが好ましい。なお、ここで列挙したアミノ酸残基について置換変異後の好ましいアミノ酸の種類を開示したが、上述と同様に、置換変異後のアミノ酸の種類は、性質の類似したアミノ酸を分類した上記8つのグループに基づいて適宜選択することができる。
また、特に、5重変異型ギ酸脱水素酵素としては、73番目のグリシンのアラニンへの置換変異(アミノ酸の一文字表記を利用してG73Aと略記する。以下同様)、V99I、V153I、H155Q及びN298Yを有する変異型ギ酸脱水素酵素を挙げることができる。これらG73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素は、野生型のギ酸脱水素酵素と同等の比活性を有しながらも、上述のように定義される優れた耐久性を示すことができる。
さらに、5重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、H155Q及びH287Qを有する変異型ギ酸脱水素酵素を挙げることができる。これらG73A、V99I、V153I、H155Q及びH287Qを有する5重変異型ギ酸脱水素酵素もまた、優れた耐久性を示すことができる。
さらにまた、5重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、H155K及びN298Yを有する変異型ギ酸脱水素酵素を挙げることができる。これらG73A、V99I、V153I、H155K及びN298Yを有する5重変異型ギ酸脱水素酵素もまた、優れた耐久性を示すことができる。なお、後述の実施例に示すように、G73A、V99I、V153I、H155K及びN298Yを有する5重変異型ギ酸脱水素酵素と、G73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素とを耐久性の点で比較すると、G73A、V99I、V153I、H155K及びN298Yを有する5重変異型ギ酸脱水素酵素の方が優れた耐久性を示す。したがって、155番目のヒスチジンの置換変異後のアミノ酸は、グルタミンよりもリシンが好ましいことが理解できる。
さらにまた、5重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、Q159L及びN298Yを有する変異型ギ酸脱水素酵素、V99I、V153I、H155Q、H287Q及びN298Yを有する変異型ギ酸脱水素酵素を挙げることができる。これら5重変異型ギ酸脱水素酵素についても、G73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素と同等に野生型のギ酸脱水素酵素と比較して優れた耐久性を示すことができる。
さらにまた、5重変異型ギ酸脱水素酵素としては、E60V、G73A、V99I、V153I及びH155Qを有する変異型ギ酸脱水素酵素、E60V、V99I、V153I、H155Q及びN298Yを有する変異型ギ酸脱水素酵素を挙げることができる。これら5重変異型ギ酸脱水素酵素についても、野生型のギ酸脱水素酵素と比較して優れた耐久性を示すことができる。
一方、6重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素に対して、更にM239L、K82R、H287Q、E60V、I31L、I136V及びE293Dから選ばれる1つの置換変異を導入したものであることが好ましい。これら6重変異型ギ酸脱水素酵素は、G73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素と比較してより優れた耐久性を示すことができる。また、G73A、V99I、V153I、H155Q、M239L及びN298Yを有する6重変異型ギ酸脱水素酵素、E60V、G73A、V99I、V153I、H155Q及びN298Yを有する6重変異型ギ酸脱水素酵素並びにG73A、V99I、V153I、H155K、M239L及びN298Yを有する6重変異型ギ酸脱水素酵素は、上述のように耐久性に優れるばかりでなく、野生型のギ酸脱水素酵素と比較して優れた比活性を示すことができる。
また、6重変異型ギ酸脱水素酵素としては、上述したG73A、V99I、V153I、Q159L及びN298Yを有する5重変異型ギ酸脱水素酵素に対してM239Lを導入したものを挙げることができる。このM239Lを導入することにより、上述したG73A、V99I、V153I、Q159L及びN298Yを有する5重変異型ギ酸脱水素酵素と同等の耐久性を維持しながら、比活性を向上させることができる。
さらに、6重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、H155Q及びN298Yを有する変5重異型ギ酸脱水素酵素に対してH287Pを導入したものを挙げることができる。しかし、上述のようにG73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素に対してH287Qを導入した6重変異型ギ酸脱水素酵素と比較すると、比活性が低下していることが認められる。したがって、287番目のヒスチジンの置換変異後のアミノ酸は、プロリンよりもグルタミンが好ましいことが理解できる。
さらにまた、6重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素に対して、R50G又はN343Rを導入したものを挙げることができる。これら6重変異型ギ酸脱水素酵素についても、G73A、V99I、V153I、H155Q及びN298Yを有する5重変異型ギ酸脱水素酵素と同等の耐久性を示すことができる。
さらにまた、6重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、H155K及びN298Yを有する5重変異型ギ酸脱水素酵素に対して、K82R又はQ159Lを導入したものを挙げることができる。これら6重変異型ギ酸脱水素酵素については、G73A、V99I、V153I、H155K及びN298Yを有する5重変異型ギ酸脱水素酵素と比較して優れた耐久性を示すことができる。
さらにまた、6重変異型ギ酸脱水素酵素としては、I64V、G73A、V99I、V153I、H155Q及びN298Yを有する変異型ギ酸脱水素酵素、G73A、V99I、V153I、H155Q、Q159N及びN298Yを有する変異型ギ酸脱水素酵素を挙げることができる。これら5重変異型ギ酸脱水素酵素についても、野生型のギ酸脱水素酵素と比較して優れた耐久性を示すことができる。
一方、7重変異型ギ酸脱水素酵素としては、上述したG73A、V99I、V153I、H155Q、M239L及びN298Yを有する6重変異型ギ酸脱水素酵素に対して、K82Rを導入したものを挙げることができる。この7重変異型ギ酸脱水素酵素は、上述したG73A、V99I、V153I、H155Q、M239L及びN298Yを有する6重変異型ギ酸脱水素酵素と比較してより優れた耐久性を示すと共に、優れた比活性を示すことができる。
また、7重変異型ギ酸脱水素酵素としては、G73A、K82R、V99I、V153I、H155Q、H287Q及びN298Yを有するもの、G73A、V99I、V153I、H155Q、Q159L、M239L及びN298Yを有するもの、G73A、V99I、V153I、H155Q、Q159L、H287Q及びN298Yを有するものを挙げることができる。これら7重変異型ギ酸脱水素酵素についても、野生型のギ酸脱水素酵素と比較して優れた耐久性を示すことができる。
さらに、7重変異型ギ酸脱水素酵素としては、G73A、V99I、V153I、H155Q、D286F、H287P及びN298Yを有するもの、G73A、V99I、V153I、H155Q、D286F、H287P及びN298Yを有するものを挙げることができる。これら7重変異型ギ酸脱水素酵素についても、野生型のギ酸脱水素酵素と比較して優れた耐久性を示すことができる。
アカカビ由来のギ酸脱水素酵素の製造
上述した本発明に係る変異型ギ酸脱水素酵素のなかでも、アカカビ由来のギ酸脱水素酵素に由来する変異型ギ酸脱水素酵素は、置換変異前あっても非常に高い比活性を示す。したがって、アカカビ由来のギ酸脱水素酵素に由来する変異型ギ酸脱水素酵素は、他の生物由来のギ酸脱水素酵素由来の変異型ギ酸脱水素酵素と比較して耐久性及び/又は比活性が際だって高いため好ましい。
上述した本発明に係る変異型ギ酸脱水素酵素のなかでも、アカカビ由来のギ酸脱水素酵素に由来する変異型ギ酸脱水素酵素は、置換変異前あっても非常に高い比活性を示す。したがって、アカカビ由来のギ酸脱水素酵素に由来する変異型ギ酸脱水素酵素は、他の生物由来のギ酸脱水素酵素由来の変異型ギ酸脱水素酵素と比較して耐久性及び/又は比活性が際だって高いため好ましい。
ここで、アカカビ由来のギ酸脱水素酵素は、従来公知のタンパク質の製造方法では取得することができない。したがって、アカカビ由来の変異型ギ酸脱水素酵素についても、従来公知のタンパク質の製造方法で取得することが困難であり、以下に説明する手法に準じて製造することができる。すなわち、アカカビ由来のギ酸脱水素酵素の製造方法は、アカカビ由来のギ酸脱水素酵素遺伝子を誘導型プロモーターの制御下に配置したベクターを導入した宿主を準備する。そして当該宿主を培養し、対数増殖期を過ぎた時にギ酸脱水素酵素遺伝子の発現を誘導する。次に、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度で培養することでギ酸脱水素酵素を宿主内に発現させる。
アカカビ由来のギ酸脱水素酵素の製造方法において、誘導型プロモーターとしては、特に限定されず、従来公知のものを使用することができる。例えば、上記の宿主として大腸菌を使用する場合には、イソプロピル-β-チオガラクトピラノシド(IPTG)の存在下に転写活性を示す誘導型プロモーターを使用することができる。このようなプロモーターの例としては、Trpプロモーター、Lacプロモーター、Trcプロモーター及びTacプロモーターを挙げることができる。また、IPTG以外の誘導物質の存在下に転写活性を示す他のプロモーターや、培地成分及び温度等の培養条件に応じて転写活性を示す他のプロモーターも、誘導型プロモーターとして使用することができる。
また、アカカビ由来のギ酸脱水素酵素の製造方法において、ベクターとしては、上記の宿主内で複製可能なものであれば特に限定されず、如何なるベクターをも使用することができる。例えば、上記宿主として大腸菌を使用する場合には、ベクターとしてはプラスミドベクター、ファージベクターのいずれであっても良い。具体的なベクターとしては、pCDFシリーズ、pRSFシリーズ、pETシリーズ等を例示列挙することができる。
さらに宿主としては、発現ベクターに組み込まれたプロモーターから転写可能な宿主であれば特に限定されないが、例えば、発現ベクターがpET(T7プロモーター)系の場合には大腸菌BL21(DE3)を使用することができる。上述したベクターを宿主に導入する手法としては、一般的に形質転換法として知られる各種の手法を適用することができる。具体的な手法としては、例えば、リン酸カルシウム法、エレクトロポレーション法、リポフェクション法等を適用することができる。
特にアカカビ由来のギ酸脱水素酵素の製造方法においては、ベクターを導入した宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導する。ギ酸脱水素酵素遺伝子の発現を誘導する前において、宿主の培養条件としては、何ら限定されず、例えば当該宿主の生育至適温度や生育至適pHを勘案して適宜、設定すればよい。しかし、培養を継続しながら宿主の増殖を観察し、いわゆる対数増殖期を過ぎた時点で以下の条件を満足するような培養条件に変更する。すなわち、条件1としてギ酸脱水素酵素遺伝子の発現を誘導し、条件2として宿主の生育至適温度よりも低く且つ宿主が生存可能な温度で培養する。
ここで、対数増殖期を過ぎた時とは、横軸に培養時間及び縦軸に細胞数の対数をとった増殖曲線において、所定の傾きの略直線で表される部分から、接線の傾きが低下し始める時点を意味する。なお、培養曲線は培地中のOD600nmを測定することによって作製することができる。また、ギ酸脱水素酵素遺伝子の発現を誘導する際には、対数増殖期を過ぎて定常期に入ってからが好ましい。ここで定常期とは、上述した増殖曲線の接線の傾きがほぼ0となる期間である。
また、宿主の生育至適温度とは、宿主毎に異なる温度範囲として公知である、例えば、大腸菌B株を宿主とした場合、生育至適温度は37℃である。例えば、大腸菌B株を宿主とした場合、生育可能な温度は15~37℃である。したがって、大腸菌B株を宿主とした場合、宿主の生育至適温度よりも低く且つ宿主が生存可能な温度としては15~37℃を意味する。特に、大腸菌B株を宿主とした場合、対数増殖期を過ぎた時に約20℃にして培養を継続することが好ましい。
宿主の対数増殖期を過ぎたときに上記温度範囲に設定することでギ酸脱水素酵素遺伝子が発現し、宿主内に非常に高い比活性を示すギ酸脱水素酵素が生成することとなる。培養後、目的のギ酸脱水素酵素が宿主内に生産されるため菌体又は細胞を破砕し、粗酵素懸濁液を調製する。この粗酵素懸濁液には、非常に高い比活性を示すギ酸脱水素酵素が含まれる。したがって、得られた粗酵素懸濁液をそのまま利用することができる。なお、得られた粗酵素懸濁液からギ酸脱水素酵素を単離精製することもできる。このとき、蛋白質の単離精製に用いられる一般的な生化学的方法、例えば硫酸アンモニウム沈殿、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等を単独で又は適宜組み合わせて用いることができる。単離精製されたギ酸脱水素酵素は、所定のpHの緩衝液等に懸濁された状態で利用することができる。
変異型ギ酸脱水素酵素の利用形態
以上で説明した変異型ギ酸脱水素酵素は、変異導入前のギ酸脱水素酵素と比較して抜群に高い耐久性を示すため、従来公知のギ酸脱水素酵素が使用されている反応系のいずれにも優れた代替物として利用することができる。例えば、変異型ギ酸脱水素酵素の利用形態としてはNADHの再生系を挙げることができる。NADHは種々の酵素反応で利用されNAD+へと変換される。NADHは、例えば、化学工業や製薬工業の分野において光学異性体を生物的に合成する際の補酵素として利用される。NADHの再生系とは、反応系に残存するNAD+を還元してNADHとし、NADHを回収して再び上記酵素反応に利用することを意味する。上述した変異型ギ酸脱水素酵素をギ酸及びNAD+を含む反応系に作用させることによって、当該NAD+を還元してNADHを合成することができる。
以上で説明した変異型ギ酸脱水素酵素は、変異導入前のギ酸脱水素酵素と比較して抜群に高い耐久性を示すため、従来公知のギ酸脱水素酵素が使用されている反応系のいずれにも優れた代替物として利用することができる。例えば、変異型ギ酸脱水素酵素の利用形態としてはNADHの再生系を挙げることができる。NADHは種々の酵素反応で利用されNAD+へと変換される。NADHは、例えば、化学工業や製薬工業の分野において光学異性体を生物的に合成する際の補酵素として利用される。NADHの再生系とは、反応系に残存するNAD+を還元してNADHとし、NADHを回収して再び上記酵素反応に利用することを意味する。上述した変異型ギ酸脱水素酵素をギ酸及びNAD+を含む反応系に作用させることによって、当該NAD+を還元してNADHを合成することができる。
以上のように変異型ギ酸脱水素酵素をNADHの再生系に利用することによって、反応系に含まれるNAD+からNADHを効率よく製造することができる。特に、変異型ギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して非常に優れた耐久性を示すため、変異導入前のギ酸脱水素酵素と比較して長時間に亘って高い活性を維持することができる。このため、変異型ギ酸脱水素酵素を利用することによって、従来公知のギ酸脱水素酵素の使用した場合と比較してNADHの生産性を大幅に向上させることができる。
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
〔実施例1〕
本実施例ではアカカビ由来のギ酸脱水素酵素遺伝子をクローニングし、当該遺伝子に対してランダム変異を導入し、得られた変異遺伝子に由来するギ酸脱水素酵素における耐久性及び比活性を評価した。
本実施例ではアカカビ由来のギ酸脱水素酵素遺伝子をクローニングし、当該遺伝子に対してランダム変異を導入し、得られた変異遺伝子に由来するギ酸脱水素酵素における耐久性及び比活性を評価した。
(1)試薬
試薬は、特に記載の無い場合ナカライテスク社製のものを使用した。
試薬は、特に記載の無い場合ナカライテスク社製のものを使用した。
・リン酸カリウム緩衝液(KPB) pH7.5
<SolutionA>0.5M KH2PO4 13.6g/200ml
<SolutionB>0.5M K2HPO4 26.13g/300ml
0.5M KPB pH7.5はSolutionAとSolutionBを次の割合で混合した。
<SolutionA>0.5M KH2PO4 13.6g/200ml
<SolutionB>0.5M K2HPO4 26.13g/300ml
0.5M KPB pH7.5はSolutionAとSolutionBを次の割合で混合した。
[SolutionA 16ml]+[SolutionB 84ml]⇒100ml
・EcoPro T7 system(Novagen社製)
Lysate、Methionine
・1.62M ギ酸ナトリウム(ギ酸Na)
5.5g/50ml 0.5M KPB pH7.5、調製後0.22μmのフィルター(ミリポア社製)で滅菌した。
・EcoPro T7 system(Novagen社製)
Lysate、Methionine
・1.62M ギ酸ナトリウム(ギ酸Na)
5.5g/50ml 0.5M KPB pH7.5、調製後0.22μmのフィルター(ミリポア社製)で滅菌した。
・16.2mM NAD
581mg/50ml 0.5M KPB pH7.5、調製後0.22μmのフィルター(ミリポア社製)で滅菌した。
581mg/50ml 0.5M KPB pH7.5、調製後0.22μmのフィルター(ミリポア社製)で滅菌した。
・mPMS(DOJINDO社製)
methoxy PMS 0.5mg/mlとなるよう蒸留水で調製した。
methoxy PMS 0.5mg/mlとなるよう蒸留水で調製した。
・WST1(DOJINDO社製)
8mg/mlに蒸留水で調製した。
8mg/mlに蒸留水で調製した。
・PD培地
Potato dextrose broth(Difco社製) 24g/L。pH7に調整後オートクレーブして使用した。
Potato dextrose broth(Difco社製) 24g/L。pH7に調整後オートクレーブして使用した。
・LB培地
LB Broth(Difco社製) 20g/L。オートクレーブして使用した。使用前にアンピシリン(SIGMA社製)を50μg/mlとなるように添加した。
LB Broth(Difco社製) 20g/L。オートクレーブして使用した。使用前にアンピシリン(SIGMA社製)を50μg/mlとなるように添加した。
・100mM MgCl2
MgCl2・6H2O 2.03g/100ml。オートクレーブして使用した。
MgCl2・6H2O 2.03g/100ml。オートクレーブして使用した。
・ExTaq Polymerase[5U/μl](タカラバイオ社製)
10XBuffer
・KOD-Plus-:KOD-Plus-Polymerase [1U/μl] (TOYOBO社製)
25mM MgSO4、2mM dNTP、10×Buffer
・Pyrobest DNA polymerase(タカラバイオ社製)
・Triton X-100
・100mM dATP、dCTP、dGTP、dTTP(タカラバイオ社製)
・RNeasy Plant Mini Kit(QIAGEN社製)
・RNA PCR Kit(タカラバイオ社製)
・MinElute Gel Extraction Kit(QIAGEN社製)
・MinElute PCR Purification Kit(QIAGEN社製)
・BigDyeTerminator v3.1(ABI社製)
・dH2O:DNase/RNase Free Distilled Water(Invitrogen社製)
・制限酵素NdeI/EcoRI(タカラバイオ社製)
・DNA Ligation kit ver2.1、solutionI(タカラバイオ社製)
・JM109 Competent Cells(タカラバイオ社製)
・pET-23b(+) vector (Novagen社製)
・pT7 Blue T-vector(Novagen製)
・KOD-Plus-Mutagenesis Kit (TOYOBO社製)
(2)Gibberella zeae FDH遺伝子のクローニング
(2-1) 微生物株
Gibberella zeaeは独立行政法人製品評価技術基盤機構の関連機関であるNITE Biological Resource Center(以下NBRCと記載)に保存されている株(NBRC No. 4474)を購入し、指定される方法で復元したものをPD(Potato Dextrose)培地を用いて培養した。
10XBuffer
・KOD-Plus-:KOD-Plus-Polymerase [1U/μl] (TOYOBO社製)
25mM MgSO4、2mM dNTP、10×Buffer
・Pyrobest DNA polymerase(タカラバイオ社製)
・Triton X-100
・100mM dATP、dCTP、dGTP、dTTP(タカラバイオ社製)
・RNeasy Plant Mini Kit(QIAGEN社製)
・RNA PCR Kit(タカラバイオ社製)
・MinElute Gel Extraction Kit(QIAGEN社製)
・MinElute PCR Purification Kit(QIAGEN社製)
・BigDyeTerminator v3.1(ABI社製)
・dH2O:DNase/RNase Free Distilled Water(Invitrogen社製)
・制限酵素NdeI/EcoRI(タカラバイオ社製)
・DNA Ligation kit ver2.1、solutionI(タカラバイオ社製)
・JM109 Competent Cells(タカラバイオ社製)
・pET-23b(+) vector (Novagen社製)
・pT7 Blue T-vector(Novagen製)
・KOD-Plus-Mutagenesis Kit (TOYOBO社製)
(2)Gibberella zeae FDH遺伝子のクローニング
(2-1) 微生物株
Gibberella zeaeは独立行政法人製品評価技術基盤機構の関連機関であるNITE Biological Resource Center(以下NBRCと記載)に保存されている株(NBRC No. 4474)を購入し、指定される方法で復元したものをPD(Potato Dextrose)培地を用いて培養した。
(2-2) ギ酸脱水素酵素遺伝子の単離
(2-2-1) ギ酸脱水素酵素遺伝子の増幅
2-1の方法で培養して得た菌体をRNeasy Plant Mini Kit(QIAGEN社製)を用いてTotal RNA(mRNA、rRNA及びtRNA等を含む)を調製した。まず、RNA PCR Kit(タカラバイオ社製)を用いてTotal RNAを鋳型としたcDNA合成をおこなった。表1に反応液組成を示す。
(2-2-1) ギ酸脱水素酵素遺伝子の増幅
2-1の方法で培養して得た菌体をRNeasy Plant Mini Kit(QIAGEN社製)を用いてTotal RNA(mRNA、rRNA及びtRNA等を含む)を調製した。まず、RNA PCR Kit(タカラバイオ社製)を用いてTotal RNAを鋳型としたcDNA合成をおこなった。表1に反応液組成を示す。

上記組成の反応液を用いて、50℃で2時間、その後99℃で5分間、その後4℃とする反応サイクルでcDNAの合成反応を行った。
次に、合成されたcDNAを鋳型とし、Pyrobest DNA polymeraseを用いてPCRをおこなった。反応液50μl中の成分組成を表2に示す。

上記組成の反応性を用いて、95℃で1分、その後、(95℃で30秒、60℃で30秒及び72℃で1分を1サイクルとして25サイクル繰り返し、その後72℃で10分、その後4℃とする反応サイクルでPCRを行った。なお、本PCRにおいては、一対のプライマーとしてGib FDH1-F-NdeI(forward):CGC CAT ATG GTC AAG GTT CTT GCA GTT C(配列番号13)及びGib FDH1-R(reverse):CTA TTT CTT CTC ACG CTG ACC(配列番号14)を使用した。
(2-2-2) ギ酸脱水素酵素遺伝子のクローニング及び構造解析
アガロースゲル電気泳動により得られた各種PCR産物のサイズを確認後、MinElute Gel Extraction Kit(QIAGEN製)を用いてアガロースゲルより精製したPCR産物をpT7 Blue T-vector(Novagen製)、JM109 competent cells(タカラバイオ社製)を用いてサブクローニングした(GzFDH/pT7)(図2参照)。単離したギ酸脱水素酵素遺伝子配列はデータベース上に公開されている配列(Genbank No. XP_386303)とアミノ酸レベルで100%の一致を示していた。
アガロースゲル電気泳動により得られた各種PCR産物のサイズを確認後、MinElute Gel Extraction Kit(QIAGEN製)を用いてアガロースゲルより精製したPCR産物をpT7 Blue T-vector(Novagen製)、JM109 competent cells(タカラバイオ社製)を用いてサブクローニングした(GzFDH/pT7)(図2参照)。単離したギ酸脱水素酵素遺伝子配列はデータベース上に公開されている配列(Genbank No. XP_386303)とアミノ酸レベルで100%の一致を示していた。
(2-2-3) ギ酸脱水素酵素遺伝子発現用ベクターの作製
2-2-2で作製されたプラスミド(FDH/pT7)を制限酵素NdeI/EcoRIで処理した。表3に反応液組成を示す。制限酵素処理の反応条件としては37℃で2時間とした。
2-2-2で作製されたプラスミド(FDH/pT7)を制限酵素NdeI/EcoRIで処理した。表3に反応液組成を示す。制限酵素処理の反応条件としては37℃で2時間とした。

反応後の溶液を0.8%アガロースゲル電気泳動に供し、NdeI/EcoRI断片としてベクターから切り出されたギ酸脱水素酵素遺伝子(約1.1kb)をMinElute Gel Extraction Kit(QIAGEN製)を用いて精製した。次にFDH/pT7と同様に制限酵素処理した遺伝子発現用ベクターpET23b(+)(Novagen社製)のNdeI/EcoRI部位にDNA Ligation kit Ver.2.1(タカラバイオ社製)を用いて上記ギ酸脱水素酵素遺伝子断片を導入後、JM109 competent cells(タカラバイオ社製)によるサブクローニングを行った(FDH/pET23b(+))(図2参照)。
(3)Site-directed mutagensisによる変異型ギ酸脱水素酵素の作製
(3-1)多重変異型ギ酸脱水素酵素
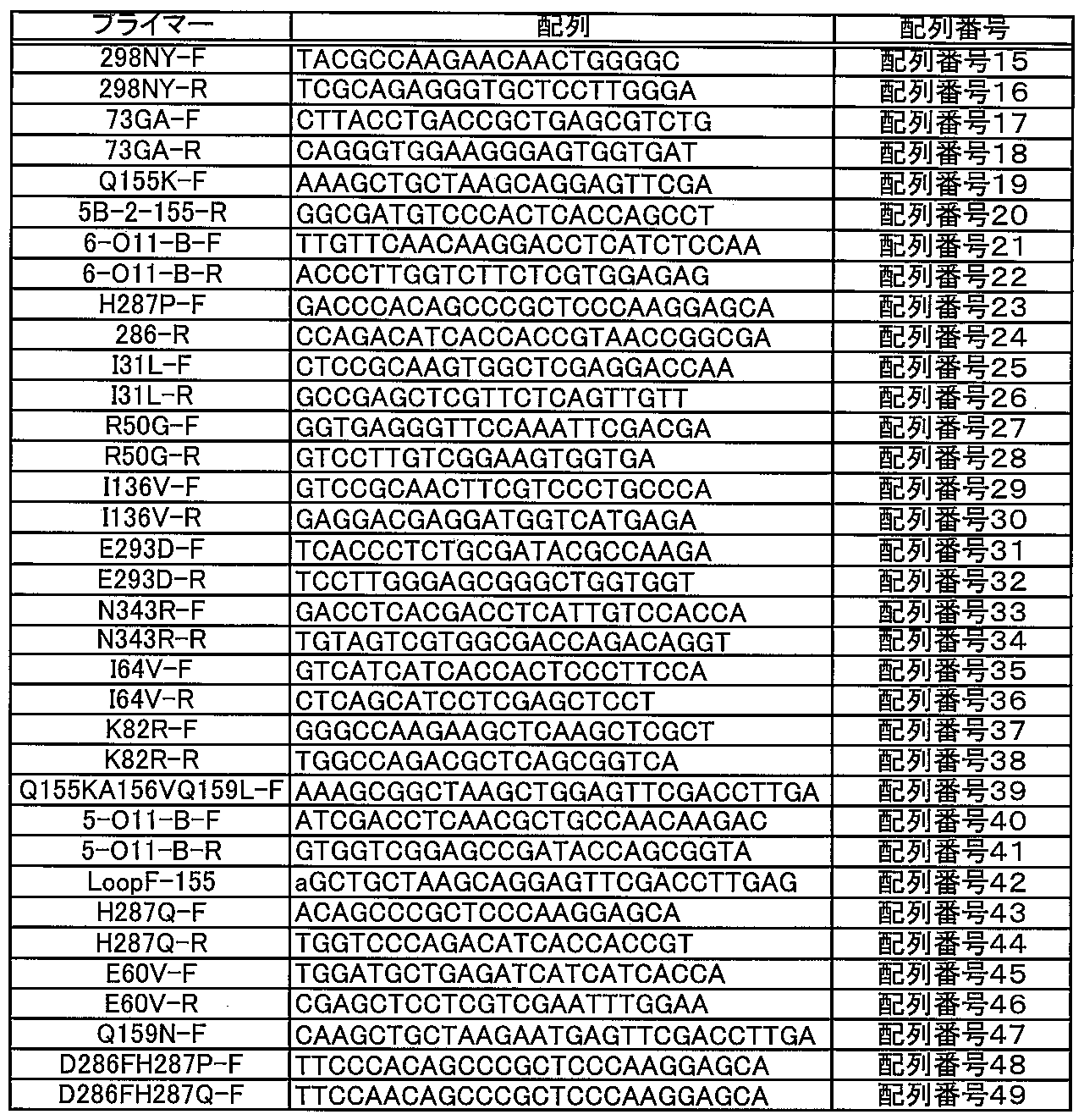
KOD-Plus-Mutagenesis Kitを用いて、プロトコールに従いFDHに部位特異的に変異を導入した。作製した変異体、作製に用いた鋳型/プライマーの組み合わせを表4に示す。また使用したプライマーの配列を表5に示す。
(3-1)多重変異型ギ酸脱水素酵素
KOD-Plus-Mutagenesis Kitを用いて、プロトコールに従いFDHに部位特異的に変異を導入した。作製した変異体、作製に用いた鋳型/プライマーの組み合わせを表4に示す。また使用したプライマーの配列を表5に示す。

なお、表4における鋳型の欄に記載したWTは野性型のFDHを意味し、例えば、V99IはFDHにおける99番目のバリンをイソロイシンに置換した変異型FDHを意味する。
具体的には、先ず野性型のFDHに対してV99I変異を導入した単独変異型ギ酸脱水素酵素遺伝子を作製し、これにV153I/H155Q変異若しくはV153I/Q159L変異を導入して3重変異型ギ酸脱水素酵素遺伝子を作製し、そして3重変異型ギ酸脱水素酵素遺伝子に対してN298Y変異、G73A変異、H287Q変異若しくはE60V変異を導入することで4重変異型ギ酸脱水素酵素を作製した。
以上により作製した4重変異型ギ酸脱水素酵素のうち表6に示す3種類の4重変異型ギ酸脱水素酵素遺伝子を後述の(4)組換え体FDHの性状評価に使用した。
また、同様にして5重~7重変異型ギ酸脱水素酵素遺伝子を作製し、表7に示す5重~7重変異型ギ酸脱水素酵素遺伝子を後述の(4)組換え体FDHの性状評価に使用した。
(4)組換え体FDHの性状評価
(4-1)大腸菌S30による組換え体FDHの合成
(4-1-1)鋳型の調製
前項(3-1)で構築したプラスミドを鋳型としてPCRを行った。この増幅産物を翻訳反応の鋳型として用いた。PCRでは、一対のプライマーとしてSingle-F及びSingle-R1を使用した。PCRの反応液組成を表8に示した。
(4-1)大腸菌S30による組換え体FDHの合成
(4-1-1)鋳型の調製
前項(3-1)で構築したプラスミドを鋳型としてPCRを行った。この増幅産物を翻訳反応の鋳型として用いた。PCRでは、一対のプライマーとしてSingle-F及びSingle-R1を使用した。PCRの反応液組成を表8に示した。
Single-F:5’-CGA TCC CGC GAA ATT AAT ACG ACT-3’(配列番号50)
Single-R1:5’-TCC GGA TAT AGT TCC TCC TTT CAG-3’ (配列番号51)
Single-R1:5’-TCC GGA TAT AGT TCC TCC TTT CAG-3’ (配列番号51)

上記組成の反応液を用いて、94℃で2分の後、94℃で15秒及び68℃で1分30秒を1サイクルとして30サイクル行い、その後、68℃で2分、その後4℃とする反応サイクルでPCRを行った。PCRにより増幅された約1.4kbpの断片をMinElute PCR Purification Kitで精製し翻訳反応に使用した。
(4-1-2) 無細胞翻訳反応
4-1-1で精製したDNA断片を鋳型としてEcoPro T7 system(Novagen社製)を用い翻訳反応を行った。方法はキットのプロトコールに従った。
4-1-1で精製したDNA断片を鋳型としてEcoPro T7 system(Novagen社製)を用い翻訳反応を行った。方法はキットのプロトコールに従った。
(4-2)S30中での耐久性評価(加速試験)
S30(大腸菌粗抽出液)中における変異酵素の耐久性を加速試験により評価した。手順を以下に示す。4-1-2で調製した翻訳産物を1.8μlずつPCRチューブに分注し、サーマルサイクラーで52℃、56℃又は57℃で加熱した。加熱開始から0、25及び50分ごとにチューブを取り出し氷につけて冷却した。その後、表9に示す組成の活性測定用試薬のチューブに98μlずつ添加した。その後、サーマルサイクラー37℃で加熱(30分程度)し、チューブを取り出し氷につけて反応を停止させた。
S30(大腸菌粗抽出液)中における変異酵素の耐久性を加速試験により評価した。手順を以下に示す。4-1-2で調製した翻訳産物を1.8μlずつPCRチューブに分注し、サーマルサイクラーで52℃、56℃又は57℃で加熱した。加熱開始から0、25及び50分ごとにチューブを取り出し氷につけて冷却した。その後、表9に示す組成の活性測定用試薬のチューブに98μlずつ添加した。その後、サーマルサイクラー37℃で加熱(30分程度)し、チューブを取り出し氷につけて反応を停止させた。

なお、FDH触媒によるギ酸分解反応は次式で示される。
HCOO- + NAD+ → CO2 + NADH
ここに電子伝達物質であるMethoxy PMS(mPMS)と、酸化還元発色指示薬であるWST1(共にDOJINDO社製)を加えることで次式のように反応が進むため、波長438nmの吸光度で黄色ホルマザンを測定することで、ギ酸分解量の定量が可能となる。なお黄色ホルマザンの吸光係数はNADHの約6倍であり、NADHの直接測定よりも高感度での定量が可能となる。
ここに電子伝達物質であるMethoxy PMS(mPMS)と、酸化還元発色指示薬であるWST1(共にDOJINDO社製)を加えることで次式のように反応が進むため、波長438nmの吸光度で黄色ホルマザンを測定することで、ギ酸分解量の定量が可能となる。なお黄色ホルマザンの吸光係数はNADHの約6倍であり、NADHの直接測定よりも高感度での定量が可能となる。
NADH + mPMS → NAD+ + mPMS(還元型)
mPMS(還元型) + WST1 → mPMS + 黄色ホルマザン(37000/M・cm、438nm)
そして、プレートリーダー(Spectrafluor Plus:TECAN社製)で吸光度430nmを測定することで黄色ホルマザンを定量し、4-1-2で調整した複数のアミノ酸変異を有するFDHの活性を評価することができる。
mPMS(還元型) + WST1 → mPMS + 黄色ホルマザン(37000/M・cm、438nm)
そして、プレートリーダー(Spectrafluor Plus:TECAN社製)で吸光度430nmを測定することで黄色ホルマザンを定量し、4-1-2で調整した複数のアミノ酸変異を有するFDHの活性を評価することができる。
(4-3)大腸菌での組換え体FDHの過剰発現と精製
(4-3-1)発現誘導
G.zeae由来FDH(野生型および変異体)の発現は以下のように行った。使用菌株としては前項(3-1)で構築した組換え体プラスミド5B-2を導入した大腸菌BL21(DE3)株と野生型FDHを導入した株を使用した。LB培地 5mlを使用して37℃で22時間の前培養を行った。前培養の培養液が1.5%となるように、LB培地 500mlに植菌し、 37℃で4時間(O.D.600:3~)の本培養を行った。また、タンパク質の発現を誘導する誘導培養として、終濃度1mMとなるようにIPTGを添加し、20℃で16時間(O.D.600:3.5~4.1)培養した。
(4-3-1)発現誘導
G.zeae由来FDH(野生型および変異体)の発現は以下のように行った。使用菌株としては前項(3-1)で構築した組換え体プラスミド5B-2を導入した大腸菌BL21(DE3)株と野生型FDHを導入した株を使用した。LB培地 5mlを使用して37℃で22時間の前培養を行った。前培養の培養液が1.5%となるように、LB培地 500mlに植菌し、 37℃で4時間(O.D.600:3~)の本培養を行った。また、タンパク質の発現を誘導する誘導培養として、終濃度1mMとなるようにIPTGを添加し、20℃で16時間(O.D.600:3.5~4.1)培養した。
培養後、培養液を一旦氷につけて冷やした後に集菌した。培養液を500mlチューブ2本に分けて、5200rpmで10分間、4℃にて遠心分離した。上清を捨てて、菌体にそれぞれ60mlの10mM KPB(pH7.5)を加えて懸濁し、120ml分の懸濁液を50mlマルチビーズショッカー用専用破砕チューブ(安井器械株式会社製)3本に分けて、5600rpmで5分間、4℃にて再度遠心分離を行い、菌体を得た。
(4-3-2) 発現誘導菌体の破砕
(4-3-1)で得た菌体を冷却しながら破砕し、粗抽出液を調製した。集菌後の湿菌体重量を測定し、10mM KPB(pH7.5)を10倍量(湿菌体重量1gに対し10mlの割合)、マルチビーズショッカー専用グラスビーズ0.1mm(安井器械株式会社製)をKPBと等量(KPB10mlに対して10gの割合)を添加した。破砕にはマルチビーズショッカー(安井器械株式会社製)を用いた。
(4-3-1)で得た菌体を冷却しながら破砕し、粗抽出液を調製した。集菌後の湿菌体重量を測定し、10mM KPB(pH7.5)を10倍量(湿菌体重量1gに対し10mlの割合)、マルチビーズショッカー専用グラスビーズ0.1mm(安井器械株式会社製)をKPBと等量(KPB10mlに対して10gの割合)を添加した。破砕にはマルチビーズショッカー(安井器械株式会社製)を用いた。
粗抽出液は手順に従って調整した。先ず、6000rpmで120秒間振とうし、インターバル60秒間の後、再び同条件で振とうする工程を3回繰り返した。その後、6000rpmで20分間、4℃にて遠心分離し、上清を回収した。その後、6000rpmで10分間、4℃にて遠心分離し上清を0.45μmのフィルター (ミリポア社製)でろ過することで、粗抽出液を調製した。
(4-3-3)イオン交換カラム
前項4-3-2で調製した粗抽出液をHiTrap Q FFカラム 5ml(GEヘルスケアバイオサイエンス社製)により分画した。手順を以下に示す。先ず、10mM KPB(pH7.5) 25mlを流速1ml/minでカラムに供給して平衡化を行った。次に、粗抽出液を流速1ml/minで供給し、タンパク質成分を結合させた。次に、10mM KPB(pH7.5) 25mlを流速1ml/minでカラムに供給し、洗浄を行った。次に、10mM KPB(pH7.5)をベースとしてNaCl(10ml, 0~200mM)のグラジエントを流速0.5ml/minでカラムに供給し、溶出を行った。なお、1つのフラクションが0.5mlとなるように分画した。最後に、10mM KPB(pH7.5)及び500mM NaCl 15mlを流速1ml/min でカラムに供給し、洗浄を行った。
前項4-3-2で調製した粗抽出液をHiTrap Q FFカラム 5ml(GEヘルスケアバイオサイエンス社製)により分画した。手順を以下に示す。先ず、10mM KPB(pH7.5) 25mlを流速1ml/minでカラムに供給して平衡化を行った。次に、粗抽出液を流速1ml/minで供給し、タンパク質成分を結合させた。次に、10mM KPB(pH7.5) 25mlを流速1ml/minでカラムに供給し、洗浄を行った。次に、10mM KPB(pH7.5)をベースとしてNaCl(10ml, 0~200mM)のグラジエントを流速0.5ml/minでカラムに供給し、溶出を行った。なお、1つのフラクションが0.5mlとなるように分画した。最後に、10mM KPB(pH7.5)及び500mM NaCl 15mlを流速1ml/min でカラムに供給し、洗浄を行った。
溶出ステップで得たフラクションのうちFDH活性のピークが認められたフラクションをハイドロキシアパタイトカラムによる二次精製のサンプルとした。
(4-3-4)ハイドロキシアパタイトカラム
前項4-3-3で得たFDH活性含有フラクションを用いて二次精製を行った。各フラクションを一つにまとめ、Amicon Ultra-15(30kDaカット)(ミリポア社製)を用いて5000rpmで4℃にて30分間遠心分離し、脱塩後10mM KPB(pH7.2)で15ml程度に希釈した。二次精製にはEcono-Pac CHT-IIカラム 5ml(BIO-RAD社製)を使用した。手順を以下に示す。
前項4-3-3で得たFDH活性含有フラクションを用いて二次精製を行った。各フラクションを一つにまとめ、Amicon Ultra-15(30kDaカット)(ミリポア社製)を用いて5000rpmで4℃にて30分間遠心分離し、脱塩後10mM KPB(pH7.2)で15ml程度に希釈した。二次精製にはEcono-Pac CHT-IIカラム 5ml(BIO-RAD社製)を使用した。手順を以下に示す。
先ず、1mM KPB(pH7.2) 30mlを流速1ml/minでカラムに供給し、平衡化を行った。次に、脱塩した後のサンプルを流速0.5ml/minでカラムに供給し(2循環させた)、タンパク質成分を結合させた。次に、1mM KPB(pH7.2) 10mlを流速1ml/minでカラムに供給し、洗浄を行った。次に、1mM~10mM KPB(pH6.8)各10mlを流速0.5ml/minでカラムに供給し、溶出を行った。なお、1つのフラクションが0.5mlとなるように分画(計49フラクション)した。最後に、500mM KPB(pH6.8)5mlを流速1ml/minでカラムに供給し、洗浄を行った。
(4-3-5)精製タンパク質の濃度測定
Protein Assay試薬を用いたブラッドフォード法を使って、精製後のタンパク濃度定量を行った。次に、その値をもとに、野生型FDH及び変異型FDH(5B-2)のそれぞれについて、タンパク濃度が0.067mg/mlとなるように1mM MOPS(pH7.5)で調整した。
Protein Assay試薬を用いたブラッドフォード法を使って、精製後のタンパク濃度定量を行った。次に、その値をもとに、野生型FDH及び変異型FDH(5B-2)のそれぞれについて、タンパク濃度が0.067mg/mlとなるように1mM MOPS(pH7.5)で調整した。
(4-4)FDH変異体の耐久性測定
野生型FDH及び変異型FDH(5B-2)のそれぞれについて0.067mg/mlに調整した酵素溶液を0.7M MOPS pH7.5、10mg/ml BSA及び0.3Mトレハロースとなるように0.2mlチューブ(BIO-RAD社製)に5μlずつ分注し、サーマルサイクラー(BIO-RAD社製)37℃で500時間後の残存活性を測定した。
野生型FDH及び変異型FDH(5B-2)のそれぞれについて0.067mg/mlに調整した酵素溶液を0.7M MOPS pH7.5、10mg/ml BSA及び0.3Mトレハロースとなるように0.2mlチューブ(BIO-RAD社製)に5μlずつ分注し、サーマルサイクラー(BIO-RAD社製)37℃で500時間後の残存活性を測定した。
(5)結果のまとめ
前記(4-4)で行った耐久性試験の結果を図3に示す。図3から分かるように、プラスミド名5B-2に示される5重変異型ギ酸脱水素酵素(G73A、V99I、V153I、H155Q及びN298Y)は、野生型ギ酸脱水素酵素がほぼ完全に失活するような時間が経過した後においても80%以上の活性を維持できることが明らかとなった。このように、プラスミド名5B-2に示される5重変異型ギ酸脱水素酵素は、極めて優れた耐久性を有する酵素といえる。
前記(4-4)で行った耐久性試験の結果を図3に示す。図3から分かるように、プラスミド名5B-2に示される5重変異型ギ酸脱水素酵素(G73A、V99I、V153I、H155Q及びN298Y)は、野生型ギ酸脱水素酵素がほぼ完全に失活するような時間が経過した後においても80%以上の活性を維持できることが明らかとなった。このように、プラスミド名5B-2に示される5重変異型ギ酸脱水素酵素は、極めて優れた耐久性を有する酵素といえる。
また、前記(3-1)で取得した3種類の4重変異型ギ酸脱水素酵素について、前記(4-2)で行った耐久性加速試験の結果を図4に示す。また、これら3種類の4重変異型ギ酸脱水素酵素及び野生型のギ酸脱水素酵素について比活性を測定した結果を図5に示す。図5に示すように、上記(3-1)で取得した3種類の4重変異型ギ酸脱水素酵素については、野生型とほぼ同等の比活性を示すことが分かった。これに対して、図4に示すように、上記(3-1)で取得した3種類の4重変異型ギ酸脱水素酵素については、野生型と比較すると耐久性が大幅に向上していることが分かった。
上記(3-1)で取得した5重変異型ギ酸脱水素酵素、6重変異型ギ酸脱水素酵素及び7重変異型ギ酸脱水素酵素並びに野生型ギ酸脱水素酵素について、耐久性の加速試験結果を図6~14に示す。また、上記(3-1)で取得した5重変異型ギ酸脱水素酵素、6重変異型ギ酸脱水素酵素及び7重変異型ギ酸脱水素酵素並びに野生型ギ酸脱水素酵素について、比活性を測定した結果を図15及び16に示す。なお、図15及び16は、野生型のギ酸脱水素酵素の比活性を1としたときの相対値として示している。また、図15は、野生型のギ酸脱水素酵素と同等の比活性を示す変異型ギ酸脱水素酵素を纏めて示している。また、図16は、主として野生型のギ酸脱水素酵素よりやや劣る比活性を示す変異型ギ酸脱水素酵素を示している。
図6~9に示すように、プラスミド名5B-2、5B-4、5C-2、6B-1、6B-11、6B-2、6B-3、6B-5、6B-7、6B-8、6C-1及び7B-3に示される変異型ギ酸脱水素酵素は、野生型のギ酸脱水素酵素と同等の比活性を示しながらも、優れた耐久性を示すことが明らかとなった。
また、図10~12に示すように、5A-2、5B-5、6A-1、6B-4、6B-6、6B-9、6C-2、6D-1、7B-4、7D-1及び7D-2に示される変異型ギ酸脱水素酵素は、野生型のギ酸脱水素酵素と比較するとやや比活性が劣るものの、優れた耐久性を示すことが明らかとなった。
さらに、図13及び14に示すように、プラスミド名5B-6、5B-7、6B-10、6B-12、7B-1及び7B-2に示される変異型ギ酸脱水素酵素は、上述した変異型ギ酸脱水素酵素と比較すると、比活性及び/又は耐久性の点で劣るものの野生型のギ酸脱水素酵素と比較すると優れた耐久性を示すことが明らかとなった。
Claims (7)
- 配列番号2に示すアミノ酸配列からなるギ酸脱水素酵素における、
A)99番目のバリン、153番目のバリン、155番目のヒスチジン及び298番目のアスパラギンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列
B)73番目のグリシン、99番目のバリン、153番目のバリン及び155番目のヒスチジンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列、及び
C)73番目のグリシン、99番目のバリン、153番目のバリン及び159番目のグルタミンに相当するアミノ酸残基が他のアミノ酸に置換したアミノ酸配列
のうちいずれか1つのアミノ酸配列を含み、耐久性が向上した変異型ギ酸脱水素酵素。 - 上記A)のアミノ酸配列に対して、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、73番目のグリシン、82番目のリシン、136番目のイソロイシン、159番目のグルタミン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸及び343番目のアスパラギンからなる群から選ばれる1~3つのアミノ酸に相当するアミノ酸残基が他のアミノ酸に更に置換したアミノ酸配列を含むことを特徴とする請求項1記載の変異型ギ酸脱水素酵素。
- 上記B)のアミノ酸配列に対して、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、82番目のリシン、136番目のイソロイシン、159番目のグルタミン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸、298番目のアスパラギン及び343番目のアスパラギンからなる群から選ばれる1~3つのアミノ酸に相当するアミノ酸残基が他のアミノ酸に更に置換したアミノ酸配列を含むことを特徴とする請求項1記載の変異型ギ酸脱水素酵素。
- 上記C)のアミノ酸配列に対して、31番目のイソロイシン、50番目のアルギニン、60番目のグルタミン酸、64番目のイソロイシン、82番目のリシン、136番目のイソロイシン、155番目のヒスチジン、239番目のメチオニン、286番目のアスパラギン酸、287番目のヒスチジン、293番目のグルタミン酸、298番目のアスパラギン及び343番目のアスパラギンからなる群から選ばれる1~3つのアミノ酸に相当するアミノ酸残基が他のアミノ酸に更に置換したアミノ酸配列を含むことを特徴とする請求項1記載の変異型ギ酸脱水素酵素。
- 上記31番目のイソロイシンに相当するアミノ酸の置換後のアミノ酸がロイシンであり、上記50番目のアルギニンに相当するアミノ酸の置換後のアミノ酸がグリシンであり、上記60番目のグルタミン酸に相当するアミノ酸の置換後のアミノ酸がバリンであり、上記64番目のイソロイシンに相当するアミノ酸の置換後のアミノ酸がバリンであり、上記73番目のグリシンに相当するアミノ酸の置換後のアミノ酸がアラニンであり、上記82番目のリシンに相当するアミノ酸の置換後のアミノ酸がアルギニンであり、上記99番目のバリンに相当するアミノ酸の置換後のアミノ酸がイソロイシンであり、上記136番目のイソロイシンに相当するアミノ酸の置換後のアミノ酸がバリン、上記153番目のバリンに相当するアミノ酸の置換後のアミノ酸がイソロイシンであり、上記155番目のヒスチジンに相当するアミノ酸の置換後のアミノ酸がグルタミン又はリシンであり、上記159番目のグルタミンに相当するアミノ酸の置換後のアミノ酸がロイシン又はアスパラギンであり、上記239番目のメチオニンに相当するアミノ酸の置換後のアミノ酸がロイシンであり、上記286番目のアスパラギン酸に相当するアミノ酸の置換後のアミノ酸がフェニルアラニンであり、上記287番目のヒスチジンに相当するアミノ酸の置換後のアミノ酸がグルタミン又はプロリンであり、上記293番目のグルタミン酸に相当するアミノ酸の置換後のアミノ酸がアスパラギン酸であり、上記298番目のアスパラギンに相当するアミノ酸の置換後のアミノ酸がチロシンであり、上記343番目のアスパラギンに相当するアミノ酸の置換後のアミノ酸がアルギニンであることを特徴とする請求項1乃至4いずれか一項記載の変異型ギ酸脱水素酵素。
- 請求項1乃至5いずれか一項記載の変異型ギ酸脱水素酵素をコードする遺伝子。
- ギ酸及びNAD+を含む反応系に請求項1乃至5いずれか一項記載の変異型ギ酸脱水素酵素を作用させる、
NADHの製造方法。
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PCT/JP2009/063743 WO2011016102A1 (ja) | 2009-08-03 | 2009-08-03 | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 |
| US13/388,943 US8481294B2 (en) | 2009-08-03 | 2009-08-03 | Mutant formate dehydrogenase, gene encoding the same, and method for producing NADH |
| EP09848035.3A EP2471914B1 (en) | 2009-08-03 | 2009-08-03 | Mutant formate dehydrogenase, gene encoding same, and method for producing nadh |
| JP2011525702A JP5605361B2 (ja) | 2009-08-03 | 2009-08-03 | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PCT/JP2009/063743 WO2011016102A1 (ja) | 2009-08-03 | 2009-08-03 | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2011016102A1 true WO2011016102A1 (ja) | 2011-02-10 |
Family
ID=43544024
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2009/063743 WO2011016102A1 (ja) | 2009-08-03 | 2009-08-03 | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US8481294B2 (ja) |
| EP (1) | EP2471914B1 (ja) |
| JP (1) | JP5605361B2 (ja) |
| WO (1) | WO2011016102A1 (ja) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104342406A (zh) * | 2013-07-26 | 2015-02-11 | 南京朗恩生物科技有限公司 | 热稳定性增强的甲酸脱氢酶突变体及其制备方法 |
| JPWO2013051685A1 (ja) * | 2011-10-07 | 2015-03-30 | 味の素株式会社 | 変異型γ−グルタミルトランスフェラーゼ、及び、γ−グルタミルバリルグリシン又はその塩の製造法 |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN107109443B (zh) * | 2014-11-06 | 2021-11-02 | 庆尚大学校产学协力团 | 阿洛酮糖的制备方法 |
| CN106479988B (zh) * | 2016-11-08 | 2019-08-06 | 江南大学 | 一种酶活和稳定性提高的甲酸脱氢酶突变体及其构建方法 |
| CN117603924B (zh) * | 2023-04-25 | 2024-05-17 | 杭州力文所生物科技有限公司 | 蛋白可溶性表达提高的甲酸脱氢酶突变体及其应用 |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH1023896A (ja) | 1996-05-07 | 1998-01-27 | Unitika Ltd | 組換えプラスミド、それにより形質転換された大腸菌、その培養物及びそれを用いたアミノ酸又はその誘導体の製造方法 |
| JP2002233395A (ja) | 2001-02-06 | 2002-08-20 | Mitsubishi Rayon Co Ltd | 補酵素nadhの再生方法 |
| JP2003180383A (ja) | 2001-09-21 | 2003-07-02 | Degussa Ag | カンジダボイジニイからのギ酸デヒドロゲナーゼの新規の変異体 |
| JP2008100448A (ja) | 2006-10-20 | 2008-05-01 | Toyoda Gosei Co Ltd | ウエザストリップ |
Family Cites Families (16)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS63313580A (ja) | 1987-06-15 | 1988-12-21 | Sagami Chem Res Center | 蟻酸脱水素酵素 |
| JPH06102019B2 (ja) | 1989-07-31 | 1994-12-14 | 工業技術院長 | ギ酸脱水素酵素及びその製造方法 |
| DE19753350A1 (de) | 1997-12-03 | 1999-06-10 | Degussa | Neue Mutanten der Formiatdehydrogenase aus Candida boidinii, neue Gensequenzen diese codierend sowie Verwendung der neuen Formiatdehydrogenasen |
| JP2000069971A (ja) | 1998-08-28 | 2000-03-07 | Tadayuki Imanaka | 新規な耐熱性蟻酸脱水素酵素 |
| JP2000078970A (ja) | 1998-09-04 | 2000-03-21 | Unitika Ltd | ギ酸脱水素酵素及びその製造方法 |
| JP2000245471A (ja) | 1999-03-01 | 2000-09-12 | Unitika Ltd | ギ酸脱水素酵素遺伝子、それを含有する組換えベクター、その組換えベクターを含有する形質転換体及びその形質転換体を用いたギ酸脱水素酵素の製造方法 |
| JP2003339390A (ja) | 1999-03-04 | 2003-12-02 | Nara Institute Of Science & Technology | 蟻酸デヒドロゲナーゼポリペプチド、当該ポリペプチドをコード化するdnaフラグメント |
| JP3448609B2 (ja) | 1999-03-04 | 2003-09-22 | 奈良先端科学技術大学院大学長 | 植物の光酸化障害を回避させる方法 |
| JP4287144B2 (ja) | 2000-12-04 | 2009-07-01 | 株式会社カネカ | 新規ギ酸脱水素酵素及びその製造方法 |
| US20040115691A1 (en) * | 2002-07-10 | 2004-06-17 | Rozzell J. David | Mutants of enzymes and methods for their use |
| JP4116349B2 (ja) | 2002-07-10 | 2008-07-09 | 株式会社カネカ | 補酵素依存型を改変したギ酸脱水素酵素 |
| DE10327454A1 (de) | 2003-06-18 | 2005-01-20 | Juelich Enzyme Products Gmbh | Oxidoreduktase aus Pichia capsulata |
| JP5332277B2 (ja) | 2008-04-08 | 2013-11-06 | トヨタ自動車株式会社 | Nadhの製造方法及びギ酸脱水素酵素の製造方法 |
| JP2010041971A (ja) * | 2008-08-14 | 2010-02-25 | Toyota Motor Corp | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 |
| JP5381113B2 (ja) * | 2009-01-16 | 2014-01-08 | トヨタ自動車株式会社 | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 |
| JP5201218B2 (ja) * | 2009-06-15 | 2013-06-05 | トヨタ自動車株式会社 | 無細胞タンパク質合成溶液、無細胞タンパク質合成キット及びタンパク質合成方法 |
-
2009
- 2009-08-03 WO PCT/JP2009/063743 patent/WO2011016102A1/ja active Application Filing
- 2009-08-03 EP EP09848035.3A patent/EP2471914B1/en not_active Not-in-force
- 2009-08-03 JP JP2011525702A patent/JP5605361B2/ja not_active Expired - Fee Related
- 2009-08-03 US US13/388,943 patent/US8481294B2/en active Active
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH1023896A (ja) | 1996-05-07 | 1998-01-27 | Unitika Ltd | 組換えプラスミド、それにより形質転換された大腸菌、その培養物及びそれを用いたアミノ酸又はその誘導体の製造方法 |
| JP2002233395A (ja) | 2001-02-06 | 2002-08-20 | Mitsubishi Rayon Co Ltd | 補酵素nadhの再生方法 |
| JP2003180383A (ja) | 2001-09-21 | 2003-07-02 | Degussa Ag | カンジダボイジニイからのギ酸デヒドロゲナーゼの新規の変異体 |
| JP2008100448A (ja) | 2006-10-20 | 2008-05-01 | Toyoda Gosei Co Ltd | ウエザストリップ |
Non-Patent Citations (18)
| Title |
|---|
| "Amino Acid-Peptide-Protein 5.1 Amino acid", KAGAKU-DOJIN PUBLISHING COMPANY, INC., article "McKee Biochemistry (Biochemistry: the molecular basis of life" |
| BABA, T. ET AL., LANCET, vol. 359, 2002, pages 1819 - 1827 |
| BIOCHEMISTRY (MOSCOW, vol. 69, no. 11, 2004, pages 1252 - 1267 |
| BIOKHIMIYA, vol. 69, no. 11, 2004, pages 1537 - 1554 |
| BIOMOLECULAR ENGINEERING, vol. 23, 2006, pages 98 - 110 |
| CAZALET ET AL., NAT. GENET, vol. 36, 2004, pages 1165 - 1173 |
| CHIEN ET AL., SCIENCE, vol. 305, 1966, pages 2004 |
| HENIKOFF S.; HENIKOFF J.G.: "Amino-acid substitution matrices from protein blocks", PROC. NATL. ACAD. SCI. U.S.A., vol. 89, 1992, pages 10915 - 10919, XP002599751, DOI: doi:10.1073/pnas.89.22.10915 |
| HWANG ET AL., MOL. BIOL. CELL, vol. 14, 2003, pages 2314 - 2326 |
| J. SAMBROOK ET AL.: "Molecular Cloning, A Laboratory Manual", 1989, COLD SPRING HARBOR LABORATORY |
| LARSSON ET AL., NAT. GENET, vol. 37, 2005, pages 153 - 159 |
| LI ET AL., PROC. NATL. ACAD. SCI. U.S.A., vol. 102, 2005, pages 12344 - 12349 |
| LOFTUS ET AL., SCIENCE, vol. 307, 2005, pages 1321 - 1324 |
| PARKHILL ET AL., NAT. GENET, vol. 35, 2003, pages 32 - 40 |
| POPOV, V.O. ET AL.: "NAD+-dependent formate dehydrogenase", BIOCHEM. J., vol. 301, 1994, pages 625 - 643, XP002909477 * |
| ROJKOVA, A.M. ET AL.: "Bacterial formate dehydrogenase. Increasing the enzyme thermal stability by hydrophobization of alpha-helices", FEES LETTERS, vol. 445, 1999, pages 183 - 188, XP004259872 * |
| See also references of EP2471914A4 |
| TISHKOV, V.I. ET AL.: "Protein engineering of formate dehydrogenase", BIOMOL. ENG., vol. 23, 2006, pages 89 - 110, XP028041903 * |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPWO2013051685A1 (ja) * | 2011-10-07 | 2015-03-30 | 味の素株式会社 | 変異型γ−グルタミルトランスフェラーゼ、及び、γ−グルタミルバリルグリシン又はその塩の製造法 |
| US9677106B2 (en) | 2011-10-07 | 2017-06-13 | Ajinomoto Co., Inc. | Method for producing gamma-glutamylvalylglycine or a salt thereof |
| CN104342406A (zh) * | 2013-07-26 | 2015-02-11 | 南京朗恩生物科技有限公司 | 热稳定性增强的甲酸脱氢酶突变体及其制备方法 |
| CN104342406B (zh) * | 2013-07-26 | 2017-03-08 | 南京朗恩生物科技有限公司 | 热稳定性增强的甲酸脱氢酶突变体及其制备方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2011016102A1 (ja) | 2013-01-10 |
| US20120129222A1 (en) | 2012-05-24 |
| EP2471914A1 (en) | 2012-07-04 |
| US8481294B2 (en) | 2013-07-09 |
| EP2471914A4 (en) | 2013-03-20 |
| EP2471914B1 (en) | 2014-10-08 |
| JP5605361B2 (ja) | 2014-10-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Hoelsch et al. | Engineering of formate dehydrogenase: synergistic effect of mutations affecting cofactor specificity and chemical stability | |
| JP6185484B2 (ja) | バシラス由来の新規のメタノールデヒドロゲナーゼ酵素 | |
| Zhang et al. | A new NAD (H)-dependent meso-2, 3-butanediol dehydrogenase from an industrially potential strain Serratia marcescens H30 | |
| JP5605361B2 (ja) | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 | |
| Wang et al. | Characterization of a stereospecific acetoin (diacetyl) reductase from Rhodococcus erythropolis WZ010 and its application for the synthesis of (2 S, 3 S)-2, 3-butanediol | |
| Fedorov et al. | 1-Aminocyclopropane-1-carboxylate (ACC) deaminases from Methylobacterium radiotolerans and Methylobacterium nodulans with higher specificity for ACC | |
| Tang et al. | Molecular cloning and characterization of a malic enzyme gene from the oleaginous yeast Lipomyces starkeyi | |
| JP5867586B2 (ja) | 炭化水素合成酵素遺伝子及びその利用 | |
| Itoh et al. | PCR-based amplification and heterologous expression of Pseudomonas alcohol dehydrogenase genes from the soil metagenome for biocatalysis | |
| US20220348974A1 (en) | Biotin synthases for efficient production of biotin | |
| Machielsen et al. | Laboratory evolution of Pyrococcus furiosus alcohol dehydrogenase to improve the production of (2S, 5S)-hexanediol at moderate temperatures | |
| Matsuo et al. | Isocitrate dehydrogenase isozymes from a psychrotrophic bacterium, Pseudomonas psychrophila | |
| Ying et al. | Characterization of an allylic/benzyl alcohol dehydrogenase from Yokenella sp. strain WZY002, an organism potentially useful for the synthesis of α, β-unsaturated alcohols from allylic aldehydes and ketones | |
| Yoshida et al. | Three inositol dehydrogenases involved in utilization and interconversion of inositol stereoisomers in a thermophile, Geobacillus kaustophilus HTA426 | |
| Patel et al. | Substrate specificity of the Bacillus licheniformis lyxose isomerase YdaE and its application in in vitro catalysis for bioproduction of lyxose and glucose by two-step isomerization | |
| Neveling et al. | Purification of the pyruvate dehydrogenase multienzyme complex of Zymomonas mobilis and identification and sequence analysis of the corresponding genes | |
| Sasaki et al. | Lactobacillus reuteri ATCC 53608 mdh gene cloning and recombinant mannitol dehydrogenase characterization | |
| JP5381113B2 (ja) | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 | |
| Erb et al. | Phenylacetate metabolism in thermophiles: characterization of phenylacetate-CoA ligase, the initial enzyme of the hybrid pathway in Thermus thermophilus | |
| JP2010041971A (ja) | 変異型ギ酸脱水素酵素、これをコードする遺伝子及びnadhの製造方法 | |
| JP5332277B2 (ja) | Nadhの製造方法及びギ酸脱水素酵素の製造方法 | |
| CN106795511B (zh) | 氧化酶、编码该酶的多核苷酸、以及它们的应用 | |
| WO2013002277A1 (ja) | 酵素機能改変方法及びその変異体 | |
| KR20190097250A (ko) | 신규한 효소를 사용한 메틸글리옥살의 히드록시아세톤으로의 전환 및 그의 적용 | |
| Si et al. | Cloning and overexpression of ketopantoic acid reductase gene from Stenotrophomonas maltophilia and its application to stereospecific production of D-pantoic acid |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 09848035 Country of ref document: EP Kind code of ref document: A1 |
|
| DPE1 | Request for preliminary examination filed after expiration of 19th month from priority date (pct application filed from 20040101) | ||
| WWE | Wipo information: entry into national phase |
Ref document number: 2011525702 Country of ref document: JP |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 13388943 Country of ref document: US |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2009848035 Country of ref document: EP |