KR20170138543A - 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 - Google Patents
메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 Download PDFInfo
- Publication number
- KR20170138543A KR20170138543A KR1020177033598A KR20177033598A KR20170138543A KR 20170138543 A KR20170138543 A KR 20170138543A KR 1020177033598 A KR1020177033598 A KR 1020177033598A KR 20177033598 A KR20177033598 A KR 20177033598A KR 20170138543 A KR20170138543 A KR 20170138543A
- Authority
- KR
- South Korea
- Prior art keywords
- nucleic acid
- yeast cell
- sequence
- dna
- leu
- Prior art date
Links
Images










Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/80—Vectors or expression systems specially adapted for eukaryotic hosts for fungi
- C12N15/81—Vectors or expression systems specially adapted for eukaryotic hosts for fungi for yeasts
- C12N15/815—Vectors or expression systems specially adapted for eukaryotic hosts for fungi for yeasts for yeasts other than Saccharomyces
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/37—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from fungi
- C07K14/39—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from fungi from yeasts
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/415—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from plants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/795—Porphyrin- or corrin-ring-containing peptides
- C07K14/805—Haemoglobins; Myoglobins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/14—Fungi; Culture media therefor
- C12N1/16—Yeasts; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/635—Externally inducible repressor mediated regulation of gene expression, e.g. tetR inducible by tetracyline
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/67—General methods for enhancing the expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/80—Vectors or expression systems specially adapted for eukaryotic hosts for fungi
- C12N15/81—Vectors or expression systems specially adapted for eukaryotic hosts for fungi for yeasts
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02E—REDUCTION OF GREENHOUSE GAS [GHG] EMISSIONS, RELATED TO ENERGY GENERATION, TRANSMISSION OR DISTRIBUTION
- Y02E50/00—Technologies for the production of fuel of non-fossil origin
- Y02E50/10—Biofuels, e.g. bio-diesel
Abstract
메틸영양성 효모를 유전적으로 조작하기 위한 방법 및 물질이 제공된다.
Description
일반적으로 본 개시내용은 메틸영양성 효모를 유전적으로 조작하기 위한 DNA 구축물 및 이러한 DNA 구축물을 사용하는 방법에 관한 것이다.
메틸영양성 효모 예컨대 피키아 파스토리스(Pichia pastoris)가 재조합 단백질의 발현에 통상적으로 사용된다. 메틸영양성 효모에서 하나 이상의 폴리펩티드를 효율적으로 발현시키는데 사용될 수 있는 구축물이 본원에서 제공된다.
본 개시내용은 AOX1 프로모터로부터 전사 활성화제인 Mxr1을 과다발현하는 피. 파스토리스(P. pastoris) 균주를 사용하여 또한 AOX1 프로모터로부터 발현되는 트랜스진의 발현을 증가시키는 것을 개시하며, 이는 하나 이상의 단백질의 재조합 생산을 유의하게 개선한다. 추가적으로, AOX1 프로모터로부터의 Mxr1의 발현은 억제성 탄소원이 고갈될 때, 일반적으로 절대적인 유도제인 메탄올의 부재 하에 AOX1 프로모터로부터의 다른 트랜스진의 발현을 허용하는 양성 피드백 루프를 생성시킨다. Mxr1의 발현은 생산되는 단백질의 양의 유의한 증가를 초래한다.
한 측면에서, 재조합 핵산 분자를 포함하는 메틸영양성 효모 세포가 제공된다. 전형적으로 이러한 재조합 핵산 분자는 적어도 하나의 메탄올-유도성 프로모터 요소에 작동가능하게 연결된 전사 활성화제를 코딩하는 외인성 핵산을 포함한다. 대표적인 메틸영양성 효모는 칸디다(Candida), 한세눌라(Hansenula), 피키아(Pichia) 및 토룰롭시스(Torulopsis) 속의 것일 수 있다. 대표적인 메틸영양성 효모는 피키아 파스토리스이다.
일부 실시양태에서, 재조합 핵산 분자는 메틸영양성 효모 세포의 게놈 내로 안정적으로 통합된다. 일부 실시양태에서, 재조합 핵산 분자는 복제-적격 플라스미드로부터 염색체외적으로 발현된다.
일부 실시양태에서, 전사 활성화제를 코딩하는 외인성 핵산은 피키아 파스토리스로부터의 Mxr1 서열, 한세눌라 폴리모르파(Hansenula polymorpha)로부터의 Adr1 서열, 칸디다 보이디니이(Candida boidinii)로부터의 Trm1 서열, 및 칸디다 보이디니이로부터의 Trm2 서열을 포함한다. 전사 활성화제를 코딩하는 대표적인 핵산이 DQ395124에서 제시된다. 대표적인 전사 활성화제는 ABD57365에 제시된 아미노산 서열을 갖는다.
일부 실시양태에서, 적어도 하나의 메탄올-유도성 프로모터 요소는 피키아 파스토리스로부터의 알콜 옥시다제 1 (AOX1) 프로모터 요소, 칸디다 보이디니이로부터의 AOD1 프로모터 요소, 한세눌라 폴리모르파로부터의 MOX 프로모터 요소, 피키아 메타놀리카(Pichia methanolica)로부터의 MOD1 프로모터 요소, 피키아 파스토리스로부터의 DHAS 프로모터 요소, 피키아 파스토리스로부터의 FLD1 프로모터 요소, 또는 피키아 파스토리스로부터의 PEX8 프로모터 요소이다.
일부 실시양태에서, 메틸영양성 효모 세포는 적어도 하나의 메탄올-유도성 프로모터 요소에 작동가능하게 연결된 폴리펩티드를 코딩하는 적어도 하나의 이종성 핵산을 포함하는 핵산 분자를 추가로 포함한다. 일부 실시양태에서, 적어도 하나의 이종성 핵산은 철 보조인자 예컨대 헴의 생합성에서 수반되는 적어도 하나의 폴리펩티드 (예를 들어, ALA 신타제, ALA 데히드라타제, 포르포빌리노겐 데아미나제, UPG III 신타제, UPG III 데카르복실라제, CPG 옥시다제, PPG 옥시다제, 및/또는 페로킬라타제)를 코딩한다. 일부 실시양태에서, 철 보조인자 생합성에서 수반되는 폴리펩티드 중 하나 이상이 적어도 하나의 메탄올-유도성 프로모터 요소에 연결된다.
또 다른 측면에서, 세포에서 이종성 폴리펩티드를 발현시키는 방법이 제공된다. 전형적으로 이러한 방법은 본원에 기술된 바와 같은 메틸영양성 효모 세포를 제공하는 단계; 재조합 핵산 분자를 메틸영양성 효모 세포 내로 도입하며, 이러한 재조합 핵산 분자는 적어도 하나의 피키아 파스토리스 알콜 옥시다제 1 (AOX1) 프로모터 요소에 작동가능하게 연결된 폴리펩티드를 코딩하는 적어도 하나의 이종성 핵산을 포함하는 것인 단계; 및 세포를 재조합 핵산 분자의 발현에 적합한 조건 하에 배양하며, 이에 의해 이종성 폴리펩티드를 발현시키는 단계를 포함한다.
일부 실시양태에서, 세포가 배양되는 조건은 철 또는 그의 제약상 또는 대사상 허용되는 염의 첨가를 포함한다. 일부 실시양태에서, 도입 단계는 형질도입, 전기천공, 바이오리스틱 입자 전달 또는 화학적 형질전환과 같은 기술을 포함한다. 일부 실시양태에서, 배양 단계는 세포를 메탄올의 존재 하에 배양하는 것을 포함한다.
또 다른 측면에서, 활성화시키는 프로모터에 작동가능하게 연결된 전사 활성화제를 포함하는 재조합 생물이 제공된다. 일부 실시양태에서, 이러한 프로모터에 작동가능하게 연결된 폴리펩티드를 발현하는 재조합 생물이 제공된다. 또 다른 측면에서, 본원에 기술된 바와 같이 유도제의 첨가 없이 유도성 프로모터로부터 폴리펩티드를 발현시키는 방법이 제공된다.
달리 정의되지 않는 한, 본원에서 사용된 모든 기술 용어 및 과학 용어는 사안 방법 및 조성물이 속하는 분야의 통상의 기술자가가 통상적으로 이해하는 바와 동일한 의미를 갖는다. 본원에 기술된 것과 유사하거나 등가인 방법 및 물질이 사안 방법 및 조성물의 실행 또는 테스트에서 사용될 수 있지만, 적합한 방법 및 물질이 하기에서 기술된다. 추가적으로, 물질, 방법 및 예는 예시적일 뿐이고, 제한적인 것으로 의도되지 않는다. 본원에서 언급된 모든 발표물, 특허 출원, 특허 및 다른 참고문헌은 전문이 참조로 포함된다.
도 1은 헴 생합성 경로에서 수반되는 단계를 도시하는 개략도이다.
도 2는 생산 균주 MXY0183의 구축에서 사용된 플라스미드의 개략도이다.
도 3은 모 균주 Bg11로부터의 생산 균주 MXY0183 및 MXY0207의 생성을 나타내는 개략도이다.
도 4는 플라스미드 pGAB 및 pMx354를 나타내는 개략도이다.
도 5는 모 균주 Bg11로부터의 항생제 선택이 없는 생산 균주 MXY0291 및 MXY0338의 생성을 나타내는 개략도이다.
도 6은 생산 균주 MXY0291을 제조하기 위해 공동-형질전환에 의해 도입된 Mxr1 및 LegH 변이체 3을 함유하는 선형 DNA 조각의 개략도이다.
도 7은 피키아의 천연 비-pAOX1 구성적 프로모터의 제어 하에 LegH를 발현하는 선형 구축물을 나타내는 개략도이다.
도 8은 균주 MXY0183과 연관된 표현형 변화를 나타내는 사진이다. 유도 시작 (0 hr) 및 유도 72 hr 후의 진탕 플라스크가 제시된다. 1, MXY0051; 2, MXY0118; 3, MXY0183.
도 9는 변형된 피. 파스토리스 균주로부터의 LegH 생산을 나타낸다. 패널 A는 진탕 플라스크에서 성장된 피. 파스토리스 균주로부터의 용해물을 나타내는 SDS 겔이다: 51, MXY0051; 118, MXY0118; 183, MXY0183. 패널 B는 균주 MXY0118, MXY0183, 및 MXY0207로부터의 LegH 생산을 비교하는 표이다.
도 10은 균주 MXY0206으로의 실험으로부터의 데이터를 나타낸다. 패널 A는 억제성 탄소원에서의 48 hr의 성장 후의 균주 MXY0183 (왼쪽) 및 MXY0206 (오른쪽)의 진탕 플라스크 배양물의 사진이다. 패널 B는 BMY 배지에서의 48 hr의 성장 후의 균주 MXY0183 (왼쪽) 및 MXY0206 (오른쪽)의 진탕 플라스크 배양물로부터의 세포 펠릿의 사진이다. 패널 C는 (임의의 유도제의 부재 하에서의) 헴-로딩된 LegH의 상대적인 수율을 나타내는 그래프이다.
도 11은 2 L 발효기 탱크에서 메탄올과 글리세롤 또는 메탄올과 글루코스의 존재 하에서 성장되었을 때의 본원에 기술된 균주의 상대적인 수율을 나타내는 요약 표이다.
도 2는 생산 균주 MXY0183의 구축에서 사용된 플라스미드의 개략도이다.
도 3은 모 균주 Bg11로부터의 생산 균주 MXY0183 및 MXY0207의 생성을 나타내는 개략도이다.
도 4는 플라스미드 pGAB 및 pMx354를 나타내는 개략도이다.
도 5는 모 균주 Bg11로부터의 항생제 선택이 없는 생산 균주 MXY0291 및 MXY0338의 생성을 나타내는 개략도이다.
도 6은 생산 균주 MXY0291을 제조하기 위해 공동-형질전환에 의해 도입된 Mxr1 및 LegH 변이체 3을 함유하는 선형 DNA 조각의 개략도이다.
도 7은 피키아의 천연 비-pAOX1 구성적 프로모터의 제어 하에 LegH를 발현하는 선형 구축물을 나타내는 개략도이다.
도 8은 균주 MXY0183과 연관된 표현형 변화를 나타내는 사진이다. 유도 시작 (0 hr) 및 유도 72 hr 후의 진탕 플라스크가 제시된다. 1, MXY0051; 2, MXY0118; 3, MXY0183.
도 9는 변형된 피. 파스토리스 균주로부터의 LegH 생산을 나타낸다. 패널 A는 진탕 플라스크에서 성장된 피. 파스토리스 균주로부터의 용해물을 나타내는 SDS 겔이다: 51, MXY0051; 118, MXY0118; 183, MXY0183. 패널 B는 균주 MXY0118, MXY0183, 및 MXY0207로부터의 LegH 생산을 비교하는 표이다.
도 10은 균주 MXY0206으로의 실험으로부터의 데이터를 나타낸다. 패널 A는 억제성 탄소원에서의 48 hr의 성장 후의 균주 MXY0183 (왼쪽) 및 MXY0206 (오른쪽)의 진탕 플라스크 배양물의 사진이다. 패널 B는 BMY 배지에서의 48 hr의 성장 후의 균주 MXY0183 (왼쪽) 및 MXY0206 (오른쪽)의 진탕 플라스크 배양물로부터의 세포 펠릿의 사진이다. 패널 C는 (임의의 유도제의 부재 하에서의) 헴-로딩된 LegH의 상대적인 수율을 나타내는 그래프이다.
도 11은 2 L 발효기 탱크에서 메탄올과 글리세롤 또는 메탄올과 글루코스의 존재 하에서 성장되었을 때의 본원에 기술된 균주의 상대적인 수율을 나타내는 요약 표이다.
폴리펩티드의 재조합 발현을 증가시키도록 세포를 유전적으로 조작하는 것을 허용하는 핵산 구축물이 본원에서 제공된다. 일부 실시양태에서, 유도 분자의 부재 하에 유도성 프로모터로부터의 폴리펩티드의 재조합 발현을 증가시키도록 세포를 유전적으로 조작하는 것을 허용하는 핵산 구축물이 본원에서 제공된다. 어떠한 특정 메커니즘에도 제한되지 않으면서, 본원에 기술된 방법은 전사 활성화제의 낮은 수준의 천연 발현이 전사 활성화제에 작동가능하게 연결된 프로모터를 유도하는 양성 피드백 루프를 생성시킨다. 이는 전사 활성화제 뿐만 아니라 동일한 유도성 프로모터에 작동가능하게 연결된 하나 이상의 표적 폴리펩티드의 발현 증가에 이른다.
메틸영양성 효모 세포를 유전적으로 조작하는 것을 허용하는 핵산 구축물이 본원에서 제공된다. 본원에서 피키아 종 (즉, 피. 파스토리스)을 사용하여 방법이 예시되지만, 피키아 속의 다른 종, 또는 칸디다, 한세눌라, 피키아 및 토룰롭시스 속 중 임의의 것으로부터의 종을 사용할 수 있다.
메틸영양성 효모 세포를 유전적으로 조작하는 것은 세포 내로 재조합 핵산 분자를 도입하는 것을 전형적으로 포함한다. 본원에 기술된 바와 같이, 재조합 핵산 분자는 적어도 하나의 유도성 프로모터 요소에 작동가능하게 연결된 전사 활성화제를 코딩하는 외인성 핵산을 전형적으로 포함한다.
본원에 기술된 방법에서 사용되는 재조합 핵산 분자는 전형적으로 DNA지만, 적절한 환경 하에 RNA 분자를 사용할 수 있다. 본원에서 사용된 바와 같이, "외인성"은 외부 공급원으로부터 세포의 게놈 내로 도입된 임의의 핵산 서열을 지칭하고, 여기서 외부 공급원은 동일하거나 상이한 생물 또는 합성으로 생성된 핵산일 수 있다. 예를 들어, 외인성 핵산은 하나의 미생물 (예를 들어, 하나의 메틸영양성 효모 속 또는 종)으로부터의 핵산이 상이한 속 또는 종의 메틸영양성 효모 내로 도입된 핵산일 수 있지만, 외인성 핵산은 상응하는 천연 핵산 서열이 존재함에도 불구하고 추가적인 카피로서 메틸영양성 효모 내로 재조합으로 도입된 메틸영양성 효모로부터의 핵산일 수도 있다. 예를 들어, 피. 파스토리스는 Mxr1 전사 활성화제를 코딩하는 내인성 핵산을 함유한다; 추가적인 피. 파스토리스 Mxr1 핵산 (예를 들어, 피. 파스토리스 내로 재조합으로 도입됨) 또는 내인성 피. 파스토리스 Mxr1 핵산을 변형시키는 것이 외인성으로 간주된다.
전사 활성화제, 및 전사 활성화제를 코딩하는 핵산 (예를 들어, 전사 활성화제를 코딩하는 외인성 핵산)이 관련 기술분야에 공지되어 있다. 예를 들어, 피키아 파스토리스로부터의 전사 활성화제는 Mxr1 서열이지만, 적합한 전사 활성화제를 한세눌라 폴리모르파 (Adr1 서열; 예를 들어, 핵산 서열은 진뱅크(GenBank) 수탁 번호 AEOI02000005의 염기 858873 내지 862352, 아미노산 서열은 진뱅크 수탁 번호 ESX01253 참조) 및 칸디다 보이디니이 (Trm1 서열; 예를 들어, 핵산 서열은 진뱅크 수탁 번호 AB365355, 아미노산 서열은 진뱅크 수탁 번호 BAF99700을 참조한다; Trm2 서열; 예를 들어, 핵산 서열은 진뱅크 수탁 번호 AB548760, 아미노산 서열은 진뱅크 수탁 번호 BAJ07608 참조)에서 또한 발견할 수 있다. 대표적인 피. 파스토리스 Mxr1 핵산 서열을, 예를 들어, 진뱅크 수탁 번호 DQ395124에서 확인할 수 있는 한편, 대표적인 피. 파스토리스 Mxr1 폴리펩티드 서열을, 예를 들어, 진뱅크 수탁 번호 ABD57365에서 확인할 수 있다.
전사 활성화제 예컨대 Mxr1은 정상적으로는 낮은 수준으로 발현될 수 있다. 따라서, 외인성 핵산 (즉, 전사 활성화제)를 유도성인 프로모터의 제어 하에 놓는 것이 바람하다. 본원에서 사용된 바와 같이, "작동가능하게 연결된"은 프로모터 또는 다른 발현 요소(들)가 핵산 코딩 서열에 대해 이러한 핵산의 발현을 지시하거나 조절하는 방식으로 (예를 들어, 인-프레임(in-frame)으로) 위치하는 것을 의미한다.
메틸영양성 효모를 유전적으로 조작할 때 사용할 수 있는 다수의 유도성 프로모터가 있다. 예를 들어, 메탄올-유도성 프로모터, 또는 이로부터의 프로모터 요소를 사용할 수 있다. 관련 기술분야에 메탄올 유도성 프로모터가 공지되어 있다. 예를 들어, 피. 파스토리스로부터의 통상적으로 사용되는 메탄올-유도성 프로모터는 메탄올에 반응하여 강하게 전사되는 알콜 옥시다제 1 (AOX1) 유전자로부터의 프로모터 또는 그의 일부분이다. 그러나, 칸디다 보이디니이로부터의 알콜 옥시다제 (AOD1) 프로모터 (예를 들어, 진뱅크 수탁 번호 YSAAOD1A 참조), 한세눌라 폴리모르파로부터의 알콜 옥시다제 (MOX) 프로모터 (예를 들어, 진뱅크 수탁 번호 X02425 참조), 피키아 메타놀리카로부터의 MOD1 또는 MOD2 프로모터 (예를 들어, 문헌 [Raymond et al., 1998, Yeast, 14:11-23]; 및 [Nakagawa et al., 1999, Yeast, 15:1223-30] 참조), 피. 파스토리스로부터의 DHAS 프로모터 (예를 들어, 진뱅크 수탁 번호 FJ752551 참조) 또는 이로부터의 프로모터 요소, 피키아 파스토리스로부터의 포름알데히드 데히드로게나제 (FLD1) 프로모터 (예를 들어, 진뱅크 수탁 번호 AF066054 참조), 또는 피. 파스토리스로부터의 PEX8 프로모터 (예를 들어, 문헌 [Kranthi et al., 2010, Yeast, 27:705-11] 참조)를 비제한적으로 포함하여, 다른 메탄올-유도성 프로모터 또는 이로부터의 프로모터 요소를 사용할 수 있다. 일부 실시양태에서, 전사 활성화제는 피키아 파스토리스로부터의 Mit1 서열 (예를 들어, 진뱅크 수탁 번호 CAY70887 참조)이다. 이들 프로모터 모두가 메탄올에 의해 유도되는 것으로 공지되어 있다.
통상의 기술자는 본원에 기술된 재조합 핵산 분자가 메틸영양성 효모 세포의 게놈 내로 안정적으로 통합될 수 있거나, 또는 복제-적격 플라스미드로부터 염색체외적으로 발현될 수 있다는 것을 이해할 것이다. 둘 다를 달성하는 방법이 관련 기술분야에 널리 공지되어 있고, 일상적으로 사용된다.
본원에서 증명된 바와 같이, 피키아 내의 메탄올-조절 전사 활성화제가 AOX1 프로모터에 결합할 수 있고, Mxr1과 협동으로 작용하여 AOX1 프로모터로부터의 전사를 활성화시킬 수 있다. 일부 실시양태에서, 2개의 메탄올-조절 전사 활성화제 (예를 들어, Mxr1 및 Mit1)가 메탄올 유도성 프로모터 요소에 작동가능하게 연결될 수 있다.
본원에 기술된 바와 같은 재조합 핵산 분자를 포함하는 균주를 메틸영양성 효모 세포 내에서 제2 재조합 핵산 분자를 조절하는데 (예를 들어, 과발현시키는데) 사용할 수 있다. 제2 재조합 핵산 분자는, 예를 들어, 하나 이상의 관심 폴리펩티드를 코딩하는 하나 이상의 이종성 핵산을 포함할 수 있다. 전사 활성화제를 코딩하는 외인성 핵산과 유사하게, 이종성 핵산은 게놈에 대해 천연이 아니거나 생물의 게놈 내에 있지 않은 임의의 핵산 서열을 지칭한다 (예를 들어, 이종성 핵산은 하나의 미생물 (예를 들어, 하나의 메틸영양성 효모 속 또는 종)으로부터의 핵산이 상이한 속 또는 종의 메틸영양성 효모 내로 도입된 핵산일 수 있음).
단순히 예로서, 하나 이상의 관심 폴리펩티드를 코딩하는 이종성 핵산은 헴-보조인자 생합성에서 수반되는 핵산일 수 있다. 피키아 파스토리스 게놈의 서열로부터 결정되고 주해된 바와 같은 헴 생합성에서 수반되는 8개의 상이한 효소를 코딩하는 핵산이 본원에서 예시된다. 예를 들어, ALA 신타제, ALA 데히드라타제, 포르포빌리노겐 데아미나제, UPG III 신타제, UPG III 데카르복실라제, CPG 옥시다제, PPG 옥시다제, 및 페로킬라타제를 코딩하는 이종성 핵산이 본원에 기술된 메틸영양성 효모 균주에서 발현될 수 있다. 하나 초과의 이종성 핵산 (예를 들어, 트랜스진)을 함유하도록 메틸영양성 효모를 유전적으로 조작하기 위해, 메탄올-유도성 및 구성적 프로모터 또는 이로부터의 요소의 조합물을 조합하여 이러한 핵산의 발현을 추가로 증가시킬 수 있다.
사카로미세스 세레비시아에(Saccharomyces cerevisiae)에서의 기존의 연구에서, ALA 데히드라타제 및 포르포빌리노겐 데아미나제가 헴 생합성에서의 속도 제한 효소로서 확인되었다 (예를 들어, 문헌 [Hoffman et al., 2003, Biochem. Biophys. Res. Commun., 310(4):1247-53] 참조). 그러나, 글리세르알데히드-3-포스페이트 데히드로게나제 (GAP) 프로모터로부터의 피. 파스토리스에서의 개별적인 헴 효소의 이종성 발현은 헴 보조인자를 함유하는 재조합 단백질의 발현과 연관된 한계를 극복하지 못했다 (문헌 [Krainer et al., 2015, Microb. Cell Fact., 13;14:4] 참조). 본원에 기술된 바와 같이, 피. 파스토리스에서의 재조합 헴 함유 단백질의 고도로 효율적인 발현이 메탄올-유도성 프로모터로부터 전체 헴 생합성 경로를 공동-발현시킴으로써 달성되었지만, 헴 생합성 경로에서 수반되는 유전자 중 하나 이상이 하나 이상의 구성적 프로모터로부터 발현될 수 있다는 것이 이해될 것이다.
철-보조인자 생합성에서 수반되는 효소에 더하여, 식물 헤모글로빈을 포함하는 글로빈 단백질 패밀리 (Pfam 데이터베이스 내의 PF00042)의 구성원을 코딩하는 핵산이 존재할 수 있다는 것이 이해될 것이다. 본원에서의 실시예에서, 대두 레그헤모글로빈 (LegH)을 코딩하는 핵산이 존재한다. LegH는 철 보조인자인 헴에 결합하는 단백질이고, 이는 415 nm에서의 특징적인 흡수 및 독특한 적색을 초래한다. LegH 단백질 (일명 LGB2)은 대두의 뿌리혹에서 자연적으로 발견되고 (예를 들어, 유니프롯KB(UniprotKB) 수탁 번호 P02236 참조), 본원에서 사용된 핵산 서열은 피. 파스토리스에서의 발현에 대해 코돈이 최적화되었다. 예를 들어, WO 2014/110539 및 WO 2014/110532를 참조한다.
대안적으로, 관심 폴리펩티드를 코딩하는 이종성 핵산은, 예를 들어 비제한적으로, 데히드린, 피타제, 프로테아제, 카탈라제, 리파제, 퍼옥시다제, 아밀라제, 트랜스글루타미나제, 옥시도리덕타제, 트랜스퍼라제, 히드롤라제, 리아제, 이소머라제, 리가제, 또는 임의의 이러한 폴리펩티드에 대한 항체일 수 있다. 다른 실시양태에서, 이종성 핵산은 소형 분자, 예컨대 에탄올, 락트산, 부탄올, 아디프산 또는 숙신산의 생산을 위한 경로에서 수반되는 하나 이상의 효소를 코딩할 수 있다.
전사 활성화제를 코딩하는 외인성 핵산과 유사하게, 관심 폴리펩티드를 코딩하는 이종성 핵산이 유도성 프로모터 요소 (예를 들어, 메탄올-유도성 프로모터 요소)에 작동가능하게 연결될 수 있거나, 또는 관심 폴리펩티드를 코딩하는 이종성 핵산이 구성적 프로모터 또는 구성적 프로모터 요소에 작동가능하게 연결될 수 있다. 유도성 프로모터 및 이로부터의 요소는 상기에 논의되어 있다. 구성적 프로모터 및 구성적 프로모터 요소는 관련 기술분야에 공지되어 있다. 예를 들어, 피. 파스토리스로부터의 통상적으로 사용되는 구성적 프로모터는 구성적 방식으로 강하게 전사되는 전사 신장 인자 EF-1α 유전자 (TEF1)로부터의 프로모터 또는 그의 일부분이다. 그러나, 피. 파스토리스로부터의 글리세르알데히드-3-포스페이트 데히드로게나제 (GAPDH) 프로모터 (예를 들어, 진뱅크 수탁 번호 U62648.1 참조), 피. 파스토리스로부터의 잠재적 글리코실 포스파티딜 이노시톨 (GPI)-앵커 단백질인 GCW14p (PAS_chr1-4_0586)로부터의 프로모터 (예를 들어, 진뱅크 수탁 번호 XM_002490678 참조), 또는 피. 파스토리스로부터의 3-포스포글리세레이트 키나제 유전자 (PGK1)로부터의 프로모터 (예를 들어, 진뱅크 수탁 번호 AY288296 참조)를 비제한적으로 포함하여, 다른 구성적 프로모터 또는 이로부터의 프로모터 요소를 사용할 수 있다.
본원에 기술된 재조합 핵산 분자와 유사하게, 제2 재조합 핵산 분자는 메틸영양성 효모 세포의 게놈 내로 안정적으로 통합될 수 있거나, 또는 복제-적격 플라스미드로부터 염색체외적으로 발현될 수 있다.
통상의 기술자는 유도성 (예를 들어, 메탄올-유도성) 및 구성적 프로모터 (또는 이로부터의 프로모터 요소)의 조합물을 조합하여 이에 작동가능하게 연결된 임의의 핵산의 발현을 추가로 증가시킬 수 있다는 것을 이해할 것이다.
통상의 기술자는 프로모터 요소에 작동가능하게 연결된 관심 폴리펩티드를 코딩하는 이종성 핵산이 본원에 기술된 재조합 핵산 분자로부터 분리될 수 있거나, 또는 본원에 기술된 재조합 핵산 분자 내에 함유된 프로모터 요소에 작동가능하게 연결된 전사 활성화제를 코딩하는 외인성 핵산에 인접할 수 있다는 것을 이해할 것이다. 통상의 기술자는 제2 핵산 분자가 본원에 기술된 재조합 핵산 분자에 인접하면, 단일 프로모터 또는 이로부터의 프로모터 요소가 양쪽 또는 모든 유전자 (예를 들어, 전사 활성화제를 코딩하는 외인성 핵산, 뿐만 아니라 관심 폴리펩티드(들)를 코딩하는 하나 이상의 이종성 핵산)의 전사를 구동시키는데 사용될 수 있다는 것을 또한 이해할 것이다.
핵산을 메틸영양성 효모 세포 내로 도입하는 방법이 관련 기술분야에 공지되어 있고, 비제한적으로, 형질도입, 전기천공, 바이오리스틱 입자 전달, 및 화학적 형질전환을 포함한다.
추가적으로, 메틸영양성 효모 세포를 배양하는 방법이 관련 기술분야에 공지되어 있다. 예를 들어, 문헌 [Pichia Protocols, Methods In Molecular Biology, 389, Cregg, Ed., 2007, 2nd Ed., Humana Press, Inc.]을 참조한다. 일부 환경 하에, 메탄올을 배양 배지에 도입 또는 첨가하는 것이 바람직할 수 있지만, 본원에서 증명된 바와 같이, 하나 이상의 관심 폴리펩티드의 높은 수준의 효율적인 발현을 수득하기 위해 메탄올이 요구되지 않는다. 일부 환경 하에 (예를 들어, 철-보조인자 생합성에서 수반되는 효소(들)를 코딩하는 하나 이상의 핵산이 발현되는 경우), 배양 배지에 철 또는 그의 제약상 또는 대사상 허용되는 (또는 GRAS) 염을 보충하는 것이 바람직할 수 있다.
피키아 균주는 단독 탄소원으로서의 메탄올 상에서 성장할 수 있다. 알콜 옥시다제의 작용에 의해 메탄올을 포름알데히드로 전환하는 것에 의해 메탄올 이용이 시작된다. 메틸영양성 효모인 피키아 파스토리스는 알콜 옥시다제에 대한 2개의 유전자인 AOX1 및 AOX2를 함유한다. 알콜 옥시다제 활성이 감소된 균주 ("메탄올 이용 느림" 또는 MutS 균주)는 종종 알콜 옥시다제 활성이 감소되지 않은 균주보다 AOX1 프로모터로부터 발현된 재조합 단백질을 더 많이 생산한다. 양쪽 AOX 유전자에서 돌연변이되고 알콜 옥시다제 활성이 완전히 결여된 균주는 메탄올을 대사할 수 없지만, 여전히 메탄올에 의해 AOX1 프로모터로부터의 발현에 대해 유도될 수 있다. 이들 균주는 다른 탄소원을 성장에 이용하는 능력을 유지하지만, 여전히 메탄올 첨가 시 AOX1 프로모터로부터 이종성 단백질을 발현한다. 이들 균주는 메탄올을 대사하지 않기 때문에 ("메탄올 이용 마이너스" 또는 Mut- 균주), 훨씬 더 적은 메탄올이 단백질 발현 유도에 요구되고, 이러한 돌연변이를 보유하는 균주는 대규모 발효에서 메탄올 공급에 관련된 문제를 피한다. 예를 들어, 문헌 [Chiruvolu et al., 1997, Enzyme Microb. Technol., 21:277-83]을 참조한다. Mut- 균주에서의 AOX1 프로모터로부터의 LegH 발현이 LegH 수율을 크게 개선한 것으로 본원에서 결정되었다. 따라서, AOX1 유전자 및 AOX2 유전자 양쪽 모두에서 돌연변이가 있는 메틸영양성 효모가 본원에 기술된 방법에서 사용될 수 있다.
관심 단백질, 또는 하나 이상의 관심 단백질을 포함하는 복합체 (예를 들어, 헴-결합된 LegH, 데히드린, 피타제, 프로테아제, 카탈라제, 리파제, 퍼옥시다제, 아밀라제, 트랜스글루타미나제, 옥시도리덕타제, 트랜스퍼라제, 히드롤라제, 리아제, 이소머라제, 리가제, 또는 항체)를 이러한 효모 세포로부터 정제할 수 있다. 폴리펩티드를 정제하는 방법이 관련 기술분야에 공짖되어 있다. 본원에서 사용된 바와 같이, "정제된" 폴리펩티드는 이에 천연적으로 동반되는 세포 성분으로부터 분리 또는 정제된 폴리펩티드이다. 전형적으로, 폴리펩티드는 자신이 천연적으로 회합되는 폴리펩티드 또는 천연 발생 분자가 건조 중량 기준으로 적어도 70% (예를 들어, 적어도 75%, 80%, 85%, 90%, 95%, 또는 99%) 없으면 "정제된" 것으로 간주된다. 화학적으로 합성된 폴리펩티드는 이에 천연적으로 동반되는 성분으로부터 선천적으로 분리되기 때문에, 합성 폴리펩티드는 "정제된" 것이다.
본원에서 사용된 바와 같이, 핵산은 DNA 및 RNA를 포함할 수 있고, 하나 이상의 뉴클레오티드 유사체 또는 백본 변형을 함유하는 핵산을 포함한다. 핵산은 단일 가닥 또는 이중 가닥일 수 있고, 이는 일반적으로 그의 의도되는 용도에 좌우된다. 소정의 서열과 상이한 핵산 및 폴리펩티드가 또한 제공된다. 핵산 및 폴리펩티드는 소정의 핵산 또는 폴리펩티드 서열에 대한 적어도 50%의 서열 동일성 (예를 들어, 적어도 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 또는 99%의 서열 동일성)을 가질 수 있다.
퍼센트 서열 동일성의 계산에서, 2개의 서열을 정렬하고, 2개의 서열 사이의 동일하게 매칭되는 뉴클레오티드 또는 아미노산 잔기의 수를 결정한다. 동일한 매칭의 수를 정렬된 영역의 길이 (즉, 정렬된 뉴클레오티드 또는 아미노산 잔기의 수)로 나누고 100을 곱하여 퍼센트 서열 동일성 값에 도달한다. 정렬된 영역의 길이가 하나의 서열 또는 둘 다의 서열의 일부분에서 최단 서열의 전장 크기까지일 수 있다는 것이 이해될 것이다. 단일 서열이 하나 초과의 다른 서열과 정렬될 수 있고, 따라서 각각의 정렬된 영역에 걸쳐 상이한 퍼센트 서열 동일성 값을 가질 수 있다는 것이 또한 이해될 것이다.
퍼센트 서열 동일성을 결정하기 위해 2종 이상의 서열을 정렬하는 것은 컴퓨터 프로그램 ClustalW 및 디폴트 파라미터를 사용하여 수행할 수 있고, 이는 핵산 또는 폴리펩티드 서열의 정렬이 이들의 전체 길이에 걸쳐 수행되는 것을 허용한다 (전체 정렬). 문헌 [Chenna et al., 2003, Nucleic Acids Res., 31(13):3497-500]. ClustalW는 질의 서열과 하나 이상의 대상 서열 사이의 최상의 매칭을 계산하고, 동일성, 유사성 및 차이가 결정될 수 있도록 이들을 정렬한다. 서열 정렬을 최대화하도록 하나 이상의 잔기의 갭이 질의 서열, 대상 서열, 또는 양쪽 내로 삽입될 수 있다. 핵산 서열을 신속하게 쌍으로 정렬하기 위해, 디폴트 파라미터를 사용할 수 있고 (즉, 워드 크기: 2; 윈도우 크기: 4; 채점 방법: 백분율; 상부 대각선의 수: 4; 및 갭 페널티: 5); 다중 핵산 서열의 정렬을 위해, 하기 파라미터를 사용할 수 있다: 갭 개방 페널티: 10.0; 갭 연장 페널티: 5.0; 및 가중치 변환: 예. 폴리펩티드 서열을 신속하게 쌍으로 정렬하기 위해, 하기 파라미터를 사용할 수 있다: 워드 크기: 1; 윈도우 크기: 5; 채점 방법: 백분율; 상부 대각선의 수: 5; 및 갭 페널티: 3. 폴리펩티드 서열의 다중 정렬을 위해, 하기 파라미터를 사용할 수 있다: 가중치 행렬: 블로썸; 갭 개방 페널티: 10.0; 갭 연장 페널티: 0.05; 친수성 갭: 켜짐; 친수성 잔기: Gly, Pro, Ser, Asn, Asp, Gln, Glu, Arg, 및 Lys; 및 잔기-특이적 갭 페널티: 켜짐. 예를 들어, 월드와이드웹 상의 베일러 의과 대학의 서치 런처(Baylor College of Medicine Search Launcher) 웹사이트 또는 유럽 생물정보학 연구소(European Bioinformatics Institute) 웹사이트에서 ClustalW을 실행할 수 있다.
변화를 핵산 분자 내로 도입함으로써, 코딩된 폴리펩티드의 아미노산 서열의 변화를 일으킬 수 있다. 예를 들어, 돌연변이유발 (예를 들어, 부위-지정 돌연변이유발, PCR-매개 돌연변이유발)을 사용하여 또는 변화가 있는 핵산 분자를 화학적으로 합성함으로써, 핵산 코딩 서열 내로 변화를 도입할 수 있다. 이러한 핵산 변화는 하나 이상의 아미노산 잔기에서의 보존적 및/또는 비-보존적 아미노산 치환에 이를 수 있다. "보존적 아미노산 치환"은 하나의 아미노산 잔기가 유사한 측쇄가 있는 상이한 아미노산 잔기로 교체되는 치환이고 (예를 들어, 문헌 [Dayhoff et al., 1978, Atlas of Protein Sequence and Structure, 5(Suppl. 3):345-352]을 참조하고, 이는 아미노산 치환에 대한 빈도 표를 제공함), 비-보존적 치환은 아미노산 잔기가 유사한 측쇄가 없는 아미노산 잔기로 교체되는 치환이다. 발현 (예를 들어, 전사 및/또는 번역) 증가, 더 엄격한 조절, 탈조절, 이화대사산물 억제의 상실, 변형된 특이성, 분비, 열안정성, 용매 안정성, 산화 안정성, 프로테아제 저항성, 촉매 활성 및/또는 색상을 비제한적으로 포함하는 하나 이상의 성질을 개선하도록 핵산 및/또는 폴리펩티드 서열이 본원에 기술된 바와 같이 변형될 수 있다.
본원에서 사용된 바와 같이, "단리된" 핵산 분자는 단리된 핵산 분자가 유래되는 생물의 게놈 (예를 들어, PCR 또는 제한 엔도뉴클레아제 소화에 의해 생산된 cDNA 또는 게놈 DNA 단편) 내에서 천연적으로 핵산의 한쪽 또는 양쪽 끝부분의 측면에 있는 서열이 없는 핵산 분자이다. 일반적으로 이러한 단리된 핵산 분자는 조작 편의를 위해 또는 하기에 더욱 상세하게 논의된 융합 핵산 분자를 생성시키기 위해 벡터 (예를 들어, 클로닝 벡터, 또는 발현 벡터) 내로 도입된다. 추가적으로, 단리된 핵산 분자는 조작된 핵산 분자 예컨대 재조합 또는 합성 핵산 분자를 포함할 수 있다.
관련 기술분야에서 일상적인 기술을 사용하여 핵산을 단리할 수 있다. 예를 들어, 재조합 핵산 기술, 및/또는 폴리머라제 연쇄 반응 (PCR)을 비제한적으로 포함하는 임의의 방법을 사용하여 핵산을 단리할 수 있다. 일반적인 PCR 기술이, 예를 들어, 문헌 [PCR Primer: A Laboratory Manual, Dieffenbach & Dveksler, Eds., Cold Spring Harbor Laboratory Press, 1995]에 기술되어 있다. 재조합 핵산 기술은, 예를 들어, 제한 효소 소화 및 라이게이션을 포함하고, 이를 사용하여 핵산을 단리할 수 있다. 단리된 핵산은 단일 핵산 분자로서 또는 일련의 올리고뉴클레오티드로서 화학적으로 합성될 수도 있다.
공지된 방법 예컨대 DEAE 이온 교환, 겔 여과, 및 히드록시아파타이트 크로마토그래피에 의해 천연 공급원 (예를 들어, 생물학적 샘플)으로부터 폴리펩티드를 정제할 수 있다. 또한, 예를 들어, 발현 벡터 내의 핵산을 발현시킴으로써 폴리펩티드를 정제할 수 있다. 추가적으로, 정제된 폴리펩티드를 화학적 합성에 의해 수득할 수 있다. 임의의 적절한 방법, 예를 들어, 칼럼 크로마토그래피, 폴리아크릴아미드 겔 전기영동, 또는 HPLC 분석을 사용하여 폴리펩티드 순도의 정도를 측정할 수 있다.
핵산 (예를 들어, 폴리펩티드를 코딩하는 핵산)을 함유하는 구축물 또는 벡터가 또한 제공된다. 발현 구축물 또는 벡터를 포함하는 구축물 또는 벡터는 시판되거나, 또는 관련 기술분야에서 일상적인 재조합 DNA 기술에 의해 생산될 수 있다. 핵산을 함유하는 구축물 또는 벡터는 이러한 핵산에 작동가능하게 연결된 발현 요소를 가질 수 있고, 선택성 마커 (예를 들어, 항생제 저항성 유전자)를 코딩하는 서열과 같은 서열을 추가로 포함할 수 있다. 핵산을 함유하는 구축물 또는 벡터는 키메라 또는 융합 폴리펩티드 (즉, 폴리펩티드의 N-말단 또는 C-말단 중 하나에 있을 수 있는 이종성 폴리펩티드에 작동적으로 연결된 폴리펩티드)를 코딩할 수 있다. 대표적인 이종성 폴리펩티드는 코딩된 폴리펩티드의 정제에서 사용될 수 있는 것 (예를 들어, 6xHis 태그, 글루타티온 S-트랜스퍼라제 (GST))이다.
발현 요소는 핵산 코딩 서열의 발현을 지시 및 조절하는 핵산 서열을 포함한다. 발현 요소의 한 예는 프로모터 서열이다. 발현 요소는 인트론, 인핸서 서열, 반응 요소, 또는 핵산 발현을 조정하는 유도성 요소를 또한 포함할 수 있다. 발현 요소는 박테리아, 효모, 곤충, 포유동물 또는 바이러스 기원의 것일 수 있고, 벡터는 기원이 상이한 요소의 조합물을 함유할 수 있다.
본원에 기술된 바와 같은 벡터는 숙주 세포 내로 도입될 수 있다. 본원에서 사용된 바와 같이, "숙주 세포"는 핵산이 도입된 특정 세포를 지칭하고, 벡터를 보유하는 이러한 세포의 자손을 또한 포함한다. 숙주 세포는 임의의 원핵생물 또는 진핵생물 세포일 수 있다. 예를 들어, 핵산이 박테리아 세포 예컨대 이. 콜라이(E. coli), 또는 곤충 세포, 효모 또는 포유동물 세포 (예컨대 차이니즈 햄스터 난소 세포 (CHO) 또는 COS 세포)에서 발현될 수 있다. 다른 적합한 숙주 세포가 관련 기술분야의 통상의 기술자에게 공지되어 있다. 생체 내 및 시험관 내 둘 다에서 핵산을 숙주 세포 내로 도입하는 다수의 방법이 관련 기술분야의 통상의 기술자에게 널리 공지되어 있고, 전기천공, 인산칼슘 침전, 폴리에틸렌 글리콜 (PEG) 형질전환, 열 충격, 리포펙션(lipofection), 미세주입, 및 바이러스-매개 핵산 전달을 비제한적으로 포함한다.
수많은 증폭 기술 (예를 들어, 문헌 [PCR Primer: A Laboratory Manual, 1995, Dieffenbach & Dveksler, Eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY]; 및 미국 특허 번호 4,683,195; 4,683,202; 4,800,159; 및 4,965,188 참조)을 적절한 올리고뉴클레오티드 (예를 들어, 프라이머) 쌍과 함께 사용하여 핵산을 검출할 수 있다. 원본 PCR에 대한 다수의 변형이 개발되었고, 핵산을 검출하는데 사용될 수 있다.
혼성화를 사용하여 핵산을 검출할 수도 있다. 핵산들 사이의 혼성화가 문헌 [Sambrook et al., 1989, Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY]의 섹션 7.37-7.57, 9.47-9.57, 11.7-11.8, 및 11.45-11.57에 상세하게 논의되어 있다. 뉴클레오티드 약 100개 미만의 올리고뉴클레오티드 프로브에 대한 적합한 서던 블롯 조건이 문헌 [Sambrook et al.]에 개시되어 있다 (섹션 11.45-11.46). 길이가 뉴클레오티드 100개 미만인 서열과 제2 서열 사이의 Tm을 섹션 11.46에서 제공된 식을 사용하여 계산할 수 있다. 문헌 [Sambrook et al.]에는 뉴클레오티드 약 100개 초과의 올리고뉴클레오티드 프로브에 대한 서던 블롯 조건이 추가적으로 개시되어 있다 (섹션 9.47-9.54 참조). 길이가 뉴클레오티드 100개 초과인 서열과 제2 서열 사이의 Tm을 문헌 [Sambrook et al.]의 섹션 9.50-9.51에서 제공된 식을 사용하여 계산할 수 있다.
핵산을 함유하는 막이 예비혼성화 및 혼성화되는 조건, 뿐만 아니라 핵산을 함유하는 막을 세정하여 과량의 비-특이적으로 결합된 프로브를 제거하는 조건이 혼성화의 엄격성에서 중요한 역할을 할 수 있다. 이러한 혼성화 및 세정을, 적절한 경우에, 중도 또는 고도의 엄격성 조건 하에 수행할 수 있다. 예를 들어, 세정 용액 내의 염 농도를 감소시킴으로써 및/또는 세정이 수행되는 온도를 증가시킴으로써 세정 조건을 더욱 엄격하게 만들 수 있다. 단순한 예로서, 고도의 엄격성 조건은 65℃에서 0.2X SSC에서 막을 세정하는 것을 전형적으로 포함한다.
추가적으로, 혼성화의 양을 해석하는 것이, 예를 들어, 표지된 올리고뉴클레오티드 프로브의 특이적 활성, 프로브가 혼성화된 주형 핵산 상의 프로브-결합 부위의 수, 및 자가방사선사진 또는 다른 검출 매질에 대한 노출량에 의해 영향을 받을 수 있다. 관련 기술분야의 통상의 기술자는 수많은 혼성화 및 세정 조건을 고정된 표적 핵산에 대한 프로브 핵산 분자의 혼성화를 검사하는데 사용할 수 있지만, 동일한 혼성화, 세정, 및 노출 조건 하에 표적 핵산에 대한 프로브의 혼성화를 검사하는 것이 더욱 중요하다는 것을 쉽게 이해할 것이다. 바람직하게는, 표적 핵산이 동일한 막 상에 있다.
한 핵산에 대한 혼성화가 또 다른 핵산에 대한 혼성화보다 적어도 5배 (예를 들어, 적어도 6배, 7배, 8배, 9배, 10배, 20배, 50배, 또는 100배) 더 크면 핵산 분자가 한 핵산에 혼성화하지만 또 다른 핵산에는 혼성화하지 않는 것으로 간주된다. 혼성화의 양을 직접적으로 막 상에서 또는 자가방사선사진으로부터, 예를 들어, 포스포이미저(PhosphorImager) 또는 덴서토미터(Densitometer) (몰레큘러 다이나믹스(Molecular Dynamics), 캘리포니아주 서니베일)를 사용하여 정량할 수 있다.
항체를 사용하여 폴리펩티드를 검출할 수 있다. 항체를 사용하여 폴리펩티드를 검출하는 기술은 효소 결합 면역흡착 검정법 (ELISA), 웨스턴 블롯, 면역침전 및 면역형광을 포함한다. 항체는 폴리클로날 또는 모노클로날일 수 있다. 폴리펩티드에 대한 특이적 결합 친화성이 있는 항체를 관련 기술분야에 널리 공지된 방법을 사용하여 생성시킬 수 있다. 이러한 항체를 관련 기술분야에 공지된 방법을 사용하여 고체 지지체 예컨대 미량역가 플레이트에 부착시킬 수 있다. 폴리펩티드의 존재 하에, 항체-폴리펩티드 복합체가 형성된다.
일반적으로 검출 (예를 들어, 증폭 생성물, 혼성화 복합체, 또는 폴리펩티드의 검출)은 검출가능한 표지를 사용하여 달성된다. 용어 "표지"는 직접적인 표지, 뿐만 아니라 간접적인 표지의 사용을 포괄하도록 의도된다. 검출가능한 표지는 효소, 치환기, 형광 물질, 발광 물질, 생물발광 물질 및 방사성 물질을 포함한다.
선택용 서열이 결여된 (즉, 선택성 마커가 결여된) 균주를 생성시키는데 사용될 수 있는 방법이 본원에서 기술된다. 이들 방법은 원형 플라스미드 DNA 벡터 및 선형 DNA 서열을 사용하는 것을 포함하고, 여기서 원형 플라스미드 DNA 벡터는 선택 마커 및 DNA 복제 기원 (일명 자가 복제 서열 (ARS))을 함유하고, 선형 DNA 서열은 동종 재조합에 의한 피키아 게놈 내로의 통합을 위한 서열을 함유한다. 선형 DNA 분자는 하나 이상의 관심 단백질, 예컨대 비제한적으로 헴-결합된 LegH, 데히드린, 피타제, 프로테아제, 카탈라제, 리파제, 퍼옥시다제, 아밀라제, 트랜스글루타미나제, 옥시도리덕타제, 트랜스퍼라제, 히드롤라제, 리아제, 이소머라제, 리가제, 소형 분자, 예컨대 에탄올, 락트산, 부탄올, 아디프산 또는 숙신산의 생산을 위한 경로에서 수반되는 하나 이상의 효소, 또는 임의의 이러한 단백질에 대한 항체를 코딩하는 핵산 서열을 추가적으로 포함할 수 있다.
피키아 세포를 양쪽 DNA 분자로 형질전환시킬 수 있고, 원형 플라스미드 상의 선택성 마커의 존재에 의해 형질전환체를 선택할 수 있다. 그 후, 예를 들어, PCR을 사용하여, 게놈 내로의 선형 DNA 분자의 통합에 대해 형질전환체를 스크리닝할 수 있다. 마커가 없는 선형 DNA 분자가 올바르게 통합된 형질전환체가 확인되면, 세포를 원형 플라스미드에 대한 선택의 부재 하에 성장시킬 수 있다. 선택의 부재 하에 마커-보유 플라스미드가 안정적으로 유지되지 않기 때문에, 선택이 완화된 후에 이러한 플라스미드가 상실되고, 종종 매우 신속하게 상실된다. 생성된 균주는 통합된 선형 DNA를 선택용 이종성 서열의 부재 하에 보유한다. 따라서, 이러한 접근법은 재조합 단백질 수율에 영향을 거의 미치지 않거나 미치지 않으면서 선택성 마커 (예를 들어, 이종성 선택 마커)가 결여된 피키아 균주를 구축하는데 사용될 수 있다.
본 발명에 따라, 관련 기술분야의 기술 내의 통상적인 분자생물학, 미생물, 생화학, 및 재조합 DNA 기술을 사용할 수 있다. 이러한 기술은 문헌에 충분하게 설명되어 있다. 본 발명이 하기 실시예에서 추가로 기술될 것이고, 이는 특허청구범위에 기술된 사안의 방법 및 조성물의 범주를 제한하지 않는다.
실시예
파트
A. 물질 및 방법
실시예
1 -
폴리머라제
연쇄 반응
퓨젼 하이-피델리티(Phusion Hi-fidelity) DNA 폴리머라제 (뉴 잉글랜드 바이오랩스(New England Biolabs))를 사용하여 게놈 DNA 또는 플라스미드 DNA 주형으로부터 관심 유전자를 증폭시켰다. 간략하게, 1-2 U의 퓨젼 DNA 폴리머라제의 존재 하에 각각 0.6 μM의 정방향 및 역방향 프라이머를 10-50 ng의 주형 DNA 및 400 μM의 뉴클레오티드 믹스와 함께 인큐베이션한다. 반응 조건은 하기와 같았다:

실시예
2 -
라이게이션에
의한 플라스미드 구축
50-100 ng의 제한 효소로 소화된 플라스미드 및 3X 몰 과량의 PCR로 증폭된 삽입물을 T4 DNA 리가제 (뉴 잉글랜드 바이오랩스)의 존재 하에 인큐베이션하였다. 16℃에서 2 hr 초과 동안 라이게이션을 수행하였다. 2 μl의 라이게이션 반응물을 DH10B 전기천공적격 이. 콜라이 세포 내로 형질전환시켰다.
실시예
3 - 이.
콜라이
일렉트로맥스
(
ElectroMax
)
DH10B
T1 파지-저항성 적격 세포 내로의 형질전환
1.5-2 μl의 라이게이션 혼합물을 1 mm 갭 큐벳 (바이오래드(BioRad), 카탈로그 # 165-2089)을 사용하여 1.7 kV로 설정된 마이크로펄서(MicroPulser) (바이오래드)를 사용한 전기천공에 의해 20 μl의 일렉트로맥스 DH10B T1 파지-저항성 적격 세포 (인비트로젠(Invitrogen), 카탈로그 # 12033-015) 내로 형질전환시켰다; 펄스 후, 1 ml SOC를 세포에 첨가하고, 200 rpm으로 진탕시키면서 세포를 37℃에서 1시간 동안 인큐베이션하였다. 10 μl의 회수 혼합물을 100 μg/ml의 농도로 앰피실린을 함유하는 LB 한천 플레이트 상에 플레이팅하였다. 플레이트를 37℃에서 인큐베이션하였다.
실시예
4 - 피.
파스토리스
내로의 형질전환을 위한 플라스미드 DNA의 선형화
플라스미드 DNA를 1-4시간 동안 37℃에서 1x 컷스마트 버퍼(CutSmart Buffer) 내의 PmeI 제한 엔도뉴클레아제 (뉴 잉글랜드 바이오랩스, 카탈로그 # R0560L)로 또는 1-4시간 동안 50℃에서 1x 컷스마트 버퍼 내의 SfiI 제한 엔도뉴클레아제 (뉴 잉글랜드 바이오랩스, 카탈로그 # R0123L)로 소화시켰다. 선형화된 플라스미드를 자이모클린 겔 DNA 회수 키트(Zymoclean Gel DNA Recovery Kit) (자이모 리서치(Zymo Research), 카탈로그 # D4002)를 사용하여 0.8% 아가로스 겔로부터 겔 정제하였다. DNA를 20 μl H2O에 용출시켰다.
실시예
5 - 피.
파스토리스
형질전환-적격 세포의 제조
선택된 피. 파스토리스 균주를 25 ml YPD 배지에서 지수 성장 중기 (~2 OD)까지 성장시켰다. 930 x g로 15분 동안 원심분리하여 세포를 수집하였다. 세포 펠릿을 80% YPD 및 200 mM HEPES, pH 6.8의 용액 2 ml에 재현탁시켰다. 75 μl의 1 M DTT를 첨가하였다. 재현탁된 세포 펠릿을 100 rpm으로 30℃에서 25분 동안 혼합시켰다. 40 ml 부피의 빙냉 멸균수를 현탁액에 첨가하고, 1125 x g로 15분 동안 원심분리하여 세포를 수집하고, 얼음 상에 놓았다. 2회의 추가적인 세정 단계에 대해 이전과 같이 세포 펠릿을 40 ml 빙냉수에 재현탁시키고 수집하였다. 그 후, 세포 펠릿을 20 ml의 빙냉 1 M 소르비톨에 재현탁시키고, 이전과 같이 원심분리에 의해 수집하였다. 최종 세포 펠릿을 0.3 ml 빙냉, 멸균 소르비톨에 현탁시키고, 분취하고, -80℃에서 동결시켰다.
실시예
6 - 피.
파스토리스
내로의 형질전환
30-100 ng의 선형화된 플라스미드 DNA를 1 mm 갭 진펄서(GenePulser) 큐벳 (바이오래드)과 1.15 kV로 설정된 진펄서 (바이오래드)를 사용하여 30 μl의 전기천공적격 피. 파스토리스 세포 내로 형질전환시켰다. 1 ml의 YPD/1 M 소르비톨을 1:1 비로 세포에 첨가하여 혼합시켰다. 100 rpm으로 진탕하면서 세포를 30℃에서 3시간 동안 인큐베이션하였다. 100 μl의 회수 혼합물을 적절한 항생제를 함유하는 YPD 플레이트 상에 플레이팅하고, 나머지 세포를 적절한 항생제가 있는 YPD 플레이트 상에 플레이팅하였다. 플레이트를 30℃에서 48시간 동안 인큐베이션하였다. 1차 형질전환 플레이트를 적절한 항생제가 있는 YPD 플레이트 상에 스트리킹하고, 플레이트를 48시간 동안 30℃에서 인큐베이션하였다. 개별적인 클론을 항생제가 있는 YPD 플레이트 상에 패칭하고, 이러한 패치를 염색체 내로의 통합을 증명하기 위해 콜로니 PCR 또는 gDNA 프렙을 행하는데 또는 추가 분석을 위해 균주를 진탕 플라스크에서 성장시키는데 사용하였다.
실시예
7 -
LegH
생산을 위한 진탕 플라스크에서의 배양물 성장
신선한 패치로부터의 균주를 성장 배지 BMGY (0.75% 글리세롤이 보충된 BMY) 내로 접종하고, 200 rpm으로 진팅시키면서 30℃에서 철야로 성장시켰다. 다음날, 철야 배양물을 0.1 mM 암모늄 Fe(III) 시트레이트가 보충된 BMMY 배지 (BMY+ 1% 메탄올)로 희석함으로써 메탄올로 LegH 발현을 유도하였다. 배양물을 0.5-0.7의 OD600으로 성장시켰다. 소포제를 0.01%의 최종 농도로 첨가하였다. 배양물을 총 72시간 동안 성장시켰다; 진탕 플라스크 부피의 1/10의 10x BMMY 배지 (BMY+10% 메탄올)를 첨가함으로써 배양물에 24시간 마다 메탄올을 보충하였다. 72시간의 성장 유도 후에 세포를 원심분리로 수확하였다.
실시예
8 - 진탕 플라스크 배지
10 g의 효모 추출물 및 20 g 소이톤을 790 ml 물에 용해시킴으로써 BMY 배지를 제조하였다. 혼합물을 오토클레이빙으로 멸균하고, 실온으로 냉각시켰다. 100 ml의 1 M 인산칼륨 완충제 (pH 6.0) 및 100 ml의 10X 아미노산 무함유 효모 질소 베이스 (100 mL당 13.4 g의 YNB 분말; 시그마-알드리치(Sigma-Aldrich))를 필터 멸균하고 (0.2 μm 구멍 크기의 PES), 배지에 첨가하였다. pH 조정이 요구되지 않는다.
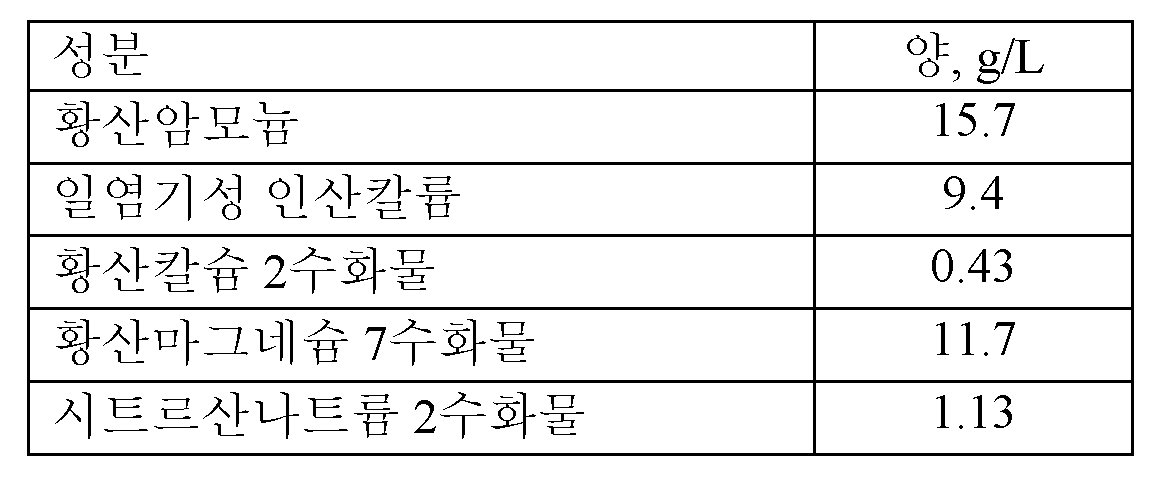
BMY 배지 성분

최종 성분을 물에 용해시키고, 오토클레이빙하여 멸균하였다.
진탕 플라스크용 저-오스몰농도 배지
실시예
9 - 발효 배지 및 공급원료
하기 지시된 성분을 용해시키고, 물로 부피를 조정하였다. 성분은 FCC 식품 등급 또는 등가물이었다. 오토클레이빙에 의해, 정치 증기에 의해, 또는 등가물로 배지를 멸균하였다.
95 g/L 글리세롤 함유 발효용 저-오스몰농도 배지

멸균 후, 배지를 실온으로 냉각되게 하고, 하기를 첨가하였다:

미량 금속 PTM1 용액은 분말 믹스로서 선라이즈 사이언스(Sunrise Science) (카탈로그 번호 4052-A-B-1L)로부터 입수가능하다. 파우치 A 및 파우치 B를 950 mL 물에서 혼합하고, 5 mL 황산을 첨가하였다. 혼합 시 약간의 침전이 예상된다; 혼합물을 필터 멸균하고 (0.2 μm 구멍 크기의 PES), 4℃의 암실에서 보관하였다.
비타민 용액 레시피

대안적으로, 미량 금속 PTM1을 하기와 같이 제조할 수 있다:

성분을 함께 혼합하고, 필터 멸균하고, 실온에서 보관한다. 17.5 g의 앰버펌(AmberFerm) 4000을 320 mL 물 내로 혼합하고 교반하여 용해시킴으로써 글리세롤 공급원료 믹스를 제조하였다. 물-앰버펌 혼합물을 850 g의 글리세롤에 첨가하고, 격렬하게 교반함으로써 잘 혼합시켰다. 공급원료 믹스를 오토클레이빙으로 멸균하였다.
글리세롤 공급원료 용액

12 mL/L의 PTM1 용액이 보충된 99-100% 메탄올을 사용하여 메탄올 공급원료를 제조하였다.
실시예
10 - 실험실 규모의 고-산소 전달 발효를 위한 프로토콜
시드 진탕 플라스크 프로토콜
무균성 생물안전 후드에서, 저-오스몰농도 배지 및 BMY를 9:1 저-오스몰농도:BMY 비로 혼합하였다. 12.5 g/L 농도의 글리세롤을 배지에 첨가하였다. USP 식품 등급 글리세롤/글리세린 (50% v/v (63% w/w) 글리세롤/물 용액 내의 99.7% 순도)을 사용하였고, 오토클레이빙하여 멸균하였다. 시그마 204 또는 등가의 소포제를 0.25 mL/L의 농도로 배지에 첨가하였다. 글리세롤 시드 바이알을 회수하고, 70% IPA 또는 에탄올을 외부에 분무하고, 실온에서 약 5분 동안 생물안전 후드 내부에서 해동시켰다. 배플형 진탕 플라스크에 글리세롤 시드 바이알을 접종하였다; 1 L의 진탕 플라스크 배지마다 1 mL의 접종물 바이알을 사용하였다. 진탕 (1" 쓰로와 함께 200 RPM)하면서 배양물을 24시간 동안 30℃에서 성장시켰다. 1:10 내지 1:5의 실제 배지 부피 : 공칭 진탕 플라스크 부피 비를 사용하였다. 2.8 L 공칭 부피 플라스크와 250 내지 500 mL의 배지가 일상적으로 성공적으로 사용되었다. 24시간의 성장 후 600 nm에서의 OD를 측정하였다; OD가 15 이상이면, 배양물을 발효기에 접종하는데 사용하였다. OD가 15 미만이면, 배양물을 1-2시간 더 성장시킨 후, OD를 다시 측정하였다. 15 내지 30시간 후에 OD가 15에 도달하지 않았으면, 시드 플라스크를 실패한 것으로 간주하였다.
발효 프로토콜
발효 배지 및 공급원료를 본원에 기술된 바와 같이 제조하였다. 초기 부피는 최대 발효기 부피의 약 40%이어야 하고, 예를 들어, 발효기의 최대 작업 부피가 10 L이면, 4 L이어야 한다. 이는 발효가 끝날 때 공정이 최대 작업 부피에 접근할 것이기 때문이다. 10% 접종물-발효기 비로 발효기에 진탕 플라스크 시드를 접종하고, 예를 들어, 4 L의 초기 배지가 발효기 내에 존재하면, 발효기에 약 0.4 L의 진탕 플라스크 시드를 접종한다. 이러한 시점의 발효기 내의 총부피를 T0 부피로 지칭하고, 예를 들어, 이러한 대표적인 예에서는 4.4 L이다. 공정 제어는 하기를 포함한다: 30℃ 온도; 20% 포화 설정점을 유지하도록 교반-통기 케스케이드에 의해 용존 산소를 제어함; 및 28% NH4OH 첨가를 통해 pH를 제어하고, 설정점은 공정의 단계에 좌우됨.
뱃치 단계 (접종에서 글리세롤 고갈까지이고, DO 급증에 의해 신호됨): 용존 산소에 대한 PID 제어의 반응성에 따라, 세포에서 배지 내에 존재하는 글리세롤이 고갈될 때 강한 DO 급증 또는 교반-통기 속도의 빠른 하락 또는 둘 다의 조합이 관찰될 수 있다. 이것이 발생할 때 페드-뱃치 단계가 시작된다. 뱃치 단계의 기간은 대략 20시간이지만, 24시간까지 허용가능한 것으로 간주된다. pH 설정점은 5.0이다. 뱃치 단계 끝의 습식 세포 중량은 대략 220 g/L일 것이다.
페드 - 뱃치 단계: T0 부피를 기초로 12-14 g/L/hr의 순수 글리세롤에 도달하도록 글리세롤 공급을 개시한다. 대략 350 g/L의 습식 세포 중량에 도달했을 때까지 공급속도를 유지하였고, 이는 약 7-10시간이 걸릴 것이다. pH 설정점은 5.0이다.
전이 단계: 샘플을 취한 후 전이 단계를 시작한다. 발효 브로쓰에서 1-2 g/L 메탄올 농도에 도달했을 때까지, T0 부피를 기초로 1 g/L/hr의 순수 메탄올에 도달하도록 메탄올 공급을 개시하였다. 브로쓰 내의 0.25-1 g/L의 메탄올 농도를 유지하도록 나머지 발효 동안 메탄올 공급 속도를 조정하였다. 2시간 과정에 걸쳐 선형으로, T0 부피를 기초로 12-14 g/L/hr에서 8-9 g/L/hr의 순수 글리세롤로 글리세롤 공급 속도를 감소시켰다. 20분 마다 공급 속도를 단계식으로 감소시키는 것이 또한 허용가능할 것이다. pH 설정점을 3.5로 변화시키고, 자연적으로 새로운 설정점으로 맞춰지도록 발효를 허용하였다 (즉, 산을 첨가하지 않음).
생산 단계 (글리세롤 공급의 경사 감소 종료에서 발효 종료까지): pH 설정점은 3.5였다. 발효 브로쓰에서 0.25-1 g/L의 메탄올 농도를 유지하였다. 글리세롤의 공급 속도를 T0 부피를 기초로 8-9 g/L/hr의 순수 글리세롤로 유지하였다. 대략 12시간마다 샘플을 취했다. 4℃에서 4000 내지 7000 RCF로 샘플을 스피닝하고, 상청액을 경사분리하였다. 상청액을 별개의 튜브에 저장하였다. 각각의 시점의 5 mL의 상청액 샘플 3개 및 펠릿을 -80℃에서 동결시켰다. 용기에 대한 최대 통기 및 교반 속도에서도 생산 동안의 15-20% DO를 유지할 수 없으면, 글리세롤 공급 속도를 T0 부피를 기초로 5 g/L/hr의 순수 글리세롤까지 저하시킬 수 있다. 접종 60시간 후에 발효가 종료되었다. 1000 L 규모에서, 수확 공정은 공급원료 및 통기를 중단하고, 브로쓰를 8℃로 냉각하고, 샤플스 또는 디스크 스택 원심분리를 사용하여 페이스트를 농축하는 것으로 이루어졌다. 수확은 일반적으로 약 5-10시간이 걸리고, 생성물 품질의 검출가능한 손실을 초래하지 않는다. 실험실 규모의 경우, 3x5 mL 샘플에 더하여, 추가적인 50 mL 샘플을 종료 시에 수집하는 것이 충분하다. 습식 세포 중량은 >450 g/L이었고, 스피닝된 펠릿은 더 하얀색으로 보이는 유도 전 샘플로부터의 스피닝된 샘플과 대조적으로 분홍색으로 보였다. 약 6-12시간의 유도 후에 브로쓰의 색이 백색에서 더욱 뚜렷한 분홍색으로 변하기 시작하였다.
파트
B. 생산 균주 구축
생산 균주
MXY0183
실시예
11 - 헴 생합성 경로의 각각의 효소의
pGAN
또는
pGAZ
통합 벡터 내로의
클로닝
pGAN (nat 선택 마커가 있음) 및 pGAZ (제오신 선택 마커가 있음)를 바이오그래매틱스, 인크.(Biogrammatics, Inc.) (캘리포니아주 칼스배드)로부터 구입하였다. 각각의 유전자는 AOX1 프로모터의 제어 하에 놓였고, FDH 종결인자가 각각의 유전자의 정지 코돈 직후에 놓였다. 헴 생합성 경로 내의 유전자를 야생형 피. 파스토리스 균주로부터 PCR 증폭시키거나 또는 이전 구축물로부터 서브클로닝하였다.
헴 생합성에서 수반되는 효소를 포함하는 헴 생합성 경로가 도 1에서 제시된다. 헴 생합성 동안 생산되는 중간체가 상자에서 제시되고, 각각의 단계를 촉매하는 효소가 오른쪽에 제시된다. 에스. 세레비시아에(S. cerevisiae)에서 나타난 바와 같은 속도 제한 효소 단계가 밑줄이 그어져 제시된다.
ALA 신타제, ALA 데히드라타제, UPGIII 신타제, UPGIII 데카르복실라제, CPG 옥시다제 및 PPG 옥시다제 유전자를 제한 엔도뉴클레아제인 BsaI에 의한 인식을 위한 부위를 함유하는 프라이머로 PCR 증폭시켰다. 엘림바이오팜(ElimBiopharm)에 의해 올리고뉴클레오티드가 합성되었다.
퓨전 하이-피델리티 DNA 폴리머라제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0530L)를 사용하여 게놈 DNA로부터 유전자를 증폭시켰다. PCR 생성물을 수득하고, DNA 클린&컨센트레이터(DNA Clean&Concentrator)-5 (카탈로그 # D4004)를 사용하여 정제하고, DNA를 25 μl의 H2O에 용출시켰다. 벡터 DNA, pGAZ 및 pGAN, 및 PCR 생성물을 50 μl 반응 부피로 37℃에서 BsaI (뉴 잉글랜드 바이오랩스, 카탈로그 # R0535S)로 소화시켰다.
선형화된 벡터 및 소화된 PCR 생성물을 자이모클린 겔 DNA 회수 키트 (자이모 리서치 카탈로그 # D4002)를 사용하여 0.8% 아가로스 겔로부터 정제하였다. DNA를 20 μl H2O에 용출시켰다. T4 DNA 리가제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0202S)를 사용하여 철야로 16℃에서 10 μl로 라이게이션 반응을 설정하였다.
PBD 및 페로킬라타제 유전자를 이전에 구축된 플라스미드로부터 서브클로닝하였다: pJAZ_PBD를 37℃에서 5분 동안 1x 패스트 다이제스트(Fast Digest) 완충제 내의 BstBI(Bsp119I) (써모사이언티픽(ThermoScientific), FD0124) 및 NotI (써모사이언티픽, FD0596)로 소화시켰다. pJAZ_Ferroch를 37℃에서 5분 동안 1x 패스트 다이제스트 완충제 내의 MfeI (MunI, 써모사이언티픽, FD0753) 및 NotI (써모사이언티픽, FD0596)로 소화시켰다.
소화된 생성물을 자이모클린 겔 DNA 회수 키트 (자이모 리서치 카탈로그 # D4002)를 사용하여 0.8% 아가로스 겔로부터 정제하였다. DNA를 20 μl H2O에 용출시켰다.
T4 DNA 리가제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0202S)를 사용하여 철야로 16℃에서 10 μl로 라이게이션 반응을 설정하였다.
1.5 μl의 라이게이션 혼합물을 1.7 kV로 설정된 마이크로펄서 (바이오래드)를 사용한 전기천공에 의해 20 μl의 일렉트로맥스 DH10B T1 파지-저항성 적격 세포 (인비트로젠, 카탈로그 # 12033-015) 내로 형질전환시켰다; 200 rpm으로 진탕시키면서 세포를 37℃에서 1시간 동안 1 ml SOC 내에서 인큐베이션하였다. 10 μl의 회수 혼합물을 100 μg/ml의 농도로 앰피실린을 함유하는 LB 한천 플레이트 상에 플레이팅하였다. 플레이트를 철야로 37℃에서 인큐베이션하였다. 삽입물의 존재에 대해 콜로니 PCR에 의해 콜로니를 스크리닝하였다. 유전자의 서열을 확인하였다.

실시예
12 - 피.
파스토리스
mutS
게놈 내로의 통합을 위한 플라스미드 상에서의 헴 생합성 유전자의 조립
전체 카세트인 "프로모터-유전자-종결인자"를 피키아 게놈 내로의 통합을 위한 플라스미드를 조립하기 위해 제한 엔도뉴클레아제에 대한 부위를 함유하는 프라이머로 PCR 증폭시켰다.
pGAN
플라스미드 상에서의
Paox1
_UPS_
FDHterm
-
Paox1
_
UPD
_
FDHterm
-
Paox1
_CPO_
FDH
term.의 조립 (pMx327)
pGAN-CPG옥시다제 (pMx312)를 벡터로 사용하여 UPS 및 UPD 카세트를 클로닝하였다. UPG III 신타제 카세트를 상응하게 제한 엔도뉴클레아제에 대한 NheI 및 SphI 인식 부위를 함유하는 AOX1 프로모터 / FDH1 종결인자에 대한 프라이머로 pMx310으로부터 PCR 증폭시켰다:

UPG III 데카르복실라제 카세트를 상응하게 제한 엔도뉴클레아제에 대한 SphI 및 AgeI 인식 부위를 함유하는 AOX1 프로모터 / FDH1 종결인자에 대한 프라이머로 pMx311로부터 PCR 증폭시켰다:
퓨전 하이-피델리티 DNA 폴리머라제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0530L)를 사용하여 플라스미드로부터 DNA를 증폭시켰다.
수득된 PCR 생성물을 DNA 클린&컨센트레이터-5 (자이모 리서치, 카탈로그 # D4004)를 사용하여 정제하고, DNA를 25 μl의 H2O에 용출시켰다.
벡터로 지정된 pGAN-CPG옥시다제 (pMx312)를 1x 컷스마트 버퍼에서 NheI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3131S) 및 AgeI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3552S)로 철야로 37℃에서 소화시켰다.
UPG III 신타제 카세트 PCR 생성물을 1x 컷스마트 버퍼에서 NheI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3131S) 및 SphI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3182S)로 철야로 37℃에서 소화시켰다.
UPG III 데카르복실라제 카세트 PCR 생성물을 1x 컷스마트 버퍼에서 SphI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3182S) 및 AgeI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3552S)로 철야로 37℃에서 소화시켰다.
소화된 벡터 및 PCR 생성물을 자이모클린 겔 DNA 회수 키트 (자이모 리서치, 카탈로그 # D4002)를 사용하여 0.8% 아가로스 겔로부터 겔 정제하였다. DNA를 20 μl H2O에 용출시켰다.
NheI-SphI로 소화된 UPG III 신타제 카세트, SphI-AgeI로 소화된 UPG III 데카르복실라제 카세트 및 NheI-AgeI로 소화된 벡터 사이의 3-방식 라이게이션을 T4 DNA 리가제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0202S)를 사용하여 철야로 16℃에서 10 μl로 설정하였다.
1.5 μl의 라이게이션 혼합물을 1.7 kV로 설정된 마이크로펄서 (바이오래드)를 사용한 전기천공에 의해 20 μl의 일렉트로맥스 DH10B T1 파지-저항성 적격 세포 (인비트로젠, 카탈로그 # 12033-015) 내로 형질전환시켰다; 200 rpm으로 진탕시키면서 세포를 37℃에서 1시간 동안 1 ml SOC 내에서 인큐베이션하였다. 10 μl의 회수 혼합물을 100 μg/ml의 농도로 앰피실린을 함유하는 LB 한천 플레이트 상에 플레이팅하였다. 플레이트를 철야로 37℃에서 인큐베이션하였다. 삽입물의 존재에 대해 콜로니 PCR에 의해 콜로니를 스크리닝하였다. 벡터와 삽입물 사이의 접합부의 서열을 확인하였다.
Paox1
_
ALA신타제
_
FDH1term
.-
Paox1
_
PPG옥시다제
_
FDH1term
-
Paox1
_
Fc
_
FDH1term
.-Paox1_PBD_
FDH1term
카세트 (
pMx330
)의 조립
a. 유전자-카세트의 PCR 증폭:
ALA신타제 카세트를 상응하게 제한 엔도뉴클레아제에 대한 NheI 및 XhoI 인식 부위를 함유하는 AOX1 프로모터 / FDH1 종결인자에 대한 프라이머로 pMx310으로부터 PCR 증폭시켰다:

PPG옥시다제 카세트를 상응하게 제한 엔도뉴클레아제에 대한 XhoI 및 AflII 인식 부위를 함유하는 AOX1 프로모터 / FDH1 종결인자에 대한 프라이머로 pMx313으로부터 PCR 증폭시켰다:

페로킬라타제 카세트를 상응하게 제한 엔도뉴클레아제에 대한 AflII 및 AgeI 인식 부위를 함유하는 AOX1 프로모터 / FDH1 종결인자에 대한 프라이머로 pMx323으로부터 PCR 증폭시켰다:

G418 마커를 하기 프라이머를 사용하여 바이오그래매틱스로부터 구입한 pJAG 플라스미드로부터 PCR 증폭시켰다:

퓨전 하이-피델리티 DNA 폴리머라제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0530L)를 사용하여 플라스미드로부터 DNA를 증폭시켰다. PCR 생성물을 수득하고, DNA 클린&컨센트레이터-5 (자이모 리서치, 카탈로그 # D4004)를 사용하여 정제하고, DNA를 25 μl의 H2O에 용출시켰다.
b. 벡터 제조
벡터로 지정된 pGAZ-PBD (pMx321)를 1x 컷스마트 버퍼에서 NheI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3131S) 및 XhoI (뉴 잉글랜드 바이오랩스, 카탈로그 # R0146S)로 철야로 37℃에서 소화시켰다.
pGAZ-ALAsyn.-PBD (pMx328)를 1x NE버퍼3.1에서 MluI (뉴 잉글랜드 바이오랩스, 카탈로그 # R0198S) 및 BbvCI (뉴 잉글랜드 바이오랩스, 카탈로그 # R0601S)로 철야로 37℃에서 소화시켰다.
pGAG-ALAsyn-PBD (pMx332)를 1x 컷스마트 버퍼에서 XhoI (뉴 잉글랜드 바이오랩스, 카탈로그 # R0146S) 및 AgeI-HF (뉴 잉글랜드 바이오랩스, 카탈로그 # R3552S)로 철야로 37℃에서 소화시켰다.
c. 중간체 구축물 제조 및 최종 카세트 조립
소화된 벡터 및 PCR 생성물을 자이모클린 겔 DNA 회수 키트 (자이모 리서치, 카탈로그 # D4002)를 사용하여 0.8% 아가로스 겔로부터 겔 정제하였다. DNA를 20 μl의 H2O에 용출시켰다.
NheI-XhoI 제한 엔도뉴클레아제로 소화된 pGAZ-PBD (pMx321) 벡터와 동일한 효소로 소화된 ALA신타제 카세트 PCR 생성물을 T4 DNA 리가제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0202S)를 사용하여 철야로 16℃에서 10 μl 반응으로 라이게이션시켜, pGAZ-ALAsyn.-PBD 플라스미드 (pMx328)가 산출되었다.
XhoI 및 AgeI-HF 제한 엔도뉴클레아제로 소화된 pGAG-ALAsyn-PBD (pMx332)와 XhoI, AflII 및 AflII, AgeI-HF 제한 엔도뉴클레아제로 상응하게 소화된 PPG옥시다제 카세트 및 페로킬라타제 카세트 PCR 생성물을 T4 DNA 리가제 (뉴 잉글랜드 바이오랩스, 카탈로그 # M0202S)를 사용하여 3-방식 라이게이션 반응으로 라이게이션시켜, pGAG-ALA신타제_PPG옥시다제_Fc_PBD (pMx330)가 산출되었다.
1.5 μl의 라이게이션 혼합물을 1.7 kV로 설정된 마이크로펄서 (바이오래드)를 사용한 전기천공에 의해 20 μl의 일렉트로맥스 DH10B T1 파지-저항성 적격 세포 (인비트로젠, 카탈로그 # 12033-015) 내로 형질전환시켰다; 200 rpm으로 진탕시키면서 세포를 37℃에서 1시간 동안 1 ml SOC 내에서 인큐베이션하였다. 10 μl의 회수 혼합물을 100 μg/ml의 농도로 앰피실린을 함유하는 LB 한천 플레이트 상에 플레이팅하였다. 플레이트를 철야로 37℃에서 인큐베이션하였다. 삽입물의 존재에 대해 콜로니 PCR에 의해 콜로니를 스크리닝하였다. 벡터와 삽입물 사이의 접합부의 서열을 확인하였다.

실시예
13 - 유전자 카세트가 있는 선형화된 플라스미드의 피.
파스토리스
Bg11
게놈 내로의 통합
생산 균주인 MXY0183을 생성시키는데 사용된 플라스미드가 도 2에서 제시된다. 생산 균주인 MXY0183에 이른 변형을 만드는데 취해진 단계가 도 3에서 도시된다.
피. 파스토리스 Bg11 게놈 내로 도입될 제1 효소는 ALAD였다. pAOX1-구동 ALAD를 함유하는 플라스미드 (pMX229, 도 2i 및 도 3)를 PmeI 제한 효소 (뉴 잉글랜드 바이오랩스)를 사용하여 선형화시켰다. 선형화된 플라스미드를 기술된 바와 같이 0.8% 아가로스 겔로부터 정제하고, 천연 AOX1 유전자좌에서의 상동 재조합을 사용하여 피. 파스토리스 내로 형질전환시켜, 균주 MXY099가 생성되었다 (도 3).
pMX282 (도 2ii 및 도 3)로 지정된 pAOX1 프로모터 제어 하의 대두 LegH 유전자 (피. 파스토리스에 대해 최적화된 서열; 서열식별번호(SEQ ID NO): 3)의 2개의 카피를 함유하는 플라스미드를 SfiI 제한 효소를 사용하여 선형화시켰다. 선형화된 플라스미드를 기술된 바와 같이 0.8% 아가로스 겔로부터 정제하고, ALAD를 함유하는 피. 파스토리스 균주 내로 형질전환시켜, 균주 MXY0118이 생성되었다 (도 3). qPCR을 사용하여, 균주 MXY0118이 LegH 유전자의 여러 카피를 함유하였음이 결정되었으며, 이는 재조합 시점의 플라스미드 pMX282의 콘카테머화로 인해서일 것이다.
AOX1 프로모터의 제어 하의 우로포르피리노겐 III 신타제 (UPS), 우로포르피리노겐 III 데카르복실라제 (UPD) 및 코프로포르피리노겐 III 옥시다제 (CPO) (각각 단계 4, 5 및 6을 촉매하는 효소)를 코딩하는 유전자를 함유하는 플라스미드 pMX327 (도 2iii 및 도 3)을 SfiI 제한 엔도뉴클레아제로 선형화시키고, MXY0118 내로 도입하여, 균주 MXY0170이 산출되었다 (도 3).
피. 파스토리스 게놈으로부터의 ALA 신타제 (ALAS), 프로토포르피린 III 옥시다제 (PPO), 페로킬라타제 (FC) 및 포르포빌리노겐 데아미나제 (PBD) (각각 단계 1, 7, 8 및 3을 촉매하는 효소)를 코딩하는 유전자를 플라스미드 pMX330 (도 2iv 및 도 3) 상에서 조립시켰다. pMX330을 SfiI 제한 엔도뉴클레아제로 선형화하고, MXY0170 내로 형질전환시켜, 균주 MXY0183이 생성되었다 (도 3). MXY0183의 유전자형을 PCR 및 qPCR을 사용하여 확인하였다.
생산 균주
MXY0207
실시예
14 -
pGAB
발현 벡터의 구축
pGAZ 벡터 (바이오그래매틱스, 인크., 캘리포니아주 칼스배드) 내의 제오신 저항성 유전자의 오픈 리딩 프레임을 아스페르길루스 테레우스(Aspergillus terreus)로부터의 블라스티시딘 S 데아미나제 (BSD) 유전자로부터의 오픈 리딩 프레임으로 교체함으로써 pGAB 발현 벡터 (도 4A)를 구축하였고, 이는 플라스미드를 보유하는 형질전환체를 항생제 블라스티시딘 S로 선택하는 것을 허용한다.
본원에 기술된 바와 같이 하이-피델리티 폴리머라제 연쇄 반응을 사용하여 올리고뉴클레오티드 프라이머 MxO0476 및 MxO0477을 사용하여 BSD 오픈 리딩 프레임을 상업적으로 합성된 DNA 분자로부터 증폭시켰다.

BSD PCR 생성물을 1x TBE 완충제 (89 mM 트리스(Tris), 89 mM 붕산, 2 mM EDTA, pH 8.3)에서 1% 아가로스 겔 상에서의 겔 전기영동으로 정제하고, SYBR 세이프(SYBR Safe) DNA 겔 염색 (라이프 테크놀러지즈(Life Technologies), 캘리포니아주 칼스배드)를 사용하여 가시화하였다. 원하는 DNA 단편을 아가로스 겔로부터 절제하고, DNA를 자이모클린 겔 DNA 회수 키트 (자이모 리서치, 캘리포니아주 어바인)를 사용하여 회수하였다.
정제된 BSD PCR 생성물 및 pGAZ 벡터를 1x NE버퍼 3.1 (100 mM NaCl, 50 mM 트리스-HCl, 10 mM MgCl2, 100 μg/ml BSA, pH 7.9 @ 25℃)에서 각각 10 유닛의 MluI 및 BbvCI 제한 엔도뉴클레아제 (뉴 잉글랜드 바이오랩스, 매사추세츠주 입스위치)로 1시간 동안 37℃에서 소화시켰다. 소화된 DNA 생성물을 상기 기술된 바와 같이 겔 전기영동으로 회수하였다.
정제된 MluI 및 BbvCI-소화 BSD 생성물 및 pGAZ 벡터를 20 μl 반응으로 1x T4 DNA 리가제 반응 완충제 (50 mM 트리스-HCl, 10 mM MgCl2, 1 mM ATP, 10 mM DTT, pH 7.5 @ 25℃)에서 16℃에서 2시간 동안 20 μl 반응으로 400 유닛의 T4 DNA 리가제 (뉴 잉글랜드 바이오랩스)와 함께 인큐베이션하였다. 전기천공적격 이. 콜라이 DH10B 세포를 2 μl의 라이게이션 반응물로 형질전환시키고, 항생제 저항성 형질전환체를 100 μg/μl 앰피실린이 보충된 LSB 한천 플레이트 상에서 선택하였다.
실시예
15 -
Mxr1
발현 벡터의 구축
Mxr1 오픈 리딩 프레임을 pGAB 벡터 내로 도입함으로써 Mxr1 발현 벡터인 pMx354를 구축하였다 (도 4B). 번역 개시부가 피키아 파스토리스로부터의 메탄올-유도성 알콜 옥시다제 1 (AOX1) 프로모터의 바로 하류에 있고 번역 정지 신호에 피. 파스토리스 FDH1 유전자로부터의 전사 종결인자 서열이 바로 이어지면서 Mxr1 오픈 리딩 프레임이 pGAB 내로 삽입되었다.
Mxr1 단백질을 코딩하는 오픈 리딩 프레임을 바이오그래매틱스, 인크. (캘리포니아주 칼스배드)로부터 수득된 피키아 파스토리스 균주 Bg11 MutS로부터 단리된 게놈 DNA로부터 증폭시켰다. Mxr1 오픈 리딩 프레임을 프라이머 MxO0495 (TTT TGC GGC CGC ATG AGC AAT CTA CCC CCA ACT TTT G (서열식별번호: 45)) 및 MxO0496 (AAA AGC GGC CGC CTA GAC ACC ACC ATC TAG TCG GTT (서열식별번호: 46))으로 피. 파스토리스 게놈 DNA로부터 증폭시켰고, 이들은 측면의 NotI 제한 엔도뉴클레아제 인식 부위를 부가하였다. 본원에 기술된 바와 같은 폴리머라제 연쇄 반응을 사용하여 증폭을 달성하였다.
증폭된 Mxr1 PCR 생성물 및 pGAB 벡터를 1x NE버퍼 3.1 (100 mM NaCl, 50 mM 트리스-HCl, 10 mM MgCl2, 100 μg/ml BSA, pH 7.9 @ 25℃)에서 10 유닛의 NotI 제한 엔도뉴클레아제 (뉴 잉글랜드 바이오랩스)로 1시간 동안 37℃에서 소화시켰다. 소화 후, NotI-소화 pMx352 벡터를 1x 앤탁틱(Antarctic) 포스파타제 완충제 (50 mM 비스-트리스-프로판-HCl, 1 mM MgCl2, 0.1 mM ZnCl2, pH 6 @ 25℃)에서 37℃에서 15분 동안 5 유닛의 앤탁틱 포스파타제 (뉴 잉글랜드 바이오랩스)로 처리하였다.
정제된 NotI-소화 Mxr1 단편 및 pMx352 벡터를 1x TBE 완충제 (89 mM 트리스, 89 mM 붕산, 2 mM EDTA, pH 8.3)에서 1% 아가로스 겔 상에서의 전기영동으로 분리하고, SYBR 세이프 DNA 겔 염색 (라이프 테크놀러지즈, 캘리포니아주 칼스배드)을 사용하여 가시화하였다. 원하는 DNA 단편을 아가로스 겔로부터 절제하고, DNA를 자이모클린 겔 DNA 회수 키트 (자이모 리서치, 캘리포니아주 어바인)를 사용하여 회수하였다.
Mxr1 오픈 리딩 프레임을 함유하는 NotI-소화 단편을 라이게이션에 의해 AOX1 프로모터의 바로 하류의 NotI 부위에서 pGAB 내로 도입하였다. 137 ng의 Mxr1 오픈 리딩 프레임을 코딩하는 NotI-소화 DNA 및 60 ng의 NotI-소화, 포스파타제-처리 pMx352를 함유하는 혼합물을 20 μl 반응으로 1x T4 DNA 리가제 반응 완충제 (50 mM 트리스-HCl, 10 mM MgCl2, 1 mM ATP, 10 mM DTT, pH 7.5 @ 25℃)에서 16℃에서 2시간 동안 20 μl 반응으로 400 유닛의 T4 DNA 리가제 (뉴 잉글랜드 바이오랩스)와 함께 인큐베이션하였다. 전기천공적격 이. 콜라이 DH10B 세포를 2 μl의 라이게이션 반응물로 형질전환시키고, 항생제 저항성 형질전환체를 100 μg/μl 앰피실린이 보충된 LSB 한천 플레이트 상에서 선택하였다. 플레이트를 37℃에서 철야로 인큐베이션하였다. 프라이머 MxO0495 및 MxO0496을 사용하여 PCR에 의해 삽입물의 존재에 대해 콜로니를 스크리닝하였다. 최종 벡터의 서열을 DNA 시퀀싱으로 확인하였다.
클로닝 동안, 6개의 추가적인 아미노산을 Mxr1의 N-말단에서 도입하였다. Mxr1 오픈 리딩 프레임이 "핵산 서열" 섹션에서 제시되고, 여기서 클로닝으로부터의 잔여 아미노산이 밑줄이 그어져 제시된다. N-말단에 추가적인 6-아미노산이 있는 Mxr1 서열을 함유하는 피키아 생산 균주 및 야생형 Mxr1 (즉, N-말단에 추가적인 6 아미노산이 없음)을 함유하는 피키아 균주가 발효 탱크에서 구별가능하지 않았다.
실시예
16 - 천연
Mxr1
발현 벡터의 구축
pMX354로 지정된, pAOX1 프로모터의 제어 하의 Mxr1 전사 조절인자 유전자를 함유하는 플라스미드를 PCR 증폭용 주형으로서 사용하였다. AOX1 프로모터의 3' 끝부분, LegH 오픈 리딩 프레임, 및 AOX1 종결인자를 하기 제시된 프라이머 MxO0617 및 MxO0647을 사용하여 pMX354로부터 증폭시켰다. AOX1 종결인자, 링커 및 AOX1 프로모터의 5' 끝부분을 프라이머 MxO0618 및 MxO0646을 사용하여 pMX382로부터 증폭시켰다.

PCR 생성물을 수득하고, DNA 클린&컨센트레이터-5를 사용하여 정제하고, DNA를 12 μl의 H2O에 용출시켰다. 그 후, 정제된 PCR 생성물을 조합하고, 프라이머 MxO0617 및 MxO0618을 사용하는 후속 라운드의 PCR 증폭을 위한 주형으로서 사용하였다. 생성된 PCR 생성물은 AOX1 프로모터의 3' 끝부분에 이어지는 Mxr1 오픈 리딩 프레임, AOX1 종결인자, 짧은 링커 서열, 및 AOX1 프로모터의 5' 끝부분으로 구성되었다. PCR 생성물을 수득하고, 본원에 기술된 바와 같이 정제하였다. 정제된 PCR 생성물을 제로 블런트® TOPO® PCR 클로닝 키트(Zero Blunt® TOPO® PCR Cloning Kit) (인비트로젠, 카탈로그 # K2800-20)를 사용하여 pCR™-블런트(Blunt) II-TOPO® 벡터 내로 클로닝하여, pMX402 벡터가 생성되었다.
실시예
17 - 피.
파스토리스
균주
MXY0206
및
MXY0207의
구축
pMx354 Mxr1 발현 벡터 (도 4B)를 DNA 형질전환에 의해 MXY0183 균주 내로 도입하였다 (도 3).
1시간 동안 37℃에서 1x NE버퍼 4 (50 mM 아세트산칼륨, 20 mM 트리스-아세테이트, 10 mM 아세트산마그네슘, 1 mM DTT, pH 7.9@25℃)에서 20 유닛의 PmeI 제한 엔도뉴클레아제 (뉴 잉글랜드 바이오랩스)로 소화시킴으로써 pMx354 벡터 (1.5 μg)를 AOX1 프로모터 서열 내의 독특한 PmeI 부위에서 선형화시켰다.
PmeI-소화 pMX354 벡터를 상기 기술된 바와 같이 겔 전기영동으로 정제하고 자이모클린 겔 DNA 회수 키트를 사용하여 회수하였다. 선형화된 pMX354 벡터를 형질전환에 의해 균주 MXY0183 내로 도입하고, 블라스티시딘-함유 배지 상에서 선택하였다. 2개의 독립적인 클론이 형질전환으로부터 수득되었고, 이들을 MXY0206 및 MXY0207로 지정하였다. 이들 균주 내의 AOX1 프로모터의 제어 하의 Mxr1의 추가적인 카피의 존재를 PCR로 확인하였다.
생산 균주
MXY0291
실시예
18 - 균주
MXY0213
및
MXY0260의
구축
도 5는 헴 생합성 경로의 7개의 효소를 함유하는, 항생제 마커가 없는 균주 MXY0213을 구축하기 위해 취해진 단계를 나타낸다. 각각의 끝부분에서 pAOX1 프로모터에 대한 상동성이 있는 pAOX1 하의 변이체 Mxr1 (N 말단의 6개의 추가 아미노산)을 함유하는 선형 DNA 조각을 공동-형질전환을 사용하여 도입하였다 (도 5 및 도 6i). 동시에 이러한 선형 Mxr1 발현 카세트를 형질전환에 의해 pIL75 플라스미드와 함께 피키아 균주 MXY213 내로 도입하였다. pIL75 벡터는 형질전환된 세포의 게놈 내로의 통합 없이 플라스미드 벡터를 유지하는 것을 허용하는 panARS 자가 복제 서열 (Liachko & Dunham, 2014, FEMS Yeast Res., 14:364-7), 및 항생제 G418로 형질전환체를 선택하기 위한 kanMX 마커를 보유한다. 형질전환된 세포를 pIL75 플라스미드 상의 kanMX 마커의 존재에 대해 G418이 보충된 배지 상에서 선택하였다. pIL75 플라스미드를 받아들였고 또한 Mxr1 발현 카세트가 정확하게 통합된 형질전환체에 대해 콜로니 PCR에 의해 피키아 형질전환체를 스크리닝하였다.
실시예
19 -
LegH
발현 카세트를
피키아
내로 도입하기 위한 공동-형질전환
pMX399로 지정된, pAOX1 프로모터의 제어 하의 대두 LegH 유전자의 상이한 피키아 파스토리스-코돈 최적화 변이체 (변이체 3; 서열식별번호: 5)를 함유하는 플라스미드를 유전자의 PCR 증폭을 위한 주형의 공급원으로서 사용하였다. TOPO 클로닝 플라스미드 pMX401로부터의 백본을 PCR 증폭시켰다. 삽입물 및 벡터를 깁슨(Gibson) 어셈블리 (NEB의 깁슨 어셈블리 키트)를 사용하여 조립하여, 플라스미드 pMX422를 생성시켰다. 이러한 플라스미드를 하기 제시된 프라이머 MxO0617 및 MxO0618을 사용하는 후속 라운드의 PCR 증폭을 위한 주형으로서 사용하였다.

생성된 PCR 생성물은, 5'에서 3' 방향으로, AOX1 프로모터의 3' 끝부분에 이어지는 LegH 변이체 3 오픈 리딩 프레임, AOX1 종결인자, 짧은 링커 서열, 및 AOX1 프로모터의 5' 끝부분으로 구성되었다 (도 6ii). PCR 생성물을 수득하고, 본원에 기술된 바와 같이 아가로스 겔 전기영동으로 정제하였다.
LegH 발현 카세트가 게놈 내로 통합된 형질전환체를 PCR로 스크리닝하고, qPCR을 사용하여 LegH 유전자 카피수에 대해 특성화하였다.
실시예
20 - 형질전환체에서의 선택
마커를
보유하는 플라스미드 벡터의 교정
대두 LegH 발현 카세트가 정확하게 통합된 것으로 콜로니 PCR에 의해 나타났고 카피수가 높은 것으로 qPCR에 의해 나타난 클론에서, G418 상에서의 선택에 요구되는 pIL75 플라스미드를 항생제에 대한 선택을 완화함으로써 제거하였다. 형질전환체를 G418 항생제가 결여된 배지 상에 단일 콜로니에 대해 스트리킹하였다. 선택의 부재 하에 panARS 플라스미드는 안정적으로 유지되지 않기 때문에, 이러한 조건 하에 pIL75가 형질전환된 세포로부터 신속하게 상실되었다. 생성된 피키아 균주 MXY0291은 MXY0207과 유사한 카피수로 LegH 발현을 위한 서열을 함유하지만, 선택용 이종성 서열이 결여된다.
생산 균주
MXY0330
,
MXY0333
, 및
MXY0338
실시예
21 - 균주
MXY0306의
구축
균주 MXY0291의 유전자형 PCR에서, 이러한 균주의 구축 동안 CPG옥시다제 코딩 서열의 일부분이 결실되었음이 밝혀졌다. 말단절단 카피의 교체에 의해 전장 CPG옥시다제 코딩 영역을 복원하였다. 간략하게, pAOX1 프로모터 및 전장 CPG옥시다제 코딩 영역을 함유하는 선형 DNA 단편을 하기 제시된 프라이머 MxO0866 및 MxO0867을 사용하여 플라스미드 pMX312로부터 PCR 증폭에 의해 생성시켰다.
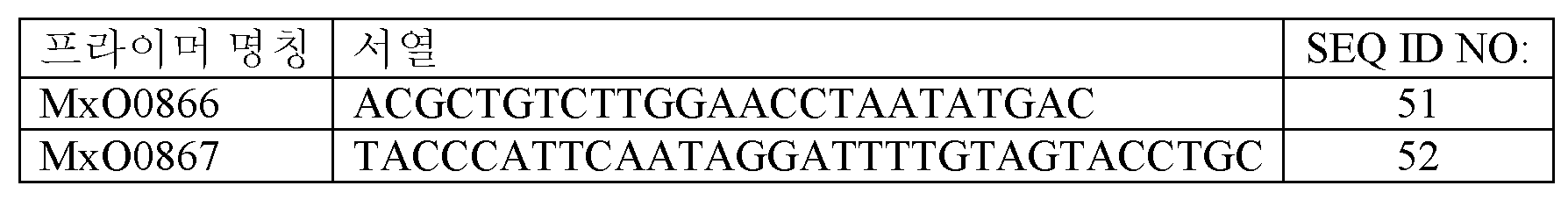
선형 pAOX1-CPG옥시다제 DNA 단편을 pIL75 플라스미드와의 공동-형질전환에 의해 균주 MXY0291 내로 도입하였다. G418을 함유하는 배지 상에서 형질전환체를 선택한 후, PCR에 의해 전장 CPG옥시다제 코딩 영역의 존재에 대해 스크리닝하였다. 전장 CPG옥시다제를 함유하는 단리물을 확인하고, 이어서 상기 기술된 바와 같이 G418 상에서의 선택에 요구되는 플라스미드 벡터를 교정하였다. 이러한 균주를 MXY0306로 지정하였다 (도 5 참조).
실시예
22 -
하이브리드
프로모터 균주를 위한 선형 구축물
LegH 변이체 3을 본원에 지시된 3개의 피키아 파스토리스의 천연 구성적 프로모터 각각의 지시 하에 발현시켰다. 선형 구축물이 도 7에 제시되고, 이는 프로모터의 3' 절반, 이어지는 LegH var3, 이어지는 FDH1 전사 종결인자를 함유하였다. 여기에 아시비아 고시피이(Ashbya gossypii)로부터의 pTEF 프로모터, 아스페르길루스 니둘란스(Aspergillus nidulans)로부터의 아세트아미다제 유전자 (amdS) 및 아시비아 고시피이로부터의 TEF 종결인자를 함유하는 항생제 저항성 카세트가 바로 이어졌다. 마지막으로, 구축물은 프로모터의 5' 절반을 함유하였다. 이러한 선형 카세트를 하기 표에 열거된 올리고뉴클레오티드 프라이머를 사용하여 증폭시켜, 천연 피키아 게놈 내의 각각의 프로모터에 상동성인 5' 및 3' 끝부분 상의 수백개의 염기쌍을 함유하는 구축물을 생성시켰다.
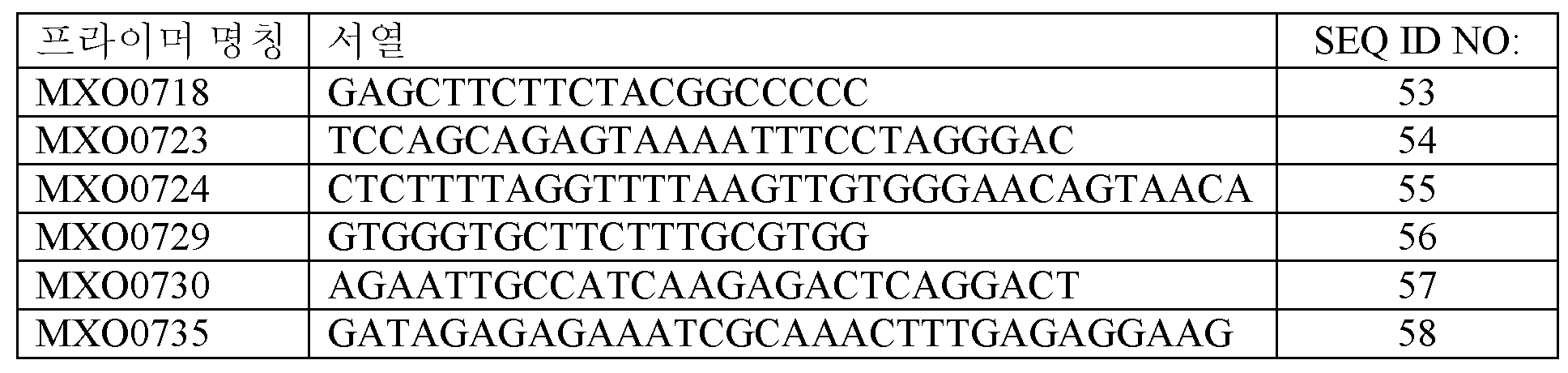
선형 구축물을 증폭시키는데 사용된 프라이머
적격 MXY0306 세포를 각각의 선형 카세트로 형질전환시키고, amdS 선택 카세트를 함유하는 형질전환체를 아세트아미드를 단독 질소원으로서 함유하는 한천 플레이트 상에서 성장하는 능력을 기초로 선택하였다. 이들 균주를 정제하고, 단리하고, 구성적 프로모터의 제어 하의 LegH의 존재를 PCR로 증명하였다 (도 5).
실시예
23 - 핵산 서열
Mxr1 핵산 서열 (밑줄친 뉴클레오티드는 클로닝 동안 도입된 N-말단의 6개의 아미노산을 코딩함) (서열식별번호: 1)


Mxr1 단백질 서열 (클로닝 동안 도입된 N-말단의 6개의 아미노산을 밑줄침) (서열식별번호: 2)

피키아 파스토리스-코돈-최적화 LegH 핵산 서열 (서열식별번호: 3)

피키아 파스토리스-코돈-최적화 LegH 아미노산 서열 (서열식별번호: 4)

피키아 파스토리스-코돈-최적화 LegH 변이체 3 핵산 서열 (서열식별번호: 5)

피키아 파스토리스-코돈-최적화 LegH 변이체 3 아미노산 서열 (서열식별번호: 6)

피키아 파스토리스 pAOX1 프로모터 (서열식별번호: 7)
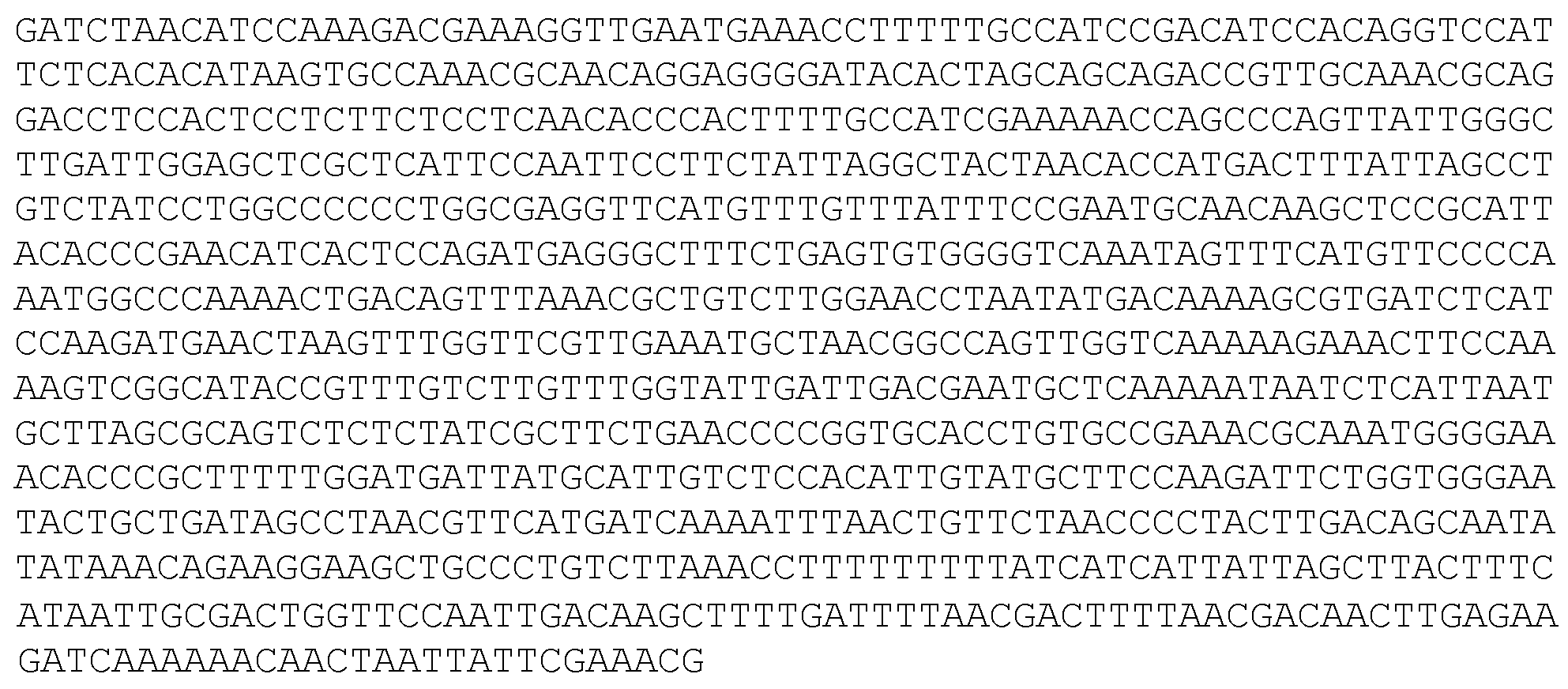
피키아 파스토리스 pGAP 프로모터 (서열식별번호: 8)

피키아 파스토리스 pGCW14 프로모터 (서열식별번호: 9)

피키아 파스토리스 pTEF1 프로모터 (서열식별번호: 10)

헴 생합성 효소 1- ALA 신타제 (서열식별번호: 11)

헴 생합성 효소 2- ALA 데히드라타제 (서열식별번호: 12)
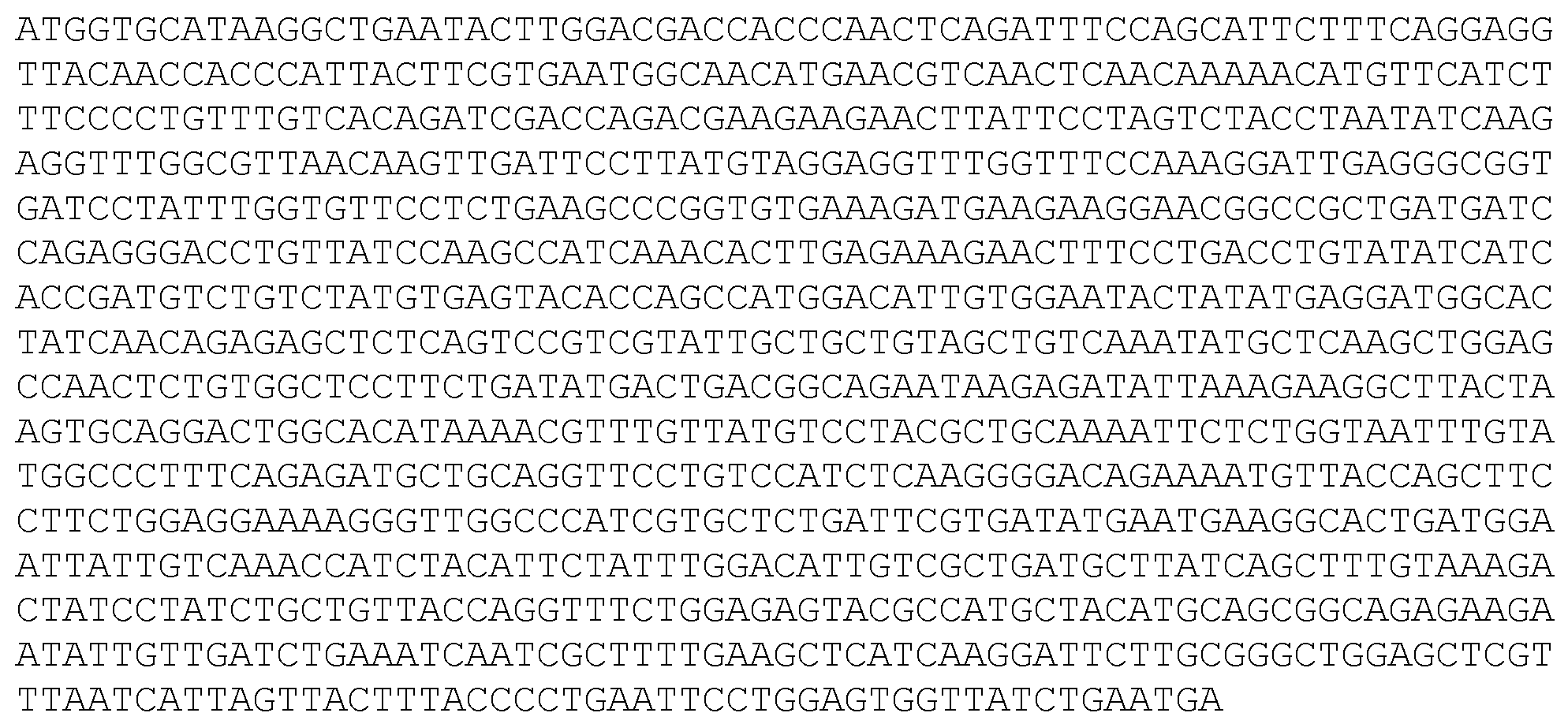
헴 생합성 효소 3- 포르포빌리노겐 데아미나제 (서열식별번호: 13)

헴 생합성 효소 4- 우로포르피리노겐 III 신타제 (서열식별번호: 14)
헴 생합성 효소 5- 우로포르피리노겐 III 데카르복실라제 (서열식별번호: 15)

헴 생합성 효소 6- 코프로포르피리노겐 III 옥시다제 (서열식별번호: 16)

헴 생합성 효소 7- 프로토포르피리노겐 옥시다제 (서열식별번호: 17)

헴 생합성 효소 8- 페로킬라타제 (서열식별번호: 18)

파트
C. 결과 및 논의
실시예
24 - 균주
MXY0183의
특성화
균주 MXY0183에 대한 최적 성장 조건은 3.0 내지 6.0의 표적 pH 및 28-35℃의 온도를 포함한다. LegH 단백질을 생산하기 위해, 균주 MXY0183은 6일의 기간 동안 살아 있어야 하고 호기성으로 성장 중이어야 한다.
균주 MXY0183과 연관된 유전자의 발현은 균주에 대한 표현형 변화를 초래하였다. 도 8은 유도 시작 (0 hr) 및 유도 72 hr 후의 진탕 플라스크의 사진을 나타낸다. #1로 지정된 플라스크는 숙주 균주인 MXY0051을 함유한다. #2 및 #3으로 지정된 플라스크는 각각 중간체 균주 (즉, MXY0118이고, LegH 유전자의 >10개의 카피 및 헴 생합성 경로로부터의 ALA 데히드라타제를 함유함) 및 생산 균주 (즉, MXY0183이고, LegH 유전자의 >10개의 카피 및 헴 생합성 경로로부터의 8개의 효소를 함유함) 중 하나를 함유한다. 72시간 후의 플라스크 #3에서의 특징적인 적색은 헴-결합된 LegH의 생산을 증명한다.
진탕 플라스크에서의 성장 후, 상기 지시된 피. 파스토리스 균주 MXY0051, MXY0118, 및 MXY0183을 용해시키고, 단백질을 SDS 겔 상에 러닝시켰다 (도 9A). 화살표는 LegH 단백질의 위치를 나타낸다. 균주 MXY0183 및 균주 MXY0118에서의 LegH 생산의 비교가 도 9B에서 제시되고, 이는 MXY0183 균주에 의한 LegH 단백질 의 헴 로딩의 효율을 증명한다.
실시예
25 - 균주
MXY0207의
특성화
그 후, 헴 생합성에서 수반되는 8개의 효소를 코딩하는 유전자의 존재 하에 전사 활성화제인 Mxr1을 과발현시키는 것의 이점을 결정하기 위해 실험을 수행하였다. LegH 서열의 >10개의 카피 및 헴 생합성에서 수반되는 8개의 효소를 코딩하는 유전자를 함유하는 균주 MXY0183, 및 LegH 서열의 >10개의 카피, 헴 생합성에서 수반되는 8개의 효소를 코딩하는 유전자, 및 Mxr1 전사 활성화제를 함유하는 자매 균주 MXY0206 및 MXY0207을 이들 균주에 대한 억제성 탄소원인 글리세롤의 존재 하에 진탕 플라스크 배양물에서 성장시켰다. 48 hr 후의 진탕 플라스크 배양물의 사진이 도 10A에서 제시되고, 추가적인 탄소원 없이 48시간 동안 BMY 배지에서 성장된 세포로부터의 펠릿의 사진이 도 10B에서 제시된다; 이들 실험은 AOX1 프로모터의 제어 하의 트랜스진 (예를 들어 헴 효소)의 유의한 발현이 Mxr1이 AOX1 프로모터로부터 또한 발현되는 균주의 성장 배지에서 억제성 탄소원이 소비될 때 유도성 탄소원의 부재 하에 발생한다는 것을 증명하였다. 진탕 플라스크 배양물이 유도제의 부재 하에 성장되었을 때의 헴-로딩된 LegH의 상대적인 수율이 도 10C에서 제시된다. 이들 실험은 재조합 헴-로딩된 단백질의 유의한 생산이 Mxr1 발현이 AOX1 프로모터에 의해 또한 구동되는 피키아 균주에서 메탄올 유도의 부재 하에 AOX1 프로모터-구동 트랜스진으로부터 달성된다는 것을 증명한다.
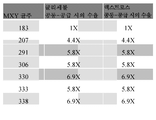
선택된 균주를 2 L 발효기 탱크에서 성장시키고, LegH 및 헴-로딩된 LegH의 상대적인 수율을 결정하였다 (도 11). 균주 MXY0183에 비교하여, MXY0207 균주가 더욱더 많은 LegH를 생산하였고, 매우 효과적으로 LegH 단백질에 헴-로딩하기 위한 충분한 헴을 생산할 수 있었다.
실시예
26 - 균주
MXY0291의
특성화
상기 실시예 18-20에서 기술된 바와 같이, 균주 MXY0291은 항생제 저항성 유전자가 없으면서 MXY0207가 LegH를 생산하는 능력을 재현하도록 구축되었다. 균주 MXY0291이 LegH var3의 ~ 16개의 카피, Mxr1, 및 8개의 헴 생합성 효소 중 7개를 함유한 것으로 결정되었다. 2 L 발효기 탱크에서 성장시켰을 때, 이러한 균주는 MXY0207에 비교하여 개선된 LegH 수율을 나타냈다. 이러한 개선은 메탄올 / 글리세롤 및 메탄올 / 덱스트로스 (D-글루코스)를 함유하는 유도 배지 둘 다에서 나타났다 (도 11).
실시예
27 -
하이브리드
프로모터 균주의 특성화
프로모터 pAOX1의 제어 하의 LegH의 여러 카피, 모든 헴 생합성 효소, 및 전사 인자 Mxr1를 이미 함유하는 균주 (상기에서 MXY0291로 지칭됨)에서, 대두 레그헤모글로빈 (LegH)의 추가적인 카피를 3개의 상이한 구성적 프로모터인 pGAP, pGCW14 및 pTEF1 하에 발현시켰다. 덱스트로스 (즉, D-글루코스)의 존재 하에 메탄올로 유도되었을 때, 구성적 프로모터 및 pAOX1가 LegH의 발현을 구동시키는 반면, pAOX1 프로모터만 헴 효소의 발현을 구동시킨다. 이는 이전의 균주 MXY0291에 비교하여 LegH 수율에서의 추가적인 개선에 이른다 (도 11).
사안 방법 및 조성물이 다수의 상이한 측면과 연계되어 본원에서 기술되었지만, 다양한 측면에 대한 상기 설명은 사안 방법 및 조성물을 예시하도록, 그리고 그의 범주를 제한하지 않도록 의도된다는 것을 이해하여야 한다. 다른 측면, 장점 및 변형이 하기 특허청구범위의 범주 내에 속한다.
개시된 방법 및 조성물에 사용될 수 있거나, 이와 연계되어 사용될 수 있거나, 이에 대비하여 사용될 수 있거나, 또는 그의 생성물인 방법 및 조성물이 개시된다. 이들 물질 및 다른 물질이 본원에서 개시되고, 이러한 방법 및 조성물의 조합, 부분집합, 상호작용, 군 등이 개시되는 것으로 이해된다. 즉, 이들 조성물 및 방법의 각각의 다양한 개별적 및 총괄적 조합 및 치환에 대한 구체적인 언급이 명시적으로 개시되지 않을 수 있지만, 각각이 본원에서 구체적으로 구상되고 기술된다. 예를 들어, 특정한 사안 조성물 또는 특정한 방법이 개시 및 논의되고, 다수의 조성물 또는 방법이 논의되면, 구체적으로 달리 대조적으로 지시되지 않는 한 조성물 및 방법의 각각의 모든 조합 및 치환이 구체적으로 구상된다. 유사하게, 이들의 임의의 부분집합 또는 조합이 또한 구체적으로 구상되고 개시된다.
SEQUENCE LISTING
<110> Impossible Foods Inc.
<120> EXPRESSION CONSTRUCTS AND METHODS OF GENETICALLY ENGINEERING
METHYLOTROPHIC YEAST
<130> 38767-0048WO1
<140> PCT/US2016/031797
<141> 2016-05-11
<150> 62/313,491
<151> 2016-03-25
<150> 62/236,506
<151> 2015-10-02
<160> 58
<170> PatentIn version 3.5
<210> 1
<211> 3486
<212> DNA
<213> Komagataella phaffii
<400> 1
atgcgagacc gcggccgcat gagcaatcta cccccaactt ttggttccac tagacaatct 60
ccagaagacc aatcacctcc cgtgcccaag gagctgtcat tcaatgggac cacaccctca 120
ggaaagctac gcttatttgt ctgtcagaca tgtactcgag catttgctcg tcaggaacac 180
ttgaaacgac acgaaaggtc tcacaccaag gagaaacctt tcagctgcgg catttgttct 240
cgtaaattca gccgtcgaga tctgttattg agacatgccc aaaaactgca cagcaactgc 300
tctgatgcgg ccataacaag actaaggcgc aaggcaactc gtcggtcttc taatgccgcg 360
ggttccatat ctggttctac tccggtgaca acgccaaata ctatgggtac gcccgaagat 420
ggcgagaaac gaaaagttca gaaactggcc ggccgccggg actcaaatga acagaaactg 480
caactgcaac aacaacatct acagcaacaa ccacagttgc aataccaaca atctcttaag 540
cagcatgaaa atcaagtcca gcagcctgat caagatccat tgatatcccc gagaatgcaa 600
ttattcaatg attccaacca tcacgtaaac aatttgtttg atcttggact aagaagagct 660
tccttctccg ccgttagtgg aaataattat gcccattatg tgaataattt tcaacaagat 720
gcctcttcta ccaatccaaa tcaagattca aataatgccg aatttgagaa tattgaattt 780
tctaccccac aaatgatgcc cgttgaagat gctgaaactt ggatgaacaa catgggtcca 840
attccgaact tctctctcga tgtgaacagg aacattggtg atagctttac agatatacaa 900
cacaagaatt cagagcctat tatatccgaa ccgcccaagg acaccgctcc aaacgacaag 960
aagttgaatg gctactcttt ttacgaagcc cccatcaagc cattagaatc cctattttct 1020
gtcaggaata caaagagaaa caagtataaa acaaatgacg actctccaga caccgtggat 1080
aataactccg caccggctgc taataccatt caagaacttg agtcttcttt gaatgcatcc 1140
aagaattttt gcttgccaac tggttattcc ttctatggta atttggacca acagactttc 1200
tctaacacgt tatcatgcac ttcttctaat gccacaattt cgcccattct actcgataac 1260
tccattaata ataactccac tagtgacgtg agaccagaat ttagaacaca aagtgtcacc 1320
tctgaaatga gtcaagcccc tccccctcct caaaaaaaca actcgaaata ttccaccgaa 1380
gttcttttta ccagcaacat gcggtcgttt attcactacg ctctttccaa gtatcctttt 1440
attggtgtgc ccactccaac tcttccggag aacgaaagac taaatgaata tgctgattca 1500
ttcaccaacc gtttcttaaa tcattatcct ttcatacatg tcacgattct caaagaatac 1560
tcccttttca aggcaatttt agatgagaat gagtcgacta agaactggga aaataatcag 1620
ttttacttag agaaccaacg aatatcaatt gtttgtcttc ctcttttggt ggctacgata 1680
ggtgcagtac tatcaaacaa caaaaaggat gcttcgaatt tatacgaagc ttcaaggcgt 1740
tgtattcatg tttacttaga ttccaggaaa aagataccca cttccttgtc cgcaaataac 1800
aatgactctc cactttggct aattcaatcc ctgacgttat ctgttatgta tgggttattt 1860
gcggacaatg acattagttt gaatgtcgtg atcagacaag ttaacgcact taattctctg 1920
gtcaagactt cgggcctgaa taggacctca attatagatc ttttcaacat caacaaacct 1980
ttggataatg aactctggaa tcaattcgtg aaaatagagt ccaccgtaag gacaatccac 2040
acgatttttc aaatcagttc caacttaagc gccttgtaca atattattcc atcgttgaaa 2100
attgatgacc taatgattac tctaccagtt cccacaacac tttggcaagc tgattctttt 2160
gtgaaattca aaagtctaag ttacggaaat cagatccctt ttcaatatac aagagtacta 2220
cagaatttga ttgattacaa tcagccattg agcgatggaa aatttttgta tgaaaaccat 2280
gtaagtgagt ttggactcat atgcctacag aatggtctac accaatacag ctatttccaa 2340
aaattgactg ctgtcaataa cagagaagat gcgctattca caaaggttgt taattcactt 2400
cacagttggg ataggatgat ttcgaattct gatttgtttc caaagaagat atatcagcag 2460
agttgcttga ttttggactc aaagttgctt aataatttcc tgattgtcaa gagctcattg 2520
aaagtttcga ccggagacgt tagttctttg aataagttaa aagaaaacgt gtggcttaaa 2580
aactggaatc aagtgtgtgc tatctattat aacagcttca tgaacattcc tgctcccagt 2640
attcaaaaga agtacaatga catagagttt gtggatgaca tgattaattt gagtctaatc 2700
atcatcaaga ttatgaaact cattttctat aacaatgtca aagacaatta tgaggatgaa 2760
aatgacttca aattgcaaga gttaaattta acatttgaca attttgatga gaaaatatcc 2820
ttgaatttga caatattatt cgatatattt ttgatgatct acaagataat taccaattac 2880
gaaaagttta tgaagatcaa acacaagttt aattactaca attctaattc gaatataagc 2940
ttcttgcatc atttcgaact ctcctcggtt atcaataaca cccaaatgaa ccagaatgat 3000
tatatgaaaa cagatattga tgaaaagctt gatcagcttt tccacatcta tcaaacattt 3060
ttccggctgt atctggattt agaaaagttt atgaagttca aattcaacta tcatgacttt 3120
gagacagagt tttcaagtct ctcaatatcc aatatactga acactcatgc tgcttctaac 3180
aatgacacaa atgctgctga tgctatgaat gccaaggatg aaaaaatatc tcccacaact 3240
ttgaatagcg tattacttgc tgatgaagga aatgaaaatt ccggtcgtaa taacgattca 3300
gaccgcctgt tcatgctgaa cgagctaatt aattttgaag taggtttgaa atttctcaag 3360
ataggtgagt cattttttga tttcttgtat gagaataact acaagttcat ccacttcaaa 3420
aacttaaatg acggaatgtt ccacatcagg atatacctag aaaaccgact agatggtggt 3480
gtctag 3486
<210> 2
<211> 1161
<212> PRT
<213> Komagataella phaffii
<400> 2
Met Arg Asp Arg Gly Arg Met Ser Asn Leu Pro Pro Thr Phe Gly Ser
1 5 10 15
Thr Arg Gln Ser Pro Glu Asp Gln Ser Pro Pro Val Pro Lys Glu Leu
20 25 30
Ser Phe Asn Gly Thr Thr Pro Ser Gly Lys Leu Arg Leu Phe Val Cys
35 40 45
Gln Thr Cys Thr Arg Ala Phe Ala Arg Gln Glu His Leu Lys Arg His
50 55 60
Glu Arg Ser His Thr Lys Glu Lys Pro Phe Ser Cys Gly Ile Cys Ser
65 70 75 80
Arg Lys Phe Ser Arg Arg Asp Leu Leu Leu Arg His Ala Gln Lys Leu
85 90 95
His Ser Asn Cys Ser Asp Ala Ala Ile Thr Arg Leu Arg Arg Lys Ala
100 105 110
Thr Arg Arg Ser Ser Asn Ala Ala Gly Ser Ile Ser Gly Ser Thr Pro
115 120 125
Val Thr Thr Pro Asn Thr Met Gly Thr Pro Glu Asp Gly Glu Lys Arg
130 135 140
Lys Val Gln Lys Leu Ala Gly Arg Arg Asp Ser Asn Glu Gln Lys Leu
145 150 155 160
Gln Leu Gln Gln Gln His Leu Gln Gln Gln Pro Gln Leu Gln Tyr Gln
165 170 175
Gln Ser Leu Lys Gln His Glu Asn Gln Val Gln Gln Pro Asp Gln Asp
180 185 190
Pro Leu Ile Ser Pro Arg Met Gln Leu Phe Asn Asp Ser Asn His His
195 200 205
Val Asn Asn Leu Phe Asp Leu Gly Leu Arg Arg Ala Ser Phe Ser Ala
210 215 220
Val Ser Gly Asn Asn Tyr Ala His Tyr Val Asn Asn Phe Gln Gln Asp
225 230 235 240
Ala Ser Ser Thr Asn Pro Asn Gln Asp Ser Asn Asn Ala Glu Phe Glu
245 250 255
Asn Ile Glu Phe Ser Thr Pro Gln Met Met Pro Val Glu Asp Ala Glu
260 265 270
Thr Trp Met Asn Asn Met Gly Pro Ile Pro Asn Phe Ser Leu Asp Val
275 280 285
Asn Arg Asn Ile Gly Asp Ser Phe Thr Asp Ile Gln His Lys Asn Ser
290 295 300
Glu Pro Ile Ile Ser Glu Pro Pro Lys Asp Thr Ala Pro Asn Asp Lys
305 310 315 320
Lys Leu Asn Gly Tyr Ser Phe Tyr Glu Ala Pro Ile Lys Pro Leu Glu
325 330 335
Ser Leu Phe Ser Val Arg Asn Thr Lys Arg Asn Lys Tyr Lys Thr Asn
340 345 350
Asp Asp Ser Pro Asp Thr Val Asp Asn Asn Ser Ala Pro Ala Ala Asn
355 360 365
Thr Ile Gln Glu Leu Glu Ser Ser Leu Asn Ala Ser Lys Asn Phe Cys
370 375 380
Leu Pro Thr Gly Tyr Ser Phe Tyr Gly Asn Leu Asp Gln Gln Thr Phe
385 390 395 400
Ser Asn Thr Leu Ser Cys Thr Ser Ser Asn Ala Thr Ile Ser Pro Ile
405 410 415
Leu Leu Asp Asn Ser Ile Asn Asn Asn Ser Thr Ser Asp Val Arg Pro
420 425 430
Glu Phe Arg Thr Gln Ser Val Thr Ser Glu Met Ser Gln Ala Pro Pro
435 440 445
Pro Pro Gln Lys Asn Asn Ser Lys Tyr Ser Thr Glu Val Leu Phe Thr
450 455 460
Ser Asn Met Arg Ser Phe Ile His Tyr Ala Leu Ser Lys Tyr Pro Phe
465 470 475 480
Ile Gly Val Pro Thr Pro Thr Leu Pro Glu Asn Glu Arg Leu Asn Glu
485 490 495
Tyr Ala Asp Ser Phe Thr Asn Arg Phe Leu Asn His Tyr Pro Phe Ile
500 505 510
His Val Thr Ile Leu Lys Glu Tyr Ser Leu Phe Lys Ala Ile Leu Asp
515 520 525
Glu Asn Glu Ser Thr Lys Asn Trp Glu Asn Asn Gln Phe Tyr Leu Glu
530 535 540
Asn Gln Arg Ile Ser Ile Val Cys Leu Pro Leu Leu Val Ala Thr Ile
545 550 555 560
Gly Ala Val Leu Ser Asn Asn Lys Lys Asp Ala Ser Asn Leu Tyr Glu
565 570 575
Ala Ser Arg Arg Cys Ile His Val Tyr Leu Asp Ser Arg Lys Lys Ile
580 585 590
Pro Thr Ser Leu Ser Ala Asn Asn Asn Asp Ser Pro Leu Trp Leu Ile
595 600 605
Gln Ser Leu Thr Leu Ser Val Met Tyr Gly Leu Phe Ala Asp Asn Asp
610 615 620
Ile Ser Leu Asn Val Val Ile Arg Gln Val Asn Ala Leu Asn Ser Leu
625 630 635 640
Val Lys Thr Ser Gly Leu Asn Arg Thr Ser Ile Ile Asp Leu Phe Asn
645 650 655
Ile Asn Lys Pro Leu Asp Asn Glu Leu Trp Asn Gln Phe Val Lys Ile
660 665 670
Glu Ser Thr Val Arg Thr Ile His Thr Ile Phe Gln Ile Ser Ser Asn
675 680 685
Leu Ser Ala Leu Tyr Asn Ile Ile Pro Ser Leu Lys Ile Asp Asp Leu
690 695 700
Met Ile Thr Leu Pro Val Pro Thr Thr Leu Trp Gln Ala Asp Ser Phe
705 710 715 720
Val Lys Phe Lys Ser Leu Ser Tyr Gly Asn Gln Ile Pro Phe Gln Tyr
725 730 735
Thr Arg Val Leu Gln Asn Leu Ile Asp Tyr Asn Gln Pro Leu Ser Asp
740 745 750
Gly Lys Phe Leu Tyr Glu Asn His Val Ser Glu Phe Gly Leu Ile Cys
755 760 765
Leu Gln Asn Gly Leu His Gln Tyr Ser Tyr Phe Gln Lys Leu Thr Ala
770 775 780
Val Asn Asn Arg Glu Asp Ala Leu Phe Thr Lys Val Val Asn Ser Leu
785 790 795 800
His Ser Trp Asp Arg Met Ile Ser Asn Ser Asp Leu Phe Pro Lys Lys
805 810 815
Ile Tyr Gln Gln Ser Cys Leu Ile Leu Asp Ser Lys Leu Leu Asn Asn
820 825 830
Phe Leu Ile Val Lys Ser Ser Leu Lys Val Ser Thr Gly Asp Val Ser
835 840 845
Ser Leu Asn Lys Leu Lys Glu Asn Val Trp Leu Lys Asn Trp Asn Gln
850 855 860
Val Cys Ala Ile Tyr Tyr Asn Ser Phe Met Asn Ile Pro Ala Pro Ser
865 870 875 880
Ile Gln Lys Lys Tyr Asn Asp Ile Glu Phe Val Asp Asp Met Ile Asn
885 890 895
Leu Ser Leu Ile Ile Ile Lys Ile Met Lys Leu Ile Phe Tyr Asn Asn
900 905 910
Val Lys Asp Asn Tyr Glu Asp Glu Asn Asp Phe Lys Leu Gln Glu Leu
915 920 925
Asn Leu Thr Phe Asp Asn Phe Asp Glu Lys Ile Ser Leu Asn Leu Thr
930 935 940
Ile Leu Phe Asp Ile Phe Leu Met Ile Tyr Lys Ile Ile Thr Asn Tyr
945 950 955 960
Glu Lys Phe Met Lys Ile Lys His Lys Phe Asn Tyr Tyr Asn Ser Asn
965 970 975
Ser Asn Ile Ser Phe Leu His His Phe Glu Leu Ser Ser Val Ile Asn
980 985 990
Asn Thr Gln Met Asn Gln Asn Asp Tyr Met Lys Thr Asp Ile Asp Glu
995 1000 1005
Lys Leu Asp Gln Leu Phe His Ile Tyr Gln Thr Phe Phe Arg Leu
1010 1015 1020
Tyr Leu Asp Leu Glu Lys Phe Met Lys Phe Lys Phe Asn Tyr His
1025 1030 1035
Asp Phe Glu Thr Glu Phe Ser Ser Leu Ser Ile Ser Asn Ile Leu
1040 1045 1050
Asn Thr His Ala Ala Ser Asn Asn Asp Thr Asn Ala Ala Asp Ala
1055 1060 1065
Met Asn Ala Lys Asp Glu Lys Ile Ser Pro Thr Thr Leu Asn Ser
1070 1075 1080
Val Leu Leu Ala Asp Glu Gly Asn Glu Asn Ser Gly Arg Asn Asn
1085 1090 1095
Asp Ser Asp Arg Leu Phe Met Leu Asn Glu Leu Ile Asn Phe Glu
1100 1105 1110
Val Gly Leu Lys Phe Leu Lys Ile Gly Glu Ser Phe Phe Asp Phe
1115 1120 1125
Leu Tyr Glu Asn Asn Tyr Lys Phe Ile His Phe Lys Asn Leu Asn
1130 1135 1140
Asp Gly Met Phe His Ile Arg Ile Tyr Leu Glu Asn Arg Leu Asp
1145 1150 1155
Gly Gly Val
1160
<210> 3
<211> 438
<212> DNA
<213> Artificial Sequence
<220>
<223> Glycine max leghemoglobin codon-optimized for Pichia pastoris
<400> 3
atgggtgctt tcaccgagaa gcaggaagca cttgtttcct cttcgttcga agcttttaag 60
gctaacatcc ctcaatactc tgttgtgttt tacacgtcca ttctagaaaa agctcctgct 120
gccaaggacc tcttctcttt tctgtccaac ggtgtagatc catccaatcc caaattaaca 180
ggtcacgctg agaaattgtt cggtttagtc agagatagcg ctggacaatt gaaagcaaat 240
ggtactgtgg ttgctgatgc tgccttgggc agcatccatg cacagaaggc aattacagac 300
ccacaatttg ttgttgtgaa ggaagctctg cttaaaacta taaaggaagc cgtcggagac 360
aaatggagtg acgagttgtc atcagcttgg gaggtagctt atgatgagtt ggccgcagca 420
atcaaaaagg cattctaa 438
<210> 4
<211> 145
<212> PRT
<213> Artificial Sequence
<220>
<223> Glycine max leghemoglobin codon-optimized for Pichia pastoris
<400> 4
Met Gly Ala Phe Thr Glu Lys Gln Glu Ala Leu Val Ser Ser Ser Phe
1 5 10 15
Glu Ala Phe Lys Ala Asn Ile Pro Gln Tyr Ser Val Val Phe Tyr Thr
20 25 30
Ser Ile Leu Glu Lys Ala Pro Ala Ala Lys Asp Leu Phe Ser Phe Leu
35 40 45
Ser Asn Gly Val Asp Pro Ser Asn Pro Lys Leu Thr Gly His Ala Glu
50 55 60
Lys Leu Phe Gly Leu Val Arg Asp Ser Ala Gly Gln Leu Lys Ala Asn
65 70 75 80
Gly Thr Val Val Ala Asp Ala Ala Leu Gly Ser Ile His Ala Gln Lys
85 90 95
Ala Ile Thr Asp Pro Gln Phe Val Val Val Lys Glu Ala Leu Leu Lys
100 105 110
Thr Ile Lys Glu Ala Val Gly Asp Lys Trp Ser Asp Glu Leu Ser Ser
115 120 125
Ala Trp Glu Val Ala Tyr Asp Glu Leu Ala Ala Ala Ile Lys Lys Ala
130 135 140
Phe
145
<210> 5
<211> 438
<212> DNA
<213> Artificial Sequence
<220>
<223> Glycine max leghemoglobin codon-optimized for Pichia pastoris
<400> 5
atgggtgcat ttacagaaaa acaagaggct ttagtatcct catcttttga agctttcaaa 60
gccaatattc ctcaatactc cgttgttttc tatacgtcca ttttggaaaa ggctccagca 120
gctaaggacc ttttctcttt cttgtcgaac ggcgtggatc cctcaaatcc taagctgact 180
ggtcacgccg agaagctttt tggtttggtc agagacagcg ccggacagct gaaagctaac 240
ggtacagttg tggcagatgc tgccttggga tctatacatg cacaaaaggc tatcaccgac 300
ccacagtttg tggttgtaaa agaggctcta ctcaaaacta tcaaggaagc agttggtgac 360
aaatggagcg atgaattgtc cagtgcatgg gaggtcgctt acgatgagtt agctgctgca 420
atcaaaaagg ctttctaa 438
<210> 6
<211> 145
<212> PRT
<213> Artificial Sequence
<220>
<223> Glycine max leghemoglobin codon-optimized for Pichia pastoris
<400> 6
Met Gly Ala Phe Thr Glu Lys Gln Glu Ala Leu Val Ser Ser Ser Phe
1 5 10 15
Glu Ala Phe Lys Ala Asn Ile Pro Gln Tyr Ser Val Val Phe Tyr Thr
20 25 30
Ser Ile Leu Glu Lys Ala Pro Ala Ala Lys Asp Leu Phe Ser Phe Leu
35 40 45
Ser Asn Gly Val Asp Pro Ser Asn Pro Lys Leu Thr Gly His Ala Glu
50 55 60
Lys Leu Phe Gly Leu Val Arg Asp Ser Ala Gly Gln Leu Lys Ala Asn
65 70 75 80
Gly Thr Val Val Ala Asp Ala Ala Leu Gly Ser Ile His Ala Gln Lys
85 90 95
Ala Ile Thr Asp Pro Gln Phe Val Val Val Lys Glu Ala Leu Leu Lys
100 105 110
Thr Ile Lys Glu Ala Val Gly Asp Lys Trp Ser Asp Glu Leu Ser Ser
115 120 125
Ala Trp Glu Val Ala Tyr Asp Glu Leu Ala Ala Ala Ile Lys Lys Ala
130 135 140
Phe
145
<210> 7
<211> 939
<212> DNA
<213> Pichia pastoris
<400> 7
gatctaacat ccaaagacga aaggttgaat gaaacctttt tgccatccga catccacagg 60
tccattctca cacataagtg ccaaacgcaa caggagggga tacactagca gcagaccgtt 120
gcaaacgcag gacctccact cctcttctcc tcaacaccca cttttgccat cgaaaaacca 180
gcccagttat tgggcttgat tggagctcgc tcattccaat tccttctatt aggctactaa 240
caccatgact ttattagcct gtctatcctg gcccccctgg cgaggttcat gtttgtttat 300
ttccgaatgc aacaagctcc gcattacacc cgaacatcac tccagatgag ggctttctga 360
gtgtggggtc aaatagtttc atgttcccca aatggcccaa aactgacagt ttaaacgctg 420
tcttggaacc taatatgaca aaagcgtgat ctcatccaag atgaactaag tttggttcgt 480
tgaaatgcta acggccagtt ggtcaaaaag aaacttccaa aagtcggcat accgtttgtc 540
ttgtttggta ttgattgacg aatgctcaaa aataatctca ttaatgctta gcgcagtctc 600
tctatcgctt ctgaaccccg gtgcacctgt gccgaaacgc aaatggggaa acacccgctt 660
tttggatgat tatgcattgt ctccacattg tatgcttcca agattctggt gggaatactg 720
ctgatagcct aacgttcatg atcaaaattt aactgttcta acccctactt gacagcaata 780
tataaacaga aggaagctgc cctgtcttaa accttttttt ttatcatcat tattagctta 840
ctttcataat tgcgactggt tccaattgac aagcttttga ttttaacgac ttttaacgac 900
aacttgagaa gatcaaaaaa caactaatta ttcgaaacg 939
<210> 8
<211> 558
<212> DNA
<213> Pichia pastoris
<400> 8
cgactattat cgatcaatga aatccatcaa gattgaaatc ttaaaattgc ccctttcact 60
tgacaggatc cttttttgta gaaatgtctt ggtgtcctcg tccaatcagg tagccatctc 120
tgaaatatct ggctccgttg caactccgaa cgacctgctg gcaacgtaaa attctccggg 180
gtaaaactta aatgtggagt aatggaacca gaaacgtctc ttcccttctc tctccttcca 240
ccgcccgtta ccgtccctag gaaattttac tctgctggag agcttcttct acggccccct 300
tgcagcaatg ctcttcccag cattacgttg cgggtaaaac ggaggtcgtg tacccgacct 360
agcagcccag ggatggaaaa gtcccggccg tcgctggcaa taatagcggg cggacgcatg 420
tcatgagatt attggaaacc accagaatcg aatataaaag gcgaacacct ttcccaattt 480
tggtttctcc tgacccaaag actttaaatt taatttattt gtccctattt caatcaattg 540
aacaactatc aaaacacg 558
<210> 9
<211> 822
<212> DNA
<213> Pichia pastoris
<400> 9
caggtgaacc cacctaacta tttttaactg ggatccagtg agctcgctgg gtgaaagcca 60
accatctttt gtttcgggga accgtgctcg ccccgtaaag ttaatttttt tttcccgcgc 120
agctttaatc tttcggcaga gaaggcgttt tcatcgtagc gtgggaacag aataatcagt 180
tcatgtgcta tacaggcaca tggcagcagt cactattttg ctttttaacc ttaaagtcgt 240
tcatcaatca ttaactgacc aatcagattt tttgcatttg ccacttatct aaaaatactt 300
ttgtatctcg cagatacgtt cagtggtttc caggacaaca cccaaaaaaa ggtatcaatg 360
ccactaggca gtcggtttta tttttggtca cccacgcaaa gaagcaccca cctcttttag 420
gttttaagtt gtgggaacag taacaccgcc tagagcttca ggaaaaacca gtacctgtga 480
ccgcaattca ccatgatgca gaatgttaat ttaaacgagt gccaaatcaa gatttcaaca 540
gacaaatcaa tcgatccata gttacccatt ccagcctttt cgtcgtcgag cctgcttcat 600
tcctgcctca ggtgcataac tttgcatgaa aagtccagat tagggcagat tttgagttta 660
aaataggaaa tataaacaaa tataccgcga aaaaggtttg tttatagctt ttcgcctggt 720
gccgtacggt ataaatacat actctcctcc cccccctggt tctctttttc ttttgttact 780
tacattttac cgttccgtca ctcgcttcac tcaacaacaa aa 822
<210> 10
<211> 424
<212> DNA
<213> Pichia pastoris
<400> 10
ataactgtcg cctcttttat ctgccgcact gcatgaggtg tccccttagt gggaaagagt 60
actgagccaa ccctggagga cagcaaggga aaaataccta caacttgctt cataatggtc 120
gtaaaaacaa tccttgtcgg atataagtgt tgtagactgt cccttatcct ctgcgatgtt 180
cttcctctca aagtttgcga tttctctcta tcagaattgc catcaagaga ctcaggacta 240
atttcgcagt cccacacgca ctcgtacatg attggctgaa atttccctaa agaatttctt 300
tttcacgaaa attttttttt tacacaagat tttcagcaga tataaaatgg agagcaggac 360
ctccgctgtg actcttcttt tttttctttt attctcacta catacatttt agttattcgc 420
caac 424
<210> 11
<211> 1683
<212> DNA
<213> Pichia pastoris
<400> 11
atggagtttg tcgcccgtca gtccatgaat gcctgtccct ttgtcaggtc aacttctacc 60
caccatttga agaagttggc agcaaacagt tctctagctg ctactgctag tcattgtccc 120
gtggttggcc ctgctctcca acagcagaga tactactctc aaccttccaa gccagcccaa 180
gcccaaacct ccgacattgc tactgggatc aagaaggatg tttctccgat ccgtatggac 240
tctaatgaaa ccgcctttga ttacaatgga atgtatgagt ctgatcttgc gaataaacgt 300
aaagataact cgtatcgtta tttcaataac atcaaccgtc tagccaagga gtttcccaag 360
gcacatcgcc agaccgaaga tgacaaggtg accgtctggt gctctaacga ctacttagga 420
atgggtaggc atcctgagat tatcaaaacc atgaaggcta ccatggacaa gtacggttcc 480
ggagcaggag gaactaggaa cattgcaggt cataaccacg ccgctatcaa tttggaaagc 540
gagttggctt gcttgaacaa gaaggaagcg gctctggtgt tttcatcatg tttcatagct 600
aacgatgcaa tcatctcgtt gttgggacaa aaaatcaaaa atttggtcat tttctctgac 660
cagtcgaatc atgcttccat gatattgggt gtgcgtaact ccaaagcgaa gaagcacatc 720
ttcaagcaca acaatttgaa ggatctggag tcgcagttag ctcagtaccc caagtcgact 780
cctaaactga tcgccttcga gtcagtttac tctatgtgtg gatctgtggc tcccattgag 840
aagatttgcg atttggctaa aaggtacggt gccctcacct tcttggatga agttcatgct 900
gttggaatgt atggtcctca tggacagggt gtagctgagc atttggactt tgatctgcat 960
ttacagtctg gaatcgccag tcctagcgtg gtggacaaac gcaccatatt ggatcgtgtc 1020
gacatgatta ctggtacttg cggaaagtca tttggtactg ttggaggtta cgttgctggt 1080
agtgccaacc taattgattg gttaagatcc tatgcgccag gtttcatttt cactaccaca 1140
cttcctcctg ctatcatggc tggtacagcc acttctgttc gtattgttag ggccgacatt 1200
gaggcccgta tcaagcaaca gcttaatact cgctacgtca aagactcatt tgaaaacctt 1260
ggtattccag tcattccaaa cccaagtcac attgttcctg ttctagttgg aaatgctgca 1320
gatgccaaga aggcatccga tatgttaatg aacaaacacc gtatttatgt tcaagctatt 1380
aactacccta ctgtgcctgt cggtgaagaa cgactaagga ttactcctac tccaggtcat 1440
ggaaaggaga tttgtgacca gctgatcagc gctgtcgacg atgtttttac tgagcttaat 1500
ttaccaagaa tcaacaaatg gcagtcccaa ggtggtcatt gcggtgttgg tgatgctaat 1560
tacgtaccag aacccaatct gtggactcag gaccagctca gcttgacaaa ccaagacttg 1620
cactccaatg tgcacaaccc agtgattgag cagatcgaaa cctcatcagg agtcagattg 1680
tag 1683
<210> 12
<211> 1026
<212> DNA
<213> Pichia pastoris
<400> 12
atggtgcata aggctgaata cttggacgac cacccaactc agatttccag cattctttca 60
ggaggttaca accacccatt acttcgtgaa tggcaacatg aacgtcaact caacaaaaac 120
atgttcatct ttcccctgtt tgtcacagat cgaccagacg aagaagaact tattcctagt 180
ctacctaata tcaagaggtt tggcgttaac aagttgattc cttatgtagg aggtttggtt 240
tccaaaggat tgagggcggt gatcctattt ggtgttcctc tgaagcccgg tgtgaaagat 300
gaagaaggaa cggccgctga tgatccagag ggacctgtta tccaagccat caaacacttg 360
agaaagaact ttcctgacct gtatatcatc accgatgtct gtctatgtga gtacaccagc 420
catggacatt gtggaatact atatgaggat ggcactatca acagagagct ctcagtccgt 480
cgtattgctg ctgtagctgt caaatatgct caagctggag ccaactctgt ggctccttct 540
gatatgactg acggcagaat aagagatatt aaagaaggct tactaagtgc aggactggca 600
cataaaacgt ttgttatgtc ctacgctgca aaattctctg gtaatttgta tggccctttc 660
agagatgctg caggttcctg tccatctcaa ggggacagaa aatgttacca gcttccttct 720
ggaggaaaag ggttggccca tcgtgctctg attcgtgata tgaatgaagg cactgatgga 780
attattgtca aaccatctac attctatttg gacattgtcg ctgatgctta tcagctttgt 840
aaagactatc ctatctgctg ttaccaggtt tctggagagt acgccatgct acatgcagcg 900
gcagagaaga atattgttga tctgaaatca atcgcttttg aagctcatca aggattcttg 960
cgggctggag ctcgtttaat cattagttac tttacccctg aattcctgga gtggttatct 1020
gaatga 1026
<210> 13
<211> 1035
<212> DNA
<213> Pichia pastoris
<400> 13
atgaaccaaa tcgaacagag cggacccatt gattgcagtt ccttgaaatt ggggtcccga 60
aagtccgctc tggctataat ccaggcagaa atcgtccgcc aattgatatt gaaagaatac 120
cctgaattgg agacgaagtt ggtcagtgtg tccaccctgg gggaccaagt ccagaataaa 180
gcacttttca cgtttggagg aaaatctttg tggaccaaag aacttgagat gttgttgttg 240
gagagtgtgg gaggatttga ccaaatagac atgattgtac actcgttgaa agacatgcca 300
actcatttac cagacgaatt tgagctgggt tgcattattg aaagagaaga ccctagagac 360
gctttggtcg tgcaagatgg tttatcttac aagtcattgg ccgaccttcc agagggagct 420
gtggtcggta cgtcttcggt tagaagatcg gctcaactac tgaagaattt ccctcatctg 480
aaattcaaat ctgttagagg aaaccttcag accagactaa gaaaattaga tgatccagat 540
tccgagtact gctgtctcct ccttgcagca gccggtttaa tcaggacagg cttacaacac 600
agaatttcaa tgtatttgaa cgacgatgtg atgtaccact ccgtcggaca aggagcatta 660
ggagtagaga tcagaaaagg tgaccaattc atgaaaaata tctgtgaaaa gattgggcat 720
agaaccacca cccttcgttg tcttgcagag agagcactgc tgagatatct agagggaggc 780
tgctcggtgc caattggggt ctccactatt tatagcgagg atacgaagga acttaccatg 840
aactccctag tcgtcagttg taacggtcgt gactcggtaa cagaatcaat gactgaagtc 900
gtgactactg aagagcaagc tgaagatttc ggtgaaaggc tggcccagaa gctcatagat 960
caaggtgcga aacgcattct tgacgagatc aacttcaaca agatcaaaga gattaaggaa 1020
gagggtttac attaa 1035
<210> 14
<211> 798
<212> DNA
<213> Pichia pastoris
<400> 14
atgccaaaag ccattcttct gaagaataaa actacaccga aggatcctta tctggagaac 60
ttcgtaagta gtggctactc gaccgatttc gtaccacttt tagatcatat tcacatggag 120
aaatctgaga tcatcgcatt tctcaagact gactactttt tgcataaaac tttggcgttt 180
attattacgt cccaaagagc tgtagaaatg ctgaatgagt gtatgcaaat actgagacgt 240
actgatcctg aaattacaca aatcatctat agtaaacctg tctatacagt tggccctgcc 300
acctacagaa tacttgcgga tgctggcttc gtggatctac gaggcggaga taaggcagga 360
aacggatcca ttctagccca gataattttg aatgatgaca tttacactgg aattgaagat 420
tctgacaagc atataacgtt tttcacggga gaaacaagga gagacataat tcccaaatgt 480
ttactctcta acaactttca actttacgaa aagattgtct acaagactct tcctagggat 540
gatatcgtga ctagattcaa gtctgccgtt gacagcatcg accaatcgca aagaagttcc 600
agttgggtgg tcttcttttc gcctcaagga acagaggaca ttgtaacgta tcttcaacac 660
accaaagacc aatttaatat tgcatctatc gggccaacca cagaaaaata ccttctaagc 720
aaaaacctga aaccaaaagt tgtggcacct aagccagagc ctatctcttt actattgtct 780
atacaaaaag tgcactaa 798
<210> 15
<211> 1086
<212> DNA
<213> Pichia pastoris
<400> 15
atgagtagat ttccagaact gaagaatgac cttattttaa gggcagctcg tggtgaaaaa 60
gttgaacgtc ccccaatatg gattatgaga caggccggaa gatatcttcc ggagtaccat 120
gaggtcaaag gaggtaggga cttctttgaa acttgcaggg atgctgagat tgcttctgaa 180
attactatcc agccgattac gcattttgac ggtctgatcg atgcagctat tatcttcagt 240
gatatcttgg tgattcctca agctatgggc atggaagtta agatggtgga caaagttggc 300
ccacagttcc ccaatccgct aagaaaaccg tctgacttgg atcatttgaa aaaagacgtt 360
gacgttttga aggaactcga ttgggccttc aaagctatct cattgaccag aaaaaaactc 420
aatggacgag tgcctttgct tggattttgt ggtgctcctt ggactctact ggtttatatg 480
actgaaggag gcggtaccaa gatgtttcga tttgcaaaag agtggatcta caagtttacc 540
aaggaatctc atcaattact ccaacagatc actgacgttg cagttgaatt cttagctcag 600
caagttgttg caggtgccca aatgttacaa gtttttgaat cttggggcgg tgaattgggg 660
cctgatgaat tcgatgagtt ttctttgcct tatttgagac agatttcctc taaacttccc 720
ctgaggttga aggaacttgg aatcacagag aatgttccca taactgtctt tgctaaaggc 780
tcttggtacg ccttggagca attgtgcgac agtggttatg atgttgtctc gttggattgg 840
ttattccgtc caagtgatgc tgtccagatt gctaacggaa gaatcgcatt gcaaggtaat 900
cttgaccctg gaaccatgta cggctccaaa gaaaccattt ccaagaaagt ggacaaaatg 960
atcaagggtt ttggtggagg aaagcaaaac tacataatta attttggaca cggcactcat 1020
ccattcatgg atccagaaca gatcagatgg ttcttacaag aatgtcatcg cattggatct 1080
caatag 1086
<210> 16
<211> 978
<212> DNA
<213> Pichia pastoris
<400> 16
atggccatcg actctgatat caatctaagc tctcccaatg attccatccg tcaaaggatg 60
ttcgagctta tccagcggaa gcaactcgaa attgtcgctg cattggaggc aattgaagga 120
aacgatacca aatttcgttc tgattcttgg gaaagaggag ccgaaggtgg aggaggaaga 180
tctatgctta ttcaagatgg aagagtgttt gaaaaggctg gtgtaaatat ttccaaggtt 240
catggcgtat tgcctcctca agctgtgagc cagatgagaa atgaccactc caagctagat 300
ctgcctgcgg gaacctctct gaagttcttt gcctgtgggc tttcgttggt cattcatccc 360
cataatcccc atgctccaac tacccatctg aattatcgct acttcgaaac ttgggatgaa 420
actggaaagc ctcacacctg gtggtttggg ggcggtgctg atttaacgcc ttcgtacctg 480
tatcccgagg atgccaagca attccatcaa gcccataagg atgccctgga caaacacgat 540
gttagcttgt acccgagatt caaaaagtgg tgtgatgaat actttctgat caaacatcga 600
aatgaaacta gaggtattgg gggtattttc tttgatgatt ttgacgagtt tgatgctgag 660
aggtccctga agttggttga agattgtttc aatgctttct tggaatctta tcccgctatc 720
actcgaaaaa ggatggacac cccttcaact gatgctgaga agaactggca acaaattaga 780
agaggaagat atgtcgaatt caacttagta ttggatagag gtactcaatt tggtttgaga 840
acgcctggat ctcgtgttga aagtattttg atgtcgttgc caagaacagc tggttgggtc 900
tatgatcatc atccagagcc tggctccaga gaagaggagt tattgcaggt actacaaaat 960
cctattgaat gggtatga 978
<210> 17
<211> 1683
<212> DNA
<213> Pichia pastoris
<400> 17
atgctgaaaa gtcttgcacc aaattcctca attgccgttt taggttcagg gatatctgga 60
ttgactttca gctttttttt gaatcggttg cgtcccgatg ttaagatcca tatctttgaa 120
aaatccaagc aggttggagg atggatcaga tcagaagagc atgaaacttt tcattttgaa 180
aagggaccca gaactttgag aggcacaaat acgggtacct tgatgttgtt ggatcttctt 240
accaagatag gagcaaatga caaggtcctg ggactgcaca aagattctct tgctaataaa 300
aagtatctgt tgtccccgtt ctcagatgtt cacggaaaca acgcaaagct tcttcaagtg 360
ccacaggatt tcagctcttt tgtaaagttc atgtttgacc cgttgtctaa ggatctcatt 420
ctcggtcttt tgaaagaacc atggcaacca aaattaaagt attcagatga gtcggttgac 480
cattttttca acagaagatt tgctaccaaa ctatcagaga atatcgtcag cgcaattgtg 540
catggaatct atgcgggcga cgtgaagaag ttaagtgtga aagccatctt ccctaggctc 600
cctgagatgg aacaggaaag tggctctatt ataaggtata tgatcgccca atacaggaca 660
aaaaagaacg tcaaacaaaa agttgaccct tttttggcag attatgaaaa attgatcggt 720
acatctttga gtttcaaaaa tatttctttg tttctgaaaa actttcccat gctgagtttt 780
cagggtggac tacagaaact tcccatctca ttgaagaacc atttatcaca gattgaaaac 840
atcaagtttc attttgacag caaaatcaaa aacattgctt tggagagcgg taaggtggca 900
ttgactgacc atgatcaggt ttatcttgtt gaccatgtga gatctaccat taataccaac 960
gaattggcca aaatcatttc acccgttgtt ccaagttcta ctaagaaaaa atccgttttc 1020
aaatccaaag cgaatggccc agggctggtc aaatgtttga gctggctaca ctatacaaat 1080
atactaatgt gcaacattta tatacctaag cacgtctcaa aatctatcac cggatttgga 1140
tacttggttc ctcgatcaat gtcttctcag gcatccaaac ttctcggtgt catatttgac 1200
tcagacatcg agactgcaat gactcctaat tttacagagg ccaacattac ggcgataaac 1260
agtaactctg catctcccaa gcaactccaa aagttttctg accaattcgt caataatgat 1320
ctccctaaat acaccaagtt gacgctaatg cttggaggtc attatctcaa gtcggaggca 1380
gacatgcccg gttccgcaga gagtaaacat gctgtcaagg cgattctgtc aaatcacctg 1440
aatattgatc tagatgagtt tgcatctttg ccagacttca agatggaaat caccaagatc 1500
cccaactgca ttccccaata tgaagttggg tatcttgatc tcaagagaaa ggttcagaat 1560
gcagcctcca aagagttcaa cgaccaaata agttttggag gcatggcatt tggtgatggt 1620
gtggggatcc ctgactgtgt ccagaatgca ttcaaagatt cggctaccct cagtggcatt 1680
taa 1683
<210> 18
<211> 1125
<212> DNA
<213> Pichia pastoris
<400> 18
atgcttaacc gtcgtttcca atctaccgtg tcctcgagtc tgaacaaggg cactggaata 60
gtgttcatga atatgggtgg tccctccact gtcaaggaaa cctatgactt tttatttcgt 120
cttttctcgg acggagattt aatcccgttt ggcagatttc agaacatcct ggcccgcttc 180
attgcaagta gaagaacacc caaaattgaa tcctactaca aagctatcgg aggtgggtct 240
cctatccgaa agtggtctga ataccagagt tctaaactat gtgaaaaatt agacattatc 300
agtccacaat cggctcctca taagccttat gttgccttca gatacgctaa tcctctcact 360
gaagatactt tacaaaagat gaaaaatgat ggaattacta aggccattgc cttttctcaa 420
tatccgcaat ttagttattc aaccaccgga tcatcgatta acgaacttta caggcaatcg 480
aaaattttgg accctgatca atctattaaa tggacagtta tagatcgctg gcctgaccac 540
ccagccttag ttaaaacttt cgcagctcat atcaaagata ctctaaacag attcaaaact 600
gaaaatggac tgactgacac aaaagacgtc gtcctccaat tcagtgctca ttctttacca 660
atggatattg tcaataaagg agattcgtat cctgcagaag tcgcagcgag tgtctttgcc 720
attatgaaag aacttaactt ctcaaatcct tataaattaa cctggcaatc acaggttggc 780
ccaaagcctt ggctgggtgc tcaaactgaa aaaattacca agcagctagc atccagtgat 840
gttcctggag tcgttttggt tcctattgcc tttacctctg atcatattga aactctccat 900
gaactggata ttgaactgat tcaagaacta cctaatcctt caaaagtaaa gcgagttgaa 960
tcgttgaacg gagaccaaac tttcattgac tccttggcag aactagtgaa gagtcacatt 1020
gattcgaagg ttgtattttc caaccagttg ccattggatt ccatgctggg agtagtgtca 1080
gataattccc tcacagatcc aaaagagttt ttcagagccc attga 1125
<210> 19
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 19
gagggtctcg gatggagttt gtcgcccgtc 30
<210> 20
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 20
gagggtctcg attacaatct gactcctgat gagg 34
<210> 21
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 21
gagggtctcg gatggtgcat aaggctgaat acttg 35
<210> 22
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 22
gagggtctcg attattcaga taaccactcc agg 33
<210> 23
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 23
gagggtctcg gatgccaaaa gccattcttc tgaag 35
<210> 24
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 24
gagggtctcg attagtgcac tttttgtata gac 33
<210> 25
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 25
gagggtctcg gatgagtaga tttccagaac tgaag 35
<210> 26
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 26
gagggtctcg attattgaga tccaatgcg 29
<210> 27
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 27
gagggtctcg gatggccatc gactctgata tc 32
<210> 28
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 28
gagggtctcg attataccca ttcaatagga t 31
<210> 29
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 29
gagggtctcg gatgctgaaa agtcttgcac caaa 34
<210> 30
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 30
gagggtctcg attaaatgcc actgagggta gc 32
<210> 31
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 31
caatcgctag catccaacat ccaaagacga aagg 34
<210> 32
<211> 42
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 32
ggatagcatg caccttatca agatagctag aaatagaaat gg 42
<210> 33
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 33
caatagcatg caacatccaa agacgaaagg ttgaatg 37
<210> 34
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 34
catggtaccg gtaccttatc aagatagcta gaaatagaaa tgg 43
<210> 35
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 35
caatcgctag catccaacat ccaaagacga aagg 34
<210> 36
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 36
gatattgctc gagaccttat caagatagct agaaatagaa atg 43
<210> 37
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 37
caatctcgag aacatccaaa gacgaaaggt tg 32
<210> 38
<211> 44
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 38
caaccatttc tatttctagc tatcttgata aggtcttaag tcca 44
<210> 39
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 39
catggtaccg gtaccttatc aagatagcta gaaatagaaa tgg 43
<210> 40
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 40
ttacttaagt ccaacatcca aagacgaaag gttg 34
<210> 41
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 41
tcacagacgc gttgaattgt cc 22
<210> 42
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 42
ttgctcctca gcttagaaga actcgtccaa catcaagtg 39
<210> 43
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 43
ttagtcttgc tcctcagctt agcc 24
<210> 44
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 44
tcacagacgc gttgaattgt cc 22
<210> 45
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 45
ttttgcggcc gcatgagcaa tctaccccca acttttg 37
<210> 46
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 46
aaaagcggcc gcctagacac caccatctag tcggtt 36
<210> 47
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 47
actagatggt ggtgtctagt caagaggatg tcagaatgcc atttg 45
<210> 48
<211> 46
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 48
tctgacatcc tcttgactag acaccaccat ctagtcggtt ttctag 46
<210> 49
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 49
aaacgctgtc ttggaaccta atatgac 27
<210> 50
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 50
aaactgtcag ttttgggcca tttg 24
<210> 51
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 51
acgctgtctt ggaacctaat atgac 25
<210> 52
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 52
tacccattca ataggatttt gtagtacctg c 31
<210> 53
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 53
gagcttcttc tacggccccc 20
<210> 54
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 54
tccagcagag taaaatttcc tagggac 27
<210> 55
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 55
ctcttttagg ttttaagttg tgggaacagt aaca 34
<210> 56
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 56
gtgggtgctt ctttgcgtgg 20
<210> 57
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 57
agaattgcca tcaagagact caggact 27
<210> 58
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic oligonucleotide
<400> 58
gatagagaga aatcgcaaac tttgagagga ag 32
Claims (38)
- 재조합 핵산 분자를 포함하며, 여기서 재조합 핵산 분자는 적어도 하나의 메탄올-유도성 프로모터 요소에 작동가능하게 연결된 전사 활성화제를 코딩하는 외인성 핵산을 포함하는 것인 메틸영양성 효모 세포.
- 제1항에 있어서, 메틸영양성 효모 세포가 칸디다(Candida), 한세눌라(Hansenula), 피키아(Pichia) 및 토룰롭시스(Torulopsis)로 이루어진 군으로부터 선택된 효모의 것인 효모 세포.
- 제1항 또는 제2항에 있어서, 메틸영양성 효모 세포가 피키아 효모의 것인 효모 세포.
- 제1항 또는 제2항에 있어서, 세포가 피키아 세포인 효모 세포.
- 제1항 내지 제4항 중 어느 한 항에 있어서, 세포가 피키아 파스토리스(Pichia pastoris) 세포인 효모 세포.
- 제1항 내지 제5항 중 어느 한 항에 있어서, 재조합 핵산 분자가 메틸영양성 효모 세포의 게놈 내로 안정적으로 통합된 것인 효모 세포.
- 제1항 내지 제5항 중 어느 한 항에 있어서, 재조합 핵산 분자가 복제-적격 플라스미드로부터 염색체외적으로 발현되는 것인 효모 세포.
- 제1항 내지 제7항 중 어느 한 항에 있어서, 전사 활성화제를 코딩하는 외인성 핵산이 피키아 파스토리스로부터의 Mxr1 서열, 한세눌라 폴리모르파(Hansenula polymorpha)로부터의 Adr1 서열, 칸디다 보이디니이(Candida boidinii)로부터의 Trm1 서열, 또는 칸디다 보이디니이로부터의 Trm2 서열을 포함하는 것인 효모 세포.
- 제1항 내지 제8항 중 어느 한 항에 있어서, 전사 활성화제를 코딩하는 외인성 핵산이 피키아 파스토리스로부터의 Mxr1 서열을 포함하는 것인 효모 세포.
- 제1항 내지 제9항 중 어느 한 항에 있어서, 전사 활성화제가 진뱅크(GenBank) 수탁 번호 ABD57365에 제시된 서열을 포함하는 것인 효모 세포.
- 제10항에 있어서, 전사 활성화제가 진뱅크 수탁 번호 DQ395124에 제시된 핵산 서열에 의해 코딩되는 것인 효모 세포.
- 제1항 내지 제11항 중 어느 한 항에 있어서, 적어도 하나의 메탄올-유도성 프로모터 요소가 피키아 파스토리스로부터의 알콜 옥시다제 1 (AOX1) 프로모터 요소, 칸디다 보이디니이로부터의 AOD1 프로모터 요소, 한세눌라 폴리모르파로부터의 MOX 프로모터 요소, 피키아 메타놀리카(Pichia methanolica)로부터의 MOD1 프로모터 요소, 피키아 파스토리스로부터의 DHAS 프로모터 요소, 피키아 파스토리스로부터의 FLD1 프로모터 요소, 및 피키아 파스토리스로부터의 PEX8 프로모터 요소로 이루어진 군으로부터 선택되는 것인 효모 세포.
- 제1항 내지 제11항 중 어느 한 항에 있어서, 적어도 하나의 메탄올-유도성 프로모터 요소가 피키아 파스토리스로부터의 알콜 옥시다제 1 (AOX1) 프로모터 요소인 효모 세포.
- 제1항 내지 제13항 중 어느 한 항에 있어서, 적어도 하나의 메탄올-유도성 프로모터 요소에 작동가능하게 연결된 폴리펩티드를 코딩하는 적어도 하나의 이종성 핵산을 포함하는 핵산 분자를 추가로 포함하는 효모 세포.
- 제14항에 있어서, 이종성 핵산이 철 보조인자의 생합성에서 수반되는 적어도 하나의 폴리펩티드를 코딩하는 것인 효모 세포.
- 제15항에 있어서, 철 보조인자가 헴인 효모 세포.
- 제16항에 있어서, 헴 생합성에서 수반되는 적어도 하나의 폴리펩티드가 ALA 신타제, ALA 데히드라타제, 포르포빌리노겐 데아미나제, UPG III 신타제, UPG III 데카르복실라제, CPG 옥시다제, PPG 옥시다제, 및 페로킬라타제로 이루어진 군으로부터 선택되는 것인 효모 세포.
- 제16항에 있어서, 헴 생합성에서 수반되는 적어도 하나의 폴리펩티드가 ALA 신타제, ALA 데히드라타제, 포르포빌리노겐 데아미나제, UPG III 신타제, UPG III 데카르복실라제, CPG 옥시다제, PPG 옥시다제, 및 페로킬라타제로 이루어진 군으로부터 선택된 2종 이상의 폴리펩티드를 포함하는 것인 효모 세포.
- 제1항 내지 제18항 중 어느 한 항에 있어서, 선택용 이종성 서열이 결여된 효모 세포.
- 제1항 내지 제19항 중 어느 한 항의 효모 세포를 제공하는 단계;
재조합 핵산 분자를 메틸영양성 효모 세포 내로 도입하며, 이러한 재조합 핵산 분자는 적어도 하나의 피키아 파스토리스 알콜 옥시다제 1 (AOX1) 프로모터 요소에 작동가능하게 연결된 폴리펩티드를 코딩하는 적어도 하나의 이종성 핵산을 포함하는 것인 단계; 및
세포를 재조합 핵산 분자의 발현에 적합한 조건 하에 배양하며,
이에 의해 이종성 폴리펩티드를 발현시키는 단계
를 포함하는, 세포에서 이종성 폴리펩티드를 발현시키는 방법. - 제20항에 있어서, 이종성 폴리펩티드가 글로빈 패밀리 PF00042의 구성원인 방법.
- 제20항에 있어서, 이종성 폴리펩티드가 식물 헤모글로빈인 방법.
- 제22항에 있어서, 식물 헤모글로빈이 레그헤모글로빈인 방법.
- 제23항에 있어서, 레그헤모글로빈이 서열식별번호: 4 및 서열식별번호: 6으로 이루어진 군으로부터 선택된 서열을 갖는 것인 방법.
- 제20항 내지 제24항 중 어느 한 항에 있어서, 도입 단계가 형질도입, 전기천공, 바이오리스틱 입자 전달, 및 화학적 형질전환으로 이루어진 군으로부터 선택된 기술을 포함하는 것인 방법.
- 제20항 내지 제25항 중 어느 한 항에 있어서, 세포가 배양되는 조건이 철 또는 그의 제약상 또는 대사상 허용되는 염의 첨가를 포함하는 것인 방법.
- 제20항 내지 제26항 중 어느 한 항에 있어서, 배양 단계가 세포를 메탄올의 존재 하에 배양하는 것을 포함하는 것인 방법.
- 제20항 내지 제26항 중 어느 한 항에 있어서, 배양 단계가 세포를 메탄올의 부재 하에 배양하는 것을 포함하는 것인 방법.
- 제20항 내지 제28항 중 어느 한 항에 있어서, 배양 단계가 세포를 선택의 부재 하에 배양하는 것을 포함하는 것인 방법.
- 재조합 핵산 분자를 포함하며, 여기서 재조합 핵산 분자는
피. 파스토리스로부터의 Mxr1 전사 활성화제 서열을 코딩하는 핵산;
글로빈 패밀리 (PF00042)의 구성원을 코딩하는 핵산; 및
헴 생합성에서 수반되는 적어도 하나의 폴리펩티드를 코딩하는 핵산
을 포함하는 것인 메틸영양성 피키아 효모 세포. - 제30항에 있어서, Mxr1 전사 활성화제 서열을 코딩하는 핵산이 메탄올-유도성 프로모터 요소에 작동가능하게 연결된 것인 효모 세포.
- 제30항에 있어서, 글로빈 패밀리의 구성원이 레그헤모글로빈인 방법.
- 제30항 내지 제32항 중 어느 한 항에 있어서, 글로빈 패밀리의 구성원을 코딩하는 핵산이 메탄올-유도성 프로모터 요소에 작동가능하게 연결된 것인 효모 세포.
- 제30항 내지 제32항 중 어느 한 항에 있어서, 글로빈 패밀리의 구성원을 코딩하는 핵산이 구성적 프로모터 요소에 작동가능하게 연결된 것인 효모 세포.
- 제30항 내지 제34항 중 어느 한 항에 있어서, 헴 생합성에서 수반되는 적어도 하나의 폴리펩티드를 코딩하는 핵산이 메탄올-유도성 프로모터 요소에 작동가능하게 연결된 것인 효모 세포.
- 제30항 내지 제34항 중 어느 한 항에 있어서, 헴 생합성에서 수반되는 적어도 하나의 폴리펩티드를 코딩하는 핵산이 구성적 프로모터 요소에 작동가능하게 연결된 것인 효모 세포.
- 제30항, 제31항, 제33항 및 제35항 중 어느 한 항에 있어서, 메탄올-유도성 프로모터 요소가 피. 파스토리스로부터의 알콜 옥시다제 I (AOX1) 프로모터 요소인 효모 세포.
- 제30항, 제34항 및 제36항 중 어느 한 항에 있어서, 구성적 프로모터 요소가 피. 파스토리스로부터의 전사 신장 인자 EF-1α 유전자 (TEF1) 프로모터 요소인 효모 세포.
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020207026647A KR102504198B1 (ko) | 2015-05-11 | 2016-05-11 | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 |
Applications Claiming Priority (17)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201562159899P | 2015-05-11 | 2015-05-11 | |
| US62/159,899 | 2015-05-11 | ||
| US201562183074P | 2015-06-22 | 2015-06-22 | |
| US62/183,074 | 2015-06-22 | ||
| US201562185921P | 2015-06-29 | 2015-06-29 | |
| US62/185,921 | 2015-06-29 | ||
| US201562203052P | 2015-08-10 | 2015-08-10 | |
| US62/203,052 | 2015-08-10 | ||
| US201562220366P | 2015-09-18 | 2015-09-18 | |
| US62/220,366 | 2015-09-18 | ||
| US201562222388P | 2015-09-23 | 2015-09-23 | |
| US62/222,388 | 2015-09-23 | ||
| US201562236506P | 2015-10-02 | 2015-10-02 | |
| US62/236,506 | 2015-10-02 | ||
| US201662313491P | 2016-03-25 | 2016-03-25 | |
| US62/313,491 | 2016-03-25 | ||
| PCT/US2016/031797 WO2016183163A1 (en) | 2015-05-11 | 2016-05-11 | Expression constructs and methods of genetically engineering methylotrophic yeast |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020207026647A Division KR102504198B1 (ko) | 2015-05-11 | 2016-05-11 | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| KR20170138543A true KR20170138543A (ko) | 2017-12-15 |
| KR102229968B1 KR102229968B1 (ko) | 2021-03-22 |
Family
ID=57249545
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020207026647A KR102504198B1 (ko) | 2015-05-11 | 2016-05-11 | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 |
| KR1020177033598A KR102229968B1 (ko) | 2015-05-11 | 2016-05-11 | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 |
| KR1020237006299A KR20230031993A (ko) | 2015-05-11 | 2016-05-11 | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020207026647A KR102504198B1 (ko) | 2015-05-11 | 2016-05-11 | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020237006299A KR20230031993A (ko) | 2015-05-11 | 2016-05-11 | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 |
Country Status (21)
| Country | Link |
|---|---|
| US (5) | US10689656B2 (ko) |
| EP (2) | EP3294762B1 (ko) |
| JP (4) | JP6831797B2 (ko) |
| KR (3) | KR102504198B1 (ko) |
| CN (1) | CN107614516A (ko) |
| AU (3) | AU2016262494B2 (ko) |
| BR (1) | BR112017024123A2 (ko) |
| CA (1) | CA2985641A1 (ko) |
| DK (1) | DK3294762T3 (ko) |
| ES (1) | ES2908463T3 (ko) |
| HK (1) | HK1252605A1 (ko) |
| HR (1) | HRP20220299T1 (ko) |
| HU (1) | HUE058104T2 (ko) |
| LT (1) | LT3294762T (ko) |
| MX (3) | MX2017014469A (ko) |
| NZ (1) | NZ737094A (ko) |
| PL (1) | PL3294762T3 (ko) |
| PT (1) | PT3294762T (ko) |
| RS (1) | RS63099B1 (ko) |
| SI (1) | SI3294762T1 (ko) |
| WO (1) | WO2016183163A1 (ko) |
Families Citing this family (25)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| HRP20220299T1 (hr) | 2015-05-11 | 2022-05-13 | Impossible Foods Inc. | Ekspresijski konstrukti i postupci za genetsku modifikaciju metilotrofnog kvasca |
| US10988515B2 (en) * | 2017-01-13 | 2021-04-27 | Bolt Threads, Inc. | Elastomeric proteins |
| WO2018140654A1 (en) * | 2017-01-25 | 2018-08-02 | Altius Institute For Biomedical Sciences | Methods and platform of designing genetic editing tools |
| EP3684389A4 (en) | 2017-09-21 | 2021-06-16 | Dana Farber Cancer Institute, Inc. | INSULATION, PRESERVATION, COMPOSITIONS AND USES OF EXTRACTS FROM JUSTICIA PLANTS |
| US11051532B2 (en) | 2017-09-22 | 2021-07-06 | Impossible Foods Inc. | Methods for purifying protein |
| US10221403B1 (en) * | 2018-04-25 | 2019-03-05 | Life Rainbow Biotech Co., Ltd. | Method of preparing zearalenone hydrolase |
| CN108913732B (zh) * | 2018-07-31 | 2022-02-08 | 华东理工大学 | 一种莫纳可林j异源生产的方法及应用 |
| CN111286520B (zh) * | 2018-12-10 | 2021-05-07 | 上海凯赛生物技术股份有限公司 | 用于发酵生产l-赖氨酸的重组dna、菌株及其应用 |
| CN109929769B (zh) * | 2019-03-05 | 2021-07-23 | 天津科技大学 | 一种利用fld1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株及构建方法 |
| CN109749948A (zh) * | 2019-03-05 | 2019-05-14 | 天津科技大学 | 一种利用tef1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株及构建方法 |
| US11427932B2 (en) | 2019-04-17 | 2022-08-30 | Impossible Foods Inc. | Materials and methods for protein production |
| MX2021012851A (es) | 2019-04-25 | 2022-02-10 | Impossible Foods Inc | Cepas y métodos para la producción de proteínas que contienen hemo. |
| WO2021108502A1 (en) | 2019-11-27 | 2021-06-03 | Evelo Biosciences, Inc. | Methods and compositions for culturing hemoglobin-dependent bacteria |
| EP4090746A4 (en) * | 2020-01-10 | 2024-01-24 | Intron Biotechnology Inc | METHOD FOR PRODUCING SOY LEGHHEMOGLOBIN USING ESCHERICHIA COLI |
| US10894812B1 (en) | 2020-09-30 | 2021-01-19 | Alpine Roads, Inc. | Recombinant milk proteins |
| EP4222167A1 (en) | 2020-09-30 | 2023-08-09 | Nobell Foods, Inc. | Recombinant milk proteins and food compositions comprising the same |
| US10947552B1 (en) | 2020-09-30 | 2021-03-16 | Alpine Roads, Inc. | Recombinant fusion proteins for producing milk proteins in plants |
| WO2022072833A2 (en) | 2020-10-02 | 2022-04-07 | Impossible Foods Inc. | Expression constructs and methods of genetically engineering cells |
| WO2022072846A2 (en) | 2020-10-02 | 2022-04-07 | Impossible Foods Inc. | Transgenic plants with altered fatty acid profiles and upregulated heme biosynthesis |
| WO2022117729A1 (en) * | 2020-12-02 | 2022-06-09 | Planeat Foods Pte. Ltd. | Method of producing globin polypeptide recombinantly and meat substitute food product |
| WO2022251166A2 (en) | 2021-05-25 | 2022-12-01 | Evelo Biosciences, Inc. | Bacterial compositions comprising soy hemoglobin |
| AU2022369299A1 (en) | 2021-10-19 | 2024-03-14 | Eat Scifi Inc. | Plant base/animal cell hybrid meat substitute |
| WO2023114395A1 (en) * | 2021-12-16 | 2023-06-22 | Impossible Foods Inc. | A genetic factor to increase expression of recombinant proteins |
| US11931687B2 (en) | 2022-02-15 | 2024-03-19 | Streamline Innovations, Inc. | Gas purification apparatus and method |
| KR20230164436A (ko) | 2022-05-25 | 2023-12-04 | 한화솔루션 주식회사 | 야생형 효모 균주를 이용한 헴 고생산 방법 |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2010516268A (ja) * | 2007-01-26 | 2010-05-20 | ノボザイムス アクティーゼルスカブ | ピキアにおけるメタノール誘導性プロモーターからのメタノール非依存的誘導の方法 |
| WO2014008729A1 (zh) * | 2012-07-12 | 2014-01-16 | 华东理工大学 | 消除甲醇诱导型启动子单一甲醇碳源依赖性的方法 |
| WO2014110532A2 (en) * | 2013-01-11 | 2014-07-17 | Maraxi, Inc. | Methods and compositions for affecting the flavor and aroma profile of consumables |
Family Cites Families (38)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4683202A (en) | 1985-03-28 | 1987-07-28 | Cetus Corporation | Process for amplifying nucleic acid sequences |
| US4965188A (en) | 1986-08-22 | 1990-10-23 | Cetus Corporation | Process for amplifying, detecting, and/or cloning nucleic acid sequences using a thermostable enzyme |
| US4683195A (en) | 1986-01-30 | 1987-07-28 | Cetus Corporation | Process for amplifying, detecting, and/or-cloning nucleic acid sequences |
| DK162399C (da) | 1986-01-28 | 1992-03-23 | Danisco | Fremgangsmaade til ekspression af gener i baelgplanteceller, dna-fragment, rekombineret dna-fragment samt plasmid til brug ved udoevelsen af fremgangsmaaden |
| US4800159A (en) | 1986-02-07 | 1989-01-24 | Cetus Corporation | Process for amplifying, detecting, and/or cloning nucleic acid sequences |
| US6946587B1 (en) | 1990-01-22 | 2005-09-20 | Dekalb Genetics Corporation | Method for preparing fertile transgenic corn plants |
| US5204253A (en) | 1990-05-29 | 1993-04-20 | E. I. Du Pont De Nemours And Company | Method and apparatus for introducing biological substances into living cells |
| US5753465A (en) | 1994-08-30 | 1998-05-19 | Carnegie Mellon University | Unmodified recombinant human adult hemoglobin production |
| US5538800A (en) | 1994-09-29 | 1996-07-23 | At&T Corp. | Magnetoresistive oxide material and articles comprising the material |
| FR2736930B1 (fr) | 1995-07-17 | 1997-09-19 | Biocem | Procede de production, par des cellules vegetales, de proteines heminiques, proteines ainsi obtenues et produits contenant ces proteines |
| US5824511A (en) | 1995-08-01 | 1998-10-20 | University Technology Corporation | Method for enhancing the production of hemoproteins |
| DE69728878T2 (de) | 1996-06-10 | 2005-04-07 | Novozymes Biotech, Inc., Davis | Verfahren zur vermehrung der hemoproteinherstellung in filamentösen fungi |
| JP3794760B2 (ja) | 1996-07-30 | 2006-07-12 | 三井化学株式会社 | フィターゼ生産酵母 |
| US5959187A (en) | 1996-09-26 | 1999-09-28 | Bailey; James E. | Expression of oxygen-binding proteins in plants |
| JPH10117776A (ja) | 1996-10-22 | 1998-05-12 | Japan Tobacco Inc | インディカイネの形質転換方法 |
| US7803765B2 (en) | 1999-05-05 | 2010-09-28 | Phylogica Limited | Methods of constructing biodiverse gene fragment libraries and biological modulators isolated therefrom |
| EP1356060B1 (en) | 2000-09-18 | 2005-12-14 | Genencor International, Inc. | Twin-arginine translocation in bacillus |
| US8021695B2 (en) | 2002-02-15 | 2011-09-20 | Arch Personal Care Products, L.P. | Personal care composition containing leghemoglobin |
| BR0317650A (pt) | 2002-12-23 | 2005-12-06 | Max Planck Gesellschaft | Planta transformada, sequência de nucleotìdeo, estrutura de gene, vetor, processo para modificar o teor de reserva de armazenagem em plantas, e, uso de uma planta |
| CA2525493C (en) | 2003-04-11 | 2018-10-30 | Ventria Bioscience | Human blood proteins expressed in oryza seeds |
| KR100660539B1 (ko) | 2004-07-29 | 2006-12-22 | 삼성전자주식회사 | 자기 기억 소자 및 그 형성 방법 |
| AU2006283077B2 (en) | 2005-08-24 | 2012-06-28 | The Scripps Research Institute | Translation Enhancer-Element dependent vector systems |
| JP2008017733A (ja) | 2006-07-11 | 2008-01-31 | Tokyo Univ Of Agriculture | 異種タンパク質の発現方法及び異種タンパク質の発現方法により得られたタンパク質 |
| JP2008017774A (ja) * | 2006-07-13 | 2008-01-31 | Nipro Corp | ピキア酵母を用いた組換えヒトヘモグロビンの製造 |
| US20110265221A1 (en) | 2007-07-10 | 2011-10-27 | Monsanto Technology Llc | Transgenic plants with enhanced agronomic traits |
| EP2058398A1 (en) | 2007-11-09 | 2009-05-13 | Nipro Corporation | Production of recombinant human hemoglobin using pichia yeast |
| US9085766B2 (en) | 2010-05-21 | 2015-07-21 | Cornell University | Methods of producing recombinant heme-binding proteins and uses thereof |
| AU2011287293B2 (en) | 2010-08-02 | 2015-02-26 | Cellectis S.A. | Method for targeted genomic events in algae |
| WO2012083424A1 (en) | 2010-12-21 | 2012-06-28 | The University Of Western Ontario | Novel amino acid linker sequences for ligand immobilization |
| WO2012102171A1 (ja) | 2011-01-27 | 2012-08-02 | 株式会社カネカ | 形質転換用酵母およびタンパク質の製造方法 |
| AU2012281069B2 (en) | 2011-07-12 | 2016-10-13 | Impossible Foods Inc. | Methods and compositions for consumables |
| WO2014045621A1 (ja) | 2012-09-19 | 2014-03-27 | 国立大学法人大阪大学 | 肺炎球菌表面タンパク質aを含む肺炎球菌ワクチン |
| DK3044320T3 (da) | 2013-09-11 | 2020-03-16 | Impossible Foods Inc | Sekretion af hæmholdige polypeptider |
| US10172380B2 (en) | 2014-03-31 | 2019-01-08 | Impossible Foods Inc. | Ground meat replicas |
| HRP20220299T1 (hr) | 2015-05-11 | 2022-05-13 | Impossible Foods Inc. | Ekspresijski konstrukti i postupci za genetsku modifikaciju metilotrofnog kvasca |
| US20190292217A1 (en) | 2016-12-02 | 2019-09-26 | Impossible Foods Inc. | Transgenic plants with upregulated heme biosynthesis |
| WO2018102656A1 (en) | 2016-12-02 | 2018-06-07 | City Of Hope | Producing recombinant products in contained systems |
| MX2021012851A (es) | 2019-04-25 | 2022-02-10 | Impossible Foods Inc | Cepas y métodos para la producción de proteínas que contienen hemo. |
-
2016
- 2016-05-11 HR HRP20220299TT patent/HRP20220299T1/hr unknown
- 2016-05-11 ES ES16793420T patent/ES2908463T3/es active Active
- 2016-05-11 HU HUE16793420A patent/HUE058104T2/hu unknown
- 2016-05-11 KR KR1020207026647A patent/KR102504198B1/ko active IP Right Grant
- 2016-05-11 JP JP2017558948A patent/JP6831797B2/ja active Active
- 2016-05-11 WO PCT/US2016/031797 patent/WO2016183163A1/en active Application Filing
- 2016-05-11 CN CN201680029580.4A patent/CN107614516A/zh active Pending
- 2016-05-11 US US15/573,300 patent/US10689656B2/en active Active
- 2016-05-11 SI SI201631500T patent/SI3294762T1/sl unknown
- 2016-05-11 EP EP16793420.7A patent/EP3294762B1/en active Active
- 2016-05-11 DK DK16793420.7T patent/DK3294762T3/da active
- 2016-05-11 EP EP22151183.5A patent/EP4039695A1/en active Pending
- 2016-05-11 RS RS20220256A patent/RS63099B1/sr unknown
- 2016-05-11 AU AU2016262494A patent/AU2016262494B2/en active Active
- 2016-05-11 BR BR112017024123-4A patent/BR112017024123A2/pt active Search and Examination
- 2016-05-11 MX MX2017014469A patent/MX2017014469A/es unknown
- 2016-05-11 KR KR1020177033598A patent/KR102229968B1/ko active IP Right Grant
- 2016-05-11 NZ NZ737094A patent/NZ737094A/en unknown
- 2016-05-11 CA CA2985641A patent/CA2985641A1/en active Pending
- 2016-05-11 LT LTEPPCT/US2016/031797T patent/LT3294762T/lt unknown
- 2016-05-11 PL PL16793420T patent/PL3294762T3/pl unknown
- 2016-05-11 PT PT167934207T patent/PT3294762T/pt unknown
- 2016-05-11 KR KR1020237006299A patent/KR20230031993A/ko not_active Application Discontinuation
-
2017
- 2017-08-16 US US15/678,891 patent/US9938327B2/en active Active
- 2017-08-16 US US15/678,342 patent/US10273492B2/en active Active
- 2017-11-10 MX MX2022013540A patent/MX2022013540A/es unknown
- 2017-11-10 MX MX2022006513A patent/MX2022006513A/es unknown
-
2018
- 2018-09-06 US US16/123,121 patent/US11319544B2/en active Active
- 2018-09-17 HK HK18111904.2A patent/HK1252605A1/zh unknown
-
2019
- 2019-10-17 AU AU2019250216A patent/AU2019250216B2/en active Active
-
2020
- 2020-02-03 JP JP2020016038A patent/JP7308160B2/ja active Active
-
2021
- 2021-09-30 JP JP2021161491A patent/JP2022025068A/ja active Pending
-
2022
- 2022-01-25 US US17/584,028 patent/US20220290166A1/en active Pending
- 2022-03-03 AU AU2022201465A patent/AU2022201465A1/en active Pending
- 2022-11-04 JP JP2022177137A patent/JP2023022042A/ja active Pending
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2010516268A (ja) * | 2007-01-26 | 2010-05-20 | ノボザイムス アクティーゼルスカブ | ピキアにおけるメタノール誘導性プロモーターからのメタノール非依存的誘導の方法 |
| WO2014008729A1 (zh) * | 2012-07-12 | 2014-01-16 | 华东理工大学 | 消除甲醇诱导型启动子单一甲醇碳源依赖性的方法 |
| WO2014110532A2 (en) * | 2013-01-11 | 2014-07-17 | Maraxi, Inc. | Methods and compositions for affecting the flavor and aroma profile of consumables |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR102229968B1 (ko) | 메틸영양성 효모를 유전적으로 조작하는 발현 구축물 및 방법 | |
| DK2588616T3 (en) | PROCEDURE FOR MAKING A RELATIONSHIP OF INTEREST | |
| EP2106447B1 (en) | Method for methanol independent induction from methanol inducible promoters in pichia | |
| US11427932B2 (en) | Materials and methods for protein production | |
| Cannio et al. | Thermoadaptation of a mesophilic hygromycin B phosphotransferase by directed evolution in hyperthermophilic Archaea: selection of a stable genetic marker for DNA transfer into Sulfolobus solfataricus | |
| CN110938649A (zh) | 一种提高外源蛋白表达量的蛋白合成体系及其应用方法 | |
| JP2018533932A (ja) | 新規エピソームプラスミドベクター | |
| CN115247135A (zh) | 强化高尔基体至胞外蛋白运输过程提高毕赤酵母胞外葡萄糖氧化酶产量 | |
| JP2003210173A (ja) | 新規プロモーター | |
| JP2007049963A (ja) | 新規酵母プロモーター |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| E902 | Notification of reason for refusal | ||
| E902 | Notification of reason for refusal | ||
| E701 | Decision to grant or registration of patent right | ||
| A107 | Divisional application of patent | ||
| GRNT | Written decision to grant |