KR20150132522A - 주요 전하 아이소폼이 풍부한 항체 조성물을 포함하는 신규 의약품 - Google Patents
주요 전하 아이소폼이 풍부한 항체 조성물을 포함하는 신규 의약품 Download PDFInfo
- Publication number
- KR20150132522A KR20150132522A KR1020157029822A KR20157029822A KR20150132522A KR 20150132522 A KR20150132522 A KR 20150132522A KR 1020157029822 A KR1020157029822 A KR 1020157029822A KR 20157029822 A KR20157029822 A KR 20157029822A KR 20150132522 A KR20150132522 A KR 20150132522A
- Authority
- KR
- South Korea
- Prior art keywords
- antibody
- composition
- monoclonal antibody
- antibody composition
- isoforms
- Prior art date
Links
Images




Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
- A61K39/39591—Stabilisation, fragmentation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2896—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against molecules with a "CD"-designation, not provided for elsewhere
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/06—Immunosuppressants, e.g. drugs for graft rejection
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K1/00—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length
- C07K1/14—Extraction; Separation; Purification
- C07K1/16—Extraction; Separation; Purification by chromatography
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/24—Immunoglobulins specific features characterized by taxonomic origin containing regions, domains or residues from different species, e.g. chimeric, humanized or veneered
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
- C07K2317/732—Antibody-dependent cellular cytotoxicity [ADCC]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
- C07K2317/734—Complement-dependent cytotoxicity [CDC]
Abstract
본 발명은 ADCC를 통하여 타겟 세포를 파괴하는 메카니즘을 포함하는 항체 치료법의 기술 분야에 속한다. 본 발명은, 항체 조성물에 원래 존재하는 서로 다른 전하 아이소폼들(charge isoforms)을 크로마토그래피로 분획하고 크로마토그램의 주 피크(main peak)에 해당하는 하나 이상의 크로마토그래프 분획물들을 조합하여 얻어진, 의약품 용도로 사용하기 위한, 정제된 항체 조성물에 관한 것이며, 여기서그렇게 하여 얻어진 상기 모노클론 항체 조성물은 상기 주 피크가 풍부하고, 상기 피크는 얻어진 조성물의 크로마토그램의 적어도 85%를 나타낸다.
Description
본 발명은 ADCC를 통하여 타겟 세포를 파괴하는 메카니즘을 포함하는 항체 치료법의 기술 분야에 속한다. 본 발명은, 항체 조성물에 원래 존재하는 서로 다른 전하 아이소폼들(charge isoforms)을 크로마토그래피로 분획하고 크로마토그램의 주 피크(main peak)에 해당하는 하나 이상의 크로마토그래프 분획물들을 조합하여 얻어진, 의약품 용도로 사용하기 위한, 정제된 항체 조성물에 관한 것이며, 여기서그렇게 하여 얻어진 상기 모노클론 항체 조성물은 상기 주 피크가 풍부하고, 상기 피크는 얻어진 조성물의 크로마토그램의 적어도 85%를 나타낸다.
지난 수 십년 동안, 암, Rh 마이너스(-) 임산부의 동종 면역(allo-immunization)의 예방, 감염성 질환, 염증성 질환 및 뚜렷한 자가면역 질환의 다양한 치료법 분야에서 항체들, 종종 모노클론 항체들을 이용한 수동(passive) 면역요법 치료가 활발하게 개발되어 왔다.
항체를 이용한 수동 면역요법 치료가 오늘날 치료 효과를 나타내고 있지만, 관찰된 임상 반응 수준은 여전히 불충분하고, 따라서 2차 효과(secondary effects)를 제한하기 위하여, 임상 반응을 증가시킬 수 있고 보다 적은 용량의 투여를 가능하게 하는, 보다 효과적인 항체 조성물이 요구된다.
임의의 생물학적 제품처럼, 항체 조성물은 천연의 이종에서 얻어진다. 실제로, 치료법에 사용되는 항체 조성물은 생물학적 시스템(세포, 유전자변형 동물 또는 식물)에서 생산되고, 상기 생물학적 시스템에서 일반적으로 단백질들 및 따라서 특히 항체들은, 다수의 번역 후 변형(post-translation modification)에 종속되고(효소적 변형 또는 분해), 이것은 하나의 항체 분자를 다른 것으로 변화시킬 것이고, 이에 의해 생산된 항체 조성물 내에 마이크로-이질성(micro-heterogeneity)을 산출할 것이다.
항체들은 4개의 폴리펩티드 쇄로 이루어진 당단백질들이다: 다양한 다수의 이황화결합 및 비공유성 상호작용과 관련된, 2개의 일반적으로 동일한 중쇄(소위 "중(heavy)"쇄를 위한 "H"쇄)와 2개의 일반적으로 동일한 경쇄(소위 "경(light)"쇄를 위한 "L"쇄). 이들 쇄는 Y-형 구조를 형성하고, 중쇄는 Y의 줄기에 기여하고 Y의 각 가지(arm)의 절반에 기여하고, 경쇄는 Y의 각 가지의 절반에 기여한다. 각 경쇄는 불변 영역(CL)와 가변 영역(VL)으로 구성된다; 중쇄는 항체의 아이소타이프(isotype)에 따라서 하나의 가변 단편(VH)과 3 또는 4개의 불변 단편(CH1 내지 CH3 또는 CH4)으로 구성된다(IgGs는 3개의 불변 단편들 CH1 내지 CH3을 포함한다). 경쇄의 결합(VL+CL) 및 중쇄의 VH와 CH1 영역의 결합은 단편 Fab를 형성하고, 결합된 도메인 VL과 VH는 항원의 인식을 책임진다. 2개의 중쇄들의 불변 영역(CH2 및 CH3) 또는 (CH2 내지 CH4)은 불변 Fc 단편을 형성한다.
항체들은 다음의 번역 후 변형에 종속되는 것으로 알려져 있다: 중쇄 또는 경쇄의 말단 변형, Fc 부분(그리고 선택적으로 Fcbs)의 글리코실화, 탈아미드화, 이성질화, 산화, 파쇄(fragmentation) 및 응집(Vlasak 등, 2008 참조).
대부분의 번역 후 변형은, 직접적으로 하전된 그룹의 수를 변형시키거나 또는 간접적으로 구조적 변형을 도입함으로써-그것들 자체가 하전된 잔기(residues)의 국소 분획을 변형하거나 그것들의 pKa를 변화시킴-, 항체의 표면 전하 특성의 변경을 가져온다. 따라서, 이러한 모든 변형들은 또한 동일 항체에 상이한 전하들을 갖고, 구별된 많은 등전점들(isoelectric points, pl)을 갖는, 마이크로-이질성을 산출하고, 이에 따라 아이소폼들이 항체 조성물 내에 공존한다(Vlasak 등, 2008 참조).
번역 후 변형 중에서, 항체의 불변 영역 Fc의 글리코실화는 항체의 많은 생물학적 특징들에 강력한 영향을 미치는 것으로 현재 잘 알려져 있다: 생체 내 반감기(Wright 등, 1994 참조), ADCC 반응을 유도하는 능력(항체-의존성 세포독성 세포 반응, Satoh 등, 2006, Presta 등, 2006, 참조), CDC 반응(보체-의존성 세포독성 반응, Wright 등, 1994, Presta 등, 2006, 참조) 등. 특히, 푸코실화된 글리칸 형태에서 항체 조성물의 함량은 현재 생체 내 ADCC 반응을 유도하기 위하여 조성물의 능력에 매우 강하게 영향을 미치는 것으로 알려져 있다.
반대로, 많은 문헌들은 모노클론 항체의 상업적 집단(batch)의 재현성 및 품질을 입증하기 위하여 항체 조성물에 존재하는 전하 아이소폼에 특징을 부여하는 것을 목적으로 하지만, 항체 조성물 내에서 동일한 항체의 많은 뚜렷한 전하 아이소폼의 존재를 초래하는 다른 번역 후 변형이 지금까지 생체 내 항체들의 생물학적 특징에 거의 또는 전혀 영향을 주지 않는 것으로 여겨지고 있다. 따라서, 선행 기술에서 전하 아이소폼과 관련하여 항체의 상업적 집단의 품질을 추적하는데 필수불가결한 것으로 일반적으로 간주되지만, 이러한 추적은 제품의 품질의 순수 추적으로 간주되고, 특정 전하 아이소폼이 매우 풍부한, 항체 조성물의 정제된 분획을 이용하는 것에 대한 제시는 전혀 없었다. 실제로, 항체 조성물의 적어도 특정 생물학적 특징에 대해 상당한 효과를 나타내지 않는 경우, 제조 방법을 복잡하게 만들고 수율을 감소시키기 위하여, 전체 조성물을 사용하지 않을 이유는 없었다. 이제, 상기에서 언급된 것처럼, 글리코실화를 제외하고, 항체 조성물 내에서 항체의 많은 뚜렷한 전하 아이소폼의 존재를 초래하는 다른 번역 후 변형은 지금까지 항체들의 생물학적 특징들을 변경시키지 않는 것으로 간주되었다.
몇몇 전하 아이소폼의 발생을 초래하는 변형(modification)들 중 하나는 항체의 중쇄의 C-말단 라이신의 효소적 절단이다. 항체가 카르복시펩티다아제를 발현하는 세포에서 생산되자마자, 이러한 절단은 항체 분자들에 따라 상이한 수준으로 발생한다. 라이신의 곁사슬 때문에 C-말단 라이신의 존재는 다소 염기성(basic nature)을 부여한다. 그러므로 중쇄 모두 또는 어느 하나 위의 절단은 좀더 산성 아이소폼을 생성한다. 일반적으로 중쇄 위에 0, 1, 또는 2 C-말단 라이신을 갖는 아이소폼이 존재하고, 이에 따라 약간 상이한 pl을 갖는 3개의 아이소폼을 생성한다(Vlasak 등, 2008 참조). 이러한 특정 변형에서, 안테스 등(Antes et al., 2007)은 혈청의 존재 또는 부재시에 생산된 암의 수동 면역요법에서 사용되는 인간화된 모노클론 항-루이스-YIGN311 항체의 집단(batches)의 등전점 맞춤(isoelectric focusing, IEF)에 의한 분석을 개시한다. 저자들은 혈청의 존재 또는 부재시에 생산된 항체 조성물의 전하 아이소폼의 프로파일이 상이하고, 혈청의 부재시에 생산된 조성물이 혈청의 존재시에 생산된 것보다 항체의 중쇄의 C-말단 라이신의 효소적 절단에 의해 영향을 덜 받는다는 것을 보여준다. (보체를 통한) CDC 반응을 유도하는 양 조성물의 개별 능력에 대한 이러한 변형의 효과 분석은 이러한 변형들과 관련된 어떠한 유의적인 효과도 없다는 것을 보여주었다.
항체 조성물 내의 몇몇 전하 아이소폼의 생성을 초래하는 또 하나의 형태의 변형은 N-말단 글루타민 또는 글루탐산 잔기의 사이클리제이션(cyclisation)이고, 이것은 파이로글루타메이트(pE) 그룹의 형성을 가져오고 이에 따라 좀더 산성의 아이소폼이 생성된다. 이러한 변형은 다른 수준에서, 전체 항체 조성물에서, 전신적으로(systematically) 발생하지만, 항체의 기능적 특징에 영향을 미칠 수 있는 것으로 간주되지 않는다(Vlasak 등, 2008 참조).
항체 조성물 내에서 몇몇 전하 아이소폼의 생성을 초래하는 또 하나의 형태의 변형은, 좀더 산성의 아이소폼을 생성하는, 특히 라이신 잔기 위의, 공유 부가물(covalent adduct)의 형성 및 주목할만한 당화반응(glycation) 현상(비효소적 당의 부가)의 형성이다. 이러한 변형 형태는 또한 항체의 기능적 특징에 영향을 미칠수 없는 것으로 간주된다(Vlasak 등, 2008 참조).
항체 조성물 내에서 몇몇 전하 아이소폼의 생성을 초래하는 다른 통상의 형태의 변형은, 좀더 산성의 아이소폼을 생성하는, 아스파리긴 잔기의 탈아미드화 및 아스파르트산 잔기의 이성질화이다. 항체의 불변 부분에서, 탈아미드화 현상에 민감한 아스파라긴 잔기들은, FcRn 수용체와의 결합 자리 및 FcγR 수용체와의 결합 자리에서 떨어져 있는, CH3 도메인에 위치된다. 따라서 이러한 변형들 일반적으로 항체의 기능적 특징에 영향을 미칠수 없는 것으로 간주된다(Vlasak 등, 2008 참조).
카일 등(Khawli et al., 2010)과 간드히 등(Gandhi et at., 2011)은 양이온 교환 수지를 이용하는 크로마토그래피 기술를 이용하여, 수동 면역요법에서 사용된 모노클론 항체 조성물의 주요, 산성 및 염기성 아이소폼의 분리를 기술한다; 몇몇 아이소폼의 존재를 초래하는 번역 후 변형의 분석; 뿐만 아니라 3개의 정제된 분획물들(산성, 주요 및 염기성 분획물들)의 특정 기능적 특징들 및 약물 동역학적 특징들의 연구. 양자의 경우에, 천연 조성물의 크로마토그램은 항상 주(major) 피크를 보이고, 산성 아이소폼을 포함하는 피크와 염기성 아이소폼을 포함하는 피크로 둘러싸인다. 확인된 번역 후 변형은 주목할만하게 특정 이황화 결합의 감소(Khawli 등, 2010), 당화반응(Khawli 등, 2010; Gandhi 등, 2011), 탈아미드화(Khawli 등, 2010; Gandhi 등, 2011), 중쇄의 C-말단 라이신의 절단(Khawli 등, 2010; Gandhi 등, 2011), 응집물의 존재(Gandhi 등, 2011), 산화 현상(Gandhi 등, 2011)을 포함한다. 약물 동역학 특징들의 분석(생체 내 FcRn 결합 및 시험 Khawli 등, 2010)은 3개의 시험 정제 분획물에서 어떠한 유의적인 거동의 차이가 나타나지 않았다. 양 문헌에서, 반응기 세포(effector cell)의 부재시에 항체에 특이적인 항원을 발현하는 세포주의 증식을 시험관 내(in vitro) 억제하는 3개의 정제 분획의 능력이, 또한 시험되었다. 이러한 시험은 항원에 결합할 수 있는 능력 및 세포자살(apoptosis)을 유도할 수 있는 능력을 나타낼 가능성을 보여준다. 양 문헌에서 산성 분획이 매우 조금 낮은 용량(capability)을 갖지만, 결과들은 유의적이지 않았고 따라서 3개의 정제 분획들 사이에서 유의적인 차이가 관찰되지 않았다. 더욱이, 이들 분획들의 분리 이전에, 주요 아이소폼이 풍부한 분획은 전체 항체 조성물과 비교하여 능력을 향상시키지 않았다.
또한, 다른 문헌들은 항체의 특정 전하 아이소폼을 어떻게 분석 및/또는 분리하는지 기술하지만, 상이한 아이소폼들의 반응기(effector) 특징들을 비교하지는 않는다. 따라서, EP1308456 및 WO2004/024866은 정제 전 및 후의 조성물의 반응기(effector) 특징들에 대한 시험 없이, 모노클론 항체 조성물의 산성 변형체를 제거하는 것을 목적으로 하는 크로마토그래피 방법을 개시한다. 또한, WO2011/009623은 정제 전 및 후의 조성물의 반응기 특징들에 대한 시험 없이, 모노클론 항체 조성물의 산성 변형체 또는 염기성 변형체를 억제하는 것을 목적으로 하는 크로마토그래피 방법을 개시한다. 추가로, 상기 문헌에 기술된 방법은 단일 형태의 변형체의 억제만이 가능하고 산성 변형체들의 제거에만 실질적으로 적용된다.
따라서, 항체들의 기능성 특징들에 영향을 미치는 것으로 알려진, 글리코실화는 제외하고, (주요 아이소폼을 산출하는 상이한 변형들로부터) 몇몇 전하 아이소폼을 생성하는 다른 번역 후 변형과 관련된 선행 기술에서 가능한 요소들은, 이들 변형이 항체의 기능적 특징들에 어떠한 영향도 미치지 않는다는 것을 제시한다. 그러나, 본 발명자들은 놀랍게도, 항체 조성물의 주요 전하 아이소폼이 풍부한, 크로마토그래피에 의해 정제된 분획이 이 수용체를 발현하는 반응기 세포에 의하여 CD16 수용체를 통하여 반응기 반응을 유도하는 능력이 상당히 더 크다는 것을 발견하였다. 따라서, 항체 조성물의 주요 전하 아이소폼 내에서 풍부한 정제된 분획이 더 강한 ADCC 반응과 생체 내 더 강한 CDC 반응을 유도할 가능성을 부여하고, 따라서 임상 반응을 증가시키고 및/또는 투여 용량을 감소시켜서, 2차 효과를 제한할 수 있다.
따라서 본 발명은 다음을 포함하는 방법에 의해 얻어질 수 있는, 모노클론 항체 조성물에 관한 것이다:
a) 비-인간 유전자 변형 동물 또는 유전자 변형 식물로부터, 세포 클론 유래 모노클론 항체 조성물을 제조하는 단계,
b) 단계 a)에서 얻어진 조성물을 크로마토그래피에 의해 분획하는 단계, 및
c) 단계 b)에서 얻어지고 크로마토그램의 주 피크에 대응하는, 하나 또는 수 개의 크로마토그래피 분획물을 조합하는 단계, 여기서 이렇게 하여 얻어진 모노클론 항체 조성물은 상기 주 피크가 풍부하게 되고, 의약품 용도를 위하여, 상기 주 피크는 단계 c)에서 얻어진 조성물의 크로마토그램 중 적어도 85%, 유리하게는 적어도 86%, 적어도 87%, 적어도 88%, 적어도 89%, 보다 유리하게는 적어도 90%, 적어도 91%, 적어도 92%, 적어도 93%, 적어도 94%, 또는 심지어 적어도 95%, 적어도 96%, 적어도 97%, 적어도 98%, 적어도 98.5%, 적어도 99%, 또는 적어도 99.5%를 나타낸다.
유리하게는, 단계 b)는 표준 이온 교환 크로마토그래피, 크로마토포커싱, 또는 소수성 상호작용 크로마토그래피에 의하여 단계 a)에서 얻어진 조성물을 분획하여 이루어진다.
유리하게는, 이온 교환 크로마토그래피는 다음의 용출(elution) 수단 중 하나를 사용한다.
·이온 힘 구배; 및/또는
· pH 구배; 또는
· 치환 분자(displacement molecule).
유리하게는, 의약품 용도의 이러한 조성물에서, 조성물에 존재하는 항체의 중쇄 중 적어도 95%, 바람직하게는 적어도 96%, 적어도 97%, 적어도 98%, 또는 심지어 적어도 98.5%, 적어도 99%, 또는 적어도 99.5%는 어떠한 C-말단 라이신 잔기도 포함하지 않는다.
본 발명은 또한 모노클론 항체 조성물에 관한 것으로서, 의약품으로서의 그것의 용도를 위하여, 조성물에 존재하는 항체의 중쇄 중 적어도 95%, 바람직하게는 적어도 96%, 적어도 97%, 적어도 98%, 또는 심지어 적어도 98.5%, 적어도 99%, 또는 적어도 99.5%는 어떠한 C-말단 라이신 잔기도 포함하지 않는다.
본 발명에 따른 의약품 용도를 위한 조성물에서, 항체는 유리하게는 건강한 공여 세포에 존재하는 비-유비쿼터스 항원(non-ubiquitous antigen), 암 세포의 항원, 또는 병원체에 감염된 세포의 항원을 향한다.
특히, 아래의 실시 형태들이 바람직하다:
- 항체는 항-Rh(D) 항체이고 조성물은 Rh 마이너스(-) 개체에서 동종 면역(allo-immunization)을 예방하기 위한 것이다.
- 항체는 암 세포의 항원을 향하고 조성물은 암을 치료하기 위한 것이다.
- 항체는 병원체에 감염된 세포의 항원을 향하고 조성물은 상기 병원 유기체에 의한 감염을 치료하기 위한 것이다.
- 항체는 면역 세포의 항원을 향하고 조성물은 자가면역 질환을 치료하기 위한 것이다.
바람직한 실시형태에서, 본 발명에 따른 의약품 용도의 조성물에서, 항체는, FcγRⅢ 수용체와의 결합 및 FcγRⅢ 수용체를 통한 반응기 특징(effector property)을 증가시키는 Fc 단편의 변형을 포함한다. 본 발명에 따른 의약품 용도의 조성물은 주목할 만하게 FcγRⅢ 수용체와의 결합 및/또는 낮은 푸코스(fucose) 함량을 증가시키는 Fc 단편에서의 돌연변이를 포함할 수 있다. 특히, 바람직하게는, 조성물에 존재하는 항체들은, 65% 미만의 푸코스 함량을 가지면서, Fc 단편의 N-글리코실화 부위에 바이안테나리(biantennary) 형태의 글리칸 구조체를 갖는다.
바람직한 실시형태에서, 본 발명에 따른 의약품 용도의 조성물에서, 항체는, 단백질 C1q와의 결합 및 보체를 통한 반응기 특징들을 증가시키는 Fc 단편의 변형을 포함한다.
본 발명은 또한, FcγRⅢ 수용체(CD16)를 발현하는 면역계의 반응기 세포에 의하여 상기 항원을 발현하는 타겟 세포들의 항체(ADCC)에 의존하여, 세포독성을 유도하기 위한, 정해진 항체에 대한 모노클론 항체 조성물의 능력을 증가시키는 크로마토그래피 분획 단계의 사용에 관한 것이다.
본 발명은 또한 보체에 의하여 상기 항원을 발현하는 타겟 세포들의 보체-의존적 세포독성(CDC)를 유도하기 위한, 정해진 항체에 대한(directed against) 모노클론 항체 조성물의 능력을 증가시키는 크로마토그래피 분획 단계의 사용에 관한 것이다.
도 1. 감소하는 pH 구배(2개의 버퍼-버퍼 A(디에탄올아민 25 mM), 버퍼 B(폴리버퍼 96 + 파말라이트(pharmalyte) 8-10.5)-를 이용하여 9.5 내지 8.0)에 의한 용출물(elution)을 이용한, 항-CD20 항체 조성물의 크로마토포커싱(음이온 교환 수지 (GE Life Science사의 컬럼 MonoTM P)에 의한 3개의 분리물에서 얻은 크로마토그램들. 항체 조성물은 탈염되었고, 20 mg을 컬럼에 주입하였다. 2 ml 분획물이 수집되었다. 분획물들 33 내지 50이 분석을 위해 수집되었다.
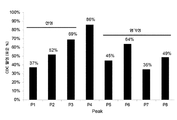
도 2. 양이온 교환 크로마토그래피(A와 동일한 칼럼 및 용출물)에 의한 11개의 분리물에 대응하는 11개의 크로마토그램의 중첩. 분획물 F1 내지 F20을 수집하였고, 피크 마다 그룹화하였다: P1(산성, F1 내지 F3), P2(산성, F4 및 F5), P3(산성, F6), P4(주 피크, F7 내지 F10), P5(염기성, F11), P6(염기성, F12 내지 F14), P7(염기성, F15 내지 F17), 및 P8(염기성, F18 내지 F20).
도 3. (A) CEX에 의해 정제된 항-CD20 항체 조성물의 크로마토그램, (B) 정제 이전의 항-CD20 항체 조성물의 크로마토그램. 정제(A) 이전에 크로마토그램의 주 피크에 대응하는 분획물 1 내지 20을 조립하여 형성된 조성물의 크로마토그램. 다양한 피크들의 백분율을 나타낸다.
도 4. 양이온 교환 크로마토그래피에 의해 정제된 분획물들의 CD16(Biacore)과의 결합이다. 각 샘플의 CD16과의 결합은 비교(reference) 샘플의 CD16과의 결합의 백분율로 표시된다.
도 5. (A) 크로마토포커싱에 의해 또는 (B) 양이온 교환 크로마토그래피에 의해 정제된 분획물들의 CD16 활성. 각 샘플의 CD16 활성(CD16 쥬르카트(Jurkat) 세포에 의한 IL-2의 분비)은 비교 샘플의 CD16 활성의 백분율로 표시된다.
도 6. 양이온 교환 크로마토그래피에 의해 정제된 분획물들의 보체-의존성 세포 독성(CDC). 각 샘플의 CDC 반응은 비교 샘플의 CDC 반응의 백분율로 표시된다.
도 2. 양이온 교환 크로마토그래피(A와 동일한 칼럼 및 용출물)에 의한 11개의 분리물에 대응하는 11개의 크로마토그램의 중첩. 분획물 F1 내지 F20을 수집하였고, 피크 마다 그룹화하였다: P1(산성, F1 내지 F3), P2(산성, F4 및 F5), P3(산성, F6), P4(주 피크, F7 내지 F10), P5(염기성, F11), P6(염기성, F12 내지 F14), P7(염기성, F15 내지 F17), 및 P8(염기성, F18 내지 F20).
도 3. (A) CEX에 의해 정제된 항-CD20 항체 조성물의 크로마토그램, (B) 정제 이전의 항-CD20 항체 조성물의 크로마토그램. 정제(A) 이전에 크로마토그램의 주 피크에 대응하는 분획물 1 내지 20을 조립하여 형성된 조성물의 크로마토그램. 다양한 피크들의 백분율을 나타낸다.
도 4. 양이온 교환 크로마토그래피에 의해 정제된 분획물들의 CD16(Biacore)과의 결합이다. 각 샘플의 CD16과의 결합은 비교(reference) 샘플의 CD16과의 결합의 백분율로 표시된다.
도 5. (A) 크로마토포커싱에 의해 또는 (B) 양이온 교환 크로마토그래피에 의해 정제된 분획물들의 CD16 활성. 각 샘플의 CD16 활성(CD16 쥬르카트(Jurkat) 세포에 의한 IL-2의 분비)은 비교 샘플의 CD16 활성의 백분율로 표시된다.
도 6. 양이온 교환 크로마토그래피에 의해 정제된 분획물들의 보체-의존성 세포 독성(CDC). 각 샘플의 CDC 반응은 비교 샘플의 CDC 반응의 백분율로 표시된다.
따라서, 본 발명은 다음을 포함하는 방법에 의해 얻어질 수 있는, 모노클론 항체 조성물에 관한 것이다:
a) 비-인간 유전자 변형 동물 또는 유전자 변형 식물로부터, 세포 클론 유래 모노클론 항체 조성물을 제조하는 단계,
b) 단계 a)에서 얻어진 조성물을 크로마토그래피에 의해 분획하는 단계, 및
c) 단계 b)에서 얻어지고 크로마토그램의 주 피크에 대응하는, 하나 또는 수 개의 크로마토그래피 분획물을 조합하는 단계,
그리고, 최종 얻어진 모노클론 항체 조성물은 상기 주 피크가 풍부하고, 의약품 용도를 위하여, 상기 주 피크는 단계 c)에서 얻어진 조성물의 크로마토그램 중 적어도 85%, 유리하게는 적어도 86%, 적어도 87%, 적어도 88%, 적어도 89%, 보다 유리하게는 적어도 90%, 적어도 91%, 적어도 92%, 적어도 93%, 적어도 94%, 또는 심지어 적어도 95%, 적어도 96%, 적어도 97%, 적어도 98%, 적어도 98.5%, 적어도 99%, 또는 적어도 99.5%를 나타낸다.
단계 a)에서, 모노클론 항체 조성물은 유전자 변형 동물 또는 유전자 변형 식물로부터, 세포 클론에서 제조된다.
"항체" 또는 "면역글로불린"은 특정 항원과 결합하기 위한 적어도 하나의 도메인과 FcR 수용체와 결합할 수 있는 Fc 단편을 포함하는 불변 영역을 포함하는 분자를 의미한다. 인간 및 마우스와 같은, 대부분의 포유동물에서, 항체는 4개의 폴리펩타이드 쇄로 구성된다: 분자에 대한 유연성을 보장하는 다양한 수의 이황화 결합을 통해 함께 연결된 2개의 중쇄와 2개의 경쇄. 각 경쇄는 불변 영역(CL)과 가변 영역(VL)으로 이루어진다; 중쇄는 항체의 아이소타입에 따라서 1개의 가변 영역(VH)과 3 또는 4개의 불변 영역(CH1 내지 CH3 또는 CH1 내지 CH4)로 이루어진다. 낙타나 라마와 같은, 일부 희귀 포유동물에서, 항체들은 2개의 중쇄 만으로 이루어지고, 중쇄 각각은 가변 영역(VH)과 불변 영역을 포함한다.
가변 영역들은 항원의 인식에 참여하지만, 불변 영역들은 항체의 생물학적, 약물 역동학 및 반응기 특징들에 참여한다. 서열이 하나의 항체와 다른 항체에서 매우 상이한, 가변 영역들과는 달리, 불변 영역들은 하나의 항체와 다른 항체에서 매우 비슷한 하나의 아미노산 서열, 종(species) 및 아이소타입의 특징에 의해 특징화되고, 선택적으로 몇몇 체세포 돌연변이를 갖는다. Fc 단편은 선천적으로 영역 CH1을 제외한 중쇄의 불변 영역으로 이루어지고, 즉, 더 낮은 경계선 영역으로 이루어지고 불변 영역 CH2 및 CH3 또는 CH2 내지 CH4(아이소타입에 따라서)으로 이루어진다. 인간 IgG1에서, 완전한 Fc 단편은, 226 위치의 시스테인 잔기(C226)에서 시작하는 중쇄의 C-말단 부분으로 이루어지고, Fc 단편에서 아미노산 잔기의 숫자는 에덜만 등(Edelman et al., 1969) 및 카바트 등(Kabat et al., 1991)이 기술한 인덱스 EU의 모든 현재 기재에 있다. 면역글로불린의 다른 형태의 대응하는 Fc 단편들은 서열 정열에 의하여 통상의 기술자에 의해 쉽게 확인될 수 있다.
Fc 단편은, 2개의 중쇄 각각에, 297 위치의 아스파라긴 잔기(Asn 297)에 결합된 N-글리칸의 존재와 함께 CH2 영역에서 글리코실화된다.
Fc에 위치하는, 다음의 결합 영역들은 항체의 생물학적 특성에서 중요하다:
- FcRn 수용체와 결합하기 위한 영역, 항체의 약물 역동학적 특징(생체 내 반감기)에 참여함:
상이한 데이터는, CH2 및 CH3 도메인의 경계면에 위치하는 특정 잔기들이 FcRn 수용체와의 결합에 참여하는 것을 제시한다.
- 보체 C1q의 단백질과 결합하는 영역(domain), CDC 반응에 참여함("보체-의존성 세포독성"에 대하여): CH2 영역에 위치함;
- FcR 수용체와 결합하는 영역, 식세포 작용(phagocytosis)의 반응 또는 ADCC("항체-의존성 세포 독성"에 대하여) 형태에 참여함: CH2 영역에 위치함.
본 발명의 의미에서, 항체의 Fc 단편은 상기에서 기술된 것처럼, 천연일 수 있고, 또는 그것이 FcR 수용체(IgGs를 위한 FcγR 수용체)와 결합하기 위한 기능성 영역, 바람직하게는 수용체 FcRn과 결합하기 위한 기능성 영역을 포함하는 경우라면, 그 밖의 다양한 방법으로 변형될 수 있다. Fc 단편이 수용체 FcR(IgGs를 위한 수용체 FcγR)와 결합하기 위한 기능성 영역, 바람직하게는 수용체 FcRn과 결합하기 위한 기능성 영역을 포함하는 경우라면, 변형은 Fc 단편의 특정 부위의 결실을 포함할 수 있다. 항체가 수용체 FcR와 결합하기 위한 기능성 영역, 바람직하게는 수용체 FcRn과 결합하기 위한 기능성 영역을 포함하는 경우라면, 변형은 또한 항체의 생물학적 특징들에 영향을 미칠 수 있는 다양한 아미노산의 치환을 포함할 수 있다. 특히, 항체가 IgG인 경우, 수용체 FcγRⅢ(CD16)과의 결합을 향상시키기 위한 돌연변이들을 포함할 수 있고, WO00/42072, 쉴드 등(Shields et al., 2001), 라자 등(Lazar et al., 2006), WO2004/029207, WO/2004063351, WO2004/074455에 기술된 바와 같다. 수용체 FcRn과의 결합을 향상시키기 위해 허용되는 돌연변이들 및 그에 따른 생체 내 반감기가 또한 존재하고, 쉴드 등(Shields et al., 2001), 달 아쿠어 등(Dall'Acqua et al., 2002), 힌톤 등(Hinton et al., 2004), 달 아쿠어 등(Dall'Acqua et al., 2006(a)), WO00/42072, WO02/060919A2, WO2010/045193, 또는 WO2010/106180A2에 기술된 바와 같다. 다른 돌연변이들, 예를 들어 보체의 단백질과의 결합을 감소 또는 증가시키기 위한 것과 그에 따른 CDC 반응은 존재 또는 부존재할 수 있다(WO99/51642, WO2004074455A2, Idusogie et al., 2001, Dall'Acqua et al., 2006(b), 및 Moore et al., 2010, 참조).
"모노클론 항체" 또는 "모노클론 항체 조성물"은 동일하고 특별한 항원 특이성을 갖는 항체 분자들을 포함하는 조성물을 의미한다. 상기 조성물에 존재하는 항체 분자들은 그것들의 번역 후 변형과 관련하여, 특히 그것들의 글리코실화 구조 또는 그것들의 등전점과 관련하여 다양할 수 있지만, 그것들 모두는 동일한 중쇄 서열 및 경쇄 서열에 의해 인코딩되고 그에 따라, 임의의 번역 후 변형 이전에, 동일한 단백질 서열을 갖는다. 번역 후 변형(예를 들어 중쇄의 C-말단 라이신의 절단, 아스파라긴 잔기들의 탈아미드화 및/또는 아스파레이트 잔기들의 이성질화)과 관련된, 단백질 서열에서의 특정 차이들은, 그럼에도 불구하고 조성물에 존재하는 다양한 항체 분자들 사이에서 존재할 수 있다.
본 발명의 범위 내에서 의약품으로 사용되는 조성물에 존재하는 모노클론 항체는 바람직하게는 잡종의(chimeric), 인간화된, 또는 인간일 수 있다. 실제로, 이것은 투여된 항체에 대하여 환자의 면역 반응을 회피할 가능성을 부여한다.
"키메라" 항체는, 특정 종의 항체에서 유래된 천연 가변 영역(경쇄 및 중쇄)을 포함하는 항체를 가리키는 의미이고, 상기 특정 종과 이종인 종의 항체의 경쇄 및 중쇄의 불변 영역들과 관련된다. 유리하게도, 본 발명에 따른 의약품 용도를 위한 모노클론 항체 조성물이 키메라 모노클론 항체를 포함한다면, 키메라 모노클론 항체는 인간 불변 영역을 포함한다. 비-인간 항체에서 출발하여, 키메라 항체는 통상의 기술자에게 잘 알려진 유전자 재조합 기술을 사용하여 제조될 수 있다. 예를 들면, 키메라 항체는 중쇄와 경쇄에 대하여, 프로모터와 비-인간 항체의 가변 영역을 코딩하는 서열, 및 인간 항체의 불변 영역을 코딩하는 서열을 포함하는 재조합 DNA를 클로닝하여 제조될 수 있다. 키메라 항체들을 제조하기 위한 방법으로서, 예를 들어 문헌[Verhoeyn et al., 1988]을 참조할 수 있다.
"인간화된" 항체는, 비-인간 기원의 항체에서 유래된 CDR 영역을 함유하는 항체를 가리키는 의미이고, 항체 분자의 다른 부분은 하나의(또는 몇 개의) 인간 항체들에서 유래된다. 또한, 백본 부분(FR로 불림)의 잔기들 중 일부는 결합 친화성을 유지하도록 변형될 수 있다(Jones et al., 1986; Verhoeyen et al., 1988; Riechmann et al., 1988). 본 발명에 따른 인간화된 항체들은, 알마그로 등(Almagro et al., 2008)의 리뷰에 요약된 바와 같이, "CDR 그래프팅", "재포장(resurfacing)", 초인간화(SuperHumanization), "인간 스트링 함량(Human string content)", "FR 도서관", "안내된 선택(Guided selection)", "FR 셔플링" 및 "휴머니어링(Humaneering)" 기술과 같이 당업자에게 공지된 기술에 의해 제조될 수 있다.
"인간" 항체는, 전체 서열이 인간 기원인, 즉, 코딩 서열이 항체들을 코딩하는 인간 유전자들의 재조합에 의해 생산되는 항체를 의미한다. 실제로, 이제 유전자 변형 동물(예, 마우스)를 제조하는 것이 가능하고, 면역화(immunization)하면서, 내생적 면역글로불린 생산의 부재시에 인간 항체들의 전체 리스트를 생산하는 것이 가능하다(Jakobovits et al., 1993(a) 및 (b); Bruggermann et al., 1993; 및 Duchosal et al.,1 992, U.S. 특허 5,591,669, 5,598,369, 5,545,806, 5,545,807, 6,150,584, 참조). 인간 항체들은 또한 파아지 디스플레이뱅크에서 획득할 수 있다(Hoogenboom et al., 1991; Marks et al., 1991; Vaughan et al., 1996). 항체들은, 그것들의 불변 영역의 특성에 따라서 수 개의 아이소타입일 수 있다: 불변 영역 γ, α, μ, ε 및 δ 각각은 IgG, IgA, IgM, IgE 및 IgD 면역글로불린에 대응한다. 유리하게는, 본 발명의 범위 내의 의약품으로 사용되는 조성물에 존재하는 모노클론 항체는 IgG 아이소타입이다. 실제로, 이 아이소타입은 대다수의 개체(인간)에서 ADCC("항체 의존성 세포성 세포 독성") 활성을 발생시키는 능력을 보여준다. γ 불변 영역은 수 개의 서브 타입들-γ1, γ2, γ3, 인간 보체에 결합하는 특이성을 갖는 이 세 개의 불변 영역들과 γ4-을 포함하고, 그에 따라 서브-아이소타입 IgG1, IgG2, IgG3, 및 IgG4를 생성한다. 유리하게는, 본 발명의 범위 내의 의약품으로 사용되는 조성물에 존재하는 모노클론 항체는 아이소타입 IgG1 또는 IgG3, 바람직하게는 IgG1이다.
모노클론 항체의 조성물은 세포클론, 비-인간 유전자변형 동물 또는 유전자변형 식물에 의해, 당업자에게 공지된 기술에 의하여 제조될 수 있다.
주목할 만하게, 조성물을 생산하는 세포 클론들은 3가지의 주요 기술에 의해 얻어질 수 있다:
1) 무한증식 라인(immortalized line)과 관심있는 항체를 생산하는 림프구 B의 융합에 의한 융합세포(hybridoma)를 획득하는 것,
2) 엡스타인-바 바이러스(Epstein-Barr virus, EBV)에 의하여 관심있는 항체를 생산하는 림프구 B를 무한증식하는 것,
3) (일반적으로 융합세포 또는 무한증식 림프구 B로부터) 관심있는 항체를 코딩하는 서열들을 분리하는 것, 항체의 중쇄 및 경쇄를 코딩하는 서열을 발현하는 하나 또는 수 개의 벡터(들)에 클로닝하는 것, 세포주를 발현 벡터(들)에 의해 변형(transform)시키는 것 및 상이한 획득한 세포 클론들을 분리하는 것. 항체의 중쇄 및 경쇄의 발현 벡터는 항체의 중쇄 및 경쇄를 인코딩하는 서열을 발현하기 위하여 요구되는 요소들을 포함하고, 특히, 프로모터, 전사 개시 코돈, 종결 서열 및 전사를 조절하기 위한 적절한 서열을 포함한다. 이 요소들은 발현에 사용되는 호스트에 따라서 변하며, 자신의 일반 상식을 고려하여 당업자가 쉽게 선택한다. 벡터는 주목할 만하게 플라스미드 또는 바이러스이다. 변형(transformation) 기술은 또한 당업자에게 잘 알려져 있다.
항체의 중쇄 및 경쇄를 인코딩하는 서열들의 하나 또는 수 개의 발현 벡터들에 의한 세포주들의 변형은, 특히 키메라 또는 인간화된 항체들을 획득하기 위하여, 가장 일반적으로 사용된다.
변형된 세포주는 바람직하게는 진핵세포 기원이고, 특히 곤충, 식물, 이스트 또는 포유동물 세포들에서 선택될 수 있다. 항체 조성물은 이어서 적절한 조건 하에서 호스트 세포들을 배양하여 제조할 수 있다. 항체들을 제조하기 위한 적절한 세포주들은 특히 다음에서 선택된 세포주들을 포함한다: SP2/0; YB2/0; IR983F; 인간 골수종(human myeloma) Namalwa; PERC6; CHO 세포주, 특히 CHO-K-1, CHO-Lec10, CHO-Lec1, CHO-Lec13, CHO Pro-5, CHO dhfr-, 또는 유전자 FUT8 및/또는 유전자 GMD를 인코딩하는 2개의 대립유전자(alleles)를 제거한 CHO 세포주; Wil-2; 주르카트(Jurkat); 베로(Vero); MoIt-4; COS-7; 293-HEK; BHK; K6H6; NSO; SP2/0-Ag 14, P3X63Ag8.653, 오리 배아 세포주(duck embryo cell line) EB66®(Vivalis); 및 랫트 헤파토마 세포주(rat hepatoma lines) H4-II-E(DSM ACC3129), H4-II-Es (DSM ACC3130)(WO2012/041768, 참조). 바람직한 실시형태에서, 항체는 다음의 세포주들 중 하나에서 제조된다: YB2/0; 유전자 FUT8 및/또는 유전자 GMD를 인코딩하는 2개의 대립유전자(alleles)를 제거한 CHO 세포주; 오리 배아 세포주 EB66® (Vivalis); 및 랫트 헤파토마 세포주 H4-II-E (DSM ACC3129), H4-II-Es (DSM ACC3130). 바람직한 실시형태에서, 항체는 YB2/0(ATCC CRL-1662)에서 제조된다.
대안으로, 항체 조성물은 비-인간 유전자 변형 동물에서 생산될 수 있다.
비-인간 유전자 변형 동물은 수정란에 관심있는 유전자(들)(여기에서 항체의 중쇄 및 경쇄를 코딩하는 재배열 유전자들)을 직접 주입하여 얻을 수 있다(Gordon et al., 1980). 비-인간 유전자 변형 동물은 또한 관심있는 유전자(들)(여기에서 항체의 중쇄 및 경쇄를 코딩하는 재배열 유전자들)을 배아 줄기 세포에 도입하고 키메라 응집 방법 또는 키메라 주입 방법에 의하여 동물을 제조하여 얻을 수 있다(Manipulating the Mouse Embryo, A Laboratory Manual, Second edition, Cold Spring Harbor Laboratory Press (1994); Gene Targeting, A Practical Approach, IRL Press at Oxford University Press (1993), 참조). 비-인간 유전자변형 동물은 또한 클로닝 기술에 의해 얻어질 수 있는데, 상기 클로닝 기술에서는 관심있는 유전자(들)(여기에서 항체의 중쇄 및 경쇄를 코딩하는 재배열 유전자들)이 도입되어 있는 핵이, 핵이 적출된 난자로 이식된다(Ryan et al., 1997; Cibelli et al., 1998, WO0026357A2). 관심있는 항체를 생산하는 비-인간 유전자변형 동물은 상기 방법들에 의하여 제조될 수 있다. 이어서 항체는 유전자변형 동물에 축적될 수 있고, 특히 동물의 젖이나 알(egg)에서 수확될 수 있다. 비-인간 유전자변형 동물의 젖(milk)에서 항체를 생산하기 위한, 방법들이 특히 WO9004036A1, WO9517085A1, WO0126455A1, WO2004050847A2, WO2005033281A2, WO2007048077A2에 기술되어 있다. 관심있는 단백질들을 젖에서 정제하는 방법들 역시 공지되어 있다(WO0126455A1, WO2007106078A2, 참조). 관심있는 비-인간 유전자변형 동물들은 주목할 만하게도 쥐, 토끼, 랫트, 염소, 소(특히 젖소) 및 가금류(특히 닭)을 포함한다.
항체 조성물은 유전자변형 식물에서 제조될 수 있다. 많은 항체들이 이미 유전자변형 식물에서 제조되었고, 관심있는 항체를 발현하는 유전자변형 식물을 획득하고 항체들을 회수하는데 요구되는 기술들은 당업자에게 잘 알려져 있다(Stoger et al., 2002, Fisher et al., 2003, Ma et al., 2003, Schillberg et al., 2005, 참조). 또한 천연 인간 항체들(자일로스 없음)의 글리코실화와 밀접한 글리코실화를 획득하기 위하여 식물에서 얻어진 글리코실화에 영향을 미치는 것이 가능하고, 그리고 예를 들어 추가의 약간의 푸코실화의 경우에는 소 간섭 RNA(small interfering RNA)에 의해 가능하다(Forthal et al., 2010).
본 발명에 따른 의약품으로의 사용을 위한 모노클론 항체 조성물을 얻기 위해 허용되는 방법의 단계 b)에서, 단계 a)에서 얻어진 조성물에 존재하는 항체들의 상이한 전하 아이소폼들이 크로마토그래피에 의해서 단계 a)에서 얻어진 조성물을 분획하여 분리된다.
도입부에서 설명한 것처럼, 비-인간 유전자변형 동물 또는 유전자 변형 식물인, 세포 클론에 의해 생산된 임의의 모노클론 항체 조성물은, 특정 갯수의 전하 아이소폼 또는 동일 모노클론 항체의 변이체(variant)의 존재를 특징으로 한다. 이러한 상이한 전하 아이소폼 또는 변이체의 존재는, 직접적으로 전하 그룹(charge group)의 수를 변경하는 것에 의하여, 또는 간접적으로 구조적 변형을 도입하는 것에 의하여 - 그것 자체가 하전된 잔기의 국소(local) 분포를 변경시키거나 pKa를 변경시킨다-, 항체의 표면 전하 특성의 변경을 가져오는 번역 후 변형의 존재와 관련된다. 전하 아이소폼 또는 변이체 각각은, 등전점(pl, 추가로 등전점 수소 포텐셜이라 함(pHI))을 특징으로 하고, 등전점은, 이 분자의 전체 전하(global charge)가 제로인, pH(수소 포텐셜)에 상응하고, 즉, 분자가 전기적으로 중성(쌍성 이온 형태 또는 혼합된 이온)인 pH에 상응한다. 따라서, 정해진 pH에서, 모노클론 항체의 상이한 전하 아이소폼 또는 변이체는 가변성 순(net) 전하를 갖는데, 그것들의 pl이 음의 전하를 갖는 pH 미만이고(분자는 염기성 배지로 자신의 양성자를 산출하는 경향이 있다), 그것들의 pl은 중성인 pH와 동일하고, 그것들의 pl은 양의 전하를 갖는 pH보다 더 크다(분자는 양성자를 보유하거나 산성 배지에서 양성자 중 일부를 포획하는 경향이 있다). 모노클론 항체의 상이한 전하 아이소폼 또는 변이체는, 변이체 각각에 존재하는 번역 후 변형의 빈도에 따라서, 가변성 부분에 존재한다. 모노클론 항체 조성물은 일반적으로 주요 변이체 또는 아이소폼을 포함하고, 그것들의 pl이 주요 아이소폼의 pl보다 작은 지 큰 지에 따라서, 다수의 소위 산성 또는 염기성 변이체 또는 아이소폼을 수반한다. 항체, 생산 방식 및 항체가 이미 거쳤을 수 있는 정제 단계에 따라서, 산성 아이소폼, 주 피크 및 염기성 아이소 폼의 비율(이온 교환 크로마토그래피의 크로마토그램에서 계산됨)은, 일반적으로 대략 다음의 값에서 변한다: 10 내지 30%의 산성 아이소폼, 50 내지 75%의 주 피크, 및 8 내지 20%의 염기성 아이소폼(Farnan et al., 2009, Rea et al., 2011, Rea et al., 2012, Khawli et al., 2010, Zhang et al., 2011, WO2011/009623, 및 EP1308456, 참조).
정해진 pH에서의 pl 및 순 전하 측면의 차이 때문에, 정해진 항체 조성물에 존재하는 항체들의 전하 아이소폼들은 다른 크로마토그래피 장비로 분리될 수 있다.
크로마토그래피는 화학 물질들(액체 또는 기체 균질 혼합물)을 분리하는 기술로서, 러닝 이동상 및 정지상(또는 고정상) 사이의 거동의 차이에 기반한 것이다. 크로마토그래피 방법은 사용되는 상(phase)들의 특성에 따라서 또는 분리에 적용되는 현상의 특성에 따라서 분류될 수 있다.
본 발명의 실시형태에서, 단계 b)의 분획은 이온 교환 크로마토그래피에 의하여 달성될 수 있다. 실제로 이것은 동일한 단백질의 전하 아이소폼을 분리할 수 있다. 이온(음이온 또는 양이온) 교환 크로마토그래피에서, 상이한 성분의 분리를 가능하게 하는 파라미터는 그것들의 순(net) 전하이다.
먼저, 항체 조성물을 이온 교환 수지에 적재한다. 이를 위하여, 양성(음이온 교환 크로마토그래피) 또는 음성(양이온 교환 크로마토그래피) 하전된 수지(고정 또는 정지 상)가 사용된다. 수지의 이온과 반대의 전하를 갖는 분자들이 수지에 유지/고정될 것이다.
당업자에게 공지된, 강하거나 약하고, 관심있는 항체 조성물의 분리를 위해 적절한, 양이온 또는 음이온 교환 수지의 임의의 형태가 사용될 수 있다. 그것의 단백질 서열에 따라서, 항체 조성물의 평균 등전점(isoelectric point, pl)은 일반적으로 5 내지 9에서 변하고, 종종 대부분이 7 내지 9에서 변한다. 8 이상의 pl에서, 양이온 교환 수지가 이용된다. 반대로, 6 이하의 pl에서 음이온 교환 수지가 이용된다. 6 내지 8에 포함되는 pl에서, 이온(양이온 또는 음이온) 교환 수지의 양 형태가 시험될 수 있다. 따라서, 양이온 교환 크로마토그래피(음으로 하전된 수지) 이후에 이온 힘 구배로 용출하는 것이 대부분 종종 이용된다고 할지라도, 특정 경우에 음이온 교환 크로마토그래피(양으로 하전된 수지)를 이용하는 것도 가능하다.
이온 교환 수지는 일반적으로, 그 위에서 양으로 하전된 그룹으로 그래프트되거나(음이온 교환 수지) 음으로 하전된 그룹으로 그래프트된(양이온 교환 수지), 가교된 폴리머 또는 겔로 이루어진다. 가교된 폴리머 또는 겔은 특히 덱스트란(예:Sephadex®), 아가로즈(예:Sepharose®), 셀룰로즈, 메타크릴레이트 폴리머(예:Fratogel®), 비닐 폴리머(예:Fractoprep®) 예를 들어 폴리(스티렌 디비닐벤젠)(예:Monobeads™; Source™; Bio Mab NP-5 또는 NP-10; SepaxAntibodix™ NP1.7, NP3, NP5 및 NP10)에서 선택될 수 있다.
겔은 바람직하게는 10 내지 200 μm에 포함되는 평균 직경을 갖는, 비즈 형태일 수 있다.
양이온 교환 수지에서, 음으로 하전된 그룹은 가교된 폴리머, 예를 들어 설포프로필(SP), 메틸 설포네이트(S) 또는 카르복시메틸(CM) 형태의 그룹에 그래프트된다.
음이온 교환 수지에서, 양으로 하전된 그룹은 가교된 폴리머, 예를 들어 4차 암모늄 형태(Q), 특히 4차 아미노에틸(QAE), 디에틸아미노아텔(DEAE), 디메틸아미노에틸(DMAE), 트리메틸아미노에틸(TMAE), 또는 디메틸아미노프로필(ANX)의 그룹에 그래프트된다.
본 발명의 범위 내에서 사용될 수 있는 양이온 교환 수지들은 수지들 Source™ 15S 또는 30S, Mono-S(GE Life Sciences사에서 판매됨); ProPac® WCX(특히, ProPac® WCX-10), ProPac® SCX(특히, ProPac® SCX-10 또는 SCX-20), ProSwift WCX, MAbPac® SCX(특히, MAbPac® SCX-10)(Dionex사에서 판매됨); Bio Mab(특히, Bio Mab NP-5 또는 NP-10, Agilent사에서 판매됨), PL-SCX(Agilent사에서 판매됨); SepaxAntibodix™(특히, SepaxAntibodix™ NP1.7, NP3, NP5 및 NP10)(Sepax사에서 판매됨)(Farnan et al.,2009, Khawli et al., 2010, Gandhi et al.,2011, Zhang et al.,2011, Rea et al., 2011 및 McAtee et al., 2012, 참조)를 포함한다. 또한, 본 발명의 범위 내에서 사용될 수 있는 음이온 교환 수지는 수지들 Source™ 15Q 또는 30Q, Mono™-Q(GE Life Sciences사에서 판매됨); ProPac® WAX(특히, ProPac® WAX-10), ProPac® SAX(특히, ProPac® SAX-10)(Dionex사에서 판매됨)을 포함한다.
일단 항체 조성물이 이온 교환 수지에 적재되면, 상이한 용출 방법들이 전하 아이소폼들을 분리하기 위하여 사용될 수 있다.
고정된 분자의 용출은 주목할 만하게 수지의 이온과 반대되는 전하를 갖는 이온을 포함하는 용출 버퍼(이동 상)를 사용하여 달성될 수 있고, 그것은 수지에 의해 보유된 전하들과 상호작용하기 위하여 고정된 분자들과 경쟁하게 될 것이다. 강한 이온 농도를 갖는 버퍼를 직접 사용하거나(모든 분자들을 한꺼번에 용출하기 위하여), 또는 반대로 이온 농도를 서서히 증가시키는 것(이어서 이것은 이온 힘 구배라고 한다)이 가능하고, 이것은 수지와의 정전기적 상호작용의 힘에 따라서, 상이한 분자들을 성공적으로 떼어낼 가능성을 부여한다. 실제로, 이 마지막 시나리오에서, 하나는 낮은 이온 농도, 다른 하나는 강한 이온 농도를 갖는 2개의 버퍼 용액이 사용된다. 2개의 구동 펌프가, 시간 경과에 따라 변화하는 비율(강한 이온 농도 용액의 비율이 서서히 증가함)에 따라서, 이들 용액 양자를 끌어올리고 혼합한다. 이러한 혼합의 생성물이 컬럼에서 사용된다. 이 장비로 항체 조성물의 전하 아이소폼을 분리하기 위한 특정 방법들의 예시들이 문헌[Gandhi et al., 2011.Rea et al., 2012]에 기술되어 있고, 또한 이 장비의 원리, 및 컬럼, 버퍼 및 항체들의 전하 아이소폼 또는 변이체를 분리하기 위한 작동 파라미터를 적절하게 선택하는 방법이 기술된다(447~451페이지 섹션 7 참조).
대안의 이온 교환 수지 크로마토그래피에서, 용출은 이온 힘 구배가 아니라, pH 구배로 달성된다. 실제로, 다수의 이온화 그룹들이 pH에 민감한다. 증가하는 pH 구배를 이용하면(즉, pH를 증가시켜서), 산성 그룹(음으로 하전된)의 이온화는 쉽게 되고(favored), 염기성 그룹(양으로 하전된)의 이온화는 쉽게 되지 않는다. pH를 증가시키는 것에 의하여, 따라서, 순(net) 음 전하의 발생이 pH 민감성 이온화 그룹을 갖는 분자들에 대하여 쉽게 된다. 따라서 증가하는 pH 구배는 또한 음으로 하전된 수지(양이온 교환체)에 고정된 항체 조성물의 전하 아이소폼의 분리를 가능하게 한다. 감소하는 pH 구배를 이용하면(즉, pH를 감소시켜서), 염기성 그룹(양으로 하전된)의 이온화가 쉽게 되고 산성 그룹(음으로 하전된)의 이온화가 쉽게 되지 않는다. pH를 감소시키는 것에 의하여, pH 민감성 이온화 그룹을 갖는 분자들에 대한 순(net) 양이온 전하의 발생이 쉽게 된다. 그러므로 감소하는 pH 구배는 또한 양으로 하전된 수지(음이온 교환체)에 고정된 항체 조성물의 전하 아이소폼을 분리할 가능성을 부여한다. pH 구배에 의한 용출을 이용하여 이온 교환 크로마토그래피에 의해 항체들의 전하 아이소폼들을 분리하는 특정 방법들의 예시는 문헌[Farnan et al., 2009 및 Rea et al., 2011.Rea et al.,2012]에 개시되어 있고 또한 이 장비의 원리, 및 컬럼, 버퍼 및 항체들의 전하 아이소폼 또는 변이체를 분리하기 위한 작동 파라미터를 적절하게 선택하는 방법이 기술된다(451~452페이지 섹션 8 참조). 실시예 1은 또한 양이온 교환 크로마토그래피와 증가하는 pH 구배를 이용한 용출에 의해 항체 조성물의 전하 아이소폼을 분리하는 것을 기술한다.
이온 교환 크로마토그래피의 다른 대안에서, 용출은 또한 문헌[Rea et al., 2012, 페이지 453 섹션 9 참조)에 기술된 것처럼 이온 힘 구배 및 pH 구배(소위 "하이브리드" 용출)를 조합하여 달성될 수 있다.
이온 교환 크로마토그래피의 또 다른 대안에서, 여기에서 "치환 이온 교환 크로마토그래피(displacement ion exchange chromatography)"이라 하고, 항체 조성물의 전하 아이소폼의 분리를 가능하게 하고, 이온(음이온 또는 양이온) 교환체 수지는 또한 고정 또는 정지 상으로 사용되지만, 용출은 이온 힘 및/또는 pH 구배가 아니라, 치환 분자, 즉 크로마토그래피 수지에 대하여 강한 친화성을 갖는 분자에 의해 이루어지고, 그것은 미리 수지에 고정되어 있는 항체 분자들과 수지 상에서의 결합에 대해 경쟁할 것이고, 이에 따라 치환 분자보다 수지에 대하여 더 낮은 친화성을 갖는 항체 분자들이 치환될 것이다. 따라서, 항체 분자들은 치환 분자 물결(wave)에 의해 컬럼을 따라 이동하도록 힘을 받게 될 것이다. 후자(latter)가 컬럼을 횡단할 때, 새로운 평형이 설정되고, 여기서 항체 분자들이, 유용하게 남아있는 수지와 결합하는 자리에 대하여 서로 경쟁하게 된다. 이러한 역동적인 평형(balancing) 공정 동안, 상이한 항체들의 전하 변이체들 또는 아이소폼들이 이온 교환 수지에 대한 그것들의 더 높은 또는 더 낮은 친화성에 따라 분리된다. 이러한 크로마토그래피 분리 방법의 원리와, 항체 조성물의 전하 아이소폼들을 분리하기 위한 응용에 요구되는 수지, 버퍼, 재료들이 문헌[Khawli et al., 2010, Zhang et al., 2011, 및 McAtee et al., 2012]에 특히 기술되어 있다.
이온 교환 크로마토그래피의 이들 상이한 용출 모드들에서, 임의의 적절한 용출(pH 또는 이온 힘 구배) 또는 치환 버퍼는 선택된 칼럼에 따라서 사용될 수 있다. 수지들 및 관련 버퍼들에 대한 예시들이 문헌[Farnan et al.-2009, Khawli et al., 2010, Gandhi et al., 2011, Zhang et al., 2011, Rea et al., 2011 및 McAtee et al., 2012]에 기술되어 있다.
항체 조성물의 전하 아이소폼들의 분리를 허용하는 다른 크로마토그래피 기술은 크로마토포커싱이다. 이 기술에서, 단백질들이 그것들의 등전점(pl)에 따라 분리된다. 이 기술은 특정 수지(고정 또는 정지 상)의 연관성 및 특정한 양쪽성 버퍼의 사용에 기초한다. 주목할 만하게, 선형 pH 구배를 획득하는 것은 분리를 위해 사용되는 pH의 전체 범위에 대하여 동등한 버퍼 용량을 요구하고, 따라서, 특별히 이 적용을 위하여 설계된 버퍼와 하전된 버퍼 아민들로 치환된 수지들이 요구된다.
분리의 원리는 다음과 같다: 크로마토포커싱 수지는 요구되는 최대 pH보다 약간 높은 pH에서 개시 버퍼로 평형을 맞춘다. 용출 버퍼(요구되는 최저 pH로 조절됨)를 칼럼에 통과시키고 수지와 단백질의 아민들로 적정을 시작한다. 용출 버퍼가 서서히 칼럼을 통과하면, pH는 감소되고 하향 이동 pH 구배가 발생한다. 컬럼 상에 용출 버퍼의 제1 부피를 통과시킨 후 샘플이 컬럼에 적용될 수 있다. 샘플의 단백질들이 컬럼에 도입되자마자 샘플의 단백질들이 적정된다(pH의 조정). 그것들의 pl 위의 pH인 단백질들이 음으로 하전되고 칼럼의 상부 근처에 유지된다(양으로 하전된 아민 그룹들과 결합함으로써). pH가 그것들의 pl보다 더 큰 영역을 이루기 이전에, 그것들의 pl 아래의 pH인 단백질들이 버퍼 흐름을 갖고 칼럼을 따라 이동하기 시작한다. 이것이 분리 공정의 시작이다.
서서히, pH가 칼럼의 상부에서 계속 감소하기 때문에(pH 구배의 시간-의존적 전하), pl이 새로운 pH보다 더 큰 임의의 단백질이 양으로 하전될 것이고, 양으로 하전된 아민 그룹에 의해 밀리게 되고 용출 버퍼를 이용하여 칼럼을 따라 이동하기 시작하고, 그 이동은 pH 구배의 이동 보다 더 빠르다. 서서히 단백질이 칼럼을 따라 이동하면, pH는 증가한다. 단백질은 pH가 그것의 pl보다 더 높은 영역에 이르는 경우, 단백질은 다시 음으로 하전되고 다시 칼럼과 결합한다. 단백질은, 이동성 pH 구배가 그것의 pl 아래의 국소 pH를 감소시킬 때까지, 단백질이 다시 양으로 하전되고 다시 이동을 시작하는 때에는 결합한 그대로 남아있다. 이 과정은, 그것의 pl에 인접한 pH에서 단백질이 칼럼에서 용출될 때까지 반복된다.
이 장비의 이름은 기술의 맞춤(focusing) 효과에서 유래한다. 실제로, pH 낮춤 구배에서, 단백질은 3개의 전하 상에 존재할 수 있다: 양성, 음성 및 중성. 또한, 크로마토포커싱에서, pH 구배가 발달하기 때문에 그리고 단백질이 칼럼의 상이한 pH 영역들을 통과하여 이동하기 때문에, 단백질의 전하 상태는 연속적으로 서서히 변화한다. 영역의 후면에서 분자들은 이 동일 영역의 전면의 분자들보다 보다 빠르게 이동할 것이고, 서서히 단백질들의 좁은 밴드들을 증가시키면서 형성하고, 각 밴드는 동일 pl을 갖는 하나 또는 수 개의 단백질들에 대응한다.
그러므로, 크로마토포커싱에서, pH 구배가 생길 때 상이한 pl을 갖는 단백질들이 상이한 속도로 칼럼을 통과하여 서서히 이동하고, 양으로 하전된 버퍼 아민 그룹들을 갖는 수지와 연속적으로 결합하고 해리하는 한편, 서서히 좁은 밴드로 포커싱되고 최종 용출된다. 최대 pl을 갖는 단백질들이 먼저 용출되는 한편, 최저 pl을 갖는 단백질들이 마지막으로 용출될 것이다.
크로마토포커싱에 의한 분리에 사용되는 수지는 표준 수지(상기에서 기술된 가교된 폴리머 또는 겔, 바람직하게는 상기에서 기술된 비즈와 같은 것), 특히 폴리(스티렌 디비닐벤젠) 또는 가교된 아가로즈 형에 기초하고, 가교된 아가로즈 형은 양으로 하전된 버퍼 아민 그룹들을 그래프팅시킨 것을 특징으로 한다. 이들 양으로 하전된 버퍼 아민 그룹들은 특히 2차, 3차 및/또는 4차 아민 그룹들이다. 크로마토포커싱에 유용한 수지들의 예시는 GE Life Sciences사 또는 GE Healthcare사에서 판매되는, Mono™-P 칼럼(2차, 3차 및/또는 4차 아민 그룹으로 그래프트된, 가교 폴리(스티렌 디비닐벤젠)), PBE 94 및 PBE 118(에테르 결합을 통해 단당류와 결합된 2차, 3차 및/또는 4차 아민 그룹으로 그래프트된 가교 6% 아가로즈 수지)를 포함한다. Mono™-P 칼럼 및 PBE 94 칼럼은 pH 9 내지 pH 4에서의 분리에 적절한 반면에, 칼럼 PBE 118은 pH 9 초과에서 시작하는 pH 구배를 이용한 분리에 적절한다. Mono™-P 및 PBE 94 칼럼, 그리고 특히 칼럼 Mono™-P가, 바람직하다.
사용되는 개시 버퍼는 상이한 pH(HCl 형 산, 아세트산 또는 아미노디아세트산의 첨가)에서 특히 디에탄올아민 용액, 트리스(Tris) 용액, 트리에틴올아민 용액, 비스-트리스 용액, 트리에틸아민 용액, 에탄올아민 용액, 이미다졸 용액, 히스티딘 용액, 또는 피페라진 용액에 기초할 수 있다.
사용된 용출 양쪽성(amphoteric) 버퍼는 특히 폴리버퍼 74(pH 범위: 7-4, Mono™-P 및 PBE 94 칼럼에서), 폴리버퍼 96(pH 범위: 9-6, Mono™-P 및 PBE 94 칼럼에서), 및 파말라이트(Pharmalyte) pH 8-10.5 (pH 범위: 11-8, PBE 118 칼럼에서)를 포함한다.
사용 및 이들 버퍼의 선택에 대한 특별한 지시 사항은 이들 칼럼들의 제조사에서 이용할 수 있다.
항체 조성물의 전하 아이소폼의 분리를 가능하게 하는 또 다른 크로마토그래피 기술은 소수성 상호작용 크로마토그래피이다.
따라서, 바람직하게는, 본 발명에 따른 의약품으로 사용하기 위한 모노클론 항체 조성물을 얻는 것을 허용하는 방법의 단계 b)에서, 단계 a)의 분획은 다음의 크로마토그래피 기술들 중 하나에 의해 수행된다:
· 이온 힘 구배에 의한 용출을 이용한 이온(음이온 또는 양이온) 교환 크로마토그래피(이온 힘 구배를 이용한 이온 교환 크로마토그래피),
· (양이온 교환의 경우에는 증가하는, 음이온 교환의 경우에는 감소하는) pH 구배에 의한 용출을 이용한 이온(음이온 또는 양이온) 교환 크로마토그래피(pH 구배를 이용한 이온 교환 크로마토그래피),
· 이온 힘 및 pH 구배에 의한 용출을 이용한 이온(음이온 또는 양이온) 교환 크로마토그래피(하이브리드 이온 교환 크로마토그래피),
· 치환 분자에 의한 용출을 이용한 이온(음이온 또는 양이온) 교환 크로마토그래피(치환 이온 교환 크로마토그래피),
· 크로마토포커싱, 및
· 소수성 상호작용 크로마토그래피.
유리하게는, 본 발명에 따른 의약품으로 사용하기 위한 모노클론 항체 조성물을 얻는 것을 허용하는 방법의 단계 b)에서, 단계 a)의 분획은 다음의 크로마토그래피 장비들 중 하나에 의해 수행된다:
· 이온 교환 크로마토그래피(용출 모드와 무관하게), 특히 pH 구배를 이용한 이온 교환 크로마토그래피, 및
· 크로마토포커싱.
특히, 본 발명자들은 두 개의 상이한 장비들을 이용하여 모노클론 항체 조성물의 전하 아이소폼들 또는 변이체들을 분리할 수 있었고, 그것들은 본 발명의 범위 내에 사용될 수 있다:
· 크로마토포커싱: 감소하는 pH 구배(두 개의 버퍼를 사용하여 9.5 내지 8.0)에 의한 용출을 이용하는 칼럼 Mono™ P(GE Life Sciences): 버퍼 A(디에탄올아민 25 mM), 버퍼 B(폴리버퍼 96 + 파말라이트 8-10.5));
· 증가하는 pH 구배에 의한 용출을 이용한 양이온 교환 크로마토그래피(컬럼 SCX, MabPac, Dionex)(버퍼 A: 20 mM NaH2PO4, 60 mM NaCl(pH 6); 버퍼 B: 20 mM Na2HPO4, 60 mM NaCl (pH10), 구배: 60분 이내에 10% 내지 60%의 버퍼 B).
전하 아이소폼들의 분리를 허용하는 크로마토그래피 기술에 의해 얻어진 항체 조성물의 크로마토그램은 주요 전하 아이소폼 뿐만 아니라 주요 아이소폼에 인접한(즉, 주요 아이소폼과 비교하여 많은 변형을 갖지 않고 이에 따라 특정 pH에서 pl 및 순 전하가 주요 아이소폼의 그것과 인접한) 다른 아이소폼을 포함하는 주 피크를 항상 포함하고, 한편으로는, 주요 아이소폼과 비교하여 그것의 pl이 열등한, 소위 "산성" 아이소폼과, 주요 아이소폼과 비교하여 그것의 pl이 우수한, 소위 "염기성" 아이소폼을 포함하는 소수 피크들로 둘러싸인다(도 1 및 2 참조).
사용된 크로마토그래피 기술에 따라서, 상이한 아이소폼들이 크로마토그램에 나타나고 다음의 순서로 용출된다:
· 용출 모드(이온 힘 구배, pH 구배, pH 및 이온 힘 구배에 의한 용출, 또는 치환 분자에 의한 용출)에 상관없이, 양이온 교환 크로마토그래피(음으로 하전된 수지)의 사용: 산성 아이소폼(주요 아이소폼보다 적게 양(+)으로 하전됨)이 먼저 용출되고, 이어서 주요 아이소폼이, 그 이후에 염기성 아이소폼(주요 아이소폼보다 좀더 양(+)으로 하전됨)이 용출된다(Khawli et al., 2010의 도 1, Rea et al, 2012의 도 3; Farnan et al., 2009의 도 1 및 Rea et al., 2011의 도 1; Zhang et al., 2011의 도 1 및 McAtee et al., 2012의 도 8.10.2; 및 본 발명의 도 2 참조);
· 용출 모드(이온 힘 구배, pH 구배, pH 및 이온 힘 구배에 의한 용출, 또는 치환 분자에 의한 용출)에 상관없이, 음이온 교환 크로마토그래피(양으로 하전된 수지)의 사용: 염기성 아이소폼(주요 아이소폼보다 적게 음(-)으로 하전됨)이 먼저 용출되고, 이어서 주요 아이소폼이, 그 이후에 산성 아이소폼(주요 아이소폼보다 좀더 음(-)으로 하전됨)이 용출된다;
· 크로마토포커싱: 염기성 아이소폼이 먼저 용출되고, 이어서 주요 아이소폼, 그 이후에 산성 아이소폼이 용출된다(본 발명의 도 1 참조).
세포 클론, 비-인간 유전자변형 동물 또는 유전자변형 식물에 의해 생산된 항체 조성물 내에 존재하는 항체의 전하 아이소폼 또는 변이체들은 또한 크로마토그래피 이외의 기술을 이용하여 분리될 수 있다. 그러나, 이들 장비가 전하 아이소폼들 및 변이체들을 분석하거나 특성화하는 목적에 매우 유용하다면, 그것들은 허용가능한 양으로 이들 아이소폼을 분리하는 것을 허용하지 않고 따라서 제조 목적으로 잘 사용되지 않는다.
이러한 다른 장비들 중에서, 특히 등전점 포커싱(isoelectric focusing)("등전점 포커싱"을 "IEF"라고 하고, 또한 전기포커싱(electrofocusing)이라고 한다)이 언급될 수 있다.
등전점 포커싱(IEF)의 기본 원리는 전기장에 종속된 단백질들이 움직일 수 있는 pH 구배를 겔(선택적으로 모세관에 포함된)에서 발생시키는 것이다. 단백질들은 전기장에서 움직일 것이다. 단백질들의 pl에 상당하는 pH에 도달하면, 그것들의 순 전하가 0일 것이므로 그것들은 고정될 것이다. 이러한 방식으로, 그것들의 pl에 따라서 제제(preparation)의 단백질을 분리할 수 있다. 특정 수의 양 또는 음의 이온화 그룹들(아민, 카르복실 또는 설페이트)를 갖고 특정 완충 능력을 갖는 고분자 전해질을 이용하여 이러한 pH 구배를 생성할 수 있다. 이들 분자는 양쪽성 물질(ampholyte)로 불린다. 이들 양쪽성 물질이 음극(anode)에서 강산의 용액에 의하여 그리고 양극(cathod)에서 강염기의 용액에 의해 제한되는 전기장에 속하게 되면, 그것들은 이동할 것이고 그것들의 pl 순서대로 분포될 것이다. 그것들의 완충 용량은 그것들 주위에 그것들의 pl과 동일한 소(small) pH 영역을 유지하는데 기여할 것이다. 따라서, 특정 pH 범위를 커버하는 pl을 갖는 일련의 양쪽성 물질 각각은, 연속적인 pH 구배를 발생시킨다. 이 시스템에서 소량의 단백질들이 이동하게 되면, 그것의 형성 후 또는 형성 동안, 그것들은 또한 이동하고 그것들의 pl에서 고정될 것이다.
겔에 대해 불활성 매트릭스로서, 아가로즈, 아크릴아마이드 또는 보다 드물게는 덱스트란을 사용할 수 있고, 여기에서 pH 구배가 형성될 것이다. 폴리아크릴아마이드 겔은 가장 흔히 사용된다. pl만이 이동에 영향을 미쳐야 하기 때문에, 아크릴아마이드의 농축물이 사용되어야 하고, 이를 위해 다공도(porosity)는 작은 단백질에 비하여 큰 단백질의 속도를 늦출 것이지만, 그것은 취급이 용이해지도록 충분한 고형물이다. 5~6% 겔이 일반적으로 적절하다.
음극의 버퍼는 강산이고, 일반적으로 인산이다. 양극에서, 강염기가 사용되고, 종종 트리에탄올아민이다.
양쪽성 물질은 중합 이전에 겔을 제조하기 위하여 혼합물에 포함된다. 고분자 전해질(polyelectrolytes) 물질인, 이들 분자는 전기장에서 움직이고 자신의 pl의 순서로 서로의 다음에 위치된다. 많은 회사들이 매우 좁거나 매우 넓은 pH 범위를 커버하는 양쪽성 물질들의 혼합물을 대량으로 만든다: 암폴린(Ampholine)®(특히 Sigma Aldrich사에 의해 판매되는 Ampholine® pH 6/8 및 Ampholine® pH 7/9), 파말라이트(Pharmalyte)®(특히 Sigma Aldrich 사 및 GE Healthcare, Life Sciences사에 의해 판매되는 Pharmalyte® pH 8/10.5), 바이오라이트(BioLite)® (특히 Bio-Rad사에 의해 판매되는 BioLite® pH 6/8, BioLite® pH 7/9 및 BioLite® pH 8/10), 줌(Zoom)®(특히 Life technologies/Invitrogen사에 의해 판매되는 Zoom® pH 6/9), 서바라이트(Servalyt)™ (특히 Serva사에 의해 판매되는 Servalyt™ pH 6/8, Servalyt™ pH 6/9, Servalyt™ pH 7/9), 시누라이트(SinuLyte)™(특히 Sinus사에 의해 판매되는 SinuLyte™ pH 6/8, SinuLyte™ pH 6/9, SinuLyte™ pH 7/9, SinuLyte™ pH 8/10) 등. 양 전극에 전압이 가해지면, 양쪽성 물질 각각은 그것의 등전점에서 가능한 멀리 이동할 것이고 그곳에 고정될 것이다. 다양한 pH 진폭을 갖는 구배들이 다양한 양쪽성 물질들을 조합하여 발생될 수 있다. 특히, 항체 조성물에서 전하 아이소폼들의 분석을 위하여, 구배들은, 항체의 평균 pl에 중심을 둔 작은 pH 범위 상에서, 상이한 아이소폼들의 pl 범위에 대응하면서(예를 들면 pH 6 내지 pH 8 또는 pH 7 내지 pH 9), 양쪽성 물질 각각의 사이에서, 매우 작은 간격(예, 0.1 pH 유닛)을 갖도록 생성될 것이고, 이것은 상이한 전하 아이소폼들의 매우 정밀한 분리를 가능하게 한다.
분석되는 항체 조성물은 겔의 중합 후에 추가되거나 중합 전 혼합물에 직접 추가될 수 있다. 항체들이 양쪽성 물질들보다 더 크기 때문에, 항체들은 훨씬 더 천천히 이동하고 따라서 양쪽성 물질들은 항체들의 실질적인 이동 전에 그것들의 pl에서 상당히 안정화될 수 있다. 이동 시간은 중요하지 않다(not critical). 실제로 항체들이 그것의 pl에서 유지되는 지점에서 고정될 경우 항체들은 겔을 벗어날 위험이 없다. 양쪽성 물질이 적절히 이동할 시간을 갖도록 그리고 항체들이 그것들의 pl을 유지하기 위한 시간을 갖도록 이동만은 충분히 긴 시간 동안 지속되어야 한다. 2 mA에서, 요구되는 시간은 약 1 시간으로 추정된다.
이동 후에, 겔은 항체 조성물에 존재하는 상이한 전하 아이소폼들을 분석하기 위하여 염색된다. 염색은 표준 전기영동에서 사용되는 임의의 통상의 기술에 의해 이루어질 수 있다. 그러나, 양쪽성 물질들은 염색될 수 있기 때문에 양쪽성 물질들은 겔로부터 제거되어야 한다. 그러므로, 일반적으로 염색은 5% 내지 10% 트리클로로아세트산 욕(bath)에 침지하거나 부위(site)에 항체들을 고정하는 동안 그것들을 겔 밖으로(out of gel) 확산시켜서 수행될 수 있다.
정해진 pl을 갖는 마커들의 사용은 상이한 전하 아이소폼들의 pl을 상당히 특이적으로 측정하는 것을 가능하게 한다.
염색 후에, 분석된 조성물에서 전체 아이소폼들에 대한 IEF에서 분리된 전하 아이소폼 각각의 비율은 이미지 분석 소프트웨어 패키지, 예를 들어 Bio-Rad사에 의해 판매되는, 소프트웨어 패키지 Quantity One®에 의해 정량화될 수 있다.
항체 조성물에 존재하는 전하 아이소폼을 분리하는데 매우 정확하고 민감하다고 하더라도, 등전점 포커싱 장비는 분리된 아이소폼들을 쉽게 수확할 수 없고 따라서 상이한 아이소폼들을 제조 분리할 목적보다는 일반적인 분석과 정량화의 목적으로 일반적으로 사용된다.
방법의 단계 c)에서, 의약품으로 사용되기 위한, 본 발명에 따른, 관심있는 조성물은, 단계 b)에서 얻은 하나 또는 수 개의 크로마토그래피 분획물들을 조합하여 얻어지고, 이것들은 크로마토그램의 주 피크에 대응하고, 최종 획득된 모노클론 항체 조성물은 상기 주 피크가 풍부하고, 상기 주 피크는 단계 c)에서 얻은 조성물의 크로마토그램의 적어도 85%, 유리하게는 적어도 86%, 적어도 87%, 적어도 88%, 적어도 89%, 보다 유리하게는 적어도 90%, 적어도 91%, 적어도 92%, 적어도 93%, 적어도 94%, 또는 심지어 적어도 95%, 적어도 96%, 적어도 97%, 적어도 98%, 적어도 98.5%, 적어도 99%, 또는 적어도 99.5%를 나타낸다.
유리하게는, 본 발명에 따른 의약품으로서 사용하기 위한 조성물에서, 조성물에 존재하는 항체들의 중쇄 중 적어도 95%, 유리하게는 적어도 96%, 적어도 97%, 적어도 98%, 또는 심지어 적어도 98.5%, 적어도 99%, 또는 적어도 99.5%가 어떠한 C-말단 라이신 잔기도 포함하지 않는다.
본 발명은 또한 모노클론 항체 조성물에 관련되고, 여기서 조성물에 존재하는 항체들의 중쇄 중 적어도 95%, 유리하게는 적어도 96%, 적어도 97%, 적어도 98%, 또는 심지어 적어도 98.5%, 적어도 99%, 또는 적어도 99.5%가, 의약품으로서의 그것의 용도에 대하여, 어떠한 C-말단 라이신 잔기도 포함하지 않는다. 실제로 조성물에 존재하는 항체들의 염기성 아이소폼들은 C-말단 라이신 잔기를 갖는 적어도 하나의 중쇄를 갖는다. 그러므로 이러한 조성물은 전적으로 주요 아이소폼 및 산성 아이소폼을 포함한다. 염기성 아이소폼이 FcγRⅢ를 통한 그리고 보체를 통한 반응기 효과에 대하여 매우 활성이 아니고(실시예 참조) 정제 전에 약 8 내지 20%를 나타내기 때문에(크로마토그래피에 의해 측정된 것처럼), 이러한 조성물은, 염기성 아이소폼들을 배제(exclusion)하기 전에, 총 조성물보다 FcγRⅢ를 통해 더 강한 ADCC와 더 강한 반응 CDC를 유도할 수 있다. 이러한 조성물은 상기에서 기술된 것처럼 크로마토그래피 분리를 통해 얻을 수 있지만, 수집된 분획물들은 이 경우에 산성 및 주요 아이소폼들의 그것에 상응한다.
상기에서 기술된 방법에 의해 얻을 수 있고 의약품으로서 사용하기 위한 항체 조성물은 모노클론 항체들로 치료될 수 있는 임의의 질병에 사용될 수 있고, 특히 ADCC에 의해 또는 CDC에 의해 타겟 세포들을 파괴하는 것이 치료에 유용한 경우에 사용될 수 있다.
오늘날, ADCC는 암을 치료하려는 항체들에 의한 수동적 면역요법 치료의 임상적 효율에 대한 필수적인 메카니즘이고(Wallace et al., 1994; Velders et al., 1998; Cartron et al., 2002; Ianello et al., 2005; Weiner et al., 2010), Rh 마이너스(-) 임산부의 동종 면역을 예방하기 위한 필수적인 메카니즘이다(Beliard et al., 2008). 또한, ADCC 반응도 바이러스(Ahmad et al., 1996, Miao et al.,2009), 박테리아(Albrecht et al., 2007; Casadevall et al., 2002), 및 기생충(Zeitlin et al., 2000)에 대한 항-감염 반응에서 중요한 역할을 하는 것으로 알려져 있다. 또한, 자가면역질환의 측면에서, 새로운 치료법은 예를 들어 B 림프구 또는 T 림프구와 같은, 공격을 책임지는 면역 세포들을 제거하는 것을 목적으로 하고, 이에 따라 ADCC가 매우 중요한 역할을 한다(Edwards et al., 2006; Chan et al., 2010).
CDC 반응은 또한 다양한 질병에서, 특히 암의 치료에서 중요한 것으로 알려져 있다.
따라서, 본 발명에 따른 의약품으로서 사용하기 위한 조성물에서, 항체는 유리하게도 건강한 공여 세포들, 암세포의 항원, 병원체에 의해 감염된 세포의 항원, 또는 면역 세포의 항원에 존재하는 비-유비쿼터스 항원에 대하여 지향한다.
특히, 다음의 실시형태들이 바람직하다:
- 항체는 항-Rh(D) 항체(주목할만하게 롤두맙(Roledumab), 아토롤리무맙(Atorolimumab) 또는 모롤리무맙(Morolimumab), 특히 롤두맙)이고, 조성물은 Rh 마이너스(-) 개인에서 동종 면역을 예방하기 위한 것이다.
- 항체는 암의 항원을 향하고 조성물은 암을 치료하기 위한 것이다.
- 항체는 병원체에 의해 감염된 세포의 항원을 향하고 조성물은 상기 병원체에 의한 감염을 치료하기 위한 것이다.
- 항체는 면역 세포의 항원을 향하고 조성물은 자가면역 질환을 치료하기 위한 것이다.
암 치료의 측면에서, 항체들은 다음의 항원들을 특히 향할 수 있다: CD20, Her2/neu, CD52, EGFR, EPCAM, CCR4, CTLA-4(CD152), CD19, CD22, CD3, CD30, CD33, CD4, CD40, CD51(Integrin alpha-V), CD80, CEA, FR-alpha, GD2, GD3, HLA-DR, IGF1R(CD221), 포스파티딜세린(phosphatidylserine), SLAMF7 (CD319), TRAIL-R1, TRAIL-R2.
보다 구체적으로는, 그것들의 치료학적 관심에 대해 알려져 있는 특정(항원/암) 쌍(pairs)을 아래 표 1에 나타내었다(이 항원 특이성의 항체들은 언급된 암의 치료를 위해 적어도 1개국에서 승인되었거나, 임상 시험이 진행 중이다).
| 항원 | 이 항원에 대한 항체의 예시 | 이 항원 특이적 항체로 치료될 수 있는 암(들) |
| CD20 | 리툭시맙(rituximab), 오파투무맙(ofatumumab), 오크렐리주맙(Ocrelizumab), 토시투모맙(Tositumomab), 벨투주맙(Veltuzumab), 유블리툭시맙(Ublituximab) |
혈액암, 특히: 비-호지킨 림프종, B-세포 림프종, 만성 골수성 백혈병, 소포성 림프종 |
| Her2/neu | 트라스투주맙(Trastuzumab), 퍼투주맙(Pertuzumab) | 고형 암, 특히: 유방암, 비-소세포성 폐암, 췌장암, 전립선암, 난소암 |
| CD52 | 알렘투주맙(alemtuzumab) | 혈액학적 암, 특히: 만성 림프구성 백혈병, 만성 골수성 백혈병, 피부 또는 말초 T-세포 림프종 |
| EGFR | 세툭시맙(Cetuximab), 파니투무맙(panitumumab), 푸툭시맙(Futuximab), 임가투주맙(Imgatuzumab), 마투주맙(Matuzumab), 넥시투무맙(Necitumumab), 니모투주맙(Nimotuzumab), 잘루투무맙(Zalutumumab) | 고형 종양, 특히: 대장암, 두부암 및 경부암, 폐암, 식도암, 위암, 신경아교종, 퇴화된 성상세포종, 교아종 |
| EPCAM | 에드레콜로맙(Edrecolomab), 아데카투무맙(Adecatumumab), 솔리토맙(Solitomab), | 고형 암, 특히: 대장암, 전립선암, 유방암 |
| CCR4 | 모가물리주맙(Mogamulizumab) | 혈액학적 암, 특히: 성인 T-세포 백혈병/림프종 |
| CTLA-4 (CD152의 이름으로 공지됨) | 이필리무맙(Ipilimumab), 트레멜리무맙(Tremelimumab), | 고형 종양, 특히: 흑색종, 전립선암, 방광암 |
| CD19 | 블리나투모맙(Blinatumomab)(CD19 및 CD3 양자를 타겟함) | 혈액암, 특히: 비-호지킨 림프종, 급성 림프아구 백혈병, 폐암, 소화관암 |
| CD22 | 에프라투주맙(Epratuzumab) | 혈액암, 특히: B-세포암 |
| CD3 | 오텔릭시주맙(Otelixizumab), 테플리주맙(Teplizumab), 비실리주맙(Visilizumab) | 혈액암, 특히: 다발성 골수종 |
| CD30 | 이라투무맙(Iratumumab) | 혈액암, 특히: 비-호지킨 림프종 |
| CD33 | 린투주맙(Lintuzumab) | 혈액암, 특히: 급성 골수성 백혈병, 골수이형성 증후군 |
| CD4 | 세델리주맙(Cedelizumab), 클렌노릭시맙(Clenoliximab), 프릴릭시맙(Priliximab), 자놀리무맙(Zanolimumab) | 흑색종, 피부 또는 말초 T-세포 림프종 |
| CD40 | 닥세투주맙(Dacetuzumab), 루카투무맙(Lucatumumab), 테넬릭시맙(Teneliximab) | 혈액암, 특히: 비-호지킨 림프종, 호지킨 림프종, 다발성 골수종 |
| CD51 (인테그린 알파-V) | 인테투무맙(Intetumumab) | 고형 종양 |
| CD80 | 갈릭시맙(Galiximab) | 혈액암, 특히: B-세포 림프종 |
| CEA | 라베투주맙(Labetuzumab) | 고형 종양, 특히: 대장암 |
| FR-알파 | 팔레투주맙(Farletuzumab) | 난소암 |
| 강글리오시드 GD2 | 3F8, TRBS07 | 신경아세포종, 흑색종 |
| 강글리오시드 GD3 | 에크로멕시맙(Ecromeximab), 미투모맙(Mitumomab) | 흑색종, 소세포성 폐암 |
| HLA-DR | 아폴리주맙(Apolizumab) | 혈액암 |
| IGF1R(CD221) | 식수투무맙(Cixutumumab), 피지투무맙(Figitumumab), 로바투무맙(Robatumumab), 가니투맙(Ganitumab) | 고형 종양, 특히: 비-소세포성 폐암, 부신피질 암종, 췌장암 |
| 포스파티딜세린 | 바비툭시맙(Bavituximab) | 고형 종양, 특히: 유방암, 비-소세포성 폐암 |
| SLAMF7(CD319) | 엘로투주맙(Elotuzumab) | 다발성 골수종 |
| TRAIL-R1 | 마파투무맙(Mapatumumab) | 고형 종양, 특히: 비-소세포성 폐암, 대장암; 비-호지킨 림프종 |
| TRAIL-R2 | 코나투무맙(Conatumumab), 렉사투무맙(Lexatumumab), 티가투주맙(Tigatuzumab) | 고형 종양, 특히: 유방암, 췌장암, 대장암, 비-소세포성 폐암, 난소암 |
병원 유기체에 의한 감염을 치료하는 측면에서, 항체들은 다음의 항원들을 특히 향할 수 있다: 클로스트리듐 디피실(Clostridium difficile )의 항원들, 스타 필로코커스 아우레우스(Staphylococcus aureus )의 항원들(특히 ClfA 및 지질타이코산), 거대세포바이러스의 항원들(특히 당단백질 B), 에스케리키아 콜리(Escherichia coli)의 항원(특히 시가양 독소, 단위 IIB 하), 호흡기세포융합바이러스의 항원(특히 단백질 F), B형 간염 바이러스의 항원, 인플루엔자 A 바이러스의 항원(특히 헤마글루티닌), 혈청형 IATS O11의 슈도모나스 에루지노사(Pseudomonas aeruginosa)의 항원, 광견병 바이러스의 항원(특히 당단백질), 포스파티딜세린.
보다 구체적으로, 그것들의 치료학적 관심에 대해 알려져 있는 특정(항원/감염 질병) 쌍은 아래의 표 2에 나타내었다(이 항원 특이적 항체들은 언급된 감염 질환을 치료하기 위해 적어도 하나의 국가에서 승인되었거나, 임상 시험이 진행 중이다).
| 항원 | 이 항원에 대한 항체의 예시 | 이 항원에 특이적 항체로 치료될 수 있는 감염 질병(들) |
| 클로스트리듐 디피실의 항원 | 악토주맙(Actoxumab), 베즐로톡주맙(Bezlotoxumab) | 클로스트리듐 디피실 감염 |
| 스타필로코커스 아우레우스의 ClfA 항원 | 테피바주맙(Tefibazumab) | 스타필로코커스 아우레우스 감염 |
| 거대세포 바이러스의 항원 | 세비루맙(Sevirumab) | 거대세포바이러스에 의한 감염 |
| 거대세포 바이러스의 당단백질 B | 레가비루맙(Regavirumab) | 거대세포바이러스에 의한 감염 |
| 시가양 독소, 에스케리키아 콜리의 서브 유닛 IIB | 울톡사주맙(Urtoxazumab) | 에스케리키아 콜리, 혈청형 O121에 의한 감염 |
| 호흡기세포융합 바이러스의 단백질 F | 팔리비주맙(Palivizumab), 모타비주맙(Motavizumab) | 호흡기세포융합 바이러스에 의한 감염 |
| B형 간염 바이러스의 표면 항원 | 엑스비비루맙(Exbivirumab), 리비비루맙(Libivirumab) | B형 간염 바이러스에 의한 감염 |
| B형 간염 바이러스의 표면 항원 | 투비루맙(Tuvirumab) | B형 간염 바이러스에 의한 감염 |
| 인플루엔자 A 바이러스의 헤마글루티닌 | CR6261 | 인플루엔자, 특히 스페니쉬 인플루엔자 및 H5N1 |
| 스타필로코커스 아우레우스의 지질타이코산 | 파기박시맙(Pagibaximab) | 스타필로코커스 아우레우스 감염, 스타필로코커스 아우레우스 에 의한 패혈성 쇼크 |
| 포스파티딜세린 | 바비툭시맙(Bavituximab) | 간염 C 바이러스, 인플루엔자 A 및 B, HIV 1 및 2, 풍진, 호흡기세포융합 바이러스, 피친데 바이러스에 의한 감염 |
| 슈도모나스 에루지 노사 혈청형 IATS O11의 항원 | 파노바쿠맙(Panobacumab) | 슈도모나스 에루지노사에 의한 감염 |
| 광견병 바이러스의 당단백질 | 포라비루맙(Foravirumab), 라피비루맙(Rafivirumab) | 광견병 바이러스의 감염 |
자가면역 질환을 치료하는 측면에서, 항체들은 다음의 항원들을 특히 향할 수 있다:CD20, CD52, CD25, CD2, CD22, CD3, 및 CD4.
보다 구체적으로, 그것들의 치료학적 관심에 대해 알려져 있는 특정 (항원/자가면역 질환) 쌍을 아래의 표 3에 나타내었다(이 항원 특이적 항체들은 언급된 자가면역 질환을 치료하기 위해 적어도 하나의 국가에서 승인되었거나, 임상 시험이 진행 중이다).
| 항원 | 이 항원에 대한 항체의 예시 | 이 항원 특이적 항체로 치료될 수 있는 자가면역질병(들) |
| CD20 | 리툭시맙(rituximab), 오파투무맙(ofatumumab), 오크렐리주맙(Ocrelizumab), 토시투모맙(Tositumomab), 벨투주맙(Veltuzumab), 유블리툭시맙(Ublituximab) |
류마티스 관절염, 혈소판감소성 자반병, 홍반성 낭창, 다발성 경화증 |
| CD52 | 알렘투주맙(alemtuzumab) | 다발성 경화증 |
| CD25 | 다실주맙(Daclizumab), 바실릭시맙(Basiliximab), 이놀리모맙(Inolimomab) | 포도막염, 다발성 경화증, 건선, 제1형 당뇨병, 궤양성 대장염 |
| CD2 | 시플리주맙(Siplizumab) | 건선 |
| CD22 | 에프라투주맙(Siplizumab) | 홍반성 낭창 |
| CD3 | 오텔릭시주맙(Otelixizumab), 테플리주맙(Teplizumab), 비실리주맙(Visilizumab) | 제1형 당뇨병, 궤양성 대장염, 크론병 |
| CD4 | 세델리주맙(Cedelizumab), 클렌노릭시맙(Clenoliximab), 프릴릭시맙(Priliximab), 자놀리무맙(Zanolimumab) | 류마티스 관절염, 크론병, 다발성 경화증, 건선 |
본 발명에 따른 의약품으로서 사용되기 위한 항체 조성물들은 특히 ADCC 반응을 의미하는 치료법을 위한 것이고, 이것은 상기에서 상세히 설명한 것처럼 많은 시나리오들을 포함한다. 따라서, 이들 항체는 또한 FcγRⅢ 수용체를 통해 생체 내 ADCC 반응을 유도하기 위한 기타 수단들에 의해, 가능한 강하게, 최적화되는 것이 유리하다. 따라서, 유리한 실시형태에서, 본 발명에 따른 의약품으로 사용하기 위한 조성물 내에서, 항체는 FcγRⅢ 수용체에 대한 그것의 결합과 그것의 FcγRⅢ 수용체를 통한 반응기 특징을 향상시키는 Fc 단편의 변형을 포함한다.
두 개의 주요 수단들은 당장은(for the momnet) FcγRⅢ 수용체를 통해 ADCC 활성을 최적화하기 위해 기술되었다:
- Fc 단편의 특정 아미노산 잔기들에서 적어도 하나의 돌연변이 삽입, 특히 WO00/42072, [Shields 등, 2001], [Lazar 등, 2006], WO2004/029207, WO/2004063351, WO2004/074455에 기술됨.
- Fc 단편의 중쇄 각각의 Asn297 잔기에 부착된 N-글리칸의 특성의 최적화.
따라서, 유리한 실시형태에서, 본 발명에 따른 의약품으로서 사용되기 위한 조성물은 모노클론 항체를 포함하고, 이것의 서열은, WO00/42072, [Shields 등, 2001], [Lazar 등, 2006], WO2004/029207, WO/2004063351, WO2004/074455에 기술된 것처럼, FcγRⅢ 수용체에 대한 결합을 향상시키는 Fc 단편의 적어도 하나의 아미노산 잔기가 변형되었다.
특히, Fc의 다음의 위치에서의 돌연변이는 FcγRⅢ 수용체에 대한 친화성과 이 수용체를 통해 ADCC를 유도하는 능력을 증가시킨다는 것에 대해 기술되어 있다: 219, 222, 224, 239, 247, 256, 267, 270, 283, 280, 286, 290, 294, 295, 296, 298, 300, 320, 326, 330, 332, 333, 334, 335, 339, 360, 377, 396.
보다 구체적으로, 다음의 치환체들은 FcγRⅢ 수용체에 대한 친화성과 이 수용체를 통해 ADCC를 유도하는 능력을 증가시키는 것을 허용한다는 것에 대해 기술되어 있다: S219Y; K222N; H224L; L234E, L234Y, L234V; L235D, L235S, L235Y, L235I; S239D, S239T; V240I, V240M; P247L; T256A, T256N; V264I, V264T; V266I; S267A; D270E; D280A, D280K, D280H, D280N, D280T, D280Q, D280Y; V282M; E283Q; N286S; K290A, K290Q, K290S, K290E, K290G, K290D, K290P, K290N, K290T, K290S, K290V, K290T, K290Y; E294N; Q295K; Y296W; S298A, S298N, S298V, S298D, S298E; Y300I, Y300L; K320M, K320Q, K320E; N325T; K326S, K326N, K326Q, K326D, K326E; A330K, A330L, A330Y, A330I; I332E, I332D; E333A, E333Q, E333D; K334A, K334N, K334Q, K334S, K334E, K334D, K334M, K334Y, K334H, K334V, K334L, K334I; T335E, T335K; A339T; K360A; F372Y; I377F; V379M; P396H, P396L; D401V.
관심있는 돌연변이의 조합들은 다음을 포함한다: E333A/K334A, T256A/S298A, S298A/E333A, S298A/K334A, S298A/E333A/K334A, S267A/D280A(WO00/42072), S239D/I332E, S239D/I332E/A330L(Lazar et al., 2006), V264I/I332E, S298A/I332E, S239E/I332E, S239Q/I332E, S239D/I332D, S239D/I332E, S239D/I332N, S239D/I332Q, S239E/I332D, S239E/I332N, S239N/I332E, S239Q/I332D, A330Y/I332E, V264I/A330Y/I332E, A330L/I332E, V264I/A330L/I332E, S239E/V264I/I332E, S239E/V264I/A330Y/I332E, S239D/A330Y/I332E, S239N/A330Y/I332E, S239D/A330L/I332E, S239N/A330L/I332E, V264I/S298A/I332E, S239D/S298A/I332E, S239N/S298A/I332E, S239D/V264I/I332E(WO2004/029207).
대안으로 또는 추가로, 본 발명에 따른 의약품으로서 사용되기 위한 모노클론 항체 조성물은 낮은 푸코스 함량을 포함한다. "푸코스 함량"은 각 항체의 각 중쇄의 Fc 단편의 Asn297 잔기에 부착된 N-글리칸 내에 푸코실화된 형태의 백분율을 의미한다. "낮은 푸코스 함량"은 65% 이하의 푸코스 함량을 의미한다. 실제로, 항체 조성물의 푸코스 함량은 FcγRⅢ 수용체를 통해 강한 ADCC 반응을 유도하는 이 조성물의 능력에 중요한 역할을 하는 것으로 오늘날 알려졌다. 유리하게는, 푸코스 함량은 65% 이하, 바람직하게는 60%, 55% 또는 50% 이하, 또는 심지어 45%, 40%, 35%, 30%, 25% 또는 20% 이하이다. 그러나, 푸코스 함량이 0이 되는 것이 필수적이지 않고, 예를 들어 5%, 10%, 15% 또는 20% 이상일 수 있다. 예를 들어, 푸코스 함량은 5 내지 65%, 5 내지 60%, 5 내지 55%, 5 내지 50%, 5 내지 45%, 5 내지 40%, 5 내지 35%, 5 내지 30%, 5 내지 25%, 5 내지 20%, 10 내지 65%, 10 내지 60%, 10 내지 55%, 10 내지 50%, 10 내지 45%, 10 내지 40%, 10 내지 35%, 10 내지 30%, 10 내지 25%, 10 내지 20%, 15 내지 65%, 15 내지 60%, 15 내지 55%, 15 내지 50%, 15 내지 45%, 15 내지 40%, 15 내지 35%, 15 내지 30%, 15 내지 25%, 15 내지 20%, 20 내지 65%, 20 내지 60%, 20 내지 55%, 20 내지 50%, 20 내지 45%, 20 내지 40%, 20 내지 35%, 20 내지 30%, 20 내지 25%로 이루어질 수 있다.
그것들이 낮은 푸코스 함량을 갖는다면, 항체 조성물은 또한 글리코실화의 다른 타입(바이안테너리 복합체 타입의 N-글리칸의 경우 N-아세틸글루코사민(GlcNAc) 잔기 또는 갈락토스 잔기를 이분하는 다양한 비율을 갖는, 올리고만노즈 또는 바이안테너리(biantennary) 복합체 타입의 N-글리칸)을 가질 수 있다. 따라서, 올리고만노즈 타입의 N-글리칸은 다른 글리코실화 억제제들이 존재시 배양에 의해 얻어질 것이고, 억제제들의 예는 α1,2-만노시다제 I(예, 데옥시만노지리마이신(Deoxymannojirimycin) 또는 "DMM") 또는 α-글루코시다제(예, 카스타노스퍼민 또는 "Cs"); 또는 CHO Lec 1 주(line)에서 항체를 제조하는 또 다른 것이다. 유전자변형 염소의 젖의 생산은 또한 주요 N-글리칸이 올리고만노즈 형태인 항체들을 얻게 하고, 이것은 하나 또는 두 개의 갈락토즈를 갖는 푸코실화된 바이안테너리 복합체 형태를 갖고, 임의의 GlcNAc을 이분화하지 않고, 시알릴화하지 않는다(G1F 또는 G2F)(WO2007048077A2 참조). 바이안테너리 복합체 형태의 N-글리칸들은 대부분의 포유동물 세포뿐 아니라 박테리아, 효모균 또는 식물에서 얻어질 수 있지만, 이것의 글리코실화 조직(machinery)은 변형되었다. 푸코스 함량을 제한하기 위하여, Fc 단편에 결합된 GlcNAc 상에 푸코스의 첨가를 담당하는 효소 FUT8(1,6-푸코실트랜스퍼라제)의 낮은 활성을 천연적으로 갖는 세포주들; 예를 들어 세포주는 YB2/0, 오리 배아 세포주 EB66®, 또는 랫트 헤파토마 세포주 H4-II-E(DSM ACC3129), H4-II-Es(DSM ACC3130);가 사용될 수 있다. 다른 유전자에 대해 돌연변이된 세포주들 및 낮은 푸코스 함량을 야기하는 낮은(sub) 발현 또는 과-발현의 세포주들은 CHO Lec13 세포주인, GDP-푸코스의 합성을 감소시키는 CHO 세포주의 돌연변이와 같이, 사용될 수도 있다. 그것은 또한 관심있는 세포주를 선택하는 것 및 N-글리칸들의 푸코실화 경로에 관여하는 단백질의 발현을 감소시키거나 발현되지 않게 하는 것(특히 간섭 RNA들을 사용하거나 또는 관심있는 단백질을 발현하는 유전자의 돌연변이 또는 결실에 의해)을 가능하게 한다(특히 FUT8, Yamane-Ohnuki et al., 2004 참조; 또한 GMD, GDP-푸코스의 이동에 관여하는 유전자, Kanda et al., 2007 참조). 또 다른 대안으로, 관심있는 세포주를 선택하는 것과 단백질 GnTⅢ(β(1,4)-N-아세틸글루코사민트랜스퍼라제 Ⅲ)와 같은, N-글리칸의 푸코실화를 어떻게든 간섭하는 단백질을 과발현시키는 것이 있다. 특히, 약간 푸코실화된 N-글리칸들을 갖는 항체들이 특히 다음에 의해 얻어졌다:
· YB2/0(EP1176195A1, WO01/77181, Shinkawa et al., 2003 참조), CHO Lec13(Shields et al., 2002 참조), EB66®(Olivier et al.,2010), 또는 랫트 헤파토마 세포주 H4-II-E(DSM ACC3129), H4-II-Es(DSM ACC3130)(WO2012/041768 참조) 내에서 생산.
·FUT8를 향하는 소(small) 간섭 RNAs의 존재에서 야생형 CHO 세포주(Mori et al., 2004, Suzuki et al., 2007, Cardarelli et al., 2009, Cardarelli et al., 2010, Herbst et al., 2010), 또는 GMD(골지체에서 GDP-푸코스의 이동체를 코딩하는 유전자, Imai-Nishiya et al., 2007 참조) 내에서 생산
· 1,6-푸코실트랜스퍼라제를 인코딩하는 유전자 FUT8의 2개의 대립유전자를 제거한 CHO 세포주에서(Yamane-Ohnuki et al., 2004) 또는 골지체에서 GDP-푸코스의 이동체를 인코딩하는 유전자 GMD의 양 대립유전자를 제거한 CHO 세포주에서(Kanda et al., 2007) 생산
· 유전자 변형에 의해 효소 GnTⅢ(β(1,4)-N-아세틸글루코사민트랜스퍼라제 Ⅲ)를 인코딩하는 유전자를 과-발현시킨 CHO 세포주 내에서 생산(Umana et al., 1999). 적은 푸코실화뿐 아니라, 얻어진 N-글리칸은 GlcNAc를 이분화하는 강한 함량을 특징으로 한다.
· 소 간섭 RNAs를 사용하여 β1,2-자이로스 및 α1,3-푸코스 잔기 함량들의 강한 감소를 갖는, 유전자 변형 식물들(N. 벤타미아나(Benthamiana)) 내에서 생산(Forthal et al., 2010).
올리고만노즈 타입의 N-글리칸은 바이안테너리 복합체 타입의 N-글리칸과 비교하여 생체내에서 감소된 반감기를 갖는다. 그 결과, 유리하게는, 조성물에 존재하는 항체들은 상기 기술한 것처럼, 낮은 푸코스 함량을 갖는 바이안테너리 복합체 타입의 Fc 단편 글리칸 구조의 N-글리코실화 자리를 갖는다.
특히, 상기 한정된 바와 같이, 모노클론 항체 조성물은 60% 초과의 G0+G1+G0F+G1F 폼들의 함량과 낮은 푸코스 함량을 가질 수 있다. 또한, 상기 한정된 바와 같이, 이것은 65% 초과의 G0+G1+G0F+G1F의 함량과 낮은 푸코스 함량을 가질 수 있다. 또한, 상기 한정된 바와 같이, 이것은 70% 초과의 G0+G1+G0F+G1F의 함량과 낮은 푸코스 함량을 가질 수 있다. 또한, 상기 한정된 바와 같이, 이것은 75% 초과의 G0+G1+G0F+G1F의 함량과 낮은 푸코스 함량을 가질 수 있다. 또한, 상기 기술한 바와 같이, 이것은 80% 초과의 G0+G1+G0F+G1F 폼들의 함량과 낮은 푸코스 함량을 가질 수 있다. 이것은 또한, 60%, 65%, 70%, 75% 또는 80% 초과의 G0+G1+G0F+G1F 폼들의 함량과 50% 미만의 G0F+G1F 폼들의 함량을 가질 수 있다. G0, G1, G0F 및 G1F 폼들은 아래와 같다:

이러한 항체 조성물들은 특히 YB2/0에서, CHO Lec13에서, FUT8 또는 GMD을 향하는 소 간섭 RNAs의 존재시 배양된 야생형 CHO 세포주에서, 1,6-푸코실트랜스퍼라제를 인코딩하는 유전자 FUT8의 양 대립유전자 또는 골지체에서 GDP-푸코스의 이동체를 인코딩하는 유전자 GMD의 양 대립유전자를 제거한 CHO 세포주에서 생산되어 얻어질 수 있다.
본 발명에 따른 의약품으로서 사용되기 위한 항체 조성물들은 CDC 반응에 관여하는 치료법을 위한 것일 수도 있다. 따라서, 이들 항체가 가능한 강하게 단백질 C1q를 통해 생체 내에서 CDC 반응을 유도하는 다른 수단에 의해 최적화되는 것은, 추가적으로 또는 대안으로, FcγRⅢ를 통한 활성을 증가시키는 변형에 유리하다. 따라서, 유리한 실시형태에서, 본 발명에 따른 의약품으로서 사용되기 위한 조성물에서, 항체는 보체를 통해 단백질 C1q에 결합하는 것과 반응기 특성들을 강화하는 Fc 단편의 변형을 포함한다.
이러한 돌연변이는 다음의 문헌들에 특히 기술되어 있다: WO2004074455A2, [Idusogie et al., 2001], [Dall'Acqua et al., 2006(b)], 및 [Moore et al., 2010].
본 발명은 또한 FcγRⅢ(CD16) 수용체를 발현하는 면역 시스템의 반응기 세포들에 의해 상기 항체를 발현하는 타겟 세포들의 항체-의존적 세포성 세포독성(ADCC)을 유도하는 정해진 항체를 향하는 모노클론 항체 조성물의 능력을 증가시키기 위해 크로마토그래피 분획 단계를 사용에 관한 것이다.
최종 얻어진 조성물은 FcγRⅢ(CD16) 수용체를 발현하는 면역 시스템의 반응기 세포들, 특히 자연 살해 세포(또는 NK 세포)에 의해 관심있는 항원을 발현하는 타겟 세포의 ADCC를 유도하는 능력을 향상시킨다. 바람직하게는, 주 피크의 아이소폼들이 풍부한 조성물과 분획되기 전 조성물에서 얻어진 ADCC 수준의 비 R은 다음의 식으로 정의되고:

이것은 적어도 1.15(적어도 15%의 ADCC 수준의 증가에 대응함); 유리하게는 적어도 1.16; 적어도 1.17; 적어도 1.18; 적어도 1.19 보다 유리하게는 적어도 1.20; 적어도 1.25; 적어도 1.30; 적어도 1.35; 적어도 1.40; 적어도 1.45; 또는 심지어 적어도 1.50(적어도 50%의 ADCC 수준의 증가에 대응함)이다.
또한, 본 발명은 정해진 항체를 향하는 모노클론 항체 조성물의 보체에 의해 상기 항원을 발현하는 타겟 세포들의 보체-의존성 세포독성(CDC)를 유도하는 능력을 증가시키는 크로마토그래피 분획 단계의 사용에 관한 것이다.
최종 얻어진 조성물은 관심있는 항원을 발현하는 타겟 세포들의 보체에 의해 세포용해를 유도하는 능력을 향상시킨다. 바람직하게는, 주 피크의 아이소폼들이 풍부한 조성물에서 얻어진 CDC 수준과 분획되기 전 조성물에서 얻어진 CDC 수준의 비 R은 다음의 식으로 정의되고:

이것은 적어도 1.15(적어도 15%의 CDC 수준의 증가에 대응함); 유리하게는 적어도 1.16; 적어도 1.17; 적어도 1.18; 적어도 1.19 보다 유리하게는 적어도 1.20; 적어도 1.25; 적어도 1.30; 적어도 1.35; 적어도 1.40; 적어도 1.45; 또는 심지어 적어도 1.50(적어도 50%의 CDC 수준의 증가에 대응함)이다.
상기의 양자의 사용에서, 크로마토그래피 분획 단계는 본 발명에 따른 의약품으로서 사용되기 위한 주요 아이소폼이 풍부한 항체 조성물을 얻기 위해 상기 기술된 임의의 방법으로 수행될 수 있다. 특히, 분획은 다음의 크로마토그래피 기술들 중 하나에 의해 수행될 수 있다:
· 용출 모드(이온 힘 구배, pH 구배, pH 및 이온 힘 구배, 치환 분자)와 관련없는, 이온 교환 크로마토그래피;
· 크로마토포커싱;
· 소수성 상호작용 크로마토그래피.
CD16을 발현하는 반응기 세포들을 통해 ADCC 또는 CDC 반응 능력을 증가시키는 목적을 수행하기 위한 이러한 크로마토그래피의 분획 단계를 위한 모노클론 항체 조성물은 상기 기술된 임의의 모노클론 항체 조성물일 수 있다. 특히, 조성물 내에 존재하는 모노클론 항체는 인간, 인간화된 것 또는 키메라일 수 있다.
타겟 세포들이 암 세포들인 경우, 이것은 또한 임의의 타입의 항원, 특히 상기 기술된 것들을 향할 수 있고, 항체는 암 세포 항원, 특히 암의 치료 측면에서 상기 기술된 항원들 중 하나를 향할 수 있다. 타겟 세포들이 병원체에 감염된 세포들인 경우, 항체는 감염된 세포들의 항원, 특히 감염 질병의 치료 측면에서 상기 기술된 항원들 중 하나를 향할 수 있다. 타겟 세포들이 자가면역 질병의 발생과 관련된 면역 세포들인 경우, 항체는 면역 세포들의 항원, 특히 자가면역 질병의 치료 측면에서 상기 기술된 항원들 중 하나를 향할 수 있다.
크로마토그래피 분획 단계(단계 a) 후 크로마토그램의 주 피크에 대응하여 얻어진 크로마토그래프 분획물들을 조합하는 단계(단계 b)가 따르고, 최종 얻어진 모노클론 항체 조성물은 상기 주 피크가 풍부하고, 상기 피크는 단계 b)에서 얻어진 조성물의 크로마토그램의 적어도 85%를 나타낸다(분획 및 관심있는 크로마토그래프 분획물의 조합 이후).
다음의 실시예들은 본 발명의 예시에 해당한다.
실시예
실시예
1. 항-
CD20
항체 조성물의 전하
아이소폼들의
정제된
분획물들의
제조,
아이소폼들과
그들의 반응기 특성들의 특징
장비 및 방법
항-CD 항체 조성물
모든 분리와 분석은 클론 YB2/0에 의해 생산된 항-CD20 항체 조성물의 배치에서 수행하였다.
크로마토포커싱에
의한 전하
아이소폼들의
분리
동일한 항체 조성물의 전하 아이소폼의 3개의 제제 분리는 크로마토포커싱으로 수행하였다.
음이온 교환 수지 Mono P 5/200 GL을 사용하였다. 20 mg의 탈염된 단백질을 각 분리시 주입하였다. 용출은 다음의 2개의 버퍼들을 사용하여, pH 구배를 감소시키며(pH 9.5에서 8.0) 수행하였다:
- 버퍼 A : 디에탄올아민 25 mM,
- 버퍼 B : 폴리버퍼 96 + 파말라이트 8-10.5.
분리의 용출물은 2mL 분획물에 수집되었다. 관심있는 분획물들은 33 내지 50 분획물이다.
3개 분리의 분획물들은 분석을 위해 농축되었다.
분리 1(S1)은 분획물들을 여과시켜 살균하기 위해 특정 농도가 되게 하였다:
- 1mL의 부피를 얻기 위한 아미콘 울트라 10kDa(Amicon Ultra 10kDa) 상에 분획물을 농축
- 여과물을 살균
- 농축물을 측정하기 위해 분취물과 활성을 측정하기 위해 분취물을 샘플링.
양이온 교환 크로마토그래피(
CEX
)에 의한 전하
아이소폼들의
분리
전하 아이소폼들의 11개의 분리는 pH 구배를 증가시켜 용출시킨 양이온 교환 크로마토그래피(CEX)에 의해 이루어졌다.
양이온 교환 수지 SCX(MabPac SCX 10.4x250 mm, Dionex)는 30℃에서 사용되었다. 용출은 다음의 2개의 버퍼들을 사용하여, pH 구배를 증가시키며(pH 6에서 10) 수행하였다:
- 버퍼 A : 20 mM NaH2PO4, 60 mM NaCl (pH 6)
- 버퍼 B : 20 mM NaH2PO4, 60 mM NaCl (pH 10).
구배는 다음의 방법으로 얻어졌다: 60분 동안 10% 내지 60%의 버퍼 B.
분리의 용출물은 분획물에서 수집되었다. 관심있는 분획물들은 분획물 1 내지 20이다.
BIACORE
에 의해
CD16
수용체에 대한 결합의 분석
Biacore T100 시스템(GEHealthcare)의 SPR("표면 플라즈몬 공명(Surface plasmon resonance)") 장치를 사용하여 수용체 CD16a에 결합하는 항체 조성물의 능력을 측정하는 방법을 개발하였다. 가용성 수용체 CD16a는 아민 커플링을 사용하여 검출 칩 상에 고정되었다. 유세포(flow cell)는 항체에 사용되고, 배경 노이즈를 제거하기 위해 다른 유세포는 자유롭게 방치된다. 항체들은 세개의 농축물들에 주입되고, 동역학 파라미터들은 결합 단계(association phase)와 분리 단계(dissociation phase) 양자의 결합 비를 농도 각각에서 산출하기 위해 추정되었다. 공명 단위(RU)로 나타내는 SPR 신호는 수용체에서 항체의 결합과 분리를 나타낸다.
CD16
을 통한 반응기 세포들의 활성화(
CD16
주르카트
시험)
CD16 수용체(FcγRⅢ)를 통한 반응기 세포들의 반응을 유도하는, 상이한 전하 아이소폼들을 포함하는, 크로마토포커싱과 양이온 교환 크로마토그래피(CEX)에 의해 분리된 다양한 분획물들의 능력을 시험하였다.
사용된 시험은 다음과 같다:
항체들은 WIL2-S 세포들(양(+) CD20 타겟 세포들)과 CD16 주르카트 세포(반응기 세포들)(유전자형 CD16FF)로 인큐베이트시켰다. CD16 주르카트 세포들에서 분비된 사이토카인(IL2)의 양은 ELISA로 측정하였다.
보다 구체적으로, 96-웰 플레이트에서 다음을 혼합하였다:
· 항체 : IMDM 5% FCS 내 0.156 내지 10 ng/ml 범위의 희석액 50μl
· IMDM 5% FCS 내 40ng/ml에서 희석액의 PMA 50μl(즉, 2 ng PMA/50μl)
· WIL2-S 세포들 : IMDM 5% FCS 내 6x105/ml에서 50μl(즉, 30x103 cells/50μl)
· CD16 주르카트 : IMDM 5% FCS 내 10x106/ml에서 50μl(즉, 500x103 cells/50μl)
2개의 대조군이 사용되었다: 음성 대조군은 어떠한 타겟 세포도 없고, 양성 대조군은 최대 활성을 갖는다:
어떠한 세포도 없는 음성 대조군은: 웰 당 다음을 첨가하였다:
· 10x106 cells/ml의 50μl의 CD16 주르카트 세포들(즉, 500x103 cells/50μl)
· 40 ng/ml의 50μl의 PMA(즉, 2ng PMA/50μl)
· 최고 농도에서 50μl의 항체
· 50μl의 IMDM 배지 + 5% FCS
최대 활성 양성 대조군은: 웰 당 다음을 첨가하였다:
· 10x106 cells/ml에서 50μl의 CD16 주르카트 세포들(즉, 500x103 cells/50μl)
· 40 ng/ml에서 50μl의 PMA(즉, 2ng PMA/50μl)
· 5μg/ml에서 50μl의 이오노마이신
· 50μl의 IMDM 배지 + 5% SVF
약하게 교반하고 하룻밤 동안 37℃ ± 0.5℃에서 인큐베이트한다.
1분 동안 125g의 세포들을 붓는다.
상청액 160μl를 둥근 바닥을 가진 96-웰 플레이트로 이동시킨다.
1분 동안 125g의 세포들을 다시 붓는다.
상청액 안에 IL-2를 넣는다. 450 nm에서 기록한다.
각 샘플의 CD16 활성(IL-2의 분비)을 비교 샘플의 CD16의 백분율로 나타낸다.
보체
-의존성 세포독성(
CDC
)
타겟 세포들 Wil2-S을 10%의 FCS를 갖는 보충되지 않은(de-complemented) IMDM 배지(배지 I10)에서 배양하였다. 플라스크 F175에 0.2 106 cells/ml를 갖는 배지 100 ml를 일주일에 2회 이식하였다. 시험은 24시간 내지 72시간 동안 이식된 세포들에서 수행되었고, 보충되지 않은 배지 IMDM+5% FCS(배지 I5)에서 1.106 cells/ml으로 다시 취하였다.
인간 혈청(순혈종의 사람(full blood)의 응집원에 의해 얻은 인간 혈청 AB)을 사용할 때 하루 해동하였다. 해동은 +4℃에서 수행하였다. 해동한 후, 혈청을 배지 I5에서 ½까지 희석하였다.
CellTiter-Blue®(Promega)를 -20℃에서 저장하였고, 사용 전에 해동하기 위해 실온에 방치하였다.
I5 배지에서 연구될 항체들의 농도를 1μg/ml로 조정하였다.
U형 바닥을 갖는 96-웰 플레이트에서 다음을 첨가하였다:
· 50μl의 타겟 세포(1.106 cells/ml의 Wil2S)
· 50μl의 테스트할 항체
· 50μl의 ½ 희석된 인간 혈청
세포들을 1.106C/ml까지 조정한 후 플레이트에 직접 침전시키고(deposited) 37℃에 두었다.
세포들을 5분 동안 인큐베이트시키고, 샘플들을 혈청을 침전시키기 전에 37℃에서 교반하였다.
2개의 대조군이 만들어졌다: 어떠한 세포들도 포함하지 않는 대조군(C-)과 항체를 갖는 대조군(AC-). 손실된 성분들을 I5 배지로 대체하였다.
그것들을 37℃에서 2시간 동안 교반하면서 인큐베이트시켰다. 그 다음 30μl의 CellTiter-Blue®를 각 웰에 첨가하였고, 첨가시 역 피펫팅(reverse pipetting)으로 균질화를 수행하였고, 인큐베이션은 37℃에서 3시간 동안 그리고 교반하면서 30분 동안 수행하였다.
인큐베이션이 끝났을 때, 정지하고 25μl의 3% SDS를 첨가하여 반응물을 안정화시키면서 기록을 다음날로 연기하였다. 그 다음 플레이트를 실온에 두었다.
인큐베이션이 끝났을 때 또는 다음날, 플레이트를 2분 동안 125g에서 원심분리하였다. 100μl의 각 웰을 샘플링하고, 그 다음 플레이트를 평평하게 유지하면서 투명한 바닥을 갖는 검은 광학 플레이트에 분획하였다.
플레이트의 기록을 다음의 파라미터들을 갖는 형광 리더기로 수행하였다:
· 여기 : 530/25 nm
· 방출 : 590/20 nm
· 플레이트의 바닥을 통한 기록(FOND)
· 간격 시간 : 20 μs
· 플래쉬의 수 : 25
· 증가(Gain) : 어떠한 항체도 함유하지 않는 대조군 웰에서 취한 웰로부터 계산함(세포+혈청+I5 배지)
· 오비탈 모드에서 15초 동안 강도 2로 교반
질량 분석법에 의한
아이소폼들의
특성
크로마토포커싱 또는 양이온 교환 크로마토그래피(CEX)로 분리된 다양한 분획물들에 존재하는 다양한 전하 아이소폼들을 Chevreux-2011에 기술된 바와 같은 질량 분석법으로 분석하였다.
이 방법은 박테리아 프로테아제 시스테인(IdeS, 스트렙토코커스 피오게네스(Streptococcus pyogenes )의 면역글로불린을 분해하는 효소)를 사용하는 것을 포함하고, 이것은 구체적으로 경계 영역 하에서 IgGs를 절단하고, 중쇄를 각각 VH-CH1 및 CH2-CH3 영역을 포함하는 25 kDa의 2개의 단편으로 절단한다. 단편들을 아세토니트릴 구배의 액체 크로마토그래피로 분리하였고, 다음의 과정에 따라 질량 분석기로 분석하였다:
크로마토포커싱 또는 CEX로 정제한 100μg의 단편을 동결 건조하였고, 20μl의 분해 버퍼(50 mM NaH2PO2 및 150 mM NaCl, pH 6.30)로 재용해시켰고, 100 IU의 IdeS 효소를 효소 키트(FabRICATOR Kit, Genovis, Lund, Sweden)의 지시사항에 따라 첨가하였다. 가수분해를 향상시키기 위해 제제를 37℃에서 1시간 동안 50W 전원의 마이크로파 보조기(CEM Discover System, CEM, Matthews, NC, USA)로 인큐베이트시켰다. 그 다음, 25μl의 변성 버퍼(denaturing buffer)(8M 우레아 및 0.4M의 NH4HCO3, pH 8.0)를 첨가하였고, 이어서 250mM의 디티오트레이톨(dithiothreitol: DTT) 용액 5μl을 첨가하였다. 단백질을 완전히 환원(reduction)시키기 위해 샘플을 50℃에서 20분 동안 50W 전원의 마이크로파 보조기로 인큐베이트시켰고, 그 다음 이것을 액체크로마토그래피-질량 분석법(LC-MS)으로 분석하였다.
20μg의 양에 대응하는 반응 혼합물의 분취물을 350μl/min의 흐름에서 70℃로 평형을 이룬 역상 ProSphere C4 컬럼(150 x 2.1 mM, 5μm, Alltech) 상에 주입하였다. 역상 크로마토그래피를 울트라-수행 액체 크로마토그래피 시스템(UPLC, Acquity UPLC, Waters, Milford, MA, USA)을 사용하여 수행하였다. 이동 상 A로서 0.1%의 트리플루오로아세트산(TFA)과 이동상 B로서 0.1% TFA를 포함하는 아세토니트릴을 사용하여 구배를 생성하였다. 5분 동안 10% B의 등용매 용출(isocratic elution)한 후, 5분 동안 B를 27%까지 증가시킨 후 추가 10분 동안 40%까지 증가시켰다. 그 다음 컬럼을 90%의 B로 3분 동안 세척하고 10%의 B로 2분 동안 재-평형시켜, 전체 시간 25분을 제공하였다.
그 다음 500 내지 3,000 m/z의 양이온 모드를 작동시키고 수지의 제조업자에 의해 기술된 공정에 따라 눈금이 매겨진 질량 분석기 QSTAR(QSTAR XL, Applied Biosystems, Toronto, Canada)로 용출된 종들을 분석하였다.
결과
크로마토포커싱에
의한 전하
아이소폼들의
분리
3개의 분리의 크로마토그램은 도 1에 나타내었고, 이것은 그것들이 완벽하게 겹쳐질 수 있고, 따라서 크로마토포커싱에 의한 분리 방법의 재현성을 설명한다. 음이온 교환 수지의 사용과 pH 구배의 감소 때문에 염기성 아이소폼들이 먼저 용출되고, 이어서 주요 아이소폼, 그리고 다음으로 산성 아이소폼들이 용출된다.
분획물들 33 내지 50을 그들의 생화학적 특성과 반응기 특성들의 이후 분석에 따라 수집하였다.
양이온 교환 크로마토그래피(
CEX
)에 의한 전하
아이소폼들의
분리
전하 아이소폼들의 양이온 교환 크로마토그래피(CEX)에 의한 11개의 분리의 크로마토그램들은 도 2에 나타내었다. 양이온 교환 수지의 사용으로 인하여, 산성 아이소폼들이 먼저 용출되고, 이어서 주요 아이소폼, 그리고 다음으로 염기성 아이소폼들이 용출된다.
피크 4(P4, 주 피크)를 정제의 효율을 체크하기 위해 CEX로 재분석하였다. CEX로 분리하기 전과 후에 얻어진 산성, 주요 및 염기성 아이소폼들의 백분율을 도 3과 아래의 표 4에 나타내었고, 주 피크의 정제의 효율을 명확히 나타내었다.
| 샘플 | 산성 폼들 | 주 피크 | 염기성 폼들 |
| 분리 전 | 12.6 % | 58.7 % | 28.7 % |
| 분리 후 | 3.5 % | 93.4 % | 3.1 % |
BIACORE
에 의한
CD16
수용체에 대한 결합의 분석
상이한 전하 아이소폼들을 포함하는, 양이온 교환 크로마토그래피로 분리한 다양한 분획물들의, 수용체 CD16과 결합하는 능력을 시험하였다.
결과들은 도 4에 나타내었고, 산성 폼들(P1 내지 P3)과 피크 7(P7)은 친화성의 손실이 나타났지만, 다른 염기성 폼들(P6, P8)은 친화성의 손실이 나타나지 않았다.
CD16
을 통한 반응기 세포들의 활성화(
CD16
주르카트
시험)
상이한 전하 아이소폼들을 포함하는, 크로마토포커싱 및 양이온 교환 크로마토그래피(CEX)로 분리한 다양한 분획물들의, CD16 수용체(FcγRⅢ)를 통해 반응기 세포들의 반응을 유도하는 능력을 시험하였다.
결과는 도 5(CEX)와 아래의 표 5(크로마토포커싱에 의한 분리)에 나타내었다.
각각의 경우, 주요 아이소폼에 대응하는 분획물이 산성 또는 염기성 아이소폼들을 포함하는 분획물들보다 유의적으로 더 큰 CD16 주르카트 세포들의 활성화를 유도한다는 것이 관찰되었다.
따라서, CD16을 통해 반응기 세포들을 활성화시키는 다양한 전하 아이소폼들의 능력은 유의적으로 변하고, 주요 아이소폼은 CD16을 발현하는 반응기 세포들을 활성화시키는 다른 아이소폼들과 비교할 때 유의적으로 향상된 능력을 갖는다.
항체 조성물이 존재하는 경우 수용체 CD16으로 감염된 주르카트 세포들에 의한 IL-2의 분비량을 측정하는, 상기 기술된 테스트는 CD16을 발현하는 반응기 세포들에 의해 ADCC를 유도하는 항체 조성물의 능력을 나타내는 것을 보여준다(WO2004/024768). 따라서, 아래의 표 4 내지 6에 나타낸 결과들은 정제 전 크로마토포커싱 또는 CEX의 주 피크에 대응하는 정제된 분획물들이 다른 아이소폼들, 및 CD16을 발현하는 반응기 세포들을 통해 ADCC를 유도하는 모든 아이소폼들을 포함하는 전체 조성물과 비교하면 유의적으로 향상된 능력을 갖는다.
| 샘플 | CD16 주르카트 활성 (비교 조성물에 대한 %) |
| 비교 조성물 | 100% |
| 분리 전 전체 조성물 | 80% |
| F36 (염기성 아이소폼) | 62% |
| F39 (주요 아이소폼) | 96% |
| F43 (산성 아이소폼) | 59% |
| F44 (산성 아이소폼) | 71% |
| F48 (산성 아이소폼) | 27% |
| F49 (산성 아이소폼) | 38% |
| F50 (산성 아이소폼) | 17% |
보체
-의존성 세포독성(
CDC
)
다른 전하 아이소폼들을 포함하는, 양이온 교환 크로마토그래피(CEX)로 분리된 다양한 분획물의 보체-의존성 세포독성 반응(CDC)을 유도하는 능력을 측정하였다.
결과는 도 6에 나타내었고, 주 피크(P4)의 활성 손실과 비교하면 산성 폼들(P1: 37%, P2: 52%, 및 P3: 69%)과 염기성 폼들(P5: 45%, P6=K1: 64%, P7: 35%, 및 P8=K2: 49%)에서 강한 활성 손실이 나타난다. 따라서 주요 아이소폼에 대응하는 피크(P4)는 산성 또는 염기성 아이소폼들을 포함하는 분획물들보다 유의적으로 큰 CDC 반응을 유도한다.
질량 분석법에 의한
아이소폼들의
특성화
N-말단 라이신을 갖거나 갖지 않는 중쇄의 백분율을 특성화하기 위해, 크로마토포커싱으로 정제된 분획물들과 CEX로 정제된 분획물들을 LC-MS로 분석하였다.
분리 전 주 피크에 대응하는 크로마토포커싱으로 정제된 분획물들과 CEX로 정제된 분획물들에 대해, 95%를 초과하는 중쇄들이 어떠한 C-말단 라이신도 포함하지 않는 것으로 분석되었다.
결론
상기에 나타낸 결과는 이온 교환 크로마토그래피(CEX)에 의해 또는 크로마토포커싱에 의해 분리된 주 피크에 대응하는 항체 조성물의 전하 아이소폼들이 수용체 FcγRⅢ(CD16)을 통해 및 또한 보체를 통해 반응기 세포들을 활성화하는 동일한 항체 조성물의 산성 또는 염기성 아이소폼들보다 유의적으로 큰 능력을 가지는 것을 나타낸다.
따라서, 이 주 피크에 대응하는 정제된 분획물들의 사용은 ADCC 또는 CDC 반응의 중요한 역할을 하는 모노클론 항체들에 의한 치료되는 병리학의 범위 내, 예를 들어 특히 동종 면역을 예방하거나, 또는 암, 감염 질병, 자가면역 질병의 치료 내에서 CD16(ADCC, 사이토카인의 분비)를 통해 반응기 특성을 더 증가시킨다.
참고 문헌
Ahmad et al. FASEB J. 1996. 10:258-266.
Albrecht MT, et al. INFECTION AND IMMUNITY, Nov. 2007, p. 5425-5433.
Almagro et al. Frontiers in Bioscience 13, 1619-1633, January 1, 2008.
Antes B, et al. J Chromatogr B Analyt Technol Biomed Life Sci. 2007 Jun 1;852(1-2):250-6.
Beliard R et al. Br J Haematol. 2008 Apr;141(1):109-19.
Bruggermann et al., Year in Immuno., 7:33 (1993);
Cardarelli et al. Clin Cancer Res 2009 April 28;15:3376-3383,
Cardarelli et al. Cancer Immunol Immunother. 2010. 59. 257-265,
Cartron G, et al. Blood. 2002 Feb 1;99(3):754-8.
Casadevall A. et al. Emerg Infect Dis. 2002 Aug;8(8):833-41;
Chan AC, et al. Nat Rev Immunol. 2010 May;10(5):301-16.
Chevreux G et al. Anal Biochem. 2011 Aug 15;415(2):212-4;
Cibelli et al., 1998 Science, 280: 1256-1258
Dall'Acqua et al. 2002, J lmmunol.;169:5171 -80.
Dall'Acqua et al. 2006, J. Biol. Chem.;281:23514-24. (a)
Dall'Acqua et al. J Immunol 2006; 177:1129-1138. (b)
Duchosal et al. Nature 355:258 (1992)
Edelman, G.M. et al., Proc. Natl. Acad. USA, 63, 78-85 (1969).
Edwards JC, et al. Nat Rev Immunol. 2006 May;6(5):394-403.
EP 1 176 195 A1,
EP 1 308 456,
EP 1 829 961,
Farnan D, Moreno GT. Anal Chem. 2009 Nov 1;81(21):8846-57.
Fisher R, et al. Vaccine 21 (2003) 820-825.
Forthal et al., J Immunol 2010;185;6876-6882,
Gandhi S, et al. Pharm Res. 2012 Jan;29(1):209-24.
Gene Targeting, A Practical Approach, IRL Press at Oxford University Press (1993)
Gordon et al., 1980 Proc Natl Acad Sci U S A.;77:7380-4
Herbst R. et al. J Pharmacol Exp Ther. 2010 Oct;335(1):213-22,
Hinton et al. 2004, J Biol Chem.;279:6213-6.
Hoogenboom et al., J. Mol. Biol., 227:381 (1991);
Ianello A, et al. Cancer and Metastasis Reviews 24: 487-499, 2005.
Idusogie EE et al. J Immunol. 2001; 166:2571-5.
Imai-Nishiya et al., BMC Biotechnology 2007, 7:84;
Jakobovits et al., Proc. Natl. Acad. Sci. USA. 90:2551 (1993) (a);
Jakobovits et al., Nature, 362:255-258 (1993) (b);
Jones et al. Nature, 321: 522-525, 1986;
Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991).
Kanda Y et al., Journal of Biotechnology 130 (2007) 300-310,
Khawli LA, et al. MAbs. 2010 Nov-Dec;2(6):613-24.
Lazar, G. A., et al. Proc Natl Acad Sci U S A. 103(11): 4005-10.
Ma JK, et al. Nat Rev Genet. 2003 Oct;4(10):794-805.
Manipulating the Mouse Embryo, A Laboratory Manual, Second edition, Cold Spring Harbor Laboratory Press (1994);
Marks et al., J. Mol. Biol., 222:581- 5 597 (1991);
McAtee CP et al. Curr Protoc Protein Sci. 2012 Aug;Chapter 8:Unit 8.10,
Miao C, et al. Journal of General Virology (2009), 90, 1119-1123.
Moore GL. et al. mAbs 2:2, 181-189; March/April, 2010;
Mori K, et al. Biotechnol Bioeng. 2004 Dec 30;88(7):901-8,
Olivier S. et al. MAbs. 2010 Jul-Aug; 2(4): 405-415,
Presta LG. Adv Drug Deliv Rev. 2006 Aug 7;58(5-6):640-56.
Rea JC, et al. J Pharm Biomed Anal. 2011 Jan 25;54(2):317-23.
Rea Jennifer C. Innovations in Biotechnology. InTech. February 17, 2012. Chapter 19.
Riechmann et al. Nature, 332: 323-327, 1988,
Ryan et al., 1997 Science; 278: 873 - 876;
Satoh M, et al. Expert Opin Biol Ther. 2006 Nov;6(11):1161-73.
Schillberg S, et al. Vaccine 23 (2005) 1764-1769.
Shields RL, et al. J Biol Chem. 2001 Mar 2;276(9):6591-604.
Stoger E, et al. Molecular Breeding 9: 149-158, 2002.
Suzuki et al. Clin Cancer Res 2007 March 15;13:1875-1882,
Umana et al. Nat Biotechnol. 1999 Feb;17(2):176-80,
US 5,591,669,
US 5,598,369,
US 5,545,806,
US 5,545,807,
US 6,150,584
Vaughan et al. Nature Biotech 14:309 (1996)
Velders MP et al., Br J Cancer. 1998 Aug;78(4):478-83.
Verhoeyn et al. BioEssays, 8: 74, 1988,
Verhoeyen et al. Science, 239: 1534-1536, 1988;
Vlasak J, Ionescu R. Curr Pharm Biotechnol. 2008 Dec;9(6):468-81.
Wallace PK et al., J Leukoc Biol. 1994 Jun;55(6):816-26.
Weiner LM, et al. Nat Rev Immunol. 2010 May;10(5):317-27.
WO9004036A1,
WO9517085A1,
WO9951642,
WO0026357A2,
WO00/42072,
WO01/77181,
WO01/26455A1,
WO02/060919A2,
WO2004/024768,
WO2004/024866,
WO2004/029207,
WO2004/050847A2,
WO2004/063351
WO2004/074455
WO2005/033281A2,
WO2007/048077A2
WO2007/106078A2,
WO2010/045193,
WO2010/106180A2,
WO2011/009623,
WO2012/041768A1
Wright A, Morrison SL. J Exp Med. 1994 Sep 1;180(3):1087-96,
Yamane-Ohnuki N. et al. Biotechnol Bioeng. 2004 Sep 5;87(5):614-22,
Zeitlin L. et al. Microbes Infect. 2000 May;2(6):701-8,
Zhang T. et al. J Chromatogr A. 2011 Aug 5;1218(31):5079-86.
Claims (16)
- 모노클론 항체 조성물로서,
a) 세포 클론, 비-인간 유전자 변형 동물 또는 유전자 변형 식물로부터 모노클론 항체 조성물을 제조하는 단계,
b) 단계 a)에서 얻어진 조성물을 크로마토그래피에 의해 분획하는 단계, 및
c) 단계 b)에서 얻어지고 크로마토그램의 주 피크에 대응하는, 하나 또는 수 개의 크로마토그래피 분획물을 조합하는 단계를 포함하는 방법에 의해 얻을 수 있으며, 상기 방법으로 얻어진 모노클론 항체 조성물은 상기 주 피크가 풍부하고, 상기 주 피크는 단계 c)에서 얻은 조성물의 크로마토그램 중 적어도 85%를 나타내는, 의약품으로서 사용하기 위한 모노클론 항체 조성물. - 제1항에 있어서, 상기 단계 b)의 분획은 이온 교환 크로마토그래피, 크로마토포커싱, 또는 소수성 상호작용 크로마토그래피에 의하여 수행되는 것을 특징으로 하는, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제2항에 있어서, 이온 교환 크로마토그래피는 다음의 용출 수단 중 하나를 이용하는 것을 특징으로 하는, 의약품으로서 사용하기 위한 모노클론 항체 조성물:
·이온 힘 구배; 및/또는
· pH 구배; 또는
· 치환 분자. - 제1항 내지 제3항 중 어느 한 항에 있어서, 조성물에 존재하는 항체의 중쇄 중 적어도 95%는 어떠한 C-말단 라이신 잔기도 포함하지 않는 것을 특징으로 하는, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 조성물에 존재하는 항체의 중쇄 중 적어도 95%는 어떠한 C-말단 라이신 잔기도 포함하지 않는, 약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제1항 내지 제5항 중 어느 한 항에 있어서, 상기 항체는 건강한 공여 세포에 존재하는 비-유비쿼터스 항원(non-ubiquitous antigen), 암 세포의 항원, 병원체에 감염된 세포의 항원, 또는 면역 세포의 항원에 대한 것인, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제1항 내지 제6항 중 어느 한 항에 있어서, 상기 항체는 항-Rh(D) 항체이고 상기 조성물은 Rh 마이너스(-) 개체에서 동종 면역을 예방하기 위한 것인, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제1항 내지 제6항 중 어느 한 항에 있어서, 상기 항체는 암 세포의 항원을 향하고 상기 조성물은 암을 치료하기 위한 것인, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제1항 내지 제6항 중 어느 한 항에 있어서, 상기 항체는 병원체에 감염된 세포의 항원을 향하고 상기 조성물은 상기 병원체에 의한 감염을 치료하기 위한 것인, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제1항 내지 제6항 중 어느 한 항에 있어서, 상기 항체는 면역 세포의 항원을 향하고 상기 조성물은 자가면역 질환을 치료하기 위한 것인, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제1항 내지 제10항 중 어느 한 항에 있어서, 상기 항체는 FcγRⅢ 수용체와의 결합 및 FcγRⅢ 수용체를 통한 반응기 특징을 증가시키는 Fc 단편의 변형을 포함하는, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제11항에 있어서, 상기 항체는 Fc 단편의 특정 아미노산 잔기에 적어도 하나의 돌연변이를 포함하는, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제11항 또는 제12항에 있어서, 65% 이하의 푸코스 함량을 포함하는, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- 제1항 내지 제10항 중 어느 한 항에 있어서, 상기 항체는 단백질 C1q와의 결합 및 보체를 통한 반응기 특징들을 증가시키는 Fc 단편의 변형을 포함하는, 의약품으로서 사용하기 위한 모노클론 항체 조성물.
- FcγRⅢ(CD16) 수용체를 발현하는 면역계의 반응기 세포들에 의하여 상기 항원을 발현하는 타겟 세포들의 항체 의존적 세포성 세포독성(ADCC)을 유도하기 위한, 정해진 항체에 대한 모노클론 항체 조성물의 능력을 증가시키기 위한 크로마토그래피 분획 단계의 사용.
- 보체에 의하여 항원을 발현하는 타겟 세포들의 보체 의존적 세포독성(CDC)을 유도하기 위한, 정해진 항체에 대한 모노클론 항체 조성물의 능력을 증가시키기 위한 크로마토그래피 분획 단계의 사용.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR1352360A FR3003171B1 (fr) | 2013-03-15 | 2013-03-15 | Nouveaux medicaments comprenant une composition d'anticorps enrichie en isoforme de charge majoritaire |
| FR1352360 | 2013-03-15 | ||
| PCT/EP2014/055179 WO2014140322A1 (fr) | 2013-03-15 | 2014-03-14 | Nouveaux médicaments comprenant une composition d'anticorps enrichie en isoforme de charge majoritaire |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20150132522A true KR20150132522A (ko) | 2015-11-25 |
Family
ID=48771615
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020157029822A KR20150132522A (ko) | 2013-03-15 | 2014-03-14 | 주요 전하 아이소폼이 풍부한 항체 조성물을 포함하는 신규 의약품 |
Country Status (11)
| Country | Link |
|---|---|
| US (1) | US20160046722A1 (ko) |
| EP (1) | EP2968533A1 (ko) |
| JP (1) | JP2016512216A (ko) |
| KR (1) | KR20150132522A (ko) |
| CN (1) | CN105163758B (ko) |
| AU (1) | AU2014230134A1 (ko) |
| BR (1) | BR112015023209A8 (ko) |
| CA (1) | CA2907358A1 (ko) |
| FR (1) | FR3003171B1 (ko) |
| MX (1) | MX2015012812A (ko) |
| WO (1) | WO2014140322A1 (ko) |
Families Citing this family (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US10323081B2 (en) * | 2011-07-06 | 2019-06-18 | Genmag A/S | Modulation of complement-dependent cytotoxicity through modifications of the C-terminus of antibody heavy chains |
| WO2017192093A1 (en) | 2016-05-04 | 2017-11-09 | Delaval Holding Ab | A cartridge for a teatcup, and a teatcup |
| FR3053688A1 (fr) | 2016-07-06 | 2018-01-12 | Laboratoire Francais Du Fractionnement Et Des Biotechnologies | Mutants fc a activite fonctionnelle amelioree |
| EP3498293A1 (en) | 2017-12-15 | 2019-06-19 | Institut National De La Sante Et De La Recherche Medicale (Inserm) | Treatment of monogenic diseases with an anti-cd45rc antibody |
| EP3626265A1 (en) | 2018-09-21 | 2020-03-25 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Anti-human cd45rc antibodies and uses thereof |
| AU2020211728A1 (en) | 2019-01-23 | 2021-08-12 | Encefa | CD31 competitors and uses thereof |
| CN113874720A (zh) * | 2019-05-23 | 2021-12-31 | 瑞泽恩制药公司 | 抗体结构域特异性电荷变异体的表征分析 |
| CN114829407A (zh) * | 2019-09-23 | 2022-07-29 | 南开大学 | 利用哺乳动物展示筛选FcγR特异性结合Fc |
| WO2023041717A1 (en) | 2021-09-16 | 2023-03-23 | Aboleris Pharma | Anti-human cd45rc binding domains and uses thereof |
| CN114236010A (zh) * | 2021-12-18 | 2022-03-25 | 苏州莱奥生物技术有限公司 | 一种生物活性药物的药代动力学分析方法 |
Family Cites Families (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| HUP0104865A3 (en) * | 1999-01-15 | 2004-07-28 | Genentech Inc | Polypeptide variants with altered effector function |
| FR2807767B1 (fr) * | 2000-04-12 | 2005-01-14 | Lab Francais Du Fractionnement | Anticorps monoclonaux anti-d |
| FR2844513B1 (fr) * | 2002-09-13 | 2007-08-03 | Lab Francais Du Fractionnement | Anticorps pour adcc et induisant la production de cytokines. |
| BRPI0314814C1 (pt) * | 2002-09-27 | 2021-07-27 | Xencor Inc | anticorpo compreendendo uma variante de fc |
| SI1558648T1 (sl) * | 2002-10-17 | 2012-05-31 | Genmab As | Človeška monoklonalna protitelesa proti CD |
| EP2368578A1 (en) * | 2003-01-09 | 2011-09-28 | Macrogenics, Inc. | Identification and engineering of antibodies with variant Fc regions and methods of using same |
| FR2861078B1 (fr) * | 2003-10-16 | 2007-09-21 | Lab Francais Du Fractionnement | NOUVELLES IgG3 UTILES POUR STIMULER LA PHAGOCYTOSE |
| SG10201912554TA (en) * | 2005-03-23 | 2020-02-27 | Genmab As | Antibodies against cd38 for treatment of multiple myeloma |
| US10323081B2 (en) * | 2011-07-06 | 2019-06-18 | Genmag A/S | Modulation of complement-dependent cytotoxicity through modifications of the C-terminus of antibody heavy chains |
-
2013
- 2013-03-15 FR FR1352360A patent/FR3003171B1/fr not_active Expired - Fee Related
-
2014
- 2014-03-14 AU AU2014230134A patent/AU2014230134A1/en not_active Abandoned
- 2014-03-14 MX MX2015012812A patent/MX2015012812A/es unknown
- 2014-03-14 BR BR112015023209A patent/BR112015023209A8/pt not_active Application Discontinuation
- 2014-03-14 CA CA2907358A patent/CA2907358A1/fr not_active Abandoned
- 2014-03-14 WO PCT/EP2014/055179 patent/WO2014140322A1/fr active Application Filing
- 2014-03-14 EP EP14709977.4A patent/EP2968533A1/fr not_active Withdrawn
- 2014-03-14 JP JP2015562236A patent/JP2016512216A/ja active Pending
- 2014-03-14 KR KR1020157029822A patent/KR20150132522A/ko not_active Application Discontinuation
- 2014-03-14 CN CN201480024699.3A patent/CN105163758B/zh not_active Expired - Fee Related
- 2014-03-14 US US14/776,725 patent/US20160046722A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| MX2015012812A (es) | 2016-05-09 |
| CA2907358A1 (fr) | 2014-09-18 |
| BR112015023209A8 (pt) | 2018-01-23 |
| JP2016512216A (ja) | 2016-04-25 |
| CN105163758A (zh) | 2015-12-16 |
| EP2968533A1 (fr) | 2016-01-20 |
| AU2014230134A1 (en) | 2015-10-29 |
| BR112015023209A2 (pt) | 2017-07-18 |
| US20160046722A1 (en) | 2016-02-18 |
| CN105163758B (zh) | 2017-11-17 |
| FR3003171A1 (fr) | 2014-09-19 |
| FR3003171B1 (fr) | 2015-04-10 |
| WO2014140322A1 (fr) | 2014-09-18 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR20150132522A (ko) | 주요 전하 아이소폼이 풍부한 항체 조성물을 포함하는 신규 의약품 | |
| TWI732176B (zh) | 全人源的抗b細胞成熟抗原(bcma)單鏈抗體及其應用 | |
| Jefferis | Recombinant antibody therapeutics: the impact of glycosylation on mechanisms of action | |
| CN111712258A (zh) | 抗trem2抗体和相关方法 | |
| JP2021534196A (ja) | 抗tigit抗体 | |
| US20200277385A1 (en) | Anti-CD303 Monoclonal Antibodies | |
| MX2013010210A (es) | Lineas de celulas de bajo nivel de fucosa y usos de las mismas. | |
| CN115942976A (zh) | 掩蔽的il-2细胞因子和其切割产物 | |
| CN115734806A (zh) | 掩蔽的il-12细胞因子和其切割产物 | |
| US8470320B2 (en) | Humanised antibodies with anti-tumour activity | |
| JP2022514734A (ja) | 抗trem2抗体の使用方法 | |
| Wang et al. | The interplay of protein engineering and glycoengineering to fine‐tune antibody glycosylation and its impact on effector functions | |
| KR20210047316A (ko) | 비푸코실화된 항체 및 그의 제조법 | |
| AU2010314798B2 (en) | Treatment of cancer involving mutated KRAS or BRAF genes | |
| EP3178848A1 (en) | Type ii anti-cd20 antibody for reducing formation of anti-drug antibodies | |
| JP2016504337A (ja) | イオン交換クロマトグラフィーを使用して高マンノースグリコフォームのレベルを制御する方法 | |
| KR102651432B1 (ko) | 비푸코실화된 항체 및 그의 제조법 | |
| US20190002567A1 (en) | New use of an anti-cd303 transmembrane protein antibody | |
| TW201309728A (zh) | 具高抗體介導細胞毒殺作用之抗cd20抗體在治療瓦氏症中的用途 | |
| RU2778053C2 (ru) | Антитела к lag3 | |
| EP2660324A1 (en) | Production of secreted therapeutic antibodies in microalgae | |
| CN117715940A (zh) | 抗trem-1抗体 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WITN | Application deemed withdrawn, e.g. because no request for examination was filed or no examination fee was paid |