JP6970974B2 - 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ - Google Patents
様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ Download PDFInfo
- Publication number
- JP6970974B2 JP6970974B2 JP2018511114A JP2018511114A JP6970974B2 JP 6970974 B2 JP6970974 B2 JP 6970974B2 JP 2018511114 A JP2018511114 A JP 2018511114A JP 2018511114 A JP2018511114 A JP 2018511114A JP 6970974 B2 JP6970974 B2 JP 6970974B2
- Authority
- JP
- Japan
- Prior art keywords
- cancer
- peptide
- cell
- cells
- tumor
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 108090000765 processed proteins & peptides Proteins 0.000 title claims description 320
- 206010028980 Neoplasm Diseases 0.000 title claims description 315
- 238000009169 immunotherapy Methods 0.000 title description 32
- 210000004027 cell Anatomy 0.000 claims description 179
- 201000011510 cancer Diseases 0.000 claims description 132
- 108090000623 proteins and genes Proteins 0.000 claims description 110
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 claims description 90
- 108091008874 T cell receptors Proteins 0.000 claims description 89
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 80
- 238000011282 treatment Methods 0.000 claims description 74
- 239000000427 antigen Substances 0.000 claims description 68
- 108091007433 antigens Proteins 0.000 claims description 66
- 102000036639 antigens Human genes 0.000 claims description 66
- 102000004169 proteins and genes Human genes 0.000 claims description 56
- 208000005718 Stomach Neoplasms Diseases 0.000 claims description 52
- 208000000236 Prostatic Neoplasms Diseases 0.000 claims description 51
- 206010060862 Prostate cancer Diseases 0.000 claims description 49
- 206010017758 gastric cancer Diseases 0.000 claims description 49
- 201000011549 stomach cancer Diseases 0.000 claims description 48
- 206010061535 Ovarian neoplasm Diseases 0.000 claims description 43
- 238000000034 method Methods 0.000 claims description 42
- 206010033128 Ovarian cancer Diseases 0.000 claims description 41
- 206010061902 Pancreatic neoplasm Diseases 0.000 claims description 35
- 208000002154 non-small cell lung carcinoma Diseases 0.000 claims description 34
- 208000029729 tumor suppressor gene on chromosome 11 Diseases 0.000 claims description 33
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 claims description 31
- 201000002528 pancreatic cancer Diseases 0.000 claims description 31
- 208000008443 pancreatic carcinoma Diseases 0.000 claims description 31
- 150000007523 nucleic acids Chemical class 0.000 claims description 30
- 210000004443 dendritic cell Anatomy 0.000 claims description 29
- 206010005003 Bladder cancer Diseases 0.000 claims description 28
- 230000002018 overexpression Effects 0.000 claims description 28
- 206010041067 Small cell lung cancer Diseases 0.000 claims description 27
- 208000007097 Urinary Bladder Neoplasms Diseases 0.000 claims description 27
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 27
- 201000001441 melanoma Diseases 0.000 claims description 27
- 208000000587 small cell lung carcinoma Diseases 0.000 claims description 27
- 102000039446 nucleic acids Human genes 0.000 claims description 26
- 108020004707 nucleic acids Proteins 0.000 claims description 26
- 201000005112 urinary bladder cancer Diseases 0.000 claims description 26
- 238000000338 in vitro Methods 0.000 claims description 23
- 230000006870 function Effects 0.000 claims description 22
- 208000031261 Acute myeloid leukaemia Diseases 0.000 claims description 21
- 208000000461 Esophageal Neoplasms Diseases 0.000 claims description 20
- 206010030155 Oesophageal carcinoma Diseases 0.000 claims description 20
- 201000004101 esophageal cancer Diseases 0.000 claims description 20
- 208000033776 Myeloid Acute Leukemia Diseases 0.000 claims description 19
- 208000015914 Non-Hodgkin lymphomas Diseases 0.000 claims description 19
- 210000000612 antigen-presenting cell Anatomy 0.000 claims description 18
- 208000006990 cholangiocarcinoma Diseases 0.000 claims description 18
- 208000002495 Uterine Neoplasms Diseases 0.000 claims description 17
- 239000003814 drug Substances 0.000 claims description 17
- 150000003839 salts Chemical class 0.000 claims description 17
- 229940079593 drug Drugs 0.000 claims description 16
- 206010046766 uterine cancer Diseases 0.000 claims description 16
- 206010004593 Bile duct cancer Diseases 0.000 claims description 15
- 208000026900 bile duct neoplasm Diseases 0.000 claims description 15
- 239000003795 chemical substances by application Substances 0.000 claims description 14
- 208000024313 Testicular Neoplasms Diseases 0.000 claims description 13
- 210000000941 bile Anatomy 0.000 claims description 13
- 239000003446 ligand Substances 0.000 claims description 13
- 206010057644 Testis cancer Diseases 0.000 claims description 12
- 210000003236 esophagogastric junction Anatomy 0.000 claims description 12
- 239000013604 expression vector Substances 0.000 claims description 12
- 201000003120 testicular cancer Diseases 0.000 claims description 12
- 108010075704 HLA-A Antigens Proteins 0.000 claims description 11
- 239000012636 effector Substances 0.000 claims description 10
- 102000008949 Histocompatibility Antigens Class I Human genes 0.000 claims description 9
- 238000003745 diagnosis Methods 0.000 claims description 9
- 238000004519 manufacturing process Methods 0.000 claims description 9
- 239000012528 membrane Substances 0.000 claims description 7
- 230000004927 fusion Effects 0.000 claims description 6
- 108010088652 Histocompatibility Antigens Class I Proteins 0.000 claims description 4
- 239000003053 toxin Substances 0.000 claims description 4
- 231100000765 toxin Toxicity 0.000 claims description 4
- 108700012359 toxins Proteins 0.000 claims description 4
- 210000000013 bile duct Anatomy 0.000 claims description 3
- 238000012258 culturing Methods 0.000 claims description 3
- 230000003308 immunostimulating effect Effects 0.000 claims description 3
- 239000008194 pharmaceutical composition Substances 0.000 claims description 3
- 230000003213 activating effect Effects 0.000 claims description 2
- 239000001963 growth medium Substances 0.000 claims description 2
- 108091023037 Aptamer Proteins 0.000 claims 1
- 102000011786 HLA-A Antigens Human genes 0.000 claims 1
- 239000004480 active ingredient Substances 0.000 claims 1
- 239000003937 drug carrier Substances 0.000 claims 1
- 230000003278 mimic effect Effects 0.000 claims 1
- 102000004196 processed proteins & peptides Human genes 0.000 description 150
- 230000014509 gene expression Effects 0.000 description 117
- 206010006187 Breast cancer Diseases 0.000 description 70
- 208000026310 Breast neoplasm Diseases 0.000 description 70
- 210000001519 tissue Anatomy 0.000 description 61
- 235000018102 proteins Nutrition 0.000 description 55
- 235000001014 amino acid Nutrition 0.000 description 54
- 206010009944 Colon cancer Diseases 0.000 description 53
- 229940024606 amino acid Drugs 0.000 description 53
- 150000001413 amino acids Chemical class 0.000 description 53
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 51
- 210000004881 tumor cell Anatomy 0.000 description 47
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 45
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 45
- 230000004083 survival effect Effects 0.000 description 40
- 230000027455 binding Effects 0.000 description 36
- 238000013459 approach Methods 0.000 description 33
- 208000010839 B-cell chronic lymphocytic leukemia Diseases 0.000 description 31
- 230000001965 increasing effect Effects 0.000 description 31
- 229920001184 polypeptide Polymers 0.000 description 31
- 206010073071 hepatocellular carcinoma Diseases 0.000 description 29
- 231100000844 hepatocellular carcinoma Toxicity 0.000 description 29
- 208000000102 Squamous Cell Carcinoma of Head and Neck Diseases 0.000 description 28
- 229960005486 vaccine Drugs 0.000 description 28
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 27
- 208000031422 Lymphocytic Chronic B-Cell Leukemia Diseases 0.000 description 25
- 206010027476 Metastases Diseases 0.000 description 25
- 208000032852 chronic lymphocytic leukemia Diseases 0.000 description 25
- 208000005017 glioblastoma Diseases 0.000 description 25
- 208000006265 Renal cell carcinoma Diseases 0.000 description 24
- 230000009401 metastasis Effects 0.000 description 24
- 201000010099 disease Diseases 0.000 description 23
- 230000018109 developmental process Effects 0.000 description 22
- 239000012071 phase Substances 0.000 description 22
- 238000006467 substitution reaction Methods 0.000 description 22
- 238000002255 vaccination Methods 0.000 description 22
- 238000011161 development Methods 0.000 description 21
- 230000028993 immune response Effects 0.000 description 21
- 208000020816 lung neoplasm Diseases 0.000 description 21
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 20
- 230000000694 effects Effects 0.000 description 20
- 230000008595 infiltration Effects 0.000 description 19
- 238000001764 infiltration Methods 0.000 description 19
- 230000005867 T cell response Effects 0.000 description 18
- 238000002512 chemotherapy Methods 0.000 description 18
- 201000000459 head and neck squamous cell carcinoma Diseases 0.000 description 18
- 201000005202 lung cancer Diseases 0.000 description 18
- 230000035772 mutation Effects 0.000 description 18
- 238000002560 therapeutic procedure Methods 0.000 description 18
- 206010008342 Cervix carcinoma Diseases 0.000 description 17
- 208000032612 Glial tumor Diseases 0.000 description 17
- 206010018338 Glioma Diseases 0.000 description 17
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 description 17
- 125000000539 amino acid group Chemical group 0.000 description 17
- 239000000090 biomarker Substances 0.000 description 17
- 201000010881 cervical cancer Diseases 0.000 description 17
- -1 9-fluorenylmethyloxycarbonyl (Fmoc) group Chemical group 0.000 description 16
- 102000043131 MHC class II family Human genes 0.000 description 16
- 108091054438 MHC class II family Proteins 0.000 description 16
- 108020004414 DNA Proteins 0.000 description 15
- 101000607318 Homo sapiens UL16-binding protein 3 Proteins 0.000 description 15
- 108091054437 MHC class I family Proteins 0.000 description 15
- 102100040011 UL16-binding protein 3 Human genes 0.000 description 15
- 229960005386 ipilimumab Drugs 0.000 description 15
- 230000037361 pathway Effects 0.000 description 15
- 238000010837 poor prognosis Methods 0.000 description 15
- 208000015347 renal cell adenocarcinoma Diseases 0.000 description 15
- 239000012634 fragment Substances 0.000 description 14
- 230000001225 therapeutic effect Effects 0.000 description 14
- 239000000047 product Substances 0.000 description 13
- 238000004393 prognosis Methods 0.000 description 13
- 102000005962 receptors Human genes 0.000 description 13
- 108020003175 receptors Proteins 0.000 description 13
- 230000004044 response Effects 0.000 description 13
- 230000011664 signaling Effects 0.000 description 13
- 230000008685 targeting Effects 0.000 description 13
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 12
- 102100038449 Claudin-6 Human genes 0.000 description 12
- 102100035300 Cystine/glutamate transporter Human genes 0.000 description 12
- 101000882898 Homo sapiens Claudin-6 Proteins 0.000 description 12
- 108091006241 SLC7A11 Proteins 0.000 description 12
- 102000001742 Tumor Suppressor Proteins Human genes 0.000 description 12
- 108010040002 Tumor Suppressor Proteins Proteins 0.000 description 12
- 231100000504 carcinogenesis Toxicity 0.000 description 12
- 238000001727 in vivo Methods 0.000 description 12
- 208000032839 leukemia Diseases 0.000 description 12
- 229940023041 peptide vaccine Drugs 0.000 description 12
- 230000035755 proliferation Effects 0.000 description 12
- 239000000523 sample Substances 0.000 description 12
- 230000015572 biosynthetic process Effects 0.000 description 11
- 230000003247 decreasing effect Effects 0.000 description 11
- 230000004069 differentiation Effects 0.000 description 11
- 239000013598 vector Substances 0.000 description 11
- 108700028369 Alleles Proteins 0.000 description 10
- 208000005623 Carcinogenesis Diseases 0.000 description 10
- 102100025064 Cellular tumor antigen p53 Human genes 0.000 description 10
- 108010019670 Chimeric Antigen Receptors Proteins 0.000 description 10
- 102100028972 HLA class I histocompatibility antigen, A alpha chain Human genes 0.000 description 10
- 102000043129 MHC class I family Human genes 0.000 description 10
- 108010038807 Oligopeptides Proteins 0.000 description 10
- 102000015636 Oligopeptides Human genes 0.000 description 10
- 108050003627 Wnt Proteins 0.000 description 10
- 230000036952 cancer formation Effects 0.000 description 10
- 208000029742 colonic neoplasm Diseases 0.000 description 10
- 230000002163 immunogen Effects 0.000 description 10
- 230000001394 metastastic effect Effects 0.000 description 10
- 206010061289 metastatic neoplasm Diseases 0.000 description 10
- 230000004048 modification Effects 0.000 description 10
- 238000012986 modification Methods 0.000 description 10
- 102100026097 Claudin-9 Human genes 0.000 description 9
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 9
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 9
- 101000721661 Homo sapiens Cellular tumor antigen p53 Proteins 0.000 description 9
- 101000912661 Homo sapiens Claudin-9 Proteins 0.000 description 9
- 102000013814 Wnt Human genes 0.000 description 9
- 230000005975 antitumor immune response Effects 0.000 description 9
- 230000006907 apoptotic process Effects 0.000 description 9
- 230000000711 cancerogenic effect Effects 0.000 description 9
- 230000004663 cell proliferation Effects 0.000 description 9
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 9
- 230000003828 downregulation Effects 0.000 description 9
- 230000005764 inhibitory process Effects 0.000 description 9
- 210000002540 macrophage Anatomy 0.000 description 9
- 239000003550 marker Substances 0.000 description 9
- 230000011987 methylation Effects 0.000 description 9
- 238000007069 methylation reaction Methods 0.000 description 9
- 102000040430 polynucleotide Human genes 0.000 description 9
- 108091033319 polynucleotide Proteins 0.000 description 9
- 239000002157 polynucleotide Substances 0.000 description 9
- 230000019491 signal transduction Effects 0.000 description 9
- 102100024090 Aldo-keto reductase family 1 member C3 Human genes 0.000 description 8
- 108060000903 Beta-catenin Proteins 0.000 description 8
- 102000015735 Beta-catenin Human genes 0.000 description 8
- 101001006776 Homo sapiens Kinesin-like protein KIFC1 Proteins 0.000 description 8
- 208000022361 Human papillomavirus infectious disease Diseases 0.000 description 8
- 102100027942 Kinesin-like protein KIFC1 Human genes 0.000 description 8
- 206010061309 Neoplasm progression Diseases 0.000 description 8
- 108010065942 Prostaglandin-F synthase Proteins 0.000 description 8
- DTQVDTLACAAQTR-UHFFFAOYSA-N Trifluoroacetic acid Chemical compound OC(=O)C(F)(F)F DTQVDTLACAAQTR-UHFFFAOYSA-N 0.000 description 8
- 238000004458 analytical method Methods 0.000 description 8
- 231100000315 carcinogenic Toxicity 0.000 description 8
- 230000000875 corresponding effect Effects 0.000 description 8
- 230000001024 immunotherapeutic effect Effects 0.000 description 8
- 238000004895 liquid chromatography mass spectrometry Methods 0.000 description 8
- 208000014018 liver neoplasm Diseases 0.000 description 8
- 230000001404 mediated effect Effects 0.000 description 8
- 201000002740 oral squamous cell carcinoma Diseases 0.000 description 8
- 210000001550 testis Anatomy 0.000 description 8
- 238000012546 transfer Methods 0.000 description 8
- 230000005751 tumor progression Effects 0.000 description 8
- 201000009030 Carcinoma Diseases 0.000 description 7
- 102000004127 Cytokines Human genes 0.000 description 7
- 108090000695 Cytokines Proteins 0.000 description 7
- 102100034523 Histone H4 Human genes 0.000 description 7
- 101001067880 Homo sapiens Histone H4 Proteins 0.000 description 7
- 101001130243 Homo sapiens RAD51-associated protein 1 Proteins 0.000 description 7
- 102100037850 Interferon gamma Human genes 0.000 description 7
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 7
- 208000007433 Lymphatic Metastasis Diseases 0.000 description 7
- 206010025323 Lymphomas Diseases 0.000 description 7
- 241000699666 Mus <mouse, genus> Species 0.000 description 7
- 241000699670 Mus sp. Species 0.000 description 7
- 108091028043 Nucleic acid sequence Proteins 0.000 description 7
- 102100031535 RAD51-associated protein 1 Human genes 0.000 description 7
- 102100027974 Semaphorin-3A Human genes 0.000 description 7
- 108010090319 Semaphorin-3A Proteins 0.000 description 7
- 230000004913 activation Effects 0.000 description 7
- 210000000481 breast Anatomy 0.000 description 7
- 239000011575 calcium Substances 0.000 description 7
- 239000003153 chemical reaction reagent Substances 0.000 description 7
- 230000002596 correlated effect Effects 0.000 description 7
- 229940029030 dendritic cell vaccine Drugs 0.000 description 7
- 210000002443 helper t lymphocyte Anatomy 0.000 description 7
- 210000002865 immune cell Anatomy 0.000 description 7
- 210000000987 immune system Anatomy 0.000 description 7
- 230000003993 interaction Effects 0.000 description 7
- 210000004072 lung Anatomy 0.000 description 7
- 238000013508 migration Methods 0.000 description 7
- 239000000203 mixture Substances 0.000 description 7
- 230000036961 partial effect Effects 0.000 description 7
- 238000012545 processing Methods 0.000 description 7
- 230000001629 suppression Effects 0.000 description 7
- 238000013518 transcription Methods 0.000 description 7
- 230000035897 transcription Effects 0.000 description 7
- 210000003171 tumor-infiltrating lymphocyte Anatomy 0.000 description 7
- 102100028080 ATPase family AAA domain-containing protein 5 Human genes 0.000 description 6
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 6
- 102100032187 Androgen receptor Human genes 0.000 description 6
- 229940045513 CTLA4 antagonist Drugs 0.000 description 6
- 108010022366 Carcinoembryonic Antigen Proteins 0.000 description 6
- 102100025475 Carcinoembryonic antigen-related cell adhesion molecule 5 Human genes 0.000 description 6
- 102100038423 Claudin-3 Human genes 0.000 description 6
- 108091026890 Coding region Proteins 0.000 description 6
- 102100032952 Condensin complex subunit 3 Human genes 0.000 description 6
- RTZKZFJDLAIYFH-UHFFFAOYSA-N Diethyl ether Chemical compound CCOCC RTZKZFJDLAIYFH-UHFFFAOYSA-N 0.000 description 6
- 101000789829 Homo sapiens ATPase family AAA domain-containing protein 5 Proteins 0.000 description 6
- 101000882908 Homo sapiens Claudin-3 Proteins 0.000 description 6
- 101000942622 Homo sapiens Condensin complex subunit 3 Proteins 0.000 description 6
- 101001050567 Homo sapiens Kinesin-like protein KIF2C Proteins 0.000 description 6
- 101000662909 Homo sapiens T cell receptor beta constant 1 Proteins 0.000 description 6
- 101000662902 Homo sapiens T cell receptor beta constant 2 Proteins 0.000 description 6
- 101001121409 Homo sapiens Transcription factor Ovo-like 2 Proteins 0.000 description 6
- 229940076838 Immune checkpoint inhibitor Drugs 0.000 description 6
- 108010002350 Interleukin-2 Proteins 0.000 description 6
- 102000000588 Interleukin-2 Human genes 0.000 description 6
- 208000008839 Kidney Neoplasms Diseases 0.000 description 6
- 102100023424 Kinesin-like protein KIF2C Human genes 0.000 description 6
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 6
- 206010038389 Renal cancer Diseases 0.000 description 6
- 102100037272 T cell receptor beta constant 1 Human genes 0.000 description 6
- 102100037298 T cell receptor beta constant 2 Human genes 0.000 description 6
- 102100026385 Transcription factor Ovo-like 2 Human genes 0.000 description 6
- 102100027881 Tumor protein 63 Human genes 0.000 description 6
- 101710140697 Tumor protein 63 Proteins 0.000 description 6
- 230000002411 adverse Effects 0.000 description 6
- 108010080146 androgen receptors Proteins 0.000 description 6
- 230000000259 anti-tumor effect Effects 0.000 description 6
- 238000002619 cancer immunotherapy Methods 0.000 description 6
- 239000013592 cell lysate Substances 0.000 description 6
- 239000000306 component Substances 0.000 description 6
- 235000018417 cysteine Nutrition 0.000 description 6
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 6
- SDUQYLNIPVEERB-QPPQHZFASA-N gemcitabine Chemical compound O=C1N=C(N)C=CN1[C@H]1C(F)(F)[C@H](O)[C@@H](CO)O1 SDUQYLNIPVEERB-QPPQHZFASA-N 0.000 description 6
- 229960005277 gemcitabine Drugs 0.000 description 6
- 239000012274 immune-checkpoint protein inhibitor Substances 0.000 description 6
- 239000003112 inhibitor Substances 0.000 description 6
- 230000002401 inhibitory effect Effects 0.000 description 6
- 201000010982 kidney cancer Diseases 0.000 description 6
- 201000007270 liver cancer Diseases 0.000 description 6
- 230000003211 malignant effect Effects 0.000 description 6
- 230000005012 migration Effects 0.000 description 6
- 239000000178 monomer Substances 0.000 description 6
- 210000000822 natural killer cell Anatomy 0.000 description 6
- 102000054765 polymorphisms of proteins Human genes 0.000 description 6
- 238000001959 radiotherapy Methods 0.000 description 6
- 230000002829 reductive effect Effects 0.000 description 6
- 230000001105 regulatory effect Effects 0.000 description 6
- 239000007790 solid phase Substances 0.000 description 6
- 238000011272 standard treatment Methods 0.000 description 6
- 239000000126 substance Substances 0.000 description 6
- 238000001356 surgical procedure Methods 0.000 description 6
- 230000004614 tumor growth Effects 0.000 description 6
- 208000003174 Brain Neoplasms Diseases 0.000 description 5
- 102100038447 Claudin-4 Human genes 0.000 description 5
- 102100024230 Dendritic cell-specific transmembrane protein Human genes 0.000 description 5
- 102000001301 EGF receptor Human genes 0.000 description 5
- 108060006698 EGF receptor Proteins 0.000 description 5
- 101000882890 Homo sapiens Claudin-4 Proteins 0.000 description 5
- 101000832060 Homo sapiens Dendritic cell-specific transmembrane protein Proteins 0.000 description 5
- 101001130401 Homo sapiens E3 ubiquitin-protein ligase RAD18 Proteins 0.000 description 5
- 101000971521 Homo sapiens Kinetochore scaffold 1 Proteins 0.000 description 5
- 101001005725 Homo sapiens Melanoma-associated antigen 10 Proteins 0.000 description 5
- 101000983140 Homo sapiens Protein associated with UVRAG as autophagy enhancer Proteins 0.000 description 5
- 101000716994 Homo sapiens Suppressor APC domain-containing protein 2 Proteins 0.000 description 5
- 101000795350 Homo sapiens Tripartite motif-containing protein 59 Proteins 0.000 description 5
- 108010074328 Interferon-gamma Proteins 0.000 description 5
- 102100021464 Kinetochore scaffold 1 Human genes 0.000 description 5
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 5
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 5
- 239000004472 Lysine Substances 0.000 description 5
- 102100025049 Melanoma-associated antigen 10 Human genes 0.000 description 5
- 206010035226 Plasma cell myeloma Diseases 0.000 description 5
- 102100026827 Protein associated with UVRAG as autophagy enhancer Human genes 0.000 description 5
- 102000001170 RAD18 Human genes 0.000 description 5
- 102100020923 Suppressor APC domain-containing protein 2 Human genes 0.000 description 5
- 208000006842 Tonsillar Neoplasms Diseases 0.000 description 5
- 102100029717 Tripartite motif-containing protein 59 Human genes 0.000 description 5
- 239000002671 adjuvant Substances 0.000 description 5
- 230000008901 benefit Effects 0.000 description 5
- 230000033228 biological regulation Effects 0.000 description 5
- 201000006612 cervical squamous cell carcinoma Diseases 0.000 description 5
- 238000006243 chemical reaction Methods 0.000 description 5
- 230000001419 dependent effect Effects 0.000 description 5
- 238000001514 detection method Methods 0.000 description 5
- 229940011871 estrogen Drugs 0.000 description 5
- 239000000262 estrogen Substances 0.000 description 5
- 230000002496 gastric effect Effects 0.000 description 5
- 230000012010 growth Effects 0.000 description 5
- 230000036039 immunity Effects 0.000 description 5
- 230000001939 inductive effect Effects 0.000 description 5
- 230000000977 initiatory effect Effects 0.000 description 5
- 238000002955 isolation Methods 0.000 description 5
- 230000003902 lesion Effects 0.000 description 5
- 210000004185 liver Anatomy 0.000 description 5
- 238000004949 mass spectrometry Methods 0.000 description 5
- 230000007246 mechanism Effects 0.000 description 5
- 210000002752 melanocyte Anatomy 0.000 description 5
- 210000004379 membrane Anatomy 0.000 description 5
- 230000002611 ovarian Effects 0.000 description 5
- 238000010647 peptide synthesis reaction Methods 0.000 description 5
- 238000002360 preparation method Methods 0.000 description 5
- 238000002271 resection Methods 0.000 description 5
- 230000028327 secretion Effects 0.000 description 5
- 239000011734 sodium Substances 0.000 description 5
- 238000003786 synthesis reaction Methods 0.000 description 5
- 230000003827 upregulation Effects 0.000 description 5
- 230000003612 virological effect Effects 0.000 description 5
- NHJVRSWLHSJWIN-UHFFFAOYSA-N 2,4,6-trinitrobenzenesulfonic acid Chemical compound OS(=O)(=O)C1=C([N+]([O-])=O)C=C([N+]([O-])=O)C=C1[N+]([O-])=O NHJVRSWLHSJWIN-UHFFFAOYSA-N 0.000 description 4
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 4
- 101100326430 Caenorhabditis elegans bub-1 gene Proteins 0.000 description 4
- 229910020366 ClO 4 Inorganic materials 0.000 description 4
- 108020004705 Codon Proteins 0.000 description 4
- 102100027995 Collagenase 3 Human genes 0.000 description 4
- 102000053602 DNA Human genes 0.000 description 4
- 102100036218 DNA replication complex GINS protein PSF2 Human genes 0.000 description 4
- AOJJSUZBOXZQNB-TZSSRYMLSA-N Doxorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)[C@H](O)[C@H](C)O1 AOJJSUZBOXZQNB-TZSSRYMLSA-N 0.000 description 4
- 206010014759 Endometrial neoplasm Diseases 0.000 description 4
- 102100041006 Forkhead box protein J1 Human genes 0.000 description 4
- 102100039545 Homeobox protein Hox-D11 Human genes 0.000 description 4
- 101000736065 Homo sapiens DNA replication complex GINS protein PSF2 Proteins 0.000 description 4
- 101000892910 Homo sapiens Forkhead box protein J1 Proteins 0.000 description 4
- 101000962591 Homo sapiens Homeobox protein Hox-D11 Proteins 0.000 description 4
- 101001044927 Homo sapiens Insulin-like growth factor-binding protein 3 Proteins 0.000 description 4
- 101001078133 Homo sapiens Integrin alpha-2 Proteins 0.000 description 4
- 101001091231 Homo sapiens Kinesin-like protein KIF18A Proteins 0.000 description 4
- 101000877861 Homo sapiens Protein FAM83B Proteins 0.000 description 4
- 101001062098 Homo sapiens RNA-binding protein 14 Proteins 0.000 description 4
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 4
- 101000577874 Homo sapiens Stromelysin-2 Proteins 0.000 description 4
- 108091008026 Inhibitory immune checkpoint proteins Proteins 0.000 description 4
- 102000037984 Inhibitory immune checkpoint proteins Human genes 0.000 description 4
- 108090000723 Insulin-Like Growth Factor I Proteins 0.000 description 4
- 102100022708 Insulin-like growth factor-binding protein 3 Human genes 0.000 description 4
- 102100025305 Integrin alpha-2 Human genes 0.000 description 4
- 229940126262 KIF18A Drugs 0.000 description 4
- 102100034895 Kinesin-like protein KIF18A Human genes 0.000 description 4
- 208000001894 Nasopharyngeal Neoplasms Diseases 0.000 description 4
- 206010061306 Nasopharyngeal cancer Diseases 0.000 description 4
- 206010048757 Oncocytoma Diseases 0.000 description 4
- 208000002063 Oxyphilic Adenoma Diseases 0.000 description 4
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Chemical compound OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 4
- 108010004729 Phycoerythrin Proteins 0.000 description 4
- 102100035443 Protein FAM83B Human genes 0.000 description 4
- 108091008611 Protein Kinase B Proteins 0.000 description 4
- 102100030624 Proton myo-inositol cotransporter Human genes 0.000 description 4
- 102100029250 RNA-binding protein 14 Human genes 0.000 description 4
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 4
- 208000015634 Rectal Neoplasms Diseases 0.000 description 4
- 108091006309 SLC2A13 Proteins 0.000 description 4
- 108091006682 SLCO5A1 Proteins 0.000 description 4
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 4
- 102100021990 Solute carrier organic anion transporter family member 5A1 Human genes 0.000 description 4
- 208000036765 Squamous cell carcinoma of the esophagus Diseases 0.000 description 4
- 102100028848 Stromelysin-2 Human genes 0.000 description 4
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 4
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 4
- 241000700605 Viruses Species 0.000 description 4
- 208000008383 Wilms tumor Diseases 0.000 description 4
- 150000001408 amides Chemical group 0.000 description 4
- 230000003321 amplification Effects 0.000 description 4
- RDOXTESZEPMUJZ-UHFFFAOYSA-N anisole Chemical compound COC1=CC=CC=C1 RDOXTESZEPMUJZ-UHFFFAOYSA-N 0.000 description 4
- 239000002246 antineoplastic agent Substances 0.000 description 4
- 230000001640 apoptogenic effect Effects 0.000 description 4
- 235000009582 asparagine Nutrition 0.000 description 4
- 229960001230 asparagine Drugs 0.000 description 4
- 210000003719 b-lymphocyte Anatomy 0.000 description 4
- 210000004369 blood Anatomy 0.000 description 4
- 239000008280 blood Substances 0.000 description 4
- 210000004899 c-terminal region Anatomy 0.000 description 4
- 230000005907 cancer growth Effects 0.000 description 4
- 229940022399 cancer vaccine Drugs 0.000 description 4
- 125000002915 carbonyl group Chemical group [*:2]C([*:1])=O 0.000 description 4
- 230000012292 cell migration Effects 0.000 description 4
- 230000021615 conjugation Effects 0.000 description 4
- 230000002074 deregulated effect Effects 0.000 description 4
- 230000007705 epithelial mesenchymal transition Effects 0.000 description 4
- 208000007276 esophageal squamous cell carcinoma Diseases 0.000 description 4
- 201000006585 gastric adenocarcinoma Diseases 0.000 description 4
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 4
- 210000003714 granulocyte Anatomy 0.000 description 4
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 4
- JYGXADMDTFJGBT-VWUMJDOOSA-N hydrocortisone Chemical compound O=C1CC[C@]2(C)[C@H]3[C@@H](O)C[C@](C)([C@@](CC4)(O)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 JYGXADMDTFJGBT-VWUMJDOOSA-N 0.000 description 4
- 230000002209 hydrophobic effect Effects 0.000 description 4
- 230000006058 immune tolerance Effects 0.000 description 4
- 230000005847 immunogenicity Effects 0.000 description 4
- 230000006698 induction Effects 0.000 description 4
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 4
- 230000002601 intratumoral effect Effects 0.000 description 4
- JXDYKVIHCLTXOP-UHFFFAOYSA-N isatin Chemical compound C1=CC=C2C(=O)C(=O)NC2=C1 JXDYKVIHCLTXOP-UHFFFAOYSA-N 0.000 description 4
- 210000001616 monocyte Anatomy 0.000 description 4
- 229960003301 nivolumab Drugs 0.000 description 4
- 238000003199 nucleic acid amplification method Methods 0.000 description 4
- 239000002773 nucleotide Substances 0.000 description 4
- 125000003729 nucleotide group Chemical group 0.000 description 4
- 210000000056 organ Anatomy 0.000 description 4
- 210000005259 peripheral blood Anatomy 0.000 description 4
- 239000011886 peripheral blood Substances 0.000 description 4
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 4
- 230000008569 process Effects 0.000 description 4
- 230000002062 proliferating effect Effects 0.000 description 4
- 230000001737 promoting effect Effects 0.000 description 4
- 125000006239 protecting group Chemical group 0.000 description 4
- 238000000746 purification Methods 0.000 description 4
- 238000011002 quantification Methods 0.000 description 4
- 239000002516 radical scavenger Substances 0.000 description 4
- 206010038038 rectal cancer Diseases 0.000 description 4
- 201000001275 rectum cancer Diseases 0.000 description 4
- 230000022983 regulation of cell cycle Effects 0.000 description 4
- 239000011347 resin Substances 0.000 description 4
- 229920005989 resin Polymers 0.000 description 4
- 230000002381 testicular Effects 0.000 description 4
- 238000012360 testing method Methods 0.000 description 4
- 230000005748 tumor development Effects 0.000 description 4
- 230000002792 vascular Effects 0.000 description 4
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 4
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 3
- MZOFCQQQCNRIBI-VMXHOPILSA-N (3s)-4-[[(2s)-1-[[(2s)-1-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-methyl-1-oxopentan-2-yl]amino]-5-(diaminomethylideneamino)-1-oxopentan-2-yl]amino]-3-[[2-[[(2s)-2,6-diaminohexanoyl]amino]acetyl]amino]-4-oxobutanoic acid Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@@H](N)CCCCN MZOFCQQQCNRIBI-VMXHOPILSA-N 0.000 description 3
- 102100036962 5'-3' exoribonuclease 1 Human genes 0.000 description 3
- 108010085238 Actins Proteins 0.000 description 3
- 208000024893 Acute lymphoblastic leukemia Diseases 0.000 description 3
- 208000014697 Acute lymphocytic leukaemia Diseases 0.000 description 3
- 102100022476 Adenosylhomocysteinase 3 Human genes 0.000 description 3
- 102100036799 Adhesion G-protein coupled receptor V1 Human genes 0.000 description 3
- 102100022724 Alpha-1,4-N-acetylglucosaminyltransferase Human genes 0.000 description 3
- 102100023635 Alpha-fetoprotein Human genes 0.000 description 3
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 3
- 108700020463 BRCA1 Proteins 0.000 description 3
- 102000036365 BRCA1 Human genes 0.000 description 3
- 101150072950 BRCA1 gene Proteins 0.000 description 3
- 102100021663 Baculoviral IAP repeat-containing protein 5 Human genes 0.000 description 3
- UHOVQNZJYSORNB-UHFFFAOYSA-N Benzene Chemical compound C1=CC=CC=C1 UHOVQNZJYSORNB-UHFFFAOYSA-N 0.000 description 3
- 102100036305 C-C chemokine receptor type 8 Human genes 0.000 description 3
- 102000015347 COP1 Human genes 0.000 description 3
- 108060001826 COP1 Proteins 0.000 description 3
- 102100025570 Cancer/testis antigen 1 Human genes 0.000 description 3
- 102100025832 Centromere-associated protein E Human genes 0.000 description 3
- 102100039537 Claudin-14 Human genes 0.000 description 3
- 108050005238 Collagenase 3 Proteins 0.000 description 3
- 102100032202 Cornulin Human genes 0.000 description 3
- CMSMOCZEIVJLDB-UHFFFAOYSA-N Cyclophosphamide Chemical compound ClCCN(CCCl)P1(=O)NCCCO1 CMSMOCZEIVJLDB-UHFFFAOYSA-N 0.000 description 3
- 241000701022 Cytomegalovirus Species 0.000 description 3
- 102100022204 DNA-dependent protein kinase catalytic subunit Human genes 0.000 description 3
- 102100032297 Dynein axonemal heavy chain 17 Human genes 0.000 description 3
- 206010014733 Endometrial cancer Diseases 0.000 description 3
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 3
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 3
- 241000282412 Homo Species 0.000 description 3
- 101000804879 Homo sapiens 5'-3' exoribonuclease 1 Proteins 0.000 description 3
- 101000822527 Homo sapiens Adenosylhomocysteinase 3 Proteins 0.000 description 3
- 101000928167 Homo sapiens Adhesion G-protein coupled receptor V1 Proteins 0.000 description 3
- 101000823037 Homo sapiens Alpha-1,4-N-acetylglucosaminyltransferase Proteins 0.000 description 3
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 3
- 101000716063 Homo sapiens C-C chemokine receptor type 8 Proteins 0.000 description 3
- 101000856237 Homo sapiens Cancer/testis antigen 1 Proteins 0.000 description 3
- 101000914247 Homo sapiens Centromere-associated protein E Proteins 0.000 description 3
- 101000888570 Homo sapiens Claudin-14 Proteins 0.000 description 3
- 101000920981 Homo sapiens Cornulin Proteins 0.000 description 3
- 101000619536 Homo sapiens DNA-dependent protein kinase catalytic subunit Proteins 0.000 description 3
- 101001016203 Homo sapiens Dynein axonemal heavy chain 17 Proteins 0.000 description 3
- 101001050275 Homo sapiens Keratin, type I cuticular Ha1 Proteins 0.000 description 3
- 101001007039 Homo sapiens Keratin, type I cuticular Ha5 Proteins 0.000 description 3
- 101001027621 Homo sapiens Kinesin-like protein KIF20A Proteins 0.000 description 3
- 101001027602 Homo sapiens Kinesin-like protein KIF26B Proteins 0.000 description 3
- 101001121408 Homo sapiens L-amino-acid oxidase Proteins 0.000 description 3
- 101000874532 Homo sapiens Lactosylceramide 1,3-N-acetyl-beta-D-glucosaminyltransferase Proteins 0.000 description 3
- 101001005719 Homo sapiens Melanoma-associated antigen 3 Proteins 0.000 description 3
- 101000653374 Homo sapiens Methylcytosine dioxygenase TET2 Proteins 0.000 description 3
- 101000972278 Homo sapiens Mucin-6 Proteins 0.000 description 3
- 101000934338 Homo sapiens Myeloid cell surface antigen CD33 Proteins 0.000 description 3
- 101000766826 Homo sapiens Protein CIP2A Proteins 0.000 description 3
- 101000804792 Homo sapiens Protein Wnt-5a Proteins 0.000 description 3
- 101000994788 Homo sapiens Ras GTPase-activating-like protein IQGAP3 Proteins 0.000 description 3
- 101001094545 Homo sapiens Retrotransposon-like protein 1 Proteins 0.000 description 3
- 101000914484 Homo sapiens T-lymphocyte activation antigen CD80 Proteins 0.000 description 3
- 101000771618 Homo sapiens WD repeat-containing protein 62 Proteins 0.000 description 3
- 101000744897 Homo sapiens Zinc finger homeobox protein 4 Proteins 0.000 description 3
- 102100027735 Hyaluronan mediated motility receptor Human genes 0.000 description 3
- 102000037982 Immune checkpoint proteins Human genes 0.000 description 3
- 108091008036 Immune checkpoint proteins Proteins 0.000 description 3
- 108010055717 JNK Mitogen-Activated Protein Kinases Proteins 0.000 description 3
- 102100023131 Keratin, type I cuticular Ha1 Human genes 0.000 description 3
- 102100028354 Keratin, type I cuticular Ha5 Human genes 0.000 description 3
- 102100037694 Kinesin-like protein KIF20A Human genes 0.000 description 3
- 102100037692 Kinesin-like protein KIF26B Human genes 0.000 description 3
- 102100026388 L-amino-acid oxidase Human genes 0.000 description 3
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 3
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 3
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 3
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 3
- 102100035655 Lactosylceramide 1,3-N-acetyl-beta-D-glucosaminyltransferase Human genes 0.000 description 3
- GQYIWUVLTXOXAJ-UHFFFAOYSA-N Lomustine Chemical compound ClCCN(N=O)C(=O)NC1CCCCC1 GQYIWUVLTXOXAJ-UHFFFAOYSA-N 0.000 description 3
- 108091054455 MAP kinase family Proteins 0.000 description 3
- 102000043136 MAP kinase family Human genes 0.000 description 3
- 102100028389 Melanoma antigen recognized by T-cells 1 Human genes 0.000 description 3
- 108050008953 Melanoma-associated antigen Proteins 0.000 description 3
- 102100025082 Melanoma-associated antigen 3 Human genes 0.000 description 3
- 206010027480 Metastatic malignant melanoma Diseases 0.000 description 3
- 241001465754 Metazoa Species 0.000 description 3
- 102100030803 Methylcytosine dioxygenase TET2 Human genes 0.000 description 3
- 102100034256 Mucin-1 Human genes 0.000 description 3
- 102100022493 Mucin-6 Human genes 0.000 description 3
- 208000034578 Multiple myelomas Diseases 0.000 description 3
- 101100519207 Mus musculus Pdcd1 gene Proteins 0.000 description 3
- 102100038895 Myc proto-oncogene protein Human genes 0.000 description 3
- 101710135898 Myc proto-oncogene protein Proteins 0.000 description 3
- 102100025243 Myeloid cell surface antigen CD33 Human genes 0.000 description 3
- NWIBSHFKIJFRCO-WUDYKRTCSA-N Mytomycin Chemical compound C1N2C(C(C(C)=C(N)C3=O)=O)=C3[C@@H](COC(N)=O)[C@@]2(OC)[C@@H]2[C@H]1N2 NWIBSHFKIJFRCO-WUDYKRTCSA-N 0.000 description 3
- 206010029260 Neuroblastoma Diseases 0.000 description 3
- 208000007571 Ovarian Epithelial Carcinoma Diseases 0.000 description 3
- 102000038030 PI3Ks Human genes 0.000 description 3
- 108091007960 PI3Ks Proteins 0.000 description 3
- 208000006664 Precursor Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 description 3
- 102000007066 Prostate-Specific Antigen Human genes 0.000 description 3
- 108010072866 Prostate-Specific Antigen Proteins 0.000 description 3
- 102100028634 Protein CIP2A Human genes 0.000 description 3
- 102100034417 Ras GTPase-activating-like protein IQGAP3 Human genes 0.000 description 3
- 108091006531 SLC28A3 Proteins 0.000 description 3
- 102100023085 Serine/threonine-protein kinase mTOR Human genes 0.000 description 3
- 102100021470 Solute carrier family 28 member 3 Human genes 0.000 description 3
- 208000034254 Squamous cell carcinoma of the cervix uteri Diseases 0.000 description 3
- 108010002687 Survivin Proteins 0.000 description 3
- 102100027222 T-lymphocyte activation antigen CD80 Human genes 0.000 description 3
- 102100040254 TBC1 domain family member 7 Human genes 0.000 description 3
- 108010065917 TOR Serine-Threonine Kinases Proteins 0.000 description 3
- 108010017842 Telomerase Proteins 0.000 description 3
- 208000003721 Triple Negative Breast Neoplasms Diseases 0.000 description 3
- 108700025716 Tumor Suppressor Genes Proteins 0.000 description 3
- 102000044209 Tumor Suppressor Genes Human genes 0.000 description 3
- 108010067390 Viral Proteins Proteins 0.000 description 3
- 102100029478 WD repeat-containing protein 62 Human genes 0.000 description 3
- 230000004156 Wnt signaling pathway Effects 0.000 description 3
- 102000043366 Wnt-5a Human genes 0.000 description 3
- 102100039968 Zinc finger homeobox protein 4 Human genes 0.000 description 3
- 230000001594 aberrant effect Effects 0.000 description 3
- 230000000735 allogeneic effect Effects 0.000 description 3
- 230000002424 anti-apoptotic effect Effects 0.000 description 3
- 230000001093 anti-cancer Effects 0.000 description 3
- 230000005809 anti-tumor immunity Effects 0.000 description 3
- 230000000890 antigenic effect Effects 0.000 description 3
- 230000009286 beneficial effect Effects 0.000 description 3
- 229960000397 bevacizumab Drugs 0.000 description 3
- 230000022131 cell cycle Effects 0.000 description 3
- 230000007910 cell fusion Effects 0.000 description 3
- 230000010261 cell growth Effects 0.000 description 3
- 238000002659 cell therapy Methods 0.000 description 3
- 229940030156 cell vaccine Drugs 0.000 description 3
- 210000003793 centrosome Anatomy 0.000 description 3
- 238000007385 chemical modification Methods 0.000 description 3
- 210000000349 chromosome Anatomy 0.000 description 3
- 230000001684 chronic effect Effects 0.000 description 3
- DQLATGHUWYMOKM-UHFFFAOYSA-L cisplatin Chemical compound N[Pt](N)(Cl)Cl DQLATGHUWYMOKM-UHFFFAOYSA-L 0.000 description 3
- 229960004316 cisplatin Drugs 0.000 description 3
- 239000002299 complementary DNA Substances 0.000 description 3
- 239000000470 constituent Substances 0.000 description 3
- 238000009109 curative therapy Methods 0.000 description 3
- 229960004397 cyclophosphamide Drugs 0.000 description 3
- 229940127089 cytotoxic agent Drugs 0.000 description 3
- 230000034994 death Effects 0.000 description 3
- 231100000517 death Toxicity 0.000 description 3
- 230000007547 defect Effects 0.000 description 3
- 230000002950 deficient Effects 0.000 description 3
- 208000035475 disorder Diseases 0.000 description 3
- 102000052116 epidermal growth factor receptor activity proteins Human genes 0.000 description 3
- 108700015053 epidermal growth factor receptor activity proteins Proteins 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- 229960000390 fludarabine Drugs 0.000 description 3
- GIUYCYHIANZCFB-FJFJXFQQSA-N fludarabine phosphate Chemical compound C1=NC=2C(N)=NC(F)=NC=2N1[C@@H]1O[C@H](COP(O)(O)=O)[C@@H](O)[C@@H]1O GIUYCYHIANZCFB-FJFJXFQQSA-N 0.000 description 3
- 102000037865 fusion proteins Human genes 0.000 description 3
- 108020001507 fusion proteins Proteins 0.000 description 3
- 230000030279 gene silencing Effects 0.000 description 3
- 230000002068 genetic effect Effects 0.000 description 3
- 230000013595 glycosylation Effects 0.000 description 3
- 208000002672 hepatitis B Diseases 0.000 description 3
- 239000000833 heterodimer Substances 0.000 description 3
- 210000004408 hybridoma Anatomy 0.000 description 3
- 230000003053 immunization Effects 0.000 description 3
- 239000002955 immunomodulating agent Substances 0.000 description 3
- 229940121354 immunomodulator Drugs 0.000 description 3
- 230000001976 improved effect Effects 0.000 description 3
- 230000006872 improvement Effects 0.000 description 3
- 229960003130 interferon gamma Drugs 0.000 description 3
- 201000007450 intrahepatic cholangiocarcinoma Diseases 0.000 description 3
- 238000011835 investigation Methods 0.000 description 3
- 210000000265 leukocyte Anatomy 0.000 description 3
- 230000021633 leukocyte mediated immunity Effects 0.000 description 3
- 210000004698 lymphocyte Anatomy 0.000 description 3
- 239000006166 lysate Substances 0.000 description 3
- 208000021039 metastatic melanoma Diseases 0.000 description 3
- 230000004899 motility Effects 0.000 description 3
- 210000003205 muscle Anatomy 0.000 description 3
- YOHYSYJDKVYCJI-UHFFFAOYSA-N n-[3-[[6-[3-(trifluoromethyl)anilino]pyrimidin-4-yl]amino]phenyl]cyclopropanecarboxamide Chemical compound FC(F)(F)C1=CC=CC(NC=2N=CN=C(NC=3C=C(NC(=O)C4CC4)C=CC=3)C=2)=C1 YOHYSYJDKVYCJI-UHFFFAOYSA-N 0.000 description 3
- 230000001613 neoplastic effect Effects 0.000 description 3
- 201000008026 nephroblastoma Diseases 0.000 description 3
- 210000004789 organ system Anatomy 0.000 description 3
- 210000002997 osteoclast Anatomy 0.000 description 3
- 201000008968 osteosarcoma Diseases 0.000 description 3
- 201000008129 pancreatic ductal adenocarcinoma Diseases 0.000 description 3
- 230000026731 phosphorylation Effects 0.000 description 3
- 238000006366 phosphorylation reaction Methods 0.000 description 3
- 230000001817 pituitary effect Effects 0.000 description 3
- 229920001223 polyethylene glycol Polymers 0.000 description 3
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 3
- 239000000092 prognostic biomarker Substances 0.000 description 3
- 230000005855 radiation Effects 0.000 description 3
- 238000011127 radiochemotherapy Methods 0.000 description 3
- 230000009467 reduction Effects 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 238000004007 reversed phase HPLC Methods 0.000 description 3
- 229960004641 rituximab Drugs 0.000 description 3
- 230000003248 secreting effect Effects 0.000 description 3
- 210000000130 stem cell Anatomy 0.000 description 3
- 230000000638 stimulation Effects 0.000 description 3
- 238000002626 targeted therapy Methods 0.000 description 3
- 230000002103 transcriptional effect Effects 0.000 description 3
- 230000009466 transformation Effects 0.000 description 3
- 230000005945 translocation Effects 0.000 description 3
- 208000022679 triple-negative breast carcinoma Diseases 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- DHBXNPKRAUYBTH-UHFFFAOYSA-N 1,1-ethanedithiol Chemical compound CC(S)S DHBXNPKRAUYBTH-UHFFFAOYSA-N 0.000 description 2
- ASOKPJOREAFHNY-UHFFFAOYSA-N 1-Hydroxybenzotriazole Chemical compound C1=CC=C2N(O)N=NC2=C1 ASOKPJOREAFHNY-UHFFFAOYSA-N 0.000 description 2
- DURPTKYDGMDSBL-UHFFFAOYSA-N 1-butoxybutane Chemical compound CCCCOCCCC DURPTKYDGMDSBL-UHFFFAOYSA-N 0.000 description 2
- VOXZDWNPVJITMN-ZBRFXRBCSA-N 17β-estradiol Chemical compound OC1=CC=C2[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CCC2=C1 VOXZDWNPVJITMN-ZBRFXRBCSA-N 0.000 description 2
- VUCNQOPCYRJCGQ-UHFFFAOYSA-N 2-[4-(hydroxymethyl)phenoxy]acetic acid Chemical class OCC1=CC=C(OCC(O)=O)C=C1 VUCNQOPCYRJCGQ-UHFFFAOYSA-N 0.000 description 2
- WHNPOQXWAMXPTA-UHFFFAOYSA-N 3-methylbut-2-enamide Chemical compound CC(C)=CC(N)=O WHNPOQXWAMXPTA-UHFFFAOYSA-N 0.000 description 2
- JVJFIQYAHPMBBX-UHFFFAOYSA-N 4-hydroxynonenal Chemical compound CCCCCC(O)C=CC=O JVJFIQYAHPMBBX-UHFFFAOYSA-N 0.000 description 2
- 108091007504 ADAM10 Proteins 0.000 description 2
- 102000007469 Actins Human genes 0.000 description 2
- 208000010507 Adenocarcinoma of Lung Diseases 0.000 description 2
- 208000036764 Adenocarcinoma of the esophagus Diseases 0.000 description 2
- 230000007730 Akt signaling Effects 0.000 description 2
- 102100026446 Aldo-keto reductase family 1 member C1 Human genes 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- 108010074708 B7-H1 Antigen Proteins 0.000 description 2
- 241000193830 Bacillus <bacterium> Species 0.000 description 2
- 102100027314 Beta-2-microglobulin Human genes 0.000 description 2
- 208000019838 Blood disease Diseases 0.000 description 2
- 208000006386 Bone Resorption Diseases 0.000 description 2
- 102100028622 Brefeldin A-inhibited guanine nucleotide-exchange protein 3 Human genes 0.000 description 2
- 101710149863 C-C chemokine receptor type 4 Proteins 0.000 description 2
- 125000001433 C-terminal amino-acid group Chemical group 0.000 description 2
- 102100040841 C-type lectin domain family 5 member A Human genes 0.000 description 2
- 102100032976 CCR4-NOT transcription complex subunit 6 Human genes 0.000 description 2
- 210000003359 CD4-positive helper T lymphocyte Anatomy 0.000 description 2
- 108010084313 CD58 Antigens Proteins 0.000 description 2
- 102000008203 CTLA-4 Antigen Human genes 0.000 description 2
- 108010021064 CTLA-4 Antigen Proteins 0.000 description 2
- 101100283604 Caenorhabditis elegans pigk-1 gene Proteins 0.000 description 2
- 208000000419 Chronic Hepatitis B Diseases 0.000 description 2
- 241000193468 Clostridium perfringens Species 0.000 description 2
- 102000000529 Costimulatory and Inhibitory T-Cell Receptors Human genes 0.000 description 2
- 108010041504 Costimulatory and Inhibitory T-Cell Receptors Proteins 0.000 description 2
- 102100021906 Cyclin-O Human genes 0.000 description 2
- 102100033270 Cyclin-dependent kinase inhibitor 1 Human genes 0.000 description 2
- 102100039077 Cytosolic 10-formyltetrahydrofolate dehydrogenase Human genes 0.000 description 2
- 108010041986 DNA Vaccines Proteins 0.000 description 2
- 229940021995 DNA vaccine Drugs 0.000 description 2
- QOSSAOTZNIDXMA-UHFFFAOYSA-N Dicylcohexylcarbodiimide Chemical compound C1CCCCC1N=C=NC1CCCCC1 QOSSAOTZNIDXMA-UHFFFAOYSA-N 0.000 description 2
- 102100039673 Disintegrin and metalloproteinase domain-containing protein 10 Human genes 0.000 description 2
- 206010059866 Drug resistance Diseases 0.000 description 2
- 102100024284 Dynein axonemal assembly factor 10 Human genes 0.000 description 2
- 102100032245 Dynein axonemal heavy chain 2 Human genes 0.000 description 2
- 101710146739 Enterotoxin Proteins 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- 102100026406 G/T mismatch-specific thymine DNA glycosylase Human genes 0.000 description 2
- 102100037488 G2 and S phase-expressed protein 1 Human genes 0.000 description 2
- 102100030708 GTPase KRas Human genes 0.000 description 2
- 229940032072 GVAX vaccine Drugs 0.000 description 2
- 206010061968 Gastric neoplasm Diseases 0.000 description 2
- 102000009465 Growth Factor Receptors Human genes 0.000 description 2
- 108010009202 Growth Factor Receptors Proteins 0.000 description 2
- 102100028976 HLA class I histocompatibility antigen, B alpha chain Human genes 0.000 description 2
- 108010058607 HLA-B Antigens Proteins 0.000 description 2
- 102000006354 HLA-DR Antigens Human genes 0.000 description 2
- 108010058597 HLA-DR Antigens Proteins 0.000 description 2
- 206010019043 Hair follicle tumour benign Diseases 0.000 description 2
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 2
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 2
- 206010019695 Hepatic neoplasm Diseases 0.000 description 2
- 208000028782 Hereditary disease Diseases 0.000 description 2
- 102100027703 Heterogeneous nuclear ribonucleoprotein H2 Human genes 0.000 description 2
- 108010033040 Histones Proteins 0.000 description 2
- 101000718028 Homo sapiens Aldo-keto reductase family 1 member C1 Proteins 0.000 description 2
- 101000695868 Homo sapiens Brefeldin A-inhibited guanine nucleotide-exchange protein 3 Proteins 0.000 description 2
- 101000749314 Homo sapiens C-type lectin domain family 5 member A Proteins 0.000 description 2
- 101000897441 Homo sapiens Cyclin-O Proteins 0.000 description 2
- 101000959030 Homo sapiens Cytosolic 10-formyltetrahydrofolate dehydrogenase Proteins 0.000 description 2
- 101000831170 Homo sapiens Dynein axonemal assembly factor 10 Proteins 0.000 description 2
- 101001016199 Homo sapiens Dynein axonemal heavy chain 2 Proteins 0.000 description 2
- 101001026457 Homo sapiens G2 and S phase-expressed protein 1 Proteins 0.000 description 2
- 101000584612 Homo sapiens GTPase KRas Proteins 0.000 description 2
- 101001081143 Homo sapiens Heterogeneous nuclear ribonucleoprotein H2 Proteins 0.000 description 2
- 101001076297 Homo sapiens IGF-like family receptor 1 Proteins 0.000 description 2
- 101000635408 Homo sapiens Inactive N-acetylated-alpha-linked acidic dipeptidase-like protein 2 Proteins 0.000 description 2
- 101001003169 Homo sapiens Insulin-like growth factor-binding protein-like 1 Proteins 0.000 description 2
- 101000998810 Homo sapiens Insulin-like peptide INSL6 Proteins 0.000 description 2
- 101000599940 Homo sapiens Interferon gamma Proteins 0.000 description 2
- 101000619643 Homo sapiens Ligand-dependent nuclear receptor-interacting factor 1 Proteins 0.000 description 2
- 101001017764 Homo sapiens Lipopolysaccharide-responsive and beige-like anchor protein Proteins 0.000 description 2
- 101000636209 Homo sapiens Matrix-remodeling-associated protein 5 Proteins 0.000 description 2
- 101001120864 Homo sapiens Meckelin Proteins 0.000 description 2
- 101000578784 Homo sapiens Melanoma antigen recognized by T-cells 1 Proteins 0.000 description 2
- 101001036675 Homo sapiens Melanoma-associated antigen B6 Proteins 0.000 description 2
- 101001055106 Homo sapiens Metastasis-associated in colon cancer protein 1 Proteins 0.000 description 2
- 101001133056 Homo sapiens Mucin-1 Proteins 0.000 description 2
- 101000972276 Homo sapiens Mucin-5B Proteins 0.000 description 2
- 101001097889 Homo sapiens Platelet-activating factor acetylhydrolase Proteins 0.000 description 2
- 101001064853 Homo sapiens Polyunsaturated fatty acid lipoxygenase ALOX15 Proteins 0.000 description 2
- 101000944004 Homo sapiens Potassium channel subfamily U member 1 Proteins 0.000 description 2
- 101001103597 Homo sapiens RING finger protein 24 Proteins 0.000 description 2
- 101000633778 Homo sapiens SLAM family member 5 Proteins 0.000 description 2
- 101000798076 Homo sapiens T cell receptor delta constant Proteins 0.000 description 2
- 101000891627 Homo sapiens TBC1 domain family member 7 Proteins 0.000 description 2
- 101001095487 Homo sapiens Telomere-associated protein RIF1 Proteins 0.000 description 2
- 101000666331 Homo sapiens Teneurin-4 Proteins 0.000 description 2
- 101000638427 Homo sapiens Tonsoku-like protein Proteins 0.000 description 2
- 101000667110 Homo sapiens Vacuolar protein sorting-associated protein 13B Proteins 0.000 description 2
- 101000818532 Homo sapiens Zinc finger and BTB domain-containing protein 20 Proteins 0.000 description 2
- 101000723920 Homo sapiens Zinc finger protein 40 Proteins 0.000 description 2
- MHAJPDPJQMAIIY-UHFFFAOYSA-N Hydrogen peroxide Chemical compound OO MHAJPDPJQMAIIY-UHFFFAOYSA-N 0.000 description 2
- 206010020880 Hypertrophy Diseases 0.000 description 2
- 102100025958 IGF-like family receptor 1 Human genes 0.000 description 2
- 102100031009 Inactive N-acetylated-alpha-linked acidic dipeptidase-like protein 2 Human genes 0.000 description 2
- 206010061218 Inflammation Diseases 0.000 description 2
- 102000004877 Insulin Human genes 0.000 description 2
- 108090001061 Insulin Proteins 0.000 description 2
- 102000004218 Insulin-Like Growth Factor I Human genes 0.000 description 2
- 102000014429 Insulin-like growth factor Human genes 0.000 description 2
- 102100020781 Insulin-like growth factor-binding protein-like 1 Human genes 0.000 description 2
- 102100033235 Insulin-like peptide INSL6 Human genes 0.000 description 2
- 108010064593 Intercellular Adhesion Molecule-1 Proteins 0.000 description 2
- 102100037877 Intercellular adhesion molecule 1 Human genes 0.000 description 2
- 102000006992 Interferon-alpha Human genes 0.000 description 2
- 108010047761 Interferon-alpha Proteins 0.000 description 2
- 102000013462 Interleukin-12 Human genes 0.000 description 2
- 108010065805 Interleukin-12 Proteins 0.000 description 2
- 102000019145 JUN kinase activity proteins Human genes 0.000 description 2
- 208000004527 Joubert syndrome 6 Diseases 0.000 description 2
- 150000008575 L-amino acids Chemical class 0.000 description 2
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 2
- 241000713666 Lentivirus Species 0.000 description 2
- 102100033353 Lipopolysaccharide-responsive and beige-like anchor protein Human genes 0.000 description 2
- 108700001646 MCC Genes Proteins 0.000 description 2
- 101150067806 MCC gene Proteins 0.000 description 2
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 2
- 102100030776 Matrix-remodeling-associated protein 5 Human genes 0.000 description 2
- 102100026047 Meckelin Human genes 0.000 description 2
- 102000000440 Melanoma-associated antigen Human genes 0.000 description 2
- 102100039483 Melanoma-associated antigen B6 Human genes 0.000 description 2
- 208000024556 Mendelian disease Diseases 0.000 description 2
- 102100026892 Metastasis-associated in colon cancer protein 1 Human genes 0.000 description 2
- AIJULSRZWUXGPQ-UHFFFAOYSA-N Methylglyoxal Chemical compound CC(=O)C=O AIJULSRZWUXGPQ-UHFFFAOYSA-N 0.000 description 2
- 108090000744 Mitogen-Activated Protein Kinase Kinases Proteins 0.000 description 2
- 102000004232 Mitogen-Activated Protein Kinase Kinases Human genes 0.000 description 2
- YXOLAZRVSSWPPT-UHFFFAOYSA-N Morin Chemical compound OC1=CC(O)=CC=C1C1=C(O)C(=O)C2=C(O)C=C(O)C=C2O1 YXOLAZRVSSWPPT-UHFFFAOYSA-N 0.000 description 2
- 102100022494 Mucin-5B Human genes 0.000 description 2
- ZDZOTLJHXYCWBA-VCVYQWHSSA-N N-debenzoyl-N-(tert-butoxycarbonyl)-10-deacetyltaxol Chemical compound O([C@H]1[C@H]2[C@@](C([C@H](O)C3=C(C)[C@@H](OC(=O)[C@H](O)[C@@H](NC(=O)OC(C)(C)C)C=4C=CC=CC=4)C[C@]1(O)C3(C)C)=O)(C)[C@@H](O)C[C@H]1OC[C@]12OC(=O)C)C(=O)C1=CC=CC=C1 ZDZOTLJHXYCWBA-VCVYQWHSSA-N 0.000 description 2
- 125000001429 N-terminal alpha-amino-acid group Chemical group 0.000 description 2
- 108050006691 NEDD4-binding protein 2 Proteins 0.000 description 2
- 102100036542 NEDD4-binding protein 2 Human genes 0.000 description 2
- 101710104492 NUP210 Proteins 0.000 description 2
- 208000034176 Neoplasms, Germ Cell and Embryonal Diseases 0.000 description 2
- 102100035570 Nuclear pore membrane glycoprotein 210 Human genes 0.000 description 2
- 206010030137 Oesophageal adenocarcinoma Diseases 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 108010067244 Origin Recognition Complex Proteins 0.000 description 2
- 102100040591 Origin recognition complex subunit 1 Human genes 0.000 description 2
- 108010011536 PTEN Phosphohydrolase Proteins 0.000 description 2
- 102000014160 PTEN Phosphohydrolase Human genes 0.000 description 2
- 206010033701 Papillary thyroid cancer Diseases 0.000 description 2
- 102000002508 Peptide Elongation Factors Human genes 0.000 description 2
- 108010068204 Peptide Elongation Factors Proteins 0.000 description 2
- 102000011755 Phosphoglycerate Kinase Human genes 0.000 description 2
- 102100037518 Platelet-activating factor acetylhydrolase Human genes 0.000 description 2
- 239000004952 Polyamide Substances 0.000 description 2
- 102100031950 Polyunsaturated fatty acid lipoxygenase ALOX15 Human genes 0.000 description 2
- 102100033525 Potassium channel subfamily U member 1 Human genes 0.000 description 2
- 102100024216 Programmed cell death 1 ligand 1 Human genes 0.000 description 2
- 102100033810 RAC-alpha serine/threonine-protein kinase Human genes 0.000 description 2
- 102100033479 RAF proto-oncogene serine/threonine-protein kinase Human genes 0.000 description 2
- 102100039516 RING finger protein 24 Human genes 0.000 description 2
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 2
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 2
- 102100029216 SLAM family member 5 Human genes 0.000 description 2
- 108091006478 SLC25A43 Proteins 0.000 description 2
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 2
- 208000000453 Skin Neoplasms Diseases 0.000 description 2
- 102100032413 Solute carrier family 25 member 43 Human genes 0.000 description 2
- 241000713880 Spleen focus-forming virus Species 0.000 description 2
- 206010041848 Squamous cell carcinoma of the cervix Diseases 0.000 description 2
- 230000006044 T cell activation Effects 0.000 description 2
- 102100032272 T cell receptor delta constant Human genes 0.000 description 2
- 108700042076 T-Cell Receptor alpha Genes Proteins 0.000 description 2
- 108700042077 T-Cell Receptor beta Genes Proteins 0.000 description 2
- 208000000389 T-cell leukemia Diseases 0.000 description 2
- 208000028530 T-cell lymphoblastic leukemia/lymphoma Diseases 0.000 description 2
- 102100038123 Teneurin-4 Human genes 0.000 description 2
- MUMGGOZAMZWBJJ-DYKIIFRCSA-N Testostosterone Chemical compound O=C1CC[C@]2(C)[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CCC2=C1 MUMGGOZAMZWBJJ-DYKIIFRCSA-N 0.000 description 2
- 101001099217 Thermotoga maritima (strain ATCC 43589 / DSM 3109 / JCM 10099 / NBRC 100826 / MSB8) Triosephosphate isomerase Proteins 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 239000004473 Threonine Substances 0.000 description 2
- 206010062129 Tongue neoplasm Diseases 0.000 description 2
- 102100031224 Tonsoku-like protein Human genes 0.000 description 2
- 102000040945 Transcription factor Human genes 0.000 description 2
- 108091023040 Transcription factor Proteins 0.000 description 2
- 101710150448 Transcriptional regulator Myc Proteins 0.000 description 2
- DTQVDTLACAAQTR-UHFFFAOYSA-M Trifluoroacetate Chemical compound [O-]C(=O)C(F)(F)F DTQVDTLACAAQTR-UHFFFAOYSA-M 0.000 description 2
- 102100033579 Trophoblast glycoprotein Human genes 0.000 description 2
- 206010046799 Uterine leiomyosarcoma Diseases 0.000 description 2
- 102100039113 Vacuolar protein sorting-associated protein 13B Human genes 0.000 description 2
- 102000009524 Vascular Endothelial Growth Factor A Human genes 0.000 description 2
- 108010073929 Vascular Endothelial Growth Factor A Proteins 0.000 description 2
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 2
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 2
- 102100021146 Zinc finger and BTB domain-containing protein 20 Human genes 0.000 description 2
- 102100028440 Zinc finger protein 40 Human genes 0.000 description 2
- 239000000654 additive Substances 0.000 description 2
- 230000000996 additive effect Effects 0.000 description 2
- 208000009956 adenocarcinoma Diseases 0.000 description 2
- 125000001931 aliphatic group Chemical group 0.000 description 2
- 150000003862 amino acid derivatives Chemical class 0.000 description 2
- 239000003098 androgen Substances 0.000 description 2
- 150000008064 anhydrides Chemical class 0.000 description 2
- 230000006023 anti-tumor response Effects 0.000 description 2
- 239000008346 aqueous phase Substances 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- 125000000637 arginyl group Chemical group N[C@@H](CCCNC(N)=N)C(=O)* 0.000 description 2
- 235000003704 aspartic acid Nutrition 0.000 description 2
- 230000001363 autoimmune Effects 0.000 description 2
- 230000033590 base-excision repair Effects 0.000 description 2
- 108010081355 beta 2-Microglobulin Proteins 0.000 description 2
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 2
- 230000000903 blocking effect Effects 0.000 description 2
- 210000001185 bone marrow Anatomy 0.000 description 2
- 230000024279 bone resorption Effects 0.000 description 2
- 201000003163 breast adenoma Diseases 0.000 description 2
- 125000004744 butyloxycarbonyl group Chemical group 0.000 description 2
- LYQFWZFBNBDLEO-UHFFFAOYSA-M caesium bromide Chemical compound [Br-].[Cs+] LYQFWZFBNBDLEO-UHFFFAOYSA-M 0.000 description 2
- AIYUHDOJVYHVIT-UHFFFAOYSA-M caesium chloride Chemical compound [Cl-].[Cs+] AIYUHDOJVYHVIT-UHFFFAOYSA-M 0.000 description 2
- 238000009566 cancer vaccine Methods 0.000 description 2
- 230000021235 carbamoylation Effects 0.000 description 2
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 2
- 230000015556 catabolic process Effects 0.000 description 2
- 230000006369 cell cycle progression Effects 0.000 description 2
- 230000030833 cell death Effects 0.000 description 2
- 210000000170 cell membrane Anatomy 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 230000036755 cellular response Effects 0.000 description 2
- 229960005395 cetuximab Drugs 0.000 description 2
- 230000002759 chromosomal effect Effects 0.000 description 2
- 230000001886 ciliary effect Effects 0.000 description 2
- 238000003776 cleavage reaction Methods 0.000 description 2
- 210000001072 colon Anatomy 0.000 description 2
- 238000002648 combination therapy Methods 0.000 description 2
- 230000000295 complement effect Effects 0.000 description 2
- 230000001054 cortical effect Effects 0.000 description 2
- 238000004132 cross linking Methods 0.000 description 2
- 239000003431 cross linking reagent Substances 0.000 description 2
- YPHMISFOHDHNIV-FSZOTQKASA-N cycloheximide Chemical compound C1[C@@H](C)C[C@H](C)C(=O)[C@@H]1[C@H](O)CC1CC(=O)NC(=O)C1 YPHMISFOHDHNIV-FSZOTQKASA-N 0.000 description 2
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 2
- 210000004405 cytokine-induced killer cell Anatomy 0.000 description 2
- 231100000433 cytotoxic Toxicity 0.000 description 2
- 230000001472 cytotoxic effect Effects 0.000 description 2
- 230000006378 damage Effects 0.000 description 2
- 238000007405 data analysis Methods 0.000 description 2
- 230000007812 deficiency Effects 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- 239000005547 deoxyribonucleotide Substances 0.000 description 2
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 2
- 238000010511 deprotection reaction Methods 0.000 description 2
- OQALFHMKVSJFRR-UHFFFAOYSA-N dityrosine Chemical compound OC(=O)C(N)CC1=CC=C(O)C(C=2C(=CC=C(CC(N)C(O)=O)C=2)O)=C1 OQALFHMKVSJFRR-UHFFFAOYSA-N 0.000 description 2
- 229960003668 docetaxel Drugs 0.000 description 2
- 229960004679 doxorubicin Drugs 0.000 description 2
- 229950009791 durvalumab Drugs 0.000 description 2
- 230000004064 dysfunction Effects 0.000 description 2
- 230000008482 dysregulation Effects 0.000 description 2
- 230000013020 embryo development Effects 0.000 description 2
- 230000002708 enhancing effect Effects 0.000 description 2
- 231100000655 enterotoxin Toxicity 0.000 description 2
- 239000000147 enterotoxin Substances 0.000 description 2
- 229940088598 enzyme Drugs 0.000 description 2
- 208000028653 esophageal adenocarcinoma Diseases 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 238000004108 freeze drying Methods 0.000 description 2
- 125000000524 functional group Chemical group 0.000 description 2
- 210000001156 gastric mucosa Anatomy 0.000 description 2
- 238000001415 gene therapy Methods 0.000 description 2
- 235000013922 glutamic acid Nutrition 0.000 description 2
- 239000004220 glutamic acid Substances 0.000 description 2
- 238000006206 glycosylation reaction Methods 0.000 description 2
- 238000000227 grinding Methods 0.000 description 2
- 239000003102 growth factor Substances 0.000 description 2
- 210000003780 hair follicle Anatomy 0.000 description 2
- 201000010536 head and neck cancer Diseases 0.000 description 2
- 208000014829 head and neck neoplasm Diseases 0.000 description 2
- 208000014951 hematologic disease Diseases 0.000 description 2
- 208000018706 hematopoietic system disease Diseases 0.000 description 2
- 208000006359 hepatoblastoma Diseases 0.000 description 2
- 210000003494 hepatocyte Anatomy 0.000 description 2
- 102000040060 histone H4 family Human genes 0.000 description 2
- 108091069671 histone H4 family Proteins 0.000 description 2
- 108010003425 hyaluronan-mediated motility receptor Proteins 0.000 description 2
- 229960000890 hydrocortisone Drugs 0.000 description 2
- 125000001165 hydrophobic group Chemical group 0.000 description 2
- 238000004191 hydrophobic interaction chromatography Methods 0.000 description 2
- 230000005746 immune checkpoint blockade Effects 0.000 description 2
- 238000002649 immunization Methods 0.000 description 2
- 230000002584 immunomodulator Effects 0.000 description 2
- 230000001506 immunosuppresive effect Effects 0.000 description 2
- 230000004054 inflammatory process Effects 0.000 description 2
- 238000001802 infusion Methods 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 229940125396 insulin Drugs 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 230000000968 intestinal effect Effects 0.000 description 2
- 230000003834 intracellular effect Effects 0.000 description 2
- 108010028930 invariant chain Proteins 0.000 description 2
- PGLTVOMIXTUURA-UHFFFAOYSA-N iodoacetamide Chemical compound NC(=O)CI PGLTVOMIXTUURA-UHFFFAOYSA-N 0.000 description 2
- 238000004255 ion exchange chromatography Methods 0.000 description 2
- 150000002500 ions Chemical class 0.000 description 2
- 229960004942 lenalidomide Drugs 0.000 description 2
- GOTYRUGSSMKFNF-UHFFFAOYSA-N lenalidomide Chemical compound C1C=2C(N)=CC=CC=2C(=O)N1C1CCC(=O)NC1=O GOTYRUGSSMKFNF-UHFFFAOYSA-N 0.000 description 2
- AMXOYNBUYSYVKV-UHFFFAOYSA-M lithium bromide Chemical compound [Li+].[Br-] AMXOYNBUYSYVKV-UHFFFAOYSA-M 0.000 description 2
- KWGKDLIKAYFUFQ-UHFFFAOYSA-M lithium chloride Chemical compound [Li+].[Cl-] KWGKDLIKAYFUFQ-UHFFFAOYSA-M 0.000 description 2
- 230000007774 longterm Effects 0.000 description 2
- 201000005296 lung carcinoma Diseases 0.000 description 2
- 210000001165 lymph node Anatomy 0.000 description 2
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 2
- 238000012423 maintenance Methods 0.000 description 2
- 230000036210 malignancy Effects 0.000 description 2
- 201000010893 malignant breast melanoma Diseases 0.000 description 2
- 239000011159 matrix material Substances 0.000 description 2
- 229960000485 methotrexate Drugs 0.000 description 2
- UZKWTJUDCOPSNM-UHFFFAOYSA-N methoxybenzene Substances CCCCOC=C UZKWTJUDCOPSNM-UHFFFAOYSA-N 0.000 description 2
- ZCQGVFNHUATAJY-UHFFFAOYSA-N methyl 2-[methyl(prop-2-enoyl)amino]acetate Chemical compound COC(=O)CN(C)C(=O)C=C ZCQGVFNHUATAJY-UHFFFAOYSA-N 0.000 description 2
- 230000002438 mitochondrial effect Effects 0.000 description 2
- 229960004857 mitomycin Drugs 0.000 description 2
- 238000011242 molecular targeted therapy Methods 0.000 description 2
- UXOUKMQIEVGVLY-UHFFFAOYSA-N morin Natural products OC1=CC(O)=CC(C2=C(C(=O)C3=C(O)C=C(O)C=C3O2)O)=C1 UXOUKMQIEVGVLY-UHFFFAOYSA-N 0.000 description 2
- 235000007708 morin Nutrition 0.000 description 2
- 201000000050 myeloid neoplasm Diseases 0.000 description 2
- CMWYAOXYQATXSI-UHFFFAOYSA-N n,n-dimethylformamide;piperidine Chemical compound CN(C)C=O.C1CCNCC1 CMWYAOXYQATXSI-UHFFFAOYSA-N 0.000 description 2
- 210000005170 neoplastic cell Anatomy 0.000 description 2
- 230000000926 neurological effect Effects 0.000 description 2
- FEMOMIGRRWSMCU-UHFFFAOYSA-N ninhydrin Chemical compound C1=CC=C2C(=O)C(O)(O)C(=O)C2=C1 FEMOMIGRRWSMCU-UHFFFAOYSA-N 0.000 description 2
- 238000010606 normalization Methods 0.000 description 2
- 239000000101 novel biomarker Substances 0.000 description 2
- 238000011275 oncology therapy Methods 0.000 description 2
- 206010030983 oral lichen planus Diseases 0.000 description 2
- 230000008520 organization Effects 0.000 description 2
- 229960002621 pembrolizumab Drugs 0.000 description 2
- 238000009520 phase I clinical trial Methods 0.000 description 2
- OJUGVDODNPJEEC-UHFFFAOYSA-N phenylglyoxal Chemical compound O=CC(=O)C1=CC=CC=C1 OJUGVDODNPJEEC-UHFFFAOYSA-N 0.000 description 2
- 208000001095 pilomatrixoma Diseases 0.000 description 2
- 210000004180 plasmocyte Anatomy 0.000 description 2
- 229920002401 polyacrylamide Polymers 0.000 description 2
- 229920002647 polyamide Polymers 0.000 description 2
- 230000004481 post-translational protein modification Effects 0.000 description 2
- 102000003998 progesterone receptors Human genes 0.000 description 2
- 108090000468 progesterone receptors Proteins 0.000 description 2
- 230000000750 progressive effect Effects 0.000 description 2
- 230000002035 prolonged effect Effects 0.000 description 2
- 210000002307 prostate Anatomy 0.000 description 2
- 230000009145 protein modification Effects 0.000 description 2
- 229940023143 protein vaccine Drugs 0.000 description 2
- 108010014186 ras Proteins Proteins 0.000 description 2
- 102000016914 ras Proteins Human genes 0.000 description 2
- 230000007420 reactivation Effects 0.000 description 2
- 238000001953 recrystallisation Methods 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 230000000717 retained effect Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 230000007017 scission Effects 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- DCKVNWZUADLDEH-UHFFFAOYSA-N sec-butyl acetate Chemical compound CCC(C)OC(C)=O DCKVNWZUADLDEH-UHFFFAOYSA-N 0.000 description 2
- 238000000926 separation method Methods 0.000 description 2
- 210000002966 serum Anatomy 0.000 description 2
- 238000001542 size-exclusion chromatography Methods 0.000 description 2
- 201000000849 skin cancer Diseases 0.000 description 2
- JHJLBTNAGRQEKS-UHFFFAOYSA-M sodium bromide Chemical compound [Na+].[Br-] JHJLBTNAGRQEKS-UHFFFAOYSA-M 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- 239000006104 solid solution Substances 0.000 description 2
- 230000009870 specific binding Effects 0.000 description 2
- 230000003595 spectral effect Effects 0.000 description 2
- 238000004611 spectroscopical analysis Methods 0.000 description 2
- 206010041823 squamous cell carcinoma Diseases 0.000 description 2
- 238000010561 standard procedure Methods 0.000 description 2
- 230000004936 stimulating effect Effects 0.000 description 2
- 238000010998 test method Methods 0.000 description 2
- 208000030045 thyroid gland papillary carcinoma Diseases 0.000 description 2
- 231100000331 toxic Toxicity 0.000 description 2
- 230000002588 toxic effect Effects 0.000 description 2
- 230000001052 transient effect Effects 0.000 description 2
- 230000007704 transition Effects 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- 238000011269 treatment regimen Methods 0.000 description 2
- 125000002221 trityl group Chemical group [H]C1=C([H])C([H])=C([H])C([H])=C1C([*])(C1=C(C(=C(C(=C1[H])[H])[H])[H])[H])C1=C([H])C([H])=C([H])C([H])=C1[H] 0.000 description 2
- 229940030325 tumor cell vaccine Drugs 0.000 description 2
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 2
- 230000034512 ubiquitination Effects 0.000 description 2
- 238000010798 ubiquitination Methods 0.000 description 2
- 210000003932 urinary bladder Anatomy 0.000 description 2
- 210000003741 urothelium Anatomy 0.000 description 2
- 238000007738 vacuum evaporation Methods 0.000 description 2
- DNXHEGUUPJUMQT-UHFFFAOYSA-N (+)-estrone Natural products OC1=CC=C2C3CCC(C)(C(CC4)=O)C4C3CCC2=C1 DNXHEGUUPJUMQT-UHFFFAOYSA-N 0.000 description 1
- FDKXTQMXEQVLRF-ZHACJKMWSA-N (E)-dacarbazine Chemical compound CN(C)\N=N\c1[nH]cnc1C(N)=O FDKXTQMXEQVLRF-ZHACJKMWSA-N 0.000 description 1
- LMDZBCPBFSXMTL-UHFFFAOYSA-N 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide Chemical compound CCN=C=NCCCN(C)C LMDZBCPBFSXMTL-UHFFFAOYSA-N 0.000 description 1
- KFDPCYZHENQOBV-UHFFFAOYSA-N 2-(bromomethyl)-4-nitrophenol Chemical compound OC1=CC=C([N+]([O-])=O)C=C1CBr KFDPCYZHENQOBV-UHFFFAOYSA-N 0.000 description 1
- IZQAUUVBKYXMET-UHFFFAOYSA-N 2-bromoethanamine Chemical compound NCCBr IZQAUUVBKYXMET-UHFFFAOYSA-N 0.000 description 1
- JQPFYXFVUKHERX-UHFFFAOYSA-N 2-hydroxy-2-cyclohexen-1-one Natural products OC1=CCCCC1=O JQPFYXFVUKHERX-UHFFFAOYSA-N 0.000 description 1
- 102100022644 26S proteasome regulatory subunit 4 Human genes 0.000 description 1
- AOJJSUZBOXZQNB-VTZDEGQISA-N 4'-epidoxorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)[C@@H](O)[C@H](C)O1 AOJJSUZBOXZQNB-VTZDEGQISA-N 0.000 description 1
- 102100033793 ALK tyrosine kinase receptor Human genes 0.000 description 1
- 101710168331 ALK tyrosine kinase receptor Proteins 0.000 description 1
- 102000000872 ATM Human genes 0.000 description 1
- 102100028161 ATP-binding cassette sub-family C member 2 Human genes 0.000 description 1
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 1
- 102000015693 Actin Depolymerizing Factors Human genes 0.000 description 1
- 108010038798 Actin Depolymerizing Factors Proteins 0.000 description 1
- 102100022900 Actin, cytoplasmic 1 Human genes 0.000 description 1
- 102100030891 Actin-associated protein FAM107A Human genes 0.000 description 1
- 206010000830 Acute leukaemia Diseases 0.000 description 1
- 206010052747 Adenocarcinoma pancreas Diseases 0.000 description 1
- 208000003200 Adenoma Diseases 0.000 description 1
- 208000009888 Adrenocortical Adenoma Diseases 0.000 description 1
- 206010073478 Anaplastic large-cell lymphoma Diseases 0.000 description 1
- 101710145634 Antigen 1 Proteins 0.000 description 1
- 102100021569 Apoptosis regulator Bcl-2 Human genes 0.000 description 1
- 102100023180 Armadillo repeat-containing protein 5 Human genes 0.000 description 1
- 244000186140 Asperula odorata Species 0.000 description 1
- 206010003571 Astrocytoma Diseases 0.000 description 1
- 241000501754 Astronotus ocellatus Species 0.000 description 1
- 108010004586 Ataxia Telangiectasia Mutated Proteins Proteins 0.000 description 1
- 206010003694 Atrophy Diseases 0.000 description 1
- 238000012935 Averaging Methods 0.000 description 1
- 102000019260 B-Cell Antigen Receptors Human genes 0.000 description 1
- 108010012919 B-Cell Antigen Receptors Proteins 0.000 description 1
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 1
- 208000032800 BCR-ABL1 positive blast phase chronic myelogenous leukemia Diseases 0.000 description 1
- 208000032791 BCR-ABL1 positive chronic myelogenous leukemia Diseases 0.000 description 1
- BXTVQNYQYUTQAZ-UHFFFAOYSA-N BNPS-skatole Chemical compound N=1C2=CC=CC=C2C(C)(Br)C=1SC1=CC=CC=C1[N+]([O-])=O BXTVQNYQYUTQAZ-UHFFFAOYSA-N 0.000 description 1
- 241001289141 Babr Species 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 208000023514 Barrett esophagus Diseases 0.000 description 1
- 208000023665 Barrett oesophagus Diseases 0.000 description 1
- 206010004146 Basal cell carcinoma Diseases 0.000 description 1
- 101150008012 Bcl2l1 gene Proteins 0.000 description 1
- 208000004860 Blast Crisis Diseases 0.000 description 1
- 206010006049 Bovine Tuberculosis Diseases 0.000 description 1
- 206010055113 Breast cancer metastatic Diseases 0.000 description 1
- 102100024217 CAMPATH-1 antigen Human genes 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 101150013553 CD40 gene Proteins 0.000 description 1
- 102100032937 CD40 ligand Human genes 0.000 description 1
- 108010065524 CD52 Antigen Proteins 0.000 description 1
- 210000001266 CD8-positive T-lymphocyte Anatomy 0.000 description 1
- 210000001239 CD8-positive, alpha-beta cytotoxic T lymphocyte Anatomy 0.000 description 1
- 102100033787 CMP-sialic acid transporter Human genes 0.000 description 1
- 102000012000 CXCR4 Receptors Human genes 0.000 description 1
- 108010061299 CXCR4 Receptors Proteins 0.000 description 1
- 101100220616 Caenorhabditis elegans chk-2 gene Proteins 0.000 description 1
- 101100314454 Caenorhabditis elegans tra-1 gene Proteins 0.000 description 1
- 102100036431 Calcineurin subunit B type 1 Human genes 0.000 description 1
- 241000178270 Canarypox virus Species 0.000 description 1
- 102100026550 Caspase-9 Human genes 0.000 description 1
- 108090000566 Caspase-9 Proteins 0.000 description 1
- 102000011068 Cdc42 Human genes 0.000 description 1
- 108050001278 Cdc42 Proteins 0.000 description 1
- 229940123587 Cell cycle inhibitor Drugs 0.000 description 1
- 206010007953 Central nervous system lymphoma Diseases 0.000 description 1
- VEXZGXHMUGYJMC-UHFFFAOYSA-M Chloride anion Chemical compound [Cl-] VEXZGXHMUGYJMC-UHFFFAOYSA-M 0.000 description 1
- 102100034330 Chromaffin granule amine transporter Human genes 0.000 description 1
- 108010077544 Chromatin Proteins 0.000 description 1
- 208000037051 Chromosomal Instability Diseases 0.000 description 1
- 206010061764 Chromosomal deletion Diseases 0.000 description 1
- 208000031639 Chromosome Deletion Diseases 0.000 description 1
- 208000010833 Chronic myeloid leukaemia Diseases 0.000 description 1
- 108090000317 Chymotrypsin Proteins 0.000 description 1
- 208000030808 Clear cell renal carcinoma Diseases 0.000 description 1
- 206010053138 Congenital aplastic anaemia Diseases 0.000 description 1
- 206010062759 Congenital dyskeratosis Diseases 0.000 description 1
- 108091029523 CpG island Proteins 0.000 description 1
- JPVYNHNXODAKFH-UHFFFAOYSA-N Cu2+ Chemical compound [Cu+2] JPVYNHNXODAKFH-UHFFFAOYSA-N 0.000 description 1
- 102000005636 Cyclic AMP Response Element-Binding Protein Human genes 0.000 description 1
- 108010045171 Cyclic AMP Response Element-Binding Protein Proteins 0.000 description 1
- 108010058546 Cyclin D1 Proteins 0.000 description 1
- 108010025464 Cyclin-Dependent Kinase 4 Proteins 0.000 description 1
- 108010016788 Cyclin-Dependent Kinase Inhibitor p21 Proteins 0.000 description 1
- 102000000578 Cyclin-Dependent Kinase Inhibitor p21 Human genes 0.000 description 1
- 102100036252 Cyclin-dependent kinase 4 Human genes 0.000 description 1
- 108010037462 Cyclooxygenase 2 Proteins 0.000 description 1
- 150000008574 D-amino acids Chemical class 0.000 description 1
- 102100033462 DENN domain-containing protein 1B Human genes 0.000 description 1
- 102100024810 DNA (cytosine-5)-methyltransferase 3B Human genes 0.000 description 1
- 101710123222 DNA (cytosine-5)-methyltransferase 3B Proteins 0.000 description 1
- 230000005778 DNA damage Effects 0.000 description 1
- 231100000277 DNA damage Toxicity 0.000 description 1
- 230000033616 DNA repair Effects 0.000 description 1
- 230000006820 DNA synthesis Effects 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 108010049207 Death Domain Receptors Proteins 0.000 description 1
- 102000009058 Death Domain Receptors Human genes 0.000 description 1
- 206010073135 Dedifferentiated liposarcoma Diseases 0.000 description 1
- 102100023933 Deoxyuridine 5'-triphosphate nucleotidohydrolase, mitochondrial Human genes 0.000 description 1
- QSJXEFYPDANLFS-UHFFFAOYSA-N Diacetyl Chemical compound CC(=O)C(C)=O QSJXEFYPDANLFS-UHFFFAOYSA-N 0.000 description 1
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 1
- 206010061818 Disease progression Diseases 0.000 description 1
- 102100036109 Dual specificity protein kinase TTK Human genes 0.000 description 1
- 208000000471 Dysplastic Nevus Syndrome Diseases 0.000 description 1
- 206010062805 Dysplastic naevus Diseases 0.000 description 1
- 108050002772 E3 ubiquitin-protein ligase Mdm2 Proteins 0.000 description 1
- 102000012199 E3 ubiquitin-protein ligase Mdm2 Human genes 0.000 description 1
- 102100026245 E3 ubiquitin-protein ligase RNF43 Human genes 0.000 description 1
- 101710109241 E3 ubiquitin-protein ligase RNF43 Proteins 0.000 description 1
- 101150029707 ERBB2 gene Proteins 0.000 description 1
- 102100039578 ETS translocation variant 4 Human genes 0.000 description 1
- 201000009273 Endometriosis Diseases 0.000 description 1
- 102100031780 Endonuclease Human genes 0.000 description 1
- 108010042407 Endonucleases Proteins 0.000 description 1
- 102000005593 Endopeptidases Human genes 0.000 description 1
- 108010059378 Endopeptidases Proteins 0.000 description 1
- HTIJFSOGRVMCQR-UHFFFAOYSA-N Epirubicin Natural products COc1cccc2C(=O)c3c(O)c4CC(O)(CC(OC5CC(N)C(=O)C(C)O5)c4c(O)c3C(=O)c12)C(=O)CO HTIJFSOGRVMCQR-UHFFFAOYSA-N 0.000 description 1
- 102000007594 Estrogen Receptor alpha Human genes 0.000 description 1
- 108010007005 Estrogen Receptor alpha Proteins 0.000 description 1
- DNXHEGUUPJUMQT-CBZIJGRNSA-N Estrone Chemical compound OC1=CC=C2[C@H]3CC[C@](C)(C(CC4)=O)[C@@H]4[C@@H]3CCC2=C1 DNXHEGUUPJUMQT-CBZIJGRNSA-N 0.000 description 1
- 108010007457 Extracellular Signal-Regulated MAP Kinases Proteins 0.000 description 1
- 201000004939 Fanconi anemia Diseases 0.000 description 1
- 102000017177 Fibromodulin Human genes 0.000 description 1
- 108010013996 Fibromodulin Proteins 0.000 description 1
- 240000008168 Ficus benjamina Species 0.000 description 1
- 108010009306 Forkhead Box Protein O1 Proteins 0.000 description 1
- 241000700662 Fowlpox virus Species 0.000 description 1
- 102100039818 Frizzled-5 Human genes 0.000 description 1
- 101710140951 Frizzled-5 Proteins 0.000 description 1
- 102100024165 G1/S-specific cyclin-D1 Human genes 0.000 description 1
- 102100038904 GPI inositol-deacylase Human genes 0.000 description 1
- 102100036482 GTP-binding protein 10 Human genes 0.000 description 1
- 235000008526 Galium odoratum Nutrition 0.000 description 1
- 208000022072 Gallbladder Neoplasms Diseases 0.000 description 1
- 241001663880 Gammaretrovirus Species 0.000 description 1
- 102400000921 Gastrin Human genes 0.000 description 1
- 108010052343 Gastrins Proteins 0.000 description 1
- 229920002148 Gellan gum Polymers 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 208000034951 Genetic Translocation Diseases 0.000 description 1
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 1
- 108060005986 Granzyme Proteins 0.000 description 1
- 102000001398 Granzyme Human genes 0.000 description 1
- 102100028971 HLA class I histocompatibility antigen, C alpha chain Human genes 0.000 description 1
- 102100031547 HLA class II histocompatibility antigen, DO alpha chain Human genes 0.000 description 1
- 108010052199 HLA-C Antigens Proteins 0.000 description 1
- 108010037031 Hair-Specific Keratins Proteins 0.000 description 1
- 102000011733 Hair-Specific Keratins Human genes 0.000 description 1
- 102100031561 Hamartin Human genes 0.000 description 1
- 206010019375 Helicobacter infections Diseases 0.000 description 1
- 208000002250 Hematologic Neoplasms Diseases 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 108010027412 Histocompatibility Antigens Class II Proteins 0.000 description 1
- 102000018713 Histocompatibility Antigens Class II Human genes 0.000 description 1
- 102100030649 Histone H2B type 1-J Human genes 0.000 description 1
- 102100039869 Histone H2B type F-S Human genes 0.000 description 1
- 102100033068 Histone acetyltransferase KAT7 Human genes 0.000 description 1
- 108090000246 Histone acetyltransferases Proteins 0.000 description 1
- 102000003893 Histone acetyltransferases Human genes 0.000 description 1
- 208000017604 Hodgkin disease Diseases 0.000 description 1
- 208000021519 Hodgkin lymphoma Diseases 0.000 description 1
- 208000010747 Hodgkins lymphoma Diseases 0.000 description 1
- 101000619137 Homo sapiens 26S proteasome regulatory subunit 4 Proteins 0.000 description 1
- 101001063917 Homo sapiens Actin-associated protein FAM107A Proteins 0.000 description 1
- 101000971171 Homo sapiens Apoptosis regulator Bcl-2 Proteins 0.000 description 1
- 101000684964 Homo sapiens Armadillo repeat-containing protein 5 Proteins 0.000 description 1
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 1
- 101000714321 Homo sapiens Calcineurin subunit B type 1 Proteins 0.000 description 1
- 101000793825 Homo sapiens Capping protein, Arp2/3 and myosin-I linker protein 2 Proteins 0.000 description 1
- 101000577887 Homo sapiens Collagenase 3 Proteins 0.000 description 1
- 101000944380 Homo sapiens Cyclin-dependent kinase inhibitor 1 Proteins 0.000 description 1
- 101000870914 Homo sapiens DENN domain-containing protein 1B Proteins 0.000 description 1
- 101000659223 Homo sapiens Dual specificity protein kinase TTK Proteins 0.000 description 1
- 101000813747 Homo sapiens ETS translocation variant 4 Proteins 0.000 description 1
- 101001099051 Homo sapiens GPI inositol-deacylase Proteins 0.000 description 1
- 101001071647 Homo sapiens GTP-binding protein 10 Proteins 0.000 description 1
- 101000866278 Homo sapiens HLA class II histocompatibility antigen, DO alpha chain Proteins 0.000 description 1
- 101000795643 Homo sapiens Hamartin Proteins 0.000 description 1
- 101001084678 Homo sapiens Histone H2B type 1-J Proteins 0.000 description 1
- 101001035372 Homo sapiens Histone H2B type F-S Proteins 0.000 description 1
- 101000944166 Homo sapiens Histone acetyltransferase KAT7 Proteins 0.000 description 1
- 101001046870 Homo sapiens Hypoxia-inducible factor 1-alpha Proteins 0.000 description 1
- 101000599951 Homo sapiens Insulin-like growth factor I Proteins 0.000 description 1
- 101100020228 Homo sapiens KLHL31 gene Proteins 0.000 description 1
- 101001027143 Homo sapiens Kelch domain-containing protein 7B Proteins 0.000 description 1
- 101001004623 Homo sapiens Lactase-like protein Proteins 0.000 description 1
- 101000777628 Homo sapiens Leukocyte antigen CD37 Proteins 0.000 description 1
- 101000608935 Homo sapiens Leukosialin Proteins 0.000 description 1
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 description 1
- 101000990902 Homo sapiens Matrix metalloproteinase-9 Proteins 0.000 description 1
- 101000801539 Homo sapiens Mitochondrial import receptor subunit TOM34 Proteins 0.000 description 1
- 101000972282 Homo sapiens Mucin-5AC Proteins 0.000 description 1
- 101000979342 Homo sapiens Nuclear factor NF-kappa-B p105 subunit Proteins 0.000 description 1
- 101000986765 Homo sapiens Oxytocin receptor Proteins 0.000 description 1
- 101000923340 Homo sapiens Phospholipid-transporting ATPase VB Proteins 0.000 description 1
- 101000701518 Homo sapiens Probable phospholipid-transporting ATPase IM Proteins 0.000 description 1
- 101001129654 Homo sapiens Prohibitin-2 Proteins 0.000 description 1
- 101000977257 Homo sapiens Protein MMS22-like Proteins 0.000 description 1
- 101000598778 Homo sapiens Protein OSCP1 Proteins 0.000 description 1
- 101000893100 Homo sapiens Protein fantom Proteins 0.000 description 1
- 101000687459 Homo sapiens REST corepressor 3 Proteins 0.000 description 1
- 101000743242 Homo sapiens RNA-binding protein 4 Proteins 0.000 description 1
- 101001092185 Homo sapiens Regulator of cell cycle RGCC Proteins 0.000 description 1
- 101100094993 Homo sapiens SAPCD2 gene Proteins 0.000 description 1
- 101000904787 Homo sapiens Serine/threonine-protein kinase ATR Proteins 0.000 description 1
- 101000711475 Homo sapiens Serpin B10 Proteins 0.000 description 1
- 101000873927 Homo sapiens Squamous cell carcinoma antigen recognized by T-cells 3 Proteins 0.000 description 1
- 101000820589 Homo sapiens Succinate-hydroxymethylglutarate CoA-transferase Proteins 0.000 description 1
- 101100480755 Homo sapiens TBC1D7 gene Proteins 0.000 description 1
- 101000976959 Homo sapiens Transcription factor 4 Proteins 0.000 description 1
- 101000596771 Homo sapiens Transcription factor 7-like 2 Proteins 0.000 description 1
- 101000800498 Homo sapiens Transketolase-like protein 1 Proteins 0.000 description 1
- 101000667104 Homo sapiens Vacuolar protein sorting-associated protein 13C Proteins 0.000 description 1
- 101000941898 Homo sapiens Volume-regulated anion channel subunit LRRC8E Proteins 0.000 description 1
- 101000955409 Homo sapiens Xyloside xylosyltransferase 1 Proteins 0.000 description 1
- 101100268070 Homo sapiens ZBTB20 gene Proteins 0.000 description 1
- 101000788678 Homo sapiens Zinc finger MYM-type protein 1 Proteins 0.000 description 1
- 101000976574 Homo sapiens Zinc finger protein 131 Proteins 0.000 description 1
- 101000633054 Homo sapiens Zinc finger protein SNAI2 Proteins 0.000 description 1
- 101000873780 Homo sapiens m7GpppN-mRNA hydrolase Proteins 0.000 description 1
- 206010071119 Hormone-dependent prostate cancer Diseases 0.000 description 1
- 241000714260 Human T-lymphotropic virus 1 Species 0.000 description 1
- 241000701806 Human papillomavirus Species 0.000 description 1
- 208000037147 Hypercalcaemia Diseases 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- 102100022875 Hypoxia-inducible factor 1-alpha Human genes 0.000 description 1
- 101150106931 IFNG gene Proteins 0.000 description 1
- 206010061598 Immunodeficiency Diseases 0.000 description 1
- 108060003951 Immunoglobulin Proteins 0.000 description 1
- 208000026350 Inborn Genetic disease Diseases 0.000 description 1
- 102100037852 Insulin-like growth factor I Human genes 0.000 description 1
- 102000003777 Interleukin-1 beta Human genes 0.000 description 1
- 108090000193 Interleukin-1 beta Proteins 0.000 description 1
- 102000007482 Interleukin-13 Receptor alpha2 Subunit Human genes 0.000 description 1
- 108010085418 Interleukin-13 Receptor alpha2 Subunit Proteins 0.000 description 1
- 108090001005 Interleukin-6 Proteins 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- 102100037648 Kelch domain-containing protein 7B Human genes 0.000 description 1
- 102000010638 Kinesin Human genes 0.000 description 1
- 108010063296 Kinesin Proteins 0.000 description 1
- 101150048929 Kl gene Proteins 0.000 description 1
- 108010062028 L-BLP25 Proteins 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- 239000005411 L01XE02 - Gefitinib Substances 0.000 description 1
- 239000002138 L01XE21 - Regorafenib Substances 0.000 description 1
- 102100025640 Lactase-like protein Human genes 0.000 description 1
- 108010000851 Laminin Receptors Proteins 0.000 description 1
- 102000002297 Laminin Receptors Human genes 0.000 description 1
- 208000032004 Large-Cell Anaplastic Lymphoma Diseases 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 102100031586 Leukocyte antigen CD37 Human genes 0.000 description 1
- 102100039564 Leukosialin Human genes 0.000 description 1
- 229910018119 Li 3 PO 4 Inorganic materials 0.000 description 1
- 229910013684 LiClO 4 Inorganic materials 0.000 description 1
- 229910013553 LiNO Inorganic materials 0.000 description 1
- 102100029185 Low affinity immunoglobulin gamma Fc region receptor III-B Human genes 0.000 description 1
- 206010050017 Lung cancer metastatic Diseases 0.000 description 1
- 229910052765 Lutetium Inorganic materials 0.000 description 1
- 102000019149 MAP kinase activity proteins Human genes 0.000 description 1
- 108040008097 MAP kinase activity proteins Proteins 0.000 description 1
- 108010010995 MART-1 Antigen Proteins 0.000 description 1
- 108700041567 MDR Genes Proteins 0.000 description 1
- 229940124647 MEK inhibitor Drugs 0.000 description 1
- PEEHTFAAVSWFBL-UHFFFAOYSA-N Maleimide Chemical compound O=C1NC(=O)C=C1 PEEHTFAAVSWFBL-UHFFFAOYSA-N 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 102100030412 Matrix metalloproteinase-9 Human genes 0.000 description 1
- 201000008643 Meckel syndrome Diseases 0.000 description 1
- 208000000172 Medulloblastoma Diseases 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 206010027406 Mesothelioma Diseases 0.000 description 1
- 206010059282 Metastases to central nervous system Diseases 0.000 description 1
- 108091028695 MiR-224 Proteins 0.000 description 1
- 208000032818 Microsatellite Instability Diseases 0.000 description 1
- 102000029749 Microtubule Human genes 0.000 description 1
- 108091022875 Microtubule Proteins 0.000 description 1
- 108091027766 Mir-143 Proteins 0.000 description 1
- 102100033583 Mitochondrial import receptor subunit TOM34 Human genes 0.000 description 1
- 102100024193 Mitogen-activated protein kinase 1 Human genes 0.000 description 1
- 229930192392 Mitomycin Natural products 0.000 description 1
- 102100034085 Monocarboxylate transporter 14 Human genes 0.000 description 1
- 108010008707 Mucin-1 Proteins 0.000 description 1
- 102100022496 Mucin-5AC Human genes 0.000 description 1
- 108010066419 Multidrug Resistance-Associated Protein 2 Proteins 0.000 description 1
- 241000699660 Mus musculus Species 0.000 description 1
- 101000794231 Mus musculus Mitotic checkpoint serine/threonine-protein kinase BUB1 beta Proteins 0.000 description 1
- 101100523539 Mus musculus Raf1 gene Proteins 0.000 description 1
- 101100096028 Mus musculus Smok1 gene Proteins 0.000 description 1
- 108010021466 Mutant Proteins Proteins 0.000 description 1
- 102000008300 Mutant Proteins Human genes 0.000 description 1
- 241000186359 Mycobacterium Species 0.000 description 1
- 241000186366 Mycobacterium bovis Species 0.000 description 1
- 208000033761 Myelogenous Chronic BCR-ABL Positive Leukemia Diseases 0.000 description 1
- 208000004304 Myofibromatosis Diseases 0.000 description 1
- LPPJGTSPIBSYQO-YLNOTJRMSA-N N-Acetyl-S-(1,2-dichlorovinyl)-cysteine Chemical compound CC(=O)N[C@H](C(O)=O)CS\C(Cl)=C/Cl LPPJGTSPIBSYQO-YLNOTJRMSA-N 0.000 description 1
- VIHYIVKEECZGOU-UHFFFAOYSA-N N-acetylimidazole Chemical compound CC(=O)N1C=CN=C1 VIHYIVKEECZGOU-UHFFFAOYSA-N 0.000 description 1
- PCLIMKBDDGJMGD-UHFFFAOYSA-N N-bromosuccinimide Chemical compound BrN1C(=O)CCC1=O PCLIMKBDDGJMGD-UHFFFAOYSA-N 0.000 description 1
- 108010057466 NF-kappa B Proteins 0.000 description 1
- 102000003945 NF-kappa B Human genes 0.000 description 1
- 208000009905 Neurofibromatoses Diseases 0.000 description 1
- 108010094052 Nik related kinase Proteins 0.000 description 1
- 102100034340 Nik-related protein kinase Human genes 0.000 description 1
- 102100023050 Nuclear factor NF-kappa-B p105 subunit Human genes 0.000 description 1
- 206010061534 Oesophageal squamous cell carcinoma Diseases 0.000 description 1
- 208000035327 Oestrogen receptor positive breast cancer Diseases 0.000 description 1
- 206010031112 Oropharyngeal squamous cell carcinoma Diseases 0.000 description 1
- 102000016978 Orphan receptors Human genes 0.000 description 1
- 108070000031 Orphan receptors Proteins 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 102100028139 Oxytocin receptor Human genes 0.000 description 1
- 102000042685 PI3/PI4-kinase family Human genes 0.000 description 1
- 108091081974 PI3/PI4-kinase family Proteins 0.000 description 1
- 239000012828 PI3K inhibitor Substances 0.000 description 1
- 229930012538 Paclitaxel Natural products 0.000 description 1
- 208000009608 Papillomavirus Infections Diseases 0.000 description 1
- 241001645095 Parisis Species 0.000 description 1
- BUQLXKSONWUQAC-UHFFFAOYSA-N Parthenolide Natural products CC1C2OC(=O)C(=C)C2CCC(=C/CCC1(C)O)C BUQLXKSONWUQAC-UHFFFAOYSA-N 0.000 description 1
- KHGNFPUMBJSZSM-UHFFFAOYSA-N Perforine Natural products COC1=C2CCC(O)C(CCC(C)(C)O)(OC)C2=NC2=C1C=CO2 KHGNFPUMBJSZSM-UHFFFAOYSA-N 0.000 description 1
- 102000017794 Perilipin-2 Human genes 0.000 description 1
- 108010067163 Perilipin-2 Proteins 0.000 description 1
- 102100032666 Phospholipid-transporting ATPase VB Human genes 0.000 description 1
- 208000007913 Pituitary Neoplasms Diseases 0.000 description 1
- 108010022233 Plasminogen Activator Inhibitor 1 Proteins 0.000 description 1
- 102100039418 Plasminogen activator inhibitor 1 Human genes 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 101710093543 Probable non-specific lipid-transfer protein Proteins 0.000 description 1
- 102100030468 Probable phospholipid-transporting ATPase IM Human genes 0.000 description 1
- 102100031156 Prohibitin-2 Human genes 0.000 description 1
- 102000003946 Prolactin Human genes 0.000 description 1
- 108010057464 Prolactin Proteins 0.000 description 1
- 102100038280 Prostaglandin G/H synthase 2 Human genes 0.000 description 1
- 102000001253 Protein Kinase Human genes 0.000 description 1
- 102100023475 Protein MMS22-like Human genes 0.000 description 1
- 102100037778 Protein OSCP1 Human genes 0.000 description 1
- 102000006478 Protein Phosphatase 2 Human genes 0.000 description 1
- 108010058956 Protein Phosphatase 2 Proteins 0.000 description 1
- 101710149109 Protein Vpx Proteins 0.000 description 1
- 102100040970 Protein fantom Human genes 0.000 description 1
- 102000004022 Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000412 Protein-Tyrosine Kinases Proteins 0.000 description 1
- 102000007568 Proto-Oncogene Proteins c-fos Human genes 0.000 description 1
- 108010071563 Proto-Oncogene Proteins c-fos Proteins 0.000 description 1
- 101710141955 RAF proto-oncogene serine/threonine-protein kinase Proteins 0.000 description 1
- 101150083042 RBM14 gene Proteins 0.000 description 1
- 102100024871 REST corepressor 3 Human genes 0.000 description 1
- 238000012228 RNA interference-mediated gene silencing Methods 0.000 description 1
- 102100038153 RNA-binding protein 4 Human genes 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 102100035542 Regulator of cell cycle RGCC Human genes 0.000 description 1
- 102100035123 Retrotransposon-like protein 1 Human genes 0.000 description 1
- 108010034782 Ribosomal Protein S6 Kinases Proteins 0.000 description 1
- 102000009738 Ribosomal Protein S6 Kinases Human genes 0.000 description 1
- 102100028029 SCL-interrupting locus protein Human genes 0.000 description 1
- 101150077807 SERPINB10 gene Proteins 0.000 description 1
- 108091006627 SLC12A9 Proteins 0.000 description 1
- 108091006771 SLC16A14 Proteins 0.000 description 1
- 108091006772 SLC18A1 Proteins 0.000 description 1
- 108091006172 SLC21 Proteins 0.000 description 1
- 108091006540 SLC35A1 Proteins 0.000 description 1
- 101100100680 Schizosaccharomyces pombe (strain 972 / ATCC 24843) trp4 gene Proteins 0.000 description 1
- 229940124639 Selective inhibitor Drugs 0.000 description 1
- 102100023921 Serine/threonine-protein kinase ATR Human genes 0.000 description 1
- 102100034012 Serpin B10 Human genes 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- 102100035748 Squamous cell carcinoma antigen recognized by T-cells 3 Human genes 0.000 description 1
- 206010041849 Squamous cell carcinoma of the hypopharynx Diseases 0.000 description 1
- 206010041865 Squamous cell carcinoma of the tongue Diseases 0.000 description 1
- 102100021652 Succinate-hydroxymethylglutarate CoA-transferase Human genes 0.000 description 1
- 108700005078 Synthetic Genes Proteins 0.000 description 1
- 102220468097 T cell receptor beta constant 1_C33A_mutation Human genes 0.000 description 1
- 102000003622 TRPC4 Human genes 0.000 description 1
- 206010043276 Teratoma Diseases 0.000 description 1
- 208000033133 Testicular seminomatous germ cell tumor Diseases 0.000 description 1
- NYTOUQBROMCLBJ-UHFFFAOYSA-N Tetranitromethane Chemical compound [O-][N+](=O)C([N+]([O-])=O)([N+]([O-])=O)[N+]([O-])=O NYTOUQBROMCLBJ-UHFFFAOYSA-N 0.000 description 1
- 210000000068 Th17 cell Anatomy 0.000 description 1
- 102000019347 Tob1 Human genes 0.000 description 1
- 108700038377 Tob1 Proteins 0.000 description 1
- 101150041680 Tob1 gene Proteins 0.000 description 1
- 206010044002 Tonsil cancer Diseases 0.000 description 1
- 108020004566 Transfer RNA Proteins 0.000 description 1
- 102100033108 Transketolase-like protein 1 Human genes 0.000 description 1
- 101150099990 Trpc4 gene Proteins 0.000 description 1
- 108090000631 Trypsin Proteins 0.000 description 1
- 102000004142 Trypsin Human genes 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 108090000704 Tubulin Proteins 0.000 description 1
- 102000004243 Tubulin Human genes 0.000 description 1
- 108060008683 Tumor Necrosis Factor Receptor Proteins 0.000 description 1
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 1
- 102100039094 Tyrosinase Human genes 0.000 description 1
- 108060008724 Tyrosinase Proteins 0.000 description 1
- 108090000848 Ubiquitin Proteins 0.000 description 1
- 102000044159 Ubiquitin Human genes 0.000 description 1
- 201000005969 Uveal melanoma Diseases 0.000 description 1
- 108091008605 VEGF receptors Proteins 0.000 description 1
- 241000700618 Vaccinia virus Species 0.000 description 1
- 102100039112 Vacuolar protein sorting-associated protein 13C Human genes 0.000 description 1
- 102000009484 Vascular Endothelial Growth Factor Receptors Human genes 0.000 description 1
- 102100032675 Volume-regulated anion channel subunit LRRC8E Human genes 0.000 description 1
- 101150109862 WNT-5A gene Proteins 0.000 description 1
- 208000000260 Warts Diseases 0.000 description 1
- 108700025700 Wilms Tumor Genes Proteins 0.000 description 1
- 208000026448 Wilms tumor 1 Diseases 0.000 description 1
- 102100022748 Wilms tumor protein Human genes 0.000 description 1
- 101710127857 Wilms tumor protein Proteins 0.000 description 1
- 241001492404 Woodchuck hepatitis virus Species 0.000 description 1
- 102100039027 Xyloside xylosyltransferase 1 Human genes 0.000 description 1
- 101150020014 ZBTB20 gene Proteins 0.000 description 1
- 102100039102 ZW10 interactor Human genes 0.000 description 1
- 101710177447 ZW10 interactor Proteins 0.000 description 1
- 102100025100 Zinc finger MYM-type protein 1 Human genes 0.000 description 1
- 102100023571 Zinc finger protein 131 Human genes 0.000 description 1
- 102100029570 Zinc finger protein SNAI2 Human genes 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 230000002159 abnormal effect Effects 0.000 description 1
- 230000035508 accumulation Effects 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- 159000000021 acetate salts Chemical class 0.000 description 1
- 150000001242 acetic acid derivatives Chemical class 0.000 description 1
- 125000002777 acetyl group Chemical group [H]C([H])([H])C(*)=O 0.000 description 1
- 230000021736 acetylation Effects 0.000 description 1
- 238000006640 acetylation reaction Methods 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 238000005903 acid hydrolysis reaction Methods 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 229940125666 actinium-225 Drugs 0.000 description 1
- QQINRWTZWGJFDB-YPZZEJLDSA-N actinium-225 Chemical compound [225Ac] QQINRWTZWGJFDB-YPZZEJLDSA-N 0.000 description 1
- 239000008186 active pharmaceutical agent Substances 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 230000010933 acylation Effects 0.000 description 1
- 238000005917 acylation reaction Methods 0.000 description 1
- 208000025084 adenoid cystic breast carcinoma Diseases 0.000 description 1
- 230000001919 adrenal effect Effects 0.000 description 1
- 229960002833 aflibercept Drugs 0.000 description 1
- 108010081667 aflibercept Proteins 0.000 description 1
- 230000032683 aging Effects 0.000 description 1
- 239000000556 agonist Substances 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 229960000548 alemtuzumab Drugs 0.000 description 1
- 230000029936 alkylation Effects 0.000 description 1
- 238000005804 alkylation reaction Methods 0.000 description 1
- 239000013566 allergen Substances 0.000 description 1
- 208000030961 allergic reaction Diseases 0.000 description 1
- 108010026331 alpha-Fetoproteins Proteins 0.000 description 1
- 125000003368 amide group Chemical group 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 238000009167 androgen deprivation therapy Methods 0.000 description 1
- AEMFNILZOJDQLW-QAGGRKNESA-N androst-4-ene-3,17-dione Chemical compound O=C1CC[C@]2(C)[C@H]3CC[C@](C)(C(CC4)=O)[C@@H]4[C@@H]3CCC2=C1 AEMFNILZOJDQLW-QAGGRKNESA-N 0.000 description 1
- AEMFNILZOJDQLW-UHFFFAOYSA-N androstenedione Natural products O=C1CCC2(C)C3CCC(C)(C(CC4)=O)C4C3CCC2=C1 AEMFNILZOJDQLW-UHFFFAOYSA-N 0.000 description 1
- 230000033115 angiogenesis Effects 0.000 description 1
- 239000004037 angiogenesis inhibitor Substances 0.000 description 1
- 230000001772 anti-angiogenic effect Effects 0.000 description 1
- 239000000611 antibody drug conjugate Substances 0.000 description 1
- 230000005875 antibody response Effects 0.000 description 1
- 229940049595 antibody-drug conjugate Drugs 0.000 description 1
- 230000030741 antigen processing and presentation Effects 0.000 description 1
- 230000014102 antigen processing and presentation of exogenous peptide antigen via MHC class I Effects 0.000 description 1
- 230000005775 apoptotic pathway Effects 0.000 description 1
- 230000005756 apoptotic signaling Effects 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 238000012098 association analyses Methods 0.000 description 1
- 210000001130 astrocyte Anatomy 0.000 description 1
- 101150051494 atr gene Proteins 0.000 description 1
- 230000037444 atrophy Effects 0.000 description 1
- 230000002238 attenuated effect Effects 0.000 description 1
- 230000003305 autocrine Effects 0.000 description 1
- 230000006472 autoimmune response Effects 0.000 description 1
- 229940030547 autologous tumor cell vaccine Drugs 0.000 description 1
- 239000002774 b raf kinase inhibitor Substances 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 1
- 230000002146 bilateral effect Effects 0.000 description 1
- 238000002306 biochemical method Methods 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 230000005540 biological transmission Effects 0.000 description 1
- 238000001574 biopsy Methods 0.000 description 1
- 238000001815 biotherapy Methods 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 210000004204 blood vessel Anatomy 0.000 description 1
- 210000000988 bone and bone Anatomy 0.000 description 1
- 210000002798 bone marrow cell Anatomy 0.000 description 1
- 201000008274 breast adenocarcinoma Diseases 0.000 description 1
- 201000007389 breast adenoid cystic carcinoma Diseases 0.000 description 1
- 201000008275 breast carcinoma Diseases 0.000 description 1
- 208000024119 breast tumor luminal A or B Diseases 0.000 description 1
- 230000009702 cancer cell proliferation Effects 0.000 description 1
- 238000005251 capillar electrophoresis Methods 0.000 description 1
- 150000001720 carbohydrates Chemical class 0.000 description 1
- 231100000357 carcinogen Toxicity 0.000 description 1
- 239000003183 carcinogenic agent Substances 0.000 description 1
- 239000000969 carrier Substances 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 229960000419 catumaxomab Drugs 0.000 description 1
- 230000021164 cell adhesion Effects 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 230000025084 cell cycle arrest Effects 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 230000005859 cell recognition Effects 0.000 description 1
- 230000009134 cell regulation Effects 0.000 description 1
- 230000010307 cell transformation Effects 0.000 description 1
- 230000003833 cell viability Effects 0.000 description 1
- 210000002421 cell wall Anatomy 0.000 description 1
- 230000010001 cellular homeostasis Effects 0.000 description 1
- 210000003169 central nervous system Anatomy 0.000 description 1
- 210000004718 centriole Anatomy 0.000 description 1
- 210000002230 centromere Anatomy 0.000 description 1
- QUPYHCHUQVNFJW-UHFFFAOYSA-M cesium;thiocyanate Chemical compound [Cs+].[S-]C#N QUPYHCHUQVNFJW-UHFFFAOYSA-M 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- AOXOCDRNSPFDPE-UKEONUMOSA-N chembl413654 Chemical compound C([C@H](C(=O)NCC(=O)N[C@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@H](CCSC)C(=O)N[C@H](CC(O)=O)C(=O)N[C@H](CC=1C=CC=CC=1)C(N)=O)NC(=O)[C@@H](C)NC(=O)[C@@H](CCC(O)=O)NC(=O)[C@@H](CCC(O)=O)NC(=O)[C@@H](CCC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H]1N(CCC1)C(=O)CNC(=O)[C@@H](N)CCC(O)=O)C1=CC=C(O)C=C1 AOXOCDRNSPFDPE-UKEONUMOSA-N 0.000 description 1
- 230000002925 chemical effect Effects 0.000 description 1
- 125000003636 chemical group Chemical group 0.000 description 1
- 230000000973 chemotherapeutic effect Effects 0.000 description 1
- 208000018805 childhood acute lymphoblastic leukemia Diseases 0.000 description 1
- 201000011633 childhood acute lymphocytic leukemia Diseases 0.000 description 1
- 208000011654 childhood malignant neoplasm Diseases 0.000 description 1
- 229960004630 chlorambucil Drugs 0.000 description 1
- JCKYGMPEJWAADB-UHFFFAOYSA-N chlorambucil Chemical compound OC(=O)CCCC1=CC=C(N(CCCl)CCCl)C=C1 JCKYGMPEJWAADB-UHFFFAOYSA-N 0.000 description 1
- VDQQXEISLMTGAB-UHFFFAOYSA-N chloramine T Chemical compound [Na+].CC1=CC=C(S(=O)(=O)[N-]Cl)C=C1 VDQQXEISLMTGAB-UHFFFAOYSA-N 0.000 description 1
- 150000003841 chloride salts Chemical class 0.000 description 1
- 210000004252 chorionic villi Anatomy 0.000 description 1
- 210000003483 chromatin Anatomy 0.000 description 1
- 229960002376 chymotrypsin Drugs 0.000 description 1
- 210000004081 cilia Anatomy 0.000 description 1
- 206010073251 clear cell renal cell carcinoma Diseases 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 239000003245 coal Substances 0.000 description 1
- 201000010897 colon adenocarcinoma Diseases 0.000 description 1
- 230000000112 colonic effect Effects 0.000 description 1
- 210000004922 colonic epithelial cell Anatomy 0.000 description 1
- 230000005757 colony formation Effects 0.000 description 1
- 201000002758 colorectal adenoma Diseases 0.000 description 1
- 230000004732 colorectal carcinogenesis Effects 0.000 description 1
- 238000009096 combination chemotherapy Methods 0.000 description 1
- 230000000052 comparative effect Effects 0.000 description 1
- 239000002131 composite material Substances 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 239000012141 concentrate Substances 0.000 description 1
- 238000004624 confocal microscopy Methods 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 239000010949 copper Substances 0.000 description 1
- 229910001431 copper ion Inorganic materials 0.000 description 1
- 230000009260 cross reactivity Effects 0.000 description 1
- XLJMAIOERFSOGZ-UHFFFAOYSA-M cyanate Chemical compound [O-]C#N XLJMAIOERFSOGZ-UHFFFAOYSA-M 0.000 description 1
- OILAIQUEIWYQPH-UHFFFAOYSA-N cyclohexane-1,2-dione Chemical compound O=C1CCCCC1=O OILAIQUEIWYQPH-UHFFFAOYSA-N 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 230000001461 cytolytic effect Effects 0.000 description 1
- 210000004292 cytoskeleton Anatomy 0.000 description 1
- 210000000172 cytosol Anatomy 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- GYOZYWVXFNDGLU-XLPZGREQSA-N dTMP Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](COP(O)(O)=O)[C@@H](O)C1 GYOZYWVXFNDGLU-XLPZGREQSA-N 0.000 description 1
- 108010011219 dUTP pyrophosphatase Proteins 0.000 description 1
- 229960002465 dabrafenib Drugs 0.000 description 1
- BFSMGDJOXZAERB-UHFFFAOYSA-N dabrafenib Chemical compound S1C(C(C)(C)C)=NC(C=2C(=C(NS(=O)(=O)C=3C(=CC=CC=3F)F)C=CC=2)F)=C1C1=CC=NC(N)=N1 BFSMGDJOXZAERB-UHFFFAOYSA-N 0.000 description 1
- 229960003901 dacarbazine Drugs 0.000 description 1
- 229950007409 dacetuzumab Drugs 0.000 description 1
- 125000001295 dansyl group Chemical group [H]C1=C([H])C(N(C([H])([H])[H])C([H])([H])[H])=C2C([H])=C([H])C([H])=C(C2=C1[H])S(*)(=O)=O 0.000 description 1
- 230000007850 degeneration Effects 0.000 description 1
- 230000000593 degrading effect Effects 0.000 description 1
- 230000003111 delayed effect Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 230000003831 deregulation Effects 0.000 description 1
- 125000004386 diacrylate group Chemical group 0.000 description 1
- 125000004989 dicarbonyl group Chemical group 0.000 description 1
- FFYPMLJYZAEMQB-UHFFFAOYSA-N diethyl pyrocarbonate Chemical compound CCOC(=O)OC(=O)OCC FFYPMLJYZAEMQB-UHFFFAOYSA-N 0.000 description 1
- 208000030315 diffuse gastric adenocarcinoma Diseases 0.000 description 1
- 230000007646 directional migration Effects 0.000 description 1
- 230000005750 disease progression Effects 0.000 description 1
- 230000009433 disease-worsening effect Effects 0.000 description 1
- 230000005782 double-strand break Effects 0.000 description 1
- 230000034431 double-strand break repair via homologous recombination Effects 0.000 description 1
- 238000013399 early diagnosis Methods 0.000 description 1
- 230000000678 effect on lipid Effects 0.000 description 1
- 210000003162 effector t lymphocyte Anatomy 0.000 description 1
- 238000001962 electrophoresis Methods 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 238000001437 electrospray ionisation time-of-flight quadrupole detection Methods 0.000 description 1
- 238000009261 endocrine therapy Methods 0.000 description 1
- 229940034984 endocrine therapy antineoplastic and immunomodulating agent Drugs 0.000 description 1
- 230000012202 endocytosis Effects 0.000 description 1
- 201000003908 endometrial adenocarcinoma Diseases 0.000 description 1
- 208000029382 endometrium adenocarcinoma Diseases 0.000 description 1
- 230000003511 endothelial effect Effects 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 229960001904 epirubicin Drugs 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- 230000009786 epithelial differentiation Effects 0.000 description 1
- 208000010932 epithelial neoplasm Diseases 0.000 description 1
- 230000004743 esophageal carcinogenesis Effects 0.000 description 1
- 229960005309 estradiol Drugs 0.000 description 1
- 102000015694 estrogen receptors Human genes 0.000 description 1
- 108010038795 estrogen receptors Proteins 0.000 description 1
- 201000007280 estrogen-receptor negative breast cancer Diseases 0.000 description 1
- 201000007281 estrogen-receptor positive breast cancer Diseases 0.000 description 1
- 229960003399 estrone Drugs 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 230000017188 evasion or tolerance of host immune response Effects 0.000 description 1
- 210000001808 exosome Anatomy 0.000 description 1
- 201000001343 fallopian tube carcinoma Diseases 0.000 description 1
- 229950008454 favipiravir Drugs 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 230000001605 fetal effect Effects 0.000 description 1
- 210000002950 fibroblast Anatomy 0.000 description 1
- 230000003176 fibrotic effect Effects 0.000 description 1
- 239000012467 final product Substances 0.000 description 1
- 238000009093 first-line therapy Methods 0.000 description 1
- 102000006815 folate receptor Human genes 0.000 description 1
- 108020005243 folate receptor Proteins 0.000 description 1
- 239000011724 folic acid Substances 0.000 description 1
- 229960000304 folic acid Drugs 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 231100000221 frame shift mutation induction Toxicity 0.000 description 1
- 230000037433 frameshift Effects 0.000 description 1
- 239000012520 frozen sample Substances 0.000 description 1
- 201000008396 gallbladder adenocarcinoma Diseases 0.000 description 1
- 201000010175 gallbladder cancer Diseases 0.000 description 1
- 150000002270 gangliosides Chemical class 0.000 description 1
- 229960002584 gefitinib Drugs 0.000 description 1
- XGALLCVXEZPNRQ-UHFFFAOYSA-N gefitinib Chemical compound C=12C=C(OCCCN3CCOCC3)C(OC)=CC2=NC=NC=1NC1=CC=C(F)C(Cl)=C1 XGALLCVXEZPNRQ-UHFFFAOYSA-N 0.000 description 1
- 230000005861 gene abnormality Effects 0.000 description 1
- 230000004547 gene signature Effects 0.000 description 1
- 230000009368 gene silencing by RNA Effects 0.000 description 1
- 102000054766 genetic haplotypes Human genes 0.000 description 1
- 230000007614 genetic variation Effects 0.000 description 1
- 239000003862 glucocorticoid Substances 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-L glutamate group Chemical group N[C@@H](CCC(=O)[O-])C(=O)[O-] WHUUTDBJXJRKMK-VKHMYHEASA-L 0.000 description 1
- 125000000291 glutamic acid group Chemical group N[C@@H](CCC(O)=O)C(=O)* 0.000 description 1
- 210000001173 gonocyte Anatomy 0.000 description 1
- 159000000011 group IA salts Chemical class 0.000 description 1
- 230000009036 growth inhibition Effects 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 230000002489 hematologic effect Effects 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 229920000140 heteropolymer Polymers 0.000 description 1
- 125000000487 histidyl group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C([H])=N1 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 210000005260 human cell Anatomy 0.000 description 1
- 229940084986 human chorionic gonadotropin Drugs 0.000 description 1
- 239000000017 hydrogel Substances 0.000 description 1
- 230000000148 hypercalcaemia Effects 0.000 description 1
- 208000030915 hypercalcemia disease Diseases 0.000 description 1
- 230000006607 hypermethylation Effects 0.000 description 1
- 238000009802 hysterectomy Methods 0.000 description 1
- 230000002519 immonomodulatory effect Effects 0.000 description 1
- 230000007124 immune defense Effects 0.000 description 1
- 229940127121 immunoconjugate Drugs 0.000 description 1
- 102000018358 immunoglobulin Human genes 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 238000011532 immunohistochemical staining Methods 0.000 description 1
- 229960003444 immunosuppressant agent Drugs 0.000 description 1
- 230000001861 immunosuppressant effect Effects 0.000 description 1
- 239000003018 immunosuppressive agent Substances 0.000 description 1
- 229940032219 immunotherapy vaccine Drugs 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 238000011355 in situ vaccination Methods 0.000 description 1
- 230000000415 inactivating effect Effects 0.000 description 1
- 201000011489 infantile myofibromatosis Diseases 0.000 description 1
- 230000028709 inflammatory response Effects 0.000 description 1
- 238000011221 initial treatment Methods 0.000 description 1
- 238000011081 inoculation Methods 0.000 description 1
- 230000002452 interceptive effect Effects 0.000 description 1
- 229940079322 interferon Drugs 0.000 description 1
- 230000037041 intracellular level Effects 0.000 description 1
- 230000010189 intracellular transport Effects 0.000 description 1
- JDNTWHVOXJZDSN-UHFFFAOYSA-N iodoacetic acid Chemical compound OC(=O)CI JDNTWHVOXJZDSN-UHFFFAOYSA-N 0.000 description 1
- 230000005865 ionizing radiation Effects 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 125000000741 isoleucyl group Chemical group [H]N([H])C(C(C([H])([H])[H])C([H])([H])C([H])([H])[H])C(=O)O* 0.000 description 1
- 210000002510 keratinocyte Anatomy 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 210000003292 kidney cell Anatomy 0.000 description 1
- 229940043355 kinase inhibitor Drugs 0.000 description 1
- 231100000518 lethal Toxicity 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 229950001237 lilotomab Drugs 0.000 description 1
- 206010024627 liposarcoma Diseases 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 229960002247 lomustine Drugs 0.000 description 1
- 201000005249 lung adenocarcinoma Diseases 0.000 description 1
- 230000004758 lung carcinogenesis Effects 0.000 description 1
- 201000005243 lung squamous cell carcinoma Diseases 0.000 description 1
- 208000037841 lung tumor Diseases 0.000 description 1
- OHSVLFRHMCKCQY-UHFFFAOYSA-N lutetium atom Chemical compound [Lu] OHSVLFRHMCKCQY-UHFFFAOYSA-N 0.000 description 1
- 210000002751 lymph Anatomy 0.000 description 1
- 208000022766 lymph node neoplasm Diseases 0.000 description 1
- 230000035168 lymphangiogenesis Effects 0.000 description 1
- 230000001926 lymphatic effect Effects 0.000 description 1
- 102100035860 m7GpppN-mRNA hydrolase Human genes 0.000 description 1
- 102000033952 mRNA binding proteins Human genes 0.000 description 1
- 108091000373 mRNA binding proteins Proteins 0.000 description 1
- 229940124302 mTOR inhibitor Drugs 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 239000003628 mammalian target of rapamycin inhibitor Substances 0.000 description 1
- 238000007726 management method Methods 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 238000001840 matrix-assisted laser desorption--ionisation time-of-flight mass spectrometry Methods 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000008099 melanin synthesis Effects 0.000 description 1
- 206010027191 meningioma Diseases 0.000 description 1
- 150000002730 mercury Chemical class 0.000 description 1
- 230000004060 metabolic process Effects 0.000 description 1
- 239000002207 metabolite Substances 0.000 description 1
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 1
- 108091031479 miR-204 stem-loop Proteins 0.000 description 1
- 108091032382 miR-204-1 stem-loop Proteins 0.000 description 1
- 108091085803 miR-204-2 stem-loop Proteins 0.000 description 1
- 108091089766 miR-204-3 stem-loop Proteins 0.000 description 1
- 108091073500 miR-204-4 stem-loop Proteins 0.000 description 1
- 108091053626 miR-204-5 stem-loop Proteins 0.000 description 1
- 238000002493 microarray Methods 0.000 description 1
- 238000012737 microarray-based gene expression Methods 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 201000011591 microinvasive gastric cancer Diseases 0.000 description 1
- 210000004688 microtubule Anatomy 0.000 description 1
- 230000027829 mitochondrial depolarization Effects 0.000 description 1
- 239000002829 mitogen activated protein kinase inhibitor Substances 0.000 description 1
- 230000017205 mitotic cell cycle checkpoint Effects 0.000 description 1
- 230000008600 mitotic progression Effects 0.000 description 1
- 230000003990 molecular pathway Effects 0.000 description 1
- 238000002625 monoclonal antibody therapy Methods 0.000 description 1
- 210000005087 mononuclear cell Anatomy 0.000 description 1
- 210000004877 mucosa Anatomy 0.000 description 1
- 238000012243 multiplex automated genomic engineering Methods 0.000 description 1
- 208000025113 myeloid leukemia Diseases 0.000 description 1
- AMCGLRWKUQPNKD-MRXNPFEDSA-N n-[(1r)-1-(3-cyclopentyloxyphenyl)ethyl]-3-[(2,4-dioxopyrimidin-1-yl)methoxy]propane-1-sulfonamide Chemical compound N([C@H](C)C=1C=C(OC2CCCC2)C=CC=1)S(=O)(=O)CCCOCN1C=CC(=O)NC1=O AMCGLRWKUQPNKD-MRXNPFEDSA-N 0.000 description 1
- 230000014399 negative regulation of angiogenesis Effects 0.000 description 1
- 230000017066 negative regulation of growth Effects 0.000 description 1
- 238000011227 neoadjuvant chemotherapy Methods 0.000 description 1
- 230000009826 neoplastic cell growth Effects 0.000 description 1
- 230000010309 neoplastic transformation Effects 0.000 description 1
- 230000004770 neurodegeneration Effects 0.000 description 1
- 201000011519 neuroendocrine tumor Diseases 0.000 description 1
- 201000004931 neurofibromatosis Diseases 0.000 description 1
- 208000029974 neurofibrosarcoma Diseases 0.000 description 1
- 210000000440 neutrophil Anatomy 0.000 description 1
- 208000004649 neutrophil actin dysfunction Diseases 0.000 description 1
- 229950010203 nimotuzumab Drugs 0.000 description 1
- 230000000683 nonmetastatic effect Effects 0.000 description 1
- 238000002414 normal-phase solid-phase extraction Methods 0.000 description 1
- 230000004942 nuclear accumulation Effects 0.000 description 1
- 108010054452 nuclear pore complex protein 98 Proteins 0.000 description 1
- 230000005937 nuclear translocation Effects 0.000 description 1
- 230000000269 nucleophilic effect Effects 0.000 description 1
- 230000030648 nucleus localization Effects 0.000 description 1
- 238000011580 nude mouse model Methods 0.000 description 1
- 229960003347 obinutuzumab Drugs 0.000 description 1
- 229960002450 ofatumumab Drugs 0.000 description 1
- 230000000771 oncological effect Effects 0.000 description 1
- 208000010655 oral cavity squamous cell carcinoma Diseases 0.000 description 1
- 208000022698 oropharynx squamous cell carcinoma Diseases 0.000 description 1
- 201000003733 ovarian melanoma Diseases 0.000 description 1
- 230000003647 oxidation Effects 0.000 description 1
- 238000007254 oxidation reaction Methods 0.000 description 1
- 229960001592 paclitaxel Drugs 0.000 description 1
- 201000002094 pancreatic adenocarcinoma Diseases 0.000 description 1
- 208000021010 pancreatic neuroendocrine tumor Diseases 0.000 description 1
- 208000003154 papilloma Diseases 0.000 description 1
- KTEXNACQROZXEV-PVLRGYAZSA-N parthenolide Chemical compound C1CC(/C)=C/CC[C@@]2(C)O[C@@H]2[C@H]2OC(=O)C(=C)[C@@H]21 KTEXNACQROZXEV-PVLRGYAZSA-N 0.000 description 1
- 229940069510 parthenolide Drugs 0.000 description 1
- 244000052769 pathogen Species 0.000 description 1
- 230000001717 pathogenic effect Effects 0.000 description 1
- 230000007918 pathogenicity Effects 0.000 description 1
- 230000001575 pathological effect Effects 0.000 description 1
- 230000009745 pathological pathway Effects 0.000 description 1
- 230000016446 peptide cross-linking Effects 0.000 description 1
- 229940125667 peptide vaccine candidate Drugs 0.000 description 1
- 229930192851 perforin Natural products 0.000 description 1
- 210000004976 peripheral blood cell Anatomy 0.000 description 1
- 210000005105 peripheral blood lymphocyte Anatomy 0.000 description 1
- 230000002093 peripheral effect Effects 0.000 description 1
- 230000002085 persistent effect Effects 0.000 description 1
- 239000005426 pharmaceutical component Substances 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- 238000009521 phase II clinical trial Methods 0.000 description 1
- 238000009522 phase III clinical trial Methods 0.000 description 1
- 239000003757 phosphotransferase inhibitor Substances 0.000 description 1
- 238000002428 photodynamic therapy Methods 0.000 description 1
- 210000002826 placenta Anatomy 0.000 description 1
- 239000013612 plasmid Substances 0.000 description 1
- 150000003057 platinum Chemical class 0.000 description 1
- 201000010065 polycystic ovary syndrome Diseases 0.000 description 1
- 238000006116 polymerization reaction Methods 0.000 description 1
- 208000015768 polyposis Diseases 0.000 description 1
- 230000008092 positive effect Effects 0.000 description 1
- 230000023603 positive regulation of transcription initiation, DNA-dependent Effects 0.000 description 1
- 238000011248 postoperative chemotherapy Methods 0.000 description 1
- 230000012357 postreplication repair Effects 0.000 description 1
- 230000001124 posttranscriptional effect Effects 0.000 description 1
- GKKCIDNWFBPDBW-UHFFFAOYSA-M potassium cyanate Chemical compound [K]OC#N GKKCIDNWFBPDBW-UHFFFAOYSA-M 0.000 description 1
- ZNNZYHKDIALBAK-UHFFFAOYSA-M potassium thiocyanate Chemical compound [K+].[S-]C#N ZNNZYHKDIALBAK-UHFFFAOYSA-M 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000004321 preservation Methods 0.000 description 1
- 208000016800 primary central nervous system lymphoma Diseases 0.000 description 1
- CPTBDICYNRMXFX-UHFFFAOYSA-N procarbazine Chemical compound CNNCC1=CC=C(C(=O)NC(C)C)C=C1 CPTBDICYNRMXFX-UHFFFAOYSA-N 0.000 description 1
- 229960000624 procarbazine Drugs 0.000 description 1
- 201000007282 progesterone-receptor negative breast cancer Diseases 0.000 description 1
- 230000002250 progressing effect Effects 0.000 description 1
- 230000000770 proinflammatory effect Effects 0.000 description 1
- 229940097325 prolactin Drugs 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 201000005825 prostate adenocarcinoma Diseases 0.000 description 1
- 208000023958 prostate neoplasm Diseases 0.000 description 1
- 108060006633 protein kinase Proteins 0.000 description 1
- 239000003197 protein kinase B inhibitor Substances 0.000 description 1
- 230000007026 protein scission Effects 0.000 description 1
- 230000016434 protein splicing Effects 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 230000006337 proteolytic cleavage Effects 0.000 description 1
- 238000004445 quantitative analysis Methods 0.000 description 1
- 238000009801 radical cystectomy Methods 0.000 description 1
- 238000007347 radical substitution reaction Methods 0.000 description 1
- 150000003254 radicals Chemical group 0.000 description 1
- 239000000376 reactant Substances 0.000 description 1
- 230000009257 reactivity Effects 0.000 description 1
- 230000008707 rearrangement Effects 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000000306 recurrent effect Effects 0.000 description 1
- 230000008929 regeneration Effects 0.000 description 1
- 238000011069 regeneration method Methods 0.000 description 1
- 230000007261 regionalization Effects 0.000 description 1
- 229960004836 regorafenib Drugs 0.000 description 1
- FNHKPVJBJVTLMP-UHFFFAOYSA-N regorafenib Chemical compound C1=NC(C(=O)NC)=CC(OC=2C=C(F)C(NC(=O)NC=3C=C(C(Cl)=CC=3)C(F)(F)F)=CC=2)=C1 FNHKPVJBJVTLMP-UHFFFAOYSA-N 0.000 description 1
- 230000001373 regressive effect Effects 0.000 description 1
- 230000006884 regulation of angiogenesis Effects 0.000 description 1
- 210000005084 renal tissue Anatomy 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 230000028617 response to DNA damage stimulus Effects 0.000 description 1
- 201000009410 rhabdomyosarcoma Diseases 0.000 description 1
- 102220259646 rs764841552 Human genes 0.000 description 1
- 210000003079 salivary gland Anatomy 0.000 description 1
- 201000007416 salivary gland adenoid cystic carcinoma Diseases 0.000 description 1
- 210000004706 scrotum Anatomy 0.000 description 1
- 238000009094 second-line therapy Methods 0.000 description 1
- 238000011519 second-line treatment Methods 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 230000001235 sensitizing effect Effects 0.000 description 1
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 1
- 108091006024 signal transducing proteins Proteins 0.000 description 1
- 102000034285 signal transducing proteins Human genes 0.000 description 1
- 238000011125 single therapy Methods 0.000 description 1
- 238000009097 single-agent therapy Methods 0.000 description 1
- 229940074386 skatole Drugs 0.000 description 1
- 210000002363 skeletal muscle cell Anatomy 0.000 description 1
- 201000010153 skin papilloma Diseases 0.000 description 1
- 201000010106 skin squamous cell carcinoma Diseases 0.000 description 1
- 210000000813 small intestine Anatomy 0.000 description 1
- 201000010329 small intestine neuroendocrine neoplasm Diseases 0.000 description 1
- VGTPCRGMBIAPIM-UHFFFAOYSA-M sodium thiocyanate Chemical compound [Na+].[S-]C#N VGTPCRGMBIAPIM-UHFFFAOYSA-M 0.000 description 1
- 238000010532 solid phase synthesis reaction Methods 0.000 description 1
- 239000000243 solution Substances 0.000 description 1
- 210000001082 somatic cell Anatomy 0.000 description 1
- 230000020347 spindle assembly Effects 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 108010088201 squamous cell carcinoma-related antigen Proteins 0.000 description 1
- 210000004085 squamous epithelial cell Anatomy 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 238000011255 standard chemotherapy Methods 0.000 description 1
- 239000007858 starting material Substances 0.000 description 1
- 238000011476 stem cell transplantation Methods 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 206010042863 synovial sarcoma Diseases 0.000 description 1
- 238000011521 systemic chemotherapy Methods 0.000 description 1
- 229940037128 systemic glucocorticoids Drugs 0.000 description 1
- RCINICONZNJXQF-MZXODVADSA-N taxol Chemical compound O([C@@H]1[C@@]2(C[C@@H](C(C)=C(C2(C)C)[C@H](C([C@]2(C)[C@@H](O)C[C@H]3OC[C@]3([C@H]21)OC(C)=O)=O)OC(=O)C)OC(=O)[C@H](O)[C@@H](NC(=O)C=1C=CC=CC=1)C=1C=CC=CC=1)O)C(=O)C1=CC=CC=C1 RCINICONZNJXQF-MZXODVADSA-N 0.000 description 1
- 208000024662 testicular seminoma Diseases 0.000 description 1
- 229960003604 testosterone Drugs 0.000 description 1
- 238000004809 thin layer chromatography Methods 0.000 description 1
- 125000003396 thiol group Chemical group [H]S* 0.000 description 1
- 210000001541 thymus gland Anatomy 0.000 description 1
- 201000006134 tongue cancer Diseases 0.000 description 1
- 201000002743 tongue squamous cell carcinoma Diseases 0.000 description 1
- 208000025093 tonsil carcinoma Diseases 0.000 description 1
- 229960001479 tosylchloramide sodium Drugs 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 229960004066 trametinib Drugs 0.000 description 1
- LIRYPHYGHXZJBZ-UHFFFAOYSA-N trametinib Chemical compound CC(=O)NC1=CC=CC(N2C(N(C3CC3)C(=O)C3=C(NC=4C(=CC(I)=CC=4)F)N(C)C(=O)C(C)=C32)=O)=C1 LIRYPHYGHXZJBZ-UHFFFAOYSA-N 0.000 description 1
- 238000001890 transfection Methods 0.000 description 1
- 206010044412 transitional cell carcinoma Diseases 0.000 description 1
- 102000035160 transmembrane proteins Human genes 0.000 description 1
- 108091005703 transmembrane proteins Proteins 0.000 description 1
- 229960000575 trastuzumab Drugs 0.000 description 1
- 229950007217 tremelimumab Drugs 0.000 description 1
- 239000012588 trypsin Substances 0.000 description 1
- 239000000107 tumor biomarker Substances 0.000 description 1
- 230000005740 tumor formation Effects 0.000 description 1
- 230000005851 tumor immunogenicity Effects 0.000 description 1
- 102000003298 tumor necrosis factor receptor Human genes 0.000 description 1
- 208000025444 tumor of salivary gland Diseases 0.000 description 1
- 230000005760 tumorsuppression Effects 0.000 description 1
- 229940121358 tyrosine kinase inhibitor Drugs 0.000 description 1
- 239000005483 tyrosine kinase inhibitor Substances 0.000 description 1
- 150000004917 tyrosine kinase inhibitor derivatives Chemical class 0.000 description 1
- 125000001493 tyrosinyl group Chemical group [H]OC1=C([H])C([H])=C(C([H])=C1[H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 208000024719 uterine cervix neoplasm Diseases 0.000 description 1
- 229960000653 valrubicin Drugs 0.000 description 1
- ZOCKGBMQLCSHFP-KQRAQHLDSA-N valrubicin Chemical compound O([C@H]1C[C@](CC2=C(O)C=3C(=O)C4=CC=CC(OC)=C4C(=O)C=3C(O)=C21)(O)C(=O)COC(=O)CCCC)[C@H]1C[C@H](NC(=O)C(F)(F)F)[C@H](O)[C@H](C)O1 ZOCKGBMQLCSHFP-KQRAQHLDSA-N 0.000 description 1
- 239000003981 vehicle Substances 0.000 description 1
- 229960003862 vemurafenib Drugs 0.000 description 1
- GPXBXXGIAQBQNI-UHFFFAOYSA-N vemurafenib Chemical compound CCCS(=O)(=O)NC1=CC=C(F)C(C(=O)C=2C3=CC(=CN=C3NC=2)C=2C=CC(Cl)=CC=2)=C1F GPXBXXGIAQBQNI-UHFFFAOYSA-N 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- OGWKCGZFUXNPDA-XQKSVPLYSA-N vincristine Chemical compound C([N@]1C[C@@H](C[C@]2(C(=O)OC)C=3C(=CC4=C([C@]56[C@H]([C@@]([C@H](OC(C)=O)[C@]7(CC)C=CCN([C@H]67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)C[C@@](C1)(O)CC)CC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-XQKSVPLYSA-N 0.000 description 1
- OGWKCGZFUXNPDA-UHFFFAOYSA-N vincristine Natural products C1C(CC)(O)CC(CC2(C(=O)OC)C=3C(=CC4=C(C56C(C(C(OC(C)=O)C7(CC)C=CCN(C67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)CN1CCC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-UHFFFAOYSA-N 0.000 description 1
- 229960004528 vincristine Drugs 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 229940023147 viral vector vaccine Drugs 0.000 description 1
- 230000009278 visceral effect Effects 0.000 description 1
Images




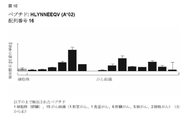


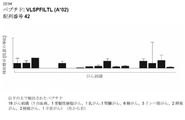



























Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4748—Tumour specific antigens; Tumour rejection antigen precursors [TRAP], e.g. MAGE
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
- A61K35/14—Blood; Artificial blood
- A61K35/17—Lymphocytes; B-cells; T-cells; Natural killer cells; Interferon-activated or cytokine-activated lymphocytes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/04—Peptides having up to 20 amino acids in a fully defined sequence; Derivatives thereof
- A61K38/08—Peptides having 5 to 11 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/0005—Vertebrate antigens
- A61K39/0011—Cancer antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/385—Haptens or antigens, bound to carriers
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
- A61K39/39533—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals
- A61K39/39558—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals against tumor tissues, cells, antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/10—Cellular immunotherapy characterised by the cell type used
- A61K40/11—T-cells, e.g. tumour infiltrating lymphocytes [TIL] or regulatory T [Treg] cells; Lymphokine-activated killer [LAK] cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/40—Cellular immunotherapy characterised by antigens that are targeted or presented by cells of the immune system
- A61K40/41—Vertebrate antigens
- A61K40/42—Cancer antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/7051—T-cell receptor (TcR)-CD3 complex
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70539—MHC-molecules, e.g. HLA-molecules
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2833—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against MHC-molecules, e.g. HLA-molecules
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/30—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants from tumour cells
- C07K16/3076—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants from tumour cells against structure-related tumour-associated moieties
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/32—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against translation products of oncogenes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/06—Linear peptides containing only normal peptide links having 5 to 11 amino acids
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/08—Linear peptides containing only normal peptide links having 12 to 20 amino acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/115—Aptamers, i.e. nucleic acids binding a target molecule specifically and with high affinity without hybridising therewith ; Nucleic acids binding to non-nucleic acids, e.g. aptamers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0636—T lymphocytes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6876—Nucleic acid products used in the analysis of nucleic acids, e.g. primers or probes
- C12Q1/6883—Nucleic acid products used in the analysis of nucleic acids, e.g. primers or probes for diseases caused by alterations of genetic material
- C12Q1/6886—Nucleic acid products used in the analysis of nucleic acids, e.g. primers or probes for diseases caused by alterations of genetic material for cancer
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/574—Immunoassay; Biospecific binding assay; Materials therefor for cancer
- G01N33/57484—Immunoassay; Biospecific binding assay; Materials therefor for cancer involving compounds serving as markers for tumor, cancer, neoplasia, e.g. cellular determinants, receptors, heat shock/stress proteins, A-protein, oligosaccharides, metabolites
- G01N33/57492—Immunoassay; Biospecific binding assay; Materials therefor for cancer involving compounds serving as markers for tumor, cancer, neoplasia, e.g. cellular determinants, receptors, heat shock/stress proteins, A-protein, oligosaccharides, metabolites involving compounds localized on the membrane of tumor or cancer cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/515—Animal cells
- A61K2039/5158—Antigen-pulsed cells, e.g. T-cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/40—Fusion polypeptide containing a tag for immunodetection, or an epitope for immunisation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/16—Aptamers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/50—Cell markers; Cell surface determinants
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2600/00—Oligonucleotides characterized by their use
- C12Q2600/106—Pharmacogenomics, i.e. genetic variability in individual responses to drugs and drug metabolism
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2600/00—Oligonucleotides characterized by their use
- C12Q2600/156—Polymorphic or mutational markers
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/435—Assays involving biological materials from specific organisms or of a specific nature from animals; from humans
- G01N2333/705—Assays involving receptors, cell surface antigens or cell surface determinants
- G01N2333/70503—Immunoglobulin superfamily, e.g. VCAMs, PECAM, LFA-3
- G01N2333/70539—MHC-molecules, e.g. HLA-molecules
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Immunology (AREA)
- General Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Medicinal Chemistry (AREA)
- Biomedical Technology (AREA)
- Zoology (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biophysics (AREA)
- Biotechnology (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Cell Biology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Epidemiology (AREA)
- Microbiology (AREA)
- Pharmacology & Pharmacy (AREA)
- General Engineering & Computer Science (AREA)
- Oncology (AREA)
- Hematology (AREA)
- Gastroenterology & Hepatology (AREA)
- Physics & Mathematics (AREA)
- Toxicology (AREA)
- Mycology (AREA)
- Analytical Chemistry (AREA)
- Pathology (AREA)
- Plant Pathology (AREA)
- Urology & Nephrology (AREA)
- Hospice & Palliative Care (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
Description
2012年には、1410万件の新規がん症例、3260万人のがん患者(診断の5年以内)、および820万件のがん死亡が世界的に推定された(Ferlay et al.,2013;Bray et al.,2013)。
CLLは現在治療可能ではないが、多くの患者は、遅い疾患進行または症状悪化のみを示す。患者が治療の早期開始の恩恵を受けることはないので、最初のアプローチは「経過観察」である(Richards et al.,1999)。症候性のまたは迅速に進行する疾患がある患者に対しては、いくつかの治療選択肢が利用できる。これらとしては、化学療法;標的療法;モノクローナル抗体、キメラ抗原受容体(CAR)、および能動的免疫療法のような免疫ベース治療法;および幹細胞移植が挙げられる。
活性免疫療法としては、遺伝子治療、全修飾腫瘍細胞ワクチン、DCベースのワクチン、および腫瘍関連抗原(TAA)由来ペプチドワクチンのストラテジーが挙げられる。
結腸直腸がん(CRC)の病期次第で、異なる標準的治療法が、結腸および直腸がんに対して利用できる。標準手順としては、外科手術、放射線療法、化学療法、およびCRC標的療法が挙げられる(Berman et al.,2015a;Berman et al.,2015b)。
食道がんの一次治療ストラテジーは、腫瘍病期と部位、組織型、および患者の医学的状態に左右される。扁平上皮がん患者の小さなサブグループを除いて、手術のみでは十分でない。
胃がん(GC)は、粘膜層を覆う細胞内で始まり、増殖するにつれて外層を通して広がる。外科手術は胃がんの一次治療法であり、唯一の根治的治療法である。進行したGCに対する現行の薬剤投与計画は有効性に劣り、低い5年生存率をもたらす。免疫療法は、GC患者の生存率を改善する代替アプローチであるかもしれない。腫瘍関連リンパ球およびサイトカイン誘導キラー細胞の養子免疫伝達;HER2/neu、MAGE−3または血管内皮成長因子受容体1および2を標的とするペプチドベースのワクチン;およびHER2/neuを標的とする樹状細胞ベースのワクチンが、臨床GC治験において有望な結果を示した。免疫チェックポイント阻害および遺伝子操作T細胞は、追加的な治療の選択肢に相当するかもしれず、それは現在前臨床および臨床試験で評価中である(Matsueda and Graham,2014)。
神経膠芽腫(WHOグレードIV)に対する療の選択肢は、非常に限られている。ドイツ神経学会によって発表された指針によれば、若年患者の標準的治療法には、腫瘍の切除または生検、限局性放射線療法、およびテモゾロミドまたはCCNU/ロムスチンによる、またはプロカルバジンとCCNUおよびビンクリスチン(PCV)との併用による、化学療法が含まれる。米国、カナダ、スイスでは、ベバシズマブ(抗VEGF抗体)による治療も、再発治療のために承認されている(Leitlinien fur Diagnostik und Therapie in der Neurologie,2014)。
疾病管理は、診断時の腫瘍段階、および肝臓の全体的な状態に左右される。手術が治療選択肢ではない場合、異なるその他の治療法が利用できる。
黒色腫の標準的治療は、周囲の健常組織を含めた完全外科的切除である。切除が完全ではない、または全く不可能な場合、患者は一次放射線療法を受け、これは進行期のインターフェロンα投与と組み合わせされ得る(ステージIIB/CおよびIIIA−C)。
治療選択肢は、がん型(小細胞または非小細胞)および病期によって決定され、外科手術、放射線療法、化学療法、およびベバシズマブやエルロチニブやゲフィチニブなどの標的生物学的療法が挙げられる
外科的切除は、初期ならびに進行期卵巣がんにおける一次治療である術後化学療法の適応対象でない非常に低悪性度の卵巣がん(IA期、グレード1)を除いて、外科的除去には、白金類似体による全身化学療法が続く。
膵臓がん患者のための治療の選択肢は、非常に限られている。効果的治療のための1つの大きな問題は、診断時における典型的に進行した腫瘍病期である。さらに、膵臓がんは化学療法薬に対してかなり抵抗性であり、それは、緻密で乏血管性の線維形成性腫瘍間質によって引き起こされるかもしれない。
前立腺がんのための治療ストラテジーは、主にがん病期に左右される。樹状細胞ベースのワクチンであるシプロイセルTは、FDAによって承認された最初の抗がんワクチンであった。CRPCがある患者の生存期間に対するその好ましい効果のために、さらなる免疫療法の開発に向けた多大な努力がなされている。ワクチン接種ストラテジーについては、ペプチドワクチンである前立腺特異的抗原(PSA)−TRICOM、個別化ペプチドワクチンであるPPV、DNAワクチンであるpTVG−HP、およびGM−CSF発現全細胞ワクチンであるGVAXが、異なる臨床試験で有望な結果を示した。さらに、シプロイセルT以外の樹状細胞ベースのワクチン、すなわちBPX−101およびDCVAC/Paは、前立腺がん患者において臨床的奏効を引き起こすことが示された。イピリムマブおよびニボルマブのような免疫チェックポイント阻害物質が、単剤療法として、ならびにアンドロゲン奪取療法、局所放射線療法、PSA−TRICOM、およびGVAXをはじめとするその他の治療法との併用で、現在臨床試験で評価されている。第II相試験で進行を有意に遅延させ、無憎悪生存期間を延長させた免疫調節物質タスキニモドは、現在第III相試験でさらに調査されている。もう一つの免疫修飾物質であるレナリドミドは、初期臨床試験で有望な効果をもたらしたが、第III相試験では生存率を改善できなかった。これらの期待外れな結果にもかかわらず、さらなるレナリドミド治験が継続されている(Quinn et al.,2015)。
初期治療は、最も一般的には、罹患した腎臓の部分的または完全除去のどちらかであり、依然として根治的治療の主流である(Rini et al.,2008)。RCCの既知の免疫原性(immunogenity)は、進行したRCCにおける免疫療法およびがんワクチンの使用を支持する基礎に相当する。リンパ球PD−1発現と、RCCの進行期、グレード、および予後との間の興味深い相関、ならびにRCC腫瘍細胞による選択的PD−L1発現と、より芳しくない臨床転帰との潜在的関連性は、単独での、または抗血管新生薬またはその他の免疫療法アプローチと併用される、RCC治療のための新規抗PD−1/PD−L1薬の開発をもたらした(Massari et al.,2015)。進行したRCCでは、TRIST試験と称される第III相がんワクチン治験が、第一選択標準的治療に追加された、TroVax(ポックスウイルスベクターと共に腫瘍関連抗原5T4を使用したワクチン)が、局所的進行またはmRCCがある患者の生存期間を延長するかどうかを評価する。いずれの群でも生存期間中央値に達しておらず、399人の患者(54%)が試験に残っているが、データの解析は、TroVaxが免疫学的に活性であり、5T4特異的抗体応答の強度と改善された生存期間との間に相関があることの双方を実証する、以前の臨床結果を裏付けている。さらにRCCにおいて過剰発現されるエピトープを使用してペプチドワクチンを検索する、いくつかの研究がある。
SCLCの治療および予後は、診断されたがん病期に大きく左右される。免疫療法は、過度に調査されたがん療法の分野に相当する。SCLCの治療には、様々なアプローチが点在する。アプローチの1つは、天然ヒト免疫抑制剤であるCTLA−4のブロックを標的とする。CTLA−4の阻害は、がんと戦うために免疫系を増強することを意図する。近年、SCLCの治療のための有望な免疫チェックポイント阻害剤の開発が始まっている。もう一つのアプローチは抗がんワクチンに基づき、それは現在臨床試験においてSCLC治療のために利用できる(American Cancer Society,2015;National Cancer Institute,2015)。
AMLの1つの治療選択肢は、CD33を抗体薬物コンジュゲート(抗CD33+カレキアマイシン、SGN−CD33a、抗CD33+アクチニウム−225)、二重特異性抗体(CD33+CD3(AMG330)またはCD33+CD16の認識)およびキメラ抗原受容体(CAR)で標的化することである(Estey,2014)。
NHLの治療は、組織型および病期に左右され(National Cancer Institute,2015):リンパ腫患者では、自然腫瘍退縮が観察され得る。したがって、能動免疫療法が治療選択肢である(Palomba,2012)。
子宮内膜がんの治療は、病期に依存的する。子宮内膜(endometrical)がんの大部分は、通常は診断時に子宮体に限定されて、子宮摘出によって治癒され得る、高度から中程度に分化した類内膜腺がんを含んでなる(World Cancer Report,2014)。
胆管がんは診断が困難なため、大部分が進行期に同定されている。胆管がんは治療が困難であり、通常は致死性である。
膀胱がんの標準的治療法としては、外科手術、放射線療法、化学療法、および免疫療法が挙げられる。
頭頸部扁平上皮がん(HNSCC)は、疫学、病因、および治療において差異を有する不均一腫瘍である(Economopoulou et al.,2016)。
a)がん精巣抗原:T細胞によって認識され得る初めて同定されたTAAはこのクラスに属し、元々はがん精巣(CT)抗原と称されたが、それは、そのメンバーが組織学的に異なるヒト腫瘍において発現し、正常組織では精巣の精母細胞/精原細胞のみに存在し、時として胎盤に存在するためであった。精巣の細胞は、クラスIおよびII HLA分子を発現しないので、これらの抗原は正常組織のT細胞によって認識され得ず、したがって免疫学的に腫瘍特異的と見なされる。CT抗原の周知の例は、MAGEファミリーメンバーおよびNY−ESO−1である。
b)分化抗原:これらのTAAは、腫瘍と、それから腫瘍が生じる正常組織との間で共有される。既知の分化抗原のほとんどは、黒色腫および正常メラノサイトに見いだされる。これらのメラノサイト系関連タンパク質の多くは、メラニン生合成に関与し、したがって腫瘍特異的でないが、それでもなおがん免疫療法のために広く利用されている。例としては、黒色腫に対するチロシナーゼとMelan−A/MART−1、または前立腺がんに対するPSAが挙げられるが、これに限定されるものではない。
c)過剰発現TAA:広範に発現されるTAAをエンコードする遺伝子は、組織学的に異なる型の腫瘍において検出され、多数の正常組織においても概してより低い発現レベルで検出されている。正常組織によってプロセスされて潜在的に提示さるエピトープの多くは、T細胞認識の閾値レベル未満であり得る一方で、腫瘍細胞におけるそれらの過剰発現は、以前確立された免疫寛容を破壊することにより、抗がん応答を始動し得る。このクラスのTAAの顕著な例は、Her−2/neu、サバイビン、テロメラーゼまたはWT1である。
d)腫瘍特異的抗原:これらのユニークなTAAは、正常な遺伝子(β−カテニン、CDK4など)の変異から生じる。これらの分子変化のいくつかは、腫瘍性形質転換および/または進行に関連している。腫瘍特異的抗原は、通常、正常組織に対する自己免疫反応のリスクなしに、強力な免疫応答を誘導できる。他方、これらのTAAは、ほとんどの場合、その上でそれらが同定されたまさにその腫瘍のみと関係があり、通常は、多くの個々の腫瘍間で共有されない。腫瘍特異的(関連)イソ型を有するタンパク質では、ペプチドの腫瘍特異性(または関連性)はまた、ペプチドが腫瘍(関連)エクソンに由来する場合に生じてもよい。
e)異常な翻訳後修飾から生じるTAA:このようなTAAは、特異的でなく腫瘍において過剰発現もされないタンパク質から生じてもよいが、それでもなお、腫瘍において主に活性である翻訳後プロセスによって腫瘍関連になる。このクラスの例は、腫瘍にMUC1のような新規エピトープをもたらす改変グリコシル化パターン、または腫瘍特異的であってもなくてもよい分解中のタンパク質スプライシングのような事象から生じる。
f)オンコウイルスタンパク質:これらのTAAはウイルスタンパク質であり、それらは発がん過程において重要な役割を果たしてもよく、外来性である(ヒト由来でない)ため、それらはT細胞応答を誘起し得る。このようなタンパク質の例は、子宮頸がんにおいて発現されるヒト乳頭腫16型ウイルスタンパク質E6およびE7である。

Bub1発現は、リンパ腫、乳がん、胃がん、および前立腺がんのサブセットにおいて増加する。Bub1過剰発現は、臨床予後不良と相関する(Ricke and van Deursen,2011)。Bub1突然変異は、染色体不安定性を示す結腸直腸がん腫において見いだされ得る(Williams et al.,2007)。
同一性百分率=100[1−(C/R)]
式中、Cは、参照配列と比較される配列との間のアライメント長にわたる、参照配列と比較配列の間の差異の数であり、
(i)比較配列中に対応する整列塩基またはアミノ酸を有しない、参照配列中の各塩基またはアミノ酸、および
(ii)参照配列中の各ギャップ、および
(iii)比較配列中の整列塩基またはアミノ酸と異なる、参照配列中の各整列塩基またはアミノ酸が差異を構成して、
(iiii)アライメントは、整合配列の1位から開始しなくてはならず;
Rは、比較配列とのアライメント長にわたる参照配列中の塩基またはアミノ酸の数であり、参照配列中に生じる任意のギャップもまた、塩基またはアミノ酸として数えられる。
(a)溶液中のまたは凍結乾燥形態の上述の医薬組成物を含有する容器;
(b)任意選択的に、凍結乾燥製剤のための希釈剤または再構成溶液を含有する第2の容器;および
(c)任意選択的に、(i)溶液の使用、または(ii)凍結乾燥製剤の再構成および/または使用のための取扱説明書
を含んでなるキットをさらに目的とする。
1.悪性物質からのHLAリガンドを質量分析法によって同定した
2.ゲノム規模メッセンジャーリボ核酸(mRNA)発現解析を使用して、一連の正常器官および組織と比較して悪性組織中の遺伝子過剰発現を同定した
3.同定されたHLAリガンドを遺伝子発現データと比較した。好ましくは、ステップ2で検出されたような選択的に発現されまたは過剰発現される遺伝子によってコードされる、腫瘍組織上で過剰提示されまたは選択的に提示されるペプチドが、多重ペプチドワクチンのための適切なTUMAP候補と見なされた。
4.同定されたペプチドのTUMAPとしての妥当性を支持する追加的な証拠を同定するために、文献調査を実施した
5.mRNAレベルでの過剰発現の関連性をステップ3からの選択されたTUMAPの腫瘍組織上における再検出と、健常組織における検出の欠如(またはまれな)検出によって確認した。
6.選択されたペプチドによる生体内T細胞応答の誘導が可能かどうかを評価するために、健常ドナーならびにがん患者からのヒトT細胞を使用して、生体外免疫原性アッセイを実施した。
細胞表面に提示される腫瘍関連ペプチドの同定および定量化
組織サンプル
患者の腫瘍組織は、外科手術または検死解剖に与えられた、全ての患者の告知に基づく同意書意のもとに得られた。組織は切除の直後に衝撃凍結されて、TUMAPの単離まで−70℃未満で保存された。
ショック凍結組織サンプルからのHLAペプチド貯留は、HLA−A*02特異的抗体BB7.2、HLA−A、−B、−C特異的抗体W6/32、HLAクラスII特異的抗体L243、CNBr−活性化セファロース、酸処理、および限外濾過を用いて、わずかな修正を加えて、既報のプロトコル(Falk et al.,1991;Seeger et al.,1999)に従って、固体組織からの免疫沈降によって得られた。
得られたHLAペプチド貯留は、逆相クロマトグラフィー(nanoAcquity UPL C system、Waters)によってそれらの疎水性に従って分離し、ESI源を装着したLTQ−velosおよびfusion hybrid質量分光計(ThermoElectron)内で溶出ペプチドを分析した。ペプチド貯留は、毎分400nLの流速を適用して、1.7μm C18逆相材料(Waters)で充填された分析用融合シリカマイクロキャピラリーカラム(75μm内径×250mm)上に直接挿入した。引き続いて、毎分300nLの流速で10%から33%へのBの二段階180分間二成分勾配を用いて、ペプチドを分離した。勾配は、溶媒A(水中の0.1%ギ酸)および溶媒B(アセトニトリル中の0.1%ギ酸)から構成された。nanoESI源への導入には、金被覆ガラス毛管(PicoTip,New Objective)を使用した。LTQ−Orbitrap質量分光計は、TOP5ストラテジーを使用してデータ依存モードで操作した。手短に述べると、Orbitrap(R=30000)内の高質量精度の完全スキャンでスキャンサイクルを開始し、これもまたOrbitrap(R=7500)内の5種の最も豊富な前駆イオンのMS/MSスキャンがそれに続き、以前選択されたイオンは動的に排除された。タンデム質量スペクトルは、SEQUESTおよび追加的な手動調節によって解釈した。同定されたペプチド配列は、生成された天然ペプチド断片化パターンと、配列が同一の合成参照ペプチドの断片化パターンとの比較によって確認した。
本発明のペプチドをコードする遺伝子発現プロファイリング
正常細胞と比較した腫瘍細胞上のペプチドの過剰提示または特異的提示は、免疫療法におけるその有用性にとって十分であり、いくつかのペプチドは、それらの起源タンパク質が正常組織にもまた存在するにもかかわらず、腫瘍特異的である。それでもなお、mRNA発現プロファイリングは、免疫療法のためのペプチド標的の選択において、安全性のレベルを高めることができる。特に、親和性成熟TCRのような安全性リスクが高い治療選択肢では、理想的な標的ペプチドは、腫瘍に特有であり正常組織には見られないタンパク質から誘導され、遺伝子発現の高い腫瘍対正常比は、治療濃度域を示唆する。さらに、ペプチド提示についてまだ分析されていない腫瘍実体における起源遺伝子の過剰発現は、特定のペプチドが各実体において重要であってもよいことを示唆する。
外科的に除去された組織標本は、告知に基づく同意書が各患者から入手された後に、上述の通り提供された(実施例1を参照されたい)。腫瘍組織標本を手術直後にスナップ凍結し、その後、液体窒素下で乳鉢と乳棒を用いて均質化した。TRI試薬(Ambion,Darmstadt,Germany)を使用して、これらのサンプルから全RNAを調製し、RNeasy(QIAGEN,Hilden,Germany)による精製がそれに続き;どちらの方法も製造業者のプロトコルに従って実施した。
腫瘍および正常組織RNAサンプルの遺伝子発現解析は、CeGaT(Tubingen,Germany)によって、次世代配列決定(RNAseq)によって実施した。簡単に述べると、配列決定ライブラリーは、RNA断片化、cDNA転換、および配列決定アダプターの付加を含む、Illumina HiSeq v4試薬キットを使用して、販売業者(Illumina Inc.,San Diego,CA,USA)のプロトコルに従って作製される。複数のサンプルに由来するライブラリーは等モル混合され、Illumina HiSeq 2500配列決定装置上で、製造会社の使用説明書に従って配列決定され、50bpのシングルエンドリードが生成された。処理された読み取りは、STARソフトウェアを使用して、ヒトゲノム(GRCh38)にマッピングされる。発現データは、ensembl配列データベース(Ensembl77)の注釈に基づいて、RPKM(100万個のマッピングされた読み取り当たりキロベース当たり読み取り、ソフトウェアCufflinksによって生成される)として転写物レベルで、そしてエクソンレベルで(全読み取り、ソフトウェアBedtoolsによって生成される)提供される。エクソン読み取りは、エクソン長さおよびアライメントサイズについて正規化されて、RPKM値が得られる。
MHCクラスI提示ペプチドの生体外免疫原性
本発明のTUMAPの免疫原性に関する情報を得るために、本発明者らは、ペプチド/MHC複合体および抗CD28抗体を負荷した人工抗原提示細胞(aAPC)によるCD8+T細胞の反復刺激に基づく、生体外T細胞プライミングアッセイを用いて研究を実施した。このようにして、本発明者らは、本発明のHLA−A*02:01拘束性TUMAPの免疫原性を示し得て、これらのペプチドが、それに対するCD01+前駆T細胞がヒトに存在する、T細胞エピトープであることを実証した(表9)。
ペプチドMHC複合体(pMHC)および抗CD28抗体を負荷した、人工抗原提示細胞による生体外刺激を実施するために、本発明者らは、最初に、告知に基づく同意後に、University clinics Mannheim,Germanyから得られた健常ドナーのCD8ミクロビーズ(Miltenyi Biotec,Bergisch−Gladbach,Germany)を使用した正の選択を通じて、新鮮HLA−A*02白血球除去生成物からCD8+T細胞を単離した。
HLAクラスIペプチドを試験するために、ペプチド特異的T細胞株の生成によって生体外免疫原性が実証され得た。本発明の2種のペプチドの、TUMAP特異的多量体染色後の例示的フローサイトメトリー結果は、対応する陰性対照と共に図3に示される。本発明の5つのペプチドからの結果を表12Aに要約する。本発明の7種のペプチドに関する、TUMAP特異的多量体染色後における例示的フローサイトメトリー結果は、対応する陰性対照と共に図4および図5に示される。本発明からの74種のペプチドの結果は、表12Bに要約される。
ペプチドの合成
Fmocストラテジーを使用した標準的な十分に確立された固相ペプチド合成を使用して、全てのペプチドを合成した。個々のペプチドのアイデンティティーおよび純度は、質量分析および分析用RP−HPLCによって判定された。ペプチドは、純度>50%の白色から灰白色の凍結乾燥物(トリフルオロ酢酸塩)として得られた。全てのTUMAPは、好ましくはトリフルオロ酢酸塩または酢酸塩として投与され、その他の塩形態もまた可能である。
MHC結合アッセイ
本発明によるT細胞ベースの治療法のための候補ペプチドを、それらのMHC結合能力(親和性)についてさらに試験した。個々のペプチド−MHC複合体は、UVリガンド交換によって生成され、UV感受性ペプチドはUV照射に際して切断されて、分析される目的ペプチドで交換された。ペプチド受容性MHC分子と効果的に結合して安定化し得るペプチド候補のみが、MHC複合体の分離を防止する。交換反応の収率を判定するために、安定化MHC複合体の軽鎖(β2m)の検出に基づくELISAを実施した。アッセイは、Rodenko et al.(Rodenko et al.,2006)に一般的に記載されるようにして実施した。
細胞表面に提示される腫瘍関連ペプチドの絶対定量化
抗体および/またはTCRなどのバインダーの生成は、骨の折れる工程であり、いくつかの選択された標的に対してのみ実施されてもよい。腫瘍関連および特異的ペプチドの場合、選択基準としては、提示の排他性および細胞表面に提示されるペプチドの密度が挙げられるが、これに限定されない。実施例1に記載されるペプチドの単離および相対定量化に加えて、本発明者らは、記載される細胞当たりの絶対的ペプチドコピー数も分析された。固形腫瘍サンプル中の細胞当たりのTUMAPコピーの定量化は、単離されたTUMAPの絶対定量化、TUMAP単離の効率、および分析される組織サンプルの細胞計数を必要とする。
質量分析によるペプチドの正確な定量化のために、内標準法を使用して各ペプチドの検量線を作成した。内標準は各ペプチドの二重同位体標識変異型であり、すなわち、2つの同位体標識アミノ酸がTUMAP合成に含まれた。それは、腫瘍関連ペプチドとはその質量異なるのみであるが、他の物理化学的性質に差異を示さない(Anderson et al.,2012)。内標準を各MSサンプルに添加して、全てのMSシグナルを内標準のMSシグナルに対して正規化し、MS実験間の可能な技術的変動を平準化した。
あらゆるタンパク質精製処理と同様に、組織サンプルからのタンパク質の単離には、目的タンパク質のいくらかの損失が伴う。TUMAP単離の効率を判定するために、絶対定量化のために選択された全てのTUMAPについて、ペプチド/MHC複合体が生成された。添加されたものを天然ペプチド/MHC複合体から識別できるように、TUMAPの単一同位体標識バージョンが使用され、すなわち、1つの同位体標識アミノ酸がTUMAP合成に含まれた。これらの複合体は、新鮮に調製された組織溶解産物に、すなわち、TUMAP単離手順の可能な限り早い時点で添加され、次に、以下の親和性精製において、天然ペプチド/MHC複合体のように捕捉された。したがって単一標識TUMAPの回収率を測定することで、個々の天然TUMAPの単離効率に関する結論が可能になる。
絶対ペプチド定量化に供した組織サンプルの細胞数を測定するために、本発明者らは、DNA含量分析を適用した。この方法は、異なる起点の幅広いサンプルに、最も重要なことには、冷凍サンプルに適用できる(Alcoser et al.,2011;Forsey and Chaudhuri,2009;Silva et al.,2013)。ペプチド単離プロトコル中に、組織サンプルを均質溶解産物に処理して、それから小さな溶解産物アリコートを取り出す。アリコートを3つに分割し、それからDNAを単離する(QiaAmpDNAMiniKit,Qiagen,Hilden,Germany)。蛍光ベースのDNA定量化アッセイ(Qubit dsDNA HS Assay Kit,Life Technologies,Darmstadt,Germany)を使用して、少なくとも2つの反復試験において、各DNA単離からの全DNA含有量を定量化する。
前述の実験のデータを用いて、本発明者らは、サンプルの全ペプチド量を総細胞数で除算して、それに続いて単離効率により除算することで、細胞当たりのTUMAPコピー数を算出した。選択されたたペプチドのコピー細胞数は、表15に示される。
Aalto, Y. et al., Leukemia 15 (2001): 1721-1728
Abaan, O. D. et al., Cancer Res 73 (2013): 4372-4382
Accardi, L. et al., Int.J Cancer 134 (2014): 2742-2747
Adams, D. J. et al., Mol.Cell Biol 25 (2005): 779-788
Agha-Hosseini, F. et al., Med.J Islam Repub.Iran 29 (2015): 218
Agostini, M. et al., Oncotarget. 6 (2015): 32561-32574
Akiyama, Y. et al., Oncol.Rep. 31 (2014): 1683-1690
Al-haidari, A. A. et al., Int.J Colorectal Dis. 28 (2013): 1479-1487
Alcoser, S. Y. et al., BMC.Biotechnol. 11 (2011): 124
Allison, J. P. et al., Science 270 (1995): 932-933
Alonso, C. N. et al., Leuk.Res. 36 (2012): 704-708
Amaro, A. et al., Cancer Metastasis Rev 33 (2014): 657-671
American Cancer Society, (2015), www.cancer.org
Ammirante, M. et al., Nature 464 (2010): 302-305
Ampie, L. et al., Front Oncol. 5 (2015): 12
An, C. H. et al., Hum.Pathol. 43 (2012): 40-47
Andersen, R. S. et al., Nat.Protoc. 7 (2012): 891-902
Anderson, N. L. et al., J Proteome.Res 11 (2012): 1868-1878
Appay, V. et al., Eur.J Immunol. 36 (2006): 1805-1814
Arai, E. et al., Int.J Cancer 137 (2015): 2589-2606
Armitage, J. O., Blood 110 (2007): 29-36
Armstrong, C. M. et al., Am.J Clin Exp.Urol. 3 (2015): 64-76
Asahara, S. et al., J Transl.Med. 11 (2013): 291
Atcheson, E. et al., Biosci.Rep. 31 (2011): 371-379
Avigan, D. et al., Clin Cancer Res. 10 (2004): 4699-4708
Azevedo, R. et al., J Control Release 214 (2015): 40-61
Baek, J. M. et al., Biochem.Biophys.Res Commun. 461 (2015): 334-341
Baker, M. et al., PLoS.One. 8 (2013): e62516
Banchereau, J. et al., Cell 106 (2001): 271-274
Bankovic, J. et al., Lung Cancer 67 (2010): 151-159
Barlin, J. N. et al., Neoplasia. 17 (2015): 183-189
Batliner, J. et al., Mol.Immunol. 48 (2011): 714-719
Battistella, M. et al., J Cutan.Pathol. 41 (2014): 427-436
Beatty, G. et al., J Immunol 166 (2001): 2276-2282
Becker, M. A. et al., Mol.Cancer Ther. 14 (2015): 973-981
Beggs, J. D., Nature 275 (1978): 104-109
Benada, J. et al., Biomolecules. 5 (2015): 1912-1937
Benjamini, Y. et al., Journal of the Royal Statistical Society.Series B (Methodological), Vol.57 (1995): 289-300
Bentz, S. et al., Digestion 88 (2013): 182-192
Berard, A. R. et al., Proteomics. 15 (2015): 2113-2135
Berman, R. S. et al., National Cancer Institute: PDQ(R) Colon Cancer Treatment (2015a)
Berman, R. S. et al., National Cancer Institute: PDQ(R) Rectal Cancer Treatment (2015b)
Berndt, S. I. et al., Nat Commun. 6 (2015): 6889
Bie, L. et al., PLoS.One. 6 (2011): e25631
Bill, K. L. et al., Lab Invest (2015)
Binsky-Ehrenreich, I. et al., Oncogene 33 (2014): 1006-1016
Black, J. D. et al., Toxins.(Basel) 7 (2015): 1116-1125
Bo, H. et al., BMC.Cancer 13 (2013): 496
Bockelman, C. et al., Cancer Biol Ther. 13 (2012): 289-295
Boeva, V. et al., PLoS.One. 8 (2013): e72182
Bogdanov, K. V. et al., Tsitologiia 50 (2008): 590-596
Bogni, A. et al., Leukemia 20 (2006): 239-246
Boldt, H. B. et al., Endocrinology 152 (2011): 1470-1478
Bormann, F. et al., Mol.Genet.Genomics 286 (2011): 279-291
Boulter, J. M. et al., Protein Eng 16 (2003): 707-711
Braumuller, H. et al., Nature (2013)
Bray, F. et al., Int J Cancer 132 (2013): 1133-1145
Brenner, S. et al., Cancer Lett. 356 (2015): 517-524
Bridgewater, J. et al., J Hepatol. 60 (2014): 1268-1289
Brocker, E. B. et al., Int.J Cancer 41 (1988): 562-567
Brossart, P. et al., Blood 90 (1997): 1594-1599
Bruckdorfer, T. et al., Curr.Pharm.Biotechnol. 5 (2004): 29-43
Bryant, N. L. et al., J Neurooncol. 101 (2011): 179-188
Burgess, A. W. et al., Exp.Cell Res 317 (2011): 2748-2758
Butler, J. E. et al., J Immunol. 182 (2009): 6600-6609
Butterfield, L. H. et al., Clin Cancer Res 12 (2006): 2817-2825
Butterfield, L. H. et al., Clin Cancer Res 9 (2003): 5902-5908
Byrd, J. C. et al., N.Engl.J Med. 369 (2013): 32-42
Byrns, M. C. et al., J Steroid Biochem.Mol.Biol 125 (2011): 95-104
Cai, C. J. et al., Sichuan.Da.Xue.Xue.Bao.Yi.Xue.Ban. 41 (2010): 941-945
Camoes, M. J. et al., PLoS.One. 7 (2012): e49819
Cao, S. et al., J Virol. 89 (2015): 713-729
Cao, W. et al., J Biol Chem 282 (2007): 18922-18928
Carballido, E. et al., Cancer Control 19 (2012): 54-67
Carbonnelle-Puscian, A. et al., Leukemia 23 (2009): 952-960
Card, K. F. et al., Cancer Immunol Immunother. 53 (2004): 345-357
Carlsten, M. et al., Cancer Res 67 (2007): 1317-1325
Carr, J. C. et al., Surgery 152 (2012): 998-1007
Carr, J. C. et al., Ann.Surg.Oncol 20 Suppl 3 (2013): S739-S746
Cassoni, P. et al., J Neuroendocrinol. 16 (2004): 362-364
Catellani, S. et al., Blood 109 (2007): 2078-2085
Cavard, C. et al., J Pathol. 218 (2009): 201-209
Chae, Y. K. et al., Oncotarget. 6 (2015): 37117-37134
Chang, Y. S. et al., Cancer Chemother.Pharmacol. 59 (2007): 561-574
Chanock, S. J. et al., Hum.Immunol. 65 (2004): 1211-1223
Chapiro, J. et al., Radiol.Med. 119 (2014): 476-482
Che, J. et al., Tumour.Biol 36 (2015): 6559-6568
Chen, H. S. et al., Zhonghua Gan Zang.Bing.Za Zhi. 11 (2003): 145-148
Chen, H. W. et al., Mol.Carcinog 52 (2013): 647-659
Chen, J. et al., Cancer Chemother.Pharmacol. 75 (2015): 1217-1227
Chen, R. S. et al., Oncogene 28 (2009): 599-609
Chen, W. L. et al., BMC.Cancer 12 (2012): 273
Chen, Y. et al., Am.J Physiol Lung Cell Mol.Physiol 306 (2014): L797-L807
Cheong, S. C. et al., Oral Oncol 45 (2009): 712-719
Chinwalla, V. et al., Oncogene 22 (2003): 1400-1410
Chisholm, K. M. et al., PLoS.One. 7 (2012): e30748
Choi, H. H. et al., Oncotarget. 6 (2015a): 19721-19734
Choi, H. H. et al., Oncotarget. 6 (2015b): 11779-11793
Chudnovsky, Y. et al., Cell Rep. 6 (2014): 313-324
Cicek, M. et al., PLoS.One. 6 (2011): e17522
Cipriano, R. et al., Oncotarget. 4 (2013): 729-738
Cipriano, R. et al., Mol.Cancer Res 12 (2014): 1156-1165
Cohen, C. J. et al., J Mol Recognit. 16 (2003a): 324-332
Cohen, C. J. et al., J Immunol 170 (2003b): 4349-4361
Cohen, S. N. et al., Proc.Natl.Acad.Sci.U.S.A 69 (1972): 2110-2114
Cohen, Y. et al., Hematology. 19 (2014): 286-292
Coligan, J. E. et al., Current Protocols in Protein Science (1995)
Colombetti, S. et al., J Immunol. 176 (2006): 2730-2738
Coosemans, A. et al., Anticancer Res 33 (2013): 5495-5500
Cotterchio, M. et al., PLoS.One. 10 (2015): e0125273
Counter, C. M. et al., Blood 85 (1995): 2315-2320
Courtial, N. et al., FASEB J 26 (2012): 523-532
Crawford, H. C. et al., Curr.Pharm.Des 15 (2009): 2288-2299
Cribier, B. et al., Br.J Dermatol. 144 (2001): 977-982
Cui, D. et al., Oncogene 33 (2014): 2225-2235
Dahlman, K. B. et al., PLoS.One. 7 (2012): e34414
Dai, X. et al., J Virol. 88 (2014): 12694-12702
de Kruijf, E. M. et al., BMC.Cancer 12 (2012): 24
De, S. et al., Cancer Res 69 (2009): 8035-8042
Dedes, K. J. et al., Sci.Transl.Med. 2 (2010): 53ra75
Dengjel, J. et al., Clin Cancer Res 12 (2006): 4163-4170
Denkberg, G. et al., J Immunol 171 (2003): 2197-2207
Dhanoa, B. S. et al., Hum.Genomics 7 (2013): 13
Ding, M. et al., Oncotarget. 6 (2015): 7686-7700
Donnard, E. et al., Oncotarget. 5 (2014): 9199-9213
Drayton, R. M. et al., Clin Cancer Res 20 (2014): 1990-2000
Drutskaya, M. S. et al., IUBMB.Life 62 (2010): 283-289
Du, C. et al., Gastric.Cancer 18 (2015): 516-525
Du, H. et al., Protein Pept.Lett. 16 (2009): 486-489
Duffy, M. J. et al., Clin Cancer Res 15 (2009): 1140-1144
Dufour, C. et al., Cancer 118 (2012): 3812-3821
Economopoulou, P. et al., Ann.Transl.Med. 4 (2016): 173
Ehlken, H. et al., Int.J Cancer 108 (2004): 307-313
Eichhorst, B. F. et al., Blood 107 (2006): 885-891
Eijsink, J. J. et al., Int.J Cancer 130 (2012): 1861-1869
Eisele, G. et al., Brain 129 (2006): 2416-2425
Elbelt, U. et al., J Clin Endocrinol.Metab 100 (2015): E119-E128
Elsnerova, K. et al., Oncol Rep. (2016)
Emens, L. A., Expert.Rev.Anticancer Ther. 12 (2012): 1597-1611
Engelmann, J. C. et al., PLoS.Comput.Biol 11 (2015): e1004293
Enguita-German, M. et al., World J Hepatol. 6 (2014): 716-737
Er, T. K. et al., J Mol.Med.(Berl) (2016)
Eruslanov, E. et al., Clin.Cancer Res. 19 (2013): 1670-1680
Espiard, S. et al., Endocrinol.Metab Clin North Am. 44 (2015): 311-334
Estey, E. H., Am.J Hematol. 89 (2014): 1063-1081
Etcheverry, A. et al., BMC.Genomics 11 (2010): 701
Faget, J. et al., Oncoimmunology 2 (2013): e23185
Falk, K. et al., Nature 351 (1991): 290-296
Fang, M. et al., Mol.Cell Biol 33 (2013): 2635-2647
Fang, Y. et al., Tumour.Biol 33 (2012): 2299-2306
Farrell, A. S. et al., Mol.Cancer Res 12 (2014): 924-939
Ferlay et al., GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No.11 [Internet], (2013), http://globocan.iarc.fr
Fernandez-Calotti, P. X. et al., Haematologica 97 (2012): 943-951
Fevre-Montange, M. et al., J Neuropathol.Exp.Neurol. 65 (2006): 675-684
Finocchiaro, G. et al., Ann.Transl.Med. 3 (2015): 83
Fiorito, V. et al., Biochim.Biophys.Acta 1839 (2014): 259-264
Fokas, E. et al., Cell Death.Dis. 3 (2012): e441
Fong, L. et al., Proc.Natl.Acad.Sci.U.S.A 98 (2001): 8809-8814
Ford-Hutchinson, A. W., Eicosanoids 4 (1991): 65-74
Forsey, R. W. et al., Biotechnol.Lett. 31 (2009): 819-823
Fremont, S. et al., EMBO Rep. 14 (2013): 364-372
Fritz, P. et al., Pathol.Res Pract. 208 (2012): 203-209
Fuge, O. et al., Res Rep.Urol. 7 (2015): 65-79
Fujita, H. et al., J Histochem.Cytochem. 63 (2015): 217-227
Fukuyama, R. et al., Oncogene 27 (2008): 6044-6055
Furman, R. R. et al., N.Engl.J Med. 370 (2014): 997-1007
Furukawa, T. et al., Sci.Rep. 1 (2011): 161
Gabrielson, M. et al., Biochem.Biophys.Res Commun. 469 (2016): 1090-1096
Gabrielson, M. et al., Oncol Rep. 29 (2013): 1268-1274
Gabrilovich, D. I. et al., Nat Med. 2 (1996): 1096-1103
Galazis, N. et al., Gynecol.Endocrinol. 29 (2013): 638-644
Gandhi, A. V. et al., Ann Surg.Oncol 20 Suppl 3 (2013): S636-S643
Gao, M. et al., Diagn.Pathol. 8 (2013): 205
Garbe, C. et al., J Invest Dermatol. 100 (1993): 239S-244S
Garcia-Irigoyen, O. et al., Hepatology 62 (2015): 166-178
Gattinoni, L. et al., Nat Rev.Immunol 6 (2006): 383-393
Gazy, I. et al., Mutat.Res Rev Mutat.Res 763 (2015): 267-279
Gelsi-Boyer, V. et al., Mol.Cancer Res 3 (2005): 655-667
Ghosh, A. et al., Int.J Biol Sci. 12 (2016): 30-41
Giannopoulos, K. et al., Leukemia 24 (2010): 798-805
Giannopoulos, K. et al., Int.J Oncol 29 (2006): 95-103
Gnjatic, S. et al., Proc Natl.Acad.Sci.U.S.A 100 (2003): 8862-8867
Godkin, A. et al., Int.Immunol 9 (1997): 905-911
Goede, V. et al., N.Engl.J Med. 370 (2014): 1101-1110
Gonda, T. J. et al., Expert.Opin.Biol Ther. 8 (2008): 713-717
Goni, M. H. et al., Anticancer Res 13 (1993): 1155-1160
Granziero, L. et al., Blood 97 (2001): 2777-2783
Green, J. et al., Cochrane.Database.Syst.Rev (2005): CD002225
Green, M. R. et al., Molecular Cloning, A Laboratory Manual 4th (2012)
Greenfield, E. A., Antibodies: A Laboratory Manual 2nd (2014)
Grimm, M. et al., J Transl.Med. 12 (2014): 208
Grinberg-Rashi, H. et al., Clin Cancer Res 15 (2009): 1755-1761
Grivas, P. D. et al., Semin.Cancer Biol 35 (2015): 125-132
Gruel, N. et al., Breast Cancer Res 16 (2014): R46
Gunawardana, C. et al., Br.J Haematol. 142 (2008): 606-609
Guo, P. et al., Onco.Targets.Ther. 8 (2015a): 73-79
Guo, T. et al., Int.J Cancer (2016)
Guo, Z. et al., Tumour.Biol 36 (2015b): 3583-3589
Guo, Z. et al., Tumour.Biol 36 (2015c): 4777-4783
Guyonnet, Duperat, V et al., Biochem.J 305 ( Pt 1) (1995): 211-219
Hallek, Michael et al., ASH Annual Meeting Abstracts 112 (2008): 325
Halon, A. et al., Arch.Gynecol.Obstet. 287 (2013): 563-570
Handkiewicz-Junak, D. et al., Eur.J Nucl.Med.Mol.Imaging (2016)
Hapgood, G. et al., Blood 126 (2015): 17-25
Harig, S. et al., Blood 98 (2001): 2999-3005
Hayette, S. et al., Oncogene 19 (2000): 4446-4450
He, H. et al., Diagn.Mol.Pathol. 21 (2012): 143-149
He, M. et al., J Dig.Dis. 12 (2011): 393-400
Heerma van Voss, M. R. et al., Histopathology 65 (2014): 814-827
Heishima, K. et al., PLoS.One. 10 (2015): e0137361
Hill, S. J. et al., Genes Dev. 28 (2014): 1957-1975
Hinrichs, C. S. et al., Nat.Biotechnol. 31 (2013): 999-1008
Hirahata, M. et al., Cancer Med. (2016)
Hirano, Y. et al., Genes Cells 11 (2006): 1295-1304
Hlavac, V. et al., Medicine (Baltimore) 93 (2014): e255
Holla, S. et al., Mol.Cancer 13 (2014): 210
Holtl, L. et al., Clin.Cancer Res. 8 (2002): 3369-3376
Hong, L. et al., Hum.Pathol. 45 (2014): 2423-2429
Honore, B. et al., Exp.Cell Res 294 (2004): 199-209
Horig, H. et al., Cancer Immunol Immunother. 49 (2000): 504-514
Hu, X. T. et al., Zhonghua Zhong.Liu Za Zhi. 30 (2008): 515-518
Hu, X. T. et al., Oncol Rep. 22 (2009): 1247-1252
Huang, P. Y. et al., Leuk.Lymphoma 55 (2014): 2085-2092
Huang, Y. et al., Clin Epigenetics. 8 (2016): 9
Huang, Y. et al., PLoS.One. 8 (2013a): e82519
Huang, Y. et al., Cell Biosci. 3 (2013b): 16
Huang, Y. X. et al., Nan.Fang Yi.Ke.Da.Xue.Xue.Bao. 29 (2009): 1329-1332
Hubertus, J. et al., Oncol Rep. 25 (2011): 817-823
Huisman, C. et al., Mol.Ther. (2015)
Huisman, C. et al., Mol.Oncol 7 (2013): 669-679
Hung, C. F. et al., Immunol.Rev 222 (2008): 43-69
Hus, I. et al., Oncol Rep. 20 (2008): 443-451
Hussein, S. et al., Sci.Rep. 5 (2015): 15752
Huu, N. T. et al., FEBS J 282 (2015): 4727-4746
Hwang, M. L. et al., J Immunol. 179 (2007): 5829-5838
Ihn, H. J. et al., Exp.Biol Med.(Maywood.) 240 (2015): 1690-1697
Ilm, K. et al., Mol.Cancer 14 (2015): 38
Imai, K. et al., Br.J Cancer 104 (2011): 300-307
Inoue, K. et al., Subcell.Biochem. 85 (2014): 17-40
Ishida, T. et al., Leukemia 20 (2006): 2162-2168
Ishizone, S. et al., Cancer Sci. 97 (2006): 119-126
Iunusova, N. V. et al., Izv.Akad.Nauk Ser.Biol (2014): 448-455
Iunusova, N. V. et al., Izv.Akad.Nauk Ser.Biol (2013): 284-291
Iwakawa, R. et al., Carcinogenesis 36 (2015): 616-621
Jager, D. et al., Cancer Res 60 (2000): 3584-3591
Jaiswal, A. S. et al., Bioorg.Med.Chem Lett. 24 (2014): 4850-4853
Januchowski, R. et al., Biomed.Pharmacother. 67 (2013): 240-245
Januchowski, R. et al., Biomed.Pharmacother. 68 (2014): 447-453
Jelinek, J. et al., PLoS.One. 6 (2011): e22110
Jenne, D. E. et al., Am.J Hum.Genet. 69 (2001): 516-527
Jiang, H. et al., Int.J Mol.Med. 35 (2015a): 1374-1380
Jiang, H. et al., Exp.Ther.Med. 8 (2014a): 769-774
Jiang, H. N. et al., PLoS.One. 8 (2013): e67637
Jiang, L. et al., Cell Cycle 14 (2015b): 2881-2885
Jiang, L. et al., Oncotarget. 5 (2014b): 7663-7676
Jiang, Y. et al., Mol.Cell 53 (2014c): 75-87
Jiao, X. L. et al., Eur.Rev Med.Pharmacol.Sci. 18 (2014): 509-515
Johnson, M. A. et al., Growth Horm.IGF.Res 24 (2014): 164-173
Jones, R. T. et al., Urol.Clin North Am. 43 (2016): 77-86
Ju, W. et al., Oncol.Res. 18 (2009): 47-56
Jung, G. et al., Proc Natl Acad Sci U S A 84 (1987): 4611-4615
Junttila, M. R. et al., Cell Cycle 7 (2008): 592-596
Kachakova, D. et al., J BUON. 18 (2013): 660-668
Kadeh, H. et al., Asian Pac.J Cancer Prev. 16 (2015): 6609-6613
Kalikin, L. M. et al., Genomics 57 (1999): 36-42
Kalos, M. et al., Sci.Transl.Med. 3 (2011): 95ra73
Kang, Y. K. et al., Cancer Res 68 (2008): 7887-7896
Kanthan, R. et al., J Oncol 2015 (2015): 967472
Kanzaki, H. et al., Oncol Rep. 18 (2007): 1171-1175
Kanzaki, H. et al., J Cancer Res Clin Oncol 134 (2008): 211-217
Kanzawa, M. et al., Pathobiology 80 (2013): 235-244
Karim, H. et al., Biochem.Biophys.Res Commun. 411 (2011): 156-161
Karrman, K. et al., Br.J Haematol. 144 (2009): 546-551
Kasiappan, R. et al., Mol.Cancer 9 (2010): 311
Katkoori, V. R. et al., PLoS.One. 7 (2012): e30020
Kato, S. et al., Int.J Oncol 29 (2006): 33-40
Kaufman, H. L. et al., Clin Cancer Res 14 (2008): 4843-4849
Kayser, G. et al., Pathology 43 (2011): 719-724
Kelavkar, U. et al., Curr.Urol.Rep. 3 (2002): 207-214
Kelavkar, U. P. et al., Prostaglandins Other Lipid Mediat. 82 (2007): 185-197
Khanna, A. et al., Int.J Cancer 138 (2016): 525-532
Khanna, A. et al., Cancer Res 73 (2013): 6548-6553
Khatamianfar, V. et al., BMJ Open. 2 (2012)
Kibbe, A. H., Handbook of Pharmaceutical Excipients rd (2000)
Kim, D. S. et al., J Proteome.Res 9 (2010a): 3710-3719
Kim, H. S. et al., Korean J Intern.Med. 25 (2010b): 399-407
Kim, J. et al., J Biol Chem 286 (2011): 43294-43300
Kim, J. H. et al., J Prev.Med.Public Health 49 (2016): 61-68
Kim, J. W. et al., Cancer Sci. 100 (2009): 1468-1478
Kim, J. Y. et al., BMB.Rep. 47 (2014a): 451-456
Kim, K. et al., Mol.Cancer Res 6 (2008): 426-434
Kim, S. M. et al., Int.J Cancer 134 (2014b): 114-124
Kim, Y. D. et al., Int.J Mol.Med. 29 (2012): 656-662
Kindla, J. et al., Cancer Biol Ther. 11 (2011): 584-591
Kirschner, L. S. et al., Horm.Cancer 7 (2016): 9-16
Kitchen, M. O. et al., Epigenetics. 11 (2016): 237-246
Kiyomitsu, T. et al., Mol.Cell Biol 31 (2011): 998-1011
Kleylein-Sohn, J. et al., J Cell Sci. 125 (2012): 5391-5402
Klopfleisch, R. et al., J Proteome.Res 9 (2010): 6380-6391
Knollman, H. et al., Ther.Adv.Urol. 7 (2015a): 312-330
Knollman, H. et al., Ther.Adv.Urol. 7 (2015b): 312-330
Kocer, B. et al., Pathol.Int. 52 (2002): 470-477
Kohnz, R. A. et al., ACS Chem Biol 10 (2015): 1624-1630
Kohonen-Corish, M. R. et al., Oncogene 26 (2007): 4435-4441
Koido, S. et al., World J Gastroenterol. 19 (2013): 8531-8542
Kong, D. S. et al., Oncotarget. (2016)
Krackhardt, A. M. et al., Blood 100 (2002): 2123-2131
Krieg, A. M., Nat Rev.Drug Discov. 5 (2006): 471-484
Kronenberger, K. et al., J Immunother. 31 (2008): 723-730
Krupenko, S. A. et al., Cell Growth Differ. 13 (2002): 227-236
Kubota, T. et al., Cell Cycle 12 (2013): 2570-2579
Kuchenbaecker, K. B. et al., Nat Genet. 47 (2015): 164-171
Kuefer, M. U. et al., Oncogene 22 (2003): 1418-1424
Kumar, A. et al., Cell Biochem.Biophys. 67 (2013): 837-851
Kumar, R. et al., DNA Repair (Amst) 15 (2014): 54-59
Kunimoto, K. et al., J Cell Physiol 220 (2009): 621-631
Kuwada, M. et al., Cancer Lett. 369 (2015): 212-221
Landi, D. et al., Cancer 118 (2012): 4670-4680
Lanier, M. H. et al., Mol.Biol Cell 26 (2015): 4577-4588
Lee, D. G. et al., Curr.Cancer Drug Targets. 11 (2011): 966-975
Lee, J. H. et al., Ann.Surg. 249 (2009a): 933-941
Lee, K. Y. et al., Yonsei Med.J 50 (2009b): 60-67
Lee, M. A. et al., BMC.Cancer 14 (2014a): 125
Lee, S. Y. et al., Eur.J Cancer 50 (2014b): 698-705
Lee, W. C. et al., J Immunother. 28 (2005): 496-504
Lei, N. et al., Oncol Rep. 32 (2014): 1689-1694
Leitlinie Endometriumkarzinom, 032/034, (2008)
Leitlinie Magenkarzinom, 032-009OL, (2012)
Leitlinien f?r Diagnostik und Therapie in der Neurologie, 030/099, (2014)
Leonetti, M. D. et al., Proc.Natl.Acad.Sci.U.S.A 109 (2012): 19274-19279
Leung, J. et al., Immune.Netw. 14 (2014): 265-276
Li, J. et al., Mol.Biol Rep. 41 (2014): 8071-8079
Li, J. et al., Zhongguo Fei.Ai.Za Zhi. 18 (2015a): 16-22
Li, J. et al., Tumour.Biol (2016)
Li, J. F. et al., Zhonghua Wei Chang Wai Ke.Za Zhi. 15 (2012a): 388-391
Li, L. et al., Pharmacogenet.Genomics 22 (2012b): 105-116
Li, W. Q. et al., Carcinogenesis 34 (2013): 1536-1542
Li, Y. et al., Cancer Biol Ther. 16 (2015b): 1316-1322
Li, Y. et al., Cancer Epidemiol. 39 (2015c): 8-13
Li, Y. F. et al., Int.J Biol Sci. 8 (2012c): 1168-1177
Liang, Y. C. et al., Oncotarget. 6 (2015): 38046-38060
Liao, W. et al., Oncotarget. 5 (2014): 10271-10279
Liddy, N. et al., Nat Med. 18 (2012): 980-987
Lin, C. et al., Oncotarget. 6 (2015): 8434-8453
Lin, J. C. et al., RNA. 20 (2014): 1621-1631
Lin, Y. W. et al., Eur.J Cancer 45 (2009): 2041-2049
Lin, Z. et al., Diagn.Pathol. 8 (2013): 133
Lindqvist, B. M. et al., Epigenetics. 7 (2012): 300-306
Linhares, N. D. et al., Eur.J Med.Genet. 57 (2014): 643-648
Linher-Melville, K. et al., Mol.Cell Biochem. 405 (2015): 205-221
Linkov, F. et al., Eur.Cytokine Netw. 20 (2009): 21-26
Listerman, I. et al., Cancer Res 73 (2013): 2817-2828
Liu, C. et al., Int.J Clin Exp.Pathol. 8 (2015): 7446-7449
Liu, L. et al., Biochem.J 451 (2013a): 55-60
Liu, M. et al., Asian Pac.J Cancer Prev. 14 (2013b): 6281-6286
Liu, Q. et al., Med.Oncol 31 (2014a): 882
Liu, T. et al., DNA Repair (Amst) 11 (2012): 131-138
Liu, W. J. et al., Leuk.Lymphoma 55 (2014b): 2691-2698
Liu, X. et al., Mol.Biol Rep. 41 (2014c): 7471-7478
Ljunggren, H. G. et al., J Exp.Med. 162 (1985): 1745-1759
Lleonart, M. E. et al., Oncol Rep. 16 (2006): 603-608
Llovet, J. M. et al., N.Engl.J Med. 359 (2008): 378-390
Lobito, A. A. et al., J Biol Chem 286 (2011): 18969-18981
Loddo, M. et al., J Pathol. 233 (2014): 344-356
Lollini, P. L. et al., Int.J Cancer 55 (1993): 320-329
Longenecker, B. M. et al., Ann N.Y.Acad.Sci. 690 (1993): 276-291
Lonsdale, J., Nat.Genet. 45 (2013): 580-585
Lu, G. et al., Cancer Cell 26 (2014): 222-234
Lucas, S. et al., Int.J Cancer 87 (2000): 55-60
Luhrig, S. et al., Cell Div. 8 (2013): 3
Luis, Espinoza J. et al., Cancer Sci. 104 (2013): 657-662
Lukas, T. J. et al., Proc.Natl.Acad.Sci.U.S.A 78 (1981): 2791-2795
Lukka, H. et al., Clin Oncol (R Coll.Radiol.) 14 (2002): 203-212
Luna, B. et al., Mol.Neurobiol. 52 (2015): 1341-1363
Lundblad, R. L., Chemical Reagents for Protein Modification 3rd (2004)
Ma, J. et al., Pathol.Oncol Res 19 (2013a): 821-832
Ma, L. D. et al., Zhongguo Shi Yan.Xue.Ye.Xue.Za Zhi. 21 (2013b): 1429-1434
Ma, T. et al., Zhonghua Yi.Xue.Za Zhi. 94 (2014): 3005-3007
Maggioni, A. et al., Protein Expr.Purif. 101 (2014): 165-171
Mahomed, F., Oral Oncol 47 (2011): 797-803
Mantel, A. et al., Exp.Dermatol. 23 (2014): 573-578
Mantia-Smaldone, G. M. et al., Hum.Vaccin.Immunother. 8 (2012): 1179-1191
Marchio, C. et al., J Clin Pathol. 63 (2010): 220-228
Marechal, R. et al., Clin Cancer Res 15 (2009): 2913-2919
Marine, J. C., Nat Rev Cancer 12 (2012): 455-464
Markus, M. A. et al., Genomics 107 (2016): 138-144
Marten, A. et al., Cancer Immunol.Immunother. 51 (2002): 637-644
Martin, R. W. et al., Cancer Res 67 (2007): 9658-9665
Martinez, I. et al., Eur.J Cancer 43 (2007): 415-432
Marzec, K. A. et al., Biomed.Res Int. 2015 (2015): 638526
Mason, C. C. et al., Leukemia (2015)
Massari, F. et al., Cancer Treat.Rev. 41 (2015): 114-121
Massoner, P. et al., PLoS.One. 8 (2013): e55207
Matsueda, S. et al., World J Gastroenterol. 20 (2014): 1657-1666
Matsuura, N. et al., Nihon Rinsho 53 (1995): 1643-1647
Maus, M. V. et al., Blood 123 (2014): 2625-2635
Mayr, C. et al., Exp.Hematol. 34 (2006): 44-53
Mayr, C. et al., Blood 105 (2005): 1566-1573
McGilvray, R. W. et al., Int.J Cancer 127 (2010): 1412-1420
Medeiros, A. C. et al., Cancer Epidemiol.Biomarkers Prev. 3 (1994): 331-333
Mehta, J. et al., PLoS.One. 10 (2015): e0120622
Mei, J. Z. et al., Nan.Fang Yi.Ke.Da.Xue.Xue.Bao. 27 (2007): 887-889
Mencia, N. et al., Biochem.Pharmacol. 82 (2011): 1572-1582
Mendoza-Maldonado, R. et al., PLoS.One. 5 (2010): e13720
Meziere, C. et al., J Immunol 159 (1997): 3230-3237
Migliorini, D. et al., J Clin Invest 121 (2011): 1329-1343
Milutin, Gasperov N. et al., PLoS.One. 10 (2015): e0129452
Missero, C. et al., Exp.Dermatol. 23 (2014): 143-146
Miyagi, Y. et al., Clin Cancer Res 7 (2001): 3950-3962
Miyamoto, K. et al., Int.J Cancer 116 (2005): 407-414
Mohanraj, L. et al., Recent Pat Anticancer Drug Discov 6 (2011): 166-177
Mohelnikova-Duchonova, B. et al., Cancer Chemother.Pharmacol. 72 (2013): 669-682
Molina, J. R. et al., Mayo Clin Proc. 83 (2008): 584-594
Morgan, R. A. et al., Science 314 (2006): 126-129
Mori, M. et al., Transplantation 64 (1997): 1017-1027
Morin, P. J., Cancer Res 65 (2005): 9603-9606
Morita, T. et al., Int.J Cancer 109 (2004): 525-532
Mortara, L. et al., Clin Cancer Res. 12 (2006): 3435-3443
Moser, J. J. et al., J Neurosci.Res 85 (2007): 3619-3631
Mou, X. et al., Sci.Rep. 4 (2014): 6138
Moulton, H. M. et al., Clin Cancer Res 8 (2002): 2044-2051
Mueller, L. N. et al., J Proteome.Res 7 (2008): 51-61
Mueller, L. N. et al., Proteomics. 7 (2007): 3470-3480
Mukhopadhyay, P. et al., Biochim.Biophys.Acta 1815 (2011): 224-240
Muller, M. R. et al., Blood 103 (2004): 1763-1769
Mumberg, D. et al., Proc.Natl.Acad.Sci.U.S.A 96 (1999): 8633-8638
Nagashio, R. et al., Sci.Rep. 5 (2015): 8649
Naito, T. et al., J Biol Chem 290 (2015): 15004-15017
Nakajima, H. et al., Cancer Sci. 105 (2014): 1093-1099
Nakano, K. et al., Exp.Cell Res 287 (2003): 219-227
Nakarai, C. et al., Clin Exp.Med. 15 (2015): 333-341
Nakashima, A. et al., Biochem.Biophys.Res Commun. 361 (2007): 218-223
National Cancer Institute, (5-6-2015), www.cancer.gov
Nguyen, M. H. et al., Int.J Oncol 41 (2012): 1285-1296
Ni, I. B. et al., Hematol.Rep. 4 (2012): e19
Ni, L. et al., J Cell Biochem. 106 (2009): 920-928
Nobusawa, S. et al., Brain Tumor Pathol. 31 (2014): 229-233
O'Brien, S. et al., Lancet Oncol 15 (2014): 48-58
O'Geen, H. et al., PLoS.Genet. 3 (2007): e89
Obama, K. et al., Clin Cancer Res 14 (2008): 1333-1339
Oehler, V. G. et al., Blood 114 (2009): 3292-3298
Ogasawara, N. et al., J Biochem. 149 (2011): 321-330
Ogbomo, H. et al., Neoplasia. 10 (2008): 1402-1410
Ogiso, Y. et al., Cancer Res 62 (2002): 5008-5012
Oh, Y. et al., J Biol.Chem 287 (2012): 17517-17529
Okabe, N. et al., Int.J Oncol 46 (2015): 999-1006
Okuno, K. et al., Exp.Ther Med. 2 (2011): 73-79
Olkhanud, P. B. et al., Cancer Res 69 (2009): 5996-6004
Olszewski-Hamilton, U. et al., Biomark.Cancer 3 (2011): 31-40
Orentas, R. J. et al., Front Oncol 2 (2012): 194
Orzol, P. et al., Histol.Histopathol. 30 (2015): 503-521
Ouyang, M. et al., BMC.Cancer 15 (2015): 132
Ozawa, H. et al., Ann.Surg.Oncol 17 (2010): 2341-2348
Ozeki, N. et al., Int.J Mol.Sci. 17 (2016)
Palma, M. et al., Cancer Immunol Immunother. 57 (2008): 1705-1710
Palmer, D. H. et al., Hepatology 49 (2009): 124-132
Palomba, M. L., Curr.Oncol Rep. 14 (2012): 433-440
Pan, J. et al., Leuk.Res 36 (2012): 889-894
Pannu, V. et al., Oncotarget. 6 (2015): 6076-6091
Parikh, R. A. et al., Genes Chromosomes.Cancer 53 (2014): 25-37
Parikh, S. A. et al., Blood 118 (2011): 2062-2068
Parisi, M. A., Am.J Med.Genet.C.Semin.Med.Genet. 151C (2009): 326-340
Park, E. et al., Mol.Cell 50 (2013): 908-918
Park, M. J. et al., Immunol.Invest 40 (2011): 367-382
Park, Y. R. et al., Cancer Genomics Proteomics. 13 (2016): 83-90
Parplys, A. C. et al., DNA Repair (Amst) 24 (2014): 87-97
Pasmant, E. et al., Mol.Med. 17 (2011): 79-87
Patil, A. A. et al., Oncotarget. 5 (2014): 6414-6424
Pattabiraman, D. R. et al., Leukemia 27 (2013): 269-277
Pawar, S. et al., J Ovarian.Res 7 (2014): 53
Payne, S. R. et al., Prostate 69 (2009): 1257-1269
Peng, B. et al., Mol.Biosyst. 11 (2015): 105-114
Pequeux, C. et al., Cancer Res 62 (2002): 4623-4629
Perrais, M. et al., J Biol Chem 276 (2001): 15386-15396
Petrini, I., Ann.Transl.Med. 3 (2015): 82
Phan, G. Q. et al., Cancer Control 20 (2013): 289-297
Phe, V. et al., BJU.Int. 104 (2009): 896-901
Piasecka, D. et al., Postepy Biochem. 61 (2015): 198-206
Pinheiro, J. et al., nlme: Linear and Nonlinear Mixed Effects Models (http://CRAN.R-project.org/packe=nlme) (2015)
Plebanski, M. et al., Eur.J Immunol 25 (1995): 1783-1787
Porta, C. et al., Virology 202 (1994): 949-955
Porter, D. L. et al., N.Engl.J Med. 365 (2011): 725-733
Potapenko, I. O. et al., Mol.Oncol 9 (2015): 861-876
Przybyl, J. et al., Int.J Biochem.Cell Biol 53 (2014): 505-513
Qian, M. X. et al., Cell 153 (2013): 1012-1024
Qiu, J. et al., Leukemia 17 (2003): 1891-1900
Quinn, D. I. et al., Urol.Oncol. (2015)
Qureshi, R. et al., Cancer Lett. 356 (2015): 321-331
Rainer, J. et al., Mol.Endocrinol. 26 (2012): 178-193
Raja, S. B. et al., J Cell Sci. 125 (2012): 703-713
Rajadhyaksha, A. M. et al., Am.J Hum.Genet. 87 (2010): 643-654
Rajkumar, T. et al., BMC.Cancer 11 (2011): 80
Rakic, M. et al., Hepatobiliary.Surg.Nutr. 3 (2014): 221-226
Rammensee, H. G. et al., Immunogenetics 50 (1999): 213-219
Ramsay, R. G. et al., Expert.Opin.Ther.Targets. 7 (2003): 235-248
RefSeq, The NCBI handbook [Internet], Chapter 18, (2002), http://www.ncbi.nlm.nih.gov/books/NBK21091/
Reid-Lombardo, K. M. et al., Cancer Epidemiol.Biomarkers Prev. 20 (2011): 1251-1254
Reinisch, W. et al., J Immunother. 25 (2002): 489-499
Reinmuth, N. et al., Dtsch.Med.Wochenschr. 140 (2015): 329-333
Relogio, A. et al., PLoS.Genet. 10 (2014): e1004338
Rendon-Huerta, E. et al., J Gastrointest.Cancer 41 (2010): 52-59
Resende, C. et al., Helicobacter. 16 Suppl 1 (2011): 38-44
Richards, S. et al., J Natl.Cancer Inst. 91 (1999): 861-868
Ricke, R. M. et al., Cell Cycle 10 (2011): 3645-3651
Rincon, R. et al., Oncotarget. 6 (2015): 4299-4314
Rini, B. I. et al., Curr.Opin.Oncol. 20 (2008): 300-306
Rini, B. I. et al., Cancer 107 (2006): 67-74
Riordan, J. D. et al., PLoS.Genet. 9 (2013): e1003441
Ritter, A. et al., Cell Cycle 14 (2015): 3755-3767
Robak, T. et al., Expert.Opin.Biol.Ther 14 (2014): 651-661
Roca, H. et al., PLoS.One. 8 (2013): e76773
Rock, K. L. et al., Science 249 (1990): 918-921
Rodenko, B. et al., Nat Protoc. 1 (2006): 1120-1132
Rodini, C. O. et al., Int.J Oncol 40 (2012): 1180-1188
Rodriguez, F. J. et al., J Neuropathol.Exp.Neurol. 67 (2008): 1194-1204
Romanuik, T. L. et al., BMC.Med.Genomics 3 (2010): 43
Ronchi, C. L. et al., Neoplasia. 14 (2012): 206-218
Rouanne, M. et al., Crit Rev Oncol Hematol. 98 (2016): 106-115
Rucki, A. A. et al., World J Gastroenterol. 20 (2014): 2237-2246
Rudland, P. S. et al., Am.J Pathol. 176 (2010): 2935-2947
Rutkowski, M. J. et al., Mol Cancer Res 8 (2010): 1453-1465
Ryu, B. et al., PLoS.One. 2 (2007): e594
S3-Leitlinie Exokrines Pankreaskarzinom, 032-010OL, (2013)
S3-Leitlinie Lungenkarzinom, 020/007, (2011)
S3-Leitlinie maligne Ovarialtumore, 032-035OL, (2013)
S3-Leitlinie Mammakarzinom, 032-045OL, (2012)
S3-Leitlinie Melanom, 032-024OL, (2013)
S3-Leitlinie Prostatakarzinom, 043/022OL, (2014)
S3-Leitlinie Zervixkarzinom, 032/033OL, (2014)
Sadeque, A. et al., BMC.Med.Genomics 5 (2012): 59
Saeki, M. et al., PLoS.One. 8 (2013): e67326
Safarpour, D. et al., Arch.Pathol.Lab Med. 139 (2015): 612-617
Saiki, R. K. et al., Science 239 (1988): 487-491
Salim, H. et al., Genes Chromosomes.Cancer 52 (2013): 895-911
Salman, B. et al., Oncoimmunology. 2 (2013): e26662
Sandoval, J. et al., J Clin Oncol 31 (2013): 4140-4147
Sangro, B. et al., J Clin Oncol 22 (2004): 1389-1397
Sankaranarayanan, P. et al., PLoS.One. 10 (2015): e0121396
Santarlasci, V. et al., Eur.J Immunol. 44 (2014): 654-661
Sarma, S. N. et al., Environ.Toxicol.Pharmacol. 32 (2011): 285-295
Sasao, T. et al., Reproduction. 128 (2004): 709-716
Satija, Y. K. et al., Int.J Cancer 133 (2013): 2759-2768
Sato, N. et al., Genes Chromosomes.Cancer 49 (2010): 353-367
Savaskan, N. E. et al., Ann.Anat. 192 (2010): 309-313
Savaskan, N. E. et al., Curr.Neuropharmacol. 13 (2015): 258-265
Sawada, G. et al., Oncol Rep. 30 (2013): 1971-1975
Schetelig, J. et al., J Clin Oncol 26 (2008): 5094-5100
Scheurer, B. et al., Immunopharmacology 38 (1997): 167-175
Schmidt, S. M. et al., Cancer Res 64 (2004): 1164-1170
Schreiber, M. et al., J Biol Chem 273 (1998): 3509-3516
Seeger, F. H. et al., Immunogenetics 49 (1999): 571-576
Seidl, C. et al., Invest New Drugs 28 (2010): 49-60
Seppanen, M. et al., Acta Obstet.Gynecol.Scand. 87 (2008): 902-909
Shareef, M. M. et al., Arab.J Gastroenterol. 16 (2015): 105-112
Sharma, R. K. et al., Clin Exp.Metastasis 33 (2016): 263-275
Sharpe, D. J. et al., Oncotarget. 5 (2014): 8803-8815
Shen, C. et al., Cancer Res 73 (2013): 3393-3401
Shen, Y. et al., Oncotarget. 6 (2015a): 20396-20403
Shen, Y. et al., Cancer Cell Microenviron. 2 (2015b)
Sherman, F. et al., Laboratory Course Manual for Methods in Yeast Genetics (1986)
Sherman, S. K. et al., Surgery 154 (2013): 1206-1213
Shi, M. et al., World J Gastroenterol. 10 (2004): 1146-1151
Shi, Z. et al., Tumour.Biol 36 (2015): 8519-8529
Shimizu, F. et al., Lab Invest 83 (2003): 187-197
Shioji, G. et al., J Hum.Genet. 50 (2005): 507-515
Showel, M. M. et al., F1000Prime.Rep. 6 (2014): 96
Siegel, S. et al., Blood 102 (2003): 4416-4423
Siew, Y. Y. et al., Int.Immunol. 27 (2015): 621-632
Silva, L. P. et al., Anal.Chem. 85 (2013): 9536-9542
Silvestris, F. et al., Adv.Exp.Med.Biol 714 (2011): 113-128
Singh, V. et al., Curr.Cancer Drug Targets. 13 (2013): 379-399
Singh-Jasuja, H. et al., Cancer Immunol.Immunother. 53 (2004): 187-195
Skawran, B. et al., Mod.Pathol. 21 (2008): 505-516
Small, E. J. et al., J Clin Oncol. 24 (2006): 3089-3094
Smetsers, S. et al., Fam.Cancer 11 (2012): 661-665
Smith, P. et al., Clin Cancer Res 13 (2007): 4061-4068
Sohrabi, A. et al., Asian Pac.J Cancer Prev. 15 (2014): 6745-6748
Song, H. R. et al., Mol.Carcinog 52 Suppl 1 (2013): E155-E160
Sonora, C. et al., J Histochem.Cytochem. 54 (2006): 289-299
Spaner, D. E. et al., Cancer Immunol Immunother. 54 (2005): 635-646
Srivastava, N. et al., Cancer Manag.Res. 6 (2014): 279-289
Stacey, S. N. et al., Nat Commun. 6 (2015): 6825
Stahl, M. et al., Ann.Oncol. 24 Suppl 6 (2013): vi51-vi56
Stangel, D. et al., J Surg.Res 197 (2015): 91-100
Stein, U., Expert.Opin.Ther.Targets. 17 (2013): 1039-1052
Steinberg, R. L. et al., Urol.Oncol (2016a)
Steinberg, R. L. et al., Urol.Oncol (2016b)
Steinway, S. N. et al., PLoS.One. 10 (2015): e0128159
Stenman, G. et al., Cell Cycle 9 (2010): 2986-2995
Stevanovic, S. et al., J Clin Oncol 33 (2015): 1543-1550
Stintzing, S., F1000Prime.Rep. 6 (2014): 108
Stratakis, C. A. et al., DNA Seq. 9 (1998): 227-230
Struyf, S. et al., Am.J Pathol. 163 (2003): 2065-2075
Sturm, M. et al., BMC.Bioinformatics. 9 (2008): 163
Su, Z. et al., Cancer Res. 63 (2003): 2127-2133
Subhash, V. V. et al., BMC.Cancer 15 (2015): 550
Sui, Y. et al., Oncogene 26 (2007): 822-835
Sukocheva, O. A. et al., World J Gastroenterol. 21 (2015): 6146-6156
Sun, S. et al., Gene 584 (2016): 90-96
Sun, W. et al., World J Gastroenterol. 19 (2013): 2913-2920
Sutherland, C. L. et al., Blood 108 (2006): 1313-1319
Suzuki, N. et al., J Orthop.Res 32 (2014): 915-922
Tabares-Seisdedos, R. et al., Mol.Psychiatry 14 (2009): 563-589
Takahashi, M. et al., Int.J Oncol 27 (2005): 1483-1487
Takatsu, H. et al., J Biol Chem 286 (2011): 38159-38167
Takayama, M. A. et al., Genes Cells 5 (2000a): 481-490
Takayama, T. et al., Cancer 68 (1991): 2391-2396
Takayama, T. et al., Lancet 356 (2000b): 802-807
Taketani, T. et al., Cancer Res 62 (2002): 33-37
Tang, C. et al., Int.J Clin Exp.Pathol. 7 (2014): 4782-4794
Tatenhorst, L. et al., J Neuropathol.Exp.Neurol. 63 (2004): 210-222
Taverniti, V. et al., Nucleic Acids Res 43 (2015): 482-492
Taylor, M. et al., Breast Cancer Res 9 (2007): R46
Terabayashi, T. et al., PLoS.One. 7 (2012): e39714
Teufel, R. et al., Cell Mol Life Sci. 62 (2005): 1755-1762
Thakkar, J. P. et al., Cancer Epidemiol.Biomarkers Prev. 23 (2014): 1985-1996
Tian, Y. et al., Diagn.Pathol. 9 (2014): 42
Ting, L. et al., DNA Repair (Amst) 9 (2010): 1241-1248
Toda, M. et al., Meta Gene 2 (2014): 686-693
Toomey, P. G. et al., Cancer Control 20 (2013): 32-42
Torelli, G. F. et al., Haematologica 99 (2014): 1248-1254
Tran, E. et al., Science 344 (2014): 641-645
Tsujikawa, T. et al., Int.J Cancer 132 (2013): 2755-2766
Tumova, L. et al., Mol.Cancer Ther. 13 (2014): 812-822
Urata, Y. N. et al., Sci.Rep. 5 (2015): 13676
Ushiku, T. et al., Histopathology 61 (2012): 1043-1056
Utrera, R. et al., EMBO J 17 (1998): 5015-5025
Vainio, P. et al., Am.J Pathol. 178 (2011a): 525-536
Vainio, P. et al., Oncotarget. 2 (2011b): 1176-1190
Valque, H. et al., PLoS.One. 7 (2012): e46699
van de Klundert, M. A. et al., PLoS.One. 7 (2012): e48940
Van Ginkel, P. R. et al., Biochim.Biophys.Acta 1448 (1998): 290-297
van Muijen, G. N. et al., Recent Results Cancer Res 139 (1995): 105-122
Van, Seuningen, I et al., Biochem.J 348 Pt 3 (2000): 675-686
Vater, I. et al., Leukemia 29 (2015): 677-685
Ventela, S. et al., Oncotarget. 6 (2015): 144-158
Verreman, K. et al., Biochem.J 439 (2011): 469-477
Vici, P. et al., J Exp.Clin Cancer Res 33 (2014): 29
Von Hoff, D. D. et al., N.Engl.J Med. 369 (2013): 1691-1703
von Rundstedt, F. C. et al., Transl.Androl Urol. 4 (2015): 244-253
Wallrapp, C. et al., Ann.Oncol 10 Suppl 4 (1999): 64-68
Walsh, M. D. et al., Mod.Pathol. 26 (2013): 1642-1656
Walter, S. et al., J Immunol 171 (2003): 4974-4978
Walter, S. et al., Nat Med. 18 (2012): 1254-1261
Walton, E. L. et al., Biology.(Basel) 3 (2014): 578-605
Wan, W. et al., World J Surg.Oncol 12 (2014): 185
Wang, C. et al., Nucleic Acids Res 43 (2015a): 4893-4908
Wang, C. Q. et al., Oral Surg.Oral Med.Oral Pathol.Oral Radiol. 117 (2014a): 353-360
Wang, D. et al., Chin Med.Sci.J 14 (1999a): 107-111
Wang, G. et al., Tumour.Biol 36 (2015b): 1055-1065
Wang, G. H. et al., Oncol Lett. 5 (2013a): 544-548
Wang, H. et al., Carcinogenesis 30 (2009a): 1314-1319
Wang, J. et al., Ann.Surg.Oncol 22 (2015c): 685-692
Wang, J. et al., J Exp.Clin Cancer Res 34 (2015d): 13
Wang, J. W. et al., Oncogene 23 (2004): 4089-4097
Wang, L. et al., J Cutan.Pathol. 42 (2015e): 361-367
Wang, L. et al., Mol.Biol Rep. 38 (2011a): 229-236
Wang, L. et al., Diagn.Pathol. 8 (2013b): 190
Wang, N. et al., Arch.Gynecol.Obstet. 283 (2011b): 103-108
Wang, Q. et al., Cell 138 (2009b): 245-256
Wang, Q. et al., BMC.Cancer 11 (2011c): 271
Wang, Q. et al., Onco.Targets.Ther. 8 (2015f): 1971-1977
Wang, Q. et al., PLoS.One. 8 (2013c): e61640
Wang, R. et al., Mol.Cell Biochem. 405 (2015g): 97-104
Wang, S. et al., J Cell Sci. 120 (2007): 567-577
Wang, W. Z. et al., J Exp.Clin Cancer Res 29 (2010): 140
Wang, X. W. et al., Gut Liver 8 (2014b): 487-494
Wang, X. Z. et al., Oncogene 18 (1999b): 5718-5721
Wang, Y. et al., Cancer Cell 26 (2014c): 374-389
Wang, Y. P. et al., Ai.Zheng. 27 (2008): 243-248
Wang, Z. et al., J Cancer Res Clin Oncol 141 (2015h): 1353-1361
Wang, Z. et al., Oncotarget. (2016)
Wang, Z. et al., Glycobiology 22 (2012): 930-938
Watanabe, N. et al., J Biol Chem 278 (2003): 26102-26110
Watts, C. A. et al., Chem Biol 20 (2013): 1399-1410
Wells, J. et al., J Biol Chem 284 (2009): 29125-29135
Weng, Y. R. et al., Carcinogenesis 35 (2014): 1389-1398
Wheler, J. J. et al., BMC.Cancer 15 (2015): 442
Whitaker, H. C. et al., Oncogene 33 (2014): 5274-5287
Wierda, W. G. et al., Blood 118 (2011): 5126-5129
Wierinckx, A. et al., Endocr.Relat Cancer 14 (2007): 887-900
Wilhelm, S. M. et al., Cancer Res 64 (2004): 7099-7109
Willcox, B. E. et al., Protein Sci. 8 (1999): 2418-2423
Williams, G. L. et al., Cell Cycle 6 (2007): 1699-1704
Wilson, P. M. et al., Nat Rev.Clin Oncol 11 (2014): 282-298
Wilzen, A. et al., Int.J Oncol 34 (2009): 697-705
Wittig, B. et al., Hum.Gene Ther. 12 (2001): 267-278
Wlcek, K. et al., Cancer Biol Ther. 11 (2011): 801-811
Wong, R. P. et al., Pigment Cell Melanoma Res 25 (2012): 213-218
World Cancer Report, (2014)
World Health Organization, (2014), http://www.who.int/en/
Wu, J. et al., ACS Chem Biol 8 (2013): 2201-2208
Wu, Y. et al., Cancer Lett. 356 (2015): 646-655
Xie, B. et al., Pathol.Oncol Res 19 (2013): 611-617
Xie, C. et al., Biochem.Biophys.Res Commun. 445 (2014): 263-268
Xiong, D. et al., Carcinogenesis 33 (2012): 1797-1805
Xu, X. et al., Oncogene 26 (2007): 7371-7379
Xu, X. et al., J Biol Chem 289 (2014): 8881-8890
Xue, J. H. et al., Acta Pharmacol.Sin. 32 (2011): 1019-1024
Yamada, T. et al., Br.J Cancer 108 (2013): 2495-2504
Yamashita, J. et al., Acta Derm.Venereol. 92 (2012): 593-597
Yamazoe, S. et al., J Exp.Clin Cancer Res 29 (2010): 53
Yan-Chun, L. et al., Appl.Immunohistochem.Mol.Morphol. (2015)
Yan-Fang, T. et al., PLoS.One. 10 (2015): e0126566
Yang, J. J. et al., Haematologica 99 (2014a): e11-e13
Yang, L. et al., J Biol Chem 291 (2016): 3905-3917
Yang, L. et al., PLoS.One. 10 (2015a): e0133896
Yang, T. T. et al., Sci.Rep. 5 (2015b): 14096
Yang, Y. et al., Oncol Lett. 9 (2015c): 1833-1838
Yang, Y. et al., PLoS.One. 9 (2014b): e97578
Yang, Y. M. et al., Cancer Sci. 102 (2011): 1264-1271
Yao, Y. et al., Cell Physiol Biochem. 35 (2015): 983-996
Ye, B. G. et al., Oncotarget. (2016)
Yeh, I. et al., Nat.Commun. 6 (2015): 7174
Yeh, S. et al., Proc.Natl.Acad.Sci.U.S.A 97 (2000): 11256-11261
Yonezawa, S. et al., Pathol.Int. 49 (1999): 45-54
Yoshimaru, T. et al., Nat Commun. 4 (2013): 2443
Yoshimaru, T. et al., Sci.Rep. 4 (2014): 7355
Young, A. et al., BMC.Cancer 14 (2014): 808
Yu, C. J. et al., Int.J Cancer 69 (1996): 457-465
Yu, H. et al., Nat Chem Biol 11 (2015a): 847-854
Yu, T. et al., Cell Res 24 (2014): 1214-1230
Yu, X. et al., Tumour.Biol 36 (2015b): 967-972
Yuan, M. et al., Oncotarget. 5 (2014): 2820-2826
Zaganjor, E. et al., Proc.Natl.Acad.Sci.U.S.A 111 (2014): 10568-10573
Zamuner, F. T. et al., Mol.Cancer Ther. 14 (2015): 828-834
Zaremba, S. et al., Cancer Res. 57 (1997): 4570-4577
Zavala-Zendejas, V. E. et al., Cancer Invest 29 (2011): 1-11
Zekri, A. R. et al., BMC.Res Notes 1 (2008): 106
Zeng, B. et al., Curr.Cancer Drug Targets. 13 (2013a): 103-116
Zeng, S. et al., Eur.J Cancer 49 (2013b): 3752-3762
Zeng, X. et al., Ai.Zheng. 26 (2007): 1080-1084
Zeng, X. X. et al., Eur.Rev Med.Pharmacol.Sci. 19 (2015): 4353-4361
Zhan, W. et al., PLoS.One. 10 (2015): e0142596
Zhang, B. et al., J Huazhong.Univ Sci.Technolog.Med.Sci. 30 (2010a): 322-325
Zhang, G. et al., BMC.Cancer 14 (2014): 310
Zhang, J. et al., Theor.Biol Med.Model. 9 (2012a): 53
Zhang, Q. et al., Zhongguo Fei.Ai.Za Zhi. 13 (2010b): 612-616
Zhang, W. et al., Clin Cancer Res 7 (2001): 822s-829s
Zhang, W. et al., Biochem.J (2016)
Zhang, X. et al., EMBO J 30 (2011): 2177-2189
Zhang, X. et al., Med.Oncol 32 (2015): 148
Zhang, X. et al., Int.J Med.Sci. 10 (2013a): 1795-1804
Zhang, Y. et al., Gene 497 (2012b): 93-97
Zhang, Y. et al., J Ovarian.Res 6 (2013b): 55
Zhao, J. et al., Int.J Med.Sci. 11 (2014a): 1089-1097
Zhao, J. G. et al., FEBS Lett. 588 (2014b): 4536-4542
Zhen, T. et al., Oncotarget. 5 (2014): 3756-3769
Zheng, M. et al., Breast Cancer Res Treat. 148 (2014): 423-436
Zheng, M. Z. et al., J Transl.Med. 5 (2007): 36
Zhong, M. et al., Mol.Cancer Res 8 (2010): 1164-1172
Zhong, T. et al., Biomed.Pharmacother. 69 (2015): 317-325
Zhou, X. et al., J Cancer Res Clin Oncol 141 (2015): 961-969
Zhou, Y. et al., Front Biosci.(Landmark.Ed) 16 (2011): 1109-1131
Zhou, Z. et al., Gastroenterology 147 (2014): 1043-1054
Zhu, H. H. et al., Asian Pac.J Trop.Med. 7 (2014): 488-491
Zhu, J. et al., Int.J Clin Exp.Pathol. 8 (2015a): 9479-9486
Zhu, P. et al., Oncol Lett. 10 (2015b): 1487-1494
Zighelboim, I. et al., J Clin Oncol 27 (2009): 3091-3096
Zimmerman, K. M. et al., Mol.Cancer Res 11 (2013): 370-380
Zocchi, M. R. et al., Blood 119 (2012): 1479-1489
Zou, J. X. et al., Mol.Cancer Res 12 (2014): 539-549
Zou, T. T. et al., Oncogene 21 (2002): 4855-4862
Claims (13)
- 配列番号241に示されるアミノ酸配列からなるペプチドまたはその薬学的に許容可能な塩を含む、がんの診断および/または治療において使用するための薬剤であって、前記ペプチドがHLA−A*24に結合する、薬剤。
- 前記ペプチドが、非ペプチド結合を含む、請求項1に記載の薬剤。
- 請求項1に記載の薬剤をエンコードする核酸、または前記核酸が異種プロモーター配列と結合する、核酸。
- 請求項3に記載の核酸を発現できる、発現ベクター。
- 請求項1に記載の薬剤、請求項3に記載の核酸または請求項4に記載の発現ベクターを含んでなる組換え宿主細胞であって、前記宿主細胞が樹状細胞若しくは抗原提示細胞である、組換え宿主細胞。
- 請求項1に記載の薬剤を提示し、または請求項3に記載の核酸を発現し、または請求項4に記載の発現ベクターを発現する、請求項5に記載の宿主細胞を培養するステップと、前記薬剤を前記宿主細胞またはその培養液から単離するステップとを含んでなる、請求項1に記載の薬剤を製造する方法。
- T細胞を、適切な抗原提示細胞の表面に、または抗原提示細胞を模倣する人工コンストラクトの表面に発現される抗原負荷ヒトクラスI MHC分子に、前記T細胞を抗原特異的様式で活性化するのに十分な時間にわたり、生体外で接触させるステップを含んでなり、前記抗原が、請求項1または2に記載の薬剤である、活性化Tリンパ球を製造するインビトロ法。
- 請求項1または2に記載の薬剤を提示する細胞を選択的に認識する、請求項7に記載の方法によって製造される活性化Tリンパ球。
- MHC分子と結合している請求項1または2に記載の薬剤を特異的に認識する抗体、又は請求項1または2に記載の薬剤を特異的に認識する抗体、または前記抗体が可溶性または膜結合抗体である、モノクローナル抗体。
- がんが、配列番号241に示されるアミノ酸配列からなるペプチドが由来するタンパク質の過剰発現を示す、神経膠芽腫、乳がん、結腸直腸がん、腎細胞がん、慢性リンパ球性白血病、肝細胞がん、非小細胞肺がんおよび小細胞肺がん、非ホジキンリンパ腫、急性骨髄性白血病、卵巣がん、膵臓がん、前立腺がん、胃食道接合部がんを含む食道がん、胆嚢がんおよび胆管がん、黒色腫、胃がん、精巣がん、膀胱がん、または子宮がんおよびその他の腫瘍の群から選択される、請求項1に記載の薬剤。
- T細胞受容体(TCR)、または可溶性または膜結合T細胞受容体であって、HLAリガンドと反応性であり、
前記リガンドが請求項1または2に記載の薬剤、またはMHC分子と結合している請求項1または2に記載の薬剤の一部である、T細胞受容体、または
前記HLAリガンドと反応性であるT細胞受容体が可溶性分子として提供される、または
前記HLAリガンドと反応性であるT細胞受容体が免疫刺激ドメインもしくは毒素を含むさらなるエフェクター機能を保有する、T細胞受容体。 - 以下のa)〜g)から選択される少なくとも1つの活性成分、及び薬学的に許容できる担体を含んでなる、がんの診断および/または治療において使用するための医薬組成物;
a)配列番号241に示されるアミノ酸配列からなるペプチド;
b)a)に記載のペプチドまたはMHC分子と結合している前記ペプチドに反応性のT細胞受容体;
c)a)〜b)のいずれかをコードする核酸、または前記核酸を含んでなる発現ベクター;
d)c)の発現ベクターを含んでなる宿主細胞;
e)T細胞を、抗原特異的様式でT細胞を活性化するのに十分な時間にわたり、適切な抗原提示細胞の表面に発現されるa)に記載のペプチドと生体外で接触させるステップを含んでなる方法によって得られる、活性化Tリンパ球;
f)a)に記載のペプチドまたはMHC分子と結合している前記ペプチドを特異的に認識する抗体、または可溶性T細胞受容体、または免疫活性化ドメインまたは毒素との融合によって潜在的に修飾される、a)に記載のペプチドまたはMHC分子と結合している前記ペプチドを特異的に認識する抗体、または可溶性T細胞受容体;
g)配列番号241に示されるアミノ酸配列からなるペプチドを認識し、またはMHC分子と結合している配列番号241に示されるアミノ酸配列からなるペプチドを認識する、アプタマー。 - がんが、配列番号241に示されるアミノ酸配列からなるペプチドの過剰発現を示す、神経膠芽腫、乳がん、結腸直腸がん、腎細胞がん、慢性リンパ球性白血病、肝細胞がん、非小細胞肺がんおよび小細胞肺がん、非ホジキンリンパ腫、急性骨髄性白血病、卵巣がん、膵臓がん、前立腺がん、胃食道接合部がんを含む食道がん、胆嚢がんおよび胆管がん、黒色腫、胃がん、精巣がん、膀胱がん、または子宮がんおよびその他の腫瘍の群から選択される、請求項12に記載の医薬組成物。
Priority Applications (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2021072989A JP7506026B2 (ja) | 2015-08-28 | 2021-04-23 | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
| JP2024095596A JP2024123066A (ja) | 2015-08-28 | 2024-06-13 | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201562211276P | 2015-08-28 | 2015-08-28 | |
| GB1515321.6 | 2015-08-28 | ||
| US62/211,276 | 2015-08-28 | ||
| GBGB1515321.6A GB201515321D0 (en) | 2015-08-28 | 2015-08-28 | Novel peptides, combination of peptides and scaffolds for use in immunotherapeutic treatment of various cancers |
| PCT/EP2016/070146 WO2017036936A1 (en) | 2015-08-28 | 2016-08-26 | Novel peptides, combination of peptides and scaffolds for use in immunotherapeutic treatment of various cancers |
Related Child Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2018142058A Division JP6742370B2 (ja) | 2015-08-28 | 2018-07-30 | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
| JP2021072989A Division JP7506026B2 (ja) | 2015-08-28 | 2021-04-23 | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2018532382A JP2018532382A (ja) | 2018-11-08 |
| JP2018532382A5 JP2018532382A5 (ja) | 2019-10-03 |
| JP6970974B2 true JP6970974B2 (ja) | 2021-11-24 |
Family
ID=54326487
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2018511114A Active JP6970974B2 (ja) | 2015-08-28 | 2016-08-26 | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
| JP2021072989A Active JP7506026B2 (ja) | 2015-08-28 | 2021-04-23 | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021072989A Active JP7506026B2 (ja) | 2015-08-28 | 2021-04-23 | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
Country Status (18)
| Country | Link |
|---|---|
| US (7) | US10525114B2 (ja) |
| EP (2) | EP3461494A1 (ja) |
| JP (2) | JP6970974B2 (ja) |
| KR (3) | KR20180038550A (ja) |
| CN (1) | CN107921111B (ja) |
| AU (3) | AU2016313684B2 (ja) |
| BR (1) | BR112018003460A2 (ja) |
| CA (2) | CA2996963A1 (ja) |
| CR (6) | CR20180174A (ja) |
| EA (2) | EA201890560A1 (ja) |
| GB (1) | GB201515321D0 (ja) |
| MA (7) | MA43458A1 (ja) |
| MX (1) | MX2018002480A (ja) |
| PE (2) | PE20181517A1 (ja) |
| SG (3) | SG10201807588TA (ja) |
| TW (15) | TWI722662B (ja) |
| UA (1) | UA124577C2 (ja) |
| WO (1) | WO2017036936A1 (ja) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2021118721A (ja) * | 2015-08-28 | 2021-08-12 | イマティクス バイオテクノロジーズ ゲーエムベーハー | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
Families Citing this family (25)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB201507030D0 (en) | 2015-04-24 | 2015-06-10 | Immatics Biotechnologies Gmbh | Immunotherapy against lung cancers, in particular NSCLC |
| WO2017106638A1 (en) | 2015-12-16 | 2017-06-22 | Gritstone Oncology, Inc. | Neoantigen identification, manufacture, and use |
| JP7075125B2 (ja) | 2016-05-25 | 2022-05-25 | イマティクス バイオテクノロジーズ ゲーエムベーハー | 標的としてのおよび胆嚢がんおよび胆管がんおよびその他のがんに対する免疫療法で使用するための新規ペプチド、ペプチド組み合わせ |
| GB201609193D0 (en) | 2016-05-25 | 2016-07-06 | Immatics Biotechnologies Gmbh | Novel peptides, combination of peptides as targets for use in immunotherapy against gallbladder cancer and cholangiocarcinoma and other cancers |
| DE102016123893A1 (de) | 2016-12-08 | 2018-06-14 | Immatics Biotechnologies Gmbh | T-Zellrezeptoren mit verbesserter Bindung |
| KR102379955B1 (ko) | 2016-12-08 | 2022-03-29 | 이매틱스 바이오테크놀로지스 게엠베하 | 짝짓기가 향상된 t 세포 수용체 |
| CN108250286B (zh) * | 2016-12-28 | 2021-06-08 | 香雪生命科学技术(广东)有限公司 | 源自于pasd1的肿瘤抗原短肽 |
| WO2018189152A2 (en) | 2017-04-10 | 2018-10-18 | Immatics Biotechnologies Gmbh | Peptides and combination of peptides for use in immunotherapy against leukemias and other cancers |
| SG10202100326SA (en) | 2017-04-10 | 2021-02-25 | Immatics Biotechnologies Gmbh | Peptides and combination of peptides for use in immunotherapy against leukemias and other cancers |
| DE102017115966A1 (de) | 2017-07-14 | 2019-01-17 | Immatics Biotechnologies Gmbh | Polypeptidmolekül mit verbesserter zweifacher Spezifität |
| SI3652215T1 (sl) | 2017-07-14 | 2021-08-31 | Immatics Biotechnologies Gmbh | Izboljšana polipeptidna molekula z dvojno specifičnostjo |
| WO2019068007A1 (en) * | 2017-09-28 | 2019-04-04 | Immpact-Bio Ltd. | UNIVERSAL PLATFORM FOR PREPARING AN INHIBITORY CHIMERIC ANTIGEN RECEPTOR (ICAR) |
| ES3029080T3 (en) | 2017-10-10 | 2025-06-23 | Seattle Project Corp | Neoantigen identification using hotspots |
| CN111630602A (zh) | 2017-11-22 | 2020-09-04 | 磨石肿瘤生物技术公司 | 减少新抗原的接合表位呈递 |
| DE102018107224A1 (de) * | 2018-02-21 | 2019-08-22 | Immatics Biotechnologies Gmbh | Peptide und Kombinationen von Peptiden nicht-kanonischen Ursprungs zur Verwendung in der Immuntherapie gegen verschiedene Krebsarten |
| TW202016131A (zh) * | 2018-05-16 | 2020-05-01 | 德商英麥提克生物技術股份有限公司 | 用於抗癌免疫治療的肽 |
| CN109467596B (zh) * | 2018-11-12 | 2021-11-09 | 湖北省农业科学院畜牧兽医研究所 | 转录因子sp1在调控猪rtl1基因表达中的应用 |
| CN109738929B (zh) * | 2018-12-03 | 2022-10-21 | 中国辐射防护研究院 | 一种基于居民食物最大消费量的剂量估算方法及系统 |
| JPWO2020230792A1 (ja) * | 2019-05-13 | 2020-11-19 | ||
| CN110123453B (zh) * | 2019-05-31 | 2021-07-23 | 东北大学 | 一种基于无标记增强现实的手术导航系统 |
| WO2021207826A1 (en) * | 2020-04-14 | 2021-10-21 | Université de Montréal | Novel tumor-specific antigens for acute lymphoblastic leukemia (all) and uses thereof |
| CN111910000B (zh) * | 2020-07-02 | 2022-07-01 | 中山大学肿瘤防治中心(中山大学附属肿瘤医院、中山大学肿瘤研究所) | 一种预测鼻咽癌预后的肿瘤微环境成分标志物组合及系统 |
| WO2022225852A1 (en) * | 2021-04-19 | 2022-10-27 | Board Of Regents, The University Of Texas System | Methods and compositions comprising mhc class i peptides |
| CN115607573B (zh) * | 2022-12-16 | 2023-05-23 | 北京大学第三医院(北京大学第三临床医学院) | 一种用于调节杀伤性t细胞活性的方法、药物及其应用 |
| WO2025015234A2 (en) * | 2023-07-13 | 2025-01-16 | Ltz Therapeutics Inc. | Multispecific antibodies and uses thereof |
Family Cites Families (41)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0832125A4 (en) * | 1995-06-05 | 1998-12-30 | Human Genome Sciences Inc | HUMAN G PROTEIN RECEPTOR HGBER32 |
| WO1999045954A1 (en) * | 1998-03-13 | 1999-09-16 | Epimmune, Inc. | Hla-binding peptides and their uses |
| US7342108B2 (en) * | 1998-06-25 | 2008-03-11 | The General Hospital Corporation | De novo DNA cytosine methyltransferase genes, polypeptides and uses thereof |
| JP4315301B2 (ja) * | 1998-10-30 | 2009-08-19 | 独立行政法人科学技術振興機構 | ヒトH37タンパク質と、このタンパク質をコードする cDNA |
| US6680197B2 (en) * | 1998-12-28 | 2004-01-20 | Corixa Corporation | Compositions and methods for the therapy and diagnosis of breast cancer |
| CA2311201A1 (en) * | 1999-08-05 | 2001-02-05 | Genset S.A. | Ests and encoded human proteins |
| US20040002068A1 (en) * | 2000-03-01 | 2004-01-01 | Corixa Corporation | Compositions and methods for the detection, diagnosis and therapy of hematological malignancies |
| US6436703B1 (en) | 2000-03-31 | 2002-08-20 | Hyseq, Inc. | Nucleic acids and polypeptides |
| GB0119473D0 (en) * | 2001-08-09 | 2001-10-03 | Astrazeneca | Compounds |
| WO2003016523A1 (en) * | 2001-08-21 | 2003-02-27 | Millennium Pharmaceuticals, Inc. | Screen for cdc7 inhibitors |
| US7193069B2 (en) * | 2002-03-22 | 2007-03-20 | Research Association For Biotechnology | Full-length cDNA |
| DE10316701A1 (de) | 2003-04-09 | 2004-11-04 | Hinzmann, Bernd, Dr. | Humane Nukleinsäuresequenzen aus Bronchialkarzinomen |
| EP1622934A2 (en) * | 2003-05-05 | 2006-02-08 | Pharmacia Italia S.p.A. | Truncated forms of human dbf4, complexes with their interacting partners and methods for identification of inhibitors thereof |
| EP1892306A3 (en) | 2003-10-06 | 2008-06-11 | Bayer HealthCare AG | Methods and kits for investigating cancer |
| JP4721903B2 (ja) * | 2004-01-29 | 2011-07-13 | 大日本住友製薬株式会社 | 新規腫瘍抗原蛋白質及びその利用 |
| WO2009008414A1 (ja) * | 2007-07-10 | 2009-01-15 | Shionogi & Co., Ltd. | Mmp13に対する中和活性を有するモノクローナル抗体 |
| RS54147B1 (en) * | 2007-07-27 | 2015-12-31 | Immatics Biotechnologies Gmbh | Novel immunotherapy against brain tumors |
| ES2555282T3 (es) | 2007-07-27 | 2015-12-30 | Immatics Biotechnologies Gmbh | Nuevos epítopos inmunogénicos para inmunoterapia |
| AU2008311869B2 (en) * | 2007-10-17 | 2013-11-21 | The Board Of Trustees Of The Leland Stanford Junior University | Method and composition for crystallizing G protein-coupled receptors |
| WO2009123568A1 (en) | 2008-03-31 | 2009-10-08 | Agency For Science, Technology & Research | High efficiency linear transmitter |
| TW201000115A (en) * | 2008-06-11 | 2010-01-01 | Oncotherapy Science Inc | IQGAP3 epitope peptides and vaccines containing the same |
| US20110318380A1 (en) * | 2008-10-01 | 2011-12-29 | Dako Denmark A/S | MHC Multimers in Cancer Vaccines and Immune Monitoring |
| SI2172211T1 (sl) | 2008-10-01 | 2015-03-31 | Immatics Biotechnologies Gmbh | Sestavek s tumorjem povezanih peptidov in zadevnih cepiv proti raku za zdravljenje glioblastoma (GBM) in drugih vrst raka |
| ITBO20090075A1 (it) * | 2009-02-13 | 2010-08-14 | Magneti Marelli Spa | Macchina elettrica con singolo statore e due rotori tra loro indipendenti e veicolo stradale provvisto di tale macchina elettrica |
| GB0911566D0 (en) * | 2009-07-03 | 2009-08-12 | Immunocore Ltd | T cell receptors |
| GB201004551D0 (en) * | 2010-03-19 | 2010-05-05 | Immatics Biotechnologies Gmbh | NOvel immunotherapy against several tumors including gastrointestinal and gastric cancer |
| US8710014B2 (en) * | 2010-10-08 | 2014-04-29 | Proteapex Therapeutics Llc | Compositions and methods for inhibition of MMP13:MMP-substrate interactions |
| MX350200B (es) * | 2011-02-01 | 2017-08-30 | Genmab As | Anticuerpos humanos y conjugados de anticuerpo-farmaco contra cd74. |
| CA2848162C (en) | 2011-09-12 | 2023-03-14 | Creatics Llc | Non-invasive methods of detecting target molecules |
| EP2788012A4 (en) | 2011-11-11 | 2015-08-05 | Cedars Sinai Medical Center | COMPOSITIONS AND METHODS OF TREATING NURSE DISEASES |
| WO2013151667A1 (en) | 2012-04-02 | 2013-10-10 | modeRNA Therapeutics | Modified polynucleotides |
| US9254311B2 (en) * | 2012-04-02 | 2016-02-09 | Moderna Therapeutics, Inc. | Modified polynucleotides for the production of proteins |
| EP2867252B1 (en) * | 2012-08-15 | 2019-12-04 | Université de Montréal | Method for identifying novel minor histocompatibility antigens |
| DK3636665T3 (da) * | 2012-09-14 | 2022-10-03 | Us Health | T-cellereceptorer, som genkender mhc klasse ii-begrænset mage-a3 |
| WO2014160275A2 (en) | 2013-03-14 | 2014-10-02 | Battell Memorial Institute | Biomarkers for liver fibrosis |
| TWI777198B (zh) | 2013-08-05 | 2022-09-11 | 德商伊瑪提克斯生物科技有限公司 | 新穎肽類,細胞及其用於治療多種腫瘤的用途,其製造方法及包含其等之醫藥組成物(七) |
| WO2015092710A1 (en) * | 2013-12-19 | 2015-06-25 | Glaxosmithkline Biologicals, S.A. | Contralateral co-administration of vaccines |
| GB201515321D0 (en) * | 2015-08-28 | 2015-10-14 | Immatics Biotechnologies Gmbh | Novel peptides, combination of peptides and scaffolds for use in immunotherapeutic treatment of various cancers |
| WO2017184590A1 (en) * | 2016-04-18 | 2017-10-26 | The Broad Institute Inc. | Improved hla epitope prediction |
| TW202016131A (zh) * | 2018-05-16 | 2020-05-01 | 德商英麥提克生物技術股份有限公司 | 用於抗癌免疫治療的肽 |
| US11945850B2 (en) * | 2018-09-17 | 2024-04-02 | Immatics Biotechnologies Gmbh | B*44 restricted peptides for use in immunotherapy against cancers and related methods |
-
2015
- 2015-08-28 GB GBGB1515321.6A patent/GB201515321D0/en not_active Ceased
-
2016
- 2016-08-26 SG SG10201807588TA patent/SG10201807588TA/en unknown
- 2016-08-26 CN CN201680050119.7A patent/CN107921111B/zh active Active
- 2016-08-26 KR KR1020187007555A patent/KR20180038550A/ko not_active Ceased
- 2016-08-26 CR CR20180174A patent/CR20180174A/es unknown
- 2016-08-26 TW TW108141497A patent/TWI722662B/zh not_active IP Right Cessation
- 2016-08-26 TW TW108141498A patent/TWI722663B/zh not_active IP Right Cessation
- 2016-08-26 EA EA201890560A patent/EA201890560A1/ru unknown
- 2016-08-26 TW TW108141488A patent/TWI722654B/zh not_active IP Right Cessation
- 2016-08-26 PE PE2018001375A patent/PE20181517A1/es unknown
- 2016-08-26 TW TW108141489A patent/TWI722655B/zh not_active IP Right Cessation
- 2016-08-26 TW TW109122012A patent/TWI782290B/zh active
- 2016-08-26 MA MA43458A patent/MA43458A1/fr unknown
- 2016-08-26 TW TW107128393A patent/TWI699377B/zh not_active IP Right Cessation
- 2016-08-26 AU AU2016313684A patent/AU2016313684B2/en not_active Ceased
- 2016-08-26 TW TW108141490A patent/TWI722656B/zh not_active IP Right Cessation
- 2016-08-26 KR KR1020187023153A patent/KR102308798B1/ko active Active
- 2016-08-26 MA MA43460A patent/MA43460A1/fr unknown
- 2016-08-26 TW TW108141499A patent/TWI722664B/zh not_active IP Right Cessation
- 2016-08-26 MX MX2018002480A patent/MX2018002480A/es unknown
- 2016-08-26 CR CR20200493A patent/CR20200493A/es unknown
- 2016-08-26 MA MA045073A patent/MA45073A/fr unknown
- 2016-08-26 EP EP18201208.8A patent/EP3461494A1/en active Pending
- 2016-08-26 TW TW108141494A patent/TWI722659B/zh not_active IP Right Cessation
- 2016-08-26 UA UAA201808797A patent/UA124577C2/uk unknown
- 2016-08-26 CR CR20200492A patent/CR20200492A/es unknown
- 2016-08-26 MA MA43461A patent/MA43461B1/fr unknown
- 2016-08-26 CA CA2996963A patent/CA2996963A1/en active Pending
- 2016-08-26 EA EA201891874A patent/EA201891874A3/ru unknown
- 2016-08-26 TW TW108141496A patent/TWI722661B/zh not_active IP Right Cessation
- 2016-08-26 SG SG10202001831SA patent/SG10202001831SA/en unknown
- 2016-08-26 SG SG10202001830PA patent/SG10202001830PA/en unknown
- 2016-08-26 TW TW108141487A patent/TWI722653B/zh not_active IP Right Cessation
- 2016-08-26 KR KR1020217031080A patent/KR102515909B1/ko active Active
- 2016-08-26 JP JP2018511114A patent/JP6970974B2/ja active Active
- 2016-08-26 TW TW108141491A patent/TWI722657B/zh not_active IP Right Cessation
- 2016-08-26 CA CA3072922A patent/CA3072922A1/en active Pending
- 2016-08-26 CR CR20200494A patent/CR20200494A/es unknown
- 2016-08-26 TW TW108141495A patent/TWI722660B/zh not_active IP Right Cessation
- 2016-08-26 BR BR112018003460-6A patent/BR112018003460A2/pt not_active Application Discontinuation
- 2016-08-26 WO PCT/EP2016/070146 patent/WO2017036936A1/en not_active Application Discontinuation
- 2016-08-26 EP EP16762752.0A patent/EP3341014A1/en active Pending
- 2016-08-26 MA MA43459A patent/MA43459A1/fr unknown
- 2016-08-26 MA MA43457A patent/MA43457A1/fr unknown
- 2016-08-26 TW TW108141492A patent/TWI722658B/zh not_active IP Right Cessation
- 2016-08-26 MA MA042694A patent/MA42694A/fr unknown
- 2016-08-26 CR CR20180423A patent/CR20180423A/es unknown
- 2016-08-26 PE PE2018000318A patent/PE20181164A1/es unknown
- 2016-08-26 TW TW105127400A patent/TW201708246A/zh unknown
- 2016-08-26 CR CR20200495A patent/CR20200495A/es unknown
-
2018
- 2018-05-17 US US15/982,500 patent/US10525114B2/en active Active
- 2018-05-17 US US15/982,293 patent/US10576132B2/en active Active
- 2018-10-18 AU AU2018250464A patent/AU2018250464B2/en not_active Ceased
-
2019
- 2019-11-04 US US16/673,608 patent/US10568950B1/en active Active
- 2019-11-04 US US16/673,619 patent/US10695411B2/en active Active
-
2020
- 2020-03-16 AU AU2020201896A patent/AU2020201896B2/en not_active Ceased
- 2020-05-29 US US16/887,765 patent/US10898557B2/en not_active Expired - Fee Related
- 2020-06-29 US US16/915,420 patent/US10898558B2/en not_active Expired - Fee Related
- 2020-09-10 US US17/017,358 patent/US11065316B2/en active Active
-
2021
- 2021-04-23 JP JP2021072989A patent/JP7506026B2/ja active Active
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2021118721A (ja) * | 2015-08-28 | 2021-08-12 | イマティクス バイオテクノロジーズ ゲーエムベーハー | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
| JP7506026B2 (ja) | 2015-08-28 | 2024-06-25 | イマティクス バイオテクノロジーズ ゲーエムベーハー | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7506026B2 (ja) | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ | |
| JP6742370B2 (ja) | 様々ながんの免疫治療で使用するための新規ペプチド、ペプチドおよびスキャフォールド組み合わせ |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20190814 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20190814 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20190816 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20200716 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20201013 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20210126 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210423 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20210519 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210818 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20210928 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20211025 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6970974 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |