JP5710487B2 - 抗notch2抗体および使用方法 - Google Patents
抗notch2抗体および使用方法 Download PDFInfo
- Publication number
- JP5710487B2 JP5710487B2 JP2011530173A JP2011530173A JP5710487B2 JP 5710487 B2 JP5710487 B2 JP 5710487B2 JP 2011530173 A JP2011530173 A JP 2011530173A JP 2011530173 A JP2011530173 A JP 2011530173A JP 5710487 B2 JP5710487 B2 JP 5710487B2
- Authority
- JP
- Japan
- Prior art keywords
- antibody
- amino acid
- seq
- acid sequence
- hvr
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 238000000034 method Methods 0.000 title description 160
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 232
- 102000001756 Notch2 Receptor Human genes 0.000 claims description 198
- 108010029751 Notch2 Receptor Proteins 0.000 claims description 197
- 102000005650 Notch Receptors Human genes 0.000 claims description 73
- 108010070047 Notch Receptors Proteins 0.000 claims description 73
- 206010028980 Neoplasm Diseases 0.000 claims description 72
- 230000000694 effects Effects 0.000 claims description 66
- 230000014509 gene expression Effects 0.000 claims description 63
- 239000003814 drug Substances 0.000 claims description 60
- 239000012634 fragment Substances 0.000 claims description 59
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 46
- 201000010099 disease Diseases 0.000 claims description 45
- 102000008394 Immunoglobulin Fragments Human genes 0.000 claims description 40
- 108010021625 Immunoglobulin Fragments Proteins 0.000 claims description 40
- 201000011510 cancer Diseases 0.000 claims description 38
- 238000011282 treatment Methods 0.000 claims description 35
- 238000004519 manufacturing process Methods 0.000 claims description 34
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 25
- 230000001965 increasing effect Effects 0.000 claims description 25
- 101000577199 Homo sapiens Neurogenic locus notch homolog protein 2 Proteins 0.000 claims description 18
- 239000003112 inhibitor Substances 0.000 claims description 16
- 101100348842 Mus musculus Notch2 gene Proteins 0.000 claims description 14
- 201000001441 melanoma Diseases 0.000 claims description 14
- 230000036210 malignancy Effects 0.000 claims description 13
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 10
- 102000046883 human NOTCH2 Human genes 0.000 claims description 8
- 238000005734 heterodimerization reaction Methods 0.000 claims description 4
- 101100074828 Caenorhabditis elegans lin-12 gene Proteins 0.000 claims description 3
- 239000013642 negative control Substances 0.000 claims description 3
- 210000004027 cell Anatomy 0.000 description 229
- 230000027455 binding Effects 0.000 description 169
- 108090000623 proteins and genes Proteins 0.000 description 118
- 239000000427 antigen Substances 0.000 description 96
- 108091007433 antigens Proteins 0.000 description 95
- 102000036639 antigens Human genes 0.000 description 95
- 108090000765 processed proteins & peptides Proteins 0.000 description 71
- 102000004169 proteins and genes Human genes 0.000 description 70
- 102000004196 processed proteins & peptides Human genes 0.000 description 62
- 235000018102 proteins Nutrition 0.000 description 61
- 229920001184 polypeptide Polymers 0.000 description 56
- 108010029755 Notch1 Receptor Proteins 0.000 description 55
- 102000001759 Notch1 Receptor Human genes 0.000 description 55
- 235000001014 amino acid Nutrition 0.000 description 55
- 239000013598 vector Substances 0.000 description 51
- 238000003556 assay Methods 0.000 description 47
- -1 1 μM in CmpE Chemical compound 0.000 description 39
- 229940024606 amino acid Drugs 0.000 description 39
- 238000006467 substitution reaction Methods 0.000 description 38
- 108060003951 Immunoglobulin Proteins 0.000 description 37
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 description 37
- 229940079593 drug Drugs 0.000 description 37
- 102000018358 immunoglobulin Human genes 0.000 description 37
- 230000011664 signaling Effects 0.000 description 37
- 125000005647 linker group Chemical group 0.000 description 36
- 150000007523 nucleic acids Chemical class 0.000 description 36
- 108020004414 DNA Proteins 0.000 description 35
- 150000001413 amino acids Chemical group 0.000 description 35
- 102000039446 nucleic acids Human genes 0.000 description 35
- 108020004707 nucleic acids Proteins 0.000 description 35
- 210000004408 hybridoma Anatomy 0.000 description 34
- 239000000203 mixture Substances 0.000 description 31
- 239000003446 ligand Substances 0.000 description 30
- 241000699670 Mus sp. Species 0.000 description 29
- 238000001727 in vivo Methods 0.000 description 29
- 102000005962 receptors Human genes 0.000 description 29
- 108020003175 receptors Proteins 0.000 description 29
- 108091035707 Consensus sequence Proteins 0.000 description 24
- 241001465754 Metazoa Species 0.000 description 24
- 238000000338 in vitro Methods 0.000 description 24
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 23
- 241000699666 Mus <mouse, genus> Species 0.000 description 23
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 23
- 230000006870 function Effects 0.000 description 23
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 22
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 22
- 229940127089 cytotoxic agent Drugs 0.000 description 22
- 239000000562 conjugate Substances 0.000 description 20
- 229940127121 immunoconjugate Drugs 0.000 description 20
- 230000004048 modification Effects 0.000 description 20
- 238000012986 modification Methods 0.000 description 20
- 108010087819 Fc receptors Proteins 0.000 description 19
- 102000009109 Fc receptors Human genes 0.000 description 19
- 230000004071 biological effect Effects 0.000 description 19
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 18
- 239000012636 effector Substances 0.000 description 18
- 238000003752 polymerase chain reaction Methods 0.000 description 18
- 239000003053 toxin Substances 0.000 description 18
- 231100000765 toxin Toxicity 0.000 description 18
- 108700012359 toxins Proteins 0.000 description 18
- AOJJSUZBOXZQNB-TZSSRYMLSA-N Doxorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)[C@H](O)[C@H](C)O1 AOJJSUZBOXZQNB-TZSSRYMLSA-N 0.000 description 17
- 125000003729 nucleotide group Chemical group 0.000 description 17
- 241000894007 species Species 0.000 description 17
- 239000003795 chemical substances by application Substances 0.000 description 16
- 231100000599 cytotoxic agent Toxicity 0.000 description 16
- 238000012360 testing method Methods 0.000 description 16
- 238000002965 ELISA Methods 0.000 description 15
- 241000588724 Escherichia coli Species 0.000 description 15
- 239000002246 antineoplastic agent Substances 0.000 description 15
- 239000002254 cytotoxic agent Substances 0.000 description 15
- 102000004190 Enzymes Human genes 0.000 description 14
- 108090000790 Enzymes Proteins 0.000 description 14
- 239000002253 acid Substances 0.000 description 14
- 125000000539 amino acid group Chemical group 0.000 description 14
- 238000005516 engineering process Methods 0.000 description 14
- 229940088598 enzyme Drugs 0.000 description 14
- 230000001976 improved effect Effects 0.000 description 14
- 238000003780 insertion Methods 0.000 description 14
- 230000037431 insertion Effects 0.000 description 14
- 239000002609 medium Substances 0.000 description 14
- 239000002773 nucleotide Substances 0.000 description 14
- 239000000243 solution Substances 0.000 description 14
- 241000124008 Mammalia Species 0.000 description 13
- 206010035226 Plasma cell myeloma Diseases 0.000 description 13
- 229940049595 antibody-drug conjugate Drugs 0.000 description 13
- 238000012217 deletion Methods 0.000 description 13
- 230000037430 deletion Effects 0.000 description 13
- 239000013604 expression vector Substances 0.000 description 13
- 238000002823 phage display Methods 0.000 description 13
- 239000013612 plasmid Substances 0.000 description 13
- 102000040430 polynucleotide Human genes 0.000 description 13
- 108091033319 polynucleotide Proteins 0.000 description 13
- 239000002157 polynucleotide Substances 0.000 description 13
- 229940124597 therapeutic agent Drugs 0.000 description 13
- 229920001213 Polysorbate 20 Polymers 0.000 description 12
- 239000000611 antibody drug conjugate Substances 0.000 description 12
- 230000008499 blood brain barrier function Effects 0.000 description 12
- 210000001218 blood-brain barrier Anatomy 0.000 description 12
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 12
- 238000010494 dissociation reaction Methods 0.000 description 12
- 230000005593 dissociations Effects 0.000 description 12
- 230000004927 fusion Effects 0.000 description 12
- 230000012010 growth Effects 0.000 description 12
- 230000003834 intracellular effect Effects 0.000 description 12
- 230000035772 mutation Effects 0.000 description 12
- 239000002953 phosphate buffered saline Substances 0.000 description 12
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 12
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 12
- 239000013615 primer Substances 0.000 description 12
- 239000000126 substance Substances 0.000 description 12
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 11
- 241000282412 Homo Species 0.000 description 11
- 108010076504 Protein Sorting Signals Proteins 0.000 description 11
- 241000283984 Rodentia Species 0.000 description 11
- 238000013459 approach Methods 0.000 description 11
- 238000010367 cloning Methods 0.000 description 11
- 230000002401 inhibitory effect Effects 0.000 description 11
- 230000005764 inhibitory process Effects 0.000 description 11
- 238000002360 preparation method Methods 0.000 description 11
- 230000009467 reduction Effects 0.000 description 11
- 230000002829 reductive effect Effects 0.000 description 11
- HXCHCVDVKSCDHU-PJKCJEBCSA-N s-[(2r,3s,4s,6s)-6-[[(2r,3s,4s,5r,6r)-5-[(2s,4s,5s)-5-(ethylamino)-4-methoxyoxan-2-yl]oxy-4-hydroxy-6-[[(2s,5z,9r,13e)-9-hydroxy-12-(methoxycarbonylamino)-13-[2-(methyltrisulfanyl)ethylidene]-11-oxo-2-bicyclo[7.3.1]trideca-1(12),5-dien-3,7-diynyl]oxy]-2-m Chemical compound C1[C@H](OC)[C@@H](NCC)CO[C@H]1O[C@H]1[C@H](O[C@@H]2C\3=C(NC(=O)OC)C(=O)C[C@@](C/3=C/CSSSC)(O)C#C\C=C/C#C2)O[C@H](C)[C@@H](NO[C@@H]2O[C@H](C)[C@@H](SC(=O)C=3C(=C(OC)C(O[C@H]4[C@@H]([C@H](OC)[C@@H](O)[C@H](C)O4)O)=C(I)C=3C)OC)[C@@H](O)C2)[C@@H]1O HXCHCVDVKSCDHU-PJKCJEBCSA-N 0.000 description 11
- 238000010186 staining Methods 0.000 description 11
- 210000000130 stem cell Anatomy 0.000 description 11
- 235000000346 sugar Nutrition 0.000 description 11
- 230000001225 therapeutic effect Effects 0.000 description 11
- 241000894006 Bacteria Species 0.000 description 10
- 241000700159 Rattus Species 0.000 description 10
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 10
- 229930195731 calicheamicin Natural products 0.000 description 10
- 238000004113 cell culture Methods 0.000 description 10
- 239000003153 chemical reaction reagent Substances 0.000 description 10
- 150000001875 compounds Chemical class 0.000 description 10
- 235000018417 cysteine Nutrition 0.000 description 10
- 201000000050 myeloid neoplasm Diseases 0.000 description 10
- 238000003127 radioimmunoassay Methods 0.000 description 10
- 238000011160 research Methods 0.000 description 10
- 238000012216 screening Methods 0.000 description 10
- 238000012384 transportation and delivery Methods 0.000 description 10
- 210000004881 tumor cell Anatomy 0.000 description 10
- QWPXBEHQFHACTK-KZVYIGENSA-N (10e,12e)-86-chloro-12,14,4-trihydroxy-85,14-dimethoxy-33,2,7,10-tetramethyl-15,16-dihydro-14h-7-aza-1(6,4)-oxazina-3(2,3)-oxirana-8(1,3)-benzenacyclotetradecaphane-10,12-dien-6-one Chemical compound CN1C(=O)CC(O)C2(C)OC2C(C)C(OC(=O)N2)CC2(O)C(OC)\C=C\C=C(C)\CC2=CC(OC)=C(Cl)C1=C2 QWPXBEHQFHACTK-KZVYIGENSA-N 0.000 description 9
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 9
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 9
- 241000288906 Primates Species 0.000 description 9
- 210000001744 T-lymphocyte Anatomy 0.000 description 9
- 150000001299 aldehydes Chemical class 0.000 description 9
- 230000001580 bacterial effect Effects 0.000 description 9
- 230000015572 biosynthetic process Effects 0.000 description 9
- 230000000903 blocking effect Effects 0.000 description 9
- 210000004369 blood Anatomy 0.000 description 9
- 239000008280 blood Substances 0.000 description 9
- 230000000875 corresponding effect Effects 0.000 description 9
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 9
- 229960000485 methotrexate Drugs 0.000 description 9
- 230000000269 nucleophilic effect Effects 0.000 description 9
- 238000000746 purification Methods 0.000 description 9
- 230000010076 replication Effects 0.000 description 9
- 210000001519 tissue Anatomy 0.000 description 9
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 8
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 8
- 101150113031 Jag1 gene Proteins 0.000 description 8
- NKANXQFJJICGDU-QPLCGJKRSA-N Tamoxifen Chemical compound C=1C=CC=CC=1C(/CC)=C(C=1C=CC(OCCN(C)C)=CC=1)/C1=CC=CC=C1 NKANXQFJJICGDU-QPLCGJKRSA-N 0.000 description 8
- 101150117115 V gene Proteins 0.000 description 8
- 241000700605 Viruses Species 0.000 description 8
- 230000004913 activation Effects 0.000 description 8
- 238000007792 addition Methods 0.000 description 8
- 108010044540 auristatin Proteins 0.000 description 8
- 239000011324 bead Substances 0.000 description 8
- 238000006243 chemical reaction Methods 0.000 description 8
- 238000003776 cleavage reaction Methods 0.000 description 8
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 8
- 125000000151 cysteine group Chemical class N[C@@H](CS)C(=O)* 0.000 description 8
- 231100000433 cytotoxic Toxicity 0.000 description 8
- 230000001472 cytotoxic effect Effects 0.000 description 8
- 150000002148 esters Chemical class 0.000 description 8
- 108020001507 fusion proteins Proteins 0.000 description 8
- 102000037865 fusion proteins Human genes 0.000 description 8
- 230000016784 immunoglobulin production Effects 0.000 description 8
- 238000002347 injection Methods 0.000 description 8
- 239000007924 injection Substances 0.000 description 8
- 238000005259 measurement Methods 0.000 description 8
- 230000008569 process Effects 0.000 description 8
- 230000007017 scission Effects 0.000 description 8
- 210000000813 small intestine Anatomy 0.000 description 8
- JJAHTWIKCUJRDK-UHFFFAOYSA-N succinimidyl 4-(N-maleimidomethyl)cyclohexane-1-carboxylate Chemical compound C1CC(CN2C(C=CC2=O)=O)CCC1C(=O)ON1C(=O)CCC1=O JJAHTWIKCUJRDK-UHFFFAOYSA-N 0.000 description 8
- 206010006187 Breast cancer Diseases 0.000 description 7
- 208000026310 Breast neoplasm Diseases 0.000 description 7
- 102100024746 Dihydrofolate reductase Human genes 0.000 description 7
- 101000978766 Homo sapiens Neurogenic locus notch homolog protein 1 Proteins 0.000 description 7
- 108010073807 IgG Receptors Proteins 0.000 description 7
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 7
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 7
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 7
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 7
- 108091034117 Oligonucleotide Proteins 0.000 description 7
- 238000001042 affinity chromatography Methods 0.000 description 7
- 239000003242 anti bacterial agent Substances 0.000 description 7
- 229940088710 antibiotic agent Drugs 0.000 description 7
- 239000000872 buffer Substances 0.000 description 7
- 210000004899 c-terminal region Anatomy 0.000 description 7
- 230000000295 complement effect Effects 0.000 description 7
- 230000021615 conjugation Effects 0.000 description 7
- 108020001096 dihydrofolate reductase Proteins 0.000 description 7
- AMRJKAQTDDKMCE-UHFFFAOYSA-N dolastatin Chemical compound CC(C)C(N(C)C)C(=O)NC(C(C)C)C(=O)N(C)C(C(C)C)C(OC)CC(=O)N1CCCC1C(OC)C(C)C(=O)NC(C=1SC=CN=1)CC1=CC=CC=C1 AMRJKAQTDDKMCE-UHFFFAOYSA-N 0.000 description 7
- 229930188854 dolastatin Natural products 0.000 description 7
- 239000003623 enhancer Substances 0.000 description 7
- 239000001963 growth medium Substances 0.000 description 7
- 230000000968 intestinal effect Effects 0.000 description 7
- 210000004698 lymphocyte Anatomy 0.000 description 7
- 150000002482 oligosaccharides Chemical class 0.000 description 7
- 229920000642 polymer Polymers 0.000 description 7
- 230000002285 radioactive effect Effects 0.000 description 7
- 238000010188 recombinant method Methods 0.000 description 7
- 238000012552 review Methods 0.000 description 7
- 150000003839 salts Chemical class 0.000 description 7
- 239000000523 sample Substances 0.000 description 7
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 description 7
- RCINICONZNJXQF-MZXODVADSA-N taxol Chemical compound O([C@@H]1[C@@]2(C[C@@H](C(C)=C(C2(C)C)[C@H](C([C@]2(C)[C@@H](O)C[C@H]3OC[C@]3([C@H]21)OC(C)=O)=O)OC(=O)C)OC(=O)[C@H](O)[C@@H](NC(=O)C=1C=CC=CC=1)C=1C=CC=CC=1)O)C(=O)C1=CC=CC=C1 RCINICONZNJXQF-MZXODVADSA-N 0.000 description 7
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 6
- 208000010839 B-cell chronic lymphocytic leukemia Diseases 0.000 description 6
- 108091026890 Coding region Proteins 0.000 description 6
- 206010009944 Colon cancer Diseases 0.000 description 6
- 241000196324 Embryophyta Species 0.000 description 6
- WZUVPPKBWHMQCE-UHFFFAOYSA-N Haematoxylin Chemical compound C12=CC(O)=C(O)C=C2CC2(O)C1C1=CC=C(O)C(O)=C1OC2 WZUVPPKBWHMQCE-UHFFFAOYSA-N 0.000 description 6
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 6
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 description 6
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 description 6
- 102100029185 Low affinity immunoglobulin gamma Fc region receptor III-B Human genes 0.000 description 6
- 208000031422 Lymphocytic Chronic B-Cell Leukemia Diseases 0.000 description 6
- ZDZOTLJHXYCWBA-VCVYQWHSSA-N N-debenzoyl-N-(tert-butoxycarbonyl)-10-deacetyltaxol Chemical compound O([C@H]1[C@H]2[C@@](C([C@H](O)C3=C(C)[C@@H](OC(=O)[C@H](O)[C@@H](NC(=O)OC(C)(C)C)C=4C=CC=CC=4)C[C@]1(O)C3(C)C)=O)(C)[C@@H](O)C[C@H]1OC[C@]12OC(=O)C)C(=O)C1=CC=CC=C1 ZDZOTLJHXYCWBA-VCVYQWHSSA-N 0.000 description 6
- 108010039491 Ricin Proteins 0.000 description 6
- 108010003723 Single-Domain Antibodies Proteins 0.000 description 6
- 230000009824 affinity maturation Effects 0.000 description 6
- 235000004279 alanine Nutrition 0.000 description 6
- 238000004458 analytical method Methods 0.000 description 6
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 6
- 208000032852 chronic lymphocytic leukemia Diseases 0.000 description 6
- 238000011198 co-culture assay Methods 0.000 description 6
- 230000003013 cytotoxicity Effects 0.000 description 6
- 231100000135 cytotoxicity Toxicity 0.000 description 6
- 238000001514 detection method Methods 0.000 description 6
- 230000018109 developmental process Effects 0.000 description 6
- 238000003745 diagnosis Methods 0.000 description 6
- 229960004679 doxorubicin Drugs 0.000 description 6
- 238000009472 formulation Methods 0.000 description 6
- 230000013595 glycosylation Effects 0.000 description 6
- 238000006206 glycosylation reaction Methods 0.000 description 6
- 102000045609 human NOTCH1 Human genes 0.000 description 6
- 230000003053 immunization Effects 0.000 description 6
- 239000000463 material Substances 0.000 description 6
- 229920001542 oligosaccharide Polymers 0.000 description 6
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 6
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical compound FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 6
- 210000001525 retina Anatomy 0.000 description 6
- 208000024891 symptom Diseases 0.000 description 6
- 238000002560 therapeutic procedure Methods 0.000 description 6
- 238000013518 transcription Methods 0.000 description 6
- 230000035897 transcription Effects 0.000 description 6
- 238000001890 transfection Methods 0.000 description 6
- 238000012546 transfer Methods 0.000 description 6
- 229960005486 vaccine Drugs 0.000 description 6
- 229960004528 vincristine Drugs 0.000 description 6
- OGWKCGZFUXNPDA-XQKSVPLYSA-N vincristine Chemical compound C([N@]1C[C@@H](C[C@]2(C(=O)OC)C=3C(=CC4=C([C@]56[C@H]([C@@]([C@H](OC(C)=O)[C@]7(CC)C=CCN([C@H]67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)C[C@@](C1)(O)CC)CC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-XQKSVPLYSA-N 0.000 description 6
- OGWKCGZFUXNPDA-UHFFFAOYSA-N vincristine Natural products C1C(CC)(O)CC(CC2(C(=O)OC)C=3C(=CC4=C(C56C(C(C(OC(C)=O)C7(CC)C=CCN(C67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)CN1CCC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-UHFFFAOYSA-N 0.000 description 6
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 5
- SGKRLCUYIXIAHR-AKNGSSGZSA-N (4s,4ar,5s,5ar,6r,12ar)-4-(dimethylamino)-1,5,10,11,12a-pentahydroxy-6-methyl-3,12-dioxo-4a,5,5a,6-tetrahydro-4h-tetracene-2-carboxamide Chemical compound C1=CC=C2[C@H](C)[C@@H]([C@H](O)[C@@H]3[C@](C(O)=C(C(N)=O)C(=O)[C@H]3N(C)C)(O)C3=O)C3=C(O)C2=C1O SGKRLCUYIXIAHR-AKNGSSGZSA-N 0.000 description 5
- STQGQHZAVUOBTE-UHFFFAOYSA-N 7-Cyan-hept-2t-en-4,6-diinsaeure Natural products C1=2C(O)=C3C(=O)C=4C(OC)=CC=CC=4C(=O)C3=C(O)C=2CC(O)(C(C)=O)CC1OC1CC(N)C(O)C(C)O1 STQGQHZAVUOBTE-UHFFFAOYSA-N 0.000 description 5
- DWJXYEABWRJFSP-XOBRGWDASA-N DAPT Chemical compound N([C@@H](C)C(=O)N[C@H](C(=O)OC(C)(C)C)C=1C=CC=CC=1)C(=O)CC1=CC(F)=CC(F)=C1 DWJXYEABWRJFSP-XOBRGWDASA-N 0.000 description 5
- 108090000331 Firefly luciferases Proteins 0.000 description 5
- GHASVSINZRGABV-UHFFFAOYSA-N Fluorouracil Chemical compound FC1=CNC(=O)NC1=O GHASVSINZRGABV-UHFFFAOYSA-N 0.000 description 5
- 241000233866 Fungi Species 0.000 description 5
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 5
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 5
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 5
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 5
- QWPXBEHQFHACTK-UHFFFAOYSA-N Maytansinol Natural products CN1C(=O)CC(O)C2(C)OC2C(C)C(OC(=O)N2)CC2(O)C(OC)C=CC=C(C)CC2=CC(OC)=C(Cl)C1=C2 QWPXBEHQFHACTK-UHFFFAOYSA-N 0.000 description 5
- NWIBSHFKIJFRCO-WUDYKRTCSA-N Mytomycin Chemical compound C1N2C(C(C(C)=C(N)C3=O)=O)=C3[C@@H](COC(N)=O)[C@@]2(OC)[C@@H]2[C@H]1N2 NWIBSHFKIJFRCO-WUDYKRTCSA-N 0.000 description 5
- 229930012538 Paclitaxel Natural products 0.000 description 5
- 239000002202 Polyethylene glycol Substances 0.000 description 5
- 150000007513 acids Chemical class 0.000 description 5
- 239000000556 agonist Substances 0.000 description 5
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 5
- 230000003321 amplification Effects 0.000 description 5
- 239000002585 base Substances 0.000 description 5
- 230000001588 bifunctional effect Effects 0.000 description 5
- 210000004556 brain Anatomy 0.000 description 5
- 150000001720 carbohydrates Chemical class 0.000 description 5
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 5
- 238000004587 chromatography analysis Methods 0.000 description 5
- 238000010276 construction Methods 0.000 description 5
- STQGQHZAVUOBTE-VGBVRHCVSA-N daunorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(C)=O)[C@H]1C[C@H](N)[C@H](O)[C@H](C)O1 STQGQHZAVUOBTE-VGBVRHCVSA-N 0.000 description 5
- 238000011161 development Methods 0.000 description 5
- 239000000539 dimer Substances 0.000 description 5
- 229960003722 doxycycline Drugs 0.000 description 5
- 238000011977 dual antiplatelet therapy Methods 0.000 description 5
- 102000015694 estrogen receptors Human genes 0.000 description 5
- 108010038795 estrogen receptors Proteins 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- 229960002949 fluorouracil Drugs 0.000 description 5
- 238000001415 gene therapy Methods 0.000 description 5
- 229940088597 hormone Drugs 0.000 description 5
- 239000005556 hormone Substances 0.000 description 5
- 238000002649 immunization Methods 0.000 description 5
- 229940072221 immunoglobulins Drugs 0.000 description 5
- 150000002632 lipids Chemical class 0.000 description 5
- 210000004962 mammalian cell Anatomy 0.000 description 5
- 230000001404 mediated effect Effects 0.000 description 5
- DNIAPMSPPWPWGF-UHFFFAOYSA-N monopropylene glycol Natural products CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 5
- 238000002703 mutagenesis Methods 0.000 description 5
- 231100000350 mutagenesis Toxicity 0.000 description 5
- 238000003199 nucleic acid amplification method Methods 0.000 description 5
- 229960001592 paclitaxel Drugs 0.000 description 5
- 239000002245 particle Substances 0.000 description 5
- 229920001223 polyethylene glycol Polymers 0.000 description 5
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 5
- 238000012545 processing Methods 0.000 description 5
- 239000000047 product Substances 0.000 description 5
- 238000000159 protein binding assay Methods 0.000 description 5
- 230000004044 response Effects 0.000 description 5
- 230000028327 secretion Effects 0.000 description 5
- 210000002966 serum Anatomy 0.000 description 5
- 230000019491 signal transduction Effects 0.000 description 5
- 238000001179 sorption measurement Methods 0.000 description 5
- 239000000758 substrate Substances 0.000 description 5
- 238000003786 synthesis reaction Methods 0.000 description 5
- 241000701161 unidentified adenovirus Species 0.000 description 5
- VPFUWHKTPYPNGT-UHFFFAOYSA-N 3-(3,4-dihydroxyphenyl)-1-(5-hydroxy-2,2-dimethylchromen-6-yl)propan-1-one Chemical compound OC1=C2C=CC(C)(C)OC2=CC=C1C(=O)CCC1=CC=C(O)C(O)=C1 VPFUWHKTPYPNGT-UHFFFAOYSA-N 0.000 description 4
- 101710132601 Capsid protein Proteins 0.000 description 4
- 101710094648 Coat protein Proteins 0.000 description 4
- 238000000116 DAPI staining Methods 0.000 description 4
- 102000009123 Fibrin Human genes 0.000 description 4
- 108010073385 Fibrin Proteins 0.000 description 4
- BWGVNKXGVNDBDI-UHFFFAOYSA-N Fibrin monomer Chemical compound CNC(=O)CNC(=O)CN BWGVNKXGVNDBDI-UHFFFAOYSA-N 0.000 description 4
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 4
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 4
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 4
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 4
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 4
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 4
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 4
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 4
- 206010025323 Lymphomas Diseases 0.000 description 4
- 239000004472 Lysine Substances 0.000 description 4
- 101710125418 Major capsid protein Proteins 0.000 description 4
- 108090000157 Metallothionein Proteins 0.000 description 4
- OVRNDRQMDRJTHS-UHFFFAOYSA-N N-acelyl-D-glucosamine Natural products CC(=O)NC1C(O)OC(CO)C(O)C1O OVRNDRQMDRJTHS-UHFFFAOYSA-N 0.000 description 4
- MBLBDJOUHNCFQT-LXGUWJNJSA-N N-acetylglucosamine Natural products CC(=O)N[C@@H](C=O)[C@@H](O)[C@H](O)[C@H](O)CO MBLBDJOUHNCFQT-LXGUWJNJSA-N 0.000 description 4
- 208000015914 Non-Hodgkin lymphomas Diseases 0.000 description 4
- 108091028043 Nucleic acid sequence Proteins 0.000 description 4
- 101710141454 Nucleoprotein Proteins 0.000 description 4
- 108700020796 Oncogene Proteins 0.000 description 4
- 229910019142 PO4 Inorganic materials 0.000 description 4
- 102000035195 Peptidases Human genes 0.000 description 4
- 108091005804 Peptidases Proteins 0.000 description 4
- 102100024616 Platelet endothelial cell adhesion molecule Human genes 0.000 description 4
- 101710083689 Probable capsid protein Proteins 0.000 description 4
- 239000004365 Protease Substances 0.000 description 4
- 108020004511 Recombinant DNA Proteins 0.000 description 4
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 4
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 4
- 108010090804 Streptavidin Proteins 0.000 description 4
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 4
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 4
- VWQVUPCCIRVNHF-OUBTZVSYSA-N Yttrium-90 Chemical compound [90Y] VWQVUPCCIRVNHF-OUBTZVSYSA-N 0.000 description 4
- 239000002671 adjuvant Substances 0.000 description 4
- WQZGKKKJIJFFOK-PHYPRBDBSA-N alpha-D-galactose Chemical compound OC[C@H]1O[C@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-PHYPRBDBSA-N 0.000 description 4
- 230000004075 alteration Effects 0.000 description 4
- 150000001412 amines Chemical class 0.000 description 4
- 125000003277 amino group Chemical group 0.000 description 4
- 238000010171 animal model Methods 0.000 description 4
- 239000005557 antagonist Substances 0.000 description 4
- 230000008901 benefit Effects 0.000 description 4
- 239000012472 biological sample Substances 0.000 description 4
- 230000011712 cell development Effects 0.000 description 4
- 230000024245 cell differentiation Effects 0.000 description 4
- 230000010261 cell growth Effects 0.000 description 4
- 238000012512 characterization method Methods 0.000 description 4
- 239000002738 chelating agent Substances 0.000 description 4
- 229960002173 citrulline Drugs 0.000 description 4
- 239000011248 coating agent Substances 0.000 description 4
- 238000000576 coating method Methods 0.000 description 4
- 208000029742 colonic neoplasm Diseases 0.000 description 4
- 238000002648 combination therapy Methods 0.000 description 4
- 239000002299 complementary DNA Substances 0.000 description 4
- 238000004590 computer program Methods 0.000 description 4
- 239000007822 coupling agent Substances 0.000 description 4
- 239000013078 crystal Substances 0.000 description 4
- 238000012258 culturing Methods 0.000 description 4
- 230000006378 damage Effects 0.000 description 4
- 230000002950 deficient Effects 0.000 description 4
- 230000001419 dependent effect Effects 0.000 description 4
- 238000006471 dimerization reaction Methods 0.000 description 4
- 229960003668 docetaxel Drugs 0.000 description 4
- 210000002889 endothelial cell Anatomy 0.000 description 4
- 229950003499 fibrin Drugs 0.000 description 4
- 210000004602 germ cell Anatomy 0.000 description 4
- 102000005396 glutamine synthetase Human genes 0.000 description 4
- 108020002326 glutamine synthetase Proteins 0.000 description 4
- 210000002175 goblet cell Anatomy 0.000 description 4
- 239000003966 growth inhibitor Substances 0.000 description 4
- 230000001900 immune effect Effects 0.000 description 4
- 230000002163 immunogen Effects 0.000 description 4
- 230000001939 inductive effect Effects 0.000 description 4
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 4
- 230000003993 interaction Effects 0.000 description 4
- 238000002955 isolation Methods 0.000 description 4
- 210000003292 kidney cell Anatomy 0.000 description 4
- 239000002502 liposome Substances 0.000 description 4
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 4
- 239000003550 marker Substances 0.000 description 4
- 210000000822 natural killer cell Anatomy 0.000 description 4
- 108010068617 neonatal Fc receptor Proteins 0.000 description 4
- 230000002265 prevention Effects 0.000 description 4
- 230000000069 prophylactic effect Effects 0.000 description 4
- 230000001105 regulatory effect Effects 0.000 description 4
- 235000004400 serine Nutrition 0.000 description 4
- JQWHASGSAFIOCM-UHFFFAOYSA-M sodium periodate Chemical compound [Na+].[O-]I(=O)(=O)=O JQWHASGSAFIOCM-UHFFFAOYSA-M 0.000 description 4
- 229960001603 tamoxifen Drugs 0.000 description 4
- 125000003396 thiol group Chemical group [H]S* 0.000 description 4
- CNHYKKNIIGEXAY-UHFFFAOYSA-N thiolan-2-imine Chemical compound N=C1CCCS1 CNHYKKNIIGEXAY-UHFFFAOYSA-N 0.000 description 4
- 150000003573 thiols Chemical class 0.000 description 4
- 230000009466 transformation Effects 0.000 description 4
- 230000009261 transgenic effect Effects 0.000 description 4
- 239000003981 vehicle Substances 0.000 description 4
- JXLYSJRDGCGARV-CFWMRBGOSA-N vinblastine Chemical compound C([C@H](C[C@]1(C(=O)OC)C=2C(=CC3=C([C@]45[C@H]([C@@]([C@H](OC(C)=O)[C@]6(CC)C=CCN([C@H]56)CC4)(O)C(=O)OC)N3C)C=2)OC)C[C@@](C2)(O)CC)N2CCC2=C1NC1=CC=CC=C21 JXLYSJRDGCGARV-CFWMRBGOSA-N 0.000 description 4
- 230000003612 virological effect Effects 0.000 description 4
- JWDFQMWEFLOOED-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 3-(pyridin-2-yldisulfanyl)propanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCSSC1=CC=CC=N1 JWDFQMWEFLOOED-UHFFFAOYSA-N 0.000 description 3
- BQWBEDSJTMWJAE-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-[(2-iodoacetyl)amino]benzoate Chemical compound C1=CC(NC(=O)CI)=CC=C1C(=O)ON1C(=O)CCC1=O BQWBEDSJTMWJAE-UHFFFAOYSA-N 0.000 description 3
- IEUUDEWWMRQUDS-UHFFFAOYSA-N (6-azaniumylidene-1,6-dimethoxyhexylidene)azanium;dichloride Chemical compound Cl.Cl.COC(=N)CCCCC(=N)OC IEUUDEWWMRQUDS-UHFFFAOYSA-N 0.000 description 3
- YBBNVCVOACOHIG-UHFFFAOYSA-N 2,2-diamino-1,4-bis(4-azidophenyl)-3-butylbutane-1,4-dione Chemical compound C=1C=C(N=[N+]=[N-])C=CC=1C(=O)C(N)(N)C(CCCC)C(=O)C1=CC=C(N=[N+]=[N-])C=C1 YBBNVCVOACOHIG-UHFFFAOYSA-N 0.000 description 3
- RTQWWZBSTRGEAV-PKHIMPSTSA-N 2-[[(2s)-2-[bis(carboxymethyl)amino]-3-[4-(methylcarbamoylamino)phenyl]propyl]-[2-[bis(carboxymethyl)amino]propyl]amino]acetic acid Chemical compound CNC(=O)NC1=CC=C(C[C@@H](CN(CC(C)N(CC(O)=O)CC(O)=O)CC(O)=O)N(CC(O)=O)CC(O)=O)C=C1 RTQWWZBSTRGEAV-PKHIMPSTSA-N 0.000 description 3
- CSCPPACGZOOCGX-UHFFFAOYSA-N Acetone Chemical compound CC(C)=O CSCPPACGZOOCGX-UHFFFAOYSA-N 0.000 description 3
- 108010088751 Albumins Proteins 0.000 description 3
- 102000009027 Albumins Human genes 0.000 description 3
- 108090001008 Avidin Proteins 0.000 description 3
- UHOVQNZJYSORNB-UHFFFAOYSA-N Benzene Chemical compound C1=CC=CC=C1 UHOVQNZJYSORNB-UHFFFAOYSA-N 0.000 description 3
- 241000283707 Capra Species 0.000 description 3
- DLGOEMSEDOSKAD-UHFFFAOYSA-N Carmustine Chemical compound ClCCNC(=O)N(N=O)CCCl DLGOEMSEDOSKAD-UHFFFAOYSA-N 0.000 description 3
- 102000000844 Cell Surface Receptors Human genes 0.000 description 3
- 108010001857 Cell Surface Receptors Proteins 0.000 description 3
- 241000282693 Cercopithecidae Species 0.000 description 3
- 206010008342 Cervix carcinoma Diseases 0.000 description 3
- 108020004705 Codon Proteins 0.000 description 3
- 241000699800 Cricetinae Species 0.000 description 3
- 241000699802 Cricetulus griseus Species 0.000 description 3
- UHDGCWIWMRVCDJ-CCXZUQQUSA-N Cytarabine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@@H](O)[C@H](O)[C@@H](CO)O1 UHDGCWIWMRVCDJ-CCXZUQQUSA-N 0.000 description 3
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Natural products CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 3
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 3
- 102000016607 Diphtheria Toxin Human genes 0.000 description 3
- 108010053187 Diphtheria Toxin Proteins 0.000 description 3
- 241000255925 Diptera Species 0.000 description 3
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 3
- 241000206602 Eukaryota Species 0.000 description 3
- PNNNRSAQSRJVSB-SLPGGIOYSA-N Fucose Natural products C[C@H](O)[C@@H](O)[C@H](O)[C@H](O)C=O PNNNRSAQSRJVSB-SLPGGIOYSA-N 0.000 description 3
- 229940125373 Gamma-Secretase Inhibitor Drugs 0.000 description 3
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- 241000238631 Hexapoda Species 0.000 description 3
- 208000017604 Hodgkin disease Diseases 0.000 description 3
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 3
- 102100029098 Hypoxanthine-guanine phosphoribosyltransferase Human genes 0.000 description 3
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 description 3
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 description 3
- 241000235649 Kluyveromyces Species 0.000 description 3
- 241001138401 Kluyveromyces lactis Species 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 3
- SHZGCJCMOBCMKK-DHVFOXMCSA-N L-fucopyranose Chemical compound C[C@@H]1OC(O)[C@@H](O)[C@H](O)[C@@H]1O SHZGCJCMOBCMKK-DHVFOXMCSA-N 0.000 description 3
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 3
- 208000031671 Large B-Cell Diffuse Lymphoma Diseases 0.000 description 3
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 3
- 229930126263 Maytansine Natural products 0.000 description 3
- 102000003792 Metallothionein Human genes 0.000 description 3
- 102000029749 Microtubule Human genes 0.000 description 3
- 108091022875 Microtubule Proteins 0.000 description 3
- 208000034578 Multiple myelomas Diseases 0.000 description 3
- 241000699660 Mus musculus Species 0.000 description 3
- 101100294417 Mus musculus Notch1 gene Proteins 0.000 description 3
- NQTADLQHYWFPDB-UHFFFAOYSA-N N-Hydroxysuccinimide Chemical compound ON1C(=O)CCC1=O NQTADLQHYWFPDB-UHFFFAOYSA-N 0.000 description 3
- OVRNDRQMDRJTHS-RTRLPJTCSA-N N-acetyl-D-glucosamine Chemical compound CC(=O)N[C@H]1C(O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-RTRLPJTCSA-N 0.000 description 3
- 229930193140 Neomycin Natural products 0.000 description 3
- 241000283973 Oryctolagus cuniculus Species 0.000 description 3
- 102000004316 Oxidoreductases Human genes 0.000 description 3
- 108090000854 Oxidoreductases Proteins 0.000 description 3
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 3
- 208000009052 Precursor T-Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 description 3
- 208000017414 Precursor T-cell acute lymphoblastic leukemia Diseases 0.000 description 3
- 241000589517 Pseudomonas aeruginosa Species 0.000 description 3
- 108010052090 Renilla Luciferases Proteins 0.000 description 3
- IIDJRNMFWXDHID-UHFFFAOYSA-N Risedronic acid Chemical compound OP(=O)(O)C(P(O)(O)=O)(O)CC1=CC=CN=C1 IIDJRNMFWXDHID-UHFFFAOYSA-N 0.000 description 3
- 230000018199 S phase Effects 0.000 description 3
- 241000282887 Suidae Species 0.000 description 3
- 208000029052 T-cell acute lymphoblastic leukemia Diseases 0.000 description 3
- 101150006914 TRP1 gene Proteins 0.000 description 3
- 239000004473 Threonine Substances 0.000 description 3
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 description 3
- JXLYSJRDGCGARV-WWYNWVTFSA-N Vinblastine Natural products O=C(O[C@H]1[C@](O)(C(=O)OC)[C@@H]2N(C)c3c(cc(c(OC)c3)[C@]3(C(=O)OC)c4[nH]c5c(c4CCN4C[C@](O)(CC)C[C@H](C3)C4)cccc5)[C@@]32[C@H]2[C@@]1(CC)C=CCN2CC3)C JXLYSJRDGCGARV-WWYNWVTFSA-N 0.000 description 3
- 241000607479 Yersinia pestis Species 0.000 description 3
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 3
- 125000000217 alkyl group Chemical group 0.000 description 3
- 210000004102 animal cell Anatomy 0.000 description 3
- 239000003886 aromatase inhibitor Substances 0.000 description 3
- 229940046844 aromatase inhibitors Drugs 0.000 description 3
- 125000003118 aryl group Chemical group 0.000 description 3
- 125000004429 atom Chemical group 0.000 description 3
- 239000011230 binding agent Substances 0.000 description 3
- 229960002685 biotin Drugs 0.000 description 3
- 235000020958 biotin Nutrition 0.000 description 3
- 239000011616 biotin Substances 0.000 description 3
- 210000000170 cell membrane Anatomy 0.000 description 3
- 238000005119 centrifugation Methods 0.000 description 3
- 201000010881 cervical cancer Diseases 0.000 description 3
- 229960004630 chlorambucil Drugs 0.000 description 3
- JCKYGMPEJWAADB-UHFFFAOYSA-N chlorambucil Chemical compound OC(=O)CCCC1=CC=C(N(CCCl)CCCl)C=C1 JCKYGMPEJWAADB-UHFFFAOYSA-N 0.000 description 3
- 230000024203 complement activation Effects 0.000 description 3
- 239000000356 contaminant Substances 0.000 description 3
- 229920001577 copolymer Polymers 0.000 description 3
- 238000004132 cross linking Methods 0.000 description 3
- 230000001086 cytosolic effect Effects 0.000 description 3
- 238000002784 cytotoxicity assay Methods 0.000 description 3
- 231100000263 cytotoxicity test Toxicity 0.000 description 3
- 229960000975 daunorubicin Drugs 0.000 description 3
- 230000007423 decrease Effects 0.000 description 3
- CYQFCXCEBYINGO-IAGOWNOFSA-N delta1-THC Chemical compound C1=C(C)CC[C@H]2C(C)(C)OC3=CC(CCCCC)=CC(O)=C3[C@@H]21 CYQFCXCEBYINGO-IAGOWNOFSA-N 0.000 description 3
- 238000001212 derivatisation Methods 0.000 description 3
- 229960003957 dexamethasone Drugs 0.000 description 3
- UREBDLICKHMUKA-CXSFZGCWSA-N dexamethasone Chemical compound C1CC2=CC(=O)C=C[C@]2(C)[C@]2(F)[C@@H]1[C@@H]1C[C@@H](C)[C@@](C(=O)CO)(O)[C@@]1(C)C[C@@H]2O UREBDLICKHMUKA-CXSFZGCWSA-N 0.000 description 3
- 206010012818 diffuse large B-cell lymphoma Diseases 0.000 description 3
- 230000029087 digestion Effects 0.000 description 3
- 125000005442 diisocyanate group Chemical group 0.000 description 3
- ZWIBGKZDAWNIFC-UHFFFAOYSA-N disuccinimidyl suberate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCCC(=O)ON1C(=O)CCC1=O ZWIBGKZDAWNIFC-UHFFFAOYSA-N 0.000 description 3
- VJJPUSNTGOMMGY-MRVIYFEKSA-N etoposide Chemical compound COC1=C(O)C(OC)=CC([C@@H]2C3=CC=4OCOC=4C=C3[C@@H](O[C@H]3[C@@H]([C@@H](O)[C@@H]4O[C@H](C)OC[C@H]4O3)O)[C@@H]3[C@@H]2C(OC3)=O)=C1 VJJPUSNTGOMMGY-MRVIYFEKSA-N 0.000 description 3
- 150000002222 fluorine compounds Chemical class 0.000 description 3
- 229930182830 galactose Natural products 0.000 description 3
- 230000036541 health Effects 0.000 description 3
- 229960001001 ibritumomab tiuxetan Drugs 0.000 description 3
- 210000002865 immune cell Anatomy 0.000 description 3
- 238000003018 immunoassay Methods 0.000 description 3
- 238000002991 immunohistochemical analysis Methods 0.000 description 3
- 238000003364 immunohistochemistry Methods 0.000 description 3
- 238000001114 immunoprecipitation Methods 0.000 description 3
- 239000002596 immunotoxin Substances 0.000 description 3
- 230000006872 improvement Effects 0.000 description 3
- 238000011534 incubation Methods 0.000 description 3
- 230000006698 induction Effects 0.000 description 3
- 208000015181 infectious disease Diseases 0.000 description 3
- 238000001802 infusion Methods 0.000 description 3
- 238000007912 intraperitoneal administration Methods 0.000 description 3
- UWKQSNNFCGGAFS-XIFFEERXSA-N irinotecan Chemical compound C1=C2C(CC)=C3CN(C(C4=C([C@@](C(=O)OC4)(O)CC)C=4)=O)C=4C3=NC2=CC=C1OC(=O)N(CC1)CCC1N1CCCCC1 UWKQSNNFCGGAFS-XIFFEERXSA-N 0.000 description 3
- 150000002576 ketones Chemical class 0.000 description 3
- 210000002429 large intestine Anatomy 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 238000013507 mapping Methods 0.000 description 3
- WKPWGQKGSOKKOO-RSFHAFMBSA-N maytansine Chemical compound CO[C@@H]([C@@]1(O)C[C@](OC(=O)N1)([C@H]([C@@H]1O[C@@]1(C)[C@@H](OC(=O)[C@H](C)N(C)C(C)=O)CC(=O)N1C)C)[H])\C=C\C=C(C)\CC2=CC(OC)=C(Cl)C1=C2 WKPWGQKGSOKKOO-RSFHAFMBSA-N 0.000 description 3
- 229960004961 mechlorethamine Drugs 0.000 description 3
- 210000004379 membrane Anatomy 0.000 description 3
- 239000012528 membrane Substances 0.000 description 3
- GLVAUDGFNGKCSF-UHFFFAOYSA-N mercaptopurine Chemical compound S=C1NC=NC2=C1NC=N2 GLVAUDGFNGKCSF-UHFFFAOYSA-N 0.000 description 3
- 229960005558 mertansine Drugs 0.000 description 3
- ANZJBCHSOXCCRQ-FKUXLPTCSA-N mertansine Chemical compound CO[C@@H]([C@@]1(O)C[C@H](OC(=O)N1)[C@@H](C)[C@@H]1O[C@@]1(C)[C@@H](OC(=O)[C@H](C)N(C)C(=O)CCS)CC(=O)N1C)\C=C\C=C(C)\CC2=CC(OC)=C(Cl)C1=C2 ANZJBCHSOXCCRQ-FKUXLPTCSA-N 0.000 description 3
- 108020004999 messenger RNA Proteins 0.000 description 3
- 229910052751 metal Inorganic materials 0.000 description 3
- 239000002184 metal Substances 0.000 description 3
- 150000002739 metals Chemical class 0.000 description 3
- MYWUZJCMWCOHBA-VIFPVBQESA-N methamphetamine Chemical compound CN[C@@H](C)CC1=CC=CC=C1 MYWUZJCMWCOHBA-VIFPVBQESA-N 0.000 description 3
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 3
- 244000005700 microbiome Species 0.000 description 3
- 210000004688 microtubule Anatomy 0.000 description 3
- 229960004857 mitomycin Drugs 0.000 description 3
- KKZJGLLVHKMTCM-UHFFFAOYSA-N mitoxantrone Chemical compound O=C1C2=C(O)C=CC(O)=C2C(=O)C2=C1C(NCCNCCO)=CC=C2NCCNCCO KKZJGLLVHKMTCM-UHFFFAOYSA-N 0.000 description 3
- 210000001616 monocyte Anatomy 0.000 description 3
- 229960004927 neomycin Drugs 0.000 description 3
- 210000005155 neural progenitor cell Anatomy 0.000 description 3
- 229910052757 nitrogen Inorganic materials 0.000 description 3
- 238000011275 oncology therapy Methods 0.000 description 3
- 210000001672 ovary Anatomy 0.000 description 3
- 238000004091 panning Methods 0.000 description 3
- 230000036961 partial effect Effects 0.000 description 3
- 230000037361 pathway Effects 0.000 description 3
- 210000005105 peripheral blood lymphocyte Anatomy 0.000 description 3
- 230000002093 peripheral effect Effects 0.000 description 3
- 210000001322 periplasm Anatomy 0.000 description 3
- 102000013415 peroxidase activity proteins Human genes 0.000 description 3
- 108040007629 peroxidase activity proteins Proteins 0.000 description 3
- 235000021317 phosphate Nutrition 0.000 description 3
- 239000010452 phosphate Substances 0.000 description 3
- 230000003389 potentiating effect Effects 0.000 description 3
- GZUITABIAKMVPG-UHFFFAOYSA-N raloxifene Chemical compound C1=CC(O)=CC=C1C1=C(C(=O)C=2C=CC(OCCN3CCCCC3)=CC=2)C2=CC=C(O)C=C2S1 GZUITABIAKMVPG-UHFFFAOYSA-N 0.000 description 3
- 238000003259 recombinant expression Methods 0.000 description 3
- 230000006798 recombination Effects 0.000 description 3
- 238000005215 recombination Methods 0.000 description 3
- 238000011084 recovery Methods 0.000 description 3
- QFJCIRLUMZQUOT-HPLJOQBZSA-N sirolimus Chemical compound C1C[C@@H](O)[C@H](OC)C[C@@H]1C[C@@H](C)[C@H]1OC(=O)[C@@H]2CCCCN2C(=O)C(=O)[C@](O)(O2)[C@H](C)CC[C@H]2C[C@H](OC)/C(C)=C/C=C/C=C/[C@@H](C)C[C@@H](C)C(=O)[C@H](OC)[C@H](O)/C(C)=C/[C@@H](C)C(=O)C1 QFJCIRLUMZQUOT-HPLJOQBZSA-N 0.000 description 3
- 150000003384 small molecules Chemical class 0.000 description 3
- 239000007790 solid phase Substances 0.000 description 3
- 238000007920 subcutaneous administration Methods 0.000 description 3
- 150000008163 sugars Chemical class 0.000 description 3
- 239000006228 supernatant Substances 0.000 description 3
- 230000008685 targeting Effects 0.000 description 3
- 210000001541 thymus gland Anatomy 0.000 description 3
- 230000005030 transcription termination Effects 0.000 description 3
- 238000011830 transgenic mouse model Methods 0.000 description 3
- 229930013292 trichothecene Natural products 0.000 description 3
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 3
- 241001515965 unidentified phage Species 0.000 description 3
- 241001430294 unidentified retrovirus Species 0.000 description 3
- 238000011144 upstream manufacturing Methods 0.000 description 3
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 description 3
- 229960003048 vinblastine Drugs 0.000 description 3
- UGGWPQSBPIFKDZ-KOTLKJBCSA-N vindesine Chemical compound C([C@@H](C[C@]1(C(=O)OC)C=2C(=CC3=C([C@]45[C@H]([C@@]([C@H](O)[C@]6(CC)C=CCN([C@H]56)CC4)(O)C(N)=O)N3C)C=2)OC)C[C@@](C2)(O)CC)N2CCC2=C1N=C1[C]2C=CC=C1 UGGWPQSBPIFKDZ-KOTLKJBCSA-N 0.000 description 3
- 229960004355 vindesine Drugs 0.000 description 3
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 3
- 230000003442 weekly effect Effects 0.000 description 3
- XRASPMIURGNCCH-UHFFFAOYSA-N zoledronic acid Chemical compound OP(=O)(O)C(P(O)(O)=O)(O)CN1C=CN=C1 XRASPMIURGNCCH-UHFFFAOYSA-N 0.000 description 3
- WOWDZACBATWTAU-FEFUEGSOSA-N (2s)-2-[[(2s)-2-(dimethylamino)-3-methylbutanoyl]amino]-n-[(3r,4s,5s)-1-[(2s)-2-[(1r,2r)-3-[[(1s,2r)-1-hydroxy-1-phenylpropan-2-yl]amino]-1-methoxy-2-methyl-3-oxopropyl]pyrrolidin-1-yl]-3-methoxy-5-methyl-1-oxoheptan-4-yl]-n,3-dimethylbutanamide Chemical compound CC(C)[C@H](N(C)C)C(=O)N[C@@H](C(C)C)C(=O)N(C)[C@@H]([C@@H](C)CC)[C@H](OC)CC(=O)N1CCC[C@H]1[C@H](OC)[C@@H](C)C(=O)N[C@H](C)[C@@H](O)C1=CC=CC=C1 WOWDZACBATWTAU-FEFUEGSOSA-N 0.000 description 2
- AGGWFDNPHKLBBV-YUMQZZPRSA-N (2s)-2-[[(2s)-2-amino-3-methylbutanoyl]amino]-5-(carbamoylamino)pentanoic acid Chemical compound CC(C)[C@H](N)C(=O)N[C@H](C(O)=O)CCCNC(N)=O AGGWFDNPHKLBBV-YUMQZZPRSA-N 0.000 description 2
- FDKXTQMXEQVLRF-ZHACJKMWSA-N (E)-dacarbazine Chemical compound CN(C)\N=N\c1[nH]cnc1C(N)=O FDKXTQMXEQVLRF-ZHACJKMWSA-N 0.000 description 2
- IAKHMKGGTNLKSZ-INIZCTEOSA-N (S)-colchicine Chemical compound C1([C@@H](NC(C)=O)CC2)=CC(=O)C(OC)=CC=C1C1=C2C=C(OC)C(OC)=C1OC IAKHMKGGTNLKSZ-INIZCTEOSA-N 0.000 description 2
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 2
- GZCWLCBFPRFLKL-UHFFFAOYSA-N 1-prop-2-ynoxypropan-2-ol Chemical compound CC(O)COCC#C GZCWLCBFPRFLKL-UHFFFAOYSA-N 0.000 description 2
- MQLACMBJVPINKE-UHFFFAOYSA-N 10-[(3-hydroxy-4-methoxyphenyl)methylidene]anthracen-9-one Chemical compound C1=C(O)C(OC)=CC=C1C=C1C2=CC=CC=C2C(=O)C2=CC=CC=C21 MQLACMBJVPINKE-UHFFFAOYSA-N 0.000 description 2
- HZAXFHJVJLSVMW-UHFFFAOYSA-N 2-Aminoethan-1-ol Chemical compound NCCO HZAXFHJVJLSVMW-UHFFFAOYSA-N 0.000 description 2
- FZDFGHZZPBUTGP-UHFFFAOYSA-N 2-[[2-[bis(carboxymethyl)amino]-3-(4-isothiocyanatophenyl)propyl]-[2-[bis(carboxymethyl)amino]propyl]amino]acetic acid Chemical compound OC(=O)CN(CC(O)=O)C(C)CN(CC(O)=O)CC(N(CC(O)=O)CC(O)=O)CC1=CC=C(N=C=S)C=C1 FZDFGHZZPBUTGP-UHFFFAOYSA-N 0.000 description 2
- FBUTXZSKZCQABC-UHFFFAOYSA-N 2-amino-1-methyl-7h-purine-6-thione Chemical compound S=C1N(C)C(N)=NC2=C1NC=N2 FBUTXZSKZCQABC-UHFFFAOYSA-N 0.000 description 2
- HNXQXTQTPAJEJL-UHFFFAOYSA-N 2-aminopteridin-4-ol Chemical compound C1=CN=C2NC(N)=NC(=O)C2=N1 HNXQXTQTPAJEJL-UHFFFAOYSA-N 0.000 description 2
- NDMPLJNOPCLANR-UHFFFAOYSA-N 3,4-dihydroxy-15-(4-hydroxy-18-methoxycarbonyl-5,18-seco-ibogamin-18-yl)-16-methoxy-1-methyl-6,7-didehydro-aspidospermidine-3-carboxylic acid methyl ester Natural products C1C(CC)(O)CC(CC2(C(=O)OC)C=3C(=CC4=C(C56C(C(C(O)C7(CC)C=CCN(C67)CC5)(O)C(=O)OC)N4C)C=3)OC)CN1CCC1=C2NC2=CC=CC=C12 NDMPLJNOPCLANR-UHFFFAOYSA-N 0.000 description 2
- FPQQSJJWHUJYPU-UHFFFAOYSA-N 3-(dimethylamino)propyliminomethylidene-ethylazanium;chloride Chemical compound Cl.CCN=C=NCCCN(C)C FPQQSJJWHUJYPU-UHFFFAOYSA-N 0.000 description 2
- OSJPPGNTCRNQQC-UWTATZPHSA-N 3-phospho-D-glyceric acid Chemical compound OC(=O)[C@H](O)COP(O)(O)=O OSJPPGNTCRNQQC-UWTATZPHSA-N 0.000 description 2
- FWBHETKCLVMNFS-UHFFFAOYSA-N 4',6-Diamino-2-phenylindol Chemical compound C1=CC(C(=N)N)=CC=C1C1=CC2=CC=C(C(N)=N)C=C2N1 FWBHETKCLVMNFS-UHFFFAOYSA-N 0.000 description 2
- GANZODCWZFAEGN-UHFFFAOYSA-N 5-mercapto-2-nitro-benzoic acid Chemical compound OC(=O)C1=CC(S)=CC=C1[N+]([O-])=O GANZODCWZFAEGN-UHFFFAOYSA-N 0.000 description 2
- VVIAGPKUTFNRDU-UHFFFAOYSA-N 6S-folinic acid Natural products C1NC=2NC(N)=NC(=O)C=2N(C=O)C1CNC1=CC=C(C(=O)NC(CCC(O)=O)C(O)=O)C=C1 VVIAGPKUTFNRDU-UHFFFAOYSA-N 0.000 description 2
- 108010066676 Abrin Proteins 0.000 description 2
- 108010051457 Acid Phosphatase Proteins 0.000 description 2
- 102000013563 Acid Phosphatase Human genes 0.000 description 2
- 229920000936 Agarose Polymers 0.000 description 2
- OMNVYXHOSHNURL-WPRPVWTQSA-N Ala-Phe Chemical compound C[C@H](N)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 OMNVYXHOSHNURL-WPRPVWTQSA-N 0.000 description 2
- OGSPWJRAVKPPFI-UHFFFAOYSA-N Alendronic Acid Chemical compound NCCCC(O)(P(O)(O)=O)P(O)(O)=O OGSPWJRAVKPPFI-UHFFFAOYSA-N 0.000 description 2
- 244000036975 Ambrosia artemisiifolia Species 0.000 description 2
- 235000003129 Ambrosia artemisiifolia var elatior Nutrition 0.000 description 2
- 101710154825 Aminoglycoside 3'-phosphotransferase Proteins 0.000 description 2
- BFYIZQONLCFLEV-DAELLWKTSA-N Aromasine Chemical compound O=C1C=C[C@]2(C)[C@H]3CC[C@](C)(C(CC4)=O)[C@@H]4[C@@H]3CC(=C)C2=C1 BFYIZQONLCFLEV-DAELLWKTSA-N 0.000 description 2
- 206010003445 Ascites Diseases 0.000 description 2
- 241000228212 Aspergillus Species 0.000 description 2
- 101000669426 Aspergillus restrictus Ribonuclease mitogillin Proteins 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- MLDQJTXFUGDVEO-UHFFFAOYSA-N BAY-43-9006 Chemical compound C1=NC(C(=O)NC)=CC(OC=2C=CC(NC(=O)NC=3C=C(C(Cl)=CC=3)C(F)(F)F)=CC=2)=C1 MLDQJTXFUGDVEO-UHFFFAOYSA-N 0.000 description 2
- 241000194108 Bacillus licheniformis Species 0.000 description 2
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 2
- 108010006654 Bleomycin Proteins 0.000 description 2
- 208000031648 Body Weight Changes Diseases 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- 241000701822 Bovine papillomavirus Species 0.000 description 2
- 208000003174 Brain Neoplasms Diseases 0.000 description 2
- BHPQYMZQTOCNFJ-UHFFFAOYSA-N Calcium cation Chemical compound [Ca+2] BHPQYMZQTOCNFJ-UHFFFAOYSA-N 0.000 description 2
- 241000282465 Canis Species 0.000 description 2
- 101710158575 Cap-specific mRNA (nucleoside-2'-O-)-methyltransferase Proteins 0.000 description 2
- GAGWJHPBXLXJQN-UORFTKCHSA-N Capecitabine Chemical compound C1=C(F)C(NC(=O)OCCCCC)=NC(=O)N1[C@H]1[C@H](O)[C@H](O)[C@@H](C)O1 GAGWJHPBXLXJQN-UORFTKCHSA-N 0.000 description 2
- OKTJSMMVPCPJKN-NJFSPNSNSA-N Carbon-14 Chemical compound [14C] OKTJSMMVPCPJKN-NJFSPNSNSA-N 0.000 description 2
- 108091006146 Channels Proteins 0.000 description 2
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 2
- 108700032819 Croton tiglium crotin II Proteins 0.000 description 2
- XZMCDFZZKTWFGF-UHFFFAOYSA-N Cyanamide Chemical compound NC#N XZMCDFZZKTWFGF-UHFFFAOYSA-N 0.000 description 2
- CMSMOCZEIVJLDB-UHFFFAOYSA-N Cyclophosphamide Chemical compound ClCCN(CCCl)P1(=O)NCCCO1 CMSMOCZEIVJLDB-UHFFFAOYSA-N 0.000 description 2
- 241000701022 Cytomegalovirus Species 0.000 description 2
- SRBFZHDQGSBBOR-IOVATXLUSA-N D-xylopyranose Chemical compound O[C@@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-IOVATXLUSA-N 0.000 description 2
- 101150074155 DHFR gene Proteins 0.000 description 2
- 101150011813 DLL1 gene Proteins 0.000 description 2
- 102000053602 DNA Human genes 0.000 description 2
- 230000004568 DNA-binding Effects 0.000 description 2
- 108010092160 Dactinomycin Proteins 0.000 description 2
- WEAHRLBPCANXCN-UHFFFAOYSA-N Daunomycin Natural products CCC1(O)CC(OC2CC(N)C(O)C(C)O2)c3cc4C(=O)c5c(OC)cccc5C(=O)c4c(O)c3C1 WEAHRLBPCANXCN-UHFFFAOYSA-N 0.000 description 2
- 108010053770 Deoxyribonucleases Proteins 0.000 description 2
- 102000016911 Deoxyribonucleases Human genes 0.000 description 2
- 241000702421 Dependoparvovirus Species 0.000 description 2
- 229920002307 Dextran Polymers 0.000 description 2
- RTZKZFJDLAIYFH-UHFFFAOYSA-N Diethyl ether Chemical compound CCOCC RTZKZFJDLAIYFH-UHFFFAOYSA-N 0.000 description 2
- 108010016626 Dipeptides Proteins 0.000 description 2
- ZQZFYGIXNQKOAV-OCEACIFDSA-N Droloxifene Chemical compound C=1C=CC=CC=1C(/CC)=C(C=1C=C(O)C=CC=1)\C1=CC=C(OCCN(C)C)C=C1 ZQZFYGIXNQKOAV-OCEACIFDSA-N 0.000 description 2
- 241000255581 Drosophila <fruit fly, genus> Species 0.000 description 2
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 2
- 206010058314 Dysplasia Diseases 0.000 description 2
- 241000283086 Equidae Species 0.000 description 2
- PIICEJLVQHRZGT-UHFFFAOYSA-N Ethylenediamine Chemical compound NCCN PIICEJLVQHRZGT-UHFFFAOYSA-N 0.000 description 2
- 101710082714 Exotoxin A Proteins 0.000 description 2
- 108010021468 Fc gamma receptor IIA Proteins 0.000 description 2
- 108010021472 Fc gamma receptor IIB Proteins 0.000 description 2
- 241000724791 Filamentous phage Species 0.000 description 2
- PXGOKWXKJXAPGV-UHFFFAOYSA-N Fluorine Chemical compound FF PXGOKWXKJXAPGV-UHFFFAOYSA-N 0.000 description 2
- VWUXBMIQPBEWFH-WCCTWKNTSA-N Fulvestrant Chemical compound OC1=CC=C2[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3[C@H](CCCCCCCCCS(=O)CCCC(F)(F)C(F)(F)F)CC2=C1 VWUXBMIQPBEWFH-WCCTWKNTSA-N 0.000 description 2
- 101150074355 GS gene Proteins 0.000 description 2
- 108010015133 Galactose oxidase Proteins 0.000 description 2
- 108700004714 Gelonium multiflorum GEL Proteins 0.000 description 2
- 108010073178 Glucan 1,4-alpha-Glucosidase Proteins 0.000 description 2
- 102100022624 Glucoamylase Human genes 0.000 description 2
- 108010070600 Glucose-6-phosphate isomerase Proteins 0.000 description 2
- 102000010437 HD domains Human genes 0.000 description 2
- 108050001906 HD domains Proteins 0.000 description 2
- 208000009889 Herpes Simplex Diseases 0.000 description 2
- 208000010747 Hodgkins lymphoma Diseases 0.000 description 2
- 101100118545 Holotrichia diomphalia EGF-like gene Proteins 0.000 description 2
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 2
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 2
- 102000004877 Insulin Human genes 0.000 description 2
- 108090001061 Insulin Proteins 0.000 description 2
- ZCYVEMRRCGMTRW-AHCXROLUSA-N Iodine-123 Chemical compound [123I] ZCYVEMRRCGMTRW-AHCXROLUSA-N 0.000 description 2
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 2
- 101150056261 Jag2 gene Proteins 0.000 description 2
- 102000049556 Jagged-1 Human genes 0.000 description 2
- 108700003486 Jagged-1 Proteins 0.000 description 2
- 102000049546 Jagged-2 Human genes 0.000 description 2
- 241000235058 Komagataella pastoris Species 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- 239000002147 L01XE04 - Sunitinib Substances 0.000 description 2
- 241000254158 Lampyridae Species 0.000 description 2
- 108090001090 Lectins Proteins 0.000 description 2
- 102000004856 Lectins Human genes 0.000 description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 2
- 108010000817 Leuprolide Proteins 0.000 description 2
- 102100029205 Low affinity immunoglobulin gamma Fc region receptor II-b Human genes 0.000 description 2
- 230000027311 M phase Effects 0.000 description 2
- 108010014251 Muramidase Proteins 0.000 description 2
- 102000016943 Muramidase Human genes 0.000 description 2
- 108010062010 N-Acetylmuramoyl-L-alanine Amidase Proteins 0.000 description 2
- 238000005481 NMR spectroscopy Methods 0.000 description 2
- 230000004989 O-glycosylation Effects 0.000 description 2
- 206010061902 Pancreatic neoplasm Diseases 0.000 description 2
- 108090000526 Papain Proteins 0.000 description 2
- 206010057249 Phagocytosis Diseases 0.000 description 2
- 101100413173 Phytolacca americana PAP2 gene Proteins 0.000 description 2
- 239000004372 Polyvinyl alcohol Substances 0.000 description 2
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 2
- NBBJYMSMWIIQGU-UHFFFAOYSA-N Propionic aldehyde Chemical compound CCC=O NBBJYMSMWIIQGU-UHFFFAOYSA-N 0.000 description 2
- 239000012980 RPMI-1640 medium Substances 0.000 description 2
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 2
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 2
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 2
- 108700008625 Reporter Genes Proteins 0.000 description 2
- 229920002684 Sepharose Polymers 0.000 description 2
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 2
- CDBYLPFSWZWCQE-UHFFFAOYSA-L Sodium Carbonate Chemical compound [Na+].[Na+].[O-]C([O-])=O CDBYLPFSWZWCQE-UHFFFAOYSA-L 0.000 description 2
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical compound [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 2
- 241000256251 Spodoptera frugiperda Species 0.000 description 2
- CYQFCXCEBYINGO-UHFFFAOYSA-N THC Natural products C1=C(C)CCC2C(C)(C)OC3=CC(CCCCC)=CC(O)=C3C21 CYQFCXCEBYINGO-UHFFFAOYSA-N 0.000 description 2
- NAVMQTYZDKMPEU-UHFFFAOYSA-N Targretin Chemical compound CC1=CC(C(CCC2(C)C)(C)C)=C2C=C1C(=C)C1=CC=C(C(O)=O)C=C1 NAVMQTYZDKMPEU-UHFFFAOYSA-N 0.000 description 2
- 229940123237 Taxane Drugs 0.000 description 2
- FOCVUCIESVLUNU-UHFFFAOYSA-N Thiotepa Chemical compound C1CN1P(N1CC1)(=S)N1CC1 FOCVUCIESVLUNU-UHFFFAOYSA-N 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 102000006601 Thymidine Kinase Human genes 0.000 description 2
- 108020004440 Thymidine kinase Proteins 0.000 description 2
- DKJJVAGXPKPDRL-UHFFFAOYSA-N Tiludronic acid Chemical compound OP(O)(=O)C(P(O)(O)=O)SC1=CC=C(Cl)C=C1 DKJJVAGXPKPDRL-UHFFFAOYSA-N 0.000 description 2
- 102000004338 Transferrin Human genes 0.000 description 2
- 108090000901 Transferrin Proteins 0.000 description 2
- 108700019146 Transgenes Proteins 0.000 description 2
- 239000007983 Tris buffer Substances 0.000 description 2
- 239000013504 Triton X-100 Substances 0.000 description 2
- 229920004890 Triton X-100 Polymers 0.000 description 2
- 102000004243 Tubulin Human genes 0.000 description 2
- 108090000704 Tubulin Proteins 0.000 description 2
- DRTQHJPVMGBUCF-XVFCMESISA-N Uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-XVFCMESISA-N 0.000 description 2
- 240000001866 Vernicia fordii Species 0.000 description 2
- 241000863480 Vinca Species 0.000 description 2
- 229940122803 Vinca alkaloid Drugs 0.000 description 2
- IXKSXJFAGXLQOQ-XISFHERQSA-N WHWLQLKPGQPMY Chemical compound C([C@@H](C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)NC(=O)[C@@H](N)CC=1C2=CC=CC=C2NC=1)C1=CNC=N1 IXKSXJFAGXLQOQ-XISFHERQSA-N 0.000 description 2
- 238000012452 Xenomouse strains Methods 0.000 description 2
- IEDXPSOJFSVCKU-HOKPPMCLSA-N [4-[[(2S)-5-(carbamoylamino)-2-[[(2S)-2-[6-(2,5-dioxopyrrolidin-1-yl)hexanoylamino]-3-methylbutanoyl]amino]pentanoyl]amino]phenyl]methyl N-[(2S)-1-[[(2S)-1-[[(3R,4S,5S)-1-[(2S)-2-[(1R,2R)-3-[[(1S,2R)-1-hydroxy-1-phenylpropan-2-yl]amino]-1-methoxy-2-methyl-3-oxopropyl]pyrrolidin-1-yl]-3-methoxy-5-methyl-1-oxoheptan-4-yl]-methylamino]-3-methyl-1-oxobutan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]-N-methylcarbamate Chemical compound CC[C@H](C)[C@@H]([C@@H](CC(=O)N1CCC[C@H]1[C@H](OC)[C@@H](C)C(=O)N[C@H](C)[C@@H](O)c1ccccc1)OC)N(C)C(=O)[C@@H](NC(=O)[C@H](C(C)C)N(C)C(=O)OCc1ccc(NC(=O)[C@H](CCCNC(N)=O)NC(=O)[C@@H](NC(=O)CCCCCN2C(=O)CCC2=O)C(C)C)cc1)C(C)C IEDXPSOJFSVCKU-HOKPPMCLSA-N 0.000 description 2
- 230000002159 abnormal effect Effects 0.000 description 2
- DZBUGLKDJFMEHC-UHFFFAOYSA-N acridine Chemical compound C1=CC=CC2=CC3=CC=CC=C3N=C21 DZBUGLKDJFMEHC-UHFFFAOYSA-N 0.000 description 2
- RJURFGZVJUQBHK-UHFFFAOYSA-N actinomycin D Natural products CC1OC(=O)C(C(C)C)N(C)C(=O)CN(C)C(=O)C2CCCN2C(=O)C(C(C)C)NC(=O)C1NC(=O)C1=C(N)C(=O)C(C)=C2OC(C(C)=CC=C3C(=O)NC4C(=O)NC(C(N5CCCC5C(=O)N(C)CC(=O)N(C)C(C(C)C)C(=O)OC4C)=O)C(C)C)=C3N=C21 RJURFGZVJUQBHK-UHFFFAOYSA-N 0.000 description 2
- 230000003213 activating effect Effects 0.000 description 2
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 2
- 229940009456 adriamycin Drugs 0.000 description 2
- 238000012867 alanine scanning Methods 0.000 description 2
- 108010011559 alanylphenylalanine Proteins 0.000 description 2
- 229940100198 alkylating agent Drugs 0.000 description 2
- 239000002168 alkylating agent Substances 0.000 description 2
- SHGAZHPCJJPHSC-YCNIQYBTSA-N all-trans-retinoic acid Chemical compound OC(=O)\C=C(/C)\C=C\C=C(/C)\C=C\C1=C(C)CCCC1(C)C SHGAZHPCJJPHSC-YCNIQYBTSA-N 0.000 description 2
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 description 2
- 108010001818 alpha-sarcin Proteins 0.000 description 2
- YBBLVLTVTVSKRW-UHFFFAOYSA-N anastrozole Chemical compound N#CC(C)(C)C1=CC(C(C)(C#N)C)=CC(CN2N=CN=C2)=C1 YBBLVLTVTVSKRW-UHFFFAOYSA-N 0.000 description 2
- 230000033115 angiogenesis Effects 0.000 description 2
- 235000003484 annual ragweed Nutrition 0.000 description 2
- 230000001093 anti-cancer Effects 0.000 description 2
- 230000000259 anti-tumor effect Effects 0.000 description 2
- 238000011091 antibody purification Methods 0.000 description 2
- 230000005875 antibody response Effects 0.000 description 2
- 102000025171 antigen binding proteins Human genes 0.000 description 2
- 108091000831 antigen binding proteins Proteins 0.000 description 2
- 229940041181 antineoplastic drug Drugs 0.000 description 2
- 238000003782 apoptosis assay Methods 0.000 description 2
- 230000006907 apoptotic process Effects 0.000 description 2
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 2
- 239000008228 bacteriostatic water for injection Substances 0.000 description 2
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 2
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 2
- QZPQTZZNNJUOLS-UHFFFAOYSA-N beta-lapachone Chemical compound C12=CC=CC=C2C(=O)C(=O)C2=C1OC(C)(C)CC2 QZPQTZZNNJUOLS-UHFFFAOYSA-N 0.000 description 2
- 229960001561 bleomycin Drugs 0.000 description 2
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 2
- 230000004579 body weight change Effects 0.000 description 2
- 229910052796 boron Inorganic materials 0.000 description 2
- GXJABQQUPOEUTA-RDJZCZTQSA-N bortezomib Chemical compound C([C@@H](C(=O)N[C@@H](CC(C)C)B(O)O)NC(=O)C=1N=CC=NC=1)C1=CC=CC=C1 GXJABQQUPOEUTA-RDJZCZTQSA-N 0.000 description 2
- 235000006263 bur ragweed Nutrition 0.000 description 2
- 229910001424 calcium ion Inorganic materials 0.000 description 2
- 244000309466 calf Species 0.000 description 2
- 230000005907 cancer growth Effects 0.000 description 2
- 235000014633 carbohydrates Nutrition 0.000 description 2
- 230000004663 cell proliferation Effects 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 230000002759 chromosomal effect Effects 0.000 description 2
- 229960004316 cisplatin Drugs 0.000 description 2
- DQLATGHUWYMOKM-UHFFFAOYSA-L cisplatin Chemical compound N[Pt](N)(Cl)Cl DQLATGHUWYMOKM-UHFFFAOYSA-L 0.000 description 2
- ACSIXWWBWUQEHA-UHFFFAOYSA-N clodronic acid Chemical compound OP(O)(=O)C(Cl)(Cl)P(O)(O)=O ACSIXWWBWUQEHA-UHFFFAOYSA-N 0.000 description 2
- 238000003501 co-culture Methods 0.000 description 2
- 235000003488 common ragweed Nutrition 0.000 description 2
- 230000002860 competitive effect Effects 0.000 description 2
- 239000003636 conditioned culture medium Substances 0.000 description 2
- 238000007796 conventional method Methods 0.000 description 2
- 238000010168 coupling process Methods 0.000 description 2
- COFJBSXICYYSKG-OAUVCNBTSA-N cph2u7dndy Chemical compound OS(O)(=O)=O.C([C@H](C[C@]1(C(=O)OC)C=2C(=CC3=C([C@]45[C@H]([C@@]([C@H](O)[C@]6(CC)C=CCN([C@H]56)CC4)(O)C(N)=O)N3C)C=2)OC)C[C@@](C2)(O)CC)N2CCC2=C1NC1=CC=CC=C21 COFJBSXICYYSKG-OAUVCNBTSA-N 0.000 description 2
- PSNOPSMXOBPNNV-VVCTWANISA-N cryptophycin 1 Chemical compound C1=C(Cl)C(OC)=CC=C1C[C@@H]1C(=O)NC[C@@H](C)C(=O)O[C@@H](CC(C)C)C(=O)O[C@H]([C@H](C)[C@@H]2[C@H](O2)C=2C=CC=CC=2)C/C=C/C(=O)N1 PSNOPSMXOBPNNV-VVCTWANISA-N 0.000 description 2
- PSNOPSMXOBPNNV-UHFFFAOYSA-N cryptophycin-327 Natural products C1=C(Cl)C(OC)=CC=C1CC1C(=O)NCC(C)C(=O)OC(CC(C)C)C(=O)OC(C(C)C2C(O2)C=2C=CC=CC=2)CC=CC(=O)N1 PSNOPSMXOBPNNV-UHFFFAOYSA-N 0.000 description 2
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 2
- 229960003901 dacarbazine Drugs 0.000 description 2
- 230000003247 decreasing effect Effects 0.000 description 2
- 239000000032 diagnostic agent Substances 0.000 description 2
- 229940039227 diagnostic agent Drugs 0.000 description 2
- 238000000502 dialysis Methods 0.000 description 2
- 229930191339 dianthin Natural products 0.000 description 2
- 206010013023 diphtheria Diseases 0.000 description 2
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 2
- 150000004662 dithiols Chemical class 0.000 description 2
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 2
- 230000003828 downregulation Effects 0.000 description 2
- 229950004203 droloxifene Drugs 0.000 description 2
- 229940120655 eloxatin Drugs 0.000 description 2
- 238000010828 elution Methods 0.000 description 2
- 230000009764 endothelial cell sprouting Effects 0.000 description 2
- 108010028531 enomycin Proteins 0.000 description 2
- 230000002255 enzymatic effect Effects 0.000 description 2
- 229930013356 epothilone Natural products 0.000 description 2
- 150000003883 epothilone derivatives Chemical class 0.000 description 2
- 229960005420 etoposide Drugs 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 238000000855 fermentation Methods 0.000 description 2
- 230000004151 fermentation Effects 0.000 description 2
- 210000003754 fetus Anatomy 0.000 description 2
- 210000002950 fibroblast Anatomy 0.000 description 2
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 2
- 239000007850 fluorescent dye Substances 0.000 description 2
- OVBPIULPVIDEAO-LBPRGKRZSA-N folic acid Chemical compound C=1N=C2NC(N)=NC(=O)C2=NC=1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 OVBPIULPVIDEAO-LBPRGKRZSA-N 0.000 description 2
- 235000008191 folinic acid Nutrition 0.000 description 2
- 239000011672 folinic acid Substances 0.000 description 2
- VVIAGPKUTFNRDU-ABLWVSNPSA-N folinic acid Chemical compound C1NC=2NC(N)=NC(=O)C=2N(C=O)C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 VVIAGPKUTFNRDU-ABLWVSNPSA-N 0.000 description 2
- YAKWPXVTIGTRJH-UHFFFAOYSA-N fotemustine Chemical compound CCOP(=O)(OCC)C(C)NC(=O)N(CCCl)N=O YAKWPXVTIGTRJH-UHFFFAOYSA-N 0.000 description 2
- 229960004783 fotemustine Drugs 0.000 description 2
- 238000013467 fragmentation Methods 0.000 description 2
- 238000006062 fragmentation reaction Methods 0.000 description 2
- 230000033581 fucosylation Effects 0.000 description 2
- 230000002538 fungal effect Effects 0.000 description 2
- CHPZKNULDCNCBW-UHFFFAOYSA-N gallium nitrate Chemical compound [Ga+3].[O-][N+]([O-])=O.[O-][N+]([O-])=O.[O-][N+]([O-])=O CHPZKNULDCNCBW-UHFFFAOYSA-N 0.000 description 2
- 239000003540 gamma secretase inhibitor Substances 0.000 description 2
- 238000001502 gel electrophoresis Methods 0.000 description 2
- 229960003297 gemtuzumab ozogamicin Drugs 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- XKUKSGPZAADMRA-UHFFFAOYSA-N glycyl-glycyl-glycine Chemical compound NCC(=O)NCC(=O)NCC(O)=O XKUKSGPZAADMRA-UHFFFAOYSA-N 0.000 description 2
- 150000004820 halides Chemical class 0.000 description 2
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 2
- 210000003494 hepatocyte Anatomy 0.000 description 2
- 238000004128 high performance liquid chromatography Methods 0.000 description 2
- 238000007489 histopathology method Methods 0.000 description 2
- 229920001519 homopolymer Polymers 0.000 description 2
- 229940042795 hydrazides for tuberculosis treatment Drugs 0.000 description 2
- 229910052739 hydrogen Inorganic materials 0.000 description 2
- 239000001257 hydrogen Substances 0.000 description 2
- NPZTUJOABDZTLV-UHFFFAOYSA-N hydroxybenzotriazole Substances O=C1C=CC=C2NNN=C12 NPZTUJOABDZTLV-UHFFFAOYSA-N 0.000 description 2
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 2
- 229960001101 ifosfamide Drugs 0.000 description 2
- HOMGKSMUEGBAAB-UHFFFAOYSA-N ifosfamide Chemical compound ClCCNP1(=O)OCCCN1CCCl HOMGKSMUEGBAAB-UHFFFAOYSA-N 0.000 description 2
- 230000036039 immunity Effects 0.000 description 2
- 230000005847 immunogenicity Effects 0.000 description 2
- 239000003547 immunosorbent Substances 0.000 description 2
- 230000002637 immunotoxin Effects 0.000 description 2
- 229940051026 immunotoxin Drugs 0.000 description 2
- 231100000608 immunotoxin Toxicity 0.000 description 2
- 230000008676 import Effects 0.000 description 2
- APFVFJFRJDLVQX-AHCXROLUSA-N indium-111 Chemical compound [111In] APFVFJFRJDLVQX-AHCXROLUSA-N 0.000 description 2
- 108091008042 inhibitory receptors Proteins 0.000 description 2
- 229940125396 insulin Drugs 0.000 description 2
- 238000001990 intravenous administration Methods 0.000 description 2
- 210000003734 kidney Anatomy 0.000 description 2
- 238000002372 labelling Methods 0.000 description 2
- HPJKCIUCZWXJDR-UHFFFAOYSA-N letrozole Chemical compound C1=CC(C#N)=CC=C1C(N1N=CN=C1)C1=CC=C(C#N)C=C1 HPJKCIUCZWXJDR-UHFFFAOYSA-N 0.000 description 2
- 229960001691 leucovorin Drugs 0.000 description 2
- 210000000265 leukocyte Anatomy 0.000 description 2
- GFIJNRVAKGFPGQ-LIJARHBVSA-N leuprolide Chemical compound CCNC(=O)[C@@H]1CCCN1C(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](CC(C)C)NC(=O)[C@@H](NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H]1NC(=O)CC1)CC1=CC=C(O)C=C1 GFIJNRVAKGFPGQ-LIJARHBVSA-N 0.000 description 2
- 229960004338 leuprorelin Drugs 0.000 description 2
- 238000011068 loading method Methods 0.000 description 2
- DHMTURDWPRKSOA-RUZDIDTESA-N lonafarnib Chemical compound C1CN(C(=O)N)CCC1CC(=O)N1CCC([C@@H]2C3=C(Br)C=C(Cl)C=C3CCC3=CC(Br)=CN=C32)CC1 DHMTURDWPRKSOA-RUZDIDTESA-N 0.000 description 2
- 229960000274 lysozyme Drugs 0.000 description 2
- 239000004325 lysozyme Substances 0.000 description 2
- 235000010335 lysozyme Nutrition 0.000 description 2
- 125000005439 maleimidyl group Chemical group C1(C=CC(N1*)=O)=O 0.000 description 2
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 description 2
- 230000008774 maternal effect Effects 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- RQZAXGRLVPAYTJ-GQFGMJRRSA-N megestrol acetate Chemical compound C1=C(C)C2=CC(=O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@@](C(C)=O)(OC(=O)C)[C@@]1(C)CC2 RQZAXGRLVPAYTJ-GQFGMJRRSA-N 0.000 description 2
- 229960004296 megestrol acetate Drugs 0.000 description 2
- 229960001924 melphalan Drugs 0.000 description 2
- SGDBTWWWUNNDEQ-LBPRGKRZSA-N melphalan Chemical compound OC(=O)[C@@H](N)CC1=CC=C(N(CCCl)CCCl)C=C1 SGDBTWWWUNNDEQ-LBPRGKRZSA-N 0.000 description 2
- 229960001428 mercaptopurine Drugs 0.000 description 2
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 2
- HPNSFSBZBAHARI-UHFFFAOYSA-N micophenolic acid Natural products OC1=C(CC=C(C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-UHFFFAOYSA-N 0.000 description 2
- 229960001156 mitoxantrone Drugs 0.000 description 2
- 108010010621 modeccin Proteins 0.000 description 2
- 229940035032 monophosphoryl lipid a Drugs 0.000 description 2
- 238000010172 mouse model Methods 0.000 description 2
- 229960000951 mycophenolic acid Drugs 0.000 description 2
- HPNSFSBZBAHARI-RUDMXATFSA-N mycophenolic acid Chemical compound OC1=C(C\C=C(/C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-RUDMXATFSA-N 0.000 description 2
- LBWFXVZLPYTWQI-IPOVEDGCSA-N n-[2-(diethylamino)ethyl]-5-[(z)-(5-fluoro-2-oxo-1h-indol-3-ylidene)methyl]-2,4-dimethyl-1h-pyrrole-3-carboxamide;(2s)-2-hydroxybutanedioic acid Chemical compound OC(=O)[C@@H](O)CC(O)=O.CCN(CC)CCNC(=O)C1=C(C)NC(\C=C/2C3=CC(F)=CC=C3NC\2=O)=C1C LBWFXVZLPYTWQI-IPOVEDGCSA-N 0.000 description 2
- 230000007935 neutral effect Effects 0.000 description 2
- 108010087904 neutravidin Proteins 0.000 description 2
- 210000000440 neutrophil Anatomy 0.000 description 2
- 230000001293 nucleolytic effect Effects 0.000 description 2
- 239000002777 nucleoside Substances 0.000 description 2
- 235000015097 nutrients Nutrition 0.000 description 2
- 229960001756 oxaliplatin Drugs 0.000 description 2
- DWAFYCQODLXJNR-BNTLRKBRSA-L oxaliplatin Chemical compound O1C(=O)C(=O)O[Pt]11N[C@@H]2CCCC[C@H]2N1 DWAFYCQODLXJNR-BNTLRKBRSA-L 0.000 description 2
- 230000001590 oxidative effect Effects 0.000 description 2
- WRUUGTRCQOWXEG-UHFFFAOYSA-N pamidronate Chemical compound NCCC(O)(P(O)(O)=O)P(O)(O)=O WRUUGTRCQOWXEG-UHFFFAOYSA-N 0.000 description 2
- 210000000496 pancreas Anatomy 0.000 description 2
- 201000002528 pancreatic cancer Diseases 0.000 description 2
- 208000008443 pancreatic carcinoma Diseases 0.000 description 2
- 229940055729 papain Drugs 0.000 description 2
- 235000019834 papain Nutrition 0.000 description 2
- 230000001575 pathological effect Effects 0.000 description 2
- 238000010647 peptide synthesis reaction Methods 0.000 description 2
- 125000001151 peptidyl group Chemical group 0.000 description 2
- 230000008782 phagocytosis Effects 0.000 description 2
- 239000008194 pharmaceutical composition Substances 0.000 description 2
- 108010076042 phenomycin Proteins 0.000 description 2
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 2
- 235000011007 phosphoric acid Nutrition 0.000 description 2
- 238000000053 physical method Methods 0.000 description 2
- BASFCYQUMIYNBI-UHFFFAOYSA-N platinum Chemical compound [Pt] BASFCYQUMIYNBI-UHFFFAOYSA-N 0.000 description 2
- 229920002451 polyvinyl alcohol Polymers 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 208000017426 precursor B-cell acute lymphoblastic leukemia Diseases 0.000 description 2
- 230000002062 proliferating effect Effects 0.000 description 2
- 230000035755 proliferation Effects 0.000 description 2
- 235000013930 proline Nutrition 0.000 description 2
- 235000019833 protease Nutrition 0.000 description 2
- 125000006239 protecting group Chemical group 0.000 description 2
- 230000017854 proteolysis Effects 0.000 description 2
- 230000006337 proteolytic cleavage Effects 0.000 description 2
- ZCCUUQDIBDJBTK-UHFFFAOYSA-N psoralen Chemical compound C1=C2OC(=O)C=CC2=CC2=C1OC=C2 ZCCUUQDIBDJBTK-UHFFFAOYSA-N 0.000 description 2
- 230000005180 public health Effects 0.000 description 2
- RXWNCPJZOCPEPQ-NVWDDTSBSA-N puromycin Chemical compound C1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CO RXWNCPJZOCPEPQ-NVWDDTSBSA-N 0.000 description 2
- 230000005855 radiation Effects 0.000 description 2
- 235000009736 ragweed Nutrition 0.000 description 2
- 229960004622 raloxifene Drugs 0.000 description 2
- 238000002708 random mutagenesis Methods 0.000 description 2
- ZAHRKKWIAAJSAO-UHFFFAOYSA-N rapamycin Natural products COCC(O)C(=C/C(C)C(=O)CC(OC(=O)C1CCCCN1C(=O)C(=O)C2(O)OC(CC(OC)C(=CC=CC=CC(C)CC(C)C(=O)C)C)CCC2C)C(C)CC3CCC(O)C(C3)OC)C ZAHRKKWIAAJSAO-UHFFFAOYSA-N 0.000 description 2
- 229930002330 retinoic acid Natural products 0.000 description 2
- RODXRVNMMDRFIK-UHFFFAOYSA-N scopoletin Chemical compound C1=CC(=O)OC2=C1C=C(OC)C(O)=C2 RODXRVNMMDRFIK-UHFFFAOYSA-N 0.000 description 2
- 230000003248 secreting effect Effects 0.000 description 2
- 238000010187 selection method Methods 0.000 description 2
- 229940095743 selective estrogen receptor modulator Drugs 0.000 description 2
- 239000000333 selective estrogen receptor modulator Substances 0.000 description 2
- 230000035945 sensitivity Effects 0.000 description 2
- 238000000926 separation method Methods 0.000 description 2
- 229960002930 sirolimus Drugs 0.000 description 2
- 239000001632 sodium acetate Substances 0.000 description 2
- 235000017281 sodium acetate Nutrition 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- 125000006850 spacer group Chemical group 0.000 description 2
- 230000009870 specific binding Effects 0.000 description 2
- 210000000952 spleen Anatomy 0.000 description 2
- 210000004989 spleen cell Anatomy 0.000 description 2
- 238000010561 standard procedure Methods 0.000 description 2
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 2
- PVYJZLYGTZKPJE-UHFFFAOYSA-N streptonigrin Chemical compound C=1C=C2C(=O)C(OC)=C(N)C(=O)C2=NC=1C(C=1N)=NC(C(O)=O)=C(C)C=1C1=CC=C(OC)C(OC)=C1O PVYJZLYGTZKPJE-UHFFFAOYSA-N 0.000 description 2
- 229960001052 streptozocin Drugs 0.000 description 2
- ZSJLQEPLLKMAKR-GKHCUFPYSA-N streptozocin Chemical compound O=NN(C)C(=O)N[C@H]1[C@@H](O)O[C@H](CO)[C@@H](O)[C@@H]1O ZSJLQEPLLKMAKR-GKHCUFPYSA-N 0.000 description 2
- 230000001629 suppression Effects 0.000 description 2
- 125000000101 thioether group Chemical group 0.000 description 2
- 229960001196 thiotepa Drugs 0.000 description 2
- 125000000341 threoninyl group Chemical group [H]OC([H])(C([H])([H])[H])C([H])(N([H])[H])C(*)=O 0.000 description 2
- 229940104230 thymidine Drugs 0.000 description 2
- WYWHKKSPHMUBEB-UHFFFAOYSA-N tioguanine Chemical compound N1C(N)=NC(=S)C2=C1N=CN2 WYWHKKSPHMUBEB-UHFFFAOYSA-N 0.000 description 2
- UCFGDBYHRUNTLO-QHCPKHFHSA-N topotecan Chemical compound C1=C(O)C(CN(C)C)=C2C=C(CN3C4=CC5=C(C3=O)COC(=O)[C@]5(O)CC)C4=NC2=C1 UCFGDBYHRUNTLO-QHCPKHFHSA-N 0.000 description 2
- 231100000331 toxic Toxicity 0.000 description 2
- 230000002588 toxic effect Effects 0.000 description 2
- 239000012581 transferrin Substances 0.000 description 2
- 102000035160 transmembrane proteins Human genes 0.000 description 2
- 108091005703 transmembrane proteins Proteins 0.000 description 2
- 150000003327 trichothecene derivatives Chemical class 0.000 description 2
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 2
- 230000004614 tumor growth Effects 0.000 description 2
- 238000000108 ultra-filtration Methods 0.000 description 2
- 239000013603 viral vector Substances 0.000 description 2
- 238000005406 washing Methods 0.000 description 2
- 229920003169 water-soluble polymer Polymers 0.000 description 2
- 238000001262 western blot Methods 0.000 description 2
- 229960004276 zoledronic acid Drugs 0.000 description 2
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 description 1
- DNXHEGUUPJUMQT-UHFFFAOYSA-N (+)-estrone Natural products OC1=CC=C2C3CCC(C)(C(CC4)=O)C4C3CCC2=C1 DNXHEGUUPJUMQT-UHFFFAOYSA-N 0.000 description 1
- NNJPGOLRFBJNIW-HNNXBMFYSA-N (-)-demecolcine Chemical compound C1=C(OC)C(=O)C=C2[C@@H](NC)CCC3=CC(OC)=C(OC)C(OC)=C3C2=C1 NNJPGOLRFBJNIW-HNNXBMFYSA-N 0.000 description 1
- DVWJFTGEISXVSH-CWVFEVJCSA-N (1R,3S,5S,7Z,11R,12S,13Z,15Z,17Z,19Z,21R,23S,24R,25S)-21-[(2R,3S,4S,5S,6R)-4-amino-3,5-dihydroxy-6-methyloxan-2-yl]oxy-12-ethyl-1,3,5,25-tetrahydroxy-11-methyl-9-oxo-10,27-dioxabicyclo[21.3.1]heptacosa-7,13,15,17,19-pentaene-24-carboxylic acid Chemical compound CC[C@H]1\C=C/C=C\C=C/C=C\[C@@H](C[C@@H]2O[C@@](O)(C[C@H](O)[C@H]2C(O)=O)C[C@@H](O)C[C@@H](O)C\C=C/C(=O)O[C@@H]1C)O[C@@H]1O[C@H](C)[C@@H](O)[C@H](N)[C@@H]1O DVWJFTGEISXVSH-CWVFEVJCSA-N 0.000 description 1
- JKHVDAUOODACDU-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 3-(2,5-dioxopyrrol-1-yl)propanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCN1C(=O)C=CC1=O JKHVDAUOODACDU-UHFFFAOYSA-N 0.000 description 1
- PVGATNRYUYNBHO-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-(2,5-dioxopyrrol-1-yl)butanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCN1C(=O)C=CC1=O PVGATNRYUYNBHO-UHFFFAOYSA-N 0.000 description 1
- PMJWDPGOWBRILU-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-[4-(2,5-dioxopyrrol-1-yl)phenyl]butanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCC(C=C1)=CC=C1N1C(=O)C=CC1=O PMJWDPGOWBRILU-UHFFFAOYSA-N 0.000 description 1
- VLARLSIGSPVYHX-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 6-(2,5-dioxopyrrol-1-yl)hexanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCN1C(=O)C=CC1=O VLARLSIGSPVYHX-UHFFFAOYSA-N 0.000 description 1
- WCMOHMXWOOBVMZ-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 6-[3-(2,5-dioxopyrrol-1-yl)propanoylamino]hexanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCNC(=O)CCN1C(=O)C=CC1=O WCMOHMXWOOBVMZ-UHFFFAOYSA-N 0.000 description 1
- IHVODYOQUSEYJJ-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 6-[[4-[(2,5-dioxopyrrol-1-yl)methyl]cyclohexanecarbonyl]amino]hexanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCNC(=O)C(CC1)CCC1CN1C(=O)C=CC1=O IHVODYOQUSEYJJ-UHFFFAOYSA-N 0.000 description 1
- DIGQNXIGRZPYDK-WKSCXVIASA-N (2R)-6-amino-2-[[2-[[(2S)-2-[[2-[[(2R)-2-[[(2S)-2-[[(2R,3S)-2-[[2-[[(2S)-2-[[2-[[(2S)-2-[[(2S)-2-[[(2R)-2-[[(2S,3S)-2-[[(2R)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[2-[[(2S)-2-[[(2R)-2-[[2-[[2-[[2-[(2-amino-1-hydroxyethylidene)amino]-3-carboxy-1-hydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1,5-dihydroxy-5-iminopentylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]hexanoic acid Chemical compound C[C@@H]([C@@H](C(=N[C@@H](CS)C(=N[C@@H](C)C(=N[C@@H](CO)C(=NCC(=N[C@@H](CCC(=N)O)C(=NC(CS)C(=N[C@H]([C@H](C)O)C(=N[C@H](CS)C(=N[C@H](CO)C(=NCC(=N[C@H](CS)C(=NCC(=N[C@H](CCCCN)C(=O)O)O)O)O)O)O)O)O)O)O)O)O)O)O)N=C([C@H](CS)N=C([C@H](CO)N=C([C@H](CO)N=C([C@H](C)N=C(CN=C([C@H](CO)N=C([C@H](CS)N=C(CN=C(C(CS)N=C(C(CC(=O)O)N=C(CN)O)O)O)O)O)O)O)O)O)O)O)O DIGQNXIGRZPYDK-WKSCXVIASA-N 0.000 description 1
- LNNDRFNNTDYHIO-OMYILHBOSA-N (2S)-1-[(2S)-2-[[(2S)-2-[2-[(3R,6S)-6-[[(2S)-2-[[(2R)-2-[[(2R)-2-[[(2R)-2-acetamido-3-naphthalen-2-ylpropanoyl]amino]-3-(4-chlorophenyl)propanoyl]amino]-3-pyridin-3-ylpropanoyl]amino]-3-hydroxypropanoyl]-methylamino]-1-amino-7-(4-hydroxyphenyl)-1,4,5-trioxoheptan-3-yl]hydrazinyl]-4-methylpentanoyl]amino]-6-(propan-2-ylamino)hexanoyl]-N-[(2R)-1-amino-1-oxopropan-2-yl]pyrrolidine-2-carboxamide Chemical compound CC(C)C[C@H](NN[C@H](CC(N)=O)C(=O)C(=O)[C@H](Cc1ccc(O)cc1)N(C)C(=O)[C@H](CO)NC(=O)[C@@H](Cc1cccnc1)NC(=O)[C@@H](Cc1ccc(Cl)cc1)NC(=O)[C@@H](Cc1ccc2ccccc2c1)NC(C)=O)C(=O)N[C@@H](CCCCNC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@H](C)C(N)=O LNNDRFNNTDYHIO-OMYILHBOSA-N 0.000 description 1
- WDQLRUYAYXDIFW-RWKIJVEZSA-N (2r,3r,4s,5r,6r)-4-[(2s,3r,4s,5r,6r)-3,5-dihydroxy-4-[(2r,3r,4s,5s,6r)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-6-[[(2r,3r,4s,5s,6r)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxymethyl]oxan-2-yl]oxy-6-(hydroxymethyl)oxane-2,3,5-triol Chemical compound O[C@@H]1[C@@H](CO)O[C@@H](O)[C@H](O)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)[C@H](O)[C@@H](CO[C@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)O1 WDQLRUYAYXDIFW-RWKIJVEZSA-N 0.000 description 1
- RIWLPSIAFBLILR-WVNGMBSFSA-N (2s)-1-[(2s)-2-[[(2s,3s)-2-[[(2s)-2-[[(2s,3r)-2-[[(2r,3s)-2-[[(2s)-2-[[2-[[2-[acetyl(methyl)amino]acetyl]amino]acetyl]amino]-3-methylbutanoyl]amino]-3-methylpentanoyl]amino]-3-hydroxybutanoyl]amino]pentanoyl]amino]-3-methylpentanoyl]amino]-5-(diaminomethy Chemical compound CC(=O)N(C)CC(=O)NCC(=O)N[C@@H](C(C)C)C(=O)N[C@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N1CCC[C@H]1C(=O)NCC RIWLPSIAFBLILR-WVNGMBSFSA-N 0.000 description 1
- YXTKHLHCVFUPPT-YYFJYKOTSA-N (2s)-2-[[4-[(2-amino-5-formyl-4-oxo-1,6,7,8-tetrahydropteridin-6-yl)methylamino]benzoyl]amino]pentanedioic acid;(1r,2r)-1,2-dimethanidylcyclohexane;5-fluoro-1h-pyrimidine-2,4-dione;oxalic acid;platinum(2+) Chemical compound [Pt+2].OC(=O)C(O)=O.[CH2-][C@@H]1CCCC[C@H]1[CH2-].FC1=CNC(=O)NC1=O.C1NC=2NC(N)=NC(=O)C=2N(C=O)C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 YXTKHLHCVFUPPT-YYFJYKOTSA-N 0.000 description 1
- FLWWDYNPWOSLEO-HQVZTVAUSA-N (2s)-2-[[4-[1-(2-amino-4-oxo-1h-pteridin-6-yl)ethyl-methylamino]benzoyl]amino]pentanedioic acid Chemical compound C=1N=C2NC(N)=NC(=O)C2=NC=1C(C)N(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FLWWDYNPWOSLEO-HQVZTVAUSA-N 0.000 description 1
- KYBXNPIASYUWLN-WUCPZUCCSA-N (2s)-5-hydroxypyrrolidine-2-carboxylic acid Chemical compound OC1CC[C@@H](C(O)=O)N1 KYBXNPIASYUWLN-WUCPZUCCSA-N 0.000 description 1
- CGMTUJFWROPELF-YPAAEMCBSA-N (3E,5S)-5-[(2S)-butan-2-yl]-3-(1-hydroxyethylidene)pyrrolidine-2,4-dione Chemical compound CC[C@H](C)[C@@H]1NC(=O)\C(=C(/C)O)C1=O CGMTUJFWROPELF-YPAAEMCBSA-N 0.000 description 1
- XRBSKUSTLXISAB-XVVDYKMHSA-N (5r,6r,7r,8r)-8-hydroxy-7-(hydroxymethyl)-5-(3,4,5-trimethoxyphenyl)-5,6,7,8-tetrahydrobenzo[f][1,3]benzodioxole-6-carboxylic acid Chemical compound COC1=C(OC)C(OC)=CC([C@@H]2C3=CC=4OCOC=4C=C3[C@H](O)[C@@H](CO)[C@@H]2C(O)=O)=C1 XRBSKUSTLXISAB-XVVDYKMHSA-N 0.000 description 1
- INAUWOVKEZHHDM-PEDBPRJASA-N (7s,9s)-6,9,11-trihydroxy-9-(2-hydroxyacetyl)-7-[(2r,4s,5s,6s)-5-hydroxy-6-methyl-4-morpholin-4-yloxan-2-yl]oxy-4-methoxy-8,10-dihydro-7h-tetracene-5,12-dione;hydrochloride Chemical compound Cl.N1([C@H]2C[C@@H](O[C@@H](C)[C@H]2O)O[C@H]2C[C@@](O)(CC=3C(O)=C4C(=O)C=5C=CC=C(C=5C(=O)C4=C(O)C=32)OC)C(=O)CO)CCOCC1 INAUWOVKEZHHDM-PEDBPRJASA-N 0.000 description 1
- NOPNWHSMQOXAEI-PUCKCBAPSA-N (7s,9s)-7-[(2r,4s,5s,6s)-4-(2,3-dihydropyrrol-1-yl)-5-hydroxy-6-methyloxan-2-yl]oxy-6,9,11-trihydroxy-9-(2-hydroxyacetyl)-4-methoxy-8,10-dihydro-7h-tetracene-5,12-dione Chemical compound N1([C@H]2C[C@@H](O[C@@H](C)[C@H]2O)O[C@H]2C[C@@](O)(CC=3C(O)=C4C(=O)C=5C=CC=C(C=5C(=O)C4=C(O)C=32)OC)C(=O)CO)CCC=C1 NOPNWHSMQOXAEI-PUCKCBAPSA-N 0.000 description 1
- FPVKHBSQESCIEP-UHFFFAOYSA-N (8S)-3-(2-deoxy-beta-D-erythro-pentofuranosyl)-3,6,7,8-tetrahydroimidazo[4,5-d][1,3]diazepin-8-ol Natural products C1C(O)C(CO)OC1N1C(NC=NCC2O)=C2N=C1 FPVKHBSQESCIEP-UHFFFAOYSA-N 0.000 description 1
- IEXUMDBQLIVNHZ-YOUGDJEHSA-N (8s,11r,13r,14s,17s)-11-[4-(dimethylamino)phenyl]-17-hydroxy-17-(3-hydroxypropyl)-13-methyl-1,2,6,7,8,11,12,14,15,16-decahydrocyclopenta[a]phenanthren-3-one Chemical compound C1=CC(N(C)C)=CC=C1[C@@H]1C2=C3CCC(=O)C=C3CC[C@H]2[C@H](CC[C@]2(O)CCCO)[C@@]2(C)C1 IEXUMDBQLIVNHZ-YOUGDJEHSA-N 0.000 description 1
- LKJPYSCBVHEWIU-KRWDZBQOSA-N (R)-bicalutamide Chemical compound C([C@@](O)(C)C(=O)NC=1C=C(C(C#N)=CC=1)C(F)(F)F)S(=O)(=O)C1=CC=C(F)C=C1 LKJPYSCBVHEWIU-KRWDZBQOSA-N 0.000 description 1
- AGNGYMCLFWQVGX-AGFFZDDWSA-N (e)-1-[(2s)-2-amino-2-carboxyethoxy]-2-diazonioethenolate Chemical compound OC(=O)[C@@H](N)CO\C([O-])=C\[N+]#N AGNGYMCLFWQVGX-AGFFZDDWSA-N 0.000 description 1
- NWUYHJFMYQTDRP-UHFFFAOYSA-N 1,2-bis(ethenyl)benzene;1-ethenyl-2-ethylbenzene;styrene Chemical compound C=CC1=CC=CC=C1.CCC1=CC=CC=C1C=C.C=CC1=CC=CC=C1C=C NWUYHJFMYQTDRP-UHFFFAOYSA-N 0.000 description 1
- FJQZXCPWAGYPSD-UHFFFAOYSA-N 1,3,4,6-tetrachloro-3a,6a-diphenylimidazo[4,5-d]imidazole-2,5-dione Chemical compound ClN1C(=O)N(Cl)C2(C=3C=CC=CC=3)N(Cl)C(=O)N(Cl)C12C1=CC=CC=C1 FJQZXCPWAGYPSD-UHFFFAOYSA-N 0.000 description 1
- WDCYWAQPCXBPJA-UHFFFAOYSA-N 1,3-dinitrobenzene Chemical compound [O-][N+](=O)C1=CC=CC([N+]([O-])=O)=C1 WDCYWAQPCXBPJA-UHFFFAOYSA-N 0.000 description 1
- WNXJIVFYUVYPPR-UHFFFAOYSA-N 1,3-dioxolane Chemical compound C1COCO1 WNXJIVFYUVYPPR-UHFFFAOYSA-N 0.000 description 1
- VILFTWLXLYIEMV-UHFFFAOYSA-N 1,5-difluoro-2,4-dinitrobenzene Chemical compound [O-][N+](=O)C1=CC([N+]([O-])=O)=C(F)C=C1F VILFTWLXLYIEMV-UHFFFAOYSA-N 0.000 description 1
- DIYPCWKHSODVAP-UHFFFAOYSA-N 1-[3-(2,5-dioxopyrrol-1-yl)benzoyl]oxy-2,5-dioxopyrrolidine-3-sulfonic acid Chemical compound O=C1C(S(=O)(=O)O)CC(=O)N1OC(=O)C1=CC=CC(N2C(C=CC2=O)=O)=C1 DIYPCWKHSODVAP-UHFFFAOYSA-N 0.000 description 1
- CULQNACJHGHAER-UHFFFAOYSA-N 1-[4-[(2-iodoacetyl)amino]benzoyl]oxy-2,5-dioxopyrrolidine-3-sulfonic acid Chemical compound O=C1C(S(=O)(=O)O)CC(=O)N1OC(=O)C1=CC=C(NC(=O)CI)C=C1 CULQNACJHGHAER-UHFFFAOYSA-N 0.000 description 1
- LOTKRQAVGJMPNV-UHFFFAOYSA-N 1-fluoro-2,4-dinitrobenzene Chemical compound [O-][N+](=O)C1=CC=C(F)C([N+]([O-])=O)=C1 LOTKRQAVGJMPNV-UHFFFAOYSA-N 0.000 description 1
- PNDPGZBMCMUPRI-HVTJNCQCSA-N 10043-66-0 Chemical compound [131I][131I] PNDPGZBMCMUPRI-HVTJNCQCSA-N 0.000 description 1
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 1
- LDGWQMRUWMSZIU-LQDDAWAPSA-M 2,3-bis[(z)-octadec-9-enoxy]propyl-trimethylazanium;chloride Chemical compound [Cl-].CCCCCCCC\C=C/CCCCCCCCOCC(C[N+](C)(C)C)OCCCCCCCC\C=C/CCCCCCCC LDGWQMRUWMSZIU-LQDDAWAPSA-M 0.000 description 1
- BTOTXLJHDSNXMW-POYBYMJQSA-N 2,3-dideoxyuridine Chemical compound O1[C@H](CO)CC[C@@H]1N1C(=O)NC(=O)C=C1 BTOTXLJHDSNXMW-POYBYMJQSA-N 0.000 description 1
- KGLPWQKSKUVKMJ-UHFFFAOYSA-N 2,3-dihydrophthalazine-1,4-dione Chemical class C1=CC=C2C(=O)NNC(=O)C2=C1 KGLPWQKSKUVKMJ-UHFFFAOYSA-N 0.000 description 1
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 1
- QCXJFISCRQIYID-IAEPZHFASA-N 2-amino-1-n-[(3s,6s,7r,10s,16s)-3-[(2s)-butan-2-yl]-7,11,14-trimethyl-2,5,9,12,15-pentaoxo-10-propan-2-yl-8-oxa-1,4,11,14-tetrazabicyclo[14.3.0]nonadecan-6-yl]-4,6-dimethyl-3-oxo-9-n-[(3s,6s,7r,10s,16s)-7,11,14-trimethyl-2,5,9,12,15-pentaoxo-3,10-di(propa Chemical compound C[C@H]1OC(=O)[C@H](C(C)C)N(C)C(=O)CN(C)C(=O)[C@@H]2CCCN2C(=O)[C@H](C(C)C)NC(=O)[C@H]1NC(=O)C1=C(N=C2C(C(=O)N[C@@H]3C(=O)N[C@H](C(N4CCC[C@H]4C(=O)N(C)CC(=O)N(C)[C@@H](C(C)C)C(=O)O[C@@H]3C)=O)[C@@H](C)CC)=C(N)C(=O)C(C)=C2O2)C2=C(C)C=C1 QCXJFISCRQIYID-IAEPZHFASA-N 0.000 description 1
- 125000000979 2-amino-2-oxoethyl group Chemical group [H]C([*])([H])C(=O)N([H])[H] 0.000 description 1
- BMUXBWLKTHLRQC-UHFFFAOYSA-N 2-azanylethanoic acid Chemical compound NCC(O)=O.NCC(O)=O.NCC(O)=O BMUXBWLKTHLRQC-UHFFFAOYSA-N 0.000 description 1
- FDAYLTPAFBGXAB-UHFFFAOYSA-N 2-chloro-n,n-bis(2-chloroethyl)ethanamine Chemical compound ClCCN(CCCl)CCCl FDAYLTPAFBGXAB-UHFFFAOYSA-N 0.000 description 1
- VNBAOSVONFJBKP-UHFFFAOYSA-N 2-chloro-n,n-bis(2-chloroethyl)propan-1-amine;hydrochloride Chemical compound Cl.CC(Cl)CN(CCCl)CCCl VNBAOSVONFJBKP-UHFFFAOYSA-N 0.000 description 1
- YIMDLWDNDGKDTJ-QLKYHASDSA-N 3'-deamino-3'-(3-cyanomorpholin-4-yl)doxorubicin Chemical compound N1([C@H]2C[C@@H](O[C@@H](C)[C@H]2O)O[C@H]2C[C@@](O)(CC=3C(O)=C4C(=O)C=5C=CC=C(C=5C(=O)C4=C(O)C=32)OC)C(=O)CO)CCOCC1C#N YIMDLWDNDGKDTJ-QLKYHASDSA-N 0.000 description 1
- UAIUNKRWKOVEES-UHFFFAOYSA-N 3,3',5,5'-tetramethylbenzidine Chemical compound CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 UAIUNKRWKOVEES-UHFFFAOYSA-N 0.000 description 1
- PWMYMKOUNYTVQN-UHFFFAOYSA-N 3-(8,8-diethyl-2-aza-8-germaspiro[4.5]decan-2-yl)-n,n-dimethylpropan-1-amine Chemical compound C1C[Ge](CC)(CC)CCC11CN(CCCN(C)C)CC1 PWMYMKOUNYTVQN-UHFFFAOYSA-N 0.000 description 1
- QGJZLNKBHJESQX-UHFFFAOYSA-N 3-Epi-Betulin-Saeure Natural products C1CC(O)C(C)(C)C2CCC3(C)C4(C)CCC5(C(O)=O)CCC(C(=C)C)C5C4CCC3C21C QGJZLNKBHJESQX-UHFFFAOYSA-N 0.000 description 1
- CLOUCVRNYSHRCF-UHFFFAOYSA-N 3beta-Hydroxy-20(29)-Lupen-3,27-oic acid Natural products C1CC(O)C(C)(C)C2CCC3(C)C4(C(O)=O)CCC5(C)CCC(C(=C)C)C5C4CCC3C21C CLOUCVRNYSHRCF-UHFFFAOYSA-N 0.000 description 1
- VXGRJERITKFWPL-UHFFFAOYSA-N 4',5'-Dihydropsoralen Natural products C1=C2OC(=O)C=CC2=CC2=C1OCC2 VXGRJERITKFWPL-UHFFFAOYSA-N 0.000 description 1
- AOJJSUZBOXZQNB-VTZDEGQISA-N 4'-epidoxorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)[C@@H](O)[C@H](C)O1 AOJJSUZBOXZQNB-VTZDEGQISA-N 0.000 description 1
- CLPFFLWZZBQMAO-UHFFFAOYSA-N 4-(5,6,7,8-tetrahydroimidazo[1,5-a]pyridin-5-yl)benzonitrile Chemical compound C1=CC(C#N)=CC=C1C1N2C=NC=C2CCC1 CLPFFLWZZBQMAO-UHFFFAOYSA-N 0.000 description 1
- AKJHMTWEGVYYSE-AIRMAKDCSA-N 4-HPR Chemical compound C=1C=C(O)C=CC=1NC(=O)/C=C(\C)/C=C/C=C(C)C=CC1=C(C)CCCC1(C)C AKJHMTWEGVYYSE-AIRMAKDCSA-N 0.000 description 1
- XZKIHKMTEMTJQX-UHFFFAOYSA-N 4-Nitrophenyl Phosphate Chemical compound OP(O)(=O)OC1=CC=C([N+]([O-])=O)C=C1 XZKIHKMTEMTJQX-UHFFFAOYSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- DODQJNMQWMSYGS-QPLCGJKRSA-N 4-[(z)-1-[4-[2-(dimethylamino)ethoxy]phenyl]-1-phenylbut-1-en-2-yl]phenol Chemical compound C=1C=C(O)C=CC=1C(/CC)=C(C=1C=CC(OCCN(C)C)=CC=1)/C1=CC=CC=C1 DODQJNMQWMSYGS-QPLCGJKRSA-N 0.000 description 1
- ZMRMMAOBSFSXLN-UHFFFAOYSA-N 4-[4-(2,5-dioxopyrrol-1-yl)phenyl]butanehydrazide Chemical compound C1=CC(CCCC(=O)NN)=CC=C1N1C(=O)C=CC1=O ZMRMMAOBSFSXLN-UHFFFAOYSA-N 0.000 description 1
- TVZGACDUOSZQKY-LBPRGKRZSA-N 4-aminofolic acid Chemical compound C1=NC2=NC(N)=NC(N)=C2N=C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 TVZGACDUOSZQKY-LBPRGKRZSA-N 0.000 description 1
- 101710169336 5'-deoxyadenosine deaminase Proteins 0.000 description 1
- IDPUKCWIGUEADI-UHFFFAOYSA-N 5-[bis(2-chloroethyl)amino]uracil Chemical compound ClCCN(CCCl)C1=CNC(=O)NC1=O IDPUKCWIGUEADI-UHFFFAOYSA-N 0.000 description 1
- NMUSYJAQQFHJEW-KVTDHHQDSA-N 5-azacytidine Chemical compound O=C1N=C(N)N=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 NMUSYJAQQFHJEW-KVTDHHQDSA-N 0.000 description 1
- 229940117976 5-hydroxylysine Drugs 0.000 description 1
- WYXSYVWAUAUWLD-SHUUEZRQSA-N 6-azauridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=N1 WYXSYVWAUAUWLD-SHUUEZRQSA-N 0.000 description 1
- 229960005538 6-diazo-5-oxo-L-norleucine Drugs 0.000 description 1
- YCWQAMGASJSUIP-YFKPBYRVSA-N 6-diazo-5-oxo-L-norleucine Chemical compound OC(=O)[C@@H](N)CCC(=O)C=[N+]=[N-] YCWQAMGASJSUIP-YFKPBYRVSA-N 0.000 description 1
- 102100031126 6-phosphogluconolactonase Human genes 0.000 description 1
- 108010029731 6-phosphogluconolactonase Proteins 0.000 description 1
- CJIJXIFQYOPWTF-UHFFFAOYSA-N 7-hydroxycoumarin Natural products O1C(=O)C=CC2=CC(O)=CC=C21 CJIJXIFQYOPWTF-UHFFFAOYSA-N 0.000 description 1
- ZGXJTSGNIOSYLO-UHFFFAOYSA-N 88755TAZ87 Chemical compound NCC(=O)CCC(O)=O ZGXJTSGNIOSYLO-UHFFFAOYSA-N 0.000 description 1
- FUXVKZWTXQUGMW-FQEVSTJZSA-N 9-Aminocamptothecin Chemical compound C1=CC(N)=C2C=C(CN3C4=CC5=C(C3=O)COC(=O)[C@]5(O)CC)C4=NC2=C1 FUXVKZWTXQUGMW-FQEVSTJZSA-N 0.000 description 1
- HDZZVAMISRMYHH-UHFFFAOYSA-N 9beta-Ribofuranosyl-7-deazaadenin Natural products C1=CC=2C(N)=NC=NC=2N1C1OC(CO)C(O)C1O HDZZVAMISRMYHH-UHFFFAOYSA-N 0.000 description 1
- 108091007505 ADAM17 Proteins 0.000 description 1
- 208000002008 AIDS-Related Lymphoma Diseases 0.000 description 1
- 102000007469 Actins Human genes 0.000 description 1
- 108010085238 Actins Proteins 0.000 description 1
- 208000030090 Acute Disease Diseases 0.000 description 1
- 208000024893 Acute lymphoblastic leukemia Diseases 0.000 description 1
- 208000014697 Acute lymphocytic leukaemia Diseases 0.000 description 1
- 208000031261 Acute myeloid leukaemia Diseases 0.000 description 1
- 102100036664 Adenosine deaminase Human genes 0.000 description 1
- 241000256118 Aedes aegypti Species 0.000 description 1
- 241000256173 Aedes albopictus Species 0.000 description 1
- 108010012934 Albumin-Bound Paclitaxel Proteins 0.000 description 1
- 101710187573 Alcohol dehydrogenase 2 Proteins 0.000 description 1
- 101710133776 Alcohol dehydrogenase class-3 Proteins 0.000 description 1
- 102100021266 Alpha-(1,6)-fucosyltransferase Human genes 0.000 description 1
- 102100023635 Alpha-fetoprotein Human genes 0.000 description 1
- CEIZFXOZIQNICU-UHFFFAOYSA-N Alternaria alternata Crofton-weed toxin Natural products CCC(C)C1NC(=O)C(C(C)=O)=C1O CEIZFXOZIQNICU-UHFFFAOYSA-N 0.000 description 1
- QGZKDVFQNNGYKY-OUBTZVSYSA-N Ammonia-15N Chemical compound [15NH3] QGZKDVFQNNGYKY-OUBTZVSYSA-N 0.000 description 1
- 108010049777 Ankyrins Proteins 0.000 description 1
- 102000008102 Ankyrins Human genes 0.000 description 1
- 108090000672 Annexin A5 Proteins 0.000 description 1
- 102000004121 Annexin A5 Human genes 0.000 description 1
- 108700031308 Antennapedia Homeodomain Proteins 0.000 description 1
- 108020000948 Antisense Oligonucleotides Proteins 0.000 description 1
- 102100029470 Apolipoprotein E Human genes 0.000 description 1
- 101710095339 Apolipoprotein E Proteins 0.000 description 1
- 241000209524 Araceae Species 0.000 description 1
- 208000002109 Argyria Diseases 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 240000006439 Aspergillus oryzae Species 0.000 description 1
- 235000002247 Aspergillus oryzae Nutrition 0.000 description 1
- 101710192393 Attachment protein G3P Proteins 0.000 description 1
- 241001203868 Autographa californica Species 0.000 description 1
- 241000713842 Avian sarcoma virus Species 0.000 description 1
- 108091008875 B cell receptors Proteins 0.000 description 1
- 208000036170 B-Cell Marginal Zone Lymphoma Diseases 0.000 description 1
- 208000012526 B-cell neoplasm Diseases 0.000 description 1
- 230000003844 B-cell-activation Effects 0.000 description 1
- 208000025321 B-lymphoblastic leukemia/lymphoma Diseases 0.000 description 1
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 1
- 239000012664 BCL-2-inhibitor Substances 0.000 description 1
- 244000063299 Bacillus subtilis Species 0.000 description 1
- 235000014469 Bacillus subtilis Nutrition 0.000 description 1
- 108090000363 Bacterial Luciferases Proteins 0.000 description 1
- 231100000699 Bacterial toxin Toxicity 0.000 description 1
- 229940123711 Bcl2 inhibitor Drugs 0.000 description 1
- VGGGPCQERPFHOB-MCIONIFRSA-N Bestatin Chemical compound CC(C)C[C@H](C(O)=O)NC(=O)[C@@H](O)[C@H](N)CC1=CC=CC=C1 VGGGPCQERPFHOB-MCIONIFRSA-N 0.000 description 1
- 102100026189 Beta-galactosidase Human genes 0.000 description 1
- DIZWSDNSTNAYHK-XGWVBXMLSA-N Betulinic acid Natural products CC(=C)[C@@H]1C[C@H]([C@H]2CC[C@]3(C)[C@H](CC[C@@H]4[C@@]5(C)CC[C@H](O)C(C)(C)[C@@H]5CC[C@@]34C)[C@@H]12)C(=O)O DIZWSDNSTNAYHK-XGWVBXMLSA-N 0.000 description 1
- 229940122361 Bisphosphonate Drugs 0.000 description 1
- 241000409811 Bombyx mori nucleopolyhedrovirus Species 0.000 description 1
- ZOXJGFHDIHLPTG-UHFFFAOYSA-N Boron Chemical compound [B] ZOXJGFHDIHLPTG-UHFFFAOYSA-N 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 101800004538 Bradykinin Proteins 0.000 description 1
- 102400000967 Bradykinin Human genes 0.000 description 1
- 102100026008 Breakpoint cluster region protein Human genes 0.000 description 1
- 208000011691 Burkitt lymphomas Diseases 0.000 description 1
- 108010037003 Buserelin Proteins 0.000 description 1
- COVZYZSDYWQREU-UHFFFAOYSA-N Busulfan Chemical compound CS(=O)(=O)OCCCCOS(C)(=O)=O COVZYZSDYWQREU-UHFFFAOYSA-N 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- 241000222120 Candida <Saccharomycetales> Species 0.000 description 1
- 241000222122 Candida albicans Species 0.000 description 1
- 241000282421 Canidae Species 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- GAGWJHPBXLXJQN-UHFFFAOYSA-N Capecitabine Natural products C1=C(F)C(NC(=O)OCCCCC)=NC(=O)N1C1C(O)C(O)C(C)O1 GAGWJHPBXLXJQN-UHFFFAOYSA-N 0.000 description 1
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 1
- OKTJSMMVPCPJKN-OUBTZVSYSA-N Carbon-13 Chemical compound [13C] OKTJSMMVPCPJKN-OUBTZVSYSA-N 0.000 description 1
- 229920002134 Carboxymethyl cellulose Polymers 0.000 description 1
- 201000009030 Carcinoma Diseases 0.000 description 1
- AOCCBINRVIKJHY-UHFFFAOYSA-N Carmofur Chemical compound CCCCCCNC(=O)N1C=C(F)C(=O)NC1=O AOCCBINRVIKJHY-UHFFFAOYSA-N 0.000 description 1
- 102000005600 Cathepsins Human genes 0.000 description 1
- 108010084457 Cathepsins Proteins 0.000 description 1
- JWBOIMRXGHLCPP-UHFFFAOYSA-N Chloditan Chemical compound C=1C=CC=C(Cl)C=1C(C(Cl)Cl)C1=CC=C(Cl)C=C1 JWBOIMRXGHLCPP-UHFFFAOYSA-N 0.000 description 1
- XCDXSSFOJZZGQC-UHFFFAOYSA-N Chlornaphazine Chemical compound C1=CC=CC2=CC(N(CCCl)CCCl)=CC=C21 XCDXSSFOJZZGQC-UHFFFAOYSA-N 0.000 description 1
- 241000282552 Chlorocebus aethiops Species 0.000 description 1
- MKQWTWSXVILIKJ-LXGUWJNJSA-N Chlorozotocin Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](C=O)NC(=O)N(N=O)CCCl MKQWTWSXVILIKJ-LXGUWJNJSA-N 0.000 description 1
- 208000017667 Chronic Disease Diseases 0.000 description 1
- 108090000746 Chymosin Proteins 0.000 description 1
- 102100032768 Complement receptor type 2 Human genes 0.000 description 1
- 108060003955 Contactin Proteins 0.000 description 1
- 102000018361 Contactin Human genes 0.000 description 1
- 229920000742 Cotton Polymers 0.000 description 1
- 239000004971 Cross linker Substances 0.000 description 1
- 229930188224 Cryptophycin Natural products 0.000 description 1
- 101710112752 Cytotoxin Proteins 0.000 description 1
- IGXWBGJHJZYPQS-SSDOTTSWSA-N D-Luciferin Chemical compound OC(=O)[C@H]1CSC(C=2SC3=CC=C(O)C=C3N=2)=N1 IGXWBGJHJZYPQS-SSDOTTSWSA-N 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- QNAYBMKLOCPYGJ-UWTATZPHSA-N D-alanine Chemical compound C[C@@H](N)C(O)=O QNAYBMKLOCPYGJ-UWTATZPHSA-N 0.000 description 1
- QNAYBMKLOCPYGJ-UHFFFAOYSA-N D-alpha-Ala Natural products CC([NH3+])C([O-])=O QNAYBMKLOCPYGJ-UHFFFAOYSA-N 0.000 description 1
- MNQZXJOMYWMBOU-VKHMYHEASA-N D-glyceraldehyde Chemical compound OC[C@@H](O)C=O MNQZXJOMYWMBOU-VKHMYHEASA-N 0.000 description 1
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 1
- HMFHBZSHGGEWLO-SOOFDHNKSA-N D-ribofuranose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H]1O HMFHBZSHGGEWLO-SOOFDHNKSA-N 0.000 description 1
- 239000012624 DNA alkylating agent Substances 0.000 description 1
- 230000007035 DNA breakage Effects 0.000 description 1
- 239000003155 DNA primer Substances 0.000 description 1
- 229940124087 DNA topoisomerase II inhibitor Drugs 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 102220469628 Decapping and exoribonuclease protein_S28T_mutation Human genes 0.000 description 1
- CYCGRDQQIOGCKX-UHFFFAOYSA-N Dehydro-luciferin Natural products OC(=O)C1=CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 CYCGRDQQIOGCKX-UHFFFAOYSA-N 0.000 description 1
- 101710088194 Dehydrogenase Proteins 0.000 description 1
- 102100026662 Delta and Notch-like epidermal growth factor-related receptor Human genes 0.000 description 1
- XXGMIHXASFDFSM-UHFFFAOYSA-N Delta9-tetrahydrocannabinol Natural products CCCCCc1cc2OC(C)(C)C3CCC(=CC3c2c(O)c1O)C XXGMIHXASFDFSM-UHFFFAOYSA-N 0.000 description 1
- NNJPGOLRFBJNIW-UHFFFAOYSA-N Demecolcine Natural products C1=C(OC)C(=O)C=C2C(NC)CCC3=CC(OC)=C(OC)C(OC)=C3C2=C1 NNJPGOLRFBJNIW-UHFFFAOYSA-N 0.000 description 1
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 1
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 1
- AUGQEEXBDZWUJY-ZLJUKNTDSA-N Diacetoxyscirpenol Chemical compound C([C@]12[C@]3(C)[C@H](OC(C)=O)[C@@H](O)[C@H]1O[C@@H]1C=C(C)CC[C@@]13COC(=O)C)O2 AUGQEEXBDZWUJY-ZLJUKNTDSA-N 0.000 description 1
- AUGQEEXBDZWUJY-UHFFFAOYSA-N Diacetoxyscirpenol Natural products CC(=O)OCC12CCC(C)=CC1OC1C(O)C(OC(C)=O)C2(C)C11CO1 AUGQEEXBDZWUJY-UHFFFAOYSA-N 0.000 description 1
- XEHFSYYAGCUKEN-UHFFFAOYSA-N Dihydroscopoletin Natural products C1CC(=O)OC2=C1C=C(OC)C(O)=C2 XEHFSYYAGCUKEN-UHFFFAOYSA-N 0.000 description 1
- 108090000204 Dipeptidase 1 Proteins 0.000 description 1
- 206010061818 Disease progression Diseases 0.000 description 1
- 102100031111 Disintegrin and metalloproteinase domain-containing protein 17 Human genes 0.000 description 1
- CYQFCXCEBYINGO-DLBZAZTESA-N Dronabinol Natural products C1=C(C)CC[C@H]2C(C)(C)OC3=CC(CCCCC)=CC(O)=C3[C@H]21 CYQFCXCEBYINGO-DLBZAZTESA-N 0.000 description 1
- 241000255601 Drosophila melanogaster Species 0.000 description 1
- 206010059866 Drug resistance Diseases 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 102000001301 EGF receptor Human genes 0.000 description 1
- 108060006698 EGF receptor Proteins 0.000 description 1
- 238000012286 ELISA Assay Methods 0.000 description 1
- 241000257465 Echinoidea Species 0.000 description 1
- AFMYMMXSQGUCBK-UHFFFAOYSA-N Endynamicin A Natural products C1#CC=CC#CC2NC(C=3C(=O)C4=C(O)C=CC(O)=C4C(=O)C=3C(O)=C3)=C3C34OC32C(C)C(C(O)=O)=C(OC)C41 AFMYMMXSQGUCBK-UHFFFAOYSA-N 0.000 description 1
- SAMRUMKYXPVKPA-VFKOLLTISA-N Enocitabine Chemical compound O=C1N=C(NC(=O)CCCCCCCCCCCCCCCCCCCCC)C=CN1[C@H]1[C@@H](O)[C@H](O)[C@@H](CO)O1 SAMRUMKYXPVKPA-VFKOLLTISA-N 0.000 description 1
- 241000588914 Enterobacter Species 0.000 description 1
- 241000588921 Enterobacteriaceae Species 0.000 description 1
- 101710181478 Envelope glycoprotein GP350 Proteins 0.000 description 1
- 102400001368 Epidermal growth factor Human genes 0.000 description 1
- 101800003838 Epidermal growth factor Proteins 0.000 description 1
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 1
- HTIJFSOGRVMCQR-UHFFFAOYSA-N Epirubicin Natural products COc1cccc2C(=O)c3c(O)c4CC(O)(CC(OC5CC(N)C(=O)C(C)O5)c4c(O)c3C(=O)c12)C(=O)CO HTIJFSOGRVMCQR-UHFFFAOYSA-N 0.000 description 1
- OBMLHUPNRURLOK-XGRAFVIBSA-N Epitiostanol Chemical compound C1[C@@H]2S[C@@H]2C[C@]2(C)[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CC[C@H]21 OBMLHUPNRURLOK-XGRAFVIBSA-N 0.000 description 1
- 241000588698 Erwinia Species 0.000 description 1
- 241000588722 Escherichia Species 0.000 description 1
- 241001522878 Escherichia coli B Species 0.000 description 1
- 241001302584 Escherichia coli str. K-12 substr. W3110 Species 0.000 description 1
- 229930189413 Esperamicin Natural products 0.000 description 1
- 108090000371 Esterases Proteins 0.000 description 1
- JOYRKODLDBILNP-UHFFFAOYSA-N Ethyl urethane Chemical compound CCOC(N)=O JOYRKODLDBILNP-UHFFFAOYSA-N 0.000 description 1
- DBVJJBKOTRCVKF-UHFFFAOYSA-N Etidronic acid Chemical compound OP(=O)(O)C(O)(C)P(O)(O)=O DBVJJBKOTRCVKF-UHFFFAOYSA-N 0.000 description 1
- 206010073306 Exposure to radiation Diseases 0.000 description 1
- 241000282324 Felis Species 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- BJGNCJDXODQBOB-UHFFFAOYSA-N Fivefly Luciferin Natural products OC(=O)C1CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 BJGNCJDXODQBOB-UHFFFAOYSA-N 0.000 description 1
- 230000037057 G1 phase arrest Effects 0.000 description 1
- 101710198884 GATA-type zinc finger protein 1 Proteins 0.000 description 1
- 108700012941 GNRH1 Proteins 0.000 description 1
- 229910052688 Gadolinium Inorganic materials 0.000 description 1
- JRZJKWGQFNTSRN-UHFFFAOYSA-N Geldanamycin Natural products C1C(C)CC(OC)C(O)C(C)C=C(C)C(OC(N)=O)C(OC)CCC=C(C)C(=O)NC2=CC(=O)C(OC)=C1C2=O JRZJKWGQFNTSRN-UHFFFAOYSA-N 0.000 description 1
- 108700023863 Gene Components Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 208000034951 Genetic Translocation Diseases 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 102000034615 Glial cell line-derived neurotrophic factor Human genes 0.000 description 1
- 108091010837 Glial cell line-derived neurotrophic factor Proteins 0.000 description 1
- DTHNMHAUYICORS-KTKZVXAJSA-N Glucagon-like peptide 1 Chemical compound C([C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(N)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@H](CCC(N)=O)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@@H](NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC=1N=CNC=1)[C@@H](C)O)[C@@H](C)O)C(C)C)C1=CC=CC=C1 DTHNMHAUYICORS-KTKZVXAJSA-N 0.000 description 1
- 102400000322 Glucagon-like peptide 1 Human genes 0.000 description 1
- 102000030595 Glucokinase Human genes 0.000 description 1
- 108010021582 Glucokinase Proteins 0.000 description 1
- 239000004366 Glucose oxidase Substances 0.000 description 1
- 108010015776 Glucose oxidase Proteins 0.000 description 1
- 102100031132 Glucose-6-phosphate isomerase Human genes 0.000 description 1
- 102000005731 Glucose-6-phosphate isomerase Human genes 0.000 description 1
- 108010018962 Glucosephosphate Dehydrogenase Proteins 0.000 description 1
- 102100041003 Glutamate carboxypeptidase 2 Human genes 0.000 description 1
- 235000010469 Glycine max Nutrition 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 239000000579 Gonadotropin-Releasing Hormone Substances 0.000 description 1
- BLCLNMBMMGCOAS-URPVMXJPSA-N Goserelin Chemical compound C([C@@H](C(=O)N[C@H](COC(C)(C)C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N1[C@@H](CCC1)C(=O)NNC(N)=O)NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H]1NC(=O)CC1)C1=CC=C(O)C=C1 BLCLNMBMMGCOAS-URPVMXJPSA-N 0.000 description 1
- 108010069236 Goserelin Proteins 0.000 description 1
- 241000219146 Gossypium Species 0.000 description 1
- 102100021186 Granulysin Human genes 0.000 description 1
- QXZGBUJJYSLZLT-UHFFFAOYSA-N H-Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg-OH Natural products NC(N)=NCCCC(N)C(=O)N1CCCC1C(=O)N1C(C(=O)NCC(=O)NC(CC=2C=CC=CC=2)C(=O)NC(CO)C(=O)N2C(CCC2)C(=O)NC(CC=2C=CC=CC=2)C(=O)NC(CCCN=C(N)N)C(O)=O)CCC1 QXZGBUJJYSLZLT-UHFFFAOYSA-N 0.000 description 1
- 239000007995 HEPES buffer Substances 0.000 description 1
- 101150092640 HES1 gene Proteins 0.000 description 1
- 208000031886 HIV Infections Diseases 0.000 description 1
- 208000037357 HIV infectious disease Diseases 0.000 description 1
- 229940121710 HMGCoA reductase inhibitor Drugs 0.000 description 1
- 101100082540 Haemophilus influenzae (strain ATCC 51907 / DSM 11121 / KW20 / Rd) pcp gene Proteins 0.000 description 1
- 102220525298 Heat shock 70 kDa protein 6_S93I_mutation Human genes 0.000 description 1
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 1
- 241000700721 Hepatitis B virus Species 0.000 description 1
- 102000005548 Hexokinase Human genes 0.000 description 1
- 108700040460 Hexokinases Proteins 0.000 description 1
- 101000819490 Homo sapiens Alpha-(1,6)-fucosyltransferase Proteins 0.000 description 1
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 1
- 101000941929 Homo sapiens Complement receptor type 2 Proteins 0.000 description 1
- 101001054266 Homo sapiens Delta and Notch-like epidermal growth factor-related receptor Proteins 0.000 description 1
- 101000892862 Homo sapiens Glutamate carboxypeptidase 2 Proteins 0.000 description 1
- 101001040751 Homo sapiens Granulysin Proteins 0.000 description 1
- 101000843556 Homo sapiens Transcription factor HES-1 Proteins 0.000 description 1
- 101000851376 Homo sapiens Tumor necrosis factor receptor superfamily member 8 Proteins 0.000 description 1
- 108091006905 Human Serum Albumin Proteins 0.000 description 1
- 102000008100 Human Serum Albumin Human genes 0.000 description 1
- 241000701109 Human adenovirus 2 Species 0.000 description 1
- 241000700588 Human alphaherpesvirus 1 Species 0.000 description 1
- 241000701024 Human betaherpesvirus 5 Species 0.000 description 1
- 101100273566 Humulus lupulus CCL10 gene Proteins 0.000 description 1
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 1
- VSNHCAURESNICA-UHFFFAOYSA-N Hydroxyurea Chemical compound NC(=O)NO VSNHCAURESNICA-UHFFFAOYSA-N 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 1
- MPBVHIBUJCELCL-UHFFFAOYSA-N Ibandronate Chemical compound CCCCCN(C)CCC(O)(P(O)(O)=O)P(O)(O)=O MPBVHIBUJCELCL-UHFFFAOYSA-N 0.000 description 1
- XDXDZDZNSLXDNA-TZNDIEGXSA-N Idarubicin Chemical compound C1[C@H](N)[C@H](O)[C@H](C)O[C@H]1O[C@@H]1C2=C(O)C(C(=O)C3=CC=CC=C3C3=O)=C3C(O)=C2C[C@@](O)(C(C)=O)C1 XDXDZDZNSLXDNA-TZNDIEGXSA-N 0.000 description 1
- XDXDZDZNSLXDNA-UHFFFAOYSA-N Idarubicin Natural products C1C(N)C(O)C(C)OC1OC1C2=C(O)C(C(=O)C3=CC=CC=C3C3=O)=C3C(O)=C2CC(O)(C(C)=O)C1 XDXDZDZNSLXDNA-UHFFFAOYSA-N 0.000 description 1
- JJKOTMDDZAJTGQ-DQSJHHFOSA-N Idoxifene Chemical compound C=1C=CC=CC=1C(/CC)=C(C=1C=CC(OCCN2CCCC2)=CC=1)/C1=CC=C(I)C=C1 JJKOTMDDZAJTGQ-DQSJHHFOSA-N 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 206010053574 Immunoblastic lymphoma Diseases 0.000 description 1
- 102000009786 Immunoglobulin Constant Regions Human genes 0.000 description 1
- 108010009817 Immunoglobulin Constant Regions Proteins 0.000 description 1
- 108010041012 Integrin alpha4 Proteins 0.000 description 1
- 102000003996 Interferon-beta Human genes 0.000 description 1
- 108090000467 Interferon-beta Proteins 0.000 description 1
- 108700003489 Jagged-2 Proteins 0.000 description 1
- 241000588748 Klebsiella Species 0.000 description 1
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 1
- RHGKLRLOHDJJDR-BYPYZUCNSA-N L-citrulline Chemical compound NC(=O)NCCC[C@H]([NH3+])C([O-])=O RHGKLRLOHDJJDR-BYPYZUCNSA-N 0.000 description 1
- SXTAYKAGBXMACB-DPVSGNNYSA-N L-methionine sulfoximine Chemical compound CS(=N)(=O)CC[C@H](N)C(O)=O SXTAYKAGBXMACB-DPVSGNNYSA-N 0.000 description 1
- LRQKBLKVPFOOQJ-YFKPBYRVSA-N L-norleucine Chemical compound CCCC[C@H]([NH3+])C([O-])=O LRQKBLKVPFOOQJ-YFKPBYRVSA-N 0.000 description 1
- 239000005511 L01XE05 - Sorafenib Substances 0.000 description 1
- 102000007330 LDL Lipoproteins Human genes 0.000 description 1
- 108010007622 LDL Lipoproteins Proteins 0.000 description 1
- 101150108666 LPG3 gene Proteins 0.000 description 1
- 108010023244 Lactoperoxidase Proteins 0.000 description 1
- 102100038609 Lactoperoxidase Human genes 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 229920001491 Lentinan Polymers 0.000 description 1
- 102220498649 Leucine-rich repeat and IQ domain-containing protein 1_I29N_mutation Human genes 0.000 description 1
- GQYIWUVLTXOXAJ-UHFFFAOYSA-N Lomustine Chemical compound ClCCN(N=O)C(=O)NC1CCCCC1 GQYIWUVLTXOXAJ-UHFFFAOYSA-N 0.000 description 1
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- DDWFXDSYGUXRAY-UHFFFAOYSA-N Luciferin Natural products CCc1c(C)c(CC2NC(=O)C(=C2C=C)C)[nH]c1Cc3[nH]c4C(=C5/NC(CC(=O)O)C(C)C5CC(=O)O)CC(=O)c4c3C DDWFXDSYGUXRAY-UHFFFAOYSA-N 0.000 description 1
- 235000007688 Lycopersicon esculentum Nutrition 0.000 description 1
- 206010025312 Lymphoma AIDS related Diseases 0.000 description 1
- 241000282553 Macaca Species 0.000 description 1
- 241000282567 Macaca fascicularis Species 0.000 description 1
- 241000282560 Macaca mulatta Species 0.000 description 1
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 1
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 1
- PWHULOQIROXLJO-UHFFFAOYSA-N Manganese Chemical compound [Mn] PWHULOQIROXLJO-UHFFFAOYSA-N 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 102220490832 Mannosyl-oligosaccharide glucosidase_Y32F_mutation Human genes 0.000 description 1
- 208000025205 Mantle-Cell Lymphoma Diseases 0.000 description 1
- VJRAUFKOOPNFIQ-UHFFFAOYSA-N Marcellomycin Natural products C12=C(O)C=3C(=O)C4=C(O)C=CC(O)=C4C(=O)C=3C=C2C(C(=O)OC)C(CC)(O)CC1OC(OC1C)CC(N(C)C)C1OC(OC1C)CC(O)C1OC1CC(O)C(O)C(C)O1 VJRAUFKOOPNFIQ-UHFFFAOYSA-N 0.000 description 1
- 241001441512 Maytenus serrata Species 0.000 description 1
- 240000004658 Medicago sativa Species 0.000 description 1
- 235000017587 Medicago sativa ssp. sativa Nutrition 0.000 description 1
- 241000219828 Medicago truncatula Species 0.000 description 1
- 108010006035 Metalloproteases Proteins 0.000 description 1
- 102000005741 Metalloproteases Human genes 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 229930192392 Mitomycin Natural products 0.000 description 1
- 241000218984 Momordica Species 0.000 description 1
- 244000302512 Momordica charantia Species 0.000 description 1
- 235000009811 Momordica charantia Nutrition 0.000 description 1
- 241001529936 Murinae Species 0.000 description 1
- 208000033776 Myeloid Acute Leukemia Diseases 0.000 description 1
- FSVCELGFZIQNCK-UHFFFAOYSA-N N,N-bis(2-hydroxyethyl)glycine Chemical compound OCCN(CCO)CC(O)=O FSVCELGFZIQNCK-UHFFFAOYSA-N 0.000 description 1
- OVRNDRQMDRJTHS-CBQIKETKSA-N N-Acetyl-D-Galactosamine Chemical compound CC(=O)N[C@H]1[C@@H](O)O[C@H](CO)[C@H](O)[C@@H]1O OVRNDRQMDRJTHS-CBQIKETKSA-N 0.000 description 1
- OVBPIULPVIDEAO-UHFFFAOYSA-N N-Pteroyl-L-glutaminsaeure Natural products C=1N=C2NC(N)=NC(=O)C2=NC=1CNC1=CC=C(C(=O)NC(CCC(O)=O)C(O)=O)C=C1 OVBPIULPVIDEAO-UHFFFAOYSA-N 0.000 description 1
- GXCLVBGFBYZDAG-UHFFFAOYSA-N N-[2-(1H-indol-3-yl)ethyl]-N-methylprop-2-en-1-amine Chemical compound CN(CCC1=CNC2=C1C=CC=C2)CC=C GXCLVBGFBYZDAG-UHFFFAOYSA-N 0.000 description 1
- MBLBDJOUHNCFQT-UHFFFAOYSA-N N-acetyl-D-galactosamine Natural products CC(=O)NC(C=O)C(O)C(O)C(O)CO MBLBDJOUHNCFQT-UHFFFAOYSA-N 0.000 description 1
- OVRNDRQMDRJTHS-FMDGEEDCSA-N N-acetyl-beta-D-glucosamine Chemical compound CC(=O)N[C@H]1[C@H](O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-FMDGEEDCSA-N 0.000 description 1
- 230000004988 N-glycosylation Effects 0.000 description 1
- 108010072915 NAc-Sar-Gly-Val-(d-allo-Ile)-Thr-Nva-Ile-Arg-ProNEt Proteins 0.000 description 1
- RHGKLRLOHDJJDR-UHFFFAOYSA-N Ndelta-carbamoyl-DL-ornithine Natural products OC(=O)C(N)CCCNC(N)=O RHGKLRLOHDJJDR-UHFFFAOYSA-N 0.000 description 1
- 241000244206 Nematoda Species 0.000 description 1
- 108700037638 Neurogenic locus notch homolog protein 1 Proteins 0.000 description 1
- 244000061176 Nicotiana tabacum Species 0.000 description 1
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 1
- KGTDRFCXGRULNK-UHFFFAOYSA-N Nogalamycin Natural products COC1C(OC)(C)C(OC)C(C)OC1OC1C2=C(O)C(C(=O)C3=C(O)C=C4C5(C)OC(C(C(C5O)N(C)C)O)OC4=C3C3=O)=C3C=C2C(C(=O)OC)C(C)(O)C1 KGTDRFCXGRULNK-UHFFFAOYSA-N 0.000 description 1
- 230000005913 Notch signaling pathway Effects 0.000 description 1
- 101710163270 Nuclease Proteins 0.000 description 1
- 108010038807 Oligopeptides Proteins 0.000 description 1
- 102000015636 Oligopeptides Human genes 0.000 description 1
- 229930187135 Olivomycin Natural products 0.000 description 1
- 102100021079 Ornithine decarboxylase Human genes 0.000 description 1
- 108700005126 Ornithine decarboxylases Proteins 0.000 description 1
- 101100081884 Oryza sativa subsp. japonica OSA15 gene Proteins 0.000 description 1
- 239000007977 PBT buffer Substances 0.000 description 1
- 238000012408 PCR amplification Methods 0.000 description 1
- 101150082245 PSAG gene Proteins 0.000 description 1
- 241000609499 Palicourea Species 0.000 description 1
- 241000282577 Pan troglodytes Species 0.000 description 1
- 108010067372 Pancreatic elastase Proteins 0.000 description 1
- 102000016387 Pancreatic elastase Human genes 0.000 description 1
- 206010033661 Pancytopenia Diseases 0.000 description 1
- 241000282520 Papio Species 0.000 description 1
- 229930040373 Paraformaldehyde Natural products 0.000 description 1
- 208000018737 Parkinson disease Diseases 0.000 description 1
- 229930182555 Penicillin Natural products 0.000 description 1
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 1
- 108010087702 Penicillinase Proteins 0.000 description 1
- 235000002233 Penicillium roqueforti Nutrition 0.000 description 1
- 108010057150 Peplomycin Proteins 0.000 description 1
- 108090000284 Pepsin A Proteins 0.000 description 1
- 102000057297 Pepsin A Human genes 0.000 description 1
- 102000007079 Peptide Fragments Human genes 0.000 description 1
- 108010033276 Peptide Fragments Proteins 0.000 description 1
- 240000007377 Petunia x hybrida Species 0.000 description 1
- 102000001105 Phosphofructokinases Human genes 0.000 description 1
- 108010069341 Phosphofructokinases Proteins 0.000 description 1
- 102000012288 Phosphopyruvate Hydratase Human genes 0.000 description 1
- 108010022181 Phosphopyruvate Hydratase Proteins 0.000 description 1
- ABLZXFCXXLZCGV-UHFFFAOYSA-N Phosphorous acid Chemical group OP(O)=O ABLZXFCXXLZCGV-UHFFFAOYSA-N 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- KMSKQZKKOZQFFG-HSUXVGOQSA-N Pirarubicin Chemical compound O([C@H]1[C@@H](N)C[C@@H](O[C@H]1C)O[C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1CCCCO1 KMSKQZKKOZQFFG-HSUXVGOQSA-N 0.000 description 1
- 231100000742 Plant toxin Toxicity 0.000 description 1
- 208000007452 Plasmacytoma Diseases 0.000 description 1
- 241000276498 Pollachius virens Species 0.000 description 1
- 229920000805 Polyaspartic acid Polymers 0.000 description 1
- 239000004698 Polyethylene Substances 0.000 description 1
- 241001505332 Polyomavirus sp. Species 0.000 description 1
- 102000004257 Potassium Channel Human genes 0.000 description 1
- 208000006664 Precursor Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 description 1
- HFVNWDWLWUCIHC-GUPDPFMOSA-N Prednimustine Chemical compound O=C([C@@]1(O)CC[C@H]2[C@H]3[C@@H]([C@]4(C=CC(=O)C=C4CC3)C)[C@@H](O)C[C@@]21C)COC(=O)CCCC1=CC=C(N(CCCl)CCCl)C=C1 HFVNWDWLWUCIHC-GUPDPFMOSA-N 0.000 description 1
- 108010050254 Presenilins Proteins 0.000 description 1
- 102000015499 Presenilins Human genes 0.000 description 1
- 108010009341 Protein Serine-Threonine Kinases Proteins 0.000 description 1
- 102000009516 Protein Serine-Threonine Kinases Human genes 0.000 description 1
- 108700037966 Protein jagged-1 Proteins 0.000 description 1
- 101710170213 Protein jagged-2 Proteins 0.000 description 1
- 102100024924 Protein kinase C alpha type Human genes 0.000 description 1
- 101710109947 Protein kinase C alpha type Proteins 0.000 description 1
- 241000588769 Proteus <enterobacteria> Species 0.000 description 1
- 241000589516 Pseudomonas Species 0.000 description 1
- 101100084022 Pseudomonas aeruginosa (strain ATCC 15692 / DSM 22644 / CIP 104116 / JCM 14847 / LMG 12228 / 1C / PRS 101 / PAO1) lapA gene Proteins 0.000 description 1
- 108010011939 Pyruvate Decarboxylase Proteins 0.000 description 1
- 102000013009 Pyruvate Kinase Human genes 0.000 description 1
- 108020005115 Pyruvate Kinase Proteins 0.000 description 1
- 102000004879 Racemases and epimerases Human genes 0.000 description 1
- 108090001066 Racemases and epimerases Proteins 0.000 description 1
- AHHFEZNOXOZZQA-ZEBDFXRSSA-N Ranimustine Chemical compound CO[C@H]1O[C@H](CNC(=O)N(CCCl)N=O)[C@@H](O)[C@H](O)[C@H]1O AHHFEZNOXOZZQA-ZEBDFXRSSA-N 0.000 description 1
- 241000700157 Rattus norvegicus Species 0.000 description 1
- 101100016884 Rattus norvegicus Hes1 gene Proteins 0.000 description 1
- 208000015634 Rectal Neoplasms Diseases 0.000 description 1
- 241000242739 Renilla Species 0.000 description 1
- 206010057190 Respiratory tract infections Diseases 0.000 description 1
- 108091027981 Response element Proteins 0.000 description 1
- 108010083644 Ribonucleases Proteins 0.000 description 1
- 102000006382 Ribonucleases Human genes 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- PYMYPHUHKUWMLA-LMVFSUKVSA-N Ribose Natural products OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PYMYPHUHKUWMLA-LMVFSUKVSA-N 0.000 description 1
- 241000714474 Rous sarcoma virus Species 0.000 description 1
- 241000235070 Saccharomyces Species 0.000 description 1
- 241000607142 Salmonella Species 0.000 description 1
- 241000293869 Salmonella enterica subsp. enterica serovar Typhimurium Species 0.000 description 1
- 241000277331 Salmonidae Species 0.000 description 1
- 108010084592 Saporins Proteins 0.000 description 1
- 229920002305 Schizophyllan Polymers 0.000 description 1
- 241000235347 Schizosaccharomyces pombe Species 0.000 description 1
- 241000311088 Schwanniomyces Species 0.000 description 1
- 241001123650 Schwanniomyces occidentalis Species 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- 102000011842 Serrate-Jagged Proteins Human genes 0.000 description 1
- 108010036039 Serrate-Jagged Proteins Proteins 0.000 description 1
- 241000607720 Serratia Species 0.000 description 1
- 241000607715 Serratia marcescens Species 0.000 description 1
- 241000607768 Shigella Species 0.000 description 1
- 241000700584 Simplexvirus Species 0.000 description 1
- 240000003768 Solanum lycopersicum Species 0.000 description 1
- 244000061456 Solanum tuberosum Species 0.000 description 1
- 235000002595 Solanum tuberosum Nutrition 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- BXFOFFBJRFZBQZ-QYWOHJEZSA-N T-2 toxin Chemical compound C([C@@]12[C@]3(C)[C@H](OC(C)=O)[C@@H](O)[C@H]1O[C@H]1[C@]3(COC(C)=O)C[C@@H](C(=C1)C)OC(=O)CC(C)C)O2 BXFOFFBJRFZBQZ-QYWOHJEZSA-N 0.000 description 1
- 241001116500 Taxus Species 0.000 description 1
- 241001116498 Taxus baccata Species 0.000 description 1
- WFWLQNSHRPWKFK-UHFFFAOYSA-N Tegafur Chemical compound O=C1NC(=O)C(F)=CN1C1OCCC1 WFWLQNSHRPWKFK-UHFFFAOYSA-N 0.000 description 1
- CBPNZQVSJQDFBE-FUXHJELOSA-N Temsirolimus Chemical compound C1C[C@@H](OC(=O)C(C)(CO)CO)[C@H](OC)C[C@@H]1C[C@@H](C)[C@H]1OC(=O)[C@@H]2CCCCN2C(=O)C(=O)[C@](O)(O2)[C@H](C)CC[C@H]2C[C@H](OC)/C(C)=C/C=C/C=C/[C@@H](C)C[C@@H](C)C(=O)[C@H](OC)[C@H](O)/C(C)=C/[C@@H](C)C(=O)C1 CBPNZQVSJQDFBE-FUXHJELOSA-N 0.000 description 1
- CGMTUJFWROPELF-UHFFFAOYSA-N Tenuazonic acid Natural products CCC(C)C1NC(=O)C(=C(C)/O)C1=O CGMTUJFWROPELF-UHFFFAOYSA-N 0.000 description 1
- 239000004098 Tetracycline Substances 0.000 description 1
- 238000012338 Therapeutic targeting Methods 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-N Thiophosphoric acid Chemical class OP(O)(S)=O RYYWUUFWQRZTIU-UHFFFAOYSA-N 0.000 description 1
- 108091036066 Three prime untranslated region Proteins 0.000 description 1
- 101710183280 Topoisomerase Proteins 0.000 description 1
- 239000000365 Topoisomerase I Inhibitor Substances 0.000 description 1
- 239000000317 Topoisomerase II Inhibitor Substances 0.000 description 1
- IWEQQRMGNVVKQW-OQKDUQJOSA-N Toremifene citrate Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O.C1=CC(OCCN(C)C)=CC=C1C(\C=1C=CC=CC=1)=C(\CCCl)C1=CC=CC=C1 IWEQQRMGNVVKQW-OQKDUQJOSA-N 0.000 description 1
- 101710120037 Toxin CcdB Proteins 0.000 description 1
- 108700009124 Transcription Initiation Site Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102100030798 Transcription factor HES-1 Human genes 0.000 description 1
- 102000007238 Transferrin Receptors Human genes 0.000 description 1
- 108010033576 Transferrin Receptors Proteins 0.000 description 1
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 description 1
- 241000223259 Trichoderma Species 0.000 description 1
- UMILHIMHKXVDGH-UHFFFAOYSA-N Triethylene glycol diglycidyl ether Chemical compound C1OC1COCCOCCOCCOCC1CO1 UMILHIMHKXVDGH-UHFFFAOYSA-N 0.000 description 1
- FYAMXEPQQLNQDM-UHFFFAOYSA-N Tris(1-aziridinyl)phosphine oxide Chemical compound C1CN1P(N1CC1)(=O)N1CC1 FYAMXEPQQLNQDM-UHFFFAOYSA-N 0.000 description 1
- 102100036857 Tumor necrosis factor receptor superfamily member 8 Human genes 0.000 description 1
- 108010092464 Urate Oxidase Proteins 0.000 description 1
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Natural products NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 1
- 201000005969 Uveal melanoma Diseases 0.000 description 1
- 206010046865 Vaccinia virus infection Diseases 0.000 description 1
- 244000000188 Vaccinium ovalifolium Species 0.000 description 1
- 108020005202 Viral DNA Proteins 0.000 description 1
- 241000235013 Yarrowia Species 0.000 description 1
- 240000008042 Zea mays Species 0.000 description 1
- 235000005824 Zea mays ssp. parviglumis Nutrition 0.000 description 1
- 235000002017 Zea mays subsp mays Nutrition 0.000 description 1
- ZYVSOIYQKUDENJ-ASUJBHBQSA-N [(2R,3R,4R,6R)-6-[[(6S,7S)-6-[(2S,4R,5R,6R)-4-[(2R,4R,5R,6R)-4-[(2S,4S,5S,6S)-5-acetyloxy-4-hydroxy-4,6-dimethyloxan-2-yl]oxy-5-hydroxy-6-methyloxan-2-yl]oxy-5-hydroxy-6-methyloxan-2-yl]oxy-7-[(3S,4R)-3,4-dihydroxy-1-methoxy-2-oxopentyl]-4,10-dihydroxy-3-methyl-5-oxo-7,8-dihydro-6H-anthracen-2-yl]oxy]-4-[(2R,4R,5R,6R)-4-hydroxy-5-methoxy-6-methyloxan-2-yl]oxy-2-methyloxan-3-yl] acetate Chemical class COC([C@@H]1Cc2cc3cc(O[C@@H]4C[C@@H](O[C@@H]5C[C@@H](O)[C@@H](OC)[C@@H](C)O5)[C@H](OC(C)=O)[C@@H](C)O4)c(C)c(O)c3c(O)c2C(=O)[C@H]1O[C@H]1C[C@@H](O[C@@H]2C[C@@H](O[C@H]3C[C@](C)(O)[C@@H](OC(C)=O)[C@H](C)O3)[C@H](O)[C@@H](C)O2)[C@H](O)[C@@H](C)O1)C(=O)[C@@H](O)[C@@H](C)O ZYVSOIYQKUDENJ-ASUJBHBQSA-N 0.000 description 1
- SPJCRMJCFSJKDE-ZWBUGVOYSA-N [(3s,8s,9s,10r,13r,14s,17r)-10,13-dimethyl-17-[(2r)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1h-cyclopenta[a]phenanthren-3-yl] 2-[4-[bis(2-chloroethyl)amino]phenyl]acetate Chemical compound O([C@@H]1CC2=CC[C@H]3[C@@H]4CC[C@@H]([C@]4(CC[C@@H]3[C@@]2(C)CC1)C)[C@H](C)CCCC(C)C)C(=O)CC1=CC=C(N(CCCl)CCCl)C=C1 SPJCRMJCFSJKDE-ZWBUGVOYSA-N 0.000 description 1
- HIHOWBSBBDRPDW-PTHRTHQKSA-N [(3s,8s,9s,10r,13r,14s,17r)-10,13-dimethyl-17-[(2r)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1h-cyclopenta[a]phenanthren-3-yl] n-[2-(dimethylamino)ethyl]carbamate Chemical compound C1C=C2C[C@@H](OC(=O)NCCN(C)C)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HIHOWBSBBDRPDW-PTHRTHQKSA-N 0.000 description 1
- IFJUINDAXYAPTO-UUBSBJJBSA-N [(8r,9s,13s,14s,17s)-17-[2-[4-[4-[bis(2-chloroethyl)amino]phenyl]butanoyloxy]acetyl]oxy-13-methyl-6,7,8,9,11,12,14,15,16,17-decahydrocyclopenta[a]phenanthren-3-yl] benzoate Chemical compound C([C@@H]1[C@@H](C2=CC=3)CC[C@]4([C@H]1CC[C@@H]4OC(=O)COC(=O)CCCC=1C=CC(=CC=1)N(CCCl)CCCl)C)CC2=CC=3OC(=O)C1=CC=CC=C1 IFJUINDAXYAPTO-UUBSBJJBSA-N 0.000 description 1
- IHGLINDYFMDHJG-UHFFFAOYSA-N [2-(4-methoxyphenyl)-3,4-dihydronaphthalen-1-yl]-[4-(2-pyrrolidin-1-ylethoxy)phenyl]methanone Chemical compound C1=CC(OC)=CC=C1C(CCC1=CC=CC=C11)=C1C(=O)C(C=C1)=CC=C1OCCN1CCCC1 IHGLINDYFMDHJG-UHFFFAOYSA-N 0.000 description 1
- 108010023617 abarelix Proteins 0.000 description 1
- 229960002184 abarelix Drugs 0.000 description 1
- 229940028652 abraxane Drugs 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 229930183665 actinomycin Natural products 0.000 description 1
- RJURFGZVJUQBHK-IIXSONLDSA-N actinomycin D Chemical compound C[C@H]1OC(=O)[C@H](C(C)C)N(C)C(=O)CN(C)C(=O)[C@@H]2CCCN2C(=O)[C@@H](C(C)C)NC(=O)[C@H]1NC(=O)C1=C(N)C(=O)C(C)=C2OC(C(C)=CC=C3C(=O)N[C@@H]4C(=O)N[C@@H](C(N5CCC[C@H]5C(=O)N(C)CC(=O)N(C)[C@@H](C(C)C)C(=O)O[C@@H]4C)=O)C(C)C)=C3N=C21 RJURFGZVJUQBHK-IIXSONLDSA-N 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 229940037127 actonel Drugs 0.000 description 1
- 125000002015 acyclic group Chemical group 0.000 description 1
- 230000002730 additional effect Effects 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 239000003463 adsorbent Substances 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 230000002776 aggregation Effects 0.000 description 1
- 238000004220 aggregation Methods 0.000 description 1
- 238000013019 agitation Methods 0.000 description 1
- 229940062527 alendronate Drugs 0.000 description 1
- 239000003513 alkali Substances 0.000 description 1
- 125000003342 alkenyl group Chemical group 0.000 description 1
- 229940045714 alkyl sulfonate alkylating agent Drugs 0.000 description 1
- 150000008052 alkyl sulfonates Chemical class 0.000 description 1
- 208000026935 allergic disease Diseases 0.000 description 1
- 230000007815 allergy Effects 0.000 description 1
- HMFHBZSHGGEWLO-UHFFFAOYSA-N alpha-D-Furanose-Ribose Natural products OCC1OC(O)C(O)C1O HMFHBZSHGGEWLO-UHFFFAOYSA-N 0.000 description 1
- 108010026331 alpha-Fetoproteins Proteins 0.000 description 1
- 229960000473 altretamine Drugs 0.000 description 1
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 1
- 229960003437 aminoglutethimide Drugs 0.000 description 1
- ROBVIMPUHSLWNV-UHFFFAOYSA-N aminoglutethimide Chemical compound C=1C=C(N)C=CC=1C1(CC)CCC(=O)NC1=O ROBVIMPUHSLWNV-UHFFFAOYSA-N 0.000 description 1
- 229940126574 aminoglycoside antibiotic Drugs 0.000 description 1
- 239000002647 aminoglycoside antibiotic agent Substances 0.000 description 1
- 229960002749 aminolevulinic acid Drugs 0.000 description 1
- 229960003896 aminopterin Drugs 0.000 description 1
- UMGDCJDMYOKAJW-UHFFFAOYSA-N aminothiocarboxamide Natural products NC(N)=S UMGDCJDMYOKAJW-UHFFFAOYSA-N 0.000 description 1
- 238000003453 ammonium sulfate precipitation method Methods 0.000 description 1
- APKFDSVGJQXUKY-INPOYWNPSA-N amphotericin B Chemical compound O[C@H]1[C@@H](N)[C@H](O)[C@@H](C)O[C@H]1O[C@H]1/C=C/C=C/C=C/C=C/C=C/C=C/C=C/[C@H](C)[C@@H](O)[C@@H](C)[C@H](C)OC(=O)C[C@H](O)C[C@H](O)CC[C@@H](O)[C@H](O)C[C@H](O)C[C@](O)(C[C@H](O)[C@H]2C(O)=O)O[C@H]2C1 APKFDSVGJQXUKY-INPOYWNPSA-N 0.000 description 1
- 229960000723 ampicillin Drugs 0.000 description 1
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 1
- 229960001220 amsacrine Drugs 0.000 description 1
- XCPGHVQEEXUHNC-UHFFFAOYSA-N amsacrine Chemical compound COC1=CC(NS(C)(=O)=O)=CC=C1NC1=C(C=CC=C2)C2=NC2=CC=CC=C12 XCPGHVQEEXUHNC-UHFFFAOYSA-N 0.000 description 1
- 229960002932 anastrozole Drugs 0.000 description 1
- BBDAGFIXKZCXAH-CCXZUQQUSA-N ancitabine Chemical compound N=C1C=CN2[C@@H]3O[C@H](CO)[C@@H](O)[C@@H]3OC2=N1 BBDAGFIXKZCXAH-CCXZUQQUSA-N 0.000 description 1
- 229950000242 ancitabine Drugs 0.000 description 1
- 239000003098 androgen Substances 0.000 description 1
- 229940030486 androgens Drugs 0.000 description 1
- 230000002491 angiogenic effect Effects 0.000 description 1
- 239000003957 anion exchange resin Substances 0.000 description 1
- 150000001450 anions Chemical class 0.000 description 1
- 230000008485 antagonism Effects 0.000 description 1
- 230000002280 anti-androgenic effect Effects 0.000 description 1
- 230000003432 anti-folate effect Effects 0.000 description 1
- 230000000843 anti-fungal effect Effects 0.000 description 1
- 230000001130 anti-lysozyme effect Effects 0.000 description 1
- 230000000340 anti-metabolite Effects 0.000 description 1
- 230000002421 anti-septic effect Effects 0.000 description 1
- 239000000051 antiandrogen Substances 0.000 description 1
- 229940030495 antiandrogen sex hormone and modulator of the genital system Drugs 0.000 description 1
- 210000000628 antibody-producing cell Anatomy 0.000 description 1
- 229940127074 antifolate Drugs 0.000 description 1
- 238000010913 antigen-directed enzyme pro-drug therapy Methods 0.000 description 1
- 230000007503 antigenic stimulation Effects 0.000 description 1
- 239000013059 antihormonal agent Substances 0.000 description 1
- 229940100197 antimetabolite Drugs 0.000 description 1
- 239000002256 antimetabolite Substances 0.000 description 1
- 229940045686 antimetabolites antineoplastic purine analogs Drugs 0.000 description 1
- 229940045687 antimetabolites folic acid analogs Drugs 0.000 description 1
- 229940045713 antineoplastic alkylating drug ethylene imines Drugs 0.000 description 1
- 229940045688 antineoplastic antimetabolites pyrimidine analogues Drugs 0.000 description 1
- 239000003418 antiprogestin Substances 0.000 description 1
- 239000000074 antisense oligonucleotide Substances 0.000 description 1
- 238000012230 antisense oligonucleotides Methods 0.000 description 1
- 230000001640 apoptogenic effect Effects 0.000 description 1
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 1
- 150000008209 arabinosides Chemical class 0.000 description 1
- 238000000149 argon plasma sintering Methods 0.000 description 1
- 229940078010 arimidex Drugs 0.000 description 1
- 229940087620 aromasin Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 125000000613 asparagine group Chemical group N[C@@H](CC(N)=O)C(=O)* 0.000 description 1
- 208000006673 asthma Diseases 0.000 description 1
- 230000002238 attenuated effect Effects 0.000 description 1
- 229960002756 azacitidine Drugs 0.000 description 1
- 229950011321 azaserine Drugs 0.000 description 1
- 150000001541 aziridines Chemical class 0.000 description 1
- 239000000688 bacterial toxin Substances 0.000 description 1
- 238000013320 baculovirus expression vector system Methods 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- WPYMKLBDIGXBTP-UHFFFAOYSA-N benzoic acid Chemical compound OC(=O)C1=CC=CC=C1 WPYMKLBDIGXBTP-UHFFFAOYSA-N 0.000 description 1
- 108010051210 beta-Fructofuranosidase Proteins 0.000 description 1
- 108010005774 beta-Galactosidase Proteins 0.000 description 1
- DRTQHJPVMGBUCF-PSQAKQOGSA-N beta-L-uridine Natural products O[C@H]1[C@@H](O)[C@H](CO)O[C@@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-PSQAKQOGSA-N 0.000 description 1
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 1
- 102000006635 beta-lactamase Human genes 0.000 description 1
- GUBGYTABKSRVRQ-QUYVBRFLSA-N beta-maltose Chemical compound OC[C@H]1O[C@H](O[C@H]2[C@H](O)[C@@H](O)[C@H](O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@@H]1O GUBGYTABKSRVRQ-QUYVBRFLSA-N 0.000 description 1
- QGJZLNKBHJESQX-FZFNOLFKSA-N betulinic acid Chemical compound C1C[C@H](O)C(C)(C)[C@@H]2CC[C@@]3(C)[C@]4(C)CC[C@@]5(C(O)=O)CC[C@@H](C(=C)C)[C@@H]5[C@H]4CC[C@@H]3[C@]21C QGJZLNKBHJESQX-FZFNOLFKSA-N 0.000 description 1
- 229960002938 bexarotene Drugs 0.000 description 1
- 229960000997 bicalutamide Drugs 0.000 description 1
- 239000007998 bicine buffer Substances 0.000 description 1
- 238000013357 binding ELISA Methods 0.000 description 1
- 238000002306 biochemical method Methods 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 229960000074 biopharmaceutical Drugs 0.000 description 1
- 238000005460 biophysical method Methods 0.000 description 1
- 229950008548 bisantrene Drugs 0.000 description 1
- 150000004663 bisphosphonates Chemical class 0.000 description 1
- 230000037396 body weight Effects 0.000 description 1
- 229960001467 bortezomib Drugs 0.000 description 1
- 108010006025 bovine growth hormone Proteins 0.000 description 1
- 239000012888 bovine serum Substances 0.000 description 1
- 229940098773 bovine serum albumin Drugs 0.000 description 1
- QXZGBUJJYSLZLT-FDISYFBBSA-N bradykinin Chemical compound NC(=N)NCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(=O)NCC(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@@H](CO)C(=O)N2[C@@H](CCC2)C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)CCC1 QXZGBUJJYSLZLT-FDISYFBBSA-N 0.000 description 1
- 210000000481 breast Anatomy 0.000 description 1
- 229960005520 bryostatin Drugs 0.000 description 1
- MJQUEDHRCUIRLF-TVIXENOKSA-N bryostatin 1 Chemical compound C([C@@H]1CC(/[C@@H]([C@@](C(C)(C)/C=C/2)(O)O1)OC(=O)/C=C/C=C/CCC)=C\C(=O)OC)[C@H]([C@@H](C)O)OC(=O)C[C@H](O)C[C@@H](O1)C[C@H](OC(C)=O)C(C)(C)[C@]1(O)C[C@@H]1C\C(=C\C(=O)OC)C[C@H]\2O1 MJQUEDHRCUIRLF-TVIXENOKSA-N 0.000 description 1
- MUIWQCKLQMOUAT-AKUNNTHJSA-N bryostatin 20 Natural products COC(=O)C=C1C[C@@]2(C)C[C@]3(O)O[C@](C)(C[C@@H](O)CC(=O)O[C@](C)(C[C@@]4(C)O[C@](O)(CC5=CC(=O)O[C@]45C)C(C)(C)C=C[C@@](C)(C1)O2)[C@@H](C)O)C[C@H](OC(=O)C(C)(C)C)C3(C)C MUIWQCKLQMOUAT-AKUNNTHJSA-N 0.000 description 1
- CUWODFFVMXJOKD-UVLQAERKSA-N buserelin Chemical compound CCNC(=O)[C@@H]1CCCN1C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](COC(C)(C)C)NC(=O)[C@@H](NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H]1NC(=O)CC1)CC1=CC=C(O)C=C1 CUWODFFVMXJOKD-UVLQAERKSA-N 0.000 description 1
- 229960002719 buserelin Drugs 0.000 description 1
- 229960002092 busulfan Drugs 0.000 description 1
- ZTQSAGDEMFDKMZ-UHFFFAOYSA-N butyric aldehyde Natural products CCCC=O ZTQSAGDEMFDKMZ-UHFFFAOYSA-N 0.000 description 1
- 102220401484 c.164C>A Human genes 0.000 description 1
- 108700002839 cactinomycin Proteins 0.000 description 1
- 229950009908 cactinomycin Drugs 0.000 description 1
- 239000011575 calcium Substances 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- IVFYLRMMHVYGJH-PVPPCFLZSA-N calusterone Chemical compound C1C[C@]2(C)[C@](O)(C)CC[C@H]2[C@@H]2[C@@H](C)CC3=CC(=O)CC[C@]3(C)[C@H]21 IVFYLRMMHVYGJH-PVPPCFLZSA-N 0.000 description 1
- 229950009823 calusterone Drugs 0.000 description 1
- 229940088954 camptosar Drugs 0.000 description 1
- 229950007296 cantuzumab mertansine Drugs 0.000 description 1
- 229960004117 capecitabine Drugs 0.000 description 1
- 150000004657 carbamic acid derivatives Chemical class 0.000 description 1
- OWIUPIRUAQMTTK-UHFFFAOYSA-N carbazic acid Chemical class NNC(O)=O OWIUPIRUAQMTTK-UHFFFAOYSA-N 0.000 description 1
- 229960003669 carbenicillin Drugs 0.000 description 1
- FPPNZSSZRUTDAP-UWFZAAFLSA-N carbenicillin Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)C(C(O)=O)C1=CC=CC=C1 FPPNZSSZRUTDAP-UWFZAAFLSA-N 0.000 description 1
- 125000004432 carbon atom Chemical group C* 0.000 description 1
- 229910002091 carbon monoxide Inorganic materials 0.000 description 1
- 239000002041 carbon nanotube Substances 0.000 description 1
- 229910021393 carbon nanotube Inorganic materials 0.000 description 1
- 125000002915 carbonyl group Chemical group [*:2]C([*:1])=O 0.000 description 1
- YAYRGNWWLMLWJE-UHFFFAOYSA-L carboplatin Chemical compound O=C1O[Pt](N)(N)OC(=O)C11CCC1 YAYRGNWWLMLWJE-UHFFFAOYSA-L 0.000 description 1
- 229960004562 carboplatin Drugs 0.000 description 1
- 239000001768 carboxy methyl cellulose Substances 0.000 description 1
- 235000010948 carboxy methyl cellulose Nutrition 0.000 description 1
- 239000008112 carboxymethyl-cellulose Substances 0.000 description 1
- XREUEWVEMYWFFA-CSKJXFQVSA-N carminomycin Chemical compound C1[C@H](N)[C@H](O)[C@H](C)O[C@H]1O[C@@H]1C2=C(O)C(C(=O)C3=C(O)C=CC=C3C3=O)=C3C(O)=C2C[C@@](O)(C(C)=O)C1 XREUEWVEMYWFFA-CSKJXFQVSA-N 0.000 description 1
- 229930188550 carminomycin Natural products 0.000 description 1
- XREUEWVEMYWFFA-UHFFFAOYSA-N carminomycin I Natural products C1C(N)C(O)C(C)OC1OC1C2=C(O)C(C(=O)C3=C(O)C=CC=C3C3=O)=C3C(O)=C2CC(O)(C(C)=O)C1 XREUEWVEMYWFFA-UHFFFAOYSA-N 0.000 description 1
- 229960003261 carmofur Drugs 0.000 description 1
- 229960005243 carmustine Drugs 0.000 description 1
- 239000000969 carrier Substances 0.000 description 1
- 229950001725 carubicin Drugs 0.000 description 1
- 108010047060 carzinophilin Proteins 0.000 description 1
- 238000012219 cassette mutagenesis Methods 0.000 description 1
- 239000003729 cation exchange resin Substances 0.000 description 1
- 229960000590 celecoxib Drugs 0.000 description 1
- RZEKVGVHFLEQIL-UHFFFAOYSA-N celecoxib Chemical compound C1=CC(C)=CC=C1C1=CC(C(F)(F)F)=NN1C1=CC=C(S(N)(=O)=O)C=C1 RZEKVGVHFLEQIL-UHFFFAOYSA-N 0.000 description 1
- 230000006369 cell cycle progression Effects 0.000 description 1
- 230000030833 cell death Effects 0.000 description 1
- 230000032823 cell division Effects 0.000 description 1
- 239000002771 cell marker Substances 0.000 description 1
- 230000005754 cellular signaling Effects 0.000 description 1
- 238000007385 chemical modification Methods 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 239000003638 chemical reducing agent Substances 0.000 description 1
- 229940044683 chemotherapy drug Drugs 0.000 description 1
- 229950008249 chlornaphazine Drugs 0.000 description 1
- 229960001480 chlorozotocin Drugs 0.000 description 1
- 238000011098 chromatofocusing Methods 0.000 description 1
- 239000013611 chromosomal DNA Substances 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 229940080701 chymosin Drugs 0.000 description 1
- 235000013477 citrulline Nutrition 0.000 description 1
- 229960002286 clodronic acid Drugs 0.000 description 1
- HJKBJIYDJLVSAO-UHFFFAOYSA-L clodronic acid disodium salt Chemical compound [Na+].[Na+].OP([O-])(=O)C(Cl)(Cl)P(O)([O-])=O HJKBJIYDJLVSAO-UHFFFAOYSA-L 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- 230000004186 co-expression Effects 0.000 description 1
- 229960001338 colchicine Drugs 0.000 description 1
- 210000001072 colon Anatomy 0.000 description 1
- 230000000112 colonic effect Effects 0.000 description 1
- 238000012875 competitive assay Methods 0.000 description 1
- 230000009137 competitive binding Effects 0.000 description 1
- 230000004154 complement system Effects 0.000 description 1
- 230000009918 complex formation Effects 0.000 description 1
- 238000010668 complexation reaction Methods 0.000 description 1
- 239000008139 complexing agent Substances 0.000 description 1
- JNGZXGGOCLZBFB-IVCQMTBJSA-N compound E Chemical compound N([C@@H](C)C(=O)N[C@@H]1C(N(C)C2=CC=CC=C2C(C=2C=CC=CC=2)=N1)=O)C(=O)CC1=CC(F)=CC(F)=C1 JNGZXGGOCLZBFB-IVCQMTBJSA-N 0.000 description 1
- 230000001268 conjugating effect Effects 0.000 description 1
- 239000013068 control sample Substances 0.000 description 1
- 239000005289 controlled pore glass Substances 0.000 description 1
- 235000005822 corn Nutrition 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- 229940111134 coxibs Drugs 0.000 description 1
- 239000003431 cross linking reagent Substances 0.000 description 1
- 108010006226 cryptophycin Proteins 0.000 description 1
- 108010089438 cryptophycin 1 Proteins 0.000 description 1
- 238000005520 cutting process Methods 0.000 description 1
- 125000000392 cycloalkenyl group Chemical group 0.000 description 1
- 125000000753 cycloalkyl group Chemical group 0.000 description 1
- 239000003255 cyclooxygenase 2 inhibitor Substances 0.000 description 1
- 229960004397 cyclophosphamide Drugs 0.000 description 1
- UFULAYFCSOUIOV-UHFFFAOYSA-N cysteamine Chemical compound NCCS UFULAYFCSOUIOV-UHFFFAOYSA-N 0.000 description 1
- 229960000684 cytarabine Drugs 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 208000024389 cytopenia Diseases 0.000 description 1
- 210000005220 cytoplasmic tail Anatomy 0.000 description 1
- 229940104302 cytosine Drugs 0.000 description 1
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 1
- 239000002619 cytotoxin Substances 0.000 description 1
- 229960000640 dactinomycin Drugs 0.000 description 1
- 125000001295 dansyl group Chemical group [H]C1=C([H])C(N(C([H])([H])[H])C([H])([H])[H])=C2C([H])=C([H])C([H])=C(C2=C1[H])S(*)(=O)=O 0.000 description 1
- 230000007547 defect Effects 0.000 description 1
- 230000000593 degrading effect Effects 0.000 description 1
- 230000002939 deleterious effect Effects 0.000 description 1
- 108700041286 delta Proteins 0.000 description 1
- YSMODUONRAFBET-UHFFFAOYSA-N delta-DL-hydroxylysine Natural products NCC(O)CCC(N)C(O)=O YSMODUONRAFBET-UHFFFAOYSA-N 0.000 description 1
- 229960005052 demecolcine Drugs 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 239000003599 detergent Substances 0.000 description 1
- 230000001627 detrimental effect Effects 0.000 description 1
- 238000002405 diagnostic procedure Methods 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- WVYXNIXAMZOZFK-UHFFFAOYSA-N diaziquone Chemical compound O=C1C(NC(=O)OCC)=C(N2CC2)C(=O)C(NC(=O)OCC)=C1N1CC1 WVYXNIXAMZOZFK-UHFFFAOYSA-N 0.000 description 1
- 229950002389 diaziquone Drugs 0.000 description 1
- 239000012954 diazonium Substances 0.000 description 1
- PZXJOHSZQAEJFE-UHFFFAOYSA-N dihydrobetulinic acid Natural products C1CC(O)C(C)(C)C2CCC3(C)C4(C)CCC5(C(O)=O)CCC(C(C)C)C5C4CCC3C21C PZXJOHSZQAEJFE-UHFFFAOYSA-N 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- MCWXGJITAZMZEV-UHFFFAOYSA-N dimethoate Chemical compound CNC(=O)CSP(=S)(OC)OC MCWXGJITAZMZEV-UHFFFAOYSA-N 0.000 description 1
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 1
- 229940042396 direct acting antivirals thiosemicarbazones Drugs 0.000 description 1
- 230000006806 disease prevention Effects 0.000 description 1
- 230000005750 disease progression Effects 0.000 description 1
- 208000035475 disorder Diseases 0.000 description 1
- 125000002228 disulfide group Chemical group 0.000 description 1
- 150000002019 disulfides Chemical class 0.000 description 1
- NAGJZTKCGNOGPW-UHFFFAOYSA-N dithiophosphoric acid Chemical class OP(O)(S)=S NAGJZTKCGNOGPW-UHFFFAOYSA-N 0.000 description 1
- AFOSIXZFDONLBT-UHFFFAOYSA-N divinyl sulfone Chemical compound C=CS(=O)(=O)C=C AFOSIXZFDONLBT-UHFFFAOYSA-N 0.000 description 1
- 101150109170 dll4 gene Proteins 0.000 description 1
- 239000003534 dna topoisomerase inhibitor Substances 0.000 description 1
- 229940115080 doxil Drugs 0.000 description 1
- NOTIQUSPUUHHEH-UXOVVSIBSA-N dromostanolone propionate Chemical compound C([C@@H]1CC2)C(=O)[C@H](C)C[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H](OC(=O)CC)[C@@]2(C)CC1 NOTIQUSPUUHHEH-UXOVVSIBSA-N 0.000 description 1
- 229960004242 dronabinol Drugs 0.000 description 1
- 229950004683 drostanolone propionate Drugs 0.000 description 1
- 238000012377 drug delivery Methods 0.000 description 1
- 238000009510 drug design Methods 0.000 description 1
- 238000009509 drug development Methods 0.000 description 1
- 238000002651 drug therapy Methods 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 229960005501 duocarmycin Drugs 0.000 description 1
- VQNATVDKACXKTF-XELLLNAOSA-N duocarmycin Chemical compound COC1=C(OC)C(OC)=C2NC(C(=O)N3C4=CC(=O)C5=C([C@@]64C[C@@H]6C3)C=C(N5)C(=O)OC)=CC2=C1 VQNATVDKACXKTF-XELLLNAOSA-N 0.000 description 1
- 229930184221 duocarmycin Natural products 0.000 description 1
- 239000000975 dye Substances 0.000 description 1
- AFMYMMXSQGUCBK-AKMKHHNQSA-N dynemicin a Chemical compound C1#C\C=C/C#C[C@@H]2NC(C=3C(=O)C4=C(O)C=CC(O)=C4C(=O)C=3C(O)=C3)=C3[C@@]34O[C@]32[C@@H](C)C(C(O)=O)=C(OC)[C@H]41 AFMYMMXSQGUCBK-AKMKHHNQSA-N 0.000 description 1
- FSIRXIHZBIXHKT-MHTVFEQDSA-N edatrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CC(CC)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FSIRXIHZBIXHKT-MHTVFEQDSA-N 0.000 description 1
- 229950006700 edatrexate Drugs 0.000 description 1
- 230000002500 effect on skin Effects 0.000 description 1
- VLCYCQAOQCDTCN-UHFFFAOYSA-N eflornithine Chemical compound NCCCC(N)(C(F)F)C(O)=O VLCYCQAOQCDTCN-UHFFFAOYSA-N 0.000 description 1
- 229960002759 eflornithine Drugs 0.000 description 1
- 229940121647 egfr inhibitor Drugs 0.000 description 1
- 238000001962 electrophoresis Methods 0.000 description 1
- 239000012149 elution buffer Substances 0.000 description 1
- 230000002124 endocrine Effects 0.000 description 1
- 238000009261 endocrine therapy Methods 0.000 description 1
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 1
- 210000003038 endothelium Anatomy 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 229950011487 enocitabine Drugs 0.000 description 1
- 230000006862 enzymatic digestion Effects 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- YQGOJNYOYNNSMM-UHFFFAOYSA-N eosin Chemical compound [Na+].OC(=O)C1=CC=CC=C1C1=C2C=C(Br)C(=O)C(Br)=C2OC2=C(Br)C(O)=C(Br)C=C21 YQGOJNYOYNNSMM-UHFFFAOYSA-N 0.000 description 1
- 229940116977 epidermal growth factor Drugs 0.000 description 1
- YJGVMLPVUAXIQN-UHFFFAOYSA-N epipodophyllotoxin Natural products COC1=C(OC)C(OC)=CC(C2C3=CC=4OCOC=4C=C3C(O)C3C2C(OC3)=O)=C1 YJGVMLPVUAXIQN-UHFFFAOYSA-N 0.000 description 1
- 229960001904 epirubicin Drugs 0.000 description 1
- 208000010932 epithelial neoplasm Diseases 0.000 description 1
- 229950002973 epitiostanol Drugs 0.000 description 1
- YSMODUONRAFBET-UHNVWZDZSA-N erythro-5-hydroxy-L-lysine Chemical compound NC[C@H](O)CC[C@H](N)C(O)=O YSMODUONRAFBET-UHNVWZDZSA-N 0.000 description 1
- ITSGNOIFAJAQHJ-BMFNZSJVSA-N esorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)C[C@H](C)O1 ITSGNOIFAJAQHJ-BMFNZSJVSA-N 0.000 description 1
- 229950002017 esorubicin Drugs 0.000 description 1
- LJQQFQHBKUKHIS-WJHRIEJJSA-N esperamicin Chemical compound O1CC(NC(C)C)C(OC)CC1OC1C(O)C(NOC2OC(C)C(SC)C(O)C2)C(C)OC1OC1C(\C2=C/CSSSC)=C(NC(=O)OC)C(=O)C(OC3OC(C)C(O)C(OC(=O)C=4C(=CC(OC)=C(OC)C=4)NC(=O)C(=C)OC)C3)C2(O)C#C\C=C/C#C1 LJQQFQHBKUKHIS-WJHRIEJJSA-N 0.000 description 1
- 229960001842 estramustine Drugs 0.000 description 1
- FRPJXPJMRWBBIH-RBRWEJTLSA-N estramustine Chemical compound ClCCN(CCCl)C(=O)OC1=CC=C2[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CCC2=C1 FRPJXPJMRWBBIH-RBRWEJTLSA-N 0.000 description 1
- 229940011871 estrogen Drugs 0.000 description 1
- 239000000262 estrogen Substances 0.000 description 1
- JKKFKPJIXZFSSB-CBZIJGRNSA-N estrone 3-sulfate Chemical compound OS(=O)(=O)OC1=CC=C2[C@H]3CC[C@](C)(C(CC4)=O)[C@@H]4[C@@H]3CCC2=C1 JKKFKPJIXZFSSB-CBZIJGRNSA-N 0.000 description 1
- 238000012869 ethanol precipitation Methods 0.000 description 1
- QCYAXXZCQKMTMO-QFIPXVFZSA-N ethyl (2s)-2-[(2-bromo-3-oxospiro[3.5]non-1-en-1-yl)amino]-3-[4-(2,7-naphthyridin-1-ylamino)phenyl]propanoate Chemical compound N([C@@H](CC=1C=CC(NC=2C3=CN=CC=C3C=CN=2)=CC=1)C(=O)OCC)C1=C(Br)C(=O)C11CCCCC1 QCYAXXZCQKMTMO-QFIPXVFZSA-N 0.000 description 1
- 229960005237 etoglucid Drugs 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 229940085363 evista Drugs 0.000 description 1
- 230000005284 excitation Effects 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 229960000255 exemestane Drugs 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 229950011548 fadrozole Drugs 0.000 description 1
- 229940043168 fareston Drugs 0.000 description 1
- 229940087861 faslodex Drugs 0.000 description 1
- 229940087476 femara Drugs 0.000 description 1
- 229950003662 fenretinide Drugs 0.000 description 1
- 238000000684 flow cytometry Methods 0.000 description 1
- 229960000961 floxuridine Drugs 0.000 description 1
- ODKNJVUHOIMIIZ-RRKCRQDMSA-N floxuridine Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(F)=C1 ODKNJVUHOIMIIZ-RRKCRQDMSA-N 0.000 description 1
- 229960000390 fludarabine Drugs 0.000 description 1
- GIUYCYHIANZCFB-FJFJXFQQSA-N fludarabine phosphate Chemical compound C1=NC=2C(N)=NC(F)=NC=2N1[C@@H]1O[C@H](COP(O)(O)=O)[C@@H](O)[C@@H]1O GIUYCYHIANZCFB-FJFJXFQQSA-N 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 1
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 1
- YLRFCQOZQXIBAB-RBZZARIASA-N fluoxymesterone Chemical compound C1CC2=CC(=O)CC[C@]2(C)[C@]2(F)[C@@H]1[C@@H]1CC[C@](C)(O)[C@@]1(C)C[C@@H]2O YLRFCQOZQXIBAB-RBZZARIASA-N 0.000 description 1
- 229960001751 fluoxymesterone Drugs 0.000 description 1
- MKXKFYHWDHIYRV-UHFFFAOYSA-N flutamide Chemical compound CC(C)C(=O)NC1=CC=C([N+]([O-])=O)C(C(F)(F)F)=C1 MKXKFYHWDHIYRV-UHFFFAOYSA-N 0.000 description 1
- 229960002074 flutamide Drugs 0.000 description 1
- 235000019152 folic acid Nutrition 0.000 description 1
- 239000011724 folic acid Substances 0.000 description 1
- 229960000304 folic acid Drugs 0.000 description 1
- 239000004052 folic acid antagonist Substances 0.000 description 1
- 150000002224 folic acids Chemical class 0.000 description 1
- 230000003325 follicular Effects 0.000 description 1
- 229960004421 formestane Drugs 0.000 description 1
- OSVMTWJCGUFAOD-KZQROQTASA-N formestane Chemical compound O=C1CC[C@]2(C)[C@H]3CC[C@](C)(C(CC4)=O)[C@@H]4[C@@H]3CCC2=C1O OSVMTWJCGUFAOD-KZQROQTASA-N 0.000 description 1
- 229940001490 fosamax Drugs 0.000 description 1
- 238000005194 fractionation Methods 0.000 description 1
- 229960002258 fulvestrant Drugs 0.000 description 1
- 230000005714 functional activity Effects 0.000 description 1
- 125000000524 functional group Chemical group 0.000 description 1
- UIWYJDYFSGRHKR-UHFFFAOYSA-N gadolinium atom Chemical compound [Gd] UIWYJDYFSGRHKR-UHFFFAOYSA-N 0.000 description 1
- 229940044658 gallium nitrate Drugs 0.000 description 1
- 210000001035 gastrointestinal tract Anatomy 0.000 description 1
- QTQAWLPCGQOSGP-GBTDJJJQSA-N geldanamycin Chemical compound N1C(=O)\C(C)=C/C=C\[C@@H](OC)[C@H](OC(N)=O)\C(C)=C/[C@@H](C)[C@@H](O)[C@H](OC)C[C@@H](C)CC2=C(OC)C(=O)C=C1C2=O QTQAWLPCGQOSGP-GBTDJJJQSA-N 0.000 description 1
- 229960005277 gemcitabine Drugs 0.000 description 1
- SDUQYLNIPVEERB-QPPQHZFASA-N gemcitabine Chemical compound O=C1N=C(N)C=CN1[C@H]1C(F)(F)[C@H](O)[C@@H](CO)O1 SDUQYLNIPVEERB-QPPQHZFASA-N 0.000 description 1
- 229940020967 gemzar Drugs 0.000 description 1
- 238000003500 gene array Methods 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 239000011521 glass Substances 0.000 description 1
- 229940084910 gliadel Drugs 0.000 description 1
- 108060003196 globin Proteins 0.000 description 1
- 102000018146 globin Human genes 0.000 description 1
- 239000003862 glucocorticoid Substances 0.000 description 1
- 229940116332 glucose oxidase Drugs 0.000 description 1
- 235000019420 glucose oxidase Nutrition 0.000 description 1
- 150000004676 glycans Chemical class 0.000 description 1
- 102000006602 glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 description 1
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 1
- 230000002414 glycolytic effect Effects 0.000 description 1
- 229930182470 glycoside Natural products 0.000 description 1
- 108010067216 glycyl-glycyl-glycine Proteins 0.000 description 1
- 229960002913 goserelin Drugs 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 201000009277 hairy cell leukemia Diseases 0.000 description 1
- 230000009931 harmful effect Effects 0.000 description 1
- 210000003128 head Anatomy 0.000 description 1
- 208000014829 head and neck neoplasm Diseases 0.000 description 1
- 108010067006 heat stable toxin (E coli) Proteins 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 125000000623 heterocyclic group Chemical group 0.000 description 1
- UUVWYPNAQBNQJQ-UHFFFAOYSA-N hexamethylmelamine Chemical compound CN(C)C1=NC(N(C)C)=NC(N(C)C)=N1 UUVWYPNAQBNQJQ-UHFFFAOYSA-N 0.000 description 1
- 230000001744 histochemical effect Effects 0.000 description 1
- 230000013632 homeostatic process Effects 0.000 description 1
- 210000005260 human cell Anatomy 0.000 description 1
- 208000033519 human immunodeficiency virus infectious disease Diseases 0.000 description 1
- 210000004754 hybrid cell Anatomy 0.000 description 1
- 229940088013 hycamtin Drugs 0.000 description 1
- 150000002429 hydrazines Chemical class 0.000 description 1
- 150000002431 hydrogen Chemical class 0.000 description 1
- 230000007062 hydrolysis Effects 0.000 description 1
- 238000006460 hydrolysis reaction Methods 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 238000004191 hydrophobic interaction chromatography Methods 0.000 description 1
- 125000002349 hydroxyamino group Chemical group [H]ON([H])[*] 0.000 description 1
- 229960001330 hydroxycarbamide Drugs 0.000 description 1
- 229910052588 hydroxylapatite Inorganic materials 0.000 description 1
- 238000012872 hydroxylapatite chromatography Methods 0.000 description 1
- 125000004029 hydroxymethyl group Chemical group [H]OC([H])([H])* 0.000 description 1
- 239000002471 hydroxymethylglutaryl coenzyme A reductase inhibitor Substances 0.000 description 1
- 229940015872 ibandronate Drugs 0.000 description 1
- 229960000908 idarubicin Drugs 0.000 description 1
- 229950002248 idoxifene Drugs 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 150000002463 imidates Chemical class 0.000 description 1
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Substances C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 1
- 230000014726 immortalization of host cell Effects 0.000 description 1
- 230000028993 immune response Effects 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 238000002513 implantation Methods 0.000 description 1
- DBIGHPPNXATHOF-UHFFFAOYSA-N improsulfan Chemical compound CS(=O)(=O)OCCCNCCCOS(C)(=O)=O DBIGHPPNXATHOF-UHFFFAOYSA-N 0.000 description 1
- 229950008097 improsulfan Drugs 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 238000005462 in vivo assay Methods 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 229940055742 indium-111 Drugs 0.000 description 1
- 230000002458 infectious effect Effects 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 150000002484 inorganic compounds Chemical class 0.000 description 1
- 229910010272 inorganic material Inorganic materials 0.000 description 1
- 239000000138 intercalating agent Substances 0.000 description 1
- 210000000936 intestine Anatomy 0.000 description 1
- 238000001361 intraarterial administration Methods 0.000 description 1
- 230000007154 intracellular accumulation Effects 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000010253 intravenous injection Methods 0.000 description 1
- 239000001573 invertase Substances 0.000 description 1
- 235000011073 invertase Nutrition 0.000 description 1
- 238000005342 ion exchange Methods 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 229960004768 irinotecan Drugs 0.000 description 1
- 229910052742 iron Inorganic materials 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- 125000000468 ketone group Chemical group 0.000 description 1
- 230000002147 killing effect Effects 0.000 description 1
- 229940043355 kinase inhibitor Drugs 0.000 description 1
- 150000002596 lactones Chemical class 0.000 description 1
- 229940057428 lactoperoxidase Drugs 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 238000011031 large-scale manufacturing process Methods 0.000 description 1
- 229940115286 lentinan Drugs 0.000 description 1
- 229960003881 letrozole Drugs 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- RGLRXNKKBLIBQS-XNHQSDQCSA-N leuprolide acetate Chemical compound CC(O)=O.CCNC(=O)[C@@H]1CCCN1C(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](CC(C)C)NC(=O)[C@@H](NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H]1NC(=O)CC1)CC1=CC=C(O)C=C1 RGLRXNKKBLIBQS-XNHQSDQCSA-N 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 239000007791 liquid phase Substances 0.000 description 1
- 201000007270 liver cancer Diseases 0.000 description 1
- 208000014018 liver neoplasm Diseases 0.000 description 1
- 244000144972 livestock Species 0.000 description 1
- 229960002247 lomustine Drugs 0.000 description 1
- 229950001750 lonafarnib Drugs 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 101150074251 lpp gene Proteins 0.000 description 1
- 238000003670 luciferase enzyme activity assay Methods 0.000 description 1
- 210000004072 lung Anatomy 0.000 description 1
- 210000005265 lung cell Anatomy 0.000 description 1
- 108010078259 luprolide acetate gel depot Proteins 0.000 description 1
- 229940087857 lupron Drugs 0.000 description 1
- RVFGKBWWUQOIOU-NDEPHWFRSA-N lurtotecan Chemical compound O=C([C@]1(O)CC)OCC(C(N2CC3=4)=O)=C1C=C2C3=NC1=CC=2OCCOC=2C=C1C=4CN1CCN(C)CC1 RVFGKBWWUQOIOU-NDEPHWFRSA-N 0.000 description 1
- 229950002654 lurtotecan Drugs 0.000 description 1
- 230000002535 lyotropic effect Effects 0.000 description 1
- 230000002934 lysing effect Effects 0.000 description 1
- 230000002101 lytic effect Effects 0.000 description 1
- 150000002671 lyxoses Chemical class 0.000 description 1
- 229920002521 macromolecule Polymers 0.000 description 1
- 210000002540 macrophage Anatomy 0.000 description 1
- 239000011777 magnesium Substances 0.000 description 1
- 229910052749 magnesium Inorganic materials 0.000 description 1
- 238000002595 magnetic resonance imaging Methods 0.000 description 1
- 238000012423 maintenance Methods 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 208000026037 malignant tumor of neck Diseases 0.000 description 1
- 229910052748 manganese Inorganic materials 0.000 description 1
- 239000011572 manganese Substances 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- MQXVYODZCMMZEM-ZYUZMQFOSA-N mannomustine Chemical compound ClCCNC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CNCCCl MQXVYODZCMMZEM-ZYUZMQFOSA-N 0.000 description 1
- 229950008612 mannomustine Drugs 0.000 description 1
- 201000007924 marginal zone B-cell lymphoma Diseases 0.000 description 1
- 210000003826 marginal zone b cell Anatomy 0.000 description 1
- 208000021937 marginal zone lymphoma Diseases 0.000 description 1
- 229940099262 marinol Drugs 0.000 description 1
- 241001515942 marmosets Species 0.000 description 1
- 238000004949 mass spectrometry Methods 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- HAWPXGHAZFHHAD-UHFFFAOYSA-N mechlorethamine Chemical compound ClCCN(C)CCCl HAWPXGHAZFHHAD-UHFFFAOYSA-N 0.000 description 1
- PSGAAPLEWMOORI-PEINSRQWSA-N medroxyprogesterone acetate Chemical compound C([C@@]12C)CC(=O)C=C1[C@@H](C)C[C@@H]1[C@@H]2CC[C@]2(C)[C@@](OC(C)=O)(C(C)=O)CC[C@H]21 PSGAAPLEWMOORI-PEINSRQWSA-N 0.000 description 1
- 229960002985 medroxyprogesterone acetate Drugs 0.000 description 1
- 210000002752 melanocyte Anatomy 0.000 description 1
- 229960003151 mercaptamine Drugs 0.000 description 1
- 230000004060 metabolic process Effects 0.000 description 1
- 230000009401 metastasis Effects 0.000 description 1
- VJRAUFKOOPNFIQ-TVEKBUMESA-N methyl (1r,2r,4s)-4-[(2r,4s,5s,6s)-5-[(2s,4s,5s,6s)-5-[(2s,4s,5s,6s)-4,5-dihydroxy-6-methyloxan-2-yl]oxy-4-hydroxy-6-methyloxan-2-yl]oxy-4-(dimethylamino)-6-methyloxan-2-yl]oxy-2-ethyl-2,5,7,10-tetrahydroxy-6,11-dioxo-3,4-dihydro-1h-tetracene-1-carboxylat Chemical compound O([C@H]1[C@@H](O)C[C@@H](O[C@H]1C)O[C@H]1[C@H](C[C@@H](O[C@H]1C)O[C@H]1C[C@]([C@@H](C2=CC=3C(=O)C4=C(O)C=CC(O)=C4C(=O)C=3C(O)=C21)C(=O)OC)(O)CC)N(C)C)[C@H]1C[C@H](O)[C@H](O)[C@H](C)O1 VJRAUFKOOPNFIQ-TVEKBUMESA-N 0.000 description 1
- YACKEPLHDIMKIO-UHFFFAOYSA-N methylphosphonic acid Chemical compound CP(O)(O)=O YACKEPLHDIMKIO-UHFFFAOYSA-N 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 108010029942 microperoxidase Proteins 0.000 description 1
- 230000008880 microtubule cytoskeleton organization Effects 0.000 description 1
- 230000003278 mimic effect Effects 0.000 description 1
- 229960003539 mitoguazone Drugs 0.000 description 1
- MXWHMTNPTTVWDM-NXOFHUPFSA-N mitoguazone Chemical compound NC(N)=N\N=C(/C)\C=N\N=C(N)N MXWHMTNPTTVWDM-NXOFHUPFSA-N 0.000 description 1
- VFKZTMPDYBFSTM-GUCUJZIJSA-N mitolactol Chemical compound BrC[C@H](O)[C@@H](O)[C@@H](O)[C@H](O)CBr VFKZTMPDYBFSTM-GUCUJZIJSA-N 0.000 description 1
- 229950010913 mitolactol Drugs 0.000 description 1
- 230000011278 mitosis Effects 0.000 description 1
- 229960000350 mitotane Drugs 0.000 description 1
- 230000000394 mitotic effect Effects 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 230000004001 molecular interaction Effects 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 239000012120 mounting media Substances 0.000 description 1
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 1
- 231100000219 mutagenic Toxicity 0.000 description 1
- 230000003505 mutagenic effect Effects 0.000 description 1
- GNOLWGAJQVLBSM-UHFFFAOYSA-N n,n,5,7-tetramethyl-1,2,3,4-tetrahydronaphthalen-1-amine Chemical compound C1=C(C)C=C2C(N(C)C)CCCC2=C1C GNOLWGAJQVLBSM-UHFFFAOYSA-N 0.000 description 1
- NJSMWLQOCQIOPE-OCHFTUDZSA-N n-[(e)-[10-[(e)-(4,5-dihydro-1h-imidazol-2-ylhydrazinylidene)methyl]anthracen-9-yl]methylideneamino]-4,5-dihydro-1h-imidazol-2-amine Chemical compound N1CCN=C1N\N=C\C(C1=CC=CC=C11)=C(C=CC=C2)C2=C1\C=N\NC1=NCCN1 NJSMWLQOCQIOPE-OCHFTUDZSA-N 0.000 description 1
- 229950006780 n-acetylglucosamine Drugs 0.000 description 1
- 239000002105 nanoparticle Substances 0.000 description 1
- 229930014626 natural product Natural products 0.000 description 1
- 229940086322 navelbine Drugs 0.000 description 1
- 210000003739 neck Anatomy 0.000 description 1
- QZGIWPZCWHMVQL-UIYAJPBUSA-N neocarzinostatin chromophore Chemical compound O1[C@H](C)[C@H](O)[C@H](O)[C@@H](NC)[C@H]1O[C@@H]1C/2=C/C#C[C@H]3O[C@@]3([C@@H]3OC(=O)OC3)C#CC\2=C[C@H]1OC(=O)C1=C(O)C=CC2=C(C)C=C(OC)C=C12 QZGIWPZCWHMVQL-UIYAJPBUSA-N 0.000 description 1
- MQYXUWHLBZFQQO-UHFFFAOYSA-N nepehinol Natural products C1CC(O)C(C)(C)C2CCC3(C)C4(C)CCC5(C)CCC(C(=C)C)C5C4CCC3C21C MQYXUWHLBZFQQO-UHFFFAOYSA-N 0.000 description 1
- 208000015122 neurodegenerative disease Diseases 0.000 description 1
- 230000004766 neurogenesis Effects 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- 102000046701 nicastrin Human genes 0.000 description 1
- 108700022821 nicastrin Proteins 0.000 description 1
- XWXYUMMDTVBTOU-UHFFFAOYSA-N nilutamide Chemical compound O=C1C(C)(C)NC(=O)N1C1=CC=C([N+]([O-])=O)C(C(F)(F)F)=C1 XWXYUMMDTVBTOU-UHFFFAOYSA-N 0.000 description 1
- 229960002653 nilutamide Drugs 0.000 description 1
- 229960001420 nimustine Drugs 0.000 description 1
- VFEDRRNHLBGPNN-UHFFFAOYSA-N nimustine Chemical compound CC1=NC=C(CNC(=O)N(CCCl)N=O)C(N)=N1 VFEDRRNHLBGPNN-UHFFFAOYSA-N 0.000 description 1
- YMVWGSQGCWCDGW-UHFFFAOYSA-N nitracrine Chemical compound C1=CC([N+]([O-])=O)=C2C(NCCCN(C)C)=C(C=CC=C3)C3=NC2=C1 YMVWGSQGCWCDGW-UHFFFAOYSA-N 0.000 description 1
- 229950008607 nitracrine Drugs 0.000 description 1
- KGTDRFCXGRULNK-JYOBTZKQSA-N nogalamycin Chemical compound CO[C@@H]1[C@@](OC)(C)[C@@H](OC)[C@H](C)O[C@H]1O[C@@H]1C2=C(O)C(C(=O)C3=C(O)C=C4[C@@]5(C)O[C@H]([C@H]([C@@H]([C@H]5O)N(C)C)O)OC4=C3C3=O)=C3C=C2[C@@H](C(=O)OC)[C@@](C)(O)C1 KGTDRFCXGRULNK-JYOBTZKQSA-N 0.000 description 1
- 229950009266 nogalamycin Drugs 0.000 description 1
- 229940085033 nolvadex Drugs 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 239000012038 nucleophile Substances 0.000 description 1
- 150000003833 nucleoside derivatives Chemical class 0.000 description 1
- 238000011580 nude mouse model Methods 0.000 description 1
- 229960000435 oblimersen Drugs 0.000 description 1
- MIMNFCVQODTQDP-NDLVEFNKSA-N oblimersen Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](COP(S)(=O)O[C@@H]2[C@H](O[C@H](C2)N2C3=NC=NC(N)=C3N=C2)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C3=C(C(NC(N)=N3)=O)N=C2)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C3=C(C(NC(N)=N3)=O)N=C2)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(NC(=O)C(C)=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C3=C(C(NC(N)=N3)=O)N=C2)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C3=C(C(NC(N)=N3)=O)N=C2)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C3=NC=NC(N)=C3N=C2)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(NC(=O)C(C)=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(N=C(N)C=C2)=O)COP(O)(=S)O[C@@H]2[C@H](O[C@H](C2)N2C(NC(=O)C(C)=C2)=O)CO)[C@@H](O)C1 MIMNFCVQODTQDP-NDLVEFNKSA-N 0.000 description 1
- 238000002515 oligonucleotide synthesis Methods 0.000 description 1
- CZDBNBLGZNWKMC-MWQNXGTOSA-N olivomycin Chemical class O([C@@H]1C[C@@H](O[C@H](C)[C@@H]1O)OC=1C=C2C=C3C[C@H]([C@@H](C(=O)C3=C(O)C2=C(O)C=1)O[C@H]1O[C@@H](C)[C@H](O)[C@@H](OC2O[C@@H](C)[C@H](O)[C@@H](O)C2)C1)[C@H](OC)C(=O)[C@@H](O)[C@@H](C)O)[C@H]1C[C@H](O)[C@H](OC)[C@H](C)O1 CZDBNBLGZNWKMC-MWQNXGTOSA-N 0.000 description 1
- 229950011093 onapristone Drugs 0.000 description 1
- 230000003287 optical effect Effects 0.000 description 1
- 239000003960 organic solvent Substances 0.000 description 1
- 230000003204 osmotic effect Effects 0.000 description 1
- 229940043515 other immunoglobulins in atc Drugs 0.000 description 1
- 230000002018 overexpression Effects 0.000 description 1
- 239000007800 oxidant agent Substances 0.000 description 1
- 150000002923 oximes Chemical class 0.000 description 1
- 229960003978 pamidronic acid Drugs 0.000 description 1
- 229920002866 paraformaldehyde Polymers 0.000 description 1
- 230000005298 paramagnetic effect Effects 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 229940046159 pegylated liposomal doxorubicin Drugs 0.000 description 1
- 229940049954 penicillin Drugs 0.000 description 1
- 229950009506 penicillinase Drugs 0.000 description 1
- XYJRXVWERLGGKC-UHFFFAOYSA-D pentacalcium;hydroxide;triphosphate Chemical compound [OH-].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O XYJRXVWERLGGKC-UHFFFAOYSA-D 0.000 description 1
- 229960002340 pentostatin Drugs 0.000 description 1
- FPVKHBSQESCIEP-JQCXWYLXSA-N pentostatin Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(N=CNC[C@H]2O)=C2N=C1 FPVKHBSQESCIEP-JQCXWYLXSA-N 0.000 description 1
- QIMGFXOHTOXMQP-GFAGFCTOSA-N peplomycin Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCCN[C@@H](C)C=1C=CC=CC=1)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1NC=NC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C QIMGFXOHTOXMQP-GFAGFCTOSA-N 0.000 description 1
- 229950003180 peplomycin Drugs 0.000 description 1
- 229940111202 pepsin Drugs 0.000 description 1
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 1
- 230000010412 perfusion Effects 0.000 description 1
- SZFPYBIJACMNJV-UHFFFAOYSA-N perifosine Chemical compound CCCCCCCCCCCCCCCCCCOP([O-])(=O)OC1CC[N+](C)(C)CC1 SZFPYBIJACMNJV-UHFFFAOYSA-N 0.000 description 1
- 229950010632 perifosine Drugs 0.000 description 1
- KHIWWQKSHDUIBK-UHFFFAOYSA-N periodic acid Chemical compound OI(=O)(=O)=O KHIWWQKSHDUIBK-UHFFFAOYSA-N 0.000 description 1
- 230000035699 permeability Effects 0.000 description 1
- 230000008823 permeabilization Effects 0.000 description 1
- 239000012071 phase Substances 0.000 description 1
- 101150009573 phoA gene Proteins 0.000 description 1
- 125000002467 phosphate group Chemical group [H]OP(=O)(O[H])O[*] 0.000 description 1
- 150000004713 phosphodiesters Chemical class 0.000 description 1
- 150000003013 phosphoric acid derivatives Chemical group 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- 239000003757 phosphotransferase inhibitor Substances 0.000 description 1
- 230000004962 physiological condition Effects 0.000 description 1
- 230000035790 physiological processes and functions Effects 0.000 description 1
- 229960000952 pipobroman Drugs 0.000 description 1
- NJBFOOCLYDNZJN-UHFFFAOYSA-N pipobroman Chemical compound BrCCC(=O)N1CCN(C(=O)CCBr)CC1 NJBFOOCLYDNZJN-UHFFFAOYSA-N 0.000 description 1
- 229960001221 pirarubicin Drugs 0.000 description 1
- 229960004403 pixantrone Drugs 0.000 description 1
- PEZPMAYDXJQYRV-UHFFFAOYSA-N pixantrone Chemical compound O=C1C2=CN=CC=C2C(=O)C2=C1C(NCCN)=CC=C2NCCN PEZPMAYDXJQYRV-UHFFFAOYSA-N 0.000 description 1
- 239000003123 plant toxin Substances 0.000 description 1
- 229920003023 plastic Polymers 0.000 description 1
- 239000004033 plastic Substances 0.000 description 1
- 229910052697 platinum Inorganic materials 0.000 description 1
- 229960001237 podophyllotoxin Drugs 0.000 description 1
- YJGVMLPVUAXIQN-XVVDYKMHSA-N podophyllotoxin Chemical compound COC1=C(OC)C(OC)=CC([C@@H]2C3=CC=4OCOC=4C=C3[C@H](O)[C@@H]3[C@@H]2C(OC3)=O)=C1 YJGVMLPVUAXIQN-XVVDYKMHSA-N 0.000 description 1
- YVCVYCSAAZQOJI-UHFFFAOYSA-N podophyllotoxin Natural products COC1=C(O)C(OC)=CC(C2C3=CC=4OCOC=4C=C3C(O)C3C2C(OC3)=O)=C1 YVCVYCSAAZQOJI-UHFFFAOYSA-N 0.000 description 1
- 229920000191 poly(N-vinyl pyrrolidone) Polymers 0.000 description 1
- 229920001481 poly(stearyl methacrylate) Polymers 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 108010054442 polyalanine Proteins 0.000 description 1
- 108010064470 polyaspartate Proteins 0.000 description 1
- 229920005862 polyol Polymers 0.000 description 1
- 150000003077 polyols Chemical class 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 1
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 1
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 1
- 230000023603 positive regulation of transcription initiation, DNA-dependent Effects 0.000 description 1
- 108020001213 potassium channel Proteins 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 229960004694 prednimustine Drugs 0.000 description 1
- 229960005205 prednisolone Drugs 0.000 description 1
- OIGNJSKKLXVSLS-VWUMJDOOSA-N prednisolone Chemical compound O=C1C=C[C@]2(C)[C@H]3[C@@H](O)C[C@](C)([C@@](CC4)(O)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 OIGNJSKKLXVSLS-VWUMJDOOSA-N 0.000 description 1
- 229960004618 prednisone Drugs 0.000 description 1
- XOFYZVNMUHMLCC-ZPOLXVRWSA-N prednisone Chemical compound O=C1C=C[C@]2(C)[C@H]3C(=O)C[C@](C)([C@@](CC4)(O)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 XOFYZVNMUHMLCC-ZPOLXVRWSA-N 0.000 description 1
- 229940063238 premarin Drugs 0.000 description 1
- 238000011809 primate model Methods 0.000 description 1
- CPTBDICYNRMXFX-UHFFFAOYSA-N procarbazine Chemical compound CNNCC1=CC=C(C(=O)NC(C)C)C=C1 CPTBDICYNRMXFX-UHFFFAOYSA-N 0.000 description 1
- 229960000624 procarbazine Drugs 0.000 description 1
- 239000000583 progesterone congener Substances 0.000 description 1
- 230000003623 progesteronic effect Effects 0.000 description 1
- 238000004393 prognosis Methods 0.000 description 1
- 230000005522 programmed cell death Effects 0.000 description 1
- 230000000750 progressive effect Effects 0.000 description 1
- 230000002035 prolonged effect Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 210000002307 prostate Anatomy 0.000 description 1
- 235000019419 proteases Nutrition 0.000 description 1
- 239000003528 protein farnesyltransferase inhibitor Substances 0.000 description 1
- 239000003531 protein hydrolysate Substances 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- WOLQREOUPKZMEX-UHFFFAOYSA-N pteroyltriglutamic acid Chemical compound C=1N=C2NC(N)=NC(=O)C2=NC=1CNC1=CC=C(C(=O)NC(CCC(=O)NC(CCC(=O)NC(CCC(O)=O)C(O)=O)C(O)=O)C(O)=O)C=C1 WOLQREOUPKZMEX-UHFFFAOYSA-N 0.000 description 1
- 150000003212 purines Chemical class 0.000 description 1
- 229950010131 puromycin Drugs 0.000 description 1
- NHKZQYJGWDGXMY-UHFFFAOYSA-N pyridin-2-ylsulfanyl pentanoate Chemical compound CCCCC(=O)OSC1=CC=CC=N1 NHKZQYJGWDGXMY-UHFFFAOYSA-N 0.000 description 1
- 150000003230 pyrimidines Chemical class 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 238000010791 quenching Methods 0.000 description 1
- 230000000171 quenching effect Effects 0.000 description 1
- 150000003254 radicals Chemical class 0.000 description 1
- 238000007420 radioactive assay Methods 0.000 description 1
- 238000001959 radiotherapy Methods 0.000 description 1
- 229920005604 random copolymer Polymers 0.000 description 1
- 229960002185 ranimustine Drugs 0.000 description 1
- 229940099538 rapamune Drugs 0.000 description 1
- 229910052761 rare earth metal Inorganic materials 0.000 description 1
- 150000002910 rare earth metals Chemical class 0.000 description 1
- BMKDZUISNHGIBY-UHFFFAOYSA-N razoxane Chemical compound C1C(=O)NC(=O)CN1C(C)CN1CC(=O)NC(=O)C1 BMKDZUISNHGIBY-UHFFFAOYSA-N 0.000 description 1
- 229960000460 razoxane Drugs 0.000 description 1
- 230000008707 rearrangement Effects 0.000 description 1
- 210000003370 receptor cell Anatomy 0.000 description 1
- 206010038038 rectal cancer Diseases 0.000 description 1
- 210000000664 rectum Anatomy 0.000 description 1
- 201000001275 rectum cancer Diseases 0.000 description 1
- 230000022983 regulation of cell cycle Effects 0.000 description 1
- 230000003252 repetitive effect Effects 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 239000011347 resin Substances 0.000 description 1
- 229920005989 resin Polymers 0.000 description 1
- 230000004043 responsiveness Effects 0.000 description 1
- 230000000284 resting effect Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 201000006845 reticulosarcoma Diseases 0.000 description 1
- 208000029922 reticulum cell sarcoma Diseases 0.000 description 1
- 230000002207 retinal effect Effects 0.000 description 1
- 238000004007 reversed phase HPLC Methods 0.000 description 1
- PYWVYCXTNDRMGF-UHFFFAOYSA-N rhodamine B Chemical compound [Cl-].C=12C=CC(=[N+](CC)CC)C=C2OC2=CC(N(CC)CC)=CC=C2C=1C1=CC=CC=C1C(O)=O PYWVYCXTNDRMGF-UHFFFAOYSA-N 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 229940089617 risedronate Drugs 0.000 description 1
- 238000011808 rodent model Methods 0.000 description 1
- NSFWWJIQIKBZMJ-PAGWOCKZSA-N roridin a Chemical compound C([C@@]12[C@]3(C)[C@H]4C[C@H]1O[C@@H]1C=C(C)CC[C@@]13COC(=O)[C@@H](O)[C@H](C)CCO[C@H](\C=C\C=C/C(=O)O4)[C@H](O)C)O2 NSFWWJIQIKBZMJ-PAGWOCKZSA-N 0.000 description 1
- 102200048328 rs1801270 Human genes 0.000 description 1
- 102200015995 rs28931574 Human genes 0.000 description 1
- 102220083620 rs371904636 Human genes 0.000 description 1
- 102220141469 rs543096490 Human genes 0.000 description 1
- 102220040412 rs587778307 Human genes 0.000 description 1
- 102220062178 rs786202417 Human genes 0.000 description 1
- 102220122065 rs886042999 Human genes 0.000 description 1
- VHXNKPBCCMUMSW-FQEVSTJZSA-N rubitecan Chemical compound C1=CC([N+]([O-])=O)=C2C=C(CN3C4=CC5=C(C3=O)COC(=O)[C@]5(O)CC)C4=NC2=C1 VHXNKPBCCMUMSW-FQEVSTJZSA-N 0.000 description 1
- DWZAJFZEYZIHPO-UHFFFAOYSA-N santin Natural products C1=CC(OC)=CC=C1C1=C(OC)C(=O)C2=C(O)C(OC)=C(O)C=C2O1 DWZAJFZEYZIHPO-UHFFFAOYSA-N 0.000 description 1
- 238000013391 scatchard analysis Methods 0.000 description 1
- FWYIBGHGBOVPNL-UHFFFAOYSA-N scopoletin Natural products COC=1C=C2C=CC(OC2=C(C1)O)=O FWYIBGHGBOVPNL-UHFFFAOYSA-N 0.000 description 1
- 238000002864 sequence alignment Methods 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 238000013207 serial dilution Methods 0.000 description 1
- 125000003607 serino group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C(O[H])([H])[H] 0.000 description 1
- 210000000717 sertoli cell Anatomy 0.000 description 1
- 230000035939 shock Effects 0.000 description 1
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 1
- 239000000377 silicon dioxide Substances 0.000 description 1
- 229940112726 skelid Drugs 0.000 description 1
- 210000003491 skin Anatomy 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- PTLRDCMBXHILCL-UHFFFAOYSA-M sodium arsenite Chemical compound [Na+].[O-][As]=O PTLRDCMBXHILCL-UHFFFAOYSA-M 0.000 description 1
- 229910000029 sodium carbonate Inorganic materials 0.000 description 1
- 239000001509 sodium citrate Substances 0.000 description 1
- NLJMYIDDQXHKNR-UHFFFAOYSA-K sodium citrate Chemical compound O.O.[Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O NLJMYIDDQXHKNR-UHFFFAOYSA-K 0.000 description 1
- MKNJJMHQBYVHRS-UHFFFAOYSA-M sodium;1-[11-(2,5-dioxopyrrol-1-yl)undecanoyloxy]-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].O=C1C(S(=O)(=O)[O-])CC(=O)N1OC(=O)CCCCCCCCCCN1C(=O)C=CC1=O MKNJJMHQBYVHRS-UHFFFAOYSA-M 0.000 description 1
- ULARYIUTHAWJMU-UHFFFAOYSA-M sodium;1-[4-(2,5-dioxopyrrol-1-yl)butanoyloxy]-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].O=C1C(S(=O)(=O)[O-])CC(=O)N1OC(=O)CCCN1C(=O)C=CC1=O ULARYIUTHAWJMU-UHFFFAOYSA-M 0.000 description 1
- VUFNRPJNRFOTGK-UHFFFAOYSA-M sodium;1-[4-[(2,5-dioxopyrrol-1-yl)methyl]cyclohexanecarbonyl]oxy-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].O=C1C(S(=O)(=O)[O-])CC(=O)N1OC(=O)C1CCC(CN2C(C=CC2=O)=O)CC1 VUFNRPJNRFOTGK-UHFFFAOYSA-M 0.000 description 1
- MIDXXTLMKGZDPV-UHFFFAOYSA-M sodium;1-[6-(2,5-dioxopyrrol-1-yl)hexanoyloxy]-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].O=C1C(S(=O)(=O)[O-])CC(=O)N1OC(=O)CCCCCN1C(=O)C=CC1=O MIDXXTLMKGZDPV-UHFFFAOYSA-M 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 230000000392 somatic effect Effects 0.000 description 1
- 229960003787 sorafenib Drugs 0.000 description 1
- 238000009987 spinning Methods 0.000 description 1
- 229950006315 spirogermanium Drugs 0.000 description 1
- 210000004988 splenocyte Anatomy 0.000 description 1
- 238000012453 sprague-dawley rat model Methods 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 230000003637 steroidlike Effects 0.000 description 1
- 150000003431 steroids Chemical class 0.000 description 1
- 230000000638 stimulation Effects 0.000 description 1
- 210000002784 stomach Anatomy 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- 238000010254 subcutaneous injection Methods 0.000 description 1
- 239000007929 subcutaneous injection Substances 0.000 description 1
- 125000001424 substituent group Chemical group 0.000 description 1
- WINHZLLDWRZWRT-ATVHPVEESA-N sunitinib Chemical compound CCN(CC)CCNC(=O)C1=C(C)NC(\C=C/2C3=CC(F)=CC=C3NC\2=O)=C1C WINHZLLDWRZWRT-ATVHPVEESA-N 0.000 description 1
- 229960001796 sunitinib Drugs 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 229940034785 sutent Drugs 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 238000001308 synthesis method Methods 0.000 description 1
- 230000009897 systematic effect Effects 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 238000012353 t test Methods 0.000 description 1
- FQZYTYWMLGAPFJ-OQKDUQJOSA-N tamoxifen citrate Chemical compound [H+].[H+].[H+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O.C=1C=CC=CC=1C(/CC)=C(C=1C=CC(OCCN(C)C)=CC=1)/C1=CC=CC=C1 FQZYTYWMLGAPFJ-OQKDUQJOSA-N 0.000 description 1
- 229940099419 targretin Drugs 0.000 description 1
- RCINICONZNJXQF-XAZOAEDWSA-N taxol® Chemical compound O([C@@H]1[C@@]2(CC(C(C)=C(C2(C)C)[C@H](C([C@]2(C)[C@@H](O)C[C@H]3OC[C@]3(C21)OC(C)=O)=O)OC(=O)C)OC(=O)[C@H](O)[C@@H](NC(=O)C=1C=CC=CC=1)C=1C=CC=CC=1)O)C(=O)C1=CC=CC=C1 RCINICONZNJXQF-XAZOAEDWSA-N 0.000 description 1
- 229940063683 taxotere Drugs 0.000 description 1
- 229960001674 tegafur Drugs 0.000 description 1
- WFWLQNSHRPWKFK-ZCFIWIBFSA-N tegafur Chemical compound O=C1NC(=O)C(F)=CN1[C@@H]1OCCC1 WFWLQNSHRPWKFK-ZCFIWIBFSA-N 0.000 description 1
- 229960000235 temsirolimus Drugs 0.000 description 1
- NRUKOCRGYNPUPR-QBPJDGROSA-N teniposide Chemical compound COC1=C(O)C(OC)=CC([C@@H]2C3=CC=4OCOC=4C=C3[C@@H](O[C@H]3[C@@H]([C@@H](O)[C@@H]4O[C@@H](OC[C@H]4O3)C=3SC=CC=3)O)[C@@H]3[C@@H]2C(OC3)=O)=C1 NRUKOCRGYNPUPR-QBPJDGROSA-N 0.000 description 1
- 229960001278 teniposide Drugs 0.000 description 1
- 229960002180 tetracycline Drugs 0.000 description 1
- 229930101283 tetracycline Natural products 0.000 description 1
- 235000019364 tetracycline Nutrition 0.000 description 1
- 150000003522 tetracyclines Chemical class 0.000 description 1
- 229930183279 tetramycin Natural products 0.000 description 1
- 229940126585 therapeutic drug Drugs 0.000 description 1
- 150000003584 thiosemicarbazones Chemical class 0.000 description 1
- 229940019375 tiludronate Drugs 0.000 description 1
- 229960003087 tioguanine Drugs 0.000 description 1
- RUELTTOHQODFPA-UHFFFAOYSA-N toluene 2,6-diisocyanate Chemical compound CC1=C(N=C=O)C=CC=C1N=C=O RUELTTOHQODFPA-UHFFFAOYSA-N 0.000 description 1
- 229940044693 topoisomerase inhibitor Drugs 0.000 description 1
- 229960000303 topotecan Drugs 0.000 description 1
- 229960005026 toremifene Drugs 0.000 description 1
- XFCLJVABOIYOMF-QPLCGJKRSA-N toremifene Chemical compound C1=CC(OCCN(C)C)=CC=C1C(\C=1C=CC=CC=1)=C(\CCCl)C1=CC=CC=C1 XFCLJVABOIYOMF-QPLCGJKRSA-N 0.000 description 1
- 230000007888 toxin activity Effects 0.000 description 1
- 239000011573 trace mineral Substances 0.000 description 1
- 235000013619 trace mineral Nutrition 0.000 description 1
- 238000003146 transient transfection Methods 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- 102000027257 transmembrane receptors Human genes 0.000 description 1
- 108091008578 transmembrane receptors Proteins 0.000 description 1
- 229950001353 tretamine Drugs 0.000 description 1
- IUCJMVBFZDHPDX-UHFFFAOYSA-N tretamine Chemical compound C1CN1C1=NC(N2CC2)=NC(N2CC2)=N1 IUCJMVBFZDHPDX-UHFFFAOYSA-N 0.000 description 1
- 229960001727 tretinoin Drugs 0.000 description 1
- PXSOHRWMIRDKMP-UHFFFAOYSA-N triaziquone Chemical compound O=C1C(N2CC2)=C(N2CC2)C(=O)C=C1N1CC1 PXSOHRWMIRDKMP-UHFFFAOYSA-N 0.000 description 1
- 229960004560 triaziquone Drugs 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- LZAJKCZTKKKZNT-PMNGPLLRSA-N trichothecene Chemical compound C12([C@@]3(CC[C@H]2OC2C=C(CCC23C)C)C)CO1 LZAJKCZTKKKZNT-PMNGPLLRSA-N 0.000 description 1
- 150000005671 trienes Chemical class 0.000 description 1
- 229960001670 trilostane Drugs 0.000 description 1
- KVJXBPDAXMEYOA-CXANFOAXSA-N trilostane Chemical compound OC1=C(C#N)C[C@]2(C)[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CC[C@@]32O[C@@H]31 KVJXBPDAXMEYOA-CXANFOAXSA-N 0.000 description 1
- NOYPYLRCIDNJJB-UHFFFAOYSA-N trimetrexate Chemical compound COC1=C(OC)C(OC)=CC(NCC=2C(=C3C(N)=NC(N)=NC3=CC=2)C)=C1 NOYPYLRCIDNJJB-UHFFFAOYSA-N 0.000 description 1
- 229960001099 trimetrexate Drugs 0.000 description 1
- 229950000212 trioxifene Drugs 0.000 description 1
- 229960000875 trofosfamide Drugs 0.000 description 1
- UMKFEPPTGMDVMI-UHFFFAOYSA-N trofosfamide Chemical compound ClCCN(CCCl)P1(=O)OCCCN1CCCl UMKFEPPTGMDVMI-UHFFFAOYSA-N 0.000 description 1
- HDZZVAMISRMYHH-LITAXDCLSA-N tubercidin Chemical compound C1=CC=2C(N)=NC=NC=2N1[C@@H]1O[C@@H](CO)[C@H](O)[C@H]1O HDZZVAMISRMYHH-LITAXDCLSA-N 0.000 description 1
- 230000005747 tumor angiogenesis Effects 0.000 description 1
- 230000005740 tumor formation Effects 0.000 description 1
- 238000013414 tumor xenograft model Methods 0.000 description 1
- 230000007306 turnover Effects 0.000 description 1
- 108010087967 type I signal peptidase Proteins 0.000 description 1
- 229940121358 tyrosine kinase inhibitor Drugs 0.000 description 1
- 239000005483 tyrosine kinase inhibitor Substances 0.000 description 1
- 150000004917 tyrosine kinase inhibitor derivatives Chemical class 0.000 description 1
- 229950009811 ubenimex Drugs 0.000 description 1
- 238000002604 ultrasonography Methods 0.000 description 1
- HFTAFOQKODTIJY-UHFFFAOYSA-N umbelliferone Natural products Cc1cc2C=CC(=O)Oc2cc1OCC=CC(C)(C)O HFTAFOQKODTIJY-UHFFFAOYSA-N 0.000 description 1
- ORHBXUUXSCNDEV-UHFFFAOYSA-N umbelliferone Chemical compound C1=CC(=O)OC2=CC(O)=CC=C21 ORHBXUUXSCNDEV-UHFFFAOYSA-N 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- DRTQHJPVMGBUCF-UHFFFAOYSA-N uracil arabinoside Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-UHFFFAOYSA-N 0.000 description 1
- 229960001055 uracil mustard Drugs 0.000 description 1
- 229940045145 uridine Drugs 0.000 description 1
- 208000007089 vaccinia Diseases 0.000 description 1
- 229940099039 velcade Drugs 0.000 description 1
- 230000007332 vesicle formation Effects 0.000 description 1
- GBABOYUKABKIAF-IELIFDKJSA-N vinorelbine Chemical compound C1N(CC=2C3=CC=CC=C3NC=22)CC(CC)=C[C@H]1C[C@]2(C(=O)OC)C1=CC([C@]23[C@H]([C@@]([C@H](OC(C)=O)[C@]4(CC)C=CCN([C@H]34)CC2)(O)C(=O)OC)N2C)=C2C=C1OC GBABOYUKABKIAF-IELIFDKJSA-N 0.000 description 1
- 229960002066 vinorelbine Drugs 0.000 description 1
- CILBMBUYJCWATM-PYGJLNRPSA-N vinorelbine ditartrate Chemical compound OC(=O)[C@H](O)[C@@H](O)C(O)=O.OC(=O)[C@H](O)[C@@H](O)C(O)=O.C1N(CC=2C3=CC=CC=C3NC=22)CC(CC)=C[C@H]1C[C@]2(C(=O)OC)C1=CC([C@]23[C@H]([C@@]([C@H](OC(C)=O)[C@]4(CC)C=CCN([C@H]34)CC2)(O)C(=O)OC)N2C)=C2C=C1OC CILBMBUYJCWATM-PYGJLNRPSA-N 0.000 description 1
- 210000002845 virion Anatomy 0.000 description 1
- 235000012431 wafers Nutrition 0.000 description 1
- XLYOFNOQVPJJNP-OUBTZVSYSA-N water-17o Chemical compound [17OH2] XLYOFNOQVPJJNP-OUBTZVSYSA-N 0.000 description 1
- 229940053867 xeloda Drugs 0.000 description 1
- 150000003742 xyloses Chemical class 0.000 description 1
- 229950009268 zinostatin Drugs 0.000 description 1
- 229940002005 zometa Drugs 0.000 description 1
- 229960000641 zorubicin Drugs 0.000 description 1
- FBTUMDXHSRTGRV-ALTNURHMSA-N zorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(\C)=N\NC(=O)C=1C=CC=CC=1)[C@H]1C[C@H](N)[C@H](O)[C@H](C)O1 FBTUMDXHSRTGRV-ALTNURHMSA-N 0.000 description 1
Images














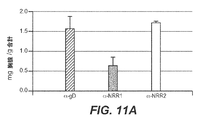






























Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/30—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants from tumour cells
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/30—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants from tumour cells
- C07K16/3053—Skin, nerves, brain
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/55—Fab or Fab'
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/567—Framework region [FR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
Description
本出願は、2008年10月1日に出願した米国仮特許出願番号61/101917の優先権を主張する。その開示は出典明記によって完全に本明細書において援用される。
本発明は、概して分子生物学の分野に関する。特に、本発明は、抗Notch2ネガティブ制御領域(NRR)抗体含む抗Notch抗体とその使用に関する。抗Notch1 NRR抗体とその使用方法も提供される。
Notch2 NTMは、細胞外領域(S2切断部位を内部に持つ)、膜貫通部分(S3切断部位を内部に持つ)、及びRAMドメイン、6つのアンキリンリピート、トランスアクチベーションドメイン及びカルボキシ末端PEST配列を含む大きな細胞内部分を含む。ECN及びNTMサブユニットの安定な結合は、ECNのカルボキシ終末端(HD−Nと称される)及びNTMの細胞外アミノ末端(HD−Cと称される)を含むヘテロ二量体化ドメイン(HD)に依存している。リガンド誘導性の活性化の前に、Notchは、3つのLNRとHDドメインを含むネガティブ制御領域(NRR)によって静止立体構造に維持される。Notch2 NRRの結晶構造は、Gordon et al., (2007) Nature Structural & Molecular Biology 14:295-300, 2007において報告される。
ECNサブユニットへのNotchリガンドの結合により、制御された膜内タンパク質分解により生じる2つの連続的なタンパク質分解性切断が開始する。部位S2のメタロプロテアーゼ(ADAM17)による第一切断によって、Notch膜貫通サブユニットの、原形質膜の内側の小葉の近くの部位S3での第二切断が起こりやすくなる。プレセニリンとニカストリンを含む多タンパク質複合体により触媒される部位S3切断は、Notch膜貫通サブユニットの細胞内部分を遊離し、核に転位させ、標的遺伝子の転写を活性化する。(Notchのタンパク分解性切断の概要については、Sisodia et al., Nat. Rev. Neurosci. 3:281-290, 2002を参照のこと)。
特許出願及び出版物を含め本明細書において引用するすべての文献は、出典明記によってすべてが援用される。
一態様では、Notch2 NRRに結合するモノクローナル抗体が提供される。一実施態様では、抗体は、Notch2活性を阻害する。他の実施態様では、抗体は、Notch2以外のNotchファミリに有意に結合しない。他の実施態様では、抗体は、マウスNotch2 NRRおよびヒトNotch2 NRRに結合する。他の実施態様では、抗体は、10nM以下のKdでNotch2 NRRに結合する。
さらなる実施態様では、Notch2 NRRに結合するモノクローナル抗体であって、
(a) 配列番号:3のコンセンサス配列と一致するアミノ酸配列を含むHVR-H1;
(b) 配列番号:4のアミノ酸配列を含むHVR-H2;
(c) 配列番号:5のアミノ酸配列を含むHVR-H3;
(d) 配列番号:10のコンセンサス配列に一致するアミノ酸配列を含むHVR-L1;
(e) 配列番号:14のコンセンサス配列に一致するアミノ酸配列を含むHVR-L2;及び、
(f) 配列番号:19のコンセンサス配列に一致するアミノ酸配列を含むHVR-L3
を含む抗体が提供される。
そのような実施態様では、抗体は、配列番号:1−2から選択されるアミノ酸配列を含むHVR-H1;配列番号:6−9から選択されるアミノ酸配列を含むHVR-L1;配列番号:11−13から選択されるアミノ酸配列を含むHVR-L2;そして、配列番号:15−18から選択されるアミノ酸配列を含むHVR-L3を含む。そのような実施態様では、HVR-H1は配列番号:1のアミノ酸配列を含み、HVR-L1は配列番号:6のアミノ酸配列を含み、HVR-L2は配列番号:11のアミノ酸配列を含み、そして、HVR-L3は配列番号:15のアミノ酸配列を含む。他の実施態様では、HVR-H1は配列番号:2のアミノ酸配列を含み、HVR-L1は配列番号:7のアミノ酸配列を含み、HVR-L2は配列番号:11のアミノ酸配列を含み、そして、HVR-L3は配列番号:16のアミノ酸配列を含む。他の実施態様では、HVR-H1は配列番号:2のアミノ酸配列を含み、HVR-L1は、配列番号:8のアミノ酸配列を含み、HVR-L2は配列番号:12のアミノ酸配列を含み、そして、HVR-L3は配列番号:17のアミノ酸配列を含む。他の実施態様では、HVR-H1は配列番号:2のアミノ酸配列を含み、HVR-L1は配列番号:9のアミノ酸配列を含み、HVR-L2は配列番号:13のアミノ酸配列を含み、そして、HVR-L3は配列番号:18のアミノ酸配列を含む。上記何れかの実施態様では、抗体は、ヒトVHアクセプター2フレームワークおよびヒトVLκサブグループIコンセンサスフレームワークから選択される少なくとも一のフレームワークをさらに含む。
他の態様では、抗体D、抗体D-1、抗体D-2又は抗体D-3から選択した抗体と同じエピトープに結合する単離された抗体が、提供される。他の態様では、Notch2のLNR−AドメインおよびHD−Cドメインに結合する単離された抗体が、提供される。
他の態様では、抗Notch2 NRR抗体は、Fab、Fab'-SH、Fv、scFv又は(Fab')2の断片から選択される抗体断片である。他の態様では、抗Notch2 NRR抗体は、ヒト化、キメラ又はヒトの抗体である。
上記何れかの実施態様は単独でも組み合わせてもよい。
他の態様では、Notch1の発現又は活性の増加と関連する疾患の治療方法であって、必要とする被検体に、抗Notch1 NRR抗体と、デキサメサゾンおよびタモキシフェンから選択した治療薬の有効量を同時に投与することを含み、該治療薬が抗体によって生じる変化した腸管細胞の分化を低減する方法が、提供される。そのような実施態様では、疾患はT細胞悪性腫瘍である。
本発明は、Notchに結合する単離された抗体、および例えばNotchの発現又は活性と関連する疾患の診断又は治療のための同抗体の使用方法を提供する。
本明細書中に記載又は引用される技術及び手順は、一般に、十分に理解されるものであり、当業者によって、従来の方法論を用いて共通して実施されるものである。その例として、以下の文献に記載される方法論が広く利用されている。
Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.;Current Protocols in Molecular Biology (F. M. Ausubel, et al. eds., (2003));the series Methods in Enzymology (Academic Press, Inc.): PCR 2: A Practical Approach (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) Antibodies, A Laboratory Manual, and Animal Cell Culture (R. I. Freshney, ed. (1987));Oligonucleotide Synthesis (M. J. Gait, ed., 1984);Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J. E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R. I. Freshney), ed., 1987);Introduction to Cell and Tissue Culture (J. P. Mather and P. E. Roberts, 1998) Plenum Press;Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J. B. Griffiths, and D. G. Newell, eds., 1993-8) J. Wiley and Sons;Handbook of Experimental Immunology (D. M. Weir and C. C. Blackwell, eds.);Gene Transfer Vectors for Mammalian Cells (J. M. Miller and M. P. Calos, eds., 1987);PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994);Current Protocols in Immunology (J. E. Coligan et al., eds., 1991);Short Protocols in Molecular Biology (Wiley and Sons, 1999);Immunobiology (C. A. Janeway and P. Travers, 1997);Antibodies (P. Finch, 1997);Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989);Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000);Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999);The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995);及び、Cancer: Principles and Practice of Oncology (V. T. DeVita et al., eds., J.B. Lippincott Company, 1993)。
本出願を理解するために、以下の定義を適用し、必要な場合はいつでも、単数で使用した用語には複数形も含まれ、その逆もそうである。以下に示すいずれかの定義が出典明記によって本明細書中に援用されるいずれかの文献と矛盾する場合、以下の定義を調節してよい。
「単離された」抗体とは、その自然環境の成分から同定され分離され及び/又は回収されたものである。その自然環境の汚染成分とは、抗体の研究、診断又は治療的な使用を妨害する物質であり、酵素、ホルモン、及び他のタンパク質様又は非タンパク質様溶質が含まれる。ある実施態様では、タンパク質は、(1)例えばローリー法で測定した場合95%を超える抗体、ある実施態様では99重量%を超えるまで、(2)例えばスピニングカップシークエネーターを使用することにより、少なくとも15のN末端あるいは内部アミノ酸配列の残基を得るのに充分なほど、あるいは、(3)例えばクーマシーブルーあるいは銀染色を用いた非還元あるいは還元条件下でのSDS-PAGEにより均一になるまで十分なほど精製される。抗体の自然環境の少なくとも一の成分が存在しないため、単離された抗体には、組換え細胞内のインサイツでの抗体が含まれる。しかしながら、通常は、単離された抗体は少なくとも一の精製工程により調製される。
「抗Notch2 NRR抗体」又は「Notch2 NRRに結合する抗体」なる用語は、十分な親和性でNotch2 NRRを結合することができ、Notch2を標的とする診断及び/又は治療上の薬剤として有用である抗体を指す。好ましくは、関係のないNotch以外のタンパク質に対する抗Notch2 NRR抗体の結合の程度は、例えば、ラジオイムノアッセイ(RIA)によって測定するところの、Notch2 NRRへの抗体の結合のおよそ10%未満である。ある実施態様では、Notch2 NRRに結合する抗体は、1μM以下、100nM以下、10nM以下、1nM以下、又は0.1nM以下の解離定数(Kd)を有する。ある実施態様では、抗Notch2 NRR抗体は、異なる種、例えば齧歯動物(マウス、ラット)および霊長類のNotch間で保存されるNotchのエピトープに結合する。
「可変」という用語は、可変ドメインのある部位が、抗体の中で配列が広範囲に異なっており、その特定の抗原に対する各特定の抗体の結合性及び特異性に使用されているという事実を意味する。しかしながら、可変性は抗体の可変ドメインにわたって一様には分布していない。軽鎖及び重鎖の可変ドメインの両方の高頻度可変領域(HVR)と呼ばれる3つのセグメントに濃縮される。可変ドメインのより高度に保持された部分はフレームワーク領域(FR)と呼ばれる。天然の重鎖及び軽鎖の可変ドメインは、βシート構造を結合し、ある場合にはその一部を形成するループ結合を形成する、3つのHVRにより連結されたβシート配置を主にとる4つのFR領域をそれぞれ含んでいる。各鎖のHVRは、FRによって近接して結合され、他の鎖のHVRと共に、抗体の抗原結合部位の形成に寄与している(Kabatら, Sequence of Proteins ofImmunological Interest, 5th Ed. National Institutes of Health, BEthesda, MD. (1991))。定常ドメインは、抗体の抗原への結合に直接関連しているものではないが、種々のエフェクター機能、例えばADCCへの抗体の関与を示す。
その重鎖の定常ドメインのアミノ酸配列に基づいて、抗体(免疫グロブリン)は異なるクラスが割り当てられる。免疫グロブリンには5つの主なクラスがある:IgA、IgD、IgE、IgG及びIgM、更にそれらは、IgG1、IgG2、IgG3、IgG4、IgA1、及びIgA2等のサブクラス(アイソタイプ)に分かれる。免疫グロブリンの異なるクラスに対応する重鎖定常ドメインはそれぞれα、δ、ε、γ、及びμと呼ばれる。免疫グロブリンの異なるクラスのサブユニット構造及び三次元立体配位はよく知られており、例えば、Abbas等.Cellular and Mol. Immunology, 第4版(2000)(W.B. Saunders, Co., 2000)に概説されている。抗体は、抗体と1以上の他のタンパク質又はペプチドとの共有結合性又は非共有結合性の会合により形成された融合大分子の一部であり得る。
本明細書中の「ネイキッド抗体」は、細胞障害性部分又は放射性標識にコンジュゲートしていない抗体である。
「抗体断片」は、好ましくは抗原結合領域を含む、インタクトな抗体の一部を含む。抗体断片の例には、Fab、Fab'、F(ab')2およびFv断片;ダイアボディ;線形抗体;単鎖抗体分子;及び抗体断片から形成される多特異性抗体が含まれる。
「Fv」は、完全な抗原結合部位を含む最小抗体断片である。ある実施態様では、二本鎖Fv種は、堅固な非共有結合をなした一つの重鎖及び一つの軽鎖可変ドメインの二量体からなる。一本鎖Fv(scFv)種では、柔軟なペプチドリンカーによって1の重鎖及び1の軽鎖可変ドメインは共有結合性に連鎖することができ、よって軽鎖及び重鎖は、二本鎖Fv種におけるものと類似の「二量体」構造に連結することができる。この配置において、各可変ドメインの3つのHVRは相互に作用してVH-VL二量体表面に抗原結合部位を形成する。集合的に、6つのHVRが抗体に抗原結合特異性を付与する。しかし、単一の可変ドメイン(又は抗原に対して特異的な3つのHVRのみを含むFvの半分)でさえ、全結合部位よりも親和性が低くなるが、抗原を認識して結合する能力を有している。
「一本鎖Fv」又は「scFv」抗体断片は、抗体のVH及びVLドメインを含み、これらのドメインは単一のポリペプチド鎖に存在する。通常、scFvポリペプチドはVH及びVLドメイン間にポリペプチドリンカーを更に含み、それはscFvが抗原結合に望まれる構造を形成するのを可能にする。scFvの概説については、The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg及びMoore編, Springer-Verlag, New York, pp. 269-315 (1994)のPluckthunを参照のこと。
「モノクローナル」との形容は、抗体の、実質的に均一な抗体の集団から得られたものであるという特性を示し、抗体を何か特定の方法で生産しなければならないことを意味するものではない。例えば、本発明に従って使用されるモノクローナル抗体は様々な技術によって作製することができ、それらの技術には、例えば、ハイブリドーマ法(例えば、Kohler and Milstein, Nature, 256:495-97 (1975);Hongo等, Hybridoma, 14 (3): 253-260 (1995);Harlow等, Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988);Hammerling等: Monoclonal Antibodies and T-Cell hybridomas 563-681 (Elsevier, N. Y., 1981))、組換えDNA法(例えば、米国特許第4816567号参照)、ファージディスプレイ技術(例えば、Clackson等, Nature, 352:624-628 (1991);Marks等, J. Mol. Biol. 222:581-597 (1992);Sidhu等, J. Mol. Biol. 338(2): 299-310 (2004);Lee等, J. Mol. Biol. 340(5);1073-1093 (2004);Fellouse, Proc. Natl. Acad. Sci. USA 101(34):12467-12472 (2004);及びLee等, J. Immounol. Methods 284(1-2): 119-132 (2004))、並びに、ヒト免疫グロブリン座位の一部又は全部、或るいはヒト免疫グロブリン配列をコードする遺伝子を有する動物にヒト又はヒト様抗体を生成する技術(例えば、国際公開第98/24893号;国際公開第96/34096号;国際公開第96/33735号;国際公開第91/10741号;Jakobovits等, Proc. Natl. Acad. Sci. USA 90: 2551 (1993);Jakobovits等, Nature 362: 255-258 (1993);Bruggemann等, Year in Immunol. 7:33 (1993);米国特許第5545807号;同第5545806号;同第5569825号;同第5625126号;同第5633425号;同第5661016号; Marks等, Bio.Technology 10: 779-783 (1992);Lonberg等, Nature 368: 856-859 (1994);Morrison, Nature 368: 812-813 (1994);Fishwild等, Nature Biotechnol. 14: 845-851 (1996);Neuberger, Nature Biotechnol. 14: 826 (1996)及びLonberg及びHuszar, Intern. Rev. Immunol. 13: 65-93 (1995)参照)が含まれる。
多数のHVRの描写が使用され、ここに含まれる。カバット相補性決定領域(CDR)は配列変化に基づいており、最も一般的に使用されている(Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991))。Chothiaは、代わりに構造的ループの位置に言及している(Chothia and Lesk J. Mol. Biol. 196:901-917 (1987))。AbM HVRは、カバットHVRとChothia構造的ループの間の妥協を表し、Oxford MolecularのAbM抗体モデリングソフトウェアにより使用される。「接触」HVRは、利用できる複合体結晶構造の分析に基づく。これらHVRのそれぞれからの残基を以下に示す。
ループ カバット AbM Chothia 接触
L1 L24-L34 L24-L34 L26-L32 L30-L36
L2 L50-L56 L50-L56 L50-L52 L46-L55
L3 L89-L97 L89-L97 L91-L96 L89-L96
H1 H31-H35B H26-H35B H26-H32 H30-H35B (カバット番号付け)
H1 H31-H35 H26-H35 H26-H32 H30-H35 (Chothia番号付け)
H2 H50-H65 H50-H58 H53-H55 H47-H58
H3 H95-H102 H95-H102 H96-H101 H93-H101
「フレームワーク」又は「FR」残基は、ここで定義する高頻度可変領域残基以外の可変ドメイン残基である。
カバット番号付けシステムは一般に、可変ドメイン内の残基を指す場合に用いられる(軽鎖のおよそ残基1−107及び重鎖の残基1−113)(例えばKabat et al., Sequences of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991))。「EU番号付けシステム」又は「EUインデックス」は一般に、イムノグロブリン重鎖定常領域を指す場合に用いられる(例えば上掲のKabat et al.において報告されたEUインデックス)。「カバットにおけるEUインデックス」はヒトIgG1 EU抗体の残基番号付けを指す。本明細書中で特に明記しない限り、抗体の可変ドメイン内の残基番号の参照は、カバット番号付けシステムによって番号付けする残基を意味する。本明細書中で特に明記しない限り、抗体の定常ドメイン内の残基番号の参照は、EU番号付けシステムによる残基番号付けを意味する(例えば、米国特許出願2008/0181888A1のEU番号付けに関する図を参照のこと)。
「ブロック(遮断)」抗体又は「アンタゴニスト」抗体は、結合する抗原の生物学的活性を阻害するか又は低減するものである。あるブロック抗体又はアンタゴニスト抗体は、抗原の生物学的活性を実質的ないしは完全に阻害する。
本明細書中で用いる「アゴニスト抗体」は、対象のポリペプチドの機能的な活性の少なくとも一を部分的ないし完全に模倣する抗体である。
「アポトーシスを誘発する」抗体とは、アネキシンVの結合、DNAの断片化、細胞の縮み、小胞体の拡張、細胞断片化、及び/又は膜小胞の形成(アポトーシス性本体と呼ばれる)といった、標準的なアポトーシスアッセイにより決定される、プログラムされた細胞死を誘発するものである。
「Fc領域」なる用語は、天然配列Fc領域及び変異形Fc領域を含む、免疫グロブリン重鎖のC末端領域を定義するために使用される。免疫グロブリン重鎖のFc領域の境界は変化するかも知れないが、通常、ヒトIgG重鎖Fc領域はCys226の位置又はPro230からの位置のアミノ酸残基からFc領域のカルボキシル末端まで伸長すると定義される。Fc領域のC末端リジン(EU番号付けシステムによれば残基447)は、例えば、抗体の産生又は精製中に、又は抗体の重鎖をコードする核酸を組み換え遺伝子操作することによって取り除かれてもよい。したがって、インタクト抗体の組成物は、すべてのK447残基が除去された抗体群、K447残基が除去されていない抗体群、及びK447残基を有する抗体と有さない抗体の混合を含む抗体群を含みうる。
「機能的Fc領域」は、天然配列Fc領域の「エフェクター機能」を有する。例示的「エフェクター機能」には、C1q結合、補体依存性細胞障害作用(CDC)、Fcレセプター結合、抗体依存性細胞媒介性細胞障害作用(ADCC)、食作用、細胞表面レセプター(例えばB細胞レセプター;BCR)の下方制御などが含まれる。そのようなエフェクター機能は、通常、Fc領域が結合ドメイン(例えば、抗体可変ドメイン)と組み合わさることを必要とし、例えば本明細書中の定義に開示される様々なアッセイを使用して評価される。
「変異体Fc領域」は、少なくとも1つのアミノ酸修飾、好ましくは一又は複数のアミノ酸置換により、天然配列のFc領域とは異なるアミノ酸配列を包含するものである。好ましくは、変異体Fc領域は、天然配列のFc領域もしくは親ポリペプチドのFc領域と比較した場合、少なくとも1つのアミノ酸置換、例えば、天然配列のFc領域又は親のポリペプチドのFC領域におよそ1からおよそ10のアミノ酸置換、好ましくはおよそ1からおよそ5のアミノ酸置換を有する。本明細書中の変異体Fc領域は、好ましくは、天然配列のFc領域及び/又は親ポリペプチドのFc領域と、少なくともおよそ80%の同一性を有するか、最も好ましくは少なくともおよそ90%の配列同一性を、より好ましくは少なくともおよそ95%の配列又はそれ以上の同一性を有するものであろう。
インビボでのヒトFcRnへの結合とヒトFcRn高親和性結合ポリペプチドの血清半減期は、例えばヒトFcRnを発現するトランスジェニックマウス又は形質転換されたヒト細胞株、又は変異体Fc領域を有するポリペプチドを投与された霊長類動物においてアッセイすることができる。国際公開公報00/42072(Presta) にFcRへの結合を向上又は減弱させた抗体変異型が述べられている。例としてShields等, J. Biol. Chem. 9(2): 6591-6604 (2001)も参照のこと。
他の実施態様によると、〜10反応単位(RU)の固定した抗原CM5チップを用いて25℃のBIAcore(登録商標)-2000又はBIAcore(登録商標)-3000(BIAcore, Inc., Piscataway, NJ)にて表面プラズモン共鳴アッセイを行ってKd又はKd値を測定する。簡単に言うと、カルボキシメチル化デキストランバイオセンサーチップ(CM5, BIAcore Inc.)を、提供者の指示書に従ってN-エチル-N'-(3-ジメチルアミノプロピル)-カルボジイミド塩酸塩(EDC)及びN-ヒドロキシスクシニミド(NHS)で活性化した。抗原を10mM 酢酸ナトリウム(pH4.8)で5μg/ml(〜0.2μM)に希釈し、結合したタンパク質の反応単位(RU)がおよそ10になるように5μl/分の流速で注入した。抗原の注入後、反応しない群をブロックするために1Mのエタノールアミンを注入した。動力学的な測定のために、2倍の段階希釈したFab(0.78nMから500nM)を25℃、およそ25μl/分の流速で0.05%Tween20TM界面活性剤(PBST)を含むPBSに注入した。会合及び解離のセンサーグラムを同時にフィットさせることによる単純一対一ラングミュア結合モデル(simple one-to-one Langmuir binding model) (BIAcore Evaluationソフトウェアバージョン3.2)を用いて、会合速度(kon)と解離速度(koff)を算出した。平衡解離定数(Kd)をkoff/kon比として算出した。例として、Chen, Y.,等, (1999) J. Mol Biol 293:865-881を参照。上記の表面プラスモン共鳴アッセイによる結合速度が106M−1S−1を上回る場合、分光計、例えば、流動停止を備えた分光光度計(stop-flow equipped spectrophometer)(Aviv Instruments)又は撹拌キュベットを備えた8000シリーズSLM-Aminco分光光度計(ThermoSpectronic)で測定される、漸増濃度の抗原の存在下にて、PBS(pH7.2)、25℃の、20nMの抗抗原抗体(Fab型)の蛍光放出強度(励起=295nm;放出=340nm、帯域通過=16nm)における増加又は減少を測定する蛍光消光技術を用いて結合速度を測定することができる。
また、本発明の「結合速度」又は「会合の速度」又は「会合速度」又は「kon」は、〜10反応単位(RU)の固定した抗原CM5チップを用いて25℃のBIAcore(登録商標)-2000又はBIAcore(登録商標)-3000(BIAcore, Inc., Piscataway, NJ)を用いた前述と同じ表面プラズモン共鳴アッセイにて測定される。
ここで用いる「実質的に低減する」又は「実質的に異なる」という句は、当業者が2つの数値(一般に、分子に関連するもの、及び参照/比較分子に関連する他のもの)の差異に、該値(例えばKd値)によって測定される生物学的性質上統計学的に有意であると認められるほど、2つの数値が有意に異なっていることを意味する。前記2つの値間の差異は、参照/比較分子の値の、例えば約10%より大きく、約20%より大きく、約30%より大きく、約40%より大きく、及び/又は約50%より大きい。
「VLサブグループIコンセンサスフレームワーク」は、上掲のKabat等の可変軽鎖κサブグループIのアミノ酸配列から得られたコンセンサス配列を含んでなる。一実施態様では、VHサブグループIコンセンサスフレームワークアミノ酸配列は、以下の配列:(配列番号:60)−L1−(配列番号:61)−L2−(配列番号:62)−L3−(配列番号:63)の配列のそれぞれの少なくとも一部又は全部を含む。
「単離された」核酸分子は、例えば天然の環境に通常付随している少なくとも一の他の核酸分子から分離された核酸分子である。さらに、単離された核酸分子は、例えば、核酸分子を通常発現するが、その核酸分子がその天然の染色体位置と異なる染色体位置にあるか又は染色体外に存在する、細胞に含まれる核酸分子を含む。
ここで使用される「オリゴヌクレオチド」とは、短く、一般的に単鎖であり、また必ずしもそうではないが、一般的に約200未満のヌクレオチド長さの、一般的に合成のポリヌクレオチドを意味する。「オリゴヌクレオチド」及び「ポリヌクレオチド」なる用語は、相互に排他的なものではない。ポリヌクレオチドについての上述した記載はオリゴヌクレオチドと等しく、十分に適用可能である。
本明細書において使われるように、特に明記しない限り、「Notch1」なる用語は、霊長類(例えばヒト)および齧歯動物(例えばマウスおよびラット)のような哺乳動物を含む任意の脊椎動物の供与源由来の任意の天然のNotch1を指す。この用語は、「完全長」、プロセシングされていないNotch1並びに細胞内のプロセシングから生じるNotch1の任意の形態を包含する。また、この用語は、Notch1の天然に生じる変異体、例えばスプライス変異体又は対立遺伝子変異体を包含する。
本明細書において使われるように、特に明記しない限り、「Notch2」なる用語は、霊長類(例えばヒト)および齧歯動物(例えばマウスおよびラット)のような哺乳動物を含む任意の脊椎動物の供与源由来の任意の天然のNotch2を指す。この用語は、「完全長」、プロセシングされていないNotch2並びに細胞内のプロセシングから生じるNotch2の任意の形態を包含する。また、この用語は、Notch2の天然に生じる変異体、例えばスプライス変異体又は対立遺伝子変異体を包含する。
「Notch2 NRR」なる用語は、3つのLNRモジュール(LNR−A、LNR−BおよびLNR−C)およびHDドメイン(HD−NおよびHD−C)からなるNotch2の領域を指す。例示的なヒトおよびマウスのNotch2 NRR配列は、図18(それぞれ配列番号:28および29)に示される。Notch2 NRRは、例えばS1でのNotch2のプロセシングにより生じる非共有結合断片、並びに単一の近接するポリペプチド配列からなってよい。例として、そして、図18に示すように、ヒトNotch2 NRRは、ヒトNotch2(配列番号:75)のアミノ酸1422−1677、あるいは、配列番号:75のアミノ酸1609−1677に非共有的に結合した配列番号:75のアミノ酸1422−1608からなってよい。
分率X/Yの100倍
ここで、Xは配列アラインメントプログラムALIGN-2のA及びBのプログラムアラインメントによって同一であると一致したスコアのアミノ酸残基の数であり、YはBの全アミノ酸残基数である。アミノ酸配列Aの長さがアミノ酸配列Bの長さと異なる場合、AのBに対する%アミノ酸配列同一性は、BのAに対する%アミノ酸配列同一性とは異なることは理解されるであろう。特に断らない限りは、ここでの全ての%アミノ酸配列同一性値は、直ぐ上のパラグラフに示したようにALIGN-2コンピュータプログラムを用いて得られる。
これには、哺乳動物の問題の疾患の素因になる病的状態を含み、慢性及び急性の疾患が含まれる。本明細書において治療される疾患の非限定的な例は、癌などの状態等である。
「B細胞新生物」とも称される「B細胞悪性腫瘍」には、ホジキン病(例えばリンパ球優性ホジキン病(LPHD);非ホジキンリンパ腫(NHL);濾胞性中心細胞リンパ腫(FCC);急性リンパ性白血病(ALL、B−ALLとも称される);前駆体B−リンパ芽球性白血病;慢性リンパ球性白血病(CLL、B−CLLとも称される);ヘアリー細胞白血病;周辺帯B細胞リンパ腫(結節性、MALTおよび脾臓タイプを含む);多発性骨髄腫(形質細胞腫、プラズマ細胞骨髄腫);小さい非切れ込み型細胞リンパ腫(例えばバーキットリンパ腫);大細胞リンパ腫(びまん性大細胞(B細胞)、びまん性混合細胞および免疫芽球性リンパ腫を含む);マントル細胞リンパ腫;小リンパ球性リンパ腫;AIDS関連のリンパ腫およびワルデンシュトレームマクログロブリン血症が含まれるがこれらに限定されない。緩徐進行性リンパ腫は遅生育の不治の疾患であり、平均的な患者は多くの寛解と再燃の期間を経て6〜10年生き延びる。悪性のリンパ腫は、高悪性度及びいくつかの分類の中悪性度のNHLを含む、即育型のリンパ腫である。
「メラノーマ」なる用語は、初めに皮膚に見られるが、眼(ブドウ膜メラノーマ)又は腸にも見られるメラニン細胞の悪性腫瘍を指す。
「個体」、「被検体」又は「患者」は脊椎動物である。ある実施態様では、脊椎動物は哺乳動物である。哺乳動物には、限定するものではないが、家畜動物(ウシなど)、スポーツ用動物、愛玩動物(ネコ、イヌ及びウマ)、霊長類、マウス及びラットが含まれる。ある実施態様では、哺乳動物はヒトである。
「無菌の」製剤は、防腐性であるか、又は生きているすべての微生物およびそれらの胞子を含まない。
「有効量」とは所望の治療又は予防結果を達成するために必要な用量及び時間での効果的な量を意味する。
本発明の物質/分子の「治療的有効量」は、例えば個体の疾病ステージ、年齢、性別、及び体重、並びに個体に所望の応答を誘発する物質/分子の能力などの因子に従って変わりうる。また、治療的有効量は物質/分子の任意の毒性又は有害な効果よりも治療的に恩恵のある効果が上回る量を包含する。「予防的有効量」とは所望の予防結果を達成するために必要な用量及び時間での効果的な量を意味する。典型的には必ずではないが、予防的用量は疾患の初期ステージ又はその前の患者に用いるので、予防的有効量は治療的有効量よりも少ないであろう。
「毒素」は、細胞の成長又は増殖に対して有害な作用を有することができる任意の物質である。
本発明は、Notch結合薬剤、例えば抗体ないしその断片の同定にある程度基づく。特定の実施態様では、本発明は、Notch2ネガティブ制御領域(NRR)結合剤、例えばNotch2 NRR及びその断片を結合する単離された抗体に関する。このような抗体は、例えば、Notch2の発現および/または活性(例えば発現又は活性の増加)と関係する疾患の診断又は治療に有用である。特定の実施態様では、抗Notch2 NRR抗体は、癌、例えばB細胞悪性腫瘍およびメラノーマの診断又は治療に有用である。したがって、本発明は、Notch2 NRR結合に関連した方法、組成物、キットおよび製造品を提供する。
1.例示的なファージライブラリ由来の抗体
実施例Bに記載のように、ファージライブラリから得られた例示的なモノクローナル抗体は、本明細書において提供される。Notch2 NRRに結合する「抗体D」と称する抗体を単離した。その抗体は、抗体D-1、抗体D-2および抗体D-3と称する抗体を生成するために親和性成熟した。抗体D、抗体D-1、抗体D-2および抗体D-3の重鎖及び軽鎖の高頻度可変領域(HVR)は、図1および2に示す。抗体D、抗体D-1、抗体D-2および抗体D-3の重鎖および軽鎖の可変ドメインの配列は、図3および4に示す。さらに、抗Notch2 NRR抗体の実施態様は以下の通りに提供する。
一態様では、Notch2 NRRに特異的に結合する抗体であって、
(a) 配列番号:3のコンセンサス配列と一致するアミノ酸配列を含むHVR-H1;
(b) 配列番号:4のアミノ酸配列を含むHVR-H2;
(c) 配列番号:5のアミノ酸配列を含むHVR-H3;
(d) 配列番号:10のコンセンサス配列に一致するアミノ酸配列を含むHVR-L1;
(e) 配列番号:14のコンセンサス配列に一致するアミノ酸配列を含むHVR-L2;及び、
(f) 配列番号:19のコンセンサス配列に一致するアミノ酸配列を含むHVR-L3
から選択される少なくとも1、2、3、4、5又は6のHVRを含む抗体が提供される。
更なる態様では、抗体は、配列番号:5のアミノ酸配列を含むHVR-H3と、上記の(a)、(b)、(d)、(e)および(f)から選択される少なくとも1、2、3、4又は5のHVRとを含む。更なる態様では、抗体は上記の(a)、(b)、(c)、(d)、(e)および(f)を含む。(a)、(d)、(e)および(f)について、以下の何れか一又は複数の実施態様が考慮される。HVR-H1は、配列番号:1−2から選択されるアミノ酸配列を含む。HVR-L1は、配列番号:6−9から選択されるアミノ酸配列を含む。HVR-L2は、配列番号:11−13から選択されるアミノ酸配列を含む。そして、HVR−L3は、配列番号:15−18から選択されるアミノ酸配列を含む。
他の態様では、Notch2 NRRに特異的に結合する抗体であって、配列番号:10のコンセンサス配列に一致するアミノ酸配列を含むHVR-L1、配列番号:14のコンセンサス配列に一致するアミノ酸配列を含むHVR-L2、および配列番号:19のコンセンサス配列に一致するアミノ酸配列を含むHVR-L3を含む抗体が提供される。以下の実施態様は何れかの組み合わせて考慮される。HVR-L1は、配列番号:6−9から選択されるアミノ酸配列を含む。HVR-L2は、配列番号:11−13から選択されるアミノ酸配列を含む。そして、HVR−L3は、配列番号:15−18から選択されるアミノ酸配列を含む。一実施態様では、Notch2 NRRに結合する抗体は、配列番号:6のアミノ酸配列を含むHVR-L1;配列番号:11のアミノ酸配列を含むHVR-L2;及び配列番号:15のアミノ酸配列を含むHVR-L3を含む。他の実施態様では、Notch2 NRRに結合する抗体は、配列番号:7のアミノ酸配列を含むHVR-L1;配列番号:11のアミノ酸配列を含むHVR-L2;及び配列番号:16のアミノ酸配列を含むHVR-L3を含む。他の実施態様では、Notch2 NRRに結合する抗体は、配列番号:8のアミノ酸配列を含むHVR-L1;配列番号:12のアミノ酸配列を含むHVR-L2;及び配列番号:17のアミノ酸配列を含むHVR-L3を含む。他の実施態様では、Notch2 NRRに結合する抗体は、配列番号:9のアミノ酸配列を含むHVR-L1;配列番号:13のアミノ酸配列を含むHVR-L2;及び配列番号:18のアミノ酸配列を含むHVR-L3を含む。
(a) 配列番号:1のアミノ酸配列を含むHVR-H1;
(b) 配列番号:4のアミノ酸配列を含むHVR-H2;
(c) 配列番号:5のアミノ酸配列を含むHVR-H3;
(d) 配列番号:6のアミノ酸配列を含むHVR-L1;
(e) 配列番号:11のアミノ酸配列を含むHVR-L2;及び
(f) 配列番号:15のアミノ酸配列を含むHVR-L3
を含む抗体が提供される。
他の実施態様では、Notch2 NRRに特異的に結合する抗体であって、
(a) 配列番号:2のアミノ酸配列を含むHVR-H1;
(b) 配列番号:4のアミノ酸配列を含むHVR-H2;
(c) 配列番号:5のアミノ酸配列を含むHVR-H3;
(d) 配列番号:7のアミノ酸配列を含むHVR-L1;
(e) 配列番号:11のアミノ酸配列を含むHVR-L2;及び
(f) 配列番号:16のアミノ酸配列を含むHVR-L3
を含む抗体が提供される。
他の実施態様では、Notch2 NRRに特異的に結合する抗体であって、
(a) 配列番号:2のアミノ酸配列を含むHVR-H1;
(b) 配列番号:4のアミノ酸配列を含むHVR-H2;
(c) 配列番号:5のアミノ酸配列を含むHVR-H3;
(d) 配列番号:8のアミノ酸配列を含むHVR-L1;
(e) 配列番号:12のアミノ酸配列を含むHVR-L2;及び
(f) 配列番号:17のアミノ酸配列を含むHVR-L3
を含む抗体が提供される。
他の実施態様では、Notch2 NRRに特異的に結合する抗体であって、
(a) 配列番号:2のアミノ酸配列を含むHVR-H1;
(b) 配列番号:4のアミノ酸配列を含むHVR-H2;
(c) 配列番号:5のアミノ酸配列を含むHVR-H3;
(d) 配列番号:9のアミノ酸配列を含むHVR-L1;
(e) 配列番号:13のアミノ酸配列を含むHVR-L2;及び
(f) 配列番号:18のアミノ酸配列を含むHVR-L3
を含む抗体が提供される。
ある実施態様では、抗Notch2 NRR抗体は親和性成熟している。例えば、示したHVR位置(カバット番号による)内に以下の何れか一又は複数の置換が何れかの組合せでなされていてよい。
- HVR-H1(配列番号:1)において:S28T;T30S;
- HVR-L1(配列番号:6)において:S28N;I29N又はV;S30R又はK;S31R;Y32F
- HVR-L2(配列番号:11)において:G50R;S53I又はT;A55E
- HVR-L3(配列番号:15)において:S93I又はR;L96W又はH
本明細書中に開示する特定の抗体、すなわち抗体D並びに抗体Dの親和性成熟した形態(D-1、D-2及びD-3)は、更に親和性成熟されてもよい。したがって、本明細書中に記載の何れかの抗体の親和性成熟形態が提供される。
ある実施態様では、上記何れかのHVR配列を有する抗Notch2 NRR抗体は、図6Aおよび6Bに示す、ヒト可変軽鎖(VL)コンセンサスフレームワーク配列を更に含んでよい。一実施態様では、VLコンセンサスフレームワーク配列は、例えば図6Aおよび6Bに示す、ヒトVLκサブグループIコンセンサスフレームワーク(κv1)配列を含む。他の実施態様では、VLフレームワークコンセンサス配列は、図7又は8に示すように、huMAb4D5-8のFR1−FR4を含む。特定の実施態様では、VLフレームワークコンセンサス配列は、配列番号:60、61、62および63の配列を含む。
ある実施態様では、上記何れかの抗体をコードするポリヌクレオチドが提供される。一実施態様では、ポリヌクレオチドを含むベクターが提供される。一実施態様では、ベクターを含む宿主細胞が提供される。一実施態様では、宿主細胞は真核生物のものである。一実施態様では、宿主細胞はCHO細胞である。一実施態様では、抗Notch2 NRR抗体の製造方法であって、抗体をコードするポリヌクレオチドの発現に適する条件下で宿主細胞を培養し、抗体を単離することを含む方法が提供される。
一実施態様では、本発明は、Notch2活性を阻害する単離された抗Notch2 NRR抗体を提供する。例えば、本発明の抗体は、任意の生物学上関連するNotch2シグナル伝達経路の破壊を含むNotch2シグナル伝達の一又は複数の態様を調整しうる。
さらなる態様では、本発明は、100nM以下のKdでNotch2 NRRを結合する単離された抗Notch2 NRR抗体を提供する。ある実施態様では、抗Notch2 NRR抗体は、10nM以下、1nM以下、又は0.1nM以下のKdでNotch2 NRRを結合する。本明細書中の実施例に記載のように、例示的なファージ抗体D-3は5nMのKdで結合する。当分野で十分に確立されているので、リガンドのそのレセプターに対する結合親和性は様々なアッセイの何れかを用いて決定され、様々な定量的値で表されうる。したがって、一実施態様では、結合親和性は、Kd値として表され、(例えば、結合力効果を最小限にすることによって)内因性の結合親和性を反映する。一般に、そして、好ましくは、無細胞又は細胞関連のいずれの環境であっても、結合親和性はインビトロで測定される。例えばBiacore、ラジオイムノアッセイ(RIA)およびELISAを含め、本明細書中に記載のものを含め、当分野で公知の多くのアッセイの何れかを用いて、結合親和性測定値を得ることができる。
他の実施態様では、抗本明細書中に示す抗体と同じエピトープに結合する単離された抗体が提供される。一実施態様では、抗体D、抗体D-1、抗体D-2および抗体D-3から選択した抗体と同じエピトープに結合する単離された抗Notch2 NRR抗体が提供される。他の実施態様では、本発明は、抗体D、抗体D-1、抗体D-2および抗体D-3から選択される抗体と競合する抗Notch2 NRR抗体を提供する。他の実施態様では、Notch2のLNR−AドメインおよびHD−Cドメインから選択される少なくとも一つのドメインに結合する単離された抗体が提供される。そのような実施態様では、抗体は、LNR−AドメインおよびHD−Cドメインの両方に結合する。他の実施態様では、抗体は、さらにLNR-Bおよび/またはHD−Nドメインに結合する。これらのドメインは図18に記載する。
抗Notch2 NRR抗体の前記実施態様は、単独で、または何れかの組合せで存在してもよい。さらに、本明細書に記載の何れかの抗Notch2 NRR抗体は、以下の形質の何れか一又は複数を保持してよい。ある実施態様では、抗Notch2 NRR抗体はモノクローナル抗体である。ある実施態様では、抗Notch2 NRR抗体は、Fab、Fab'-SH、Fv、scFv又は(Fab')2断片から選択される抗体断片である。他の実施態様では、抗Notch2 NRR抗体は、ヒト化、ヒト又はキメラの抗体である。
本発明は抗体断片を包含する。抗体断片は伝統的な手段、例えば酵素的消化によって、あるいは組換え法によって生産することができる。ある状況下では、抗体全体よりも、抗体断片を用いることに利点がある。より小さなサイズの断片によって迅速なクリアランスが可能となり、固形腫瘍への接近の改良につながり得る。ある種の抗体断片の概説については、Hudson等 (2003) Nat. Med. 9:129-134を参照のこと。
本発明は、ヒト化抗体を含む。非ヒト抗体をヒト化するための様々な方法が従来技術に既知である。例えば、ヒト化抗体は、非ヒトのソースからそれに導入された一以上のアミノ酸残基を有することができる。これらの非ヒトアミノ酸残基は、しばしば「移入」残基と呼ばれ、一般に「移入」可変ドメインに由来する。ヒト化は、基本的にヒト抗体の該当する配列を高頻度可変領域配列で置換することにより、Winter及び共同研究者(Jones等(1986)Nature 321:522-525;Riechmann等(1988)Nature, 332:323-327;Verhoeyen等(1988)Science 239:1534-1536)の方法に従って実施される。よって、このような「ヒト化」抗体は、インタクトなヒト可変ドメインより実質的に少ない分が非ヒト種由来の対応する配列で置換されたキメラ抗体(米国特許第4816567号)である。実際には、ヒト化抗体は典型的には幾つかの高頻度可変領域残基が、及び場合によっては幾つかのFR残基が齧歯類抗体の類似する部位由来の残基によって置換されたヒト抗体である。
本発明のヒト抗体は、上記のように、ヒト由来のファージディスプレイライブラリーから選択したFvクローン可変ドメイン配列を既知のヒト定常ドメイン配列と組み合わせることによって構築することができる。あるいは、本発明のヒトモノクローナル抗体は、ハイブリドーマ法によって作製することができる。ヒトモノクローナル抗体の生産のためのヒトミエローマ及びマウス-ヒトヘテロミエローマ細胞株は、例えば、Kozbor, J. Immunol. 133, 3001(1984);Brodeur等, Monoclonal Antibody Production Techniques and Applications, pp.51-63(Marcel Dekker, Inc., New York, 1987);及びBoerner 等, J. Immunol., 147: 86 (1991)に記載されている。
二重特異性抗体は、少なくとも2つの異なるエピトープに対して結合特異性を有するモノクローナル抗体である。ある実施態様では、二重特異性抗体はヒト抗体又はヒト化抗体である。ある実施態様では、結合特異性の一つはNotch2に対するものであり、他方は任意の他の抗原に対するものである。ある実施態様では、二重特異性抗体は、Notch2の2つの異なるエピトープに結合しうる。ある実施態様では、二重特異性抗体はNotch2を発現する細胞に細胞傷害剤を局在化するためにも使用されうる。これらの抗体はNotch2結合アーム及び細胞傷害剤、例えば、サポリン(saporin)、抗インターフェロン-α、ビンカアルカロイド、リシンA鎖、メトトレキセート又は放射性同位体ハプテンと結合するアームを有する。二重特異性抗体は完全長抗体又は抗体断片(例えばF(ab')2二重特異性抗体)として調製することができる。
多価抗体は、抗体が結合する抗原を発現する細胞により、二価抗体よりも早くインターナリゼーション(及び/又は異化)されうる。本発明の抗体は、3又はそれ以上の結合部位を有する多価抗体(IgMクラス以外のもの)であり得(例えば四価抗体)、抗体のポリペプチド鎖をコードする核酸の組換え発現により容易に生成することができる。多価抗体は二量化ドメインと3又はそれ以上の抗原結合部位を有する。ある実施態様では、二量化ドメインはFc領域又はヒンジ領域を有する(又はそれらからなる)。このシナリオでは、抗体はFc領域と、Fc領域のアミノ末端に3又はそれ以上の抗原結合部位を有しているであろう。ある実施態様では、多価抗体は3から約8の抗原結合部位を有する(又はそれらからなる)。かかる一実施態様では、多価抗体は4つの抗原結合部位を有する(又はそれらからなる)。多価抗体は少なくとも1つのポリペプチド鎖(例えば2つのポリペプチド鎖)を有し、ポリペプチド鎖は2又はそれ以上の可変ドメインを有する。例えば、ポリペプチド鎖はVD1-(X1)n-VD2-(X2)n-Fcを有し、ここでVD1は第1の可変ドメインであり、VD2は第2の可変ドメインであり、FcはFc領域のポリペプチド鎖の一つであり、X1及びX2はアミノ酸又はポリペプチドを表し、nは0又は1である。例えば、ポリペプチド鎖は、VH-CH1-可動性リンカー-VH-CH1-Fc領域鎖;又はVH-CH1-VH-CH1-Fc領域鎖を有し得る。ここでの多価抗体は、少なくとも2つ(例えば4つ)の軽鎖可変ドメインポリペプチドをさらに有する。ここでの多価抗体は、例えば約2から約8の軽鎖可変ドメインポリペプチドを含む。ここで考えられる軽鎖可変ドメインポリペプチドは軽鎖可変ドメインを含み、場合によってはCLドメインを更に含む。
ある実施態様では、本発明の抗体は単一ドメイン抗体である。単一ドメイン抗体は、抗体の重鎖可変ドメインの全て又は一部又は軽鎖可変ドメインの全て又は一部を含む単一ポリペプチド鎖である。ある実施態様では、単一ドメイン抗体はヒト単一ドメイン抗体(Domantis, Inc., Waltham, MA; 例えば米国特許第6248516B1号を参照)。一実施態様では、単一ドメイン抗体は抗体の重鎖可変ドメインの全て又は一部からなる。
ある実施態様では、ここに記載された抗体のアミノ酸配列の修飾を考える。例えば、抗体の結合親和性及び/又は生物学的特性を向上することができれば望ましい。抗体のアミノ酸配列変異体は、抗体の核酸に適切なヌクレオチド変化を導入して、又はペプチド合成により調製されうる。そのような修飾は、抗体のアミノ酸配列内の残基の、例えば、欠失型、及び/又は挿入及び/又は置換を含む。最終構成物が所望する特徴を有していれば、欠失、挿入及び置換をどのように組合せてもよい。アミノ酸変化は、配列ができるときに被検体の抗体アミノ酸配列中に導入されうる。

(1)無極性:Ala(A),Val(V),Leu(L),Ile(I),Pro(P),Phe(F),Trp(W),Met(M)
(2)無電荷極性:Gly(G),Ser(S),Thr(T),Cys(C),Tyr(Y),Asn(N),Gln(Q)
(3)酸性:Asp(D),Glu(E)
(4)塩基性:Lys(K),Arg(R),His(H)
別法では、天然に生じる残基は共通の側鎖特性に基づいて群に分けることができる:
(1)疎水性:ノルロイシン、Met、Ala、Val、Leu、Ile;
(2)中性の親水性:Cys、Ser、Thr、Asn、Gln;
(3)酸性:Asp、Glu;
(4)塩基性:His、Lys、Arg;
(5)鎖配向に影響する残基:Gly、Pro;及び
(6)芳香族:Trp、Tyr、Phe。
本発明の抗体は当該分野において知られ直ぐに利用できる更なる非タンパク質性部分を含むように更に修飾することができる。好ましくは、抗体の誘導体化に適した部分は水溶性ポリマーである。水溶性ポリマーの非限定的な例には、限定されるものではないが、ポリエチレングリコール(PEG)、エチレングリコール/プロピレングリコールのコポリマー、カルボキシメチルセルロース、デキストラン、ポリビニルアルコール、ポリビニルピロリドン、ポリ-1,3-ジオキソラン、ポリ-1,3,6-トリオキサン、エチレン/無水マレイン酸コポリマー、ポリアミノ酸(ホモポリマーかランダムコポリマー)、及びデキストラン又はポリ(n-ビニルピロリドン)ポリエチレングリコール、プロピレングリコールホモポリマー、プロリプロピレンオキシド/エチレンオキシドコポリマー、ポリオキシエチレン化ポリオール(例えばグリセロール)、ポリビニルアルコール、及びそれらの混合物が含まれる。ポリエチレングリコールプロピオンアルデヒドは水中におけるその安定性のために製造の際に有利であろう。ポリマーは任意の分子量であってよく、分枝状でも非分枝状でもよい。抗体に結合するポリマーの数は変化してもよく、一を超えるポリマーが結合する場合、それらは同じでも異なった分子でもよい。一般に、誘導体化に使用されるポリマーの数及び/又はタイプは、限定されるものではないが、その抗体誘導体が定まった条件下での治療に使用されるかどうか、改善される抗体の特定の性質又は機能を含む考慮事項に基づいて決定することができる。
1.ハイブリドーマベース法
本発明のモノクローナル抗体は、Kohler等, Nature, 256:495 (1975)により最初に記述され、更に例えばHongo等, Hybridoma, 14 (3): 253-260 (1995), Harlow等, Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2版 1988); Hammerling等, Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N.Y., 1981)、及びNi, Xiandai Mianyixue, 26(4):265-268 (2006)でヒト-ヒトハイブリドーマに関するものた記載されたハイブリドーマ法を使用して作製することができる。更なる方法には、例えばハイブリドーマ細胞株からのモノクローナルヒト天然IgM抗体の生産に関する米国特許第7189826号に記載されているものが含まれる。ヒトハイブリドーマ技術(トリオーマ技術)はVollmers及びBrandlein, Histology and Histopathology, 20(3):927-937 (2005)並びに Vollmers及びBrandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3):185-91 (2005)に記載されている。
本発明の抗体は、所望の活性又は活性群を持つ抗体をスクリーニングするためにコンビナトリアルライブラリーを使用することによって作製されうる。例えば、ファージディスプレイライブラリーを作製し、所望される結合特性を有する抗体のためにかかるライブラリーをスクリーニングするために様々な方法が知られている。かかる方法は一般にHoogenboom等 Methods in Molecular Biology 178:1-37 (O’Brien等編, Human Press, Totowa, NJ, 2001)に記載されている。例えば、興味ある抗体を生産する一方法は、Lee等, J. Mol. Biol. (2004), 340(5):1073-93に記載されたファージ抗体ライブラリーを使用することによる。
抗体はまた組換え法を使用して生産することができる。抗Notch2抗体の組換え生産のために、抗体をコードする核酸が単離され、更なるクローニング(DNAの増幅)又は発現のために、複製可能なベクター中に挿入される。抗体をコードするDNAは直ぐに単離され、常套的な手法を用いて(例えば、抗体の重鎖及び軽鎖をコードする遺伝子に特異的に結合可能なオリゴヌクレオチドを使用することによって)配列決定される。多くのベクターが利用可能である。ベクター成分には、一般に、これらに制限されるものではないが、次のものの一又は複数が含まれる:シグナル配列、複製開始点、一又は複数のマーカー遺伝子、エンハンサーエレメント、プロモーター、及び転写終結配列である。
本発明の抗体は直接的に組換え手法によって生産されるだけではなく、シグナル配列あるいは成熟タンパク質あるいはポリペプチドのN末端に特異的切断部位を有する他のポリペプチドである異種性ポリペプチドとの融合ペプチドとしても生産される。好ましく選択された異種シグナル配列は宿主細胞によって認識され加工される(すなわち、シグナルペプチダーゼによって切断される)ものである。天然抗体シグナル配列を認識せずプロセシングしない原核生物宿主細胞に対して、シグナル配列は、例えばアルカリホスファターゼ、ペニシリナーゼ、lppあるいは熱安定なエンテロトキシンIIリーダーの群から選択される原核生物シグナル配列により置換される。酵母での分泌に対して、天然シグナル配列は、例えば酵母インベルターゼリーダー、α因子リーダー(酵母菌属(Saccharomyces)及びクルイベロマイシス(Kluyveromyces)α因子リーダーを含む)、又は酸ホスフォターゼリーダー、白体(C.albicans)グルコアミラーゼリーダー、又は国際公開第90/13646号に記載されているシグナルにより置換されうる。哺乳動物細胞での発現においては、哺乳動物のシグナル配列並びにウイルス分泌リーダー、例えば単純ヘルペスgDシグナルが利用できる。
発現及びクローニングベクターは共に一又は複数の選択された宿主細胞においてベクターの複製を可能にする核酸配列を含む。一般に、クローニングベクターにおいて、この配列は宿主染色体DNAとは独立にベクターが複製することを可能にするものであり、複製開始点又は自律的複製配列を含む。そのような配列は多くの細菌、酵母及びウイルスに対してよく知られている。プラスミドpBR322に由来する複製開始点は大部分のグラム陰性細菌に好適であり、2μプラスミド開始点は酵母に適しており、様々なウイルス開始点(SV40、ポリオーマ、アデノウイルス、VSV又はBPV)は哺乳動物細胞におけるクローニングベクターに有用である。一般には、哺乳動物の発現ベクターには複製開始点成分は不要である(SV40開始点が典型的にはただ初期プロモーターを有しているために用いられる)。
発現及びクローニングベクターは、典型的には、選択可能マーカーとも称される選択遺伝子を含む。典型的な選択遺伝子は、(a)アンピシリン、ネオマイシン、メトトレキセートあるいはテトラサイクリンのような抗生物質あるいは他の毒素に耐性を与え、(b)栄養要求性欠陥を補い、又は(c)例えばバシリに対する遺伝子コードD-アラニンラセマーゼのような、複合培地から得られない重要な栄養素を供給するタンパク質をコードする。
発現及びクローニングベクターは一般に宿主生物体によって認識され抗体核酸に作用可能に結合しているプロモーターを含む。原核生物宿主での使用に好適なプロモーターは、phoAプロモーター、βラクタマーゼ及びラクトースプロモーター系、アルカリホスファターゼ、トリプトファン(trp)プロモーター系、及びハイブリッドプロモーター、例えばtacプロモーターを含む。しかし、他の既知の細菌プロモーターも好適である。細菌系で使用するプロモータもまた抗体をコードするDNAと作用可能に結合したシャイン・ダルガーノ(S.D.)配列を含むであろう。
より高等の真核生物によるこの発明の抗体をコードしているDNAの転写は、ベクター中にエンハンサー配列を挿入することによってしばしば増強される。哺乳動物遺伝子由来の多くのエンハンサー配列が現在知られている(グロビン、エラスターゼ、アルブミン、α-フェトプロテイン及びインスリン)。しかしながら、典型的には、真核細胞ウィルス由来のエンハンサーが用いられるであろう。例としては、複製起点の後期側のSV40エンハンサー(100−270塩基対)、サイトメガロウィルス初期プロモーターエンハンサー、複製起点の後期側のポリオーマエンハンサー及びアデノウィルスエンハンサーが含まれる。真核生物プロモーターの活性化のための増強要素については、Yaniv, Nature, 297:17-18 (1982)もまた参照のこと。エンハンサーは、抗体コード配列の5'又は3'位でベクター中にスプライシングされうるが、好ましくはプロモーターから5'位に位置している。
真核生物宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、又は他の多細胞生物由来の有核細胞)に用いられる発現ベクターは、また転写の終結及びmRNAの安定化に必要な配列を含む。このような配列は、真核生物又はウィルスのDNA又はcDNAの5'、時には3'の非翻訳領域から一般に取得できる。これらの領域は、抗体をコードしているmRNAの非翻訳部分にポリアデニル化断片として転写されるヌクレオチドセグメントを含む。一つの有用な転写終結成分はウシ成長ホルモンポリアデニル化領域である。国際公開第94/11026号とそこに開示された発現ベクターを参照のこと。
ここに記載のベクター中のDNAをクローニングあるいは発現させるために適切な宿主細胞は、上述の原核生物、酵母、又は高等真核生物細胞である。この目的にとって適切な原核生物は、限定するものではないが、真正細菌、例えばグラム陰性又はグラム陽性生物体、例えばエシェリチアのような腸内菌科、例えば大腸菌、エンテロバクター、エルウィニア(Erwinia)、クレブシエラ、プロテウス、サルモネラ、例えばネズミチフス菌、セラチア属、例えばセラチア・マルセスキャンス及び赤痢菌属、並びに桿菌、例えば枯草菌及びバシリ・リチェフォルミス(licheniformis)(例えば、1989年4月12日に公開されたDD266710に開示されたバシリ・リチェニフォルミス41P)、シュードモナス属、例えば緑膿菌及びストレプトマイセス属を含む。一つの好適な大腸菌クローニング宿主は大腸菌294(ATCC31446)であるが、他の大腸菌B、大腸菌X1776(ATCC31537)及び大腸菌W3110(ATCC27325)のような株も好適である。これらの例は限定するものではなく例示的なものである。
この発明の抗体を産生するために用いられる宿主細胞は種々の培地において培養することができる。市販培地の例としては、ハム(Ham)のF10(シグマ)、最小必須培地((MEM),(シグマ)、RPMI-1640(シグマ)及びダルベッコの改良イーグル培地((DMEM),シグマ)が宿主細胞の培養に好適である。また、Ham等, Meth. Enz. 58:44 (1979), Barnes等, Anal. Biochem. 102:255 (1980), 米国特許第4767704号;同4657866号;同4927762号;同4560655号;又は同5122469号;国際公開第90/03430号;国際公開第87/00195号;又は米国再発行特許第30985号に記載された何れの培地も宿主細胞に対する培地として使用できる。これらの培地には何れもホルモン及び/又は他の増殖因子(例えばインシュリン、トランスフェリン、又は表皮成長因子)、塩類(例えば、塩化ナトリウム、カルシウム、マグネシウム及びリン酸塩)、バッファー(例えばHEPES)、ヌクレオチド(例えばアデノシン及びチミジン)、抗生物質(例えば、GENTAMYCINTM薬)、微量元素(最終濃度がマイクロモル範囲で通常存在する無機化合物として定義される)及びグルコース又は等価なエネルギー源を必要に応じて補充することができる。任意の他の必要な補充物質もまた当業者に知られている適当な濃度で含むことができる。培養条件、例えば温度、pH等々は、発現のために選ばれた宿主細胞について過去に用いられているものであり、当業者には明らかであろう。
組換え技術を用いる場合、抗体は細胞内、細胞膜周辺腔に生産され、又は培地内に直接分泌される。抗体が細胞内に生産された場合、第1の工程として、宿主細胞か溶解された断片の何れにしても、粒子状の細片が、例えば遠心分離又は限外濾過によって除去される。Carter等, Bio/Technology 10: 163-167 (1992)は、大腸菌の細胞膜周辺腔に分泌された抗体の単離方法を記載している。簡単に述べると、細胞ペーストを、酢酸ナトリウム(pH3.5)、EDTA、及びフェニルメチルスルホニルフルオリド(PMSF)の存在下で約30分間解凍する。細胞細片は遠心分離で除去できる。抗体が培地に分泌された場合は、そのような発現系からの上清を、一般的には先ず市販のタンパク質濃縮フィルター、例えばAmicon又はPelliconの限外濾過装置を用いて濃縮する。PMSFなどのプロテアーゼ阻害剤を上記の任意の工程に含めて、タンパク質分解を阻害してもよく、また抗生物質を含めて外来性の汚染物の増殖を防止してもよい。
本発明は、また化学療法剤、薬剤、成長阻害剤、毒素(例えば、細菌、糸状菌、植物又は動物由来の酵素活性性毒素、又はその断片)、又は放射性同位体(すなわち放射性コンジュゲート)などの一又は複数の細胞傷害剤にコンジュゲートした抗体を含んでなる、免疫コンジュゲート(交換可能に「抗体-薬剤コンジュゲート」又は「ADC」とも称される)を提供する。
ある実施態様では、免疫コンジュゲートは、一又は複数のメイタンシノイド分子にコンジュゲートした抗体(完全長又は断片)を含んでなる。
ある実施態様では、免疫コンジュゲートは、ドラスタチン又はドロスタチンペプチジル類似体及び誘導体、アウリスタチン(auristatin) (米国特許第5635483号;同第5780588号)にコンジュゲートした抗体を含んでなる。ドラスタチン及びアウリスタチンは、微小管動態、GTP加水分解及び核と細胞の分割を妨げ(Woyke 等 (2001) Antimicrob. Agents and Chemother. 45(12): 3580-3584)、抗癌活性(米国特許第5663149号)及び抗真菌性活性(Pettit 等 (1998) Antimicrob. Agents Chemother. 42:2961-2965)を有することが示されている。ドラスタチン又はアウリスタチン薬剤成分は、ペプチジル薬剤分子のN(アミノ)末端又はC(カルボキシル)末端により抗体に接着しうる(国際公開第02/088172号)。
他の実施態様では、免疫コンジュゲートは、一又は複数のカリケアマイシン分子と結合した抗体を含んでなる。抗生物質のカリケアマイシンファミリーはサブピコモル濃度で二重鎖DNA破壊を生じることができる。カリケアマイシンファミリーのコンジュゲートの調製については、米国特許第5712374号、同5714586号、同5739116号、同5767285号、同5770701号、同5770710号、同5773001号、同5877296号(全て、American Cyanamid Company)を参照のこと。使用可能なカリケアマイシンの構造類似体には、限定するものではないが、γ1I、α2I、α3I、N-アセチル-γ1I、PSAG及びθI1(Hinman等, Cancer Research, 53:3336-3342(1993)、Lode等 Cancer Research, 58:2925-2928(1998)及び上述したAmerican Cyanamidの米国特許)が含まれる。抗体が結合可能な他の抗腫瘍剤は、葉酸代謝拮抗薬であるQFAである。カリケアマイシン及びQFAは双方共、細胞内に作用部位を有し、原形質膜を容易に通過しない。よって、抗体媒介性インターナリゼーションによるこれらの薬剤の細胞への取込により、細胞傷害効果が大きく向上する。
抗体と結合可能な他の抗腫瘍剤には、BCNU、ストレプトゾイシン、ビンクリスチン及び5-フルオロウラシル、米国特許第5053394号、同5770710号に記載されており、集合的にLL-E33288複合体として知られている薬剤ファミリー、並びにエスペラマイシン(esperamicine)(米国特許第5877296号)が含まれる。
抗体薬剤コンジュゲート(ADC)において、抗体(Ab)はリンカー(L)を介して、一又は複数の薬剤部分(D)、例えば1抗体につき約1から約20の薬剤部分にコンジュゲートされる。式IのADCは、(1)抗体の求核基を二価のリンカー試薬と反応させて共有結合を介してAb-Lを形成した後、薬剤部分Dと反応させること;及び(2)薬剤部分の求核基を二価のリンカー試薬と反応させて共有結合を介してD-Lを形成した後、抗体の求核基と反応させることを含み、当業者に知られている有機化学反応、状態及び試薬を用いる幾つかの手段によって調製されうる。ADCを調製するための更なる方法はここに記載される。
Ab−(L−D)p I
1.診断方法と検出方法
一態様では、本発明の抗体は、生体試料中のNotch2の存在を検出するために有用である。本明細書中で用いる「検出する」なる用語は、定量的又は定性的な検出を包含する。ある実施態様では、生体試料は、細胞又は組織、例えば癌性組織を含む。
一態様では、本発明は、生体試料中のNotch2の存在を検出する方法を提供する。ある実施態様では、前記方法は、Notch2への抗Notch2抗体の結合に許容される条件下で抗Notch2抗体(例えば抗Notch2 NRR抗体)と生体試料を接触させ、抗Notch2抗体とNotch2との間に複合体が形成されるか否かを検出することを含む。
ある実施態様では、抗体は標識される。標識には、限定するものではないが、直接検出される標識又は分子(例えば、蛍光、発色、高電子密度、化学発光、及び放射性標識)、並びに、例えば酵素反応又は分子相互作用によって間接的に検出される酵素ないしはリガンドなどの分子が含まれる。例示的な標識には、限定するものではないが、放射性同位体である32P、14C、125I、3H及び131I、フルオロフォア、例えば希有土類キレート又はフルオレセイン及びその誘導体、ローダミン及びその誘導体、ダンシル、ウンベリフェロン、ルシフェラーゼ、例えばホタルルシフェラーゼ及び細菌ルシフェラーゼ(米国特許第4737456号)、ルシフェリン、2,3-ジヒドロフタルアジネジオン(dihydrophthalazinediones)、西洋わさびペルオキシダーゼ(HRP)、アルカリホスファターゼ、β-ガラクトシダーゼ、グルコアミラーゼ、リソチーム、サッカライドオキシダーゼ、例えばグルコースオキシダーゼ、ガラクトースオキシダーゼ及びグルコース-6-リン酸デヒドロゲナーゼ、複素環のオキシダーゼ、例えばウリカーゼ及びキサンチンオキシダーゼ、HRP、ラクトペルオキシダーゼ又はマイクロペルオキシダーゼなどの色素前駆体を酸化するために水素ペルオキシダーゼを用いる酵素とカップリングさせたもの、ビオチン/アビジン、スピン標識、バクテリオファージ標識、安定した遊離基などが含まれる。
診断ないし検出の上記いずれかの実施態様は、抗Notch2抗体の代わりに、ないしは抗Notch2抗体に加えて本発明のイムノコンジュゲートを用いて行われうることは理解される。
本発明の抗体は、例えばインビトロ、エクスビボ及びインビボの治療方法において用いられうる。一態様では、本発明は、インビボ又はインビトロでのNotch2活性、すなわちNotch2シグナル伝達の阻害方法であって、Notch2への抗体の結合に許容される条件下で細胞を本発明の抗Notch2 NRR抗体にさらすことを含む方法を提供する。ある実施態様では、細胞は、癌細胞、例えば癌性のB細胞又はメラノーマ細胞である。一実施態様では、本発明の抗Notch2 NRR抗体がNotch2活性を阻害するために用いられてよく、前記方法は、Notch2活性が阻害されるように、Notch2を本発明の抗Notch2 NRR抗体にさらすことを含む。
本発明の抗Notch2 NRR抗体は、例えば、Notch2の発現及び/又は活性と関係する疾患の治療、例えばNotch2の発現又は活性の増加と関係する疾患、又はNotch2の発現又は活性が病態に寄与する疾患の治療のために用いられてよい。「Notch2の発現又は活性の増加と関係する疾患」は、Notch2発現又は活性が正常より有意に高い疾患を指す。
更なる態様では、本発明の抗体を用いて、癌、特に癌幹細胞を含む固形腫瘍を治療するか又は予防する。癌幹細胞は、増殖して更なる癌幹細胞並びに他の腫瘍細胞群を引き起こすことが可能である。Al-Hajj et al., Proc. Nat'l Acad. Sci. USA 100: 3983-3988 (2003)を参照。癌幹細胞は、癌の再発および薬物治療に対する癌の耐性に寄与すると考えられている。Notchレセプターは、癌幹細胞マーカーとして、そして、固形腫瘍の形成及び再発の原因である癌幹細胞を取り除くための標的として同定された。国際公開2008/091641を参照。このような固形腫瘍には、胸部、大腸、膵臓、前立腺、肺、頭部および頸部、直腸、結腸直腸の癌が含まれるがこれらに限定されない。
本発明の抗体は、治療において単独で又は他の組成物と組み合わせて用いられてよい。例えば、本発明の抗体は、少なくとも一の更なる治療薬および/またはアジュバントと同時に投与されてよい。ある実施態様では、抗Notch1抗体(例えば抗Notch1 NRR抗体)は、別の方法でこのような抗Notch1抗体によって誘導されうる腸管の細胞分化の変化を阻害するかまたは低減する他の治療薬と共に投与される。ある実施態様では、他の治療薬は、デキサメサゾン又はタモキシフェンである。このような併用療法は、例えば腫瘍関連の血管形成および癌を含む血管形成性疾患を治療する際に有用であろう。ある実施態様では、抗Notch2抗体(例えば抗Notch2 NRR抗体)は、更なる治療薬、例えば化学療法剤と共に投与される。このような併用療法は、例えば、B細胞悪性腫瘍およびメラノーマといった癌を治療する際に有用である。
上述のかかる併用療法には、併用投与(2以上の薬剤が同一又は別個の製剤中に含まれる場合)、及び本発明の抗体の投与が、他の治療剤及び/又はアジュバントの投与の前、同時、及び/又は後に、本発明の抗体の投与がなされうる別個の投与が含まれる。また、本発明の抗体は放射線療法と組み合わせて用いられてよい。
一実施態様では、本発明の抗体は、Notch2の発現および/または活性の増加と関係する疾患を患っている個体においてNotch2を結合させる方法に用いられ、この方法は個体においてNotch2が結合されるように、個体に抗体を投与することを含む。一実施態様では、Notch2はヒトNotch2であり、個体はヒト個体である。あるいは、個体は、本発明の抗体が結合するNotch2を発現する哺乳動物であってよい。また更に、個体は、Notch2が(例えば、Notch2の投与によって、または、Notch2をコードする導入遺伝子の発現によって)導入された哺乳動物であってもよい。
本発明の抗体(及び任意の更なる治療薬又はアジュバント)は、非経口的、皮下、腹膜内、肺内、鼻腔内、そして、必要に応じて局所の治療のために、病巣内投与を含む任意の好適な手段によって投与することができる。非経口注入には、筋肉内、静脈内、動脈内、腹膜内、又は皮下的な投与を含む。加えて、抗体を、特に抗体の用量を減少して、パルス注入によって好適に投与する。投与が短期のものであるか長期のものであるかにある程度依存して、任意の好適な経路、例えば、静脈内又は皮下注射などの注射によって投与することができる。
ある実施態様では、核酸(場合によってベクターに含まれている)は患者の細胞内へインビボ及びエクスビボの方法によって導入されてよい。インビボ運搬の一例では、患者の例えば治療的介入を必要とする部位に核酸が直接注入される。インビボ運搬の更なる例では、核酸は、ウイルスベクター(例えば、アデノウイルス、単純ヘルペスIウイルス、又はアデノ関連ウイルス)、及び脂質ベースの系(例えば、遺伝子の脂質媒介移入に有用な脂質は、DOTMA、DOPE、及びDC-Cholである)による形質移入を用いて細胞内に導入される。遺伝子マーキング及び遺伝子治療プロトコールの概説については、Anderson et al., Science, 256:808-813 (1992)、及び国際公開93/25673及びそこに引用された参考文献を参照のこと。エクスビボ治療の例では、患者の細胞が取り除かれ、核酸がこれら単離した細胞内に導入され、変更した細胞が直接又は、例えば、患者内に着床される多孔性膜内に被包して患者に投与される(例として米国特許第4892538号および同第5283187号を参照)。核酸のエクスビボ運搬のために一般的に用いられるベクターはレトロウイルスである。
標的細胞への抗体の移入は、当分野で公知の他の方法によって増強することができる。例えば、HIV Tat又はアンテナペディアホメオドメインタンパク質由来の配列のような特定の配列は、細胞膜全体に異種タンパク質の効率的な取り込みを導くことができる。例としてChen et al., Proc. Natl. Acad. Sci. USA (1999), 96:4325-4329を参照。
血液脳関門に抗体を搬送する物理的方法には、限定するものではないが、血液脳関門を完全に包囲すること、又は血液脳関門に開口部を形成することが含まれる。包囲法には、限定するものではないが、脳への直接注入(例えば、Papanastassiou等, Gene Therapy 9: 398-406 (2002))、間質性注入/対流強化送達(例えば、Bobo等, Proc. Natl. Acad. Sci. USA 91: 2076-2080 (1994)参照)、及び脳への送達装置の移植(例えば、Gill等, Nature Med. 9: 589-595 (2003);及びGliadel WafersTM, Guildford Pharmaceutical参照)が含まれる。関門に開口を形成する方法には、限定するものではないが、超音波(例えば、米国特許出願公開第2004/0038086号参照)、浸透圧(例えば、高浸透圧性マンニトールの投与による(Neuwelt, E. A., Implication of the Blood-Brain Barrier and its Manipulation, Vols 1 & 2, Plenum Press, N.Y. (1989)))、例えば、ブラジキニン又はパーミアライザーA−7による透過性化(例えば、米国特許第5112596号、同第5268164号、同第5506206号、及び同第5686416号参照)、及び抗体をコードする遺伝子を含むベクターによる血液脳関門にまたがるニューロンの形質移入(例えば、米国特許出願公開第2003/0083299号)が含まれる。
血液脳関門を越えて抗体を搬送する幹細胞に基づく方法は、対象の抗体を発現するように神経前駆細胞(NPC)を遺伝的に操作し、次いで、治療する個体の脳に幹細胞を移植することを伴う。詳細なオンライン文献であるBehrstock等 (2005) Gene Ther. 15 Dec. 2005(神経栄養因子GDNFを発現するように遺伝子操作したNPCが齧歯類及び霊長類のモデルの脳に移植した場合にパーキンソン病の症状を低減したことを報告している)を参照。
本発明の抗体は、医学的実用性に合わせた様式で調製し、1回分に分けて、投与される。ここで考慮する要因は、治療する特定の疾患、治療する特定の哺乳動物、個々の患者の臨床状態、疾患の原因、薬剤の運搬部位、投与の方法、投与の日程計画、及び医師が知る他の因子を含む。必要ではないが場合によっては、問題の疾患を予防するか又は治療するために一般に用いられる一つ以上の作用剤と抗体とを調製する。そのような他の作用剤の有効量は、製剤中の抗体の量、疾患の型又は治療、及び上記の他の因子に依存する。これらは、一般的に、ここに記載されるものと同じ用量及び投与経路で、又はここに記載される用量の1〜99%で、或いは経験的/臨床的に適切と判断される任意の用量及任意の投与経路で、用いられる。
上記いずれかの治療方法は、抗Notch2抗体の代わりに、ないしは抗Notch2抗体に加えて本発明のイムノコンジュゲートを用いて行われうることは理解される。
本発明の抗Notch2 NRR抗体は、当分野で公知の様々なアッセイによって物理的/化学的な特性及び/又は生物学的活性に特徴がありうる。
a)活性アッセイ
一態様では、生物学的活性を有する抗Notch2抗体(例えば抗Notch2 NRR抗体)を同定するためのアッセイが提供される。生物学的活性には、例えば、Notch2活性、例えばNotch2シグナル伝達の阻害又は低減を含みうる。また、このようなインビボ及び/又はインビトロで生物学的活性を有する抗体が提供される。
ある実施態様では、本発明の抗Notch2 NRR抗体は、周辺帯B細胞の生成を阻害するその能力について試験される。例示的なアッセイは実施例に示す。特定の他の実施態様では、本発明の抗体は、Notch2シグナル伝達に応答するリポーター遺伝子の発現を阻害するその能力について試験される。例示的なアッセイは実施例に示す。
一態様では、本発明の抗体は、例えばELISA、ウエスタンブロットなどといった公知の方法によって、その抗原結合活性について試験される他の態様では、競合アッセイを用いて、本発明の抗体と競合する抗体(例えばモノクローナル抗体)を同定してよい。一実施態様では、抗体は、Notch2への結合について、抗体D、抗体D-1、抗体D-2又は抗体D-3と競合する。そのような実施態様では、競合抗体は、抗体D、抗体D-1、抗体D-2又は抗体D-3が結合する同じエピトープ(例えば線形又は立体構造のエピトープ)に結合する。他の実施態様では、競合抗体は、実施例B(5)に記載のように抗NRR1が結合する同じエピトープ(例えば線形又は立体構造のエピトープ)、すなわちNotch1 NRRのLNR−A、LNR−BおよびHD−Cドメインを含むエピトープに結合する。例示的な競合アッセイには、限定するものではないが、Harlow and Lane (1988) Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY)に挙げられるものなどの常套的アッセイが含まれる。抗体が結合するエピトープをマッピングするための詳細な例示的な方法は、Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ)のMorris (1996) "Epitope Mapping Protocols,"に示される。2つの抗体のそれぞれが50%以上他の結合をブロックする場合、これらの抗体は同じエピトープに結合すると言える。
一態様では、本発明の抗体はさらに、限定するものではないが、N末端配列決定法、アミノ酸分析、非変性サイズ排除高圧液体クロマトグラフィ(HPLC)、質量分析、イオン交換クロマトグラフィ及びパパイン消化を含む、一連のアッセイによって特徴付けることができる。
上記いずれかのアッセイは、抗Notch2抗体の代わりに、ないしは抗Notch2抗体に加えて本発明のイムノコンジュゲートを用いて行われうることは理解される。
本発明の他の態様では、上記の疾患の治療、予防及び/又は診断に有用な物質を含む製造品が提供される。該製造品は容器と該容器上又は該容器に付随するラベル又はパッケージ挿入物を具備する。好適な容器には、例えば、ビン、バイアル、シリンジ等々が含まれる。容器は、様々な材料、例えばガラス又はプラスチックから形成されうる。容器は、症状を治療、予防及び/又は診断するために有効な組成物単独又は他の組成物と組み合わせる組成物を収容し、滅菌アクセスポートを有しうる(例えば、容器は皮下注射針が貫通可能なストッパーを有するバイアル又は静脈内投与溶液バッグでありうる)。組成物中の少なくとも一つの活性剤は本発明の抗体ないしイムノコンジュゲートである。ラベル又はパッケージ挿入物は、組成物が選択された症状の治療に使用されることを示す。更に、製造品は、(a)本発明の抗体ないしイムノコンジュゲートを含有する組成物を中に収容する第一の容器;と(b)更なる細胞障害剤又はそれ以外の治療薬を含有する組成物を中に収容する第二の容器とを含みうる。本発明のこの実施態様における製造品は、組成物を特定の症状の治療に使用することができることを示しているパッケージ挿入物を更に含む。あるいは、もしくは付加的に、製造品は、薬学的に許容されるバッファー、例えば注射用の静菌水(BWFI)、リン酸緩衝生理食塩水、リンガー液及びデキストロース溶液を含む第二の(又は第三の)容器を更に具備してもよい。更に、他のバッファー、希釈剤、フィルター、針、及びシリンジを含む、商業上及び使用者の見地から望ましい他の材料を含んでもよい。
1.抗Notch2 NRR抗体の生成
抗Notch2 NRR抗体を同定するためのライブラリの分類とスクリーニング
選択された相補性決定領域(H1、H2、H3、L3)に人工的な多様性を有し、ヒトIgGレパートリの天然の多様性を疑似するヒトファージ抗体ライブラリをパニングのために用いた。Fab断片は、M13バクテリオファージ粒子の表面上に二価で表出された。(Lee et al., J. Mol. Biol. 340:1073-1093 (2004)を参照)。Notch2 NRR断片は、バキュロウイルス発現ベクターシステム又は293T細胞を使用して、エピトープタグ(FLAG又は6×His)に融合した分泌タンパク質として発現され、親和性クロマトグラフィを用いて90%より多い純度に精製され、光散乱を用いて凝集の欠如について試験した。Notch2 NRR抗原の配列は以下の通りである。
4ラウンドのパニングの後、有意な濃縮が観察された。VHおよびVH/VLライブラリ分類からそれぞれ96のクローンを拾い上げ、それらがヒトとマウス両方のNotch2 NRRに特異的に結合したが否かを決定した。これらのクローンの可変領域をPCR配列決定し、特有の配列クローンを同定した。
ファージ抗体の親和性は、点競合ELISAを用いて層別化した。さらに、競合ファージ-結合ELISAを使用してファージ抗体IC50値を決定した。ヒト及びマウスのNotch2 NRRを結合する特有のファージ抗体を選択し、インビトロ細胞アッセイでの評価のために完全長IgGに再編成した。
対象のクローンを、LPG3およびLPG4ベクターのそれぞれに別々のクローンのVL及びVH領域をクローニングし、哺乳類のCHO細胞において過渡的に発現させ、プロテインAカラムにて精製することによって、再編成した。
ファジミドpW0703(ファジミドpV0350-2b由来(Lee et al., J. Mol. Biol 340, 1073-1093 (2004))、M13バクテリオファージの表面上に一価のFabを表出しているもの)を、親和性成熟のためのVH/VLライブラリからの抗体Dの軽鎖(VL)及び重鎖(VH)可変ドメインを移植するためのライブラリテンプレートとして用いた。次いで、停止コドンをライブラリテンプレートのCDR-L3に組み込んだ。70−10−10−10混合の野生型ヌクレオチドを好む(favoring)塩基により害を及ぼすオリゴヌクレオチド方策を用いることによって、選択した位置でおよそ50%の変異率を導入したソフトランダム化方策を用いた。、突然変異誘発DNAを、により合成した(Gallop et al., Journal of Medicinal Chemistry 37:1233-1251 (1994))。具体的には、CDR-L1の位置28−32、CDR-L2の位置50及び53−55、CDR-L3の位置91−94及び96、CDR-H1の28−35、CDR-H2の50−58、CDR-H3の95−100の残基を標的とした。また、L1/L2/L3、L3/H1/H2およびL3/H3のソフトランダム化CDRループの組合せを有する3つの異なるライブラリを構築した。
親和性改善選別のために、ファージライブラリは、第1ラウンドではプレート分類に、その後5回のラウンドでは溶液分類を行った。ライブラリは、それぞれヒト及びマウスのNotch2 NRRに対して分類した。プレート分類の第1ラウンドでは、3つのライブラリを、標的コートプレート(NUNC Maxisorpプレート)に対して別々に分類し、ファージインプットは、室温、2時間で、1%BSAおよび0.05%Tween20中でおよそ3O.D/mlであった。後に続く溶液分類のラウンドでは、プレート分類の第1ラウンドにて増殖した1O.D./mlのファージを、50nMのビオチン化標的タンパク質(この濃度は親のクローンファージIC50値を基にしている)を含む、1%Superblock(Pierce Biotechnology)と0.05%Tween20を含む100μlのバッファ中で室温で30分間インキュベートした。さらに、混合物を1%Superblockにて10倍に希釈し、100μl/ウェルをニュートラビジンコートウェルに添加し(5μg/ml)、室温で15分間、ゆっくりと撹拌しながら、ビオチン化標的をファージに結合させた。ウェルをPBS−0.05%Tween20にて10回洗浄した。バックグラウンド結合を決定するために、ビオチン化していない標的を有するファージを含むコントロールウェルをニュートラビジンコートプレート上で捕獲した。結合したファージを0.1N HClにて20分間溶出し、1/10量の1M トリスpH11にて中和し、力価を測定して、次のラウンドのために増殖させた。次に、後続のラウンドの溶液分類はともに、選択ストリンジェンシーを高めた2つの方法により実施した。その一つ目は、10nMから0.5nMにビオチン化標的タンパク質濃度を減少することによるオンレート(on-rate)選択であり、その二つ目は、弱い結合物を完全に除くために過剰量(100〜500倍以上)の非ビオチン化標的タンパク質を添加することによるオフレート(off-rate)選択である。また、ファージインプットはバックグラウンドファージ結合を低くするために低くした(0.1〜0.5O.D/ml)。
第6ラウンド分類からコロニーを拾い、96ウェルプレート(Falcon)中で50μg/mlカルベニシリンと1E10/mlのKO7を含む150μl/ウェルの2YT培地にて37℃で終夜生育させた。同じプレートから、XL-1感染親ファージのコロニーをコントロールとして拾った。96ウェルNunc Maxisorpプレートを、100μl/ウェルのヒト及びマウスのNotch2 NRRタンパク質(1μg/ml)を含むPBSにて、4℃で終夜又は室温で2時間かけてコートした。65μlの1% BSAにて30分間、そして40μlの1% Tween20にてさらに30分間、プレートをブロックした。
ファージ上清を、10nMの標的タンパク質を含む又は含まないELISA(酵素結合吸着アッセイ)バッファ(0.5%BSAと0.05%Tween20を含むPBS)にて1:5に希釈して全量を100μlとし、Fプレート(NUNC)にて室温で少なくとも1時間インキュベートした。標的タンパク質を含む又は含まない75μlの混合物を、標的タンパク質コートプレートに並んで移した。プレートを15分間緩やかに振とうし、標的タンパク質コートプレートに結合していないファージを捕獲させた。プレートを、PBS−0.05%Tween20にて少なくとも5回洗浄した。結合は、ELISAバッファ(1:5000)に西洋ワサビペルオキシダーゼ(HRP)結合抗M13抗体を加えて定量し、室温で30分間インキュベートした。PBS−0.05%Tween20にて少なくとも5回プレートを洗浄した。次に、3,3',5,5'-テトラメチルベンジジン(TMB)ペルオキシダーゼ基質とペルオキシダーゼ溶液B(H2O2)((Kirkegaard-Perry Laboratories (Gaithersburg, MD))を1:1の比率で含む100μl/ウェルの溶液をウェルに加えて、室温にて5分間インキュベートした。各ウェルに100μlの1Mリン酸(H3PO4)を加えることによって反応を止め、室温で5分間インキュベートした。各ウェルの黄色の450nmのOD(光学密度)を、標準的なELISAプレート読み込み器を用いて定量した。OD減少(%)は、以下の方程式によって算出した。
OD450nm減少(%)=[(競合物質を含むウェルのOD450nm)/(競合物質を含まないウェルのOD450nm)]*100
親ファージのウェルのOD450nm減少(%)(100%)と比較して、ヒト及びマウスの標的について50%より低いOD450nm減少(%)を有するクローンを配列分析に選択した。ファージ調製のために特有のクローンを選択し、親クローンと比較することによって、ヒト及びマウスのNotch2 NRRに対する結合親和性(ファージIC50)を決定した。最も親和性を改善したクローンをヒトIgG1に再編成し、哺乳動物細胞に発現させた。
抗Notch2 NRR抗体の結合親和性は、BIAcoreTM-3000装置を用いた表面プラスモン共鳴(SRP)によって測定した。抗Notch2 NRRヒトIgGを、CM5センサーチップ上にコートしたマウス抗ヒトIgGによって捕獲し、およそ200反応単位(RU)を達成した。動態測定のために、ヒトおよびマウスのNotch2 NRRの二倍段階希釈物(3.9nM〜500nM)を、30μl/分の流速、25℃で、PBTバッファ(0.05%のTween20を含むPBS)に注入した。会合速度(kon)および解離速度(koff)は、単一の1対1ラングミュア結合モデル(BIAcore評価ソフトウェアバージョン3.2)を使用して算出した。平衡解離定数(KD)はkoff/konの比率として算出した。
特定の抗Notch1 NRR抗体の生成および特徴づけは先に述べた。米国特許出願公開番号US2009/0081238A1を参照。
抗Flag抗体を用いて、黒色壁のある高い結合プレート(Greiner)において4つすべてのNotchレセプターから、Flagタグ付加NRRタンパク質を捕獲した。NRR抗体およびE25コントロール抗体はNRRタンパク質に結合し、PBSにて洗い流した。アルカリホスフェートコンジュゲート抗ヒト(H+L)二次抗体(Jackson ImmunoResearch)を用いて、結合した抗NRRおよびE25を検出した。結合していない二次抗体を洗い流し、APはPNPP基質(Pierce)にて検出した。
発現プラスミドは、Notch1又はNotch2の細胞外ドメインおよび膜貫通ドメインがN末端にmycタグ付加され、ミニマルCMV最初期プロモータの上流のテトラマイシン反応エレメント(TRE)の制御下で発現されるように作製した。これらのプラスミドはCHO-K1細胞(Clonetech)に安定して形質移入され、結合して形質移入したプラスミド内のTREを活性化するドキシサイクリンの存在下でトランス活性化因子タンパク質を発現するように操作した。1mg/mlのドキシサイクリン又はベヒクルにて48時間細胞を処理し、次いで、抗NRR、抗myc(9E10, Millipore)又はE25コントロール抗体とヤギ抗ヒトAlexa488又はAlexa647二次抗体(Invitrogen)とで染色した。染色の後、細胞はFACScalibur機(BD Biosciences)にて実行し、15000イベントを回収し、範囲ゲートを描いて、陽性及び陰性に染色した集団を評価した。
Notch1を安定して形質移入したか又は他のNotchレセプターを含むプラスミドを過渡的に形質移入したNIH−3T3細胞に、Notch反応TP−1(12×CSL)ホタルルシフェラーゼレポータと、形質移入効率のコントロールのために構成的に活性なウミシイタケルシフェラーゼレポータ(pRL-CMV、Promega)とを同時に形質移入した。細胞は、6時間から終夜の形質移入から回復させる。抗体の処理とリガンドを安定して形質移入したNIH-3T3細胞を用いてレセプター細胞を刺激する。20時間後に、ホタルおよびウミシイタケのルシフェラーゼを、二重Gloルシフェラーゼアッセイシステム(Promega)にて測定する。ホタルシグナルを形質移入効率のためのコントロールのウミシイタケシグナルにて除して、各条件に対して8つの複製物を分析した。平均および標準偏差を算出し、値を規準化して、リガンドを形質移入していないNIH-3T3細胞にて刺激した同時培養物について値を算出した。
Notch1およびNotch2 NRR配列は、NRR −LNR−A;LNR−B;LNR−C;HD−N;及びHD−Cの5つのドメイン間の接合での特有の制限部位により設計され、Blue Heron Technology (Bothell, WA)によって製造された。操作した制限部位を用いて2つの合成配列間の様々なドメインを交換することにより、キメラNRRタンパク質を生成した。キメラクローンは、変更したpUC19ベクターにアッセンブリし、発現のためにSEAP含有ベクターであるpCSC.APへ移した。APタグ付加NRRタンパク質は、293T細胞の形質移入によって生産した。72時間の培養物の後、馴化培地を回収し、遠心分離法によって浄化し、Phospha-Lightシステム(Applied Biosystems, Bedford, MA)を用いてAP活性について試験した。各試料のAP活性を規準化し、ELISAを行ってキメラNRRタンパク質を結合するα-NRR1又はα-NRR2の能力を試験した。ELISAは以下の通りに実行した。α-NRR1又はα-NRR2を、96ウェルフォーマットのプレート結合a−Fc抗体によって捕獲させ、終夜馴化培地にて培養した。プレートを洗浄して結合していないタンパク質を除去し、捕獲したタンパク質はPhospha-Lightシステムを使用してAP活性について試験した。結果として生じたAP活性を用いて、特定のキメラタンパク質を結合する各α-NRR抗体の能力を定量化し、結合のための重要なエピトープを推定した。
HUVEC細胞は、既に記載されているように(Nakatsu et al., 2003)、フィブリンゲルビーズ上で増殖させた。Cytodex3ビーズ(Amersham Pharmacia Biotech)は、2mlのEGM-2培地(Clonetics)のビーズにつき350−400のHUVECにてコートした。およそ200のHUVECコートビーズを、12ウェル組織培養プレートの1ウェルのフィブリン凝塊に包埋した。6×104のデトロイト551線維芽細胞を、5ug/mlの抗NRR1、5ug/mlの抗DLL4、1uMのDBZ又はベヒクルコントロールの何れかを含有する培地と凝塊の上にプレートした。培地は2日おきに交換した。次いで、7〜9日後に、フィブリンゲルを4%のパラホルムアルデヒドにて固定し、内皮細胞を、検出のためにFITCコンジュゲート二次抗体を用いて抗CD31(R&D Systems AF806)にて染色した。
CD1新生仔に、10mg/kgのブタクサコントロール又は20mg/kgのNotch1をP1およびP3に腹腔内投与した。P5に眼を採取し、4%PFAにて固定した。網膜を切り出し、1時間ブロックした後(5%BSA、0.5%トリトンX-100)、1Mのクエン酸ナトリウムにて10分間処理した。次いで、網膜は、ビオチン化したイソレクチンB4(Sigma)およびKi67抗体(NeoMarkers)と共に4℃で終夜インキュベートした。その後網膜を洗浄し、ストレプトアビジンAlexa488およびヤギ抗ウサギCy3を含む1%BSA、0.5%トリトンX-100にて染色した。網膜を平らにマウントし、落射蛍光顕微鏡を使用して撮像した。
ヒト腫瘍細胞株Calu-6およびHM7を、10%v/v FBS、1%v/vペニシリン/ストレプトマイシン、2mMのL−Glnおよび1μg/mlファンギゾンTM(InvitrogenTM, CA)を添加したハムのF12、低グルコースDMEM1:1中で生育させた。細胞を、95%空気/5%CO2の空気下で37℃でインキュベートした。マウス異種移植片実験のために、腫瘍細胞を、1×108又は1×109細胞/mlの濃度で懸濁し、Balb-cヌードマウス(Harlan Sprague Dawley, IN)の背面の脇側に皮下投与した(100μl/マウス)。腫瘍が400mm3の体積に達したときに、0日目のコントロールとして、コホートをランダムに選択した(n=10)。残りのマウスは10匹のマウス群に分け、ファージ抗体の抗Notch1を10mg/kgの用量で週に2回腹腔内投与した。移植した腫瘍は、記述したように長軸及び垂直軸に沿って週2回測定した。腫瘍を測定した各日に、各マウスの腫瘍体積を算出し、各コントロール抗体(抗ブタクサ)、抗VEGF群からの腫瘍体積平均を、JMPTM統計分析システム(ウィンドウズ用バージョン5.1;SAS Institute, Cary, NC)において実行するP<0.05レベルのダネットt検定によって、抗Notch1処理マウスの腫瘍体積平均と比較した。腫瘍体積が2000mm3に達したときに、マウスを屠殺した。腫瘍は、クリオスタットにて7ミクロンに切り、アセトンにて固定し、DAPI(Invitrogen)、ハムスター抗CD31(Serotech, Inc.)及び抗ハムスターCy3二次(Jackson Immunolabs)にて染色した。スライドは、蛍光色素マウンティング培地(Dako)にてマウントした。画像は、ツァイス社製Axioskop2顕微鏡にて撮像し、DAPI染色およびCy3染色の領域についてImageJにて分析した。Cy3染色はDAPI染色の領域にて除して、存在する細胞数に対して規準化し、値は抗ブタクサ処理した腫瘍に対して規準化した。すべての動物実験は、Institutional Animal Care and Use Committeeが承認したプロトコールに従って実施した。
12週齢のBalb-cマウス(Jackson)に、5mg/kgの抗gD、抗NRR1又は抗NRR2を週2回、2週間腹腔内投与した。既に記載されているように(Saito, et al., Immunity, Vol. 18, 675-685, May 2003)、マウスから脾細胞を単離し、B220-PerCP、CD23-PE及びCD21-FITC(BD Biosciences)にてFACSのために染色した。
6匹の6つの群の10週齢のC57BL/6マウス(Jackson)に、MCTを1日おきに、30umol/kgのDBZを1日おきに、10、2あるいは0.4mg/kgの抗NRR1又は10mg/kgの抗gDコントロール(HSV−1)を0、2および6日目に、あるいは示したように、投与した。ホルマリン固定しパラフィン包埋したマウス小腸組織を3ミクロン厚で切断した。製造業者(PolyScientific)によって推奨されるように、腸管細胞種の組織化学的識別はアルシアンブルーにて実行した。抗Ki67染色のために、切片を、Target Retrieval溶液(S1700、DAKO)にて前処理し、ウサギ抗Ki67(1:200、クローンSP6、Neomarkers)と共にインキュベートした。7.5μg/ml(Vector labs)の二次ヤギ抗ウサギは、ベクタステインABC Eliteキット(Vector labs)にて検出した。Ki67染色した切片は、マイヤーのヘマトキシリンにて対比染色した。抗リゾチーム染色のために、切片を、CC1mエピトープ回復条件、OmniMap-ウサギ検出およびベンタナ社のヘマトキシリンII/ブルーイング対比染色を使用したディスカバリーXTプラットフォーム(Ventana Medical Systems, Inc.)にて処理した。HES-1染色のために、抗ラットHES-1(クローンNM1、MBL, International)の後にTSA−HRPを行った。
1.Notch1及びNotch2ネガティブ調節領域を特異的に標的とする合成抗体の生成
ヒトおよびマウスにおけるNotch1およびNotch2のシグナル伝達の独立した拮抗作用を可能にするために、発明者等は、ファージディスプレイを使用してNRRを標的とし、合成のヒト抗体のために以下の性質により選択した。第一に、各抗体を、その同族の標的であるNotch1又はNotch2を強力に阻害するが、他のNotchレセプターは阻害しないように選別した。第二に、抗体を、ヒトの標的配列への結合について選別し、治療的ターゲッティングを可能にした。第三に、インビボで全身的にNotch1又はNotch2を阻害する生物学的結果を発見するため、そしてマウスモデルにおけるこれらレセプターの機能を調べるために、抗体を、オルソログなマウス配列への結合について同時に選別した。第四に、ヒト抗抗体免疫応答を防止するために、相補性決定領域を、ヒト免疫グロブリンフレームワークにクローニングした。
この手法の後、発明者等は、それぞれ所望の性質を有する抗Notch1 NRR抗体および抗Notch2 NRR抗体を成功裏に単離した。これらの抗体は、γ-分泌酵素インヒビターといった複数のNotchレセプターを標的とする「パンNotch」インヒビターよりもいくつかの利点を有する。これらの抗体は個々のNotchレセプターを区別するので、それら個々のレセプターの異なる機能を分析することができる。さらに、パンNotchインヒビターは、Notch1およびNotch2を二重阻害するため、インビボで望ましくない副作用を示しうるので、Notch1又はNotch2に対して選択的である抗体は治療のためにはより魅力的な候補となりうる。上記の手法に従って単離した抗Notch1 NRRおよび抗Notch2 NRR抗体は、更に詳細に以下に記述し、特徴を示す。
抗Notch2 NRR抗体の単離および特徴づけをは本明細書において記述する。抗Notch2 NRR抗体を単離し、「抗体D」と称する。その抗体を上記の通りに親和性成熟し、抗体D-1、抗体D-2および抗体D-3とした。図1は、抗体D、抗体D-1、抗体D-2および抗体D-3のHVR-H1、HVR-H2およびHVR-H3配列を示す。図2は、抗体D、抗体D-1、抗体D-2および抗体D-3のHVR-L1、HVR-L2およびHVR-L3配列を示す。図3は、抗体D、抗体D-1、抗体D-2および抗体D-3の重鎖可変領域配列を示す。図4は、抗体D、抗体D-1、抗体D-2および抗体D-3の軽鎖可変領域配列を示す。便宜上、抗体D-3は、本明細書中で「抗NRR2」と称する。
抗NRR抗体がNotchシグナル伝達に作用したかどうかを評価するために、発明者等はまず、NotchリガンドとしてJag1を、又はNotchレセプターとしてNotch1(図10A)ないしはNotch2(図10B)を発現するように操作したNIH-3T3細胞株を用いて共培養アッセイを行った。強力なNotchレポーターシグナル(ホタルルシフェラーゼ)がリガンド発現細胞およびレセプター発現細胞の両方の存在に依存していたので、このアッセイはNotchシグナル伝達を正確に反映しており(図10A及びB、−Jag1を+Jag1に比較する)、GSI(γ-分泌酵素インヒビター)が含まれる場合には、レポーターレベルはバックグラウンドにまで下がった(図10AおよびB、DMSOをDAPTと比較する)。
抗NRR1の漸増量の添加によりNotch1細胞のシグナル伝達が阻害され、80から400ng/mlの抗体濃度で完全な阻害が観察された(図10A)。コントロール抗体は、Notch1以外の抗原に対する他の多くの抗体(データは示さない)のように、シグナル伝達を阻害しなかった(図10A、α-gDをα-NRR1の力価に比較する)。この阻害活性が抗NRR1結合を反映していたかどうかを更に試験するために、発明者等は、精製したNRR1抗原の添加が、シグナル伝達細胞に発現されるNotch1への抗体結合に競合することによって、抗NRR1抗体の阻害濃度(80ng/ml)の存在下でシグナル伝達を救うか否かを調べた。NRR2ではなくNRR1抗原の添加により(他のアッセイにおいて活性を示し、適切にホールドされたもの;図10Bを参照)、コントロールレベルにまでシグナル伝達を修復した(図10A、80ng/mlのα-NRR1+NRR1を80ng/mlのα-NRR1単独および80ng/mlのα-NRR1+NRR2と比較する)。コントロールとして、NRR1及びNRR2抗原はいずれもコントロール抗体を含有した共培養物におけるシグナル伝達に作用しなかった。これは、NRR断片自体が、Notchシグナル伝達に対して直接効果を示さないことを裏付けるものである(図10A、α-gD+NRR1及びα-gD+NRR2)。抗NRR2活性をアッセイするためにNotch2発現細胞を使用して、発明者等は、Notch2シグナル伝達の抗NRR2阻害について類似の結果を観察した(図10B)。両抗体は、発明者等が試験したNotchリガンドのすべて、すなわちJag1、Jag2、Dll1およびDll4により誘導されるシグナル伝達を阻害した(図16およびデータは示さない)。まとめると、これらの結果は、抗NRR1及び抗NRR2はそれぞれNotch1およびNotch2からのシグナル伝達の強力なパラログ特異的なインヒビターである。
Notchシグナル伝達が直接腫瘍形成に作用することを示した1つのメカニズムは、T−ALLにおけるNotch1シグナル伝達の突然変異の活性化によるものである。特に、T−ALLにおけるNotch1活性化突然変異は2つの広いカテゴリーに分類される。(1)PESTドメインを切断することによりICDを安定化し、リガンド依存的な様式でシグナル伝達を増加するもの、及び(2)NRRを不安定化することによりADAMを切断し、リガンドに非依存的な様式でシグナル伝達を刺激するもの。抗NRR1がこのような変異レセプターによるシグナル伝達を無効にするか否かを試験するために、発明者等は、Notch1-WT(Notch1野生型)、Notch1−PEST(PESTドメインを欠くNotch1)又はNotch1-L1594Pを発現するように共培養アッセイを変更した(NRRのヘテロ二量体化(HD)ドメインのL1594P突然変異を保有するNotch1)。野生型Notch1シグナル伝達について観察されたように(図10C、上パネル)、抗NRR1は、Notch1−ΔPESTおよびNotch1-L1594Pによるリガンド依存性及びリガンド非依存性のシグナル伝達を完全に阻害した(図10C、中央および下パネル)。ゆえに、抗NRR1は、抗体が標的とする同じドメイン内にあり、不安定化するL1594P突然変異を含む変異レセプターの両種類によるシグナル伝達を中和(アンタゴナイズ)する。
抗NRR1および抗NRR2がインビボでレセプター特異的なインヒビターとして機能するか否かを決定するために、発明者等は、抗体がどのように、遺伝学的技術がNotch1又はNotch2の役割を確立する細胞運命決定に作用するかについて調査した。具体的には、Notch1又はNotch2の条件的不活性化により既に、Notch1はリンパ系発達の間にB細胞に対するT細胞の運命を決定するために重要であるのに対して、Notch2は脾臓周辺帯B(MZB)細胞を生成するために必要であることが示された。T細胞発達におけるNotch1の重要な機能と一致して、発明者等は、抗NRR2ではなく抗NRR1にてマウスを処置すると、胸腺重量が有意に低減したことを発見した(図11AおよびB)。同様に、抗NRR2でなく抗NRR1は、胸腺の細胞の総数を激減させ(図11B)、CD4+/CD8+二重陽性T細胞の生成をほとんど完全に阻害した(図11C)。
対照的に、MZB細胞を調べると、抗体効果は逆転していた。抗NRR2による処置によりMZB細胞がほとんど除去され(6.61+/−0.25のコントロール値と比較して0.97+/−0.45)、精製した組み換えリンホトキシン−βレセプター(LTβR)-Fc融合タンパク質(ポジティブコントロールとして用いた)(3.48+/−0.06)よりも更に集団を激減した(図11D)。抗NRR1が抗gDコントロール抗体(6.61+/−0.25;図11D)と比較してMZB集団を有意に低減しなかったので(6.00+/−0.44)、この阻害作用は抗NRR2に特異的であった。まとめると、これらのインビボ研究は、両抗体が細胞運命決定の生物学上有意な変化を誘導しうるそれぞれの標的の強力なインヒビターであることを示す。さらに、抗NRR1はMZB細胞でなくTの生成に作用するのに対して、抗NRR2はT細胞でなくMZBの生成に作用し、各々の抗体はインビボでその意図する標的に特異的に機能する。
抗NRR1および抗NRR2の結合特異性を解明し、抗体アンタゴニスト活性に機構的な洞察をするために、発明者等は、キメラNRRタンパク質のバッテリーに対する両抗体の結合を試験した。具体的には、発明者等は、Notch1とNotch2との間の各NRRサブドメイン(LNR−A、LNR−B、LNR−C、HD−NおよびHD−C)を交換することによってキメラNRRを作製し、簡易な検出のためにアルカリホスファターゼに融合させた分泌タンパク質のようにこれらキメラを発現させ、抗NRR1又は抗NRR2への各キメラNRRの規準化した量の結合を検定した。発明者等が発現させた26のキメラNRRのうち、17は分泌タンパク質のように効率的に検出され(図12A)、7は僅かに検出され、2はバックグラウンドを上回って検出されなかった(図17)ことから、すべてではないがほとんどのキメラNRRが発現され、適切にホールドされ、分泌されたことが示唆された。NRR2の単一LNR−AドメインをNRR1主鎖へ交換することにより抗NRR1の結合が破壊されたことは(図12A、キメラBC.HdおよびNRR2のLNR−Aを有する他のすべて)、NRR1のLNR−Aサブドメインとの抗NRR1の接触は結合に必須であることを示す。対照的に、NRR2のLNR-Cおよび/またはHD−NドメインをNRR1に交換することが抗NRR1結合に作用しなかったことは(図12A、キメラAB.Hd、ABC.HcおよびAB.Hc)、これらドメインはいずれも結合特異性に必須であることを示す。最後に、AB.Hcは、(ABキメラへの弱い結合が観察されたが)完全な抗NRR1結合を以前として持つNRR1サブドメインを最も少なく持つキメラであったことから、ほとんどそしておそらくすべての抗NRR1結合特異性に必要な接触はLNR−A、LNR-BおよびHD−Cドメインに含まれることが示唆された。
また、この構造により、Notch2よりもNotch1に対するFab特異性の基礎が明らかになる。Notch1およびNotch2のNRRドメイン間におよそ45%の配列同一性があるにもかかわらず、界面に関与する大部分の残基は、2つのタンパク質間で異なっている。例えば、Fabを結合する際に少なくとも25%の溶媒アクセス表面領域を覆う21のNRR残基のうち、Notch1およびNotch2において6つしか同一ではない。これら6つの残基は、Fabエピトープの4分の1未満の表面積でしかなく、エピトープの全体にわたって分布している(図12Dおよび18)。
インビボでの個々のレセプターの機能的な重要性を定めるためにNotchレセプター特異的抗体を利用し始めるために、発明者等はまず、抗NRR1にてNotch1シグナル伝達を選択的にブロックすると哺乳類の脈管形成が破壊するか否かを調べた。数多くの報告は、Notch経路が血管内皮性増殖因子(VEGF)の下流で機能し、チップ(tip)細胞とストーク(stalk)細胞の間の内皮細胞運命選択を制御することによって脈管形成に重要な役割を果たすことを示している。Dll4特異的ブロック抗体を含む様々な遺伝的及び生化学的手法を用いた実験は、2つのNotchレセプターであるNotch1およびNotch4は脈管形成においてDll4のレセプターとして関与していたが、Dll4をNotch血管形成シグナル伝達に関与する重要なNotchリガンドとして定めた。
発明者等はまず、Notch1の選択的な阻害がインビトロの内皮出芽に作用するか否かを試験した。ヒト皮膚線維芽細胞と共培養したヒト臍帯静脈内皮細胞(HUVEC)は、内腔様構造の出芽を生成する。先の研究と同様に、発明者等は、DBZを使用したパンNotch阻害、γ分泌酵素のインヒビター並びにDll4の選択的抗体ブロックにより内皮細胞出芽の数と長さが増加したことがわかった。重要なことに、抗NRR1を使用してNotch1シグナル伝達を選択的にブロックすることにより類似の表現型が生成した(図13Aおよび13B)。
抗NRR1を使用したNotch1の選択的ブロックが腫瘍成長を阻害するために十分か否かを決定するために、発明者等は、臨床前腫瘍モデルを使用し、抗NRR1、抗VEGF又は抗ブタクサ抗体による処置の後に定着させた腫瘍の成長を比較した。Calu6およびHM7の両モデルにおいて、抗NRR1にて処置した群は、試験した初めの時点、初回投与の3〜4日後でさえ、コントロール群と比較して腫瘍のサイズが有意に減少していた(図13A−C)。Calu6モデルでは、抗NRR1群はおよそ250mm3から100mm3未満に腫瘍サイズが減少しており、これは抗VEGF群に観察された減少と類似していた。これに反して、抗ブタクサコントロール群の腫瘍は400mm3より大きいサイズに成長していた(図14A)。発明者等は、HM7モデルを強力に成長させて類似の結果を観察した。抗NRR1処置により、12〜13日間にわたって腫瘍サイズが静止したまま(図14B)又はほぼ静止したまま(図14C)であり、これは、コントロール群に観察された250mm3から900mm3への6日間の成長と比較して劇的に異なっている(図14Bおよび5C)。10および20mg/kgの群は2.5および5mg/kgの群と比較して強い阻害傾向を示したが(図14C)、発明者等は、20mg/kgから2.5mg/kgの抗NRR1用量濃度を用いても腫瘍成長に対する同様な阻害を観察した。
複数のNotchレセプターを阻害するパンNotchインヒビターであるγ-分泌酵素インヒビター(GSI)により、体重損失および腸管のゴブレット細胞異形成が生じる。これは、Notchが腸管の陰窩始原細胞の増殖を維持し、分泌細胞運命への分化を妨げることによって細胞運命を決定する際に働く役割を反映する。(van Es et al., Nature 435:959-963 (2005)を参照)。更に、条件的Notchノックアウトマウスを使用した遺伝子の研究は、Notch1およびNotch2の破壊が腸管のゴブレット細胞異形成を引き起こすために必要であることを示唆する。(Riccio et al., EMBO Rep. 9: 377-383 (2008)を参照)。発明者等は、抗NRR1および抗NRR2を用いて、体重損失および腸管の細胞分化に対する、Notch1又はNotch2、あるいはその両方の選択的な阻害の効果を決定した。
抗NRR1にて処置したマウスは、抗体投薬クールの間、総体重の軽度の減少を示した。抗NRR1および抗NRR2を直接比較する第一実験では、発明者等は、抗NRR1により総体重の5%が減少したのに対して、抗NRR2並びに抗gDおよび抗LTβR-Fcコントロール群では体重の増加が観察された(図15A)。第二の実験では、抗NRR1又は抗NRR2をそれぞれ用いたNotch1又はNotch2の選択的な阻害は、体重に対してほとんどあるいはまったく効果がなかった。実験の結果を図24Aに示す。ここでは、マウスに、矢印で示される日に5mg/kgの抗NRR1(正方形)、抗NRR2(三角形)又はコントロール抗体(「α-gD」、菱形)を投与した。しかしながら、抗NRR1および抗NRR2の両方にて処置したマウス(図24Aの「X」)は、7日目という早い時期に開始時の体重のほぼ20%を失った。
図23は、デキサメサゾンが抗NRR1によって生じた腸管の表現型を少なくとも部分的にレスキューすることを示す。抗体は、以下の通りに雌NCR.ヌードマウス(1群につき3匹マウス)の腹膜内(IP)に投与した。
グループA) ベヒクルの組合せ、MCT、1日1回、4mg/kgの抗gDコントロール抗体、3日おき(図に示さない);
グループB) デキサメサゾン、90mg/kg、毎日(図に示さない);
グループC) 抗NRR1、4mg/kg、3日おき;
グループD) デキサメサゾン、90mg/kg、毎日および抗NRR1、4mg/kg、3日おき。
ベヒクルは、0.5%(w/v) ヒドロキシプロピルメチルセルロース(Methocel E4M)および0.1%(w/v)Tween80を含む水(MCT)であり、前もって不都合な臨床効果を引き起こさないことを示した。
9日目に腸を採取し、抗Ki67(増殖マーカー)およびアルシアンブルー(ムチン用)にて染色した。この実験は、デキサメサゾンと抗NRR1の組合せにて処置したマウスは、抗NRR1単独で処置したマウスと比較して軽度の腸管の表現型を現したことから、デキサメサゾンが抗NRR1が誘導する分化の変化から腸を保護することを示唆した。
抗NRR1及び抗NRR2の、メラノーマ細胞株の成長に対する効果を調査した。ヒトメラノーマ細胞株SK23およびLOX−IMVIを、Jagged-1リガンド(R&D Systems, Minneapolis, MN)でコートしない(−jag-1)又はJagged-1リガンドでコートした(+jag-1)96ウェルプレートに、低密度(SK23では4000細胞/ウェル、LOX−IMVIでは2500細胞/ウェル)で2%FBSを含む培地に播いた。抗NRR1(20mg/ml)、抗NRR2(20mg/ml)又はγ−分泌酵素インヒビター(GSI) DAPT(5μM)を培養物に加え、その後1日おきに再添加した。等しい体積のDMSOをベヒクルコントロールとし、等しい濃度の抗gD HuB6ファージ抗体をアイソタイプコントロール抗体とした。プレートに播いて6日後に、Cell Titer Gloアッセイ(Promega, Madison, WI)を行った。結果を図25Aおよび25Bに示す(抗NRR1および抗NRR2を「抗N1」および「抗N2」とする)。図25Aおよび25Bでは、細胞生存度(y軸)を、DMSOベヒクル、−jag-1コントロールウェルからの値と比較して表した。両メラノーマ細胞株は、抗NRR1処置でなく、GSI処置(特にJag1リガンドの存在下)および抗NRR2処置に応答して生存度の減少を示した。
びまん性大細胞型B細胞リンパ腫(DLBCL)は、最も一般的なタイプの非ホジキンリンパ腫である。DLBCL患者の癌細胞の最近の研究は、患者のおよそ8%がNotch2にPESTドメイン突然変異を保因していることを示した。この変異は、リガンド活性化後のNotch2シグナル伝達を延長することが予測されるものである。(Lee et al., Cancer Sci. 100: 920-926, 2009を参照)。しかしながら、DLBCL細胞がNotch2阻害にどのように応答しているのかはわかっていない。したがって、発明者等は、5つのDLBCL細胞株に対する抗NRR2の効果を調査した。図26の右に列挙するDLBCL株を、384ウェルプレートのウェルに数通り3日間成長させた。細胞培養物には示した濃度の抗NRR2を含め(図26および27では「抗Notch2」と示す)、これは10μg/mlで開始した3倍段階希釈物に対応している。成長は、Cell Titer Gloアッセイ(Promega)を使用して評価し、アイソタイプコントロール抗体による処置と比較して細胞生存度の割合としてプロットした。図26のデータポイントは、示した抗体濃度での各細胞株についての各々のウェルから得た平均値を表す。図26に示すように、細胞株の一つ、「DB」の増殖は、抗NRR2による処置によって強く阻害された。図27は経時的なDB細胞株の増殖を示す。DB株は12ウェルプレートのウェル中で生育させ、培養物を、示した濃度のDMSO、γ−分泌酵素インヒビターDAPT、抗gDアイソタイプコントロール抗体又は抗NRR2にて処置した。成長は、播種して培養物を処置した2、3および5日後に生きている細胞(Vi-CELL, Beckman Coulter, Fullerton, CA)を計数することによって評価した。値は、3つの別々の培養物からの測定値の平均±標準偏差を表す。図27は、抗NRR2がインビトロでDB DLBCL細胞株の成長を抑制することを示す。
Claims (25)
- Notch2ネガティブ制御領域(NRR)のLin12/Notchリピート(LNR)−Aドメインおよびヘテロ二量体化ドメイン(HD)−Cドメインに結合し、Notch2活性を阻害するモノクローナル抗体であって、ここで、該抗体は、10nM以下のKdでマウスNotch2 NRRおよびヒトNotch2 NRRに結合し、
配列番号:1−2から選択されるアミノ酸配列を含むHVR-H1;
配列番号:4のアミノ酸配列を含むHVR-H2;
配列番号:5のアミノ酸配列を含むHVR-H3;
配列番号:6−9から選択されるアミノ酸配列を含むHVR-L1;
配列番号:11−13から選択されるアミノ酸配列を含むHVR-L2;及び、
配列番号:15−18から選択されるアミノ酸配列を含むHVR-L3
を含む、モノクローナル抗体。 - Notch2以外のNotchファミリに有意に結合しない、請求項1に記載の抗体。
- HVR-H1が配列番号:1のアミノ酸配列を含み、HVR-L1が配列番号:6のアミノ酸配列を含み、HVR-L2が配列番号:11のアミノ酸配列を含み、そして、HVR-L3が配列番号:15のアミノ酸配列を含む、請求項1に記載の抗体。
- HVR-H1が配列番号:2のアミノ酸配列を含み、HVR-L1が配列番号:7のアミノ酸配列を含み、HVR-L2が配列番号:11のアミノ酸配列を含み、そして、HVR-L3が配列番号:16のアミノ酸配列を含む、請求項1に記載の抗体。
- HVR-H1が配列番号:2のアミノ酸配列を含み、HVR-L1が配列番号:8のアミノ酸配列を含み、HVR-L2が配列番号:12のアミノ酸配列を含み、そして、HVR-L3が配列番号:17のアミノ酸配列を含む、請求項1に記載の抗体。
- HVR-H1が配列番号:2のアミノ酸配列を含み、HVR-L1が配列番号:9のアミノ酸配列を含み、HVR-L2が配列番号:13のアミノ酸配列を含み、そして、HVR-L3が配列番号:18のアミノ酸配列を含む、請求項1に記載の抗体。
- ヒトVHアクセプター2フレームワークおよびヒトVLκサブグループIコンセンサスフレームワークから選択される少なくとも一のフレームワークをさらに含む、請求項1に記載の抗体。
- 配列番号:20−21から選択されるアミノ酸配列に少なくとも90%の配列同一性を有する重鎖可変ドメインと、配列番号:22−25から選択されるアミノ酸配列に少なくとも90%の配列同一性を有する軽鎖可変ドメインとを含む、請求項1に記載の抗体。
- 配列番号:20のアミノ酸配列に少なくとも90%の配列同一性を有する重鎖可変ドメインと、配列番号:22のアミノ酸配列に少なくとも90%の配列同一性を有する軽鎖可変ドメインとを含む、請求項8に記載の抗体。
- 重鎖可変ドメインが配列番号:20のアミノ酸配列を含み、軽鎖可変ドメインが配列番号:22のアミノ酸配列を含む、請求項9に記載の抗体。
- 配列番号:21のアミノ酸配列に少なくとも90%の配列同一性を有する重鎖可変ドメインと、配列番号:23−25から選択されるアミノ酸配列に少なくとも90%の配列同一性を有する軽鎖可変ドメインとを含む、請求項8に記載の抗体。
- 重鎖可変ドメインが配列番号:21のアミノ酸配列を含み、軽鎖可変ドメインが配列番号:23−25から選択されるアミノ酸配列を含む、請求項11に記載の抗体。
- Fab、Fab'-SH、Fv、scFv又は(Fab')2の断片から選択される抗体断片である、請求項1から12の何れか一に記載の抗体。
- ヒト、ヒト化、又はキメラである、請求項1から13の何れか一に記載の抗体。
- 請求項1から12の何れか一の抗体を含む、Notch2活性の阻害剤。
- Notch2の発現又は活性の増加と関連する疾患を治療するための医薬であって、請求項1から12の何れか一に記載の抗体の有効量を含有してなる医薬。
- B細胞悪性腫瘍を治療するための医薬であって、請求項1から12の何れか一に記載の抗体の有効量を含有してなる医薬。
- メラノーマを治療するための医薬であって、請求項1から12の何れか一に記載の抗体の有効量を含有してなる医薬。
- Notch2の発現又は活性の増加と関係する疾患を治療するための医薬を製造するための、請求項1から12の何れか一に記載の抗体の使用。
- B細胞悪性腫瘍を治療するための医薬を製造するための、請求項1から12の何れか一に記載の抗体の使用。
- メラノーマを治療するための医薬を製造するための、請求項1から12の何れか一に記載の抗体の使用。
- 医薬として使用するための請求項1から12の何れか一に記載の抗体。
- Notch2の発現又は活性の増加と関係する疾患の治療に使用するための、請求項1から12の何れか一に記載の抗体。
- B細胞悪性腫瘍の治療に使用するための、請求項1から12の何れか一に記載の抗体。
- メラノーマの治療に使用するための、請求項1から12の何れか一に記載の抗体。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US10191708P | 2008-10-01 | 2008-10-01 | |
| US61/101,917 | 2008-10-01 | ||
| PCT/US2009/059028 WO2010039832A1 (en) | 2008-10-01 | 2009-09-30 | Anti-notch2 antibodies and methods of use |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015042089A Division JP2015145379A (ja) | 2008-10-01 | 2015-03-04 | 抗notch2抗体および使用方法 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2012504634A JP2012504634A (ja) | 2012-02-23 |
| JP2012504634A5 JP2012504634A5 (ja) | 2012-11-15 |
| JP5710487B2 true JP5710487B2 (ja) | 2015-04-30 |
Family
ID=41349057
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2011530173A Active JP5710487B2 (ja) | 2008-10-01 | 2009-09-30 | 抗notch2抗体および使用方法 |
| JP2015042089A Pending JP2015145379A (ja) | 2008-10-01 | 2015-03-04 | 抗notch2抗体および使用方法 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015042089A Pending JP2015145379A (ja) | 2008-10-01 | 2015-03-04 | 抗notch2抗体および使用方法 |
Country Status (18)
| Country | Link |
|---|---|
| US (5) | US8404239B2 (ja) |
| EP (1) | EP2334331B1 (ja) |
| JP (2) | JP5710487B2 (ja) |
| KR (1) | KR101712214B1 (ja) |
| CN (1) | CN102170909B (ja) |
| AR (1) | AR073717A1 (ja) |
| AU (1) | AU2009298569A1 (ja) |
| BR (1) | BRPI0913827A2 (ja) |
| CA (1) | CA2736429A1 (ja) |
| CL (1) | CL2011000711A1 (ja) |
| ES (1) | ES2541726T3 (ja) |
| IL (1) | IL211623A0 (ja) |
| MX (1) | MX2011002928A (ja) |
| PE (1) | PE20120080A1 (ja) |
| RU (1) | RU2580029C2 (ja) |
| TW (1) | TW201018484A (ja) |
| WO (1) | WO2010039832A1 (ja) |
| ZA (1) | ZA201101651B (ja) |
Families Citing this family (18)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7919092B2 (en) | 2006-06-13 | 2011-04-05 | Oncomed Pharmaceuticals, Inc. | Antibodies to notch receptors |
| US8088617B2 (en) | 2007-01-24 | 2012-01-03 | Oncomed Pharmaceuticals, Inc. | Antibodies that bind the glutamate ligand binding region of Notch1 |
| SG192467A1 (en) | 2008-07-08 | 2013-08-30 | Oncomed Pharm Inc | Notch1 receptor binding agents and methods of use thereof |
| US9132189B2 (en) | 2008-07-08 | 2015-09-15 | Oncomed Pharmaceuticals, Inc. | Notch1 binding agents and methods of use thereof |
| CA2736429A1 (en) * | 2008-10-01 | 2010-04-08 | Genentech, Inc. | Anti-notch2 antibodies and methods of use |
| EP2621950A4 (en) | 2010-09-27 | 2015-09-02 | Janssen Biotech Inc | ANTIBODIES FOR BINDING HUMAN COLLAGEN II |
| US8906649B2 (en) | 2010-09-27 | 2014-12-09 | Janssen Biotech, Inc. | Antibodies binding human collagen II |
| US9663573B2 (en) * | 2011-10-05 | 2017-05-30 | Genentech, Inc. | Methods of treating liver conditions using Notch2 antagonists |
| JP2015517529A (ja) * | 2012-05-16 | 2015-06-22 | オンコメッド ファーマシューティカルズ インコーポレイテッド | Notch2/3抗体によって癌を治療するための方法 |
| WO2014028446A1 (en) | 2012-08-13 | 2014-02-20 | Genentech, Inc. | Anti-jagged anitbodies and methods of use |
| EP2970468B1 (en) | 2013-03-13 | 2021-07-07 | Novartis AG | Notch2 binding molecules for treating respiratory diseases |
| KR20150128707A (ko) * | 2013-03-15 | 2015-11-18 | 제넨테크, 인크. | 간암의 진단 및 치료를 위한 조성물 및 방법 |
| SG10201808259TA (en) | 2014-02-12 | 2018-10-30 | Genentech Inc | Anti-jagged1 antibodies and methods of use |
| RU2017103289A (ru) | 2014-07-11 | 2018-08-14 | Дженентек, Инк. | Ингибирование пути notch |
| WO2017053939A1 (en) * | 2015-09-26 | 2017-03-30 | Kung, Hsing-Jien | Method and composition for treatment of hair loss |
| GB201521447D0 (en) * | 2015-12-04 | 2016-01-20 | Ucb Biopharma Sprl | De novo antibody design |
| US10858443B2 (en) | 2017-05-31 | 2020-12-08 | Trustees Of Boston University | Synthetic notch protein for modulating gene expression |
| PE20231300A1 (es) * | 2020-07-17 | 2023-08-24 | Genentech Inc | Anticuerpos anti-notch2 y metodos de uso |
Family Cites Families (62)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3896111A (en) | 1973-02-20 | 1975-07-22 | Research Corp | Ansa macrolides |
| US4151042A (en) | 1977-03-31 | 1979-04-24 | Takeda Chemical Industries, Ltd. | Method for producing maytansinol and its derivatives |
| US4137230A (en) | 1977-11-14 | 1979-01-30 | Takeda Chemical Industries, Ltd. | Method for the production of maytansinoids |
| US4307016A (en) | 1978-03-24 | 1981-12-22 | Takeda Chemical Industries, Ltd. | Demethyl maytansinoids |
| US4265814A (en) | 1978-03-24 | 1981-05-05 | Takeda Chemical Industries | Matansinol 3-n-hexadecanoate |
| JPS5562090A (en) | 1978-10-27 | 1980-05-10 | Takeda Chem Ind Ltd | Novel maytansinoid compound and its preparation |
| JPS55164687A (en) | 1979-06-11 | 1980-12-22 | Takeda Chem Ind Ltd | Novel maytansinoid compound and its preparation |
| US4256746A (en) | 1978-11-14 | 1981-03-17 | Takeda Chemical Industries | Dechloromaytansinoids, their pharmaceutical compositions and method of use |
| JPS5566585A (en) | 1978-11-14 | 1980-05-20 | Takeda Chem Ind Ltd | Novel maytansinoid compound and its preparation |
| JPS55102583A (en) | 1979-01-31 | 1980-08-05 | Takeda Chem Ind Ltd | 20-acyloxy-20-demethylmaytansinoid compound |
| JPS55162791A (en) | 1979-06-05 | 1980-12-18 | Takeda Chem Ind Ltd | Antibiotic c-15003pnd and its preparation |
| JPS55164685A (en) | 1979-06-08 | 1980-12-22 | Takeda Chem Ind Ltd | Novel maytansinoid compound and its preparation |
| JPS55164686A (en) | 1979-06-11 | 1980-12-22 | Takeda Chem Ind Ltd | Novel maytansinoid compound and its preparation |
| US4309428A (en) | 1979-07-30 | 1982-01-05 | Takeda Chemical Industries, Ltd. | Maytansinoids |
| JPS5645483A (en) | 1979-09-19 | 1981-04-25 | Takeda Chem Ind Ltd | C-15003phm and its preparation |
| JPS5645485A (en) | 1979-09-21 | 1981-04-25 | Takeda Chem Ind Ltd | Production of c-15003pnd |
| EP0028683A1 (en) | 1979-09-21 | 1981-05-20 | Takeda Chemical Industries, Ltd. | Antibiotic C-15003 PHO and production thereof |
| WO1982001188A1 (en) | 1980-10-08 | 1982-04-15 | Takeda Chemical Industries Ltd | 4,5-deoxymaytansinoide compounds and process for preparing same |
| US4450254A (en) | 1980-11-03 | 1984-05-22 | Standard Oil Company | Impact improvement of high nitrile resins |
| US4315929A (en) | 1981-01-27 | 1982-02-16 | The United States Of America As Represented By The Secretary Of Agriculture | Method of controlling the European corn borer with trewiasine |
| US4313946A (en) | 1981-01-27 | 1982-02-02 | The United States Of America As Represented By The Secretary Of Agriculture | Chemotherapeutically active maytansinoids from Trewia nudiflora |
| JPS57192389A (en) | 1981-05-20 | 1982-11-26 | Takeda Chem Ind Ltd | Novel maytansinoid |
| US5053394A (en) | 1988-09-21 | 1991-10-01 | American Cyanamid Company | Targeted forms of methyltrithio antitumor agents |
| US5770701A (en) | 1987-10-30 | 1998-06-23 | American Cyanamid Company | Process for preparing targeted forms of methyltrithio antitumor agents |
| US5606040A (en) | 1987-10-30 | 1997-02-25 | American Cyanamid Company | Antitumor and antibacterial substituted disulfide derivatives prepared from compounds possessing a methyl-trithio group |
| CA2026147C (en) | 1989-10-25 | 2006-02-07 | Ravi J. Chari | Cytotoxic agents comprising maytansinoids and their therapeutic use |
| US5208020A (en) | 1989-10-25 | 1993-05-04 | Immunogen Inc. | Cytotoxic agents comprising maytansinoids and their therapeutic use |
| IE20030749A1 (en) | 1991-05-03 | 2003-11-12 | Indiana University Foundation | Human notch and delta binding domains in torporythmic proteins, and methods based thereon |
| US5786158A (en) | 1992-04-30 | 1998-07-28 | Yale University | Therapeutic and diagnostic methods and compositions based on notch proteins and nucleic acids |
| US20050112121A1 (en) | 1992-04-30 | 2005-05-26 | Yale University | Therapeutic and diagnostic methods and compositions based on notch proteins and nucleic acids |
| WO1994011026A2 (en) | 1992-11-13 | 1994-05-26 | Idec Pharmaceuticals Corporation | Therapeutic application of chimeric and radiolabeled antibodies to human b lymphocyte restricted differentiation antigen for treatment of b cell lymphoma |
| US5635483A (en) | 1992-12-03 | 1997-06-03 | Arizona Board Of Regents Acting On Behalf Of Arizona State University | Tumor inhibiting tetrapeptide bearing modified phenethyl amides |
| US5780588A (en) | 1993-01-26 | 1998-07-14 | Arizona Board Of Regents | Elucidation and synthesis of selected pentapeptides |
| US5773001A (en) | 1994-06-03 | 1998-06-30 | American Cyanamid Company | Conjugates of methyltrithio antitumor agents and intermediates for their synthesis |
| US5663149A (en) | 1994-12-13 | 1997-09-02 | Arizona Board Of Regents Acting On Behalf Of Arizona State University | Human cancer inhibitory pentapeptide heterocyclic and halophenyl amides |
| US5731168A (en) | 1995-03-01 | 1998-03-24 | Genentech, Inc. | Method for making heteromultimeric polypeptides |
| US5712374A (en) | 1995-06-07 | 1998-01-27 | American Cyanamid Company | Method for the preparation of substantiallly monomeric calicheamicin derivative/carrier conjugates |
| US5714586A (en) | 1995-06-07 | 1998-02-03 | American Cyanamid Company | Methods for the preparation of monomeric calicheamicin derivative/carrier conjugates |
| US5780300A (en) | 1995-09-29 | 1998-07-14 | Yale University | Manipulation of non-terminally differentiated cells using the notch pathway |
| US20050158859A1 (en) | 1995-09-29 | 2005-07-21 | Yale University | Manipulation of non-terminally differentiated cells using the Notch pathway |
| US6436650B1 (en) | 1997-07-23 | 2002-08-20 | Yale University | Activated forms of notch and methods based thereon |
| US6692919B1 (en) | 1997-07-23 | 2004-02-17 | Yale University | Activated forms of notch and methods based thereon |
| CA2343963A1 (en) | 1998-10-02 | 2000-04-13 | The Government Of The United States Of America As Represented By The Secretary, Dept. Of Health And Human Services, The National Institutes Of Health | Apotosis inducing agents and methods |
| US8084258B2 (en) | 1999-07-12 | 2011-12-27 | University Of Basel | Manipulation of tissue of organ type using the notch pathway |
| CA2422538C (en) | 2000-09-22 | 2012-06-19 | Genentech, Inc. | Notch receptor agonists and uses |
| EP1356048A2 (en) | 2000-12-04 | 2003-10-29 | Argonex Pharmaceuticals | Cytotoxic t-lymphocyte-inducing immunogens for prevention, treatment, and diagnosis of cancer |
| US6884869B2 (en) | 2001-04-30 | 2005-04-26 | Seattle Genetics, Inc. | Pentapeptide compounds and uses related thereto |
| US7755007B2 (en) | 2003-04-17 | 2010-07-13 | K&H Manufacturing, Inc | Heated pet mat |
| WO2005074633A2 (en) | 2004-02-03 | 2005-08-18 | The Regents Of The University Of Michigan | Compositions and methods for characterizing, regulating, diagnosing, and treating cancer |
| US20070077245A1 (en) | 2004-08-04 | 2007-04-05 | The Brigham And Women's Hospital, Inc. | NOTCH mutations leading to increased receptor signaling |
| WO2006053063A2 (en) | 2004-11-05 | 2006-05-18 | The Regents Of The University Of California | Notch-1 assay to detect neurodegenerative diseases |
| WO2006068822A1 (en) | 2004-12-20 | 2006-06-29 | Genentech, Inc. | Notch receptor agonists and uses |
| CA2630839C (en) | 2005-12-16 | 2017-01-17 | Regeneron Pharmaceuticals, Inc. | Therapeutic methods for inhibiting tumor growth with dll4 antagonists |
| US20080241150A1 (en) * | 2006-05-15 | 2008-10-02 | The Brigham And Women's Hospital, Inc. | Functional negative regulatory domain sequences from human NOTCH1 and 2 and isolated LNR domains from human NOTCH1 |
| US7919092B2 (en) * | 2006-06-13 | 2011-04-05 | Oncomed Pharmaceuticals, Inc. | Antibodies to notch receptors |
| WO2008051797A2 (en) | 2006-10-19 | 2008-05-02 | Genentech, Inc. | Anti-notch3 agonist antibodies and their use in the treatment of notch3-related diseases |
| WO2008076960A2 (en) | 2006-12-18 | 2008-06-26 | Genentech, Inc. | Antagonist anti-notch3 antibodies and their use in the prevention and treatment of notch3-related diseases |
| US8088617B2 (en) * | 2007-01-24 | 2012-01-03 | Oncomed Pharmaceuticals, Inc. | Antibodies that bind the glutamate ligand binding region of Notch1 |
| PE20090321A1 (es) | 2007-06-04 | 2009-04-20 | Genentech Inc | Anticuerpos anti-notch1 nrr, metodo de preparacion y composicion farmaceutica |
| SG192467A1 (en) | 2008-07-08 | 2013-08-30 | Oncomed Pharm Inc | Notch1 receptor binding agents and methods of use thereof |
| CA2736429A1 (en) * | 2008-10-01 | 2010-04-08 | Genentech, Inc. | Anti-notch2 antibodies and methods of use |
| PE20121494A1 (es) * | 2009-06-18 | 2012-11-01 | Pfizer | Anticuerpos anti notch-1 |
-
2009
- 2009-09-30 CA CA 2736429 patent/CA2736429A1/en not_active Abandoned
- 2009-09-30 JP JP2011530173A patent/JP5710487B2/ja active Active
- 2009-09-30 BR BRPI0913827-7A patent/BRPI0913827A2/pt not_active IP Right Cessation
- 2009-09-30 AU AU2009298569A patent/AU2009298569A1/en not_active Abandoned
- 2009-09-30 RU RU2011117237/10A patent/RU2580029C2/ru not_active IP Right Cessation
- 2009-09-30 ES ES09793181.0T patent/ES2541726T3/es active Active
- 2009-09-30 MX MX2011002928A patent/MX2011002928A/es active IP Right Grant
- 2009-09-30 EP EP20090793181 patent/EP2334331B1/en active Active
- 2009-09-30 PE PE2011000807A patent/PE20120080A1/es not_active Application Discontinuation
- 2009-09-30 CN CN200980139202.1A patent/CN102170909B/zh active Active
- 2009-09-30 AR ARP090103776 patent/AR073717A1/es unknown
- 2009-09-30 TW TW98133312A patent/TW201018484A/zh unknown
- 2009-09-30 WO PCT/US2009/059028 patent/WO2010039832A1/en active Application Filing
- 2009-09-30 US US12/570,267 patent/US8404239B2/en active Active
- 2009-09-30 KR KR1020117007489A patent/KR101712214B1/ko active IP Right Grant
-
2011
- 2011-03-03 ZA ZA2011/01651A patent/ZA201101651B/en unknown
- 2011-03-08 IL IL211623A patent/IL211623A0/en unknown
- 2011-03-31 CL CL2011000711A patent/CL2011000711A1/es unknown
-
2013
- 2013-02-22 US US13/773,843 patent/US20140037643A1/en not_active Abandoned
- 2013-10-11 US US14/051,699 patent/US20140314782A1/en not_active Abandoned
-
2014
- 2014-06-06 US US14/298,488 patent/US20170204175A9/en not_active Abandoned
-
2015
- 2015-03-04 JP JP2015042089A patent/JP2015145379A/ja active Pending
-
2017
- 2017-07-21 US US15/656,967 patent/US20180057589A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| BRPI0913827A2 (pt) | 2020-12-29 |
| PE20120080A1 (es) | 2012-02-17 |
| US20150104461A1 (en) | 2015-04-16 |
| JP2012504634A (ja) | 2012-02-23 |
| US20140314782A1 (en) | 2014-10-23 |
| CA2736429A1 (en) | 2010-04-08 |
| TW201018484A (en) | 2010-05-16 |
| AU2009298569A1 (en) | 2010-04-08 |
| ES2541726T3 (es) | 2015-07-23 |
| IL211623A0 (en) | 2011-05-31 |
| MX2011002928A (es) | 2011-04-11 |
| US20140037643A1 (en) | 2014-02-06 |
| KR101712214B1 (ko) | 2017-03-03 |
| KR20110061600A (ko) | 2011-06-09 |
| US8404239B2 (en) | 2013-03-26 |
| RU2011117237A (ru) | 2012-11-10 |
| EP2334331B1 (en) | 2015-04-29 |
| US20180057589A1 (en) | 2018-03-01 |
| EP2334331A1 (en) | 2011-06-22 |
| US20100080808A1 (en) | 2010-04-01 |
| ZA201101651B (en) | 2012-06-27 |
| RU2580029C2 (ru) | 2016-04-10 |
| CL2011000711A1 (es) | 2012-07-20 |
| CN102170909A (zh) | 2011-08-31 |
| AR073717A1 (es) | 2010-11-24 |
| JP2015145379A (ja) | 2015-08-13 |
| US20170204175A9 (en) | 2017-07-20 |
| WO2010039832A1 (en) | 2010-04-08 |
| CN102170909B (zh) | 2015-11-25 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5710487B2 (ja) | 抗notch2抗体および使用方法 | |
| JP5955828B2 (ja) | 抗vegf抗体 | |
| JP5383905B2 (ja) | ヒト化抗egfl7抗体とその使用方法 | |
| JP2018082713A (ja) | 抗notch1 nrr抗体とその使用方法 | |
| US9096686B2 (en) | E518A/K mutant smoothened and methods of using the same | |
| WO2011014457A1 (en) | Combination treatments | |
| JP5363972B2 (ja) | Egfl7に対する抗体とその使用方法 | |
| US20110171231A1 (en) | ANTI-PlGF ANTIBODIES AND METHODS USING SAME | |
| AU2013204108B2 (en) | Anti-VEGF antibodies | |
| AU2015252098A1 (en) | Mutant smoothened and methods of using the same | |
| AU2015200714A1 (en) | Anti-VEGF antibodies |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20120928 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20120928 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20140325 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140623 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140630 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140723 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140730 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140821 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140828 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20140924 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20150203 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20150304 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 5710487 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |