発明の概要
本発明は、部分的に、1つまたは複数(例えば、1、2、3、4、5等)の組換え部位(例えば、1つまたは複数のatt部位、1つまたは複数のlox部位等)および/または1つまたは複数(例えば、1、2、3、4、5等)のトポイソメラーゼ認識部位(例えば、1つまたは複数のIA型トポイソメラーゼ、IB型トポイソメラーゼ、II型トポイソメラーゼ等)を含む核酸分子、ならびにトポイソメラーゼ(例えば、部位特異的トポイソメラーゼ)で切断された核酸分子に関する。本発明は1つまたは複数の組換え部位および/または1つまたは複数のトポイソメラーゼを含む核酸分子にも関する。本発明はより具体的には、少なくとも1つの組換え部位を含む少なくとも1つの第1の核酸分子と、少なくとも1つのトポイソメラーゼ認識部位および/または少なくとも1つのトポイソメラーゼを含む少なくとも1つの第2の核酸分子の連結または結合に関する。これらの少なくとも第1および第2の分子を結合すると、(1) 少なくとも1つの組換え部位、および(2)少なくとも1つのトポイソメラーゼ認識部位および/または少なくとも1つのトポイソメラーゼを含む少なくとも1つの第3の(またはキメラ)分子が生成する。これらの核酸分子は、線状または閉鎖環状(例えば、弛緩型、スーパーコイル等)であり得る。そのような組換え部位、トポイソメラーゼ認識部位、およびトポイソメラーゼは、核酸分子の末端または末端付近、および/または核酸分子内部を含め、本発明の任意の数の核酸分子上の任意の位置に存在し得る。さらに、同一または異なる組換え部位、トポイソメラーゼ認識部位、および/またはトポイソメラーゼの任意の組み合わせが、本発明にしたがって使用できる。
本発明は、部分的に、核酸分子が(1)少なくとも1つ(例えば、1、2、3、4、5、6、7、8等)の組換え部位、および(2)少なくとも1つ(例えば、1、2、3、4、5、6、7、8等)のトポイソメラーゼ(例えば、共有結合トポイソメラーゼ)または少なくとも1つ(例えば、1、2、3、4、5、6、7、8等)のトポイソメラーゼ認識部位を含む、核酸分子(例えば、反応液)を含む、核酸分子および組成物を含む。特定の態様では、上述の核酸分子のトポイソメラーゼまたはトポイソメラーゼ認識部位、ならびに組換え部位は、内部、または1つまたは両方の末端または末端付近に存在し得る。例えば、少なくとも1つのトポイソメラーゼまたは少なくとも1つのトポイソメラーゼ認識部位のうちの1つまたは複数(例えば、1、2、3、4、5、6、7、8等)、ならびに少なくとも1つの認識部位のうちの1つまたは複数は、1つの5'端もしくはその付近、1つの3'端もしくはその付近、両方の5'端もしくはその付近、両方の3'端もしくはその付近、1つの5'端および1つの3'端もしくはその付近、1つの5'端および両方の3'端もしくはその付近、または1つの3'端および両方の5'端もしくはその付近に存在し得る。さらに本発明は、本発明の核酸分子および組成物の調製および使用方法も提供する。
特定の局面では、本発明は(1)種々の型のトポイソメラーゼ(例えば、IA型トポイソメラーゼ、IB型トポイソメラーゼ、II型トポイソメラーゼ等)が結合(例えば、共有結合)し、および/または(2)種々の型のトポイソメラーゼによって認識される2つまたはそれ以上のトポイソメラーゼ認識部位を含む核酸分子、ならびにそのような核酸分子を含む組成物の調製および使用方法も提供する。多くの態様では、これらの核酸分子はさらに1つまたは複数(例えば、1、2、3、4、5、6、7、8等)の組換え部位も含む。
さらに本発明は、少なくとも1つの核酸セグメントが少なくとも1つのトポイソメラーゼまたはトポイソメラーゼ認識部位および/または1つまたは複数の組換え部位を含む、2つまたはそれ以上の核酸セグメントの結合方法を提供する。さらに、本発明の方法で使用される核酸セグメントが、同一または異なる核酸セグメント上で複数(例えば、2、3、4、5、6、7、8等)のトポイソメラーゼを含む場合には、これらのトポイソメラーゼは同一または異なる型のトポイソメラーゼでよい。同様に、本発明の方法で使用される核酸セグメントが、同一または異なる核酸セグメント上で、複数のトポイソメラーゼ認識部位を含む場合には、これらのトポイソメラーゼ認識部位は同一または異なる型のトポイソメラーゼによって認識され得る。さらに、本発明の方法で使用される核酸セグメントが1つまたは複数の組換え部位を含む場合には、これらの組換え部位は同一または異なる核酸セグメント上の1つまたは複数の組換え部位と組換えできる。したがって、本発明は任意の1つのトポイソメラーゼまたはトポイソメラーゼ認識部位を利用した方法を用いた核酸セグメントの結合方法を提供する。さらに本発明は、(1)任意の組み合わせのトポイソメラーゼまたはトポイソメラーゼ認識部位、および/または(2)任意の組み合わせの組換え部位を利用した方法を用いた、核酸セグメントの結合方法を提供する。本発明はまた、上述の方法で産生される核酸分子、ならびにこれらの分子およびこれらの分子を含む組成物の利用も提供する。
一般に、本発明は部分的には、異なる機能的および構造的要素を含む任意の数(例えば、2、3、4、5、6、7、8、9、10等)の核酸セグメントを結合する方法を提供する。したがって、本発明は部分的には、核酸分子産物に異なる性質を与える任意の数(例えば、2、3、4、5、6、7、8、9、10等)の核酸セグメントを結合させる方法を提供する。多くの場合、本発明の方法によって、結合された個々の核酸セグメントの性質および/または要素の間に機能的な相互作用がある(例えば、発現制御配列およびオープンリーティングフレームの間の機能的な相互作用/連結)、核酸分子が形成される。産物分子に与えられる(1)機能的および構造的要素、ならびに(2)性質の例は、マルチクローニング部位(例えば、少なくとも2つの制限エンドヌクレアーゼ切断部位を含む核酸領域)、パッケージングシグナル(例えば、アデノウイルスパッケージングシグナル、アルファウイルスパッケージングシグナル等)、制限エンドヌクレアーゼ切断部位、オープンリーディングフレーム(例えば、インテインコード配列、アフィニティ精製タグコード配列等)、発現制御配列(例えば、プロモーター、オペレーター等)などが含まれるが、これらに限定されるわけではない。核酸セグメントから産物核酸分子に与えられ得る別の要素および性質は、本明細書の別の部分に記載されている。本発明は、上述の方法で産生された核酸分子、ならびにこれらの核酸分子およびこれらの核酸分子を含む組成物の使用も提供する。
さらに本発明は、部分的には、少なくとも1つ(例えば、1、2、3、4、5、6、7、8等)の核酸セグメントが1つまたは複数(例えば、1、2、3、4、5、6、7、8等)のトポイソメラーゼおよび/または1つまたは複数のトポイソメラーゼ認識部位を含み、少なくとも1つの核酸セグメントが1つまたは複数の組換え部位を含む、2つまたはそれ以上(例えば、2、3、4、5、6、7、8等)の核酸セグメントの結合方法も含む。特定の態様では、本発明は1方の核酸セグメントが1つまたは複数のトポイソメラーゼまたはトポイソメラーゼ認識部位を含むが組換え部位は含まず、他方の核酸セグメントが1つまたは複数の組換え部位を含むがトポイソメラーゼもトポイソメラーゼ認識部位も含まない、少なくとも2つ(例えば、2、3、4、5、6、7、8等)の核酸分子の結合方法(例えば、組換えを用いた方法および/または1つまたは複数のトポイソメラーゼが仲介する方法)を提供する。従って、本発明の方法は、核酸セグメントの結合によって、結合したまたはキメラの核酸分子を調製するために使用できるが、ここで産物核酸分子は(1)1つまたは複数(例えば、1、2、3、4、5、6、7、8等)のトポイソメラーゼおよび/または1つまたは複数(例えば、1、2、3、4、5、6、7、8等)のトポイソメラーゼ認識部位、および(2)1つまたは複数(例えば、1、2、3、4、5、6、7、8等)の組換え部位を含む。さらに本発明は、そのような方法によって調製された核酸分子、そのような核酸分子を含む組成物、およびそのような核酸分子の使用方法も提供する。
本発明は本明細書に記載される、1つまたは複数の核酸セグメントおよび/または核酸分子を含む組成物も提供する。そのような組成物は、1つまたは複数の他の核酸分子(組換え部位、トポイソメラーゼ認識部位、トポイソメラーゼ等を含みうる)、1つまたは複数のヌクレオチド、1つまたは複数のポリメラーゼ、1つまたは複数の逆転写酵素、1つまたは複数の組み換えタンパク質、1つまたは複数のトポイソメラーゼ、1つまたは複数の緩衝液および/または塩、1つまたは複数の固相支持体、1つまたは複数のポリアミン、1つまたは複数のベクター、1つまたは複数の制限酵素などからなる群より選択される1つまたはいくつかの他の成分を含みうる。例えば、本発明の組成物は、少なくとも1つのトポイソメラーゼ認識部位、および核酸セグメントの少なくとも1つのトポイソメラーゼ認識部位を少なくとも1つ認識する少なくとも1つのトポイソメラーゼを含む核酸セグメントを含む混合液(例えば、反応液)を含むが、これらに限定されるわけではない。さらに本発明の組成物は、(1)少なくとも1つのトポイソメラーゼ認識部位、または少なくとも1つのトポイソメラーゼが結合(例えば、共有結合)した少なくとも1つの核酸セグメント、および(2)1つまたは複数の他の成分を含む、少なくとも1つの核酸セグメントを含む。そのような他の成分の例には、トポイソメラーゼ;1つまたは複数のトポイソメラーゼまたはトポイソメラーゼ認識部位を含むまたは含まない他の核酸セグメント;ポリアミン(例えば、スペルミン、スペルミジン等);水;その他が含まれるが、これらに限定されるわけではない。本発明の組成物中に存在する核酸セグメントは、さらに1つまたは複数の組換え部位および/または1つまたは複数のレコンビナーゼを含みうる。
本発明の方法によりまたは本発明の方法と組み合わせて産生された核酸分子またはセグメント、および本発明の核酸分子またはそのセグメントは、本明細書に具体的に記載された分子またはセグメント、および本明細書に具体的に記載された分子またはセグメントに実質的な配列同一性を持つ分子またはセグメントも含む。特定の分子またはセグメントに「実質的な配列同一性」を持つ分子またはセグメントとは、その分子またはセグメントが、所定の(または「参照」)分子またはセグメントに対して、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも約90%、91%、92%、93%、94%、95%、96%、97%、98%、または99%同一であることを意味する。参照核酸分子またはセグメントに対して例えば、少なくとも65%「同一」なヌクレオチド配列を持つ核酸分子またはセグメントとは、その核酸分子またはセグメントが参照ヌクレオチド配列の100ヌクレオチドごとに最高35の点変異を含みうるという以外は、その核酸分子またはセグメントのヌクレオチド配列が参照配列と同一であることを意味する。すなわち、参照ヌクレオチド配列に少なくとも65%同一なヌクレオチド配列を持つポリヌクレオチドを入手するためには、参照配列のヌクレオチドの最高5%までが欠失または他のヌクレオチドで置換されるか、または参照ヌクレオチド中の総ヌクレオチド数の最高35%までのヌクレオチドが参照配列に挿入できる。参照配列のこれらの変異は、参照ヌクレオチド配列の5'もしくは3'端位置(もしくは両方)、またはこれらの末端位置の間の任意の場所に、参照配列のヌクレオチドの間に個々に散在するか、または参照配列内で1つまたは複数の連続するグループとして、存在し得る。
実際問題として、任意の特定の核酸配列分子またはセグメントが特定の参照分子またはセグメントと少なくとも約65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、または99%同一であるかどうかは、通常はFASTA(ドイツ、ハイデルベルク)、BLAST(ワシントンDC)、または2つの配列の間の相同性の最高のセグメントを見つけるために、局所相同性アルゴリズム(SmithおよびWaterman、Advances in Applied Mathematics 2:482-489 (1981))を使用するBESTFIT(Wisconsin Sequence Analysis Package、Unix用バージョン8、Genetics Computer Group、University Research Park、575 Science Drive、Madison、WI 53711)のような既知のコンピュータプログラムを用いて決定できる。FASTA、BLAST、BESTFITまたは任意の他の配列整列プログラムを用いて、特定の配列が、本発明による参照配列に例えば65%同一性を持つかどうかを決定するときには、参照ヌクレオチド配列の全長にわたって同一性のパーセンテージが計算され、参照配列中のヌクレオチドの総数の最高35%までのギャップが認められるように、パラメーターを設定する。
トポイソメラーゼ(例えば、部位特異的トポイソメラーゼ)で切断された核酸分子は、核酸分子のリン酸基にトポイソメラーゼ分子がさらに共有結合していることが多い。本発明は、上述および本明細書に記載される核酸分子の調製方法ならびにそのような分子を用いた組換え方法を含む。
特定の態様では、本発明の核酸分子はベクターである。別の態様では、本発明は本発明の核酸分子を含む宿主細胞、および、例えば発現産物(例えば、タンパク質、ポリペプチド、抗原、抗原決定基、エピトープ等、またはその断片)を産生するためにそのような宿主細胞を作製および使用する方法を含む。
特定の態様では、本発明の核酸分子は、組換え部位の間に1つまたは複数(例えば1、2、3、4、5等)のトポイソメラーゼ認識部位が存在するような、2つまたはそれ以上の組換え部位を含む。別の特定の態様では、本発明の核酸分子は、2つまたはそれ以上のトポイソメラーゼ認識部位の間に、1つまたは複数(例えば1、2、3、4、5等)の組換え部位が存在するような、2つまたはそれ以上のトポイソメラーゼ認識部位を含む。
別の特定の態様では、本発明の核酸分子は、2つの組換え部位の間に2つのトポイソメラーゼ認識部位が存在するような、2つの組換え部位を含む。したがって、そのような分子がトポイソメラーゼ認識部位の間で切断されて線状になれば、得られた線状分子のトポイソメラーゼ認識部位は、組換え部位に対して遠位に(すなわち、直線分子の2つの末端に近いほうに)存在することになる。したがって、本発明は1つまたは複数の組換え部位および1つまたは複数のトポイソメラーゼ認識部位を含む、線状核酸分子を提供する。特定の態様では、その1つまたは複数のトポイソメラーゼ認識部位は、1つまたは複数の組換え部位に対して遠位に存在する。そのような分子の例は、以下の実施例8に示されている。
第1の核酸の組換え部位およびトポイソメラーゼ認識部位の位置は、トポイソメラーゼを介してこの分子が第2の核酸分子へ連結すると、第2の核酸分子が2つまたはそれ以上の組換え部位の間に来るようなものでよい。例えば、線状の第1の核酸分子は、各末端またはその付近に1つの組換え部位を持ち、さらに2つの組換え部位の1つの遠位側にトポイソメラーゼ認識部位を含む場合がある。そのような場合には、線状の第1の核酸分子をトポイソメラーゼとインキュベートすると、第1の核酸分子にトポイソメラーゼが共有結合し、ここでトポイソメラーゼが第1の核酸分子の末端またはその付近で、隣の/最も近い組換え部位の遠位側になるように設計できる。第1の核酸分子のこの末端は、平滑末端であるか、5'または3'のいずれかに突出があってもよい。適当な第2の核酸分子(例えば、第1の核酸分子のトポイソメラーゼ修飾末端の少なくとも1つの鎖に配列相補性を持つ分子)とインキュベートすると、第2の核酸分子の1つの末端の片方または両方の鎖が、第1の核酸分子の1つの末端の片方または両方の鎖に共有結合できる。さらに、環状核酸分子が望ましい場合には、第2の核酸分子の第2の末端を、トポイソメラーゼ、リガーゼ、または他の方法によって、第1の核酸分子の第2の末端に結合できる。上述の過程の結果、2つの組換え部位の間に位置した核酸挿入物を含む核酸分子が作製される。関連する過程の具体的な例は、以下の実施例8に示されている。トポイソメラーゼを用いて核酸分子を共有結合する方法は、本明細書の別の箇所でさらに詳しく説明されている。
核酸挿入物が1つまたは複数の組換え部位の間に挿入されたら、この挿入物および隣接核酸は、組換えクローニングによって他の核酸分子に移すことができる。したがって、本発明は上述および本明細書の他の部分に記載される核酸分子の作製方法も提供する。
本発明の核酸分子中に存在する組換え部位とトポイソメラーゼ認識部位の間のヌクレオチド数で示される距離は、その分子が使用される特定の適用例によって異なるが、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、20、25、30、40、50、60、80、100、150、200、300、500、700、900、1000等、またはそれ以上のヌクレオチドでよい。さらに、本発明の核酸分子中に存在する組換え部位とトポイソメラーゼ認識部位の間のヌクレオチド数で示される距離は、以下の範囲内でありうる:0〜10ヌクレオチド、10〜30ヌクレオチド、20〜50ヌクレオチド、40〜80ヌクレオチド、70〜100ヌクレオチド、90〜200ヌクレオチド、120〜400ヌクレオチド、200〜400ヌクレオチド、200〜1000ヌクレオチド、200〜2000ヌクレオチド等。
本発明は、一般に本発明のセグメントまたは分子を2つまたはそれ以上(例えば、3またはそれ以上、4またはそれ以上、5またはそれ以上等)結合または連結する材料および方法も提供する。1つの局面では、そのような分子が結合されるために、セグメントまたは分子のうちの少なくとも1つが、少なくとも1つの組換え部位を含み、かつセグメントまたは分子のうちの少なくとも1つが、少なくとも1つのトポイソメラーゼ認識部位を含みうる。本発明による複数の核酸分子を結合する方法は、インビボまたはインビトロで実行できる。したがって、本発明は新規または固有の配列の組み合わせを作製する方法およびそのような方法によって作製された配列にも関する。本発明の方法によって作製された核酸分子は、当業者に周知の任意の目的のために使用できる。1つの局面では、本発明の方法によって結合される核酸分子またはセグメントのうちの少なくとも1つ(および多くの場合は2つまたはそれ以上)は、少なくとも1つ、および好ましくは少なくとも2つの組換え部位を含むが、各分子は複数(例えば、3またはそれ以上、4またはそれ以上、5またはそれ以上)の組換え部位を含みうる。別の局面では核酸分子は少なくとも1つのトポイソメラーゼ認識部位および/または少なくとも1つのトポイソメラーゼを含む場合がある。さらに別の局面では、分子は、(1)少なくとも1つの組換え部位、および(2)少なくとも1つのトポイソメラーゼ認識部位および/または少なくとも1つのトポイソメラーゼを含みうる。そのような組換え部位およびトポイソメラーゼ認識部位(同一または異なる場合がある)は、各核酸分子またはセグメント中で様々な位置にあり、本発明で使用される核酸は様々なサイズで、環状、スーパーコイル、線状等を含め種々の形状をとりうる。本発明で使用される核酸分子は、1つもしくは複数のベクター、またはその分子が宿主細胞中でベクターとして機能できるようにする1つもしくは複数の配列(複製開始点など)も含む場合がある。1つの局面では、本発明で使用する核酸分子またはセグメントは、分子の少なくとも1つの末端またはその付近に少なくとも1つの組換え部位を持つ線状分子で、好ましくは分子の両方の末端またはその付近に少なくとも1つの組換え部位を含む。別の局面では、対象核酸分子上に複数の組換え部位が存在する場合には、そのような部位はその分子上で互いに組換えをしないまたは実質的に組換えない。この態様では、対応する結合パートナー組換え部位は、好ましくは、本発明の方法によって連結または結合する1つまたは複数の他の核酸分子上にある。例えば、本発明で使用される第1の核酸分子は、少なくとも第1および第2の組換え部位を含み、第2の核酸分子は少なくとも第3および第4の組換え部位を含みうるが、第1と第2の部位は互いに組換えず、第3と第4の部位は互いに組換えをしないが、第1と第3および/または第2と第4の部位は組換えを行いうる。
本発明の方法で結合する核酸分子(例えば、「開始分子」)は、開始分子の全体または一部を含む1つまたは複数のハイブリッド分子(例えば、「産物核酸分子」)を産生するために使用できる。開始分子は、任意の供給源に由来する、または任意の方法で産生された、任意の核酸分子でよい。そのような分子は、天然の供給源(任意の動物または非動物由来の細胞、組織、および器官など)、または非天然(例えば、核酸誘導体)または合成によるものでよい。本発明で使用するセグメントまたは分子は、これらに限定されるわけではないが、PCRのような増幅、天然源からの単離、化学合成、より大きな核酸分子(ゲノムまたはcDNA)の切断または制限消化、転写、逆転写などを含む、当業者に周知の任意の方法で産生でき、また組換え部位および/またはトポイソメラーゼ認識部位および/またはトポイソメラーゼは、組換え部位および/またはトポイソメラーゼ認識部位および/またはトポイソメラーゼを含むアダプターの連結、組換え部位および/またはトポイソメラーゼ認識部位および/またはトポイソメラーゼを含むプライマーを用いた増幅または核酸合成、組換え部位および/またはトポイソメラーゼ認識部位および/またはトポイソメラーゼを含む、核酸分子(例えば、トランスポゾンまたは組込み配列)の挿入または組込み等を含む当業者に周知の任意の方法でそのような分子に付加できる。1つの局面では、本発明で使用する核酸分子は、核酸ライブラリまたはcDNAライブラリのような分子群である。
本発明に記載されるような方法を用いて組換えによって核酸分子が結合されたら、その後これらの核酸分子は、やはり本明細書に記載されるトポイソメラーゼを介する結合法および/または組換えを介する結合法を用いて、他の核酸分子と結合することができる。
本発明で使用する組換え部位は、組み換えタンパク質によって触媒または促進される組換え反応に関係する核酸分子上の任意の認識配列でよい。複数の組換え部位を利用する本発明の態様では、そのような組換え部位は同一でも異なっていても良く、互いに組換えする、または互いに組換えしない、もしくは互いに実質的に組換えしない場合がある。本発明で考慮される組換え部位は、野生型の変異型、誘導体、もしくは変種、または天然に存在する組換え部位も含む。好ましい組換え部位の修飾には、組換えを増強するものが含まれるが、ここで増強とは実質的に、以下からなる群から選択される:(i)組込み型組換えの促進;(ii)切除型組換えの促進;(iii)宿主因子の要求性の緩和;(iv)コインテグレートまたは産物の形成の効率の増加;および(v)コインテグレートまたは産物の形成の特異性の増加。好ましい修飾には、組換え特異性を増強するもの、1つまたは複数の終止コドンを除去するもの、および/またはヘアピンの形成を防ぐものが含まれる。修飾された組換え部位を通って翻訳または転写が起こったときに、転写または翻訳産物(例えば、mRNAまたはタンパク質)に所望のアミノ酸の変化を起こすように、組換え部位に所望の修飾をすることもできる。本発明にしたがって使用できる組換え部位には、att部位、frt部位、dif部位、psi部位、cer部位、およびlox部位、またはその変異体、誘導体、および変種(またはその組み合わせ)が含まれる。本発明で考慮される組換え部位には、そのような組換え部位の一部も含まれる。
各開始核酸分子は、1つまたは複数の組換え部位および/または1つまたは複数のトポイソメラーゼ認識部位および/または1つまたは複数のトポイソメラーゼに加えて、プライマー部位としての使用に適した配列(例えば、配列決定プライマーまたは増幅プライマーがそこにハイブリダイズして核酸合成、増幅、または配列決定が開始する配列)、転写もしくは翻訳シグナル、またはプロモーターおよび/またはオペレーター、リボソーム結合部位、トポイソメラーゼ認識配列(もしくは部位)、Kozak配列、および開始コドンのような調節配列、終止コドンのような転写および/または翻訳終止シグナル(1つまたは複数のサプレッサーtRNA分子によって最適に抑制できる)、tRNA(例えば、サプレッサーtRNA)、複製開始点、選択可能マーカー、およびGST、GUS、GFP、オープンリーティングフレーム(orf)配列のような融合タンパク質を作製するために使用できる遺伝子または遺伝子の一部(例えば、N末端またはC末端)、および相同組換え(例えば、遺伝子ターゲティング)で使用するための配列を含め、種々の分子生物学技術で望まれるまたは使用される、関心対象の任意の他の配列を含むがこれらに限定されるわけではない、種々の配列(またはその組み合わせ)を含みうる。
本発明は2つまたはそれ以上(例えば、2、3、4、5、6、7、8、9、10等)の核酸分子(本明細書では「ヌクレオチド配列」と呼ばれる場合があるが同等である)、例えば、2本鎖(「ds」)または1本鎖(「ss」)核酸分子、に少なくとも1つ(例えば、1、2、3、4、5、6、7、8、9、10等)のトポイソメラーゼを接触させることによって、共有結合した組換え核酸分子を作製する方法にも関する。本明細書で言及される全ての核酸分子またはヌクレオチド配列、例えば本明細書で開示される方法、組成物、およびキットで使用または作製されるものは、その分子または配列が本明細書で特にssおよび/またはdsと述べられているかいないかに限らず、ssまたはdsの核酸分子またはヌクレオチド配列であることは、当業者には理解されるだろう。
1つのそのような局面では、本発明の方法によりそのような核酸配列を所望の方向および/または順序で結合でき、これは必要ならばさらに、例えばインビトロまたはインビボで行われる転写またはトランスフェクション手順、翻訳反応または他のタンパク質発現手順、組換え反応等を含む、種々の分析または手順で操作または使用できる。別の局面では、同一または異なる核酸配列のうち3もしくはそれ以上、4もしくはそれ以上、5もしくはそれ以上等、またはその集団またはライブラリは、本発明の方法にしたがって結合できる。さらに別の局面では、本発明の方法を用いて単一の核酸分子の各末端を結合して、共有結合で閉鎖したまたはスーパーコイルの分子を作製できる。
結合する核酸配列は、任意の供給源に由来でき、cDNA、ゲノムDNA、ベクター、オリゴヌクレオチド等のような、天然および化学合成もしくは組換えで合成した核酸分子でよい。さらに、核酸配列は、遺伝子調節要素、複製開始点、スプライス部位、ポリアデニル化部位、例えば、タグ配列、検出可能または選択可能なマーカー、細胞局在ドメイン、または他のペプチドまたはポリペプチド等をコードし得るオープンリーディングフレームのような、機能的配列をを1つまたは複数含んでもよいが、含まなくてもよい。本発明は、同一または異なる任意の数の核酸配列を、必要ならば事前に決められた順序もしくは方向またはその両方で、結合することができる。
結合する核酸分子(例えば、dsまたはss核酸分子)は、例えば1本鎖、2本鎖、線状、環状、またはスーパーコイルのような任意の形状で良く、結合する各核酸分子はトポイソメラーゼの基質であるか、そのような基質になるように修飾できるという点で、部分的に特徴づけられる。トポイソメラーゼは、核酸分子の少なくとも1つの鎖を、好ましくはリン酸ジエステル結合により、別の核酸分子の少なくとも1つの鎖に結合できる、任意のトポイソメラーゼでよい。トポイソメラーゼは、部位特異的トポイソメラーゼでも、緩和した特異性を持っても良く、好ましくは切断を行なう場所またはその付近で、核酸分子の1つの鎖と安定な複合体(例えば、共有結合複合体)を形成する。
本発明の方法は、一般に、結合する核酸分子(例えば、dsまたはss核酸分子)とトポイソメラーゼを、1つの核酸分子の末端の両方の鎖が、少なくとも1つ(例えば、1、2、3、4、5、6、7、8、9、10等)の他の核酸分子の末端の両方の鎖に連結するような条件下で、接触させて行なう。したがって本発明の方法は、共有結合した組換え核酸分子(1本鎖または2本鎖)を産生するが、この分子は基質の核酸分子が連結した部位でニックを含まない。本発明は、そのような方法によって調製された組換え核酸分子も提供する。本発明の特定の局面では、そのような組換え核酸分子は、さらに1つまたは複数の組換え部位を含む。
本発明の方法は、成分の種々の組み合わせを用いて実行できる。例えば、本方法は共有結合する2つまたはそれ以上の基質核酸分子(例えば、ss核酸分子またはds核酸分子)と少なくとも1つのトポイソメラーゼを接触させることによって実行できるが、ここでトポイソメラーゼは核酸分子の一方または両方の鎖を切断し、切断部位の端のヌクレオチドと安定な複合体を作る。その後、トポイソメラーゼの結合した末端またはトポイソメラーゼの結合した核酸分子は、互いに接触し、基質核酸分子の各鎖が連結し、それにより1つまたは複数の共有結合した組換え核酸分子が生成する。好ましくは、トポイソメラーゼは各連結部位で、ホスホジエステル結合の形成を仲介する。本方法はまた、2つまたはそれ以上のトポイソメラーゼの結合核酸分子を、単独もしくは過剰のトポイソメラーゼの存在下で接触させる、または1つまたは複数のトポイソメラーゼ結合核酸分子(ssまたはds)に、トポイソメラーゼ切断部位を含む1つまたは複数の核酸分子(やはりssまたはds)およびトポイソメラーゼを接触させても実行できる。また本発明は、そのような方法で調製された組換え核酸分子も提供する。本発明の特定の局面では、そのような組換え核酸分子は、さらに1つまたは複数の組換え部位を含む。種々の態様では、トポイソメラーゼは比較的緩和した特異性を持ち、種々のヌクレオチド配列に結合しこれを切断できるか、トポイソメラーゼは部位特異的トポイソメラーゼで、特異的なヌクレオチド配列に結合しこれを切断する。トポイソメラーゼはまた、ds核酸分子の一方の鎖を切断するI型トポイソメラーゼであるか、ds核酸分子の両方の鎖を切断するII型トポイソメラーゼである。トポイソメラーゼがII型トポイソメラーゼの場合には、切断により線状ds核酸分子が産生され、その一方または両方の末端にトポイソメラーゼが結合している。そのような場合の特定の局面では、ds核酸分子の中で、結合したトポイソメラーゼを含む鎖に対して相補的な鎖は、突出した配列を形成する。
本発明の方法を実行する利点は、トポイソメラーゼによる連結反応が非常に迅速に、広い温度範囲で起きることである。さらなる利点は、本発明の方法によって作製される組換え核酸分子は、2つの核酸分子が結合した部位でニックを含まないという点である。したがって、共有結合した組換え核酸分子は、例えばポリメラーゼ連鎖反応(PCR)のような増幅反応の基質として、その後の手順にそのまま使用できる。
例として、本発明の方法は1)第1の末端および第2の末端を持ち、第1の末端または第2の末端または両方の3'端またはその付近にトポイソメラーゼ認識部位を持つ、第1の核酸分子(ssまたはdsでありうる);2)第1の末端および第2の末端を持ち、第1の末端または第2の末端または両方の3'端またはその付近にトポイソメラーゼ認識部位を持つ、少なくとも第2の核酸分子(やはりssまたはdsでありうる);ならびに3)部位特異的トポイソメラーゼを、全ての成分が接触し、トポイソメラーゼがその活性を示すことができるような条件下で接触させることによって実行できる。トポイソメラーゼ認識配列を含む鎖に相補的な鎖は、5'ヒドロキシル基を持つ可能性があり、トポイソメラーゼによる切断後に、さらに5'突出配列を有しうる。
本発明の方法はまた、1)第1の末端および第2の末端を持つ核酸分子(ssまたはdsでありうる)で、第1の末端および第2の末端の両方が、3'端またはその付近にトポイソメラーゼ認識部位を持つ分子、および2)部位特異的トポイソメラーゼを、成分が接触し、トポイソメラーゼがその活性を示すことができるような条件下で接触させても実行できる。例えば、トポイソメラーゼは、VacciniaトポイソメラーゼまたはS. cerevisiaeトポイソメラーゼのようなIB型トポイソメラーゼでよい。そのような方法は、共有結合した閉鎖環状またはスーパーコイルds核酸分子の調製手段を提供する。
本発明の方法は1)第1の末端および第2の末端を持ち、第1の末端または第2の末端または両方の5'端またはその付近にトポイソメラーゼ認識部位を持つ、第1の核酸分子(ssまたはdsでありうる);2)第1の末端および第2の末端を持ち、第1の末端または第2の末端または両方の5'端またはその付近にトポイソメラーゼ認識部位を持つ少なくとも第2の核酸分子(やはりssまたはdsでありうる);ならびに3)少なくとも1つの部位特異的トポイソメラーゼを、全ての成分が接触し、少なくとも1つのトポイソメラーゼがその活性を示すことができるような条件下で接触させることによって実行できる。例えば、トポイソメラーゼは大腸菌トポイソメラーゼIまたはトポイソメラーゼIIIのようなIA型トポイソメラーゼ、または真核細胞のトポイソメラーゼIIIでよい。核酸分子の切断後、トポイソメラーゼは好ましくは5'端に安定に結合している。トポイソメラーゼ認識部位または結合したトポイソメラーゼを含む末端の3'端は、3'ヒドロキシル基を含むか、3'ヒドロキシル基を含むように修飾できる。トポイソメラーゼによる切断後、切断された核酸分子は、3'突出配列を有しうる。
本明細書に例示した方法は、2つまたはそれ以上の部位特異的トポイソメラーゼを用いて実行でき、第1、第2、または他の核酸基質は、1つの末端の3'端または5'端に、2つまたはそれ以上のトポイソメラーゼのうちの1つのトポイソメラーゼ認識部位を対応して持つ。2つまたはそれ以上のトポイソメラーゼおよび対応するトポイソメラーゼ認識部位を利用すると、事前に決められた順序、方向、またはその組み合わせで、核酸分子(ssまたはdsでありうる)を結合することが容易にできる。したがって、本発明の方法は1つのトポイソメラーゼを用いて例示されているが、2つまたはそれ以上のトポイソメラーゼを用いて本方法が同様に実行できることが認識されるだろう。場合によっては、少なくとも1つのトポイソメラーゼの使用と言及されていても、他に記載がないかぎり、基質核酸分子が適当なトポイソメラーゼ認識部位を含むかぎり、1つ、2つ、3つ、またはそれ以上のトポイソメラーゼを用いて本方法は実行できる。トポイソメラーゼ結合核酸基質についても、トポイソメラーゼは同一または異なるという点で、同様に考えられる。
別の態様では、本発明の方法は、1)第1の末端または第2の末端または両方の末端の3'端またはその付近にトポイソメラーゼ認識部位、5'端またはその付近にトポイソメラーゼ認識部位を持つ、第1および第2の末端を持つ第1の核酸分子(ssまたはdsでありうる);2)第1の末端および第2の末端を持つ少なくとも第2の核酸分子(やはりssまたはdsでありうる);ならびに3)少なくとも2つ(例えば、2、3、4、5、6、7、8、9、10等)の部位特異的トポイソメラーゼを、全ての成分が接触し、各々のトポイソメラーゼがその効果を示すことのできる条件下で接触させることによって実行できる。基質の第1の核酸分子の末端をトポイソメラーゼが切断すると、一方または両方の末端の5'端または3'端は突出配列を含むか、平滑末端であるか、または一方の末端が突出を含み、第2の末端が平滑末端を含みうる。突出配列が存在する場合には、一般に第2の(または他の)核酸分子の突出配列に十分な相補性を持ち、2つの分子が互いに特異的にハイブリダイゼーションできる。
本発明のトポイソメラーゼを介する結合方法によって核酸分子が結合したら、得られた核酸分子は、本明細書の別の箇所に記載されるような組換え反応に使用できる。
そのような態様で有用な異なるトポイソメラーゼの数は、第1の核酸分子が第1の末端もしくは第2の末端のみにトポイソメラーゼ認識部位を持つのか、または両方の末端にトポイソメラーゼ認識部位を含むのか、および核酸分子が両方の末端にトポイソメラーゼ認識部位を含む場合には、少なくとも3'認識配列または5'認識配列が異なるかどうかに、部分的に依存する。また、少なくとも第2の核酸分子の1つまたはそれ以上も、その第1の末端または第2の末端または両方の末端で、3'端もしくはその付近にトポイソメラーゼ認識部位、および/または5'端もしくはその付近にトポイソメラーゼ認識部位を含み、他の核酸分子の3'端もしくは5'端もしくは両方またはその付近のトポイソメラーゼ認識部位は、第1の核酸分子のトポイソメラーゼ認識部位と同一または異なるようにして、本発明は実行できる。したがって、異なるトポイソメラーゼの数は、さらに本発明による方法によって結合される異なる基質核酸分子の数にも依存する。
部位特異的トポイソメラーゼを用いて本発明の方法を実行する利点は、第1の核酸分子、第2の核酸分子、およびさらに1つまたは複数の核酸分子(ssまたはdsでありうる)が、事前に決められた方向で共有結合できることである。さらなる利点は、所望の共有結合組換え核酸分子の末端に特異的なプライマーを用いて、増幅反応を実行することによって、インビトロで機能的な産物が選択できることである。したがって、本発明の方法にしたがって作製した共有結合した組換え核酸分子(ssまたはdsでありうる)は、例えば、細胞のトランスフェクションのため、または増幅(例えば、PCR)、組換え反応(例えば、本明細書に記載されたような組換え反応)、インビトロ転写反応、もしくはカップリングした転写/翻訳反応の実行の鋳型として、さらなる手順に直接使用できる。したがって、共有結合した組換え核酸分子は、さらに操作を行なわなくても、種々の目的のために有用である。
本発明の1つの局面では、本発明の方法で使用される第1の核酸分子および他の核酸分子は、核酸分子の少なくとも第1の集団、例えば、cDNAライブラリまたは合成オリゴヌクレオチドのコンビナトリアルライブラリのようなコンビナトリアルライブラリに由来する可能性があり、さらに本発明の方法で使用される第2の核酸分子および他の核酸は、核酸分子の少なくとも第2の集団に由来しうる。そのような方法では、第1の核酸分子を第2の核酸分子に結合すると、共有結合した組換え核酸分子(ssまたはdsでありうる)のコンビナトリアル集団を作製する方法が提供される。そのような方法にしたがって、1つまたは複数の標的核酸分子を、その集団の組換え核酸分子に結合して、さらなる集団を産生することもできる。コンビナトリアル分子のそのような集団は、例えば、タンパク質発現および望ましい特性を持つ融合タンパク質のスクリーニングによって、さらに操作または解析できる。
1つの態様では、本発明の方法は、本発明の方法に使用される第1の核酸分子(ssまたはdsでありうる)および他の核酸分子が、例えば単離された遺伝子のcDNAまたはコード配列のようなオープンリーディングフレームを含み、さらに第2の核酸分子(ssまたはdsでありうる)がプロモーターのような調節要素を含み、これがコード配列の5'末端に機能的に共有結合されてコード配列がそれから転写されるようにして、実行できる。本発明の方法で使用される第2の核酸分子、および他の核酸も、互いに機能的に連結した例えばプロモーター(例えば、GAL4プロモーター)、オペレーター(例えば、tetオペレーター、ガラクトースオペロンオペレーター、lacオペロンオペレーター等)、内部のリボソーム進入部位、およびATG開始メチオニンコドンのような、2つまたはそれ以上の調節要素を含む可能性があり、これは本発明の方法にしたがってコード配列を含む第1の核酸分子の5'端に機能的に共有結合できる。そのような方法は、コード配列の3'端に機能的に共有結合できる、例えばポリアデニル化シグナルを含む第3の核酸分子(ssまたはdsでありうる)を接触させることをさらに含みうる。そのような方法は、機能的ユニットとして転写、翻訳またはその両方ができる発現可能核酸分子の作製のために有用である。さらに/または、例えばエピトープタグのような検出可能マーカーをコードする核酸分子は、本発明の方法にしたがって第1または第2の(または他の)核酸分子に機能的に結合できる。そのようなコンストラクトの中で、ヌクレオチド配列を望ましい方向で持つ組換え核酸分子(ssまたはdsでありうる)の作製は、例えば、トポイソメラーゼによって共有結合される核酸分子の末端に、相補的な5'突出配列を入れることによって、容易にできる。
別の態様では、本発明の方法は、本発明の方法で使用される少なくとも第1の核酸分子または少なくとも第2の核酸分子、ならびに他の核酸が、例えば、cDNAライブラリ、ヌクレオチド配列のコンビナトリアルライブラリ、またはヌクレオチド配列の変化させた集団のような複数のヌクレオチド配列のうちの1つであるように、実行できる。別の態様では、本発明の方法はさらに、作製された共有結合したds組換え核酸分子(例えば、一方または両方の鎖で共有結合した組換え核酸分子)にPCRプライマー対を接触させて、共有結合した組換え核酸分子の全体または一部を増幅することを含む。大量の産物の作製に加えて、増幅反応は、特に共有結合する核酸分子が相補的な突出配列を含んでいる場合に、所望の共有結合したds組換え核酸分子を含むコンストラクトに選択性を持つことができる。したがって、本発明の方法は、ハイスループット分析に適したインビトロ選択方法を提供する。
本発明の方法は、1)第1の末端または第2の末端または両方で、3'端に共有結合したトポイソメラーゼを持つ(「トポイソメラーゼ結合」)、第1の末端および第2の末端を持つ第1の核酸分子(ssまたはdsでありうる);ならびに2)少なくとも第2のトポイソメラーゼ結合核酸分子(ssまたはdsでありうる)を接触させることでも、例示できる。好ましくは、トポイソメラーゼ結合核酸分子は、結合したトポイソメラーゼを含む末端に5'ヒドロキシル基を含むが、5'ヒドロキシル基はホスファターゼを用いても作製できる。本発明の方法は、第1の核酸分子および第2の核酸分子のみを用いても実行できるが、必要ならば各々定義されたようなヌクレオチド配列を持つ第3、第4、またはそれ以上の核酸分子(ssまたはdsでありうる)を含めてもよい。第1または第2の(または他の)核酸分子は、独立してヌクレオチド配列の一方または両方の末端の3'端にトポイソメラーゼが共有結合していても良く、他に記載が無いかぎり、第1および第2の(または他の)核酸分子は同一でも異なっていてもよい。特定のそのような局面では、本明細書に記載される方法で使用される核酸分子のうちの少なくとも1つは、少なくとも1つの組換え部位を含む。さらに、上述の方法によって作製される核酸分子は、本明細書の別の箇所に記載されるような組換え反応に使用できる。
本発明の方法は、さらに1)第1の末端または第2の末端または両方で、5'端に共有結合したトポイソメラーゼを持つ(すなわちトポイソメラーゼ結合5'端)、第1の末端および第2の末端を持つ第1の核酸分子(ssまたはdsでありうる);ならびに2)少なくとも1つのトポイソメラーゼ結合5'端を含む少なくとも第2のトポイソメラーゼ結合核酸分子(ssまたはdsでありうる)を接触させることでも、例示できる。トポイソメラーゼ結合核酸分子は、結合したトポイソメラーゼを含む末端に3'ヒドロキシル基を含み得るが、3'ヒドロキシル基はホスファターゼを用いても作製できる。本明細書に開示されるそのような方法は、第1の核酸分子および第2の核酸分子のみを用いても実行できるが、必要ならば、少なくとも1つのトポイソメラーゼ結合5'端を含め、各々定義されたようなヌクレオチド配列を持つ第3、第4、またはそれ以上の核酸分子(ssまたはdsでありうる)を含めてもよい。第1または第2の(または他の)核酸分子は、独立して核酸分子の一方または両方の末端の5'端にトポイソメラーゼが共有結合していても良く、他に記載が無いかぎり、第1および第2の(または他の)核酸分子は同一でも異なっていてもよい。特定のそのような局面では、本明細書に記載される方法で使用される核酸分子のうちの少なくとも1つは、少なくとも1つの組換え部位を含む。さらに、上記および本発明の別の箇所に記載される方法によって作製される核酸分子は、本明細書の別の箇所に記載されるような組換え反応にも使用できる。
本発明の方法は、さらに1)第1の末端または第2の末端または両方で、5'端に共有結合した第1のトポイソメラーゼを持ち、第1の末端または第2の末端または両方で、3'端に共有結合した第2のトポイソメラーゼを持つ(すなわち、一方または両方の末端が、トポイソメラーゼ結合5'端およびトポイソメラーゼ結合3'端を含む)、第1の末端および第2の末端を持つ第1の核酸分子;ならびに2)好ましくはトポイソメラーゼを含む第1の核酸分子の末端に共有結合する末端の5'端および3'端にヒドロキシル基を持つまたは持つようにできる少なくとも第2の核酸分子を接触させることでも、例示できる。本方法は、それぞれトポイソメラーゼ結合3'端またはトポイソメラーゼ結合5'端を含む末端の5'端または3'端のいずれかが、トポイソメラーゼ認識部位を含み、トポイソメラーゼ認識部位に関してその活性を示すことのできるトポイソメラーゼを、その成分に接触させることを含むようにも実行できる。特定のそのような局面では、本明細書に記載される方法で使用される核酸分子のうちの少なくとも1つは、少なくとも1つの組換え部位を含む。さらに、上記および本発明の別の箇所に記載される方法によって作製される核酸分子は、本明細書の別の箇所に記載されるような組換え反応にも使用できる。
本発明のそのような方法は、第1の核酸分子および第2の核酸分子のみを用いても実行できるが、必要ならば第3、第4、またはそれ以上の核酸分子を含めてもよいが、ここで核酸分子は第1の核酸分子、第2の核酸分子、またはその組み合わせについて定義された通りである。第1または第2の(または他の)核酸分子は、独立して、第2の末端(すなわち、定義されていない末端)の5'端、3'端、または5'と3'端の両方に共有結合したトポイソメラーゼを持つことができるが、持たなくてもよい。さらにこれらの核酸分子のうちの1つまたは複数は、さらに1つまたは複数の組換え部位を持つ場合がある。他に記載が無いかぎり、第1および第2の(または他の)核酸分子は同一でも異なっていてもよい。
本発明はさらに1)プライマー対の少なくとも1つのプライマーはトポイソメラーゼ認識部位および選択的に1つまたは複数の組換え部位の相補配列をコードするPCRプライマー対を用いた、第1の核酸分子の一部の増幅段階であって、第1の末端または第2の末端または両方が3'端またはその付近にトポイソメラーゼ認識部位を持つような、第1の末端および第2の末端を持つ増幅された第1の核酸分子が作製される増幅段階;ならびに2) a)増幅された第1の核酸分子、b) 第1の末端または第2の末端または両方が、3'端またはその付近にトポイソメラーゼ認識部位またはその切断産物を持ち、同じ末端の5'端にヒドロキシル基を持つまたは持つようにできる、第1の末端および第2の末端を持つ少なくとも第2の核酸分子;およびc)部位特異的トポイソメラーゼを、トポイソメラーゼ認識部位を持つ増幅された第1の核酸分子の末端およびトポイソメラーゼ認識部位を持つ少なくとも第2の核酸分子の末端をトポイソメラーゼが切断でき、かつその連結活性を発揮できるような条件下で、接触させる段階により共有結合したds組換え核酸分子を作製する方法にも関する。トポイソメラーゼ認識部位の相補配列をコードするPCRプライマーは、その5'端にヒドロキシル基を持つか、そのプライマーを用いて作製された増幅された第1の核酸分子にホスファターゼを接触させてその5'端にヒドロキシル基を形成できる。トポイソメラーゼ認識部位の相補配列をコードするPCRプライマーは、プライマーを用いて増幅された第1の核酸分子を部位特異的トポイソメラーゼが切断すると、核酸分子は5'突出配列を持ち、これが本発明の方法にしたがって第1の核酸分子に共有結合される第2の(または他の)核酸分子の5'突出配列に相補的になるようなヌクレオチド配列を、その5'端に有しうる。特定の局面では、本明細書に記載される方法で使用される核酸分子の少なくとも1つは、少なくとも1つの組換え部位を持つ。さらに、上述および本明細書の他の部分に記載された方法で作製された核酸分子は、本明細書の他の部分で記載されるような、組換え反応にも使用できる。
本発明はさらに1)プライマー対の少なくとも1つのプライマーはトポイソメラーゼ認識部位および選択的に1つまたは複数の組換え部位をコードするPCRプライマー対を用いた、第1の核酸分子の一部の増幅段階であって、第1の末端または第2の末端または両方が5'端またはその付近にトポイソメラーゼ認識部位を持つような、第1の末端および第2の末端を持つ増幅された第1の核酸分子が作製される増幅段階;ならびに2) a)増幅された第1の核酸分子、b) 第1の末端または第2の末端または両方が、5'端またはその付近にトポイソメラーゼ認識部位を持ち、同じ末端の3'端にヒドロキシル基を持つまたは持つようにできる、第1の末端および第2の末端を持つ少なくとも第2の核酸分子;およびc)少なくとも1つの部位特異的トポイソメラーゼを、トポイソメラーゼ認識部位を持つ増幅された第1の核酸分子の末端およびトポイソメラーゼ認識部位を持つ少なくとも第2の核酸分子の末端を少なくとも1つのトポイソメラーゼが切断でき、かつその連結活性を発揮できるような条件下で、接触させる段階により、共有結合したds組換え核酸分子を作製する方法にも関する。増幅された第1の核酸分子は、一般にトポイソメラーゼ認識部位を含む末端の3'端にヒドロキシル基を持つか、そのような3'ヒドロキシル基を持つように修飾できる。トポイソメラーゼ認識部位をコードするPCRプライマーは、増幅された第1の核酸分子を部位特異的トポイソメラーゼが切断すると、核酸分子は3'突出配列を持ち、これが本発明の方法にしたがって第1の核酸分子が共有結合される第2の(または他の)核酸分子の3'突出配列に相補的になるようなヌクレオチド配列を、その5'端(すなわちトポイソメラーゼ認識部位に対して5')に有しうる。特定の局面では、本明細書に記載される方法で使用される核酸分子の少なくとも1つは、少なくとも1つの組換え部位を持つ。さらに、上述および本明細書の他の部分に記載された方法で作製された核酸分子は、本明細書の他の部分で記載されるような、組換え反応にも使用できる。
本発明はさらに1)プライマー対の少なくとも1つのプライマーはトポイソメラーゼ認識部位、トポイソメラーゼ認識部位に相補的なヌクレオチド配列、および選択的に組換え部位を含むPCRプライマー対を用いた、第1の核酸分子の一部の増幅段階であって、増幅された第1の核酸分子が第1の末端または第2の末端または両方の5'末端またはその付近にトポイソメラーゼ認識部位を、3'端またはその付近にトポイソメラーゼ認識部位を持つような、第1の末端および第2の末端を持つ増幅された第1の核酸分子が作製される増幅段階;ならびに2) a)増幅された第1の核酸分子、b) 第2の核酸分子が、第1の末端または第2の末端または両方に、5'ヒドロキシル基および3'ヒドロキシル基を持つまたは持つようにできる、第1の末端および第2の末端を持つ少なくとも第2の核酸分子;およびc)少なくとも2つの部位特異的トポイソメラーゼを、i)増幅された第1の核酸分子の末端の5'端またはその付近のトポイソメラーゼ認識部位を少なくとも1つのトポイソメラーゼが切断でき、かつii) 少なくとも1つのトポイソメラーゼが、増幅された第1の核酸分子の末端の3'端またはその付近のトポイソメラーゼ認識部位を切断でき、かつその連結活性を発揮できるような条件下で、接触させる段階により共有結合したds組換え核酸分子を作製する方法にも関する。したがって本発明は、一方または両方の末端に、5'端またはその付近にトポイソメラーゼ認識部位、および3'端またはその付近でトポイソメラーゼ認識部位を含む、核酸分子を提供する。さらに、本発明は5'端、または3'端、または両方にトポイソメラーゼが結合した核酸分子を提供する。特定の局面では、本明細書に記載される方法で使用される核酸分子の少なくとも1つは、少なくとも1つの組換え部位を持つ。さらに、上述および本明細書の他の部分に記載された方法で作製された核酸分子は、本明細書の他の部分で記載されるような、組換え反応にも使用できる。
本発明はさらに、1つまたは複数のIA型部位特異的トポイソメラーゼの少なくとも1つの認識部位、1つまたは複数のIB型部位特異的トポイソメラーゼの認識部位に相補的な少なくとも1つのヌクレオチド配列、および選択的に、少なくとも1つの組換え部位を含む、オリゴヌクレオチドに関する。そのようなオリゴヌクレオチドは、例えば、プライマー伸長反応のプライマーとして、またはPCRのような増幅反応を行なうためのプライマーの1つとして、有用である。本明細書ではオリゴヌクレオチドプライマーと呼ばれるそのようなオリゴヌクレオチドは、プライマー対の1つである可能性があり、これは一方の末端の5'端またはその付近にIA型トポイソメラーゼ認識部位、および同じ末端の3'端またはその付近にIB型トポイソメラーゼ認識部位を含む、ds核酸増幅産物を作製するために有用であり得る。さらにオリゴヌクレオチドプライマーは、例えば、制限エンドヌクレアーゼ認識部位、ペプチドタグ、および必要ならば、1つまたは複数の別のIA型またはIB型トポイソメラーゼ認識部位のような関心対象の任意の他のヌクレオチド配列またはペプチドをコードする(またはこれに相補的な)ヌクレオチド配列を含み得るため、本発明の方法の実施に、1つまたは複数の好都合のまたは容易に入手できるトポイソメラーゼが選択できる。オリゴヌクレオチドプライマーはさらに、増幅された第1の核酸分子が部位特異的トポイソメラーゼに切断されると、本発明の方法にしたがって第1の核酸分子が共有結合される第2の(または他の)核酸分子の、3'または5'突出配列に相補的な、それぞれ3'または5'突出配列を核酸分子が含むようになるヌクレオチド配列を、その5'端、すなわちIAトポイソメラーゼ認識部位またはIBトポイソメラーゼ認識部位に相補的なヌクレオチド配列の5'側に、含み得る。または、増幅された核酸分子が切断されると、平滑末端のトポイソメラーゼ結合核酸分子が生成するように、オリゴヌクレオチドプライマーを設計することもできる。
さらに本発明は、少なくとも1つのトポイソメラーゼ認識部位、またはそれに相補的なヌクレオチド配列、および少なくとも1つの組換え部位を含む、オリゴヌクレオチドにも関する。そのようなオリゴヌクレオチドは、例えばプライマー対の1つとして、上述のように使用できる。
本発明のオリゴヌクレオチドは、15〜20、15〜30、15〜50、20〜30、20〜50、30〜40、30〜50、30〜80、30〜100、40〜50、40〜70、40〜80、40〜100、50〜60、50〜80、50〜100、15〜80、15〜100、または20〜100(など)のヌクレオチド長であることが多い。
本発明はまたプライマー対も提供し、これは、増幅反応においてプライマーの1つがフォワードプライマーとして有用であり、もう一つのプライマーはリバースプライマーとして有用である、上記で定義された少なくとも1つのオリゴヌクレオチドプライマーを含む。そのようなプライマー対の第2のプライマーは、IA型トポイソメラーゼ認識部位、IB型トポイソメラーゼ認識部位に相補的なヌクレオチド配列、またはその両方を、必須ではなく含み得るが、関心対象の認知の他のヌクレオチド配列および/または少なくとも1つの組換え部位を含み得る。1つの態様では、プライマー対は本発明のオリゴヌクレオチドプライマーを2つ含み、その1つのオリゴヌクレオチドプライマーはフォワードプライマーとして有用で、第2のオリゴヌクレオチドプライマーはリバースプライマーとして有用である。そのようなプライマー対は、例えば、端のIA型またはIB型または両方のトポイソメラーゼ認識部位が同一または異なるような、両方の末端の両方の端にトポイソメラーゼ認識部位を持つ、および/または1つまたは複数の組換え部位を持つ、核酸分子増幅産物を作製するために有用である。
したがって、本発明はさらに、第1の末端または第2の末端または両方の末端の、5'端またはその付近にIA型トポイソメラーゼ認識部位を含み、3'端またはその付近にIB型トポイソメラーゼ認識部位を含む、第1の末端および第2の末端を持つ核酸分子に関する。また、本発明は、核酸分子が、必要ならば一方または両方の末端に、IA型トポイソメラーゼまたはIB型トポイソメラーゼまたは両方が安定に結合したトポイソメラーゼ結合分子であること以外は、上記に定義された核酸分子を提供する。これらの核酸分子は、さらに1つまたは複数の組換え部位を含みうる。
1つの態様では、本発明の方法で使用される第1の核酸分子および他の核酸分子は、ポリペプチド(例えば、インテインを持つポリペプチドでありうる)、アンチセンスヌクレオチド配列、干渉RNA(すなわちRNAi)分子、リボザイム、転移RNA(すなわち、これらに限定されるわけではないが、サプレッサーtRNAを含むtRNA)、3重鎖ヌクレオチド配列等の様な分子をコードする発現可能なヌクレオチド配列を含み、第2の(または他の)核酸分子は、プロモーター(例えば、GAL4オペレーター)、オペレーター(例えば、tetオペレーター、ガラクトースオペロンオペレーター、lacオペロンオペレーター等)、エンハンサー、サイレンサー、翻訳開始部位、もしくはポリアデニル化シグナルのような転写調節要素を含むか、または開始メチオニン、終止コドン、細胞の区画化ドメイン、相同性ドメイン等のような翻訳調節要素をコードするか、または機能的に連結したその組み合わせを含む。増幅された第1の核酸分子について調製された増幅された第2の(または他の)核酸分子であり得る、第2の(または他の)核酸分子および本発明で使用される他の核酸は、1つまたは複数のマルチクローニング部位(MCS)、検出可能な標識、例えば、酵素、酵素の基質、蛍光化合物、発光化合物、化学発光化合物、放射性核種、常磁性化合物、およびビオチンも含み得る。または、オリゴヌクレオチドタグ、または例えばポリヒスチジンタグ、V5エピトープ、またはmycエピトープのようなペプチドタグであるタグを含み得る。
別の態様では、本発明の方法はポリペプチド(例えば、インテインを含むポリペプチド)、またはそのドメインをコードする第1の核酸分子、および転写活性化ドメインまたはDNA結合ドメインをコードする第2の(または他の)核酸分子を用いて行われる。そのような方法は、ツーハイブリッドアッセイシステム、特にハイスループットなツーハイブリッドアッセイを行なうために有用なキメラポリペプチドをコードする、共有結合したds組換え核酸分子の作製に使用できる。別の態様では、第1の核酸分子は複数のヌクレオチド配列を含み、これはcDNAライブラリ、ヌクレオチド配列のコンビナトリアルライブラリ、変化させたヌクレオチド配列の集団等であり得る。
本発明の方法は、標的ゲノムDNA配列へ、部位特異的挿入を行なうために有用な、共有結合したds組換え核酸分子を作製するための手段を提供する。標的ゲノムDNA配列は、任意のゲノム配列、特に遺伝子、および好ましくはヌクレオチド配列の一部または全体が既知の遺伝子であり得る。本方法は、2組のPCRプライマー対および1つの核酸分子を利用して実行できる。核酸分子は第1の末端および第2の末端を持ち、ポリペプチド、例えば、選択可能なマーカーをコードするが、核酸分子は各末端の3'端にトポイソメラーゼ認識部位またはその切断産物を含み、選択的に、各末端の5'端にヒドロキシル基を含み、好ましくは5'端は互いに異なる突出配列を含む。同様に、核酸分子は一方または両方の末端の5'端またはその付近にトポイソメラーゼ認識部位またはその切断産物を含み、選択的に3'端の一方または両方は同一または好ましくは互いに異なる突出配列を含む。または核酸分子の一方または両方の5'端および3'端は各々トポイソメラーゼ認識部位またはその切断産物を含み得る(図11参照)。特定の局面では、本明細書に記載される方法で使用される核酸分子のうちの少なくとも1つは、少なくとも1つの組換え部位を含む。さらに、上記または本明細書の他の部分に記載される方法で生成する核酸分子は、本明細書の他の部分に記載されるような組換え反応にも使用できる。
2組のPCRプライマー対は一般に、Taqポリメラーゼのような適当なDNAポリメラーゼおよび増幅する配列を含む鋳型の存在下では、プライマーが、ポリペプチドの挿入のために標的部位(例えば、選択可能マーカー)の上流(および隣接する)および下流の(および隣接する)ゲノムDNA配列の部分を増幅するように、選択される。PCRプライマー対のセットは、増幅産物が、実行する本発明の特定の方法に適当な、5'端、または3'端、または両方またはその付近を含め、少なくとも選択可能マーカーに共有結合する末端に、トポイソメラーゼ認識部位を含むように設計される。したがって、第1のPCRプライマー対は、例えば、1) 5'から3'方向に、増幅産物が共有結合される選択可能マーカーの末端の5'突出配列に相補的なヌクレオチド配列、トポイソメラーゼ認識部位に相補的なヌクレオチド配列、標的ゲノムDNA配列の3'配列に相補的なヌクレオチド配列を含む第1のプライマー;および2)第1のプライマーが相補的な3'配列の上流の標的ゲノムDNAのヌクレオチド配列を含む第2のプライマーを含み得る。第2のPCRプライマー対は、1) 5'から3'方向に、共有結合される選択可能マーカーの末端の5'突出配列に相補的なヌクレオチド配列、トポイソメラーゼ認識部位に相補的なヌクレオチド配列、および標的ゲノムDNA配列の5'配列のヌクレオチド配列を含む第1のプライマー、ただし標的ゲノムDNAの5'配列は第1のPCRプライマー対の第1のプライマーが相補的である標的ゲノムDNAの3'配列の下流にある;および2)第1のプライマーに含まれる標的ゲノムDNAの5'配列の下流にある、標的ゲノムDNAの3'配列に相補的な、ヌクレオチド配列を含む第2のプライマーを含む。
選択可能マーカーを含む核酸分子、PCR増幅産物、および少なくとも1つのトポイソメラーゼが接触すると、本発明の方法にしたがって共有結合したds組換え核酸分子が生成する。生成したds組換え核酸分子は、ゲノムにおいて相同組換えを行なうため、例えば、細胞中の遺伝子の機能をノックアウトする、または生成したds組換え核酸分子を含む細胞に新規の表現型を与えるために、有用である。本方法はさらに、生成した組換え核酸分子が、そのゲノム中で安定に維持されているトランスフェニック非ヒト生物を産生するためにも使用できる。
本発明は、本発明の方法にしたがって調製された組成物、および本方法の実施に有用な組成物にも関する。そのような組成物は、本発明の方法で使用された1つまたは複数の反応物および/または本発明の方法にしたがって産生された1つまたは複数のds組換え核酸を含み得る。そのような組成物は、例えば、1つまたは複数のトポイソメラーゼ認識部位を持つ1つまたは複数の核酸分子;1つまたは複数のトポイソメラーゼ結合核酸分子;1つまたは複数の組換え部位を含む1つまたは複数の核酸分子;プライマーを用いて調製された増幅産物の一方または両方の末端の1つまたは両方の端にトポイソメラーゼ認識部位を含む核酸分子を調製するために有用な1つまたは複数のプライマー;1つまたは複数のトポイソメラーゼ;例えば、タグ、マーカー、調節要素等をコードするヌクレオチド配列を含む1つまたは複数の基質核酸分子;本発明の方法にしたがって産生された共有結合した1つまたは複数のds組換え核酸分子;本明細書に開示される核酸分子、プライマー、または組換え核酸分子を含むまたは含むために有用な1つまたは複数の細胞;プライマー伸長または増幅反応を実行するための1つまたは複数のポリメラーゼ;1つまたは複数の反応緩衝液等を含み得る。1つの態様では、本発明の組成物は、2つまたはそれ以上の異なるトポイソメラーゼ結合核酸分子および/または2つまたはそれ以上の異なる組換え部位を含む。組成物はさらに少なくとも1つのトポイソメラーゼを含み得る。本発明の組成物は、部位特異的トポイソメラーゼおよび共有結合したds組換え核酸分子も含み得るが、組換え核酸分子は各鎖に部位特異的トポイソメラーゼの認識部位を少なくとも1つ含み、1つの鎖のトポイソメラーゼ認識部位は相補鎖のトポイソメラーゼ認識部位から約100ヌクレオチド以内、一般には約5、10、20、または30ヌクレオチド以内にある。
本発明の方法で産生された産物分子は、開始分子またはセグメント、分子上の組換え部位の位置、および部位の組換え順序によって、開始分子(またはその一部)の任意の組み合わせを含むことができ、任意のサイズおよび任意の形状(例えば、環状、線状、スーパーコイルなど)であり得る。
本発明の任意の産物分子は、さらに任意の数の標準的な分子生物学的手法またはその組み合わせ(インビトロまたはインビボ)において、操作、解析、または使用できる。これらの手法には、配列決定、増幅、核酸合成、タンパク質またはペプチドの発現(例えば、融合タンパク質発現、抗体発現、ホルモン発現など)、タンパク質間相互作用(ツーハイブリッド解析または逆ツーハイブリッド解析)相同組換えまたは遺伝子ターゲティング、およびコンビナトリアルライブラリ解析および操作が含まれる。また本発明は、1つまたは複数のベクターへの本発明の核酸分子のクローニング(好ましくは組換えによる)、または特定の機能的ベクター配列(例えば、複製開始点)を付加することによる、本発明の核酸分子のベクターへの変換にも関する。1つの局面では、組換えおよび/またはトポイソメラーゼを介する結合は、インビトロで行われ、さらなる操作または分析は、インビトロで直接行われる。したがって、さらなる解析および操作は、本発明の分子を宿主細胞に導入するおよび/または宿主細胞で維持される能力によって制限されない。したがって、宿主細胞を経た後またはインビボで直接に(宿主細胞中)インビトロ分析または操作を行なうこともできるが、インビトロで直接本発明の分子をさらに操作または分析することによって、時間を節約し収率を高めることができる。
本発明による核酸合成段階は、以下の段階を含んでもよい:
(a)関心対象の核酸分子または鋳型を、1つまたは複数のプライマーおよび1つまたは複数のヌクレオチドと混合して、反応液を作る段階;ならびに
(b)分子または鋳型の全体または一部に相補的な核酸分子を合成するために十分な条件下で、反応液をインキュベートする段階。
合成された分子は、その後、第1の合成された分子の全体または一部に相補的な核酸分子をさらに合成するための鋳型として使用できる。したがって、2本鎖核酸分子(例えば、DNA)が調製できる。好ましくは、そのような第2の合成段階は、第1の核酸分子の全体または一部に相補的な第2の核酸分子を合成するために十分な条件下で、1つまたは複数のプライマーおよび1つまたは複数のヌクレオチドの存在下で行われる。通常は、1つまたは複数の核酸分子の合成は、1つまたは複数のポリメラーゼ(好ましくは耐熱性または中温性のDNAポリメラーゼ)の存在下で行われるが、逆転写酵素もそのような合成反応に使用できる。したがって、さらなる核酸分子の合成の鋳型として使用される核酸分子は、RNA、mRNA、DNA、または非天然もしくは誘導体の核酸分子であり得る。本発明による核酸合成は、プライマー部位を含む開始核酸分子を使用することによって、産物分子に1つまたは複数のプライマー部位を組み込むことによって、促進されうる。したがって、本発明の方法によって、開始分子中のプライマー部位の位置、および産物分子における開始分子の付加の順序によって、産物分子中に1つまたはいくつかの所望の位置にプライマー部位を付加することができる。
本発明による配列決定段階は、以下の段階を含んでもよい:
(a)配列決定する核酸分子を、1つまたは複数のプライマー、1つまたは複数のヌクレオチド、および1つまたは複数の停止剤と混合して反応液を作製する段階;
(b)配列決定をする分子の全体または一部に相補的な分子群を合成するために十分がな条件下で該反応液をインキュベートする段階;ならびに
(c)該分子群を分離して、配列決定する分子の全体または一部のヌクレオチド配列を決定する段階。
そのような配列決定段階は、好ましくは1つまたは複数のポリメラーゼ(例えば、DNAポリメラーゼおよび/または逆転写酵素)および1つまたは複数のプライマーの存在下で行われる。配列決定のための好ましい停止剤には、ジデオキシヌクレオチドのような誘導体ヌクレオチド(ddATP、ddTTP、ddGTP、ddCTPおよびその誘導体)が含まれる。本発明による核酸配列決定は、プライマー部位を含む開始核酸分子を使用することによって、産物分子に1つまたは複数の配列決定プライマー部位を組み込むことによって、促進できる。したがって、本発明の方法によって、開始分子中のプライマー部位の位置、および産物分子における開始分子の付加の順序によって、産物分子中に1つまたはいくつかの所望の位置に配列決定プライマー部位を付加することができる。
本発明によるタンパク質発現段階は、以下の段階を含んでもよい:
(a)1つまたは複数の発現シグナルを含む、発現する核酸分子を入手する段階;および
(b)該発現シグナルの制御下にある核酸分子の全体または一部を発現して、該分子またはその一部にコードされるペプチドまたはタンパク質を産生する段階。
この場合、発現シグナルは発現される配列に機能的に連結しているといわれる。発現されるタンパク質またはペプチドは、好ましくは宿主細胞中(インビボ)で発現されるが、当技術分野で周知の技法を用いてインビトロで発現してもよい。タンパク質またはペプチドの発現後、タンパク質またはペプチド産物は、選択的に単離または精製してもよい。さらに、発現されたタンパク質またはペプチドは、ツーハイブリッド相互作用、タンパク質機能分析、およびアゴニスト/アンタゴニスト-タンパク質相互作用(例えば、薬剤、化合物、または他のペプチドによるタンパク質機能の刺激または阻害)を含む種々のタンパク質解析技法で使用できる。本発明および特に本発明のコンビナトリアル分子の発現によって産生された新規かつ固有のハイブリッドタンパク質またはペプチド(例えば、融合タンパク質)は、一般に治療薬に有用でありうる。本発明によるタンパク質発現は、1つまたは複数の転写または翻訳シグナル、または調節配列、開始コドン、終止シグナル、スプライスドナー/受容体配列(例えば、イントロン配列)などを含む開始核酸分子を使用することにより、産物分子にこのような配列を組み込むことによって、促進できる。したがって、本発明の方法によって、開始分子中の発現配列の位置、および産物分子における開始分子の付加の順序によって、産物分子中に1つまたはいくつかの所望の位置に発現配列を付加することができる。
本発明による相同組換えは、以下の段階を含んでもよい:
(a)1つまたは複数の組換え部位および/または1つまたは複数のトポイソメラーゼ認識部位を含む本発明の少なくとも1つの第1の核酸分子(好ましくは産物分子)と、少なくとも1つの標的核酸分子とを混合する段階であって、該第1の分子と標的分子は、1つまたは複数の相同配列を持つ段階;ならびに
(b)該第1の分子および標的核酸分子に相同組換えによる組換えを起こさせる段階。相同組換えで使用できる核酸コンストラクトの1つの例は、図37に示されている。本発明はさらに、相同組換えに使用できる核酸分子の調製方法、そのような方法によって調製された核酸分子、および本発明の方法に係る相同組換えを受けた細胞も含む。
そのような相同組換えは、インビトロで行われる場合もあるが、好ましくはインビボ(例えば、宿主細胞中)で行われる。好ましくは、相同組換えによって、組換え部位を含む本発明の核酸分子(第1の核酸分子)の全体または一部が、相同配列を含む標的核酸分子の1つまたは複数の部位に移動する。そのような相同組換えの選択は、陽性選択または陰性選択(例えば、選択可能マーカーを使用)を用いて、所望の産物を選択および/または望ましくない産物以外を選択することによって、促進できる。1つの好ましい局面では、本発明の核酸分子は、少なくとも1つの選択可能マーカーおよび標的分子に相同な配列を少なくとも2つ含む。好ましくは、第1の分子は、少なくとも1つの選択可能マーカーを挟む少なくとも2つの相同配列を含む。
本発明はこのように、関心対象の配列または遺伝子、特に動物、植物、ヒト、昆虫、細菌等の中の遺伝子または配列、またはそのような宿主または宿主細胞中のウイルスのような外来物質の配列を、ノックアウトまたは変異する(または既存の配列を変化させる、例えば変異配列を野生型配列に変換する)ために使用できる遺伝子ターゲティング核酸分子またはベクターの作製を容易にする。そのような遺伝子ターゲティングは、好ましくは、そのような宿主細胞のゲノムにおけるターゲティング配列を含む。そのような遺伝子ターゲティングは、インビトロまたはインビボで行なえる。したがって、1つの好ましい局面では、本発明は、以下の段階を含む配列または遺伝子のターゲティングまたは変異方法に関する:
(a)1つまたは複数の組換え部位および/または1つまたは複数のトポイソメラーゼ認識部位(および好ましくは1つまたは複数の選択可能マーカー)を含む本発明の少なくとも1つの核酸分子を入手する段階であって、該分子は関心対象の標的遺伝子または配列に相同な1つまたは複数の配列を含む(該1つまたは複数の相同配列は好ましくは本発明の分子上の1つまたは複数の選択可能マーカーに隣接する)段階;ならびに
(b)関心対象の該標的配列または遺伝子と、本発明の該分子の間で1つまたは複数の部位で相同組換えが起きるために十分な条件下で、関心対象の1つまたは複数の標的遺伝子または配列を、本発明の該分子と接触させる段階であって、それにより本発明の分子の全体または一部が、標的配列または遺伝子内に挿入される段階。
そのような配列によって通常発現される発現産物(通常はタンパク質またはペプチド)が産生されない、またはより高いもしくは低いレベルで産生される、または活性の上昇もしくは低下、または不活性なもしくは部分的に不活性な発現産物をもたらしうる改変されたタンパク質配列を持つ程度に産生されるように、そのようなターゲティング法は、配列または標的遺伝子の欠失、不活化、または部分的不活化を引き起こす。本発明の分子上に好ましくは存在する選択可能マーカーは、相同組換えイベントが成功した候補(例えば宿主細胞)の選択を容易にする。したがって、本発明は本発明のターゲティング法によって産生された修飾された遺伝子または配列を含む、宿主細胞、組織、器官、および動物(例えば、トランスジェニック動物)を産生する方法を提供する。修飾された配列または遺伝子は、好ましくは本発明の分子によって提供される少なくとも1つの組換え部位および/または少なくとも1つの選択可能マーカーを含む。
したがって、より具体的には本発明は、以下の段階を含む、配列または遺伝子のターゲティングまたは変異方法に関する:
(a)1つまたは複数の組換え部位、関心対象の標的遺伝子または配列に相同な1つまたは複数の配列が隣接する少なくとも1つの選択可能マーカー、および選択的に、1つまたは複数のトポイソメラーゼ認識部位を含む、本発明の少なくとも1つの核酸分子を入手する段階;
(b)関心対象の該標的配列または遺伝子と該分子の間で1つまたは複数の部位で相同組換えが起きるために十分な条件下で、関心対象の1つまたは複数の標的遺伝子または配列を該分子に接触させる段階であって、これにより本発明の分子の全体または一部が、標的配列または遺伝子中に挿入される(および好ましくは少なくとも1つの選択可能マーカーおよび/または少なくとも1つの組換え部位が挿入される)段階;ならびに
(c)選択的に、本発明の分子の全体または一部を含む該配列もしくは遺伝子、または本発明の該分子の全体または一部を含む該遺伝子または配列を含む宿主細胞を選択する段階。
本発明の別の局面では、本発明にしたがって標的配列に挿入された組換え部位は、標的配列に挿入された分子の全部または一部を切り出すまたは除去するために使用できる。したがって、本発明はそのような配列のインビトロまたはインビボでの除去を可能にし、標的遺伝子または配列の再活性化を可能にすることがある。いくつかの態様では、上述のようにして挿入された変化を含む配列の同定および単離後に、本発明の分子上に存在する選択可能マーカーを除去できる。
また本発明は、本発明の開始または産物核酸分子を、1つもしくは複数のベクターにクローニングする方法、または本発明の産物分子を1つもしくは複数のベクターに変換する方法も提供する。1つの局面では、開始分子を組換えて、1つまたは複数の産物分子を作製し、そのような産物分子を1つまたは複数のベクターにクローニングする(好ましくは組換えによる)。別の局面では、いくつかの開始分子がベクター内で結合するように開始分子を1つまたは複数のベクターに直接にクローニングし、本発明の産物分子を含むベクターを作製する。別の局面では、開始分子をベクター内で結合しないように(すなわち、開始分子がベクター配列で分離されるように)1つまたは複数のベクターに直接にクローニングする。さらに別の局面では、産物分子と開始分子の組み合わせを、1つまたは複数のベクターに任意の順序でクローニングし、元の開始分子および産物分子の組み合わせから得られる新しい産物分子を含むベクターを作製する。
したがって、本発明は、以下の段階を含むクローニング法に関する:
(a)1つまたは複数の組換え部位および/または1つまたは複数のトポイソメラーゼ認識部位を含む、本発明の少なくとも1つの核酸分子を入手する段階;ならびに
(b)該分子の全体または一部を1つまたは複数のベクターに移動する段階。さらに本発明はそのような方法で調製されたベクター、これらのベクターを含む組成物、およびこれらのベクターを用いる方法も含む。
そのようなベクターは多くの場合1つまたは複数の組換え部位および/または1つまたは複数のトポイソメラーゼ認識部位を含み、この分子のそのようなベクターへの移動は、好ましくはベクター上の1つまたは複数の部位と、本発明の分子上の1つまたは複数の部位の間の組換えによって、行われる。別の局面では、本発明の産物分子は、必要なベクター配列(例えば、複製開始点)を含むことによって、ベクターとして機能するように変換できる。したがって、本発明にしたがって、そのようなベクター配列は、そのような配列を含む開始分子を使用して、産物分子に組み込むことができる。そのようなベクター配列は、開始分子中のその配列の位置および産物分子中の開始分子の付加の順序によって、産物分子中の1つまたはいくつかの所望の位置に付加することができる。ベクター配列を含む産物分子は、線状であるか、産物分子内の組換え部位の組換えもしくはトポイソメラーゼを介する結合反応によって、環状もしくはスーパーコイル型に変換できる。そのような産物分子の環化は、産物分子の両方の末端またはその付近の組換え部位の組換えによって行われることが多い。
本発明で使用されるベクター配列は、1つまたは複数の配列決定または増幅プライマー部位、1つまたは複数のマルチクローニング部位、1つまたは複数の選択可能マーカー(例えば、毒性遺伝子、抗生物質抵抗性遺伝子、選択可能マーカー等)、1つまたは複数の転写または翻訳部位またはシグナル、1つまたは複数の転写または翻訳終結部位、1つまたは複数のトポイソメラーゼ認識部位、1つまたは複数のトポイソメラーゼ、1つまたは複数の複製開始点、1つまたは複数の組換え部位(またはその一部)等を含む、1つまたはいくつかの要素および/または機能的配列および/または部位(またはその組み合わせ)を含み得る。本発明で使用されるベクター配列はまた、本明細書に記載されるようにして所望の融合タンパク質の発現を可能にするように抑制できる、終止コドンも含み得る。したがって、本発明にしたがって、1つまたは複数のそのような要素、機能的配列、および/または部位を本発明の任意の核酸分子に導入するために、ベクター配列が使用でき、そのような配列は、そのようなベクターにクローニングされた任意のそのような核酸分子を操作または解析するために使用できる。例えば、ベクターによって提供されるプライマー部位(好ましくはそのようなベクター中でクローニングされた挿入物の両側に存在する)は、ベクター中にクローニングされた産物分子の全体または一部の配列決定または増幅を可能にする。また、ベクターに含まれる転写または調節配列は、ベクターにクローニングされた産物分子の全体または一部によってコードされるペプチド、ポリペプチド、またはタンパク質の発現を可能にする。同様に、ベクターが提供する遺伝子、遺伝子の一部、または配列タグ(GUS、GST、GFP、Hisタグ、エピトープタグ等)は、ベクター中にクローニングされた産物分子との遺伝子融合の集団の作製を可能にするが、そのようなベクターにクローニングされた産物配列と組み合わせて、ベクターによって提供される配列タグによてコードされるいくつかのペプチド、ポリペプチド、またはタンパク質融合の産生を可能にする。そのような遺伝子、遺伝子の一部、または配列タグは、選択的に抑制された終止コドンと組み合わせて使用して、ベクターにクローニングされた関心対象の配列、およびベクターの提供する遺伝子またはタグ配列によってコードされる融合タンパク質の、制御された発現を可能にする。コンストラクト中では、ベクターは1つまたは複数の組換え部位、1つまたは複数の終止コドン、および1つまたは複数のタグ配列を含み得る。いくつかの態様では、タグ配列は組換え部位に隣接しうる。選択的に、関心対象の遺伝子にタグ配列を制御して付加するために、タグの配列または組換え部位の配列中に終止コドンを組み込むことができる。この種の態様では、関心対象の遺伝子は、関心対象の遺伝子のタグとコード配列が同じ読み枠になるように、ベクター中に挿入できる。関心対象の遺伝子には、終止コドンが抑制されない場合には天然のN端から遺伝子の発現ができるように、翻訳開始シグナル、例えば、シャインダルガーノ配列、コザック配列、および/またはIRES配列を提供してもよい。関心対象の遺伝子にはまた、コード配列の3'端に終止コドンを提供してもよい。いくつかの態様では、関心対象の遺伝子のN末端およびC末端の両方に、タグ配列が提供され得る。または、N末端のタグ配列に終止コドンを提供し、関心対象の遺伝子に終止コドンを提供し、C末端のタグに終止コドンを提供することもできる。終止コドンは、同一でも異なっていてもよい。いくつかの態様では、N末端タグの終止コドンは、関心対象の遺伝子の終止コドンとは異なる。この種の態様では、一方または両方の終止コドンに対応するサプレッサーtRNAを提供してもよい。両方が提供されるときには、サプレッサーtRNAの各々は、同一のベクター、異なるベクター、または宿主細胞のゲノムによって、独立して提供され得る。サプレッサーtRNAは、ともに同一の方法で提供される必要はなく、例えば、1つは関心対象の遺伝子を含むベクター上で提供され、他方は宿主細胞ゲノムで提供されてもよい。このように、本発明の1つのそのような局面における核酸分子は、2つのコード領域を分離する抑制可能な終止コドンを含むことができる。発現シグナル(例えば、プロモーター)の位置によって、サプレッサーtRNAの発現によって終止コドンが抑制され、それによって融合ペプチド、例えば発現されるタンパク質のNおよび/またはC末端にアフィニティタグ配列を持つ融合タンパク質が、産生される。終止コドンが抑制されないと、Nおよび/またはC末端のタグ配列のない、関心対象の配列の発現が行われる。したがって、本発明は、必要に応じた、融合タンパク質の制御された発現のための、関心対象の遺伝子または配列(例えば、1つまたは複数のオープンリーディングフレームまたは「orf」)を含むベクターの、組換えによる効率的な構築を可能にする。好ましくは、1つまたは複数のベクターにクローニングされる本発明の開始分子または産物分子は、少なくとも1つのオープンリーディングフレーム(orf)を含む。そのような開始分子または産物分子は、機能的配列(例えば、プライマー部位、転写または翻訳部位またはシグナル、終結部位(例えば、選択的に抑制され得る終止コドン)、複製開始点等)も含む可能性があり、好ましくは転写調節配列を含む遺伝子発現調節配列および内部リボソーム進入部位(IRES)として機能する配列を含む。好ましくは、開始または産物分子および/またはベクターのうちの少なくとも1つは、プロモーターとして機能する配列を含む。そのような開始または産物分子および/またはベクターは、転写終結配列、選択可能マーカー、制限酵素認識部位等も含みうる。
いくつかの態様では、ベクターは同一の選択可能マーカーを2コピー含み、この各々には組換え部位および/またはトポイソメラーゼ認識部位が隣接している。他の態様では、ベクターは各々2つの組換え部位に隣接する2つの異なる選択可能マーカーを含む。いくつかの態様では、1つまたは複数の選択可能マーカーは陰性選択マーカーである。
特定の局面では、本発明は、少なくとも1つの第1および第2の組換え部位を含む少なくとも1つの第1の核酸分子、および少なくとも1つの第3および第4の組換え部位を含む少なくとも1つの第2の核酸分子を提供する段階であって、第1または第2の組換え部位のいずれかは、第3または第4の組換え部位のいずれかと組換えできる段階、2つの核酸分子が1つまたは複数の産物核酸分子に組み込まれるように組換え反応を行なう段階、および1つまたは複数のベクター中に産物核酸分子をクローニングする段階を含むクローニング法を提供する。特定のそのような態様では、組換え部位は第1および/または第2の核酸分子を挟む。さらに、クローニング段階は、多くの場合1つまたは複数の組換え部位を含むベクターへの産物分子の組換え反応によって行われる。1つの局面では、クローニング段階は、第1の組換え反応で反応しなかった、産物核酸分子中の部位と、未反応の部位と組換えできる組換え部位を持つベクターの間の組換え反応を含む。
いくつかの態様では、組換え部位および/またはトポイソメラーゼ認識部位は、通常の結合手法を用いて関心対象の分子に結合できる。例えば、同一または異なる1つまたは複数の反応性の官能基を含むように、組換え部位および/またはトポイソメラーゼ認識部位を含むオリゴヌクレオチドを合成できる。適当な反応性の官能基には、アミン基、エポキシ基、ビニル基、チオール基等が含まれるが、これらに限定されるわけではない。1つまたは複数の反応性の官能基を含むオリゴヌクレオチドの合成は、当技術分野における日常的作業である。合成されたら、1つまたは複数の反応性の官能基を含むオリゴヌクレオチドは、関心対象の分子または化合物上の1つまたは複数の反応性の基に結合できる。オリゴヌクレオチドは1つまたは複数の反応性の官能基を1つまたは複数の反応性の官能基と反応させることによって、直接結合できる。いくつかの態様では、結合は、オリゴヌクレオチド上に存在する1つまたは複数の反応性の官能基、および関心対象の分子上に存在する1つまたは複数の反応性の基と反応できる、適当な結合基を用いて行われる。別の態様では、直接の結合および結合基を用いた結合の両方が用いられる。当業者は、オリゴヌクレオチド上の反応性の官能基は、関心対象の分子および/または化合物上の反応性の官能基と同一または異なることが理解できるだろう。オリゴヌクレオチドの関心対象の分子への結合のための適当な試薬および手法は、ヘルマンソン(Hermanson)、「生物接合体技術(Bioconjugate techniques)」、アカデミックプレス(Academic Press)社、サンディエゴ(San Diego)、CA、1996に記載されている。
本発明は、本発明の方法を実施するための組成物、およびそのような組成物を含むキット、ならびに本発明の方法を実施する際に生成する組成物にも関する。
本発明の組成物、方法、およびキットは、ファージλ部位特異的組換えシステムを用いて調製および実施できる。さらに、そのような組成物、方法、およびキットは、GATEWAY(商標)組換えクローニングシステムおよび/またはTOPO(登録商標)クローニングシステムおよび/またはpENTR Directional TOPO(登録商標)クローニングシステムを用いても調製および実施でき、これらはインビトロゲン社(Invitrogen Corporation)(Carlsbad、California)から市販されている。
別の局面では、本発明は1つまたは複数(例えば、1、2、3、4、5等)の組換え部位および/または1つまたは複数(例えば、1、2、3、4、5等)のトポイソメラーゼ認識部位を含む単離された核酸分子を提供する。本発明の1つのそのような分子は、1つのトポイソメラーゼ認識部位を挟む2つまたはそれ以上の組換え部位を含む。本発明の別のそのような分子は、各組換え部位がトポイソメラーゼ認識部位に隣接するような、2つまたはそれ以上の組換え部位および2つまたはそれ以上のトポイソメラーゼ認識部位を含む。本発明のこの局面による核酸分子は、線状、環状であるか、またはコイル、スーパーコイル等のような種々の形状および構造のうちのいずれかをとり得る。本発明のこの局面による核酸分子において都合よく利用される組換え部位には、att部位(attB部位、attP部位、attL部位、attR部位等を含むが、これらに限定されるわけではない)、lox部位(loxP部位、loxP511部位等を含むが、これらに限定されるわけではない)、psi部位、dif部位、cer部位、frt部位、ならびに組換えを受ける能力を保持したこれらの組換え部位の変異体、変種、および誘導体が含まれるが、これらに限定されるわけではない。本発明のこの局面の核酸分子中で都合よく利用されるトポイソメラーゼ認識部位は、好ましくはI型トポイソメラーゼ(IA型トポイソメラーゼ(大腸菌トポイソメラーゼI、大腸菌トポイソメラーゼIII、真核生物トポイソメラーゼII、アルシエール(archeal)リバースジャイレース、酵母トポイソメラーゼIII、ショウジョウバエトポイソメラーゼIII、ヒトトポイソメラーゼIII、肺炎球菌トポイソメラーゼIII、およびプラスミドRP4のtraEタンパク質)のような)、およびIB型トポイソメラーゼ(真核生物核I型トポイソメラーゼおよびポックスウイルス(ワクシニアウイルス、ショープ線維腫ウイルス、ORFウイルス、鶏痘ウイルス、伝染性軟肬腫ウイルス、およびアムサクタ・ムーレイ(Amsacta moorei)昆虫ポックスウイルスから単離または産生された)を含むが、これらに限定されるわけではない)、およびII型トポイソメラーゼ(細菌ジャイレース、細菌DNAトポイソメラーゼIV、真核生物DNAトポイソメラーゼII(コウシ胸腺II型トポイソメラーゼなど)、およびT11ファージにコードされるDNAトポイソメラーゼを含むが、これらに限定されるわけではない)によって認識および結合される。
本発明はそのような単離された核酸分子を含むベクター(発現ベクターの場合もある)も提供する。本発明のこの局面による代表的なベクターには、pcDNAGW-DT(sc)、pENTR-DT(sc)、pcDNA-DEST41、pENTR/D-TOPO、pENTR/SD/D-TOPO、pcDNA3.2/V5/GWD-TOPO、およびpcDNA6.2/V5/GWD-TOPOが含まれるが、これらに限定されるわけではない。本発明は、本発明のそのような単離核酸分子またはベクターを含む宿主細胞も提供する。
関連する局面では、本発明は核酸分子のクローニングのインビトロ法を提供する。本発明のこの局面による方法は、以下の段階を含む1つまたは複数の段階を含み得る:
(a)クローニングする核酸分子(特定の態様ではPCR産物のような線状分子(平滑末端を持つまたは持たない場合がある)であり、選択的に1つまたは複数の遺伝子またはオープンリーディングフレームを含み得る)を入手する段階;
(b)クローニングする核酸分子を、少なくとも1つの第1の組換え部位に隣接する少なくとも1つの第1のトポイソメラーゼ認識部位、および少なくとも1つの第2の組換え部位を含むベクター(発現ベクターの場合がある)とインビトロで混合する段階であって、第1と第2の組換え部位は互いにはおよび少なくとも1つのトポイソメラーゼとは組換えをしない段階;ならびに
(c)クローニングする核酸分子が第1および第2のトポイソメラーゼ認識部位の間でベクターに挿入される条件下で反応液をインキュベートし、それによって第1および第2の組換え部位の間にその核酸分子が存在する第1の産物分子を産生する段階。さらに本発明は、上述の方法によって調製された核酸分子も含む。
本発明のこの局面による方法は、例えば、互いに組換えをしない少なくとも第3および第4の組換え部位を含む少なくとも1つのベクターを、第1と第3の間、第2と第4の組換え部位の間での組換えに有利な条件下で、第1の産物分子に接触させ、それにより少なくとも第2の産物分子を産生する段階を含む、1つまたは複数の別の段階を含むことがある。本発明にしたがって、これらの方法によって産生された第1および/または第2の産物分子は、宿主細胞に挿入できる。本発明のこの局面で使用されるベクターは、選択可能マーカー、クローニング部位、制限酵素部位、プロモーター、オペロン、複製開始点、および遺伝子または部分的遺伝子(例えば、遺伝子断片または要素)からなる群より選択される少なくとも1つのさらなる核酸配列を含み得る。
本発明のこの局面の方法で使用される組換え部位およびトポイソメラーゼ認識部位は、本明細書の別の箇所に記載されるものを含むが、これらに限定されるわけではない。特定の方法では、第2の産物核酸分子およびベクターは、Cre、Int、IHF、Xis、Fis、Hin、Gin、Cin、Tn3リゾルバーゼ、TndX、XerC、またはXerDを含むがこれらに限定されるわけではない、少なくとも1つの組み換えタンパク質の存在下で結合される。
本発明は本発明のこれらの単離された核酸分子を含むキットも提供するが、これは選択的に、1つまたは複数のトポイソメラーゼ、1つまたは複数の組み換えタンパク質、1つまたは複数のベクター、1つまたは複数のポリメラーゼ活性を持つポリペプチド、および1つまたは複数の宿主細胞からなる群より選択される1つまたは複数の他の成分も含み得る。
本発明の他の好ましい態様は、当技術分野における知識、本発明の以下の図面および説明、ならびに添付の特許請求の範囲を考慮すると、当業者には明らかであると考えられる。
発明の詳細な説明
定義
以下の説明において、組換え核酸技術に使用される数多くの用語は広範に使用される。このような用語を与える範囲を含む、明細書および特許請求の範囲の理解を明確にし、かつ常に一定にするために、以下の定義を提供する。
遺伝子:
本明細書において使用する遺伝子は、ポリペプチド、タンパク質または機能的RNA(例えば、リボザイム、tRNA、rRNA、mRNA等)の発現に必要な情報を含む核酸配列である。それは、プロモーターおよび構造遺伝子オープンリーディングフレーム配列(orf)、ならびにタンパク質の発現に関与する他の配列を含む。
構造遺伝子:
本明細書において使用する構造遺伝子は、メッセンジャーRNAに転写され、次いで、特定のポリペプチドを特徴とするアミノ酸配列に翻訳される核酸配列をいう。
宿主:
本明細書において使用する宿主は、複製可能な発現ベクター、クローニングベクターまたは任意の核酸分子のレシピエントである、任意の原核生物または真核生物である。核酸分子は、構造遺伝子、(プロモーター、エンハンサー、リプレッサー等などの)転写調節配列および/または複製開始点を含んでもよいが、これらに限定されない。本明細書において使用する「宿主」、「宿主細胞」、「組換え宿主」、および「組換え宿主細胞」は交換して使用することができる。このような宿主の例は、マニアティス(Maniatis)ら、分子クローニング:実験マニュアル(Molecular Cloning: A Laboratory Manual)、コールドスプリングハーバーラボラトリー(Cold Spring Harbor Laboratory)、コールドスプリングハーバー、ニューヨーク(1982)参照。
転写調節配列:
本明細書において使用する転写調節配列は、1つまたは複数の構造遺伝子のメッセンジャーRNAへの転写を調節するように作用をする、核酸分子に任意の配置または構造で含有される機能的なヌクレオチド鎖である。転写調節配列の例には、プロモーター、オペレーター、エンハンサー、リプレッサー等が含まれるが、これらに限定されない。転写調節配列は、機能的RNA(例えば、リボザイム、tRNA、rRNA、mRNA等)をコードする核酸分子の転写を調節することもできる。
プロモーター:
本明細書において使用するプロモーターは転写調節配列の一例であり、具体的には、開始コドンの近位に位置する遺伝子の5'側-領域と一般的記載される核酸配列である。隣接する核酸セグメントの転写はプロモーター領域から開始される。抑制性プロモーターの転写速度は、抑制剤に応答して低下する。誘導プロモーターの転写速度は、誘導剤に応答して増加する。構成的プロモーターの転写速度は特に調節されないが、一般的な代謝条件の影響下で変更しうる。
挿入物:
本明細書において使用する挿入物は、大型の核酸分子の一部である、望ましい核酸セグメントである。
標的核酸分子:
本明細書において使用する標的核酸分子は、好ましくは本発明の化合物および方法の使用により作動する核酸の、関心対象の核酸セグメントである。このような標的核酸分子は、好ましくは、遺伝子または遺伝子の一部を1つまたは複数含む。
挿入物ドナー:
本明細書において使用する挿入物ドナーは、挿入物を保有する本発明の2つの親核酸分子(例えば、RNAまたはDNA)の一方である。挿入物ドナー分子は、組換え部位の両側に隣接する挿入物を含む。本発明の一態様において、挿入物ドナーは選択的に超コイル状態の環状核酸分子であり、組換えシグナルの外側にクローニングベクター配列をさらに含む(図1参照)。挿入物ドナーを作製するために挿入物集団または核酸セグメント集団を使用する場合には、挿入物ドナー集団が生じ、本発明により使用することができる。
産物:
本明細書において使用する産物は、組換えクローニング過程中の第2の組換え事象後に作製される、A配列およびD配列を含む望ましい娘分子の1つである(図1参照)。産物は、クローニングまたはサブクローニングされた核酸を含む。本発明によると、挿入物ドナー集団を使用する場合には、得られる産物分子集団は挿入物ドナーの挿入物集団の全てまたは一部を含有し、好ましくは挿入物ドナーの元の分子の代表的な集団を含む。
認識配列:
本明細書において使用する認識配列(本明細書において「認識部位」と二者択一的にまたは等価にいわれる)は、タンパク質、化合物DNAまたはRNA分子(例えば、制限エンドヌクレアーゼ、トポイソメラーゼ、修飾メチラーゼまたはリコンビナーゼ)が認識し、結合する特定の配列である。本発明において、認識配列とは通常、(リコンビナーゼ認識部位と二者択一的に呼ばれることもある)組換え部位またはトポイソメラーゼ認識部位をいう。例えば、Creリコンビナーゼの認識配列は、8塩基対のコア配列に隣接する13塩基対の2つの逆方向反復(リコンビナーゼ結合部位として働く)を含む、34塩基対配列であるloxPである。サウアー(Sauer, B)、「バイオテクノロジーの最新の見解(Current Opinion in Biotechnology) 」5:521-527(1994)の図1参照。このような認識配列の他の例は、リコンビナーゼ酵素によって認識されるattB、attP、attLおよびattR配列である(インテグラーゼattBは9塩基対のコア型Int結合部位2つと7塩基対の重複領域を含む、約25塩基対の配列である。attPは、コア型Int結合部位およびアーム型Int結合部位ならびに補助的なタンパク質組み込み宿主因子(IHF)、FISおよび除去酵素(Xis)の部位を含む約240塩基対の配列である。ランディ(Landy)、「バイオテクノロジーの最新の見解(Current Opinion in Biotechnology)」3: 699-707(1993)参照。このような部位は、本発明の方法における産物の産生を増大するために、本発明により操作することもできる。組換え反応を不可逆的にするために、このような操作部位がP1またはH1ドメインを欠損する場合には、これらの部位のドメインが何らかの方法で改変されていることを示すために、このような部位をattR'またはattP'と命名することができる。トポイソメラーゼ認識部位の例には、大腸菌(E. coli)トポイソメラーゼIII(I型トポイソメラーゼ)によって認識される配列5'-GCAACTT-3'、ワクシニアウイルスDNAトポイソメラーゼIを含む、ほとんどのポックスウイルストポイソメラーゼによって特異的に結合されるトポイソメラーゼ認識部位である配列5'-(C/T)CCTT-3'、および本明細書の他の個所で考察されている当技術分野において既知の他の配列が含まれるが、これらに限定されない。
組換えタンパク質:
本明細書において使用する組換えタンパク質は、切り出しまたは組み込みタンパク質、酵素、補助因子または1つもしくは複数の組換え部位に関与する組換え反応を含み、野生型タンパク質(Landy、「バイオテクノロジーの最新の見解(Current Opinion in Biotechnology) 」3: 699-707(1993)参照)または突然変異体、誘導体(例えば、組換えタンパク質配列またはそれらの断片を含む融合タンパク質)、断片およびそれらの変種であってもよい、関連するタンパク質を含む。
組換え部位:
本明細書において使用する組換え部位は、組換えタンパク質による組み込み/組換え反応に関与する核酸分子上の認識配列である。組換え部位は、組み込みまたは組換えの最初の段階において、部位特異的な組換えタンパク質によって認識され結合される関与中の核酸分子上の、核酸の別個の部分またはセグメントである。例えば、Creリコンビナーゼの組換え部位は、8塩基対のコア配列に隣接する13塩基対の2つの逆方向反復(リコンビナーゼ結合部位として働く)を含む34塩基対配列であるloxPである。サウアー(Sauer, B)、Curr. Opin. Biotech. 5:521-527(1994)の図1参照。認識配列の他の例には、本明細書に記載するattB、attP、attLおよびattR配列ならびに認識タンパク質(Intならびに補助的なタンパク質組み込み宿主因子(IHF)、FISおよび除去酵素(Xis)によって認識される、それらの突然変異体、断片、変種および誘導体が含まれる。ランディ(Landy)、Curr. Opin. Biotech. 3: 699-707(1993)参照。
組換えクローニング:
本明細書において使用する組換えクローニングは、(その内容が完全に参照として本明細書に組み入れられる)米国特許第5,888,732号、同第6,143,557号、同第6,171,861号、同第6,270,969号および同第6,277,608号に記載されているものなどの方法ならびに本明細書にも記載されている方法であり、それによって核酸分子セグメントまたはこのような分子集団は、インビトロまたはインビボにおいて交換、挿入、取替え、置換または改変される。好ましくは、このようなクローニング方法はインビトロにおける方法である。
抑制カセット:
本明細書において使用するリプレッションカセットは、サブクローニングベクターに存在するリプレッサーまたは選択マーカーを含む核酸セグメントである。
選択マーカー:
本明細書において使用する選択マーカーは、多くは特定の条件下でそれを含む分子(例えば、レプリコン)または細胞を含むようにまたは含まないように選択することができる核酸セグメントである。これらのマーカーは、RNA、ペプチドもしくはタンパク質の産生などであるが、これらに限定されない作用をコードすることができるか、またはRNA、ペプチド、タンパク質、無機および有機化合物もしくは組成物等の結合部位を提供することができる。選択マーカーの例には、(1)毒性化合物(例えば、抗生物質)に対する耐性を提供する産物をコードする核酸セグメント、(2)レシピエント細胞において欠損している産物をコードする核酸セグメント(例えば、tRNA遺伝子、栄養要求性マーカー)、(3)遺伝産物の活性を抑制する産物をコードする核酸セグメント、(4)容易に同定することができる産物をコードする核酸セグメント(例えば、ガラクトシダーゼ、緑色蛍光タンパク質(GFP)および細胞表面タンパク質などの表現型マーカー)、(5)細胞の生存および/または機能に有害な産物に結合する核酸セグメント、(6)上記の1番〜5番に記載する核酸セグメントのいずれかの活性を阻害する核酸セグメント(例えば、アンチセンスオリゴヌクレオチド)、(7)基質を修飾する産物に結合する核酸セグメント(例えば、制限エンドヌクレアーゼ)、(8)望ましい分子を単離または同定するために使用できる核酸セグメント(例えば、特定のタンパク質結合部位)、(9)非機能的であってもよい特定のヌクレオチド配列をコードする核酸セグメント(例えば、分子の亜集団のPCR増幅のため)、(10)欠損していると、特定の化合物に対する耐性または感受性を直接または間接的に示す核酸セグメント、および/または(11)レシピエント細胞において毒性である産物をコードする核酸セグメントが含まれるが、これらに限定されない。
選択スキーム:
本明細書において使用する選択スキームは、導入クローンもしくはベクター、目的ベクター、ドナーベクター、発現クローンまたはベクター、任意の中間体(例えば、融合体もしくはレプリコン)および/または副産物を含む混合物からの、望ましい産物の選択、濃縮または同定を可能にする任意の方法である。1つの好ましい態様の選択スキームは、組換えクローニング中に結合されるまたは結合されない少なくとも2つの成分を有する。一方の成分は選択マーカーである。他方の成分は、選択マーカーのインビトロもしくはインビボにおける発現、または選択マーカーを保有するプラスミドを保有する細胞(または例えば、レプリコンのような核酸分子)の生存を制御する。一般に、この制御要素は選択マーカーのリプレッサーまたはインデューサーであるが、選択マーカーの発現または活性を制御する他の手段を使用してもよい。リプレッサーまたはアクチベーターを使用するかどうかは、当業者に容易に理解されるように、マーカーが陽性または陰性選択用であるかどうかおよび種々の核酸セグメントの正確な配列に依存している。いくつかの好ましい態様において、選択スキームにより、1つまたは複数の望ましい産物だけが選択または濃縮される。本明細書に規定するように、望ましい核酸分子を求めて選択する段階は、(a)望ましい核酸分子の存在を求めて選択または濃縮する段階、および(b)望ましい核酸分子でない核酸分子の存在を含まないように選択または濃縮する段階を含む。
一態様において、(逆に実施することができる)選択スキームは3つの形態のうちの1つを取り、図1に関して考察されている。第一に、選択マーカーおよびリプレッサーを用いて本明細書において例示されているように、セグメントDを有し、セグメントCを欠損する分子を選択する。第二は、セグメントCを有する分子を除外し、セグメントDを有する分子を含むように選択する。第2の形態の可能な態様は、インビトロにおける反応産物を導入する細胞に対して毒性の遺伝子を保有する、核酸セグメントを有してもよい。毒性遺伝子は、毒性遺伝産物(毒性タンパク質またはRNA)として発現される核酸であってもよく、またはそれ自体単独で毒性であってもよい(後者の場合には、毒性遺伝子は、「遺伝的特徴」という古典的な定義を有すると理解される)。
このような毒性遺伝産物の例は当技術分野において既知であり、制限エンドヌクレアーゼ(例えば、DpnI)、アポトーシス関連遺伝子(例えば、ASK1またはbcl-2/ced-9ファミリーのメンバー)、ヒト免疫不全ウイルス(HIV)の遺伝子を含むレトロウイルス遺伝子、NP-1などのデフェンシン、逆方向反復または対になった回文配列の核酸配列、(X174またはバクテリオファージT4の遺伝子などのバクテリオファージ溶解遺伝子、rpsLなどの抗生物質感受性遺伝子、pheSなどの抗菌感受性遺伝子、プラスミドキラー遺伝子、GATA-1などの、細菌に毒性の遺伝産物を産生する真核細胞の転写ベクター遺伝子、kicB、ccdB、(X174E(Liu, Q.ら、Curr. Biol. 8:1300-1309(1998))のように、抑制機能が存在しない場合に宿主を殺す遺伝子、ならびにレプリコンの安定性および/または複製に負に影響する他の遺伝子が含まれるが、これらに限定されない。または、毒性遺伝子は、インビトロにおいて選択可能で、例えば制限部位であってもよい。
誘導プロモーターに機能的に結合する制限エンドヌクレアーゼをコードする多数の遺伝子は既知であり、本発明に使用することができる。例えば、米国特許第4,960,707号(DpnIおよびDpnII)、同第5,000,333号、同第5,082,784号および同第5,192,675号(KpnI)、同第5,147,800号(NgoAIIIおよびNgoAI)、同第5,179,015号(FspIおよびHaeIII)、同第5,200,333号(HaeIIおよびTaqI)、同第5,248,605号(HpaII)、同第5,312,746号ClaI)、同第5,231,021号および同第5,304,480号(XhoIおよびXhoII)、同第5,334,526号(AluI)、同第5,470,740号(NsiI)、同第5,534,428号(SstI/SacI)、同第5,202,248号(NcoI)、同第5,139,942号(NdeI)および同第5,098,839号(PacI)参照。また、Wilson, G. G.、Nucl. Acids Res. 19: 2539-2566(1991)、およびLunnen, K. D.ら、Gene 74: 25-32(1988)も参照。
第二の形態において、セグメントDは選択マーカーを保有する。毒性遺伝子は、ベクタードナー、融合体および副産物分子を保有する形質転換体を排除すると思われるが、産物を含む細胞を含むように選択し、挿入物ドナーだけを保有する細胞を含まないように選択するために、選択マーカーを使用することができる。
第三の形態は、同一分子にセグメントAおよびDをシス形態で有する細胞を選択するが、両方のセグメントを異なる分子にトランス形態で有する細胞を選択しない。これは、2つの不活性な断片に分割され、セグメントAおよびDにそれぞれ1つずつある選択マーカーによって具体化されうる。
セグメントが組換え事象によって接合される場合には、それらが機能的な選択マーカーを再構成するように、断片は組換え部位に対して配列される。例えば、組換え事象はプロモーターに構造核酸分子(例えば、遺伝子)を接続することができ、構造核酸分子の2つの断片を接続することができ、または生存に必要なヘテロダイマー遺伝産物をコードする核酸分子を接続することができ、またはレプリコンの一部を接続することができる。
部位特異的リコンビナーゼ:
本明細書において使用する部位特異的リコンビナーゼは、典型的には、少なくとも以下の4つの活性(またはその組み合わせ)を有する種類のリコンビナーゼである:(1)1つまたは2つの特異的な核酸配列の認識、(2)前記配列の切断、(3)鎖交換に関与するトポイソメラーゼ活性および(4)切断された核酸鎖を再閉するリガーゼ活性。サウアー (Sauer, B)、「バイオテクノロジーの最新の見解Current Opinions in Biotechnology」5: 521-527(1994)参照。保存的な部位特異的な組換えは、両方のパートナーに対する高度な特異性による相同組換えおよび転位によって識別される。鎖交換機序は、DNA合成が存在しない場合には、特異的な核酸配列の切断および再接続に関与する(Landy, A.(1989) Ann. Rev. Biochem. 58: 913-949)。
ベクター:
本明細書において使用するベクターは、有用な生物学的または生化学的特性を挿入物に提供する核酸分子(好ましくは、DNA)である。例として、プラスミド、ファージ、自己複製配列(ARS)、セントロメア、および複製可能かまたはインビトロもしくは宿主細胞において複製されうる、または宿主細胞内の望ましい位置に望ましい核酸セグメントを搬送することができる他の配列が含まれる。ベクターは、ベクターの生物学的必須機能を損失することなく、測定可能な様式で配列を切断することができ、複製およびクローニングを生じさせるために、核酸断片をスプライシングすることができる1つまたは複数の制限エンドヌクレアーゼ認識部位を有することができる。ベクターは、例えばPCRのプライマー部位、転写および/または翻訳開始および/または調節部位、組換えシグナル、レプリコン、選択マーカー等をさらに提供することができる。明らかに、組換え、転位または制限酵素の使用を必要としない望ましい核酸断片を挿入する方法(PCR断片のUDGクローニング(全体が参照として本明細書に組み入れられる、米国特許第5,334,575号)、TA Cloning(登録商標)ブランドPCRクローニング(Invitrogen Corporation、カリフォルニア州カールスバッド)(直接ライゲーションクローニングとしても既知)等などであるが、これらに限定されない)を利用して、本発明により使用されるクローニングベクターに断片をクローニングすることができる。クローニングベクターは、クローニングベクターで形質転換した細胞を同定する際に使用するのに適切な1つまたは複数の選択マーカーをさらに含有してもよい。
サブクローニングベクター:
本明細書において使用するサブクローニングベクターは、好ましくは適当なレプリコンを含む、環状または直鎖状核酸分子を含むクローニングベクターである。本発明において、サブクローニングベクター(図1のセグメントD)は、最終的な産物に組み込まれて、クローニングされた核酸挿入物に対して作用するかまたはそれと共に作用することが望ましい、機能的および/または調節的構成要素を含有してもよい(図1のセグメントA)。サブクローニングベクターは、選択マーカー(好ましくはDNA)も含有してもよい。
ベクタードナー:
本明細書において使用するベクタードナーは、望ましい産物の一部となる核酸ベクターを含む核酸セグメントを保有する、本発明の2つの親核酸分子(例えば、RNAまたはDNA)の一方である。ベクタードナーは、組換え部位が隣接するサブクローニングベクターD(または、挿入物ドナーがクローニングベクターを含有しない場合には、クローニングベクターと呼ぶことができる)およびセグメントCを含む(図1参照)。セグメントCおよび/またはDは、選択スキームについて上記するように、望ましい産物である娘分子の選択に貢献する要素を含むことができる。組換えシグナルは同じであっても、または異なってもよく、同じまたは異なるリコンビナーゼによる作用を受けることができる。また、ベクタードナーは直鎖状であっても、または環状であってもよい。
プライマー:
本明細書において使用するプライマーは、核酸分子(例えば、DNA分子)の増幅または重合過程中にヌクレオチドモノマーの共有結合によって伸長される一本鎖または二本鎖オリゴヌクレオチドである。一局面において、プライマーは配列決定用プライマー(例えば、普遍的配列決定用プライマー)であってもよい。別の局面において、プライマーは組換え部位またはその一部を含んでもよい。
鋳型:
本明細書において使用する鋳型は、増幅、合成または配列決定される二本鎖または一本鎖核酸分子である。二本鎖DNA分子の場合には、第1および第2の鎖を形成するための鎖の変性は、好ましくは、これらの分子が増幅、合成もしくは配列決定される前に実施されるか、または二本鎖分子を鋳型として直接使用することができる。一本鎖鋳型では、鋳型の少なくとも一部に相補的なプライマーを適当な条件下でハイブリダイゼーションし、ポリメラーゼ活性(例えば、DNAポリメラーゼおよび/または逆転写酵素)を有する1つまたは複数のポリペプチドが、鋳型の全てまたは一部に相補的な分子を合成することができる。または、二本鎖鋳型では、1つまたは複数の転写調節配列(例えば、1つまたは複数のプロモーター)を1つまたは複数のポリメラーゼと併用使用して、鋳型のすべてまたは一部に相補的な核酸分子を作製することができる。本発明により新規に合成された分子は、元の鋳型と同じかまたはそれより短い鎖長であってもよい。新規に合成される分子の合成または伸長過程中のミスマッチの組み込みまたは鎖の翻訳スリップにより、ミスマッチのある塩基対が1つまたはいくつか作製される。従って、合成された分子は鋳型に正確に相補的である必要はない。また、核酸鋳型集団を合成または増幅過程中に使用して、元の鋳型集団を典型的に代表する核酸分子集団を作製することができる。
組み込み:
本明細書において使用する組み込みは、核酸(例えば、DNA)分子またはプライマーの一部になることを意味する。
ライブラリ:
本明細書において使用するライブラリは核酸分子(環状または直鎖状)の集合である。一態様において、ライブラリは、通常の起源生物、器官、組織または細胞由来であってもそうでなくてもよい、複数(すなわち、2つまたはそれ以上)の核酸分子を含んでもよい。別の態様において、ライブラリは生物の核酸含量の全てもしくは一部もしくはかなりの部分の代表であるか(「ゲノム」ライブラリ)、または細胞、組織、器官もしくは生物において発現される核酸分子(それから誘導されるcDNAライブラリまたはセグメント)の全てもしくは一部もしくはかなりの部分を代表する核酸分子の組である。ライブラリは新規合成によって作製されるランダム配列、1つまたは複数の配列の突然変異等を含んでもよい。このようなライブラリは1つまたは複数のベクターに含有されても、されなくてもよい。
増幅:
本明細書において使用する増幅は、ポリメラーゼ活性を有する1つまたは複数のポリペプチド(例えば、1つもしくは複数の核酸ポリメラーゼまたは1つもしくは複数の逆転写酵素)を使用することにより、数多くのコピー数のヌクレオチド配列を増加する、任意のインビトロ方法である。核酸増幅により、ヌクレオチドがDNAおよび/またはRNA分子またはプライマーに組み込まれて、鋳型に相補的な新規核酸分子が形成される。形成された核酸分子およびその鋳型を鋳型として使用して、追加の核酸分子を合成することができる。本明細書において使用する1回の増幅反応は、多数回の核酸複製からなってもよい。DNA増幅反応は、例えばポリメラーゼ連鎖反応(PCR)を含む。1回のPCR反応は、DNA分子の5〜100サイクルの変性および合成からなってもよい。
ヌクレオチド:
本明細書において使用するヌクレオチドは塩基-糖-リン酸塩の組み合わせである。ヌクレオチドは、核酸分子(DNAおよびRNA)のモノマー単位である。ヌクレオチドという用語は、リボヌクレオシド三リン酸ATP、UTP、CTG、GTPおよびdATP、dCTP、dITP、dUTP、dGTP、dTTPなどのデオキシリボヌクレオシド三リン酸またはそれらの誘導体を含む。このような誘導体には、例えば[(S]dATP、7-デアザ-dGTPおよび7-デアザ-dATPが含まれる。本明細書において使用するヌクレオチドという用語は、ジデオキシリボヌクレオシド三リン酸(ddNTP)およびそれらの誘導体も意味する。ジデオキシリボヌクレオシド三リン酸の例示的な例には、ddATP、ddCTP、ddGTP、ddITPおよびddTTPが含まれるが、これらに限定されない。本発明によると、「ヌクレオチド」は未標識でも、または既知の技法により検出できるように標識されてもよい。検出可能な標識には、例えば放射性同位体、蛍光標識、化学発光標識、生物発光標識および酵素標識が含まれる。
核酸分子:
本明細書において使用する核酸分子は、全長のポリペプチドまたは任意の鎖長の断片をコードしても、または非コード領域であってもよい、任意の鎖長の連続するヌクレオチド(リボNTP、dNTPもしくはddNTPまたはそれらの組み合わせ)の配列である。本明細書において使用する「核酸分子」および「ポリヌクレオチド」という用語は交換して使用することができる。
オリゴヌクレオチド:
本明細書において使用するオリゴヌクレオチドは、1つのヌクレオチドのペントースの3'側位置と隣接するヌクレオチドのペントースの5'側位置との間の、ホスホジエステル結合によって接続される、共有結合したヌクレオチド配列を含む合成または天然分子である。
ポリペプチド:
本明細書において使用するポリペプチドは任意の鎖長の連続するアミノ酸配列である。本明細書において使用する「ペプチド」、「オリゴペプチド」または「タンパク質」という用語は「ポリペプチド」という用語に交換して使用することができる。
ハイブリダイゼーション:
本明細書において使用するハイブリダイゼーションおよびハイブリダイゼーションするという用語は、2つの相補的な一本鎖核酸分子(RNAおよび/またはDNA)が塩基対形成して、二本鎖分子を形成することをいう。本明細書において使用するように、2つの核酸分子はハイブリダイゼーションすることができるが、塩基対形成は完全に相補的ではない。従って、ミスマッチ塩基は2つの核酸分子のハイブリダイゼーションを妨害しないが、ただし、当技術分野において既知の適当な条件を使用する。いくつかの局面において、ハイブリダイゼーションは「ストリンジェントな条件」下であると言われる。本明細書において使用する「ストリンジェントな条件」は、50%ホルムアミド、5×SSC(150 mM NaCl、15 mMクエン酸三ナトリウム)、50 mM リン酸ナトリウム(pH 7.6)、5×デンハルト液、10%硫酸デキストラン、および変性させて切断したサケ精子20g/mlを含む溶液中で、42℃において一晩インキュベーションし、その後フィルターを0.1×SSC中で約65℃において洗浄することを意味する。
本明細書において使用する組換え核酸技術、ならびに分子生物学および細胞生物学の分野において使用される他の用語は、適用可能な技術分野の当業者に一般に理解されると考えられる。
概説
本発明は、核酸または他の分子および/または化合物(またはそれらの組み合わせ)の2つまたはそれ以上のセグメントまたは分子を組換えおよび/またはトポイソメラーゼ媒介により接続する方法、組成物、ならびにキットに関する。本発明はまた、接続されたこのような核酸または他の分子および/または化合物を、好ましくは(組換えタンパク質認識配列、トポイソメラーゼ認識配列等を含んでもよい)組換え部位またはそれらの一部を介して、1つまたは複数の支持体または構造物に接着する段階に関する。従って、本発明は一般に、1つまたは複数のトポイソメラーゼ認識部位および/または1つもしくは複数の組換え部位またはそれらの一部を含む核酸リンカーを介して、任意の数の核酸または他の分子および/または化合物を接続する段階に関する。本発明により作製された接続産物は、出発物質に応じて、任意の数の同じもしくは異なる核酸または他の分子、および/または化合物を含んでもよい。このような出発物質には、任意の核酸(またはペプチド核酸(PNA)などの誘導体)、化合物、検出できるように標識された分子(蛍光分子および化学発光分子)、薬剤、ペプチドまたはタンパク質、脂質、炭水化物および1つまたは複数の組換え部位またはその一部を含む、他の分子および/または化合物が含まれるが、これらに限定されない。本発明によるこのような組換え部位の組換えおよび/またはトポイソメラーゼ媒介性接続反応により、任意の数または組み合わせのこのような出発分子および/または化合物を接続して、本発明の接続産物を作製することができる。また、本発明の接続産物のある部分または成分の欠失または置換は、組換えにより実施することができる。
いくつかの態様において、本発明の組換えクローニング方法および/またはトポイソメラーゼ媒介性接続方法などにより、接続セグメントをベクターなどの異なる核酸分子に挿入することができる。従っていくつかの態様において、本発明は、組換え反応および/またはトポイソメラーゼ媒介性接続反応により2つまたはそれ以上の核酸セグメントを組み合わせ、接続した2つまたはそれ以上のセグメントを、組換えクローニングによりベクターに挿入することによって、核酸分子(RNAまたはDNA)を構築する段階に関する。接続した核酸分子を、組換え反応により追加の核酸分子とさらに組み合わせる態様では、2つの組換え事象、すなわちセグメントの接続とセグメントのベクターへの挿入の時期は重大ではない。すなわち、2つまたはそれ以上の核酸セグメントが、ベクターに挿入される前に一体として接続されるかどうか、または例えば、各セグメントの1つの組換え部位がベクターの組換え部位と最初に反応し、その後核酸セグメントの組換え部位が互いに反応してセグメントを接続するかどうかは、本発明にとって重大ではない。さらに、核酸セグメントはベクター内の任意の1箇所またはいくつかの位置にクローニングされてもよく、互いに隣接して挿入される必要はないが、いくつかの態様では、ベクター内の2つまたはそれ以上のこのようなセグメントの接続が好ましい。本発明によると、組換えクローニングにより、組み合わせられた核酸セグメントを含む分子(好ましくはベクター)を効率的に選択および同定することができる。従って、関心対象の2つまたはそれ以上の核酸セグメントを組み合わせて、選択的に、組み合わせた核酸分子をさらに操作するのに適切な1つのベクターに挿入することができる。
別の態様において、それぞれが少なくとも1つの(例えば、1、2、3、4、5、6、7、8等)の組換え部位と、選択的に、少なくとも1つの(例えば、1、2、3、4、5、6、7、8等)トポイソメラーゼ認識部位を含む、少なくとも2つの(例えば、2、3、4、5、6、7、8等)核酸セグメントに適切な組換えタンパク質および/またはトポイソメラーゼを接触させて、分子内の組換え部位の位置に応じて、2つの分子の全てまたは一部を接続する。このようなある種の態様では、少なくとも2つの組換え部位を含む核酸分子などにおいて、2つの組換え部位の少なくとも一方が、分子のトポイソメラーゼ認識部位の各末端に隣接する。別の認識部位(例えば、別の組換え部位またはトポイソメラーゼ認識部位)に「隣接する」組換え部位(またはトポイソメラーゼ認識部位)は、2つの部位が互いから約20ヌクレオチド以内にあるか、または互いから約20、19、18、17、16、15、14、13、12、11、10、9、8、7、6、5、4、3、2、1または0ヌクレオチド以内にあることを意味する。個々の核酸セグメントはそれぞれ、プライマー部位として使用するのに適切な配列(配列決定プライマーまたは増幅プライマーなどのプライマーがハイブリダイゼーションして、核酸合成、増幅または配列決定することができる配列)、プロモーター、リボソーム結合部位、Kozak配列および開始コドンなどの転写もしくは翻訳シグナル、または調節配列、停止コドンなどの停止シグナル、複製開始点、組換え部位(またはそれらの一部)、トポイソメラーゼ認識部位(またはそれらの一部)、選択マーカーおよびGST、GUS、GFP、6ヒスチジン、エピトープハプテン等などのタンパク質融合物を作製するための遺伝子または遺伝子の一部(例えば、N末端またはカルボキシ末端)ならびにそれらの組み合わせを含むが、これらに限定されない種々の配列を含んでもよい。このようなセグメントをクローニングするために使用されるベクターは、これらの機能的な配列(例えば、プロモーター、プライマー部位等)を含んでもよい。このような配列を含むセグメントを組み合わせ、選択的には1つまたは複数のベクターへの配列のクローニング後に、標的配列の配列決定または増幅(すなわち、組み込み配列によって組入れられるプライマー部位の少なくとも1つを使用することによる)、標的配列の突然変異(すなわち、標的配列内または標的配列に対する挿入、欠損または置換による)および標的配列またはその一部からのタンパク質発現(すなわち、セグメントおよび/またはベクターによって含有される翻訳および/または転写シグナルの発現による)を含む種々の方法で、分子を操作することができる。
本発明はまた、開示されている組換えクローニング方法を使用したコンビナトリアルライブラリの作製に関する。従って、接続された核酸セグメントの1つまたは複数は核酸ライブラリを含むことができる。このようなライブラリは、例えばペプチド、ポリペプチドまたはタンパク質配列をコードする配列の順列に対応する核酸配列を含むことができる。順列は1つの配列からなる別の核酸セグメントに接続できるか、または2つのセグメントの接続により、2つのペプチド、ポリペプチドまたはタンパク質配列の全ての順列の可能な組み合わせを全て示すライブラリを作製することができるように、第2の核酸セグメントもまた、別のペプチド、ポリペプチドまたはタンパク質配列の順列に対応するライブラリであってもよい。コンビナトリアルライブラリの用途の数多くの例は当技術分野において既知である。例えば、それらの全てが具体的に参照として本明細書に組み入れられる、Waterhouseら、Nucleic Acids Research、1993年、21巻、9号、2265〜2266ページ、Tsurushitaら、Gene、1996、第172巻、第1号、59〜63ジ、Persson、Int Rev Immunol、1993、10: 2〜3 153〜63、Chanockら、Infect Agents Dis 1993年6月2: 3 118〜31、Burioniら、Res Virol 1997年5月〜4月 148: 2 161〜4、Leung、Thromb Haemost 1995年7月74: 1 373〜6、Sandhu、Crit Rev Biotechnol 1992、12: 5〜6 437〜62、ならびに米国特許第5,733,743号、同第5,871,907号、および同第5,858,657号を参照。
組換え部位
本発明に使用する組換え部位は、組換え反応の基質として働くことができる任意の核酸配列であってよい。このような組換え部位は野生型もしくは天然型組換え部位または改変もしくは突然変異組換え部位であってもよい。本発明に使用する組換え部位の例には、λファージ組換え部位(attP、attB、attLおよびattR、ならびにそれらの突然変異体または誘導体など)ならびにphi80、P22、P2、186、P4およびP1(loxPおよびloxP511などのlox部位を含む)などの他のバクテリオファージの組換え部位が含まれるが、これらに限定されない。新規突然変異att部位(例えば、attB 1〜10、attP 1〜10、attR 1〜10およびattL 1〜10)は、具体的に参照として本明細書に組み入れられる、1999年5月28日に提出された、以前の特許出願第60/136,744号に記載されている。固有の特性を有する他の組換え部位(すなわち、第1の部位に対応する部位を組換え、かつ異なる特異性を有する第2の部位を組換えない)は当業者に既知であり、本発明を実施するために使用することができる。
これらの系に対応する組換えタンパク質を、示した組換えと共に本発明により使用することができる。本発明に使用するための組換え部位および組換えタンパク質を提供する他の系には、出芽酵母(Saccharomyces cerevisiae)のFLP/ERT系、リゾルバーゼファミリー(例えば、(Tn3リゾルバーゼ、Hin、GinおよびCin)およびIS231ならびに他のバシラス・スリンジエンシス(Bacillus thuringiensis)転位要素が含まれる。本発明に使用するための他の適切な組換え系には、大腸菌(E. coli)のXerCおよびXerDリコンビナーゼならびにpsi、difおよびcer組換え部位が含まれる。他の適切な組換え部位は、具体的に参照として本明細書に組み入れられる、エレッジ(Elledge)およびリウ(Liu)に付与された米国特許第5,851808号に見出すことができる。本発明に使用するための好ましい組換えタンパク質および突然変異または改変組換え部位には、そのすべての開示内容が全体として参照として本明細書に組み入れられる、米国特許第5,888,732号、同第6,171,861号、同第6,143,557号、同第6,270,969号および同第6,277,608号、ならびに共通所有の同時係属米国特許出願第09/438,358号(1999年11月12日提出)、同第09/517,466号(2000年3月2日提出)、同第09/695,065号(2000年10月25日提出)および同第09/732,914号(2000年12月11日提出)に記載されているものならびにインビトロジェンコーポレーション(Invitrogen Corporation、Carlsbad、CA)製のGATEWAY(商標)クローニング技術に関連するものが含まれる。
トポイソメラーゼクローニング
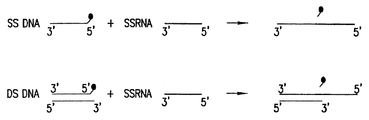
本発明はまた、2つまたはそれ以上のヌクレオチド配列から組換え核酸分子を作製するために、1つまたは複数のトポイソメラーゼを使用する方法に関する。第1の局面において、本発明は、共有結合で一本鎖に接続された二本鎖組換え核酸分子を作製する方法を提供する。このような方法は、一方の鎖が共有結合されるが、両方の鎖は共有結合されないように、第1および少なくとも1つの第2のヌクレオチド配列に少なくとも1つ(例えば、1、2、3、4、5、6、7、8、9、10等)のトポイソメラーゼ(例えば、IA型、IB型および/またはII型トポイソメラーゼ)を接続することに関する(例えば、図11参照)。第2の局面において、本発明は、両方の鎖に共有結合された二本鎖組換え核酸分子を作製する方法を提供する。このような方法は、リガンドの末端が両方の鎖に共有結合される(すなわち、二本鎖組換え核酸分子は、末端がライゲーションされている位置にニックを含有しない。例えば、図5参照)ように、第1および少なくとも1つの第2のヌクレオチド配列に少なくとも1つのトポイソメラーゼを接続することに関する。第3の局面において、本発明は、1つの鎖に共有結合されている組換え核酸分子を作製する方法であって、本発明の方法により接続された基質ヌクレオチド配列は、第2(またはそれ以上)の一本鎖ヌクレオチド配列または核酸分子に共有結合することができる少なくとも1つの一本鎖ヌクレオチド配列を含む方法を提供する(例えば、図15参照)。
1つの鎖に共有結合された二本鎖組換え核酸分子を作製する方法は、第2のヌクレオチド配列を第1のヌクレオチド配列に共有結合させられるように、5'端または3'端に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)またはその切断産物を有する第1の核酸分子に、第2(または他の)核酸分子、および選択的にトポイソメラーゼ(例えば、IA型、IB型および/またはII型トポイソメラーゼ)を接触させることによって実施することができる。本明細書に開示するように、本発明の方法は、任意の数のヌクレオチド配列、典型的には、ヌクレオチド配列の少なくとも1つが一方または両方の5'端に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ)またはその切断産物を有する核酸分子を使用して、実施することができる(例えば、図11A〜図11F参照)。
両方の鎖に共有結合された二本鎖組換え核酸分子を作製する方法は、例えば第1の末端および第2の末端を有する第1の核酸分子(ここで第1の末端もしくは第2の末端または両方の末端において、第1の核酸分子は3'端または3'端付近にトポイソメラーゼ認識部位(またはその切断産物)を有する第1の核酸分子);第1の末端および第2の末端を有する少なくとも1つの第2の核酸分子(ここで、第1の末端もしくは第2の末端または両方の末端において、少なくとも第2の二本鎖ヌクレオチド配列は、3'端または3'端付近にトポイソメラーゼ認識部位(またはその切断産物)を有する);少なくとも1つの部位特異的トポイソメラーゼ(IA型および/またはIB型トポイソメラーゼ)を、全ての成分が接触し、トポイソメラーゼがその活性を発揮できる条件下において接触させることによって実施することができる。本発明のこの局面の方法により作製された共有結合二本鎖組換え核酸は、どちらの鎖の核酸分子が接続されている位置にもニックがないことを一部の特徴とする。一態様において、本発明の方法は、共有結合される対象の2つの末端の3'端または5'端に、トポイソメラーゼ認識部位またはその切断産物をそれぞれ有する、第1の核酸分子と第2(または他の)核酸分子を接触させることによって実施される。別の態様において、本発明の方法は、少なくとも一方の末端の5'端および3'端に、トポイソメラーゼ認識部位またはその切断産物を有する第1の核酸分子と、認識部位を含む第1の核酸分子の末端に接続される末端に3'側ヒドロキシル基および5'側ヒドロキシル基を有する第2(または他の)核酸分子を接触させることによって実施される。本明細書に開示されるように、本発明の方法は、端(termini)および末端(ends)の種々の組み合わせを有する任意の数の核酸分子を使用して実施することができる(例えば、図12A〜12D)。
トポイソメラーゼは、二本鎖核酸分子の一本鎖を切断するI型(IA型およびIB型トポイソメラーゼを含む)および核酸分子の両方の鎖を切断するII型トポイソメラーゼ(ジャイレース)として分類される。IA型およびIB型トポイソメラーゼは1本鎖の核酸を切断する。IA型トポイソメラーゼによる核酸分子の切断により切断部位に5'側リン酸塩および3'側ヒドロキシルが生じ、IA型トポイソメラーゼが切断された鎖の5'端に共有結合する。これと比較すると、IB型トポイソメラーゼによる核酸分子の切断は切断部位に3'側リン酸塩および5'側ヒドロキシルを生じ、IB型トポイソメラーゼは切断された鎖の3'端に共有結合する。本明細書に開示するように、I型およびII型トポイソメラーゼならびにそれらの触媒ドメインおよび突然変異体は、本発明の方法により両方の鎖に共有結合された二本鎖組換え核酸分子を作製するのに有用である。
IA型トポイソメラーゼには、大腸菌(E. coli)トポイソメラーゼI、大腸菌(E. coli)トポイソメラーゼIII、真核細胞トポイソメラーゼII、始生型(archeal)型逆方向のジャイレース、酵母トポイソメラーゼIII、ショウジョウバエトポイソメラーゼIII、ヒトトポイソメラーゼIII、肺炎球菌(Streptococcus pneumoniae)トポイソメラーゼIII、および他のIA型トポイソメラーゼを含むもの等が含まれる(そのそれぞれが参照として本明細書に組み入れられる、Berger、Biochim. Biophys. Acta 1400: 3-18、1988、DiGateおよびMarians、J. Biol. Chem. 264: 17924-17930、1989、KimおよびWang、J. Biol. Chem. 267: 17178-17185、1992、Wilsonら、J. Biol. Chem. 275: 1533-1540、2000、Hanaiら、Proc. Natl. Acad. Sci., USA 93: 3653-3657、1996、米国特許第6,277,620号参照)。配列5'-GCAACTT-3'を認識し、結合し、切断するIA型トポイソメラーゼである大腸菌(E. coli)トポイソメラーゼIIIは、本発明の方法に特に有用となりうる(参照として本明細書に組み入れられる、Zhangら、J. Biol. Chem. 270: 23700-23705、1995)。プラスミドRP4のtraEタンパク質である相同物は、リ(Li)ら、J. Biol. Chem. 272: 19582-19587(1997)に記載されており、本発明を実施する際にも使用することができる。DNA-タンパク質付加物は、酵素が5'側-チミジン残基に共有結合し、2つのチミジン残基の間を切断して形成される。
IB型トポイソメラーゼには、全ての真核細胞に存在する核のI型トポイソメラーゼならびにワクシニアウイルスおよび他の細胞ポックスウイルスによってコードされるものが含まれる(参照として本明細書に組み入れられる、Chengら、Cell 92: 841-850、1998)。真核細胞IB型トポイソメラーゼは、酵母、ショウジョウバエおよびヒト細胞を含む哺乳類細胞において発現されるものによって例示される(そのそれぞれが参照として本明細書に組み入れられる、CaronおよびWang、Adv. Pharmacol. 29B: 271-297、1994、Guptaら、Biochim. Biophys. Acta 1262: 1-14、1995参照、Berger上記、1998年も参照)。ウイルスのIB型トポイソメラーゼは脊椎動物ポックスウイルス(ワクシニア、ショープ線維腫ウイルス、伝染性膿疱性皮膚炎ウイルスおよび伝染性軟肬腫ウイルス)および昆虫ポックスウイルス(Amsacta moorei entomopoxvirus)によって産生されるものによって例示される(そのそれぞれが参照として本明細書に組み入れられる、Shuman、Biochim. Biophys. Acta 1400: 321-337、1998、Petersenら、Virology 230: 197-206、1997、ShumanおよびPrescott、Proc. Natl. Acad. Sci., USA 84: 7478-7482、1987、Shuman、J. Biol. Chem. 269: 32678-32684、1994、米国特許第5,766,891号、PCT/US95/16099、PCT/US98/12372参照、またChengら、上記、1998年も参照)。
II型トポイソメラーゼには、例えば細菌ジャイレース、細菌DNAトポイソメラーゼIV、真核細胞DNAトポイソメラーゼIIおよびT偶数系ファージがコードするDNAトポイソメラーゼが含まれる(そのそれぞれが参照として本明細書に組み入れられる、RocaおよびWang、Cell 71:833-840、1992、Wang、J.Biol.Chem. 266: 6659-6662、1991、Berger、上記、1998年参照)。IB型トポイソメラーゼと同様に、II型トポイソメラーゼは切断作用およびライゲーション作用の両方を有する。また、IB型トポイソメラーゼと同様に、基質核酸分子は、II型トポイソメラーゼが一本鎖の切断部位と共有結合を形成することができるように作製することができる。例えば、仔ウシ胸腺II型トポイソメラーゼは、5'末端から3ヌクレオチドの位置の陥凹した5'側トポイソメラーゼ認識部位を含む基質核酸分子を切断し、切断部位の5'側の3つのヌクレオチド配列を解離し、核酸分子の5'端にトポイソメラーゼを共有結合することができる(Andersonら、上記、1991年)。さらに、このようなII型トポイソメラーゼが結合した核酸分子に、3'側ヒドロキシル基を有する第2のヌクレオチド配列を接触させると、II型トポイソメラーゼは配列をライゲーションすることができ、次いで組換え核酸分子から放出される。このように、II型トポイソメラーゼも本発明の方法を実施するのに有用である。
IA型、IB型およびII型トポイソメラーゼを含む特定のトポイソメラーゼファミリーは、それぞれファミリーの他のメンバーと共通の構造的特徴を共有することをトポイソメラーゼの構造分析は示している(Berger、上記、1998年)。また、構造は、特に触媒ドメインでは保存性が高いことを種々のIB型トポイソメラーゼの配列分析は示している(Shuman、上記、1998年、Chengら、上記、1998年、Petersonら、上記、1997)。例えば、314個のアミノ酸のワクシニアトポイソメラーゼの81位〜314位のアミノ酸を含むドメインは、他のIB型トポイソメラーゼと実質的な相同性を共有しており、単離されたドメインは全長のトポイソメラーゼと本質的に同じ活性を有するが、単離されたドメインは代謝回転速度が遅く、認識部位への結合親和性が低い(Shuman、上記、1998年、Chengら、上記、1998年)。また、アミノ末端ドメイン(アミノ酸残基70位〜72位)が突然変異されている突然変異体のワクシニアトポイソメラーゼは、全長のトポイソメラーゼと同一の特性を示す(Chengら、上記、1998年)。実際、ワクシニアIB型トポイソメラーゼの突然変異分析は、トポイソメラーゼの活性に影響を与えることなく、突然変異させることができる多数のアミノ酸残基を明らかにしており、活性に必要ないくつかのアミノ酸を同定している(Shuman、上記、1998年)。ワクシニアトポイソメラーゼ触媒ドメインおよび他のIB型トポイソメラーゼ間に共有される高い相同性ならびにワクシニアトポイソメラーゼの詳細な突然変異分析を考慮すると、IB型トポイソメラーゼおよび種々のアミノ酸突然変異を有するIB型トポイソメラーゼの単離された触媒ドメインは、本発明の方法に使用できると認められるであろう。
種々のトポイソメラーゼはある範囲の配列特異性を示す。例えば、II型トポイソメラーゼは種々の配列に結合することができるが、特異性の高い認識部位を切断する(参照として本明細書に組み入れられる、Andersenら、J.Biol.Chem.266: 9203-9210、1991)。これと比較すると、IB型トポイソメラーゼには、特異的なヌクレオチド配列に結合して切断する部位特異的トポイソメラーゼが含まれる(「トポイソメラーゼ認識部位」)。例えば、IB型トポイソメラーゼのようなトポイソメラーゼによる核酸分子の切断の結果、ホスホジエステル結合のエネルギーは、トポイソメラーゼの特定のチロシン残基とトポイソメラーゼ認識部位の3'側ヌクレオチド間にホスホチロシル結合が形成されることにより保存される。トポイソメラーゼ切断部位が核酸分子の3'端に近い場合には、下流の配列(切断部位の3'側)が解離して、新たに形成された3'側末端にトポイソメラーゼが共有結合した核酸分子が生じる(図29参照)。
1つの鎖に共有結合した二本鎖組換え核酸分子を作製する本発明の方法は、1) 第1の末端および第2の末端を有する第1の核酸分子であって、第1もしくは第2の末端または両方の5'端または5'端付近に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)を有し、選択的に、1つまたは複数の組換え部位を含む、第1の核酸分子;2)第1の末端および第2の末端を有するか、または第1の末端および第2の末端を有するように作製することができる、少なくとも1つの第2の核酸分子;3)少なくとも1つの(例えば、1、2、3、4、5、6、7、8、9、10等)部位特異的トポイソメラーゼ(例えば、IA型またはIB型トポイソメラーゼ認識部位)を、全ての成分が接触し、少なくとも1つのトポイソメラーゼがその活性を発揮できる条件下において接触させることによって実施することができる。例えば、トポイソメラーゼは、大腸菌(E. coli)トポイソメラーゼIなどのIA型トポイソメラーゼ、大腸菌(E. coli)トポイソメラーゼIIIまたは真核細胞トポイソメラーゼIIIであってもよい。核酸分子が切断されると、トポイソメラーゼは好ましくは、5'端に安定に結合される。トポイソメラーゼによる切断の結果、切断された核酸分子は3'側突出配列を含むことが多くある。核酸分子が上記の方法により接続されると、得られた分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。
1つの鎖に共有結合された二本鎖組換え核酸分子を作製する本発明の方法は、任意の組み合わせの末端が接続され、接続される末端の1つの鎖が共有結合され、他方の鎖は共有結合されないが、ニックを有するように実施することができる。例えば、第1の核酸分子は、ATG開始コドンが第1の末端または第1の末端付近にあり、ポリAシグナルが第2の末端または第2の末端付近にコードされるコード配列を含むことができ、第2の核酸分子は、コード配列の上流に位置づけられると機能し、第1の末端が第2の末端の上流にあるプロモーター要素を含むことができ、部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)が第1の核酸分子の第1の末端の5'端またはその付近にあり、トポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ)が第1の核酸分子の第1の末端の5'端を第2の核酸分子の第1の核酸分子の3'端に共有結合して二本鎖組換え核酸分子を作製し、ポリペプチドをコード配列から発現することができる条件下で、接触を実施する方法を実施することができる。または、トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)が第1の核酸分子の第2の末端の5'端またはその付近にあり、トポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ認識部位)が第1の核酸分子の第2の末端の5'端を第2の核酸分子の第1の末端の3'端に共有結合して、アンチセンス分子を発現させることができる二本鎖組換え核酸分子を作製する条件下で接触を実施する方法を実施することができる。核酸分子が上記の方法によって接続されたら、得られた分子を本明細書の別の箇所に記載するような組換え反応に使用することができる。
上記の第1の核酸分子および第2の核酸分子を使用する別の例として、トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)が第1の核酸分子の第1の末端および第2の末端の5'端または5'端付近にあり、IA型トポイソメラーゼが、第1の核酸分子の第1の末端の5'端を第2の核酸分子の第1の末端の3'端に共有結合させ、第1の核酸分子の第2の末端の5'側を第2の核酸分子の第2の末端の3'端に接続させる条件下において、接触を実施する方法を実施することができる。このように、本発明の方法によって作製される二本鎖組換え核酸分子は環状化され、鎖がトポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ)によって共有結合された位置の反対側の各鎖にニックを含む。さらに、第2の核酸分子のプロモーターが第1の核酸分子の発現を開始することができる。一態様において、環状化された二本鎖組換え核酸分子はベクターを含む。核酸分子が上記の方法によって接続されたら、得られた分子を本明細書の別の箇所に記載するような組換え反応に使用することができる。
上記の第1の核酸分子および第2の核酸分子を使用する別の例として、トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)が第1の核酸分子の第1の末端および第2の末端の5'端または5'端付近にあり、トポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ)が、第1の核酸分子の第1の末端の5'端を第2の核酸分子の第1の末端の3'端に共有結合させ、第1の核酸分子の第2の末端の5'側を第2の核酸分子の第1の末端の3'端に接続させる条件下において接触を実施する方法を実施することができる。このように、本発明の方法によって作製される二本鎖組換え核酸分子は環状化され、鎖がトポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ認識部位)によって共有結合された位置の反対側の各鎖にニックを含む。さらに、第2の核酸分子のプロモーターがアンチセンス配列の発現を開始することができる。一態様において、環状化された二本鎖組換え核酸分子はベクターを含む。核酸分子が上記の方法によって接続されたら、得られた分子を本明細書の別の箇所に記載するような組換え反応に使用することができる。
本明細書に開示するように、第1の核酸分子および少なくとも1つの第2の核酸分子に関与する、1つの鎖に共有結合された二本鎖組換え核酸分子を作製する方法はさらに、1つの鎖に共有結合された二本鎖組換え核酸分子を増幅する段階を含む。増幅反応は、二本鎖組換え核酸分子に増幅反応プライマー対を接触させることによって実施することができ、対の第1のプライマーは共有結合されている鎖の第1または第2の核酸分子の一方の末端または末端付近に結合し、他方の核酸分子に対して増幅反応を開始させて、ヌクレオチド配列が二本鎖組換え核酸分子のニックの入った鎖と同一の第1の伸長産物を作製することができ、対の第2のプライマーは、第1のプライマーの存在下において第1の伸長産物の、典型的には3'端または3'端付近に結合することができ、共有結合された鎖および伸長産物(またはそれから作製される複数の伸長産物)を鋳型として使用して増幅産物を作製することができる。例えば、本発明の方法は、IA型トポイソメラーゼ認識部位が第1の核酸分子の第1の末端または末端付近にあるように実施することができ、本発明の方法は、さらに二本鎖組換え核酸分子に増幅反応プライマー対を接触させる段階であって、順方向プライマーは第1の核酸分子の第2の末端または末端付近に結合することができ、逆方向プライマーは第2の核酸分子の第2の末端の少なくとも一部に相補的なヌクレオチド配列に結合することができる段階、および二本鎖組換え核酸分子を増幅する段階を含む。第1の核酸分子はコード領域を含んでもよく、第2の核酸分子は調節要素を含んでもよい。核酸分子が上記の方法によって接続されたら、得られた分子を本明細書の別の箇所に記載するような組換え反応に使用することができる。
1つの鎖に共有結合した二本鎖組換え核酸分子を作製する本発明の方法は、1) 第1の末端および第2の末端を有し、第1の核酸分子が第1の末端もしくは第2の末端または両方の5'端または5'端付近に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)を有する第1の核酸分子、2)第1の末端および第2の末端を有する、または第1の末端および第2の末端を有するように作製することができる少なくとも1つの第2の核酸分子、3)末端がそれぞれ5'端および3'端をさらに含む第1の末端および第2の末端を有する、または第1の末端および第2の末端を有するように作製することができる少なくとも1つの第3の核酸分子、ならびに4)少なくとも1つの(例えば、1、2、3、4、5、6、7、8、9、10等)部位特異的トポイソメラーゼ(例えば、IA型またはIB型トポイソメラーゼ認識部位)を、全ての成分が接触し、少なくとも1つのトポイソメラーゼがその活性を発揮できる条件下において接触させることによって実施することができる。例えば、トポイソメラーゼは、大腸菌(E. coli)トポイソメラーゼIなどのIA型トポイソメラーゼ、大腸菌(E. coli)トポイソメラーゼIIIまたは真核細胞トポイソメラーゼIIIであってもよい。核酸分子が切断されると、トポイソメラーゼは、好ましくは、5'端に安定に結合される。好ましくは、トポイソメラーゼによる切断の結果、切断された核酸分子は3'側突出配列を含む。核酸分子が上記の方法により接続されると、得られた分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。
部位特異的トポイソメラーゼ認識部位(例えば、IA型またはIB型トポイソメラーゼ認識部位)またはその切断産物を有する第1の核酸分子、少なくとも1つの第2の核酸分子および少なくとも1つの第3の核酸分子に関与する、1つの鎖に共有結合された二本鎖組換え核酸分子を作製する本発明の方法は、任意の組み合わせの末端が接続され、接続される末端にある1つの鎖が共有結合され、1つの鎖にはニックが入るように実施することができる。この態様によると、末端のいずれかはIA型、II型またはIB型トポイソメラーゼ認識部位を有してもよいか、またはその切断部位を含んでもよいが、ただし、第1の二本鎖組換えヌクレオチド分子はトポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)またはその切断産物を5'端または5'端付近に有し、ただ1つのトポイソメラーゼまたはトポイソメラーゼ認識部位が、接続される末端に存在する。例えば、第1の核酸分子が第1および第2の末端または第1および第2の各末端付近に部位特異的IA型トポイソメラーゼ認識部位を含む場合には、本発明の方法は、第1の核酸分子および第2の核酸分子に、各末端が5'端および3'端をさらに含む第1の末端および第2の末端を有するまたは第1の末端および第2の末端を有するように作製することができる、少なくとも1つの第3の核酸分子を、トポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ)が第1の核酸分子の第1の末端の5'端に第2のヌクレオチド配列の第1の末端の3'端を共有結合させ、第1の核酸分子の第2の末端の5'端に第3のヌクレオチド配列の第1の末端の3'端を共有結合することができるような条件下において、接触させる段階をさらに含んでもよい。本発明のこのような方法を実施するために、末端およびトポイソメラーゼ認識部位またはそれらの切断産物の他の組み合わせを使用できることが認識される。核酸分子が上記の方法によって接続されると、得られた分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。
本発明の方法はまた、第1の核酸分子および第2の核酸分子に、各末端が5'端および3'端をさらに含む第1の末端および第2の末端を含み、第1の末端もしくは第2の末端、または第1の末端および第2の末端の両方またはその付近にIB型トポイソメラーゼ認識部位を含む、少なくとも1つ第3の核酸分子と少なくとも1つの(例えば、1、2、3、4、5、6、7、8、9、10等)IB型トポイソメラーゼを、IB型トポイソメラーゼが第3の核酸分子の第1の末端または第2の末端の3'端を、第2の核酸分子の第1の末端または第2の末端の5'端に共有結合させられる条件下において接触させることによって実施することができる。このような方法では、第3の核酸分子が第1の末端の3'端または3'端付近にIB型トポイソメラーゼ認識部位を含む場合には、接触させる段階は、IB型トポイソメラーゼが第3の核酸分子の第1の末端の3'端を第2の核酸分子の第1の末端の5'端に共有結合させることができる条件下において、実施することができる。本発明のこのような方法を実施するために、末端およびトポイソメラーゼ認識部位またはそれらの切断産物の他の組み合わせを使用できることが認識される。核酸分子が上記の方法によって接続されると、得られた分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。
別の態様において、1つの鎖に共有結合した二本鎖組換え核酸分子を作製する本発明の方法は、1) 第1の末端および第2の末端を有し、第1の核酸分子が末端の5'端または5'端付近に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)、および他方の末端の3'端または3'側付近にIB型トポイソメラーゼ認識部位を有する、第1の核酸分子、2)第1の末端および第2の末端を有する、または第1の末端および第2の末端を有するように作製することができる、少なくとも1つの第2の核酸分子、3)少なくとも1つの(例えば、1、2、3、4、5、6、7、8、9、10等)部位特異的トポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ認識部位)、ならびに4)少なくとも1つの(例えば、1、2、3、4、5、6、7、8、9、10等)IB型トポイソメラーゼを、全ての成分が接触し、少なくとも1つのトポイソメラーゼがその活性を発揮できる条件下において、接触させることによって実施することができる。例えば、認識部位が5'端または5'端付近にあるトポイソメラーゼは、大腸菌(E. coli)トポイソメラーゼIなどのIA型トポイソメラーゼ、大腸菌(E. coli)トポイソメラーゼIIIまたは真核細胞トポイソメラーゼIIIであってもよい。核酸分子が切断されると、IA型トポイソメラーゼは、好ましくは、5'端に安定に結合され、IB型トポイソメラーゼは、好ましくは、3'端に安定に結合される。好ましくは、トポイソメラーゼによる切断の結果、切断された核酸分子は3'側突出配列および5'側突出配列を含む。本発明の方法は、二本鎖組換え核酸分子にDNAリガーゼを接触させ、それによって両方の鎖に共有結合した二本鎖組換え核酸分子を作製する段階をさらに含んでもよい。核酸分子が上記の方法により接続されると、得られた分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。
第1の核酸分子、第2の核酸分子および少なくとも1つの第3の核酸分子を接触させることにより、1つの鎖に共有結合した二本鎖組換え核酸分子を作製する方法は、二本鎖組換え核酸分子、好ましくは共有結合された鎖を増幅する段階をさらに含んでもよい。増幅反応は、二本鎖組換え核酸分子に増幅反応プライマー対を接触させることによって実施することができ、対の第1のプライマーは共有結合されている鎖の第1または第2の核酸分子の一方の末端または末端付近に結合し、他方の核酸分子に対して増幅反応を開始させて、共有結合された鎖と相補的な第1の伸長産物を作製することができ、対の第2のプライマーは、第1のプライマーの存在下において第1の伸長産物の、典型的には3'端または3'端付近に選択的に結合することができ、共有結合された鎖および伸長産物(またはそれから作製される複数の伸長産物)を鋳型として使用して増幅産物を作製することができる。本発明の方法は、トポイソメラーゼ認識部位(例えば、IA型またはIB型トポイソメラーゼ認識部位)が第1の核酸分子の第1の末端または末端付近にあるように実施することができ、本発明の方法はさらに、二本鎖組換え核酸分子に増幅反応プライマー対を接触させる段階であって、順方向プライマーは第1の核酸分子の第2の末端または末端付近に結合することができ、逆方向プライマーは第3の核酸分子の少なくとも一部に相補的なヌクレオチド配列に結合することができる段階、および二本鎖組換え核酸分子を増幅する段階を含む。第1の核酸分子はコード領域を含んでもよく、第3の核酸分子は調節要素を含んでもよい。さらに、接続される末端は相補的な突出配列を有してもよい。核酸分子が上記の方法によって接続されたら、得られた分子を本明細書の別の箇所に記載するような組換え反応に使用することができる。
1つの鎖に共有結合され、選択的に、1つまたは複数の認識部位を含む二本鎖組換え核酸分子を作製する、開示された方法の代表的な態様は、図11A〜図11Fに例示されている。図11Aにおいて、核酸分子の1つには一方の末端の5'端にトポイソメラーゼが接続されており、3'側突出を有するこの分子に、実質的に相補的な3'側突出を有する第2の核酸分子を適切な条件下において接触させると、3'側突出を含むヌクレオチドはハイブリダイゼーションすることができ、トポイソメラーゼはライゲーションを触媒することができる。図11Bは、一方のヌクレオチド配列の2つの異なる末端の5'端および3'端にトポイソメラーゼ分子が接続した第1の核酸分子を示しさらに、第1の核酸分子が他の2つのヌクレオチド配列に接続して、ニックの全くない1つの鎖とニックが2つある別の鎖を有する核酸分子を作製することを示す。図11Cは、一方の末端の5'端にトポイソメラーゼ分子が接続した第1の核酸分子、および一方の末端の5'端にトポイソメラーゼ分子が接続した第2の核酸分子を示しさらに、第1および第2の核酸分子が他の1つのヌクレオチド配列に接続して、ニックのない1つの鎖とニックが2つある別の鎖を有する核酸分子を作製することを示す。図11Dにおいて、接続される核酸分子の1つは、両方の末端の5'端に部位特異的IA型トポイソメラーゼが接続されており、ヌクレオチド配列を接触させると、相補的な3'側突出はハイブリダイゼーションすることができ、トポイソメラーゼはライゲーションを触媒する。図11Eは、5'端にIA型トポイソメラーゼをトポイソメラーゼ付加した1つの核酸分子および接続される反対側の鎖の3'側にIB型トポイソメラーゼをトポイソメラーゼ付加した別の核酸分子を使用し、ヌクレオチド配列を接触させると、相補的な3'側突出がハイブリダイゼーションすることができ、トポイソメラーゼがライゲーションを触媒する、3つの核酸分子を接続する別の例を示す。図11Fは、この場合には、5'端にトポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ)および反対側の鎖の3'側にIB型トポイソメラーゼをトポイソメラーゼ付加した1つの核酸分子を使用し、ヌクレオチド配列を適切な条件下において接触させると、相補的な3'側突出がハイブリダイゼーションすることができ、トポイソメラーゼがライゲーションを触媒する、3つの核酸分子を接続する別の例を例示する。核酸分子が上記の方法によって接続されたら、得られた分子を本明細書の別の箇所に記載するような組換え反応に使用することができる。
図11A〜図11Fに記載された例は、平滑末端を有するように接続されているものの反対側の核酸分子の末端を示し、3'側突出配列を有するように接続されていることを示す。しかしながら、基質の核酸分子は、末端が例えば、互いにライゲーションされて環状分子を形成することができるように、または適当な末端を有する他の核酸分子にライゲーションできるように、平滑および/または相補的である末端またはそれらの組み合わせを含む任意の末端および突出を適宜有してもよい。従って、図11A〜図11Fに示す平滑末端の1つまたは複数が、どちらかがチミジン残基などの単一ヌクレオチドを構成するか、または同じもしくは異なってもよい、多数のヌクレオチド(例えば、2、3、4、5、6、7、8、9、10、11、12、13、14ヌクレオチド等)を構成することができる、5'側突出または3'側突出を含むヌクレオチド配列で置換されていてもよい。開示されている方法のある種の態様において、第1の核酸分子は接続される平滑末端を有し、第2の核酸分子は、部位特異的トポイソメラーゼ(例えば、IA型またはIB型トポイソメラーゼ)により接続される末端に、平滑末端を含むものに対して相補的な配列を含む突出を含み、それによって接続反応の末端を適切に位置づける手段として鎖侵入を促進する。
図11A〜図11Cに例示するように、本発明のこの局面の方法を使用して作製される二本鎖組換え核酸分子は、接続される末端に1つの鎖(両方の鎖ではない)が共有結合されているものを含む(すなわち、これらの方法を使用して作製される二本鎖組換え核酸分子は、2つの末端が接続されている各位置にニックを有する)。ポリメラーゼを使用して、共有結合されている鎖を最初に複製することによって二本鎖組換え核酸分子を複製することができるという点において、これらの態様は特に有利である。例えば、PCRなどの増幅反応を実施するのに有用なポリメラーゼなどの熱安定なポリメラーゼは、共有結合された鎖を複製するために使用することができるが、ニックを有する鎖は複製の適切な鋳型にならない。
本発明はまた、作製された産物が両方の鎖にライゲーションされており、従ってニックを有しないように、2つの異なる核酸分子の末端または同一核酸分子の2つの末端を、共有結合によりライゲーションする方法を提供する。本発明のこの局面の代表的な態様を図12に例示する。例えば、図12Aにおいて、核酸分子の一方には、その3'端および5'端にトポイソメラーゼ分子が接続されており、5'側突出を有するこの分子に、適切な条件下において、実質的に相補的な5'側突出を有する第2の核酸分子を接触させると、5'側突出を含むヌクレオチドはハイブリダイゼーションすることができ、トポイソメラーゼは核酸分子の両方のライゲーションを触媒することができる。図12Bにおいて、接続される核酸分子の各末端には3'端にトポイソメラーゼ分子が接続されており、ヌクレオチド配列を適切な条件下において接触させると、5'側突出を含むヌクレオチドがハイブリダイゼーションすることができ、トポイソメラーゼはライゲーションを触媒する(接続される各核酸分子が、接続される末端の5'端にトポイソメラーゼが接続されている、図12Cと比較すること)。図12Dは、両方の末端の両側の末端にトポイソメラーゼが負荷されている核酸分子を介して、3つの核酸分子を接続することを例示している。図11と同様に、図12A〜12Dに記載する例は、平滑末端を有するとき接続されない核酸分子の末端を示す。しかし、図11に関して考察されているように、図12に例示する方法に使用される基質核酸分子は、末端が互いにライゲーションして、例えば環状分子を形成することができるように、または適当な末端、平滑末端、5'側突出、3'側突出等を適宜有する他の核酸分子にライゲーションすることができるように、トポイソメラーゼ付加末端を含む任意の末端を適宜有してもよい。核酸分子が上記の方法によって接続されたら、得られた分子を本明細書の別の箇所に記載するような組換え反応に使用することができる。
共有結合されたトポイソメラーゼは、触媒することができるライゲーション反応を触媒する以外に、逆反応、例えばIB型トポイソメラーゼがホスホチロシル結合により接続される認識配列の3'側ヌクレオチドと、切断前は核酸分子の5'端を含み、切断後に遊離5'側ヒドロキシ基を有するヌクレオチド配列との再連結を触媒することもできる。このように、組み合わせ核酸分子を作製するためにIB型トポイソメラーゼを使用する方法が開発されている。例えば、結合されたIB型トポイソメラーゼを有するクローニングベクターが開発されており、市販品を購入することができる(Invitrogen Corp.、カリフォルニア州ラジョラ)。このようなクローニングは、直鎖状である場合には、各3'端に共有結合されたIB型トポイソメラーゼを有する(「トポイソメラーゼ付加」)。このようなベクターにクローニングされるcDNAライブラリ、または制限断片または切断されたゲノムDNA配列を含むものなどのヌクレオチド配列を、例えばホスファターゼで処理して、5'側ヒドロキシ末端を作製し、次いでトポイソメラーゼが、ヌクレオチド配列のヒドロキシル基を有する5'端と、共有結合したトポイソメラーゼを有するベクターの3'端をライゲーションすることができる条件下において、直鎖状トポイソメラーゼ付加ベクターに付加される。5'端を含む、作製されたPCR増幅産物などのヌクレオチド配列は、迅速な接続反応(室温において約5分)でトポイソメラーゼ付加ベクターにクローニングすることができる。トポイソメラーゼ接続反応に固有の、迅速な接続および広い温度範囲により、トポイソメラーゼ付加ベクターを使用することは、一般に自動システムを使用して実施されるハイスループット用途にとって理想的である。
II型トポイソメラーゼは、一般に、組み合わせ核酸分子を作製するため、またはクローニング手法に使用されていないが、上記のIB型トポイソメラーゼは種々の手法に使用される。本明細書に開示するように、IA型トポイソメラーゼは、IB型トポイソメラーゼについて記載されているものと同様の種々の手法に使用することができる。しかし、2つまたはそれ以上のヌクレオチド配列を接続するためにIB型トポイソメラーゼを使用する、以前に記載されている方法には、接続されたトポイソメラーゼは、接続されている鎖の3'端と、5'側ヒドロキシル基を有する第2の鎖との接続しか行わないという欠点がある。トポイソメラーゼは相補鎖を接続することができないので、作製される核酸分子はニックを有する。ニックは一般に細胞内で分解されるので、このようなニックの存在は、宿主細胞のトランスフェクションのための組換え分子の使用を妨げないが、二本鎖核酸分子にこのようなニックが存在すると組換え分子の直接使用の大きな制約となる。例えば、プライマー伸長反応はニックのところで停止するので、ニックを有する核酸分子鎖はPCRで増幅することができない。従って、一般に、両方の鎖に共有結合され、PCRなどのその後の操作に有用である二本鎖組換え核酸分子を得るためには、以前に記載されている方法によりトポイソメラーゼを使用して作製される核酸構築物は、例えばDNAリガーゼで処理しなければならない。
以前に記載された、核酸構築物を作製するための方法もまた一般に、特に3つ以上のヌクレオチド配列を接続する場合には数多くの段階を必要とし、配列を所定の方向にライゲーションしなければならない場合にはさらに数多くの段階を必要とした。例えば、一般に、接続されるヌクレオチド配列を逐次的にライゲーションして中間構築物を作製し、そのそれぞれをクローニングし、宿主細胞内で増幅し、単離し、特徴づけなければならない。次のヌクレオチド配列をライゲーションできるように、適切な配列を有する十分な量および形態の構築物を単離しなければならず、適切な構築物を同定するために、クローニング、増幅、単離および特徴づけ過程を再度実施しなければならない。明らかに、接続される異なるヌクレオチド配列の数が増加すると、実施しなければならない本質的に反復的な手法の数も増加し、それにより高価で手間を要し、時間のかかる過程となる。
本明細書に開示するように、両方の鎖に共有結合された二本鎖組換え核酸分子を作製するための本発明の方法の利点は、両方の鎖に共有結合された機能的な二本鎖組換え核酸分子を得るために、別個のライゲーション反応を実施する必要がないことである(図8および12参照)。また、数多くの異なる核酸分子を所定の方向に共有結合する場合には、その後の段階に進む前に、中間構築物をクローニング、特徴づけおよび単離する必要がないように、本発明のこの局面の方法を実施することができる(実施例1.B参照)。このように、本発明のこの局面の方法は、以前の既知の方法を使用したとき考えられるよりはるかに迅速で、実質的に低価格で、両方の鎖に共有結合した二本鎖組換え核酸分子を作製する手段を提供する。
追加の利点として、作製される構築物は、二本鎖ヌクレオチド配列が接続されている部位にニックがないので、作製後の両方の鎖に共有結合された二本鎖組換え核酸分子は、さらなる手法、例えばPCR増幅手法などの伸長もしくはプライマーに関与する特定の手法または他の転写もしくは翻訳手法に直接使用できるということである。本明細書に開示するように、1つの鎖に共有結合された二本鎖組換え核酸分子を作製するための本発明の方法は、ある種の態様でも、作製後の二本鎖組換え核酸分子は、さらなる手法、例えばPCR増幅手法などの伸長もしくはプライマーに関与する特定の手法または他の転写もしくは翻訳手法に直接使用できる形態であるという点で有利である。その理由は、ある種の態様では、作製後の二本鎖組換え核酸分子は、二本鎖ヌクレオチド配列が接続された部位にニックがない一本鎖を有するからである。
「ヌクレオチド配列」または「核酸分子」という用語は、本明細書において、別個の核酸分子をいうために使用される。このように使用される場合には、「ヌクレオチド配列」という用語は、組成物中の成分または本発明の方法に使用される成分を明確に識別できるように、単に便宜上使用される。従って、例えば本発明の方法では、組換え「核酸分子」産物を作製するために使用される反応体(基質)に対応する「核酸分子」に言及される。
本発明のある種の方法は、一般に、本明細書において、ワクシニアトポイソメラーゼなどのIB型トポイソメラーゼ、またはIA型トポイソメラーゼの使用に言及して例示されている。しかし、本発明の方法は、単に成分をそれなりに調整することによって、例示されているもの以外のトポイソメラーゼを使用しても実施することができることが認識されている。例えば、以下にさらに詳細に記載されているように、トポイソメラーゼ認識部位に相補的なヌクレオチド配列を少なくとも一部を含むPCRプライマーを使用して、直鎖状核酸分子の一方または両方の3'端にIB型トポイソメラーゼ認識部位を組入れる方法が開示されている。これと比較すると、IA型、または望ましい場合には、II型トポイソメラーゼのトポイソメラーゼ認識部位は、認識部位を有するPCRプライマーを使用して核酸分子に組入れることができる。
部位特異的IB型トポイソメラーゼにより核酸分子を切断すると、共有結合したトポイソメラーゼを有するものに相補的な5'側突出配列が、同じ末端に形成される。さらに、本明細書に開示されるように、IB型トポイソメラーゼ認識部位を核酸分子に組入れることができ、トポイソメラーゼによる核酸分子の切断の結果、規定された所定の配列を有する5'側突出配列を、相補鎖内に作製することができるPCRプライマーを設計することができる。このように、本発明の方法は、成分である核酸分子が所定の方向に機能的に接続された、二本鎖組換え核酸分子を作製する方法に容易に適合させることができる。本発明の開示を考慮すると、IA型トポイソメラーゼ認識部位を、異なる配列ライブラリを含む核酸分子に組入れることができるように、また望ましい場合には、部位特異的トポイソメラーゼによる切断の結果、3'側突出配列を作製するように、PCRプライマーを設計することもできることが認識される。
本明細書に開示するように、両方の鎖に共有結合した二本鎖組換え核酸分子を作製する方法は、共有結合される各核酸分子の末端または末端付近にトポイソメラーゼを提供することによって、以前から既知の方法を拡張している。例えば、IB型トポイソメラーゼに関しては、本発明の方法は、接続される各直鎖状核酸分子の3'側または3'側付近にトポイソメラーゼ認識部位またはその切断産物(すなわち、共有結合したIB型トポイソメラーゼ)を提供する。本明細書において使用する「トポイソメラーゼ認識部位」という用語は、部位特異的トポイソメラーゼによって認識され、結合される規定されたヌクレオチド配列を意味する。例えば、ヌクレオチド配列5’-(C/T)CCTT-3’は、ワクシニアウイルスDNAトポイソメラーゼを含むほとんどのポックスウイルストポイソメラーゼによって特異的に結合され、認識部位の最も3'端のチミジンの下流の鎖を切断して、5’-(C/T)CCTT-PO4-TOPOを含むヌクレオチド配列、すなわち、トポイソメラーゼのチロシン残基を介して3'側リン酸塩に共有結合したトポイソメラーゼの複合体を作製することができるトポイソメラーゼ認識部位である(それぞれが参照として本明細書に組み入れられる、Shuman、J.Biol.Chem. 226: 11372-11379、1991、SekiguchiおよびShuman、Nucl. Acids Res. 22: 5360-5365、1994参照、また米国特許第5,766,891号、PCT/US95/16099号、PCT/US98/12372号参照)。これと比較すると、ヌクレオチド配列5'-GCAACTT-3'は、IA型大腸菌(E. coli)トポイソメラーゼIIIの認識部位である。
核酸分子の一方または両方の末端の5'端もしくは3'端または両方に共有結合したトポイソメラーゼを有するものを含むトポイソメラーゼ付加核酸分子は、数多くの方法のいずれかにより作製することができる。いくつかの場合に適当な条件下において、I型トポイソメラーゼは一本鎖ヌクレオチド配列を切断することができる。例えば、IA型トポイソメラーゼである、大腸菌(E. coli)トポイソメラーゼIのアミノ末端67kDaドメインを含むドメインは、トポイソメラーゼ認識部位を有する一本鎖ヌクレオチド配列を切断することができる。条件が、トポイソメラーゼが一本鎖ヌクレオチド配列を切断することができるようである場合には、核酸分子の一方の末端の5'端および3'端にトポイソメラーゼ認識部位を有する核酸分子の切断は平行に実施することができる。または、トポイソメラーゼの一方または両方が認識および切断のための核酸分子を必要とする場合には、反応は逐次的に実施され、トポイソメラーゼ認識部位のより末端側(遠位側)が最初に切断され、次いで二本鎖の状態でいる内側(近位)部位が切断される。例えば、一方の末端の5'端または5'側付近に大腸菌(E. coli)トポイソメラーゼIII認識部位を有し、同じ末端の3'端または3'端付近にワクシニアIB型トポイソメラーゼ認識部位を含有し、IB型認識部位がIA型認識部位より末端近くにある核酸分子をワクシニアトポイソメラーゼとインキュベーションして、IB型トポイソメラーゼ付加核酸分子を作製し、次いで大腸菌(E. coli)トポイソメラーゼとインキュベーションして、5'端にIA型トポイソメラーゼが結合し、3'端にIB型トポイソメラーゼが結合した核酸分子を作製することができる。従って、本発明は、少なくとも一方の末端の一方または両方の末端に接続されたトポイソメラーゼを含む核酸分子を作製する方法を含み、さらにこのようなトポイソメラーゼ付加核酸分子を提供する。
本明細書において使用する「切断産物」という用語は、トポイソメラーゼ認識部位に言及して使用する場合には、一般にトポイソメラーゼ認識部位においてトポイソメラーゼにより切断されており、IA型もしくはII型トポイソメラーゼの場合にはトポイソメラーゼ認識部位の5'端ヌクレオチドの5'側リン酸基に共有結合されたトポイソメラーゼの複合体、またはIB型トポイソメラーゼの場合には、トポイソメラーゼ認識部位の3'端ヌクレオチドの3'側基に共有結合されたトポイソメラーゼの複合体を含むヌクレオチド配列をいう。トポイソメラーゼが共有結合されたトポイソメラーゼ切断型核酸分子を含むこのような複合体は、本明細書において「トポイソメラーゼ活性化」または「トポイソメラーゼ付加」ヌクレオチド配列という。トポイソメラーゼが認識部位の核酸分子を切断して、共有結合する、未切断トポイソメラーゼ認識部位およびトポイソメラーゼを含む核酸分子のように、トポイソメラーゼ活性化核酸分子を本発明の方法に使用することができる。
両方の鎖に共有結合された二本鎖組換え核酸分子を作製する方法の一態様において、IB型トポイソメラーゼが存在する場合には、各ヌクレオチド配列が切断されて、共有結合されたトポイソメラーゼを有する3'端を作製するように、トポイソメラーゼ認識部位は、接続される各ヌクレオチド配列の3'端または3末端付近に存在する(図8参照)。共有結合されるヌクレオチド配列は、トポイソメラーゼ認識部位を有するものと同じ末端に5'側ヒドロキシル基を有しても、または5'側ヒドロキシル基がホスファターゼを使用して作製されてもよい。このようなヌクレオチド配列が接触すると、部位特異的トポイソメラーゼは、3'側リン酸を有する各鎖をそれぞれの5'側ヒドロキシル基にライゲーションし、直鎖状、環状または正もしくは負の超らせん核酸分子として作製することができる、両方の鎖に共有結合された二本鎖組換え核酸分子を作製することができる。
好ましくは、本発明のある種の局面の方法によりIB型トポイソメラーゼによって接続されるヌクレオチド配列の末端の5'端は、望ましい場合には、所定の方向配向を含む、ヌクレオチド配列の最初の結合を促進することができる、相補的な5'側突出配列を有する。または、本発明のある種の局面の方法によりIB型トポイソメラーゼにより接続されるヌクレオチド配列の末端の5'側は、望ましい場合には、所定の方向配向を含む、鎖侵入によりヌクレオチド配列の最初の結合を促進するために、配列の一方が5'側突出配列を有し、他方のヌクレオチド配列が5'端の平滑末端に相補鎖を有する相補的な5'側配列を有する。「5'側突出」または「5'側突出配列」という用語は、本明細書において、核酸分子の相補鎖の末端を越えて5'側方向に伸長する核酸分子の鎖をいうために使用される。便利なことに、5'側突出は、IB型トポイソメラーゼによる核酸分子の部位特異的な切断の結果として作製することができる(実施例1)。
好ましくは、本発明のある種の局面の方法によりIA型トポイソメラーゼによって接続されるヌクレオチド配列の末端の3'端は、望ましい場合には、所定の方向配向を含む、ヌクレオチド配列の最初の結合を促進することができる、相補的な3'側突出配列を有する。または、本発明のある種の局面の方法によりトポイソメラーゼ(例えば、IA型またはII型トポイソメラーゼ)により接続されるヌクレオチド配列の末端の3'側は、望ましい場合には、所定の方向配向を含む、鎖侵入によりヌクレオチド配列の最初の結合を促進するために、配列の一方が3'側突出配列を有し、他方のヌクレオチド配列が3'端の平滑末端に相補鎖を有する相補的な3'側配列を有する。「3'側突出」または「3'側突出配列」という用語は、本明細書において、核酸分子の相補鎖の末端を越えて5'側方向に伸長する核酸分子の鎖をいうために使用される。便利なことに、3'側突出は、IA型またはII型トポイソメラーゼによる切断の結果として作製することができる。
3'側または5'側突出配列は任意の配列を有してもよいが、一般に、配列は、本発明の方法により1つの核酸分子の所定の末端を第2のヌクレオチド配列の所定の末端にライゲーションすることができるように選択される(図9C、実施例1.B.参照)。このように、3'側または5'側突出は回文的であってもよいが、回文的突出を有する核酸分子は互いに結合して、所定の配向の2つまたはそれ以上の核酸分子を含む両方の鎖に共有結合された二本鎖組換え核酸分子の収率を低下させるため、3'側または5'側突出は一般的に回文的ではない。例えば、図9Aに示す核酸分子の5'側突出配列は回文であるので、例えばAGCT突出を介する第1のCMV要素と第2のCMV要素の結合は、AGCT突出を介するCMV要素とGFP要素の結合と同じくらい可能性がある。このように、図9Cに示す要素を使用してこのような構築物を作製する効率と比較すると、5'側から3'側の順序で、CMV要素、GFP要素およびBGH要素を含む機能的に共有結合した構築物を含む構築物を作製する効率は低下すると思われる。図9Bに示す要素は、望ましい構築物を作製するためには、示しているGFP要素の1つの末端およびBGH要素の末端に回文的突出を有し、従って、図9Cの要素より効率が低いと思われるが、図9Aのものより効率がよいと思われる。
本発明の方法およびキットに使用するヌクレオチド配列は、トポイソメラーゼ切断後の再連結を防ぐために架橋用ホスホロチオエートを含むように設計することができる。例えば、トポイソメラーゼが大腸菌(E. coli)トポイソメラーゼIIIである場合には、GCAACTT切断/認識配列の2つのチミジンの間に架橋用ホスホロチオエートを組入れることができる。切断される場合には、切断された(clipped)配列は3'側-OHの代わりに3'側-SHを有し、従って再連結を防ぐ(Burginら、Nucl. Acids Res. 23: 2973-2979、1995参照)。
本発明の一局面の方法またはキットに有用な核酸分子は、末端の3'端または5'端にトポイソメラーゼ認識部位を有するように、PCRなどの増幅方法により増幅することができる。さらに、増幅された核酸分子の切断の結果、切断された核酸分子が一方または両方の末端に5'側または3'側突出を有するように、PCRに使用される1つまたは両方のプライマーを設計することができる。一態様において、第1の核酸分子の5'側突出配列が第2(または他の)核酸分子の5'側突出配列に相補的であり、それによって好ましくは所定の配向のヌクレオチド配列の結合を促進するようにPCRプライマーを設計し、その結果それらは本発明の方法により共有結合することができる。本発明によると、接続される異なる核酸分子のための独自の突出配列を設計することによって、任意の数の核酸分子を望ましい順序および/または配向で接続することができる。
PCRは、本発明の方法に関して2つの方法で使用されることが認識されるべきである。一局面において、望ましい核酸分子、例えばエピトープ標識または細胞画分化ドメインなどの関心対象の、転写もしくは翻訳調節要素またはコード配列をコードする核酸分子に特定の特徴を与えるようにPCRプライマーを設計する。この局面では、増幅の結果、核酸分子が適宜、一方または両方の末端にトポイソメラーゼ認識部位を有するように、PCRプライマーを設計することができる。本明細書に開示するように、部位特異的トポイソメラーゼによる増幅産物の切断の結果、切断された核酸分子がトポイソメラーゼ切断末端に5'側または3'側突出配列を有するように、PCRプライマーは追加の配列を含んでもよい。5'端に結合し、切断するトポイソメラーゼに関与する本発明の一態様において(例えば、IA型トポイソメラーゼに関与する態様)、トポイソメラーゼ切断後の再連結を遮断することができ、トポイソメラーゼ付加増幅産物の作製を助けることができる架橋用ホスホロチオエート結合(上記参照)を有するようにPCRプライマーを設計することができる。
PCRを使用して作製される突出配列は、PCR反応の人為的結果として作製される一塩基突出を含んでもよい。例えば、校正機能を持たず、本質的な末端トランスフェラーゼ活性を有するTaqなどのポリメラーゼが通常使用され、各末端に鋳型由来でない1つの3'側A突出を有するPCR産物を作製する。本発明の方法を使用して、これらの増幅産物を、T/Aクローニング反応では、ベクター(おのおの参照として本明細書に組み入れられる、米国特許第5,487,993号および同第5,856,144号参照)であってもよい3'側一塩基T突出または3'側一塩基dU突出を一方または両方の末端に有するトポイソメラーゼ付加核酸分子を接続することができる。
PCRはまた、本発明の方法によって作製される、1つまたは両方の鎖に共有結合された共有結合型二本鎖組換え核酸分子を増幅するために使用される。例えば、図13に例示するように、本発明の方法は、プロモーターを含むヌクレオチド配列、コード配列を含むヌクレオチド配列、およびポリアデニル化シグナルを含むヌクレオチド配列を含む3つの基質核酸分子から、発現可能な二本鎖組換え核酸分子を作製することができる。二本鎖組換え核酸分子の作製は、接続される二本鎖ヌクレオチド配列の末端に相補的な3'側(または5'側)突出配列を組入れることによって促進することができる。例えば、発現可能な二本鎖組換え核酸分子は、第1の末端の5'端にIA型トポイソメラーゼおよび第2の末端の3'端にIB型トポイソメラーゼを有する第1の核酸分子に、第2の核酸分子および第3の二本鎖ヌクレオチド配列を接触させることによって作製することができる。プロモーターの上流であるプロモーターを含むヌクレオチド配列の一部に特異的な第1のプライマーおよびシグナルの下流であるポリアデニル化シグナルを含むヌクレオチド配列の一部に特異的な第2のプライマーを有するPCRプライマー対を設計することによって、プロモーター、コード配列およびポリアデニル化シグナルを適切な(所定の)配向で含む全長の機能的な二本鎖組換え核酸分子だけが増幅される。特に、例えばコード配列に接続したプロモーターだけを有する部分的な反応産物およびニックを有する反応産物は増幅されない。従って、本発明の方法に有用であるように核酸分子を特異的に設計するため、および望ましい成分および特性を有する反応産物だけを特異的に増幅するためにPCRを使用することができる。
本明細書において使用する「共有結合された」という用語は、二本鎖組換え核酸分子に言及して使用される場合には、核酸分子が、トポイソメラーゼ媒介接続により両方の鎖にライゲーションされる少なくとも2つの核酸分子から作製されることを意味する。例えば、共有結合される核酸分子の一方に共有結合されるトポイソメラーゼは、他方の核酸分子に共有結合されるトポイソメラーゼと同じであっても、異なってもよいことが認識されるべきである。従って、ワクシニアトポイソメラーゼが一方の核酸分子に共有結合され、別のポックスウイルスまたは真核細胞核IB型トポイソメラーゼが他方の鎖に結合されてもよい。しかし、一般に、トポイソメラーゼは、異なる場合には、同じファミリー、例えばIA型またはIB型またはII型のメンバーであるが、トポイソメラーゼが、例えば5'側リン酸に共有結合され、相補的な3'側突出を作製する場合には、トポイソメラーゼは、例えば異なるファミリー、例えばIA型とII型由来であってもよい。
「共有結合される」という用語はまた、1つの鎖にライゲーションされる少なくとも2つのヌクレオチド配列から作製される一本鎖または二本鎖核酸分子に言及して本明細書において使用される。例えば、トポイソメラーゼが、結合する第1の核酸分子の5'端を、第2の核酸分子の3'端に共有結合することができる条件下において、5'端または5'端付近に結合された1つのトポイソメラーゼを含む第1のトポイソメラーゼ付加核酸分子が第2の二本鎖ヌクレオチド配列と接触する場合に作製される二本鎖組換え核酸分子は、1つの鎖に共有結合された二本鎖組換え核酸分子を作製することができる。
一態様において、本発明の方法により作製される両方の鎖に共有結合された二本鎖組換え核酸分子は、どちらかの鎖の2つのヌクレオチド配列がライゲーションされる部位にニックがないが、分子の別の箇所にニックを有することがある。1つの鎖に共有結合された二本鎖組換え核酸分子を作製する方法では、少なくとも末端が相補鎖に接続された位置にニックを有する二本鎖組換え核酸分子が作製される。このニックのある二本鎖組換え核酸分子は、当技術分野において既知であるように、ニックのある二本鎖組換え核酸分子を細胞に導入することによって、またはリガーゼを使用するなどのライゲーション反応を二本鎖組換え核酸分子に実施することによって、両方の鎖に共有結合された二本鎖組換え核酸分子に変換することができる。
「組換え」という用語は、本発明の方法により、少なくとも2つのヌクレオチド配列を接続することによって作製される核酸分子を言及するために本明細書において使用される。このように、本発明に含まれる二本鎖組換え核酸分子は、天然、例えば減数分裂中に作製されうる核酸分子とは識別される。例えば、本発明のある種の局面の方法により作製される、両方の鎖に共有結合された二本鎖組換え核酸分子は、核酸分子が接続された部位または部位付近に、相補鎖のそれぞれに1つずつ存在する2つのトポイソメラーゼ認識部位の存在によって同定することができる。
本発明の方法は、第1の末端および第2の末端を有し、第1の核酸分子が、第1の末端もしくは第2の末端または両方において、3'端または3'端付近にトポイソメラーゼ認識部位またはその切断産物を有し、同じ末端の5'端にヒドロキシル基を有する(または、例えばホスファターゼによって有するように作製することができる)第1の核酸分子と、第1の末端および第2の末端を有し、少なくとも第2の核酸分子が、第1の末端もしくは第2の末端または両方において、3'端または3'端付近にトポイソメラーゼ認識部位またはその切断産物を有し、同じ末端の5'端にヒドロキシル基を有する(または、有するように作製することができる)少なくとも1つの第2の核酸分子と、トポイソメラーゼを、全ての成分が接触し、トポイソメラーゼがその活性を発揮できる条件下において接触させることによって実施することができる。適宜、トポイソメラーゼと第1および第2(または他の)核酸分子を接触させ、切断されると、各ヌクレオチド配列は、接触の結果、第1および少なくとも第2のヌクレオチド配列が両方の鎖に共有結合されるように、切断部位の3'端に共有結合されたトポイソメラーゼを含み、5'端にヒドロキシル基を有する、または有してもよい。従って、本発明は、このような方法によって作製される、両方の鎖に共有結合された二本鎖組換え核酸分子を提供する。
本明細書において使用する「その部位または部位付近」は、ヌクレオチド配列の3'側(IB型)または5'側(IA型またはII型)に対するトポイソメラーゼ認識部位の近位性に言及して使用する場合には、部位が、それぞれ、3'端または5'端から約1〜100ヌクレオチド以内にある、一般に末端から約1〜20ヌクレオチド以内にある、特にそれぞれの末端から約2〜12ヌクレオチド以内にあることを意味する。トポイソメラーゼ認識部位を末端の約10〜15ヌクレオチド以内に位置づける利点は、トポイソメラーゼによる切断の結果、切断部位の下流の配列部分が、共有結合されたトポイソメラーゼを有する残りのヌクレオチド配列から自然に分離することができるということである(一般に「自殺切断(suicide cleavage)」と言われる、例えばShuman、上記、1991年、Andersenら、上記、1991年参照)。トポイソメラーゼ認識部位が末端からの約12〜15ヌクレオチドより大きい場合には、例えばトポイソメラーゼ切断部位を含む二本鎖の一部の融解温度より高い温度におけるインキュベーション段階を提供することによって、反応条件を改良することによって、配列の残りから分離するように切断部位の上流または下流のヌクレオチド配列を誘導することができる。
例えば、一方または両方の末端から約2〜15ヌクレオチドのところにIB型トポイソメラーゼ認識部位を含むように、第1または第2の(または他の)核酸分子を構築する追加の利点は、部位特異的トポイソメラーゼによる核酸分子の切断により、5'側突出が作製されることである。2〜15ヌクレオチドを含むと思われるこのような5'側突出配列を、本明細書に開示されているPCR方法を使用して、望ましい任意の配列を有するように、それぞれ、設計することができる。従って本発明の方法により、切断された第1の核酸分子が、選択された第2の(または他の)核酸分子に共有結合される場合、および選択された配列が5'側突出配列を有する場合には、2つ(または2つまたはそれ以上)の配列が5'側突出の相補性により所定の配向で共有結合されるように、第1の核酸分子の5'側突出は、選択された第2の(または他の)二本鎖配列に相補的であるように設計することができる。上記に考察されるように、例えばIA型またはII型トポイソメラーゼによる切断の結果作製される3'側突出配列に関して同様の方法を使用することができる。
本明細書において使用する「第1の末端」および「第2の末端」を有するヌクレオチド配列への言及は、ヌクレオチド配列が直鎖状であることを意味する。基質核酸分子は、超らせんを含んで、直鎖状または環状であってもよいが、1つまたは複数のトポイソメラーゼによる切断の結果として、直鎖状トポイソメラーゼ付加核酸分子が一般に作製される。例えば、各鎖が切断され、介在する配列が分離し、トポイソメラーゼが各末端に共有結合した直鎖状核酸分子を作製するように、互いから約100ヌクレオチド以内で、相補鎖内に、好ましくは互いから約20ヌクレオチド以内で、相補鎖内に2つのIB型トポイソメラーゼを有する環状核酸分子に部位特異的IB型トポイソメラーゼを接触させることができる。
核酸分子の第1の末端または第2の末端への言及は、ヌクレオチド配列の任意の特定の配向を意味する意図ではなく、互いに対する末端の相対的な重要性を意味する意図ではないことが認識されるべきである。第1の末端および第2の末端を有するヌクレオチド配列が二本鎖ヌクレオチド配列である場合には、各末端は5'端および3'端を有する。従って、例えば第1の末端であっても、第2の末端であってもよい同じ末端の、3'端にトポイソメラーゼ認識部位を有し、5'端にヒドロキシル基を有するヌクレオチド配列に本明細書において言及される。
本発明の方法は、第1の核酸分子および第2の核酸分子だけを使用して実施しても、または適宜第3、第4またはそれ以上の核酸分子をさらに含んでもよい。一般に、このようなヌクレオチド配列はそれぞれ、3'端または5'端の少なくとも一方またはその付近にトポイソメラーゼ認識部位またはその切断産物を有し、同じ末端の5'端にヒドロキシル基を有しても、またはホスファターゼを使用してヒドロキシル基を作製してもよい。第2のヌクレオチド配列に接続される末端または末端付近に、ヌクレオチド配列がトポイソメラーゼ認識部位を持たない場合には、例えばトポイソメラーゼ認識部位の相補鎖を含むプライマーを使用した配列のPCR増幅によって、本明細書に開示する方法を使用して、トポイソメラーゼ認識部位をヌクレオチド配列に組入れることができる。
「第1のヌクレオチド配列」、「第2のヌクレオチド配列」、「第3のヌクレオチド配列」等という用語は、いくつかのヌクレオチド配列のどれが言及されているかを示す手段を提供するためだけに本明細書において使用される。従って、特定のヌクレオチド配列に関して具体的に規定される任意の特徴がない場合には、「第1」、「第2」、「第3」等という用語は、ヌクレオチド配列またはヌクレオチド配列の集団もしくは多数のヌクレオチド配列に言及して使用される場合には、ヌクレオチド配列に関する任意の特定の順序、重要性または他の情報を示すことを意図していない。従って、例示されている方法が、例えば増幅産物が一方または両方の末端にトポイソメラーゼ認識部位を有するように、PCRを使用して第1の核酸分子を増幅することに言及している場合には、第2の(または他の)核酸分子も同様にこのように増幅することができることが認識される。
「少なくとも第2のヌクレオチド配列」という用語は、第1のヌクレオチド配列以外の1つまたは複数のヌクレオチド配列を意味するために本明細書において使用される。従って、この用語は第2のヌクレオチド配列だけまたは第2のヌクレオチド配列と第3のヌクレオチド配列(またはそれ以上)を言及することができる。このように、「第2の(または他の)ヌクレオチド配列」または「第2(および他の)ヌクレオチド配列」という用語は、「少なくとも第2のヌクレオチド配列」という用語が第2、第3またはそれ以上のヌクレオチド配列を言及することができるということを認識して本明細書において使用される。特に記載しない限り、「少なくとも第2のヌクレオチド配列」という用語の意味に含まれるヌクレオチド配列は、第1のヌクレオチド配列と同じまたは実質的に同じであってもよいことが認識されるべきである。例えば、本発明の方法を使用して、第1および第2の核酸分子を共有結合することができるように、トポイソメラーゼによる切断の結果作製される相補的な5'側突出配列を有することを除いて、第1および第2の核酸分子は同じであってもよい。このように、本発明の方法を使用して、選択的に、例えば調節要素などの第3の核酸分子が点在してもよく、共有結合された配列を所定の方向配向、例えばそれぞれ互いに対して5'側から3'側への配向で有してもよい直鎖状の第1および第2の核酸分子を作製することができる。
本明細書に開示するように、本発明の方法は、所定の方向配向で2つまたはそれ以上の二本鎖ヌクレオチドを共有結合する手段を提供する。「方向配向」または「所定の方向配向」または「所定の配向」という用語は、2つまたはそれ以上のヌクレオチド配列を特定の順序で共有結合することを言及するために本明細書において使用される。従って、本発明の方法は、例えば機能的な発現可能な二本鎖組換え核酸分子を作製するために、コード配列の上流のプロモーター調節要素を共有結合し、コード領域の下流のポリアデニル化シグナルを共有結合するための手段を提供するか、またはそれらがインフレームにおいて転写および翻訳されて融合ポリペプチドを作製することができるように、2つのコード配列を共有結合するための手段を提供する。
本発明の方法はまた、第1の末端および第2の末端を有し、第1の末端もしくは第2の末端または両方において、第1の核酸分子は、IB型トポイソメラーゼが3'端(トポイソメラーゼ付加している)に共有結合しており、同じ末端の5'端にヒドロキシル基を有する(または有するように作製することができる)第1の核酸分子と、同じ末端の5'端にヒドロキシル基を有する(または有するように作製することができる)少なくとも第2のIB型トポイソメラーゼ付加核酸分子を接触させることによって実施することができる。トポイソメラーゼおよび5'側ヒドロキシル基を有する末端においてトポイソメラーゼ活性化した第1のヌクレオチド配列と、少なくとも第2のヌクレオチド配列を接触させる結果、ホスホジエステル結合が各鎖に形成され、それによって両方の鎖に接続された二本鎖組換え核酸分子を形成する。
本発明は、2つまたはそれ以上の(例えば、2、3、4、5、6、7等)ヌクレオチド配列を接続する方法であって、接続された二本鎖組換え核酸分子は1つの鎖に共有結合されているが、両方の鎖に共有結合されていない(すなわち、二本鎖組換え核酸分子は、2つの末端が接続されて、二本鎖組換え核酸分子を作製した1つの鎖の各位置にニックを有する)方法をさらに提供する。さらに、ヌクレオチド配列の1つまたは複数は、1つまたは複数の組換え部位を含むことができる。例示目的のために図11Aに示す略図を使用すると、本発明は、第1の末端と第2の末端を有し、第1の末端もしくは第2の末端または両方の末端において、部位特異的IA型トポイソメラーゼが5'端に共有結合されている第1の核酸分子と、トポイソメラーゼが少なくとも一方の末端のどちらの端にも共有結合されていない第2の核酸分子を接触させる段階を含む、少なくとも2つのヌクレオチド配列を接続する方法を含む。さらに、第2のヌクレオチド配列は、典型的には、第1の核酸分子に接続される末端の3'端にヒドロキシル基を有する。多数の例において、接続される2つのヌクレオチド配列は、ハイブリダイゼーションを可能にするのに十分な配列相補性を有する3'側または5'側突出を有する。関連する態様において、上記の第1および第2の核酸分子は、同じ核酸分子の第1および第2の末端であってもよい。従って、2つの末端を接続すると、環状分子が形成される。上記の方法で核酸分子が接続されると、得られる分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。本発明は、本発明の方法によって作製される核酸分子と、このような核酸分子を含む組成物と、このような核酸分子を使用する方法とをさらに含む。
例示目的のために図11Bに示す略図を使用すると、本発明は、3つ以上のヌクレオチド配列を接続する方法を含む。本発明の任意の数の変種が考えられるが、3つのヌクレオチド配列は、一方の末端の5'端および3'端またはその付近にトポイソメラーゼを有し、選択的に、1つまたは複数の認識部位を有するリンカー分子を使用することによって接続することができる。3つのヌクレオチド配列の接続の結果、接続部にニックのない第1の鎖と接続部にニックのある第2の鎖を有する一塩基配列が形成される。この方法は、3つのヌクレオチド配列を接続するために一トポイソメラーゼ修飾分子を使用する利点を有する。上記の方法で核酸分子が接続されると、得られる分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。本発明は、本発明の方法によって作製される核酸分子と、このような核酸分子を含む組成物と、このような核酸分子を使用する方法とをさらに含む。
本発明は、2つまたはそれ以上の(例えば、2、3、4、5、6、7等)核酸分子の両方の鎖を共有結合する方法をさらに提供する。例示の目的のために図12Aに示す略図を使用すると、本発明は、第1の末端と第2の末端を有し、第1の末端もしくは第2の末端または両方の末端において、3'端および5'端にそれぞれ1つのトポイソメラーゼが共有結合されている2つのトポイソメラーゼ(例えば、IA型およびIB型トポイソメラーゼ)を有する第1の末端と、少なくとも一方の末端のどちらの端にもトポイソメラーゼが共有結合されていない第2の核酸分子を接触させる段階を含む、少なくとも2つのヌクレオチド配列を接続する方法を含む。さらに、第2のヌクレオチド配列は、第1の核酸分子に接続される末端の5'端および3'端にヒドロキシル基を有することが多い。多くの例において、接続される2つのヌクレオチド配列は、ハイブリダイゼーションを可能にするのに十分な配列相補性を有する3'側または5'側突出、および選択的に、1つまたは複数の組換え部位を有する。関連する態様において、上記の第1および第2の核酸分子は、同じ核酸分子の第1および第2の末端であってもよい。従って、2つの末端が接続されると、環状分子が形成される。上記の方法で核酸分子が接続されると、得られる分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。本発明は、本発明の方法によって作製される核酸分子と、このような核酸分子を含む組成物と、このような核酸分子を使用する方法とをさらに含む。
例示目的のために図5Dに示す略図を使用すると、本発明は、3つ以上のヌクレオチド配列を接続するための方法を含む。本発明の任意の数の変種が考えられるが、3つのヌクレオチド配列は、各末端の5'端および3'端または端付近にトポイソメラーゼを有し、選択的に、1つまたは複数の認識部位を有するリンカー分子を使用することによって接続することができる。従って、3つのヌクレオチド配列の接続の結果、接続部にニックのない一塩基配列が形成される。この方法は、3つのヌクレオチド配列を接続するために一トポイソメラーゼ修飾分子を使用する利点を有する。上記の方法で核酸分子が接続されると、得られる分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。本発明は、本発明の方法によって作製される核酸分子と、このような核酸分子を含む組成物と、このような核酸分子を使用する方法とをさらに含む。
本発明は、1つの試験管または多数の試験管内で実施することができるトポイソメラーゼ媒介性接続反応および組換え反応を実施する方法をさらに提供する。例えば、トポイソメラーゼ媒介性接続反応および組換え反応を実施するのに必要な成分全てを1つの試験管内で組み合わせて、両方の反応を本質的に同時に行うことができる。1つの試験管または多数の試験管内で実施することができるトポイソメラーゼ/組換え反応の例を図35〜40に示す。従って、特定の態様において、本発明は、(1)1つまたは複数の核酸分子または核酸分子の2つの末端をトポイソメラーゼ媒介性反応により互いに接続する反応、ならびに(2)1つまたは複数の組換え部位が1つまたは複数の他の組換え部位との組換えを受ける一試験管反応を提供する。任意の数のトポイソメラーゼ媒介性接続反応および/または組換え反応を本発明の方法において実施することができる。特定の態様において、本発明の反応混合物中の1つまたは複数の核酸分子は、(1)1つまたは複数の組換え部位と、(2)1つまたは複数のトポイソメラーゼまたは1つまたは複数のトポイソメラーゼ認識部位を有する。
以下の実施例9に説明するように、ある種の例において、トポイソメラーゼは、特定の組換え反応を阻害することが見出されている。このような例では、トポイソメラーゼ媒介性接続反応を受けた核酸分子を、反応混合物に存在するトポイソメラーゼから分離し、ついで組換え反応の基質として使用することができる。このような例では、トポイソメラーゼ媒介性接続反応および組換え反応は別個の試験管内で行われることが多い。トポイソメラーゼ媒介性接続反応の産物をトポイソメラーゼから分離することができる方法の例には、フェノール/クロロホルム抽出、典型的には、その後の核酸の沈殿(例えば、エタノール沈殿)およびクロマトグラフィー(例えば、カラムクロマトグラフィー)が含まれるが、これらに限定されない。
または、反応混合物中に存在するトポイソメラーゼは、例えば加熱することによって(例えば、約65℃で約60分、約70℃で約60分、約75℃で約60分、約70℃で約40分、約75℃で約40分、約80℃で約40分、約80℃で約30分、約85℃で約20分、約90℃で約15分、約95℃で約5分または約99℃で約1分加熱する)またはプロテアーゼ(例えば、プロテイナーゼK)を使用することによって不活性化することができる。この例では、一般に、トポイソメラーゼ媒介性接続反応および組換え反応を同じ試験管内で行うことが可能である。
一試験管反応の具体的な態様では、それぞれが1つまたは複数のトポイソメラーゼまたはトポイソメラーゼ認識部位を含む2つまたはそれ以上の核酸セグメントを、トポイソメラーゼ媒介性接続反応(例えば、トポイソメラーゼ媒介性接続反応)を使用して互いに接続する。その後、試験管を約85℃までの温度に約20分間加熱し、1つまたは複数のリコンビナーゼを添加する。さらに、2つまたはそれ以上の核酸セグメントの1つまたは複数が組換え部位を含まない場合、または追加の核酸セグメントによる組換えが望ましい場合には、1つまたは複数の組換え部位を含む核酸セグメントを添加することができる。典型的には、試験管内に存在する組換え部位は、互いに組換えることができるものである。
一試験管反応の他の具体的な態様では、2つまたはそれ以上の核酸セグメントが、1つまたは複数のリコンビナーゼにより触媒される組換えを受ける。組換えが生じたら、トポイソメラーゼを試験管に添加して、核酸セグメントのトポイソメラーゼ媒介性接続を促進する。上記のように、追加の核酸セグメントを、選択的に、トポイソメラーゼと共に反応混合物に添加することができる。さらに、1つまたは複数のトポイソメラーゼが接続される核酸セグメントを反応混合物に添加する場合には、追加のトポイソメラーゼを添加する必要がないことが多い。従って、特定の態様において、トポイソメラーゼ修飾核セグメントを上記の反応混合物に添加することができ、特定の反応条件に応じて、追加のトポイソメラーゼを添加しても、しなくてもよい。
本発明はまた、1つまたは複数の(例えば、1、2、3、4、5、6等)マルチクローニング部位を有する核酸分子を作製する方法を提供する。例えば、本発明の方法に使用する1つまたは複数の核酸セグメントは、1つまたは複数のマルチクローニング部位を含むことができる。別の例として、本発明の方法により、核酸分子を作製するために使用する核酸セグメントにマルチクローニング部位を付加するか、または1つもしくは複数のマルチクローニング部位を有するリンカーを接続することにより、本発明の方法によって作製される核酸分子にマルチクローニング部位を付加してもよい。関連する局面において、本発明は、1つまたは複数のマルチクローニング部位を有する、本発明の方法により作製される核酸分子、および本発明の方法により作製される核酸分子を修飾するための、1つまたは複数のこのようなマルチクローニング部位の用途を含む。本発明はまた、上記の方法により作製される核酸分子、ならびにこれらの分子およびこれらの分子を含む組成物の用途を提供する。
ウイルスベクター
本発明はさらに、ウイルス核酸領域を有する核酸分子を作製する方法、ならびにこのような方法で作製される核酸分子およびこれらの核酸分子を含む組成物を提供する。
アデノウイルスは、例えば遺伝子治療に使用できるウイルスベクターである。アデノウイルスは気道上皮に遺伝子を送達するための特に興味深い搬送体であり、このようなベクターの用途は本発明の範囲内に含まれる。アデノウイルスは通常気道上皮に感染し、軽度な疾患を生じる。アデノウイルスに基づく送達系の他の標的は、肝臓、中枢神経系、内皮細胞および筋肉である。アデノウイルスは、分割していない細胞に感染することができるという利点を持つ。コザルスキー(Kozarsky)およびウィルソン(Wilson)、Current Opinion in Genetics and Development 3: 499-503(1993)は、アデノウイルスに基づいた遺伝子治療の総説を示している。ブウト(Bout)ら、Human Gene Therapy 5: 3-10(1994)は、アカゲザルの気道上皮に遺伝子を送達するためのアデノウイルスベクターの用途を証明した。遺伝子治療にアデノウイルスを使用する他の例は、全ての開示全体が参照として本明細書に組み入れられる、ローゼンフェルド(Rosenfeld)ら、Science 252: 431-434(1991)、ローゼンフェルド(Rosenfeld)ら、Cell 68: 143-155(1992)、マストランゲリ(mastrangeli)ら、J. Clin. Invest. 91: 225-234(1993)、PCT国際公開公報第94/12649号および国際公開公報第96/17053号、米国特許第5,998,205号ならびにワン(Wang)ら、Gene Therapy 2: 775-783(1995)に見出すことができる。
アデノ関連ウイルス(AAV)およびヘルペスウイルス、ならびにこれらのウイルスから作製されるベクターも、遺伝子治療に使用するために提案されている(Walshら、1993、Proc. Soc. Exp. Biol. Med. 204: 289-300、米国特許第5,436,146号、Wagstaffら、Gene Ther. 5: 1566-70(1998))。ヘルペスウイルスベクターは、遺伝子発現が神経細胞で望まれる用途にとって、特に有用である。
従って、本発明は、ウイルスベクター(例えば、アデノウイルスベクター、アルファウイルスベクター、ヘルペスウイルスベクター、アデノ関連ウイルスベクター等)の1つまたは複数の機能的特性を有する、核酸分子を作製する方法を含む。特定の態様において、本発明の方法は、核酸セグメントの1つまたは複数が、産物の核酸分子に、ウイルスベクターとして機能する能力(例えば、特定の宿主細胞において複製する能力、ウイルス粒子内にパッケージングされる能力等)を与える領域を有する核酸セグメントの接続を含む。
特定の態様において、本発明は、アデノウイルスの配列を含む少なくとも1つの(例えば、1、2、3、4等)核酸セグメントを、1つまたは複数の他の核酸セグメントに接続することによって、アデノウイルスベクターを作製する方法を含む。アデノウイルスベクターおよびアデノウイルスベクターを作製するために使用できる核酸セグメントの具体的な例は、全体の開示内容が参照として本明細書に組み入れられる、米国特許第5,932,210号、同第6,136,594号および同第6,303,362号に開示されている。本発明の方法によって作製されるアデノウイルスベクターは複製能力があっても、複製欠損であってもよい。
アデノウイルスベクターの一例は、アデノウイルスの核酸を含む核酸セグメントを、1つまたは複数の他の核酸セグメントに接続することによって作製することができる。例えば、複製欠損アデノウイルスベクターが望ましい場合には、アデノウイルスの核酸は以下の領域の1つまたは複数の、全てまたは一部の欠損を有する:E1a領域、E1b領域および/またはE3領域。これらの領域に欠損を有するアデノウイルスベクターは、例えば米国特許第6,136,594号に記載されている。本発明はさらに、本発明の方法により作製されるアデノウイルスベクター、ならびにこれらのベクターおよびこれらのベクターを含む、組成物の用途を含む。本発明の方法により作製されるアデノウイルスベクターの用途の一例は、核酸セグメントを哺乳類(例えば、ヒト)の細胞へ送達することを含む。従って本発明は、遺伝子治療プロトコールに使用するのに適切なベクターを作製する方法を提供する。典型的には、このようなベクターは複製欠損である。
具体的な態様において、本発明のアデノウイルスベクターは、以下の領域の1つまたは複数の、全てまたは一部の欠損であることを除いて、実質的にアデノウイルスゲノム全体を含む:E1a領域、E1b領域および/またはE3領域。さらに別の具体的な態様において、アデノウイルス以外の核酸がE1a領域、E1b領域および/またはE3領域の1つまたは複数に存在してもよい。
特定の態様において、本発明の方法により作製されるアデノウイルスベクターは、大腸菌(E. coli)などの原核細胞内でのベクターの増幅を可能にする、複製起点および/または選択マーカーを少なくとも1つ有する。
アデノ関連ウイルスおよびヘルペスウイルスベクターは、上記のものと同様の本発明の方法により作製することができる。従って本発明は、このようなベクターを作製する方法、ならびにこれらの方法によって作製されるベクター、これらのベクターおよびこれらのベクターを含む組成物の用途をさらに提供する。
本発明は、アルファウイルスベクター(例えば、シンドビスウイルスベクター、セムリキ森林熱ウイルスベクター、ロスリバーウイルスベクター、ベネズエラウマ脳炎ウイルスベクター、西部ウマ脳炎ウイルスベクター、東部ウマ脳脊髄炎ウイルスベクター等)を作製する方法、ならびにこのような方法によって作製されるアルファウイルスベクター、これらのアルファウイルスベクターおよびこれらのアルファウイルスベクターを含む組成物を使用する方法をさらに提供する。
特定の態様において、本発明は、アルファウイルスの配列を含む少なくとも1つ核酸セグメントを、1つまたは複数の他の核酸セグメントに接続することによって、アルファウイルスベクターを作製する方法を含む。アルファウイルスベクターおよびアルファウイルスベクターを作製するために使用できる核酸の具体的な例は、全体の開示内容が参照として本明細書に組み入れられる、米国特許第5,739,026号および同第6,224,879号、GibcoBRL社の取り扱い説明書10179-018号、SFV遺伝子発現系(SFV Gene Expression System)およびインビトロジェン(Invitrogen)社のシンドビス発現系マニュアル、カタログ番号K750-01(バージョンE)に記載されている。
具体的な態様において、アルファウイルスベクターを作製するために本発明の方法において使用されるアルファウイルスベクター配列は、以下の成分の1つまたは複数のを含む:(アルファウイルス起源であっても、なくてもよい)1つまたは複数のパッケージングシグナル、1つまたは複数のサブゲノムプロモーターおよび/または1つまたは複数の非構造タンパク質(例えば、nsp1、nsp2、nsp3、nsp4等)をコードする核酸。
本発明のアルファウイルスベクターは、DNAまたはRNA分子として細胞に導入することができる。DNA形態のこのようなベクターを細胞に導入する場合には、発現制御配列(例えば、誘導性、抑制性または構成性発現制御配列)を使用して、1つまたは複数の非構造タンパク質を翻訳することができるRNA分子を作製することができる。具体的な態様において、これらの非構造タンパク質は、DNA形態のアルファウイルスベクターから作製される転写物の全てまたは一部に対応するRNA分子を増幅する、RNA依存的RNAポリメラーゼを形成する。従って、これらの非構造タンパク質は、RNA鋳型からのRNA分子の追加のコピーの作製を触媒して、RNAを増幅することができる。さらに、高レベルの発現が望まれる核酸セグメントをサブゲノムプロモーターに機能的に結合して、核酸セグメントに対応する高レベルのRNAを作製することができる。
例示的な一態様において、本発明の方法によって作製されるアルファウイルスベクターは、誘導性プロモーターが、シンドビスウイルス起源ではない核酸セグメントに機能的に結合したシンドビスウイルス、およびシンドビスサブゲノムプロモーターのnsp1、nsp2、nsp3およびnsp4をコードするRNA分子の転写を誘導するDNAを含む。本発明はまた、本発明の方法によって作製されるアルファウイルスベクター、このようなアルファウイルスベクターを使用する方法、およびこのようなアルファウイルスベクターを含む組成物を提供する。
本発明はさらに、核酸セグメントの1つまたは複数が、1つまたは複数の(例えば、1、2、3、4等)ウイルスパッケージングシグナル(例えば、上記に言及したウイルス由来の1つまたは複数のパッケージングシグナル)を有する核酸セグメントを接続する方法を提供する。これらのパッケージングシグナルは、本発明の方法によって作製される核酸分子のパッケージングを誘導するために使用することができる。パッケージングされた核酸分子を作製する1つの方法は、ウイルス様粒子の作製に適切なタンパク質を発現するパッケージング細胞系統に本発明の核酸分子を導入または発現することによる。本発明はさらに、本発明のパッケージングされた核酸分子、本発明のパッケージングされた核酸分子を作製する方法および本発明のパッケージングされた核酸分子を含む組成物を提供する。
本発明はまた、組成物および本発明の方法を実施するのに有用な成分を含む、このような組成物を含むキットを提供する。一局面において、本発明の組成物は、本発明の方法の一段階を特徴とする単離された成分を含む。例えば、本発明の組成物は、同じまたは異なるトポイソメラーゼ付加核酸分子の2つまたはそれ以上を含むことができる。本明細書において使用する「異なる」という用語は、本発明の組成物の核酸分子に言及して使用する場合には、核酸分子が最適に配列されたとき互いに95%未満の配列の同一性を有し、一般に90%未満の配列の同一性を有し、かつ通常70%未満の配列の同一性を有することを意味する。従って、例えば互いの多型変種である点においてのみ異なる核酸分子、または異なる5'側または3'側突出配列を単に有する核酸分子は、本発明の組成物の目的のためには、「異なる」と考えられない。これと比較すると、異なる核酸分子は、ポリペプチドをコードする第1の配列および調節要素を含む第2の配列、または第1のポリペプチドをコードする第1の配列および非相同ポリペプチドをコードする第2の配列によって例示される。
本発明の組成物が3つ以上の異なる単離された核酸分子または3つ以上の異なるトポイソメラーゼ付加核酸分子を含む場合には、核酸分子はそれぞれ互いに異なる。すなわち、それらは全て互いと異なる。しかし、核酸分子のそれぞれ、例えば第1の核酸分子と言及される配列は、一般に、互いに同一または実質的に同一であるこのようなヌクレオチド配列集団を含むことが認識されている。従って、「異なる」という用語は、例えば第1の核酸分子(または第1の核酸分子の集団)と第2の(および他の)核酸分子を比較する際に使用されることが明白になるはずである。2つまたはそれ以上の異なるトポイソメラーゼ付加核酸分子を含む組成物はさらに、トポイソメラーゼを含むことができる。本発明の組成物の成分を含むこのような核酸分子の例は本明細書に開示されており、例えばコード配列、転写調節要素、翻訳調節要素、エピトープ標識または抗生物質耐性遺伝子などの検出または選択マーカーをコードする要素、細胞画分化ドメインまたはシグナルペプチドなどのポリペプチドドメインをコードする要素等を含む。
本明細書において使用する「単離された」という用語は、言及されている分子が、天然に存在するもの以外の形態で存在することを意味する。一般に、単離されたヌクレオチド配列は、例えば細胞のゲノムの一部ではない任意のヌクレオチド配列であっても、または通常にヌクレオチド配列を含む細胞から物理的に分離した、任意のヌクレオチド配列であってもよい。本発明の種々の組成物は、単離された核酸分子の混合物を含むことが認識されるべきである。このように、「単離された」という用語のみが、天然の状態から分子を単離することに関して使用されるが、その分子が唯一の成分であることは示していないことが理解される。
本発明の組成物は、それぞれが一方または両方の末端または末端付近にトポイソメラーゼ認識部位を有する、2つの異なる核酸分子、およびトポイソメラーゼ認識部位において核酸分子に結合して切断することができる、部位特異的トポイソメラーゼを含むことができる。選択的に、異なる核酸分子の少なくとも1つはトポイソメラーゼ付加核酸分子であってもよい。好ましくは、トポイソメラーゼ付加核酸分子に共有結合したトポイソメラーゼは、組成物中のトポイソメラーゼと同じファミリーであってもよい。
種々の組み合わせの成分を本発明の方法に使用することができる。例えば、本発明の方法は、選択的に、1つまたは複数の組換え部位を含む、トポイソメラーゼ活性化された第1の核酸分子および第1および第2の末端を有し、第1の末端もしくは第2の末端または両方において、3'端または3'端付近にトポイソメラーゼ認識部位を有し、同じ末端の5'端にヒドロキシル基を有する第2の核酸分子を接触させることによって実施することができる。接続される一方または両方の末端の5'端が5'側リン酸基を有する場合には、ホスファターゼに反応混合物の成分を接触することもできる。このような接触の結果、トポイソメラーゼは第2のヌクレオチド配列を切断してトポイソメラーゼ活性化された第2の核酸分子を作製することができ、ホスファターゼは、適宜同じ末端に5'側ヒドロキシル基を作製し、第2の核酸分子はトポイソメラーゼ活性化された第1の核酸分子に共有結合することができる。このように、本発明の組成物は、本発明の方法を実施するのに有用な種々の組み合わせの成分のいずれかを含むことができることが認識される。上記の方法で核酸分子が接続されると、得られる分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。本発明は、本発明の方法によって作製される核酸分子と、このような核酸分子を含む組成物、およびこのような核酸分子を使用する方法をさらに含む。
一般に、両方の鎖に共有結合された二本鎖組換え核酸分子を作製する本発明の方法は、第1および第2の配列がそれぞれ、接続される末端に、トポイソメラーゼ認識部位、例えば5'-(C/T)CCTT-3'を有する、第1の核酸分子に第2の核酸分子を接触させることによって、両方の鎖に共有結合された二本鎖組換え核酸分子を作製することができるという傾向に基づいている(Shuman、上記、1991年、米国特許第5,776,891号)。切断の結果、部位特異的トポイソメラーゼは3'端に共有結合する。切断されたヌクレオチド配列も、結合されたトポイソメラーゼと同じ末端に5'側ヒドロキシル基を有し、2つのヌクレオチド配列の末端が結合する場合には、各3'端のトポイソメラーゼは、結合するヌクレオチド配列の5'側ヒドロキシル基にその末端を共有結合することができる(図1参照)。
本明細書において使用される「全ての成分が接触する条件下において」第1のヌクレオチド配列と少なくとも第2のヌクレオチド配列を接触させるという言及は、反応条件は、ヌクレオチド配列のトポイソメラーゼ切断末端にとって、トポイソメラーゼがその酵素活性を発揮し、第1のヌクレオチド配列の3'端または5'端を第2のヌクレオチド配列の、それぞれ5'端または3'端に共有結合するように十分接近するのに適当であることを意味する。反応温度、イオン強度、pH等を含むこのような条件の例は本明細書に開示されており、例えばトポイソメラーゼ切断の結果作製される末端の特定の5'側突出配列に必要な他の適当な条件は、当技術分野において既知であるように、実験的またはヌクレオチド配列の特定のハイブリダイゼーションの条件を予測する式を使用して決定することができる(例えば、それぞれが参照として本明細書に組み入れられる、Sambrookら、「分子クローニング:実験マニュアル(Molecular Cloning: A Laboratory manual)」(Cold Spring Harbor Laboratory Press 1989)、Ausubelら、「分子生物学の最新プロトコール(Current Protocols in Molecular Biology)」、John Wiley and Sons、メリーランド州バルチモア(1987年および1995年までの補遺)参照)。
一態様において、本発明の方法は、1つまたは複数の調節要素をコード配列と推定されるものに機能的に接続することによって、cDNAまたは単離されたゲノムDNA配列のオープンリーディングを発現可能にする手段を提供する。従って、オープンリーディングフレームを含む第1の核酸分子は、部位特異的トポイソメラーゼによる切断の結果、一方または両方の末端が規定された5'側または3'側突出を有するか、または平滑であるように、一方または両方の末端にトポイソメラーゼ認識部位および選択的に、適宜1つまたは複数の組換え部位を有する増幅された第1の核酸分子を作製するプライマー対を使用するPCRによって増幅することができる。増幅された第1の核酸分子の両方の末端がこのように構築される場合には、5'側または3'側突出配列は、必ずしもではないが、一般に互いに異なる。次いで、本発明の方法により第2のヌクレオチド配列がコード配列の5'端に機能的に共有結合されるように、増幅された第1の核酸分子に、プロモーターなどの望ましい調節要素ならびにある種の態様では、(a)トポイソメラーゼ認識部位を伴う、1つまたは複数のトポイソメラーゼ認識部位、および/または(b)1つまたは複数の組換え部位を、組換えを促進する条件下において接触させることができる。
このような方法では、第2の(または他の)核酸分子はまた、2つまたはそれ以上の調節要素、例えばプロモーター、配列内リボソーム進入部位、およびATG開始メチオニンコドン等、または関心対象の他の配列、例えば互いに機能的に結合して、コード配列を含む第1の核酸分子の5'端に機能的に結合することができるエピトープ標識をコードする配列を含んでもよい。このような方法は、例えばコード配列の3'端に、本発明の方法により機能的に結合し、それによって発現可能な二本鎖組換え核酸分子を作製することができるポリアデニル化シグナルを含む、第3の核酸分子を接触させる段階をさらに含んでもよい。このように、本発明の方法は、機能的単位として転写、翻訳または両方を行うことができる機能的な二本鎖組換え核酸分子を作製する手段を提供する。本明細書において開示するように、部位特異的トポイソメラーゼによって接続される核酸分子の末端に、トポイソメラーゼ切断によって作製される相補的な5'側または3'側突出配列を導入すると、構築物中に望ましい方向の配向のヌクレオチド配列を有する二本鎖組換え核酸分子の作製が容易になる。
別の態様において、本発明の方法は、第1の核酸分子または第2の(または他の)核酸分子またはそれらの組み合わせが複数のヌクレオチド配列の1つであるように実施される。本明細書において使用する「複数」という用語は、第1または少なくとも第2のヌクレオチド配列に言及して使用する場合には、それらのヌクレオチド配列が関連するが異なることを意味する。本発明の目的のためには、複数の中の各ヌクレオチド配列が、1つまたは複数の末端および/または少なくとも1つの組換え部位に、少なくとも1つのトポイソメラーゼ認識部位またはその切断された形態を有するという点において、複数のヌクレオチド配列が「関連」している。さらに、複数のヌクレオチド配列は、例えばcDNAライブラリ、ヌクレオチド配列のコンビナトリアルライブラリ、ヌクレオチド配列の多様な集団等を含んでもよいという点において、複数のヌクレオチド配列は「異なる」。cDNAライブラリ、コンビナトリアルライブラリ、ヌクレオチド配列の多様な集団を含むライブラリ等を作製する方法は、当技術分野において既知である(例えば、それぞれ参照として本明細書に組み入れられる、米国特許第5,837,500号、米国特許第5,622,699号、米国特許第5,206,347号、ScottおよびSmith、Science 249: 386-390、1922、Marklandら、Gene 109: 13-19,1991、O'Connellら、Proc. Natl. Acad. Sci., USA 93:5883-5887、1996、TuerkおよびGold、Science 249: 505-510、1990、Goldら、Ann. Rev. Biochem. 64: 763-797、1995参照)。
本発明はさらに、プライマー対の少なくとも1つプライマーがトポイソメラーゼ認識部位またはその相補物をコードするPCRプライマー対、および選択的に、1つまたは複数の組換え部位を使用して第1のヌクレオチド配列の一部を増幅し、それによって第1の末端および第2の末端を有し、第1の末端もしくは第2の末端または両方が3'端および/または5'端にトポイソメラーゼ認識部位を有する第1の核酸分子を作製し;第1の核酸分子;第1の末端および第2の末端を有し、第1の末端もしくは第2の末端または両方が3'端および/または5'端にトポイソメラーゼ認識部位またはその切断産物を有する、少なくとも第2の核酸分子;ならびにトポイソメラーゼを接触させることによって、両方の鎖に共有結合された二本鎖組換え核酸分子を作製する方法を提供する(図1参照)。トポイソメラーゼ認識部位を有する第1の核酸分子の末端と、トポイソメラーゼ認識部位を有する少なくとも第2の核酸分子の末端とが結合することができる条件下において接触させる場合には、両方の鎖に共有結合された二本鎖組換え核酸分子が作製される。上記の方法で核酸分子が接続されると、得られる分子は、本明細書の別の箇所に記載されているような組換え反応に使用することができる。本発明は、本発明の方法によって作製される核酸分子、このような核酸分子を含む組成物、およびこのような核酸分子を使用する方法とをさらに含む。
本明細書に開示するように、1つまたは複数のトポイソメラーゼ認識部位、および選択的に、1つまたは複数の組換え部位を増幅された核酸分子の一方または両方の末端に組入れるように設計されたプライマーを使用するPCR方法は、本発明の方法に有用な核酸分子を作製するための便利な手段を提供する。ある種の態様において、プライマー対のプライマーの少なくとも1つは、トポイソメラーゼ認識部位に相補的なヌクレオチド配列および増幅される標的核酸分子の3'端に相補的なヌクレオチド配列を3'側から5'側方向に含むように設計される(すなわち、標的特異的部位)。また、プライマーは、部位特異的トポイソメラーゼによる増幅産物の切断の結果、望ましい5'側突出を形成するトポイソメラーゼ認識部位の相補物の5'側位置に、任意の鎖長(一般に約1〜100ヌクレオチド、通常約2〜20ヌクレオチド、特に約4〜12ヌクレオチド)の望ましいヌクレオチド配列を有してもよい。PCRプライマー対の第2のプライマーは増幅されるヌクレオチド配列の望ましい配列に相補的であってもよく、トポイソメラーゼ認識部位の相補物、部位特異的トポイソメラーゼによる切断の結果5'側突出を形成すると思われる配列、または任意の他の配列を適宜含んでもよい。
このようなプライマーは、例えばatt部位、lox部位等などの部位特異的な組入れ認識部位を含む、関心対象の任意の他の配列を含んでもよく、または上記に考察するように、トポイソメラーゼ認識部位を関心対象のこのような配列を含む核酸分子に組入れるためだけに使用してもよい。本発明の方法により作製され、att部位またはlox部位などの部位特異的な組入れ認識部位を有する二本鎖組換え核酸分子は、必要な組入れ部位、例えばatt部位またはlox部位をそれぞれ有し、例えばλIntおよびIHFタンパク質またはCreリコンビナーゼベクターのような、部位特異的な事象に必要な適当な酵素と接触する結果、遺伝子座等などの望ましい遺伝子座に特異的に組入れることができる。本発明の方法による両方の鎖に共有結合した二本鎖組換え核酸分子に、例えばattBまたはattP配列を組入れると、GATEWAY(商標)クローニングシステム(Invitrogen Corp.、カリフォルニア州ラジョラ)を使用して核酸分子を都合よく操作することができる。
一態様において、本発明の方法により作製される構築物は、PCR反応または他の増幅反応によってさらに増幅される。構築物は少なくとも1つの鎖に共有結合されるので、本発明の方法により作製される二本鎖組換え核酸分子の直接PCRが可能である。このように、PCRを使用して大量の構築物を作製することができる。さらに重要なことには、上記に示すようにPCRは、部分的な反応産物を得ることなく、本発明の方法により作製される望ましい産物だけを入手するインビトロ選択方法を提供する。例えば本発明の方法を使用して、プロモーターを含む第1の核酸分子、コード領域を含む第2の核酸分子、およびポリアデニル化シグナルを含む第3の核酸分子を、5'側から3'側方向に機能的に結合して含む、両方の鎖に共有結合した二本鎖組換え核酸分子を作製することができる。
本明細書に開示するように、所定の配向を有する構築物は、接続される核酸分子の末端に相補的な5'側突出配列を含ませることによって作製することができる。第1の核酸分子に相補的で、プロモーター配列の上流の第1のプライマー、および第3の核酸分子に相補的で、ポリアデニル化シグナルの下流の第2のプライマーを含むPCRプライマー対を選択することによって、プロモーター、コード領域およびポリアデニル化シグナルを含む機能的増幅産物を作製することができる。一方、第1または第2のプライマーは、それぞれ、部分的な産物にハイブリダイゼーションしないので、第1の核酸分子または第3の二本鎖ヌクレオチドを欠損する部分的な反応産物は増幅されない。また、第2の核酸分子を欠損する構築物は、第1および第3の核酸分子の5'側突出配列の相補性を欠損するため、作製されないと思われる。このように本発明の方法は、両方の鎖に共有結合された望ましい機能的な二本鎖組換え核酸分子を入手する手段を提供する。
このような方法におけるPCRの用途はさらに、関心対象の構築物を同定するために、本発明の方法により作製される大多数の核酸分子をスクリーニングする手段を提供する。自動ハイスループット分析にPCRを使用する方法は日常的で、既知であるので、本発明の方法はハイスループット系に使用するために容易に適合させられることが認識される。このような系を使用すると、大多数の構築物を並行してスクリーニングすることができ、部分的または不完全な反応産物を同定して廃棄し、それによってさらに別の実験で構築物を特徴づけ、構築物の機能を調査しなければならないと思われる時間および費用の浪費を防ぐことができる。
本発明の方法は、分子生物学分野に対して広い用途がある。以下にさらに詳細に考察するように、例えば本発明の方法を使用して、DNAまたはRNAプローブを標識することができ、指向的クローニングを実施することができ(実施例1.B参照)、センスまたはアンチセンスRNA分子を作製することができ(実施例2.A参照)、ツーハイブリッドアッセイを実施するためのベイト(bait)またはプレイ(prey)構築物を作製することができ(実施例2.C.参照)、直鎖状発現要素を作製することができ(実施例2.Aおよび2.B参照)およびインビトロにおける転写/翻訳共役アッセイに有用な構築物を作製することができる(実施例2.B参照)。例えば、両方の鎖に共有結合された二本鎖組換え核酸分子を作製する方法は、オープンリーディングフレームに結合したプロモーターまたは他の調節要素などの2つまたはそれ以上のヌクレオチド配列を含む直鎖状核酸分子からなる、直鎖状発現要素(LEE)を作製する手段を提供する(実施例1参照)。LEEは効率的に細胞にトランスフェクトし、それによってベクターに発現要素をクローニングする必要性を回避することが報告されている(SykesおよびJohnston、Nat. Biotechnol. 17: 355-359、1999)。LEEの成分は共有結合されなくても、ライゲーション反応により共有結合されてもよい。共有結合されないLEEの作製には、デオキシウリジン残基を有するPCRプライマーを使用して、各ヌクレオチド配列成分を増幅し、次いでPCR産物をウラシル-DNAグリコシラーゼで処理して、ハイブリダイゼーションすることができる突出末端を作製することが必要である。しかし、共有結合されていないこのようなLEEを使用するトランスフェクションの効率は変わりやすく、共有結合されているLEEの効率よりかなり低いこともある(SykesおよびJohnston、上記、1999年)。さらに、このようなLEEはPCR増幅の鋳型として好適ではない。その理由は、プライマー伸長反応は鋳型のニックを越えて進行することができず、従って停止し、不完全な反応産物を作製するからである。
本発明の方法は、共有結合されたLEEを作製するための、直接的かつ簡単な手段を提供し、それによってLEEを作製するために以前に記載されている、不便な追加の段階を必要とせず、共有結合されていないLEEを使用したときに観察されるトランスフェクション効率の変動を少なくする。例えば、関心対象のオープンリーディングフレームをコードする第1の核酸分子は、一方または両方の末端にトポイソメラーゼ認識部位またはその切断産物を有するように、本明細書において開示されているPCRによって増幅することができる。さらに、増幅された第1の核酸分子の部位特異的トポイソメラーゼによる切断の結果、所定の望ましい5'側突出配列を有するように、PCRプライマーを設計することができる。第2のヌクレオチド配列(適宜、第3またはそれ以上)は、トポイソメラーゼ認識部位またはその切断産物を有する以外に、調節要素、例えばプロモーター、エンハンサー、サイレンサー、スプライシング受容部位、翻訳開始部位、リボソーム認識部位もしくは配列内リボソーム進入部位、ポリアデニル化シグナル、開始メチオニンコドンもしくは停止コドンを含むもしくはコードしてもよく、またはエピトープ標識もしくは細胞画分化ドメインなどの任意の他の望ましい配列をコードしてもよい。好ましくは、第1の核酸分子に共有結合された第2の(または他の)核酸分子は、接続される第1の核酸分子の末端の5'側突出と相補的な、5'側突出配列を有する。トポイソメラーゼの存在下においてこのようなヌクレオチド配列と接触させると、プロモーターは、例えばオープンリーディングフレームの5'端に機能的に共有結合することができ、ポリアデニル化シグナルはオープンリーディングフレームの3'端に機能的に共有結合して、共有結合した機能的なLEEを作製することができる(実施例1参照)。
本発明に有用な調節要素の例は本明細書に開示されており、転写調節要素、翻訳調節要素、ヌクレオチド配列またはポリペプチドの細胞内への(細胞からの)輸送または局在化を促進する要素、検出可能な表現型を与える要素等を含む。転写調節要素には、例えばサイトメガロウイルス、モロニー白血病ウイルスおよびヘルペスウイルス由来のもの、ならびにメタロチオネイン、骨格アクチン、ホスホエノールピルビン酸カルボキシラーゼ、ホスホグリセリン酸、ジヒドロ葉酸還元酵素およびチミジンキナーゼをコードする遺伝子由来のものなどのプロモーター、ならびにラウス肉腫ウイルスLTRなどのウイルス末端反復配列(LTR)などのプロモーターおよびオペレーター、免疫グロブリンエンハンサーなどの構成的に活性であってもよいエンハンサー、またはSV40エンハンサーなどの誘導性であってもよいエンハンサー等が含まれる。例えば、メタロチオネインプロモーターは、銅、ニッケルまたはカドミウムイオンなどの金属イオンに暴露すると、高レベルの発現に誘導することができる、構成的に活性なプロモーターである。これと比較すると、テトラサイクリン(tet)誘導性プロモーターは、テトラサイクリンまたはテトラサイクリン類似物に暴露されると誘導されるが、そうでないと不活性であるプロモーターの一例である。転写調節要素は組織特異的調節要素、例えばコードされる産物の発現が、患者の筋肉細胞または培養中の細胞の混合集団、例えば器官培養物中の筋肉細胞に限定されるような、筋肉細胞特異的調節要素であってもよい。例えば、筋肉クレアチンキナーゼプロモーター(参照として本明細書に組み入れられる、Sternbergら、Mol. Cell. Biol. 8: 2896-2909、1988)およびミオシン軽鎖エンハンサー/プロモーター(参照として本明細書に組み入れられる、Donoghueら、Proc. Natl. Acad. Sci., USA 88: 5847-5851、1991)を含む筋肉細胞特異的調節要素は、当技術分野において既知である。細胞の特定の発育段階にだけ発現される他の組織特異的プロモーターおよび調節要素は、当技術分野において既知である。
別の態様において、本発明に使用される、または本発明を実施することによって作製されるヌクレオチド配列に含有される調節要素は、1つまたは複数のオペレーターであってもよい。数多くのオペレーターが当技術分野において既知である。本発明に使用するのに適切なオペレーターの一例は大腸菌(E. coli)のトリプトファンオペロンのトリプトファンオペレーターである。トリプトファンリプレッサーは、2分子のトリプトファンに結合している場合には、大腸菌(E. coli)トリプトファンオペレーターに結合し、プロモーターに対して適切に位置づけられている場合には、転写を遮断する。本発明に使用するのに適切なオペレーターの別の例は大腸菌(E. coli)テトラサイクリンオペロンのオペレーターである。大腸菌(E. coli)のテトラサイクリン耐性系の成分は真核細胞中で機能することも見出されており、遺伝子発現を調節するために使用されている。例えば、テトラサイクリンが存在しない場合にテトラサイクリンオペレーターに結合し、遺伝子転写を抑制するテトラサイクリンリプレッサーは、テトラサイクリンオペレーター配列を含むプロモーターからの転写を抑制するのに十分に高い濃度で、植物細胞中で発現されている(Gatzら、Plants 2:397-404(1992))。テトラサイクリンに調節された発現系は、例えば全体の開示内容が参照として本明細書に組み入れられる、米国特許第5,789,156号に記載されている。本発明に使用することができるオペレーターの別の例には、大腸菌(E. coli)のLacオペレーターおよびモリブデン酸塩輸送オペレーター/プロモーター系が含まれる(例えば、Croninら、Genes Dev. 15: 1461-1467(2001)およびGrundenら、J. Biol. Chem.、274: 24308-24315(1999)参照)。
従って、特定の態様において本発明は、原核または真核細胞において発現を調節するために使用することができる、1つまたは複数のオペレーターを含む核酸分子を作製する方法を提供する。当業者が認識しているように、オペレーターを含む核酸分子が、インビボまたはインビトロにおいて転写装置が存在する条件下に置かれている場合には、発現の調節は、リプレッサーおよび適当なリプレッサーがオペレーターに結合するのを促進する、1つまたは複数の代謝物を核酸分子に接触させることによって調節されることが多い。従って、本発明はさらに、オペレーターの機能を調節するリプレッサーをコードする核酸分子を作製する方法、ならびにこれらの方法によって作製される核酸分子、これらの分子を含む組成物、およびこれらの分子および組成物の用途を提供する。
本発明の方法により構築物を作製する際に有用な調節要素または他の要素は種々の方法で得ることができる。特に、要素の多くは市販のベクターに含まれ、それらから単離することができ、例えば本明細書に開示するPCR方法を使用して、一方または両方の末端にトポイソメラーゼ認識部位を有するように改変することができる。また、本明細書において有用な要素の配列または要素をコードする配列は一般に既知であり、文献に開示されている。多くの場合、要素、例えば多数の転写および翻訳調節要素ならびに細胞画分化ドメインは配列が比較的短く、従って要素または要素をコードするヌクレオチド配列を化学的合成しやすい。従って、一態様において、本発明の方法により二本鎖組換え核酸分子を作製する際に有用な本発明の組成物を含む要素、または本発明のキットに含まれる要素は化学的に合成することができ、要素の一方または両方の末端にトポイソメラーゼ認識部位を有するように、さらに部位特異的トポイソメラーゼによる切断により突出配列を有するように適宜合成することができる。
トポイソメラーゼ付加ベクターは以下の方法で作製することができる(Genome Res. 9: 383-392、1999):ベクターは、「付着末端」を残す制限酵素で直鎖状にされる。T4 DNAリガーゼなどのリガーゼを使用して、アダプターオリゴヌクレオチドを直鎖状DNAの両方の末端および両方の鎖にライゲーションする。アダプターオリゴヌクレオチドは、トポイソメラーゼによって切断され、ベクターの各3'端に共有結合型のトポイソメラーゼ-DNA複合体を捕捉するように、5'-CCCTT-3'ワクシニアI型トポイソメラーゼ認識配列を有し、位置づける。次いで、適合されたベクターを、ベクターの各末端の「トポイソメラーゼ部位」を完成させる、精製したワクシニアトポイソメラーゼおよびアニーリングオリゴヌクレオチドと共にインキュベーションする。アニーリングオリゴヌクレオチドは、5'-CCCTT-3'を含むオリゴヌクレオチドの、最後のTの反対側の「下の(bottom)」鎖に裂け目またはニックを残す作用をする。トポイソメラーゼ切断部位の「下流」にあるオリゴヌクレオチドアダプター断片(「脱離基」)はトポイソメラーゼ切断の結果放出され、トポイソメラーゼ-ベクター精製過程中に除去される。「脱離基」からの5'側ヒドロキシルが存在しない場合には、トポイソメラーゼはDNA末端との共有結合複合体に捕捉されて、トポイソメラーゼ付加ベクターを作製する。
核酸分子が本発明の方法により共有結合される場合には、作製される組換え核酸分子が望ましい構造を有し、望ましい機能を発揮するか、または望ましい発現産物をコードするように、ヌクレオチド配列は一般に、機能的に結合される。本明細書において使用する「機能的に結合する」という用語は、2つまたはそれ以上のヌクレオチド配列が、一方もしくは両方の配列またはそれらの組み合わせに起因する機能を発揮する単位として作用するように、互いに位置づけられることを意味する。「機能的に共有結合する」という用語は、本明細書において、一方または両方の鎖に共有結合された二本鎖組換え核酸分子を作製するために、本発明の方法により作製される機能的に結合されたヌクレオチド配列を言及するために本明細書において使用される。例えば、通常細胞のゲノムに結合している、オープンリーディングフレームを発現する方法と同様に、プロモーターがオープンリーディングフレームに調節作用を与えるように、オープンリーディングフレームを有するヌクレオチド配列をプロモーターに機能的に結合することができる。同様に、転写および翻訳の結果、キメラ融合ポリペプチドが作製されるように、オープンリーディングフレームを含む2つまたはそれ以上のヌクレオチド配列を、インフレームで機能的に結合することができる。
本発明の方法により作製される、一方または両方の鎖に共有結合された二本鎖組換え核酸分子は一般に直鎖状であるが、作製される構築物は環状二本鎖組換え核酸分子であってもよい。さらに、環状二本鎖組換え核酸分子がベクターの特徴を有し、例えば原核宿主細胞、真核宿主細胞または両方における複製に必要な調節要素を有し、抗生物質耐性等を与えるポリペプチドをコードするヌクレオチド配列を有することができるように、環状二本鎖組換え核酸分子を作製することができる。このような方法の利点は、本発明の方法により環状化された、作製された二本鎖組換え核酸分子を、構築物が増幅される適当な宿主細胞に形質転換またはトランスフェクトすることができることである。従って、本発明の方法により作製される大量の直鎖状二本鎖組換え核酸分子を作製するために使用できるPCRなどのインビトロにおける方法以外に、本発明の方法により作製される大量の環状産物を得るために宿主細胞を使用するインビボ方法を使用することができる。本発明によるトポイソメラーゼ認識部位を含む、細菌の複製起点、抗生物質耐性遺伝子等を含むこのような要素は、本明細書に開示する本発明のキットに含む有用な成分であってもよい。
一方または両方の鎖に共有結合される直鎖状二本鎖組換え核酸分子は、全て既知であり、市販品を購入することができるプラスミドベクターまたはバクテリオファージ、バキュロウイルス、レトロウイルス、レンチウイルス、アデノウイルス、ワクシニアウイルス、セムリキ森林熱ウイルスおよびアデノ関連ウイルスなどのウイルスベクターであってもよいベクターにクローニングすることもできることが認識されるべきである(Promega、ウィスコンシン州マディソン、Stratagene、カリフォルニア州ラジョラ、GIBCO/BRL、メリーランド州ゲイサースバーグ)。望ましい場合には、一方または両方の末端にトポイソメラーゼ認識部位またはその切断産物を有するように、例えばPCR方法を使用して本発明の方法によりベクターを環状化し、改変してもよく、または当業者により構築されてもよい(一般に、それぞれが参照として本明細書に組み入れられる、Meth. Enzymol. 第185巻、Goeddel編(Academic Press, Inc.、1990年)、Jolly、Canc. Gene Ther. 1: 51-64、1994、Flotte、J. Bioenerg. Biomemb. 25: 37-42、1993、Kirshenbaumら、J. Clin. Invest. 92: 381-387、1993参照)。
細胞、特に被験者の細胞に導入される、一方または両方の鎖に共有結合された二本鎖組換え核酸分子を作製する目的のために、本発明の方法が実施される場合には、ウイルス発現ベクターは特に有用となりうる。ウイルスベクターは、それらが比較的高い効率で宿主細胞に感染することができ、特異的な細胞種に感染することができ、宿主の特定の細胞に感染するように改変することができるという利点を有する。
ウイルスベクターは特定の宿主系に使用するために開発されており、例えば昆虫細胞に感染するバキュロウイルスベクター、レトロウイルスベクター、ヒト免疫不全ウイルス(HIV)に基づいたものなどの他のレンチウイルスベクター、哺乳類細胞に感染するアデノウイルスベクター、アデノ関連ウイルス(AAV)ベクター、ヘルペスウイルスベクター、ワクシニアウイルスベクター等が含まれる(それぞれ参照として本明細書に組み入れられる、MillerおよびRosman、BioTechniques 7: 980-990、1992、Andersonら、Nature 392: 25-30 補遺、1998、VermaおよびSomia、Nature 389: 239-242、1997、Wilson、New Engl. J. Med. 334: 1185-1187(1996)参照)。例えば、HIVに基づいたウイルスベクターは、T細胞に感染させるために使用することができ、アデノウイルスに基づいたウイルスベクターは、例えば気道上皮細胞に感染させるために使用することができ、ヘルペスウイルスに基づいたウイルスベクターは、神経細胞に感染させるために使用することができる。AAVベクターなどの他のベクターは広い宿主細胞範囲を持つことができ、従って、種々の細胞種に感染させるために使用することができるが、受容体媒介事象による標的特異性を変更するために特異的な受容体またはリガンドを用いてベクターを改変することもできる。
本発明の方法は、キメラポリペプチドをコードする核酸が作製されるように、オープンリーディングフレームを有する第1の核酸分子を、オープンリーディングフレームを有する第2の(および他の)核酸分子に機能的に共有結合するために使用することができる。キメラポリペプチドは、2つの(またはそれ以上の)コードされるペプチド(またはポリペプチド)が1つの産物に翻訳されている、すなわち、ペプチドがペプチド結合により共有結合されている融合ポリペプチドを含む。例えば、第1の核酸分子は、原形質膜局在化ドメイン、核局在化シグナル、ミトコンドリア膜局在化シグナル、小胞体局在化シグナル等などの細胞画分化ドメインまたは結合されたペプチドの細胞内転位を促進する、ヒト免疫不全ウイルスTATタンパク質ドメインなどのタンパク質産生ドメインをコードすることができる(それぞれ参照として本明細書に組み入れられる、Schwarzeら、Science 285: 1569-1572、1999、Derossiら、J. Biol. Chem.、271: 18188、1996、Hancockら、EMBO J. 10: 4033-4039、1991、Bussら、Mol. Cell. Biol. 8: 3960-3963、1988、米国特許第5,776,689号参照)。このようなドメインは、本発明の方法により共有結合されている、第2の核酸分子によってコードされたドメインおよびポリペプチドを含む融合ポリペプチドを、細胞内の特定の画分に標的化するために有用であり、または細胞からの分泌もしくは細胞内への流入に有用となりうる。このように本発明は、キメラポリペプチドをコードする、両方の鎖に共有結合された二本鎖組換え核酸分子を作製する手段を提供する。
本発明の方法により作製される核酸分子から発現される融合ポリペプチドは、発現された融合ポリペプチドを検出、単離等することができるように、検出可能な標識またはタグの特徴を有するペプチドも含むことができる。例えば、トポイソメラーゼ認識部位またはその切断産物を有する核酸は、アルカリ性ホスファターゼ、-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、ルシフェラーゼもしくは他の酵素などの酵素をコードしてもよく、またはポリヒスチジン配列(例えば、ヘキサヒスチジン)、V5エピトープ、c-mycエピトープ、ヘマグルチニンAエピトープ、FLAGエピトープ等などのペプチド標識をコードしてもよい。検出可能な標識を含む融合ポリペプチドの発現は、例えばルシフェラーゼを含む融合ポリペプチドにルシフェリンを添加する結果生じる発光を検出することによって、またはポリヒスチジン標識を含む融合ポリペプチドへのニッケルイオンの結合を検出することによって、適当な試薬を使用して検出することができる。同様に、標識を含む融合ポリペプチドの単離は、例えば抗c-mycエピトープ抗体が結合したカラムに、mycエピトープを含む融合ポリペプチドを通過させ、次いで結合した融合ポリペプチドを溶出することによって、またはニッケルイオンもしくはコバルトイオン親和性カラムにポリヒスチジン標識を含む融合ポリペプチドを通過させ、結合したポリペプチドを溶出することによって実施することができる。このような融合ポリペプチドを検出または単離する方法は、選択した検出可能な標識またはタグに基づいて、当業者に既知である(例えば、それぞれ参照として本明細書に組み入れられる、Hoppら、BioTechnology 6: 1204、1988、米国特許第5,011,912号参照)。
本発明の方法は、ヌクレオチド配列がプローブとして有用であるように、ヌクレオチド配列を化合物または小型の有機もしくは無機部分で標識するために使用することもできる。例えば、本発明の方法により共有結合された二本鎖組換え核酸分子を作製すると、作製された核酸分子が標識されているように、トポイソメラーゼ認識部位またはその切断産物を3'端に有する核酸分子を、アビジンまたはストレプトアビジンを使用して検出することができるビオチン、蛍光化合物(例えば、Cy3、Cy5、Fam、フルオロレセインまたはローダミン)、放射性核種(例えば、硫黄-35、テクネシウム-99、リン-32またはトリチウム)、常磁性スピン標識(例えば、炭素-13)、化学発光化合物等などの検出可能な部分に結合することができる。ヌクレオチド配列をこのような部分で検出できるように標識する方法は、当技術分野において既知である(例えば、参照として本明細書に組み入れられる、Hermanson、「生物接合技法(Bioconjugate Techniques)」(Academic Press 1996年)参照)。さらに、本発明により作製される二本鎖核酸分子の捕捉を可能にするために、検出可能な標識を使用することができる。最後に、検出可能な標識、例えばビオチンは、第1の核酸分子のトポイソメラーゼ付加末端を、第2の核酸分子の標識された末端にライゲーションするのを遮断するために使用することができ、それによって第2の核酸分子の未標識の末端へのライゲーションを誘導する方法を提供することができる。細胞画分化ドメインをコードするヌクレオチド配列または検出可能な標識もしくはタグを含むか、または転写もしくは翻訳調節要素を含む、本明細書に開示されているかまたはさもなければ当技術分野において既知のこのような要素は、本明細書に開示するキットの有用な成分となりうる。
本発明の方法は、例えばツーハイブリッドアッセイを実施するのに有用なキメラポリペプチドをコードする、二本鎖組換え核酸分子を便利に作製する手段を提供する。このような方法では、第1の核酸分子は、1つまたは複数の他のポリペプチドと特異的に相互作用する能力を有すると思われるか、またはその能力について検討される、ポリペプチドまたはその関連するドメインをコードする。第1の核酸分子は、一方または両方の末端にトポイソメラーゼ認識部位および適宜5'側突出配列を有するように、本明細書において開示するように改変される。第1の核酸分子が本発明の方法により共有結合される第2の核酸分子は、転写活性化ドメインまたはDNA結合ドメインをコードすることができ(実施例2.C)、トポイソメラーゼ認識部位、またはその切断産物、および結合される第1の核酸分子の末端の5'側突出配列に相補的な5'側突出配列を有する。トポイソメラーゼとの接触の結果、ヌクレオチド配列がトポイソメラーゼ付加されていない場合には、ツーハイブリッドアッセイを実施するのに有用な第1のハイブリッド(例えば、それぞれ参照として本明細書に組み入れられる、FieldsおよびSong、Nature 340: 245-246、1989、米国特許第5,283,173号、Fearonら、Proc. Natl. Acad. Sci., USA 89: 7958-7962、1992、Chienら、Proc. Natl. Acad. Sci., USA 88: 9578-9582、1991、Young、Biol. Reprod. 58: 302-311(1998)参照)、または逆ツーハイブリッドアッセイ法などの改良型のツーハイブリッドアッセイ法(参照として本明細書に組み入れられる、LeannaおよびHannink、Nucl. Acids Res. 24: 3341-3347、1996)、抑制されたトランス活性化因子系(参照として本明細書に組み入れられる、米国特許第5,885,779号)、タンパク質補充(recruitment)系(参照として本明細書に組み入れられる、米国特許第5,776,689号)等が作製される。第1のハイブリッドタンパク質のポリペプチドまたはそのドメインと相互作用する能力について試験される、複数のポリペプチドを含んでもよい、第2のハイブリッドタンパク質を作製するために、同様の方法を使用する。
同様に、キメラタンパク質を作製するこのような方法は、接続される末端の少なくとも1つの5'端に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)またはその切断産物を含み、トポイソメラーゼによる切断の結果、相補的な3'側突出をさらに含むことができる、第1および第2の核酸分子を使用して、1つの鎖に共有結合された二本鎖組換え核酸分子を作製する本発明の方法により実施することができる。
同様に、キメラタンパク質を作製するこのような方法は、接続される末端の少なくとも5'端に部位特異的トポイソメラーゼ認識部位またはその切断産物を含み、トポイソメラーゼによる切断の結果、相補的な3'側突出をさらに含むことができるか、または第1もしくは第2の核酸分子の一方が少なくとも一方の末端の5'端および3'端にトポイソメラーゼ認識部位またはその切断産物を含み、他方の核酸分子が接続される末端に3'側ヒドロキシル基および5'側ヒドロキシル基を有し、トポイソメラーゼによる切断の結果、トポイソメラーゼ付加核酸分子が相補的である5'側もしくは3'側突出を有し、接続される他方の核酸分子の末端の、それぞれ、5'側もしくは3'側突出または平滑末端へのハイブリダイゼーションを促進する、第1および第2の核酸分子を使用して、両方の鎖に共有結合された二本鎖組換え核酸分子を作製する本発明の方法により実施することができる。
別の態様において、本発明はまた、トポイソメラーゼ媒介クローニングの容易さおよび効率を伴って、DNA断片をクローニングベクターまたは発現ベクターに指向的に挿入する方法を提供する。本発明はまた、望ましい配向でクローニングされた挿入物を同定するのに必要な、手間のかかるスクリーニング過程を少なくするので、現在のクローニング系を上回る利点を有する。本発明のこの局面は、最も簡単な形態では、1つのトポイソメラーゼ分子が両方の3'末端に共有結合された直鎖状発現ベクターからなる。直鎖状ベクターの少なくとも一方の末端は5'側一本鎖突出を有するが、対向する末端は平滑であっても、T/Aクローニングの一塩基3'側T伸長部を有しても、第2の5'側一本鎖突出配列を有してもよい。これらの一本鎖配列突出は本明細書において「SSS」といわれ、任意の都合の良い配列からなってもよい。
本発明のこの局面によるトポイソメラーゼ付加クローニングベクターの構築は、例えば(pDONRベクター(図32参照)またはpDESTベクター(図33参照)であってもよい)ベクターのエンドヌクレアーゼ消化、次に合成オリゴヌクレオチドの相補的なアニーリング、およびワクシニアトポイソメラーゼIによるヘテロニ重鎖の部位特異的切断によって実施することができる。任意の適合性エンドヌクレアーゼによるベクターの消化は、特異的な付着末端を作製する。特注のオリゴヌクレオチドはこれらの付着末端にアニーリングすることができ、トポイソメラーゼIの修飾後、特注末端のベクターを形成する配列を有する(図32および33参照)。SSSの配列および鎖長は、使用者の希望に基づいて変わる。
本発明のこの局面により提供されるTOPO SSSベクターの一用途において、ベクターに挿入されるDNA断片はPCR産物である。特注プライマーによるPCR増幅後、挿入部位の一方または両方の末端にSSSを有するトポイソメラーゼI付加クローニングベクターに、産物を指向的に挿入することができる。所定のプライマー対の少なくとも一方のプライマーが5'末端に追加の配列を有するように、特注プライマーを設計することができる。付加される配列は、ベクターの一本鎖突出の配列に相補的であるように設計することができる。ベクターの5'側一本鎖突出とPCR産物の5'末端との間の相補性が、PCR産物のトポイソメラーゼ媒介ベクターへの指向的な挿入を媒介する。具体的には、ベクターの一方の末端とPCR産物の一方の末端だけが相補的なSSS領域を有するので、産物の挿入は指向的になる。トポイソメラーゼIは、PCR産物のベクターへのライゲーションを触媒する。
本発明のこの局面は、トポイソメラーゼが付加した(SSS)、突出している一本鎖片のDNAを有する、改変されたクローニングベクター、または「TOPO SSSベクター」を提供する。改変されたベクターは、その後の発現のためにPCR増幅されたオープンリーディングフレーム(ORF)、またはそれ以外に適切なオープンリーディングフレーム(ORF)の指向的な挿入を可能にし、トポイソメラーゼ媒介クローニングの効率を利用する。
上記のように、トポイソメラーゼは、DNA鎖の切断および再接続によりDNAの形態的状態を改変するクラスの酵素である(参照として本明細書に組み入れられる、Shumanら、米国特許第5,766,891号)。ワクシニアウイルスは、二本鎖DNAに部位特異的に一本鎖にニックを入れ、5'側ヒドロキシルにより再連結することができる、314アミノ酸I型トポイソメラーゼ酵素をコードする。部位特異的I型トポイソメラーゼには、ポックスウイルストポイソメラーゼなどのウイルストポイソメラーゼが含まれるが、これらに限定されない。ポックスウイルストポイソメラーゼの例には、ショープ線維腫ウイルスおよびORFウイルスが含まれる。他の部位特異的トポイソメラーゼは当業者に既知であり、本発明を実施するために使用することができる。
シュマン(Shuman)は、ワクシニアトポイソメラーゼは二本鎖DNAに結合し、一本鎖のホスホジエステル骨格を切断すると同時に、高いレベルの配列特異性を示すことを教示している。切断は、切断されやすい鎖のコンセンサスペンタピリミジン要素5'-(C/T)CCTT-3'または関連した配列で生じる。一態様において、切断されやすい結合は、二本鎖DNAの3'末端から2〜12 bpの範囲内に位置する。別の態様において、ワクシニアトポイソメラーゼによる切断可能な複合体形成は、切断部位の上流の二本鎖の6ヌクレオチドおよび下流の2ヌクレオチドを必要とする。ワクシニアトポイソメラーゼ切断可能な例には、+6/-6二本鎖
(配列番号:29)
、+8/-4二本鎖
(配列番号:30)
、+10/-2二本鎖
(配列番号:31)
、+11/-1二本鎖
(配列番号:32)
が含まれるが、これらに限定されない。
他の部位特異的I型トポイソメラーゼの例は当技術分野において既知である。これらの酵素は、出芽酵母(Saccharomyces cerevisiae)、サッカロミセス・ポンベ(Saccharomyces pombe)、およびテトラヒメナ(Tetrahymena)を含むが、これらに限定されない多数の生物によってコードされるが、これらの種のトポイソメラーゼI酵素は、ワクシニアよりコンセンサス配列に対する特異性が低い。それぞれ、(Lynn, R. M.、Bjornsti, M.、Caron, P. R.およびWang、J. C. (1989)ペプチド配列決定および部位特異的突然変異は、チロシン-727を出芽酵母(Saccharomyces cerevisiae)DNAトポイソメラーゼIの活性部位チロシンとして同定する(Peptide sequencing and site-directed mutagenesis identify tyrosine-727 as the active tyrosine of Saccharomyces cerevisiae DNA topoisomerase I)、Proc. Natl. Acad. Sci., USA 86: 3559-3563)、(Eng, W.、Pandit, S. D.、およびDternglanz, R.、(1989) 真核細胞DNAトポイソメラーゼIの活性部位チロシンのマッピング(Mapping of the active site tyrosine of eukaryotic DNA topoisomerase I)、J. Biol. Chem.、264: 13373-13376)および(Busk, H..、Thomsen, B.、Bonven, B. J.、Nielsen, O. F.、およびWestergaard, O. (1987)トポイソメラーゼIのヘキサデカマー認識を有する超らせんDNAの優先的な弛緩(Preferential relaxation of supercoiled DNA containing a hexadecameric recognition for topoisomerase I)、Nature、327: 638-640)。
本発明のこの局面に関して、本明細書において使用するドナーという用語は、3'末端付近に5'-CCCTT切断部位を有する二本鎖DNAを示し、アクセプターという用語は、5'側-OH端を有する二本鎖DNAを示す。トポイソメラーゼによる共有結合により活性化されると、ドナーは、SSS相補性を有するアクセプターまで移動する。
本発明のこの局面によると、トポイソメラーゼ修飾ベクターは、DNAセグメントの指向的な挿入を促進するために、少なくとも1つの5'側一本鎖突出配列を有するようにさらに適合させられる。好ましい態様において、クローニングされるセグメントは、結果として生じる組換えベクターから発現されるオープンリーディングフレーム(ORF)を構成するPCR産物である。ORFを増幅するために使用されるプライマーは、プライマー対の少なくとも1つプライマーが5'末端に追加の配列を有するように設計される。この配列は、本発明のトポイソメラーゼ修飾ベクターに存在する、5'側一本鎖突出の配列に相補的であるように設計される。
本発明のこの局面に従う、これらに限定されるわけではないが、特定の好ましい態様は、以下の実施例5〜8に詳細に記載されている。
本発明の方法を使用して集合される核酸分子は、任意の数の目的のために、直接使用されても、または増幅して使用されてもよい。図34を参照すると、本発明の方法を使用して集合される核酸セグメントは、任意の数の方法によって作製することができる。例えば、これらのセグメントは、当技術分野において既知の任意の方法によって得ることができる。核酸セグメントが、本発明の方法を使用した集成に適切な1つまたは複数(例えば、1、2、3、4等)の端および/または領域を持たない場合には、このような末端および/または領域を付加することができる。適切な端および/または領域は、例えばPCRを使用して核酸を増幅することによって、または1つもしくは複数(例えば、1、2、3、4等)のアダプターリンカー(1つまたは複数のトポイソメラーゼ認識部位を有するアダプターリンカー)を付加することによって付加することができる。次いで、本明細書の別の箇所に記載されている本発明の方法を使用して、適切な端および/または領域を有する核酸セグメントを集合させることができる。
図34に示すように、一旦集合されると、接続された核酸セグメントは(例えば、インビボまたはインビトロにおいて)増幅して、多くは本明細書の別の箇所に記載されている任意の数の方法または過程に使用することができる。または、集合された核酸セグメントを、インビトロにおける転写/翻訳、組換えクローニングなどの用途または細胞を形質転換もしくはトランスフェクトするために直接使用することができる。従って本発明は、核酸を製造するための多用途の組成物および方法を提供する。
本発明は、トポイソメラーゼおよび組換えを使用して核酸分子を接続するための組成物および方法を提供する。本発明の特定の態様において、核酸分子は1つまたは複数(例えば、1、2、3、4、5、6、7、8、9、10等)の組換え反応を受け、次いで、1つまたは複数(例えば、1、2、3、4等)のトポイソメラーゼによって触媒される鎖の共有結合に関与する方法によって、1つまたは複数(例えば、1、2、3、4、5、6、7、8、9、10等)の他の核酸分子に接続される。他の態様において、核酸分子は、1つまたは複数(例えば、1、2、3、4等)のトポイソメラーゼによって触媒される鎖の共有結合に関与する方法によって他の核酸分子に接続され、次いで1つまたは複数(例えば、1、2、3、4、5、6、7、8、9、10等)の組換え反応を受ける。当業者が認識しているように、本発明は、核酸分子のトポイソメラーゼ媒介接続または組換え反応の任意の特定の順序に縛られない。従って、一般に本発明は、組換え反応およびトポイソメラーゼの使用による核酸セグメントの接続を共に実施するための、組成物と方法とに関する。
従って、本発明はまた、本発明の方法および組成物により使用するためのアダプター-リンカー分子を提供する。本発明により提供され、本発明に関連して使用することができるアダプターリンカーは、トポイソメラーゼ部位および組換え部位の両方を有することができる。本発明の方法の一例は図35に略図で示されている。図35は、一塩基組換え部位を有するトポイソメラーゼ適合核酸セグメント(「アダプターリンカー」)を、挿入物と呼ばれる別の核酸セグメントに接続することを伴う過程を示す。これら2つの核酸セグメントは、本明細書に記載する任意のトポイソメラーゼ媒介過程によって接続することができる。
本発明のアダプターリンカーは、(1)1つまたは複数の組換え部位および/または(2)1つもしくは複数のトポイソメラーゼ認識部位または1つもしくは複数のトポイソメラーゼを含むことができる。特定の態様において、アダプターリンカーの1つまたは複数の組換え部位の少なくとも1つは、1つもしくは複数のトポイソメラーゼ認識部位または1つもしくは複数のトポイソメラーゼの少なくとも1つから、0、1、2、3、4、5、6、7、8、9、10、15または20ヌクレオチド以内に配置される。具体的な態様において、本発明のアダプターリンカーに存在する組換え部位はattL、attB、attPまたはattL組換え部位である。別の具体的な態様において、トポイソメラーゼ認識部位認識は、IB型トポイソメラーゼ、IA型トポイソメラーゼもしくはII型トポイソメラーゼの認識部位であり、またはトポイソメラーゼはIB型トポイソメラーゼ、IA型トポイソメラーゼもしくはII型トポイソメラーゼである。また、トポイソメラーゼ認識部位またはトポイソメラーゼは、組換えの結果、特定の組換え部位が産物分子に結合しているように、組換え部位に対して、本発明のアダプターリンカー内に配置することができる。例えば、リンカーが核酸分子に接続し、組換えが生じたとき、attBまたはattP部位のどちらかが、アダプターリンカーが接続した核酸分子に作製されるように、トポイソメラーゼ認識部位をアダプターリンカーのattL部位のどちらかの末端に配置することができる。従って、核酸分子にライゲーションし、組換えの結果、任意の数の種類の組換え部位が核酸分子産物に存在するように、組換え部位に対して位置づけられたトポイソメラーゼ認識部位および/またはトポイソメラーゼをアダプターリンカーは有することができる。
本発明はさらに、同じまたは異なる特異性を有する組換え部位を有するアダプターリンカーを使用して、任意の数の核酸セグメントを接続する方法、ならびに同じまたは異なる特異性を有する組換え部位を有するアダプターリンカー、およびこのようなアダプターリンカーを有するキットを提供する。例えば、attL1およびattL3部位がセグメントAの末端に存在し、attR3およびattR4部位がセグメントBの末端に存在し、attL4およびattL2部位がセグメントCの末端に存在するように、セグメントA、BおよびCと呼ばれる3つの別個のPCR産物をアダプターリンカーに接続することができる。従って、端または端付近にattR1およびattR2組換え部位を有する直鎖状ベクターによる組換えの結果、3つのPCR産物は全て互いに接続され、ベクターに挿入されて、環状核酸分子を作製する。上記の任意の数の変種が考えられ、本発明の範囲内に含まれる。
本発明はさらに、(1)同じもしくは異なる特異性を有する1つもしくは複数の組換え部位、および/または(2)1つもしくは複数のトポイソメラーゼもしくはトポイソメラーゼ認識部位を有する、2つもしくはそれ以上(例えば、2、3、4、5、6、7 8 、9等)のアダプターリンカーのセット、これらのアダプターリンカーセットを使用して1つまたは複数の組換え部位を有する核酸分子を作製する方法、ならびにこのようなアダプターリンカーセットまたはこれらのセットの個別のメンバーを含む組成物、これらのセットの1つまたは複数のアダプターリンカーで適合された核酸分子、およびこれらの核酸分子を使用する方法を提供する。
トポイソメラーゼにより媒介された集合後、1つまたは複数(例えば、1、2、3、4等)の適切な組換え部位を有する別の核酸セグメントを用いて集合された核酸分子を組換えることができる。図35に示す組換え部位はattL1およびattR1部位であるが、任意の適切な組換え部位を使用することができる(例えば、lox部位、attR部位、attL部位、attB部位、attP部位等)。別の適切な組換え部位は本明細書の別の箇所に記載されている。
従って、本発明は、トポイソメラーゼ認識部位および互いに組換えをする組換え部位を使用して、核酸分子を作製する方法を含む。本発明はまた、本発明の方法によって作製される分子および本発明の方法に使用される分子、ならびに本明細書に記載する方法によって作製される核酸分子を使用する方法を含む。
本発明はさらに、多数(例えば、2、3、4、5、6、7、8、9、10等)の組換え部位およびトポイソメラーゼ認識部位を使用して核酸分子を作製する方法、ならびにこのような方法によって作製される核酸分子およびこのような方法に使用される核酸分子を含む。さらに、これらの組換え部位は、多数(例えば、2、3、4、5、6、7、8、9、10等)の特異性を有することができる。また、トポイソメラーゼ認識部位は、これらの端を異なる核酸セグメントに接続する端を作製するように設計することができる。例えば、これらの末端は、トポイソメラーゼによる切断の結果、異なる「付着末端」を作製するように設計することができる。
上記に記載する方法の別の例を図36に示す。図36は、2つの核酸セグメントが、核酸セグメントの末端の鎖のトポイソメラーゼ媒介共有結合に関与する方法を使用して接続される方法を示す。次いで、結果として生じる核酸分子は組換えを受け、(1)複製起点を有する核酸セグメントに接続されているトポイソメラーゼ集合核酸分子、および(2)プロモーターによる負の選択マーカー(例えば、ccdB遺伝子)の置換が生じる。次いで、組換えた核酸産物を、両方の末端がトポイソメラーゼ適合され、正の選択マーカーを有する核酸セグメントに接続する。この最後の段階により、核酸分子は環状化する。
図36に示す環状核酸最終産物を、本明細書の別の箇所に記載されているような原核(例えば、細菌)細胞または真核(例えば、酵母、植物、動物(ヒトなどの哺乳類を含む)細胞であってもよい宿主細胞に導入することができる。さらに、この最終産物を含む細胞を、正および負の選択を使用して選択することができる。従って、例えば負の選択マーカーがプロモーターで置換されていない核酸分子を獲得した細胞は含まれないように選択される。本発明はさらに、任意の数の段階および成分が変更されている、図35および図36に記載するものと同様の方法および組成物を含む。変更されてもよい段階および成分の例は本明細書の別の箇所に記載されている。本発明はさらに、上記の方法によって作製される核酸分子を使用する方法を含む。
当業者が認識しているように、図35および36に示すものなどの方法に使用される核酸セグメントは任意の数の異なる要素を有してもよい。例えば、正の選択マーカーは図36に示すプロモーターに置換してもよい。さらに、図35に示す挿入物は、任意の数の機能を有する核酸を含むことができる。特に、挿入物が、転写される領域を含む場合には、転写物は、翻訳が行われていない状況で機能を果たすmRNAまたはRNAであってもよい。翻訳が行われていない状況で機能を果たすRNAの例には、トランスファーRNA(例えば、サプレッサーtRNA)、アンチセンスRNA、リボソームRNAおよびリボザイムが含まれる。また、本発明の方法によって接続および/または組換えられた核酸セグメントの2つまたはそれ以上は、1つまたは複数(例えば、1、2、3、4、5、6、7等)のオープンリーディングフレームの全てまたは一部を有することができる。このような例では、転写および翻訳により1つまたは複数の融合タンパク質が作製されるように、核酸セグメントを互いに接続することができる。本発明の方法に使用することができる別の核酸要素は本明細書の別の箇所に記載されている。
図35に示す過程の最終産物などの核酸分子が本発明の方法によって作製されたら、選択的に、核酸分子を1つまたは複数(例えば、1、2、3、4等)の他の核酸分子に接続しても、または末端を互いに接続することによって環状化してもよい。さらに、3つ以上の核酸分子を本発明の方法によって互いに接続する場合には、種々の中間分子または最終産物の末端を互いに接続して、これらの分子を環状化することができる。
本発明の方法は、相同組換えを実施するための組成物および方法、ならびに遺伝子組換え動物を産生する組成物および方法を提供する。外因性DNA構築物と同族染色体配列間の相同組換えによる遺伝子ターゲティングにより、ゲノムの所定の部位における正確な改変を行うことができる。遺伝子ターゲティングは、例えばマウス胚性幹(ES)細胞において十分に確立されており、大多数のマウス遺伝子の改変を実施するために使用されている(例えば、Brandonら、Curr. Biol. 5: 625-634、758-765、873-881(1995)参照)。遺伝子ターゲティングは体細胞においても実施することができる(例えば、Itzakiら、Nat. Genet. 15:258-265(1997)参照)。次いで、相同組換えによる遺伝子ターゲティングによって改変された細胞を当技術分野において既知の方法で操作して、遺伝子組換え動物を樹立することができる。
相同組換え用途に使用することができる本発明の組成物の一例は、図37に示す最終産物の核酸分子である。図37はさらに、このような組成物を作製するための方法の一例を示す。特に、図37は、トポイソメラーゼ適合された核酸セグメントの、トポイソメラーゼ適合されていない核酸セグメントへの接続を示す。この例では、核酸の最終産物の設計者が染色体内に組入れようとしている、挿入物と本明細書において呼ぶ核酸セグメントに、2つの組換え部位の間に位置づけられた(1)正の選択マーカーおよび(2)負の選択マーカーを有する領域が隣接している。次いで、組換えを使用して、2つの負の選択マーカーを、最終産物を組入れる染色体領域に相同性を有する核酸と置換する(図37の標識した「HR1」および「HR2」)。
本発明を実施する際に使用する相同性領域は、核酸分子を組入れる細胞の染色体によって変わる。さらに、多くの例において、相同性領域は、特定の生物の細胞への導入を促進するように選択される。このような生物は単細胞生物(例えば、酵母、原生動物等)または多細胞生物(例えば、植物、動物等)であってもよい。
従って、本発明は、相同組換えを実施するための核酸分子および組成物、ならびにこれらの分子および組成物に関与する相同組換えにより作製される細胞を提供する。本発明の方法は、多数の核酸セグメントの接続に使用することができる。例えば、図38は、トポイソメラーゼを使用して4つの核酸セグメントを接続して、末端付近に組換え部位(標識された「L1」および「L2」)が配置された直鎖状核酸分子を作製する略図を示す。第1段階では、attL1組換え部位およびattL2組換え部位を有するトポイソメラーゼ適合核酸セグメントを、トポイソメラーゼを使用して2つの他の核酸セグメントに接続する。この特定の例では、互いに接続される端の各鎖はトポイソメラーゼ分子に共有結合される。従って、核酸鎖のトポイソメラーゼ媒介性接続の結果、接続点にニックは存在しない。第2の段階において、LR CLONASE(商標)の存在下において、attLおよびattR組換え部位間の組換えを可能にする条件下において、トポイソメラーゼ集合された核酸セグメントに、複製起点(標識された「ori」)、正の選択マーカー(標識された「PM」)、attR1組換え部位およびattR2組換え部位を接触させる。特定のこのような方法において、例えばTOPO-適合ベクターを室温(例えば、約20〜20℃)において約5〜30分(好ましくは約10分)間、1つまたは複数の核酸セグメント(例えば、1つまたは複数のPCR産物)と共にインキュベーションし、約80℃において約20分間インキュベーションして反応物を熱処理し、次いで、LR反応のインキュベーション時間を約3時間に延長することを除いて、製造業者の指示(Invitrogen Corporation)により反応混合物を標準的なLR反応に使用する。組換えにより、attB1およびattB2組換え部位によって起点および選択マーカーから分離された種々の出発核酸セグメントを含む、環状核酸分子が形成される。当業者が認識しているように、任意の適切な組換え部位を、この図に示すatt組換え部位の代わりに使用してもよい。従って、本発明はまた、このような核酸を含む組成物、このような核酸を作製するために使用される組成物、ならびに本明細書の別の箇所に記載されている本発明の組換えおよびトポイソメラーゼ媒介性接続における、このような核酸および組成物の用途を提供する。
本発明はさらに、第2の核酸分子と1つまたは複数の相同性領域を有する第1の核酸分子を使用して、第2の核酸分子の核酸を第1の核酸分子に挿入するクローニング反応を実施するのに適切な核酸分子を提供する。本発明はさらに、このようなクローニング反応を実施するための組成物および方法を提供する。
上記の方法の一例は、全体の開示内容が参照として本明細書に組み入れられる、PCT国際公開公報第01/04288号に記載されているRecE/Tである。典型的には、RecE/Tクローニングでは、直鎖状の第1の核酸分子(例えば、ベクター)を、(1)「第2の核酸分子」と本明細書において呼ばれる、細胞(例えば、プラスミド、細菌人工染色体、天然染色体等)に存在する核酸分子の、2つの別個の隣接領域と相同性を有する端の領域(例えば、鎖長が約20〜約30、約20〜約40、約20〜約50、約30〜約40、約40〜約50、約40〜約60、約40〜約80、約50〜約90等である核酸領域)、(2)選択マーカーおよび(3)複製起点を有する細胞に導入する。鎖状の第1の核酸分子は、一般に、環状になる場合に複製する。さらに、第1の核酸分子は、典型的には、第2の核酸分子による組換えを受け、相同性領域間に介在している第2の核酸分子から核酸を獲得した場合に環状になる。このような態様では、第1の核酸分子の相同性領域は、典型的には、第2の核酸分子と比較したとき、逆方向である。一般に、組換えが生じる細胞はRecE/TまたはRedAlpha/Betaなどのリコンビナーゼを発現するものである。従って本発明は、一部には、RecE/Tクローニングを実施する方法、このような方法によって作製される核酸分子、ならびにこのような核酸分子を含む組成物、このような核酸分子および組成物を使用する方法を提供する。
RecE/T方法の改良法を使用して、数多くの異なる最終産物を作製することができる。例えば、相同性領域が種々の方法で配列されている場合には、(1)第2の核酸分子に挿入するか、または(2)第2の核酸分子から核酸を欠損させるように、第1の核酸分子を設計することができる。典型的には、第2の核酸分子を第2の核酸分子に挿入することが望ましい場合には、第1の核酸分子の相同性領域は、第2の核酸分子の相同性領域に対して同じ方向である。さらに、第2の核酸分子から核酸を欠損させることが望ましい場合には、第1の核酸分子の相同性領域は、一般に、第2の核酸分子の相同性領域に対して逆方向である。また、第1の核酸分子を第2の核酸分子に挿入することが望ましい場合には、典型的には、第1の核酸分子は複製起点がない。本発明は、上記の方法を実施する方法を提供する。本発明はまた、上記の方法に使用するための核酸分子および組成物を提供する。
本発明はまた、トポイソメラーゼおよび組換え部位を使用して、1段階の方法で2つの核酸セグメントを接続して、環状核酸分子を作製することができる。この態様の一例を図39に図示し、核酸セグメントの1つは、attL1組換え部位(標識された「L1」)、プロモーター(標識された「P」)および一方の末端に共有結合されたトポイソメラーゼを有する。他の核酸セグメントは、attR1組換え部位(標識された「R1」)、オープンリーディングフレーム(標識された「ORF」)、複製起点(標識された「ORI」)、正の選択マーカー(標識された「PM」)および一方の末端に共有結合されたトポイソメラーゼ分子を有する。従って、LR CLONASE(商標)の存在下において、attLおよびattR組換え部位間の組換えならびに核酸鎖のトポイソメラーゼ媒介接続を可能にする条件下において、これら2つの核酸セグメントを互いに接触させると、示す構造を有する環状分子が形成される。特定のこのような方法において、例えばTOPO-適合ベクターを室温(例えば、約20〜20℃)において約5〜30分(好ましくは約10分)間、1つまたは複数の核酸セグメント(例えば、1つまたは複数のPCR産物)と共にインキュベーションし、約80℃において約20分インキュベーションして反応物を熱処理し、次いで、LR反応のインキュベーション時間を約3時間に延長することを除いて、製造業者の指示(Invitrogen Corporation)により反応混合物を標準的なLR反応に使用する。当業者が認識しているように、任意の適切な組換え部位を、この図に示すatt組換え部位の代わりに使用してもよい。
本発明はまた、トポイソメラーゼ媒介方法を使用して、2つの核酸セグメントを接続して、環状核酸セグメントを作製することもできる。本発明のこの局面の一態様の略図を図40に例示する。図40に示すように、環状分子は、attL1組換え部位とattL2組換え部位(標識された「L1」および「L2」)との間に位置づけられたオープンリーディングフレーム(標識された「ORF」)を有する。次いで、トポイソメラーゼ集合された産物は、attR1およびattR2組換え部位を有する別の環状分子による組換えを受けて、attB1組換え部位とattB2組換え部位との間に位置づけられたオープンリーディングフレームを有する第3の環状核酸分子を作製する。さらに、オープンリーディングフレームはプロモーターに機能的に結合される。従って、この方法によって作製される第1の核酸分子は発現構築物である。当業者が認識しているように、任意の適切な組換え部位を、この図に示すatt組換え部位の代わりに使用してもよい。
本明細書に開示するように、第1の核酸分子は、例えばcDNAライブラリ、ヌクレオチド配列のコンビナトリアルライブラリまたは種々のヌクレオチド配列集団のような複数のヌクレオチド配列の1つであってもよい。このように、本発明の方法の特に有用な態様は、ポリペプチド集団間に生じるタンパク質-タンパク質相互作用を同定するためにハイスループットツーハイブリッドアッセイ法を実施するための、キメラポリペプチドをコードする組換えポリペプチドを作製する段階にある(それぞれ参照として本明細書に組み入れられる、米国特許第6,057,101号および米国特許第6,083,693号参照)。このような方法では、ポリペプチドをコードする2つの集団(複数)のヌクレオチド配列を検討し、この複数性は、関連しているが異なる数個のヌクレオチド配列から何万ものこのような配列からなる複雑さを有する。例えば、PCRプライマー対の少なくとも一方のプライマーが(a)少なくとも1つのトポイソメラーゼ認識部位もしくはその相補鎖、または(b) 少なくとも1つの組換え部位を含むPCRプライマー対を使用して、各ヌクレオチド配列を複数に増幅するために本発明の方法を実施することによって、それぞれが(a)少なくとも1つのトポイソメラーゼ認識部位を含む増幅された複数のヌクレオチド配列に、少なくとも1つのトポイソメラーゼおよび少なくとも1つのトポイソメラーゼ認識部位を有し、転写活性化ドメインまたはDNA結合ドメインをコードするヌクレオチド配列を接触させる、またはそれぞれが(b)少なくとも1つの組換え部位を含む増幅された複数のヌクレオチド配列に、少なくとも1つのトポイソメラーゼおよび少なくとも1つの組換え部位を有し、転写活性化ドメインまたはDNA結合ドメインをコードするヌクレオチド配列を接触させることによって、キメラベイトポリペプチド集団およびキメラプレイポリペプチド集団をコードする共有結合された組換えポリペプチドを作製することができる。
本発明の方法を実施する際には、第1の核酸分子はまた、例えばリボプローブ、アンチセンスヌクレオチド配列、リボザイムまたは3本鎖ヌクレオチド配列として機能することができるリボ核酸(RNA)分子をコードしてもよく、または第1のヌクレオチド配列からRNAを発現するのに有用な調節要素をコードしてもよい(実施例2.A)。例えば、大量のRNAを作製することが望ましい場合には、本発明の方法を実施するための第2の核酸分子成分は、T7、T3またはSP6 RNAポリメラーゼプロモーターなどのRNAポリメラーゼプロモーターを含むことができる。RNA分子が細胞内で発現される場合、例えばアンチセンス分子が哺乳類細胞内で発現される場合には、第2の(または他の)核酸分子は、哺乳類細胞内で活性であるプロモーター、特に標的細胞においてだけ活性である組織特異的プロモーターを含むことができる。さらに、RNA分子が、例えばインビトロにおける転写/翻訳反応において翻訳される場合には、第1のヌクレオチド配列または第2の(もしくは他の)ヌクレオチド配列は、適当な翻訳調節要素を含むことができる(実施例2.B)。
本発明の方法はまた、インビボにおいて遺伝子のサイレンシングを可能にする構築物を作製するために使用することができる。遺伝子をサイレンシングする方法は、RNA干渉(RNAi)と呼ばれる二本鎖RNAの作製に関与する(例えば、Metteら、EMBO J.、19: 5194-5201(2000)参照)。RNAiが機能すると考えられる機序は、フジョセ(Fjose)ら、Biotechnol. Annu. Rev. 7: 31-57(2001)に総説が示されているが、特異的なRNA分子の分解を誘導する二本鎖RNAiの能力に基づいていると思われる。この機序は、リボヌクレアーゼを相同なRNA標的(例えば、mRNA標的)に誘導する短いRNAへと、二本鎖RNAを変換することに関与すると報告されている。本発明の方法は、RNAiなどの分子を作製するために数多くの方法において使用することができる。従って、本発明の核酸分子の発現産物は、遺伝子発現をサイレンシングするために使用することができる。
RNAiを作製するように設計された核酸分子の一例は、両方の鎖に対応するRNAが2つの別個の転写物としてまたは同転写物の一部として作製されるように、核酸セグメントが1つまたは複数のプロモーターに接続している分子である。例えば核酸分子は、介在する核酸セグメントによって、オープンリーディングフレームの2つのコピーに、異なる方向へ転写を誘導する2つのプロモーターが接続される、本発明の方法を使用して作製されてもよい。従って、プロモーターの一方はセンス鎖mRNAの転写を誘導し、他方のプロモーターはアンチセンスmRNAの転写を誘導する。RNAiを作製するために使用してもよい核酸分子の別の例は、各末端においてオープンリーディングフレームの反対方向への転写を誘導するプロモーターが隣接しているものである。第3の例として、一方の端に「スナップバック」領域(例えば、鎖長が6、7、8、9、10等のヌクレオチドである領域)を有するRNAをコードする核酸分子から、二本鎖RNAを作製することができる。従って、この種類のRNA転写物は一方の端または端付近にヘアピンターンを形成する。このようなRNA分子を、適当な条件下において、RNA依存的RNAポリメラーゼの存在下においてインキュベーションすると、ヘアピンによって作製される二本鎖領域を使用して、第2の鎖の合成を刺激して二本鎖RNA分子を作製することができる。
上記の核酸分子などのRNAiを作製するように設計された核酸セグメントは、全長の遺伝子またはオープンリーディングフレームに対応する必要はない。例えば、核酸セグメントがORFの全てもしくは一部に対応するか、またはORFの全てもしくは一部に対応しないRNA分子をコードする場合には、セグメントはORFの一部(例えば、ORFの5'末端または3'末端の約15、約16、約17、約18、約19、約20、約21、約22、約23、約24、約25、約26、約27、約28、約29、約30、約40、約50、約60等のヌクレオチド)に対応するだけでよい。
従って、特定の態様において、本発明は、少なくとも3つのセグメントを含む核酸分子を作製する方法を提供する。いくつかの態様において、これらのセグメントの少なくとも2つは、少なくとも1つの領域の配列の同一性を共有する(例えば、ヌクレオチド鎖長が少なくとも約15、少なくとも約16、少なくとも約17、少なくとも約18、少なくとも約19、少なくとも約20、少なくとも約21、少なくとも約22、少なくとも約23、少なくとも約24、少なくとも約25、少なくとも約26、少なくとも約27、少なくとも約28、少なくとも約29、少なくとも約30、少なくとも約40、少なくとも約50、少なくとも約60、少なくとも約70、少なくとも約80、少なくとも約90、少なくとも約100のヌクレオチド等)。他の態様において、(例えば、センスおよびアンチセンス転写物を作製するために)1つの核酸セグメントに、分子の内側部分の反対方向の転写を与えることができる領域が隣接している。本発明はさらに、本発明の方法によって作製される核酸分子、および遺伝子発現を阻害または特定のRNA分子の分解を促進するためのこのような分子の用途を提供する。
本発明はまた、アンチセンスRNA(例えば、アンチセンスmRNA)を発現するために使用できる、核酸分子を作製するための方法を提供する。RNAiに使用できる核酸分子の作製について上記したものと同様の方法を使用してもよいが、典型的にはアンチセンス鎖だけが、アンチセンスRNAを作製するために使用できる本発明の方法によって作製される分子に転写される。
関連する態様において、センスRNAまたはアンチセンスRNAの転写を誘導するプロモーターは構成的(例えば、CMVプロモーター、SV40プロモーター等)、誘導性(例えば、メタロチオネインプロモーター等)または抑制性であってもよい。従って、例えばセンスRNAおよびアンチセンスRNAの転写を誘導するために2つの異なる誘導性プロモーターを使用することができる。このような例において、センスRNA、アンチセンスRNAまたはセンスRNA、およびアンチセンスRNAの作製を誘導するためにプロモーター活性化を使用することができる。さらに、作製されるセンスRNAおよび/またはアンチセンスRNAの量は、例えばプロモーターの漸増的誘導および/または抑制を使用することによって関連させることができる。
RNAiおよびアンチセンスRNAなどの化合物の用途に関与する遺伝子サイレンシング方法は、例えば遺伝子の機能を同定するために特に有用である。さらに具体的には、遺伝子サイレンシング方法は、細胞または生物において1つまたは複数の遺伝子の発現を低下または防止するために使用することができる。次いで、遺伝子機能の選択的な阻害に関連する表現型発現を使用して、「サイレンシングされた」遺伝子に役割を割り当てることができる。一例として、チャン(Chuang)ら、Proc. Natl. Acad. Sci. (USA) 97: 4985-4990(2000)は、RNAiのインビボにおける産生はシロイヌナズナ(Arabidopsis thaliana)における遺伝子活性を変更することができることを証明している。従って、本発明は、RNAiおよびアンチセンスRNAの発現を含む、細胞および組織における核酸分子の発現を調節する方法を提供する。本発明はさらに、DNA分子の一本鎖または両方の鎖に対応するRNAを作製するために使用できる、核酸分子を作製する方法を提供する。
従って本発明は、センスRNAおよび/またはアンチセンスRNAの発現を含む、核酸分子のインビボ(例えば、細胞および組織内)および/またはインビトロにおける発現を調節する方法を提供する。本発明はさらに、核酸分子(例えば、DNA分子)の一方または両方の鎖に対応するRNAを作製するために使用できる、核酸分子を作製する方法を提供する。本発明はまた、上記の方法を実施するための組成物、および上記方法によって作製される核酸分子(例えば、RNAおよびDNA分子)を提供する。
本発明はまた、リボザイムに関与する遺伝子サイレンシングのための化合物および方法に関する。特に、本発明は、1)標的遺伝子に対応するアンチセンスRNAおよび2)RNAを切断する1つまたは複数のリボザイム(例えば、ハンマーヘッドリボザイム、ヘアピンリボザイム、デルタリボザイム、テトラヒメナ(Tetrahymena)L-21リボザイム等)を含む、アンチセンスRNA/リボザイム融合物を提供する。このような融合物を発現するベクター、このようなベクターを作製する方法、および遺伝子発現を抑制するためにこのようなベクターを使用する方法が、本発明によりさらに提供されている。
転写物のアンチセンスRNA部分は、特定のmRNA分子にハイブリダイゼーションするように設計することができるので、リボザイムに融合したアンチセンス分子の発現を、例えば細胞内の特定のRNA分子を切断するために使用することができる。さらに、ハイブリダイゼーションしているRNA分子を切断するために、転写物のリボザイム部分を設計することができる。例えば、リボザイムは、二本鎖RNAを切断するものであってもよい(例えば、テトラヒメナ(Tetrahymena)L-21リボザイム)。
本発明の方法は、標的DNA配列に部位特異的に挿入することができる、発現可能な二本鎖組換え核酸分子を作製するのに特に有用となりうる。標的DNA配列は任意のDNA配列であってもよく、特にゲノムDNA配列であってもよく、好ましくはヌクレオチド配列の一部または全てが既知である遺伝子であってもよい。本発明の方法は、第1の末端および第2の末端を有し、ポリペプチド、例えば選択マーカーをコードし、各末端の3'端に少なくとも1つのトポイソメラーゼ認識部、位および/もしくは少なくとも1つの組換え部位またはその切断産物、ならびに選択的に、各末端の5'端にヒドロキシル基を含み、好ましくは、5'端は、互いに異なる5'側突出配列を含む第1の核酸分子を使用して、本発明の方法により第1の核酸分子を第1および第2のPCR増幅産物に共有結合することによって実施することができる。第1および第2の増幅産物は、構築物が挿入される部位の上流および下流の配列から作製され、各増幅産物は、少なくとも1つのトポイソメラーゼ認識部位および選択的に、少なくとも1つの組換え部位、好ましくは部位特異的トポイソメラーゼとの接触により作製される5'側突出配列を有する。好ましくは、それぞれを第1の核酸分子の所定の末端に接続することができるように、第1および第2の増幅産物は異なる5'側突出配列を有する。このような方法は同様に、一方または両方の末端の5'端に少なくとも1つのトポイソメラーゼ認識部位、および選択的に、少なくとも1つの組換え部位またはその切断産物を含み、トポイソメラーゼによる切断の結果、トポイソメラーゼ付加分子が、トポイソメラーゼを含む一方または両方の末端に3'側突出を含む二本鎖増幅産物を使用して実施することができる。また本発明の方法は、一方または両方の末端の5'端および3'端またはその付近に、トポイソメラーゼ認識部位および選択的に、組換え部位またはその切断産物を含み、トポイソメラーゼによる切断の結果、好ましくは、トポイソメラーゼを含む一方または両方の末端に5'側または3'側突出を有する二本鎖増幅産物を使用して実施することができる。上記の方法によって核酸分子が接続されると、得られる分子は本明細書の別の箇所に記載されているような組換え反応に使用することができる。
第1および第2の増幅産物は、2セットのPCRプライマー対を使用して作製することができる。2セットのPCRプライマー対は、Taqポリメラーゼなどの適当なポリメラーゼおよび増幅される配列を含む鋳型の存在下において、選択マーカーの挿入部位の上流側に隣接する標的DNA配列の部分、および下流側に隣接する標的DNA配列の部分を、プライマーが増幅するように選択することができる。また、PCRプライマー対のセットは、増幅産物がトポイソメラーゼ認識部位を有し、部位特異的トポイソメラーゼによる切断により、選択マーカーに共有結合される末端に5'側突出配列を有するように設計することができる。このように、第1のPCRプライマー対は、1)増幅産物が共有結合される選択マーカーの末端の5'側突出配列に相補的なヌクレオチド配列、トポイソメラーゼ認識部位に相補的なヌクレオチド配列、および挿入部位の上流の標的DNA配列の3'側配列に相補的なヌクレオチド配列を、5'側から3'側の方向に含む第1のプライマー、ならびに2)第1のプライマーが相補的である3'側配列の上流、すなわち、挿入部位の下流の標的ゲノムDNAのヌクレオチド配列を含む第2のプライマーを含む。第2のPCRプライマー対は、1)共有結合される選択マーカーの末端の5'側突出配列に相補的なヌクレオチド配列、トポイソメラーゼ認識部位に相補的なヌクレオチド配列、および標的DNA配列の5'側配列のヌクレオチド配列を5'側から3'側に含み、標的ゲノムDNAの5'側配列は、第1のPCRプライマー対の第1のプライマーが相補的である、標的DNA配列の3'側配列の下流であり、第2のプライマー対の第2のプライマーは、第1のプライマーに含まれる標的ゲノムDNAの5'側配列の下流である、標的DNA配列の3'側配列に相補的なヌクレオチド配列を含む第1のプライマーを含む。当業者は、標的ゲノムDNAに相補的なプライマーの配列は、標的DNAの配列に基づいて選択されることを認識している。これらのプライマーはさらに、1つまたは複数の組換え部位を含むことができる。
選択マーカー、第1および第2の増幅産物、ならびにトポイソメラーゼ(分子がトポイソメラーゼ付加されていない場合)を含む核酸分子を接触させると、両方の鎖に共有結合された二本鎖組換え核酸分子が本発明の方法により作製される。作製された二本鎖組換え核酸分子は、標的ゲノムDNAの上流および下流配列に特異的なPCRプライマーを使用して適宜さらに増幅することができ、機能的な構築物だけが確実に増幅される。作製された二本鎖組換え核酸分子は、例えば細胞の遺伝子の機能をノックアウトするか、または作製された組換え核酸分子を含む細胞に新規表現型を与えるために、ゲノムに相同組換えを実施するのに有用である。本発明の方法は、作製された二本鎖組換え核酸分子がゲノムに安定に維持されている遺伝子組換え非ヒト生物を産生するために、さらに使用することができる。
本発明の方法はまた、複数の核酸分子のそれぞれを含む、関心対象の核酸分子の一方または両方の末端に、アダプターまたはリンカー配列を共有結合するのに有用である。例えば、第1の核酸分子の両方の末端にリンカーを配置することが望ましい場合には、本発明の方法は、一方または両方の3'端または5'端にトポイソメラーゼ認識部位またはその切断産物を有し、両方の5'端のヒドロキシル基、および1つまたは複数の組換え部位を含むことができる第1の核酸分子;それぞれが適当な3'端または5'端にトポイソメラーゼ認識部位またはその切断産物を含むことができ、同じ末端の5'側ヒドロキシル基および1つまたは複数の組換え部位を適宜含むことができる、第2の核酸分子;ならびに少なくとも第3の二本鎖ヌクレオチド配列を、トポイソメラーゼに接触させることによって実施することができる。適当な端は、リンカーが、少なくとも一方の鎖の第1のヌクレオチド配列に共有結合される末端である。一態様において、一方または両方のリンカー配列は、リンカーが共有結合する第1の核酸分子の末端の5'端の配列に相補的である突出配列を有し、それによって、適切な(所定の)方向でのヌクレオチド配列の最初の結合を促進する(例えば、図2および実施例1.B参照)。このような方法を実施する際に、第2のおよび少なくとも第3のヌクレオチド配列を含むリンカー配列は同じであっても、異なってもよい。
図14は、配列の一方の末端の5'端に結合したトポイソメラーゼ(例えば、IA型トポイソメラーゼ)を有し、同じ末端が3'側突出をさらに含む核酸分子を作製する方法の一例を示す(図14の(4)参照)。段階Aにおいて、トポイソメラーゼが修飾するヌクレオチド配列を、「付着末端」を作製する制限酵素で消化する。次いで、制限酵素処理したヌクレオチド配列を段階Bにおいて、5'端に接続したトポイソメラーゼおよびリガーゼ(例えば、T4 DNAリガーゼなどのDNAリガーゼ)を有する、直鎖状の一本鎖ヌクレオチド配列を接触させる。直鎖状の一本鎖ヌクレオチド配列はまた、2つの分子がハイブリダイゼーションするように、制限酵素によって作製される「付着」末端との十分な配列相補性を共有する領域を3'端に有する。従って、段階Bにおいて、2つのヌクレオチド配列が互いにライゲーションされる。段階Cにおいて、第2段階の産物を、段階Bにおいて作製された直鎖状の一本鎖核酸分子の一部との配列相補性を共有する第3のヌクレオチド配列、およびリガーゼに接触させる。(4)に示す段階Cの産物は、一方の末端の5'端に接続されたトポイソメラーゼおよび同じ末端に、3'側突出を有する核酸分子である。例示した方法の数多くの変種が本発明の範囲内であることが認識される。例えば、一方の末端の3'端に接続されたトポイソメラーゼを含む、またはトポイソメラーゼが接続される末端に5'側突出を有するか、もしくは平滑末端である核酸分子を作製するために、同様の方法を実施することができる。別の例では、図14において番号3と標識したヌクレオチド配列は以下の方法で作製することができる:核酸分子を制限酵素で消化して、IA型トポイソメラーゼ認識部位を含む一本鎖5'側突出を有する核酸分子を作製することができる。次いで、一本鎖突出を有する核酸分子にIA型トポイソメラーゼを接触させて、IA型トポイソメラーゼ付加核酸分子を作製することができる。
図15は、一本鎖または二本鎖DNAが一本鎖RNAに共有結合されている本発明の2つの態様を示す。一本鎖DNAが一本鎖RNAに接続される場合には、リボヌクレオチド配列の3'末端はデオキシリボヌクレオチド配列の5'末端に共有結合される。二本鎖DNAが一本鎖RNAに接続される場合には、リボヌクレオチド配列の3'端は、2つの分子がハイブリダイゼーションするように、デオキシリボヌクレオチド配列の3'側突出との十分な配列相補性を共有する。上記のように、リボヌクレオチド配列の3'末端もデオキシリボヌクレオチド配列の5'末端に共有結合される。認識されているように、上記の数多くの変種は本発明の範囲内である。例えば、RNA分子は二本鎖であってもよい。別の例において、ヌクレオチド配列の全てがデオキシリボヌクレオチド配列であっても、および/または1つまたは複数の組換え部位を含んでもよい。
本発明は、各末端が5'端および3'端を含み、ベクターが第1の末端、第2の末端または第1の末端および第2の末端の両方の5'端またはその付近に部位特異的IA型トポイソメラーゼ認識部位を含む、第1の末端および第2の末端を有するかまたは有するように作製することができる、二本鎖組換え核酸分子を提供する。二本鎖組換え核酸分子は、IA型トポイソメラーゼ認識部位を含まない末端の3'端またはその付近に、IB型トポイソメラーゼ認識部位をさらに含むことができる。二本鎖組換え核酸分子はベクターであってもよい。
本発明は、各末端が5'端および3'端を有し、部位特異的IA型トポイソメラーゼが第1の末端、第2の末端、または第1の末端および第2の末端の両方の5'端に結合している、第1の末端および第2の末端を有するトポイソメラーゼ付加した二本鎖組換え核酸分子をさらに提供する。例えば、トポイソメラーゼ付加した二本鎖組換え核酸分子は、第1および第2の末端のそれぞれの5'端に結合されたIA型トポイソメラーゼを含むことができる。トポイソメラーゼ付加した核酸二本鎖組換え核酸分子は、IA型トポイソメラーゼが結合していない末端の3'端に結合したIB型トポイソメラーゼを含むことができる。トポイソメラーゼ付加した二本鎖組換え核酸分子はベクターであってもよい。
キット
本発明はまた、本発明の方法を都合よく実施するのに有用な成分を含むキットを提供する。一態様において、本発明のキットは、ポリペプチド、特に選択マーカーをコードし、各末端にトポイソメラーゼ認識部位を有する第1の核酸分子を含む。好ましくは、第1のヌクレオチド配列はトポイソメラーゼ活性化ヌクレオチド配列を含む。さらに好ましくは、トポイソメラーゼ付加した第1のヌクレオチド配列は各末端に5'側突出配列を含み、最も好ましくは、5'側突出配列は互いに異なる。選択的に、5'端のそれぞれは5'側ヒドロキシル基を含む。
またキットは、上流もしくは下流調節要素または他の要素であってもよく、一方または両方の末端にトポイソメラーゼ認識部位を有する調節要素を含む、少なくとも1つのヌクレオチド配列(またはその相補物)を含むことができる。好ましくは、キットは、それぞれが、異なる調節要素または他の要素、例えば標識もしくは他の検出可能な分子または細胞画分化ドメインをコードする配列を含む、複数の核酸分子を含む。異なる要素は異なる種類の特定の調節要素、例えば構成プロモーター、誘導プロモーターおよび組織特異的プロモーターであっても、または例えば、転写および翻訳調節要素、エピトープ標識等を含む異なる種類の要素であってもよい。このような核酸分子はトポイソメラーゼ活性化することができ、特に選択マーカーなどのポリペプチドが1つまたは複数の細胞種においてインビトロにおいて発現可能であるように、所定の方向で要素を機能的に共有結合するのを促進する、5'側突出または3'側突出を有することができる。
キットはまた、キットの成分を使用して作製された、一方または両方の鎖に共有結合された望ましい二本鎖組換え核酸分子を増幅するために、第1および第2のプライマーを含むプライマー対を選択し、使用することができるように、第1および第2のプライマーを含むプライマーを含むことができる。例えば、プライマーは、作製される二本鎖組換え核酸分子の5'末端に一般に位置づけられる要素に相補的な第1のプライマー、例えばプロモーター要素を含む核酸分子の部分、および作製される二本鎖組換え核酸分子の3'末端に一般に位置づけられる要素に相補的な第2のプライマー、例えば転写終了部位を含むまたはエピトープ標識をコードする核酸分子の部分を含むことができる。両方の鎖に共有結合された二本鎖組換え核酸分子を作製するためのキットから選択される要素に応じて、全長の機能的構築物を増幅するために、適当な第1および第2のプライマーを選択し、使用することができる。
別の態様において、本発明のキットは、それぞれが1つもしくは複数の組換え部位を有することができる、および/または一方もしくは両方の末端がトポイソメラーゼ活性化され、それぞれが5'側突出配列もしくは3'側突出配列またはそれらの組み合わせを有することができる、複数の異なる要素を含む。5'側または3'側突出配列は特定の要素に固有であってもよく、または複数の関連する要素、例えば複数の異なるプロモーター要素に共通であってもよい。好ましくは、要素の5'側突出配列は、1つまたは複数の要素が有用な機能を提供するように、機能的に共有結合されうるように設計され、コザック配列を含む要素および翻訳開始部位を含む要素は、要素が本発明の方法により機能的に共有結合されうるように、相補的な5'側突出を有することができる。
キットの複数の要素は、転写または翻訳調節要素、細菌、昆虫、酵母または哺乳類宿主細胞におけるヌクレオチド配列の複製に必要な要素、制限エンドヌクレアーゼまたはリコンビナーゼなどの部位特異的核酸結合タンパク質のための認識配列を含む要素、エピトープ標識または薬剤耐性遺伝子などの発現可能な産物をコードする要素等を含む、任意の要素を含むことができる。このように、本発明のキットは、例えば本発明の方法により作製される構築物が導入または発現される特定の細胞に応じて選択することができる、便利な異なる要素源を提供する。キットはまた、キットの要素を使用して作製された、一方または両方の鎖に共有結合された二本鎖組換え核酸分子を増幅するために上記のように組み合わせることができる、第1および第2のプライマーを含むPCRプライマーを含むことができる。選択的に、キットは、少なくとも一方の鎖に共有結合されるのに有用な量の部位特異的トポイソメラーゼ、選択的に、トポイソメラーゼ活性化された核酸分子またはトポイソメラーゼ認識部位を含むヌクレオチド配列であってもよい、第2の(または他の)核酸分子に対するトポイソメラーゼ認識部位を含む第1の核酸分子をさらに含む。
さらに別の態様において、本発明のキットは、選択マーカーをコードし、各末端にトポイソメラーゼ認識部位および/または組換え部位を有する第1の核酸分子;本発明の方法による所定の方向の第1の核酸分子に、一方または両方の鎖に共有結合することができる第1および第2の増幅産物を作製することができる、第1および第2のPCRプライマー対を含む。作製されたこのような構築物は細胞に導入することができ、部位特異的に相同組換えによって細胞のゲノムに組入れることができ、安定に維持され、細胞内で異種ポリペプチドを発現または標的遺伝子機能をノックアウトすることができる。ノックアウトされる標的遺伝子は、例えば配列の少なくとも一部が既知であるか、または容易に決定することができ、その機能が破壊されることが望ましい任意の遺伝子、例えば発癌遺伝子、アポトーシスに関与する遺伝子、セリン/スレオニンもしくはチロシンキナーゼをコードする遺伝子、または任意の他の遺伝子であってもよい。
両方の鎖に共有結合された二本鎖組換え核酸分子を作製するのに有用な本発明のキットの第1のPCRプライマー対は、共有結合される核酸分子の5'側突出配列に相補的なヌクレオチド配列(例えば、選択マーカーをコードする核酸分子の末端)、トポイソメラーゼ認識部位、ならびに/または組換え部位に相補的なヌクレオチド配列および標的DNA配列の3'側配列に相補的なヌクレオチド配列を5'側から3'側の方向に含む第1のプライマーを含む。第1のPCRプライマー対はまた、第1のプライマーが相補的である3'側配列の上流の標的DNA配列のヌクレオチド配列を含む第2のプライマーを含む。
両方の鎖に共有結合された二本鎖組換え核酸分子を作製するのに有用なキットの第2のPCRプライマー対は、共有結合される核酸分子の5'側突出配列に相補的なヌクレオチド配列、トポイソメラーゼ認識部位、ならびに/または組換え部位に相補的なヌクレオチド配列および標的DNA配列の5'側配列を5'側から3'側の方向に含み、標的遺伝子の5'側配列が、第1のプライマー対の第1のプライマーが相補的である標的DNA配列の3'側配列の下流にある、第1のプライマーを含む。第2のPCRプライマー対はまた、第1のプライマーに含有される標的DNA配列の5'側配列の下流である標的遺伝子の、3'側配列に相補的なヌクレオチド配列を含む第2のプライマーを含む。
別の態様において、両方の鎖に共有結合された二本鎖組換え核酸分子を作製するのに有用な本発明のキットは、転写活性化ドメインをコードし、3'端にトポイソメラーゼ認識部位またはその切断産物を含む第1の核酸分子と、DNA結合ドメインをコードし、3'端にトポイソメラーゼ認識部位またはその切断産物を含む第2の核酸分子を含む。部位特異的トポイソメラーゼによる切断の結果、第1もしくは第2の核酸分子は5'側突出を有することができ、または両方の配列が、同じであっても、互いに異なってもよい5'側突出を有することができる。核酸分子が5'側突出を有する場合には、突出は、第1または第2の核酸分子が本発明の方法により共有結合される核酸分子に一般に相補的である。キットはまた、一方または両方の3'端にトポイソメラーゼ認識部位またはその切断産物および選択的に、同じ末端にヒドロキシル基を含むことができる1つまたは対のアダプター、リンカー等を含むことができる。このようなアダプター、リンカー等は、それらが、上記の2つの核酸分子の一方または他方およびキットの一部に相補的である5'側突出を含むように選択される。
同様に、本発明のキットは、一方または両方の5'端にトポイソメラーゼ認識部位および/または組換え部位またはその切断産物および選択的に、同じ末端にヒドロキシル基を含む1つまたは対のアダプター、リンカー等を含むことができる。このようなアダプター、リンカー等は、上記の2つの核酸分子の一方または他方およびキットの一部に相補的である3'側突出を有するように選択される。また、キットは、一方または両方の5'側および/または3'端にトポイソメラーゼ認識部位またはその切断産物および選択的に、同じ末端にヒドロキシル基を含む1つまたは対のアダプター、リンカー等を含むことができる。
アダプター、リンカー等は、一般に、それらが本明細書に開示する2つの核酸分子の一方または他方およびキットの一部に相補的である、5'側および/または3'側突出を含むように選択される。このようなアダプター、リンカー等は、キットにより提供される第1または第2の核酸分子の一方または他方に共有結合される核酸分子の末端に接続することができ、それによってツーハイブリッドアッセイに有用なベイトおよびプレイポリペプチドをコードするキメラポリペプチドの構築を促進する。このようなキットはまた、トポイソメラーゼ認識部位またはその切断産物を含む増幅された複数のヌクレオチド配列を作製するために使用できる、PCRプライマーまたはプライマー対を含むことができる(表1および実施例1参照)。
1つの鎖に共有結合された二本鎖組換え核酸分子を作製するために使用できる本発明のキットのPCRプライマー対は、結合される核酸分子の5'側突出配列(例えば、選択マーカーをコードする核酸分子の末端)、トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)および選択的に、標的DNA配列の5'側配列に相補的なヌクレオチド配列を5'側から3'側の方向で含む第1のプライマーを含むことができる。PCRプライマー対はまた、第1のプライマーが相補的である5'側配列の下流の標的DNA配列のヌクレオチド配列を含む第2のプライマーを含む。
別の態様において、本発明のキットは、転写活性化ドメインをコードし、5'端に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)および選択的に、組換え部位またはその切断産物を含む第1の核酸分子、ならびにDNA結合ドメインをコードし、5'端に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)またはその切断産物を含む第2の核酸分子を含む。部位特異的トポイソメラーゼによる切断の結果、第1または第2の核酸分子は3'側突出を有することができ、または両方の配列は、同じであっても、互いに異なってもよい3'側突出を有することができる。核酸分子が3'側突出を有する場合には、突出は、一般に、第1または第2の核酸分子が本発明の方法により接続される核酸分子に相補的である。キットはまた、一方または両方の5'端に部位特異的トポイソメラーゼ認識部位(例えば、IA型またはII型トポイソメラーゼ認識部位)および選択的に、組換え部位またはその切断産物を含み、キットの2つの核酸分子の一方または他方に相補的である5'側突出を有することができる、1つまたは対のアダプター、リンカー等を含むことができる。
1つまたは両方の鎖に共有結合され、本発明の方法により作製される二本鎖組換え核酸分子は、例えば細胞内でポリペプチドを発現させるため、異常な状態を診断または治療するため等を含む種々の目的に使用することができる。このように、本発明は、1つまたは複数の細胞においてポリペプチドを発現することによって、またはアンチセンス分子を発現する等によって、異常な状態を治療するのに有用となりうる医薬を提供する。このような二本鎖組換え核酸分子は、エキソビボにおいて細胞と接触させることによって細胞に提供することができ、次いで細胞を被験者に投与することができ、このような方法はまた、このような投与の前に二本鎖組換え核酸分子を含む細胞を選択および/または増殖しても、または被験者に直接提供してもよい。生存している被験者への投与のためには、1つまたは両方の鎖に共有結合された二本鎖組換え核酸分子は、一般に、被験者に投与するのに適切な組成物に製剤化される。従って、本発明は、1つまたは両方の鎖に共有結合され、本発明の方法により作製された二本鎖組換え核酸分子を含む組成物を提供する。本明細書に開示するこのような核酸分子は、病的な状態に罹患している被験者を治療するための医薬として有用である。
投与のための組成物は、一般に、当技術分野において既知の薬学的に許容されうる1つまたは複数の担体を使用して製剤化され、例えば水もしくは緩衝生理食塩液などの水性溶液またはグリコール、グリセロール、オリーブオイルなどの油もしくは注射用有機エステルなどの他の溶媒または担体を含む。薬学的に許容されうる担体は、例えば結合物を安定化または結合物の吸収を増加する作用をする、生理学的に許容されうる化合物を含むことができる。生理学的に許容されうるこのような化合物には、例えばブドウ糖、ショ糖もしくはデキストランなどの炭水化物、アスコルビン酸もしくはグルタチオンなどの抗酸化剤、キレート剤、低分子量タンパク質または他の安定化剤もしくは賦形剤が含まれる。当業者は、生理学的に許容されうる化合物を含む薬学的に許容されうる担体の選択は、例えば経口または静脈内および注射、挿管または当技術分野において既知の他のこのような方法によるなどの非経口であってもよい、組成物の投与経路に依存することを周知である。本発明の組成物はまた、診断試薬、栄養補給物質、毒素または例えば、癌化学療法剤のような治療薬などの第2の試薬を含むこともできる。
一方または両方の鎖に共有結合された二本鎖組換え核酸分子は、水中油型エマルジョン、マイクロエマルジョン、ミセル、混合ミセル、リポソーム、マイクロスフェアまたは他のポリマー基質などの封入材料内に組入れることができる(例えば、それぞれ参照として本明細書に組み入れられる、Gregoriadis、Liposome Technology、第1巻(CRC Press、フロリダ州ボッカラトン、1984年)、Fraleyら、Trends Biochem. Sci.、6: 77(1981)参照)。例えば、リン脂質または他の脂質からなるリポソームは、製造および投与が比較的簡単である、無毒性で、生理学的に許容可能で、代謝可能な担体である。「ステルス」リポソーム(例えば、それぞれ参照として本明細書に組み入れられる、米国特許第5,882,679号、同第5,395,619号および同第5,225,212号参照)は、薬学的組成物を調製するのに特に有用なこのような封入材料の一例であり、他の「隠蔽型(masked)」リポソームも同様に使用することができ、このようなリポソームは、核酸分子が循環液中に存在する時間を延長する。陽イオンリポソームは、例えば特異的な受容体またはリガンドで修飾することもできる(参照として本明細書に組み入れられる、Morishitaら、J. Clin. Invest.、91: 2580-2585(1993))。核酸分子は、アデノウイルスポリリジン複合体と複合体を形成することによって、細胞に導入することもできる(例えば、参照として本明細書に組み入れられる、Michaelら、J. Biol. Chem.、268: 6866-6869(1993)参照)。このような組成物は、エキソビボを含むインビボまたはインビトロにおいて核酸分子を細胞に導入し、核酸分子を含む細胞を被験者に投与するのに特に有用となりうる(参照として本明細書に組み入れられる、米国特許第5,399,346号参照)。本発明の方法により作製された核酸分子はまた、遺伝子銃方法を使用して細胞に導入することもできる(例えば、SykesおよびJohnston、上記、1999年参照)。
宿主細胞
本発明はまた、本発明の核酸分子またはベクター、特に本明細書において詳細に記載する核酸分子、およびベクターの1つまたは複数を含む宿主細胞に関する。本発明のこの局面により使用できる代表的な宿主細胞には、細菌細胞、酵母細胞、植物細胞および動物細胞が含まれるが、これらに限定されない。好ましい細菌宿主細胞にはエシェリキア(Escerichia)種細胞(特に、大腸菌(E. coli)細胞、最も特には大腸菌(E. coli)株DH10B、Stbl2、DH5(DB3、DB3.1(好ましくは大腸菌(E. coli)LIBRARY EFFICIENCY(登録商標) DB3.1(商標)コンピテント細胞、Invitrogen Corporation、カリフォルニア州カールスバッド)、DB4およびDB5(開示内容が全体として参照として本明細書に組み入れられる、2000年3月2日提出の米国特許出願第09/518,188号)、バシラス(Bacillus)種細胞(特に、枯草菌(B. sutilis)およびB. メガテリウム(B. megaterium)細胞)、ストレプトマイセス(Streptomyces)種細胞、エルウィニア(Erwinia)種細胞、クレブシエラ(Klebsiella)種細胞、セラチア(Serratia)種細胞(特にS. マルセッセンス(S. marcessans)細胞)、シュードモナス(Pseudomonas)種細胞(特に、緑膿菌(P. aeruginosa)細胞)およびサルモネラ(Salmonella)種細胞(特に、S. ティフィムリウム(S. typhimurium)およびS. ティフィ(S. typhi)細胞)が含まれる。好ましい動物宿主細胞には、昆虫細胞(最も特には、キイロショウジョウバエ(Drosophila melanogaster)細胞、(ヨトウガ(Spodoptera frugiperda)Sf9およびSf21細胞ならびに(キャベツシャクトリムシハイファイブ(Trichoplusa High-Five)細胞)、線虫細胞(特に、線虫(C. elegans)細胞)、鳥類細胞、両生類細胞(特に、アフリカツメガエル(Xenopus laevis)細胞)、爬虫類細胞ならびに哺乳類細胞(最も特には、NIH3T3、CHO、COS、VERO、BHKおよびヒト細胞)が含まれる。好ましい酵母宿主細胞には、出芽酵母(Saccharomyces cerevisiae)細胞およびピキア・パストリス(Pichia pastoris)細胞が含まれる。これらおよび他の適切な宿主細胞は、例えばインビトロジェンコーポレーション(Invitrogen Corporation)(カリフォルニア州カールスバッド)、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(バージニア州マナサス)およびアグリカルチャル・リサーチ・カルチャー・コレクション(Agricultural Research Culture Collection)(NRRL、イリノイ州ペオリア)から購入できる。
本発明の核酸分子および/またはベクターの1つまたは複数を含む宿主細胞を産生するために、本明細書に記載する宿主細胞に本発明の核酸分子および/またはベクターを導入する方法は当業者に既知である。例えば、感染、形質導入、エレクトロポレーション、トランスフェクションおよび形質転換という既知の技法を使用して、本発明の核酸分子および/またはベクターを宿主細胞に導入することができる。本発明の核酸分子および/またはベクターは、単独または他の核酸分子および/またはベクターおよび/またはタンパク質、ペプチドまたはRNAと併用して導入してもよい。または、本発明の核酸分子および/またはベクターは、リン酸カルシウム沈殿などの沈殿物としてまたは脂質との複合体の形態で宿主細胞に導入することができる。エレクトロポレーションも、本発明の核酸分子および/またはベクターを宿主に導入するために使用することができる。同様に、このような分子を、大腸菌(E. coli)などの化学的適格細胞に導入することができる。ベクターがウイルスである場合には、インビトロにおいてパッケージングしても、またはパッケージング用細胞に導入してもよく、パッケージングしたウイルスを細胞に形質導入することができる。このように、本発明のこの局面により本発明の核酸分子および/またはベクターを細胞に導入するのに適切な多種多様な技法は既知であり、当業者にとって日常的である。このような技法は、例えばこれらの技法を詳細に記載した多数の実験マニュアルを例示的なものであり、関連する開示内容に関して全体の内容が参照として本明細書に組み入れられる、サムブルック(Sambrook, J.)ら、「分子クローニング、実験マニュアル(Molecular Cloning:A Laboratory Manual)」、第2版、ニューヨーク州コールドスプリングハーバー、Cold Spring Harbor Laboratory Prees、16.30〜16.55(1989年)、ワトソン(Watson, J. D.)ら、「組換えDNA(Recombinant DNA)」、第2版、ニューヨーク、W. H. Freeman and Co.、213〜234(1992年)、およびウィナッカー(Winnacker, E.-L.,)、「遺伝子からクローンまで(From Genes to Clones)」、ニューヨーク、VCH Publishers(1987年)に十分に概説されている。
ポリメラーゼ
本発明に使用するポリメラーゼには、ポリメラーゼ(DNAおよびRNAポリメラーゼ)および逆転写酵素が含まれるが、これらに限定されない。DNAポリメラーゼには、テルムス・サーモフィラス(Thermus thermophilus)(Tth)DNAポリメラーゼ、テルムス・アクアティカス(Thermus aquaticus)(Taq)DNAポリメラーゼ、テルモトガ・ネオポリタナ(Thermotoga neopolitana)(Tne)DNAポリメラーゼ、テルモトガ・マリチマ(Thermotoga maritima)(Tma)DNAポリメラーゼ、テルモコッカス・リトラリス(Thermococcus litoralis)(TliまたはVENT(商標))DNAポリメラーゼ、ピロコッカス・フリオサス(Pyrococcus furiosus)(Pfu)DNAポリメラーゼ、DEEPVENT(商標)DNAポリメラーゼ、ピロコッカス・ウーゼィ(Pyrococcus woosii)(Pwo)DNAポリメラーゼ、ピロコッカス(Pyrococcus)種KOD2(KOD)DNAポリメラーゼ、バシラス・ステアロサーモフィラス(Bacillus sterothermophilus)(Bst)DNAポリメラーゼ、バシラス・カルドフィラス(Bacillus caldophilus)(Bca)DNAポリメラーゼ、スルホロブス・アシドカルダリウス(Sulfolobus acidocaldarius)(Sac)DNAポリメラーゼ、テルモプラズマ・アシドフィラム(Thermoplasma acidophilum)(Tac)DNAポリメラーゼ、テルムス・フラブス(Thermus flavus)(Tfl/Tub)DNAポリメラーゼ、テルムスルーバー(Thermus ruber)(Tru)DNAポリメラーゼ、テルムス・ブロキアヌス(Thermus brockianus)(DYNAZYME(商標))DNAポリメラーゼ、メタノバクテリウム・サーモオートトロフィカム(Methanobacterium thermoautotrophicum)(Mth)DNAポリメラーゼ、マイコバクテリウム(mycobacterium)DNAポリメラーゼ(Mtb、Mlep)、大腸菌(E. coli) pol I DNAポリメラーゼ、T5 DNA ポリメラーゼ、T7 DNA ポリメラーゼおよび一般にpol I型DNAポリメラーゼおよびそれらの突然変異体、変種および誘導体が含まれるが、これらに限定されない。
本発明に使用する核酸ポリメラーゼは中温性であっても、好熱性であってもよいが、好ましくは、好熱性である。好ましい中温性DNAポリメラーゼには、Pol IファミリーのDNAポリメラーゼ(およびそれぞれのクレノウフラグメント)が挙げられ、いずれも、大腸菌(E. coli)、H. インフルエンゼ(H. influenzae)、D. ラジオデュランス(D. radiodurans)、H. ピロリ(H. pylori)、(C. aurantiacus)、R. プロワツェキィ(R. prowazekii)、梅毒トレポネーマ(T. pallidum)、シネコシスキス(Synechocystis)種、枯草菌(B. sutilis)、L. ラクティス(L. lactis)、肺炎連鎖球菌(S. pneumoniae)、M. ツベルクローシス(M. tuberculosis)、らい菌(M. leprae)、M. スメグマチス(M. smegmatis) などの生物、バクテリオファージL5、phi-C31、T7、T3、T5、SP01、SP02、出芽酵母(S. cerevisiae)MIP-1ならびに真核細胞の線虫(C. elegans)およびキイロショウジョウバエ(D. melanogaster)(Astatke, M.ら、1998、J. Mol. Biol. 278、147-165)から単離するでき、任意の起源から単離したpol III型DNAポリメラーゼならびにそれらの突然変異体、誘導体または変種であってもよい。本発明の方法および組成物に使用できる好ましい好熱性DNAポリメラーゼには、Taq、Tne、Tma、Pfu、KOD、Tfl、Tth、Stoffelフラグメント、VENT(商標)およびDEEPVENT(商標)DNAポリメラーゼならびにそれらの突然変異体、変種および誘導体が含まれる(米国特許第5,436,149号、米国特許第4,889,818号、米国特許第4,965,188号、米国特許第5,079,352号、米国特許第5,614,365号、米国特許第5,374,553号、米国特許第5,270,179号、米国特許第5,047,342号、米国特許第5,512,462号、国際公開公報第92/06188号、国際公開公報第92/06200号、国際公開公報第96/10640号、国際公開公報第97/09451号、Barnes, W. M.、Gene 112: 29-35(1992)、Lawyer, F. C.ら、PCR Meth. Appl. 2: 275-287(1993)、Flaman, J.-Mら、Nucl. Acids Res. 22(15): 3259-3260(1994))。
本発明に使用するための逆転写酵素には、逆転写酵素活性を有する任意の酵素が含まれる。このような酵素には、レトロウイルス逆転写酵素、レトロトランスポゾン逆転写酵素、B型肝炎ウイルス逆転写酵素、カリフラワーモザイクウイルス逆転写酵素、細菌逆転写酵素、Tth DNAポリメラーゼ、Taq DNAポリメラーゼ(Saiki, R. K.ら、Science 239: 487-491(1988)、米国特許第4,889,818号および同第4,965,188号)、Tne DNAポリメラーゼ(国際公開公報第96/10640号および国際公開公報第97/09451号)、Tma DNAポリメラーゼ(米国特許第5,374,533号)ならびにそれらの突然変異体、変種または誘導体(例えば、国際公開公報第97/09451号および国際公開公報第98/47912号参照)が含まれるが、これらに限定されない。本発明に使用される好ましい酵素は、RNase H活性が低下、実質的に低下または排除されたものが含まれる。「RNase H活性が実質的に低下された」酵素は、その酵素が、対応する野生型または野生型モロニーマウス白血病ウイルス(M-MLV)、トリ骨髄芽球症ウイルス(AMV)もしくはラウス肉腫ウイルス(RSV)逆転写酵素などのRNase H+酵素のRNase H活性の約20%未満、さらに好ましくは約15%未満、約10%未満、約5%未満、最も好ましくは約2%未満であることを意味する。任意の酵素のRNase H活性は、例えばその全ての開示内容が全体として参照として本明細書に組み入れられる、米国特許第5,244,797号、コテヴィクツ(Kotewicz, M. L.)ら、Nucl. Acids Res. 16: 265(1988)およびゲラード(Gerard, G. F.)ら、FOCUS 14(5): 91(1992)に記載されているものなどの種々のアッセイによって測定することができる。本発明に使用する特に好ましいポリペプチドには、M-MLV H逆転写酵素、RSV H-逆転写酵素、AMV H-逆転写酵素、RAV(ラウス関連ウイルス)H-逆転写酵素、MAV(骨髄芽球症関連ウイルス)H-逆転写酵素およびHIV H-逆転写酵素、が含まれるが、これらに限定されない(米国特許第5,244,797号および国際公開公報第98/47912号参照)。しかし、リボ核酸分子からDNA分子を作製することができる(すなわち、逆転写酵素活性を有する)任意の酵素を、本発明の組成物、方法およびキットに等価に使用することができるが当業者によって理解されている。
本発明に使用するポリメラーゼ活性を有する酵素は、例えばインビトロジェンコーポレーション(Invitrogen Corporation)(カリフォルニア州カールスバッド)、パーキン-エルマー(Perkin-Elmer)(ニュージャージー州ブランチバーグ)、ニューイングランドバイオラボズ(New England BioLabs)(マサチューセッツ州ベバーリー)またはベーリンガーマンハイムバイオケミカルズ(Boehringer Mannheim Biochemicals)(インディアナ州インディアナポリス)から購入することができる。本発明に使用する逆転写酵素活性を有する酵素は、例えばインビトロジェンコーポレーション(Invitrogen Corporation)(カリフォルニア州カールスバッド)、ファルマシア(Pharmacia)(ニュージャージー州ピスカッタウェイ)、シグマ(Sigma)(ミズーリ州セントルイス)またはベーリンガーマンハイムバイオケミカルズ(Boehringer Mannheim Biochemicals)(インディアナ州インディアナポリス)から購入することができる。または、ポリメラーゼまたはポリメラーゼ活性を有する逆転写酵素は、当業者に既知の、天然のタンパク質を単離および精製するための標準的な手法により、天然のウイルスまたは細菌起源から単離することができる(例えば、Houts, G. E.ら、J. Virol. 29: 517(1979)参照)。また、このようなポリメラーゼ/逆転写酵素は、当業者に既知の組換えDNA技法によって作製することができる(例えば、Kotewicz, M. L.ら、Nucl. Acids Res. 16: 265(1988)、米国特許第5,244,797号、国際公開公報第98/47912号、Soltis, D. A.、およびSkalka, A. M.、Proc. Natl. Acad. Sci. USA 85: 3372-3376(1988)参照)。ポリメラーゼ活性および逆転写酵素活性を有する酵素の例は、本願に記載されているもののいずれかを含んでもよい。
核酸合成、増幅および配列決定方法
本発明は、DNA(cDNAを含む)およびRNA分子などの核酸分子の合成に関与する任意の方法を組み合わせて使用することができる。このような方法には、核酸合成方法、核酸増幅方法および核酸配列決定方法が含まれるが、これらに限定されない。このような方法は本発明に使用する分子(例えば、出発分子)を作製するために使用することができ、または本発明によって作製された分子もしくはベクターをさらに操作するために使用することができる。
本発明のこの局面による核酸合成方法は1つまたは複数の段階を含んでもよい。例えば、本発明は、(a)核酸鋳型(例えば、本発明の核酸分子またはベクター)に1つまたは複数のプライマーおよびポリメラーゼまたは逆転写酵素活性を有する1つまたは複数の酵素を混合して、混合物を形成する段階、ならびに(b)鋳型の全てまたは一部に相補的な第1の核酸分子を作製するのに十分な条件下において、混合物をインキュベーションする段階を含む、核酸分子を合成するための方法を提供する。本発明のこの局面によると、核酸鋳型はcDNA分子もしくはライブラリなどのDNA分子またはmRNA分子などのRNA分子であってもよい。pH、温度、イオン強度およびインキュベーション時間などの合成を可能にするのに十分な条件は、当業者が最適化することができる。望ましい場合には、合成過程中または後に、組換え部位をこのような合成分子に付加することができる(例えば、1997年10月24日提出の米国特許仮出願第60/065,930号に基づいた、米国特許出願第09/177,387号参照)。
本発明によると、標的または鋳型核酸分子またはライブラリは、種々の細胞、組織、器官または生物などの天然の起源から得られる核酸分子から調製することができる。核酸分子源として使用できる細胞は、原核細胞(エシェリキア(Escerichia)属、バシラス(Bacillus)属、セラチア(Serratia)属、サルモネラ(Salmonella)属、スタヒロコッカス(Staphylococcus)属、ストレプトコッカス(Streptococcus)属、クロストリジウム(Clostridium)属、クラミジア(Chlamydia)属、ナイセリア(Neisseria)属、マイコプラズマ(Mycoplasma)属、ボレリア(Borrelia)属、レジオネラ(Legionella)属、シュードモナス(Pseudomonas)属、マイコバクテリウム(Mycobacterium)属、ヘリコバクター(Helicobacter)属、エルウィニア(Erwinia)属、アグロバクテリウム(Agrobacterium)属、リゾビウム(Rhizobium)属およびストレプトマイセス(Streptomyces)属の種のものを含む、細菌細胞)または真核細胞(真菌(特に酵母)、植物、原生動物および他の寄生虫ならびに昆虫(特に、ドロソフィラ(Drosophila)種細胞)、線虫(特に、線虫(Caenorhabditis elegans)細胞)および哺乳類(特にヒト細胞)を含む動物を含む)であってもよい。
当然のことであるが、有利に使用することができる他の核酸合成技法は当業者に容易に明らかになる。
本発明の他の局面において、本発明は、核酸分子を増幅または配列決定する方法と併用して使用することができる。本発明のこの局面による核酸増幅方法は、一段階(例えば、一段階TR-PCR)または二段階(例えば、二段階RT-PCR)逆転写酵素-増幅反応として一般に当技術分野において既知の方法において、逆転写酵素活性を有する1つまたは複数のポリペプチドの使用を含むことができる。長い核酸分子(すなわち、鎖長が約3〜5 Kbより長い)の増幅のためには、国際公開公報第98/06736号および国際公開公報第95/16028号に記載されているように、DNAポリメラーゼの組み合わせを使用することができる
本発明による増幅方法は、1つまたは複数の段階を含むことができる。例えば、本発明は、(a)ポリメラーゼ活性を有する1つまたは複数の酵素に、1つまたは複数の核酸鋳型を混合する段階、および(b)ポリメラーゼ活性を有する酵素に、鋳型の全てまたは一部に相補的な1つまたは複数を増幅させるのに十分な条件下において混合物をインキュベーションする段階とを含む、核酸分子を増幅する方法を提供する。本発明はまた、このような方法によって増幅される核酸分子を提供する。望ましい場合には、増幅過程中または後に、組換え部位を増幅されたこのような分子に付加することができる(例えば、1997年10月24日提出の米国特許仮出願第60/065,930号に基づいた、1998年10月23日提出の米国特許出願第09/177,387号参照)。
核酸分子または断片を増幅および合成するための一般的な方法は当業者に既知である(例えば、米国特許第4,683,195号、同第4,683,202号および同第4,800,159号、Innis, M.A.ら編、「PCRプロトコール:方法と応用指針(PCR Protocols: A Guide to Methods and Application)」、カリフォルニア州サンディエゴ:Academic Press, Inc. (1990年)、Griffin, H. G.およびGriffin, A. M.編、「PCR技術:最新技術(PCR Technology: Current Innovations)」、フロリダ州ボカラトン:CRC Press(1994年)参照)。例えば、本発明により使用できる増幅方法には、PCR(米国特許第4,683,195号および同第4,683,202号)、鎖置換増幅(Strand Displacement Amplification)(SDA、米国特許第5,455,166号、欧州特許第0 684 315号)および核酸配列に基づいた増幅(Nucleic Acid Sequence-Based Amplification)(NASBA、米国特許第5,409,818号、欧州特許第0 329 822号)が含まれる。
典型的には、これらの増幅方法は、(a)1つまたは複数のプライマー配列の存在下においてポリメラーゼ活性を有する、1つまたは複数の酵素に核酸試料を混合する段階、および(b)好ましくは、PCRまたは等価な自動増幅技法によって、核酸試料を増幅して、増幅された核酸断片の集合体を作成する段階を含む。
本発明の方法による増幅または合成後、使用または特徴づけのために増幅または合成した核酸断片を単離することができる。この段階は、通常、増幅または合成した核酸断片をサイズまたはゲル電気泳動、キャピラリー電気泳動、クロマトグラフィー(サイジング(分粒)、アフィニティーおよびイムノクロマトグラフィー)、密度勾配遠心分離および免疫吸着を含む任意の物理的もしくは生化学的手段によって分離することによって実施することができる。ゲル電気泳動による核酸断片の分離は、多数の核酸断片を感度高く分離する迅速で、再現性の高い手段を提供し、いくつかの核酸試料中の断片を直接同時に比較することができるので、特に好ましい。別の好ましい態様において、本発明の方法によって増幅または合成した3つの断片または任意の核酸断片を単離し、特徴づけるために、この方法を拡張することができる。従って、本発明はまた、本発明の増幅または合成方法によって作製した単離された核酸分子に関する。
この態様において、増幅または合成した核酸断片は、電気泳動溶出または物理的切断などの標準的な技法により、同定(上記参照)のために使用したゲルから取り出される。単離した独自の核酸断片は、種々の原核細胞(細菌細胞)または真核細胞(酵母、植物またはヒトおよび他の哺乳類を含む動物細胞)のトランスフェクションまたは形質転換に適切な、発現ベクターを含む標準的なベクターに挿入することができる。または、本発明の方法によって作製した核酸分子は、例えば配列決定(すなわち、核酸断片のヌクレオチド配列を決定する)、以下に記載する方法および当技術分野において標準的である他の方法によってさらに特徴づけることができる(例えば、DNA配列決定方法に関する、米国特許第4,962,022号および同第5,498,523号参照)。
本発明による核酸配列決定方法は1つまたは複数の段階を含むことができる。例えば、本発明は、(a)ポリメラーゼ活性を持つ酵素に、配列決定する核酸分子、1つまたは複数のヌクレオチドおよび1つまたは複数の停止剤(ジデオキシヌクレオチドなど)を混合して、混合物を形成する段階、(b)配列決定する分子の全てまたは一部に相補的な分子の集団を合成するのに十分な条件下において混合物をインキュベーションする段階、ならびに(c)配列決定する分子の全てまたは一部のヌクレオチド配列を決定するために、集団を分離する段階を含む、核酸分子を配列決定する方法と組み合わせることができる。
使用することができる核酸配列決定技法には、米国特許第4,962,022号および米国特許第5,498,523号に開示されているものなどのジデオキシ配列決定方法が含まれる。
キット
別の局面において、本発明は、本発明と併用して使用することができるキットを提供する。本発明のキットは任意の数の成分を含むことができるが、典型的には、少なくとも2つの成分を含む。本発明のこの局面によるキットは、本発明の1つまたは複数の核酸分子またはベクター、1つまたは複数のプライマー、本発明の分子および/または化合物、本発明の支持体、1つまたは複数のポリメラーゼ、1つまたは複数の逆転写酵素、1つまたは複数の組換えタンパク質(または本発明の方法を実施するための他の酵素)、1つまたは複数のトポイソメラーゼ、1つまたは複数の緩衝液、1つまたは複数の界面活性剤、1つまたは複数の制限エンドヌクレアーゼ、1つまたは複数のヌクレオチド、1つまたは複数の停止剤(例えば、ddNTPs)、1つまたは複数のトランスフェクション剤、ピロホスファターゼ等からなる群から選択される、1つまたは複数の成分を含むことができる、1つまたは複数の容器を含むことができる。本発明のキットはまた、本発明の方法を実施するための取扱い説明書を含んでもよい。
例えば、本発明のキットは、(1)1つまたは複数(例えば、1、2、3、4、5、6、7、8、9、10等)の組換え部位、および/または1つまたは複数(例えば、1、2、3、4、5、6、7、8、9、10等)のトポイソメラーゼ認識部位を含む第1の核酸分子、ならびに(2)本明細書に記載する方法を使用して、第1の核酸分子(nucleic molecule)を別の核酸分子に共有結合する取扱い説明書を含むことができる。特定の態様において、取扱い説明書は、一方または両方の鎖に2つまたはそれ以上の核酸分子を接続する方法を記載している。関連する態様において、第1の核酸分子は、キットに加えられる前に、トポイソメラーゼ付加される。
本発明の追加のキットは、例えばキットの成分の精度または適合度を試験するのに有用となりうる、1つまたは複数の対照ヌクレオチド配列を含んでもよい、1つまたは複数のトポイソメラーゼ付加した核酸分子物質、1つまたは複数のトポイソメラーゼ、1つまたは複数のリコンビナーゼ(または組換えタンパク質)、1つまたは複数のリコンビナーゼ(または組換えタンパク質)を含む1つまたは複数の組成物、少なくとも1つのトポイソメラーゼ認識部位および/または少なくとも1つの組換え部位を含むことができる1つまたは複数のプライマー、少なくとも1つのトポイソメラーゼ認識部位および/または少なくとも1つの組換え部位または少なくとも1つのトポイソメラーゼ認識部位および少なくとも1つのトポイソメラーゼ認識部位に相補的な、少なくとも1つのヌクレオチド配列の両方に相補的なヌクレオチド配列、キットの核酸分子またはキットを使用して作製される核酸分子を含むことができるまたは含むのに有用となりうる1つまたは複数の細胞、キットを使用して方法を実施するための1つまたは複数の試薬、ポリマー、緩衝液等、キットを使用して方法を実施するための取扱い説明書等を含むことができる。
別の局面において、本発明のキットは、第1の末端および第2の末端を有し、発現されるポリペプチド、例えば選択マーカーをコードし、一方または両方の末端の3'端にトポイソメラーゼ認識部位または切断産物を含む核酸分子を含むことができる。選択的に、核酸分子は、他方の末端、すなわち、トポイソメラーゼ認識部位を含有しない、またはトポイソメラーゼ付加されていない末端の一方または両方の5'端にヒドロキシル基を含む。さらに一方または両方の5'端は、互いに異なる突出配列を含むことができる。本発明のキットは、第1の末端および第2の末端を有し、発現されるポリペプチド、例えば選択マーカーをコードし、一方または両方の末端の5'端にトポイソメラーゼ認識部位または切断産物を含む核酸分子を含むことができる。選択的に、核酸分子は、一方または両方の末端の3'端にヒドロキシル基を含有し、一方または両方の3'端は、互いに異なる突出配列を含むことができる。また、本発明のキットは、第1の末端および第2の末端を有し、発現されるポリペプチド、例えば選択マーカーをコードし、一方または両方の末端の5'端および3'端にトポイソメラーゼ認識部位または切断産物を含む核酸分子を含むことができる。このように、本発明のキットは、1つまたは複数のトポイソメラーゼ認識部位またはトポイソメラーゼ付加核酸分子を含むこのような核酸分子の種々の組み合わせのいずれかを含むことができることが認識されるべきである。
本発明のキットはまた、少なくとも第1の末端の3'端に調節要素または他のヌクレオチド配列、例えばコード配列およびトポイソメラーゼ認識部位および/または組換え部位またはその切断産物ならびに選択的に、認識部位を含む末端の5'端にヒドロキシル基を含む核酸分子、または少なくとも第1の末端の5'端にトポイソメラーゼ認識部位またはその切断産物および選択的に、認識部位を含む末端の3'端にヒドロキシル基を含む核酸分子、または少なくとも第1の末端の5'端および3'端にトポイソメラーゼ認識部位を含む核酸分子を含むことができる。ある種の態様において、キットは、種々の上流の調節要素、種々の下流の調節要素、要素およびそれらの組み合わせを含む分子を検出または同定するのに有用な種々の要素を含むことができる。例えば、キットは、構成的または誘導的に活性であるおよび小数種類の細胞または多数の異なる種類の細胞において活性である種々の遺伝子プロモーター要素、配列内リボソーム進入部位などのリボソーム結合を可能にする要素、コザック(Kozak)配列または開始メチオニンをコードする要素等を含むことができる。さらに、またはあるいは、キットはポリアデニル化シグナル配列、転写または翻訳等を停止するような配列のような種々の下流調節要素を含むことができる。同様にキットは、エピトープ標識等などの検出可能なマーカーをコードする要素を含むこともできる。本発明のある種のこのような局面では、キットは、それぞれが少なくとも1つのトポイソメラーゼ認識部位および/または少なくとも1つの組換え部位を含む、種々のこのような要素を含む。特定の他のこのような局面では、これらの要素は、互いにまたは本発明の方法により選択マーカーなどのポリペプチドをコードする核酸分子に機能的に共有結合することができるように、突出配列を有することができる。
選択的に、キットは、キットに含まれる種々の要素の1つを含む構築物を増幅することができる要素特異的プライマーを含む。キットがこのようなプライマーを含む場合には、調節要素または他の要素を含む核酸分子は、プライマーによって特異的に認識され、調節要素によりプライマーが伸長し、調節要素を含むプライマーを伸長することができるヌクレオチド配列を有する。特にキットは、例えばキットの特定の5'側調節要素および特定の3'側調節要素を含む構築物を増幅するプライマー対を作製するように組み合わせることができる、要素特異的な順方向および逆方向プライマーを含むことができる。このようなプライマー対は、本発明の方法により作製された共有結合した機能的な望ましい二本鎖核酸分子を選択的に増幅することができるが、部分的な反応産物を増幅しない。
別の態様において、本発明のキットは、第1の末端および第2の末端を有し、一方または両方の3'端またはその付近にトポイソメラーゼ認識部位またはその切断産物および/または組換え部位を有し、転写活性化ドメインをコードする第1の核酸分子、ならびに第1の末端および第2の末端を有し、一方または両方の3'端またはその付近にトポイソメラーゼ認識部位またはその切断産物を有し、DNA結合ドメインをコードする第2の核酸分子を含むか、または第1の末端および第2の末端を有し、一方または両方の5'端またはその付近にトポイソメラーゼ認識部位またはその切断産物および/または組換え部位を有し、転写活性化ドメインをコードする第1の核酸分子、および一方または両方の5'端またはその付近にトポイソメラーゼ認識部位またはその切断産物および/または組換え部位を有し、DNA結合ドメインをコードする第2の核酸分子を含む。本発明のキットはまた、第1の末端および第2の末端を有し、転写活性化ドメインをコードする第1の核酸分子と、第1の末端および第2の末端を有し、DNA結合ドメインをコードする第2の核酸分子を有し、少なくとも第1の核酸分子または第2の核酸分子が少なくとも一方の末端の5'側または末端付近および3'端またはその付近にトポイソメラーゼ認識部位またはその切断産物を有し、他方の二本鎖ヌクレオチドは、認識部位を含む核酸分子の末端に共有結合される末端に3'側ヒドロキシルおよび5'側ヒドロキシルを有する。このようなキットは、例えばツーハイブリッドアッセイを実施するためのキメラポリペプチドをコードする共有結合された二本鎖組換え核酸分子を作製するのに有用である。キットは、ヌクレオチド配列を増幅して、第1または第2の核酸分子に共有結合することができ、プライマー対の少なくとも1つのプライマーはトポイソメラーゼ認識部位、トポイソメラーゼ認識部位の相補物または両方を含む、プライマー対をさらに含むことができる。好ましくは、このようなプライマー対を使用して作製される増幅産物は、部位特異的トポイソメラーゼによる切断後、共有結合される第1または第2の核酸分子に相補的な3'側または5'側突出配列を含む。このようなキットは、キットの第1または第2のヌクレオチド配列を含み、ツーハイブリッドアッセイを実施するのに有用なキメラポリペプチドをコードする組換えポリヌクレオチドの作製を促進することができる。
本発明はまた、本発明の方法を実施するため、特に本発明の産物の核酸分子を作製する際に使用するための追加のキットに関する。本発明はまた、本発明の方法により相同組換え(特に遺伝子ターゲティング)を実施するためのキットに関する。本発明のこのようなキットはまた、本発明の方法により作製された組換え部位を含む分子、および/または化合物をさらに操作するためのさらに別の成分を含むことができる。本発明のキットは、本発明の1つまたは複数の核酸分子(特に、1つまたは複数の組換え部位および選択的に、1つまたは複数の反応性官能部分を含む出発物質)、本発明の1つまたは複数の分子および/または化合物、本発明の1つまたは複数の支持体および/または本発明の1つまたは複数のベクターを含むことができる。このようなキットは、選択的に、1つまたは複数の宿主細胞、1つまたは複数のヌクレオチド、1つまたは複数のポリメラーゼおよび/または逆転写酵素、1つまたは複数の適切な緩衝液、1つまたは複数のプライマー、1つまたは複数の停止剤、コンビナトリアルライブラリを作製するための1つまたは複数の分子集団、および1つまたは複数のコンビナトリアルライブラリからなる群から選択される、追加の成分を1つまたは複数含むことができる。
別の態様において、本発明のキットは、ポリペプチド、特に選択マーカーをコードし、各末端にトポイソメラーゼ認識部位を有する第1の核酸分子を含む。特定の好ましいこのような態様において、第1の核酸分子は環状分子(例えば、プラスミド、ベクター等)であり、分子の1つまたは複数の、好ましくは2つまたはそれ以上のトポイソメラーゼ認識部位に隣接している、少なくとも1つの組換え部位を含み、さらに好ましくは少なくとも2つの組換え部位を含む。好ましくは、第1のヌクレオチド配列はトポイソメラーゼ活性化ヌクレオチド配列を含む。さらに好ましくは、トポイソメラーゼ付加した第1のヌクレオチド配列は、各末端に5'側突出配列を含み、最も好ましくは、5'側突出配列は互いに異なる。選択的に、5'端のそれぞれは5'側ヒドロキシル基を含む。
本発明のこの局面によるキットはまた、1つまたは複数のトポイソメラーゼ認識部位を有し、選択的に、一方または両方の末端に1つまたは複数の組換え部位を有する、上流または下流の調節要素または他の要素であってもよい調節要素を含む、少なくとも1つのヌクレオチド配列を含むことができる。好ましくは、キットは、それぞれが異なる調節要素または他の要素、例えば標識もしくは他の検出可能な分子または細胞画分化ドメインをコードする配列を含む、複数の核酸分子を含む。異なる要素は、異なる種類の特定の調節要素、例えば構成的もしくは誘導的プロモーターもしくは組織特異的プロモーターであっても、または例えば、転写および翻訳調節要素、エピトープ標識等を含む異なる種類の要素であってもよい。このような核酸分子はトポイソメラーゼ活性化することができ、特に選択マーカーなどのポリペプチドがインビトロまたは1つもしくは複数の細胞種において発現可能であるように、要素を所定の方向で機能的に共有結合するのを促進する5'側突出配列を有することができる。
このようなキットはまた、第1および第2のプライマーを含むプライマー対が選択され、キットの成分を使用して作製される共有結合された望ましい二本鎖組換え核酸分子を増幅するために使用できるように、第1および第2のプライマーを含むプライマーを含むことができる。例えば、プライマーは、作製された二本鎖組換え核酸分子の5'端、例えばプロモーター要素を含む核酸分子の部分に一般に位置づけられている要素に相補的である第1のプライマー、ならびに作製された二本鎖組換え核酸分子の3'端に、例えば転写停止部位を含むまたはエピトープ標識を含む核酸分子の部分に一般に位置づけられている要素に相補的である第2のプライマーを含むことができる。共有結合された二本鎖組換え核酸分子を作製するためのキットから選択される要素に応じて、適当な第1および第2のプライマーを選択し、全長の機能的構築物を増幅するために使用することができる。
別の態様において、本発明のキットは、それぞれは一方または両方の末端がトポイソメラーゼ活性化されてもよく、それぞれは5'側突出配列を含むことができる複数の異なる要素を含む。5'側突出配列は特定の要素に固有であっても、または複数の関連する要素、例えば複数の異なるプロモーター要素に共通であってもよい。好ましくは、要素の5'側突出配列は、1つまたは複数の要素が機能的に共有結合されて、有用な機能、例えばコザック(Kozak)配列を含む要素および本発明の方法により要素を機能的に共有結合することができるように、相補的な5'側突出を有する翻訳開始部位を含む要素を提供することができるように設計される。
キットの複数の要素は、転写または翻訳調節要素、細菌、昆虫、酵母または哺乳類宿主細胞内でのヌクレオチド配列の複製に必要な要素、制限エンドヌクレアーゼまたはリコンビナーゼなどの部位特異的核酸結合タンパク質の認識配列を含む要素、エピトープ標識または薬剤耐性遺伝子などの発現可能な産物をコードする要素等を含む、任意の要素を含むことができる。このように、本発明のキットは、例えば本発明の方法により作製された構築物を導入または発現させる特定の細胞に応じて選択することができる、便利な異なる要素源を提供する。キットはまた、第1および第2のプライマーを含み、上記のように組み合わせて、キットの要素を使用して作製される共有結合された二本鎖組換え核酸分子を増幅することができるPCRプライマーを含むことができる。選択的に、キットは、トポイソメラーゼ活性化核酸分子であっても、またはトポイソメラーゼ認識部位を含むヌクレオチド配列であってもよい、トポイソメラーゼ認識部位を含む第1の核酸分子を第2の(または他の)核酸分子に共有結合するのに有用な量の1つまたは複数のトポイソメラーゼ(例えば、1つまたは複数の部位特異的トポイソメラーゼ)および/または1つまたは複数のリコンビナーゼ(またはリコンビナーゼタンパク質)をさらに含む。
さらに別の態様において、本発明のキットは、選択マーカーをコードし、各末端にトポイソメラーゼ認識部位を有する第1の核酸分子、ならびに本発明の方法により所定の方向で第1の核酸分子に共有結合することができる第1および第2の増幅産物を作製することができる、第1および第2のPCRプライマー対を含む。作製されたこのような構築物は細胞に導入することができ、部位特異的に相同組換えによって細胞のゲノムに組入れることができ、そこで安定に維持され、細胞内で異種ポリペプチドを発現するか、または標的遺伝子機能をノックアウトすることができる。ノックアウトされる標的遺伝子は、例えば配列の少なくとも一部が既知であり、容易に決定することができ、機能を破壊することが望ましい任意の遺伝子、例えば発癌遺伝子、アポトーシスに関与する遺伝子、セリン/スレオニンもしくはチロシンキナーゼをコードする遺伝子または任意の他の遺伝子であってもよい。
本発明のキットの第1のPCRプライマーは、共有結合される核酸分子の5'側突出配列に相補的なヌクレオチド配列(例えば、選択マーカーをコードする核酸分子の末端)、トポイソメラーゼ認識部位および/または組換え部位に相補的なヌクレオチド配列および標的DNA配列の3'側配列に相補的なヌクレオチド配列を5'側から3'側の方向に含む第1のプライマーを含む。第1のPCRプライマー対はまた、第1のプライマーが相補的である3'側配列の上流の標的DNA配列のヌクレオチド配列を含む第2のプライマーを含む。
本発明のキットの第2のPCRプライマー対は、共有結合される核酸分子の5'側突出配列に相補的なヌクレオチド配列、トポイソメラーゼ認識部位に相補的なヌクレオチド配列および選択的に、組換え部位に相補的なヌクレオチド配列および標的DNA配列の5'側配列のヌクレオチド配列を5'側から3'側に含み、第1のプライマー対の第1のプライマーが相補的である標的DNA配列の、3'側配列の下流に標的遺伝子の5'側配列を有する第1のプライマーを含む。第2のPCRプライマー対はまた、第1のプライマーに含有される標的DNA配列の5'側配列の下流にある標的遺伝子の、3'側配列に相補的なヌクレオチド配列を含む第2のプライマーを含む。
別の態様において、本発明のキットは、転写活性化ドメインをコードし、3'端またはその付近にトポイソメラーゼ認識部位またはその切断産物を含む第1の核酸分子、ならびにDNA結合ドメインをコードし、3'端またはその付近にトポイソメラーゼ認識部位および選択的に、組換え部位またはその切断産物を含む第2の核酸分子を含む。部位特異的トポイソメラーゼによる切断の結果、第1または第2の核酸分子は5'側突出を有しても、または両方の配列が、同じまたは互いに異なる5'側突出を有してもよい。核酸分子が5'側突出を有する場合には、突出は一般に、第1または第2の核酸分子が本発明の方法により共有結合される核酸分子に相補的である。
キットはまた、一方または両方の3'端にトポイソメラーゼ認識部位および選択的に、組換え部位またはその切断産物を含み、選択的に、同じ末端にヒドロキシル基を含む1つまたは対のアダプター、リンカー等を含むことができる。このようなアダプター、リンカー等は、上記の2つの核酸分子の一方または他方およびキットの一部に相補的である5'側突出を有するように選択される。同様にキットはまた、一方または両方の5'端にトポイソメラーゼ認識部位および選択的に、組換え部位またはその切断産物を含み、選択的に、同じ末端にヒドロキシル基を含む1つまたは対のアダプター、リンカー等を含むことができる。このようなアダプター、リンカー等は、上記の2つの核酸分子の一方または他方およびキットの一部に相補的である3'側突出を有するように選択される。また、キットは、両方の5'側および/または3'端またはその付近にトポイソメラーゼ認識部位またはその切断産物を含み、選択的に、同じ末端にヒドロキシル基を含む1つまたは対のアダプター、リンカー等を含むことができる。このようなアダプター、リンカー等は、上記の2つの核酸分子の一方または他方およびキットの一部に相補的である5'側および/または3'側突出を有するように選択される。このようなアダプター、リンカー等を、キットに提供されている第1または第2の核酸分子の一方または他方に共有結合する核酸分子の末端に接続することができ、ツーハイブリッドアッセイに有用なベイトおよびプレイポリペプチドをコードする、キメラポリペプチドの構築を促進することができる。このようなキットはまた、トポイソメラーゼ認識部位またはその切断産物を含む増幅された複数のヌクレオチド配列を作製するために使用できる、PCRプライマーまたはプライマー対を含むことができる。本発明のこの局面による追加のキットは、選択的に、1つまたは複数のトポイソメラーゼ、1つまたは複数の組換えタンパク質、1つまたは複数のベクター、ポリメラーゼ活性を有する1つまたは複数のポリペプチドおよび1つまたは複数の宿主細胞などの追加の成分を、1つまたは複数含むことができる。
本明細書に記載する方法および用途の他の適切な改良および適合は、当業者に既知の情報を考慮して、本発明の説明から容易に明らかになり、本発明の範囲または本発明の任意の態様から逸脱することなく行うことができることは、関連分野の当業者に理解される。本発明を詳細に説明したが、例示的な目的のためだけに含まれ、本発明を限定する意図のものではない、以下の実施例を参照することによって、本発明はさらに明白に理解される。
実施例
実施例1
トポイソメラーゼを使用した共有結合二本鎖組換え核酸分子の構築
本実施例は、トポイソメラーゼを使用して共有結合二本鎖(ds)組換え核酸分子を産生することができることを示す。
A. 方法
特記しない限り、以下の方法を使用して試験を行った。PCRを、10ngプラスミド(鋳型)、各プライマー100ng、Taq DNAポリメラーゼ(Sigma)2.5U、10×PCR緩衝液5μl、およびdNTP(各200μl)4μlを含む反応物50μl中で行った。94℃で4分間の最初の変性を行い、その後94℃(45秒間)での変性、55℃(45秒間)のプライマーアニーリング、および72℃(標的配列1kbあたり1分)の伸長を30サイクルしてPCRを行った。サイクル後、反応物を、72℃(10分間)でインキュベートし、4℃に置いた。
各増幅エレメント(PCRで作製または合成)50ng〜100ng、500mM Tris(pH7.5)0.5μl、およびトポイソメラーゼ0.5μgを含む5μl中でトポイソメラーゼ連結反応を行った。反応物を室温で5分間インキュベートし、1〜2μlのTopo連結産物を、線状断片作製に使用した。
Topo結合産物(鋳型)1μl〜2μl、各プライマー100ng、Taq DNAポリメラーゼ(Sigma)2.5U、10×PCR緩衝液5μl、dNTP(各200μM)4μlを含む反応物50μl中でPCRによる線状断片作製を行った。上記のようにPCRを行った。
得られた線状断片を、SNAP Miniprepキット(Invitrogen)を使用して、製造者が記載のように精製した。本質的に、PCR産物100μlを、結合緩衝液300μl、イソプロパノール750μlと混合し、混合物を、SNAP Miniprep Column/Collection Tubeに入れ、7,000rpmで30秒間遠心分離した。カラムを、700μlの洗浄緩衝液で洗浄し、7,000rpmで30秒間遠心分離し、その後1×Final Wash900μlで洗浄し、7,000rpmで30秒間遠心分離した。次いで、カラムを7,000rpmでさらに30秒間遠心分離して、全ての残存液体を除去した。水(30μl〜50μl)を添加し、カラムを7,000rpmで30秒間遠心分離して、精製DNAを溶出させた。DNA濃度を、分光光度法で決定した。
B. トポイソメラーゼ結合線状核酸分子の作製
緑色蛍光タンパク質(GFP;図9A〜Cを参照のこと)のコード配列へのエレメントの定方向付加が試験されるようにPCRプライマーを設計した。CMVプロモーター(約700bp)およびBGHポリアデニル化シグナル配列(約380bp)を、pCMV/myc/nucプラスミド鋳型から増幅し、GFPエレメント(約700bp)を、図9Dに記載のプライマーを使用してpcDNA3.1/GFPプラスミド鋳型(Invitrogen)から増幅した。得られた増幅産物を、上記のようにトポイソメラーゼを使用して結合し、ライゲーション反応物の一部を、プライマーF6945(配列番号:11)およびF6948(配列番号:15)を使用したPCRの鋳型として使用して、全構築物(CMV+GFP+BGH;約1,700bp)を増幅した。さらに、ライゲーション混合物5μlを、プロテイナーゼKにて37℃で30分間処理して、任意の結合トポイソメラーゼを除去し、3%〜8%NuPAGE Tris-酢酸ゲルでの電気泳動に供して、ライゲーション産物を試験した。
回文突出配列(図9Aまたは図9B)を有するエレメントを使用して組換え核酸分子を作製した場合は正確なサイズ(1.7kb)の少量のライゲーション産物のみが認められ、非回文突出(図9C)を有するエレメントを使用すると大量の所望の産物が作製された。これらの結果は、所定の方向で機能的に連結されたヌクレオチド配列を含む両鎖に共有結合したds組換え核酸分子の作製効率が突出配列の性質に関連することを示す。特に、回文領域を欠く突出配列の選択により、両鎖に共有結合した所望のds組換え核酸分子が有効に産生される一方で、突出中の回文配列の存在により、意図する産物以外のライゲーション産物が形成されて所望の産物の作製効率が減少する。
実施例2
トポイソメラーゼ作製ds組換え核酸分子機能の特徴付け
本実施例は、本発明の方法により両鎖に共有結合した機能的ds組換え核酸分子の作製手段が得られることを示す。
A. TOPOライゲーション構築物からのセンスおよびアンチセンスmRNAの発現
関心対象の遺伝子に隣接する機能的上流および下流エレメントを含むds組換え核酸分子の作製能力を、T7プロモーター配列またはT3プロモーター配列のいずれかを含む2つの合成エレメントを使用して試験した。合成オリゴヌクレオチド対のアニーリングによってエレメントを作製した。T7リンカーを、等モル量のT7topオリゴヌクレオチド(F9304;配列番号:20)およびT7bottomオリゴヌクレオチド(F9305;配列番号:21)の混合によって作製した(図9D)。T3リンカーを、等モル量のT3topオリゴヌクレオチド(F9661;配列番号:23)およびT7bottomオリゴヌクレオチド(F9662;配列番号:24)の混合によって作製した(図9D)。混合物を、熱湯中で5分間加熱し、その後室温に冷却した。両エレメントを、片方の末端にトポイソメラーゼ認識部位を含むように設計した。
GFP遺伝子を、GFPプライマーF8418(配列番号:17)およびF8420(配列番号:18;図9D;図9Cも参照のこと)を使用して増幅した。非精製GFP PCR産物(2μl)を、50ngのT7リンカーおよび50ngのT3リンカーと混合し、トポイソメラーゼを添加し、室温で5分間TOPO結合反応を進行させた。T7配列およびT3配列のプライマーを使用した50μlのPCR反応の鋳型として2μlの結合反応物を使用した。
増幅後、PCR反応物4μlアリコートを、インビトロ転写の鋳型として使用した。製造者の説明にしたがって、Promega RiboProbe In vitro転写系キットを使用して反応を行った。T7RNAポリメラーゼあるいはT3 RNAポリメラーゼ(最終体積20μl)を使用して、37℃で60分間反応を進行させた。インビトロ転写反応物のアリコートをRNaseあるいはDNaseで消化し、その後非消化および消化試料を2%TBEゲルの電気泳動に供した。非消化試料中に推定サイズの所定のバンド(センス方向あるいはアンチセンス方向のいずれか)が認められた。DNaseで処理した試料では産物バンドの減少が認められなかった。試料をRNaseで処理した際に産物バンドが消滅し、これにより、産物がRNAであることが示された。これらの結果は、本発明の方法に従ってトポイソメラーゼを使用して所定の方向で両鎖に共有結合したds組換え核酸分子を作製できること、およびこのような核酸分子からRNA転写物を発現させることができることを示す。
B. TOPOライゲーション構築物からの翻訳産物の発現
トポイソメラーゼライゲーションポリヌクレオチドのインビトロ転写/翻訳共役を補助する能力を試験した。本発明の方法に従って1kb、2kb、あるいは3kbのlacZ PCR産物へのT7プロモーター(+コザック配列)を含むエレメントの結合によってds組換え核酸分子を作製した。作製した2種類の産物2μlを、PCR増幅反応の鋳型(プライマー、配列番号:25〜28;図9D)として使用した。増幅反応物の非精製アリコート(3μl)を、製造者の説明書にしたがってPCR DNAキット用のTNT T7 Quickを使用した転写/翻訳共役の鋳型として使用した(Promega)。
各反応から2μlアリコートを、Tris-グリシンゲル(Novex)での電気泳動によって分離し、オートラジオグラフィーによって視覚化し、予想サイズに移動したタンパク質産物が明らかとなった。これらの結果は、本発明の方法を使用してインビトロ転写/翻訳共役によるポリペプチド発現のための鋳型として有用な両鎖に共有結合したds組換え核酸分子を産生することができることを示す。
C. ツーハイブリッドアッセイ用のTOPOライゲーション産物の作製
ツーハイブリッドアッセイにより、インビボでのタンパク質-タンパク質間相互作用の強力な検出法が得られる。これらのアッセイは、多数の真核生物転写アクチベーターが2つの物理的および機能的に異なるドメイン(特定のDNA配列に結合するDNA結合ドメインおよび基本的転写機構と相互作用する転写活性化ドメインを含む)からなるという事実に基づく。トランス活性化ドメインのDNA結合ドメインとの会合により、機能的RNAポリメラーゼII複合体の構築を促進することができるので、例えば、検出可能なレポーター遺伝子の転写活性化が可能となる(FieldおよびSong、Nature、340、pp245〜246、1989)。第1のタンパク質であるXがDNA結合ドメイン(例えば、GAL4結合ドメイン)と融合し、第2のタンパク質であるY(Xと同一であっても異なっていてもよい)がトランス活性化ドメイン(例えば、VP16ドメイン)と融合する場合、タンパク質XおよびYの相互作用をGAL4プロモーターを有するレポーター遺伝子の転写の検出によって同定することができる。
本発明の方法の哺乳動物ツーハイブリッドアッセイのための融合タンパク質を発現するための線状構築物の作製能力を試験した。PCRを使用して、適切な末端にトポイソメラーゼ部位を含むGAL4(F10779およびF12667プライマー;それぞれ配列番号:1および3)、VP16(F10779およびF12668プライマー;それぞれ配列番号:1および5)、p53(F12669およびF12505プライマー;それぞれ配列番号:8および4)、T抗原(F12670およびF12505プライマー;それぞれ配列番号:9および4)、およびSV40pA(F12016およびF561プライマー;それぞれ配列番号:6および7)エレメントを作製した。トポイソメラーゼを使用して、共有結合した二本鎖構築物GAL4+p53+SV40pAおよびVP16+T抗原+SV40pAを作製し、得られたライゲーション産物をPCR増幅用鋳型として使用した。
精製GAL4+p53+SV40pAおよびVP16+T抗原+SV40pAPCR構築物を、lacZレポーター遺伝子(pGene/lacZプラスミド;Invitrogen)と共にCHO細胞(6ウェルプレート、1×105細胞/ウェル)に同時トランスフェクトした。並行実験では、非精製構築物を含むPCR反応混合物の使用と同様に、発現構築物を含むプラスミドベクターの使用を試験した。挿入物を含まないGAL4+pAおよびVP16+pA(負の対照)またはp53+VP16(正の対照)を使用して対照反応を行った。トランスフェクションの48時間後に細胞を溶解し、レポーター遺伝子活性をβ-ガラクトシダーゼアッセイキットを使用して測定した。
陽性対照(図10、試料3)ならびにレポーター遺伝子、線状GAL4+p53+SV40pA、およびVP16+T抗原+SV40pA構築物で同時トランスフェクトした試料(図10、試料4)で高レベルのレポーター遺伝子活性が検出された。構築物のプラスミドバージョンを使用した場合(図10、試料1)、低レベルの活性(しかし、陰性対照の活性よりは高い;試料5、6、8、および9)が検出された。非精製PCR作製プレイおよびベイト構築物(試料7)と同時トランスフェクトした試料でも低レベルの活性が認められた。これらの結果は、本発明の方法を使用してツーハイブリッドアッセイに有用な構築物を調製することができることを示す。
実施例3
定方向Topo負荷ゲートウェイベクターの産生および使用
緒言
トポイソメラーゼおよびGATEWAY(商標)組換えクローニングテクノロジーの組み合わせとして、定方向Topo負荷ゲートウェイベクターを開発した。これらのツールは、ドナーベクターへの組換え前にPCR増幅ORFのいずれかの末端へのattB部位(25塩基対)の付加の軽減によってゲートウェイシステムへの侵入を容易にする。代わりに、4塩基のtag認識配列(CACC)をORFの5'末端に付加し、PCR産物を定方向TOPOクローニングしてエントリー適合性発現ベクターまたはゲートウェイ適合性発現ベクターを作製する(図8および9を参照のこと)。
本実施例では、全部で3つのTopo-ゲートウェイベクターおよび1つのデスティネーションベクターを作製した。2つのTopoエントリーベクターを産生した。(1)attL部位間に定方向クローニングしたORFを任意のN末端融合原核生物DESTベクターおよび全ての真核生物DESTベクターに導入したpENTR/D-TOPO(登録商標)(図22)および(2)原核生物リボソーム結合部位(シャイン・ダルガルノ)の下流にORFを定方向TOPOクローニングしたpENTR/SD/D-TOPO(登録商標)(図23)。この様式でクローニングした遺伝子を、N末端タグを含まない原核生物DESTベクターに導入し、細菌で発現させて天然のN末端を含むタンパク質を産生することができる。
1つの定方向Topoゲートウェイ哺乳動物発現ベクター(pcDNA/GWD-TOPO(登録商標))も構築した(図19)。このベクターにより、pcDNA3.1誘導体へのORFの定方向クローニングが可能である。このベクターにクローニングされたORFは、CMVプロモーターの調節下にて哺乳動物細胞で発現する。クローニングORFは、ベクター中のattB部位に隣接し、BPクロナーゼおよびLRクロナーゼ反応を介してゲートウェイ系周囲に移動する。このベクターはまた、哺乳動物細胞系列での安定なクローン選択のためのC末端V5タグ、TKポリアデニル化シグナル、およびネオマイシン(G418)耐性マーカーをコードする。最後に、ゲートウェイデスティネーションベクターを、ccdBおよびクロラムフェニコール耐性カセットの導入によりpcDNA/GWD-TOPO(登録商標)から構築した。
これらのTopoゲートウェイエントリーベクターおよびTopoゲートウェイ発現ベクターは、研究者がプライマーへのattB部位の付加を行うことなくPCR増幅遺伝子を直接クローニングしてBPクロナーゼ反応を行うことによりゲートウェイ系に容易に侵入される。
材料と方法
pcDNA/GWD-TOPO(登録商標)の構築
pcDNA3.1attBのマルチクローニング部位(BGHポリアデニル化シグナルを有する初期バージョン)の第1の置換によってpcDNA/GWD-TOPO(登録商標)を構築した。BsrGI(MCSに隣接する各att部位内で切断する)での親ベクターの消化および新規のMCSをコードする二本鎖オリゴヌクレオチドの挿入によってこれを行った(図18)。適切な挿入を確認した後すぐに、V5/HisタグおよびBGHポリアデニル化シグナルを、V5タグに置換し、その後3つの終止コドン(TAG、TGA、TAA)および単純ヘルペスウイルス由来のチミジンキナーゼ(TK)ポリアデニル化シグナルに置換した。AscIおよびAvrIIでのベクターの消化、ベクター断片の精製、および三重ライゲーションにおける新規の配列をコードする2つの断片の挿入によってこれを行った(図19)。
pcDNA-DEST40の構築
pDONR221でのBPクロナーゼ反応を介してpcDNA/GWD-TOPO(登録商標)からpcDNA-DEST40を作製した。pDONR221を、適切な緩衝液中でpcDNAGW-DT(sc)およびBPクロナーゼ(Invitrogen Corporation;Carlsbad、CA)と組み合わせた。標準的なプロトコールに従って反応物をインキュベートし、カナマイシンプレート上で形質転換体を選択した。産物(ccdB、ccdA、およびクロラムフェニコール耐性遺伝子に隣接するattP部位を含むpcDNAデスティネーションベクター)を、培地を含むアンピシリン/クロラムフェニコールにて選択した(図20を参照のこと)。
pENTR/D-TOPO(登録商標)(sc)の構築
pcDNA/GWD-TOPO(登録商標)(sc)でのBP組換えによるattP部位間への適応配列カセットの付加によってpDONOR221を改変して、pENTR/D-TOPO(登録商標)(sc)を作製した(図21)。pDONR221を、適切な緩衝液中でpcDNA/GWD-TOPO(登録商標)(sc)およびBPクロナーゼと組み合わせた。pENTR/D-TOPO(登録商標)(sc)の形質転換および増殖にDH10BsbcC細胞を使用する以外は、標準的なプロトコールに従って反応物をインキュベートした。この細胞系列は、極めて近接してヘアピン構造(例えば、attL部位)を保有するプラスミドを維持可能な変異を有する。このプラスミドは、選択培地でのTop10細胞の成長を補助しなかった。
pENTR/D-TOPO(登録商標)およびpENTR/SD/D-TOPO(登録商標)の作製
NotI、AscI、およびXhoIでの段階的消化法(sequential digestion)によってベクターpENTR/D-TOPO(登録商標)(sc)に定方向をもってTOPOを付加し、15℃で一晩pENTR/SD/D-TOPO(登録商標)には定方向TOPOアダプターTopo-D71、Topo-D72、Topo-D75、およびTopo-D76またはpENTR/D-TOPO(登録商標)にはTopo D-73、Topo-D74、Topo-D75、およびTopo-D76でライゲートした(図26を参照のこと)。アダプター付加ベクター(adapted vector)を、室温でのイソプロパノール沈殿によって遊離のオリゴヌクレオチドから分離した。共通のアニーリングオリゴTopo D-70、T4キナーゼ、および組換えワクシニアトポイソメラーゼIを加えることにより精製され、アダプター付加ベクターにTOPOを負荷した。37℃で15分間のインキュベーション後、負荷ベクターを、アガロースゲル電気泳動(NB JC-12、2001-035、pg.3)または25Q MacroPrepカラム(BioRad)(NB2000-0342、pg.45)のいずれかによって精製した。定方向TOPOクローニング効率を、1ngの精製ベクターの5ngの定方向(CACC)750bp被験挿入物との室温で5分間のインキュベーションによってアッセイした。Top10ケミカルコンピテント細胞を、2μlのクローニング反応物で形質転換し、抗生物質選択としてカナマイシンを含むLBプレート上で成長させた。
Topo-ゲートウェイクローニングおよび遺伝子発現
これらのベクターのTopoクローニング、ゲートウェイクローニング、およびタンパク質産生を補助する能力を試験するために、ヒトHLAクラスIをコードする遺伝子(アクセッション番号D32129)を、4塩基CACCタグがATG開始コドンのすぐ上流にその5'末端が組み込まれたプライマーを使用したPCRによって増幅した。このPCR産物を、pENTR/D-TOPO(登録商標)およびpENTR/SD/D-TOPO(登録商標)の両方にクローニングした。各HLA反応由来の10個のクローンを、コロニー定方向PCR反応(d-PCR)で使用した。本実験では、クローンを、T7プライマー(5'側のattL 1部位に結合する)および129方向逆プライマー(HLAの3'末端に特異的)を使用して増幅した。
HLA遺伝子に加えて、クロラムフェニコールアセチルトランスフェラーゼ(CAT)の遺伝子を同様に増幅し、2つのエントリーベクターにクローニングした。ミニプレップおよび消化分析後、各反応由来の1つのクローンを単離し、M13正方向およびM13逆方向プライマーを使用して配列決定した。全てのエントリークローンを配列決定によって確認し、pcDNA/GW DEST40(pENTR-D-TOPO(登録商標)クローン)またはpET DEST42(pENTR/SD/D-TOPO(登録商標)クローン)とのL/Rクロナーゼ反応によって組換えた。NcoI(定方向アダプター付加ORF, caCCATGGの5'末端に部位が出現する)およびNotI(データ示さず)での消化によって陽性クローンを確認した。得られたpcDNA-DEST40(HLAおよびCAT)およびpcDNA/GW/D-TOPO(登録商標)(HLAおよびCAT)構築物を使用して、COS細胞をトランスフェクトした。リポフェクタミン2000、8μgのDNA、および至適な緩衝液を使用して、細胞をトランスフェクトした。細胞に反応物を5時間適用し、培地を交換した。37℃で一晩のインキュベーション後、細胞を採取し、溶解し、標準的な手順を使用して4%〜20%のTris-グリシンゲルにて電気泳動した。電気泳動後、タンパク質をニトロセルロース膜に移し、ブロックし、V5-HRP抗体およびECL検出で探索した。
各pET DEST42反応由来の1つの陽性クローンを使用して、BL21(DE3)細胞を形質転換し、LB/Amp中で一晩成長させた。次いで、培地を同一の培地で25倍に希釈し、O.D.(600nm)=0.5まで成長させ、その時点で組換えタンパク質の発現を最終濃度1mMのIPTGの添加によって誘導した。培養物を37℃で3時間成長させた後、細胞を遠心分離によって採取した。細胞ペレットのアリコートをNuPage変性試料緩衝液中でボイルし、4%〜12%のNuPageポリアクリルアミドゲルにて電気泳動し、SafeStain(商標)(Invitrogen)を使用して染色した。pET DEST42ベクター中の被験遺伝子発現の陽性対照として、HLAおよびCAT遺伝子を、pET100CATおよびHLA(dTopo、attB部位なし)に直接TOPOクローニングした。これらの構築物を使用して、BL21(DE3)大腸菌細胞をトランスフェクトし、対数期まで成長させ、上記のようにIPTGで誘導した。
結果と考察
pENTR-dTopoおよびpENTR/SD-dTopoにおけるHLAおよびCATクローンの定方向クローニング効率
pENTR/D-TOPO(登録商標)およびpENTR/SD/D-TOPO(登録商標)にクローニングしたHLA ORFが確実に正確な方向であるように定方向PCR反応を設計した。10個のコロニーを各Topoクローニング形質転換物から選択し、「材料と方法」に記載のようにPCR反応物に直接置いた。試験した10個のpENTR/SD-HLAクローンのうちの8個が正確な方向であり、10個のpENTR-HLAクローンのうちの9個が正確であった。約10%〜15%の挿入物バックグラウンドを示さないゲル精製ベクター(データ示さず)を使用してこれらの試験を行った。
あるいは、CATクローンの制限分析を行った。クローンを単離し、NcoIおよびAscIでDNAを消化した。正確に配向したCATクローンの2つのNcoI部位の1つが、Kozac定方向アダプター付加配列およびCAT遺伝子の第1の2つのコドン(caCCATGG)の一部として各ORFの5'末端に出現する。AscIは、ベクター中のORFの3'末端に存在する。正確に配向したクローンは、2つのNcoI部位(一方は5'末端であり、他方は内部である)を有し、AscIでの二重消化後に500bpおよび150bpの断片が得られる。CAT ORFは、その3'末端で至適なタグ配列に対して1塩基対のミスマッチを含む配列CGCCをコードする。この密接な相同性により、ほんの50%の効率(8個のクローンのうち4個、データ示さず)で定方向クローニングするためのCAT PCR産物が得られた。
エントリークローンの配列決定
DESTベクターへの組換えおよびその後の発現のために選択された各エントリークローンを両末端から配列決定して、アダプターおよびORFが正確にライゲーションされたことを確認した。M13正方向プライマーおよびM13逆方向プライマーを使用し、反応物をABI 3700キャピラリーシークエンサーでの配列決定のためにResGenに送った。これらの反応物から、最小で600塩基の読み取り可能な配列が得られた。attL部位による反応の進行につれてシグナルがいくらか喪失するが、この手順を使用したこの時点で有意なシグナルが残存することが明白である(データ示さず)。
COS細胞におけるHLAおよびCATの発現
pcDNA/GW/D-TOPO(登録商標)およびpcDNA-DEST40由来の発現を、これらの構築物中の標的遺伝子としてのHLAおよびCATでのCOS細胞のトランスフェクションによって試験した。採取した溶解物を、V5抗体を使用したウェスタンブロットによってV5タグ化組換えタンパク質を探索した。図27に示したデータは、Topoクローニング(図27、レーン3およびレーン6)またはpENTR/D-TOPO(登録商標)からのLRクロナーゼ導入後(図27、レーン2およびレーン5)に遺伝子が直接クローニングされるかのいずれかでHLAおよびCAT遺伝子がこれらのベクター中で発現することを示す。
HLAおよびCATの細菌性発現
pENTR/SD/D-TOPO(登録商標)にクローニングしたCATおよびHLA遺伝子を、LRクロナーゼ反応を介してpDEST-42(pET、C末端V5/His)に導入した。図28に示した結果により、attB部位に隣接するかしないCAT遺伝子が細菌中で発現されることが示唆される(図28、レーン6とレーン7を比較のこと)。pENTR/SD/D-TOPO(登録商標)由来のpET DESTベクターに導入後にCAT遺伝子が大腸菌で十分に発現するという所見により、Topo-ゲートウェイ系を使用したORFのクローニングおよび発現のためのこの系の有用性が実証される。
興味深いことに、pDEST42(attB部位に隣接)にクローニングしたHLAは、2つの独立した実験でBL21(DE3)細胞で発現できなかった(図28、レーン3およびレーン4)。上記のように、同一のエントリークローン由来のHLA遺伝子は、哺乳動物DESTベクターに組換えられた場合にCOS細胞中で十分に発現した。さらに、attB部位に隣接する場合にpET系がHLA遺伝子発現を補助することができないという事実により、少なくとも細菌中でのゲートウェイ系を使用した発現における遺伝子特異的バリエーションが存在し得る。この結果に関連し得る1つの事実は、対照ベクター(pET100d-Topo)から発現したHLAがゲル中で異常に泳動されることであった(推定41kDaの代わりに30kDa)。このヒトタンパク質は、任意の場合に細菌中で発現することができず、attB部位の添加によって発現における問題がさらに深刻になり得る。この所見の理解にはより多くの実験が必要である。
結論として、本発明者らは、2つの新規Topoゲートウェイエントリーベクター(一方は新規のTopoゲートウェイ発現ベクターであり、他方はこれから得られる新規のDESTベクター)の構築および試験を記載した。全体で、Topoクローニングの容易さおよび効率ならびにゲートウェイ系の多能性を組み合わせた新規のツールにより、最小の費用および手間で多数の異なる状況において多数の遺伝子をクローニングおよび発現可能である。
実施例4
トポイソメラーゼクローニングの別法
本発明の1つの好ましい別の態様では、TOPO SSSベクターを、市販のクローニングベクターを最初に得ることによって作製する。このようなベクターの1つは、pUni/V5-HisバージョンA(Invitrogen Corp.、Carlsbad、CA)(独自に設計したエレメントを含む環状スーパーコイルベクター)である。これらのエレメントには、真核生物宿主中でのmRNA安定性を増加させるためのBGHポリアデニル化配列、T7転写終結領域、R6Kg DNA複製起点、カナマイシン耐性遺伝子、および抗生物質耐性選択のためのプロモーターが含まれる。さらに、pUni/V5-HisバージョンAは、一連の制限エンドヌクレアーゼ認識部位をコードする合成DNA配列であるマルチクローニング部位を含む。これらの部位を、特定の位置でのベクターへのDNAのクローニング用に操作する。また、ベクターのマルチクローニング部位内で、エンドヌクレアーゼ認識部位の5'へのloxP部位の挿入により、種々の他の発現ベクター(Echo(商標)Cloning System、Invitrogen Corporation、Carlsbad、CA)へのCreリコンビナーゼ媒介融合が促進される。抗V5抗体を使用した発現融合タンパク質の容易な検出に最適なC末端V5エピトープタグが存在する。最適なC末端ポリヒスチジン(6×His)タグもまた、発現タンパク質を迅速に精製および検出することができるように存在する。loxP部位由来の細菌リボソーム結合部位下流により、大腸菌での転写開始が可能になる。エレメントのこの組み合わせは、pUni/V5-HisバージョンAクローニングベクターに特異的であるが、多数の類似のクローニングおよび発現ベクターが市販されているか、配列および当該分野で周知の方法によって構築することができる。pUni/V5-HisバージョンAは、2.2kbの二本鎖プラスミドである。
pUni/V5-HisバージョンA由来のトポイソメラーゼI負荷クローニングベクターの構築を、ベクターのエンドヌクレアーゼ消化、合成オリゴヌクレオチドの相補的アニーリング、およびワクシニアトポイソメラーゼIによるヘテロ二重鎖の部位特異的切断によって行う。SacIおよびEcoRIは、pUni/V5-HisバージョンA内に存在する多数の制限エンドヌクレアーゼ部位の2つである。対応する制限酵素SacIおよびEcoRIでのpUni/V5-HisバージョンAの消化により、ベクター上の付着末端(5'-AGCT-3'および5'-AATT-3')が遊離する。これらの酵素は、New England Biolabs(Beverly、MA、カタログ番号RO156S(SacI)およびカタログ番号RO101S(EcoRI))を含む多数の販売者から容易に入手可能である。消化pUni/V5-HisバージョンAは、イソプロパノール沈殿を使用して消化断片から容易に分離される。これらおよび他のDNA消化および単離方法は、当業者に周知である(Sambrook,J.、Fritsch,E. F.、およびT. Maniatis.、1989、「Molecular Cloning, A Laboratory Manual.」、第2版、Cold Spring Harbor Laboratory Press、p. 5.28〜5.32)。
次いで、精製および消化されたベクターを、2つの特異的オリゴヌクレオチドアダプターおよびT4DNAリガーゼとインキュベートする。アダプターは、ベクターのSacIおよびEcoRI末端に適合する末端を含むオリゴヌクレオチド二重鎖である。当業者には、異なる制限酵素部位を有する他のベクターのために適切な配列を含む他のアダプターオリゴヌクレオチドを作製できることが容易に理解できるだろう。T4 DNAリガーゼとのインキュベーション後、ライゲーションしたアダプターを含むベクターを、イソプロパノールを使用して精製する。TOPO D1およびTOPO D2のアニーリング由来のアダプター二重鎖は、一方の末端に一本鎖EcoRI突出を有し、他方の末端に12ヌクレオチドの一本鎖突出を有する。
第1のアダプターオリゴヌクレオチド(TOPO D1)は、EcoRI付着末端(3'-TTAA-5')に相補性を示す。さらに、TOPO D1は3'末端の16bp上流に存在するトポイソメラーゼ・コンセンサス・ペンタピリミジン・エレメント5'-CCCTTを含むさらなる24bpを有する。TOPO D1アダプターオリゴの残存する配列およびサイズを変化させることができ、研究者の特定の要求に適合するように改変することができる。本発明のこの好ましい態様の1つのこのような局面によれば、
(配列番号:33)は、使用したアダプターの全長配列である。
第2のアダプターオリゴヌクレオチド(TOPO D2)は、TOPO D1に完全な相補性を示さなければならない。TOPO D2は、EcoRI付着フラップの5'を直接補って、線状ベクターの下の鎖(bottom strand)を伸長させる。さらに、TOPO D2は、定方向クローニングに必要なSSSである配列3'-GTGGを含む。この態様では、SSSを、mRNA上のリボソーム結合効率の増加によって真核細胞中のORFの発現を補助することが公知のKozac配列を補うために選択するが、配列および長さは各使用者の特定の要求を満たすように高度に変化させることができる。TOPO D2の完全な配列は、
(配列番号:34)である。上記と類似して、オリゴヌクレオチドTOPO D4およびTOPO D5のアニーリングに起因するアダプター二重鎖は、一方の末端に一本鎖SacI突出を有し、他方の末端に12ヌクレオチドの一本鎖突出を有する。
第3のアダプターオリゴヌクレオチド(TOPO D5)は、SacI付着末端3'-TCGA-5'に相補性を示す。TOPO D1と同様、TOPO D5は、一本鎖突出を作製するさらなる塩基を有する。長さおよび配列は、使用者の要求に基づいて変化し得る。本態様では、TOPO D5'の配列は、
(配列番号:35)である。
第4のアダプターオリゴヌクレオチド(TOPO D4)は、TOPO D5に完全な相補性を示し、線状化ベクターの上の鎖にわたるSacIの付着フラップの5'に直接的な相補性を示す。TOPO D4はまた、トポイソメラーゼコンセンサス配列5'-CCCTTを含む。TOPO D4アダプターオリゴの残存する配列およびサイズは変化させることができ、特定の要求に適合させるために改変することができる。本態様では、TOPO D4配列は、さらなる12bpの一本鎖突出を含む
(配列番号:36)である。
これらのアダプターオリゴヌクレオチドを、ホスホラミダイド法を含む任意の多数の技術を使用して化学合成することができる(Caruthers,M. H.、Barone,A. D.、Beaucage,S. L.、Dodds,D. R.、Fisher,E. F.、McBride,L. J.、Matteucci,M.、Stabinsky,Z.、およびTang,J. Y.、1987、「Chemical Synthesis of Deoxyoligonucleotides」、Methods Enzymol.、154、pp287〜313)。このオリゴの化学合成方法および他の方法は、当業者に周知である。
精製され消化されたベクターおよびアダプターオリゴヌクレオチドの相補アニーリングを、T4 DNAリガーゼの存在下でのDNAのインキュベーションによって行う。典型的なライゲーション反応を、リガーゼおよび適切な反応緩衝液の存在下での適切なDNA断片とのクローニングベクターのインキュベーションによって行う。ライゲーション反応用の緩衝液は、反応のためのエネルギーを提供するためのATP、ジチオスレイトールのような還元剤、およびTris-HClのようなpH安定剤を含むべきである。クローニングベクターおよびDNA断片の濃度比は各個々の反応に依存し、その決定手順は、文献に豊富に記載されている(例えば、「Protocols and Appications Guide」、1991、Promega Corporation、Madison、WI、p.45を参照のこと)。T4リガーゼは、インキュベーション中の隣接する5'リン酸塩と3'ヒドロキシ末端との間のホスホジエステル結合の形成を触媒する。付着末端のライゲーションを12℃〜15℃で30分間で行うことができ、平滑末端のライゲーションには室温で4時間〜16時間必要である(Ausubel,F.M.、Brent,R.、Kingston,R.E.、Moore,D.D.、Seidman,J.G.、Smith,J.A.、Struhl,K.、1992、第2版、「Short Protocols in Molecular Biology」、John Wiley & Sons,Inc.、New York、NY、pp.3.14〜3.37)が、各試験によりパラメーター範囲は変化する。本態様では、精製消化pUni/V5-HisバージョンAおよびアダプターオリゴを、T4リガーゼおよび適切な緩衝液の存在下、12.5℃で16時間インキュベートした。得られた線状化およびアダプター付加ベクターは、塩基対相補性およびT4リガーゼ触媒ホスホジエステル結合によってアダプターオリゴヌクレオチドに結合した精製クローニングベクターを含む。
トポイソメラーゼでのアダプター付加ベクターの有効な改変には、TOPO D1およびTOPO D4の一本鎖突出上に二本鎖DNAを作製するためのアニーリングオリゴの付加が必要である。ワクシニアトポイソメラーゼIは、最初に二本鎖DNAに非共有結合する。次いで、コンセンサス・ペンタピリミジン配列5'-CCCTTに局在して共有結合するまで二重鎖に沿って酵素が核酸し、トポイソメラーゼアダプター付加複合体が形成される(Shumanら、米国特許第5,766,891号を参照のこと)。切断されにくい鎖中のホスホジエステル結合により切断時の遊離基の解離を防止するので、アダプター付加ベクターとアニーリングオリゴとの間のホスホジエステル結合の形成を防止するためのアダプター付加ベクターの改変は、DNAリガーゼの非存在下でおこる。
アニーリングオリゴヌクレオチド(TOPO D3)は、TOPO D1およびTOPO D4の一本鎖DNA突出に相補性を示さなければならない。本態様では、突出は共に以下の配列
(配列番号:37)を共有する。したがって、TOPO D3は、アダプターオリゴの一本鎖突出およびさらなる3bpの突出3'-AAC-5'に完全に相補的な以下の配列
(配列番号:38)を有する。
トポイソメラーゼの存在下でのアニーリングオリゴとのアダプター付加ベクターのインキュベーションにより、トポイソメラーゼが共有結合することができる二本鎖DNAが作製される。結合したトポイソメラーゼは、5'-CCCTT認識モチーフが存在するまで分散機構の促進によって二本鎖DNAを検索する。モチーフの切断しやすい鎖の3'のホスホジエステル骨格の切断は、好ましいオリゴヌクレオチド切断配列5-CCCTTの3'リン原子への求核攻撃を介して触媒され、3'ホスホチロシル結合によって酵素へDNAが共有結合する(Shuman,S.、Kane,E. M.、Morham,S. G.、1989、Proc. Natl. Acad. Sci. USA、86、pp9793〜9796を参照のこと)。切断しやすい鎖の切断により、3'末端アダプターオリゴ、5'-CCCTTモチーフ、およびアニーリングオリゴTOPO D3を含む二本鎖遊離基が作製される。ホスホチロシル結合の5'ヒドロキシル媒介攻撃を介してベクターのトポイソメラーゼ改変末端に遊離基を再び連結させる(religate)することができるが、遊離基がもはやベクターに共有結合しない場合はこの反応は好ましくない。キナーゼが遊離基の5'ヒドロキシルをリン酸化して結合を防止することができるので、切断/再ライゲーション反応物へのT4ポリヌクレオチドキナーゼおよびATPの添加により、トラップされたトポイソメラーゼが蓄積する方に平衡がさらにシフトする(Ausubel,F.M.、Brent,R.、Kingston,R.E.、Moore,D.D.、Seidman,J.G.、Smith,J.A.、Struhl,K.、1992、第2版、「Short protocols in Molecular Biology」、John Wiley & Sons,Inc.、New York、NY、pp.3.14〜3. 30)。得られた線状化ベクターは、TOPO D4/D3遊離基由来の平滑末端およびTOPO D1/D3遊離基由来のSSS保有末端を含む。両線状クローニングベクターの末端にトポイソメラーゼを負荷し、受容体分子を迅速且つ有効な定方向トポイソメラーゼ媒介挿入を行うことができる。
上記の例でトポイソメラーゼ改変定方向クローニングベクターを形成するためのpUni/V5-HisバージョンAの改変を詳述しているが、当業者はこれらの方法を任意のプラスミド、コスミド、ウイルス、または他のDNAに適用させる方法を理解できるだろう。この例は配列5'-GGTG-3'を含む5'一本鎖突出を含むベクターを示しているが、当業者はアダプター二重鎖およびアニーリングオリゴヌクレオチドのデザインを所与のベクターの一方または両方の末端で任意の配列または長さの突出に特別に設計することができることにも留意すべきである。
具体的には、任意のプラスミド、コスミド、ウイルス、または他のDNAを改変して、任意の都合の良い配列および長さのSSSを含めることができる。これらは基本的段階で、DNAを線状化することが知られる処理にベクターを最初に供する。一般的手順には、制限消化およびトポイソメラーゼIIでの処理が含まれるが、これらに限定されない。線状化後、特別なSSSを添加する。上記の例では、相補オリゴヌクレオチドを制限消化の付着末端に付加して所望のSSSを獲得するが、SSS形成オリゴヌクレオチドを、T4平滑末端ライゲーションによっても付加することができる。SSS配列を、トポイソメラーゼI媒介一本鎖ニックによって露呈する。その後、このSSSを使用して、1つあるいは複数の相補SSSを含むPCR産物を定方向に挿入することができる。
同様に、トポイソメラーゼの改変を任意の二本鎖プラスミド、コスミド、ウイルス、または他のDNA片に適用することができる。二本鎖DNAへのトポイソメラーゼIの結合方法は、当該分野で周知である(Shumanら、米国特許第5,766,891号を参照のこと)。二本鎖DNA片へのトポイソメラーゼの有利な配置を、トポイソメラーゼI配列の組み込みによって決定する(Shumanら、米国特許第5,766,891号を参照のこと)。トポイソメラーゼIは、二本鎖DNAに結合し、切断しやすい鎖に切れ目を入れて所定の一本鎖突出配列を出現させ、正確なSSS媒介配向で入ってきたPCR産物をライゲーションする。
実施例5
特別なトポイソメラーゼI適合化ベクターの産生
本発明のこの局面の別のプラスミドへの適用の例として、pCR2.1(Invitrogen Corporation;Carlsbad、CA)を、特別な(custom)一本鎖配列を有するトポイソメラーゼI順応ベクターを作製するように改変した。
プラスミドpCR2.1は、3.9kbのT/Aクローニングベクターである。この内でこのベクターの配列は、多数の固有に設計したエレメントである。これらのエレメントには、f1起点、ColE1起点、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、LacZ-α断片、および組換えプラスミドの青白選択が可能なLacZ-α断片内に存在するマルチクローニング配列が含まれる。pCR2.1のマルチクローニング配列は、以下を含む。多数の制限酵素部位(HindIII、SpeI、およびEcoRIが含まれるがこれらに限定されない)、M13正方向プライマーおよびM13逆方向プライマー、ならびにT7 DNAポリメラーゼプロモーター。
特別な一本鎖配列を保有するトポイソメラーゼI負荷ベクターの構築は、エンドヌクレアーゼ消化、その後の合成オリゴヌクレオチドのアニーリング、およびワクシニアトポイソメラーゼIによるヘテロ二重鎖の部位特異的切断からなる。制限酵素HindIII、SpeI、およびEcoRIでのpCR2.1の消化により、ベクター上にHindIIIおよびEcoRI付着末端が遊離する。HindIII切断部位由来のpCR2.1下流の解離断片を、SpeIでさらに切断してそのサイズを減少させる。断片サイズの減少により、イソプロパノール沈殿によってより小さな消化片から消化ベクターを精製する。これらの酵素は、New England Biolabs(Beverly、MA、カタログ番号RO104S(HindIII)、RO133S(SpeI)、およびRO101S(EcoRI))を含む多数の販売者から容易に入手可能である。DNAの消化および単離方法は、当業者に周知である(Sambrook,J.、Fritsch,E. F.、およびT. Maniatis.、1989、「Molecular Cloning, A Laboratory Manual.」、第2版、Cold Spring Harbor Laboratory Press、p. 5. 28〜5. 32)。
精製した消化ベクターを、4つのアダプターオリゴヌクレオチドおよびT4DNAリガーゼとインキュベートする。これらのアダプターオリゴヌクレオチドを、HindIII付着末端、EcoRI付着末端、または互いに相補的なように設計する。T4 DNAリガーゼとのインキュベーション後、アダプター付加ベクターを、イソプロパノールを使用して精製する。
第1のアダプターオリゴヌクレオチド(TOPO H)は、HindIII付着末端3'-TCGA-5'に相補的である。さらに、TOPO Hは、3'末端の19bp上流に存在するトポイソメラーゼコンセンサスペンタピリミジンエレメント5'-CCCTTを含むさらなる24bpを有する。TOPO Hアダプターオリゴの残存する配列およびサイズを変化させることができ、研究者の特定の要求に適合するように改変することができる。本態様では、
(配列番号:39)は、使用したアダプターの全長配列である。
第2のアダプターオリゴヌクレオチド(TOPO 16)は、TOPO Hに完全な相補性を示さなければならない。TOPO 16は、HindIII付着末端の5'を直接補って線状ベクターの下の鎖を伸長させる。さらに、TOPO 16は、定方向クローニング用に選択された一本鎖配列である配列3'-TAAGを含む。TOPO 16の完全な配列は、
(配列番号:40)である。
第3のアダプターオリゴヌクレオチド(TOPO 1)は、EcoRI付着末端3'-TTAA-5'に相補性を示す。TOP Hと同様、TOPO 1は、3'末端の12bp上流に存在するトポイソメラーゼIコンセンサス配列CCCTTを含むさらなる塩基を有する。TOPO1の長さおよび配列は、使用者の要求に基づいて変化し得る。本態様では、TOPO 1'の配列は、
(配列番号:41)である。
第4のアダプターヌクレオチド(TOPO 2)は、TOPO 1に完全な相補性を示し、線状化ベクターの上の鎖を伸長させるEcoRIの付着末端の5'を直接補う。本態様では、TOPO 2配列は、3'-GCGGGAA-5'である。
精製され消化されたベクターおよびアダプターオリゴヌクレオチドの相補アニーリングを、T4 DNAリガーゼの存在下でのDNAのインキュベーションによって行う。T4リガーゼは、インキュベーション中での隣接する5'リン酸と3'ヒドロキシル末端との間のホスホジエステル結合の形成を触媒する。本態様では、精製した消化pCR2.1およびアダプターオリゴをT4リガーゼおよび適切な緩衝液の存在下、12.5℃で16時間インキュベートした。得られた線状化アダプター付加ベクターは、塩基対相補性およびT4リガーゼ触媒ホスホジエステル結合を介してアダプターオリゴヌクレオチドに結合した精製クローニングベクターを含む。ライゲーション技術は、文献に豊富に記載されている(Ausubel,F. M. ら、1992、第2版、「Short protocols in Molecular Biology」、John Wiley & Sons,Inc.、New York、NY、p. 3.14〜3. 37)。
トポイソメラーゼでのアダプター付加ベクターの負荷には、TOPO HおよびTOPO 1の一本鎖突出上の二本鎖DNAを作製するためのアニーリングオリゴヌクレオチドを付加する必要がある。切断されない鎖中のホスホジエステル結合により切断時の遊離基の解離が防止されるので、DNAリガーゼの存在下でアダプター付加ベクターが負荷されて、アダプター付加ベクターとアニーリングオリゴとの間のホスホジエステル結合の形成が防止される。
アニーリングオリゴヌクレオチド(TOPO 17)は、TOPO Hの一本鎖DNA突出に相補性を示さなければならない。本態様では、突出は配列5'-CGATAGTG-3'を有する。したがって、TOPO 17は、アダプターオリゴの一本鎖突出に完全に相補的な配列3'-GCTATCAC-5'を有する。
アニーリングオリゴヌクレオチド(TOPO 3)は、TOPO 1の一本鎖DNA突出に相補性を示さなければならない。本態様では、突出は以下の配列
(配列番号:42)を有する。したがって、TOPO 3は、アダプターオリゴの一本鎖突出およびさらなる3bpの突出5'-CAA-3'に完全に相補的な以下の配列
(配列番号:43)を有する。
トポイソメラーゼの存在下でのアニーリングオリゴとアダプター付加ベクターのインキュベーションにより、トポイソメラーゼが非共有結合することができる二本鎖DNAが作製される。結合したトポイソメラーゼは、5'-CCCTT認識モチーフが存在するまで分散機構の促進によって二本鎖DNAを検索する。モチーフの切断しやすい鎖の3'のホスホジエステル骨格の切断により、3'ホスホチロシル結合によって酵素にDNAが共有結合される(Shuman,S. ら、1989、Proc. Natl. Acad. Sci. U.S.A.、86、pp9793〜9796を参照のこと)。切断しやすい鎖の切断により、3'末端アダプターオリゴ、5'-CCCTTモチーフ、および相補アニーリングオリゴヌクレオチドを含む二本鎖遊離基が作製される。トポイソメラーゼによっても触媒されるホスホチロシル結合の5'ヒドロキシル媒介攻撃によってトポイソメラーゼアダプター付加ベクターに遊離基を再ライゲーションすることができる。平衡反応物へのT4ポリヌクレオチドキナーゼの添加により、遊離基の5'ヒドロキシルのキナーゼ媒介リン酸化を介して反作用を防止する(Ausubel,F. M. ら、1992、第2版、「Short protocols in Molecular Biology」、John Wiley & Sons,Inc.、New York、NY、pp.3.14〜3. 30)。得られた線状化ベクターは、TOPO 1/3遊離基由来の平滑末端およびTOPO H/17遊離基由来の一本鎖配列を含む。両線状クローニングベクターの末端にトポイソメラーゼを負荷し、受容体分子を迅速且つ有効な定方向トポイソメラーゼ媒介挿入を行うことができる。
実施例6
トポイソメラーゼを使用した定方向クローニング
本発明のこの局面はまた、DNAの定方向クローニング法を提供する。このような方法では、pUni/V5-HisバージョンAから構築したTOPO SSSベクターを、GeneStorm Expression Ready Clones(Invitrogen Corporation、Carlsbad、CA)の定方向挿入に使用した。ベクターに付加した一本鎖はORF発現を増強することが公知のKozak配列に相同性を示すので、改変pUniベクターをこれらのORFのクローニングのために選択した。しかし、上記のように、任意のプラスミド、コスミド、ウイルス、または他のDNAを必要な一本鎖配列を有するように改変することができることに留意されたい。同様に、任意のDNA断片を、任意のベクターSSSに相同な配列を有するように改変することができる。関心対象の点として、SSS配列により定方向クローニング効率を得ることができる。例えば、GC含量の低いSSSほどアニーリング安定性が低く、クローニングすべきDNA断片の両末端に対する相補性が高いSSSもまたこれらのDNA挿入物を方向付ける能力を低下させる。したがって、SSS配列を、これらおよび類似の問題を回避するように慎重に設計すべきである。
本発明のこの局面は、本発明に従って構築したベクターへのPCR産物の定方向挿入で特に有用である。所望の挿入物のPCR増幅では、TOPO SSSベクターに定方向クローニングすべき挿入物の同定配列に相補的なようにPCRプライマーを設計する。DNAコード鎖の上流に結合するように設計したプライマーを、その5'末端上でさらなるベクターのSSS相補性配列で改変する。得られたPCR産物は、TOPO SSSクローニングベクターへのSSS媒介定方向挿入を行い、その後の産物を発現する相補配列を有する。
1つのこのような態様は、SSSを含む5'オリゴヌクレオチドプライマーでドナー二重鎖DNA分子を増幅するPCRによってドナー二重鎖DNA基質にSSS部位を移入する段階を含む。所望の領域の公知の外側の領域に相補的なオリゴヌクレオチドプライマーの設計によって、DNA領域のPCR増幅を行う。好ましい態様では、DNAの二本鎖領域のコード鎖に相同性を示すプライマーは、TOPO SSSクローニングベクターのSSSに対してヌクレオチド相補性を示すさらなる配列を有する。
ハイスループット様式で本発明を使用して、本発明者らは、TOPO SESSベクターへの定方向クローニングのためにGeneStorm発現系(Invitrogen Corporation、Carlsbad、CA)の82個の公知のORFを選択したが、各使用者が所望する任意のDNA配列を選択することができる。これらの各ORFについて、コード鎖および非コード鎖に相補的なプライマーを設計する。改変pUni/V5-HisバージョンA(実施例4に記載のTOPO SSSベクター)に定方向様式でPCR産物をクローニングするために、所与のプライマー対を含むように改変された所与の1つのプライマー対を、ベクター内に含まれるSSSに相補的なヌクレオチド配列を含むように改変した。本実施例では、コードプライマーは、TOPO SSSクローニングベクターの「SSS」(3'-GTGG-5')に相補的な付加配列5'-CACC-3'を含んでいた。上記ORFのその各プライマーを使用したPCR増幅により、その5'末端にSSSを有する二本鎖DNA断片が産生される。本PCR増幅ではpfuポリメラーゼを使用したが、PCR反応をpfuなどの非好熱性ポリメラーゼまたはtaqのような好熱性ポリメラーゼのいずれかを使用して行うことができ、その後平滑末端作製段階によって非鋳型ヌクレオチドを除去し、これらの酵素がPCR産物の末端で遊離することが周知である。
本実施例では、0.1μgの各プライマーを、ORFを含む0.05μgのDNAを含む全部で50μlのPCR反応混合物と組み合わせた。プライマーおよびベクターに加え、反応混合物は、水、PCR緩衝液の塩、10mM dNTP、および1.25単位のpfuポリメラーゼも含んでいた。サーマルサイクリング温度は次の通りであった。94℃での最初の変性;94℃での変性、55℃でのプライマーアニーリング、および72℃での伸長(各1分間)を25サイクル;および72℃で15分間の伸長で終了。これらのパラメーターは、増幅される各DNA断片によって変化し、当業者に周知の方法を使用して種々の長さおよび組成物の断片に最適化することができる(Ausubel,F.M.、Brent,R.、Kingston,R.E.、Moore,D.D.、Seidman,J.G.、Smith,J.A.、Struhl,K.、1992、第2版、「Short protocols in Molecular Biology」、John Wiley & Sons,Inc.、New York、NY、pp. 15.3〜15.4)。3'突出の平滑末端への変換技術もまた、当業者に公知である(「Protocols and Appications Guide」、1991、Promega Corporation、Madison、WI、pp. 43〜44)。
SSS相補配列を含むPCR増幅ドナー二重鎖DNAの改変pUni/V5-HisバージョンATOPO SSSベクターとのインキュベーションにより、ドナーDNAの定方向クローニングを行う。例えば、GeneStormクローン集団(Invitrogen Corporation、Carlsbad、CA)由来の82個のORFを、SSSアダプター付加プライマーを使用して増幅した。記載の改変プライマー対を使用した82個のGeneStorm ORFの増幅により、その5'末端にSSS相補配列を有するPCR産物が得られた。ORF PCR産物を、滅菌水または塩溶液中で10ngのTOPO SSSクローニングベクターと組み合わせる。反応物を穏やかに混合し、室温(22℃〜23℃)で5分間インキュベートする。5分後、反応物を氷上に置き、OneShot(登録商標)化学的形質転換またはエレクトロポレーション(Invitrogen Corporation、Carlsbad、CA、それぞれカタログ番号C4040-10およびC4040-50)(Invitrogen TOPOクローニングプロトコール、Invitrogen Corporation、Carlsbad、CA)に供した。トポイソメラーゼは再結合反応の触媒によってベクターおよび産物の隣接する鎖に結合した(図17)。したがって、その5'末端でSSSを使用して構築したDNA断片は、TOPO SSSクローニングベクターに高い効率で正確に挿入された。
形質転換宿主細胞の複数のコロニーの配列決定によって90%を超える効率で5'SSSを含むDNA断片の定方向挿入が行われる。本実施例では、GeneStorm ORFを含むTOPO SSSクローニングベクターを、形質転換コンピテント大腸菌宿主細胞とインキュベートした。74個の形質転換反応物では、選択した8個のコロニーのうち少なくとも7個でTOPO SSSクローニングベクターへのORFの定方向クローニングが行われ、これらのクローニング反応物の59個が選択した8個全てでコロニーで定方向を有していた。全定方向クローニングスコアは、656のうち609であり、定方向挿入は、選択したクローンの93%を超えて存在した(表1参照のこと)。
(表1) TOPO SSSクローニングベクターを使用したORFの定方向クローニング
実施例7
レポーター遺伝子の定方向クローニング
類似の例では、上記の改変pCR2.1 TOPO SSSベクターを使用して、レポーター分子である緑色蛍光タンパク質(GFP)をコードする遺伝子をコードするPCR作製ORFを、lacZ(ベクター中に存在する断片)にてインフレームで定方向クローニングを行った。GFP遺伝子増幅に使用したプライマーは、不可欠なSSS相補配列5'-ATTC-3'および翻訳開始メチオニンの周知の配列5'-ATG-3'を含んでいた。上記の必要なクローニング段階を使用して、PCR増幅GFPをベクターに挿入し、形質転換細胞を固体寒天プレートで成長させた。成長したコロニーは、正確に挿入されたPCR産物を示した(表2を参照のこと)。
(表2) 改変pCR2.1TOPO SSSクローニングベクターへのGFPのインフレームおよび定方向挿入
これらのデータは、現在のクローニング技術よりも実質的に改良されているので、本発明はハイスループット技術と高度に適合するクローニングである。定方向クローニング効率が90%を超える場合、使用者は、各クローニングDNA断片の2つのコロニーのみを必要とする。従って、96ウェルプレートでは、定方向挿入についてで48個の異なるクローンをスクリーニングすることができ、これは現在のクローニング技術の400%倍を超える。本発明の使用により、多数のハイスループット遺伝子発現操作が簡素化され、現在の費用の一部でこれを行うことができる。
実施例8
エントリーベクターへの平滑末端PCR産物の定方向トポイソメラーゼクローニング
概説
さらなる態様では、本発明の組成物、キット、および方法を効率的な5分クローニングストラテジー(「TOPO(登録商標)クローニング」、Invitorogen Corporation、Carlsbad、CA)と組み合わせて、本発明の組み合わせクローニング系(例えば、Invitorogen Corporation、Carlsbad、CAのGATEWAY(登録商標)システム)に入れるためにベクターに平滑末端PCR産物を定方向クローニングする。本発明のこのクローニングストラテジーを使用して、リガーゼ、その後のPCR手順、または制限酵素を使用せずに90%を超える効率で平滑末端PCR産物が定方向クローニングされる。
関心対象のGATEWAY(商標)デスティネーションベクターでの組換え後のPCR産物の最適な発現のために、任意の適切な発現ベクターを使用することができる。例として、以下の多数の利点を有する市販のpENTR定方向TOPO(登録商標)ベクターが含まれる(Invitorogen Corporation、Carlsbad、CA)が、これらに限定されない。
これらのpENTR/D-TOPO(登録商標)およびpENTR/SD/D-TOPO(登録商標)ベクターを、GATEWAY(商標)系へのエントリーのための平滑末端PCR産物の迅速な定方向TOPO(登録商標)クローニングを容易にするように設計する。これらのベクターの特徴には、以下が含まれる。
・GATEWAY(商標)デスティネーションベクターでのエントリークローンの部位特異的組換え用のattL1およびattL2部位;
・平滑末端PCR産物の迅速且つ有効な定方向クローニング用の定方向TOPO(登録商標)クローニング部位;
・大腸菌中での関心対象のPCR産物の基本的発現を防止するためのrrnB転写終結末端;
・大腸菌での選択用のカナマイシン耐性遺伝子
・大腸菌中でのプラスミドの高コピー複製および維持のためのpUC起点;
・原核生物系でのPCRの有効な翻訳のためのT7遺伝子10翻訳エンハンサーおよびリボソーム結合部位(pENTR/SD/D-TOPO(登録商標)のみ)。
本発明のGATEWAY(商標)組換えクローニング系と組み合わせてpENTR定方向TOPO(登録商標)ベクターの使用を使用して、平滑末端PCR産物中に含まれる関心対象の遺伝子を、以下のいくつかの簡単な段階によって容易に発現することができる。
1. 平滑末端PCR産物を、上記のpENTR TOPO(登録商標)ベクターの1つにクローニングして(本明細書記載の「TOPO(登録商標)クローニング」手順のトポイソメラーゼを使用する)、エントリークローンを作製する。
2. このエントリークローンと選択したGATEWAY(商標)デスティネーションベクター(本明細書記載のものなど)との間での組換え反応によって発現構築物を作製する。
3. 発現構築物を、適切な宿主細胞(例えば、細菌、哺乳動物、酵母、昆虫、または他の適切な宿主細胞。選択は上記の発現構築物の産物について選択される特異的デスティネーションベクターに依存する)に移入し、PCR産物(発現構築物に含まれる)の関心対象の遺伝子によってコードされる組換えタンパク質を、特定の宿主細胞系に適切な発現条件を使用して発現する。
定方向TOPO(登録商標)クローニング
ワクシニアウイルス由来のトポイソメラーゼIは、特定の部位(CCCTT)で二重鎖DNAに結合し、一方の鎖中のホスホジエステル骨格を切断する(Shuman、1991)。破壊されたホスホジエステル骨格由来のエネルギーは、切断された鎖の3'リン酸とトポイソメラーゼIのチロシン残基(Tyr-274)との間の共有結合の形成によって変換される。元の切断鎖の5'ヒドロキシルによってDNAと酵素との間のホスホ-チロシン結合を攻撃して反作用させてトポイソメラーゼを放出することができる(Shuman、1994)。TOPO(登録商標)クローニングは、PCR産物の効率的なクローニングにこの反応を利用している。
得られたDNAへの3'一本鎖末端(突出)の付加によって、TOPO(登録商標)オリゴヌクレオチドを使用した二本鎖DNAの定方向結合を行う(ChengおよびShuman、2000)。この一本鎖突出は、TOPO(登録商標)負荷DNA断片の5'末端と同一である。本発明により、TOPO(登録商標)負荷DNAへの4つのヌクレオチド突出配列の付加および「全ベクター」形式へのアダプター付加によってこのアプローチを改変した。
この系では、PCR産物を、正方向プライマーの4塩基(CACC)の付加によって定方向クローニングを行う。クローニングベクター中の突出(GTGG)は、PCR産物の5'末端に侵入し、付加した塩基とアニーリングされ、正確な方向でPCR産物を安定化する。90%以上の効率で挿入物を正確な方向でクローニングすることができる。
方法
PCR産物の設計
関心対象の遺伝子を増幅するためのPCRプライマーの設計は、発現に重要である。使用するpENTR TOPO(登録商標)ベクターに依存して、PCRプライマーの設計の際にはいくつかのの以下の検討材料に留意しなければならない。
・定方向クローニングを容易にするために必要な配列;
・PCR産物の適切な翻訳開始に必要な配列;および
・GATEWAY(商標)デスティネーションベクターでのエントリークローンの組換え後にN末端またはC末端とPCR産物がインフレームで融合されるかどうか。
正方向PCRプライマー設計のガイドライン
正方向PCRプライマーを設計する場合、以下の点を考慮しなければならない。
定方向クローニングを可能にするために、正方向PCRプライマーMUSTは、プライマーの5'末端に配列CACCを含まなければならない。4つのヌクレオチドCACCは、各pENTR TOPO(登録商標)ベクター中の突出配列GTGGと塩基対合する。
PCR産物が哺乳動物細胞中で発現される場合(エントリークローンのGATEWAY(商標)デスティネーションベクターでの組換え後)、関心対象の配列は、適切な翻訳開始用のATG開始コドンを含むKozak翻訳開始配列を含まなければならない(Kozak、1987;Kozak、1991;Kozak、1990)。Kozakコンセンサス配列の例は、(G/A)NNATGGである。他の配列が可能であるが、-3位のGまたはAおよび+4位のGは、機能で最も重要である(太字で示す)。ATG開始コドンに下線を引く。注釈:関心対象の配列がKozak配列中に開始コドンを含まない場合、正方向PCRプライマーを、プライマーの5'末端にKozak配列を含むように設計することができる(以下を参照のこと)。
PCR産物がN末端融合タグを使用することなく原核細胞中で発現される場合(GATEWAY(商標)デスティネーションベクターでのエントリークローンの組換え後)、PCR産物を、pENTR/SD/D-TOPO(登録商標)エントリーベクターにTOPO(登録商標)クローニングすべきである。上記のように、pENTR/SD/D-TOPO(登録商標)は、大腸菌中でのPCR産物の有効な翻訳を可能にするためにT7遺伝子10翻訳エンハンサーおよびリボソーム結合部位(RBS)を含む。適切な翻訳の任意選択的なスペーシングを確実にするために、PCR産物のATG開始コドンが定方向クローニングに必要なCACCに直接後続するように正方向PCRプライマーを設計すべきである(以下を参照のこと)。
正方向プライマー設計の例
以下は、理論上のタンパク質のN末端のDNA配列および対応する正方向PCRプライマーの推定配列である。ATG開始コドンに下線を引いている。
DNA配列
(配列番号:44):
推定正方向PCRプライマー
(配列番号:45):
正方向PCRプライマーを上記のように設計する場合、(a)ATG開始コドンは、哺乳動物細胞でPCR産物が適切に翻訳を開始させるKozak配列(箱で囲んだ配列参照のこと)に含まれ(5'CACC突出に後続するPCR産物の最初の3つの塩基対が機能的コドンを構築することに留意)、および(b)ATG開始コドンは、原核細胞中でPCR産物を適切に翻訳させるRBS(pENTR/SD/D-TOPO(登録商標)のみ)から適切な間隔がある。
逆方向プライマー設計のガイドライン
逆方向PCRプライマーを設計する場合、以下の点を考慮する。それぞれpENTR/D-TOPO(登録商標)およびpENTR/SD/D-TOPO(登録商標)のTOPO(登録商標)クローニング部位の線図については図26および図27を参照のこと。
PCR産物が高い効率で定方向クローニングされることを確実にするために、逆方向PCRプライマーは5'末端で突出配列GTGGに相補的であってはならない。1つの塩基対のミスマッチにより、定方向クローニング効率が90%から50%に減少し、ORFが反対方向にクローニングされる可能性が増大し得る(以下の「例A」を参照のこと)。2つの塩基対のミスマッチ由来の反対方向のPCR産物のクローニングの証拠は認められなかった。
PCR産物がインフレームでC末端タグと融合する場合(GATEWAY(商標)デスティネーションベクターでのエントリークローンの組換え後)、逆方向PCRプライマーを関心対象の遺伝子中の天然の終止コドンを除去するように設計すべきである(以下の「例B」を参照のこと)。
PCR産物がC末端タグと融合しない場合(GATEWAY(商標)デスティネーションベクターとのエントリークローンの組換え後)、終止コドンを含む天然の配列が逆方向プライマーに含まれるか、終止コドンが逆方向プライマー結合部位から上流であることを確実にすべきである(以下の「例B」を参照のこと)。
逆方向プライマー設計の例A
以下は、理論上のタンパク質のC末端の配列である。タンパク質はC末端タグにインフレームで融合されるべきである(GATEWAY(商標)デスティネーションベクターでのエントリークローンの組換え後)。終止コドンに下線を引く。
DNA配列
(配列番号:46):
1つの解決法は、終止コドンのすぐ上流から始まるが最後の2つのコドンは4bpの突出配列と同一であるGTGG(以下の下線部分)を含むように逆方向PCRプライマーを設計することである。結果として、PCR産物を反対方向にクローニングする可能性が高まり、4bpの突出配列に逆方向プライマーは相補的になり得る。この条件を回避すべきである。
DNA配列
(配列番号:46):
推定逆方向PCRプライマー配列
(配列番号:47):
別の解決法は、終止コドンのすぐ下流とハイブリダイズするが依然としてORFのC末端を含むように逆方向プライマーを設計することである。終止コドンはグリシン、アラニン、またはリジンなどの害の無いアミノ酸のコドンと置換されることに留意されたい。
逆方向プライマー設計の例B
以下は、理論上のタンパク質のC末端配列
(配列番号:48)である。終止コドンに下線を引いている。
ORFをインフレームでC末端タグ(組換え後のデスティネーションベクターから得た)と融合するために、最後のコドン(TGC)に相同なヌクレオチドからの開始によって終止コドンを除去し、上流に繋げる。逆方向プライマー
(配列番号:49)は以下である。
これは、終止コドンを使用せずにC末端を増幅し、ORFをインフレームでC末端タグに結合させる。ORFをインフレームでC末端タグに結合することが望ましくない場合、終止コドンを含むように逆方向プライマー
(配列番号:50)を単純に設計する。
重要:pENTR TOPO(登録商標)ベクターが平滑末端のPCR産物を受け入れることに留意すべきである。pENTR TOPO(登録商標)ベクターへのライゲーションを妨げるので、PCRのために5'リン酸をプライマーに付加すべきではない。さらに、オリゴヌクレオチドが長い場合(30ヌクレオチド超)に使用前にゲル精製することが推奨される。
平滑末端PCR産物の産生
上記のガイダンスに従って一旦PCRストラテジーが選択されてプライマーが合成されると、平滑末端PCR産物を産生することができる。任意の熱安定性校正ポリメラーゼ(PCR用のThermalAce(商標)、PLATINUM(登録商標)、Pfx、Pfu、またはVent(登録商標)を含む)を、この目的に使用することができる。平滑末端PCR産物を産生するために、ポリメラーゼ製造の説明書および推奨に従うべきである。個別の一本鎖PCR産物を産生するためにPCR条件を最適にすることが重要である。下記の方法に従ったPCR断片のゲル精製もまた推奨される。
PCR産物の産生
PCRによって増幅産物を産生するために、以下のガイダンスを使用して25μlまたは50μlのPCR反応混合物を準備する。
使用するDNAポリメラーゼについての製造者の説明に従う。
プライマーおよび鋳型に適切なサイクリングパラメーターを使用する。
全てのPCR産物が完全に伸長していることを確実にするために7分間〜30分間の最終伸長を行う。
サイクリング後、チューブを氷上に置くか、-20℃2週間保存すべきである。
PCR産物のチェック
PCR産物の質および量を評価するために、5μl〜10μlを各PCR反応物から取り出し、以下のためにアガロースゲル電気泳動によって分析する。
正確なサイズの個別の一本鎖バンドの存在
個別の一本鎖バンドが存在しない場合、選択したポリメラーゼでのPCR反応の最適化について製造者の説明書を参照する。あるいは、所望の産物を、ゲル精製することができる(以下を参照のこと)。
PCR産物濃度の評価
TOPO(登録商標)クローニングのために、最も高いクローニング効率を得るためにPCR産物:TOPO(登録商標)ベクター=5:1の分子比が推奨されている。例えば、20ngの500bpのPCR産物または10ngの100bpのPCR産物を、TOPO(登録商標)クローニング反応に使用することができる。TOPO(登録商標)クローニングを行う前にPCR産物の濃度を調整する必要があり得る。
注釈:ThermalAce(商標)ポリメラーゼを平滑末端PCR産物の産生に使用する場合、ThermalAce(商標)は他の校正ポリメラーゼよりも高い収率を得ることができることに留意すべきである。0.5kb〜1.0kbの範囲でPCR産物を作製する場合、一般に、TOPO(登録商標)クローニング反応を行う前に1×ThermalAce(商標)緩衝液でPCR反応物を5倍に希釈する。1.0kbを超えるPCR産物には、希釈する必要はない。
TOPO(登録商標)反応の準備
緒言
一旦所望のPCR産物が産生されると、これをpENTR TOPO(登録商標)ベクターにTOPO(登録商標)クローニングし、TOP10大腸菌に組換えベクターを形質転換する準備ができる。最高の結果を得ることを確実にするために必要とするもの全てを準備および用意することが有用である。本発明者らは、開始前に「TOPO(登録商標)クローニング反応の準備」(10頁〜11頁)および「OneShot(登録商標)TOP10コンピテント細胞の形質転換」(12頁〜14頁)というタイトルの章を読むことを提案する。TOPO(登録商標)クローニングを行うのが始めてである場合、試料を使用して22頁〜24頁の対照反応を並行して行う。
HTP形式(以下を参照のこと)でTOPO(登録商標)クローニングを行う場合、Bulk TOP10細胞(500反応キット)またはMultiShot(商標)TOP10細胞(480反応キット)を使用してTOP10大腸菌を形質転換することができる。使用するキットによって、15頁〜16頁または17頁〜18頁のTOPO(登録商標)クローニングおよび形質転換プロトコールを参照のこと。
注釈:最近の実験により、TOPO(登録商標)クローニング反応物に塩(200mM NaCl、10mM MgCl2)を添加すると形質転換体の数が増加し得ることが示されている。これらの結果から、TOPO(登録商標)クローニング反応物に塩を添加することを推奨する。この関心対象のストック塩溶液がキットに含まれている。TOPO(登録商標)クローニング反応物に添加する塩の量は、ケミカルコンピテント細胞(添付)またはエレクトロコンピテント細胞(注文情報についてはxページを参照のこと)のいずれを形質転換する予定であるかによって変化することに留意されたい。この理由のために、2つの異なるTOPO(登録商標)クローニング反応物を使用すれば最良の結果が得られる。以下の情報を慎重に読むこと。
ケミカルコンピテント大腸菌細胞の形質転換
TOPO(登録商標)クローニングおよびケミカルコンピテント大腸菌への形質転換のために、TOPO(登録商標)クローニング反応物中での塩化ナトリウムおよび塩化マグネシウムの添加(最終濃度が200mM NaCL、10mM MgCl2)により、長期間コロニー数が増加する。塩溶液(1.2M NaCl、0.06M MgCl2)を使用して、TOPO(登録商標)クローニング反応物のNaClおよびMgCl2を推奨濃度に調整する。
エレクトロコンピテント大腸菌の形質転換
TOPO(登録商標)クローニングおよびエレクトロコンピテント大腸菌への形質転換のために、TOPO(登録商標)クローニング反応物に塩を含めることができるが、エレクトロポレーションの際のアーク放電を防止するために塩の量を50mM NaCl、2.5mM MgCl2に減少させなければならない。TOPO(登録商標)クローニング反応物への都合の良い添加のために塩溶液を水で4倍希釈して300mM NaCl、15mM MgCl2溶液を調製する。
TOPO(登録商標)クローニング反応の準備
以下の表は、ケミカルコンピテントOneShot(登録商標)TOPO大腸菌(添付)またはエレクトロコンピテント大腸菌のいずれかへの最終的な形質転換用にどのようにしてTOPO(登録商標)クローニング反応物(6μl)を準備するかを記載している。必要なTOPO(登録商標)クローニング反応物の最適化に関するさらなる情報を、21頁に見出すことができる。ThermalAceポリメラーゼを使用してPCR産物を作製する場合、実施前にPCR反応物を希釈する必要があり得ることに留意のこと(9頁を参照のこと)。
注釈:TOPO(登録商標)ベクター溶液の青色は正常であり、これを使用して溶液を視覚化する。
(表3) TOPO(登録商標)クローニング反応混合物の準備
*全試薬を終了まで-20℃で保存する。塩溶液および水を室温または4℃で保存することができる。
TOPO(登録商標)クローニング反応の実施
反応物を穏やかに混合し、室温(22℃〜23℃)で5分間インキュベートする。
注釈:ほとんどの適用では、5分間で分析用の多数のコロニーが得られる。必要に応じて、TOPO11クローニング反応の長さを30秒から30分間に変化させることができる。PCR産物の通常のサブクローニングでは、30秒間で十分であり得る。PCR産物が大きい(1kb超)か、PCR産物にTOPO(登録商標)クローニングを行う場合、反応時間を延ばすほどコロニー収率を増加させることができる。
反応物を氷上に置き、次の頁の「One Shot7 TOP10コンピテント細胞の形質転換」に進む。注釈:TOPO7クローニング反応物を-20℃で一晩保存することができる。
One Shot(登録商標) TOP10コンピテント細胞の形質転換
緒言
一旦TOPO(登録商標)クローニング反応を行うと、pENTR TOPO(登録商標)構築物がコンピテント大腸菌に形質転換される。形質転換を容易にするために20反応キットと共にOne Shot(登録商標)TOP10ケミカルコンピテント大腸菌(ボックス2)が含まれるが、エレクトロコンピテント細胞を形質転換することもできる(注文情報についてはxページを参照のこと)。ケミカルコンピテントまたはエレクトロコンピテント大腸菌の形質転換プロトコールを、この節に記載されている。
使用者が用意する材料
一般的な微生物用備品(すなわち、プレート、スプレッダー)に加えて、以下の試薬および装置が必要である。
(a)42℃の水浴(またはキュベット(オプション)を具備するエレクトーポレーター)
(b)50μg/mlのカナマイシンを含むLBプレート(各形質転換について2つ)
(c)37℃の振盪および非振盪インキュベーター。
挿入物の存在についての青白スクリーニングを行わない。ほとんどの形質転換体は、正確な方向でクローニングされた関心対象のPCR産物を含む組換えプラスミドを含む。キットには方向およびオープンリーディングフレームを確認するためにマルチクローニング部位中の挿入物を越えて配列決定するための配列決定プライマーが含まれる。
形質転換の準備
各形質転換のために、コンピテント細胞用の1つのバイアルと2つの選択プレートが必要である。
水浴を42℃に平衡化するか(化学的形質転換用)、エレクトロコンピテント大腸菌を使用する場合、エレクトロポレーターを準備する。
エレクトロポレーションのために、少量の4倍塩溶液を希釈して、希釈塩溶液(例えば、滅菌水150μlに塩溶液5μlを添加する)を調製する。
Box2由来のSOC培地を含むバイアルを室温に加温する。
50μg/mlのカナマイシンを含むLBプレートを37℃で30分間加温する。
各形質転換についてBox2由来のOne Shot(登録商標)TOP10細胞の1バイアルを氷上で解凍する。
重要:一般に、定方向TOPO(登録商標)クローニングによりコロニーの収量は従来の二方向TOPO TAクローニング(登録商標)の1/5から1/10であることに留意されたい。750bpの被験挿入物を定方向TOPO(登録商標)クローニングする場合、22〜23頁のプロトコールを使用して1800個〜3000個のコロニーが得られる。全コロニー数がより少ないにもかかわらず、90%を超えるコロニーが正確な方向でPCR挿入物を含むプラスミドを含む。
One Shot(登録商標)TOP10化学的形質転換プロトコール
1. TOPO(登録商標)クローニング反応、工程2、11頁由来の2μlのTOPO(登録商標)クローニング反応物を、One Shot(登録商標)TOP10ケミカルコンピテント大腸菌のバイアルに添加し、おだやかに混合する。ピペッターを上下して混合しないこと。
2. 氷上で5分間〜30分間インキュベートする。
注釈:氷上で長時間のインキュベーションは形質転換効率を最小にする。インキュベーション時間は、使用者の判断による。
3. 振盪せずに42℃で30秒間細胞に熱ショックを与える。
4. その直後にチューブを氷上に移す。
5. 250μlの室温のSOC培地を添加する。
6. チューブに固く栓をし、チューブを37℃で30分間水平方向に振盪する(200rpm)。
7. 予め加温した選択プレート上で各形質転換由来の50μl〜200μlを画線し、37℃で一晩インキュベートする。少なくとも1つのプレートで確実に十分なコロニー間隔が得られるように2つの異なる体積を播種することを推奨する。
8. 有効なTOPO7クローニング反応により数百個のコロニーを得ることができる。分析のために約5コロニーを選択する(「形質転換体の分析」(19頁)を参照のこと)。
エレクトロポレーションによる形質転換
アーク放電を回避するために、エレクトロポレーションにはエレクトロコンピテント細胞のみを使用する。エレクトロポレーションにはOne Shot(登録商標)TOP10ケミカルコンピテント細胞を使用しないこと。
1. TOPO7クローニング反応、工程2、11頁由来のTOPO7クローニング反応物2μlを、エレクトロコンピテント大腸菌50μlを含む0.1cmのキュベットに添加し、おだやかに混合する。ピペッターを上下して混合しないこと。泡の形成を避けること。
2. プロトコールおよびエレクトロポレーターを使用して試料をエレクトロポレーションする。
注釈:アーク放電の問題が発生した場合、以下を参照のこと。
3. その直後に室温のSOC培地250μlを添加する。
4. 溶液を15mlのスナップキャップチューブ(すなわち、Falcon)に移し、37℃で少なくとも1時間振盪して、カナマイシン耐性遺伝子を発現させる。
5. 予め加温した選択プレート上で各形質転換由来の20μl〜100μlを広げ、37℃で一晩インキュベートする。少量でも確実に広がるように、SOC20μlを添加する。少なくとも1つのプレートで確実に十分なコロニー間隔が得られるように2つの異なる体積を播種することを推奨する。
6. 有効なTOPO7クローニング反応により数百個のコロニーを得ることができる。分析のために約5コロニーを選択する(「Analyzing Transformants」(19頁)を参照のこと)。
TOPO(登録商標)クローニング反応への希釈塩溶液の添加により、TOPO(登録商標)クローニング反応物中のNaClおよびMgCl2の最終濃度がそれぞれ50mMおよび2.5mMとなる。エレクトロポレーション中の試料のアーク放電を防止するために、細胞体積は、50μlと80μlとの間(0.1cmキュベット)または100μl〜200μl(0. 2cmキュベット)であるべきである。
形質転換中にアーク放電が認められた場合、以下の提案のうちの1つを試す。
エレクトロポレーターの充電で通常使用する電圧を10%減少させる。
100ohmへの付加抵抗の減少によってパルス長を減少させる。
TOPO(登録商標)クローニング反応をエタノール沈殿させ、エレクトロポレーション前に水に再懸濁する。
ハイスループット用途
480および500反応用のpENTRおよびpENTR/SD定方向TOPO(登録商標)クローニングキットを、ハイスループット(HTP)適用の使用にGATEWAY(商標)エントリークローンを産生するように設計する。これらのキットでは、pENTR TOPO(登録商標)ベクターがバラで提供されており、2つの形式のケミカルコンピテントTOP10大腸菌を選択する。
TOPO7クローニング反応物を含む96ウェルプレートへ無菌の溝から細胞を簡単に移せるように5mlアリコートで細胞がバラで提供されている(カタログ番号K2400-500およびK2420-500)。
TOPO7クローニング反応物を細胞に添加するために96ウェルプレート(12ウェルのストリップウェル)の予め等分した細胞を提供している(Invitrogen Corporation、Carlsbad、CA;カタログ番号K2400-480およびK2420-480)。
HTP TOPO(登録商標)クローニングおよびバルク細胞での形質転換
説明
このプロトコールでは、TOPO(登録商標)クローニング反応を、96ウェルのU底ポリスチレンプレート(Costar、カタログ番号3366、330μl/ウェル)に準備し、TOP10コンピテント細胞を、分注のために溝に入れる。
開始前
96ウェル金属加熱ブロック(VWR、カタログ番号13259-260)を、ブロックが冷却するまで氷上で冷却する。
SOCのバイアルを室温にする。
96ウェル金属ブロックを含む熱ブロックまたはサーマルサイクラーを42℃に予備加熱する。
注釈:水浴を使用することができるが、細胞が汚染しないように注意すること。
・1チューブ(5ml)のTOP10ケミカルコンピテント大腸菌を氷上で解凍する(30分〜60分)。
・50μg/mlのカナマイシンを含むLB寒天プレートを37℃に加温する。細胞の形質転換効率を試験するためにpUC19対照を含める場合、50〜100μg/mlのアンピシリンを含むLB寒天プレートが必要である。
対照:便宜上、TOPO(登録商標)クローニングおよび形質転換反応を試験するために50μlアリコートのコンピテント細胞を使用する。さらに、内部対照としてpUC19プラスミドを含めることができる(以下の手順を参照のこと)。
手順
1. 以下のように6μlのTOPO(登録商標)クローニング反応物を準備する。対照としてpUC19を含める場合、2〜3ウェルを空にしておく。
PCR産物 1μl
塩溶液 1μl
滅菌水 3μl
pENTR TOPO(登録商標)ベクター 1μl
最終体積 6μl
2. 室温で5分間〜10分間インキュベートする。
3.96ウェルプレートを冷却したブロックに5分間置く。
4. pUC19を含める場合、1μl(10pg)のプラスミドを2〜3個の空のウェルに添加する。
5. 解凍したTOP10大腸菌を滅菌した溝に注ぎ、直ちに45μl/ウェルで分注する。ピペットを穏やかに1〜2分間上下して混合する。
6. プレートをParafilm(登録商標)で被覆し、これを冷却ブロック上で20分間インキュベートする。
7. プレートを予備加温熱ブロックまたはサーモサイクラーのいずれかに移し、細胞を42℃で30秒間熱ショックを与える。
8. プレートを冷却ブロックに戻し、プレートを冷却ブロックに完全に接触させることを確実にするために押しつける。1分間インキュベートする。
9. Parafilm(登録商標)を除去し、150μL/ウェルのSOCを添加する。
10. プレートを再度被覆し、プレートを37℃で1時間インキュベートする。
注釈:穏やかな振盪(125RPM)が最適である。
11. 各ウェル由来の50μg/mlカナマイシン50μLを含むLB寒天プレートに播種する。pUC19のために、形質転換混合物10μl+SOC20μlを、100μg/mlのアンピシリンを含むLBプレートに播種する。37℃で一晩インキュベートする。
12. 翌日、5個〜10個のコロニーを選択し、所望のように処理する。
多すぎるコロニー
得られたコロニーが多すぎる場合、播種する細菌培養物の量を減少させる、および/またはさらなるSOCでの形質転換物を希釈する。
HTP TOPOクローニングおよびMultiShot(商標)細胞での形質転換
説明
このプロトコールでは、TOPO(登録商標)クローニング反応を、96ウェルプレートに準備し、2μlを、ウェルあたり15μlのケミカルコンピテントTOP10大腸菌を含む96ウェルのMultiShot(商標)プレートの各ウェルに移す。
開始前
・96ウェル金属加熱ブロック(VWR、カタログ番号13259-260)を、ブロックが冷却するまで氷上で冷却する。
・SOCのバイアルを室温にする。
・50μg/mlのカナマイシンを含むLB寒天プレートを37℃に加温する。細胞の形質転換効率を試験するためにpUC19対照を含める場合、50〜100μg/mlのアンピシリンを含むLB寒天プレートが必要である。
・96ウェル金属ブロックを含む熱ブロックまたはサーマルサイクラーを42℃に予備加熱する。
・注釈:水浴を使用することができるが、細胞が汚染しないように注意すること。
・サーモサイクラーを使用する場合、42℃の温度を保持するように機械をプラグラムする。
対照:TOPO(登録商標)クローニング反応物および形質転換物を試験するために、1列(12ウェル)にTOP10細胞を含む試験プレートが含まれる。さらに、内部対照としてpUC19プラスミドを含めることができる(以下の手順を参照のこと)。
手順
1. 96ウェルプレートでは、各ウェル中に以下の6μlのTOPO(登録商標)クローニング反応物を準備する。
PCR産物 1μl
塩溶液 1μl
滅菌水 3μl
pENTR TOPO(登録商標)ベクター 1μl
最終体積 6μl
2. 室温で5〜10分間インキュベートする。
3. 96ウェルプレートを冷却したブロックに5分間置く。
4. フリーザーからケミカルコンピテントTOP10大腸菌の96ウェルMultiShot(商標)プレートを取り出し、これを第2の冷却ブロックに置く。細胞を30秒以内に解凍すべきである。
5. アルミニウムホイルシールを慎重に取り除く。
6. マルチチャネルピペットを使用して、細胞を含む96ウェルプレートの各ウェルに2μlの各TOPO(登録商標)クローニング反応物(約3.3ng)を添加する。一定の結果を得るために体積を約2μlに維持する。pUC19対照のために、1μl(10pg)のDNAを添加する。
7. 細胞を添付のプラスチック製の蓋で覆い、冷却ブロック中で20分間細胞およびDNAをインキュベートする。
8. プレートを予備加温熱ブロックまたはサーモサイクラーのいずれかに移し、細胞を42℃で30秒間熱ショックを与える。
9. 細胞プレートを冷却ブロックに戻し、プレートをブロックに押し付け、プレートを1分間冷却する。
10. プラスチック製の蓋を除去し、90μlのSOCを各ウェルに添加する。
11. プレートを蓋で覆い、プレートを37℃で1時間インキュベートする。注釈:穏やかな振盪(125RPM)が最適である。
12. 各ウェル由来の50μg/mlカナマイシン100μLを含むLB寒天プレートに播種する。pUC19対照のために、形質転換混合物10μl+SOC20μlを、100μg/mlアンピシリンを含むLBプレートに播種する。37℃で一晩インキュベートする。
注釈:得られたコロニーが多すぎる場合、播種する細胞の量を減少させるか、細胞への反応物の添加前にTOPO(登録商標)クローニング反応物を滅菌水もしくはTE緩衝液で希釈することができる。
形質転換体の分析
陽性クローンの分析
1.5コロニーを選択し、これらを50〜100μg/mlのカナマイシンを含むLBまたはSOB培地で一晩培養する。
2. 選択した方法を使用してプラスミドDNAを単離する。自動化または手動の配列決定用の非常に純度の高いプラスミドDNAが必要な場合、S.N.A.P.J. MidiPrepキット(カタログ番号K1910-01)の使用を推奨する。
3. 挿入物の存在および正確な方向を確認するために制限分析を使用してプラスミドを分析する。一度ベクターを切断し、一度挿入物を切断する制限酵素または酵素の組み合わせを使用する。
配列決定
遺伝子が正確な方向でクローニングされていることを確認するために、構築物を配列決定することができる。挿入物の配列決定を補助するために、キットにはM13正方向(-20)プライマーおよびM13逆方向プライマーが含まれている。Invitrogen(注文情報についてはx頁を参照のこと)のM13正方向(-20)プライマーおよびM13逆方向プライマーもまた個別に利用可能である。
重要:Invitrogen CorporationウェブサイトからpENTR/D-TOPO(登録商標)またはpENTR/SD/D-TOPO(登録商標)の配列をダウンロードする場合(本明細書中図26および図27に記載を参照のこと)、突出配列(GTGG)がCACCと既にハイブリダイズされていることに留意すること。DNA配列分析プログラムにより、相補配列を含まない突出を得ることが可能である。
PCRによる形質転換の分析
PCRを使用して陽性形質転換体を分析することができる。PCRプライマーのために、M13正方向(-20)プライマーまたはM13逆方向プライマーと挿入物内でハイブリダイズするプライマーとの組み合わせを使用する。増幅条件を決定しなければならない。初めてこの技術を使用する場合、並行して制限分析を行うことを推奨する。鋳型のミスプライミングまたは鋳型の汚染が原因の産物が得られるかもしれない。
便宜上、以下のプロトコールを提供する。他のプロトコールも適切である。
1. PCR緩衝液、dTNP、プライマー、およびTaqポリメラーゼからなるPCRカクテルを調製する。反応物の体積は20μlを使用する。分析されるコロニー数を増殖させる(例えば5)。
2.5コロニーを選択し、これらをそれぞれ20μlのPCRカクテルに再懸濁する(さらなる分析のためにコロニーを保存するためのパッチプレートを作製すること)。
3. 反応物を94℃で10分間インキュベートして、細胞を溶解し、ヌクレアーゼを不活化する。
4. 20サイクル〜30サイクル増幅させる。
5. 最後の伸長のために、72℃で10分間インキュベートする。-4℃で保存する。
6. アガロースゲル電気泳動によって視覚化する。
重要:形質転換体または正確な挿入物の獲得に関して問題が生じる場合、22頁〜24頁に記載の対照反応を行う。これらの反応により、実験修正の一助となる。
長期保存
一旦正確なクローンが同定されると、確実にコロニーが精製され、長期保存用のグリセロールストックを作製する。-20℃でプラスミドDNAを保存することを推奨する。
1. 50μg/mlのカナマイシンを含むLBプレート上に1コロニーの元のコロニーを画線する。
2. 1コロニーを単離し、50μg/mlのカナマイシンを含む1〜2mlのLB中でインキュベートする。
3. 培養物が静止期に達するまで成長させる。
4. 0.85mlの培養物を0.15mlの滅菌グリセロールと混合し、凍結バイアルに移す。
5. -80℃で保存する。
デスティネーションベクターでのエントリー構築物の組換え
一旦エントリークローンが得られると、pENTR TOPO(登録商標)構築物を選択した任意のGATEWAY(商標)デスティネーションベクターで形質転換して、発現クローンを作製することができる。この「LR」組換え反応を、CLONASE(商標)(組換えタンパク質のカクテル)で媒介する。LR CLONASE(商標)酵素混合物は、Invitrogen Corporation(Carlsbad、CA)から市販されている。このような一定の方法では、例えば、TOPOアダプター付加ベクターを、1つまたは複数の核酸セグメント(例えば、1つまたは複数のPCR産物)と室温(例えば、約20℃〜20℃)で約5分間〜30分間(好ましくは、約10分間)インキュベートし、反応物を約80℃で約20分間のインキュベーションによって熱ショックを与え、LR反応物のインキュベーション時間を約3時間に増加する以外は、反応混合物を、製造者の説明書(Invitrogen Corporation)にしたがって、標準的なLR反応で使用する。
TOPO(登録商標)クローニング反応の最適化
クローニングプロセスの高速化
効率の高いTOPO(登録商標)クローニングによりクローニングプロセスが簡素化する。PCR産物を日常的にクローニングし、このプロセスを高速化したい場合、以下を考慮する。
・TOPO(登録商標)クローニング反応を5分間の代わりに30秒間だけインキュベートする。
最多数のコロニーを得ることができないが、高いTOPO(登録商標)クローニング効率が得られた場合、ほとんどの形質転換体は挿入物を含む。
・ケミカルコンピテント細胞に3glのTOPO(登録商標)クローニング反応物を添加し、氷上の5分間だけインキュベートする。
インキュベーション時間の増加(30分間)により、形質転換は有効に改良されない。
より多くの形質転換体の獲得
巨大なPCR産物、有毒遺伝子、またはPCR産物のクローニングプールをTOPO(登録商標)クローニングする場合、所望するクローンを得るためにより多数の形質転換体が必要である。コロニー数を増加させるために、
・塩補足TOPO(登録商標)クローニング反応物を5分間の代わりに20〜30分間インキュベートする。
塩補足TOPO(登録商標)クローニング反応物のインキュベーション時間の増加により、より多数の分子をライゲーション可能であり、形質転換効率が増加する。塩の添加により、PCR産物のライゲーションおよびDNAの解離後のトポイソメラーゼIへのDNAの再結合およびニッキングが防止されるようである。
・TOPO7クローニング反応で使用したPCR産物の量を最大コロニー産出量について滴定する。
希釈PCR産物のクローニング
希釈PCR産物をクローニングするために、以下を行うことができる。
・PCR産物の量を増加させる。
・TOPO(登録商標)クローニング反応物を20〜30分間インキュベートする。
・PCR産物を濃縮する。
対照反応の実施
緒言
20個の反応キットを初めて使用する場合、結果の評価を支援するために以下の対照TOPO(登録商標)クローニング反応を実施することを推奨する。対照反応は、キットに含まれる試薬を使用してPCR産物を産生することと、この産物をTOPO(登録商標)クローニング反応に直接使用することを含む。
開始前
各形質転換のために、50μg/mlのカナマイシンを含む2つのLBプレートを調製する。
対照PCR産物の産生
対照PCR産物を増幅させるために熱安定性校正ポリメラーゼおよび適切な緩衝液を使用する。使用するポリメラーゼの製造者の説明書に従う。
1. 750bpの対照PCR産物を産生するために、以下の50μlのPCR反応物を調製する。
対照DNA鋳型(100ng) 1μl
10×PCR緩衝液(酵素に適切なもの) 5μl
dNTP混合物 0.5μl
対照PCRプライマー(各0.1μg/μl) 1μl
滅菌水 41.5μl
熱安定性ポリメラーゼ(1〜2.5単位/μl) 1μl
全体積 50μl
2. 70μl(1滴)の鉱物油を重層する。
3. 以下のサイクリングパラメータを使用して増幅する。
4. 反応物から10・1を除去し、アガロースゲル電気泳動によって分析する。分離した750bpのバンドを、次の頁の視覚可能な対照TOPO7クローニング反応に供するべきである。
対照TOPO(登録商標)クローニング反応
先の頁で産生した対照PCR産物およびTOPO(登録商標)ベクターであるpENTRを使用して、6plのTOPO(登録商標)クローニング反応を下記のように準備する。
1. 対照TOPO(登録商標)クローニング反応物を準備する。
2. 室温で5分間インキュベーションし、氷上に置く。
3. 3μlの各反応物を別のバイアルのOne Shot(登録商標)TOP10細胞中で形質転換する(13頁)。
4. 100μl〜200μlの各形質転換混合物を、50μl/mlのカナマイシンを含むLBプレートに広げる。少なくとも1つのプレートで確実に十分なコロニー間隔が得られるように2つの異なる体積を播種することを推奨する。
5. 37℃で一晩インキュベートする。
結果の分析
ベクター+PCR挿入物反応由来の数百のコロニーが得られるはずである。形質転換を分析するために、プラスミドDNAを単離し、下記の適切な制限酵素で消化する。以下の表は、正確な方向または逆方向にクローニングされた挿入物について調査すべき消化パターンを列挙している(ジャネット:以下に記入せよ)。
90%を超えるコロニーが正確な方向で750bpの挿入物を含む。ベクターのみの反応では比較的少数のコロニーが産生されべきである。
形質転換対照
One Shot(登録商標)TOP10コンピテント細胞の形質転換効率をチェックするためにpUC19プラスミドを含む。13頁のプロトコールを使用して、1バイアルのOne Shot(登録商標)TOP10細胞を10pgのpUC19で形質転換する。形質転換混合物10μl+SOC20μlを、100μg/mlのアンピシリンを含むLBプレートに播種する。形質転換効率は、1×109cfu/μgのDNAのはずである。
クローニング効率に影響を与える因子
より低いクローニング効率により以下の利点が得られることに留意されたい。これらのほとんどは容易に正確であるが、巨大な挿入物をクローニングする場合、90%の定方向クローニング効率を期待することはできない。
PCR産物のゲル精製
緒言
スメア、多段バンド、プライマー-二量体産物、または巨大PCR産物(3kb超)には、ゲル精製が必要であり得る。PCR産物を精製したい場合、非常に慎重に全てのヌクレアーゼ汚染を除去する。DNA断片を単離するかオリゴヌクレオチドを除去するプロトコールは多数存在する。最も一般的なプロトコールについては、「Current Protocols in Molecular Biology」、ユニット2. 6(Ausubelら、1994)を参照のこと。3つの簡単なプロトコールが以下に示す。
注釈:PCR産物を精製してクローニング効率を増加させることができる(例えば、PCR産物が非常に希薄である場合)。1つのバンドになるようにPCRを最適化することもできる(9頁の「平滑末端PCR産物の産生」を参照のこと)。
S.N.A.P.(商標)ゲル精製キットの使用
Invitrogenから市販されているS.N.A.P. (商標)ゲル精製キット(カタログ番号K1999-25)により、通常のアガロースゲルからPCR産物を迅速に精製可能である。
1. 増幅反応物を1%〜5%の通常のTAEアガロースゲルで電気泳動する。(注釈:アガロースゲルの調製にTBEを使用してはならない。下記のようにホウ酸塩がヨウ化ナトリウム段階を妨害する)
2. PCR産物を含むゲルスライスを切断し、2倍体積の6Mヨウ化ナトリウム溶液中、65℃で溶解する。
3. 1.5倍体積の結合緩衝液を添加する。
4. 段階3由来の溶液(時間で1mlに過ぎない)をS.N.A.P. (商標)カラムにロードする。微量遠心分離機にて3000×gで1分間遠心分離し、上清を捨てる。
5. 段階3由来の溶液が残存している場合、段階4を繰り返す。
6. 900μlの最終洗浄緩衝液を添加する。
7. 微量遠心分離機にて最高速度で1分間遠心分離し、タイル流動物を捨てる。
8. 段階7を繰り返す。
9. 精製PCR産物を40μlのTEまたは滅菌水に溶出する。TOPO(登録商標)クローニング反応物に4μlを使用し、11頁に記載のように進行させる。
迅速S.N.A.P.(商標)法
さらにより簡単な方法はPCR産物を含むゲルスライスを単純に切断し、S.N.A.P.(商標)カラムベッドの上に置き、最高速度で10秒間遠心分離することである。TOPO(登録商標)クローニング反応物に1μl〜2μlの流動物を使用する(11頁)。最良の結果のためには、できるだけ確実にゲルスライスを小さくする。
低融点アガロース法
低融点アガロースを使用したい場合、以下の手順を使用する。ゲル精製によりPCR産物が希釈され、クローニング効率が潜在的に損失することに留意されたい。
1. 低融点アガロースゲル(0.8〜1.2%)を含むTAE緩衝液でPCR反応物をできるだけ電気泳動する。
2. 関心対象のバンドを視覚化し、バンドを切り出す。
3. ゲルスライスを微量遠心管に入れ、チューブをゲルスライスが融解するまで65℃でインキュベートする。
4. チューブを37℃にし、アガロース融解を維持する。
5. 11頁に記載のようにTOPO(登録商標)クローニング反応物にPCR産物を含む4μlの融解アガロースを添加する。
6. TOPO(登録商標)クローニング反応物を37℃で5分間〜10分間インキュベートする。これは、アガロースの融解を維持するためである。
7. 13頁の方法を使用してOneShot(登録商標)TOP10細胞に2μl〜4μlを直接形質転換する。
注釈:PCR産物の精製によってクローニング効率が減少し得る。1つのバンドが得られるようにPCRを至適化することができる。
実施例9
GATEWAY(商標)ベクターを使用したTOPO連結反応物の反応条件の最適化
GATEWAYベクターと組み合わせてTOPOクローニング手順を使用するために、組み合わせた反応物の最適条件を調査した。これらの研究の実施で、いくつかの問題に取り組んだ。
十分なBP反応用の鋳型およびTOPO反応成分によるBP反応の阻害
これらの課題に取り組むために、本明細書記載のTOPOツールを使用して、attB1+CAT+attB2鋳型を作製した。二次PCRを行い、研究対象の試験に十分な鋳型を作製し、産物を使用してBP反応を行った。各処理段階で以下の反応条件を使用した。
BP反応後、混合物をケミカルコンピテント大腸菌細胞(例えば、TOP10;Invitrogen Corporation)に化学的に形質転換し、細胞を播種して組換え効率を決定した。
結果
これらの結果は、TOPOツールによりその後のBP反応用の鋳型が作製されることを示す。さらに、これらの結果は、TOPO連結がその後のBP反応を阻害することを示す。
BP反応に対するattB1およびattB2の存在の効果
研究のこの部分では、その後のBP反応に対する反応混合物中の過剰なattB1およびattB2アダプターの存在の効果を試験した。この問題に取り組むために、異なる量のattB1およびattB2アダプターを鋳型(attB1+CAT+attB2、20ng)に添加し、標準的な条件下で(室温で60分間)BP反応を行った。BP反応後、混合物をケミカルコンピテント大腸菌細胞(例えば、TOP10;Invitrogen Corporation)に化学的に形質転換し、細胞を播種して組換え効率を決定した。
結果:
これらの結果は、過剰なattB1アダプターおよびattB2アダプターの存在は認められた形質転換効率に有意な影響を与えないことを示し、これは、BP反応が反応混合物中でattB1およびattB2の存在によって有意に影響を受けないことを示す。
TOPO連結反応物由来のインヒビターの除去
BP反応物中の産物の使用前にTOPO連結反応物由来のインヒビターの最適な除去方法に取り組むために、種々の処理を評価した。室温で5分間インキュベートした以下の反応混合物を使用してTOPO連結反応を行った。
TOPO連結反応後、BP反応を行う前に反応混合物の7個の異なる試料を以下の1つの条件下で処理した。
(1)1つの反応に0. 6%SDS+3mM EDTA 1μlを37℃で15分間添加した。
(2)4つの反応に0. 6%SDS+3mM EDTA 4μlを37℃で15分間添加し、SNAPを水20μlに精製する。
(3)4つの反応に0. 6%SDS+3mM EDTA 4μl+プロテイナーゼK(2μg/μl)1μlを37℃で15分間添加し、SNAPを水20μlに精製する。
(4)1つの反応に2.5M NaCl 0.8μlを37℃で17分間添加する。
(5)4つの反応に2.5M NaCl 3.2μlを37℃で15分間添加し、SNAPを水20μlに精製する。
(6)4つの反応に2.5M NaCl 3.2μlおよび2μl/μlプロテイナーゼK 1μlを37℃で15分間添加し、SNAPを水20μlに精製する(陽性対照;使用した鋳型 0. 8ng)。
(7)(陰性対照;使用した鋳型なし)。
無塩緩衝液を使用して室温で60分間BP反応を行った。非精製混合物のために、BP反応物10μlあたりTOPO連結反応混合物 1μlを使用した。精製混合物のために、BP反応物10μlあたりTOPO連結反応混合物 5.5μlを使用した。BP反応後、混合物をケミカルコンピテント大腸菌細胞(例えば、TOP10;Invitrogen Corporation)に化学的に形質転換し、細胞を播種して組換え効率を決定した。
結果
これらの結果は、以下のことを示す:(1)精製にはBP反応を効率的に行う必要はないこと;(2)その後BP反応効率を最大にするためにTOPO連結反応後にプロテイナーゼKでの反応混合物の処理は必要ないこと;および(3)反応混合物のSDS処理およびNaCl処理により同一の形質転換効率が得られること(したがって、BP反応の際に同一の結果が得られる)。
BP反応温度の最適化
TOPO連結後のBP反応の最適反応温度を決定するために、種々の温度下で行うBP反応の鋳型としてattB1+CAT+attB2 PCR産物を使用した。BP反応後、混合物をケミカルコンピテント大腸菌細胞(例えば、TOP10;Invitrogen Corporation)に化学的に形質転換し、細胞を播種して組換え効率を決定した。
結果
これらの結果は、BP反応には室温(約20℃〜25℃)が最適な温度であることを示す。
attB1:挿入物:attB2のモル比の最適化
BP反応でのattB1、挿入物、およびattB2鋳型の最適なモル比を決定するために、これらの鋳型を種々のモル比で混合し、上記の最適条件下でBP反応を行った。BP反応後、混合物をケミカルコンピテント大腸菌細胞(例えば、TOP10;Invitrogen Corporation)に化学的に形質転換し、細胞を播種して組換え効率を決定した。
結果
これらの結果は、1:1:1のattB1:挿入物:attB2の比がBP反応に最適であることを示す。
BP反応に対する塩の効果の決定
BP反応溶液中の塩の存在が組換え効率に影響を与えるかどうかを決定するために、無塩緩衝液または塩を含む標準的なBP反応緩衝液中でBP反応を行った。
結果(形成コロニー数)
これらの結果は、BP反応において反応緩衝液中の塩の有無は組換え効率に影響を与えないことを示す。
最適なTOPO連結反応数の決定
次の一連の実験では、1つのTOPO連結反応で精製後のBP反応のための最適な組換え効率が得るのに十分であるかという疑問を試験した。1つのTOPO連結反応を、以下の反応混合物を使用して行った。
attB1およびattB2(各20ng/μl) 0.5μl
CAT(100ng/μl) 1.7μl
500mM Tris 0.5μl
トポイソメラーゼ(1μl/μl) 1μl
dH2O 最終体積を5μlにするのに十分な量
反応混合物を37℃で15分間インキュベートし、0. 6%SDS+3mM EDTA 1μlを添加し、混合物を37℃で15分間インキュベートし、SNAPカラム(上記)を使用して水20μlに精製した。以下のように、このTOPO連結反応産物を使用してBP反応を行った。
標準的なBP反応緩衝液 2μl
pDONR22(300ng/μl) 0.5μl
TOPO連結産物(上記) 5.5μl
BPクロナーゼ 2μl
反応混合物を室温で60分間インキュベートし、2μg/μlのプロテイナーゼK 1μlを添加し、混合物を37℃で15分間インキュベートし、75℃で15分間インキュベートした。反応混合物 4μlをケミカルコンピテント大腸菌細胞(例えば、TOP10;Invitrogen Corporation)に化学的に形質転換し、細胞を播種して組換え効率を決定した。
結果(形成コロニー数)
これらの結果は、TOPO連結反応により、有効なBP反応十分な鋳型が得られることを示す。
精製方法の最適化
SNAP精製カラム(Invitrogen Corporation)またはCONCERT精製システム(Invitrogen Corporation)がTOPO連結後BP反応用の異なる最適な精製鋳型が得られるかどうかを決定するために実験を行った。TOPO連結反応後にいくつかの試料をSNAPカラムを使用して精製し、他の試料をCONCERTプラスミド精製システムを使用して精製すること以外は、上記のようにTOPO連結反応およびBP反応を行った。次いで、精製試料は標準的なBP反応で使用し、次いで、反応混合物を化学的形質転換またはエレクトロポレーションによる形質転換に使用した。形質転換後、細胞を播種して組換え効率を決定した。
結果(形成対照数)
これらの結果は、SNAPおよびCONCERT精製システムは、TOPO連結反応後のBP反応用の精製鋳型が得られるように十分に作用することを示す。
最適な条件
上記の実験結果に基づいて、組み合わせTOPO連結Gateway反応の至適条件が以下であると決定した。
(1)TOPO連結反応
(a)1:1モル比のattB/挿入物(反応体積5μl)
(b)37℃で15分間インキュベートする。
(c)0. 6%SDS+3mM EDTA 1μlを添加し、37℃で15分間インキュベートする。
(d)SNAPカラムまたはCONCERTシステムを使用して20μlのdH2Oに精製する。
(2)BP反応
(a)以下の反応混合物を調製する。
(i)精製TOPO連結産物 5.5μl;
(ii)標準的なBP反応緩衝液 2μl;
(iii)pDONR222(30μg/μl) 0.5μl;
(iv)BPクロナーゼ 2μl;
(b)反応混合物を室温で60分間インキュベートする。
(c)2μg/μlのプロテイナーゼK 1μlを添加する;
(d)37℃で15分間インキュベートする;
(e)75℃で15分間インキュベートする;
(3)形質転換
(a)BP反応由来の反応混合物2μl〜4μlを使用し、化学的形質転換またはエレクトロポレーションを行う。
これらの最適化条件の効率を示すために、TOPO連結およびその後のBP反応に供した種々のサイズのCAT挿入物およびlacZ挿入物を使用して実験を行い、その後形質転換およびプレーティングを行った。
結果
化学的形質転換
電気的形質転換
これらの結果をあわせると、上記の条件は種々のサイズの挿入物における組み合わせTOPO連結ゲートウェイ反応に最適であることが示される。
実施例10
二次PCR法を使用しない哺乳動物発現カセットの構築
関心対象のエレメントおよび遺伝子の調製
以下のプライマー組(以下の表4を参照のこと)および鋳型を、関心対象のエレメントおよび遺伝子のPCR増幅に使用した。
(A)プライマー組:配列番号:51および配列番号:52;鋳型:pcDNA 4/TetO。
PCR産物:5'エレメント。
(B)プライマー組:配列番号:53および配列番号:54;鋳型:pcDNA 3.2/V5。
PCR産物:3'エレメント。
(C)プライマー組:配列番号:55および配列番号:56;鋳型:pcDNA 3.1/CAT。
PCR産物:CAT挿入物。
プラチナTaq DNAポリメラーゼハイフィデリティ(Invitrogen Corporation;Carlsbad、CA)を、PCRに使用した。PCR条件を以下に示した。
94℃で4分間(1サイクル)
94℃で30秒間→55℃で30秒間→68℃で1分間(30サイクル)
68℃で10分間(1サイクル)
4℃(終了)
以下の条件を使用してPCRで作製された断片を精製した。
試薬:SNAP MiniPrepキット(Invitrogen Corporation)
段階
(1)50μlのPCR産物を150μlの結合緩衝液と混合する。十分に混合する。
(2)350μlのイソプロパノールを添加する。十分に混合する。
(3)試料をSNAP MiniPrepカラムにロードする。
(4)14000rpmで1分間遠心分離する。カラム流出物を捨てる。
(5)500μlの洗浄緩衝液を添加し、14000rpmで1分間遠心分離する。カラム流出物を捨てる。
(6)700μlの1×最終洗浄緩衝液を添加し、14000rpmで1分間遠心分離する。カラム流出物を捨てる。
(7)14000rpmで1分間の遠心分離によってカラムを乾燥させる。
(8)カラムを新規の遠心管に移す。50μlのdH2Oをカラムに添加する。室温で2分間〜5分間インキュベートする。14000rpmで1分間遠心分離する。流出物を回収する。
(9)260nmのUV吸収によってDNA濃度を測定する。
TOPO連結試薬
二次PCRを使用した発現カセットの産生のために、以下の結合条件を使用した。
5'エレメント(700bp) 75ng
3'エレメント(350bp) 35ng
500mM Tris(pH7.5) 0.5μl
トポイソメラーゼ(1μg/μl) 0.5μl
CAT挿入物(700bp) 150ng
dH2O 最終体積が5μlになる量
反応を室温で5分間〜15分間行った。配列番号:51および配列番号:54のプライマー組を使用した第2ラウンドのPCRの鋳型として、反応物の半分を使用した。PCR条件は、伸長時間が2分間であること以外は、上記と同一である。PCR後、上記のようにDNAを精製した。精製DNAを、トランスフェクションに使用した。
二次PCRを行わないで発現カセットを産生するために、以下の結合条件を使用した。
5'エレメント(700bp) 510ng
3'エレメント(350bp) 230ng
500mM Tris(pH7.5) 1.5μl
トポイソメラーゼ(1μg/μl) 3μl
CAT挿入物(700bp) 450ng
dH2O 最終体積が15μlになる量
反応を37℃で15分間行った。最終濃度50μg/mlのプロテイナーゼKを添加し、混合物を37℃で10分間インキュベートした。処理DNAを、トランスフェクションのために用意した。
遺伝子発現研究
3つの細胞系列(懸濁液TRex-CHO、接着TRex-CHO、および接着TRex-293細胞系列)を、これらの発現カセットを試験するための細胞系列として使用した。標準的な細胞培養法を使用した。24ウェル細胞培養プレートを使用した。トランスフェクション試薬としてリポフェクタミン2000を使用した。24時間のトランスフェクション後、1μg/mlの最終濃度のテトラサイクリンを添加した。対照実験のために、テトラサイクリンを添加しなかった。溶解前に細胞をさらに24時間インキュベートした。タンパク質の移行のためにウェスタンブロットを使用し、検出には抗V5または抗CAT抗体を使用した。
結果と考察
本研究の目的は、二次PCRを使用することなく発現カセットを作製することができるかを示すことであった。本研究で、二次PCR段階を使用して産生した発現カセットから作製した発現データと二次PCR段階を使用しないで産生した発現カセットを使用して得た発現データとを比較した。二次PCRを使用して産生した発現カセットのために、約1.2μg/ウェルのDNAを24ウェルプレート形式へのトランスフェクションに使用した。二次PCRを使用せずに産生した発現カセットのために、1つの結合反応由来の産物を使用した(約1.2μg/ウェル)。検出データは、二次PCR段階を使用することなく本発明の方法を使用して機能的発現カセットを産生することができることを示した(図30)。
実施例11
Topoツール法を使用したゲートウェイ適合カセットの作製
アダプターの調製
等量の配列番号:57および配列番号:58(上記の表4を参照のこと)を、40mM NaClと混合し、混合物を95℃で5分間変性させ、室温にゆっくり冷却してattB1アダプターを形成した。等量の配列番号:59および配列番号:60(上記の表4を参照のこと)を、40mM NaClと混合し、混合物を95℃で5分間変性させ、室温にゆっくり冷却してattB2アダプターを形成した。
TOPO連結
実施例10に記載のようにCAT挿入物を作製した。結合条件を上記のように最適化した(実施例9および10を参照のこと)。
attB1アダプター(40bp) 10ng
attB2アダプター(40bp) 10ng
500mM Tris(pH7.5) 0.5μl
トポイソメラーゼ(1μg/μl) 1μl
CAT挿入物(700bp) 170ng
dH2O 最終体積が5μlになる量
反応を37℃で15分間行った。それぞれ最終濃度0.1%および0.5mMの最終濃度のSDSおよびEDTAを添加した。混合物を37℃で15分間インキュベートした。
精製
処理した混合物に水(15μl)を添加した。SNAP MiniPrepキット(Invitrogen)を使用してDNAを精製した。
段階
(1)処理した産物を結合緩衝液 60μlと混合する。十分に混合する。
(2)イソプロパノール 140μlを添加する。十分に混合する。
(3)試料をSNAP MiniPrepカラムにロードする。
(4)14000rpmで1分間遠心分離する。カラム流出物を捨てる。
(5)洗浄緩衝液500μlを添加し、14000rpmで1分間遠心分離する。カラム流出物を捨てる。
(6)1×最終洗浄緩衝液 700μlを添加し、14000rpmで1分間遠心分離する。カラム流出物を捨てる。
(7)14000rpmで1分間の遠心分離によってカラムを乾燥させる。
(8)カラムを新規の遠心管に移す。dH2O 20μlをカラムに添加する。室温で2分間〜5分間インキュベートする。14000rpmで1分間遠心分離する。流出物を回収する。
BP反応
BP反応緩衝液 2μl
精製産物 5.5μl
pDONR22(300ng/μl) 0.5μl
BPクロナーゼ 2μl
反応混合物を室温で60分間インキュベートし、1μlのプロテイナーゼK(2μl/μl)を添加した。混合物を37℃で15分間インキュベートし、その後75℃で15分間インキュベートして酵素を不活化した。
形質転換
処理した混合物をTOP10コンピテント細胞(化学的)に形質転換するか、ElectroMaxコンピテント細胞にエレクトロポレーションした。細胞をLP-カナマイシンプレートに播種し、37℃で一晩インキュベートする。コロニーを計数する。挿入物を確実にこれらのコロニー中に存在させるために、コロニーPCRを行うためのプライマー組(配列番号:61および配列番号:62)を設計した。挿入物が存在する場合、約700bpのバンドのPCR産物が産生され、しかし挿入物が存在しない場合、PCR産物のバンドは2.2kbのサイズである。
結果と考察
本研究では、TOPOツールの付着末端を使用して産生したPCR産物がattB1およびattB2に直接結合することができることを示すことを所望した。結合した産物をBP組換え反応に直接使用して、GATEWAY(商標)エントリークローンを作製することができる(表5)。
挿入物を確実にこれらのコロニー中に存在させるために、18個の陽性コロニーおよび2個の陰性コロニーを選択してPCRを行った。PCRの結果は、チェックした18個のコロニー全てに正確なサイズの産物が存在することを示した(図31)。
本発明を理解を明確にすることを目的として例示および例によって幾らか詳細に説明しており、本発明またはその特定の態様の範囲に影響を与えることなく広範および等価な範囲の条件、処方物、および他のパラメーター内での本発明の修正または変更によって本発明を行うことができること、およびこのような修正形態または変更形態が添付の特許請求の範囲の範囲内に含まれることが意図されることが当業者に明白である。
以下の同一所有権者の同時係属中の米国特許出願の全体が本明細書中で参考として組み入れられる。2000年12月8日提出の米国特許仮出願番号第60/254,510号;2000年12月11日提出の米国特許出願番号第09/732,914号、2001年3月21日提出の米国特許仮出願番号第60/291,972号;2001年9月14日提出の米国特許仮出願番号第60/318,902号;および2001年9月28日提出の米国特許仮出願番号第60/326,092号。
本明細書記載の全ての刊行物、特許、および特許出願は、本発明に属する当業者の技術レベルの指標である。各刊行物、特許、または特許出願が特異的且つ個別に参考として組み入れられるかのような程度に本明細書中で参考として組み入れられる。